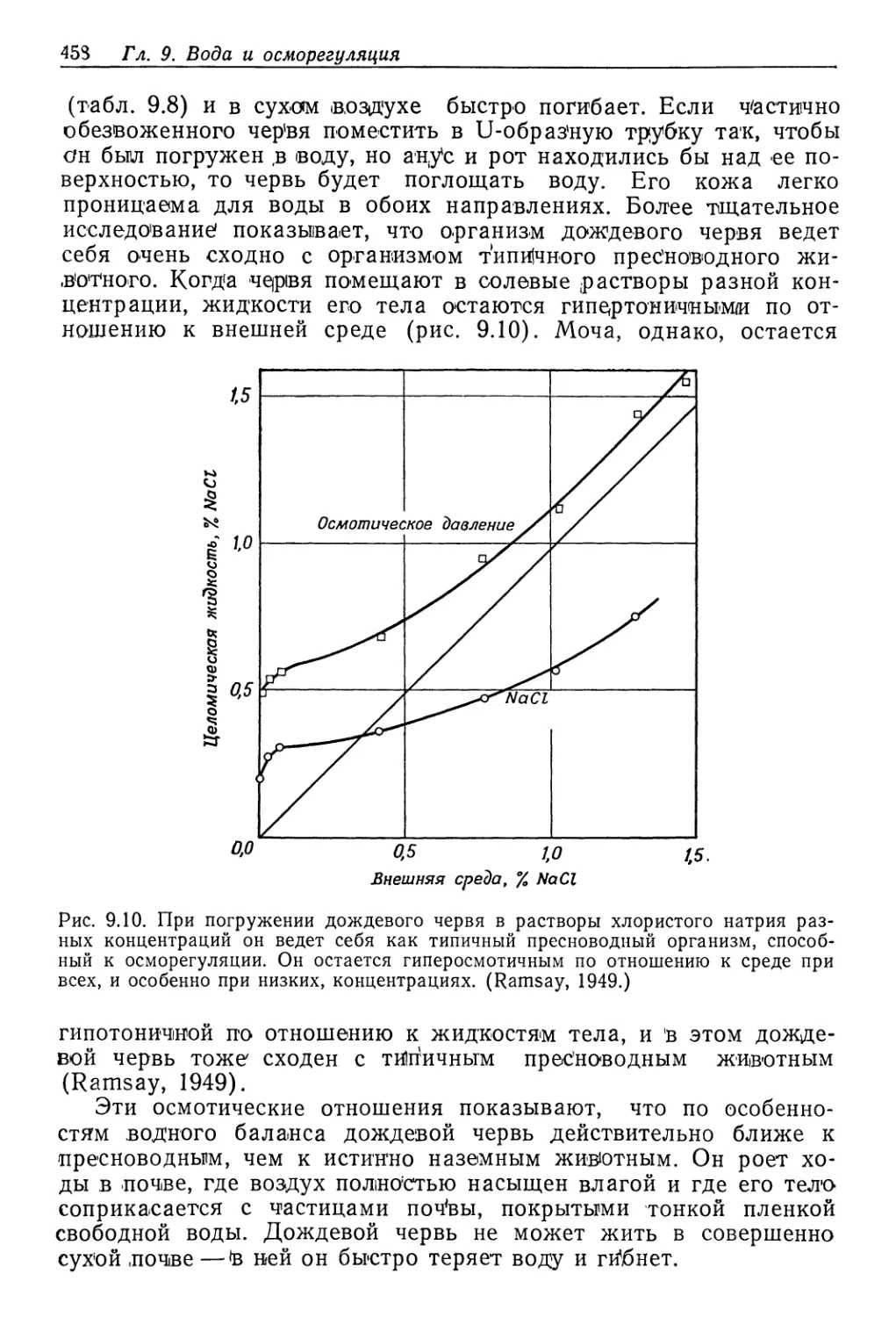
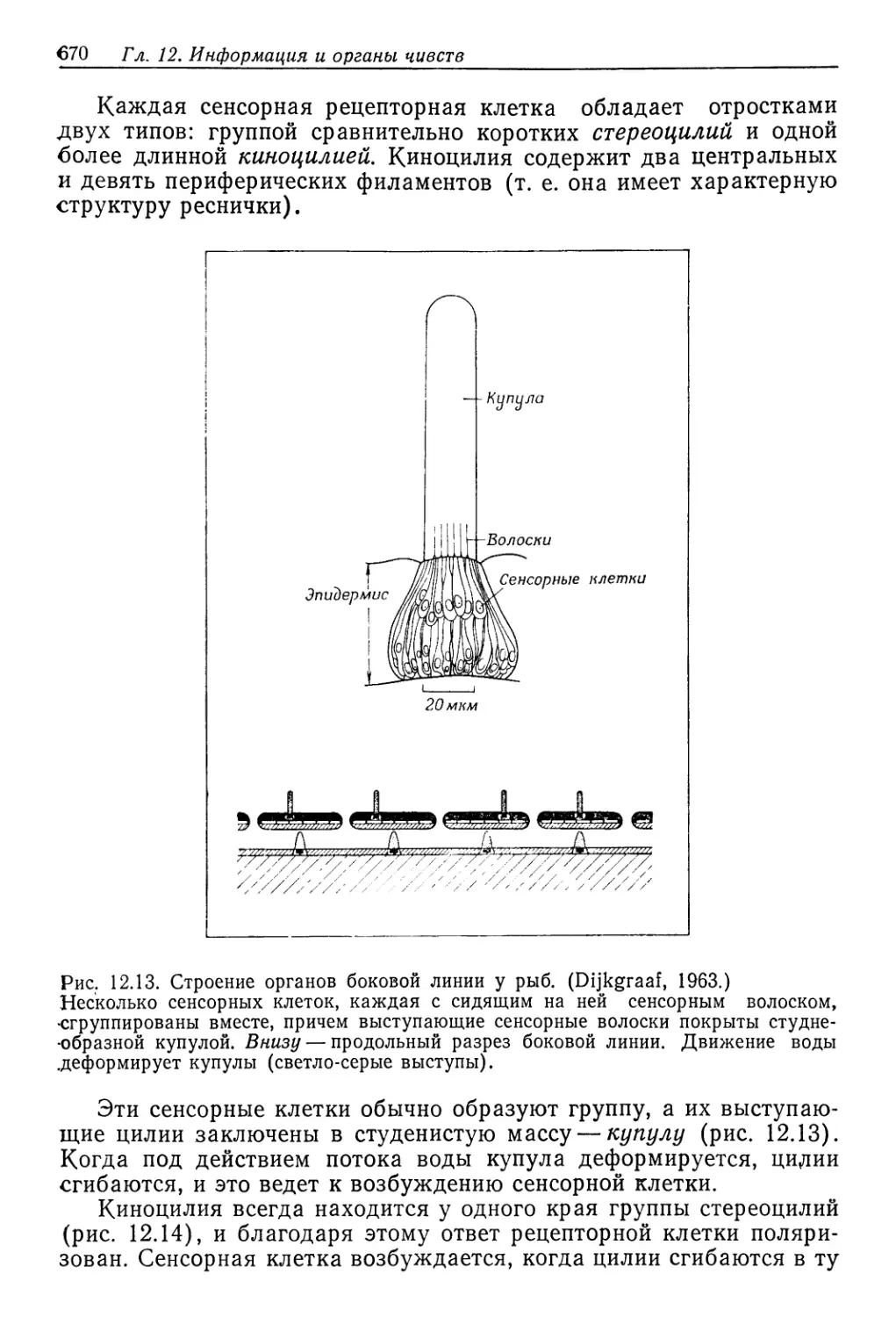
/




















































































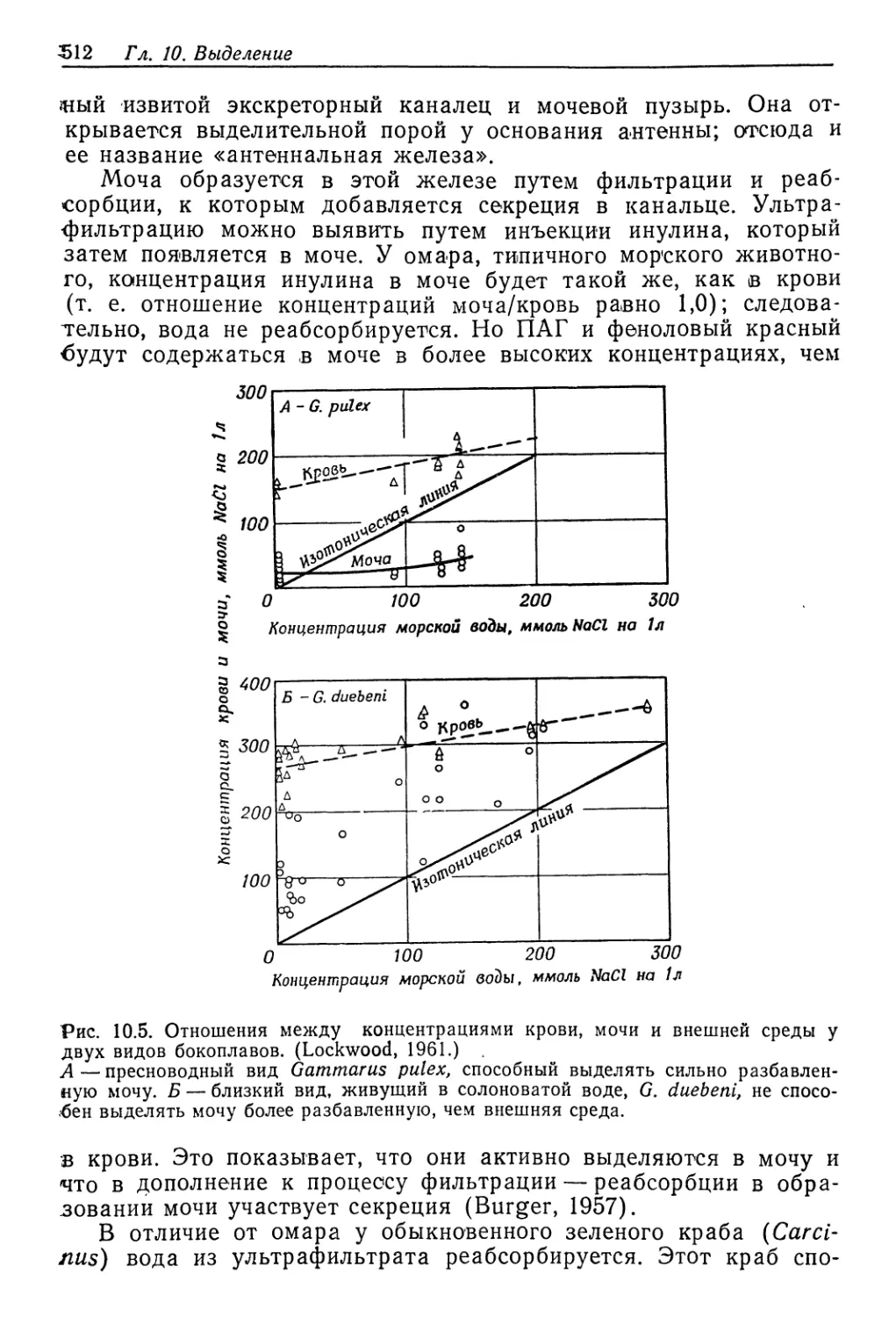

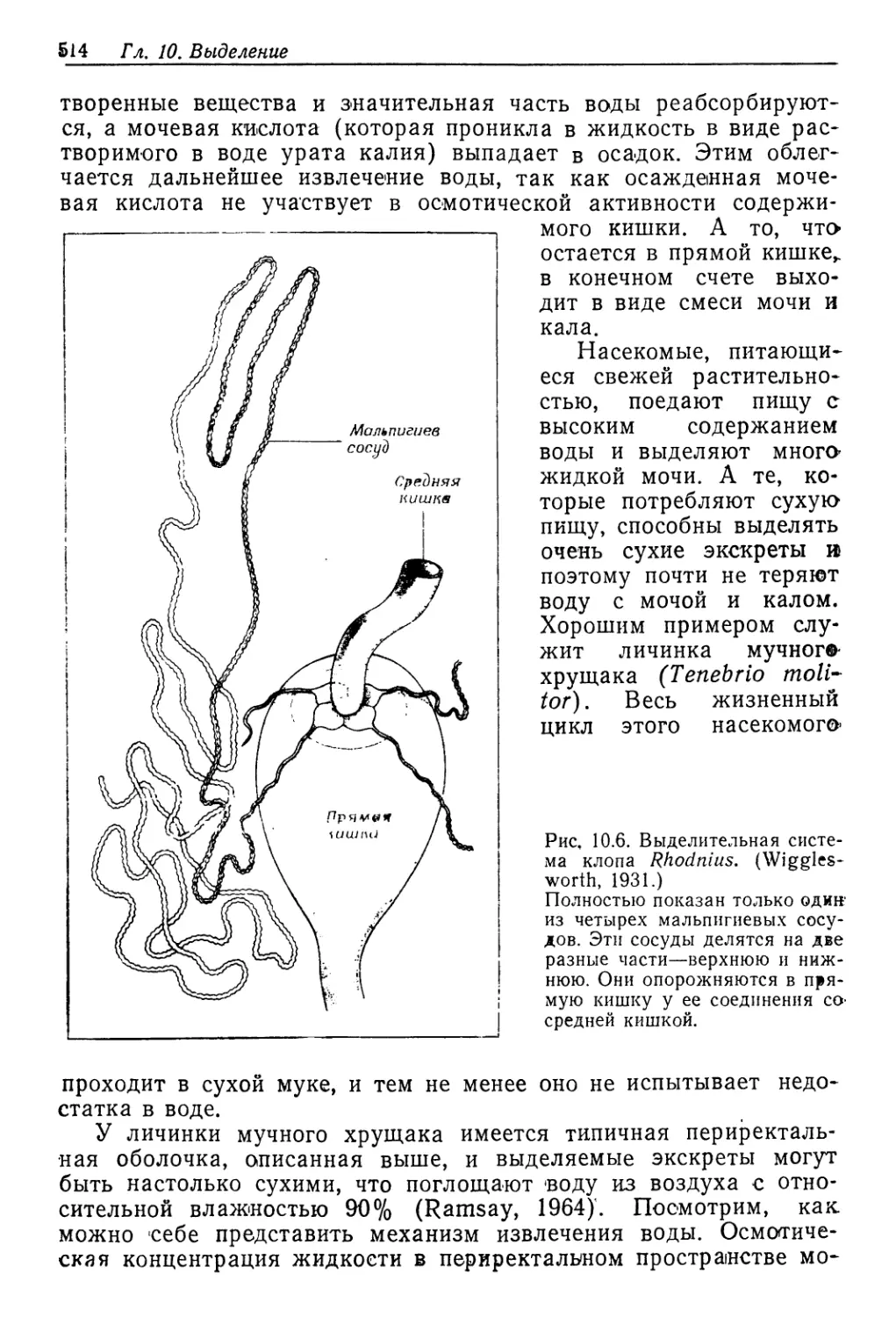




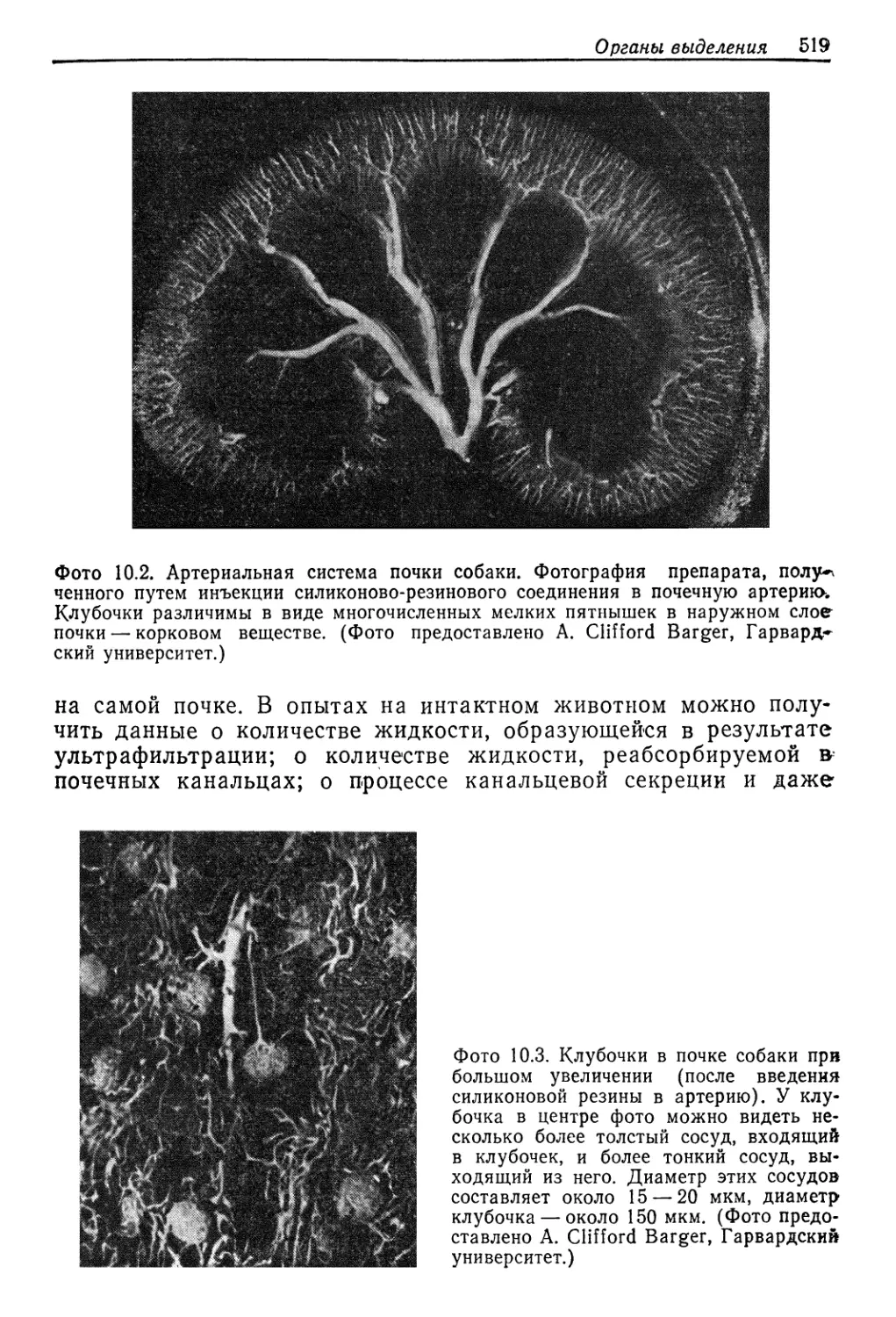



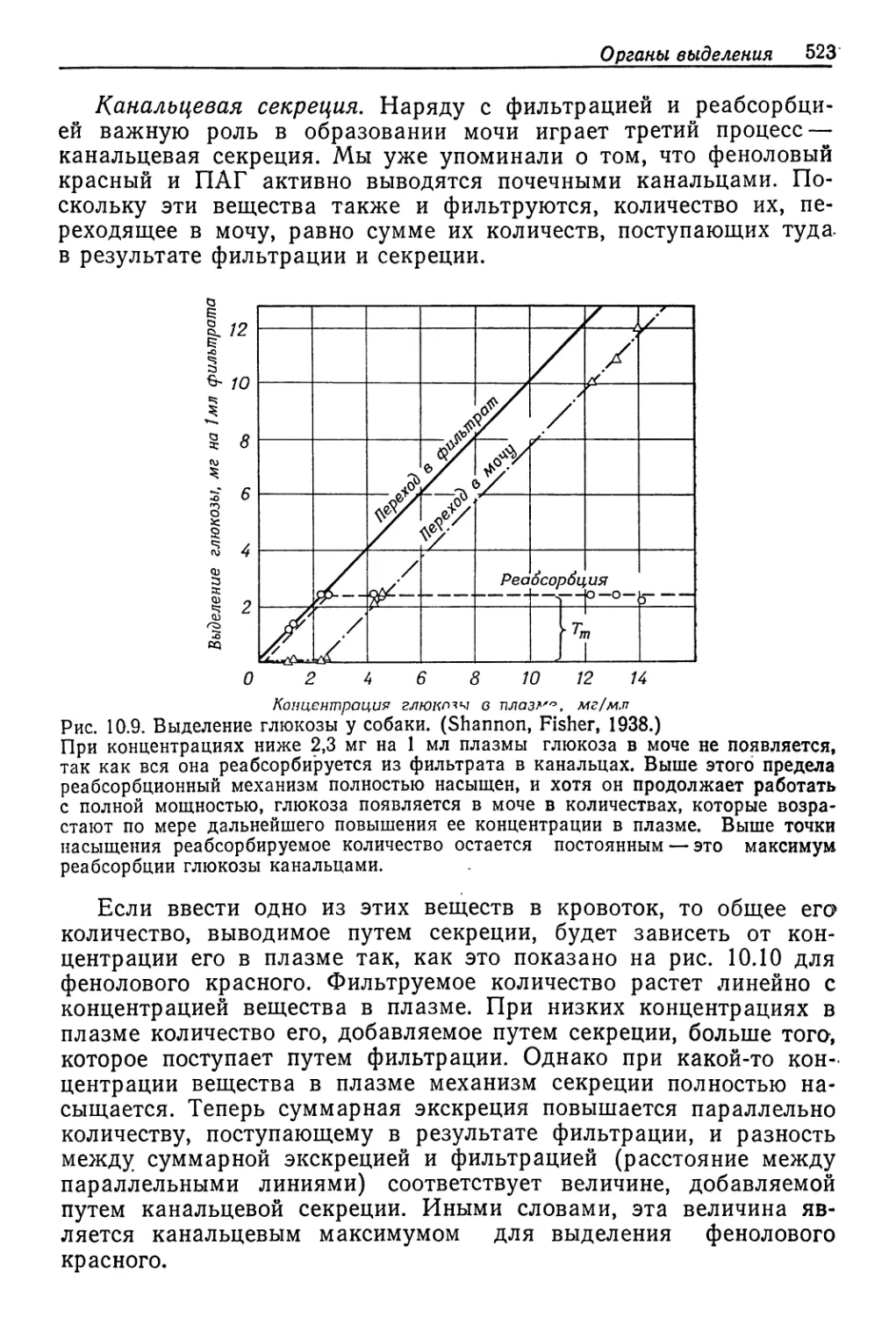





















































































































































































































































Author: Шмидт-Ниельсен К.
Tags: физиология животных биофизика, биохимия и физиология животных и человека биология зоология
Year: 1982




Text
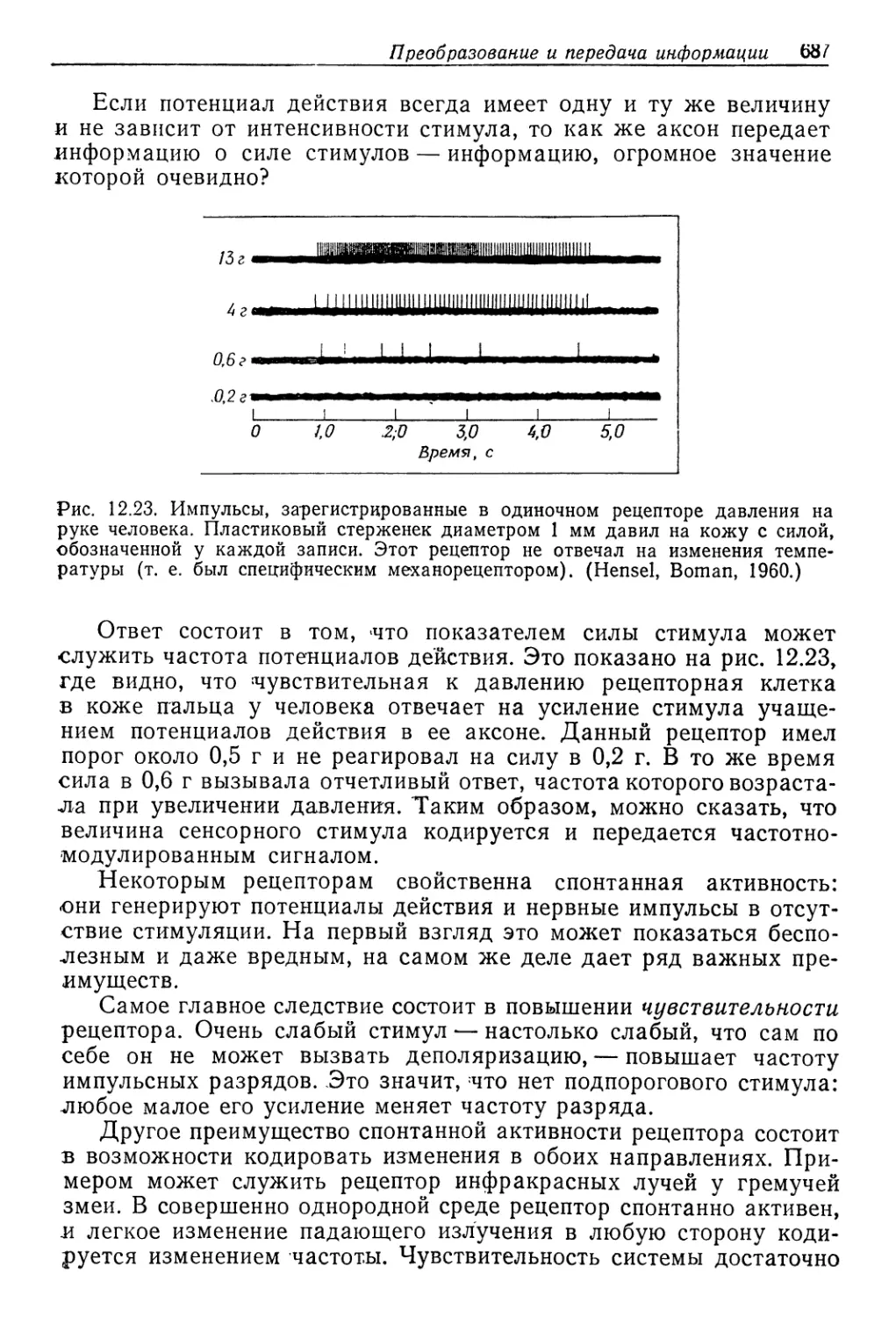
b’i
К. Шмидт-Ниельсен
ФИЗИОЛОГИЯ
животных
ПРИСПОСОБЛЕНИЕ И СРЕДА
ci
Second edition
Knut Schmidt-Nielsen
James B. Duke Professor of Physiologyr
Department of Zoology, Duke-
University
ANIMAL
PHYSIOLOGY
ADAPTATION AND ENVIRONMENT
Cambridge University Press
Cambridge London New York Melbourne
К. Шмидт-Ниельсен
ФИЗИОЛОГИЯ
животных
ПРИСПОСОБЛЕНИЕ И СРЕДА
Книга вторая
Перевод с английского
канд. биол. наук М. Д. ГРОЗДОВОЙ
и
канд. физ.-мат. наук Г. И. РОЖКОВОЙ
под редакцией
акад. Е. М. КРЕПСА
Москва «Мир» 1982
ББК 28.901
Ш 73
УДК 591.1
Шмидт-Ниельсен К.
ТТТ 73 Физиология животных: Приспособление и среда, Книга 2:
Пер. с англ./Перевод Гроздовой М. Д., Рожковой Г. И.; Под
ред. и с предисл. Е. М. Крепса. — М.: Мир, 1982. — 384 с., ил.
Книга известного американского физиолога посвящена вопросам сравнительной
физиологии, рассматриваемым в экологическом аспекте. Русское издание выходит в
двух книгах. Во вторую книгу вошли следующие разделы: вода; движение, информа-
ция и интеграция. В приложении дана система классификации животных.
Предназначена для физиологов, зоологов, экологов, гидробиологов, специалистов
по охране окружающей среды, для преподавателей, аспирантов и студентов универ-
ситетов, педагогических, сельскохозяйственных и рыботехнических институтов.
21008—177
Шп../пп оо 177-82, ч. 1 2007020000
Urrl tv 1j —OZ
ББК 28.901
591.4
Редакция литературы по биологии
© Cambridge University Press 1975, 1979
© Перевод на русский язык, «Мир», 1982
ЧАСТЬ ЧЕТВЕРТАЯ
ВОДА
ГЛАВА 9
ВОДА И ОСМОРЕГУЛЯЦИЯ
В самом общем виде живой организм можно описать как вод-
ный раствор, заключенный в оболочку — поверхность тела. Объем
организма и концентрация растворенных веществ должны сохра-
няться постоянными в довольно узких пределах, так как для оп-
тимального функционирования организма требуется совершенно
определенный и относительно неизменный состав жидкостей те-
ла. Значительные отклонения от нормального состава обычно не-
совместимы с жизнью.
Перед животными стоит задача поддерживать надлежащие
концентрации растворенных веществ в жидкостях тела, несмотря
на то что они почти всегда отличаются от соответствующих кон-
центраций во внешней среде. Разница концентраций стремится
выравняться, нарушая требуемое постоянство внутренней среды.
Животные могут све'сти к минимуму возникающие трудности,
уменьшив градиенты или проницаемость; и действительно, они
применяют оба этих способа. Тем не менее всегда происходит не-
которая диффузионная утечка, и постоянство внутренней среды
не может сохраняться, если организм не создает противоток, в
точности равный этой утечке. Такой протйвоток требует затраты
энергии.
Задачи поддержания постоянных концентраций воды и рас-
творенных в ней веществ меняются в зависимости от окружаю-
щей среды, и они совершенно различны в морокой воде, в прес-
ной воде и на суше. Поэтому удобно рассмотреть эти условия
жизни по отдельности, проанализировать основные физиологиче-
ские проблемы в каждой среде и выяснить, как их решают раз-
ные животные.
В этой главе речь пойдет сначала о водных, а затем о назем-
ных животных.
422 Гл. 9. Вода и осморегуляция
ВОДНАЯ СРЕДА
Прежде чем рассматривать физиологические проблемы, свой-
ственные той или иной среде, полезно ознакомиться с ее наибо-
лее важными физическими и химическими -свойствами.
Больше двух третей (71%) поверхности Земли покрыто водой,
главным образом океаном; общее количество пресной воды в озе-
рах и реках составляет меньше 1% от площади и 0,01% от объе-
ма морской воды (Sverdrup et al., 1942; Hutchinson, 1957, 1967).
На суше жизнь сосредоточена в тонкой пленке на поверхности
земли и непосредственно под ней и над ней; в воде же организмы
живут не только на дне, но и во всей толще водных масс до океан-
ких глубин более 10 000 м.
Всякая вода содержит растворенные вещества — соли, газы,
небольшие количества органических соединений, разные примеси;
первостепенное физиологическое значение имеет температура
воды. В этой главе речь пойдет главным образом о растворен-
ных солях; значение растворенных газов было рассмотрено в
гл. 1, влияние температуры — в гл. 7 и 8. Краткий обзор свойств
растворов, осмотического давления и связанных с ними вопросов
сделан в Приложении Д.
Морская вода содержит 3,5% солей (т. е. в 1 л морской воды
содержится 35 г солей)1. Ионы представлены главным образом
натрием и хлором; в значительных количествах присутствуют
также магний, сульфат и кальций (табл. 9.1). Общая концентра-
ция солей несколько варьцрует в зависимости от географическо-
го положения. Например, в Средиземном море содержание со-
лей доходит почти до 4%, так как сильное испарение не компен-
сируется достаточным притоком пресной воды из рек. В других
местах, особенно в прибрежных районах, содержание солей не-
сколько ниже, чем в открытом океане, но относительные количе-
ства растворенных ионов остаются почти постоянными и очень
близки к соотношениям, указанным в табл. 9.1.
В пресной воде в отличие от морской содержание растворен-
ных веществ сильно варьирует. Очень небольшие количества со-
лей есть даже в дождевой воде, но ее состав сильно изменяется
по мере того, как вода течет по поверхности земли и проникает
в почву. Источником солей в дождевой воде служит море; брыз-
ги из океана испаряются и частицы соли уносятся токами возду-
ха, иногда далеко в глубь суши, и там (с дождем падают на зем-
лю. Если вода течет по твердой, нерастворимой породе, напри-
мер по граниту, в нее почти ничего не переходит, и такую воду
называют мягкой. Если же вода просачивается в пористый из-
вестняк, она растворяет сравнительно много солей кальция, и
1 Нельзя точно определить количество растворенных солей, выпаривая воду
и взвешивая сухой остаток, так как условия высушивания влияют на содержание
кристаллизационной воды, а также на вес бикарбонатов и карбонатов.
Водная среда 423
Таблица 9.1
Состав морской воды. Кроме перечисленных ионов,
морская вода содержит небольшие количества
практически всех элементов, встречающихся на Земле.
(Potts, Раггу, 1964)
Ион Количество на I л морс- кой воды Количество на 1 кг воды!)
ммоль г ммоль г
Натрий 470,2 10,813 475,4 10,933
Магний 53,57 1,303 54,17 1,317
Кальций 10,23 0,410 10,34 0,414
Калий 9,96 0,389 10,07 0,394
Хлор 548,3 19,440 554,4 19,658
Сульфат 28,25 2,713 28,56 2,744
Бикарбонат 2,34 0,143 2,37 0,145
’) В термодинамике концентрации вычисляются на 1 кг воды и
обозначаются как моляльность раствора. Краткие сведения по фи-
зике растворов даны в Приложении Д.
тогда ее называют жесткой. Общее содержание солей в .пресной
воде колеблется от величин менее 0,1 ммоль/л до более чем
10 ммоль/л, а относительные количества разных ионов могут
варьировать в очень широких пределах. Это имеет физиологиче-
ское значение, особенно в тех /случаях, когда главными ионами
являются магний и сульфат. Состав разных типов мяг’кой и
жесткой воды приведен в табл. 9.2.
Таблица 9.2
Типичный состав мягкой, жёсткой и материковой соленой воды
в миллимолях на 1 кг воды; последовательность ионов та же,
что и в табл. 9.1. (Пересчитано по данным Livingstone, 1963)
Ион Мягкая озерная вода!) Речная вода2) Жесткая речная водаЗ) Соленая вода!) Вода Мертвого моря5)
Натрий 0,17 0,39 6,13 640 840
Магний 0,15 0,21 0,66 6 2302
Кальций 0,22 0,52 5,01 32 583
Калий — 0,04 0,11 16 152
Хлор 0,03 0,23 13,44 630 6662
Сульфат 0,09 0,21 1,40 54 8,4
Бикарбонат 0,43 1,11 1,39 3 Следы
‘) Озеро Ниписсинг, Онтарио.
2) Средний состав североамериканских рек.
3) Река Тускароэс, штат Огайо.
4) Бэд Уотер («Гнилая вода») в Долине Смерти, штат Калифорния.
5) Мертвое море, Израиль. Эта вода содержит 118 ммоль брома на
1 кг Н2О.
424 Гл. 9. Вода и осморегуляция _
Природная дождевая .вода имеет .слегка кислую реакции. Это
обусловлено тем, что в ней растворяемся атмосферная двуокись
углерода, и pH в таких случаях достигает .приблизительно 5,6.
Впрочем, на северо-востоке США pH дождевой воды часто быва-
ет ниже — всего лишь 4. Это объясняют тем, что сернистый газ
и закись азота, выделяющиеся при сгорании ископаемого топли-
ва, образуют сильные кислоты, которые снижают pH осад'ков
(Galloway et al., 1976).
Высокая кислотность дождя встречается не только в США.
На большие области Скандинавии, лежащие с .подветренной сто-
роны от промышленных районов Центральной Европы, пролива-
ется кислая дождевая вода, которая создает серьезные проблемы
для /жизни животных, а в некоторых местах сильно вредит попу-
ляциям рыб. Это вредное влияние особенно заметно в Норвегии,
где численность лососей в южных частях страны быстро падает,
хотя в более отдаленных районах такого снижения не наблюда-
ется. Точно так же во многих озерах южной Норвегии пропала
кумжа, и число таких озер быстро увеличивается.
Почему Норвегия особенно страдает от кислой реакции дож-
девой воды, в то время как в других областях, расположенных
ближе к промышленным районам, этого не отмечается? Дело
здесь в том, что во многих частях южной Норвегии подстилаю-
щая порода состоит из гранита, очень устойчивого против вы-
ветривания. В результате вода в реках и,озерах чрезвычайно
мягкая и практически не содержит минеральных веществ, кото-
рые нейтрализовали бы кислоты.
Степень кислотности, которую .может выно|сить та или иная
рыба, связана не только с pH — очень важно также .содержание
растворенных солей. Кислота, вероятно, подавляет активное по-
глощение натрия, поскольку в Экспериментах показано, что этот
процесс тормозится высокой кислотностью воды (Maetz et al.,
1976). Это согласуется с данными о том, что рыба исчезает сна-
чала из озер с чрезвычайно низким содержанием растворенных
солей и что она гибнет особенно сильно весной, во время
снеготаяния, когда с потоками талой воды в практически незабу-
ференную воду внезапно устремляется волна кислоты (Leivestad,
Muniz, 1976).
В некоторых местах вода внутренних водоемов содержит
очень много солей. Большое Соленое озеро в штате Юта насы-
щено хлористым натрием, который кристаллизуется на его бере-
гах. Мертвое море в Израиле тоже насыщено, но .здесь преобла-
дают ионы магния и хлора, а сернокислый кальций кристалли-
зуется. В Большом Соленом озере не может жить никакая рыба,
но некоторые животные здесь процветают, например рачок Arte-
mia. Насыщенный солевой раствор Мертвого моря имеет иной
состав. В нем нет ни высших животных, ни растений, выживают
только микроорганизмы. Вода некоторых источников содержит
Водная среда 425
необычайно много солей, но такие местообитания хотя и пред-
ставляют интерес, однако по сравнению с большими массами
морской и пресной воды имеют мало значения.
Солоноватые воды встречаются в прибрежных районах, где
морская вода смешивается с пресной. В устье крупной реки пре-
сная вода разбавляет океанскую воду на большом расстоянии,
а при сильных приливах соленая вода заходит далеко в реку.
В {Этой области — в речном эстуарии — степень .солено'сти быстро
меняется с приливным диплом — часто от почти пресной до почти
неразбавленной морокой воды. В больших замкнутых простран-
ствах, как, например, в Балтийском море, дело обстоит иначе.
Соленость Балтики у западных берегов Швеции составляет около
3% и постепенно уменьшается до 0,5% и ниже в самой северной
части. В этой обширной области географический -градиент /соле-
ности относительно устойчив и существует почти незаметный пе-
реход от солоноватой воды к дресной.
Трудно точно сказать, еде морская вода становился солонова-
той и где; на другом конце шкалы, очень разбавленную солоно-
ватую воду практически можно уже считать пресной. Согласно
общепринятому определению, солоноватой называют воду с со-
леностью от 3,0 до 0,05%. Солоноватая вода имеет большое био-
логическое значение. Она создает .барьер для распространения
многих морских животных, с одной стороны, и .пресноводных —
с другой, а также образует интересный переход между морской
и пресноводной средами обйтания. Но в географическом смысле
значение солоноватой воды невелико, так как она покрывает
меньше 1% земной поверхности.
ОПРЕДЕЛЕНИЯ
Некоторые водные животные переносят большие .колебания
концентрации солей в воде; их называют эвригалинными (от
греческого эурис — широкий -и галос— соль). Другие животные
обладают ограниченной толерантностью к изменениям концент-
рации солей; их называют стеногалинными (от греческого сте-
нде— узкий). Морское животное, которое может проникать в .со-
лоноватые воды и выживать в них — эвригалинное. В высшей
степени эвригалинное животное может даже находиться более
или менее долго в пресной воде. ЭвригалИнными называют так-
же тех пресноводных животных, которые переносят значительное
повышение солености воды. Стеногалинный организм, морской
или пресноводный, может переносить изменение концентрации со-
лей в воде только в узких пределах.
Между эвригалинными и стеногалинными животными нет
резкой грани, и нет общепринятого определения, которое позво-
ляло бы отнести данное животное к той или другой группе.
426 Гл. 9. Вода и осморегуляция
У большинства морских беспозвоночных тканевая жидкость
имеет такое же .осмотическое давление, как и морская вода; они
изоосмотичны со средой .своего обитания (от греческого изос —
равный). На изменения концентрации среды животное может
реагировать одним из двух способов. Первый состоит в изменении
концентрации жидкостей тела в соответствии с концентрацией сре-
ды — животное остается изоосмотичным со средой; таких живот-
ных называют осмоконформерами. При втором способе живот-
ное поддерживает или регулирует свою осмотическую концентра-
цию несмотря на изменение среды; таких животных называют
осморегуляторами. Например, морокой краб, сохраняющий высо-
кую концентрацию солей в жидкостях тела после того, как его
перенесли в разбавленную солоноватую воду, .является типичным
осморегулятором.
Концентрации отдельных веществ, растворенных в жидкостях
тела животного, обычно значительно отличаются от их концент-
раций во внешней среде, даже если животное изоосмотично со
средой. Эти различия, как правило, тщательно регулируются, что
носит название ионной регуляции. .В той или иной степени ионная
регуляция, по-видимому, осуществляется у всех живых организ-
мов, как у осморегуляторов, так и у осмоконформеров.
У пресноводных животных жидкости тела имеют более высо-
кую осмотическую концентрацию, чем окружающая среда; та-
ких животных называют гиперосмотичными. Если же в организ-
ме осмотическая концентрация ниже, чем в среде (как, напри-
мер, у морских костистых рыб), то животное называют гипоос-
мотичным.
Концентрацию растворенного вещества обычно выражают в
единицах молярности — ’в молях (моль) на литр раствора.
В биологическом контексте часто удобно пользоваться единицей
миллимоль (ммоль). Например, концентрация раствора
0,5 моль/л равна 500 ммоль/л.
Осмотическую концентрацию раствора можно (Выражать как
осмолярность (в осмолях на литр). Осмолярность раствора зави-
сит от числа растворенных частиц; ее можно определить, не зная,
какие именно вещества содержатся в растворе. Осмолярность
раствора неэлектролита (например, сахарозы или мочевины)
равна молярной концентрации. У раствора .электролита (напри-
мер, хлористого натрия, который в растворе диссоциирует на
Na+ и С1~) осмотическая концентрация выше, чем молярная.
Степень диссоциации электролита зависит от его концентрации
и от взаимодействия его с другими ионами. Общую осмотиче-
скую концентрацию определить нетрудно. В биологических ис-
следованиях для этого чаще всего измеряют понижение точки
замерзания или давление паров раствора. При рассмотрении осмо-
регуляции у животных в большинстве случаев значение имеет
осмотическая концентрация, а не подробный перечень растворен-
ных веществ.
Водные беспозвоночные 421
Осмотическая концентрация обычной морской воды, содержа-
щей в одном литре около 470 ммоль натрия, около 550 ммоль
хлора и значительные количества двухвалентных ионов (магния
и сульфата), составляет около 1000 мосмоль на литр.
Термин изотонический применяемся в ином смысле. Мы гово-
рим, что живая клетка изотонична с данным раствором, если
она и не набухает, и не сморщивается в нем. Например, .эритро-
циты млекопитающих, взвешенные в растворе хлористого натрия
концентрацией 150 ммоль/л (около 0,9%), сохраняют свои раз-
меры, форму и объем. Но в изоосмотическом растворе мочевины
(0,3 моль/л) они быстро набухают и лопаются. Раствюр мочеви-
ны хотя и изоосмотичен, но не изотоничен. Растворенная моче-
вина быстро проникает сквозь мембрану эритроцита, так что
концентрации ее внутри и вне клеТки уравниваются, но электро-
литы не выходят из клетки, и клетка ведет себя так, как если
бы она находилась в дистиллированной воде. Вследствие разни-
цы осмотических концентраций вода входит внутрь, и клетка на-
бухает и лопается. Изоосмотичность определяется в понятиях фи-
зической химии, а изотоничность — это описательное слово, ха-
рактеризующее поведение клеток в данном расТворе.
ВОДНЫЕ БЕСПОЗВОНОЧНЫЕ
МОРСКИЕ ЖИВОТНЫЕ
У большинства морских беспозвоночных осмотическая кон-
центрация жидкостей тела такая же, как у окружающей мор-
ской воды; эти животные — осмоконформеры. Этим устраня-
ется большое физиологическое «затруднение», связанное с ос-
мосом: таким животным не приходится решать проблем, связан-
ных с осмотическим движением воды. •
Хотя, как правил'о, морские беспозвоночные являются о’смо-
конформерами, это не значит, что состав веществ, растворенных
в жидкости их тела, такой же, как в морской воде. Напротив,
имеются характерные различия, которые животные должны со-
хранять, для чего требуется активное регулирование концентра-
ции ионов.
КОНЦЕНТРАЦИЯ ИОНОВ В ЖИДКОСТЯХ ТЕЛА
Концентрации наиболее важных ионов в крови некоторых
беспозвоночных приведены в табл. 9.3. У одних животных эти
концентрации сходны с концентрациями в морской воде, у дру-
гих существенно отличаются. НапрИ'мер, у некоторых беспозво-
ночных концентрация магния такая же, как в морской воде, а у
других она гораздо ниже. То же самое относится к сульфату.
428 Гл. 9. Вода и осморегуляция
Таблица 9.3
Концентрации важнейших ионов (в миллимолях на 1 кг воды)
в морской воде и в жидкостях тела некоторых морских животных.
(Potts, Parry, 1964)
Na Mg Са К Cl SO4 Белок (г/л)
Морская вода 478,3 54,5 10,5 10,1 558,4 28,-8 —
Медуза Aurelia 474 53,0 10,0 10,7 580 15,8 0,7
Полихета Aphrodite 476 , 54,6 10,5 10,5 557 26,5 0,2
Морской еж Echinus 474 53,5 10,6 10,1 557 28,7 0,3
Мидия (Mytilus) 474 52,6 11,9 12,0 553 28,9 1,6
Кальмар Loligo 456 55,4 10,6 22,2 578 8,1 150
Равноногий рак Ligia 566 20,2 34,9 13,3 629 4,0 ——
Краб Maia 488 44,1 13,6 12,4 554 14,5 —•
Зеленый краб (Carcinus) 531 19,5 13,3 12,3 557 16,5 60
Норвежский омар (Nephrops) 541 9,3 11,9 7,8 552 19,8 33
Миксина (Myxine) 537 18,0 5,9 9,1 542 6,3 67
Такие различия могут поддерживаться только при условии,
что поверхность тела, в'ключая тонкую поверхностную оболочку
жабр, относительно непроницаема для данного иона. Но неко-
торые кол’ичества этих ионов все равно проникают внутрь, так
как ни одно животное не бывает полностью непроницаемым и вся
поглощаемая пища содержит растворенные вещества. Поэтому
животные должны обладать механизмом, который удалял бы
одни ионы, а концентрацию других удерживал на уровне более
высоком, чем ® воде. Регулируемое выведение растворенных ве-
ществ — одна из главных функций почек и иных выделителЫных
органов.
Некоторые из ,животных, перечисленных в табл. 9.3, поддер-
живают концентрацию сульфата на уровне вдвое ниже, чем в
морской воде. Такая большая разница явно указывает на за-
трудненное поглощение этого иона и на его выведение — иными
словами, на активную регуляцию его содержания в организме.
Если же внутренняя концентрация лишь ненамного отличается
от .концентрации в морской воде, то в таких случаях труднее ре;
шить, объясняется ли эта разница регулированием. В частности,
на распределение ионов по обе стороны полупроницаемой мем-
браны значительное влияние (именуемое эффектом Доннана, см.
Приложение Д) оказывают белки. Поэтому различие в ионных
концентрациях не обязательно указывает на активную регуля-
цию содержания данного иона.
Английский исследователь Дж. Д. Робертсон .(Robertson,
1957) с помощью .простого .приема обошел трудности, ,с которы-
ми Связана точная оценка роли белков и эффекта Доннана в
распределении различных ионов. Он .помещал .пробу в полупро-
Водные беспозвоночные 429
ницаемый целлофановый мешочек, который затем погружал в
морскую воду. Поскольку соли и вода проходят через целлофан,
а бел’йи не проходят, |концентрации ионов .внутри {мешочка в со-
стоянии равновесия /будут несколько иными, чем в морской «воде;
эта разница создается доннановоки’м (Эффектом белков. Такая
процедура называется диализом. Концентрация данного иона
внутри «мешочка служит «нулевым уровнем», и концентрацию то-
го же иона в теле животного выражают в процентах от ^этой ве-
личины. Если есть измеримая разница между обе’имй величина-
ми, то она должна быть результатом активного регулирования
концентрации данного иона. Некоторые результаты таких опы-
тов представлены в табл. 9.4.
Таблица 9.4
Регуляция ионов у некоторых морских беспозвоночных.
Концентрации в плазме или целомической жидкости, выраженные в процентах
от концентрации в жидкости тела, диализованной против морской воды.
(Robertson, 1957)
Животное Na Mg Са К С1 so4
Кишечнополостные
Aurelia aurita 99 97 96 106 104 47
Иглокожие
Marthasterias glacialis 100 98 101 111 101 100
Оболочники
Salpa maxima 100 95 96 113 102 65
Кольчатые черви
Arenicola marina 100 100 100 104 100 92
Сипункулиды
Phascolosoma vulgare 104 69 104 НО 99 91
Членистоногие
Maia squinado 100 81 122 125 102 66
Dromia vulgaris 97 99 84 120 103 53
Carcinus maenas НО 34 108 118 104 61
Pachygrapsus marmoratus^ 94 24 92 95 87 46
Nephrops norvegicus ИЗ 17 124 77 99 69
Моллюски
Pecten maximus 100 97 103 130 100 97
Neptunea antiqua 101 101 102 114 101 98
Sepia officinalis 93 98 91 205 105 22
’) Этот грапсоидный краб — единственное из упомянутых здесь животных, гипоосмотич-
ное по отношению к морской воде концентрации в морской воде). (концентрация ионов у него составляет 86°/о, от
У иглокож'их не обнаружено сколько-нибудь значительной ре-
гуляции какого-либо иона. Кишечнополостное Aurelia 'регулиру-
ет только концентрацию сульфата, которая удерживается на
430 Гл. 9. Вода и осморегуляция
уровне гораздо более низком, чем в морской воде. Для этого
животного низ'кая концентрация сульфата прямо связана с про-
блемой плавучести: исключение тяжелого иона сульфата пони-
жает плотность медузы, предотвращая такИм образом ее погру-
жение на глубину ((см. дл. 11).
У членистоногих, представленных в табл. 9.4, отмечается ин-
тересная особенность: у •видо'в, способных быСтро двигаться, уро-
вень магния в плазме низок. Краб Pachygrapsus и сходный
с омаром Nephrops — животные активные и быстро передвигаю-
щиеся; напротив, краб Maia движется /медленно, и концентрация
магния у него высокая. Магний подавляет нервно-мышечную пе-
редачу, и можно было /бы сделать поспешное заключение, что
высокая концентрация магния у этих ракообразных действи-
тельно .связана с низкИм уровнем их активности. Однако тот
факт, чгго у каракатицы (Sepia), которая быстро движется и хо-
рошо плавает, содержание магния такое же высокое, как у гре-
бешка (Pecten), заставляет сомневаться в наличии причинно-
следственной связи между .концентрацией магния и активностью.
ВНУТРИКЛЕТОЧНЫЕ КОНЦЕНТРАЦИИ И РЕГУЛЯЦИЯ ОБЪЕМА КЛЕТКИ
До сих пор мы обсуждали соотношения между окружающей
водой и концентрацией растворенных веществ в крови и других
жидкостях тела. Что касается концентраций ионов внутри -кле-
ток, то они обычно весьма отличны от внеклеточных; например,,
концентрации натрия и хлора внутри клеток обыкновенно ниже,
чем снаружи, а для калия соотношение обычно обратное. Одна-
ко, несмотря на различия в концентрациях отдельных ионов,
клетки изоосмотичны с окружающей тканевой жвд'костью и
кровью. -
Чтобы получить представление о проблемах регуляции объе-
ма клетки, можно принять, чТо одна и та же клеточная мембра-
на проницаема для веды, но непроницаема для растворенных в
ней вещеСтв. Изменение внеклеточной концентрации должно*
вызывать соответствующее изменение объема клетки. При пони-
жении внешней концентрации клетка будет доглощать воду и.
набухать, а при ее повышении вода будет выходить из клетки и
клетка будеТ сморщиваться. В действительности, од'нако, у
большинства животных объем клетки регулируется так, что по-
сле начального отклонения она возвращается ,к исходному объе-
му, который в дальнейшем остаемся относительно постоянным.
Теперь твердо установлено, что важную роль в регуляции
объема клетки при изменении солености среды играет внутрикле-
точная концентрация свободных аминокислот. При повышении
или понижении солености воды содержание аминокислот в клет-
ке возрастает или падает, и клетки, таким образом, остаются
изотоничными с окружающей их средой.
Водные беспозвоночные 431
Была высказана мысль, чгго понижение концентрации амино-
кислот могло бы осуществля'ться путем синтеза белка, а ее вос-
становление— путем расщепления белков. По другому предполо-
жению, при воздействии на животное низкой солености внутри-
клеточные аминокислоты выходят из клетки через растянутые
вначале клеточные мембраны вместе с осмотически связанной
с ними водой, и таким .образом прежний объем ,клетки восста-
навливается. Согласно этой гипотезе, которая подкрепляется ис-
следованиями на сердечной мышце моллюсков, регуляция объе-
ма клеток связана с изменениями проницаемости клеточной
мембраны, на которую в свою очередь влияет внеклеточная ос-
мотическая концентрация (Pierce, Greenberg, 1973).
Изменения, происходящие в клетках позвоночных при адап-
тации к изменениям солености, сходны с темп, которые наблю-
даются у беспозвоночных. При повышении или понижении соле-
ности среды в концентрациях внутриклеточных аминокислот про-
исходят соответствующие сдвиги, которые нужны для
поддержания .постоянного объема клетки (Fugelli, Zachariassen,
1976). Таким образом, механизмы регуляции клеточного объема
у позвоночных и беспозвоночных в принципе сходны.
ЖИВОТНЫЕ В ПРЕСНОЙ И В СОЛОНОВАТОЙ ВОДЕ
Если перевести .разных морских животных в несколько раз-
бавленную морскую воду, скажем с .80% ее обычной солености,
большинство из них .может выжить. До если через некоторое
время исследовать жидкости их тела, то окажется, что живот-
ные приспособились к разбавлению среды и у них установились
новые, более низкие .концентрации ионов. Осмоконформеры (на-
пример, морские звезды и устрицы) приобретают такую же ос-
мотиче’скую концентрацию, как и разбавленная среда, хотя со-
держание отдельных ионов у них по-прежнему отличается от
содержания их в окружающей воде. Между тем осморегуляторы
более или менее успешно сопротивляются разбавлению их внут-
ренней среды и остаются гиперосмотичными.
Таким образом, морские животные, проникающие в солоно-
ватую воду, делятся на два типа — на пассивных осмоконфор-
меров и активных осморегуляторов. Такой осмо конфо'рмер, как
устрица, выносит значительное понижение солености; он также
ослабляет воздействие периодического разбавления воды в
эстуарии, держа свою раковину закрытой. Но в течение более
длительных дер’иодов активные регуляторы лучше выдерживают
колебания солености.
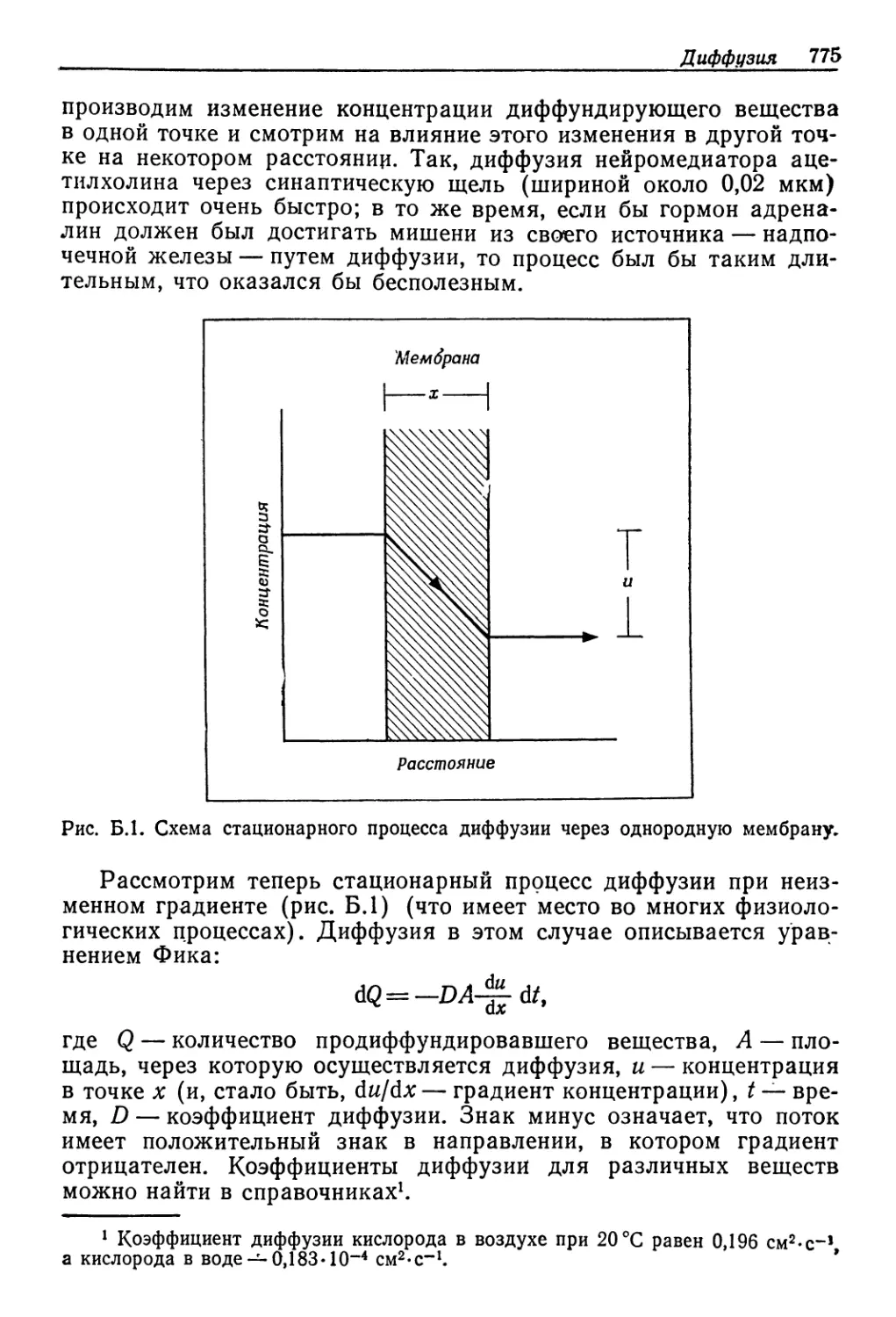
Посмотрим, как они реагируют на разбавление мюрс'кой во-
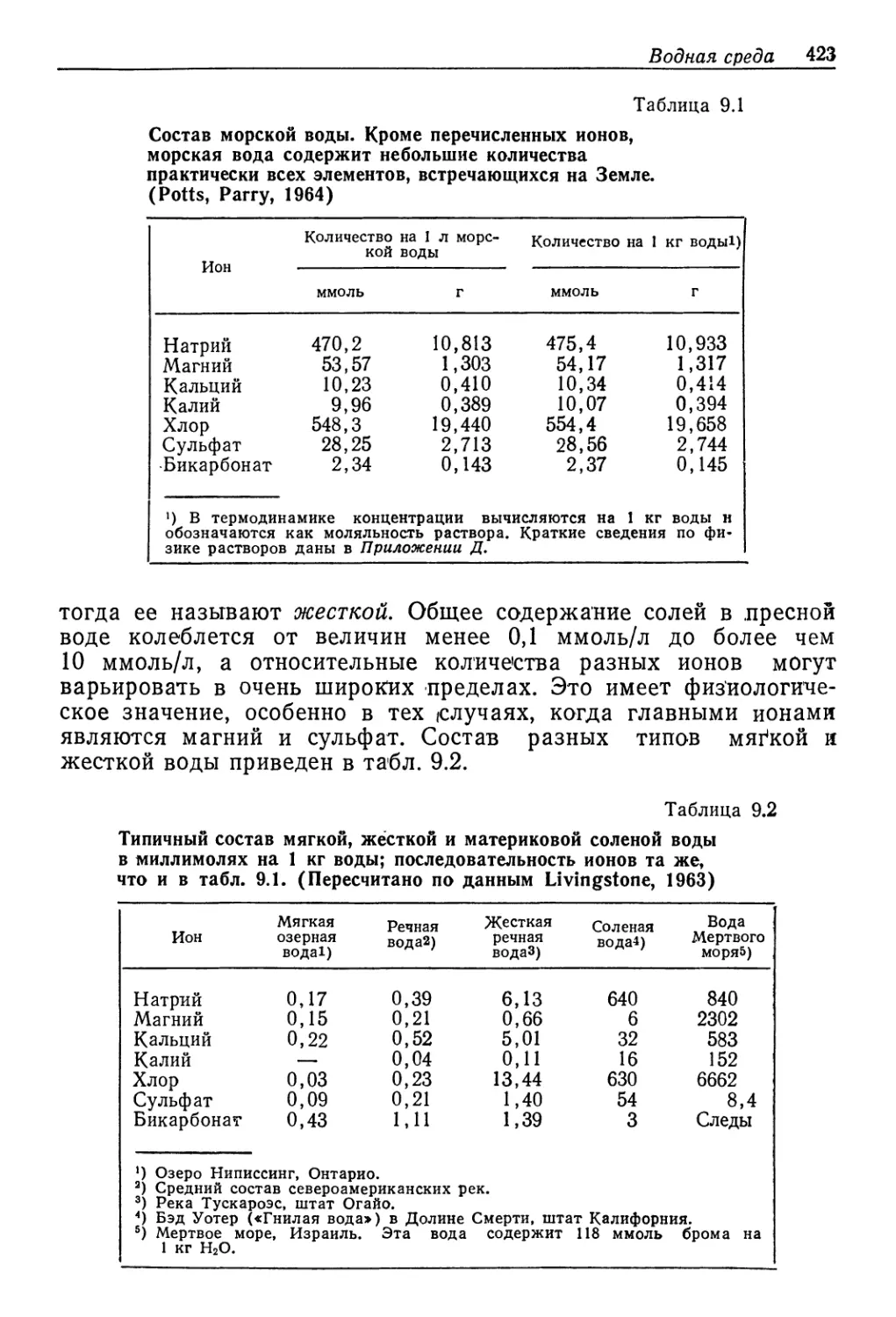
ды. На рис. 9.1 по'казано соотношение концентраций в жидко-
стях тела и в воде для ряда осморегуляторов. Даже для хоро-
ших регуляторов .существуют некоторые ограничения. Примерам
432 Гл. 9. Вода и осморегуляция
служ’ит европейский краб Carcinus, который не выж*ивает в со-
лоноватой воде, если ее концентрация составляет менее одной
трети концентрации нормальной морской воды. Предел разведе-
ния, переносимого этим крабом и многими другими соло'новато-
водными формами, варьирует .в зависимости от географических
условий. Краб Carcinus из Северного моря гораздо менее толе-
100 200 300 400 500 600
Концентрация среды, ммоль/л
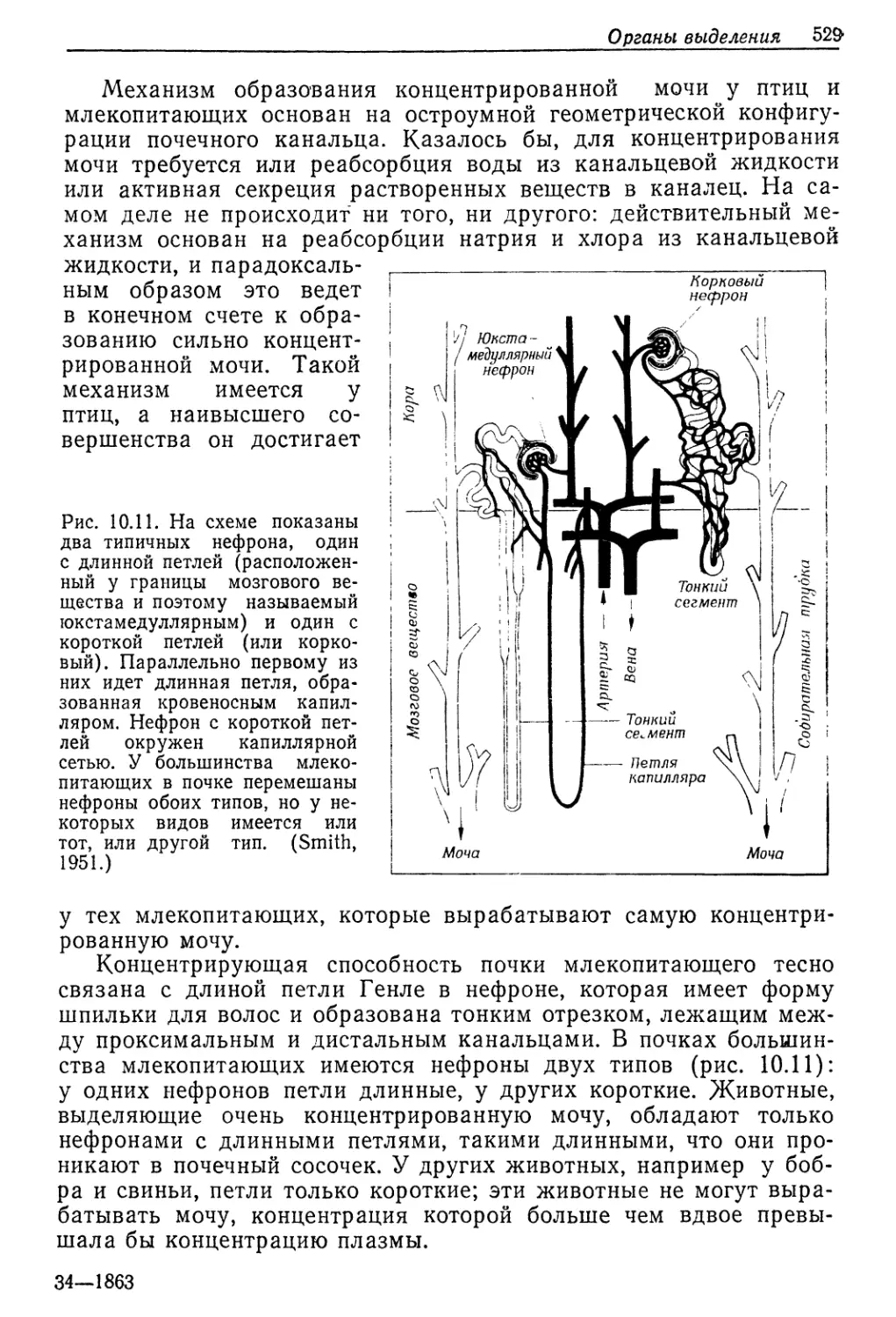
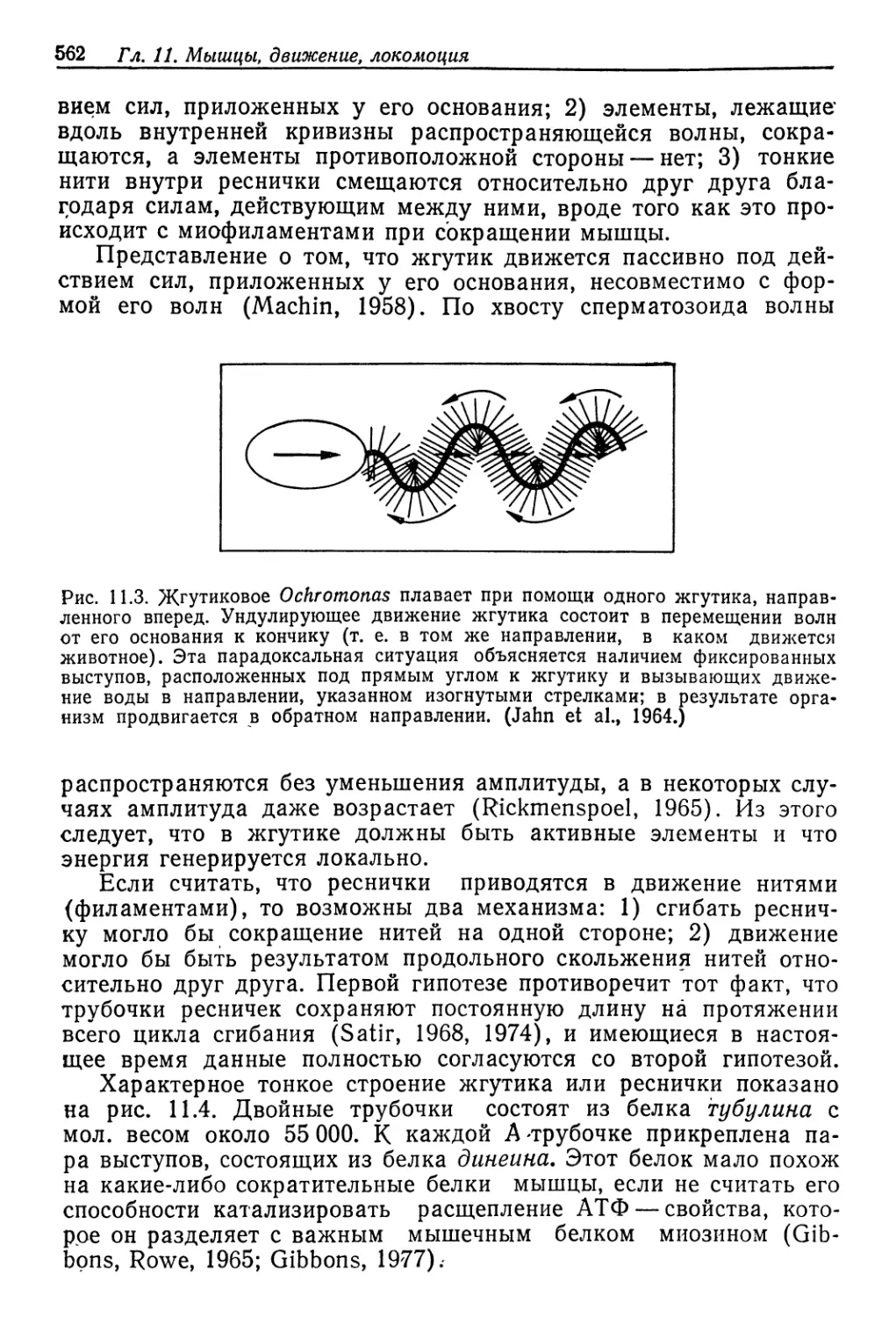
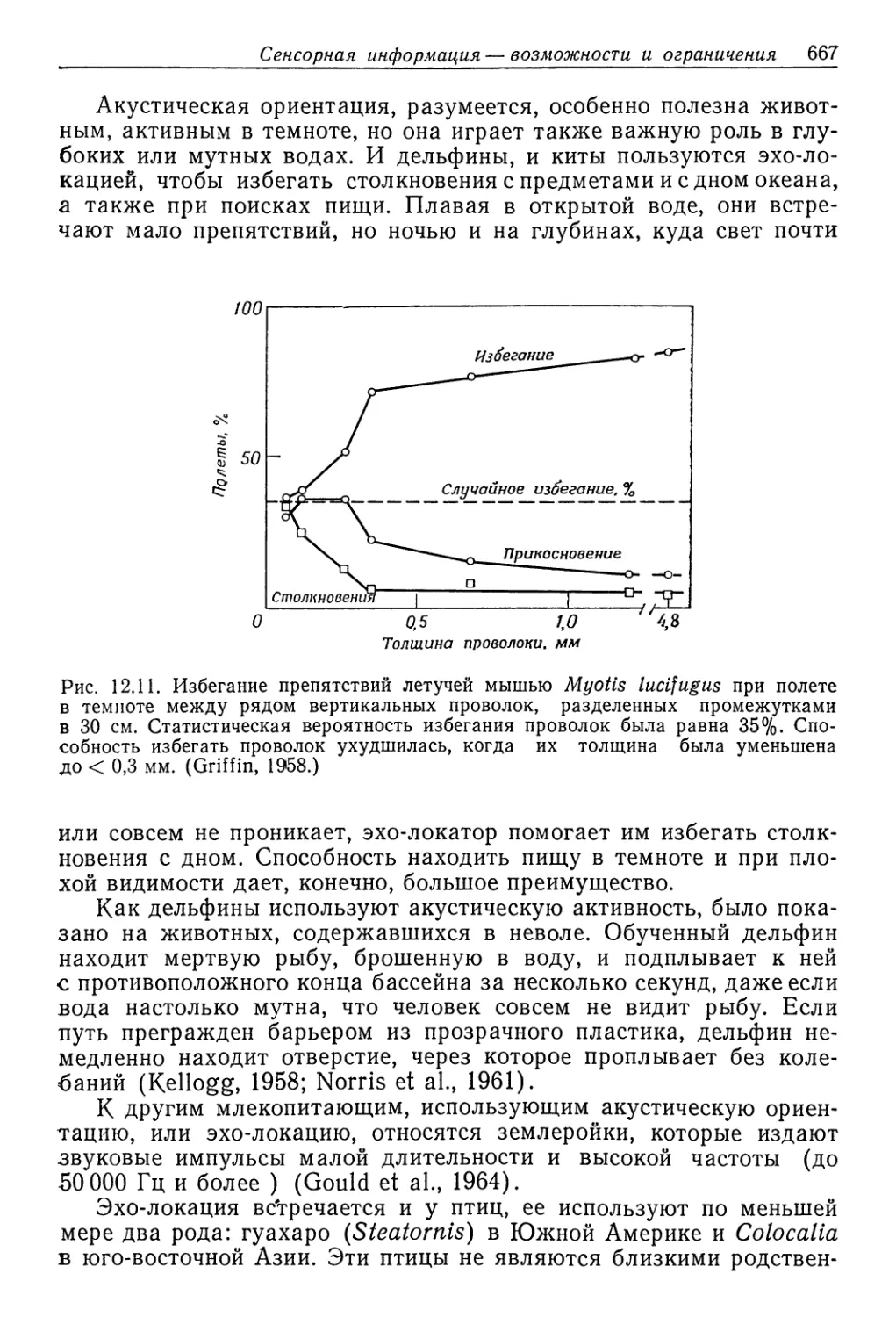
Рис. 9.1. Соотношение между концентрациями жидкостей в теле и во внешней
среде у разных животных, обитающих в солоноватой воде. Концентрация нераз-
веденной морской воды указана стрелкой. Линия, идущая по диагонали, соот-
ветствует одинаковым концентрациям жидкости тела и среды. (Beadle, 1943.)
рантен к разбавлению воды, чем такой же краб из Балтийского
мюря, где концентрация ксолей в норме гораздо ниже, чем в от-
крытом море.
Другой краб — ’китайский Eriocheir — выдерживает гораздо
большие разведения и даже способен проникать в црес'ные во-
ды. Для размножения он должен вернуться ,в мере и поэтому не
может прочно обосноваться и проводить весь свой жизненный
цикл в пре'сной воде.,
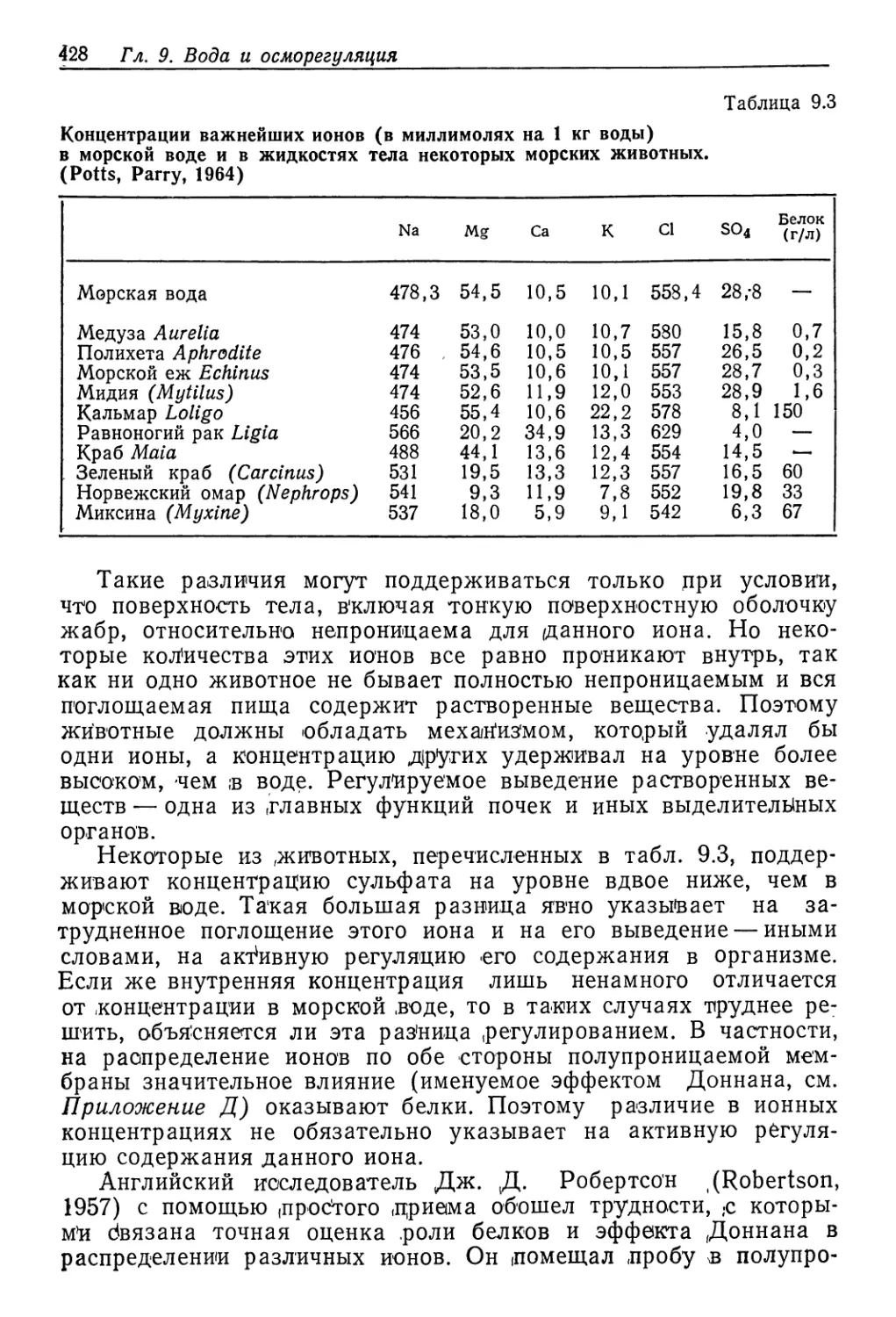
В' принципе пресноводные животные ведут себя в . осмотиче-
ском отношении сходню с преуспевающими осморегуляторами в
солоноватой воде, но концентрации, которые они цоддерж'ивают
в жидкостях тела, сильно различаются ,(!рис. 9.2). Речной рак
Potamobius поддерживает осмотическую концентрацию на уров-
не около 500 мосмоль на 1 л, а у дрес'новодного моллюска без-
зубки (Anodonta) она составляет ^менее од'ной десятой этой ве-
Водные беспозвоночные 43$
личины — в'сего лишь около 50 мосмоль на 1 л. При этом без-
зубка все еще является гиперосмотичной, и не найдено ни одно-
го пресноводного животного, у которого концентр ац'ия жидко-
стей тела могла бы снизиться до уровня окружающей воды.
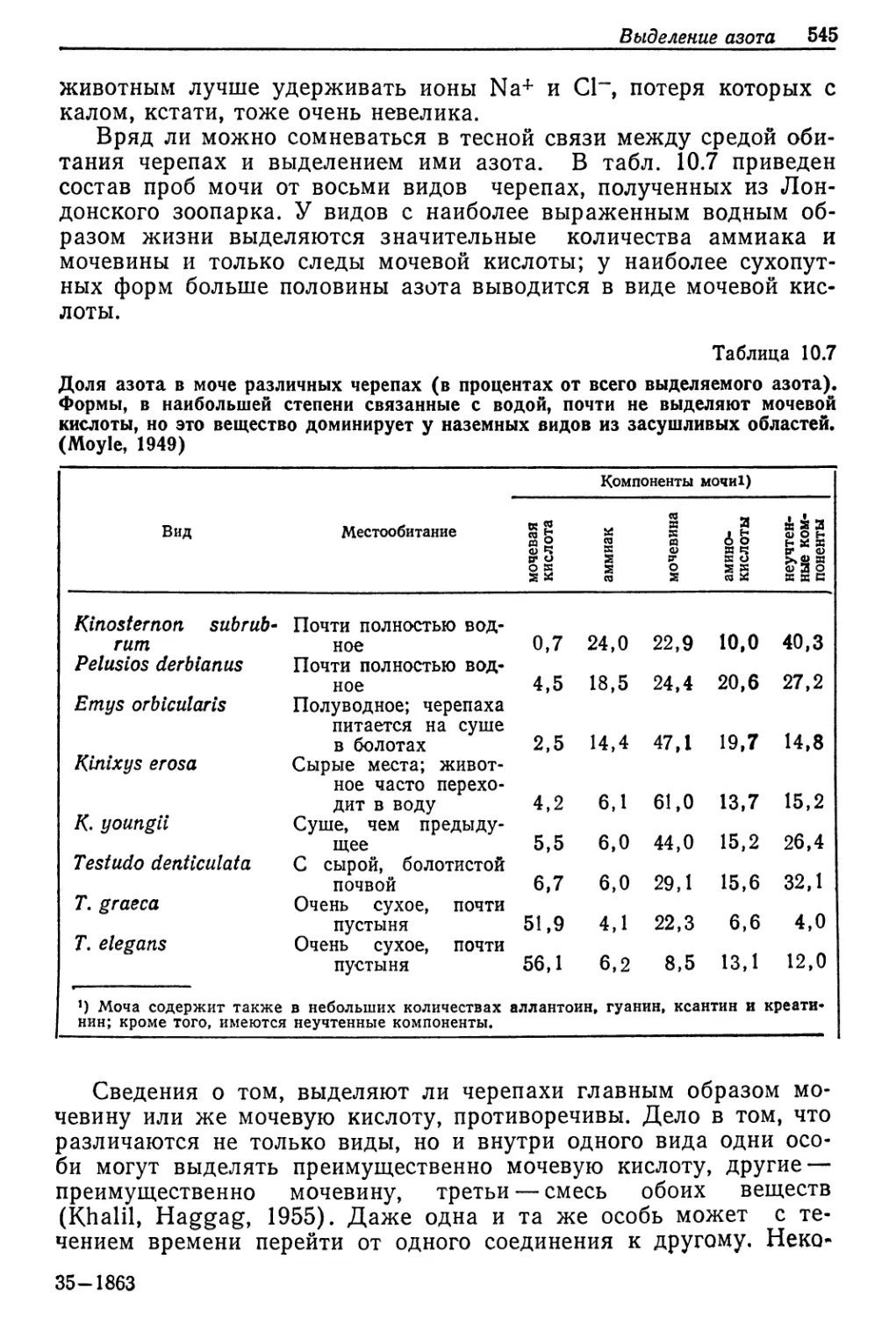
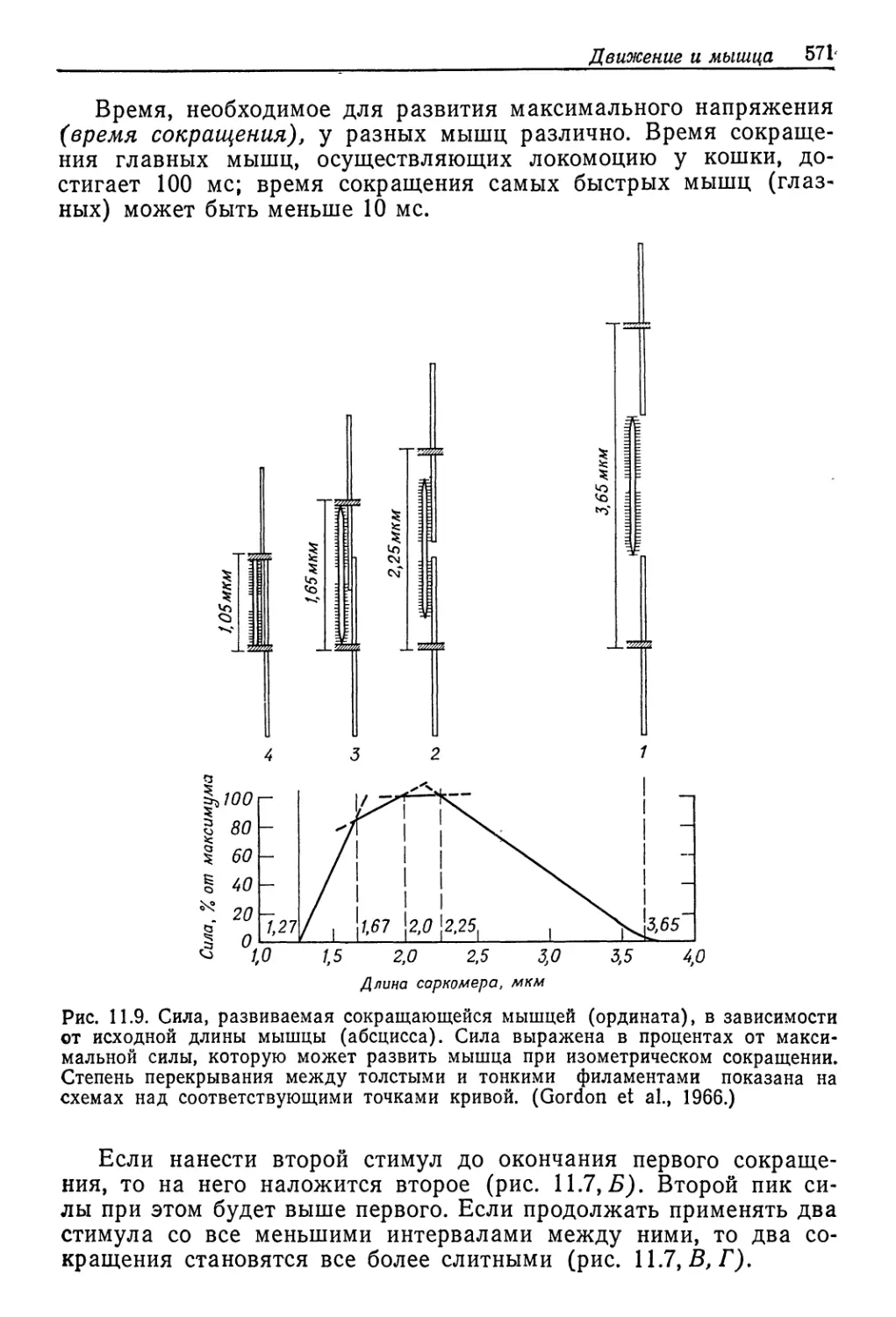
Концентрация среды, ммоль/л
Рис. 9.2. Соотношение между концентрациями жидкостей тела и внешней среды
у разных пресноводных животных. Концентрация неразведенной морской воды
указана стрелкой. Линия, идущая по диагонали, соответствует одинаковым кон-
центрациям жидкости тела и среды. (Beadle, 1943.)
Большинство главных типов животных имеет представителей
как в море, так и в .пресных водах, хотя дислю морских Пидов
гораздо больше. Иглокожие и головоногие моллюски (осьминоги
и кальмары) в цресной воде совсем не встречаются.
МЕХАНИЗМЫ ОСМОРЕГУЛЯЦИИ
Когда животное гиперосмоти|чцо по отношению к окружаю-
щей среде, оно сталкивается с двумя физиологическими «труд-
ностями»: 1) вода стремится проникнуть внутрь тела из-за бо-
лее высокой концентрации жидкостей .организма и 2) растворен-
ные вещества стремятся выходить наружу, так как их внутрен-
няя концентрация выше, а входящая вода должна выводиться,
и она уносит с собой некоторое количество этих веществ. Эти
трудности можно было бы уменьшить, сделав все поверхности
тела очень мало проницаемыми, однако не известно ни одного
животного с совсем непроницаемыми покровами. По крайней ме“
ре дыхательные поверхности должны быть тонкими и достаточ-
но обширными, чтобы могла происходить необходимая диффу-
28-1863
434 Гл. 9. Вода и осморегуляция
зая газов, и эти поверхности обычно служат местом наибольшей
утечки растворенных ‘веществ и наибольшего проникновения
.воды.
Как может ж'ивотное 'Кюмпенсйровать 1потерю растворенных
веществ? Оно могл*о бы (получать необходимые ионы с пищей, и
при очень малой проницаемости покровов этого было бы доста-
точно. Однако у многих пресноводных животных проницаемость
не столь мала, и тем ‘не менее даже не получая пищи, они под-
держивают нормальные концеНтрацйи ’ио'нов. Это достигается
путем прямого поглощения ионов из ^среДы.
Проще все(го показать это, поместив животное, например реч-
ного рака, 'в проточную или часто сменяемую Дистиллированную
воду. Это вызовет постепенную утечку солей организма, и кон-
центрация их снизится, скажем, с 500 до 450 мосмоль/л, что рак
легко переносит. Если теперь снова поместить рака в обычную
пресную воду, концентрация солей в ёго кр'ови снова Возрастет,
хотя в пресной воде о’на 'в 100 раз меньше, чем в крови (напри-
мер, 5 мосмоль/л).
Поскольку в этом опыте рак извлекает ионы из разбавленно-
го раствора и перемещает >их в область гораздо большей кон-
центрации— в кровь, ионы передвигаются прот'ив концентраци-
онного градиента. Такой перенос называют активным транспор-
том1.
Не всегда известно, какие органы осуществляют активное по-
глощение ионов. Было высказано предположение, что у некото-
рых животных таким органом служит вся поверхность тела, но
при этом исходили главным образом из того, что не найдено
каких-то определенных органов, которым можно было бы припи-
сать такую роль. В ютн'ошении других животных, особенно рако-
образных и насекомых, весьма вероятно, <что общие покровы те-
ла не участвуют в активном транспорте. Есть убедительные дан-
ные о том, что у ракообразных органами активного транспорта
ионов служат жабры, а у водных лич’инок насекомых за погло-
щение ионов ответственны определенные придатки, а именно
«анальные жабры». Осморегуляция не вступает 'в конфликт с ды-
ханием; обе функции может выполнять один и тот же орган, на-
пример жабры у пресноводных крабов и речных раков. Но
«анальные жабры» лич’инок насекомых, вероятно, не несут Дыха-
тельной функции и служат только органами осморегуляцИи.
Активный транспорт требует .затраты энергии, и было бы ин-
тересно знать, насколько возрастает эта потребность при переме-
щении животного в более разбавленную среду. При перемеще-
1 Активный транспорт определяют как перенос против электрохимического
градиента, и как таковой он требует затраты энергии. Его часто называют пере-
носом «в гору» в отличие от пассивной диффузии «под гору», в направлении
концентрационного градиента.
Водные беспозвоночные 435-
нии краба Carcinus, хорошо переносящего солоноватую воду, из-
морской (воды во всё более разбавленные растворы, потребление
кислорода у непо заметно возрастает. Если поместить его *в мор-
скую воду, разведенную в четыре раза, потребление кислорода
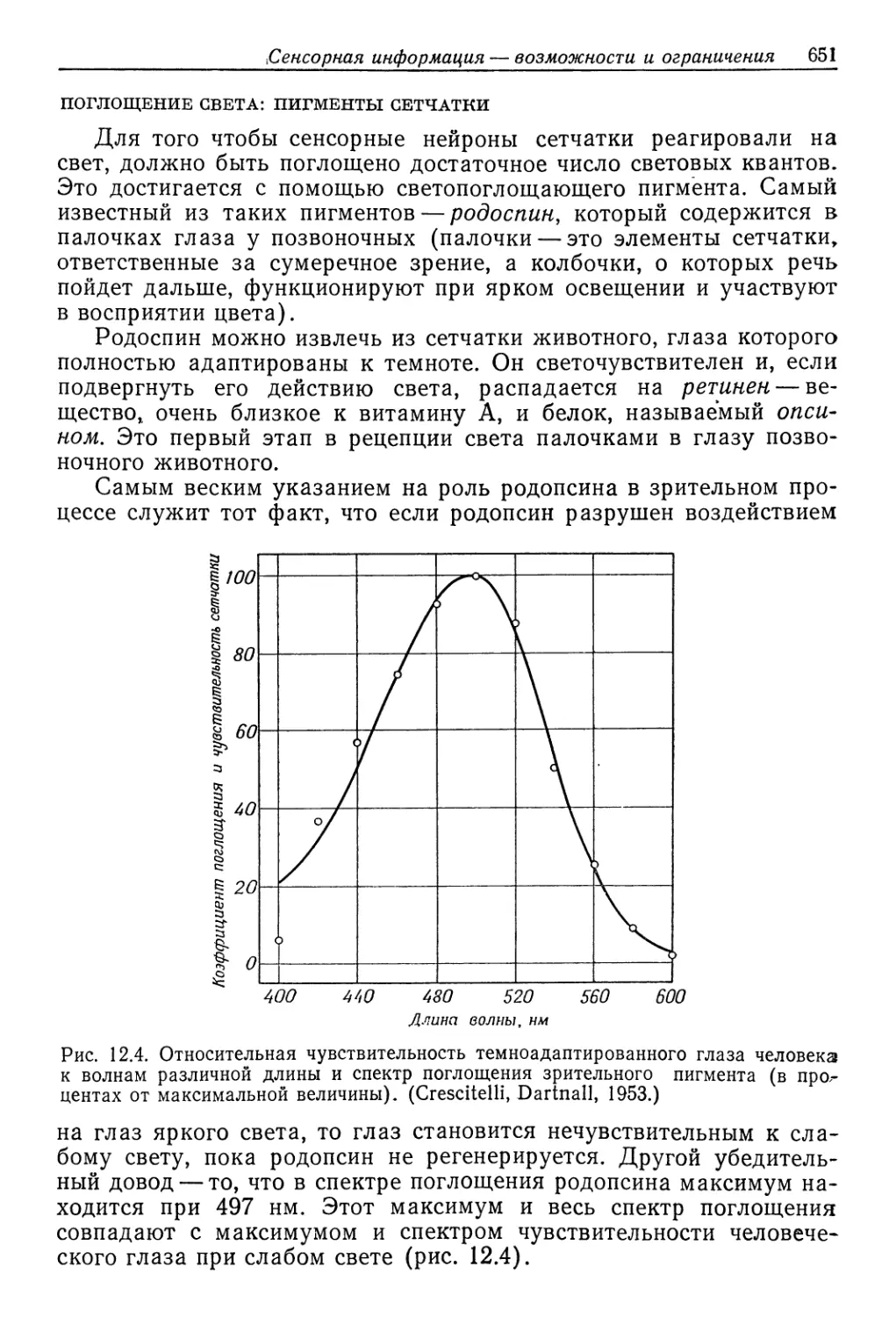
возрастет приблизительно на 50%. Легко можно было бы заклю-
чить, что это увеличение объясняется работой активного переноса,
ионов, которую приходится осуществлять в разбавленной мор-
окой воде. Между тем другой 'краб—Eriocheir — сохраняет одну
и ту же интенсивность метаболизма в морской, солоноватой и
пресной воде. Поэтому он, видимо, не 'тратит измеримого количе-
ства дополнительной энергии для осморегуляции даже в пресной
воде. Такая разница была бы объяснима, если бы покровы у
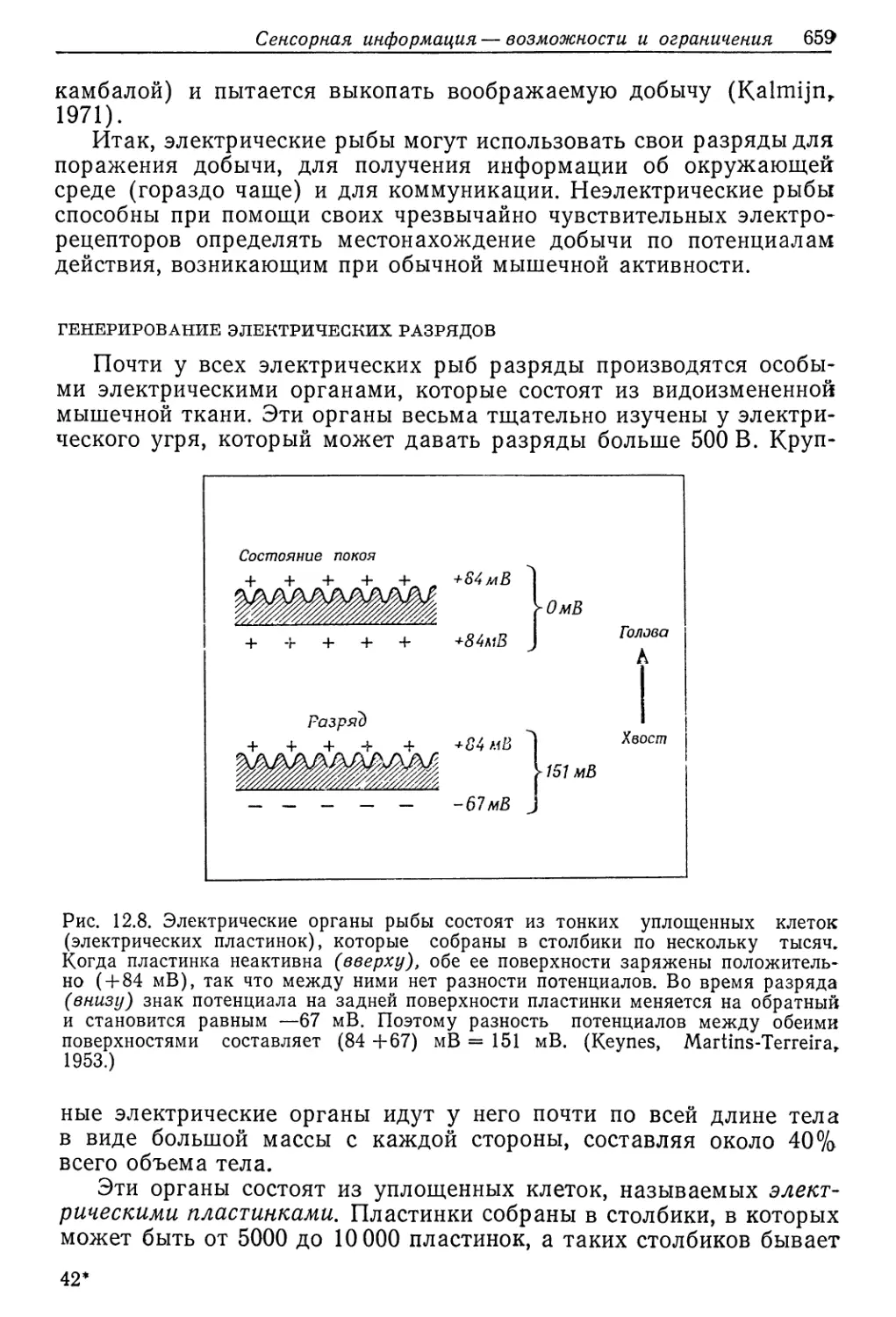
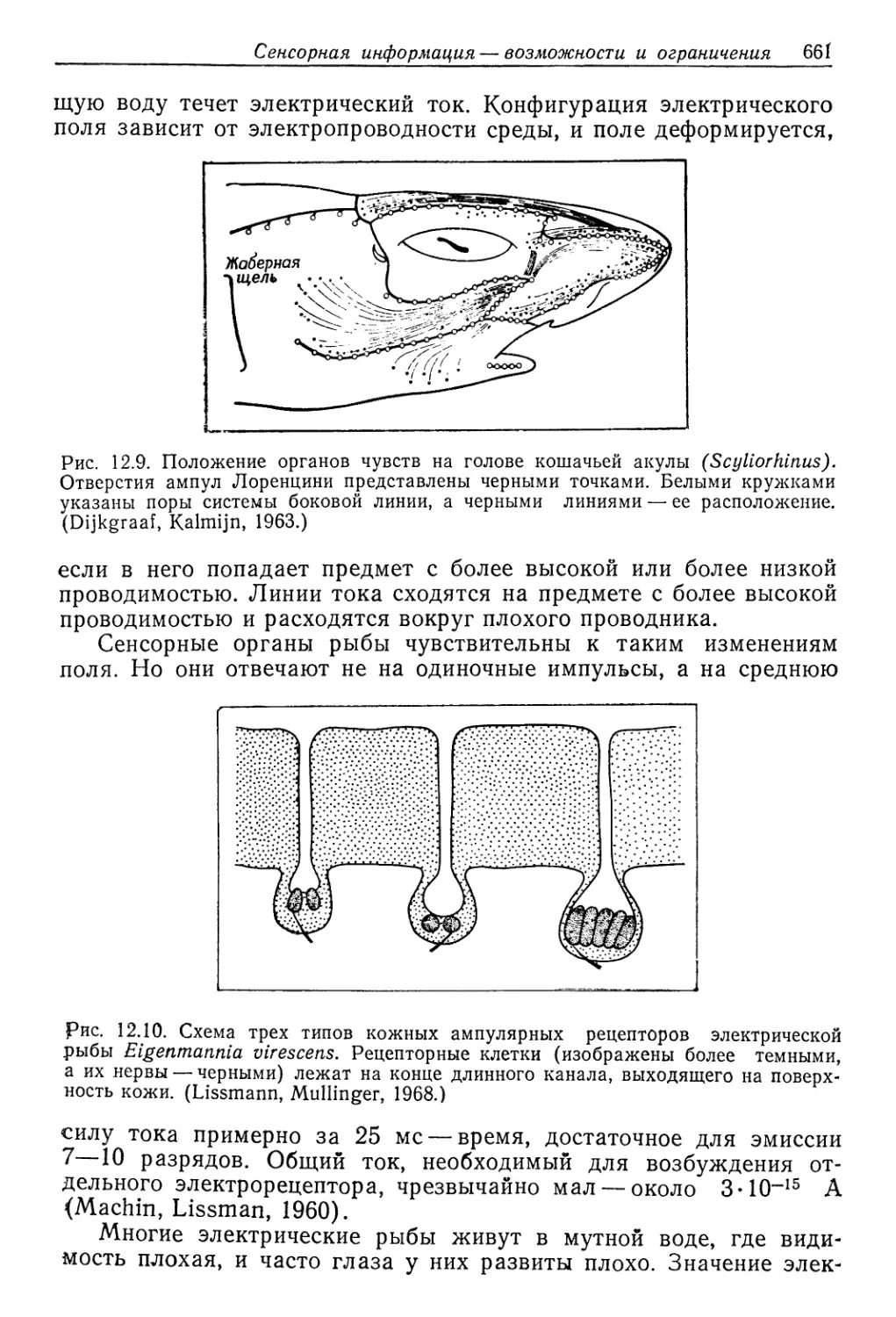
этого краба были менее про’ницаемым'и, чем у Carcinus, но в этом
отношении между ними 'нет резкой разницы. Поэтому измерение-
сдвигов потребления 'О2 не дает надежной информации о коли-
честве энергии, затрачиваемой на осморегуляЦию (Potts, Parry,.
1964).
Однако эту энергию можно вычислить на основе термодина-
мических соображений. Если нам известно, сколько растворен-
ных веществ теряется за данный отрезо'к времени, то мы знаем,,
сколько их должно быть поглощено из окружающей среды для*
сохранения постоянства внутренней среды животного. Необходи-
мая осмотическая работа (Т^осм) зависит от 'концентрации рас-
творенных веществ во внешней среде (Сер) и в кров'и (СКР), а
работу по перемещению 1 моля дает следующее уравнение (где-
R — универсальная газовая постоянная, а Т — абсолютная тем-
пература):
Госм=ЯТ1п
ьср
В нашей модели мы примем, что поверхность животного про-
ницаема для воды, но непроницаема для растворенных веществ..
В гипотонической среде вода входит внутрь вследствие осмоса,
и такое же количество воды должно быть выведено *в виде моти.
Приток воды определяется проницаемостью животного, и поэто-
му объем .мочи «служит мер-ой -этой проницаемости. Некоторые-
растворенные вещества вывюдятся с мочой, и общее 'количество»
потерянных веществ равно произведению объема мочи на ее кон-
центрацию УмХСм.
Теперь мы видцм, что осмотическая работа зависит от прони-
цаемости ж'ивотного и потери растворенных веществ:
°ср
Некоторые животные выделяют мочу, изотоничную крови..
Многие другие реабсорбируют из мочи ряд веществ и выделяют
гипотоничную мочу. В последнем случае потеря растворенных
28*
436 Гл. 9. Вода и осморегуляция
веществ меньше, но требуется работа для их реабсорбции, что-
бы моча стала гипотон'ичной. Это усложняет вычисление мини-
мума необходимой осмотической работы, но расчеты показыва-
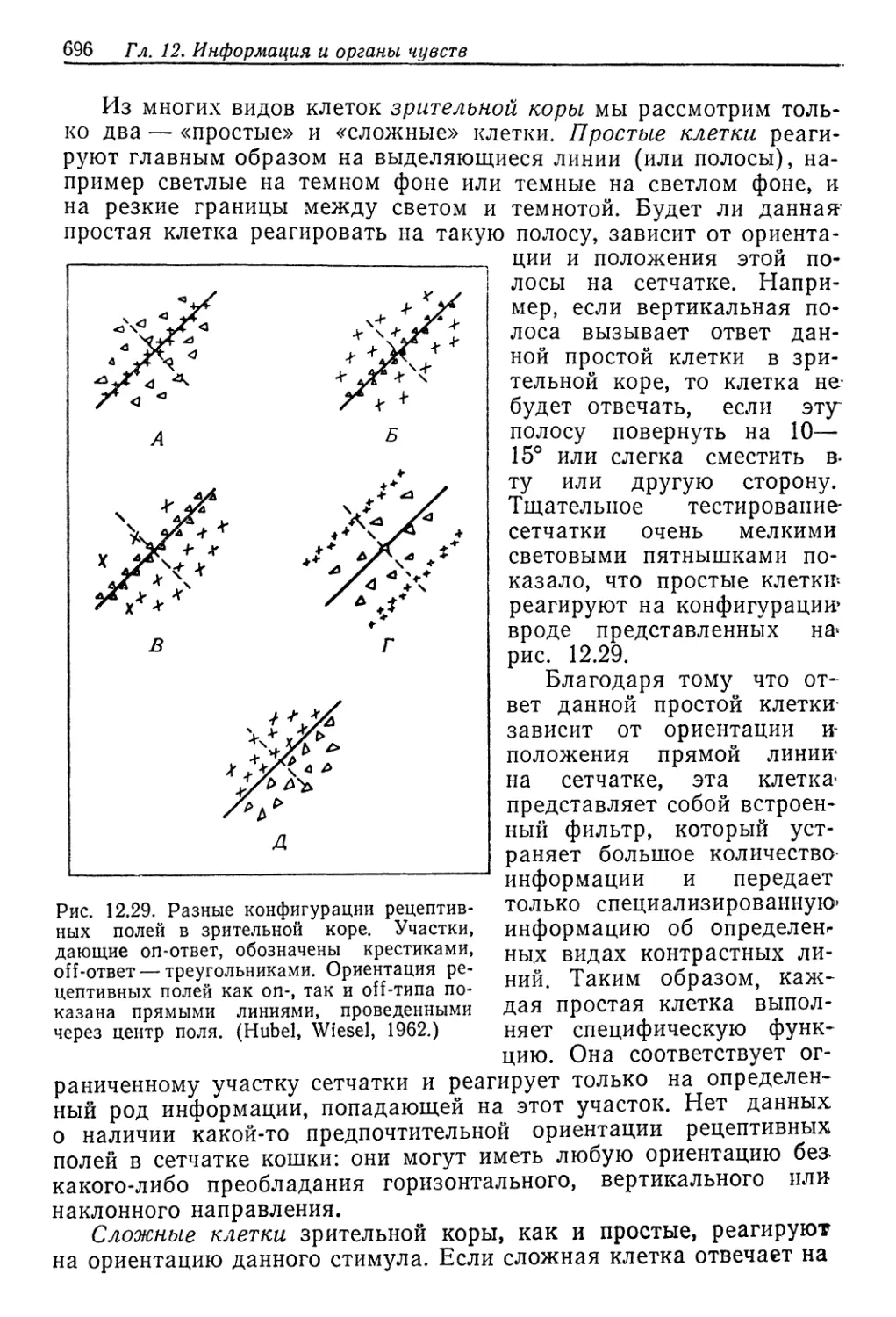
ют, что реабсорбция растворенных веществ из мочи значительно
снижает затрату энергии.
В табл. 9.5 приведены оценки минимальной осмотиче'ской ра-
боты, необходимой для осморегуляции в прёснбй воде для трех
Таблица 9.5
Метаболические затраты на осморегуляцию у трех животных весом 60 г
л пресной воде с концентрацией растворенных веществ 6 ммоль/л.
Эти затраты (столбец г) вычислены с использованием данных о концентрации
веществ в крови и моче (столбцы а и б) и об объеме мочи (столбец в),
В последнем столбце (е) вычисленная осмотическая работа сравнивается
с наблюдаемой интенсивностью метаболизма. (Potts, 1954)
Животное (а) Концентра- ция раство- ренных веществ в крови, ммоль/л (б) Концентра- ция раство- ренных веществ в моче, ммоль/л (в) Объем мочи, мл/ч (2) Осмоти- ческая работа, кал/ч (б) Интенсив- ность ме- таболизма, кал/ч (е) Осмоти- ческая ра- бота, % от интенсив- ности ме- таболизма
Краб Eriocheir 320 320 о,1 0,076 14 0,5
Речной рак 420 124 0,1 0,037 10 0,3
Беззубка 42 24 0,5 0,015 1 ,2 1,3
видов животных. Для (вычисления осмотической работы исполь-
зованы данные о концентрации крови и ю концентрации и объе-
ме 1МЮЧИ.
Первое животное — краб Eriocheir — представляет собой мор-
ское животное, легко проникающее в пресную воду. Концентра-
ция растворенных веществ в его моче такая же, как <в крювИ, что
весьма обычно для морских беспозвоночных. Осмотическая ра-
бота, необходимая для возмещения утраченных растворенных ве-
ществ путем их поглощения 'из разбавленной внешней среды,
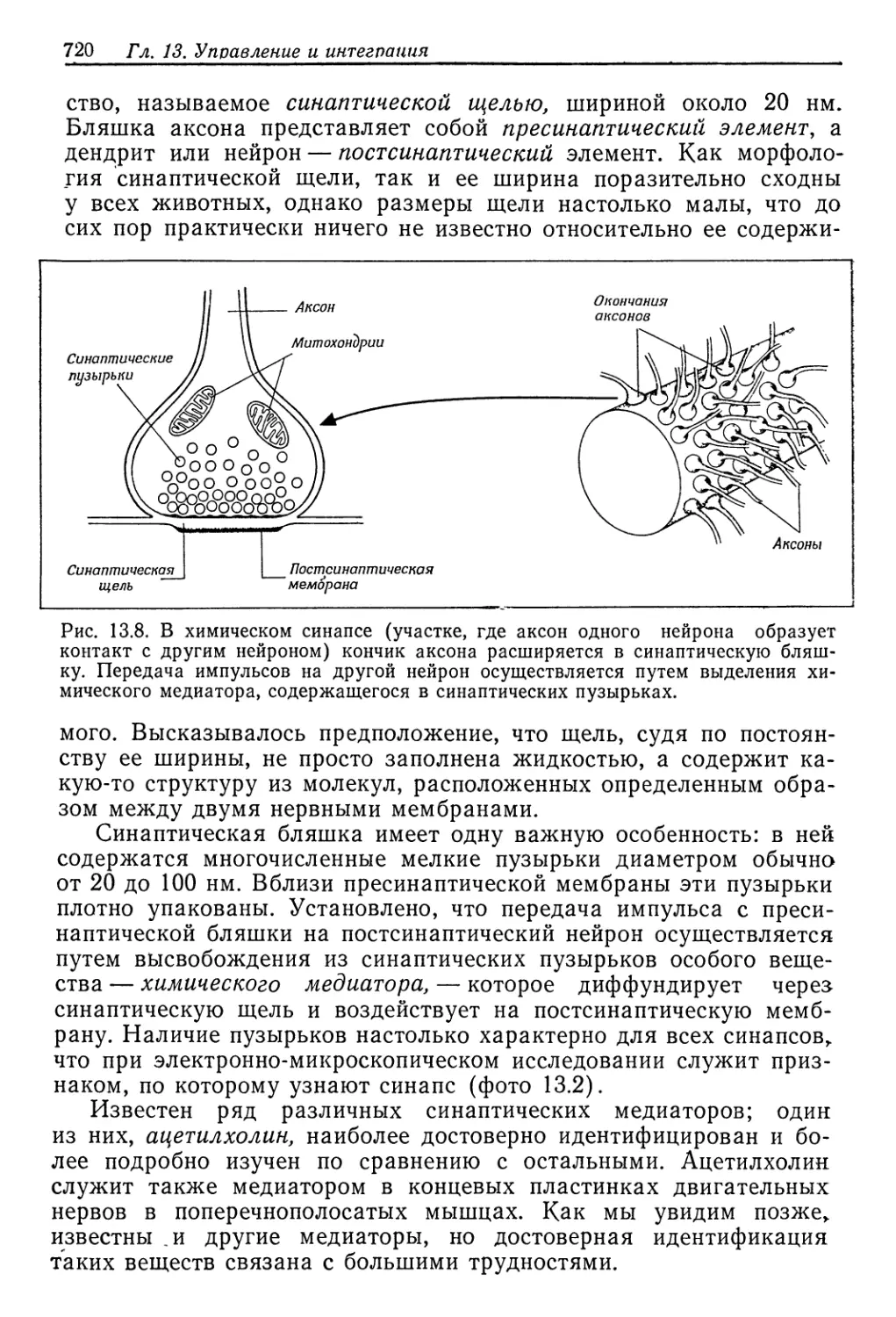
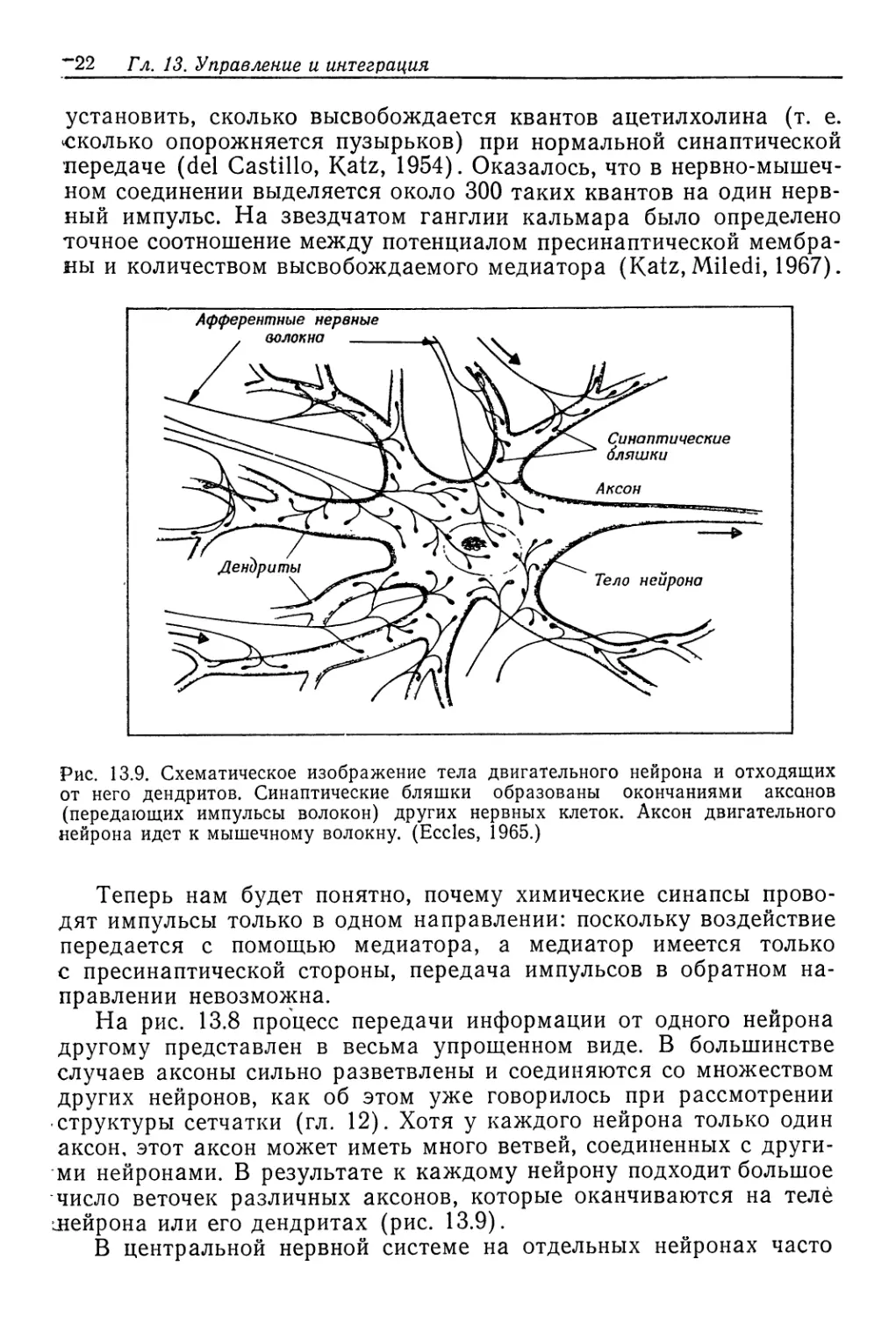
весьма мала—‘0,5% от интенсивности метаболизма.
Речной рак (Potamobius fluviatilis)—животное пресновод-
ное. Он выделяет разбавленную мочу, концентрация которой в
три раза меньше концентрации крови. Объем мочи такой же,
как у Eriocheir, и поскольку концентрация крови выше, прони-
цаемость должна быть несколько ниже. Но наибольшая эконо-
мия достигается путем обратного извлечения растворенных ве-
ществ из мочи, и в результате общая осмотическая работа при-
мерно вдвое меньше, чем у Eriocheir. У рака юна составляет
.лишь 0,3% от интенсивности метаболизма.
У третьего животного — пресноводного двустворчатого мол-
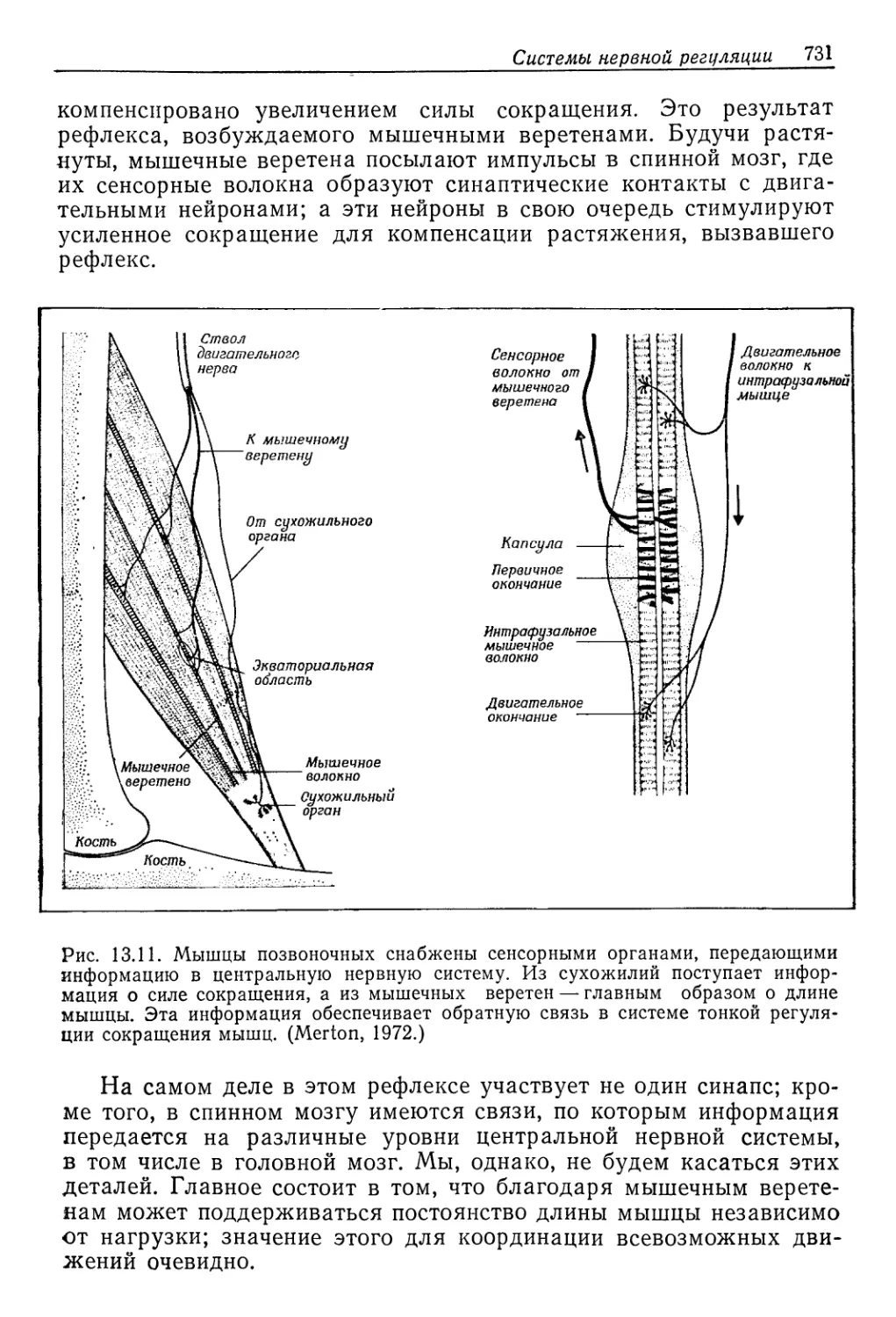
люска беззубки (Anodonta cygnea) — концентрация растворен-
ных веществ в крови очень невелика, она в десять раз меньше,
Водные беспозвоночные 437
чем у рака. Беззубка тоже выделяет разбавленную мочу, при-
мерно ‘в два раза менее концентрированную, чем кровь. Одна'ко
объем мочи большой, что указывает на выоокую проницаемость
из-за большой поверхности соприкосновения мягких тканей с на-
ружной средой. Высокая проницаемость делает для моллюс’ка
фактически обязательной очень низкую внутреннюю концентра-
цию; если бы она была такой же, как у рака, то осмотический
приток воды и, следовательно, объем мочи 'были бы в 10 раз
больше. Это означало бы соответствующее увеличение количе-
ства растворенных веществ, подлежащих возмещению из внеш-
ней среды. Но прирост необходимой работы был бы гораздо
больше, так как приходилось бы переносить ионы против вдеся-
теро большей концентрации их в крови. Поэтому для животного
с высокой проницаемостью и низким уровнем метаболизма важ-
но, чтобы концентрация крови была как можно меньшей.
Хотя стратегия осморегуляции у животных, представленных
в табл. 9.5, не одинакова, у всех у них минимальная потребность
в энергии для этой цели соответствует лишь малой доле всего
метаболизма. На самом деле затраты выше, так как описывае-
мый процесс не обладает 100%-ной эффективностью. Тем не ме-
нее мы должны заключить, что большое повышение интенсивно-
сти обмена, наблюдаемое у некоторых животных при переходе
в разбавленную среду, не может служить прямым показателем
повышенной потребности в энергии для осморегуляции.
ЖИВОТНЫЕ В СОЛОНОВОДНОЙ СРЕДЕ: ГИПОРЕГУЛЯЦИЯ
Креветка Palaemonetes и ее близкий родственник Leander
отличаются одной важной особенностью от Цсех животных, пред-
ставленных на рис. 9.1. В обычной, неразведенной морской воде
они гипотоничны (т. е. жидкости их тела имеют меньшую осмо-
тическую концентрацию, чем наружная среда), и это требует
активной осморегуляЦии. Такого рода гипорегуляция весьма не-
обычна для морских беспозвоночных, и поэтому принято считать,
что эти две креветки принадлежат к группе, обитавшей раньше
в пресной воде: в море они переселились вторично и сохранили
внутреннюю концентрацию ниже концентрации морской воды.
Вода некоторых соленых водоемов содержит гораздо больше
солей, чем морская вода, и в таких экстремальных условиях ги-
порегуляция имеет еще большее значение. Вероятно, наиболее
изученным примером может служить рачок артемия (Artemia),
который в огромных количествах водится в соленых озерах и в
прибрежных бассейнах, где добывают соль путем выпаривания
морской воды.
Хотя артемия не выживает в пресной воде, она приспосабли-
вается к разным условиям — от воды с соленостью в 10 раз
меньшей чем в море, до кристаллизующегося раствора, содер-
438 Гл. 9. Вода и осморегуляция
жащего 300 г соли на 1 л. В .разбавленной морской воде артемия
гипертонична по отношению к ней и ведет .себя как обитатель
солоноватых вод. При больших 'концентрациях соли артемия —
превосходный гипорегулятор; в концентрированном рассоле она
продолжает удерживать осмотическое давление жидкостей свое-
го тела на уровне ненамного выше, чем 7ю осмотического дав-
ления среды (рис. 9.3).
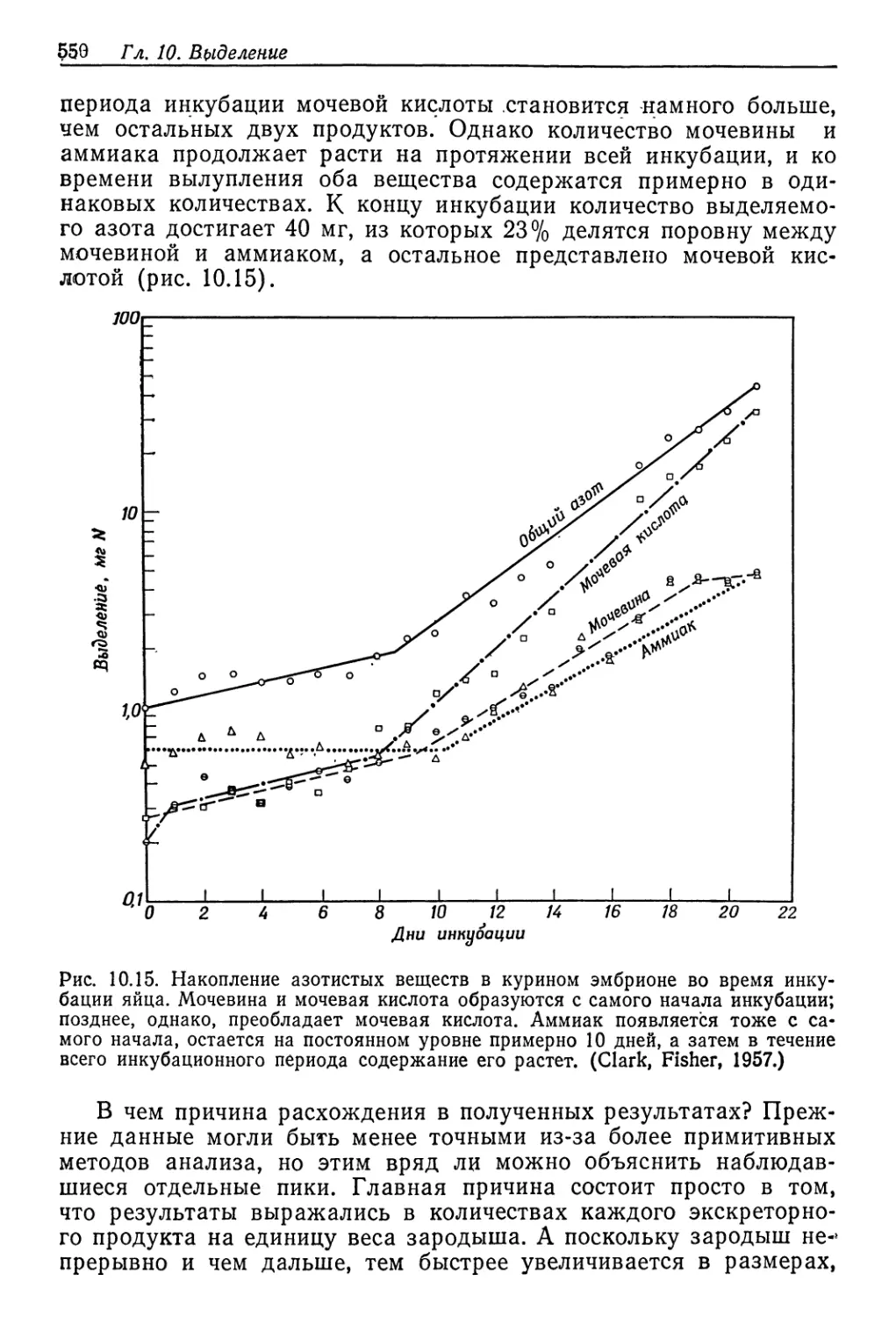
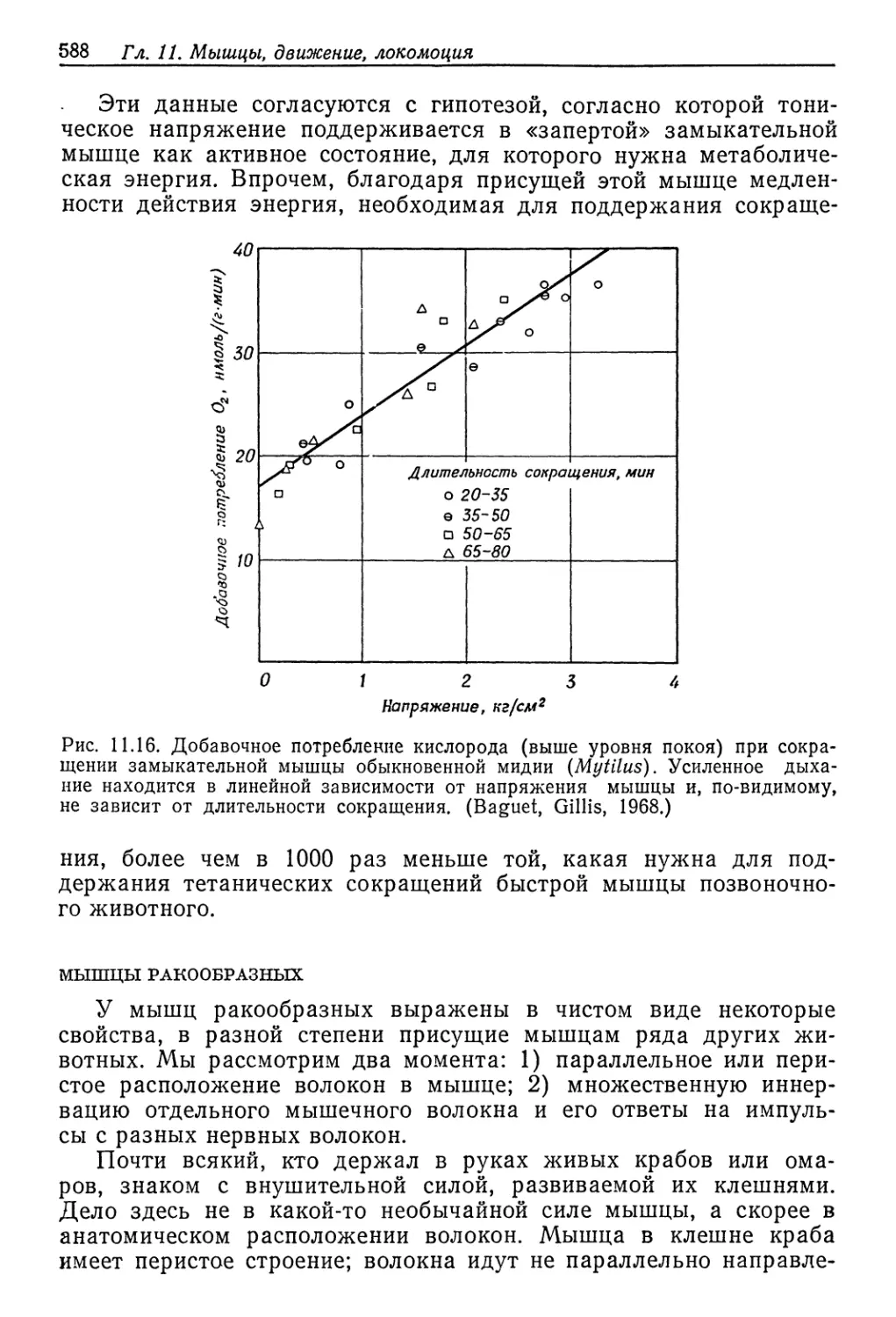
Осмотическое давление внешней среды t % Na Cl
Рис. 9.3. Осмотическое давление жидкостей тела артемии в сильно концентриро-
ванных растворах NaCl. Линия с крутым наклоном в начале графика соответ-
ствует равным концентрациям жидкости тела и среды. (Croghan, 1958а.)
Артемия сохраняет низкую осмотическую концентрацию не
потому, что она совершенно непроницаема для воды и ионов, а
благодаря активной регуляц'ии. Она непрерывно заглатывает
окружающую жидкость, и в содержимом кишки осмотическое
давление заметно выше, чем в гемолимфе (крови). Но хотя это
так, .концентрации натрия и хлора в кишечной жидкости значи-
тельно ниже, чем в гемолимфе (Croghan, 1958b). Это значит, что.
натрий и хлор удаляются из кишки путем их активного всасы-
вания, .а удаление этих ионов из организма для поддержания их
концентрации в крови на относительно низком уровне должно
происходить где-то в другом месте. Весьма вероятно, что главную
роль в этом процессе играет жаберный эпителий.
У личинок артемии активное выведение натрия, по-видимому,.
происходит в другом месте, а именно в «шейном органе»—‘спе-
циализированной структуре на дорсальной поверхности тела
(Conte et al., 1972).
Водные беспозвоночные 439
Личинки некоторых в'идов комаров одинаково хорошо себя
чувствуют как в пресных водоемах, так и в соленой воде, в не-
сколько раз более концентрированной, чем их гемолимфа, и да-
же выдерживают соленость в три раза большую, чем в морской
веде. Еще более примечательно то, что личинки комаров Aedes
campestris могут отлично существовать и в щелочных соленых
озерах, где из солей преобладает бикарбонат натрия, а pH вы-
ше 10.
При низкой солености воды личин>ки Aedes гиперосмотичны
по отношению к среде, а при больших концентрациях соли они
гиподсмотичны. Эти личинки могут адаптироваться к концент-
рациям в пределах 500-кратных различий, причем концентрации
главных ионов в их гемолимфе изменяются не более чем вдвое.
На повышение внешней солености личинки Aedes реагируют
увеличением в несколько раз количества выпиваемой жидкости.
По-видимому, это единственная возможность получать воду для
Компенсации ее утечки в концентрированную среду. Однако питье
означает также поглощение большого количества растворенных
ионов. Избыточная соль выделяется через мальпигиевы сосуды
и заднюю кишку (подробнее см. в ,гл. 10). Так называемые
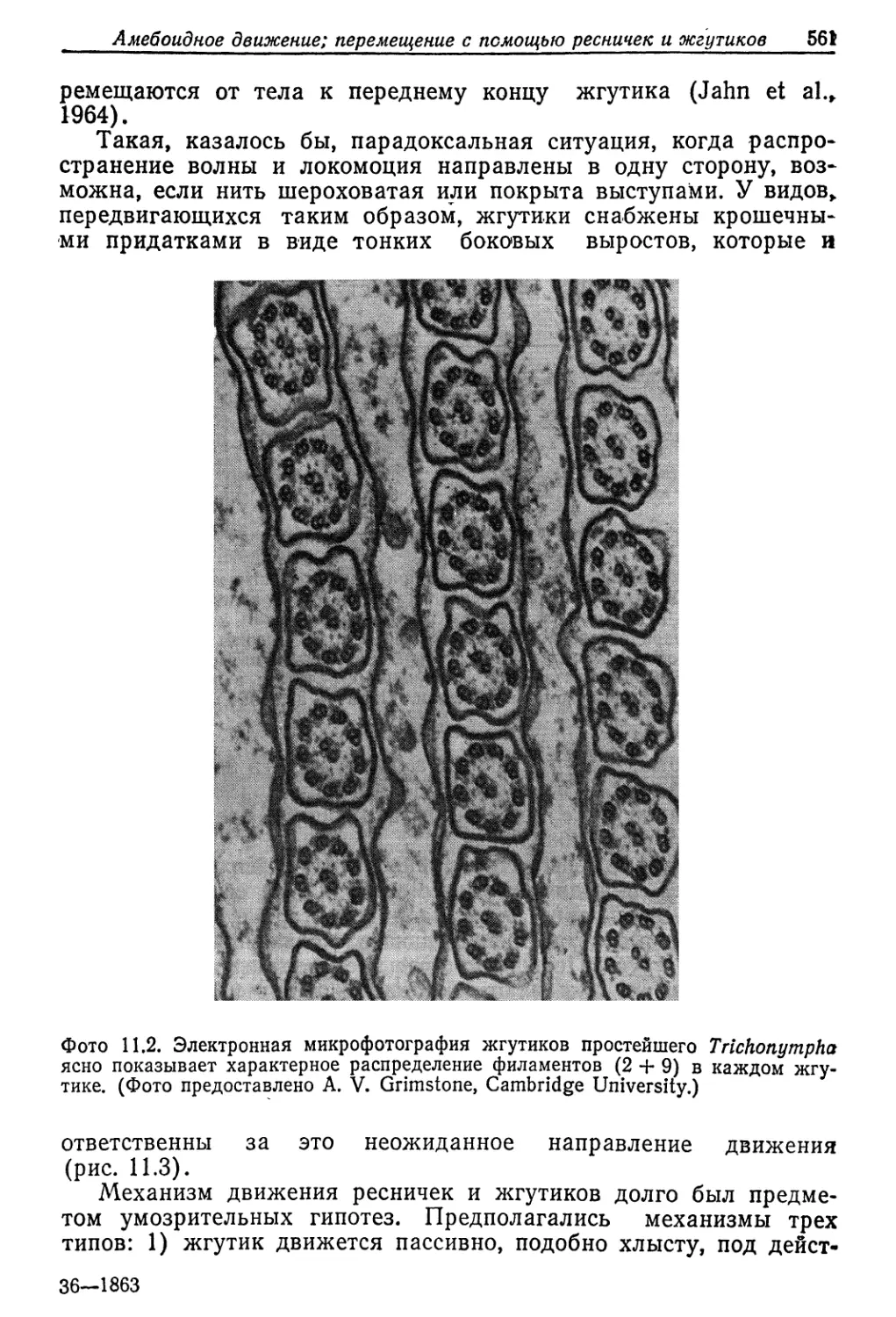
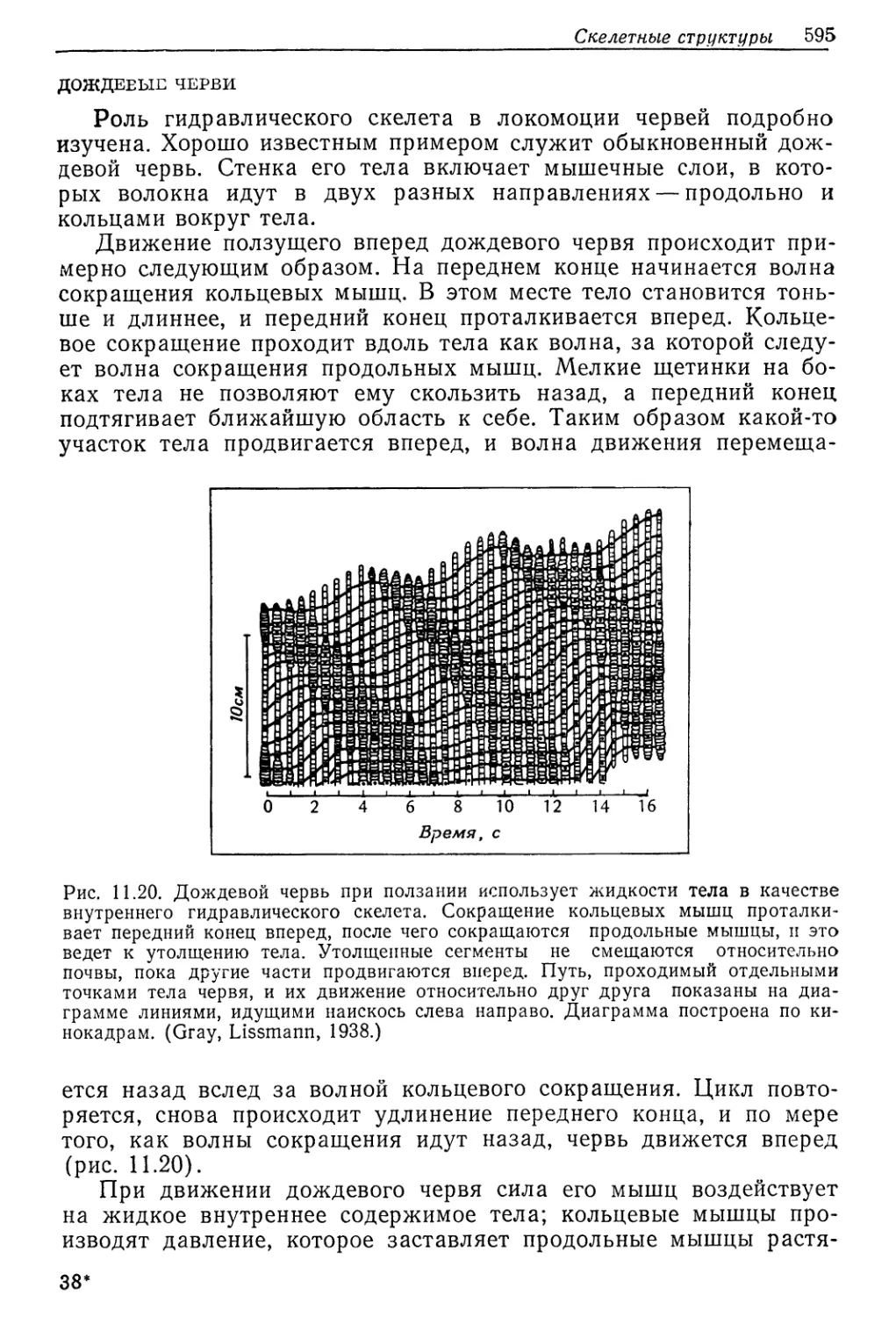
Рис. 9.4. Суточный обмен ионов и воды у личинки Aedes campestris весом 8 мг,
•адаптировавшейся к гиперосмотической щелочной среде. (Phillips et al., 1977.)
анальные папиллы, которые в разбавленной среде служат глав-
ным местом активного поглощения ионов, по-видимому, не игра-
ют роли в выведении избыточной соли.
На рис. 9.4 представлен общий ионный баланс у личинки Ae-
des в щелочной воде. За сутки личинка весом 8 мг выпивает
440 Гл. 9, Вода и осморегуляция
2,4 мкл 'воды, что больше одной трети общего количества воды
в ее теле (6,5 мкл). Количество натрия, поглощаемого за сутки
(1,2 мкмоль), огромно—оно больше того, которое’ содержится
в организме (0,96 мкмоль). Весь этот поглощаемый натрий, а
также небольшое его количество, поступающее через поверх-
ность тела (0,1 м’Кмоль), выводится из организма. На это выве-
дение затрачивается лишь часть (1,8 мкл) воды, поглощаёмой
организмом (2,4 мкл), так что ее остается достаточно для ком-
пенсации потерь от диффузии в концентрированную среду через
поверхность тела (0,2 мкл) и анальные папиллы (0,4 мкл). Та-
ким образом, гипотоничная личинка способна осуществлять ос-
мюрегуляцию и оставаться в равновесии с сильно концентриро-
ванным раствором.
В целом для 'беспозвоночных гипорегуляция—скорее Исклю-
чение, чем правило. .Перейдем теперь к позвоночным: мы уви-
дим, что осмотическая гипорегуляция распространена у них го-
раздо шире, хотя все же это не универсальный способ, так как
есть и другие пути решения осмотических проблем.
ВОДНЫЕ ПОЗВОНОЧНЫЕ
Этот раздел будет посвящен прежде всего рыбам и амфиби-
ям. Никто не стал бы отрицать, чТо киты и морские черепахи то-
же водные животные, но о них речь пойдет в другом контексте.
Они происходят от наземных .предков и дышат воздухом, поэто-
му удобнее рассматривать их как наземных животных, обитаю-
щих в среде, где нет пресной воды.
Основные стратегии, используемые водными позвоночными,
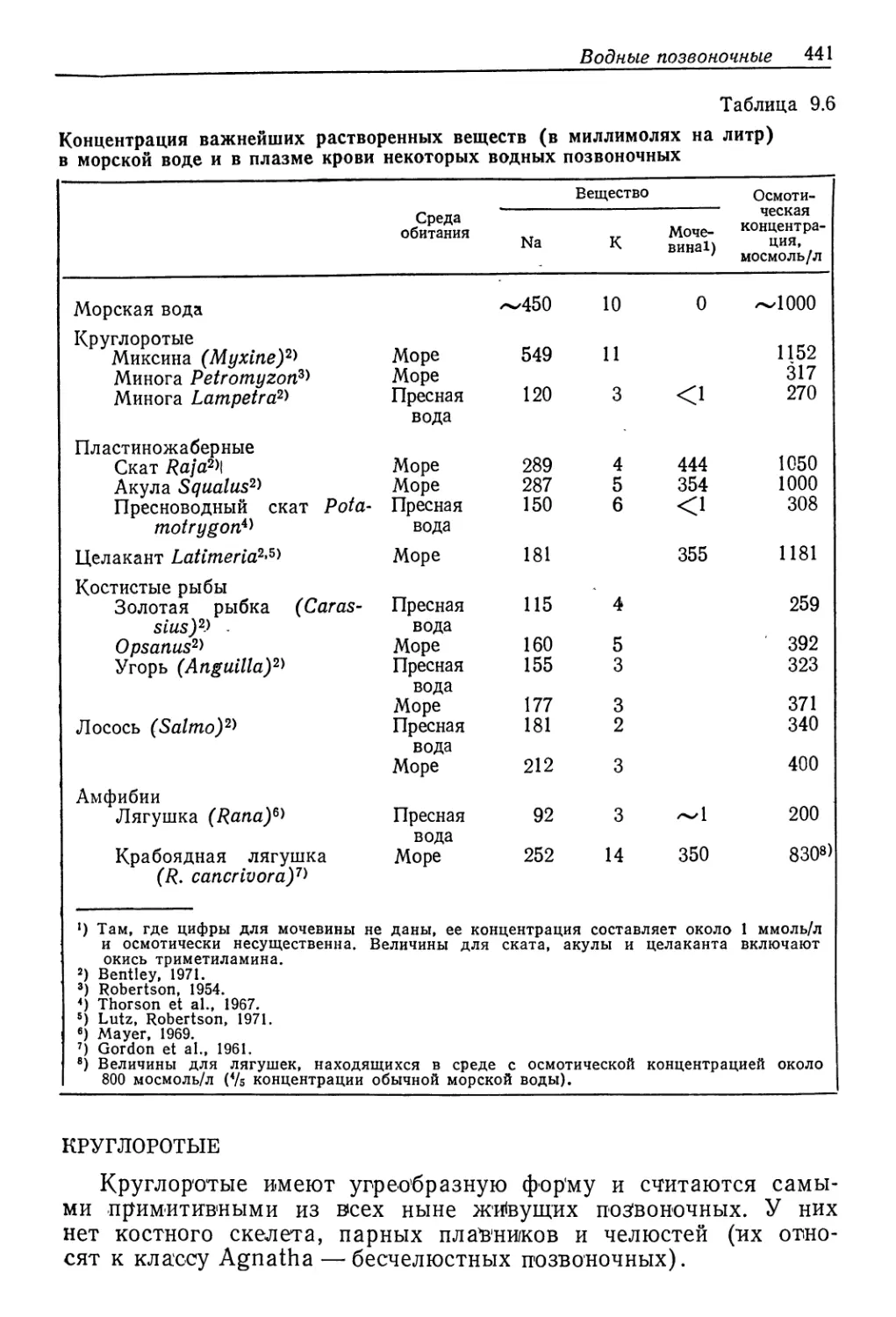
будут понятны из рассмотрения табл. 9.6. В ней приведены при-
меры как морских, так и пресноводных позвоночных. Морские
представители четко делятся на две группы: те, у кого осмоти-
ческие концентрации такие же, как в морс*кой воде, или немного
выше (миксины, пластиножаберные, Latimeria и крабоядная ля-
гушка), и те, у кого они примерно в три раза ниже, чем в мор-
ской воде (миноги, костистые рыбы). Для первой группы сохра-
нение водного баланса не составляет серьезной проблемы, так
как при равенстве внутренней и внешней концентраций осмоти-
ческого тока воды нет. Напротив, явно гипооомютичным «живот-
ным постоянно угрожает утечка воды в осмотически более кон-
центрированную среду. Таким образом, осмотические проблемы
и способы их решения у разных морских позвоночных совершен-
но различны. С другой стороны, у всех пресноводных позвоноч-
ных концентрация солей в жидкостях тела всего лишь в 3—4 ра-
за меньше, чем в морской воде; поэтому они гиперосмотичны по
отношению к среде и в принципе сходны с пресноводными беспоз-
воночными.
Водные позвоночные 441
Таблица 9.6
Концентрация важнейших растворенных веществ (в миллимолях на литр)
в морской воде и в плазме крови некоторых водных позвоночных
Среда обитания Вещество Осмоти- ческая концентра- ция, мосмоль/л
Na К Моче- вина!)
Морская вода ~450 10 0 ~1000
Круглоротые
Миксина (Myxine)2' Море 549 11 1152
Минога Petromyzon6' Море 317
Минога Lampetra2' Пресная 120 3 <1 270
вода
Пластиножаберные
Скат Raja2'\ Море 289 4 444 1050
Акула Squalus2' Море 287 5 354 1000
Пресноводный скат Pota- Пресная 150 6 <1 308
motrygon*' вода
Целакант Latimeria2'6' Море 181 355 1181
Костистые рыбы
Золотая рыбка (С ar as - Пресная 115 4 259
sius)2) - вода
Opsanus2' Море 160 5 392
Угорь (Anguilla)2' Пресная 155 3 323
вода
Море 177 3 371
Лосось (Sal то)2' Пресная 181 2 340
вода
Море 212 3 400
Амфибии
Лягушка (Rana)6' Пресная 92 3 —1 200
Крабоядная лягушка о U Да Море 252 14 350 8308)
(R. cancrivora)7'
9 Там, где цифры для мочевины не даны, ее концентрация составляет около 1 ммоль/л
и осмотически несущественна. Величины для ската, акулы и целаканта включают
окись триметиламина.
2) Bentley, 1971.
3) Robertson, 1954.
4) Thorson et al., 1967.
s) Lutz, Robertson, 1971.
6) Mayer, 1969.
7) Gordon et al., 1961.
8) Величины для лягушек, находящихся в среде с осмотической концентрацией около
800 мосмоль/л (Vs концентрации обычной морской воды).
КРУГЛОРОТЫЕ
Круглоротые имеют угрео'бразную фор'му и считаются самы-
ми прим-итив'ными из Bicex ныне живущих позвоночных. У них
нет костного скелета, парных плавников и челюстей (их отно-
сят к классу Agnatha— бесчелюстных позвоночных).
442 Гл, 9. Вода и осморегуляция
Имеются две группы круглорстых: миноги и миксины. Миноги
живут и в море, и в пресной воде; мийсины — только морские
стеногалинные животные. Интересно, что миноги и миксины раз*
решили проблему жизни в морской воде по-разному. Из всех
истинных позвоночных только у миксин концентрации солей в-
жидкостях тела сходны с концентрациями их в морской воде;
нормальная концентрация натрия в крови миксин даже немного-
выше, чем в окружающей среде. Тем не менее микс'ины в значи-
тельной степени способны к ионной регуляции, хотя, будучи изо-
осмотичными и обладая высокими концентрациями солей, они
ведут себя в осмотическом отношении как -беспозвоночные.
За исключением миКсин, у всех морских позвоночных кон*
центрации солей в жидкостях тела намного ниже, чем (вю внеш*
ней среде. На этот факт ссылались как на аргумент в пользу
того, что позвоночные вначале появились в прёсной воде и лишь
позднее расселились в море. Круглоротые во многих отношениях
сходны с предками современных позвоночных, и знакомство с их
анатомией имело большое значение для интерпретации ископае-
мых форм позвоночных и для понимания их ранней эволюции.
Тот факт, что миксины с их высокой концентрацией солей от*
личаются в этом отношении от остальных позвоночных, означа-
ет, что теория пресноводного происхождения всех позвоночных
не подтверждается физиологическими данными: низкая конце'нт*
рация солей свойственна не всем позвоночным. Однако совре*
менные физиологические признаки не могут служить аргумен*
тами в вопросах эволюции, так как в общем физиологическая
адаптация осуществляется легче, чем морфологические измене-
ния. Поэтому анатомическая структура и ископаемые остатки
имеют большее значение для эволюционных гипотез, чем физио-
логические данные.
Представители второй группы круглоротых — миноги —
встречаются и в пресных водах, и в море, ню даже морская ми-
нога (Petromyzon marinas) относится к анадромным формам1 и
поднимается для нереста в реки.
У миног — и пресноводных, и морских — осмотические кон-
центрации приблизительно в три или четыре раза ниже,"чем у
морской воды. Их главная осмотическая проблема сходна с той»
которая стоит перед костистыми рыбами, как морскими, так и
пресноводным^. Эти проблемы будут подробно рассмотрены поз*
же в этой главе.
1 Рыб, которые для икрометания поднимаются из моря в пресные воды, на*
зывают анадромными (от греч. ana — вверх и dromein — бежать). Хорошо извест*
ными примерами служат лососи и шэд. Катадромными (от греч. kata — вниз>
называют рыб, живущих в пресной воде и спускающихся для нереста в море.
Обыкновенный угорь — катадромная рыба: он развивается до взрослого состоя*
ния в пресной воде, а для размножения спускается в море.
Водные позвоночные 443
МОРСКИЕ ПЛАСТИНОЖАБЕРНЫЕ
Пластиножаберные — акулы и скаты — почти все без исклю-
чения морские животные. Они разрешали осмотическую пробле-
му жизни в море очень интересным способом. Подобию большин-
ству позвоночных, они поддерживают концентрации солей в ж:ид-
костях своего тела на уровне приблизительно в три раза ниже,
чем в морской воде, но при этом сохраняют осмотическое равно-
весие. Это достигается путем добавления к жидкостям больших
количеств органических веществ, главным образом мючйвмны,
вследствие чего общая осмотическая концентрация крови равна
или несколько выше концентрации морской воды (табл. 9.6).
Помимо мочевины осмотически важным органическим веще-
ством в крови пластиножаберных является окись триметиламина.
,NH2
О=С^
\nh2
Мочевина
СН3
I
Нзс—N=O
СН3
Окись триметил-
амина
Мочевина представляет собой конечный продукт белкового
обмена у млекопитающих и некоторых других позвоночных;
у млекопитающих Она выводится с мочой, но у акулы почка ак-
тивно реабсорбирует мочевину, которая, таким образом, остает-
ся в .крови. Окись ^риметиламина встречается у многих мордких
организмов, но ее (Происхождение и обмен еще мало изучены.
Трудно сказать, получают л*и ее акулы с пищей или же она обра-
зуется в их организме.
Содержание мочевины в крови .морских пластиножаберных
в сто с лишним раз больше, чем у млекопитающих, и другие по-
звоночные не могли |бы переносить таких высоких концентраций.
У пластиножаберных мочевина является нормальным компонен-
том всех жидкостей тела, и без ее высокой концентрации ткани
не могут должным образом функционировать. Изолированное
сердце акулы продолжает нормально сокращаться в течение не-
скольких часов, если перфузировать его солевым раствором,
сходным .по ионному составу с кровью и содержащим в высокой
концентрации мочевину. Если же мочевину удалить, состояние
сердца быстро ухудшается и оно перестает биться.
Хотя пластиножа'берные разрешили осмотическую проблему
жизни в море путем поддержания изоосмотичности, они все же
способны к широкой регуляции ионного состава своих жидко-
стей. Например, концентрация натрия удерживается на уровне
примерно вд|вое ниже, чем в морской воде. Это означает, что
натрий будет стремиться диффундировать в организм акулы из-
вне, главным образом через тонкий жаберный эпителий; кроме
того, некоторое количество натрия поступает с пищей. Посколь-
444 Гл. 9. Вода и осморегуляция
ку концентрация натр'ия стремится повыситься, а ее нужно
удерживать на низком уровне, излишний натрий должен выво-
диться наружу.
Часть натрия выводится через почку, но, вероятно, более важ-
ную роль играет особый орган — ректальная железа. .Эта неболь-
шая железа открывается через проток в задний отдел кишечни-
ка— прямую кишку. Она выделяет жидкость с высокой концент-
рацией натрия и хлора, даже несколько более высокой, чем в
морской воде. Например, у акул, находившихся в морской воде
с концентрацией натрия 440 ммоль/л, содержание натрия в сек-
рете ректа'льной железы достигало 500—560 ммоль/л (Burger,
Hess, 1960).
Однако выведение солей у пластиножаберных нельзя пол-
ностью объяснить функцией ректальной железы. Если у колючей
акулы (Squalus acanthias) удалить ректальную железу, то кон-
центрация ионов в плазме все еще сможет оставаться на обыч-
ном уровне, т. е. примерно вдвое ниже, чем в морской воде. По-
скольку жабры немного проницаемы для солей, концентрации
ионо>в в крови постепенно повышались бы, если бы не было дру-
гих путей их выведения. Главную «роль в выделении натрия, по-
видимому, в'се же играет почка; пока не известно, происходит
ли активное выведение ионов из крови пластиножаб^рных также
и через жабры.
Тот факт, -что пластиножаберные находятся почти в осмоти-
ческом равновесии с морской водой, снимает проблему большой
осмотической утечки воды (проблему, которая очень важна для
морских когтистых рыб). Пластиножаберные не нуждаются в
питье морской воды и тем самым избегают поглощения боль-
шого количества натрия.
Однако интересно то, что концентрация растворенных ве-
ществ в крови пластиножаберных рыб обычно немного выше, чем
в морской воде. Это вызывает небольшой осмотический приток
воды через жабры. Таким путем рыба медленно поглощает во-
ду, которая используется для образования мочи и секрета рек-
тальной железы. Поскольку избыток осмотической концентрации
следует приписать мочевине, удержание мочевины можно счи-
тать изящным решением трудной осмотической задачи: оно’ по-
зволяет сохранять низкую концентрацию солей организму, оби-
тающему в море.
ПРЕСНОВОДНЫЕ ПЛАСТИНОЖАБЕРНЫЕ
Подавляющее большинство властиножаберных живет в мо-
ре, но некоторые из них заходят в реки и озера, а отдельные ви-
ды постоянно живут в пресной воде. Даже среди тех пластино-
жаберных, которые считаются типично морскими, есть виды с
поразительной толерантностью к низкой солености внешней ере-
Водные позвоночные 445-
ды. Б некоторых частях света и акулы, и скаты заходят в ре'ки
и оказываются вполне приспособленными к пресной воде. Хо-
рошо известный пример—существование акулы Carcharhinus
leucas в озере Никарагуа. Раньше считали, .что эта акула живет
только в озере, но последние данные показывают, что она мор-
фологически неотличима от соответствующей морской формы и
может свободно переходить в море (Thorson et al., 1966).
Четыре вида пласТиножаберных рыб, обнаруженные в реке
Перак в Малайзии, вероятно, не живут все время в пресной во-
де, а регулярно -заходят в ,нее из моря. Концентрация кров'и у них
ниже, чем у чисто морских форм; в частности, содержание моче-
вины в их кров'и втрое ниже, чем у морских акул, хотя оно все
же гораздо выше, чем у других позвоночных.
Низкий уровень растворенных веществ в крови облегчает за-
дачу осморегуляции, так как осмотический приток воды умень-
шен, а более низкие концентрации солей поддерживать легче.
При меньшем осмотическом притоке воды меньшее количество
ее должно выводиться почками. А поскольку моча неизбежно
содержит в'се-таки растворенные вещества, уменьшение ее коли-
чества в свою очередь снижает потер’и солей. Разумеется, трудно
сказать, я!вляется ли пониженная концентрация крови первич-
ным приспособлением или же это просто результат усиленного
поглощения воды и сопутствующей потери растворенных ве-
ществ с мочой (Smith, 1931).
Одна пластиножаберная рыба — амазонский речной скат Ро-
tamotrygon—постоянно живет в пресной воде. Э'тот скат часто
Таблица 9.7
Концентрации растворенных веществ
в сыворотке крови амазонского ската.
Они примерно такие же, как
у костистых рыб. Хотя скат
относится к пластиножаберным,
мочевина в жидкостях его тела
практически отсутствует.
(Thorson et al., 1967)
Растворенное вещество Количество на 1 л
Натрий 150
Калий 5,9
Кальций 3,6
Магний 1,8
Хлор 149
Мочевина 0,5
Окись триметил- 0
амина
ммоль
Осмоляльность 308 мосмоль
Белок 18 г
446 Гл. 9. Вода и осморегуляция
встречается в осушительных каналах системы рек Амазонки и
Ориноко более чем в 4000 .км от океана. Он, не выживаем в мор-
ской воде даже в том случае, если переход осуществляется пу-
тем постепенного повышения концентрации солей (Pang et al.,
1972). Средний состав его крови (табл. 9.7) свидетельствует
о полной адаптации к пресной воде; мочевины в крови так же
мало, как у пресноводных костистых рыб.
Самая поразительная особенность — это низкая концентра-
ция мочевины; она ниже, чем у большинства млекопитающих.
Ясно, что удержание мочевины не является физиологической не-
обходимостью для всех пластиножаберных рыб. Этот интерес-
ный факт опять-таки показывает, что физиологические функции
гораздо больше подвержены изменениям, чем большинство ана-
томических структур, и что физиологические сходства и разли-
чия не могут служить надежной основой для выводов относи-
тельно эволюции.
ЦЕЛАКАНТ
До 1938 г. считалось, что группа так называемых кистеперых
рыб (Crossoptenygii) вымерла более 75 млн. лет назад, так как
ее представители совершенно не встречаются среди более позд-
них ископаемых находок. В филогенетическом отношении они
отстоят очень далеко от современных рыб, близки к двоякоды-
шащим рыбам и являются предками амфибий. В 1938 г. всемир-
ную научную сенсацию вызвал выловленный у берегов юго-во-
сточной Африки экземпляр целаканта, получившего название
Latimeria. Это была крупная рыба длиной более 1,5 м, весом
больше 50 кг, но она плохо сохранилась, и поэтому не удалось
получить подробных сведений об ее анатомии.
После интенсивных поисков около Мадагаскара было пойма-
но еще несколько живых экземпляров, и хотя ни один из них не
прожил достаточно долго для того, чтобы можПо было подверг-
нуть его физиологическому исследованию, известно, что цела-
кант разрешил свои проблемы осморегуляции так .же, как пла-
стиножаберные рыбы. Данные, приведенные в табл. 9.6, полу-
чены .на замороженном экземпляре латимерии; .высокое содер-
жание мочевины ставит ее в один ряд с пла'стиножаберными.
Дополнительные анализы подтвердили факт ' высокой кон-
центрации мочевины и выявили также высокое содержание оки-
си триметиламина в крови (>100 ммоль/л) и в мышцах
(>200 ммоль/л) (Lutz, Robertson, 1971). Цифры, приведенные
для концентрации натр'ия в плазме, пожалуй, занижены, так как
замораживание и оттаивание приводят к обмену натрием и ка-
лием между плазмой крови и эритроцитам^: натрия в плазме
становится меньше, а калия больше (концентрация калия дейст-
вительно оказалась необычно высокой — 51 ммоль/л) (Pickford,
Grant, 1967).
Водные позвоночные 447
КОСТИСТЫЕ РЫБЫ
Костистые рыбы поддерживают свои осмотические концент-
рации на уровне примерно в три или четыре раза ниже, чем в
морской воде (ом. табл. 9.6). В целом цифры для морс'ких и
пресноводных рыб лежат в одних и тех же пределах, хотя у
морских имеется тенденция к несколько более высоким концент-
рациям. Некоторые рыбы переносят изменение солености в ши-
роком диапазоне и мигрируют из морской '.воды в солоноватую и;
пресную и обратно.
Эти миграции часто связаны с жизненным циклом; например,
лососевые размножаются в пресной воде, их молодь скатывает-
ся в море и, достигнув зрелости, возвращается в пресную воду
для нереста. У обыкновенного угря мы находим обратную кар-
тину: личинки выводятся в море, затем перемещаются с морски-
ми течениями и достигают прибрежных зон, откуда заходят в.
пресные воды, а перед наступлением зрелости угорь возвраща-
ется в море для размножения. Переход из одной среды в дру-
гую требует глубоких изменений в осморегуляторных процессах.
МОРСКИЕ КОСТИСТЫЕ РЫБЫ
Морские рыбы гипоосмотичны, и им постоянно угрожает
утечка воды из организма в более концентрированную морскую’
воду, так как поверхности их тела, особенно обширная поверх-
ность жабр, несколько проницаемы для воды. Эти рыбы долж-
ны как-то возмещать неизбежную осмотическую потерю воды,,
и для этого пьют морскую воду.
Хоть питье компенсирует потерю воды, вместе с водой йз ки-
шечного тракта .всасываются большие количества солей. Кон-
центрация солей в организме возрастает, и встает задача удале-
ния излишней соли. Для того чтобы после питья морской воды в
теле задерживалась только вода, соли должны выводиться в:
концентрации более высокой, чем их концентрация в поступа-
ющей воде. Почка костистой рыбы не может служить для этой
цели, так как она не способна сделать мочу более Концейгриро-
ванной, чем кровь.
Поэтому избыток солей должен выводиться каким-то другим:
органом. Для этого служат жабры, которые, тайим образом, вы-
полняют двойную функцию, участвуя и в осморегуляции, и в га-
зообмене. Секреция соли через жаберный эпителий должна пред-
ставлять собой активный транспорт, так как она направлена от
более низкой концентрации (в крови) к более высокой ('во внеш-
ней среде).
Основные аспе'кты осморегуляции у морских костистых рыб’
суммированы на рис. 9.5. На верхней схеме показано движение-
воды: вода теряется осмотически через мембрану жабр и ,с мо-
чой. Для возмещения ее утеЧки рыба пьет морскую воду, вме-
448 Гл. 9. Вода и осморегуляция
сте с которюй из кишечника всасываются и соли. На нижней схе-
ме показано движение солей, поступающих в организм при
Литье .морокой воды. Двойная стрелка у жабр означает выведе-
ние натрия и хлора путем активного транспорта. Выделение
этих ионов с мочой имеет второстепенное значение, так как у
костистых рыб моча обычно более разбавленная, чем жидкости
Морские костистые рыбы
Соли из
морской
воды *
Mg2*, SO^-
fe моче)
Рис. 9.5. У морской костистой ры-
бы осмотическое давление жидко-
стей тела ниже, чем у воды, в ко-
торой она живет. Вследствие бо-
лее высокой осмотической кон-
центрации среды рыба непрерыв-
но теряет воду (верхняя схема),
главным образом через тонкие
жаберные мембраны. Кроме того,
вода выходит с мочой. Для воз-
мещения этих потерь рыба в зна-
чительных количествах пьет мор-
скую воду. Содержащиеся в этой
воде натрий и хлор всасываются
в кишечнике и выводятся через
жабры путем активного, транспор-
та (двойная стрелка на нижней
схеме); магний и сульфат выде-
ляются через почку.
тела. Однако пбчка играет важную роль в выделении двухна-
.лентных ионов .магНия и сульфата, которые составляют пример-
но одну десятую солей морской воды. ЭГи ионы не выводятся
через жабры, которые, по-видимому, активно выделяют только
натрий и хлор.
Хотя морские рыбы пьют воду, измерение количества выпи-
ваемой воды показало, что с нею поглощается лишь небольшая
доля поступающего натрия, а основной его приток происходит в
другом мес!те— по^вид’имому, в жабрах, обладающих некоторой
проницаемостью. Независимо от того, поступает ли натрий через
всю .поверхность тела или через жабры, ясно, что у рыб, адап-
тированных ,к морской воде, покровы относительно проницаемы
для ионов, а у рыб, адаптированных к пресной воде, относитель-
но непроницаемы для них (Motais, Maetz, 1965).
Изменения проницаемости для натрия и хлора, происходя-
щие при адаптации к разной солености, изучались на рыбке Fun-
dulus heteroclitus, которая легко адаптируется и к пресной, и к
морской воде. Проницаемость снижается через несколько минут
после перевода в пресную воду, но повышение ее при возвраще-
нии в морскую воду занимает много часов (Potts, Evans, 1967).
Преимущество низкой проницаемости для ионов в пресной
воде очевидно, но трудно понять, в чем преимущество более вы-
сокой проницаемости в морской воде. Морские рыбы должны
Водные позвоночные 449
совершать работу, чтобы поддерживать стационарное осмотиче-
ское состояние в морской воде< и малая проницаемость, оче-
видно, должна была бы уменьшать количество необходимой ра-
боты. Рыбе требуется несколько часов, чтобы вернуться к высо-
кой проницаемости в морской воде, и можно только гадать,
почему она не сохраняет постоянно низкую проницаемость, кото-
рая, по-видимому, лежит в пределах ее физиологических воз-
можностей.
Мало вероятно, что'бы весь жаберный эпителий участвовал
в транспорте ионов; последний скорее всего осуществляется осо-
быми крупными клетками, получившими название хлоридных
клеток. До недавнего времени было не я^сно, транспортируется
ли ион хлора активно, а за ним пассивно следует натрий, или
же активно транспортируется ион натрия, а за ним пассивно
следует хлор. КлеТки были названы хлоридными, хотя функция
их не была точно известна (Keys, Willmer, 1932). Но теперь вы-
ясняется, что это наз‘ва’ние, видимо, дано правильно, так как у
угрей, помещенных в морскую во'ду, ион хлора выводиться пу*тем
активного Транспорта (Maetz, Campanini, 1966). Разность потен-
циалов по обе Стороны жаберной мембраны указывает на актив-
ный транспорт хлора, но натрий не всегда находится в пассив-
ном равновесии и тоже может переноситься активно (House,
1963).
ПРЕСНОВОДНЫЕ КОСТИСТЫЕ РЫБЫ
Осмотические условия для рыб в пресной воде примерно та-
ковы же, что и для пресноводных беспозвоночных. Осмотическая
концентрация в крови—порядка 300 мо'смоль/л—гораздо вы-
ше, чем в окружающей пресной воде.
Общая схема осморегуляции у пресноводных костистых рыб
показана на рис. 9.6. Главную проблему создает осмотический
приток воды. Важ'ную роль в этом притоке играют жабры ввиду
их большой поверхности и сравнительно высокой проницаемо-
сти; кожа имеет меньшее значение. Излишек воды выводится
В виде мочи; моча эта очень жидкая и вырабатывается в коли-
чествах, составляющих за сут*ки до одной трети веса тела. .Хотя
она содержит, вероятно, всего лишь 2—.10 ммоль/л растворен-
ных веществ, из-за большого объема мочи происходит значи-
тельная утечка этих веществ, которая должна быть восполнена.
Жабры также в некоторой степени проницаемы для ионов, по-
тёря которых тоже должна быть возмещена их поглощением.
Некоторые растворённые вещества поступают с пищей, но в
основном они поглощаются путем активного транспорта в жаб-
рах. Это удалось показать, поместив рыбу в перегороженную
камеру, где можно отдельно исследовать голову и остальное те-
ло (р'и|с. 9.7). В таких опытах активное поглощение ионов прои-с-
29—1863
450 Гл. 9. Вода и осморегуляция
ходит только в передней части .камеры; из этого следует, что ко-
жа не участвует в нем: за этот процесс ответственны только
жабры.
Пресноводные костистые рыбы
Вода
Моча
Растворенные вещества
Рис. 9.6. У пресноводной костистой ры-
бы осмотическая концентрация жидко-
стей тела выше, чем у внешней среды,
и поэтому в организм все время осмоти-
чески поступает вода, главным образом
через жабры (верхняя схема). Излишек
воды удаляется в виде мочи. Потеря
растворенных веществ через жабры и
с мочой возмещается главным образом
путем активного поглощения их в жаб-
рах (жирная стрелка на нижней схе-
ме).
Рис. 9.7. Поместив рыбы в камеру, где
передняя и задняя части разделены ре-
зиновой перепонкой, можно показать,,
что кожа не участвует в активном по-
глощении ионов: оно происходит только
в жабрах. (Krogh, 1937.)
КАТАДРОМНЫЕ И АНАДРОМНЫЕ РЫБЫ
Большинство костистых рыб обладают лишь ограниченной
способностью переходить из пресной воды в море и обратно; они
сравнительно стеногал'и'нны. Но, как уже было сказано, у миног,
лососей и угрей такие миграции составляют часть нормального
жизненного цикла (Koch, 1968); при этом резко изменяются тре-
бования, предъявляемые к осмореруляторным механизмам.
При переходе угря из пресной воды в морскую осмотическая
потеря воды достигает 4% веса тела за 10 ч (Keys, 1933). Если
лишить угря возможности пиТь морскую воду, введя ему в пи-
щевод раздутый баллончик, он будет непрерывно терять воду и
че’рез несколько дней погибнет от обезвоживания. Но если угорь
может пить, то он вскоре наминает заглатывать морскую воду,
Водные позвоночные 451
потеря в весе прекращается и через день-два наступает равно-
весное состояние. Если же, наоборот, перевести угря из морской
воды в пресную, то .вначале он будет прибавлять в весе, но по-
том усилится образование мочи и через один-два дня также на-
ступит равновесие.
При переходе угря из пресной воды в морскую или обратно
меняется не только направление осмотического тока воды, но
для достижения равновесия и компенсации избытка или потери
растворенных веществ должно изменяться также направление
активного транспорта в жабрах. Как происходит это изменение,
неизвестно, хотя предполагается участие эндокринных механиз-
мов. Неизвестно также, осуществляют л/л транспорт в двух на-
правлениях разные популяци'и клеток, из которых та или другая
активируется при надобности. Вторая возможность состоит в
том, что меняется на обратную полярность транспортного меха-
низма во всех действующих клётках. На этот вопрос пока дет
ответа.
Судя по имеющимся данным, перемена направления транс-
порта в отдельной клетке мало вероятна. Среди многочисленных
органов и типов кле'ток, участвующих в каком-либо активном
транспорте, нельзя назвать ни одного, который определенно был
бы способен к такой функциональной инверсии. Кожа лягушки,
которая, подобно жабрам рыбы, может активно поглощать ионы
из разведенных растворов в пресной воде, по*Видимому, не спо-
собна изменять направление транспорта у того единственного
вида, который обитает в морской воде,—у крабоядной лягушки
(подробнее см. в следующем разделе).
АМФИБИИ
Большинство а'мфибий—водные или полуводные животные.
Они откладывают яйца в воду, и личинки их живут в воде и ды-
шат жабрами. Во время метаморфоза многие (но не все) амфи-
бии переходят на легочное дыхание. Некоторые хвостатые ам-
фибии и во взрослом состоянии сохраняют жабры и остаются
полностью водными животными; большинство лягушек, однако,
выходят на сушу, хотя обычно они живут поблизости от воды
или влажных мест.
Недавно в ,Африке и в Южной Америке было изучено не-
сколько видо'в атиНич'ных лягушек, которые отлично чувствуют
себя в очень сухих местообитаниях и весьма устойчивы к потере
воды путем испарения. Ниже в этой плаве будут описаны их не-
обычные физиологические особенности.
ПРЕСНОВОДНЫЕ АМФИБИИ
В отношении осморегуляции амфибии очень сходны с кости-
стыми рыбами. Практически все они — пресноводные животные,
и у взрослой амфибии главным органом осморегуляци’и служит
29*
452 Гл. 9. Вода и осморегуляция
кожа. Когда животное находится в воде, происходит осмотиче-
ское поглощение воды, которая вно’вь выводится в виде очень
жидкой мочи. Однако теряется некоторое количество растворен-
ных веществ — как с мочой, так и через кожу. Эти потери воз-
мещает активный захват соли из сильно разведенной среды, ^1е-
Рис. 9.8. Аппарат для измерения транспорта натрия в изолированном лоскуте ко-
жи лягушки. (Ussing, Zerahn, 1951.)
Кожа разделяет две камеры с раствором Рингера, и транспорт натрия через
кожу создает разность потенциалов (напряжение) между обеими ее сторонами.
Если теперь пропускать ток в направлении, противоположном разности потен-
циалов, создаваемой транспортом натрия, то сила тока, при которой разность
потенциалов исчезнет, будет прямой мерой транспорта натрия через кожу.
А и А'— агаровые мостики, соединяющие растворы с каломельными электрода-
ми; Б и Б' — агаровые мостики для электрической связи с внешними источника-
ми напряжения.
хан1из(м Tjpa’HonojpiTa локализован у взрослого животного в коже,
и кожа лягушки стала общеизвестной моделью для изучения ак-
тивного транспорта ионов.
Куски лягушачьей ко'ж!и легко вырезать и ,использовать в ка-
честве мембраны, разделяющей две камеры, (которые заполня-
ются жидкостями с разными концентрациями. Анализируя из-
менения, происходящие ,в обеих камерах, можно изучать транс-
портную функцию кож!и (рис. ,9.8). Такие изолированные кусочки
кожи переживают в тече'ние многих часов. Этот аппарат для
исследования процессов активного транспорта ’первоначально
был сконструирован Уссингом, и его называют камерой Уссинга.
Когда кожа лягушки разделяет в такой камере два солеВых
раствора1 одинакового состава, между внутренней и внешней
1 Раствор соли должен иметь такую же величину pH и осмотическую кон-
центрацию, как кровь, и содержать приблизительно в тех же концентрациях
важнейшие ионы — Na+, К+, Са2+ и С1“. Такой сбалансированный солевой рас-
твор называют раствором Рингера — по имени английского физиолога, который
установил, что для переживания изолированного сердца лягушки необходимы
определенные количественные соотношения между этими ионами.
Водные позвоночные 4&3
сторонами кожи быстро создается разность потенциалов около
50 мВ. Внутренняя сторона заряжена положительно, поэтому
предполагают, что разность потенциалов обусловлена активным
переносом внутрь положительных ионов натрия. Когда разность
потенциалов установилась, ионы хлора .проходят через кожу пу-
тем Диффузий, ускоряемой электрическим полем. Накоплено-
огромное количестве данных в пользу такой интерпретаци/и. На
активный характер транспорта ясно указывают возникающий по-
тенциал и тот факт, что ингибиторы метаболизма (напрУмер,
циаУид) подавляют как образование этого потенциала, так и
перенос ионов.
Путем приложения к коже внешней разности потенциалов
той же величины, но с обратным знаком можно привести кож_-
ный потенциал к нулю. Ток, нужный для удержания потенциала
на нуле, должен быть равен току, генерируемому транспортом
натрия через кожу. Поэтому такой ток, называемый закорачива-
ющим, служит прямой мерой переноса натр'ия внутрь. Такой ме-
тод стал весьма ценным приемом измерения активного транспор-
та ионов и во многих других системах.
ЛЯГУШКА, ОБИТАЮЩАЯ В СОЛЕНОЙ ВОДЕ
Лягушки и хвостатые амфибии обычно жи'вут только в прес-
ной воде, а в морской воде они через несколько часов погиба-
ют. Единственное исключение — обитающая в юго-восточной
Азии крабоядная лягуш1ка (Rana cancrivora). Эта маленькая,
обычного вида лягушка ж,и1вет в прибрежных мангровых боло-
тах, где в поисках пищи плавает в неразбавленной морской воде.
Если лягушке нужно сохранять в морокой воде сравнительно
низкую концентрацию солей, характерную для поз’воночных, то
для решения этой задачи у нее ес'ть два пути. Оди’н из них (ис-
пользуемый морскими костистыми рыбами) состоит в том, что-
бы противодействовать «осмотической потере воды и компенси-
ровать диффузию солей внутрь через кожу. Другой способ
(свойственный морским пластиножаберным рыбам) заключает-
ся в накоплении мочевины и поддержании осмотического равно-
весия между жидкостями тела и внешней средой, ч’то снимает
проблему осмотической потери воды. Лягушка, обитающая в со-
леной воде, использует тот же способ, что и пласгНиножабер-
ные,—к жидкостям тела добавляются большие количества мо-
чевины, так что его концентрация может достигать 480 ммоль/л
(Gordon et al., 1961).
Такая стратегия представляется удачной. Кожа амфибий от-
носительно проницаема для воды, и поэтому им проще поддер-
живать такую же осмотическую концентрацию, как во внешней
среде, и устранять осмотическую потерю воды. Для того чтобы
устранить потерю воды путем одного лишь повышения внутрен-
454 Гл. 9. Вода и осморегуляция
ней концентрац'ии соли, лягушка должна была бы обладать то-
лерантностью к солям, исключительной среди позвоночных (кро-
ме миксин). А если бы она прибегла к способу, 'используемому
костистыми рыбами, и оставалась пипоосмОтичной, ее солевой
баланс еще больше нарушался бы из-за необходимости шить со-
леную воду.
Крабоядная лягушка, помещенная в морскую воду, не будет
совершенно изооамотична с окружающей средой; подобно аку-
лам, она остается слегка гиперосмотичной. В результате проис-
ходит медленный приток воды, полезный потому, что он нужен
для о-бразо'вания мочи. Получать воду таким образом, несомнен-
но, выгоднее, чем (пиТь морскую воду, что неизбежно повысило
бы поступление в организ'м солей.
У крабоядной лягушки, как и у пластиножаберных, мочевина
является осмотически важным веществом, а не просто ркскре-
том. Кроме того, она необходима для нормального сокращения
мышц, которое без нее быстро нарушается (Thesleff, Schmidt-
Nielsen, 1962). Поскольку мочевина нужна для нормальной жизни
этого животного, она должна задерживаться в организме, а не
выводиться с мочой. У акул это происходит благодаря активной
реабсорбции в почечных канальцах (см. гл. ДО). Но у крабояд-
ной лягушки м(о'чев1ина накапливается гла’вным образом вследст-
вие уменьшения объема мочи, когда лягушка находится в мор-
ской воде. По-видимому, мочевина не реабсорбируется активно,
так как концентрация ее в моче всегда немного выше, чем в
плазме (Schmidt-Nielsen, Lee, 1962).
Головастики крабоядной лягушки еще более толерантны к
высокой солености, чем взрослые животные. Но способ о'сморе-
гуляции у них такой, как у косТистых рыб, и, следовательно,
иной, чем у пластиножаберных и у взрослых лягушек (Gordon,
Tucker, 1965).
Хотя и головастики, »и взро’слые крабояд’ные лягушки очень
хорошо переносят морскую воду, они все-таки нуждаются и в
пресной воде, поскольку для оплодотворения яиц и для мета-
морфоза во взрослую форму им требуется относительно низкая
концентрация солей в воде. Из-за частых ливней в тропиках Око'-
ло берега легко образуются временные пресные водоемы, так
что икрометание может происходить в несоленой воде. Голова-
стик -хорошо переносит соль, но метаморфоз не начинается, пока
соленость остается высокой, и организм проходит эту критиче-
скую стадию только после того, как сильный дождь- разбавит
воду.
Хотя для размножения крабоядной лягушки нужна пресная
вода, толерантность к морокой воде позволяет ей использовать
богатую .прибрежную тропическую среду, недоступную всем ос-
тальным амфибиям.
Наземная среда 455
НАЗЕМНАЯ СРЕДА
Самое большое физиологическое преимущество жизни на су-
ше состоит в доступности кислорода; наибольшей угрозой для
наземной жизни является опасность высыхания. Успешный пе-
реход к наземной ж!изни в широком масштабе произошел толь/ко
у двух типов животных — у членистоногих и позвоночных, ко-
торые. Ж1и|вут и благоденствуют даже в самых сухих и жар'ких
местообитаниях. Кроме того, некоторые улитки тоже жчйвут на1,
суше и являются подлинно назем'ными животными; некоторые
из них встречаются даже в пустынях. Очевидно, что угроза вы-
сыхания представляет собой серьезную преграду для выхода на
сушу, так как многим «наземным» животным, не относящимся к
указанным типам, нужны достаточно влажные местообитания,
и эти организмы являются наземными только в формальном
смысле сло^ва. Например, дождевому червю совершенно необхо-
дима влажная среда, и он быстро- погибает от обезвоживания,
если даже ненадолго попадает на открытый воздух. Он мало
устойчив к потере воды и ищет для себя микросреду, где воз-
дух и пю'чва влажные (у него Имеются соответствующие пове-
денческие реакции). Другим примером могут служить лягушки
и улитки, которых мы можем объединить по признаку влажной
Кожи, быстро испаряющей воду.
Однако, прежде чем о'бсуждать данные о конкретных живот-
ных, надо кое-что знать о физических факторах, влияющих на
испарение. Рассказав об этом, мы сможем перейти сначала к не-
которым наземным животным с влажной кожей, жизнь которых
сильно завидит от наличия воды, а затем к членистоногим и на-
земным позвоночным.
ИСПАРЕНИЕ
Мы знаем, что испарение с открытой водной поверхности
возрастает с температурой и ч*го в сухом воздухе оно и|дет быст-
рее, чем во влажном. Для того чтобы понять, как идет испаре-
ние с поверхности тела животного, нужно иметь более точное
•представление о физических законах, управляющих скоростью
перехода воды из жидкой фазы в газообразную.
Давление водя'ных паров над открытой водной поверхностью
быстро увеличив'ается ,с повышением температуры (рис. 9.9).
Для некоторой ориентировки нетрудно запомнить, что при тем-
пературе тела млекопитающего (38 °C) давление водяных паров
(50 мм рт. ст.) приблизительно вдвое больше, чем при комнат-
ной температуре 25 °C (24 мм рт. ст.) и в десять с лишним раз
выше, чем при 0°С (4,6 мм р(т. ст.).
Если воздух уже содержит какое-то количество водяных па-
ров, то разность между их давлением на поверхности воды и в
456 Гл. 9. Вода и осморегуляция
воздухе будет меньше, и соответственно слабее будет сила, за-
ставляющая водяные пары диффундировать из насыщенного1 (по-
граничного сдоя у водной поверхности в воздух.
В качестве весьма приближенной меры этой силы, вызываю-
щей испарение, многие исследователи .используют дефицит на-
сыщения. Он выражается разностью между давлением водяных
паров у открытой водной поверхности (при данной температуре)
и в воздухе. Если относительная влажность воздуха равна 50%,
то дефицит насыщения возрастает с температурой как разность
между двумя кривыми на р(и'с. 9.9.
Рис. 9.9. Давление водяных паров над свободной водной поверхностью быстро
возрастает с повышением температуры (белые кружки, жирная линия). При тем-
пературе тела млекопитающих (38 °C) оно приблизительно в два раза больше,
чем при комнатной температуре (25°C), а при более высокой температуре растет
все круче.
Дефицит насыщения оказался полезным во многих экологи-
ческих исследованиях, касавшихся как животных, так и расте-
ний, но теоретически такое его использование не вполне правиль-
но, так как это не единственный физический фактор, определя-
ющий скорость Испарения воды (Ramsey, 1935; Edney, 1957).
Во-первых, на эту скорость очень сильно влияет движение воз-
духа, т. е. свободная или принудительная конвекция. Второй
фактор, который при некоторых обстоятельствах играет важную
роль, — это охлаждение той поверхности, ,с которой происходит
испарение. Поэтому испаряющая поверхность может быть хо-
лоднее, чем остальные части системы, причем разница темпера-
тур зависит от скорости испарения, а также от переноса, тепла
из других участков, т. е. от параметра, который неред'ко трудно
бывает измерить. Далее, диффузия водяных паров в воздух
ускоряется с повышением темпе'раТуры. И наконец, диффузия
растет с понижением барометрического давления. Хотя каждая
из этих переменных является сложной функцией, которая в свою
очередь влияет на все другие переменные, математическое мо-
Животные с влажной кожей 457
дел(И/ро1вание меже*? оказаться чрезвычайно эффективным спосо-
бом анализа обмена воды ,в наземных условиях (Tracy, 1976).
Для животных в естественной ср'еде одной йд са'мых важных
•переменных является подвижность воздуха. При ветре (а в при-
роде воздух практически никогда не бывает совершенно непо-
движным) воздушный слой, прилагающий к поверхности, быстро
обновляется и испарение усиливается. Мы ощущаем это как ох-
лаждение, когда влажную кон^у обдувает ветер или по'ток воз-
духа от вентилятора. Охлаждение поверхности, вызываемое ис-
парением, изменяет также плотность прилегающего воздуха и
создает конвекционные токи. Сила их на горизонтальной и вер-
тикальной поверхностях различна, и в результате при прочих
равных физических условиях испарение с влажной поверхности
меняется в зависимости от ее ориентации. На'коне'ц, испарение
зависит еще от кривизны поверхности.
Из-за этих сложностей трудно точно описать переменные, оп-
ределяющие скорость испарения влаги. Дефицит насыщения
остается самым полезным приближением, каким мы располага-
ем, и, относя определенную скорость испарения к дефициту на-
сыщения, мы полуЧаем достаточно хорошую основу для сравне-
ний. Но следует иметь в виду, что результаты, получаемые для
животных разной величины и формы при равных, казалось бы,
условиях, не дают точной основы для сопоставления, и, в част-
ности, их нельзя .прямо сравнивать с испаре'нием на горизон-
тальной поверхности открытого, водного пространства.
ЖИВОТНЫЕ С ВЛАЖНОЙ КОЖЕЙ
Рассмотрим два крайних случая. С влажной поверхности те-
ла ж/йвотного идет интенсивное испарение, и потеря воды опре-
деляется главным образом выходом водяных паров в окружа-
ющий воздух (диффузия при участии конвекции). В противопо-
ложном случае (например; при испарении с сухой кутикулы на-
секомого) наибольшим препятствием служит сама поверхность,
и всякое изменение в проницаемости этой преграды сильно влия-
ет на интенсивность испарения. В таком случае изменения в де-
фиците насыщения, кон'векцин и т. д. не имеют большого значе-
ния, и иногда ими даже можно пренебречь. Эти два случая мо-
гут быть описаны как «система, ограниченная паром» (vapor-
limited syste’m), в которой испарение лимитируется переносом
воды в воздухе, и «система, ограниченная оболочкой» (membra-
ne-limited system), в которой главным препятствием для испаре-
ния служит мембрана (Beament, ,1961).
ДОЖДЕВЫЕ ЧЕРВИ
Дождевой червь, помещенный в сухой воздух, быстро теряет
в весе в результате испарения влаги. Он отдает много воды
453 Гл. 9. Вода и осморегуляция
(табл. 9.8) и в сухом .воздухе быстро погибает. Если частично
обезвоженного чер'вя поместить в U-образ'ную трубку так, чтобы
он был погружен ,в воду, но ан.ус и рот находились бы над ее по-
верхностью, то червь будет поглощать воду. Его кожа легко
проницаема для воды в обоих направлениях. Более тщательное
исследование' показывает, что организм дождевого червя ведет
себя очень сходно с организмом типичного пресноводного жи-
вотного. Когда че)рвя помещают в солевые растворы разной кон-
центрации, жидкости его тела остаются гипертоничными по от-
ношению к внешней среде (рис. 9.10). Моча, однако, остается
Рис. 9.10. При погружении дождевого червя в растворы хлористого натрия раз-
ных концентраций он ведет себя как типичный пресноводный организм, способ-
ный к осморегуляции. Он остается гиперосмотичным по отношению к среде при
всех, и особенно при низких, концентрациях. (Ramsay, 1949.)
гипотоничной по отношению к жидкостям тела, и в этом дожде-
вой червь тоже сходен с типичным пресноводным животным
(Ramsay, 1949).
Эти осмотические отношения показывают, что по особенно-
стям водного баланса дождевой червь действительно ближе к
пресноводным, чем к истинно наземным животным. Он роет хо-
ды в почве, где воздух полностью насыщен влагой и где его тело
соприкасается с частицами поч'вы, покрытыми тонкой пленкой
свободной воды. Дождевой червь не может жить в совершенно
сухой почве—<в ней он бьгстро теряет воду и гибнет.
Животные с влажной кожей 459
Таблица 9.8
Испарение воды с поверхности тела
различных животных при комнатной
температуре. Цифры показывают
порядок величины; точные значения
меняются в зависимости от условий
опыта. Все величины выражены в
микрограммах воды, испаряющейся
за 1 ч с 1 см2 поверхности тела при
дефиците насыщения 1 мм рт. ст.
(О 13 кПа) (Schmidt-Nielsen, 1969)
Дождевой червь 400
Лягушка 300
Саламандра 600
Садовая улитка, активная 870
Садовая улитка, неактивная 39
Человек (не потеющий) 48
Крыса 46
Игуана 10
Мучной червь 6
ЛЯГУШКИ И ДРУГИЕ АМФИБИИ
У лягушки, даже пойманной на суше, кожа влажная и про-
хладная. Интенсивность испарения воды с ее кожи примерно та-
кая же, как у дождевого червя (табл. 9.8), а двукратное разли-
чие между лягуш(кой и саламандрой по причинам, изложенным
выше, несущественно. У обеих скорость испарения того же по-
рядка, что и при испарении с открытой водной поверхности, так
что кожа амфибий, по-видимому, не служит сколько-нибудь зна-
чительным препятствием. Поэтому взрослые амфибии, которые
живут на суше и дышат воздухом, обычно встречаются о'коло
воды и в сырых местах, где испарение невелико. При погруже-
нии в воду они в осмотическом отношении ведут себя как ти-
пичные пресноводные животные.
Хотя амфибии обладают влажной кожей и обычно держат-
ся поблизости ют водоемов, некоторые из них, как это ни пара-
доксально, хорошо приспособились к жизни в пустыне. Несколь-
ко видов лягушек обитают в засушливых пустынных областях
внутренней Австралии. Они прячутся в норах на глубине не-
скольких футов, где проводят в спячке долгие периоды засухи.
Поскольку размножение1 их происходит в воде, ойи производят
потомство только после дождей, создающих временные водоемы.
Во время дождя эти лягушки выходят из нор, восстанавлива-
ют нормальное содержание воды в своем теле и откладывают
яйца; из яиц чрезвычайно быстро выводятся головастики, кото-
рые проходят метаморфоз до того, как высохнут лужи. Когда
лягушка снова впадает в спячку, ее мочевой пузырь бывает на-
полнен Ж1ИДКЮЙ мочой, которая играет в'ажную роль в 'водном?
460 Гл. 9. Вода и осморегуляция
балансе. У некоторых лягушек вес этой мочи составляем до 30%
общего веса тела. Эта моча ’чрезвычайно жидкая, и ее осмоти-
ческая концентрация соответствует менее чем 0,1% NaCl. Моча
служит основным запасом воды и постепенно расходуется во
вре'мя спячки (Ruibal, 1962).
Сообщают, что Австралийские аборигены используют пустын-
ную лягушку Chiroleptes как источник питьевой воды, поскольку
она так раздувается от жидкой моч!и, чМо становится «похожа
на шишковатый теннисный мяч» (Buxton, 1923). Когда запас во-
ды в мочевом пузыре иссякает, дальнейшее обезвоживание ве-
дет к концентрированию жидкостей тела и прогрессирующей де-
гидратации крови и тканей.
Лягушки, проводящие период засухи в глубоких норах, сход-
ны в этом отношении с двоя ко дышащими рыбами, которые зары-
ваются в ил. Когда озеро выСыхает, рыба внедряется в грунт и
остается там в сухом коконе, из которого к твердой, как камень,
поверхности ведет дыхательный канал. В таком состоянии рыба
может прожить несколько лет и для сохранения минимального
уровня метаболизма постепенно расходует белки своего тела.
Б организме не Хватает воды для выделения мочевины, образу-
ющейся при белковом обмене, и концентрация мочевины в жид-
кости тела может дойти до 500 ммоль/л (Smith, 1959).
Идет ли испарение с кожи у пустынной лягушки менее ин-
тенсивно, чем у других амфибий? При измерении в сходных ус-
ловиях ('включая температуру и влажность воздуха) разница в
испарении оказывается незначительной, и она не позволяет объ-
ясни/ть столь большое' различие в среде обитания. Утверждали,
что лягушки теряют воду быстрее, чем жабы, у которых кожа
более толстая и ороговевшая, но, по-видимому, это не так (Bent-
ley, 1966). Поэтому уменьшение испарения с кожи, очевидно,
не играет важной роли в адаптации амфибий к наземной
жизни.
Однако существует несколько поразительных исключений.
Южноафриканская лягушка Chiromantis не нуждается во влаж-
ной среде, чтобы избегать быстрого обезвоживания. Помещен-
ная в сухую атмосферу, она теряет путем испарения во много
раз меньше воды, чем другие лягушки; по этому признаку она
фактически очень близка к реп*тил1иям (рис. 9.11). У некоторых
южноамериканских лягушек рода Phyllomedusa испарение тоже
составляет лишь около V20 величины, характерной для других
лягушек (Shoemaker, McClanahan, 1975). В коже Phyllomedusa
имеются особые железы, выделяющее восковидный секрет, ко-
торый, по-видимому, делает кожу водонепроницаемой: поверх-
ность кожи становится настолько гидрофобной, что капельки во-
ды не растекаются по ней, а ска!ты/ваются с нее (Blaylock et al.,
1976). У Chiromantis нет желез, выделяющих воск, и ее кожа
непроницаема для воды благодаря какому-то иному механизму.
Животные с влажной кожей 461
Эти необычные лягушки обладают еще 'одной физиологиче-
ской особенностью, резко отличающей их от остальных лягушек.
Они выделяют не мочевину, как все другие амфибии, а моче'вую
кислоту, что характерно для рептилий. Подробнее об этом будет
сказано в гл. 10 в связи с проблемой выделения азота.
Наличие у некоторых амфибий физиологических черт, прису-
щ'их рептилиям, служит иллюстрацией того, о чем уже говори-
Рис. 9.1. У лягушки Chiromantis при 25 °C и относительной влажности 30%
испарение воды идет очень медленно — так же, как у рептилий (Chamaeleo). По-
сле 6 дней пребывания на воздухе потеря веса составляет у нее лишь несколько
граммов. Другие амфибии быстро теряют в весе. Жаба (Bufo) теряет около
40% веса тела и погибает через 2 дня, обыкновенная лягушка (Rana) гибнет че-
рез день, а морская амфибия Xenopus — меньше чем через 10 ч. (Loveridge,
1970.)
лось выше. Физиологические -признаки, как правило, гораздо
пластичнее и легче .адаптируются к условиям среды, чем морфо-
логические; поэтому они менее пригодны для установления эво-
люционных отношений.
462 Гл. 9. Вода и осморегуляция
УЛИТКИ
Улитки и слизни обладают влажной кожей, с поверхности
•которой интенсивно идет испарение (табл. 9.8). Для голых слиз-
ней необходима влажная среда, и они активны преимуществен-
но после дождя и ночью, когда относительная влажность высо-
ка. В остальное время они Скрываются в такой микросреде, где
влажность достаточна для них. В отличие от слизней улитки но-
сят на себе водонепроницаемую раковину, в которую могут пря-
таться.
Было высказано предположение, что слизь, покрывающая те-
ло улитки, препятствует испарению влаги, но это трудно понять.
Слизь, снятая с животного, испаряет воду практически так же
быстро, как свободная водная поверхность, пока эта слизь не
высохнет полностью. Однако этот вопрос не вполне ясен, так
как, если слегка прикоснуться к находящейся в докое улитке, из
слизистых желез »в ее коже выделяется свежая слизь и испаре-
ние усиливается (Ma'chin, 1964).
Сухопутная улитка, находящаяся в неактивном состоянии
или спячке, превосходно защищена от .потери воды. Она уходит
в свою раковину и закрывает отверстие эпифрагмой—«мембра-
ной, которая состоит из высохшей слизи, а иногда содержа так-
же много кристаллического карбоната кальция.
Благодаря очень малой потере воды неактивной улиткой,
ушедшей в раковину, закрытую эпифрагмой, некоторые виды вы-
живают в жарких и сухих пустынных местностях. Одна из таких
улито(к, Sphincterochila, обитает в пустынях Ближнего Востока.
Раковины с этими улитками' в состоянии спячки .можно увиде’ть
на поверхности почвы в середине лета, ничем не прикрытые от
воздействия солнца и жары. Улитки становятся активными по-
сле дождей, которые выпадают в зимние месяцы, с ноября по
март; в это время улитки питаются и размножаются.
На протяжении долгого засушливого сезона улитки пребы-
вают в состоянии спячки. В этот период потеря воды у Sphinc-
terochila составляет меньше 0,5 -мг в день, а так как в организ-
ме животного имеется около 1,5 г воды, оно может прожить не-
сколько лет и не подвергнуться сильному обезвоживанию. Само
тело улитки (без известковой раковины) содержит больше 80%
воды. Такое высокое содержание воды сохраняемся даже в са-
мое жаркое и сухое время года. Это означает, что улиТка не
расходует постепенно свои водные запасы: проблема ре’шается
здесь г/утем необычайного снижения потерь воды в период спяч-
ки (Schmidt-Nielsen et al., 1971).
ЧЛЕНИСТОНОГИЕ
Самые преуспевающие наземные животные относятся .к типу
членистоногих, который включает насекомых, ракообразных и
Членистоногие 463
некоторых других животных, обладающих членистом жестким
наружным скелетом. Насекомые по числу видов сста1вили дале-
ко позади «все остальные группы животных— описано около
миллиона различных видов насекомых.
Насекомые и паукообразные (пауки, клещи, бкорпионы и
др.) — преимущественно наземные животные, и они очень хоро-
шо приспособлены к жизни на суше и воздушному дыханию.
Только .немногие Из них вторично переселились в пресную воду,
а морских форм в этих двух классах дочти нет. Наземными яв-
ляются еще два класса членистоногих: дВупарноногие (Diplopo-
da) и губоногие , (Chilopo’da). В отл!ичие от .них большинство ра-
кообразных— водные животные, так же как и меростомовые
(Xiphosura), предста»вленные живым ископаемым — мечехво-
стом (Limulus).
РАКООБРАЗНЫЕ
Хотя большинство ракообразных живет в воде, значительное
число их, включая некоторых десятиногих (крабов и раков), от-
носится к наземным животным—даже в такой мере наземным,
что в воде они погибают. Однако преимущественно они -сосредо-
точены во влажных местообитаниях и по соседству с водой. В то
же время некоторые равноногие (Isopoda), например мокрицы,
живут только на суше, а» отдельные виды даже в жарких, сухих
пустынях.
Крабы обычны в литоральной зоне, а ,в тропиках многие из
них постоянно живут выше верхней границы прилива. Но им
нужна влажная среда, и часто их норы расположены там, где
на дне остаются лужи воды. Более сухопутные виды почти пол-
ностью эмансипированы от воды, но для размножения все 'кра-
бы возвращаются в воду.
ПоЛуназемные и наземные крабы, как правило, обладают
превосходной осморегуляц)ией. В зависимости от условий и сво-
их потребностей они одинаково эффективно осуществляют и ги-
пер-, и г.ипорегуляцию. Интересная особенность некоторых сухо-
путных крабов (Cardiosoma и Gecarcinus) —То, что они могут
вбирать воду из сырого песка и влажного субстрата даже в от-
сутствие видимых количеств свободной воды. Таким образом
они могут выживать в течение многих месяцев без прямого до-
ступа к воде (Gross et al., 1966; Bliss, 1966).
Краб-привидение (Ocypode quadrata}, снующий, как нередко
можно видеть, по песчаному берегу, долж-е'н часто погружаться
в морскую воду. Хорошо известные манящие крабы (Uca), жи-
вущие на побережьях всех тропических морей .и даже в умерен-
ных зонах, роют глубокие норы на берегу около или выше верх-
ней границы при'лива, но им необходимо часто возвращаться в
воду.
464 Гл, 9, Вода и осморегуляция
Обыкновенные наземные равноногие рачки, называемые мок-
рицами, чаще всего встречаются во влажных местах подальше
от открытых участков. В течение дня они прячутся, а передви-
гаются ночью, когда относительная влажность повышена. Для
размножения эти ра/кообразные совершенно не нуждаются в
свободной воде и, таким образом, я'вляются истинно наземными
животными.
Хотя мокрицы большей частью живут в микросреде с высо-
кой влажностью, иногда они приходят в соприкосновение с бо-
лее сухйи воздухом; тогда они. теряют воду Вследствие испаре-
ния и должны ее возмещать. Как правило, мокрицы питаются
влажными разлагающимися растительными остатками, которые,
вероятно, служат для них обычным источником воды. Однако
установлено, что некоторые равно'нопие могут поглощать воду
путем питья, а также через анус. Если их содержать на влаж-
ном гипсе, они даже способны поглощать воду с его поверхности
(Edney, 1954).
По сравнению с кутикулой насекомых кутикула равноногих
раков сравнительно проницаема для воды. Главная причина это-
го различия состоит, по-видимому, в том, что кутикула насеко-
мого покрыта тонким -слоем воска, который сильно уменьшает
потерю воды; все попытки обнаружить восковой слой у мокриц
оказались безуспешными (Edney, 1954).
Казалось бы, наземных ракообразных вряд л И можно найти
в пустыне, а между тем мокрица Hemilepistus очень часто 'встре-
чается в такого рода местах. Эти животные продолжают ды-
шать жабрами и выживают благодаря тому, что выкапывают
узкие вертикальные ходы глубиной около 30 см, где проводят
жаркие часы суток. Измерения температуры и влажности пока-
зали, что их убежища гораздо прохладнее, чем поверхность
грунта в пустыне, и, что особенно важно, там высока относи-
тельная влажность — около 95%. Кутикула Hemilepistus не-
сколько менее проницаема для воды, чем у других мо<криц, но
главным средством адаптации этого животного к условиям пу-
стыни, по-видимому, служит его поведение (Edney, 1956).
НАСЕКОМЫЕ И ПАУКООБРАЗНЫЕ
Если многочисленность означает преуспевание, то насекомые,
бесспорно, самые преуспевающие животные на Земле. Различ-
ных видов насекомых больше, чем видов всех остальных живот-
ных вместе взятых, и, кроме того, популяции многих насеко{мых
чрезвычайно велики. Они живут почти во всех мыслимых ме’сто-'
обитаниях на суше и в пресной воде; только полярные области
и океан, по-видимому, относительно свободны от них. Сухость,
условия пустыни и отсутствие -открытой воды, как видно, не слу-
жат преградой для этих животных. Платяная мбль, отлично су-
Членистоногие 465
ществующая на шерстяной одежде, или мучной хрущак, прово-
дящий .всю свою жизнь от яйца до и(маго в сухой муке, живут,
развиваются и благоденствуют в среде, практически лишенной
свободной воды. Тем не менее, поскольку взрослое насекомое
почти на две трети состоит из воды, эти организмы должны об-
ладать необыкновенной способностью удерживать воду и сокра-
щать ее потери.
В'виду значительного сходства между насекомыми и пауко-
образными эти два класса будут рассмотрены вместе.
ВОДНЫЙ БАЛАНС: ПРИБЫЛЬ И ПОТЕРИ
Для сохранения водного баланса организма все посери во-
ды за определенное время должны уравновешиваться ее равной
прибылью.
Водный баланс складывается из следующих компонентов:
Потеря воды
Испарение
с поверхности тела
с поверхности дыхательных органов
Кал
Моча
Другие (специфические) выделения
Прибыль воды
Питье
Поглощение через поверхность тела
из воды
из воздуха
Вода в пище
Вода—продукт окисления (метаболическая вода).
Задача поддержания баланса решается разными путями. Во-
да может быть в избытке или ее может не хватать, и поэтому
физиологические механизмы должны справляться с обеими си-
туациями. Например, пресноводное насекомое заглатывает мно-
го воды вместе с пищей, и в то же время вода поступает в ор-
ганизм путам осмоса. Избыток воды должен быть удален, и эту
задачу обычно выполняет ренальный орган или почка, которые
для этого должны вырабатывать большее количество жидкой
мочи. Напротив, наземное насекомое, обитающее в сухой среде,
получает очень мало воды, и все потери должны быть сведены
к Такому уровню, чтобы в сумме они в конечном счете не превы-
шали общее поступление воды в организм.
Любой организм выдерживает некоторые отклонения от
среднего содержания в нем воды, причем одни животные в
этом отношении выносливее других. Млекопитающие, напри-
30—1863
-466 Гл. 9, Вода и осморегуляция
мер, обычно переносят потерю 10% воды, хотя цри этом состоя-
ние организма существенно ухудшается; потеря 15 или 20% во-
ды для большинства из них, вероятно, гибельна. Многие низшие
организмы выдерживают большие потери. Например, некоторые
лягушки могут переносить потерю 40% воды, но лишь очень
немногие животные выживают при потере половины воды1.
Когда содержание воды <в организме выражено в процентах
веса тела, данные не всегда легко интерпретировать. Напрцмер,
количество жира у животных сильно варьирует, а жировая ткань
содержит сравнительно мало воды — около 10% или меньше.
Поэтому у животного с большим запасом жира общее содержа-
ние воды в процентах ниже, чем у тощего животного, хотя сте-
пень гидратации мышц и внутренних органов в обоих случаях
совершенно одинакова. Если же содержание воды выражать в
процентах ма'ссы тела за вычетом жира, величины обычно
варьируют гораздо меньше. Недостаток этого способа — то, что
здесь нужно определять содержание жира, а это сильно услож-
няет дело.
Обычай выражать содержание воды в процентах веса тела
может привести к неверным выводам и в другом отношении, что
лучше всего объяснить на примере. Возьмем насекомое, в ко-
тором содержится 75% ,воды. Если оно потеряет ровно половину
этой воды, то будет содержать ее 60%. Такой результат может
показаться неожиданным, но из рис. 9.12 видно, что расчет пра-
вилен. На первый взгляд снижение содержания воды с 75 до
60% не кажется чрезмерным, хотя в действительности потеряна
половина всей воды. Таких подча1с обманчивых результатов про-
центного выражения можно избежать, пользуясь абсолютными
цифрами. Есл'и в приведенном примере насекомое весило 100 мг,
то вначале в нем содержалось 75 мг воды, а после обезвожива-
ния— 37,5 мг; теперь сразу ясно, что оно потеряло половину
воды.
ПОТЕРЯ ВОДЫ В РЕЗУЛЬТАТЕ ИСПАРЕНИЯ
Вода испаряется как с поверхности органов дыхания, так
и со всей поверхности тела, но иногда в опыте трудно опреде-
1 Есть два способа определения общего содержания воды в организме. Его
можно измерить, убив животное, полностью высушив его тело (например, в су-
шильном шкафу) и определив потерю веса. При таком «деструктивном» способе
какое-то число животных используется для установления нормы . (контроль),
а другие животные — для опытов. Недеструктивным является изотопный метод.
В организм вводят небольшой объем воды, меченной изотопом (дейтерием или
тритием). После равномерного распределения меченой воды во всей воде орга-
низма берут пробу крови или другой жидкости тела и определяют в ней кон-
центрацию метки. После этого нетрудно вычислить объем воды, в котором рас-
пределилась изотопная метка. Такой метод позволяет использовать одно и то же
животное для повторных определений.
Членистоногие 467
лить эти две переменные по отдельности. Органы дыхания на-
секомых и позвоночных сильно различаются по структуре и
функции. У позвоночных выстилка легких всегда влажная, вы-
дыхаемый воздух насыщен водяными парами и потеря воды че-
рез дыхательные пути существенна. Между тем у насекомых ор-
ганы дыхания состоят из трубочек, выстланных хитиновой кути-
Рис. 9.12. Как показывает диаграмма, если содержание воды в организме выра-
жать в процентах веса тела, то это может привести к ошибочным заключениям.
Если первоначально содержание воды составляло 75% веса тела (А) и ровно
половина этой воды была потеряна, то ее содержание в обезвоженном организме
составит 60% (Б). Таким образом, потеря воды в действительности весьма зна-
чительная, покажется не очень большой.
кулой, и только их самые тонкие веточки относительно прони-
цаемы для воды. Тем не менее насекомые теряют через трахей-
ную систему значительное для них количество воды, и возмож-
ные способы снижения этих потерь были рассмотрены при спи-
сании дыхания насекомых в гл. 2.
Вся поверхность тела насекомого покрыта жесткой, сухой ку-
тикулой, и легко можно было бы подумать, что кутикула пол-
ностью непроницаема для воды. В действительности это- не так.
Существуют огромные различия в испарении влаги между вида-
30*
468 Гл. 9. Вода й осморегуляция
ми, обычно живущими во влажной среде, и теми, которые оби-
тают в совершенно сухих местах 1(табл. 9.9). Испарение у вод-
Таблица 9.9
Испарение воды с поверхности тела
насекомых и паукообразных. Потеря влаги
через органы дыхания не включена. Все
данные представлены в микрограммах воды,
испаряющейся за 1 ч с 1 см2 поверхности
тела при дефиците насыщения 1 мм рт. ст.
(0,13 кПа). (Schmidt-Nielsen, 1969)
Личинки толстоножек (Bibio) 900
Таракан Periplaneta 49
Пустынная саранча (Schistocerca) 22
Муха цеце (Glossina) 13
Мучной червь (личинка Tenebrio) 6
Мучной клещ (Acarus) 2
Клещ Dermacentor 0,8
ных личинок толстоножек в воздухе составляет величину того
же порядка, что и у животного с влажной Дожей (см. 'табл. '9.8).
Это значит, что кутикула почти не препятствует Испарению во-
ды. В то же время у насекомых, живущих в очень сухих местах,
проницаемость покровов может быть ниже, чем у любых других
животных, сведениями о которых мы располагаем.
Кутикула насекомого — очень сложное образование, состоя-
щее из нескольких слоев. Жесткий хитин сам по себе не столь
уж непроницаем для воды; испарению препятствует главным об-
разом покрывающий его тонкий восковой слой. Если соскрести
его абразивом (например, частицами глинозема), испарение
сильно возрастает, и насекомые, которые в норме могут жить
в очень сухом материале, например в хранящемся зерне, будут
гибнуть ют обезвоживания (Wigglesworth, 1945). Это представ-
ляет известный практический интерес, так как абразивы приме-
няются в 'качестве нетоксичных инсектицидов, которые затем до-
вольно легко можно отделить от зерна благодаря разнице в
плотности. Подобное действие оказывают не только острые час-
тицы, но и такие вещества, как сухая распыленная глина, веро-
ятно потому, что они тоже разрушают структуру воскового слоя
Испарение воды с кутикулы насекомых возрастает с повыше-
нием температуры (риС. 9.13). Оно увеличивается гораздо боль-
ше, чем давление водяных паров, и это, по-видимому,, связано
с физическими изменениями в восковом слое. Если откладывать
проницаемость против температуры поверхности насекомого, а
не температуры воздуха, то на полученной кривой будет резкий
перелом. Соответствующая температура для разных видов раз-
лична. Были исследованы пробы воскового слоя, и оказалось,
что, его физические свойства резко изменяются при той же тем-
Членистоногие 469
пературе, при которой сразу возрастает потеря воды (Beament,
1959).
Для членистоногих восковой слой не является обязательным
условием жизни на суше. Как мы уже упоминали, 'многие Isopo-
da (равноногие раки) приспособлены к наземному существова-
Рис. 9.13. Потеря воды через кутикулу таракана возрастает с повышением тем-
пературы воздуха (треугольники). Если полученные данные выразить как функ-
цию температуры поверхности тела (кружочки), то при температуре около 30°C
на кривой появится резкий перелом, который соответствует точке резкого изме-
нения физических свойств покрывающего поверхность воска. (Beament, 1958.)
нию, а некоторые из них живут даже в .пустынях. Воск на их 'ку-
тикуле не обнаружен, и у них потеря воды путем испарения не-
посредственно 'Связана с дефицитом насыщения в атмосфере и в
остальном независима от температуры. У насекомых, напротив,
испарение меняется в зависимости от температуры даже при по-
стоянном дефиците давления водяных ларов.
ПОТЕРЯ ВОДЫ С КАЛОМ И МОЧОЙ
Насекомые выбрасывают экскременты и мочу через одно и
то же отверстие — анус. Моча образуется в мальпигиевых сосу-
дах, которые открываются в задний отдел кишки. Жидкая моча
и фекальные массы из кишечника поступают в прямую кишку,
тде происходит реабсорбция воды. Это будет подробнее рассмот-
рено в гл. 10.
НАКОПЛЕНИЕ ЭКСКРЕТОВ
Если экскреторные продукты не выводятся с мочой, а сохра-
няются в теле, то вода не расходуется на их выделение. Так как
мочевая кислота очень мало растворима, ее депонирование в те-
ле— вполне осуществимый подход к решению задачи «экскре-
470 Гл. 9. Вода и осморегуляция
ции», и у насекомых отложение мочевой кислоты в разных час-
тях тела, по-ви[димому, очень распространено. Например, у не-
которых видов тараканов количество мочевой кислоты может
составлять до 10% общего сухого веса тела.
У многих насекомых мочевая кислота откладывается в жиро-
вом теле, у других она накапливается в кутикуле. Мочевая кис-
лота, отложенная в кутикуле, вероятно, никогда больше не вы-
свобождается. Но в жировом теле положение может быть ины*м.
Возможно, что мочевая кКслота служит хранилищем (депо) азо-
та, откуда он в период нехватки мобилизуется и используется в
процессах метаболизма; однако еще нет достаточных данных о
существовании такой функции.
«АКТИВНЫЙ» ТРАНСПОРТ ВОДЫ
Насекомое, обитающее в очень сухой среде (например, муч-
ной червь), извлекает воду из содержимого прямой кишки, пока
фекальные шарики не станут сов'сем сухими. Такое извлечение
воды, казалось бы, должно быть Активным процессом, так как
вода перемещается из области высокой осмотической концентра-
ции в кишке к более низкой концентрации в крови. Однако вви-
ду сложного строения прямой кишки и окружающих ее структур
возможны и другие гипотезы. Вообще в животных системах
большую часть переноса воды против градиента можно объяс-
Рис. 9.14. Модель Керрана, демонстрирующая переход воды против осмотическо-
го градиента из камеры А в камеру В. Во время опыта выводная трубка из
камеры Б закрыта. Подробности см. в тексте. (Curran, Macintosh, 1962.)
нить первичным транспортом какого-то растворенного вещества,
за которым пассивно следует вода благодаря осмотическим си-
лам, действие которых объясняет теория трех компартментов, пред-
ложенная Керраном (Curran, 1960).
Гипотеза Керрана станет понятной, если обратиться к искус-
ственной модели, представленной на рис. 9.14. Здесь изображен
Членистоногие 471
цилиндр, состоящий Из трех отделений. Отделения А и Б разде-
лены целлофановой мембраной, которая легко проницаема для
воды, но не пропускает сахарозу. Отделения Б и В разделены
пористой стеклянной пластинкой (годится и лист фильтроваль-
ной бумаги, если для него будет подходящая механическая опо-
ра). Сначала наполним отделение А 0,1 М раствором сахарозы,
Рис. 9.15. Модель транспорта воды через эпителиальную клетку, состоящая из
трех отделений. Стрелками указан активный транспорт Na+. Площадь, покрытая
точками, соответствует отделению Б на рис. 9.14.
отделение Б — 0,5 М сахарозой и отделение В — дистиллирован-
ной водой. Концентрированный раствор в Б тотчас же будет
осмотически отсасывать поду из Л и В, но если не допускать уве-
личения объема жидкости в 5, то она потечет через пористую
пластинку «в отделение В. Поэтому в конечном итоге вода будет
перемещаться из А в В, несмотря на то что осмотическая кон-
центрация в А выше, чем в В. Движение воды против суммарно-
го осмотического градиента, разумеется, не может длиться бес-
конечно, так как различия между концентрациями постепенно
исчезнут и система перестанет действовать. Но если как-нибудь
можно будет поддерживать высокую концентрацию растворен-
ных веществ в отделении Б, движение воды не прекратится.
Модель Керрана позволяет объяснить движение воды через
различные биологические мембраны. На рис. 9.15 схематически
представлена типичная эпителиальная клетка, мембрана ‘кото-
рой на стороне, обращенной к тканевой жидкости, образует глу-
бокие впячивания. Активный транспорт натрия создает в поло-
-сти впячивания высокую осмотическую концентрацию. Тогда под
472 Гл. 9. Вода и осморегуляция
действием осмотических сил вода диффундирует в эту полость,
и повысившееся гидростатическое давление вызывает ток жидко-
сти из отверстия в тканевую жидкость. В результате происходит
общее движение воды с наружной стороны через эпителий в тка-
невую жидкость. Для этого перемещения воды, которое может
идти против суммарного осмотического градиента от наружной
стороны клетки к тканевой жидкости, первичной движущей си-
лой служит активный транспорт натрия.
Возможно, что это весьма распространенный механизм пере-
носа воды. Для многих эпителиев характерны длинные, узкие
межклеточные или внутриклеточные пространства, и имеющиеся
экспериментальные данные говорят в пользу этой гипотезы (Dia-
mond, 1962; Diamond, Bossert, 1967).
Реабсорбция воды из задней кишки насекомых изучалась на
тараканах, и результаты согласуются с гипотезой трех компарт-
ментов. Из ректальных подушечек были взяты пробы жидкости
объемом всего лишь 0,1 нл (0,0001 мм3). Здесь жидкость более
концентрированная, чем в просвете прямой кишки, а кровь ме-
нее концентрированная. Таким образом, эта жидкость соответ-
ствует отделению Б в модели Керрана (Wall, Oschman, 1970).
питье воды
Наиболее- очевидный способ получения воды—-питье. Разу-
меется, свободная вода доступна всем пресноводным насекомым
и другим насекомым во многих других условиях, например при
наличии росы или дождя. Однако большинству из них свободная
вода доступна только временами и с нерегулярными промежут-
ками, а многие насекомые обитают в сухих местах, где никогда
не бывает свободной воды, и они должны получать ее из дру-
гих источников.
ВОДА в ПИЩЕ
Растительноядные насекомые получают большие количества
воды с пищей, так как свежий растительный материал содержит
много воды. В сочных плодах, листьях и т. п. ее может быть
больше 90%, но даже и самый сухой растительный материал со-
держит некоторое количество свободной воды. Сухие зерна и се-
мена, мука, шерсть и другие, казалось бы, совершенно сухие ма-
териалы, питаясь которыми на'секомые могут расти и ' размно-
жаться, иногда содержат 5—10% свободной воды.
Когда в пище воды много, задача состоит в том, чтобы вы-
вести ее избыток из организма. Это составляет нормальную
функцию органов выделения, которые тогда вырабатывают боль-
шое количество жидкой мочи. Напротив, в сухих условиях цель
Членистоногие 473
состоит в выведении экскретов с минимальной потерей воды и в
выработке такой концентрированной мочи, на (какую только спо-
собен экскреторный механизм.
МЕТАБОЛИЧЕСКАЯ ВОДА
У животных, обитающих в безводной среде, самым значи-
тельным источником влаги служит вода, образующаяся при
окислении органических (веществ. Ее можно называть водой
окисления, но принят также термин «метаболическая вода».
Всем известно, что при горении органических веществ образу-
ется вода; можно видеть, как она конденсируется на наружной
стороне холодной кастрюли, поставленной на газ, или капает с
выхлопной трубы автомашины холодным утром. Реакцию окис-
ления глюкозы можно записать следующим образом:
CgHjgOg -J- 60g -► 6СО2 6Н2О.
(180 г) (192 г) (264 г) (108 г)
В общем итоге можно считать, что вода образуется в резуль-
тате окисления водорода. При окислении компонентов пищи ко-
личество воды зависит от количества водорода, содержащегося
в данном пищевом продукте. Из приведенного уравнения легко
вычислить, что 1 г глюкозы дает 0,60 г воды, и никакими физио-
логическимЩ ухищрениями нельзя получить воды больше, чем
следует из этого уравнения.
При метаболизме более сложных углеводов, например крах-
мала, воды образуется немного меньше, потому что эти вещест-
ва содержат меньше водорода. Так, окисление крахмала про-
исходит следующим образом:
(СбН10О5)п + пО2 -> 6пСО2 + 5пН2О.
В этом случае образуется 0,56 г воды на 1 г окисленного
крахмала (табл. 9.10).
Окисление жира дает воды больше, чем ее получается из
углеводов, — около 1,07 г на 1 г жира; эта цифра слегка варьи-
рует в зависимости от состава жира и степени насыщенности
жирных кислот. Таким образом, при ок'ислёнии жира образуется
примерно вдвое больше воды, чем при окислении крахмала. Од-
нако в некоторых отношениях эта цифра обманчива, поскольку
жир дает также больше энергии на 1 г (9,4 ккал, тогда как
крахмал дает 4,2 ккал). Поэтому при данной интенсивности об-
мена животное использует жир в количестве вдвое меньшем и
соответственно уменьшается образование воды, так что крахмал
Имеет в этом отношении небольшое преимущество перед жиром.
Это видно из последнего столбца табл. 9.10, где указано количе-
ство образующейся воды на единицу доставляемой питательным
веществом энергии.
474 Г л. 9. Вода и осморегуляция
Таблица 9.10
Количество воды, образующееся при
окислении различных пищевых веществ.
(King, 1957; Schmidt-Nielsen, , 1964)
Вещество Образова- ние воды, г НоО на 1 г Метаболи- ческая энергия, ккал/г Образова- ние воды, г НгО/ккал
Крахмал 0,56 4,2 0,13
Жир Белок (при вы- делении мо- 1,07 9,4 0,11
чевины) Белок (при вы- делении мо- чевой кис- 0,39 4,3 0,09
лоты) 0,50 4,4 0,11
Белковый обмен несколько более сложен, так как азот белка
образует экскреты, содержащие некоторое количество водорода;
этот водород выводится и не окисляется в воду. Выход метабо-
лической воды зависит от природы конечного продукта белково-
го обмена. Если это мочевина, то образуется 0,39 г воды на 1 г
белка; если же конечный продукт — 'мочевая кислота (как у на-
секомых), то тогда вода образуется в большем количестве.
Мочевина (CH4ON2) содержит два атома водорода на один
атом азота, а мочевая кислота (C5H4O3N4) —только один атом
водорода на атом азота, т. е. вдвое меньше. Поэтому в случае
выделения мочевой кислоты выход метаболической воды возра-
стает до 0,50 г на 1 г белка. Таким образом, чтобы точно рас-
считать количество метаболической воды, надо знать не только
точные количества и состав окисляемых пищевых веществ, но и
природу конечного продукта белкового обмена. Но это важно
лишь тогда, когда требуются очень точные сведения, так как
различия в количестве воды, образующейся при окислении раз-
ных пищевых веществ, относительно невелики (табл. 9.10, по-
следний столбец).
ПОГЛОЩЕНИЕ ВОДЫ ЧЕРЕЗ ПОВЕРХНОСТЬ ТЕЛА
У водных насекомых вода поступает внутрь через поверх-
ность тела так же, как у других пресноводных животных: более
высокая осмотическая концентрация веществ, растворенных в
жидкостях тела, вызывает осмотический приток воды. Для вод-
ных насекомых задача состоит в том, чтобы выводить лиш'нюю
воду, и, как уже упоминалось, э'то делает почка или соответст-
вующий выделительный орган.
Членистоногие 475
Но у наземных насекомых дело обстоит иначе; им часто не-
достает воды. Очень интересное явление наблюдается у некото-
рых насекомых и паукообразных: они способны поглощать во-
дяные пары прямо из атмосферного воздуха. Первое сообщение
об этом 'было встречено с недоверием, так как чрезвычайно
трудно представить себе какой-либо правдоподобный механизм
извлечения воды прямо из воздуха. Однако ранние данные были
подтверждены рядом компетентных исследователей. Хорошо до-
кументированные примеры приведены в табл. 9.11. Таблица со-
Таблица 9.11
Насекомые, способные извлекать воду
из атмосферного воздуха. Второй столбец —
предельная относительная влажность,
ниже которой чистая прибавка количества воды
уже невозможна
Предель-
ная отно-
Животное сительная
влажность,
%
Клещ Ornithodorus 94
Клещ Ixodes 92
Личинка мучного хрущака (Tenebrio) 90
Клещ Echinolaelaps 90
Пустынный таракан (Arenivaga) 83
Прямокрылое Chortophaga 82
Мучной клещ (Acarus) 70
Блоха (Xenopsylla) 50
Чешуйница Thermobia 45
держит также сведения о самой низкой относительной влажно-
сти, при которой для каждого животного возможно такое погло-
щение водяных паров.
Особенности такого механизма поглощения воды лучше все-
го описать на примере, для чего мы воспользуемся пустынным
тараканом Arenivaga. Он извлекает воду из атмосферы только
после частичного обезвоживания организма. Если содержать та-
ракана в очень сухом воздухе, пока он не потеряет, скажем, 10%
своего веса, то при последующем помещении его в атмосферу с
относительной влажностью выше 83% он начнет поглощать вла-
гу. Этот процесс будет продолжаться до полной регидратации
организма, а затем прекратится (иными словами, поглощение
воды регулируется в точном соответствии с потребностью в ней).
На рис. 9.16 показано понижение и последующее повышение со-
держания воды в организме пустынного таракана после обезво-
живания и последующей регидратации.
Мы видели, что атмосферная влажность, ниже которой по-
глощение воды невозможно, для разных видов различна (табл.
476 Гл. 9. Вода и осморегуляция
9.11). Этот специфический для каждого вида минимум не зави-
сит от температуры; значит, он определяется относительной
влажностью, а не дефицитом давления водяных паров (т. е. этот
процесс, по-ви'димому, сходен с гигроскопическим эффектом).
Это обстоятельство трудно понять, и любая приемлемая гипоте-
за должна удовлетворительно объяснить его.
Рис. 9.16. Соотношение воды и сухого вещества у пустынного таракана Areniva-
ga до и после обезвоживания в сухом воздухе и последующей регидратации пу-
тем поглощения водяных паров из воздуха при относительной влажности 95%.
Насекомых с исходным весом около 250 мг в течение опыта не кормили, и по-
этому они теряли сухое вещество. В результате процент воды в конце опыта
был выше (73,2%), чем в начале (67,2%). По оси ординат все значения выра-
жены в процентах от исходного веса. (Edney, 1966.)
Одна из гипотез состоит в том, что температурные микрогра-
диенты внутри тела животного ведут к конденсации водяного
пара. Для конденсации из атмосферы с относительной влаж-
ностью 90% требуется градиент больше 2°, что вряд ли мыслимо
в опытах, проводимых в условиях постоянной температуры. При
относительной влажности 50%—едва ли не самой низкой, при
которой у того или иного вида отмечено поглощение воды, —
для того, чтобы произошла конденсация, температура воздуха
должна понизиться с 25 до 14 °C. Очевидно, что в опытах, про-
водимых при контролируемой постоянной температуре, такие пе-
репады исключены (Edney, 1966).
Анатомическую локализацию поглощения водяного пара
установить трудно. Казалось бы, несложно определить, участву-
ет ли здесь дыхательная система, так как сравнительно легко
закупорить воском отверстия трахей. У некоторых насекомых
поглощение воды при этом прекращается, но у клеща Ixodes оно
Наземные позвоночные МТ
продолжается и после закрытия дыхалец. У тех видов, у кото-
рых поглощение воды прекращается, это могло бы объясняться
вторичным эффектом отключения дыхательной системы: оно ве-
дет к аноксии, а аноксия сама по себе приостанавливает извле-
чение водяных napote.
Многие неясности частично связаны с малыми размерами не-
которых подопытных организмов (домовый пыльный клещ ве-
сит 0,01 мг), но имеющиеся
данные указывают на то, что
нет единого механизма для
всех животных, способных
поглощать водяные пары. У
клещей это поглощение, веро-
ятно, происходит через рото-
вое отверстие, а у личинки
мучного хрущака и чешуйни-
цы — почти наверное через
Фото 9.1. Клещ Dermatophagoides fa-
rinae способен извлекать водяные па-
ры из воздуха. В очень сухом возду-
хе он обезвоживается, но если отно-
сительная влажность превышает 70%,
клещ может поглощать воду прямо
из атмосферы. На верхнем снимке
показан обезвоженный клещ, кото-
рый весит 8 мкг (0,008 мг), на ниж-
нем снимке — клещ, вес которого воз-
рос до 13 мкг в результате поглоще-
ния воды из воздуха. Длина живот-
ного составляет около 0,4 — 0,5 мм.
(Фото предоставлены L. С. Arlien,
Wright State University, Дэйтон, штат
Огайо.)
прямую кишку (Noble-Nesbitt, 1977). У пустынного таракана
Arenivava этот механизм связан с хорошо выраженной пузырча-
той структурой в гипофаринксе, которая выдвигается и становит-
ся видимои в периоды активного поглощения воды (O’Donnell,
1977).
НАЗЕМНЫЕ ПОЗВОНОЧНЫЕ
РЕПТИЛИИ
Современные рептилии делятся на четыре основных отряда.
Из них крокодилы всегда связаны с водой, а представители
остальных трех отрядов — змеи, ящерицы и черепахи — хорошо
478 Гл. 9. Вода и осморегуляция
приспособлены к жизни в сухих местах, но -среди них имеются
также годные или полуводные животные. Все водные рептилии
обладают легкими, дышат воздухом и несомненно произошли от
наземных предков.
Кожа у рептилий сухая, чешуйчатая и считается непроницае-
мой для воды. Рассмотрим испарение -влаги с кожи у некоторых
рептилий (табл. 9.12). Чтобы легче было сравнивать, используем
Таблица 9.12
Испарение воды с поверхности тела рептилий при
23—25 °C. Все данные выражены в микрограммах
воды на 1 см2 поверхности тела при дефиците
насыщения в 1 мм рт. ст. (0,13 кПа).
(Schmidt-Nielsen, 1969)
Кайман (Caiman) 65
Уж (Natrix) 41
Черепаха Pseudemys 24
Черепаха Теггарепе 11
Игуана (Iguana) 10
Змея Pit ио phis 9
Ящерица Sauromalus 3
Пустынная черепаха (Gopherus) 3
те же единицы измерения, что и раньше (табл. 9.8 и 9.9). Можно
видеть, что испарение с кожи у рептилий, обитающих в сухой
среде, во много раз меньше, чем у водных рептилий, и даже у
этих последних (если содержать их на воздухе) оно на порядок
меньше, чем у таких животных с влажной кожей, как лягушки
(ом. табл. 9.8).
Можно было бы думать, что у рептилий с влажной поверх-
ностью дыхательных путей испаряется гораздо больше воды,
чем с сухой кожи, но на самом деле это не так. Доля участия
сухой кожи в общем испарении всегда вдвое или больше превы-
шает долю участия дыхательного аппарата (рис. 9.17). Если со-
держать ужа (Natrix) на воздухе, то почти 90% всей воды бу-
дет испаряться с поверхности тела. Сходные соотношения имеют
место у черепах и ящериц: и у них кожа играет более важную
роль б потере воды, чем дыхательные пути. Даже у пустынной
ящерицы Sauromalus две трети общего испарения приходится
на долю кожи и только одна треть — на долю дыхательных пу-
тей.
Испарение тесно коррелирует со средой обитания. Чем суше
обычная для животного среда, тем слабее испарение. У пустын-
ной гремучей змеи общее испарение с поверхности тела и дыха-
тельных путей составляет менее 0,5% от веса тела в сут^ки, и при
таком его уровне змея, вероятно, могла бы прожить без воды 2
пли 3 месяца. А если бы она находилась под землей в норе или
Наземные позвоночные 479?
Рис. 9.17. У рептилий испарение с сухой кожи превышает потерю влаги с поверх-
ности дыхательных путей и составляет от 66 -до 88% всего испарения. В скоб-
ках после названия каждого животного указана суточная величина общего
испарения (с кожи и из дыхательных путей) в граммах воды на 100 г веса
тела. Она зависит от местообитания и у ужа более чем в 10 раз превышает испа-
рение у пустынной черепахи. Все наблюдения проведены в сухом воздухе при'
23 °C. Д.п. — дыхательные пути. (Schmidt-Nielsen, 1969.)
ходе, где влажность выше, то, несомненно, могла бы прожить
еще дольше.
Помимо потерь в результате испарения вода расходуется еще
при образовании мочи. В качестве конечного продукта белкового'
обмена рептилии выделяют главным образом мо-чевую кислоту,,
а поскольку это соединение чрезвычайно мало растворимо, для
489 Гл. 9. Вода и осморегуляция
его выведения требуется очень мало воды. Связь между экскре-
цией азота и водным обменом будет (подробнее рассмотрена в
гл. 10, посвященной выделению и выделительным органам.
Перед морскими рептилиями стоит особая проблема, потому
что вода, в которой они живут, и значительная часть их пищи
содержат много соли. Как это влияет .на процессы выделения,
будет рассмотрено позже в настоящей главе.
Для возмещения потерь воды рептилиям, разумеется, так же
необходимо получать воду, как и другим животным. Нет ника-
ких указаний на то, чтобы рептилии способны были поглощать
влагу из атмосферы, как это делают некоторые насекомые. По-
этому в отсутствие питьевой воды они должны получать всю
воду из таких источ|нико'в, как пища и окислительные процессы.
ПТИЦЫ II МЛЕКОПИТАЮЩИЕ
До сих пор мы рассматривали водный баланс, не касаясь
проблем терморегуляции; однако некоторые животные, в особен-
ности птицы и млекопитающие, используют воду для того, чтобы
•охлаждать свое тело (в жаркой среде. Человек и ряд других мле-
копитающих потеют, многие млекопитающие, включая собак, а
также птицы резко учащают дыхание, и усиленное испарение
предотвращает перегрев. Этот аспект расходования воды был
рассмотрен в гл. 8 и здесь обсуждаться уже не будет.
Млекопитающие таких размеров, как грызуны, удобны для
.анализа основных сторон водного баланса, так как они не при-
бегают к учащенному дыханию, а кожных желез у них слишком
мало для того, чтобы они могли играть существенную роль в
терморегуляции. Это дает возможность наследовать у них ос-
новные, неизбежные потери в'о'ды — испарение и выведение с мо-
чой и калом — без учета ее больших и весьма изменчивых коли-
честв, затрачиваемых на терморегуляцию. Тогда можно увидеть,
покрывают ли имеющиеся источники воды потребность в ней.
В большинстве пустынь свободная вода появляется только во
время редких дождей; и тем не менее птицы и мелкие грызуны
обитают во многих пустынях, где дождь выпадает раз в год или
еще реже. Эти животные должны получать всю воду из пищи,
так как роса играет, по-видимому, минимальную роль, по край-
ней мере в водном балансе вы!сших позвоночных. /Многие живот-
ные получают воду в избытке из зеленых листьев, стеблей, пло-
дов, корней и клубней; хищники получают много воды в виде
жидкостей тела своих жертв, ткани которых могут содержать
50—80% воды.
Некоторые грызуны пустынь действительно используют тако-
го рода влажную пищу; другие, однако, питаются главным обра-
зом сухими семенами и другим сухим растительным материалом
и получают свободную воду лишь в минимальных количествах.
Наземные позвоночные 481
Хорошо известным' примером могут служить кенгуровые крысы
(фото 9.2) и пероцнаты, во множестве обитающие в североаме-
риканских пустынях. Они способны (неопределенно долгое время
обходиться сухой пищей и ничего не пить. Поэтому рассмотре-
ние их водного баланса позволит раскрыть механизмы, благо-
даря которым млекопитающее может жить и поддерживать вод-
ное равновесие, казалось бы, совсем без поступления воды в ор-
ганизм извне.
Фото 9.2. Кенгуровая крыса (Dipodomys spectabilis). Этот грызун часто встре-
чается в пустынях Северной Америки. Он не пьет воды и питается главным об-
разом семенами и другими сухими растительными материалами. Благодаря эко-
номному расходу воды при образовании мочи и малому испарению через
дыхательные пути крыса обходится в основном водой, образующейся при окисли-
тельных метаболических процессах. (К. Schmidt-Nielsen.)
Кенгуровая крыса—не особенно «сухое» животное: в ее теле
содержится воды столько же, сколько и у других млекопитаю-
щих (около 66%). Даже если в течение недель и месяцев она
питалась только сухим ячменем или овсом, содержание воды в
ее организме остается неизменным. При этом вес тела остается
прежним или даже увеличивается. Это показывает, что живот-
ное, питаясь сухой пищей, поддерживает свой водный баланс;
потери воды не превышают ее поступления.
Данные по суммарному водному обмену кенгуровой крысы
приведены в табл. 9.13. Они относятся к промежутку, в течение
которого животное съедает и перерабатывает в организме 100 г
ячменя. Крыса весом около 35 г съедает это количество пример-
но за месяц. Впрочем, точный срок не имеет значения, так как
баланс был бы совершенно таким же и при большей интенсив-
ности метаболизма, когда животное поедало бы то же количе-
ство пищи, скажем, за две недели.
В отношении источников воды мы находим, что 54 г воды
образуются при окислении пищи. Эту цифру можно вычислить
из состава зерна. Использовав величины образования метаболиче-
31—1863
482 Гл. 9. Вода и осморегуляция
Таблица 9.13
Общий водный баланс кенгуровой крысы,
уравновешивающий потери и поступление воды за
время, в течение которого животное потребляет
и метаболизирует 100 г ячменя (обычно около
месяца). Температура воздуха 25 °C;
относительная влажность 20%.
(Schmidt-Nielsen, 1964)
Поступление воды мл Потери воды мл
Метаболическая
вода 54,0 Моча 13,5
Вода, полученная
извне 6,0 Кал 2,6
Испарение 43,9
Общий приход ВО- Общая по-
ДЫ 60,0 теря воды 60,0
ской воды при окислении крахмала, белка и жира (см. табл.
9.10), мы получим цифру, приведенную в табл. 9.13. Зермо со-
держит также нем-ного свободной воды, точное количество кото-
рой зависит от влажности воздуха. При 20%-ной влажности
100 г ячменя содержат 6 г воды, что дает общее (поступление во-
ды в размере 60 г. Этого долж|но хватить на все нужды живот-
ного.
Что касается потерь воды, то мы видим, что около четверти
ее идет на образование мочи. В количественном отношении
главным экскретов является мочевина, которая получается из
белков зерна. Это вещество должно быть выведено из организма
(нет указаний на то, чтобы оно откладывалось, подобно тому
как это происходит у насекомых), и чем более концентрирован-
ной будет моча, тем меньше расход воды. Почки кенгуровой
Крысы обладают поразительной способностью концентрировать,
мочу — они делают это намного эффективнее, чем почки боль-
шинства млекопитающих, не живущих в пустыне (см. табл.
10.2). Если мы знаем, сколько белка содержимся в 100 г ячменя,
мы можем вычислить, сколько мочевцны должно быть выведено
и сколько для этого нужно воды. Количество воды, теряемой
с калом, можно определить, собрав его и измерив содержание в
нем воды.
Самый главный путь потери воды — это испарение.. В табл.
9.13 поступление и потеря воды точно уравновешивают друг
друга. При более низкой влажности воздуха поступление воды
слегка уменьшится, так как в зерне будет меньше свободной ?в<У-
ды, и в то же время в более сухом воздухе больше воды будет
испаряться в дыхательных путях. Поэтому в очень сухой атмо-
сфере кенгуровая крыса не сможет сохранять водный баланс.
Наземные позвоночные 483
В более влажном воздухе — при относительной влажности выше
20%—с поверхности дыхательных путей испаряется меньше во-
ды, так как ее больше во вдыхаемом воздухе, и животное легко
поддерживает водный баланс. В природных условиях кенгуро-
вые крысы много времени проводят в своих подземных норах,
где влажность воздуха несколько выше, чем над поверхностью
грунта пустьцни, и это помогает сохранять водное равновесие.
Интенсивность испарения с поверхности дыхательных путей
зависит от того, сколько воздуха поступает в легкие (от дыха-
тельного объема). Поскольку выдыхаемый воздух всегда насы-
щен водяными парам’и, количество воды, уже содержащейся во
вдыхаемом воздухе, определяет, сколько добавочной воды нуж-
но для насыщения воздуха; нет данных о том, чтобы какое-ли-
бо млекопитающее выдыхало воздух, не полностью насыщенный
водяными парами.
Дыхательный объем зависит от потребления кислорода (от
которого зависит также количество образующейся метаболиче-
ской воды). Млекопитающие обычно извлекают около 5 мл кис-
лорода из 100 мл альвеолярного воздуха, прежде чем выдохнут
его. Если бы можно было увеличить извлечение кислорода, то
соответственно снизились бы дыхательный объем (при данном
потреблении кислорода) и количество испаряемой воды. Этот
путь уменьшения потери воды при дыхании, по-видимому, не
используется ни одним млекопитающим. Для того чтобы в лег-
ких извлекалось больше кислорода, гемоглобин крови должен
обладать большим сродством к нему, но кровь кенгуровой кры-
сы не отличается в этом отношении от крови других грызунов.
Повышенное извлечение кислорода повело бы также к увеличе-
нию содержания СО2 в крови, что отразилось бы на основном
кислотно-щелочном -равновесии, но у данного животного не об-
наружено подобных отклонений от общей «нормы» для млеко-
питающих.
Однако испарение значительно снижается благодаря тому,
что выдыхаемый воздух холоднее внутренних областей тела. Хо-
тя воздух в легких имеет температуру тела и насыщен влагой,
проходя через нос, он охлаждается. Механизм этого процесса
•очень прост (рис. 9.18). При вдохе стенки дыхательных путей
отдают тепло проходящему по ним воздуху; температура стенок
понижается и в результате испарения может упасть ниже тем-
пературы вдыхаемого воздуха. При выдохе, когда теплый воздух
из легких проходит около охлажденных поверхностей, он охлаж-
дается, а вода конденсирует1ся на стенках. Степень охлаждения
(т. е. окончательная температура выдыхаемого воздуха) варьи-
рует в зависимости от температуры и влажности вдыхаемого
воздуха.
Насколько важно это охлаждение для баланса воды? Его
значение, разумеется, изменяется в зависимости от состояния
31*
484 Гл. 9. Вода и осморегуляция
воздуха, которым дышит кенгуровая крыса. Возьмем вполне
возможный пример, когда температура воздуха равна 30 °C, а
относительная влажность—25%. При вдыхании такого воздуха
в легкие он нагревается до температуры тела (38°C) и в ре-
зультате насыщения водяными парами содержит теперь 46 мг
Рис. 9.18. Модель теплообмена в носовых ходах. (Schmidt-Nielsen, 1972.)
Окружающий воздух насыщен влагой, температура его 28 °C, температура те-
ла 38 °C. Проходя по носовым ходам (слева), вдыхаемый воздух согревается
и поглощает водяные пары; в легкие он попадает уже насыщенным влагой и
нагретым до 38 °C. При выдохе (справа) воздух течет вдоль прохладных стенок
и отдает тепло, а водяные пары конденсируются. К моменту завершения тепло-
вого и водного обмена температура выдыхаемого воздуха снижается до 28°С„
и он остается насыщенным.
Н2О на 1 л. При выходе этот воздух охлаждается до 27°C, и, хо-
тя он по-прежнему насыщен, теперь в нем вдвое меньше воды
(половина воды задерживается в носовых ходах и идет на
увлажнение воздуха при следующем вдохе). Но «следует обра-
тить внимание на то, что выдыхаемый воздух насыщен и всегда
содержит больше водяных .паров, чем вдыхаемый; иными слова-
ми, нос не служит системой для извлечения воды из воздуха, и в
дыхательных путях в итоге всегда происходит потеря воды.
Теплообмен в носовых ходах характерен не только для кен-
гуровых крыс — он имеет место у всех животных и является не-
избежным следствием теплообмена между воздухом и стенками
носовой полости; этот теплообмен полнее, когда ходы, узкие и
поверхность их велика, как это бывает у мелких грызунов.
У птиц с их более короткими и широкими носовыми ходами
степень охлаждения меньше (рис. 9.19). В сухом воздухе у кен-
гуровой крысы температура выдыхаемого воздуха ниже, чем
вдыхаемого; у птиц температура выдыхаемого воздуха выше
(хотя он тоже заметно охлаждается). У человека, у которого
Морские позвоночные, дышащие воздухом ' 485"
носовые ходы гораздо шире, теплообмен очень неполный и обыч-
но температура выдыхаемого воздуха только на несколько гра-
дусов ниже температуры тела. Поэтому у человека и других
млекопитающих теплообмен в носу и связанное с ним сбереже-
ние воды имеют меньшее значение, и с выдыхаемым воздухом
теряется больше воды.
10 15 20 25 30 35
Температура окружающего воздуха, °C
Рис. 9.19. Температура выдыхаемого воздуха у семи видов птиц, измеренная в
сухом воздухе. (Schmidt-Nielsen et al., 1970.)
Температура тела птиц была между 40 и 41 °C. Две из них, кактусовый крапив-
ник и волнистый попугайчик, живут в пустыне, но по степени охлаждения возду-
ха (и, следовательно, по возмещению воды) не отличаются от птиц, обитающих
в других местностях. Менее сильное охлаждение выдыхаемого воздуха у утки
связано с большими размерами ее дыхательных путей.
МОРСКИЕ ПОЗВОНОЧНЫЕ, ДЫШАЩИЕ ВОЗДУХОМ
Высшие позвоночные — рептилии, птицы и млекопитающие —
это наземные животные, и, как мы видели, некоторые из них
превосходно чувствуют себя в самых сухих пустынях мира. Но
несколько групп наземных позвоночных вторично проникли в
море, оставаясь воздуходышащими. В отношении проблем, свя-
занных с водой ,и солью, они, по существу, являются наземными
животными и по сравнению с рыбами физиологически намного
больше изолированы от окружающей мор'ской воды. В отличие
от рыб, у которых жабры относительно проницаемы для воды,
высшие морские позвоночные дышат легкими и таким образом
избегают осмотической проблемы, возникающей при тесном со-
486 Гл. 9. Вода и осморегуляция
прикосновении большой поверхности жабр .с морской водой.
Они отличаются физиологически от своих наземных родичей
прежде всего тем, что вынуждены пйть только морскую воду,
и тем, что значительная часть их пищи содержит много соли.
Воды в море достаточно; трудность состоит в том, что вся
она содержит около 35 г соли на 1 л при осмотической концент-
рации около 1000 мосмоль. Большая доля дищи также содержит
много соли, хотя по ионному составу она часто существенно от-
личается от морской воды. Поедаемые растения и беспозвоноч-
ные животные создают в общем такую же осмотическую про-
блему, как используемая для питья морская вода. Как мы ви-
дели, морские костистые рыбы содержат гораздо ^меньше соли,
и для животных, питающихся главным образом рыбой, пробле-
ма, связанная с солью, менее серьезна, чем для тех, которые пи-
таются растениями или беспозвоночными. Так или иначе, все
они должны выделять конечные продукты белкового обмена; у мле-
копитающих это главным образом мочевина, а у птиц и репти-
лий— мочевая кислота.
Когда позвоночное животное пьет морскую воду, оно погло-
щает соли и концентрация их в жидкостях тела возрастает.
Если только соли не выводятся в объеме воды, меньшем, чем
объем выпитой воды, чистой прибыли воды быть не может. Ины-
ми словами, соли должны выделяться в растворе не менее кон-
центрированном, чем морская вода; в противном случае орга-
низм будет все больше и больше обез’вожи1ваться.
Почки рептилий не могут вырабатывать мочу более концент-
рированную, чем жидкости тела, а у птиц обычно образуется
моча, концентрация которой не более чем вдвое превышает кон-
центрацию крови. Поскольку концентрация жидкостей тела со-
ставляет у птиц около 300—400 мосмоль/л, моча не может до-
стичь концентрации морской воды (1000 мосмоль/л). Поэтому
почки не способны создавать концентрацию, достаточную для
того, чтобы эти животные могли пить морскую воду и есть пи-
щу с высоким содержанием соли, а если они это делают, то, зна-
чит, обладают другими средствами для выведения соли.
МОРСКИЕ РЕПТИЛИИ
Морских представителей имеют три отряда рептилий — че-
репахи, ящерицы и змеи. Некоторые морские змеи совсем не
связаны с сушей, даже во время размножения, так как они жи-
вородящие, никогда не покидают море и на суше были бы со-
вершенно беспомощны. Морские черепахи большую часть жиз-
ни проводят в открытом океане, но для размножения возвра-
щаются на пеЬчаные тропические побережья; на сушу выходят
только самки для откладки яиц, а самцы никогда не ступают
на землю, после того как, вылупившись, перебрались ц море.
Морские позвоночные, дышащие воздухом 487
Морские ящерицы больше связаны с сушей. Примером мо-
жет служить галапагосская морская игуана Amblyrhynchus cris~
lotus. Она живет в полосе прибоя на Галапагосских островах, ла-
зает по скалам и питается только водорослями. Четвертый от-
ряд современных рептилий, крокодилы, по-видимому, не имеет
истинно морских представителей. Живущий в соленой воде Сго-
codylus porosus связан главным образом с эстуариями; он пита-
ется преимущественно рыбой и, вероятно, не может долго суще-
ствовать в настоящей морской среде.
Почки рептилий не приспособлены для выведения избыточ-
ной соли, и она выводится солевыделяющими (или просто соле-
выми) железами, расположенными в голове. Солевые железы
вырабатывают сильно концентрированную жидкость, которая со-
держит главным образом натрий и хлор в концентрациях, значи-
тельно более высоких, чем в морской воде. Эти железы не
функционируют непрерывно, как почка; они выделяют свой сек-
рет только время от времени в ответ на солевую нагрузку, по-
вышающую концентрации солей в плазме. Подобные железы
есть и у морских птиц, у которых они подробно изучены.
У морской ящерицы солевые железы изливают свой секрет в
переднюю часть носовой полости, в которой имеется гребень, не
позволяющий жидкости стекать назад и быть проглоченной.
Иногда при резком выдохе жидкость выбрасывается из ноздрей
в виде мелких брызг. Галапагосская игуана питается только во-
дорослями, которые близки по содержанию солей к морской во-
де. Поэтому животному необходим механизм для выделения
солей в высокой концентрации (Schmidt-Nielsen, Fange,
1958).
Морские черепахи, как растительноядные, так и хищные, об-
ладают крупными выделяющими соль железами, расположенны-
ми в орбитах обоих глаз. Проток железы открывается в заднем
углу орбиты, и черепаха, получившая солевую нагрузку, пла-
чет поистине солеными слезами. (Человеческие слезы, которые,
как всем известно, имеют соленый вкус, изоосмотичны с плаз-
мой крови. Поэтому слезные железы у человека не играют осо-
бой роли в выведении соли.)
Морские змеи тоже выделяют соленую жидкость при соле-
вой нагрузке и обладают солевыми железами, открывающимися
в ротовую полость, откуда выделяемая жидкость выводится на-
ружу (Dunson, 1968). Морские змеи — близкие родственники
кобр и очень ядовиты, что несколько затормозило физиологиче-
ское изучение их солевого обмена, имеющего ряд интересных ас-
пектов.
Хотя морские рептилии обладают механизмом выведения со-
ли в форме очень концентрированной жидкости, остается нере-
шенным вопрос, действительно ли многие из них пьют воду в
значительных количествах?
488 Гл. 9. Вода и осморегуляция
МОРСКИЕ ПТИЦЫ
Среди птиц немало обитателей моря, но большинство из них
живет на море и над ним, а не в нем «самом. Многие обитают
на берегу, но есть и истинно пелагические птицы. Молод'ой аль-
батрос, вылупившийся на тихоокеанском острове, проводит три
или четыре года над окрытым океаном, прежде чем вернуться
в места гнездовий. Пингвин, наиболее приспособленная морская
птица, утратил способность летать. Он отличный пловец, далеко
продвинувшийся в своей эволюции по направлению к полностью
водному образу жизни. Тем не менее в физиологическом смысле
пингвин остался в основном наземным, воздуходышащим живот-
ным, размножающимся |на суше. И когда говорят, что император-
ский пингвин даже не выводит птенцов на земле, то это лишь иг-
ра слов, потому что он высиживает свое яйцо на льду в течение
долгой темной антарктической зимы.
У всех морских птиц имеются парные носовые солевые желе-
зы, соединенные протоком с носовой полостью (Fange et al.,
1958). Носовые железы есть и у большинства наземных птиц, но
они очень небольшие; у морских же Птиц они во всех без исклю-
чения случаях сильно развиты. Чаще всего они расположены на
верхней части черепа, над орбитой каждого глаза, в неглубокой
впадине в кости (рис. 9.20). У тех птиц, которые регулярно по-
требляют с пищей много соли или получают для питья солевые
растворы, железы увеличиваются в размерах и становятся боль-
ше нормальных (Schmidt-Nielsen, Kim, 1964).
Рис. 9.20. У чайки железы, выделяющие соль, расположены на верхушке головы
над каждым глазом, в небольших углублениях в кости. (Schmidt-Nielsen, 1963.)
Солевые железы начинают функционировать только в ответ на
осмотическую нагрузку (например, после потребления морской
воды или соленой пищи). В остальное время они бездействуют
й этим отличаются от почки, которая вырабатывает мочу непре-
рывно. Выделяемая жидкость имеет простой состав и содержит
главным образом натрий и хлор в довольно постоянных концент-
Морские позвоночные, дышащие воздухом 489
рациях. В этом отношении солевая железа тоже резко отличает-
ся от почки, которая в широких пределах из/мёняет концентрации
и относительные количества выделяемых компонентов. Кроме то-
го, моча содержит разнообразные органические вещества, тогда
как в секрете солевой железы имеются лишь следы неэлектроли-
тов (Schmidt-Nielsen, 1960).
Хотя концентрация соли в выделяемой Жидкости неизменно
высока, в этом отношении имеются характерные видовые разли-
чия, связанные с нормальной экологией птиц и способами их пи-
тания. Например, баклан, живущий на побережье и питающийся
рыбой, выделяет жидкость с относительно низким содержанием
соли — около 500—600 ммоль натрия на 1 л (табл. 9.14). У сереб-
Таблица 9.14
Концентрация натрия в носовых выделениях
различных птиц. Концентрация хлора в каждой
данной пробе почти равна концентрации натрия;
другие ионы содержатся лишь в небольших
количествах. (Schmidt-Nielsen, 1960)
Вид Концентрация натрия, ммоль/л
Кряква 400—600.
Баклан 500—600
Американский черный водорез 550—700
Пеликан бурый 600—750
Чайка серебристая 600—800
Клуша 700—900
Пингвин Гумбольдта 725—Й50
Кайра 750—850
Альбатрос черноногий 800—900
Качурка северная 900—1100
ристой чайки, которая поедает больше беспозвоночных и саот-
вётственно потребляет больше соли, концентрация натрия состав-
ляет 600—800 ммоль/л. У северной качурки, проводящей много
времени в открытом океане и питающейся планктонными ракооб-
разными, концентрация натрия в носовой жидкости достигает
1100 ммоль/л.
Солевая железа обладает поразительной способностью выде-
лять соль. В одном опыте, в котором чайка (клуша) получила
морскую воду в количестве, равном почти одной десятой веса ее
тела (что для человека составило бы 7 д), весь избыток соли
был выведен примерно за три часа. Детали, этого -опыта приве-
дены в табл. 9.15, Птица вёсила 1420 г и ей. было введено через
желудочный зонд 134 мл морокой воды. Общий объем жидкости,
выведенной за последующие три часа (131,5 мл.), был почти ра-
вен объему введенной воды, причем (меньщ,е половины (56,3 мд)
49ft Гл. 9. Вода и осморегуляция
было выведено солевой железой, а остальное (75,2 мл) — через
клоаку. Однако количество натрия, выделенного носовой железой,
было приблизительно в 10 раз больше, чем то, которое содержа-
лось в экскретах из клоаки (моча, смешанная с кало'м).
Рассмотрение табл. 9.15 показывает, что объем выделений из
носа был наибольшим в течение второго часа, после чего он по-
Таблица 9.15
Выделение из носа и клоаки у клуши в течение 175 мин после введения
Ь желудок морской воды в количестве, составлявшем почти одну десятую веса
fee тела/(Schmidt-Nielsen, 1960)
Время, мин Выделения нз носа Выделения из клоаки
Объем, мл Концентрация натрия, ммоль/л Количество натрия, ммоль Объем, мл Концентрация натрия, ммоль/л Количество натрия, ммоль
15 2,2 798 1,7 5,8 38 0,28
40 10,9 756 8,2 14,6 71 1,04
70 14,2 780 11,1 25,0 80 2,00
100 16,1 776 12,5 12,5 61 0,76
130 6,8 799 5,4 6,2 33 0,21
169 4,1 800 3,3 7,3 10 0,07
175 2,0 780 1,5 3,8 12 0,05
Всего 56,3 43,7 75,2 4,41
степенно уменьшался. Концентрация носовых выделений была по-
стоянной, несмотря на изменения в скорости секреции. Из клоаки
было больше выделений (главным образом моча), чем из носа,
но концентрация соли была здесь совсем низкой. Почка птицы
может вырабатывать мочу с максимальной концентрацией нат-
рия около 300 ммоль/л, но в этом опыте концентрация была во
много раз меньше. Кроме того, в течение опыта концентрация
надрия в моче колебалась примерно в 8-кратных пределах, что
Обычно для почки, но резко контрастирует с очень постоянной
его концентрацией в носовом секрете.
Солевая железа выделяет удивительно много жидкости, осо-
бенно если учесть очень большую осмотическую работу, произ-
водимую при образовании концентрированного солевого раство-
ра. В расчете на 1 кг веса тела количество этой жидкости пример-
но вдвое больше количества мочи, выделяемой почками человека
при максимальном водном диурезе (табл. 9.16). При пересчете
-на 1 г веса железы получается еще более резкое различие. Один
Грамм солевой железы чайки может продуцировать 0,6 мл жид-
кости в минуту, а почки человека при максимальном диурезе вы-
рабатывают только одну двадцатую этого количества. У почки,
когда она выделяет концентрированную мочу (концентрация ко-
торой‘-все же вдйое меньше, чем у секрета солевой железы чай-
Морские позвоночные, дышащие воздухом 491
ки), Скорость выделения может снизиться до 1% скорости При
максимальном водном диурезе. Совершенно очевидно, что соле-
вая железа — одна из самых эффективных среди известных си-
стем ионного транспорта.
Жидкость, выделяемая солевой железой морокой птицы, всег-
да содержит главным образом натрий и очень мало калия. Отно-
шение между этими двумя ионами составляет приблизительно
Таблица 9.16
Секреция жидкости солевой железой серебристой
чайки и почками человека при максимальном
водном диурезе
Скорость секреции Солевая железа Почки человека
На 1 кг веса тела (мл/мин) 0,5 0,24
На 1 г веса железы (мл/мин) 0,6 0,03
30:1, и если увеличить количество калия в пище чайки, то отно-
шение Na/K сильно не изменится (Schmidt-Nielsen, 1965). У мор-
ских черепах и морских змей отношение Na/K тоже высокое, но
у морской игуаны оно ниже, а относительное количество калия,,
выделяемого солевой железой, выше (табл. 9.17). Это легко по-
нять: морская игуана питается главным образом морскими водо-
ро/слями, а растения вообще содержат много калия. Поэтому со-
держание калия в пище морской игуаны относительно велико и
соответственно велика потребность в его выведении.
Таблица 9.17
Концентрация натрия и калия (в миллимолях на литр)
в морской воде и в секрете носовой солевой железы.
Отношение Na+/K+ велико у морских видов и мало
у наземных растительноядных животных
Na+ K+ Na+/K+
Морская вода 470 10 47
Серебристая чайка (Larus)^ 718 24 30
Морская черепаха Lepidochelys2'* 713 29 25
Морская змея Pelamis2^ 607 28 24
Морская игуана (Amblyrhynchus) 1434 235 6,7
Ложная игуана (Ctenosaura) 9 Schmidt-Nielsen, 1960. 2) Dunson, 1969. s) Templeton, 1967. 78 527 0,15
492 Гл. 9. Вода и осморегуляция
Типичная сухопутная ящерица из засушливой местности, лож-
ная игуана (Ctenosaura), выделяет носовой секрет с очень высо-
кой концентрацией калия и низкой концентрацией натрия, что
дает отношение Na/K, равное 0,15.
Наземные рептилии отличаются от родственных морских форм
еще одной особенностью солевой железы: состав ее секрета ме-
няется в зависимости от характера солевой нагрузки. Если сухо-
путная рептилия получает натриевую нагрузку, в секрете воз-
растает содержание натрия; если же она получает калиевую на-
грузку, то секрет содержит преимущественно калий. У наземных
рептилий состав выделяемой жидкости регулируется в соответ-
ствии с надобностью, а солевые железы морских птиц и реп-
тилий, по-видимому, весьма специализированы для выделения
натрия и хлора.
МОРСКИЕ МЛЕКОПИТАЮЩИЕ
Представители трех отрядов млекопитающих — ластоногие,
киты и сирены — являются исключительно морскими животны-
ми в том смысле, что практически всю свою жизнь они проводят
в море. Тюлени ненадолго возвращаются на сушу для родов и
выкармливания детенышей, а киты и сирены (ламантин и дю-
гонь) даже приносят потомство в воде.
Пища этих млекопитающих очень различна по солевому со-
ставу; Тюлени и киты плотоядны и питаются рыбой, некоторыми
крупными беспозвоночными и морским планктоном. Те из них,
кто питается рыбой, потребляют пищу с довольно низким содер-
жанием соли (меньше 1%), но с относительно высоким содер-
жанием белка. Антарктический крабоядный тюлень и морж (пи-
щу его составляют Двустворчатые моллюски и другие придон-
ные организмы) питаются животными, изоосмотичными по
отношению к морской воде. Пищу беззубых китов составляют
планктонные ракообразные с высоким содержанием соли, харак-
терным для морских беспозвоночных. Если вместе с пищей за-
глатывается морская вода, то это еще увеличивает солевую
нагрузку. Ламантины и дюгони — травоядные животные и пита-
ются растениями, которые находятся в осмотическом равно-
весии с морской водой; поэтому они тоже поглощают много
соли.
Обладают ли морские млекопитающие ка’ким-нибудь физио-
логическим механизмом, который соответствовал бы солевым
железам птиц и рептилий? Они в этом не нуждаются, так как
почки китов и тюленей могут выделять мочу более концентри-
рованную, чем морская вода. Максимальная концентрация хло-
ра, обнаруженная в моче кита, составила 820 м*моль/л (Krogh,
1939). рто намного больше, чем в морской воде (около
535 ммФ.ль/л).,_и поэтому для кита, -потребляющего пищу с вы-
Морские позвоночные, дышащие воздухом 493
соким содержанием соли или даже морскую воду, выведение со-
ли не должно составлять трудности.
Хорошо известно, что для человека морская вода токсична
и что потерпевшие кораблекрушение в открытом море только
ускоряют обезвоживание организма, если пьют морскую воду.
В табл. 9.18 показано влияние питья морокой веды на водный
баланс человека и кита. Кит может выпить один литр морской
воды и при этом, после того как соли будут выведены, приобре-
сти около трети литра чистой воды. Почка человека менее эф-
фективна; максимальная концентрация соли в его моче ниже,
чем в морской воде, и если человек выпьет литр морской воды,
то в итоге чистая потеря воды неизбежно составит у него около
одной трети литра, и он будет в худшем положении, чем если
бы вовсе не пил. Кроме того, обезвоживанию организма способ-
ствуют большие количества магния и сульфата в морской воде,
которые действуют как слабительное и вызывают понос, тем са-
мым увеличивая потерю воды.
Таблица 9.18
Влияние 1 л морской воды, введенной через рот, на водный баланс у человека
и у кита
Введенная вода Образовавшаяся моча Водный баланс: Пополнение или потеря воды, мл
Объем, мл Концентрация, ммоль/л Объем, мл Концентрация, ммоль/л
Человек 1000 535 1350 400 —350
Кит 1000 535 650 820 -ро50
Еще нет достаточно надежных сведений о том, поглощают ли
тюлени и киты сколько-нибудь значительное количество «морской
воды путем питья или случайного заглатывания с пищей. Из-
вестно, однако, что ластоногие, видимо, не нуждаются в питье
морской воды. Калифорнийских морских львов (Zalophus call-
fornianus) содержали в неволе и кормили одной только рыбой;
даже не имея доступа к воде в течение 45 дней, эти животные
оставались совершенно нормальными, и их водный баланс был
положительным (Pilson, 1970). Интересно отметить, что и при
питании беспозвоночными животными, например кальмарами,
состояние морских львов, по-1видимому, остается таким же хо-
рошим.
Хотя кровь кальмара находится в осмотическом равновесии
с морской ведой и содержит много соли, осмотическая концент-
рация внутриклеточных жидкостей в значительной части -обу-
словлена присутствием органических веществ. Поэтому кальма-
ры и многие другие беспозвоночные при их поедании фактически
создают меньшую солевую нагрузку, чем можно было бы ожи-
494 Гл. 9. Вода и осморегуляция
дать, исходя только из осмотических концентраций жидкостей
их тела.
В отношении водного баланса у млекопитающих есть одно
важное отличие от птиц и рептилий: самки выкармливают сво-
их детенышей молоком, для образования которого нужно много
воды. Сократить эту потерю воды могла бы выработка более
концентрированного молока. Давно известно, что в молоке тю-
леней и китов очень высок процент жира, а содержание белка
выше, чем в молоке коровы (табл. 9.19). Обычно это объясняли
Состав и энергетическая ценность молока млекопитающих
Таблица 9.19
Животное Состав, г на 100 г молока Энергетичес- кая ценность, ккал на 1 г Н2О
н2о Жиры Белки Углеводы
Корова1) 87,3 3,7 3,3 4,8 0,8
Человек1) 87,6 3,8 1,2 7,0 0,8
Гренландский тюлень2) 45,3 42,7 10,5 0 9,9
Тюлень Уэддела среднее значение3) 43,6 42,2 (14,1)4) 10,5
крайнее значение3) 27,2 57,9 (19,5)4) — 23
’) Коп, Cowie, 1961. 2) Sivertsen, 1935. 3) Kooyman, Drabek, 1968. 4) Цифры относятся к общему количеству нежировых твердых веществ, но фактически
это в основном белки, так как : молоко тюленей почти не содержит углеводов.
тем, что их детеныши должны быстро развиваться и в особен-
ности получать большое количество жира для отложения его
под кожей, где' он служит теплоизоляцией.
Но высокое содержание жира в тюленьем молоке можно
рассматривать и в связи с ограниченностью водных ресурсов
матери. У тюленя Уэддела, изученного лучше других ластоно-
гих, содержание жира в молоке в период лактации постепенно
растет, а содержание воды соответственно снижается. Наи-
большее содержание жира, найденное в молоке этого тюленя,
составляло 57,9% (почти вдвое больше, чем в сбитых, сливках),
а содержание воды в той же пробе — всего лишь 7,2% (что на-
много меньше, чем в обычном постном мя1се, где оно равно
65%). Таким образом, тюлени выкармливают свое потомство
при минимальном расходе воды. На каждый граМхМ истраченной
воды они передают детенышам в 10 раз больше питательных^
веществ, чем сухопутные млекопитающие (последний столбе'ц в
табл. 9.19). С этими данными согласуется тот факт, что самка
тюленя Уэддела в неволе не страдает от обезвоживания во вре-
мя лактации (Kooyman, Drabek, 1968).
Литература 495
* *
*
При рассмотрении осморегуляции мы снова и снова убеж-
дались в том, какую важную роль играют органы выделения в
поддержании относительного постоянства (стационарного со-
стояния) внутренних концентраций солей и содержания воды в
живых организмах. Перейдем теперь к более подробному опи-
санию органов выделения и их функций.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Beadle L. С. (1943). Osmotic regulation and the faunas of inland waters, Biol.
Rev., 18, 172—183.
Beament J. W. L. (1958). The effect of temperature on the waterproofing mecha-
nism of an insect, J. Exp. Biol., 35, 494—519.
Beament J. W. L. (1959). The waterproofing mechanisms of arthropods. 1. The
effect of temperature on cuticle permeability in terrestrial insects and ticks,
J. Exp. Biol.» 36, 391—422.
Beament J. W. L. (1961). The water relations of insect cuticle, Biol. Rev., 36, 281—
320.
Bentley P. J. (1966). Adaptations of amphibia to arid environments, Science, 152,
619—623.
Bentley P. J. (1971). Endocrines and Osmoregulation: A Comparative Account of
the Regulation of Water and Salt in Vertebrates, New York. Springer-Verlag,
300 pp.
Blaylock L. A., Ruibal R., Platt-Aloia K. (1976). Skin structure and wiping beha-
vior of Phyllomedusine frogs. Copeia, 1976, 283—295.
Bliss D. E. (1966). Water balance in the land crab, Gecarcinus lateralis, during
the intermolt cycle, Am. Zool., 6, 197—212.
Burger J. W., Hess W. N. (1960). Function of the rectal gland in the spiny dogfish,
Science, 131, 670—671.
Buxton P, A. (1923). Animal Life in Deserts: A Study of the Fauna in Relation to
the Environment, London, Arnold. 176 pp.
Conte F. P., Hootman S. R., Harris P. J. (1972). Neck organ of Artemia salina
nauplii: A larval salt gland, J. Comp. Physiol., 80, 239—246.
Croghan P. C. (1958a). The osmotic and ionic regulation of Artemia salina (L.),
J. Exp. Biol., 35, 219—233.
Croghan P. C. (1958b). The mechanism of osmotic regulation in Artemia salina (L.):
The physiology of the gut, J. Exp. Biol., 35, 243—249.
Curran P. F, (1960). Na, Cl, and water transport by rat ileum in vitro, J. Gen.
Physiol., 43, 1137—1148.
Curran P. F.> Macintosh J. R. (1962). A model system for biological water trans-
port, Nature, Lond., 193, 347—348.
Diamond J. M. (1962). The mechanism of water transport by the gall-bladder,
J. Physiol., 161, 503—527.
Diamond J. M.. Bossert W. H. (1967). Standing-gradient osmotic flow, J. Gen.
Physiol., 50. 2061—2083.
Dunson W A. (1968). Salt gland secretion in the pelagic sea snake Pelamis,
Am. J. Physiol., 215, 1512—1517.
Dunson W. A. (1969). Electrolyte excretion by the salt gland of the Galapagos
marine iguana, Am. J. Physiol., 216, 995—1002.
Edney E. B. (1954). Woodlice and the land habitat, Biol. Rev., 29. 185—219.
Edney E. B. (1956). The micro-climate in which woodlice live, Proc. 10th Int,
Congr. Entomol., 2, 709—712.
496 Гл, 9. Вода и осморегуляция
Edney Е. В. (1957). The Water Relations of Terrestrial Arthropods, Cambridge.
Cambridge University Press, 109 pp.
Edney E. B. (1966). Absorption of water vapour from unsaturated air by Areni-
vaga sp. (Polyphagidae, Dictyoptera), Comp. Biochem. Physiol., 19, 387—
408.
Fdnge R., Schmidt-Nielsen K-> Osaki H. (1958). The salt gland of the herring gull,
Biol. Bull., 115, 162—171.
Fugelli K., Zachariassen К. E. (1976). The distribution of taurine, gamma-amino-
butyric acid and inorganic ions between plasma and erythrocytes in flounder
(Platichthys flesus) at different plasma osmolalities» Comp. Biochem. Physiol.,
55A, 173—177.
Galloway J. N., Likens G. E., Edgerton E. S. (1976). Acid precipitation in the
northeastern United States : pH and acidity, Science, 194, 722—724.
Gordon M. S.. Schmidt-Nielsen K., Kelly H. M. (1961). Osmotic regulation in the
crab-eating frog (Rana cancrivora), J. Exp. Biol., 38, 659—678.
Gordon M. S., Tucker V. A. (1965). Osmotic regulation in the tadpoles of the crab-
eating frog (Rana cancrivora), J. Exp. Biol., 42, 437—445.
Gross W. J., Lasiewski R. C., Dennis M., Rudy P., Jr. (1966). Salt and water balan-
ce in selected crabs of Madagascar. Comp. Biochem. Physiol.,* 17, 641 —
660.
House C. R. (1963). Osmotic regulation in the brackish water teleost Blennius
pholis, J. Exp. Biol.» 40, 87—104.
Hutchinson G. E. (1957). A Treatise on Limnology, vol. 1, Geography, Physics and
Chemistry, New York, J. Wiley, 1015 pp. '
Hutchinson G. E. (1967). A Treatise on Limnology, vol. II, Introduction to Lake
Biology and the Limnoplankton. New York, Wiley, 1115 pp.
Keys A. B. (1933). The mechanism of adaptation to varying salinity in the com-
mon eel and the general problem of osmotic regulation in fishes, Prog. R. Soc.
Lond., B, 112, 184—199.
Keys A. B., Willmer E. N. (1932). «Chloride secreting cells» in the gills of fishes,
with special reference to the common eel, J. Physiol., 76, 368—378.
King J. R. (1957). Comments on the theory of indirect calorimetry as applied to
birds, Northwest Sci., 31, 155—169.
Koch H. J. A. (1968). Migration. In: Perspectives in Endocrinology: Hormones in
the Lives of Lower Vertebrates (E. J. W. Barrington and С. B. Jorgensen,
eds.), pp. 305—349, London, Academic Press.
Kon S.K., Cowie A. T. (1961). Milk: The Mammary Gland and Its Secretion,
vol. II, New York, Academic Press, 423 pp.
Kooyman G. L., Drabek С. M. (1968). Observations on milk, blood and urine con-
stituents of the Weddell seal, Physiol. Zool., 41, 187—194.
Krogh A. (1937). Osmotic regulation in fresh water fishes by active absorption
of chloride ions, Z. Vergl. Physiol., 24, 656—666.
Krogh A. (1939). Osmotic Regulation in Aquatic Animals, Cambridge, Cambridge
' University Press, 242 pp. Reprinted by Dover Publications, New York,
1965.
Leivestad H., Muniz I. P. (1976). Fish kill at low pH in a Norwegian river, Natu-
re, Lond., 259, 391—392.
Livingstone D. A. (1963). Chemical composition of rivers and lakes. In: Data of
Geochemistry, 6th ed. (M. Fleischer, ed.), Geological Survey Professional Pa-
per 440, Chapter G, 64 pp., Washington, D. C., GPO.
Loveridge J. P. (1970). Observations on nitrogenous excretion and water relations
of Chiromantis xerampelina (Amphibia, Anura), Arnoldia, 5, J—6.
.Lutz P. L., Robertson J. D. (1971). Osmotic constituents of the coelacanth Latime-
ria chalumnae Smith, Biol. Bulk, 141, 553—560.
Machin J. (1964). The evaporation of water from Helix aspersa L. The nature of
the evaporating surface, J. Exp. Biol., 41, 759—769.
Maetz J., Campanini G. (1966). Potentiels transepitheliaux de la branchie d’an-
guille in vivo en eau douce et en eau de mer, J. Physiol.. (Paris), 58,> 248.
Литература 49Г
Maetz J., Payan P.> de Renzis G, (1976). Controversial aspects of ionic uptake in-
freshwater animals. In: Perspectives in Experimental Biology, vol. 1, Zoology'
(P. S. Davies, ed.), pp. 77—92, Oxford, Pergamon Press.'
Mayer N. (1969). Adaptation de Rana esculenta a des milieux varies: Etude Spe-
ciale de Fexcretion renale de 1’eau et des electrolytes au cours des changements-
de milieux, Comp. Biochem. Physiol., 29, 27—50.
Motais R.> Maetz J. (1965). Comparaison des echanges de sodium chez un teleosteen.
euryhalin (le flet) et un teleosteen stenohalin (le serran) en eau de mer: Impor-
tance relative du tube digestif et de la branchie dans ces echanges, C. R. Acad.
Sci. Paris, 261, 532—535.
Noble-Nesbitt J. (1977). Absorption of water vapour by Thermobia domestica and’
other insects. In: Comparative Physiology: Water, Ions and Fluid Mechanics-
(K. Schmidt-Nielsen, L. Bolis and S. H. P. Maddrell, eds.), pp. 53—66, Lon-
don, Cambridge University Press.
O'Donnell M. J. (1977). Site of water vapor absorption in the desert cockroach,.
Arenivaga investigata, Proc. Natl. Acad. Sci. USA, 74, 1757—1760.
Pang P. К. T.. Griffith R. W., Kahn N. (1972). Electrolyte regulation in the fresh
water stingrays (Potamotrygonidae), Fed. Proc., 31, 344.
Phillips J. E., Bradley T. J., Maddrell S. H. P. (1977). Mechanisms of ionic and;
osmotic regulation in saline-water mosquito larvae. In: Comparative Physiolo-
gy: Water, Ions and Fluid Mechanics (K. Schmidt-Nielsen, L. Bolis and'.
S. H. P. Maddrell, eds.), pp. 151—171, London, Cambridge University Press.
Pickford G. E.. Grant F. B. (1967). Serum osmolality in the coelacanth, Latimeriar
chalumnae: Urea retention and ion regulation, Science, 155, 568—570.
Pierce S. K., Jr., Greenberg M. J. (1973). The initiation and control of free amino-
acid regulation of cell volume in salinity-stressed marine bivalves, J. Exp. Biol.,.
59, 435—440.
Pilson M. E. Q. (1970). Water balance in sea lions, Physiol. Zool., 43, 257—
269.
Potts W. T. W. (1954). The energetics of osmotic regulation in brackish- and fresh-
water animals, J. Exp. Biol., 31, 618—630. *
Potts W. T. W.. Evans D. H. (1967). Sodium and chloride balance in the killifish
Fundulus heieroclitus, Biol. Bull., 133, 411—425.
Potts W. T. W., Parry G. (1964). Osmotic and Ionic Regulation in Animals, Oxford
Pergamon Press, 423 pp.
Ramsay J. A. (1935). Methods of measuring the evaporation of water from animals,
J. Exp. Biol., 12, 355—372.
Ramsay J. A. (1949). The osmotic relations of the earthworm, J. Exp. Biol., 26;
46—56.
Robertson J. D. (1954). The chemical composition of the blood of some aquatic chor-
dates, including members of the Tunicata, Cyclostomata and Osteichthyes,.
J. Exp. Biol., 31, 424—442.
Robertson J. D. (1957). Osmotic and ionic regulation in aquatic invertebrates. In:
Recent Advances in Invertebrate Physiology (В. T. Scheer, ed.), pp. 229—246,
Eugene University of Oregon Press.
Ruibal R. (1962). The adaptive value of bladder water in the toad, Bufo с о gnat us
Physiol. Zool., 35, 218—223.
Schmidt-Nielsen K. (1960). The salt-secreting gland of marine birds, Circulation,,
21, 955—967.
Schmidt-Nielsen K. (1963). Osmotic regulation in higher vertebrates. Harvey Leet.,
58, 53—95.
Schmidt-Nielsen K. (1964). Desert Animals: Physiological Problems of Heat and’
Water, Oxford, Clarendon Press, 277 pp. To be reprinted by Dover Publications,
New York.
Schmidt-Nielsen K. (1965). Physiology of salt glands. In: Funktionelle und mor-
phologische Organisation der Zelle: Sekretion und Exkretion, pp. 269—288,.
Berlin, Springer-Verlag.
Schmidt-Nielsen K. (1969). The neglected interface: The biology of water as a li-
quid gas system, Q. Rev. Biophys., 2, 283—304. •
32—1863
498 Гл. 9. Вода и осморегцляция
Schmidt-Nielsen К. (1972). How Animals Work, Cambridge, Cambridge University
Press, 114 pp.
Schmidt-Nielsen K-, Fange R. (1958). Salt glands in marine reptiles, Nature, Lond.,
182, 783—785.
Schmidt-Nielsen K., Hainsworth F. R.. Murrish D. E. (1970). Countercurrent heat
exchange in the respiratory passages: Effect on water and heat balance, Respir.
Physiol., 9, 263—276.
Schmidt-Nielsen K-, Kim Y. T. (1964). The effect of salt intake on the size and
function of the salt gland of ducks, Auk., 81, 160—172.
Schmidt-Nielsen K., Lee P. (1962). Kidney function in the crab-eating frog (Rana
cancrivora), J. Exp. Biol., 39, 167—177.
Schmidt-Nielsen K., Taylor C. R., Shkolnik A. (1971). Desert snails: Problems of
heat, water and food, J. Exp. Biol., 55, 385—398.
Shoemaker V. H., McClanahan L. L., Jr. (1975). Exaporative water loss, nitrogen
excretion and osmoregulation in Phyllomedusine frogs, J. Comp. Physiol., 100,
331—345.
Sivertsen E. (1935). Ueber die chemische Zusammensetzung von Robbenmilch, Ma-
gazin for Naturvidenskaberne, 75, 183—185.
Smith H. (1931). The absorption and excretion of water and salts by the elasmo-
branch fishes. 1. Fresh water elasmobranchs, Am. J. Physiol.. 98, 279—
295
Smith H. W. (1959). From Fish to Philosopher. Summit, N. J., CIBA, 304 pp.
Reprinted (1961), Garden City, N. Y., Doubleday.
Sverdrup H. U., Johnson M. W., Fleming R. H. (1942). The Oceans: Their Physics,
Chemistry, and General Biology, Englewood Cliffs, N. J., Prentice-Hall,
1087 pp.
Templeton J. R. (1967). Nasal salt gland excretion and adjustment to sodium
loading in the lizard, Ctenosaura pectinata, Copeia, 1967, 136—140.
Thesleff S., Schmidt-Nielsen K. (1962). Osmotic tolerance of the muscles of the
crab-eating frog Rana cancrivora, J. Cell Comp. Physiol., 59, 31—34.
Thorson T. B., Cowan С. M., Watson D. E. (1967). Potamotrygon spp.: Elasmo-
branchs with low urea content. Science, 158, 375—377.
Thorson T. B., Watson D. E., Cowan С. M. (1966). The status of the freshwater
shark of Lake Nicaragua, Copeia, 1966, 385—402.
Tracy C. R. (1976). A model of the dynamic exchanges of water and energy bet-
ween a terrestrial amphibian and its environment, Ecol. Monogr., 46, 293—
326.
Ussing H. H., Zerahn K. (1951). Active transport of sodium as the source of
electric current in the shortcircuited isolated frog skin, Acta Physiol. Scand.,
23, 110—127.
Wall B. J.. Oschman J. L. (1970). Water and solute uptake by rectal pads of Pe-
riplaneta americana, Am. J. Physiol., 218. 1208—1215.
Wigglesworth V. B. (1945). Transpiration through the cuticle of insects, J. Exp.
Biol., 21, 97—114.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Bayly I. A. E. (1972). Salinity tolerance and osmotic behavior of animals in athalas-
sic^ saline and marine hypersaline waters, Annu. Rev. Ecol. Syst., 3, 233—
Bentley P. J. (1971). Endocrines and Osmoregulation: A Comparative Account of
the Regulation of Water and Salt in Vertebrates, New York, Springer-Verlag,
300 pp.
Bentley P. J. (1976). Osmoregulation. In: Biology of the Reptilia, vol. 5, Physiolo-
gy A (C. Gans and W. R. Dawson, eds.). pp. 365—412, London, Academic
Press.
Burton R. F. (1973). The significance of ionic concentrations in the internal media
of animals, Biol. Rev., 48, 195—231.
Литература 49$>
Dunson W. А. (1975). Salt and water balance in sea snakes. In: The Biology of
Sea Snakes (W. A. Dunson, ed.). pp. 329—353, Baltimore, University Park
Press.
Edney E. B. (1977). Water Balance in Land Arthropods, Berlin. Springer-Verlag,
282 pp.
Gupta B. L., Mor eton R. B., Oschman J. L., Wall В. J. (eds.) (1977). Transport of
Ions and Water in Animals, London» Academic Press.
Keynes R. C. (1971). A discussion on active transport of salts and water in living,
tissues, Philos. Trans. R. Soc. Lond. (Biol.), 262, 83—342.
Kirschner L. B., et al. (1970). Refresher course on ionic regulation in organisms,
Am. Zool., 10, 329—436.
Krogh A. (1965). Osmotic Regulation in Aquatic Animals, New York, Dover,v
242 pp. Republication of 1st ed. (1939).
Maetz J., Bornancin M. (1975). Biochemical and biophysical aspects of salt excre-
tion by chloride cells in teleosts, Fortschr. Zool., 23, 322—362.
Peaker M., Linzell J. L. (1975). Salt Glands in Birds and Reptiles, Cambridge,
Cambridge University Press, 307 pp.
Potts W. T. W., Parry G. (1964). Osmotic and Ionic Regulation in Animals, Oxford,
Pergamon Press, 423 pp.
Schmidt-Nielsen К.» Bolis L., Maddrell S. H. P. (eds.) (1978). Comparative Phy-
siology: Water, Ions and Fluid Mechanics, Cambridge, Cambridge University
Press, 360 pp.
Shoemaker V. H., Nagy K. A. (1977). Osmoregulation in amphibians and reptiles^
Annu. Rev. Physiol., 39, 449—471.
Smith H. W. (1961). From Fish to Philosopher, Garden City, N. Y., Doubleday.
32/
ГЛАВА 10
ВЫДЕЛЕНИЕ
В предыдущей главе было показано, что органы выделения
играют важную роль в осморетуляции и поддержании водного
баланса и концентраций растворенных веществ. В настоящей
главе речь пойдет о том, как работает выделительная (экскре-
торная) система.
Органы выделения выполняют много функций, и все они
связаны с сохранением постоянства внутренней среды организ-
ма. Для поддержания постоянства состава требуется одно ос-
новное условие: поступление любого вещества в организм долж-
но быть уравновешено удалением такого же его количества.
А для этого в свою очередь нужно, чтобы экскреторные пути об-
ладали переменной мощностью, приспособленной для выделения
тщательно контролируемых количеств каждого из несчетного
•числа различных веще’ств. Главными функциями выделительных
систем можно считать следующие:
1) поддержание надлежащих концентраций отдельных ионов
(Na+, К+, С1-, Са2+, Н+ и др.);
2) поддержание надлежащего объема тела (содержания в
нем воды);
3) поддержание осмотических концентраций (что вытекает
из задач 1 и 2);
4) выведение конечных продуктов обмена (например, моче-
вины, мочевой кислоты и т. п.);
5) выведение посторонних веществ и/или продуктов их об-
мена.
Первые три функции уже рассматривались в главе 9. Чет-
вертая касается выделяемых продуктов обмена. Одним из глав-
ных конечных продуктов метаболической активности является
окись углерода, и почти вся она выводится через органы дыха-
ния; большая часть остальных конечных продуктов обмена уда-
ляется экскреторными органами. Эти органы выводят также
множество посторонних веществ — или в неизмененном виде,
или же с некоторыми изменениями, которые делают их -безвред-
ными (детоксикация) или легче выводимыми.
Таким образом, главная роль экскреторных (выделитель-
ных) органов состоит в удалении из организма точно регулируе-
мых количеств избыточных веществ, что помогает поддерживать
стационарное состояние при всех тех воздействиях, которые мог-
ли бы его нарушить.
Органы выделения 501
Органы выделения очень разнообразны, но в принципе су-
ществуют только два основных процесса, ответственных за об-
разование выделяемой жидкости: ультрафильтрация и активный
транспорт.
При ультрафильтрации жидкость под давлением проходит
через полупроницаемую мембрану, которая задерживает белки
и другие крупные молекулы, но пропускает воду и низкомолеку-
лярные растворенные вещества (ем. Приложение Д).
Активный транспорт представляет собой движение растворен-
ных веществ против электрохимического градиента благодаря
процессам, требующим затраты метаболической энергии. Если
эти вещества переносятся из внутренней среды животного в про-
свет экскреторного органа или органеллы, то это называется
активной секрецией. Если же перенос идет в обратном направ-
лении, то речь идет об активной реабсорбции.
В тех выделительных органах, где первоначальная жидкость
образуется путем ультрафильтрации, она по мере прохождения
по каналам экскреторного органа видоизменяется: одни вещест-
ва удаляются из фильтрата путем активной реабсорбции, дру-
гие добавляются к нему в результате активной секреции. Таким
образом, фильтрационную систему часто, если не всегда, допол-
няют процессы активного транспорта.
Выделительные органы и органеллы весьма разнообразны
по своей морфологической структуре и анатомическому располо-
жению, но все же их можно подразделить на сравнительно не-
большое число функциональных типов. Одни органы являются
универсальными, неспециализированными, и их можно считать
почками в обобщенном смысле этого слова, а продукт их экс-
креции— 'мочой. Другие экскреторные органы играют более
специальную роль, выполняя одну определенную функцию. Не-
сколько примеров помогут лучше уяснить это различие:
Универсальные [выделительные ърганы
Сократительные (пульсирующие) вакуоли простейших
Нефридии беспозвоночных
Мальпигиевы сосуды насекомых
Почки позвоночных
Специализированные выделительные органы
Жабры (у ракообразных и рыб)
Ректальные железы (у пла-стиножаберных рыб)
Солевые железы (у рептилий и птиц)
Печень (у позвоночных)
Из универсальных экскреторных органов лучше всего изуче-
на почка. Она подробно исследована, и функция ее понятна. Со-
кратительная вакуоль составляет только ча1сть клетки (т. е. яв-
ляется органеллой), и она так мала, что исследование образую-
502 Гл. 10. Выделение
щейся в ней жидкости весьма затруднено; тем не менее о ее
функции кое-что известно. Мальпигиевы со'суды насекомых тща-
тельно изучались у нескольких видов, и работа их стала вполне
понятной. Выделительные органы других беспозвоночных, одна-
ко, гораздо меньше исследо’ваны, и существуют лишь приблизи-
тельные представления о том, как функционируют .некоторые из
них. В отношении большинства этих органов неизвестно даже,
каков начальный этап образования выделяемой жидкости —
ультрафильтрация это или же секреция.
Некоторые специализированные экскреторные органы были
рассмотрены в предыдущей главе, посвященной осморегуляции.
Ничего, однако, не было сказано о печени позвоночных. Одна из^
ее выделительных функций состоит в следующем. Эритроциты,
достигнув предела своей нормальной продолжительности жизни
(у человека околю 100 дней), выводятся из кровотока и разру-
шаются; порфириновая группа молекулы гемоглобина преобра-
зуется печенью в соединения, называемые желчными пигмента-
ми, которые переходят в желчь, попадают в кишечник и в конце
концов удаляются с калом. Таким образом, в дополнение ко
многим другим своим функциям печень служит экскреторным
органом специально для выведения порфирина. Она играет так-
же важную роль в метаболизме и детоксикации разнообразных
чужеродных веществ и мюжет, кроме того, выполнять другие, ме-
нее важные экскреторные функции.
ОРГАНЫ ВЫДЕЛЕНИЯ
Функцию разных выделительных органов легче будет рас-
сматривать, если предварительно разделить их на группы или
типы1. Основные типы и их распределение в животном мире
приблизительно следующие:
Экскреторных органов не обнаружено
Кишечнополостные
Иглокожие
Сократительные вакуоли
Простейшие
Губки
1 Эта функциональная группировка широко используется, но она не совпа-
дает с морфологической классификацией экскреторных органов по Гудричу
(Goodrich, 1945), основанной на том, произошел ли орган из трубки, которая
развивается снаружи внутрь (нефридий) или же изнутри (из целома) наружу
(целомодукт). Выделительные органы моллюсков, членистоногих и позвоночных
образуются из целомодуктов; у остальных беспозвоночных (кроме простейших,
губок, кишечнополостных и иглокожих) они представляют собой нефридии.
Гудрич различает также разнообразные смешанные типы. У насекомых экскре-
торные органы — мальпигиевы сосуды — возникают из заднего отдела кишки
и не укладываются в классификацию Гудрича.
Органы выделения 503
Нефридиальные 'органы
'Протонефридий (замкнутый)
Плоские черви
Круглые черви
Метанефридий (незамкнутый)
Кольчатые черви
Нефрид'ий
Моллюски
Антеннальная железа (зеленая железа)
Ракообразные
Мальпигиевы сосуды
Насекомые
Почки
Позвоночные
У кишечнополостных и иглокожих специальных выделитель-
ных органов не найдено. Это удивительно, так как пресновод-
ные кишечнополостные явно гипертоничны по отношению к сре-
де, и в их тело, несомненно, поступает вода осмотическим путем.
Как удаляется излишек воды, неизвестно. Что касается иглоко-
жих, то проблемы осморегуляции для них не существует, так
как в пресной воде они не встречаются, а морские формы всег-
да ИЗОТ01НИЧНЫ с морской водой.
СОКРАТИТЕЛЬНЫЕ ВАКУОЛИ
Сократительными вакуолями обладают две группы живот-
ных— простейшие и губки. По-видимому, такие вакуоли есть у
всех пресноводных простейших. Не столь ясно, имеются ли они
у всех морских форм, но они обнаружены по крайней мере у не-
которых морских инфузорий. Наличие сократительных вакуолей
у пресноводных губок раньше подвергалось сомнению, но теперь
доказано бесспорно (Jepps, 1947).
Так как пресноводные формы всегда гиперосмотичны по от-
ношению к среде и поверхность их проницаема для воды, им по-
стоянно приходится выводить из организма воду. Они должны
не только удалять излишек воды, но также возмещать утрачен-
ные растворенные вещества, вероятно, путем активного погло-
щения солей из внешней среды. Определение водной проницае-
мости крупной амебы Chaos chaos показало, что вычисленный
осмотический приток воды хорошо согласуется с наблюдаемой
скоростью выведения жидкости сократительной вакуолью. Этим
подтверждается широко распространенное мнение, что главная
504 Гл. 10. Выделение
функция сократительной вакуоли состоит в осморегуляции и ре-
гуляции объема клетки (L^vtrup, Pigon, 1951).
Наблюдая сократительную вакуоль у пресноводных простей-
ших под микроскопом, можно видеть непрерывные циклические
изменения. Вакуоль набирает воду и постепенно увеличивается
в объеме, пока не достигнет критических размеров. Тогда она
внезапно выбрасывает свое содержимое наружу и уменьшается
Рис. 10.1. Сократительная вакуоль Amoeba proieus ограничена мембраной и
окружена слоем мелких пузырьков, которые наполнены жидкостью и, по-видимо-
му, опорожняются в вакуоль. Вокруг этой структуры лежит слой митохондрий^
которые, вероятно, доставляют энергию для секреторного процесса. (Mercer,.
1959.)
в объеме, после чего снова начинает увеличиваться, и цикл по-
вторяется.
Просвет сократительной вакуоли у амебы окружен одиноч-
ной тонкой мембраной. К этой мембране снаружи прилегает
толстый (0,5—2 мкм) слой плотно упакованных мелких пу-
зырьков диаметром от 0,02 до 0,2 мкм. Вокруг этого слоя мел-
ких пузырьков лежит слой митохондрий, которые, по-видимому,,
доставляют энергйю для осмотической работы, создающей гипо-
тоничность содержимого вакуоли (рис. 10.1). Судя по электрон-
ным микрофотографиям, пузырьки опорожняются в сократитель-
ную вакуоль в результате слияния мембран.
Роль сократительной вакуоли в осморегуляции хорошо про-
демонстрирована у эвригалинной амебы Amoeba lacerata. Эта
амеба исходно является пресноводным организмом, но облада-
ет высокой толерантностью к соли и даже может адаптировать-
ся к 50%-ной морской воде. Скорость опорожнения ее сократи-
Органы выделения 505
тельной вакуоли при адаптации к разным концентрациям солей
находится в обратной зависимости от осмотической концентра-
ции среды (рис. 10.2).
По-видимому, сократительные вакуоли удаляют воду с той
же скоростью, с какой происходит ее осмотический приток, так.
как по мере повышения концентрации среды количество посту-
Рис. 10.2. Скорость выведения жидкости сократительной вакуолью Amoeba la-
cerata в зависимости от концентрации внешней среды (выраженной в процентах
от концентрации морской воды). Амебы исследовались в том же растворе, в ка-
ком были выращены. (Hopkins, 1946.)
пающей воды снижается. В морской среде, где, как надо пола-
гать, внутренняя и внешняя осмотические концентрации почти
одинаковы, сократительные вакуоли (у тех форм, у которых их
наблюдали) опорожняются очень медленно. В этих случаях
приходится предположить, что они не служат в первую очередь
для осморегуляции, а выполняют другие экскреторные функции.
Если у пресноводных простейших главная функция сократи-
тельной вакуоли состоит в удалении воды, то ее содержимое
должно быть гипотоничным по отношению к остальной части
клетки. Так и обстоит дело в действительности. В микроскопи-
ческих пробах жидкости, взятых из сократительной вакуоли,
осмотическая концентрация примерно в три раза ниже, чем в
цитоплазме, но в несколько раз выше, чем во внешней среде
(В. Schmidt-Nielsen, Schrauger, 1963).
506 Гл. 10. Выделение
Сократительная вакуоль может удалять гипотоничную жид-
кость и служить для выведения воды. Но из-за того, что выво-
димая жидкость обладает более высокой осмотической концент-
рацией, чем внешняя среда, происходит непрерывная потеря рас-
творенных веществ, и из этого следует, что амеба должна быть
способна поглощать нужные ей веще!ст'ва, вероятно, путем их ак-
тивного переноса прямо из внешней среды.
Каким образом вакуоль может увеличиваться в объеме и в
то же время содержать жидкость менее концентрированную, чем
цитоплазма? Здесь возможны разные объяснения. Согласно од-
ному из них, происходит активный транспорт воды в вакуоль.
Но по ряду причин такая гипотеза мало правдоподобна. Другая
возможность состоит в том, что вначале вакуоль содержит изо-
тоничную жидкость, из которой осмотически активные вещества
извлекаются, перед тем как жидкость будет выведена наружу.
Но такое предположение противоречит данным о том, что жид-
кость гипотонична и состав ее относительно постоянен на про-
тяжении всего периода увеличения вакуоДи.
Сведения о составе вакуолярной жидкости позволяют нам
предположить третий механизм. Как видно из табл. 10.1, осмо-
Таблица 10.1
Концентрация веществ, растворенных
в цитоплазме и в сократительной вакуоли
пресноводной амебы. Средний объем вакуоли
составлял около 0,2 нл. (Riddick, 1968)
Концентрация В наруж- ной среде В цито- плазме В вакуоли Отноше- ние ва- куоль/ци- топлазма
Осмотическая,
мосмоль/л <2 117 51 <0,49
Na+, ммоль/л 0,2 0,60 19,9 33
К+, ммоль/л 0,1 31 4,6 0,15
тическая концентрация жидкости в вакуоли примерно вдвое ни-
же, чем в цитоплазме, -но более чем в 25 раз превышает кон-
центрацию в наружной среде. Содержание натрия в жидкости
вакуоли относительно велико — оно в 3 раза выше, чем в цито-
плазме. В то же время калия в вакуоли сравнительно мало, его
концентрация здесь существенно ниже, чем в цитоплазме. В сум-
ме натрий и калий в вакуолярной жидкости составляют около
25 ммоль/л, а если анионом является С1“, то эти три иона обес-
печивают почти всю осмотическую концентрацию жидко'сти
(51 мо^смоль/л).
Органы выделения 50?
Наиболее вероятен следующий механизм образования сокра-
тительной вакуоли. Окружающие ее мелкие пузырьки вначале
наполняются жидкостью, изотоничной по отношению к цитоплаз-
ме. Затем пузырьки путем активного транспорта накачивают в
эту жидкость натрий и удаляют калий — таким образом, что
удаление калия превышает накопление натрия. Мембрана пу-
зырьков должна быть относительно непроницаема для воды, что-
бы в пузырьке могла образоваться жидкость, гипотоничная по
отношению к цитоплазме. Если затем эти гипотоничные пузырь-
ки будут сливаться и опорожняться в сократительную вакуоль,
как па это указывают электронные микрофотографии, то ваку-
оль будет вместилищем вырабатываемой пузырьками жидкости.
Энергия для осмотической работы поставляется слоем митохонд-
рий, смежным с пузырьками. Поскольку активность сократи-
тельной вакуоли приводит к непрерывной потере натрия, необхо-
димо предположить, что эта потеря возмещается активным за-
хватом натрия поверхностью клетки (Riddick, 1968).
ВЫДЕЛИТЕЛЬНЫЕ ОРГАНЫ БЕСПОЗВОНОЧНЫХ
ПРОТОНЕФРИДИИ И МЕТАНЕФРИДИИ
Истинные экскреторные органы имеются только у животных
с двусторонней симметрией тела. Самая обычная форма таких
органов, широко распространенная у беспозвоночных, — это
простая или разветвленная трубочка, которая открывается нару-
жу через отверстие (нефрндиальную пору). Эти трубочки делят-
ся на два основных тина: протонефридии, у которых внутренний
конец замкнут и оканчивается слепо, и метанефридии, которые
соединяются с полостью тела воронкообразной структурой, назы-
ваемой нефридиостомом или нефростомом.
Протонефридии встречаются главным образом у животных,
не имеющих истинной полости тела (целома). У животного мо-
жет быть два или более протонефридия, которые часто образу-
ют обширные разветвления. Их слепые концы расширяются в
виде луковицы, в полость которой выступают одна или несколь-
ко длинных ресничек (рис. 10.3). Если ресничка одна, терми-
нальную клетку называют соленоцитам; если же ре'сничек много
(иногда несколько десятков), то тогда эта структура называет-
ся пламенной клеткой, так как пучок ресничек с его волнооб-
разными движениями несколько напоминает мерцающее пламя
свечи (фото 10.1). Функциональные различия между пламенны-
ми клетками и соленоцитами не установлены.
Для метанефридиев характерно отсутствие разветвлений, и
внутренние их концы открываются воронками в целомиче'с'кую
полость. Метанефридии бывают только у животных, обладаю-
щих целомом, и притом не у всех; некоторые животные с цело-
508 Гл. 10. Выделение
мом имеют протонефридии, которые обычно характерны для
ацеломических и псевдоцеломических форм (рис. 10.4).
Функция метанефридия (часто называемого просто нефри-
дием) достаточно ясна, главным образом благодаря исследова-
ниям Дж. Рамсея (Ramsay, 1949), которому удалось взять мель-
чайшие пробы жидкости из разных частей нефридии дождевого
червя. Полученные данные говорят в пользу следующего пред-
ставления. Жидкость из
целома попадает в нефри-
дий через воронкообразный
нефростом, и по мере ее
прохождения по сильно из-
витому протоку состав ее
изменяется. Когда жидкость
входит в нефридий, она изо-
тонична, но в концевых
участках органа из нее из-
влекаются соли, так что
наружу выводится жидкая
моча (значение жидкой мо-
чи для осморегуляции у
дождевого червя рассмотре-
но в гл. 9). Поэтому нефри-
дий работает как фильт-
рационно - реабсорбционная
Рис. 10.3. У планарии выделитель-
ная система сильно разветвленная.
Выделяемая жидкость вначале об-
разуется в соленоцитах или пла-
менных клетках, а затем проходит
по протокам нефридиев и выво-
дится через выделительные поры.
почка, в которой жидкость сначала образуется путем ультрафиль-
трации, а затем видоизменяется при прохождении по мочевому
канальцу.
Как функционирует протонефридий, не столь ясно, так как
принято считать, что слепой конец его не пригоден для ультра-
фильтрации. Кроме того, этот конец расположен в ткани между
клетками тела и в отличие от метанефридия не соприкасается
с целомической жидкостью. Функция соленоцитов и пламенных
клеток не ясна, хотя было высказано предположение, что непре-
рывное биение ресничек может создавать отрицательное давле-
ние, достаточное для ультрафильтрации. Из-за малого размера
Органы выделения 509»
соответствующих структур трудно было бы получить прямое под-
тверждение этой гипотезы.
Однако есть данные, что протонефридий коловратки Asplan-
chna функционирует на основе фильтрации и реабсорбции.
Жидкости тела Asplanchna гипертоничны по отношению к внеш-
ней среде, но коловратка выделяет жидкую мочу; если переме-
стить животное в более разбавленную среду, образуется еще
более жидкая моча. Из этого, во всяком случае, следует, что
протонефридий участвует в осморегуляции и выделении воды
(Braun et al., 1966).
Фото 10.1. Пламенная клетка в экскреторном органе личинки печеночной двуустки
(Fasciola hepatica). (Фото G. Ciimmel, Freie Universitat, Зап. Берлин.) На фо-
то (А) на поперечном срезе видно, что «пламя» (Пл) представляет собой плот-
ный пучок ресничек. Как показано также на схематических рисунках (Б и В),
пламя окружено тонкой стенкой (Ст) из палочек. Первичная моча, вероятно,,
образуется путем ультрафильтрации через эту стенку.
Самый важный вопрос относительно протонефридия заклю-
чается в том, происходит ли в нем ультрафильтрация. Обычно,,
чтобы выявить наличие этого процесса, в организм вводят ину-
лин и смотрят, появится ли он в моче. Инулин (о котором еще-
будет речь позже, в связи с почкой позвоночных животных) —
это растворимый полисахарид с мол. весом около 5000. Он не-
претерпевает в организме каких-либо превращений и может пе-
реходить в мочу только в случае ее образования путем фильт-
рации. Он никогда не выводится посредством клеточного транс-
ЗЛО Гл. 10. Выделение
порта или секреции, так как инертен по отношению ко всем из-
вестным процессам активного транспорта.
Введенный в организм Asplanchna, инулин действительно по-
является в моче, что указывает на процесс фильтрации. Кроме
того, концентрация инули-
на в моче Asplanchna вы-
ше, чем в жидкости тела;
это означает, что вода реаб-
сорбируется перед выходом
.мочи наружу. Стенка пла-
менной клетки у Asplanchna
чрезвычайно тонка и, по-ви-
димому, пригодна для уль-
трафильтрации (Pontin,
1964).
Понятно, что другие
протонефридии могут функ-
ционировать по-иному, но
поскольку у Asplanchna
Рис. 10.4. Схема строения перед-
него конца тела примитивного
кольчатого червя (в продольном
разрезе). (Goodrich, 1945.)
Показано отношение нефридиев и
целомодуктов с сегментарными
целомическими полостями (Ц).
Слева — протонефридии (Пн) с
соленоцитами (Сол); справа — ме-
танефридии (Мн) с воронкообраз-
ными нефростомами (Нет). Про-
дукты гонад (Г) выводятся через
цсломодукты (Цд) у нефростома.
продемонстрирована начальная фильтрация, функциональные раз-
личия между разными типами нефридиев представляются сейчас
менее существенными.
почки моллюсков
Важнейшие группы моллюсков — это головоногие (осьминоги
и кальмары), двустворчатые (устрицы и др.) и брюхоногие
(улитки). Осьминоги и кальмары — чисто мор'ские животные, но
двустворчатые моллюски и улитки встречаются как в морской,
так и в пресной воде, а среди улиток имеются также и наземные
виды. Почечная функция изучена у представителей всех этих
Органы, выделения 511
групп достаточно хорошо. У всех первоначальная жидкость об-
разуется путем ультрафильтрации крови. В этой жидкости
растворены те же вещества, что и в крови, за исключением бел-
ков, и практически в тех же концентрациях1. Поэтому ультра-
фильтрат содержит не только то, что подлежит выделению, но
и такие ценные вещества, как глюкоза и аминокислоты. Было-’
бы большим расточительством изливать ультрафильтрат нару-
жу, если бы эти ценные соединения не реабсорбировались пе-
ред выведением мочи.
Помимо ультрафильтрации, за которой следует избиратель-
ная реабсорбция, происходит также активная секреция некото-
рых веществ, которые добавляются к моче в определенных отде-
лах почки. Двумя веществами, активно выделяемыми в почках
разных типов (в том числе и у позвоночных), являются пара-
аминогиппуровая кислота (ПАГ) и краситель феноловый крас-
ный (фенолсульфофталеин). Последний имеет то преимущество,
что благодаря своей окраске он хорошо виден, так что его кон-
центрацию легко измерить. Феноло'вый красный и ПАГ снова
будут упомянуты, когда пойдет речь о почке позвоночного, так
как они играют важную роль в выявлении секреторных процес-
сов.
У моллюсков есть одна своеобразная особенность: функция
обеих почек не всегда одинакова. Например, получены следую-
щие данные о морском ушке {Haltons'). Начальная ультрафильт-
рация происходит в обеих почках, поскольку инулин появляется
в моче из той и другой почки и концентрация его в крови, в пе-
рикардиальной жидкости и в окончательной моче из обеих по-
чек одна и та же. Это означает, что объем мочи равен объему
фильтрата и что вода не реабсорбируется в сколько-нибудь за-
метных количествах.
Естественно, что у морского животного, которое изотонично
по отношению к морской воде и поэтому не должно решать важ-
ной задачи регуляции содержания воды, вода в почках не реаб-
сорбируется. Однако нужно регулировать ионный состав жидко-
стей и выделять продукты обмена, и в этом отношении две поч-
ки Haliotis функционально различны, так как ПАГ и феноловый
красный активно секретируются в основном правой почкой, а ре-
абсорбция глюкозы происходит, по-видимому, главным образом
в левой почке (Harrison, 1962).
АНТЕННАЛЬНАЯ ЖЕЛЕЗА РАКООБРАЗНЫХ
У ракообразных органом выделения служат антеннальные,.
или зеленые, железы. Эти парные железы расположены в голо-
ве. Каждая железа состоит из мешка, за которым следуют длин-
1 Небольшие различия между концентрациями в крови и ультрафильтрате
объясняются доннановским эффектом белков крови (см. Приложение Д).
3512 Гл. 10. Выделение
<ный извитой экскреторный каналец и мочевой пузырь. Она от-
крывается выделительной порой у основания антенны; отсюда и
ее название «антеннальная железа».
Моча образуется в этой железе путем фильтрации и реаб-
сорбции, к которым добавляется секреция в канальце. Ультра-
фильтрацию можно выявить путем инъекции инулина, который
затем появляется в моче. У омара, типичного морского животно-
го, концентрация инулина в моче будет такой же, как в крови
(т. е. отношение концентраций моча/кровь равно 1,0); следова-
тельно, вода не реабсорбируется. Но ПАГ и феноловый красный
будут содержаться в моче в более высоких концентрациях, чем
Концентрация морской воды, ммоль NaCl на 1л
Рис. 10.5. Отношения между концентрациями крови, мочи и внешней среды у
двух видов бокоплавов. (Lockwood, 1961.)
А — пресноводный вид Gatnmarus pulex, способный выделять сильно разбавлен-
ную мочу. Б — близкий вид, живущий в солоноватой воде, G. duebeni, не спосо-
бен выделять мочу более разбавленную, чем внешняя среда.
в крови. Это показывает, что они активно выделяются в мочу и
что в дополнение к процессу фильтрации — реабсорбции в обра-
зовании мочи участвует секреция (Burger, 1957).
В отличие от омара у обыкновенного зеленого краба (Carci-
nus) вода из ультрафильтрата реабсорбируется. Этот краб спо-
Органы выделения 513
собен к хорошей осморегуляции и может проникать в солонова-
тую воду. Концентрация инулина в его моче может быть в не-
сколько раз выше, чем в крови, что должно быть следствием
реабсорбции воды из исходного фильтрата. Движение воды ско-
рее всего объясняется активной реабсорбцией натрия, за кото-
рой следует пассивная реабсорбция воды. В результате образу-
ется моча с более низкой концентрацией натрия, чем в крови, и
это создает очевидное преимущество для животного, которое
проникает в менее соленую среду, где ему трудно сохранять вы-
сокие концентрации солей в крови (Riegel, Lockwood, 1961).
У пресноводного бокоплава Gammarus pulex концентрация
мочи не только очень низка, но может быть даже ниже концент-
рации внешней среды (рис. 10.5). У его близкого родственника
G. duebeni, обитающего в солоноватой воде и переносящего прес-
ную, моча по своей концентрации ближе к концентрации крови.
Этот вид неспособен вырабатывать очень разбавленную мочу и
в пресной воде не может успешно конкурировать с G. pulex,
МАЛЬПИГИЕВЫ СОСУДЫ НАСЕКОМЫХ
Для многих выделительных органов беспозвоночных еще не
решен вопрос о том, образуется ли первичная моча путем ульт-
рафильтрации. Но для экскреторных органов насекомых ответ,
по-видимому, определенно отрицательный. Он основан на том
факте, что инулин, который считается самым надежным показа-
телем ультрафильтрации, не переходит у насекомых в мочу, а
это говорит об отсутствии ультрафильтрации (Ramsay, Riegel,
1961). Как же в таком случае образуется у насекомых моча?
Их выделительная система состоит из трубочек, называемых
мальпигиевыми сосудами, которых может быть от двух до не-
скольких сотен. Каждый сосуд открывается в кишечник на гра-
нице между средней и задней кишкой; другой его конец слепой
и у большинства насекомых лежит в гемоцеле (рис. 10.6). Но
у некоторых насекомых, особенно у жуков, питающихся сухими
веществами (например, у личинок мучного хрущака Tenebrio),
имеется специальное приспособление, которое обусловливает по-
разительную способность извлекать воду из экскрементов. У них
слепой конец мальпигиева сосуда тесно связан с прямой киш-
кой и вся эта структура окружена оболочкой (периректальной
мембраной). Образуемое ею пространство заполнено перирек-
тальной жидкостью, которая окружает и мальпигиев сосуд, и
эпителий прямой кишки, но отделена от общей гемолимфы.
Мальпигиев сосуд функционирует следующим образом.
В просвет сосуда активно транспортируется калий, а за ним
пассивно следует вода под влиянием осмотических сил. В ре-
зультате этого в просвете накапливается много жидкости, бога-
той калием, и отсюда она переходит в заднюю кишку. Там рас-
33—1863
514 Гл. 10. Выделение
творенные вещества и значительная часть воды реабсорбируют-
ся, а мочевая кислота (которая проникла в жидкость в виде рас-
творимого в воде урата калия) выпадает в осадок. Этим облег-
чается дальнейшее извлечение воды, так как осажденная моче-
вая кислота не участвует в осмотической активности содержи-
мого кишки. А то, что*
остается в прямой кишке,,
в конечном счете выхо-
дит в виде смеси мочи и
кала.
Насекомые, питающи-
еся свежей растительно-
стью, поедают пищу с
высоким содержанием
воды и выделяют многа
жидкой мочи. А те, ко-
торые потребляют сухую
пищу, способны выделять
очень сухие экскреты и
поэтому почти не теряют
воду с мочой и калом.
Хорошим примером слу-
жит личинка мучного
хрущака (Tenebrio moli-
tor). Весь жизненный
цикл этого насекомого
Рис, 10.6. Выделительная систе-
ма клопа Rhodnius. (Wiggles-
worth, 1931.)
Полностью показан только один
из четырех мальпигиевых сосу-
дов. Эти сосуды делятся на две
разные части—верхнюю и ниж-
нюю. Они опорожняются в пря-
мую кишку у ее соединения со-
средней кишкой.
проходит в сухой муке, и тем не менее оно не испытывает недо-
статка в воде.
У личинки мучного хрущака имеется типичная периректаль-
ная оболочка, описанная выше, и выделяемые экскреты могут
быть настолько сухими, что поглощают воду из воздуха с отно-
сительной влажностью 90% (Ramsay, 1964)'. Посмотрим, как.
можно себе представить механизм извлечения воды. Осмотиче-
ская концентрация жидкости в периректальном пространстве мо-
Органы выделения 515
жет быть очень высокой [понижение точки замерзания (ЛТ) до-
стигает 8°]—намного больше концентрации гемолимфы (где
ДТ составляет от 0,7 до 1,4°). Разница более выражена при со-
держании мучного хрущака в среде с очень низкой влажностью.
Возможны два способа функционирования ректального комп-
лекса у личинки мучного хрущака: 1) активный транспорт воды
Рис. 10.7. Модель движения воды в стенке прямой кишки у таракана Periplane-
ta atnericana. (Oschman, Wall, 1969.)
Движение растворенных веществ показано черными стрелками, движение водь! —
светлыми стрелками. Эти перемещения представлены раздельно лишь для ясно-
сти, на самом деле они происходят в одном и том же внутриклеточном1 синусе.
Растворенные вещества активно накачиваются во внутриклеточный синус как из
просвета кишки, так и из жидкости, втекающей в субэпителиальный синус.
Высокая осмотическая концентрация во внутриклеточном синусе вызывает осмо-
тический приток воды, что приводит к переходу воды и солей в субэпителиаль-
ный синус. Эта система похожа на модель Керрана с тремя отделениями
(рис. 9.14).
из просвета этого комплекса в гемолимфу или 2) активный
транспорт какого-то растворенного вещества (предположительно
хлористого калия) из гемолимфы в периректальное пространст-
во, повышающий концентрацию периректальной жидкости и тем
самым в свою очередь приводящий к осмотическому удалению
воды из прямой кишки (Grimstone et al., 1968).
Второй из этих механизмов реабсорбции воды в принципе
идентичен тому, который постулируется в теории трех ком,парт-
ментов Кёррана (гл. 9). При изучении тараканов были получены
дополнительные данные в пользу этой гипотезы. Если таракан
имеет возможность пить воду, он выделяет разбавленную мочу;
если же лишить его воды, у него будут образовываться сухие фе-
кальные шарики и содержимое его прямой кишки будет сильно
гиперосмотично по отношению к гемолимфе. Подробный анализ
33*
516 Гл. 10. Выделений
осмотических концентраций в ректальном комплексе позволил
предположить механизм реабсорбции воды, схематически пред-
ставленный на рис. 10.7.
Таким образом, следует заключить, что, выделительная си-
стема насекомых, по-видимому, работает без всякой начальной
ультрафильтрации, что она основана на первичной секреции ка-
лия в мальпигиевы сосуды, за которой следует пассивное пере-
мещение воды, и что вода вместе с растворенными веществами
извлекается обратно в задней кишке и ректальном комплексе.
Получено достаточно данных в пользу того, что движение воды
основано на первичном транспорте растворенных веществ.
ПОЧКИ позвоночных животных
Почки всех позвоночных — рыб, амфибий, рептилий, птиц и
млекопитающих — сходны в том отношении, что они функциони-
руют по принципу фильтрации — реабсорбции, к которому1 до-
бавляется секреция в канальцах. Лишь у нескольких костистых
рыб! мы находим отклонение от этого общего плана: у них нет
механизма ультрафильтрации и почка работает полностью- па
секреторному типу.
Каковы преимущества и недостатки фильтрационного меха-
низма? Первичный ультрафильтрат содержит все компоненты
крови, кроме таких высокомолекулярных веществ, как белки.
Многие соединения, переходящие в фильтрат, представляют цен-
ность ДЛЯ' организма и не должны теряться. Поэтому нужен ме-
ханизм реабсорбции для сохранения таких веществ, как глюко-
за, аминокислоты и витамины.
Фильтрационно-реабсорбционная почка способна . перераба-
тывать , большие количества жидкости, и при этом часто
больше”99% фильтрата реабсорбируется и меньше 1% выводит-
ся в виде мочи. Может показаться, что выгоднее было бы иметь
почку, работающую только на основе канальцевой секреции, так
как это сэкономило бы много энергии, необходимой для реаб-
сорбции. Известно, что в эволюционном смысле такое решение
осуществимо, так как некоторые костистые рыбы обходятся без.
ультрафильтрации; у насекомых, как мы видели, этого механиз-
ма тоже нет.
Но существует одно уникальное следствие ультрафильтрации:
любое перешедшее в фильтрат вещество остается в моче, если
только оно не реабсорбируется. Встречаясь с «новыми» вещест-
вами, подлежащйми выведению, организм при помощи фильтра-
ционной почки удаляет их, не нуждаясь в создании специаль-
ного секреторного .механизма для каждого нового вещества, с
которым оно может столкнуться. Это дает организму горазда
большую свободу обследования новых местообитаний, изменения
пищевого рациона и,т. п. В этом смысле секреторная почка со-
Органы выделения 517
здает гораздо больше ограничений. Это согласуется с тем фак-
том, что среди позвоночных животных почками секреторного ти-
па обладает лишь несколько видов морских рыб, которые живут
в стабильной, «консервативной» среде.
Все позвоночные могут продуцировать мочу, изотоничную ил»
гипотоничную по отношению к крови, но только у птиц и млеко-
питающих возможно образование мочи более концентрирован-
ной, чем жидкости тела. В пресной воде разбавленная моча слу-
жит для выведения излишней воды без потери растворенных ве-
ществ. В морской воде животные не могут использовать разбав-
ленную или изотоничную мочу для удаления излишка солей, и у
них имеются вспомогательные органы для выделения солей (на-
пример, жабры, ректальные железы, солевые железы). Морские
млекопитающие обладают почками с исключительной концент-
рирующей способностью и решают свои солевые проблемы путем
почечной экскреции. У наземных млекопитающих способность к
выработке концентрированной мочи тоже очень важна для их
водного баланса. То, что птицы и рептилии выделяют мочевую
кислоту (а не мочевину), позволяет им выделять полутвердую
мочу, для которой требуется лишь умеренное количество воды
(см. раздел «Выделение азота» ниже в этой главе).
СТРОЕНИЕ ПОЧЕК
Чтобы понять, как почка работает, надо знать ее строение
(рис. 10.8). Почки у всех позвоночных состоят из множества
элементов — нефронов. У мелких рыб их может быть всего не-
сколько десятков, у крупных млекопитающих — несколько мил-
лионов в каждой почке. Каждый нефрон начинается мальпигие-
вым тельцем, в котором происходит ультрафильтрация плазмы
крови. В тельце входит небольшая артерия, которая там распа-
дается на пучок или клубок капилляров — почечный клубочек.
Через стенки клубочковых капилляров жидкость вытесняется
под давлением крови; эта жидкость — клубочковый фильтрат —
поступает в каналец, ведущий от мальпигиева тельца. В каналь-
це эта жидкость изменяется в результате процессов реабсорбции
и секреции и образует окончательную мочу.
Почечный каналец можно подразделить на две части: прок-
симальный каналец, в котором реабсорбируются многие раство-
ренные вещества, такие, как соли и глюкоза, а также вода, и
дистальный каналец, в котором продолжается процесс превра^
щения канальцевой жидкости в мочу. Дистальные канальцы со-
единяются и образуют собирательные трубки, из которых моча
поступает в почечную лоханку. Отсюда она проходит через мо-
четочник в мочевой пузырь, а из него время от времени излива-
ется наружу. В нефроне млекопитающих проксимальный и ди-
стальный отделы канальца разделены характерным узким сег-
518 Гл. 10. Выделение
ментом, образующим петлю наподобие шпильки для волос —
так называемую петлю Генле. Подобный сегмент, хотя и не так
хорошо развитый, имеется в почке у птиц. Именно в этой части
нефрона происходит образование мочи, более концентрирован-
Рис. 10.8. Схема почки млекопитающего.
Почка состоит из большого числа (до нескольких миллионов) отдельных нефро-
нов. На схеме слева представлен только один нефрон; он показан в увеличенном
виде справа. Наружный слой почки, кора, содержит мальпигиевы тельца и прок-
симальные и дистальные извитые канальцы. Капиллярная сеть в мальпигиевом
тельце называется клубочком. Внутренняя часть почки — мозговое вещество —
содержит петли Генле и собирательные трубки.
Моча первоначально образуется путем ультрафильтрации в мальпигиевых тель-
цах, а затем фильтрат видоизменяется и сильно уменьшается в объеме по мере
прохождения по извитому канальцу; из канальцев моча попадает в собиратель-
ные трубки, которые выводят ее в почечную лоханку, откуда она проходит через
мочеточник в мочевой пузырь.
ной, чем плазма крови. Рыбы, амфибии и рептилии не имеют та-
кой петли и не могут вырабатывать мочу более высокой кон-
центрации, чем плазма.
КАК ИССЛЕДУЮТ ПОЧЕЧНУЮ ФУНКЦИЮ
Существующие методы исследования почечной функции дают
поразительно много сведений без прямого экспериментирования
Органы выделения 519
Фото 10.2. Артериальная система почки собаки. Фотография препарата, полу*\
ченного путем инъекции силиконово-резинового соединения в почечную артерию,
Клубочки различимы в виде многочисленных мелких пятнышек в наружном слое
почки — корковом веществе. (Фото предоставлено A. Clifford Barger, Гарвард*
ский университет.)
на самой почке. В опытах на интактном животном можно полу-
чить данные о количестве жидкости, образующейся в результате
ультрафильтрации; о количестве жидкости, реабсорбируемой в
почечных канальцах; о процессе канальцевой секреции и даже
Фото 10.3. Клубочки в почке собаки при
большом увеличении (после введения
силиконовой резины в артерию). У клу-
бочка в центре фото можно видеть не-
сколько более толстый сосуд, входящий
в клубочек, и более тонкий сосуд, вы-
ходящий из него. Диаметр этих сосудов
составляет около 15 — 20 мкм, диаметр
клубочка — около 150 мкм. (Фото предо-
ставлено A. Clifford Barger, Гарвардский
университет.)
520 Гл. 10. Выделение
об объеме кровотока в почке. Эти методы широко применяются
и в клинике — без ненужного риска и дискомфорта — для изуче-
ния функции почек у человека. Поэтому следует уяснить себе
принципы, на которых основаны такие исследования.
Много сведений о функции почки получено и путем прямого
экспериментирования на этом органе. В особенности это отно-
сится -к механизму формирования концентрированной мочи. При
помощи методов так называемой микропункции, при которой в
разные сегменты нефрона вводят тонкие пипетки и извлекают
ничтожные количества жидкости, получена информация о рабо-
те различных отделов нефрона. Такие методы требуют хирурги-
ческого вмешательства и неприменимы для исследований на че-
ловеке.
Фильтрация. Ультрафильтрация в мальпигиевом тельце про-
исходит благодаря кровяному давлению, которое проталкивает
жидкость через тонкие стенки капилляров. Поскольку эти стен-
ки не пропускают белков, последние остаются в плазме, а с во-
дой фильтруются только низкомолекулярные вещества.
Ультрафильтрация возможна только в том случае, если кро-
вяное давление выше осмотического давления белков крови
(коллоидного осмотического давления). Если у собаки постепен-
но сдавливать зажимом почечную артерию и таким образом по-
нижать кровяное давление в почке, образование мочи прекра-
тится, когда кровяное давление в почке снизится до уровня кол-
лоидного осмотического давления. Много лет назад это показал
Старлинг (Starling, 1899). Если снова постепенно ослаблять за-
эцим, чтобы кровяное давление в капиллярах повышалось,
фильтрация возобновится, как только кровяное давление превы-
сит коллоидное осмотическое давление, и чем больше будет раз-
ница между ними, тем быстрее пойдет фильтрация.
В почке млекопитающего мальпигиевы тельца заложены глу-
боко, а в почке амфибии видны на ее поверхности. Крупному
американскому специалисту по физиологии почек Ричардсу (Ri-
chards, 1935) удалось осуществить введение микропипеток в
мальпигиевы тельца амфибий, и он показал, что концентрации
С1~, глюкозы, мочевины и фосфата, а также общее осмотическое
давление в фильтрате те же, что и в крови, если не считать не-
больших различий, обусловленных доннановским эффектом бел-
ков. Эти опыты, по существу, доказали, что первичная моча дей-
ствительно представляет собой ультрафильтрат'Крови.
Общее количество жидкости, образующейся в результате
ультрафильтрации, можно определить при помощи инъекции по-
лисахарида инулина, не обнажая почки и не прибегая к каким-
либо другим грубым вмешательствам. Молекулярный вес инули-
на около 5000, и поэтому он проходит через капиллярную стен-
ку, которая задерживает только вещества с молекулярным ве-
сом, превышающим примерно 70 000. Инулин не подвергается
Органы выделения 521
никаким превращениям, а также не реабсорбируется и не секре-
тируется почечными канальцами; поэтому весь инулин, находи-
мый в окончательной моче, попадает в нее только в результате
фильтрации.
Этот факт можно выразить простым уравнением:
Инулин в фильтрате = Инулин в моче.
Количество инулина в фильтрате равно объему фильтрата
(1/ф), умноженному на концентрацию инулина в фильтрате
(Сф); точно так же количество инулина в моче равно объему мо-
чи (Vm), умноженному на концентрацию инулина «в моче (См).
Оба этих количества равны между собой, и поэтому получаем:
Уф-сф=км.см
или
I/ _ VM’CM
Сф •
Три величины в правой части уравнения определить нетруд-
но. Ум и См находят, измеряя объем мочи за данный период вре-
мени и определяя содержание в ней инулина. Величина Сф рав-
на концентрации инулина в плазме крови, и ее можно устано-
вить, исследовав пробу крови. В результате мы получаем все дан-
ные, необходимые для вычисления скорости клубочковой 'фильт*
рации (Уф).
Рассмотрим пример. Предположим, что измеренная скорость
тока мочи составляет у человека 1,3 мл/мин и что концентрация
инулина в моче равна 2%, а в плазме — 0,02%. Это дает ско-
рость клубочковой фильтрации, равную 130 мл/мин, что является
для человека нормой.
Теперь видно, что для определения скорости фильтрации у
интактного животного достаточно 1) инъецировать подходящее
количество инулина1, 2) собрать мочу за известный период вре-
мени и измерить ее объем и 3) определить концентрацию инули-
на в моче и в плазме. Оказывается, что у человека скорость
фильтрации довольно постоянна — около 130 мл/мин, а колеба-
ния объема мочи объясняются главным образом изменением ко-
личества реабсорбируемой воды. У многих животных, особенно у
низших позвоночных, скорость фильтрации может значительно
варьировать, а у лягушки фильтрация иногда полностью прекра-
щается на долгое время без всякого вреда для организма.
1 Обычно инулин вводят непрерывно в течение всего опыта. Благодаря этому
концентрация его в плазме в Период наблюдения остается постоянной, а не сни-
жается.
522 Гл. 10. Выделение
Скорость фильтрации у человека равна 130 мл, или 0,13 л, в
минуту; таким образом, за час образуются 7,8 л фильтрата, что
примерно вдвое больше всего объема плазмы крови у человека,
и из этого объема удаляются продукты, подлежащие экскреции.
Если бы глюкоза, аминокислоты и другие важные вещества,
содержащиеся в фильтрате, терялись, то процесс фильтрации бы-
стро лишил бы организм этих ценных соединений; но они сохра-
няются благодаря канальцевой реабсорбции.
Канальцевая реабсорбция. Одним из важных веществ, реаб-
сорбируемых в проксимальном канальце, является глюкоза, и она
может служить примером, иллюстрирующим общий принцип ка-
нальцевой реабсорбции. Обратное всасывание глюкозы происхо-
дит путем активного транспорта, и в норме реабсорбируется вся
перешедшая в фильтрат глюкоза.
Если содержание глюкозы в плазме выше нормального уров-
ня, то реабсорбционный механизм должен справляться с этим
повышенным ее количеством, а если оно превышает мощность
транспортного механизма, то реабсорбируется не вся глюкоза и
некоторая часть ее остается в моче. Так происходит у больных са-
харным диабетом; концентрация глюкозы в их крови часто под-
нимается выше нормы и когда ее содержание в фильтрате чрез-
мерно возрастает, глюкоза появляется в моче.
Эта ситуация представлена на рис. 10.9. Глюкоза фильтрует-
ся непрерывно, и так как скорость фильтрации постоянна, ее ко-
личество в фильтрате растет линейно с увеличением ее концентра-
ции в плазме. При низких уровнях глюкозы в плазме из фильтра-
та реабсорбируется вся глюкоза, и она совсем не появляется в
моче. Такова нормальная ситуация, поскольку обычно глюкоза
плазмы остается на уровне около 1 мг/мл. Если ее содержание в
плазме повышено, но не более чем до пороговой величины око-
ло 2,3 мг/мл, вся глюкоза, поступающая в фильтрат, продолжает
реабсорбироваться, и в моче ее нет. Но если ее количество в
плазме превысит пороговый уровень, она уже не будет реабсорби-
роваться полностью и часть ее будет оставаться в моче. Иными
словами, в какой-то момент достигается предельная мощность
реабсорбционного механизма. Соответствующий уровень часто
называют почечным порогом, или канальцевым максимумом
{Км), Для глюкозы. Выше этого порога в моче появляется все
больше и больше глюкозы (рис. 10.9). Линии для глюкозы в
фильтрате и для глюкозы в моче идут параллельно; постоянство
расстояния между ними говорит о постоянстве канальцевого мак-
симума.
Таким же образом можно определить канальцевый максимум
для любого другого реабсорбируемого вещества: повышают его
концентрацию в плазме до тех пор, пока мощность реабсорбци-
онного механизма не будет превышена и вещество не начнет по-
являться в моче.
Органы выделения 523
Канальцевая секреция. Наряду с фильтрацией и реабсорбци-
ей важную роль в образовании мочи играет третий процесс —
канальцевая секреция. Мы уже упоминали о том, что феноловый
красный и ПАГ активно выводятся почечными канальцами. По-
скольку эти вещества также и фильтруются, количество их, пе-
реходящее в мочу, равно сумме их количеств, поступающих туда
в результате фильтрации и секреции.
Концентрация глюкгнч в плаз^, мг/м.п
Рис. 10.9. Выделение глюкозы у собаки. (Shannon, Fisher, 1938.)
При концентрациях ниже 2,3 мг на 1 мл плазмы глюкоза в моче не появляется,
так как вся она реабсорбируется из фильтрата в канальцах. Выше этого предела
реабсорбционный механизм полностью насыщен, и хотя он продолжает работать
с полной мощностью, глюкоза появляется в моче в количествах, которые возра-
стают по мере дальнейшего повышения ее концентрации в плазме. Выше точки
насыщения реабсорбируемое количество остается постоянным — это максимум
реабсорбции глюкозы канальцами.
Если ввести одно из этих веществ в кровоток, то общее его
количество, выводимое путем секреции, будет зависеть от кон-
центрации его в плазме так, как это показано на рис. 10.10 для
фенолового красного. Фильтруемое количество растет линейно с
концентрацией вещества в плазме. При низких концентрациях в
плазме количество его, добавляемое путем секреции, больше того,
которое поступает путем фильтрации. Однако при какой-то кон-
центрации вещества в плазме механизм секреции полностью на-
сыщается. Теперь суммарная экскреция повышается параллельно
количеству, поступающему в результате фильтрации, и разность
между суммарной экскрецией и фильтрацией (расстояние между
параллельными линиями) соответствует величине, добавляемой
путем канальцевой секреции. Иными словами, эта величина яв-
ляется канальцевым максимумом для выделения фенолового
красного.
524 Гл. 10. Выделение
Другие посторонние для организма вещества тоже выводятся
путем канальцевой секреции. Часто это происходит с фенольны-
ми соединениями и продуктами их детоксикации. К хорошо изве-
стным веществам, быстро удаляемым путем канальцевой секре-
ции, относится пенициллин; этот его недостаток компенсируют
повторными инъекциями больших количеств.
Рис. 10.10. Выделение фенолового красного у лягушки-быка. (Forster, 1940.)
Количество его в фильтрате возрастает пропорционально концентрации в плаз-
ме. Кроме того, феноловый красный добавляется к моче путем активного транс-
порта в канальцах. Количество, добавляемое секрецией в канальцах, при кон-
центрациях выше 0,05 мг на 1 мл плазмы остается постоянным, и это говорит
отом, что достигнут максимум возможной экскреции фенолового красного.
‘ Почечный кровоток. Процесс канальцевой секреции можно
очень интересным способом использовать для измерения крово-
тока в почке. Для этого нужно взять вещество, которое выводит-
ся путем канальцевой секреции с такой легкостью, что кровь,
прошедшая через почку, полностью очищается от него. Одно из
таких' веществ — /шрааминогиппуровая кислота (ПАГ), которая
выводится так быстро, что совершенно отсутствует в крови, вы-
ходящей из почки (если только не превышен канальцевый макси-
мум). В этом методе используется то обстоятельство, что почка
млекопитающего получает кровь только из почечной артерии; вся
кровь проходит сначала по клубочковым капиллярам, а затем по
капиллярам, окружающим почечные канальцы. Производят
инъекцию некоторого количества ПАГ, а затем определяют,
сколько ее появляется в моче за данное время. Из концентрации
ПАГ в плазме крови можно вычислить, какой объем плазмы дол-
жен был пройти через почку за этот период. У человека ток плаз-
мы через почку в норме составляет около 0,7 л/мин. Если гема-
токрит равен 45%, то это соответствует почечному кровотоку,
Органы выделения 525
равному 1,25 л/мин. Последняя величина составляет от У4 до Vs
всего минутного объема сердца в покое (5—6 л/мин), что гово-
рит об удивительно мощном кровоснабжении почек у млекопита-
ющих.
Такой метод неприменим для определения притока артериаль-
ной крови к почкам низших позвоночных, так как система крово-
снабжения почек у .них иная: почки получают кровь из двух ис-
точников. Помимо крови из почечной артерии в них поступает
также венозная кровь из задней части тела (по так называемой
воротной системе почек). Значение воротного почечного кровооб-
ращения не ясно. Может быть, дело в том, что при его наличии
образование мочи может продолжаться даже в том случае, если
прекратится клубочковая фильтрация. У лягушки, например,
фильтрация может приостановиться, а феноловый красный, по-
ступающий в почку по воротной системе, по-прежнему будет вы-
водиться путем канальцевой секреции.
ВЫДЕЛЕНИЕ У РЫБ
У большинства костистых рыб почки состоят из нефронов с
типичными для позвоночных свойствами. Пресноводные рыбы ги-
пертоничны по отношению к среде, и излишек воды, поступающий
в организм под действием осмотического градиента, удаляется в
форме гипотоничной мочи.
У морских костистых рыб, которые испытывают дефицит воды,
так как они гипотоничны по отношению к среде, образуется лишь
небольшое количество мочи. Они не могут вырабатывать кон-
центрированную мочу, и излишек солей выводится жабрами.
Для компенсации осмотической потери воды эти рыбы пьют мор-
скую воду, и одна из важных функций почек состоит в выведе-
нии двухвалентных ионов, содержащихся в морской воде, — маг-
ния и сульфата.
У некоторых морских костистых рыб почки лишены клубочков.
Это относится к морскому черту (Lophius piscatorius), рыбе Ор-
sanus tau и некоторым морским иглам (рода Syngnathus). Эти
рыбы сыграли большую роль в изучении почечной функции, так
как у них можно исследовать процессы секреции в такой почке,
где фильтрация невозможна. Если вводить им инулин, он не по-
является в моче (введение инулина служит превосходным функ-
циональным тестом на механизм фильтрации).
Среди рыб, не имеющих клубочков и тем самым лишенных на-
чального процесса фильтрации, есть несколько антарктических
родов. Для таких рыб отсутствие фильтрационного процесса име-
ет большой смысл. Как уже говорилось в гл. 7, многие антаркти-
ческие рыбы избегают замерзания благодаря гликопротеидным
антифризам. Молекулярный вес этих веществ варьирует в преде-
лах от нескольких тысяч до 30 000. Поскольку клубочек задержи-
526 Гл, 10. Выделение
вает белки с молекулярным весом около 70 000, но пропускает
менее крупные молекулы, при наличии фильтрации гликопротеи.-
ды быстро терялись бы, если бы только почка не обладала соот-
ветствующим реабсорбирующим механизмом. Так или иначе, ре-
абсорбция гликопротеидов потребовала бы значительной затраты
энергии, в чем, однако, нет нужды, так как эти вещества просто»
не могут переходить в канальцевую жидкость. Поэтому бесклу-
бочковая почка, очевидно, очень выгодна для тех рыб, которые
используют антифризы для защиты от повреждения, связанного
с образованием льда (Dobbs et al., 1974).
Примечательно, что у некоторых пресноводных рыб-игл, по-
видимому, тоже нет клубочков, но эти рыбы и функция их. почек
еще не изучены достаточно подробно. У пресноводной рыбы
должно происходить значительное осмотическое поглощение во-
ды, и трудно представить себе, каким образом нефильтрующая
почка удаляла бы избыточную воду. Очевидно, что этот вопрос
требует дальнейшего изучения.
В почках пластиножаберных имеются клубочки, и, как мы уже
знаем, этим рыбам нужна мочевина для поддержания осмотиче-
ского баланса. Поскольку молекула мочевины невелика (мол.
вес 60), в клубочке она проходит в фильтрат, но реабсорбируется
в канальцах. Это значит, что в почках пластиножаберных имеется
специальный механизм активного транспорта мочевины. Детали
этого механизма будут рассмотрены позже в настоящей главе.
ВЫДЕЛЕНИЕ У АМФИБИЙ
Первые несомненные данные о начальной ультрафильтрации
в почке были получены на лягушках. Это связано с тем, что у
амфибий многие клубочки довольно крупны и лежат так близко
к поверхности почки, что можно ввести в них тонкую микропипет-
ку и извлечь очень малый объем жидкости для анализа. Еще од-
но преимущество экспериментов на холоднокровных животных
состоит в том, что их почки хорошо функционируют при комнат-
ной температуре в отличие от почки млекопитающего, для кото-
рой необходимо тщательно поддерживать температуру тела.
Большинство амфибий живет в пресной воде или около нее.
Когда они находятся в воде, в организм осмотическим путем по-
ступает вода, и для ее удаления они выделяют большое количе-
ство очень разбавленной мочи. Неизбежная потеря натрия ком-
пенсируется активным поглощением его через кожу из среды, со-
держащей его в очень низкой концентрации.
В почке лягушки для выведения мочевины используется не-
только клубочковая фильтрация, но и канальцевая секреция
(Marshall, 1933). Это дает некоторые преимущества по следую-
щей причине. Когда лягушка находится в сухом воздухе, она
сокращает образование мочи (потерю воды), снижая скорость
Органы выделения 527
клубочковой фильтрации и повышая реабсорбцию воды в каналь-
цах, но при этом выделение мочевины может оставаться на вы-
соком уровне. Из общего количества мочевины, выводимого поч-
кой, может фактически фильтроваться всего лишь седьмая часть,
л остальное добавляется к моче путем канальцевой секреции
{В. Schmidt-Nielsen, Forster, 1954). Это означает, что выделение
мочевины будет продолжаться на высоком уровне даже при поч-
ти полном прекращении фильтрации в клубочках. Таким образом,
в отличие от почки пластиножаберной рыбы, где мочевина актив-
но реабсорбируется, у амфибий она активно выводится.
В главе 9, посвященной осморегуляции, говорилось о том, что
одна из амфибий — крабоядная лягушка (Rana cancrivora), оби-
тающая в юго-восточной Азии, живет и отыскивает пищу в соле-
ной морской воде. Эта лягушка, подобно пластиножаберным ры-
бам, удерживает в крови много мочевины, что обеспечивает осмо-
тическое равновесие с окружающей морокой водой, но в то же
время сохраняет относительно низкую концентрацию солей, ха-
рактерную для позвоночных животных в целом. Однако крабо-
ядная лягушка отличается от акул в отношении почечного меха-
низма, позволяющего удерживать мочевину.
Почка крабоядной лягушки не реабсорбирует мочевину, но
когда животное находится в морской воде, в организм поступает
очень мало воды, и очень медленный ток мочи в сочетании с пре-
кращением секреции мочевины снижает потери этой последней.
Если же поместить крабоядную лягушку в разбавленную мор-
скую воду, то осмотический приток воды усилится, объем мочи
увеличится и потеря мочевины возрастет. Тем самым снизится
осмотическая разница между жидкостями тела и внешней сре-
дой и в результате уменьшится осмотический приток воды
(Schmidt-Nielsen, Lee, 1962).
ВЫДЕЛЕНИЕ У РЕПТИЛИЙ
Почка рептилий соответствует схеме, типичной для позвоноч-
ных. Она вырабатывает разбавленную или изотоническую мочу,
но не может продуцировать мочу более концентрированную, чем
плазма крови. Это соответствует потребностям пресноводных реп-
тилий (например, крокодилов и пресноводных черепах), так как
разбавленная моча служит для выведения излишней воды.
Морские рептилии сталкиваются с противоположной пробле-
мой— с нехваткой воды и избытком солей. Их почки не справля-
ются с излишками солей, которые выводятся носовыми или ор-
битальными солевыми железами.
Наземные рептилии, обитающие в сухих местах, где воды не
хватает, выделяют азотистые отходы в форме мочевой кислоты,
которая чрезвычайно мало растворима и поэтому выпадает в мо-
че в осадок. В результате моча выходит в виде пасты или полу-
твердого шарика, для чего требуется очень мало воды.
528 Гл. 10. Выделение
Выделение мочевой кислоты дает еще одно преимущество. Не
только она, но и ее натриевая и калиевая соли очень мало рас-
творимы. При включении Na+ и К+ в фракцию мочи, выпадаю-
щую в осадок, эти важные ионы могут выводиться без дополни-
тельного расхода воды, необходимого при их экскреции почкой
млекопитающего или же солевой железой птицы (McNabb,
McNabb, 1975).
У многих наземных пресмыкающихся имеются носовые соле-
вые железы, через которые выводится излишек натрия и калия,
но у пустынных рептилий, лишенных таких желез (например, у
Amphibolurus), и у наземных змей выделение мочевой кислоты
служит механизмом для удаления солей с минимальной потерей
воды (Minnich, 1972).
КОНЦЕНТРИРОВАНИЕ МОЧИ У ПТИЦ И МЛЕКОПИТАЮЩИХ
Из всех позвоночных только у птиц и млекопитающих может
образовываться моча, гиперосмотичная по отношению к плазме
крови. У птиц эта способность не очень выражена; осмотическая
концентрация их мочи максимум в два раза больше, чем плазмы.
Между тем почка млекопитающего способна вырабатывать мочу
с концентрацией, которая можетбытьв25 раз выше, чем у плаз-
мы (McMillen, Lee, 1969). Самая концентрированная моча обра-
зуется у обитателей пустыни; а такие животные, как бобры, ко-
торым вода всегда доступна в избытке, обладают лишь умерен-
ной концентрационной способностью (табл. 10.2).
Таблица 10.2
Максимальная концентрирующая способность
почки у разных млекопитающих связана с их
обычной средой обитания: у пустынных животных
моча наиболее концентрированная, у пресновод-
ных животных — наименее концентрированная
Максимальная осмотическая Животное концентрация мочи, осмоль/л Отношение концентра- ции мо- ча/плазма
Бобр1) 0,52 2
Свинья1) 1,1 3
Человек2) 1,4 4
Белая крыса2) 2,9 9
Кошка2) 3,1 10
Кенгуровая крыса2) 5,5 14
Гофер (sand rat)2) 6,3 Австралийская прыга- 17
ющая мышь3) 9,4 1) В. Schmidt-Nielsen, O’Dell, 1961. 2) К. Schmidt-Nielsen, 1964. 3) MacMillen, Lee, 1967. 25
Органы выделения 529
Механизм образования концентрированной мочи у птиц и
млекопитающих основан на остроумной геометрической конфигу-
рации почечного канальца. Казалось бы, для концентрирования
мочи требуется или реабсорбция воды из канальцевой жидкости
или активная секреция растворенных веществ в каналец. На са-
мом деле не происходит ни того, ни другого: действительный ме-
ханизм основан на реабсорбции натрия и хлора из канальцевой
жидкости, и парадоксаль-
ным образом это ведет
в конечном счете к обра-
зованию сильно концент-
рированной мочи. Такой
механизм имеется у
птиц, а наивысшего со-
вершенства он достигает
Рис. 10.11. На схеме показаны
два типичных нефрона, один
с длинной петлей (расположен-
ный у границы мозгового ве-
щества и поэтому называемый
юкстамедуллярным) и один с
короткой петлей (или корко-
вый). Параллельно первому из
них идет длинная петля, обра-
зованная кровеносным капил-
ляром. Нефрон с короткой пет-
лей окружен капиллярной
сетью. У большинства млеко-
питающих в почке перемешаны
нефроны обоих типов, но у не-
которых видов имеется или
тот, или другой тип. (Smith,
1951.)
у тех млекопитающих, которые вырабатывают самую концентри-
рованную мочу.
Концентрирующая способность почки млекопитающего тесно
связана с длиной петли Генле в нефроне, которая имеет форму
шпильки для волос и образована тонким отрезком, лежащим меж-
ду проксимальным и дистальным канальцами. В почках большин-
ства млекопитающих имеются нефроны двух типов (рис. 10.11):
у одних нефронов петли длинные, у других короткие. Животные,
выделяющие очень концентрированную мочу, обладают только
нефронами с длинными петлями, такими длинными, что они про-
никают в почечный сосочек. У других животных, например у боб-
ра и свиньи, петли только короткие; эти животные не могут выра-
батывать мочу, концентрация которой больше чем вдвое превы-
шала бы концентрацию плазмы.
34—1863
530 Гл. 10. Выделение
Роль петли в образовании концентрированной мочи была вы-
яснена швейцарским исследователем Вирцем и его сотрудниками,
а затем другие европейские и американские ученые получили до-
полнительные данные. Вирц полагал, что первичным фактором
является активный транспорт натрия из почечного канальца, но
более поздние данные говорят о том, что таким фактором, воз-
можно, служит хлоридный насос. Для понимания концентрирую-
Рис. 10.12. Схема концентрирую-
щего механизма петли Генле в
почке млекопитающего. (Кокко,
Tisher, 1976.)
Активный транспорт иона С1~ по-
казан жирными стрелками, пассив-
ный переход воды и мочевины —
тонкими стрелками. М — мочевина,
РВ — растворенные вещества; зна-
ки «плюс» и «минус» означают
соответственно проницаемость или
непроницаемость данного участка
для данного вещества, ± означает
малую проницаемость.
щего механизма эта разница мало существенна: какой бы из
двух ионов ни переносился активно, за ним следует соответству-
ющий противоион, поэтому можно говорить о первичном актив-
ном транспорте NaCl.
Механизм образования концентрированной мочи легче всего
понять, если обратиться к рис. 10.12 (Kokko, Tisher, 1976). Местом
активной реабсорбции хлористого натрия служит восходящее ко-
лено петли Генле, в особенности толстый отрезок восходящего
колена. В этом участке С1~ переносится в интерстиций, сопро-
вождаемый своим противоионом Na+. Далее следует рассмотреть
нисходящее колено петли Генле, проницаемое для воды, но не
проницаемое для растворенных в ней веществ. Поскольку хло-
ридный насос повысил концентрацию интерстициальной жидкости,
вода отсасывается из нисходящего колена петли, что делает со-
держимое канальца более концентрированным. Таким образом,
канальцевая жидкость, проходящая через поворот петли, стала
более концентрированной и теперь подвергается действию насоса:
отсасывается еще некоторое количество С1~, дополнительно уда-
ляется вода и т. д. В результате одиночный эффект ионного на-
соса умножается по мере того, как все больше воды выходит из
Органы выделения 53i
петли и все более концентрированная жидкость поступает в ее
восходящую часть.
Следующий вопрос: что происходит с водой? Важную роль
здесь играют капиллярные петли, идущие параллельно петлям
Генле. Эти капилляры, называемые vasa recta, вполне проницае-
мы для воды и растворенных в ней веществ. Поэтому в их со-
держимом концентрация растворенных веществ у поворота петли
повышена так же, как в канальцевой жидкости и интерстиции. Но
стенка капилляра непроницаема для белков, и поэтому коллоид-
ное осмотическое давление белков плазмы, как и в других капил-
лярах, втягивает воду в кровяное русло. Отсюда излишек воды
переносится в венозную систему. В общем итоге картина такова:
если почка должна извлекать из крови концентрированную жид-
кость (т. е. мочу), то оттекающая венозная кровь неизбежно
должна быть более разбавленной, чем притекающая артериаль-
ная кровь.
Далее следует рассмотреть дистальную часть канальца. Из
жидкости, поступающей из толстого восходящего сегмента в ди-
стальный каналец, удаляется хлористый натрий, и здесь его кон-
центрация ниже, чем в плазме. Удаленный в толстом сегменте
NaCl накапливается в интерстиции вокруг петли, и здесь кон-
центрация его высока.
При входе в собирательную трубку разбавленная канальце-
вая жидкость бедна солью, но содержит мочевину, которая игра-
ет в этой системе важную роль. Нисходящая собирательная труб-
ка проницаема для воды и мочевины. Вследствие высокой осмо-
тической концентрации в интерстиции вода выходит из собира-
тельной трубки, мочевина концентрируется и тоже диффундирует
наружу. Эта мочевина значительно повышает интерстициальную
осмотическую концентрацию. Когда канальцевая жидкость, про-
текающая в нисходящей собирательной трубке, достигает осмоти-
ческого равновесия с концентрированной жидкостью в интерсти-
ции мозгового вещества, она превращается в мочу с высокой кон-
центрацией растворенных веществ. Высокая концентрация в моз-
говом веществе создается благодаря мочевине, поступающей из
собирательных трубок и соли, переходящей из петель Генле, а
пассивная диффузия воды из собирательной трубки служит ме-
ханизмом образования концентрированной мочи.
Эта модель концентрирования мочи основана на высокоизби-
рательной проницаемости различных отделов нефрона. Несмотря
на очень малую толщину канальцев, удалось осуществить микро-
перфузию изолированных отрезков петли Генле, и найденные ве-
личины проницаемости согласуются с требованиями модели
(Кокко, 1974).
Наконец, нужно ответить на последний вопрос: как образуется
разбавленная моча? При водном диурезе канальцевая жидкость,
переходящая в собирательные трубки из дистального извитого
34*
532 Гл. 10. Выделение
канальца, остается разбавленной. Это обусловлено пониженной
водопроницаемостью собирательных трубок, благодаря которой
вода не удаляется. Изменения проницаемости для воды регули-
руются антидиуретическим гормоном (АДГ). В его отсутствие
собирательные трубки непроницаемы для воды, и образуется раз-
бавленная моча. В присутствии АДГ они водопроницаемы, вода
выходит из них и образуется концентрированная моча.
Жидкости в нисходящем и восходящем коленах текут в про-
тивоположных направлениях, т. е. создается противоток. В любом
месте петли разность концентраций между восходящим и нисхо-
дящим коленами не так велика, но на всем протяжении петли
эти небольшие различия складываются (имеет место эффект ум-
ножения). Поэтому вся система называется противоточной умно-
жающей системой. Конечная концентрация, достижимая в такой
системе, определяется длиной петли: чем длиннее петля, тем вы-
ше возможная концентрация мочи. Поэтому у каждого данного
вида животных максимальная концентрация мочи в высокой сте-
пени коррелирует с относительной длиной петли.
Почка птицы отличается от почки млекопитающего тем, что у
одних нефронов имеются петли, а у других нет. По-видимому,
максимальная концентрирующая способность почки у птицы за-
висит от численного соотношения между нефронами с петлями и
без них, а не с максимальной длиной петель, как у млекопитаю-
щих (Poulson, 1965).
Многие птицы живут в относительно сухих местах, питаются
сухой пищей (например, 'семенами) -и имеют лишь ограниченный
доступ ж воде. Естественно возникает вопрос: почему у них почки
не обладают такой же концентрирующей способностью, как у мле-
копитающих? Вероятно, это связано с тем, что в качестве конеч-
ного продукта азотистого обмена птицы выделяют мочевую кис-
лоту.
Если бы почка птицы извлекала воду в гораздо большей сте-
пени, чем это происходит в действительности, то весьма мало рас-
творимая мочевая кислота осаждалась бы в канальцах и закупо-
ривала их, если бы в них не оставалось достаточно жидкости, ко-
торая вымывала бы осадок. Хотя моча у птицы содержит осаж-
денную мочевую кислоту, в нефронах и мочеточнике она должна
оставаться сравнительно жидкой, пока не попадет в клоаку —
последний отдел кишечника, где моча и фекальные массы накап-
ливаются и откуда время от времени выбрасываются через об-
щее наружное отверстие. После того как моча попала В клоаку,
из нее еще может быть извлечено дополнительное количество
воды.
роль клолки
Когда моча птицы или рептилии попадает в клоаку, она жид-
кая или полужидкая. Но если животному нужно сберегать воду,
Органы выделения 533
моча выводится наружу в виде полутвердой пасты или катышка
(это значит, что вода удалена из нее). Такая реабсорбция воды
может, по крайней мере частично, происходить вне клоаки, так
-как показано, что моча иногда переходит из клоаки в нижний
отдел кишечника, где соли и вода всасываются обратно (Nechay
ct al., 1968; Skadhauge, 1967, 1976).
Каков механизм всасывания воды из содержимого клоаки?
Это может быть перенос, сопряженный с транспортом растворен-
ных веществ, или, по крайней мере у ящериц, пассивный процесс,
обусловленный -коллоидным осмотическим давлением белков плаз-
мы. В клоаке ящерицы вода поглощается до тех пор, пока сила,
необходимая для извлечения еще большего количества воды, не
’Сравняется с коллоидным осмотическим давлением белков плазмы;
тогда всасывание воды прекращается. Если же осмотическое дав-
ление белков плазмы уравновесить, введя в клоаку раствор бел-
ка с такой же коллоидной осмотической концентрацией, что и в
плазме, то вода не будет реабсорбироваться. По-видимому, яще-
рицам для образования в клоаке полусухой мочи достаточно пас-
сивного осмотического удаления воды. Но не исключено, что в
клоаке происходит также активный транспорт натрия или дру-
гих растворенных веществ и сопряженная с ним реабсорбция во-
ды (Murrish, Schmidt-Nielsen, 1970).
Когда соль реабсорбируется из клоаки вместе с водой, может
возникнуть трудность с удалением избытка соли. Однако ящерицы
(и многие птицы) выделяют избыточную соль через носовые же-
лезы. Таким образом, ящерицы, обитающие в сухих местах, мо-
гут использовать почки главным образом для экскреции мочевой
кислоты и теряют при этом минимум воды, а носовые железы
служат для выведения солей в сильно концентрированном раство-
ре, также лишь с небольшой потерей воды (Schmidt-Nielsen et
al., 1963).
РЕГУЛИРОВАНИЕ КОНЦЕНТРАЦИИ И ОБЪЕМА МОЧИ
Если животное находится в калорийном равновесии, вес его
тела, а значит, и его объем остаются удивительно постоянными.
Важную роль в поддержании этого постоянства играет точная ре-
гуляция скорости образования мочи. Общий объем мочи зависит
от потребности в выведении воды. Если вода поступает в орга-
низм в избытке, выводится большой объем разбавленной мочи;
-если же организму воды не хватает, объем мочи уменьшается, а
концентрация возрастает до возможного максимума.
У человека возможны более чем 100-кратные различия в коли-
честве выделяемой мочи. Если он лишен воды, образование мочи
может снизиться до 10 мл/ч, но дальше снижаться не будет, пока
положение не станет критическим и не наступит почечная недо-
статочность. Когда же, наоборот, вода потребляется в большом
количестве, образование мочи может превысить 1 л/ч.
534 Гл. 10. Выделение
Можно было бы подумать, что при усиленном питье выделе-
ние мочи возрастет еще больше, но это невозможно. Максималь-
ный объем мочи определяется количеством фильтрата, которое у
человека весьма постоянно и не возрастает, когда мы больше
пьем. В проксимальном канальце около 85% профильтровавшейся'
воды неизменно реабсорбируются, а остальные 15% составляют
максимальный возможный объем выделяемой мочи. Причина об-
ратного всасывания воды в проксимальном канальце состоит в
том, что здесь реабсорбируются важнейшие растворенные веще-
ства, такие, как глюкоза и хлористый натрий. По мере их уда-
ления содержимое канальца становится гипоосмотичным, и в ре-
зультате этого вода пассивно следует за растворенными вещест-
вами сквозь стенку канальца. Таким образом, в проксимальном
канальце реабсорбция воды является прямым и неизбежным ре-
зультатом всасывания глюкозы и натрия; это так называемая
облигатная реабсорбция воды.
Дистальный механизм (петля Генле и собирательная трубка),
как уже говорилось, ответствен за образование концентрирован-
ной мочи. Процессом концентрирования управляет ант ид иу ретине-
ский гормон (АДГ), который теперь обычно называют вазопрес-
сином. В отсутствие этого гормона, вырыбатываемого задней до-
лей гипофиза, образуется большой объем сильно разбавленной
мочи. Под влиянием вазопрессина реабсорбция воды усиливается,
объем мочи уменьшается и концентрация ее возрастает.
Вазопрессин влияет на проницаемость собирательных трубок.
Без него их проницаемость для воды мала, и разбавленная жид-
кость, перейдя в собирательную трубку из дистального канальца,
уже не теряет больше воды даже при высокой осмотической кон-
центрации в окружающих тканях. С увеличением количества ва-
зопрессина проницаемость собирательных трубок возрастает.
Благодаря высокой осмотической концентрации в ткани сосочка
(создаваемой умножающим противоточным механизмом) вода
осмотически адсорбируется из жидкости собирательных трубок,
и в них остается лишь небольшой объем концентрированной мочи.
Это представление о роли собирательных трубок было прове-
рено в опытах на крысах путем введения нитевидных полиэтиле-
новых капилляров через кончик сосочка и извлечения проб на
разных уровнях собирательной трубки (Hilger et al., 1958). Ког-
да в крови имеется вазопрессин, моча собирательных трубок нахо-
дится в осмотическом равновесии с жидкостью в петле Генле,
а при отсутствии вазопрессина этого равновесия нет из-за пони-
женной проницаемости собирательной трубки (Gottschalk, 1961).
Может показаться парадоксальным, что гормон, вызывающий
образование концентрированной мочи, действует путем повыше-
ния проницаемости собирательных трубок. Другой парадоксаль-
ный момент состоит в том, что транспортный механизм, ответст-
венный за концентрирование мочи, основан на извлечении натрия
Выделение азота 535
и хлора из канальцевой жидкости, а не на их добавлении. Но в
конце концов именно их реабсорбция в сочетании с повышением
проницаемости собирательных трубок под влиянием вазопресси-
на приводит к тому, что окончательная моча достигает максималь-
ной концентрации.
Интересно, что эффект вазопрессина, состоящий в повышении
проницаемости собирательной трубки, сходен с его воздействием
на кожу лягушки — модель, которую часто используют при изуче-
нии транспорта солей. В коже лягушки механизм переноса нат-
рия действует в том же направлении, что и натриевый насос в
почечном канальце — возвращает натрий в организм. В обеих си-
стемах вазопрессин повышает проницаемость для воды. В почке
млекопитающего он сберегает воду, а в коже лягушки повышен-
ная проницаемость создает усиленный осмотический приток воды.
ВЫДЕЛЕНИЕ АЗОТА
Большая часть поедаемой животными пищи содержит три
главных питательных компонента: углеводы, жиры и белки (а
также небольшие количества нуклеиновых кислот). В процессе
метаболизма из углеводов и жиров образуются двуокись углеро-
да и вода как единственные конечные продукты окисления. Бел-
ки и нуклеиновые кислоты тоже дают углекислоту и воду, но, кро-
ме того, химически связанный азот этих пищевых веществ вклю-
чается в ряд относительно простых азотсодержащих продуктов,
подлежащих экскреции. Из них наибольший интерес представля-
ют три соединения — аммиак, мочевина и мочевая кислота.
Аммиак Мочевина Мочевая кислота
(кето-форма)
NH3 CON2H4 C5H4O3N4
С=О
/Н ш Hl/ 'c-NH
/ / 2 । у ч
n-h о=с( ;с=о
\н чШ2 о=С C-NH
В процессе превращений аминокислот аминогруппа (—NH2)
удаляется путем дезаминирования и образует аммиак (NH3).
Многие животные, в том числе водные безпозвоночные, выде-
ляют аммиак в неизмененном состоянии, другие перед выделением
синтезируют из него мочевину, а третьи — мочевую кислоту. Азо-
тистые соединения, содержащиеся в нуклеиновых кислотах, — это
пурины или пиримидины, которые могут выводиться в виде од-
ного из продуктов расщепления, указанных в табл. 10.3. Основ-
ная масса азотистых экскретов образуется из белка (аминокис-
лот) путем дезаминирования.
536 Гл. 10. Выделение
Таблица 10.3
Конечные продукты метаболизма основных групп пищевых
веществ. Аммиак, образующийся при белковом обмене,
выделяется в неизмененном виде, или же из него могут
синтезироваться другие азотсодержащие продукты; пурины из
нуклеиновых кислот выделяются как таковые или в виде
любого из многочисленных продуктов расщепления, включая
аммиак
Пищевые вещества Конечные продукты
Углеводы —>СО2Ч-Н2О
Жиры —>СО2+Н2О
^Мочевина
Белки —^NH3("
^Мочевая кислота
Нуклеиновые кислоты—^Пурины + Пиримидины
I I
Мочевая кислота [З-Аминокислоты
! 1
Аллантоин NH3
I
Аллантоиновая
кислота
I
Мочевина
!
NH3
Кроме того, выделяются и другие азотсодержащие продукты,
из которых одни происходят из пищи, а другие образуются в про-
цессе метаболизма. Например, моча позвоночных в норме со-
держит небольшое количество креатинина. Это вещество обычно
вырабатывается мышцами и представляет особый интерес, так
как его можно применять при исследовании почечной функции
подобно инулину. Он образуется в организме с относительно пог
стоянкой скоростью, и наблюдение за его экскрецией позволяет
изучать работу почек, не вводя посторонних веществ. По-видимо^-
му, креатинин — это продукт обмена высокоэнергетического фос-
фатного соединения фосфокреатина.
Другая группа азотистых соединений, часто находимых в мо^
че, образуется в процессах детоксикации. Например, бензойная!
кислота (входящая в состав некоторых растительных продуктов)1,,
соединяясь с аминокислотой глицином, образует гиппуровую кис-
лоту (от греч. hippos — лошадь, так как впервые она была обна*-
ружена в моче лошади). Другие фенольные соединения обезвре-
живаются подобным же образом и затем выводятся с мочож Гип-
Выделение азота 537
пуровая кислота и близкая к ней гшрааминогиппуровая кислота
играют важную роль в исследованиях по физиологии почек, так
как легко секретируются канальцами, и их используют для изме-
рения почечного кровотока, как уже было описано выше.
Существенным аспектом теории азотистого обмена является
предположение, что в метаболизме животных никогда не участ-
вует газообразный азот. Всеми признано, что ассимилировать мо-
лекулярный азот (N2) и превращать его в химические соединения
способны только азотфиксирующие бактерии и что только у де-
Главные азотистые экскреты у разных групп животных
Таблица 10.4
Животные Главный конечный про- дукт белкового обмена Среда обитания взрослого живот- ного Окружающая сре- да в эмбриональ- ный период
Водные беспозвоноч- Аммиак Водная Водная
ные
Костистые рыбы Аммиак, некоторое количество моче- вины » »
Пластиножаберные Мочевина
Крокодилы Аммиак, некоторое количество моче- вой кислоты Полуводная Клейдоическое яйцо0
Личинки амфибий Аммиак Водная Водная
Взрослые амфибии Мочевина Полуводная »
Млекопитающие » Наземная »
Черепахи Мочевина п мочевая кислота » Клейдоическое яйцо0
Насекомые Мочевая кислота »
Наземные брюхоно- » » »
гие моллюски
Ящерицы » »
Змеи » » »
Птицы » » *) Роль клейдоических яиц рассматривается ниже в настоящей главе.
нитрифицирующих бактерий имеются метаболические пути, веду-
щие к образованию молекулярного газообразного азота1.
Форма выведения белкового азота — в виде аммиака, мочеви-
ны или же мочевой кислоты — тесно связана с условиями жизни
животного и наличием воды (табл. 10.4). Аммиак весьма токси-
чен даже в очень малых концентрациях, поэтому он должен бы-
1 Убедительные данные показывают, однако, что это не всегда верно. Неко-
торые исследователи (например, Costa et al., 1968, 1974) сообщают об образова-
нии газообразного азота у млекопитающих, получающих большие количества
белка. Эти сведения должны изменить некоторые из наших представлений о бел-
ковом обмене и конечных азотистых продуктах.
538 Гл. 10. Выделение
стро удаляться либо путем выведения во внешнюю среду, либо
путем превращения в менее токсичные вещества (мочевину или
мочевую кислоту).
АММИАК
У большинства водных беспозвоночных конечным продуктом
белкового обмена является аммиак. Благодаря его легкой раство-
римости и небольшому молекулярному весу он диффундирует
чрезвычайно быстро. Значительная его часть может быть выделе-
на через любую поверхность, соприкасающуюся с водой, — не обя-
зательно через почку. У костистых рыб большая часть азота вы-
водится в форме аммиака через жабры. У карпа и золотой рыб-
ки жабры выделяют в 6—10 раз больше азота, чем почки, и толь-
ко 1'0% его составляет мочевина; остальные 90% выводятся в ви-
де аммиака (Smith, 1929).
МОЧЕВИНА
Мочевина легко растворима в воде и обладает довольно малой
токсичностью. Синтез мочевины у высших животных был изучен
знаменитым биохимиком Гансом Кребсом — тем самым ученым,
по имени которого был назван цикл окислительного энергетиче-
ского обмена (цикл трикарбоновых кислот, или цикл Кребса).
При синтезе мочевины аммиак и двуокись углерода, конденси-
руясь с фосфатом, образуют карбамоилфосфат, который затем ис-
пользуется для синтеза цитруллина из**орнитина, как показано на
рис. 10.13. После этого добавляется еще одна молекула аммиака
из аспарагиновой кислоты, и это ведет к образованию аминокис-
лоты аргинина. В присутствии фермента аргиназы аргинин рас-
падается на мочевину и орнитин. Из орнитина синтезируется но-
вая молекула цитруллина, и весь цикл повторяется; поэтому весь
этот путь превращений называют орнитиновым циклом синтеза
мочевины. Наличие аргиназы у животного говорит о его способ-
ности вырабатывать мочевину и часто указывает на то, что мо-
чевина является у него главным азотистым экскретом. Но это не
обязательно так, поскольку возможно наличие аргиназы и при
отсутствии всего цикла.
МОЧЕВИНА У ПОЗВОНОЧНЫХ
Позвоночные животные, выделяющие главным образом, мочевину
и обладающие для ее синтеза ферментами орнитинового цикла,
представлены на рис. 10.14. Некоторое количество мочеви-
ны выделяют костистые рыбы, а у пластиножаберных, амфибий и
млекопитающих это главный азотистый экскрет. У пластиножа-
берных (акул и скатов), а также у крабоядной лягушки и цела-
канта Latimeria мочевина задерживается в организме, играет
Выделение азота 539
важную роль в осморегуляции и поэтому является ценным про-
дуктом обмена. У пластиножаберных мочевина фильтруется в по-
чечном клубочке, но ввиду ее значения для осморегуляции она не
должна теряться с мочой; поэтому она возвращается в результате
активной реабсорбции в канальцах. У амфибий дело обстоит ина-
Рис. 10,13. Мочевина синтезируется из аммиака и двуокиси углерода путем кон-
денсации с аминокислотой орнитином. На нескольких следующих этапах обра-
зуется аргинин, от которого под действием фермента аргиназы отщепляется
мочевина; при этом аргинин превращается в орнитин, снова поступающий в цикл.
Ф — фермент, переносящий карбамоильную группу.
че. Мочевина фильтруется, и, кроме того, значительное количест-
во ее добавляется к моче путем активной секреции в канальцах.
Таким образом, и у пластиножаберных, и у амфибий имеет место
активный канальцевый транспорт мочевины, но он идет у этих
групп в разных направлениях. Очевидно, насосные механизмы
здесь метаболически не идентичны, поскольку опыты с рядом
близких друг к другу производных мочевины дают в обеих груп-
пах животных различные результаты (табл. 10.5). Это превосход-
ный пример того, как одна и та же физиологическая функция воз-
никает в двух группах независимо, причем для достижения одной
и той же цели (в данном случае для активного транспорта мо-
чевины) не обязательно используются одинаковые механизмы.
У крабоядной лягушки, которая тоже сохраняет мочевину для
осморегуляции, активной реабсорбции этого вещества в каналь-
540 Гл. 10. Выделение
цах не обнаружено (Schmidt-Nielsen, Lee, 1962). Моча образует-
ся у нее медленно, и почечные канальцы весьма проницаемы для
мочевины. Поэтому мочевина диффундирует из канальцевой жид-
МОЧЕВАЯ
МОЧЕВИНА
КИСЛОТА
АММИАК
i
!
I
!
Рис. 10.14. Выделение азота на разных этапах филогенеза позвоночных. Линиям»
окружены группы животных, которые выделяют соответственно аммиак, мочеви-
ну и мочевую кислоту в качестве основного экскрета. (В. Schmidt-Nielsen*
1972.)
кости обратно в кровь и оказывается в моче приблизительно в
той же концентрации, что и в крови. Таким образом, лишь неболь-
шие количества ее теряются с мочой.
Если у обычных лягушек происходит активная канальцевая
секреция мочевины, то почему крабоядная лягушка не использу-
Выделение азота 541
Таблица 10.5
Мочевина активно транспортируется почечным
канальцем акулы (активная реабсорбция)
и лягушки (активная секреция). Но с тремя
другими близкими веществами результаты
получаются у этих двух видов животных
совершенно различными. Это говорит о том, что
клеточный механизм транспорта в их почках
различен. (В. Schmidt-Nielsen, Rabinovitz, 1964)
Вещество Транспорт у акулы Транспорт у ля- гушки
Мочевина Активный Активный
Метилмочевина Активный Нет
Тиомочевина Нет Активный
Ацетамид Активный Нет
ет такой насос, попросту поменяв его направление на обратное?
На этот вопрос ответить нелегко, но, по-видимому, направление-
активного транспорта — консервативная физиологическая функ-
ция, изменить которую непросто. Как мы уже видели, и, в коже
лягушки, и в почке млекопитающего сохраняется направление
активного транспорта хлористого натрия извне внутрь организ-
ма. Но в почке млекопитающего направленный внутрь обратный
транспорт NaCl из канальцевой жидкости в организм использу-
ется в умножающей противоточной системе таким образом, что-
конечным результатом тем не менее оказывается концентриро-
ванная моча.
Обычное представление о выделении мочевины почкой млеко-
питающего состоит в том, что мочевина фильтруется в клубочке,,
а затем пассивно проходит по канальцам, хотя некоторая ее доля
благодаря высокой способности к диффузии пассивно диффунди-
рует обратно в кровь. Имеются, однако, убедительные доказа-
тельства того, что мочевина служит важным элементом умно-
жающей противоточной системы и что способ выделения моче-
вины является существенным элементом функции почки у мле-
копитающих.
МОЧЕВИНА И МЕТАМОРФОЗ У АМФИБИЙ
Головастики лягушек и жаб выделяют главным образом ам-
миак; взрослые животные выделяют мочевину. У лягушки (Rana
temporaria), жабы (Bufo bufo), тритона (Triturus vulgaris) и дру-
гих амфибий при метаморфозе происходит четкий переход от
выделения аммиака к экскреции мочевины. Однако южноафри-
канская шпорцевая лягушка (Xenopus), которая и во взрослом со-
стоянии остается в воде, продолжает выделять аммиак и на этой:
стадии (табл. 10.6).
542 Гл. 10. Выделение
Переход к выделению мочевины во время метаморфоза у по-
луназемных амфибий связан с заметным повышением активности
всех ферментов орнитинового цикла в печени (Brown et al.,
1959).
Таблица 10.6
Выделение аммиака у наземной жабы Bufo bufo и
у полностью водной бесхвостой амфибии Xenopus taevls.
Цифры указывают выделение свободного аммиака в процентах
от общего количества выделяемых аммиака и мочевины
на разных стадиях развития. (Munro, 1953)
Стадия Bufo Xenopus
Без задних конечностей 85
Задние конечности развились на три четверти 80
Задние конечности функционируют 85 83
Передние конечности свободные, хвост сохраняется 50 81
Хвост атрофируется 36
Без хвоста 20 77
Взрослая 15 81
Интересно, что особи водной амфибии Xenopus, извлеченные
на несколько недель из воды, накапливают в крови и тканях мо-
чевину. Накопление мочевины можно вызвать, поместив животных
в 0,9%-ный раствор NaCl. Когда взрослых особей содержали вне
воды, но в сыром мху (чтобы избежать обезвоживания), концент-
рация мочевины в крови увеличивалась в 10—20. раз и достигала
почти 100 ммоль/л. После возвращения животных в воду избы-
точная мочевина выделялась (Balinsky et al., 1961).
У группы особей Xenopus, которые в естественных условиях
переживали летнюю засуху в иле около высохшего пруда, кон-
центрация мочевины также была повышена в 15—20 раз. Среди
ферментов, участвующих в синтезе мочевины, количество карба-
моилфосфатсинтетазы, ответственной за первый этап синтеза (см.
рис. 10.13), возросло приблизительно в шесть раз, но активность
остальных ферментов цикла не изменилась. Возможно, что син-
тез карбамоилфосфата является этапом, лимитирующим скорость
синтеза мочевины, и увеличение количества этого фермента, ве-
роятно, удерживает аммиак плазмы на низком уровне, когда жи-
вотные находятся вне воды (Balinsky et al., 1967).
МОЧЕВИНА У ДВОЯКОДЫШАЩИХ РЫБ
У африканской двоякодышащей рыбы Protopterus происходят
совершенно такие же изменения, как у амфибий. В обычных ус-
ловиях, когда такая рыба живет в воде, она выделяет много ам-
Выделение азота 54$
миака (и некоторое количество мочевины), но когда в период
засухи она находится в коконе в засохшем иле, то все азотистые
отходы превращаются у нее в мочевину, которая накапливается в
крови, где ее концентрация к концу трехлетнего пребывания ры-
бы в коконе может достигать 3% (500 ммоль/л) (Smith, 1959).
В печени африканской двоякодышащей рыбы обнаружены все
пять ферментов орнитинового цикла (Janssens, Cohen, 1966).
Уровни двух ферментов, лимитирующих скорость синтеза мочеви-
ны, сходны у этой рыбы и у головастика лягушки Rana catesbeia-
па и значительно ниже уровней, найденных у взрослых лягушек.
Это согласуется с фактом преимущественного выделения аммиа-
ка двоякодышащей рыбой, когда она находится в воде. Однако*
было вычислено, что того количества ферментов орнитинового
цикла, которое содержится в печени двоякодышащей рыбы, не
находящейся в спячке, достаточно, чтобы обеспечить накопление
мочевины, фактически наблюдаемое во время спячки (^orster^
Goldstein, 1966).
У австралийской двоякодышащей рыбы Neoceratodus концент-
рации ферментов орнитинового цикла невелики, что согласуется
с образом жизни этой рыбы: она пользуется легким только как
добавочным органом дыхания и может лишь недолго выживать
на воздухе (о дыхании двоякодышащих рыб см. гл. 2). Синтез
мочевины в срезах печени австралийской двоякодышащей рыбы
идет в сто раз медленнее, чем у африканской. Это опять-таки со-
гласуется с чисто водным образом жизни первой из них (Gold-
stein et al., 1967).
МОЧЕВАЯ КИСЛОТА
Выделение мочевой кислоты преобладает у насекомых, назем-
ных улиток, большинства рептилий и у птиц. Все это — типична
наземные животные, и образование у них мочевой кислоты мож-
но рассматривать как эффективное приспособление, сберегающее
воду при жизни на суше. Поскольку мочевая кислота и ее соли
очень плохо растворимы в воде (растворимость ее составляет
около 6 мг на 1 л воды), реабсорбция воды из мочи ведет к вы-
падению мочевой кислоты и ее солей в осадок.
МОЧЕВАЯ КИСЛОТА У ПТИЦ И НАСЕКОМЫХ
' Полутвердая белая часть птичьего помета представляет собой
мочу и состоит главным образом из мочевой кислоты; для выве-
дения азотистых экскретов птицы расходуют очень мало воды.
У некоторых насекомых уменьшение потерь воды с мочой зашло
так далеко, что они вообще не выделяют мочевую кислоту, а от-
кладывают ее в разных частях организма, главным образом в
544 Гл. 10. Выделение
жировом теле. Поэтому для удаления конечных азотистых про-
дуктов таким формам вода совсем не нужна (Kilby, 1963).
Было высказано предположение, что использование мочевой
кислоты в качестве главного экскрета дает птицам еще одно пре-
имущество. Поскольку для образования мочи им нужно мало во-
ды, выделение мочевой кислоты, согласно этому предположению,
уменьшает вес тела у летающих птиц. Но это соображение не
убедительно, так как птицы, имеющие доступ к воде (как пресно-
водные, так и морские), часто выделяют большие количества
жидкой мочи.
КЛЕЙД0ИЧЕСКОЕ ЯЙЦО
Джозеф Нидхем предположил, что разница между теми позво-
ночными, которые вырабатывают мочевину (млекопитающие и
амфибии), и теми, которые продуцируют мочевую кислоту (реп-
тилии и птицы), связана прежде всего со способом размножения.
Яйцо амфибии развивается в воде, а эмбрион млекопитающего —
в жидкой среде в матке, где отходы метаболизма попадают в
кровь матери. С другой стороны, эмбриональное развитие репти-
лий и птиц происходит в замкнутом, так называемом клейдоиче-
ском1 яйце, которое обменивается с внешней средой только газа-
ми, а все экскреты остаются внутри скорлупы. В клейдоическом
яйце запас воды очень невелик, а аммиак, разумеется, слишком
токсичен, чтобы эмбрион мог выносить его присутствие в боль-
ших количествах. Если бы вырабатывалась мочевина, она оста-
валась бы в яйце и накапливалась в растворенном состоянии.
Между тем мочевая кислота может выпадать в осадок и тем са-
мым по существу элиминироваться; это и происходит, когда она
откладывается в виде кристаллов в аллантоисе, который, таким
образом, служит эмбриональным мочевым пузырем.
МОЧЕВАЯ КИСЛОТА У РЕПТИЛИЙ
Ящерицы и змеи выделяют главным образом мочевую кисло-
ту; многие черепахи выделяют смесь мочевой кислоты и мочеви-
ны, а крокодилы — главным образом аммиак (Cragg et al.,
1961). Это соответствует общему представлению, что способ
экскреции азота тесно связан с количеством доступной воды в
окружающей среде.
Крокодилы и аллигаторы выделяют аммиак в моче, где глав-
ным катионом является. NH4+, а главным анионом — НСО3“ (Co-
ulson et al., 1950; Coulson, Hernandez, 1955). Возможно, что при-
сутствие в моче указанных ионов помогает этим пресноводным
1 По-гречески kleisto — замкнутый, от kleis — ключ.
Выделение азота 545
животным лучше удерживать ионы Na+ и С1~ потеря которых с
калом, кстати, тоже очень невелика.
Вряд ли можно сомневаться в тесной связи между средой оби-
тания черепах и выделением ими азота. В табл. 10.7 приведен
состав проб мочи от восьми видов черепах, полученных из Лон-
донского зоопарка. У видов с наиболее выраженным водным об-
разом жизни выделяются значительные количества аммиака и
мочевины и только следы мочевой кислоты; у наиболее сухопут-
ных форм больше половины азота выводится в виде мочевой кис-
лоты.
Таблица 10.7
Доля азота в моче различных черепах (в процентах от всего выделяемого азота).
Формы, в наибольшей степени связанные с водой, почти не выделяют мочевой
кислоты, но это вещество доминирует у наземных видов из засушливых областей.
(Moyle, 1949)
Компоненты мочи!)
Вид Местообитание мочевая кислота аммиак мочевина амино- кислоты неучтен- ные ком- поненты
Kinosternon subrub- rum Почти полностью вод- ное 0,7 24,0 22,9 10,0 40,3
Pelusios derbianus Почти полностью вод- ное 4,5 18,5 24,4 20,6 27,2
Etnys orbicularis Полуводное; черепаха питается на суше в болотах 2,5 14,4 47,1 19,7 14,8
Kinixys erosa Сырые места; живот- ное часто перехо- дит в воду 4,2 6,1 61,0 13,7 15,2
K. youngii Суше, чем предыду- щее 5,5 6,0 44,0 15,2 26,4
Testudo denticulata С сырой, болотистой почвой 6,7 6,0 29,1 15,6 32,1
T. graeca Очень сухое, почти пустыня 51,9 4,1 22,3 6,6 4,0
T. elegans Очень сухое, почти пустыня 56,1 6,2 8,5 13,1 12,0
’) Моча содержит также в небольших количествах аллантоин, гуанин, ксантин и креати-
нин; кроме того, имеются неучтенные компоненты.
Сведения о том, выделяют ли черепахи главным образом мо-
чевину или же мочевую кислоту, противоречивы. Дело в том, что
различаются не только виды, но и внутри одного вида одни осо-
би могут выделять преимущественно мочевую кислоту, другие —
преимущественно мочевину, третьи — смесь обоих веществ
(Khalil, Haggag, 1955). Даже одна и та же особь может с те-
чением времени перейти от одного соединения к другому. Неко-
35- 1863
546 Гл. 10. Выделение
торое количество выпавшей в осадок мочевой кислоты задержи-
вается в клоаке, а жидкая часть мочи выводится наружу; это де-
лает ненадежным определение образующейся мочевой кислоты
путем анализа одной или нескольких проб мочи: при неполном
опорожнении клоаки могут получиться совсем низкие цифры, а
при такой ее эвакуации, когда выходит осадок, накопившийся за
некоторое время, мочевой кислоты окажется слишком много.
У черепахи Testudo mauritanica переход от мочевины к моче-
вой кислоте и обратно находится, по-видимому, в прямой зави-
симости от температуры и содержания воды в организме. Выде-
ление мочевой кислоты возрастает при неблагоприятном водном
балансе, но механизм, управляющий этим сдвигом биохимической
активности, неясен.
ДВЕ НЕОБЫЧНЫЕ ЛЯГУШКИ 1
В главе 9 мы уже упоминали, что африканская лягушка Chi-
romantis xerampelina теряет воду через кожу очень медленно,,
примерно с той же скоростью, что и рептилии. Она сходна с реп-
тилиями и тем, что выделяет в основном мочевую кислоту, а не
мочевину, как это обычно свойственно взрослым амфибиям. Это
сенсационный факт, так как он противоречит общепринятому
представлению о выделении азота у амфибий. Точность этого со-
общения не вызывает сомнений, так как мочевую кислоту опреде-
ляли в моче Chiromantis специфичным для этого вещества фер-
ментативным методом, и было установлено, что она составляет
до 60—75% сухого веса мочи (Loveridge, 1970).
Южноафриканская лягушка Phyllomedusa sauvagii в этом от-
ношении тоже сходна с рептилиями. Потеря воды через кожу со-
ставляет у нее величину того же порядка, что и у рептилий с их
сухими покровами, а моча содержит много полутвердого осадка
урата (Shoemaker et al., 1972). В форме урата у Phyllomedusa
выделяется 80% общего азота, и повышенное потребление воды
не изменяет интенсивности образования урата. Этот вид продол-
жает выделять главным образом мочевую кислоту даже при из-
бытке воды. Когда лягушка нуждается в сохранении воды, экск-
реция мочевой кислоты (вместо мочевины) приобретает очень
большое значение. Вычислено, что если бы экскреторным продук-
том у этой лягушки была мочевина, то для образования мочи ей
потребовалось бы около 60 мл воды в день на 1 кг веса тела.
А между тем, благодаря тому что Р. sauvagii выделяет мочевую
кислоту, она теряет с мочой всего лишь 3,8 мл воды в день на
1 кг веса тела (Shoemaker, McClanahan, 1975).
1 Два описанных здесь вида лягушек обитают в сухих, полупустынных местно-
стях. — Прим. ред.
Выделение азота 547
АММИАК И ПОЧЕЧНАЯ ФУНКЦИЯ
Из всего сказанного выше может создаться впечатление, что
аммиак выделяют главным образом водные животные, но это не
совсем верно. Аммиак в норме содержится и в моче наземных жи-
вотных, где он служит для регуляции pH мочи. Если моча стано-
вится кислой из-за выделения кислых продуктов обмена, для
.нейтрализации добавляется аммиак.
Избыток кислоты обычно образуется при белковом обмене, так
как конечным продуктом окисления серусодержащей аминокис-
лоты цистеина является серная кислота. Чем кислее моча, тем
больше добавляется аммиака. Аммиак, используемый для нейтра-
лизации кислой мочи, образуется в почках из аминокислоты глут-
амина. Почки содержат глутаминазу, и она имеется здесь спе-
циально для выработки аммиака. Поэтому аммиак в моче млеко-
питающего прямо не связан с тем аммиаком, который образуется
в печени при дезаминировании аминокислот, и в этом смысле его
не следует рассматривать как нормальный конечный продукт бел-
кового обмена.
НУКЛЕИНОВЫЕ КИСЛОТЫ И ВЫДЕЛЕНИЕ АЗОТА
Нуклеиновые кислоты содержат две группы азотистых соеди-
нений: пурины (аденин и гуанин) и пиримидины (цитозин и ти-
мин). У некоторых животных пурины выделяются в виде моче-
вой кислоты (которая и сама является пурином); у других жи-
вотных пуриновая структура расщепляется до ряда промежуточ-
ных соединений или до аммиака, причем любое из этих веществ
может выводиться из организма.
Метаболическое расщепление пуринов и выделение его конеч?
ных продуктов изучены не так тщательно, как обмен белкового
азота. Важнейшие данные приведены в табл. 10.8. У птиц, назем-
ных рептилий и насекомых пурины расщепляются до мочевой кис-
лоты и последняя выводится из организма. Это те животные, у
которых из аминного азота синтезируется мочевая кислота; оче-
видно, что для животного было бы бессмысленно синтезировать
мочевую кислоту и в то же время обладать механизмами ее раз-
ложения. Поэтому нельзя ожидать дальнейшего распада пури-
нов у тех животных, у которых мочевая кислота — конечный про-
дукт белкового обмена.
Среди млекопитающих человек, высшие обезьяны и далмат-
ский дог составляют особую группу: они выделяют мочевую кис-
лоту, тогда как остальные млекопитающие выделяют аллантоин.
Аллантоин образуется из мочевой кислоты путем одного превра-
щения в присутствии фермента уриказы. У человека и высших
обезьян нет этого фермента. Из-за своей малой растворимости
мочевая кислота иногда откладывается в организме человека, вы-
зывая припухлость суставов и очень мучительное заболевание —
35*
548 Гл. 10. Выделение
Таблица 10.8
Азотистые конечные продукты пуринового обмена у разных животных.
(Keilin, 1959)
Животные Конечный продукт
Птицы \ Рептилии (наземные) | Насекомые Человек, высшие обезьяны, далматский дог J > Мочевая кислота
Млекопитающие (за исключением перечисленных выше) Брюхоногие моллюски Двукрылые > Аллантоин
Некоторые костистые рыбы Аллантоиновая кислота
Амфибии 1 Костистые рыбы | Пластиножаберные Пластинчатожаберные моллюски (пресноводные) J |> Мочевина
Пластинчатожаберные моллюски (морские) 1 Большинство остальных водных беспозвоночных j • Аммиак
подагру. Если бы у человека сохранился фермент уриказа, по-
дагры не существовало бы.
Хотя далматский дог выделяет гораздо больше мочевой кис-
лоты, чем другие собаки, это не следствие какого-то дефекта ме-
таболизма. Печень всякой собаки содержит уриказу и вырабаты-
вает некоторое количество аллантоина. Но у далматского дога
имеется почечный дефект, препятствующий канальцевой реаб-
сорбции мочевой кислоты (которая происходит у других млеко-
питающих, в том числе у человека); поэтому у дога мочевая кис-
лота теряется с мочой быстрее, чем перерабатывается печенью в
аллантоин (Yu et al., 1960). Немало данных говорит о том, что
мочевая кислота у далматского дога не только фильтруется в
клубочке, но и экскретируется путем активного транспорта в ка-
нальцах (Keilin, 1959).
Пурины аденин и гуанин сходны по своей структуре с мочевой
кислотой: они содержат одно шестичленное кольцо и одно пяти-
членное. Но пиримидины (цитозин и тимин) представляют собой
одиночные шестичленные кольца, содержащие два атома азота.
У высших позвоночных пиримидины расщепляются путем разры-
ва этого кольца с образованием одной молекулы аммиака и одной
молекулы р-аминокислоты. Последняя метаболизируется затем по
обычной схеме дезаминирования.
Самая поразительная черта обмена нуклеиновых кислот со-
стоит в том, что «высшие» животные, перечисленные в начале
табл. 10.8, полностью лишены ферментов, нужных для расщепле-
Выделение азота 549
ния пуринов. Среди «низших» животных мы находим все боль-
шее усложнение биохимических и ферментных систем, осуществ-
ляющих дальнейшее расщепление пуринов, так что самые «низ-
шие» формы обладают наиболее полным ферментным аппаратом.
ДРУГИЕ АЗОТИСТЫЕ СОЕДИНЕНИЯ
У пауков главным экскретом является гуанин. По-видимому,
он синтезируется из аминного азота, хотя весь путь его образова-
ния неизвестен. Некоторые пауки, в том числе птицеядные таран-
тулы, после приема пищи выделяют более 90% всего азота в фор-
ме гуанина (Peschen, 1939). У обыкновенного садового паука
Epeira diadema идентификация гуанина была подтверждена весь-
ма специфическим ферментативным методом (Vajropala, 1935).
Гуанин довольно часто встречается и у разнообразных других
животных. Например, серебристый блеск рыбьей чешуи обуслов-
лен отложением кристаллов гуанина. Садовая улитка Helix вы-
деляет гуанин, но лишь в пределах около 20% общего количест-
ва экскретируемых пуринов, а остальные 80% составляет мочевая
кислота. Возможно, что эта фракция — продукт обмена нуклеино-
вых кислот, а мочевая кислота образуется в результате белкового
обмена.
Аминокислоты не занимают важного места среди продуктов
азотистого обмена, но в небольших количествах они содержатся
в моче многих животных. Казалось бы, животному выгоднее дез-
аминировать аминокислоту, выделять аммиак обычным путем и
использовать образующуюся органическую кислоту в энергетиче-
ском обмене. Но поскольку аминокислоты играют лишь незначи-
тельную роль в выделении азота, этот вопрос не будет здесь об-
суждаться.
ТЕОРИЯ РЕКАПИТУЛЯЦИИ
Обычно считали, что выделение азота у развивающегося ку-
риного эмбриона изменяется во времени и проходит через ряд пи-
ков: вначале основным продуктом является аммиак, затем моче-
вина и, наконец, мочевая кислота. Предполагалось, что такое раз-
витие рекапитулирует этапы эволюции, которая у птиц заканчи-
вается выделением мочевой кислоты. Как сообщалось, образова-
ние аммиака у куриного зародыша достигает максимума через
4 дня, мочевины — через 9 дней и мочевой кислоты — через
11 дней после начала инкубации (Baldwin, 1949).
Более новые работы говорят о том, что выделение азота у ку-
риного эмбриона резко отличается от этой ранее описанной кар-,
тины (Clark, Fischer, 1957). Все три главных экскреторных про-
дукта— аммиак, мочевина и мочевая кислота — образуются и при-
сутствуют с самого начала эмбрионального развития. К концу
550 Гл. 10. Выделение
периода инкубации мочевой кислоты становится намного больше,
чем остальных двух продуктов. Однако количество мочевины и
аммиака продолжает расти на протяжении всей инкубации, и ко
времени вылупления оба вещества содержатся примерно в оди-
наковых количествах. К концу инкубации количество выделяемо-
го азота достигает 40 мг, из которых 23% делятся поровну между
Рис. 10.15. Накопление азотистых веществ в курином эмбрионе во время инку-
бации яйца. Мочевина и мочевая кислота образуются с самого начала инкубации;
позднее, однако, преобладает мочевая кислота. Аммиак появляется тоже с са-
мого начала, остается на постоянном уровне примерно 10 дней, а затем в течение
всего инкубационного периода содержание его растет. (Clark, Fisher, 1957.)
В чем причина расхождения в полученных результатах? Преж-
ние данные могли быть менее точными из-за более примитивных
методов анализа, но этим вряд ли можно объяснить наблюдав-
шиеся отдельные пики. Главная причина состоит просто в том,
что результаты выражались в количествах каждого экскреторно-
го продукта на единицу веса зародыша. А поскольку зародыш не-*
прерывно и чем дальше, тем быстрее увеличивается в размерах,
Выделение азота 551
то при делении количества каждого вещества на вес эмбриона
создается искусственный пик.
На самом деле все три экскреторных продукта присутствуют
с самого начала и на протяжении эмбрионального развития их
становится постепенно все больше, но после 10-го дня инкубации
количество аммиака возрастает незначительно. Мочевина, выра-
батываемая зародышем, синтезируется не из азота аминокислот
в цикле орнитина, а в результате воздействия аргиназы на арги-
нин (Eakin, Fisher, 1958). Таким образом, ни образование ам-
миака, ни синтез мочевины в курином эмбрионе не подтверждают
представление о том, что онтогенез биохимических механизмов
повторяет эволюционную историю выделения азота.
* * *
Мы рассмотрели разнообразные органы выделения и описали
их общие особенности. Эти органы удаляют отходы метаболизма,
помогают поддерживать нужные концентрации солей и других
растворенных веществ и регулируют содержание воды в организ-
ме, тщательно сохраняя воду, если ее в организме мало, и выводя
ее избыточные количества.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Baldwin Е. (1949). An Introduction to Comparative Biochemistry, Cambridge,
Cambridge University Press, 164 pp.
Balinsky J. B., Choritz E, L., Coe C. G. L., van der Schans G. S. (1967). Amino
acid metabolism and urea synthesis in naturally aestivating Xenopus laevis,
Comp. Biochem. Physiol., 22, 59—68.
Balinsky J. B., Cragg M. M., Baldwin E. (1961). The adaptation of amphibian
waste nitrogen excretion to dehydration, Comp. Biochem. Physiol., 3, 236—
244.
Braun G., Kummel G., Mangos J. A. (1966). Studies on the ultrastructure and
function of a primitive excretory organ, the protonephridium of the Rotifer
Asplanchna priodonta, Pfliigers Arch., 289, 141—154.
Brown G. W. Jr., Brown W. R., Cohen P. P. (1959). Comparative biochemistry of
urea synthesis. 2. Levels of urea cycle enzymes in metamorphosing Rana ca-
tesbeiana tadpoles, J. Biol. Chem., 234, 1775—1780.
Burger J. W. (1957). The general form of excretion in the lobster, Homarus, Biol.
Bull, 113, 207—223.
Clark H., Fischer D. (1957). A reconsideration of nitrogen excretion by the chick
embryo, J. Exp. Zool, 136, 1—15.
Costa G., Kerins M. E., Kantor F., Griffith K., Cummings W. B. (1974). Conversion
of protein nitrogen into gaseous catabolities by the chick embryo, Proc. Natl.
Acad. Sci. USA, 71, 451—454.
Costa G., Ullrich L., Kantor F., Holland J. F. (1968). Production of elemental
nitrogen by certain mammals including man, Nature, Lond, 218, 546—
551.
Coulson R. A., Hernandez T. (1955). Renal excretion of carbon dioxide and ammo-
nia by the alligator, Proc. Soc. Exp. Biol. Med.. 88, 682—687.
Coulson R. A., Hernandez T., Brazda F. G. (1950). Biochemical studies on the alli-
gator, Proc. Soc. Exp. Biol. Med, 73, 203—206.
552 Гл. 10. Выделение
Cragg М. М, Balinsky J. В., Baldwin Е. (1961). A comparative study of nitrogen
excretion in some amphibia and reptiles, Comp. Biochem. Physiol., 3, 227—
235.
Dobbs G. H., Ill, Lin Y., De Vries A, L. (1974). Aglomerularism in antarctic fish,
Science, 185, 793—794.
Eakin R. E, Fisher J. R. (1958). Patterns of nitrogen excretion in developing chick
embryos. In: The Chemical Basis of Development (W. D. McElroy and
B. Glass, eds.), pp. 514—522, Baltimore, Johns Hopkins Press.
Forster R. P. (1940). A renal clearance analysis of phenol red elimination in the
frog, J. Cell Comp Physiol., 16, 113—122.
Forster R. P.. Goldstein L. (1966). Urea synthesis in the lungfish: Relative impor-
tance of purine and ornithine cycle pathways, Science, 153, 1650—1652.
Goldstein L., Janssens P. A., Forster R. P. (1967). Lungfish Neoceratodus forsteri:
Activities of ornithine-urea cycle and enzymes, Science, 157, 316—317.
Goodrich E. S. (1945). The study of nephridia and genital ducts since 1895,
Q. J. Microsc. Sci., 86, 113—392.
Gottschalk C. W. (1961). Micropuncture studies of tubular function in the mamma-
lian kidney, Physiologist, 4, 35—55.
Grimstone A. V., Mullinger A. M., Ramsay J. A. (1968). Further studies on the rectal
complex of the mealworm, Tenebrio molitor, L. (Coleoptera, Tenebrionidae),
Philos. Trans. R. Soc. Lond., B, 253, 343—382.
Harrison F. M. (1962). Some excretory processes in the abalone, Haliotis rufescens,
J. Exp. Biol., 39, 179—192.
Hilger H. H„ Klumper J. D., Ullrich K. J. (1958). Wasserruckresorption und lonen-
transport durch die Sammelrohrzellen der Saugetierniere (Mikroanalytische Un-
tersuchungen), Pflugers Arch., 267, 217—237.
Hopkins D. L. (1946). The contractile vacuole and the adjustment to changing
concentration in fresh-water amoebae, Biol. Bull., 90, 158—176.
Janssens P. A., Cohen P. P. (1966). Ornithine-urea cycle enzymes in the African
lungfish, Protopterus aethiopicus, Science, 152, 358—359.
Jepps M. W. (1947). Contribution to the study of the sponges, Proc. R. Soc.
Lond., B, 134, 408—417.
Keilin J. (1959). The biological significance of uric acid and guanine excretion,
Biol. Rev., 34, 265—296.
Khalil F., Hag gag G. (1955). Ureotelism and uricotelism in tortoises, J. Exp. Zool.,
130, 423—432.
Kilby B. A. (1963). The biochemistry of the insect fat body, Adv. Insect Physiol.,
1,111—174.
Kokko J. P. (1974). Membrane characteristics governing salt and water transport
in the loop of Henle, Fed. Proc., 33, 25—30.
Kokko J. P., Tisher С. C. (1976). Water movement across nephron segments invol-
ved with the countercurrent multiplication system, Kidney Int., 10, 64—
81.
Lockwood A. P. M. (1961). The urine of Gammarus duebeni and G. pulex. J. Exp.
Biol., 38, 647—658.
Loveridge J. P. (1970^. Observations on nitrogenous excretion and water relations
of Chiromantis xerampelina (Amphibia, Anura), Arnoldia, 5, 1—6.
Lovtrup S., Pigon A. (1951). Diffusion and active transport of water in the amoeba
Chaos chaos L., C. R. Trav. Lab. Carlsberg (Ser. Chim.), 28, 1—36.
MacMillen R. E., Lee A. K. (1967). Australian desert mice: Independence of exoge-
nous water, Science, 158, 383—385.
MacMillen R. E., Lee A. K. (1969). Water metabolism of Australian hopping mice,
• Comp. Biochem. Physiol., 28, 493—514.
Marshall E. K., Jr.. (1933). The secretion of urea in the frog, J. Cell. Comp. Phy-
siol., 2, 349—353.
McNabb R. A., McNabb F. M. (1975). Urate excretion by the avian kidney,
Comp. Biochem; Physiol., 51A, 253—258.
Mercer E. H. (1959). An electron microscopic study of Amoeba proteus. Proc. R.
Soc. Lond., B, 150, 216—232.*
Литература 553
Minnich J. E. (1972). Excretion of urate salts by reptiles, Comp. Biochem. Physiol.»
41A, 535—549.
Moyle V. (1949). Nitrogenous excretion in chelonian reptiles, Biochem. J., 44, 581—
584.
Munro A, F. (1953). The ammonia and urea excretion of different species of Amphi-
bia during their development and metamorphosis, Biochem. J., 54, 29—
36.
Murrish D. E., Schmidt-Nielsen K. (1970). Water transport in the cloaca of lizards:
Active or passive? Science, 170, 324—326.
Nechay B. R., Boyarsky S., Catacutan-Labay P. (1968). Rapid migration of urine
into intestine of chickens, Comp. Biochem. Physiol., 26, 369—370.
Needham J. (1931). Chemical Embryology, vol. I, pp. 1—614; vol. II, pp. 615—1254;
vol. Ill, pp. 1255—1724, Cambridge, Cambridge University Press.
Oschman J. L., Wall B. J. (1969). The structure of the rectal pads of Periplanet a
americana L. with regard to fluid transport, J. Morphol., 127, 475—510.
Peschen К. E, (1939). Untersuchungen uber das Vorkommen und den Stoffwechsel
des Guanins im Tierreich, Zool. Jahrb., 59, 429—462.
Pontin R. M. (1964). A comparative account of the protonephridia of Asplanchna
(Rotifera) with special reference to the flame bulbs, Proc. Zool. Soc. bond.,
142, 511—525.
Poulson T. L. (1965). Countercurrent multipliers in avian kidneys, Science, 148,
389—391.
Ramsay J, A. (1949). The site of formation of hypotonic urine in the nephridium,
J. Exp. Biol., 26, 65—75.
Ramsay J. A. (1964). The rectal complex of the mealworm Tenebrio molitor, L.
(Coleoptera, Tenebrionidae), Philos. Trans. R. Soc. bond., B, 248, 279—
314.
Ramsay J. A., Riegel J. A. (1961). Excretion of inulin by Malpighian tubules, Na-
ture, bond., 191, 1115.
Richards A. N. (1935). Urine formation in the amphibian kidney, Harvey beet., 30,
93—118.
Riddick D. H. (1968). Contractile vacuole in the amoeba, Pelomyxa carolinensis,
Am. J. Physiol., 215, 736—740.
Riegel J. A., Lockwood A. P. M. (1961). The role of antennal gland in the osmo-
tic and ionic regulation of Carcinus maenas, J. Exp. Biol., 38, 491—499.
Schmidt-Nielsen B. (1972). Mechanisms of urea excretion by the vertebrate kidney.
In: Nitrogen Metabolism and the Environment (J. W. Campbell and b. Gold-
stein, eds.), pp. 79—103, bondon, Academic Press.
Schmidt-Nielsen B., Forster R. P. (1954). The effect of dehydration and low tem-
perature on renal function in the bullfrog, J. Cell. Comp. Physiol., 44, 233—
246.
Schmidt-Nielsen B., O'Dell R. (1961). Structure and concentrating mechanism in
the mammalian kidney, Am. J. Physiol., 200, 1119—1124.
Schmidt-Nielsen B., Rabinowitz L. (1964). Methylurea and acetamide: Active reab-
sorption by elasmobranch renal tubules, Science, 146, 1587—1588.
Schmidt-Nielsen B., Schrauger C. R. (1963). Amoeba proteus: Studying the contra-
ctile vacuole by micropuncture, Science, 139, 606—607.
Schmidt-Nielsen K. (1964). Desert Animals: Physiological Problems of Heat and
Water, Oxford, Clarendon Press, 277 pp. To be reprinted by Dover Publications,
New York.
Schmidt-Nielsen K., Borut A., Lee P., Crawford E, C., Jr. (1963). Nasal salt excre-
tion and the possible function of the cloaca in water conservation, Science, 142,
1300—1301.
Schmidt-Nielsen K., Lee P. (1962). Kidney function in the crab-eating frog (Rana
cancrivora), J. Exp. Biol., 39, 167—177.
Shannon J. A., Fisher S. (1938). The renal tubular reabsorption of glucose in the
normal dog, Am. J. Physiol., 122, 766—774.
Shoemaker V. H., Balding D., Ruibal R. (1972). Uricotelism and low evaporative
water loss in a South American frog, Science, 175, 1018—1020.
554 Гл. 10. Выделение
Shoemaker V. Н., McClanahan L. L., Jr. (1975). Evaporative water loss, nitrogen
excretion and osmoregulation in Phyllomedusine frogs, J. Comp. Physiol., 100,
331—345.
Skadhauge E. (1967). In vivo perfusion studies of the cloacal water and electroly-
te resorption in the fowl (Gallus domesticus), Comp. Biochem. Physiol., 23,
483—501.
Skadhauge E. (1976). Cloacal absorption of urine in birds, Comp. Biochem. Phy-
siol., 55A, 93—98.
Smith H. W. (1929). The excretion of ammonia and urea by the gills of fish,
J. Biol. Chem., 81, 727—742.
Smith H. W. (1951). The Kidney: Structure and Function in Health and Disease,
New York, Oxford University Press, 1049 pp.
Smith H. W. (1959). From Fish to Philosopher. Summit, N. J., CIBA, 304 pp. Reprin-
ted (1961), Garden City, N. Y. Doubleday.
Starling E. H. (1899). The glomerular functions of the kidney, J. Physiol., 24, 317—
330.
Vajropala K. (1935). Guanine in the excreta of arachnids, Nature, bond., 136,
145.
Wigglesworth V. B. (1931). The physiology of excretion in a blood-sucking insect,
Rhodnius prolixus (Hemiptera, Reduviidae). 2. Anatomy and histology of the
excretory system, J. Exp. Biol., 8, 428—442.
Yu T. F., Berger L., Kupfer S., Gutman A. B. (1960). Tubular secretion of urate
in the dog. Am. J. Physiol., 199, 1199—1204.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Cochran D. G. (1975). Excretion in insects. In: Insect Biochemistry and Function
(D. J. Candy and B. A. Kilby, eds.), pp. 179—281, New York, Wiley.
Dantzler W. H. (1976). Renal function (with special emphasis on nitrogen excre-
tion). In: Biology of the Reptilia, vol. 5, Physiology A (C. Gans and W. R. Daw-
son, eds.), pp. 447—503, London, Academic Press.
Forster R. P. (1973). Comparative vertebrate physiology and renal concepts. In:
Handbook of Physiology, Sect. 8, Renal Physiology (J. Orloff, R. W. Berliner
and S. R. Geiger, eds.), pp. 161—184, Washington, D. C., American Physiologi-
cal Society.
Kirschner L: B? (1967). Comparative physiology: Invertebrate excretory organs,
Annu. Rev. Physiol., 29, 169—196.
Maddrell S. H. P. (1971). The mechanism of insect excretory systems, Adv. In-
sect. Physiol., 8, 199—331.
Orloff J., Berliner R. W., Geiger S. R. (eds.) (1973). Handbook of Physiology,
Sect. 8, Renal Physiology, Washington, D. C., American Physiological Society,
1082 pp.
Peaker M., Linzell J. L. (1975). Salt Glands in Birds and Reptiles, Cambridge,
Cambridge University Press, 307 pp.
Riegel J. A. (1972). Comparative Physiology of Renal Excretion, Edinburgh, Oli-
ver and Boyd, 204 pp.
Smith H. W. (1951). The Kidney: Structure and Function in Health and Disease,
New York, Oxford University Press, 1049 pp.
Wall В. J., Oschman J. L. (1975). Structure and function of the rectum in insects,
Fortschr. Zool., 23, (2/3), 193—222.
Wessing A. (ed.) (1975). Excretion, Fortschr. Zool., 23, 1—362.
ЧАСТЬ ПЯТАЯ
ДВИЖЕНИЕ, ИНФОРМАЦИОННЫЕ
ПРОЦЕССЫ, ИНТЕГРАЦИЯ
ГЛАВА 11
МЫШЦЫ, ДВИЖЕНИЕ, ЛОКОМОЦИЯ
Большинство животных, даже те, которые прикреплены к суб-
страту и никогда не перемещаются (как, например, губки и ко-
ралловые полипы), совершают много движений. Эти движения
служат не только для локомоции (т. е. перемещения в пространст-
ве), но и для многих других целей. Сюда относятся такие разно-
образные формы активности, как проталкивание крови в сосу-
дистой системе, продвижение пищи по пищеварительному трак-
ту и перемещение внешней среды — смена воздуха в легких или
воды около жабр.
Используется лишь несколько основных механизмов движения,
но зато применение их очень многообразно. Поэтому мы сосредо-,
точимся на этих основных механизмах и не будем подробно опи-
сывать разнообразные устройства, существующие у различных
животных. Три главных механизма — это амебоидное, ресничное
и мышечное движение.
Амебоидное движение получило свое название от способа пе-.
ремещения амебы — одноклеточного организма, описанного во
всех учебниках биологии. Этот вид локомоции связан с сущест-
венным изменением формы клетки, токами цитоплазмы и актив-
ностью псевдоподий. Внутренний механизм такого движения наи-
менее понятен, хотя он и привлекал к себе много внимания.
Передвижение с помощью ресничек характерно для инфузорий,
например туфельки (Paramecium), но реснички встречаются у
представителей всех типов животных и служат разнообразным-
целям. Например, дыхательные пути у позвоночных, дышащих
воздухом, выстланы ресничными клетками, которые передвигают
посторонние частицы, попавшие на их поверхность, и таким обра-
зом медленно удаляют их. Реснички используются также для со-
здания токов жидкости во внутренних структурах или во внешней
среде, например в каналах амбулакральной системы иглокожих
555
556 Гл. 11. Мышцы, движение, локомоция
и в воде, омывающей жабры пластинчатожаберных моллюсков.
Сперматозоиды почти у всех животных подвижны и в большинст-
ве случаев передвигаются при помощи хвоста, который функцио-
нирует в принципе сходно с ресничкой.
Однако подавляющее большинство движений связано у жи-
вотных с использованием мышц, которые во всем животном мире
обладают одним общим свойством — способностью развивать силу
путем укорочения (сокращения).
Мы уже рассматривали использование мышц для проталкива-
ния жидкостей с помощью насосов, как перистальтических (про-
движение пищи по кишечнику), так и клапанных (сердце). В этой
главе речь пойдет о механизмах локомоции, из которых самый
главный и самый распространенный — это опять-таки использо-
вание мышц. В главе 6, посвященной энергетическому обмену,
была рассмотрена потребность в энергии для целей локомоции;
в конце настоящей главы будут описаны способы, с помощью ко-
торых некоторые водные животные снижают затрату мышечной
энергии.
АМЕБОИДНОЕ ДВИЖЕНИЕ; ПЕРЕМЕЩЕНИЕ
С ПОМОЩЬЮ РЕСНИЧЕК И ЖГУТИКОВ
АМЕБОИДНОЕ ДВИЖЕНИЕ
Движение, называемое амебоидным, характерно для некоторых
простейших, для определенных клеток слизистых грибов и для
лейкоцитов позвоночных животных. Движение этих клеток свя-
зано с токами цитоплазмы, изменением формы клетки и вытяги-
ванием псевдоподий. Эти явления легко наблюдать под микроско-
пом, но механизмы такого движения недостаточно ясны.
Когда амеба движется, ее цитоплазма переливается во вновь
образующиеся выступы клетки (псевдоподии), которые постепен-
но вытягиваются на том месте, где перед этим начала формиро-
ваться лишь маленькая псевдоподия. По мере перемещения клет-
ки на ее переднем крае образуются новые псевдоподии, а задние
выступы втягиваются.
У амебы самый наружный слой — эктоплазма — состоит из не-
сколько менее жидкого, гелеобразного вещества. При образова-
нии псевдоподии в нее перетекает более жидкая эндоплазма, а на
поверхности образуется новая эктоплазма. При этом в 'задней
части продвигающейся клетки эктоплазматический гель должен
превращаться в более жидкий эндоплазматический золь.
Какая сила заставляет эндоплазму течь? Было высказано
предположение, что сокращение вещества на заднем конце аме-
бы толкает эндоплазму вперед и выжимает ее в вытягивающуюся
псевдоподию; меньшая вязкость некоторых участков эндоплазмы
Амебоидное движение; перемещение с помощью ресничек и жгутиков 557
облегчает ее течение, и движущей силой служит незначительная
.разность давлений.
Гипотезе, согласно которой главную роль в вытягивании псев-
доподий играет гидростатическое давление, противоречат экспери-
ментальные данные. Если всосать псевдоподию амебы в капил-
ляр, в котором давление понижено, то это не препятствует обра-
зованию других псевдоподий, как следовало бы ожидать исходя
•из этой гипотезы (Allen et al., 1971). Если бы течение эндоплаз-
мы объяснялось градиентом давления вдоль линии тока, то под
слиянием обратного градиента оно переменило бы направление
на обратное.
Амебоидное движение имеет очевидное сходство с движением
цитоплазмы (циклозом), которое часто наблюдается во всех ви-
дах клеток, как растительных, так и животных, и играет важ-
ную роль в процессах внутриклеточного транспорта. В последние
годы представление о том, как возникают внутриклеточные дви-
жущие силы, радикально изменилось благодаря тому, что два
важных белка, ответственных за сокращение мышц, — актин и
миозин — найдены во всех эукариотических клетках (т. е. в клет-
ках всех живых организмов, кроме бактерий и сине-зеленых во-
дорослей). Актины из самых разных источников поразительно
сходны по своей структуре и образуют в клетке тонкие нити (фи-
ламенты). Поэтому можно думать, что как цитоплазматические
токи, так и процессы образования псевдоподий обусловлены тем
же в своей основе механизмом, что и мышечное сокращение.
Миозины из разных клеток различаются в большей степени, чем
актины, но все они способны обратимо присоединяться к акти-
новым нитям и все катализируют гидролиз аденозинтрифосфата
(АТФ)—универсального источника энергии для движения (Pol-
lard, 1977).
Наличие у амеб системы, генерирующей силу, помогает по-
нять, как может возникнуть амебоидное движение. Но дальней-
шая экспериментальная работа будет нелегкой, так как мы не
умеем непосредственно измерять действующие здесь механиче-
ские силы — отчасти из-за малой величины амеб, а отчасти из-за
отсутствия механических структур, доступных для прямого мани-
пулирования.
ДВИЖЕНИЕ С ПОМОЩЬЮ РЕСНИЧЕК И ЖГУТИКОВ
Реснички и жгутики имеют сходную внутреннюю структуру;
они различаются лишь характером биения. Жгутик, как и хвост
сперматозоида, совершает симметричные волнообразные колеба-
ния, которые распространяются вдоль него. В отличие от жгутика
ресничка ударяет асимметрично, быстро, рывком в одну сторону,
а затем производит медленное обратное движение, в результате
которого возвращается в исходное положение (рис. 11.1). При
558 Гл, 11. Мышцы, движение, локомоция
движении жгутика вода отталкивается параллельно его продоль-
ной оси, а при движении реснички перемещается параллельно по-
верхности, несущей на себе реснички (рис; 11.2).
Клетка обычно обладает одним или всего лишь немногими
жгутиками; на ресничной клетке, например на клетке парамеции,
может быть несколько тысяч ресничек, равномерно распределен-
Рис. 11.1. Биение ресничек у гребневика Pleurobrachia. (Sleigh, 1968.)
Схема составлена по данным киносъемки, произведенной при 17 °C. Положение
одной реснички показано через интервалы в 5 мс. Рабочее движение занимает
10 мс, а возвратное — 50 мс. Затем ресничка остается в исходном положении
в течение 20 мс, пока на 65-й миллисекунде не начнется следующее биение.
ных по ее поверхности. Однако трудно четко разграничить жгу-
тики и реснички: их внутреннее строение идентично, движения
возникают в самих органеллах, и нередко встречаются промежу-
точные формы движения.
«Жгутики» бактерий совершенно иные. Они более тонкие
(диаметр их около 0,02 мкм в отличие от 0,25 мкм у истинных
жгутиков и ресничек), короткие и относительно жесткие, их пово-
рачивают силы, действующие у основания жгутика, где он при-
креплен к клетке (Berg, Anderson, 1973).
Реснички и жгутики есть у многих простейших и имеют пер-
востепенное значение для локомоции. Сперматозоиды множества
Амебоидное движение; перемещение с помощью ресничек и жгутиков 559
животных плавают при помощи жгутиков. Покрытые ресничками
жабры и щупальца, часто встречающиеся у беспозвоночных, осу-
ществляют две главные функции: дыхательный газообмен и про-
цеживание воды, для захвата пищевых частиц. Реснички распро-
странены и у более высокоорганизованных животных, у которых
они служат, например, для передвижения жидкости по трубкам
Рис. 11.2. Типичное биение жгутика (слева) прогоняет воду параллельно его
главной оси (стрелка). Биение реснички (справа) прогоняет воду параллельно
поверхности, кк которой ресничка прикреплена. (Sleigh, 1974.)
в репродуктивных и выделительных системах (в нефридиях коль-
чатых червей и т. п.). У млекопитающих ресничные эпителии по-
могают переносу различных материалов по внутренним поверх-
ностям, например передвижению слизи в дыхательных путях или
яйца по яйцеводу.
Реснички встречаются у животных всех типов. Прежде насеко-
мых считали исключением, так как у них нет функционирующих
ресничек. Однако видоизмененные ресничные структуры, опозна-
ваемые по характерному расположению внутренних филаментов
(схема 9+2), найдены у насекомых в глазах, а также в боль-
шинстве других органов чувств у животных почти всех типов.
Важнейшие исключения — глаза некоторых беспозвоночных и
вкусовые рецепторы позвоночных. Во многих случаях, когда в ор-
гане чувств находят ресничную структуру, нелегко бывает устано-
вить ее значение для фото- или хеморецепции. Эти вопросы труд-
ны для разрешения, и только исследование их на широкой срав-
нительной основе, по-видимому, сможет привести к приемлемым
обобщениям (Barber, 1974).
560 Гл. 1L Мышцы, движение, локомоция
Как показала электронная микроскопия, реснички и жгутики
обладают одинаковым строением: пара филаментов в центре
окружена еще девятью тонкими нитями. Такое расположение по
схеме 94-2 характерно для всех групп животных от простейших
до позвоночных и свойственно подавляющему большинству спер-
Фото 11.1. Координированное биение ресничек у инфузории Paramecium cauda-
turn создает видимость волн, пробегающих по ее поверхности. Длина животного
около 150 мкм. (Фото предоставлено R. L. Hammersmith, Indiana University.)
матозоидов. Эта комбинация 94-2 почти универсальна, хотя и не
обязательна, так как известны исключения: бывают сперматозои-
ды с тремя центральными филаментами, с одним или совсем без
них (Blum, Lubbiner, 1973).
Реснички, используемые для движения, способны действовать
только в водной среде и поэтому встречаются только на поверх-
ностях клеток, обращенных к жидкости или покрытых жидкой
пленкой, например слизью.
Интуитивно можно было бы ожидать, что организм, обладаю-
щий одним жгутиком, например сперматозоид, будет плыть в
сторону, противоположную направлению волны, бегущей по жгу-
тику. Это действительно так в случае гладкой волнообразно дви-
жущейся нити: она действительно будет толкать организм в про-
тивоположную сторону. Поэтому кажется удивительным, что не-
которые жгутиковые движутся при помощи длинного жгута, ко-
торый бьет спереди в направлении движения, так что волны пе-
Амебоидное движение; перемещение с помощью ресничек и жгутиков 561
ремещаются от тела к переднему концу жгутика (Jahn et al.»
1964).
Такая, казалось бы, парадоксальная ситуация, когда распро-
странение волны и локомоция направлены в одну сторону, воз-
можна, если нить шероховатая или покрыта выступами. У видов,
передвигающихся таким образом, жгутики снабжены крошечны-
ми придатками в виде тонких боковых выростов, которые и
Фото 11.2. Электронная микрофотография жгутиков простейшего Trichonympha
ясно показывает характерное распределение филаментов (2 + 9) в каждом жгу-
тике. (Фото предоставлено А. V. Grimstone, Cambridge University.)
ответственны за это неожиданное направление движения
(рис. 11.3).
Механизм движения ресничек и жгутиков долго был предме-
том умозрительных гипотез. Предполагались механизмы трех
типов: 1) жгутик движется пассивно, подобно хлысту, под дейст-
36—1863
562 Гл. 11. Мышцы, движение, локомоция
вием сил, приложенных у его основания; 2) элементы, лежащие’
вдоль внутренней кривизны распространяющейся волны, сокра-
щаются, а элементы противоположной стороны — нет; 3) тонкие
нити внутри реснички смещаются относительно друг друга бла-
годаря силам, действующим между ними, вроде того как это про-
исходит с миофиламентами при сокращении мышцы.
Представление о том, что жгутик движется пассивно под дей-
ствием сил, приложенных у его основания, несовместимо с фор-
мой его волн (Machin, 1958). По хвосту сперматозоида волны
Рис. 11.3. Жгутиковое Ochromonas плавает при помощи одного жгутика, направ-
ленного вперед. Ундулирующее движение жгутика состоит в перемещении волн
от его основания к кончику (т. е. в том же направлении, в каком движется
животное). Эта парадоксальная ситуация объясняется наличием фиксированных
выступов, расположенных под прямым углом к жгутику и вызывающих движе-
ние воды в направлении, указанном изогнутыми стрелками; в результате орга-
низм продвигается в обратном направлении. (Jahn et al., 1964.)
распространяются без уменьшения амплитуды, а в некоторых слу-
чаях амплитуда даже возрастает (Rickmenspoel, 1965). Из этого
следует, что в жгутике должны быть активные элементы и что
энергия генерируется локально.
Если считать, что реснички приводятся в движение нитями
(филаментами), то возможны два механизма: 1) сгибать реснич-
ку могло бы сокращение нитей на одной стороне; 2) движение
могло бы быть результатом продольного скольжения нитей отно-
сительно друг друга. Первой гипотезе противоречит тот факт, что
трубочки ресничек сохраняют постоянную длину на протяжении
всего цикла сгибания (Satir, 1968, 1974), и имеющиеся в настоя-
щее время данные полностью согласуются со второй гипотезой.
Характерное тонкое строение жгутика или реснички показано
на рис. 11.4. Двойные трубочки состоят из белка тубулина с
мол. весом около 55 000. К каждой А «трубочке прикреплена па-
ра выступов, состоящих из белка динеина. Этот белок мало похож
на какие-либо сократительные белки мышцы, если не считать его
способности катализировать расщепление АТФ — свойства, кото-
рое он разделяет с важным мышечным белком миозином (Gib-
bons, Rowe, 1965; Gibbons, 1977).'
Амебоидное движение, перемещение с помощью ресничек и жгутиков 563
Жгутик сгибается, когда динеиновые выступы* прикрепляются
к соседней трубочке В, вызывая активные скользящие движения
за счет энергии АТФ. Процесс этот сходен со скольжением фи-
ламентов в мышце, но каковы молекулярные перемещения при
движении жгутика, еще точно не выяснено.
Можно создать экспериментальную модель ресничного аппа-
рата парамеции и изучать на этой модели регуляцию движения
ресничек. Клетку обрабатывают детергентом, что ведет к разру-
Рис. 11.4. Схема внутреннего строения жгутика или реснички (в поперечном раз-
резе). Двойные трубочки состоят из белка тубулина. К А-трубочкам прикреп-
лены выступы, содержащие белок динеин. (Brokaw, Gibbons, 1975.)
шению клеточной мембраны, тогда как функция ресничного ап-
парата сохраняется. После этого инфузорию можно реактивиро-
вать, и она будет плавать в растворе АТФ, содержащем Йоны
магния. Можно показать, что направление, в котором бьют рес-
нички, определяется концентрацией ионов кальция (Naitoh, Ка-
neko, 1972). Когда живая парамеция встречает препятствие, она
отступает и плывет назад, так как направление биений меняется
на обратное; эта реакция начинается с повышения проницаемости
клетки для ионов кальция (Eckert, 1972). Как мы увидим позже,
ионы кальция играют важную роль в управлении многими про-
цессами, в том числе мышечным сокращением.
Суммарный коэффициент полезного действия при превращении
метаболической энергии в механическую у плывущего спермато-
зоида, по-видимому, не меньше 19% и, вероятно, ближе к 25% (Rick-
menspoel et al, 1969). Эта величина поразительно сходна с ве*
личиной к. п. д. для мышц, совершающих внешнюю работу;
36*
564 Гл. 11. Мышцы, движение, локомоция
РАЗМЕРЫ ТЕЛА И МЕХАНИЗМ ЛОКОМОЦИИ
Мелкие организмы используют для плавания реснички или
жгутики; более крупные животные передвигаются с помощью
мышц (рис. 11.5). Для такого различия есть серьезные основания.
Одно из них чисто геометрическое. Мелкие организмы обладают
гораздо большей поверхностью по отношению к своему объему
я при движении ресничек в прилегающем слое воды продвигают-
Мышцы
Реснички (гребные пластинки)
Эукариотические жгутики
Бактериальные жгутики
___I_____I_____I_____I I I I I
100 нм 1мкм Юмкм ЮОмкм 1мм 10мм 100 мм 1м
Длина тела
Рис. 11.5. Размеры тела у организмов, которые для плавания пользуются жгу-
тиками, ресничками или мышцами. Соответствующие диапазоны размеров в зна-
чительной степени перекрываются; некоторые организмы, использующие мышцы,
меньше, чем самые крупные из тех животных, которые используют жгутики,
а диапазон для ресничных организмов перекрывает обе эти группы. Гребные
пластинки представляют собой группы ресничек в форме гребешка, которые
служат гребневикам для плавания. (Sleigh, 1977.)
ся вперед, используя вязкость воды для получения необходимого
толчка. Но представьте себе кита, покрытого ресничками, кото-
рый должен будет отталкиваться ими от слоя воды толщиной
лишь в долю миллиметра. По отношению к массе животного по-
лучаемый при этом толчок был бы неизмеримо мал.
Другая причина связана с законами гидродинамики. У очень
.небольшого, медленно плавающего организма движения всецело
зависят от сил вязкости — как это было бы, например, при пла-
вании человека в сиропе. У крупного животного, напротив, пла-
вание зависит от инерционных сил — от толчка, создаваемого
ускорением массы воды в противоположном направлении. Для
этого животное пользуется органами с достаточно большой по-
верхностью, такими, как плавники и ласты, а также мышцами,
чтобы развивать силу.
Теперь мы перейдем к рассмотрению функции мышц — не
только при локомоции, но и при всех разнообразных движениях,
в основе которых лежит создание силы сокращающейся мышцей.
Движение и мышца 565
ДВИЖЕНИЕ И МЫШЦА
СТРОЕНИЕ МЫШЦ
Насколько известно, биохимический механизм сокращения во
всех мышцах один и тот же. Механическую часть составляют два
белка — актин и миозин, а непосредственным источником энер-
гии для сокращения служит АТФ. Однако детали устройства
сильно варьируют, и в соответствии с этим удобно подразделять
мышцы на ряд типов. Эта классификация основана главным об-
разом на данных о мышцах позвоночных животных по той про-
стой причине, что они лучше изучены, чем мышцы беспозвоноч-
ных.
Наиболее широкое подразделение связано с наличием или
отсутствием у мышцы правильной поперечной исчерченности, ко-
торая видна в обычный световой микроскоп. Скелетные мышцы и
сердечная мышца позвоночных поперечнополосатые; мышцы внут-
ренних органов — в стенках мочевого пузыря, кишечника, крове-
носных сосудов, матки и т. д. — гладкие. Хотя сердечная мышца
и имеет поперечную исчерченность, ее часто выделяют в особый
тип, так как она отличается от скелетных мышц характерными
свойствами, из которых функционально самое важное то, что со-
кращение этой мышцы, начавшись, распространяется по всему
сердцу.
Скелетные мышцы обычно называют произвольными, так как
мышцы конечностей и туловища находятся под контролем воли.
Это не значит, что мы всегда отдаем себе отчет в наших движе-
ниях или сознательно управляем ими; напротив, локомоция, ды-
хание и т. п. происходят без осознания того, какие мышцы при
этом действуют. Произвольная мышца всегда обладает поперечной
исчерченностью, но сам термин «произвольная» не очень удачен
и применим в лучшем случае только к мышцам человека.
Гладкая мускулатура у позвоночных почти всегда находится
в стенках полых внутренних органов; она не контролируется во-
лей, и ее называют непроизвольной. Обычно гладкие мышцы со-
кращаются гораздо медленнее поперечнополосатых, и в норме мы
совершенно не ощущаем, какова степень их сокращения в стен-
ках наших кровеносных сосудов, желудка, кишечника и т. д.
ПОПЕРЕЧНОПОЛОСАТЫЕ МЫШЦЫ
Организация поперечнополосатой мышцы схематически пока-
зана на рис. 11.6. Мышца состоит из множества параллельных
мышечных волокон диаметром от 0,01 до 0,1 мм, длина которых,
однако, может достигать нескольких сантиметров. Эти волокна
в свою очередь состоят из более тонких мышечных фибрилл.
Фибриллы обладают характерной поперечной исчерченностью,
обусловленной так называемыми линиями (дисками) Z, которые
566 Гл. 11. Мышцы, движение, локомоция
повторяются через совершенно правильные промежутки величи*
ной около 2,5 мкм в расслабленной мышце. От тонкого диска Z
в обе стороны тянутся очень тонкие миофиламенты, (нити), а £
промежутки между их концевыми участками входят несколько
более толстые миофиламенты, благодаря чему между линиями Z
образуются менее выраженные полосы; вид этих полос изменяется
Рис. 11.6. Схема строения поперечнополосатой мышцы позвоночного. Вся мышца
состоит из волокон, которые в световом микроскопе выглядят поперечно исчер-
ченными. Волокна состоят из фибрилл, у которых имеются более темные и более
светлые полосы. Электронный микроскоп показывает, что эти полосы создаются
периодическим правильным расположением толстых и тонких филаментов.
в зависимости от степени сокращения мышцы. Диаметр тонких
филаментов около 0,005 мкм, толстых — приблизительно вдвое'
больше, т. е. около 0,01 мкм. Длина толстых филаментов при-
мерно 1,5 мкм. Тонкие филаменты несколько варьируют по длине
и часто занимают по обе стороны диска Z от 2 до 2,6 мкм (или
половину этого расстояния, если измерять от диска Z в одну сто-
рону до кончика нити). Расположение филаментов оказалось'
весьма регулярным и упорядоченным.
В поперечнополосатой мышце толстые филаменты состоят из
миозина, а тонкие — из актина. Те и другие соединены между
собой системой молекулярных поперечных связей, и когда мыш-
ца сокращается и укорачивается, эти поперечные связи перестраи-
ваются таким образом, что толстые филаменты скользят между
тонкими и расстояние между дисками Z уменьшается (Huxley,
1969).
Эта модель строения мышцы хорошо подтверждается элект-
ронными микрофотографиями миофиламентов на разных стадиях
сокращения. Ни толстые, ни тонкие филаменты не изменяют при
сокращении своей длины, но ясно видно, что они движутся отно-
сительно друг друга. Однако еще больше в пользу модели со
Движение и мышца 567
скользящими нитями говорит тот факт, что сила, развиваемая
мышцей, находится в прямой зависимости от степени перекрыва-
ния этих нитей (с. 571—572).
Фото 11.3. Электронная микрофотография участка волокна летательной мышцы
водяного клопа Lethocerus cordofanus. Можно видеть очень правильное располо-
жение толстых и тонких миофиламентов. Две темные полосы, пересекающие во-
локно, — диски Z; расстояние между ними, составляющее около 3 мкм, опреде-
ляет длину одного саркомера. Более темные тельца вверху слева и внизу спра-
ва — митохондрии. (Фото предоставлено В. М. Luke, Oxford University.)
СЕРДЕЧНАЯ МЫШЦА И ГЛАДКАЯ МУСКУЛАТУРА
Сердечная мышца, подобно скелетным мышцам, поперечно ис-
черчена, но обладает иными свойствами, что связано с ее струк-
турой. Ее волокна не расположены параллельным пучком, а вет-
вятся и соединены между собой в единую сеть. Из этой струк-
туры вытекает важное следствие: когда в одном участке сердеч-
ной мышцы начинается сокращение, оно быстро распространяется
по всей мышечной массе. Другая важная особенность сердечной
мышцы — то, что за сокращением тотчас же следует расслабле-
ние. В результате здесь не бывает продолжительного сокращения,
подобного длительному напряжению скелетной мышцы. Эти два
свойства весьма существенны для нормальных ритмических сокра-
щений сердца.
Гладкая мышца лишена поперечной исчерченности, характер-
ной для скелетных мышц, но сокращение ее связано с теми же
568 Гл. 11. Мышцы, движение, локомоция
белками, т. е. актином и миозином, и с использованием энергии
АТФ. Отсутствие поперечной исчерченности, по-видимому, объ-
ясняется тем, что в гладкой мышце нет упорядоченного располо-
жения толстых и тонких филаментов.
Гладкие мышцы изучены не так основательно, как попереч-
нополосатые. Это обусловлено несколькими причинами. Одна из
них — то, что волокна гладкой мышцы часто перемежаются с
соединительнотканными волокнами. Другая причина состоит в
том, что волокна гладкой мышцы не образуют правильных па-
раллельных пучков, которые легко было бы изолировать и изу-
чать. Кроме того, гладкомышечные клетки гораздо мельче; длина
их часто составляет лишь долю миллиметра. И наконец, глад-
кая мышца сокращается во много раз медленнее, чем поперечно-
полосатая; по существу, все гладкие мышцы не способны к бы-
строму сокращению.
ВЕЩЕСТВА, СЛУЖАЩИЕ ДЛЯ ЗАПАСАНИЯ ЭНЕРГИИ
Непосредственным источником энергии для мышечного сокра-
щения служит аденозинтрифосфат (АТФ)—главный источник
энергии почти для всех биологических процессов, требующих ее
затраты. АТФ представляет собой единственное вещество, кото-
рое мышечные белки используют непосредственно. Когда конце-
вая фосфатная группа этого соединения отщепляется, высокая
энергия разрываемой химической связи становится доступной для
использования в механизме мышечного сокращения.
Несмотря на свое большое значение, АТФ содержится в мыш-
це в очень малых количествах. Всего его запаса может хватить
не больше чем на десяток быстрых сокращений; отсюда следует,
что этот запас должен очень быстро пополняться, иначе мышца
скоро перестанет работать. Источником для этого служит другое
органическое фосфатное соединение — креатинфосфат, который
присутствует в мышце в больших количествах. Его фосфатная
группа переносится на аденозиндифосфат (АДФ), и таким обра-
зом восстанавливается запас АТФ.
Но со временем и запас креатинфосфата нужно будет попол-
нить, и для этого используется первичный источник мышечной
энергии — окисление углеводов или жирных кислот. Углеводное
«топливо» запасается в мышце в форме гликогена, количество
которого часто достигает 0,5—2% влажного веса мышцы и кото-
рый может дать, вероятно, в сотню раз больше энергии, чем весь
имеющийся креатинфосфат. Даже при недостатке кислорода гли-
коген все еще может давать энергию, расщепляясь до молочной
кислоты, но выход энергии в этом случае невелик — он составляет
лишь около 7% энергии, получаемой при полном окислении (см.
гл. 6).
Движение и мышца 569
Креатинфосфат обнаружен в мышцах всех исследованных по-
звоночных, но явно отсутствует в мышцах некоторых беспозво-
ночных. У этих животных найдены другие органические фосфаты;
один из них — аргининфосфат, который никогда не встречается
в мышцах позвоночных.
Все эти различные органические фосфаты доставляют энер-
гию при отщеплении фосфатной группы, и высокая энергия соот-
ветствующих связей становится доступной для нуждающихся в
ней процессов. Тот факт, что креатинфосфат используется только
в нескольких группах беспозвоночных, но повсеместно присутст-
вует у позвоночных, навел на мысль, что по биохимическому сход-
ству между этими группами беспозвоночных и позвоночными мож-
но судить, от какого типа первых произошли вторые. Однако со-
временные данные говорят скорее о том, что распространение
аргинин- и креатинфосфата среди беспозвоночных очень нерегу-
лярно и не связано с классификацией этих животных. В частно-
сти, картина распространения креатинфосфата, по-видимому, не
такова, чтобы она могла служить ключом к эволюции позвоноч-
ных.
МЫШЕЧНОЕ СОКРАЩЕНИЕ
В физиологии мышц сокращением называют определенное со-
стояние механической активности. Оно может сопровождаться
укорочением мышцы, но если концы мышцы прочно закреплены и
укорочение невозможно, то и тогда активное состояние обознача-
ют термином «сокращение». В этом случае развивается сила, воз-
действующая на точки прикрепления мышцы; поскольку длина
мышцы при этом не меняется, такое сокращение называют изо-
метрическим. Если же к одному концу мышцы прикреплен груз,
который она в состоянии поднять, то при сокращении она уко-
рачивается; и так как нагрузка здесь остается с начала до кон-
ца постоянной, такое сокращение называют изотоническим.
Фактически ни одно мышечное движение не бывает ни чисто
изометрическим, ни чисто изотоническим: обычно во время сокра-
щения изменяются и длина мышцы, и нагрузка на нее. Например,
когда рука поднимает груз, то и длина сокращающихся мышц, и
нагрузка на них изменяются непрерывно, так как меняются и
действие рычага, и угол приложения силы.
Хотя чисто изометрическое или чисто изотоническое сокраще-
ние в реальной мышечной деятельности организма не осуществ-
ляется, при изучении изолированных мышц и их функции удобно
различать эти два вида сокращения. Изометрические сокращения
позволяют получать много сведений о силах, развиваемых мыш-
цами, а по изотоническим можно легко вычислить величину со-
вершаемой работы. Поэтому в лаборатории обычно выбирается
первый или второй режим в зависимости от задачи исследования.
570 Гл. 1L Мышцы, движение, локомоция
СИЛА
Сокращение мышцы можно вызвать, нанеся на нее одиноч-
ный короткий электрический импульс, и если при этом не давать
ей укоротиться (изометрическое сокращение), то развиваемая ею
Рис. 11.7. Сила, развиваемая при изометрическом сокращении икроножной мыш-
цей кошки: А — после одного стимула, Б, В п Г — после двух стимулов, нано-
симых с уменьшающимися интервалами. Отметки времени через каждые 20 мс.
Температура 34,5 °C. (Cooper, Eccles, 1930.)
сила будет соответствовать кривой, приведенной на рис. 11.7, Л.
Непосредственно после стимуляции отмечается очень короткий
интервал (латентный период) в несколько миллисекунд перед на-
чалом сокращения. Затем развиваемая мышцей сила быстро ра-
Рис. 11.8. Сила, развиваемая при изометрическом сокращении глазодвигатель-
ной мышцей кошки в ответ на многократную стимуляцию: I — с частотой 125
в секунду (интервал 8 мс), II — с частотой 210 в секунду. Во втором случае
происходит почти полное слияние в тетанус. Температура 36 °C. (Cooper, Eccles,
стет до максимума, а потом снова снижается, но несколько более
медленно. Такое одиночное сокращение в ответ на одиночный сти-
мул называется фазическим.
Движение и мышца 57Г
Время, необходимое для развития максимального напряжения
(время сокращения), у разных мышц различно. Время сокраще-
ния главных мышц, осуществляющих локомоцию у кошки, до-
стигает 100 мс; время сокращения самых быстрых мышц (глаз-
ных) может быть меньше 10 мс.
Длина саркомера, мкм
Рис. 11.9. Сила, развиваемая сокращающейся мышцей (ордината), в зависимости
от исходной длины мышцы (абсцисса). Сила выражена в процентах от макси-
мальной силы, которую может развить мышца при изометрическом сокращении.
Степень перекрывания между толстыми и тонкими филаментами показана на
схемах над соответствующими точками кривой. (Gordon et al., 1966.)
Если нанести второй стимул до окончания первого сокраще-
ния, то на него наложится второе (рис. 11.7, Д). Второй пик си-
лы при этом будет выше первого. Если продолжать применять два
стимула со все меньшими интервалами между ними, то два со-
кращения становятся все более слитными (рис. 11.7, ДД).
572 Гл. 11. Мышцы, движение, локомоция
Многократная стимуляция с интервалами достаточной дли-
тельности будет вызывать ряд отдельных сокращений, но если
интервалы укорачивать, сокращения начнут сливаться и в конце
концов возникает одно длительное сокращение (рис. 11.8,//), на-
зываемое тетанусом.
Максимальная сила, развиваемая при тетанусе, зависит от ис-
ходной длины мышцы. Когда мышцу заставляют сокращаться изо-
метрически (фиксировав ее концы), она будет развивать меньшую
силу, если расстояние между концами мышцы будет меньше или
больше, чем ее длина в покое. Максимум силы достигается, если
мышце придают длину, равную ее длине в состоянии покоя.
Это обстоятельство тесно связано с положением филаментов по
отношению друг к другу (рис. 11.9). Если вначале мышца рас-
тянута настолько, что тонкие и толстые филаменты не перекры-
ваются (положение /), то никакой силы не возникает. Это согла-
суется с полной неспособностью филаментов к образованию по-
перечных мостиков в этом положении. Но при меньшей степени
растяжения мышцы по мере того, как перекрывание филаментов
увеличивается, сила растет. Она максимальна при такой длине
мышцы, когда поперечные мостики на всем протяжении толстых
филаментов соприкасаются с тонкими (положение 2). Если же
перед началом сокращения мышца еще короче, то сила снова
будет уменьшаться и быстро упадет до нуля, когда концы тол-
стых филаментов придут в соприкосновение с дисками Z (поло-
жения 3 и 4).
ЧТО ЗАСТАВЛЯЕТ ФИЛАМЕНТЫ СКОЛЬЗИТЬ?
Молекулярные события, происходящие при мышечном сокра-
щении, ' изучены лучше большинства других процессов в живом
организме. Для их описания следует рассмотреть структуру тол-
стых и тонких филаментов.
Толстые филаменты состоят из белка миозина. Каждая мо-
лекула миозина похожа на тонкую палочку с круглой «голов-
кой». Один толстый филамент содержит несколько сот молекул
миозина, расположенных как показано на рис. 11.10. При этом
их головки в каждой половине филамента направлены в сторону
его ближайшего конца; в результате посередине остается глад-
кая зона, а в остальных частях всюду выступают головки.
Тонкие филаменты построены сложнее, они состоят из трех
важных белков. Остов филамента образует актин — глобулярный
белок с относительно небольшими молекулами, которые образу-
ют здесь двойную четкообразную спираль. Каждая молекула ак-
тина слегка асимметрична, и в каждом филаменте все эти моле-
кулы ориентированы в одну сторону. Такая направленность иг-
рает важную роль в сокращении мышцы.
Движение и мышца 57Э
Вторым важным белком тонкого филамента является тропо-
миозин; его длинные и тонкие молекулы соединены своими кон-
цами и образуют очень тонкую нитевидную структуру, которая»
лежит в бороздках двойной спирали из молекул актина. Одна
молекула тропомиозина по своей длине соответствует цепочке из»
семи молекул актина. Тонкий филамент имеет длину около 1 мкм-
и содержит около 400 молекул актина и 60 молекул тропомиозина.
Рис. 11.10. Тонкий филамент (вверху) представляет собой двойную спираль и»
глобулярного белка актина. В желобке между тяжами актина лежат две тонкие
нити из белка тропомиозина; кроме того, на каждые семь молекул актина имеет-
ся по одной молекуле тропонина. Толстый филамент (внизу) состоит из палочко-
образного белка миозина с расширенными «головками», которые выступают в>
сторону актиновых тяжей.
К каждой молекуле тропомиозина прикреплена молекула
третьего белка — тропонина. Этот белок связывает кальций и иг-
рает ключевую роль в инициации сокращения. Связав эти ионы,,
он претерпевает конформационное изменение, необходимое для
взаимодействия между головками миозина в толстых нитях и ак-
тином тонких нитей (Squire, 1975).
Взаимодействие между толстыми и тонкими филаментами со-
стоит в циклическом замыкании и разрыве поперечных мостиков
между теми и другими. Миозиновые головки прикрепляются к
тонким филаментам под определенным углом; они претерпевают
конформационное изменение, в результате которого этот угол из-
меняется, и мостики тянут тонкую нить вдоль толстой (Huxley,.
1973). Поперечные мостики на противоположных сторонах от
гладкой зоны в середине толстого филамента поворачиваются в-
разные стороны и притягивают друг к другу концы тонких фи-
ламентов, уменьшая тем самым расстояние между дисками Z.
Это сближение дисков Z и приводит к укорочению мышцы.
Чтобы произошло значительное укорочение, каждый попереч-
ный мостик должен много раз прикрепиться, повернуться, отъеди-
574 Гл. 11. Мышцы, движение, локомоция
ниться и снова прикрепиться в другой, более дальней точке тон-
кого филамента. Каждый цикл прикрепления требует затраты
энергии в форме АТФ. Молекула АТФ присоединяется на время к
головке миозина, образуя с нею активный комплекс, который мо-
жет прикрепиться к актиновому тяжу при наличии подходящего
места, не блокированного тонкой нитью тропомиозина (Weber,
Murray, 1973).
Роль ионов Са2+. Тропомиозин лежит в канавках актиновых
филаментов, где он препятствует их взаимодействию с головками
миозина. Регулирующий белок тропонин обладает высоким срод-
ством к кальцию. Сразу же после стимуляции мышцы концентра-
ция ионов кальция в мышечном волокне резко возрастает; ионы
кальция связываются с тропонином, который в свою очередь пре-
терпевает конформационное изменение, позволяющее тропомио-
зину сойти со своей блокирующей позиции на актиновых тяжах.
Повышение концентрации ионов кальция тотчас после стиму-
ляции было продемонстрировано с помощью белка экворина.
Этот белок, который можно выделить из одной светящейся меду-
зы, испускает свет в присутствии ионов кальция. После инъекции
экворина в мышечные волокна гигантского усоногого рака Bala-
nus nubilis электрическое раздражение этих волокон вызывает их
слабое свечение, что говорит о повышении в них концентрации
ионов кальция. Измерение этой эмиссии света служит чувстви?
тельным индикатором очень быстрых и кратковременных измене-
ний внутриклеточной концентрации Са2+ (Ridgway, Ashley, 1967).
КАК «ЗАПУСКАЕТСЯ» СОКРАЩЕНИЕ?
В норме мышечное сокращение инициируется, когда к нервно-
мышечному соединению (двигательной концевой пластинке) при-
ходит нервный импульс. Импульс быстро распространяется в фор-
ме электрической деполяризации по поверхностной мембране мы-
шечного волокна (сарколемме), на короткий момент аннулируя
обычный поверхностный потенциал около 60 мВ. Эта деполяриза-
ция почти мгновенно передается внутрь мышечной клетки и вы-
зывает ее сокращение.
Как мы увидим позже, сигнал, идущий от концевой пластин-
ки к мышечному волокну, состоит в диффузии химического пере-
датчика ацетилхолина через узкую щель между двигательной
концевой пластинкой и сарколеммой. Но расстояние от поверхно-
сти до внутренних частей мышечного волокна так велико, что
если бы сигнализация и здесь была основана на диффузии како-
го-либо вещества, то импульс не мог бы передаваться ко всем
филаментам одновременно. Одна из классических проблем мы-
шечной физиологии состояла в том, чтобы объяснить, каким обра-
зом эта передача может осуществляться столь быстро, что все
филаменты сокращаются одновременно.
Движение и мышца 575
Ответ на этот вопрос дает ультраструктура мышечного во-
локна. Сарколемма (наружная мембрана мышечной клетки) со-
единена со сложной системой поперечных трубочек, которые про-
низывают мышечную клетку, как правило, около дисков Z, — так
называемой Т-системой (рис. 11.11). Импульс, пришедший к дви-
гательной концевой пластинке, вызывает деполяризацию сарко-
Рис. 11.11. Система Т-трубочек и саркоплазматический ретикулум, окружающий
миофибриллы в волокне поперечнополосатой мышцы. (Peachey, 1965.)
леммы; и этот процесс деполяризации продолжается в Т-системе,
запуская дальнейшие этапы сокращения.
Нужно упомянуть еще одну важную структуру, а именно си-
стему уплощенных пузырьков, или саркоплазматический ретику-
лум, который окружает внутреннюю область мышечного волокна
наподобие муфты (рис. 11.11).
Теперь мы знакомы со структурной основой быстрой передачи
сигнала всему мышечному волокну. При распространении им-
пульса по сарколемме и Т-системе происходящее изменение пе-
Зт76 Гл. И. Мышцы, движение, локомоция
редается на мембрану саркоплазматического ретикулума и вызы-
вает повышение ее проницаемости. В результате ионы кальция, в
состоянии покоя замкнутые в полостях саркоплазматического ре-
тикулума, теперь могут выйти оттуда (рис. 11.12). Внезапное по-
явление кальция приводит к изменению конфигурации тропони-
на, и теперь становится возможным взаимодействие миозина с
актином, лежащее в основе сокращения. Концентрация кальция,
тического ретикулума.
необходимая для активирования сократительного процесса, со-
ставляет не более 0,01 ммоль/л. Освободившийся кальций немед-
ленно вновь захватывается саркоплазматическим ретикулумом и
остается здесь до прихода следующего импульса; в расслаблен-
ной скелетной мышце концентрация кальция, как полагают, рав-
на всего лишь 0,0002 ммоль/л.
Теперь мы можем составить перечень важнейших событий, с
которыми связано мышечное сокращение (табл. 11.1).
СИЛА И РАБОТА
Если во время стимуляции мышце дана возможность свободно
сокращаться, она укоротится, скажем, на одну треть. Но если
прикрепить к ней груз, укорочение будет меньше. Если груз по-
степенно увеличивать, мышца сможет поднимать его на все мень-
шую высоту, пока груз не окажется настолько тяжелым, что мыш-
ца вовсе не поднимет его. Таким образом, мы получим кривую
зависимости укорочения от силы. Подобная кривая представлена
на рис. 11.13; в данном случае при нагрузке чуть меньше 100 г
укорочение оказалось уже невозможным.
Движение и мышца 577
Таблица ПЛ
Последовательность событий при стимуляции и сокращении мышцы
Стимуляция
1. Деполяризуется сарколемма
2. Деполяризуется Т-система
3. Ионы Са2+ выходят из саркоплазматического ретикулума
4. Ионы Са2+ диффундируют к тонким филаментам
Сокращение
5. Ионы Са2+ связываются с тропонином
6. Комплекс тропонин — Са2+ снимает блокаду актина тропомиозином
7. Головки толстого филамента (содержащие комплекс миозин —АТФ)
образуют поперечные мостики к актиновому тяжу
8. Поперечные мостики поворачиваются при гидролизе АТФ и образо-
вании свободного АДФ
Расслабление
9. Ионы Са2+ отделяются от тонких филаментов мембранами саркоплаз-
матического ретикулума
10. Ионы Са2+ диффундируют от тонких филаментов к саркоплазматиче-
скому ретикулуму
11. Са2+ высвобождается из комплекса тропонин — Са2+
12. Тропонин дает возможность тропомиозину вернуться в блокирующее
положение
13. Миозин-актиновые поперечные мостики разрываются
14. В головках толстых филаментов снова образуется комплекс АТФ —
миозин
Намного ли различаются по силе мышцы разных животных?
Когда муравей тащит в челюстях зерно, которое весит больше не-
го самого, создается впечатление, что его мышцы должны быть
необычайно сильными. Объективное сравнение мышечной силы
показывает, что это не так. При сравнении разных мышц нуж-
но, разумеется, учитывать различие в их величине.
Сила, развиваемая мышцей данного типа, находится в прямой
зависимости от площади ее поперечного сечения, а не от ее дли-
ны. Поэтому можно сравнивать весьма различные мышцы, если
относить развиваемую ими силу к их поперечному сечению, как
это сделано в табл. 11.2 для мышц разных животных. Хотя мно-
гие мышцы слабее, чем приведенные в таблице, верхний предел,
по-видимому, примерно одинаков для всех мышц независимо от
их принадлежности тому или иному животному, а именно около
4—6 кг на 1 см2 площади поперечного сечения.
Тот факт, что сила сокращения одинакова у такого множества
разнообразных мышц, может показаться неожиданным, но его
легко понять, если вспомнить о принципиальном единстве самого
сократительного механизма. Любое мышечное сокращение основа-
но на скольжении актиновых и миозиновых филаментов, энергию
для которого доставляет АТФ. Трудно было бы ожидать, что этот
механизм можно улучшить и обеспечить большую силу на еди-
37—1863
578 Гл. 11. Мышцы, движение, локомоция
ницу площади поперечного сечения, так как максимальная сила
зависит от числа миофиламентов, которые помещаются на данной
площади, а это число в свою очередь зависит от величины белко-
вых молекул, образующих филаменты1.
Рис. 11.13. Изотоническое сокращение портняжной мышцы жабы Bufo mar in us.
Укорочение мышцы падает до нуля при увеличении нагрузки до 97 г. Работа,
совершаемая при сокращении, возрастает до максимума при нагрузке около 40%
от максимальной, а при дальнейшем увеличении нагрузки снова снижается.
(С любезного разрешения Frans Jobsis.)
Когда мышца поднимает груз, совершаемая ею внешняя рабо-
та равна произведению веса этого груза на длину его пути. Это
показывает дугообразная кривая на рис. 11.13. Когда нагрузка
равна нулю, внешняя работа тоже равна нулю, хотя мышца уко-
рачивается максимально. В другом крайнем случае, когда груз
так велик, что совсем не может быть поднят, внешняя работа
также равна нулю. Между этими крайними точками внешняя ра-
бота достигает максимума, когда груз составляет около 40% наи-
большего веса, который мышца в состоянии поднять, а ее укоро-
чение составляет около трети максимального укорочения.
Очевидно, что работа, которую мышца способна совершить,
1 Диаметр толстых и тонких филаментов лимитирует возможную плотность
их расположения. Но можно представить себе увеличение силы путем удлине-
ния зоны перекрывания тонких и толстых филаментов (т. е. путем увеличения
расстояния между дисками Z и удлинения самих филаментов). Это дало бы
большее число поперечных мостиков на один филамент и тем самым увеличило
бы силу. Но такой путь, возможно, неосуществим из-за предела прочности фи-
ламентов, которые должны были бы выдерживать возросшую растягивающую
силу.
Движение и мышца 579
Таблица 11.2
Максимальная сила сокращения мышц у разных животных
Животное Мышца Сила сок- ращения , кг/см2
Кольчатый червь Areni- cold^ Стенка тела 3
Двустворчатый моллюск Anodonta2'* Двустворчатый моллюск Mytilus^ Мышца-замыкатель 5
Передний ретрактор бис- суса 4,5
Осьминог (Octopus')^ Ретрактор воронки 5,1
Насекомое Locusta^ Мышцы задней пары ног 4,7
Насекомое Decticus^ Сгибатель голени 5,9
Насекомое Drosophila^ Летательные мышцы 5
Лягушка (Ranay) Передняя большеберцо- вая мышца (20 °C) 4,4
Кролик8) Скелетные мышцы (37 °C) 5,0
Человек9) *) Trueman, 1966. 2) Weber, Portzehl, 1954. 3) Abbott, Lowy, 1953. 4) Lowy, Millman, 1962. Б) Wigglesworth, 1972. 6) Roeder, 1953. 7) Casella, 1950. 8) Wilson, 1972. 9) Haxton, 1944. Икроножная мышца (произвольное сокра- щение) 4,2
зависит от ее величины. Рассмотрим две мышцы одинаковых раз-
меров, развивающие одинаковую предельную силу на единицу
площади поперечного сечения. Если они укоротятся в равной сте-
пени, то произведут одинаковую работу. А теперь рассмотрим две
мышцы одного поперечного сечения, но разной длины — одну
вдвое длиннее другой, причем обе укорачиваются на одну и ту же
долю их длины покоя, скажем на одну треть. Максимальная ра-
бота их будет находиться в прямой зависимости от их первона-
чальной длины (т. е. вдвое больше у первой, чем у второй), хотя
сила сокращения у них одинаковая (одно и то же поперечное се-
чение).
Можно продолжить это рассуждение и сказать, что, поскольку
работа есть произведение силы на расстояние, а объем мышцы
пропорционален произведению поперечного сечения на длину, ра-
бота, которую мышца может совершить при своем сокращении,
должна быть прямо пропорциональна ее объему. Разумеется, из
этого общего правила возможны исключения, но его все же по-
лезно помнить.
Что касается мощности (работы в единицу времени), то здесь
положение совершенно иное. Поскольку время сокращения «бы-
37*
580 Гл. 11. Мышцы, движение, локомоция
строй» мышцы короче, ее мощность будет больше, чем у «медлен-
ной» мышцы. При сравнении мышц слона и землеройки можно
ожидать, что 1) у этих мышц приблизительно одинаковая сила
на единицу площади поперечного сечения; 2) они способны уко-
рачиваться приблизительно на одну и ту же долю длины покоя;
3) работа, совершаемая при сокращении 1 г мышцы, одна и та
же у обоих животных. Однако благодаря тому, что у землеройки
мышцы сокращаются гораздо быстрее (т. е. за меньшее время),
мощность на 1 г мышечной ткани у землеройки соответственно
гораздо больше, чем у слона. Именно это отражают различия в
интенсивности метаболизма у разных животных (см. гл. 6), ко-
торую обычно выражают в единицах мощности (работа или энер-
гия в единицу времени) и которая намного выше у мелких жи-
вотных.
КАК ИСПОЛЬЗУЮТСЯ МЫШЦЫ
Способы использования мышц у разных животных весьма раз-
личны (и прежде всего зависят от функции данной мышцы).
В самом деле, к летательным мышцам насекомого, сокращающим-
ся несколько сот раз в секунду,предъявляются совсем иные тре-
бования, нежели к мышце, которая замыкает раковину моллюска
и остается иногда в сокращенном состоянии несколько часов. Опи-
сать, каким образом мышца выполняет разные задачи, можно*
лучше всего, рассмотрев несколько характерных типов мышц и
их видоизменения, связанные со специфическими потребностями.
БЫСТРЫЕ И МЕДЛЕННЫЕ МЫШЦЫ ПОЗВОНОЧНЫХ
Поперечнополосатая мышца позвоночного состоит из мышеч-
ных волокон, которые разделяются по меньшей мере на два типа»
часто называемых «быстрыми» и «медленными» волокнами. Эти
термины легко могут привести к путанице, и сейчас принято на-
зывать быстрые волокна фазическими, а медленные — тонически-
ми. Обычно эти два вида волокон различаются по содержанию
миоглобина. Фазические волокна, в которых меньше миоглобина,
называли также бледными или белыми, а тонические, с. большим
количеством миоглобина — красными. Мышца может состоять
только из быстрых волокон, только из медленных или из тех и
Других.
Различие между фазическими (быстрыми) и тоническими
(медленными) волокнами было впервые установлено у лягушки.
Главная функциональная разница между ними состоит в том, что
первые используются при быстрых движениях, а вторые поддер-
живают длительное, но слабое напряжение. Фазическая система
связана с крупными нервными волокнами (толщиной от 10 до
20 мкм) со скоростью проведения от 8 до 40 м/с, которые вызы-
Движение и мышца 581
вают быстрые сократительные реакции. Эта фазическая система
используется, например, при прыжках. Тоническая (медленная)
система иннервируется тонкими нервными волокнами (толщиной
около 5 мкм), которые проводят импульсы со скоростью от 2 до
8 м/с и вызывают более медленные, градуальные мышечные со-
кращения, сопровождаемые нераспространяющимися мышечными
потенциалами малой амплитуды и большой длительности. Тони-
ческая система осуществляет, например, поддержание позы (Kuf-
fler, Williams, 1953а).
Портняжная мышца лягушки, идущая вдоль бедра и участ-
вующая главным образом в прыжках, состоит только из фазиче-
ских волокон. Нерв, идущий к этой мышце, содержит крупные,
быстропроводящие нервные волокна, из которых каждое образу-
ет контакт с мышечными волокнами посредством одиночной тер-
минали (концевой пластинки). Фазические волокна реагируют по
типу «всё или ничего»: когда стимул превышает некоторую мини-
мальную (пороговую) величину, волокно отвечает полным сокра-
щением и дальнейшее усиление стимула уже не приводит к уве-
личению реакции. Иными словами, мышечное волокно или совсем
не реагирует, или отвечает максимальной реакцией. Это и назы-
вается реакцией типа «всё или ничего».
Тоническая мышца обладает иными свойствами. Ее волокна,
по своему поведению во многих отношениях несходные с фази-
ческими, иннервируются нервными волокнами малого диаметра,
концевые контакты которых распределены по всей длине мышеч-
ного волокна. В отличие от фазических мышечных волокон с их
быстрым сокращением у тонических волокон реакция не обяза-
тельно распространяется по всему волокну в виде полного ответа
типа «всё или ничего», а при многократном раздражении проис-
ходит суммация. Ответ на одиночный стимул невелик, при по-
вторной же стимуляции напряжение растет с увеличением ее ча-
стоты. Таким образом, для того чтобы напряжение тонической
мышцы значительно возросло, нужна многократная стимуляция
тонких нервов. Учащение стимуляции ускоряет прирост напряже-
ния. После сокращения медленное мышечное волокно расслаб-
ляется в 50—100 раз медленнее, чем фазическое. Реакция тако-
го типа играет важную роль в регулировании общей позы у ля-
гушки (Kuffler, Williams, 1953 b).
Для раздельного изучения функции мышечных волокон раз-
ных типов нет необходимости искать мышцы с волокнами только
одного типа. Наблюдаемые различия между мышцами, состоящи-
ми только из фазических и только из тонических волокон, могут
быть результатом просто разной локализации и разного назначе-
ния тех и других мышц. Поэтому лучше исследовать волокна раз-
ного типа в одной и той же мышце.
Для этой цели можно воспользоваться икроножной мышцей
кошки, которая идет от колена к пяточному сухожилию и слу-
582 Гл. 11. Мышцы, движение, локомоция
жит для разгибания лапы в голеностопном суставе. Она содер-
жит мышечные волокна даже не двух, а трех разных функцио-
нальных типов. Это можно показать следующим образом. Через
введенный микроэлектрод стимулируют одиночный мотонейрон в
соответствующем участке спинного мозга. Это вызывает актива-
цию только одной двигательной единицы (которая состоит из
0 2 4 6 15
50
Долго не утомляющиеся
ъ 20
ь
§
10
1
О
Рис. 11.14. Икроножная мышца кошки состоит из волокон трех типов, реагирую-
щих на стимуляцию (40 раз в секунду) тремя различными способами. Одни
быстро сокращаются и быстро утомляются (вверху)} другие быстро сокраща-
ются и мало утомляемы (в середине), третьи медленно сокращаются и не утом-
ляются (внизу). (Burke et al., 1971.)
Движение и мышца §83
многих мышечных волокон, совместно иннервируемых одним ак-
соном). При такой активации происходит сокращение волокон од-
ного из трех разных типов в зависимости от того, какой именно
нейрон стимулирован.
На рис. 11.14 показана реакция трех типов волокон: 1) быст-
ро сокращающегося и быстро утомляющегося; 2) быстро сокра-
щающегося и мало подверженного утомлению и 3) медленно со-
кращающегося и не утомляющегося даже1При длительной стимуля-
ции. У медленно сокращающихся (тонических) волокон время
сокращения в два с лишним раза больше, чем у быстро сокра-
щающихся и быстро утомляющихся (фазических) волокон
(табл. 11.3). Еще поразительнее разница в развиваемой силе.
Быстрые волокна развивают силу в 10 раз большую, чем медлен-
ные. Расплатой за это, очевидно, и служит более быстрое утом-
ление этих волокон.
Таблица 11.3
Время сокращения и максимальная
сила, развиваемая при тетаническом
сокращении мышечными волокнами
трех разных функциональных типов
в икроножной мышце кошки.
(Burke et al., 1971)
Тип волокон
3 S3 к
\о 3\о ч
Время быстрого
сокращения,
мс 34 40 73
Сила при тета-
нусе, г 60 20 5
Принадлежность мышцы к фазическому или же тоническому
типу не обязательно является постоянным врожденным призна-
ком. Это свойство, по-видимому, каким-то образом зависит от
способа использования мышцы. Изменяя место прикрепления су-
хожилий у мышц задней ноги кролика, удавалось придать тони-
ческой мышце свойства, более близкие к свойствам фазических
мышц.
У новорожденного котенка все мышцы, по-видимому, одина-
ково медленные, и дифференциация их происходит, когда они на-
чинают функционировать. Путем перерезки нервов, идущих к
двум мышцам, и их последующего перекрестного соединения с
этими мышцами было показано, что при соединении нервных во-
локон от быстрых (фазических) мотонейронов с медленной (тони-
584 Гл. 11. Мышцы, движение, локомоция
ческой) мышцей эта мышца превращается в быструю (фазиче-
скую). Подобным же образом медленные (тонические) мотонейро-
ны превращают фазическую мышцу в тоническую (Buller et al.,
1960). Впрочем, выяснилось, что перекрестная иннервация не
полностью переводит мышцу из одного типа в другой, а создает
некую смешанную форму (Buller, Lewis, 1964).
В мышцах рыб тоже есть фазические и тонические волокна.
У пелагических рыб, например скумбрии и тунца, которые непре-
рывно плавают с относительно малой скоростью, волокна двух
типов обособлены в разных мышечных массах, резко различаю-
щихся по внешнему виду. Тоническая мускулатура имеет темно-
красную окраску из-за высокой концентрации миоглобина. Ее
слой залегает вдоль боковой линии и уходит вглубь до позвоноч-
ника. Непрерывное плавание всецело осуществляется с помощью
этой красной мускулатуры; вся масса белой мускулатуры (фази-
ческого типа) служит резервной силой для коротких вспышек
быстрой двигательной активности (рис. 11.15).
Рис. 11.15. У полосатого тунца (А)
красная (тоническая) плаватель-
ная мускулатура, используемая
при длительном плавании, распо-
ложена вдоль боковой линии меж-
ду кожой и позвоночником (на
схеме Б слегка затушевана).
Остальная обширная масса белой
(быстрой) мускулатуры исполь-
зуется при быстрых бросках.
(Rayner, Kennan, 1967.)
Плавательные мышцы акул тоже подразделяются на два по-
добных типа, причем тонические волокна образуют менее 20%
мышечной массы. И у них тонические волокна используются при
обычном передвижении, а фазические — при энергичных бросках,
например для преследования добычи. Фазические мышцы спо-
собны развивать высокую анаэробную мощность, чего и следует
Движение и мышца 585
ожидать при максимальной активности, и соответственно содер-
жат много гликогена. После усиленной активности количество
гликогена в быстрых мышцах заметно уменьшается. Между тем
в течение долгих периодов медленного плавания уровень глико-
гена в них остается постоянным; это говорит о том, что при
обычном плавании фазическая мускулатура не используется (Bo-
ne, 1966).
ГЛАДКИЕ МЫШЦЫ
В отличие от поперечнополосатых мышц (у позвоночных это
скелетные и сердечная мышцы) мышцы внутренних органов глад-
кие (иначе их называют неисчерченными или непроизвольными).
У позвоночных животных мускулатура этого типа имеется в стен-
ках желудка, кишечника, мочевого пузыря, мочеточников, матки,
бронхов, кровеносных сосудов и т. д. Гладкие мышцы встречаются
также у множества беспозвоночных, но у них они не образуют
отчетливых функциональных групп.
Гладкая мускулатура позвоночного животного иннервируется
двумя группами нервов вегетативной (автономной) нервной си-
стемы. Одна группа стимулирует сокращение, вторая тормозит
(иначе говоря, эти две группы являются антагонистами). Гладкие
мышцы отличаются от произвольных тем, что гладкомышечные
слои, находящиеся главным образом в стенках полых органов,
содержат множество нервных волокон и нервных клеток. Это за-
трудняло проведение экспериментов с изолированными гладкими
мышцами, так как при нанесении стимула трудно отделить его
прямое воздействие на мышечные клетки от действия на нервные
клетки. Еще одна трудность при изучении гладких мышц связана
с тем, что их волокна не образуют хорошо оформленных пучков;
они здесь тоньше и короче, чем в поперечнополосатой мышце,
и часто идут во многих направлениях.
Для сокращения гладкой мышцы не требуется стимуляции ее
нервами. Она способна к спонтанным ритмическим сокращениям,
которые могут сильно варьировать и по частоте, и по интенсив-
ности. Заложенные в мышце нейроны и нервные волокна иногда
существенно влияют на эту активность, но детали такого взаимо-
действия еще предстоит выяснить.
Внезапное растяжение гладкой мышцы немедленно вызывает
сокращение. Поэтому за растяжением полого органа часто сле-
дует сокращение. Но изолированный лоскут гладкой мускулату-
ры ведет себя несколько иначе. Под действием ’ незначительной
нагрузки он медленно растягивается и может при одной и той же
нагрузке принимать в разное время разную длину. Таким обра-
зом, гладкая мышца не имеет определенной длины покоя.
Для гладкой мускулатуры позвоночных типична медленная
реакция. Ее самое характерное свойство — то, что она способна
585 Гл. 11. Мышцы, движение, локомоция
поддерживать сокращение долгое время с очень малой затратой
энергии. Способность находиться в состоянии слабого сокраще-
ния, или тонуса, при самой разной длине очень существенна для
экономии энергии при длительном напряжении, поскольку энерге-
тические затраты на его поддержание, как правило, находятся в
обратной зависимости от быстроты сокращения (Riiegg, 1971).
В гладкой мышце поперечной исчерченности не видно потому,
что миофиламенты, которые, вероятно, в ней имеются, не рас-
положены так регулярно, чтобы получались видимые полосы.
Гладкие мышцы позвоночных содержат тонкие (актиновые) фи-
ламенты, но, хотя в ней есть также миозин, до сих пор не удава-
лось обнаружить толстых филаментов вроде тех, какие имеются
в поперечнополосатой мышце.
Многие мышцы беспозвоночных животных гладкие. Один из
их видов, который будет описан ниже, — это медленная замыка-
тельная мышца двустворчатых моллюсков; она не только закры-
вает створки раковины, но и может оставаться сокращенной ча-
сы и дни. Однако не все гладкие мышцы моллюсков сокращаются
медленно; мускулатура мантии осьминога и кальмара, ответствен-
ная за реактивное движение этих животных в воде, сокращается
быстро. Ее полное сокращение у интактного животного длится
0,1 или 0,2 с, что для гладкой мышцы действительно очень мало
(Trueman, Packard, 1968).
По строению и составу гладкие мышцы у беспозвоночных не-
сколько иные, чем у позвоночных. Например, гладкая мышца
моллюска содержит белок тропомиозин А, который больше нигде
не встречается. Тем не менее основной механизм сокращения,
по-видимому, всегда связан со взаимным скольжением миофила-
ментов, происходящим одинаково и в гладких, и в поперечнопо-
лосатых мышцах всех животных.
ЗАМЫКАТЕЛЬНАЯ МЫШЦА МОЛЛЮСКОВ
Двустворчатые моллюски защищаются от врагов, закрывая
свою раковину. Одна или несколько мышц-аддукторов смыкают
створки и удерживают их в таком положении против пружиняще-
го действия замка. У видов, обитающих в литоральной зоне, ра-
ковина во время отлива должна быть плотно закрыта, и сокра-
щение мышцы длится часами; при этом в закрытой раковине, ве-
роятно, создаются полностью анаэробные условия. Если на мол-
люска нападает морская звезда, она присасывается своими ам-
булакральными ножками к обеим створкам раковины и пытается
разомкнуть их; замыкательный механизм помогает моллюску вы-
держать это испытание на выносливость.
У некоторых, но не у всех, видов замыкательные мышцы раз-
делены на две части; одна состоит из гладких, а вторая — из по-
перечнополосатых волокон. Функционально они различаются тем,.
Движение и мышца 587
что поперечнополосатая часть сокращается быстро, и ее называ-
ют быстрой или фазической; неисчерченная часть мышцы сокра-
щается гораздо медленнее и называется медленной или то-
нической. Длительное сокращение, позволяющее долго удер-
живать створки раковины закрытыми, производится главным
образом медленной частью замыкательной мышцы (Hoyle,
1964).
Высказано предположение, что замыкательные мышцы, уко-
ротившись, переходят в состояние «зацепления» (catch). Это
означает, что сокращение поддерживается без дальнейшей за-
траты энергии — иными словами, фаза расслабления невероятно
растянута и может длиться часами. Вопрос о том, каким обра-
зом замыкательная мышца сохраняет такое состояние, разрешен
только отчасти, и соответствующий механизм еще во многом не-
ясен.
Гипотезы, объясняющие это состояние мышцы, в основном под-
разделяются на две группы. В одних предполагается изменение
на молекулярном уровне, которое препятствует разрыву попереч-
ных мостиков, образовавшихся между миофиламентами. Согласно
другим гипотезам, требуется многократная реактивация сократи-
тельного аппарата нервной системой или внутренними мышечны-
ми потенциалами действия.
Из мышц замыкательного типа лучше всего изучен передний
ретрактор биссуса обыкновенной мидии (Mytilus edulis), содер-
жащий только гладкие волокна. Его сокращение можно вызывать
электрическими стимулами, но в зависимости от характера стиму-
ла либо через несколько секунд наступает полное расслабление,
либо сокращение длится минутами и часами (Johnson, Twarog,
1960).
Поддерживать состояние напряжения без затраты энергии
было бы особенно выгодно для животного в анаэробных услови-
ях. Потребление кислорода передним ретрактором биссуса слиш-
ком мало для того, чтобы его можно было обнаружить в виде
повышенного потребления О2 целым животным, отчасти потому,
что трудно исключить участие анаэробных процессов и такие ис-
точники энергии, как высокоэнергетические фосфатные соедине-
ния.
Потребление кислорода изолированным передним ретрактором
биссуса медленно снижается во времени — приблизительно так
же, как уменьшается напряжение мышцы. Во время сокращения
этой мышцы расход кислорода падает при уменьшении внешней
нагрузки на мышцу и снова растет, если нагрузку опять увели-
чить путем растяжения мышцы до ее исходной длины. Это под-
тверждает, что потребление кислорода мышцей в состоянии «за-
цепления» определяется ее напряжением. Измерение дополни-
тельного расхода О2 (выше уровня покоя) выявляет его линей-
ную зависимость от напряжения мышцы (рис. 11.16).
588 Гл. И. Мышцы, движение, локомоция
Эти данные согласуются с гипотезой, согласно которой тони-
ческое напряжение поддерживается в «запертой» замыкательной
мышце как активное состояние, для которого нужна метаболиче-
ская энергия. Впрочем, благодаря присущей этой мышце медлен-
ности действия энергия, необходимая для поддержания сокраще-
Рис. 11.16. Добавочное потребление кислорода (выше уровня покоя) при сокра-
щении замыкательной мышцы обыкновенной мидии (Mytilus). Усиленное дыха-
ние находится в линейной зависимости от напряжения мышцы и, по-видимому,
не зависит от длительности сокращения. (Baguet, Gillis, 1968.)
ния, более чем в 1000 раз меньше той, какая нужна для под-
держания тетанических сокращений быстрой мышцы позвоночно-
го животного.
МЫШЦЫ РАКООБРАЗНЫХ
У мышц ракообразных выражены в чистом виде некоторые
свойства, в разной степени присущие мышцам ряда других жи-
вотных. Мы рассмотрим два момента: 1) параллельное или пери-
стое расположение волокон в мышце; 2) множественную иннер-
вацию отдельного мышечного волокна и его ответы на импуль-
сы с разных нервных волокон.
Почти всякий, кто держал в руках живых крабов или ома-
ров, знаком с внушительной силой, развиваемой их клешнями.
Дело здесь не в какой-то необычайной силе мышцы, а скорее в
анатомическом расположении волокон. Мышца в клешне краба
имеет перистое строение; волокна идут не параллельно направле-
Движение и мышца 589
нию создаваемой ими внешней силы, а под углом, что дает им
большое механическое преимущество (рис. 11.17).
В замкнутом внутреннем пространстве клешни перистая мыш-
ца данного объема может иметь более короткие волокна и боль-
шее их число, чем мышца с параллельными волокнами. Увеличе-
ние силы благодаря такому механическому устройству достига-
Рис. 11.17. Расположение мышц в клешне краба или омара создает для них ме-
ханическое преимущество. Перистое строение позволяет также мышце утолщаться
во время сокращения, что иначе было бы невозможно в ограниченном простран-
стве клешни. (Alexander, 1968.) '
ется за счет расстояния, на которое может быть передвинута точ-
ка прикрепления мышцы. Аддуктор клешни краба благодаря пе-
ристому строению способен развивать силу приблизительно вдвое
большую, чем если бы он состоял из параллельных волокон (Ale-
xander, 1968).
Второе преимущество перистой мышцы в клешне краба свя-
зано с ограниченным пространством, в котором она работает.
Когда сокращается мышца с параллельными волокнами, ее попе-
речное сечение увеличивается, и в ^кестком панцире клешни это
создавало бы трудности. Между тем перистая мышца благодаря
своему строению может почти целиком заполнять полость клеш-
ни, так как при сокращении волокок угол, под которым они ле-
жат, изменяется таким образом, что создается пространство для
их утолщения. Для мышцы с параллельными волокнами в огра-
ниченном пространстве это невозможно.
Перистая мышца может совершать лишь такую же работу, что
и мышца равного объема с параллельными волокнами, так как в
590 Гл. 11. Мышцы, движение, локомоция
противовес большей силе сокращения уменьшается укорочение.
Следует отметить, что перистые мышцы есть не только у рако-
образных; они достаточно часто встречаются и у позвоночных.
Наибольший интерес в мышце ракообразного представляет ее
множественная иннервация: к одному мышечному волокну могут
подходить два и больше нервных волокон. Кроме множественной
иннервации, вызывающей сокращение мышцы, многие мышцы
волокна
Медленное
Тормозное
Ракообразные
Мышечные
Фазическое Тоническое
Позвоночные
Рис. 11.18. Мышца ракообразного иннервируется несколькими нейронами, нерв-
ные волокна широко ветвятся и снабжают своими окончаниями всю мышцу
(вверху слева). Мышца позвоночного иннервируется множеством спинномозговых
нейронов, каждый из которых снабжает нервными окончаниями сравнительно
малое число мышечных волокон (вверху справа). Мышечное волокно ракообраз-
ного может иннервироваться тремя разными нервными волокнами: одним для
быстрого, вторым — для медленного сокращения и третьим — для торможения
(внизу слева).
членистоногих снабжены также тормозными нервами, стимуляция
которых вызывает расслабление мышцы, если она находилась в
сокращенном состоянии.
Другая особенность ракообразных состоит в том, что вся мыш-
ца часто иннервируется всего лишь несколькими или даже од-
ним аксоном. Благодаря этому она действует как единое целое,
а градации ее ответов создаются изменением нервной импульса-
ции в сочетании с балансом между возбуждающими и тормозны-
ми импульсами. Поэтому одна и та же мышца в зависимости от
получаемой импульсации может действовать то как медленная,
то как быстрая.
Различие между иннервацией мышцы позвоночного и ракооб-
разного показано на рис. 11.18. Как уже говорилось, самая пора-
зительная особенность мышцы ракообразного — то, что она ин-
нервируется двумя или большим числом разных нейронов. Часто
к ней подходят одновременно быстрое возбудительное волокно,
медленное возбудительное и, наконец, тормозное. Во многих
мышцах большинство волокон иннервируется быстрым нервом,
а меньшее их число — медленным. В сочетании с тормозными во-
Движение и мышца 591
локнами это создает возможность градации сокращений в широ-
ких пределах. Вторая особенность состоит в том, что вся мышца
обычно иннервируется очень малым числом центральных нейро-
нов, и соответственно к ней подходит очень мало аксонов, кото-
рые ветвятся и идут ко всем волокнам мышцы.
Напротив, мышца позвоночного животного иннервируется мно-
жеством центральных нейронов, и каждое нервное волокно под-
ходит лишь к небольшому числу волокон всей мышцы. В быстрой
(фазической) мышце позвоночного градация сокращения зависит
от числа аксонов, по которым^ в данный момент приходят воз-
буждающие импульсы к соответствующим мелким группахМ мы-
шечных волокон. В мышцах позвоночных торможение отсутству-
ет; у них оно происходит на уровне центральных нейронов, а не
на уровне самой мышцы (Hoyle, 1957; Wiersma, 1961).
ЛЕТАТЕЛЬНАЯ МУСКУЛАТУРА НАСЕКОМЫХ
У многих насекомых — стрекоз, бабочек, саранчи — взмахи
крыльев повторяются со сравнительно малой частотой, и каж-
дое сокращение летательных мышц происходит в ответ на нерв-
ный импульс. Поскольку сокращение синхронизировано с нервным
импульсом, мышцы такого типа называются синхронными.
У многих мелких насекомых — пчел, мух, комаров — крылья
совершают от 100 до 1000 и более взмахов в секунду (Sotavalta,
1953). Такая частота слишком высока, чтобы для каждого сокра-
щения к мышце мог прийти отдельный нервный импульс, оказать
свое действие и угаснуть до прихода следующего импульса. По-
этому для таких частых сокращений мышцьг должны возбуж-
даться иным механизмом. Эти быстрые мышцы тоже снабжены
нервами, но нервные импульсы приходят к ник с меньшей ча-
стотой, чем происходят сокращения, и такие мышцы называются
асинхронными. Мы находим их у представителей четырех отрядов
насекомых: мух и комаров (Diptera), ос и пчел (Hymenoptera),
жуков (Coleoptera) и некоторых клопов (Hemiptera).
Первый удивительный факт состоит в том, что мышцы в груди
(тораксе) такого насекомого с быстрыми крыльями прикреплены
вовсе не к крыльям, а к стенке груди. Они образуют две группы,
которые для удобства можно назвать вертикальной и горизон-
тальной. Первичный результат мышечного сокращения состоит в
изменении формы груди, и поскольку укорочение мышцы при этом
очень невелико (лишь несколько процентов ее длины), оно мо-
жет происходить очень быстро. Но самая важная особенность
асинхронной мышцы — то, что она сокращается в ответ на рас-
тяжение.
При сокращении вертикальных мышц грудь мгновенно, са
«щелчком», изменяет свою форму, и в результате этого с сокра-
щающихся вертикальных мышц внезапно. снимается напряжение
592 Гл. И. Мышцы', движение, локомоция
(рис. 11.19). Но это внезапное изменение формы груди ведет к
растяжению горизонтальной группы летательных мышц, которое
в свою очередь служит стимулом к их сокращению. Сокращение
горизонтальных мышц изменяет форму груди в противоположном
направлении, и она опять-таки «щелчком» возвращается в ис-
Рис. 11.19. У некоторых насекомых летательные мышцы образуют вместе с эла-
стичной грудью осцил/ирующую систему, в которой частота взмахов может быть
Намного больше част/ты нервных импульсов. Подробности см. в тексте.
ходное состояние, снимая напряжение с горизонтальных мышц.
Это ведет к j/астяжению вертикальных мышц, которые реагируют
на него новь/м сокращением, и т. д.
Эластичная грудь с ее мышцами представляет собой механи-
ческий осциллятор, который благодаря притоку энергии от мышц
совершает колебания, не затухающие под действием сил трения.
Разумеется, наибольшая из этих демпфирующих сил связана с
сопротивлением, которое встречают движения крыльев, толкаю-
щие насекомое вперед, и большая часть мышечной работы тра-
тится на преодоление этой силы. Полет продолжается несмотря
на то,/что нервные импульсы поступают с интервалами, соответ-
ствующими нескольким мышечным сокращениям. В некоторых
случаях на каждый нервный импульс было зарегистрировано по
40 взмахов крыльев. Однако нервные импульсы нужны и для
инициации полета, и для поддержания длительной работы крыль-
ев (Pringle,. 1949; 1957).
Скелетные структуры 59£
СКЕЛЕТНЫЕ СТРУКТУРЫ
Для того чтобы мышцы могли использовать свою силу, они
должны быть прикреплены к какой-нибудь механической струк-
туре; они бесполезны для животного, если ничего не тянут. Со-
ответствующей механической структурой для передачи силы слу-
жит скелет. Обычно он представляет собой жесткое образование
с сочленениями, хотя жидкость тоже может быть использована
для передачи силы — подобно тому, как гидравлическая система
используется в автомобиле. Существуют следующие основные ти-
пы скелетных структур:
Жесткий скелет
Внутренний скелет, или эндоскелет (у позвоночных)
Наружный скелет, или экзоскелет (у насекомых, ракообраз-
ных)
Гидравлический скелет
Жидкость+мягкие стенки (у червей, осьминогов, морских
звезд)
Жидкость+твердые части (в ногах пауков)
Позвоночные обладают твердым внутренним скелетом — ко-
стями; насекомое или ракообразное использует свою жесткую ку-
тикулу в качестве наружного скелета, У некоторых животных (на-
пример, у дождевых червей) для передачи силы служат внутрен-
ние жидкости; это гидравлические скелеты.
ЖЕСТКИЕ СКЕЛЕТЫ
У позвоночного животного большая часть мягких тканей окру-
жает скелет снаружи. Наилучшее ли это устройство? У членисто-
ногих скелет образует поверхность тела, и все мягкие части, та-
ким образом, хорошо защищены. Не лучше ли такое устройство?
Сравним оба вида структур. В силу простой механики полая
цилиндрическая трубка выдерживает гораздо больший вес, не про-
гибаясь, чем сплошной цилиндрический стержень, сделанный из
такого же количества материала. Поэтому, когда для животного
возможный вес скелета лимитирован, выгоднее построить из дан-
ного материала трубку. Именно такую стратегию избрали члени-
стоногие, и в этом отношении они имеют преимущество перед
позвоночными.
Почему же в таком случае позвоночные не используют подоб-
ное устройство? Во-первых, при постоянном весе материала жест-
кость полого цилиндра по мере увеличения его радиуса очень
быстро уменьшается, и в конце концов стенки будут такими тон-
кими, что вся структура сомнется. Во избежание прогиба гипо-
тетического экзоскелета очень крупного животного пришлось бы
увеличить его толщину, а тем самым и вес. У экзоскелета есть
38—1863
>594 Гл. И. Мышцы, движение, локомоция
еще один механический недостаток: даже если он достаточно про-
чен для того, чтобы служить опорой, он очень чувствителен к
ударам — возникает опасность прогиба или пробоины. Эта опас-
ность, несущественная для таких мелких животных, как насеко-
мые, быстро возрастает с увеличением размеров тела.
Еще одна трудность при наличии экзоскелета связана с ро-
стом животного. Большинство взрослых насекомых не растет; в
течение всей жизни они сохраняют одни и те же размеры и же-
сткую форму, а на стадии растущей личинки снабжены более
мягкой кутиколой, способной растягиваться. Но и такую кути-
кулу личинке приходится время от времени сбрасывать, чтобы в
достаточной степени увеличиваться в размерах.
Водные членистоногие, периодически линяющие, растут в те
периоды, когда старый экзоскелет сброшен, а новый еще мягок и
растяжим. Если не считать беззащитности и невозможности дви-
гаться во время этого периода, механическая невыгода такой вре-
менной утраты жесткости для водного животного не слишком ве-
лика, так как вес его тела поддерживается водой. Для наземного
животного эта проблема была бы невероятно трудной: тело лю-
бого сколько-нибудь крупного животного «расплылось» бы под
собственной тяжестью, если бы ему пришлось проходить процесс
линьки.
Тем не менее некоторые защитные достоинства экзоскелета
все же используются и у позвоночных. Такой жизненно важный
орган, как головной мозг, заключен в жесткую костную структу-
ру. Однако черепная коробка не составляет части локомоторного
скелета.
Одно из преимуществ экзоскелета заключается в его упругих
свойствах. Эластическая деформация скелетных элементов со-
здает запас энергии, используемый для локомоции; примером
могут служить полет насекомых (рассмотренный выше) и прыж-
ки блох (см. ниже).
ГИДРАВЛИЧЕСКИЕ СКЕЛЕТЫ
Гидравлические скелеты в одном отношении существенно от-
личаются от жестких. Жесткий скелет состоит из элементов, кото-
рые обычно подвижны относительно некоторой точки (сочлене-
ния). Мышцы прикреплены по обе стороны от этой точки, и раз-
виваемая ими сила передается через жесткий элемент и действует
в каком-то другом месте. А для того, чтобы мог работать* гидрав-
лический скелет, жидкость должна быть заключена в ограничен-
ное пространство и мышечная сила должна использоваться для
создания давления в этой жидкости (т. е. мышца обычно окружа-
ет пространство с жидкостью кольцевыми слоями). В этом случае
мышца ни к чему не прикреплена; мышечное кольцо тянет толь-
ко само себя.
Скелетные структуры 595
ДОЖДЕВЫЕ ЧЕРВИ
Роль гидравлического скелета в локомоции червей подробно
изучена. Хорошо известным примером служит обыкновенный дож-
девой червь. Стенка его тела включает мышечные слои, в кото-
рых волокна идут в двух разных направлениях — продольно и
кольцами вокруг тела.
Движение ползущего вперед дождевого червя происходит при-
мерно следующим образом. На переднем конце начинается волна
сокращения кольцевых мышц. В этом месте тело становится тонь-
ше и длиннее, и передний конец проталкивается вперед. Кольце-
вое сокращение проходит вдоль тела как волна, за которой следу-
ет волна сокращения продольных мышц. Мелкие щетинки на бо-
ках тела не позволяют ему скользить назад, а передний конец
подтягивает ближайшую область к себе. Таким образом какой-то
участок тела продвигается вперед, и волна движения перемеща-
Рис. 11.20. Дождевой червь при ползании использует жидкости тела в качестве
внутреннего гидравлического скелета. Сокращение кольцевых мышц проталки-
вает передний конец вперед, после чего сокращаются продольные мышцы, и это
ведет к утолщению тела. Утолщенные сегменты не смещаются относительно
почвы, пока другие части продвигаются вперед. Путь, проходимый отдельными
точками тела червя, и их движение относительно друг друга показаны на диа-
грамме линиями, идущими наискось слева направо. Диаграмма построена по ки-
нокадрам. (Gray, Lissmann, 1938.)
ется назад вслед за волной кольцевого сокращения. Цикл повто-
ряется, снова происходит удлинение переднего конца, и по мере
того, как волны сокращения идут назад, червь движется вперед
(рис. 11.20).
При движении дождевого червя сила его мышц воздействует
на жидкое внутреннее содержимое тела; кольцевые мышцы про-
изводят давление, которое заставляет продольные мышцы растя-
38
596 Гл. 11. Мышцы, движение, локомоция
гиваться, а когда в свою очередь сокращаются продольные, дав-
ление растягивает кольцевые мышцы, и червь утолщается.
Тело дождевого червя разделено на части поперечными пере-
городками. Это очень помогает локомоции, так как делает раз-
личные участки относительно независимыми. У многих червей, на-
пример у пескожила (Arenicola), таких перегородок нет (Seymo-
ur, 1971). Одна из невыгод их отсутствия состоит в том, что в слу-
чае ранения жидкость тела вытекает и червь практически не спо-
собен двигаться. И наоборот, как известно почти каждому, если
разрезать пополам дождевого червя, то обе части передвигаются
почти так же, как целый червь.
ноги ПАУКОВ
Пауки не могут выпрямлять ноги при помощи мышц, так как
у них нет мышц-разгибателей. Для сгибания ног служат мышцы-
сгибатели, но как ноги разгибаются? Есть две возможности: ис-
пользование упругих сил и гидравлических сил.
Здесь могли бы действовать эластичные шарнирные суставы,
устроенные так, чтобы ноги всегда выпрямлялись сами при рас-
слаблении сгибателей. Это было бы похоже на шарнирное устрой-
ство у двустворчатых моллюсков, у которых раковины раскрыва-
ются при расслаблении замыкательной мышцы. Но такую воз-
можность исключает одно простое наблюдение: суставы изолиро-
ванной ноги паука сохраняют приданное им положение в боль-
шом диапазоне углов сгибания.
Пауки используют второй возможный механизм — гидравли-
ческий. Это можно доказать несколькими способами. Проще все-
го отделить ногу и сдавить ее около места разреза пинцетом; тог-
да отверстие будет зажато, давление внутренней жидкости повы-
сится и в результате ножка разогнется. Кроме того, измерения по-
казывают, что существует прямая зависимость между внутрен-
ним давлением в ноге и углом поворота ее суставов. Далее, если
паук ранен и при этом потерял много крови, он совершенно не
в состоянии разгибать ноги, которые остаются поджатыми к те-
лу. Кровяное давление у пауков поразительно высокое — до
400 мм рт. ст., или около 0,5 атм (~50 кПа). Оно очень сильно
отличается от кровяного давления у других членистоногих, кото-
рое обычно равно лишь нескольким миллиметрам ртутного стол-
ба (Раггу, Brown, 1959а).
Некоторые мелкие пауки могут прыгать на жертву с расстоя-
ния более 10 см (фото 11.4). Для прыжка они производят вне-
запное разгибание одной пары ножек. Могут ли они использо-
вать для этого очень быстрого движения гидравлическое давле-
ние?
Сам по себе прыжок всецело осуществляется за счет резкого
распрямления последней (четвертой) пары ножек. Крутящий мо-
мент, необходимый для прыжка, можно вычислить из длины прыж-
Скелетные структуры 597
ка; оказалось, что нужные гидравлические силы, которые у пры-
гающего паука трудно измерить непосредственно (он весит всего
лишь около 10 мг), лежат в пределах величин кровяного давле-
ния, часто встречающихся у пауков. Рапидная съемка прыгаю-
Фото 11.4. У прыгающего паука Sitticus pubescens, как и у других пауков, ноги
лишены мышц-разгибателей. Когда паук прыгает на добычу, он пользуется зад-
ней парой ножек, которые выпрямляются под действием давления крови. (Фото
предоставлено G. A. Parry, Cambridge University.)
щих пауков подтверждает предположение об участии в прыжке
кровяного давления, так как в момент прыжка мельчайшие ще-
тинки на ногах выпрямляются совершенно так же, как при по-
вышении давления внутренней жидкости; но механизм этого по,-
вышения с точностью не известен (Parry, Brown, 1956 b).
БИОМЕХАНИКА ЛОКОМОЦИИ
Главное назначение скелетных мышц — доставлять механиче-
скую энергию для передвижения животных. Животные ходят,
бегают и прыгают, летают по воздуху и плавают в воде. Если не
598 Гл. 11. Мышцы, движение, локомоция
говорить о перемещении с помощью псевдоподий, ресничек и
жгутиков, а также о небольшом числе необычных способов пе-
редвижения, в локомоции животных всегда участвуют мышцы.
В главе 6 была рассмотрена потребность в энергии для передви-
жения, а сейчас речь пойдет о механических аспектах этой про-
блемы.
БЕГ
Животное, бегущее по горизонтальной поверхности, затрачи-
вает энергию в связи с тремя главными обстоятельствами:
1. При движении в воздушной среде приходится преодолевать
сопротивление воздуха.
2. Когда животное бежит, его центр тяжести непрерывно при-
поднимается и снова опускается. При каждом подъеме соверша-
ется работа, увеличивающая потенциальную энергию тела, и
большая часть энергии при опускании центра тяжести теряется.
3. Во время бега движение конечностей все время то ускоря-
ется, то вновь замедляется. Для ускорения, т. е. увеличения кине-
тической энергии, требуется работа; для замедления — тоже.
Затрата механической энергии на преодоление трения в су-
ставах очень мала, но некоторое ее количество расходуется на
усиленные дыхательные движения и дополнительную работу
сердца. Однако и это количество очень невелико по сравнению с
тем, которое требуется для механических движений тела.
Сопротивление воздуха можно вычислить на основании зако-
нов аэродинамики или найти путем прямых измерений давления
ветра на объекты, сходные по форме и величине с бегущим жи-
вотным. Сопротивление можно также оценить по повышению по-
требления кислорода во время бега при разной скорости ветра.
Результаты показывают, что для людей затраты энергии на пре-
одоление сопротивления воздуха при беге по треку на среднюю
дистанцию составляет до 7,5% общего расхода энергии. Можно
думать, что сопротивление воздуха возрастает приблизительно
пропорционально квадрату скорости, и при спринте такая затра-
та может составить 13% всего расхода энергии на бег. При бе-
ге с малой скоростью или ходьбе сопротивление воздуха ничтож-
но. Установлено, что при беге позади другого бегуна на расстоя-
нии 1 м от него сопротивления воздуха практически нет (Pugh,
1971). Для других млекопитающих, кроме тех, которые бегают
с исключительно большой скоростью, сопротивление воздуха не
имеет существенного значения для общего расхода энергии.
Трением о твердый субстрат тоже можно пренебречь (боль-
шинство из нас знает по собственному опыту, что совсем другое
дело — ходьба по рыхлому песку). Таким образом, работа, со-
вершаемая против внешней среды (преодоление сопротивления
воздуха и трения о почву) составляет лишь небольшую долю об-
Биомеханика локомоции 599
щих затрат, и поэтому следует заключить, что большая часть
энергии, расходуемой на бег, рассеивается внутри тела в форме
тепла. (В гл. 6 мы упоминали, что с полетом птицы дело обстоит
иначе, так как у нее, вероятно, 25% энергии расходуется на ра-
боту, совершаемую против внешней среды.)
БЕГ В ГОРУ
При беге вверх требуется дополнительная энергия. Тело, дви-
жущееся с подъемом вверх, приобретает потенциальную энергию,
которая равна произведению его веса на вертикальную состав-
ляющую его пути. Поэтому можно предсказать, что энергия, не-
обходимая для перемещения единицы веса тела на единицу вер-
тикального пути, должна быть одинаковой для животных самой
различной величины.
Подъем 1 кг веса тела на 1 м вертикали увеличивает его по-
тенциальную энергию на 1 кг/м (что равно 9,8 Дж, или 2,34 кал,
и соответствует потреблению 0,49 мл О2). Если коэффициент по-
лезного действия мышц у крупных и мелких животных одинаков и
равен, скажем, 20%, то они использовали бы в пять раз боль-
ше О2. Это предполагаемое сходство относительных метаболиче-
ских затрат на вертикальное движение в достаточной мере под-
тверждено многими исследованиями на животных разной величи-
ны— от мыши до лошади.
Такое сходство затрат на вертикальное движение ведет к од-
ному интересному следствию: маленькому животному должно
быть гораздо легче бежать вверх, чем крупному. Почему?
Ответ прост. Нормальная удельная интенсивность метаболиз-
ма у 30-граммовой мыши примерно в 15 раз больше, чем у 1000-
килограммовой лошади. Поскольку вертикальная составляющая
передвижения единицы веса тела для обоих животных од-
на и та же, относительное усиление обмена, связанное с подъе-
мом, у мыши должно быть в пятнадцать раз меньше, чем у ло-
шади.
Этот удивительный вывод экспериментально был проверен
Тейлором и др. (Taylor et al., 1972). Мышей обучали бегать по
тредбану с наклоном в 15°, т. е. с весьма крутым подъемом. Ре-
зультаты не показали статистически значимого различия в по-
треблении кислорода между мышами, бегущими вверх, по гори-
зонтали и вниз. Между тем у шимпанзе (с весом тела 17 кг) по-
требление О2 при подъеме почти удваивалось по сравнению с бе-
гом по горизонтали. А для лошади это увеличение должно быть
*еще больше.
Тейлор пришел к выводу, что вертикальное передвижение со
^скоростью 2 км/ч требует прироста потребления кислорода на
.23% (едва заметного) у мыши, на 189% (почти втрое) у шим-
панзе и на 630% у лошади (как при тяжелой работе). Это объ-
600 Гл. 11. Мышцы, движение, локомоция
ясняет ту легкость, с которой белка бегает вверх и вниз по стволу
дерева без видимого усилия. Такому маленькому животному поч-
ти безразлично, вверх бежать или вниз.
ЭНЕРГИЯ, СБЕРЕГАЕМАЯ УПРУГОСТЬЮ
Теннисный мяч, упавший на твердую поверхность, подпрыги-
вает много раз, но высота отскока постепенно снижается. Когда
мяч ударяется о поверхность, его кинетическая энергия расходу-
ется на упругую деформацию, а затем снова освобождается, и
мяч подскакивает. Пока он летит вверх, его кинетическая энергия
переходит в потенциальную, но лишь для того, чтобы снова пе-
рейти в кинетическую при снижении. Энергия постепенно теря-
ется из-за трения о воздух, а также внутреннего трения во время
эластической деформации мяча. Однако небольшого притока энер-
гии, такого, который лишь едва превысил бы потери от трения,
достаточно, чтобы мяч продолжал подскакивать.
Есть ли здесь какое-либо сходство с локомоцией животного?
Как мы видели, много энергии уходит на ускорение и замедление
движений конечностей и на перемещение центра тяжести в поле
земного тяготения. Если бы какие-то упругие элементы могли
действовать как пружины и запасать часть кинетической энергии,
то это дало бы большую экономию.
Эластичные элементы, требующие рассмотрения, — это сухо-
жилия и мышцы. На первый взгляд может показаться странным,
что мышцы можно считать упругими телами, поскольку их глав-
ная функция — сокращаться и развивать силу. Но посмотрим на
бегущего человека в тот момент, когда его нога касается земли.
Если бы мышцы ног были вялыми и расслабленными, человек
упал бы; очевидно, они должны сопротивляться растяжению и,
следовательно, быть сокращены еще до интересующего нас мо-
мента. Кинетическая энергия тела создает силу для растяжения
мышц, а они сопротивляются, что требует работы. Может ли ка-
кая-то доля кинетической энергии поглощаться упругими эле-
ментами и затем освобождаться?
Наиболее важные исследования по этому вопросу провела
группа итальянских ученых в Милане, среди них Р. Маргариа и
Дж. Каванья, и полученные ими результаты очень много дали
для понимания энергетики локомоции.
Изменения потенциальной и кинетической энергии во время
бега и величину сил, действующих на опору, можно изучать с
помощью силовой платформы — устройства в принципе очень про-
стого. Человек или животное бежит по твердой платформе изве-
стной массы, подвешенной на очень тугих пружинах, и соответст-
вующие датчики регистрируют малейшие движения платформы в
трех измерениях. Зарегистрированные движения используются за-
тем для вычисления сил и их направления в течение всего период
Биомеханика локомоции 601
да, когда нога соприкасается с платформой. Из этих данных мож-
но вычислить всю механическую работу, необходимую для одного
шага. Оказалось, что работа на 1 км пути не зависит от скорости
бега, а общая механическая работа бегущего человека составля-
ет около 0,4—0,5 ккал/(кг-км) (Cavagna, 1964).
Одновременное измерение потребления кислорода показыва-
ет расход метаболической энергии. Расчет коэффициента полезного
действия (отношения полезной работы к затрате энергии) дает
величину около 50%. Такая цифра кажется совершенно непомер-
ной, так как все имеющиеся данные говорят о том, что к. п.д.
мышц при превращении химической энергии во внешнюю механи-
ческую работу не может превышать 25%. Но это расхождение
исчезает, если половина механической работы, зарегистрирован-
ной с помощью силовой платформы, совершалась за счет накоп-
ленной кинетической энергии, освобождавшейся при упругой от-
даче растянутых мышц ноги. Вывод о том, что запасание энер-
гии в форме упругой деформации играет важную роль в локомо-
ции человека и животных, был полностью подтвержден последую-
щими работами.
Каждый легко может сам убедиться в том, что упругая отда-
ча существенно сберегает энергию. Попробуйте из положения
стоя приседать, сильно сгибая колени; вы увидите, что поднимать-
ся гораздо легче, если выпрямлять ноги немедленно, чем если за-
держиваться на секунду в положении с согнутыми коленями,
прежде чем снова выпрямиться. При сгибании колен мышцы-раз-
гибатели напряжены и контролируют движение вниз. Если они
начнут тотчас снова сокращаться, то поможет упругая отдача; ес-
ли же позволить им расслабиться до подъема туловища, такой
отдачи не будет. Но реальна ли кажущаяся разница в усилии?
Чтобы выяснить это объективно, можно измерять потребление
кислорода. Когда упражнения со сгибанием колен выполняются
на платформе, можно точно измерить работу, совершаемую при
выпрямлении ног, и она окажется одинаковой при обоих упраж-
нениях. Разницы трудно было бы и ожидать, поскольку при каж-
дом распрямлении испытуемый поднимает одну и ту же массу
(свое тело) на одну и ту же высоту, и измерения подтверждают
этот вывод.
В серии опытов испытуемые совершали ровно 20 приседаний
в минуту, но в одном случае выпрямляясь немедленно после сги-
бания колен, а в другом — после полуторасекундной задержки,
причем общее число приседаний в минуту оставалось одним и тем
же (Thys et al., 1972). Результаты подтвердили субъективное впе-
чатление (табл. 11.4). Когда использовалась упругая отдача, по-
требление кислорода снижалось на 22%. Совершаемую механи-
ческую работу измеряли на платформе, что давало возможность
вычислить коэффициент полезного действия, который поразитель-
ным образом возрастал на 37%.
6^2 Гл. 11. Мышцы, движение, локомоция
Таблица 11.4
Потребление кислорода и механи-
ческий коэффициент полезного дей-
ствия при приседании. В одном
случае испытуемые делали короткую
паузу, сидя на корточках; в другом
случае использовали упругую отдачу,
немедленно выпрямляясь. (Thys et al.,
1972)
Потребление Коэффициент
кислорода, л/мин полезного действия
Без отдачи 1,89 0,19
С отдачей 1,49 0,26
Разность -22% 4-37%
Другое сходное упражнение состоит в стоянии на цыпочках с
выпрямленными неподвижными коленями и многократных неболь-
ших подскоках на обеих ногах. В этом упражнении эластическая
энергия накапливается в сокращенных икроножных мышцах, когда
они растягиваются. Таким образом, только часть положительной
работы по подъему тела совершается за счет энергии химических
процессов, происходящих в мышцах; другая часть производится за
счет эластической энергии, запасаемой в сокращенных мышцах.
Эта последняя составляет 60% всей энергии, и только 40% дают
обменные процессы (Thys et al., 1975).
Тот факт, что при упражнениях и локомоции мышцы часто ак-
тивно сокращаются сразу же после того, как они были растянуты
внешней силой, чрезвычайно важен. Опыты на изолированных
мышцах показывают, что при укорочении после растяжения мышца
развивает большую силу, чем в том случае, когда при той же ско-
рости и длине она укорачивается после изометрического сокраще-
ния. Согласно имеющимся данным, помимо эластических характе-
ристик мышцы возможны также некоторые изменения сократитель-
ного процесса, так как во время сокращения после растяжения из-
меняются показатели силы и скорости (Cavagna et al., 1968; Ca-
vagna, Citterio, 1974).
Иной подход применен при исследовании животных. Обученная
немецкая овчарка совершала с разбега длинный прыжок на три
метра через ряд барьеров. В том месте, где собака отрывалась от
опоры, была установлена силовая платформа, которая давала ин-
формацию о силах, развиваемых во время прыжка, и их направле-
нии. Нас будет интересовать здесь совместная работа икроножной
и подошвенной мышц — единственных мышц, сила которых дейст-
вует на голеностопный сустав (рис. 11.21) (Alexander, 1974). Эти
мышцы должны развивать силу в течение всего времени соприкос-
Биомеханика локомоции 603
новения задних лап с опорой. Из силы, действующей на платфор-
му, с учетом отношения плеч рычага, которым служит кость у го-
леностопного сустава, можно вычислить общую силу, развиваемую
обеими мышцами.
Рис. 11.21. Контурные изображения собаки, совершающей с разбега длинный
прыжок с силовой платформы. Цифры обозначают кинокадры, снятые с часто-
той 56 кадров в секунду. Стрелка на каждом контуре показывает направление
и величину результирующей силы для данного кадра. Точками помечены суставы,
использованные при вычислении силы. Справа внизу — контур скелета задней
ноги в момент отрыва, показывающий положение икроножной и подошвенной
мышц. (Alexander, 1974.)
На рис. 11.22 показаны изменения силы во время приземления
и при отрыве от опоры. На этом графике сила, равиваемая сов-
местно икроножной и подошвенной мышцами, отложена против из-
менения длины этих мышц, вычисленного по кинокадрам, снятым
во время прыжка. Числа на графике соответствуют кадрам пленки,
начиная с какого-то момента после соприкосновения задних лап с
платформой. Кадры 15 и 16 соответствуют периоду прироста силы
по мере растяжения мышц; затем мышцы снова укорачиваются
(17—20), и сила падает до нуля, когда задние ноги отрываются от
платформы.
Эта запись чрезвычайно интересна. При растяжении мышц над
ними совершается работа, равная силе, умноженной на расстояние.
В фазу отрыва от платформы эта работа возмещается, и поскольку
путь остается в точности тем же, работа (силахрасстояние) во
время отдачи должна быть точно такой же, как работа, произве*
денная над мышцей при растяжении. Таким образом, икроножная
604 Гл. 11. Мышцы, движение, локомоция
и подошвенная мышцы с их сухожилиями вели себя как пассивные
упругие тела. По степени растяжения сухожилий и их эластическим
свойствам (сила известна по данным регистрации) можно вычис-
лить, что большая часть энергии упругости, вероятно, накаплива-
ется в сухожилиях.
Рис. 11.22. Сила, развиваемая совместно икроножной и подошвенной мышцами
при прыжке собаки весом 35 кг, отнесенная к изменениям длины мышц. Цифры
соответствуют номерам кадров, как на рис. 11.21. Сила в 1 кН равна 102 кгс;
следовательно, максимальная сила приблизительно в 2 кН (кадр 16) составляет
около 200 кгс. (Alexander, 1974.)
Если эти мышцы не совершают работы, то что же переносит со-
баку через барьеры? Только частично этот перенос можно отнести
за счет энергии, накопленной во время пробега и освобождаемой
при упругой отдаче. Во время соприкосновения задних ног с поч-
вой укорачиваются и совершают работу мышцы-разгибатели, на-
ходящиеся на бедре. Масса этих мышц значительна, и они выпол-
няют немалую долю работы при длинном прыжке (Alexander,
1974).
Кенгуру прыгают больше других животных; что мы можем уз-
нать из этих прыжков? Когда кенгуру движутся медленно, исполь-
Биомеханика локомоции 605
зуя все четыре ноги и хвост, потребление кислорода у них круто
возрастает с увеличением скорости (рис. 11.23). Но при больших
скоростях, когда животные прыгают только на задних ногах, при
Длина шага, м Число шагов, мин-} Потребление кислорода, мл/(г-ч)
Рис. 11.23. Записи, полученные на двух кенгуру (весом 18 и 28 кг), движущихся
по тредбану. А — потребление кислорода; Б — частота шагов; В — длина шага.
Левая часть каждого графика относится к движению на всех четырех ногах
при участии хвоста («пятиногое движение»). Правая часть относится к прыж-
кам на мощных задних ногах, причем тяжелый хвост используется как балансир.
Пунктирная линия на графике А — потребление кислорода млекопитающим весом
18 кг, бегущим на четырех ногах. (Dawson, Taylor, 1973.)
606 Гл. 11. Мышцы, движение, локомоция
увеличении скорости от 8 до 22 км/ч (т. е. почти втрое) потребле-
ние кислорода больше не растет (Dawson, Taylor, 1973).
В связи с вопросом о роли эластичности интересно рассмотреть
частоту прыжков у кенгуру. Несмотря на трехкратное увеличение
скорости, число прыжков в минуту сильно не меняется; скорость
возрастает почти исключительно благодаря удлинению каждого
прыжка.
Что это значит? Если продолжительность одного прыжка не из-
меняется, то не должна изменяться и его высота, а только длина.
Иными словами, требуется увеличение не вертикальной силы,
я только вектора, направленного вперед, — для удлинения прыжка.
Более длинный прыжок означает увеличение кинетической энергии,
которой обладает тело животного в момент соприкосновения с зем-
лей, и если значительная часть этой кинетической энергии может
быть поглощена упругими элементами, то это даст большую эко-
номию энергии, нужной для увеличения скорости.
Измерения на силовых платформах и вычисление сил в длинном
ахилловом сухожилии кенгуру говорят о том, что экономия, свя-
занная с накоплением энергии эластичным сухожилием, достигает
40% той положительной работы, которую мышцы ног должны со-
вершать во время соприкосновения животного с землей (Alexander,
Vernon, 1975).
ЕЩЕ О ПРЫЖКАХ
Блохи и кузнечики совершают прыжки, которые иной раз в
50 раз превышают длину их тела, что дало повод для досужих вы-
числений: как далеко мог бы прыгнуть человек при таком же со-
отношении? В Северной Америке он запрыгнул бы на верхушку
небоскреба Эмпайр Стейт Билдинг, а во Франции — выше Эйфеле-
вой башни. Но лучше рассмотреть этот вопрос реалистично.
Предположим, что кузнечик весит 1 г и может с места прыгнуть
на 50 см — обе эти цифры вполне реальны. Но представим себе
более крупное животное такого же строения, линейные размеры
которого ровно в 10 раз больше. Какова будет в соответствующем
масштабе ожидаемая высота его прыжка? Масса этого животного
в 1000 раз больше, чем у кузнечика, но поперечное сечение его ска-
кательных мышц (которое определяет возможную силу мышцы)
увеличится только в 100 раз. Поэтому на единицу своей массы
крупное животное будет обладать лишь одной десятой силы для
ускорения тела.
Поскольку ускорение равно частному от деления силы на мас-
су, при отрыве от опоры оно тоже будет в десять раз меньшим,
чем у мелкого животного. Но поскольку все линейные размеры уве-
личены в 10 раз, ускорение будет продолжаться на всем «пути от-
рыва», который тоже в 10 раз больше, чем у мелкого животного
Поэтому в конечном итоге скорость при отрыве одинакова у обоих
Биомеханика локомоции 607
животных. После этого движение замедляется силой земного тяго-
тения. Поскольку и кинетическая энергия при отрыве, и замедляю-
щая сила земного притяжения пропорциональны массе тела, это
замедление позволит обоим животным подняться на одинаковую
высоту. (Здесь мы не учли дополнительную потерю скорости из-за
сопротивления воздуха, которое для такого малого объекта, как
блоха, становится существенным.)
Теперь можно сделать вывод, что если мышцы мелких и круп-
ных животных развивают одинаковую силу на единицу поперечного
сечения, то мелкое и крупное животные одинакового строения спо-
собны прыгать точно на одну и ту же высоту.
Рассмотрим теперь реальные прыжки разных животных
(табл. 11.5). (Речь будет идти только о прыжках с места, так как
при прыжке с разбега для увеличения высоты используется кине-
тическая энергия тела. Поэтому мы не учитываем мировые рекор-
ды по прыжкам в высоту для человека или для бегущей лошади,
в обоих случаях составляющие около 2 м.) При прыжке с места
человек может взять высоту около 1,6 м. Но его центр тяжести
поднимается на меньшую высоту, так как вначале он находится не
на уровне земли, а приблизительно на уровне 1 м (см. примечание
к табл. 11.5). Таким образом, человек и кузнечик смещают свой
центр тяжести вверх примерно на одно и то же расстояние, хотя
по своей массе они различаются в 10 000—100 000 раз.
Таблица 11.5
Механические характеристики прыжков с места у насекомых и человека.
Высота прыжков — величина одного и того же порядка, хотя массы тела
различаются более чем в 100 миллионов раз (Блоха так мала, что для нее имеет
значение сопротивление воздуха, и для достижения данной высоты исходное
ускорение у нее должно быть больше вычисленной величины 320 g)
Блоха PulexX} Жук-щел- кун Athousl) Саранча Schist осег- ccfi) Человек (Ното)1)
Масса тела 0,49 мг 40 МГ 3 Г 70 кг
Высота прыжка (А), см 20 30 59 60
Путь ускорения, см Скорость при отрыве (u = y/i-2g), 0,075 0,077 4 40
см/с Время ускорения (время до отрыва) 190 240 340 343
(/=2s/u), с 0,00079 0,00064 0,00235 0,233
Ускорение (а=и//), см/с2 Ускорение относительно ускорения си- 241 000 374 000 14 500 1471
лы тяжести, g ’) Bennet-Clark, Lucey, 1967. 2) Evans, 1972, 1973. 3) Hoyle, 1955; Alexander, 1968. 245 382 15 1,5
4) Оценка, основанная на том, что центр тяжести в начале прыжка опускается до 60 см
над землей, до момента отрыва ускоряется на пути 40 см (до высоты 100 см), , а затем
поднимается до высоты 160 см. Мировой рекорд по 165 см над землей. прыжкам с места составляет около
ФОБ Гл. 11. Мышцы, движение, локомоция
Разумеется, в действительности животные геометрически не
подобны друг другу. И тем не менее удивительно, насколько ре-
корды по прыжкам сходны у разных животных. То, что прыжки
человека и других млекопитающих не пропорциональны линейным
размерам тела и что это вообще было бы невозможно, объясняется
простыми законами физики.
Вывод о том, что сходные животные независимо от размеров
тела должны быть в состоянии прыгать на одинаковую высоту,
можно сформулировать еще следующим образом. Примем, что
мышцы, участвующие в прыжке, составляют одну и ту же часть
всей массы тела. Сила мышцы пропорциональна площади ее по-
перечного сечения, а укорочение пропорционально исходной длине.
Поперечное сечение, умноженное на длину, дает объем мышцы,
л затрата энергии на одно сокращение измеряется произведением
•силы на расстояние. Поэтому затрата энергии пропорциональна
мышечной массе, а значит, и массе тела. Поскольку при прыжке
происходит только одно сокращение участвующих в нем мышц, ра-
бота, производимая во время отрыва и используемая для ускоре-
ния,— одна и та же по отношению к массе тела. Из этого следует,
что все геометрически подобные друг другу животные с различной
массой должны прыгать на одну и ту же высоту, если только их
мышцы сокращаются с одинаковой силой.
Галаго. Если сказанное выше верно, то каким же образом не-
которые животные прыгают гораздо выше? Рекорд по прыжкам
<с места, пожалуй, принадлежит малому галаго — небольшому
тропическому примату, который весит около 250 г. В хорошо конт-
ролируемых условиях галаго прыгнул на 2,25 м (Hall-Craggs,
1965). Это в три раза выше прыжка с места у человека. Если
только мышцы галаго не обладают большей силой на 1 см2 попе-
речного сечения (что мало вероятно), такое достижение можно
объяснить только большей мышечной массой (большей энергией,
получаемой от сокращения при отрыве) и, возможно, более благо-
приятной в механическом отношении «конструкцией» конечностей.
Галаго действительно обладает крупными прыгательными мыш-
цами, составляющими почти 10% массы тела (Alexander, 1968),
т. е. почти вдвое больше, чем у человека. Если принять, что соче-
тание всех механических преимуществ у высокоспециализирован-
ного прыгающего животного может дать 50 %-ное увеличение
прыжка, то в комбинации с удвоением массы прыгательныхмышц
это даст трехкратное увеличение прыжка — это отнюдь не завы-
шенная приблизительная оценка. Такой прыжок с места внуши-
тельнее, чем прыжок прямокрылого, но следует помнить', что гала-
го— теплокровное животное, весьма приспособленное к длинным
прыжкам в своем естественном местообитании — в джунглях.
Блоха. Помимо того, что животные геометрически не совсем
подобны друг другу, мы сталкиваемся еще с одной трудностью.
Чем меньше животное, тем короче путь при отрыве, а поскольку
Биомеханика локомоции 609
скорость отрыва должна быть одинаковой для всех, то небольшое
животное должно гораздо сильнее ускорять массу своего тела. По-
скольку времени для отрыва намного меньше, мощность мышцы
соответственно должна быть больше (т. е. мышца должна сокра-
щаться очень быстро).
Для животного такой величины, как блоха, время отрыва со-
ставляет меньше 1 мс, а путь, на котором происходит ускорение,
Рис. 11.24. Жук-щелкун Athous haemorroidalis, лежащий на спине и готовый
к прыжку, с выступом, установленным на краю ямки в задней части тела.
(Evans, 1972.)
равен всего лишь 0,75 мм. Среднее ускорение во время отрыва пре-
вышает 2000 м/с2, что приблизительно соответствует 200 g
(Rothschild et al., 1972).
Но мышцы просто неспособны сокращаться так быстро. Как же
тогда блоха вообще может прыгать? Она использует принцип ка-
тапульты и запасает энергию в кусочке упругого материала (рези-
лина) у основания задних ног. Резилин — белок, очень сходный
по своим механическим свойствам с каучуком (Weis-Fogh, 1960;
Anderson, Weis-Fogh, 1964). Для сжатия резилина используются
относительно медленные мышцы, а когда включается освобождаю-
щий механизм, резилин возвращает всю энергию с к.п.д. около
100%. Таким образом, упругая отдача действует примерно так, как
в рогатке, и сообщает блохе необходимое большое ускорение.
Жук-щелкун, Если положить жука-щелкуна на спину на глад-
кую твердую поверхность, то вначале он барахтается, пытаясь пе-
ревернуться, но ноги его не находят опоры, и он перестает двигать-
ся. Затем он вдруг подпрыгивает в воздух и при этом слышится
щелчок. Если он снова упадет на спину, то продолжает щелкать,
пока не перевернется. Этот механизм прыжков без участия ног
подбрасывает жука на высоту до 30 см, причем ускорение при от-
рыве может достигать почти 400 g.
Механизм этот прост. На первом грудном сегменте имеется вы-
ступ, направленный назад, и он частично входит в ямку во втором
сегменте (рис. 11.24). Когда этот выступ упирается в край ямки,
он не дает крупной прыгательной мышце в протораксе откидывать
переднюю половину тела вверх. Поэтому напряжение сокращаю-
щейся мышцы растет, и она действует как пружина: когда выступ
39-1863
6*0 Гл. И, Мышцы, движение, локомоция
в конце концов соскальзывает, тело жука со щелчком сгибается,
и накопленная в мышце энергия создает мгновенную силу, нужную
для отрыва. Тщательные измерения показывают, что во время
щелчка центр тяжести жука поднимается приблизительно на 0,6—
0,7 мм всего лишь за 0,5 мс.
Жук-щелкун, как и блоха, слишком мал для того, чтобы его
мышцы могли придать его телу нужное ускорение за короткое
время. Половины миллисекунды недостаточно для сокращения
мышцы, и наилучший способ быстро получить энергию состоит в
том, чтобы запасти ее в эластичной структуре (Evans, 1973).
ПОЛЕТ И ПЛАВАНИЕ
Воздух и вода представляют собой текучие среды, и перед жи-
вотными, которые движутся в них, стоят одни и те же проблемы.
Под животным нет твердого субстрата, который поддерживал бы
его вес, и силы, приводящие тело в движение, действуют против
среды, которая буквально ускользает. Плотность и вязкость у воз-
духа гораздо меньше, чем у воды, и это различие имеет существен-
ное значение для локомоции животных в этих двух средах, но есть
много гидродинамических принципов, общих для обеих сред.
Судя по числу летающих форм, полет является чрезвычайно
выгодным способом передвижения. Примерно из миллиона совре-
менных видов животных более трех четвертей — летающие насе-
комые. Способность летать возникала независимо по меньшей мере
четыре раза: у насекомых, у вымерших крупных рептилий (птеро-
завров), у птиц и у млекопитающих (летучих мышей). Размеры
активно летающих животных варьируют в пределах приблизитель-
но от 1 мкг у очень мелких насекомых до более 10 кг у самых
крупных летающих птиц.
Сила, необходимая для продвижения животного в воздухе по
прямому пути, состоит из двух компонент — подъемной силы, рав-
ной весу тела, и тяги, т. е. силы, которая толкает тело вперед, пре-
одолевая сопротивление воздуха. У летающих животных подъемная
сила и тяга создаются одними и теми же поверхностями. Как мы
увидим позже, для животных, плавающих в воде, это не всегда так.
Основную силу для полета птиц создают крупные грудные мыш-
цы, которые у всех птиц составляют около 15% массы тела незави-
симо от его размеров. Мышцы, поднимающие крыло при каждом
взмахе, составляют только около 7ю всей массы летательных
мышц и поэтому не могут вносить существенного вклада в ту силу,
которая нужна для продвижения вперед. Существуют, конечно, не-
большие отклонения от общего правила, но обычно у птиц двига-
тель составляет по весу одну и ту же долю всего «летательного
аппарата» независимо от его размеров (Greenewalt, 1975).
Колибри отличаются от остальных птиц: масса летательных
мышц составляет у них 25—30% массы тела. Это находится в со-
Биомеханика локомоции 611
ответствии с тем, что для «зависания» в воздухе на одном месте
нужна большая затрата энергии, чем при машущем полете вперед
(Weis-Fogh, 1973). Кроме того, мышцы, ответственные за подъем
крыльев, составляют здесь около трети всей массы летательных
мышц. Это указывает на то, что как источник энергии для поддер-
жания тела в воздухе существенное значение имеют оба движения
крыла — и вверх, и вниз, а поскольку птица остается на одном мес-
те, вся энергия тратится на создание подъемной силы. Этот вывод
подтверждается аэродинамическим анализом движений крыльев у
колибри (Stolpe, Zimmer, 1939).
Поразительно, что, несмотря на сложность гидродинамической
теории полета, полет птиц и насекомых удалось хорошо понять
(Pennycuick, 1969; Tucker, 1973; Weis-Fogh, 1973; 1976; Lighthill,
1974). Мощность, необходимая для полета птицы, является чрезвы-
чайно сложной функцией величины и формы тела, скорости движе-
ния, размеров и формы крыльев. Кроме того, точный анализ ослож-
няется непрерывным изменением формы крыла и скорости его
движения при каждом взмахе. Согласно аэродинамической теории
(Lighthill, 1974), эта мощность возрастает пропорционально весу
тела с показателем степени 1,17. Анализ, проведенный Таккером
с учетом эмпирических измерений, привел к результатам, которые
в упрощенной форме говорят о том, что требуемая сила должна
быть пропорциональна весу птицы (Tucker, 1973). Для дальнейших
рассуждений не столь существенно, будем ли мы брать вес тела в
степени 1,17 или же 1,0.
Известно, что у птицы метаболизм покоя возрастает пропорцио-
нально массе тела с показателем степени 0,75 (с. 276). Можно
принять (хотя это достоверно не известно), что максимальная ста-
бильная мощность находится в такой же зависимости от размеров
тела. Это просто означает, что возможная мощность растет с уве-
личением размеров птицы не так быстро, как мощность, необходи-
мая для полета. Это показано на рис. 11.25, где кривая мощности,
требуемой для полета, построена так, что она пересекает кривую
максимальной возможной мощности при весе тела 10 кг — пример-
но таком, как у самых крупных летающих птиц. Следовательно,
еще более крупная птица, например страус (вес около 100 кг),
даже если бы она по своей анатомии была приспособлена к полету,
не могла бы мобилизовать столько энергии, сколько нужно для
стабильного машущего полета по горизонтали.
Интересно сравнить эти сведения с подробностями полета че-
ловека на аппарате весом 32 кг, приводимом в движение мышеч-
ной силой, когда хорошо тренированный спортсмен, весивший 62 кг,
завоевал знаменитую премию Кремера. Вращая пропеллер с по-
мощью цепной передачи велосипедного типа, он успешно пролетел
намеченным курсом почти 2000 м приблизительно за 7 мин (Hirst,
1977). Это было рекордное достижение тренированного человека в
идеальных условиях; подобные полеты вряд ли могут быть намного
39*
612 Гл. 11. Мышцы, движение, локомоция
более длительными. Однако наши представления о мощностях, не-
обходимых для машущего полета и полета с помощью пропеллера^
хорошо согласуются с данными о действительных полетах. И в са-
мом деле, пропеллер представляет собой просто такое приспособ-
ление, в котором поверхность, создающая горизонтальную тягу,,
отделена от поверхности, ответственной за подъемную силу.
Вес тела, кг
Рис. 11.25. Мощность, нужная для полета птицы, по сравнению с максимальной
возможной мощностью. Хорошо знакомое соотношение между метаболизмом
покоя и весом тела представлено в виде линии регрессии с наклоном 0,75 (I).
Линия регрессии для принятой максимальной мощности (в 15 раз большей, чем
в покое) идет параллельно на уровне в 15 раз более высоком (II). Вычисленная
сила, нужная для полета, прямо пропорциональна весу тела, и соответствующая
линия регрессии с наклоном 1,0 (III) проведена так, что она пересекает линию
максимальной мощности при весе тела 10 кг — приблизительной величине самых
крупных летающих птиц.
Плавание в принципе не очень отличается от полета по воздуху,
если не считать того, что большая плотность воды уменьшает по-
требность в подъемной силе. В самом деле, некоторые плавающие
животные обладают той же плотностью, что и вода, и подъемная
сила им вообще не нужна; вся необходимая для плавания энергия
может быть использована для создания горизонтальной тяги. Су-
щественное значение имеет большая плотность и вязкость воды;
рыбы не плавают с такой быстротой, с какой летают птицы.
Для таких крупных животных, как рыбы, сопротивление движе-
нию растет приблизительно пропорционально квадрату скорости,
что сильно ограничивает возможную скорость (Lighthill, 1974). Мы
Плавучесть 613
уже видели, как некоторые из самых крупных и быстрых хищных
рыб приобрели способность поддерживать в своей «силовой уста-
новке»— мышечной массе — высокую температуру для получения
максимальной мощности.
Как мог бы организм сравняться по плотности с водой, приоб-
рести тем самым нейтральную плавучесть и избежать затраты энер-
гии на создание подъемной силы? Эта проблема решается самыми
разными способами, к рассмотрению которых мы переходим.
ПЛАВУЧЕСТЬ
Если плывущее животное тяжелее воды, то оно расходует часть
энергии на то, чтобы держаться в воде на определенном уровне,
и только часть — на продвижение вперед. Если бы животное об-
ладало той же плотностью, что и вода (т. е. нейтральной плаву-
честью), у него оставалось бы больше энергии для передвижения.
Для крупных животных проблема плавучести важнее, чем для
очень мелких. Мелкие организмы обладают относительно большой
поверхностью, и часто у них имеются разного рода выросты, за-
медляющие их погружение на дно; самые мелкие взвешены в воде,
как пылинки в воздухе, и проходят таким образом весь свой жиз-
ненный цикл. У крупных организмов по механическим причинам
не может быть таких больших выступов, чтобы они могли тормо-
зить погружение, и такие организмы должны поэтому применять
другие средства.
Единственный способ избежать погружения на дно, не требую-
щий никаких движений (т. е. затраты энергии), — это снижение
плотности, или удельного веса. Можно назвать пять возможных
способов уменьшения тенденции к погружению и приближения к
нейтральной плавучести:
1) уменьшение количества тяжелых веществ (например, карбо-
ната или фосфата кальция);
2) замена тяжелых ионов (например, Mg2+ и SO42“) ионами
Na+, Cl" или даже более легкими (Н+ и NH4+);
3) удаление ионов без их замены (т. е. разбавление жидкостей
организма);
4) увеличение количества веществ более легких, чем вода, глав-
ным образом жиров и масел;
5) использование полостей, наполненных газом (например, пла-
вательных пузырей у рыб).
Рассмотрим эти пять возможностей, положительные и отрица-
тельные стороны и ограничения каждой из них. Поскольку морская
вода тяжелее (удельный вес 1,026), чем пресная (удельный вес
1,000), животному легче достичь нейтральной плавучести в море;
некоторые из способов решения этой задачи в море вообще непри-
годны для обитателей пресных вод.
614 Гл. И, Мышцы, движение, локомоция
УМЕНЬШЕНИЕ КОЛИЧЕСТВА ТЯЖЕЛЫХ ВЕЩЕСТВ
У многих животных скелет состоит из очень тяжелых веществ,
часто из солей кальция — карбоната или фосфата. Карбонат каль-
ция обычно встречается у беспозвоночных (раковины двустворча-
тых моллюсков и улиток, кораллы и др.), а фосфат кальция — ос-
новной компонент костей позвоночных животных, где он содержит-
ся в форме минерала апатита. У ряда организмов (например, у
некоторых губок) скелет состоит из кремнезема; другие (многие чле-
нистоногие) используют хитин, в чистом виде или вместе с карбо-
натом кальция. Об относительном влиянии этих веществ на общий
вес животного в воде можно судить по их удельному весу
(табл. 11.6).
Таблица 11.6
Удельный вес морской воды, пресной
воды и веществ, часто входящих в
состав скелета у водных организмов.
(Denton, 1961)
Вещество Удельный вес
Морская вода 1,026
Пресная вода 1,000
Карбонат кальция
Арагонит 2,9
Кальцит 2,7
Фосфат кальция
Апатит 3,2
Хитин 1,2
Легко видеть, что уменьшение количества солей кальция может
намного снизить вес животного. Поэтому у пелагических голожа-
берных улиток нет раковин, а у заднежаберных улиток, которые
живут на дне, раковины хорошо развиты. Плавающие кишечно-
полостные (например, медузы) совершенно лишены тяжелого ске-
лета из карбоната кальция, характерного для многих сидячих ки-
шечнополостных, например коралловых полипов.
Интересно отметить, что представители одной группы пелагиче-
ских организмов — плавающие крабы — не пошли далеко по пути
уменьшения массы своего тяжелого экзоскелета (состоящего глав-
ным образом из карбоната кальция). Это относится и к обыкно-
венному голубому крабу (Callinectes), которому, чтобы не идти ко
дну, приходится непрерывно плавать.
Отказ от механических опорных структур, разумеется, представ-
ляет значительную невыгоду. Поэтому такое решение чаще встре-
чается у относительно мелких животных. Вместо этого возможна
замена тяжелых материалов для построения скелета более легки-
ми. Например, у обыкновенного кальмара скелет из карбоната
Плавучесть 615
кальция, который имеется у родственной ему каракатицы, заменен
более легкой структурой из хитина (так называемое перо каль-
мара).
ЗАМЕНА ТЯЖЕЛЫХ ИОНОВ БОЛЕЕ ЛЕГКИМИ
Пожалуй, самые лучшие примеры такого решения проблемы
встречаются у растений. Две многоклеточные водоросли, Valonia
и Halicystis, демонстрируют, чего можно достичь, изменив свой ион-
ный состав. Обе они обладают почти нейтральной плавучестью:
первая лишь очень медленно опускается в морской воде, а у вто-
рой даже слегка положительная плавучесть. У обеих в изобилии
имеется клеточный сок, изотоничный морской воде, но иного со-
става (табл. 11.7). Обе почти полностью лишены тяжелого иона
сульфата. Valonia, кроме того, не содержит магния, который име-
ется у Halicystis в концентрации втрое меньшей, чем в морской
воде. Другого важного двухвалентного иона — кальция — в обеих
водорослях тоже гораздо меньше, чем в морской воде.
Таблица 11.7
Ионный состав морской воды и сока двух
многоклеточных водорослей, обладающих
приблизительно нейтральной плавучестью.
Ионные молекулярные концентрации даны
относительно концентрации хлора (С1~ = 100).
(Gross, Zeuthen, 1948)
Морская вода Сок Valonia Сок Halicystis
С1- 100 100 100
Na+ 85,87 15,08 92,4
К+ 2,15 86,24 1,01
Са2+ 2,05 0,285 1,33
Mg24* 9,74 Следы 2,77
So42- 6,26 Следы Следы
Удельный вес 1,0277 1,0290 1,0250
То же для искусст- венного раствора1) 1,0285 1,0285 1,0252
’) Раствор с указанными выше ионными концентрациями.
Из одновалентных ионов К+ тяжелее, чемНа+. Valonia, которая
чуть тяжелее морской воды, накапливает в клеточном соке глав-
ным образом К+, как это вообще происходит у растений. Но Hali-
cystis, которая немного легче морской воды, исключила из своего
состава большую часть калия и использует в качестве самого глав-
ного катиона более легкий натрий.
Динофлагелляты Noctiluca miliaris — широко распространенные
организмы, вызывающие свечение моря. Они немного меньше 1 мм
616 Гл. 11. Мышцы, движение, локомоция
в диаметре и обычно скапливаются на поверхности, так как замет-
но легче морской воды. При механическом раздражении они испус-
кают вспышки света, которые видны ночью как фосфоресценция
моря. Их свечение было предметом многих исследований, но здесь
мы рассмотрим только вопрос об их плавучести.
Состав клеточного сока Noctiluca приведен в табл. 11.8. Этот
организм изотоничен морской воде и обязан своей положительной
плавучестью более низкой концентрации ионов Са2+, Mg2+, и в осо-
бенности тяжелого иона SO42”. Кроме того, он накапливает ион
аммония, который легче двух самых распространенных одновалент-
ных катионов — Na+ и К+.
Таблица 11.8
Ионный состав морской воды
и клеточного сока Noctiluca —
обычного источника фосфоресценции
в море. Этот организм достигает
положительной плавучести, исключая
тяжелые ионы и заменяя их главным
образом NH4+. (Kesseler, 1966)
В соке, В морской
мэкв/л воде, мэкв/л
Na+ 414 418
К+ 34 8,8
NH4+ 58,5 О
Са2^ 9,5 18,5
Mg2+ 15 95
Cl- 496,5 498,9
SO42“ Следы 46,6
Н2РО4“ 13 Следы
pH 4,35 8,2
Ион водорода (Н+) намного легче всех других катионов. Было
высказано предположение, что положительная плавучесть могла
бы достигаться повышением концентрации ионов водорода за счет
других катионов, но это неосуществимо. Хотя клеточный сок Nocti-
luca довольно кислый (т. е. концентрация ионов Н+ в нем высока),
легко видеть, что это не может заметно сказываться на удельном
весе.
Если принять, что величина pH сока Noctiluca равна 4,0, то кон-
центрация ионов водорода в нем составляет 10~4 моль/л, т. е.
0,1 ммоль/л и, следовательно, эти ионы могут заменить не больше
Убооо доли всех катионов. Чтобы ионы Н+ заменили существенную
часть катионов, pH должно быть значительно ниже 2, что соответ-
ствует концентрации Н+ больше 10 ммоль/л. Такая высокая кис-
лотность не обнаружена ни в одной клетке; она встречается только
в немногих кислых секретах, например в желудочном соке позво-
Плавцчесто 617
ночных и в слюне некоторых морских улиток, которые пользуются
ею при просверливании раковин своих жертв.
Принцип замены тяжелых ионов легкими могут с таким же ус-
пехом использовать и крупные организмы. Поразительным приме-
ром служит группа глубоководных кальмаров сем. Cranchidae.
У этих животных наполненный жидкостью целом очень велик и
занимает около двух третей всего объема тела. Если его вскрыть
и выпустить жидкость, животное потеряет свою нейтральную пла-
вучесть и пойдет ко дну. Удельный вес этой жидкости около 1,010
(удельный вес морской воды 1,026), она осмотически изотонична
морской воде и имеет pH 5,2. Концентрация NH4+в целомической
жидкости составляет около 480 мэкв/л, а концентрация натрия —
80 мэкв/л. Хлор здесь остается почти единственным анионом; тя-
желый ион SO42~ исключен (Denton, 1960; Denton et al., 1969).
Высокая концентрация ионов аммония объясняется просто. Це-
ломическая жидкость слегка кислая, и аммиак — нормальный
конечный продукт белкового обмена у большинства водных живот-
ных— диффундирует в эту жидкость и задерживается в ней. Пони-
жать pH здесь мог бы натриевый насос, который удаляет из цело-
мической жидкости ионы Na+, — его действие создавало бы избы-
ток анионов и тем самым повышало бы концентрацию ионов
водорода и, значит, увеличивало бы кислотность. Кислая жидкость
служила бы ловушкой для аммиака. Вычислено, что если аммиак
является у Cranchidae единственным конечным продуктом белково-
го обмена, то для сохранения нейтральной плавучести должно
удерживаться около 40% всего аммиака, образующегося в течение
жизни животного (Denton, 1971).
Способ, используемый кальмарами сем. Cranchidae, вполне эф-
фективно обеспечивает нейтральную плавучесть. Однако он связан
с некоторыми структурными неудобствами, так как две трети всего
объема животного занимает жидкость (т. е. объем жидкости, кото-
рую животное носит в себе, в два раза превышает объем собствен-
но тела).
ГИПОТОНИЧНОСТЬ
Еще одним подходом к снижению веса могло бы быть удаление
некоторых ионов без их замены. Конечно, это создало бы осмоти-
ческие проблемы, так как разбавленный раствор стал бы гипото-
ничным по отношению к морской воде. Как правило, беспозвоноч-
ные находятся в осмотическом равновесии с морской водой, и,
по-видимому, ни одно из этих животных не использовало гипото-
ничность как средство увеличения плавучести.
Однако у костистых рыб концентрации ионов гораздо ниже, чем
в-морской воде, и это помогает им снизить свой вес в воде. Правда,
значение этого фактора невелико, так как в скелете и- мышцах
костистых рыб содержатся существенные количества относительно
618 Гл. 11. Мышцы, движение, локомоция
тяжелых веществ. Впрочем, у многих глубоководных форм, у кото-
рых скелет и мышцы сильно редуцированы, гипотоничность значи-
тельно помогает уравновешивать сравнительно малые количества
тяжелых компонентов.
ЖИРЫ И МАСЛА
Во многих планктонных организмах довольно велико содержа-
ние жиров. Разумеется, жир служит обычной формой хранения за-
пасов энергии как у животных, так и у многих растений, но жиры
и масла играют также важную роль в плавучести. Планктонные
растения часто откладывают жир, а не более тяжелый крахмал,
который у растений является обычным резервным веществом. Так,
например, все диатомовые водоросли откладывают только жир.
У пластиножаберных рыб — в отличие от костистых — никогда
не бывает плавательного пузыря. Но у них большая печень, осо-
бенно у тех акул, которые являются хорошими пловцами. Печень
может составлять у них около одной пятой массы тела, а у кости-
стых рыб — обычно около 1—2%.
Изучение акулы Etmopterus spinax показало, что в среднем
17% веса ее тела приходится на печень, которая на 75% состоит
из жира (S. Schmidt-Nielsen et al., 1934). (В отлйчие от этого пе-
чень млекопитающего содержит около 5% жира.) Половину жира
в печени акулы составляет сквален — интересный ненасыщенный
углеводород (С30Н70), название которого происходит от названия
акулы Squalus.
Большая часть жиров и масел имеет удельный вес около 0,90—
0,92, но у сквалена он ниже и равен 0,86. Разница невелика, но
для плавучести в морской воде эффект данного объема сквалена
приблизительно на 50% больше, чем эффект равного объема более
обычного жира или масла. В этом ли причина накопления огром-
ных количеств сквалена в печени пластиножаберных рыб, сказать
трудно; так или иначе, его наличие, несомненно, помогает рыбе
приблизиться к нейтральной плавучести. Он особенно эффективен
потому, что пластиножаберные в отличие от костистых рыб обла-
дают сравнительно легким хрящевым скелетом, который не отяго-
щают тяжелые фосфаты кальция.
Интересно, что у скатов, живущих на дне, печень меньше и со-
держание жира в ней ниже. У пяти видов ската Raja печень со-
ставляет в среднем 7,53% массы тела и в ней меньше 50% жира.
В этом есть смысл, так как скаты живут около дна и не относятся
к очень хорошим пловцам.
Нужно отметить, что сравнительно низкие концентрации ионов
в жидкостях тела пластиножаберных по отношению к морской
воде не способствуют существенно их плавучести. Они изоосмотич-
ны с морской водой, разница в концентрации солей компенсирует-
ся в основном мочевиной, а изоосмотичные растворы мочевины
Плавучесть 619
очень ненамного легче морской воды. У окиси триметиламина, ко-
торая тоже содержится в жидкостях тела этих рыб, плотность за-
метно меньше, чем у мочевины, и это вещество играет некоторую,
хотя и небольшую, роль в плавучести.
У ряда глубоководных костистых рыб плавательный пузырь
наполнен не воздухом, а жиром. Это относится к некоторым видам
Gonostoma и Cyclothone (Marshall, 1960), но поскольку в жировых
плавательных пузырях других рыб много холестерина (до 49%),
который имеет больший удельный вес (1,067), чем морская вода
(1,026), роль наполненного жиром пузыря в регуляции плавучести
сомнительна (Phleger, 1971).
КАМЕРЫ, НАПОЛНЕННЫЕ ГАЗОМ
По сравнению с водой газ имеет очень низкую плотность, и даже
относительно малый его объем превосходно решает проблему пла-
вучести. Но это решение имеет некоторые ограничения и недостат-
ки. Многое зависит от того, окружено ли газовое пространство мяг-
кой тканью или жесткими стенками.
ПУЗЫРИ С МЯГКИМИ СТЕНКАМИ
Представим себе наполненный газом резиновый баллон, пол-
ностью погруженный в жидкость. Чем глубже его опускают, тем
сильнее давит на него снаружи вода, а поскольку давление газа
внутри соответствует давлению воды снаружи, баллон все больше
сжимается.
Давление воды возрастает приблизительно на 1 атм с каждыми
10 метрами глубины, и для'организма, живущего на значительной
глубине, это создает свои проблемы. Рассмотрим организм, оби-
тающий на глубине 1000 м, где давление воды составляет около
100 атм. Создание и сохранение объема газа при таком давлении
связано с тремя трудностями: 1) газ приходится выделять в по-
лость пузыря против давления в 100 атм; 2) количество газа, не-
обходимое для того, чтобы наполнить тот же объем при таком дав-
лении, в 100 раз больше, чем у поверхности воды; и 3) при таком
высоком давлении трудно предотвратить потерю газа путем диффу-
зии. Напряжение газов в воде на любой глубине океана обычно
мало отличается от их напряжения на поверхности, где имеет ме-
сто равновесие с 0,2 атм кислорода и 0,8 атм азота. Газы для пу-
зыря извлекаются из воды и путем секреции транспортируются в
его полость против высокого давления, причем должна быть пре-
дотвращена их утечка в результате диффузии.
Мягкие стенки пузыря создают еще одну трудность: его объем
меняется, когда животное движется в воде вверх и вниз. Допустим,
что газа в пузыре достаточно, чтобы животное находилось в состоя-
нии нейтральной плавучести на данной глубине. Но если оно опу-
620 Гл. 11. Мышцы, движение, локомоция
стится ниже, то газ сожмется, животное станет тяжелее и утратит
-плавучесть. Если же оно поднимется выше, то газ расширится и по-
ложительная плавучесть возрастет. Это может быть опасно; если
только животное не опустится активно снова до нейтрального уров-
ня, газ будет расширяться все быстрее и быстрее, по мере того как
животное поднимается со все
большей скоростью. Если оно
не обладает механизмом для
выпускания газа, то, утратив
контроль над своим подъе-
мом, оно может даже лопнуть
от расширения газа.
Если же содержащее газ
пространство заключено в
жесткие стенки, то ограниче-
ния для вертикальных пере-
движений будут менее строги-
ми, но стенки должны обла-
дать достаточной механиче-
ской прочностью, чтобы вы-
держивать изменения давле-
ния. Однако если придать
стенкам большую прочность,
то они станут тяжелее, а это
противоречит назначению га-
зового поплавка (пневмато-
фора) — сделать организм
более легким. Как мы увидим,
Фото 11.5. Физалия (Physalia pelagi-
са). Пневматофор этого колониально-
го кишечнополостного наполнен газом,
значительную долю которого состав-
ляет окись углерода. Длина пневма-
тофора может превышать 20 см,
а длина щупалец, вызывающих чрез-
вычайно болезненные ожоги, дости-
гает более 10 м. (Фото предоставле-
но Ch. Е. Lane, University of Miami.)
механическая прочность пневматофора налагает ограничения на
вертикальное распространение каракатицы (Sepia), у которой
плавучесть обеспечивают- жесткие структуры, наполненные газом.
Плавательный пузырь рыб, несомненно, представляет собой наи-
лучший из известных типов газовых поплавков. Этот принцип ис-
пользуется также небольшим числом других организмов, например
колониальными сифонофорами (представители кишечнополост-
Плав учесть 621
ных). Одна из них, Physalia, часто встречается вдоль атлантиче-
ского побережья Северной Америки (фото 11.5). Всю колонию
поддерживает «поплавок» — наполненная газом пузыревидная ка-
мера длиной нередко более 10 см. Она плавает на поверхности
океана и, увлекаемая ветром, тянет за собой всю колонию.
Поскольку газовый пузырь физалии не погружен в воду, а дер-
жится на поверхности, внутреннее давление в нем практически
равно атмосферному. Это упрощает задачу наполнения его газом.
Однако состав этого газа весьма удивителен: газ содержит до 15%
окиси углерода, концентрация кислорода меньше атмосферной,
а содержание двуокиси углерода ничтожно; остальное—азот. Газ
вырабатывается газовой железой, которая использует для выра-
ботки окиси углерода аминокислоту серин (CH2OHCHNH2COOH)
(Wittenberg, 1960).
У других колониальных сифонофор тоже имеются пневматофо-
ры, содержащие окись углерода. У одной из них, Nanomia bijuga,
пневматофоры очень небольшие, объемом от 0,14 до 0,46 мкл . и с
концентрацией окиси углерода от 77 до 93% (Pickwell et al., 1964).
Окись углерода, конечно, очень токсична для человека и многих
других высших животных, и в природе встречаются только ее
следы. Она найдена в полых стеблях некоторых морских водорос-
лей, где может составлять несколько процентов всего газа. В сов-
еем малых количествах окись углерода содержится в выдыхаемом
воздухе у людей, даже у некурящих. Она не вырабатывается в ки-
шечнике бактериями, а образуется в печени при метаболическом
.распаде гемоглобина (Ludwig et al., 1957). Источником выдыхае-
мой окиси углерода не является загрязненный воздух, поскольку
антарктический тюлень Уэддела (который, надо полагать, дышит
совершенно чистым воздухом) тоже выдыхает ее в измеримых ко-
личествах (Pugh, 1959).
Наличием пневматофоров у планктонных организмов, возможно,
объясняется загадочное явление, называемое глубинным рассеи-
вающим слоем. Когда на судах измеряют глубину с помощью эхо-
лота, звуковые волны часто отражаются не только от дна, но так-
же от какого-то слоя в толще воды, который тем самым создает
ложное представление о расстоянии до дна. Этот слой, который
часто находится днем и ночью на разной глубине, и называют глу-
бинным рассеивающим слоем. Многие годы причина такого отра-
жения звука была неизвестна, но теперь почти с уверенностью
можно сказать, что его вызывают косяки рыб с плавательными
пузырями или пелагические сифонофоры с пневматофорами.
Пузыри сифонофор имеют отверстие, и при подъеме в воде газ
может выходить из них. Простой опыт показал, что эти организмы
способны также и к секреции газа (Pickwell et al., 1964).Животных
помещали в 10-миллилитровый шприц и вытягиванием поршня
создавали в шприце разрежение. В результате газ выходил из
пневматофоров и они становились тяжелее воды, но уже через час
622 Гл. 11. Мышцы, движение, локомоция
снова плавали на поверхности. Теперь они были наполнены глав-
ным образом окисью углерода, подобно пневматофорам физалий.
Ночью глубинный рассеивающий слой часто смещается по на-
правлению к поверхности океана. Поскольку сифонофоры при
подъеме выделяют газ, они, вероятно, выпускают пузырьки, и этим
можно объяснить то, что интенсивность отражения звука от рас-
сеивающего слоя повышается по мере его подъема.
Если, поднимаясь, животные выпускают газ, то надо полагать,
что такое же его количество вновь образуется у них при спуске на
глубину. Однако они могли бы избежать многократной секреции
и освобождения газа, если бы сохраняли постоянное его количество
и справлялись с изменениями его объема при вертикальной миг-
рации. Это осуществимо, если животное находится в состоянии
нейтральной плавучести на более высоком уровне, но вряд ли воз-
можно, если нейтральный уровень лежит глубже.
Рассмотрим пример. Предположим, что объем плавательного
пузыря рыбы (при нейтральной плавучести) равен 5% объема
тела. Утренний спуск с глубины 50 м на глубину 300 м уменьшит
объем газа в пять раз, т. е. до 1 % объема тела. При этом пла-
вучесть станет отрицательной, но рыба сможет держаться на новом
уровне, если будет немного плавать. Но если рыба, пузырь которой
занимает 5% ее объема, находится в состоянии нейтральной пла-
вучести на глубине 300 м, то при подъеме до уровня 50 м объем ее
плавательного пузыря увеличится в пять раз (если только газ не
будет выходить). Пятикратное расширение пузыря создало бы
серьезную опасность потери управления и подъема до самой по-
верхности, где при давлении 1 атм плавательный пузырь катастро-
фически увеличился бы в 30 раз по сравнению с первоначальным
объемом! Если организмы глубинного рассеивающего слоя пере-
мещаются по вертикали, не выпуская газа, то надо полагать, что
они находятся в состоянии нейтральной плавучести не на нижнем,
а на верхнем уровне.
Об изменении объема можно кое-что узнать, не прибегая к пря-
мому наблюдению. Глубинный рассеивающий слой особенно хо-
рошо отражает звук на частотах, соответствующих резонансной ча-
стоте газовых пузырьков, которая в свою очередь зависит от их
объема и от давления в них. Если известно давление (глубину
можно прямо определить по показаниям эхолота) и если принять,
что резонансная частота соответствует наилучшему отражению
звука, то можно вычислить величину пузырьков. Если теперь слой
движется вверх, то есть две возможности: либо газ будет выходить
и пузырьки сохранят постоянный объем, либо газ останется в пу-
зырьках, они расширятся и давление в них изменится. Если пу-
зырек сохраняет постоянный объем, то его резонансная частота
должна изменяться в соответствии с Р 2 (где Р—гидростатическое
давление); если же объем меняется с давлением, а масса газа оста-
ется постоянной, то резонансная частота будет изменяться в соот-
Плавучесть 623
ветствии с Р5/6. Записи звука сложны и часто их трудно интерпре-
тировать, но создается впечатление, что у организмов глубинного,
рассеивающего слоя могут реализоваться обе эти возможности
(Hersey et al., 1962).
ПНЕВМАТОФОРЫ С ЖЕСТКИМИ СТЕНКАМИ
Сложности, связанные с изменениями объема, а значит, и пла-
вучести при переходе на другую глубину, исчезают, если газ за-
ключен в полость с жесткими стенками. Такое решение задачи мы
находим у каракатицы {Sepia officinalis)—родственницы кальма-
ра и осьминога, которая часто встречается в верхних слоях при-
брежных водных масс. Почти одну десятую объема ее.тела занима-
ет рудиментарная раковина — обызвествленная внутренняя струк-
тура, хорошо знакомая любителям птиц, которые используют ее
как источник кальция для своих питомцев (рис. 11.26). Роль ее
изучал английский исследователь Дентон (Denton, Gilpin-Brown,
1959, 1961).
Рис. 11.26. Схема строения каракатицы — морского головоногого моллюска, ис-
пользующего наполненную газом жесткую структуру (рудимент раковины) для
создания нейтральной плавучести в воде. Задняя часть раковины (зачерненный
участок) наполнена жидкостью. В морской воде такой поплавок снижает на 4%
вес животного, измеренный в воздухе, и таким образом уравновешивает избы-
точный вес остальных тканей. (Denton, Gilpin-Brown, 1961.)
Раковина занимает 9,3% объема тела, а ее удельный вес сразу
после извлечения лежит в пределах от 0,57 до 0,64. Она создает
подъемную силу, достаточную для придания животному нейтраль-
ной плавучести в морской воде.
624 Гл. 11. Мышцы, движение, локомоция
Раковина каракатицы обладает своеобразным пластинчатым
строением. Она состоит из тонких слоев карбоната кальция, ук-
репленных хитином и соединенных многочисленными «распорками»
длиной около 2/з мм из того же материала (фото 11.6). У совсем
взрослого животного таких параллельных слоев может быть около
100, а пространство между ними в основном заполнено газом.
Фото 11.6. Структура раковины каракатицы. Это наполненное газом образование
состоит из карбоната кальция. Оно построено из тонких слоев этого вещества,
разделенных промежутками 2/з мм и поддерживаемых столбиками из того же
материала. (Фото предоставлено М. L. Blankenship, Duke University.)
Если эту рудиментарную раковину извлечь из животного и про-
колоть под водой, газ не будет выходить из нее. Это значит, что
он не находится в ней под давлением. Если же раковину разда-
вить, то газ выходит из нее и его можно собрать для анализа. Он
состоит из азота под давлением в 0,8 атм и небольшого количества
кислорода.
Плавичесть 625*-
Около каудальной поверхности раковины между обызвествлен-
ными слоями находится немного жидкости. Ее можно собрать под.
парафиновым маслом, подвергнув раковину воздействию вакуума.
Жидкость содержит главным образом хлористый натрий; она гипо-
осмотична по отношению к морской воде и жидкостям тела (кото-
рые изоосмотичны с морской водой). Поэтому более высокое осмо-
тическое давление вне раковины стремится извлечь из нее воду.
Осмотическому отсасыванию воды противодействует гидростатиче-
ское давление, которое стремится вдавить воду обратно. Макси-
мальная сила, с которой отсасывается вода, ограничена разностью*
концентраций. Если бы из внутренней жидкости были извлечены
все ионы, то разность концентраций между этой жидкостью и
кровью составила бы около 1,1 осмоль, что соответствует давлению
24 атм. Иными словами, если внешнее гидростатическое давление
превысит 24 атм, жидкость будет вдавливаться в раковину, не-
смотря на осмотические силы, действующие в обратном направле-
нии. На глубине около 240 м давление равно 24 атм, и на большей
глубине извлечь жидкость из раковины осмотическим путем невоз-
можно. Но каракатицы живут в верхних слоях воды и, вероятно,
не спускаются ниже примерно 200 м. Поэтому осмотических сил
достаточно, чтобы вода не вдавливалась внутрь раковины.
Давление газа внутри раковины равно около 0,8 атм, и она
должна быть достаточно прочной, чтобы выдерживать гидростати-
ческое давление окружающей воды. Так оно и есть на самом деле:
если покрыть раковину тонким пластиковым мешочком (чтобы
в нее не могла проникать вода) и подвергнуть ее высокому дав-
лению, она выдержит больше 25 атм, прежде чем вода ее раз-
давит.
Преимущество жесткой системы состоит, разумеется, в том, что-
на плавучесть практически не влияют изменения глубины. Это дает
каракатице большую свободу передвижения вверх и вниз без ка-
кого-либо регуляторного изменения механизма плавучести. Поми-
мо того, что раковина делает каракатицу относительно независи-
мой от глубины, она еще служит ей скелетом.
Механизм заполнения многокамерной раковины кораблика
(Nautilus) в принципе такой же, как у каракатицы (рис. 11.27). По>
мере своего роста этот моллюск добавляет к своей раковине одну
за другой новые воздушные камеры. Вновь построенная камера
(ближайшая к животному) вначале наполнена жидкостью, в кото-
рой преобладает хлористый натрий. Натрий удаляется из жидкости
путем активного транспорта; поэтому вода вытягивается осмотиче-
ски, как и у каракатицы, а в камеру на место воды диффундирует
газ. Давление проникшего таким образом азота достигает 0,8 атм,
т. е. того же напряжения, что в морской воде и в жидкостях тела
животного. С кислородом дело обстоит иначе, так как животное
потребляет его, и поэтому напряжение Ог в жидкостях тела ниже
атмосферного. Давление газа в раковине всегда составляет около*.
40—1863
626 Гл. 11. Мышцы, движение, локомоция
0,9 атм независимо от того, на какой глубине поймано животное.
Так же, как и у каракатицы, у Nautilus свободу вертикального
передвижения ограничивает только предельное давление, которое
может выдержать его жесткая раковина.
Итак, мы рассмотрели примеры различных механизмов, исполь-
зуемых животными для улучшения плавучести. Каждый из них
Рис. 11.27. Многокамерная раковина Nautilus macrotnphalus в ее естественном
положении (в разрезе). Камеры наполнены газом, и по мере роста животного
добавляются новые камеры. Каждая новая камера вначале наполнена жид-
костью, которая затем удаляется осмотическим путем и замещается газом. В са-
мых новых камерах сохраняется немного жидкости. (Denton, Gilpin-Brown,
1966.)
имеет свои выгодные и невыгодные стороны; они резюмированы
в табл. 11.9. Очевидно, что газовый поплавок представляет собой
одно из лучших приспособлений для создания подъемной силы и
имеет также ряд других достоинств. Но если газ заключен в мяг-
кую структуру, такое устройство обладает одним большим недо-
статком: оно чрезвычайно чувствительно к изменениям давления.
У костистых рыб этот механизм используется в форме наполненно-
го газом плавательного пузыря с мягкими стенками, о котором речь
пойдет ниже.
Плавучесть &Т
Таблица 11.9
Достоинства и недостатки различных способов улучшения плавучести водных
организмов (плохие качества указаны знаком минус; неплохие, хорошие и
превосходные — одним, двумя или тремя плюсами соответственно)
Эффективность
плавучести
Затрата энергии
Редукция тяжелых
структур О
Замена тяжелых ио-
нов 200
Жир 50
Сквален 35
Пневматофор с мягки-
ми стенками 5
Пневматофор с жест-
кими стенками 10
+ — +++ —
++ ++ +++ + +
+++ +++
Не требуется
Все время требуется
Вначале большие за-
траты, в дальней-
шем не требуется
Вначале большие за-
траты, в дальней-
шем не требуется
— +++ Все время требуется
+++ +++ ++
4—|—(- Вероятно, требуется
ПЛАВАТЕЛЬНЫЙ ПУЗЫРЬ РЫБ
У многих костистых рыб имеется плавательный пузырь, кото-
рый создает подъемную силу, необходимую для нейтральной пла-
вучести. Наличие у той или иной рыбы плавательного пузыря
больше зависит от ее образа жизни, чем от систематического поло-
жения. У некоторых придонных рыб плавательного пузыря нет,
и это представляется понятным, так как нейтральная плавучесть
не дает особых выгод рыбе, обитающей на дне. У многих пелаги-
ческих форм и рыб, живущих у поверхности воды, плавательный
пузырь имеется, а у других таких форм он отсутствует. Его нет,
например, у атлантической скумбрии. Для быстрого и сильного
хищника это составляет определенное преимущество, поскольку,
как мы видели, пузырь с мягкими стенками существенно ограничи-
вает свободу передвижения вверх и вниз в толще воды. Ни у одной
пластиножаберной рыбы плавательного пузыря нет.
Плавательный пузырь представляет собой более или менее уд-
линенный мешок с мягкими стенками, лежащий в брюшной поло-
сти под самым позвоночником (рис. 11.28; фото 11.7). Форма его
сильно варьирует, но объем у разных видов довольно постоянный:
пузырь занимает около 5% объема тела у морских рыб и 7% у
пресноводных. Это объясняется тем, что удельный вес рыбы без
плавательного пузыря равен примерно 1,07; поэтому объем газа,
40
л628 Гл. 11. Мышцы, движение, локомоция
-придающий нейтральную плавучесть в пресной воде, составляет
около 7% объема тела, а в морской воде (уд. вес 1,026) около 5%.
Плавательный пузырь может придать рыбе идеальную ней-
тральную плавучесть, и тогда ей не нужно тратить энергию, чтобы
держаться на одном уровне. Однако это связано с серьезным не-
'Рис. 11.28. Плавательный пузырь поме-
щается у рыбы в брюшной полости под
самым позвоночником. Форма пузырей
сильно варьирует, хотя их относитель-
ный объем обычно мало различается у
разных видов рыб.
Фото 11.7. Рентгенограмма рыбы
Chelmon rostratus. Плавательный
пузырь хорошо просматривается
как светлый участок под самым
позвоночником. Длина этого эк-
земпляра около 10 см. (Фото
предоставлено J. Lundberg, Duke
University.)
удобством: рыба находится в состоянии равновесия только на од-
ной определенной глубине. Если она погружается глубже, возрос-
шее давление воды сжимает ее плавательный пузырь, плавучесть
уменьшается и рыбе приходится активно плавать, чтобы не опус-
титься еще ниже. Это не очень серьезная проблема, так как даже
при почти полном сдавлении пузыря рыба не будет намного тяже-
лее воды.
Но если рыба поднимется выше уровня нейтральной плавуче-
сти, положение может стать катастрофическим. Давление снизит-
ся, плавательный пузырь расширится и подъемная сила возрастет.
Рыбе придется теперь активно опускаться, иначе она будет под-
ниматься все выше и в результате все большего расширения пузы-
ря в конце концов потеряет управление.
У некоторых рыб плавательный пузырь соединен с пищеводом,
что позволяет газу выходить, но у других рыб он совершенно зам-
кнут. Если рыбу с замкнутым пузырем поднять с большой глуби-
ны, она появится на поверхности с раздувшимся, вывернутым че-
рез рот или даже разорванным плавательным пузырем.
В эмбриогенезе плавательный пузырь образуется как выпячива-
ние пищеварительного тракта. Те рыбы, у которых связь пузыря
.с пищеводОхМ сохраняется, называются открытопузырными. Такие
Плавучесть 629
рыбы могут наполнять пузырь, заглатывая воздух у поверхности
воды. Но для глубоководных рыб этот способ непрактичен или
даже невозможен, так как им приходилось бы набирать в пузырь
у поверхности очень большой объем воздуха, чтобы создать ней-
тральную плавучесть на глубине, где давление во много раз
'больше.
У многих рыб соединительный канал дегенерирует и пузырь не
-сообщается с внешней средой. У таких рыб, называемых закрыто-
пузырными, газы для плавательного пузыря должны поступать из
крови и выделяться в пузырь под давлением, равным давлению
на той глубине, где эти рыбы живут. Рыбы с плавательными пузы-
рями были добыты и с глубины больше 4000 м. Поскольку стенка
пузыря мягкая, газ в нем на такой глубине должен находиться под
давлением более 400 атм. А это создает проблемы, о которых го-
ворилось выше в настоящей главе.
В плавательных пузырях содержатся те же газы, что и в атмо-
сфере (кислород, азот и двуокись углерода), но в иных соотноше-
ниях. Если удалить из пузыря некоторое количество газа, проколов
его иглой от шприца, то рыба должна будет возместить потерю пу-
тем секреции. У большинства рыб вновь выделенный газ богат кис-
лородом (табл. 11.10).
Таблица 11.10
Состав газа, выделяемого газовой
железой трески. Железу покрывали
тонкой пластиковой пленкой и
стимулировали; затем собирали
пузырьки размером около 0,5 мкл и
анализировали газ. (Scholander, 1956)
В среднем
о2, % СО2, % n2, %
81,1 8,6 10,3
70,3 5,0 24,7
76,6 15,8 7,6
49,5 6,9 43,6
61,1 6,8 32,1
66,5 5,9 27,6
30,1 13,9 56,0
69,2 7,2 23,6
63,1 8,8 28,1
Но так обстоит дело не у всех рыб; например, сиг (Coregonus
albus) наполняет свой плавательный пузырь практически чистым
азотом. Поскольку он живет в озерах на глубине порядка 100 м,
азот должен секретироваться под давлением около 10 атм. Газо-
образный азот чрезвычайно инертен, и трудно было понять, каким
630 Гл. 11. Мышцы, движение, локомоция
образом возможна секреция инертного газа. Кроме того, настоя-
щие инертные газы (такие, как аргон) в пузыре тоже сконцентри-
рованы и находятся в таком же соотношении с азотом, как и в ат-
мосфере (Wittenberg, 1958). Это говорит о том, что механизм сек-
реции не связан с химическими реакциями; он должен иметь физи-
ческую природу.
Даже если газ в плавательном пузыре богат кислородом, кон-
центрация азота в нем все же может быть выше, чем в окружаю-
щей воде. Предположим, что рыба извлечена с глубины 500 м и
что в пробе газа из ее пузыря обнаружено 80% О2 и 20% N2
(а также следы двуокиси углерода). На той глубине, где была
поймана рыба, газы в пузыре находились под общим давлением
50 атм, так что их парциальные давления составляли 40 атм для
кислорода и 10 атм для азота. Таким образом, по сравнению с на-
пряжениями газов в окружающей воде — 0,2 атм О2 и 0,8 атм N2 —
в плавательном пузыре напряжение кислорода в 200 раз, а азота
в 12,5 раз больше. Прежде чем рассмотреть, каким образом воз-
можна секреция газов при столь высоких давлениях, надо обра-
титься к более простому вопросу о том, как они удерживаются в
плавательном пузыре и не уходят из него.
КАК СОХРАНЯЕТСЯ ГАЗ В ПЛАВАТЕЛЬНОМ ПУЗЫРЕ?
Как всегда, для того чтобы понять функцию, надо знать кое-что
о структуре. В стенке плавательного пузыря у большинства рыб
имеется газовая железа, заметная благодаря своему ярко-красно-
му цвету. Если бы газовая железа снабжалась обычной артериаль-
ной кровью, которая поступает из жабр и поэтому уравновешена
в отношении газов с наружной водой, то кровь растворяла бы газы
плавательного пузыря и уносила их с собой. Как можно это предот-
вратить?
Кровь проходит в газовую железу через своеобразную структу-
ру, называемую rete mirabile (мн. ч. retia mirabilia), т. е. чудесным
сплетением. Форма этого сплетения и газовой железы у разных ви-
дов различна, но общий принцип устройства состоит в следующем.
Перед тем как подойти к газовой железе, артерия распадается на
огромное множество прямых параллельных капилляров, которые
перед входом в газовую железу снова соединяются в один сосуд
(рис. 11.29). Вена, идущая от газовой железы, тоже разветвляется
на такое же число параллельных капилляров, которые проходят
между артериальными капиллярами, а затем вновь образуют одну
общую вену.
Посмотрим, каким образом эта структура способствует удержа-
нию в плавательном пузыре газов, которые там уже находятся.
Такая система аналогична противоточному теплообменнику в плав-
нике кита, но здесь она имеет дело не с теплом, а с растворенными
газами. Предположим, что в плавательном пузыре находится газ
Плавучесть 631
под давлением 100 атм. Венозная кровь, покидая пузырь, содержит
растворенные газы под этим же высоким давлением. Когда она
входит в венозные капилляры, от крови артериальных капилляров
ее отделяют только тончайшие стенки тех и других сосудов. Поэто-
му газы диффундируют из венозной крови в артериальную. По
мере прохождения по венозным капиллярам кровь теряет все
Рис. 11.29. Схема кровоснабжения плавательного пузыря рыбы. Кровь подходит
•к пузырю или по пучку параллельных капилляров (rete mirabile), который снаб-
жает кровью газовую железу, или через сосуд, идущий к задней части пузыря,
где тонкие разветвления в его стенке могут служить для быстрого поглощения
газов. Во время секреции газа сосуды, где происходит его поглощение, закрыты
и не пропускают крови.
больше и больше газа вследствие диффузии в артериальные ка-
пилляры. В конце венозных капилляров, перед выходом крови из
сплетения, окружающие артериальные капилляры содержат кровь,
которая идет прямо из жабр и еще не обогащена дополнительным
количеством газа. Поэтому венозная кровь может терять газы, пока
не наступит диффузионное равновесие с притекающей артериаль-
ной кровью (т. е. пока в венозной крови, выходящей из сплетения,
не исчезнет избыток газа по сравнению с артериальной кровью).
Таким образом, rete mirabile служит ловушкой, которая удер-
живает газы в плавательном пузыре и препятствует их утечке в
кровоток. Это типичный противоточный обменник, работа которого
определяется пассивной диффузией между двумя потоками жидко-
сти, идущими в противоположных направлениях. Обмену газами
способствуют большая поверхность (т. е. многочисленность капил-
ляров), короткий диффузионный путь (т. е. малая толщина стенки
капилляров, составляющая всего лишь долю микрометра) и длина
капилляров (которая очень велика).
632 Гл. 11. Мбиицы, движение, локомоция ~
Некоторые измерения весьма информативны в этом отношении.
Сплетение от обыкновенного угря весило около 65 мг и состояло
примерно из 100 000 артериальных капилляров и такого же числа
венозных капилляров. Капилляры имели длину около 4 мм, что в
сумме составляет для каждого вида капилляров общую длину
400 м. Их диаметр был, как у большинства капилляров, около 7—
10 мкм, и простой расчет показывает, что вся поверхность капил-
ляров одного типа (венозных или артериальных) превышала
100 см2, и все это в объеме капли воды (Krogh, 1929). Таким об-
разом, газообмен между артериальной и венозной кровью мог про-
исходить на весьма значительной площади.
Капилляры в сплетении плавательного пузыря необычайно длин-
ны, их длина часто достигает нескольких миллиметров, тогда как
у мышечных капилляров, относящихся к самым длинным во всем
теле, она составляет лишь около 0,5 мм. Существует отчетливая
корреляция между длиной капилляров чудесного сплетения и дав-
лением газа в плавательном пузыре. Те рыбы, которые живут на
самых больших глубинах и выделяют в пузырь газы под наиболее
высокими давлениями, обладают самыми длинными капиллярами;
рекорд в этом отношении принадлежит глубоководной рыбе Basso-
zetas taenia, у которой капилляры чудесного сплетения достигают
огромной длины в 25 мм (Marshall, 1960).
Эффективность такого устройства оценил Шоландер (Scholan-
der, 1954), который нашел, что его размеры были бы достаточны
для поддержания в плавательном пузыре давлений выше 4000 атм.
Рыбы, обладающие плавательным пузырем, почти наверное не
встречаются на глубинах больше 5000 м (500 атм); из этого следу-
ет, что чудесное сплетение может надежно предотвращать утечку
газа из пузыря с оттекающей от него кровью. Поскольку работа
этого приспособления зависит только от диффузии и не требует
затраты энергии, оно представляет собой весьма удачное решение
проблемы.
Другой путь возможной утечки газа из плавательного пузыря —
диффузия через его стенку. Поскольку общее давление газа в пу-
зыре почти равно гидростатическому давлению воды, где нахо-
дится рыба, парциальные давления неизбежно выше, чем в окру-
жающей воде. Обычно парциальное давление азота в воде (и в
крови) равно 0,8 атм. Парциальное давление кислорода в воде,
а значит, и в крови, как правило, не превышает 0,2 атм. В отличие
от этого в плавательном пузыре парциальные давления могут из-
меряться сотнями атмосфер.
Содержание воды в стенке плавательного пузыря такое же, как
в других тканях, и поэтому можно было бы ожидать, что кислород
будет легко диффундировать наружу. Между тем его диффузион-
ная способность оказывается неожиданно низкой. Вероятно, это*
объясняется наличием слоев очень тонких пластинок кристалличе-
ского гуанина (толщина каждого слоя всего лишь около 0,02 мкм);.
Плавучесть 633
Эти тонкие слои образуют барьер для диффузии газов из плава-
тельного пузыря и тем самым уменьшают потребность в метаболи-
ческой энергии для секреции газов в пузырь с целью поддержания
его объема (Lapennas, Schmidt-Nielsen, 1977).
КАК ПРОИСХОДИТ СЕКРЕЦИЯ ГАЗА В ПЛАВАТЕЛЬНЫЙ ПУЗЫРЬ?
Задача выделения газа в плавательный пузырь против высокого
давления гораздо сложнее задачи удержания его там после того,
как он туда попал. Рассмотрим сначала секрецию кислорода, по-
скольку он чаще всех других газов выделяется в плавательный пу-
зырь. Представим себе пузырь, который уже содержит кислород
под высоким давлением, скажем 100 атм (что соответствует глуби-
не 1000 м), и предположим, что против этого огромного давления
должно быть выделено еще дополнительное количество газа. По-
скольку напряжение кислорода в артериальной крови, притекаю-
щей из жабр, в лучшем случае равно его напряжению в окружаю-
щей воде, артериальное напряжение кислорода не превышает
0,2 атм.
Рассмотрим теперь содержание кислорода в крови, а не его на-
пряжение. Если кислород переходит из крови в плавательный пу-
зырь, то это значит, что выходящая из чудесного сплетения веноз-
ная кровь должна содержать меньше Ог, чем притекающая арте-
риальная кровь. Возьмем произвольный пример: пусть артериаль-
ная кровь содержит, скажем, 10 мл Ог на 100 мл крови, а венозная,
когда она вытекает из сети, содержит 9 мл О2 на 100 мл. На
рис. 11.30, где для простоты вся сеть представлена одной петлей,
эти два числа указаны у левого края схемы. Разность между арте-
риальной и венозной кровью, т. е. 1 мл О2 на 100 мл крови, — это
тот кислород, который переходит в плавательный пузырь.
Теперь предположим, что газовая железа начинает вырабаты-
вать молочную кислоту (что действительно происходит при секре-
ции газа). Молочная кислота поступает в кровь и снижает ее
сродство к кислороду. У рыб это действие кислоты выражено го-
раздо сильнее, чем у млекопитающих, и называется эффектом Рута.
Молочная кислота вытесняет кислород из гемоглобина, и поэтому
-напряжение кислорода в крови, оттекающей от плавательного пу-
зыря, повышено (рис. 11.31). Теперь в венозном капилляре оно
выше, чем в артериальном, и кислород диффундирует в этот по-
следний. Так происходит, пока молочная кислота добавляется к
венозной крови, которая, таким образом, выходит из сети с мень-
шим количеством кислорода, чем в притекающей артериальной
крови. Благодаря противотоку молочная кислота тоже задержива-
ется в петле, что усиливает ее действие.
В этой системе важно четко различать количество и напряже-
ние. Количество кислорода в венозной крови, вытекающей из чу-
десного сплетения, должно быть меньше, чем в поступающей арте-
634 Гл. 11. Мышцы, движение, локомоция
риальной крови, но для того, чтобы кислород диффундировал из
венозных капилляров в артериальные, его напряжение должно
быть выше в венозных капиллярах. Это более высокое напряжение
создается под влиянием кислоты, которую вырабатывает газовая
железа. Так кислород непрерывно возвращается в артериальный
капилляр и постепенно накапливается в петле, где его концентра-
Рис. 11.30. Схема противоточной умножающей системы в плавательном пузыре
рыбы. Многочисленные капилляры чудесного сплетения представлены на схеме
одной петлей. По мере того как газовая железа вырабатывает молочную кислоту,
напряжение кислорода в венозных капиллярах растет и газ диффундирует в
артериальные капилляры, оставаясь, таким образом, в петле. Оттекающая веноз-
ная кровь содержит меньше кислорода, чем притекающая артериальная.
ция может стать очень высокой. Как работает сама газовая желе-
за, детально не известно, но описанная система объясняет, каким
образом в крови, поступающей в железу, кислород накапливается
в очень высоких концентрациях.
Описанный механизм получил экспериментальное подтвержде-
ние. Норвежскому исследователю Дж. Стену (Steen, 1963) уда-
лось взять очень малые пробы крови из артерии и вены rete mira-
bile угря во время секреции кислорода. И действительно, оказа-
лось, что содержание кислорода в венозной крови было понижено,
а его напряжение повышено в результате образования молочной
кислоты и двуокиси углерода в газовой железе.
Теперь мы видим, что во время секреции rete mirabile пред-
ставляет собой противоточную умножающую систему. Небольшая
разница в напряжении кислорода между венозной и артериальной
кровью умножается на протяжении ее капилляров. Следовательно,
чем длиннее капилляры, тем сильнее эффект умножения. Это опять-
таки согласуется с фактом гораздо большей длины капилляров чу-
десного сплетения у рыб, живущих на большей глубине. Умножение
требует затраты энергии, как любой процесс переноса веществ
Плавучесть 635
против градиента; в данном случае необходимую движущую силу
создает молочная кислота, выделяемая газовой железой.
Описанная система основана на использовании кислоты, кото-
рая понижает сродство гемоглобина к кислороду. Однако на газы
крови может влиять не только кислота; любое растворенное в кро-
ви вещество понижает растворимость в ней газов. Это можно было
7.5
6. 1-6.2
f 6.8—“ .0 Золотист ской с ый мор- зкунъ
100
60
20
Давление 0%, агпм
Рис. 11.31. На кривых кислородной диссоциации для крови рыбы виден выражен-
ный эффект кислотности, называемый эффектом Рута. Верхняя кривая на каж-
дом графике — для необработанной крови при 4 °C, нижняя кривая — для крови
с молочной кислотой, добавленной в количестве, создающем указанную величи-
ну pH. Полоска у оси абсцисс показывает, на каких глубинах обитает рыба.
Обратите внимание, что давление О2 дано по оси абсцисс в атмосферах.
,бы назвать эффектом высаливания. Таким образом, и молочная
кислота наряду с эффектом Рута оказывает также высаливающее
действие на газы крови — не только на кислород, но и на другие
газы, включая азот.
636 Гл. 11. Мышцы, движение, локомоция
Поскольку добавление растворимого вещества понижает раство-
римость азота, чудесное сплетение действует как умножитель не
только в отношении кислорода, но и в отношении азота, и теперь
понятно, каким образом в плавательном пузыре могут накапли-
ваться инертные газы. До того как был понят эффект высалива-
ния, очень трудно было объяснить, как может происходить секре-
ция какого-либо инертного газа. На такую возможность еще в
1934 году указал Генри Кох (Koch), но на нее почти не обратили
внимания до конца 60-х годов; к этому времени накопилось доста-
точно экспериментальных данных, позволивших ясно понять то, ка-
ким образом чудесное сплетение выполняет функцию противоточ-
ной умножающей системы.
Но эффект высаливания, по-видимому, не так действен, как эф-
фект Рута. Тщательные расчеты показывают, что эффект высали-
вания от повышения в крови содержания солей на 0,02 моль/л со-
здает концентрацию азота, соответствующую 25 атм, тогда как
благодаря эффекту Рута добавление молочной кислоты в коли-
честве всего лишь 0,005 моль/л (такая концентрация имеет место-
в период секреции газа у угрей) могло бы создать давление О2
около 3000 атм (Kuhn et al., 1963).
Еще одно открытие, по-видимому, окончательно разрешает за-
гадку. Когда в кровь рыбы добавляется кислота и кислород вытес-
няется из гемоглобина, этот эффект называют off-реакцией Рута^
Обратный процесс, когда кислород снова связывается с гемоглоби-
ном при повышении pH, называют оп-эффектом Рута. Установлен
интересный факт, а именно, что скорости off- и on-эффектов Рута
совершенно различны. Период полуосуществления для off-реакции
равен 50 мс (при 23°C), а для on-реакции это будет величина по-
рядка 10—20 с. Когда кислота диффундирует в артериальные ка-
пилляры сплетения, кровь в них быстро отдает свой кислород.
Но когда та же самая кровь течет по венозным капиллярам и мо-
лочная кислота диффундирует из нее в артериальные капилляры,,
кислород вступает в соединение с гемоглобином не сразу — он не-
которое время остается несвязанным. Иными словами, сохраняется
высокое напряжение кислорода. Таким образом, медленное осу-
ществление on-эффекта Рута позволяет большой доле гемоглобина
выходить из венозных капилляров в неоксигенированном виде,,
хотя напряжение кислорода может быть все еще весьма высоким
(Berg, Steen, 1968).
Если функция rete mirabile как противоточной умножающей
системы ясна, то функция секреторного эпителия самой газовой же-
лезы изучена недостаточно. Один из нерешенных вопросов состоит
в том, каким образом газ переходит из капилляров в газовую фазу
самого пузыря. Роль секреторного эпителия в этом процессе не
раскрыта.
* я»
*
Плавучесть 63Т
Мы видели, как движение животного осуществляется при помо-
щи различных механизмов, создающих силу. Лежащий в их основе
принцип — скольжение смежных микрофиламентов относительно
друг друга — по-видимому, является универсальным механизмом
создания силы. У всех животных, кроме самых мелких, для локо-
моции используются силы, приложенные к элементам скелета —
либо гидравлического, либо твердого, который может быть внут-
ренним (например, у позвоночных) или наружным (например,,
у членистоногих). В локомоторных адаптациях животных выявля-
ется много интересных принципов биомеханики, в том числе различ-
ные способы создания нейтральной плавучести, благодаря которой,
животные могут оставаться на определенном уровне в воде без за-
траты энергии.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Abbott В. С., Lowy J. (1953). Mechanical properties of Mytilus muscle, J. Physiol.,
120, 50P.
Alexander R. McN. (1968). Animal Mechanics, London, Sidgwick and Jackson,
346 pp.
Alexander R. McN. (1974). The mechanics of jumping by a dog (Canis familiaris),
J. Zool., Lond., 173, 549—573.
Alexander R. McN., Vernon A. (1975). The mechanics of hopping by kangaroos
(Macropodidae), J. Zool., Lond., 177, 265—303.
Allen R. D., Francis D., Zeh R. (1971). Direct test of the positive pressure gradient
theory of pseudopod extension and retraction in amoebae, Science, 174, 1237—
1240.
Andersen S. 0., Weis-Fogh T. (1964). Resilin: A rubber-like protein in arthropod
cuticle, Adv. Insect. Physiol., 2, 1—65.
Baguet F., Gillis J. M. (1968). Energy cost of tonic contraction of a lamellibranch-
catch muscle, J. Physiol., 198, 127—143.
Barber V. C. (1974). Cilia in sense organs. In: Cilia and Flagella (M. A. Sleigh,,
ed.), pp.403—433, London, Academic Press.
Bennet-Clark H. C., Lucey E. C. A. (1967). The jump of the flea: A study of the
energetics and a model of the mechanism, J. Exp. Biol.. 47, 59—76.
Berg H. C., Anderson R. A. (1973). Bacteria swim by rotating their flagellar fila-
ments, Nature, Lond., 245, 380—382.
Berg T., Steen J. B. (1968). The mechanism of oxygen concentration in the swim-
bladder of the eel, J. Physiol., 195, 631—638.
Blum J. J., Lubliner J. (1973). Biophysics of flagellar motility, Annu. Rev. Bio-
phys. Biomed. Engin., 2, 181—219.
Bone Q. (1966). On the function of the two types of myotomal muscle fibre in
elasmobranch fish, J. Mar. Biol. Assoc. UK, 46, 321—349.
Brokaw C. J., Gibbons I. R. (1975). Mechanisms of movement in flagella and cilia.
In: Swimming and Flying in Nature, vol. 1 (T. Y.-T. Wu, C. J. Brokaw and
C. Brennen, eds.), pp. 89—126, New York, Plenum Press.
Buller A. J., Eccles J. C., Eccles R. M. (1960). Interactions between motoneurons
and muscles in respect to the characteristic speeds of their responses, J. Phy-
siol., 150, 417—439.
Buller A. J., Lewis D. M. (1964). The rate of rise of tension in isometric tetani of
cross-innervated mammalian skeletal muscles, J. Physiol., 170, 67P — 68P.
Burke R. E., Levine D. N., Zajack F. E., Ill, Tasiris P., Engel W. K. (1971). Mam-
malian motor units: Physiological-histochemical correlation in three types in
cat gastrocnemius, Science, 174, 709—712.
638 Гл. И. Мышцы, движение, локомоция
Casella С. (1950). Tensile force in total striated muscle, isolated fibre and sarco-
lemma, Acta Physiol. Scand., 21, 380—401.
Cavagna G. A., Citterio G. (1974). Effect of stretching on the elastic characteri-
stics and the contractile component of frog striated muscle, J. Physiol., 239,
1—14.
Cavagna G. A., Dusman B., Margaria R. (1968). Positive work done by a previously
stretched muscle, J. Appl. Physiol., 24, 21—32.
Cavagna G. A., Saibene F, P., Margaria R. (1964). Mechanical work in running,
J. Appl. Physiol., 19, 249—256.
Cooper S., Eccles J, C, (1930). The isometric responses of mammalian muscles,
- J. Physiol., 69, 377—385.
Dawson T. J., Taylor C. R. (1973). Energetic cost of locomotion in kangaroos, Na-
ture, bond., 246, 313—314.
Denton E. (1960). The buoyancy of marine animals, Sci. Am., 203, 119—128.
Denton E. J. (1961). The buoyancy of fish and cephalopods, Prog. Biophys. Bio-
phys. Chem., 11, 178—234.
Denton E. J. (1971). Examples of the use of active transport of salts and water to
give buoyancy in the sea. Pilos, Trans. R. Soc. bond., B, 262, 277—287.
Denton E. J., Gilpin-Brown J. B. (1959). On the buoyancy of the cuttlefish, Nature,
bond., 184, 1330—1332.
Denton E. J., Gilpin-Brown J. B. (1961). The distribution of gas and liquid within
the cuttlebone, J. Mar. Biol. Assoc. UK, 41, 365—381.
Denton E. J., Gilpin-Brown J. B. (1966). On the buoyancy of the pearly Nautilus,
J. Mar. Biol. Assoc. UK, 46, 723—759.
Denton E. J., Gilpin-Brown J. B., Shaw T. L. (1969). A buoyancy mechanism found
in cranchid squid, Proc. R. Soc. bond., B, 174, 271—279.
Eckert R. (1972). Bioelectric control of ciliary activity, Science, 176, 473—481.
Evans M. E. G. (1972). The jump of the click beetle (Coleoptera: Elateridae):
A preliminary study, J. Zool., bond., 167, 319—336.
Evans M. E. G. (1973). The jump of the click beetle (Coleoptera: Elateridae):
Energetics and mechanics, J. Zool., bond., 169, 181—194.
Gibbons I. R. (1977). Structure and function of flagellar microtubules. In: Interna-
tional Cell Biology (B. R. Brinkley and K. R. Porter, eds.), pp. 348—357, New
York, Rockefeller University Press.
Gibbons I. R., Rowe A. J. (1965). Dynein: A protein with adenosine triphosphatase
activity from cilia, Science, 149, 424—426.
Gordon A. M., Huxley A. F., Julian F. J. (1966). The variation in isometric tension
with sarcomere length in vertebrate muscle fibres, J. Physiol., 184, 170—
192.
Gray J., Lissmann H. W. (1938). Studies in animal locomotion. 7. bocomotory
reflexes in the earthworm, J. Exp. Biol., 15, 506—517.
Greenewalt С. H. (1975). The flight of birds: The significant dimensions, their de-
parture from the requirements for dimensional similarity and the effect on flight
aerodynamics of that departure, Trans. Am. Philos. Soc. (new series), 65 (4),
1—67.
Gross F., Zeuthen E. (1948). The buoyancy of plankton diatoms: A problem of cell
physiology, Proc. R. Soc. bond., B, 135, 382—389.
Hall-Craggs E. С. B. (1965). An analysis of the jump of the lesser galago (Galago
senegalensis), J. Zool., 147, 20—29.
Haxton H. A. (1944). Absolute muscle force in the ankle flexors of man, J. Phy-
siol., 103, 267—273.
Hersey J. B., Backus R. H., Hellwig J. (1962). Sound-scattering spectra of deep
scattering layers in the western North Atlantic Ocean, Deep Sea Res., 8, 196—
210.
Hirst M. (1977). America’s man-powered prizewinner, Flight Int., 112, 1253—
1256.
Hoyle G. (1955). Neuromuscular mechanisms of a locust skeletal muscle, Proc. R,
Soc. bond., B. 143, 343—367.
Литература 639
Hoyle G. (1957). Comparative Physiology of the Nervous Control of Muscular
Contraction, Cambridge, Cambridge University Press, 147 pp.
Hoyle G. (1964). Muscle and neuromuscular physiology. In: Physiology of Mollusca,
vol. ) (К. M. Wilbur and С. M. Yonge, eds.), pp. 313—351, New York, Acade-
mic Press.
Huxley H. E. (1969). The mechanisms of muscular contraction, Science, 164, 1356—
1366.
Huxley H. E. (1973). Muscular contraction and cell motility, Nature Lond., 243,
445—449.
Jahn T. L.f Landman M. D., Fonseca J. R. (1964). The mechanism of locomotion
of flagellates. 2. Function of the mastigonemes of Ochromonas, J. Protozool.,
11, 291—296.
Johnson W. H.f Twarog В. M. (1960). The basis for prolonged contractions in mob
luscan muscles, J. Gen. Physiol., 43, 941—960.
Kesseler H, (1966). Beitrag zur Kenntnis der chemischen und physikalischen Eigen-
schaften des Zellsaftes von Noctiluca miliaris, Veroff. Inst. Meeresforsch. Bre-
merhaven, 2, 357—368.
Koch H. (1934). L’emission de gaz dans la vesicule gazeuse des poissons, Rev.
Quest. Sci., 26, 385—409.
Krogh A. (1929). The Anatomy and Physiology of Capillaries, 2nd ed., New Haven,
Conn., Yale University Press, 422 pp.
Kuffer S. W., Williams E. M. V. (1953a). Small nerve junctional potentials: The
distribution of small motor nerves to frog skeletal muscle and the membrane
characteristics of the fibres they innervate, J. Physiol., 121, 289—317.
Kuffler S. W., Williams E. M. V. (1953b). Properties of the «slow» skeletal muscle
fibres of the frog, J. Physiol., 121, 318—340.
Kuhn W., Ramel A., Kuhn H. J., Marti E, (1963). The filling mechanism of the
swimbladder: Generation of high gas pressures through hairpin countercurrent
multiplication, Experientia, 19,497—511.
Lapennas G. N., Schmidt-Nielsen K. (1977). Swimbladder permeability to oxygen,
J. Exp. Biol., 67, 175—196.
Lighthill J. (1974). Aerodynamic Aspects of Animal Flight. Fluid Sceince Lecture,
British Hydromechanics Research Association, 30pp.
Lowy J., Millman В. M. (1962). Mechanical properties of smooth muscles of cepha-
lopod molluscs, J. Physiol., 160, 353—363.
Ludwig G. D., Blakemore W. S., Drabkin D. L. (1957). Production of carbon mono-
xide and bile pigment by haemin oxidation, Biochem. J., 66, 38P.
Machin К. E. (1958). Wave propagation along flagella, J. Exp. Biol., 35, 796—
806.
Marshall N. B, (1960). Swimbladder structure of deepsea fishes in relation to their
• systematics and biology, Discovery Rep., 31, 1—122.
Naitoh Y., Kaneko H. (1972). Reactivated triton-extracted models of paramecium:
Modification of ciliary movement by calcium ion, Science, 176, 523—524.
Parry D. A., Brown R. H, J, (1959a). The hydraulic mechanism of the spider leg,
J. Exp. Biol., 36, 423—433.
Parry D. A., Brown R. H. J. (1959b). The jumping mechanism of salticid spiders,
J. Exp. Biol., 36, 654—664.
Peachey L. D. (1965). The sarcoplasmic reticulum and transverse tubules of the
frog’s sartorius, J. Cell. Biol., 25, 209—231.
Pennycuick C. J. (1969). The mechanics of bird migration, Ibis, 111, 525—556.
Phleger C. F. (1971). Pressure effects on cholesterol and lipid synthesis by the
swimbladder of an abyssal Coryphaenoides species, Am. Zool., 11, 559—
Pickwell G. V., Barham E. G., Wilson J. W. (1964). Carbon monoxide production by
a bathypelagic siphonophore, Science, 144, 860—862.
Pollard T. D. (1977). Cytoplasmic contractile proteins. In: International Cell Biolo-
gy (B. R. Brinkley and K. R. Porter, eds.), pp. 378—387, New York, Rockefel-
ler University Press.
640 Гл. 11. Мышцы, движение, локомоция
Pringle L W. S. (1949). The excitation and contraction of the flight muscles of in-
sects, J. Physiol., 108, 226—232.
Pringle J. W. S. (1957). Insect Flight, Cambridge, Cambridge University Press,
132 pp.
Pugh L. G. С. E. (1959). Carbon monoxide content of the blood and other observa-
tions on Weddell seals, Nature, Lond., 183, 74—76.
Pugh L. G. С. E. (1971). The influence of wind resistance in running and walking
and the mechanical efficiency of work against horizontal or vertical forces,
J. Physiol., 213, 255—276.
Rayner M. D., Kennan M. J, (1967). Role of red and white muscles in the swim-
ming of the skipjack tuna, Nature, Lond., 214, 392—393.
Ridgway E. B., Ashley С. C. (1967). Calcium transients in single muscle fibers,
Biochem. Biophys. Res. Commun., 29, 229—234.
Rikmenspoel R. (1965). The tail movement of bull spermatozoa: Observations and
model calculations, Biophys. J., 5, 365—392.
Rikmenspoel R., Sinton S., Janick J. J. (1969). Energy conversion in bull sperm
flagella, J. Gen. Physiol., 54, 782—805.
Roeder K. D. (1953). Insect Physiology, New York, Wiley, 1100 pp.
Ruegg J. C. (1971). Smooth muscle tone, Physiol. Rev., 51, 201—248.
Satir P. (1968). Studies on cilia. 3. Further studies on the cilium tip and a «sliding
filament» model of ciliary motility, J. Cell. Biol., 39, 77—94.
Satir P. (1974). The present status of the sliding microtubule model of ciliar mo-
tion. In: Cilia and Flagella (M. A. Sleigh, ed.), pp. 131—142, London, Acade-
mic Press.
Schmidt-Nielsen S., Flood A., Stene J. (1934). On the size of the liver of some
gristly fishes, their content of fat and vitamin A, Kongelige Norske Videnska-
bers Selskab Forhandlinger, 7, 47—50.
Scholander P. F. (1954). Secretion of gases against high pressures in the swimblad-
der of deep sea fishes. 2. The rete mirabile, Biol. Bull., 107, 260—277.
Scholander P. F. (1956). Observations on the gas gland in living fish, J. Cell.
Comp. Physiol., 48, 523—528.
Scholander P. F., van Dam L. (1954). Secretion of gases against high pressures in
the swimbladder of deep sea fishes. 1. Oxygen dissociation in blood, Biol. Bull.,
107, 247—259.
Seymour M. K. (1971). Burrowing behaviour in the European lugworm Aren't col a
marina (Polychaeta: Arenicolidae), J. Zool., 164, 93—132.
Sleigh M. A. (1968). Paterns of ciliary beating, Symp. Soc. Exp. Biol., 22, 131—
150.
Sleigh M. A. (1974). Cilia and Flagella, London, Academic Press, 500 pp.
Sleigh M. A. (1977). Fluid propulsion by cilia and flagella. In: Comparative Phy-
siology: Water, Ions and Fluid Mechanics (K. Schmidt-Nielsen, L. Bolis and
S. H. P. Maddrell, eds.), pp. 255—265, Cambridge, Cambridge University Press.
Sotavalta O. (1953). Recordings of high wing-stroke and thoracic vibration fre-
quency in some midges, Biol. Bull., 104, 439—444.
Squire J. M. (1975). Muscle filament structure and muscle contraction, Annu. Rev.
Biophys. Bioeng., 4, 137—163.
Steen J. B. (1963). The physiology of the swimbladder in the eel Anguilla vulga-
ris. 3. The mechanism of gas secretion, Acta Physiol. Scand., 59, 221—
241.
Stolpe M., Zimmer K. (1939). Der Schwirrflug des Kolibri im Zeitlupenfilm, J. Or-
nithol., 87, 136—155.
Taylor C. R., Caldwell S. L., Rowntree V. J. (1972). Running up and down hills:
Some consequences of size, Science, 178, 1096—1097.
Thys H., Cavagna G. A., Margaria R. (1975). The role played by elasticity in an
exercise involving movements of small amplitude, Pfliigers. Arch., 354, 281—
Phys H., Faraggiana T., Margaria R. (1972). Utilization of muscle elasticity in
exercise, J. Appl. Physiol., 32, 491—494.
Литература 641
Trueman Е. R. (1966). Observations on the burrowing of Arenicola marina (L.),
J. Exp. Biol., 44, 93—118.
Trueman E. R., Packard A. (1968). Motor performances of some cephalopods,
J. Exp. Biol., 49, 495—507.
Tucker V. A. (1973). Bird metabolism during flight: Evaluation of a theory, J. Exp.
Biol., 58, 689—709.
Weber A., Murray J. M. (1973). Molecular control mechanisms in muscle contra-
ction, Physiol. Rev., 53, 612—673.
Weber H. H., Portzehl H. (1954). The transference of the muscle energy in the
contraction cycle, Prog. Biophys. Biophys. Chem., 4, 60—111.
Weis-Fogh T. (1960). A rubber-like protein in insect cuticle, J. Exp. Biol., 37, 889—
907.
Weis-Fogh T. (1973). Quick estimates of flight fitness in hovering animals, inclu-
ding novel mechanisms for lift production, J. Exp. Biol., 59, 169—230.
Weis-Fogh T. (1976). Energetics and aerodynamics of flapping flight: A synthesis.
In: Insect Flight (R. C. Rainey, ed.), pp. 48—72, New York, Wiley.
Wiersma C. A. G. (1961). The neuromuscular system. In: The Physiology of Crusta-
cea, vol. II (T. H. Waterman, ed.), pp. 191—240, New York, Academic
Press.
Wigglesworth V. B. (1972). The Principles of Insect Physiology, 7th ed., London,
Chapman and Hall, 827 pp.
Wilson J. A. (1972). Principles of Animal Physiology, New York, Macmillan.
Wittenberg J. B. (1958). The secretion of inert gas into the swimbladder of fish,
J. Gen. Physiol., 41, 783—804.
Wittenberg J. B. (1960). The source of carbon monoxide in the float of the Portu-
guese man-of-war, Physalia physalis L., J. Exp. Biol., 37, 698—705.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Alexander R. McN. (1968). Animal Mechanics, Seattle, University of Washington
Press, 346 pp. {Имеется перевод: Александер P. Биомеханика. — M.: Мир,
1970.]
Blake J. R., Sleigh M. A. (1974). Mechanics of ciliary locomotion, Biol. Rev., 49,
85—125.
Biilbring E., Brading A. F., Jones A. W., Tomita T. (1970). Smooth Muscle, London,
Arnold, 676 pp.
Carlson F. D., Wilkie D. R. (1974). Muscle Physiology, Englewood Cliffs, N. J.,
Prentice-Hall, 170 pp.
Close R. J. (1972). Dynamic properties of mammalian skeletal muscles, Physiol.
Rev., 52, 129—197.
Cold Spring Harbor Laboratory (1973). The mechanisms of muscle contraction,
Cold Spring Harbor Symp. Quant. Biol., 37, 706 pp.
Gray J. (1968). Animal Locomotion, London, Weidenfeld and Nicolson, 479 pp.
Hill A. V. (1965). Trails and Trials in Physiology: A Bibliography 1909—1964;
with Reviews of Certain Topics and Methods and a Reconnaissance for Further
Research, London, Edward Arnold, 374 pp.
Hubbard J. I. (1973). Microphysiology of vertebrate neuromuscular transmission,
Physiol. Rev., 53, 674—723.
Karpovich P. V., Sinning W. E. (1971). Physiology of Muscular Activity, 7th ed.,
Philadelphia, Saunders, 374 pp.
Lighthill M /. (1969). Hydromechanics of acquatic animal propulsion, Annu. Rev.
Fluid Meeh., 1, 413—446.
Margaria R. (1976). Biomechanics and Energetics of Muscular Exercise, London,
Oxford University Press, 146 pp.
Nachtigall W. (1974). Locomotion: Mechanics and hydrodynamics of swimming in
aquatic insects. In: The Physiology of Insecta, 2nd ed., vol. 3 (M. Rockstein,
ed.), pp. 381—432, New York, Academic Press.
41—1863
642 Гл. 11, Мышцы, движение, локомоция
Pedley Т. J. (ed.) (1977). Scale Effects in Animal Locomotion, London, Academic
Press, 545 pp.
Rainey R. C. (ed.) (1970). Insect Flight, New York, Wiley, 287 pp.
Sleigh M. A. (ed.) (1974). Cilia and Flagella, London, Academic Press, 500 pp.
Squire J. M. (1975). Muscle filament structure and muscle contraction, Annu. Rev.
Biophys. Bioeng., 4, 137—163.
Wainwright S. A., Biggs W, D., Currey J. D„ Gosline J. M. (1976). Mechanical
Design in Organisms, New York, Wiley, 423 pp.
Westfall J. A., et al. (1973). Symposium on invertebrate neuromuscular systems,
Am. Zool., 13, 233—445.
Wu T. Y.-T., Brokaw C. J., Brennen C. (eds.) (1975). Swimming and Flying in Na-
ture, vols. 1 and 2, New York, Plenum Press, 1005 pp.
ГЛАВА 12
ИНФОРМАЦИЯ И ОРГАНЫ ЧУВСТВ
В предшествующих главах были рассмотрены некоторые аспек-
ты внешней среды, существенные для животных, — кислород, пища,
температура, вода. Настоящая глава посвящена вопросу о том, как
животные получают информацию об окружающей их среде и как
они эту информацию используют. Нас будет интересовать: 1) какая
информация может быть получена и 2) как она перерабатывается
и передается в центральную нервную систему. В следующей, по-
следней, главе мы ознакомимся с тем, как используется информа-
ция, поступающая извне и из самого организма, и как регулируют-
ся и интегрируются различные физиологические функции.
Практически все животные нуждаются в информации об окру-
жающей их среде — как в поисках пищи и особей другого пола,
так и при избегании хищников. Они должны находить в ней свой
путь и оценивать ее важные свойства (например, температуру).
Большую часть информации о среде организм получает с по-
мощью специализированных органов чувств. По традиции эти ор-
ганы разделяют на экстероцепторы, отвечающие на внешние стиму-
лы, например на свет и звук, и проприоцепторы, реагирующие на
внутреннюю информацию, например о положении конечностей. Та-
кое деление не имеет большого внутреннего смысла и в лучшем
случае просто удобно. Другая традиционная классификация исхо-
дит из пяти наиболее очевидных чувств человека, к которым отно-
сятся зрение, слух, вкус, обоняние и осязание. В действительности
наш сенсорный аппарат далеко не так ограничен.
Список внешних стимулов, на которые реагируют по крайней
мере некоторые животные, достаточно велик (табл. 12.1). Разные
категории в этом списке не имеют четких границ, и разделение их
несколько произвольно. Но все же ради удобства мы будем при-
держиваться этой последовательности, когда будем рассматривать
возможности и ограничения при использовании разных видов до-
ступной информации. Эта информация естественным образом рас-
падается на три основные категории: информация об электромаг-
нитной и тепловой энергии, о механической энергии и механических
•силах и о химических агентах.
В данном контексте не имеет значения вопрос о том, может ли
информация попадать в центральную нервную систему, минуя ор-
ганы чувств (экстрасенсорное, или внечувственное, восприятие).
В настоящей главе речь пойдет об измеримых физических величи-
нах, которые можно опознавать, описывать и изменять контроли-
41*
644 Гл. 12. Информация и органы чувств
Таблица 12.1
Внешние стимулы, имеющие значение для сенсорного
восприятия
I
Электромагнитная и тепловая энергия
Свет
Инфракрасное излучение
Тепловая энергия: «тепло», «холод»
Электрическая энергия
Магнитная энергия
Механическая энергия и сила
Звук и его отражение (при эхо-локации)
Прикосновение и вибрация
Давление
Сила тяжести
Инерция
Химические агенты
Вкус
Запах
Влажность
руемым образом. Хотя некоторые сенсорные механизмы, возможно,
еще неизвестны (это касается, например, реакции на магнитные
поля), но само слово «экстрасенсорный» по определению значит,
что никакая сенсорная структура в нем не участвует.
Большое разнообразие стимулов и поразительные структурные
различия между органами могут создать впечатление запутанности,
пока не станет ясно, что общая последовательность событий во всех:
(Свет)
(Глаз)
( Клетки
сетчатки)
(Зрительный
нерв)
( Зрительная
кора)
Рис. 12.1. Упрощенная схема последовательности событий при сенсорном вос-
приятии. В качестве примера взято зрительное восприятие, хотя у высших жи-
вотных его организация гораздо сложнее, чем показано на схеме.
случаях сходна (рис. 12.1). Внешний стимул воздействует на вспо-
могательную структуру, которая или очень сложна, как глаз и ухо,
или гораздо проще, как, например, тактильные рецепторы в коже.
Через вспомогательные структуры внешний стимул доходит до од-
ного или нескольких сенсорных нейронов. В разных органах чувств
эти нейроны обладают многими общими свойствами, но различа-
Сенсорная информация — возможности и ограничения 645
ются тем, что каждый их тип особенно чувствителен к одному спе-
цифическому виду стимулов. Сенсорные нейроны сетчатки высоко-
чувствительны к свету, нейроны уха — к вибрациям в пределах ча-
стот, которые мы обозначаем как «звук», и т. д. Однако все сенсор-
ные нейроны реагируют одинаково; они преобразуют стимул в
нервные импульсы, которые по соответствующим сенсорным нер-
вам передаются в центральную нервную систему.
Теперь мы приходим к тому интригующему факту, что все нерв-
ные импульсы, проводимые по разным сенсорным нервам, имеют
в основе своей одну и ту же природу. Например, зрительный нерв
проводит нервные импульсы такого же рода, что и слуховой нерв,
а идентификация первоначального стимула происходит в централь-
ной нервной системе. При искусственной стимуляции слухового
нерва вызываемые ею нервные импульсы воспринимаются мозгом
как «звук», а искусственное раздражение зрительного нерва вос-
принимается как «свет». Большинству из нас это знакомо по соб-
ственному опыту: мы знаем, что механическое давление на глазное
яблоко воспринимается как свет, а от резкого удара по глазу
«из глаз сыплются искры», хотя никакого света при этом не
было.
Возвращаясь к схеме, представленной на рис. 12.1, мы скажем,
что Сенсорные нейроны служат преобразователями, которые полу-
чают информацию извне и кодируют ее в форме импульсов, идущих
по сенсорным нервам1. В центральной нервной системе эти нервные
сигналы снова декодируются и соответствующая информация ин-
тегрируется и используется. Наибольший интерес представляют
преобразование и кодирование информации, с одной стороны, и ее
декодирование и переработка — с другой. Сенсорные нервы служат
линиями передачи, и в нашем контексте это наименее интересная
часть системы.
В этой главе мы сначала рассмотрим, какие виды информации
могут быть получены, а также возможности и ограничения, при-
сущие различным ее видам. Сенсорная информация кодируется и
передается в виде нервных сигналов и снова декодируется в цент-
ральной нервной системе. Поразительно то, что все эти различные
операции производятся нервными клетками, которые действуют в
основе своей одним и тем же способом, основанным на изменении
разности электрических потенциалов по обе стороны клеточной
мембраны. Поэтому мы познакомимся с мембранным потенциалом
нервных клеток и рассмотрим, как генерируются и передаются им-
1 Преобразователь в общем смысле этого слова представляет собой прибор,
который превращает, или «переводит», энергию, поступающую в одной форме,
в другую форму. Например, микрофон — это преобразователь, который переводит
звуковую энергию в электрическую. Преобразователь часто нуждается в допол-
нительном источнике энергии и, таким образом, является, строго говоря, регуля-
тором расхода энергии. При этом на выходе уровень энергии часто бывает выше,
чем на входе (т. е. преобразователь действует также в качестве усилителя).
646 Гл. 12. Информация и органы чувств
пульсы. Затем можно будет вернуться к вопросу о том, как все
богатство поступающей информации просматривается, сортирует-
ся и перерабатывается.
СЕНСОРНАЯ ИНФОРМАЦИЯ -
ВОЗМОЖНОСТИ И ОГРАНИЧЕНИЯ
ОПРЕДЕЛЕНИЕ НАПРАВЛЕНИЯ И РАССТОЯНИЯ
Информация о направлении, откуда поступает стимул, имеет
первостепенное значение, ибо она позволяет животному в зависи-
мости от того, что ему нужно, приблизиться к источнику стимуля-
ции или удалиться от него. Без знания направления это невоз-
можно. Расстояние тоже играет важную роль, хотя обычно не само
по себе, а в сочетании с данными о направлении. Для стимулов
той или иной природы существуют строгие физические ограничения
в отношении того, что вообще возможно в области их восприятия
и чему могут способствовать вспомогательные структуры.
Самым дирекциональным (направленным) чувством является
зрение; у большинства животных это, вероятно, самое главное из
отдельных видов чувств. Причина направленности зрения заключа-
ется, конечно, в том, что лучи света распространяются прямолиней-
но. Кроме информации о местонахождении различных объектов,
зрение дает много сведений об их природе. Хотя зрение — самое
«направленное» чувство, информацию о направлении дают и другие
органы чувств, но с некоторыми неизбежными ограничениями.
В принципе существуют лишь три способа получения информа-
ции о направлении:
1. Сам сенсорный орган дирекционален; это значит, что генери-
руемые им сигналы различны в зависимости от того, повернут ли
он к источнику стимула или от него (как, например, глаз).
2. Сравниваются сигналы, получаемые парой одинаковых сен-
сорных органов. Таким образом можно получить информацию о
направлении источника звука с помощью двух ушей, хотя сам по
себе орган слуха не очень дирекционален.
3. Сигналы сравниваются в пространстве и времени; это дела-
ется путем последовательной оценки их состояния в ряде точек
пространства.
Информацию о расстоянии получить труднее. Здесь первое ме-
сто опять-таки принадлежит зрению; расстояние оценивается с по-
мощью двух глаз путем учета различий между изображениями на
обеих сетчатках. Большинство других органов чувств способно
лишь грубо оценивать расстояния по ослаблению сигналов. Впро-
чем, как мы увидим позже, некоторые слуховые механизмы ис-
пользуются для чрезвычайно точной информации как о направле-
нии, так и о расстоянии.
Сенсорная информация — возможности и ограничения
647
СВЕТ И ЗРЕНИЕ
Зрение, несомненно, способно давать более детальную инфор-
мацию, чем любое другое чувство. При помощи соответствую-
щих вспомогательных структур (глаза) свет создает подробное
изображение окружающей среды как ближней, так и отдален-
ной.
Информация, получаемая с помощью света, зависит от различий:
в его интенсивности, так как равномерно освещенное пространствен
не содержит никакой информации, представляющей интерес. Много
дополнительной информации несут различия в длине волны, или
цвете. Наконец, для животных, обладающих соответствующими
вспомогательными органами, весьма информативной может быть
также плоскость поляризации света. Скорость распространения
света так велика (300 000 км/с), что существенной для животных
задержки не происходит. Это значит, что изменения в окружающей
среде воспринимаются мгновенно, если не считать времени, идуще-
го на преобразование сигнала и передачу его в центральную нерв-
ную систему. Сравните это, например, с тем, как медленно воспри-
нимается изменение обонятельного сигнала.
ГЛАЗА, СОЗДАЮЩИЕ ИЗОБРАЖЕНИЕ
Чувствительность к свету распространена чрезвычайно широко;
ею обладают даже многие одноклеточные организмы. У многокле-
точных животных эта чувствительность обычно сосредоточена в
определенных местах — зрительных пятнах. Если эти пятна защи-
щены от света с одной стороны, то они чувствительны и к его на-
правлению.
У более высокоорганизованных животных светочувствительные
структуры все усложняются и в своей окончательной форме пред-
ставляют собой превосходные приборы, создающие изображение,
т. е. глаза. Такие хорошо развитые глаза имеются у представите-
лей четырех типов животных: у червей, моллюсков, членистоногих
и позвоночных. Глаза различны по строению и развитию, и надо
полагать, что они развились в этих группах животных независимо
друг от друга.
Глаза, создающие изображение, бывают двух типов: фасеточ-
ные, как у насекомых, или с одной линзой, подобные фотокамере,
как у позвоночных (рис. 12.2). У фасеточного глаза разрешающая
способность одна и та же независимо от того, далеко находится
предмет или совсем близко, но глаз с одной линзой (хрусталиком)
должен обладать каким-то фокусирующим приспособлением, чтобы
работать одинаково хорошо на разных расстояниях. Интересно,
что у тех немногих червей, у которых глаза хорошо развиты и
создают изображение, встречаются и фасеточные, и хрусталиковые
(камерные) глаза.
Рис. 12.2. Глаза позвоночных и некоторых беспозвоночных имеют хрусталик и
действуют по тому же принципу, что и фотокамера. Насекомые обладают слож-
ными глазами, в которых изображение создается комбинацией отдельных эле-
ментов. На рисунке показано строение глаза у млекопитающего (А), осьмино-
га (Б), насекомого (В) и кольчатого червя (Г).
Сенсорная информация — возможности и ограничения 649
ДИАПАЗОН ЧУВСТВИТЕЛЬНОСТИ
Те длины волн, которые человек воспринимает как свет, лежат
в очень узком участке — от 380 до 760 нм — того широкого спек-
тра электромагнитного излучения, который охватывает область от
чрезвычайно коротковолновых гамма-лучей до длинных радиоволн
(рис. 12.3).
Видимый
свет
Космиче- Гамма- Рентгенов-
ские лучи личи ские лучи лыпрафиоле!
Ишрракра- Диапазон Телееи- радиоволны
сныч свет радара
1пм
1 нт
1мм
1мкм
1м
1км
ФСзжок
10s
103 1 10'3 10'6
Энергия, ккал на 1моль квантов
1О'Э
Рис. 12.3. Спектр электромагнитных излучений от самых коротких космических
лучей до длинных радиоволн. Как свет воспринимается только узкая полоса
приблизительно от 380 до 760 нм. Энергия, которую несет каждый квант электро-
магнитного излучения, возрастает в 10 раз на каждое десятикратное укорочение
волны. Ф — фиолетовый, С — синий, 3 — зеленый, Ж — желтый, О — оранжевый,
К — красный.
Зрительная чувствительность у всех других животных лежит в
том же или очень близком диапазоне длин волн. Удивительно, что
не только животные, но и растения реагируют на свет в этом же
диапазоне. Это относится и к фотосинтезу, и к ростовым реакциям
на свет (фототропизму) у растений. Причина такого универсаль-
ного значения очень узкой полосы электромагнитного спектра
проста. Энергия, которую несет каждый квант излучения, обрат-
но пропорциональна длине волны. Поэтому энергия кванта, соот-
ветствующего более длинным волнам, недостаточна, чтобы вы-
звать заметный фотохимический эффект, а более короткие волны
(ультрафиолетовые и короче) несут столько энергии, что оказы-
вают на органические вещества разрушительное действие. Уни-
версальное биологическое использование того, что мы называем
«светом», — это результат исключительной «пригодности» именно
этих длин волн.
Их диапазон не в точности один и тот же для всех животных.
Например, зрение насекомых заходит в ближнюю ультрафиолето-
вую часть спектра, т. е. в область несколько более коротких волн,
чем зрение позвоночных (фото 12.1). Это видно из способности
медоносных пчел отличать любой спектральный цвет между 313 и-
615 нм от белого света — способности, которая не зависит от ин-;
тенсивности света и, следовательно, должна быть основана на раз^
659 Гл. 12. Информация и органы чувств
личении длин волны («цветовое» зрение) (Kuhn, 1927). Сетчатка
млекопитающих тоже была бы чувствительна к ультрафиолетовым
лучам, но они не доходят до нее главным образом из-за некоторой
желтизны хрусталика, который действует как фильтр.
Это сужение полосы вос-
принимаемых длин волн,
пожалуй, имеет свои преи-
мущества. Если линза (хру-
сталик) состоит из одно-
родного материала, она
преломляет коротковолно-
вое излучение сильнее, чем
длинноволновое. Это зна-
чит, что лучи с разной дли-
ной волны не могут быть
сфокусированы одновремен-
но. В оптических приборах
это осложнение называется
хроматической аберрацией
и корригируется примене-
нием сложных линз, состоя-
щих из нескольких элемен-
тов с разными показателя-
Фото 12.1. Зрение насекомых. То,
что из раскраски цветка воспри-
нимает пчела, поразительно отли-
чается от того, что видит глаз че-
ловека. Человеческому глазу цве-
ток болотной калужницы (Caltha
palustris) представляется однород-
ным ярко-желтым (вверху); на
снимке в ультрафиолете (внизу)
выявляется резкий рисунок, кото-
рый служит указателем нектара,
направляющим пчел к источнику
пищи. (Фото предоставлено Т. Eis-
ner, Cornell Uniwersity.)
ми преломления. Для глаза, который не обладает цветовой кор-
рекцией, самый простой способ уменьшить трудность одновремен-
ного фокусирования волн разной длины состоит в сужении поло-
сы тех длин волн, которые могут попасть в него. Поэтому лучше
всего устранить самые короткие волны, для которых ошибка ре-
фракции имеет наибольшую величину. Для насекомых с их нефо-
кусирующими сложными глазами это несущественно, так как раз-
решение определяется угловым расстоянием между отдельными
элементами глаза.
Сенсорная информация — возможности и ограничения 651
ПОГЛОЩЕНИЕ СВЕТА: ПИГМЕНТЫ СЕТЧАТКИ
Для того чтобы сенсорные нейроны сетчатки реагировали на
свет, должно быть поглощено достаточное число световых квантов.
Это достигается с помощью светопоглощающего пигмента. Самый
известный из таких пигментов — родоспин, который содержится в
палочках глаза у позвоночных (палочки — это элементы сетчатки,
ответственные за сумеречное зрение, а колбочки, о которых речь
пойдет дальше, функционируют при ярком освещении и участвуют
в восприятии цвета).
Родоспин можно извлечь из сетчатки животного, глаза которого
полностью адаптированы к темноте. Он светочувствителен и, если
подвергнуть его действию света, распадается на ретинен — ве-
щество,, очень близкое к витамину А, и белок, называемый опси-
ном. Это первый этап в рецепции света палочками в глазу позво-
ночного животного.
Самым веским указанием на роль родопсина в зрительном про-
цессе служит тот факт, что если родопсин разрушен воздействием
Рис. 12.4. Относительная чувствительность темноадаптированного глаза человека
к волнам различной длины и спектр поглощения зрительного пигмента (в про,-
центах от максимальной величины). (Crescitelli, Dartnail, 1953.)
на глаз яркого света, то глаз становится нечувствительным к сла-
бому свету, пока родопсин не регенерируется. Другой убедитель-
ный довод — то, что в спектре поглощения родопсина максимум на-
ходится при 497 нм. Этот максимум и весь спектр поглощения
совпадают с максимумом и спектром чувствительности человече-
ского глаза при слабом свете (рис. 12.4).
652 Гл. 12. Информация и органы чувств
Труднее было установить роль пигментов в восприятии света
колбочками, прежде всего из-за тех сложностей, с которыми связа-
но извлечение соответствующих пигментов. Давно уже было ясно,
что для восприятия цвета недостаточно одного пигмента и что на-
личием трех пигментов с пиками поглощения в синей, зеленой и
красной частях спектра можно объяснить различение всех видимых
нами цветов (трихроматическая теория зрения).
Трудности были преодолены путем измерения спектров погло-
щения непосредственно в человеческом глазу. Проводя с помощью
остроумных приборов измерения на отдельных колбочках, удалось
показать, что имеются три вида колбочек: чувствительные к сине-
му свету — с максимумом поглощения около 450 нм, к зеленому —
с максимумом поглощения около 525 нм и к красному — с макси-
мумом поглощения при 555 нм. По-видимому, с колбочками этих
трех типов и связано цветовое зрение человека на ярком свету
.(Brown, Wald, 1964).
Спектральная чувствительность человеческого глаза согласует-
ся с трехпигментной теорией и измеренными спектрами поглоще-
ния. Кроме того, все, что известно о дефектах цветового зрения
у человека (часто называемых цветовой слепотой), находит удов-
летворительное объяснение на основе свойств тех же-трех пигмен-
тов (Wald, 1964; Rushton, 1972).
ПОЛЯРИЗОВАННЫЙ СВЕТ
Одно свойство света, не воспринимаемое человеком, — это пло-
скость поляризации1. По этой причине информация, которую несет
поляризованный свет, и ее значение для животных оставались не-
замеченными, пока немецкий зоолог Карл Фриш не обнаружил,
что медоносные пчелы пользуются плоскостью поляризации как по-
казателем направления.
Фриш изучал общение между пчелами и стремился выяснить,
каким образом пчела, нашедшая хороший источник пищи, передает
информацию об этом другим пчелам. Обычно пчелы указывают на-
правление к источнику пищи относительно направления на солнце
(т. е. пользуются солнцем как компасом), и они обладают внутрен-
1 В поляризованном свете все колебания распространяющихся волн (луча)
происходят в одной плоскости, перпендикулярной лучу. В обычном неполяри-
зованном свете колебания совершаются по всем направлениям (но тоже перпен-
дикулярно лучу). Неполяризованный свет может стать поляризованным, отра-
зившись от воды и других неметаллических поверхностей или пройдя через
определенные вещества. Плоскость поляризации может нести информацию, ко-
торой не содержит неполяризованный свет. По-видимому, в природе самая важная
информация, которую животные получают таким путем, связана с тем, что свет
голубого неба поляризован, причем поляризация в любой точке неба зависит от
ее положения относительно солнца. Это косвенным образом дает сведения о по-
ложении солнца, чрезвычайно важные для ориентации и навигации животных.
Сенсорная информация — возможности и ограничения 653
ними часами, которые «компенсируют» перемещение солнца по
небосклону.
Оказалось, однако, что пчелы ориентируются правильно даже
тогда, когда солнца не видно, — достаточно, чтобы был виден не-
большой кусок голубого неба. Свет голубого неба поляризован
настолько, что дает информацию об истинном положении солнца,
и пчелы это используют. Один из способов показать их зависи-
мость от плоскости поляризации света состоит в том, чтобы отра-
зить свет зеркалом. Это заставляет пчел ориентироваться в сторо-
ну, противоположную верному направлению, как и следовало ожи-
дать при повороте плоскости поляризации в результате отражения
в зеркале. Точно так же с помощью фильтров, изменяющих на-
правление поляризации, пчелам можно навязать любое отклонение
от верной ориентации (von Frisch, 1948).
После того как было показано значение поляризованного света
для пчел, установили, что и другие насекомые, как правило, чув-
ствительны к нему. Неожиданным оказалось то, что и некоторые
водные животные чувствительны к поляризации света. Впервые та-
кое наблюдение было сделано на мечехвосте (Limulus) (Water-
man, 1950). Многие другие водные животные, в том числе осьмино-
ги и рыбы, тоже обладают такой чувствительностью (Waterman,
Forward, 1970).
Значительное количество данных показывает, что способность
определять плоскость поляризации света играет важную роль в
ориентации и навигации не только сухопутных, но и многих водных
животных (Waterman, Horch, 1966).
ТЕМПЕРАТУРА
Мы склонны думать, что мы ощущаем температуру окружаю-
щей среды, и наше поведение, казалось бы, нередко подтверждает
это. Но на самом деле ощущается не окружающая температура,
а температура кожи на глубине соответствующих рецепторов.
Нашу неспособность судить о температуре воздуха хорошо демон-
стрируют электрические сушилки для рук. Сушилка выпускает
струю горячего воздуха (температурой до 100°C), и пока кожа
влажная, воздух кажется умеренно теплым, но как только она вы-
сыхает, воздушная струя становится болезненно горячей.
Тепло и холод воспринимаются в коже разными рецепторами.
Это можно показать, исследуя активность отдельных нервных во-
локон, идущих от кожных рецепторов, чувствительных к темпера-
туре. Волокна от рецептора, чувствительного к теплу, неактивны,
пока температура не поднимется выше определенной точки, начи-
ная с которой активность растет приблизительно пропорционально
повышению температуры, пока не будет достигнут некоторый верх-
ний предел, выше которого активность снова снижается.
654 Гл, 12. Информация и органы чувств
Именно это показывают записи, получаемые от кожи мошонки
у крысы (рис. 12.5). Тепловые рецепторы «молчат» примерно до
температуры 37 °C. Выше этой точки их активность быстро растет,
а в области выше 43 °C снова падает. Ниже 37 °C тепловые рецеп-
торы молчат, а ответ холодовых рецепторов усиливается, опять-таки
Рис. 12.5. Кривые ответов, полученных от одиночных тепловых и холодовых ре-
цепторов в коже мошонки у крыс. (Iggo, 1969.)
приблизительно пропорционально падению температуры. Ниже оп-
ределенного уровня их активность уменьшается и в конце концов
падает до нуля.
В узком нейтральном диапазоне, который соответствует нор-
мальной температуре кожи мошонки у крысы в состоянии теплово-
го равновесия, наблюдается низкий уровень активности как тепло-
вых, так и холодовых рецепторов. Поэтому даже незначительный
сдвиг в ту или другую сторону воспринимается быстро и точно.
Это делает возможной чувствительную систему контроля, основан-
ную на получении точной информации о небольших изменениях
температуры кожи, а значит, и об изменениях теплообмена между
кожей и окружающей средой.
ИНФРАКРАСНОЕ ИЗЛУЧЕНИЕ
Те кожные терморецепторы, о которых шла речь выше, чувстви-
тельны к локальной температуре кожи. Они информируют о внеш-
ней среде только косвенно, так как температура кожи определяет-
ся балансом между притоком тепла изнутри тела и отдачей его пу-
тем теплопроводности и излучения в окружающую среду. Эти
рецепторы обнаруживают излучение только по его влиянию на темпе-
ратуру кожи; таким образом мы можем, например, ощущать излу-
чение от горячей печки или какого-нибудь другого горячего пред-
мета. До известной степени можно также ощущать потерю тепла
Сенсорная информация — возможности и ограничения 655
•с излучением. Например, холодная кирпичная стена дома может
вызвать определенное ощущение «холода»; мы чувствуем, что хо-
лод идет от стены, и часто воспринимаем это как «поток холод-
ного воздуха», хотя совершенно очевидно, что никакой воздух че-
рез стену не проходит.
Некоторые животные воспринимают инфракрасное излучение
непосредственно: они обладают особыми органами чувств, реаги-
рующими на радиацию этого типа. Так называемые лицевые ямки
на голове у некоторых змей представляют собой такие специализи-
рованные рецепторы инфракрасных лучей.
Когда гремучая змея нападает, то направление броска опре-
деляется инфракрасным излучением, исходящим от ее жертвы. Гре-
мучая змея бросается только на>. теплокровных животных, а если
добыча мертва и тело ее имеет температуру окружающего воздуха,
змея не нападает. Однако ослепленная змея делает точный бросок
на перемещаемую мертвую крысу при условии, что эта крыса теп-
лее воздуха. Слепая змея не может руководствоваться зрением. Не
направляет ее и обоняние, так как она схватывает даже передви-
гаемую электрическую лампочку, завернутую в ткань. Очевидно,
в определении локализации теплых предметов участвует лицевая
ямка. Все змеи, обладающие лицевыми ямками, питаются преиму-
щественно теплокровной добычей, и это тоже говорит в пользу
того, что эти органы чувствительны к инфракрасному излучению.
У гремучей змеи ямки лежат по одной с каждой стороны головы
между ноздрей и глазом; они обильно иннервированы, что само по
себе наводит на мысль о сенсорной функции этого органа.
Чувствительность лицевой ямки исследовали, регистрируя ак-
тивность в идущем от нее нерве. Разнообразные стимулы — звук,
вибрация или свет умеренной интенсивности (с отфильтрованной
инфракрасной частью спектра) —не оказывают заметного влияния
на активность нерва. Но если к голове приближают предмет,
температура которого отличается от температуры среды, актив-
ность нерва резко изменяется независимо от температуры возду-
ха между предметом и головой (Bullock, Cowles 1952; Barret et al.,
1970).
Как происходит рецепция инфракрасного излучения? Ямка по-
крыта тонкой прозрачной перепонкой (рис. 12.6), и было высказа-
но предположение, что повышение температуры в камере за пере-
понкой вызывает расширение газа; в результате перепонка дефор-
мируется, и эту деформацию воспринимает соответствующий рецеп-
тор. Такая гипотеза весьма мало правдоподобна, так как разрез в
перепонке, открывающий доступ в камеру наружному воздуху, не
снижает чувствительности. Такой результат несовместим с гипоте-
зой о том, что ощущается изменение давления.
Остается рассмотреть две другие возможности. Либо инфра-
красное излучение поглощается специфическим веществом, анало-
гичным светочувствительным пигментам глаза, либо лицевая ямка
656 Гл. 12. Информация и органы чувств
Наружная Перепонка
Внутренняя номера
Рис. 12.6. Органы лицевой ямки на голове
гремучей змеи представляют собой специа-
лизированные рецепторы инфракрасного
излучения, которые служат для обнаруже-
ния теплокровной добычи. Две ямки, на-
правленные вперед, лежат на обеих сторо-
нах между глазом и ноздрей. Ямка состоит
из наружной и внутренней камер, разде-
ленных тонкой перепонкой. (Gamow, Har-
ris, 1973.)
чувствительна к легкому повышению температуры, вызываемому
поглощением инфракрасных лучей.
Инфракрасное излучение, испускаемое телом млекопитающего,
имеет пик в области 10 000 нм, и малая энергия кванта при этой
длине волны делает какое-либо фотохимическое изменение пигмен-
та совершенно невероятным. Чистое инфракрасное излучение мо-
жет быть получено от лазера, и в опытах с таким излучением из-
вестной длины волны были
получены веские данные в
пользу того, что механизм
рецепции в лицевой ямке
чисто тепловой (Harris, Ga-
mow, 1971).
Могут ли лицевые ямки
служить для стереоскопиче-
ского восприятия инфра-
красного источника подобно
тому, как наши два глаза ис-
пользуются для стереоско-
пического зрения? Это ка-
жется весьма вероятным —
на это указывает не толь-
ко точность броска на жерт-
ву, но и данные об актив-
ности головного мозга. Ког-
да инфракрасное излучение
падает на лицевую ямку,
электрические потенциалы
можно зарегистрировать в
tectum opticum — той части
головного мозга, с которой
связан зрительный нерв.
Это интересно само по себе:
хотя нерв, отходящий от лицевой ямки (ветвь тройничного нерва),
идет совершенно отдельно от зрительного нерва, по-видимому,
один и тот же отдел мозга перерабатывает и зрительную инфор-
мацию, и сигналы от рецепторов инфракрасного излучения.
Многие нейроны в tectum opticum отвечают на раздражение ли-
цевой ямки противоположной стороны головы. Это напоминает то,
что происходит с информацией от глаз: перекрест зрительного нер-
ва (хиазма) необходим при бинокулярном зрении для оценки рас-
стояния. По-видимому, таким же образом интегрируется и оцени-
вается информация от двух лицевых ямок; этот вывод согласуется
с данными об изменениях нейронной активности в tectum opticum
при перемещении инфракрасного источника. Ответ становится осо-
бенно заметным при таком положении источника, когда излучение
падает одновременно на обе ямки. Поэтому можно думать, что
Сенсорная информация — возможности и ограничения 65 7
лицевые ямки действительно обеспечивают стереоскопическое вос-
приятие и этим значительно способствуют точной локализации до-
бычи (Goris, Terashima, 1973).
Мало вероятно, чтобы специализированные органы, чувстви-
тельные к инфракрасным лучам, имели значение для многих жи-
вотных. В частности, почти невозможно представить себе, чтобы
подобный орган был полезен для водных животных, отчасти из-за
очень малой проницаемости воды для инфракрасного излучения,
а отчасти потому, что прямой контакт поверхности тела со средой,
обладающей высокой теплопроводностью и теплоемкостью, делал
бы невозможным восприятие небольших количеств тепла, прино-
симых излучением.
«ЖИВОТНОЕ ЭЛЕКТРИЧЕСТВО»
Как известно, некоторые рыбы способны производить сильные
электрические разряды. Народы древности — греки и египтяне —
знали и писали о мощных «ударах», наносимых электрическим ска-
том (Torpedo) и электрическим сомом (Malapterurus), но это яв-
ление представлялось совершенно загадочным, пока не стала из-
вестна природа электричества.
Мощные электрические разряды служат очевидным целям на-
падения и защиты. Самые сильные разряды производит южноаме-
риканский электрический угорь (Electrophorus)} они достигают
500—600 В и настолько сильны, что убивают других рыб, а воз-
можно, и животных величиной с человека. Такой сильный разряд
способны производить лишь несколько видов рыб, притом не очень
близких между собой: одни относятся к костистым, другие — к
пластиножаберным; они встречаются как в пресных водах, так и
в море.
Когда электрических рыб изучили более детально, стало ясно,
что даже слабые разряды — настолько слабые, что они не оказы-
вают прямого действия на других рыб, — служат разнообразным
полезным целям. Очевидно, что очень слабые разряды не могут ог-
лушить жертву и бесполезны как орудие нападения или защиты.
Тем не менее теперь выяснилось, что они могут использоваться для
получения информации об окружающей среде. Электрические раз-
ряды особенно пригодны для отличения неэлектропроводных пред-
метов от хороших проводников, каковы, например, другие живот-
ные. Пресноводное животное, скажем рыба, обладает гораздо
большей электропроводностью (благодаря содержанию солей в
жидкостях его тела), чем плохо проводящая ток вода. Слабые
электрические разряды могут быть использованы для ориентации
в мутной воде, а также для обнаружения хищников и добычи. Кро-
ме того, они служат для коммуникации между особями, и в некото-
рых случаях рыба отвечает половым поведением только на элек-
трические сигналы особей другого пола (Black-Cleworth, 1970; Hop-
kins, 1972).
658 Гл. 12. Информация и органы, чувств
Наконец, недавно стало известно, что многие рыбы, не способ-
ные сами подавать электрические сигналы, весьма чувствительны
к слабым электрическим полям, возникающим при обычной мы-
шечной активности других организмов. Таким образом акулы и
скаты могут обнаруживать других рыб, даже если те неподвижны.
Голодная акула Scyliorhinus реагирует даже на камбалу, которая
Рис. 12.7. Пищевые реакции небольшой акулы Scyliorhinus canicula.
А. Когда голодная акула проплывает около камбалы, полностью зарывшейся
в песок, она обнаруживает ее и немедленно нападает. Б. Чтобы исключить обоня-
тельные сигналы, камбалу накрывают агаровой камерой, которую перфузируют
водой, выходящей на некотором расстоянии (прерывистая стрелка). Тем не ме-
нее акула бросается точно в то место, где находится жертва. В. Когда в агаро-
вую камеру положены куски рыбы, акула ищет пищу там, где вытекает пер-
фузат. Г. Если покрыть агаровую камеру пластиковой пленкой, сигнал стано-
вится слабее и акула проплывает мимо, не замечая его. Д. Искусственное элек-
трическое поле той же величины, что и создаваемое дыхательными движениями
камбалы, возбуждает акулу, которая немедленно атакует «добычу». (Kalmijn,
1971.)
спокойно лежит на дне аквариума, полностью закрытая песком.
Если акула проплывает на расстоянии менее 15 см от нее, она вне-
запно поворачивается к укрывшейся жертве, удаляет песок, всасы-
вая его, а затем выбрасывая, и схватывает добычу (рис. 12.7).
Тщательные исследования показали, что эта способность обна-
руживать спрятанную камбалу не связана с обонянием. По-види-
мому, акула отвечает на очень слабые электрические потенциалы,
создаваемые дыхательными движениями укрытого животного, так
как она реагирует на искусственный электрический разряд в 4 мкА
(того же порядка величины, что и разряд, производимый живой
Сенсорная информация — возможности и ограничения 659
камбалой) и пытается выкопать воображаемую добычу (Kalmijnr
1971).
Итак, электрические рыбы могут использовать свои разряды для
поражения добычи, для получения информации об окружающей
среде (гораздо чаще) и для коммуникации. Неэлектрические рыбы
способны при помощи своих чрезвычайно чувствительных электро-
рецепторов определять местонахождение добычи по потенциалам
действия, возникающим при обычной мышечной активности.
ГЕНЕРИРОВАНИЕ ЭЛЕКТРИЧЕСКИХ РАЗРЯДОВ
Почти у всех электрических рыб разряды производятся особы-
ми электрическими органами, которые состоят из видоизмененной
мышечной ткани. Эти органы весьма тщательно изучены у электри-
ческого угря, который может давать разряды больше 500 В. Круп-
Состояние покоя
+84 мВ
»0мВ
+84мВ
Голова
А
Хбос/п
Рис. 12.8. Электрические органы рыбы состоят из тонких уплощенных клеток
(электрических пластинок), которые собраны в столбики по нескольку тысяч.
Когда пластинка неактивна (вверху), обе ее поверхности заряжены положитель-
но ( + 84 мВ), так что между ними нет разности потенциалов. Во время разряда
(внизу) знак потенциала на задней поверхности пластинки меняется на обратный
и становится равным —67 мВ. Поэтому разность потенциалов между обеими
поверхностями составляет (84 +67) мВ — 151 мВ. (Keynes, Martins-Terreira.
1953.)
ные электрические органы идут у него почти по всей длине тела
в виде большой массы с каждой стороны, составляя около 40%
всего объема тела.
Эти органы состоят из уплощенных клеток, называемых элект-
рическими пластинками. Пластинки собраны в столбики, в которых
может быть от 5000 до 10 000 пластинок, а таких столбиков бывает
42
^660 Гл. 12. Информация и органы чувств
около 70 на каждой стороне тела. Две поверхности каждой элек-
трической пластинки заметно отличаются друг от друга: одна по-
верхность снабжена густой сетью нервных окончаний, а другая
изрезана глубокими складками и извилинами.
Когда орган находится в состоянии покоя, обе поверхности пла-
стинок заряжены положительно по отношению к внутренности
пластинки, что создает потенциал +84 мВ (рис. 12.8). Поэтому
разность потенциалов между внешними поверхностями обеих мем-
бран равна нулю. Но при разряде знак потенциала иннервирован-
ной поверхности меняется на обратный и общая разность потен-
циалов между обеими сторонами одной пластинки достигает около
150 мВ. Ввиду последовательного расположения электрических
пластинок напряжение суммируется, как при последовательном
соединении ряда батарей. Поскольку таким образом соединено не-
сколько тысяч электропластинок, электрический угорь может со-
здать напряжение в несколько сотен вольт. (Из-за внутренних по-
терь напряжение не достигает полностью величины, получаемой
при идеальном последовательном соединении пластинок.)
Расположение электрических пластинок параллельными столби-
ками усиливает ток. У электрического угря он может быть равен
около 1 А. Гигантский электрический скат (Torpedo} обитает в
морской воде, сопротивление которой гораздо ниже, чем у пресной
воды. Напряжение у ската меньше (около 50 В), но поскольку
каждый электрический орган состоит приблизительно из 2000 па-
раллельно включенных столбиков, суммарный ток получается не-
обычайно сильным — до 50 А. Ток в 50 А при 50 В соответствует
при разряде мощности 2500 Вт (Keynes, Martins-Ferreira, 1953).
.ЭЛЕКТРОРЕЦЕПТОРЫ
У большинства рыб в коже имеется множество сенсорных орга-
нов, расположенных главным образом на голове и вдоль боковой
.линии. Один вид таких органов — ампулы Лоренцини — имеет ха-
рактерную форму маленьких бутылочек. Эти ампулы находятся на
поверхности головы у всех морских пластиножаберных рыб
(рис. 12.9), и подобные им рецепторы есть у многих костистых рыб.
Соответствующие перерезки нервов и поведенческие опыты пока-
зали, что ампулы Лоренцини у акулы Scyliorhinus представляют
собой электрорецепторы (Dijkgraaf, Kalmijn, 1963). У электриче-
ских рыб такие рецепторы делятся на несколько разных типов;
чаще всего они снабжены каналом, который идет от поверхности
цожи к расширенной ампуле, лежащей в глубине кожи (рис. 12.10).
Электрорецепция детально изучена у африканской электриче-
ской рыбы Gymnarchus niloticus (Machin, Lissman, 1960). Эта
рыба, имеющая только латинское название, разряжается непре-
рывным потоком импульсов с частотой около 300—400 в секунду.
При каждом разряде кончик ее хвоста на мгновение становится
электроотрицательным по отношению к голове, и через окружаю-
Сенсорная информация — возможности и ограничения 661
щую воду течет электрический ток. Конфигурация электрического
поля зависит от электропроводности среды, и поле деформируется,
Рис. 12.9. Положение органов чувств на голове кошачьей акулы (Scyliorhinus).
Отверстия ампул Лоренцини представлены черными точками. Белыми кружками
указаны поры системы боковой линии, а черными линиями — ее расположение.
(Dijkgraaf, Kalmijn, 1963.)
если в него попадает предмет с более высокой или более низкой
проводимостью. Линии тока сходятся на предмете с более высокой
проводимостью и расходятся вокруг плохого проводника.
Сенсорные органы рыбы чувствительны к таким изменениям
поля. Но они отвечают не на одиночные импульсы, а на среднюю
рис. 12.10. Схема трех типов кожных ампулярных рецепторов электрической
рыбы Eigenmannia virescens. Рецепторные клетки (изображены более темными,
а их нервы — черными) лежат на конце длинного канала, выходящего на поверх-
ность кожи. (Lissmann, Mullmger, 1968.)
силу тока примерно за 25 мс — время, достаточное для эмиссии
7—10 разрядов. Общий ток, необходимый для возбуждения от-
дельного электрорецептора, чрезвычайно мал — около 3-10"15 А
(Machin, Lissman, 1960).
Многие электрические рыбы живут в мутной воде, где види-
мость плохая, и часто глаза у них развиты плохо. Значение элек-
662 Гл. 12. Информация и органы чувств
трорецепции, позволяющей сканировать окружение и находить
добычу, в таких условиях очевидно. Такое чувство имеет то преиму-
щество перед зрением, что эта система не зависит от суточного све-
тового цикла, а недостаток ее — ограниченная дальность восприя-
тия.
Как обстоит дело с интерференцией сигналов? Если на одном и
том же пространстве живет много электрических рыб, то как они
не путают собственные сигналы с сигналами других особей? Если
электрическую рыбу подвергнуть действию искусственных электри-
ческих импульсов той же частоты, что и ее собственные, то она
реагирует тем, что уводит их частоту от частоты искусственного
сигнала. По-видимому, это позволяет рыбе избегать смешения сво-
их сигналов с сигналами от других рыб. Такой вывод подтвержда-
ется тем, что в природе электрическая рыба нередко меняет часто-
ту своих импульсов в случае приближения другой рыбы с такой
же частотой (Bullock et al., 1972).
РЕЦЕПЦИЯ МАГНИТНЫХ ПОЛЕЙ
Теперь уже нет сомнения в том, что многие животные реагиру-
ют на магнитные поля, хотя бесчисленные эксперименты с искус-
ственными магнитами не дали ясных результатов. Но тот факт,
что магнитное поле оказывает на животное какое-то действие, еще
не означает, что в природе это животное воспринимает магнитное
поле или использует его.
По-видимому, одна из причин, почему многие опыты не выяви-
ли никакого эффекта, состоит в том, что экспериментаторы приме-
няли очень сильные магнитные поля. Между тем поля такой же
величины, как естественное магнитное поле Земли, систематически
влияют на направление движения брюхоногих моллюсков Nassa-
rius и планарий Dugesia. При существенном ослаблении искусст-
венного магнитного поля реакция тоже становится минимальной.
Видимо, эта реакция (ориентация «по компасу») великолепно при-
способлена к силе естественного геомагнетизма, и этим может объ-
ясняться неудача множества ранних опытов (Brown et al., 1964).
Многие другие беспозвоночные тоже реагируют на слабые маг-
нитные поля, но значение их реакций часто трудно определить.
Можно полагать, что если реакция не носит просто случайного ха-
рактера, она существенна для выбора направления и ориентиров-
ки, как это установлено для медоносных пчел. Когда пчелы сооб-
щают другим рабочим пчелам о направлении к источнику пищи,
то на виляющий танец, который они исполняют на вертикальном
соте, влияет магнитное поле земли (Lindauer, Martin, 1968).
Роль магнитного поля в ориентации птиц, очевидно, не вызыва-
ет сомнений. Когда голуби направляются домой из незнакомых
мест, они пользуются для получения навигационной информации
множеством разных признаков. Одним из ориентиров служит солн-
Сенсорная информация — возможности и ограничения 663
це, по которому определяются страны света. Голуби успешно доби-
раются домой даже в пасмурные дни, однако они теряют ориента-
цию, если к ним прикрепить маленькие магниты, тогда как конт-
рольные птицы с такими же, но ненамагниченными кусочками ме-
талла не теряют направления. По-видимому, магнитные сигналы —
это лишь один из видов информации, которыми пользуется голубь,
и поэтому они не позволяют полностью объяснить превосходные
ориентационные способности этих птиц (Keeton, 1971).
В лабораторных экспериментах, где легче вычленять и контро-
лировать отдельные элементы внешней среды, действие магнитных
полей можно исследовать с большей точностью. У европейской за-
рянки (Erithacus rubecula) весной, а затем снова осенью наступает
период миграционного беспокойства. Посаженная в круглую клет-
ку, она стремится двигаться в том направлении, которое было бы
для нее естественным направлением миграции в данное время года.
Предпочитаемое направление полета сохраняется в наглухо за-
крытом помещении, лишенном каких-либо зрительных ориентиров
или других известных сигналов, которые позволили бы птице пра-
вильно ориентироваться. Однако, если экспериментальную клетку
поместить в стальную камеру, которая создает полную защиту от
магнитных полей, птицы перестают находить естественное мигра-
ционное направление (Fromme, 1961). Кроме того, искусственно
«создаваемые магнитные поля влияют на выбор направления. По-
саженные в клетку овсянки Passerina суапеа во время своего миг-
рационного периода в соответствии с естественным геомагнитным
полем ориентируются на север. Если же горизонтальную состав-
ляющую магнитного поля искусственно отклонить на 120° по часо-
вой стрелке, птицы соответственно начинают ориентироваться на
востоко-юго-восток (Emlen et al., 1976). Имеются также сообще-
ния, что на свободных птиц, совершающих перелет в ночное время,
влияют естественные возмущения геомагнитного поля (Moore,
1977).
По-видимому, магнитный компас этих птиц нечувствителен к
полярности поля, но наклон магнитных линий осевого направле-
ния используется для получения информации о полярности север —
;юг (Wiltschko, Wiltschko, 1972).
Что именно в организме реагирует на магнитные поля, совер-
шенно неизвестно. Нет никаких сведений о сенсорных органах, ко-
торые могли бы быть ответственны за «магнитное чувство», и нет
даже убедительных гипотез о его возможных механизмах.
ЗВУК И СЛУХ
При попытке определить звук мы легко попадаем в порочный
круг. Мы знаем, что звук представляет собой регулярные волны
«сжатия, которые мы воспринимаем ушами; а ухо в свою очередь
.можно определить как орган, чувствительный к звуку. Так как вол-
664 Гл. 12. Информация и органы чувств
ны сжатия могут распространяться в воздухе, в воде и в твердых
телах, нельзя ограничить значение слова «слух» только восприя-
тием звука в воздухе.
При описании человеческого слуха мы не встречаемся с боль-
шими концептуальными трудностями. Наши уши чувствительны к
регулярным воздушным волнам сжатия в пределах примерно от
40 до 20 000 Гц (колебаний в секунду). Но собака воспринимает
и более высокие частоты, совершенно неслышимые для людей, —
до 30 000 или 40 000 Гц. Летучие мыши воспринимают даже часто-
ты до 100 000 Гц, и мы продолжаем считать это слухом. Для этого
есть две основательные причины. Одна из них заключается в том,
что уши у собак и летучих мышей очень сходны с нашими. Другая
причина — то, что диапазон воспринимаемых частот непрерывен;
просто наше ухо устроено так, что оно нечувствительно к очень вы-
соким частотам, которые слышат многие другие животные.
С другой стороны, мы знаем, что волны сжатия при частотах
ниже тех, которые мы воспринимаем как звук, часто могут воспри-
ниматься как вибрация (т. е. мы все еще чувствительны к частотам
ниже тех, которые мы считаем звуком).
Когда рыба «слышит» шаги на берегу, действительно ли это
слух? Вибрация от шагов передается через почву и воду, и эти
низкочастотные колебания, вероятно, воспринимаются сенсорными
органами боковой линии, а не уха. Как в этом случае нужно го-
ворить— что рыба «слышит» или что она «чувствует» приближаю-
щиеся шаги? Вообще мы обычно говорим о слухе, когда животные
обладают специально развитыми органами, чувствительными к
тому, что мы считаем звуком, но очевидно, что это не лучший спо-
соб определять понятие «слух».
Определение слуха становится гораздо труднее применительно
к беспозвоночным животным, у которых органы, воспринимающие
вибрации, весьма отличны от уха позвоночных и во многих случаях
вообще пока не идентифицированы.
Какого рода информацию можно извлечь из звуковых волн? Че-
ловек может воспринимать звуковые волны и их изменения во
времени (ритмы и т. п.), определять их интенсивность, различать
разные частоты (высоту тона) и производить очень сложный час-
тотный анализ. Он также способен определять направление, откуда
приходит звук. Кроме того, некоторые животные пользуются зву-
ками, которые они сами издают, для получения подробной инфор-
мации о физической структуре окружающей среды.
Сложность частотного анализа, к которому способен, человек,
поразительна, если принять во внимание, что то, что он слышит,—
это, собственно говоря, не что иное, как микроскопические переме-
щения барабанной перепонки внутрь и наружу. Несмотря на такую
простоту физической основы слуха, среднее музыкальное ухо раз-
личает отдельные инструменты в оркестре, где множество их зву-
чит одновременно.
Сенсорная информация — возможности и ограничения 665
Насекомые издают и воспринимают весьма разнообразные зву-
ки, широко используя их для общения, часто с особями другого пола.
Органы слуха у насекомых различны по своему строению и нахо-
дятся в разных частях тела. По-видимому, слуховые органы насе-
комых нечувствительны к высоте тона так, как чувствительно наше
ухо; вместо этого информация передается главным образом в виде
изменений интенсивности звука, его длительности и временной
•структуры (ритма).
Записи звуков, производимых самцами насекомых для привле-
чения самок, можно проигрывать с такими искажениями частот,
что они станут неузнаваемы для человеческого уха и тем не менее
будут привлекать самку. Смысл сообщения передается, по-видимо-
му, не высотой и качеством звука, а формой или ритмом (паттер-
ном) звуковых импульсов. Звуковой паттерн у насекомых относи-
тельно фиксирован и весьма видоспецифичен. Человеческое ухо по
своим врожденным свойствам непригодно для восприятия важных
особенностей звуковых сигналов насекомых, но с тех пор, как по-
явилась электронная аппаратура для регистрации и анализа зву-
ков, стало возможным транскрибировать «песни» насекомых в ви-
димые фигуры, которые мы лучше понимаем и можем оценивать
более осмысленно.
ИНФОРМАЦИЯ О НАПРАВЛЕНИИ
Оценка направления на источник звука зависит главным обра-
зом от наличия двух ушей, раздвинутых в пространстве. Некоторая
информация извлекается благодаря тому, что сигналы, приходящие
к двум ушам, часто различаются по интенсивности, но большее
значение имеет разница во времени прихода звука от его источ-
ника.
Направление сложного сигнала определяется путем одновремен-
ного сравнения интенсивности, частотного спектра, фазы и времени
прихода сигналов к двум ушам. Сравнивать их должна централь-
ная нервная система, которая должна также выделять значимый
сигнал из окружающего шума (Erulkar, 1972).
Человек локализует звук гораздо менее точно, чем такие хищ-
ники, как кошки или совы, которые обычно охотятся в темноте. На-
пример, сова сипуха способна локализовать добычу в полной тем-
ноте, пользуясь только слухом, с ошибкой меньше 1° как в верти-
кальной, так и в горизонтальной плоскостях. Эту точную локали-
зацию источника звука она производит по частотам выше 5000 Гц,
обладающим большей направленностью, чем более низкие звуки
(Payne, 1971).
ЭХО-ЛОКАЦИЯ
Совы определяют местоположение своей жертвы по звукам, ис-
ходящим от нее самой, но некоторые позвоночные извлекают
666 Гл. 12, Информация и органы чувств
информацию из слабых отражений (эхо) звуков, которые они же
сами и издают. Такая эхо-локация (биосонар) особенно хорошо
развита у летучих мышей, но существует также у китов, дельфи-
нов, землероек и некоторых птиц.
Давно известно, что летучие мыши могут свободно летать в
совершенно темном помещении независимо от того, открыты ли их
слабо развитые глаза. Одна из наиболее изученных летучих мы-
шей— большой бурый кожан (Eptesicus fuscus). Летая в темноте,
он издает звуковые импульсы, неслышимые для человека, так как
частота их составляет примерно от 25 000 до 50 000 Гц. Каждый
импульс, или «щелчок», длится около 10—15 мс, и такие импульсы
могут повторяться около пяти раз в секунду. Получая информацию
об окружающей среде с помощью эхо, летучая мышь способна из-
бегать всяких препятствий и даже пролетать между проволоками,
натянутыми в помещении.
Если летучая мышь приближается к особенно «интересному»
предмету, например к летящему насекомому, число звуковых щелч-
ков возрастает, а длительность каждого щелчка уменьшается, так
что частота их может достигать 200 в секунду, а отдельный щел-
чок длится меньше 1 мс.
Одно из преимуществ использования высокочастотного звука
(с короткой длиной волны) состоит в том, что его направленность
гораздо точнее, чем у низкочастотного звука, который рассеивает-
ся слишком широко и дает диффузные отражения, не позволяющие
точно засекать положение предметов. Кроме того, звуковые волны
отражаются только от объектов, имеющих определенную мини-
мальную величину по отношению к длине волн. Чем короче волны,
тем меньше может быть предмет, дающий отчетливое отражение,
и поэтому самые высокие частоты позволяют обнаруживать самые
мелкие предметы.
Поразительное умение летучей мыши не натыкаться на тонкие
проволоки можно испытать в длинном помещении, где животное
имеет возможность летать взад и вперед. Если середину помеще-
ния разделить вертикальными проволоками, натянутыми с проме-
жутками в 30 см, то для летучей мыши, не имеющей о них никакой
информации, вероятность не наткнуться на проволоку составит
35%. Мышь успешно избегает проволок толщиной в несколько
миллиметров, но если их толщина будет меньше 0,3 мм, то живот-
ное при полете будет чаще наталкиваться на них или касаться их.
Если проволоки очень тонки — 0,07 мм, — число столкновений и
прикосновений растет, а число удачных пролетов снижается до
случайной величины 35%, такой же, как в опытах, где летучих
мышей делают на время глухими, затыкая им уши (рис. 12.11)
Самые короткие волны, которые посылала летучая мышь, ис-
пользованная в этих опытах (бурая ночница), — около 3 мм
(100 000 Гц). Таким образом, животное обнаруживает предметы
диаметром не более чем около Vio длины волны.
Сенсорная информация — возможности и ограничения 667
Акустическая ориентация, разумеется, особенно полезна живот-
ным, активным в темноте, но она играет также важную роль в глу-
боких или мутных водах. И дельфины, и киты пользуются эхо-ло-
кацией, чтобы избегать столкновения с предметами и с дном океана,
а также при поисках пищи. Плавая в открытой воде, они встре-
чают мало препятствий, но ночью и на глубинах, куда свет почти
Рис. 12.11. Избегание препятствий летучей мышью Myotis lucifugus при полете
в темноте между рядом вертикальных проволок, разделенных промежутками
в 30 см. Статистическая вероятность избегания проволок была равна 35%. Спо-
собность избегать проволок ухудшилась, когда их толщина была уменьшена
до < 0,3 мм. (Griffin, 1958.)
или совсем не проникает, эхо-локатор помогает им избегать столк-
новения с дном. Способность находить пищу в темноте и при пло-
хой видимости дает, конечно, большое преимущество.
Как дельфины используют акустическую активность, было пока-
зано на животных, содержавшихся в неволе. Обученный дельфин
находит мертвую рыбу, брошенную в воду, и подплывает к ней
с противоположного конца бассейна за несколько секунд, даже если
вода настолько мутна, что человек совсем не видит рыбу. Если
путь прегражден барьером из прозрачного пластика, дельфин не-
медленно находит отверстие, через которое проплывает без коле-
баний (Kellogg, 1958; Norris et al., 1961).
К другим млекопитающим, использующим акустическую ориен-
тацию, или эхо-локацию, относятся землеройки, которые издают
звуковые импульсы малой длительности и высокой частоты (до
50 000 Гц и более ) (Gould et al., 1964).
Эхо-локация встречается и у птиц, ее используют по меньшей
мере два рода: гуахаро (Steatornis) в Южной Америке и Colocalia
в юго-восточной Азии. Эти птицы не являются близкими родствен-
€68 Гл. 12. Информация и органы чувств
никами, но обе живут и устраивают гнезда в глубоких пещерах.
Более известны гуахаро, которые свободно летают в темных пеще-
рах, не наталкиваясь на стены и другие препятствия. Система эхо-
локации у них в принципе такая же, как у летучих мышей, но с
той разницей, что издаваемые ими звуки имеют частоту всего
лишь 7000 Гц. Поэтому их хорошо слышит человек, и их сравни-
вали с треском пишущей машинки. Если этим птицам заткнуть
уши, они теряют способность ориентироваться в темноте, но все еще
летают по светлому помещению, очевидно пользуясь для ориента-
ции глазами (Griffin, 1953, 1954).
ДРУГИЕ ВИДЫ МЕХАНОРЕЦЕПЦИИ
Как мы видели, зрение и слух дают большое количество подроб-
ной информации об окружающей среде — слух, как выяснилось,
гораздо больше, чем можно было думать. Нет сомнений, что те
животные, которые пользуются эхо-локацией, получают о своем
непосредственном окружении трехмерное впечатление, до некото-
рой степени аналогичное тому, которое дает людям зрение, но с
меньшей дальностью восприятия.
Другие органы чувств дают свою дополнительную информацию,
но большей частью менее обширную. Сюда относятся различные
виды механорецепции, которые еще плохо изучены.
Чувствительность к прикосновению, которая по существу обна-
руживает деформацию кожной поверхности, широко распростра-
нена и дает сведения о предметах, непосредственно соприкасаю-
щихся с телом. С нею сходно по своей природе вибрационное чув-
ство. К той же категории относятся ощущения, доставляемые
боковой линией рыб. Боковая линия, по существу, очень сложный
орган, который служит множеству целей, в том числе воспринима-
ет быстроту тока воды вдоль тела, что дает рыбе информацию о
скорости ее собственного движения (Cahh, 1967).
У различных водных беспозвоночных обнаружена чувствитель-
ность к гидростатическому давлению. Некоторые из них очень чув-
ствительны к малым изменениям давления, но механизм рецепции
остается неясным. Нетрудно понять, как животное с заполненной
газом полостью внутри могло бы воспринимать изменения давле-
ния, так как возникающее при этом изменение формы или объема
полости легко регистрировалось бы механорецепторами. Но боль-
шинство планктонных организмов, реагирующих на давление, не
имеет газовых полостей, которые позволили бы объяснить их вы-
сокую чувствительность к малым изменениям этого фактора (En-
right, 1963).
Чувствительность к силе земного притяжения чрезвычайно рас-
пространена и дает информацию о верхе и низе. Более сложные
организмы имеют рецепторы гравитации, называемые статоциста-
Сенсорная информация — возможности и ограничения 669
ми, которые все устроены по одному принципу (рис. 12.12). Тяже-
лое тельце — статолит, часто состоящий из карбоната кальция,—
опирается на сенсорные волоски. При любом смещении относитель-
но направления силы тяжести статолит стимулирует пучки ресни-
чек на других сенсорных клетках, которые посылают информацию
в центральную нервную систему.
Подобным же образом воспринимается угловое ускорение..
Жидкость внутри соответствующей структуры приходит в движе-
Рис. 12.12. Статоцист пелагической улитки. Внутри статоциста на чувствительных
сенсорных клетках лежит статолит — тяжелое тельце из карбоната кальция.
Клетки реагируют на изменения в положении статолита, когда животное ме-
няет свое положение в гравитационном поле. (Tschachotin, 1908.)
ние и отклоняет реснички клеток в направлении, которое зависит
от направления ускорения. У позвоночных орган, чувствительный
к угловому ускорению, находится во внутреннем ухе и состоит из
трех полукружных каналов, лежащих в трех плоскостях под пря-
мыми углами друг к другу. Они заполнены жидкостью, и при лю-
бом угловом движении эта жидкость в силу инерции стимулирует
сенсорные клетки в плоскости движения. Поскольку каналы ориен-
тированы в трех плоскостях, воспринимается любое угловое ус-
корение.
Все рецепторные клетки, имеющие отношение к слуху, к вос-
приятию положения и равновесия и к рецепции движения воды бо-
ковой линией рыб, принадлежат к одному и тому же основному
типу. У этих клеток имеются отростки, сходные с ресничками, —
цилии, и смещение этих отростков вызывает соответствующую сен-
сорную реакцию. Рассмотрим в качестве примера типичную рецеп-
торную клетку в боковой линии рыбы — в системе, которая служит
для восприятия движения тела относительно окружающей воды.
€70 Гл. 12. Информация и органы чивств
Каждая сенсорная рецепторная клетка обладает отростками
двух типов: группой сравнительно коротких стереоцилий и одной
более длинной киноцилией. Киноцилия содержит два центральных
и девять периферических филаментов (т. е. она имеет характерную
структуру реснички).
Рис. 12.13. Строение органов боковой линии у рыб. (Dijkgraaf, 1963.)
Несколько сенсорных клеток, каждая с сидящим на ней сенсорным волоском,
•сгруппированы вместе, причем выступающие сенсорные волоски покрыты студне-
образной купулой. Внизу — продольный разрез боковой линии. Движение воды
.деформирует купулы (светло-серые выступы).
Эти сенсорные клетки обычно образуют группу, а их выступаю-
щие цилии заключены в студенистую массу — купулу (рис. 12.13).
Когда под действием потока воды купула деформируется, цилии
сгибаются, и это ведет к возбуждению сенсорной клетки.
Киноцилия всегда находится у одного края группы стереоцилий
(рис. 12.14), и благодаря этому ответ рецепторной клетки поляри-
зован. Сенсорная клетка возбуждается, когда цилии сгибаются в ту
Сенсорная информация — возможности и ограничения 671
сторону, где расположена киноцилия. В боковой линии соседние
волосковые клетки ориентированы в противоположных направлен
ниях; поэтому одни из них возбуждаются движением воды в одном
направлении, а другие — движением в обратном направлении.
Рис. 12.14. Обычные сенсорные клетки механорецепторов — так называемые волос-
ковые клетки — несут на себе одну более длинную киноцилию, сидящую сбоку
от группы более коротких стереоцилий. Деформация выступающих цилий вызы-
вает в сенсорной клетке потенциалы действия. (Flock» 1971.)
ХИМИЧЕСКАЯ ЧУВСТВИТЕЛЬНОСТЬ — ВКУС И ОБОНЯНИЕ
Для человека ощущение вкуса связано с веществами, которые
соприкасаются со слизистой рта, а запах относится к газообразным
веществам, попадающим в нос с воздухом. Такое деление весьма
субъективно, так как вкусовая чувствительность, локализованная
во рту, выделяет только четыре основных качества: соленый, слад-
кий, кислый и горький вкус. Когда мы говорим о «вкусе» пищи,
сюда включается и информация, получаемая через нос1. Когда мы
1 Текстура пищи тоже составляет важный компонент ее «вкуса»; после гомо-
генизации хорошо знакомых пищевых продуктов их часто бывает очень трудно
распознать.
€72 Гл. 12. Информация и органы чувств
распространяем термины «вкус» и «запах» на сенсорную сферу
других позвоночных, дышащих воздухом, то аналогия эта оправда-
на, так как их органы чувств по своему строению очень сходны
с нашими.
Для водных животных различие между вкусом и запахом уже
теряет смысл. Так, например, некоторые рыбы обладают невероят-
.Рис. 12.15. У сома вкусовые почки разбросаны по всему телу. Каждое черное
.пятнышко соответствует сотне вкусовых почек. Их более высокая концентрация
на губах и усиках показана соответственно мелкими точками и сплошным зачер-
нением. Вверху — вид сбоку, внизу — спинная (слева) и брюшная (справа) по-
верхности тела. (Atema, 1971.)
но тонкой химической чувствительностью, но как мы скажем —
чувствуют они «запах» или же «вкус» воды? Говоря простым язы-
ком, часто удобно употреблять слово «запах» для предметов, нахо-
дящихся на расстоянии, а «вкус» — для веществ, непосредственно
соприкасающихся с животным, а именно с его ртом. Однако у со-
ма хеморецепторы расположены по всему телу, а между тем мы
считаем естественным называть эти рецепторы вкусовыми почками
(рис. 12.15).
Хеморецепторы рыб чрезвычайно чувствительны к аминокисло-
там, и это безусловно важно при поисках пищи. Электрофизиоло-
гическая регистрация ответов хеморецепторов на усах сомика
(Ictalurus punctatus) выявляет различную чувствительность к раз-
ным аминокислотам — их пороговые концентрации варьируют в
пределах от 10“9 до 10~и моль/л (от 1 до 100 мкг/л). Эти невероят-
Сенсорная информация — возможности и ограничения 673
но малые концентрации являются самыми низкими из электрофи-
зиологических порогов, установленных для вкусовых рецепторов
позвоночных (Caprio, 1975)<
Химическая чувствительность не может быть использована для
сканирования окружающей среды, подобно тому как используются
свет и звук, так как химические вещества, прежде чем вызвать
ощущение, должны попасть на сенсорный орган животного или
путем диффузии, или вместе с перемещением среды. С точки зре-
ния физиологии свет доходит до рецептора мгновенно, а звук —
быстро, но все же с такой задержкой, которая сама по себе несет
ценную информацию (в случае пространственного слуха и при эхо-
локации). Ограничения для использования химической чувстви-
тельности гораздо серьезнее, и для получения детальной информа-
ции здесь обычно нужны длительные периоды времени, часто в
сочетании с передвижением самого животного.
Самая полезная информация, получаемая при помощи органов
химического чувства, связана с нахождением пищи или особей дру-
гого пола, а также с обнаружением врагов. Эти органы надо счи-
тать прежде всего полезными настораживающими или предупреж-
дающими приборами, которые позволяют сосредоточить усилия
других органов чувств.
Химическая чувствительность того или иного рода, по-видимо-
му, распространена среди животных повсеместно; даже очень при-
митивные организмы, включая простейших, реагируют на химиче-
ские стимулы. Очевидно, что для амебы представляет интерес,
съедобна ли данная частица и соответственно должна ли она быть
поглощена. Однако у самых простых организмов нет специальных
хеморецепторных органов. Наивысшего развития такие органы
достигают у позвоночных и членистоногих.
В наличии химической чувствительности у менее высокооргани-
зованных животных легко убедиться, наблюдая за их поведением,
но практически все имеющиеся данные о физиологических процес-
сах в хеморецепторах получены на членистоногих и позвоночных.
Это связано с тем, что у этих животных хеморецепторные органы
образуют отдельные структуры, которые, особенно у насекомых,
легко доступны и очень удобны для электрофизиологических иссле-
дований.
ВКУС И ОБОНЯНИЕ У НАСЕКОМЫХ
Разделим вкус и обоняние в соответствии с тем, какая среда
несет химический стимул, — примем, что для вкуса такой средой
служит вода, а для обоняния воздух. Тогда можно будет сказать,
что вкусовые и обонятельные рецепторы у насекомого являются
четко обособленными структурами. Они могут находиться почти
всюду: на ротовых частях, на антеннах, на лапках. Органы вкуса
обычно представляют собой щетинки или волоски длиной в долю
43-1863
674 Гл. 12. Информация и органы чувств
Фото 12.2. Голова плодовой муш-
ки Drosophila с многочисленными
сенсорными волосками. Большие
выпуклые участки по обеим сторо-
нам головы — два сложных глаза.
Внизу снимка видны ротовые ча-
сти с сидящими на них сенсорны-
ми волосками. (Фото предоставле-
но R. Falk, Hebrew University,
Иерусалим.)
миллиметра, с отверстием на верхушке и с одним или несколькими
сенсорными нейронами у основания. Обонятельные органы могут
быть подобны волоскам или щетинкам, а иногда это ямки или
тонкостенные выступы самой разной формы.
Для органов вкуса обычно требуется гораздо более высокая
концентрация стимулирующих молекул, чем для обонятельных ре-
цепторов. Эти последние в некото-
рых случаях невероятна чувстви-
тельны к специфическим пахучим
веществам, играющим важную роль
в поисках пищи, добычи и особей
другого пола.
Вещества, которые служат для
нахождения особей того же вида, а
также для передачи разного рода
информации у колониальных насе-
комых, называются феромонами
(от греческого pherein — нести).
Феромоны составляют химический
язык, который играет важную роль
при прокладывании путей, узнава-
нии особей из того же гнезда, мар-
кировке расположения источников
пищи, подаче сигналов тревоги
и т. д.
Сенсорные волоски на хоботке
падальной мухи Phormia regina
служат превосходной эксперимен-
тальной моделью. Отделив голову
мухи и прикрепив ее к тонкой мик-
ропипетке, наполненной солевым
раствором, создают одновременно опору для нее и электрическую
связь с ее внутренней частью. Проколов микроэлектродом стен-
ку сенсорного волоска на некотором расстоянии от его кончика,
можно регистрировать электрическую активность в волоске. За-
тем при помощи микроманипулятора можно привести в контакт
с открытым кончиком волоска тончайшую стеклянную трубочку,
наполненную тестируемым раствором, и если этот раствор содер-
жит надлежащее стимулирующее вещество, он вызывает электри-
ческую активность, регистрируемую микроэлектродом (Morita,
Yamashita, 1959).
Типичный вкусовой волосок (рис. 12.16) может иметь у осно-
вания пять сенсорных нейронов. Один из них — всегда механоре-
цептор, чувствительный к отклонению волоска; из остальных один
обычно чувствителен к сахару, один к воде и один или два к солям
и различным другим веществам (Hodgson, 1961). Отводя электри-
ческую активность прямо от вкусового волоска, можно определить,
Сенсорная информация — возможности и ограничения 675
какие вещества стимулируют рецептор и какая концентрация для
этого нужна.
В отличие от вкусовых рецепторов, чувствительных лишь к
«очень узкому кругу веществ, обонятельные рецепторы насекомых
очень чувствительны к весьма разнообразным стимулам. Они
сложнее вкусовых рецепторов и часто содержат большое число
<енсорных нейронов.
Рис. 12.16. Хоботок падальной мухи несет на себе множество сенсорных волосков,
которые воспринимают различные вещества и, таким образом, служат вкусовыми
рецепторами. У основания полого волоска лежат несколько хеморецепторных ней-
ронов, каждый из которых более или менее специализирован в отношении опре-
деленной группы веществ, и один тактильный рецептор.
Обонятельные рецепторы часто (но не всегда) бывают распо-
ложены на антеннах. Хорошо известна способность самцов неко-
торых ночных бабочек находить самок того же вида даже на рас-
стоянии нескольких километров. Самка выделяет половой аттрак-
тант, а самец снабжен гигантскими антеннами, которые играют
главную роль в чувствительности к половому аттрактанту в неве-
роятно малых концентрациях (рис. 12.17). У самца сатурнии Telea
polyphemus на каждой антенне сидит около 70 000 сенсорных орга-
нов, содержащих около 150 000 сенсорных клеток. У самки же ан-
тенны гораздо меньше, и на каждой только около 14 000 сенсорных
органов и 35 000 сенсорных клеток (Boeckh et al., 1960). Около
двух третей сенсорных клеток у самца специализированы для ре-
цепции полового аттрактанта самки. Благодаря своей форме антен-
ны действуют наподобие гребней, как бы прочесывая воздух в по-
43*
676 Гл. 12. Информация и органы чувств
Рис. 12.17. Крупные антенны самца бабочки Telea polyphemus представляют со-
бой сенсорные органы, высокочувствительные к половому аттрактанту самки. Их
форма приспособлена для контакта с возможно большим объемом воздуха.
Фото 12.3. Большие антенны самца ночной бабочки Antherаеа permyi служат ему
для того, чтобы находить самку того же вида по запаху полового аттрактанта^
который она выделяет. (Фото предоставлено М. V. Williams, Harvard’ Univer-
sity.)
исках нужных молекул. При помощи полового аттрактанта
с радиоактивной меткой было показано, что из воздуха, проходя-
щего через антенны, улавливается больше 25% молекул этого ве-
щества (Schneider et al., 1968).
Опыты с самцами тутового шелкопряда (Bombyx) позволили
определить, сколько примерно нужно молекул, чтобы вызвать у на-
секомого реакцию. Если воздух содержит одну молекулу полового-
аттрактанта на 1015 других молекул и движется со скоростью
60 см/с в течение всего лишь двух секунд, то самец уже реаги-
Преобразование и передача информации 677
рует. Число молекул, попадающих на антенны, близко к тому, при
котором каждый из 10 000 рецепторов за 2 с получает максимум
одну молекулу аттрактанта (Schneider, 1969).
Невероятно, чтобы самец, чувствующий присутствие самки на
расстоянии нескольких километров, мог использовать градиенты
концентраций аттрактанта в воздухе, чтобы установить, в каком
направлении она находится. Типичное поведение самца состоит
в том, что он взлетает против ветра и таким образом попадает на
верное направление. Затем он летит по воздушному следу аттрак-
танта, пересекая его характерными зигзагами. Животные, идущие
по пахучему следу, обычно поступают так: когда они сходят со
следа и теряют его, они поворачивают обратно и тем самым снова
попадают на след. Таким образом, отклонение автоматически ве-
дет к возвращению на след, и животное многократно пересекает
его по извилистой линии. Когда же оно оказывается в непосредст-
венной близости к цели, то может окончательно локализовать ее
по градиентам концентрации.
Для того чтобы локализацию химического сигнала (такого, как
половой аттрактант) можно было определить на большом расстоя-
нии, нужны относительно устойчивые молекулы. А у веществ, слу-
жащих сигналами тревоги, которые действуют недолго и в ограни-
ченном участке, молекула должна быть относительно нестойкой во
избежание длительных остаточных эффектов, которые могли бы
внести путаницу. Если пахучее вещество нестойко и быстро раз-
рушается, то оно создает крутые градиенты около источника, по-
ложение которого поэтому легче фиксировать.
ПРЕОБРАЗОВАНИЕ И ПЕРЕДАЧА ИНФОРМАЦИИ
Сенсорная информация, доставляемая светом, звуком и т. п.,
должна быть преобразована в нервные импульсы, поступающие
в центральную нервную систему, которая интегрирует и использу-
ет эту информацию, направляя на периферию соответствующие
ответы. Предыдущий раздел был посвящен вопросу о природе сен-
сорной информации и о том, как ее получают органы чувств.
Теперь следует рассмотреть, как стимул действует на рецепторные
органы, как он преобразуется и кодируется нервными сигналами
и в какой форме информация передается в центральную нервную
систему.
Для того, чтобы это понять, надо знать, как работает нервная
клетка. Ввиду поразительного единообразия, присущего функции
нервных клеток, выбор животного здесь сравнительно безразличен.
Улитка или кальмар, лягушка или рыба — все эти объекты одина-
ково пригодны для иллюстрации принципов, лежащих в основе
функционирования нервов и органов чувств.
Рассмотрим теперь нервную клетку, природу нервных импуль-
сов, кодирование и декодирование сенсорной информации. В гла-
678 Гл. 12. Информация и органы чувств
ве 13 мы вернемся к нервной функции и подробно ознакомимся
<с тем, как импульсы передаются от одной нервной клетки к дру-
гой.
СТРОЕНИЕ НЕРВНЫХ КЛЕТОК
У всех животных нервная система состоит из множества отдель-
ных нервных клеток (нейронов). Нейроны могут быть весьма раз-
личной формы и величины, но обладают рядом важных общих
особенностей (рис. 12.18). Они состоят из тела клетки, которое
содержит ядро, и множества отходящих от тела тонких волокон.
Каждый нейрон обычно имеет одно длинное волокно (аксон), кото-
Рис. 12.18. Нейроны могут быть разной формы, но каждый нейрон имеет тело и
несколько длинных отростков. На этих схематических рисунках изображены: мо-
тонейрон членистоногого (А), спинномозговой сенсорный нейрон млекопитающе-
го (Б), биполярный нейрон из сетчатки позвоночного (В), нейрон из нервной се-
ти кишечнополостного (Г) и корзинчатая клетка из мозжечка млекопитающе-
го (Д). (Aidley, 1971.)
рое у крупных животных может достигать в длину нескольких мет-
ров, и много коротких волокон (дендритов), которые сильно вет-
вятся и большей частью имеют длину менее 1 мм. У позвоночных
тело нейрона обычно очень небольшое, его диаметр часто меньше
0,1 мм, а толщина волокна меньше 0,01 мм. Вся клетка, включая
все волокна, одета тонкой оболочкой (нервной мембраной). Слож-
Преобразование и передача информации 679
ная нервная система содержит огромное число нейронов. У чело-
века, например, один только головной мозг содержит около
10 000 000 000 нервных клеток.
Длинные волокна — аксоны — служат основными проводящими
линиями в теле. То, что обычно называют нервом или нервным
стволом, состоит из сотен или тысяч аксонов, каждый из которых
отходит от своего нейрона. В нерве нет клеточных тел — они на-
ходятся в центральной нервной системе, в особых скоплениях, на-
зываемых ганглиями, и в сенсорных органах.
Те места, где нервные клетки и их отростки вступают в кон-
такт с другими нервными клетками, называются синапсами. Один
нейрон через свои синапсы может соединяться с сотнями других
нейронов. Самая важная особенность синапса состоит в том, что
он функционирует как одностороннее передаточное устройство.
Импульс может передаваться только в одном направлении — от
аксона к следующей клетке, а не в противоположную сторону (под-
робности см. в гл. 13). В результате проведение импульсов по аксо-
ну в любом участке нервной системы происходит только в одном
направлении.
Сам по себе аксон отлично может проводить импульсы в обе
стороны, но в целостной нервной системе всякое проведение в лю-
бом аксоне идет всегда в одном направлении. Однако в нервном
стволе одни волокна могут проводить импульсы в одну сторону,
а другие — в другую. Например, в нерве, идущем к определенной
мышце, импульсы, вызывающие в ней сокращение, передаются из
центральной нервной системы к мышце, а в то же самое время
сенсорная информация от мышцы проходит по другим аксонам об*
ратно в центральную нервную систему.
КАК РАБОТАЕТ НЕРВНАЯ КЛЕТКА?
Уже почти два столетия известно, что передача нервных им-
пульсов и мышечная активность связаны с электрическими явле-
ниями. Но детального понимания природы этих явлений удалось
достичь только в последние десятилетия. Чрезвычайно важным
событием было открытие того, что некоторые нервные волокна
кальмара состоят из одного аксона гигантских размеров. Большин-
ство аксонов имеют диаметр от 1 мкм и менее до примерно 10 мкм;
диаметр же гигантских аксонов достигает около 1000 мкм
(рис. 12.19). Благодаря их исключительной толщине на них можно
проводить много экспериментов, неосуществимых на тонких аксо-
нах. Однако общие принципы, установленные в исследованиях на
гигантских аксонах, распространяются и на функцию других ней-
ронов, а также некоторых тканей иного типа, например мышц.
Для нормального нейрона, включая его аксон, характерна раз-
ность потенциалов между внутренней и наружной сторонами его
мембраны. В неактивном нейроне эта разность потенциалов назы-
68) Гл. 12. Информация и органы чувств
вается потенциалом покоя. В гигантском аксоне кальмара потен-
циал покоя составляет около —70 мВ ( + с внутренней стороны).
Во время активности он может изменяться до +55 мВ (+ снару-
жи), что дает общее изменение, т. е. потенциал действия, в 125 мВ.
Рис. 12.19. Мантия обыкновенного кальмара снабжена нервными волокнами
исключительно большой толщины. Эти так называемые гигантские аксоны быстро
проводят возбуждение и служат для того, чтобы вызывать почти одновременное
сокращение всей мускулатуры мантии. (Keynes, 1958.)
МЕМБРАННЫЙ ПОТЕНЦИАЛ ПОКОЯ
Самые важные сведения, которые нужно рассмотреть в связи
с мембранным потенциалом, касаются концентрации ионов внутри
и снаружи аксона и проницаемости мембраны аксона для этих
ионов. Внутри аксона — в аксоплазме — сравнительно высока
концентрация калия и низка концентрация натрия; в наружной
жидкости, наоборот, мало калия и много натрия (табл. 12.2). Хлор
распределен в соответствии с доннановским равновесием и не бу-
дет играть большой роли в наших рассуждениях.
Таблица 12.2
Концентрация (в миллимолях на литр)
важнейших ионов в аксоплазме гигантских
аксонов кальмара, в крови кальмара и в морской
воде. (В основном по данным Hodgkin, Huxley,
1952)
Ион Аксоплазма Кровь Морская вода
Калий 400 20 10
Натрий 59 450 470
Хлор 100 570 550
Кальций1) 0,3-Ю“3 10 10
Магний 10 55 54
’) Концентрация ионизированного кальция в аксо-
плазме точно (Baker et al., не определена, 1971). но она очень низка
Преобразование и передача информации 681
Мембрана нерва в состоянии покоя проницаема для ионов ка-
лия, а ее проницаемость для натрия так мала, что ею можно пре-
небречь. Поэтому ионы калия стремятся диффундировать из аксо-
на наружу, в область более низкой их концентрации, создавая тем
самым электрический потенциал, отрицательный внутри аксона и
положительный снаружи. Отрицательный потенциал с внутренней
стороны мембраны противодействует выходу положительных ионов
наружу, и в состоянии равновесия этот потенциал в точности ком-
пенсирует действие концентрационного градиента, вытесняющего
калий наружу. Величина мембранного потенциала зависит от кон-
центрации калия внутри и снаружи и может быть вычислена на
основе законов физической химии.
Разность потенциалов (£), создаваемая неравными концентра-
циями ионов по обе стороны проницаемой мембраны, описывается
уравнением Нернста (см. Приложение Д). Введя в уравнение соот-
ветствующие численные значения констант, получаем (при 2О°С)з
E=581og-^-,
где Со и G — соответственно наружная и внутренняя концентрации
рассматриваемого иона.
Отношение концентрации калия в крови к концентрации его в
аксоплазме гигантского аксона равно 1 : 20, а вычисленный потен-
циал покоя должен быть равен —75 мВ (внутренняя сторона элек-
троотрицательна по отношению к наружной). Мембранный потен-
циал можно измерить, введя в аксон тонкий микроэлектрод. На-
блюдаемый потенциал покоя очень близок к предсказанному, но
вследствие небольшой утечки натрия он обычно на несколько
милливольт меньше, т. е. около —70 мВ.
Принципиальную правильность вывода, что мембранный потен-
циал обусловлен калиевым градиентом, можно проверить, измерив
этот трансмембранный градиент. При повышении или понижении
концентрации калия в наружной среде трансмембранный потен-
циал изменяется именно так, как предсказывает уравнение Нерн-
ста. Если уравнять наружную концентрацию калия с внутренней,
то трансмембранный потенциал в соответствии с предсказанием
падает до нуля (Hodgkin, Keynes, 1955).
Самый убедительный опыт, показывающий, что главным эле-
ментом является нервная мембрана, а аксоплазма непосредственно
не играет роли, состоит в удалении всего содержимого аксона и
замене его искусственным солевым раствором (рис. 12.20) (Baker
et al., 1961, 1962). Такой перфузируемый аксон ведет себя порази-
тельно сходно с нормальным аксоном; он сохраняет возбудимость
в течение многих часов и способен дать несколько сот тысяч потен-
циалов действия.
Эти изящные эксперименты, возможные благодаря величине
гигантских аксонов, показывают две важные вещи. Во-первых, для
682 Гл. 12. Информация и органы чувств
генерации мембранных потенциалов аксоплазма не нужна; ее мож-
но заменить искусственным солевым раствором. Во-вторых, если
изменить внутреннюю концентрацию калия, мембранный потенциал
изменится так, как предсказывает уравнение Нернста, и если внут-
ренняя концентрация калия сравняется с внешней, то потенциал
покоя исчезнет. Мембранный потенциал даже изменит свой знак
Рис. 12.20. Из гигантского аксона можно выдавить аксоплазму и заменить ее
искусственным раствором желаемого состава. Потенциал действия, отводимый от
такого аксона, заполненного изотоническим раствором сульфата калия (а), прак-
тически неотличим от импульса в интактном аксоне, наполненном естественной
аксоплазмой (б). (Baker et al., 1962.)
на обратный, если сделать внутреннюю концентрацию калия ниже
внешней (Baker et al., 1961, 1962). Однако изменение концентрации
натрия или хлора не вызовет в потенциале покоя таких изменений.
ПОТЕНЦИАЛ ДЕЙСТВИЯ
Теперь нам нужно будет рассмотреть потенциал действия.
Когда речь шла о потенциале покоя, было сделано допущение, что
проницаемость мембраны для натрия равна нулю, и было пока-
зано, что вследствие проницаемости для калия внутренний потен-
циал должен быть отрицательным. Поэтому положение таково, что
и высокая внешняя концентрация натрия, и отрицательный внут-
ренний потенциал способствовали бы переходу Na+ в аксон. Отвле-
чемся пока от калия и предположим, что в мембране что-то вре-
менно изменяется и на мгновение делает ее весьма проницаемой
для ионов натрия. Как концентрационный градиент, так и мемб-
ранный потенциал втягивают натрий внутрь, делая внутреннюю
Преобразование и передача информации 683
сторону электроположительной, пока потенциал (теперь положи-*
тельный) не воспрепятствует дальнейшему поглощению ионов нат-
рия. В соответствии с уравнением Нернста, потенциал будет равен
+ 55 мВ (+• внутри). Таким образом, мембранный потенциал
изменился с —70 мВ (калиевый потенциал) до +55 мВ (натрие-
вый потенциал). Иными словами, мембранный потенциал аксона
может сдвинуться на 125 мВ вследствие одного только изменения
относительных проницаемостей для ионов натрия и калия.
Если мгновение спустя мембрана вновь станет непроницаемой
для натрия, то проницаемость для калия приведет к восстановле-
нию потенциала покоя. Как мы увидим, натриевая проницаемость
действительно меняется именно таким образом.
За время краткого изменения проницаемости в клетку проник
кает некоторое количество натрия, который во избежание посте-
пенной инактивации системы должен выводиться обратно. Это
происходит путем его активного переноса изнутри наружу. Количе-
ство натрия, вошедшего во время потенциала действия в аксон,
можно вычислить на основании теоретических соображений и из-
мерить опытным путем. Эксперимент, представленный на рис. 12.21,
показывает, что происходит при погружении гигантского аксона в
морскую воду, содержащую радиоактивные ионы натрия. Когда
многократная стимуляция аксона вызывает потенциалы действия,
ионы натрия входят в него и внутренняя жидкость становится ра-
диоактивной. После прекращения стимуляции и возвращения аксо-
на в состояние покоя натрий медленно выводится наружу. Чтобы
показать, что это осуществляется путем активного транспорта,
натриевый насос «отключали» с помощью ингибитора динитрофе-
нола; после этого выход натрия прекращался и его внутренняя
концентрация оставалась постоянной. При новой стимуляции аксо-
на натрий опять входил в него; это еще раз показало, что потен-
циалы действия и проникновение в аксон натрия связаны друг
с другом.
Количество натрия, входящего в аксон при одном потенциале
действия, чрезвычайно мало. Гигантские аксоны, в которых на-
триевый насос инактивирован динитрофенолом или цианидом,
сохраняют нормальную возбудимость и выдают потенциалы дей-
ствия в течение нескольких часов. В одном случае такой гигант-
ский аксон продолжал проводить импульсы с частотой 50 в секун-
ду в течение целых 70 мин (т. е. всего 210 000 импульсов), прежде
чем система «разрядилась» в результате накопления натрия с внут-
ренней стороны мембраны (Hodgkin, Keynes, 1955).
Пока мы не принимали во внимание ион хлора. Распределение
ионов С1~ изменяется в зависимости от электрического потенциала
в соответствии с уравнением Нернста, и для наших целей нет нуж-
ды рассматривать их передвижение. Действие их невелико и, что
самое главное, проницаемость для хлора во время потенциала дей-
ствия не меняется.
684 Гл. 12. Информация и органы чувств
Итак, мы установили, что мембранный потенциал аксона — как
в покое, так и в состоянии активности — определяется проницае-
мостью мембраны и что величину его можно вычислить исходя из
определенных основных принципов. Избирательно меняя проницае-
мость для натрия и калия, мембранный потенциал можно сместить
в любую точку между —75 мВ и 4-55 мВ.
Рис'. 12.21. Поглощение метки аксоном кальмара из морской воды, содержащей
радиоактивный’ натрий. В аксон, стимулированный в течение 5 мин и дававший
потенциалы действия с частотой 50 в секунду, вошло значительное количество
цатрия., После прекращения стимуляции концентрация натрия в аксоне посте-
пенно снижается по мере его активного выведения наружу. В случае отравления
натриевого насоса ингибитором динитрофенолом (ДНФ) содержание натрия
остается постоянным. Повторная стимуляция аксона вызывает новый приток
натрия; а после, удаления ингибитора натрий снова выводится путем активного
транспорта. (Hodgkin, Keynes, 1955.)
Проницаемости изменяются чрезвычайно быстро, в течение мик-
росекунд. Экспериментальные трудности, связанные с измерением
ионных токов, которые проходят через мембрану за такое короткое
£ремя, разрешаются с помощью приема, называемого фиксацией
йбтенциала. Удерживая мембранный потенциал на желаемом пот
стоянном уровне («фиксируя» его), можно измерить ток, переноси-
мый через мембрану каждым ионом. Таким образом можно выяс-
нить зависимость проницаемости мембраны для ионов от мембран-
ного потенциала.
Преобразование и передача информации 685
Самым важным результатом таких экспериментов, проведенных
в основном английскими исследователями Ходжкином, Хаксли и
Катцем, явилось определение ионной проницаемости мембраны как
•функции трансмембранного потенциала. Уменьшение потенциала
покоя (деполяризация) повышает проницаемость мембраны для
,*4Г
Рис. 12.22. Вычисленные изменения натриевой и калиевой проводимости во время
распространяющегося потенциала действия в гигантском аксоне кальмара. Основ-
ной прирост натриевой проводимости происходит в восходящей фазе потенциала
действия; повышение калиевой проводимости идет медленно и служит для вос-
становления потенциала покоя после снижения натриевой проводимости. (Hodg-
kin, Huxley, 1952.)
натрия. Такое изменение усиливает приток натрия внутрь, что еще
^больше снижает мембранный потенциал, а это в свою очередь
еще больше повышает проницаемость для натрия и т. д. Таким об-
разом, здесь возникает положительная обратная связь (см. гл. 13),
которая за чрезвычайно короткое время доводит проницаемость
дЛя натрия до максимума.* Однако полная деполяризация' в свою
очередь инактивирует систему, которая позволяет йону~ натрия
686 Гл. 12. Информация и органы чувств
проходить через мембрану, и таким образом восстанавливается ее*
исходная низкая проницаемость (Hodgkin, Huxley, 1952).
Деполяризация мембраны изменяет также проницаемость ее
для калия, но не так быстро. Если бы проницаемость-для обоих,
ионов возрастала одновременно, то вместо потенциала действия
происходило бы смещение мембранного потенциала до некоторого»
промежуточного уровня. Но так как проницаемость для калия из-
меняется уже после того, как она снизилась для натрия, конечный:
эффект состоит в восстановлении исходного состояния и возврате'
потенциала к его первоначальному значению (так называемая:
реполяризация мембраны) (рис. 12.22).
НЕРВНЫЕ ИМПУЛЬСЫ
До сих пор речь шла только о местных процессах в данной:
точке нервной мембраны. Но теперь мы располагаем сведениями,,
нужными для понимания того, каким образом потенциал действия
распространяется по аксону (т. е. как проводится по аксону нерв-
ный импульс).
Мы видели, что местная деполяризация нервной мембраны уси-
ливает приток натрия внутрь и в результате положительной обрат-
ной связи это приводит к потенциалу действия. Участок мембраны,,
лежащий в непосредственной близости, на может сохранить полг-
ностью свой потенциал покоя и отчасти деполяризуется; натриевая
проводимость возрастает, внутрь входит еще больше натрия и т. д.,.
пока и здесь не возникнет полный потенциал действия. Поэтому
деполяризация переходит с одного участка на другой и весь про-
цесс перемещается по аксону в виде быстро распространяющейся”
волны потенциала действия.
Очень слабый стимул не вызывает потенциала действия: для:
этого нужен стимул определенной минимальной (пороговой) силы..
Ниже этого порога потенциала действия нет, но он появится, если’
стимул превысит пороговый уровень. Однако на- величину потен-
циала действия сила стимула не влияет. Если даже мы удвоим или
утроим силу надпорогового раздражителя, потенциал действия'
будет оставаться прежним. Объясняется это,, конечно, тем, что его
величину определяют концентрации ионов по оба стороны мембра-
ны. Поэтому стимул либо вызовет полный потенциал действия,,
либо совсем не вызовет его. В физиологии такая реакция назы-
вается реакцией типа всё или ничего.
Как только мембрана аксона деполяризовалась и возник по-
тенциал действия, он быстро перемещается по аксону в виде рас-
пространяющегося импульса. Поскольку потенциал действия опре-
деляется местными ионными концентрациями, он распространяется’
по аксону, не изменяя своей амплитуды. Это называется бездекре*
ментным проведением и составляет главную особенность аксона;,
отличающую его, например, от пассивного проводника электриче-
ских сигналов.
Преобразование и передача информации
Если потенциал действия всегда имеет одну и ту же величину
и не зависит от интенсивности стимула, то как же аксон передает
информацию о силе стимулов — информацию, огромное значение
которой очевидно?
Рис. 12.23. Импульсы, зарегистрированные в одиночном рецепторе давления на
руке человека. Пластиковый стерженек диаметром 1 мм давил на кожу с силой,
обозначенной у каждой записи. Этот рецептор не отвечал на изменения темпе-
ратуры (т. е. был специфическим механорецептором). (Hensel, Boman, 1960.)
Ответ состоит в том, <что показателем силы стимула может
-служить частота потенциалов действия. Это показано на рис. 12.23,
где видно, что чувствительная к давлению рецепторная клетка
в коже пальца у человека отвечает на усиление стимула учаще-
нием потенциалов действия в ее аксоне. Данный рецептор имел
порог около 0,5 г и не реагировал на силу в 0,2 г. В то же время
сила в 0,6 г вызывала отчетливый ответ, частота которого возраста-
ла при увеличении давления. Таким образом, можно сказать, что
величина сенсорного стимула кодируется и передается частотно-
модулированным сигналом.
Некоторым рецепторам свойственна спонтанная активность:
-они генерируют потенциалы действия и нервные импульсы в отсут-
ствие стимуляции. На первый взгляд это может показаться беспо-
лезным и даже вредным, на самом же деле дает ряд важных пре-
имуществ.
Самое главное следствие состоит в повышении чувствительности
рецептора. Очень слабый стимул — настолько слабый, что сам по
себе он не может вызвать деполяризацию, — повышает частоту
импульсных разрядов. Это значит, что нет подпорогового стимула:
любое малое его усиление меняет частоту разряда.
Другое преимущество спонтанной активности рецептора состоит
в возможности кодировать изменения в обоих направлениях. При-
мером может служить рецептор инфракрасных лучей у гремучей
змеи. В совершенно однородной среде рецептор спонтанно активен,
я легкое изменение падающего излучения в любую сторону коди-
руется изменением частоты. Чувствительность системы достаточно
688 Гл. 12. Информация и органы чувств
высока для того, чтобы рецептор реагировал всякий раз, когда
перед змеей внезапно окажется участок поверхности с температу-
рой на 0,1° выше или ниже, чем в предыдущий момент (Bullocks
Diecke, 1956). Таким образом, спонтанная активность не только
обеспечивает высокую чувствительность, но и позволяет опреде-
лять, в каком направлении изменился стимул.
Итак, мы установили некоторые основные принципы сенсорной
физиологии. Информация передается по чувствительным нервам
в виде потенциалов действия; во всех таких нервах потенциалы
действия имеют одну и ту же природу; величина этих потенциалов
постоянна и не зависит от интенсивности стимула; и наконец, ин-
формация о силе стимула кодируется частотой потенциалов дей-
ствия. Теперь, можно перейти к вопросу о дальнейшей переработке
сенсорной информации.
СОРТИРОВКА И ПЕРЕРАБОТКА СЕНСОРНОЙ ИНФОРМАЦИИ
Если бы вся поступающая в сенсорные органы информация пе-
редавалась в центральную нервную систему, то мозг получал бы
чудовищную массу сигналов, с которой, вероятно, было бы совер-
шенно невозможно справиться. Но прежде чем сигналы пройдут
дальше, происходит их немалая предварительная обработка и
фильтрация; она начинается на уровне сенсорных нейронов и про-
должается на нескольких уровнях на пути к головному мозгу.
Фильтрующие сети избирательно пропускают только часть полу-
чаемой ими информации; кроме того, они выполняют некоторые
этапы переработки, которые улучшают организацию данных, пере-
даваемых на высшие уровни. Рассмотрим эти принципы на приме-
ре обработки зрительной информации.
Несмотря на особую сложность зрительной системы, перера-
ботка сигналов в ней изучена лучше, чем в других сложных сен-
сорных системах. Это объясняется несколькими причинами. Одна
из них — то, что здесь легко придавать искусственным стимулам
(световым) то или иное направление, распределять их во времени
и дозировать с большой точностью. Вторая причина состоит в том,,
что в качестве экспериментального объекта использовались глаза
членистоногих: строение их сложного глаза обусловливает легкий
доступ к отдельным сенсорным элементам. Впрочем, даже сложная
переработка сигналов в глазу позвоночных изучена поразительно
хорошо.
ЛАТЕРАЛЬНОЕ ТОРМОЖЕНИЕ
Мечехвост (Lirnulus) обладает сложным глазом, в котором лег-
ко стимулировать отдельные рецепторные клетки тонким лучом
света. Можно также отводить импульсы от того волокна зритель-
ного нерва, которое соединено с данным рецептором. Оказывается^
что сигналы в аксоне представляют стимул не полностью; картина
Преобразование и передача информации 689’
этих сигналов зависит также от количества света, падающего на
другие рецепторные клетки. Это объясняется тем, что каждый зри-
тельный рецептор связан со своими соседями и тормозит их актив-
ность. Благодаря этой особенности, называемой латеральным тор-
можением, усиливается контраст между количествами света, па-
дающими на два смежных рецептора.
Здесь, пожалуй, требуется дополнительное разъяснение. На
рис. 12.24 приведены записи активности двух соседних рецепторных
клеток в глазу мечехвоста. На верхней записи видны разделенные
равными промежутками импульсы, возникающие при стимуляции
одного рецептора (Л). Частота разряда составляла 35 импульсов
в секунду. Когда подвергался стимуляции также и соседний рецеп-
тор (Б), частота разрядов в А упала до 29 (средйяя запись).
Клетка Б также испытывала влияние своего соседа Д, поскольку
после прекращения стимуляции А частота разряда Б возросла-
с 23 до 31 импульса в секунду (нижняя запись). Таким образом^
между этими двумя клетками имело место взаимное латеральное-
торможение (Hartline, Ratliff, 1957).
Эффект латерального торможения поясняет схема на рис. 12.25.
Рассмотрим 12 клеток, стимулируемых равномерным светом двух
интенсивностей, причем резкий переход от одной к другой лежит
между клетками 6 и 7. В этой системе клетки 2, 3, 4 и 5 находятся
рядом с ярко освещенными клетками, и поэтому они испытывают ла-
теральное торможение. Но клетка 6 не подвергается в той же сте-
пени латеральному торможению от клетки 7, которая лежит в сла-
бо освещенной зоне. Поэтому частота ее разряда выше. Далее идут
слабо освещенные клетки, из которых на рецепторы 8, 9, 10 и 11
латеральное торможение действует в равной степени. Клетка 7
тормозится своим ярко освещенным соседом, рецептором 6, и по-
этому разряжается реже, чем остальные слабо освещенные рецеп-
торы. Конечный результат состоит в том, что переход от ярко
освещенных рецепторов к слабо освещенным подчеркнут: сигналы
в зрительном нерве дают верную картину края, но с обостренным
контрастом между двумя зонами.
Прежде чем перейти к глазу позвоночного, надо подчеркнуты
еще один аспект зрительной рецепции. Замечено, что у гребешка
(Pecten), у которого по краю мантии сидят хорошо развитые гла-
за, формирующие изображение, сетчатка состоит из двух слоев.
Для одного слоя, как и следовало ожидать, свет служит нормаль-
ным стимулом. Но второй слой ведет себя совсем иначе: он не
отвечает на усиление освещения, а чувствителен только к его
ослаблению. Этот феномен часто встречается и в глазах позвоноч-
ных, и мы скоро вернемся к нему. Легко можно представить себе,
насколько он важен. Когда на животное внезапно падает тень, это-
часто означает приближение хищника, и в таком случае ослабле-
ние стимуляции гораздо важнее для животного, чем любая другая
информация.
44—1863
A IIIIIIIIHIIIIIIIIIIIIIIIIIMIIIIIIIIIIIIIIIIIIIII
Б Oc-1
A IllllllllllUlIllIjllllllilllllllllJIllllllllllll 29c-’
6 IIIIIIIIHIIIIIIIIIIOHIIHIIIII
А Ос'1
Б i|i||i|WllllllllllllllllllllllllillllHNHO
I_____________I____________I____________I
0,5 1,0 1,5
Время, c
Рис. 12.24. Потенциалы действия, отводившиеся одновременно от двух смежных
омматидиев А и Б в сложном глазу мечехвоста (Limulus). Частота разрядов
указана справа около каждой записи. Вверху: освещен один только оммати-
дий А. В середине: освещены А и Б, что приводит к латеральному торможению
(см. текст). Внизу: освещен только омматидий Б, (Hartline, Ratliff, 1957.)
Рис. 12.25. Схема, показывающая, что латеральное торможение создает усилен-
ный краевой эффект. Подробности см. в тексте.
Преобразование и передача информации 691
Мы познакомились с двумя сравнительно простыми, но важны-
ми аспектами зрительных систем: усилением краевого эффекта и
значением темных зон в зрительном поле.
ПЕРЕРАБОТКА ИНФОРМАЦИИ
Глаз лягушки. Сетчатка лягушки не имеет центральной ямки1,,
и палочки и колбочки распределены в ней равномерно, так что
строение сетчатки всюду более или менее одинаково. Фоторецепто-
ры соединены с нейронами разного типа, лежащими в сетчатке на
разных уровнях.
Здесь мы рассмотрим только те нейроны, которые называются
ганглиозными клетками и аксоны которых образуют зрительный
нерв. Ганглиозных клеток около полумиллиона — это число при-
мерно соответствует числу волокон в зрительном нерве (Maturana^
1959). Но рецепторных клеток (палочек и колбочек) в сетчатке
больше миллиона. Ясно, что зрительный нерв не может переда-
вать полностью, от точки к точке, изображение, падающее на сет-
чатку; аналогия между сетчаткой и пленкой в фотоаппарате годит-
ся не во всем, так как перед посылкой сигналов в мозг должна
произойти некоторая их сортировка или обработка.
Зрительный нерв содержит пять разных видов волокон, беру-
щих начало от ганглиозных клеток. Они были четко идентифициро-
ваны, так как они по-разному отвечают на определенные виды*
стимуляции сетчатки. Одни волокна реагируют только на начало
освещения; они называются оп-волокнами. Другие отвечают только^
на окончание световой стимуляции и носят название off-волокон.
Волокна третьего типа отвечают и на начало, -и на конец освеще-
ния; их называют on-off-волокнами. Они дают выраженную реак-
цию на движение линии в зрительном поле, поэтому их можно
было бы также назвать детекторами движущегося края.
Далее, имеются волокна, которые отвечают на резкую границу
в поле зрения, как неподвижную, так и перемещающуюся, и эти
детекторы края четко отличаются от on-ff-волокон. Наконец, есть,
волокна, которые можно было бы назвать детекторами мух, так
как они реагируют на мелкие темные движущиеся объекты и не
отвечают на крупные и темные, а также на неподвижные объекты.
Последние две категории не отвечают на изменения общей осве-
щенности; даже включение или выключение света не действует на'
них (рис. 12.26).
Теперь мы видим, что сетчатка лягушки в значительной степе-
ни способна анализировать зрительные сигналы перед тем, каю
1 В глазу млекопитающего небольшая центральная зона сетчатки, fovea cen-
tralis, содержит только колбочки. Именно эта зона обеспечивает самое острое'
зрение на сильном свету, но из-за отсутствия палочек она нечувствительна к су-
меречному освещению.
44*
*692 Гл. 12. Информация и органы чувств
информация поступает в зрительный нерв, и если рассматривать
эту информацию с учетом того, что именно в ней может быть важ-
но для лягушки, то такая система окажется весьма целесообраз-
ной. С точки зрения лягушки насекомые, которыми она питается,
Рис. 12.26. У лягушки переработка и отбор зрительной информации в значитель-
ной части происходят в сетчатке глаза. Разные волокна зрительного нерва пере-
дают различные виды информации. Некоторые волокна не отвечают на движение
крупных предметов (А), на движение всего поля зрения (Б) или на включение
и выключение света. Они реагируют на мелкие движущиеся предметы, если эти
^предметы темнее фона (В), а если светлее, ,то не реагируют (Г). Ответ не зави-
хит от общего уровня освещения (Д), а движение предметов с размытыми краями
.не вызывает реакции (Е). (Bullock, Horridge, 1965.)
Преобразоание и передача информации 693
относятся к самым существенным в ее жизни предметам. Поэтому
важен всякий маленький темный объект, особенно если он движет-
ся, и лягушка устроена так, что реагирует специфическим образом
на этот стимул благодаря волокнам — «детекторам мух». Мелкие
неподвижные объекты — пятна, тени или камушки — не представ-
ляют особого интереса, и информация о них не доходит до той
части системы, которая предназначена для быстрой реакции на
живую добычу. Поэтому самая важная переработка сигналов, свя-
занных с пищей, происходит еще в сетчатке и передается в цент-
ральную нервную систему в виде специализированной информации
(Lettvin et al., 1961). Мы не будем обсуждать подробно остальные
типы детекторных систем сетчатки, а перейдем к зрительной систе-
ме млекопитающих.
Глаз млекопитающего. У кошки и многих других млекопитаю-
щих в сетчатке глаза имеется около 100 млн. рецепторных клеток.
Зрительный же нерв состоит примерно из 1 млн. аксонов. Из этого
сразу же видно, что мозг не может получать информацию отдель-
но от каждой рецепторной клетки; прежде чем информация посту-
пит в центральную нервную систему, происходит ее значительная
сортировка и переработка.
Несмотря на сложность того, что происходит в зрительной сис-
теме млекопитающих, она основательно изучена. Несколько упро-
щая дело, можно сказать, что передача информации происходит
на шести уровнях; три из них находятся в сетчатке глаза, один —
в латеральном коленчатом теле и два — в зрительной коре боль-
шого мозга (рис. 12.27).
Проследим эти уровни и рассмотрим переработку, которой под-
вергается информация до того, как она поступит в зрительную ко-
ру, соединенную в свою очередь с другими частями центральной
нервной системы.
Светочувствительные рецепторные нейроны в сетчатке
(рис. 12.28) связаны со слоем нервных клеток, называемых бипо-
лярными1, а биполярные клетки (биполяры) соединены далее со
слоем ганглиозных клеток сетчатки, аксоны которых образуют
зрительный нерв. Связи между этими тремя типами клеток очень
сложны. Одна рецепторная клетка может быть соединена с не-
сколькими биполярами, а несколько рецепторных клеток — с
одним и тем же биполяром. То же самое относится к связям меж-
ду биполярами и ганглиозными клетками. Поскольку' на каждую
ганглиозную клетку приходится больше 100 рецепторных, она
должна, очевидно, получать информацию от множества рецепто-
ров. Все вместе рецепторные клетки такой группы составляют ре-
цептивное поле, т. е. тот участок сетчатки, с которым связана дан-
ная ганглиозная клетка.
1 Для простоты рассмотрения мы не будем говорить об амакринных и го-
ризонтальных клетках.
694 Гл. 12. Информация и органы чувств
Не входя в детали, вспомним, что клетки сетчатки обладают
on- и off-свойствами и что синаптические связи бывают возбуж-
дающими и тормозными (более подробно о возбуждении и тормо-
жении см. в гл. 13).
Рис. 12.27. Вид человеческого мозга снизу: схематически показаны главные пути
от глаза к зрительной коре.
Одна из самых поразительных особенностей ганглиозных кле-
ток — то, что они и в отсутствие какой бы то ни было стимуляции
посылают в зрительный нерв импульсы с постоянной частотой око-
ло 20—30 в секунду. Еще удивительнее тот факт, что освещение
всей сетчатки не оказывает значительного влияния на число им-
пульсов, генерируемых ганглиозными клетками. Но если на одно
рецептивное поле падает небольшое световое пятно, оно оказывает
на соответствующую ганглиозную клетку очень заметное, хотя и
весьма сложное действие.
Как показал Куффлер (Kuffler, 1953), рецептивные поля со-
стоят из двух концентрических зон — одной с on- и другой с of {-ре-
цепторами. Рецептивные поля делятся на два типа: поля с оп-цент-
рами, в которых центральная область состоит из оп-рецепторов и
окружена кольцом off-рецепторов, и поля с off-центрами, где цент-
ральная зона содержит off-рецепторы и окружена кольцом оп-ре-
цепторов. Если небольшое световое пятнышко падает на середину
поля с on-центром, активность соответствующей ганглиозной клет-
Преобразование и передача информации 695
ки усиливается. Освещение двумя пятнами двух соседних участков
в оп-области усиливает ответ, но если одно из пятен попадает на
on-область, а другое — на окружающую off-зону, то ответ сильно
уменьшается. Это позволяет понять, почему равномерное освеще-
ние всей сетчатки вызывает лишь очень слабый ответ: различные
юп- и off-зоны в значительной степени нейтрализуют друг друга.
Рис. 12.28. Упрощенная схема связей между рецепторными (R), биполярными (В)
и ганглиозными (G) клетками в сетчатке глаза. Показаны также горизонталь-
ные (Н), амакринные (А) клетки и одна глиальная, или опорная, клетка (GI).
(Рисунок предоставлен J. Е. Dowling. Harvard University.)
Значение этой предварительной «оценки» или обработки сигна-
лов очевидно. Регистрация и передача в головной мозг сведений об
общей интенсивности равномерного освещения не представляет
особого интереса; гораздо важнее детали границ между светом и
темнотой. Мы имеем здесь случай избирательного уничтожения
информации, производимого в глазу. Глаз отбирает ту информа-
цию, которую следует передать по зрительному нерву, и таким об-
разом понижает информационную нагрузку на мозг.
Четвертый уровень переработки зрительной информации нахо-
дится в головном мозгу, в латеральном коленчатом теле. Его
клетки соответствуют рецептивным полям сетчатки; они обладают
некоторыми свойствами, общими с ганглиозными клетками, и функ-
ция их состоит в усилении контраста между небольшими световы-
ми пятнами и уровнями диффузного освещения. Но для наших
целей достаточно рассматривать коленчатое тело просто как пе-
редаточную станцию, состоящую из синаптических связей между
зрительными нервами и зрительной корой большого мозга.
696 Гл. 12. Информация и органы чувств
Рис. 12.29. Разные конфигурации рецептив-
ных полей в зрительной коре. Участки,
дающие on-ответ, обозначены крестиками,
off-ответ — треугольниками. Ориентация ре-
цептивных полей как on-, так и off-типа по-
казана прямыми линиями, проведенными
через центр поля. (Hubei, Wiesel, 1962.)
Из многих видов клеток зрительной коры мы рассмотрим толь-
ко два — «простые» и «сложные» клетки. Простые клетки реаги-
руют главным образом на выделяющиеся линии (или полосы), на-
пример светлые на темном фоне или темные на светлом фоне, и
на резкие границы между светом и темнотой. Будет ли данная*
простая клетка реагировать на такую полосу, зависит от ориента-
ции и положения этой по-
лосы на сетчатке. Напри-
мер, если вертикальная по-
лоса вызывает ответ дан-
ной простой клетки в зри-
тельной коре, то клетка He-
будет отвечать, если эту
полосу повернуть на 10—
15° или слегка сместить в-
ту или другую сторону.
Тщательное тестирование-
сетчатки очень мелкими
световыми пятнышками по-
казало, что простые клетки*
реагируют на конфигурации*
вроде представленных на*
рис. 12.29.
Благодаря тому что от-
вет данной простой клетки
зависит от ориентации и
положения прямой линии*
на сетчатке, эта клетка*
представляет собой встроен-
ный фильтр, который уст-
раняет большое количестве-
информации и передает
только специализированную’
информацию об определен^
ных видах контрастных ли-
ний. Таким образом, каж-
дая простая клетка выпол-
няет специфическую функ-
цию. Она соответствует ог-
раниченному участку сетчатки и реагирует только на определен-
ный род информации, попадающей на этот участок. Нет данных
о наличии какой-то предпочтительной ориентации рецептивных
полей в сетчатке кошки: они могут иметь любую ориентацию беа
какого-либо преобладания горизонтального, вертикального или
наклонного направления.
Сложные клетки зрительной коры, как и простые, реагируют
на ориентацию данного стимула. Если сложная клетка отвечает на
Л итератора 697
определенную ориентацию светлой полосы на сетчатке, то поворот
Зтой полосы приводит к прекращению активности клетки. Если же
полоса передвигается по сетчатке, не изменяя направления, актив-
ность не прекращается. Это легче всего объяснить, предположив,
что сложная клетка получает информацию от множества простых
клеток, которые все относятся к одному и тому же общему типу
с одинаковой ориентацией полей. Для этого требуется огромная
сложность связей в зрительной коре, но все имеющиеся данные
указывают на то, что клетки действительно расположены такими
анатомически четко очерченными группами.
Теперь можно резюмировать основные этапы переработки зри-
тельной информации в глазу млекопитающего. Рецепторы могут
быть on- или off-типа. Они не посылают информацию в централь-
ную нервную систему по отдельности,, а организованы в поля, ко-
торые легко перерабатывают информацию не об общем уровне
освещения, .а о контрасте между светом и темнотой. Это проис-
ходит при помощи двух уровней передающих нейронов в сетчатке,
которые выбирают наиболее важные сведения и отбрасывают ме-
нее существенные.
Отобранная информация передается в коленчатое тело, которое
•акцентирует данные о контрасте, получаемые от рецептивных по-
лей с on-центром и с off-центром. И наконец, зрительная кора
реорганизует информацию от коленчатого тела так, что для даль-
нейшей переработки отбираются линии и контуры. Здесь первый
этап (в простых клетках) касается ориентации линий; затем ин-
формация конвергирует на сложные клетки, от которых передается
другим частям головного мозга для интеграции с общей активно-
стью организма.
Процессы предварительной обработки и сортировки сенсорных
сигналов, происходящие перед их передачей в центральную нерв-
ную систему, устраняют большое количество несущественной ин-
формации и сильно уменьшают нагрузку на управляющие системы,
ответственные за адекватные реакции. Теперь мы перейдем к во-
просу о том, как работают эти системы и как они координируют
многочисленные функции в организме животного.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Aidley D. J. (1971). The Physiology of Excitable Cells, Cambridge, Cambridge Uni-
versity Press, 468 pp.
Atema J. (1971). Structures and functions of the sense of taste in the catfish
(Ictalurus natalis), Brain Behav. EvoL, 4, 273—294.
Baker P. F., Hodgkin A. L., Ridgway E. B. (1971). Depolarization and calcium entry
in squid giant axons, J. Physiol., 218, 709—755.
Baker P. F., Hodgkin A. L., Shaw T. I. (1961). Replacement of the protoplasm of
a giant nerve fibre with artificial solutions, Nature, Lond., 190, 885—887.
Baker P. F., Hodgkin A. L., Shaw T. I. (1962). Replacement of the axoplasm of
giant nerve fibres with artificial solutions, J. Physiol., 164, 330—354.
698 Гл. 12. Информация и органы чувств
Barrett R., Mader son Р. F. A., Meszler R. M. (1970). The pit organs of snakes.
In: Biology of the Reptilia, vol. II (C. Gans and T. S. Parsons, eds.), pp. 277—
300, London, Academic Press.
Black-Clew or th P. (1970). The role of electrical discharges in the nonproductive
social behaviour of Gymnotus carapo (Gymnotidae, Pisces), Anim. Behav. Mo-
nogr., 3, 1—77.
Boeckh J., Kaissling K.-E., Schneider D. (1960). Sensillen und. Bau der Antennen-
geissel von Telea polyphemus (Vergleiche mit weiteren Saturniden: Antheraea^
Platysamia und Philosamia), Zool. Jahrb., Abt. Anat., 78, 559—584.
Brown F. A., Jr., Barnwell F. H., Webb H. M. (1964). Adaptation of the magneto-
receptive mechanism of mud-snails to geomagnetic strength, Biol. Bull., 127,
221—231.
Brown P. K., Wald G. (1964). Visual pigments in single rods and cones of the
human retina, Science, 144, 45—52.
Bullock T. H., Cowles R. B. (1952). Physiology of an infrared receptor: The facial
pit of pit vipers, Science, 115, 541—543.
Bullock T. H., Diecke F. P. J. (1956). Properties of an infra-red receptor, J. Phy-
siol., 134. 47—87.
Bullock T. H., Hamstr a R. H., Jr., Scheich H. (1972). The jamming avoidance
response of high frequency electric fish. 1. General features, J. Comp. Physiol.,.
77, 1—22.
Bullock T. H., Horridge G. A. (1965). Structure and Function in the Nervous
System of Invertebrates, vols 1 and 2, San Francisco, Freeman, 1719 pp.
Cahn P. H. (ed.) (1967). Lateral Line Detectors, Bloomington, Indiana Univer-
sity Press, 496 pp.
Caprio J. (1975). High sensitivity of catfish taste receptors to amino acids, Comp.
Biochem. Physiol., 52A, 247—251.
Crescitelli P., Dartnall H. J. A. (1953). Human visual purple, Nature, Lond., 172».
195—197.
Dijkgraaf S. (1963). The functioning and significance of the lateral-line organs^
Biol. Rev., 38, 51—105.
Dijkgraaf S., Kalmijn A. J. (1963). Untersuchungen fiber die Funktion der Loren-
zinischen Ampullen an Haifischen, Z. Physiol., 47, 438—456.
Emlen S. T., Wiltschko W.t Demong N. J., Wiltschko- R., Bergman S. (1976). Mag-
netic direction finding: Evidence for its use in migratory indigo buntings.
Science, 193, 505—508.
Enright J. T. (1963). Estimates of the compressibility of some marine crustaceans,
Limnol. Oceangr., 8, 382—387.
Erulkar S. D. (1972). Comparative aspects of spatial localization of sound, Phy-
siol. Rev., 52, 237—360.
Flock A. (1971). Sensory transduction in hair cells. In: Handbook of Sensory Phy-
siology, vol. 1, Principles of Receptor Physiology (W. R. Lowenstein, ed.),
pp. 396—441, Berlin, Springer-Verlag.
Von Frisch K. (1948). Geldste und ungeldste Ratsel der Bienensprache, Naturwis-
senschaften, 35, 38—43.
Fromme H. G. (1961). Untersuchungen fiber das Orientierungsvermogen nachtlich
ziehender Kleinvogel (Erithacus rubecula, Sylvia communis), 7. Tierpsychol.,
18, 205—220.
Gamow R. I., Harris J. F. (1973). The infrared receptors of snakes, Sci. Am., 228,
94—101.
Goris R. C., Terashima S. (1973). Central response to infra-red stimulation of the
pit receptors in a crotaline snake, Trimeresurus flavoviridis, J. Exp. Biol., 58',
59—76.
Gould E., Negus N. C., Novick A. (1964). Evidence for echo location in shrews^
J. Exp. Zool., 156, 19—39.
Griffin D. R. (1953). Acoustic orientation in the oil bird, Steatornis, Proc. NatL
Acad. Sci. USA, 39, 884—893.
Griffin D. R. (1954). Bird sonar, Sci. Am., 190, 78—83.
Литература 699
Griffin D. R. (1958). Listening in the Dark: The Acoustic Orientation of Bats and
Men, New Haven, Conn., Yale University Press, 413 pp.
Harris J. F., Gamow R. I. (1971). Snake infrared receptors: Thermal or photoche-
mical mechanism? Science, 172, 1252—1253.
Hartline H. K., Ratliff F. (1957). Inhibitory interaction of receptor units in the eye
of Limulus, J. Gen. Physiol., 40, 357—376.
Hensel H., Boman К. K. A. (1960). Afferent impulses in cutaneous sensory nerves
in human subjects, J. Neurophysiol., 23, 564—578.
Hodgkin A. L., Huxley A. F. (1952). The dual effect of membrane potential on so-
dium conductance in the giant axon of Loligo, J. Physiol., 116, 497—506.
Hodgkin A. L., Keynes R. D. (1955). Active transport of cations in giant axons
from Sepia and Loligo, J. Physiol., 128, 28—60.
Hodgson E. S. (1961). Taste receptors, Sci. Am., 204, 125—144.
Hopkins C. D. (1972). Sex differences in electric signaling in an electric fish, Scien-
ce, 176, 1035—1037.
Hubei D. H.t Wiesel T. N. (1962). Receptive fields, binocular interaction and fun-
ctional architecture in the cat’s visual cortex, J. Physiol., 160, 106—154.
Jggo A. (1969). Cutaneous thermoreceptors in primates and sub-primates, J. Phy-
siol., 200, 403—430.
Hal mi ]'n A. J. (1971). The electric sense of sharks and rays, J. Exp. Biol., 55, 371—
383.
.Keeton W. T. (1971). Magnets interfere with pigeon homing, Proc. Natl. Acad. Sci.
USA, 68, 102—106.
.Kellogg W. N. (1958). Echo ranging in the porpoise, Science, 128, 982—988.
Keynes R. D. (1958). The nerve impulse and the squid, Sci. Am., 199, 83—90.
Keynes R. D., Martins-Ferreira H. (1953). Membrane potentials in the electroplates
of the electric eel, J. Physiol., 119, 315—351.
Kuffler S. W. (1953). Discharge patterns and functional organization of mammalian
retina, J. Neurophysiol., 16, 37—68.
Kiihn A. (1927). Ueber den Farbensinn der Bienen, Z. vergl. Physiol., 5, 762—
800.
Lettvin J. Y., Maturana H. R., Pitts W. H, McCulloch W. S. (1961). Two remarks
on the visual system of the frog. In: Sensory Communication (W. A. Ro-
senblith. ed.), pp. 575—776, New York, M. LT. Press and Wiley. [Имеется пе-
ревод: Дж. Леттвин, X. Матурана, У. Пите, У. Мак-Каллох. Два замечания
по поводу зрительной системы лягушки. — В кн.: Теория связи в сенсорных
системах. — М.: Мир, 1964, с. 416—433.]
Lindauer М., Martin Н. (1968). Die Schwereorientierung der Bienen unter dem
Einfluss des Erdmagnetfeldes, Z. Phylsiol., 60, 219—243.
Lissmann H. W., Mullinger A. M. (1968). Organization of ampullary electric re-
ceptors in Gymnotidae (Pisces), Proc. R. Soc. Lond., B, 169, 345—378.
Machin К. E., Lissmann H. W. (1960). The mode of operation of the electric re-
ceptors in Gymnarchus niloticus, J. Exp. Biol., 37, 801—811.
Maturana H R. (1959). Number of fibers in the optic nerve and the number of
ganglion cells in the retina of anurans, Nature, Lond., 183, 1406—1407.
Moore F. R. (1977). Geomagnetic disturbance of the orientation of nocturnally
migrating birds, Science, 196, 682—684.
Morita H, Yamashita S. (1959). Generator potential of insect chemoreceptors,
Science, 130, 922.
Norris K. S., Prescott J. H, Asa-Dorian P. V., Perkins P. (1961). An experimental
demonstration of echo-location behavior in the porpoise, Tursiops truncatus
(Montagu), Biol. Bull., 120, 163—176.
Payne R. S. (1971). Acoustic location of prey by barn owls, J. Exp. Biol., 54, 535—
573.
Rushton W. A. H. (1972). Pigments and signals in colour vision, J. Physiol., 220,
1—31P.
Schneider D. (1969). Insect olfaction: Deciphering system for chemical messages,
Science, 163, 1031—1037.
700 Гл. 12, Информация и органы чувств
Schneider D., Kasang G., Kais sling K.-E. (1968). Bestimmung der RiechschwelTe
von Bombyx mori mit Tritium-markiertem Bombykol, Naturwissenschaften, 55,
395.
Tschachoiin S. (1908). Die Statocyste der Heteropoden, Z. Wissensch. Zool., 90»
343_______422.
Wald G. (1964). The receptors of human color vision, Science, 145, 1007—1016.
Waterman T. H. (1950). A light polarization analyzer in the compound eye of Lb
tnulus, Science, 111, 252—254.
Waterman T. H., Forward R. B., Jr. (1970). Field evidence for polarized light sen*
sitivity in the fish Zenarchopterus, Nature, Lond., 228, 85—87.
Waterman T. H., Horch K. W. (1966). Mechanism of polarized light perception»
Science, 154, 467—475.
Wiltschko W., Wiltschko R. (1972). Magnetic compass of European robins, Science»
176, 62—64.
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Aidley D. J. (1971). The Physiology of Excitable Cells, Cambridge, Cambridge Uni-
versity Press, 468 pp.
Autrum H. et al. (editorial board) (1971—1978). Handbook of Sensory Physiology;
vol. 1, Principles of Receptor Physiology; vol. 2, Somatosensory System; vol. 3»
Interoceptors; vol. 4, Chemical Senses; vol. 5, Auditory System; vol. 6, Vestibu-
lar System; vol. 7, Photochemistry of Vision; vol. 8, Perception; vol. 9, Deve-
lopment of Sensory Systems, New York, Springer-Verlag.
Bench R. J., Pye A., Fye J. D. (eds.) (1975). Sound Reception in Mammals, London»
Academic Press, 340 pp.
Bennett M. V. L. (1970). Comparative physiology: Electric organs, Annu. Rev.
Physiol., 32, 471—528.
Catton W. T. (1970). Mechanoreceptor function, Physiol. Rev., 50, 297—318.
Cold Spring Harbor Laboratory (1965). Sensory receptors, Cold Spring Harbor
Symp. Quant. Biol., 30, 1—649.
Davson H. (1972). The Physiology of the Eye, 3rd ed., New York, Academic Press»
643 pp.
Daw N. W. (1973). Neurophysiology of color vision, Physiol. Rev., 53, 571 —
611.
de Vries H. (1956). Physical aspects of the sense organs, Prog. Biophys., 6, 207—
264.
Erulkar S. D. (1972). Comparative aspects of spatial localization of sound, Phy-
siol. Rev., 52, 237—360.
Griffin D. R. (1958). Listening in the Dark: The Acoustic Orientation of Bats and
Men, New Haven, Conn., Yale University Press, 413 pp.
Hodgkin A. L. (1964). The ionic basis of nervous conduction, Science, 145, 1148—
1154.
Horridge G. A. (ed.) (1975). The Compound Eye and Vision of Insects, Oxford»
Clarendon Press, 595 pp.
Huxley A. F. (1964). Excitation and conduction in nerve: Quantitative analysis»
Science, 145, 1154—1159.
Roeder K. D. (1967). Nerve Cells and Insects Behavior, rev. ed., Cambridge, Mass.»
Harvard University Press, 238 pp.
Rushton W. A. H. (1965). Chemical basis of colour vision and' colour blindness»
Nature, Lond., 206, 1087—1091.
Sales G., Pye D. (1974). Ultrasonic Communication by Animals^ London, Chapman
and Hall, 281 pp.
ГЛАВА 13
УПРАВЛЕНИЕ И ИНТЕГРАЦИЯ
Ранее мы уже неоднократно упоминали о регуляции и контро-
ле (управлении), но не касались деталей этих процессов. «Регули-
ровать» — значит приводить к какому-то желаемому уровню (или
поддерживать на этом уровне) ту или иную переменную: количе-
ство, концентрацию, скорость и т. д. Возьмем, например, регуляцию
дыхания. Нам кажется само собой разумеющимся, что дыхание
должно обеспечивать доставку кислорода с той скоростью, с кото-
рой он используется в организме (т. е. скорости поглощения кис-
лорода в легких и его использования в тканях должны быть коор-
динированы). Подобным же образом все физиологические процессы
подлежат контролю, регулированию и интеграции.
Интеграция означает объединение частей в целое. В физиоло-
гии этим словом описывают процессы такого управления всеми
функциональными элементами, которое объединяет их в слаженно
функционирующий организм, так что ни один процесс не протекает
стихийно, с собственной, не зависящей от остальных элементов
скоростью.
Существуют разные способы регулирования физиологических
функций. Мы видели, например, что двуокись углерода играет*
важную роль в регуляции дыхания у животных, дышащих возду-
хом, однако она не влияет непосредственно на дыхательные мыш-
цы. У млекопитающих сокращение диафрагмы при вдохе контроли-
руется нервом, идущим от дыхательного центра в головном мозгу»
а этот центр чувствителен к уровню СО2 в крови. В результате
дыхательные движения находятся под контролем нервной системы.
Рассмотрим другой случай. Выделение пищеварительного сока
поджелудочной железой усиливается под действием гормона (сек-
ретина), но, кроме того, поджелудочная железа снабжена нервами»
которые тоже стимулируют секрецию. Некоторые процессы, напри-
мер высвобождение из печени накопленного сахара, видимо, нахо-
дятся в основном под эндокринным контролем. Очевидно, регу-
ляция физиологических процессов может быть как нервной, так и
гормональной; обе управляющие системы могут функционировать
слаженным образом, и иногда трудно определить, где кончается
влияние одной и начинает действовать другая.
Хотя сферы влияния этих двух регулирующих систем во многом
перекрываются, между ними есть два существенных различия:
первое из них касается быстроты действия, второе — величины
объекта (мишени).
702 Гл. 13. Управление и интеграция
Быстрые реакции, например сокращение скелетных мышц, обес-
печиваются при помощи нервов. Скорость проведения нервных
импульсов достигает около 100 м/с, так что время передачи сигна-
ла по нерву измеряется миллисекундами. Для осуществления гор-
мональной регуляции необходимо, чтобы гормон дошел до органа-
мишени. Быстроту переноса гормона лимитирует скорость передви-
жения крови, которая его переносит. Поэтому минимальное время
реакции измеряется здесь секундами.
Второе важное различие между нервной и гормональной регу-
ляцией состоит в том, что воздействие со стороны нервной системы
может быть точно локализовано, тогда как гормоны нередко ока-
зывают более диффузное влияние. Одиночный аксон двигательного
-нерва соединяется лишь с ограниченным числом мышечных воло-
кон; это позволяет локально воздействовать на отдельную мышцу,
не затрагивая остальных; возможно даже воздействие только на
часть волокон одной мышцы. В отличие от этого гормоны влияют
на все чувствительные к ним клетки, до которых они доходят с то-
ком крови; объектами гормонального воздействия оказываются
целые органы и системы органов. Например, норадреналин стиму-
лирует выделение сахара из всей печени. Величина эффекта, одна-
ко, может регулироваться путем изменения количества гормона,
поступающего в кровь.
Как правило, гормоны регулируют такие процессы, которые
изменяются медленно (например, секрецию пищеварительных со-
ков, концентрацию и объем мочи, экскрецию натрия), а также
некоторые очень медленные процессы, например развитие гонад
шли рост тела. Однако, как уже упоминалось, нельзя провести рез-
кой грани между нервной и гормональной регуляцией; нервная
система не только участвует в регулировании функции эндокрин-
ных желез, но и играет важную роль в выработке гормонов.
РЕГУЛЯЦИЯ И ТЕОРИЯ УПРАВЛЕНИЯ
Изучение регуляторных механизмов, ответственных за поддер-
.жание стационарных состояний в живых организмах, составляет
важный раздел физиологии. Полезно будет рассмотреть работу
этих механизмов в свете элементарной теории управления.
Возьмем такой пример: у птиц и млекопитающих температура
внутренних областей тела поддерживается на практически постоян-
ном уровне несмотря на большие колебания температуры воздуха
ш теплопродукции самого организма. Понятно, что эта регуляция
температуры тела обеспечивается с помощью сложной системы.
ТЕОРЕТИЧЕСКИЕ СООБРАЖЕНИЯ
В технике управляющие системы играют настолько важную
роль, что теория и вопросы конструирования таких систем состав-
ляют самостоятельную дисциплину, имеющую собственную терми-
Регуляция и теория управления 703
нологию и теоретические подходы. Для физиолога здесь важны два
аспекта: 1) множество расплывчатых и туманных старых терминов
заменено точными, четко определенными понятиями, и 2) благода-
ря развитию теоретического подхода к этому вопросу удалось
более точно определить компоненты физиологических механизмов
и лучше понять взаимоотношения между ними.
ОБРАТНАЯ СВЯЗЬ
Рассмотрим хорошо всем знакомое регулирующее устройство —
термостат, поддерживающий заданную температуру в помещении
или в водяной бане. Действие этой, регуляторной системы (как и
всех других) направлено на регулируемую переменную (в данном
случае температуру), кото-
рая поддерживается на
уровне, более или менее
близком к заданному. Фак-
тическое значение регули-
руемой переменной сравни-
вается с заданным значени-
ем. Это производится с по-
мощью детектора ошибок;
последний подает сигнал,
который включает тот или
иной корректирующий ме-
ханизм; в результате в ре-
гулируемую систему вно-
сится необходимая поправ-
ка.
Если нам нужно поддер-
живать «постоянную» тем-
пературу в водяной бане,
то мы можем использовать
устройство, изображенное
на рис. 13.1; действие его
Рис. 13.1. Регулирование температуры во-
дяной бани (схема). Подробности см. в
тексте.
практически самоочевидно.
Оператор А может по-
высить температуру бани,
включив электрический по-
догреватель. Термометр сообщает ему, какова температура воды.
А уже получил от руководящего лица В инструкцию о том, како-
ва должна быть температура воды. Если она упала ниже задан-
ного уровня, А включает подогрев. Глядя на термометр, он опре-
деляет, когда температура достигнет желаемого значения, и тог-
да отключает подогреватель.
Оператора А можно заменить автоматическим устройством, а
именно термостатом вроде тех, которые регулируют систему цент-
704 Гл, 13. Управление и интеграция
рального отопления в домах. Всю систему можно представить
блок-схемой, приведенной на рис. 13.2, А. Если речь идет о темпе-
ратуре воды, то информация о выходном параметре системы пере-
дается обратно в термостат, в результате чего производятся необ-
ходимые действия для коррекции любого отклонения от заданного
уровня температуры воды. Такой принцип называется обратной
Рис. 13.2. Схемы регулирующих систем. А. Система, представленная на рис. 13.1.
Б. Система, описанная в терминах, принятых в теории управления.
связью; этот термин употребляют в том случае, когда определен-
ный параметр, регистрируемый на выходе системы, сравнивается
>с его заданным значением. В приведенном примере повышение
температуры воды корректируется путем уменьшения нагрева. Это
пример отрицательной обратной связи; так называют ситуацию,
когда отклонение измеряемого параметра в ту или другую сторону
включает корректирующее действие, противоположное по направ-
лению.
Для дальнейшего обсуждения полезно несколько расширить
приведенную схему и ввести стандартные термины, используемые
в теории управляющих систем (рис. 13.2,5).
Типичная отрицательная обратная связь представляет собой
.замкнутую систему управления, что ясно видно на схеме. Сигнал
относительно регулируемой переменной поступает обратно на вход
системы, так что получается замкнутый контур. Для полноты кар-
тины следует упомянуть также о незамкнутых системах, хотя они
имеют меньшее значение в физиологической регуляции. Предста-
вим себе, что котельная дома обеспечивается топливом в зависимо-
сти от погоды: чем ниже температура воздуха, тем больше посту-
пает топлива. Можно так тщательно отработать эту систему, что
при определенном снижении температуры будет прибавляться ров-
но столько топлива, сколько нужно для поддержания температуры
в помещениях на постоянном уровне. В этом примере входной
Регуляция и теория управления '705
переменной будет наружная температура, а выходной — количество
тепла, вырабатываемого котельной. Однако при появлении возму-
щающего фактора (например, сильного ветра) может оказаться,
что дом охлаждается быстрее обычного, а приток* тепла остается
прежним, и тогда температура в помещении понизится. В такой
системе нет обратной связи; это незамкнутая система регулиро-
вания.
Отрицательная обратная связь уменьшает расхождение между
заданным и фактическим значениями переменной; хороший пример
•тому — термостат. Перейдем теперь к положительной обратной
связи. Существует ли она? Играет ли она какую-либо роль в био-
логии?
Мой коллега Стивен Фогель предложил в качестве примера по-
ложительной обратной связи следующую ситуацию. Представим
себе, что у мужа и жены есть отдельные одеяла с электроподогре-
вом, причем каждое одеяло снабжено терморегулятором, устроен-
ным по принципу отрицательной обратной связи. Предположим,
что муж предпочитает прохладное одеяло, а жена более теплое.
Вообразим себе теперь, что они нечаянно обменялись термостати-
рующнми приставками. Тогда муж устанавливает «свой» термостат
на любимую им низкую температуру; в результате одеяло жены
оказывается слишком холодным, и она устанавливает «свой» тер-
мостат на более высокую температуру. Вскоре муж находит свое
одеяло чересчур теплым и еще больше понижает заданный уровень
на «своем» термостате, тогда как жена крутит ручку «своего»
регулятора в противоположном направлении. Это пример положи-
тельной обратной связи, где отклонение от заданного значения при-
водит к еще большему отклонению.
Очевидно, что положительная обратная связь не годится для
целей регулирования, так как она приводит систему в одно из
экстремальных состояний. Тем не менее в биологии встречаются
ситуации, когда положительная обратная связь оказывается полез-
ной.
Отрицательная обратная связь позволяет поддерживать стацио-
нарное состояние; положительная же обратная связь ведет к уско-
ряющемуся переходу системы в какое-то экстремальное состояние.
Мы уже обсуждали пример положительной обратной связи — гене-
рирование нервного импульса. Действительно, уменьшение потен-
циала нервной мембраны приводит к повышению ее проницаемости
для ионов натрия; в свою очередь переход натрия внутрь еще боль-
ше снижает мембранный потенциал, и так далее до тех пор, пока
не разовьется полный потенциал действия. Таким образом, поло-
жительная обратная связь может усилить слабый сигнал и обеспе-
чить полную реакцию на него.
Положительная обратная связь полезна также при синхрониза-
ции событий. Известный пример — положительная обратная
связь при спаривании. При встрече подходящих партнеров переход
45—1863
706 Гл. 13. Управление и интеграция
к спариванию облегчается благодаря положительной обратной
связи; по мере ухаживания взаимные эмоциональные реакции уси-
ливают сексуальное влечение; в конце концов такая обратная
связь приводит к спариванию и обеспечивает завершение этого
акта.
ПРИНЦИПЫ РЕГУЛЯЦИИ: «ВСЕ ИЛИ НИЧЕГО», ПРОПОРЦИОНАЛЬНОЕ И
ИНТЕГРАЛЬНОЕ РЕГУЛИРОВАНИЕ, РЕГУЛИРОВАНИЕ ПО ПРОИЗВОДНОЙ
Регуляция температуры в термостате осуществляется по прин-
ципу «всё или ничего». Подогрев либо включен, либо выключен,
так что тепло подается с перерывами. Это неизбежно ведет к ко-
лебаниям температуры. Для включения подогрева температура
должна несколько упасть по сравнению с заданной. Точно так же
и отключение подогрева произойдет только тогда, когда темпера-
тура несколько превысит заданный уровень. Величину отклонений
можно уменьшить путем повышения чувствительности системы, но
полностью исключить колебания в системе, работающей по принци-
пу «всё или ничего», невозможно.
Большей стабильности регулируемой переменной можно достичь
с помощью систем другого типа. Большое значение имеет принцип
пропорциональной регуляции; его лучше всего можно пояснить на
примере механической системы (рис. 13.3). Представим себе, что
Рис. 13.3. Схема простейшей системы пропорциональной регуляции; система обес-
печивает соответствие притока воды в бак ее оттоку.
уровень воды в баке регулируется с помощью поплавка; это дает
возможность осуществлять непрерывную регуляцию вместо преры-
вистой. Если скорость вытекания воды из бака почему-либо возрас-
тет, то уровень воды снижается и в результате открывается кла-
пан, что ускоряет приток воды. Чем больше понижается уровень
воды в баке, тем больше возрастает ее приток. Если же оттоку
воды что-то препятствует, то уровень ее повышается и это ведет
к закрытию клапана. В такой системе регуляция осуществляется
непрерывно и степень регуляторного воздействия пропорциональна
Регуляция и теория управления 707
отклонению от заданного уровня. (Под «пропорциональностью»
обычно понимают линейную зависимость между величинами на
входе и выходе, отсчитываемыми от нуля; однако на практике этот
термин часто применяют к системам, не вполне отвечающим при-
веденному строгому определению.)
Пропорциональный регулятор обладает одной интересной и
весьма важной особенностью. Допустим, что система находится
в стационарном состоянии, соответствующем заданному уровню.
Если произойдет какое-либо возмущение (например, изменение в
-оттоке воды), то система быстро достигнет нового стационарно-
го состояния, однако возвращение к первоначальному задан-
ному уровню оказывается невозможным. Последнее требует разъ-
яснения.
Возьмем исходное стационарное состояние, при котором вода
в баке находилась на первоначально заданном уровне. Теперь
уменьшим отток воды и посмотрим, какова будет компенсаторная
реакция. Очевидно, что поплавок поднимется и уменьшит приток
воды. Новое стационарное состояние установится при более высо-
ком уровне воды, и поплавок должен будет находиться на этом
новом уровне, чтобы обеспечить равенство притока замедленному
-оттоку (т. е. новый уровень представляет собой отклонение от пер-
воначально заданного). Регулирующее воздействие не может вер-
нуть уровень воды к исходному (сделать отклонение равным ну-
лю), так как в этом случае и поплавок оказался бы в исходном
положении, в котором приток не соответствует уменьшенному отто-
ку. Представим себе далее, что в систему введено другое возмуще-
ние, а именно появился второй ввод для воды. В этом случае уро-
вень воды вновь повысится и в результате приток уменьшится, но
новый стационарный уровень опять-таки будет выше исходного.
К физиологическим системам, работающим по принципу про-
порционального регулирования, относится регуляция дыхания (вен-
тиляции легких) у млекопитающих по уровню СОг в крови. Когда
у млекопитающего в покое Рсо2 в артериальной крови составляет
40 мм рт. ст., мы можем считать это «заданным уровнем» арте-
риального РСо2. При повышении содержания СО2 во вдыхаемом
воздухе РСо2 в артериальной крови возрастет, что приведет к уве-
личению вентиляции легких, причем степень этого увеличения бу-
дет находиться в прямой зависимости от повышения РСо2 (гл. 2)1.
1 На самом деле регуляция дыхания более сложна. Мы знаем, что при фи-
зической работе образование СО2 возрастает; если бы регуляция осуществлялась
по принципу простой пропорциональности, то при нагрузке уровень артериального
Рсо2 как сигнала, вызывающего увеличение вентиляции, был бы слегка повышен.
В действительности же происходит обратный сдвиг: артериальное Рсо2 при на-
грузке несколько снижается. Это нередко объясняют как результат изменения
«заданного уровня» при физической нагрузке. На самом же деле причина со-
стоит в том, что дыхательный центр получает и интегрирует целый ряд различ-
ных сигналов, в том числе нервные импульсы от работающих мышц.
45*
708 Гл. 13. Управление и интеграция
Как мы видели, стационарное состояние с нулевым отклонением'
от исходного уровня не может быть достигнуто с помощью просто-
го пропорционального регулирования. Однако это может быть
осуществлено путем интегрального регулирования. В этом случае’
на вход регуляторной системы поступает общее отклонение за
какой-то промежуток времени. В результате сигнал на выходе
системы будет пропорционален интегралу по времени от величины
входного сигнала (т. е. входу здесь пропорционален не выход, а
скорость изменения выхода). Если в такой системе появляется воз-
мущающий фактор, который остается постоянным, то отклонение-
выхода будет со временем стремиться к нулю. Как бы велико ни
было возмущение, если оно постоянно, то по механизму интеграль-
ной регуляции может быть достигнуто стационарное состояние'
с нулевым отклонением от заданного.
Есть еще один тип регулирования — так называемая регуляция
по производной (или регуляция по скорости), в которой для оцен-
ки ожидаемой величины нужного корректирующего воздействия'
используется временная производная величины сигнала, т. е. ско-
рость изменения сигнала. Поскольку такая система регуляции реа-
гирует только на изменение сигнала, а не на сигнал как таковой,,
она эффективна только в сочетании с другими системами. В фи-
зиологии такое сочетание имеет особую ценность, так как обеспе-
чивает необходимое регулирующее воздействие при кратковремен-
ных изменениях регулируемой переменной.
ФИЗИОЛОГИЧЕСКИЕ МЕХАНИЗМЫ
Вернемся к'регуляции температуры тела. Мы сравнивали ее-
с регуляцией температуры воды в термостате по принципу отрица-
тельной обратной связи. По тому же принципу может регулиро-
ваться температура в домах с центральным отоплением; летом,,
если очень жарко, к этому можно добавить еще систему охлажде-
ния (тогда мы получим кондиционер). Кондиционер действует та-
ким образом, что при чрезмерном повышении температуры он на-
чинает охлаждать помещение, уменьшая тем самым отклонение от
заданного уровня (т. е. здесь тоже имеется отрицательная обрат-
ная связь). Таким образом, как обогрев, так и охлаждение поме-
щения регулируются по принципу отрицательной обратной связи.
Аналогия с регуляцией температуры тела очевидна. Если тем-
пература тела начинает падать, теплопродукция организма возра-
стает — главным образом в результате непроизвольных мышечных
сокращений (дрожь). Если же из-за тепловой нагрузки, внешней
или внутренней, температура тела повышается, организм охлаж-
дается с помощью потоотделения или испарения влаги в верхних:
дыхательных путях (учащенное дыхание). Таким образом, и согре-
вание, и охлаждение тела осуществляются с помощью отрицатель-
ных обратных связей.
Регуляция и теория управления
709
Центр терморегуляции находится в головном мозгу, в гипота-
ламусе. Это можно показать различными способами. Например,
при локальном повышении температуры крови в сонной артерии
собака начинает учащенно дышать. Отсюда видно, что регуляция
происходит в голове, поскольку в результате учащенного дыхания
животное теряет слишком много тепла и его внутренняя темпера-
тура снижается. И наоборот, если температуру крови в сонной
артерии понизить, собака начнет дрожать, в результате чего внут-
ренняя температура тела будет повышаться. Точную локализацию,
центра терморегуляции можно определить путем обогрева или
охлаждения отдельных участков гипоталамуса.
Центр терморегуляции можно рассматривать как термостат,
а «нормальную» температуру тела — как заданный уровень. Одна-
ко система не так проста, поскольку в ней нет постоянного задан-
ного уровня и она имеет много входов. Начнем с того, что темпера-
тура тела колеблется на протяжении суток, даже если внешняя
температура и собственная теплопродукция остаются неизменными.
Это означает, что заданный уровень подвержен суточным цикли-
ческим колебаниям. Кроме того, как мы уже знаем, при физиче-
ской нагрузке установка уровня внутренней температуры тела
сдвигается в сторону повышения по сравнению с покоем. Среди
многих сигналов, поступающих в центр терморегуляции, важную
информацию доставляют сигналы от холодовых и тепловых рецеп-
торов кожи; дополнительную информацию несут и многие другие
сигналы, в частности температура артериальной крови, приходя-
щей к гипоталамусу.
Как уже упоминалось, температура тела регулируется как
путем согревания, так и путем охлаждения организма (т. е. регу-
ляторные воздействия могут быть направлены и в ту, и в другую
сторону). В этом состоит важный принцип физиологической регу-
ляции; он используется гораздо чаще, чем может показаться на
первый взгляд.
Еще одним примером может служить сердце. Сердечной мышце
присуща способность к ритмическим сокращениям. Сокращение
начинается в синусном узле и распространяется по всей сердечной
мышце, после чего наступает расслабление, а затем новое сокра-
щение. Частота сокращений регулируется двумя нервами: ускоряю-
щим, который учащает ритм сердца, и замедляющим (ветвь блуж-
дающего нерва), который снижает частоту сокращений. Таким об-
разом, частота сердечных сокращений зависит от баланса между
двумя антагонистическими нервными воздействиями — стимули-
рующим и тормозящим.
Как мы увидим, баланс между стимуляцией и торможением —
явление очень обычное в системах физиологической регуляции.
Для того чтобы лучше понять этот общий принцип, полезно позна-
комиться с работой нервных клеток и с тем, как они передают воз-
буждающие и тормозные импульсы.
710 Гл, 13. Управление и интеграция
СИСТЕМЫ НЕРВНОЙ РЕГУЛЯЦИИ
Основными элементами нервной системы любого типа являются
нервные клетки, или нейроны. Две важнейшие в функциональном
отношении части нейрона — это длинный нитевидный отросток, на-
зываемый аксоном, и участок соединения между клетками — си-
напс. У каждого нейрона только один аксон, но число синаптиче-
ских соединений, образуемых нейроном, может достигать нескольких
•сотен и даже тысяч, и это имеет огромное значение для инте-
грации нервной деятельности. Аксоны функционируют как провод-
ники, а синапсы — как очень сложные включающие или выклю-
чающие устройства.
Сначала мы ознакомимся с рядом важных физиологических
характеристик аксона, а затем перейдем к вопросу о том, как си-
напсы обрабатывают информацию, получаемую нейронами. Далее
в качестве примера интеграции мы рассмотрим, каким образом
одновременные возбуждающие и тормозные влияния со стороны
центральной нервной системы обеспечивают управление мышцами
конечности в процессе локомоции.
АКСОНЫ
ПОТЕНЦИАЛЫ ДЕЙСТВИЯ
Нервный импульс, или потенциал действия, уже был рассмот-
рен в гл. 12. Он представляет собой быстрое кратковременное из-
менение мембранной проницаемости, сопровождающееся передви-
жением ионов и изменением мембранного потенциала. Величины
потенциала покоя и потенциала действия очень сходны у всех
представителей животного царства, хотя и существуют небольшие
различия. Мембранный потенциал покоя как аксона, так и тела
нервной клетки обычно составляет 60—90 мВ (+ снаружи).
Потенциал действия, который представляет собой локальное
падение мембранного потенциала, варьирует обычно в пределах от
—80 до —120 мВ по отношению к потенциалу покоя.
Различия в величине потенциалов не связаны с размерами тела,
размерами аксона или принадлежностью животного к позвоночным
или беспозвоночным. Длительность потенциала действия значи-
тельно более изменчива (табл. 13.1).
СКОРОСТЬ ПРОВЕДЕНИЯ
Совсем иначе обстоит дело со скоростью проведения: она изме-
няется в громадном диапазоне от нерва к нерву и от животного
к животному. Несколько типичных примеров приведено в табл. 13.2.
Двигательные нервы позвоночных характеризуются значительно
большей скоростью проведения, чем обычные двигательные нервы
беспозвоночных. Это не связано с тем, что у беспозвоночных более
Системы нервной регуляции
711
Таблица 13.1
Потенциалы покоя и потенциалы действия нейронов различных животных.
(Bullock, Horridge, 1965)
Животное Волокно или клетка Потенциал покоя, мВ Пик потенциала действия, мВ Длитель- 1 ность по- тенциала действия, мс
Кальмар Loligo Гигантский аксон 60 120 0,75
Дождевой червь Lumbri- CUS Медианное гигантское волокно 70 100 1,0
Рак Cambarus Медианное гигантское волокно 90 145 2
Таракан Periplaneta Гигантское волокно 70 80—104 0,4
Краб Carcinus Аксон диаметром 30 мкм из нерва конечности 71—94 116—153 1,0
Лягушка (Rana) Аксон седалищного нерва 60—80 100—130 1,0
Голожаберный моллюск Aplysia Нейрон висцерального ганглия 40—60 80—120 10
Улитка Onchidium Нейрон висцерального ганглия 60—70 80—100 9
Рак Cambarus Рецептор растяжения 70—80 80—90 2,5
Рыба-иглобрюх (Sphae- roides) Супрамедуллярная клетка 50—80 80—110 3
Жаба (Bufo) Нейрон дорсального корешка 50—80 80—125 2,8
Жаба (Bufo) Спинальный мотоней- рон 40—60 40—84 2
Кролик (Oryctolagus) Симпатический нейрон 65-82 75—103 4—7
Кошка (Felis) Спинальный мотоней- рон 55-80 80—110 1-1,5
низкая температура тела, поскольку у холоднокровных позвоноч-
ных скорость проведения также достаточно велика. Если соответ-
ствующие величины для холоднокровных позвоночных пересчитать
на температуру тела млекопитающих (температурный коэффи-
циент Qio для нервных волокон равен 1,8), то полученные значения
будут соответствовать скоростям проведения у млекопитающих.
Для беспозвоночных результат был бы иным. Однако у некоторых
беспозвоночных имеются немногочисленные быстропроводящие
аксоны — так называемые гигантские аксоны, которые значительно
крупнее обычных и могут достигать почти 1 мм в диаметре. Ско-
рость проведения в них примерно в 10 раз больше, чем в обычных
аксонах того же животного.
Биологическое значение высокой скорости проведения очевидно.
Она обеспечивает быструю двигательную реакцию, используемую
чаще всего для спасения от врага. Одну из самых быстрых реак-
ций такого типа можно наблюдать у таракана, которому требует-
712 Гл. 13. Управление и интеграция
Таблица 13.2
Скорость проведения (м/с) в нервах
различных животных. (По данным
Bullock, Horridge, 1965)
Животное Обычные двигательные нервы Гиган- тские аксоны
Позвоночные
Кошка 30—120
Змея 10—35
Лягушка 7—30
Рыба 3—36
Беспозвоночные
Таракан 2 10
Кальмар 4 35
Дождевой червь 0,6 30
Краб 4
Улитка 0,8
Актиния 0,1
ся всего 25 мс для того, чтобы отреагировать на струю воздуха,
направленную на кончик брюшка (где имеются рецепторные во-
лоски). Другим примером может служить кальмар, у которого ги-
гантские аксоны проходят вдоль всей мантии. Благодаря быстрому
проведению импульсов по этим аксонам вся мускулатура мантии
может сокращаться почти одновременно, а это очень важно при
реактивном движении за счет отдачи выбрасываемой струи воды.
Если бы импульсы доходили до более отдаленных участков ман-
тии с заметным запозданием, то и сокращение распространялось
бы постепенно и не было бы быстрого, мощного выброса водяной
струи. Дождевому червю, который влажным утром наполовину
высовывается из своего подземного хода, гигантские волокна поз-
воляют почти мгновенно спрятаться в землю при механическом
раздражении; эта реакция составляет резкий контраст с тем срав-
нительно медленным передвижением, которое вообще характерно
для дождевого червя.
Как правило, существует прямая зависимость между скоростью
проведения и диаметром аксона (Hodgkin, 1954). Исходя из тео-
рии проводимости кабеля можно предсказать, что скорость прове-
дения (и) должна быть примерно пропорциональна корню квад-
ратному из диаметра волокна (d):
Множество наблюдений говорит о том, что эта формула приб-
лиженно верна, но величина константы k у разных животных раз-
лична. Приведенное уравнение означает, что для нервного волокна
Системы нервной регуляции 713
данного типа 10-кратное увеличение скорости проведения возмож-
но при 100-кратном увеличении его диаметра. Упомянутые выше
гигантские аксоны могут быть толще обычных аксонов того же
животного в 50 и даже 100 раз, так что характерные для них вы-
сокие скорости проведения соответствуют теоретическому расчету.
МИЕЛИНИЗИРОВАННЫЕ ВОЛОКНА ПОЗВОНОЧНЫХ
У позвоночных нет гигантских аксонов; и тем не менее двига-
тельные нервы этих животных обладают очень большой скоростью
проведения — наибольшей, какую только можно встретить в жи-
Рис. 13.4. Схема строения миелинизированного нервного волокна. Миелиновая
оболочка образуется из опорной (шванновской) клетки, которая обрастает нерв
по спирали до тех пор, пока он не будет окружен многослойной оболочкой, со-
стоящей из мембран этой клетки, как показано внизу на поперечных разрезах.
(Aidley, 1971.)
вотном царстве. То, что нервные волокна млекопитающих, будучи
очень тонкими, способны к быстрому проведению, обусловлено
особенностями их структуры. Весь аксон у этих животных одет
тонкой оболочкой из жироподобного вещества миелина; эта обо-
714 Гл. 13, Управление и интеграция
лочка через короткие интервалы прерывается, обнажая мембрану
нерва. Эти открытые участки называют перехватами Ранвье\ рас-
стояние между перехватами составляет от доли миллиметра до
нескольких миллиметров (рис. 13.4). Миелиновая оболочка обра-
зуется из глиальных (опорных) клеток, которые формируют много-
слойную обкладку таким образом, что их цитоплазма исчезает, а
многочисленные слои плазматических мембран остаются.
Фото 13.1. Миелинизированный нерв. На этом поперечном срезе зрительного нер-
ва крысы видно, что каждый аксон окружен слоями миелина. В зрительном нерве
содержится несколько сотен тысяч таких аксонов; диаметр каждого аксона
около 1 мкм. (Фото предоставлено A. Peters, Boston University Medical Cen-
ter.)
Рассмотрим проведение импульса по миелинизированному ак-
сону. В области перехвата потенциал действия ничем не отличает-
ся от любого другого потенциала действия (т. е. в этом месте про-
исходит локальная деполяризация мембраны). Это означает, что
данный перехват по отношению к соседнему становится электроот-
рицательным (так как обычный положительный заряд мембраны
Системы нервной регуляции 715
здесь исчез). В результате между этим и соседними перехватами
тотчас же возникает электрический ток, достаточный для того,
чтобы вызвать деполяризацию и появление потенциала действия
в соседнем перехвате. Это происходит практически мгновенно —
потенциал действия как будто перескакивает от одного перехвата
к другому. Деполяризация в области перехвата развивается не так
Рис. 13.5. Скорость проведения импульсов в миелинизированных волокнах кошки
как функция диаметра волокна. (Hursh, 1939.)
уж быстро, но, едва возникнув, она тотчас вызывает деполяриза-
цию следующего перехвата и т. д. Быстрая передача возбуждения
от перехвата к перехвату носит название сальтаторного проведения
(от лат. saltare — танцевать, прыгать); поскольку оно осуществля-
ется почти без задержки, потенциал действия быстро пробегает
вдоль всего волокна, несмотря на малую толщину аксона, при
которой обычным способом импульс распространялся бы очень
медленно.
Скорость проведения в миелинизированном аксоне лягушки
определяется следующими параметрами. При диаметре аксона
10 мкм скорость проведения составляет 20 м/с, а время задержки
в каждом перехвате — примерно 0,06 мс. Поскольку у лягушки
в аксоне такого калибра расстояния между перехватами составля-
ют около 1,6 мм, нетрудно убедиться, что большая часть времени
проведения расходуется на задержки в перехватах; затрата вре-
мени на передачу от перехвата к перехвату крайне мала (Tasaki,
1959).
Этот вывод был подтвержден изящным экспериментом с охлаж-
дением нерва. При охлаждении скорость передачи импульса замед-
ляется. Но если будет охлажден только участок между двумя пере-
хватами, то время распространения импульса между ними почти
не изменится. Этот результат согласуется с гипотезой о том, что
716 Гл. 13, Управление и интеграция
проведение между перехватами представляет собой электрический
феномен, а изменение потенциала в перехватах не отличается по
своей природе от потенциала действия в немиелинизированных
аксонах (Hodler et al., 1951).
Скорость проведения в миелинизированных аксонах все же за-
висит от их диаметра, но здесь она прямо пропорциональна диа-
Рис. 13.6. Скорость проведения импульсов в миелинизированных и немиелинизи-
рованных волокнах по-разному зависит от диаметра волокон. Вследствие этого
при диаметре меньше определенной величины миелинизированное волокно про-
водит импульсы медленнее, чем немиелинизированное того же диаметра. Пунктир-
ная линия получена путем экстраполяции данных, приведенных на рис. 13.5:
сплошная линия — теоретическая кривая, рассчитанная по максимальным скоро-
стям проведения в немиелинизированных С-волокнах кошки. (Rushton, 1951.)
метру (рис. 13.5). В основе такой зависимости лежит, по-видимому,
то обстоятельство, что расстояние между перехватами тем больше,
чем толще волокно. Поскольку проведение замедляется именно в
перехватах, при меньшем их числе потенциал действия распростра-
няется по нервному волокну быстрее (Rushton, 1951).
Из того, что в миелинизированных волокнах скорость пропор-
циональна диаметру волокна, а в немиелинизированных — квад-
ратному корню из диаметра, вытекает интересное следствие: при
очень малых диаметрах (менее 1 мкм) миелинизированные волок-
на проводят импульсы медленнее, чем немиелинизированные
(рис.- 13.6). С этим-согласуется тот факт, что у животных диаметр
миелинизированных волокон не бывает, видимо, меньше 1 мкм.
Системы нервной регуляции 717
У так называемых С-волокон в симпатической нервной системе нет
.миелиновой оболочки; самые крупные и «быстрые» С-волокна име-
ют диаметр 1,1 мкм и скорость проведения 2,3 м/с. Соответствую-
.щая точка на рис. 13.6 отмечена кружком.
Наибольшее преимущество миелинизированных волокон связано
с их малой толщиной, благодаря которой в очень сложной нервной
системе с высокой скоростью проведения импульсов проводники
могут не занимать слишком много места. Представим себе, что
мы хотим в 10 раз увеличить скорость проведения в немиелинизи-
рованном волокне. Это потребует 100-кратного увеличения диамет-
ра волокна, а объем нерва на единицу длины возрастет в
10 000 раз. Очевидно, что если бы большая скорость достигалась
таким путем, то нервный ствол, состоящий из сотен и тысяч немие-
.линизированных аксонов, достигал бы невероятного размера. Пред-
ставим себе, какой величины был бы зрительный нерв, если бы
•свойственная ему высокая скорость проведения была достигнута
без миелинизации. У человека толщина этого нерва 3 мм; но если
бы он состоял из волокон, лишенных миелиновой оболочки, но
проводящих импульсы с той же скоростью, то он должен был бы
иметь толщину 30 см!
МИЕЛИНИЗИРОВАННЫЕ НЕРВНЫЕ ВОЛОКНА БЕСПОЗВОНОЧНЫХ
Как мы видели, высокая скорость нервного проведения у поз-
воночных обусловлена наличием миелиновой оболочки и сальта-
торной передачей импульсов. У многих беспозвоночных быстрое про-
ведение достигается увеличением диаметра нервного волокна (они
используют гигантские аксоны в случаях, когда быстрое проведе-
ние жизненно необходимо, например при реакциях бегства). Одна-
ко дело этим не ограничивается.
У беспозвоночных встречаются два рода структурных модифи-
каций аксона, направленных на ускорение проведения. Во-первых,
как уже говорилось, существуют гигантские аксоны, в которых
быстрота прямо определяется их кабельными свойствами (Hodgkin,
1954). Во-вторых, некоторые нервные волокна беспозвоночных
имеют многослойную оболочку, поразительно сходную с миелино-
вой оболочкой нервов позвоночных. Такие покрытые «миелином»
волокна были обнаружены у насекомых, дождевых червей, крабов
и креветок.
Скорость проведения в таких волокнах намного выше, чем
в других аксонах сходного калибра у беспозвоночных. В волокнах
креветки Palaemonetes vulgaris, имеющих диаметр 26 мкм, ско-
рость проведения достигает 18—23 м/с (Holmes et al., 1941), т. е.
импульсы распространяются гораздо быстрее, чем в таких же
тонких волокнах других беспозвоночных, хотя и медленнее, чем в
миелинизированных аксонах большинства позвоночных.
« ли
i л. id. Управление и интеграция
Оболочки аксонов у креветки несколько отличаются от миели-
новых оболочек позвоночных (рис. 13.7). Слои расположены более
рыхло, между внутренним слоем оболочки и аксоном имеется меж-
клеточное пространство. Поэтому число мембранных слоев в обо-
лочке данной толщины значительно меньше, чем в миелиновой обо-
лочке той же толщины у позвоночного (Heuser, Doggenweiler»
1966).
Рис. 13.7. Поперечный срез аксона креветки, одетого многослойной миелиноподоб-
ной оболочкой. (Heuser, Doggenweiler, 1966.)
Второе отличие связано с тем, что промежутки между слоями
образуют резервуар для ионов. Это увеличивает емкостное сопро-
тивление оболочки нерва и тем самым влияет на кабельные свой-
ства волокон, что позволяет объяснить меньшую скорость проведе-
ния в аксонах креветки по сравнению с миелинизированными аксо-
нами позвоночных. Кроме того, расстояние между перехватами
Ранвье в нервах креветки, видимо, короче, чем у позвоночных, и
поскольку наибольшая задержка в проведении импульсов происхо-
дит именно в перехватах, увеличение числа перехватов должно
замедлять проведение. Однако структурное сходство с миелинизи-
рованными нервами позвоночных все же поразительно, и скорость
проведения для волокон данного диаметра очень велика.
Системы нервной регуляции 719
СИНАПС: ВОЗБУЖДЕНИЕ, ТОРМОЖЕНИЕ И ИНТЕГРАЦИЯ
Сигналы, идущие по аксонам, передаются другим нейронам
через синапсы. Передача в синапсах может осуществляться двумя
способами — электрическим и химическим. Электрическая переда-
ча встречается реже, чем химическая, однако она обладает такими
свойствами, что в определенных случаях оказывается биологически
выгодной.
СИНАПСЫ С ЭЛЕКТРИЧЕСКОЙ ПЕРЕДАЧЕЙ
В «электрических» синапсах кончик аксона одного нейрона на-
столько сближен с другим нейроном, что расстояние между двумя
мембранами не превышает 2 нм. Это пространство образует учас-
ток с малым сопротивлением, обеспечивающий прохождение тока
из окончания аксона в другую клетку. Помимо пути с малым со-
противлением между двумя нейронами для электрической передачи
необходимо также высокое сопротивление, которое предотвращало
бы утечку тока в области контакта.
Электрическая передача происходит без измеримой задержки,
которая характерна для химической передачи. Это имеет большое
значение для функции определенных синапсов как позвоночных, так
и беспозвоночных.
Впервые электрические синапсы были обнаружены в брюшном
нервном тяже речного рака (Furshpan, Potter, 1959), а позже они
были описаны у ряда членистоногих, кольчатых червей и мол-
люсков. У рыб электрическая передача имеет место в синапсах
определенных крупных нервных клеток, известных как маутнеров-
ские нейроны, где быстрота передачи играет важную роль в реак-
ции бегства (см. далее в этой главе); возможно, что электрическая
передача не так редко встречается в центральной нервной системе
позвоночных.
Во многих электрических синапсах передача легко идет в обоих
направлениях; в других синапсах область контакта пропускает ток
только от пресинаптического нейрона к постсинаптическому, но не
в обратном направлении. Таким образом, некоторые, но не все
электрические синапсы обладают свойствами выпрямителей: они
допускают лишь одностороннюю передачу импульсов. В этом отно-
шении они сходны с химическими синапсами.
СИНАПСЫ С ХИМИЧЕСКОЙ ПЕРЕДАЧЕЙ
Для того чтобы понять, как функционируют «химические» си-
напсы, необходимо ознакомиться с их структурой (рис. 13.8). Кон-
чик аксона расширяется и образует бляшку, которая вступает в
контакт с дендритом или телом другого нейрона. Аксон не слива-
ется со вторым нейроном — между ними остается узкое простран-
720 Гл. 13. Управление и интеграция
ство, называемое синаптической щелью, шириной около 20 нм.
Бляшка аксона представляет собой пресинаптический элемент, а
дендрит или нейрон — постсинаптический элемент. Как морфоло-
гия синаптической щели, так и ее ширина поразительно сходны
у всех животных, однако размеры щели настолько малы, что до
сих пор практически ничего не известно относительно ее содержи-
Рис. 13.8. В химическом синапсе (участке, где аксон одного нейрона образует
контакт с другим нейроном) кончик аксона расширяется в синаптическую бляш-
ку. Передача импульсов на другой нейрон осуществляется путем выделения хи-
мического медиатора, содержащегося в синаптических пузырьках.
мого. Высказывалось предположение, что щель, судя по постоян-
ству ее ширины, не просто заполнена жидкостью, а содержит ка-
кую-то структуру из молекул, расположенных определенным обра-
зом между двумя нервными мембранами.
Синаптическая бляшка имеет одну важную особенность: в ней
содержатся многочисленные мелкие пузырьки диаметром обычно
от 20 до 100 нм. Вблизи пресинаптической мембраны эти пузырьки
плотно упакованы. Установлено, что передача импульса с преси-
наптической бляшки на постсинаптический нейрон осуществляется
путем высвобождения из синаптических пузырьков особого веще-
ства — химического медиатора, — которое диффундирует черев
синаптическую щель и воздействует на постсинаптическую мемб-
рану. Наличие пузырьков настолько характерно для всех синапсов,,
что при электронно-микроскопическом исследовании служит приз-
наком, по которому узнают синапс (фото 13.2).
Известен ряд различных синаптических медиаторов; один
из них, ацетилхолин, наиболее достоверно идентифицирован и бо-
лее подробно изучен по сравнению с остальными. Ацетилхолин
служит также медиатором в концевых пластинках двигательных
нервов в поперечнополосатых мышцах. Как мы увидим позже»
известны .и другие медиаторы, но достоверная идентификация
таких веществ связана с большими трудностями.
Системы нервной регуляции 72>
Ацетилхолин высвобождается из пресинаптических бляшек не-
большими дискретными порциями, или «квантами», соответственно»
числу опорожнившихся пузырьков. Для его высвобождения необ-
ходимы ионы кальция; в среде с пониженной концентрацией каль-
ция медиатор выделяется в малом количестве или не выделяется
вовсе. Опыты с использованием различных концентраций кальция
позволили детально изучить механизм выделения ацетилхолина и
Фото 13.2. Синапс в продолговатом мозгу золотой рыбки. Нервное окончание
(большой прямоугольный участок, идущий от середины электронной микрофото-
графии вправо вверх) содержит многочисленные синаптические пузырьки (СП)
и одну митохондрию (М). Синаптическая щель (Щ) отделяет пресинаптическую
мембрану нервного окончания от тела постсинаптической клетки (которое за-
полняет всю левую нижнюю часть снимка). Диаметр синаптических пузырьков
40—50 нм. (Фото предоставлено D. Robertson, Duke University.)
46—1863
~22 Гл. 13. Управление и интеграция
установить, сколько высвобождается квантов ацетилхолина (т. е.
сколько опорожняется пузырьков) при нормальной синаптической
передаче (del Castillo, Katz, 1954). Оказалось, что в нервно-мышеч-
ном соединении выделяется около 300 таких квантов на один нерв-
ный импульс. На звездчатом ганглии кальмара было определено
точное соотношение между потенциалом пресинаптической мембра-
ны и количеством высвобождаемого медиатора (Katz, Miledi, 1967).
Рис. 13.9. Схематическое изображение тела двигательного нейрона и отходящих
от него дендритов. Синаптические бляшки образованы окончаниями аксонов
(передающих импульсы волокон) других нервных клеток. Аксон двигательного
нейрона идет к мышечному волокну. (Eccles, 1965.)
Теперь нам будет понятно, почему химические синапсы прово-
дят импульсы только в одном направлении: поскольку воздействие
передается с помощью медиатора, а медиатор имеется только
с пресинаптической стороны, передача импульсов в обратном на-
правлении невозможна.
На рис. 13.8 процесс передачи информации от одного нейрона
другому представлен в весьма упрощенном виде. В большинстве
случаев аксоны сильно разветвлены и соединяются со множеством
других нейронов, как об этом уже говорилось при рассмотрении
• структуры сетчатки (гл. 12). Хотя у каждого нейрона только один
аксон, этот аксон может иметь много ветвей, соединенных с други-
ми нейронами. В результате к каждому нейрону подходит большое
число веточек различных аксонов, которые оканчиваются на теле
нейрона или его дендритах (рис. 13.9).
В центральной нервной системе на отдельных нейронах часто
Системы нервной регуляции 723
бывают сотни синапсов, а двигательные нейроны спинного мозгам
позвоночных могут иметь более 1000 входных синапсов. Некоторые
специализированные нейроны так плотно покрыты синаптическими:
бляшками, что число связей, образуемых отдельной нервной клет-
кой, оценивается примерно в 10 000. Этот факт в сочетании с дан-
ными об огромном числе нейронов (в мозгу человека около 1010)
дает представление о степени сложности центральной нервной си-
стемы. Следует, однако, подчеркнуть, что связи между нейронами
Фото 13.3. Синаптические бляшки. На этой микрофотографии, полученной с по-
мощью сканирующего электронного микроскопа, видны многочисленные синап-
тические бляшки, контактирующие с поверхностью одного из нейронов в нервной
системе морского моллюска Aplysia. Диаметр каждой дисковидной бляшки
около 1 мкм. (Фото предоставлено R. Lewis, University of California, Berkeley.)
отнюдь не случайны; напротив, они высокоспецифичны и формиру-
ют в центральной нервной системе проводящие пути, функциони-
рующие с высокой точностью. Эти пути были подробнейшим обра-
зом изучены и описаны нейроанатомами и нейрофизиологами.
46’
724 Гл. 13. Управление и интеграция
ЛОСТСИНАПТИЧЕСКИЕ ПОТЕНЦИАЛЫ
В понимании процессов синаптической передачи был достигнут
‘большой прогресс в 50-х годах, когда появился метод отведения
электрической активности от отдельных нейронов. Для этого ис-
пользуют тончайшие стеклянные пипетки с диаметром кончика ме-
нее 1 мкм. Такую пипетку заполняют солевым раствором (обычно
концентрированным раствором КС1), превращая ее в микроэлек-
трод, с помощью которого можно регистрировать любое изменение
разности потенциалов между кончиком электрода и обычной «зем-
лей». Когда такой микроэлектрод медленно перемещают и вводят
в нейрон, то вначале регистрируется нулевой потенциал (по отно-
шению к «земле»), но в момент прокола мембраны нервной клетки
потенциал становится отрицательным. Мембрана как бы смыкается
вокруг стекла, и функция нейрона, по всей видимости, не нару-
шается; это позволяет записывать активность клетки в течение
нескольких часов. Используя пипетки не с одним, а с двумя или
несколькими каналами, удается даже вводить ничтожные количе-
ства различных веществ в те участки, где регистрируют потенциал.
С помощью этого метода можно исследовать процессы, происхо-
дящие в синапсах. Когда импульс доходит до пресинаптической
мембраны, наступает задержка длительностью в доли миллисекун-
ды, после чего потенциал постсинаптической мембраны изменяется.
Этот сдвиг называют постсинаптическим потенциалом (ПСП)
(рис. 13.10). ПСП сначала быстро возрастает, а затем угасает, но
это происходит гораздо медленнее.
Постсинаптические потенциалы отличаются от потенциалов
действия в двух важных отношениях: они имеют, во-первых, значи-
тельно меньшую амплитуду и, во-вторых, большую (иногда в 10—
100 раз) длительность. Эти особенности ведут к двум важным
следствиям: с одной стороны, единичный ПСП всегда (или почти
всегда) оказывается недостаточным для возбуждения потенциала
действия в постсинаптическом нейроне; с другой стороны, большая
длительность ПСП создает возможность значительного взаимодей-
ствия нескольких ПСП в одном и том же нейроне (как в одном
синапсе, так и в соседних синапсах одного нейрона). Поскольку
взаимодействие постсинаптических потенциалов играет очень боль-
шую роль, мы рассмотрим этот вопрос подробнее.
Что вызывает появление ПСП? Мы будем считать, что ПСП
возникает в результате выделения вещества-медиатора из преси-
наптической мембраны и что задержка в появлении ПСП соответ-
ствует времени диффузии медиатора через синаптическую щель.
Из нейромедиаторов лучше всего изучен ацетилхолин; его роль
передатчика в синапсах и нервно-мышечных соединениях не вызы-
вает сомнений. В синапсах величина ПСП, по-видимому, находится
в прямой зависимости от количества выделившегося ацетилхолина.
Однако система .не могла бы работать, если бы не происходило
'быстрого удаления ацетилхолина; в противном случае ацетилхолин
Системы нервной регуляции 725
постепенно накапливался бы и поддерживал постоянный ПСП.
В синапсах всегда имеется фермент ацетилхолинэстераза, который
гидролизует ацетилхолин и таким образом удаляет его. По мере
исчезновения медиатора ПСП постепенно затухает.
Рис. 13.10. Потенциал действия, дойдя до терминальной бляшки (А), вызывает
появление потенциала на постсинаптической мембране (Б); этот постсинаптиче-
ский потенциал меньше по величине, но более длителен. (Stevens, 1966.)
Суммация. Если очередной импульс придет к синапсу еще до
того, как исчезнет предыдущий ПСП, то количество медиатора в
синапсе увеличится и новый ПСП будет соответственно больше.
Иными словами, произойдет суммация двух импульсов; поскольку
она происходит во времени, ее называют временной суммацией.
Представим себе, что импульсы приходят к синапсу непрерывно
о постоянной частотой. При этом постсинаптические потенциалы
суммируются и образуется общий ПСП, величина которого находит-
ся в прямой зависимости от частоты поступления импульсов. Если
эта частота возрастет, ПСП увеличится; новый ПСП установится
на таком стационарном уровне, при котором скорость распада ме-
диатора равна скорости его выделения из окончания аксона.
Таким образом, величина ПСП непосредственно отражает частоту
импульсации в аксоне; другими словами, ПСП является частотно-
зависимым потенциалом. Это позволяет понять, каким образом се-_
726 Гл. 13. Управление и интеграция
рия потенциалов действия, возникающих по принципу «всё или
ничего» и потому равных по величине, может быть использована
для передачи информации относительно переменного сигнала. На-
пример, усиление стимуляции какого-то сенсорного нейрона коди-
руется и передается на аксон этого нейрона в виде повышения
частоты потенциалов действия; на уровне синапса происходит рас-
шифровка этого кода, так как величина ПСП находится в прямой
зависимости от величины исходного сигнала.
Вернемся к постсинаптическому нейрону и вспомним, что на его
поверхности имеется множество синапсов. Распространение ПСП,
возникающего в каком-то одном синапсе, не ограничивается только
областью, лежащей непосредственно под окончанием аксона, а за-
хватывает также прилежащие участки; при этом величина ПСП
убывает с увеличением расстояния от места его возникновения.
Таким образом, вокруг синапса, где возник ПСП, мембранный по-
тенциал несколько изменяется, и если к соседнему синапсу тоже
придет импульс, то новый ПСП суммируется с уже имеющимся
потенциалом. Этот вид суммации называется пространственной
суммацией.
Таким образом, различные входные сигналы, поступающие в
нейрон через разные синапсы, влияют друг на друга и могут интег-
рироваться по принципу как временной, так и пространственной
суммации. Эти два типа суммации составляют основу интегратив-
ной деятельности каждого нейрона, а тем самым и всей нервной
системы.
Прежде чем перейти к дальнейшему обсуждению интегратив-
ных процессов, рассмотрим, какое влияние оказывают постсинап-
тические потенциалы на нейрон. Как мы уже говорили, ПСП обыч-
но слишком слаб для того, чтобы вызвать в постсинаптическом
нейроне потенциал действия. Существует, однако, такой участок
нейрона — аксонный бугорок (у основания аксона),— где при до-
статочной деполяризации мембраны возникает потенциал действия.
Если к различным синапсам одного нейрона приходит много им-
пульсов, то в результате сочетания пространственной и временной
суммации может возникнуть достаточно большой ПСП, способный
распространиться вплоть до основания аксона, деполяризовать его
и таким образом вызвать появление потенциала действия в аксоне.
ВОЗБУЖДЕНИЕ И ТОРМОЖЕНИЕ
До сих пор мы не касались вопроса о полярности ПСП. Дело
в том, что ПСП может либо уменьшать, либо увеличивать нормаль-
ный мембранный потенциал. Если ПСП слегка снижает потенциал
постсинаптической мембраны (т. е. деполяризует ее), то этот сдвиг
совпадает по направлению с тем, который происходит при развитии
потенциала действия. Такой ПСП называют возбуждающим пост-
синаптическим потенциалом (ВПСП). Если же ПСП увеличивает
Системы нервной регуляции 727
нормальный мембранный потенциал (т. е. создает гиперполяриза-
цию), то это противоположно тому, что ведет к возникновению
импульса. Такой ПСП называют тормозным постсинаптическим
потенциалом (ТПСП), так как он затрудняет генерацию потенциа-
ла действия.
Имеющиеся данные говорят о том, что по характеру ПСП каж-
дый синапс принадлежит либо к одному, либо к другому типу и
все окончания какого-то одного аксона индуцируют один и тот же
тип ПСП. Каждый данный синапс является только тормозным или
только возбуждающим, и это его свойство неизменно.
За исключением полярности, оба типа постсинаптических потен-
циалов по своей природе одинаковы. Различие лишь в том, что
ВПСП преобразуют поток поступающих импульсов в деполяриза-
цию постсинаптической мембраны, а ТПСП — в гиперполяриза-
цию. Один и тот же нейрон по-разному отвечает на поступающую
информацию в зависимости от того, какую информацию он полу-
чил перед этим или получает в данный момент из других источни-
ков. В тех случаях, например, когда уже произошла частичная де-
поляризация мембраны нейрона, дополнительный возбуждающий
потенциал может оказаться достаточным для того, чтобы в аксон-
ном бугорке возник потенциал действия; в других случаях такой
же ВПСП окажется слишком слабым для этого.
Все это позволяет понять, каким образом в единичном нейроне
может осуществляться сложная интеграция сигналов, поступающих
из разных источников. Хотя каждый синапс передает информацию
только в одном направлении, наличие возбуждающих и тормозных
синапсов в сочетании с явлениями пространственной и временной
суммации обеспечивает широкие возможности интеграции на уров-
не одного нейрона, что делает нейрон функциональным элементом
нервной системы.
ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
Следует рассмотреть еще одну особенность нейронов. Не вда-
ваясь в детали, укажем на существование такого процесса, как
пресинаптическое торможение. Суть этого важного явления состоит
в следующем. Окончание аксона может оказаться в течение неко-
торого времени слегка гиперполяризованным или же слегка депо-
ляризованным (при отсутствии потенциала действия). Если сюда
придет импульс в период небольшой деполяризации, то под дейст-
вием этого импульса выделится меньшее количество медиатора и
ПСП будет соответственно более слабым. Поскольку уменьшение
ПСП в этом случае обусловлено изменением в окончании аксона,
т. е. перед синапсом, такой тип торможения называется пресинап-
тическим.
Вопрос о том, пре- или постсинаптическим является торможе-
ние, может показаться несущественным, но это не так. Дело в том,
728 Гл. 13. Управление и интеграция
что постсинаптическое торможение может уменьшить ПСП, возни-
кающие у многих входных синапсов данного нейрона; таким обра-
зом, этот вид торможения не избирателен. Пресинаптическое тор-
можение, напротив, обладает высокой избирательностью, так как
воздействует на сигналы, приходящие в один определенный синапс.
Другая особенность пресинаптического торможения — то, что оно
не является итогом простого сложения зарядов на мембране (как
постсинаптическое торможение), а влияет на передачу каждого
импульса в более длинном ряду поступающих сигналов. Поэтому
благодаря пресинаптическому торможению возрастает как специ-
фичность, так и сложность процессов интеграции на уровне одного
нейрона.
РЕАКЦИЯ БЕГСТВА: ДВИЖЕНИЕ ХВОСТА
Нормальная реакция бегства у костистых рыб выражается в
том, что рыба делает резкое движение хвостом и затем уплывает
с помощью обычных волнообразных движений. Этой реакцией
управляют две большие и весьма примечательные нервные клетки,
расположенные в головном мозгу и называемые маутнеровскими
клетками. Их роль состоит в интеграции сигналов от органов
чувств и мозга и передаче ее двигательным нервам, управляющим
мускулатурой тела.
Маутнеровские клетки имеют многочисленные входы, многие
из которых представляют собой электрические, а не химические
синапсы. Главное выходное устройство — это большой аксон, отхо-
дящий от каждой маутнеровской клетки; эти аксоны перекрещива-
ются, так что каждый из них переходит на противоположную сто-
рону тела и, проходя вдоль спинного мозга, образует синапсы
с мотонейронами, управляющими основной плавательной мускула-
турой (Diamond, 1971).
Важное значение маутнеровских клеток состоит в том, что они
реагируют на информацию, поступающую от органов чувств, осо-
бенно на сигналы о механических возмущениях в воде. В ответ
маутнеровская клетка посылает по аксону единичный импульс, ко-
торый переходит на противоположную сторону тела и вызывает
быстрое, энергичное сокращение мышц (быстрый взмах хвоста и
последующие плавательные движения). Эту реакцию легко наблю-
дать у аквариумных рыб, постучав по стенке аквариума.
Что произойдет, если в обеих маутнеровских клетках импульсы
возникнут одновременно? При одновременном сокращении мышц
на обеих сторонах тела никакого движения не будет. Здесь-то и
выступает со всей наглядностью роль тормозных входов. Посколь-
ку сигналы от органов чувств практически никогда не бывают пол-
ностью симметричны по времени и амплитуде, одна маутнеровская
клетка всегда активируется чуть-чуть раньше другой. Клетки свя-
Системы нервной регуляции 729
ваны между собой так, что каждая из них оказывает тормозящее
влияние на другую; в результате если одна из них генерирует им-
пульс, то вторая оказывается заторможенной. Благодаря наличию
электрических синапсов передача воздействий происходит почти
мгновенно. В химической передаче это было бы связано с задерж-
кой, в результате которой торможение второй маутнеровской клет-
ки могло бы наступить недостаточно быстро и генерацию импуль-
са не удалось бы предотвратить.
Преимущество электрической передачи — ее быстрота; здесь
нет задержки, свойственной химическим синапсам. Именно в эконо-
мии времени состоит значение этого механизма быстрой передачи
сигналов в нервной системе. Как только возбудилась одна маутне-
ровская клетка, она почти мгновенно тормозит другую и таким
образом предотвращает одновременное сокращение симметричных
мышц, которое парализовало бы рыбу.
За исключением тех случаев, когда быстрота имеет жизненно
важное значение, химическая передача представляется более со-
вершенной. Основное достоинство ее состоит, по-видимому, в дли-
тельности изменений, происходящих в синапсах (возбуждающих
или тормозных); благодаря действию химического медиатора про-
цесс интеграции может охватывать значительно большие отрезки
времени. Преобладание химических синапсов в нервной системе
само по себе указывает на какое-то их функциональное преимуще-
ство; кажется вполне вероятным, что оно состоит в возможности
более сложных взаимодействий между последовательными сигна-
лами.
РЕГУЛЯЦИЯ МЫШЕЧНОЙ АКТИВНОСТИ
Как уже отмечалось в главе 11, у ракообразных мышцы нахо-
дятся под контролем небольшого числа аксонов (нередко 3—5).
Каждый аксон посылает веточки ко всем (или почти всем) волок-
нам, составляющим мышцу. В отличие от этого у позвоночных
мышцы иннервируются множеством аксонов, каждый из которых
посылает ветви лишь к небольшому числу мышечных волокон (об-
разующих двигательную единицу), В связи с таким различием в
иннервации у ракообразных мышцы находятся в основном под
периферическим контролем, а у позвоночных — под контролем
чрезвычайно сложной центральной нервной системы.
Причину этого нетрудно понять. Миелинизированные волокна
позвоночных позволяют быстро передавать импульсы без чрезмер-
ного увеличения диаметра аксона, и поэтому нерв средней толщи-
ны может обеспечить достаточно большое число связей. У ракооб-
разных же для ускорения передачи импульсов требуется
значительное утолщение нервного волокна; поэтому тонкая цент-
ральная регуляция мышечных сокращений, возможная лишь при
большом числе волокон в двигательном нерве, у них просто немые-
730 Гл. 13. Управление и интеграция
лима. В главе 11 уже говорилось о том, каким образом в этом
случае все же достигается точная регуляция работы мышц, иннер-
вируемых не более чем 2—3 возбуждающими и 1—2 тормозящими
аксонами. Посмотрим теперь, как происходит управление мышца-
ми у позвоночных.
мышцы позвоночных
Сокращение мышц у позвоночных полностью находится под
центральным контролем; у них нет необходимости в иннервации
мышц тормозными аксонами, модулирующими степень сокраще-
ния1. Мышечная система позвоночных работает под управлением
центрального «компьютера», получающего обильную информацию»
из различных источников, в том числе прямо от самих мышц; эта
информация от мышц создает обратную связь и имеет очень боль-
шое значение. На примере мышечной системы мы увидим, какого*
совершенства удается достигнуть с помощью механизмов централь-
ной регуляции и интеграции.
Нерв, идущий к мышце позвоночного, содержит волокна, стиму-
лирующие сокращение; кроме того, в нем имеются другие волокна,,
которые передают сенсорную информацию от мышцы в централь-
ную нервную систему. Сенсорные волокна бывают двух основных ти-
пов (рис. 13.11). Одни идут от миниатюрных сенсорных структур,
расположенных в сухожилиях (от сухожильных органов); дру-
гие — от специализированных мышечных волокон, так называемых
мышечных веретен. Сухожильные органы воспринимают, по-види-
мому, степень растяжения сухожилия при сокращении мышцы щ
таким образом, посылают информацию о силе этого сокращения.
Мышечные веретена, которые мы рассмотрим подробнее, доставля-
ют информацию о длине мышцы, но, как мы увидим далее, особен-
но важное значение имеет то, как эта информация используется
для регуляции сокращения мышцы по принципу обратной связи.
Мышечные веретена — очень сложно устроенные органы чувств;
возможно, что в этом отношении они уступают только глазу и уху.
Рассмотрим крайне упрощенную схему (рис. 13.12). При по-
стоянной нагрузке человек легко может поддерживать постоянную-
силу сокращения мышцы. Если неожиданно увеличить нагрузку, то
произойдет растяжение мышцы, которое почти мгновенно будет
1 Хотя скелетные мышцы позвоночных лишены, по-видимому, тормозной ин-
нервации, к другим видам мышц позвоночных это не относится. Поперечнополо-
сатая мышца сердца имеет двойную иннервацию, как стимулирующую, так и
тормозную. Все гладкие мышцы регулируются как адренэргическими, так и хо-
линэргическими нервными волокнами вегетативной нервной системы. На некото-
рые гладкие мышцы (например, в бронхах) адренэргические волокна оказывают
тормозное действие, на другие (в стенках сосудов) — стимулирующее. Поскольку
холинэргические волокна всегда антагонистичны по отношению к адренэргиче-
ским, они тоже могут оказывать либо тормозное, либо стимулирующее дей-
ствие.
Системы нервной регуляции
731
компенсировано увеличением силы сокращения. Это результат
рефлекса, возбуждаемого мышечными веретенами. Будучи растя-
нуты, мышечные веретена посылают импульсы в спинной мозг, где
их сенсорные волокна образуют синаптические контакты с двига-
тельными нейронами; а эти нейроны в свою очередь стимулируют
усиленное сокращение для компенсации растяжения, вызвавшего
рефлекс.
Капсула
Первичное
окончание
Сенсорное
волокно от
мышечного
веретена
Интрафузальное
мышечное
волокно
Двигательное
окончание ~
Рис. 13.11. Мышцы позвоночных снабжены сенсорными органами, передающими
информацию в центральную нервную систему. Из сухожилий поступает инфор-
мация о силе сокращения, а из мышечных веретен — главным образом о длине
мышцы. Эта информация обеспечивает обратную связь в системе тонкой регуля-
ции сокращения мышц. (Merton, 1972.)
На самом деле в этом рефлексе участвует не один синапс; кро-
ме того, в спинном мозгу имеются связи, по которым информация
передается на различные уровни центральной нервной системы,
в том числе в головной мозг. Мы, однако, не будем касаться этих
деталей. Главное состоит в том, что благодаря мышечным верете-
нам может поддерживаться постоянство длины мышцы независимо
ст нагрузки; значение этого для координации всевозможных дви-
жений очевидно.
732 Гл. 13. Управление и интеграция,
Мышечные веретена имеют еще одну важную особенность: они
сами способны к сокращению, но их иннервируют особые двига-
тельные волокна, отличные от тех аксонов, которые вызывают со-
кращение обычных мышечных волокон. Необходимо ознакомиться
со структурой мышечных веретен (рис. 13.11). Они представляют
Рис. 13.12. Обратная связь в регуляции мышечного сокращения. (Merton^
1972.)
Постоянная нагрузка на мышцу уравновешивается постоянной силой сокраще-
ния (А). Внезапное увеличение нагрузки (Б) вызывает растяжение мышцы; тот-
час же из мышечных веретен начинают поступать сигналы в спинной мозг; это
приводит к возбуждению дополнительных двигательных нейронов, они посылают
импульсы назад по двигательному нерву, и в результате сокращение усиливается
в соответствии с возросшей нагрузкой (В).
собой пучки из нескольких видоизмененных мышечных волокон,,
называемых интрафузалъными волокнами (от лат. fusus — верете-
но). Эти волокна способны к сокращению, но в середине волокна
(в его «экваториальном» участке) сократительный аппарат отсутст-
вует, и этот участок обвивают чувствительные нервные окончания.
Предположим, что в результате стимуляции интрафузальноё-
волокно сократится; при этом его средний участок будет растяну?
и в спинной мозг направится залп импульсов. Здесь эти импульсы
Системы нервной регуляции 733>
вызовут возбуждение мотонейронов, и в двигательном нерве воз-
никнут потенциалы действия, которые будут стимулировать сокра-
щение мышцы. Когда сокращение мышцы достигнет такого же-
уровня, как сокращение интрафузального волокна, растяжение-
среднего участка этого волокна прекратится и его рецепторы «за-
молкнут».
Эта изящная система в некоторой степени аналогична механи-
ческой сервосистеме. Например, у тяжелого грузовика руль бывает*
снабжен сервоприводом: поворот руля, производимый вручную, ве-
дет к включению мотора, который и выполняет тяжелую работу по-
разворачиванию передних колес. Сервомеханизм — это, по сущест-
ву, не что иное, как автоматическая система регуляции, действую-
щая по принципу обратной связи, где в качестве регулируемой пе-
ременной выступает положение какого-либо механического уст-
ройства. Такие системы широко используются в технике.
Описанные выше интрафузальные волокна служат остроумным*
механизмом для поддержания определенной скорости сокращения
при меняющейся нагрузке. Если сокращение данной мышцы ре-
гулируется медленным сокращением интрафузальных волокон, то
увеличение нагрузки лишь на мгновение замедлит сокращение
мышцы. В самом деле, при этом интрафузальные волокна будут*
продолжать сокращаться с прежней скоростью, а так как сокраще-
ние всей мышцы замедлится, то средний участок интрафузального
волокна подвергнется растяжению и тотчас же начнет посылать-
дополнительные импульсы в спинной мозг, что приведет к усиле-
нию сокращения. Таким образом, увеличение или уменьшение на-
грузки при мышечном сокращении автоматически компенсируется.
Для управления мышцами при движении конечностей возника-
ет необходимость в дополнительном регулирующем устройстве.
Мышцы, обеспечивающие поддержание позы, должны находиться
в состоянии небольшого сокращения; если они полностью рассла-
бятся, то тело обмякнет (как это случается при обмороке). Кроме
того, для движения конечности недостаточно того, чтобы опреде-
ленная группа мышц сократилась: необходимо, чтобы одновремен-
но расслабились мышцы-антагонисты. Это обеспечивается доволь-
но простым рефлекторным механизмом, схема которого приведена
на рис. 13.13. Предположим, стимуляция мышечного веретена в-
мышце А вызывает ее сокращение. Информация о сокращении по-
ступает в определенные нейроны спинного мозга, называемые вста-
вочными нейронами, или интернейронами1: их функция состоит а
торможении других нейронов. Связи организованы таким образом,
что эти тормозные интернейроны воздействуют на двигательные*
1 Вставочными нейронами называют такие нейроны, аксоны которых не вы-
ходят за пределы центральной нервной системы. Они бывают как тормозными,
так и возбуждающими.
734 Гл. 13. Управление и интеграция
нейроны мышцы-антагониста (5). Реципрокное действие мышц-
антагонистов, выражающееся в том, что сокращение одних сопря-
жено с расслаблением других, функционально очень выгодно; оно
не могло бы осуществиться без участия тормозных нейронов или
жакого-то другого механизма торможения.
Помимо регуляции антагонистических мышц торможение игра-
ет важную роль и в других процессах. Если, например, сокращаю-
щуюся мышцу растянуть с очень большой силой, то произойдет
Рис. 13.13. Если в движении конечности участвуют две антагонистические мыш-
цы (А и Б), то при сокращении одной из них (А) импульсы от нее поступают
в спинной мозг к тормозным интернейронам. Последние тормозят соответствую-
щие двигательные нейроны и тем самым вызывают расслабление мышцы-анта-
гониста (Б).
активация сухожильных органов и от них начнут поступать им-
пульсы к тормозным интернейронам в спинном мозгу. Это приведет
к торможению сокращения нагруженной мышцы, и в результате
ее активное сопротивление растяжению уменьшится. Этот рефлекс
предохраняет мышцы от разрыва при внезапном увеличении на-
грузки. Не будь такого защитного механизма, травмы типа растя-
жения мышц встречались бы значительно чаще.
Интеграция работы мышц при локомоции на самом деле гораз-
до сложнее, чем описано в приведенных примерах. Эта интеграция
зависит от различных команд, поступающих из мозга, и от инфор-
мации, идущей от органов чувств — глаз, ушей, кожи и т. д. Цент-
ральная нервная система представляет собой исключительно слож-
ную управляющую систему, действие которой основывается на ин-
формации и сигналах обратной связи, поступающих из множества
источников.
Обратимся теперь к эндокринной системе и к тому тесному
взаимодействию, которое происходит между эндокринной и нерв-
ной системами. Как мы увидим, центральная нервная система не
Система гормональной регуляции 735
только осуществляет управление на основе передачи и интеграции,
нервных импульсов, но и играет важную роль в выработке гор-
монов.
СИСТЕМА ГОРМОНАЛЬНОЙ РЕГУЛЯЦИИ
Под гормональным контролем находится множество физиологи-
ческих функций. Иногда этот вид регуляции называют химической
регуляцией в отличие от нервной, но этот термин неудачен и может
ввести в заблуждение. Мы уже видели, что передача нервного им-
пульса в синапсе обычно осуществляется химическим путем; а с
другой стороны, как мы увидим, нервной системе принадлежит
важная роль в самой выработке гормонов и в ее регулировании.
Кроме того, некоторые химические вещества, образующиеся в ор-
ганизме в нормальных условиях (например, двуокись углерода) г
обладают выраженным физиологическим действием и участвуют
в регуляторных процессах, однако они не являются гормонами и;
не имеют отношения к эндокринной функции.
Гормон лучше всего определить как вещество, которое выделя-
ется определенным органом или структурой и оказывает специфи-
ческое воздействие на другую структуру или функцию. Важнейшие-
гормоны позвоночных и их главные функции приведены в табл. 13.3.
До последнего времени эндокринология и биохимия гормонов:
были эмпирической областью науки, где преобладал довольно*
стандартный подход. Если возникало предположение, что какой-то*
орган продуцирует гормон, то этот орган удаляли, лишая тем
самым организм нормального источника гормона. Далее изучали
и описывали результат, и если развивающиеся симптомы можно*
было снять введением экстракта данного органа, то дальнейшая
работа состояла в очистке этого экстракта, выделении активного
компонента и, наконец, его химическом синтезе. Другой метод за-
местительной терапии состоял в трансплантации вырабатывающей
гормон ткани.
Эти общие подходы обеспечивают успех при исследовании тех.
органов, удаление которых не представляет опасности для жизни,,
например гонад. Однако многие органы, продуцирующие гормоны^
обладают и другими жизненно важными функциями, так что их.
удаление приводит к весьма серьезным последствиям. Например,,
печень и почки вырабатывают гормоны, но удаление этих органов^
быстро вызывает гибель животного по причинам, не связанным с
их гормональной функцией.
Все физиологические функции, которые регулируются в основ-
ном гормонами, удобно разделить на пять достаточно широких ка-
тегорий (табл. 13.4). Первые три категории относятся к обмену
веществ и поддержанию стационарных состояний, а две послед-
ние — к росту, развитию и размножению. Многие гормоны сходны
или идентичны у представителей всех классов позвоночных, но»
736
Важнейшие Гормоны пбзйойочйых
Таблица 13.3
Источник Гормон1) Основные функции
Гипоталамус Гормоны, стимулирующие Эти гормоны попадают через портальный кровоток в адено- или подавляющие выде- гипофиз. Подробнее см. табл. 13.6 ленпе определенных гормонов аденогипофи- зом
Гипоталамус (через ней- рогипофиз) Окситоцин Стимулирует сокращение мускулатуры матки и выделение молока Антидиуретический гормон Стимулирует реабсорбцию воды в почках
Аденогипофиз Адренокортикотропный гор- Стимулирует кору надпочечников мон (АКТГ) Тиреотропный гормон (ТТГ) Стимулирует щитовидную железу Фолликулостимулирующий Стимулирует развитие фолликулов в яичнике и семенных ка- гормон (ФСГ) нальцев в семенниках Лютеинизирующий гормон Стимулирует превращение яйцевого фолликула в желтое тело; (ЛГ) стимулирует образование прогестерона и тестостерона Пролактин Стимулирует выработку молока Меланоцитстимулирующий Стимулирует рассеяние меланина в пигментных клетках кожи гормон (МСГ) у земноводных Гормон роста Стимулирует рост (действует через печень, см. ниже)
Печень Соматомедин Стимулирует рост (опосредует действие гипофизарного гормо- на роста)
Кора надпочечников Глюкокортикоиды (корти- Регулируют обмен углеводов костерон, гидрокорти- зон и др.) Минералокортикоиды (аль- Регулируют обмен и выделение натрия достерон, дезоксикорти- костерон и др.)
47—1853 737
Кортикальные андрогены, прогестерон Стимулируют развитие вторичных половых признаков, особен- но мужских
Яичник Эстрогены Прогестерон Инициируют развитие женских вторичных половых признаков и поддерживают их; инициируют периодическое разраста- ние слизистой матки; тормозят выделение ФСГ Вместе с эстрогенами стимулирует развитие женских вторич- ных половых признаков; вызывает секреторное преобразо- вание слизистой матки; тормозит выделение ЛГ и ФСГ
Семенники Тестостерон Инициирует развитие мужских вторичных половых признаков и поддерживает их
Щитовидная железа Тироксин, трииодтиронин Кальцитонин3) Стимулируют окислительный обмен; стимулируют метаморфоз амфибий; тормозят высвобождение ТТГ Тормозит чрезмерное повышение уровня кальция в крови
Паращитовидная железа Паратгормон Повышает уровень кальция в крови
Желудок Гастрин Стимулирует секрецию желудочного сока
Двенадцатиперстная кишка Секретин Панкреозимин4) Холецистокинин4) Энтерогастрон Стимулирует секрецию сока поджелудочной железы Стимулирует секрецию ферментов поджелудочной железой Стимулирует выделение желчи из желчного пузыря Тормозит желудочную секрецию
Поджелудочная железа Инсулин Глюкагон Снижает уровень глюкозы в крови; стимулирует синтез и на- копление углеводов Повышает уровень глюкозы в крови путем мобилизации гли- когена в печени
Мозговое вещество над- почечников Адреналин, норадреналин Усиливают симпатические эффекты (расширение сосудов в мышцах, печени и легких, сужение сосудов во многих внутренних органах); повышают содержание сахара в крови
’) Термины «-тропный гормон» и «-тропин» равнозначны (например, тиреотропный гормон называют также тиреотропином. Окончание
«-тропин» (от греч. tropein — повернуть, изменить) предпочтительнее в данном случае, чем «-трофин» (от греч. trophos — кормящий)
(ср. фототрофный и гипертрофия). (Stewart, Li, 1962).
2) АДГ называют также вазопрессином, так как в высоких концентрациях он вызывает сокращение гладкой мускулатуры сосудов, что при-
водит к повышению кровяного давления.
3) У млекопитающих его вырабатывают С-клетки щитовидной железы, у низших позвоночных — ультимобранхиальные тельца.
4) Предполагается, что панкреозимин и холецистокинин идентичны друг другу.
738 Гл. 13. Управление и интеграция
Таблица 13.4
Основные категории физиологических функций, регулируемых гормонами
(и соответствующие гормоны, наиболее известные)
Пищеварение и метаболические функции
Секретин
Г астрин
Инсулин
Глюкагон
Норадреналин
Тироксин
Стероиды надпочечников
Осморегуляция, экскреция, водный и солевой обмен
Вазопрессин
Пролактин
Альдостерон
Обмен кальция
Паратгормон
Кальцитонин
Рост и морфологические изменения
Гормон роста
Андрогены коры надпочечников
Тироксин (метаморфоз амфибий)
Меланоцитстимулирующий гормон (изменение окраски у амфибий)
Репродуктивные органы и размножение
Фолликулостимулирующий гормон
Лютеинизирующий гормон
Эстрогены
Прогестерон
Пролактин
Тестостерон
некоторые обладают специфическими функциями, различными
у разных групп животных. Примером может служить пролактин,
который у млекопитающих стимулирует секрецию молока, у голу-
бей — образование «зобного молочка», а у рыб влияет на функцию
почек и осмотическую проницаемость жабр.
ХИМИЧЕСКАЯ ПРИРОДА ГОРМОНОВ ПОЗВОНОЧНЫХ
Несмотря на многочисленность гормонов и разнообразие их
функций у позвоночных, их можно разделить по химической приро-
де и структуре на три четко определенные группы: 1) производные
тирозина, 2) стероиды и 3) пептиды и белки (табл. 13.5). Эта клас-
сификация очень важна, так как со структурой гормона тесно свя-
зан механизм передачи его воздействия на клетки органа-мишени
(см. ниже в этой главе).
Системы нервной регуляции 739
Таблица 13.5
Классификация гормонов позвоночных по их структуре
и химической природе. Этой классификации
соответствует механизм действия гормона на уровне
клетки
Гормоны — производные тирозина
Катехоламины
Норадреналин
Адреналин
Тиреоидные гормоны
Тироксин
Трииодтиронин
Стероидные гормоны
Тестостерон
Эстрогены
Прогестерон
Кортикостероиды
Витамин D3
Пептидные и белковые гормоны
Пептиды
Гормоны гипоталамуса (3—14 аминокислот)
Ангиотензин (8 аминокислот)
Гастрин (17 аминокислот)
Секретин (27 аминокислот)
Глюкагон (29 аминокислот)
Кальцитонин (32 аминокислоты)
Инсулин (51 аминокислота)
Паратгормон (84 аминокислоты)
Белки с более крупными молекулами
Гормон роста
Пролактин
Лютеинизирующий гормон
Фолликулостимулирующий гормон
Тиреотропный гормон
Гормоны — производные тирозина. На рис. 13.19 приведены
структурные формулы двух широко распространенных катехола-
минов — адреналина и норадреналина. Эти два гормона отличают-
ся друг от друга только метильной группой (—СН3), которая име-
ется в адреналине и отсутствует в норадреналине.
Катехоламины образуются из аминокислоты тирозина, которая
в несколько этапов превращается в адреналин и норадреналин.
Тирозин служит также исходным материалом для синтеза тиреоид-
ных гормонов — тироксина и трииодтиронина. Эти гормоны не от-
носятся к катехоламинам, а образуют особую группу; они синте-
зируются из тироксина путем конденсации двух шестичленных
углеродных колец, в результате иодирования которых образуются
активные гормоны.
47*
740 Гл, 13. Управление и интеграция
Стероидные гормоны. Все стероидные гормоны — это производ-
ные холестерина, имеющего характерную структуру из одного-
пятичленного и трех шестичленных углеродных колец. В результа-
те модификаций этой основной структуры возникает целый ряд
важных гормонов, причем небольшие изменения структуры могут
приводить к радикальному изменению физиологического действия.
Холестерин
Тестостерон
Сравним, например, эстрадиол и тестостерон: различия в их струк-
туре очень невелики, но эффект этих двух половых гормонов диа-
метрально противоположен. К стероидам относятся также андроге-
ны, образующиеся в коре надпочечников, эстрогены, прогестерон и
такие кортикостероиды, как кортизон и альдостерон.
Пептидные и белковые гормоны. К пептидным гормонам отно-
сятся гипоталамические регуляторные гормоны, содержащие от 3
до 14 аминокислот. Тиреолиберин состоит всего лишь из трех ами-
нокислот, соматолиберин — из десяти, а соматостатин — из 14 ами-
нокислот. Ряд других пептидных и белковых гормонов приведен
в табл. 13.5.
Важнейшие гормоны, образующиеся в аденогипофизе, имеют
белковую природу и могут состоять из нескольких сотен аминокис-
лот. Некоторые из них — гликопротеиды, т. е. в дополнение к пеп-
тидной цепи содержат углеводные компоненты. Все это не очень-
крупные белки: молекулярный вес их в ряде случаев составляет
около 30 000, но не всегда можно сказать, действительно ли актив-
ный компонент, выделенный из железы, идентичен тому гормону,
который функционирует в живом организме.
ИНТЕГРАЦИЯ ЭНДОКРИННОГО И НЕРВНОГО КОНТРОЛЯ
Между эндокринными органами и центральной нервной систе-
мой существует непрерывное взаимодействие. Как мы увидим, мозг
прямо или косвенно оказывает влияние на эндокринную систему.
Но не следует забывать, что существует и обратная зависимость:
гормоны тоже оказывают большое влияние на функции централь-
ной нервной системы. Рассмотрим такой простой пример: у собак
самка во время течки положительно реагирует на ухаживание сам-
ца, хотя в другой период те же действия со стороны самца были
бы восприняты ею индифферентно или враждебно. То, что один и
Системы нервной регуляции 741
тот же сигнал вызывает совершенно разные поведенческие реак-
ции, обусловлено гормональными эффектами, и эти эффекты мож-
но воспроизвести путем инъекции соответствующих гормонов.
Центральная нервная система играет важную роль в регуляции
эндокринных функций. Как показали исследования, доминирую-
щую роль в этой регуляции играет область головного мозга, назы-
ваемая гипоталамусом. К этой регуляторной системе относятся:
также смежные с гипоталамусом участки мозга, а именно супраоп-
тическое и паравентрикулярное ядра.
СИСТЕМА ГИПОТАЛАМИЧЕСКОЙ РЕГУЛЯЦИИ
Гипоталамус расположен в основании мозга, непосредственно
над гипофизом1 и позади перекреста зрительных нервов, и образу-
ет дно третьего желудочка (рис. 13.14).
Гипоталамус осуществляет нервную регуляцию ряда функций,
в частности температуры тела и потребления воды и пищи. Как
уже говорилось, температура тела регулируется по принципу об-
ратной связи. Роль гипоталамуса в регуляций потребления пищи
можно продемонстрировать путем разрушения или электрической
стимуляции определенных его участков; если электрическое раз-
дражение правильно локализовать, оно приводит к неумеренному
потреблению пищи и ожирению. Аналогичным образом можно вы-
явить роль гипоталамуса в потреблении воды: электростимуляция
или введение небольших количеств гипертонического раствора со-
ли в определенные участки гипоталамуса заставляет животное
чрезмерно много пить. Козы, например, в этих условиях за не-
сколько минут выпивали количество воды, составлявшее 40% от
веса тела.
Гипоталамус играет важнейшую роль в эндокринной системе,
так как он регулирует функцию гипофиза, а последний называют
«главной» эндокринной железой в организме. Регуляторное воздей-
ствие на нейрогипофиз осуществляется по нервным путям, а на
аденогипофиз — через специальные кровеносные сосуды, называе-
мые воротной (или портальной) системой гипофиза.
1 Термины гипофиз (hypophysis) и питуитарная железа (pituitary) взаимо-
заменяемы. Древние греки называли этот орган гипофизом в связи с его рас-
положением под головным мозгом. Название «питуитарная железа» появилось
позже и связано с ошибочным мнением, будто эта железа выделяет носовую слизь
(лат. pituita — слизь). Термины «передняя» и «задняя» в отношении долей гипо-
физа неудачны, так как относительное положение этих двух главных частей
железы различно у разных животных. Аденогипофиз (греч. aden — железа), на-
зываемый также передней долей гипофиза, имеет железистое происхождение и
образуется путем выпячивания пищеварительной трубки. Нейрогипофиз, называе-
мый также задней долей, образуется из нервной закладки. Значительную пута-
ницу вносит термин «промежуточная доля гипофиза»; она образуется из той же
закладки, что и аденогипофиз, но ее включают вместе с нейрогипофизом в со-
став задней доли.
742 Гл. 13, Управление и интеграция
Нейрогипофиз содержит два хорошо изученных гормона — вазо-
прессин и окситоцин. Еще 50 лет назад было известно, что в ней-
рогипофизе содержатся вещества, влияющие на реабсорбцию воды
в почках и необходимые для концентрирования мочи. В настоящее
время установлено, что антидиуретический гормон (АДГ) млеко-
питающих идентичен вазопрессину, который при введении в боль-
ших, нефизиологических количествах вызывает заметное повыше-
ние. 13.14. Локализация гипофиза и гипоталамуса в головном мозгу человека и
взаимоотношения между этими органами. Функция аденогипофиза регулируется
нейрогормонами, образующимися в гипоталамусе; нейрогипофиз является нейро-
гемальным органом, который выделяет гормоны, выработанные нейронами при-
лежащих участков мозга.
Системы нервной регуляции
.743
ние кровяного давления вследствие сужения артериол. Другое ве-
щество, содержащееся в нейрогипофизе, — окситоцин — вызывает
сокращение гладкой мускулатуры матки при родах.
Вазопрессин и окситоцин — октапептиды (т. е. состоят из 8
аминокислот). Они образуются в нервных клетках, расположенных
около гипоталамуса, и транспортируются по аксонам к нервным:
окончаниям в нейрогипофиз, откуда могут переходить в кровь.
Таким образом, нейрогипофиз служит органом, накапливающим и
выделяющим гормоны (нейрогемальным органом), тогда как син-
тез этих гормонов происходит в соседних участках мозга.
В аденогипофизе, напротив, происходит синтез гормонов, а их
выделение в кровоток регулируется гипоталамусом. Эта регуляция
осуществляется с помощью гормонов, образующихся в гипоталаму-
се и попадающих в аденогипофиз через его воротную систему. Сей-
час установлено, что существуют по меньшей мере 9, а может быть
и 10, гипоталамических регулирующих гормонов, которые участву-
ют в системе управления гипофизом (табл. 13.6).
Таблица 13.6
Гипоталамические гормоны, регулирующие выделение гормонов
аденогипофизом. (Schally et al., 1973)
Гипоталамический гормон Гипофизарный гормон, выделение кото- рого стимулируется (+) или тормозится (-)
Соматолиберин Соматостатин Пролактолиберин Пролактостатин Меланолиберин Гормон роста ( + ) Гормон роста (—) Пролактин ( + ) Пролактин (—) Меланоцитстимулирующий гормон
Меланостатин Меланоцитстимулирующий гормон
Кортиколиберин Тиреолиберин Люлиберин Фоллиберин ( ) АКТГ (+) Тиреотропный гормон ( + ) Лютеинизирующий гормон ( + ) Фолликулостимулирующий гормон (+)
Три гормона аденогипофиза — гормон роста, пролактин и мела-
ноцитстимулирующий гормон — находятся под двойным гипотала-
мическим контролем, стимулирующим и тормозным. Таким образом,
выделение этих трех гормонов регулируется не с помощью простых
обратных связей, хотя обратные сигналы, несомненно, используют-
ся в процессе этой регуляции.
Выделение четырех других гормонов аденогипофиза, по-видимо-
му, регулируется обычной отрицательной обратной связью. Органа-
ми-мишенями кортикотропина (АКТГ), тиреотропина, лютеинизи-
рующего гормона (ЛГ) и фолликулостимулирующего гормона
744 Гл. 13. Управление и интеграция
(ФСГ) являются соответственно кора надпочечников, щитовидная
железа и гонады. Под влиянием стимуляции эти железы выделяют
в кровь свои гормоны (кортикостероиды, тироксин и половые сте-
роиды), которые в свою очередь тормозят секрецию аденогипофи-
зам тропных гормонов (отрицательная обратная связь). Есть вес-
кие данные в пользу того, что это торможение опосредуется через
Вазопрессин Меланоцит-
Окситоцин стимулирующий
гормон
Рис. 13.15. Схема, иллюстрирующая центральную роль гипоталамуса в эндо-
кринной регуляции. Функция нейрогипофиза регулируется по нервным путям.
Выделение ряда гормонов из аденогипофиза находится под двойным контролем
(стимуляция и торможение, обозначено ±); выделение других гормонов регули-
руется отрицательной обратной связью от органов-мишеней (показано пунктир-
ными линиями и знаками «минус»).
гипоталамус (исключение составляет тироксин, который, возможно,
действует прямо на гипофиз, образуя более короткую петлю обрат-
ной связи).
Система гипоталамической регуляции представлена на
рис. 13.15; видно, что гипоталамус играет центральную роль в эн-
докринной системе, управляя функцией аденогипофиза с помощью
ряда регулирующих гормонов. Аденогипофиз в свою очередь по
механизму отрицательной обратной связи регулирует секреторную
активность щитовидной железы, гонад и коры надпочечников; каж-
дая из трех других функций аденогипофиза регулируется двумя
гормонами — одним стимулирующим и одним тормозящим.
Системы нервной регуляции 745
В связи с центральным положением гипоталамуса возникает
вопрос о том, как регулируется функция его самого. Известно, что
гипоталамус имеет обширные нервные связи с другими частями
мозга, которые подвержены влиянию многих внешних факторов
(длины светового дня, сезона и т. д.), а также эмоциональных
моментов. Из этого можно заключить, что почти вся эндокринная
система находится под нервным контролем, осуществляемым через
«центральный командный пункт» — гипоталамус.
ЭНДОКРИННЫЕ ЖЕЛЕЗЫ,
НЕ НАХОДЯЩИЕСЯ ПОД ПРЯМЫМ КОНТРОЛЕМ ГИПОТАЛАМУСА
ЩИТОВИДНАЯ И ПАРАЩИТОВИДНАЯ ЖЕЛЕЗЫ
Паращитовидные железы, расположенные вблизи щитовидной
железы или внутри нее, вырабатывают гормон, называемый парат-
гормоном, который повышает содержание ионов кальция в крови.
Его антагонист — гормон кальцитонин, понижающий концентрацию
кальция в плазме, — выделяется С-клетками щитовидной железы
млекопитающих. Избыток паратгормона повышает концентрацию
Са2+ в плазме выше нормального уровня, причем источником каль-
ция служат главным образом кости. В результате возрастает екс-
креция кальция почками, а кости декальцинируются. Удаление па-
ращитовидных желез оказывает противоположный эффект — кон-
центрация кальция в плазме падает, что приводит к тетаническим
судорогам мышц.
Нормальная секреторная функция паращитовидных желез регу-
лируется, по-видимому, с помощью простой обратной связи: высо-
кий уровень кальция в плазме тормозит выделение паратгормона,
а низкий стимулирует его. На секрецию кальцитонина уровень
кальция в плазме оказывает прямо противоположное влияние: вы-
сокий уровень стимулирует ее, а низкий тормозит. Таким образом,
концентрация Са2+ в плазме крови зависит от соотношения между
двумя системами с отрицательной обратной связью.
ОРГАНЫ ПИЩЕВАРЕНИЯ
Различные органы желудочно-кишечного тракта выделяют гор-
моны, регулирующие секреторную активность пищеварительных
желез. Секреция этих гормонов находится под контролем главным
образом нервной системы. Кроме того, в поджелудочной железе
(точнее, в небольших скоплениях эндокринных клеток, так назы-
ваемых островках Лангерганса) образуются два очень важных гор-
мона, которые хотя и не участвуют непосредственно в пищеваре-
нии, но играют чрезвычайно важную роль в обмене углеводов.
Эти два гормона — инсулин и глюкагон.
Наиболее очевидный эффект инсулина состоит в снижении кон-
центрации глюкозы в крови в результате усиленного образования
746 Гл. 13. Управление и интеграция
и накопления гликогена в клетках (для его синтеза используётся
глюкоза, доставляемая кровью). Заболевание сахарный диабет
характеризуется недостаточным синтезом или выделением инсули-
на (может иметь место как нехватка гормона, так и резистентность
тканей к его воздействию). В результате концентрация глюкозы
в крови оказывается повышенной и глюкоза выделяется с мочой,
часто в очень больших количествах. Однако инсулин выполняет и
другие, более сложные функции в промежуточном обмене, поэтому
недостаток инсулина приводит и к ряду других серьезных послед-
ствий.
Инсулин вырабатывается в бета-клетках островков Лангерган-
са, а глюкагон — в альфа-клетках. Если уровень глюкозы в крови
поднимается выше нормы (например, после всасывания углево-
дов пищи), то происходит выделение инсулина, который стимули-
рует поглощение глюкозы мышцами и образование гликогена; в ре-
зультате концентрация сахара в крови снижается. Таким образом,
секреция инсулина регулируется с помощью отрицательной обрат-
ной £ВЯЗИ.
Если уровень глюкозы в крови оказывается слишком низким,
выделяется глюкагон; он вызывает мобилизацию глюкозы в печени
и повышает содержание ее в крови. Действие глюкагона противо-
положно действию инсулина; таким образом, нормальный уровень
глюкозы в крови поддерживается двумя гормонами-антагониста-
ми — понижающим и повышающим, причем каждый из них имеет
собственный контур отрицательной обратной связи.
МОЗГОВОЙ СЛОЙ НАДПОЧЕЧНИКОВ
Клетки мозгового вещества надпочечников имеют нервное про-
исхождение и относятся к симпатической нервной системе. Эти
клетки выделяют два близких по структуре гормона — норадрена-
лин и адреналин (см. рис. 13.19). Адреналин вызывает учащение
ритма сердца, повышает кровяное давление, а также уровень са-
хара в крови (стимулируя превращение гликогена в глюкозу), вы-
зывает расширение сосудов и увеличение кровотока в легких, сер-
дечной мышце и скелетных мышцах и в то же время сужение сосу-
дов и уменьшение кровотока в гладких мышцах, пищеварительном
тракте и коже. Эти эффекты адреналина составляют хорошо из-
вестный синдром «борьбы или бегства», который возникает при ис-
пуге, боли или ярости; они способствуют мобилизации всех физиче-
ских ресурсов организма в ответ на непосредственную опасность.
Норадреналин обладает сходным действием, и разница между
обоими гормонами в основном количественная. Например, адрена-
лин оказывает большее влияние на ритм сердца, а норадреналин
вызывает более сильное сужение сосудов в некоторых органах. Оба
гормона действуют на клетки через рецепторные участки, называе-
мые альфа-рецепторами (в основном для норадреналина) и бета-
Системы нервной регуляции 747
рецепторами (в основном для адреналина). Во многих органах
имеются как альфа-, так и бета-рецепторы; кроме того, рецепторы
того и другого типа в известной степени чувствительны к обоим
гормонам. Именно поэтому очень трудно четко описать черты
сходства и различия между адреналином и норадреналином: их
эффекты в очень большой мере перекрываются1.
КАСКАДНЫЙ ЭФФЕКТ
Регулирование метаболических функций эндокринной системой
может осуществляться путем каскадного, поэтапного усиления, что
позволяет достичь большого конечного эффекта с помощью нич-
тожных количеств гормона. Пример этого представлен на рис. 13.16,
Количество
вещества
Гипоталамус
------------Кортиколиоерин 0,1 мкг
V________
Аденогипофиз
Кортикотропин 1мкг
(АКТГ)
У________
Кора
надпочечников
-------------Кортикостероид 40мкг
V
Печень-------------Гликоген 5600 мкг
Рис. 13.16. Пример биологического усиления сигнала в эндокринной системе.
Высвобождение ничтожного количества гипоталамического гормона приводит к
отложению гликогена в печени; коэффициент усиления при этом составляет 56 000.
(По данным Bentley, 1976.)
где указаны приблизительные количества гормонов, последователь-
ное действие которых приводит к отложению гликогена. Количе-
ство гипоталамического кортиколиберина, равное всего лишь
0,1 мкг, запускает цепь событий, ведущих в конечном результате
1 Норадреналин обнаружен в нервной системе насекомых и кольчатых чер-
вей, но его, по-видимому, нет у ракообразных, моллюсков, иглокожих и пред-
ставителей ряда других типов.
748 Гл. 13. Управление и интеграция
к образованию в печени 5600 мкг гликогена. Исходное количество
кортиколиберина так мало, что коэффициент усиления здесь до-
стигает 56 000.
ВЗАИМОДЕЙСТВИЕ ГОРМОНОВ С КЛЕТКАМИ-МИШЕНЯМИ
До сих пор мы рассматривали места образования, секрецию и
транспорт гормонов, но не касались вопроса о механизме их воз-
действия на орган-мишень. Для того чтобы гормон оказывал влия-
ние на определенную мишень, но не действовал на другие органы,
к которым кровь доставляет его в той же концентрации, необходи-
мо, чтобы воспринимающие клетки имели на своей поверхности
специфические рецепторные участки, взаимодействующие с данным
гормоном, а у других клеток таких рецепторов не было.
По механизму действия на клетки-мишени все гормоны можно
разделить на две группы: одни (катехоламины и пептидные гормо-
ны) действуют, регулируя образование циклического нуклеотида
3',5'-аденозинмонофосфата (циклического АМФ, или цАМФ), а
другие (стероиды и тиреоидные гормоны) проникают внутрь клет-
ки и воздействуют непосредственно на клеточное ядро и механизм
белкового синтеза.
Роль циклического АМФ была открыта при изучении действия
адреналина на печень, где этот гормон стимулирует превращение
гликогена в глюкозу (Sutherland, 1972). В этом процессе участвует
ряд ферментов, в том числе фосфорилаза, которая действует на
этапе, лимитирующем скорость процесса в целом (рис. 13.17).
Активный фермент — фосфорилаза а — образуется из предшест-
венника (фосфорилазы &), который в присутствии киназы фосфо-
рилазы взаимодействует с аденозинтрифосфатом (АТФ). Весь про-
цесс начинается с того, что адреналин присоединяется к рецептор-
ному участку клеточной мембраны. Это ведет к активации фермента
аденилатциклазы, которая в свою очередь катализирует образо-
вание циклического АМФ из АТФ.
Циклический АМФ называют «вторым посредником» в про-
цессе реализации гормонального эффекта; цАМФ и аденилатцикла-
за были обнаружены в различных тканях позвоночных и многих
беспозвоночных, а также у бактерий (Sutherland et al., 1962). На-
чальным событием в действии всех гормонов, эффект которых
опосредован цАМФ, всегда бывает высвобождение аденилатцикла-
зы из комплекса с рецептором гормона в клеточной мембране.
Стероидные гормоны, к которым относятся мужские и женские
половые гормоны и гормоны коры надпочечников, действуют с по-
мощью иного механизма. Как мы уже знаем, гормоны влияют
только на клетки, обладающие специфическими рецепторами, и не
влияют на остальные. Эстрадиол связывается рецепторами в матке,
тестостерон — рецепторами в предстательной железе, прогесте-
рон — в яйцеводах птиц и т. д. Образовавшийся комплекс гормона
Системы нервной регуляции 749
с белком-рецептором быстро переносится в клеточное ядро, где
стимулирует или индуцирует экспрессию определенных генов (Jen-
sen et al., 1971).
Рис. 13.17. Выделение глюкозы из клеток печени под влиянием адреналина.
(Sutherland, 1972.)
Адреналин взаимодействует с рецепторным участком клеточной мембраны, в ре-
зультате чего образуется циклический АМФ («второй посредник»). Последний
представленный на схеме этап — действие фосфорилазы а — лимитирует скорость
всего процесса в целом; в этом этапе участвуют два дополнительных фермента —
фосфоглюкомутаза и глюкозо-6-фосфатаза.
При изучении рецепторов стероидных гормонов недавно был
выявлен ряд удивительных фактов. Оказалось, что стероиды, выде-
ляемые у новорожденного крысенка половыми железами, можно
обнаружить в клетках-мишенях, находящихся в определенных
участках головного мозга. Гормоны взаимодействуют с рецепто-
750 Гл. 13. Управление и интеграция
рами, имеющимися в этих участках, и вызывают формирование
нервных путей, которые будут определять тип поведения — муж-
ской или женский — у взрослой особи. Такая половая дифференци-
ровка нервных путей создает морфологическую основу для про-
явления каких-то одних типов поведения и торможения других
(McEwen, 1976). Таким образом, функция нервной системы в пе-
риод развития организма видоизменяется под влиянием гормонов;
и в то же время, как мы уже видели, центральная нервная система
служит важным регулятором эндокринных функций во всем орга-
низме.
ХИМИЧЕСКАЯ ПЕРЕДАЧА
НЕРВНЫХ СИГНАЛОВ И НЕЙРОМЕДИАТОРЫ
Действие адреналина и норадреналина, выделяющихся в кровь
из надпочечников, сходно с эффектами общей стимуляции симпати-
ческой нервной системы1. Это сходство нетрудно объяснить, если
вспомнить, что клетки мозгового слоя надпочечников имеют нерв-
ное происхождение и аналогичны крупным симпатическим ганг-
лиям, расположенным вдоль позвоночника. Сходство станет еще
более понятным, если добавить, что норадреналин служит конеч-
ным медиатором во всей симпатической системе.
Мы уже видели, что в большинстве случаев синаптическая пе-
редача осуществляется путем высвобождения медиатора ацетилхо-
лина и что последний выделяется окончаниями двигательных нер-
вов в нервно-мышечных соединениях. Действие вегетативной нерв-
ной системы также опосредовано медиаторами.
Окончания симпатических нервов выделяют норадреналин, а
окончания парасимпатических нервов — ацетилхолин. Эти два ве-
щества действуют как антагонисты: если одно стимулирует какую-
либо функцию, то другое тормозит ее. Хорошо известный пример
такого антагонизма — регуляция частоты сердечных сокращений.
Работа сердца регулируется двумя нервами. Один из них — ветвь
чревного нерва; это симпатический нерв, высвобождающий нор-
адреналин и ускоряющий ритм сердца; второй — веточка блуждаю-
щего нерва; это парасимпатический нерв, высвобождающий аце-
тилхолин и замедляющий работу сердца.
Замедление сокращений сердца под влиянием ацетилхолина
служит поразительным примером того, что реакция органа на дан-
1 Основными компонентами периферической нервной системы позвоночных
являются сенсорные нервы, двигательные нервы и вегетативная (автономная)
нервная система. Вегетативная нервная система в свою очередь подразделяется
на симпатическую и парасимпатическую системы, которые совместно регулируют
работу внутренних органов — сердца, желудка, желез, кишечника, почек и т. д.
(т. е. непроизвольные функции организма). Указанные органы снабжены одно-
временно и симпатическими, и парасимпатическими нервами, действующими как
антагонисты: одни оказывают стимулирующее действие, другие — тормозящее.
Системы нервной регуляции 751
ное вещество зависит от самого органа, а не от химической приро-
ды нейромедиатора. В самом деле, на обычную поперечнополоса-
тую мышцу ацетилхолин действует возбуждающим образом
(стимулирует сокращение), тогда как на сердце он оказывает тор-
мозящее действие (снижает частоту сокращений).
Спинной мозг
Симпатическая
система
Двигательный
нейрон
Мозговой /±\ Парасимпатиче-
ские ( V* ) ( < J 1 ская система
слои 1 АХ J V АХ J
наЪпоче ч -
ников
Нор-
АЪр Н V аЪр «А»
"°р-
«рО8Ь адР
Мышца
Рис. 13.18. Схема действия периферических нейромедиаторов у позвоночных.
Подробности см. в тексте. АХ — ацетилхолин; Адр. — адреналин; Норадр. —
норадреналин.
Основные медиаторы периферической нервной системы позво-
ночных приведены на рис. 13.18. Можно видеть, что самым распро-
страненным медиатором является ацетилхолин — он свойствен не
только окончаниям двигательных нервов и синапсам парасимпати-
ческой нервной системы, но также симпатическим ганглиям и во-
локнам, иннервирующим мозговой слой надпочечников. Таким об-
разом, вне головного и спинного мозга ацетилхолин представляет
собой универсальный медиатор передачи воздействия с нейрона на
нейрон у позвоночных. Что касается центральной нервной системы,
то здесь ситуация не столь ясна.
Несмотря на то что ацетилхолин так широко используется в си-
напсах периферической нервной системы, есть веские данные
в пользу того, что в качестве нейромедиаторов функционируют и
другие вещества, особенно в центральной нервной системе. У бес-
позвоночных важными нейромедиаторами служат соединения, отно-
сящиеся в большинстве своем к классу аминов. На рис. 13.19 при-
ведена структура некоторых из этих веществ. Недостоверность све-
дений относительно нейромедиаторов обусловлена тем, что точно
установить медиаторную функцию того или иного вещества чрезвы-
чайно трудно.
Наибольшая трудность связана с тем, что многие вещества, со-
держащиеся в организме в небольших количествах, оказывают вы-
раженное физиологическое действие, особенно на нервную систему.
752 Гл. 13. Управление и интеграция
Если какое-то вещество вызывает реакцию, сходную с реакцией
на раздражение нерва, то это еще не доказывает медиаторной
функции данного вещества. Такое доказательство должно удовле-
творять ряду требований, в частности
1) предполагаемый медиатор или его предшественник должен
содержаться в тех нейронах, из которых он высвобождается;
Рис. 13.19. Структурные формулы некоторых соединений с установленной или
предполагаемой медиаторной функцией.
2) вещество должно присутствовать во внеклеточной жидкости
около активированного синапса;
3) при воздействии на постсинаптическую структуру исследуе-
мое вещество должно воспроизводить действие медиатора;
4) должен иметься какой-то механизм удаления или инактива-
ции исследуемого вещества (ферментативная инактивация, специ-
фическое поглощение или реабсорбция).
С помощью фармакологических агентов, тормозящих синтез,
выделение, удаление или инактивацию исследуемого вещества или
его взаимодействие с мишенью, можно выяснить ряд частных во-
просов, но нельзя получить прямых доказательств медиаторной
функции вещества.
Решающее доказательство медиаторной функции состоит в де-
монстрации того факта, что под влиянием потенциала действия
данное вещество высвобождается из пресинаптического нервного
окончания в количестве, достаточном для того, чтобы вызвать соот-
ветствующий физиологический эффект в постсинаптической струк-
туре.
Получить доказательства, удовлетворяющие всем указанным
требованиям, действительно нелегко, и технические трудности
Система гормональной регуляции 753
очень часто оказываются непреодолимыми. В результате многие
работы, касающиеся предполагаемых нейромедиаторов (особенно
у беспозвоночных), остаются, по существу, незавершенными, ибо
ограничиваются в основном лишь выявлением эффекта исследуе-
мых агентов. Поэтому имеющиеся данные о нейромедиаторах не
позволяют делать обобщений. Тем не менее следует все же оста-
новиться на двух веществах—5-гидрокситриптамине и гамма-ами-
номасляной кислоте, поскольку в экспериментах на многих беспоз-
воночных неизменно обнаруживается их высокая физиологическая
активность.
Первое из этих веществ — 5-гидрокситриптамин (5-ГТ) — со-
держится в нейронах многих беспозвоночных, особенно моллюсков
и членистоногих. Оно встречается не только в нервной системе, но
его концентрация в других тканях обычно ниже. Большие количе-
ства 5-ГТ найдены в ряде животных токсинов, например в ядах ос,
моллюска Мигех и осьминога.
Сердце двустворчатых моллюсков очень чувствительно к 5-ГТ,
и есть убедительные данные о том, что это вещество служит ме-
диатором ускоряющего влияния на сердце моллюсков. Влияние
5-ГТ на кровяное давление у позвоночных (прессорное действие)
было известно еще до установления химической структуры вещест-
ва. Тогда его называли серотонином, и этот термин часто исполь-
зуют до сих пор. Предполагают, что 5-ГТ служит нейромедиатором
в центральной нервной системе позвоночных, но какова здесь его
роль в нормальных условиях, пока не ясно.
Гамма-аминомасляная кислота (ГАМК) тоже может претендо-
вать на роль нейромедиатора в центральной нервной системе поз-
воночных и почти наверняка является нейромедиатором в тормоз-
ных синапсах ракообразных. Окончания тормозных нервных воло-
кон на мышцах ракообразных высвобождают ГАМК, и эффект
торможения можно воспроизвести путем прямого воздействия этим
веществом (Otsuka et al., 1966; Takeuchi, Takeuchi, 1967).
ГАМК в довольно высоких концентрациях содержится в опреде-
ленных участках головного мозга у млекопитающих, но это еще
не доказывает ее нейромедиаторной функции. В настоящее время
проводится много исследований, направленных на выяснение роли
ГАМК и других предполагаемых нейромедиаторов в центральной
нервной системе, и особенно в головном мозгу.
Высказывались предположения, что некоторые аминокислоты,
в частности глутаминовая и аспарагиновая кислоты, тоже могут иг-
рать роль нейромедиаторов. Они встречаются в нервных тканях
как позвоночных, так и беспозвоночных, но веских доказательств
их медиаторной функции пока нет. Одна из основных трудностей
такого доказательства связана с тем, что аминокислоты всегда
присутствуют в тканях и поэтому выявление их в нейронах само по
себе не имеет особого значения. Чрезвычайно трудно также вы-
явить и специфический механизм инактивации аминокислот, свя-
48—1863
754 Гл. 13. Управление и интеграция
занный с их предполагаемой ролью нейромедиаторов, так как все
аминокислоты подвергаются превращениям в ходе обычных мета-
болических процессов.
.ДРУГИЕ ВЕЩЕСТВА,
ВОЗДЕЙСТВУЮЩИЕ НА НЕРВНЫЕ КЛЕТКИ
Становится все более очевидным, что в центральной нервной
системе образуется больше различных веществ, обладающих тем
или иным специфическим действием, чем можно было думать еще
несколько лет назад. Ряд интересных открытий был сделан при
изучении действия таких общеизвестных препаратов опия, как мор-
фин и героин. В медицине их используют в качестве болеутоляю-
щих, но они вызывают также эйфорию и, как известно, к ним мо-
жет развиваться болезненное пристрастие (наркомания).
Наиболее значительное открытие было сделано при попытке об-
наружить в мозгу специфические рецепторы опиатов; выявление
таких рецепторов создало бы основу для дальнейшего изучения
специфического действия этих лекарственных веществ: мы знали
•бы, откуда начинать поиски механизма наркотического эффекта
.(Pert, Snyder, 1973).
Рецепторы опиатов были найдены, и это открыло путь для
дальнейших исследований. Поскольку в естественных условиях в
•организме животного опиатов нет, логично было предположить, что
соответствующие рецепторы играют какую-то другую, не связан-
ную с наркотиками физиологическую роль и что должны сущест-
вовать эндогенные вещества, воздействующие на эти рецепторы
в норме. В результате исследований был обнаружен целый ряд
эндогенных веществ, участвующих в нормальном функционирова-
нии мозга и, возможно, имеющих отношение не только к снятию
боли, но и к изменениям настроения, эмоциям, а также предполо-
жительно и к некоторым типам психических расстройств.
Природные соединения, взаимодействующие с опиатными рецеп-
торами, получили название эндорфинов. К ним, в частности, отно-
сятся два пентапептида — энкефалины. Известен также ряд актив-
ных полипептидов с несколько более длинной цепью, и продолжают
выявляться все новые активные вещества. Некоторые из них со-
держат точно такую же последовательность аминокислот, как в от-
дельных фрагментах пептидных гормонов, например р-липотропи-
на, способствующего мобилизации жира из жировой ткани и со-
стоящего из 91 аминокислоты (Kosterlitz et al., 1977). Физиологи-
ческим эффектом обладали пептиды как с более длинной, так и
с более короткой цепью, и поэтому пока трудно сказать, какие
именно вещества действуют в нормальных условиях, а какие яв-
ляются предшественниками или продуктами расщепления.
Не следует думать, что эти результаты в какой-то мере завер-
шают период открытий в области нейрохимии; напротив, есть все
Процессы регулирования и интеграции у беспозвоночных 755
основания ожидать, что здесь еще будут обнаружены интересней-
шие новые принципы взаимодействий.
Перейдем теперь к примерам эндокринной регуляции у беспоз-
воночных. Роль нервной системы в выработке гормонов и регуля-
ции эндокринных функций у этих животных выражена в еще
большей степени, чем у позвоночных.
ПРОЦЕССЫ РЕГУЛИРОВАНИЯ И ИНТЕГРАЦИИ
У БЕСПОЗВОНОЧНЫХ
У наиболее высокоорганизованных беспозвоночных физиологи-
ческие функции регулируются как нервной, так и эндокринной си-
стемами. Так же как и у позвоночных, нервная система осуществля-
ет быстрое управление, необходимое для совершения действий»
связанных с бегством от врагов, добыванием пищи, спариванием
и т. д.; эндокринная же система вырабатывает гормоны, регули-
рующие более медленные процессы — рост, созревание и многие
другие метаболические функции. Нервная система играет непосред-
ственную и ведущую роль в выработке гормонов, и связь между
нервной и эндокринной системами здесь еще теснее, чем у позво-
ночных.
ФУНКЦИЯ И ЗНАЧЕНИЕ НЕРВНОЙ СИСТЕМЫ
Как мы убедились, у позвоночных центральная нервная система
координирует большое число отдельных функций, находящихся
как под нервным, так и под эндокринным контролем. Кажущийся
контраст между нервной регуляцией (обычно быстрой) и гормо-
нальной (обычно медленной вплоть до очень медленной) при более
внимательном рассмотрении исчезает. Дело не только в том, что
передача сигналов с нейрона на нейрон и с нейрона на орган-эф-
фектор происходит в большинстве случаев химическим путем, но и
в том, что центральная нервная система сама является важным
источником гормонов и в значительной части именно таким обра-
зом оказывает общее регулирующее действие на организм.
Рассматривая роль нервной системы у беспозвоночных, стоящих
на более низких ступенях организации, мы сталкиваемся с необхо-
димостью точнее определить само понятие «нервная система»;
у позвоночных значение этого термина было самоочевидным.
«Нервной системой» называют совокупность нейронов, специа-
лизированных для постоянной и организованной передачи инфор-
мации от сенсорных рецепторных участков к нейронам, от одних
нейронов к другим или от нейронов к органам-эффекторам (на-
пример, мышцам, железам и т. п.). Это определение исключает су-
ществование нервной системы у одноклеточных животных (про-
стейших), хотя функции в этих клетках-организмах могут коорди-
нироваться с помощью проводящих органелл.
48*
756 Гл. 13, Управление и интеграция
Нейрон, или нервную клетку, как правило, довольно легко рас-
познать по ее мембранному потенциалу и по способности много-
кратно генерировать потенциалы действия. Но иногда трудно бы-
вает установить, действительно ли «нейросекреторная» клетка
является модифицированным нейроном. В этом случае может по-
мочь ее локализация в непосредственной связи с нервными клетка-
ми; дополнительную информацию могут дать структурные особен-
ности, потенциалы действия, а также функция сходных образова-
ний у близких видов.
Нейроны у беспозвоночных обладают такими же свойствами,
как и у позвоночных. Они способны генерировать потенциалы дей-
ствия, которые складываются из изменений мембранной проницае-
мости и мембранного потенциала. Потенциалы действия возникают
по принципу «всё или ничего» и распространяются по нервному во-
локну, или аксону. Типичные нейроны беспозвоночных генерируют
короткие потенциалы действия длительностью от доли миллисе-
кунды до нескольких миллисекунд; такие нейроны свойственны
всем основным типам, кроме простейших и губок.
Передача информации от одного нейрона к другому происходит
через синапсы, в которых, как правило, удается выявить химичес-
кую передачу. Как и у позвоночных, мембраны двух нейронов, об-
разующих синапс, чаще всего разделены щелью, и передача осу-
ществляется путем выделения химического медиатора. Однако в
некоторых синапсах, по-видимому, происходит прямая электриче-
ская передача; в этих синапсах нервные мембраны двух клеток
соприкасаются, как это бывает и в определенных синапсах у по-
звоночных.
Типичные постсинаптические потенциалы генерируются, видимо,
у всех представителей по крайней мере четырех типов: моллюсков,
кольчатых червей, членистоногих и хордовых. Эти потенциалы мо-
гут быть возбуждающими или тормозными. Как правило, возбуж-
дающие постсинаптические потенциалы уменьшают поляризацию
мембраны (т. е. это сдвиги в сторону деполяризации), а тормозные
увеличивают мембранный потенциал. По этим общим особенно-
стям нейроны беспозвоночных и позвоночных в принципе сходны.
НЕЙРОСЕКРЕЦИЯ
У беспозвоночных специализированные нейросекреторные клет-
ки и органы были известны еще задолго до того, как выяснилось
значение этих образований у позвоночных. Насколько трудно опре-
делить и охарактеризовать, что такое нейросекреторная клетка, на-
столько же легко ее распознать на гистологическом препарате;
основной метод выявления нейросекреторных клеток до сих пор
состоит в том, что используются определенные красители и другие
вещества, обладающие особым сродством к этим клеткам.
Процессы регулирования и интеграции у беспозвоночных 757
НЕЙРОГЕМАЛЬНЫЕ ОРГАНЫ
Нейросекреторные органы представляют собой группы нейро-
нов, обладающих секреторной активностью. Отходящие от них
нервные волокна служат для транспорта секрета, и обычно их
окончания тесно связаны с сосудистой сетью. Здесь образуется
нейрогемалъный орган, в котором секрет накапливается и высво-
бождается. Нейрогемальные органы очень широко распространены,
однако нельзя с уверенностью утверждать, что они имеются при
каждом нейросекреторном органе.
Во всех типах более высокоорганизованных животных нейросек-
реторные системы играют центральную роль в функционировании
эндокринных механизмов и в его регуляции. Мы уже видели, какое
значение имеет нейросекреция в управлении эндокринными органа-
ми у позвоночных; у беспозвоночных оно, по-видимому, еще
больше.
Несколько позже в этой главе будут приведены примеры эндо-
кринной регуляции у насекомых, у которых нейросекреторным и
нейрогемальным органам принадлежит главная роль в регулятор-
ных процессах. Нейросекреторные системы сильно развиты также
у кольчатых червей и ракообразных, где они регулируют размно-
жение, обмен веществ, линьку, пигментацию и т. д.
Хотя ряд эндокринных желез как у позвоночных, так и у бес-
позвоночных не находится под прямым нейроэндокринным контро-
лем, все же нормальное функционирование большинства или даже
всех желез внутренней секреции в конечном счете зависит от регу-
ляторных воздействий со стороны нервной системы, осуществляе-
мых либо прямо по нервным путям, либо через посредство гормо-
нов. По-видимому, регулирование путем нейросекреции характер-
но для всех животных, у которых эндокринные механизмы играют
важную роль.
ГОРМОНЫ И ЭНДОКРИННАЯ ФУНКЦИЯ
Работа эндокринной системы лучше всего изучена у тех беспоз-
воночных, которые отличаются сложной морфологической органи-
зацией. Это обусловлено двумя причинами. Во-первых, высокоорга-
низованным животным, таким, например, как насекомые, необхо-
димы более сложные процессы регуляции и интеграции, чем более
примитивным формам вроде, скажем, актиний. Во-вторых, у более
сложных форм многие функции передаются специализированным
органам. Это позволяет производить эксперименты с удалением
органов, повторной их имплантацией, экстрагированием и другими
приемами, которые трудно или даже невозможно применить в опы-
тах на животных с относительно низкой организацией.
Большая часть гормонов беспозвоночных отличается от гормо-
нов позвоночных и по химической структуре, и по действию. Мно-
гие гормоны позвоночных, сильно влияющие на рост позвоночного
758 Гл. 13. Управление и интеграция
животного, развитие его половых желез, метаболические процессы
и т. п., при введении беспозвоночным не вызывают ни малейшего
эффекта.
Эндокринная функция была продемонстрирована у многих бес-
позвоночных, однако тот факт, что какая-либо вытяжка из ткани
обладает физиологическим действием, еще не доказывает, что она
содержит вещество, выполняющее роль гормона в организме дан-
ного животного. Для такого доказательства нужны более тщатель-
ные исследования, удовлетворяющие ряду требований; должно
быть, например, показано наличие исследуемого вещества в орга-
низме в нормальных условиях, установлено место его синтеза, изу-
чены последствия, к которым ведет удаление его источника или
блокада секреции; нужно выделить вещество в чистом виде и в
конце концов синтезировать его химическим путем, причем про-
дукт синтеза должен проявлять такой же эффект, как и природный
продукт, и примерно в тех же концентрациях. Ввиду сложности
такой работы эндокринная система подробно изучена лишь у не-
скольких групп беспозвоночных.
К числу животных, у которых были четко выявлены и относи-
тельно неплохо изучены органы внутренней секреции, относятся
моллюски (в особенности головоногие), кольчатые черви, ракооб-
разные, насекомые и оболочники. Все они имеют достаточно высо-
кий уровень организации, так что сложность их функций и специа-
лизация органов облегчали проведение эндокринологических иссле-
дований. Особенно подробно были изучены насекомые; это объяс-
няется тем, что они доступны, их легко собирать и выращивать,
они хорошо переносят крупные хирургические вмешательства и,
кроме того, имеют большое хозяйственное значение.
ЭНДОКРИНОЛОГИЯ НАСЕКОМЫХ
Гормоны играют весьма важную роль в физиологии насекомых,
особенно в процессах роста, линьки, окукливания и превращения во
взрослую форму. Эти процессы изучались у множества различных
видов, но если учесть, что общее число видов насекомых достигает
около миллиона, будет ясно, что доля исследованных видов очень
мала. Однако эндокринные функции у разных насекомых удиви-
тельно сходны. Например, у насекомых с неполным превращением,
которые на протяжении ряда линек постепенно превращаются во
взрослую форму, действуют те же самые гормоны, что и у насеко-
мых с полным превращением — тех, которые проходят несколько
линек на стадии личинки, затем превращаются в куколку, а из ку-
колки выходит взрослая форма (имаго). Мы рассмотрим обе груп-
пы насекомых.
В качестве примера насекомого с неполным превращением возь-
мем южноамериканского кровососущего клопа Rhodnius, родствен-
ника обыкновенного постельного клопа. Rhodnius вылупляется из
Процессы регулирования и интеграции у беспозвоночных 759
яйца в виде крошечного клопика — нимфы, которая проходит ряд
стадий (возрастов), разделенных линьками, и постепенно превра-
щается во взрослого клопа. На каждой такой стадии нимфа долж-
на напитаться кровью, для того чтобы сбросить старую кутикулу и
перейти в следующую стадию. После того как нимфа насосется
крови, проходит около 4 недель; далее она сбрасывает старую
кутикулу и, прежде чем затвердеет новая, увеличивает свои раз-
меры путем наполнения воздухом всех трахей. Теперь нимфа вновь
готова к сосанию крови, но если такой возможности не представит-
ся, она может жить без этого многие месяцы. Если же нимфа на-
сосется крови, то через 4 недели наступит следующая линька.
У клопа Rhodnius линьку стимулирует гормон экдизон (назы-
ваемый также гормоном линьки), секреция которого начинается
после сосания крови. Этот гормон выделяют проторакальные желе-
зы, расположенные в грудном отделе. Функцию этих желез в свою
очередь стимулирует гормон, выделяемый нейросекреторными клет-
ками мозга.
Взрослый клоп Rhodnius отличается от нимфы наличием крыль-
ев и зрелых гонад, а также и в некоторых других отношениях. Что
управляет развитием этих признаков? Оказалось, что отсутствие
на ранних стадиях черт, характерных для взрослого насекомого,
обусловлено действием еще одного гормона — ювенильного, кото-
рый вырабатывают небольшие группы клеток, расположенные не-
посредственно позади мозга и называемые «прилежащими телами»
(corpora allata).
Именно ювенильный гормон определяет личиночные признаки
образующейся новой кутикулы и препятствует развитию призна-
ков, характерных для взрослой формы. На всех ранних возрастных
стадиях, когда присутствует ювенильный гормон, экдизон стимули-
рует линьку, которая сопровождается увеличением размеров ним-
фы. В четвертом возрасте количество ювенильного гормона умень-
шается, а в пятом начинается образование крыльев. К моменту по-
следней линьки ювенильный гормон исчезает, и формируется взрос-
лая особь (рис. 13.20).
Посмотрим, какие факты указывают на эндокринную регуля-
цию этих событий. Если нимфу 5-го возраста обезглавить вскоре
после приема крови, то последняя линька не наступает, хотя клоп
может прожить без головы более года. Это наводит на мысль, что
гормон линьки экдизон образуется в голове. Однако если голову
удалить по прошествии определенного «критического периода», а
именно примерно на 7-й день после приема крови, то линька про-
изойдет и сформируется взрослая форма без головы. Следователь-
но, голова нужна только в начальном подготовительном периоде,
а не во время самой линьки.
Можно доказать, что это различие обусловлено определенными
эндокринными факторами; так, если кровь насекомого, обезглав-
ленного после «критического периода», ввести нимфе, обезглавлен-
760 Гл. 13. Управление и интеграция
ной до окончания этого периода, то у последней наступит линька.
Дело в том, что во время критического периода мозг насекомого
выделяет так называемый мозговой гормон1, который стимулирует
проторакальные железы. При раннем обезглавливании стимуляции
проторакальных желез не происходит; если же обезглавливание
Рис. 13.20. Кровососущий клоп Rhodnius превращается во взрослую особь, про-
ходя через пять линек. Стимулом для каждой линьки служит выделение гормона
экдизона из проторакальных желез. Во время первых четырех линек наличие
ювенильного гормона (из corpora allata) приводит к образованию новой кутикулы
личиночного типа и тем самым препятствует превращению нимфы во взрослую
форму. Во время последней линьки ювенильный гормон отсутствует, и в резуль-
тате развивается взрослая особь.
произведено после критического периода, то к этому времени моз-
говой гормон уже подействовал и больше не нужен; экдизон выде-
ляется и в надлежащее время индуцирует линьку.
Наиболее простой способ переливания крови у этих насекомых
состоит в том, что двух клопов соединяют коротким стеклянным
капилляром, который можно укрепить кусочком воска в шее обез-
главленного клопа (рис. 13.21). Если соединить таким образом
нимфу 5-го возраста, насосавшуюся крови и прошедшую критиче-
ский период, с совсем маленькой нимфой 1-го возраста, которая
1 Мозг служит источником нескольких гормонов, но в эндокринологии насе-
комых принято называть «мозговым гормоном» тот агент, который специфически
активирует проторакальную железу и стимулирует секрецию экдизона. Мозговой
гормон называют также прото ракотропным гормоном.
Процессы регулирования и интеграции у беспозвоночных 761
недавно вышла из яйца и еще ни разу не питалась, то у последней
разовьются такие признаки взрослой формы, как крылья и органы
размножения, и она примет вид карликового взрослого клопа.
Если повторить такой опыт с нимфой одного из следующих воз-
растов, то в этом случае хотя и удастся вызвать линьку, черты
взрослого животного не разовьются, так как к этому времени у
Рис. 13.21. Перенос гормонов кровью можно продемонстрировать, соединив двух
насекомых друг с другом. Слева — нимфа Rhodnius четвертого возраста, у кото-
рой удален только самый кончик головы (мозг сохранен интактным); она соеди-
нена с другой, полностью обезглавленной нимфой пятого возраста (справа)
(Wigglesworth, 1959.)
нимфы уже образуются достаточные количества ювенильного гор-
мона, который препятствует формированию признаков взрослой
особи.
Очевидно, экдизон необходим для каждой линьки, и ювениль-
ный гормон, который выделяется позднее, только видоизменяет его
действие. Поэтому если насекомое обезглавлено лишь после того,
как произошло высвобождение мозгового гормона и он стимулиро-
вал проторакальные железы, то индукция линьки имеет место; тип
линьки зависит от того, когда произведено обезглавливание — до
того, как началась секреция ювенильного гормона, или после. Тща-
тельно подобрав срок обезглавливания с учетом момента приема
насекомым крови и последующего критического периода, удалось
получить карликовых безголовых особей с морфологическими приз-
наками взрослой формы.
У насекомых с полным превращением имеют место аналогичные
события. Тщательные эндокринологические исследования были
проведены на нескольких видах бабочек, в том числе на тутовом
шелкопряде (Bombyx mort) и цекропийном шелкопряде (Нуа-
lophora cecropia; фото 13.4). Гусеница Hyalophora проходит через
четыре линьки (пять возрастов), каждая из которых запускается
мозговым гормоном, стимулирующим секрецию экдизона протора-
кальными железами.
У гусениц первых возрастов corpora allata выделяют ювениль-
ный гормон, благодаря которому насекомое после линьки остается
fl
Фото 13.4. Гусеницы цекропийного шелкопряда (Hyalophora cecropta) первого,
третьего и пятого возрастов. После пятого возраста происходит окукливание и
полный метаморфоз; в результате выводится взрослая бабочка. (Фото предостав-
лено Ch. Walcott, State University of New York, Stony Brook.)
.Личинка
(гусеница)
Куколка
Рис. 13.22. Эндокринные органы, регулирующие развитие цекропийного шелко-
пряда. Каждая линька наступает в результате действия экдизона, выделяемого
проторакальной железой. Эта железа в свою очередь находится под контролем
гормона, который вырабатывают нейросекреторные клетки в мозгу. Ювенильный
гормон, выделяемый прилежащими телами (corpora allata), обусловливает фор-
мирование кутикулы личиночного типа; при последней линьке гусеницы количе-
ство ювенильного гормона понижено, и это приводит к образованию куколки
с характерной утолщенной кутикулой; при самой последней линьке ювенильный
гормон уже полностью отсутствует и из куколки выходит взрослая бабочка (има-
го). (Schneiderman, Gilbert, 1964.)
Процессы регулирования и интеграции у беспозвоночных 763
на личиночной стадии развития; покрытая свободной складчатой
кожей, гусеница может расти и увеличиваться в размерах по мере
потребления пищи. В конце 5-го личиночного возраста секреция
ювенильного гормона прекращается, и при следующей линьке об-
разуется более твердая кутикула (т. е. происходит окукливание).
Личиночные ткани разрушаются и превращаются в структуры ку-
Рис. 13.23. При удалении corpora allata гусеница оказывается лишенной юве-
нильного гормона; в результате после линьки она превращается не в гусеницу
следующего возраста, а в куколку, из которой в дальнейшем выходит карлико-
вая взрослая особь. (Highnam, Hill, 1969.)
колки, из которой после заключительной линьки, происходящей
уже при полном отсутствии ювенильного гормона, выходит вполне
сформировавшаяся взрослая бабочка (рис. 13.22).
Для того чтобы убедиться в значении corpora allata, проще
всего удалить этот орган у гусеницы одного из ранних возрастов.
После этого линька наступает в обычные сроки (система мозг —
проторакальные железы осталась в целости), но в результате линь-
ки получается не гусеница следующего возраста, а миниатюрная
куколка, из которой впоследствии выйдет крошечная бабочка
(рис. 13.23). Удаление corpora allata — источников ювенильного
гормона — не оказывает никакого другого действия, так что кар-
ликовая бабочка ничем, кроме величины, не будет отличаться от
обычной.
Роль corpora allata можно подтвердить путем их транспланта-
ции. Если несколько таких органов от гусениц ранних возрастов
имплантировать гусенице 5-го возраста, то при очередной линьке
получится не куколка, а гусеница неестественно больших размеров.
Она будет продолжать расти, и путем пересадки ей дополнитель-
ных corpora allata можно получить гусеницу-гиганта, из которой
764 Гл. 13. Управление и интеграция
выведется гигантская бабочка. Однако у такого роста есть предел:
после искусственно полученного 7-го гусеничного возраста насеко-
мое, по-видимому, неизбежно погибает.
Следует остановиться на структуре обсуждаемых гормонов и
сравнить их с гормонами позвоночных. Экдизон представляет со-
бой стероид (Karlson, Sekeris, 1966); когда его впервые выделили
в кристаллической форме, экстракции было подвергнуто 500 кг
шелковичных червей и получено 25 мг гормона. Для установления
полной химической структуры экдизона потребовалось обрабо-
тать 1000 кг высушенных куколок (что соответствует примерно'
4 тоннам свежих куколок). Стероидная природа экдизона пред-
ставляет особый интерес в связи с тем, что стероидные гормоны
широко распространены у позвоночных.
В отличие от экдизона ювенильный гормон по своей структуре
не имеет ничего общего с гормонами позвоночных и близок к тер-
пенам. Удалось выделить большое число соединений с активностью
ювенильного гормона, и много таких соединений было синтезиро-
вано, причем некоторые из синтетических веществ оказались на-
много активнее природных. Основной природный ювенильный гор-
мон цекропийного шелкопряда имеет следующую структуру
(Roller et al., 1967):
сн3
сн3 сн2 сн3 п
I I I ?
н3с. СН^ СН, /С /С_0СНз
СН2О-СН сн2 сн сн2 сн
Синтез соединений с активностью ювенильного гормона привле-
кает большое внимание в связи с потенциальной возможностью
их использования в качестве инсектицидов. Эти соединения эффек-
тивны в чрезвычайно малых количествах, и если их воздействие
приурочить к определенной фазе развития насекомых, то этим
можно предотвратить развитие взрослых форм, а значит, и раз-
множение. Преимущество ювенильных гормонов как инсектицидов,
по сравнению с ядами, которые сейчас применяются, состоит в том,
что у насекомых менее вероятна выработка устойчивости к веще-
ствам того типа, какие необходимы для их нормального развития.
Поскольку некоторые синтетические ювенильные гормоны действу-
ют в большей степени на одних насекомых, чем на других, возмож-
но, что удастся получить инсектициды с некоторой направленно-
стью действия; это дало бы огромные преимущества по сравнению
с нынешней практикой использования высокотоксичных веществ
вроде ДДТ, которые уничтожают в равной мере и вредных, и по-
лезных насекомых. Кроме того, ювенильный гормон почти или сов-
сем не оказывает действия на многих других животных, во всяком
Процессы регулирования и интеграции у беспозвоночных 765
случае на плотоядных позвоночных; поэтому при использовании
таких веществ менее вероятны серьезные экологические послед-
ствия.
Случайное открытие природных соединений с активностью юве-
нильного гормона показало, что, по-видимому, растения уже ис-
пользуют их для защиты от насекомых. Это открытие было сделана
при следующих обстоятельствах. Чешский ученый д-р Карел Сла-
ма (Karel Slama) приехал на год в Гарвардский университет и
привез с собой для работы клопов Pyrrhocoris. В лаборатории
этих клопов кормили семенами липы и клали им бумажные по-
лотенца, чтоб насекомые могли по ним ползать; однако в Гарвар-
де нимфы 5-го возраста не превращались во взрослую форму, а
вместо этого получались необычно крупные незрелые формы 6-га
и даже 7-го возраста.
Создавалось впечатление, что клопов непреднамеренно подвер-
гали воздействию ювенильного гормона. В результате системати-
ческого сравнения условий их содержания в США и в Европе обна-
ружилось, что используемые в Гарварде бумажные полотенца со-
держали вещество, обладающее активностью ювенильного гормона
(Slama, Williams, 1965, 1966). Оказалось, что вся бумага амери-
канского производства, включая газетную, содержит это вещество.
Его не добавляли во время технологического процесса — оно уже
было в древесине тех хвойных деревьев, которые шли на изготов-
ление бумаги в США. В бумаге европейского или японского проис-
хождения этого вещества, названного «бумажным фактором», не
было. Когда его выделили, оно оказалось по химической природе
сходным с ювенильным гормоном. Это позволяет предполагать, что
многие терпены, широко распространенные у растений и особенно
свойственные вечнозеленым деревьям, служат естественными за-
щитными веществами, которые растение вырабатывает против на-
секомых-вредителей.
До сих пор мы обсуждали те гормоны насекомых, которые не-
посредственно участвуют в регуляции роста и развития; между
тем гормонами регулируются и многие другие физиологические
процессы. В качестве примера мы кратко опишем гормон, регули-
рующий экскрецию воды у одного кровососущего насекомого, у ко-
торого из-за способа питания периодически возникает большой из-
быток воды в организме.
Кровососущий клоп Rhodnius сталкивается с этой проблемой
после каждого приема пищи. Насосавшись крови, он тотчас же
выделяет большое количество разведенной мочи, освобождаясь
таким образом почти от всего избытка воды. Этот внезапный подъ-
ем данной экскреторной функции зависит от специального гормона^
стимулирующего выделение воды.
Действие этого гормона было изучено на изолированных маль-
пигиевых трубочках. Если эти органы поместить в кровь Rhodnius.
или в подходящий солевой раствор, то секреция мочи в них продол-
766 Гл.. 13. Управление и интеграция
жается. Если они помещены в кровь только что напитавшегося
клопа, то они обильно секретируют мочу — скорость ее секреции
намного больше, чем в том случае, когда трубочки омывает кровь
ют голодного насекомого. Это позволяет предположить, что в кро-
:ви содержится гормон, ответственный за стимуляцию выделения
.мочи (т. е. диуретический гормон).
Экстракты различных органов Rhodnius, известных как проду-
центы гормонов, не оказывают никакого влияния на экскрецию
мочи; эффективен только экстракт ганглиев из первого брюшного
сегмента. Эти ганглии содержат несколько групп крупных нейро-
секреторных клеток; 97% диуретической активности приходится на
долю самой задней группы этих клеток.
После того как Rhodnius насосется крови, выработка мочи воз-
растает менее чем за минуту; значит, гормон должен высвобож-
даться очень быстро — несомненно, под действием рефлекторного
механизма, запускаемого самим процессом питания или же растя-
жением брюшка поступающей пищей (Maddrell, 1963).
Приведенные примеры дают лишь некоторое общее представ-
ление о сложности гормональной регуляции у насекомых. Обраща-
ет на себя внимание важная роль нервной системы в выработке
гормонов, причем не только у насекомых, но и у других беспозво-
ночных. Число гормонов, которое удается выделить, охарактеризо-
вать химически и, наконец, синтезировать, непрерывно растет,
однако эндокринология беспозвоночных — настолько обширная
область знания, что исследования здесь будут развиваться и оста-
ваться плодотворными на протяжении многих лет.
ЦИТИРУЕМАЯ ЛИТЕРАТУРА
Aidley D. J. (1971). The Physiology of Excitable Cells, Cambridge, Cambridge Uni-
versity Press, 468 pp.
Bentley P. J. (1976). Comparative Vertebrate Endocrinology, Cambridge, Cambrid-
ge University Press, 415 pp.
Bullock T. H., Horridge G. A. (1965). Structure and Function in the Nervous
Systems of Invertebrates, vols. I and II, San Francisco, Freeman, 1719 pp.
del Castello J., Katz B. (1954). Quantal components of the end-plate potential,
J. Physiol., 124, 560—573.
Diamond J. (1971). The Mauthner cell. In: Fish Physiology, vol. 5, Sensory
Systems and Electric Organs (W. S. Hoar and D. J. Randall, eds.), pp. 265—
346, New York, Academic Press.
Eccles J. (1965). The synapse, Sci. Am., 212, 56—66.
Furshpan E. J., Potter D. D. (1959). Transmission at the giant motor synapses of
the crayfish, J. Physiol., 145, 289—325.
Heuser J. E., Doggenweiler C. F. (1966). The fine structural organization of nerve
fibers, sheaths and glial cells in the prawn, Palaemonetes vulgaris, J. Cell.
Biol., 30, 381—403.
Highnam К- C., Hill L. (1969). The Comparative Endocrinology of the Invertebra-
tes, London, Edward Arnold, 270 pp., 2 ed. (1977), 368 pp.
Hodgkin A. L. (1954). A note on conduction velocity, J. Physiol., 125, 221 —
224.
Литература 7^7
Hodler J., Stdmpfli R., Tasaki I. (1951). Ueber die Wirkung internodaler Abkiihlung:
auf die Erregungsleitung in der isolierten markhaltigen Nervenfaser des Fro*
sches, Pfliigers Arch., 253, 380—385.
Holmes W., Pumphrey R. J., Young J. Z. (1941). The structure and conductance
velocity of the medullated nerve fibres of prawns, J. Exp. Biol., 18, 50—
54.
Hursh J. B. (1939), Conduction velocity and diameter of nerve fibers, Am. J. Phy-
siol., 127, 131 — 139.
Jensen E. V., Numata M., Brecher P. L., Desombre E. R. (1971). Hormone-receptor
interaction as a guide to biochemical mechanism. In: The Biochemistry of Ste-
roid Hormone Action (Biochemical Society Symposium No. 32) (R. M. S. Smel*
lie, ed ), pp. 133—159, London, Academic Press.
Karlson P., Sekeris С. E. (1966). Ecdysone, an insect steroid hormone, and its mode
of action, Recent Prog. Horm. Res., 22, 473—502.
Katz B., Miledi R. (1967). A study of synaptic transmission in the absence of ner-
ve impulses, J. Physiol., 192, 407—436.
Kosterlitz H. W., Hughes J., Lord J. A. H., Waterfield A. A. (1977). Enkephalins,
endorphins and opiate receptors, Soc. Neurosci. Symp., 2, 291—307.
Maddrell S. H. P. (1963). Excretion in the blood-sucking bug, Rhodnius prolixus-
Stal, J. Exp. Biol., 40, 247—256.
McEwen B. S. (1976). Interactions between hormones and nerve tissue, Sci. Am.,.
235, 48—58.
Merton P. A, (1972). How we control the contraction of our muscles, Sci. Am., 226,,
30—37.
Otsuka M., Iversen L. L., Hall Z. W., Kravitz E. A. (1966). Release of gamma-
aminobutyric acid from inhibitory nerves of lobster, Proc. Natl. Acad. Sci.
USA, 56, 1110—1115.
Pert С. B., Snyder S. H. (1973). Opiate receptor: Demonstration in nervous tissue,
Science, 179, 1011—1014.
Roller H., Dahm К. H., Swedy C. C.f Trost В. M. (1967). The structure of the juve-
nile hormone, Angew. Chem. (Engl.), 6, 179—180.
Rushton W. A. H. (1951). A theory of the effects of fibre size in the medullated'
nerve, J. Physiol., 115, 101—122.
Schally A., Arimura A., Kastin A. /. (1973). Hypothalamic regulatory hormones,
Science, 179, 341—349.
Schneiderfnan H. A., Gilbert L. I. (1964). Control of growth and development im
insects, Science, 143, 325—333.
Slama K-, Williams С. M. (1965). Juvenile hormone activity for the bug Pyrrhocoris
apterus, Proc. Natl. Acad. Sci. USA, 54, 411—414.
Slama K., Williams С. M. (1966). The juvenile hormone. 5. The sensitivity of the-
bug, Pyrrhocoris apterus, to a hormonally active factor in American paperpulp,
Biol. Bull., 130, 235—246.
Stevens C. F. (1966). Neurophysiology: A Primer, New York, Wiley.
Stewart J., Li С. H. (1962). On the use of -tropin or -trophin in connection with>
anterior pituitary hormones, Science, 137, 336—337.
Sutherland E. W. (1972). Studies on the mechanism of hormone action, Science,
177, 401—408.
Sutherland E. W., Rail T. W., Menon T. (1962). Adenyl cyclase. 1. Distribution,
preparation and properties, J. Biol., Chem., 237, 1220—1227.
Takeuchi A., Takeuchi N. (1967). Anion permeability of the inhibitory post-sy-
naptic membrane of the crayfish neuromuscular junction, J. Physiol., 191, 575—
590.
Tasaki I. (1959). Conduction of the nerve impulse. In: Handbook of Physiology,,
sect. 1; Neurophysiology, vol. 1 (J. Field, H. W. Magoun and V. C. Hall,,
eds.), pp. 75—121, Washington, D. C., American Physiological Society.
Wigglesworth V. B. (1959). The Control of Growth and Form: A Study of the Epi-
dermal Cell in an Insect, Ithaca, N. Y., Cornell University Press, 140 pp.
768 Гл 13. Управление и интеграция
ДОПОЛНИТЕЛЬНАЯ ЛИТЕРАТУРА
Aidley D. J. (1971). The Physiology of Excitable Cells, Cambridge, Cambridge
University Press, 468 pp.
Barrington E. J. W. (1975). An Introduction to General and Comparative Endocri-
nology, 2nd ed., Oxford, Clarendon Press.
Barth R. H., Lester L. J. (1973). Neuro-hormonal control of sexual behavior in in-
sects, Annu. Rev. Entomol., 18, 445—472.
Benson G. K., Phillips J. G. (eds.) (1970). Hormones and the Environment, Lon-
don, Cambridge University Press, 629 pp.
Bentley P. J. (1976). Comparative Vertebrate Endocrinology, Cambridge, Cambrid-
ge University Press, 415 pp.
Bullock T. H„ Horridge G. A. (1965). Structure and Function in the Nervous
Systems of Invertebrates, vols 1 and 2, San Francisco, Freeman, 1719 pp.
Gooke L.f Lipkin M., Jr. (1972). Cellular Neurophysiology: A Source Book, New
York, Holt., 1039 pp.
Eccles J. C. (1973). The Understanding of the Brain, New York, McGraw-Hill,
238 pp.
Field J., Magoun H. W., Hall V. C. (eds.) (1959—1960). Handbook of Physiology,
sect. 1, Neurophysiology: vol. 1, pp. 1—780 (1959); vol. 2, pp. 781—1440 (I960);
vol. 3, pp.; 441—1966 (1960), Washington, D. C., American Physiological So-
ciety.
Ganong W. F., Martini L. (eds.) (1978). Frontiers in Neuroendocrinology, vol. 5,
New York, Raven Press, 416 pp.
Gershenfeld H. M. (1973). Chemical transmission in invertebrate central nervous
systems and neuromuscular junctions, Physiol. Rev., 53, 1—119.
Gilbert L. I. (ed.) (1975). The Juvenile Hormones, New York, Plenum Press.
Greep R. O., Astwood E. B. (eds.) (1972—1976). Handbook of Physiology, sect. 7,
Endocrinology, vols. 1—7, Washington, D. C., American Physiological So-
ciety.
Highnani К. C., Hill L. (1977). The Comparative Endocrinology of the Invertebra-
tes, 2nd ed., Baltimore, University Park Press, 368 pp.
Holmes R. L., Ball J. N. (1974). The Pituitary Gland: A Comparative Account,
Cambridge, Cambridge University Press, 397 pp.
Kuffler S. W., Nicholls J. G. (1976). From Neuron to Brain, Sunderland, Mass.,
Sinauer, 486 pp. [Имеется перевод: Куффлер С., Николо Дж. От нейрона к
мозгу. — М.: Мир, 1979.]
Roeder К. D. (1967). Nerve Cells and Insect Behavior, rev. ed., Cambridge, Mass.,
Harvard University Press, 238 pp.
Sutherland E. W. (1972). Studies on the mechanism of hormone action, Science,
177, 401—408.
Turner C. D., Bagnara J. T. (1976). General Endocrinology, 6th ed., Philadelphia,
Saunders, 596 pp.
Wigglesworth V. B. (1970). Insect Hormones, Edinburgh, Oliver and Boyd.
159 pp.
ПРИЛОЖЕНИЯ
ПРИЛОЖЕНИЕ А
ИЗМЕРЕНИЯ И ЕДИНИЦЫ
Цель физиологического измерения — найти значения определен-
ных физических величин. Для представления результатов измере-
ний нужны общепринятые единицы. Такие единицы, их обозначе-
ния и математические символы, связанные с их применением, стан-
дартизованы и приняты для повсеместного использования давно
учрежденным и постоянно действующим международным орга-
ном — Всеобщей палатой мер и весов (The General Conference on
Weights and Measures; Conference Generale des Poids et Mesures).
Указанная система единиц носит название Международной Систе-
мы Единиц или сокращенно — системы СИ (SI—Systeme Interna-
tional).
В этом приложении приведены 1) названия единиц и их обо-
значения, 2) краткий список коэффициентов перевода одних: еди-
ниц в другие и 3) список некоторых физических величин и реко-
мендуемых символов для их обозначения.
ЕДИНИЦЫ
В системе СИ используется семь основных единиц для семи
размерно-независимых физических величин; каждая основная
единица обозначается определенным символом, использование ко-
торого обязательно (табл. А.1).
Система СИ определяет также несколько производных единиц
(равно как и ряд вспомогательных) Хотя некоторые из них обо-
значаются специальными символами, все они выводятся из основ-
ных единиц и определяются через них. Несколько наиболее важ-
ных для физиологии единиц приведено в табл. А.2.
49—1863 769
770 Приложение А
Основные единицы
Таблица АЛ
Физическая величина .Основная единица в системе СИ Обозна- чение
Длина метр м
Масса килограмм кг
Время секунда с
Термодинамическая температу- кельвин К
ра Количество вещества моль М
Сила электрического тока ампер А
Сила света кандела кд
Об использовании этих единиц и обозначений нужно сказать
следующее.
1. Обозначения печатаются прямыми буквами. Они не являются
сокращениями и не требуют после себя точки, если только не
стоят в конце предложения.
2. В случае множественного числа обозначение не меняется.
3. Некоторые единицы названы по имени ученых (например, по
имени Вольта); такце единицы обозначают большими прямыми
буквами (В), но названия их пишут с маленькой буквы (вольт).
Часто бывает удобнее использовать более крупные или более
мелкие единицы, отличающиеся от данных в число раз, кратное 10.
Для их обозначения применяют приставки, перечисленные в
табл. А.З. Эти приставки могут добавляться к любой единице си-
стемы СИ — как основной, так и производной (однако к секундам
приставки, означающие кратные единицы, добавляются редко). Со-
четание приставки и символа рассматривается как единый символ;
составные приставки не применяются.
Производные единицы
Таблица А.2
Физическая величина Производная единица в СИ Обозна- чение Определение через основные единицы
Сила НЬЮТОН н М-КГ-С”2
Энергия джоуль Дж М2-КГ-С“2
Мощность ватт Вт М2*КГ*С“3
Давление паскаль Па М^-КГ-С”2
Вязкость пуаз П м“1*кг’С~1* 10"1
Разность электрических вольт В м2*кг-с~3*А~1
потенциалов
Электрическое сопротив- ом Ом м2-кг-с~3-А~2
ление
Электрический заряд кулон к с-А
Частота герц ' Гц С-1
Измерения и единицы 771
Десятичные доли в английском языке обычно отделяются точ-
кой (.), а во французском и немецком — запятой (,). В системе
СИ точку или запятую можно использовать только для десятичных
долей. Чтобы облегчить прочтение длинных чисел, цифры можно
группировать по три, но не следует разделять запятыми.
Пример: 212 489 143, но не 212,489,143.
Таблица А.З
Приставки для образования десятичных кратных и дольных единиц
Приставка Обозначение Множитель Приставка Обозначение Множитель
Тер а т 1012 МИЛЛИ м 10“3
Гига г 109 Микро мк ю-«
Мега м 1G6 Нано н 10“9
Кило к 103 Пико п 10~12
Деци д ю-1 Фемто ф 10“13
Санти с 10“2 Атто а Ю-18
Когда десятичный разделительный знак ставится перед первой
значащей цифрой числа, впереди этого знака всегда нужно ставить
нуль.
Пример: 0,143 872’, но не ,143 872.
С произведениями или частными от двух и более единиц опери-
руют в соответствии с математическими правилами. Заметим, что
использование более чем одного знака дроби (/) неоднозначно.
Так, например, удельную скорость поглощения кислорода можно
записать в виде мл/(кг-с) или мл-(кг-с)”1 или мл-кг^-сг1, но
не в виде мл/кг/с.
В настоящее время по ряду причин нецелесообразно выражать
результаты всех физиологических измерений через основные и про-
изводные единицы системы СИ. Это связано, в частности, с нали-
чием большого количества информации (включая широко извест-
ные таблицы физических и химических констант), в которой при-
меняются другие величины (например, калории). Еще одной при-
чиной является то, что при использовании ряда общепринятых
методов проще применять некоторые традиционные единицы (на-
пример, грамм-силу, а не единицу системы СИ — ньютон). Однако
такую практику нужно постепенно ограничивать и стараться по
возможности использовать единицы системы СИ.
НЕКОТОРЫЕ КОЭФФИЦИЕНТЫ ПЕРЕВОДА
ОДНИХ ЕДИНИЦ В ДРУГИЕ
Традиционные единицы, обычно используются в тех областях
физиологии, где речь идет об энергии (например, при обсуждении
49*
772 Приложение А
вопросов метаболизма) и давлении (например, при измерении
газообмена).
Сила
1 килограмм-сила = 9,807 Н
1 ньютон = 0,102 кг-сила
Энергия, работа (силахрасстояние)
1 кал=4,184 Дж
1 джоуль = 0,239 кал
1 литр Ог (при 0°C и 760 мм рт. ст.), когда он используется
при окислительном метаболизме, часто приравнивается к
4,8 ккал; следовательно, 1 литр 20,083 кДж
Мощность (работа в единицу времени)
1 кал-с-1=4,184 Дж-с-1=4,184 Вт
1 ккал-ч-1 = 1,163 Вт
1 ватт=0,239 кал-с-1 = 14,34 кал-мин-1 = 860,4 кал-ч-1
Давление (сила на единицу площади)
1 атм = 1,013-105Н-м-2 = 101,3 кПа
1 мм рт. ст. при 0 °C (1 торр) = 1,333-102 Н-м~2= 133,3 Па
1 кН-м-2=7,50 мм рт. ст. = 9,869-10-3 атм
Объем
1 литр определен (с 1964 г.) как единица, в точности равная
1 дм3 (0,001 м3); соответственно 1 мл = 1 см3, а 1 мкл =
= 1 мм3
-1 ^американский галлон = 3,785 литра
1-американская кварта = 0,946 литра
Длина
1 дюйм=25,4 мм
1 фут=304,8 мм=0,3048 м
1 ярд=0,914 м
1 миля =1609,3 м
1 ангстрем (А) = 10-10 м = 0,1 нм
Масса
1 фунт (avoirdupois) =0,4536 кг
1 унция = 0,0283 кг
Температура
Температурная шкала в градусах Цельсия (°C) равна темпера-
турной шкале в градусах Кельвина (К). Температура по Цельсию
определяется как превышение термодинамической температуры
над 273,15 К.
Для практических целей точку плавления льда при давлении
1 атм можно считать соответствующей 0°С; тройная точка для
льда, воды и водяного пара составляет 0,01 °C, а точка кипения
воды при давлении 1 атм— 100 °C.
Измерения и единицы 773
ФИЗИЧЕСКИЕ ВЕЛИЧИНЫ
Для представления физических- величин удобнее всего пользо-
ваться символами, особенно в тех случаях, когда соотношения
между этими величинами выражаются в форме уравнения. Приме-
ром , может служить закон Пуазейля, который гласит, что поток
жидкости Q в трубке прямо пропорционален давлению и обратно
пропорционален сопротивлению R : Q = plR.
Названия некоторых физических величин и рекомендуемые для
них обозначения приведены в табл. А.4.
Таблица А.4
Физические величины и рекомендуемые обозначения.
Одинаково часто употребляемые обозначения даны через запятую
Физическая величина Обозначение Физическая величина Обозначение
Длина 1 Время t
Высота h Скорость U, V
Радиус г Частота f
Площадь А Сила F
Объем V Работа W
Масса т Энергия Е, W
Плотность (mjV) Р Мощность Р
Давление Р Температура Т
Физическую величину следует обозначать одной буквой латин-
ского или греческого алфавита, используя при этом наклонные
буквы (курсив). Когда это необходимо, применяют индексы для
указания специфического значения символа. Например, Тт — тем-
пература тела, Ук — объем крови, Уо2—скорость поглощения кис-
лорода.
В уравнениях, описывающих физические или физиологические
соотношения, для обозначения физических величин используются
эти и подобные им символы. Обозначения единиц (м, кг, сит. д.)
никогда не следует использовать в таких уравнениях для обозна-
чения физических величин.
ЛИТЕРАТУРА
Mechtly Е. А. (1973). The International System of Units. Physical Constants and
Conversion Factors (second revision), NASA SP-9012, Washington, D. C.,
National Aeronautics and Space Administration, 21 pp.
Symbols Committee of the Royal Society (1975). Quantities, Units and Symbols,
2nd ed. London, The Royal Society, 55 pp.
\J.S. Department of Commerce (1977). The International System of Units (SI).
NBS Special Publication 330, Washington, D. C.: U. S. Government Printing
Office, 41 pp.
ПРИЛОЖЕНИЕ Б
ДИФФУЗИЯ
Диффузия — фундаментальный процесс, с помощью которого
происходит перенос вещества. Те диффузионные процессы, которые
имеют физиологическое значение, осуществляются на очень неболь-
ших расстояниях — обычно это доли миллиметра, несколько мик-
рометров и менее. Примером может служить диффузия дыхатель-
ных газов через дыхательные мембраны или диффузия ионов через
мембрану нейрона во время распространения потенциала действия.
Диффузия на более далекие расстояния не имеет физиологического
значения; транспорт здесь осуществляется путем конвекции (при-
меры— течение протоплазмы или циркуляция крови). Почему роль
диффузии велика лишь на небольших расстояниях? Дело в том,
что время диффузии возрастает пропорционально квадрату рас-
стояния.
Определим время, которое потребуется частице, чтобы продиф-
фундировать на данное расстояние. Среднее расстояние У?, на ко-
торое перемещается диффундирующая частица, зависит от средней
длины I всех этапов (шагов), из которых складывается тепловое
движение частиц (средней длины свободного пробега), и от числа
отдельных этапов п. Имеет место следующая зависимость: R = l^n.
Поскольку число отдельных шагов пропорционально времени t, мы
можем записать Rcctyt, или t<xR2. Это означает, что время диф-
фузии t возрастает пропорционально квадрату среднего расстоя-
ния R, на которое перемещается диффундирующее вещество.
Рассмотрим этот результат применительно к диффузии веществ
в тканях организма. Предположим, что мы создаем ступенчатое
увеличение концентрации кислорода в данной точке А. Молекулы
кислорода будут диффундировать в окружающие ткани и переме-
щаться в среднем на расстояние 1 мкм за 10~4 с. Для преодоле-
ния в 10 раз большего расстояния потребуется в 100 раз боль-
ше времени и т. д. Таким образом, для диффузии на расстояние
10 мкм потребуется время 10-2 с, для 1 мм— 100 с, для 10 мм —
104 с (почти три часа), а для диффузии на расстояние 1 м
необходимо время еще в 10000 раз большее, т. е._ более
3 лет. Расстояние, которое должны пройти диффундирующие моле-
кулы газа от легких человека до конечностей, составляет пример-
но 1 м; очевидно, что перенос путем одной диффузии в этом слу-
чае осуществляться не может.
Важно иметь в виду, что мы рассмотрели время, необходимое
для диффузии на определенное расстояние в том случае, когда мы
Диффузия 775
производим изменение концентрации диффундирующего вещества
в одной точке и смотрим на влияние этого изменения в другой точ-
ке на некотором расстоянии. Так, диффузия нейромедиатора аце-
тилхолина через синаптическую щель (шириной около 0,02 мкм)
происходит очень быстро; в то же время, если бы гормон адрена-
лин должен был достигать мишени из своего источника — надпо-
чечной железы — путем диффузии, то процесс был бы таким дли-
тельным, что оказался бы бесполезным.
Рис. Б.1. Схема стационарного процесса диффузии через однородную мембрану.
Рассмотрим теперь стационарный процесс диффузии при неиз-
менном градиенте (рис. Б.1) (что имеет место во многих физиоло-
гических процессах). Диффузия в этом случае описывается урав-
нением Фика:
dQ= —DA-^dt,
где Q — количество продиффундировавшего вещества, А — пло-
щадь, через которую осуществляется диффузия, и — концентрация
в точке х (и, стало быть, du/dx— градиент концентрации), t — вре-
мя, D — коэффициент диффузии. Знак минус означает, что поток
имеет положительный знак в направлении, в котором градиент
отрицателен. Коэффициенты диффузий для различных веществ
можно найти в справочниках1.
1 Коэффициент диффузии кислорода в воздухе при 20 °C равен 0,196 см2-с~\
а кислорода в воде — 0,183-10~4 см2-с“!.
776 Приложение Б
Из этого уравнения видно, что количество продиффундировав-
шего вещества пропорционально времени; это и интуитивно ясно.
Рассмотрим физиологический пример. В альвеолярном воздухе
парциальное давление кислорода равно 100 мм рт. ст., а смешан-
ная венозная кровь, поступающая в легкие, может иметь парциаль-
ное давление (напряжение) кислорода, равное 40 мм рт. ст. В нор-
ме эти концентрации остаются строго постоянными, и, следова-
тельно, кислород диффундирует из воздуха через стенки альвеол
с постоянной скоростью. Тогда очевидно, что за две минуты про-
диффундирует вдвое больше кислорода, чем за одну минуту, а за
10 минут — в 10 раз больше и т. д. Таким образом, если поддер-
живается постоянный градиент, количество продиффундировавше-
го вещества пропорционально времени.
Из уравнения Фика следует также, что при увеличении диффу-
зионного расстояния х количество продиффундировавшего веще-
ства Q уменьшается. Если х удваивается, Q уменьшается наполо-
вину, если х возрастает в 10 раз, Q снижается до одной десятой
и т. д. Таким образом, количество продиффундировавшего веще-
ства обратно пропорционально диффузионному расстоянию, но
опять-таки лишь в стационарной системе. Для этого нужно, чтобы
по одну сторону мембраны постоянно поступало диффундирующее
вещество или имелся его неиссякающий источник, а по другую
сохранялась постоянная более низкая концентрация, поддерживае-
мая путем непрерывного удаления диффундирующего вещества,
или существовал бездонный сток. Такая ситуация имеет место в
рассмотренном нами примере с легкими.
Константа диффузии. Константа диффузии К, которая была
использована для вычислений на с. 29, отличается от коэффи-
циента диффузии D, входящего в уравнение Фика. Необходимость
введения константы диффузии в физиологии возникает в связи
с изучением диффузии газов между воздухом и тканями, особенно
в легких млекопитающих. В этом случае целесообразно соотносить
скорость диффузии газа, скажем кислорода, с парциальным давле-
нием этого газа в альвеолярном воздухе. Коэффициент диффузии
использовать здесь нельзя, поскольку при этом необходимо знать
градиент концентрации du/dx в ткани, который трудно определить
непосредственно и нельзя вычислить, если неизвестна раствори-
мость данного газа в данной ткани. Использование константы диф-
фузии позволяет обойти эту трудность, связывая скорость диффу-
зии непосредственно с парциальным давлением данного газа в га-
зовой фазе.
Константа диффузии численно равна количеству газа в кубиче-
ских сантиметрах, которое диффундирует за 1 мин через 1 см2 при
градиенте давления 1 атм на 1 см. Эти единицы можно выписать
и упростить следующим образом:
-------------г-—см8 • атм-1 • мин-1.
мин-см2-атм-см-1
Диффузия 777
Константы диффузии были определены лишь для,немногих жи-
вотных тканей; значения некоторых из них приведены в табл. Б.1.
Эти константы определены эмпирически и учитывают раствори-
мость газа в исследуемой ткани. Цифры показывают, что диффу-
зия в тканях с относительно высоким содержанием воды (напри-
мер, в мышцах и соединительных тканях) протекает несколько
Таблица Б.1
Константы диффузии кислорода при 20 °C
(Krogh, 1919). Заметьте, что приведенные
величины не есть коэффициенты диффузии
К, см2.атм-1-мин—1
Воздух 11
Вода 34-1О-6
Желатина 28-10“б
Мышца 14-10~6
Соединительная ткань 11,5-10-6
Хитин1) 1,3-10”6
Резина 7,7-10“6
’) Для хитина часто приводят неверное зна-
чение константы диффузии — в 10 раз боль-
шее, чем в данной таблице. Это результат
опечатки в книге Крога (Krogh, 1941), кото-
рая была скопирована позднее многими ав-
торами.
медленнее, чем в воде, но имеет тот же порядок. С другой стороны,
скорость диффузии через кутикулу насекомых, образованную по-
лимером хитином, меньше по величине на целый порядок.
Отметим, что скорость диффузии кислорода через^ резину весь-
ма велика — она лишь примерно вдвое меньше скорости диффузии
в мышцах. Это очень важно для постановки физиологических
экспериментов, поскольку при использовании’для соединений ре-
зиновых трубок кислород может диффундировать через стенки этих
трубок; двуокись углерода благодаря своей высокой растворимости
может диффундировать через них еще быстрее. Реальная скорость
диффузии зависит от типа используемой резины или пластика;
у одних типов проницаемость весьма велика, у других — намного
меньше.
ЛИТЕРАТУРА
Krogh А. (1919). The rate of diffusion of gases through animal tissue, with some
remarks on the coefficient of invasion, J. Physiol., 52, 391—608.
Krogh A. (1968). The Comparative Physiology of Respiratory Mechanisms (reprint
ed.), New York, Dover, 172 pp.
ПРИЛОЖЕНИЕ. В
ЛОГАРИФМИЧЕСКИЕ И ПОКАЗАТЕЛЬНЫЕ УРАВНЕНИЯ
ЛОГАРИФМИЧЕСКИЕ УРАВНЕНИЯ
В общем виде логарифмическое уравнение записывается так:
У^=Ь-ха, (1)
или в логарифмической форме
log у=log & 4 a log*. (2)
Уравнение (2) показывает, что log у есть линейная функция log х,
т. е., откладывая log у как функцию log х, мы получим прямую
с наклоном а.
Пример 1. На рис. 6.12 скорость поглощения кислорода (Го2)
изображена графически, как функция массы тела Мт:
VO2 = kM^. (3)
Независимую переменную, массу тела AfT, принято откладывать по
оси абсцисс. Скорость поглощения кислорода на единицу массы
получается делением обеих частей уравнения (3) на Мт.
^=k^=kMja-". (4)
Мт Мт т ' ’
Пример 2. Рис. 6.10.
ПОКАЗАТЕЛЬНЫЕ УРАВНЕНИЯ
Показательное уравнение в общем виде записывается так:
у—Ь-ах, (5)
или в логарифмической форме
log у=log & 4-х log а. (6)
Уравнение (6) показывает, что log у является линейной функ-
цией х, т. е., откладывая log г/ в зависимости от х, мы получим
прямую. Используя полулогарифмическую . бумагу (с линейной
осью абсцисс и логарифмической осью ординат), мы получим тот
же результат при изображении зависимости у от х.
Пример. Рис. 7.4, зависимость скорости поглощения кислорода
от температуры.
ПРИЛОЖЕНИЕ Г
ТЕРМОДИНАМИЧЕСКОЕ ВЫРАЖЕНИЕ
ТЕМПЕРАТУРНЫХ ЭФФЕКТОВ
Показатель Qio (определенный в гл. 7) широко используется
в биологии, поскольку он в простой и понятной форме выражает
зависимость скорости физиологических процессов от температуры
(например, скорость потребления кислорода). Однако в физической
химии температурные эффекты выражаются иначе, на основе тер-
модинамического рассмотрения частоты соударений молекул в за-
висимости от температуры. Соответствующее уравнение в том виде,
как оно было выведено Аррениусом, выглядит так:
ит2^т1)/т2т119 (1)
где К — константа скорости, Т — термодинамическая температура
(в кельвинах), ц— энергия активации, R — универсальная газовая
постоянная, е — основание натурального логарифма.
Термодинамически корректное выражение для Qio (отношения
двух скоростей при разности температур 10 °C) можно получить,
решив уравнение (1) для при Т2—7\ = 10:
Qlo=2^ti«_==e-^ рю (Г+ЮЩ, (2)
Анализ уравнения (2) показывает, что Qio является функцией
температуры и меняется с ее изменением, а следовательно, не мо-
жет быть константой. R и jll (для каждого данного процесса) —
константы, поэтому Qio уменьшается с увеличением температуры.
Пример такого изменения Qio с температурой дан в табл. Г.1 для
ц=12 286 (значение ц выбрано так, чтобы для интервала темпера-
Таблица Г.1
Изменения Qio в интервале
температур от 0°С до 50 °C,
вычисленные для энергии активации
12 286
Tlt °C т2, °с Q10
0 10 2,22
10 20 2,10
20 30 2,00
30 40 1,91
40 50 1,84
780 Приложение Г
тур от 20 до 30°C Qio было равно 2). Если р,= 19 506, то для тем-
пературного интервала 20—30 °C Qio = 3, но по сравнению с преды-
дущим случаем Qio будет выше при более низких температурах и
ниже при более высоких температурах. Таким образом, использо-
вание Qio для описания температурных эффектов не имеет термо-
динамической основы; тем не менее Qio — очень наглядный пока-
затель, который удобен и широко используется в биологии.
ПРИЛОЖЕНИЕ Д
РАСТВОРЫ И ОСМОС
Вода — универсальный растворитель, в котором протекает по-
давляющее большинство биологических реакций и связанных с ни-
ми процессов. По этой причине следующий краткий обзор ограни-
чен лишь водными растворами. Более подробно с этими вопросами
читатель может познакомиться, обратившись к учебникам по хи-
мии и физической химии.
РАСТВОРЫ И КОНЦЕНТРАЦИИ
Одномолярный раствор — это раствор, в каждом литре которо-
го содержится 1 моль растворенного вещества. Так, например,
одномолярный раствор хлористого натрия содержит 58,45 г NaCl
на литр раствора. В биологических исследованиях обычно имеют
дело с растворами, в тысячу раз менее концентрированными, и ис-
пользуют соответствующие единицы — миллимоли. Это позволяет
обойтись при записях без большого количества нулей.
Одномоляльный раствор содержит 1 моль растворенного ве-
щества на килограмм воды. Так, например, раствор, содержащий
58,45 г NaCl на 1 кг воды, есть одномоляльный раствор. Поскольку
объем этого раствора превышает 1 л, его концентрация меньше
одномолярной.
Молярные концентрации часто используются в биологических
работах; они удобны тем, что объем данной пробы биологической
жидкости легко измерить, тогда как точное содержание воды в
ней определить весьма трудно или даже невозможно. Так, напри-
мер, измерить объем пробы крови не составляет труда, и концент-
рация натрия в ней легко определяется. В то же время связать
количество натрия с количеством воды в пробе сложнее, так как и
в плазме, и в эритроцитах содержится много других растворенных
веществ, в том числе значительное количество белков. В физиче-
ской химии часто пользуются моляльной концентрацией; это свя-
зано с рассмотрением осмотических явлений.
Иногда концентрацию измеряют в процентах (%). К сожале-
нию, это понятие весьма нечеткое и нет однозначного соглашения
относительно того, как его использовать, что часто приводит к не-
доразумениям. Что такое, например, 1%-ный раствор хлористого-
натрия? Означает ли это, что 1 г NaCl растворен в 99. г воды? Или.
под этим понимается, что в 100 мл раствора содержится 1 г NaCl?
А может быть, поскольку термин «процент» означает «сотая часть»»
782 Приложение Д
этот раствор содержит 1 г NaCl на 100 г врды? Еще более непог
нятно выражение «миллиграмм-процент» (мг%), которое по логи-
ке должно означать «миллиграммы на 100-миллиграммов». Обьпи
но, однако, оно указывает число миллиграммов вещества на 100 мл
раствора. Этой путаницы можно избежать, точно указывая едини-
цы (например, 120 мг глюкозы на 100 мл крови) или лучше — ис-
пользуя молярные концентрации.
Введение дополнительных, уточняющих слов, например «весо-
вой процент», помогает мало. Обычно (но не всегда) это выраже-
ние указывает, сколько весовых частей приходится на растворенное
вещество, если вес раствора принят за 100 частей. Соответственно
1%-ныц раствор сахарозы содержит 1 г сахарозы и 99 г воды.
Однако если взвешивать соль, содержащую кристаллизационную
воду, то и количество соли, и общее количество воды могут быть
неопределенными.
Строго говоря, термин «процент» должен означать безразмер-
ную долю. Следовательно, соотносимые величины нужно выражать
в одних и тех же единицах, которые должны сократиться при де-
лении. Заметим, что концентрацию газов можно однозначно выра-
жать в процентах. Так, например, сухой атмосферный воздух со-
держит 20,95% кислорода независимо от того, в чем мы выражаем
количество кислорода и воздуха — в числе молекул, в миллилитрах
или в единицах давления.
ОСМОС
Осмос — это движение воды между двумя растворами с разной
концентрацией в том случае, когда эти растворы разделены полу-
проницаемой мембраной (т. е. мембраной, пропускающей воду, но
непроницаемой для растворенного вещества).
Рассмотрим систему, изображенную на рис. Д.1. Молекулы воды
будут перемещаться из отсека В в отсек Л, создавая в последнем
дополнительное — осмотическое — давление. Движению воды мож-
но воспрепятствовать, приложив к поршню давление; когда это.
давление будет в точности равно осмотическому давлению, резуль-
тирующего перемещения воды не будет.
Сила л/ под действием которой перемещается вода, определя-
ется уравнением Вант-Гоффа
лУ=п£Т,
где V — объем, п — число молей растворенного вещества, R — уни-
версальная газовая постоянная, Т—абсолютная температура. Это
уравнение аналогично уравнению состояния газа где
Р — давление.
Одномоляльный раствор идеального неэлектролита имеет осмо-
тическое давление 22,4 атм при 0°С (273 К). Оно равно давлению,
Растворы и осмос - 783
оказываемому 1 молем идеального газа, который занимает объ-
ем 1 л.
Растворенные вещества снижают точку замерзания воды; так,
температура замерзания одномоляльного раствора идеального не-
электролита равна —1,86 °C. Растворенные вещества снижают
также давление пара и повышают точку кипения жидкости.
В принципе осмотичес-
кое давление раствора за-
висит только от числа при-
сутствующих в нем частиц,
но не от их размера или
природы. Следовательно,
электролит такого типа, как
хлористый натрий, который
в воде диссоциирует на ио-
ны Na+ и С1“, имеет более
высокую осмотическую кон-
центрацию, чем можно,
ожидать исходя из его мо-
ляльности.
Рис. Д.1. Когда два раствора раз-
ной концентрации разделены полу-
проницаемой мембраной (М), во-
да будет переходить из более раз-
бавленного раствора в более кон-
центрированный/ Если мембрана
жесткая, это перемещение можно
предотвратить, оказывая на пор-
шень давление, равное разности
осмотических давлений в этих
двух растворах;
К сожалению, данные по реальном^ осмотическому давлению
в растворах электролитов не совсем совпадают с результатами
расчетов, основанных на оценке ожидаемого числа ионов. Это свя-
зано с взаимодействием между положительными и отрицательными
ионами, а также'мёжду ионами и молекулами воды. Поэтому осмо-
тическое давление электролита нельзя вычислить, непосредственно
по его молярной концентрации, и нужно йводить эмпирический
осмотический коэффициент Фг
Если бы’-NaCl в растворе-полностью диссоциировал, точка* за-
мерзания его одномоляльного раствора была бы ниже, чем у воды,
на 2-1,86°C, т. е. была бы —3,72°C?На самом дел'е' этбт раствор^
замерзает при —3,38 °C. Таким; образом,; его- осмотический, коэффи-
циент райен 3,38/3,72 = 0,91. Осмотический^ коэффициент'‘зависит
от концентрации электролита и от его природы:
784 Приложение Д
ДОННАНОВСКОЕ РАВНОВЕСИЕ
Рассмотрим систему типа изображенной на рис. Д.1, но такую,
в которой разделяющая два отсека жесткая мембрана проницаема
для воды и электролитов, но не пропускает крупные заряженные
молекулы, скажем отрицательно заряженные белки (рис. Д.2).
Пусть электролитом будет хлористый натрий.
Если стенки отсеков жесткие, давление на поршень Р может
предотвратить результирующее движение воды через мембрану.
В состоянии равновесия
[Na+]AX[C1-]A= [Na+]BX[C1“]B.
Преобразуя это выражение, мы получим, что отношение коли-
чества растворенных катионов по одну сторону мембраны к коли-
честву тех же катионов по другую сторону мембраны обратно про-
порционально отношению коли-
честв анионов по обе стороны
мембраны:
[Na+]A _ [С1-]в __
[Na+]B ~ [С1-]А '
Величину г называют констан
той Доннана.
Примем, что начальные кон-
центрации NaCl по обе стороны
мембраны одинаковы и что в от-
секе А имеется также натрие-
Рис. Д.2. Осмотическая система, состоя-
щая из двух отсеков А и В, разделен-
ных жесткой мембраной, которая про-
пускает воду и мелкие ионы, но не про-
пускает крупные ионы, такие, как отри-
цательно заряженные молекулы белков.
Распределение ионов и осмотическое дав-
ление в равновесии (доннановское рав-
новесие) описаны в тексте.
вая соль белка NaPr, диссоциирующая на Na+ и Рг~. Вслед-
ствие этого суммарная концентрация Na+ в отсеке А будет больше,
чем в В, и ионы натрия начнут диффундировать в В. Это приведет
к избытку ионов Na+ по сравнению с С1_ в отсеке В, и потому
такое же количество ионов С1~ должно будет переместиться в от-
сек В. Однако этому процессу будет препятствовать результирую-
щее повышение концентрации С1“ в отсеке В по сравнению с А.
В равновесии концентрации ионов по обе стороны мембраны будут
Растворы и осмос 785
различаться и в отсеке А будет избыточный отрицательный заряд
по отношению к отсеку В. Такая равновесная ситуация с неравно-
мерным распределением диффундирующих ионов и наличием ре-
зультирующего заряда известна под названием доннановского
равновесия.
Подчеркнем, что разность потенциалов обусловлена присутст-
вием в отсеке А недиффундирующих анионов. Поскольку система
находится в равновесии, этот потенциал поддерживается без затрат
энергии и сохраняется неопределенно долго.
Указанную разность потенциалов Е можно вычислить с по-
мощью уравнения Нернста
, [Na+]A
Ь — zF Ш [Na+]B ’
где R — универсальная газовая постоянная, Т — абсолютная тем-
пература, z — заряд иона (в данном случае он равен единице),
F— число Фарадея (96 500 кулонов на моль).
Давление, приложенное к поршню на рис. Д.2, достаточное для
того, чтобы предотвратить осмотический поток воды в отсек А,
называют коллоидным осмотическим давлением. Когда давление
выше коллоидного осмотического давления, вода и электролит про-
талкиваются через мембрану, а неспособный проникать через нее
белок остается в отсеке А. Этот процесс называют ультрафильтра-
цией. Осмотическое давление ультрафильтрата равно осмотическо-
му давлению раствора в отсеке В.
ЛИТЕРАТУРА
Overbeek J. Th. G. (1956). The Donnan equilibrium, Progr. Biophys., 6, 57—
84.
Hammel H. T., Scholander P. F. (1976). Osmosis and Tensile Solvent, New York,
Springer-Verlag, 133 px>.
50— 1863
ПРИЛОЖЕНИЕ Е
ЖИВОТНОЕ ЦАРСТВО
СОКРАЩЕННАЯ КЛАССИФИКАЦИЯ
СОВРЕМЕННЫХ ЖИВОТНЫХ
ТИП PROTOZOA. Простейшие
КЛАСС FLAGELLATA. Жгутиковые
КЛАСС SARCODINA. Саркодовые. (Простейшие с псевдопо-
диями. Amoeba и др.)
КЛАСС CILIATA. Инфузории. (Paramecium и др.)
ТИП PORIFERA. Губки
ТИП COELENTERATA (или Cnidaria). Кишечнополостные (или
стрекающие)
КЛАСС HYDROZOA. Гидроидные полипы. (Hydra и др.)
КЛАСС SCYPHOZOA. Сцифомедузы
КЛАСС ANTHOZOA. (Актинии и коралловые полипы)
ТИП CTENOPHORA. Гребневики
ТИП PLATYHELMINTHES. Плоские черви
КЛАСС TURBELLARIA. (Свободноживущие плоские черви)
КЛАСС TREMATODA. Сосальщики
КЛАСС CESTODA. Ленточные черви
ТИП NEMATODA. Круглые черви. (Ascaris и др.)
ТИП MOLLUSCA. Моллюски
КЛАСС AMPHINEURA. (Хитоны)
КЛАСС GASTROPODA. Улитки, или брюхоногие моллюски
КЛАСС PELECYPODA. Двустворчатые моллюски: (Мидии
и др.)
КЛАСС CEPHALOPODA. Головоногие моллюски. Кальмары,
осьминоги, Nautilus)
ТИП ANNELIDA. Кольчатые черви
КЛАСС POLYCHAETA. (Морские черви)
КЛАСС OLIGOHAETA. (Дождевые черви и многие пресновод-
ные кольчатые черви)
КЛАСС HIRUDINEA. Пиявки
ТИП ARTHROPODA. Членистоногие
КЛАСС XIPHOSURA. Мечехвосты. (Limulus)
КЛАСС ARACHNIDA. Паукообразные. (Пауки, клещи, скор-
пионы и т. п.)
КЛАСС CRUSTACEA. Ракообразные. (Речные раки, омары,
Daphnia, Artemia и др.)
КЛАСС CHILOPODA. Губоногие Ъм™™™!
КЛАСС DIPLOPODA. Двупарноногие J (Многоножки)
КЛАСС INSECTA. Насекомые
Животное царство 787
ТИП ECHINODERMATA. Иглокожие
КЛАСС ASTEROIDEA. Морские звезды.
КЛАСС OPHIUROIDEA. Офиуры
КЛАСС ECHINOIDEA. Морские ежи
КЛАСС HOLOTHUROIDEA. Голотурии
ТИП CHORDATA. Хордовые
ПОДТИП UROCHORDATA (или Tunicata). Оболочники
ПОДТИП CEPHALOCHORDATA. (Amphioxus)
ПОДТИП VERTEBRATA. Позвоночные
КЛАСС AGNATHA. Бесчелюстные. (Миноги и миксины)
КЛАСС CHOiNDRICHTHYES. Пластиножаберные. (Акулы и
скаты)
КЛАСС OSTEICHTHYES. Костные рыбы
Подкласс Crossopterygii. Кистеперые рыбы. (Latimeria)
Подкласс Dipnoi. Двоякодышащие рыбы
Подкласс Actinopterygii. (Высшие костные рыбы)
КЛАСС AMPHIBIA. Амфибии. (Лягушки, жабы, саламандры
и др.)
КЛАСС REPTILIA. Рептилии
Отряд Chelonia. Черепахи
Отряд Crocodilia. Крокодилы
Отряд Squamata. Чешуйчатые. (Ящерицы и змеи)
КЛАСС AVES. Птицы
КЛАСС MAMMALIA. Млекопитающие
Подкласс Prototheria
Отряд Monotremata. Однопроходные. (Яйцекладущие живот-
ные — ехидна и утконос)
Подкласс Metatheria
Отряд Marsupialia. Сумчатые. (Кенгуру, опоссум и др.)
Подкласс Eutheria
Отряды: насекомоядные; (кроты, землеройки и др.); лету-,
чие мыши; приматы (лемуры, обезьяны, человек); непол-.
нозубые (ленивцы, броненосцы и др.); зайцеобразные.(кроу
лики и зайцы); грызуны; китообразные (киты и дельфины);,
хищные; ластоногие (тюлени и др.); хоботные (слоны);
сирены (ламантины и морские коровы); копытные
ЛИТЕРАТУРА
Rothschild L. (1965). A Classification of Living Animals, 2nd ed., London, Long-
mans, Green, 134 pp.
59'
ПРЕДМЕТНЫЙ УКАЗАТЕЛЬ
(Страницы до 420 — в книге 1, после 420 — в книге 2)
Артемия 437, 438
Артериальная луковица 149, 151
Артериальный конус 149, 150
Артерии 160—162
Аскорбиновая кислота 218
Аспарагиновая кислота 753
Атмосферный воздух 17
Атриовентрикулярный пучок 159
АТФ (аденозинтрифосфат) 111, 237,
238, 248, 562, 563, 565, 568, 748—
749
Аденилатциклаза 749
Аденогипофиз 741—743, 747
Аденозинмонофосфат см. АМФ
Аденозинтрифосфат см. АТФ
Адреналин 746—751
— структура 752
Азот (молекулярный) 17—18, 252—
253, 255
— выделение 532—551
— фиксация 217
Акклимация 290, 326—327
— и акклиматизация, определения
291—292, 319
Аксон 679, 710—718
Аксонный бугорок 726
Активный транспорт 434, 501, 530
-----воды 470—472
Актин 557, 565, 566
Акулы, плавучесть 618
— сердце 149
— температура тела 398, 401
— электрорецепция 658, 660, 661
Алкалоиды 226
Алкоголь 240
Аллантоин 548
Аллигатор 262
Альфа-клетки 746
Амебоидное движение 555—557
Аминокислоты 215—217
— и осморегуляция 430—431
Аммиак, выделение 538, 547
Ампулы Лоренцини 660, 661
АМФ 748—749
Амфибии, выделение 526—527
— испарение воды 459—461
— кровообращение 146, 152, 154, 155
— осморегуляция 451—454
Анадромные рыбы 442, 450—451
Анаэробный метаболизм 248—249
Антеннальная железа 511—513
Антидиуретический гормон (АДГ)
532, 534, 742
Антифризы 315—319
Аргон 17
Аутокаталитическая активация 200
Ацетилхолин 169, 720, 721, 751
— структура 752
Ацетилхолинэстераза 725
Бег, биомеханика 598—600
— энергетическая цена 283—287
Бегемот 262
Беззубка 436—437
Белка 360
Белки, калорийность 239
— переваривание 199—201
Белый медведь 359, 360, 367
Бернулли теорема 162—165
Бета-клетки 746
Биомеханика 597—613
Бишир 63
Блохи, прыжки 608—609
Бобр 262, 360
Боковая линия 669—670
Бокоплав 328
Бора эффект 109—ПО, 113, 114
Бражник 407, 408
Брожение (в рубце) 207—209
Бурый жир 395—396
Вазопрессин 534—535, 742, 744
Ванадий 223
Ваточник 230
Вены 160—162
Верблюд 378—380, 382
Викунья 115
Предметный указатель 789
Витамины 217—219
Вкус 671—677
Влажность, дефицит насыщения 456
— относительная 20
Внеклеточная вода 142, 143
Внутриклеточная вода 142, 143
Вода (в организме) 142—145
— жесткая и мягкая 422—425
— метаболическая 473—474
— морская, состав 422, 423
— поглощение из воздуха 475—476
Водный баланс млекопитающих 480—
485
----- морских 492—495
-----птиц 480, 485
-----морских 488—492
-----рептилий 477—480, 485—487
-----членистоногих 462—477
Водяной пар в воздухе 19—21
-----давление 455—456
Воздух 17—21
Воздушные мешки 71—77
Волк 360
Волнистый попугайчик, полет 288—
289
Воротная система почки 153
Воротное кровообращение 741
Воск, переваривание 201—202
Воска морские 202, 203
Временная суммация 725
«Всё или ничего» 581, 686, 706—708
Выделение 500—553
— у насекомых 513—516
Выделительные органы, типы 501—
503
Высоты (горные) 21—22, 79, 112—
115, 290—294
Вязкость 167, 168
— крови 106, 107
Vasa recta 531
Газели, терморегуляция 384—386
Газы атмосферные, растворимость
22—23
— в плавательном пузыре 631—637
— секреция 633—637
Галаго 608
Галапагосская игуана 403, 404, 492
Гамма-аминомасляная кислота
(ГАМК) 752, 753
Гемоглобин 103—116, 119—126, 129,
130
— распад 621
Гемостаз 80—182
Гемоцианин 104, 105, 121—123
Гемоэритрин 104, 105, 121
Генри закон 24
Гепарин 82
Гетеротермные организмы 299
Гибернация см. Зимняя спячка
Гигантский аксон 679—685, 711, 712
Гидравлические скелеты 593—597
5-гидрокситриптамин 752, 753
Гидролиз ферментативный 197
Гипоталамус, гормоны 741—745
Гипофииз 741—744
Глаз 647—653
— переработка информации 688—697
Гликоген 241—243
Гликозиды 227, 230
Глицерин и устойчивость к замерза-
нию 312—313
Глубинный рассеивающий слой 621
Глутаминовая кислота 752, 753
Глюкагон 745
Глюкоза, реабсорбция 522—523
Голец (американская палия) 247, 324
Головастики, кровь 112—114
Голубь 108, 190, 191, 361
Гомойотермные животные 299
Гормон роста 743, 744
Гормональная регуляция 735—755,
757—766
Гормоны беспозвоночных 757—766
— определение 735
— перечень 736—740
— химическая природа 738—741
Горная болезнь 291
Горячие источники 305
Гремучая змея 655—656
Гренландский тюлень 366
Гуанако 369
Гуанин 547
Гуахаро 667—668
Гулярное трепетание 383
Гуппи 310
Давления высокие 268—270
— низкие 21—25, 290—294
Далматский дог 547—548
Двоякодышащие рыбы, выделение
мочевины 542—543
----дыхание 61, 63, 69—71
----кровообращение 146, 151—154
Двуокись углерода (СО2), 18—19
----диффузия 26—28
----и кислородная диссоциация ге-
моглобина 109—111
----и регуляция дыхания 57—61
----при водном дыхании 131—132
----тепличный эффект 19
----транспорт в крови 127-И 29
Дельфины 365, 371, 667
Дендриты 678
Деполяризация 685
Детектор ошибок 703
Динеин 542
790 Предметный указатель
Диссоциация гемоглобина для СО2
129—131
---кислородная 108—122
Диуретический гормон 765
Дифосфоглицерат (ДФГ) ГЫ; 1'12,
115, 116
Диффузия 774—777
— кислорода 30, 122—127
— коэффициент 775—777
— облегченная 122—127
— СО2 26—28
Дождевые черви, гидравлический,
скелет 594—596
---испарение воды 457
---кровообращение 174
Доннана равновесие 784—785
Дофамин 752
Древоточец 85, 86
Дыхальца 86—88, 94—99
Дыхание в воде (опыты на собаках
и мышах) 267—268
— водное 28—40
— воздушное 43—68
— кожное 49—52, 65
— насекомых 82—99
— органы 46—47
— птиц 71—75
— птичьих яиц 80—82
— регуляция 57—61
Дыхательные пигменты 103—107
Дыхательный коэффициент (ДК) 239/
240
Ехидна 58, 339, 393
Еж 338, 388
Жаба 561
Жабры 30—40, 46, 47
— насекомых 90
Жвачные 206—209
Жгутики 557—564
Железо 221—222
Жесткая вода 423, 425
Жидкости тела 142—144
Жираф 165, 166
Жиры 239—243
— и плавучесть 618—619
— переваривание 201—203
Жук-бомбардир 231—232
Жуки-щелкуны 609—610
Закрытопузырные рыбы.629
Замерзание,, замораживание 310—319-
Замыкательная мышца» 586—587
Зеленая железа см. Антеннальная
железа
— черепаха 266
Землеройки 359, 360
Земляная белка 356
Зимняя спячка 387—397
Змеи морские 51, 266
— температура тела 404
Зоб 189
Зобное молочко 190
Золотая рыбка 246, 322—324, 721
Зрение 647—653
Зрительная информация, переработка
688—697
— кора 695—697
Иглобрюх 119, 244, 324
Иглокожие 175
Излучающая способность 349
Излучение 347—351
— тепловое 343, 375, 397—398
Изокалорический вес 242—243
Изометрическое сокращение 569, 570
Изоосмотичность 426—427
Изотоническое сокращение 569
Изотоничность 42,7
Илистый прыгун 62, 66, 263
Ильная рыба 61—63
Ингибиторы ферментов (в растениях)
228
Инертные;газы 256, 630
Инсулин 745—746
Интернейроны 733—734
Интерстициальная жидкость 142, 143
Интрафуза^ьные .волокна 732
Инулин 509, 510, 520—521
Инфракрасные лучи 347, 349. 350;’
, 398
-----рецепция 654—657
Иод 225.
Ионная регуляция, внутриклеточная
430-431
Испарение воды 351—353, 455-^-457;-
459, 482
-----и водный баланс 480, 482
-----и- терморегуляция 373—380,
, 382—387, 397—398'
— — при дыхании 483—485
Кайман 262
Калорийность питательных веществ
239—240
Калория, • определенна 239, 342 «
Кальмары 433, 493, 680
Кальций и свертывание крови 181
— роль в мышцах 574
Кальцитонин 745
Камбалд 324
Канальцевая;реабсорбция 516. 522
— секреция 5 Гб, 523—524
Капилляры 160—162
— диффузия О2 117—118
— фильтрация 168—173
Предметный указатель 791
Каракатица 430
— плавучесть 620, 623—625
Карась 248—249
Карбоангидраза 134—136
Карпозубики 307
Катадромные рыбы 443, 450—451
Каудальное сердце 148
Кашалот 254
Кенгуровая крыса 481—483
Кенгуру 383, 604—606
Кессонная болезнь 252—255
Кета 323, 324
Кефалины 754
Киви 337, 338
Киноцилия 670, 671
Кислород в атмосфере 18
— потребление 238—241
-----и размеры тела 269—279
-----при нырянии 259—266
— токсичность 255—256
Кислородной диссоциации кривая
108—122
Кислотно-щелочное равновесие 132—
134
Киты, водный баланс 492
— ныряние 254, 255
— тепловой баланс 364, 365
Клейдоические яйца 544
Клещи, поглощение воды 476—477
Клоака 532—533
Кобальт 222
Кожное дыхание 49—52, 65, 266—267
Коза, кровь 113
Коленчатые тела 695
Колибри 388, 394. 395
Коллоидное осмотическое давление
169—171
Колорадский жук 302, 303
Кольчатые черви (аннелиды), крово-
обращение 174—175
Комары 439—440
— личинки 89, 311
Конвекция 343, 346
Копрофагия 210—211
Корабельный червь 204
Кораблик 625, 626
Корова, желудок 207
Кортикотропин 747
Кошка 582—5831
Крабоядная лягушка 453—454, 527
Крабы 329, 430, 432, 463
Креатинфосфат 569
Кремний 224
Кровеносные сосуды 160—162
Кровообращение 139—182
— замкнутые й незамкнутые систе-
мы 141—142
— у беспозвоночных 174—186
— у ныряющих животных 258, 260—
263
Кровоток 157—158, 172—173
— в почках 524—525
Кровь 102
— вязкость 106—107
— объем 142—145
— плода 112—116
— свертывание 180—182
— транспорт СО2 109—111, 127—136
---О2 108—109
— форменные элементы 105—107
— pH 128—129, 132—134
Кровяное давление 166—168
Крокодилы, кровообращение 147, 152
Кролик, копрофагия 211
— молоко 191
Круглоротые, кровообращение 146,
148—151
— осморегуляция 440—442
Куница 360
Купула 670
Corpora allata 759—764
Лама 114, 115
Лангерганса островки 745
Латеральное торможение 689—691
Леггемоглобин 126
Легкие 47—48, 52—59
— объем 53—54
— птиц 72—75
Лемминг 359, 360
Ленивец 209, 373, 383
Летальная температура 305—310
Летательные мышцы, (насекомых)
. 591—592
Летучие мыши, дыхание 52, 53
---зимняя спячка 389—390
---эхо-локация 665—668
Лимфа 170—171
Липазы 203
Лихорадка 340—342
Лицевая ямка 655—656
Локомоция 555—637
— биомеханика 597—613
Лори 373
Лососи 424
Лютеинизирующий гормон1 (ЛГ) 743
Лягушка, выделение 538—541, 546
— дыхание 48—49
— зрение 69(—693
—, испарение воды 459—461
— крабоядная 453—454
— кровообращение 146
— кровь 112—114
Магнитные поля, рецепция ‘662—663
Макрель 40
792 Предметный указатель
Мальпигиевы сосуды 513—516
— тельца 517, 518. 520
Марганец 223—224
Маутнеровские клетки 728
Медведи, теплоизоляция 359, 360
Медоуказчик 202
Медузы, содержание сульфата 428—
430
Медь 223
Меланоцитстимулирующий гормон
743, 744
Менидия 324
Мертвое море 424
— пространство 54
Метаболизм анаэробный 237, 238,
248—249
— интенсивность 238—241
---и размеры тела 270—283
Метаболическая вода 474—475
Мех (шерстный покров) 350, 368—
370, 381
— как теплоизоляция 358'—361
Мечехвост 688
Миелин 713—718
Микроэлементы 219—225
Миксины, кровообращение 146, 148
— рсморегуляция 440—442
Минеральное питание 219—225
Миноги 441, 442
Минутный объем сердца 156—157
Миоглобин 105, 122, 125
— при нырянии 259, 262
Миозин 557, 565, 566, 572—573
Миофиламенты 572—576
Млекопитающие, водный баланс
480—485, 492—495
— кровообращение 145, 147, 153—
162
— легкие, объем 52—55
— ныряние 250—270
— почки 516—525, 528—532
— температура тела 339—340
Многоножки 231
Мозговой гормон (насекомых) 760
Молибден 223
Моллюски, кровообращение 175—176
— мышцы 585—588
— перегрев 307
— почки 510—511
Молоко и водный баланс 494
«Молоко» птиц 190—191
Молочная кислота 263, 264
---и плавательный пузырь 633,
635, 636
Морская вода 422, 423
Морские иглы 525
— млекопитающие 492—495
— птицы 488—492
— рептилии 486—487
Морской лев 493
— черт 525
Морфин 754
Моча, концентрирование 528—532
— образование 517—532
Мочевая кислота 543—546
— структура 535
Мочевина 443—446, 474
— выделение 538—543
— структура 535
— у акул 443. 445
— у жвачных 208
Мускульный желудок 189—190
Мухи, личинки 201
Мышечные веретена 729—735
Мышцы 565—592
— быстрые и медленные 580—585
— гладкие 565, 585—592
— поперечнополосатые 565—567
— сердечная мышца 567—569
— строение 565—567
— функция 569—592
— членистоногих 588—592
Мышьяк 224
Напряжение (газа) 25
Насекомоядные, температура тела
338—339
Насекомые водные 88—93
— водный баланс 464—477
— выделение 513—516
— гормоны 758—766
— дыхание 82—99
— кровообращение 176—178
— мышцы 591—592
— терморегуляция 405—408
Насосы (в циркуляторных системах)
- 139—140
Незаменимые аминокислоты 216
Нейрогемальные органы 757
Нейрогипофиз 741—743
Нейромедиаторы 750—754
Нейроны см. Нервные клетки
Нематоцисты 196
Нервная регуляция 710—735, 755—
756
Нервные клетки 677—680
Нернста уравнение 681, 683, 785
Нефридиальные органы 503
Нефроны 517, 518, 529, 531
Нижняя критическая температура
355—358
Низкие температуры, устойчивость к
ним 310—322
Никель 223
Норадреналин 109, 746
— структура 752
Носовые железы см. Солевые железы
Предметный указатель 793
Нуклеиновые кислоты 547—549
Ныряние 250—270
Облегченная диффузия 122—127
Обмен веществ см. Метаболизм
Обоняние 673—677
Обратная связь 703—706
Однопроходные 278—279, 338, 339
Окись углерода 621
Окситоцин 742—744
Олово 224
Омар 60, 245, 246, 430
Ондатра 262
Опиатные рецепторы 754
Опоссум 356, 388
Опсин 651
Органы чувств 643—697
Орнитиновый цикл 538, 539
Осморегуляция 426—427
— механизмы 433
— у амфибий 451—454
— у беспозвоночных 426—433, 437—
440
— у водных позвоночных 440—451
— у костистых рыб 447—451
— у круглоротых 440—442
— у пластиножаберных 441, 443—446
Осмос 781—785
Осмотический' коэффициент 783
Осьминог 433
Открытопузырные рыбы 628
Оцепенение 387—397
Падальная муха, хеморецепторы 674,
675
Панцирная щука 68—69
/Уарааминогиппуровая кислота (ПАГ)
511, 524
Паратгормон 745
Парциальное давление 25—26
Пауки, выделение 549
— гидравлический механизм движе-
ний 596—597
— прыгающие 596—597
Паукообразные, водный баланс 464—
474
— кровообращение 178
Пейсмейкер 159
Пепсин 198—200
Пептидазы 199
Перегрев, причины гибели от него
307—310
— толерантность 304—309, 319—325
Переохлаждение 310—314
Перистые мышцы 589
Перогнаты 388, 391, 392
Песец 360
Петля Генле 518, 530, 531, 534
Печеночная двуустка 509
Пилильщик хлебный 312, 313
Пингвины 190, 191, 362—364
Пирогены 340
Питание 211—225
— белковое 215—217
— минеральное 219—225
— способы 187—195
Питон 404—405
Пищеварение 195—211
— симбиотическое 202—210
Плавание 610—613
— энергетическая цена 287—290
Плавательный пузырь 627—637
Плавления точка, определение 311
Плавучесть 613—637
Пламенные клетки 507—510
Планарии 196—197, 508
Планктон 187—189
Пластиножаберные, выделение 538—
541
— сердце 149, 150
Пластрон 92
Плодовая мушка, хеморецепция 674
Плотва 39
Пневматофоры 620—621, 623—627
Поверхностное натяжение 55, 56
Поглощающая способность 348, 350
Подкожный жир 359, 365—368, 401
Пойкилотермные животные 299
Полет 610—613
— энергетическая цена 287—290
Поляризованный свет 652—653
Постсинаптические потенциалы 724—
726
Потенциал действия 680—688
— покоя 680—682
Потоотделение 375, 377, 382—387
Потребление пищи 213—214
Почечные клубочки 517—519
Почечный кровоток 524—525
Почки моллюсков 510—511
— позвоночных 503, 516—535
Пресинаптический потенциал 726—727
Пресинаптическое торможение 727—
728
Прилежащие тела (corpora allata)
759—764
«Пробуждение» от спячки 395—397
Проведение импульсов 686, 710—718
Пролактин 743, 744
Пропорциональное регулирование
706—708
Простейшие, переваривание целлюло-
зы 205—209
Пространственная суммация 726
Противотока принцип в газообмене
33—40, 78—79
794 Предметный указатель
----в плавательном пузыре 634—
635
---- В'почке 532
----в теплообмене 370—372, 398
Протонефридий 510
Проторакальная железа 760—763
Протромбин 180—181
Прыжки 596—597; 601—010
Птицы 76—80
— водный баланс 480, 485, 488—492
— дыхание 71—80
— интенсивность метаболизма 276—
279
— температура тела 339, 340
— теплоизоляция 361—362
Птицы-мыши 388
Пчелы 408
Размеры тела 279—283
----и интенсивность метаболизма
270—276
----и свойства крови ПО, 116—118
----и терморегуляция 377—382
Рак речной 60, 434, 436
Ракообразные, выделение 511—513
— испарение воды 463—464
— кровообращение 179—180
— мышцы 588—591
Реабсорбция канальцевая 516, 522
Регуляция и теория управления 702—
709
Резилин 609
Резонансная частота 384, 622
Рекапитуляции теория 549
Ректальная железа 444
Рептилии, водный баланс 477—480
— выделение 527—528
— кровообращение 147, 152—155
— морские 486—487
— солевые железы 487
Ресничное движение 557—564
Ретинен 651
Родопсин 651
Рубец 206—208
Рубнера правило 275
Рукокрылые см. Летучке мыши
Рута эффект НО, 633, 635, 636
Рыбы, антифризы и переохлаждение
315—319
— выделение 525—526
— дыхание воздухом 61—67
— кровообращение 146, 148—155
— кровь 116—120
Сальтаторное проведение 715
Саркоплазматический ретикулум 575
Сатурния 675—676
Свертывание крови 180—182
Свет 647—653
Селен 224
Сердечная мышца 567—568
Сердце 153—160
— каудальное 148
— у тюленя 260—262
— частота сокращений 155—156
Серин 621
Сиг 629
Силовая платформа 600, 603
Синапсы 719—723, 729
Синаптическая передача 724—728
— щель 720
Синильная кислота (у многоножек)
231
Синусный узел 159
Сипункулиды 121
Скап 244
Скаты 445—446, 618
Сквален 618
Скелет 593—597
Складчатозуб мексиканский 390
Скучивание 362—364
Слепая кишка 209
Слух 663—668
Слюна (жвачных) 206, 207
Собака 90, 161, 360
Сократительная вакуоль 502—507
Солевые железы 487, 490—491
Соленоциты 507—510
Солнечное излучение 347, 351, 402
Солоноватые воды 425, 431, 437
Сомик Ictalurus 119, 120, 323—326,
672—673
Старлинга закон 160
Статоцисты 669
Стеногалинные организмы 425
Стереоцилии 670, 671
Стефана — Больцмана закон 347, 350
Страус, дыхание 76
Стрижи 338
Суммация 725—726
Сумчатые, метаболизм 278—279, 383
— температура тела 339
Сурфактант 55, 56
Суслик 381—382
Сухожильные органы 730
Тараканы 475—477
Таранная вентиляция 38—40
Тауи 337
Температура, адаптация к ней 319—
330
— тела 334—342
----у верблюда 379
----у холоднокровных животных
397—409
---- толерантность к отклонениям
304—327
Предметный указатель 795
Температурные пределы (для жизни)
304—319
Температурный коэффициент (Q10)
299—304, 779
Теория трех компартментов Керрана
470—471
Тепло, перенос 342—344
Тепловое излучение 347—351
Тепловой баланс 352—353, 397
Теплоемкость 342, 343
Теплоизоляция 358—362, 364—370
Теплокровные животные 298
Теплообмен 397
Теплообменники 370—373, 398—404
Теплопроводность 343—353, 358—361,
397—398
Теплопродукция (различных органов)
334
Теплота парообразования 351—352
Термиты 205—206, 217
Термонейтральная зона 357—358, 374
Терморегуляция в жаркой среде
374—377
— в условиях холода 354—361
— при оцепенении 387—397
— у холоднокровных животных 397—
408
Терморецепция 653—657
Термофильные бактерии 305
Тестостерон 740
Тетанус 572, 583
Течение жидкостей по трубкам 162—
166
Тиреотропин 743
Тонические мышцы 580—581, 583, 584
Точка замерзания 426
Трахеи 82—89, 95—99
Триметиламина окись 443, 445
Трипсин 198—200
Тромбин 180, 181
Тропомиозин 573
Тропонин 573
Т-система 575
Тубулин 542
Тунец, мышцы 584
— температура тела 398—400
Тутовый шелкопряд 676—677
Тюлени, водный баланс 494
— дыхание 59
— ныряние 257—258
— терморегуляция 359, 360, 364—368
Тюлень Уэддела, ныряние 254, 259,
260, 262, 263
----плавучесть 621
Углеводы, калорийность 239
— переваривание 203—211
Углекислота см. Двуокись углерода
Угольная кислота 128
Угорь 447
— дыхание 47, 62, 64—68
— личинки 107
— электрический 657
Улитки 204, 462, 669
Ультрафильтрация 501, 516, 520
Ультрафиолетовые лучи 650
Упругая отдача 600—606
Уссинга аппарат 452—453
Утки, ныряние 261, 262, 264
— теплообмен в ногах 373, 374
Учащенное дыхание (для охлажде-
ния) 382—387
Фазические мышцы 580—581
Феноловый красный 511, 524
Ферментативный каскад 182
Фетальный гемоглобин 112—116
Фибрин 180, 181
Физалия 231, 620—621
Фика принцип 156
Фильтрация в капиллярах 169—170
— в почках 516, 520—522
Фитоэстрогены 230
Флоридский заяц 376
Фолликулостимулирующий гормон
(ФСГ) 743
Фосфаты органические (в эритроци-
тах) 111—112
Фтор 224
Хеморецепция 671—677
Химические медиаторы 720—723
Хлоридные клетки 449
Хлоридный насос 530—531
Хлорокруорин 104, 105
Холестерол 218
— структура 740
Хомяк, зимняя спячка 388, 396
Хром 223
цАМФ 748—749
Цекропийный шелкопряд 93—96, 723
Целакант 446
Целлюлоза, переваривание 204—211
Циклический аденозинмонофосфат см,
цАМФ
Циклическое дыхание 93—99
Циклоз 557
Цинк 223
Чайки 372, 488—491
Человек, водный баланс 378—379,
487, 490—494
— выделение 547—548
— кровообращение 158, 162, 166
— ныряние 258
— прыжки 607, 608
796 Предметный указатель
— температура тела 334—336
— теплообмен 353
Черепахи 265—266, 404, 478, 479, 487,
545, 546
Чешуйница 204—205
Членистоногие, испарение воды 430,
462—467
Чудесное сплетение (rete mirabile)
430—431, 634, 636
Шерстный покров см. Мех
Шмели 405—407
Щавелевая кислота 228
Эвригалинные организмы 425
Экдизон 759—761, 764
Эккритическая температура 402
Эктотермные животные 299
Электрические органы 657—661
Электрический сом 657
— угорь 66—68, 657
Электрорецепция 657—662
Эндокринная регуляция 735—755
Эндокринные железы беспозвоночных
757—766
— — позвоночных 745—747
Эндорфины 754
Эндотермные животные 299
Эритроциты 106, 107, 181
Эстивация 388
Эстрадиол 740
Эхо-локация 665—668
Ювенильный гормон 759—765
— — в растениях 229—230
----- структура 764
Яды растений 226—231
Яйца птиц, дыхание 80—82
Ящерицы, температура тела 386—387,
402, 403, 544
УКАЗАТЕЛЬ ЛАТИНСКИХ НАЗВАНИИ
(Страницы до 420 — в книге 1, после 420 — в книге 2)
Acantharina 220 Actinopterygii 63 Aedes 439 Aepyornis 80 Aeromonas 341 Agapema 94 Alligator 262 Amblyrhynchus 487 Ameiurus cm. Ictalurus Amia 61—63 Amoeba 504 Anabas 62 Anguilla 47, 62, 64 Anicistrus 62 Anisops 92, 93 Anodonta 432, 436 Antheraea 676 Aphelocheirus 92 Aplysia 723 Aptenodytes 362 Apteryx 337 Arapaima 62, 64 Area 105 Arenicola 104, 105, 596 Arenivaga 475 Artemia 104, 437 Asclepias 230 Asplanchna 509, 510 Astacus 60 Astropecten 104 Colius 388 Colocalia 667 Coregonus 629 Cossus 85, 86 Crocodylus 487 Crossopterygii 63 Cryptochiton 121, 122 Ctenolepisma 204, 492 Cucumaria 175 Cyclothone 619 Cyprinodon 307 Daphnia 104 Dendrostomum 121 Dermatophagoides 477 Desmognathus 50 Dipnoi 63 Dipodomys 481 Dipsosaurus 341 Drosophila 674 Dugesia 662 Dytiscus 90 Eigenmannia 661 Elecrophorus 62, 64, 66, 657 Eledone 105 Emerita 329 Epeira 549 Eptesicus 52, 390, 666 Eriocheir 432, 435, 436 Erithacus 663
Bassozetus 632 Betta 62 Birgus 47 Bombus 407 Bombyx 676, 761 Bracon 313, 314 Bradypus 209 Buenoa 92 Bufo 50, 541, 542 Etmopterus 618 Eulampis 394 Euscorpius 104 Fasciola 509 Fundulus 448 Galleria 202 Gammarus 513 Gastr ophilus 104, 105 Gillichthys 62
Callinectes 614 Calothrix 217 Caltha 650 Carassius 248 Carcharhinus 445 Carcinus 21, 432, 435, 512 Cardiosoma 47, 463 Cephas 312 Cercaertus 463 Chaenichthys 104 Chaos 503 Chelmon 628 Chelonia 266 Chiromantis 460, 461, 546 Chironomus 104—106, 120 Clarias 62 Gonostoma 619 Gymnarchus 660 Halier у pt us 104 Halicystis 615 Holocynthia 108 Helix 105, 205, 549 Hemilepistus 464 Holostei 63 Homarus 60, 105 Hoplosternum 62, 64 Hyalophora 723, 761 ktalurus 119, 120, 320—326, 672 Ixodes 476 Katsuwonis 400
798 Указатель латинских названий
Lama 369
Lampetra 105
Latimeria 440
Leander 437
Lepidosiren 62, 64, 69, 70
Lepisosteus 62, 68
Leptinotarsa 302
Limulus 104, 105, 463, 653
Lingula 104
Liolaemus 402
Lophius 525
Luidia 104
Lumbricus 104, 105, 174
Magelona 104
Maia 430
Malapterurus 657
Manduca 406
Melopsittacus 288
Menidia 324
Musca 201
Myotis 667
Mytilus 588
Myxine 105
Nassarius 662
Nautilus 625, 626
Neoceratodus 63, 64, 69, 70, 542
Nephrops 105, 430
Neurospora 104
Noctiluca 615—616
Notomastus 105
Notonecta 90, 92, 93
Ochromonas 542
Octopus 105
Ocypode 463
Oncorhynchus 323
Opsanus 246
Ophiocephalus 64
Pachygrapsus 430
Palaemonetes 437, 717
Palinurus 105
Pandalus 105
Paramecium 104, 555, 560
Pelamis 52, 266
Periophthalmus 66, 263
Peri planet a 515
Perognathus 391
Petromyzon 442
Phacoides 120
Phascolosoma 105
Phoca 366
Phormia 674
Phyllomedusa 460, 546
Physalia 231, 621
Physer 254
Pimephales 326
Pipilo 337
Plecostomus 62
Plethodontidae 49
Pleurobrachia 558
Polypedilum 305
Polypterus 63
Potamobius 432
Potamotrygon 436, 445
Protopterus 63, 64, 69, 70, 151, 542
Pseudopleuronectes 324
Pterosticus 321
Pytrhocoris 765
Raja 618
Rana 48, 320, 321, 453,526
Rhodnius 514, 760, 765, 766
Rossia 105
Ruiilus 39
Sabellidae 104
Salvelinus 247
Sauromalus 49, 386, 478
Scyliorhinus 658, 660
Sepia 430, 623
Ser pula 104
Serpulidae 104
Siphonosoma 121
Sipunculus 105
Spheroides 324
Sphincterochila 462
Spirographis 105
Spirorbis 104
Squalus 444, 618
Steatornis 667
Symbranchus 62, 66
Synechococcus 305
Syngnathus 525
Tachyglossus 58, 393
Tadarida 390
Talorchestia 328
Telea 675, 676
Tenebrio 513, 514
Teredo 204
Testudo 546
Tetrahymena 104
Tinea 36
Torpedo 657, 660
Trematomus 308, 309, 316—318
Trichomonas 205
Trichonympha 561
Triaacna 195
Iriturus 541
Tubifex 104
Valonia 615
X enopus 542, 561
Zalophus 493
ОГЛАВЛЕНИЕ
Часть четвертая. Вода
Глава 9. Вода и* осморегуляция.......................................421
Водная среда...................................................* 422
Водные беспозвоночные............................................427
Водные позвоночные...............................................440
Наземная среда...................................................455
Животные с «влажной кожей........................................457
Членистоногие....................................................462
Наземные позвоночные...........................................* 477
Морские позвоночные, дышащие воздухом............................485
Цитируемая литература................................................495
Дополнительная литература . 498
Глава 10, Выделение..................................................500
Органы выделения.................................................502
Выделение азота..................................................535
Цитируемая литература................................................551
Дополнительная литература ........................................ 555
Часть пятая. Движение, информационные процессы, интеграция
Глава 11. Мышцы, движение, локомоция..............................555
Амебоидное движение; перемещение с помощью ресничек и жгутиков 556
Движение и мышца...............................................565
Скелетные структуры............................................593
Биомеханика локомоции..........................................597
Плавучесть.....................................................613
Цитируемая литература.............................................637
Дополнительная литература ........................................ 641
Глава 12. Информация и органы чувств..............................643
Сенсорная информация — возможности и ограничения..................645
Преобразование и передача информации...........................677
Цитируемая литература.............................................697
Дополнительная литература ........................................ 705
Глава 13. Управление и интеграция.................................701
Регуляция и теория управления.................................. 7021
Системы нервной регуляции......................................715
Система гормональной регуляции.................................735
Процессы регулирования и интеграции у беспозвоночных .... 755
Цитируемая литература.............................................765
Дополнительная литература ........................................ 765
800 Оглавление
Приложения
Приложение А, Измерения и единицы..............................769
Единицы....................................................769
Некоторые коэффициенты перевода одних единиц в другие . . . 771
Физические величины........................................773
Литература.....................................................773
Приложение Б. Диффузия.........................................774
Литература.....................................................777
Приложение В. Логарифмические и показательные уравнения . . . 778
Логарифмические уравнения..................................778
Показательные уравнения....................................778
Приложение Г. Термодинамическое выражение температурных эффектов 779
Приложение Д. Растворы и осмос.................................781
Растворы и концентрации....................................781
Осмос......................................................782
Доннаиовское равновесие................................... . 784
Литература.................................................... . 785
Приложение Е. Животное царство.....................................786
Сокращенная классификация современных животных.................786
Литература.........................................................787
Предметный указатель....................................... . . 788
Указатель латинских названий ..................................... 797
Кнут Шмидт-Ниельсен
ФИЗИОЛОГИЯ ЖИВОТНЫХ:
ПРИСПОСОБЛЕНИЕ И СРЕДА
Книга 2
Ст. научи, редактор Лашкевич Ю. И.
Мл. редактор Кожанова 3. Е.
Художник Ежков В. Г.
Художественный редактор Кузнецова Л. М.
Технический редактор Борисова Н. И.
Корректор Андреева Н. В.
ИБ № ЗОН
Сдано в набор 27.07.81.
Подписано к печати 4.12.81.
Формат бОХЭО’Лб. Бумага типогр. № 1.
Гарнитура литературная, Печать высокая.
Объем 12,00 бум. л.
Усл. печ. л. 24,00. Усл. кр.-отт. 24,00.
Уч.-изд. л. 26,03. Изд. № 4/1474.
Тираж 10 060 экз. Зак. 1863. Цена 2 р. 10 к.
ИЗДАТЕЛЬСТВО «МИР».
Москва, 1-й Рижский пер., 2.
Московская типография № 11 Союзполиграфпрома
при Государственном комитете СССР по делам
издательств, полиграфии и книжной торговли.
Москва, 113105, Нагатинская ул., д. 1.