/















































































































































































































































































































































































































































Similar




Text
BE СОКОЛОВ
Систематика млекопитающих
BE. СОКОЛОВ
Систематика млекопитающих
Отряды: однопроходных, сумчатых, насекомоядных, шерстокрылов, рукокрылых, приматов, неполнозубых, ящеров
ДОПУЩЕНО МИНИСТЕРСТВОМ ВЫСШЕГО И СРЕДНЕГО СПЕЦИАЛЬНОГО ОБРАЗОВАНИЯ СССР В КАЧЕСТВЕ
УЧЕБНОГО ПОСОБИЯ ДЛЯ СТУДЕНТОВ УНИВЕРСИТЕТОВ
ИЗДАТЕЛЬСТВО
«ВЫСШАЯ ШКОЛА»
М О С К В А—1973
596.5
Ф 59
Соколов В. Е.
Ф 59 Систематика млекопитающих. Учеб, пособие для университетов. М., «Высшая школа», 1973.
432 стр. с илл.
\ /'
Книга представляет первую попытку в отечественной литературе дать таксономическую сводку современных млекопитающих, относящихся к отрядам однопроходных, сумчатых, насекомоядных, шерстокрылов, рукокрылых, приматов, неполнозубых и ящеров. Даются краткие морфологические сведения, характеризующие представителей класса млекопитающих.
Подробно приводятся диагнозы подклассов, отрядов, семейств и более кратко — родов. Дается перечень видов млекопитающих с указанием их распро-1 странения. В диагнозах указаны размеры млекопи-
тающих, форма тела, особенности волосяного покрова, строение черепа, зубная формула и некоторые другие анатомические признаки, а также экология (суточная активность, питание, размножение) и распространение.
Приводится история классификации млекопитающих со времен Аристотеля до наших дней.
В конце книги прилагается список основной литературы по систематике млекопитающих и указатель названий.
2106—091
001(01)—73 84—73
596.5
ОТ АВТОРА
В основу предлагаемой книги положен курс по систематике млекопитающих, читаемый автором для студентов кафедры зоологии позвоночных биолого-почвенного факультета Московского государственного университета.
Задачей руководства является ознакомление с систематикой ныне живущих млекопитающих и с диагностикой различных таксономических групп до родов включительно. В диагнозах рассматриваются морфологические особенности млекопитающих, их экология и распространение, а для отрядов — также и происхождение.
В связи с тем, что книга предназначена в основном для студентов, в ней не приводятся некоторые систематические категории, например трибы, а список литературы по млекопитающим не претендует на полноту. С другой стороны, поскольку настоящее руководство может оказаться полезным и для научных работников, объем диагнозов несколько расширен по сравнению с тем, который дается в университетском курсе.
В руководстве не удалось дать в одинаковой степени полное описание для ряда родов, что в большинстве случаев объясняется их слабой изученностью. Более подробно описаны млекопитающие отечественной фауны, особенно имеющие хозяйственное значение. Следует отметить, что в ряде случаев порядок перечисления видов не соответствует их филогенетическим отношениям.
С целью пояснения морфологических терминов, употребляющихся в Диагнозах, введен специальный раздел «Краткий морфологический обзор млекопитающих».
В тексте приведены списки видов млекопитающих, причем латинские названия продублированы русскими. Для многих видов это делается впервые и, вероятно, некоторые из предложенных русских названий не лучшие из возможных. Однако
важно иметь русские названия для видов млекопитающих (это сделано, например, на немецком языке—Эбле и др., Eble et al., 1954), поэтому предпринята такая попытка; в дальнейшем ряд названий может быть улучшен. К некоторым латинским названиям не удалось подобрать соответствующих русских.
Иллюстрации в основном взяты из монографий Уолкера и др. (Walker et al. 1964), Грассе (Grasse, 1955), Вебера (Weber, 1927—1928), Холла и Келсона (Hall and Kelson, 1959), Имайдзуми и Оба-ра (Imaizumi and Obara, 1966) и Кузякина (I960) Н. Н. Кондаковым изготовлены рисунки.
В работе оказали большую помощь доктор С. Андерсон, профессора В. Г. Гептнер, Б. А. Кузнецов, М. Я- Марвин, Г. А. Новиков и В. А. Попов. Пользуюсь случаем принести им глубокую благодарность.
f
ВВЕДЕНИЕ
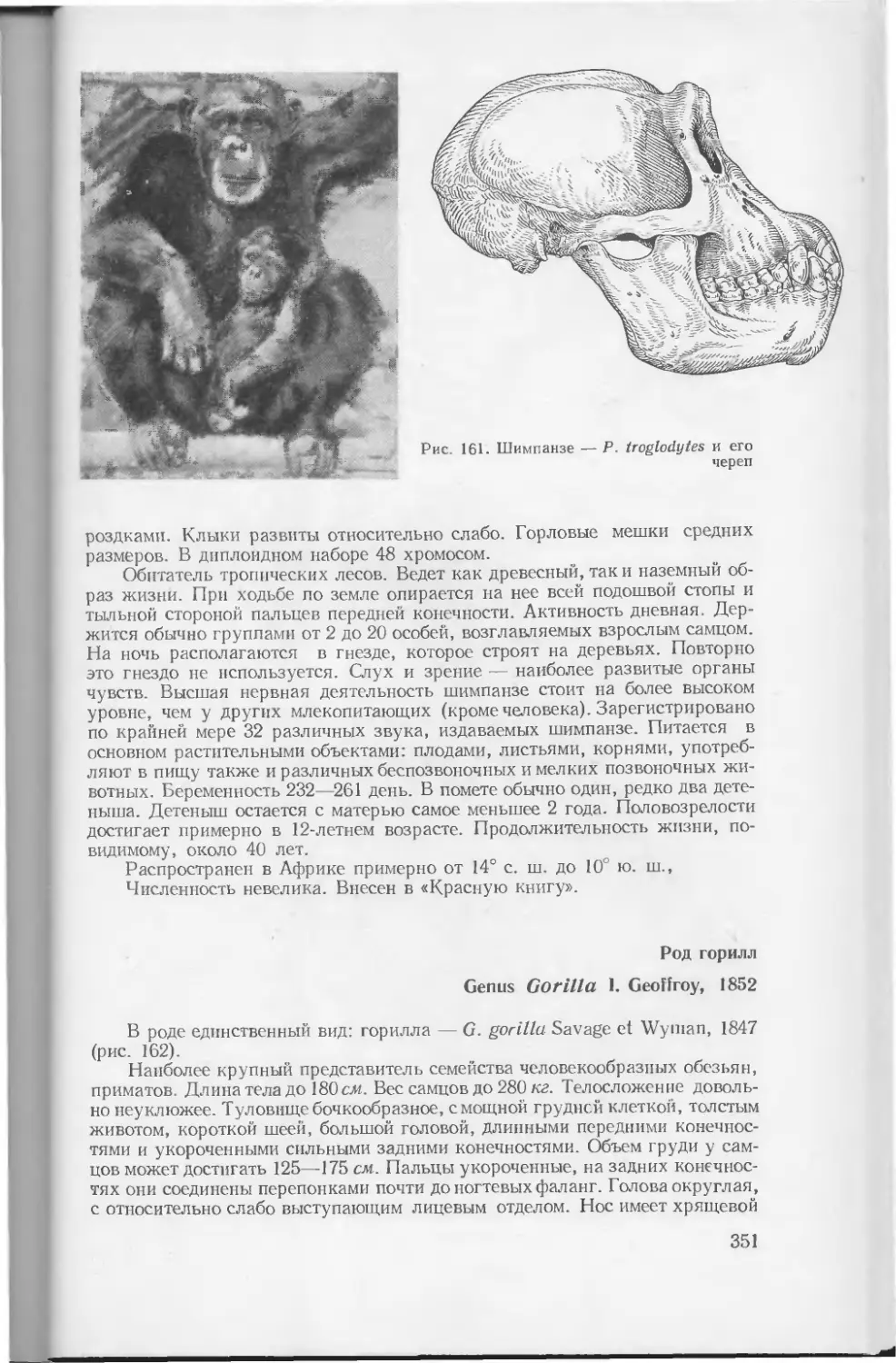
Начало классификации млекопитающих связано с именем Аристотеля (384/5—322 г. до н. э.) (рис. 1). В своем труде «История животных» Аристотель разделил всех известных ему животных на две группы: «животных без крови» и «животных, обладающих кровью», причем дополнительной особенностью последних Аристотель считает наличие спинного хребта. Животных, обладающих кровью, Аристотель делит на 5 групп: 1. Четвероногие живородные (Tetrapoda zootoka) — четвероногие, покрытые волосами. 2. Четвероногие яйцеродные (Tetrapoda ootoka, или pholidota) — большей частью яйцеродные, иногда живородные, четвероногие или безногие, часто покрытые чешуями. 3. Птицы (Ornithes) — всегда яйцеродные, крылатые, летающие, двуногие, покрытые перьями. 4. Киты (Cete) — жпвородные, дышащие легкими, безногие, водяные животные. 5. Рыбы (Ichtyes) — яйцеродные, реже живородные, чешуйчатые или голые, безногие, дышащие жабрами водяные животные. Первая группа Аристотеля соответствует млекопитающим в современном понимании. Третья и пятая группы совпадают с современными классами птиц и рыб. Земноводные и пресмыкающиеся включены во вторую группу (их нередко объединяли вместе и в значительно более позднее время). Хотя Аристотель отделил китов (четвертая группа) от млекопитающих, он не отнес их к рыбам (как это обычно делали многие более поздние исследователи), выделив в особую группу. Аристотель приводит описание многих млекопитающих: выдры, бурого медведя, лисицы, бобра, ежа, слона, двугорбого верблюда, гепарда и т. д.
В течение последующих почти 2000 лет исследования по систематике млекопитающих практически не дали ничего нового по сравнению с работами Аристотеля и в основном повторяли их. Характерными в этом отношении являются труды Конрада Геснера (Conrad’ Gesner). Крупнейший натуралист-энциклопедист, он опубликовал 5 томов по зоологии (Historia Anima-lium, 1551—1558), первый из которых посвящен млекопитающим. Перечень зверей этот ученый дал в алфавитном порядке. Описание составлено по единому плану: название животного, внешний вид, распространение, образ жизни, поведение и т. д. В отличие от Аристотеля Геснер отнес китов к рыбам. В своем сочинении Геснер дает описание сказочным животным и чудовищам, в существование которых верили многие из живших в то время ученых. Геснер был в большей степени описатель и компилятор, чем таксономист. При подготовке своей книги он использовал более 250 литературных источников (большое количество по тем временам).
Наиболее интересным исследованием, предшествовавшим работам К- Линнея и оказавшим на них влияние, являются книги Джона Рэя (John Ray) (рис. 2), в частности его книга «Synopsis Methodica Animalium Quadru-pedum et Serepentini Generis», 1963, написанная на латинском языке. Рэй разделил животных на «кровеносных» и «бескровных» (этим он сохранил деление Аристотеля). Животные «с кровью» разбиты на разделы сообразно органам дыхания. К жаберным отнесены рыбы, клеточным — все остальные. Легочные распадаются на две группы по особенностям строения сердца. У одних один желудочек — это земноводные и пресмыкающиеся в нашем по-
51
Рис. 1. Аристотель (384/5 — 322 г. до н. э.)
нимании, у^других два желудочка (птицы и млекопитающие). Животные с двумя желудочками в сердце разделены на яйцеродных (птиц) и живородных (млекопитающих). Живэродных, в свою очередь, Рэй подразделил на етвероногих и водных (китообразные). Таким образом, комплекс признаков, которыми Рэй характеризовал млекопитающих (наличие крови, легочное дыхание, два желудочка в сердце, живорождение), научно и точно определяет эту группу животных, включая такие уклоняющиеся и сомнительные для прежних систематиков группы, как летучие мыши и киты. Диагностические признаки, принятые Рэем для млекопитающих, не охватывают клоачных, но они не были известны в его время. Характеристика млекопитающих, данная Рэем, более совершенна, чем у Карла Линнея в его ранних изданиях «Systerna Naturae». Рэй первым дал достаточно четкое определение понятия вид. В книгах Рэя довольно часто можно встретить слово род, но определение рода у него остутствует. По Рэю род — скорее не таксономическая категория, а только некоторая собирательная единица, объединяющая виды в самые разнообразные по объему группы. Интересно отметить, что Рэй выделяет в своей классификации некоторые группы, которые почти полностью соответствуют ныне принятым систематическим категориям. Среди них имеется «Ungulata monochela solidipeda», включающая лошадь, осла и зебру, или «ungulata dichela», куда отнесено большинство млекопитающих, теперь называемых парнокопытными. Однако у Рэя имеются и многие ошибочно составленные группы, в частности, группа «ungulata tetrachela», объединившая носорогов, гиппопотамов, бразильского тапира и капибару.
Значение Джона Рэя в зоологии очень велико. Его можно считать наиболее крупным зоологом XVII столетия, в значительной степени наметившим пути в систематике, по которым позднее пошел Карл Линней. Если Рэй дал понятие вида, то Карл Линней (Carolus Linnaeus) (рис. 3) ввел правило номенклатуры, установил терминологию и наметил крупные таксономические подразделения млекопитающих. Первое издание его книги «Systema Naturae» появилось в 1735 г. В ней он определяет млекопитающих (не под этим названием) как животных с оволосненным телом, четырьмя ногами, самки которых живородящие и выкармливают детенышей молоком. Такой ди-
Рис. ЗЛКарл ЛиннейД1707—1778}
агноз, правда, исключает возможность присоединения к этой группе любого немлекопитающего, но зато не является всеобъемлющим и не охватывает многих млекопитающих. В 'десятом издании (1758) Линней вернулся к сущности определения млекопитающих Рэем и ввел название «Mammalia». Именно десятое издание «Systema Naturae» (1758) считают теперь отправной точкой зоологической номенклатуры вообще и номенклатуры млекопитающих в частности. Линней ввел более сложное надотрядное разделение млекопитающих, выделив в XII издании (1766) следующие группы млекопитающих: Unguicul-ata, Ungulata riMutica (последняя группа с одним отрядом Cete — киты). Unguicu-lata он подразделяет на отряд Primates, •куда входят роды людей, обезьян, лемуров, летучих мышей; отряд Bruta, включающий роды слонов, ламантинов, ленивцев, муравьедов, броненосцев; отряд
Ferae, объединивший роды тюленей,собак, кошек, виверр, куниц, медведей, опоссумов, кротов, землероек, ежей, и отряд Glires, куда вошли роды грызунов и зайцеобразных. UngulataJ разделяется Линнеем на отряд Ресога с родами верблюдов, кабарог, оленей, козлов, овец и быков и отряд Belluae с родами лошадей, гиппопотамов, свиней и носорогов.
В последующих публикациях книги Линнея, изданных Гмелиным, дается более дробное деление млекопитающих, в частности, вводится отряд Bestiae, куда включены свиньи, броненосцы, ежи, кроты, землеройки и опоссумы. Появляется большее число родсв и видов. Сбъем книги возрастает от 12 страниц в первом издании до 2400 страниц в последних. Каждая систематическая категория характеризуется короткими диагнозами.
Некоторые исследователи и до Линнея употребляли бинарную номенклатуру. Так, Джон Рэй использовал двойные названия для некоторых животных (например, Ovis strepsiceros), но как система биноминальная номенклатура была впервые использована именно Линнеем. Примечательно, что Линней вводит человека как систематическую единицу не только в класс млекопитающих, но даже в отряд приматов вместе с обезьянами, на одном уровне с последними.
В X издании своей книги Линней дает прекрасный диагноз класса млекопитающих, указывая, что млекопитающие имеют сердце с двумя желудочками и двумя предсердиями, с красной и горячей кровью; легкие, ритмически дышащие; челюсти, покрытые мышцами; самки их живородящие и секретирующие молоко; органами чувств у них служат язык, нос, глаза, уши и органы осязания; их кожный покров снабжен волосами, которые редки у тропических форм и еще более редки .у водных млекопитающих; туловище у них поддерживается четырьмя ногами, сохраняющимися и у водных форм, у которых, однако, задние конечности превращаются в хвост (это единственная ошибка во всем диагнозе, данном Линнеем).
Для диагнозов различных систематических групп Линней использовал особенности питания животных и способы добывания пищи; число зубов; характер конечностей (руки у приматов, снабженные когтями конечности у Ferae или копытами у Ресога или плавниками у Cete); способ передвижения, например лазание по деревьям у Primates, прыгание у Glires и т. д.; число и расположение сосков у самок (Primates), наличие ключицы (Primates), строение желудка (Ресога), положение ноздрей (Cete).
После Линнея появляется большое количество работ, посвященных систематике млекопитающих. Они постепенно улучшали и совершенствовали систему млекопитающих.
Бюффон и Добантон (Buffon et Daubenton) совместно опубликовали в в 1753—1767 гг. многотомное издание «Histoire des Quadrupedes», во II издании эти книги назывались «Histoire Naturelie», 1799—1805. Для каждого из описанных родов даются внешние признаки, анатомия и краткая характеристика образа жизни, а в конце перечень видов, включаемых в этот род.
Бриссон (Brisson) в книге «Regnum Animale in Classes IX. Distributum sive Synopsis Methodica», выдержавшей два издания: первое — в 1756 г. (двумя годами раньше X издания «Systema Naturae» Линнея) и второе — в 1762 г. всех животных разделяет на 9 классов. К первому он относит четвероногих с оволосненным телом и четырьмя ногами; ко второму—китообразных столым удлиненным телом, мясистыми плавниками и уплощенным горизонтальным хвостом; к остальным классам — птиц, пресмыкающихся, хрящевых рыб, настоящих рыб, ракообразных, насекомых и червей. Первый класс четвероногих Бриссон разделяет на 18 отрядов, причем названий он им не дает, проставляя только номера. Отряды Бриссон делит на секции, роды и виды. Основой диагностики родов и видов он считает число зубов. Приводит диагностические признаки для отрядов, секций, родов и видов. Более дробное деление млекопитающих, чем это сделал Линней, позволило ему избежать ряда ошибок последнего, однако и у Бриссона довольно много неестественных группировок, например в третьем отряде он объединяет слонов и моржей.
Скополи (Scopoli, 1777), следуя в большинстве случаев системе Линнея, соединяет вместе муравьедов, броненосцев и ящеров. Единственными из неполнозубых, которых Скополи не поместил с муравьедами и броненосцами, были ленивцы; их он объединил с Vespertilio, Lemur, Simia и Homo.
Эркслебен (Erxleben, 1777) вводит много новых родов и видов и приводит обзор синонимики названий млекопитающих, охватывая период от Аристотеля до 1777 г.
Блюменбах (Blumenbach) в книге «Handbuch der Naturgeschichte», 1779, затем многократно издававшейся на немецком, французском и английском языках каждый раз с новыми дополнениями (XII издание на немецком языке, 1830), выделяет 12 отрядов млекопитающих. Китообразных он рассматривает как отдельный отряд, равнозначный другим отрядам; даются краткие морфологические характеристики и географическое распространение. У Блюменбаха оказалось довольно много искусственных отрядов. Так, в шестой отряд Glires он включает многих грызунов, зайцеобразных, а также землероек, кротов, опоссума, виверр. Человека Блюменбах выделяет в особый первый отряд. Блюменбах был первым исследователем, который выделил группу промежуточного ранга между родом и отрядом, эквивалентную современным семействам. Он обособил в отдельный отряд рукокры-. лых, которых до него чаще всего соединяли с лемурами и обезьянами. В более поздних изданиях своей книги Блюменбах рассматривает утконоса в качестве отдельного рода, правда, объединяя его вместе с моржом в одну надродовую категорию.
Книга Викд‘ Азира (Vicq d‘ Azyr) «Systeme Anatomiquc des Quadrupedes», 1792 содержит описание млекопитающих на французском языке, причем автор избегает латыни даже в названиях животных и не употребляет бинарную номенклатуру. Среди млекопитающих отсутствуют китообразные, которых автор, по-видимому, считает отдельным классом. Выпал из описания и человек. Все остальные млекопитающие разделены Вик д’Азиром на 15 отрядов. Каждому из них автор дает очень краткую характеристику, в основном касающуюся внешнего вида животных.
Ж- Кювье (G. Cuvier) в «Tableau Elementaire de 1’Histoire Naturelie des Animaux», 1798, «Legons d’Anatomie Comparee», 1800, Le Regne Animal, .8
1817 уделяет значительное место классификации млекопитающих. В своем наиболее совершенном виде система млекопитающих Кювье (1817) выглядела следующим образом. Всех млекопитающих он разделил на 9 крупных категорий, в первую из которых — Bimanes отнес людей; во вторую — Quad-rumanes — обезьян и полуобезьян; в третью — Carnassiers две группы: Chiropteres рукокрылами вместе с шерстокрылами иотдельно насекомоядных (для которых Кювье вводит новый термин — Insectivores); в четвертую — Carnivores 4 группы: Plantigrades енотов, медведей, барсука, росомаху; Digitigrades различных куниц, Amphibies тюленей и моржей, Marsupiaux сумчатых;, в пятую — Rongeurs две группы: с ключицей и без ключицы, куда он отнес грызунов, а также ай-ай; в шестую — Ё dentes три группы, включающие ленивцев, броненосцев, муравьедов, трубкозубов, ящеров и однопроходных; в седьмую — Pachydermes, три группы, объединяющие слонов, носорогов, гиппопотамов, свиней, тапиров, даманов, лошадей; в восьмую— Ruminans безрогих и рогатых парнокопытных; в девятую — Cetaces две группы: ламантинов, дюгоней, морскую корову и китообразных. Кювье первым вводит понятие семейства.
Во многом типичной для своего времени работой по систематике млекопитающих явилась книга Ласепеда (Lacepede) «Tableau de Divisions, Sous-Divisions, Orderes et Genres desMammiferes», 1799. Ласепед разделяет млекопитающих на 3 раздела: четвероногих, крылатых млекопитающих и морских млекопитающих. Кроме того, он выделяет в каждом из разделов подразделы, а внутри них отряды и роды; всего он насчитывает 22 отряда. Человек не включен в систему. Объединение животных во многом ошибочно, например во втором отряде сведены вместе роды опоссумов, броненосцев и куску-сов и в третьем отряде Ласепед объединяет кенгуру и ай-ай из полуобезьян.
Де Блэнвилль (De Blainville), по-видимому, первым ввел понятие подкласса в современном смысле. В «Prodrome d’une nouvelle distribution sys-tematique du regne animale», 1816 он разделяет млекопитающих на 2 подкласса: Monodelphes, куда относит плацентарных в нашем понимании, и Didelphes, в котором объединяет сумчатых и однопроходных. Классификация, которую Де Блэнвилль употребил в своем курсе лекций в 1834 г., отличается от классификации 1816 г. тем что автор увеличил число подклассов до трех, оставив название Didelphis для сумчатых, а однопроходных выделил в особый подкласс Ornitodelphes.
Гилл (Gill) в «Оп the relations of the orders of Mammals», 1870 и «Arrangement of the Families of Mammals», 1872 следует классификации Де Блэнвилля, подразделяя.млекопитающих на 3 подкласса. Следует отметить, что в более полно составленной классификации 1872 г. Гилл вводит номенклатурные единицы Eutheria, куда помещает плацентарных и сумчатых, и Prototheria, куда относит только однопроходных. Этим Гилл подчеркивает более тесное родство между плацентарными и сумчатыми, чем между этими двумя и однопроходными.
Гексли (Huxley) в работе, посвященной эволюции позвоночных и особенно млекопитающих «Оп the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly, of the Mammalia», 1880, описывает 4 стадии, которые проходили в своей эволюции млекопитающие: Hypotheria — стадия гипотетических млекопитающих, ныне вымерших, Prototheria — стадия однопроходных, Metatheria— стадия сумчатых и, наконец, Eutheria — стадия плацентарных. По Гексли, ныне живущие эуте-рпйные звери прошли все предшествующие им три стадии. Современные однопроходные так и остановились на стадии Prototheria, а сумчатые — на стадии Metatheria. Таким образом, оригинальное понимание Гексли вышеперечисленных терминов имело чисто эволюционный смысл, но систематики взяли эти названия себе на вооружение, и в настоящее время они применяются как таксономические.
Флоуэр (Flower) в «On the arrangement of the orders and families of existing Mammalia», 1883, следуя Де Блэквиллю, делит млекопитающих на Зподкласса, однако дает им не только названия ДеБлэнвилля, но и Гексли. Из ошибочных построений системы можно отметить объединение крыланов вместе с насекомоядными в один отряд; парно- и непарнокопытных, даманов и слонов в один отряд копытных; и некоторые др. В книге, написанной совместно с Лидеккером, Флоуэр (Flower and Lydekker) «Mammals living and extinct», 1891, повторяет свою систему 1883 г.
Классификация млекопитающих, предложенная Копом (Соре) в книге «Syllabus of lectures on geology and paleontology», 1891—1898, выражает таксономические отношения между современными и ископаемыми отрядами млекопитающих. Класс млекопитающих он делит на 2 подкласса: однопроходные — Prototheria и сумчатые и плацентарные — Eutheria. Коп вводит несколько искусственных отрядов, например Bunotheria, куда он отнес ископаемых Pantotheria, Creodonta и всех насекомоядных или, например, отряд Diplarthra с подотрядами непарно-и парнокопытных и некоторые др.
Труессарт (Trouessart) в многотомном издании в 1897—1905 гг. дает полный перечень всех видов и подвидов современных и ископаемых млекопитающих. Для надвидовых категорий приведена синонимика. Указано географическое распространение.
Вебер (Weber) в «Die Saugetiere», 1904 выделяет у млекопитающих Зподкласса: Mono trema ta — однопроходные, Marsupialia — сумчатые и Мо-nodelphia — плацентарные. Во II издании, 1928 г., Вебер делит всех млекопитающих на 2 группы: 1) Prototheria, куда относит подкласс Monotrema-ta, и 2) Theria с подклассами Marsupialia и Monodelphia. Вебер выделяет 18 отрядов, большинство из которых соответствует ныне принятым отрядам за исключением 12-го отряда Subungulata, объединяющего даманов, хоботных и сирен. Отряд грызунов включает зайцеобразных и грызунов. Вебер в своей книге дает подробные морфологические диагнозы отрядов и краткие характеристики семейств и подсемейств.
Грегори (Gregory) в «The orders of mammals», 1910 приводит систему современных и вымерших млекопитающих, разделяя современных на подкласс Prototheria одним отрядом Monotremata, подкласс Theria с инфраклассом Meta theria, включающим один отряд Marsupialia и инфракласс Eutheria со всеми остальными отрядами млекопитающих. В отряд Ferae Грегори объединил ластоногими хищных, в отряд Menotyphla — тупай и прыгунчиков, в отряд Rodentia — зайцеобразных и грызунов; зубатые и усатые киты выделены в особые отряды. Грегори приводит подробные диагнозы отрядов, рассматривает их происхождение и дает исторические очерки описания отрядов. Начальный раздел его книги посвящен истории классификации класса млекопитающих.
Важной работой по классификации млекопитающих явилась книга Симпсона (Simpson) «The principles of classification and a classification of mammals», 1945. Симпсон приводит список всех отрядов, подотрядов, инфраотрядов, надсемейств, семейств, подсемейств, триб и родов млекопитающих, ныне живущих и ископаемых. Диагнозы он не дает совсем, указывая только распространение и датировку наиболее древних ископаемых остатков. Отдельно автор обсуждает эволюцию отрядов и некоторых семейств и дает исторический очерк описания различных таксономических групп. Современных млекопитающих Симпсон разделяет на два подкласса: Prototheria с одним отрядом Monotrema ta — однопроходных и Theria с двумя инфраклассами: Metatheria с одним отрядом Marsupialia — сумчатых и Eutheria со всеми остальными отрядами. Всего Симпсон выделяет 18 отрядов. Зайцеобразные отделены от грызунов и сведены в особый отряд.
Систематике млекопитающих посвящена сводка под редакцией Грассе (Grasse) «Traite de zoologie»; Tome XVII. I et II. F. Mammiferes, 1955. В двухтомной книге даны подробные диагнозы современных и ископаемых отрядов и семейств и краткие характеристики родов.
Кокрум (Cockrum) приводит в «Introduction to Mammology», 1962 специальную главу с краткими диагнозами отрядов и семейств млекопитающих, указанием на их распространение и образ жизни, а также список родов.
Двухтомная сводка млекопитающих написана группой авторов под руководством Уолкера (Walker) «Mammals of the World», 1964. В ней содержится описание всех современных отрядов и родов млекопитающих. Даются краткие морфологические характеристики и сведения по образу жизни представителей родов. Приведено большое количество оригинальных фотографий. Третий том издания включает список литературы, посвященной млекопитающим.
Диагнозы отрядов, списки родов и видов и описание некоторых видов дает Моррис (Morris) в «The mammals», 1965.
Имайдзуми и Обара (Imaizumi and Obara) запланировали дать обзор млекопитающих земного шара, но опубликовали пока только два тома, посвященных однопроходным и сумчатым (1966). В сводке, написанной на японском языке, даны характеристики современных отрядов, семейств, родов и многих видов.
Диагнозу отрядов и семейств современных млекопитающих посвящена книга Андерсона и Джонса (Anderson and Jones) «Recent mammals of the World», 1967. Даны подробные, хорошо составленные диагнозы отрядов и семейств и список родов современных млекопитающих. Указаны их распространение и образ жизни.
Описанию млекопитающих всего земного шара посвящена также работа, выполненная под руководством Гржимека (Grzimek) «Grzimeks Tierleben», 1969. Она рассчитана на широкие круги читателей. Три тома этого издания могут послужить руководством и для специалистов, так как включают описание почти всех современных млекопитающих. Следует, однако, сказать, что описание ряда групп млекопитающих, например рукокрылых, далеко не полное.
В большом числе работ описаны млекопитающие определенных зоогеографических областей или отдельных стран мира. Из наиболее значительных следует упомянуть книгу Эллермана и Моррисон-Скотта (Ellerman and Morrison-Scott) «Checklist of Palaearctic and Indian Mammals», 1946; второе издание вышло в 1966 г. Авторы приводят полный список отрядов, семейств, родов, подродов и видов млекопитающих Палеарктики и Индии, а также распространение видов и всю синонимику.
Список млекопитающих Северной Америки (до подвидов включительно) с указанием географического распространения приводят Эллиот (Elliot) в «А checklist of mammals of the North American Continent, the West Indies and the neighboring seas», 1905 и Миллер и Келлог (Miller and Kellog) в «List of North American recent mammals», 1955. Млекопитающим Северной Америки посвящено и двухтомное издание Холла и Келсена (Hall and Kel-sen) «The mammals of North America», 1959. Авторы дают диагнозы всех отрядов, семейств, родов и видов млекопитающих, населяющих Северную Америку. Приводят синонимы названий видов и их распространение. Но авторы выделяют чересчур большое число видов, подчас не заслуживающих возведения даже в ранг подвида. Так, они насчитывают 77 видов бурых медведей, тогда как на самом деле они, по-видимому, должны быть сведены в один вид.
Кабрера (Cabrera) составил список млекопитающих Южной Америки «Catalogo de los Mamiferos de America del Sur», 1, 2, 1957, 1961 с указанием их распространения.
Млекопитающим Малайзии посвящены работы Чейзена (Chasen) «А handlist of Malayasian Mammals», 1940 и Хилла (Hill) «The Robinson collection of Malayasian Mammals», 1960. Авторы приводят описание ряда новых видов и подвидов.
Тейлор (Taylor) в «Philippine land mammals», 1934 дает описание млекопитающих Филиппинских островов.
Рис. 4. Петр Симон Паллас (1741 — 1811)
По млекопитающим Австралии опубликованы работы Ле Суефа и Бар-рела (Le Souef and Burrel) «The wild animals of Australia» 1926, Иридейла и Троугтона (Iredale and Troughton) «А check-list of the mammals recorded from Australia», 1934 и Троугтона (Troughton) «Furred animals of Australia», 1947, а по Новой Гвинее и Сулавеси Лори и Хилла (Laurieand Hill) «List of land mammals of New Guinea, Celebes and adjacent islands», 1954. Фауне африканских млекопитающих посвящены исследования Аллена (Allen) «А checklist of African Mammals», 1939 и Элермана, Моррисон-Скотта и Хеймана (Ellerman, Morrison-Scott and Hayman) «Southern African mammals», 1953.
Большое число работ написано по систематике различных таксономических групп млекопитающих. (Пе
речислять их здесь не представляется возможным; основные из них приведены в списке литературы.)
История отечественной систематики млекопитающих начата академиком Петром Симоном Палласом (1741—1811) (рис. 4). В «Novae species Quadru-pedum e Glirium ordine», 1778 Паллас дает подробное описание внешних признаков многих грызунов европейской части России п Сибири, их систематического положения, внутренней анатомии, зоогеографии, причем приводит не только уже известные, нои много новых видов. Монография иллюстрирована большим числом оригинальных цветных рисунков. В 1811 г. был издан огромный труд Палласа «Zoographia Rosso-Asiatica». В связи с Отечественной войной 1812 г. атлас этого издания появился только в 1831 г. Млекопитающим посвящена первая половина I тома монографии. Паллас описал все известные ему в то время виды млекопитающих России — всего 151, многие из которых были новыми для науки. Автор приводит синонимику названий, диагноз и географическое распространение.
Академик Иван Иванович Лепехин (1840—1902) много путешествовал по европейской части России и опубликовал «Дневные записки путешествия доктора и Академии наук адьюнкта Ивана Лепехина по разным провинциям Российского государства», 1771—1804 в четырех томах. «В Дневных записках» И. И. Лепехин дает перечень видов млекопитающих тех мест, которые он посетил. Он приводит интересные сведения по образу жизнп различных млекопитающих. Однако систематике млекопитающих И. И. Лепехин уделял
немного внимания. В «Прибавлении» к «Дневным запискам» и специальных статьях им описано относительно немного видов, причем не во всех описаниях даны названия и часто отсутствуют диагнозы. И. И. Лепехин перевел на русский язык значительную часть сочинения Бюффопа «Всеобщая и частная естественная история», 1802—1804.
Академик Самуил Готлиб (Георг) Гмелин (1745—1774) путешествовал в бассейне Дона, низовьях Волги, по Кавказу и побережью Каспийского моря. Результаты путешествия опубликованы им в «Путешествии по России для исследования трех царств естества», 1771—1785. Гмелин дает описание ряда встреченных им млекопитающих, в том числе выхухоли, ушастого ежа.
Академик Иоганн Антон Гюльденштедт (1745—1781) возглавлял один из отрядов Астраханской экспедиции, организованной Петербургской Академией наук. Он обследовал верховья Волги и Дона, Северный Кавказ, до-
Рис. 5. Э. А. Эверсманн (1764—1860)
лины Куры и Риона. Особое внимание им уделено исследованию южно-русских степей.
Итоги его путешествий в обработке П. С. Палласа подведены в книге «Reisen durch Russland und im Kauk-asischen Gebirge», 1787—1791. В ней он приводит списки встреченных млекопитающих. В «Novis commentariis der Akademia» Гюльденштедт дает описание ряда новых для науки млекопитающих: крапчатого суслика, обыкновенного слепыша, перевязки.
Э. А. Эверсманн (1764—1860) (рис. 5) в известной книге «Путешествие из Оренбурга в Бухару», 1823 описал трудную и рискованную по тем временам экспедицию в Среднюю Азию. В приложении к этой книге, составленном профессором Лихтенштейном, приводится 26 видов млекопитающих, которые были отмечены во время экспедиции; 4 из них являлись новыми видами. Трехтомная «Естественная история Оренбургского края», 1840—1866 — крупнейшее описание отечественной фауны после работы Палласа.Том второй этого издания (1850) посвящен млекопитающим. Приводятся систематика, диагностические характеристики, географическое распространение и особенности образа жизни ряда видов млекопитающих.
Юлиан Симашко составил обзор известных ему отечественных млекопитающих и опубликовал «Описание всех зверей, водящихся в Империи Российской», 1851. Приведены диагнозы отрядов, семейств, родов и видов и описание образа жизни. Специальный том посвящен рисункам млекопитающих и их черепов.
Г. С. Карелин (1801—1872) много путешествовал по Оренбургскому краю, Прикаспию и Западной Сибири. В приложении к «Путешествию Г. С. Карелина по Каспийскому морю», 1883 Карелин дает список 32 видов млекопитающих с кратким указанием их географического распространения.
А. Ф. Миддендорф (1815—1894) во второй части «Путешествия на Север и Восток Сибири», 1869 дал подробное описание млекопитающих исследованных районов, их систематики и особенно образа жизни. Большое внимание Миддендорф уделяет зоогеографии.
Академик Ф. Ф. Брандт (1802—1879) написал много работ, посвященных систематике, сравнительной анатомии и зоогеографии млекопитающих. В них даны многочисленные характеристики отечественных зверей, в частности соболя, бобра, и описан ряд новых видов млекопитающих. Разработана система грызунов — они подразделяются на 3 подотряда.
Н. В. Туркин и К. А. Сатунии в книге «Звери России. Cervidac», 1902 дают перечень и синонимику родов, видов и подвидов представителей семейства оленей, их распространение и образ жизни. Большое внимание уделяется охоте и промыслу.
В конце XIX—начале XX столетия появились серьезные исследования млекопитающих Кавказа.
«Звери Кавказа» Н. Я- Динника (1847—1917) состоит из двух частей. Первая часть, 1910 включает описание китообразных и копытных, вторая, 1914 посвящена хищным. В книгах даны диагнозы семейств и описание при-
Рис. 6. С. И. Огнев (1886—1951)
знаков видов млекопитающих Кавказа и их распространение; большое внимание уделено экологии.
Первая крупная работа К. А. Сату-нина (1863—1915) «Позвоночные Московской губернии» касалась млекопитающих центра России. Затем он изучал млекопитающих Кавказа, опубликовал «Млекопитающие Кавказского края», (рукокрылые, насекомоядные, хищные), т. I, 1915. Второй том этого издания (ластоногие и часть грызунов) вышел после смерти автора в 1920 г. В систематическом порядке автор приводит описание зверей Кавказа, синонимику названий, диагнозы отрядов, семейств, родов, видов и подвидов, их географическое распространение и образ жизни. Следует отметить, что в 1914 г. К. А. Са-тунин опубликовал «Определитель млекопитающих Российской империи», вып. 1, включающий рукокрылых, насекомоядных и хищных.
Млекопитающим Сибири и Туркестана посвящены работы Н. Ф. Кащенко (1855—1935). Из его публикаций наиболее важны «Определитель млекопитающих животных Томского края», 1900 и «Обзор млекопитающих Западной Сибири и Туркестана», вып. 1. Chiroptera — Insectivora, 1905. В последней работе, помимо диагнозов видов, приведены их географическое распространение и определительные таблицы.
В эти же годы появляется книга Н. А. Смирнова (1878—1942) «Очерк русских ластоногих», 1908, в которой даны сведения по систематике ластоногих, синонимике названий, распространению, морфологическим особен-
ностям и экологии.
Огромное влияние на развитие изучения млекопитающих нашей страны оказали исследования С. И. Огнева (1886—1951) (рис. 6). Уже первая книга С. И. Огнева «Млекопитающие Московской губернии» (рукокрылые, насекомоядные, грызуны), 1913 явилась важным этапом в изучении отечественной фауны млекопитающих. Многотомное издание «Звери СССР и прилежащих стран» (с 1928 по 1950 г. вышло 7 томов) содержит описание насекомоядных, рукокрылых, хищных и ластоногих и большей части грызунов и является классическим трудом, который надолго сохранит ценность для всех исследователей млекопитающих. Через 5 лет после смерти С. И. Огнева вышел IX том издания, посвященный китообразным и написанный его учеником А. Г. Томилиным. Изложение материала едино во всех томах издания. Даются диагнозы всех видов, синонимика их названий, распространение и экология. Выделяются также подвиды млекопитающих с их синонимикой, диагнозами и географическим распространением.
Млекопитающим посвящена ценная серия, выпускаемая Зоологическим институтом Академии наук СССР, «Фауна СССР, млекопитающие». Она включает работы Б. С. Виноградова по тушканчикам (1937) и Аргиро-пуло А. И. по мышеобразным (1940); К- К. Флерова по кабаргам и оленям (1952); И. И. Соколова по копытным (1959); К. А. Гуреева по зайцеобразным (1954); И. М. Громова, Д. И. Бибикова, Н. И. Калабухова и М. Н. Мейера по наземным беличьим (1965) и Б. А. Топачевского по слепышовым (1969). Построенные по единому плану, книги этого издания содержат подробную морфологическую характеристику рассматриваемой систематической группы, сведения по экологии, филогении и полный перечень видов и подвидов с диагнозами, синонимикой названий и географическим распространением.
Важные исследования по систематике песчанок провел В. Г. Гептнер (1937), предпринявший в последние годы издание многотомной сводки «Млекопитающие Советского Союза». В этой работе приняли участие ведущие териологи СССР под редакцией В. Г. Гептнера и Н. П. Наумова. К настоящему времени выпущены том I «Копытные» (1961); том II, часть 1 «Морские коровы и хищные» (1967). Дается подробный диагноз всех отрядов, семейств, родов и видов млекопитающих Советского Союза, пересмотрены заново синонимика их названий, распространение в историческом аспекте и экология. В- И. Цалкин и В. Г. Гептнер издали «Олени СССР», 1947. В. И. Цалкин — специалист по копытным, опубликовал интересное исследование «Горные бараны Европы и Азии», 1951.
Работа С. У. Строганова «Систематика кротовых», 1948 посвящена таксономии этой группы млекопитающих, подробной диагностики каждого вида и подвида, синонимике названий и географическому распространению. Две книги того же автора из серии «Звери Сибири», 1957, 1962 дают описание насекомоядных и хищных.
Примерно в таком же плане написана книга Г. П. Адлерберга и др. «Звери Арктики», 1935. Дается описание всей фауны млекопитающих Арктики, кроме зайцеобразных и грызунов.
В монографии А. П. Кузякина «Летучие мыши», 1950 содержится описание летучих мышей отечественной фауны.
Систематическим исследованиям ластоногих посвящены работы К. К. Чапского.
В последние годы в таксономии млекопитающих развивается важное кариологическое направление. В нашей стране в этой области с успехом работают Н. Н. Воронцов и В. Н. Орлов.
Специально следует отметить важные издания определителей фауны млекопитающих СССР, способствующих развитию исследований этой группы позвоночных. К ним относятся «Определитель грызунов» Б. С. Виноградова (1933) и Б. С. Виноградова и А. И. Аргиропуло (1941); выдержавший 2 издания «Определитель млекопитающих СССР» Н. А. Бобринского, Б. А. Кузнецова и А. П. Кузякина (1944 и 1965); «Грызуны фауны СССР» Б. С. Виноградова и И. М. Громова (1952); «Хищные млекопитающие фауны СССР», Г. А. Новикова (1956) и два тома «Млекопитающие фауны СССР», составленные группой авторов (Громов И. М. и др.) из Зоологического института АН СССР под общим руководством И. И. Соколова (1963).
В последнее время появляется довольно много книг, посвященных региональной фауне млекопитающих. Среди них можно отметить книгу Б. С. Виноградова, Е. Н. Павловского и К. К. Флерова «Звери Таджикистана, их жизнь и значение для человека», 1935. Здесь дается перечень млекопитающих Таджикистана с синонимикой названий, описанием внешнего вида, указанием географического распространения и образа жизни.
Работы Б. А. Кузнецова посвящены в основном млекопитающим Средней Азии. В книге «Млекопитающие Казахстана» (1948) дается перечень млекопитающих этой республики с подробным указанием географического распространения и подвидов. Аналогично составлена книга того же автора «Звери Киргизии», 1948, в которой описание каждого вида более четко разбито на систематические заметки, распространение, места обитания, образ жизни и хозяйственное значение.
Описание насекомоядных и летучих мышей Украины приводят В. И. Абеленцев, И. Г. Пидопличко и Б. М. Попов в «Фауне Украины», т. I, 1956.
К. А. Татаринов опубликовал книгу «Звери западных областей Украины», 1956 с указанием всех видов млекопитающих зйгЙща Украины, приведением синонимики, диагноза, распространения .^‘экологии. В. Г. Гептнер в книге «Фауна позвоночных животных Бадхьйа», М956 приводит перечень млекопитающих Бадхыза, сообщает историю их описания, распространение и образ жизни. В книге И. И. Барабаш-Никифорова «Звери юго-восточной
части Черноземного центра», 1957 дается перечень млекопитающих с указанием распространения и образа жизни. Фундаментальный труд Н. К. Верещагина «Млекопитающие Кавказа», 1959 посвящен в основном истории формирования фауны Кавказа. И. Н. Сержанин выпустил 2 издания «Млекопитающие Белоруссии», 1955, 1961. Для каждого вида млекопитающих Белоруссии дается диагноз, географическое распространение и экология. Млекопитающим Волжске-Камского края посвящена книга В. А. Попова, 1960. В ней помимо перечня млекопитающих приведены данные по их морфологии, распространению, экологии, а также определительные таблицы. В 1969 г. под редакцией А. А. Слудского вышла книга «Млекопитающие Казахстана», т. I, посвященная суркам и сусликам. Приведены перечень и описание видов с указанием синонимов и подвидов, распространение и экология.
Рассчитанная на широкие круги натуралистов книга «Млекопитающие СССР» В. Е. Флинта, Ю. Д. Чугунова, В. М. Смирина содержит перечень млекопитающих СССР до вида включительно с описанием внешнего вида, географического распространения и кратких данных по образу жизни.
Большое количество монографий и статей посвящено отдельным видам млекопитающих, главным образом их экологии.
КЛАСС МЛЕКОПИТАЮЩИХ
CLASSIS MAMMALIA LINNAEUS, 1758
К классу принадлежат наиболее высоко организованные представители подтипа позвоночных. Размеры млекопитающих изменчивы: длина тела колеблется от 3,5 см у карликовой белозубки до 33 ж у голубого кита, а вес, соответственно, от 1,5 а до 120 т. Типичная форма тела млекопитающих — удлиненное туловище, опирающееся на четыре пятипалых конечности, хорошо выраженная шея, относительно небольшая голова и тонкий хвост. В зависимости от образа жизни внешний облик млекопитающих может значительно меняться.
Кожный покров млекопитающих имеет волосы (отсутствие волос— вторичное явление) и многочисленные кожные железы, сальные и потовые. Имеются млечные железы, секретом которых самки выкармливают детенышей.
Череп млекопитающих имеет два затылочных мыщелка, посредством которых он сочленяется с первым позвонком. Квадратная и сочленовная кости включаются в качестве слуховых косточек наковальни и молоточка в среднее ухо. Нижняя челюсть представлена одной зубной костью и соединяется с черепом при помощи выпуклого мыщелка, входящего в сочленовную впадину височной кости.
Зубы у большинства млекопитающих дифференцированы по форме и функции (гетеродонтные зубы) и разделяются, как правило, на резцы, клыки и щечные зубы, причем последние из них делятся на предкоренные и коренные. Зубы располагаются в особых альвеолах (текадонтная зубная система) и у многих представителей млекопитающих имеют две смены: молочную и постоянную (дифиодонтная зубная система).
Позвонки имеют плоские сочленовные поверхности (платицельный тип позвонков). Таз спереди замкнут, причем образуется симфиз лобковых костей. В стопе и кисти имеются голеностопные и предплечно-запястные сочле
нения.
Сердце у млекопитающих четырехкамерное. Сохраняется только левая дуга аорты, которая идет влево и перегибается через левый бронх. Кровь содержит безъядерные, обычно округлые, реже овальные эритроциты.
Грудная и брюшная полости разделены друг от друга грудобрюшной преградой— диафрагмой.
Головной мозг имеет хорошо развитые большие полушария и мозжечок. Большие полушария связаны между собой передней комиссурой, а в боль
шинстве случаев также и мозолистым телом.
Млекопитающие раздельнополы и рождают живых детенышей, только однопроходные откладывают яйца. Оплодотвори ’Треннее. У всех А^ётается с аллан-
высших млекопитающих серозная оболочка тоисом, дает отростки, входящие в слизис
высокая —
так называемую плаценту.
Температура тела млекопитающих п в среднем около 39° С.
и образует
Высокая организация и совершенная психика млекопитающих позволила им широко расселиться по земному шару и обитать в самых различных средах. Среди млекопитающих имеются наземные, подземные, древесные, летающие и водные формы.
По характеру питания млекопитающие могут быть разделены на узко специализированных (насекомоядных, рыбоядных, нектароядных и т. п.) и всеядных. Между этими двумя крайними типами млекопитающих можно наметить переходные формы. Для ряда видов млекопитающих характерно запасание корма.
Среди млекопитающих есть звери с дневной активностью, ночной и круглосуточной. Некоторые млекопитающие впадают в спячку на неблагоприятный период года.
Для многих млекопитающих характерны различного типа миграции, периодические и нерегулярные.
Большинство млекопитающих использует различного типа убежища для отдыха, защиты от неблагоприятных воздействий среды, выращивания потомства и т. д.
Самки млекопитающих могут приносить детенышей один раз в 2 или даже в 3 года (слоны, некоторые китообразные), один раз в году (большинство млекопитающих) или многократно (например, домовая мышь до 4—6 раз или шиншилла — до 8). Длительность беременности колеблется очень значительно: от немногих дней (у опоссума 12) до 10 и более месяцев (китообразные ) и 21—22 месяцев (слоны). Продолжительность лактации также очень изменчива у разных видов: от 10—20 дней у мелких грызунов до почти 2 лет у моржей. Число детенышей в помете может изменяться от одного (как, например, у китообразных) до 15—22 (у некоторых сумчатых, грызунов, хищных). Для огромного большинства млекопитающих характерна’забота о потомстве.
Продолжительность жизни млекопитающих весьма изменчива и колеблется от года с небольшим (некоторые грызуны) до нескольких десятков лет (приматы, хоботные).
Численность различных млекопитающих весьма неодинакова. Некоторые из них очень редки, другие, напротив, многочисленны. Численность некоторых млекопитающих может изменяться по сезонам и в различные годы.
Распространены млекопитающие всесветно: на всех материках, во всех океанах и большинстве морей земного шара. Не встречаются они лишь в центральных областях Антарктики.
Экономическое значение млекопитающих очень велико. Большинство домашних животных относятся к классу млекопитающих. Многие являются охотничьими животными. Ряд млекопитающих приносит значительный вред сельскому хозяйству, лесному хозяйству, а некоторые являются хранителями и разносчиками различных иногда очень опасных болезней.
Предками млекопитающих послужили зверозубые пресмыкающиеся (Theriodontia). Ряд ученых предполагает монофилитическое происхождение млекопитающих от высших териодонтов «иктидозавров» (Брум, Вгоош, 1932; Уатсон, Watson, 1942). Но уже в 1928 г. Симпсон высказал мнение, что по крайней мере 4 группы млекопитающих (однопроходные, многобугорчатые, триконодонты и симметродонты) произошли от териодонтов самостоятельно. Данные онтогенеза млекопитающих также свидетельствуют, по-видимому, о их монофилии (Матвеев, 1971). Эта концепция происхождения млекопитающих получила в дальнейшем широкое распространение. Она подтверждалась тем, что у зверозубых пресмыкающихся было обнаружено широкое распространение процессов параллельного формирования признаков, сходных” ^^ет^копитающими. Предположение о монофилии млекопитающих было под сомнение также и доказательством
полифилии «иктидозаврот^ (1%бмптон, 1958), которых расчленили на три-тилодонтов и диартрогнатов. А
Новейшие исследований (Татаринов, 1969) подтверждают полифилити-
ческое происхождение млекопитающих, однако не от 4—6 ветвей, как’думал Симпсон, а от 2—3. Первая из них объединяет триконодонтов, симметро-донтов, пантотериев, сумчатых и плацентарных. Вторая и третья ветви представлены, соответственно, однопроходными и многобугорчатыми. Обособление этих групп от териодонтов, по-видимому, произошло в начале триаса или даже в конце перми (Татаринов, 1969). Существует, однако, предположение о необходимости перенесения терапсид и тех пеликозавров, которые послужили предками млекопитающих, из класса пресмыкающихся в класс млекопитающих; это позволяет решить вопрос в пользу монофилии .млекопитающих (Ван Вален, Van Valen, 1960; Рид, Reed, 1960).
Таким образом, вопрос о моно- или полифилии млекопитающих окончательно не решен и требует дальнейшего исследования.
Темпы эволюции млекопитающих в различное геологическое время были не одинаковы. Наибольшее число новых семейств и отрядов млекопитающих возникло в палеоцене и эоцене. До этого времени эволюция млекопитающих шла замедленно. В юрском периоде было 6 отрядов млекопитающих: Triconodonta, Symmetrodonta, Pantotheria, Multituberculata, Docodonta, Monotremata.
Отряд Triconodonta. Максимальные размеры с домашнюю кошку. Череп удлиненный, сходный с черепом современных насекомоядных; мозговая капсула примитивная, небольших размеров. Зубы гетеродонтные, разделяющиеся на резцы, клыки, предкоренные и коренные; резцы маленькие с расширенной коронкой, клыки мощные и острые, щечные зубы островершинные; коренные с двумя или тремя буграми в одной плоскости. По характеру питания, вероятно, насекомоядные.
Отряд Symmetrodonta. Размеры небольшие, череп удлиненный. Характерны примитивные коренные зубы с 3 буграми, которые при взгляде на Жевательную поверхность зуба сверху составляют как бы вершины треугольника, причем основание этого треугольника в зубах верхней челюсти обращено кнаружи, а в нижней кнутри. Питались, по-видимому, насекомыми.
Отряд Pantotheria. Размеры мелкие, зубы гетеродонтные, верхние коренные с двумя крупными буграми, из которых один располагается с щечной стороны, а другой — со стороны языка, кроме них имеются 2 или более дополнительных бугров разных размеров; по своему строению зубы пантотериев могли служить исходным типом строения зубов сумчатых й плацентарных. Питались, по-видимому, насекомыми.
Отряд Multituberculata — отряд многобугорчатых. Максимальные размеры с сурка. По многим морфологическим признакам, а также, видимо, по образу жизни сходны с грызунами. Череп широкий и низкий, челюсти мощные. С каждой стороны в верхней челюсти 3 резца, из которых внутренний резец достигает особенно крупных размеров, а в нижней челюсти — по одному очень крупному, сходному с резцом грызунов; клыки отсутствуют, предкоренные очень изменчивы по величине и форме; коренные крупные с многочисленными бугорками на жевательной поверхности, расположенными двумя или тремя правильными рядами, проходящими спереди назад. Растительноядные млекопитающие, некоторые из них древесные.
Отряд Docodonta — гетерогенная группа примитивных млекопитающих, первоначально вследствие их сходства относимых к Pantotheria. По строению нижней челюсти, периотикума и плечевого пояса близки к однопроходным. В противоположность всем другим млекопитающим, как у некоторых зверообразных ящеров (Therapsida), задние резцы верхней челюсти расположены не на предчелюстных, а на верхнечелюстных костях (род Morganucodori).
Отряд Monotremata — однопроходные (описание см. на стр. 32).
В начале мелового периода существовали Pantotheria, Multituberculata, Symmetrodonta, Triconodonta, Monodonta. В конце мелового периода Triconodonta, Symmetrodonta, Pantotheria исчезают, но появляются 2 новые группы, по-видимому, от представителей отряда Pantotheria: сумчатых и
планцентарных млекопитающих, давших начало всей современной фауне млекопитающих (кроме однопроходных). Многобугорчатые (Multitubercu-lata) просуществовали до эоцена.
В течение всей мезозойской эры пресмыкающиеся доминировали в фауне позвоночных, в то время как млекопитающие оставались в подчиненном положении. В палеоцене происходит резкая перемена. Пресмыкающиеся теряют свое преимущественное положение, а млекопитающие становятся доминирующими на земле. В эпоху палеоцена существовало по крайней мере 16 отрядов млекопитающих: 1. Multituberculata — многобугорчатые. 2. Mar§upialia — сумчатые. 3. Insectivora — насекомоядные. 4. Primates — приматы. 5. Taeniodonta — немногочисленная группа, существовавшая в Северной Америке с нижнего палеоцена по средний эоцен; размеры довольно крупные, длина черепа до 30 см\ зубы брахиодонтные гип-селодонтные, некоторые представители имели резцы со слоем эмали лишь на
1 3
передней стороне; число резцов от — до —; клыки хорошо развиты; череп у более примитивных форм удлиненный, у остальных — укороченный; эти млекопитающие, по-видимому, обособились от насекомоядных и адаптировались к травоядному питанию.
6. Carnivora — хищные (подотряд Creodonta). 7. Condylarthra — немногочисленные примитивные растительноядные, частично всеядные копытные, имеющие сходство как с насекомоядными, так и с креодонтами; переходные формы между млекопитающими, вооруженными когтями, и копытными; существовали с нижнего палеоцена по верхний миоцен; размеры изменчивы: мелкие, величиной с ежа и крупные — с тигра; череп низкий, вытянутый, сходный с черепом хищных с сильно развитым сагиттальным гребнем. Мозг небольшой с гладкими полушариями; зубы гетеродонтные, щечные зубы , брахиодонтные, бунодонтные, буноселенодонтные или лофоселенодонтные; конечности пятипалые, первоначально стопоходящие, позже полустопохо-дящие или почти пальцеходящие. 8. Pyrotheria — малочисленная группа южноамериканских (Патагония) копытных, существовавших с нижнего эоцена до нижнего олигоцена; крупные животные величиной со слона, похожие на него, по-видимому, и своим внешним видом, в частности хоботом; зубы очень сходны с зубами древних слонов; две пары верхних и одна пара нижних резцов превращены в короткие, но сильные бивни, позади широкой диастемы (клыки отсутствуют) следуют щечные зубы с двумя поперечными гребнями на жевательной поверхности каждого из них).
9. Tillodontia — немногочисленная группа растительноядных или всеядных млекопитающих, существовавших с верхнего палеоцена по средний эоцен; размеры крупные — с медведя; череп удлиненный, низкий; зубы гетеродонтные; вторые верхний и нижний и третий верхний резцы сильно увеличены в размерах; резцы могут не иметь корней; клыки очень маленькие; щечные зубы частично брахиодонтные, частично гипселодонтные; конечности пятипалые, стопоходящие, вооруженные когтями. 10. Edentata — неполнозубые. 11. Notoungulata — многочисленные южноамериканские копытные, разнообразные по внешним признакам и размерам, существовавшие с верхнего палеоцена по плейстоцен; череп укороченный, уплощенный с широкой мозговой коробкой, массивными, широкими скуловыми дугами; зубы гетеродонтные, брахиодонтные, иногда гипселодонтные, резцы могут быть мощно развиты; в конечностях у разных видов число пальцев сокращается от 5 до 3; большинство представителей имели когти, некоторые — копыта.
12. Astrapotheria — малочисленная группа южноамериканских копытных, существовавших с палеоцена по миоцен; размеры с тапира или носорога; клыки, как правило, очень сильно развиты; задние коренные могли достигать огромных размеров; строение конечностей сходно со строением конечностей слонов. 13. Dinocerata — североамериканские и азиатские копытные, существовавшие с верхнего палеоцена по верхний эоцен; размеры от относительно мелких до крупных — с носорога; на носовых, верхнечелюстных и
теменных костях развились 3 пары массивных костных рогообразных выступа; слабо дифференцированный мозг очень мал; верхние резцы, как правило, отсутствовали, нижние имели небольшие размеры; клыки обычно сильно развиты, причем могут быть особенно удлинены; на жевательной поверхности верхних коренных зубов два сходящихся гребня; конечности пятипалые; стопа и кисть короткие, но широкие. 14. Rodentia — грызуны. 15.La-gomorpha — зайцеобразные. 16. Dermoptera — шерстокрылы.
Типичная для палеоцена фауна млекопитающих состояла из многочисленных многобугорчатых, играющих экологическую роль более поздних грызунов-, примитивных приматов, обитавших на деревьях, крупных, приспособленных для копания Taeniodonta, хищных креодонтов и наземных травоядных — Condylarthra и Pyrotheria. Все эти млекопитающие выжили в течение палеоцена, но постепенно значение их уменьшалось и, кроме некоторых приматов и креодонтов, ни один из них не дожил до конца эоцена. В эоцене происходит возникновение современной фауны млекопитающих, которая постепенно вытеснила фауну палеоцена.
Что касается места возникновения млекопитающих, то по этому поводу существуют различные мнения. В частности, Матью (Matthew, 1915) считает, что наиболее развитые представители млекопитающих должны находиться около центра происхождения, что здесь эволюция была наиболее прогрессивной и заключает на этой основе, что голарктическая область была центром эволюции млекопитающих, в то время как в тропических районах сосредотачивались примитивные формы. Другие ученые не поддержали эту точку зрения. Например, Дарлингтон (Darlington, 1957) высказал предположение о существовании центра происхождения млекопитающих в тропиках Старого Света. Его заключения основывались, в частности, на том наблюдении, что в тропиках Старого Света, особенно на Востоке, эндемиков относительно немного и есть большое количество форм, родственно связанных с фаунами окружающих областей. Исследования наиболее ранних ископаемых остатков млекопитающих показывают, что наибольшее число находок приходится на Голарктику. Однако заключение о месте происхождения млекопитающих по этому признаку вряд ли правомерно, так как этот факт может быть объяснен просто лучшей изученностью ископаемых остатков в Гол-арктике по сравнению с тропическими районами.
Класс млекопитающих подразделяют на 3 подкласса: 1. Подкласс яйцекладущих млекопитающих — Subclassis Prototheria Gill, 1872 с одним отрядом однопроходных — Monotremata Bonaparte 1838. 2. Subclassis Al-lotheria Marsch, 1880 (включает только вымерших млекопитающих)1. 3.'Подкласс живородящих млекопитающих—Subclassis Theria Parker et Haswell, 1897 с тремя инфраклассами: Infraclassis Pantotheria Simpson, 1929 (включает только вымерших млекопитающих); инфракласс сумчатых —Infraclassis Metatheria Huxsley, 1880 с одним отрядом сумчатых — Marsupialia liliger, 1811 и инфракласс плацентарных — Infraclassis Eutheria Gill, 1872.
Плацентарные включают 17 современных отрядов: насекомоядных — Insectivora Bowdich, 1821; шерстокрылов — Dermoptera liliger, 1811; рукокрылых — Chiroptera Blumenbach, 1779; приматов — Primates Linnaeus, 1758; неполнозубых — Edentata Cuvier, 1798; ящеров — Pholidota Weber, 1904; зайцеобразных — Lagomorpha Brandt, 1855; грызунов — Rodentia Bowdich, 1821, китообразных — Cetacea Brisson, 1762; хищных — Carnivora Bowdich, 1821, ластоногих — Pinnipedia liliger, 1811; трубкозубых— Tubulidenata Huxley, 1872; хоботных — Proboscidea liliger, 1811; даманов— Hyracoidea Huxley, 1869; морских коров — Sirenia liliger, 1811; непарнокопытных — Perissodactyla Owen, 1848; парнокопытных — Artio-dactyla Owen, 1848.
1 Хопсон (Hopson, 1970) предлагает рассматривать Allotheria в качестве инфракласса в подклассе Prototheria. Однопроходные сохраняются в этом же подклассе в особом инфраклассе Ornithodelphia Blainville, 1834.
КРАТКИЙ МОРФОЛОГИЧЕСКИЙ ОБЗОР МЛЕКОПИТАЮЩИХ
Цель морфологического обзора заключается в объяснении основных морфологических особенностей млекопитающих. Тело большинства млекопитающих подразделяется на голову, шею, туловище, хвост и две пары конечностей. У некоторых млекопитающих могут отсутствовать шейный отдел (китообразные), хвост (некоторые приматы, рукокрылые) или задние конечности (китообразные, сирены). Конечности большинства млекопитающих приспособлены к передвижению по земле. В зависимости от того, на какой отдел конечности опираются млекопитающие, их делят на стопоходящих (медведи), пальцестопоходящих'(кролик), пальцеходящих (большинство млекопитающих) и фалангоходящих (копытные). У некоторых млекопитающих конечности модифицированы в ласты для движения в воде (китообразные, сирены, ластоногие), в крылья для полета в воздухе (рукокрылые) или в ко-пательные лапы для движения под землей (кроты). У китообразных основным локомоторным органом служит хвост с горизонтально расположенными бесскелетными плавниками.
Кожный покров обладает волосами, имеющими в основном теплозащитную функцию, и большим количеством разнообразных желез. Волосы млекопитающих разделяются на 4 категории: вибриссы, направляющие, остевые и пуховые. Вибриссы — наиболее крупные волосы, имеющие осязательную функцию, встречаются^поодиночке или рядами на морде, а иногда на лапах и туловище с богато иннервированными волосяными влагалищами. Направляющие волосы — крупные, но значительно короче вибрисс, относительно редко 1 расположены. Остевые тоже крупные, но меньше по размерам, чем направляющие, и значительно более многочисленные, обычно имеют расширение в концевой части—-граниу. Пуховые волосы наименьшие по размсрам. ’очень тонкие, многочисленные, обеспечивают у большинства млекопитающих теплозащитные свойства меха. Как правило, волосы в коже растут пучками, причем число и расположение волос в пучке может служить'систематическим признаком. Пучки волос у многих млекопитающих располагаются определенными группами. Видоизменениями волос являются щетина и иглы, выполняющие роль пассивной защиты у дикобразов и ежей.
Обычно волосы располагаются в коже наклонно и к их сумкам подходят пучки гладких мышц, служащие для приподнимания волос. Как правило, волос состоит из сердцевинного слоя, окружающего его коркового слоя и расположенный снаружи кутикулы. Морфологическая структура волос довольно резко отличается у разных представителей млекопитающих, в частности характерные очертания клеток кутикулы волоса могут служить систематическим признаком.
У некоторых млекопитающих происходит редукция волосяного покрова, в частности у усатых китообразных остались лишь отдельные ряды вибрисс на морде. У зубатых китообразных вибриссы имеются только в эм
бриональном состоянии. Виды с почти голой кожей известны и в других отрядах млекопитающих. Отдельные участки тела могут быть лишены волос. Так, участок кожи на конце морды вокруг ноздрей обычно лишен волос (в этом случае он носит название носовое зеркало).
Окраска волосяного покрова очень разнообразна и часто является покровительственной. В определенные сезоны у млекопитающих происходит смена волосяного покрова. Обычно старые волосы выпадают и заменяются новыми.
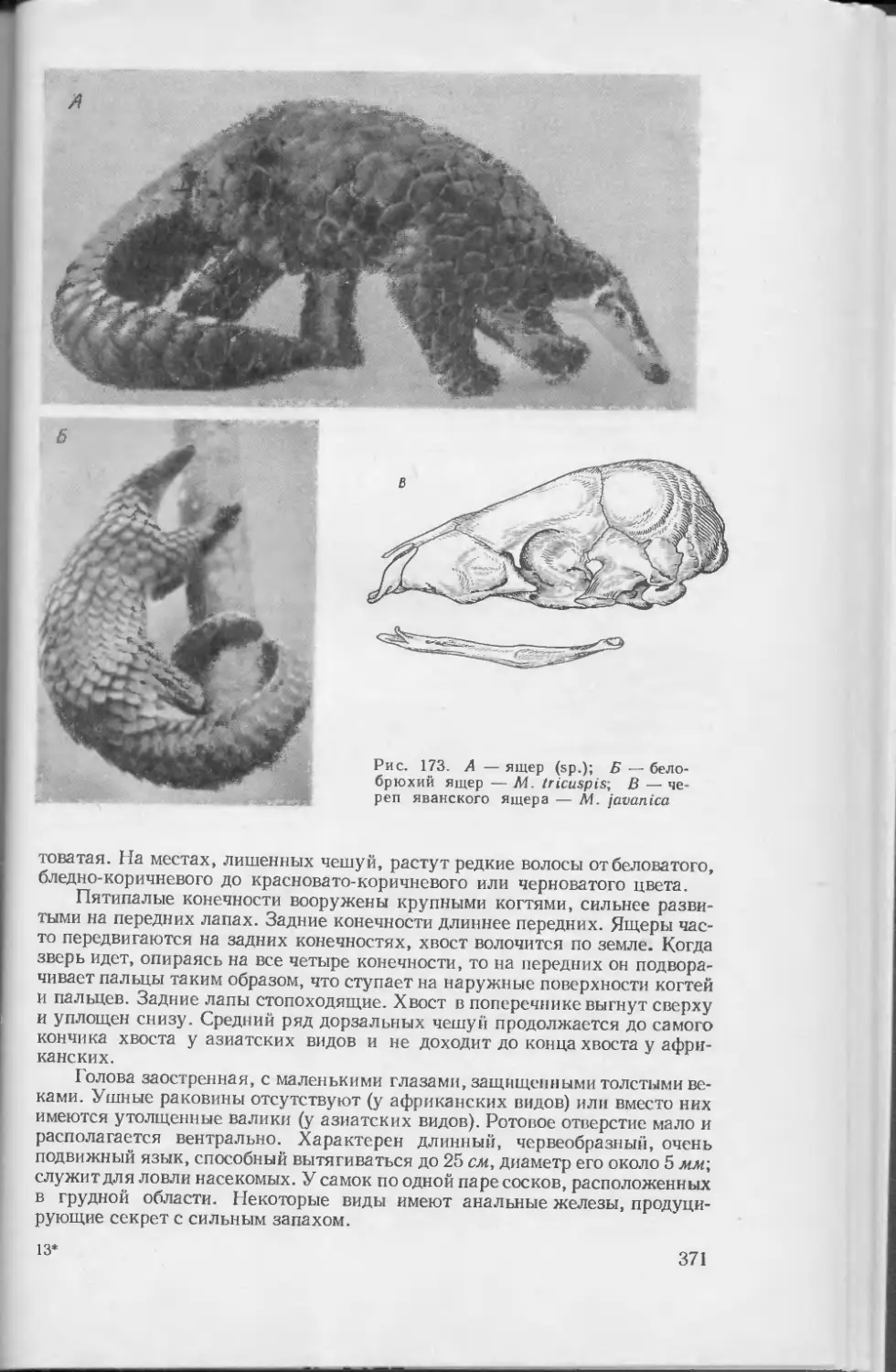
Помимо волос, у млекопитающих имеются и другие роговые образования эпидермиса: ногти, когти, копыта, рог носорога, хвостовые шпоры львов, роговые чешуи ящеров, чешуйки на хвостах и лапах многих грызунов и др. Роговой панцирь эпидермального происхождения имеется у броненосцев, причем у них под роговыми щитками располагаются еще костные щитки, являющиеся производным дермы. К роговым образованиям относятся и рога полорогих парнокопытных, а рога оленей •— производные дермы.
Млекопитающие обладают в основном двумя типами кожных желез: сальными (альвеолярными) и потовыми (трубчатыми). Протоки сальных желез, как правило, открываются в волосяные сумки, и их секрет служит жировой смазкой волос и поверхности кожи. Потовые железы открываются на поверхности кожи или в волосяные сумки. Функция потовых желез в основном терморегуляторная, а также выделительная. У некоторых млекопитающих могут отсутствовать потовые железы (например, в коже туловища грызунов) или потовые и сальные железы (китообразные). Видоизмененными потовыми железами являются млечные железы. Однако только у однопроходных они имеют трубчатое строение, все же остальные млекопитающие имеют гроздевидные железы. Наибольшего развития млечные железы достигают у самок; на поздних стадиях беременности и после родов они начинают функционировать, продуцируя молоко, которым мать вскармливает детенышей. Млечные железы, как правило, располагаются на брюшной стороне тела, и у большинства животных протоки этих желез открываются на соске; сосков нет только у однопроходных. Кроме млечных желез, у млекопитающих есть различные специфические кожные железы, представляющие собой видоизмененные потовые или сальные железы, или и те и другие. Специфические кожные железы продуцируют секрет со стойким запахом и служат для маркировки территорий, видового, индивидуального распознавания друг друга и т. д. Специфические кожные железы располагаются на подошвах лап (грызуны, хищные), между пальцами (большинство парнокопытных), перед глазами и в паху (некоторые парнокопытные), в углах рта (грызуны), около заднепроходного отверстия (грызуны, хищные). Имеются и многие другие специфические кожные железы.
Скелет разделяется на основные отделы: череп, позвоночный столб, ребра, пояса конечностей и свободные конечности.
Череп полностью окостеневает. Отдельные кости соединяются друг с другом при помощи швов, которые у некоторых млекопитающих в раннем возрасте исчезают. На наружной поверхности костей развиваются шероховатости или гребни, служащие для прикрепления мышц. Череп млекопитающих имеет узкое основание и относится к тропибазальному типу. Затылочный отдел черепа представлен одной затылочной костью (рис. 7), образовавшейся путем слияния четырех затылочных костей, ограничивающих большое затылочное отверстие. Затылочная кость несет два затылочных мыщелка, обеспечивающих подвижное сочленение черепа с первым шейным позвонком. У примитивных млекопитающих большое затылочное отверстие располагается почти под прямым углом к продольной оси черепа. У приматов с сильно развитым мозгом это отверстие перемещается книзу и находится почти в одной плоскости с основанием черепа (например, человек).
Крыша черепа образована несколькими костями. Кпереди от затылочной кости лежит непарная межтеменная кость и парные теменные. Далее расположены парные лобные кости, боковые края которых образуют нави-
сающие над глазницами надглаз-ничные отростки. Передняя часть крыши черепа занята удлиненными носовыми костями. Межтеменная кость может срастаться с теменными (жвачные парнокопытные) или с затылочной костью (хищные, приматы) или сохранять свою независимость (многие грызуны). Лобные кости' иногда срастаются в одну цельную кость (летучие мыши, приматы). В месте сочленений затылочной и теменной костей у многих млекопитающих располагается так называемый лямб-доидальный гребень. В задней части сочленения двух теменных 2 костей у многих млекопитающих располагается сагиттальный гребень. Большую часть боковой стенки мозгового черепа образует крупная височная кость; она сформировалась путем слияния чешуйчатой, каменистой и барабанной (ограничивает полость среднего уха) костей. От чешуйчатого отдела височной кости вперед отходит скуловой отросток, который соединяется со скуловой костью. Передняя часть скуловой кости прирастает к заднему краю верхнечелюстной. Образованная этими костями скуловая дуга ограничивает глазницу снаружи. У однопроходных, сумчатых, многих насекомоядных барабанная кость имеет вид коль-
Рис. 7. Череп лисицы. А — сбоку; Б — снизу:
1 — затылочная кость, 2 — большое затылочное отверстие, 3 — затылочный мыщелок, 4 — межтеменная кость, 5 — теменная кость, 6 — лобная кость 7 —• надглазничный отросток лобной кости, 8—-носовая кость, 9 — височная кость, 10 — скуловой отросток височной кости, И — скуловая кость, 12 — основная клиновидная кость, 13 —• передняя клиновидная кость, 14 —• большие крылья основной клиновидной кости, 15— малые крылья передней клиновидной кости, 16—сошник, 17 — предчелюстная кость, 18 — небный отросток предчелюстной кости, 19 — верхнечелюстная кость, 20— небный отросток верхнечелюстной кости, 21— небная кость, 22 — крыловидная кость, 23 —• слезная кость, 24 — барабанная кость, 25 — наружный слуховой проход, 26 — зубная кость, 27 — венечный отросток зубной кости, 28— место при-членения нижней челюсти, 29— резцы, 30 —клык, 31— предкоренные зубы, 32—-коренные зубы
ца. У большинства других млекопитающих она имеет вид пузыреобразного вздутия, костного слухового пузыря или барабана.
У основания мозгового черепа кпереди от затылочной кости располагается основная клиновидная кость, а впереди от нее узкая передняя клиновидная. В Стороны от основной клиновидной кости отходят отростки — большие крылья, а от передней клиновидной — малые крылья; они образуют нижнюю часть стенки глазницы. Переднюю стенку глазницы замыкает небольшая слезная кость, имеющая отверстие — слезный проток. В носовой области черепа развивается решетчатая кость со сложными пластинчатыми выростами, обонятельными раковинами. Снизу в этой области находится непарный сошник, принимающий участие в образовании носовой перегородки.
Висцеральный отдел черепа представлен несколькими костями. Спереди расположены небольшие предчелюстные кости, позади которых находятся массивные верхнечелюстные. Небные отростки этих костей вместе с небными костями образуют характерное для всех млекопитающих твердое костное небо, отграничивающее носовой проход от ротовой полости. Твердое небо скрывает лежащую впереди клиновидных решетчатую кость. К задним выступам небных костей, образующим желоб, прирастают относительно небольшие крыловидные кости. Нижняя челюсть млекопитающих представле
на только одной зубной костью. Подвижное сочленение ее с черепом осуществляется при помощи венечного отростка, соединяющегося с суставом скулового отростка височной кости. Освобожденные от функции причленения нижней челюсти квадратная и сочленовная кости у млекопитающих переходят в полость среднего уха и превращаются в слуховые косточки — наковальню и молоточек.
Подъязычный скелет состоит из толстой средней части — тела подъязычной кости и двух пар отростков, расположенных с каждой его стороны, — больших и малых рогов.
Зубная система имеет характерное строение. Зубы располагаются на предчелюстных, верхнечелюстных костях и нижней челюсти. Каждый из зубов находится в альвеоле, и по этому признаку они принадлежат к де-кадонтному типу. Большинству млекопитающих свойственна смена зубов. У детенышей зубы молочные, позднее они заменяются постоянными. Зубная система с двумя сменами принадлежит к дифиодонтному типу, а с одной сменой —• монофиодонтному. Для большинства млекопитающих характерно разделение зубов на несколько категорий, отличающихся по функции и форме. Передние зубы, или резцы (incisivi), имеют обычно долотообразную форму и служат в основном для захватывания пищи. Кзади от них располагаются крупные конические клыки (canini), удерживающие и разрывающие добычу. Щечные зубы находятся кзади от клыков и подразделяются на предкоренные (praemolares) и коренные (molares): обычно они служат для пережевывания пищи. Зубная система с различными категориями зубов называется гетеродонтной, с однородными зубами — гомодонтной.
По характеру жевательной поверхности щечные зубы могут значительно отличаться у разных млекопитающих. Наиболее примитивный характер имеют так называемые туберкулярные зубы. Эти зубы имеют на жевательной поверхности небольшие бугорки, в простейшем случае три бугорка — тритуберкулярные зубы (насекомоядные). Усложненные зубы с режущими краями называют сектор на льными, или секодонтными (у хищных). Производными туберкулярных зубов являются бунодонтные, или тупобугорчатые, зубы, обычно связанные со всеядностью; в поперечном сечении они, как правило, имеют квадратную форму и округлые низкие бугорки на жевательной поверхности (коренные зубы свиней, человека). Производными бунодонтных зубов являются селенодонтные зубы, имеющие лунчатые выемки на жевательной поверхности. Эти зубы обычно с высокими коронками и имеются у многих растительноядных животных, в частности у жвачных парнокопытных. Производными бунодонтного типа зубов являются и лофодонт-ные зубы. Бугорки жевательной поверхности этого типа зубов сливаются в поперечные гребни. Лофодонтный тип зубов распадается на билофодонтный с двумя поперечными гребнями на жевательной поверхности зуба (тапир) и полилофодонтный тип с многими поперечными гребнями (слон).
Зубы, которые не имеют корней и обладают высокой коронкой и постоянным ростом в течение жизни животного, называют гипселодонтными, а зубы с укороченной коронкой и с корнями — брахиодонтными.
Характер зубов и их число имеют большое значение в систематике млекопитающих. Для упрощения описания зубов в систематике употребляют так называемые зубные формулы. В числителе этих формул указывают число зубов верхней челюсти, в знаменателе — нижней. Сокращенно обозначают: резцы — i, клыки — с, предкоренные — р, коренные — ш. Обычно отмечают зубы одной половины челюсти; так, зубная формула обыкновенной . 3 1 4 2
лисицы: i — с — р — т— = 42.
3 13 3
Позвоночник делится на 5 отделов : шейный, грудной, поясничный, крестцовый и хвостовой. Позвонки имеют плоские сочленовные поверхности и относятся по этому признаку к платицельному типу. Шейный отдел у большинства млекопитающих содержит 7 позвонков (иногда 6—8 или 9), грудной отдел у большинства млекопитающих — от 12 до 15, число пояснич-
5
и
передняя ко-
10
Рис. 8. Плечевой пояс нечность лисицы:
1 — лопатка, 2 — гребень лопатки, 3— акромиальный отросток, 4— суставная ямка, 5 — ко-ракоидный отросток, 6 — плечевая кость, 7 — локтевая кость, 8 — лучевая кость, 9 — запястье. 10 — пястье, И — фаланги пальцев
ных позвонков изменяется от 2 до 9. У большинства млекопитающих крестцовых позвонков 4. Количество хвостовых позвонков сильно варьирует.
Ребра наибольшего развития достигают в грудной области, где каждое ребро причленяется своей головкой между двумя соседними телами позвонков, а бугорком — к нижней части поперечного отростка позвонка. В шейной области ребра рудиментарны, срастаются с телами позвонков.
Пояс передней конечности состоит из грудины, лопаток и ключиц. Грудина расчленяется на несколько отделов и заканчивается мечевидным отростком. Лопатки (рис. 8) имеют ^различную форму, гребень проходит по их верхней части и заканчивается спереди акромиальным отростком. Ключицы имеют вид относительно тонких, удлиненных костей, у некоторых млекопитающих они могут отсутствовать или быть рудиментарными. Кроме того, у однопроходных сохраняются нагрудинник, коракоид и прокоракоид.
трех отделов: плеча, предплечья и
Передняя конечность состоит из
кисти (рис. 8). В плечевом отделе имеется одна плечевая кость. Ее проксимальный отдел обладает округлой головкой, входящей в суставную ямку лопатки, а дистальный отдел — блоковидным выступом, сочленяющимся с предплечьем. Предплечье образовано локтевой и лучевой костями. Локтевая кость имеет в проксимальном отделе крупный локтевой отросток. Кисть разделяется на проксимальный отдел — запястье, промежуточный — пястье и дистальный — фаланги пальцев. В состав запястья входят кости radiale, intermedium, ulnare, centrale и пять карпальных костей. Пястье состоит из 5 пястных костей, каждый палец — из 2—3 фаланг. Первый палец многих лазящих млекопитающих может отстоять на некотором расстоянии от других пальцев — «противопоставляться» им — и образовывать таким образом хватательную лапу. Количество костей в отделах кисти варьирует; например, центральная кость (centrale) есть у грызунов и китообразных, многих неполнозубых, но отсутствует или слаборазвита у многих копытных.
Тазовый пояс состоит из двух безымянных костей. Каждая из них образована срастанием костей подвздошной, седалищной и лобковой (рис. 9). В месте схождения этих костей находится вертлужная впадина -— для сочленения с бедренной костью. У большинства млекопитающих таз закрытый: лобковые и седалищные кости правой и левой сторон срастаются друг с другом по средней линии.
Задняя конечность (рис. 10) имеет 3 отдела: бедро, голень и стопу. Бедренный отдел содержит одну кость — бедро. Проксимальный отдел бедра снабжен головкой, которая входит в вертлужную впадину таза. Бедро сочленяется с голенью коленным суставом, который защищен спереди небольшой округлой косточкой — коленной чашечкой. Голень имеет крупную большую берцовую кость и меньших размеров малую берцовую кость. Сто
па образована предплюсной, плюсной и фалангами пальцев. Предплюсна
в проксимальном отделе содержит 2 кости: внутреннюю — таранную и наружную — пяточную, направленную назад пяточным выступом. Дистальная часть предплюсны содержит 5 предплюсневых косточек. Плюсна образована 5—6 плюсневыми костями. В каждом пальце имеется до 3 фаланг.
Половой член самцов многих млекопитающих содержит кость — os
penis.
Мускулатура большинства млекопитающих наибольшее развитие получает в области конечностей и их поясов. Сильные мышцы расположены также в области шеи и спины. У средних размеров млекопитающего насчитывается 20Э—250 различных мышц. Многие млекопитающие имеют сильно развитую своеобразную подкожную мускулатуру. Характерна для млекопитающих сухожильно-мускульная диафрагма, отделяющая брюшную полость от грудной.
Головной мозг характеризуется сильным развитием передних полушарий. Поверхность полушарий у многих млекопитающих усложнена системой борозд и извилин. У примитивных млекопитающих вторичный мозговой
свод — неопаллиум развит относительно слабо. Он достигает наивысшего развития у плацентарных млекопитающих. Обонятельные доли мозга обычно хорошо развиты. По степени их развития различают макросматический мозг с сильным развитием обонятельных долей и микросматический мозг
со слабым их развитием. Китообразные имеют аносматический мозг с редуцированными обонятельными долями.
Высокого развития достигают также зрительные и “слуховые центры, сосредоточенные в среднем мозге в области четверохолмия. Кроме того, в серой коре полушария образуются вторичные ассоциативные высшие мозговые центры зрения и слуха (затылочные и височные доли полушария). Мозжечок млекопитающих сильно развит, в нем дифференцируются новые боковые доли — полушария мозжечка, которые выполняют функцию координации вторичных
Рис. 10. Задняя конечность лисицы:
1 — бедренная кость, 2 — коленная чашечка, 3 — большая берцовая кость, 4 — малая берцовая кость, 5 — предплюсна, 6 — плюсна, 7 —фаланги пальцев, 8 — таранная кость. 9 — пяточная кость
Рис. 9. Тазовый пояс лисицы (вид снизу):
1 — поцвздошная кость, 2 — седалищная кость, 3 — лобковая кость, 4 — вертлужная впадина
двигательных центров коры полушарий переднего мозга. От головного мозга млекопитающих отходят 12 пар головных нервов.
Органы обоняния большинства млекопитающих высоко развиты. Сложно устроенные обонятельные раковины распадаются на несколько групп. У некоторых млекопитающих (однопроходных, сумчатых, грызунов, копытных) около нижней стенки носовой полости располагается пара колбасовидных полостей, так называемый Якобсонов орган.
Орган слуха достигает высокого совершенства. У большинства млекопитающих есть наружное ухо, или ушная раковина, образованная хрящами и приводящаяся в движение специальной мускулатурой. Ушная раковина обрамляет наружный слуховой проход, который изнутри замкнут барабанной перепонкой. Вовнутрь от барабанной перепонки располагается полость среднего уха, в которой находятся слуховые косточки, молоточек, наковальня и стремечко, закрывающее овальное окошко внутреннего уха. Во внутреннем ухе хорошо развита улитка, к которой подходит слуховой нерв, принимающий участие в образовании кортиева органа и воспринимающего звуковые раздражения чувствующих клеток. Улитка примыкает к круглому мешочку, соединенному с овальным мешочком, от которого отходят 3 полукружных канала, расположенных во взаимно перпендикулярных плоскостях и наполненных эндолимфой с мельчайшими известковыми кристаллами. Полукружные каналы воспринимают изменение положения тела в пространстве.
Глаза имеют сложное строение. Наружная оболочка глаза — склера • образует в передней части прозрачную роговицу. Кнутри от нее располагается сосудистая оболочка, переходящая впереди в ресничное тело и радужную оболочку, которая окружает зрачок. Иногда сосудистая оболочка образует блестящий слой зеркальца (тапетум), способный в темноте отражать лучи света и создающий, впечатленце свечения глаз. Далее кнутри располагается пигментная оболочка и сетчатка с зрительными клетками. К зрачку сзади прилегает хрусталик, который прикреплен связками к мускульному ресничному телу. Впереди от хрусталика располагается передняя глазная камера с водянистой влагой, а сзади — задняя глазная камера с так называемым стекловидным телом. Глаз у большинства млекопитающих защищен двумя непрозрачными веками: верхним и нижним. Кроме того, у большинства млекопитающих имеется прозрачная мигательная перепонка. В переднем углу глаза располагается гардерова железа, выделяющая жирный смазывающий секрет, здесь же находится слезная железа. Аккомодация глаза осуществляется изменением формы хрусталика. Глаза у некоторых млекопитающих могут быть в разной степени редуцированы.
Пищеварительная система имеет целый ряд особенностей. Характерны подвижные губы, предротовая полость между губами и щеками с одной стороны и зубами — с другой. У некоторых грызунов и обезьян предротовая полость продолжается латерально и кзади в виде защечных мешков. Кнутри от зубов находится ротовая полость, величина которой у различных млекопитающих сильно варьирует. На дне ротовой полости расположен подвижный язык, богато снабженный железами; он имеет на своей поверхности различные сосочки, причем некоторые несут вкусовую функцию. Форма и функция языка могут значительно изменяться у разных млекопитающих. Помимо обычных слизистых желез, в ротовой полости находятся крупные слюнные железы: подъязычная, подчелюстная, околоушная (рис. 11) и заднеязычная. Ротовая полость с передней стороны обрамлена зубами.
Пищевод (см. рис. 11) довольно однотипен у различных млекопитающих и представляет собой проводящую пищу трубку с мускульными стенками. Внутренняя поверхность пищевода выстлана многослойным эпителием. Желудок (см. рис. 11) млекопитающих сильно варьирует у различных видов: он может быть простым, однокамерным или состоять из нескольких, иногда многих отделов, как у некоторых парнокопытных и китообразных. Обычно в желудке различают расширенную начальную часть, кардиальную, и су-
женный конец —пилорическую. В стенках желудка обычно располагаются многочисленные железы, продуцирующие желудочный сок. Кишечник млекопитающих (см. рис. 11) делится на тонкие кишки и толстые. Передняя часть тонких кишок образована V-образной двенадцатиперстной кишкой; в ее петле, как правило, располагается поджелудочная железа. На месте перехода тонких кишок в толстые находится слепая кишка, имеющая обычно червеобразный отросток. Задний отдел толстых кишок образован прямой кишкой. Длина отделов кишечника и всего кишечника в целом сильно отличается у различных млекопитающих в зависимости от их питания.
Печень (см. рис. 11) обычно имеет несколько лопастей, располагается в передней части брюшной полости и прикрепляется складкой брюшины к задней стенке диафрагмы. В печени может быть тонкостенный желчный пузырь (иногда он отсутствует), соединенный желчным протоком с полостью двенадцатиперстной кишки.
Дыхательная система (см. рис. 11) представлена легкими и дыхательными путями (гортань,
трахея, бронхи). Гортань образована хрящами — надгортанником, щитовидным, перстневидным и черпаловидными. От гортани отходит трахея, или дыхательное горло, состоящая из многочисленных неполных хрящевых колец. Трахея делится на 2 бронха, которые подходят к основанию легких. Бронхи в области легких распадаются на большое количество все уменьшающихся по калибру ответвлений — бронхиол, слепые концы которых образуют легочные альвеолы. Форма и величина легких различны у разных млекопитающих.
Кровеносная система имеет два круга кровообращения; сердце разделяется на 4 камеры — правое и левое предсердия и правый и левый желудочки. Малый круг кровообращения начинается с легочной артерии, которая отходит от правого желудочка и делится на две ветви, направляющиеся в правое и левое легкое. Легочные вены, идущие из легких, впадают в левое предсердие. Артериальная система большого круга представлена левой дугой аорты (см. рис. 11), продолжающейся в виде спинной аорты вдоль всего позвоночного столба. От дуги аорты и спинной аорты отходят многочисленные артерии к различным органам и частям тела. Некоторые артерии (или вены) распадаются на большое число мелких сосудов, связанных между собой анастомозами, из которых кровь вновь собирается в одну артерию (вену). Такая сеть мелких сосудов называется «чудесной сетью». Одна из функций «чудесной сети» заключается в накапливании — депонировании большого количества крови. Венозная кровь от головы собирается
по яремным венам, которые, сливаясь с подключичными, образуют соответственно правую и левую передние полые вены, впадающие в правое предсердие. Из задних конечностей венозная кровь собирается в подвздошные вены, которые сливаются с хвостовой и образуют непарную заднюю полую вену, направляющуюся прямо к сердцу и впадающую в правое предсердие. По пути задняя полая вена принимает ряд венозных сосудов от внутренних органов и проходит через печень (кровь из нее в сосуды печени не попадает). При выходе из печени в заднюю полую вену впадают мощные печеночные вены. Воротная система почек у млекопитающих отсутствует. Воротная система печени образована только одним сосудом — воротной веной печени, в которую вливается вена, несущая кровь от пищеварительного тракта.
Мочеполовая система. Парные бобовидные метанефрические почки, расположены в поясничной области по бокам от позвоночника (см. рис. 11). Внутреннее (мозговое) вещество почки расположено в виде пирамид, которые своими вершинами, так называемыми почечными сосочками, обращены в вогнутую сторону почки, где находится почечная лоханка с начинающимся от нее мочеточником. От почек начинаются мочеточйики, впадающие в мочевой пузырь, протоки которого открываются в мочеполовой канал.
Семенники самцов представлены овальными телами, находятся или в мошонке (скротальные семенники), или под кожей в паховой области (ин-гуинальные семенники), или, наконец, в брюшной полости (абдоминальные семенники). В передней части семенника располагается придаток семенника, от которого отходит семяпровод, впадающий в мочеполовой канал. Мочеполовой канал проходит внутри полового члена.
У парных яичников самок (см. рис.н11) расположены воронки яйцеводов, переходящие в тонкие парные яйцеводы, которые открываются в матку. Матка переходит в удлиненное влагалище/'Строение половых органов самок весьма варьирует у различных млекопитающих. Яйцеводы однопроходных переходят в раздельные матки, которые независимо друг от друга, без образования влагалища открываются в мочеполовой синус. У сумчатых часто два влагалища. Слоны, некоторые грызуны имеют двойную матку с парным отверстием во влагалище; рукокрылые, часть хищных, некоторые грызуны — двураздельную (обе матки сливаются в нижнем отделе, но перегородка между ними сохраняется на всем остальном протяжении). Двурогой маткой обладают китообразные, копытные, насекомоядные: на значительном протяжении матка имеет единую полость и лишь в верхнем отделе разделяется на два рога. Наконец, простая матка (одиночный мешкообразный орган) свойственна приматам и некоторым рукокрылым.
Процесс овуляции у самок млекопитающих может быть спонтанным — регулярно повторяющимся через определенный промежуток времени, или провоцированным — обусловленным половым актом.
После овуляции на месте лопнувшего фолликула образуется желтое тело овуляции. Если происходит оплодотворение яйцеклетки, то желтое тело овуляции превращается в особый орган внутренней секреции — желтое тело беременности. У некоторых млекопитающих яйцеклетка после оплодотворения задерживается в развитии и не имплантируется в слизистую оболочку матки. Наступает диапауза (латентный период), продолжительность которой у разных видов неодинакова.
После окончания диапаузы происходит имплантация бластоцисты в стенку матки и ее нормальное развитие. Для^эмбрионов плацентарных млекопитающих и некоторых сумчатых свойственно развитие плаценты — временного приспособления, с помощью которого зародыш может питаться за счет материнского организма. Плацента образуется срастанием аллантоиса и хориона со стенками матки.
В зависимости от расположения ворсинок на хорионе различают несколько типов плацент. Диффузная плацента имеет ворсинки, равномерно рассеянные по всему хориону (она свойственна неполнозубым, китообразным, полуобезьянам, большинству копытных). В дольчатой плаценте вор
синки собраны в группы, а группы рассеяны по всей поверхности хориона (большинство жвачных). Для кольцевой, или зональной, плаценты характерно, что ворсинки хориона собраны в широкий пояс, охватывающий поперек зародыш (некоторые хищные, хоботные, ластоногие, сирены). В дис-коидальной плаценте ворсинки собраны в группы, напоминающие по форме диск (насекомоядные, грызуны, летучие мыши, обезьяны, большинство неполнозубых). Если при родах плацента отпадает с частью оболочки матки, то такую плаценту называют отпадающей. У некоторых млекопитающих роды происходят без отпадания оболочки матки — неотпадающая плацента. к неотпадающим плацентам относятся диффузная и дольчатая, в некоторых случаях — зональная и дискоидальная, а к отпадающим — большинство зональных и дискоидальных плацент.
По характеру связи плода с маткой матери плаценты могут быть эпителиохориальными (эпителий хориона и матки соприкасается), десмохориаль-ными (ворсинки хориона проникают в железы матки, вызывают разрушение покрова желез, и эпителий хориона соприкасается с соединительной тканью слизистой матки, в которой проходят сосуды), эндотелиохориальными (ворсинки хориона вызывают разрушение слизистой оболочки матки, и эпителий ворсинок непосредственно прилегает к эпителиальным стенкам кровеносных сосудов матки) и гемохориальными (ворсинки хориона разрушают слизистую оболочку матки, в ней образуются полости, наполненные кровью матери).
Подкласс яйцекладущих млекопитающих
Subclasses Prototheria Gill, 1872
Подкласс включает единственный отряд: однопроходных —• Monotre-mata Bonaparte, 1838 (для обоих систематических групп приводится общая х а ра ктеристика).
ОТРЯД ОДНОПРОХОДНЫХ
ORDO MONOTREMATA BONAPARTE, 1838
Немногочисленная группа наиболее примитивных из ныне живущих млекопитающих. Самки откладывают 1 или 2, редко 3 яйца телолециталь-ного типа (характерно большое содержание желтка, главная масса которого помещается у одного из полюсов яйца). Вылупление детенышей из яиц происходит при помощи особого яйцевого «зуба», образующегося на небольшой яйцезубной кости (os carunculae). Выведшиеся из яиц молодые животные вскармливаются молоком. На период размножения на брюхе самки может образовываться выводковая сумка, в которой вызревает отложенное яйцо.
Размеры однопроходных невелики: длина тела 30—80 см. Имеют тяжелое телосложение, короткие стопоходящие конечности, специализированные для копания или плавания. Голова небольшая, с вытянутым «клювом», покрытым роговой оболочкой. Глаза маленькие, наружные ушные раковины едва заметны или отсутствуют вовсе. Тело покрыто грубыми волосами и колючками или мягким густым мехом. Вибриссы отсутствуют. В пяточ-
Рис. 12. Стопа: А — самца австралийской ехидны — Т. aculeatus-, Б — самца проехидны Бруийна—Z. bruijnc, В — утконоса — О. anatinus. Хорошо видна роговая шпора
Рис. 13. Череп молодого утконоса — О. anatinus. А — снизу; Б — сбоку: / — прсдч?люстная кость, 2 — предсошпик, 3 — верхнечелюстная кость, 4— носовая кость, 5—небная кость, 6 — слезное отверстие, 7 — переднелобная кость. 8— задчелоЗная кость. 9— теменная кость, /О—вэрхнезатылочная косгь, 11— боковая затылочная кость. 12 — основная затылочная кость, 13—каменистый отросток. 14 — чешуйчатая кость, 15-^ крыловидная кость. 16 — жевательная роговая пластина, 17 — крылоклиновидная кость
ной области задних конечностей имеется роговая шпора, особенно сильно развитая у самцов (рис. 12). Шпора пронизана каналом — особым протоком, связанным с так называемой голени ой железой, функция которой не совсем ясна. По-видимому, она имеет какое-то значение в размножении. Высказывается также предположение (малоубедительное), что секрет го-ленной железы ядовит и шпора служит оружием защиты. Молочные железы трубчатые. Настоящих сосков нет и выводные протоки желез открываются отдельно друг от друга на двух железистых полях брюха самки.
Череп уплощенный (рис. 13). Лицевой отдел вытянут. Хрящевой череп и соотношение костей в крыше черепа в известной степени сходны с пресмыкающимися. Крыша черепа с переднелобпыми и заднелобными костями; наличие этих костей в крыше черепа — единственный случай среди млекопитающих. Барабанная кость имеет вид уплощенного кольца, не срастающегося с черепом. Костный слуховой проход отсутствует. Молоточек и наковальня в среднем ухе срастаются между собой и имеют длинный отросток (processus folii). Слезная кость отсутствует. Скуловая кость сильно уменьшена в размерах или отсутствует. Только у однопроходных среди всех млекопитающих есть предсошник (praevomer). Межчелюстная кость имеет отросток, подобный таковому рептилий (processus ascendus); это единственный случай среди млекопитающих. Сочленовная ямка для нижней челюсти образована чешуйчатой костью. Нижняя челюсть всего с двумя слабо выраженными отростками — венечным и угловым.
Рис. 14. Плечевой пояс ехидны — Т. аси-leatus. А — спереди; Б — сбоку:
I — лопатка, 2 — ключица, 3— ребра, 4—коракоид, 5—прокоракоид, 6 — надгрудиик, 7 — сочленовная поверхность, 8 — грудина
Зубы имеются только у молодых животных или совершенно отсутствуют. Форма зубов в известной степени напоминает форму зубов мезозойских Microleptidae.
Для скелета пояса передних конечностей характерны уникальные среди млекопитающих коракоид (coracoideum) и прокоракоид (procoracoi-deum) (рис. 14). В наличии этих костей проявляется сходство плечевого пояса однопроходных с плечевым поясом рептилий. Грудина с большим надгрудинником (episternum). Ключица очень крупная. Лопатка без гребня. Плечевая кость короткая и мощная. Локтевая кость значительно длиннее лучевой. Запястье короткое и широкое. Передние и задние конечности пятипалые. Пальцы оканчиваются когтями. В тазовом поясе самцов и самок имеются так называемые сумчатые кости (ossa marsupialia), причленяющие-ся к лобковым. Функция их неясна. Симфизис тазовых костей сильно удлинен. Проксимальный отдел малой берцовой кости с большим уплощенным отростком (регопесгапоп). Позвоночный столб состоит из 7 шейных, 15—17 грудных, 2—3 поясничных, 2 крестцовых, 0—2 копчиковых и 11—20 хвостовых позвонков.
Все тело покрыто сильно развитым слоем подкожной мускулатуры (рап-niculus carnosus). Только в районе головы, хвоста, конечностей, клоаки и млечных желез подкожная мускулатура не развита. Нижняя челюсть имеет musculus detrahens, прикрепляющийся на ее внутренней стороне; это единственный случай у млекопитающих. Гортань примитивна и не обладает голосовыми связками. Мозг в общем крупный, имеет черты строения млекопитающего, однако сохраняет ряд признаков рептилий. Большие полушария с многочисленными, иногда малочисленными бороздами. Структура коры головного мозга примитивна. Обонятельные доли очень крупные. Мозжечок только частично прикрыт большими полушариями. Мозолистое тело (corpus callosum) отсутствует; оно представлено только в виде commissura dorsalis. Сильно развито обоняние. Якобсонов орган развит хорошо. Строение органов слуха примитивно. Глаза с мигательной перепонкой или без нее. Склера имеет хрящ. Сосудистая оболочка тонкая. Musculus dilatatorius и Musculus ciliaris отсутствуют. Сетчатка не имеет сосудов.
Слюнные железы маленькие или большие. Желудок простой, без пищеварительных желез, что является единственным случаем у млекопитающих.
Его функция, по-видимому, заключается в хранении пищи аналогично' функции зоба птиц. Пищеварительный тракт делится на тонкие, толстые кишки, есть слепая кишка. Открывается кишечник в клоаку, имеющуюся у обоих полов. Печень многодольчатая, с желчным пузырем.
Сердце однопроходных имеет строение, характерное для млекопитающих, однако сохраняет и некоторые рептилиеподобные признаки, заключающиеся, например в том, что правое атриовентрикулярное отверстие снабжено только одним клапаном.
Средняя температура тела ниже, чем у других млекопитающих (утконоса в среднем 32,2° С, ехидны — 31,Г С). Температура тела может варьировать между 25° и 36° С.
Мочевой пузырь, в который впадают мочеточники, открывается в клоаку. Яйцеводы впадают в клоаку порознь (нет ни влагалища, ни матки). Семенники располагаются в брюшной полости. Penis укреплен на вентральной стенке клоаки и служит только для выведения спермы.
Однопроходные обитают в лесах разного типа, в степях, поросших кустарником, на равнинах и в горах, поднимаясь до 2,5 тыс. м над уровнем моря. Они ведут полуводный (утконос) или наземный (ехидны) образ жизни; активность сумеречная и ночная; питаются насекомыми и водными беспозвоночными. Продолжительность жизни до 30 лет.
Распространены в Австралии, Тасмании, Новой Гвинее.
В отряде 2 семейства:
ехидн — Tachyglossidae Gill, 1872
утконосов — Ornithorhynchidae Burnett, 1830.
Современные однопроходные по своим признакам по сравнению со всеми другими современными млекопитающими наиболее близки к пресмыкающимся. Они, однако, не являются предками сумчатых или плацентарных млекопитающих, а представляют отдельную специализированную ветвь в эволюции млекопитающих.
Ископаемые остатки представителей отряда однопроходных известны только из Австралии. Наиболее древние находки датируются плейстоценом и существенно не отличаются от современных форм. Существуют две возможные теории, объясняющие происхождение однопроходных. Согласно одной из них, однопроходные развивались независимо и в полной изоляции от других млекопитающих, начиная с раннего периода возникновения млекопитающих, возможно, с их рептилеподобных предков. По другой теории, группа однопроходных обособилась от древних сумчатых и приобрела свои черты благодаря специализации, сохранив ряд признаков, свойственных сумчатым, подверглась дегенерации и, возможно, в известной степени возврату к формам предков (реверсии). Первая из теорий представляется более правдоподобной. Значительные отличия в морфологии между ехиднами и утконосом возникли за относительно короткий промежуток времени — начиная с верхнего эоцена. Ехидны являются вторично наземными млекопитающими, отделившимися от древних водных утконосов (Грегори, 1947).
СЕМЕЙСТВО ЕХИДН
FAMILIA TACHYGLOSSIDAE GILL, 1872
Наземные однопроходные, длина тела до 80 см. Самцы крупнее самок. Тело приземистое, шея короткая, голова с длинным цилиндрическим «клювом» (рис. 15, 16). Ротовое отверстие очень маленькое, наружные ушные раковины отсутствуют или очень небольшие. Глаза имеют всего 8—9 мм в диаметре и лишены мигательной перепонки. Хвост сильно укорочен. Короткие, но сильные конечности приспособлены для копания. Передние лапы обладают мощными изогнутыми когтями. Второй палец на задних конечностях вооружен особенно длинным когтем, которым зверь чистит свою шерсть. Кожа имеет значительную толщину. В средних и глубоких частях
Рис. 15. Австралийская ехидна — Т. аси-leatus
ее проходят пучки поперечнополосатых мышц. Потовые железы отсутствуют. Сальные железы располагаются только у волосяных сумок. В коже имеются иглы и волосы; отсутствуют иглы только на брюхе. Длина игл примерно 6—8 см, в поперечнике они округлые, снаружи гладкие, с толстым корковым слоем. Гладких мышц в коже нет. Поднятие игл, очевидно, осуществляется за счет сокращения поперечнополосатой подкожной мускулатуры. Волосы у ехидны грубые, без видимого сердцевинного слоя. Язык очень длинный, червеобразный, в длину достигает 25 см и служит для ловли насекомых. Его задняя часть покрыта роговыми зубчиками, которые в совокупности с жесткими небными складками образуют аппарат для перетирания пищи. Слюнные железы сильно развиты (особенно подчелюстная). У самок в период размножения образуется брюшная выводковая сумка, куда открываются два молочных железистых поля.
Зубы совершенно отсутствуют. Они не обнаружены даже на эмбриональных стадиях развития.
Мозг с большим количеством борозд, особенно у проехидн. Обоняние и слух развиты хорошо.
Обитатели лесных, парковых и кустарниково-степных ландшафтов, в горы поднимаются до 2.5 тыс. м над уровнем моря. Ведут одиночный образ жизни. Активны в сумеречное и ночное время. День проводят в убежищах, пустотах под камнями и корнями деревьев или в норах, которые они выкапывают сами. Копают они очень хорошо и быстро. Питаются различными насекомыми, особенно муравьями и термитами. Поедают также улиток и червей.
Размножаются раз в году. Период спаривания приходится на конец июня, июль. Откладка яиц (1 или 2, редко 3) происходит с конца августа по начало сентября. Яйцо сразу же помещается в сумку. Яйца покрыты кожистой оболочкой, достигают в длину 14—17 мм и в ширину 13—15 мм. Через 7—10 дней после откладки из яйца вылупляется детеныш, примерно 12 мм длины. Детеныш остается в сумке в течение 6—8 недель. За 43 дня детеныш увеличивает свой вес с 241 до 850 г. Питается детеныш молоком, которое выделяется на железистых полях в сумке. Молоко содержит 15— 19% жира. К выходу из сумки детеныш достигает веса примерно 400 г; глаза у него открываются примерно в возрасте 10 недель. Половая зрелость наступает в годовалом возрасте. Длительность жизни составляет около 30 лет.
Ареал охватывает Тасманию, Австралию и Новую Гвинею.
В семействе 2 рода:
ехидн — Tachyglossus liliger, 1811;
проехидн — Zaglossus Gill, 1877.
Род ехидн
Genus Tachyglossus liliger, 1811
Длина тела до 53 см. Хвост представляет собой небольшой выступ. Вес 2,5—6 кг:
Конечности укороченные, вооруженные сильными уплощенными когтями (см. рис. 12). «Клюв» составляет примерно половину длины головы, прямой или слегка изогнут кверху (см. рис. 15). Ушная раковина отсутствует. Колючки имеют желтую окраску в основании и черную на вершине, реже полностью желтую. Число хромосом в диплоидном наборе у австралийской ехидны 62—63.
Обитают в скалистых и холмистых участках, лесах и песчаных равнинах. Могут небыстро бегать; спасаясь от врага, зарываются в землю или сворачиваются в колючий шар. На холодный сезон впадают в спячку продолжительностью до 4 месяцев.
Распространены в Австралии, Новой Гвинее, Тасмании и на мелких островах в Бассовом проливе.
В роде 2 вида:
австралийская ехидна — Т. aculeatus Shaw et Nodder, 1792 (Австралия и Новая Гвинея);
тасманийская ехидна — Т. setosus Geoffroy, 1803 (Тасмания и острова в Бассовом проливе).
Род проехидн
Genus Zaglossus Gill, 1877
Проехидны крупнее ехидн, достигают длины 80 см. Длина хвоста 5—7 см. Взрослые животные имеют вес до 10 кг. Рострум («клюв») занимает ’’3 длины головы и изогнут вниз (рис. 16). Конечности, более высокие, чем у ехидн. Задние конечности пятипалые (см. рис. 12), передние трехпалые. I и V пальцы передних конечностей представлены снаружи в виде бугорков. Все пальцы вооружены когтями. Имеются небольшие наружные ушные раковины. Колючки короче, чем у ехидн, окраска от почти белой до черной.
Рис. 16. Проехидна Бартона — Z bartoni
Рис. 17. Утконос — О. anatinus
Коричневатые или черные волосы иногда полностью скрывают под собой иглы на спине. Окраска на голове более светлая, чем на туловище.
Обитают в разных местах от низменностей до высокогорья. Часто встречаются в каменистых участках. Образ жизни сходен с образом жизни ехидн.
Распространены на Новой Гвинее.
В роде 3 вида:
проехидна Бартона — Z. bartoni Thomas, 1907 (северо-восток и юг Новой Гвинеи);
проехидна Бруийна — Z. bruijni Peters et Doria, 1876 (западная часть Новой Гвинеи)
проехидна^Бубу — Z. bubuensis Laurie, 1952 (северо-восточная Новая Гвинея).
Возможно, эти 3 вида должны быть объединены в один — Z. bruijni (Ван Деузен и Джорж —• Van Deusen and George, 1969).
СЕМЕЙСТВО УТКОНОСОВ
FAMILIA ORNlTHORHyNCHIDAE BURNETT, 1830
В семействе один род: утконосов — Genus Ornithorhynchus В lumen-bach, 1800 и один вид — утконос — О. anatinus Shaw et Nodder, 1799.
Тело приземистое, достигающее в длину 30—45 см, длина хвоста 10— 15 см. Вес взрослых около 2 кг. Самцы крупнее самок. Свое название зверь получил благодаря роструму, внешне сходному с утиным клювом (рис. 17). «Клюв» покрыт мягкой резиноподобной кожей, имеет длину примерно 65 мм, ширину около 50 мм. Поверхность клюва богато иннервирована. На верхней поверхности клюва открываются носовые отверстия. Глаза находятся высоко на голове и вместе с ушными отверстиями (ушные раковины отсутствуют) располагаются с каждой стороны головы в единой впадине, которая во время ныряния зверя закрывается кожной складкой. Глаза имеют мигательную перепонку. Хвост широкий, сплюснутый в дорзо-вентральном направлении, сходный по форме с хвостом бобра. Конечности пятипалые, приспособленные как для плавания (они имеют плавательные перепонки), так и для копания. Плавательная перепонка на передних лапах выдается перед пальцами, бордюром окружает их и может подгибаться таким образом, что когти пальцев оказываются выставленными наружу, и плавательная конечность превращается в копательную (рис. 18).
Тело утконоса покрыто густым мягким мехом, обычно черно-бурым на спинной стороне и сероватым на брюшной. В коже утконоса располагаются сальные и трубчатые, слабо извитые потовые железы. Волосы двух категорий: остевые и пуховые. Растут волосы пучками и образуют группы. В каждом пучке находится по 12—20 пуховых волос, а в каждой группе — по 4—5 пучков пуховых волос, располагающихся по бокам и позади остевого. Мышцы, поднимающие волосы, отсутствуют. У основания клюва на его нижней стороне взрослые самцы имеют специфическую железу, продуцирующую секрет с мускусным запахом. Выводковой сумки у самок нет.
Зубы имеются только у молодых животных. Они закладываются в сле-.0 1 2 з ~
дующем числеы — с — р — т —. Однако прорезываются только один
верхний предкоренной, два верхних коренных и три нижних коренных зуба. Эти зубы очень непрочные и быстро стираются; их отделы, находящиеся в челюсти, рассасываются, а места, на которых они росли, сверху покрываются роговыми пластинками, которые и остаются на всю жизнь.
Характерны небольшие слюнные железы. Мозг с малым количеством борозд. Диплоидное число хромосом 70± 10 точечных микрохромосом.
Обитают утконосы по берегам больших и малых быстротекущих или медлен
Рис. 18. Передняя лапа утконоса. А — с выступающими когтями (вид снизу); Б — с расправленной плавательной перепонкой (вид сверху)
нотекущих рек или в стоячих водоемах, на равнинах и в горах до высоты 2000 м над уровнем моря. По суше утконос передвигается шагом или бегом, в воде превосходно плавает и ныряет. Обычно под водой остаются в течение 1 мин, но в случае необходимости длительность ныряния может достигать 5 мин. Питается различными мелкими водными животными, поедая ракообразных, личинок водных насекомых, червей, улиток, мелких рыб. Небольшие количества пищи могут временно помещаться в защечные мешки. По наблюдениям в неволе, количество съеденной за сутки пищи достигает примерно веса самого животного. Активны утром и вечером. Держатся поодиночке или парами. В берегах водоемов устраивают норы обычно 10 м в длину, имеющие гнездовую камеру и два входа, один из которых располагается под водой.
В период размножения самка поселяется в норе, тянущейся до 18 м в длину, и устраивает в гнездовой камере гнездо из листьев и травы. Гнездо всегда бывает влажным, что предохраняет яйца от высыхания. Период гона приходится на конец июля — начало сентября. Самка откладывает яйца в гнездо примерно месяц спустя после спаривания.Число яиц 1—2, реже 3. Яйца имеют в длину 16—18 мм и в ширину около 15 мм. После откладки яйца слипаются между собой при помощи клейкой субстанции, которая покрывает их снаружи. Самка, по-видимому, «высиживает» яйца, проводя большую часть времени свернувшись вокруг них. На этот период самка закрывает вход в нору одной или несколькими земляными пробками. Выходя наружу на короткое время, она, возвращаясь обратно, снова закрывает пробкой вход. Инкубационный период длится 9—10 дней. Детеныши появляются из яиц голые, слепые, примерно 2,5 см длиной. Самка, лежа на спине, помещает их к себе на брюхо, где спустя некоторое время они начинают питаться молоком, выступающим на железистых полях. До 2-месячного возраста терморегуляция у детенышей развита плохо. К этому времени их длина достигает примерно 23 см, глаза еще не открываются, а волосы на теле только начинают появляться. К 4 месяцам детеныши вырастают до 33 см в длину, приобретают полный меховой покров и покидают нору. Половозрелость наступает в возрасте 1 года, длительность жизни, по-видимому, составляет 10 лет.
Распространены в Восточной Австралии и Тасмании.
Утконосов усиленно промышляли из-за хорошего меха. В настоящее время добыча утконосов запрещена, и численность их несколько возросла.
Подкласс живородящих млекопитающих Subclassis theria Parker et Haswell, 1879
К подклассу принадлежат млекопитающие, самки которых рождают живых детенышей. У зародышей подавляющего большинства представителей подкласса в процессе развития образуется особый орган — плацента, который осуществляет связь между зародышем и телом матери. У самок имеется или отсутствует (у большинства) выводковая сумка. Есть вибриссы. Выводные протоки млечных желез открываются не на железистых полях, как у однопроходных, а на сосках.
В черепе передне- и заднелобных костей, а также предсошника нет. Межчелюстная кость лишена processus ascendus. Молоточек и наковальня не срастаются между собой. Слезная кость, как правило, имеется. Скуловая кость и венечный и угловой отростки нижней челюсти почти у всех представителей хорошо развиты. У большинства имеется костный слуховой проход. Барабанная кость обычно срастается с черепом.
У большинства представителей подкласса зубы гетеродонтные, дифио-донтные. В скелете туловища коракоид и прокоракоид отсутствуют. Сумчатые кости имеются лишь у немногих представителей подкласса.
Головной мозг с мозолистым телом, иногда оно не развито. Слизистая оболочка желудка содержит пищеварительные железы. Клоака отсутствует.
Температура тела, в значительно меньшей степени, чем у однопроходных, зависит от окружающей температуры.
Приспособлены к жизни в самых разнообразных условиях.
Ареал совпадает с ареалом всего класса млекопитающих.
В подклассе 2 современных инфракласса:
сумчатых — Metatheria Huxley, 1880;
плацентарных — Eutheria Gill, 1872.
Инфракласс сумчатых Infraclassis Metatheria Huxley, 1880
Инфракласс содержит единственный отряд: сумчатых — Marsupialia liliger, 1811 (для обеих систематических групп ниже приводится общая характеристика).
ОТРЯД СУМЧАТЫХ
ORDO MARSUPIALIA HUGER, 1811
К отряду принадлежат млекопитающие разного размера от небольших (длина тела плоскоголовых сумчатых мышей составляет 4—10 см) до крупных (у исполинских кенгуру от 75 до 160 см). Внешний облик сумчатых очень разнообразен: от животных кротоподобных, с укороченным валько-ватым телом на низких конечностях, до стройных и высоконогих. Хвост большинства сумчатых сильно развит, у некоторых видов хватательный. У ряда форм на конце хвоста есть обнаженный участок ороговевшей кожи. Конечности, как правило, пятипалые. Часто некоторые пальцы срастаются между собой (синдактилия). Пальцы вооружены когтями, которые иногда имеют вид ногтей. У большинства видов первый палец передней конечности не имеет когтя.
Волосяной покров обычно густой и мягкий, иногда щетинообразный. Волосы, как правило, подразделяются на направляющие, остевые и пуховые. Хорошо развиты вибриссы на морде и передних и задних конечностях. В коже имеются потовые и сальные железы.
Сумчатые обладают преанальными железами, которые обычно сильнее развиты у самцов. Млечные железы трубчатые мерокриновые, выводные протоки их открываются на сосках. Соски располагаются по бокам тела, число их сильно колеблется (от 2 до 27) не только у разных родов, но иногда и в пределах одного вида. Самки примитивных видов сумчатых имеют большее число сосков. Для самок большинства сумчатых характерна выводковая сумка на брюхе, в ней открываются соски млечных желез. Форма выводковых сумок и степень их развития неодинакова у разных видов сумчатых. Сумки могут открываться вперед или назад, иногда они представлены только боковыми складками кожи, или даже совсем отсутствуют (сумчатый муравьед).
Череп имеет различную форму — от вытянутого и низкого до короткого и высокого. Мозговой отдел черепа развит относительно слабо. Передне- и заднелобных костей нет, имеются слезная, крылоклиновидная и скуловая кости. На слезной кости открывается отверстие слезного канала. У некоторых сумчатых (опоссумы и ценолесты) костные слуховые барабаны небольшие, и костный слуховой проход отсутствует. У всех дру
гих сумчатых костные слуховые барабаны большие пли очень большие. Как правило, барабанные кости не срастаются с другими костями черепа. Угловой отросток нижней челюсти загнут внутрь.
Число и форма зубов сильно изменчивы в зависимости от характера питания. Зубы характеризуются гетеродонтностью и не полной монофио-донтностью (сменяется только по одному предкоренному зубу в каждой челюсти). Тип жевательной поверхности щечных зубов туберкулосекториальный. Число зубов изменяется от i ~-р -|-щ = 50 (опоссумы) или
. 4 1 4 — 5 4 С£? , „ . .2 1
i —---с — р ——— т~ = 50 — 56 (сумчатый муравьед) до i — с —
1 2
р ~т~ = 18 (хоботноголовый кускус).
Как правило, в нижней челюсти резцов бывает меньше, чем в верхней.
В скелете характерны сумчатые кости, которые отходят от лобкового сочленения тазовых костей и находятся в стенке брюшной полости (рис. 19). В позвоночном столбе число позвонков изменчиво: шейных 7, грудных 11— 15, поясничных 4—8, крестцовых 1—6, хвостовых 7—35. В большинстве случаев 13 грудных позвонков обладают подвижными ребрами. Только сумчатый крот имеет 6 крестцовых позвонков, у остальных сумчатых их обычно 2. Лопатка с гребнем. Всегда есть ключица, только у сумчатых барсуков она развита слабо или отсутствует. Коракоид превратился в отросток лопатки (processus coracoideus). Для лазающих сумчатых характерно подвижное сочленение между малой и большой берцовыми костями — единственный случай среди млекопитающих.
Головной мозг сумчатых примитивен и, как и у однопроходных, не имеет мозолистого тела. Обонятельные доли сильно развиты (макросматиче-ский тип). Неопаллиум развит относительно слабо и практически борозд
не имеет.
глаза редуцированы. В носовой полости в
Рис. 19. Тазовый пояс длинноногого бандикута. А — сбоку; Б — снизу;
1 — подвздошная кость, 2— седалищная кость, 3 — лоб" ковая кость, 4 — сумчатая кость
Глаза по строению не отличаются от глаз высших млекопитающих; мигательная перепонка имеется, часто она развита слабо. Склера соедини-тельнотканая, без хряща. Есть гардеровы железы. У некоторых сумчатых
большинстве случаев имеется 6 носовых раковин. Якобсонов орган развит хорошо. Молоточек и наковальня в среднем ухе часто срастаются между собой.
У некоторых сумчатых (сумчатый муравьед, хоботноголовый кускус) язык очень длинный и может далеко выдвигаться изо рта. Слюнные железы у растительноядных сумчатых сильно развиты. Иногда имеются щечные мешки. Число и расположение небных складок сильно изменчиво и имеет систематический характер. Желудок у нерастительноядных форм обычно в виде простого мешка, а у растительноядных длинный и напоминает толстый отдел ки
шечника.
У нерастительноядных сумчатых тонкий отдел кишечника, а у растительнояд-
них толстый отдел представляет наиболее длинные части кишечного тракта. Как правило, слепой отдел кишечника имеется. Печень многодольчатая, с желчным пузырем.
В кровеносной системе большинства сумчатых есть кювьеровы протоки и непарная вена (venae azygos). Температура тела ниже, чем у плацентарных млекопитающих, и может слегка изменяться в зависимости от изменений температуры окружающей среды; нормально она колеблется от 34 до 36° С.
Влагалище и матка двойные. Иногда влагалища срастаются между собой в верхних отделах, образуя непарный слепой мешок. Этот мешок своим нижним концом может достигать мочеполового синуса, прирастать к нему и прорываться в него после первых родов. В этом случае образуется третье, или непарное, влагалище (некоторые кенгуру). Плацента, за редким исключением, не развивается. Пенис имеет раздвоенную головку. Семенники располагаются в мошонке (за исключением сумчатого крота), находящейся перед пенисом. Os penis отсутствует.
У сумчатых моно- или полиэстральный цикл. Овуляция спонтанная. Хориоаллантоидная плацента имеется только у сумчатых барсуков. Длительность развития эмбрионов в половых путях самки коротка (8—42 дня). Детеныш при рождении очень мал: длина его всего 0,5—3,0 см. Сразу после рождения он самостоятельно или с помощью матери достигает сумки и прикрепляется к соску млечной железы, который охватывает ртом, причем края рта срастаются с соском. Питание молоком идет пассивно: молоко впрыскивается в рот детеныша благодаря сокращению специальной мышцы — сжимателя млечных желез (musculus compressor mammae). Для защиты от попадания молока в дыхательное горло хрящи гортани приподняты, плотно прижаты к хоанам; таким образом создается сплошной воздухопроводящий путь и детеныш может свободно дышать даже в тот момент, когда молоко из рта направляется в пищевод, обтекая по сторонам гортань. Длительность лактационного периода не менее 65 дней. В сумке детеныш остается до 250 дней.
Обитатели различных ландшафтов: открытых пространств и лесов, равнин и гор, поднимаются до 5000 м над уровнем моря. Ведут наземный, подземный, древесный (имеются планирующие формы) и полуводный образ жизни, в связи с чем у них выработались глубокие адаптации. Насекомоядные, хищные, растительноядные и всеядные. Продолжительность жиз-' ни от 5 до 25 лет.
Распространены в Австралии, на Тасмании, Новой Гвинее, включая прилегающие острова и некоторые из Больших Зондских, а также в Южной, Центральной и Северной Америке. Акклиматизированы на Новой Зеландии.
В отряде 9 семейств (71 род):
опоссумов — Didelphidae Gray, 1821 (12 родов);
хищных сумчатых — Dasyuridae Waterhouse, 1838 (13 родов);
сумчатых муравьедов — Myrmecobiidae Waterhouse, 1838 (1 род); сумчатых кротов — Notoryctidae Ogilby, 1892 (1 род);
сумчатых барсуков, или бандикутов,—Peramelidae Waterhouse, 1838 (8 родов);
ценолестов, или крысовидных опоссумов, —Caenolestidae Trouessart, 1898 (3 рода);
лазящих сумчатых, или кускусов, — Phalangeridae Thomas, 1888 (14 родов);
вомбатов — Phascolomyidae Bonaparte, 1845 (2 рода);
кенгуру — Macropodidae Owen, 1839 (17 родов).
Линней поместил выделенный им род сумчатых Didelphis в сборный отряд Bestiae. Натуралисты XVIII столетия не смогли установить различий между сумчатыми и конвергентными им плацентарными и не выделяли сумчатых в отдельную группу. Это сделал Де Блэнвилль, в 1816г., объединивший сумчатых как «Didelphis Normaux». С тех пор не возникало сомне
ний в правомочности выделения сумчатых в качестве самостоятельного отряда.
Однако до сих пор подразделения внутри отряда остаются в известной степени точно не установленными. Наиболее часто употреблялось разделение отряда сумчатых по признакам строения резцов (Грегори — Gregori, 1910) или по наличию или отсутствию срастания второго и третьего пальцев на задней лапе (Джонс, Jones, 1923—1925 гг.).
Таблица 1
Разделение сумчатых по признакам строения резцов
Подотряды Надсгмейства
Polyprotodontia — многорезцовые (резцы обеих челюстей многочисленные и примерно одинаковых размеров; клыки имеют крупные размеры) Diprotodontia — двурезцовые (два средних резца в обеих челюстях большие, остальные резцы и клыки малы или могут совершенно отсутствовать) Didelphoidea Dasyuroidea Perameloi dea Caenolestoidea Phalangeroidea
Таблица 2
Разделение сумчатых по признакам срастания второго и третьего пальцев на задних конечностях
Подотряды
Надсемейства
Didactyla — раздельнопалые (пальцы разделены)
Syndactyla — сростнопалые (пальцы сращены)
Didelphoidea Dasyuroidea Caenolestoidea Perameloi dea Phalangeroidea
Симпсон (1945) полагает, что Didelphoidea является примитивной группой сумчатых, а остальные 5 надсемейств произошли от ранних представителей этой группы. Если любые два из этих надсемейств имели общего предка, то это было настолько давно и разделение их произошло в такой короткий срок, что можно считать, что все 5 надсемейств эволюционировали независимо друг от друга. Все эти 5 надсемейств возникли примерно в одно время — вероятно, в поздний меловой период или ранний третичный, и имели с этого времени совершенно независимое развитие. Исходя из этих соображений, Симпсон (1945) оставляет 6 систематических групп сумчатых в ранге надсемейств и не объединяет их в подотряды1: надсемейство Didelphoidea (с одним сем. Didelphidae), надсемейство Dasyuroidea (с сем. Dasyuridae и Notoryctidae), надсемейство Perameloi dea (с одним сем. Peramelidae), надсемейство Caenolestoidea (с одним сем. Caenolestidae) и надсемейство Phalangeroidea (с сем. Phalangeridae, Phascolomidae, и Мас-ropodidae). Таким образом, Симпсон насчитывает в отряде сумчатых 8 современных семейств.
Некоторые систематики выделяют сумчатых муравьедов и коала в отдельные семейства (Myrmecobiidae и Phascolarctidae) и таким образом
1 Большинство современных систематиков также не разделяют отряд сумчатых на подотряды.
увеличивают число современных семейств до 9 (Бурлиер, Bourlier, 1955) или 10 (Бартон, Burton, 1962).
Следует отметить, что многие таксономические группы сумчатых не соответствуют сходным группировкам плацентарных млекопитающих. Райд (Ride, 1964) предложил новую классификацию сумчатых, в которой в пределах инфракласса Metatheria он выделяет 4 отряда вместо одного:
Ordo Marsupicarnivora Ride, 19641 Superfamilia Didelphoidea
Familia Didelphidea Superfamilia Dasyuroidea Familia Dasyuridae Familia Thylacinidae Ordo Paucituberculata Ameghino, 1894 Familia Caenolestidae Ordo Peramelina Gray, 1825 Familia Peramelidae
Ordo Diprotodonta Owen, 1866 Familia Phalangeridae Familia Vombatidae Familia Macropodidae Marsupialia Insertae sedis Familia Notoryctidae
Объединение всех сумчатых в одном отряде, по мнению Райда, не обосновано серьезными морфологическими, палеонтологическими или какими-либо другими причинами, кроме традиционной привычки. Признавая справедливыми многие, высказывания Райда, в настоящем руководстве автор все же следует большинству систематиков (в основном Хальтенорту, На1-tenorth, 1958) и сохраняет принятое ими деление сумчатых.
Наиболее древние предки сумчатых известны из ранних отложений мелового периода Северной Америки. Они принадлежат к сем. Didelphidae (вымершее подсемейство Pediomyinae с представителями Eodelphis, Pedio-mys; вымершее подсемейство Thlaeodontinae с представителями Thlaeodon, Didelphodon и др.) и имеют значительное сходство с современными опоссумами. Представители современного подсемейства Didelphinae (сем. Didelphidae) появились в палеоцене Северной Америки. В Европе (которая в третичном периоде была непосредственно связана с Северной Америкой) сумчатые (тоже Didelphidae) были распространены от эоцена вплоть до миоцена. Хотя ископаемые остатки сумчатых до сих пор еще не обнаружены в Азии, можно предполагать, что Евразия и Северная Америка — родина сумчатых. Из Азии сумчатые проникли в Австралию по существовавшему в прошлом «мосту» через Новую Гвинею (по Симпсону, 1961, сумчатые мигрировали в Новую Гвинею из Австралии). В раннем мелу образовалась Тасманийская синклиналь, отделившая Новую Зеландию от Восточной Австралии (еще до проникновения туда сумчатых). Таким образом, отсутствие сумчатых в Новой Зеландии подтверждает происхождение их из Азии. В Евразии сумчатые погибли, не выдержав конкуренции с плацентарными млекопитающими. В Австралию плацентарные млекопитающие не успели проникнуть до ее изоляции, и сумчатые, не испытывая конкуренции, образовали здесь многочисленные формы, конвергентные многим плацентарным млекопитающим.
Менее вероятно предположение о возникновении сумчатых в Южной Америке и проникновении их в Австралию по существовавшему в прошлом «мосту» через Антарктику.
1 Ниже приведены лишь современные надсемейства и семейства.
СЕМЕЙСТВО ОПОССУМОВ
FAMILIA DIDELPHIDAE GRAY, 1821
Сумчатые мелких размеров. Длина тела 7—50 см. Длина хвоста 4—55 см. Морда вытянутая и заостренная. Уши маленькие или средних размеров, неоволосненные. Хвост в большинстве случаев длинный и хватательный, обнаженный полностью или частично, иногда утолщенный в основании отложениями жира. Конечности укороченные, пятипалые; задние немного длиннее передних. Большой палец задней конечности крупных размеров, .противопоставляется другим пальцам и лишен когтя. Пальцы между собой не срастаются.
Волосяной покров в большинстве случаев короткий, густой, окраска варьирует от серой и желтовато-бурой до черной. На голове могут быть полосы (густошерстные, пушистохвостые и обыкновенные опоссумы), темные окологлазные кольца (опоссумы Чилоэ и мышевидные опоссумы) или надглазные светлые отметки (патагонские и четырехглазые опоссумы). На спине также могут быть полосы (водяные опоссумы).
Сумка некоторых представителей семейства развита хорошо и открывается вперед (обыкновенные и четырехглазые опоссумы) или назад (водяные опоссумы); у других (длиннохвостые опоссумы) она неполная (в виде двух боковых кожных складок) или совсем отсутствует (мышевидные (см. рис. 22) и голохвостые опоссумы). От 5 до 27 сосков расположены в несколько рядов или в виде кольца.
В черепе лицевой отдел удлинен и заострен,' мозговой — небольшой. Барабанные кости полукольцевидные. Скуловая дуга крупная. Зубная формула: i с -у- р т~ = 50. Зубы имеют корни. Клыки развиты хорошо. Резцы небольшие. Передние коренные зубы тритуберкулярного типа. Позвонков шейных 7, грудных 13, поясничных 6, крестцовых 2, хвостовых 19—35.
Диплоидное число хромосом 22 у обыкновенных и толстохвостых опоссумов и 14 у густошерстных и мышевидных опоссумов. Обитатели лесов разного типа, степей и полупустынь, встречаются на равнинах и в горах до 4000 м над уровнем моря. Большинство представителей семейства ведет наземный или древесный образ жизни, а водяной опоссум — полу-водный.
Активность в основном вечерняя и ночная. Большинство всеядно с преобладанием плотоядности или насекомоядности. Мышевидные, четырехглазые и водяные опоссумы моноэстральные, а обыкновенные опоссумы полиэстральные (самки приносят по 2—3 помета в год) животные. Длительность беременности 12—13 дней. В помете детенышей 4—11 (до 25). Лактационный период продолжается до 70—100 дней. После выхода из сумки детеныши часто передвигаются с матерью, удерживаясь за шерсть на ее спине. Половозрелось наступает в 6—8-месячном возрасте. Продолжительность жизни от 5 до 8 лет.
Распространены на территории от юго-восточной Канады на юг через восточные штаты США и Мексику до примерно 52° ю. ш. в Аргентине. Встречаются также на Малых Антильских островах. Обыкновенный опоссум акклиматизирован в некоторых западных районах США.
В семействе 12 родов (84 вида):
густошерстных опоссумов — Caluromys J. Allen, 1900 (3 вида);
голохвостых опоссумов — Monodelphis Burnett, 1830 (10 видов);
короткохвостых опоссумов — Minuania Cabrera, 1919 (3 вида);
опоссумов Чилоэ —Dromiciops Thomas, 1894 (1 вид);
пушистохвостых опоссумов — Glironia Thomas, 1912 (2 вида);
патагонских опоссумов — Lestodelphis Tate, 1934 (1 вид);
мышевидных опоссумов — Marmoset. Gray, 1821 (58 видов); четырехглазых опоссумов — Metachirops Matschie, 1916 (1 вид);
Рис. 20. Полосатый густошерстный опоссум — С. irrupta
длиннохвостых опоссумов — Metachirus Burmeister, 1854 (1 вид); толстохвостых опоссумов — Lutreolina Thomas, 1910 (1 вид);
обыкновенных опоссумов — Didelphis Linnaeus, 1758 (2 вида); водяных опоссумов — Chironectes liliger, 1811 (1 вид).
Род густошерстных опоссумов
Genus Caluromys J. Allen, 1900
Средних размеров опоссумы. Длина тела 22—30 см. Длина хвоста равна или больше длины тела и составляет 22—40 см. Хвост хватательный, восковании или до половины покрытый мехом (рис. 20). Волосяной покров на теле плотный и мягкий, в большинстве случаев одноцветный, серый, красный или бурый. На морде могут быть темные полосы. У полосатого густошерстного опоссума на спине проходят две Х-образно пересекающиеся черные полосы. Выводковая сумка отсутствует: имеются лишь боковые кожные складки. Сосков 7. Диплоидный набор хромосом у густошерстного опоссума 14. Ведут преимущественно древесный образ жизни. Активны вечером, ночью и ранним утром. Днем обычно прячутся в дуплах деревьев. По характеру питания склонны к всеядности. Поедают сочные плоды, семена, насекомых, мелких позвоночных и т. д. Самки приносят по 3—4 детеныша в помете. В прошлом густошерстных опоссумов добывали из-за меха.
Распространены от южной Мексики до северной Аргентины.
В роде 3 вида:
рыжий опоссум — С. laniger Desmarest, 1820 (от южной Мексики на юг до Парагвая);
желтый опоссум — С. philander Linnaeus, 1758 (северная Бразилия, Гвинея, Венесуэла, о-в Тринидад);
полосатый густошерстный опоссум — С. irrupta Sanborn, 1951 (южное Перу).
Род голохвостых опоссумов
Genus Monodelphis Burnett, 1830
Размеры мелкие. Длина тела 7—16 см. Длина хвоста 4—8 см. Глаза маленькие. Хвост обнажен почти по всей длине, не хватательный. Внешне напоминают землероек (рис. 21).
Окраска короткого, густого и довольно грубого меха от серой до красноватой, бурой и черной. Часто есть продольные темные полосы. На груди и шее самцов некоторых видов расположены специфические железы, образованные сальными и потовыми железами; они продуцируют секрет с характерным запахом. Число сосков 9—25. Сумка отсутствует.
Экология изучена слабо. Ведут наземный образ жизни. Питаются в основном мелкими грызунами и насекомыми, а также сочными плодами, семенами и падалью. Домовый голохвостый опоссум в Бразилии поселяется в жилых помещениях, где уничтожает грызунов и насекомых. В тропических областях сезонность в размножении отсутствует. Число детенышей в помете от 8 до 14. Самка носит подросших детенышей на своей спине.
Распространены от севера Аргентины до севера Перу и Панамы.
В роде 10 видов:
голохвостый опоссум — TH. henseli Thomas, 1888 (север Аргентины, Парагвай);
трехцветный опоссум — М. brevicaudata Erxleben, 1777 (от Парагвая до Венесуэлы);
трехполосый опоссум — М. americana Muller, 1776 (южная и юго-западная Бразилия);
карликовый опоссум — М. sorex Hensel, 1872 (Риу-Гранди-ду-Сул в Бразилии);
однополосый опоссум — М. unistriata Wagner, 1842 (южная Бразилия); красноголовый опоссум — М. scalops Thomas, 1888 (южная Бразилия); домовый опоссум — М. domestica Wagner, 1842 (центральная, южная и юго-западная Бразилия);
опоссум Марайо — М. maraxina Thomas, 1923 (о-в Марайо в устье Амазонки);
перуанский голохвостый опоссум — М. peruviana Osgood, 1913 (северное Перу и Боливия);
колумбийский опоссум •— М. adusta Thomas, 1897 (Колумбия и восток Панамы).
Род короткохвостых опоссумов
Genus Minuania Cabrera, 1919
Размеры небольшие. Длина тела 10—14 см. Длина хвоста 4—7см. Внешне сходны с голохвостыми опоссумами и, вероятно, могут быть отнесены к ним в качестве подрода. Волосяной покров серый, красноватый
Рис. 21. Трехцветный опоссум — М. brevicaudata
или бурый, иногда с черными продольными полосами. Выводковой сумки нет.
Наземные хищники.
Распространены от центральных и восточных районов Бразилии до западных областей Аргентины.
В роде 3 вида:
серый опоссум — М. dimidiata Wagner, 1847 (Уругвай и Риу-Гранди-ду-Сул в Бразилии);
полосатый короткохвостый опоссум — М. umbristriata Mirando-Ribei-ro, 1936 (Гояс в Бразилии);
пампасский опоссум — М. fbsteri Thomas, 1924 (восток Аргентины)..
Род опоссумов Чилоэ
Genus Drotniciops Thomas, 1894
В роде единственный вид: опоссум Чилоэ — D. australis Philippi, 1893.
Размеры мелкие. Длина тела и длина хвоста почти одинаковые—примерно по 10 см. Внешний облик мышевидный. Морда укороченная. Уши небольшие. Глаза крупные. Хвост густо покрыт волосами, кроме нижней стороны кончика хвоста, в основании утолщен. Окраска короткого, густого и шелковистого меха серо-бурая с более темными или более светлыми боками. Имеется небольшая выводковая сумка, куда открываются 4 соска.
Обитает в густых влажных лесах. Особенно часто встречается в зарослях чилийского бамбука (Chusquea sp.). Ведет древесный образ жизни. Активность в основном ночная. Делает небольшие наземные гнезда из веточек, листьев и травы. В областях ареала с значительными понижениями температуры на холодный сезон впадает в спячку. Питается в основном насекомыми и другими беспозвоночными. Период размножения приходится на весну. В помете 2—5 детенышей. Когда детеныши немного подрастут, самка помещает их в гнездо, а несколько позже они начинают передвигаться с матерью, удерживаясь на ее спине.
Распространен в центральных районах Чили от 37 до 42° ю. ш. и на о-ве Чилоэ.
Род пушистохвостых опоссумов
Genus Glironia Thomas, 1912
Размеры от мелких до средних: длина тела 16—21 см, длина хвоста 20—23 см. Внешним видом напоминают мышей. Хвост длинный и густо ово-лосненный, за исключением вентральной поверхности самого кончика. Волосяной покров густой и мягкий. Верхняя сторона тела светло- или темно-бурая, нижняя — грязно-белая. На голове имеются полосы. Конец, хвоста белый.
Обитатели влажных тропических лесов. Экология не изучена. По-видимому, ведут древесный образ жизни. Всеядны.
Распространены на юго-востоке Перу и северо-востоке Эквадора.
В роде 2 вида:
пушистохвостый опоссум — G. venusta Thomas, 1912 (юго-восток Перу);
эквадорский опоссум — G. aequatorialis Anthony, 1926 (северо-восток: Эквадора).
Род патагонских опоссумов:
Genus Lestodelphys Tate, 1934
В роде единственный вид: патагонский опоссум — L. halli Thomas, 1921.
Размеры небольшие. Длина тела 14,5 см. Длина хвоста 9,5 См. Голова укороченная, довольно широкая. Уши небольшие, округлые. Волосяной
Рис. 22. Самка карликового мышевидного опоссумы — М. murina с детенышами и череп опоссума Алстона — М. distant
покров средней длины, густой и мягкий, на спинной стороне серый, на брюшной — белый, на плечах и бедрах темные пятна. Основание хвоста сильно утолщено. Хвост покрыт короткими волосами только у основания. Щеки, надглазные пятна и пятна позади оснований ушей и кончик хвоста белые. Сосков 19.
По-видимому, населяет пампасы. Образ жизни неизвестен. Судя по строению зубной системы, является хищником.
Распространен на юго-востоке Патагонии, на юг идет дальше всех других представителей сумчатых.
Род мышевидных опоссумов
Genus Marmosa Gray, 1821
Размеры от мелких до средних. Длина тела 7—21 см. Длина хвоста 6—28 см. Самцы заметно крупнее самок. Внешний облик мыше- и крысообразный (рис. 22). Морда заостренная, глаза большие, уши средней длины, безволосые. Волосяной покров короткий, блестящий, густой и мягкий, на верхней стороне тела серый, желтоватый или темно-буро-рыжий, на нижней светлый.
Часто имеются темные окологлазные кольца. Хвост хватательный, почти полностью лишенный волос, только его основание оволоснено, безволосая часть хвоста покрыта чешуями. В хвосте откладываются запасы жира. Выводковая сумка отсутствует. Сосков 9—19, располагаются они двумя рядами или кольцом.
Обитают на равнинах и в горах, поднимаясь на высоту до 3700 м над уровнем моря. Древесные формы. Активность ночная. Как правило, держатся поодиночке. Часто строят гнезда на деревьях из листьев и веточек. Реже встречаются наземные гнезда. Питаются в основном насекомыми и плодами, а также мелкими грызунами, ящерицами и яйцами птиц. В холодных районах ареала размножаются 1—3 раза в год, а в тропических областях — в течение всего года.
Распространены от центральных районов Мексики до севера Патагонии.
В роде 49 (Тэйт, 1933) или 57 (Хальтенорт, 1958) видов. Очевидно, после ревизии рода число видов (многие из них описаны по 1 -2 экземплярам) будет сокращено. Ниже приводится перечень 58 видов (по Хальтенорту, 1958) с добавлением одного вида, описанного Петтером, 1968:
пепельный опоссум — М. cinerea Temminck, 1834 (Парагвай и восток Бразилии);
лимский опоссум — М. limae Thomas, 1920 (восток Бразилии);
буробрюхий опоссум —М. demararae Thomas, 1905 (Гвиана, Венесуэла, восточная Колумбия);
опоссум Алстона — М. alstoni Allen, 1900 (Колумбия, Коста-Рика, Никарагуа и Гондурас);
амазонский опоссум — М. domina Thomas, 1920 (среднее и нижнее течение Амазонки);
многососковый опоссум — М. constaninae Thomas, 1920 (север Бразилии, северо-запад Аргентины и восток Боливии);
опоссум Мапири — М. mapiriensis Tate, 1931 (Кордильеры в Боливии и юг Перу);
опоссум Раппоза — М. rapposa Thomas, 1899 (северо-восток Боливии и юго-восток Перу);
опоссум Германа -— М. germana Thomas, 1904 (от центральных районов Перу до севера Эквадора);
боготский опоссум — М. regina Thomas, 1898 (окрестности Боготы в Колумбии);
длинношерстный опоссум — М. phaea Thomas, 1898 (Западные Анды от юго-западной Колумбии до юго-запада Эквадора);
карликовый мышевидный опоссум (или энеева мышь) — М. marina Linnaeus, 1758 (от р. Амазонки до р. Ориноко на западе и на восток от Кордильер);
красный опоссум — М. rubra Tate, 1931 (северо-восток Эквадора);
венесуэльский опоссум — М. tyleriana Tate, 1931 (юг Венесуэлы);
перуанский мышевидный опоссум — М. quichua Thomas, 1899 (восток Перу);
светлобрюхий опоссум — М. mitis Bangs, 1898 (север Венесуэлы, север Гвианы, Малые Антильские острова, Тринидад, Гренада);
опоссум Симонса — М. simonsi Thomas, 1899 (юг Эквадора и северо-запад Перу);
роатанский опоссум—М. ruatanica Goldman, 1911 (северо-запад Южной Америки);
мексиканский опоссум — М. mexicana Merriam, 1897 (от южной Мексики до центральной Панамы);
опоссум Аллена — М. canescens Allen, 1893 (от Мексики — 25° с. ш. вдоль тихоокеанского побережья до Сальвадора);
ночной опоссум — М. noctivaga Tschudi, 1846 (от Анд на северо-востоке Эквадора до среднего течения Амазонки);
опоссум Юнга — М. yungasensis Tate, 1931 (Боливия);
белобрюхий мышевидный опоссум — М. leucastra Thomas, 1927 (север Перу);
кольцеглазый опоссум — М. ocellata Tate, 1931 (центральная Боливия);
восточный опоссум — М. incana Lund, 1841 (восток Бразилии);
щетиноплечий опоссум — М. scapulata Burmeister, 1856 (Бразилия);
тринидадский опоссум — М. carri Allen et Chapman, 1897 (о-ва Тринидад и Малые Антильские);
опоссум Кана — М. invicta Goldman, 1912 (восток Панамы);
опоссум Томаса — М. fuscata Thomas, 1896 (восток Венесуэлы и север Колумбии);
андский опоссум — М. саисае Thomas, 1900 (север Перу и север Колумбии до среднего течения Амазонки);
опоссум Пары — М. emiliae Thomas, 1909 (север Бразилии);
малолапый опоссум — М. microtarsus Wagner, 1842 (юго-восточное побережье Бразилии);
стройный опоссум — М. agilis Burmeister, 1854 (Бразилия к югу от -р. Амазонки до Кордильер);
опоссум Марика — М. marica Thomas, 1899 (север Венесуэлы);
опоссум Ундуави — М. unduaviensis Tate, 1931 (запад Боливии);
опоссум Ацерамарка —М. aceramarcae Tate, 1931 (запад Боливии);
высокогорный опоссум — М. dryas Thomas, 1898 (восток Венесуэлы);
опоссум-красавец — М. lepida Thomas, 1888 (Боливия, Перу, Эквадор, Гвиана);
опоссумЮнина — М. juniensis Tate, 1931 (центральные районы Перу);
опоссум Дамара — М. parvidens Tate, 1931 (Гайана);
чилийский опоссум — М. elegans Waterhouse, 1839 (Чили);
толстохвостый мышевидный опоссум — М. marmota Океп, 1816 (Парагвай и север Аргентины);
большеухий опоссум — М. janetta Thomas, 1926 (юг Боливии);
. короткохвостый мышевидный опоссум — Л4. pusilia Desmarest, 1805 (юг и запад Парагвая, север Аргентины);
грациозный опоссум — М. venusta Thomas, 1902 (юг Боливии, север Аргентины, Перу);
обыкновенный мышевидный опоссум — М. pallidior Thomas, 1902 (восток и юг Боливии, на восток до р. Парагвай, запад Аргентины);
сан-луисский опоссум — М. bruchi Thomas, 1921 (центральная Аргентина);
опоссум Формозы1 — М. formosa Shamel, 1930 (северо-восток Аргентины);
бархатистый опоссум — М. velutina Wagner, 1842 (юг Бразилии);
опоссум Мату-Гросу — М. meridionalis Mirando-Ribeiro, 1936 (Мату-Гросу, Бразилия);
опоссум Столле — М. stollei Mirando-Ribeiro, 1936 (Бразилия, провинция Амазонка);
опоссум Блазе — М. blasei Mirando-Ribeiro, 1936 (Гояс в Бразилии);
опоссум Морейра — М. moreirae Mirando-Ribeiro, 1936 (Бразилия);
опоссум Эрхардта — М. erhardti Mirando-Ribeiro, 1936 (Санта-Катарина в Бразилии);
опоссум Рондона — М. rondoni Mirando-Ribeiro, 1936 (Мату-Гросу в Бразилии);
опоссум Муйена —М. agricolai Mcojen, 1943 (северо-восток Бразилии);
опоссум Тейта — М. tatei Handley, 1956 (центр и запад Перу);
М. karimii Petter, 1968 (северо-восток Бразилии).
Род четырехглазых опоссумов
Genus Metachirops Matschie, 1916
В роде единственный вид: четырехглазый опоссум — М. opossum Linnaeus, 1758.
Средних размеров опоссум. Длина тела около 30 см. Длина хвоста около 35 см. Внешне напоминает крысу (рис. 23). Голова довольно крупная
Рис. 23. Четырехглазый опсссум — М. oppossum
1 Провинция Формоза в северо-восточной Аргентине.
с удлиненной, конической мордой. Глаза средней величины. Уши недлинные, округлые и голые. Хвост длинный, хватательный, в основании оволоснен, далее. почти голый, черный, в концевой четверти — бело-желтый. Волосяной покров короткий и мягкий; на спинной стороне окраска от пепельнобурой до красно-бурой, на брюшной светло-желтая. Лоб темный, почти черный, двумя светлыми треугольными пятнами над глазами. Сумка хорошо развита и открывается вперед. Сосков от 5 до 9.
Обитатель лесов, предпочитает увлажненные места около болот и рек. Ведет древесный образ жизни, может быстро бегать по земле и хорошо плавать. На деревьях строит шаровидные гнезда около 30 см в диаметре. Активность ночная. Питается мелкими млекопитающими, птицами и их яйцами, пресмыкающимися, земноводными, насекомыми, пресноводными ракообразными, улитками, земляными червями, плодами. Часто посещает фруктовые плантации. Сезонность в размножении, по-видимому, отсутствует. В помете 1—3 детенышей.
Распространен от центральной Мексики до юго-востока Бразилии и севера Аргентины.
Род длиннохвостых опоссумов
Genus Metachirus Burmeister, 1854
В роде единственный вид: длиннохвостый опоссум — М. nudicaudatus Geoffroy, 1803.
Размеры средние. Длина тела 30 см. Длина хвоста 35 см. По общему облику и окраске сходен с четырехглазым опоссумом, но отличается от него более изящным телосложением и немного более длинными конечностями. Волосяной покров короткий, густой и мягкий, на спине и боках бурый, на брюхе желтоватый или серый, морда черноватая с белыми пятнами над глазами. Хвост оволоснен только в самом основании. Сумка отсутствует, иногда на ее месте имеются боковые кожные складки. Сосков у самок 5—9. Характерно сильное развитие циркуманальной специфической кожной железы.
Обитает в густых лесах или кустарниковых зарослях на равнинах. Ведет древесный образ жизни. На деревьях строит круглое гнездо из листьев и веточек, иногда находит себе убежища под камнями или под стволами упавших деревьев. Активность ночная. Всеядный: поедает фрукты, моллюсков, насекомых, земноводных, пресмыкающихся, птиц, их яйца и мелких млекопитающих. В помете 1—3 детенышей.
Распространен от Парагвая до Коста-Рики.
Вид разделяется на 12 подвидов.
Род толстохвостых опоссумов Genus Lutreolina Thomas, 1910
В роде единственный вид: толстохвостый опоссум — L. crassicaudala Desmarest, 1804 (рис. 24).
Размеры средние. Длина тела около 30 см. Длина хвоста около 35 см. Внешний облик напоминает ласку; туловище удлиненное, конечности укороченные, уши небольшие, закругленные. Волосяной покров короткий, густой и мягкий, на спинной стороне желтоватый, желто-бурый. Основная треть хвоста покрыта волосами и утолщена (здесь откладываются запасы жира), остальная часть хвоста безволосая, черноватого цвета (однако концевая треть хвоста желто-белая). Сумка отсутствует. Сосков 9. Первые пальцы на передних и задних конечностях не полностью противопоставляются другим пальцам. Число хромосом в диплоидном наборе 22.
Рис. 24. Толстохвостый опоссум — L. crassicau.da.ta
Обитатель лесов, пампасов, берегов рек и озер. Ведет преимущественно наземный образ жизни. Активность в основном ночная. Подвижный, быстро бегающий по земле зверек. Хорошо плавает. В лесных областях находит себе убежище в дуплах деревьев. В болотистых местах устраивает круглые травяные гнезда в прибрежных зарослях. Питается мелкими млекопитающими, птицами, пресмыкающимися, рыбами и насекомыми. Поедает иногда вегетативные части растений и семена. В течение года, по-видимому, бывает два помета (первый — весной). Длительность беременности около 14 дней. В прошлом толстохвостых опоссумов добывали из-за меха.
Распространен от Гвианы до северной Патагонии.
Род обыкновенных опоссумов
Genus Didelphis Linnaeus, 1758
Размеры наиболее крупные в семействе. Длина тела 32—50 см, длина хвоста 25—55 см. Вес от 2 до 5,5 кг. Внешний вид крысообразный (рис. 25). Голова с вытянутой, заостренной мордой. Уши крупные, безволосые. Хвост хватательный, голый, покрыт волосами только у самого основания, темного цвета с белым концом. Первый палец передней лапы противопоставляется другим и не имеет когтя. Волосяной покров, в отличие от других представителей семейства, состоит из пуховых и большого числа направляющих волос. Окраска серая, черная, красноватая, иногда белая. На голове могут проходить 3 продольные темные полосы: по одной через глаза и третья между ними по средней линии головы через лоб. Самка имеет открывающуюся вперед хорошо развитую сумку; внутри нее обычно 13 сосков. Они расположены кольцом; один сосок в центре. Диплоидное число хромосом 22.
Держатся главным образом в лесах, в местах с густым кустарником и подростом. Ведут сумеречный и ночной образ жизни. День проводят в трещинах скал, дуплах, кучах хвороста, норах. Строят округлое гнездо из листьев и травы. Материал для гнезда притаскивают во рту или захватив его цепким хвостом. Хотя большую часть времени обыкновенные опоссумы проводят на земле, они хорошо лазают и по деревьям. К осени сильно жиреют и в холодные дни могут быть совершенно неактивными. В случае ранения или испуга обыкновенный опоссум может прикинуться мертвым и таким образом избежать опасности. Набор пищи очень разнообразен и состоит из многих растительных и животных объектов. Держатся поодиночке, за исключением периода размножения. Самки достигают половозре-лости уже в первый год жизни. Для них характерен полиэстральный цикл, так что они обычно имеют 2 помета ежегодно на большей части своего ареала. В период размножения длительность астрального цикла составляет
2 Д с п 3 я
К
К ш
17
Во не вы бе;
Рис. 25. Северный опоссум — D. marsupialis и его череп
28 дней. Течка длится 1 или 2 дня. Период беременности 12—13дней. Число детенышей в помете 8—18 (до 25). Детеныши остаются прикрепленными к соску матери в течение 60—70 дней, затем они покидают сумку и еще месяц проводят на теле самки, цепляясь лапами за шерсть матери, а хвостами за ее хвост, который она держит над своей спиной. В это время они продолжают питаться молоком матери, хотя и начинают принимать твердую пищу.
Распространены на большей части Северной и Южной Америки.
В роде 2 вида:
северный опоссум — D. marsupialis Linnaeus, 1758 (от южных районов Канады до севера Аргентины);
южный опоссум — D. paraquayensis Oken, 1816 (большинство областей Южной Америки).
Обыкновенные опоссумы служат лабораторными животными. В южных штатах США их добывают из-за мяса и меха.
Род водяных опоссумов
Genus Chironectes liliger, 1811
В роде единственный вид: водяной опоссум — С. minimus Zimmermann, 1780 (рис. 26).
Размеры крупные. Длина тела 35—40 см. Длина хвоста 40—45 см. Волосяной покров густой, мягкий и довольно короткий. На спинной сторо не серо-белый с черными узорчатыми пятнами. Эти пятна имеют округлые выступы, вырезанные с боков и расположенные на затылке, плечах, груди, бедрах и задних частях огузка. Брюхо белое. На голове два прямоугольных
Рис. 26. Водяной опоссум — С. minimus, его задняя лапа и череп
белых надглазничных пятна. Хвост длинный, только у основания покрыт 'черными волосами. Остальная часть хвоста голая, черного цвета. Хвост в слабой степени хватательный. Наружные ушные раковины небольшие, округлые, голые. На задних лапах между пальцами сильно развитая плавательная перепонка. Выводковая сумка хорошо развита и открывается назад. Рудиментарная сумка имеется у самцов.
Обитает по берегам различного типа водоемов. Единственный представитель отряда сумчатых, приспособленный к полуводному образу жизни. Плавает медленно. Активность ночная. Питается в основном различными мелкими водными животными, включая рыб. Поедает также и наземных животных. В Бразилии самки приносят детенышей в декабре — январе. •Самка плавает вместе с находящимися в сумке детенышами. Убежищем водяных опоссумов служат подземные норы с выходом непосредственно над уровнем воды.
Распространен от юго-запада Мексики и Гватемалы до Боливии, юга Бразилии и севера Аргентины.
СЕМЕЙСТВО ХИЩНЫХ СУМЧАТЫХ
EAMIUA DASyURIDAE WATERHOUSE, 1838
Среди других семейств отряда хищные сумчатые наиболее примитивны. К этому семейству принадлежат самые мелкие в отряде, средние или крупные и весьма разнообразные по внешнему виду и образу жизни сумчатые. Длина тела колеблется от 4—10 см у представителей рода плоскоголовых сумчатых мышей до 100—НО см у сумчатого волка. Сложение тела изме
няется от приземистого и неуклюжего до стройного, высоконогого. Форма морды от тупой до заостренной. Уши маленькие или средней высоты. В отличие от представителей семейства опоссумов, хвост у хищных сумчатых не хватательный и у большинства видов покрыт волосами по всей длине. Часто в хвосте откладывается жир, и тогда он бывает утолщен. Передние конечности пятипалые, а задние четырех- или пятипалые. Большой палец задней конечности, когда он имеется, небольших размеров и без когтя. Пальцы не срастаются между собой. Конечности стопоходящие или пальцеходящие. Задние конечности могут быть непропорционально удлинены, а передние укорочены (сумчатые тушканчики). Выводковая сумка может отсутствовать, быть слабо или хорошо развитой (в последнем случае она открывается назад). Число сосков у самок варьирует от 2 до 12 (обычно 6—8).
Волосяной покров короткий, густой и мягкий. Окраска его бурая с разными оттенками, серая, красноватая или черноватая, иногда с белыми пятнами (род пятнистых сумчатых куниц) или черными полосами (полосатая сумчатая куница и др.) на спине.
Зубная формула варьирует у различных представителей семейства от 42 до 46: i с -р р т ~. Резцы небольших размеров, клыки крупные.. Щечные зубы с тремя острыми вершинами на жевательной поверхности. Коренные особенно крупные. Позвонков шейных 7, грудных 13, поясничных 6, крестцовых 2 и хвостовых 18—25.
Мочеполовой синус и прямая кишка отделены друг от друга. Мочеточник открывается у самцов в основании пениса, а семяпровод — на его вершине. Желудок простой. Слепая кишка отсутствует. Диплоидный набор из 14 хромосом имеют следующие изученные роды: сумчатые мыши, пятнистые сумчатые куницы, узколапые сумчатые мыши и тасманийские дьяволы.
Хищные сумчатые встречаются в самых различных ландшафтах и на различной высоте от морского побережья до 4000 м над уровнем моря. Ведут наземный (большинство представителей) или древесный образ жизни, активность преимущественно ночная. Как правило, держатся поодиночке. Обычно крупные представители семейства плотоядные, мелкие — насекомоядные. Для большинства видов характерен моноэстральный тип размножения. Беременность 8—30 дней. Число детенышей 3—10. Молодые остаются в сумке в течение примерно 150 дней. Половозрелость наступает в 6—12 месяцев. Продолжительность жизни у мелких форм до 7, а у крупных до 10—12 лет.
Распространены в Австралии, Тасмании, Новой Гвинее и прилегающих мелких островах.
В семействе 13 родов (48 видов):
сумчатых мышей — Antechinus McLeay, 1841 (12 видов);
плоскоголовых сумчатых мышей — Planigale Troughton, 1928 (4 вида); сумчатых крыс — Phascogale Temminck, 1824 (3 вида);
новогвинейских сумчатых крыс — Murexia Tate et Archbold, 1937 (2 вида);
полосатых сумчатых крыс — Phascolosorex Matschie, 1916 (2 вида);
гребнехвостых сумчатых мышей—- Dasycercus Peters, 1875 (2 вида); двугребнехвостых сумчатых мышей — Dasyuroides Spencer, 1896 (1 вид); узколапых сумчатых мышей—Sminthopsis Thomas, 1887 (13 видов); сумчатых тушканчиков — Antechinomys Krefft, 1867 (2 вида);
пятнистых сумчатых куниц — Dasyurus Geoffroy, 1796 (4 вида);
полосатых сумчатых куниц — Myoictis Gray, 1858 (1 вид);
сумчатых чертей, или тасманийских дьяволов, —Sarcophilus Geoffroy St. Hilaire et F. Cuvier, 1837 (1 вид);
сумчатых волков, или тасманийских волков, — Thylacinus Temminck, 1827 (1 вид).
Рис. 27. Желтая сумчатая мышь — А. flavipes
Род сумчатых мышей
Genus Antechinus McLeay, 1841
Наименее специализированные и наиболее примитивные формы семейства хищных сумчатых. Размеры небольшие. Длина тела 7—17 см. Длина хвоста 6—18 см. Внешним видом они напоминают мышей (рис. 27). Конечности короткие, широкие. На задних лапах большой палец имеется. Волосяной покров густой, короткий или длинный, довольно грубый. Окраска спинной стороны от серой до темно-бурой. Брюшная сторона белая или серая. Хвост такой же окраски, как и спина, покрыт короткими волосами, концевой метелки или утолщения не имеет. Сумка отсутствует. У ряда видов на ее месте имеется складка кожи. Сосков 6—12. У желтой сумчатой мыши есть 2 пары анальных желез, продуцирующих секрет с мускусным запахом. Зубная формула i — с р —т — = 44 — 461. Второй верхний резец крупнее третьего и четвертого; верхний четвертый пред-коренной немного крупнее третьего, а нижний четвертый значительно меньше третьего. Диплоидное число хромосом у желтой сумчатой мыши 14.
Обитают в разнообразных лесах от густых до сильно разреженных. Ведут наземный или полудревесный (например, желтая сумчатая мышь) образ жизни. Питаются насекомыми и мелкими позвоночными, некоторые виды также нектаром или падалью. Спаривание приходится на июль — август. Детеныши рождаются в августе — сентябре от 3 до 8. Беременность 30—34 дня. При рождении детеныш имеет длину 4—5 мм, вес около 16 мг. Лактационный период продолжается около трех месяцев. Половозрелость наступает примерно в 9 месяцев.
Распространены в Австралии, Тасмании и Новой Гвинее.
В роде 12 видов:
малая сумчатая мышь — A. minimus Geoffroy, 1803 (Тасмания);
сумчатая мышь Свайнзона—A. swainsoni Waterhouse, 1840 (Новый Южный Уэльс, Виктория, Тасмания);
желтая сумчатая мышь — A. flavipes Waterhouse, 1831 (Австралия);
карликовая сумчатая мышь — A. maculatus Gould, 1851 (Новый Южный Уэльс);
пятнистоглазая сумчатая мышь — A. bellus Thomas, 1904 (северные районы Австралии);
бархатистая сумчатая мышь — A. melanurus Thomas, 1899 (Новая Гвинея);
1 Такая же зубная формула характерна и для ряда других родов: плоскоголовые сумчатые мыши; сумчатые крысы, новогвинейские сумчатые крысы, полосатые сумчатые крысы, гребнехвостые сумчатые мыши, двухгребнехвостые сумчатые крысы, узколапые сумчатые мыши и сумчатые тушканчики.
шелковистая сумчатая мышь — A. naso Jentink, 1911 (Новая Гвинея);
сумчатая мышь Вильгельмины — A. Wilhelmina Tate, 1947 (Новая Гвинея);
толстохвостая сумчатая мышь — A. macdonnelensis Spencer, 1896 (центральные районы Австралии);
сумчатая мышь Баркли — A. mimulus Thomas, 1906 (север Австралии);
крапчатая сумчатая мышь — A. apicalis Gray, 1842 (юго-запад Австралии)1;
западноавстралийская сумчатая мышь — A. rosamondae Ride, 1964 (Западная Австралия).
Некоторые исследователи (Тэйт, 1947) выделяют толстохвостую сумчатую мышь и сумчатую мышь Баркли в один род Pseudantechinus Tate, 1947, а крапчатую сумчатую мышь в другой — Parantechinus Tate, 1947. По-видимому, правильнее считать их подродами.
Род плоскоголовых сумчатых мышей
Genus Planigale Trough ton, 1928
Внешне напоминают мышей. Размеры наименьшие в семействе. Длина тела 4—10 см. Длина хвоста 5—8 см. Вес около 5 г. Самцы крупнее самок. По размерам это самые маленькие представители отряда сумчатых. Голова сильно уплощена. Уши почти голые. Хвост не утолщенный, покрыт короткими волосами с небольшой кисточкой на конце. На задних конечностях первый палец имеется. Волосяной покров мягкий и густой, на спине бурый или буро-серый, на брюхе беловатый. Сумка у самок развита хорошо и открывается назад. Сосков 10—12. Череп чрезвычайно плоский, высотой всего примерно 3—6 мм.
Обитатели скалистых или песчаных участков с травянистой растительностью. Ведут наземный образ жизни. Активность ночная. Питаются в основном насекомыми. Самка приносит от 4 до 12 детенышей (северная сумчатая мышь — в дождливый сезон с декабря по март).
Распространены в западной части Нового Южного Уэльса, северных районах Австралии, Папуа и южной части Новой Гвинеи.
В роде 4 вида:
южная сумчатая мышь — Р. tenuirostris Troughton, 1928 (северная часть Нового Южного Уэльса);
северная сумчатая мышь — Р. ingrami Thomas, 1906 (север Австралии и северная часть Квинсленда);
плоскоголовая сумчатая мышь — Р. subtilissima Lonnberg, 1913 .(возможно, она является лишь подвидом Р. ingrami, Тейт, 1947) (северо-запад Австралии);
новогвинейская сумчатая мышь — Р. novaeguineae Tate et Archbold, 1941 (юго-восток Новой Гвинеи).
Численность южной и карликовой плоскоголовых сумчатых мышей очень мала, они близки к полному исчезновению. Оба вида включены в «Красную книгу»2.
Род сумчатых крыс
Genus Phascogale Temminck, 1824
Размеры мелкие. Длина тела 12—30 см. Длина хвоста 14—22 см. Внешним видом напоминает сонь (рис. 28). Морда удлиненная. Уши крупные, тонкие и голые. Глаза крупные. Хвост длинный, его концевая половина
1 Крапчатая сумчатая мышь к настоящему времени, видимо, вымерла.
2 В «Красной книге», т. I, изданной Международным союзом охраны природы н естественных ресурсов (IUCN), приведен список видов (иногда подвидов) млекопитающих, численность которых настолько мала илн так интенсивно снижается, что есть угроза исчезновения этих видов. В книге приведены также диагнозы видов, распространение, особенности размножения, численность, меры охраны.
Рис. 28. Малая сумчатая крыса — Р. calura
покрыта удлиненными черноватыми волосами, образующими метелку, причем эти волосы могут подниматься вертикально. Пальцы вооружены сильными изогнутыми когтями, за исключением первого пальца задних конечностей. Волосяной покров' короткий и мягкий (большая и малая сумчатые крысы) или длинный и грубый (сумчатая крыса Лорентца). Спинная сторона серо-голубая, а брюшная бело-серая. Сумка почти не развита. Сосков 8—10.
Обитатели лесов различного типа, от влажных до сухих. Ведут в основном древесный образ жизни. Хорошо лазают по деревьям. Активность ночная. Гнездо ив листьев и веточек, часто устраивают в дуплах или в развилках ветвей деревьев, иногда и на земле. Хищники, питаются мелкими позвоночными (млекопитающими, птицами, пресмыкающимися) и насекомыми.
Распространены в Австралии и Новой Гвинее.
В роде 3 вида:
большая сумчатая крыса — Р. tapota-fa Meyer, 1793 (Австралия);
малая сумчатая крыса — Р. calura Gould, 1844 (юго-запад Австралии);
сумчатая крыса Лорентца —• Р. loren-tzi Jentink, 1911 (Новая Гвинея). Иногда сумчатую крысу Лорентца рас-
сматривают в качестве отдельного рода Neophascogale Stein, 1933 (Тейт, 1947).
Численность малой сумчатой крысы очень мала, и она включена в «Красную книгу».
Род новогвинейских сумчатых крыс
Genus Murexia Tate et Archbold, 1937
Мелкие хищные сумчатые. Длина тела 11—20 см. Длина хвоста 15— 18 см. Внешним видом сходны с сонями. Хвост длинный и тонкий, покрыт короткими и редкими волосами, без концевой метелки. На задних конечностях первый палец имеется. Волосяной покров короткий и густой. Спинная сторона серо-бурая или красно-бурая, иногда с широкой темной полосой по средней линии. Брюшная беловатая или светло-серая. У самок две пары сосков. Череп массивный.
Обитают в густых низменных дождевых лесах1 и поднимаются в горы до 3000 м над уровнем моря. Древесные формы. Биология изучена плохо. Питаются, очевидно, насекомыми и мелкими позвоночными.
Распространены на северо-западе и юго-востоке Новой Гвинеи и о-вах Ару.
1 Тропический дождевой лес распространен главным образом в пределах экваториальной климатической зоны (между 10° с. ш. и 10° ю. ш.), характеризующейся небольшими колебаниями температуры в течение года. Годовая сумма осадков очень велика и местами достигает нескольких метров.
В роде 2 вида:
длиннохвостая сумчатая крыса — М. longicaudata Schlegel 1866 (северо-восток и юго-восток Новой Гвинеи и о-ва Ару);
новогвинейская сумчатая крыса — М. rothschildi Tate, 1938 (юго-восток Новой Гвинеи).
Род полосатых сумчатых крыс
Genus Phascolosorex Matschie, 1916
Размеры мелкие. Длина тела 13—17 см. Длина хвоста 14—15 см. Внешне похожи на сонь. Морда удлиненная. Уши относительно короткие, округлые. Хвост по большей части своей длины покрыт короткими, грубыми и довольно редкими черными волосами; только в его основании волосы густые и мягкие. На задних конечностях первый палец имеется. Когти на пальцах (за исключением первого пальца задних конечностей, на котором коготь отсутствует) короткие. Спинная сторона серо-бурая или оранжевобурая с продольной узкой черной полосой, проходящей посредине от головы до основания хвоста, брюшная оранжевая или буро-красная. Сосков 4.
Обитают в горах, поднимаясь до 3000 м над уровнем моря. Экология не изучена. Ведут древесный образ жизни. Хищники.
Ареал охватывает горные области Новой Гвинеи.
В роде 2 вида:
оранжевобрюхая сумчатая крыса — Р. doriae Thomas, 1886; буробрюхая сумчатая крыса — Р. dorsalis Peters et Doria, 1876. Возможно, они являются лишь подвидами.
Род гребнехвостых сумчатых мышей
Genus Dasycercus Peters, 1875
Размеры небольшие: длина тела 13—15 см, длина хвоста 8—10 см. По внешнему виду несколько напоминает мышей. Уши короткие, широкие и округлые. Конечности укороченные. На задней конечности первый палец имеется. Волосяной покров мягкий и густой. На спине серый, красно-, вато-бурый, на брюхе — белый или желтый. Проксимальная часть хвоста (примерно 2/3 его длины) утолщена; здесь откладываются запасы жира. Верхняя сторона конца хвоста с удлиненными черными волосами, образующими гребень’. Сумка отсутствует; имеется лишь кольцевая складка кожи, окружающая 6—8 (иногда 4) сосков. Нижнего четвертого предкоренного зуба нет.
Обитатели пустынь и полупустынь. Активность преимущественно ночная, хотя встречаются и днем. В песчаных холмах копают неглубокие норы. Питаются насекомыми и мелкими позвоночными. Самка приносит >6—7 детенышей (в Центральной Австралии обычно с июня по сентябрь). Детеныши не покидают соски матери по крайней мере месяц.
Распространены в центральных районах Южной и Западной Австралии.
В роде 2 вида:
гребнехвостая сумчатая мышь — D. cristicauda Krefft, 1867 (центральные районы и Южная Австралия);
западная сумчатая мышь — D. blythi Waite, 1904 (Западная Австралия).
Рис. 29. Двугребнехвостая сумчатая мышь — D. by г net
Род двугребнехвостых сумчатых мышей
Genus Dasyuroides Spencer, 1896
В роде единственный вид: двугребнехвостая сумчатая мышь — D-. Ьуг-nei Spencer, 1896.
Размеры небольшие. Длина тела 14—18 см. Длина хвоста 13—14 см. Внешне сходна с крысой (рис. 29). Морда удлиненная, конусовидная. Уши средних размеров, округлые. Лапы узкие, как у узколапых сумчатых мышей. В общем напоминает гребнехвостых сумчатых мышей, но отличается от них отсутствием первого пальца на задней конечности и тем, что на конце хвоста есть гребень удлиненных черных волос не только на верхней, но и нижней стороне Основание хвоста почти не утолщено. Мех мягкий и густой. На спине и боках тела он сероватый, часто с рыжим оттенком, снизу белый; конечности тоже белые. Проксимальная половина хвоста рыжая, конец черный. Подошвы лап покрыты волосами. Выводковая сумка почти не развита. Шесть сосков расположены кольцеобразно.
Обитатель каменистых пустынь и полупустынь. Экология изучена слабо. Живет в норах. Активность ночная. Хищник, питающийся в основном насекомыми. Держатся поодиночке, иногда небольшими группами.
Ареал охватывает центральные районы и Южную Австралию.
Род узколапых сумчатых мышей
Genus Sminthopsis Thomas, 1887
Небольшие хищные сумчатые. Длина тела 7—10 см. Длина хвоста 6—20 см. Внешне сходны с мышами и крысами (рис. 30). Морда заостренная. Лапы узкие. Уши средних размеров. Хвост средней длины, иногда очень длинный, чешуйчатый, покрыт короткими грубыми волосами. У некоторых видов в основании или середине хвоста откладываются запасы жира, и в этих местах он бывает сильно утолщен. На задних конечностях первый палец имеется. Волосяной покров средней длины, густой и мягкий, на спине и боках желто-бурый или серый, на брюхе серовато-белый или белый. Конечности обычно белые. Хвост буроватый или сероватый. Часто посредине морды проходит продольная темная полоска. Сумка развита хорошо. Сосков
Рис 30 Толстохвостая узколапая сумчатая мышь — S. crassicaudata
8—10 (иногда 2 или 6). Хромосом в диплоидном наборе у толстохвостой узколапой сумчатой мыши 14.
Обитают в разнообразных ландшафтах, включая пустыни, полупустыни, кустарниковые степи и влажные леса. Ведут в основном наземный образ жизни, хотя некоторые виды хорошо лазают по деревьям. Активность ночная. Убежищами служат норы или гнезда из травы и листьев, которые зверьки устраивают на земле в густых кустах или под стволами упавших деревьев При медленном движении используют все четыре лапы, а при быстром задние. Питаются в основном насекомыми, а также мелкими позвоночными (млекопитающими, ящерицами).
Распространены в Австралии, Тасмании и о-вах Ару.
В роде 13 видов:
белоногая сумчатая мышь —5. leucopus Gray, 1842 (от юга Квинсленда на юг до Виктории и Тасмании);
малая узколапая сумчатая мышь—S. murina Waterhouse, 1838 (от юга Квинсленда до юго-запада Австралии);
сумчатая мышь Квинсленда—5. lumholtzi Iredale et Troughton, 1934 (северо-восток Квинсленда);
сумчатая мышь Томаса—S. hirtipes Thomas, 1898 (центральные районы Австралии);
песчаная сумчатая мышь—S. psammophila Spencer, 1895 (центральные районы Австралии);
полосатоголовая сумчатая мышь —S. larapinta Spencer, 1896 (Австралия);
сумчатая мышь Бёркли—S. stalkeri Thomas, 1906 (север Австралии);
сумчатая мышь Дэрби —S. froggatti Ramsay, 1887 (Западная Австралия);
длиннохвостая сумчатая мышь—S. longicaudata Spencer, 1909 (Западная Австралия);
зернистоподошвенная сумчатая мышь—S. granulipes Troughton, 1932 (юго-запад Австралии);
толстохвостая узколапая сумчатая мышь—S. crassicaudata Gould, 1884 (Австралия);
сумчатая мышь Ару —S. rufigenis Thomas, 1922 (о-ва Ару);
сумчатая мышь Лаусона—S. monticola Troughton, 1965 (Новый Южный Уэльс).
Троугтон (1965) выделяет S. crassicaudata macrura в особый вид S. тас-rura Gould, 1845, а из вида S. larapinta выделяет S. nitela Collett, 1897.
За последние 30 лет никаких сведений о длиннохвостой узколапой сумчатой мыши не имеется. Если этот вид еще не полностью исчез, то его представители сохранились в ничтожном количестве. Вид внесен в «Красную книгу».
Род сумчатых тушканчиков
Genus Antechinomys Krefft, 1867
Размеры мелкие. Длина тела 8—11 см. Длина хвоста 11—12 см. Внешне сходны с тушканчиками (рис. 31). Самцы крупнее самок. Задние конечности сильно удлинены. Передние конечности развиты достаточно хорошо. Хвост длинный, с крупным пучком темных волос на конце. Морда удлиненная и заостренная. Уши крупные, округлые на вершинах. Первый палец на задних конечностях отсутствует. Волосяной покров длинный, густой и мягкий, сероватый, снизу беловатый. По бокам головы, через глаз обычно проходит темная полоса. Помимо морды, на запястьях располагаются необычайно длинные вибриссы. В период размножения выводковая сумка, открывающаяся назад, хорошо развита. Сосков 6—8.
Обитают в основном в песчаных пустынях и полупустынях. Хищники, питаются насекомыми и мелкими позвоночными. Двигаются прыжками, при движении опираясь и на передние конечности. Активность сумеречная и ночная. День проводят в глубоких норах.
Распространены в центральных районах и на востоке Австралии. Везде малочисленны.
В роде 2 вида:
восточноавстралийский сумчатый тушканчик — A. laniger Gould, 1856 (от юга Квинсленда до северо-запада Виктории);
центральноавстралийский сумчатый тушканчик — A. spenceri Thomas, 1906 (центральные районы Австралии).
Численность восточноавстралийского сумчатого тушканчика настолько низка, что ему угрожает полное исчезновение. За последние годы он был встречен примерно всего в десяти местах на территории, ограниченной 30° и 33 ю. ш. и 146° и 148° в. д. Вид внесен в «Красную книгу».
Рис. 31. Центральноавстралийский сумчатый тушканчик — A. spenceri
Рис 32. Крапчатая сумчатая куница — D. quoll и череп пятнистохвостой сумчатой куницы — D. macu.la.tus
Род пятнистых сумчатых куниц
Genus Dasyurus Geoffrey, 1796
Размеры от мелких до средних. Длина тела 25—75 см. Длина хвоста 20—35 см. Вес может достигать 3 кг. Голова короткая и заостренная (крапчатая и чернохвостая сумчатые куницы) (рис. 32) или средней длины и притупленная (пятнистохвостая сумчатая куница). Уши небольшие (пятнистохвостая сумчатая куница) или средних размеров (крапчатая и карликовая сумчатые куницы). Хвост длинный, покрытый длинными волосами. Волосяной покров на туловище короткий или средней длины, густой и мягкий. Окраска его на спинной стороне от серо-желтой до черно-бурой с многочисленными белыми пятнами неправильной формы, на брюшной белая, серая или желтая. Первый палец на задней конечности имеется у всех видов, кроме крапчатой сумчатой куницы, а подошвенные подушки — у пятнистохвостой и карликовой сумчатых куниц. Выводковая сумка самок развивается только в период размножения и открывается назад. В остальное время она представлена кольцевой складкой кожи. Сосков б—8, зубная формула: I — с — р —/п —= 42. Первый верхний резец иногда отделен про-странством от других резцов. Клыки и коренные зубы сильно развиты. Число хромосом в диплоидном наборе 14.
Обитатели лесов и открытых равнин, встречаются в поселениях человека. Активность ночная. Убежищем в течение дня служат щели среди камней, дупла упавших деревьев, куда затаскивают сухую траву и кору. Хотя эти сумчатые наземные животные, они хорошо лазают по деревьям. Питаются животной пищей: мелкими млекопитающими, птицами, пресмыкающимися, земноводными, рыбами, моллюсками и насекомыми. У крапчатой сумчатой куницы самка обычно приносит 4—8 детенышей. Известен случай рождения у одной самки 24 детенышей. Молодые покидают соски матери в возрасте около 8 недель. Глаза открываются в 11 недель. В 15 недель они начинают есть мясо. К самостоятельной жизни переходят в 4—4,5-месячном возрасте. К этому времени они достигают веса 175 г. У пятнистохвостых сумчатых куниц рождается 4—6 детенышей; беременность около трех недель. В 4 недели длина тела детенышей достигает примерно 4 см. В 7 недель открываются глаза, и они покидают соски матери. К самостоятельной жизни переходят в возрасте 18 недель. Численность пятнистых сумчатых куниц в послед
нее время в Австралии сильно уменьшилась, но они еще довольно многочисленны в Тасмании.
Распространены в Австралии, Тасмании, Новой Гвинее.
В роде 4 вида:
крапчатая сумчатая куница —D. quoll Zimmermann, 1777 (восточная часть Нового Южного Уэльса, Виктория, Южная Австралия и Тасмания);
чернохвостая сумчатая куница—D. geoffroyi Gould, 1841 (Восточная, Северная, Юго-Западная и Южная Австралия);
пятнистохвостая сумчатая куница — D. maculatus Kerr, 1792 (Квинсленд, Новый Южный Уэльс, Виктория, юго-восток Австралии и Тасмания);
карликовая сумчатая куница—D. hallucatus Gould, 1842 (север Австралии и Новая Гвинея).
Некоторые исследователи выделяют в отдельные роды чернохвостую сумчатую куницу (Dasyurinus Matschie, 1916), пятнистохвостую сумчатую куницу (Dasyurops Matschie, 1916) и карликовую сумчатую куницу (Sata-nellus Pocock, 1926). По-видимому, правильнее считать их подродами пятнистых сумчатых куниц.
Крапчатая сумчатая куница встречается очень редко; внесена в «Красную книгу».
Род полосатых сумчатых куниц
Genus Myoictus Gray, 1858
В роде единственный вид: полосатая сумчатая куница — М. melas Muller, 1840 (рис. 33).
Размеры небольшие. Длина тела 17—21 см. Длина хвоста 14—20 см. Внешне сходна с мангустой. Голова средней длины, заостренная. Уши короткие. Хвост длинный, сверху покрытый удлиненными, а снизу короткими волосами. Окраска меха на спине красно-бурая, стремя продольными полосами (средняя полоса достигает хвоста). Голова, брюшная сторона и передние конечности желтые или красноватые. На носу может быть чер-
Рис. 33. Полосатая сумчатая куница — М. melas
Рис. 34. Сумчатый черт — S. harrisi и его череп
ная полоска. Задние конечности серо-желтые или красно-бурые. Хвост буро-черный. Выводковая сумка развита слабо. У самок по 6 сосков. Зуб-4 12 4
ная формула: i — с — р — т — = 42. Первый верхний резец отделен
от остальных резцов промежутком.
Обитает в равнинных дождевых лесах. Экология почти не изучена.
Распространение охватывает Новую Гвинею, о-ва Ару.
Род сумчатых чертей, или тасманийских дьяволов, Genus Sarcophilus Geoffroy et F. Cuvier, 1837
В роде единственный вид: сумчатый черт, или тасманийский дьявол,— S. harrisi Boitard, 1841 (рис. 34).
Размеры средние: длина тела около 50 см, длина хвоста 25 см. Тело неуклюжее, массивное. Конечности укороченные, сильные, Голова крупная с притупленной мордой. Уши небольшие. Хвост средней длины, довольно толстый в основании, к концу утончается. Его покрывают длинные волосы, которые часто вытираются, и тогда он остается почти голый. Первый палец на задней конечности отсутствует. Когти крупные. Волосяной покров на теле черный. Лапы красноватого цвета. Большое полулунное пятно на передней части груди и на крестце и небольшие круглые пятна на боках белые. Сумка у самок имеет вид серповидной складки кожи, открывающейся назад; сосков — 4. Череп массивный (рис. 34). Зубная фор-
.4124. л Tz . .
мула: i — с — р — т~ = 42. Клыки и коренные зубы особенно велики.
Число хромосом в диплоидном наборе 14.
Обитает в густом подлеске и кустарниковых зарослях. Обычно ведет ночной образ жизни. В питании широкий набор животных, поедает также падаль. Убежище находит в пустотах между камнями, под стволами упавших деревьев и т. д., где устраивает гнездо из листьев и травы. Спаривание в марте — апреле. Самка обычно приносит 4 детеныша в конце мая - начале июня. При рождении длина тела около 12 мм, в 7 недель 7 см. В 15 недель покидают соски матери, но остаются в сумке весь период лактации, около 4—5 месяцев. Половозрелость наступает, по-видимому, в 2 года. Продолжительность жизни 7—8 лет. Распространен в Тасмании.
Род сумчатых, или тасманийских, волков
Genus Thylacinus Temminck, 1827
В роде единственный вид: сумчатый, или тасманийский, волк — Т. cynocephalus Harris, 1808 (рис. 35).
Наиболее крупный представитель семейства. Длина тела 100—130 см. Длина хвоста 50—65 см. Внешне напоминает собаку. Туловище удлиненное, конечности высокие, пальцеходящие. Голова сходна с собачьей. Уши короткие, закругленные, стоячие. Хвост длинный, с короткими, прилегающими волосами. Первый палец на задних конечностях и подошвенные мозоли отсутствуют. Сумка открывается назад. Число сосков 4. Волосяной по_
Рис. 35. Сумчатый волк — Т. cynocephalus и его череп
Рис. 36. Сумчатый муравьед -М. fascialus и его череп
кров короткий, густой и грубый. На спине серо-желто-бурый с 13—19 черно-бурыми поперечными полосами, расположенными от плеч до основания хвоста, на брюшной светло-серо-желтый. Морда серая с белыми отметинами вокруг глаз и ушей. Зубная формула: i — с — р — т —= 46. Клы-3 13 4
ки и коренные сильно развиты. Характерно отсутствие сумчатых костей.
Обитатель лесистых районов. Активность ночная. Держатся поодиночке или парами, иногда семейными группами. Убежище устраивают между скалами, иногда в дуплах упавших деревьев. Питаются кенгуру, другими млекопитающими и птицами. Добычу может преследовать подолгу. Период размножения растянут. Детенышей в помете 2—4.
Прежде был широко распространен по всей Австралии. Теперь сохранился только в Тасмании. Несмотря на то, что с 1938 г. был введен запрет на добычу сумчатых волков, местное население активно их уничтожало, так как они иногда нападали на овец и домашнюю птицу. В настоящее время численность их чрезвычайно невелика, вид близок к полному исчезновению. Сумчатые волки внесены в «Красную книгу».
СЕМЕЙСТВО СУМЧАТЫХ МУРАВЬЕДОВ
FAMIL1A МУВМЕСОВНОЛЕ WATERHOUSE, 1838
В семействе единственный род: сумчатых муравьедов — Myrmecobius Waterhouse, 1836 с одним видом сумчатый муравьед — М. fasciatus Water-house, 1836 (рис. 36). Иногда сумчатых муравьедов включают в качестве подсемейства,в семейство хищных сумчатых (Андерсон и Джонс, 1967).
Размеры этого насекомоядного сумчатого невелики. Длина тела 17— 27 см. Длина хвоста 13—17 см. Вес взрослого животного от 275 до 459 г. Голова уплощенная: морда вытянутая и заостренная, рот небольшой, язык может вытягиваться изо рта по крайней мере на 10 см. Глаза большие. Уши средних размеров, остроконечные. Задняя часть тела больше, чем передняя. Длинный хвост покрыт густой шерстью, не хватательный. Лапы относительно короткие, широко расставленные. Передние конечности пяти-, задние четырехпалые. Пальцы вооружены сильными когтями.
Волосяной покров довольно длинный, жесткий ..Окраска его серовато-коричневая или красноватая с 6—12 белыми поперечными полосами, проходящими по спине. Брюхо и конечности желто-белые. На груди нахо
дится специфическая кожная железа, содержащая сальные и потовые железы. У самок 4 соска. Выводковая сумка отсутствует.
Череп низкий и уплощенный (см. рис. 36). Межглазничное пространство широкое. Заглазничные отростки хорошо развиты. Костное небо черепа
сумчатого муравьеда тянется далеко назад, значительно дальше, чем у
большинства млекопитающих. Сходную структуру неба имеют броненосцы
и ящеры. Зубная формула: i
—-— с — р ——- т — = 50 — 56. Зубы ма-3—4 1 4—5 4
ленькие, очень изменчивые по величине; щечные зубы правой и левой сторон могут иметь разную длину и ширину. Щечные зубы островершинные. Позвонков шейных 7, грудных 13, поясничных 6, крестцовых 3, хвостовых 23. Диплоидное число’хромосом 14.
Обитает в лесах, на равнинах и в горах. Ведет наземный образ жизни. В отличие от большинства представителей отряда, у сумчатых муравьедов активность в основном дневная. Ночь проводит в углублениях в земле или
в других укромных местах, где часто устраивает подстилку из листьев и травы. Сумчатые муравьеды — проворные животные, проводящие большую часть времени в поисках пищи. За исключением периода размножения, они держатся поодиночке. Охотно греются на солнце в холодные месяцы года. Основными объектами пищи служат термиты (предпочтительно) и муравьи. Другие беспозвоночные поедаются лишь случайно. В неволе сумчатый муравьед поедает от 10 до 20 тыс. термитов ежедневно. Сильными когтями передних лап зверек раскапывает почву пли сдирает кору с пня, лежащего ствола дерева. Двигая с большой скоростью длинным цилиндрическим языком, сумчатый муравьед вытаскивает термитов из разрушенных им убежищ. В Юго-Западной Австралии период спаривания длится с середины лета до ранней осени (декабрь — апрель). Детеныши рождаются летом, осенью или ранней зимой (с января по апрель — май). В помете обычно 4 детеныша. Первое время детеныши удерживаются ртом на сосках, прикрытые только шерстью матери. Подросших детенышей самка помещает в неглубокую нору, где и завершается их развитие. Половая зрелость наступает в первый год жизни.
Распространены в эвкалиптовых лесах Юго-Западной Австралии. Нуждаются в строгой охране, так как численность их сильно снижается не только вследствие прямой охоты, но главным образом из-за освоения заселенных ими территорий сельским хозяйством.
СЕМЕЙСТВО СУМЧАТЫХ КРОТОВ
FAMILIA NOTORyCTIDAE OGILBY, 1892
В семействе единственный род: сумчатых кротов — Notoryctes Stirling, 1891 (рис. 37).
По размерам и внешнему виду напоминают обыкновенного крота. Длина тела всего 15—18 см. Длина хвоста 12—26 мм. Тело неповоротливое,
Рис. 37. Сумчатый крот — Notoryctes sp. и его череп
толстое, удлиненное, шея короткая. Короткий цилиндрический хвост жесткий на ощупь, с заметными кольцевыми чешуями и ороговевшим кончиком. Голова короткая, толстая. В носовом отделе морды расположен роговой щиток желтого цвета. Ноздри небольшие, щелевидные. Ушные раковины отсутствуют — под мехом имеются лишь небольшие наружные слуховые отверстия. Недоразвитые глаза имеют всего 1 мм в диаметре, скрыты под кожей. Они не имеют хрусталика и зрачка, а зрительный нерв рудиментарен. Пятипалые конечности сумчатого крота короткие, сильные. Особенно укорочены передние конечности. Третий и четвертый пальцы передних конечностей вооружены крупными, треугольной формы когтями. Пятый палец передних конечностей имеет ноготь. Первый, второй, третий пальцы передних лап имеют по две фаланги, четвертый и пятый — по одной. Три средних пальца задних конечностей также имеют удлиненные когти, а первый и пятый пальцы —1 ногти. На первой фаланге первого пальца есть боковая костная пластинка, расширяющая подошву лап. Пальцы между собой не срастаются.
Волосяной покров мягкий, густой и короткий. Его окраска варьирует от почти белой до розовато-светло-коричневой и желтовато-красной и имеет характерный переливающийся отблеск у живых животных. Выводковая сумка небольшая, открывается назад. Складка на внутренней поверхности живота разделяет сумку неполной перегородкой на два кармана, содержащих каждый по одному соску. Самцы обладают рудиментом выводковой сумки, образованным небольшой поперечной складкой кожи. Мошонки у них нет, семенники располагаются в брюшной полости.
Череп конический, с расширенным затылочным отделом и выступающими носовыми костями (см. рис. 37). Швы между костями заднего отдела
черепа рано исчезают. Зубная формула: i
1 2 4
с — р —т —
12 4
3 — 4
3
= 40 —
— 42. Резцы, клыки и предкоренные простые, с тупыми жевательными поверхностями, небольших размеров. Верхние коренные тритуберкуляр-ные. Характерна редукция сумчатых костей и срастание между собой пяти последних шейных позвонков. Два последних крестцовых позвонка своими поперечными отростками срастаются с седалищной костью. Позвонков шейных 7, грудных 15, поясничных 4, крестцовых 6 и хвостовых 12.
Обитатели песчаных равнин и холмистых районов, поросших кустарником. Постоянных нор не делают (во всяком случае, не в период размножения), как кроты из отряда насекомоядных. Обычно они передвигаются в грунте в непосредственной близости от его поверхности. Защищенная роговым щитком голова активно участвует в копании, передние конечности отгребают почву подобно лопатам, а задними конечностями сумчатый крот отталкивает грунт позади себя. Таким образом, зверек как бы плывет в грунте, не оставляя после себя подземного туннеля. Основными объектами питания служат черви, почвенные насекомые и их личинки. Сумчатые кроты очень прожорлйвы и большую часть времени проводят в поисках пищи. Довольно часто встречаются на поверхности земли, особенно после дождей. Держатся поодиночке. О размножении сумчатых кротов известно крайне мало. Полагают, что самка в период размножения устраивает постоянную глубокую нору, в которой выращивает детенышей. В северных частях ареала самка приносит детенышей в ноябре. Продолжительность жизни 1,5 года.
Распространены на юге Австралии, в ее центральных районах и Западной Австралии. Везде немногочисленны.
В роде 2 вида:
большой сумчатый крот — N. typhlops Stirling, 1889 (Южная и Центральная Австралия);
малый сумчатый крот— N. caurinus Thomas, 1920 (Западная Австралия).
Рис. 38. Длинноносый бандикут — Р. nasuta и его череп
СЕМЕЙСТВО СУМЧАТЫХ БАРСУКОВ, ИЛИ БАНДИКУТОВ,
FAMILIA PERAMELIDAE WATERHOUSE, 1838
Сумчатые небольших размеров. Длина тела от 17 до 50 см. Длина хвоста 9—26 см. Вес может достигать 4,7 кг. Внешне несколько напоминают кроликов и крыс (рис. 38, 39, 40). Морда удлиненная и заостренная. Уши могут быть маленькими или крупными и узкими, частично или полностью безволосыми. Хвост короткий, средней длины или длинный, весь покрытый волосами, не хватательный. Задние конечности удлиненные, передние укороченные. Второй, третий и четвертый пальцы передних конечностей длинные, с мощными когтями (у свиноногих бандикутов четвертый палец короткий, без когтя), а первый и пятый — короткие, не имеющие когтей (у свиноногих бандикутов оба эти пальца отсутствуют). Для задних лап бандикутов характерна синдактилия: второй и третий пальцы срастаются между собой и раздельными остаются только когти. Соединенные вместе пальцы действуют как единый палец и предназначены для чистки меха. Наибольшим по размерам пальцем на задних ногах является четвертый; первый палец развит плохо и не имеет когтя, иногда отсутствует (свиноногие и кроличьи бандикуты). Подошва лап оволосненная или голая, имеет или не имеет 1—2 небольшие мозоли.
Волосяной покров короткий и жесткий или длинный и мягкий. На спине окраска от серой, серо-гол убой и желтой до красно- и черно-бурой, на брюхе белая, серая или желтая. На крестце иногда 2—4 поперечные темные полосы. Выводковая сумка открывается вниз и назад. Количество сосков 6—10 (у большинства 8).
Зубная формула: i —у— с -±-р — гп-^-=47— 48. Нижние резцы наклонены вперед. Крайние резцы нижней челюсти обладают двулопастной коронкой. Клыки небольшие, заостренные. Для кроличьих бандикутов характерны удлиненные клыки. Коренные с трех- или четырехугольными коронками в поперечнике, три- или квадритуберкулярные. Костные слуховые барабаны небольшие, округлые или очень крупные, грушевидные. Позвонков шейных 7, грудных 13, поясничных 6, крестцовых 1—2, хвостовых 17—23. Ключица рудиментарна или ее нет вовсе.
Единственные среди сумчатых имеют хориоаллантоидную плаценту, в которой, однако, отсутствуют сосочки. Желудок простой, слепая кишка 72
средних размеров. Диплоидное число хромосом 14 (у длинноносых и коротконосых бандикутов).
Обитатели равнин с густой травой, берегов болот и рек, густых кустарников и лесов. Осторожные, быстрые зверьки, ведущие наземный образ жизни. Активность ночная. Передвигаются по земле галопирующими прыжками на всех четырех лапах. Пища животная и растительная, но большинство насекомоядно. Преимущественно осенью самки рождают от 2 до 8 детенышей. Продолжительность жизни 8—10 лет.
Распространены в Австралии, Тасмании, Новой Гвинее и прилегающих островах.
В семействе 8 родов (19 видов):
длинноносых бандикутов — Perameles Geoffroy, 1803 (5 видов); новогвинейских бандикутов — Peroryctes Thomas, 1906 (3 вида); мышевидных бандикутов — Microperoryctes Stein, 1932 (1 вид);
колючих бандикутов — Echymipera Lesson, 1842 (3 вида);
бандикутов Керама — Rhynchomeles Thomas, 1920 (1 вид); коротконосых бандикутов Thylacis liliger, 1811 (3 вида); свиноногих бандикутов — Chaeropus Ogilby, 1838 (1 вид);
кроличьих бандикутов — Thylacomys Owen, 1838 (2 вида).
Род длинноносых бандикутов
Genus Perameles Geoffroy, 1803
Размеры от небольших до крупных7Длина тела 21—40 см. Длина хвоста 9—17 слЛ'Морда удлиненная и заостренная (см. рис. 38). Уши средней длины, заостренные. Передние конечности укорочены, задние сильно удлинены. Волосяной покров мягкий или грубый, на спине серо-оливковый или светло-оранжевый. У большинства видов на крестце проходят темные поперечные полосы. Резцов ~. Костные слуховые барабаны небольшие, полушаровидные. У грубошерстного, тасманийского и длинноносого бандикутов в диплоидном наборе 14 хромосом. ?•
Обитатели открытых ландшафтов или паркового типа лесов, а также пустынь. Активность ночная. Гнезда из листьев и ветвей устраивают на земле, иногда в норах. Питаются в основном насекомыми, но также червями, улитками, ящерицами, мелкими млекопитающими, иногда растениями. Строгой сезонности в размножении нет. Самка приносит 1—5 детенышей. Беременность около 11 дней. У детенышей длинноносого бандикута глаза открываются в возрасте 45—48 дней. На 55-й день после рождения детеныши покидают сумку, на 70-й день переходят к самостоятельной жизни. Самки тасманийского бандикута достигают половозрелости примерно в 3 месяца, самцы в 4—5 месяцев. Продолжительность жизни, видимо, около двух лет.
Распространены'в Австралии и Тасмании.
В роде 5 видов:
грубошерстный бандикут — Р. bougainvillei Quoy et Gaimard, 1824 (прежде встречался на юге и юго-западе Австралии и в Западной Австралии, теперь сохранился только на о-вах Бернье и Дорр в заливе Шарк);
пустынный бандикут— Р. eremiana Spencer, 1897 (центральные районы Австралии);
полосатый бандикут—Р. fasciata Gray, 1841 (Новый Южный Уэльс и Виктория);
тасманийский бандикут — Р. gunni Gray, 1838 (Тасмания);
длинноносый бандикут—Р. nasuta Geoffroy, 1804 (Виктория, Новый Южный Уэльс и Квинсленд).
Полосатый бандикут к настоящему времени, по-видимому, вымер, так как с 1967 г. никаких сведений о его находках не имеется. Этот вид, а также грубошерстный бандикут включены в «Красную книгу».
Род новогвинейских бандикутов
Genus Peroryctes Thomas, 1906
Размеры от небольших до крупных (наибольших в семействе). Длина тела 17—50 см. Длина хвоста 14—26 см. Морда длинная, заостренная. Уши средней длины, закругленные. Хвост покрыт прилегающими к коже воло-5
сами или голый с чешуями. Резцов-у. Костные слуховые барабаны небольшие, полушаровидные. Волосяной покров относительно мягкий и длинный, на спинной стороне серо-бурый или черно-бурый, иногда с узкой срединной полосой, на брюшной белый или оранжевый.
Обитают в густом подлеске горных лесов, поднимаясь до 3—4,500 м над уровнем моря. Экология почти не изучена.
Ареал охватывает Новую Гвинею.
В роде 3 вида:
новогвинейский бандикут — Р. raffrayanus Milne-Edwards, 1878 (северо-запад и юго-восток Новой Гвинеи);
длиннохвостый бандикут—Р. longicauda Peters et Doria, 1876 (Новая Гвинея);
папуасский бандикут — Р. papuensis Laurie, 1952 (юго-восток Новой Гвинеи).
Род мышевидных бандикутов
Genus Microperoryctes Stein, 1932
В роде единственный вид: мышевидный бандикут — М. murina Stein, 1932.
Размеры небольшие. Длина тела около 17 см. Длина хвоста около 11 см. По внешнему виду напоминает мышь. Уши средней длины, закругленные. Волосяной покров мягкий, густой, довольно короткий, на спинной стороне черный, на брюшной серо-черный. Хвост покрыт волосами. Подош-5
вы лап голые. Резцов —. Костные слуховые барабаны небольшие, полушаровидные.
Обитает в густом подлеске горных лесов. Поднимается на высоту до 2500 м над уровнем моря. Экология не изучена.
Распространен в западной части Новой Гвинеи.
Род колючих бандикутов
Genus Echymlpera Lesson, 1842
Средних и крупных размеров. Длина тела 27—45 см. Длина хвоста 7—12 см. Вес достигает 2 кг. Морда длинная, заостренная. Уши короткие, с острыми вершинами. Задние конечности сильно развиты. Подошвы лап голые. Волосяной покров на спинной стороне темно-красно-бурый, с грубыми щетиновидными волосами, часто напоминающими колючки, на брюш-4
ной желто-бурый или беловато-желтый, грубый. Резцов -у. Верхний клык обычно сильно удлинен’. Костные слуховые барабаны небольшие, полушаровидные.
Обитатели дождевых лесов. Экология не изучена. Ведут наземный образ жизни.
Распространены на Новой Гвинее с прилегающими островами.
В роде 3 вида:
колючий бандикут — Е. clara Stein, 1932 (о-в Япен и северо-запад Новой Гвинеи);
Рис. 39. Большой бандикут — Т. macrourus
плоскоиглый бандикут — Е. kalubu Fischer, 1829 (Новая Гвинея);
толстоголовый бандикут — Е. rufescens Peters et Doria, 1875 (запад Новой Гвинеи, юго-запад Папуа, о-ва Ару, Кей, п-ов Кейп-Йорк на севере
Австралии).
Род бандикутов Керама
Genus Rhynchomeles Thomas, 1920
В роде единственный вид: бандикут Корама — R. prattorum Thomas, 1920.
Размеры средние. Длина тела около 32 см. Длина хвоста около 13 см. Морда очень длинная изаостренная, безволосая. Уши овальные и короткие. Хвост почти голый, черновато-бурый. Волосяной покров завитой, мягкий, окраска его сверху и снизу одинаковая, шоколадно-бурая. На груди белое пятно. Подошвы задних лап голые. Резцов -|-. Зубы небольших размеров. Костные слуховые барабаны небольшие, полушаровидные.
Обитатели густых горных лесов. Поднимаются на высоту до 1900 м над уровнем моря. Экология не изучена.
Распространен на о-ве Керам (между Новой Гвинеей и Сулавеси).
Род коротконосых бандикутов
Genus Thylacis liliger, 1811
Размеры средние и крупные: длина тела 24—41 см, длина хвоста 9— 18 см. Вес взрослых животных обычно 1,1—1,4 кг. Морда относительно короткая и широкая (рис. 39). Уши короткие, овальные, Хвост недлинный, покрытый волосами. Мех укороченный, с отдельными выступающими длинными волосами, грубый, на спине желто-бурый или темпо-бурый, на брюхе беловатый. Подошвы задних лап голые. Сумка хорошо развита и открывается назад. Сосков 8. Число резцов у. Костные слуховые барабаны грушевидной формы. У малого и большого бандикутов 14 хромосом в диплоидном наборе.
Обитатели открытых ландшафтов и разреженных лесов типа парковых. Предпочитают близость водоемов. Активность ночная. Пищу добывают, копаясь в земле. Питаются червями, насекомыми. Устраивают длинные туннели в траве. Сгроят гнезда из листьев, прутьев и травы с примесью почвы на земле или в дуплах упавших деревьев. Вход в гнездо закрывают
изнутри. Период размножения не приурочен строго к определенному сезону, хотя, по-видимому, достигает пика в июне. Беременность 15 дней. В помете 2—8 детенышей. При рождении детеныши имеют длину тела около 14 мм. В 7-недельном возрасте они покидают соски матери.
Распространены в Австралии, на некоторых прилегающих островах — Барроу, Нуит-Архипелаг, в Тасмании и Новой Гвинее.
В роде 3 вида:
бандикут Барроу — 7?. barrowensis Thomas, 1901 (о-ва Барроу и Монте-Белло у Западной Австралии);
малый бандикут— Т. obesolus Shaw et Nodder, 1797 (Австралия, Тасмания);
большой бандикут— Т. macrourus Gould, 1842 (северная часть Нового Южного Уэльса, север Австралии, юг Новой Гвинеи).
Род свиноногих бандикутов
Genus Chaeropus Ogilby, 1838
В роде единственный вид: свиноногий бандикут — С. ecaudatus Ogilby, 1838.
Размеры небольшие. Длина тела около 25 см. Длина хвоста около 10 см. Морда укороченная, но заостренная. Уши длинные, остроконечные. Передние конечности более длинные и тонкие, чем у других представителей семейства. Первый и пятый пальцы на них отсутствуют, второй и третий сильно развиты, четвертый слабо. Когти на втором и третьем пальцах крупные. Задние конечности тонкие. Первый палец на них отсуствует, пятый едва различим, четвертый сильный и крупный с коротким тупым ногтем, второй и третий — небольших размеров. Хвост длинный и тонкий, иногда с небольшим гребнем волос на верхней стороне, серый снизу, и на боках, черный сверху. Мех грубый, но без колючек, на спине буро-серый, иногда оранжевобурый; на брюхе — белый. Число резцов -|-. Костные слуховые барабаны маленькие, полушаровидные.
Обитатели травянистых степей, саванн и полупустынь. Активность преимущественно ночная, хотя довольно часто встречаются и днем. Гнезда из травы и сухих листьев устраивает в углублениях почвы. Всеяден. Период размножения приходится на май — июнь. В помете 2 детеныша.
Прежде был распространен от запада Нового Южного Уэльса, запада Виктории, запада Квинсленда и Северной Территории до Западной Австралии. За последние годы численность снизилась настолько, что если он еще неполностью уничтожен, то близок к этому. Возможно, еще сохранился в центральных районах Австралии. Основные причины снижения численности — выпас скота и поедание бандикутов лисами.
Свиноногий бандикут включен в «Красную книгу».
Род кроличьих бандикутов
Genus Thylacomys Owen, 1838
Небольшого и крупного размера бандикуты/Длина тела 20—44 см. Длина хвоста 12—22 см. Морда длинная и конусовидная. Уши заостренные, удлиненные, сходные с ушами зайца (рис. 40)., Первый палец на задних конечностях отсутствует. Хвост довольно длинный, его концевая половина сверху покрыта длинными волосами, но самый кончик остается голым. Волосяной покров длинный, шелковистый и мягкий, на спинной стороне буро-серый или голубовато-серый, на брюхе белый. Сумка открывается 5
вниз и слегка назад. Сосков 8. Число резцов -у. Костные слуховые барабаны большие.
Рис. 40. Кроличий бандикут — Т. lagotis
Обитатели открытых ландшафтов, лесов паркового типа. Устраивает в земле поры, иногда на глубине до 1,5 м. Активность в сумерках, ночью. Хищник. Самка приносит в марте — мае 1—3 детенышей.
Распространены в Австралии. Повсюду редки.
В роде 2 вида:
кроличий бандикут — Т. lagotis Reid, 1837 (отдельные малочисленные популяции в Северной Территории на юго-западе Квинсленда, в южной части Западной Австралии и, возможно, в Южной Австралии);
малый кроличий бандикут — Т. leucura Thomas, 1887 (центральные районы Австралии).
Численность кроличьего бандикута угрожающе мала, и он включен в «Красную книгу».
СЕМЕЙСТВО ЦЕНОЛЕСТОВ, ИЛИ КРЫСОВИДНЫХ ОПОССУМОВ, FAMILIA CAENOLESTIDAE TROUESSART, 1898
Небольшие по размерам сумчатые. По внешнему виду напоминают крыс или землероек (рис. 41). Длина тела 10—13 см. Длина хвоста 6—12 см. Голова удлиненная, коническая. Глаза небольшие. Длинные вибриссы на-
ходятся на конце морды и щеках. Хвост целиком покрыт короткими, редкими, грубыми волосками. Основание хвоста иногда утолщено. Конечности пятипалые, задние лапы длиннее передних. Подошвы голые. Пальцы вооружены когтями, а первый и пятый пальцы передних лап — ногтями. Пальцы не противопоставляются друг другу и не срастаются между собой.
Волосяной покров средней высоты, плотный и мягкий, на спинной стороне буро-серый или темно-бурый, на брюшной немного светлее. Выводко
вая сумка у взрослых самок отсутствует, хотя иногда имеется у молодых. Сосков 4 или 5.
Череп вытянутый, уплощенный (см. рис. 41). Мозговая часть черепа
относительно велика. Костные слуховые барабаны полукольцевидные.
Костное небо с большими отверстиями. Зубная формула:
. 4 1
i------------с — р
3 — 4 1
1m— =46—48. Зубы брахиодонтные, секториальные. Щечные зубы в по-3 4
перечном сечении четырехугольные или треугольные. Резцы небольшие и слабые, за исключением крупного первого нижнего резца с каждой стороны. Верхние клыки с одним или двумя корнями, средней длины или длинные, слабо или сильно изогнутые. Лобковый симфизис короткий. К атлан-
' ис. 41. Эквадорский ценолест — С. fuliginosus и его череп
ту причленяются подвижные, соединенные с ним • шейные ребра. Крупная плечевая кость не пропорциональна небольшим передним конечностям. Позвонков шейных 7, грудных 13, поясничных 6, крестцовых 2, хвостовых 22—27.
Желудок разделяется на 3 отдела. Головной мозг очень примитивный, с крупными обонятельными долями и небольшим мозжечком. Куперовы железы и особенно простата очень сильно развиты.
Обитают, как правило, в лесных районах,
поднимаясь в горы до 4000 м над уровнем моря. Экология ценолестов изучена плохо. Большинство ведет наземный образ жизни. Активность вечерняя и ночная. Питаются главным образом различными беспозвоночными.
Распространение охватывает западную часть Южной Америки (Анды) от южной Венесуэлы до южного Чили.
В семействе 3 рода (3 вида);
эквадорских ценолестов — Caenolestes Thomas, 1895 (1 вид);
перуанских ценолестов Lestores Ochser, 1934 (1 вид);
чилийских ценолестов — Rhyncholestes Osgood, 1924 (1 вид).
Род эквадорских ценолестов
Genus Caenolestes Thomas, 1895
В роде единственный вид: эквадорский ценолест— С. fuliginosus Thomas, 1863 (см. рис. 41).
Длина тела 11—13 см. Длина хвоста 11—12 см. Голова удлинена. Уши относительно короткие, округлые. Глаза маленькие. Волосяной покров мягкий и густой, окраска типичная для семейства. Хвост покрыт редкими волосами. Сосков 4. Верхний клык длинный и сильный, только слегка изогнутый, с одним корнем.
Обитатели горных лесов по их верхней границе и альпийских лугов. Экология изучена плохо. Активность сумеречная и ночная. Обоняние и слух развиты хорошо, зрение слабо. Питается наземными беспозвоночными.
Распространен в Андах па юге Венесуэлы, Колумбии и Эквадора.
Род перуанских ценолестов
Genus Lestores Owen, 1934
В роде единственный вид: перуанский ценолест — L. Inca Thomas, 1917.
Длина тела примерно равна длине хвоста и составляет 10—12 см. Окраска длинного меха типичная для семейства. Сосков 4. Верхний клык короткий, имеет два корня.
Обитатель горных лесов, поднимается до 3500—4200 м над уровнем моря. Экология почти не изучена. Ведет наземный образ жизни. Активность ночная. Питается в основном насекомыми и, возможно, другими беспозвоночными.
Распространен в Андах на юге Перу.
Род чилийских ценолестов
Genus Rhyncholestes Osgood, 1924
В роде один вид: чилийский ценолест — R. raphanurus Osgood, 1924.
Длина тела 11—12 см. Длина хвоста 6—8 см. Основание хвоста иногда | утолщено за счет жировых отложений. Сосков 5. Лицевой отдел черепа I длиннее и уже, чем у представителей других родов семейства. Верхний I клык длинный, сильно изогнутый, с одним корнем. У самок он короче, I чем у самцов, и напоминает первый верхний предкоренной.
Обитатель густых лесов умеренного климата. Экология не изучена. I По-видимому, ведет наземный образ жизни. Активность ночная. Питается I мелкими беспозвоночными.
Распространен на о-ве Чилоэ и в прилегающих областях материка, на I юге Чили.
СЕМЕЙСТВО ЛАЗЯЩИХ СУМЧАТЫХ, ИЛИ КУСКУСОВ, FAMILIA PHALANGERIDAE THOMAS, 1888
Многочисленное семейство, населяющее обширную территорию. Кон-вергентны плацентарным мышам, крысам, белкам, лемурам, куницам, лисицам. Размеры от мелких до крупных. Длина тела варьирует от 6 до 82 см. Хвост обычно длинный и часто хватательный, иногда рудиментарный (у коала). Хвост бывает частично или полностью покрыт волосами. Морда удлиненная или короткая. Уши небольшие или средних размеров. Конечности пятипалые, все пальцы, за исключением первого, имеют сильно изогнутые когти, помогающие при лазании по деревьям. Лапы хватательные, первый палец противопоставляется другим (рис. 42). У некоторых родов первый и второй пальцы передней конечности противопоставляются третьему, четвертому и пятому. На задних конечностях небольшие второй и третий пальцы соединены между собой кожной перепонкой по всей своей длине. Свободными остаются только когти. У представителей трех родов: карликовых, гигантских летучих кускусов и сумчатых летяг — между передними и задними конечностями изводить планирующие прыжки.
Волосяной покров мягкий, плотный, корот-I кий, иногда длинный, | окраска его варьирует от черного, бурого и серого до желтого и белого. На теле могут быть I пятна и полосы. У кисте-1хвостых кускусов, сумчатых летяг и коала имеется грудная специфиче-I ская кожная железа; у I самцов она крупнее, чем I у самок. Сальные желе-I зы в коже многочислен-
натянута кожистая перепонка, позволяющая про-
кускусов. Слева направо: целебес-
Рис. 42. Стопа
ского — Р. celebensis и хоботноголового — Т. spen-
serae
ны. Выводковая сумка хорошо развита и открывается вперед. Сосков 2—4.
Череп широкий, уплощенный. Зубная формула изменчива:
2__з 1 1_.3 з____4
i -—— с — р -—— т -—— = 24 — 42. Щечные зубы секториальные или тупобугорчатые. Верхние резцы небольшие или сильно развитые. Нижний клык отсутствует. Позвонков шейных 7, грудных 11—13, поясничных 5—8, крестцовых 2—3 и хвостовых 8—31.
Желудок простой, слепая кишка отсутствует (хоботноголовые кускусы) или она длинная и простая (представители большинства родов), или длинная с многочисленными складками (коала, кольцехвостые кускусы, гигантские летучие кускусы).
Обитают главным образом в лесистых районах от уровня моря до примерно 4000 м над уровнем моря. Большинство — древесные формы, лишь изредка спускающиеся на землю. Лисий кузу иногда обитает на земле в безлесных местах. Лязящие сумчатые активны в сумерках и ночью. День проводят в дуплах, гнездах птиц. Медлительные животные, превосходно передвигающиеся по деревьям. Большинство растительноядно: питаются листьями, плодами, корнями и корневищами растений. Некоторые насекомоядные или всеядные. В помете 1—4 детенышей (чаще три). В году бывает 1—2 помета. Продолжительность жизни у мелких видов 6 лет, средних — 10, крупных — 12.
Ареал охватывает Австралию, Тасманию, Новую Гвинею и распространяется на запад до о-вов Сулавеси, Тимора, Керама и соседних с ними, на восток — до архипелага Бисмарка и Соломоновых островов.
Некоторые из лазящих сумчатых подверглись истреблению, в настоящее время немногочисленны и охраняются законом.
В семействе 14 родов (43 вида):
кистехвостых кускусов, или кузу — Trichosurus Lesson, 1828 (2 вида); чешуехвостых кускусов — Wyulda Alexander, 1919 (1 вид);
кускусов — Phalanger Storr, 1780 (7 видов);
полосатых кускусов — Dactylopsila Gray, 1858 (2 вида);
карликовых кускусов — Eudromicia Mjoberg, 1916 (3 вида); соневидных кускусов — Cercaertus Burmeister, 1837 (2 вида); беличьих кускусов — Gymnobelideus McCoy, 1867 (1 вид); сумчатых летяг — Petaurus Shaw et Nodder, 1791 (3 вида); перьехвостых кускусов —Distoechurus Peters, 1874 (1 вид); карликовых летучих кускусов — Acrobates Desmarest, 1817 (1 вид); хоботноголовых кускусов — Tarsipes Gervais, 1842 (1 вид);
кольцехвостых кускусов — Pseudocheirus Ogilby, 1837 (17 видов);
сумчатых медведей, или коала, — Phascolarctos De Blanville, 1816 (1 вид);
гигантских летучих кускусов — Schoinobates Lesson, 1842 (1 вид).
Род кузу, или кйстехвостых кускусов,
Genus Trichosurus Lesson, 1828
Средних и крупных размеров лазящие сумчатые. Длина тела 32—58 см. Длина хвоста 24—38 см. Тело приземистое (рис. 43). Глаза крупные. Уши средней длины, треугольные, безволосые. Хвост весь покрыт волосами, за исключением хватательной нижней поверхности на кончике. Когти на лапах длинные и сильные. Самый длинный палец четвертый. Окраска меха от серой и серо-голубой до красно-бурой и черно-бурой. Проявляется половой диморфизм: у взрослых самцов плечи рыжие, на груди имеется специфическая кожная железа, сильнее развитая у самцов. В области заднепроходного отверстия также имеется специфическая кожная железа, проду-3 1
цирующая секрет с мускусным запахом. Зубная формула; i — с —
Рис. 43, Лисий кузу — Т. vulpecula и его череп
1 _9 4
р—-—ш —=34— 36. Резцы, особенно на нижней челюсти, крупные, клыки небольшие. У кузу диплоидное число хромосом 20.
Обитатели кустарниковых зарослей и лесов. Иногда встречаются в открытых местах и даже полупустынях, в парках, пригородах и постройках человека. Ведут древесный образ жизни. Активность в основном ночная. Питаются листьями, цветками и плодами деревьев, особенно эвкалиптов, а также насекомыми. Размножаются в течение всего года, но наиболее интенсивный период размножения приходится на осень и весну, причем некоторые семьи имеют детенышей в оба эти периода. На юго-востоке Австралии самки приносят детенышей в основном в мае и июне. В помете обычно один детеныш. Беременность у лисьего кузу около 17 дней. Детеныш покидает сумку в возрасте примерно 112 дней. Затем еще в течение 1—2 месяцев он остается с матерью, которая носит его на своей спине. Половозрелость наступает в возрасте 15—18 месяцев. Максимальная продолжительность жизни 12—13 лет.
Распространены в Австралии, Тасмании и о-вах Бассова пролива. Лисий кузу был акклиматизирован в Новой Зеландии около 1900 г.
В роде 2 вида:
лисий кузу — Т. vulpecula Кегг, 1792 (ареал совпадает с ареалом рода); собачий кузу — Т. caninus Ogilby, 1836 (юг Квинсленда и Виктория).
Род чешуехвостых кускусов
Genus Wyulda Alexander, 1919
В роде единственный вид: чешуехвостый кускус — W. squamicaudata Alexander, 1919.
Размеры средние: длина тела около 40 см, длина хвоста около 30 см. Голова укороченная, широкая, с короткими, округлыми ушами. Хвост хва-4-565 81
тательный, покрыт волосами только у основания. Конечности относительно короткие, пальцы с небольшими, слабо искривленными когтями. Волосяной покров густой, короткий и мягкий, на спине серый с черноватым оттенком, голова и ноги буроватые, брюхо желтоватое. Вдоль середины спины проходит темная полоса.
Обитает преимущественно в скалах. Образ жизни не известен. Активность ночная. Питается, по-видимому, цветами и плодами.
Распространен на северо-западе Австралии.
Очень редок (известен всего по нескольким, экземплярам) и внесен в «Красную книгу».
Род кускусов
Genus Phalanger Storr, 1780
Размеры от мелких до крупных. Длина тела 24—65 см. Длина хвоста 24—60 см. Тело приземистое, голова притупленная, с небольшими округлыми ушами, лишь слегка выступающими над мехом (рис. 44). Глаза довольно крупные. Когти длинные и сильные. Хвост голый на конце (примерно х/3 длины), хватательный. Самый длинный палец на передних конечностях четвертый. Подошвы лап голые. У большинства представителей волосяной покров очень густой и мягкий, от белого, рыжего и бурого до черного. На теле могут быть белые пятна. Хорошо развитая су лка. Сосков 4. Зубная
2 3 1 2 3 4
формула: i-------с — р ------т — = 34 — 40. Клыки небольшие, тесно
2 0 2—3 4
прилегающие к резцам.
Обитатели лесов и кустарниковых зарослей. Древесные формы. Активность ночная. День проводят, свернувшись клубком в густой листве или дупле. Питаются растительной пищей (листьями, плодами) и мелкими животными (насекомыми, птицами, яйцами птиц). Самка приносит 1—2 детенышей.
Ареал охватывает п-ов Кейп-Йорк в Австралии, Новую Гвинею, о-ва Соломоновы, архипелаг Бисмарка, Молуккские, Сулавеси, Тимор и мелкие острова всей этой области.
В роде 7 видов:
пушистый кускус — Р. orientalis Pallas, 1766 (ареал почти совпадает с ареалом рода);
одноцветный кускус — Р. gyrnnotis Peters et Doria, 1875 (Новая Гвинея, о-ва Ару);
шелковистый кускус — Р vestitus Milne-Edwards, 1877 (Новая Гвинея);
1 1
। < < (
с
г
и л и п
Hl
Л|
в<
D
Рис. 44. Пятнистый кускус — Р. macula-tus
У1 В ( ТР‘ на пр cai
nej
Целебесский кускус — Р. celebensis Gray, 1858 (о-ва Сулавеси и мелкие соседние);
пятнистый кускус — Р. maculatus Desmarest, 1818 (п-ов Кейп-Йорк в Австралии, Новая Гвинея, архипелаг Бисмарка, Керам);
кускус Тейта— Р. atrimaculatus Tate, 1945 (Новая Гвинея) (возможно, является подвидом пятнистого кускуса);
медвежий кускус — Р. ursinus Temminck, 1824 (о-ва Сулавеси и прилегающие мелкие).
Один из подвидов пушистого кускуса, распространенный на п-ове Кейп-Йорк (Р. о. peninsulae), чрезвычайно редок и включен в «Красную книгу».
Род полосатых кускусов
Genus Dactyl-opsila Gray, 1858
Лазающие сумчатые небольшого и среднего размера. Длина тела 17— 32 см и длина хвоста 20—40 см. По внешнему виду похожи на белку. Уши средней величины, треугольные. Хвост длинный, густо покрыт волосами, за исключением обнаженной хватательной концевой нижней поверхности. Пальцы на лапах длинные и тонкие, особенно четвертый палец на передних конечностях, который обладает, кроме того, удлиненным и изогнутым ногтем (сходный признак у ай-ай из отряда приматов). У большого кускуса окраска меха белая или грязно-серая с 3 черными продольными полосами, средняя из которых проходит от лба до основания хвоста, а боковые — отугловрта, через глаза и уши до основания хвоста. На подбородке обычно большое черное пятно. Малый кускус имеет серо-бурый основной цвет меха, а полосы такие же, как у большого полосатого кускуса, но бурого цвета;
3 1 2__3 4
на подбородке пятна нет. Зубная формула: i ——- с — р -——/и — =
= 34 — 40. Резцы крупные и сильные.
Обитатели дождевых и разреженных лесов. Древесные формы. Активность ночная. День проводят в дуплах, где устраивают гнезда из сухих листьев. Питаются в основном личинками насекомых, которых достают из коры когтем своего длинного четвертого пальца. Поедают также мед и плоды.
Распространены на северо-востоке Квинсленда, на Новой Гвинее и некоторых прилегающих островах.
В роде 2 вида:
большой кускус — D. trivirgata Gray, 1858 (ареал совпадает с ареалом рода);
малый кускус-—D. palpator Milne-Edwards , 1888 (Папуа и северо-восток Новой Гвинеи).
Некоторые исследователи выделяют малого кускуса в отдельный род— Dactylonax Thomas, 1910.
Род карликовых кускусов
Genus Eudromicia Mjoberg, 1916
Размеры небольшие. Длина тела 7—12 см. Длина хвоста 7 14 см. Уши средней длины, голые. Хвост хватательный, покрыт волосами только восковании (рис. 45). В хвосте откладывается жир. Самый длинный палец— третий. Волосяной покров на спине имеет окраску рыжеватую или бурую, । на брюхе — белую, бело-желтую или серо-белую. Через глаза от носа могут проходить черные продольные полосы, немного не достигающие ушей. У самок по 4 соска. Зубная формула: i — с -у р -|- т— = 40. Верхний первый и второй предкоренные зубы с двумя корнями.
Рис. 45. Тасманийский кускус — Е. lepida
Обитатели горных лесов на высоте 1500—4500 м над уровнем моря. Ведут древесный образ жизни. Активность сумеречная и ночная. День проводят в дуплах или в гнездах на деревьях. Питаются насекомыми, мелкими ящерицами, семенами, сочными плодами, цветками, почками. При понижении температуры впадают в сон, который может длиться несколько дней (до 6); температура тела при этом сравнивается с окружающей температурой.
Распространены в Тасмании, Квинсленде, Новой Гвинее и некоторых прилегающих островах.
В роде 3 вида:
хвостатый кускус — Е. caudata Milne-Edwards, 1877 (Новая Гвинея и о-в Фергуссон);
карликовый кускус — Е. macrura Mjoberg, 1916 (северо-восток Квинсленда);
тасманийский кускус — Е. lepida Thomas, 1888 (Тасмания).
Род соневидных кускусов
Genus Cercaertus Burmeister, 1837
Размеры небольшие. Длина тела 8—10 см. Длина хвоста 8—11 см, вес взрослого животного 15—25 г. Внешне сходны с карликовыми кускусами, но отличаются от них меньшим числом коренных зубов. Зубная формула: 3 13 3
i — с — р — от — = 36, причем верхние первый и второй предкоренные
имеют только по одному корню. Окраска волос на спинной стороне серая, серс-бурая пли красновато-бурая, на брюшной — белая. У толстохвостого соневндного кускуса глаза окружены темными кругами. Хвост хватательный, густо покрыт волосами в основании и менее густо — к концу. У толстохвостого соневндного кускуса хвост утолщен за счет отложений жира, который расходуется во время длительного сна. Выводковая сумка хорошо развита. Сосков 4 или 6. Диплоидное число хромосом 14.
Обитатели лесов. Ведут древесный образ жизни. Активность ночная. День проводят в птичьих гнездах, дуплах, иногда просто в развилках сучьев. Могут делать собственные гнезда. Осенью сильно жиреют и зимой при понижении температуры впадают в сон. Питаются насекомыми, пауками, нектаром, сочными плодами, орехами и листьями. Самка приносит 4—5 детенышей, по-видимому, 2 раза в год.
Распространены в Австралии и Тасмании.
В роде 2 вида:
толстохвостый кускус — С. nanus Desmarest, 1818 (Тасмания, восток Австралии):;
тонкохвостый кускус — С. concinnus Gould, 1845 (юг Западной Австралии и ее центральных районов).
Род беличьих кускусов
Genus Gymnobelideus McCoy, 1867
В роде единственный вид: беличий кускус — G. leadbeateri Me Соу, 1867 (рис. 46).
Размеры мелкие. Длина тела около 14 см. Длина хвоста около 16 см. Самцы и самки одинаковы по размерам. Внешне сходны с соней-полчком. Уши средней длины, широкие, треугольные, голые. Концевая половина хвоста покрыта удлиненными волосами. Хвост не хватательный, служит лишь для баланса. Пальцы с расширенными концевыми подушечками и небольшими, но крепкими когтями. Самый длинный палец четвертый. Сумка у самок развита хорошо и содержит 4 соска. Волосяной покров на спинной стороне серый и буро-серый с черной срединной полосой, идущей от головы до крестца, и темными пятнами перед и позади глаза и позади уха, 3 13 4
на брюшной — желтый. Зубная формула: i — с — р — т — = 40.
Обитатели густых лесов, встречаются в горах на высоте около 1000 м над уровнем моря. Ведут древесный образ жизни. Активность ночная.
Рис. 46. Беличий кускус — G. leadbeate
Рис. 47. Средняя сумчатая летяга — Р. norfolcensis
Питаются насекомыми и, вероятно, нектаром. Период размножения приходится, по-видимому, на сентябрь.
Распространены на юго-востоке Виктории. До последнего времени считались вымершими. В 1961 г. обнаружена небольшая популяция беличьих кускусов.
Вид внесен в «Красную книгу».
Род сумчатых летяг
Genus Petaurus Shaw et Nodder, 1791
Лазающие сумчатые мелких и средних размеров. Длина тела 12—32 см. Длина хвоста 15—48 см. Внешним видом напоминают белку (рис. 47). Голова укороченная, но заостренная. Уши средней длины, широкие в основании, треугольные, почти голые. Хвост покрыт длинными волосами. От передней части конечности (пятого пальца) вдоль бока тела к задней конечности (к голеностопному суставу) тянется лета
тельная кожная перепонка, покрытая шерстью (на дорзальной поверхности более густой, чем на вентральной). На передних лапах самые длинные пальцы первый и пятый. Верхняя сторона тела серая, буро-серая или темно-бурая, иногда с темными полосами по бокам головы и по спине. Брюшная сторона тела белая или оранжевая. На груди имеется кожная специфическая железа. Сумка развита хорошо, в нее открываются 4 соска. Зубная формула: i с -±- р -|- т = 40. У кар-
ликовой сумчатой летяги в диплоидном наборе 22 хромосомы.
Ведут древесный образ жизни. Могут совершать дальние планирующие прыжки до 55 м длиной. Питаются насекомыми, которых могут ловить на лету, мелкими позвоночными, плодами и цветками деревьев. Активность ночная. День проводят в гнездах, которые делают из листьев в дуплах, причем материал для гнезда приносят при помощи хвоста. В помете 1—3 (обычно 2 ) детеныша. Беременность около 3 недель. Молодые животные в возрасте 4 месяцев переходят к самостоятельной жизни.
Распространены в Австралии, Тасмании, Новой Гвинее и некоторых
прилегающих островах.
В роде 3 вида:
гигантская сумчатая летяга — Р. australis Shaw et Nodder, 1791 (восток Австралии);
средняя сумчатая летяга — Р. norfolcensis Kerr, 1792 (восток Австралии);
k карликовая сумчатая летяга — Р. breviceps Waterhouse, 1839 (Новая Гвинея, восточная половина Австралии и Тасмания).
Род перьехвостых кускусов
Genus Distoechurus Peters, 1874
В роде единственный вид: перьехвостый кускус — D. pennatus Peters, 1874.
Размеры небольшие. Длина тела около 10 см. Длина хвоста 12—15 см. Морда заостроенная. Уши короткие, ширина их больше, чем высота.
Глаза крупные. Хвост длинный с густо покрытыми волосами, у основания и почти безволосой остальной частью, имеющей с каждой стороны по гребню удлиненных волос. Нижняя хватательная сторона кончика хвоста голая. Пальцы с расширенными подушечками на концевых фалангах. Самый длинный палец на передних конечностях четвертый. Когти на пальцах изогнутые, острые. Хорошо развитая сумка открывается вперед. Волосяной покров густой и мягкий, на верхней стороне тела рыжеватый, а нижней белый. Морда белая с черными продольными полосами, которые тянутся через глаза к затылку между ушами и огибают их сзади. Зубная формула: . 3 1 3 3
I — с — р — т —= 34.
2 0 2 3
Обитает в лесах горных районов на высоте до 2000 м над уровнем моря. Экология почти не изучена. Древесное животное. Активность ночная. Питается насекомыми. В помете, по-видимому, один детеныш.
Распространен в Новой Гвинее.
Род карликовых летучих кускусов
Genus Acrobates Desmarest, 1817
В роде единственный вид: карликовый летучий кускус — A. pygmaeus Shaw, 1793 (рис. 48).
Размеры наименьшие в семействе. Длина тела 6—7 см. Длина хвоста 6—9 см. Внешним видом напоминает орешниковую соню. Вдоль боков тела тянется неширокая кожная складка. Хвост длинный с гребнями удлиненных волос по бокам. Кончик хвоста снизу не имеет волос и в этой части является хватательным. Концевые фаланги пальцев с сильно расширенными подушечками. Четвертый палец на передних и задних лапах наиболее длинный и обладает особенно острым когтем. Хорошо развитая выводковая сумка. Сосков 4. Окраска верхней стороны тела изменяется от серой до бурой. Глаза окружены бурым или черным кольцом. Брюхо серо-желтое или белое. Уши покрыты редкими волосами и в основании имеют по пучку удлиненных волос.
Обитатель эвкалиптовых лесов. Ведет древесный образ жизни. Активность ночная. В дуплах устраивает гнезда из сухих листьев. Способен делать планирующие прыжки между деревьями. В холодную погоду впадает
Рис. 48. Карликовый2 летучий кускус — A. pygmaeus
Рис. 49. Хоботноголовый кускус — Т. spenserae
в сон. Держится семейными группами до 16 особей. Питается в основном насекомыми (взрослыми и их личинками), а также нектаром растений. Период размножения приходится на июль — август. В помете 2—4 детеныша.
Распространен на востоке Австралии и на небольшом безымянном острове у северного побережья Новой Гвинеи.
Иногда выделяют (Хальтенорт, 1958) еще один вид: новогвинейский карликовый летучий кускус—A. pulchellus Rotschield, 1893.
Род хоботноголовых кускусов
Genus Tarsipes Gervais et Verreaux, 1842
В роде единственный вид: хоботноголовый кускус — Т. spenserae Gray, 1842 (рис. 49).
Размеры мелкие. Длина тела 7—8 см. Длина хвоста 9—10 см. Конец морды удлинен в небольшой хоботок. Язык длинный, тонкий, способный далеко (до 2,5 см) высовываться изо рта. Глаза маленькие. Уши средней длины, округлые. Хвост хватательный, длинный и тонкий. Волосы покрывают только его основание. Метатарзальный отдел конечностей длинный и узкий. Все пальцы, кроме сросшихся второго и третьего, несущих когти,
расширены дистально и имеют ногти. Имеется своеобразная клоака, куда открываются мочеполовое и анальное отверстия. Сосков 4. Волосяной покров короткий и грубый. Спинная сторона серая, с тремя бурыми продольными полосами. Средняя полоса проходит от затылка до основания хвоста,боковые — от плеч до бедер. Бока тела бледно-красноватые, брюхо желто-белое.
2 113
Зубная формула: i — с — р — т — = 22. Коренные зубы небольших
размеров, слабые.
Обитает в зарослях кустарников и в лесах. Активность ночная. Лазающая форма. Питается насекомыми, пыльцой и нектаром цветков. День про-
водит в гнездах птиц и дуплах деревьев. Распространен на юго-западе Австралии.
Род кольцехвостых кускусов
Genus Pseudocheirus Ogilby, 1837
Многочисленная группа, включающая мелкого и среднего размера кускусов с длиной тела 19—45 см и длиной хвоста 17—37 см. Вес может достигать 1,5 кг. Внешним видом несколько сходны с куницей (рис. 50). Уши короткие, закругленные. Хвост длинный, с хватательным концом. Конец хвоста полностью или только на нижней стороне голый, обычно изогнут в виде кольца. Первый и второй пальцы передней конечности противопоставляются остальным. На передних лапах наиболее длинный палец — четвертый. Волосяной покров густой, мягкий и длинный. На спинной стороне тела окраска его изменчива: от светло-серой, серебристо-серой и зеленовато-черной до черно-бурой, часто с темными и узкими продольными штрихами, реже с тремя темными продольными полосами. На щеках часто бывают темные
Рис. 50. Восточный кускус - Р. peregrinus и его череп
полосы, вокруг глаз или ушей также могут быть темные или светлые пятна. Брюхо белое, серое или желтоватое. Хвост может быть одно-, двух- или трехцветный. Выводковая сумка развита хорошо и открывается вперед. В стыку открываются 4 соска, из которых обычно функционируют только 2.
2__3 1 3 4
Зубная формула: i ——— с — р -—— т — = 32 — 40. У восточного кускуса диплоидное число хромосом — 20.
Преимущественно древесные формы, реже наземные, иногда обитающие на скалах. Активность сумеречная и ночная. День проводят в больших гнездах, которые они строят из листьев на ветвях деревьев. Питаются листьями, цветками, плодами. Передвигаются медленно, придерживаясь й ветки хвостом. Самки приносят обычно 2 детенышей один раз в году.
Распространены в Тасмании, Австралии и Новой Гвинее.
В роде 17 видов:
восточный кускус— Р. peregrinus Boddaert, 1785 (восточное и южное побережья Австралии);
красный кускус — Р. rubidus Troughton et Le Souef, 1929 (юг Квинсленда);
кускус Виктории — Р. victoriae Matschie, 1915 (юг Нового Южного Уэльса);
тасманийский кольцехвостый кускус — Р. convoluter Океп, 1816 (Тасмания);
наземный кускус — Р. occidentalis Thomas, 1888 (юг Западной Австралии);
кускус Герберта — Р. herbertensis Collet 1884 (северо-восток Квинсленда);
пестрохвостый кускус— Р. caroli Thomas, 1921 (северо-запад Новой Гвинеи);
кускус Форбеса— Р. forbesi Thomas, 1887 (Новая Гвинея);
кускус Шлегела — Р. schlegeli Jentink, 1884 (северо-запад Новой Гвинеи);
карликовый кольцехвостый кускус — Р. pygmaeus Stein, 1932 (Новая Гвинея);
собачий кускус — Р. canescens Waterhouse 1846 (Новая Гвинея и о-в Салавати);
полосатый кускус —Р. archeri Collett, 1884 (северо-восток Квинсленда);
длинношерстный кускус — Р. albertisi Peters, 1874 (Новая Гвинея и о-в Я пен);
блестящий кускус — Р. corinnae Thomas, 1897 (Новая Гвинея);
медноволосый кускус — Р. cupreus Thomas, 1897 (Новая Гвинея);
скалистый кускус — Р. dahli Collett, 1895 (север Австралии);
лемуровидный кускус — Р. lemuroides Collett, 1884 (Квинсленд). Наземный кускус редок и включен в «Красную книгу».
Род сумчатых медведей, или коала,
Genus Phascolarctos De Blainville, 1816
В роде единственный вид: сумчатый медведь, или коала, — Р. cinereus Goldfufi, 1817 (рис. 51).
Наиболее крупный представитель семейства. Длина тела 60—82 см. Хвост снаружи незаметен. Вес взрослых особей до 16 кг. Внешним обликом сходен с медведем. Голова большая. Уши крупные, закругленные, покрытые густым мехом. Спинка носа безволосая, черная, резко отграниченная от покрытых волосами участков. Самый длинный палец на передней конечности четвертый. Первый и второй пальцы передних лап противопоставляются остальным. Волосяной покров густой и мягкий, на спинной стороне окраска изменяется от светло-серой до темно-серой, на крестце иногда беловатые пятна, брюхо беловатое. Выводковая сумка развита хорошо и открывается назад (единственный случай в семействе лазящих сумчатых). Щечные мешки имеются. Зубная формула: i -у- с -у- р -у-т ~ = 30. Число хромосом в диплоидном наборе 16.
Обитает в эвкалиптовых лесах. Ведет древесный образ жизни. Очень медлительное животное, питающееся листьями эвкалиптов определенных видов. Активность ночная. Держится поодиночке или небольшими группами. Взрослый самец обычно имеет гарем из нескольких самок. Период размножения с сентября по раннюю осень. Длительность беременности 25— 30 дней. В помете один детеныш, весит при рождении около 5,5 г. Детеныш остается в сумке в течение 6 месяцев, затем еще полгода проводит с матерью. К окончанию лактационного периода отмечено поедание молодыми коала экскрементов своих матерей. Возможно, таким путем в пищеварительный тракт молодых животных попадают от взрослых микроорганизмы, необходимые для пищеварительного процесса. Половозрелость наступает в 3—4 года. Продолжительность жизни около 20 лет.
Распространен на востоке Австралии от северной части Квинсленда до юга Виктории.
В последние годы численность коала сильно снизилась, особенно вследствие избыточного промысла. Коала добывали в большом количестве из-за превосходного меха (например, в 1924 г. из восточных районов Австралии было вывезено более 2 млн. шкур коала). В настоящее время существует запрет на добычу коала и численность их медленно возрастает.
Рис. 51. Коала— Р. cinereus и его череп
Род гигантских летучих кускусов Genus Schoinobates Lesson, 1842
В роде единственный вид: гигантский летучий кускус — S. colons Kerr, 1892.
Размеры мелкие или средние. Длина тела 30—48 см. Длина хвоста 45—47 см. Вес 1—1,5 кг. Внешний вид сходен с сумчатыми летягами. Уши большие, округлые, покрытые густым мехом. Самый длинный палец передней конечности — четвертый. Хвост длинный, весь покрыт волосами, за исключением нижней части кончика. От локтевого сустава вдоль бока тела к коленному суставу тянется летательная складка кожи, покрытая шерстью. Волосяной покров очень длинный, густой и мягкий, на спинной стороне имеет изменчивую окраску от черной или серой до светлой и белой, на брюшной грязно-серый или белый. В хорошо развитую выводковую
3 113
сумку открываются 2 соска. Зубная формула: I -—— с — р — т — =
= 28 — 30. Хромосом в диплоидном наборе 22.
Обитатель горных лесов. Ведет древесный образ жизни. Активность ночная. День проводит в дуплах деревьев. Питается листьями и почками эвкалиптов. Может совершать планирующий полет до 100 м длиной. Самка приносит одного детеныша (в Виктории в июле — августе), который остается в сумке матери, пока не достигнет возраста 4 месяца.
Распространен в прибрежных горных областях на востоке Австралии от северо-востока Квинсленда до Виктории.
СЕМЕЙСТВО ВОМБАТОВ
FAMILIA PHASCOLOMVIDAE GOLDFUSE, 1820
По внешнему облику вомбаты напоминают небольших медведей: тело толстое, короткое, приземистое и неуклюжее. Это средних размеров сумчатые с длиной тела 67—105 см и весом 15—27 кг. Хвост рудиментарен. Голова утолщенная, широкая. Нос голый или покрыт волосами. Уши средних размеров. Конечности короткие, сильные, примерно одинаковой длины, пятипалые, вооруженные большими когтями. Первый палец на задних лапах развит слабо и лишен когтя. Второй и третий пальцы на задних конечностях частично срастаются между собой. Волосяной покров грубый, без пуха у короткошерстных вомбатов и мягкий с пуховыми волосами у длинношерстных вомбатов. Выводковая сумка открывается назад. Сосков одна пара. У вомбатов найдены слабо развитые щечные мешки. Желудок простой со специализированными кардиальными железами. Диплоидное число хромосом 14.
Череп массивный, уплощенный. Зубная формула: i -5- с-5- p-LmA_ = = 24. Зубы гипселодонтные, с постоянным ростом. Резцы крупных размеров, покрыты эмалью только спереди и с боков. От щечных зубов они отделены диастемой. Предкоренные и коренные крупные, с низкими буграми на жевательной поверхности (билофодонтные). Характер жевания сходен с таковым у грызунов.
Позвонков у короткошерстных вомбатов: шейных 7, грудных 15, поясничных 4, крестцовых 4 и хвостовых 10—12, а у длинношерстных вомбатов, соответственно, 7, 13, 6, 4, 15—16. Атлас незамкнут с вентральной стороны. Ребер у короткошерстных вомбатов 15, а у длинношерстных 13.
Обитатели лесов, саванн и с-тепей в равнинных и горных областях. Преимущественно ночные звери, осторожные. Вырывают длинные и глубокие норы; описаны сложноустроенные норы вомбатов более 30 м длиной. Разрывают землю передними лапами и отбрасывают ее задними лапами, мощными резцами перекусывают попадающиеся корни. В конце норы устраивают из травы и коры гнездо. Около выхода из норы они обычно устраивают место для отдыха — лежку, где принимают солнечные ванны. К местам кормежек ведут натоптанные тропы. Питаются вомбаты в основном травой, корой и корнями высших растений, а также поедают водоросли. Способны валить крупные деревья, подобно тому, как это делают бобры. Обычно вомбаты держатся поодиночке, за исключением времени размножения. Самка приносит единственного детеныша в период с января по июль. Детеныш остается в сумке матери до декабря, когда он уже полностью покрывается шерстью.
Распространены на юго-востоке Австралии, в Тасмании и на о-ве Флиндерса в Бассовом проливе. Численность вомбатов в последние годы сильно снизилась из-за прямого уничтожения.
В семействе 2 рода (2 вида):
Рис. 52. Короткошерстный вомбат — V. иг sinus и его череп
короткошерстных, или голоносых, вомбатов — Vombatus Geoffroy St. Hilaire, 1803 (1 вид);
длинношерстных, или шерстоносых, вомбатов — Lasiorchinus Gray, 1863 (1 вид).
Род короткошерстных, или голоносых, вомбатов
Genus Vombatus Geoffroy St.-Hilaire, 1803
В роде единственный вид: короткошерстный, или голоносый, вомбат — У. ursinus Shaw, 1800 (рис. 52).
Длина тела 67—105 см. Голова большая, округлая. Спинка носа безволосая, черная. Уши с закругленной вершиной. Профиль спины приподнят в крестце, закруглен. Волосяной покров короткий и грубый. Пуховые волосы отсутствуют или развиты слабо. Окраска однообразно желто-серая или песчаная, изменяющаяся до черноватого цвета. Иногда горло и грудь более светлые. В желудке (в области малой кривизны) обнаружена особая железа, присущая также коала и бобру. Хромосом в диплоидном наборе 14.
Описание экологии см. в характеристике семейства.
Распространен в Новом Южном Уэльсе, Виктории, юго-восточной части Южной Австралии, Тасмании, на о-ве Флиндерса в Бассовом проливе.
Подвид короткошерстного вомбата, обитающий на о-ве Флиндерса Г. и. ursinus, редок и внесен в «Красную книгу».
Род длинношерстных, или шерстоносых, вомбатов Genus Lasiorchinus Gray, 1863 В роде единственный вид: длинношерстный, или шерстоносый, вомбат— L. latifrons Owen, 1845 (рис. 53) (иногда выделяют и второй вид: шерстоносого вомбата Барнарда — L. barnardi Longman, 1939).
Рис. 53. Длинношерстный вомбат — L. 1а-tifrons
Длина тела 87—102 см. Голова не столь массивна, как у короткошерстного вомбата. Спинка носа белая или бурая и покрыта волосами. Уши небольшие, треугольные. Профиль спины более прямой, чем у короткошерстного вомбата. Волосяной покров длинный и мягкий, пятнистый, буро-серый. Пуховые волосы хорошо развиты. Часто под глазами и над ними имеются белые пятна.
Описание экологии см. в характеристике семейства.
Распространен в юго-восточной части Южной Австралии, Западной Виктории, юго-западных районах Нового Южного Уэльса, центральном и южном Квинсленде. Численность длинношерстных вомбатов в последние годы сильно снизилась, и он включен в «Красную книгу».
СЕМЕЙСТВО КЕНГУРУ
FAMILIA MACROPODIDAE OWEN, 1839
Размеры очень различны. Длина тела мускусных крысиных кенгуру 25 см, а исполинских кенгуру — 160 см (наибольшая в отряде сумчатых). Длина хвоста от 15 до 105 см. Вес взрослых животных от 1,4 до 90 кг. Часто самцы крупнее самок. Голова относительно невелика, с укороченной или длинной мордой. Носовое зеркало безволосое или покрыто волосами. Уши большие или укороченные, округлые или овальные, слабо или густо покрытые волосами. Хвост обычно длинный, иногда относительно короткий, покрыт волосами по всей длине и у большинства представителей не хватательный. Он постепенно суживается от утолщенного основания к концу или остается по всей длине равной толщины (скалистые кенгуру, карликовые скалистые кенгуру), иногда с ороговевшим участком кожи на конце (когтехвостые кенгуру). У всех кенгуру, за исключением древесных, задние конечности значительно длиннее и сильнее передних. Передние конечности пятипалы, с большими или очень большими когтями. На задних конечнос
тях первый палец отсутствует (имеется только у мускусных кенгуру), небольшие второй и третий пальцы соединены между собой кожистой перепонкой, четвертый палец сильно развит и обладает мощным ногтем, пятый палец умеренной длины.
Волосяной- покров обычно густой и мягкий, окраска от черной, серой или бурой до рыжей и желтой. На спине или крестце могут быть полосы, 'тогда на морде имеются пятна. У самок хорошо развита выводковая сумка, открывающаяся вперед. Из имеющихся четырех сосков обычно функ-
ционируют только два.
Зубная система весьма изменчива, типичная формула зубов: i — с
1—2
1 — 2
4(7)
4(7)
т
= 30 — 34(44). Верхние резцы хорошо развиты.
Нижний
и верхний первые резцы особенно велики и направлены вперед. Второй и
третий нижний резцы отсутствуют, верхний клык небольшой или его нет совсем. Имеется диастема между резцами и предкоренными зубами. Предкоренные узкие, пластинчатые. Коренные широкие, квадритуберкулярные или билофодонтные. У карликового скалистого кенгуру отмечена своеобраз-
ная смена зубов, характерная такжедля слонов и сирен и не встречающаяся'у других сумчатых. Новые коренные (скрытые до этого в десне) появляются у заднего конца зубного ряда и сдвигают зубы вперед. Обычно можно видеть по 4—5 (даже 6) коренных одновременно с каждой стороны челюсти. Позвонков шейных 7, грудных 13, поясничных 6, крестцовых 2, хвостовых 21—25. Атлас обычно представляет собой неполное кольцо, незамкнутое на вентральной стороне.
Слизистая оболочка щеки от угла рта до глотки обладает железистым полем мягких сосочков. Желудок с удлиненной кардиальной железистой областью, сходной с толстым кишечником, которая вместе с эофа-гиальной бороздой обеспечивает возврат пищи в ротовую полость для повторного пережевывания. Слепая кишка имеется. Самцы обладают единым мочеполовым протоком. Диплоидное число хромосом колеблется у разных видов от 10 до 24.
Обитают в ландшафтах различного типа. Большинство ведут наземный образ жизни, но один вид древесный. У большинства кенгуру ночная активность. День проводят в травяных гнездах или в неглубоких норах. Но есть и дневные виды. Все представители семейства, за исключением мускус
ных крысиных и древесных кенгуру, передвигаются прыжками, используя только задние конечности. Наибольшего развития такой способ передвижения достигает у исполинских кенгуру. Хвост при этом служит своего рода балансиром. Скорость движения, которую могут развить крупные кенгуру, достигает 50 км/ч. Держатся преимущественно небольшими группами, состоящими из одного самца и нескольких самок.
Кенгуру растительноядны: в список их кормов входят различные виды и части растений. Однако мускусные крысиные кенгуру питаются также насекомыми и червями. Специфическая флора бактерий обеспечивает у кенгуру ферментативный пищеварительный процесс. Самки приносят одного, редко двух детенышей один раз в году (некоторые виды имеют более одного помета в течение года). Вероятно, большинству видов кенгуру свойственна задержка в имплантации — диапауза развивающейся бластоцисты. Длительность эстрального цикла от 28 до 42 дней. Беременность 27—40 дней. Мелкие по размерам кенгуру живут до 8 лет, средних размеров до 12, крупные до 16.
Область распространения охватывает Австралию, Тасманию, Новую Гвинею, некоторые прилежащие острова и архипелаг Бисмарка. Акклиматизированы в Новой Зеландии.
В случае высокой численности кенгуру могут приносить вред сельскому хозяйству, однако такие случаи редки. Некоторых кенгуру разводят на фермах. Представителей рода исполинских кенгуру добывают ради меха
(например, в Квинсленде в 1959 г. было заготовлено 584 тыс. шкур). Местное население употребляет некоторых кенгуру в пищу. В настоящее время из мяса кенгуру изготавливают консервы, имеющие спрос на внутреннем рынке и идущие на экспорт.
В семействе 17 родов (51 вид);
мускусных кенгуру — Hypsiprimnodon Ramsay, 1876 (1 вид);
широкоголовых кенгуру — Potoroops Matschie, 1916 (1 вид);
потору — Potorous Desmarest, 1804 (2 вида);
гологрудых кенгуру — Caloprymnus Thomas, 1888 (1 вид);
короткомордых кенгуру — Bettongia Gray, 1837 (3 вида);
больших крысиных кенгуру — Aepyprymnus Garrod, 1875 (1 вид);
скалистых кенгуру — Petrogale Gray, 1837 (6 видов);
когтехвостых кенгуру — Onychogalea Gray, 1841 (3 вида);
заячьих кенгуру — Lagorchestes Gould, 1841 (3 вида;);
полосатых кенгуру — Lagostrophus Thomas, 1887 (1 вид);
карликовых скалистых кенгуру — Perardorcas Thomas, 1904 (1 вид); кустарниковых кенгуру—Dorcopsis Schlegel et Miiller, 1842 (5 видов); древесных кенгуру—Dendrolagus Miiller, 1839 (7 видов);
филандеров — Thylogale Gray, 1837 (6 видов);
короткохвостых кенгуру, или куокка, —• Setonyx Lesson, 1842 (1 вид); средних кенгуру, или валлэби, — Protemnodon Owen, 1873 (6 видов); исполинских кенгуру — Macropus Shaw et Nodder, 1790 (3 вида).
Некоторые исследователи выделяют роды мускусных, широкоголовых, длинномордых, пустынных, короткомордых и больших крысиных кенгуру в отдельное семейство.
В настоящее время кенгуру нуждаются в серьезной охране, так как численность их значительно снизилась.
Род мускусных кенгуру
Genus Hypsiprymnodon Ramsay, 1876
В роде единственный вид: мускусный кенгуру — Н. moschatus Ramsay, 1876 (рис. 54).
Размеры наименьшие в семействе. Длина тела около 25 см. Длина хвоста около 15 см. Вес взрослых животных примерно 500 г. Внешний вид сходен с крысой. Голова короткая, морда заостренная. Уши средней длины, голые, слегка заостренные. Характерно, что, в отличие от других кенгуру, задние конечности мускусного кенгуру имеют хорошо выраженный подвижный первый палец, однако не противопоставляющийся другим пальцам и не имеющий когтя. Хвост относительно короткий и в отличие от других представителей семейства не имеет волос и покрыт чешуей. Волосяной покров густой и мягкий, на спинной стороне темный, красновато-серый, бока тела оранжевые, брюхо желтоватое. От зверей обоих полов сильно пахнет мускусом. Секрет с этим запахом, видимо, продуцируют какие-то кожные железы. Зубная формула: i -у- с — р 1 "2- т = 32 — 34. Первый верхний резец длинный, вдвое превышает по длине два остальных резца. Последний предкоренной, особенно в верхней челюсти, узкий, с высокой режущей коронкой, огибающей снаружи первый коренной зуб. Диплоидное число хромосом 22.
Обитает в густых зарослях кустарников, в Дождевых лесах, часто по берегам рек и озер. Активность дневная. Держится поодиночке и парами. Проворное животное, передвигающееся обычно на всех четырех лапах, но при быстром движении прыгающее только на задних. Питается насекомыми,
Рис. 54. Мускусный кенгуру — Н. moschatus и его череп
червями, корнями растений, ягодами. Размножение приходится на дождливый сезон с февраля по май.
Распространен на северо-востоке Квинсленда.
Численность мала: включен в «Красную книгу».
Род широкоголовых кенгуру
Genus Poloroops Matschie, 1916
В роде единственный вид: широкоголовый кенгуру — Р. platyops Gould, 1844.
Размеры небольшие. Длина тела около 35 см. Длина хвоста около 20см. Голова очень короткая и широкая. Уши средней длины, овальные, на вершине заостренные. Безволосый участок носового зеркала на спинку носа не распространяется. Задние конечности без первого пальца. Средний палец на передних лапах с особенно крупными когтями. Волосяной покров на спинной стороне буро-серый, на брюшной беловатый. Зубная формула: t — с — р — tn — — 341. Верхний первый резец мощно развит, значительно сильнее, чем остальные резцы. Коренные уменьшаются в размерах в направлении спереди назад.
Активность дневная. Передвигаются прыжками на задних конечностях или медленно на всех четырех лапах. Питается различными растительными объектами.
Распространен на юго-западе Западной Австралии.
1 Такая же зубная формула характерна для остальных родов крысиных кенгуру: лотору, гологрудых, короткомордых и больших.
Род потору
Genus Pot or ous Desmarest, 1804
Размеры мелкие. Длина тела около 40 см. Длина хвоста 17—23 см. Голова небольшая, узкая (рис. 55). Особенности в строении конечностей, зубной системы и т. д. сходны с таковыми широкоголовых кенгуру. Голое носовое зеркало распространяется примерно на 1,5 см на спинку носа. Волосяной покров длинный и мягкий, на спинной стороне серо-буро-красноватый (зимой черноватый), на брюшной беловатый. Диплоидное число хромосом 13 у самцов и 12 у самок.
Обитают в густых зарослях кустарника, подлеске, но могут встречаться и в степи. Активность ночная. Питаются надземными частями травянистых растений, корнями. Размножаются в течение всего года. В помете один детеныш. Беременность 38 дней. В этот период самка устраивает примитивное гнездо из травы. Детеныш остается в сумке примерно 126 дней.
Распространены в южной части Западной и Юго-восточной Австралии и Тасмании. Везде очень малочисленны.
В роде 2 вида:
трехпалый потору — Р. tridactylus Kerr, 1792 (восток Нового Южного Уэльса, Виктория, Южная Австралия и Тасмания);
потору Гильберта — Р. gilberti Gould, 1841 (юго-западные районы Западной Австралии).
По-видимому, род потору может быть объединен с родом широкоголовых кенгуру.
Подвид трехпалого потору, населяющий материк Р. t. tridactylus, к настоящему времени исчез из Южной Австралии и редок в остальных частях ареала. Внесен в «Красную книгу».
Рис. 55. Трехпалый потору— Р. tridactylus и череп Poto-rous sp.
Рис. 56. Кистехвостый кенгуру — В. lesueur и его череп
Род гологрудых кенгуру
Genus Caloprymnus Thomas, 1888
В роде единственный вид: гологрудый кенгуру — С. campestris Gould, 1843.
Размеры небольшие. Длина тела около 44 см. Длина хвоста около 36 см- Внешним видом и строением напоминает широкоголовых крысиных кенгуру. Характерна расширенная лобная часть головы и удлиненные задние конечности, которые развиты относительно сильнее, чем у других крысиных кенгуру. Передние конечности небольшие. На груди и нижней части шеи имеется участок безволосой кожи, представляющий собой, по-видимому, специфическую кожную железу. Уши длинные и более узкие, чем у других крысиных кенгуру. Хвост равномерной толщины, покрыт короткими волосами. Волосяной покров мягкий, на спинной стороне песчаного цвета, переходящего в серый, на брюшной беловатый.
Обитатель песчаных пустынь. Передвигается прыжками на задних конечностях. По сравнению с другими крысиными кенгуру в большей степени питается надземными, в меньшей— подземными частями растений. Сезонности в размножении, по-видимому, не существует.
Распространен в северной части Южной Австралии и на юго-западе Квинсленда.
В настоящее время численность этого кенгуру настолько мала, что он находится под угрозой исчезновения. Включен в «Красную книгу».
Род короткомордых кенгуру
Genus Bettongia Gray, 1837
Размеры небольшие. Длина тела 26—46 см. Длина хвоста 26—31 см. Вес около 1,5 кг. Своим обликом и строением напоминают широкомордых крысиных кенгуру. Носовое зеркало красноватого цвета. Уши укороченные и округлые (рис. 56). Хвост частично хватательный. Верхняя сторона кончика хвоста с гребнем жестких волос. Волосяной покров густой и мягкий, на спинной стороне от бурого, серо-бурого, серого и темно-серого до светло-красноватого, на брюшной — серый или белый. Копчик хвоста иногда белый. В черепе большие небные отверстия. Диплоидное число хромосом у короткомордого кенгуру 28, у тасманийского и кистехвостого — по 22.
Обитатели обычно открытых ландшафтов. Характерно, что эти кенгуру часто держатся большими группами. Строят наземные гнезда из травы. Строительный материал переносят при помощи цепкого хвоста. Кистехвостый
крысиный кенгуру выкапывает себе норы. Ведут ночной образ жизни. Питаются в основном подземными частями растений. Самка приносит потомство несколько раз в год. Беременность примерно 22 дня. Детеныш остается в сумке около 120 дней.
Распространение охватывает Тасманию и южную половину Австралии от западного до восточного побережий.
В роде 3 вида:
короткомордый кенгуру — В. gaimardi Desmarest, 1822 (юго-запад Австралии и север Квинсленда);
тасманийский кенгуру — В. cuniculus Ogilby, 1838 (Тасмания);
кистехвостый кенгуру — В. lesueur Quoy et Gaimard, 1824 (западная и центральная часть Южной Австралии).
Численность всех трех видов очень мала, и все они включены в «Красную книгу».
Род больших крысиных кенгуру
Genus Aepyprymnus Garrod, 1875 -
В роде единственный вид: большой крысиный кенгуру — A. rufescens Gray, 1837.
Размеры мелкие сравнительно с другими представителями семейства, но наибольшие среди крысиных кенгуру. Длина тела около 52 см. Длина хвоста около 38 см. Уши более широкие и округлые, чем у других крысиных кенгуру.
Две узкие полосы волос идут от спинки носа к верхней части носового зеркала. В остальном сходен с другими крысиными кенгуру. Волосяной покров длинный и довольно грубый, на спинной стороне серо-красноватый с беловатыми полосами на огузке, на брюшной белый. Задние части ушей черные.
Обитает в прибрежных ландшафтах, населяя степи, саванны и светлые леса. Держится поодиночке. Активность ночная. Строит из травы гнездо, в котором проводит дневное время (возможно, только в зимний период). Питается в основном корнями растений.
Распространен от востока Квинсленда до востока Нового Южного Уэльса.
Численность этого кенгуру сильно пострадала в результате акклиматизации лисиц.
Род скалистых кенгуру
Genus Petro gale Gray, 1837
Мелких и средних размеров: длина тела 38—80 см, длина хвоста 35— 90 см, вес от 3 до 9 кг. Голова довольно длинная (рис. 57). Носовое зеркало голое. Уши длинные. Коготь среднего пальца задней конечности короткий. Первый палец задней конечности отсутствует. Хвост длинный, одинаковой толщины от основания до кончика, полностью покрыт волосами. На конце хвоста небольшая кисть удлиненных волос. Хвост служит главным образом балансом при движении и в меньшей степени опорой при сидении. Волосяной покров мягкий, длинный и густой. Волосы спины от загривка или середины спины до затылка направлены вершинами вперед (кроме кольцехвостого кенгуру). Окраска спинной стороны красно-бурая, желто-серая, буро-серая или серая, брюшной — желто-белая, серо-белая или белая. На спине или голове могут быть темные пятна или полоски. Щеки с белыми
3 0 2 4
полосами. Зубная формула: i — с — р —т — = 32. Первый верхний резец лишь немного крупнее второго и третьего. Коренные увеличиваются 100
в направлении спереди назад. У кистехвостого кенгуру диплоидное число хромосомного набора 22.
Обитают в скалистых участках гор. Активность сумеречная. Хорошо лазают по скалам и необычайно быстро двигаются, лавируя между камнями и делая прыжки до 4 м. Питаются травянистой растительностью.
Распрост ранение охватывает почти всю Австралию.
В роде 6 видов: кистехвостый ска-
Рис. 57. Кистехвостый скалистый кенгуру — Р. penicillata
листыи кенгуру — Р. penicillata Griffith, 1827 (Австралия);
кенгуру Квинсленда — Р. inornata Gould, 1842 (Квинсленд);
кольцехвостый кенгуру — Р. xanthopus Gray, 1885 (Южная Австралия, юго-запад Квинсленда, Новый Южный Уэльс и Виктория);
короткоухий кенгуру— Р. brachyotis Gould, 1841 (северо-запад и север
Австралии);
кенгуру Лонгмана — Р. longmani Thomas, 1926 (север Австралии);
кенгуру Уилкинса — Р. wilkinsi Thomas, 1926 (север Австралии).
Подвид кистехвостого скалистого кенгуру — Р. р. penicillata и подвид кольцехвостого кенгуру Р. х. xanthopus включены в «Красную книгу».
Род когтехвостых кенгуру
Genus Onychogplea Gray, 1841
Размеры мелкие. Длина тела 38—65 см. Длина хвоста 33—65 см. Без-। волосое носовое зеркало очень мало по размерам. Коготь среднего пальца передней конечности очень длинный. Уши длинные (рис. 58). Хвост обычно равен длине тела, покрыт короткими волосами. Он утончается от основания к концу, служит опорой при сидении и балансом при прыжках. На I конце хвоста своеобразный роговой коготь неизвестной функции. Волосяной покров на спинной стороне тела серый, желто-бурый, красновато-се-। рый, темно-буро-серый, иногда с темными продольными штрихами на спине или полосками, проходящими через глаз, на брюшной — белый. Зубная 3 0(1) 2 4
формула: i — с —р — т — == 32 (34). Первый верхний резец крупный, второй меньше первого, а третий меньше второго. Верхний клык отсутствует или очень мал. У плоскокоготного кенгуру диплоидное число хромосом 20.
I Обитатели густых парковых лесов. Активность сумеречная и ночная. IОбычно держатся поодиночке. Питаются в основном корнями растений, в I меньшей степени надземными травянистыми растениями. Быстрые прыгуны. Самка приносит единственного детеныша преимущественно в мае.
Распространены с северо-западной до северо-восточной и с юго-западной | до юго-восточной Австралии. На значительной части прежнего ареала те-
перь исчезли вследствие уничтожения акклиматизированными лисицами и вытеснения домашним скотом.
В роде 3 вида:
плоскокоготиый кенгуру — О. unguifer Gould, 1840 (северо-запад Австралии и Квинсленд);
короткокоготный кенгуру— О. f гае па t a Gould, 1841 (юг Квинсленда, Виктория, Новый Южный Уэльс);
луннокоготный кенгуру — О. lunata Gould, 1840 (Южная Австралия).
Плоскокоготиый и луннокоготный кенгуру внесены в «Красную книгу».
Род заячьих кенгуру
Genus Lagorchestes Gould, 1841
Размеры мелкие. Длина тела 37—52 см. Длина хвоста 25—46 см. Самки крупнее самцов. Носовое зеркало отсутствует или оно очень мало. Уши короткие (пучкохвостый и очковый кенгуру) или средней длины (длинноухий кенгуру). Задние конечности и их пальцы особенно удлинены. Хвост утоньшается от основания к концу и служит опорой при сидении. Волосяной покров на спинной стороне буро-серый, красновато-или
i А
темно-буро-серый, на брюшной бело-серый. Зубная формула: с — р — т — = 34. Размеры резцов в верхней челюсти уменьша-
1
j г г и н
ются от первого к третьему. Верхний клык имеется; иногда он развит слабо. У пучкохвостого кенгуру диплоидное число хромосом 22.
Обитатели травянистых степей, саванн и степей с зарослями кустарников. Активность ночная. Быстрые прыгуны. Могут совершать прыжки до 2—3 м длины. Держатся поодиночке. Иногда копают норы, в которых устраивают гнездо. Питаются травянистой растительностью.
Распространены в Новом Южном Уэльсе, Южной и Западной Австралии и на Северной Территории.
В роде 3 вида:
длинноухий кенгуру — L. leporides Gould, 1841 (Новый Южный Уэльс, Южная Австралия);
пучкохвостый кенгуру — L. hirsutus Gould, 1844 (Западная Австралия);
очковый кенгуру — L. conspicillatus Gould, 1841 (Северная Территория).
Пучкохвостый кенгуру в последние годы стал редок и внесен в «Красную книгу».
Род полосатых кенгуру
Genus Lagostrophus Thomas, 1887
Вроде единственный вид: полосатый кенгуру — L. fasciatus Рёгоп et Lesueur, 1887.
Размеры мелкие: длина тела около 40 см, длина хвоста около 40 см. Морда сильно вытянута. Имеется безволосое носовое зеркало. Уши средней длины. Задние конечности и пальцы на них особенно длинные. Хвост постепенно сужается от основания к концу и служит опорой при сидении и балансом при прыжках. Окраска густого мягкого и длинного меха на спине серая с черноватыми узкими штрихами. В средней и задней частях спины располагается примерно 13 черноватых широких поперечных полос. Брюхо белое. Зубная формула: iс р т~ = 32. Верхний клык от-
сутствует. Диплоидное число хромосом 24.
Обитает в разного типа степях, зарослях кустарников, иногда около болот. Активность сумеречная и ночная. Питается различными растениями и плодами.
Распространен в центральной части Западной Австралии и на нескольких мелких островах в заливе Шарк.
Численность очень мала, и, возможно, на материке в настоящее время полностью уничтожен. Вид включен в «Красную книгу».
Род карликовых скалистых кенгуру
Genus Perardorcas Thomas, 1904
В роде единственный вид: карликовый скалистый кенгуру — Р. concin-па Gould, 1842.
Размеры мелкие. Длина тела 35—40 см. Длина хвоста 30—33 см. Голова укороченная, морда заостренная. Хвост имеет одинаковую толщину по всей длине и служит балансом при прыжках. Волосяной покров короткий, мягкий, на спинной стороне от светло-оранжевого до оранжево-серого и сероватого, на брюшной серо-белый или белый. Основание хвоста окрашено, как и тело, а терминальная его часть более темная. Зубная формула:
3 0 2 7
i — с — р — т— = 44. Верхний клык отсутствует. Коренные при старании нарастают сзади наперед (подробнее см. в описании семейства).
Обитатель кустарниковых зарослей на скалах и холмах. Экология изучена слабо; вероятно, сходна с экологией скалистых кенгуру. Питается травянистыми растениями.
Распространен на северо-западе и севере Австралии.
Род кустарниковых кенгуру
Genus Dorcopsis Schlegel et Miiller, 1842
Размеры мелкие. Длина тела 42—70 см. Длина хвоста 30—45 см. У кенгуру Маклая и горного кустарникового кенгуру размеры самцов и самок примерно одинаковы, а у остальных трех видов рода самцы примерно naVg крупнее самок. Голова узкая и заостренная. Уши маленькие и закругленные. Безволосое носовое зеркало больших размеров. Хвост длинный, постепенно утончающийся от основания к кончику; на его конце удлиненных волос нет. Хвост служит опорой тела при сидении и балансом при прыжках. У кенгуру Маклая задняя треть хвоста непокрыта волосами. У полосатого кустарникового кенгуру конец хвоста белый и почти не имеет волос; у кенгуру Гудинафа весь хвост слабо оволоснен, за исключением средней линии верхней стороны. Задние конечности сравнительно короткие. Волосяной покров на спинной стороне тела имеет окраску от серо-бурой до черно-бурой, могут быть темные пятна. Брюхо светло-буро-серое, серое или белое. Волосы на спине от загривка или середины спины и до затылка вершинами направлены вперед. Зубная формула: t
= 34., Верхний клык сравнительно крупный.
Обитают в густом подлеске низменных и дождевых или горных лесов и в кустарниковых зарослях. Питаются надземными и подземными частями травянистых растений. Активность преимущественно ночная.
Распространены в Новой Гвинее.
В роде 5 видов:
кенгуру Маклая—D. macleayi Miklouho-Maclay, 1885 (юго-восток Гвинеи);
горный кустарниковый кенгуру—D. vanheumi Thomas, 1922 (запад, центр и восток Новой Гвинеи и восток Папуа);
полосатый кустарниковый кенгуру — D. veterum Lesson, 1827 (юг Папуа и мелкие прилегающие острова);
, кенгуру Гудинафа—D. atrata Van Deusen, 1957 (о-в Гудинаф из группы о-вов Д’Антркасто);
кенгуру Хагена — D. hageni Heller, 1897 (западная часть Новой Гвинеи).
Некоторые исследователи выделяют 3 последних вида в особый род Dorcopsis Schlegel et Miiller, 1845. По-видимому, более правильно считать их объединенными в ранге подрода.
Род древесных кенгуру
Genus Dendrolagus Muller, 1839
Размеры средние. Длина тела 50—90 см. Длина хвоста 43—85 см. Голова довольно короткая и широкая. Носовое зеркало широкое. Характерно, что задние конечности лишь немного длиннее, чем большие и сильные передние конечности (рис. 59). Подошва лап очень грубая. Когти на относительно коротких пальцах мощные, сильно изогнутые. Второй и третий пальцы крупнее четвертого и пятого. Хвост очень длинный, равномерной толщины на всем своем протяжении, весь покрыт волосами. Хвост не хватательный, служит в качестве баланса и для опоры при сидении. Волосяной покров очень густой, мягкий и довольно длинный, на спине от серой,
бурой-серой до золотисто-бурой, золотисто-красной, черно-бурой и черной окраски. Волосы на спине, начиная от крестца, середины спины или загривка и до затылка, направлены вперед. Брюшная сторона белая, серо-белая, желто-белая, золотисто-желтая, красноватая или оранжево-бурая. Зубная , .3124
формула: t — с — р ~т —=
= 34. Верхний клык имеется, хотя и небольших размеров (меньше резцов).
Обитают в лесах. Ведут древесный образ жизни, однако хорошо и быстро двигаются и по земле. С дерева на дерево делают прыжки до 9 м. Могут без вреда для себя прыгать на землю с большой высоты. Питаются листвой деревьев, плодами и травянистыми растениями. Держатся небольшими группами, обычно состоящими из одного самца и нескольких самок.
Рис. 59. Кенгуру Дориа — D. dorianus и его череп
Распространены в северных районах Квинсленда и Новой Гвинее.
В роде 7 видов:
кенгуру Лумхольтца — D. lumholtzi CoWett, 1884 (северо-восток Квинсленда);
медвежий кенгуру — D- ursinus Temminck, 1836 (запад Новой Гвинеи);
кенгуру Матши—D. matschiei Forster et Rothschild, 1907 (юго-запад Новой Гвинеи);
кенгуру Гудфеллоу — D. goodfellowi Thomas, 1908 (Папуа);
кенгуру Дельта — D. deltae Troughton et Le Souef, 1936 (область Дельта в Папуа);
западный древесный кенгуру — D. spadix Troughton et Le Souef, 1936 (западное Папуа);
кенгуру Дориа — D. dorianus Ramsay, 1883 (восточное Папуа и северо-восточный Квинсленд).
Род филандеров
Genus Thylogale Gray, 1837
Мелких размеров кенгуру. Длина тела 55—70 см. Длина хвоста 30— 42 см. Уши короткие и узкие. Имеется носовое зеркало. От зеркала вверх выдается небольшой выступ голой кожи. Задние конечности относительно короткие. Волосяной покров густой и мягкий, на спинной стороне от темнооливкового до серо-бурого, темно-бурого и красно-бурого цвета. В области поясницы может проходить светлая полоса. Брюхо красно-оранжевое, серо-белое или белое. Морда иногда с черной полоской, тянущейся позади глаза к углу рта. Хвост покрыт относительно редкими волосами. Зубная формула: t — с — р ~ 32. Верхний клык отсутствует. Хромо-
сом в диплоидном наборе у краснобрюхого, красноногого и красношеего филандеров 22.
Обитают в высоких и густых зарослях травы, кустов и подлеска в лесах и в зарослях кустарников. В зарослях проделывают туннели. Держатся поодиночке. Активность ночная, днем спят в густых кустарниках. Питаются травянистыми растениями, листьями кустарников. Передвигаются на четырех или только на задних лапах. В случае опасности подают сигнал, топая задней лапой по земле.
Распространены в Австралии и на о-вах Флиндерса и Кенгуру, на мелких о-вах Бассова пролива, в Тасмании, Повой Гвинее, Архипелаге Бисмарка, о-вах Ару и Кей.
Хотя численность за последние годы существенно снизилась, остаются еще в' некоторых местах ареала довольно обычными животными. За филан-дерами охотятся из-за мяса и шкуры.
В роде 6 видов:
краснобрюхий филандер — Т. billardierri Desmarest, 1822 (Юго-Восточная Австралия, южная Виктория, Тасмания, о-ва Бассова пролива);
филандер Евгении — Т. eugenii Desmarest, 1817 (Южная, Юго-Западная и Западная Австралия с о-вами Флиндерс и Кенгуру);
белогрудый филандер — Т. parma Waterhouse, 1846 (Новый Южный Уэльс);
красношеий филандер — Т. thetis Lesson, 1828 (Новый Южный Уэльс и южный Квинсленд);
красноногий филандер — Т. stigmatica Gould, I860 (Новый Южный Уэльс и Папуа в Новой Гвинее);
новогвинейский филандер — Т. brunii Schreber, 1778 (Новая Гвинея с о-вами Ару, Кей, Архипелагом Бисмарка).
Некоторые ученые (например, Райд, Ride, 1957) присоединяют краснобрюхого и белогрудого филандеров и филандера Евгении к роду средних кенгуру. Белогрудый филандер в настоящее время очень редок, а может быть и вымер совсем. Включен в «Красную книгу». В 1870 г. белогрудый филандер был акклиматизирован на о-ве Кавау у Новой Зеландии, где численность его довольно высока.
Род короткохвостых кенгуру, или куокка,
Genus Setonyx Lesson, 1842
В роде единственный вид: короткохвостый кенгуру, или куокка, —-S. brachyurus Quoy et Gaimard, 1830.
Размеры небольшие. Длина тела около 65 см. Длина хвоста 25 см. Вес взрослых до 5 кг. Уши широкие, короткие, почти не выступающие над мехом, закругленные. Носовое'зеркало с каудально направленным безволосым участком. Хвост и задние конечности относительно короткие. Волосяной покров длинный и мягкий, на спине серо-бурый, голова
более светлая, с неясными полосами, проходящими через глаз. Брюшная сторона бело-серая. Хвост покрыт редкими волосами. Зубная формула: 3 0 2 4
i — с — р — т~ = 32. Верхний клык отсутствует. Число хромосом в диплоидном наборе 22.
Обитает в зарослях травы и кустарников в прибрежных пресноводных болотах. В густой и высокой траве устраивает длинные туннели и тропы. Держится в основном поодиночке. При быстром движении прыгает на задних лапах, при медленном движении не использует хвост в качестве опоры в отличие от большинства кенгуру. Питается листьями кустарников и травянистыми растениями. Характерно, что процесс пищеварения сходен с пищеварением жвачных парнокопытных. В желудке этого кенгуру имеется своеобразная бактериальная флора из более чем 15 видов. Период размножения приходится на зиму. Беременность около 27 дней. Самка приносит одного детеныша в год. Вес его при рождении примерно 25 г. Детеныш остается в сумке в течение 150—175 дней и к этому времени увеличивается в весе до 450 г. Задержка в развитии оплодотворенной яйцеклетки у сумчатых была впервые обнаружена у короткохвостых кенгуру. Непосредственно после рождения детеныша самка вновь спаривается, и яйцеклетка оплодотворяется. Однако имплантации и развития яйцеклетки не происходит до тех пор, пока детеныш остается в сумке. Если он погиб или изъят из сумки, яйцеклетка сразу начинает развиваться. Половозрелость наступает в 5 лет.
Распространен на юго-западе Западной Австралии и на некоторых прилегающих островах.
Род средних кенгуру, или валлэби,
Genus Protemnodon Owen, 1873
Размеры небольшие и средние. Длина тела около 60—90 см. Длина хвоста 50—85 см. Самцы на г/4—Ч3 крупнее самок. Вес 4—24 кг. Уши средней длины или длинные. Носовое зеркало имеется. Задние конечности значительно длиннее передних. На задних лапах коготь среднего пальца особенно увеличен в размере. Хвост длинный, постепенно сужающийся от основания к концу. Удлиненных волос на конце хвоста нет. Хвост служит опорой тела при сидении и балансом при прыжках. Мех довольно длинный, мягкий и густой. Окраска разнообразна у различных видов. У обыкновенного валлэби черная морда, белые щеки, через глаза проходят полосы, уши светлые с черным срединным пятном, передняя часть тела светло-серая, а задняя буро-желтая. Болотный валлэби на спинной стороне серо-бурый или оранжево-бурый, на брюшной светло-буро-желтый, щеки и подбородок белые, глазные полосы черные. Рыже-серый валлэби имеет спину серо-бурую или серую, по лбу и затылку идет срединная черноватая полоса, шея и передняя часть спины рыжие, конец морды черный. Чернополосый валлэби на спинной стороне буро-серый с темными пестринами, морда светло-серая с черными полосами, тянущимися от носа до глаз. Перчаточный валлэби имеет спинную сторону голубовато-серую, брюшную — желтоватую. Прыткий валлэби буро-желтый с большим белым полулунным пятном на бедрах. Зубная формула: i с — р т -^-=32(34). Верхний клык очень мал или совсем отсутствует. У рыжесерого валлэби в диплоидном наборе 11 хромосом у самцов и 10 у самок; у остальных представителей рода 2п=16.
Обитают в светлых и затененных лесах. Активность в основном сумеречная и ночная. Держатся поодиночке или парами, иногда небольшими группами. Питаются листьями кустарников. Средние кенгуру одни из наиболее быстрых прыгунов среди кенгуру. Беременность около 29 дней. Детеныш остается в сумке в течение примерно 250 дней.
Рис. 60. Серый кенгуру —М. gigantea и череп Macropus sp.
i j
Распространены в Тасмании с о-вами Бассова пролива, в Австралии и Новой Гвинее с некоторыми мелкими соседними островами. За этими кенгуру интенсивно охотятся из-за их превосходного меха.
В роде 6 видов:
обыкновенный валлэби — Р. canguru. Muller, 1776 (восток Австралии); болотный валлэби — Р. bicolor Desmarest, 1804 (восток Австралии); рыжесерый валлэби — Р. rufogrisea Desmarest, 1817 (Южная Австралия, Виктория, Новый Южный Уэльс, Квинсленд, о-ва Бассова пролива, Тасмания);
чернополосый валлэби — Р. dorsalis Gray, 1837 (Новый Южный Уэльс и Квинсленд);
перчаточный валлэби — Р. irma Jourdan, 1837 (Южная Австралия);
прыткий валлэби — Р. agilis Gould, 1842 (северо-запад Австралии, Квинсленд, Новая Гвинея).
Некоторые ученые, помимо перечисленных выше видов, выделяют в роде средних кенгуру: Р. bedfordi Thomas, 1900; Р. billa.rd.ieri Desmarest, 1822; Р. eugenii Desmarest, 1817; Р. greyi Waterhouse, 1846; P. parma Waterhouse, 1846. Стиртон (Stirton, 1963) выделяет из рода средних кенгуру еще 2 рода: Wallabia Trouessart, 1905 и Prionotemnus Stirton, 1955.
с
в
т
Л
Р'
Род исполинских кенгуру
Genus Macropus Shaw et Nodder, 1790
Размеры от средних до наиболее крупных среди современных сумчатых. Длина тела 75—160 см. Длина хвоста 60—105 см. Самцы на V4—*/3 крупнее самок. Носовое зеркало имеется или отсутствует. Уши очень крупные и
подвижные, могут двигаться независимо одно от другого. Хвост и задние конечности длинные и мощные (рис. 60). Хвост, толстый в основании, постепенно утончается к концу. Он служит не только балансом припрыжках, но и опорой тела при сидении, причем мускулатура его настолько сильна, что может выдержать вес всего кенгуру. Волосяной покров густой, короткий или довольно длинный, на спинной стороне тела светло-серый, темносерый, красновато-серый или буро-красный, на брюшной светлее, иногда белый. Самцы обычно темнее самок. Зубная формула:
i -у- с -у- р ~т ~ = 32. Верхний клык отсутствует. Диплоидное число хромосом у серого кенгуру 22 (у тасманийского подвида 16), у рыжего 20.
Обитают в открытых ландшафтах типа травяных степей или саванн на склонах гор и на скальных участках. Держатся небольшими группами, обычно из одного самца и нескольких самок. При быстром движении способны делать гигантские прыжки до 13,5 м длины и 3,3 м высоты и передвигаться со скоростью до 45 км!ч. Активность ночная. Днем отдыхают, иногда собираясь группами (рыжий кенгуру — сотнями). Большинство самок приносит потомство зимой. Беременность 30—40 дней. У рыжего кенгуру самка после беременности, продолжающейся 33 дня, приносит одного детеныша весом примерно 80 г. Детеныш покидает сумку в возрасте 196—240 дней, но продолжает питаться молоком матери и некоторое время после этого. Половозрелость наступает в 18—39 месяцев (в неволе быстрее, чем в диком состоянии; в засушливых условиях дольше, чем во влажном климате). Для горного исполинского кенгуру установлено достижение половозре-лости в 2 года и продолжительность жизни 15—20 лет.
Распространены в Австралии, Тасмании, на о-вах Кенгуру и Барроу.
В роде 3 вида:
горный исполинский кенгуру — М. robustus Gould, 1841 (Австралия и о-в Барроу);
серый кенгуру — М. gigantea Zimmermann, 1777 (Квинсленд, Южная Австралия, Западная Австралия, о-ва Кенгуру и Тасмания);
рыжий кенгуру — М. rufus Desmarest, 1822 (Южная Австралия, Виктория, Новый Южный Уэльс, Квинсленд, Западная Австралия).
Некоторые исследователи выделяют рыжих кенгуру в отдельный род — Megaleia Gistel, 1848. Более правильно рассматривать их в качестве под-fl рода рода исполинских кенгуру.
Инфракласс плацентарных Infraclassis Eutheria Gill, 1872
Наиболее высокоорганизованные млекопитающие. В развитии зародыша плацентарных характерно участие плаценты, связывающей его со стенками матки матери. Детеныши рождаются более или менее развитыми и способны сосать молоко. У самок нет выводковых сумок. В скелете сумчатые кости отсутствуют. Угловой отросток нижней челюсти не загибается внутрь, как у сумчатых. Зубная система дифиодонтная: резцы, клыки и предкоренные зубы имеют своих молочных предшественников. Головной мозг с хорошо развитым вторичным мозговым сводом — неопаллиумом, правая и левая половины которого соединены мозолистым телом. Температура тела у взрослых плацентарных высокая и постоянная.
Обитают на земле, в земле, в воздухе и в воде, освоили разные географические зоны и ландшафты Земного шара.
В инфраклассе 17 отрядов:
насекомоядных — Insectivora Bowdich, 1821;
шерстокрылов, или кагуанов, — Dermoptera liliger, 1811; рукокрылых — Chiroptera Blumenbach. 1779;
приматов — Primates Linnaeus, 1758;
неполнозубых — Edentata Cuvier, 1798;
ящеров — Pholidota Weber, 1904;
зайцеобразных — Lagomorpha Brandt, 1855;
грызунов — Rodentia Bowdich, 1821;
китообразных — Cetacea Brisson, 1762;
хищных — Carnivora Bowdich, 1821;
ластоногих — Pinnipedia liliger, 1811;
трубкозубых — Tubulidentata Huxley, 1872;
хоботных — Proboscidea liliger, 1811;
даманов — Hyracoidea Huxley, 1869;
сирен — Sirenia liliger, 1811;
непарнопалых — Perissodactyla Owen, 1848;
парнопалых — Artiodactyla Owen, 1848.
ОТРЯД НАСЕКОМОЯДНЫХ
ORDO INSECTIVORA BOWDICH, 1821
Плацентарные млекопитающие небольших размеров. Длина тела от 3,5 см (наименьшие размеры в классе мелкопитающих) у карликовой бело-зубки и до 44 см у большого крысиного ежа. Морда удлиненная,обычно оканчивается небольшим хоботком. Наружные ушные раковины небольшие, у некоторых представителей могут отсутствовать. Глаза маленькие, иногда находятся на разной степени редукции. Конечности четырех- или пятипалые, стопоходящие, все пальцы вооружены когтями.
Волосяной покров обычно короткий, мягкий, слабо дифференцированный; иногда тело покрыто колючками. В коже имеются сальные, примитивные потовые и специфические железы. Сосков от 2 до 12.
Череп удлинен в лицевом отделе и уплощен. Верхнечелюстные кости входят в состав стенок орбит и отделяют слезную кость от небной. Орбитальные крылья небной кости уменьшены в размерах. Скуловые кости небольших размеров или их нет совсем. Скуловые дуги развиты слабо или отсутствуют. Барабанные кости часто кольцеобразные и не образуют слуховых барабанов.
Зубы со слабо выраженной гетеродонтностью, брахиодонтные, дифио-донтные. Клыки могут иметь по два корня и внешне походить на резцы или предкоренные. Нижние коренные обычно пятивершинные, а верхние трех- или четырехвершинные. Число зубов варьирует от 26 до 48.
Лучевая и локтевая кости разделены между собой, а малая и большая берцовые часто срастаются между собой у коленного сустава. Ключица есть у всех насекомоядных, за исключением выдровой землеройки. Os penis имеется по крайней мере у кротов и тенреков.
В головном мозге сильно развиты обонятельные доли (макросматиче-ский тип). Полушария почти без борозд и извилин (у ежей, землероек и прыгунчиков имеется только Sulcus rhinal is lateralis) (Брауер и Шобер — Brauer und Schober, 1970). Желудок простой. Слепая кишка может отсутствовать. Матка у самок двурогая, плацента дискоидальная. Семенники у самцов располагаются в брюшной полости, под кожей в пахах или в мошонке перед половым органом.
Таким образом, для насекомоядных характерен ряд признаков, благодаря которым их следует считать примитивнее других плацентарных млекопитающих: небольшие размеры, стопоходящие конечности, слабогетеро-донтные зубы, туберкулярные коренные зубы, недоразвитые слуховые барабаны.
Ведут наземный, подземный, полуводный или древесный образ жизни. У большинства из них активность ночная; у некоторых круглосуточная. Питаются в основном насекомыми, хотя среди них есть и хищники. Насекомоядные— полигамы. Беременность 11—43 дня. В течение года обычно один помет, редко больше. В помете до 14 детенышей. Половозрелости достигают в возрасте от 3—4 месяцев до двух лет.
Хозяйственное значение относительно невелико. Ряд видов приносит пользу лесному и сельскому хозяйству, поедая вредных насекомых. Некоторые виды (крот) имеют промысловое значение.
Распространены всесветно, за исключением Австралии, большей части Южной Америки, Гренландии и Антарктики.
Насекомоядные —наиболее древние и примитивные среди плацентарных млекопитающих. Предки современных насекомоядных были, по-видимому, родоначальниками всех остальных плацентарных млекопитающих. Отделение насекомоядных от Pantotheria произошло, вероятно, где-то в начале мела. Наиболее древние ископаемые остатки насекомоядных известны из. верхне-меловых отложений Северной Америки (Gypsonictops).
Среди современных семейств насекомоядных, большинство из которых глубоко адаптировались к специфическим условиям существования и в связи с этим подверглись значительным изменениям, наиболее примитивным является семейство ежей. Землеройки и кроты, вероятно, отделились от ежеподобных предков примерно в конце эоцена или в начале олигоцена. Находки ископаемых остатков других современных семейств датируются миоценом (тенреки, златокроты и прыгунчики) или олигоценом (щелезубы).
Внутриотрядная систематика остается до сих пор спорной по ряду вопросов. В частности, подразделение отряда насекомоядных на подотряды. Геккель (Haeckel, 1866) предложил разделение насекомоядных на 2 подотряда: Lipotyphla и Menotyphla; к первому относятся (в современном понимании) тенреки, златокроты, ежи и землеройки, ко второму — прыгунчики и тупайи. Гилл (Gill, 1885) по строению зубов подразделил насекомоядных тоже на 2 подотряда: Zalambdodonta (тенреки и златокроты) и Dilambdodonta (ежи, землеройки, прыгунчики и тупайи). Разделения эти далеко не совершенны и искусственны, хотя некоторые современные исследователи и следуют им (например, Строганов, 1957, придерживается точки зрения Геккеля). Делались попытки вывести златокротов из отряда насекомоядных, возводя их в ранг самостоятельного отряда (Broom, 1916), что, по-видимому, неправильно. Значительную самостоятельность златокротов подчеркивают и Гейм де Бальзак и Бурлиер (Heim de Balsac et Bourliere, 1955), выделяя их в особый подотряд Chrysochloroida. Кроме того, они предлагают разделить остальных насекомоядных еще на 2 подотряда: Euinsectivora, куда ими отнесены щелезубы, тенреки, ежи, землеройки и кроты, и Menotyphla с тупайями и прыгунчиками. Эванс (Evans, 1942) также выделяет тупай и прыгунчиков в подотряд Menotyphla.
Бутлер (Butler, 1956), Мак Довел (McDowell, 1958) в отряде Lipotyphla выделяют 2 подотряда: Erinaceomorpha (с семействами Erinaceidae и Talpidae) и Soricomorpha (с семействами Tenrecidae, Chrysochloridae, Solo-nodontidae и Soricidae).
Видимо, более правильно предложение Симпсона (Simpson, 1945) не выделять подотряды в отряде насекомоядных в связи с примерно равноценной обособленностью современных семейств, если принимать, что тупайи принадлежат не к насекомоядным, а к приматам. Карлссон (Carlsson, 1909, 1922) указывает, что прыгунчики имеют более тесное отношение к ежам, а тупайи •— к приматам. Грегори (1910), Ле Грос Кларк (Le Gross Clark, 1924, 1926, 1933) отнесли тупай к приматам. Эту точку зрения разделяют Симпсон (Simpson, 1945); Нестурх, 1960; Тениус и Хофер (Thenius und Hoffer, 1960); Уолкер (Walker, 1964). По-видимому, она является наиболее правильной, и в настоящем издании тупайи отнесены к приматам, а насекомоядные не разделены на подотряды. Следует, однако, иметь в виду, что эта точка зрения не окончательна. В последнее время появляются исследования, подтверждающие необходимость отнесения тупай к насекомоядным (Ван Вален—Van Valen, 1965).
Иногда выдровых землероек (Potamogale) выделяют в особое семейство Potamogalidae (Симпсон—Simpson, 1945). Однако, по-видимому, более 112
правильно рассматривать их в качестве подсемейства семейства тенреков, тем более, что недавно описанные Micropotamogale из Африки, очевидно, служат связующим звеном между выдровыми землеройками и тенреками.
Некоторые систематики (Строганов, 1957; Гуреев, 1963; Кузякин, 1965 и др.) считают необходимым выделить выхухолей в особое семейство Desmanidae. Однако большинство исследователей (Симпсон, 1945; Гейм де Бальзак и Бурлиер, 1955; Уолкер, 1964; Андерсон и Джонс, 1967 и др.) включают их в качестве подсемейства в семейство кротов—Talpidae. Таким образом, в отряде насекомоядных выделяют 7 семейств (52 рода):
ежей — Erinaceidae Bonaparte, 1838 (7 родов);
кротов— Talpidae Gray, 1821 (12 родов);
тенреков — Tenrecidae Gray, 1821 (11 родов);
златокротов — Chrysochloridae Mivart, 1868 (5 родов);
щелезубов — Solenodontidae Dobson, 1882 (1 род);
землероек — Soricidae Gray, 1821 (21 род);
прыгунчиков — Macroscelididae Mivart, 1868 (5 родов).
СЕМЕЙСТВО ЕЖЕЙ
FAMILIA ERINACEIDAE BONAPARTE, 1838
Размеры от мелких до наиболее крупных в отряде. Длина тела от 10 (землеройковый еж) до 44 см (большой крысиный еж). Длина хвоста от 1 до 21 см. Вес большого крысиного ежа может достигать 1,1кг. Животные различного внешнего облика. Одни имеют вид типичных ежей, с неуклюжим и плотным телосложением, очень коротким хвостом, короткими, широко расставленными ногами, причем спинная сторона тела и в значительной мере бока покрыты колючими иглами. Эти виды способны сворачиваться в клубок. Менее специализированные волосатые ежи, или гимнуры, внешним видом напоминают некоторых представителей мышевидных грызунов, однако имеют более вытянутую мордочку. У всех представителей семейства морда вытянута и заострена. Глаза и ушные раковины хорошо развиты. Конечности стопоходящие, пятипалые, за исключением центральноафриканского ежа, у которого на задних конечностях по 4 пальца. Туловище покрыто редкими и тонкими волосками вперемежку с многочисленными крепкими и острыми иглами либо только волосами. Сосков 2—5 пар.
Форма черепа варьирует от узкого и удлиненного до короткого и широкого. Мозговой отдел небольших размеров. Скуловая дуга хорошо развита.
2 з 1 з 4 3
Зубная формула: i------с — р--------т — = 36 — 44. Первый верхний
3 1 2 — 4 3
резец, а иногда и первый нижний резец увеличены в размерах и имеют форму клыка. Жевательная поверхность зубов приспособлена больше к всеядности, чем к насекомоядности. В скелете туловища характерно слияние малоберцовой и большой берцовой костей. Лобковые кости имеют короткий симфиз. Позвонков шейных 7, грудных 15, поясничных 6, крестцовых 7, хвостовых 10—25.
Мочеполовое отверстие у самок открывается отдельно от заднепроходного. В мышечной системе сильно развита подкожная мускулатура, особенно у ежей. Семенники в брюшной полости.
Обитатели лесов, окультуренных районов, степей и пустынь. Активность преимущественно ночная, но есть и дневные формы. Некоторые впадают в спячку в северных частях своего ареала на холодный период года. Преимущественно всеядны, но предпочитают животные корма. В течение года один или два сезона размножения. У некоторых форм отмечена провоцированная овуляция. Длительность беременности от 34 до 49 дней (обыкновенный еж). Детенышей в помете от 1 до 7.
Распространение охватывает Африку, Евразию, на север до границы леса, Юго-Восточную Азию до Калимантана; отсутствуют на Цейлоне, Мадагаскаре и в Японии.
Рис. 61. Гимнура — Е. gymnurus и ее череп
Систематика ежей окончательно не выяснена. В частности, это касается числа и объема родов, входящих в семейство. Так, Симпсон (1945), Гейм де Бальзак и Бурлиер (1955), Андерсон и Джонс (1967) считают африканских и центральноафриканских ежей отдельными родами (Aethechinus Thomas, 1918 и Atelerix Pomel, 1848). Представляется более правомочным выделять их в качестве подродов рода ежей (Erinaceus Linnaeus, 1758), как это делают, например, Эллерман и Моррисон-Скотт (1951). Ушастых и лысых ежей часто считают (Симпсон, 1945) отдельными родами (Hemiechinus Fitzinger, 1866 и Paraechinus Trouessart, 1879), но в последнее время (особенно в отечественной литературе) их обычно объединяют в один род ушастых ежей (Громов и др., 1963) или даже присоединяют к роду ежей (Бобринский и др., 1965).
Здесь принято в семействе ежей 7 родов (15 видов):
гимнур — Eohinosorex Blainville, 1838 (1 вид);
малых гимнур — Hylomys Muller, 1839 (1 вид);
хайнаньских ежей—Neohylotnys Shaw et Wong, 1959 (1 вид);
филиппинских гимнур — Podogyninura Mearns, 1905 (1 вид); землеройковых ежей —Neotetracus Trouessart, 1909) (1 вид), ежей — Erinaceus Linnaeus, 1758 (5 видов);
ушастых ежей — Hemiechinus Fitzinger, 1866 (5 видов).
Род гимнур
Genus Echinosorex Blainville, 1838
В роде единственный вид: гимнура —Е. gymtuirus Raffles, 1821 (рис. 61).
Размеры средние и крупные (наибольшие в семействе). Длина тела 26—44 см. Длина хвоста 20—21 см. Вес самцов от 870 до 1050 г, самок — ст 1075 до 1100 г. Внешний вид несколько напоминает крысу. Голова относительно большая, с сильно вытянутой лицевой частью, сужающейся к косу. Ушные раковины средних размеров, закругленные. Глаза небольшие. На морде многочисленные длинные вибриссы. Конечности относительно короткие, пятипалые, с длинными когтями. Первый и пятый пальцы очень короткие. Пальцевые мозоли крупные. Длинный хвост покрыт чешуями, причем чешуи почти по всей длине располагаются концентрическими кругами, а у основания хвоста по диагонали. В концевой трети хвост несколько сжат с боков. Волосяной покров относительно длинный, грубый. Довольно многочисленные мягкие пуховые волосы сверху перекрыты высокими грубыми остевыми. Окраска волос на туловище обычно черная. Голова, плечи и дистальная половина хвоста белые. Вокруг глаз по черному пятну. Иногда черная полоса тянется с каждой стороны морды от глаза к уху. Самки имеют по две пары сосков: 2 грудных и 2 паховых. Имеются 2 анальные специфические кожные железы, продуцирующие секрет с сильным запахом. Зубная 3 14 3
формула: i — с — р — т — = 44.
Обитатель лесов. Часто встречается около водоемов и мангровых болот. Ведет ночной, по-видимому полуводный образ жизни. День проводит в убежищах среди камней, под корнями деревьев, в дуплах упавших стволов деревьев. Обычно держится поодиночке. Питается рыбами, лягушками, а также насекомыми. Сезонности в размножении, по-видимому, нет.
Ареал охватывает Тенассерим, Таиланд, Малайзию, Суматру и Калимантан.
Род малых крысиных ежей, или малых гимнур,
Genus Hylomys Muller, 1839
В роде единственный вид: малый крысиный еж, или малая гимнура, — Н. suillus Miiller, 1839 (рис. 62).
Рис. 62. Малая гимнура — Н. suillus
Размеры мелкие. Длина тела 10—15 см. Длина хвоста 1—3 см. Внешним видом напоминает больших крысиных ежей, но хвост короткий. Туловище удлиненное. Конечности довольно короткие. Голова крупная, с хорошо развитыми ушными раковинами с округленными вершинами. Глаза относительно крупные. Морда сильно удлинена и утоньшена в терминальном отделе. Волосяной покров густой и мягкий, на спинной стороне темно-бурый с желтоватым или сероватым оттенком, на брюшной сероватый, грязно-желтый или желтоватый. Анальные железы развиты слабо. Сосков 2 пары. Зубная формула, как и у больших крысиных ежей.
Обитатель лесов различного типа, преимущественно сильно увлажненных с густым подлеском. Ведет в основном наземный образ жизни; может лазить по кустам. Активность преимущественно ночная, но встречается и днем. Питается различными беспозвоночными, главным образом насекомыми и дождевыми червями. В размножении, по-видимому, сезонности нет.
Распространение охватывает Юньнань, Бирму, п-ов Индокитай, Таиланд, Малайзию, о-ва Тиоман, Суматру, Яву, Калимантан.
Род хайнаньских ежей
Genus Neohylomys Shaw et Wong, 1959
Возможно, этот род является лишь подродом малых гимнур. В роде единственный вид: хайнаньский еж — N. hainanensis Shaw et Wong, 1959.
Размеры небольшие. Длина тела 12—15 см. Длина хвоста 4—4,5 см. По внешнему виду сходен с малым крысиным ежом, однако общие размеры относительно больше, хвост длиннее и, в отличие от последних, у хайнаньских крысиных ежей отсутствует один нижний предкоренной зуб. Волосяной покров густой, средней длины, мягкий. На голове черновато-серый с бурым оттенком. Спина буровато-серая. Посредине спины проходит продольная, длинная, черная полоса. Брюшная сторона светло-серая или желтовато-белая. Уши, конечности и хвост почти голые, покрыты очень редкими, короткими волосами. Сосков 3 пары: пара грудных и две пары брюшных.
Экология не изучена.
Распространен на о-ве Хайнань в Южном Китае.
Род филиппинских гимнур
Genus Podogymnura Mearns, 1905
В роде единственный вид: филиппинская гимнура — Р. truei Mearns, 1905.
Размеры небольшие. Длина тела примерно 13—15 см. Длина хвоста 4—7 см. Хвост короткий, морда сильно удлиненная. Уши относительно большие. Ступни задних лап длинные. Волосяной покров длинный, довольно густой и мягкий. На спине смесь серых тонов с красновато-бурыми оттенками, на брюхе окраска беловатая с примесью бурых волос. Зубная форму-3 13 3
ла: i__ с — р — т~ — 40. Хорошо развиты длинные клыки.
Экология изучена слабо. Встречается в густых лесах, поднимаясь в горы до 2300 м над уровнем моря.
Распространены на о-ве Минданао из группы Филиппинских островов.
Включен в «Красную книгу», так как численность очень мала.
a
;o
Рис. 63. Землеройковый еж — N. sinensis и его череп
Род землеройковых ежей
Genus Neotetracus Trouessart, 1909
В роде единственный вид: землеройковый еж —- N. sinensis Trouessart, i 1909 (рис. 63).
Размеры небольшие. Длина тела 10—15 см. Длина хвоста 6—8 см.
По внешнему виду напоминает малую гимнуру, но хвост значительно более । глинный, а морда несколько короче. Ушные раковины довольно крупные, округлые. Глаза относительно большие. Волосяной покров длинный, мягкий и плотный, на спинной стороне буроватый или черный. Бока головы и шеи рыжеватые. Вдоль середины спины проходит черная полоса. Брюшная сторона красноватая или сероватая. Сосков 4. Зубная формула: 3 13 3
i — с — р — т — = 40. Верхние клыки развиты очень сильно, значительно превышая по размерам резцы.
Обитает в сырых горных лесах на высоте от 2100 до 2800 м над уровнем моря. Наземные, активность ночная. Экология изучена слабо. Питается земляными червями, а также, по-видимому, и некоторыми расти-
I тельными объектами.
Распространение охватывает Сычуань, Юньнань, Северную Бирму, п-ов Индокитай.
Род еже
Genus Erinaceus Linnaeus, 1758
I Мелких и средних размеров ежи. Длина тела 13—27 см. Длина хвоста I 1—5 см. Спинная поверхность туловища покрыта иглами, которые захо- дят и на бока (рис. 64). Между иглами располагаются тонкие, длинные, I очень редкие волосы. Набрюшной стороне тела иглы отсутствуют и замене- ны длинными и грубыми волосами. Голова относительно крупная, клино-I образная, со слабо удлиненным лицевым отделом. Ушные раковины широ-I киев основании округлые. Длина их никогда не превышает половину длины I головы. Окраска спинной стороны туловища весьма изменчива: шоко-I ладно-бурая или почти черная, иногда почти белая. Брюшная поверх-I кость обычно буроватая или сероватая. Череп несколько уплощенный в I дорзо-вентральном направлении, с расширенной мозговой коробкой, широ-I ко расставленными сильными скуловыми дугами и укороченной ростраль-I ной частью, имеющей довольно значительную ширину (рис. 64). Костные I слуховые барабаны невелики по размерам, уплощенные. Зубная формула: I 9 1 3 3
; — с — р — т — = 36. У обыкновенного ежа диплоидное число хро-I 2 2 г 2 3
I носом 48.
Обитатели различных ландшафтов. Избегают сильно заболоченных мест и сплошных массивов высокоствольных лесов. Предпочитают опушки леса, вырубки, заросли кустарников. Встречаются в лесостепи и в степи. Активность преимущественно сумеречная и ночная. На зиму обыкновенный еж устраивает наземное гнездо, собирая в кучу сухую траву и листья. Гнездо располагается под кучами валежника, под корнями деревьев. В октябре — ноябре впадает в спячку, продолжающуюся до теплых весенних дней. По характеру питания всеядны. Поедают различных беспозвоночных и позвоночных животных (мышевидных гры-
I
с
Рис. 64. Обыкновенный еж — Е. europaeus и его зунов, ящериц, лягушек, раз-череп личных насекомых, их личин-
ки), а также некоторые растительные объекты (плоды). Спаривание у обыкновенного ежа в северной части ареала приходится на весну, вскоре после пробуждения от спячки. В тропиках у представителей рода сезонности в размножении не отмечено. У обыкновенного ежа в течение года один помет. Беременность примерно 5—6 недель. Самка приносит от 3 до 8 детенышей (обычно около 4). Новорожденные обыкновенного ежа весят в среднем по 12 а и имеют хорошо заметные иголки в области головы. К 15 дням их иглистый покров уже хорошо выражен. Глаза открываются на 14—18-й день после рождения. Половозрелость наступает на 2-м году жизни. Продолжительность жизни примерно 6 лет.
Распространение охватывает Европу, Среднюю Азию, Северный и Северо-Восточный Китай, Корейский полуостров и Африку от Марокко и Ливии до Анголы. Обыкновенный еж акклиматизирован в Новой Зеландии.
г т б б п п о к X
в е м и я в в К'
н А
Систематика рода окончательно не установлена, обычно выделяют 5 видов:
обыкновенный еж — Е. europaeus Linnaeus, 1758 (Европа, Малая Азия, Западная Сибирь, северо-западный Казахстан, Амурская область, Приморский край, Северный и Северо-Восточный Китай, Корейский полуостров) ;
даурский еж — Е. dauricus Sundevall, 1842 (Забайкалье, северо-восточная часть МНР и Северный Китай);
алжирский еж — Е. algirus Duvernoy et Lerelboullett, 1842 (Северная Африка, Канарские и Болеарские острова, Юго-Западная Европа);
белобрюхий еж — Е. albiventris Wagner, 1841 (Центральная Африка); южноафриканский еж — Е. frontalis A. Smith, 1831 (Южная Африка). Иногда даурского ежа считают подвидом обыкновенного (Бобринский и др., 1965). Алжирского и южноафриканского ежей некоторые исследователи рассматривают в качестве особого рода Aerhechinus Thomas, 1918 (Симпсон, 1945 и др.), выделяя в нем 3 вида (Гржимек, 1968) или 1 вид (Гейм де Бальзак и Бурлиер, 1955 и Андерсон и Джонс, 1967). Белобрюхого ежа также иногда выделяют в особый род (Atelerix Pomel, 1848) и насчитывают в нем 6 видов (Гейм де Бальзак и Бурлиер, 1955; Гржимек, 1968).
G к; П А
1о ст за
io, рс ВИ
го/
ро. CKJ ни Уз
(Q эф! тел ро; 187
В СССР: обыкновенный еж (от северных берегов Ладожского озера на юг до Крыма и Кавказа включительно, в западных районах северного Казахстана, в Западной Сибири, в южной части Амурской области и Приморском крае) и даурский еж (Забайкалье).
Род ушастых ежей
Genus Hemiechintis Fitzinger, 1866
Размеры средние. Длина тела до 28 см. Длина хвоста 1—5 см. Иглы
е
l-о а I-и .1-м
и и и.
от
я, 1И->у-
ЭС-
[ая
:а); са).
<ий >ва-918 вид ого 1ТЫ-68).
покрывают только спинную поверхность туловища и не заходят на бока тела (рис. 65). Поверхность игл с продольными гребнями или валиками с бороздами между ними (за исключением гладкоиглого ежа). На боках и брюхе волосы мягкие и сравнительно короткие. Уши длинные, по длине превышающие половину длины головы. Окраска игл светлая, а волосяного покрова — буровато-серая или белая. В черепе характерна суженная, заостренная к концу ростральная часть. Слуховые барабаны сравнительно крупных размеров. Зубная формула, как в роде ежей. Диплоидное число хромосом ушастого и темноиглого ежей 48. i >
Обитатели степей, полупустынь и пустынь. Поднимаются в горы на высоту до 2500 м над уровнем моря. Активность сумеречная и ночная. День, как правило, проводят в норе, которую выкапывают сами, или занимают чужую. Питаются различными беспозвоночными (чаще насекомыми) и мелкими позвоночными животными (ящерицами, мышевидными грызунами и т. д.). Период размножения растянут. Самки приносят один помет в год. Детенышей в помете от 1 до 7. В северных частях ареала на зиму впадают в спячку. Основные враги: лисица, волк, бродячие собаки, филин, коршун, болотный лунь и т. д.
Распространение охватывает Юго-Восточную Европу, Переднюю, Среднюю, Центральную и Южную Азию и Северную Африку.
В роде 5 видов:
ушастый еж — Н. auritus Gmelin, 1770 (Северная Африка, Юго-Восточная Европа, Передняя и Центральная Азия, Индия, Казахстан);
афганский еж — И. mega-lotis Blyth, 1845 (Афганистан, Белуджистан и юго-западная Туркмения);
эфиопский еж—Н. aeth-iopicus Ehrenberg, 1833 (Марокко, Алжир, Судан, Аравийский полуостров, Иран);
индийский еж — И. mic-
ropus Blyth, 1846 (Индия);
темноиглый еж — Н. hy-pomelas Brandt, 1836 (Аравийский полуостров, Ирак, Афганистан Пакистан, Индия, Узбекистан, Туркмения).
Некоторые исследователи (Симпсон, 1945) выделяют эфиопского, индийского и темноиглого ежей в особый
род Paraechinus Trouessart, Рис. 65. Ушастый еж — Н. auritus и его 1879. череп
В СССР: ушастый еж (от Ростове кой области на западе до Алтая и Тувинской АССР на востоке, на север до г. Пугачева и с. Ферапонтовки, верховий р. Урал и Кулундинской степи, на юг до степей Закавказья, полупустынь, пустынь и предгорий Средней Азии включительно); афганский еж (юго-западная Туркмения); темноиглый еж (Мангышлак, Устюрт, Туркмения и Узбекистан, к северу до Самаркандской области).
СЕМЕЙСТВО КРОТОВ
FAMILIA TALPIDAE GRAY, 1821
Мелких и средних размеров насекомоядные. Длина тела примерно от 5 до 21 см. Длина хвоста от 1,5 до 20 см. Вес от 9 до 170 г. Наземные бегающие формы, подземнороющие и полуводные плавающие. Туловище, как правило, вытянутое, вальковатое, конечности укороченные, у большинства форм приспособленные к копанию, у некоторых — к плаванию. Пальцы вооружены длинными сплющенными сверху вниз когтями. Задние конечности обычно слабее передних, их ступни узкие и длинные. Голова небольшая, с удлиненной конической мордой. Глаза маленькие, у некоторых видов закрыты кожей. Наружные ушные раковины обычно отсутствуют или очень малы. Шея короткая, иногда совсем незаметная. Волосяной покров, как правило, слабо дифференцирован на категории, густой и мягкий, одноцветный, в большинстве случаев черный или черно-бурый. У водных форм волосы длинные и разделяются на категории. Сосков 3 или 4 пары.
Череп с удлиненным лицевым отделом, конической формы, со слабо развитыми гребнями. Скуловые дуги имеются, однако очень тонкие. Мозговая капсула широкая и уплощенная. Есть костные слуховые барабаны.
2__.3 1 3_-4 3
Зубная формула: I -—— с ——- р -—- т~ = 34 — 44. Резцы, клыки и предкоренные сильно изменчивы по форме. Коренные туберкулосектори-ального типа. Позвонков шейных 7, грудных 13—14, поясничных 5—6, крестцовых 5—6, хвостовых 8—27. Два крестцовых и три первых хвостовых позвонка срастаются между собой, их остистые отростки образуют сильно развитый гребень. Ключица очень короткая и широкая. Плечевая кость чрезвычайно широкая и укороченная. Ключица сочленяется непосредственно с плечевой костью — случай, уникальный для млекопитающих. Тазовые кости длинные и узкие. Таз неподвижно соединен с позвоночником. Лобковые кости не образуют вентрального симфиза. У самцов имеется os penis. Слепая кишка отсутствует.
Обитатели различных ландшафтов. Большинство видов ведет подземный образ жизни. Копают туннели на различной глубине и, как правило, питаются под землей. Некоторые виды в поисках корма периодически выходят на поверхность почвы. Ряд видов ведет наземный образ жизни, хотя убежищем для них служат норы. Наконец, есть водные формы, которые прекрасно плавают и ныряют, делают норы в берегах водоемов. Активность круглосуточная или сумеречно-ночная. Держатся обычно поодиночке. Беременность около 5 недель. Самка, по-видимому, приносит потомство раз в год. Половозрелость наступает в возрасте около года. Питаются различными беспозвоночными: наземными, подземными и водными. Некоторые виды служат объектом промысла. Поедая большое количество вредных беспозвоночных, могут приносить существенную пользу для сельского и лесного хозяйства. Некоторый вред причиняют, поедая дождевых червей.
Распространены в Северной Америке, от южной Канады до северной Мексики, в Европе и Азии, на юг до Малайи и Средиземного моря, а на север примерно до 63° с. ш.
Систематика семейства кротов имеет некоторые спорные вопросы. Ряд исследователей (Симпсон, 1945; Гейм де Бальзак и Бурлиер, 1955; Эллер-ман и Моррисон-Скотт, 1966 и др.) включает выхухолей (роды Desmana и
Galemys) в состав семейства кротов в качестве отдельного подсемейства. Некоторые систематики (Строганов, 1948, 1957; Громов и др., 1963; Бобринский и др., 1965) считают их отдельными семействами. Более правильно, видимо, объединение кротов и выхухолей в одно семейство, хотя выхухоли и являются сильно уклонившейся и высокоспециализированной группой.
Разные исследователи насчитывают в семействе кротов неодинаковое количество родов. Так, Гейм де Бальзак и Бурлиер, 1955 выделяют следующие роды: Uropsillus, Nasillus, Rhynchonax, Galemys, Desmana, Talpa, Mogera, Parascaptor, Scaptochirus, Scaptonyx, Dymecodon, Urotrichus, Neu-rotrichus, Sapanulus, Parascalops, Scapanus, Scalopus, Condylura. Симпсон (1945) и Андерсон и Джонс (1967), в отличие от этой системы, считают Rhynchonax синонимом Uropsillus, оставляя самостоятельными все остальные роды (два последних автора присоединяют к роду Talpa роды Mogera и Parascaptor). Уолкер (1964) и Эллерман и Моррисон-Скотт (1966) объединяют роды Uropsillus, Nasillus и Rhynchonax в один род Uropsillus, роды Talpa, Mogera, Parascaptor и Scaptochirus в род Talpa, а род Urotrichus и Dymecodon — в род Urotrichus. На крайней позиции в раздроблении семейства на большое число родов стоит Строганов (1948, 1957). Он выделяет роды: Rhynchonax, Nasillus, Uropsillus, Condylura, Parascalops, Scaptonyx, Urotrichus, включая Dymecodon Scapanulus, Neurotrichus, Scapanus, Scalopus, Talpa, Euroscaptor, Eoscalops, Mogera, Parascaptor, Scaptochirus (как уже сказано выше, Строганов считает выхухолей отдельным семейством).
Здесь в семействе кротов выделено 12 родов (19 видов);
землеройковых кротов — Uropsilus Milne-Edwards, 1871 (включая Nasillus и Rhynchonax) (1 вид);
выхухолей — Desmana Giildenstaedt, 1777 (1 вид);
пиренейских выхухолей — Galemys Каир, 1829 (1 вид);
кротов — Talpa Linnaeus, 1758 (включая Mogera, Asioscalops, Euroscaptor, Eoscalops, Parascaptor и Scaptochirus) (3 вида);
японских землеройковых кротов — Urotrichus Temminck, 1842 (включая Dymecodon) (2 вида);
длиннохвостых кротов—Scaptonyx Milne-Edwards, 1871 (1 вид);
американских землеройковых кротов — Neurotrichus Gunther, 1880 (1 вид);
западнокитайских кротов — Scapanulus Thomas, 1912 (1 вид); волосатохвостых кротов — Parascalops True, 1894 (1 вид);
западноамериканских кротов—Scapanus Pomel, 1848 (3 вида);
восточноамериканских кротов—Scalopus Е. Geoffroy St.-Hilaire, 1803 (3 вида);
звездорылов— Condylura liliger, 1811 (1 вид).
В СССР 2 рода: выхухолей и кротов, распространенных в лесной и лесостепной зонах европейской части СССР, Западной и Средней Сибири до юго-восточного Забайкалья, в Приморском крае.
Род землеройковых кротов
Genus Uropsilus Milne-Edwards, 1871
В роде единственный вид: землеройковый крот — U. soriceps Milne-Edwards, 1872 (рис. 66).
Некоторые авторы выделяют из рода землеройковых кротов еще один род Nasillus Thomas, 1911 с одним или двумя видами: N. gracilis Thomas, 1911 и N. investigator Thomas, 1922 (Симпсон, 1945; Андерсон и Джонс, 1967) или два рода: Nasillus Thomas, 1911 и Rhynchonax Thomas, 1911 — последний с одним видом R. andersoni Thomas, 1911 (Строганов, 1948, Гейм де Бальзак и Бурлиер, 1955). Видимо, более правильно считать указанные виды подвидами землеройкового крота, как это делают Эллерман и Моррисон-Скотт (1966).
Рис. 66. Землеройковый крот — U. soriceps и его череп
Размеры небольшие. Длина тела примерно от 6,3 до 8,8 см. Длина хвоста от 5,4 до 7,5 см. По внешнему виду сходны с землеройками. Тело относительно стройное, конечности несколько удлиненные. Длинные пальцы не соединяются между собой, снабжены слабыми, сжатыми с боков слабоизогнутыми когтями. Голова небольшая, с длинной, вытянутой в хоботок мордой. Ноздри расположены латерально. Наружные ушные раковины развиты довольно хорошо. Небольшие глаза скрыты в меху. Нижняя треть передних и задних лап, а также кисть и ступни не имеют волос, покрыты небольшими округлыми чешуйками. Хвост длинный, покрыт редкими, короткими волосами. Волосяной покров короткий, но густой и мягкий. Окраска спинной стороны варьирует от темно-бурой до темно-серой, а брюшной — от темносерой до светло-серой. Череп с расширенной мозговой коробкой и сильно удлиненной лицевой частью. Скуловая дуга сравнительно сильно развита и изогнута внутрь. Барабанные кости узкие, кольцеобразные. Зубная фор-2 1 з_____4 з
мула: i-----с — р--------т— = 34 — 38. В скелете конечностей харак-
J 1—2 13 — 4 3 г
терны тонкие плечевые кости и кости предплечья. Ключица длинная и тонкая.
Экология изучена слабо. Ведет наземный образ жизни, населяя, по-видимому, лесную подстилку. Встречается в горах на высоте 1200—4500 м над уровнем моря. Питается беспозвоночными.
Распространение охватывает Тибет, Сычуань и Юньнань.
Род выхухолей
Genus Desmana Gill dens taedt, 1777
В роде единственный вид: выхухоль — D. moschata Linnaeus, 1758 (рис. 67).
Размеры крупные. Длина тела 18—21 см. Длина хвоста 17—20 см. Туловище удлиненное, толстое, массивное. Конечности довольно короткие. Задние конечности более крупные, чем передние. Ступни широкие, кисти значительно более узкие. Между пальцами на передних и задних конечностях имеется плавательная перепонка. Когти на пальцах хорошо развиты, длинные, слабо изогнутые, сжатые с боков. По наружному краю ступней и по внутреннему и наружному краям кистей расположен гребень из жестких щетинистых волос, который увеличивает плавательную поверхность лап. Хвост длинный, резко перетянут в основании, затем расширенный и суживающийся к концу, сильно сжат с боков. Хвост покрыт короткими, редкими волосками и чешуйками. В основной трети хвоста, в основании хвоста, на вентральной поверхности располагается специфическая кожная железа. Голова крупных размеров, коническая с сильно развитым вершинным хоботком, имеющим бороздку на вентральной и дорзальной поверхности. Хоботок весьма подвижен, несколько расширен на конце в поперечном на
правлении. Шея снаружи почти незаметна. Основными локомоторными органами при движении в воде служат задние конечности и хвост. Рулями глубины являются передние конечности и, по-видимому, подвижный хоботок. Кроме того, при помощи хвоста выхухоль поворачивается в разные боковые стороны. Глаза очень невелики по размерам и имеют хорошо развитые веки. Наружных ушных раковин нет. Косо посаженные ушные отверстия имеют вид щелей длиной около 1 см, окружены небольшими кожно-хрящевыми валиками. Ушные отверстия могут смыкаться при нырянии животного. Носовые отверстия также замыкаются при нырянии при помощи особого мясистого отростка, выдающегося из верхней части носовой полости.
Волосяной покров длинный, густой и мягкий. Характерно разделение волос на категории. В отличие от большинства представителей насекомоядных волосы растут пучками. Мех на спинной стороне тела коричневый или буровато-коричневый, с серебристым налетом, на брюшной серебристо-белый или серебристо-серый. Хвост бурый. Череп несколько уплощен вдорю-вент-ральном направлении, с укороченной мозговой частью, расширенной сзади. Ростральная часть цилиндрической вытянутой формы. Скуловые дуги тонкие. Гребни на черепе хорошо развиты. На ламбдоидальном гребне имеется особый пластинчатый костный гребешок с зазубренной передней частью. Хорошо выражены костные слуховые барабаны. Межтеменная кость нале-
Рис. 67. Выхухоль — D. то-schata и ее череп
гаетна теменные. В нижней челюсти характерен сильно развитый венечный отросток. Зубная формула: t — с — р — т— = 44. Внутренняя пара верхних резцов и вторая пара нижних развиты особенно сильно. Вершины первой пары верхних резцов направлены вертикально вниз, а вершины второй пары нижних резцов вперед. Остальные резцы, клыки и передние предкоренные зубы слабо дифференцированы. Коренные зубы с широкими жевательными поверхностями, имеющими много вершинок. В скелете характерна укороченная плечевая кость, длинная и узкая лопатка, широкая и короткая рукоятка грудины. Относительно узкий таз имеет симфиз лобковых костей.
Обитает в различных пойменных старицах или по берегам медленно текущих, преимущественно тихих водоемов с наличием открытого зеркала воды и густой прибрежной и водной растительностью. Нору копает в бере-’ гах: вход делает подводный, а гнездовая камера расположена выше уровня воды. В гнездовой камере всегда имеется подстилка, для которой используются вегетативные части травянистых, преимущественно водноболотных растений. В местах с сильно меняющимся уровнем воды у выхухоли обычно бывают многоярусные норы. Активность сумеречная и ночная. Питается различными животными и растительными объектами: моллюсками, червями и насекомыми, мелкими рыбами, лягушками и их икрой, стрелолистом, тростником, камышом, рогозом. Имеет место сезонная смена объектов питания. Размножающиеся самки выхухоли встречаются на протяжении всего года. Однако наибольшая активность в размножении в летне-осенние месяцы. Наибольшее количество встреч беременных самок приходится на июнь и октябрь. Беременность примерно 40—45 дней. В помете от 1 до 5, в среднем 3—4 детенышей. Осенние пометы малочисленнее весенних. В году бывает, по-видимому, один помет. Детеныши рождаются голыми нелепыми. В возрасте около 20 дней детеныши покрываются шерстью и прозревают. Примерно в месячном возрасте переходят к самостоятельному питанию. В возрасте 1 года достигают размеров взрослых. Половозрелость, по-видимому, наступает в возрасте 1 года. Основными врагами выхухолей являются лисицы, горностай, хорь и норка. Выхухоль дает ценную шкурку с красивым и прочным мехом. Охраняется законом. Хозяйственная деятельность человека, а также прямое истребление сильно снизили численность выхухолей и значительно уменьшили ареал.
Ареал ограничивается отдельными очагами в бассейнах Дона, Волги и Урала.
Род пиренейских выхухолей
Genus Galemys Каир, 1829 »
В роде единственный вид: пиренейская выхухоль — D. pyrenaicus Geoffroy St-Hilaire, 1811.
Длина тела 11—15 см. Длина хвоста 12—15 см. Вес от 50 до 80 г. Внешним видом напоминает выхухоль, но размером меньше. Отличается от выхухоли относительно более длинным хвостом почти округлой формы в поперечнике, сжатым с боков в терминальном отделе и имеющим в концевой части гребень из жестких, коротких волос. Морда относительно более вытянута. Мех довольно длинный, густой и мягкий. На спинной стороне окраска бурая варьирует от серо-бурой до темно-бурой, на брюшной серая. Хвост беловатый. Нос и конечности темные, почти черные. Зубная формула: . з 1 4 з ..
i — с — р — т— = 44.
3 14 3
Образ жизни изучен слабо и, по-видимому, сходен с образом жизни выхухоли. Активность ночная. Держится по берегам небольших горных рек и озер на высоте 300—1200 м над уровнем моря. Питается в основном
Рис. 68. Обыкновенный крот — Т. еигораеа и его череп водными насекомыми, пресноводными ракообразными и мелкой рыбой. Часто охотится и на суше.У самцов половая активность начинается в ноябре, достигает максимума между январем и маем и постепенно затухает между августом и сентябрем. Течка у самок начинается с января. Первые беременные самки появляются в феврале, последние в июне. Самка приносит 1—5 (обычно 4) детенышей. В году бывает 2—3 помета.
Распространение охватывает Пиренейские горы между Францией и Испанией, а также горы центральной Португалии. Численность невысока и вид включен в «Красную книгу».
Род кротов
Genus Talpa Linnaeus, 1758
Размеры от мелких до крупных. Длина тела от 9 до 18 см. Длина хвоста от 1,5 до 3,4 см. Тело удлиненно вальковатое (рис. 68). Шея сильно укороченная, снаружи почти незаметная. Морда конусовидно вытянутая, тонкая, приплюснутая сверху вниз. Короткий носовой хоботок внутри имеет особую тонкую кость os praenasale и оканчивается плотной голой пластинкой. Конечности сильно укорочены, причем передние гораздо массивнее задних и имеют широкие лопатообразные ладони, вывернутые наружу. Пальцы одеты общим кожным покровом. Концы пальцев передних лап имеют когти, сплющенные сверху вниз. Конечности приспособлены к копанию в земле. Ушных раковин нет. Глаза рудиментарные и часто скрыты кожей. Хвост короткий, его длина не превышает длины головы. Волосяной покров короткий, слабо дифференцированный на категории, густой и мягкий. Волосы растут поодиночке или группами. Окраска черная, черно-бурая или светло-серо-буроватая. Характерны дополнительные линьки—до четырех в течение года. Череп с удлиненным лицевым отделом. Костные слуховые „ „ .3 1 3—4 з .п
барабаны имеются. Зубная формула: t — с р • р • m— = W — _42 — 44. Типично сильное развитие клыков верхней челюсти. В нижней челюсти клыки небольших размеров, почти не отличающиеся по форме
от резцов. Изредка нижние клыки отсутствуют. Число предкоренных у некоторых видов может изменяться. Мозговой отдел черепа широкий и уплощенный сверху вниз, без выступающих гребней. Межчелюстные кости выдаются вперед от носового отверстия. Плечевая кость очень широкая, короткая и массивная. Ключица наиболее короткая по сравнению с другими представителями семейства кротов. Диплоидное число хромосом у обыкновенного крота — 38, а у дальневосточного — 32.
Обитатели разнообразных ландшафтов, в основном леса и лесостепи. Поднимаются в горы до субальпийской и альпийской зон включительно. Предпочитают увлажненные почвы, но не заболоченные с высоким стоянием грунтовых вод. Под землей кроты роют сложную систему ходов, которые в основном делятся на 2 категории: 1) кормовые, расположенные, как правило, ближе к поверхности почвы, и 2) к гнезду, водопою и кормовым ходам. Крот по своим ходам может перемещаться на большие расстояния — до 4 км. У кротов имеется гнездовая камера с подстилкой. Активность круглосуточная. Пищу составляют различные беспозвоночные, главным образом дождевые черви. Характерна большая прожорливость кротов; без пищи зверек быстро погибает. Самка приносит в год один помет. У обыкновенного крота спаривание происходит в марте — апреле. Беременность около 40 дней. В помете 3—9, обычно около 5 детенышей. Молодые покидают гнездо в возрасте около месяца. Половозрелость наступает не ранее 5—6 месяцев. Для алтайского крота характерна задержка в развитии оплодотворенной яйцеклетки. Спаривание у них происходит с июня по август, а беременность продолжается около 9 месяцев.
Кротов добывают ради шкурки. Поедая большое количество вредных насекомых и их личинок, они приносят значительную пользу сельскому и лесному хозяйству. С другой стороны, кроты причиняют некоторый вред уничтожением земляных червей и выбрасыванием малоплодородных глубоких слоев почвы на поверхность.
В роде 3 вида:
обыкновенный крот — Т. europaea Linneaus, 1758 (Европа — на север до Южной Швеции, на юг до Испании, Италии, Сицилии и Северной Греции, на запад до Британии, на восток до Урала, Западной и Средней Сибири, Алтая и Саян);
слепой, или малый, крот — Т. caeca Savi, 1822 (Португалия, Испания, Швейцария, Италия, Югославия, Греция, Малая Азия и Кавказ);
могера — Т. micrura Hodgson, 1841 (от Советского Приморья, восточной части МНР, Корейского полуострова, о-ва Тайвань, большей части Китая до п-ова Индокитай, Таиланда, Малайзии, Бирмы, Ассама и на запад до Сиккима и Непала).
Система рода кротов окончательно не установлена, и единого мнения о количестве видов в роде не существует. Строганов (1948), предложивший наиболее дробнущ систему, выделяет следующие виды: Т. europaea Linnaeus, 1758; Т. caucasica Satinin, 1908; Т. orientalis Ognev, 1926; T. rotnana Thomas, 1902; T. coeca Savi, 1822; T. altaica Nikolsky, 1884; в роде Euroscaptor Miller, 1940 и Eoscalops Stroganov, 1941, которые мы следом за Эллерманом и Моррисоном-Скоттом, 1966 считаем синонимами Talpa, автор описывает виды: Ей. klossi Thomas, 1929; Ей. parvidens Miller, 1940; Ей. grandis Miller, 1940; Eo. longirostris Milne-Edwards, 1870; Eo. micrura Hodgson, 1840. Лей (Lay, 1965) описал новый вид крота из Западного Ирана — крот Стрита — Т. street i Lay, 1965, которого мы впредь до дальнейшего подтверждения его видового статуса относим в качестве подвида обыкновенного крота. Многие авторы полагают род Mogera Pomel, 1848 самостоятельным родом (здесь отнесен в синонимы Talpa), а Симпсон (1945), Строганов (1948) и некоторые другие исследователи выделяют также роды Parascaptor Gill, 1875 и Scaptochirus Milne-Edwards, 1867 (по предложению Эллермана и Моррисона-Скотта, 1966, мы считаем их синонимами Talpa) с видами: М. robusta Nehring, 1891; М. latouchei Thomas, 1907; М. wogura Temminck,
Рис. 69. Японский землеройковый крот — U. talpoide
1833; М. leucura Blyth, 1850; Р. leptura Thomas, 1881; Р. grandidens Stroganov, 1941;S. moschatusMilne-Edwards, 1867;S. davidianusMilne-Edwards, 1884. Систематика кротов требует дальнейшего изучения и ревизии.
В СССР 3 вида: обыкновенный крот (по всей европейской части СССР на север до Архангельска, Мезени и устья Печоры, на юг до Запорожья, Изюма, Лиски, Пензы, Вольска; кроме того, на Кавказе и в Предкавказье, на Урале, в Западной и Средней Сибири от оз. Зайсан, с. Убинского, Семипалатинска и Тары, от Сургута на Оби к востоку до р. Селенги в Забайкалье, верхней Лены и верховий бассейна Вилюя; на север до г. Дубинки и по всему бассейну Нижней Тунгуски; на юг до Алтая, Саян и Тувинской АССР включительно); слепой, или малый, крот (черноморское побережье от Новороссийска до Батуми и Западный Кавказ); могера (Приморье на Дальнем Востоке).
Род японских землеройковых кротов
Genus Urotrichus Temminck, 1842
Размеры небольшие (наименьшие в семействе). Длина тела 5—10 см. Длина хвоста 2,4—4,1 см. Внешним видом напоминают землероек (рис. 69). Передние лапы лишь слегка расширены. Имеются ушные раковины, однако небольшие и не выступают над мехом. Хвост покрыт довольно длинными и густыми волосами, иногда утолщен за счет отложения жира. Глаза под кожей. Волосяной покров густой и мягкий, темно-бурый или черный, с ме-
2 13 3
таллическим отблеском. Зубная формула: i — с — р —т — = 36. Передний верхний резец трехгранный в поперечном сечении и крупный, а следующий резец — округлый в сечении и небольшой.
Обитают в лесах, поднимаясь в горы до 2000 м над уровнем моря. Роют поверхностные ходы, но много времени проводят в поисках пищи и на поверхности почвы, могут даже вскарабкиваться на кусты. Наземная активность сумеречная и ночная. Питаются насекомыми, пауками, червями. Период размножения — апрель — май. В помете обычно 3 детеныша.
Распространение охватывает Японию (о-в Хонсю и другие острова к югу от него).
В роде 2 вида:
японский землеройковый крот— U. talpoides Temminck, 1841;
землеройковый крот Тру — U. pilirostris True, 1886.
Многие исследователи (Симпсон, 1945 и др.) выделяют землеройково-го крота Тру в особый род Dymecodon True, 1886.
Род длиннохвостых кротов
Genus Scaptonyx Milne-Edwards, 1871
В роде единственный вид: длиннохвостый крот — S. fusicaudus Milne-Edwards, 1872.
Размеры небольшие. Длина тела 6,5—9 см. Длина хвоста 2,5—3 см. Внешним обликом напоминает японских землеройковых кротов, но морда меньше вытянута в длину, а хвост относительно более длинный. Глаза не закрыты кожей. Пальцы короткие, с сильными когтями, приспособленными к копанию почвы. Наружной ушной раковины нет. Хвост покрыт редкими, короткими волосами. Волосяной покров короткий, густой и мягкий. Окраска темная, темно-бурая, почти черная. Зубная формула:
I — с — р — т~ = 42. Резцы долотообразные. Первая пара верхних резцов лишь немного крупнее остальных. Нижние резцы небольшие, однотипные. Клыки сходные предкоренными. Ключица хотя и укорочена, но в меньшей степени, чем у представителей рода кротов.
Экология изучена слабо. Обитает в горах на высоте 2100—4500 м.
Ареал охватывает северную часть Юньнаня, Сычуань и Северную Бирму.
Род американских землеройковых кротов
Genus^Neiirotrichus Gunther, 1880
В роде единственный вид: американский землеройковый крот — N. gibbsii Baird, 1857.
Размеры небольшие. Длина тела примерно 7—8,5 см. Длина хвоста 3—4 см. Вес от 9 до 11 г. Общий облик напоминает японских землеройковых кротов. Однако размеры меньше и морда менее удлинена. Глаза не скрыты под кожей, но очень малы и прикрыты мехом. Ушные раковины отсутствуют. Ноздри открываются по сторонам хоботка. Передние лапы лишь слегка расширены. Три средних пальца на передних и задних конечностях имеют особенно удлиненные когти. Довольно длинный хвост покрыт грубыми волосками и чешуйками. Волосяной покров короткий, густой и мягкий. Окраска от темно-серой до голубовато-черной, иногда с металлическим блес-3 12 3
ком. Зубная формула: i — с — р — т~ = 36. Передние верхние резцы невысокие, относительно широкие, долотообразные. Вторая и третья пары резцов наполовину меньше первой. Верхние клыки большие, крупнее первых резцов.
Обитатель лесов и зарослей кустарников. Предпочитает мягкие почвы. Поднимается в горы до высоты 2500 м над уровнем моря. Ведет подземный образ жизни. Активность дневная и ночная. Разыскивает пищу как в подстилке на поверхности почвы, так и в норах. Может вскарабкиваться невысоко на кусты, хорошо плавает. Встречается небольшими группами. Питается различными беспозвоночными: земляными червями, насекомыми, их личинками, иногда поедает некоторые растительные объекты. У одной самки в течение года может быть несколько пометов, в каждом из которых бывает от 1 до 4 молодых. Длительность беременности неизвестна. Приносит существенную пользу, поедая большое количество вредных насекомых.
Распространен по тихоокеанскому побережью Северной Америки, от южной части Британской Колумбии до Центральной Калифорнии.
Рис. 70. Волосатохвостый крот — Р. breweri и его череп
Род западнокитайских кротов
Genus Scapaniilus Thomas, 1912
В роде единственный вид: западнокитайский крот — S. oweni Thomas, 1912.
Размеры небольшие. Длина тела около 10 см. Длина хвоста примерно 3,5 см. Внешним видом напоминает японского землеройкового крота, но морда удлинена значительно меньше, кисти передних лап расширены сильнее. Когти хотя и довольно длинные, но слабые. Хвост относительно длинный и толстый, покрыт довольно густыми волосами. Волосяной покров короткий, густой и мягкий. Основная окраска серая. Зубная формула: . 2 1 3 з
i — с — р — т — = 36.
2 13 3
Экология не изучена. По-видимому, обитатель лесов. Распространение охватывает Шэньси, Ганьсу, Сычуань.
Род волосатохвостых кротов
Genus Parascalops True, 1894
В роде единственный вид: волосатохвостый крот — Р. breweri Bachman, 1842 (рис. 70).
Размеры средние. Длина тела 11—14 см. Длина хвоста примерно 2— 3,6 см. Вес взрослых 40—55 г. Внешний вид напоминает представителей рода кротов. Однако голова сильнее вытянута в лицевом отделе и более узка в этой части. Ноздри расположены по бокам и направлены несколько вверх. Наружных ушных раковин нет. Глаза очень маленькие и закрыты мехом. Кисть очень широкая, вывернутая, с сильными когтями. Хвост утолщенный, сжатый в основании, густо покрытый длинными волосами. Мех короткий, густой и мягкий, черноватый. На груди часто бывают белые пятна. Морда, хвост и конечности почти чисто белые. Сосков 4 пары. Зубная формула: i -^-с-±-р-~т~- = 44. Первая пара верхних резцов круп-нееостальных зубов и имеет долотообразую форму. В скелете конечностей
ключица развита относительно слабо, а плечевая кость очень сильно. Таз узкий. Костное соединение крестцовых позвонков с седалищными костями отсутствует.
Обитатель лесов и открытых мест. Предпочитает легкие, хорошо дренированные почвы. Распространен от уровня моря до примерно 900 м над уровнем моря. Норы имеют сложную систему, обычную для кротов. Гнездовую камеру диаметром около 15 см устраивает на глубине 25—30 см. Зимняя гнездовая камера помещается несколько глужбе, выстилается листьями и травой. Иногда кроты используют одни и те же норы в течение ряда лет. Активность круглосуточная. Ночью иногда передвигаются по поверхности земли. Питаются земляными червями, насекомыми и другими беспозвоночными. Период спаривания приходится на март — апрель. Беременность 4—6 недель. В помете от 4 до 5 детенышей.
Распространен в юго-восточной Канаде и северо-восточных районах США.
Род западноамериканских кротов
Genus Scapanus Pomel, 1848
Размеры средние и крупные. Длина тела 11—19 см. Длина хвоста 2,1— 5,5 см. Вес взрослых животных 50—170 г. Внешний вид сходен с представителями рода кротов. Морда с умеренно вытянутым хоботком. Глаза небольшие, скрытые в меху. Наружная ушная раковина отсутствует. Ноздри расположены латерально. Кисть очень широкая (ее ширина превышает длину). Пальцы не соединены перепонками. Когти сильные и длинные. Хвост довольно толстый, покрыт грубыми редкими волосками и чешуйками. Волосяной покров короткий, мягкий и густой. Окраска черно-бурая или почти черная. Отдельные индивидуумы светло-серые. Сосков 4 пары. Костные слуховые барабаны хорошо развиты. Зубная формула:
i -|- с -у- р т = 44. Таз относительно широкий. Крестцовые позвонки и седалищные кости соединены двумя костными перемычками.
Обитатели разнообразных ландшафтов, в горы поднимаются до 2700 м. Активность круглосуточная. Система подземных ходов сходна с системой у остальных кротов. Во время засушливого сезона копают особенно глубокие норы. Питаются земляными червями, насекомыми, их личинками и другими мелкими беспозвоночными. Употребляют в пищу также и некоторые растительные объекты, в частности луковицы. Период спаривания приходится на февраль —• март. Длительность беременности примерно месяц. В помете от 2 до 5 детенышей, рождаются в марте или апреле. К июню молодые переходят к самостоятельному образу жизни. Западноамериканские кроты имеют некоторое промысловое значение.
Распространение охватывает Крайний запад Северной Америки от южной части Британской Колумбии на юг до Калифорнийского залива.
В роде 3 вида:
западноамериканский крот—S. townsendii Bachman, 1839 (западная часть штатов Вашингтон и Орегон в США и северо-западная оконечность Калифорнии);
тихоокеанский крот — S. orarius True, 1896 (юго-запад Британской Колумбии, штаты Вашингтон, Орегон в США и северо-запад Калифорнии);
калифорнийский крот — S. latimanus Bachman, 1842 (штаты Калифорния, Невада и Орегон в США).
Строганов (1948) выделяет еще один вид — S. anthonyi Allen, 1893, который, по-видимому, является лишь подвидом S. latimanus.
Род восточноамериканских кротов Genus Scalopus Ё Geoffroy St.Hilaire, 1803
Размеры средние и крупные. Длина тела 11—20 см. Длина хвоста 1,8— 3,8 см. Общий облик напоминает представителей рода кротов. Отличается от западноамериканского крота более удлиненной мордой, наличием перепонки между пальцами и довольно тонким и относительно коротким хвостом. Ширина кисти превышает ее длину. Глаза закрыты кожей, наружных ушных раковин нет. Хвост почти голый. Зубная формула:
3 13 3
i — с — р —т~ — 36. Первая пара верхних резцов высокая и широкая, долотообразной формы. Остальные резцы небольшие, конические. В нижней челюсти больших размеров достигает вторая пара резцов. Слуховые костные капсулы выше, чем у западноамериканских кротов. Слуховой проход значительно короче. Таз узкий. Крестцовые позвонки соединены с седалищными костями двумя костными перемычками.
Обитатели полей, лугов, пастбищ и открытых ландшафтов. Населяют хорошо дренированные почвы. Сложная система нор и переходов сходна с .норами остальных кротов. Гнездовая камера располагается на глубине 15—20 см. Способность к копанию очень высокая: один крот в течение дня может прорыть до 31 м тоннелей, в час примерно 4,5 м. Активность круглосуточная. Питаются в основном дождевыми червями, а также различными видами насекомых и их личинками, иногда употребляют в пищу растения. Спаривание происходит весной. Беременность около 42 дней. Самка приносит от 2 до 5 детенышей и, по-видимому, имеет один помет в год. В месячном возрасте детеныши переходят к самостоятельному образу жизни. Полово-зрелости достигают в возрасте около года.
Распространение охватывает восточные штаты Соединенных Штатов Америки, и, кроме того, представители рода найдены в двух местах в Мексике: в Тамаулипас и Коауила.
В роде 3 вида:
восточноамериканский крот — S. aquaticus Linnaeus, 1758 (восточная часть США от атлантического побережья до Дакоты и Техаса и от канадской границы на юг до Флориды);
мексиканский крот—S. inflatus Jackson, 1914 (Тамаулипас в Мексике);
горный крот—S. montanus Baker, 1951 (Коауила в Мексике).
Род звездорылов
Genus Condylura liliger, 1811
Род представлен единственным видом: звездорыл — С. cristata Linnaeus, 1758 (рис. 71).
Размеры крупные. Длина тела 18—21 см. Длина хвоста 6,5—8,3 см. Вес взрослых особей 40—85 г. Сложение тела сходно с представителями рода кротов. Морда относительно укорочена и оканчивается голым овальным диском, края которого обрамлены 22 мясистыми, тонкими, хорошо иннервированными отростками, лучами, расходящимися от этого диска. Относительно крупные для крота глаза скрыты в меху. Хвост довольно длинный, покрыт редкими короткими волосками и чешуей. В хвосте к зиме откладываются запасы жира, отчего толщина его значительно увеличивается. Передние конечности хотя и имеют строение, общее для всех кротов, тем не менее развиты относительно слабее. Длина вывернутых наружу кистей равна их ширине. Волосяной покров короткий, плотный и мягкий. Окраска темно-бурая, почти черная. Зубная формула: i -|- с -J- р ~ 44.
Первая пара верхних резцов представлена относительно крупными зубами,
Рис. 71. Звездорыл — С. cristata, его носовой диск спереди и череп
изогнутыми внутрь и направленными вперед. Резцы второй верхней пары небольшие, палочкообразные, косо направленные вперед. Третья пара верхних резцов — крупные клыкообразные зубы. Верхние клыки развиты слабо и имеют коническую форму. Предкоренные зубы относительно небольшие, сжатые с боков. Нижние резцы средних размеров, долотообразные, с вершинами, направленными вперед. Нижние клыки высокие, тонкие, изогнуты назад. Таз узкий и длинный. Костные перемычки между тазовыми позвонками и седалищными костями отсутствуют.
Предпочитает сильно увлажненные заболоченные почвы. Активность круглосуточная. Одну и ту же систему ходов обычно населяют несколько звездорылов. Некоторые из ходов открываются прямо под воду. Звездорыл— прекрасный пловец и ныряльщик: при плавании работает всеми четырьмя конечностями, может плавать подо льдом. Питается различными водными насекомыми, ракообразными, мелкой рыбой, земляными червями. Часто разыскивает пищу на дне водоемов; при этом мясистые отростки морды находятся в непрерывном движении. Самка приносит от 2 до 7 детенышей в период от середины апреля до середины июня. К самостоятельному образу жизни детеныши переходят в 3-недельном возрасте. Половозрелость наступает примерно в 10 месяцев.
Распространение охватывает юго-восточную Канаду и северо-восточную часть США.
СЕМЕЙСТВО ТЕНРЕКОВ
FAMILIA TENRECIDAE GRAY, 1821
Размеры изменчивы: длина тела колеблется от 4 до 39 см, а длина хвоста — от 1 до 22 см (у некоторых видов хвост снаружи незаметен). Большинство представителей семейства обитают на Мадагаскаре и Коморских островах (по-видимому, с мелового периода) и за период своего существования образовали ряд специализированных форм. Например, обыкновенный тенрек по ряду признаков в скелете и зубах напоминает опоссумов из отряда сумчатых; малый и большой тенреки зубной системой и иглистым покровом — ежей; внешний вид длиннохвостых тенреков сходен с наземными 132
землеройками, рисовых тенреков — с кротами, болотных тенреков — с ондатрой, выдровых землероек — с выдрами. Голова с удлиненным, иногда вытянутым в хоботок лицевым отделом. Передние конечности короче задних. На передних конечностях 4 или 5 пальцев, на задних — 5.
Волосяной покров мягкий или грубый щетинистый, или тело покрыто иглами. Сосков от 2 до 12 пар (у обыкновенного тенрека— 12 пар).
Череп сильно вытянутый и узкий, не имеет перехвата в области глаз-
ниц. В связи с отсутствием скуловой кости скуловая дуга незамкнутая. Зуб-2_______________3 1 3 2-_4
ная формула: i ---с — р — т------= 32 — 40. Последний коренной
3 13 2 — 3
не вырастает до тех пор, пока не выпадет первый. Коронка коренных зубов имеет V-образную форму, иногда W-образную (у выдровой землеройки). Резцы и клыки сильно отличаются по форме.
Позвонков шейных 7, грудных 15—19, поясничных 5—6, крестцовых 2, хвостовых 7—47. Характерна сильная изменчивость числа хвостовых позвонков: от 7 у обыкновенного тенрека до 47 у длиннохвостых тенреков (больше, чем у других млекопитающих, за исключением некоторых древесных ящеров). Грудина состоит из 8 или 9 элементов. Ключица имеется у всех представителей семейства, кроме выдровой землеройки. Большая и малая берцовые кости могут быть свободны или Срастаться между собой. Тазовые кости узкие; лобковый симфиз короткий, отсутствует у выдровой землеройки. У самцов обычно os penis имеется.
Мочеполовое и анальное отверстия открываются в клоаку. Семенники располагаются в брюшной полости в области таза (длиннохвостый тенрек) или в фиброзной внутренней псевдомошонке (выдровая землеройка).
Обитают в лесах, степях, кустарниковых зарослях, болотах. В основном наземные животные, но среди них есть и полуводные (болотная, карликовые выдровые и выдровая землеройки). Активность главным образом сумеречная и ночная. День проводят в норах, дуплах упавших деревьев, под камнями. Питаются животной и растительной пищей, хотя некоторые виды в основном насекомоядные. Некоторые представители семейства (обыкновенный тенрек) впадают в спячку во время зимнего засушливого сезона. Самки рождают большое число детенышей: обыкновенный тенрек — до 16 детенышей в одном помете. Обыкновенного и большого тенреков местные жители употребляют в пищу.
Распространение охватывает Мадагаскар и Коморские острова, а также Западную и Центральную Африку. Акклиматизированы на некоторых островах Индийского океана.
В семействе 11 родов (31 вид):
обыкновенных тенреков — Tenrec Lacepede, 1799 (1 вид);
полосатых тенреков — Hemicentetes Mivart, 1871 (1 вид);
больших, или ежовых, тенреков —Setifer Froriep, 1806 (1 вид);
тенреков Фонтуанонта—Dasogale Grandidier, 1928 (1 вид);
малых тенреков — Echinops Martin, 1838 (1 вид);
рисовых тенреков — Oryzorictes Grandidier, 1870 (2 вида);
длиннохвостых тенреков — Microgale Thomas, 1882 (19 видов);
перепончатолапых, или больших, тенреков — Limnogale Forsyth Major, 1896 (1 вид);
земляных тенреков — Geogale Milne-Edwards et Grandidier, 1872 (1 вид); выдровых землероек — Potamogale Du Chaillu, 1860 (1 вид);
карликовых выдровых землероек — Micropotamogale Heim de Balsac 1954 (2 вида).
Многие систематики (Симпсон, 1945) считали, что выдровые землеройки принадлежат к особому монотипическому семейству Potamogalidae Allman, 1865. Однако описанные сравнительно недавно карликовые выдровые землеройки, по-видимому, являются связующим звеном между выдровыми землеройками и тенреками. В настоящем руководстве, следуя Кабрера, 1925; Грассе, 1955; Гут и др. (Guth et al., 1960) и Андерсон и Джонс,
Рис. 72. Обыкновенный тенрек — Т. eucaudatus и его череп
1967, выдровые землеройки и карликовые выдровые землеройки включены в семейство тенреков: можно полагать, что они должны быть объединены в особое подсемейство Potamogalinae.
Род обыкновенных тенреков
Genus Tenrec Lacepede, 1799
В роде единственный вид: обыкновенный тенрек — Т. eucaudatus Schreber, 1777 (рис. 72).
Размеры средние или крупные (наибольшие в семействе): длина тела 26—39 см, длина хвоста 1—1,6 см. Тело удлиненное. Пятипалые конечности относительно короткие (передние немного короче задних), с хорошо развитыми пальцами, вооруженными довольно длинными когтями. Первый и пятый пальцы короткие. Голова крупная, с сильно вытянутой, узкой мордой. Уши небольшие, округлые. Глаза маленькие. Бока, тело и спина покрыты грубыми волосами вперемежку с колючками. Tenpej-ш не способны сворачиваться в шар. Вдоль спины проходит гребень из удлиненных грубых волос. У молодых тенреков в середине спины имеются специализированные колючки примерно 4 см длиной, направленные вершинами вперед. Действием подкожной мускулатуры животные способны заставить эти колючки вибрировать и производить звук, меняющийся по частоте от 2 до 20 герц. Звук этот имеет коммуникативное значение; взрослые животные теряют способность производить звук. Основная окраска серовато-бурая или красновато-бурая. Некоторые особи имеют темно-бурую окраску.
Сосков 12 пар. В черепе характерны сильно развитый сагиттальный и лямбдоидальный гребни. Череп сильно удлинен. Зубная формула:
. 2 1 3 3 — 4 по лп
i — с — р — т-------= 38 — 40.
3 13 3
Обитает в различных ландшафтах Мадагаскара, за исключением юго-западных пустынных областей. Предпочитает теплые районы с песчаными почвами, поросшими кустарником, или поляны в сухих лесах, высокогорные плато. Убежище находит в дуплах упавших деревьев или под камнями. На холодный период времени года в засушливый сезон (апрель —• октябрь) впадает в спячку. Спячка проходит в норе глубиной в 1—2 м. Выход из норы животное на период спячки забивает почвой. Спячка может быть весьма продолжительной, охватывая весь сухой сезон, а иногда часть его. Температура тела в это время снижается и колеблется в пределах от 24 до 35° С, число дыханий сокращается до 30 в минуту. Активность в основном вечерняя и ночная. Питается насекомыми, моллюсками, червями и корнями растений. Период размножения, по-видимому, начинается вскоре после окончания спячки. Число детенышей 12—16. У беременных самок находили от 16 до 32 эмбрионов, из которых в дальнейшем выживают не все.
Распространен на Мадагаскаре и Коморских островах. Акклиматизирован на некоторых островах, в частности на о-ве Маврикий.
Род полосатых тенреков
Genus Hemicentetes Mivart, 1871
В роде единственный вид: полосатый тенрек — Н. semispinosusG.. Cuvier, 1798 (рис. 73).
Размеры средние. Длина тела 16—19 см. Вес 130—150 г. Хвост снаружи не заметен. Внешним видом напоминает ежа. Лапы относительно длинные, передние длиннее задних. Пальцы вооружены сильно развитыми когтями. Голова с сильно удлиненной мордой. Уши средних размеров, округлые, глаза очень маленькие. Спина и бока тела покрыты грубыми волосами вперемежку с крепкими колючками. На брюхе волосы могут быть очень
Рис. 73. Полосатый тенрек — И. semispi-nosus
грубые или мягкие. Окраска черноватая с продольными белыми или желтыми широкими полосами, на брюхе — каштаново-бурая или беловатая. Вокруг шеи позади ушей в виде воротника располагаются удлиненные колючки, которые зверек может ставить вертикально при защите. В центре спины имеется группа из 14—16 специализированных колючек на участке примерно в 1 см2. Действием подкожной мускулатуры зверек может заставить колючки тереться друг о друга и производить звук частотой 2—80 герц. Звук меняется в зависимости от поведения животного и имеет значение для внутрипопуляционной коммуникации. Череп сильно удлинен, но не имеет сагиттального и лямбдоидального гребней. Зубная формула:
3 1* 3 3
i — с — р — т~ — 40. Диплоидное число хромосом 38.
Обитает в зарослях кустарников и на опушках. Активен круглосуточно. Роет неглубокие норы. Питается различными беспозвоночными, в основном дождевыми червями. Может впадать в спячку. Беременность около 50 Дней. В помете примерно 9 детенышей. Они рождаются почти голые, весом 9,9—12,9 г, длина тела около 6 см. В возрасте 5 дней наиболее длинные колючки на затылке достигают 4 мм.
Распространен на Мадагаскаре.
Некоторые исследователи выделяют в роде еще один вид: черноголового тенрека — Н. nigriceps Gunther, 1875, которого, по-видимому, правильнее считать подвидом полосатого тенрека.
Род больших, или ежовых, тенреков
Genus Setifer Froriep, 1806
В роде единственный вид: большой, или ежовый, тенрек—S. setosus Schreber, 1777 (рис. 74).
Средних размеров тенреки. Длина тела 15—18 см. Длина хвоста 1,5— 1,6 см. Вес около 180—200 г. Внешним обликом напоминает ежа. Туловище удлинено. Голова довольно широкая, с укороченной мордой. Уши относительно короткие, округлые. Задние конечности немного длиннее передних. Пальцы с относительно короткими когтями. Тело сверху и с боков покрыто крепкими колючками длиной примерно 15 мм. Морда, конечности и брюшная сторона тела лишены колючей. Колючки имеют белые вершины. На морде характерны многочисленные довольно длинные вибриссы, достигающие 5— 7 см. Общая окраска тела беловатая, иногда черноватая. Брюхо темно-бурое или беловатое. У самок 5 пар сосков. Зубная формула:
. 2 1 3 3 ос
I — с — р — т — = 36.
2 13 3
Обитатель сухих лесов, встречается также в горах. Ведет ночной образ жизни.. В спячку, по-видимому, не впадает. В случае опасности может сворачиваться в колючий шар. Питается в основном беспозвоночными. Самка рождает в среднем 4 почти голых детеныша длиной примерно 78 мм и весом около 25 г. Местное население употребляет больших тенреков в пищу.
Распространены на Мадагаскаре.
Род тенреков Фонтуанонта
Genus Dasogale Grandidier, 1928
Род представлен единственным видом: тенреком Фонтуанонта — D. fontoynonti Grandidier, 1928.
Размеры небольшие. Длина тела около 9,5 см. Хвост снаружи виден только как небольшой пучок коротких игл. Внешне похож на маленького ежа. Лапы с удлиненными пальцами, вооруженными длинными, слабо изогнутыми когтями. Уши относительно крупные, закругленные. Спинная
часть тела и бока покрыты короткими иглами. Морда и брюшная часть тела игл не имеют. Основная окраска тела буровато-желтая. Иглы у основания белые, их средние отделы опоясаны черным кольцом, а вершинка игл снова белая. Зубная формула:
.2 1 3 2 оо
I — с — р — т — = 32.
2 13 2
Экология не изучена. Обитатель лесов.
Распространен на восточном Мадагаскаре.
Род малых тенреков
Genus Echinops Martin, 1838
В роде единственный вид: малый тенрек — Е, telfairi Martin, 1838.
Средних размеров тенрек с длиной тела около 18 см. Хвост снаружи незаметен. Внешний облик животного напоминает ежа. Спинная часть тела, верх головы и бока тела покрыты средней длины колючками. Окраска спины и боков темно-бурая, а брюшной стороны — несколько светлее. Морда серо-бурая. Зубная формула:
.2 1 3 2 оо
i — с — р —т — = 32.
2 13 2
Обитатель горных засушливых областей, хотя в общем встречается по всему Мадагаскару. Экология изучена слабо. Ведет ночной, полудревес-ный образ жизни. Питается насекомыми. Во время зимнего сезона впадает
в спячку. Число детенышей в помете от 2 до 10. Детеныши рождаются почти голые, слепые. Глаза открываются на 7—9-й день.
Распространен на Мадагаскаре.
Род рисовых тенреков
Genus Oryzorictes Grandidier, 1870
Размеры небольшие. Длина тела 8—13 см. Длина хвоста 3—5 см. Внешним видом несколько напоминают кротов. Туловище удлиненное. Конечности короткие, но сильные. Передние конечности крупнее задних. Все пальцы передних конечностей, кроме очень короткого первого (он отсутствует у четырехпалого тенрека), снабжены длинными сильными когтями, приспособленными к копанию. Лицевой отдел черепа относительно укороченный, заостренный. Уши небольшие и округлые. Глаза маленькие. Мех короткий, густой и мягкий или более грубый (у четырехпалого тенрека), на спинной стороне серо-бурый или темно-бурый, на брюшной сероватый или желто-бурый. Хвост двухцветный, как и туловище. Зубная формула: 3 13 3
i — с — р — т~ = 40. Верхние клыки длиннее верхних первых резцов.
Обитают во влажных местах, часто встречаются по бровкам рисовых полей. Экология изучена слабо. Ведут, по-видимому, подземный образ жизни. Активность ночная. На поверхности почвы показываются редко. Питаются различными беспозвоночными, в частности насекомыми и моллюсками.
Распространены по всему Мадагаскару.
В роде 2 вида:
кротовидный тенрек — О. talpoides Grandidier et Petit, 1930;
четырехпалый тенрек — О. tetradactylus Milne-Edwards et Grandidier, 1882.
Некоторые исследователи выделяют и третий вид — О. hova Grandidier, 1870.
Род длиннохвостых тенреков
Genus Mier о gale Thomas, 1882
Размеры небольшие (наименьшие в семействе). Длина тела от 4 до 13 см. Длина хвоста от Здо 16 см. Вес взрослых от 5 до 12 г. Внешним видом напоминают землероек (рис. 75). Туловище вытянутое. Конечности укороченные. Задние более длинные, чем передние. Когти на пальцах передних конечностей длиннее, чем на задних. Голова довольно крупная. Морда укороченная, остроконечная. Уши относительно высокие, широкие, округлые. Глаза небольшие. Хвост длинный (содержит до 47 позвонков), чешуйчатый, покрытый небольшими редкими волосками. У видов с особенно длинным хвостом его конец хватательный. Волосяной покров густой и мягкий. Спинная часть и бока красновато-бурые или почти черные, брюхо обычно грязножелтое, желтовато-серое. Вдоль спины изредка может проходить продоль-
3 13 3
ная темная полоса. Зубная формула: i — с — р — т— ~ 40.
Обитают в лесной зоне. Населяют обширные влажные луга с густым травостоем. Экология изучена слабо. Активность дневная и ночная.
Распространены на Мадагаскаре.
В роде 19 видов:
короткохвостый тенрек — М. brevicaudata Grandidier, 1899;
тенрек Кована — М. cowani Thomas, 1882;
тенрек Мильн-Эдвардса — М. crassipes Milne-Edwards, 1893;
тенрек Дроухарда — М. drouhardi Grandidier, 1934;
длиннохвостый тенрек — М. longicaudata Thomas, 1882;
Рис. 75. Microgale sp.
длинномордый тенрек — М. longirostris Major, 1896;
тенрек Майора — М. majori Thomas, 1918;
малый длиннохвостый тенрек — М. parvula Grandidier, 1934;
тенрек Мидонги — М. principula Thomas, 1926;
тенрек Диего-Суареза — М. prolixacaudala Grandidier, 1937;
крошечный тенрек — М. pusilia Major, 1896;
землеройковый тенрек — М. sorella Thomas, 1926;
тенрек Танала — М. taiva Major, 1896;
тенрек Томаса — М. thomasi Major, 1896;
тенрек Добсона — М. dobsoni Thomas, 1884;
генрек Талазака— М. talazacki Major, 1896;
стройный тенрек — М. gracilis Major, 1896;
тенрек Декари — М. decaryi Grandidier, 1928;
западный тенрек — М. occidentalis Grandidier et Petit, 1931;
Род перепончатолапых, или болотных, тенреков
Genus Limnogale Major, 1896
В роде единственный вид: перепончатолапый, или болотный, тенрек — L. mergulus Major, 1896 (рис. 76).
Размеры небольшие. Длина тела около 13 см. Длина хвоста примерно равна длине тела. Внешний облик несколько напоминает ондатру. Туловище удлиненное, сильное, немного приплюснутое. Голова небольшая, уплощенная, лицевой отдел относительно укорочен и широк. Уши маленькие, округлые. Глаза небольшие. Конечности пятипалые, пальцы соединены плавательной перепонкой, которая сильнее выражена на задних лапах. Хвост длинный, сильный, покрытый чешуйками и сжатый с боков в задней части. Вероятно, хвост является главным локомоторным органом во время плавания. Волосяной покров густой, довольно короткий и мягкий, на спинной стороне буроватый с примесью красноватых и черных волос, на брюшной светло-желтовато-серый.
Экология изучена слабо.
Чаще встречается на востоке Мадагаскара и в высокогорье. Ведет полу-водный образ жизни. Встречается по берегам различного типа водоемов. Питается в основном различными водными растениями, а также рыбой, лягушками, ракообразными и насекомыми. Весьма редок.
Распространен на Мадагаскаре.
Род земляных тенреков
Genus Geogale Milne-Edwards etGrandidier, 1872
В роде единственный вид: земляной тенрек — G. aurita Milne-Edwards et
Рис. 76. Перепончатолапый тенрек — L. mergulus и его череп
Grandidier, 1872.
Небольшие тенреки. Длина тела около 7 см. Длина хвоста около 3 см. Внешний вид сходен с землеройками. Тело удлиненное, конечности короткие, пятипалые, с довольно короткими когтями. Голова небольшая, с вытянутой мордой, заостренная; уши довольно крупные, округлые. Хвост длинный, покрытый редкими волосами. Волосяной покров густой
и мягкий, на спине красновато-бурый, на брюхе желтовато-белый. Зубная 3 13 3
формула: i — с — р — т~ = 38.
Экология не изучена. Может впадать в спячку.
Распространен на западном берегу Мадагаскара.
Род выдровых землероек
Genus Potamogale Du Chaillu, 1860
В роде единственный вид: выдровая землеройка — Р. velox DuChaillu, 1860 (рис. 77).
Размеры крупные. Длина тела от 29 до 35 см. Длина хвоста от 24 до 29 см. Внешним обликом напоминает небольшую выдру. Туловище удлиненное, приземистое, конечности короткие, пятипалые. Когти на пальцах короткие. Пальцы не соединены плавательной перепонкой. На передних лапах второй и третий пальцы срастаются между собой на две трети своей длины. Наружный край стопы удлинен снаружи тонкой латеральной частью.
Рис. 77. Выдровая землерой-. ка — Р. velox и ее череп
Длинный хвост сильно уплощен с боков и служит основным локомоторным органом во время плавания. Верхняя и нижняя части хвоста образуют сильно развитые кили. Голова относительно невелика с укороченной округлой мордой, сильно сплющенная сверху вниз. Уши небольшие округлые. Глаза маленькие. Вибриссы длинные, многочисленные. Волосяной покров относительно короткий, густой и мягкий, на спинной стороне темно-бурый пли черноватый, на брюшной беловатый или желтоватый. У самок одна пара паховых сосков. Семенники самцов располагаются в псевдомошонке. Зуб-3 13 3
ная формула: i — с — р — т~ = 40. Первый верхний и второй нижний резцы клыкоподобные. Первый верхний резец отделен от других зубов значительным промежутком. Тазовые кости не имеют лобкового сочленения. Ключица отсутствуют.
Обитатель берегов водоемов от уровня моря до примерно 1800 м над его уровнем. Населяет как медленно текущие мутные реки равнин, так и холодные с чистой водой быстрые горные потоки. В некоторых местах обитает по берегам небольших лесных водоемов. Как правило, ведет водный образ жизни, а в засушливый сезон — наземный. Прекрасно плавает. Активность сумеречная. День проводит в норах, вырытых в берегах. Встречается поодиночке или парами. Питается крабами, рыбой и земноводными. В неволе взрослая выдровая землеройка съедала от 15 до 20 крабов за ночь. По-видимому, определенной сезонности в размножении нет. Самка приносит 2 детенышей.
Распространен в Западной и Центральной Экваториальной Африке.
Род карликовых выдровых землероек
Micropotamogale Heim de Balsac, 1954
Размеры средние. Длина тела от 14 до 22 см. Длина хвоста от 11 до 13 см. Вес около 70 г. Тело сильно удлиненное, приземистое, конечности короткие. Плавательная перепонка на лапах хорошо развита у рувензорий-ского водяного тенрека, а у карликового водяного тенрека отсутствует. Голова относительно небольшая, с укороченной тупой мордой, несколько уплощенная. Хвост длинный, покрытый короткими волосками, немного сжатый с боков. По верхней и нижней сторонам хвоста проходят небольшие кили. Конечности пятипалые, стопоходящие. Когти короткие. Волосяной
покров довольно длинный, густой и мягкий. В черепе хорошо развиты костные слуховые барабаны.
Экология изучена слабо. Ведут полуводный образ жизни. Активность ночная. Питаются различными водными животными, в том числе рыбой.
Распространены в Западной Африке и горах Рувензори.
В роде 2 вида:
карликовая выдровая землеройка — М. lamottei Heim de Balsac, 1954 (Западная Африка от Гвинеи до Берега Слоновой кости);
рувензорийская выдровая землеройка — М. ruwenzorii de Witte et Frecnkop, 1955 (горы Рувензори и Республика Заир).
СЕМЕЙСТВО ЗЛАТОКРОТОВ
FAMILIA CHRySOCHLORIDAE MIVART, 1868
Размеры небольшие и средние. Длина тела 7,6—23,5 см. Внешним видом похожи на кротов. Тело вальковатое, цилиндрическое, шея короткая и толстая, морда конусообразно суживается к носу. Конечности короткие, передние четырехпалые (отсутствует пятый палец), задние пятипалые; два средних пальца на передних конечностях с очень крупными когтями. Хвост снаружи не виден. Глаза рудиментарные, находятся под кожей. Морда оканчивается гладкой кожистой ороговевающей снаружи подушечкой, предохраняющей морду при копании. Ушные раковины небольшие, скрытые в шерсти. Ноздри прикрыты кожистой складкой, открываются на конце морды.
Волосяной покров густой, мягкий со своеобразным металлическим оттенком красного, желтого, зеленого, бронзового или фиолетового цвета. Самки имеют одну пару брюшных и одну пару паховых сосков.
Череп конический по форме, не имеет сужения в межглазничной области. У некоторых видов имеются слуховые барабаны. Скуловой отросток верхнечелюстной кости очень велик и достигает чешуйчатой или чешуйчатой и теменной костей, образуя таким образом вторичную скуловую дугу.
3 1 3 2___3
Зубная формула: i — с — р — т—--------= 36 — 40. Резцы (особенно
первый верхний) увеличены в размерах. Второй и третий резцы, клыки и первый предкоренной сходны между собой. Второй и третий предкоренные похожи на коренные. Коронки зубов нижней челюсти вдвое выше коронок зубов верхней. Ключица очень длинная, лопатка треугольная с большим pr. acromialis. Локтевая кость с крупным локтевым отростком (pr. olecranon). Большая и малая берцовые кости срастаются между собой в проксимальном отделе. Позвонков шейных 7, грудных 19, поясничных 4, крестцовых 5, хвостовых 8—9. Тазовые кости-не имеют лобкового сочленения.
Для мышечной системы характерно сильное развитие подкожной мускулатуры. Желудок в виде удлиненного мешка. В кишечнике отсутствует слепая кишка. Сердце и легкие имеют удлиненную форму. Семенники расположены в брюшной полости.
Обитатели равнин, лесов, культурных ландшафтов. Ведут подземный образ жизни, предпочитают песчаные почвы. Копают, как обыкновенные кроты, передними лапами, но в большей степени используют при этом передний отдел морды. Некоторые виды прокладывают ходы под самой поверхностью почвы, другие копают глубже. Активны круглосуточно. Полагают, что они впадают в спячку во время наиболее холодных или засушливых месяцев. Большую часть жизни проводят под землей и появляются на ее поверхности только после дождей вслед за выползающими сюда земляными червями и другими беспозвоночными. Однако представители рода обыкновенных златокротов часто питаются на поверхности почвы главным образом почвенными беспозвоночными, а также безногими ящерицами. В Южной Африке период размножения приходится на дождливый сезон.
Самка обычно приносит двух детенышей, для которых устраивает травяное гнездо глубоко в норе.
Распространены в Африке от Камеруна, Республики Заир, Уганды и Танзании на юг до Мыса Доброй Надежды.
В семействе 5 родов (15 видов):
обыкновенных златокротов — Chrysochlor is La-cepede, 1799 (4 вида);
исполинских златокротов — Chrysospalax Gill, 1883 (2 вида);
африканских златокротов — Amblysomus Ро-mel, 1848 (7 видов);
пустынных златокротов— Eremitalpa Roberts, 1924 (1 вид);
златокротов Винтона — Cryptochloris Shortridge et Carter, 1939 (1 вид).
Рис. 78. Капский златокрот — С. asiatica и его череп. А — передняя; Б — задняя лапы
Род обыкновенных златокротов
Genus Chrysochloris Lacepede, 1799
Небольших и средних размеров златокроты. Длина тела 9—14 см (рис. 78). Коготь второго пальца передней конечности короче третьего, но длиннее когтя первого пальца. Четвертый палец представлен в виде небольшого бугорка. Волосяной покров густой и мягкий; основной цвет его желтовато-оливковый, буроватый или сероватый с зеленоватым, фиолетовым или красноватым отливом. Брюхо светлое, иногда почти белое. У самок 2 пары сосков. В черепе характерны костные слуховые барабаны. Зубная формула полная, число зубов 40.
Обитают на равнинах и в горах, поднимаясь до 2800 м над уровнем моря. Обычно роют неглубокие, проходящие непосредственно под поверхностью почвы норы. Иногда в ночное время появляются на поверхности почвы в поисках пищи. Во время дождей можно наблюдать златокротов, разыскивающих на поверхности почвы дождевых червей. Пища состоит из жуков, земляных червей и других беспозвоночных. Период размножения приходится на сезон дождей — с апреля по июль. Период лактации 2— 3 месяца.
Распространены в Капской провинции, южной Родезии, Уганде, Танзании и Республике Заир.
Систематика рода окончательно не выяснена. По данным разных ученых, род содержит от 2 до 6 видов. Здесь выделено 4 вида:
капский златокрот — С. asiatica Linnaeus, 1758 (Южная Африка);
златокрот Штульманна — С. stuhlmatmi Matschie, 1894 (Восточная Африка);
конголезский златокрот — С. congicus Thomas, 1910 (Центральная Африка);
златокрот Томаса — С. vermiculus Thomas, 1910 (Центральная Африка).
Род исполинских златокротов
Genus Chrysospalax Gill, 1883
Размеры средние и крупные (наибольшие в семействе). Длина тела 12,5—23,5 см. Когти на передних конечностях развиты относительно очень сильно. Волосяной покров более грубый, чем у других златокротов, красноватый, желтовато-бурый или темно-бурый с металлическим оттенком. У самок две пары сосков. Скуловая дуга, образованная скуловым отростком верхнечелюстной кости, тянется кверху и соединяется с лямбдовидным гребнем. Число зубов 40.
Довольно часто (особенно после дождей) охотятся на поверхности почвы. Поразительна способность этих слепых зверьков находить отверстие своей норы, куда они спасаются бегством в случае опасности. В зимние месяцы исполинский златокрот впадает в спячку.
Распространены в Южной Африке.
В роде, по-видимому, 2 вида:
грубошерстный златокрот — С. villosus A.. Smith, 1833 (Наталь, Трансвааль, восточная часть Капской провинции);
исполинский златокрот — С. trevelyani Giinther, 1875 (восточная часть Капской провинции).
Род африканских златокротов
Genus Amblysomus Pomel, 1848
Размеры небольшие. Длина тела 8—13 ши. Волосяной покров короткий, густой и мягкий, на спинной стороне обычно красноватый, буроватый, сероватый или черноватый, иногда с медно-зеленоватым оттенком. На передних лапах по 2 больших когтя. У самок 2 пары сосков. Костные слуховые 3 13 2
барабаны отсутствуют. Зубная формула: i — с — р — т— = 36. Отсутствует третий коренной зуб.
Экология изучена слабо. Обитатели горных равнин, степей и лесов. Некоторые виды копают в основном поверхностные, а другие — глубокие норы. Активны часто в дневное время. Питаются дождевыми червями, насекомыми и их личинками. Период размножения, по-видимому, приходится на дождливый сезон.
Распространены в Южно-Африканской Республике, Мозамбике, Анголе, Республике Заир, Уганде, юго-западной Танзании и в Камеруне.
В роде, по-видимому, 7 видов (по некоторым авторам 10):
желтый златокрот — A. obtusirostris Peters, 1851 (Южно-Африканская Республика, Мозамбик);
светлоносый златокрот — A. leucorhinus Huet, 1885 (Ангола, Республика Заир, Камерун);
златокрот Склатера — A. sclateri Broom, 1907 (Южно-Африканская Республика);
готтентотский златокрот — A. hottentotus A. Smith, 1829 (Южно-Африканская Республика);
златокрот Гуннинга — A. gunningi Broom, 1908 (Южно-Африканская Республика);
златокрот Фостера — A. fosteri St. Leger, 1931 (Уганда);
тропический златокрот — A. tropicalis G. Allen et Loveridge, 1927 (Танзания).
Род пустынных златокротов
Genus Eremitalpa Roberts, 1924
В роде единственный вид: пустынный златокрот — Е. granti Broom, 1907.
Размеры наименьшие в семействе. Длина тела 7,6—8,8 см. Вес тела около 15 г. Тело несколько уплощено и имеет в поперечнике очертание овала. На передних лапах три длинных когтя. На подошвах задних лап и пяточном отделе имеется утолщенная подушка. Мех длинный, густой и мягкий. Верх тела желтовато-бурый или серый. Морда беловатая. Окраска брюха изменяется от буровато-белой до бледно-рыжеватой. В черепе отсутствуют костные слуховые барабаны. Число зубов равно 40.
Обитает в песчаных почвах. Как правило, роет неглубокие норы у самой поверхности земли. Постоянных нор, по-видимому, не имеет. Изредка пустынный златокрот выходит на поверхность. Наибольшая активность отмечается в жаркие часы дня, когда основной объект пищи пустынного златокрота — безногие ящерицы — зарываются в песок, спасаясь от солнца. Кроме ящериц, пустынный златокрот, вероятно, поедает и насекомых.
Встречается в западной части Капской провинции.
Род златокротов Винтона
Genus Cryptochloris Shortridge et Carter, 1939
В роде, по-видимому, единственный вид: златокрот Винтона — С. wintoni Broom, 1907.
Мелкие златокроты. Длина тела 8—9 см. Туловище несколько уплощено и имеет в поперечнике округло-овальные очертания. На передних лапах по три сильно развитых когтя и большая внутренняя пальцевая подушка на подошве. Волосяной покров короткий, густой и мягкий, по верху тела желтовато-золотистый с фиолетовым отблеском, на брюхе золотисто-серый. На морде могут быть беловатые отметины. Число зубов 40.
Обычно копает норы непосредственно под поверхностью почвы. Населяет песчаные участки почвы. Ночью иногда выходит на поверхность. Питается безногими ящерицами (Typhlosaurus) и различными беспозвоночными.
Распространен в Капской провинции.
СЕМЕЙСТВО ЩЕЛЕЗУБОВ
FAMILIA SOLENODONTIDAE DOBSON, 1882
В семействе единственный род: шелезубов — Solenodon Brandt, 1833.
Щелезубы — крупные насекомоядные, длина тела 28—32,5 см, длина хвоста 17,5—25,5 см, вес около 1 кг. Внешним видом несколько напоминают землероек на высоких ногах (рис. 79). Удлиненное рыло имеет у гаитянского щелезуба в основании небольшую круглую кость, располагающуюся горизонтально впереди от носовых костей. Ноздри открываются по сторонам рыла. Глаза небольшие. Частично обнаженные уши немного выдаются над мехом. Конечности пятипалые; передние — стопоходящие, задние — пальцеходящие.
Волосяной покров гаитянского щелезуба грубый, а у кубинского щелезуба — мягкий и длинный. Хвост и лапы почти без волос. Конец морды голый. Окраска у гаитянского щелезуба изменяется от черноватой и темножелтоватой до красновато-коричневой, а у кубинского щелезуба она черновато-коричневая с белым или желтовато-коричневая.
Рис. 79. Гаитянский щелезуб — S. paradoxus и его череп
Кожные специфические железы под мышками и в пахах продуцируют маслянистый секрет зеленого цвета с сильным «козлиным» запахом. У самок одна пара сосков располагается в паховой области. Семенники находятся в брюшной полости.
В черепе сильно развиты сагиттальный и особенно лямбдоидальный гребни; последний нависает над затылочной костью. В области глазниц череп сильно суживается. Слезное отверстие очень крупное. Скуловая ду-
3 13 3
га отсутствует. Зубная формула: i. — с — р ~^т~^ = 40. Первый резец крупнее других. Первый резец верхней челюсти отделен от второго промежутком. На передней поверхности второго нижнего резца имеется борозда. На жевательной поверхности верхних коренных зубов 3 бугорка располагаются в виде буквы V. Позвонков шейных 7, грудных 15, поясничных 4, крестцовых 5 и хвостовых 23.
Подчелюстная слюнная железа у гаитянского щелезуба (вероятно, и кубинского щелезуба) продуцирует ядовитую слюну1. Проток железы открывается у основания глубокой бороздки второго нижнего резца. По-видимому, гаитянский щелезуб не имеет иммунитета против собственного яда, так как эти зверьки погибают даже от легких укусов, полученных во время драк между собой.
Обитатели лесов, кустарниковых зарослей. Активность в основном ночная. День проводят в норах, дуплах упавших деревьев. Пищу разыски
1 Рабб (Rabb, 1959) ставит этот факт под сомнение
вают, копаясь мордой в почве, раскапывая ее сильными лапами. Кормом служат различные беспозвоночные животные, пресмыкающиеся и растительные объекты (плоды и др.). Самка приносит трех слепых, беззубых и безволосых детенышей.
Распространены на Гаити и Кубе.
В роде 2 вида:
гаитянский щелезуб — 5. paradoxus Brandt, 1833 (Гаити); кубинский щелезуб — 5. cubanus Peters, 1861 (Куба).
Численность очень мала (особенно кубинского), и оба вида включены в «Красную книгу».
Некоторые исследователи (Кабрера, Cabrera, 1925; Холл и Келсон, Hall and Kelson, 1959 и др.) выделяют кубинского щелезуба в особый род Atopogale Cabrera, 1925. Однако различия между гаитянским и кубинским щелезубами не настолько велики, чтобы считать их родовыми признаками, поэтому такое разделение вряд ли правильно.
СЕМЕЙСТВО ЗЕМЛЕРОЕК
FAMILIA SORICIDAE GRAY, 1821
Небольших и средних размеров насекомоядные, среди которых находятся самые маленькие представители класса млекопитающих. Длина тела 3,5—18 см. Длина хвоста 1—12 см. Вес животных от 2 до 35 г. Внешним видом напоминают мышей с умеренно удлиненным туловищем, относительно короткими конечностями. Голова довольно крупная, с сильно удлиненным лицевым отделом и вытянутой в хоботок носовой частью. Глаза очень маленькие, уши разной степени развития: небольшие, скрытые в меху или довольно крупные, выступающие из него. Конечности пятипалые. Между пальцами могут быть плавательные перепонки (перепончатые белозубки) или лапы окаймлены щетинистой оторочкой (куторы). Хвост от очень короткого, не превышающего длину задней ступни, до очень длинного, более длины туловища. Половое и анальное отверстия окружены кожным валиком, образующим иногда клоаку. Волосяной покров короткий, густой и мягкий, волосы обычно не подразделяются на категории и растут поодиночке. Самки имеют от 6 до 10 сосков. Сильно развиты сальные железы. Немногочисленные потовые железы имеют мешковидную форму. На боках тела или у корня хвоста обычно имеются кожные специфические железы, продуцирующие секрет со своеобразным запахом.
Череп несколько уплощен в дорзо-вентральном направлении. Мозговой отдел расширен. Лицевая часть черепа сильно удлинена и сужена. Скуловая дуга отсутствует. Заднеглазничных отростков нет. Костные слуховые пузыри отсутствуют. Барабанная кость кольцеобразная. Некоторые кости черепа срастаются в раннем возрасте, поэтому швы между ними незаметны. Сочленовный отросток нижней челюсти с двумя отдельными суставными поверхностями, имеющими валикообразную форму. Верхняя из них сочленяется с чешуйчатой костью, а нижняя — с заднесочленовным 3 q._________________________________J ।____з з
отростком. Зубная формула: i — с —-— р —— т—= 26 — 32. Смена молочных зубов на постоянные происходит в процессе эмбрионального развития. Передняя пара верхних резцов больших размеров и выдается вперед. Они серповидно изогнуты и имеют по две вершины. Второй и третий верхние резцы, клыки и передние предкоренные зубы по форме сходны между собой и называются промежуточными зубами. Коронки верхних коренных зубов брахиодонтного типа, секториальные. Выступы на их жевательной поверхности образуют букву W. Нижние резцы крупных размеров, вытянутые вдоль продольной оси челюсти.
Позвонков в позвоночном столбе шейных 7, грудных 13—15, поясничных 5—6 (у белозубок-броненосок 11), крестцовых 4—5, число хвосто
вых позвонков сильно варьирует. Имеется тонкая слабоизогнутая ключица. Лопатки очень длинные и узкие. Плечевая кость относительно длинная и тонкая. Таз широкий. Симфиз между лобковыми костями отсутствует. Большая и малая берцовые кости срастаются в дистальных отделах. Os penis отсутствует.
Обитатели самых различных ландшафтов — от северных частей зоны тундры до тропических лесов и пустынь. Поднимаются в горы до 3500— 4000 м над уровнем моря. Большинство видов предпочитает селиться во влажных местах. Некоторые виды ведут полуводный образ жизни. Держатся поодиночке. Активны круглосуточно, наибольшая активность в сумеречные и ночные часы. В спячку не впадают. Питаются в основном различными беспозвоночными: насекомыми, их .личинками, дождевыми червями и т. д. Могут нападать и на мелких позвоночных животных: лягушек, ящериц, детенышей мышевидных грызунов. Ежесуточно землеройкам необходимо поедать большое количество пищи, в 1,5—2 раза превышающее их собственный вес. В течение года, по-видимому, 2—3 помета, в каждом из которых бывает 4—14 детенышей. Беременность 13—28 дней. Детеныши рождаются голыми, слепыми, но развиваются очень быстро и в 3—4-недельном возрасте переходят к самостоятельному образу жизни. Землеройки имеют существенное значение как истребители большого количества вредителей сельского и лесного хозяйства.
Распространение всесветное, за исключением полярных областей, Австралии и Южной Америки к югу от Эквадора, Колумбии и Венесуэлы.
Различные исследователи включают разное количество родов в семейство землероек. В настоящем руководстве принят 21 род (271 вид) землероек:
бурозубок — Sorex Linnaeus, 1758 (40 видов);
бурозубок-пигмеев — Microsorex Cones, 1877 (1 вид);
азиатских бурозубок — Soriculus Blyth, 1854 (6 видов);
цейлонских бурозубок — Podihik Deraniyagale, 1958 (1 вид);
кутор —Neomys Каир, 1829 (2 вида);
американских короткохвостых бурозубок — Blarina Gray, 1838 (2 вида);
азиатских короткохвостовых бурозубок — Blarinella Thomas, 1911 (1 вид);
короткоухих бурозубок — Cryptotis Pomel, 1848 (26 видов);
серых землероек — Notiosorex Coues, 1877 (3 вида);
белозубок — Crocidura Wagler, 1832 (148 видов);
конголезских белозубок — Paracrocidura Heim de Balsac, 1956 (1 вид); африканских гигантских белозубок —• Praesorex Thomas, 1913 (1 вид); многозубых белозубок — Sanctis Ehrenberg, 1832 (28 видов);
когтистых белозубок Келаарта — Feroculus Kelaart, 1852 (1 вид); когтистых белозубок Пирсона — Solisorex Thomas, 1924 (1 вид); горных белозубок — Surdisorex Thomas, 1906 (2 вида);
путораков — Diplomesodon Brandt, 1852 (1 вид);
кротовых белозубок — Anaurosorex Milne-Edwards, 1870 (1 вид);
азиатских водяных белозубок —• Chlmarrogale Anderson, 1877 (2 вида); тибетских водяных белозубок, или перепончатолапых белозубок — Nectogale Milne-Edwards, 1870 (1 вид);
белозубок-броненосок — Scutisorex Thomas, 1913 (2 вида).
Эта система в общем соответствует системе, предложенной Симпсоном (1945); в нее лишь добавлены 2 рода: описанные в 1956 г. конголезские бело-зубки и в 1958 г. цейлонские бурозубки, а также объединены в род многозубых белозубок мышевидные белозубки (Myosorex) и лесные белозубки (Sylvisorex).
Рис. 80. Обыкновенная бурозубка — S. araneus и ее череп
Род бурозубок
Genus Sorex Linnaeus, 1758
Землеройки мелких и средних размеров. Длина тела примерно 5—9 см. Длина хвоста 2,5—8 см. Вес 3—16 г. Хвост на всем протяжении равномерно покрыт волосами одинаковой длины (рис. 80). Пальцы на лапах вооружены тонкими слабоизогнутыми когтями. Ушные раковины небольших размеров, почти целиком скрытые в волосах. Волосяной покров густой, относительно короткий и мягкий; на спинной стороне преимущественно бурый, черно-бурый или черный, на брюшной светлый, серый или буро-серый. У некоторых видов окраска одноцветная по всему телу. Сосков 3 пары.
3 13 3
Зубная формула: i — с -— р — т— = 32. Коронки зубов имеют ко-2 0 13
ричневый или красно-коричневый цвет. В каждой половине верхней челюсти по 5 промежуточных зубов. Характерна необычайно большая изменчивость числа хромосом в диплоидном наборе (от 20 до 66).
Обитают в различных биотопах, предпочитая увлажненные места в лесах, поймы рек, захламленные берега ручьев и других водоемов. Основной пищей служат беспозвоночные: черви, насекомые, многоножки, а также их личинки. Нападают и на мелких позвоночных: лягушек, грызунов. Побочными объектами питания являются некоторые растительные объекты, например семена древесно-кустарниковых растений. Некоторые виды запасают пищу- Водяная бурозубка (5. palustris\ Америка) ведет полуводный образ жизни. Задние конечности у нее окаймлены щетинистым гребнем волос. Хорошо плавает и ныряет. Размножение у бурозубок приходится на лето. Беременность 18- 28 дней. Самка приносит от 2 до 10 детенышей. На второй неделе жизни у детенышей появляются на теле волосы, а на третьей неделе открываются глаза. На 22-й день детеныш покидает гнездо и переходит к самостоятельному образу жизни. На 25-й день после родов у самки вновь наступает течка.
Ареал очень обширен: охватывает Северное полушарие в Америке на юг до Центральной Америки, а в Европе и Азии до Израиля, Малой Азии, Кашмира и Северной Бирмы.
Различные исследователи насчитывают разное число видов в роде. До появления новых исследований по систематике бурозубок представляется наиболее правильным придерживаться системы, предложенной Холлом и Келсоном (Hall a. Kelson, 1959) и Кузякиным (1965), с небольшими изменениями. В соответствии с этой системой род бурозубок насчитывает 49 видов:
крошечная бурозубка S. minutlssimus Zimmermann, 1780 (северо-восточные районы европейской части СССР, Сибирь, северная часть МНР, Южное Приморье, о-ва Кунашир, о-в Хонсю);
бурозубка Азуми — С. hosonoi Imaizumi, 1954 (о-в Хонсю);
малая бурозубка—S. minutus Linnaeus, 1766 (большая часть Европы, лесная и лесостепная зоны Сибири, Кашмир, Палестина, Цайдам);
дальневосточная бурозубка—S. gracillimus Thomas, 1907 (Приморье, Шантарские острова, Сахалин);
кавказская малая бурозубка — S. volnuchini Ognev, 1921 (Кавказ); бухарская бурозубка—S. buchariensis Ognev, 1921 (Таджикистан); крупнозубая, или темная бурозубка, — S. daphaenodon Thomas, 1907 (Сибирь, Дальний Восток, Северо-Восточный Китай, о-ва Хоккайдо, Курильские, Сахалин);
средняя бурозубка—S. caecutiens Laxmann, 1788 (включая S. hydro-dromus Dobson, 1889 и S. cinereus Kerr, 1792) (Евразия, включая северную часть МНР, Северный Китай, Корейский полуостров, Сахалин и Японские острова, а также большая часть Северной Америки— Аляска, Канада, северо-западные, северные и северо-восточные штаты США);
японская бурозубка — S. shinto Thomas, 1905 (Япония, Сахалин, Курильские острова);
обыкновенная бурозубка — С. araneus Linnaeus, 1758 (включая S. asper Thomas, 1914) (Евразия до МНР и Китая включительно);
баргузинская бурозубка—S. isodon Turov, 1924 (Забайкалье, Амурская обл., Приморский край);
европейская длиннохвостая бурозубка — S. raddei Satunin, 1895 (Кавказ);
кавказская бурозубка—S. caucasicus Satunin, 1914 (Кавказ);
арктическая бурозубка—S. arcticus Kerr, 1792 (северные районы Северной Америки от Тихого до Атлантического океана);
сибирская бурозубка — S. sibiriensis Ognev, 1921 (Кемеровская область в Сибири);
иркутская бурозубка —S. irkutensis Ognev, 1933 (Иркутская область в Сибири);
плоскочерепная бурозубка—S. vir Gl. Allen, 1914 (Алтай, Западная и Восточная Сибирь до побережья Охотского моря и Приморья);
швейцарская бурозубка—S. gemellus Ott, 1968 (Швейцария);
когтистая бурозубка — S. unguiculatus Dobson, 1890 (Дальний Восток, Северо-Восточный Китай, Курильские острова, Корейский полуостров, Япония);
альпийская бурозубка — S. alpinus Schinz, 1837 (южная часть Западной Европы);
полосатоспинная бурозубка — S. cylindricauda Milne-Edwards, 1872 (Юньнань, Сычуань, Ганьсу, Шэньси, Северная Бирма);
гигантская бурозубка — S. pacificus Coues, 1877 (Восточная Азия и западные штаты США);
бурозубка Лиелла—S. lyilli Merriam, 1902 (Калифорния);
орегонская бурозубка — С. preblei Jackson, 1922 (штат Орегон, США); северомексиканская бурозубка—S. milleri Jackson, 1947 (северная Мексика);
длинномордая бурозубка—S. longirostris Bachman, 1837 (восточные и юго-восточные районы США);
бурозубка Вагранта—S. vagrans Baird, 1858 (Аляска, западная Канада, тихоокеанское побережье США до Калифорнии, штаты Айдахо, Монтана, Вайоминг, Юга, Колорадо, Аризона, Нью-Мехико);
украшенная бурозубка — S. ornatus Merriam, 1895 (западное и юго-западное тихоокеанское побережье США);
5. tenellus Merriam, 1895 (штаты Калифорния и Невада, США);
S. trigonirostris Jackson, 1922 (штат Орегон, США);
карликовая бурозубка — S. nanus Merriam, 1895 (штаты Юта, Колорадо, Вайоминг, Нью-Мехико, Аризона);
калифорнийская бурозубка •— S. juncensis Nelson et Goldman, 1909 (Баха Калифорния);
Санта-Каталинская бурозубка—S. willeti von Bloeker, 1942 (о-в Санта-Каталина, Калифорния);
S. sinuosus Grinnell, 1913 (о-в Гриззли, Калифорния);
бурозубка Верапаза — S. veraepacis Alston, 1877 (южная Мексика и Центральная Америка);
большезубая бурозубка —S. macrodon Merriam, 1895 (южная Мексика);
болотная бурозубка—S. palustris Richardson, 1828 (включая S. alas-canus Merriam, 1900) (юго-западная Аляска, западная, центральная и восточная Канада, запад США);
водяная бурозубка — S. bendirii Merriam, 1884 (северо-запад США);
прибыловская бурозубка — S. pribilofensis Merriam, 1895 (о-в Сент-Пол в группе островов Прибылова);
дымчатая бурозубка — S. fumeus Miller, 1895 (юго-восточная часть Канады и северо-восток США);
бурозубка Джексона — S. jacksoni Hall et Gilmore, 1932 (о-в Св. Лаврентия в Беринговом море);
бурозубка Гаспе — S. gaspensis Anthony et Godwin, 1924 (Квебек, Канада);
длиннохвостая бурозубка — S. dispar Batchelder, 1911 (юго-восток Канады и северо-восток США);
бурозубка Троубриджа — S. trowbridgii Baird, 1858 (запад США);
бурозубка Мерриама — S. merriami Dobson, 1890 (штаты: Вашингтон, Орегон, Айдахо, Монтана, Вайоминг, Невада, Юта, Колорадо, Аризона, Нью-Мехико);
бурозубка Соссюра — S. saussurei Merriam, 1892 (южная Мексика и Центральная Америка);
мексиканская длиннохвостая бурозубка—S. oreopolus Merriam, 1892 (южная Мексика);
бурозубка Склатера—S. sclateri Merriam, 1897 (южная Мексика);
санкристобальская бурозубка—S. stizodon Merriam, 1895 (южная Мексика);
В СССР: крошечная бурозубка (центральные и восточные районы европейской части СССР, большая часть Сибири, кроме Севера, Южное Приморье, Сахалин, о-в Кунашир); малая бурозубка (европейская часть СССР, запад и юг Сибири, Тянь-Шань); дальневосточная бурозубка (Приморье, о-ва Шантарские, Сахалин); кавказская малая бурозубка (Кавказ); бухарская бурозубка (хр. Петра Первого в Таджикистане); крупнозубая, или темная, бурозубка (Сибирь, Дальний Восток, Приморье, Сахалин, Курильские острова); средняя бурозубка (почти вся территория СССР, кроме южных районов европейской части СССР, Казахстана и Средней Азии); японская бурозубка (Сахалин, Курильские острова); обыкновенная бурозубка (почти вся территория СССР); баргузинская бурозубка (Забайкалье, Амурская область, Приморский край); европейская длиннохвостая бурозубка (Кавказ); кавказская бурозубка (Кавказ); сибирская бурозубка (Кемеровская обл.); иркутская бурозубка (Иркутская обл.); плоскочерепная бу-
розубка (Алтай, Западная и Восточная Сибирь до побережья Охотского моря и Приморья и от бассейна р. Нижней Тунгуски, устья р. Индигирки и Нижнеколымска к югу до южного Забайкалья и окрестностей Уссурийска в Приморье); когтистая бурозубка (Камчатка, Охотское побережье, Приморский край, Сахалин, Курильские острова); альпийская бурозубка (Карпаты) и гигантская бурозубка (Южное Приморье).
Род бурозубок-пигмеев
Genus Microsorex Coues, 1877
В роде единственный вид: бурозубка-пигмей — М. hoyi Bairn, 1858.
Размеры небольшие. Длина тела 5—6 см. Длина хвоста 2,7—3,5 см. Вес 2,3—3,5 г. Волосяной покров короткий, густой и мягкий, на спинной стороне серовато-бурый или серый. На брюшной светлее. Хвост двухцветный. Сосков 4 пары. Череп напоминает по строению череп бурозубок, но относительно более узкий и уплощенный, причем рострум более короткий и широкий. Нижняя челюсть короткая и массивная.
Обитатель различных ландшафтов. Встречается в сухих участках леса, густых зарослях травы и кустарниках. Размеры зверька настолько малы, что он может пролезать в норы крупных жуков. Хорошо лазает по кустарникам и может высоко прыгать. Активность круглосуточная. Питается различными беспозвоночными, в основном насекомыми. Характерно запасание большого количества пищи. Размножается весной и летом,. По-видимому, самка приносит более .одного помета в год, по 5—6 детенышей.
Распространен на юго-западе Аляски, большей части Канады и в северных и восточных штатах США.
Род азиатских бурозубок
Genus Soriculus Blyth, 1854
Размеры небольшие и средние. Длина тела 5—10 см. Длина хвоста примерно 4—42 см. Для большинства видов характерен удлиненный тонкий хвост. Пальцы вооружены сильными когтями. Волосяной покров короткий, плотный и мягкий, на спинной стороне красновато-бурый, темнобурый, сероватый или черноватый. На брюшной обычно светлее. Зубов 28 или 30. i
Обитают в горах во влажных участках лесов и в зарослях кустарников I на высоте 2100—2300 м над уровнем моря. ’ I
Экология почти не изучена; по-видимому, не отличается от экологии j других лесных землероек. |
Распространены в Бутане, Кумаоне, Сиккиме, Непале, Китае, Север- 1 ной Бирме, Бакбо и на Тайване.
В роде 6 видов: fl
сиккимская бурозубка—S. nigrescens Gray, 1842 (Бутан, Кумаон,
Сикким, Непал, Мишми, Северная Бирма);
S. caudatus Horsfield, 1851 (Кумаон, Сикким, Мишми, Северная Бирма, Сычуань, Юньнань, Тайвань, Бакбо);
индийская бурозубка — S. leucops Horsfield, 1855 (Непал, Сикким, Северная Бирма, Сычуань, Юньнань); I
S. hypsibius de Winton, 1899 (Юньнань и Сычуань к северу до Ганьсу • и Шэньси); I
бурозубка Заленского—S. salenskii Kastschenko, 1907 (Шэньси, Сычуань, Юньнань и Северная Бирма);
152 '
бурозубка Лове — S. lowei Osgood, 1932 (Бакбо в Индокитае).
Гейм де Бальзак и Бурлиер (1958) разделяют азиатских бурозубок на 2 рода: Soriculus и Chodsigoa.
Род цейлонских бурозубок
Genus Podihik Deraniyagale, 1958
В роде единственный вид: цейлонская бурозубка — Р. kura Deraniyagale, 1958, Цейлонские бурозубки близкородственны азиатским бурозубкам и, возможно, представляют их подрод.
Размеры мелкие. Длина тела около 4 см. Длина хвоста около 2,5 см. Характерен удлиненный и расширенный лицевой отдел. Уши крупные. Хвост, утолщенный в основании, утончается к концу и покрыт длинными редкими волосами. Ноздри вытянуты в две короткие трубочки. Волосяной покров на спинной стороне красновато-бурый. На брюшной голубоватосерый. На голове темная «шапочка». Морда красновато-серая. Конечности и уши желтые.
Экология не изучена.
Обитает в северной и центральной части Цейлона.
Род кутор
Genus Neomys Каир, 1829
Размеры мелкие и средние. Длина тела 7—10 см. Длина хвоста 4,5— 7 см. Вес 10—20 г. На задних лапах характерны удлиненные ступни. Кисти
и ступни по бокам покрыты уплощенными щетинками, которые образуют своеобразную бахрому, увеличивающую плавательную поверхность лап (рис. 81). Уши почти не выдаются над мехом. Хвост на вентральной поверхности имеет киль из удлиненных щетинистых волос. Волосяной покров очень густой, плотный, мягкий, на спинной стороне черно-бурый, бурый или светло-бурый, на брюшной белый, серо-белый или желто-белый. Сос-
ков 5 пар. Череп крупный, массивный, с расширенным мозговым отделом. Хорошо выражены сагиттальный и лямбдоидальный гребни. Зубная фор-3 12 3
мула: i — с—р—т— = 30. Вершины зубов красновато-бурые. 2 0 13
Диплоидное число хромосом у обыкновенной куторы 52.
Обитатель берегов водоемов в различных ландшафтах. Хорошо плавает и ныряет. Корм добывает в воде, но поедает его всегда на суше. Питается насекомыми, червями, моллюсками, рыбьей икрой, мальками рыб и лягушками. Активность круглосуточная, но наибольшая ночью. Убежищем служат норы, которые вырывает сама или чужие. Размножение приходится на весну и лето, в течение которых бывает по меньшей мере по 2 помета у каждой самки. Беременность, вероятно, около 24 дней. Детенышей в помете от 3 до--8. Продолжительность лактации примерно 37 дней.
Экология малой куторы почти не изучена. В отличие от обыкновенной куторы, она меньше связана с водоемами. Численность малой куторы очень невелика.
Распространение охватывает Европу, Малую Азию, Сибирь, МНР, Северный и Северо-Восточный Китай.
В роде 2 вида:
малая кутора — N. anomalus Cabrera, 1907 (Малая Азия, Испания, франция, Швейцария, Югославия, Греция, Венгрия, Италия, Австрия, Румыния, Албания, Турция, Болгария, Украина, Белоруссия);
обыкновенная кутора — N. fodiens Pennant, 1771 (Европа, Сибирь, МНР, Северный и Северо-Восточный Китай).
Рис. 81. Обыкновенная кутора—N.fodi-ens и ее череп. А — передняя; Б —задняя лапы
В СССР: малая кутора (Украина, Крым, Белоруссия); обыкновенная кутора (европейская часть от Северного Ледовитого океана на юг до Северного Кавказа, Западная Сибирь, около оз. Байкал, на восток до побережья Охотского моря, Сахалина, низовья Амура).
Род американских короткохвостых бурозубок
Genus Blarina Gray, 1838
Размеры средние. Длина тела от 7,5 до 10,5 см. Длина хвоста 1,7—3 см. Вес 15—30 г (рис. 82). Спинная сторона шиферного цвета, а брюшная — немного светлее. Имеются расхождения в зубной формуле, приводимой различными исследователями; по Холлу и Келсону (1959), общее число зубов 34: i с р -у- т; по Гейм де Бальзаку и Бурлиеру (1955), —32: по Гржимеку (1968) — 30: i-y-c-^-p-y-m —.
Подчелюстная слюнная железа секретирует ядовитую слюну, которая действует на нервную систему укушенной добычи.
Обитатели самых разнообразных ландшафтов. Обычны и многочисленны. Питаются различными беспозвоночными, мелкими позвоночными и 154
Рис. 82. Обыкновенная короткохвостая бурозубка — В. brevicauda и ее череп
растительными объектами. Могут делать небольшие запасы пищи. Наибольшая активность раннеутренняя и поздневечерняя. Беременность 17—21 день. В помете 3—9 детенышей. Период размножения растянут с ранней весны до поздней осени. Каждая самка, по-видимому, имеет несколько приплодов (обычно 2) в году. Половозрелости достигают на первом году жизни. Длительность жизни примерно 2,5 года.
Распространены в Северной Америке от Квебека до Флориды, на запад до Манитобо и Техаса.
В роде 2 вида:
обыкновенная короткохвостая бурозубка — В. brevicauda Say, 1823 (ареал совпадает с ареалом рода);
болотная короткохвостая бурозубка — В. telmalestes Merriam, 1895 (штат Виргиния, США).
Род азиатских короткохвостых бурозубок
Genus Blarlnella Thomas, 1911
В роде единственный вид: азиатская короткохвостая бурозубка — В. quadraticauda Milne-Edwards, 1872.
Размеры небольшие. Длина тела 6—7,5 см. Длина хвоста 3—4 см. Короткие уши совсем не выступают над мехом. Когти на передних и задних конечностях очень длинные. Хвост относительно короткий. Окраска спинной стороны буровато-серая с серебристым или дымчатая с серым оттенком, брюшной — несколько светлее. Конечности и хвост обычно темно-бурые.
Экология не изучена. Встречается в горах.
Ареал охватывает Сычуань, Юньнань и Ганьсу в Китае, найдена также в Северной Бирме.
Род короткоухих бурозубок
Genus Cryptotis Pomel, 1848
Размеры небольшие. Длина тела 6—9,2 см. Длина хвоста 1,2—4,2 см. Глаза очень малы. Уши не выдаются над мехом. Спинная поверхность буроватая или черноватая, брюшная — несколько светлее или такого же цве-3 12 3
та, как и спинная. Зубная формула: i — с — р — т— = 30.
Большинство видов — обитатели лесов. Один вид встречается на открытых равнинах. Обыкновенная короткоухая бурозубка ведет колониальный образ жизни. Питаются различными беспозвоночными, мелкими ящерицами, иногда растениями. Самки обыкновенной короткоухой бурозубки способны к размножению в возрасте около трех месяцев. Беременность примерно 15 дней. В помете от 3 до 6, как правило, 4—5 детенышей. Самка имеет несколько пометов в году. В южных частях своего ареала эти
землеройки размножаются, по-видимому, в течение круглого года, а на севере с марта по ноябрь.
Распространение охватывает восточные и юго-восточные штаты США, Мексику, Центральную Америку и северную часть Южной Америки. Это единственный род среди насекомоядных, распространение которого охватывает Южную Америку.
В роде 26 видов:
обыкновенная короткоухая бурозубка — С. parva Say, 1823 (восточные и юго-восточные штаты США, на запад до штатов Северная и Южная Дакота, Небраска и Канзас);
С. celatus Goodwin 1956 (южная Мексика);
стройная бурозубка — С. pergacilis Elliott, 1903 (южная Мексика);
мексиканская бурозубка — С. mexicana Coues, 1877 (южная Мексика);
широкомордая бурозубка — С. eurirhynchis Genoways et Choate, 1967 (Халиско в Мексике);
С. alticola Merriam, 1895 (Южная Мексика);
бурозубка Эндерса — С. endersi Setzer, 1950 (Панама);
С. fronatalis Miller, 1911 (южная Мексика);
сан-кристобальская бурозубка — С. griseoventris Jackson, 1933 (юж-• ная Мексика);
бурозубка Гуерреро — С. guerrerensis Jackson, 1933 (южная Мексика);
бурозубка Гудвина — С. goodwini Jackson, 1933 (Гватемала);
С. gracilis Miller, 1911 (Таламанка в Коста-Рике);
бурозубка Джексона — С. jacksoni Goodwin, 1944 (Коста-Рика);
юкатанская бурозубка — С. mayensis Merriam, 1901 (Юкатан в южной Мексике);
панамская бурозубка — С. тега Goldman, 1912 (восточная Панама); бурозубка Нельсона — С. nelsoni Merriam, 1895 (южная Мексика); сумеречная бурозубка — С. obscura Merriam, 1895 (южная Мексика); никарагуанская бурозубка — С. olivacea J. Allen, 1908 (Никарагуа); бурозубка Зетека — С. zeteki Setzer, 1950 (Панама);
С. fossor Merriam, 1895 (гора Земпоалтепек в южной Мексике);
большая короткоухая бурозубка—С. magna Merriam, 1895 (южная Мексика);
гватемальская бурозубка— С. micrura Tomes, 1862 (Гватемала);
С. soricina Merriam, 1895 (Тлалпама в южной Мексике);
черноватая бурозубка — С. nigrescens J. Allen, 1895 (Гондурас); костариканская бурозубка — С. orphila J. Allen, 1895(Коста-Рика); темная короткоухая бурозубка — С. tersus Goodwin, 1954 (Панама). Систематика короткоухих бурозубок нуждается в ревизии, и после нее число видов, по-видимому, будет сокращено.
Род серых бурозубок
Genus Notiosorex Coues, 1877
Размеры мелкие. Длина тела 5—9 см. Длина хвоста 2—5 см.* Уши довольно крупные. Хвост относительно короткий. Основная окраска тела сероватая. У самок по 3 пары сосков. Зубная формула:
. 3 1 13 оо
i — с — р — т — = 28.
2 0 1 3
Обитатели засушливых районов, часто — пустынь. Активность в основном ночная. Питаются главным образом насекомыми, а также другими беспозвоночными.
Обитают в юго-западных и юго-центральных районах США и в Мексике.
В роде 3 вида:
бурозубка Крауфорда — N. crawford i Cones, 1877 (ареал сходен с ареалом рода, кроме его крайнего юга);
пустынная бурозубка — N. gigas Merriam, 1897 (штат Халиско, Мексика);
бурозубка Филлипса — N. phillipsii Schaldach, 1966 (Оаксака, Мексика).
Род белозубой
Genus Crocidura Wagler, 1832
Размеры мелкие, средние и крупные. Длина тела от 4 до 15 см. Длина хвоста от 4 до 10 см. Уши довольно крупные, выступающие над уровнем волосяного покрова (рис. 83). Хвост покрыт короткими волосами и редкими удлиненными щетинками. Окраска бурая, коричневая, черно-бурая, серая или черная. Передние когти не удлинены. Сосков 3 пары. Зубная формула: 9 । 1__2 3
i — с — р —-—т — = 28, реже 26. Коронки зубов белые. В каждой половине верхней челюсти по 3 (иногда 4) промежуточных зуба. В диплоидном наборе у обыкновенной белозубки 42 хромосомы.
Обитатели разнообразных ландшафтов: лесов, степей, полупустынь и пустынь. Активность в основном ночная. Питаются преимущественно беспозвоночными, нападают также на лягушек и ящериц. Период размножения с марта по октябрь. Беременность около 28 дней. Ежегодно самка приносит несколько пометов. Число детенышей в помете от 3 до 7 (иногда от 1 до 10). У детенышей волосы полностью покрывают туловище на 16-й день. Глаза открываются на 13-й день. Половозрелость наступает в 2—3 ме-
сячном возрасте.
Распространены в Африке, Южной и Средней Евразии до Японии, Ти-
мора и Цейлона на востоке и юге.
Систематика белозубок разработана крайне слабо и требует существенного пересмотра. Исходя из работ Эллермана и Моррисон-Скотта (1966); Строганова (1957); Громова и др. (1963); Бобринского и др. (1965); Эллермана, Моррисон-Скотта и Хеймана (1953); Гейм де Бальзака (1956а, 19566, 1957, 1958, 1968); Гейм
де Бальзака и Эллена (1958); Лори и Хилла (Laurie and Hill, 1954); Чейзена (Chasen, 1940); Тейлора (Taylor, 1934) и некоторых других, можно выделить 148 видов белозубок:
колючая белозуб-ка—С. hispida Thomas, 1913 (Средний Андаманский остров в Бенгальском заливе);
белозубка Флауе-ра—С. floweri Dollman, 1915 (APE);
цейлонская белозубка—С. miya Phillips, 1929 (Цейлон);
малая египетская белозубка — С. religiosa Geoffroy, 1827 (АРЕ, Сомали, Уганда, Судан);
Рис. 83. Малая белозубка — С. suaveolens и ее череп
белозубка Хорсфилда — С. horsfieldi Tomes, 1856 (Цейлон, Кашмир, п-ов Индокитай, Таиланд, Северная Бирма);
малая белозубка — С. suaveolens Pallas, 1811 (южные и центральные области Западной Европы и юг и центр европейской части СССР, южный Казахстан, Средняя Азия, Прибалхашье и Джунгарский Алатау, Приморский край, Корейский полуостров, МНР, Иран, Палестина, Алжир, Марокко);
обыкновенная белозубка — С. russula Hermann, 1780 (Северная Африка, Франция, Сардиния, Корсика, Швейцария, Италия, Испания, Португалия, Бельгия, Нидерланды, ФРГ и ГДР, Польша, Кавказ, Малая Азия, Палестина, Иран, Афганистан и далее к востоку до Японии);
белобрюхая белозубка — С. leucodon Hermann, 1780 (Голландия, Бельгия, Франция, ФРГ и ГДР, Польша, Швейцария, Италия, Югославия, центральные и южные области европейской части СССР, южный Казахстан, юг Средней Азии, Западная Сибирь, Иран, юг МНР);
средиземноморская белозубка — С. caudata Miller, 1901 (Сицилия, Корсика, Балеарские острова);
белохвостая белозубка — С. pergrisea Miller, 1913 (Кашмир, Белуджистан, восточный Иран, юг Средней Азии);
серая белозубка—С. attenuata Milne-Edwards, 1872 (Китай, Хайнань, Северная Бирма, Ассам, Бутан, Сикким, Кумаон, Пенджаб, Кашмир);
С. dracula Thomas, 1912 (Юньнань, Фуцзянь, Северная Бирма, п-ов Индокитай);
большая белозубка — С. lasiura Dobson, 1890 (юг Приморского края. Корейский полуостров и южнее расположенные районы Восточной Азии);
большая египетская белозубка — С. olivieri Lesson, 1827 (APE);
красносерая белозубка — С. cyanea Duvernoy, 1838 (Южная Африка);
рыжая белозубка — С. hirta Peters, 1852 — (Ангола, южная Родезия, Замбия, Мозамбик, Южная Африка на север до Республики Заир);
черная белозубка — С. pilosa Dobson, 1890 (Южно-Африканская Республика, Мозамбик, Юго-Западная Африка, южная Родезия, Ангола, Замбия, Кения, Танзания, Эфиопия, юг Республики Заир);
белозубка Смита — С. smithi Tomas, 1895 (Южно-Африканская Республика, Юго-Западная Африка, Ангола, Замбия);
гигантская белозубка — С. flavescens Geoffroy, 1827 (Южно-Африканская Республика, Юго-Западная Африка, южная Родезия, Замбия, Ангола, большая часть Восточной Африки, за исключением Сомали, Западная Африка До Сьерра-Леоне, на север до АРЕ);
коморская белозубка — С. albicauda Peters, 1866 (Коморские острова);
белозубка Аллекса — С. allex Osgood, 1910 (Кения);
нигерийская белозубка — С. arethusa Dollman, 1915 (север Нигерии);
пустынная белозубка — С. aridula Thomas, et Hinton, 1923 (Дарфур в Судане);
семиенская белозубка — С. baileyi, Osgood, 1936 (горы Семиен в Эфиопии);
габонская белозубка — С. batesi Dollman, 1915 (Габон);
кенийская белозубка — С. beta Dollman, 1915 (центральные районы Кении);
белозубка Боливара —- С. bolivari Morales Agacino, 1934 (западная Сахара);
гамугофская белозубка — С. bottegi Thomas, 1898 (Гаму-Гофа в Эфиопии);
нижнеконголезская белозубка — С. bovei Dobson, 1887 (Республика Заир);
белозубка Бойда — С. boydi Dollman, 1915 (Республика Заир);
белозубка Бюттикофера — С. biittikoferi Jentink, 1888 (Гранд-Кейп-Маунт в Либерии);
белозубка Бутлера — С. butleri Thomas, 1911 (Бахр-эл ь-Газаль в Судане);
темная белозубка — С. caliginea Hollister, 1916 (Республика Заир);
гамбийская белозубка — С. Cinderella Thomas, 1911 (Гамбия);
С. congobelgica Hollister, 1916 (Республика Заир);
С. crenata Brosset, Dubost et de Balsac, 1965 (Габон);
асабская белозубка — C. crossiei Thomas, 1895 (нижнее течение р. Нигер в Нигерии);
дарфурская белозубка — С. darfurea Thomas et Hinton, 1923 (Дарфур в Судане);
камерунская белозубка — С. dolichura Peters, 1876 (Камерун);
суданская белозубка—С. ferruginea Heuglin, 1865 (Бахр-эль-Газаль в Судане);
белозубка Фишера — С. fischeri Pagenstecher, 1885 (Танзания);
сёверонигерийская белозубка — С. foxi Dollman, 1915 (северная Нигерия);
рыжая белозубка Сундевалла — С. fulvastra Sundevall, 1842 (Бахр-эл ь-Абьяд в Судане);
дымчатая белозубка — С. fumosa Thomas, 1904 (Эфиопия, Кения, Уганда);
бурая белозубка — С. fuscosa Thomas, 1913 (Белый Нил в Судане);
белозубка Жоффруа— С. geoffroyi J. Fischer, 1830(Мавритания);
гара-мулатская белозубка — С. glassi Heim de Balsac, 1966 (Эфиопия);
зунгерекая белозубка — С. glebula Dollman, 1915 (северная Нигерия);
стройная белозубка — С. gracilipes Peters, 1870 (Танзания);
белозубка Грассе — С. grassei Brosset, Dubost et de Balsac, 1965 (Габон);
C. hedenborgiana Sundevall, 1842 (восточный Судан);
угандская белозубка— С. hildegardeae Thomas, 1904 (Уганда, Кения, Эфиопия);
С. hindei Thomas, 1904 (Кения, Судан, Чад);
белозубка Джексона — С. jacksoni Thomas, 1904 (Кения, Республика Заир);
белозубка Ламотта — С. lamottei Heim de Balsac, 1968 (Берег Слоновой кости);
касайская белозубка — С. langi Cabrera, 1925 (Касаи в Республике Заир);
С. latona Hollister, 1916 (Республика Заир);
прибрежная белозубка — С. littoralis Heller, 1910 (оз. Альберт в Уганде);
пляшущая белозубка — С. India Hollister, 1916 (Республика Заир);
С. luluae Matschie, 1926 (р. Лулу в Республике Заир);
лунная белозубка — С. Inna Dollman, 1910 (Республика Заир);
С. lusitania Dollman, 1915 (Трарза в Мавритании);
выдровая белозубка — С. lutrella Heller, 1910 (Уганда);
белозубка Макартура — С. macarthuri St. Leger, 1934 (Кения);
лагосская белозубка — С. manni Peters, 1878 (Нигерия) (по-видимому, этот вид является подвидом С. flaoescens)\
С. marita Thomas et Hinton, 1923 (Дарфур в Судане);
белозубка Мартиенссена — С. martiensseni Neumann, 1900 (Танзания);
мавританская белозубка — С. maurisca Thomas, 1904 (Уганда, Танзания);
килиманджарская белозубка — С. топах Thomas, 1910 (Танзания);
либерийская белозубка — С. muricauda Miller, 1900 (Либерия);
темно-бурая белозубка — С. nigrofusca Matschie, 1895 (Республика Заир);
рувензорийская белозубка — С. niobe Thomas, 1906 (Рувензори в Уганде);
пахучая белозубка — С. odorata Leconte, 1857 (включая С. giffardi de Winton и С. goliath Thomas) (Габон);
итурийская белозубка — С. oritis Hollister, 1916 (р. Итури в Республике Заир);
С. parvipes Osgood, 1910 (Кения);
С. pasha Dollman, 1915 (Судан);
С. percivali Dollman, 1915 (Кения);
С. poensis Fraser, 1843 (Фернандо-По, Камерун, Гана, Гвинея, Гамбия, Нигерия);
гладкая белозубка—С. polia Hollister, 1916, (Республика Заир);
белозубка Райнея — С. raineyi Heller , 1912 (Кения);
белозубка Рузвельта-—С. roosevelti Heller, 1910 (Уганда);
занзибарская белозубка — С. sansibarica Neumann, 1900 (Занзибар); белозубка Швейтцера — С. schweitzeri Peters, 1877 (Либерия);
шелковистая белозубка — С. sericea Sundevall, 1843 (Судан);
сомалийская белозубка — С. somalica Thomas, 1895 (Сомали);
белозубка Гринвуда — С. green-woodi Heim de Balsac, 1966 (Сомали); рино-кампская белозубка — С. sururae Heller, 1910 (Уганда);
белозубка Терезы — С. theresae Heim de Balsac, 1968 (Гвинея);
белозубка Сан-Томе—С. thomensis Bocage, 1887 (о-в Сан-Томе);
ниерийская белозубка — С. ultima Dollman, 1915 (Кения);
усамборская белозубка — С. velutina Thomas, 1904 (горы Усамбора в Танзании);
сенегальская белозубка — С. viaria Geoffroy, 1834 (Сенегал);
С. voi Osgood, 1910 (Кения);
С. xantippe Osgood, 1910 (Кения);
каффская белозубка — С. zaphiri Dollman, 1915 (Каффа в Эфиопии);
луалабская белозубка — С. zimmeri Osgood, 1936 (Катанга в Республике Заир);
белозубка Блойета — С. bioyeti Milne-Edwards, 1887 (Танзания);
нимбская белозубка — С. nimbae Heim de Balsac, 1956 (Гвинея);
огненная белозубка — С. vulcani Heim de Balsac, 1956 (Камерун);
белозубка Инголдби — С. ingoldbyi Heim de Balsac, 1956 (Гана);
белозубка Эйзентраута — С. eisentrauti Heim de Balsac et alien, 1957 (Камерун);
белозубка Дуцета — C. douceti Heim de Balsac, 1958 (Либерия);
белозубка Ювената — C. jouvenetae Heim de Balsac, 1958 (Либерия);
белозубка Виммера — C. witnmeri Heim de Balsac, et Allen, 1958 (Берег Слоновой Кости);
горная белозубка — С. monticola Peters, 1870 (о-ва Ява, Калимантан, Тимор, Сумба, Флорес);
светлая белозубка — С. tenius Miller, 1839 (Тимор);
сулавесская белозубка — С. elongata Miller et Hollister, 1921 (Сулавеси);
черноватая белозубка — С. nigripes Miller et Hollister, 1921 (Сулавеси);
C. rhoditis Miller et Hollister, 1921 (Сулавеси);
львиная белозубка •— C. lea Miller et Hollister, 1921 (Сулавеси);
легкая белозубка — C. levicula Miller et Hollister, 1921 (Сулавеси);
котабатская белозубка — C. parvacauda Taylor, 1934 (о-в Котабато в группе Филиппинских островов);
С. halconus Miller, 1910 (о-в Минданао в группе Филиппинских островов);
белозубка Минданао — С. beatus Miller, 1910 (о-в Минданао в группе Филиппинских островов);
белозубка Грея — С. grayi Dobson, 1890 (о-в Лузон в группе Филиппинских островов);
белозубка Миндоро — С. mindorus Miller, 1910 (о-в Миндоро в группе Филиппинских островов);
С. grandis Miller, 1911 (о-в Минданао в группе Филиппинских островов);
палаванская белозубка — С. palawanensis Taylor, 1934 (о-в Палаван в группе Филиппинских островов);
белозубка Сула — С. edwardsianalrouessari, 1880 (о-ва Сула в группе Филиппинских, островов);
восточная белозубка — С. orientalis Jentink, 1890 (Ява);
балийская белозубка — С. baluensis Thomas, 1898 (Калимантан);
мохнатая белозубка — С. villosa Robinson et Kloss, 1918 (Суматра);
ровнохвостая белозубка — С. aequicauda Robinson et Kloss, 1918 (Малаккский полуостров и Суматра);
странная белозубка — С. paradoxura Dobson, 1886 (Суматра);
короткохвостая белозубка — С. brevicauda Jentink, 1890 (Ява);
белозубка Бартельса — С. bartelsii Jentink, 1910 (Ява);
С. brunnea Jentink, 1888 (Ява и Суматра);
С. doriae Peters, 1870 (Калимантан);
калимантанская белозубка — С. foetida Peters, 1870 (Калимантан);
белозубка Вебера — С. weberi Jentink, 1890 (Суматра);
С. lepidura Lyon, 1908 (Суматра);
банкская белозубка — С. vosmaeri Jentink, 1888 (о-в Банка у Суматры);
малайская белозубка.— С. malayana Robinson et Kloss, 1911 (Южная Азия);
белозубка Аагаарда— С. aagaardi Kloss, 1917 (Южная Азия);
дайангская белозубка — С. gravida Kloss, 1917 (о-ва Дайанга в Малаккском проливе);
кохская белозубка — С. negligens Robinson et Kloss, 1914 (о-ва Кох у побережья Малаккского полуострова);
белозубка Клосса — С. klossi Robinson, 1912 (о-в Большой Реданг у побережья Малаккского полуострова);
тиоманская белозубка — С. tionis Kloss, 1917 (о-в Тиоман у побережья Малаккского полуострова);
аорская белозубка — С. aoris Robinson, 1912 (о-в Аор у побережья Малаккского полуострова);
мапорская белозубка — С. maporensis Robinson et Kloss, 1916 (о-ва Manop в Архипелаге Риоу);
черномордая белозубка — С. melanorhyncha Jentink, 1910 (Ява);
клюворылая белозубка — С. beccarii Dobson, 1886 (Суматра);
большая яванская белозубка •— С. maxi Sody, 1936 (Ява);
малая яванская белозубка — С. minuta Otten, 1917 (Ява);
белозубка острова Рождества — С. trichura Dobson, 1888 (о-в Рождества к югу от Явы);
С. fuliginosa Bly th, 1855 (Южная Азия и Калимантан);
Никобарская белозубка — С. nicobarica Miller, 1902 (Никобарские острова);
андаманская белозубка — С. andamensis Miller, 1902 (Андаманские острова).
В СССР: малая белозубка (центральные и южные районы европейской части СССР, Казахстан, Средняя Азия и Приморский край); обыкновенная белозубка (Кавказ, Таджикистан, Киргизия); белобрюхая белозубка (южные районы европейской части СССР, Кавказ и Закавказье, юго-западная Туркмения, южный Таджикистан, южный Казахстан, юг Сибири); белохвостая белозубка (Копетдаг, Памир, Гиссарский хребет);
большая белозубка (черноморское побережье Кавказа и южные районы Приморского края).
Род конголезских белозубой
Genus Paracrocidura Heim de Balsac, 1956
В роде единственный вид: конголезская белозубка — Р. shoutedeni Heim de Balsac, 1956.
Конголезская белозубка близкородственна многозубым белозубкам и белозубкам и, возможно, является подродом одного из этих родов.
Размеры небольшие. Длина тела примерно 6,5 см. Длина хвоста около 3,5 см. Морда несколько укороченная. Уши короткие, слегка выступающие над мехом. Когти на лапах недлинные. Хвост покрыт укороченными черными волосами. Общая окраска серо-черная. Имеются подмышечные специфические кожные железы. Зубов 28.
Экология не изучена.
Распространена в Экваториальной Африке.
Род африканских гигантских белозубок
Genus Praesorex Thomas, 1913
В роде единственный вид: африканская гигантская белозубка — Р. goliath Thomas, 1906. Гейм де Бальзак и Бурлиер (1958) и некоторые другие исследователи включают гигантских белозубок в род белозубок.
Размеры крупные — наибольшие в семействе. Длина тела 15—18 см. Длина хвоста примерно 11 см. Волосяной покров очень своеобразен: длинный, грубый, довольно редкий, состоит из двух категорий волос — пуховых и остевых. Спинная сторона тела буроватая с примесью белых волос, брюшная слегка светлее. Голова темно-бурая. Хвост черноватый с белым кончиком у некоторых особей. Конечности почти черные. В проксимальной трети хвоста имеются редкие длинные волосы. Сильно развиты боковые кожные специфические железы, покрытые короткими светло-бурыми волосами.
Экология не изучена, хотя, по-видимому, мало отличается от других лесных землероек.
Распространение охватывает Западную Африку.
Род многозубых белозубок
Genus Suncus Ehrenberg, 1832
Размеры от мелких (наименьших среди млекопитающих — карликовая белозубка) до крупных: длина тела от 3,5 до 15 см. Длина хвоста от 2,5 до 10с.и. Морда удлиненная, резко суживающаяся к концу носа (рис. 84). Уши выступают над мехом. Волосяной покров короткий, очень густой и мягкий, на спине от сероватого до бурого, на брюхе беловатый или серо-беловатый. Хвост покрыт короткими волосами и редкими, длинными щетинками. У самцов этого рода на боках тела имеются сильно развитые специфические кожные железы, продуцирующие секрет с очень сильным мускусным запахом. Череп уплощен, мозговая капсула удлинена. Зубная фор-3 1 2 3
мула: i' — с -—- р — т — = 30. Коронки зубов белые. Число хромосом в диплоидном наборе у домовой белозубки 40.
Обитатели различных ландшафтов, в том числе живут рядом с человеческими постройками или в них. Активность в основном ночная. Питаются различными беспозвоночными, а также мелкими позвоночными животными. Домовая белозубка запасает пищу. В южных частях ареала, по-видимому, размножаются круглый год. Детенышей в помете от 2 до 5. Молодые землеройки часто следуют за матерью, держа зубами друг друга за хвост, а первый детеныш держится за хвост матери.
Рис. 84. Домовая многозубка S. murinus и ее череп
Распространены в Африке, на Мадагаскаре, в Южной Европе и Южной Азии — на восток до Филиппинских островов и Новой Гвинеи.
Систематика многозубых белозубок требует уточнений. Род включает ориентировочно 28 видов:
домовая многозубка — S. murinus Linnaeus, 1766 (о-ва Филиппинские, Сулавеси, Калимантан, Суматра, Ява, Бали, от Малаккского полуострова до Аннама, Юго-Восточного Китая, Тайваня, Японии, Бирмы и на запад до Кашмира, от него на юг до Цейлона, Аравийский полуостров, Израиль, АРЕ, Эфиопия);
скрытная многозубка—S. occultidens Hollister, 1913 (о-ва Негрос, Минданао в группе Филиппинских);
палаванская многозубка — S. palawanensis Taylor, 1934 (о-в Палаван в группе Филиппинских);
лусонская многозубка —S. luzoniensis Peters, 1870 (о-в Лусон в группе Филиппинских);
многозубка Хозе — S. hosei Thomas, 1893 (о-в Калимантан);
малайская многозубка—S. malayanus Kloss, 1917 (Таиланд, Малайзия);
карликовая белозубка—S. etruscus Savi, 1822 (Южная Европа, Закавказье, Туркмения, Иран, Ирак, Индия, Юньнань, Аравийский полуостров, Алжир, северная Нигерия);
S. stoliczkanus Anderson, 1877 (Индия);
многозубка Дейа—S. dayi Dobson, 1888 (южные районы Индии);
большая многозубка —S. lixus Thomas, 1898 (Замбия, Южно-Африканская Республика, Танзания, Кения);
лесная многозубка—S. varius Smuts, 1832 (Южно-Африканская Республика);
темнолапая многозубка—S. cafer Sundevall, 1846 (Южно-Африканская Республика, южная Родезия);
лазающая многозубка—S. megalura Jentink, 1888 (южная Родезия, Малави, Замбия, Ангола, Судан, Камерун, Уганда, Эфиопия, Кения);
многозубка Шаллера—S. shalleri Heim de Balsac, 1966 (Конго—Браззавиль);
угандская многозубка—S. blarina Thomas, 1906 (Уганда);
кенийская многозубка—S. infinitesimus Heller, 1912 (Кения);
занзибарская многозубка — S. leucura Matschie, 1894 (о-в Занзибар и территория между побережьем Индийского океана и озером Виктория);
мадагаскарская многозубка —S. madagascariensis Coquerel, 1848 (Мадагаскар);
многозубка Гранта —S. granti Thomas, 1907 (Уганда, Кения);
многозубка Джонстона — S. johnstoni Dobson, 1888 (Камерун и о-в Фернандо-По);
S. lunaris Thomas, 1906 (Уганда);
нигерийская многозубка—S. morio Gray, 1862 (южная Нигерия);
. камерунская многозубка—S. ollula Thomas, 1913 (Камерун);
S. oriundus Hollister, 1916 (Республика Заир);
S. preussi Matschie, 1893 (южная Нигерия);
руандская многозубка — S. ruandae Lonnberg et Gyldenstolpe, 1925 (Республика Заир);
габонская многозубка—S. reymi Brosset, Dubost et de Balsac, 1965 (Габон);
обыкновенная многозубка—S. suncoides Osgood, 1936 (Республика Заир);
Некоторые исследователи (Симпсон, 1945 и др.) разделяют род многозубых белозубок на три, выделяя лесных белозубок —Sylvisorex Thomas, 1904 и мышевидных белозубок — Myosorex Gray, 1838, в качестве самостоятельных родов. Представляется более правильной позиция Эллермана и •др. (1958), объединяющих эти три рода в один.
В СССР: карликовая белозубка (Закавказье — Ленкорань, Муган-ская степь, окрестности Нухи и Тбилиси, юго-западная Туркмения и южный Узбекистан — г. Термез).
Род когтистых белозубок Келаарта
Genus Feroculus Kelaart, 1852
В роде единственный вид: когтистая белозубка Келаарта — F. feroculus Kelaart, 1850.
Размеры средние. Длина тела 10—12 см. Длина хвоста 6—7 см. Характерны крупные передние конечности с длинными когтями. Задние конечности небольшие, с маленькими когтями. Ушные раковины выступают над мехом. Волосяной покров густой, короткий и мягкий. На спине в основном серовато-черный, на брюхе более светлый. Передние конечности почти белые, задние красновато-серые. Хвост покрыт короткими мягкими волосками и небольшим количеством длинных щетинковидных волос. Зуб-3 12 3
ная формула: i. — с — р — т — = 30.
Экология не изучена. Обитатель склонов гор. Поднимается на высоту 1800—2150 м над уровнем моря.
Встречается на Цейлоне в горных массивах центральных районов.
Род когтистых белозубок Пирсона
Genus Solisorex Thomas, 1924
В роде единственный вид: когтистая белозубка Пирсона — S. pearsoni Thomas, 1924.
Размеры средние. Длина тела 12—13 см. Длина хвоста примерно 6 — 7 см. Передние конечности крупных размеров. Пальцы с длинными сильными когтями. Уши не выступают над мехом. На хвосте нет щетинковидных волос. Волосяной покров густой, короткий и мягкий, на спинной по-
верхности темно-серовато-бурый, на брюшной немного светлее. Конечное-3 113
ти бурые. Зубная формула: t — с — Р — иг — = 28.
Обитает в горных джунглях на высоте примерно 1600 м над уровнем моря. По характеру питания, по-видимому, более хищная, нежели другие землеройки.
Распространена в горных, центральных районах Цейлона.
Род горных белозубок
Genus Surdisorex Thomas, 1906
Размеры средние. Длина тела 9—11 см. Длина хвоста 2,5—3 см. Маленькие уши не выступают над мехом. Когти на передних конечностях довольно длинные, сильные. Волосяной покров высокий, плотный и относительно грубый, бурый с седоватым оттенком. Зубная формула:
i Ас + п-Л— т—= 28 — 30.
1 2 — 3 3
Обитают в горах на высоте 2800—3300 м над уровнем моря. Экология почти не изучена. Копают сложные норы и делают туннели в лесной подстилке.
Распространены в горах Кении.
В роде 2 вида:
ниерская горная белозубка — S. norae Thomas, 1906 (Ниери в Кении).
малая горная белозубка — S. polulus Hollister, 1916 (гора Кения в Кении)..
Род путораков
Genus Diplomesodon Brandt, 1852
В роде единственный вид: пегий путорак—D. pulchellum Lichtenstein, 1823 (рис. 85).
Размеры мелкие. Длина тела 5,4—7,6 см. Длина хвоста 2—3 см. Характерны расширенные кисти и ступни конечностей. Морда относительно слабо вытянута. Уши почти не выступают над мехом. По краям ступней и кистей волосяные оторочки из жестких волос. Хвост покрыт короткими равномерной длины жесткими волосами. Окраска тела двухцветная: спина серая и буровато-серая с белым продолговатым пятном по средней линии, брюхо и Г 3 0 13
бока белые. Зубная формула: i — с — р — т — — 26.
Обитатели песчаных пустынь. Активность преимущественно сумеречная и ночная. День проводит в норах, выкапывая их или занимая старые норы грызунов. В норах имеется гнездовая камера. Питается различными беспозвоночными, а также мелкими позвоночными, главным образом яще
Рпс. 85. Пегий путорак — С. pulchellum и его череп
рицами. Самка приносит потомство, по-видимому, три раза в год. Беременность около 30 дней.
Обитатель песчаных пустынь.
Распространен в Средней Азии и Казахстане.
Род кротовых белозубок
Genus Anourosorex Milne-Edwards, 1870
В роде единственный вид: кротовая белозубка — A. squamipes Milne-Edwards, 1872.
Размеры средние. Длина тела примерно 9—11 см. Длина хвоста 1—2 см. Глаза очень маленькие. Уши скрыты в меху. Конечности укороченные, широкие, безволосые, покрытые чешуйками, с сравнительно длинными когтями. Хвост очень короткий. Волосяной покров низкий, густой и мягкий, на спинной стороне оливково-серый, на брюшной — несколько более свет-2 2 3
лый. Зубная формула: i—c-j-p—т—= 26.
Обитатель горных лесов. Живет на высоте 1500—3100 м над уровнем моря. Ведет, по-видимому, подземный образ жизни. Питается главным образом насекомыми и их личинками, а также дождевыми червями.
Распространена от Бакбо на п-ове Индокитай через Северную и Западную Бирму до Сычуаня в Китае; встречается также в горах Тайваня.
Род азиатских водяных белозубок
Genus Chlmarrogate Anderson, 1877
Средних размеров землеройки. Длина тела 8—12 см. Длина хвоста 6—10 см. Морда со вздутыми боковыми отделами в области вибрисс. Глаза очень маленькие. Ушные раковины не развиты. Пальцы по краям имеют щетинковидные гребни волос. Хвост относительно длинный. Волосяной покров густой и мягкий с удлиненными остевыми волосами, на спинной стороне сероватый, бурый или черноватый, на брюхе светло-серый или белый. Хвост одноцветный, темный или двуцветный: сверху темный, снизу светлый.
Обитают по берегам горных потоков. Ведут полуводный образ жизни. Хорошо плавают и ныряют. Питаются в основном насекомыми, водными личинками, мелкими ракообразными и рыбой.
Распространены в Японии (о-в Кюсю), Сычуани, Юньнани, Юго-Восточном Китае, Лаосе, Аннаме, Бакбо, Кашмире, Северной Бирме, на о-вах Калимантане, Суматре.
В роде 2 вида:
гималайская водяная белозубка — С. platycephala Temminck, 1842 (ареал рода, кроме Калимантана и Суматры);
калимантанская водяная землеройка — С. phaeura Thomas, 1898 (Калимантан и Суматра).
Род тибетских водяных белозубок, или перепончатолапых белозубок,
Genus Nectogale Milne-Edwards, 1870
В роде единственный вид: тибетская водяная белозубка, или перепончатолапая белозубка, —N. elegans Milne-Edwards, 1870 (рис. 86).*
Размеры средние. Длина тела 9—13 см. Длина хвоста 9—11 см. Морда еильно удлинена. Наружные ушные раковины редуцированы. Ступни задних лап удлинены и расширены. Пальцы передних и задних конечностей по краям окаймлены грубыми короткими волосами. Между пальцами име-
Рис. 86. Тибетская водяная белозубка — N. elegans
ется плавательная перепонка. Дисковидные подушечки на подошвах конечностей, по-видимому, помогают не скользить на мокрых камнях. Волосяной покров мягкий и густой, на спине серый с серебристым оттенком, на брюхе серебристо-серый, иногда желтоватый. Основной цвет хвоста черный. По бокам основной трети хвоста проходят два гребня белых волос, которые соединяются друг с другом в единый гребень, тянущийся по нижней стороне хвоста до вершины. В передней трети хвоста, кроме того, проходят два других боковых гребня волос, соединяющихся на дорзальной стороне хвоста примерно у начала его терминальной трети. Этот дорзальный гребень тянется до конца хвоста. Расположение своеобразных волосяных гребней придает поперечнику хвоста в основании четырехгранную форму, в средней части — трехгранную, в терминальной части — сжатую с боков.
Обитает по берегам горных потоков. Экология почти не изучена. Прекрасно плавает и ныряет. В берегах устраивает норы. Питается, по-видимому, водными беспозвоночными, мелкими рыбами.
Распространены в Сиккиме, Бутане, Северной Бирме, Тибете, Сычуани, Юньнани и Шэньси в Китае.
Род белозубок-броненосок
Genus Scutisorex Thomas, 1913
Размеры средние и крупные. Длина тела 12—15 см. Длина хвоста 7— 9,5 см (рис. 87). Хвост довольно длинный, без удлиненных волос. У самок 2пары паховых сосков. Волосяной покров длинный, густой и довольно гру-3 3 3
бый. Основной тон окраски серый. Зубная формула: i — с + р — т— —
1=30. Очень своеобразно строение позвоночного столба, отличающего этих землероек от всех других млекопитающих (рис. 87). Позвонков шейных 7, грудных 14, поясничных 11, крестцовых 5 и хвостовых 23. Поперечные отростки позвонков сильно разрастаются в стороны и вверх и почти по всей поверхности отдельными выростами сочленяются с передними и задними позвонками. Несмотря на такие усиленные сочленения, землеройка способна выгибать спину не только вдорзо-вентральном, но ив боковых направлениях.
Рис. 87. Конголезская белозубка-броне* носка — S. congicus
Дополнительные сочленения настолько укрепляют позвоночный столб, что, по описанию некоторых авторов, небольшая землеройка способна выдержать без вреда для себя вес взрослого человека.
Экология мало изучена. Активность круглосуточная. Питаются различными мелкими животными, а также, по-видимому, и растениями.
Встречаются в Африке: в Республике Заир и Уганде.
В роде 2 вида:
конголезская белозубка-броненоска—S. congicus Thomas, 1915 (Республика Заир);
угандская белозубка-броненоска—S. somereni Thomas, 1910 (Уганда, Республика Заир).
СЕМЕЙСТВО ПРЫГУНЧИКОВ
FAMILIA MACROSCELIDIADAE MIVART, 1868
Размеры мелкие, средние и крупные. Длина тела от 9,5 до 31 см. Хвост тонкий, длиной от 8 до 26 см (примерно 80—120% длины тела). Внешним видом напоминают тушканчиков. Морда удлиненная, тонкая, подвижная. Уши средней длины; обычно достигают глаза, если их отогнуть вперед. Задние конечности намного длиннее передних. Передние и задние конечности четырех- и пятипалые. Конечности стопоходящие или полупальцеходящие. Волосяной покров длинный, густой и мягкий. Его окраска изменяется от желтовато-бурой до буровато-черной. Подхвостовая кожная
железа, продуцирующая пахучий секрет, особенно сильно развита у самцов. Сосков 2 или 3 пары. Penis разделен на три лопасти.
Череп имеет хорошо развитый мозговой отдел.Твердое небо продолжа-
ется назад дальше коренных зубов и имеет вдоль средней линии большие парные отверстия. Скуловые дуги хорошо развиты. Костные слуховые барабаны крупные. Слезные кости очень большие. Заглазничные отростки . о л. 0 — 3 1 4,2
небольшие или отсутствуют. Зубная формула: i -с — р — т----~
3 14 2 3
=34—42. У видов с полной зубной формулой первый верхний резец
крупнее прочих, а нижние резцы имеют примерно одинаковые размеры. Верхний клык сходен с предкоренными зубами (за исключением хоботковых собачек). В ряду верхних предкоренных размер зубов увеличивается спереди назад. Задний предкоренной зуб имеет наибольшие размеры среди
остальных предкоренных и коренных зубов. Коренные зубы с бугорками на жевательной поверхности, расположенными в виде буквы W.
Позвонков шейных 7, грудных 13, поясничных 7—8, крестцовых 3, хвостовых 25—28. Таз с длинным лобковым симфизом. В отличие от других насекомоядных, имеется слепая кишка.
Обитатели равнин, поросших колючими кустарниками или травой, лесов и скальных обнажений. Активность в основном дневная, но в жаркое время переходит в ночную. Держатся поодиночке или парами, за исключением некоторых видов рода длинноухих прыгунчиков, которые живут небольшими колониями. Хотя лапы прыгунчиков плохо приспособлены для копания, эти зверьки иногда устраивают собственные норы, чаще же занимают норы грызунов. Находят также убежище под стволами упавших деревьев и под камнями. Когда прыгунчики передвигаются медленно, они используют все четыре ноги; при быстрых прыжках — только задние конечности. При этом хвост они поднимают кверху. Мелкие виды питаются главным образом муравьями и, вероятно, термитами, а также нежными ростками растений, их корнями и ягодами. Крупные виды предпочитают жуков. Самка приносит одного или двух детенышей. Они полностью покрыты волосами, глаза открыты или открываются вскоре после рождения. Сразу после рождения они способны самостоятельно передвигаться. Лактационный период короткий.
Распространены в Центральной, Южной и Восточной Африке (один вид также в Марокко и Алжире) и на о-ве Занзибар.
В семействе 4 рода (14 видов):
хоботковых собачек, или пестроспинных прыгунчиков, — Rhyncho-суоп Peters, 1847 (3 вида);
лесных прыгунчиков — Petrodromus Peters, 1846 (1 вид);
короткоухих прыгунчиков — Macroscelides A.. Smith, 1829 (1 вид); длинноухих прыгунчиков — Elephantulus Thomas, et Schwann, 1906 (9 видов).
Мак Довел (McDowell, 1958) считает, что прыгунчики, сильно отличаясь от всех насекомоядных, должны быть выделены вместе с тупайями в отдельный отряд Menotyphla или отнесены к отряду приматов. По мнению Бутлера (Butler, 1956), прыгунчики должны быть объединены в особый отряд Macroscelidea.
Род хоботковых собачек, или пестроспинных прыгунчиков,
Genus Rhynchocyon Peters, 1847
Размеры наибольшие в семействе. Длина тела от 24 до 31 см. Длина хвоста от 19 до 26 см. Конечности очень длинные, причем передние не так сильно отличаются по длине от задних, как это имеет место у других представителей семейства (рис. 88). Голова очень сильно вытянута в лицевой части. Уши относительно небольшие, круглые. Передние и задние конечности четырехпалые. Отсутствует первый палец. Волосяной покров относительно грубый, короткий. Основной тон окраски спины изменяется от рыжего до темно-бурого с тенденцией к меланизму у форм из Восточной Африки. По верху спины проходят расположенные рядами беловатые пятна. Хвост, покрытый короткими волосами, у некоторых форм бывает белым. Имеется постанальная железа. Соски брюшные. В черепе характерны относительно короткие и широкие носовые кости и очень крупные лобные. „ о Л. .0—1 1 4 2
Заглазничныеотростки развиты. Зубная формула: t —-— с — р — иг—=
=34—36. Верхние резцы отсутствуют или рудиментарны. Верхние клыки очень крупные. В крестцовом отделе позвоночника 8 позвонков.
Обитатели главным образом лесов. Активность преимущественно дневная. Держатся парами. Питаются в основном насекомыми, хо-
Рис. 88. Хоботковая собачка — Rhynchocyon sp. и ее череп
тя, вероятно, поедают мелких млекопитающих, птиц и их яйца, а также и других животных. Из листьев делают себе гнезда между стволами упавших деревьев или под ними.
Ареал охватывает Кению, Танзанию, о-в Занзибар, Замбию, Республику Заир, Малави и Мозамбик.
Систематика рода окончательно не установлена. Разные исследователи насчитывают в роде от 1 до 12 видов. По-видимому, можно выделить 3 вида: пятнистая хоботковая'собачка — R. cirnei Peters, 1847 (ареал рода, за исключением прибрежной зоны Кении, северной Танзании и Занзибара); к* хоботковая собачка Петерса — R. petersi Bocage, 1880 (Танзания, Кения, о-ва Занзибар и Мафия);
золотистая хоботковая собачка —- /?. chrysopygus Gunther, 1881 (Кения).
Некоторые исследователи считают обособленными видами также: занзибарскую хоботковую собачку — 7?. adersi Dollman, 1912; хоботковую собачку Клауда — R. claudi Thomas et Wroughton, 1907; хоботковую собачку Штульманна — 7?. stuhlmanni Matschie, 1893; хоботковую собачку Свиннертона — /?. swynnertoni Kershaw, 1923.
Род лесных прыгунчиков
Genus Petrodromus Peters, 1846
В роде 1 вид: четырехпалый прыгунчик — Р. tetradactylus Peters, 1846.
Размер средний. Длина тела от 16 до 22 см. Длина хвоста от 13 до 18 см. Общий облик типичен для семейства. Мордочка относительно короткая. Уши крупные, округлые. Задние конечности четырехпалые. Первый
палец отсутствует. Волосяной покров длинный и мягкий, на спине грязножелтый с оранжевым или желтым оттенком или бурый с рыжеватым оттенком. Над глазами имеются белые или каштаново-красные отметины. Брюшная сторона белая или охристая. Большинство особей имеет сероватые или буроватые штрихи на боках. Сосков две пары. Зубная формула:
. 3 1 4 2 .л
t — с — р — т— = 40.
3 14 2
Обитатель, как правило, кустарников и подроста в лесах, но встречается также и на каменистых участках. Гнезд не делает. Активность в основном дневная. Питается главным образом насекомыми, особенно термитами. Самка приносит в помете, по-видимому, одного детеныша.
Распространен в Африке от Республики Заир и Кении на юг до Юго-Западной Африки и Зулуленда в Южно-Африканской Республике.
Некоторые исследователи выделяют в роде в качестве отдельных видов: Р. rovumae Thomas, 1897; лесного прыгунчика — Р. sultan Thomas, 1897; прыгунчика Матши — Р. matschiei Neumann, 1900; катангского прыгунчика — Р. robustus Thomas, 1918; черного прыгунчика — Р. nigriseta Neumann,
1900.
Род короткоухих прыгунчиков
Genus Macroscelldes A. Smith, 1829
роде единственный вид: короткоухий прыгунчик — М. proboscideas 1800.
В Shaw,
Размеры небольшие (наименьшие в семействе). Длина тела 9,5—12,4 см. .Длина хвоста 9,7—13,7 см. Внешний облик типичен для семейства. Тонкая мордочка очень сильно вытянута, уши относительно короче и сильнее закруглены, чем у других родов прыгунчиков. Первый палец на задних конечностях небольших размеров и имеет коготь. Волосяной покров длинный, густой и мягкий. На верхней части тела песчано-бурый, бледно-грязно-желтый, бледно-серый, оранжево-желтый, иногда черноватый, на брюшной белый или сероватый. Сосков 3 пары. В черепе характерны очень крупные костные слуховые барабаны. Зубная формула: i с -j- р т -|- = 40. Верхний резец относительно небольших размеров.
Обитатели поросших густым кустарником равнин с песчаными почвами. Активность преимущественно дневная, причем часто и в жаркие часы дня. Убежищем служат норы, которые зверек роет обычно у корней кустарников, реже занимает чужие. Держится поодиночке или парами. Питается насекомыми, в основном муравьями и термитами, а также некоторыми растительными объектами: корнями, ягодами. Новорожденные детеныши имеют относительно очень крупные размеры, открытые глаза; тело покрыто шерстью.
Встречается в Южной Африке (Капская провинция) и в Юго-Западной Африке.
Род длинноухих прыгунчиков
Genus Elephantalus Thomas et Schwann, 1906
Размеры небольшие: длина тела 10—14 см, длина хвоста 10—16 см, вес взрослых особей примерно 25—50 г. Общий облик типичен для семейства (рис. 89). Мордочка сильно вытянутая и тонкая. Уши крупные. Задние конечности пятипалые, но первый палец очень короткий. Волосяной покров довольно длинный и мягкий, на спинной стороне серовато-желтый, серый, грязно-белый, темно-бурый; на брюшной белый, грязно-желтый или сероватый. Сосков 3 пары. Зубная формула: i -|- с -j- р т= 40. Дипло-
Рис. 89. Североафриканский прыгунчик — Е. rozeti
идное число хромосом у североафриканского прыгунчика 28, а у скалистого 14.
Обитатели различных равнинных ландшафтов и гор. Встречаются на песчаных и каменистых участках. Активность преимущественно дневная, причем не избегают и жарких часов дня. В наиболее жаркое время года иногда ведут ночной образ жизни. Держатся небольшими группами. В качестве убежища используют норы мелких грызунов, расщелины в скалах, щели между камнями, реже роют собственные норы. В некоторых местах, по видимому, живут небольшими колониями. Питаются в основном насекомыми, муравьями, термитами, в меньшей степени различными растительными объектами. Самка приносит обычно 2 (иногда 1) детенышей. Новорожденные относительно велики сравнительно с размерами матери.
Распространены почти по всей территории Африки и от Марокко и Ал жира до Южной Африки.
В роде, по-видимому, 9 видов (иногда их насчитывают 6 или 12):
североафриканский прыгунчик — Е. rozeti Duvernoy, 1830 (от Марокко до Ливии);
рыжий прыгунчик — Е. rufescens Peters, 1878 (Эфиопия, Кения, Сома ли, Судан, Танзания, Уганда);
прыгунчик Ревоала — Е. revoili Huet, 1881 (Сомали);
кустарниковый прыгунчик — Е. intufi Smith, 1836 (от Анголы и Танзании до Капской провинции);
скалистый прыгунчик— Е. rupestris Smith, 1830 (Юго-Западная Афри ка, Южно-Африканская Республика, Ботсвана и южная Родезия).
голохвостый прыгунчик — Е. myarus Thomas et Schwann, 1906 (Южно-Африканская Республика, Ботсвана, южная Родезия);
прыгунчик Эдварда — Е. edwardi Smith, 1839 (Южно-Африканская Республика);
коротконосый прыгунчик — Е. brachyrhynchus Smith, 1836 (от Республики Заир и Кении на юг до Юго-Западной Африки, Ботсваны, Южно Аф риканской Республики и Мозамбика);
темный прыгунчик — Е. fuscipes Thomas, 1894 (Центральная Африка).
Некоторые исследователи выделяют коротконосого прыгунчика в особый род Nasilio Thomas et Schwann, 1906.
Рис. 90. Шерстокрыл — Cynocephalus sp. А — нижние резцы; Б — подошва стопы
ОТРЯД ШЕРСТОКРЫЛОВ, ИЛИ КАГУАНОВ,
ORDO DERMOPTERA ILL1GER, 1811
В отряде одно семейство: шерстокрылов, или кагуанов, — Cynocepha-lidae Simpson, 1945 и единственный род шерстокрылов, или кагуанов, — Cynocephalus Boddaert, 1768.
Длина тела 36—43 см. Хвост длинный (22—27 см) и широкий. Вес 1— 1,7 кг. Голова с расширенным мозговым отделом и вытянутым лицевым несколько напоминает голову собаки (рис. 90). Уши короткие, округлые, почти полностью лишены волос и имеют розоватую окраску. Глаза крупные, карие. Передние и задние конечности длинные, примерно одинаковые по размерам. Подошвы лап широкие, голые, уплощенные, почти образующие присасывательные диски (см. рис. 90). Пальцы несколько сжаты с боков. Имеется
летательная кожная перепонка, она натянута между передними конечностями, прикрепляясь к шее позади и ниже ушей и ко вторым фалангам пальцев кисти; далее она соединяет передние и задние конечности и бока тела и идет от задних конечностей (от вторых фаланг пальцев стопы) к кончику хвоста. Заднепроходное отверстие располагается в карманообразном углублении.
Волосяной покров относительно короткий, густой и мягкий. Окраска его сильно варьирует от сероватой до коричневой. Обычно самцы имеют коричневую окраску, а самки серую. На дорзальной поверхности тела могут быть белые пятна. У самок одна пара подмышечных сосков.
Череп крупный, уплощен в дорзо-вентральном направлении. Лицевой отдел его удлинен. Мозговая коробка широкая. Скуловые дуги хорошо развиты. Имеются крупные посторбитальные отростки.
Костные слуховые барабаны уплощены. Костное небо широкое. Резцовые 2 12 3
отверстия крупные. Зубная формула: i — с — р — т— = 34 (по Ка-3 12 3
2 0 3 3 \
I — с — р — т — I. Верхние резцы уменьшены 3 1 2 3 j
брера — Cabrera, 1925,
и отделены друг от друга значительным промежутком. В нижней челюсти
первые два резца с уплощенными и расширенными коронками, которые разделены продольными разрезами (от 5 до 20), так что имеют своеобразный гребешкообразный вид (см. рис. 90). Третий нижний резец имеет пять бугорков, постепенно уменьшающихся спереди назад. Последний нижний резец и все клыки обладают каждый двумя корнями. Клыки сходны с предкоренными. Щечные зубы брахиодонтного типа. Ряды верхних коренных зубов расставлены шире, чем нижних. Позвонков шейных 7, грудных 13—14, поясничных 5—9, крестцовых 3—5, хвостовых 15—19. Ребра широкие. На грудине имеется небольшой киль. Лопатка иногда окостеневает неполностью. Ключица соединена с первым ребром. Плечевая кость длинная; лучевая — крупная, а локтевая уменьшена, однако ее локтевой отросток развит хорошо. Лобковое сочленение короткое. В задней конечности большая берцовая кость крупных размеров, а малая — небольшая и не сочленяется с пяточной костью.
Мозг макросматического типа. В каждом полушарии имеется по две продольные борозды. Язык очень крупный. Желудок простой, узкий. Длина кишечника примерно в 3 раза превышает длину тела. Тонкий отдел кишечника короче толстого. Слепая кишка длинная. Семенники находятся в мошонке, а иногда в паховых мешках. Матка двурогая.
Обитатели равнинных тропических лесов. Ведут древесный образ жизни. Способны к планирующему полету до 60 м длиной; при планировании зверьки теряют 1 м высоты на каждые 15 м полета. По земле передвигаются очень медленно. Активность ночная. Большинство исследователей сообщает, что шерстокрылы день проводят, подвесившись лапами на горизонтальной ветви спиной вниз, как ленивцы, или головой вниз на вертикальном стволе дерева. Вартон (Wharton, 1950), напротив, указывает, что надень ка-гуаны укрываются в дуплах. Передвигаясь по ветвям, кагуаны, подобно ленивцам, подвешиваются спиной вниз. Питаются листьями деревьев, плодами и семенами. Нижними резцами соскабливают мякоть листьев. Самка ежегодно приносит одного детеныша. Беременность около 60 дней. Самка таскает детеныша на себе, поперек брюха.
Местные жители на о-ве Минданао употребляют кагуанов в пищу, а на о-ве Богол используют мех.
Распространение охватывает большую часть Юго-Восточной Азии.
В роде 2 вида:
малайский шерстокрыл — С. temminckii Waterhouse, 1838 (Тенассе-рим в Южной Бирме, южная часть п-ова Индокитай, Малаккский полуостров, Суматра, Ява, Калимантан и мелкие прилегающие острова);
филиппинский шерстокрыл — С. volans. Linnaeus, 1758 (о-ва Минданао, Басилан, Самар, Лейте, Бохоль в группе Филиппинских).
Происхождение шерстокрылов остается неясным вследствие бедности найденных ископаемых остатков. По-видимому, являются древней группой, уже в палеоцене отделенной от насекомоядных. Наиболее дрейние остатки известны из позднего палеоцена и раннего эоцена Северной Америки.
ОТРЯД РУКОКРЫЛЫХ
ORDO CHIROPTERA BLUMENBACH, 1779
Размеры небольшие. Длина тела примерно от 2,8 до 40 см. Рукокрылые приспособились к полету, их передние конечности преобразовались в крылья. Эластичная кожная летательная перепонка натянута между пальцами передних конечностей, плечом, предплечьем, боками тела, задними конечностями и хвостом (если последний имеется). Иногда хвост не включен в летательную перепонку. У некоторых представителей рукокрылых хвост длинный, у других он короткий или полностью отсутствует. В передних конечностях характерно слабое развитие первого пальца и сильное удлинение остальных, причем коготь имеется только на первом пальце, иногда на втором. Задние конечности несколько вывернуты коленями в стороны. Пальцы укорочены и все снабжены когтями. Голова с удлиненной мордой, широкой ротовой щелью, небольшими глазами. Наружные ушные раковины обычно крупные. В передней их части может быть дополнительный кожный вырост— так называемый козелок.
Волосяной покров довольно густой, средней длины, мягкий, покрывающий только туловище и голову. Волосы слабо подразделяются на категории. Характерные особенности волос и их кутикулы (форма и размера клеток) могут служить таксономическим признаком на уровне семейств и родов. Кожная перепонка покрыта лишь редкими волосками. Окраска волосяного покрова обычно одноцветная, темных тонов. У многих видов на горле, лбу или летательной перепонке имеются специфические кожные железы, продуцирующие секрет с сильным мускусным запахом. У самок одна пара функционирующих сосков, расположенных кзади от подмышечных впадин или по бокам грудной области.
Череп с удлиненным или укороченным лицевым и довольно хорошо развитым мозговым отделами. Характерно раннее зарастание швов между костями черепа. Межчелюстные кости обычно недоразвиты. Общее число постоянных зубов от 20 до 38. Зубы дифиодонтные и гетеродонтные. Никогда не бывает более двух пар верхних резцов. В скелете происходит значительное изменение. Локтевая кость и часто малая берцовая рудиментарны. Лучевая кость удлинена и более или менее искривлена, длиннее плечевой. Имеется бкарпальных костей и 5 метакарпальных (первые из них значительно короче последних). В плечевой кости сильно развиты большой и малый бугры, причем первый может образовывать дополнительное сочленение с лопаткой. Первый палец передних конечностей имеет две фаланги, второй — одну, две или три фаланги или только метакарпальные кости, третий, четвертый и пятый пальцы передних конечностей обычно имеют по три фаланги (третья фаланга этих пальцев может отсутствовать). Дистальная фаланга третьего пальца хрящевая или костная. Ключица хорошо развита. Плечевой пояс
развит значительно сильнее пояса задних конечностей. Грудина, как правило, имеет небольшой киль. Первый грудной и последний шейный позвонки обычно срастаются. Большинство рукокрылых имеет 17 грудопоясничных позвонков, которые у старых особей могут частично срастаться. Обычно имеется os penis.
Из органов чувств наивысшего развития достигают слух и осязание. Зрение развито слабо. Характерен короткий пищеварительный тракт, превышающий лишь в 1,5—4 раза длину тела. Желудок, как правило, простой. Слепая кишка небольшая или отсутствует. Головной мозг с гладкими большими полушариями, не покрывающими мозжечок.
Матка простая, двурогая или переходной формы. Семенники ингуиналь-ные или абдоминальные, в период сезона размножения спускающиеся во временную мошонку. У самок некоторых видов летучих мышей функционирует только правый яичник. Плацента дискоидальная, гемохориальная.
Обитатели самых разнообразных ландшафтов — от северной границы леса на юг до пустынь включительно. Особенно многочисленны в тропиках. Активность сумеречная и ночная. День проводят в пещерах, расщелинах скал, дуплах деревьев, на чердаках и под крышами построек, некоторые — на ветвях деревьев, подвешиваясь на задних лапах вниз головой и сложив крылья. В северных частях ареала на зиму либо мигрируют на юг, либо впадают в спячку. Замечательна способность к эхолокации. Большинство питается насекомыми, ловя их на лету. Объектами пищи могут быть плоды, нектар цветков, мелкие позвоночные и даже кровь крупных млекопитающих. Некоторые рукокрылые держатся поодиночке и во время охоты, и днем в убежищах, другие во время дневок образуют крупные скопления, иногда достигающие несколько сотен особей.
Характер размножения различен у разных видов. Тропические обычно размножаются в течение всего года. Некоторые виды спариваются осенью, сперматозоиды остаются всю зиму в половых путях самок, сохраняя свою жизнеспособность. Овуляция у этих самок происходит весной, и яйцеклетка оплодотворяется перезимовавшими сперматозоидами. У других видов спаривание, овуляция и оплодотворение яйцеклетки осуществляется весной. Наконец, у некоторых американских листоносов спаривание и оплодотворение яйцеклетки происходят осенью, а затем до весны следует задержка в развитии оплодотворенной яйцеклетки. Самка приносит одного (редко двух) детеныша, которого носит во время полета на себе. Обычно детеныш цепляется передними и задними лапами за шерсть на брюхе самки, зубами за сосок. В неволе рукокрылые жили до 15—17 лет.
Большого практического значения не имеют. Многие полезны уничтожением различных вредных насекомых. Некоторые тропические формы могут быть переносчиками бешенства. Отдельные виды рукокрылых употребляются местным населением в пищу.
Широко распространены в умеренных и тропических областях обоих полушарий. Отсутствуют только на некоторых мелких удаленных островах.
В отряде 2 подотряда:
крыланов — Megachiroptera Dobson, 1875;
летучих мышей — Microchiroptera Dobson, 1875.
В подотряде крыланов всего одно семейство, а в подотряде летучих мышей — 16 семейств.
Происхождение рукокрылых изучено недостаточно. Ископаемые находки бедны. Наиболее древние остатки принадлежат к подотряду летучих мышей и могут быть отнесены к среднему миоцену как в Европе, так и в Северной Америке. Эоценовые летучие мыши имели уже хорошо развитые крылья и мало чем отличались от современных летучих мышей, что свидетельствует о том, что происхождение отряда нужно искать в значительно более ранних периодах. Предками рукокрылых, по-видимому, были примитивные насекомоядные.
Подотряд крыланов
Subordo Megachiroptera Dobson, 1875
Подотряд включает единственное семейство: крыланов — Pteropidae Gray, 1821.
Размеры мелкие, средние и крупные (наибольшие среди представителей отряда рукокрылых). Длина тела от 5 до 40 см. Хвост короткий, рудиментарный или его нет совсем. Только у длиннохвостых крыланов он относительно длинный и. имеет 10 позвонков. Наиболее крупные представители семейства (летучие лисицы) в размахе крыльев достигают 170 см. Вес тела взрослых особей от15 а у нектаро- и пыльцеядных крыланов до 900 г у наиболее крупных плодоядных видов. Второй палец всегда содержит три фаланги и сохраняет значительную независимость. Все представители семейства, за исключением родов голоспинных крыланов, пещерных крыланов, сулавесских крыланов, длиннохвостых крыланов и длинноязыких крыланов Соломоновых островов, имеют коготь на втором пальце передней конечности. Наружная ушная раковина овальная, замкнутая по внутреннему краю. Козелок отсутствует. Глаза крупные. У некоторых видов очень сильно развиты губы. Трубконосые и малые трубконосые крыланы имеют своеобразные трубчатые ноздри, открывающиеся латерально. Язык покрыт хорошо развитыми сосочками: у некоторых родов он очень длинный и может далеко высовываться изо рта. Небо обычно содержит 8 поперечных складок, которые используются жи
вотным при перетирании мякоти плодов.
Волосяной покров может быть коротким и изреженным или длинным (до Зел/) и плотным. Окраска его сильно варьирует у разных представителей, но у большинства темно-бурая. У некоторых крыланов она может быть желтой, зеленоватой, с белыми пятнами на ушах, крыльях. Характерен половой диморфизм. Он проявляется у самцов в увеличенных клыках и более яркой окраске, в более крупных размерах (пещерные крыланы, биндемы, молотоголовые крыланы, некоторые виды эполетовых крыланов), в наличии желе
зистых плечевых кожных мешков с пучками волос, растущими из этих мешков (летучие собаки, эполетовые крыланы, биндемы и карликовые эполетовые крыланы, коровьемордые крыланы, крыланы Анхиеты), очень крупными глоточными мешками (эполетовые крыланы, молотоголовые крыланы, бин-
демы).
Череп с удлиненным лицевым отделом. Костное небо сильно развито и продолжается назад дальше последних коренных зубов. Предчелюстные кости крупные. Барабанные кости кольцеобразные. Зубы по большей части приспособлены к питанию плодами. Резцы небольшие. Обычно хорошо развитые
клыки имеют даже те формы, у которых зубы имеют тенденцию к редукции.
Щечные зубы низкие, с продольной бороздкой, обычно не имеющие выступов.
Формула зубов варьирует от
2 13 2
i — с — р — т— — 34 у летучих собак 2 13 3
. 1 1 3 1 .
и летучих лисиц до t ~ с — р — т— = 24 у трубконосых и ма
лых трубконосых крыланов. Зубы слабо гетеродонтные, разделены между собой промежутками. Большой и малый бугры плечевой кости развиты слабо, и большой бугор никогда не образует дополнительного сустава с лопаткой.
Кишечник относительно длинный, длина его превышает длину тела примерно в 4 раза. Температура тела обычно высокая: от 31 до 37° С у бавеанс-кой летучей лисицы и от 34,4 до 38,6° С у ангольской летучей собаки.
Активность сумеречная и ночная, иногда и дневная. Постоянного убежища может не быть, так как крыланы часто перелетают на новые места в зависимости от того, где есть зрелые плоды. С мест дневок они могут делать дальние перелеты к местам кормежек. Крупные виды летают за 15 км и в ночь могут налетать в общей сложности 90—100 км. Крупные по размерам
формы обычно скапливаются на дневках большими группами (пальмовый крылан до 10 тыс. особей), а мелкие держатся поодиночке. День проводят на ветвях деревьев, в пещерах, под карнизами зданий. Во время отдыха голову держат перпендикулярно к длинной оси тела. Питаются в основном плодами (поедают мякоть или пьют только сок) и нектаром и пыльцой цветков. Насекомые могут служить дополнительным объектом питания некоторых крыланов. Некоторые формы мигрируют, особенно в районах, где плоды созревают периодически. Например, в южной Анголе ангольский эполето-вый крылан появляется в ноябре, а покидает эту область в начале сухого сезона. Сезонные миграции восточноавстралийской летучей лисицы известны в Восточной Австралии. Летучие собаки используют эхолокацию,для ориентации. Большинство других представителей подотряда, по-видимому, ориентируется главным образом при помощи зрения, а также обоняния. Спячки, как правило, у представителей подотряда нет.
Размножение у большинства видов, по-видимому, имеет сезонный характер. Так, в Камеруне ангольская летучая собака, молотоголовый крылан и пальмовый крылан спариваются в сентябре — ноябре, а рождают детенышей в феврале. В течение года самка приносит потомство один раз; детенышей в помете 1, реже 2. Детеныш полного роста достигает примерно в 8-ме-сячном возрасте, когда он переходит к самостоятельному образу жизни. В неволе крыланы жили до 20 лет. Крыланы хорошо приручаются и привыкают к своему хозяину. Некоторые племена употребляют крыланов в пищу. Крыланы могут наносить существенный ущерб фруктовым плантациям.
Распространены в восточном полушарии от Африки на восток до Австралии, Самоа и Каролинских островов. На некоторых островах Океании фауна млекопитающих, за исключением завезенных сюда человеком крыс, мышей и собак, представлена только крыланами. Систематика крыланов окончательно не выяснена. Число родов и объемы некоторых из них остаются спорными.
Семейство включает 41 род (146 видов):
коротконосых крыланов — Cynopterus F. Cuvier, 1825 (5 видов); темных крыланов — Thoopterus Matschie, 1899 (1 вид);
черношапочных крыланов — Chironax Andersen, 1912 (1 вид);
бурых крыланов—Dyacopterus Andersen, 1912 (1 вид);
коротконосых крыланов Лукаса — Penthetor Andersen, 1912 (1 вид); крыланов Бланфорда — Sphaerias Miller, 1906 (1 вид);
крыланов Ягора — Ptenochirus Peters, 1861 (1 вид);
крыланов Темминка — Megaerops Peters, 1865, (2 вида);
пятнокрылых крыланов — Balionycteris Matschie, 1899 (1 вид);
летучих собак, или ночных крыланов, — Rousettus Gray, 1821 (13 видов);
ошейниковых крыланов — Myonycteris Matschie, 1899 (4 вида);
летучих лисиц — Pteropus Brisson, 1762 (46 видов);
сулавесских крыланов — Neopteryx Hayman, 1946 (1 вид);
ацеродон — Acerodon Jourdan, 1837 (5 видов);
птералопекс — Pteralopex Thomas, 1888 (1 вид);
бонейя — Boneia Jentink, 1879 (1 вид);
карликовых крыланов — Aethalops Thomas, 1923 (1 вид);
крыланов Уоллэса —Styloctenium Matschie, 1899 (1 вид); голоспинных крыланов — Dobsonia Palmer, 1898 (9 видов) ; крыланов Фишера — Haplonycteris Lawrence, 1939 (1 вид);
ночных крыланов — Alionycteris Kock, 1969 (1 вид);
толстоухих крыланов — Otopterus Kock, 1969 (1 вид);
эполетовых крыланов — Epomophorus Bennett, 1836 (8 видов);
карликовых эполетовых крыланов — Mlcropteropus Matschie, 1899 (2 вида);
биндемов — Epomops Gray, 1870 (3 вида);
скотониктерис — Scotonycteris Matschie, 1894 (2 вида);
коровьемордых крыланов — Nanonycteris Matschie, 1899 (1 вид);
молотоголовых крыланов — Нypsignathus Н. Allen, 1861 (1 внд);
казиннктернс — Casinycteris Thomas, 1910 (1 вид);
крыланов Анхиеты — Pierotes Andersen, 1910 (1 вид);
соломенноцветных, или пальмовых крыланов,— Eidolon Rafinesque, 1815 (3 вида),
пещерных крыланов — Eonycteris Dobson, 1873 (5 видов);
длинноязыких крыланов — Macroglossus F- Cuvier, 1824 (З^вида);
цветочных длинноязыких крыланов — Syconycteris Matschie, 1899 (3 вида);
африканских - длинноязыких крыланов — Megaloglossus Pagenstecher, 1885 (1 вид);
темнобрюхих крыланов — Melonycteris Dobson, 1877 (1 вид):
Соломоновых длинноязыких крыланов — Nesonycteris Thomas, 1887 (1 вид);
длиннохвостых крыланов — Notopteris Gray, 1859 (1 вид);
трубконосых крыланов-—Nyctimene Borkhausen, 1797 (8 видов);
карликовых трубконосых крыланов—Paranyctimene Tate, 1942 (1 вид); крыланов-гарпий — Harpyionycteris Thomas, 1896 (1 вид).
Род коротконосых крыланов
Genus Cynopterus F. Cuvier, 1825
Размеры мелкие. Длина тела 9,5—12,7 см. Длина хвоста около 13 мм (рис, 91). Размах крыльев достигает 30—45 см. Вес взрослых животных 50— 100 г. Терминальный конец хвоста не включен в межбедреную перепонку. Ноздри сильно выдающиеся, почти трубчатые. Верхняя губа имеет посреди
не глубокую вертикальную борозду. Волосяной покров довольно длинный и плотный. Окраска его варьирует, но обычно оливково-бурая. Рострум черепа укорочен. Позади орбиты имеется небольшое отверстие. Предчелюстные кости развиты слабо, спереди соприкасаются, но не срастаются между собой. Костные слуховые барабаны крупные. Нижняя челюсть с широкими
Рис. 91. Коротконосый крылан Хорсфилда — С. horsfieldi и его череп
2 1
Верхние резцы соприкасаются друг с другом, но отделены от кой диастемой. Верхние и нижние клыки с дополнительной
2 13 1
низкими угловыми отростками. Число зубов: i — с — р — т — = 30.
клыков широ-вершинкой на внутренней стороне.
Обычными убежищами служат пещеры, заброшенные шахты, пустоты под карнизами крыш домов, изредка дупла деревьев. На Цейлоне предпочитают проводить день среди листьев пальм. Коротконосые крыланы — единственные рукокрылые Старого Света, которые иногда делают себе убежище, выкусывая центральные плоды из кисти плодов пальмы таким образом, что там образуется полость, где животные и повисают. Обычно держатся группами от 6 до 12 особей, старые самцы чаще всего поодиночке. В полете их обычно видят среди кустов и низких деревьев. За ночь они пролетают до сотни километров. Могут подниматься в горы на высоту до 2000 м над уровнем моря. Питаются преимущественно соком, в меньшей степени мякотью плодов пальм, фиговых деревьев, манговых деревьев, бананов и гуавас, а также цветками перечисленных выше деревьев и Orohylum indicum. Делают налеты на плантации. Часто уносят с собой плоды финиковых пальм и поедают их в различных местах, чем помогают их расселению. Самка приносит обычно одного детеныша. Размножение может происходить в любое время года, но массовое рождение детенышей в некоторых областях приходится в основном на март и апрель. Имеются сообщения о том, что самка может приносить 2 потомства в год.
Ареал охватывает обширную территорию от Цейлона и Индии на восток через Южный Китай и Юго-Восточную Азию до Индонезии и Филиппинских островов.
Различные исследователи по-разному трактуют систему рода коротконосых крыланов. Так, Симпсон (1945) включает в род Cynopterus следующие роды: Niadius, Thoopterus, Chironax, Dyacopterus, Penthetor. Бурлиер (1955) оставляет объединенными роды Cynopterus, Niadius, Thoopterus и Chironax, выделяя роды Dyacopterus и Penthetor. Уолкер (1964) и Андерсон и Джонс (1967) все эти роды считают осособленными друг от друга за исключением Niadius, который они присоединяют к Cynopterus. Следуя последним авторам, впредь до проведения ревизии всего подотряда крыланов, включаем в состав рода Cynopterus только род Niadius.
В роде 5 видов:
индийский коротконосый крылан — С. sphinx Vahl, 1797 (Хайнань, Индия, Цейлон, Бирма, п-ов Индокитай, Таиланд, Суматра, Ява, Бали, Тимор, Ломбок);
малайский коротконосый крылан — С. brachyotis Muller, 1838 (Кантон в Южном Китае, Цейлон, Андаманские и Никобарские острова, Тенассерим, Бирма, Ассам, Таиланд, Малаккский полуостров, Суматра, Ява, Калимантан с прилегающими островами, Сулавеси, Филиппинские острова);
коротконосый крылан Хорсфилда — С. horsfieldi Gray, 1843 (о-ва Ява, Калимантан, Суматра, Ниас и Малаккский полуостров);
цепколапый коротконосый крылан •— С. harpax Thomas et Wroughton, 1909 (Малаккский полуостров);
филиппинский коротконосый крылан — С. archipelagus Taylor, 1934 (о-в Полилло из группы Филиппинских).
Род темных крыланов
Genus Thoopterus Matschie, 1899
В роде единственный вид: темный крылан — Т. nigrescens Gray, 1870.
Длина предплечья достигает примерно 7,3 см. Хвост рудиментарный. Окраска довольно длинного волосяного покрова на теле серовато-бурая. Летательная перепонка прикрепляется на задних лапах ко второму пальцу.
В черепе постррбитальные отверстия отсутствуют. Зубная формула: 2 13 1
г — с — р —/п—= 30. Характерны сильно увеличенные в ширину коронки нижнего четвертого предкоренного и нижнего первого коренного зубов.
Экология не изучена.
Распространение охватывает о-ва Моротай из группы Молуккских и Сулавеси.
Род черношапочных крыланов
Genus Chironax Andersen, 1912
В роде единственный вид: черношапочный крылан — Ch. melanocepha-lus Temminck, 1825. Симпсон (1945) включает этого крылана в род Cy-
nopterus.
Размеры мелкие. Длина тела примерно 7,6 см. Длина предплечья около 4,3 см. Хвоста нет. Окраска волосяного покрова на голове черная, на
спине от серовато- до красновато-бурой, на брюхе светло-желтая. Череп ко-
роткий, широкий. Зубная формула:
.21 31
i — с — р — т— —
2 13 2
30.
Обитатель лесов. Питается в основном плодами фиговых пальм. Распространение охватывает п-ов Малакку, о-ва Суматру, Яву.
Род бурых крыланов
Genus Dyacopterus Andersen, 1912
В роде единственный вид: бурый крылан — D. spadiceus Thomas, 1890. Некоторые исследователи выделяют и второй вид D. brooksi Thomas, 1920. Иногда род Dyacopterus присоединяют к роду Cynopterus, (Симпсон, 1945).
Размеры мелкие. Длина тела 12,7—15,2 см, длина хвоста около 1,3 см. Длина предплечья 7,6—8,9 см. Окраска волосяного покрова черноватая на морде, желтоватая на плечах, бурая на спине и боках и грязно-беловатая на груди и брюхе. Череп короткий и широкий. В черепе имеются посторбиталь-2 12 1
ные отверстия. Зубная формула: i — с — р — т — = 28.
Экология не изучена. Встречается очень редко.
Распространен на о-вах Лусон, Калимантан, Суматра и на Малаккском полуострове.
Род коротконосых крыланов Лукаса Genus Penthet or Andersen, 1912
В роде один вид: коротконосый крылан Лукаса — Р. lucasi Dobson, 1800. Некоторые исследователи (Симпсон, 1945) включают род Penthetor в род Cynopterus.
Размеры мелкие. Длина тела около 11 см. Хвост укороченный — 0,8— 1,0ели толстый. Длина предплечья около 6см. Волосяной покров короткий, окраска его дымчато-бурая. В черепе относительно удлиненный мозго-2 13 1
вой отдел. Зубная формула: i — с — р — т — = 28.
113 2
Экология почти не изучена. День проводит в пещерах. Активным становится в сумерках. Питается плодами. Посещает фруктовые плантации.
Распространен на Малаккском полуострове, архипелаге Риоу и о-ве Калимантан.
Род крыланов Бланфорда
Genus Sphaerias Miller, 1906
В роде единственный вид: крылан Бланфорда — S. blanfordi Thomas, 1891.
Размеры мелкие. Длина тела около 6,4 см. Хвост снаружи не виден. Длина предплечья примерно 5,0 см. Межбедренная летательная перепонка уменьшена до узкой оторочки вдоль внутренней стороны бедер и верхних отделов голени. Окраска волосяного покрова грязновато-буровато-серая сверху и более светлая снизу. Лицевой отдел черепа удлинен. Число зубов: 2 13 1
i — с — р — т~ — 30. Резцы направлены вперед почти параллельно продольной оси челюсти.
Экология не изучена. Очень редок.
Распространен в Бирме и Таиланде.
Род крыланов Я гора
Genus Ptenochirus Peters, 1861
В роде единственный вид: крылан Ягора — Р. jagori Peters, 1867.
Размеры небольшие. Длина тела около 10 см. Длина хвоста примерно 1,3 см. Длина предплечья примерно 8,5 см. Окраска волосяного покрова темно-бурая сверху и более светлая снизу. Зубная формула: 2 13 1
i — с — р — т~ =28. Второй верхний резец имеет небольшие размеры.
Экология не изучена. Дневным убежищем служат в основном пещеры.
Распространение охватывает о-ва Минданао, Лусон и Миндоро в группе Филиппинских. t
Род крыланов Темминка
Genus Megaerops Peters, 1865
Размеры мелкие. Длина тела 7,6—10,2 см. Длина предплечья около 5,5 см. Хвост рудиментарный или его нет вовсе. Окраска волосяного покрова на спинной стороне желтовато-бурая или пепельно-серая, на брюшной серебристо-серая. Лицевой отдел черепа укорочен. Зубная формула: 2 13 1
i — с — р — т — = 28. Второй верхний резец небольших размеров.
Экология не изучена. Крылан Темминка на Малаккском полуострове держится в горах, не спускаясь ниже 1500 м.
Распространены во Вьетнаме, на Малаккском полуострове, о-вах Суматра и Калимантан и на о-ве Котабато из группы Филиппинских.
В роде 2 вида:
крылан Темминка — М. ecaudatus Temminck, 1837 (Вьетнам, Малаккский полуостров, о-ва Суматра, Калимантан):
крылан Ветмора — М. wetmorei Taylor, 1934 (о-в Котабато).
Некоторые исследователи включают род Megaerops в состав рода Ptenochirus.
Род пятнокрылых крыланов
Genus Balionycteris Matschie, 1899
В роде единственный вид: пятнокрылый крылан — В. maculata Matschie, 1899.
Размеры мелкие. Длина тела примерно 6,6 см. Хвост снаружи незаметен. Длина предплечья около 4,1 см. Окраска волосяного покрова на спинной 182
Рис 92. Цепкохвостая летучая собака — R. amplexicaudatus
стороне черновато-бурая, снизу сероватая. Голова черноватая. Темно-бурые крылья имеют желтые пятна. Череп укорочен, широкий, посторбиталь-Г 2 13 2
ные отверстия отсутствуют. Зубная формула: i — с — р -— т— = 30.
Экология не изучена.
Распространены на Малаккском полуострове, о-ве Калимантан и архипелаге Риоу.
Род летучие собаки, или ночные крыланы,
Genus Rousettus Gray, 1821
Размеры мелкие. Длина тела около 13 см. Хвост короткий, достигающий в длину примерно 1,4 см. Длина предплечья колеблется от 7,3 до 10,3 см. Второй палец передней конечности с хорошо развитым когтем. Ростральная часть головы вытянута не особенно сильно (рис. 92). Ноздри разделены глубокой бороздкой. Кончик хвоста свободно выдается из межбедренной перепонки. На передней части шеи по ее бокам располагаются специфические кожные железы, покрытые волосами, которые у взрослых самцов имеют рыжевато-коричневато-оливковый цвет. Окраска волосяного покрова на спинной стороне тела обычно буроватая, на брюшной несколько более светлая. Волосяной покров короткий. У некоторых видов вокруг шеи проходит своеобразный ошейник из бледных коротких волос. Лицевой отдел черепа относи-
2 13 2
тельно укорочен. Зубная формула: i с — р — т— = 34. На передней поверхности нижних резцов имеется бороздка.
Активны в основном в сумерках. Питаются соком плодов и нектаром. Способствуют переносу пыльцы с одних цветков на другие. Период спаривания в большинстве районов приходится на декабрь — март. Беременность примерно 15 недель. Самка приносит обычно одного детеныша. К самостоятельному питанию детеныши переходят в возрасте примерно трех месяцев.
Распространены на большей части Африки, Южной Азии, Малайзии и архипелагах Индийского океана на восток до Соломоновых островов.
В роде 13 видов:
египетская летучая собака — R. aegyptiacus Ё. Geoffrey, 1810 (Кипр, Израиль, Сирия, АРЕ, Судан, Эфиопия и на юг до Анголы и ЮАР);
аравийская летучая собака — R. arabicas Anderson et de Winton, 1902 (Аравийский полуостров, о-в Кешм в Персидском заливе, Западная Индия);
цепкохвостая летучая собака — R. amplexicaudatus Ё. Geoffrey, 1810 (север Таиланда и Тенассерим, п-ов Индокитай, п-ов Малакка, о-ва Суматра, Ява, Калимантан, Тимор, Флорес, Филиппинские);
летучая собака Лешенолта — R. leschenaulti Desmarest, 1820 (Индия, Непал, Бирма, Тенассерим, п-ов Индокитай, Южный Китай, Ява);
цейлонская летучая собака — R. seminudus Kelaart, 1850 (Цейлон);
ангольская летучая собака — R. angolensis Bocage, 1898 (Ангола и от Танзании и Кении до Камеруна);
кенийская летучая собака — R. kempi Thomas, 1909 (Кения);
угандская летучая собака — R. lanosus Thomas, 1906 (Уганда);
мадагаскарская летучая собака — R. Madagascar iensis Grandidier, 1928 (Мадагаскар);
летучая собака Фернандо-По — R. crypticola Cabrera, 1920 (о-в Фернандо-По);
летучая собака Смита—R. Smithii Thomas, 1908 (от Сьерра-Леоне до Южной Нигерии);
целебесская летучая собака — R. celebensis Andersen, 1907 (о-ва Сулавеси и Сангихе);
летучая собака Штреземанна — R. stresemanni Stein, 1923 (северо-запад Новой Гвинеи и о-в Япен).
Некоторые исследователи насчитывают в роде не 13, а 11 видов. Ряд исследователей (Лоренс — Lawrence, 1963) выделяют подрод Lissonycteris с тремя видами в особый род Lissonycteris Andersen, 1912.
Род ошейниковых крыланов
Genus Myonycteris Matschie, 1899
Размеры небольшие. Длина тела 11—16,5 см. Длина хвоста 0,4—1,2 см. Длина предплечья 5,5—7 см. Занимают промежуточное положение между Rousettus и Cynopterus. Морда у них короче, чем у Rousettus, но длиннее, чем у Cynopterus-, ноздри сильнее выступают, чем у Rousettus, но меньше, чем у Cynopterus. Окраска волосяного покрова спинной стороны тела светло-или темно-бурая, иногда с рыжеватым или желтоватым оттенком. У самцов на передней части шеи имеются желтовато-оранжевые или рыжевато-оливковые грубые волосы, образующие своеобразное полукольцо. Эта окрашенная полоса соответствует расположению шейной специфической кожной железы.
2 13 2
Зубная формула, как у Rousettus-. i — с — р — т— = 34, однако второй верхний и третий нижний коренные сильно редуцированы и по этому признаку ошейниковые крыланы приближаются к Cynopterus, у которых эти зубы совсем отсутствуют.
Обитатели саванн. Экология изучена слабо. День проводят на ветвях кустов, иногда на прямом солнечном свету. Держатся поодиночке.
Распространены в Африке.
В роде 4 вида:
малый ошейниковый крылан — М. torquata Dobson, 1878 (Ангола, Республика Заир, Камерун);
либерийский ошейниковый крылан — М. leptodon Andersen, 1908 (Сьерра-Леоне и Либерия);
ошейниковый крылан Вроугтона — М. wroughtoni Andersen, 1908 (Республика Заир);
Рис. 93. Летучая лисица Гулди — Р. gouldi и ее череп
ошейниковый крылан Сан-Томе — М. brachycephala Bocage, 1889 (о-в Сан-Томе в Гвинейском заливе).
Иногда вместо четырех видов в роде Myonycteris выделяют только три вида. Некоторые* исследователи присоединяют род Myonycteris к роду Pfiusettus.
Род летучих лисиц
Genus Pteropus Brisson, 1762
Размеры мелкие, средние или крупные, наибольшие среди представителей отряда (рис. 93). Длина тела примерно от 10 до 40 см. Хвоста нет (рис. 93). Межбедренная перепонка узкая. Длина предплечья изменяется примерно от 8 до 23 см. Размах крыльев может достигать 170 см, вес 900 г. Лицевой отдел головы вытянут, уши довольно короткие. Волосяной покров относительно короткий и густой, серовато-бурый или черный. Область между плечами часто бывает желтая или серовато-желтая. На втором пальце коготь хорошо развит. У некоторых видов на боковых отделах затылка или на плечах описана кожная специфическая железа в виде кармана. Череп умеренно удлинен. Предчелюстные кости хорошо развиты, спереди соприкасаются, но не срастаются друг с другом. Костные слуховые барабаны кольцевидные. Зубные формула:
i — с — p — m —= 34. Верхние резцы прилегают друг к другу, но отделены от клыков широкой диастемой. Число диплоидных хромосом у летучей лисицы Рюкю 38.
Активность сумеречная и ночная. На день собираются большими стаями в сотни и даже тысячи голов вместе. Дневки устраивают на деревьях, которые могут использоваться в течение многих лет (рис. 94). Питаются в основном соком плодов, который они получают, раздавливая мякоть плодов языком о небо. Могут разжевывать цветки, чтобы получить сок и пыльцу. Охотно пьют воду. Рождение детенышей в Индии приходится в основном на март, а в Австралии, Новой Гвинее и на Мадагаскаре обычно на октябрь. В некоторых областях представители рода не имеют определенного сезона размножения. Длительность беременности у крупных видов составляет примерно 6 месяцев. В Лондонском зоопарке лету чая лисица жил а 17 лет и 2 месяца. В некоторых областях летучие лисицы приносят большой вред фруктовым плантациям. Мясо отдельных видов употребляется местным населением в пищу.
Ареал охватывает обширную территорию — Мадагаскар, о-ва Коморские, Пемба, Индийского океана, Юго-Восточную Азию, Индонезию, о-ва Филиппинские, Марианские, Новую Гвинею, Восточную Австралию и простирается на восток до Фиджи и Самоа.
В роде 48 видов:
летучая лисица Рюкю — Р. dasymallus Temminck, 1825 (о-ва Рюкю и Тайвань);
малая летучая лисица — Р. hypomelanus Temminck, 1853 (провинция Намбо в Южном Вьетнаме, Таиланд, архипелаг Мергуи, мелкие острова в Малаккском проливе, острова к западу от Суматры, Натуна, Анамбас, мелкие у Калимантана, Сулавеси, Филиппинские, Новая Гвинея);
сибутская летучая лисица — Р. speciosus Andersen, 1908 (о-ва Сибуту, Замбоанга на о-ве Минданао; о-ва Салембу-Бесар и МатаСири в Яванском море);
марианская летучая лисица — Р. marianus Desmarest, 1822 (о-ва Марианские, Рюкю);
Рис. 94. Дневка тонганских летучих лисиц — Р. tonganus на о-вах Фиджи
никобарская летучая лисица — Р. melanotus Bly th, 1863 (о-ва Никобарские, Андаманские, Ниас, Энгано, западная Суматра, Рождества);
таиландская летучая лисица — Р. lylei Andersen, 1908 (Таиланд, провинция Намбо);
большая летучая лисица — Р. vampyrus Linnaeus, 1758 (Тенассерим,
Трунбо во Вьетнаме, п-ов Малакка, о-ва Суматра, Ява, Калимантан, филиппинские, Бали, Тимор);
бавеанская летучая лисица — Р. alectc Temminck, 1837 (о-в Бавеан и о-ва Кангеан в Яванском море);
индийская летучая лисица — Р. giganteus Briinnich, 1782 (Цейлон, Индия, Непал, Сикким, Бутан, Ассам, Манипур, Бирма);
серая летучая лисица — Р. griseus Ё. Geoffroy, 1810 (о-ва Сулавеси, Тимор, Банда);
адмиралтейская летучая лисица — Р. admiralitatum Thomas, 1894 (о-ва Адмиралтейские и Соломоновы);
тонганская летучая лисица — Р. tonganus Quoy et Gaimard, 1830 (о-ва Тонга о-в Реннелл, о-ва Соломоновы, Новые Гебриды);
седая летучая лисица — Р. caniceps Gray, 1870 (о-ва Моротай, Хальма-хера и мелкие соседние с ним, Сулавеси);
серебристая летучая лисица — Р. argentatus Gray, 1844 (Сулавеси);
черноспинная летучая лисица — Р. melanopogon Peters, 1867 (о-ва Буру, Церам и Амбоина с мелкими прилегающими, Тимор, Банда);
соломонова летучая лисица — Р. cognatus Andersen, 1908 (о-ва Сан-Кристобаль и Реннелл в группе Соломоновых);
летучая лисица Райнера — Р. rayneri Gray, 1870 (Соломоновы острова);
золотистая летучая лисица — Р. chrysoproctus Temminck, 1837 (о-в Амбоина с мелкими соседними);
ломбокская летучая лисица — Р. lombocensis Dobson, 1878 (о-ва Флорес и Алор около о-ва Тимор);
моротайская летучая лисица — Р. personatus Temminck, 1825 (о-ва Моротай, Хальмахера и мелкие соседние, Сулавеси);
летучая лисица Темминка — Р. temmincki Peters, 1867 (о-ва Амбоина, Церам, Тимор, Буру, архипелаг Бисмарка);
сулавесская летучая лисица — Р. arquatus Miller et Hollister, 1921 (о-в Сулавеси);
новогвинейская летучая лисица — Р. conspicillatus Gould, 1850 (Новая Гвинея, о-ва Хальмахера и мелкие между ним и Новой Гвинеей);
церамская летучая лисица-—Р. ocularis Peters, 1867 (о-ва Церам и Буру);
Р. neohibemicus Peters, 1876 (архипелаг Бисмарка и Новая Гвинея);
летучая лисица Ару — Р. macrotis Peters, 1867 (о-ва Ару и Новая Гвинея);
япенская летучая лисица — Р. pohlei Stein, 1933 (о-в Япен и Новая Гвинея);
летучая лисица Вуудфорда — Р. woodfordi Thomas, 1888 (Соломоновы острова и Квинсленд в Австралии);
махагская летучая лисица — Р. mahaganus Sanborn, 1931 (о-в Махага в группе Соломоновых);
беловатая летучая лисица — Р. leucopterus Temminck, 1853 (о-в Лусон в группе Филиппинских);
летучая лисица Мирнса — Р. mearnsi Hollister, 1913 (о-ва Минданао и Базилан);
карликовая летучая лисица — Р. pumilus Miller, (1911 (о-в Палмас к юго-востоку от о-ва Минданао);
лузонская летучая лисица — Р. rnirnus Andersen, 1908 (о-в Лусон и юг о-ва Сулавеси);
балутская летучая лисица — Р. balutus Hollister, 1913 (о-в Балут в группе Филиппинских);
табласская летучая лисица — Р. tablasi Taylor, 1934 (о-ва Таблас в группе Филиппинских);
летучая лисица Гулда — Р. gouldi Peters, 1867 (Квинсленд и Северная территория в Австралии);
восточноавстралийская летучая лисица — Р. poliocephalus Temminck, 1825 (восток Австралии);
альдабрская летучая лисица — Р. aldabrensis True, 1893 (о-ва Альдаб-ра к северу от Коморских);
коморская летучая лисица — Р. comorensis Nicoll, 1908 (Коморские острова);
летучая лисица Ливингстона — Р. livingstonii Gray, 1866 (Коморские острова);
маврикийская летучая лисица — Р. niger Kerr, 1792 (о-в Маврикий, Реюньон и Мадагаскар);
родригесская летучая лисица — Р. rodricensis Dobson, 1878 (о-ва Родригес в Индийском океане);
рыжая летучая лисица — Р. rufus Е. Geoffroy, 1803 (о-в Мадагаскар);
сейшельская летучая лисица — Р. seychellensis Milne-Edwards, 1877 (Сейшельские острова);
черноватая летучая лисица — Р. subniger Kerr, 1792 (о-ва Реюньон и Маврикий);
пембская летучая лисица —• Р. voeltzkowi Matschie, 1909 (о-в Пемба у побережья Танзании);
летучая лисица Остина — Р. austini Lawrence, 1945 (о-в Флорида из группы Соломоновых);
летучая лисица Жиллиарда — Р. gilliardi Van Deusen, 1969 (о-в Новая Британия из архипелага Бисмарка).
Систематика рода летучих лисиц выяснена недостаточно. Различные исследователи насчитывают в роде разное число видов (Андерсон и Джонс, 1967—примерно 65).
Род сулавесских крыланов
Genus Neopteryx Hayman, 1946
В роде единственный вид: сулавесский крылан — N. frosti Hayman, 1946.
Размеры мелкие. Длина тела примерно 10 см. Хвост отсутствует. Длина предплечья примерно 11 см. Характерно отсутствие когтя на втором пальце передней конечности, в то время как первый палец имеет хорошо развитый коготь. Летательная перепонка крыльев прикрепляется на туловище близко к средней линии спины, так что создается впечатление, что спина не покрыта волосами. Волосяной покров короткий, но густой, рыжевато-коричневый. Морда кремовато-белая или буроватая. Число зубов:
i — с—p + m— = 32.
2 1 к ' 6
Экология не изучена.
Распространен на западном Сулавеси.
Род ацеродон
Genus Acerodon Jourdan, 1837
Размеры средние. Длина тела 18—25 см. Длина предплечья 13—20 см. Размах крыльев — 1,5—1,6 м. Хвоста нет. Окраска короткого волосяного покрова варьирует. Лоб и боковые отделы головы часто имеют темно-бурый или черный цвет. Затылок может быть оранжевым или золотисто-желтым.
Плечи обычно рыжевато-бурые или каштановые. Задняя часть спины темнее передней. Брюшная сторона тела темно-бурая или черная. Череп сходен с черепом представителей рода летучих лисиц. Зубная формула:
. 2 1 3 2 „.
i — с — р — т— = 34.
2 13 3
Клыки меньше, чем у летучих лисиц, а щечные зубы больше.
Обитатели бамбуковых зарослей, болотистых лесов. На дневках иногда собираются огромными скоплениями (отмечена стая примерно в 150 тыс. особей). Питаются плодами. На кормежку могут летать за 9—16 км.
Распространены на Сулавеси и прилегающих мелких островах, а также на о-вах Тимор, Флорес и Филиппинских.
В роде 5 видов:
целебесский ацеродон — A. celebensis Peters, 1867 (о-ва Сулавеси и Сула);
ацеродон Макклота — A. Macklot i Temminck, 1837 (о-ва Тимор, Флорес, Сумбава, Сумба, Ломбок);
обыкновенный ацеродон — A. humilis Andersen, 1909 (о-ва Талауд);
дневной ацеродон — A. lucifer Elliott, 1896 (о-в Панау в группе Филиппинских);
гривастый ацеродон — A. jubatus Eschscholtz, 1831 (о-в Минданао и мелкие острова к северу от него).
Некоторые исследователи включают род Acerodon в состав рода Pteropus.
Род птералопекс
Genus Pteralopex Thomas, 1888
В роде один вид: черный птералопекс — Р. atrata Thomas, 1888. Иногда род Pteralopex присоединяют к роду Pteropus.
Размеры средние. Длина тела примерно 25 см. Длина предплечья 13— 14 см. Хвоста нет. Летательная перепонка прикрепляется к туловищу по средней линии спины. Густой и мягкий волосяной покров черного цвета. По морфологическим признакам сходны с летучими лисицами, но отличаются от них наличием бугорков на зубах. Зубная формула' .2 2 3 2 о.
i — с — р — т — =34.
2 13 3
Экология не изучена.
Распространен на Соломоновых островах.
Род бонейя
Genus Boneia Jentink, 1879
В роде единственный вид: Бонейя — В. bidens Jentinck, 1879.
Размеры небольшие. Длина тела примерно 14 см. Длина хвоста около 2,5 см. Длина предплечья примерно 9,5 см. Второй палец передней конечности имеет коготь. Кончик хвоста свободно выступает из межбедренной перепонки. Окраска волосяного покрова самцов темно-бурая на спине, рыжевато-коричневая на крестце, тускло-буроватая на брюшной стороне, золо-113 2
тистая на затылке и боках шеи. Зубная формула: i — с — р — т~ —
= 32. Характерно, что в верхней челюсти только по одному резцу с каждой стороны.
Экология не изучена.
Распространен на северном Сулавеси.
Род карликовых крыланов
Genus Aethalops Thomas, 1923
В роде единственный вид: карликовый крылан— A. alecto Thomas, 1923 (иногда выделяют и второй вид A. aequalis Allen, 1938).
Один из наиболее мелких крыланов. Длина тела от 6,5 до 7,3 см. Хвоста нет. Длина предплечья 4,4—5,1 см. Уши небольшие. Окраска волосяного покрова по большей части черная или с варьирующими оттенками темно-серого цвета. Узкая оторочка межбедренной перепонки покрыта шерстью.
Обитатель горных лесов.
Экология не изучена.
Распространен на Малаккском полуострове, Суматре и Калимантане.
Род крыланов Уоллэса
Genus Styloctenium Matschie, 1899
В роде единственный вид: крылан Уоллэса — S. wallacei Gray, 1866.
Размеры средние. Длина тела 15—18 см. Хвоста нет. Длина предплечья примерно 9—10 см. Волосяной покров довольно густой и мягкий. На спине и крестце обычно светло-серого цвета, часто с бледновато-рыжевато-бурым оттенком. Вентральная часть туловища светло-рыжевато-бурая. Голова рыжевато-бурая с белыми полосками или пятнами, что характерно для этих крыланов. Половой диморфизм в окраске не выражен. Череп по строению 2 13 2
напоминает летучих лисиц. Зубная формула: i — с — р — т— = 30.
1 1 3 2
Экология не изучена.
Обитает на Сулавеси.
Род голоспинных крыланов
Genus Dobs onia Palmer, 1898
Размеры средние. Длина тела примерно 13—20см. Длина хвоста около 1,3 см\ его кончик свободно выступает из межбедренной перепонки. Длина предплечья от 84 до 160 мм. Наиболее крупные особи могут достигать веса примерно 450 г. На втором пальце передних конечностей коготь отсутствует. Уши небольшие, заостренные. Безволосые летательные перепонки передних конечностей прикрепляются на спине вдоль хребта по его срединной части, создавая впечатление, что спина у представителей этого рода не имеет волос. Лицевой отдел черепа укорочен. Зубная формула:
i — с — р — т— = 28.
1 1 ' з 3
Экология почти не изучена. Дневки устраивают в пещерах.
Обитают на островах от Сулавеси и Молуккских до Соломоновых островов, включая Филиппинские, и п-ове Кейп - Йорк в Северо-Восточной Австралии.
В роде 9 видов:
крылан Перона — D. peroni Ё. Geoffrey, 1810 (о-ва Тимор, соседние с ним, Флорес, Бали и Сумба);
зеленый крылан — D. viridis Heude, 1896 (о-ва Кей, Амбоина и Банда);
голоспинный крылан — D. inermis Andersen, 1909 (Соломоновы острова);
хищный крылан — D. praedatrix Andersen, 1909 (о-ва Новая Британия и Новая Ирландия);
хальмахерский крылан —D. crenulata Andersen, 1909 (о-ва Хальмахер и прилегающие мелкие острова);
молуккский крылан—D. moluccensis Quoy et Gaimard, 1830 (о-ва
Буру, Амбоина, Церам, Ару, Новая Гвинея, Адмиралтейства, архипелаг Бисмарка);
сулавесский голоспинный крылан—D. exoleta Andersen, 1909 (о-в Сулавеси);
малый голоспинный крылан — D. minor Dobson, 1879 (западная Новая Гвинея);
тробрианский крылан —D. remota Cabrera, 1920 (о-ва Тробриан).
Род крыланов Фишера
Genus Haplonycteris Lawrence, 1939
В роде единственный вид: крылан Фишера — Н. fischeri Lawrence, 1939.
Размеры небольшие. Длина предплечья составляет примерно 5,1 см. Хвоста нет. Первый палец передних конечностей сильно вытянут и в длину достигает 2,5 см. Задние конечности короткие, длиной всего около 1,3 см. Окраска волосяного покрова бурая, более светлая на брюшной стороне. В зубной системе характерен всего один резец в каждой половинке челюсти. Зубная формула: i — с -i- m-|- = 28.
Экология не изучена.
Распространен на о-ве Миндоро, относящемся к Филиппинским.
Род ночных крыланов
Genus Alionycteris Kock, 1969
В роде единственный вид: ночной крылан — A. paucidentata Kock, 19S9.
Размеры мелкие: длина тела примерно 6,4 см, длина предплечья около 4,4 сж. Первый палец передних конечностей сильно развит и в длину достигает примерно 2 см. Первый и второй пальцы имеют когти. Хвост отсутствует, как и межбедренная перепонка. Волосяной покров длинный и мягкий, на спине и брюхе буровато-черный. Ноздри открываются на концах трубочек длиной 4 мм. Черноватые уши лишены волос. На небе 9 пар складок. Широкий череп с укороченным лицевым отделом. Заглазничное отверстие отсутствует. Заглазничные отростки короткие. Носовые кости короткие и уплощенные. Межглазничная область и небо широкие. Характерно умень-J 113 1
шейное количество зубов. Зубная формула: i — с — р — т~ — 26.
Экология не изучена.
Распространен на о-ве Минданао из группы Филиппинских.
Род толстоухих крыланов
Genus Otopterus Kock, 1969
В роде единственный вид: толстоухий крылан — О. cartilagonodus Kock, 1969. Размеры небольшие. Длина тела 5,5—6,7 см. Длина предплечья 4,6—4,7 см. Уши с характерно утолщенными краями розового цвета. Нос трубчатый. Хвост отсутствует. Межбедренная перепонка имеет в ширину 2,5—4,5 мм. Сверху она густо покрыта волосами. У первого и второго пальцев имеются когти. Волосяной покров длинный и мягкий, на спинной стороне черно-бурый, на брюшной светлее. Череп с укороченной лицевой частью. Заглазничного отверстия нет. Межглазничная область удлиненная, довольно узкая, имеет резко выраженные надглазничные вздутия. Носовые кости широ-
кие. Костное небо широкое. На небе 10—11 складок. Число зубов уменыпе-
. 1 1 3 1 п.
но: i — с — р — т— = 24.
1 1*3 1
Экология не изучена.
Распространен на о-ве Лусон из группы Филиппинских.
Род эполетовых крыланов
Genus Epomophorus Bennett, 1836
Морда удлиненная. Размеры средние. Длина тела от 12 до 25 см. Хвост едва заметный. Длина предплечья составляет 6,5—9,0 см. Размах крыльев у самцов примерно 51 см. Окраска волосяного покрова серовато-бурая или
рыжевато-коричневая. У обоих полов имеется по белому пятнышку у основания каждого уха. У самцов на плечах находятся большие железистые
мешки, из которых выдаются пучки длинных волос. На шее самцов расположены глоточные воздушные мешки. Характерны крупные выступающие губы, которые часто имеют своеобразные складки. В черепе лицевой отдел
2 12 1
вытянут. Зубная формула: i — с — р — т~ ~ 28.
Обитатели лесов, но могут населять также и засушливые области. На дневках собираются небольшими группами — до 50 особей вместе. Для дневок используют большие дупла, густые кустарники, плотнуюлиству деревьев. Питаются плодами фиговых деревьев, манго, гуавы и бананов. Гамбийский эполетовый крылан в Гане питается нектаром цветков Parkia clappertoniana. Могут приносить существенный вред фруктовым плантациям. Определенной сезонности в размножении не установлено. У некоторых видов (суданский крылан и крылан Валберга) отмечены местные миграции.
Распространены в Африке.
В роде 8 видов:
малый эполетовый крылан — Е. labiatus Temminck, 1837 (Замбия, Танзания, Эфиопия, Судан);
крылан Валберга — Е. wahlbergi Sundevall, 1846 (ЮАР, Мозамбик, южная Родезия, Малави, Замбия, Ангола и от Танзании и Кении до Камеруна);
эполетовый крылан — Е. crypturus Peters, 1852 (ЮАР, Мозамбик, южная Родезия, Малави, Замбия, Республика Заир).
ангольский крылан — Е. angolensis Gray, 1870 (Ангола и северо-запад Юго-Западной Африки);
суданский крылан — Е. anurus Heuglin, 1864 (Судан, Эфиопия);
гамбийский крылан — Е. gambianus Ogilby, 1835 (от Гамбии и Сенегала до Нигерии и юга Эфиопии);
крылан Пусаргуеза — Е. pousarguesi Trouessart, 1904 (Республика Заир);
крылан Неуманна — Е. netunanni Matschie, 1899 (от Камеруна до Кении и Танзании);
Симпсон (1945) объединяет в род эполетовых крыланов следующие роды: Epotnops, Hypsignathu.s,Scotonycterix, Micropteropus, Nanonycteris и Pierotes. Большинство других исследователей, в частности Бурлиер (1955), Андерсон и Джонс (1967), считают все перечисленные выше роды самостоятельными.
Род карликовых эполетовых крыланов
Genus Micropteropus Matschie, 1899
Размеры небольшие. Длина тела примерно 7,5—9,3 см. Длина предплечья 5—6 см. Хвост снаружи едва заметен. По внешнему облику сходен с предыдущим родом эполетовых крыланов, однако морда короче, уши тоже
—
относительно короче, и губы не выдаются так далеко вперед. Волосяной покров на верхней части туловища буроватый, на нижней более светлый или серый. У основания ушей имеются небольшие пучки беловатых волос. Самцы имеют специфические кожные железы в виде плечевых железистых мешков, из которых выдаются пучки удлиненных волос, напоминающих эполеты. В черепе лицевой отдел короткий и широкий. Зубная формула:
2 12 1
t — с — р — т— = 28.
2 13 2
Экология изучена слабо. На дневках не избегают прямого солнечного света и держатся поодиночке или небольшими группами. В размножении сезонности, по-видимому, нет. В бассейне р. Конго самки в большинстве случаев приносят детенышей, начиная с конца февраля, а также в нояб-[ ре или декабре.
; Распространение охватывает территорию от Анголы, юга Республики Заир и Танзании до юга Судана, Эфиопии и Сенегала.
i В роде 2 вида:
карликовый эполетовый крылан —М. pussillus Peters, 1868 (Ангола, Танзания и от Эфиопии до Гамбии);
' ангольский крылан — М. grandis Sanborn, 1950 (Ангола, юг Республи- ' ки Заир).
Некоторые исследователи (Симпсон, 1945) включают карликовых эполетовых крыланов в род эполетовых крыланов.
Род биндемов
Genus Ер о mops Gray, 1870
Размеры небольшие или средние. Длина тела 13—18 см. У большинства хвоста нет, у некоторых особей имеет в длину 1 мм. Длина предплечья 8— 10см. Окраска волосяного покрова рыжевато-коричневая, буроватая или сероватая, со многими цветовыми вариациями. У основания ушей небольшие ; беловатые пучки волос. Самцы имеют по две пары глоточных мешков, крупные железистые плечевые мешки и эполетоподобные пучки волос в области плеч. Эти пучки имеют беловатую или желтоватую окраску. Зубная формула: .21 2 1 оо
i — с — р — т — ==28.
2 1*3 2
Экология изучена слабо. Дневные убежища обычно на ветвях деревьев или кустарников. Держатся поодиночке или небольшими группами по 2— 3 особи. Питаются соком и мягкими частями плодов гуавы, бананов, фиг. Се-। зонности в размножении, по-видимому, нет.
Обитатели тропической Африки от Анголы, Замбии, Танзании, Уганды, на запад до Нигерии и Ганы, Сьерра-Леоне и Либерии.
В роде 3 вида:
крылан Фран'куета — Е. franqueti Tomes, 1860 (Ангола, Замбия и от Танзании на запад до Нигерии и Ганы);
крылан Добсона —Е. dobsoni Bocage, 1889 (Ангола, Республика Заир);
крылан Бюттикофера —Е. bilttikoferiMatschie, 1899 (Либерия и Сьерра-Леоне).
Симпсон (1945) включает род Epomops в род Epotnophortis.
Род скотониктерис
Genus Scotonycteris Matschie, 1894
Размеры небольшие. Длина тела 7—11 см (иногда до 14 см). Длина предплечья 5—7,5см. Хвост рудиментарный. Окраска волосяного покрова рыжевато-коричневая или темно-бурая на спинной стороне и более светлая на брюшной. Около уха отсутствует белое пятно, которое имеется у ряда род
ственных родов. Вместо него белое пятно между глазами или впереди, или позади глаза.
Лицевой отдел черепа укорочен. Зубная формула:
i — с — р — т— = 28.
2 1 г 3 2
Экология не изучена. Держатся, видимо, поодиночке. Дневным убежищем служат ветки деревьев.
Распространение охватывает значительную часть тропической Африки: Гану, Камерун, о-в Фернандо-По и Либерию.
В роде 2 вида:
крылан Ценкера —S. zenkeri Matschie, 1894 (Гана, Камерун, о-в Фернандо-По);
либерийский крылан—S. ophiodon Pohle, 1943 (Либерия, Гана, Камерун).
Симпсон (1945) включает скотониктерис в род эполетовых крыланов.
Род коровьемордых крыланов
Genus Nanonycteris Matschie, 1899
В роде единственный вид: коровьемордый крылан — N. veldkampii Jentink, 1888.
Размеры небольшие. Длина тела примерно 7 см. Хвост снаружи только в виде небольшого выступа. Длина предплечья 4,5—5,2 см. Голова коровье-мбрдого крылана внешне имеет отдаленное сходство с головой коровы. Волосяной покров на спине рыжевато-бурый, на брюхе кремово-буроватый. Окраска может несколько варьировать у разных особей. У обоих полов в основании уха расположено белое пятно. Взрослые самцы имеют плечевые железистые мешки с эполетообразными удлиненными пучками волос. Лицевой отдел черепа короткий и не широкий. Зубная формула:
. 2 1 2 1 «о
i — с — р — т — = 28.
2 13 2
Обитает в глухих лесах. Питается нектаром цветков. Во время кормежки остается у каждого цветка в продолжении 1—30 сек.
Распространен в Центральной Африке от побережья Гвинеи на восток до Республики Заир.
Симпсон (1945) включает коровьемордых крыланов в род эполетовых крыланов.
Род молотоголовых крыланов
Genus Hypsignathus Н. Allen, 1861
В роде единственный вид: молотоголовый крылан — Н. monstrosus Н. Allen, 1861.
Размеры средние или крупные. Длина тела примерно 25—30 см. Хвоста нет. Длина предплечья 12—14 см. Взрослые особи могут достигать веса 400—450г. Самцы крупнее самок. Размах крыльев самцов составляет примерно 97 см. Лицевая часть головы сильно удлинена и утолщена и в поперечном сечении близка к квадрату. Внешне голова несколько напоминает по форме молоток (рис. 95). Губы сильно увеличены в размерах и свисают вниз. Вокруг носа многочисленные складки. Морда покрыта своеобразными мелкими бородавками. Конец морды безволосый. Окраска волосяного покрова серовато-бурая. Грудь более светлая и вокруг шеи проходит светловатый ошейник. У основания ушей располагается по белому пятну. Железистые плечевые мешки и эполетообразные пучки волос отсутствуют как у самцов, так и у самок. У самцов пара воздушных мешков открывается по бокам глотки. Гортань очень крупная, по длине составляет примерно половину длины позвоночного столба, заполняет значительную часть грудной клетки и сдвигает
Рис. 95. Голова молотоголового крылана — Н. monstrosus
сердце и легкие назад и в бока. Голос очень громкий и своеобразный, чему способствуют, по-видимому, воздушные мешки. В черепе ростральный отдел сильно сжат с боков и имеет почти одинаковую высоту в носовой и орбитальной частях. Зубная формула:
i Л с — р —/п—= 28.
2 13 2
Обитает, как правило, в мангровых и пальмовых зарослях вдоль рек или в болотистых районах. На дневках обычно держится поодиночке или небольшими группами в листве, на деревьях, но встречается также и в пещерах. Питается соком плодов манго, бананов. Размножение, по-видимому, приурочено к определенному сезону.
Распространены в Экваториальной Африке от Уганды на запад до Гамбии, включая о-в Фернандо-По, известен в северо-восточной Анголе.
Симпсон (1945) включает молотоголового крылана в род эполетовых крыланов.
Род казиниктерис
Genus Casinycteris Thomas, 1910
В роде единственный вид: казиниктерис — С. argynnis Thomas, 1910.
Размеры небольшие. Длина тела примерно 9 см. Хвост снаружи не заметен. Длина предплечья около 6 см. Волосяной покров светло-бурый. Морда, веки, уши и крылья имеют желтоватую или ярко-оранжевую окраску. У основания ушей располагается по пучку белых волос. Позади глаз или между глазами есть белое пятно. Этот род сходен с родом скотониктерис, но отличается от него черепом с чрезвычайно укороченным костным небом и расширенным лицевым отделом. Зубная формула:
i — с — р— т— = 28.
2 13 2
Экология не изучена.
Распространен в тропической Африке от Камеруна до Республики Заир.
Род крыланов Анхиеты
Genus Pierotes Andersen, 1910
В роде единственный вид: крылан Анхиеты — Р. anchietaiSeabra, 1900.
Размеры небольшие. Длина тела примерно 9 см. Хвоста нет. Длина предплечья примерно 5 см. Размах крыльев около 34 см. Волосяной покров длинный и мягкий, на спинной стороне серовато-бурый, на брюшной значительно более светлый. У основания ушей спереди и сзади имеется по небольшому пучку белых волос. Характерно отсутствие шпоры — косточки, прикрепляющейся к пятке и поддерживающей межбедрепную перепонку. У самцов есть плечевые специфические кожные мешки, из которых выдаются пучки волос. Имеются щечные мешки. Зубы необычайно слабы. Коренные зубы имеют почти гладкую жевательную поверхность. Зубная формула: .21 3 1
1ТС 1 р з т з ~32‘
Экология не изучена. Дневное время, вероятно, проводит в густой листве деревьев. По структуре зубной системы можно предполагать, что этот крылан питается цветками или их нектаром.
Распространен в Замбии, Анголе и на юге Республики Заир.
Некоторые ученые (Симпсон, 1945), присоединяют крылана Анхиеты к роду эполетовых крыланов.
Род соломенноцветных, или пальмовых, крыланов
Genus Eidolon Rafinesque, 1815
Размеры средние. Длина тела 19—21 см. Длина хвоста от 0,5 до 2 см. Длина предплечья 11—13 см. Крылья длинные и узкие, приспособленные к дальним полетам. Размах крыльев может достигать 76 см. Второй палец передних конечностей несет коготь. Окраска волосяного покрова желтоватобурая или буроватая на спине, буровато-оливковая или буроватая на брюхе. В области шеи волосы сильнее удлинены и более мягкие, чем на туловище. Межбедренная перепонка в средней части покрыта волосами. Два или два с половиной хвостовых позвонка выступают из межбедренной перепонки. Голова с относительно заостренной мордой. Ноздри разделены глубокой бороздкой. Белые пятна у основания ушей отсутствуют. Череп с удлиненным лицевым отделом. Заглазничные отростки сильно развиты, особенно у ста-
2 13 2
рых особей. Число зубов: i — с — р — tn—= 34.
J 2 1*33
Обитатели лесов и саванн. Дневки обычно устраивают на пальмах, реже в пещерах. Держатся колониями от 20 до нескольких сотен особей; иногда собираются вместе до 10 тыс. особей. Ведут себя во время дневок шумно. Питаются в основном соком различных плодов, но могут питаться также цветками и, вероятно, молодыми проростками некоторых деревьев. Бывают активны и днем. В Камеруне и, вероятно, в Конго период спаривания приходится на сентябрь — ноябрь. Самки приносят детенышей около февраля. В помете, как правило, один детеныш.
Распространены в тропической Африке и Юго-Западной Азии.
В роде 3 вида:
аравийский крылан — Е. sabaeum Andersen, 1907 (юг Аравийского полуострова);
пальмовый крылан — Е. helvum Kerr, 1792 (встречается почти по всей Африке);
мадагаскарский крылан — Е. dupreanum Pollen, 1867 (Мадагаскар).
Род пещерных крыланов
Genus Eonycteris Dobson, 1873
Размеры небольшие. Длина тела примерно 10—12 см. Длина хвоста 1,2 — 3,0 см. Длина предплечья 6—8 см. Лицевая часть головы сильно уд-
линена и сужена. Очень подвижный язык достигает большой длины и покрыт
длинными, тонкими густо расположенными сосочками, обращенными вершиной назад. Второй палец передней конечности не имеет когтя. Зубная
.2 1 3 2 Q.
I — с — р — т— — 34.
2 1*3 3
формула:
Верхние резцы одинаковых разме
ров, простые. Нижние резцы по форме сходны с верхними, но направлены вперед и отделены друг от друга и от клыков промежутками. Клыки слабые. Окраска волосяного покрова на спинной стороне буроватая, обычно темнобурая, на брюшной более светлая. У самок бока шеи покрыты более редким мехом, в то время как у самцов эти места имеют боЛее грубые и длинные волосы, темнее по цвету, чем покрывающие голову и грудь. У Е. robustus имеются специфические кожные анальные железы бобовидной формы, рас
полагающиеся по обеим сторонам заднепроходного отверстия. Эти железы встречаются как у самцов, так и у самок.
Экология изучена слабо. Дневки обычно устраивают в пещерах, где собираются иногда по нескольку сотен особей. Питаются пыльцой и нектаром цветков.
Ареал охватывает Бирму, п-ов Индокитай, Таиланд, п-ов Малакку, о-ва Суматру, Яву, Калимантан, Сумбу, Сулавеси и Филлипинские.
В роде 5 видов:
пещерный крылан —S. spelaea Dobson, 1871 (Бирма, п-ов Индокитай, Таиланд, Малаккский полуостров, о-ва Суматра, Ява, Калимантан и Лусон из группы Филиппинских);
большой пещерный крылан — Е. major Andersen, 1910 (о-в Калимантан);
крылан Розенберга — Е. rosenbergi Jentink, 1899 (о-в Сулавеси);
лузонский крылан — Е. robusta Miller, 1913 (о-в Лусон);
длиннохвостый крылан — Е. longicauda Taylor, 1934 (о-в Лусон).
Некоторые ученые (Симпсон, 1945) выделяют,вид Е. rosenbergi в отдельный род Callynicteris Jentink, 1889.
Род длинноязыких крыланов
Genus Macroglossus F. Cuvier/1824
Размеры небольшие. Длина тела примерно 6—7 см. Хвост рудиментарен. Длина предплечья 3,7—4,9 см. На втором пальце передних конечностей хорошо развит коготь. Летательная перепонка крыльев прикрепляется низко на боках тела и к основанию четвертого пальца задних конечностей. На верхней губе глубокая вертикальная срединная борозда. Язык очень длинный, покрыт высокими густо расположенными сосочками, обращенными вершинами назад. Окраска волосяного покрова рыжевато-бурая на спинной, более светлая на брюшной стороне. В черепе лицевой отдел развит слабо, мозговой — гораздо сильнее. Заднеглазничные отростки короткие, слегка изогнутые. Нижняя челюсть очень слабая, с. длинным симфизом. Зубы отделены друг,от друга промежутками. Клыки крупные. Зубная формула: . 2 1 3 2
i — с — р — т — = 34.
2 13 3
Экология почти не изучена. День проводят на ветвях деревьев, под крышами построек, на листьях бананов. Держатся поодиночке. Питаются пыльцой и нектаром цветков.
Распространение охватывает Бирму, Таиланд, Малайзию, о-ва Суматру, Яву, Бали, Калимантан, Сулавеси, Филиппинские, Новую Гвинею, Западную Австралию, Квинсленд, архипелаг Бисмарка, о-ва Адмиралтейства, Соломоновы и некоторые прилежащие острова.
В роде 3 вида:
малый длинноязыкий крылан —-М. minimus Ё. Geoffroy, 1810 (Тенас-серим, Малаккский полуостров, о-ва Суматра, Ява, Бали и некоторые соседние мелкие);
малаккский длинноязыкий крылан — М. lagochilus Matschie, 1899 (Малаккский полуостров, о-ва Калимантан, Сулавеси, Натуна, Буру, Це-рам, Банда, Ару, Кей, Адмиралтейства, Новая Гвинея, архипелаг Бисмарка, Соломоновы и Филиппинские, Западная Австралия и северный Квинсленд);
плодоядный крылан — 714. fructivorous Taylor, 1934 (о-ва Котабато и Минданао из группы Филлипинских).
Рис. 96. Африканский длинноязыкий крылан— М. woermanni с вытянутым языком
Род цветочных длинноязыких крыланов
Genus Syconycteris Matschie, 1899
Размеры небольшие (наименьшие в семействе). Длина тела 5—6 см. Длина предплечья 4—5 см. Хвост снаружи почти незаметен. Волосяной покров на спинной стороне рыжевато-бурый или темно-бурый, на брюшной более светлый. Язык длинный, подвижный. Спинка его покрыта многочисленными высокими сосочками. Зубная формула: .2 1 3 1—2 ОА
i — с — р — т-------= 30 —
2 1 ' з 2 — 3
—34.
Экология не изучена. Питаются нектаром и пыльцой.
Обитают в Австралии, на Новой Гвинее, архипелаге Бисмарка и некоторых прилежащих островах.
В роде 3 вида:
папуасский крылан — S. crassa Thomas, 1895 (Новая Гвинея,о-ва Ару, архипелаг Бисмарка, о-в Кей, Амбоина, Церам);
тробрианский крылан—S. naias Andersen, 1911 (о-ва Тробриан);
австралийский крылан—S. australis Peters, 1867 (Квинсленд, Новый Южный Уэльс и юго-восток Новой Гвинеи).
Некоторые исследователи включают род Syconycteris в род Масго-glossus.
Род африканских длинноязыких крыланов
Genus Megatoglossus Pagenstecher, 1885
f
В роде единственный вид: африканский длинноязыкий крылан — М. woermanni Pagenstecher, 1885 (рис. 96).
Размеры мелкие. Длина тела 6,5—6,8 см. Хвоста нет. Длина предплечья 4—4,5 см. Вес примерно 16 г. Пятая метакарпальная кость короче третьей. Второй палец передней конечности вооружен когтем. Язык очень длинный и покрытый на спинке высокими многочисленными сосочками, направленными вершинами назад. Окраска волосяного покрова на спинной стороне туловища темно-бурая, на брюшной несколько более светлая. В задней части головы проходит небольшая срединная темная продольная полоска. На передней части боков шеи у самцов удлиненные волосы имеют темно-желтую или кремовую окраску. Зубная формула:
. 2 1 3 2 „.
i — с — р — т—- = 34.
2 13 3
Экология не изучена. День проводит в густой листве деревьев. Держится поодиночке.
Распространен в западной части Центральной Африки от Либерии на восток до Камеруна и на юг через Заир до Анголы.
Это единственный представитель длинноязыких крыланов, населяющих Африку.
Род темнобрюхих крыланов
Genus Melonycteris Dobson, 1877
Размеры небольшие. Длина тела около 10 см. Хвоста нет. Длина предплечья 5,7—6,3 см. Окраска волосяного покрова на спинной стороне зо-лотисто-светао-бурая, на брюшной темно-бурая, почти черная. Голова темно-бурая. Язык очень длинный, снабженный высокими многочисленными сосочками, обращенными вершинами назад. Зубная формула: . 2 1 3 2
i — с — р — т— == 34.
2 1.3 3
Экология не изучена. По-видимому, питается пыльцой и нектаром, а также соком плодов.
Распространен на Новой Гвинее, архипелаге Бисмарка, о-вах Новой Британии и Соломоновых.
В роде 2 вида:
темнобрюхий крылан —М. melanops Dobson, 1877 (Новая Гвинея, архипелаг Бисмарка, о-в Новая Британия);
флоридский темнобрюхий крылан— М. aurantius Phillips, 1966 (о-в Флорида из группы Соломоновых).
Род соломоновых длинноязыких крыланов
Genus Nesonycteris Thomas, 1887
В роде единственный вид: соломонов длинноязыкий крылан — N. wo-odfordi .Thomas, 1887.
Размеры мелкие. Длина тела примерно 10 см. Хвоста нет. Длина предплечья 5,2—5,4 см. Второй палец передней конечности не имеет когтя. Язык очень длинный, подвижный, снабжен многочисленными высокими сосочка-
ми, обращенными вершинами назад. Окраска волосяного покрова на спин-
ной стороне рыжевато-коричневая, на брюшной серовато-бурая. Голова
бледно-рыжевато-коричневая. Зубная формула:
.2 1 3 2
i — с — р —- т—=32.
1 13 з
Экология не изучена. Питается, по-видимому, нектаром и пыльцой и
соком плодов.
Распространен на Соломоновых островах.
Некоторые ученые (Андерсон и Джонс, 1967) присоединяют род Nesonycteris к роду Melonycteris.
Род длиннохвостых крыланов
Genus Notopterls Gray, 1859
В роде единственный вид: длиннохвостый крылан — N. macdonaldi Gray, 1859.
Размеры мелкие. Длина тела примерно 10 см. Хвост длинный, примерно 6 см. Длина предплечья 6—7 см. Это единственный представитель семейства крыланов с длинным хвостом, к тому же не включенным в летательную перепонку. На втором пальце передней конечности когтя нет. Летательная перепонка крыльев прикрепляется к спине по ее средней линии вдоль позвоночника. Язык очень длинный, способный далеко выдаваться вперед, покрыт многочисленными высокими сосочками, направленными вершинами назад. Окраска волосяного покрова на спинной части тела оливково-бурая или темно-бурая, на брюшной лишь немного светлее. На шее удлиненных
2 12 2
волос нет. Зубная формула: i — с — р —т — = 28.
Дневки устраивают в пещерах, иногда в дуплах деревьев. Собирается группами до нескольких сотен особей. Питается нектаром, пыльцой и соком плодов.
Распространен в Западной Полинезии (о-ва Новые Гебриды, Фиджи, Понапе из группы Каролинских и Новая Каледония).
Из всех представителей семейства крыланов в Полинезии встречаются только летучие лисицы и длиннохвостые крыланы.
Род трубконосых крыланов
Genus Nyctimene Borkhausen, 1797
Мелкие крыланы. Длина тела 7,5—12,5 см. Длина хвоста 1,7—2,5 см. Длина предплечья 5—8,6 см. Характерная особенность — вытянутые в виде трубок ноздри, направленные в стороны и достигающие в длину примерно 6 мм (рис. 97). Окраска волосяного покрова на спинной стороне темножелтовато-серая, у некоторых видов темно-желтая или кремовая; на брюшной обычно более светлая. Как правило, вдоль середины спины проходит темно-бурая полоса. Крылья и уши покрыты желтыми пятнами. Лицевой отдел черепа сильно укорочен и имеет почти одинаковую высоту по всей 113 1
своей длине. Зубная формула: i — с — р — tn— = 24. На верхних
клыках иногда имеется дополнительная вершина.
Экология изучена крайне слабо. В течение дня скрываются в листве деревьев. Держатся поодиночке. В полете издают высокий свистящий крик. Когда трубконосый крылан кричит, ноздри своеобразно выгибаются наружу и двигаются. Возможно, что они имеют определенное значение, производя ультразвуки. В неволе поедают мягкие фрукты.
Распространены от о-ва Сулавеси до о-вов Тимор, Новой Гвинеи, Квинсленда в Австралии и Соломоновых островов.
В роде 8 видов:
малый трубконосый крылан —N. minutus Andersen, 1910 (о-ва Сулаве-
си и Буру);
белобрюхий трубконосый крылан
N. albiventer Gray, 1863 (Новая Гвинея, о-ва Соломоновы, Адмиралтейства, Новая Британия, Хальмахер, Моротай, Кей, Бугенвиль);
большеголовый трубконосый крылан — N. cephalotes Pallas,
Рис. 97. Большой трубконосый крылан — Af. major и его череп
1767 (о-ва Сулавеси, Тимор, Амбоина, Буру, Церам, Новая Гвинея, Адмиралтейства);
большой трубконосый крылан — N. major Dobson, 1877 (архипелаг Бисмарка, о-ва Тробриан, Новая Гвинея, Соломоновы);
новогвинейский трубконосый крылан — N. cyclo tis Andersen, 1910 (Новая Гвинея);
трубконосый крылан Томаса —N. aello Thomas, 1900 (Новая Гвинея);
трубконосый крылан Робинсона — N. robinsoni Thomas, 1904 (северный Квинсленд в Австралии);
трионский трубконосый крылан—N. tryoni Longman, 1921 (восточный Квинсленд и Новый Южный Уэльс).
Род карликовых трубконосых крыланов
Genus Paranyctimene Tate, 1942
В роде единственный вид: карликовый трубконосый крылан — Р. raptor Tate, 1942.
Размеры небольшие. Длина тела около 8 см. Длина хвоста 1,3—2 см. Длина предплечья 4,7—5,5 см. Внешним обликом напоминает трубконосых крыланов, ноздри имеют такой же вид, однако окраска волосяного покрова на спинной стороне серовато-бурая, на брюшной темно-желтоватобурая. Продольной темной полосы вдоль спины нет. На крыльях такие же желтоватые пятна, как и у трубконосых крыланов. В отличие от последних у карликового трубконосого крылана на предкоренных и коренных зубах высокие вершины. Верхние и нижние клыки и предкоренные зубы очень высокие и тонкие. Костное небо вытянуто далеко назад. Как и у рода трубконосых крыланов, нижних резцов нет.
Экология не изучена.
Встречается в высокогорье Новой Гвинеи.
Род крыланов-гарпий
Genus Harpyionycteris Thomas, 1896
В роде единственный вид: крылан-гарпия — Н. whiteheadi Thomas, 1896 (иногда выделяют и второй вид — Н. celebensis Miller et Hollister, 1921).
Размеры небольшие. Длина тела 14—15 см. Хвоста нет. Длина предплечья примерно 9 см. Характерна очень короткая стопа и чрезвычайно узкая межбедренная перепонка, густо покрытая волосами. Окраска волосяного покрова шоколадно-бурая до темно-бурой на спине и несколько 113 2
более светлая на брюхе. Зубная формула: i’ — с — р ~т — = 30.
От всех других крыланов отличаются тем, что коренные зубы имеют от 5 до 6 бугров, нижние клыки имеют 3 вершины. Верхние резцы, верхние и нижние клыки сильнее наклонены вперед, чем соответствующие зубы других крыланов. Когда челюсти сжаты, клыки своими длинными осями направлены почти под прямым углом друг к другу.
Встречается в горах на высоте более 1000 м над уровнем моря. Экология неизвестна. День проводит, по-видимому, в пещерах. Крайне редок.
Обитает на о-вах Филиппинских и Сулавеси.
Подотряд летучих мышей
Subordo Microchiroptera Dobson, 1875
Размеры мелкие (наименьшие в отряде). Длина тела от 28 до 140 мм. Хвост может быть хорошо развит или полностью отсутствовать. Иногда хвост не включен в межбедренную перепонку. Ушная раковина незамкнута, так
что края уха не образуют сплошного кольца. В большинстве случаев есть козелок. Глаза очень маленькие. Второй палец передних конечностей целиком утратил самостоятельность, лишен концевой фаланги и когтя.
В черепе лицевой отдел укорочен. Посторбитальные отростки обычно отсутствуют или очень малы. Костное небо, как правило, не заходит дальше последних заднекоренных зубов. Обычно есть предчелюстные кости. Небные отростки предчелюстных костей развиты или их нет. Щечные зубы почти у всех представителей многобугорчатые. У плечевой кости хорошо развиты большой и малый бугры, причем большой бугор многих представителей подотряда образует дополнительный сустав с лопаткой. Кишечник относительно укороченный. Желудок простой, иногда с удлиненной кардиальной частью.
По характеру питания преимущественно насекомоядные, иногда хищные, рыбоядные, питающиеся кровью, плодоядные или нектароядные.
Распространены в тропических и умеренных областях обоих полушарий, на север и на юг до границ древесной растительности.
В подотряде 16 семейств (107 родов):
ланцетоносов, или мышехвостов, — Rhinopomatidae Miller, 1907(1 род);
мешкокрылов — Emballonuridae Dobson, 1875 (8 родов);
рыбоядных летучих мышей — Noctilionidae Gray, 1821 (1 род);
щелемордов — Nycteridae Dobson, 1875 (1 род);
ложных вампиров — Megadermatidae Gill, 1872 (3 рода);
подковоносов — Rhinolophidae Bell, 1836 (2 рода);
ложных подковоносов, или листоносов Старого Света,— Hipposide-ridae Miller, 1907 (8 родов);
американских^ листоносовPhylostomatidae Waterhouse, 1839 (44 рода);
вампиров — Desmodontidae Gill, 1866 (3 рода);
воронкоухов — Natalidae Miller, 1899 (1 род);
дымчатых летучих мышей — Furipteridae Miller, 1907 (2 рода);
американских присосконогов — Thyropteridae Miller, 1907 (1 род);
мадагаскарских присосконогов — Myzopodidae Thomas, 1904 (1 род); обыкновенных летучих мышей — Vespertilionidae Gray, 1821 (23 рода); новозеландских летучих мышей — Mystacinidae Simpson, 1945 (1 род); бульдоговых летучих мышей — Molossidae Gill, 1872 (7 родов).
Некоторые исследователи объединяют листоносов Старого Света вместе с подковоносами в одно семейство Rhinolophidae (Андерсон и Джонс, 1967). Другие (Бурлиер, 1955) считают, что вампиры должны быть включены в семейство американских листоносов.
В СССР 3 семейства: подковоносов, обыкновенных летучих мышей и бульдоговых летучих мышей (вся территория страны, на север до границы леса).
СЕМЕЙСТВО ЛАНЦЕТОНОСОВ, ИЛИ МЫШЕХВОСТОВ,
FAMILIA RHINOPOMATIDAE MILLER, 1907
В семействе единственный род: ланцетоносов, или мышехвостов,— Rhinopoma Ё. Geoffroy, 1818 (рис. 98).
Размеры мелкие и средние. Длина тела 60—80 мм. Длина хвоста около 60 мм. Длина предплечья 55—70 мм. Единственные представители подотряда летучих мышей, имеющие хвост, по длине примерно равный длине тела. В основании хвоста часто бывают отложения жира. Межбедренная перепонка очень узкая и большая часть хвоста свободно выступает из нее. Крупные уши соединены между собой кожистой складкой по верху головы. Морда со вздутыми боковыми отделами, образованными гребневидными кожными выростами. На конце морды небольшой округлый носовой листок. Ноздри открываются вперед. Второй палец передней конечности имеет две фаланги.
Рис. 98. Большой ланцетонос — R. micro-phyllum
Козелок хорошо развит. Окраска волосяного покрова серовато-бурая или темно-бурая на спинной стороне и слегка более светлая на брюшной. Морда, крестец и задняя часть брюха голые.
В черепе носовые кости расширены латерально так, что их ширина больше длины. Лобные кости небольшие и имеют впадину. Посторбитальные отростки отсутствуют. Предчелюстные кости весьма сходны с предчелюстными костями крыланов; они разделены друг с другом и не сливаются с окружающими костями. Носовая ветвь их хорошо развита, небная сильно укорочена. Костные слуховые барабаны крупные. Зубная формула: . 1 1 1 3 оо у-,
I — с — р — т — =28. Пле-2 12 3
чевая кость не имеет дополнительного сочленения с лопаткой. Седьмой шейный позвонок не сливается с первым грудным.
Плечевой и тазовый пояса и задние конечности нормальные. Имеется небольшая слепая кишка.
Обитатели пустынных и полупустынных областей. Днем держатся в пещерах, расщелинах скал и зданиях, собираются часто большими колониями. Большие мышехвосты населяют некоторые пирамиды в течение более 3000 лет. Мышехвосты часто подвешиваются не только за задние лапы головой вниз, но и за первые пальцы передних конечностей головой вверх. Питаются насекомыми. О размножении известно немного. Самки индийского мышехвоста рождают детенышей раз в год с апреля по июль.
Распространены от Рио-де-Оро в Африке на восток через всю Северную Африку и Южную Азию до Суматры.
В роде 4 вида:
большой ланцетонос, или большой мышехвост,— /?. microphyllutn Briinnich, 1782 (Аравийский полуостров, АРЕ. Израиль);
индийский мышехвост — 7?. kinneari Wroughton, 1912 (Индия);
малый ланцетонос, или малый мышехвост, — /?. hardwickei Gray, 1831 (Индия, Бирма, Таиланд, Аравийский полуостров, Иран, Израиль, АРЕ, Судан);
суматранский ланцетонос, или суматранский мышехвост, — /?. suma-trae Thomas, 1903 (о-в Суматра).
СЕМЕЙСТВО МЕШКОКРЫЛОВ
FAMILIA EMBALLONURIDAE DOBSON, 1875
Мелких и средних размеров летучие мыши. Длина тела от 37 до 100 мм. Длина хвоста от6до40лсм. Длина предплечья от 32 до 83 мм. Лицевая часть головы и губы гладкие. Носового листка нет. Уши часто соединяются между собой по верху головы. Козелок есть. Большая часть хвоста включена в межбедренную перепонку, однако кончик его свободно располагается над этой перепонкой. Таким образом, вытягивая задние конечности, мешкокры-
лы могут значительно увеличивать площадь межбедренной перепонки, а подтягиванием ног — уменьшать ее. По-видимому, это обусловливает свойственный мешкокрылам особо верткий полет с неожиданными и резкими по-
воротами.
Волосяной покров у большинства представителей семейства серый, бурый или черный, а у рода белых мешкокрылов имеет необычную для летучих мышей окраску — белую или белую с серым. Некоторые представители рода обыкновенных мешкокрылов имеют белые крылья. У некоторых видов на спине и крестце хорошо выраженные серые или белые линии. Большинство видов в области плеча на летательной перепонке имеет специфическую кожную железу в виде своеобразного железистого мешка, который открывается на верхней поверхности крыльев (см. рис. 99); у белых мешкокрылов эта железа располагается на вентральной поверхности. Мешок хорошо развит у самцов и выделяет красноватый секрет с сильным запахом. У представителей рода могильных мешкокрылов кожная специфическая железа расположена между половинками нижней челюсти.
Череп характеризуется хорошо развитыми посторбитальными отростками. Предчелюстные кости неполные, у них имеются только носовые отростки. Они никогда не срастаются друг с другом или с верхнечелюстными костями. Костное небо оканчивается на уровне последних коренных зубов или не-
много позади них.
Зубная формула изменчива: у мешкокрылов Старого 2 12 3
i 34» У американских и африканских
Света мешко-
.1 I 2 з 00
Крылов i — с — р — т — = 32, г 3 12 3
i — с — р — т — = 30. Плечевой 2 12 3
у могильных мешкокрылов и тазовый пояса нормальные.
Седьмой шейный позвонок свободный. Стопа нормальная. У плечевой кости нет дополнительного сочленения с лопаткой. Второй палец передней конечности имеет все метакарпальные кости, но нет фаланг. Третий палец имеет две фаланги.
Обитают в различных ландшафтах, тропических и субтропических областях (от дождевых лесов до пустынь). Дневными убежищами служат расщелины скал, пещеры, строения, развалины, дупла деревьев. Собираются колониями (мешкокрылы Старого Света), небольшими группами от 3 до 20 особей (хоботковые и обыкновенные мешкокрылы) или держатся поодиночке (белые мешкокрылы). Хоботковые мешкокрылы часто проводят день на вертикальной поверхности стволов деревьев недалеко от воды. Могильные
мешкокрылы днюют в расщелинах скал, дуплах деревьев или кронах кокосовых пальм поодиночке или парами. Обыкновенные и хоботковые мешкокрылы используют для ориентации ультразвуки, а также зрение. Многие представители семейства превосходные летуны. Питаются в основном насекомыми, иногда плодами. Самка приносит потомство один раз в год. В помете, как правило, один детеныш.
Распространены в обоих полушариях: в Америке от северной Мексики до южной Бразилии, в Старом Свете от Сирии на севере, до Южной Африки и Южной Австралии на юге, в Тихом океане на восток до Самоа. По-видимому, отсутствуют в Вест-Индии.
В семействе 8 родов (48 видов):
мешкокрылов Старого Света — Emballonura Temminck, 1838 (7 видов); африканских мешкокрылов — Coleura Peters, 1867 (3 вида);
хоботковых мешкокрылов — Rhynchonycteris Peters, 1867 (1 вид);
обыкновенных мешкокрылов—Saccopteryx liliger, 1811 (14 видов); могильных мешкокрылов — Taphozous Е. Geoffroy, 1818 (18 видов); белых мешкокрылов — Diclidurus Wied-Neuwied, 1820 (3 вида);
депаниктерисов — Depanycteris Thomas, 1920 (1 вид);
циттаропсов — Cyttarops Thomas, 1913 (1 вид).
’*ч®
Род мешкокрылов Старого Света
Genus Embatlonura Temminck, 1838
Размеры мелкие и средние. Длина тела примерно 38—62 мм. Длина хвоста 12—20 мм. Длина предплечья 32—50 мм. Окраска волосяного покрова обычно темно-бурая на спинной стороне и более светлая на брюшной. г 2 12 3
Зубная формула: I — с — р — т— = 34. Мешкокрылы Старого 3 12 3
Света — единственный род в семействе, имеющий два резца в верхней челюсти. Железистых мешков на крыльях нет.
День проводят в расщелинах скал и в пещерах. Питаются в основном
насекомыми, но также, по-видимому, и плодами.
Распространены на Мадагаскаре, Тенассериме, Таиланде, Малайзии, на о-вах Яве, Суматре, Калимантане, Сулавеси и на мелких прилежащих островах. Населяют также различные мелкие острова к северу и востоку от Австралии. На север идут до Филлиппинских островов, а на восток — до Соломоновых.
Род содержит 7 видов:
мадагаскарский мешкокрыл — Е. atrata Peters, 1874 (Мадагаскар);
футлярохвостый мешкокрыл — Е. monticola Temminck, 1838 (Южная Бирма, северный Таиланд, западная Малайзия, Ява, Суматра,Калимантан, Сулавеси, Филиппинские острова);
юго-восточный мешкокрыл — Е. alecto Eydoux et Gervais, 1836 (о-ва Сулавеси, Филиппинские);
черноватый мешкокрыл — Е. nigrescens Gray, 1843 (о-ва Амбоина, Буру, Новая Британия, Адмиралтейства, Соломоновы);
мешкокрыл Беккара — Е. beccarii Peters et Doria, 1880 (о-ва Новая Гвинея, Ару, Кей, Тробриан);
мешкохрыл Рафрайана —Е. raffrayana Dobson, 1879 (Новая Гвинея, Церам, Соломоновы острова);
новогвинейский мешкокрыл — Е. furax Thomas, 1911 (Новая Гвинея).
Род африканских мешкокрылов
Genus Coleura Peters, 1867
Размеры мелкие и средние. Длина тела 55—65 мм. Длина хвоста 12 — 20 мм. Длина предплечья 45—50 мм. Окраска волосяного покрова красновато-бурая или темно-бурая на спине и несколько более светлая на брюхе. По внешнему виду напоминают род мешкокрылов Старого Света, но отличаются от них тем, что в верхней челюсти всего по одному резцу в каждой половине. Кожных железистых мешков в плечевой области нет.
День проводят в пещерах, расщелинах скал, под крышами зданий. Питаются в основном насекомыми.
Распространение охватывает Восточную Африку и Сейшельские острова.
В роде 3 вида:
аденский мешкокрыл — С. gallarum Thomas, 1915 (Сомали, Судан, Республика Заир и на восток до Адена на Аравийском полуострове);
южный мешкокрыл — С. afra Peters, 1852 (Мозамбик, Ангола, Танзания, Кения, Сомали, Судан, Республика Заир, южная часть Аравийского полуострова);
сейшельский мешкокрыл — С. seychellensis Peters, 1868 (Сейшельские острова).
Рис. 99. Двулинейный мешкокрыл —S. Ы1 ineat а
Род хоботковых мешкокрылов
Genus Rhynchonycteris Peters, 1867
В роде единственный вид: хоботковый мешкокрыл — N. naso Wied-Neuwied, 1820.
Размеры маленькие. Длина тела 37—41 мм. Длина хвоста примерно 12 мм. Длина предплечья 35—41 мм. Голова с вытянутой мордочкой, которая заканчивается длинным конусообразным носом, выдающимся далеко вперед по сравнению с нижней челюстью. Бурые остевые волосы с беловатыми вершинками и шоколадно-бурые пуховые создают своеобразную желтовато-серую окраску туловища. В нижней части спины и по крестцу проходят две изогнутые полосы белого или серого цвета. Железистого мешка в области плеча нет. На предплечьях небольшие пучки волос. Череп очень мал, с удлиненным ростральным отделом. Лобная часть черепа очень низ-
1 1 1 2 3
кая. Зубная формула: i -у с — р — т~ = 32.
День проводит на ветвях деревьев или на вертикальных поверхностях камней,, иногда в свернутых листьях ложного банана. На дневках собирается группами от 10 до 40 особей. Питается мелкими насекомыми, за которыми, как правило, охотится низко над поверхностью воды.
Распространение охватывает тропическую Америку от южной Мексики до северного Перу и центральной Бразилии, включая о-в Тринидад.
Род обыкновенных мешкокрылов
Genus Saccopteryx liliger, 1811
Размеры небольшие или средние (рис. 99). Длина тела 37—62 мм. Длина хвоста 6—23 мм. Длина предплечья 35—54 мм. Уши могут быть разделены друг с другом или соединены между собой тонкой кожной перепонкой
по верху головы. Окраска волосяного покрова разнообразная: на спине может быть темно-бурой, красновато-бурой, серовато-бурой, темно-серой, а на брюхе обычно более светлой. По спине могут проходить две беловатые продольные полосы. В области плеч находятся довольно глубокие мешки со специфическими кожными железами. У белокрылого мешкокрыла крылья от предплечья с наружной стороны белые. Зубная формула:
. 1 1 2 ™ 3 QO
i — с — р — т— = 32.
3 12 3
День проводят в различных убежищах, пещерах, в расщелинах скал, в дуплах деревьев и т. п. Обычно собираются небольшими группами от 5 до 20, иногда до 50 особей. Питаются насекомыми. Самки двулинейного мешкокрыла рождают детенышей во время сезона дождей.
Распространение охватывает территорию от Мексики на юг до Колумбии, Перу и Бразилии.
В роде 14 видов:
двулинейный мешкокрыл—S. bilineata Temminck, 1838 (от южной Мексики, через Центральную Америку до Панамы включительно);
седой мешкокрыл—S. canescens Thomas, 1901 (северная Колумбия, штат Пара в Бразилии);
бразильский мешкокрыл—S. gymnura Thomas, 1901 (северо-восток Бразилии);
S. leptura Schreber, 1774 (северные районы Южной Америки);
венесуэльский мешкокрыл—S. perspicillifer Miller, 1899 (север Венесуэлы,-о-в Тринидад);
малый мешкокрыл—S. pumila Thomas, 1914 (Венесуэла и Гвиана); короткомордый мешкокрыл —S. brevirostris Wagner, 1843 (от Никара-' гуа на юг до Перу и Гвианы);
собаковидный мешкокрыл—S. macrotis Wagner, 1843 (юг Мексики, Центральная Америка, Бразилия);
мешкокрыл Капплера —S. kappleri Peters, 1867 (юг Мексики, Цент-] ральная Америка и Южная Америка от Эквадора и Перу на восток до Сан-? Паулу в Бразилии);
1 белокрылый мешкокрыл —S. leucoptera Peters, 1867 (Перу, Венесуэла, .< Гвиана, Бразилия);
; мешкокрыл Максимилиана —S. maximiliani Fischer, 1829 (юг Мексики, Центральная Америка и северо-восток Южной Америки);
s S. plicata Peters, 1867 (юг Мексики и Центральная Америка);
S. io Thomas, 1904 (юг Мексики и Гватемала);
i бурый мешкокрыл—S. infusca Thomas, 1897 (Эквадор).
Санборн (Sanborn, 1937), Холл и Келсон (1959), Андерсон и Джонс (1967) разделяют род Saccopteryx на следующие роды: Cormura Peters, 1867; Peropteryx Peters, 1867; Peronymus Peters, 1868; Centronycteris Gray, 1838; Balantiopteryx Peters, 1867. По-видимому, более правильно объединять их всех в один po&Saccopteryx, как это делают Симпсон (1945) и Бурлиер (1955).
Род могильных мешкокрылов Genus Taphozous Ё. Geoffroy, 1818
Размеры крупные, наибольшие в семействе. Длина тела до 100 мм. Длина хвоста 20—40 мм. Длина предплечья 50—83 мм. Окраска волосяного покрова на спинной стороне сероватая или различных оттенков бурая, иногда красновато-бурая, на брюшной кремовая, бледно-бурая или белая. Некоторые виды имеют беловатые пятна на теле. Железистые карманы в области плеч хорошо развиты, за исключением черномордого мешкокрыла, у которого вместо мешкообразных кожных впячиваний, имеются небольшие отверстия, открывающиеся в области плеч. Зубная формула: . 1 1 2 3 „„
I — с — р — т — = 30.
2; 1 2 3
День проводят в различных убежищах: в могильных склепах, расщелинах скал, пещерах, на деревьях. Собираются небольшими группами по 5—6 особей, иногда и больше. Впадают в спячку, перед которой накапливают большое количество жира. Являются превосходными летунами. Обычно начинают кормиться перед сумерками. Летают на высоте 60—90 м. Питаются летающими насекомыми. Некоторые виды приносят детенышей в основном с августа по октябрь, а другие, например мешкокрыл Жоффруа и длинноногий мешкокрыл, размножаются в течение всего года.
Распространение охватывает большую часть Африки, Мадагаскар и территорию к востоку через Южную Азию до Филиппинских и Соломоновых островов и Австралии.
В роде 18 видов:
южноафриканский мешкокрыл — Т. mauritianus Ё. Geoffroy, 1818 (ЮАР, Малави, Ангола, Кения,Танзания, Республика Заир, Мадагаскар);
мешкокрыл Гамильтона — Т. hamilton'i Thomas, 1920 (Судан);
мешкокрыл Гильдегарда — Т. hildegardeae, 1909 (Кения);
Т. perforates Ё. Geoffroy, 1818 (АРЕ и от Кении до Адена); сенегальский мешкокрыл — Т. senegalensis Desmarest, 1820 (Сенегал); суданский мешкокрыл — Т. Sudani Thomas, 1915 (Судан);
длинноногий мешкокрыл — Т. - longimanus Hardwicke, 1825 (Индия, Цейлон, Бирма, Тенассерим, Малайзия, Суматра, Ява, Калимантан);
чернобородый мешкокрыл — Т. melanopogon Temminck, 1841 (Малайзия, Ява, Суматра, Калимантан, Лаос, Тенассерим, Бирма, Индия, Юньнань);
мешкокрыл Теобальда — Т. theobaldi Dobson, 1872 (Тенассерим, Центральная Индия, п-ов Индокитай, Малайзия, Ява);
голобрюхий мешкокрыл — Т. nudiventris Cretzschmar, 1830 (Израиль, Аравийский полуостров, АРЕ, Судан);
Т. kachhensis Dobson, 1872 (Индия, Бирма, Малайзия, Ирак);
мешкокрыл Темминка — Т. saccolaimus Temminck, 1838 (Цейлон, Индия, Бирма, Малайзия, Суматра, Ява);
австралийский мешкокрыл —- Т. australis Gould, 1854 (п-ов Кейп-Йорк и северный Квинсленд в Австралии);
голобедрый мешкокрыл — Т. nudicluniatus De Vis, 1905 (Новая Гвинея, Соломоновы острова, северный Квинсленд);
новогвинейский могильный мешкокрыл — Т. mixtus Troughton, 1925 (Новая Гвинея);
филиппинский мешкокрыл — Т. philippinensis Waterhouse, 1845 (о-ва Лусон и Минданао в группе Филиппинских);
мешкокрыл Троугтона — Т. troughtoni Tate, 1952 (Квинсленд);
желтобрюхий мешкокрыл — Т. flaviventris Peters, 1867 (Новый Южный Уэльс, Южная Австралия).
Род белых мешкокрылов
Genus Diclidurus Wied- Neuwied, 1820
Размеры мелкие и средние. Длина тела 50—80 мм. Длина хвоста 15-25 мм. Длина предплечья 45—66 мм. Окраска волосяного покрова очень характерная: белая или белая с примесью серого. Крылья также белые. Железистого кармана на крыльях нет. Глаза относительно крупные. Уши короткие и закругленные. В том месте межбедренной перепонки, где выходит конец хвоста, находится небольшое кожное впячивание со специфическими железами. Короткий первый палец передней конечности имеет рудиментар-
112 3
ный коготь. Зубная формула: i — с — р — m— = 32.
Экология почти не изучена. Питаются, по-видимому, насекомыми.
Распространены в южной Мексике, Центральной Америке, Колумбии и Бразилии.
В роде 3 вида:
белый мешкокрыл — D. albus Wied, 1819 (Бразилия);
колумбийский мешкокрыл—D. ingens Hernandez-Camacho, 1955 (юго-восток Колумбии);
мексиканский мешкокрыл —D. virgo Thomas, 1903 (южная Мексика и Центральная Америка).
Род депаниктерис
Genus Depanycteris Thomas, 1920
В роде единственный вид: депаниктерис Изабеллы—D. Isabella Thomas, 1920.
Размеры средние. Длина тела примерно 60 мм. Длина хвоста 12 мм. Длина предплечья примерно 54 мм. Волосяной покров на спинной стороне имеет бледно-бурую ' окраску, на брюшной — желтовато-белую. Как и у белых мешкокрылов, есть специфическая кожная железа в межбедренной перепонке. Уши относительно короткие и округлые. Зубная формула: .1 1 2 3 оо
I — с — р — т — =32.
3 12 3
Экология не изучена.
Распространен в низовьях Амазонки в Бразилии.
Род циттаропс
Genus Cyttarops Thomas, (913
В роде единственный вид: циттаропс — С. alecto Thomas, 1913.
Размеры мелкие. Длина тела около 50 мм. Длина хвоста примерно 20 мм. Длина предплечья 46 мм. Окраска волосяного покрова одноцветная, дымчато-серая. Характерно строение козелка, нижняя половина наружного края которого содержит большое угольчатое образование, уникальное среди летучих мышей. Кожных специфических желез в области плеча или в области хвоста нет.
Экология не изучена.
Распространен в низовьях Амазонки в Бразилии и Гайану.
СЕМЕЙСТВО РЫБОЯДНЫХ ЛЕТУЧИХ МЫШЕЙ FAMILIA NOCTILIONIDAE GRAY, 1821
В семействе единственный род: рыбоядные летучие мыши — Noctilio Linnaeus, 1766 (рис. 100).
Среднего и крупного размера летучие мыши. Длина тела 65—132 мм. Длина предплечья 65—92 мм. Голова с вытянутой и заостренной мордой. Носового листка нет. На подбородке и передней части нижней губы имеются своеобразные складки кожи. Верхняя губа разделяется вертикальной складкой кожи, проходящей под ноздрями. Ноздри в виде коротких трубок открываются вперед и вниз. Крупные, заостренные уши не соединены друг сдругом. Козелок имеется. Хвост хорошо развит. Пяточный вырост крупный, достигает особенно большой величины у N. leporinus. У этого вида особенно длинные задние конечности, сильно развиты стопы, когти на пальцах задних конечностей крупные. По сравнению с другими представителями подотряда летучих мышей, крылья более узкие и длинные. Имеются своеобразные защечные мешки. Волосяной покров имеет изменчивую окраску: спина серовато-коричневая, желтоватая или ярко-рыжая, брюхо более светлое.
Рис. 100. Мексиканская рыбоядная летучая мышь — N. leporinus
Обычно узкая полоска желтовато-белого цвета проходит по середине спины от затылка почти до основания хвоста. По бокам от плеч до колен тянется полоса кожи, лишенная волос.
Череп не имеет посторбитальных отростков. У взрослых форм предчелюстные кости срастаются друг с другом, а также с челюстными костями; их носовые отростки необычайно длинны и хорошо развиты, а небные отростки коротки и едва заметны снизу. Костные слуховые барабаны неболь-2 113
шие. Зубная формула: I — с — р — т— = 28. Внутренний резец крупнее наружного на верхней челюсти. Имеется неясно выраженное дополнительное сочленение плечевой кости с лопаткой. Второй палец передней конечности с единственной рудиментарной фалангой. Третий и четвертый пальцы имеют по две фаланги каждый. Плечевой пояс нормальный. На грудине есть небольшой киль. Седьмой шейный позвонок не срастается с первым грудным. Малая берцовая кость имеет хрящевой верхний конец. В задней конечности очень сильно развита пяточная кость.
N. leporinus обычно находит убежище в щелях скал, пещерах, дуплах, иногда в зданиях недалеко от воды, a N. lab tails — в дуплах и зданиях. N. leporinus питается рыбой, a N. labial is — насекомыми. N. leporinus ловит рыбу в море и пресных водоемах. Летучие мыши летают низко над поверхностью воды и ловят плавающих у поверхности небольших рыб длиной 25—100 мм. Хватают их острыми длинными когтями задних лап и быстро подтягивают ко рту. Рыбу поедают на лету или относят в защечных мешках на место дневок, где рыба и съедается. Межбедренная перепонка для ловли рыбы не используется. В неволе одна рыбоядная летучая мышь съедала за ночь до 30—40 рыбок. Обычно кормятся по ночам, но иногда их можно видеть и в дневное время охотящимися вместе с пеликанами. Помимо рыб, объектами питания могут служить также и водные ракообразные. N. leporinus не способны эхолоцировать объекты под водой, но достаточно рыбке появиться над водой хотя бы на 0,5—1 мм, как летучая мышь хватает ее. Расстояние, с которого происходит обнаружение, а затем и распознавание объекта при помощи ультразвука, составляет в среднем 2 м. В помете один детеныш.
Распространение охватывает тропическую Америку на север до Кубы и Синалуа в Мексике, а на юг до северной Аргентины.
В роде 2 вида:
мексиканская рыбоядная летучая мышь — N. leporinus Linnaeus, 1758 (южная Мексика, Центральная Америка, Куба, Тринидад, Малые Антильские острова, Пуэрто-Рико, Колумбия, Венесуэла, Перу, Бразилия, Боливия, Парагвай, северо-восток Аргентины);
южная рыбоядная летучая мышь— Д'. labialis Kerr, 1792 (Центральная и Южная Америка от Никарагуа до севера Аргентины).
СЕМЕЙСТВО ЩЕЛЕМОРДОВ
FAMILIA NYCTERIDAE DOBSON, 1875
В семействе единственный род: щелеморды — Nycteris Ё. Geoffroy et Cuvier, 1795 (рис. 101).
Размеры мелкие и средние. Длина тела 45—75 мм. Длина хвоста 43— 75 мм. Длина предплечья 35—65 мм. Норда разделена продольной бороздой, от ноздрей назад по верху головы. Борозда ограничена и частично скрыта носовыми листками; позади она переходит в глубокую ямку на лбу. Нижняя губа имеет на вершине железистую поверхность. Длинные уши соединены между собой невысокой перепонкой. Есть небольшой козелок. Крылья широкие. Длинный хвост полностью включен в межбе-
Рис. 101. Голова египетского — N. thebaica и хвост яванского щелемордов — N. javanica
дренную перепонку и оканчиваются Т-образным кончиком, который служит поддержкой ее свободному краю. Этот своеобразный морфологический признак не встречается у других млекопитающих.
Волосяной покров длинный, довольно редкий, окраска его от темно- до светло-бурой и сероватой, иногда оранжевой.
В черепе посторбитальные отростки развиты слабо. Межорбитальная область черепа глубоко вогнута. Переднечелюстные кости имеют только неб-
2 113
ные отростки. Зубная формула: i — с — р — т — = 32. Бугры плечевой кости небольшие и не образуют дополнительного сочленения с лопаткой. Второй палец передней конечности содержит только метакарпальные кости и не имеет фаланг. Третий палец передней конечности с двумя фалангами. Седьмой шейный позвонок срастается с первым грудным. На грудине очень сильно развит киль. Малая берцовая кость имеется. Тазовый пояс нормальный, но необычайно укороченный и широкий.
Обитатели сухих степей с отдельными деревьями, встречаются также в дождевых лесах. День проводят в пещерах, зданиях, развалинах, норах дикобразов и трубкозубов. На дневках собираются группами до 20 особей, иногда держатся поодиночке. Голос напоминает металлический звук «тик» или «клинк». Щелеморды издают звуки высокой частоты и низкой интенсивности. Самка приносит одного детеныша, возможно 2 раза в год. Лактационный период 45—60 дней.
Ареал охватывает Африку, Аравийский полуостров, Корфу, Малак
кский полуостров, о-ва Калимантан и Яву, возможно также Сулавеси и Тимор.
В роде 21 вид:
яванский щелеморд — N. javanica Ё. Geoffroy, 1813 (Тенассерим, Малайзия, Ява, Калимантан, Тимор);
малый щелеморд — N. папа Andersen, 1912 (Ангола, Гвинея, Конго-Браззавиль, Кения);
щелеморд Бейтеса — N. arge Thomas, 1903 (Ангола, Кения, Республика ЗАИР, Камерун, Либерия);
мохнатый щелеморд — N. hispida Schreber, 1774 (Мозамбик, Ангола, Танзания, Кения и от Судана до Сенегала);
египетский щелеморд — N. thebaica Ё. Geoffroy, 1818 (о-в Корфу, Израиль, Аравийский полуостров, АРЕ, Судан, Кения, Ангола, ЮАР, южная Родезия, Малави, Юго-Западная Африка, Замбия, Мозамбик, Танзания, Эфиопия, Республика Заир).
большеухий щелеморд — N. macrotis Dobson, 1876 (Малави, Ангола, Танзания, Республика Заир и от Нигерии до Гамбии);
щелеморд Вууда — N. woodi Andersen, 1914 (южная Родезия и Замбия);
эфиопский щелеморд — N. aethiopica Dobson, 1878 (Судан);
золотистый щелеморд — N. aurita Andersen, 1912, (Сомали, Кения); авакубийскийщелеморд—N.avakubiai. Allen, 1917 (РеспубликаЗаир); гамбийский щелеморд — N. gambiensis Andersen, 1912 (Гамбия);
гигантский щелеморд — N. grandis Peters, 1865 (Гвинея);
желтый щелеморд — N. luteola Thomas, 1901 (Кения, Уганда, Занзибар);
мадагаскарский щелеморд —N. madagascariensis Grandidier, 1937 (Мадагаскар);
большой щелеморд — N. major Andersen, 1912 (от Республики Заир до Камеруна);
танзанийский щелеморд — N. marica Kershaw, 1923 (Танзания);
бледный щелеморд — N. pallida J. Allen, 1917 (от Республики Заир до Либерии);
сомалийский щелеморд — N. parisii de Beaux, 1924 (Сомали);
конголезский щелеморд — N. proximo Lonnberg et Gyldenstolpe, 1925 (Республика Заир);
N. revoilii Robin, 1881 (Сомали, Эфиопия);
щелеморд Винсона — N. vinsoni Dalquest, 1965 (Мозамбик).
Число видов в роде некоторые исследователи сокращают до 13 (Андерсон и Джонс, 1967).
СЕМЕЙСТВО ЛОЖНЫХ ВАМПИРОВ
FAMILIA MEGADERMATIDAE GILL, 1872
Средних и крупных (наибольших в подотряде) размеров летучие мыши. Длина тела 65—140 мм, длина предплечья 50—115 мм. Хвост отсутствует.
Представители рода афро-азиатских ложных вампиров—самые крупные среди подотряда летучих мышей. Морда несколько укороченная с хорошо развитым носовым листком удлиненной формы. Уши крупные, соединенные между собой по верху головы. Козелок довольно крупный, раздвоенный. Волосяной покров имеет окраску серую или бурую. У желтокрылых ложных вампиров крылья, уши и носовой листок желтовато-оранжевые. У самцов этого рода имеется на спине желтая специфическая кожная железа. Самки, по-видимому всех родов, имеют по 2 сосковидных выроста в паховой области, на которых зубами удерживаются детеныши.
В черепе посторбитальные отростки короткие или совсем отсутствуют. Предчелюстных костей нет.
О 1 1__2 3
Зубная формула: i — с — р—-—/п —=26— 28. Характерно от-
сутствие верхних резцов. Верхние клыки заметно вытянуты вперед.
Плечевая кость не имеет дополнительного сочленения с лопаткой. Второй палец передней конечности с одной фалангой, третий — с двумя. Плечевой пояс весьма специфичный; предгрудина спереди очень сильно расширена (ширина ее в 4 раза превышает длину) и срастается с первой парой ребер, с первым грудным позвонком и седьмым шейным позвонком в одно сплошное костное кольцо. Киль на грудине развит умеренно. Малая берцовая кость сильно редуцирована и занимает по своей длине лишь половину большой берцовой кости. Тазовый пояс нормальный.
День проводят в пещерах, расщелинах скал, строениях, дуплах, а некоторые виды— на ветвях деревьев и кустов. На дневки собираются группами до50—100 особей; некоторые видыдержатся поодиночке. Желтокрылый ложный вампир бывает активен и в дневное время, остальные виды только ночью. Летают, по-видимому, с закрытым ртом, производя ультразвуки через носовые отверстия. Питание разнообразно: желтокрылый ложный вампир питается комарами и мелкими мухами, афроазиатские ложные вампиры поедают насекомых, птиц, рыб, мышей, лягушек и других летучих мышей австралийский ложный вампир часто кормится различными летучими мышами. Азиатские ложные вампиры приносят потомство в мае; у желтокрылого ложного вампира, по-видимому, нет определенной периодичности в размножении, хотя большинство самок приносит потомство в апреле.
Распространение охватывает Центральную и Южную Африку, Южную и Юго-Восточную Азию, на восток до Филиппинских островов и Австралию.
В семействе 3 рода:
афроазиатских ложных вампиров —Megaderma Е. Geoffroy, 1810 (3 вида);
австралийских ложных вампиров —Macroderma Miller, 1906 (1 вид); желтокрылых ложных вампиров — Lavia Gray, 1838 (1 вид).
Род афроазиатских ложных вампиров Genus Megaderma Ё. Geoffroy, 1810
Размеры мелкие. Длина тела 65—85 мм. Длина предплечья 50—70 мм. Хвоста нет (рис. 102). Окраска волосяного покрова на спинной стороне дымчато-голубовато-серая или серовато-бурая, на брюшной — буровато-серая
или беловато-серая. Африканский ложный вампир имеет одноцветную голу-0 1 1__2 3
бовато-серую окраску. Зубная формула: i — с — р--------------т—— 26—28.
2 1 2 3
День проводят в пещерах, строениях, дуплах деревьев. На дневках собираются небольшими группами. Самцы и самки малайского ложного вампира в течение круглого года держатся вместе, а самки индийского ложного вампира перед рождением детенышей — отдельно. Период спаривания в Индии приходится на позднюю осень, обычно на ноябрь. Длительность беременности примерно 5 месяцев. Молодые рождаются перед началом дождливого сезона. Половозрелость у индийского ложного вампира наступает в 15 месяцев у самцов ив 19 — у самок. Индийский ложный вампир питается насекомыми, пауками, мелкими позвоночными (летучие мыши, грызуны, птицы, лягушки и рыбы). Когда он поедает летучих мышей, то прежде всего
высасывает кровь, а потом уже ест мясо.
Ареал охватывает восточную часть Центральной Африки от Эфиопии до Танзании, Индию, Цейлон, Бирму, Южный Китай, Малайзию, Суматру, Яву, Калимантан, Сулавеси и Филиппинские острова.
Рис. 102. Африканский ложный вампир — Л!, сог и череп малайского ложного вампира — М. spasma
В роде 3 вида:
малайский ложный вампир — М spasma Linnaeus, 1758 (Цейлон, Индия, Бирма, Тенассерим, Камбоджа, Таиланд, Малайзия, Суматра, Ява, Калимантан, Сулавеси, Филиппинские острова);
индийский ложный вампир — М. lyra Ё. Geoffroy, 1810 (Южный Китай, Индия, Бирма, Малайзия);
африканский ложный вампир — М. cor Peters, 1872 (от Эфиопии до Танзании).
Некоторые ученые выделяют африканского ложного вампира в особый род Cardioderma Peters, 1873.
Род австралийских ложных вампиров
Genus Macroderma Miller, 1906
В роде единственный вид: австралийский ложный вампир — М. gigas Dobson, 1880.
Размеры крупные (наибольшие среди подотряда летучих мышей). Длина тела 115 140 мм. Длина предплечья 105—115 мм. Снаружи хвоста не видно. Окраска волос на спине обычно белая, с серовато-бурыми вершинками, которые создают пепелы-ю-ссрый оттенок. Голова, уши, носовой листок, летательная перепонка и нижняя часть тела белые. Зубная формула: 0 1 13
i — с — р — т— — 2о.
2 1 к 2 3
День проводит в пещерах или в расщелинах скал. Обычно держится поодиночке или небольшими группами. Питается в основном мелкими летучими мышами, а также и другими мелкими млекопитающими, птицами, лягушками и насекомыми.
Обитатель тропических областей.
Распространен в Северной и Западной Австралии. Прежде населял и Южную Австралию, но в настоящее время там не встречаются в связи с резким уменьшением здесь в последние годы численности мелких насекомо
ядных летучих мышей, которыми в основном питается австралийский ложный вампир. А численность насекомоядных летучих мышей упала в свою очередь в связи с уменьшением числа насекомых, с возрастанием сухости климата.
Род желтокрылых ложных вампиров
Genus Lavia Gray, 1838
В роде единственный вид: желтокрылый ложный вампир — L. frons Ё. Geoffroy, 1810.
Размеры мелкие. Длина тела примерно до 80 мм. Длина предплечья 53—63 мм. Хвоста нет. Окраска волосяного покрова на спинной стороне серая с оттенками голубоватым, перламутровым и др.; на брюшной буроватая или оливково-зеленоватая, иногда желтая. Уши и крылья красновато-желтые. Зубная формула: i с -j- р т~~ — 26.
Обитатель лесов и открытых равнин. Часто встречается около болот, озер и рек. День проводит на ветвях деревьев и кустов, иногда в дуплах деревьев и в зданиях. Держится поодиночке или небольшими группами по 2—5 особей. Бывает активным в течение дня, не боится солнечного света. В отличие от других представителей семейства мелкими позвоночными не питается; основной пищей служат насекомые. Охотится весьма своеобразно: часто висит вниз головой на ветви, поджидая насекомого, затем слетает, хватает его и возвращается на то же место. По-видимому, определенной сезонности в размножении не имеет, хотя большинство самок приносит детенышей в апреле.
Распространен в Африке от Танзании, Кении, Уганды и Судана на запад до Сенегала. Встречается также в Замбии.
СЕМЕЙСТВО ПОДКОВОНОСОВ
FAMILIA RHINOLOPHIDAE BELL, 1836
Размеры от мелких до крупных. Длина тела от 35 до 110 мм. Длина предплечья от 30 до 75 мм. Передняя часть морды сверху имеет своеобразные кожные выросты листовидной формы, не покрытые волосами (рис. 103). Обычно среди этих кожных выростов различают «подкову», «ланцет» и «седло». «Подкова» — широкая кожная пластинка, огибающая ноздри с боков и спереди .«Ланцет» служит продолжением подковы, с заостренной вершиной, отогнутой кверху; на свободных краях «ланцета» имеется три пары неглубоких ячеек. «Седло» — небольшой гребень, расположенный между^подковой
Рис. 103. Голова и череп большого подковоноса — R. ferru.mequ.inum
и ланцетом. Ушные раковины относительно крупные, остроконечные, у некоторых видов соединяющиеся между собой кожной складкой по верху головы. Козелка нет. Крылья очень широкие. Тонкий хвост включен полностью в межбедренную перепонку .У самок в паховых областях есть кожные сосковидные выросты, они служат для прикрепления детеныша, который крепко держится за них зубами в первые дни жизни.
Череп не имеет посторбитальных отростков. Предчелюстные кости представлены только небными неокостеневающими отделами. Сильно выражены сагиттальный и ламбдоидальные гребни на черепе. Костные слуховые барабаны маленькие. Костное небо глубоко вырезано спереди и сзади; длина его меньше ширины. Зубная формула: i' — с — р ——- т — = 28 — 32. Плечевая кость не образует дополнительного сочленения с лопаткой. Второй палец передней конечности не имеет фаланг. Третий палец с двумя фалангами. Малая берцовая кость полная, но очень тонкая, нитевидная. На груди киля нет. Тазовый пояс необычный: лобковая и седалищная кости редуцированы и, таким образом, дорзальные профили безымянных костей проходят почти параллельно между собой. Все пальцы задней конечности с тремя фалангами, кроме первого пальца.Седьмой шейный и первый грудной позвонки, первое ребро, рукоятка грудины, предгрудина и, по крайней мере, вентральная половина второго ребра срастаются между собой в одно сплошное, расширенное спереди костное кольцо.
День проводят в пещерах, расщелинах скал, дуплах деревьев, под крышами домов. Некоторые виды мигрируют. Питаются насекомыми. Летают с закрытым ртом и производят через ноздри ультразвуки, при помощи которых они ориентируются и находят себе добычу. Носовые листки при этом, по-видимому, служат для придания звукам узкой направленности. Некоторые виды впадают в спячку. В помете 1—2 детеныша. Половозрелость наступает примерно с 15 месяцев. Продолжительность жизни в неволе до 15 лет.
Распространение охватывает тропические и умеренную части Восточного полушария на восток до Австралии, Новой Гвинеи, Филиппинских островов и Японии, на север до Англии, балтийского побрежья, Крыма, Кавказа и южных частей Средней Азии включительно.
В семействе 2 рода:
подковоносов — Rhinolophus Lacepede, 1799 (68 видов);
большеухих подковоносов — Rhinomegalophus Bourret, 1951 (1 вид).
Некоторые исследователи (Андерсон и Джонс, 1967) объединяют семейство листоносов Старого Света Hipposideridae и подковоносов в одно семейство Rhinolophidae. По-видимому, более правильно считать их отдельными семействами.
Род подковоносов
Genus Rhinolop has Lacepede, 1799
От мелких до крупных размеров подковоносы с длиной тела 35— 110 мм, длиной хвоста 24—43 мм и длиной предплечья 30—75 мм. Окраска меха довольно сильно варьирует от красновато-бурой до черной сверху и более светлой снизу. Волосы обыкновенно двуцветные: основания волос на спине всегда светлее вершин. Имеется сезонная изменчивость в окраске меха: в зимнее время мех более темный. У большого и южного подковоносов по 58 диплоидных хромосом, а у малого — 54.
День проводят в пещерах, в зданиях, в дуплах деревьев. Виды, населяющие умеренные зоны, на зиму впадают в спячку. Некоторые подковоносы совершают сезонные перекочевки и перелеты. Ряд видов держится поодиночке, другие — группами. Питаются главным образом насекомыми (преимущественно бабочками и двукрылыми, летающими поздно вечером и ночью), а также пауками. Часто возвращаются на места дневок, чтобы
съесть свою добычу. Могут ловить насекомых и на земле. У видов, впадающих в спячку, гон наблюдается осенью, а у невпадающих в спячку — весной. Оплодотворение яйцеклетки у тех и других происходит в весенний период. Самка приносит одного или двух детенышей с апреля по июль. Полово-зрелость, по-видимому, наступает в 2—3-летнем возрасте. Некоторые совершают сезонные кочевки и перелеты.
Распространение совпадает с ареалом семейства.
В роде 68 видов:
большой подковонос — /?. ferrumequinum Schreber, 1774 (Англия, Франция, Испания, Португалия, Италия, Швейцария, Голландия, ФРГ, ГДР, Венгрия, Греция, Корсика, Сардиния, юг европейской части СССР, Средняя Азия, Малая Азия, Ирак, Сирия, Израиль, Китай, Корейский полуостров, Япония, Кашмир, Кумаон, Непал, Сикким, Алжир, Марокко);
подковонос Жоффруа — 7?. clivosus Cretzschmar, 1828 (аравийское побережье Красного моря и африканское побережье Аденского залива, ЮАР, южная Родезия, Замбия, Малави, Юго-Западная Африка, Ангола, Танзания, о-ва Пемба и Занзибар, Кения, Республика Заир, Эфиопия, Алжир, АРЕ);
аравийский подковонос — 7?. acrotis Heuglin, 1861 (АРЕ, южная часть Аравийского полуострова, Эфиопия); ’
азиатский подковонос — 7?. affinis Horsfield, 1823 (Южный Китай, Северная Индия, Бирма, п-ов Индокитай, Малайзия, о-ва Суматра, Ява, Натуна и Анамба);
подковонос Ру — 7?. rouxi Temminck, 1835 (Цейлон, Индия, Южный Китай);
подковонос Томаса — 7?. thomasi Andersen, 1905 (Бирма, Юньнань, Бакбо);
малайский подковонос — 7?. malayanus Bonhote, 1903 (Таиланд, Бакбо);
малый подковонос — 7?. hipposideros Bechstein, 1800 (Англия, Ирландия, Франция, Испания, Португалия, Швейцария, Апеннинский полуостров, Сардиния, Корсика, Мальта, ФРГ, ГДР, Польша, Венгрия, юг европейской части СССР, Средняя Азия, Малая Азия, Иран, Кипр, Аравийский полуостров, Кашмир, Марокко, Судан, Эфиопия);
японский подковонос — R. cornutus Temminck, 1835 (Япония, Южный Китай, п-ов Индокитай, Таиланд, Индия, Бирма);
индийский подковонос — 7?. lepidus Bly th, 1844 (Сычуань, Юньнань в Китае, центральные районы, долина Ганга, Кумаон и Бенгалия в Индии, Бирма);
каштановый подковонос — 7?. subbadius Bly th, 1844 (Индия, п-ов Индокитай);
однорогий подковонос — 7?. monoceros Andersen, 1905 (Тайвань);
южный подковонос — 7?. euryale Blasius 1853 (Португалия, Испания, Франция, Апеннинский полуостров, Сардиния, Австрия, Югославия, Греция, Кавказ, Туркмения, Сирия, Израиль, Марокко, Алжир, АРЕ);
подковонос Мегели — 7?. mehelyi Matschie, 1901 (Испания, Южная Франция, Сардиния, Румыния, Закавказье);
средиземноморский подковонос — 7?. blasii Peters, 1866 (Греция, Кипр, Италия, Израиль, Закавказье, Туркмения, Малая Азия, Северная Африка, ЮАР, южная Родезия, Малави, Эфиопия);
бухарский подковонос — R. bocharicus Kastschenko et Akimow, 1917 (Средняя Азия, Афганистан);
трехлистный подковонос—R. trifoliatus Temminck, 1834 (Таиланд, Малайзия, Суматра, Ява, Калимантан и соседние мелкие острова);
большой восточный подковонос — R. luctus Temminck, 1835 (Бирма, Индия, Цейлон, Южный Китай, Малайзия, Суматра, Ява, Калимантан);
подковонос Пирсона — R. pearsoni Horsfield, 1851 (Северная Индия, Южный Китай, Бакбо); •
длинноухий подковонос — macrotis Bly th, 1844 (Сычуань и Фуцзянь
в Китае, Кумаон, Непал, п-ов Индокитай, Суматра, Филиппинские острова);
бирманский подковонос — R. coelophyllus Peters, 1867 (Бирма, Таиланд, Малайзия);
королевский подковонос — R. rex G. Allen, 1923 (Сычуань и Гуйчжоу в Китае);
подковонос Дарлинга — R. darlingi Andersen, 1905 (ЮАР, южная Родезия, Юго-Западная Африка, Малави, Ангола, Танзания);
южноафриканский подковонос — R. capensis Lichtenstein, 1823 (ЮАР, Малави);
подковонос Дента — R. denti Thomas, 1904 (ЮАР, Юго-Западная Африка, Ангола);
подковонос Свинни — R. swinnyi Gough, 1908 (ЮАР, южная Родезия);
кустарниковый подковонос — R. simulator Andersen, 1904 (ЮАР, южная Родезия, Мозамбик, Замбия, Малави);
подковонос Ландера — R. landeri Martin, 1838 (ЮАР, Мозамбик, Замбия, Малави, Республика Заир, Танзания, Кения, о-ва Занзибар, Фернандо-По);
ангольский подковонос — R. angolensis Seabra, 1898 (Ангола);
дамарский подковонос —R. fumigatus Rdppell, 1842 (ЮАР, Юго-Западная Африка, Ангола, Замбия, Танзания, о-в Пемба, Кения, Уганда, Сомали, Эфиопия);
подковонос Гильдебрандта — R. hildebrandti Peters, 1878 (ЮАР, Мозамбик, южная Родезия, Малави, Замбия, Республика Заир, Танзания, Кения, Сомали);
абасский подковонос — R. abae J. Alien, 1917 (Руспублика Заир);
ганский подковонос — R. alcyone Temminck, 1852 (Гана, Камерун);
подковонос Андрейна — R. andreinii Senna, 1905 (Эфиопия);
конголезский подковонос — R. axillaris J. Allen, 1917 (Республика Заир); ;
подковонос Брокмана — R. brockmani Thomas, 1910 (Сомали);
подковонос Декена — R. deckenii Peters, 1867 (о-в Занзибар);
подковонос Добсона — R. dobsoni Thomas, 1904 (Судан);
лисий подковонос — R. foxi Thomas, 1913 (Нигерия);
подковонос Маклауда — R. maclaudi Pousargues, 1897 (Гвинея);
угандский подковонос — R. eloquens Andersen, 1905 (Уганда);
борнейский подковонос — R. borneensis Peters, 1861 (о-ва Калимантан, Натуна, Лабуан);
R. importunus Chasen, 1939 (Ява);
яванский подковонос — R. javanicus Andersen, 1918 (Ява);
мадурский подковонос — R. madurensis Andersen, 1918 (о-в Мадура);
малайзийский подковонос — R. stheno Andersen, 1905 (Малаккский полуостров, о-ва Суматра, Ява);
сиантанский подковонос —R. nereis Andersen, 1905 (о-ва Натуна, Анам-ба, Сиантан, Бунгуран);
подковонос Робинсона — R. robinsoni Andersen, 1918 (Малаккский полуостров);
подковонос Клосса — R. classi Andersen, 1918 (восточное побережье Малаккского полуострова и мелкие прилежащие острова).
блестящий подковонос — R. refulgens Temminck, 1834 (Малаккский полуостров);
карликовый подковонос — R. pusillus Temminck, 1834 (о-ва Ява, Сиантан, Анамба);
R. acurdinatus Peters, 1871 (о-ва Ява, Бали, Суматра, Калимантан, Ниас);
филиппинский подковонос — R. philippinensis Waterhouse, 1843 (о-ва Лусон, Минданао, Калимантан, п-ов Кейп-Йорк в Австралии);
калимантанский подковонос — /?. sedulus Andersen, 1905 (Малайзия, о-в Калимантан);
лузонский подковонос — R. arcuatus Peters, 1871 (о-ва Лусон, Минданао, Миндоро, Суматра);
подковонос Крега — R. creaghi Thomas, 1896 (о-ва Калимантан, Мадура, Ява);
ломбокский подковонос — R. simplex Andersen, 1905 (о-ва Ломбок, Сумбава);
австралийский подковонос — R. megaphyllus Gray, 1834 (Новый Южный Уэльс, Квинсленд, Новая Гвинея);
кейский подковонос — R. keyensis Peters, 1871 (о-ва Кей, Хальмахера, Горам);
Целебесский подковонос — R. celebensis Andersen, 1905 (Сулавеси);
бурусский подковонос — R. toxopeusi Hinton, 1925 (о-в Буру);
восточный подковонос — R. euryotis Temminck, 1835 (о-ва Амбоина, Церам, Тимор, Халмахера, Кей, Ару, Буру, Новая Гвинея);
мохнатый подковонос — R. nirsutus Andersen, 1905 (о-в Гуимар в группе Филиппинских островов);
минданаоский подковонос —R. virgo Andersen, 1905 (о-ва Лусон, Минданао);
подковонос Андерсена — R. anderseni Cabrera, 1909 (о-ва Лусон, Палаван);
рыжеватый подковонос — R. subrufus Andersen, 1905 (о-в Лусон);
слабый подковонос — R. inops Andersen, 1905 (о-в Минданао);
подковонос Бункера — R. bunkeri Taylor, 1934 (о-ва Минданао, Котабато).
В СССР: большой подковонос (Закарпатская Украина, Крым, Кавказ, среднеазиатские республики), малый подковонос (Закарпатская Украина, Молдавия, южная Украина, Крым, Кавказ, Закавказье, южная Туркмения, Узбекистан, Таджикистан), южный подковонос (черноморское побережье Кавказа, Нагорный Карабах, юго-западная Туркмения), подковонос Мегели (Грузия, Армения, Нагорный Карабах), средиземноморский подковонос (южное Закавказье, юго-западная Туркмения, бухарский подковонос (юго-восточная Туркмения, Узбекистан, западный Таджикистан).
Род большеухих подковоносов
Genus Rhinomegalophus Bourret, 1951
В роде единственный вид: большеухий подковонос — R. paradoxolophus Bourret, 1951.
Средних размеров подковонос. Длина предплечья примерно 55 мм. Длина хвоста 24 мм. Очень крупные уши достигают длины 30 мм. Окраска волосяного покрова на спинной стороне черновато-бурая, на брюшной несколько более светлая. Структура носовых листков отличается от таковых рода подковоносов тем, что имеет сложноустроенное седло и большую придаточную лопасть.
Экология не изучена.
Представители обнаружены в пещере провинции Бакбо во Вьетнаме. Пещера расположена на уровне 1700 м над уровнем моря.
СЕМЕЙСТВО ложных ПОДКОВОНОСОВ, ИЛИ ЛИСТОНОСОВ СТАРОГО СВЕТА,
FAMILIA HIPPOSIDERIDAE MILLER, 1907
Различных размеров летучие мыши от мелких (наименьших в подотряде) до крупных. Длина тела 28—110 мм. Длина хвоста до 60 мм (у некоторых представителей нет хвоста). Длина предплечья 30—110 мм. Гигантский лис-
Рис. 104. Головы: А — персидского трилистоноса — Т. persicus; Б — золотистого листоноса —К. aurantius; В—бесхвостого листоноса — С. frithi; Г—череп обыкновенного листоноса — Hipposideros sp.
тонос с предплечьем длиной 110 мм является одним из наиболее крупных представителей подотряда летучих мышей. Если хвост имеется, он целиком включен в межбедренную перепонку. На конце морды находятся листовидные выросты кожи (рис. 104), состоящие из передней седлообразной части, иногда с небольшим дополнительным листком. Перпендикулярно к нему расположен поперечный листок, обычно разделяющийся на три отростка с заостренными вершинами. Уши короткие или длинные. У некоторых представителей семейства уши соединяются между собой по верху головы. Козелка нет. Характерно, что пальцы задних конечностей имеют только по две фаланги. У некоторых видов выражен половой диморфизм. Самки и самцы отличаются по размерам тела, носовых листков, окраской волосяного покрова.
Череп характеризуется сильным развитием лямбдоидального и сагнт-| ] J____2 з
тального гребней. Зубная формула: i’ — с — р —-—= — 30-
Седьмой шейный, первый и второй грудные позвонки, две первые пары ребер и предгрудина срастаются между собой в единое костное кольцо.
День проводят в пещерах, постройках и дуплах деревьев поодиночке, небольшими группами или довольно крупными скоплениями в несколько сотен особей. Некоторые виды впадают в спячку. Питаются насекомыми. Ориентируются при помощи ультразвуков, которые производят с закрытым ртом через ноздри.
Ареал охватывает субтропические и тропические области Африки и Южной Азии, простираясь на восток до Филиппинских островов, Соломоновых и Австралии.
В семействе 8 родов (59 видов):
обыкновенных листоносов — Hlpposideros Gray, 1831 (44 вида);
трезубценосов — Asellia Gray, 1838 (2 вида);
трезубценосов Тейта — Aselliscus Tate, 1941 (2 вида);
цоелопс Coelops Blyth, 1848 (3 вида);
африканских трезубценосов — Cloeotis Thomas, 1901 (1 вид);
воронкоухих листоносов — Paracoelops Dorst, 1947 (1 вид);
золотистых листоносов — Rhinonicteris Gray, 1847 (1 вид);
трилистоносов — Triaenops Dobson, 1871 (5 видов).
Некоторые исследователи включают семейство Hipposideridae в семейство Rhinolophidae.
Род обыкновенных листоносов
Genus Hipposideros Gray, 1831
Размеры от мелких до крупных (наибольших в семействе). Длина тела 35—110 мм. Длина хвоста 18—60 мм. Длина предплечья примерно 33 — 110 мм. Окраска волосяного покрова на спинной стороне от рыжеватой до бурой. Брюшная сторона светлее спинной. У многих видов позади носовых листков в значительном углублении расположена специфическая кожная железа, продуцирующая воскообразный секрет. У самцов эта железа более крупная. В черепе характерна расширенная посторбитальная часть. Сильно развиты скуловые дуги. Ростральный отдел черепа относительно 112 3
удлинен. Зубная формула: i — с — р — т~ = 30.
День проводят обычно в дуплах деревьев, пещерах, строениях человека, иногда в крупных норах дикобразов. Некоторые виды держатся поодиночке, другие мелкими группами, третьи собираются значительными скоплениями. Некоторые виды впадают в спячку. В поисках добычи летают довольно низко над землей. Питаются жуками, термитами и другими насекомыми. Цикады часто имеют важное значение в питании отдельных видов. И. armiger, по-видимому, разыскивает насекомых по издаваемым ими звукам. Н. commersoni в Африке поедает личинок жуков, которые находятся в диких плодах фиговых пальм. На Цейлоне большинство самок обыкновенных листоносов приносит детенышей в апреле и мае, иногда августе. На Малаккском полуострове массовое рождение детенышей приходится на ноябрь и декабрь, а в Замбии — на октябрь и ноябрь.
Распространены в Африке, на Мадагаскаре, большей части Азии и далее на восток до Филиппинских и Соломоновых островов, Австралии и о-вов Новые Гебриды. В роде 46 видов:
южноафриканский листонос — Н. cafferSundeva 11,1846 (ЮАР, Мозамбик, южная Родезия, Юго-Западная Африка, Ангола, Замбия, Малави, далее ареал простирается на север до Танзании, Уганды, Кении, Эфиопии, Республики Заир, Габона, Марокко и юго-западной части Аравии);
листонос Коммерсона — Н. commersoni Е. Geoffroy, 1813 (Юго-Западная Африка, Замбия, Малави, Мадагаскар, Мозамбик, и на север до Танзании, о-вов Занзибар и Пемба, Кении и Республики Заир);
абасский листонос — Н. и. abac J. Allen, 1917 (Республика Заир); камерунский листонос—Н. beatus Andersen, 1906 (Камерун, Либерия); ганский листонос — Н. cyclops Temminck, 1853 (Гана);
темный листонос — Н. fuliginosus Temminck, 1853 (Гвинея);
гигантский листонос — Н. gigas Wagner, 1845 (Ангола, Гамбия, Республика Заир);
эфиопский листонос — Н. megalotis Heuglin, 1861 (Эфиопия);
обыкновенный карликовый листонос — Н. nanus J. Allen, 1917 (Республика Заир);
красный листонос — Н. ruber Noak, 1893 (Танзания);
листонос Сандерсона — Н. sandersoni Sanderson, 1937 (Нигерия);
индийский листонос — Н. speoris Schneider, 1800 (Индия, Цейлон, Ява, Калимантан, Тимор);
И. larvatus Horsfield, 1823 (Хайнань, Банглодеш, Бирма, п-ов Индокитай, Малайзия, Суматра, Ява, Калимантан).
большой листонос — Н. diadema Ё. Geoffroy, 1813 (Бирма, п-ов Индокитай, Малайзия, о-ва Ява, Суматра, Калимантан, Сулавеси, Филиппинские, Новая Гвинея, Квинсленд, Соломоновы острова);
цейлонский листонос — Н. lankadiva Kelaart, 1850 (Цейлон и Индия);
двухцветный листонос — Н. bicolor Temminck, 1834 (Никобарские острова, Таиланд, острова Суматра, Ява, Цейлон, Индия, Бирма, Тенассе-рим, о-ва Тайвань, Новая Гвинея, Сулавеси, Кей, Церам, Буру, северный Квинсленд);
листонос Семона — Н. semoni Matschie, 1903 (северный Квинсленд);
далекий листонос — Н. stenotis Thomas, 1913 (Северная Австралия);
Н. ротопа Andersen, 1918 (Индия, Бирма, Южный Китай, Таиланд, Малайзия, острова к западу от Суматры, Ява, Банка);
пепельный листонос — И. cineraceus Blyth, 1853 (Пенджаб в Индии, Бирма, Таиланд, Бакбо во Вьетнаме, Малайзия, Калимантан, о-ва Анамба);
гималайский листонос — Н. armiger Hodgson, 1835 (Юго-Восточный Китай, Тайвань, о-ва Рюкю, Кумаон, Непал, Ассам, Бакбо, Малайзия);
хохлатый листонос — Н. galeritus Cantor, 1846 (Цейлон, Индия, Бирма, Малайзия, Суматра, Калимантан, северный Квинсленд);
листонос Пратта — И. pratti Thomas, 1891 (Южный Китай, Бирма, Таиланд, Бакбо, Малайзия);
шпороносный листонос — Н. calcaratus Dobson, 1877 (Новая Гвинея, архипелаг Бисмарка, Соломоновы острова);
Н. cupidus Andersen, 1918 (Новая Гвинея, о-в Япен, архипелаг Бисмарка, о-ва Табар, Соломоновы);
Н. cervinus Gould, 1863 (Квинсленд, Новая Гвинея, о-ва Япен, Ару, Кей, Тробриан, Соломоновы, архипелаг Бисмарка);
тиморский листонос — Н. crumeniferus Рёгоп, 1807 (о-в Тимор);
новогвинейский листонос — Н. muscinus Thomas et Doria, 1866 (Новая Гвинея);
листонос Уолластона — Н. wollastoni Thomas, 1913 (Новая Гвинея);
листонос Соломоновых островов — Н. dinops Andersen, 1905 (Соломоновы острова);
пеленгский листонос — Н. pelingensis Shamel, 1940 (о-в Пеленг к востоку от Сулавеси);
сулавесский листонос — Н. inexpectatus Laurie et Hill, 1954 (Сулавеси);
папуасский листонос — Н. рариа Thomas et Doria, 1866 (Новая Гвинея);
листонос Ридлея — H.ridleyi Robinson et Kloss, 1911 (Малайзия);
H. dyacorum Thomas, 1902 (Калимантан);
калимантанский листонос — Н. sabanus Thomas, 1898 (Калимантан);
малайзийский листонос — Н. nequam Andersen, 1918 (Малайзия);
листонос Дориа — Н. doriae Peters, 1871 (Калимантан);
Н. coxi Shelford, 1901 (Калимантан);
длиннохвостый листонос — Н. longicauda Peters, 1861 (о-в Ява);
минданаоский листонос — Н. obscuras Peters, 1861 (о-ва Минданао, Лусон);
увенчанный листонос — Н. coronatus Peters, 1871 (о-в Минданао);
листонос Райта — Н. wrighti Taylor, 1934 (Филиппинские острова);
малый листонос — Н. pygmaeus Waterhouse, 1843 (о-в Лусон);
лузонский листонос— Н. antricolaPeters, 1861 (о-ва Лусон, Минданао);
цветкомордый листонос — Н. omatus Thomas, 1888 (Соломоновы острова).
Некоторые исследователи (Лори и Хилл, 1954 и др.) выделяют из рода обыкновенных листоносов род цветкомордых листоносов— Anthops Thomas, 1888 с единственным видом A. ornatus.B настоящем издании, как и Симпсона (1945) и Бурлиера (1955), эти роды объединены в один род Hipposideros.
Род трезубценосов
Genus Asellia Gray, 1838
Размеры от небольших до средних. Длина тела 50—62 мм. Длина хвоста 18—25 Мм. Длина предплечья 45—58 мм. Окраска волосяного покрова сильно варьирует от желтовато-серой до оранжево-бурой. Имеет место географический морфизм окраски. Носовой листок состоит из трех частей:
передней седловидной, центральной треугольной и задней в виде трезубца. Ноздри расположены в переднем отделе. Позади этих кожных образований
есть небольшая лобная впадина со специфической кожной железой. Уши 1113
крупные, почти лишены волос. Зубная формула: I— с — р — т~ = 28.
День проводят в пещерах, под крышами зданий. На дневках иногда собираются крупными колониями.
Обитатели Северной Африки от Марокко до АРЕ и на юг до Сомали и Занзибара, а на восток до Синда и Северо-Западной Индии.
В роде 2 вида:
обыкновенный трезубценос — A. tridens Ё. Geoffroy, 1813 (Синд, Индия, Аравийский полуостров, Ирак, Израиль, АРЕ, Алжир, Марокко, на юг в Африке до Сомали и Занзибара);
эфиопский трезубценос — A. patrizii de Beaux, 1931 (Эфиопия).
Род трезубценосов Тейта
Genus Aselliscus Tate, 1941
Размеры небольшие. Длина тела примерно 38—45 мм. Длина хвоста 20—40 мм. Длина предплечья 35—42 мм. Окраска волосяного покрова одноцветно-бурая или черноватая или ярко-бурая на спине и желтоватобурая на брюхе. Верхний край носового листка разделяется на три выступа. Седло имеет два боковых листовидных выроста по краям. Лобного впячи-вания со специфической кожной железой нет. Зубная формула: .1 1 2 з оп
i — с — р — т — = 30.
2 1 Г 2 3
Экология не изучена. День проводят в пещерах. Распространены в Юго-Восточной Азии, Новой Гвинее и на островах юго-западной части Тихого океана.
В роде 2 вида:
молуккский трезубценос — A. tricuspldatus Temminck, 1835 (о-ва Амбоина, Церам, Буру, Моротай, Мисол, Кей, Новая Британия, Новая Ирландия, Муруа, Тробрнанд, Новая Гвинея, Соломоновы);
южноазиатский трезубценос — A. stoliczkana Dobson, 1871 (Южный Китай, Бирма, п-ов Индокитай, Малайзия).)
Род цоелопс
Genus Coetops Blyth, 1848
Размеры небольшие (наименьшие в семействе). Длина тела 28—43 мм. Длина предплечья 33—43 мм. Хвост очень короткий или его нет совсем. Окраска волосяного покрова на спине ярко-бурая, пепельно-бурая или черноватая, на брюхе буроватая или пепельная. Уши укороченные и закруг-
ленные (рис. 104). Боковые листоподобные выросты на морде отсутствуют. У некоторых особей на лбу расположено ^впячивание со специфической кожной железой.
Экология изучена слабо. День проводят в пещерах или в дуплах деревьев, где собираются небольшими группами.
Распространены в Южной Азии, на Яве и Филиппинских островах.
В роде 3 вида:
бесхвостый листонос — С. frithi Blyth, 1848 (Южный Китай, Тайвань, Индия, п-ов Индокитай, Ява);
листонос Робинсона — С. robinsoni Bonhote, 1908 (Малаккский полуостров);
волосатый листонос — С. hirsutus Miller, 1911 (о-в Миндоро в группе Филиппинских).
Род африканских трезубценосов
Genus Cloeotis Thomas, 1910
В роде единственный вид: африканский трезубценос — С. percivali Thomas, 1901.
Размеры небольшие. Длина тела 33—50 мм. Длина хвоста 22—33 мм. Длина предплечья 30—36 мм. Характерен относительно длинный хвост. На верхней части носового листка находятся три довольно длинных заостренных кожных выроста, имеющих вид трезубца. Уши относительно короткие и закругленные, едва возвышающиеся над уровнем меха. Окраска волосяного покрова обычно серовато-бурая, желтовато-бурая или темнобурая на спинной стороне и желтоватая на брюшной. Крылья дымчатобурые.
День проводит в пещерах, где собирается большими колониями.
Распространен в ЮАР, Кенни, южной Родезии, Замбии.
Род воронкоухих листоносов
Genus Paracoelops Dorst, 1947
В роде единственный вид: воронкоухий листонос — Р. megalotis Dorst, 1947.
Размеры небольшие. Длина тела 45 мм, длина предплечья 42 мм. Хвоста нет. Характерны крупные уши, достигающие в длину 30 мм и имеющие форму воронки, закругленной в верхних отделах. Носовой листок имеет седловидную форму и по бокам разделяется на округленные радиальные выступы. Межбедренная перепонка достигает 30 мм длины и поддерживается длинными пяточными костями. Волосы довольно длинные, особенно на
спинной стороне. Вершина головы имеет ярко-золотисто-желтую окраску,
резко контрастирующую с буроватой спиной. Брюшная сторона светло-бежевого цвета. Основание всех волос желтое. Уши бледно-бурого цвета,
крылья — темно-бурого. Зубная
, .1113
формула: i — с — р — т —
30.
Экология не изучена.
Встречается во Вьетнаме.
Род золотистых листоносов
Genus Rhinonicteris Gray, 1847
В роде единственный вид: золотистый листонос — R. aurantias Gray, 1845 (рис. 104).
Размеры небольшие. Длина тела 45—52 мм? Длина хвоста 24—30 мм.
Длина предплечья 47—50 мм. Уши не очень длинные, заостренные на вершинах. Носовой листок крупный, сложно устроенный, состоит из двух частей: нижней в виде седла, расположенной ниже ноздрей, и верхней, ограничивающей ноздри сверху. Верхняя часть имеет форму короны и обладает сложной ячеистой структурой. Кроме того, эти листки имеют закругленные выступы. Волосяной покров мягкий, шелковистый, довольно длинный. Окраска его одноцветная, золотисто-желтая, на брюшной стороне несколько светлее. Летательные перепонки желтовато-бурые.
Экология не изучена.
Распространен в Северной и Северо-Западной Австралии.
Род трилистоносов
Genus Triaenops Dobson, 1871
Размеры от небольших до средних. Длина тела 35—62 мм. Длина хвоста 20—34 мм. Длина предплечья 45—55 мм. Окраска волосяного покрова красновато-бурая или серая, иногда бледно-желтовато-бурая, почти белая. Имеет место географическая и половая изменчивость окраски. Кожные листы на конце морды обладают сложной структурой (рис. 104).
112 3
Зубная формула: i — с — р — ш — = 30.
День проводят в пещерах. Охотиться начинают ранним вечером при полном свете.
Распространены в Юго-Западной Азии, Восточной Африке и на Мадагаскаре.
В роде 5 видов:
персидский трилистонос — Т. persicus Dobson, 1871 (Иран, Аравийский полуостров, АРЕ);
африканский трилистонос — Т. afer Peters, 1877 (Кения);
мадагаскарский трилистонос — Т. furcula Trouessart, 1906 (Мадагаскар);
трилистонос Хумблота — Т. humbloti Milne-Edwards, 1881 (Мадагаскар);
красный трилистонос — Т. rufus Milne-Edwards, 1881 (Мадагаскар).
СЕМЕЙСТВО АМЕРИКАНСКИХ ЛИСТОНОСОВ
FAMILIA PHyLLOSTOMATIDAE WATERHOUSE, 1839
Размеры изменчивы: от мелких до наибольших среди американских летучих мышей. Длина тела от 40 до 135 мм. Длина хвоста от 3 до 57 мм (хвост может отсутствовать). Длина предплечья от 25 до 108 мм. Хвост может быть или длиннее, или короче межбедренной перепонки, или они могут быть равны по длине. Если хвост отсутствует, то межбедренная перепонка глубоко вырезана. Носовой листок имеется у большинства представителей семейства, но у некоторых видов он невелик (усатые, подбородколистые и голоспинные листоносы) или редуцирован (брахнфнлла, сфероннктерис, складчатомордые и цветочные листоносы) (рис. 105). Носовой листок не имеет такой сложной структуры, как у подковоносов или листоносов Старого Света. Нижняя губа часто имеет бородавки и сосочки. У усатых, подбородколистых и голоспинных листоносов сосочки имеют пластинчатовидную форму. Уши различной формы; иногда у некоторых родов сильно удлиненные. Козелок имеется. У некоторых представителей семейства ушные раковины соединены между собой по верху головы кожной перепонкой. У многих родов язык сильно удлинен, очень подвижен и покрыт по спинке длинными щетинковидными сосочками.
Рис. 105. Головы: А — меченоса — L. аи-rita; Б — L. concava с вытянутым языком; В — мексиканского длинноноса С. mexi-сапа\ Г — складчатомордого листоноса— С. senex; Д — кубинского листоноса — М. cubanus
Волосяной покров по окраске сильно изменчив: от темно-бурого до светло-оранжевого и белого. Некоторые виды имеют белые лицевые полосы и белую полосу по средине спины. У копьеносов имеется плечевая специфическая кожная железа, которая сильнее развита у самцов.
Череп не имеет посторбитальных отростков. Предчелюстные кости крупные, срастаются между собой и с верхнечелюстными костями. Зубная система сйльно изменчива. Число зубов колеблется от 26 до 34. Жевательная поверхность коренных зубов подвержена также сильной изменчивости от примитивного насекомоядного типа до полностью сходной с жевательной поверхностью зубов крыланов; иногда имеет многочисленные острые бугорки (рис. 106). Плечевой пояс нормальный. Седьмой шейный позвонок не срастается с первым грудным позвонком.
Обитают в самых различных биотопах тропиков и субтропиков: пустынях, саваннах и лесах. День проводят в самых разнообразных убежищах, от затененных до совершенно светлых: в пещерах, зданиях, дуплах деревьев, норах кроликов, листьях пальм и т. д. Некоторые держатся поодиночке, другие небольшими группами, третьи крупными скоплениями. Некоторые виды мигрируют на холодный сезон на юг (например, из юго-западных районов США). Характер питания очень разнообразен. Объектами пищи служат насекомые, мелкие позвоночные (ящерицы, птицы, летучие мыши, грызуны), плоды, нектар и пыльца (см. рис. 107). Некоторые представители семейства способствуют расселению растений, плоды которых они поедают, или опылению цветков. В помете, как правило, один детеныш. Ориентируются при помощи ультразвуковых сигналов.
Распространение охватывает Южную и Северную Америку от Бразилии и северных районов Аргентины на север до юго-запада США и Вест-Индии.
В семействе 44 рода и 126 видов:
усатых листоносов — Chilonycteris Gray, 1839 (5 видов);
голоспинных листоносов — Pteronotus Gray, 1838 (2 вида);
подбородколистых листоносов — Mortnoops Leach, 1821 (2 видя); малых большеухих листоносов Micronycteris Gray, 1866 (10 видов);
большеухих листоносов— Macrotus Gray, 1843 (1 вид);
меченосов — Lonchorhina Tomes, 1863 (1 вид);
длиннолапых листоносов — Macrophyllum Gray, 1838 (1 вид);
круглоухих листоносов— Tonatia Gray, 1827 (6 видов);
копьеносов Грея —Mi-топ Gray, 1847 (2 вида);
копьеносов — Phyllosto-mus Lacepede, 1799 (4 вида); . копьеносов Петерса — Phylloderma Peters, 1865 (2 вида);
бахромчатогубых листоносов — Trachops Gray, 1847 (1 вид);
длинноволосых ложных вампиров — Chrotopterus Peters, 1865 (1 вид);
ложных вампиров — Va-mpyrum Rafinesque, 1815 (1 вид);
длннноязыкнх листоносов— Glossophaga Ё. Geoffroy, 1818 (3 вида);
лноннктернс — Lionycte-ris Thomas, 1913 (1 вид);
копьеносов Томаса — Lo-nchophylla Thomas, 1903 (5 видов);
платалина — Platalina Thomas, 1928 (1 вид);
монофил ус — Monophy-llus Lesch, 1821 (6 видов);
длинноносых листоносов — Anoura Gray, 1838 (4 вида);
банановых листоносов — Musonycteris Schaldach et McLaughlin, 1960 (1 вид);
мексиканских длинноносое •— Choeronycteris Tschudi, 1844 (1 вид);
длинноносов Годмана — Choeroniscus Thomas, 1928 (4 вида);
склерониктерис—Scleronycteris Thomas, 1912 (1 вид);
листоносов Ундервуда — Hylonycteris Thomas, 1903 (1 вид);
листоносов Соссюра — Leptonycteris Lydekker, 1891 (1 вид);
лнхониктернс — Lichonycteris Thomas, 1895 (2 вида);
короткохвостых листоносов — Carollia Gray, 1838 (3 вида);
карликовых листоносов — Rhinophylla Peters, 1865 (1 вид);
желтоплечих листоносов — Sturnira Gray, 1842 (6 видов);
корвира — Corvira Thomas, 1915 (1 вид);
брахифилла — Brachyphylla Gray, 1834 (4 вида);
листоносов-строителей — Urodernia Peters, 1865 (1 вид);
беловолосых широконосов — Vampyrops Peters, 1865 (18 видов);
большеглазых листоносов — Chiroderma Peters, 1860 (6 видов);
белых листоносов —• Ectophylla Н. Alien, 1892 (2 вида);
фруктоядных листоносов — Artibeus Leach, 1821 (8 видов);
красных фруктоядных листоносов —Stenoderma Е. Geoffroy, 1818 (8 видов);
ипанемскнх листоносов — Pygoderma Peters, 1863 (1 вид);
складчатомордых листоносов — Centuria Gray, 1842 (1 вид);
сферониктернс—Sphaeronycteris Peters, 1882 (1 вид);
малых складчатомордов — Ametrida Gray, 1847 (2 вида);
светлых цветочных листоносов — Phyllonycteris Gundlach, 1865 (4 вида);
цветочных листоносов — Erophylla Miller, 1906 (2 вида).
Некоторые ученые (Бурлиер, 1955 и др.) включают вампиров в качестве подсемейства в семейство американские листоносы.
Род усатых листоносов
.Genus Chilonycteris Gray, 1839
Размеры небольшие (наименьшие в семействе). Длина тела от 40 до 67 мм. Длина хвоста от 20 до 30 мм. Длина предплечья от 35 до 63 мм. Хвост хорошо развит и выступает из межбедренной перепонки почти наполовину. Носового листка нет. Крылья прикрепляются по бокам тела, и вся спина равномерно покрыта волосами. Нижняя губа имеет пластинчатоподобные выросты и небольшие сосочки. Окраска волосяного покрова довольно сильно изменчива от темно-бурой или светло-серо-бурой до красноватой. Число зубов 34. Имеется третий верхний резец и третий нижний предкоренной зуб. Дополнительное сочленение плечевой кости с лопаткой несовершенно.
Обитатели тропических лесов и засушливых открытых равнин. По-видимому, предпочитают населять тропические заросли вдоль потоков. День проводят в пещерах или различного рода туннелях под землей. На дневках держатся поодиночке, реже группами, иногда вместе с другими летучими мышами. Начинают охоту ранним вечером. Обычно летают низко над землей. Питаются насекомыми,
Распространение охватывает Мексику, Центральную Америку, северную часть Южной Америки и Вест-Индию.
В роде 5 видов:
черноватый листонос — С. fuliginosa Gray, 1843 (о-ва Ямайка, Гаити, Пуэрто-Рико);
листонос Маклеа — С. macleayii Gray, 1839 (о-ва Куба, Ямайка);
листонос Торрейа — С. torrei G. Allen, 1916 (о-в Куба);
мексиканский листонос — С. personatus Wagner, 1843 (Мексика, Центральная Америка, северная и центральная Бразилия и Венесуэла);
рыжий американский листонос — С. rubiginosa Wagner, 1843 (Мексика, Центральная Америка, Венесуэла, Колумбия, о-ва Куба, Гаити и Ямайка).
Род голоспинные листоносы
Genus Pteronotus Gray, 1838
Размеры от мелких до средних. Длина тела 50—77 мм. Длина хвоста 10—26 мм. Длина предплечья 42—57 мм. Носового листка нет. Нижняя губа имеет пластинчатовидные и небольшие сосцевидные выросты. Крылья прикрепляются по средней линии спины, покрывая ее сверху и создавая впечатление, что спина не имеет волос. Окраска волосяного покрова рыжеватая или темно-бурая. Число зубов 34. Имеются верхний третий резец и нижний третий предкоренной зуб. Дополнительное сочленение плечевой кости с лопаткой выражено слабо.
День проводит в пещерах, иногда в дуплах деревьев. Часто на дневках располагаются с другими летучими мышами. Когда охотятся, обычно летают
низко над землей. Питаются насекомыми, ловя их обычно вдоль водоемов. Самки приносят детенышей по большей части в мае.
Распространены от Мексики на юг до Бразилии. Встречаются также на островах Малых Антильских и Тринидаде.
В роде 2 вида:
листонос Дэви — Р. davyi Gray, 1838 (южная Мексика, Центральная Америка, в Южной Америке от Венесуэлы до Мату-Гросу в Бразилии, о-в Тринидад и Малые Антильские острова);
голоспинный листонос — Р. suapurensis J. Allen, 1904 (Центральная Америка, Венесуэла, Колумбия, Бразилия, Гвиана).
Род подбородколистых листоносов Genus Mormoops Leach, 1821
Размеры небольшие. Длина тела 51—66 мм. Длина хвоста примерно 25—34 мм. Длина предплечья 46—56 мм. Носового листка нет. Нижняя губа имеет мясистые колышковидные выступы, подбородок — листовидные выросты. По бокам рта располагаются длинные, грубые волосы, почти скрывающие его. Вздернутый кверху короткий нос имеет борозды и гребни. Хвост хорошо развит. Окраска волосяного покрова рыжевато-бурая, темнобурая или светло-бурая на спине и грязно-буроватая на брюхе. Характерна некоторая изогнутость черепа таким образом, что нижний край большого затылочного отверстия располагается выше уровня рострума черепа. Число зубов 34. Имеются верхний третий резец и нижний третий предкорен-ной зуб.
Дневки устраивают в пещерах, часто с другими летучими мышами. Наибольшая активность отмечена в 9 ч вечера и затем в середине ночи. Летают низко над землей или водой.
Распространены от Аризоны и Техаса в США на юг через Мексику и Центральную Америку до северной части Южной Америки. Встречаются также на о-ве Тринидад, Больших Антильских островах и некоторых мелких островах у побережья Венесуэлы.
В роде 2 вида:
подбородколист Блайнвнлла — М. blainvillii Leach, 1821 (Большие Антильские острова);
подбородколистый листонос — М. megalophylla Peters, 1864 (от южных районов штатов Аризона и Техас на юг через Мексику, Центральную Америку до северных районов Южной Америки, о-ва Тринидад и мелкие у берегов Венесуэлы).
Род малых большеухих листоносов
Genus Micronycteris Gray, 1866
Размеры небольшие. Длина тела 42—52 мм. Длина хвоста около 12 мм. Длина предплечья 31—49 мм. Крупные уши соединяются между собой мало заметной складкой. У некоторых видов уши отделены друг от друга. Имеется сильно выступающий носовой листок. Конец хвоста достигает лишь середины межбедренной перепонки. Окраска волосяного покрова обычно различных оттенков бурого цвета, иногда орапжевато-красповатая или оливково-бурая на спине и буроватая или грязно-желтая на брюхе; горло может быть желтоватое или оранжевое. В черепе костные слуховые барабаны 2 12 3
небольшие. Зубная формула: t — с — р — т~ = 32.
Обитатели различных биотопов от тропических лесов до пустынь. Дневки устраивают в самых различных местах: в пещерах, строениях, развалинах, дуплах деревьев и других местах, причем часто их можно ветре-
тить в освещенных местах. Большинство видов питается насекомыми. Отмечено, что М. megalotis питался плодами гуавы.
Распространены от центральной Мексики на юг до Перу и Бразилии, а также на о-вах Тринидад, Тобаго и Гренада.
В роде 10 видов:
714. megalotis Gray, 1842 (Мексика, Центральная Америка, о-ва Тринидад, Малые Антильские и от Колумбии и Венесуэлы до южного Перу и Сан-Паулу в Бразилии);
малый большеухий листонос — М. minuta Gervais, 1855 (Бразилия, Колумбия);
волосатый большеухий листонос — М. hirsuta Peters, 1869 (от севера Колумбии до Коста-Рики и о-ва Тринидад);
М. platiceps Sanborn, 1949 (Венесуэла, Колумбия, Центральная Америка на север до Никарагуа и о-в Тринидад);
бразильский листонос — М. pusilia Sanborn, 1949 (северо-западная Бразилия);
листонос Найсфора — TH. nicefori Sanborn, 1949 (северная Колумбия);
листонос Бена — М. behni Peters, 1869 (центральная Бразилия, юг Перу и север Боливии);
TH. brachyotis Dobson, 1878 (Гвиана);
листонос Шмидта — 714. schmidtorum Sanborn, 1935 (юг Мексики, Гватемала, Гондурас);
бурый листонос — 714. sylvestris Thomas, 1896 (Центральная Америка).
Род большеухих листоносов
Genus Macrotus Gray, 1843
В роде один вид: листонос Ватерхауза — 714. waterlwusi Gray, 1843.
Размеры мелкие. Длина тела 50—69 мм. Длина хвоста 25—41 мм. Длина предплечья 45—58 мм. Характерен относительно длинный хвост, слегка выступающий из межбедренной перепонки. Голова удлинена. Ушные раковины большие, закругленные, соединяющиеся между собой по верху головы. Носовой листок крупный (около 7 мм высотой), напоминает по форме наконечник стрелы. Окраска волосяного покрова на спинной стороне буроватая или сероватая, на брюшной бурая или грязно-желтая. Костные 2 12 3
слуховые барабаны небольшие. Зубная формула: i — с — р — /л—=34.
Число диплоидных хромосом 40—46.
Обитатели обычно аридных областей. День проводят в зданиях и пещерах. Отмечены случаи миграции на зимний период к югу. Питаются насекомыми, которых собирают на земле и на растениях. Самки калифорнийского листоноса приносят детенышей обычно в июне. На дневках (в зданиях, шахтах) собираются от нескольких десятков до 400, иногда с другими летучими мышами.
Распространены от южной Невады, Аризоны и южной Калифорнии на юг до Гватемалы, а также на Багамских островах и на Больших Антильских.
Некоторые исследователи выделяют в роде, кроме листоноса Ватерхауза—М. waterlwusi Gray, 1843 (о-ва Багамские и Большие Антильские), еще 714. mexicanus Saussure, 1860 (Мексика и на юг до Гватемалы) и М. са-lifornicus Baird, 1858 (южная Невада, Аризона, южная Калифорния, Баха Калифорния и Сонора в Мексике).
Вероятно, правильнее их считать подвидами листоноса Ватерхауза (Андерсон, Anderson, 1969).
Род меченосов
’ Genus Lonchorhina Tomes, 1863
В роде единственный вид: меченос—L. aurita Tomes, 1863 (рис. 105).
Размеры небольшие. Длина тела примерно 60 мм. Длина хвоста около 57 мм. Длина предплечья около 50 мм. Характерен вертикально поднимающийся вверх заостренный к вершине длинный носовой листок. Длина его примерно равна длине наружных ушных раковин. Нижняя губа спереди имеет бороздку и небольшие вздутия по краям. Длинный хвост оканчивается на- заднем краю межбедренной перепонки. Окраска меха светло-рыжевато-бурая. Число зубов 34 (формула зубов, как у большеухих листоносов).
День проводит в пещерах. Питается, по-видимому, исключительно насекомыми.
Распространен на о-вах Багамских, Тринидад, в Венесуэле, восточной Бразилии, Центральной Америке, поднимается на север до южной Мексики.
Род длиннолапых листоносов
Genus Macrophyllum Gray, 1838
В роде единственный вид: длиннолапый листонос — М. macrophyllum Schinz, 1821.
Мелких размеров американский листонос. Длина тела 43—62 мм. ; Длина хвоста примерно 37 мм. Длина предплечья 37—45 мм. Характерная ; особенность — удлиненные задние конечности и длинный хвост, полностью включенный в межбедренную перепонку. Уши разделены друг с другом. ! Носовой листок длинный и широкий в основании. Окраска волосяного по- крова на спинной стороне черновато-бурая, на брюшной несколько более : светлая. В черепе лицевой отдел укорочен. Число зубов 34. Костные слу-I ховые барабаны маленькие.
I Как правило, держится около водоемов. День проводит в пещерах, и I зданиях.
Распространен в Сальвадоре, Гондурасе, Панаме, Колумбии, Венесуэле, Гайане, Бразилии, Перу.
Род круглоухих листоносов
Genus Tonatia Gray, 1827
Размеры мелкие и средние. Длина тела 42—82 мм. Длина хвоста 12— 21 мм. Длина предплечья 32—60 мм. Характерны довольно крупные уши, закругленные на вершинах. У одних видов уши соединены между собой у основания, у другйх раздельны. Хвост относительно короткий, достигает лишь половины межбедренной перепонки. Окраска охристая или темнобурая сверху и более светлая снизу. У листоноса Каррикера брюшная сторона белая. Костные слуховые барабаны небольшие. Зубная формула: . 2 1 2 3 о0
; । — с — р — т— — 32.
1 13 3
В День проводят в дуплах деревьев, норах кроликов.
Распространены в Гайане, Перу, Боливии, Бразилии, Колумбии. Ве-I кесуэле, Эквадоре, на о-ве Тринидад.
I В роде 6 видов:
В круглоухий листонос —• Т. bidens Spix, 1823 (Южная Америка от Ко-f лумбии и Венесуэлы до Перу, Центральная Америка на север до Коста-* Рики);
{ бразильский круглоухий листонос — Т. brasiliensis Peters, 1866 (цен-; тральная и восточная Бразилия, Венесуэла, о-в Тринидад);
листонос Каррикера — Т. carrikert J. Allen, 1910 (Венесуэла);
малый круглоухий листонос — Т. minuta Goodwin, 1942 (Эквадор, Колумбия, Перу);
никарагуасский листонос — Т. nicaraguae Goodwin, 1942 (Никарагуа);
Т. sylvicola D’Orbigny, 1835 (Гвиана, Бразилия, Боливия, Перу, Эквадор, Колумбия и Центральная Америка на север до Гондураса).
Род копьеносов Грея
Genus Mlmon Gray, 1847
Размеры мелкие и средние. Длина тела 50—75 мм. Длина хвоста 10— 25 мм.- Длина предплечья 48—57 мм. Носовой листок крупный, довольно широкий в основании, остроконечный, поднимается вертикально вверх от носовой области. Довольно крупные уши не соединяются друг с другом. На подбородке может быть довольно обширная область, которая не покрыта волосами и разделена продольным желобком. У тринидадского копьеноса на нижней губе имеется V-образная бороздка. Волосяной покров на туловище длинный и мягкий. Окраска его у копьеноса Беннетта на спине и брюхе однообразно бледно-бурая, за исключением беловатых пятен позади ушей. У тринидадского копьеноса спинная часть ярко-рыжевато-бурая или черновато-бурая; брюшная беловатая, иногда сероватая; позади ушей расположено беловатое или желтовато-оранжевое пятно. Вдоль спины проходит •срединная бледноокрашенная полоска. Иногда заднеушные пятна и срединная спинная полоса отсутствуют. Костные слуховые барабаны небольшие.
2 12 3
Зубная формула: I — с — р — m— = 30.
Обитатели лесистых районов. День проводят в дуплах деревьев или пещерах. Обычно держатся небольшими группами. Характер питания точно не установлен. Предполагается, что поедают насекомых, а также плоды. Пьют сок плодов.
Распространены от южной Мексики на юг до северной и центральной части Южной Америки. Встречаются также на о-ве Тринидад.
В роде 2 вида:
копьенос Беннетта — М. bennettii Gray, 1838 (южная Мексика, Центральная Америка, Бразилия);
тринидадский копьенос — 714. crenulatum Geoffroy, 1810 (Гвиана, Перу, Бразилия, Панама, о-в Тринидад).
Иногда род копьеносов Грея разделяют на 6 видов.
Род копьеносов
Genus Phyllo stotnus Lacepede, 1799
Средних и крупных размеров американские листоносы. Длина тела 75—130 мм. Длина хвоста 10—25 мм. Длина предплечья 55—95 мм. У наиболее крупных особей размах крыльев достигает 450 мм. Вертикально поднимающийся вверх носовой листок хорошо развит. Уши относительно крупные, заостренные, не соединяющиеся между собой. Хвост короткий, полностью включен в межбедренную перепонку и обычно не достигает се края. У самцов на горле хорошо развита мешковидная специфическая кожная железа. У самок этот горловой мешок развит слабо. Нижняя губа со своеобразной бороздкой, по краям которой расположены вздутия. Окраска темно-бурая или черновато-бурая, серая, рыжевато-бурая на спине и не-2 12 3
сколько более светлая на брюхе. Зубная формула: i— с — р — m — = 32.
Костные слуховые барабаны маленькие, уплощенные.
День проводят в пещерах или дуплах деревьев, собираясь иногда в большом количестве. В одной пещере в Панаме было найдено несколько тысяч обыкновенных копьеносов. Питание разнообразное. Бледный копьенос поедает плоды. Некоторые виды поедают нектар и пыльцу. Обыкновенный копьенос нападает на птиц, мышей, мелких летучих мышей, питается также насекомыми.
Распространены от южной Мексики на юг до Перу, Бразилии, Боливии, а также на о-вах Тринидад и Тобаго.
В роде 4 вида:
обыкновенный копьенос — Р. hastatus Pallas, 1767 (Центральная Америка на север до Гондураса, Бразилия, Гвиана, Венесуэла, Колумбия, Эквадор, Перу, Боливия);
бледный копьенос — Р. discolor Wagner, 1843 (Бразилия, Перу, Центральная Америка);
длиннолистый копьенос — P.elongatus Geoffroy, 1810 (от Эквадора и Гвианы до Мату-Гросу и южных районов Перу);
широколистый копьенос — Р. latifolius Thomas, 1901 (Гвиана).
Род копьеносов Петерса
Genus Phylloderma Peters, 1865
• Размеры средние и крупные. Длина тела 85—120 мм. Длина хвоста примерно 16 мм. Длина предплечья 67—80 мм. Большой вертикально поднимающийся носовой листок хорошо выражен. Окраска волосяного покрова темно-бурая на спине, несколько более светлая на брюхе. У самцов на горле есть мешковидная специфическая кожная железа. Зубная формула:
2 12 3
i — с — р — т — = 34. Характерны двудольчатый средний верхний резец, узкие нижние коренные зубы и наличие небольшого третьего нижнего предкоренного зуба.
Экология не изучена.
i Распространены в Гондурасе и Гвиане.
J В роде 2 вида:
1 северный копьенос — Р. septentr ionales Goodwin, 1940 (Гондурас);
I южный копьенос — Р. stenops Peters, 1865 (Гвиана).
1
э э
е
о а
Род бахромчатогубых листоносов Genus Trachops Gray, 1847
В роде единственный вид: бахромчатогубый листонос — Т. cirrhosa Spix, 1823.
Размеры средние. Длина тела 76—88 мм. Длина хвоста 12—21 мм. Длина предплечья 58—64 мм. Хвост не достигает края межбедренной перепонки. Губы покрыты характерными бородавчатовидными отростками. Уши относительно длинные, с закругленными вершинами. Волосяной покров на спинной поверхности тела имеет рыжевато-бурую окраску, на брюшной — буроватую с серым оттенком. Зубная формула:
. 2 1 2 3
i — с — р -— т — =34.
2 13 3
Экология изучена слабо. День проводит в пещерах, иногда в дуплах деревьев, собираясь небольшими группами. Есть сведения, что бахромчатогубый листонос питается мелкими ящерицами.
Распространен от южной Мексики на юг до Колумбии, Венесуэлы, Бразилии и на о-ве Тринидад.
Род длинноволосых ложных вампиров
Genus Chrotopterus Peters, 1865
В роде единственный вид: длинноволосый ложный вампир — С. auri-tus Peters, 1856.
Размеры крупные. Длина тела примерно 100—112 жж. Длина хвоста около 5 мм. Длина предплечья 75—85 жж. Губы и подбородок гладкие, за исключением небольшой бородавки в центре нижней губы и узких возвышенностей по ее краям. Хвост очень короткий, далеко не достигающий края межбедренной перепонки. Уши крупные, с закругленными вершинами. Волосяной покров длинный и мягкий. Окраска его на спине темно-бурая, на брюхе — серовато-бурая. Зубная формула: I с -у- р -|- т-|- = 32.
Экология почти не изучена. День проводит в пещерах. По характеру питания, по-видимому, хищник.
Распространен от южной Мексики до северной Аргентины и Парагвая.
Род ложных вампиров
Genus Vampyrutn Rafinesque, 1815
В роде единственный вид: ложный вампир — V. spectrum Linnaeus, 1758 (рис. 106).
Размеры крупные, наибольшие в семействе. Длина тела примерно 125— 135 мм-. Хвост отсутствует. Длина предплечья 100—108 жж. Характерна удлиненная морда с поднимающимся вертикально вверх хорошо выраженным большим носовым листком. Уши крупные. Подбородок и губы гладкие. Окраска меха рыжевато-бурая на спине и слегка более светлая на брюхе.
2 12 3
Зубная формула: i — с — р — т— = 34.
День проводит в дуплах деревьев. По характеру питания всеядный. Поедает плоды, насекомых, мелких позвоночных (птиц, млекопитающих). В неволе два ложных вампира съедали 4 мыши ежедневно.
Населяет территорию от южной Мексики на юг примерно до 14° южной широты в Перу и Бразилии и встречаются также на о-ве Тринидад.
Род длинноязыких листоносов
Genus Glossophaga Ё. Geoffroy, 1818
Размеры небольшие. Длина тела 48—65 жж. Длина хвоста около 7 жж. Длина предплечья 32—42 жж. Морда удлиненная. Имеется поднимающийся вертикально вверх небольшой носовой листок. Язык очень длинный, способный далеко вытягиваться из рта и в концевой части покрытый сверху длинными, обращенными вершинами назад нитевидными сосочками. Окраска волосяного покрова от темно-бурой до рыжевато-бурой. Скуловая дуга 2 12 3
в черепе полная (рис. 106). Зубная формула: I — с — р —т — — 34.
2 13 3
Щечные зубы узкие и удлиненные.
Обитатели лесов, а также засушливых равнин. День проводят в пещерах и постройках человека. Иногда встречаются большими группами по нескольку сотен особей. Питаются главным образом нектаром (рис. 107), соком плодов, а также насекомыми. Сезонности в размножении, по-видимо-му, нет.
Распространение охватывает северную Мексику на юг до Аргентины, а также Багамские острова, Ямайку, Тринидад и Малые Антильские.
В роде 3 вида:
длинноязыкий листонос — G. soricina Pallas, 1766 (от северной Мексики на юг до Перу, Парагвая и северной Аргентины, а также Багамские острова, Ямайка и Тринидад);
длинномордый листонос — G. longirostris Miller, 1898 (Колумбия, Венесуэла, о-ва Малые Антильские, Тринидад);
куракаосский листонос — G. elongata Miller, 1900 (о-в Куракао у побережья Венесуэлы).
Род лиониктерис
Genus Lionycteris Thomas, 1913
В роде единственный вид: лиониктерис—L. spu-relli Thomas, 1913.
Размеры мелкие. Длина тела примерно 50 мм.
Рис. 107. Длинноязыкий листонос — G. soricina питается пыльцой и нектаром цветов банана
Длина хвоста 10 мм. Длина предплечья 34—36 мм. Морда удлиненная. Носовой листок низкий и ши-
рокий. Язык очень длин-
ный, с высокими нитевидными сосочками на спинной стороне кончика. Межбедренная перепонка хорошо развита. Хвост достигает примерно половины ее длины. Окраска волосяного покрова рыжевато-бурая. Коренные
зубы .узкие, вытянутые.
Экология не изучена.
Встречается в северной части Южной Америки.
Род копьеносов Томаса
Genus Lonchophylla Thomas, 1903
Размеры небольшие. Длина тела 45— 60 мм, длина хвоста 8—10 лш, длина предплечья 30—44 лгл(. Морда вытянутая. Язык очень длинный. На спинной стороне его кончика располагаются нитевидные сосочки с вершинами, направленными назад. Короткий хвост едва достигает середины длины межбедренной перепонки. Носовой листок высокий и узкий. Окраска меха обычно темно-бурая на спине и несколько более светлая на брюхе. В черепе характерна неполная скуловая дуга. Зубная формула: .21 2 3 „
i — с — р — т — — 34.
2 13 3
День проводят в пещерах, часто вместе с другими летучими мышами. Питаются в основном нектаром, вероятно, также пыльцой, плодами и насекомыми.-Служат опылителями некоторых ночных цветков. _
Распространены в Панаме, Колумбии, Венесуэле, Бразилии, Боливии и Перу.
В роде 5 видов:
западный копьенос — L. hesperia G. Allen, 1908 (Перу);
бразильский копьенос — L. mordax Thomas, 1903 (Бразилия, Боливия);
L. concava Goldman, 1914 (Панама);
панамский копьенос — L. robusta Miller, 1912 (Панама, Колумбия); копьенос Томаса — L. thomasi J. Allen, 1904 (Венесуэла).
Род платалина
Genus Platalina Thomas, 1928
В роде единственный вид: платалина — Р. genovensium Thomas, 1928.
Размеры средние. Длина тела 72 мм. Длина хвоста около 9 мм. Длина предплечья примерно 50 мм. Морда удлиненная. Язык очень длинный, с высокими нитевидными сосочками. Коренные зубы узкие и вытянутые. Ко-.роткий хвост едва достигает середины межбедренной перепонки. Носовой листок средних размеров. Окраска волосяного покрова одноцветно-буроватая.
Характер питания, по-видимому, сходен с другими длинноязыкими листоносами.
Распространен в Перу.
Род монофиллус
Genus Monophyllus Leach, 1821
Размеры небольшие. Длина тела 52—65 мм. Длина хвоста 4—12 мм. Длина предплечья 36—42 мм. Морда вытянута, на ее конце находится поднимающийся перпендикулярно вверх средних размеров носовой листок. Язык очень длинный, покрытый впереди удлиненными нитевидными сосочками. Хвост, хотя и короткий, примерно наполовину выдается из узкой межбедренной перепонки. Окраска волосяного покрова бурая на спинной и более светлая на брюшной стороне. Зубная формула: 2 12 3
i — с — р — гг-^- = 34. Характерны очень мелкие нижние резцы. Щечные зубы узкие и удлиненные.
День проводят обычно в пещерах. Питаются соком и мякотью плодов, а также насекомыми.
Распространены в Вест-Индии.
В роде 6 видов:
ямайский листонос — М. redmani Leach, 1821 (о-в Ямайка);
кубинский листонос — М. cubanus Miller, 1902 (Куба);
пуэрто-риканский листонос — М. portoricensis Miller, 1900 (о-в Пуэрто-Рико);
барбадосский листонос — М. plethodon Miller, 1900 (о-в Барбадос); сент-люсийский листонос — М. luciae Miller, 1902 (о-в Сент-Люсия); М. frater Anthony, 1917 (о-в Пуэрто-Рико).
Некоторые исследователи (Холл и Келсон, 1959 и др.) выделяют седьмой вид: островной листонос — М. clinedophus Miller, 1900, который, по-видимому, является подвидом ямайского листоноса, или предлагают выделять в роде всего 2 вида: ямайского листоноса с подвидами ямайский, островной и пуэрто-риканский и барбадосского листоноса с подвидами барбадосский, Сент-Люсии и М. р. frater (Швартц и Джонес — Schwartz and Jones, 1967).
Род длинноносых листоносов
Genus Anoura Gray, 1838
Размеры мелкие и средние. Длина тела 50—90 лш. Хвоста нет или он достигает длины 4—7 мм. Длина предплечья 34—48 мм. Морда вытянутая, язык очень длинный с высокими нитевидными сосочками на кончике. Меж
бедренная перепонка очень узкая. Окраска волосяного покрова изменчива: одноцветно-черноватая, темно-бурая на спине или темно-бурая с серебристым оттенком на плечах и серо-бурая на брюхе. Зубная формула:
j с р /я= 32. Щечные зубы узкие и удлиненные.
Экология изучена слабо. День проводят в пещерах и в расщелинах скал. Питаются нектаром, пыльцой и, видимо, мякотью плодов.
Распространены от центральной Мексики на юг до Перу, Боливии, Бразилии. Встречаются также на о-ве Тринидад.
В роде 4 вида:
длинноносый листонос — A.caudifera Geoffroy, 1818 (Эквадор, Перу, Колумбия, Венесуэла, Гвиана, Бразилия);
листонос Жоффруа—A. geoffroyi Gray, 1838 (Венесуэла, Бразилия, Боливия, Колумбия, Эквадор, Перу, о-в Тринидад);
панамский листонос — A.cultrata Handley, 1960 (Панама);
широкомордый листонос — A. brevirostrum Carter, 1968 (Южная Америка).
Род банановых листоносов
Genus Musonycteris Schaldach et McLaughlin, 1960
• В роде единственный вид: банановый листонос — TH. harrisoni Schaldach et McLanghlin, 1960. Некоторые исследователи (Андерсен и Джонс, 1967 и др.) включают бананового листоноса в род мексиканских динно-носов — Choeronycteris Tschudi, 1844.
Размеры средние. Длина тела 70—79 жж. Длина хвоста 8—12 мм. Длина предплечья 41—43 мм. Характерна очень сильно вытянутая лицевая часть головы. Язык длинный, с высокими нитевидными сосочками в концевой части. Окраска волосяного покрова на спине светло- или темно-бурая, на брюхе светло-бурая. Щечные зубы узкие и удлиненные.
Обитатели тропических лесов. Экология почти не изучена. Питаются, по-видимому, нектаром и пыльцой.
Распространены в Мексике.
Род мексиканских длинноносое
Genus Choeronycteris Tschudi, 1844
В роде единственный вид: мексиканский длиннонос — С. mexicana Tschudi, 1844 (см., рис. 105,106).
Размеры мелкие. Длина тела 55—70 мм. Длина хвоста 10—16 жж. Длина предплечья около 44 жж. Морда сильно вытянута. Носовой листок небольшой, вертикально поднимающийся вверх на конце. Язык очень длинный с удлиненными нитевидными сосочками на кончике. Окраска волосяного покрова темно-бурая на спине и светло-бурая на брюхе. Зубная формула: 2 12 3
i —- с —— р — т — = 30. Щечные зубы узкие и удлиненные.
День проводит в пещерах и зданиях. Встречается в этих убежищах поодиночке или небольшими группами, иногда до нескольких десятков осо-• бей вместе. Предполагается возможность миграции из южной Аризоны в зимние месяцы на юг в Мексику. Питается, по-видимому, нектаром и пыльцой, мякотью и соком плодов, а также насекомыми. Опыляет некоторые растения.
Распространен от юго-западных областей США до Гватемалы.
Род длинноносое Годмана
Genus Choeroniscus Thomas, 1928
Размеры небольшие. Длина тела 50—55 мм. Длина хвоста около 12 лш. Длина предплечья 32—38 мм. Морда сильно вытянута. Язык очень длинный, с высокими нитевидными сосочками на кончике. Носовой листок не-
большой, треугольной формы. Уши маленькие. Окраска волосяного покрова темно-бурая на спине и несколько более светлая на брюхе. Щечные 2 12 3
зубы удлиненные и узкие. Зубная формула: i — с — р — т —- = 30. 0 13 3
Питаются нектаром, мякотью плодов и, по-видимому, также насеко
мыми.
Распространение охватывает территорию от Мексики на юг до Бразилии в районе Амазонки, а также о-в Тринидад.
В роде 4 вида:
длиннонос Годмана — С. godmani Thomas, 1903 (южная Мексика, Гватемала, Гондурас, Сальвадор, Никарагуа, Коста-Рика);
длиннонос инков — С. inca Thomas, 1912 (Перу);
средний длиннонос — С. intermedins J. Allen et Chapman, 1893 (Гвиана, Венесуэла, о-в Тринидад);
малый длиннонос — С. minor Peters, 1898 (Гвиана).
Род длинноносов Годмана, возможно, следует отнести в качестве подрода к роду мексиканских длинноносое.
Род склерониктерис
Genus Scleronycteris Thomas, 1912
В роде единственный вид: склерониктерис — S. ega Thomas, 1912. Вероятно, может входить в качестве подрода в род мексиканских длинноносое.
Размеры небольшие. Длина тела около 57 мм. Длина хвоста примерно 6 мм. Длина предплечья 35 мм. Морда сильно вытянута. Носовой листок небольшой. Характерен выступающий вперед и вниз подбородок. Окраска волосяного покрова на спинной стороне тела темно-бурая, на брюшной — светло-бурая. Щечные зубы удлиненные и узкие.
Экология не изучена.
Распространен в районе Амазонки в Бразилии.
Род листоносов Ундервуда
Genus Hylonycteris Thomas, 1903
В роде единственный вид: листонос Ундервуда — Н. underwoodi Thomas, 1903. Некоторые исследователи (Симпсон, 1945; Бурлиер, 1955 и др.) включают листоноса Ундервуда в состав рода мексиканских длинноносов.
Размеры небольшие. Длина тела примерно 53 мм. Длина хвоста около 6 мм. Длина предплечья 33—35 мм. Морда сильно вытянута. Язык очень длинный, снабжен в передней части высокими нитевидными сосочками. Межбедренная перепонка хорошо развита. Окраска волосяного покрова одноцветно-темно-серая или на спинной стороне темно-бурая, на брюшной более светлая. Щечные зубы узкие и вытянутые.
День проводит в пещерах, туннелях, собираясь небольшими группами. Питается в основном нектаром, мякотью плодов, а также насекомыми.
Распространен в Мексике и Коста-Рике.
Род листоносов Соссюра
Genus Leptonycteris Lydekker, 1891
В роде один вид: листонос Соссюра — L. nivalis Saussure, 1860.
Размеры средние. Длина тела 70—95 мм. Хвост очень короткий, снаружи почти незаметен, содержит всего 3 позвонка. Длина предплечья 46— 57 мм. Морда сильно вытянута. Язык очень длинный с высокими нитевидными сосочками на конце. Носовой листок вертикально поднимается вверх на конце морды, небольшой. Межбедренная перепонка очень узкая. Окраска волосяного покрова обычно рыжевато-бурая или черно-бурая на спине и светло-бурая или бурая на брюхе. Зубная формула: 2 12 2
i — с — р — т — = 30. Щечные зубы узкие и удлиненные.
День проводит в пещерах, туннелях и зданиях. Может подниматься высоко в горы, однако по большей части встречается в низинах и в засушливых зонах. Питается нектаром, пыльцой, плодами и насекомыми. Населяющие северные части ареала, пощидимому, мигрируют на зимние месяцы к югу.
Распространен от южных частей штата Аризона и штата Техаса (США) на юг до Никарагуа и Венесуэлы.
Род лихониктерис
Genus Lichonycteris Thomas, 1895
Размеры небольшие. Длина тела 50—55 мм. Длина хвоста около 8 мм. Длина предплечья примерно 33 мм. Морда сильно вытянута. На конце ее находится небольшой носовой листок треугольной формы, поднимающийся вертикально вверх. Язык очень длинный, снабженный в передней части удлиненными нитевидными сосочками. Межбедренная перепонка хорошо развита. Окраска волосяного покрова буроватая, с желтым оттенком, ино-2 13 2
гда темно-бурая. Зубная формула: i — с — р — т— = 26. Щечные зубы узкие и длинные.
Экология изучена слабо. Питаются, по-видимому, нектаром и мякотью плодов. Опыляют некоторые цветущие ночью растения.
Распространены в Бразилии, Никарагуа, Коста-Рике и Суринаме.
В роде 2 вида:
листонос Миллера — Н. degener Miller, 1931 (Бразилия в районе Амазонки);
бурый лихониктерис — L. obscura Thomas, 1895 (Суринам, Коста-Рика, Никарагуа).
Род короткохвостых листоносов
Genus Carollia Gray, 1838
Размеры небольшие. Длина тела 48—65 мм. Длина хвоста 3—14 мм. Длина предплечья 34—45 мм. На конце морды находится вертикально поднимающийся вверх средних размеров носовой листок с острой вершиной. Морда умеренно вытянута в длину. Уши средних размеров, широко расставленные по бокам головы. Окраска волосяного покрова темно-бурая, иногда светло-оранжевая. В черепе характерна неполная скуловая дуга.
2 12 3
Зубная формула: i — с — р — т— = 32.
2 12 3
День проводят в пещерах, расщелинах скал, зданиях, дуплах деревьев. Встречаются поодиночке, небольшими группами или собираются по не-.скольку тысяч особей. Питаются исключительно мякотью плодов: находят
плоды по запаху. Определенного сезона размножения, по-видимому, нет. Распространены от центральной Мексики на юг до Перу, Бразилии, Парагвая, Колумбии, Венесуэлы и северной Аргентины.
В роде 3 вида:
С. castanea Н. Allen, 1890 (южная Мексика, Центральная Америка, северные и восточные районы Южной Америки);
С. perspicittata Linnaeus, 1758 (восточные районы центральной и южной Мексики, Центральная Америка и от берегов Карибского моря в Южной Америке до юга Перу, а также о-в Тринидад);
трехцветный листонос — С. tricolor Miller, 1902 (Парагвай).
Род карликовых листоносов
Genus Rhynophylla Peters, 1865
В роде единственный вид: карликовый листонос — R. pumilio Peters, 1865.
Размеры небольшие. Длина тела около 50 мм. Хвоста нет. Длина предплечья примерно 32 мм. Уши относительно небольшие, широко расставленные. Морда средней длины. На конце ее находится поставленный вертикально вверх треугольной формы небольшой носовой листок. Межбедренная перепонка довольно хорошо развита, но имеет глубокий вырез.
Экология не изучена. По характеру питания, по-видимому, сходен с короткохвостыми листоносами.
Распространен в Бразилии, Перу и Эквадоре.
Род желтоплечих листоносов
Genus Sturnira Gray, 1842
Размеры небольшие. Длина тела 58—70 мм. Хвоста нет. Длина предплечья примерно 38—46 мм. Вес от 15 до 20 а. Уши короткие, широко расставленные, не соединяющиеся между собой. Носовой листок поднимается перпендикулярно вверх, имеет относительно небольшие размеры. Межбедренная перепонка очень узкая, покрыта волосами. Задние конечности покрыты волосами до самых когтей. Окраска волосяного покрова розовато-грязно-желтая с буроватым оттенком, темно-охристо-бурая или темно-бурая. Брюшная сторона обычно несколько светлее спинной. Спереди на плечах у самцов располагается по пучку грубых желтоватых или красноватых 2 12 3
волос. Зубная формула: I — с — р — т— = 32.
Обитатели лесных районов на равнинах и в горах. День проводят обычно в дуплах.деревьев. Питаются преимущественно плодами.
Распространены от южной Мексики на юг до Аргентины и Парагвая. Встречаются также на о-вах Ямайка и Малых Антильских.
В роде 6 видов:
желтоплечий листонос—S. lilium Geoffroy, 1810 (южная Мексика, Центральная Америка и Южная Америка от Эквадора и Гвианы до юга Перу и Ла-Платы в Аргентине, а также о-ва Ямайка, Малые Антильские);
листонос; Людовика—S. ludovici Antony, 1924 (южная Мексика, Центральная Америка, Колумбия, Венесуэла, Эквадор);
тринидадский листонос — S. tildae De la Torre, 1959 (о-в Тринидад); мохноногий листонос — S. mordax Goodwin, 1938 (Коста-Рика); доминиканский листонос —S. angeli De la Torre, 1966 (о-в Доминика); большой желтоплечий листонос —S. niagna De la Torre, 1966 (Перу); Некоторые ученые (Холл и Келсон, 1959 и др.) выделяют мохноногого листоноса в особый род — Sturnirops Goodwin, 1938.
Род корвира
Genus Corvira Thomas, 1915
В роде единственный вид: корвира — С. bidens Thomas, 1915.
Размеры средние. Длина тела 73 мм. Хвоста нет. Длина предплечья 43 мм. Задние конечности покрыты волосами. Окраска волосяного покрова на спине серо-бурая, на брюхе бурая. Пучков волос на плечах нет.
Экология неизвестна. Встречается в горах на севере Эквадора на высоте 2000 м над уровнем моря.
Возможно, этот род следует объединять с родом желтоплечих листоносов.
Род брахифилла
Genus Brachyphylla Gray, 1834
Размеры средние и крупные. Длина тела 75—118 мм. Длина предплечья 55—67 мм. Хвост снаружи незаметен. Морда укороченная. Нижняя губа имеет V-образную бороздку, ограниченную по краю небольшими бородавками. Носового листка нет. Уши небольшие, широко расставленные, не соединяются между собой. Межбедренная перепонка развита хорошо. Волосяной покров на спинной стороне имеет желтую окраску, на брюшной обычно 2 12 3
бурую. Зубная формула: i — с — р — т— = 32. Щечные зубы широкие, с крупными гребнями.
День проводят в пещерах, обычно собираясь большими стаями. Питаются в основном насекомыми, но, по-видимому, поедают также и плоды.
Распространены на о-вах Куба, Гаити и Пуэрто-Рико, в Доминиканской Республике, на о-вах Барбадос, Малых Антильских.
В роде 4 вида:
сент-винсентский листонос — В. cavernarum Gray, 1834 (о-ва Пуэрто-Рико, Малые Антильские);
малый брахифилла — В. minor Miller, 1913 (о-ва Барбадос, Малые Антильские);
эль-гуамский листонос — В. папа Miller, 1902 (о-в Куба); гаитинский листонос — В. pumila Miller, 1918 (о-в Гаити).
Род листоносов-строителей
Genus Uroderma Peters, 1865
В роде единственный вид: листонос-строитель — U. bilobatum Peters, 1866.
Размеры мелкие и средние. Длина тела 54—74 мм. Хвост снаружи не виден. Длина предплечья 39—45 мм. Носовой листок хорошо развит, состоит из двух частей: седловидной основной и возвышающейся над ней вертикальной ланцетовидно-заостренной кверху. Уши средних размеров, широко расставлены, не соединены между собой. Задний край межбедренной перепонки без волос или покрыт редкими волосами. Окраска волосяного покрова на голове и туловище серовато-бурая. Края ушей желтовато-белые. На морде располагаются 4 белые продольные полосы. Вдоль средней линии спины проходит узкая белая полоса. Зубная формула:
2 12 3
i —с —р—т—= 32. Верхние резцы двудольчатые. Число диплоидных хромосом изменчиво: 38—42—44.
День проводит в листьях деревьев. На пальмах или бананах, обкусывая листья, иногда сооружает своеобразный зонтик, защищающий от света,
ветра и дождя. Питается плодами. Самка приносит детенышей в феврале, марте или апреле (в Панаме). Наблюдается и днем.
Распространен от южной Мексики до Перу, Боливии и Бразилии, а также на о-ве Тринидад.
Род белополосых широконосое
Genus Vampyrops Peters, 1865
Размеры мелкие и средние. Длина тела 43—95 мм. Длина предплечья 30—62 мм. Хвост снаружи незаметен. Носовой листок относительно хорошо развит. Он поднимается вертикально вверх от конца морды. Уши довольно короткие, с закругленными вершинами, широко расставлены и не соединяются между собой. Межбедренная перепонка узкая. Окраска волосяного покрова обычно темно-бурая или черная, реже серовато-бурая, дымчато-серая или бледно-бурая. По морде и средней линии спины часто проходят белые полосы. На морде таких полос бывает 4. Число зубов изменчиво за счет предкоренных и коренных зубов: от 26 до 32.
День проводят в самых разнообразных местах — в пещерах, настенах каньонов, на ветвях и среди листвы деревьев, а иногда в зданиях. Обычно на дневках держатся небольшими группами. Питаются в основном плодами. Биология размножения почти не изучена.
Распространены от южной Мексики на юг до Перу, Бразилии и Парагвая, обитают также на островах Тринидад и Тобаго.
В роде 18 видов:
эквадорский широконос — V. dorsalis Thomas, 1900 (Эквадор, Перу);
V. infuscus Peters, 1880 (Бразилия, Эквадор, Перу);
линейчатый широконос — V. lineatus Geoffroy, 1810 (от Парагвая и Бразилии до Венесуэлы и Колумбии);
венесуэльский широконос — V. oratus Thomas, 1914 (север Венесуэлы);
бразильский широконос — V. recifinus Thomas, 1901 (северо-восток Бразилии);
колумбийский широконос — V. umbratus Lyon, 1902 (север Колумбии);
широконос Хеллера — V. helleri Peters, 1866 (юг Мексики);
украшенный широконос — V. vittatus Peters, 1859 (Венесуэла, северная Колумбия и на север до Панамы и Коста-Рики);
V. zarhinus Н. Allen, 1891 (север Бразилии, Эквадор, Перу);
большой широконос—V. major G. Allen, 1908 (Гватемала, Панама);
широконос Караккиоло — V. caraccioloi Thomas, 1920 (север Бразилии, Перу);
V. melissa Thomas, 1926 (Перу);
большой желтоухий широконос — V. nymphaea Thomas, 1909 (Колумбия и Панама);
малый желтоухий широконос — V. pusilia Wagner, 1843 (восток Бразилии);
эквадорский желтоухий широконос — V. thyone Thomas, 1909 (Эквадор, Колумбия, на север до Коста-Рики);
перуанский желтоухий широконос — V. venilla Thomas, 1924 (Перу); гвианский желтоухий широконос — V. brocki Peterson, 1968 (Гвиана); желтоухий широконос Добсона — V. bidens Dobson, 1878 (север Перу и запад Бразилии).
Некоторые исследователи (Кабрера, 1957 и др.) выделяют из рода белополосых широконосов в качестве отдельных родов:
Vampyrodes Thomas, 1900, Vampyressa Thomas, 1900 и Vampyriscus Thomas, 1900. По-видимому, более правильно всех их объединить в качестве особых подродов в род Vampyrops.
Род большеглазых листоносов
Genus Chiroderma Peters, 1860
Размеры мелкие и средние. Длина тела 55—77 мм. Длина предплечья 37—53 мм. Хвост отсутствует. Носовой листок довольно крупный, имеет остроконечную вершину. Уши относительно небольшие, широко расставленные, между собой не соединяющиеся, вершины округленные. Межбедренная перепонка развита хорошо. Окраска волосяного покрова на спинной стороне буроватая, на брюшной несколько более светлая. На морде проходят белые продольные полосы, и одна такая полоса располагается вдоль середины спины. Иногда полосы отсутствуют. В черепе характерно 2 12 2
отсутствие носовых костей. Зубная формула: i — с — р — т~ ~ 28.
Экология изучена слабо. Питаются, по-видимому, плодами.
Распространены от южной Мексики через Центральную Америку до северной части Южной Америки, а также на о-ве Тринидад.
В роде 6 видов:
восточнобразильский листонос — С. doriae Thomas, 1891 (восточная Бразилия);
колумбийский листонос — С. jesupi J. Allen, 1900 (Колумбия);
центральноамериканский листонос — С. isthmicum Miller, 1912 (южная Мексика, Центральная Америка);
листонос Салвина — С. salvini Dobson, 1878 (южная Мексика, Центральная Америка, Колумбия, Эквадор);
венесуэльский листонос — С. villosum Peters, 1860 (Венесуэла);
малый большеглазый листонос — С. trinitatum Goodwin, 1958 (о-в Тринидад).
Род белых листоносов
Genus Ectophylla Н. Allen, 1892
Размеры небольшие. Длина тела 35—48 мм. Длина предплечья 25— 35 мм. Хвоста нет. Носовой листок хорошо развит, довольно узкий в основании, высокий, остроконечный. Уши средних размеров, широко расставлены, не соединяются друг с другом. Межбедренная перепонка глубоко вырезана. Окраска волосяного покрова очень своеобразная, белесая или буровато-белая. Глаза окружены узким кольцом темноватых волос. Число зубов 28—30.
Экология не изучена.
Распространены в Никарагуа, Коста-Рике, Панаме, Колумбии, Гайане, Бразилии, Эквадоре, Перу, на о-ве Тринидад.
В роде 2 вида:
белый листонос — Ё. alba Н. Allen, 1892 (Гондурас и Никарагуа);
листонос Макконелла — Е. macconelli Thomas, 1901 (от Гвианы и Бразилии в районе Амазонки до Эквадора и южного Перу).
Некоторые исследователи (Бурлиер, 1955 и др.) выделяют листоноса Макконелла в отдельный род Mesophylla Thomas, 1901.
Род фруктоядных листоносов
Genus Artibeus Leach, 1821
Размеры мелкие и средние. Длина тела 53—100 мм, длина предплечья 38—76 мм. Хвоста нет. Межбедренная перепонка узкая. Носовой листок хорошо развит, остроконечный. Уши небольшие, с заостренными вершинами, широко расставлены и не соединяются друг с другом. Окраска волосяного покрова на спинной стороне сероватая, темно-бурая или почти черная,
иногда с серебристым оттенком. Брюшная сторона обычно более светлая. На морде иногда имеются 4 светлые продольные полосы. Зубная формула:
I ~ с — р = 28 — 30 — 32.
2 12 2—3
День проводят в пещерах, дуплах деревьев, среди листьев; некоторые виды сооружают для себя убежище в листьях пальм или бананов в виде зонтика, надкусывая своеобразным образом крупные листья этих деревьев. Питаются плодами различных растений; фигами, манго, авокадо, бананами и др. Характерно необычайно быстрое прохождение пищевых частиц через пищеварительный тракт — всего за 15—20 мин. Способствуют расселению ряда растений. Иногда они могут приносить существенный вред плантациям.
Распространены от Мексики по всей Центральной Америке до северной Аргентины, Парагвая и Бразилии, а также в Вестиндии.
В роде 8 видов:
мохнатый листонос — A. hirsutus Andersen, 1906 (Мексика);
ямайский фруктоядный листонос — A. jamaicensis Leach, 1821 (Бразилия, Эквадор, Колумбия, Центральная Америка, Мексика, о-ва Большие и Малые Антильские):
большой фруктоядный листонос — A. lituratus Ollers, 1818 (Бразилия, Гвиана, Венесуэла, Колумбия, Эквадор, Перу, Парагвай, Аргентина, Боливия, Центральная Америка, Мексика, Малые Антильские острова);
серый листонос — A. cinereus Gervais, 1855 (Бразилия, Колумбия, Перу, Эквадор, Центральная Америка, Мексика);
карликовый фруктоядный листонос — A. nanus Andersen, 1906 (Мексика,. Центральная Америка на юг до Коста-Рики);
южномексиканский листонос — A. turpis Andersen, 1906 (юг Мексики);
гвианский листонос-—A. concolor Peters, 1865 (Гвиана);
A. harti Thomas, 1892 (Венесуэла, Эквадор, Ц. Америка до Гондураса, о-в Тринидад).
Некоторые исследователи (Кабрера, 1957 и др.) выделяют A. harti Tomas, 1892, в особый род Enehisthenes Andersen, 1906. По-видимому, более правильно рассматривать этот вид в составе рода Artibeus.
Род красных фруктоядных листоносов
Genus Stenoderma Ё. Geoffroy, 1818
Размеры мелкие. Длина тела 48—70 мм. Длина предплечья 40—54 мм. Хвоста нет. Носовой листок хорошо развит. Уши относительно небольшие, широко расставлены, но соединяются между собой. Окраска волосяного покрова на спине обычно рыжевато-бурая, реже темно-бурая или сероватая, на брюхе более светлая. На плечах могут быть белые пятна. Число зубов 30—32.
День проводят на ветвях деревьев. Питаются, по-видимому, в основном плодами.
Распространение охватывает о-ва Малые Антильские, Гаити, Кубу, Ямайку, Пуэрто-Рико.
В роде 8 видов:
монтсерратский листонос — S. montserratensis Thomas, 1894 (о-в Монтсеррат из группы Малых Антильских);
гваделупский листонос — S. annectens Miller, 1913 (о-в Гваделупа);
доминиканский древесный листонос — S. nichollesi Thomas, 1891 (о-в Доминика);
сент-люсийский древесный листонос — S. luciae Miller, 1902 (о-в Сент-Люсия);
кубинский серпокрылый листонос — S.falcatus Gray, 1839 (о-в Куба); серпокрылый листонос — S. haitiensis J. Allen, 1908 (о-в Гаити);
фигоядный листонос—S. f lave see ns Gray, 1831 (о-в Ямайка);
красный фруктоядный листонос—S.rufum Desmarest, 1820 (о-ва Пуэрто-Рико и Виргинские).
Возможно, монтсерратский, гваделупский и сент-люсийский листоносы являются лишь подвидами доминиканского древесного листоноса (Джонс и Шварц, 1967).
Некоторые исследователи (Холл и Келсон, 1959; Андерсон и Джонс, 1967 и др.) выделяют древесных листоносов в род Ardops Miller, 1906, серпокрылых листоносов — в род Phyllops Peters, 1865, фигоядных листоносов— в род Ariteus Gray, 1838. Вероятно, более правильно считать их отдельными подродами, входящими в род Stenoderma.
Род ипанемских листоносов
Genus Pygoderma Peters, 1863
.В роде единственный вид: ипанемский листонос — Р. bilabiatum Wagner, 1843.
Размеры небольшие. Длина тела примерно 60 мм. Длина предплечья 38 мм. Хвоста нет. Морда укорочена. На середине нижней губы находится крупный бородавчатый выступ, окруженный более мелкими бугорками. Межбедренная перепонка развита довольно хорошо. Уши относительно небольшие, широкие в основании, далеко расставленные друг от друга и не соединяющиеся между собой. Окраска волосяного покрова темно-бурая, на спине почти черная, на брюхе сероватая. На плечах около крыльев по белому пятну. Для черепа характерен сильно укороченный лицевой отдел. Число зубов 28.
Экология почти не изучена. Обитатель лесных районов. Питается в основном плодами.
Распространены в восточной части Южной Америки от Гвианы до Парагвая и в Центральной Америке на север до Мексики и на о-ве Тринидад.
Род складчатомордых листоносов
Genus Centurio Gray, 1842
В роде единственный вид: складчатомордый листонос — С. senex Gray, 1842 (см. рис. 105).
Размеры небольшие. Длина тела 55—70 мм. Длина предплечья 41— 44 мм. Хвоста нет. Лицевой отдел головы сильно укорочен и весь собран в крупные кожные складки. Волос в этой части головы нет. Носовой листок отсутствует. Уши относительно небольшие, с закругленными вершинами, широко расставлены и не соединяются между собой. Во время сна морда прикрывается складками, которые в активное время располагаются ниже подбородка. На горле находятся кожные специфические железы, продуцирующие секрет с сильным запахом.Окраска волосяного покрова наспинной стороне бурая, темно-бурая или желтовато-бурая с белыми пятнами на каждом плече, на брюшной более светлая. Череп относительно высокий с округленной мозговой коробкой. Лицевой отдел сильно укорочен. Костное небо короткое. Число зубов 28 или 30.
Экология не изучена.
Распространен в южной Мексике и Центральной Америке на юг до Панамы включительно.
Род сфаерониктерис
Genus Sphaeronycteris Peters, 1882
В роде единственный вид: сфаерониктерис — S. toxophyllum Peters, 1882. Некоторые исследователи включают этот вид в род Centurio.
Размеры небольшие. Длина тела около 53 мм. Длина предплечья примерно 40 мм. Хвоста нет. Носового листка нет. На конце морды на носу
имеется небольшой вырост, напоминающий рог носорога. Морда сильно укорочена. Кожные складки на морде выражены значительно слабее, чем у складчатомордого листоноса. Окраска волосяного покрова на спинной стороне светло-бурая, а на брюшной буровато-белая.
Экология не изучена.
Распространен в Венесуэле, Колумбии и Перу.
Род малых складчатомордов
Genus Ametrida Gray, 1847
Размеры мелкие. Длина тела 35—45 мм. Длина предплечья 25—34 мм. ' Хвоста нет. Голова сильно укорочена, однако, в отличиеот двух предыдущих родов, кожные складки на лицевой части выражены слабо и имеется носовой листок. Окраска волосяного покрова на спинной стороне черновато-бурая или темно-бурая с белым пятном на каждом плече около крыльев, на брюшной светлее. Зубная формула:
.2 1 2 3
i — с — р — т— = 32.
2 12 3
Экология не изучена.
Распространен в Бразилии, Гвиане и на о-ве Тринидад.
В роде 2 вида:
бразильский малый складчатоморд — A. centurio Gray, 1847(Бразилия и о-в Тринидад);
малый складчатоморд — A. minor Н. Alien, 1894 (Гвиана).
Некоторые исследователи включают эти виды в род Centurio.
Род светлых цветочных листоносов
Genus Phyllonycteris Gundlach, 1865
Размеры средние. Длина тела 75—83 мм. Длина хвоста около 12 мм. Длина предплечья примерно 50 мм. Морда сильно удлинена и довольно узкая. Носовой листок рудиментарен. Уши средних размеров, широко расставлены, не соединены между собой. Межбедренная перепонка очень узкая, и короткий хвост выступает из нее на значительную часть своей длины. Язык очень длинный, способный далеко высовываться изо рта. Его передняя часть покрыта высокими нитевидными сосочками. Окраска волосяного покрова серовато-белая или светло-желтовато-бурая. Зубная формула:
2 12 3
i — с — р ~^т~^ ~ 32. На вентральной поверхности мозговой коробки у Р. aphylla располагается своеобразная борозда.
День проводят в пещерах, собираясь крупными группами иногда до несколько тысяч особей. Питаются соком и мякотью плодов, а также, вероятно, нектаром и пыльцой. Самки кубинского цветочного листоноса приносят детенышей преимущественно в июне.
Распространены на о-вах Куба, Ямайка, Пуэрто-Рико, Гаити.
В роде 4 вида:
пуэрториканский цветочный листонос — Р. major Anthony, 1917 (о-в Пуэрто-Рико);
гаитинский цветочный листонос — Р. obtusa Miller, 1929 (о-в Гаити); кубинский цветочный листонос — Р. poeyi Gundlach, 1861 (о-в Куба); ямайский цветочный листонос — Р. aphylla Miller, 1898 (о-в Ямайка). Пуэрториканский и гаитинский цветочные листоносы известны в настоящее время только по остаткам скелета.
Род цветочных листоносов
Genus Erophytla Miller, 1906
Размеры средние. Длина тела 65—75 мм. Длина хвоста 12—17 мм. Длина предплечья 47—55 мм. Морда сильно вытянута и относительно тонкая. Язык очень длинный, снабжен на конце высокими нитевидными сосочками. Небольшой носовой листок имеет выемку на верхнем крае или даже раздвоен. Уши не очень крупные, примерно такой же высоты, как и ширины, широко расставлены и не соединяются между собой. Межбедренная перепонка узкая и хвост на значительную часть своей длины выступает из нее. Окраска волосяного покрова на спинной стороне бледно-желтовато-бурая или темно-бурая, на брюшной более светлая. Зубная формула:
• 2 1 2 3 Qo
1 — с — р — т— = 32.
2 1'2 3
День проводят в пещерах, причем забираются в самые отдаленные темные ее уголки. Собираются большими группами по нескольку сотен особей. Питаются плодами.
Распространены на о-вах Багамских, Кубе, Ямайке, Гаити, Пуэрто-Рико.
В роде 2 вида:
бурый цветочный листонос — Е. bombifrons Miller, 1899 (о-ва Пуэрто-Рико, Гаити);
темножелтый цветочный листонос — Е. sezekorni Gundlach, 1861 (о-ва Багамские, Куба, Ямайка).
СЕМЕЙСТВО ВАМПИРОВ
FAMILIA DESMODONTIDAE GILL, 1866
Размеры средние. Длина тела 65—90 мм. Длина предплечья 50—63 мм. Хвоста нет (рис. 108). Морда короткая, конической формы. Настоящий носовой листок отсутствует. Ноздри окружены кожными выростами, представляющими редуцированный носовой листок. Уши довольно маленькие,, далеко отстоящие друг от друга, не соединяются между собой, с заостренными вершинами. Козелок имеется. Межбедренная перепонка развита относительно слабо. Окраска волосяного покрова имеет различные оттенки бу-, рого цвета. У белокрылых вампиров края крыльев и часть летательной мембраны белые.
Череп не имеет посторбитальных отростков. Зубная формула:
i — с — р —- т ——— = 20 — 26. Верхние резцы большие, клыко-* видные, направлены вперед, имеют острый режущий край, приспособленный для разрезания кожи жертвы. Клыки крупные. При сомкнутых, челюстях вершины верхних резцов помещаются в глубокой бороздке на внутренней стороне нижней челюсти позади нижних резцов. Предкоренные зубы узкие, с острыми краями. Коренные зубы отсутствуют или развиты; очень слабо. Третья фаланга третьего пальца окостеневает. Пищевод очень короткий. Кардиальная часть желудка сильно удлинена и имеет узкий просвет.
Обитают в различных ландшафтах до 2100 м над уровнем моря. Населяют открытые равнины, пустыни, горы, леса. Встречаются в зарослях, кустарников, кактусов. Держатся в засушливых местах и во влажных лесных районах. Днем вампиры находят себе убежища главным образом в пещерах, а также в дуплах деревьев, строениях, колодцах и т. д. Здесь они. встречаются поодиночке, небольшими группами, иногда крупными скоплениями до тысячи и более особей. Активность начинается только с наступлением, темноты и достигает максимума к полуночи. Обычно вампиры летают низко над землей. Испускаемые ими локационные звуки имеют малую мощность —, значительно меньшую, чем у летучих мышей, питающихся насекомыми.;
Рис. 108. Обыкновенный вампир — D. rotundus:
1 — гнщ< вед. 2 — желудок, S — заднепроходное отверстие
Питаются вампиры только свежей'кровью. Они прямо опускаются на свою жертву или садятся на землю рядом и вскарабкиваются на нее. Отыскивают участок кожи без волос или перьев: область около заднепроходного отверстия, уши у млекопитающих, гребень у курицы и т. д. Затем поверхностным укусом скусывают своими острыми зубами небольшой кусочек кожи. Это почти безболезненно для жертвы и, как правило, не беспокоит спящее животное или человека. Обычно ранки бывают 3—6 мм ширины, б—10 мм длины и 1—5 мм глубины. Всасывание крови происходит весьма своеобразно. Вампир высовывает язык, причем его боковые части заворачиваются вниз. Таким образом создается трубка, образованная нижней поверхностью языка, его боковыми частями и глубокой бороздкой, которая находится посредине нижней губы. Медленными движениями языка вперед и назад достигается вакуум в ротовой полости, заставляющий кровь течь в рот. Спинка языка остается свободной от крови. Слюна вампиров содержит анти коагулирующие ферменты, препятствующие свертыванию крови. Слюна попадает в ранку жертвы, и кровь из нес может течь, не останавливаясь в продолжение нескольких часов (до 8 ч). Процесс питья крови у вампиров обычно не длится более получаса. Вампир может выпить так много крови, что бывает не в состоянии взлететь. Нападают почти на всех теплокровных животных. В некоторых областях разведение домашнего скота становится нецелесообразным из-за нападения большого числа вампиров; сама по себе потеря крови не является серьезной опасностью, однако вампиры могут передавать бешенство и чуму рогатого скота. Кроме того, открытые раны инфицируются бактериями и паразитическими личинками насекомых, и животные часто гибнут от этих причин.
Самка приносит одного детеныша. Сезонности в размножении, как правило, нет. В течение года самка приносит более одного помета. Беременность 90—120 дней.
Распространены в тропических и субтропических областях Америки от северной Мексики на юг до центральных районов Чили, Аргентины, Уругвая, Бразилии, а также на о-ве Тринидад.
В семействе 3 рода (3 вида):
обыкновенных вампиров — Desmodus Wied-Neuwied, 1824 (1 вид);
белокрылых вампиров — Diaemus Miller, 1906 (1 вид);
мохноногих вампиров — Diphylla Spix, 1823 (1 вид).
Некоторые ученые включают вампиров в качестве подсемейства в семейство американских листоносов.
Род обыкновенных вампиров
Genus Desmodus Wied-Neuwied, 1824
В роде единственный вид: обыкновенный вампир -— D. rotundas Ceof-froy,1810 (см. рис. 108).
Длина тела 75—90 мм. Длина предплечья 50—63 мм. Характерны заостренные на вершинах уши, удлиненный первый палец передней конечности с хорошо выраженной базальной подушкой, межбедренная перепонка не покрытая волосами. Окраска волосяного покрова на спине серовато-бурая, на брюхе более светлая. Зубная формула: i -у- с -у- р -|- т -у- = 20.
Обитатель влажных и засушливых областей тропиков и субтропиков. День проводит обычно в пещерах, а также в дуплах деревьев, в шахтах, зданиях. Обычно собирается большими группами по сотне, нескольку сотен, даже по нескольку тысяч особей. Самцы и самки находятся на дневках вместе. Характер питания см. в характеристике семейства. Беременность 90— 120 дней. Сезонности в размножении нет. Самка может приносить потомство дважды в течение года. Детенышей самка, как правило, оставляет на местах дневки. Длительность жизни примерно 12 лет.
: Распространение охватывает территорию от северной Мексики на юг ' до центральных районов Чили, центральной Аргентины и Уругвая.
Род белокрылых вампиров
Genus Diaemus Miller, 1906
В роде единственный вид: белокрылый вампир — D.youngi Jentink, 1893.
Длина тела примерно 85 мм. Длина предплечья 50—66 мм. Первый палец передних конечностей довольно короткий, у его основания находится большая кожистая подушка. Окраска волосяного покрова обычно светло-бурая. Края крыльев белые, летательная перепонка между вторым и третьим пальцами также имеет белый цвет. Зубная формула: ;±с J_p — т-?- = 22.
2 12 1
Дневки белокрылых вампиров обычны в дуплах деревьев, реже в пещерах. Собираются небольшими группами по нескольку десятков особей. Предпочитает питаться кровью птиц и коз.
Распространены в тропических областях Северной и Южной Америки от Мексики, Венесуэлы и Гвианы до Перу и Бразилии, также на о-ве Тринидад.
Род мохноногих вампиров Genus Diphylla Spix, 1823
В роде единственный вид: мохноногий вампир — D. ecaudata Spix, 1823.
Длина тела 65—87 мм, длина предплечья 50—56 мм. Уши короткие, с закругленными вершинами. Первый палец передней конечности укорочен
ный, не имеет в основании кожистой подушки. Межбедренная перепонка густо покрыта волосами. Окраска волосяного покрова на спине темно-бурая или красновато-бурая, на брюхе несколько более светлая. Зубная формула:
2 112
i — с — р — т — = 26. Наружный нижний резец имеет характерную
веерообразную форму с семью выступами по режущему краю.
День проводит обычно в пещерах, где большими группами не собирается, а встречается поодиночке, иногда до 12 или 20—30 особей. Нападает преимущественно на птиц, реже на лошадей и рогатый скот. Предполагается, что мохноногий вампир имеет определенный период размножения.
Распространен в южной Мексике, Центральной Америке, Перу и Бра
зилии
СЕМЕЙСТВО ВОРОНКОУХОВ1
FAMIL1A NATALIDAE MILLER, 1899
В семействе единственный род: воронкоухи — Natalus Gray, 1838. •Воронкоухи стоят наиболее близко к семействам дымчатых летучих мышей и американских присосконогов.
Размеры мелкие. Длина тела от 35 до 55 мм. Длина хвоста от 50 до 60 мм. Длина предплечья от 27 до 41 мм. Вес обычно 4—10 г. Хвост полностью включен в межбедренную перепонку. Глаза маленькие. Уши большие, воронковидной формы, поверхность их покрыта железистыми сосочками, подобно тому, как у рода Kerivoula (см. обыкновенных летучих мышей). Козелок короткий, различной толщины, форма близка к треугольной. Морда вытянутая, носового листка не имеет (рис. 109). Ноздри овальной формы, расположены близко друг к другу, открываются рядом с краем губы. Верхняя губа часто с поперечной бороздкой, широкая, несколько вывернутая вперед и наружу. Взрослые самцы имеют на лбу под кожей специфичный орган, представленный клетками, напоминающими строением сенсорные, но, возможно, имеющие и железистые функции. Иногда этот орган снаружи не заметен. Первый палец передней конечности очень короткий
и почти полностью включен в летательную перепонку, однако коготь этого пальца развит хорошо. Пальцы задних конечностей имеют длинные, слегка искривленные когти. Волосяной покров густой, мягкий и длинный. Окраска его изменчива: может быть серой, коричневатой, желтоватой или красноватой.
Удлиненный череп не имеет посторбитальных отростков. 2 13 3
Зубная формула: i — с — р — т— = 38. Верхние резцы с каждой 3 13 3
стороны отделены пространством друг от друга и от клыков. Первые и вторые предкоренные зубы хорошо развиты.
Второй палец передней конечности имеет метакарпальную кость, но не имеет фаланг. Третий, чертвертый и пятый пальцы с фалангами. На грудине есть низкий киль. Малая берцовая кость нитевидная, развита крайне слабо, ее проксимальные отделы хрящевые. Таз нормальный, но подвздошные кости обычно раздвинуты в сторону.
Обитатели тропических равнин. День проводят в пещерах, собираясь иногда в большом числе. Питаются насекомыми. Период размножения бразильского воронкоуха приходится на засушливый сезон (в Сальвадоре и южной Мексике).
Распространены от Мексики на юг через Центральную Америку до Гвианы, Бразилии, Колумбии, Венесуэлы, а также на о-вах Тринидад и Куба.
В роде 11 видов:
мексиканский воронкоух — N. mexicanus Miller, 1902 (Мексика и Центральная Америка);
Рис. 109. Голова бразильского во ронкоуха — N. stramineus и череп мексиканского воронкоуха — N те-xicanus
большой воронкоух — N. major Miller, 1902 (о-в Гаити);
доминиканский воронкоух — N. dominicensis Shamel, 1928 (о-в Доминика);
кубинский воронкоух — N. primus Anthony, 1919 (о-в Куба),
N. lepidus Gervais, 1837 (о-в Куба);
широколапый воронкоух — N. brevimanus Miller, 1898 (о-в Провидения у берегов Никарагуа);
длинноногий воронкоух — N. tnacer Miller, 1914 (о-ва Куба, Багамские);
ямайский воронкоух—N. micropus Dobson, 1880 (о-в Ямайка);
багамский воронкоух — N. tumidifrons Miller, 1903 (Багамские о-ва);
венесуэльский воронкоух — N. tumidirostris Miller, 1906 (Венесуэла и о-в Тринидад);
бразильский воронкоух — N. straw neus Gray, 1838 (Бразилия, Гвиана, о-в Тринидад).
Некоторые исследователи (Андерсол и Джонс, 1967 и др.) уменьшают число видов воронкоухов до 4.
СЕМЕЙСТВО ДЫМЧАТЫХ ЛЕТУЧИХ МЫШЕЙ
FAMILIA FURIPTERfDAE MILLER, 1907
Размеры небольшие. Длина тела 37—58 мм. Длина хвоста 24—36 мм. Длина предплечья 30—40 мм. Крылья длинные. Первый палец передней конечности развит крайне слабо и полностью включен в летательную перепонку: только очень небольшой нефункционирующий коготь выдается над ним. Уши небольшие, с округлыми краями (рис. 110). Козелок небольшой, треугольный, с широким основанием. Ноздри округлые или треугольные, открывающиеся вниз. Носового листка нет. Длинный хвост полностью включен в межбедренную перепонку и не выступает над ее поверхностью. Верхняя губа может иметь дисковидный мясистый выступ. Волосяной покров довольно грубый, бурых или серых оттенков.
В черепе нет посторбитальных отростков. Небные отростки межчелюстных костей редуцированы до хрящевых элементов. Зубная формула:
2 12 3
i — с — р —- т— = 36. Верхние кл ыки уменьшены в величине и по высоте примерно равны предкоренным зубам. Малая берцовая кость редуцирована. Киля на грудине нет. Имеется слабо выраженное дополни-
тельное сочленение плечевой кости с лопаткой. Иногда это сочленение отсутствует. <
Обитатели засушливых равнин. Экология изучена крайне слабо. День проводят, по-видимому, в пещерах или постройках человека.
Распространены в тропической зоне и северной части Южной Америки на юг до южной Бразилии и северного Чили, а также на о-ве Тринидад.
В семействе 2 рода:
восточных дымчатых летучих мышей— Furipterus Bonaparte, 1837 (1 вид);
обыкновенных дымчатых летучих мышей — Amorphochilus Peters, 1877 (1 вид).
Род восточных дымчатых летучих мышей
Genus Furipterus Bonaparte, 1837
Рис. ПО. Голова и череп'во-сточной дымчатой летучей мыши — F. horrens
В роде единственный вид: восточная дымчатая летучая мышь — F. horrens F. Cuvier, 1828 (см. рис. ПО).
Длина тела 37—40 мм. Длина хвоста 24—36 мм. Длина предплечья 30—40 мм. Окраска волосяного покрова буровато-серая, темно-серая или серовато-голубая, иногда на брюхе более светлая. И нижняя, и верхняя поверхности межбедренной перепонки покрыты волосами.
Экология не изучена.
Распространение охватывает Панаму, Колумбию, Гвиану, Бразилию и о-в Тринидад.
Род обыкновенных дымчатых летучих мышей
Genus Amorphochilus Peters, 1877
В роде единственный вид: обыкновенная дымчатая летучая мышь — A. schnablii Peters, 1877.
Длина тела 38—58 мм. Длина хвоста примерно 30 мм. Длина предплечья 34—37 мм. На морде и губах имеются бородавчатовидные кожные выросты. Окраска волосяного покрова темно-бурая или серая.
Экология не изучена.
Распространены на западном Эквадоре, западном Перу и севере Чили.
СЕМЕЙСТВО АМЕРИКАНСКИХ ПРИСОСКОНОГОВ
FAMILIA THYROPTER1DAE MILLER, 1907
В семействе единственный род: американских присосконогов — Thy-roptera Spix, 1823.
Мелких размеров летучие мыши. Длина тела 34—52 мм. Длина хвоста 25—33 мм. Длина предплечья 27—38 мм. У основания первого пальца передних конечностей и на подошве задних конечностей имеются своеобразные округлые присасывательные диски, которые прикрепляются к коже посредством небольших ножек (рис. 111). На передних конечностях они более крупные, чем на задних. Морда удлиненная. Уши относительно небольшие, воронковидные, с козелками. Носового листка нет. Форма ноздрей округлая, над ними находятся бородавчатые выросты. Конец хвоста немного
Рис. 111. Череп и присасывательные диски на передней (А) и задней (Б) конечностях трехцветного присосконога — Т. ^tricolor
выступает из межбедренной перепонки. Третий и четвертый пальцы задней конечности сращены друг с другом. Пальцы задней конечности состоят всего из двух фаланг. Коготь на первом пальце передней конечности развит слабо. Окраска волосяного покрова на спине красновато-коричневая, на брюхе белая или бурая.
В черепе костные слуховые барабаны небольших размеров. Посторбитальных отростков в черепе нет. Предчелюстные кости полные. Зубная фор-2 13 3
мула: i — с — р — пг— = 38. Верхние резцы отделены друг от друга 3 13 3
и от клыков широкими промежутками. Предкоренные зубы хорошо развиты в верхних и нижних челюстях. Малая берцовая кость редуцирована. Имеется второе дополнительное сочленение плечевой кости с лопаткой. На грудине киля нет.
Обитатели тропических лесов. День обычно проводят в свернутых листьях бананов или других растений с крупными листьями. Здесь они прикрепляются к гладкой поверхности листа своими присосками, обычно головой вверх. Присасывательная сила присосок настолько велика, что летучая мышь может удерживаться всего на одной присоске. Обычно в убежище находится по одной летучей мыши. Питаются насекомыми. В районе Амазонки самки приносят детеныша в начале сухого сезона. При полете самка носит детеныша с собой.
Распространены в Центральной Америке, в северной части Южной Америки и на о-ве Тринидад.
В роде 2 вида:
гондурасский присосконог — Т. discifera Lichtenstein et Peters, 1854 (Центральная Америка и северная часть Южной Америки от Гвианы до Колумбии и Перу);
трехцветный присосконог — Т. tricolor Spix, 1823 (от Колумбии до Перу, Бразилии, Гвианы, Венесуэлы, а также о-в Тринидад).
СЕМЕЙСТВО МАДАГАСКАРСКИХ ПРИСОСКОНОГОВ
FAMILIA MyZOPODIDAE THOMAS, 1904
В семействе единственный род: мадагаскарские присосконоги — Му-zopoda Milne-Edwards et Grandidier, 1878 с единственным видом: мадагаскарский присосконог — М. aurita Milne-Edwards et Grandidier, 1878
Размеры небольшие. Длина тела 57 мм. Длина хвоста 48 мм. Длина предплечья 46 мм. На подошве задних конечностей и в основании первого пальца передних конечностей находятся присоски, которые расположены
Рис. 112. Мадагаскарский присосконог — М. aurita, его голова и череп: а — ушной грибовидный отросток
непосредственно в кожном покрове и в отличие от американских присоско-ногов не имеют ножек (рис. 112). Голова с укороченной мордой. Носового листка нет. Уши крупные, со своеобразным грибовидным отростком в нижней части ушной раковины (см. рис. 112). Губы широкие, верхняя губа выступает значительно дальше вперед по сравнению с нижней. Хвост примерно на1/3своей длины выступает из межбедренной перепонки. Первый палец передней конечности имеет рудиментарный коготь. На задних конечностях пальцы имеют по две фаланги. Пальцы сращены между собой.
В черепе характерен укороченный лицевой отдел и крупный, округленный мозговой. Посторбитальных отростков нет. Зубная формула:
.2 1 3 3 оо
i — с — р — т — = 38.
3 13 3
Имеется дополнительное сочленение плечевой кости с лопаткой. Киль на грудине небольшой. Малая берцовая кость рудиментарная.
Экология не изучена.
Распространены на Мадагаскаре.
СЕМЕЙСТВО ОБЫКНОВЕННЫХ ЛЕТУЧИХ МЫШЕЙ
FAMILIA VESPERTILIONIDAE GRAY, 1821
Размеры от мелких до крупных. Длина тела 32—103 мм. Длина хвоста I 1 25—75 мм. Длина предплечья 22—75 мм. Носового листка нет, за исключением австралийских гладконосов. У трубконосов и шерстокрылых трубконосов имеются трубчатые ноздри. Глаза маленькие. Уши обычно средних размеров, иногда крупные, редко небольшие, обычно отделенные друг от I 1
друга. Козелок с заостренной или округлой вершиной хорошо развит и I 1
имеет различную форму (рис. 113). Крылья разнообразны по контуру. Тонкий хвост целиком включен в межбедренную перепонку, иногда выступает из нее на длину одного или двух концевых позвонков.
Рис. 113. Голова и череп большой ночницы — М. myotis
Волосяной покров от черноватого и серого до золотисто-желтого и красноватого. У некоторых видов на подошвах задних лап или у первого пальца передних конечностей (иногда в обоих этих местах) расположены присасывательные диски. На голове или крыльях могут располагаться специфические кожные железы. У большинства видов самки имеют по два соска и только у волосатохвостых гладконосов — четыре.
В черепе предчелюстные кости разделены друг от друга и не имеют небных отростков. Посторбитальных отростков нет. Зубная форму-1113 2133
ла варьирует от i — с — р — т— = 28 до i — с — р — т— = 38.
2123 3133
3
Резцы небольшие. Число коренных зубов всегда -у; на их жевательной поверхности хорошо развиты V-образные гребни. Намечается тенденция к уменьшению числа зубов и укорочению черепа и челюстей. Плечевая кость образует хорошо развитое дополнительное сочленение с лопаткой. Третий палец передней конечности имеет две костные фаланги. На грудине имеется крупный киль. Локтевая кость и малая берцовая кости редуцированы. Седьмой шейный позвонок свободен, за исключением томопеас.
День проводят в самых разнообразных местах: в пещерах, на ветвях, в дуплах и щелях коры деревьев, в расщелинах скал, шахтах, колодцах, зданиях. Огромное большинство видов по характеру питания насекомоядные, некоторые рыбоядные. Насекомых ловят, как правило, на лету, а также на земле. Крупных насекомых могут относить на места дневки и поедать там. Некоторые виды впадают в спячку или мигрируют к югу на холодный период года. Большинство видов, обитающих в умеренных зонах, спаривается в сентябре—октябре. Однако овуляция у самок и оплодотворение яйцеклетки происходит весной, в апреле—мае. Таким образом, сперматозоиды сохраняют свою жизнеспособность в половых путях самки в течение нескольких месяцев, на время которых летучая мышь впадает в спячку. Самки приносят, как правило, одного детеныша, реже двух, иногда трех, даже четырех. Самка обычно носит детенышей во время полета на себе. Держатся поодиночке, реже парами, иногда собираются крупными колониями. Колонии бывают представлены одним видом летучих мышей, иногда несколькими.
Распространены на необычайно широкой территории, охватывающей все материки, за исключением Антарктиды. Границы ареала совпадают с границей древесной растительности.
В семействе 23 рода (298 видов):
ночниц — Myotis Каир, 1829 (66 видов);
серебристых гладконосов — Lasionycteris Peters, 1865 (1 вид);
кожанов и нетопырей— Vespertillo Linnaeus, 1758 (113 видов);
вечерниц — Nyctalus Bowdich, 1825 (7 видов);
дисконогих гладконосов — Eudiscopus Conisbee, 1953 (1 вид);
стрелоухов — Otonycteris Peters, 1859 (1 вид);
сумеречных гладконосов — Nycticeius Rafinesque, 1819 (12 видов);
желтых гладконосов — Rhogeessa Н. Allen, 1866 (4 вида);
домовых гладконосов — Scotophilus Leach, 1821 (9 видов);
выростогубых гладконосов — Chalinolobus Peters, 1866 (7 видов);
волосатохвостых гладконосов — Lasiurus Gray, 1831 (7 видов);
широкоушек — Barbastella Gray, 1821 (2 вида);
ушанов — Plecotus Geoffroy, 1818 (4 вида);
пятнистых ушанов — Euderma Н. Allen, 1892 (1 вид);
длиннокрылов Miniopterus Bonaparte, 1837 (13 видов);
трубконосов Murina Gray, 1842 (9 видов);
шерстокрылых трубконосов — Harpiocephalus Gray, 1842 (1 вид);
украшенных гладконосов — Kerivoula Gray, 1842 (24 вида);
анамигдон — Anamigdon Troughton, 1929 (1 вид);
пустынных гладконосов — Antrozous Н. Allen, 1862 (3 вида);
австралийских гладконосов — Nyctophilus Leach, 1822 (9 видов);
ламингтонских гладконосов — Lamingtona McKean et Calaby, 1968 (1 вид);
томопеас — Tomopeas Miller, 1900 (1 вид).
В СССР 8 родов: ночниц, кожанов и нетопырей, вечерниц, стрелоухов, широкоушек, ушанов, длиннокрылов и трубконосов. Распространены по всей стране на север до границы леса.
Род ночниц
Genus Myotis Каир, 1829
Размеры от мелких до крупных. Длина тела 35—85 мм. Длина хвоста 40—65 мм. Длина предплечья 29—68 мм. Уши средних размеров, причем длина значительно превышает ширину (см. рис. 113). Козелок хорошо развит. Вершина козелка сужена и направлена прямо вверх или отклонена несколько кнаружи. Крылья относительно укороченные, широкие и тупые. Хвост весь заключен в межбедренную перепонку, лишь у некоторых видов
выступает из нее на длину одного—двух концевых хвостовых позвонков. Окраска волосяного покрова в основном бурая, с различными оттенками. Брюшная сторона обычно более светлая. Ночница Ходгсона характеризует-2 13 3
ся оранжевой окраской. Зубная формула: i — с — р — т— = 38.
3 13 3
Число диплоидных хромосом — 44.
По комплексу морфологических признаков ночницы являются наиболее примитивными среди всех родов семейства.
Обитатели различных ландшафтов. День проводят в пещерах, реже в зданиях, иногда на ветвях деревьев, кустов. На месте дневок собираются поодиночке, небольшими группами, иногда крупными колониями. В северных частях своего ареала некоторые виды впадают в спячку. Полету ночниц, как правило, медленный и прямой. Питаются насекомыми. По характеру строения задних конечностей у большеногой ночницы можно предполагать, что она питается рыбой. Самки приносят детенышей весной или летом. У зимоспящих видов спаривание происходит осенью. Беременность у малой бурой ночницы 50—60 дней, у большой ночницы до 70 дней. Половозрелость наступает в годовалом возрасте. Продолжительность жизни 6—7 лет, в неволе до 18,5 лет.
Распространены необычайно широко — более широко, чем другие млекопитающие. Ареал рода совпадает с ареалом всего семейства, за исключе-
нием некоторых океанических островов. В роде 66 видов:
усатая ночница — М. mystacinus Kuhl, 1819 (от Англии, Ирландии, Швеции, Норвегии, Бельгии, Франции, Испании, Швейцарии на восток до Камчатки и Сахалина; в Восточной Европе и Западной Сибири северная граница ареала проходит примерно по 62—63° с. ш., а восточнее — несколько к югу от 60° с. ш.; южная граница ареала от Швейцарии идет через Румынию к Черному морю, Кавказу, Ирану и Афганистану, огибает с юга Гималаи и продолжается к Южному Приморью);
малая ночница — М. muricola Gray, 1846 (от Южного Сахалина, южной Якутии, окрестностей Красноярска, Алтая и Закарпатья на юг до Бирмы, Непала, Бутана, Тенассерима и Лаоса);
трехцветная ночница — М. emarginatus Geoffroy, 1806 (Голландия, Франция, ФРГ, ГДР, Швейцария, Италия, Венгрия, Греция, Малайзия, юг европейской части СССР, Средняя Азия);
сычуанская ночница — М. altarium Thomas, 1911 (Сычуань и Гуйчжоу в Китае);
гималайская ночница — М. siligorensis Horsfield, 1855 (Кумаон, Непал, Сикким, Южный Китай, Бакбо, Таиланд);
длиннохвостая ночница — М. frater G. Allen, 1923 (Юго-Восточный Китай, Корейский полуостров, Приморский край, Алтай, Таджикистан, Узбекистан);
японская ночница — М. kaguyae Imaizumi, 1956 (о-в Хонсю);
рыжая ночница — М. bocagei Peters, 1870 (Ангола, Малави, Танзания, Кения, Камерун);
южноафриканская ночница — М. tricolor Temminck, 1832 (ЮАР, Уганда, Кения, Эфиопия);
ночница Наттерера — М. nattereri Kuhl, 1818 (Англия, Ирландия, Франция, Швейцария, Испания, Италия, Голландия, Дания, Швеция, Норвегия, Польша, ФРГ, ГДР, европейская часть СССР, Средняя Азия, Южная Сибирь к востоку от Иркутска, долина Амура, Япония, Корейский полуостров, Северо-Восточный Китай);
длинноухая ночница — М. bechsteini Kuhl, 1818 (Англия, Франция, Бельгия, Голландия, Испания, Швейцария, ФРГ, ГДР, Венгрия, Швеция, Польша, Литва, Украина, Кавказ);
большая ночница — М. myotis Borkhausen, 1797 (Франция, Швейцария, Италия, Сардиния, Испания, Португалия, ФРГ, ГДР, Польша, Венгрия, Румыния, южная Швеция и на восток досоветских Карпат включитель-но, а также Иран, Афганистан, Шэньси, Сычуань, Юньнань, Фуцзянь в Китае);
остроухая ночница — М. blythi Tomes, 1857 (Испания, Швейцария, Австрия, Италия, Сардиния, Мальта, Греция, Крит, юг европейской части СССР, Алжир, Тунис, Марокко, Малая Азия, Израиль, Иран, Центральная Азия, Средняя Азия, Алтай);
сиккимская ночница —М. sicarius Thomas, 1915 (ЮАР, южная Родезия, Малавия);
ночница Велвитша — М. welwitschi Gray, 1866 (Ангола);
стройная ночница — М. formosus Hodgson, 1835 (Южный Китай, Непал, Пенджаб, Кумаон, Корейский полуостров, Тайвань, Южная Япония);
водяная ночница — М. daubentoni Kuhl, 1819 (Англия, Ирландия, Норвегия, Швеция, Франция, Швейцария, Голландия, Дания, Испания, Италия, ГДР, ФРГ, Румыния, Польша, СССР на восток до Камчатки и Сахалина; северная граница ареала идет примерно по 60° с. ш., а южная проходит через южную Украину, Нижнюю Волгу, Северный Казахстан, Алтай, северную Монголию, до Южного Приморья и Южного Сахалина, Северо-Восточный Китай, МНР, Фуклен);
длиннопалая ночница — М. capaccinii Bonaparte, 1837 (Южная Франция, Испания, Италия, Швейцария, Сардиния, Трансильвания, Болгария, низовья Амударьи, Южное Приморье, Южные Курилы, Япония, Марокко, Алжир);
пекинская ночница — М. pequinius Thomas, 1908 (окрестности Пекина, Китай); -
ночница Давида — М. davidi Peters, 1869 (окрестности Пекина, Хайнань и Гуаней в Китае);
малайзийская ночница — М. adversus Horsfield, 1824 (Малайзия, Ява, Австралия, а также, вероятно, Калимантан, Суматра, Сулавеси, Таиланд, Цейлон, Тайвань, Андаманские острова, Южная Индия);
крупнопалая ночница — М. macrodactylus Temminck, 1840 (Япония);
прудовая ночница — М. dasycneme Boie, 1825 (Голландия, Бельгия, Северная Франция, Дания, Швеция, Польша, между 49 и 63°с. ш. на восток до Енисея);
ночница' Шталькера — М. stalkeri Thomas, 1910 (о-ва Ара и Кей);
ночница 'Риккета — М. ricketti Thomas, 1894 (Китай);
ангольская железистокрылая ночница — М. seabrai Thomas, 1912 (Ангола, Юго-Западная Африка, ЮАР);
железистокрылая ночница Лесуеура — М. lesueuri Roberts, 1919 (ЮАР);
африканская ночница — М. africanus Dobson, 1875 (Габон);
ночница Гудота — М. goudotii Smith, 1834 (Мадагаскар);
ночница Скотта — М. scotti Thomas, 1927 (Эфиопия);
танзанийская ночница — М. venustus Matschie, 1899 (Танзания);
ночница Хорсфилда — М. horsfieldii Temminck, 1840 (Малайзия, о-ва Ява, Бали);
калимантанская ночница — М. lepidus Thomas, 1915 (о-в Калимантан);
ночница'Хасселта — М. hasseltii Temminck, 1840 (Малайзия, о-ва Суматра, Ява);
ночница Бартелса — М. bartelsii Temminck, 1910 (о-в Ява);
ночница Германа •— М. hermani Thomas, 1923 (о-в Суматра);
М. oreias Temminck, 1840 (Малайзия);
самарская ночница — М. rufopictus Waterhouse, 1845 (о-ва Самар, Лусон);
большеногая ночница — М. macrotarsus Waterhouse, 1845 (о-ва Лусон и Минданао);
филиппинская ночница — М. jeanni Taylor, 1934 (о-в Минданао);
минданаоская ночница — М. patriciae Taylor, 1934 (о-в Минданао);
ночница Брауна — М. browni Taylor, 1934 (о-в Минданао);
ночница Херре — М. herrei Taylor, 1934 (о-в Лусон);
малая бурая ночница — М. lucifugus Le Conte, 1831 (Аляска, Канада— кроме крайнего севера, США — кроме крайнего юга);
ночница Юма — М. yumanensis Н. Allen, 1864 (Британская Колумбия в Канаде, запад и юго-запад США, северная Мексика);
юго-восточная ночница — М. austroriparius Rhoads, 1897 (юго-восток США);
серая ночница — М. grisescens К. Howell, 1909 (юго-восток США);
пещерная ночница — М. velifer J. Allen, 1890 (юго-запад США; Мексика, Гватемала);
светлобурая ночница — М. fortidens Miller et G. Allen, 1928 (юго-запад и юг Мексики);
аризонская ночница — М. occultus Hollister, 1909 (штаты США — Аризона, Нью-Мехико, Техас, а также северная Мексика);
ночница Кина — М. Леоти Merriam, 1895 (северо-запад, центр и восток США);
монтерейская ночница — М. evotis Н. Allen, 1864 (юго-запад Канады, запад США, Мексика);
ночница Миллера —. М. milleri Elliot, 1903 (Баха Калифорния);
бахромчатая ночница — М. thysanodes Miller, 1897 (запад США, Мексика);
индианская ночница — М. sodalis Miller et G. Allen, 1928 (восток и центр США);
длинноногая ночница — М. volansH. Allen, 1866(Британская Колумбия и Альберта в Канаде; запад и юго-запад США; север Мексики и Халиско);
калифорнийская ночница М. californicus Audubon et Bachman, 1842 (Британская Колумбия в Канаде, запад США, северная Мексика);
плоскоголовая ночница — М. planiceps Baker, 1955 (штат Коахуила в Мексике);
малоногая ночница — М. subulatus Say, 1823 (юго-запад Канады, большая часть территории США, север Мексики);
черная ночница — М. nigricans Schinz, 1821 (Центральная Америка и Южная Америка от Венесуэлы и Колумбии до Бразилии, Аргентины и Перу);
парагвайская ночница — М. albescens Е. Geoffroy St. Hilaire, 1806 (почти вся Южная Америка и Центральная Америка на север до Никарагуа);
серебристая ночница — М. argentatus Dalquest et Hall, 1947 (Веракрус в Мексике);
ночница Чилоэ — М. chiloensis Waterhouse, 1838 (Бразилия, Уругвай, Эквадор, Боливия, Перу, Аргентина, Чили);
красная ночница — М. ruber Geoffroy, 1806 (от Бразилии до Парагвая, Аргентины и северного Уругвая);
курносая ночница — М. simus Thomas, 1901 (область Амазонки от Эквадора и севера Перу до штата Пара в Бразилии);
рыбоядная ночница — М. vivesi Menegaux, 1901 (Баха Калифорния, Сонора в Мексике).
Некоторые исследователи выделяют рыбоядную ночницу в род Pizonyx Miller, 1906, а железистокрылых ночниц— в род Cistugo Thomas, 1912.
В СССР: ночницы усатая (вся территория страны к северу примерно до 62—63° с. ш. в европейской части и Западной Сибири и примерно до 60°с. ш. в Восточной Сибири), малая [Южный Сахалин, Приморский и Хабаровский края, юго-восточная Сибирь к западу до Алтая (Катон-Ка-рагай, Телецкое озеро) и к северу до верхнего течения реки Лены (55° с. широты) и южной Якутии, а также Закарпатье]; трехцветная (Закарпатье, Крым, Закавказье, южная Туркмения, Узбекистан, Таджикистан); длиннохвостая (Владивосток и Иманский район в Приморском крае, Красноярская область, Алтай, юго-западный Таджикистан, окрестности Самарканда в Узбекистане); Наттерера (европейская часть СССР к северу до Эстонии, Ленинграда и южной части Кировской области и на восток до Татарской АССР, окрестности Симферополя, Дагестан, многие пункты Закавказья, северо-западный Копетдаг, р. Иркут в Восточных Саянах, ст. Тунгир в южной Якутии, долина Амура, окрестности Владивостока); длинноухая (Литва, Ольгополь на Украине, Карпаты, Кавказский заповедник, Ставрополь и Западное Закавказье); большая (области: Закарпатская, Ивано-Франковская, Львовская, Тернопольская, Черновицкая, Хмельницкая, Одесская и в Белоруссии — Брестская и Гродненская); остроухая (Молдавия, Одесская обл., Крым, Кавказ, Закавказье, Туркмения, Таджикистан, южный Узбекистан, Киргизия, южный Казахстан, северо-западный Алтай); водяная (вся территория СССР к северу до Петрозаводска, Чердыни, Тобольска, Красноярска, устья Амура; южная граница проходит через Сиваш, низовья Волги, Южный Урал, севернее пустынь и степей Средней Азии и Казахстана к оз. Зайсан и далее почти по государственной границе до Посвета); длиннопалая (Нукус в низовье Амударьи, долины рек Сучан и Шмаковка в Южном Приморье, Южные Курилы); прудовая (европейская часть и Западная Сибирь к северу примерно до 58—63 ° с. ш. и к югу до 49° с. ш.).
Индианская ночница хотя и достигает численности примерно 500 тыс. особей, однако обитает в столь узком ареале, что есть опасность ее исчезновения; поэтому она включена в «Красную книгу».
Род серебристых гладконосов
Genus Lasionycteris Peters, 1865
В роде единственный вид: серебристый гладконос — L. noctivagans Le Conte, 1831.
Размеры небольшие и средние. Длина тела 55—65 мм. Длина хвоста около 40 мм. Длина предплечья примерно 42 мм. Уши относительно короткие, их длина примерно равна ширине. Волосы, покрывающие туловище, двухцветные: основание их темно-бурое или черное, вершина серебристая. Это придает своеобразную серебристую окраску всему волосяному покрову. На брюхе серебристость выражена слабее. Характерен густой волосяной покров на верхней части основания межбедренной перепонки. Число диплоидных хромосом — 20.
Обитатель лесных районов вдоль рек, встречается в горных лесах. День проводит, как правило, в дуплах деревьев, иногда среди листвы, на ветвях деревьев, в зданиях, реже в пещерах. Держится на дневках поодиночке или небольшими группами. В некоторых частях ареала ведет оседлый образ жизни и впадает на холодный период в спячку, в других мигрирует на юг (в восточных областях США). Питается насекомыми. Самка приносит одного или двух детенышей в июне или июле. Детеныши начинают летать с 3-недельного возраста.
Распространен от восточного до западного побережий южной Канады и США.
Род кожанов и нетопырей
Genus Vespertilio Linnaeus, 1758
Размеры сильно изменчивы: мелкие, средние и крупные (наибольшие в семействе). Длина тела 35—103 мм. Длина хвоста 25—60 мм. Длина предплечья 22—72 мм. Наружные ушные раковины средних размеров (рис. 114). Козелок хорошо развит, различной формы, расширенный в средней части или булавовидный или расширенный в основной трети и постепенно суживающийся к вершине. Крылья относительно широкие. Хвост или полностью включен в межбедренную перепонку, или на 3—7лии выдается за перепонку волосяного покрова. Окраска на спине светло-бурая, темно-бурая, красновато-бурая, черная, на брюхе светло- или темно-бурая, серая, беловато-серая или белая. У кожана Флоуэра и роговэкожего кожана передние конечности и хвост покрыты небольшими ороговевшими участками. У толстопалого, яванского и тасманийского нетопырей, косолапого, малайского и калимантанского кожанов на передних конечностях около первого пальца и на стопе задних конечностей есть присасывательные подушки (см. рис. 115). Мозговая коробка у большинства видов уплощенная, лишь у примитивных нетопырей мозговая капсула довольно высокая. Сагиттальный гребень не развит. Лямбдоидальный гребень развит хорошо. Костные слуховые пузыри небольших или средних размеров. Зубная фор-2 1 1__2 3
мула: i — с —Р —~—пг —= 32 — 34. Число хромосом в диплоидном наборе 26—50.
Обитают в различных ландшафтах — лесных и открытых. День проводят в пещерах, расщелинах скал, на ветвях, среди листвы и в дуплах деревьев, в зданиях. На охоту вылетают довольно рано, до наступления сумерек. Питаются различными насекомыми. В северных частях ареала впадают в спячку или на холодный период года мигрируют к югу. Самки рождают одного-двух детенышей, иногда трех. Большинство видов приносят детенышей в мае—июле.
Распространение почти полностью соответствует ареалу семейства обыкновенных летучих мышей.
Рис. 114. Голова и череп кожановидного нетопыря — V. savii
Рис. 115. г Присоски-" на передней (Л) и задней (Б) конечностях косолапого кожана—V. pachy-pus
В роде 116 видов:
двуцветный кожан — V. murinus Linnaeus, 1758 (от Англии, Норвегии, Швеции, Дании, Франции, Швейцарии, Австрии на восток до тихоокеанского побережья СССР, северная граница проходит примерно по 60° с. ш., южная — по Черному морю, Кавказу, северо-западным Гималаям, южной Туркмении, Ирану, Кашмиру, МНР, Кашгару, Уссури, Японии);
восточный кожан — V. superans Thomas, 1890 (от Ордоса до Корейского полуострова и советского Приморья, МНР, Китай, Япония);
синдский кожан — V. nasutus Dobson, 1877 (Синд, Пенджаб, Аравийский полуостров, Иран);
кожан Уалла — V. walli Thomas, 1919 (Ирак);
толстоухий кожан — V. pachyotis Dobson, 1871 (Ассам);
северный кожанок — V. nilssoni Keyserling et Blasius, 1839 (от Норвегии, Швеции, Франции, Дании, Швейцарии, Северной Италии на восток до тихоокеанского побережья СССР, в европейской части ареала заходит за Полярный круг — на Кольском полуострове, а в Сибири — примерно до 60° с. ш., южная граница ареала на запад проходит севернее Московской и Горьковской областей, а на востоке по центральноазиатским пустыням, Кашмиру, МНР, Тибету, Северо-Восточному Китаю, Корейскому полуострову);
японский кожанок—V. japonensis Imaizumi, 1953 (о-в Хонсю);
кожанок Бобринского — V. bobrinskii Kuzyakin, 1935 (Казахстан и Средняя Азия);
серовато-желтый кожан — V. isabellinus Temminck, 1840 (Ливия, АРЕ, Аравийский полуостров);
кожан Огнева — V. ognevi Bobrinskiy, 1918 (Туркмения, Узбекистан, Казахстан, юго-западная часть МНР);
поздний кожан — V. serotinus Schreber, 1774 (от Англии, Франции, Швейцарии, Италии, Западной Африки на восток через Центральную и Южную Европу, Кавказ, Малую, Среднюю и Центральную Азию до Корейского полуострова, северная граница ареала проходит через Варшаву, Харьков, Оренбург, оз. Балхаш);
капский кожан — V. capensis Smith, 1829 (ЮАР, Мозамбик, южная Родезия, Ботсвана, Юго-Западная Африка, Ангола, Замбия, Республика Заир, Кения> Сомали);
желтоватый кожан — V. flavescens Seabra, 1900 (Ангола);
кожан южноафриканский — V. melckorum Roberts, 1919 (ЮАР);
длиннохвостый кожан — V. hottentotus Smith, 1833 (ЮАР, Юго-Западная Африка, Малави);
малый кожан — V. pusillus Leconte, 1857 (Ангола, Республика Заир, Танзания, Габон);
зулусский кожан — V. zuluensis Roberts, 1924 (ЮАР, Ботсвана, Юго-Западная Африка);
белокрылый кожан — V. tenuipinnis Peters, 1872 (Ангола, Конго-. Браззавиль, Республика Заир, Танзания, Габон);
кожан Рендалла — V. rendalli Thomas, 1889 (Малави, Мозамбик, Республика Заир, Кения, Судан и на западе Гамбии);
ангольский кожан — V. bicolor Bocage, 1889 (Ангола);
западноафриканский кожан — V. brunneus Thomas, 1880 (Западная Африка);
фараджский кожан — V. faradjius J. Allen, 1917 (Республика Заир);
гарамбский кожан —V. garambae J. Allen, 1917 (Республика Заир);
кожан Грандидье — V. grandidieri Dobson, 1876 (о-в Занзибар);
гвинейский кожан — V. guineensis Bocage, 1889 [(Гвинея (Бисау)];
кожан Хумблота — V. humbloti Milne- Edwards, 1881 (о-в Мадагаскар);
арабский кожан — V. innesi Lataste, 1887 (APE);
кожан Ловена — V. loveni Granvik, 1924 (Кения);
мадагаскарский кожан — V. matroka Thomas et Schwann, 1905 (о-в Мадагаскар);
кожан-привидение— V. phasma G. Allen, 1911 (Кения);
ганский кожан — V. platyops Thomas, 1901 (Гана);
суданский кожан — V. rectitragus Wettstein, 1916 (Судан);
угандский кожан — V. ugandae Hollister, 1916 (Уганда);
verecundus Chasen, 1940 (Малайзия);
таиландский кожан — V. demissus Thomas, 1916 (Таиланд);
большой бурый кожан — V. fuscus Palisot de Beauvois, 1796 (южная часть Канады, США, Мексика, Центральная Америка и северо-восток Южной Америки);
кожан Линна — V. lynni Shamel, 1945 (о-в Ямайка);
бразильский кожан — V. brasiliensis Desmarest, 1819 (южная Мексика, Центральная Америка, Колумбия, Эквадор, Перу, Аргентина и от Гвианы до Уругвая);
V. albigularis Peters, 1872 (юг Мексики, Гватемала, Гондурас);, панамский кожан — V. chiriquinus Thomas, 1920 (Панама);
V. diminutusOsgood, 1915 (Бразилия);
аргентинский кожан — V. fidelis Thomas, 1920 (Аргентина, Уругвай, Бразилия);
V. furinalis D’Orbigny, 1847 (Аргентина, Парагвай);
V. innoxius Gervais, 1841 (Эквадор, Перу);
чернокрылый кожан — V. melanopterus Jentink, 1904 (Гвиана, Бразилия);
перуанский кожан — V. montosus Thomas, 1920 (Перу, Боливия);
роговокожий кожан — V. notius G. Allen, 1908 (ЮАР);
кожан Флоуэра — V. floweri De Winton, 1901 (Судан);
кожан Лове — V. lowei Thomas, 1915 (Судан);
кожан Тиккела — V. tickelli Blyth, 1851 (Индия, Цейлон);
кожан Бланфорда — V. blanfordi Dobson, 1877 (Тенассерим, Малаккский полуостров);
ложный кожан — V. doriae Peters, 1968 (о-в Калимантан);
кожан Томеса — V. tomesi Thomas, 1914 (Малайзия);
косолапый кожан — V. pachypus Temminck, 1840 (Южный Китай, Бирма, Манипур, Сикким и на юг до Юго-Западной Индии, Лаос, Малайзия, о-ва Калимантан, Ява, Бали, Суматра, Лусон);
калимантанский кожан — V. robustula Thomas, 1915 (Юньнань в Китае, Лаос, Аннам, Малаккский полуостров, о-ва Суматра, Ява, Калимантан, Бали, Сулавеси, Тимор);
малайский кожан — V. malayana Chasen, 1940 (Малайзия);
кожан Молони — V. moloneyi Thomas, 1891 (Ангола, Замбия, Республика Заир, Нигерия, о-в Фернандо-По, Сьера-Леоне);
новогвинейский кожан — V. rohui Thomas, 1902 (Новая Гвинея);
южноавстралийский кожан — V. pumilus Gray, 1841 (Южная Австралия, Виктория, Новый Южный Уэльс);
больцзекрылый кожан — V. alienus Thomas, 1916 (Бразилия);
V. macrotus Poeppig, 1835 (Боливия, Аргентина, Перу, Чили);
горный кожан — V. montanus Philippi et Landbeck, 1861 (Колумбия, Эквадор, Перу, Чили, Аргентина, Уругвай);
кожан Жоффруа — V. velatus J. Geoffroy, 1824 (Бразилия);
кожан Уинтона — V. wintoni Thomas, 1901 (Ангола, Замбия, Кения);
нетопырь-карлик — V. pipistrellus Schreber, 1774 (Англия, Ирландия, Швеция, Дания, Норвегия, Франция, Швейцария, Испания, Италия и на восток через южные и центральные области европейской части СССР и Среднюю Азию до Западного Китая; известен из Японии, с Корейского полуострова, из Тайваня, Малой Азии, Ирана, Кашмира, Марокко, Экваториальной Африки);
лесной нетопырь — V. nathusii Keyserling et Blasius, 1839 (Франция, Швейцария, Испания, Италия, ФРГ, ГДР, Польша, Голландия, Дания, Венгрия и на восток до Оренбурга; на север до балтийского побережья, Ленинграда, низовий р. Вятки; на юг до Средиземного и Черного морей, Ирана и Турции, Израиля);
восточный нетопырь •— V. abramus Temminck, 1840 (от Южно-Уссурийского края и Японии до Индии, Цейлона, Малайского архипелага, Филиппинских островов и северной Австралии включительно);
нетопырь Эндо — V. endoi Imaizumi, 1959 (о-в Хонсю);
Коромандельский нетопырь — V. coromandra Gray, 1838 (Южный Китай, Хайнань, п-ов Индокитай, Бирма, Бутан, Сикким, Кумаон, п-ов Индостан, о-в Цейлон);
индийский карликовый нетопырь — V. mimus Wroughton, 1899 (Цейлон, большая часть Индии, Бирма, п-ов Индокитай);
шоколадный нетопырь — V. ajfinis Dobson, 1871 (Бирма);
серый нетопырь — V. pulveratus Peters, 1871 (Южный Китай);
тенассеримский нетопырь — V. lophurus Thomas, 1915 (Тенассерим); цейлонский нетопырь — V. ceylonicus Kelaart, 1852 (Цейлон, Индия); средиземноморский нетопырь—V. kuhli Kuhl, 1819 (Франция, ФРГ, ГДР, Швейцария, Италия, Испания, Балеарские острова, Сардиния, Греция, Крым, Кавказ, низовья Амударьи, Малая Азия, Иран, Афганистан, Израиль, Аравийский полуостров, Синд, Кашмир, АРЕ, Алжир, Марокко, Кения, ЮАР);
индийский нетопырь — V. babu Thomas, 1916 (Индия, Непал);
пустынный нетопырь — V. deserti Thomas, 1902 (Ливия);
кожановидный нетопырь — V. savii Bonaparte, 1837 (Франция, Швей
цария, Италия, Испания, Греция, Крым, Кавказ, юго-западная Туркмения, Устюрт, Западный Тянь-Шань, Южное Приморье, МНР, Сикким, Ассам, Бирма);
мадейрский нетопырь — V. maderensis Dobson, 1878 (о-в Мадейра и Канарские острова);
египетский нетопырь — V. artel Thomas, 1904 (Юг АРЕ);
черный нетопырь — V. circumdatus Temminck, 1840 (о-в Ява, Северная Бирма, Индия);
V. mordax Peters, 1866 (о-ва Ява, Цейлон, Индия);
средний нетопырь — V. annectens Dobson, 1871 (Ассам, о-в Суматра);
нетопырь Руппела — V. riippelli Fischer, 1829 (Судан, Уганда, Ангола, Ботсвана на север до АРЕ, Ирак);
нетопырь Дормера — V. dormeri Dobson, 1875 (Индия, о-в Тайвань);
большой нетопырь — V. io Thomas, 1902 (Китай);
банановый нетопырь — V. nanus Peters, 1852 (ЮАР, Мозамбик, Ангола, Замбия, Малави и далее к северу до Республики Заир, Танзания, Уганда, Кения);
малый нетопырь — V. culex Thomas, 1911 (Юго-Западная Африка, Ангола, Нигерия);
рыжий нетопырь — V. rusticus Tomes, 1861 (ЮАР, Ю.-З. Африка);
ангольский нетопырь — V. anchietai Seabra, 1900 (Ангола);
абасский нетопырь — V. abaensis J. Allen, 1917 (Республика Заир);
кенийский нетопырь — V. aero Heller, 1912 (Кения);
толстый нетопырь — V. crassulus Thomas, 1904 (Камерун);
солнечный нетопырь — V. hellos Heller, 1912 (Кения);
марасский нетопырь— V. marrensis Thomas et Hinton, 1923 (Судан);
маврский нетопырь — V. maurus Blasius, 1853 (Канарские острова); камерунский нетопырь — V. musciculus Thomas, 1913 (Камерун);
карликовый камерунский нетопырь — V. nanulus Thomas, 1904 (Камерун);
нетопырь Пагенштехера — V. pagenstecheri Noack, 1889 (Республика Заир);
нетопырь-красавец — V. pulcher Dobson, 1875 (о-в Занзибар);
нетопырь Штампфла — V. stampflii Jentinck, 1888 (Либерия);
суматранский нетопырь — V. brachypterus Temminck, 1840 (Суматра и, возможно, Ява);
тонкий нетопырь — V. tenuis Temminck, 1840 (Малайзия, о-ва Суматра, Калимантан, Ява, Минданао);
нетопырь Мюррея — V. murrayi Andrews, 1900 (о-в Рождества);
нетопырь Ридли — V. ridleyi Thomas, 1898 (Малайзия);
индонезийский нетопырь — V. imbricatus Horsfield, 1824 (о-ва Ява, Сулавеси, Бали, Суматра, Калимантан, Лусон, Малайзия);
островной нетопырь — V. angulatus Peters, 1880 (архипелаг Бисмарка, о-ва Адмиралтейства, Новая Гвинея, Соломоновы);
папуасский нетопырь — V. papuanus Peters et Doria, 1881 (Новая Гвинея, о-ва Мисол, Амбоина, Ару, Кей, Церам);
нетопырь Петерса — V. petersi Meyer, 1899 (о-в Сулавеси);
нетопырь Минахасса — V. minahassae Meyer, 1899 (о-в Сулавеси);
западный нетопырь — V. hesperus Н. Allen, 1864 (западная часть США и Мексика);
восточноамериканский нетопырь — V. subflavus F. Cuvier, 1832 (восточная часть США, восток Мексики, Гватемала, Гондурас, Никарагуа);
толстопалый нетопырь — V. tylopus Dobson, 1875 (Бирма, Малайзия, о-ва Суматра, Калимантан, возможно, до Филиппинских);
яванский нетопырь — V. javanus Chasen, 1939 (о-в Ява);
японский нетопырь — V. endoi Imaizumi, 1959 (о-в Хонсю, Япония); тасманийский нетопырь — V. tasmaniensis Gould, 1858 (Австралия и Тасмания).
Некоторые исследователи выделяют из состава рода Vespertilio следующие роды: Pipistrellus Каир, 1829, куда включают около 50 видов; Gli-schropus Dobson, 1875 с 3 видами; Eptesicus Rafinesque 1820 с 30 видами; Rhinopterus Miller, 1960 с 2 видами; Hesperoptenus Peters, 1868 с 4 видами; Tylonycteris Peters, 1872 с 3 видами; Mimetillus Thomas, 1905 с 1 видом; Phyletor Thomas, 1902 с 1 видом; Histiotis Gervais, 1855 с 3 видами; Laephotis Thomas, 1901 с 1 видом и собственно в роде Vespertilio насчитывают 2 вида. Исследования А. П. Кузякина (1950, 1965) показали, что более правильно объединять все перечисленные роды в один род Vespertilio-, по его мнению, некоторые из них не заслуживают даже подродовой категории.
В СССР: кожаны двуцветный (европейская часть к северу до Калининской и Московской областей, Кавказ, Южный Урал, Казахстан, Средняя Азия, Южная Сибирь к северу примерно до линии Новосибирск—устье Амура) и восточный (Краскино, Уссурийск в Приморье, Хабаровская область); кожаны северный (Карпаты, север Украины, Белоруссия, прибалтийские республики, центральные и северные области РСФСР к югу до Смоленской, Курской, Куйбышевской, Оренбургской, Омской и Семипалатинской областей, оз. Зайсан и далее на восток по государственной границе; на север заходит за Полярный круг на Кольском полустрове, а на Урале и Сибири распространен до 60—64° с. ш., единичные находки известны из Закавказья, Памиро-Алая и окрестностей Алма-Аты); Бобринского (окрестности Казалинска, Иргиза, Аральска, пески Малые Барсуки, Бет-пак-Дала; одиночные находки этого (?) вида в Северной Осетии и Якутии); кожаны Огнева (Туркмения, горная Бухара, Казахстан); поздний (европейская часть СССР и Казахстан к югу от линии Вильнюс—Харьков—Оренбург — оз. Балхаш); нетопырь-карлик (европейская часть СССР к северу до Эстонии, Калининской, Московской, Рязанской и Пензенской областей; Казахстан и Средняя Азия на север до Кзыл-Орды и Алма-Аты); лесной (европейская часть СССР на восток до р. Урал и к югу от линии Балтийское побережье—Ленинград—низовье р. Вятка); восточный (Южное Приморье), средиземноморский (Крым, Кавказ, низовье Амударьи); кожановидный (Крым, Кавказ, Устюрт, юго-западная Туркмения, западный Тянь-Шань, Южное Приморье).
Род вечерниц
Genus Nyctalus Bowdich, 1825
Размеры от мелких до крупных. Длина тела 100—500 мм. Длина хвоста 35—65 мм. Длина предплечья 40—70 мм. Голова крупная, с относительно укороченной лицевой частью, вздута по бокам из-за сильно развитых носовых желез, обычно желтого цвета. Уши относительно короткие, широкие, причем ширина ушной раковины равна высоте или даже превышает ее (рис. 116). Козелок имеет булавовидную форму. Крылья длинные и узкие. Хвост полностью включен в межбедренную перепонку, иногда из нее выдается лишь последний укороченный позвонок. Волосяной покров обычно густой, довольно длинный и мягкий. Окраска его от золотисто-бурого или желтовато-бурого до темно-бурого на спине и обычно светло-бурого на брюхе. Череп массивный, укороченный, широкий. Мощно развит ламбдоидаль-ный гребень. Костные слуховые барабаны довольно крупные. Зубная формула: i с р т -|- = 34. Число диплоидных хромосом у рыжей вечерницы 42.
Обитают, как правило, в лесных районах, обычно лиственных лесах. В безлесных пространствах не встречаются. День проводят в пещерах, дуплах деревьев, зданиях. Характерен быстрый полет с резкими поворотами. Питаются в основном жуками, но поедают также и других насекомых. Охотятся в ранних сумерках и на рассвете. Спариваются обычно в сентябре,
Рис. 116. Рыжая вечерница—N.noctula и ее череп
а также весной. Самки приносят одного или двух, реже трех детенышей в мае—июне. Беременность у рыжей вечерницы 50—70 дней.
Распространены в северной половине Восточного полушария от Азорских островов в Атлантическом океане по всей Евразии до Японии, Суматры, Калимантана и Филиппинских островов на востоке.
В роде 7 видов:
вечерница Жоффруа — N. joffrei Thomas, 1915 (Бирма);
азорская вечерница — N. azoreum Thomas, 1901 (Азорские острова);
малая вечерница — N. lets-leri Kuhl, 1818 (Англия, Ирландия, Голландия, Франция, Швейцария, ФРГ, ГДР, Испания, о-ва Мадейра, Польша, Румыния, европейская часть СССР на север до Московской области и Среднего Поволжья, а на юг до Кавказа; Пенджаб, Кумаон);
рыжая вечерница — N. пос-tula Schreber, 1774 (Англия, Франция, Швейцария, Испания, Италия, Норвегия, Швеция, Дания, Голландия, ФРГ, ГДР, Греция, Польша, Чехословакия, европейская часть СССР, Западная Сибирь, Тарбагатай, Семиречье, Средняя Азия, Иран, Израиль, Западный и Южный Китай, Непал, Кашмир, Бирма, Малайзия, Япония, Мозамбик, Алжир);
японская вечерница — У. furvus Imaizumi et Joshiyuki, 1968 (Япония);
гигантская вечерница — IV. lasiopterus Schreber, 1780 (Швейцария, Франция, Италия, Сицилия, европейская часть СССР);
малайзийская вечерница — N. stenopterus Dobson, 1873 (Малайзия, о-ва Бинтан, Суматра, Калимантан).
В СССР : вечерницы малая (европейская часть СССР к северу до Новгородской, Московской, Пензенской и Саратовской областей), рыжая (европейская часть СССР на север до балтийского побережья, Ленинграда, южной части Кировской области; Западная Сибирь до Алтая, Казахстан, северный Узбекистан, Киргизия); гигантская (европейская часть СССР к северу до Московской и Горьковской областей, к востоку до западных районов Оренбургской области и к югу до западного Закавказья и южного Крыма включительно).
Рис. 117. Белобрюхий стрелоух — О. hemp г ic hi и его череп
Род дисконогих гладконосов
Genus Eudiscopus Conisbee, 1953
В роде единственный вид: дисконогий гладконос — Е. denticulus Osgood, 1932.
Размеры мелкие. Длина тела 40—45 мм. Длина хвоста 39—42 мм. Длина предплечья примерно 38 мм. На подошвах задних конечностей есть крупные кожистые подушки, посредством которых летучая мышь, по-видимому, может прикрепляться к гладким вертикальным плоскостям. Окраска волосяного покрова на спинной стороне светло-бурая, на брюхе несколько более светлая. В каждой половине нижней и верхней челюстей по 3 предкоренных зуба. В черепе характерен широкий уплощенный мозговой отдел, относительно длинный и узкий лицевой.
Экология не изучена.
Распространен в Лаосе.
Род стрелоухов
Genus Otonycteris Peters, 1859
В роде единственный вид: белобрюхий стрелоух — О. hemprichi Peters, 1859.
Размеры средние. Длина тела около 73 мм. Длина хвоста 47—63 мм. Длина предплечья 57 — 67 мм. Уши очень крупные, широкие с заостренной вершиной, соединенные между собой по верху головы невысокой кожной складкой, покрытой волосами (рис. 117). Козелок относительно длинный.
Крылья очень широкие. Кончик хвоста на 5—7 мм выступает из межбедренной перепонки. Волосяной покров довольно редкий, высокий. Окраска его на спинной стороне от бледно-песчаной до темно-бурой; на брюшной обычно беловатая. Самки имеют по две пары сосков в грудной области, однако неизвестно, функционируют ли обе пары сосков. В черепе хорошо развит сагиттальный гребень. Скуловые дуги широко расставлены. Довольно сильно развиты надглазничные отростки. Костные слуховые ба-1113
рабаны крупные. Зубная формула: i — с — р — т—= 30.
Экология изучена слабо. День проводит в расщелинах скал, пещерах, иногда в жилых постройках. Собираются небольшими группами до 20 особей. В Средней Азии рождение детенышей отмечено в июне.
Распространение охватывает Северную Африку от АРЕ до Алжира, Иран, Ирак, Израиль, Аравийский полуостров, Кашмир; в СССР встре-.чается в Южной Туркмении, на Западном Памире, в Таджикистане и южном Узбекистане.
Везде малочислен.
Род сумеречных гладконосов
Genus Nycticeius Rafinesque, 1819
Размеры мелкие и средние. Длина тела 45—75 мм. Длина хвоста 30— 62 мм. Длина предплечья 28—60 мм. Уши хорошо развиты, но они далеко не такие крупные, как у стрелоухов, широко расставлены на голове и не имеют между собой кожной перепонки. Козелок есть. Волосяной покров на спинной стороне тела обычно бурый, на брюшной более светлый. У американского гладконоса диплоидное число хромосом 46.
День проводят в пещерах, дуплах, среди листвы деревьев, иногда в зданиях. Полет довольно медленный. Пустынный гладконос на холодный период года мигрирует из северной части ареала в южную. Питаются главным образом насекомыми. Размножение приходится на конец лета — начало осени. Сперва, по-видимому, сохраняется в половых протоках самки в течение зимы. Овуляция и оплодотворение яйцеклетки происходят весной. Самка приносит обычно двух детенышей в мае—июне.
Ареал очень обширен: восток Соединенных Штатов, северо-восточная Мексика, Куба, большая часть Африки, Индия, Южный Китай, Сикким, Бенгалия, Ассам, вероятно, Северная Бирма, п-ов Индокитай, а также Австралия и Новая Гвинея.
В роде 12 видов:
гладконос Шлиффена — N. schlieffeni Peters, 1859 (Аравийский полуостров, АРЕ, Судан, Эфиопия, Кения, Республика Заир и на юг до Мозамбика и Юго-Западной Африки);
пустынный гладконос — N. pallidas Dobson, 1876 (северный Синд и Пенджаб в Индии);
большеухий гладконос — N. emarginatus Dobson, 1871 (Индия);
светлокрылый гладконос — N. albofuscus Thomas, 1890 (Малави, Гамбия);
темнокрылый гладконос — N. hirundo de Winton, 1899 (Замбия, Ангола, Кения, Гана);
кенийский гладконос — N. artinii de Beaux, 1923 (Кения);
коричневый гладконос — N. cinnamomeus Wettstein, 1916 (Судан); нигерийский гладконос— N. falabae Thomas, 1915 (Судан);
гладконос Грея — N. greyi Gray, 1843 (Новая Гвинея);
гладконос Санборна — N.sanborni Troughton, 1937 (Новая Гвинея, Квинсленд);
американский гладконос — N.humeralis Rafinesque, 1818 (юго-восток США и северо-восток Мексики);
кубинский гладконос — N. cubanus Gundlach, 1861 (о-в Куба).
. Род желтых гладконосов
Genus Rhogeessa Н. Allen, 1866
Размеры мелкие. Длина тела 37—50 мм. Длина хвоста 28—48 мм. Длина предплечья 25—34 мм. Уши довольно крупные, с округлыми вершинами, высокими козелками, широко расставленные. Окраска волосяного покрова желтовато-бурая или светло-бурая на спине и слегка более свет-1113
лая на брюхе. Зубная формула: i — с — р — т~ — 30. Диплоидное число хромосом 42—44.
Обитатели низменных и прибрежных областей. День проводят преимущественно в дуплах деревьев, иногда в зданиях. На дневках образуют большие скопления. Летают медленно, довольно низко над землей. Питаются насекомыми.
Распространены от западной и восточной Мексики на юг через Центральную Америку до Венесуэлы, Колумбии и Эквадора.
В роде 4 вида:
малый желтый гладконос — /?. parvula Н. Allen, 1866 (тихоокеанское побережье Мексики и от юго-востока Мексики на юг через Центральную Америку до Колумбии и Венесуэлы);
стройный гладконос — R. gracilis Miller, 1897 (южная Мексика);
пунский гладконос — R. velilla Thomas, 1903 (о-в Пуна у побережья Эквадора);
гладконос Мейера — R. meyeri Pine, 1966 (Веракрус, Мексика).
Некоторые исследователи выделяют из рода желтых гладконосов род Baedon Miller, 1906. По-видимому, более правильно объединить их в один род, как это делает, например, Симпсон (1945).
/ Род домовых гладконосов
Genus Scotophilus Leach, 1821
Размеры средние и крупные. Длина тела 60 — 95 мм. Длина хвоста 40—65 мм. Длина предплечья 43—75 мм. Уши относительно укороченные, округлые, широко расставленные. Волосяной покров на спинной части тела желтовато-бурый, а брюшной грязновато-желтый, желтый или белый. Сильно развиты челюсти. Зубы крупные.
День проводят преимущественно в зданиях, а также в дуплах деревьев, иногда среди листвы. В колонии бывает от 20 до нескольких сотен особей. На охоту вылетают в сумерки. Отличаются быстрым и сильным полетом. Питаются различными насекомыми: жуками, термитами, ночными бабочками. Малый домовый гладконос в Индии приносит детенышей в марте. Беременность примерно 105—115 дней.
Распространение охватывает Африку, Мадагаскар, Индию, Цейлон, Сикким, Бирму, Тенассерим, Таиланд, п-ов Индокитай, Малайзию, о-ва Хайнань, Яву, Бали, Калимантан, Филиппинские, Тайвань.
В роде 9 видов:
малый домовой гладконос —S. temmincki Horsfield, 1824 (о-ва Тайвань, Хайнань, Бирма, Таиланд, Центральная часть Вьетнама, Малайзия, о-ва Ява, Бали, Калимантан, Филиппинские, Индия, Цейлон);
большой домовый гладконос—S. heathi Horsfield, 1831 (Юньнань, Хайнань, Бирма, Индия, Цейлон, Таиланд, Бакбо и, очевидно, о-в Сулавеси);
исполинский гладконос—S. gigas Dobson, 1875 (Малави, южная Родезия, Нигерия, Гана);
желтый домовый гладконос—S. nigrita Schreber, 1774 (ЮАР, Мозамбик, южная Родезия, Малави, Замбия, Юго-Западная Африка, Ангола, на север до Танзании, Кении, Республики Заир и Судана, на запад до Сенегала);
зеленый гладконос—S. viridis Peters, 1852 (ЮАР, Мозамбик, Юго-Западная Африка, Малави, Ангола, на север до Танзании);
черный гладконос—S. nigritellus De Winton, 1899 (Гамбия, Гана);
мадагаскарский гладконос — S. robustus Milne-Edwards, 1881 (Мадагаскар);
эфиопский гладконос—S. murino-flavus Heuglin, 1861 (Эфиопия, Судан);
-Целебесский гладконос—S. celebensis Sody, 1928 (о-в Сулавеси).
Род выростогубых гладконосов
Genus Chalinolobus Peters, 1866
Размеры малые и средние. Длина тела 35—67 мм. Длина хвоста 32— 60 мм. Длина предплечья 30—50 мм. Уши относительно небольшие, с округлыми вершинами. Козелок также с округлой вершиной. Уши широко расставлены. На короткой широкой морде имеются специфические кожные железы в виде небольших вздутий. Окраска волосяного покрова шоколаднобурая, темно-бурая или черная, иногда с красноватым оттенком. Вершинки волос могут быть беловатыми. На теле и на голове могут быть крупные белые пятна и полосы.
Обитатели тропических и дождевых лесов и кустарниковых саванн. День проводят в дуплах деревьев, в пещерах, шахтах и т. п., иногда среди листвы деревьев. На дневках бывают поодиночке или небольшими группами до 30—50 особей. Выростогуб Гульда, по-видимому, впадает в спячку. Питаются различными насекомыми. Охотиться начинают иногда еще при солнечном свете.
Распространение охватывает Австралию, Тасманию, Новую Зеландию, значительную часть Африки.
В роде 12 видов:
смолистый выростогуб — С. picatus Gould, 1852 (Центральная Австралия, Новая Гвинея);
выростогуб Гульда — С. gouldi Gray, 1841 (Австралия, Тасмания, о-в Норфолк);
южноавстралийский выростогуб — С. mor io Gray, 1841 (Южная Австралия и Тасмания);
уэльсский выростогуб — С. dwyeri Ryan, 1966 (Новый Южный Уэльс);
новозеландский выростогуб — С. tuberculatus Forster, 1844 (Новая Зеландия);
бабочковидный выростогуб — С. variegata Tomes, 1861 (ЮАР, Мозамбик, Юго-Западная Африка, Ангола, Малави, на север до Республики Заир, Уганды, Танзании, Судана и Ганы);
серебристый выростогуб — С. argentata Dobson, 1875 (Ангола, Танзания, Кения, Конго-Браззавиль, Камерун);
пятнистобелый выростогуб — С. alboguttatus J. Allen, 1917 (Республика Заир);
• угандский выростогуб — С. beatrix Thomas, 1901 (Камерун, Уганда); камерунский выростогуб — С. egeria Thomas, 1913 (Камерун);
конголезский выростогуб — С. humeralis J. Allen, 1917 (Республика Заир);
нигерийский выростогуб — С. poensis Gray, 1842 (Нигерия).
Род волосатохвостых гладконосов
Genus Lasiurus Gray, 1831
Размеры мелкие, средние и крупные. Длина тела 50—90 мм. Длина хвоста 40—75 мм. Длина предплечья 37—57 мм. Характерна межбедренная перепонка, сверху полностью покрытая волосами. Окраска волосяного
покрова разнообразная: красноватая, с беловатым оттенком, желтоватобурая, желтоватая или оранжевая, обычно с черноватыми или беловатыми оттенками. Число хромосом в диплоидном наборе у красного и мохнатого волосатохвостов 28.
В течение дня держатся поодиночке или мелкими группами в деревьях или кустарниках, прицепляясь задними лапами к сучьям или к стволу дерева. Иногда встречаются в дуплах деревьев. Хорошие и быстрые летуны. В Канаде и Соединенных Штатах отмечены миграции этих летучих мышей в холодный период года к югу. Летают над землей обычно на высоте 6— 15 м. Большинство видов питаются на лету. Красный волосатохвост собирает насекомых с листьев деревьев. Размножение приходится на позднее лето и раннюю осень. Род волосатохвостов уникален среди летучих мышей тем, что самки некоторых его видов приносят обычно 2 или 3, иногда по 4 детеныша. В северных частях ареала детеныши рождаются в конце мая — начале июля.
Распространение охватывает большую часть Северной и Южной Америки, включая Багамские, Большие Антильские, Галапагосские и Гавайские острова, а также Кубу.
В роде 7 видов:
красный волосатохвост — L. borealis Miiller, 1776 (юг Канады, США, кроме центральных областей, Мексика, Центральная Америка, Бразилия, Уругвай, Парагвай, Аргентина, Перу, Чили);
волосатохвост Семинола — L. seminolus Rhoads, 1895 (восток и юго-восток США и северо-восток Мексики);
волосатохвост Пфейффера — L. pfeifferi Gundlach, 1861 (о-в Куба); ямайский волосатохвост—L. degelidus Miller, 1931 (о-в Ямайка); малый волосатохвост — L. minor Miller, 1931 (о-ва Гаити и Багамские);
мохнатый волосатохвост — L. cinereus Polisot de Beauvois, 1796 (центральные и южные районы Канады, США, Мексики, Центральная и Южная Америка — от Колумбии и Венесуэлы до Аргентины и Чили);
галапагосский волосатохвост — L. brachyotis J. Allen, 1892 (Галапагосские острова).
Род широкоушек
Genus Barbastella Gray, 1821
Размеры мелкие и средние. Длина тела 43—60 мм. Длина хвоста 40— 55 мм. Длина предплечья 35—45 мм. Форма головы своеобразная. Морда сильно укороченная, тупая (рис. 118). Часто кожа между глазами и межноздревым промежутком голая. Эта область ограничена двумя большими кожными складками, которые снаружи также имеют голые участки. Относительно короткие, но очень широкие уши соединяются между собой по лбу. Козелок хорошо развит и имеет форму неправильного прямоугольного треугольника. Крылья длинные, заостренные на концах. Длинный хвост почти полностью включен в межбедренную перепонку, выступая из нее на 1— 2 мм. Волосяной покров длинный, мягкий и густой. Окраска его темнобурая или черная, вершины волос могут быть беловатыми или желтоватыми. В черепе лицевой отдел развит слабо, мозговой — сильно. Скуловые дуги слабо расставлены. Костные слуховые барабаны небольших размеров. Са-2 12 3
гиттального гребня нет. Зубная формула: i — с — р — т — = 34.
Внутренний верхний резец имеет две вершины. Диплоидное число хромосом у европейской широкоушки 32.
День проводят в пещерах, расщелинах скал, дуплах деревьев, зданиях.
Рис. 118. Голова и череп европейской широкоушки В. barbastellus
Обычно держатся поодиночке или небольшими группами. Полет довольно быстрый, с крутыми поворотами. На зиму впадают в спячку или откочевывают на юг. У европейской широкоушки самки приносят детенышей в мае—начале июня. Через месяц детеныши достигают размеров взрослых.
Ареал обширный: охватывает большую часть Европы на север до Шотландии и Норвегии и к востоку до Правобережной Украины и Крыма, встречаются на Кавказе, в Передней и Средней Азии, Гималаях, Китае, Японии и на п-ове Индокитай.
В роде 2 вида:
европейская широкоушка — В. barbastellus Schreber, 1774 (Англия, Франция, Испания, Швейцария, Италия, Норвегия, Швеция, ФРГ, ГДР, Голландия, Дания, Польша, в СССР: Литовская ССР, Беловежская Пуща, Украина, Крым, Кавказ, Закавказье);
азиатская широкоушка — В. leuco-melas Cretzschmar, 1826 (Синай, Яркенд, Южный и Центральный Китай, Северная Индия, п-ов Индокитай, Япония, в СССР: Закавказье, Восточный Кавказ, Средняя Азия).
В СССР — оба вида.
Род ушанов
Genus Plecotus Geoffroy, 1818
Размеры мелкие и средние. Длина тела 40—70 мм. Длина хвоста 35— 55 мм. Длина предплечья 35—52 мм. Укороченная морда по бокам имеет железистые вздутия. Глаза по сравнению с другими летучими мышами относительно крупных размеров. Уши очень большие (в длину достигают 40 мм) и соприкасающиеся основаниями друг с другом (рис. 119). Козелок сильно развит. Внутренняя поверхность ушей покрыта многочисленными поперечными складками. Крылья короткие и широкие. Волосяной покров довольно короткий и неровный. Окраска его на спинной стороне от бледно-палевожелтой до темно-серой или коричневато-бурой, на брюшной стороне обычно несколько светлее. Длинный хвост на 1—3 мм выступает из межбедренной перепонки. Сильно развит мозговой отдел черепа. Сагиттального гребня нет. Лямбдоидальный гребень развит слабо. Скуловые дуги расставлены узко. Костные слуховые барабаны очень большие. Зубная формула: 2 12 3
i — с — р — т — = 36. Диплоидное число хромосом у обыкновенно-
го ушана и ушана Таунсенда 32, а у аризонского ушана — 30.
День проводят в пещерах, расщелинах скал, зданиях, дуплах деревьев. В период размножения самки держатся колониями, собираясь по 20—30 и до 200 особей, а самцы поодиночке. Обыкновенный ушан в средней полосе приносит детенышей обычно в июне. Самка рождает одного детеныша. Через месяц молодые зверьки достигают размеров взрослых. У ушана Таунсенда, обитающего в Америке, спаривание происходит осенью, и сперматозоиды остаются в половых путях самки до весны. Все виды на зиму впадают в
спячку. На охоту вылетают только в полной темноте. Летают медленно. Могут, махая крыльями, держаться на одном месте. Добычу ловят как на лету, так и собирают на земле.
Ареал очень обширен, охватывая умеренные области ' вразии, Северную Африку, юго-западную Канаду, юго-восточные части Соединенных Штатов, Мексику, юго-западную часть США.
В роде 4 вида:
обыкновенный ушан—Р. attritus Linnaeus, 1758 (Ан-лия, Ирландия, Франция, Испания, Италия, Швейцария, Швеция, Норвегия, Дания, Голландия, ФРГ, Югославия, Польша, Финляндия, вся территория СССР на север до 60—62° с. ш., Канарские острова, Алжир, Тунис, АРЕ, северный Судан, Иордания, Иран, Непал, Северная Индия, МНР, Китай, Япония);
ушан Таунсенда — Р. townsendii Cooper, 1837 (Британская Колумбия в Канаде, США — в основном западные и центральные районы, Мексика);
ушан Рафинеска — Р. rafinesqut Lesson, 1827 (юго-восток США);
аризонский ушан — Р. phyllotus G. Allen, 1916 (Аризона в США и северная половина Мексики).
В СССР — обыкновенный
Рис 119 Ушан Рафинеска — Р. rafinesqut и череп обыкновенного ушана — Р. auritus
ушан.
Некоторые исследователи (Бурлиер, 1955 и др ) выделяют из рода ушанов роды Corynorhinus Н. Allen, 1865 и I dionycteris Anthony, 1923 или только один последний (Симпсон, 1945 и др.). По-видпмому, более правильно всех их объединять в один род Plecotus.
Род пятнистых ушанов
Genus Euderma Н. Allen, 1892
В роде единственный вид: пятнистый ушаи — Е. maculatum J. Allen, 1891.
Размеры средние. Длина тела 60—65 мм. Длина хвоста около 50 мм. Длина предплечья примерно 50 мм. Внешним видом несколько напоминает \шана. Уши огромные, с закругленными вершинами. Козелки крупные, довольно широкие. Окраска волосяного покрова темно-красновато-бурая
или черная с характерными белыми пятнами на плечах и в основании хвоста. На брюшной поверхности тела волосы с белыми вершинками. Уши и крылья сероватые.
Обитает от уровня моря до верхних границ леса в горах. Охотится на открытых и кустарниковых участках. Держится поодиночке. Питается мелкими насекомыми. Очень малочислен: за все время добыто 30 или 40 особей.
Встречается в западной части США и в северо-западной Мексике.
Включен в «Красную книгу».
Род длиннокрылое
Genus Miniopterus Bonaparte, 1837
Размеры мелкие и средние. Длина тела 40—63 мм. Длина хвоста 40— 67 мм. Длина предплечья 37—55 мм. Уши короткие, почти треугольной формы, слегка выступающие над густым мехом головы (рис. 120). Козелок хорошо развит, с тупой вершиной. Крылья длинные, резко суженные в вершинной части. Характерно сильное удлинение второй фаланги третьего
Рис. 120. Голова и череп обыкновенного длиннокрыла — М. schreibersi
пальца, которая почти в три раза превышает длину первой фаланги этого же пальца. Длинный хвост целиком включен в межбедренную перепонку. Волосяной покров густой, высокий. Окраска его красноватая, красновато-бурая, темно-бурая, сероватобурая или серая. У самок одна пара сосков. Череп с укороченным лицевым отделом. Мозговая капсула сильно вздута и очень крупная. Костные слуховые барабаны небольших размеров. Сагиттальный гребень хорошо выражен. Скуловые дуги слабо расставлены в стороны. Зубная формула: .21 2 3 о„
I — с — р — т — = 36.
з 13 з
Характерен сильно увеличенный киль на рукоятке грудины. Диплоидное число хромосом у обыкновенного длиннокрыла 46.
День проводят в пещерах, расщелинах скал, зданиях. На дневках собираются колониями от нескольких сотен до нескольких тысяч особей; иногда встречаются и мелкие группы не более 20 особей. Часто объединяются в колонии с другими летучими мышами, например с ночницами. На охоту вылетают рано вечером. Полет быстрый
и верткий. Питаются мелкими жуками и другими насекомыми, обычно на высоте 10—20 м над землей. Размножение в августе — сентябре. Оплодотворение происходит сразу. В тропиках беременность примерно 120 дней, в Южной Европе там, где частично впадают в спячку, происходит задержка развития эмбриона, и самки рождают детенышей только весной. В помете один детеныш.
Распространение охватывает Африку, Мадагаскар, Южную Европу, Южную Азию, Филиппины, Индонезию, Новую Гвинею с мелкими прилегающими островами, Австралию и восточную часть Новых Гебридских островов. .
В роде 13 видов:
обыкновенный длиннокрыл — М. schreibersi Kuhl, 1819 (Испания, Франция, Швейцария, Италия, Сардиния, ФРГ, ГДР, Венгрия, Польша, Греция, Крит, Болгария, в СССР: Закарпатье, Крым, Кавказ, Копетдаг; ЮАР, Мозамбик, южная Родезия, Юго-Западная Африка, Ангола, Замбия, Малави, Республика Заир, Кения, Танзания, Алжир, Иран, Индия, Цейлон, Непал, Бирма, Китай, Япония, о-ва Ява, Суматра, Калимантан, Сулавеси, Филиппинские, Новая Гвинея, Северная и Южная Австралия, Квинсленд и Новый Южный Уэльс);
• восточный длиннокрыл — М. australis Tomes, 1858 (Никобарские острова, Индия, Хайнань, о-ва Ява, Калимантан, Филиппинские, Соломоновы, Новая Гвинея, Амбоина, Кей, Ару, Церам, Пеленг);
африканский длиннокрыл — М. africanus Sanborn, 1936 (Эфиопия); камерунский длиннокрыл — М. inflatus Thomas, 1903 (Камерун); большой длиннокрыл — М. majori Thomas, 1906 (о-в Мадагаскар); мадагаскарский длиннокрыл — М. manavi Thomas, 1906 (о-в Мадагаскар);
малый длиннокрыл — М. minor Peters, 1866 (о-в Занзибар);
длиннокрыл Ньютона — М. newtoni Bocage, 1889 (о-в Сан-Томе); рыжий длиннокрыл — М. rufus Sanborn, 1936 (Республика Заир); серый длиннокрыл — М. ravus Sody, 1930 (о-в Ява);
длиннокрыл Уиткампа — М. witkampi Sody, 1930 (восточная часть Калимантана);
новогвинейский длиннокрыл — М. tristis Waterhouse, 1845 (Новая Гвинея, Филиппинские острова);
длиннокрыл Эшшольтца — М. eschscholtzii Waterhouse, 1845 (о-ва Минданао и Лусон в группе Филиппинских).
В СССР: обыкновенный длиннокрыл (Закарпатье, южный Крым, черноморское побережье Кавказа, долина Аракса, Нагорный Карабах, северо-западный Копетдаг).
Род трубконосов
Genus Murina Gray, 1842
Размеры мелкие и средние. Длина тела 33—60 мм. Длина хвоста 30— 42 мм. Длина предплечья 28—45 мм. Характерно, что ноздри открываются на вершинах своеобразных кожных трубок, основания которых соединяются, а концы направлены в стороны (рис. 121). Уши средних размеров, эллипсоидной формы, с закругленными вершинами. Козелок узкий, довольно высокий, заостренный. Крылья короткие и очень широкие. Хвост одним-двумя концевыми позвонками выступает из межбедренной перепонки. Вся верхняя поверхность межбедренной перепонки покрыта довольно густо волосами. Окраска густого и мягкого волосяного покрова на туловище одноцветная, серо-палевая, буроватая или рыжая. У самок по одной паре сосков. Череп относительно крупных размеров, с выпуклой крупной мозговой капсулой. Характерен широкий межглазничный промежуток. Скуловые дуги слабо расставлены в стороны. Гребни на черепе сильно развиты
только у крупных видов. Костные слуховые барабаны развиты слабо. Зубная формула:
. 2 1 2 3
I — с — р — т— — 34.
3 12 3
Экология изучена слабо. Часто населяют холмистые районы. Летают довольно медленно. Днем трубконосов находили в сухих листьях деревьев, в пещерах. Обычно встречаются по нескольку особей вместе.
Обитатели Восточной и Юго-Восточной Азии от верховьев Енисея и от Сахалина на юг до южных Гималаев, Северо-Восточной Индии, Цейлона, Бирмы, п-ова Индокитай, Филиппинских островов, Индонезии, Новой Гвинеи.
В роде 10 видов:
малый трубконос — М. aurata Milne-Edwards, 1872 (Уссурийский край, Сахалин, Япония, Сычуань и Юньнань в Китае, Сикким, Бирма);
большой трубконос — М. leucogaster Milne-Edwards, 1872 (Алтай, Восточная Сибирь,
Рис. 121. Голова и череп малого трубконоса — М. aurata
Япония, Китай, Северо-Восточная Индия);
японский трубконос — М. tunebrosa Mizuko, 1970 (Япония);
трубконос Хуттона — М. huttoni Peters, 1872 (Северная Индия, Западная Бирма, Лаос, Китай);
сиккимский трубконос — М. cyclotic Dobson, 1872 (Хайнань, Сикким, Бирма, Лаос, Вьетнам, Цейлон, о-в Минданао из группы Филиппинских);
серый трубконос — М. grisea Peters, 1872 (Северо-Западная Индия);
флоресский трубконос — М. florium Thomas, 1908 (о-ва Флорес, Сум-бава, Пеленг);
малайзийский трубконос — М. suilla Temminck, 1840 (Малайзия, о-ва Суматра, Калимантан, Ява);
яванский трубконос — М. balstoni, 1908 (о-в Ява);
седой трубконос — М. canescens Thomas, 1923 (о-ва Ниас и Суматра).
В СССР: малый трубконос (Уссурийский край и Южный Сахалин); большой трубконос (окрестности Красноярска, верховья Енисея, Кузнецкий Алатау, у оз. Телецкого на Алтае, Уссурийский край, Сахалин).
Род шерстокрылых трубконосов
Genus Наг piocephalus Gray, 1842
В роде единственный вид: шерстокрылый трубконос — Н. harpia Temminck, 1840.
Размеры средние. Длина тела 55—75 мм, длина хвоста 37—55 мм. Длина предплечья 40—54 мм. Уши небольшие, широко расставленные на голове, с закругленными вершинами. Козелок хорошо развит. Ноздри открываются на вершинах кожных трубок, направленных в стороны. Из межбедренной перепонки выступает лишь самый кончик хвоста. Межбедренная перепонка, задние конечности и часть крыльев покрыты мехом.
Волосяной покров туловища густой и мягкий. Его окраска на спине темнобурая, оранжево-красная, обычно с примесью серого цвета. Брюшная поверхность тела серовато-грязно-желтая.
Экология не изучена. Обычно населяет холмистые районы. Питается преимущественно жуками.
Распространен в Индии, Бирме, на п-ове Индокитай, о-вах Тайвань, Суматра, Ява и Амбоин.
Род украшенных гладконосов
Genus Kerivoula Gray, 1842
Размеры мелкие (наименьшие в семействе) и средние. Длина тела 32— 57 мм. Длина хвоста 32—55 мм. Длина предплечья 27—45 мм. Одни из наименее специализированных представителей семейства. Уши средних размеров, с заостренной вершиной. Козелки хорошо развиты, узкие и длинные. Волосяной покров длинный, мягкий, иногда завитой. Окраска его у некоторых видов очень своеобразная, например у украшенного гладконоса ярко-оранжевая или алая, крылья черные, вдоль пальцев оранжевые полосы; у дамарского украшенного гладконоса ярко-оранжево-рыжая с черной рябью, причем волосы на многих участках имеют белые вершинки. Некоторые виды этого рода имеют красновато-бурую, желтовато-бурую, оливково-бурую, темно-бурую или серую окраску. Лицевой отдел черепа низкий.
День проводят в дуплах и листьях деревьев, зданиях. Держатся на дневках поодиночке или небольшими группами до 6 особей. На охоту вылетают поздно вечером. Летают довольно медленно и низко над землей.
Распространены- в Африке, Индии и на восток по всей Южной Азии, на Филиппинских островах, в Индонезии, Новой Гвинее и Северо-Восточной Австралии.
В роде 24 вида:
украшенный гладконос — К- picta Pallas, 1767 (Южный Китай, Южная Индия, Цейлон, Малайзия, о-ва Суматра, Ява, Бали, Калимантан);
украшенный гладконос Хардвика — Д'. hardwickei Horsfield, 1824 (Южный Китай, Южная Индия, Цейлон, Бирма, Малайзия, о-ва Ява, Бали, Калимантан, Сулавеси и, вероятно, Филиппинские);
яванский украшенный гладконос — К- papillosa Temminck, 1840 (Калькутта в Индии, п-ов Индокитай, Малайзия, о-ва Суматра, Ява, Калимантан);
новогвинейский украшенный гладконос — /<. muscina Tate, 1941 (Новая Гвинея);
Д. aerosa Tomes, 1858 (по-видимому, о-в Сулавеси);
папуасский украшенный гладконос — Д. papuensis Dobson, 1878 (Новая Гвинея и Квинсленд);
сент-аигнанский украшенный гладконос — Д'. agnella Thomas, 1908 (о-в Сент-Аигнан к юго-востоку от Новой Гвинеи);
Kf.-myrella Thomas, 1914 (о-ва Адмиралтейства);
украшенный гладконос Миллера — Д. rapax Miller, 1931 (о-в Сулавеси);
карликовый украшенный гладконос — К- pusilia Thomas, 1894 (о-в Калимантан);
двуцветный украшенный гладконос — Д', bicolor Thomas, 1904 (Таиланд);
калимантанский украшенный гладконос — Д', bomb if rotis Lyon, 1911 (о-ва Калимантан, Ява);
светлый украшенный гладконос — Д. pellucida Waterhouse, 1845 (о-ва Суматра, Филиппинские);
украшенный гладконос Уайтхэда — К- whiteheadi Thomas, 1894 (о-в Лусон из группы Филиппинских);
самарский украшенный гладконос — Д'. jagorii Peters, 1866 (о-в Самар из группы Филиппинских);
малый украшенный гладконос — A. lanosa Smith, 1847 (ЮАР, Замбия, Малави, Республика Заир); *
дамарский украшенный гладконос — A. argentata Tomes, 1861 (ЮАР, Дамараленд в Юго-Западной Африке, Мозамбик, Замбия, Малави, Ангола, Республика Заир);
африканский украшенный гладконос — A. africana Dobson, 1878 (о-в Занзибар);
медноцветный гладконос — A- cuprosa Thomas, 1912 (Камерун);
эфиопский украшенный гладконос — A- eriophora Heuglin, 1877 (Эфиопия);
украшенный гладконос Гаррисона — К- harrisoni Thomas, 1901 (Эфиопия);
камерунский украшенный гладконос — A- muscilla Thomas, 1906 (Камерун);
ганский украшенный гладконос — К- phalaena Thomas, 1912 (Гана);
украшенный гладконос Смита — К- smithii Thomas, 1880 (Нигерия).
Род анамигдон
Genus Anamigdon Troughton, 1929
В роде единственный вид: анамигдон — A. solomonis Troughton, 1929. Возможно, анамигдона следует присоединить к роду украшенных гладко-носов.— Kerivoula.
Размеры мелкие. Длина тела примерно 45 мм. Длина хвоста 35 мм. Длина предплечья 38 мм. Окраска волосяного покрова на спинной части тела темная, черновато-бурая или красновато-бурая, на брюшной светлее.
Экология не изучена.
Распространен на о-ве Ровиана в группе Соломоновых островов.
Род пустынных гладконосов
Genus Antrozous Н. Allen, 1862
Размеры средние и крупные. Длина тела 60—85 мм. Длина хвоста 35—57 мм. Длина предплечья 48—60 мм. Характерны крупные уши, однако не достигающие таких размеров, как у ушанов; вершины их заострены. Козелки развиты хорошо, внутренняя поверхность имеет многочисленные поперечные складки. Уши не соединяются друг с другом и расставлены довольно широко. Ноздри открываются на передней части морды, а над ними есть небольшой седлообразный перешеек. Окраска волосяного покрова разнообразна: от- кремовой, желтоватой или светло-бурой до темно-бурой. Брюшная поверхность обычно несколько светлее спинной. Диплоидное число хромосом у бледного гладконоса 46.
День проводят в пещерах, расщелинах скал, дуплах деревьев, зданиях. В отличие от многих летучих мышей летние колонии бледного гладконоса составлены из особей обоего пола и молодых, хотя колонии из одних самок с молодыми также встречаются. Осенью эти колонии разбиваются на мелкие группы. Из северных частей ареала пустынные гладконосы зимой мигрируют на юг. Часть популяции может впадать в спячку. Бледные гладконосы обычно летают низко над землей и охотятся за насекомыми не только на лету, но и на земле. Полет относительно медленный. В секунду эта летучая мышь делает примерно 10 взмахов крыльями. Питается главным образом жуками, а также кузнечиками и скорпионами. Беременность примерно 9 недель. Самка приносит одного-двух детенышей в мае—июне. Молодые становятся способными к полету в возрасте 6—7 недель. Спаривание проис
ходит, по-видимому, в октябре, и сперма сохраняется в половых путях самок до весны, когда происходят овуляция и оплодотворение.
Распространены в западной части Северной Америки от юго-западной Канады до центральной Мексики, а также на Кубе.
В роде 3 вида:
бледный гладконос — A. paltidus Le Conte, 1856 (Британская Колумбия в Канаде, запад США, северные и центральные районы Мексики);
наяритский гладконос — A. dubiaguerciis Van Gelder, 1959 (о-ва Лас-Трес-Мариас и штат Наярит в Мексике);
гладконос Купмана — A. koopmani Orr et Silva Taboada, 1960 (о-в Куба; известен' только по черепу).
Род австралийских гладконосов
Genus Nyctophilus Leach, 1822
Размеры мелкие и средние. Длина тела 43—65 мм. Длина хвоста 35— 50 мм. Длина предплечья 35—45 мм. Уши сильно развиты, широкие и высокие в длину, достигают 25 мм, соединены между собой низкой кожной складкой. На конце морды находится небольшой носовой листок, а позади него — мясистый дискообразный вырост. Это единственный род в семействе обыкновенных летучих мышей, представители которого имеют длинные уши, соединенные в основании, и небольшой носовой листок. Окраска волосяного покрова светло-бурая, желтовато-бурая или пепельно-серая.
Экология изучена слабо. День проводят в небольших пещерах, расщелинах скал, дуплах и трещинах коры деревьев. Полет довольно медленный, порхающий. По-видимому, насекомых ловят не только на лету, но и собирают их и с поверхности земли и листьев.
Распространены на Новой Гвинее, в Австралии и на Тасмании.
В роде 9 видов:
Тиморский гладконос — A. timoriensis Ё. Geoffroy, 1806 (Юго-Западная Австралия, Новый Южный Уэльс, южный Квинсленд, Тасмания и, вероятно, о-в Тимор);
малый австралийский гладконос — N. microtis Thomas, 1888 (Новая Гвинея);
карликовый австралийский гладконос — N. microdon Laurie et Hill, 1954 (северо-восток Новой Гвинеи);
гладконос Жоффруа — N. geoffroyi Leach, 1821 (Западная, Северная, Южная и Юго-Восточная Австралия; северо-западный Квинсленд и Тасмания);
арнхемский гладконос — N. arnhemensis Jonson, 1959 (север Австралии);
североквинслендский гладконос — N. bifax Thomas, 1915 (северный Квинсленд);
североавстралийский гладконос — N. daedalus Thomas, 1915 (Северная Австралия);
гладконос Уолкера — N. walked Thomas, 1892 (Северная Австралия);
новогвинейский гладконос — N. imogene Thomas, 1914 (Новая Гвинея);
Некоторые выделяют новогвинейского гладконоса в отдельный род — Pharotis Thomas, 1914.
Род лимингтонских гладконосов
Genus Lamingtona McKean et Calaby, 1968
В роде единственный вид: ламингтонский гладконос — L. lophorhina McKean et Calaby, 1968.
Длина предплечья 3,9 — 4,0 см. На конце носа находится небольшой листок. На дорзальной поверхности морды проходит бороздка. Относитель-
но небольшие уши (длинной 1,6 — 1,9 см) не соединяются между собой. Козелок с закругленной вершиной. Хвост почти полностью заключен в межбедренную перепонку. Волосяной покров средней длины, оранжево-бурый на1 спине и брюхе. Череп и зубы сходны с черепом и зубами гладконоса Жоффруа (из рода австралийских гладконосов).
Обнаружен на горе Ламингтон на юго-востоке Папуа.
Диагноз, приведенный Мак Кином и Калеби, недостаточно полный и не содержит детальных сравнений с другими родственными родами. Возможно, при дальнейших исследованиях род ламингтонских гладконосов будет ликвидирован и в качестве вида включен в другой род (например в род австралийских гладконосов).
I I
I £ с с
Род томопеас
Genus Tomopeas Miller, 1900
В роде единственный вид: томопеас — Т. ravus Miller, 1900. Размеры небольшие. Длина тела 36—42 мм. Длина хвоста 32 мм. Длина предплечья 31—33 мм. Хвост полностью включен в межбедренную перепонку. Окраска волосяного покрова на спине бледно-бурая, на брюхе беловато-грязножелтая. Лицевая часть головы, уши и крылья черные. Последний шейный и первый грудной позвонок срастаются между собой.
Экология не изучена. Встречается высоко в горах.
Распространен в Перу.
СЕМЕЙСТВО НОВОЗЕЛАНДСКИХ ЛЕТУЧИХ МЫШЕЙ
FAMILIA MySTACINIDAE SIMPSON, 1945
Г к д н м У п
м н. та д; С1 BI В£ А
В семействе единственный род: новозеландских летучих мышеи — Mystacina Gray, 1843 и один вид: новозеландская летучая мышь — М. tu-berculata Gray, 1843.
Размеры мелкие и средние. Длина тела примерно 60—70 мм. Длина хвоста около 17 мм. Длина предплечья около 43 мм. Морда конусовидная, с рудиментарной носовой подушкой (рис. 122). Ноздри вытянуты в вертикальном направлении. Уши довольно высокие, несколько суженные; они широко расставлены по бокам головы и не соединяются между собой. Козелок длинный и заостренный. Крылья широкие, кожная летательная мем
брана утолщена у боков тела и около костей конечностей. Подошвы задних лап короткие и широкие, покрыты утолщенной бороздчатой кожей. Когти на задних и на первом пальце передних конечностей длинные, очень ост-
Рис. 122. Голова, стопа и первый палец передней конечности (виден зубец на когте) новозеландской летучей мыши —М. /uberculata
та по ве] У < ют Bai ста час TOJ нос по лет мяс HOI
ная мет лов
кор бул гре( обы зад! paci CTKI
280
рые и имеют характерный зубец на внутренней стороне. Хвост выступает над дорзальной поверхностью межбедренной перепонки. Жесткие вибриссы на морде с ложковидными вершинами. Волосяной покров очень густой, более густой по сравнению с другими летучими мышами. Окраска его на спинной стороне серовато-коричневая или бурая, на брюшной более светлая.
В черепе костные слуховые барабаны хорошо развиты. Зубная формула: 112 3
’ — с — р — т~ ~ 28. Верхние предкоренные зубы хорошо развиты.
Плечевая кость имеет дополнительное сочленение с лопаткой. Локтевая кость сильно редуцирована и составляет по своей длине лишь половину длины лучевой кости. Второй палец имеет хорошо развитые метакарпальный элемент и единственную фалангу. Плечевой пояс нормальный. Седьмой шейный позвонок не срастается с первым грудным. Таз нормальный. Малая берцовая кость развита хорошо, ее диаметр составляет примерно половину диаметра большой берцовой кости.
Обитает в лесных районах. В течение дня обычно держится небольшими группами в пещерах или дуплах деревьев. Питается преимущественно насекомыми, которых ловит на лету, а также на земле. Новозеландская летучая мышь может проворно бегать по земле и быстро карабкаться вверх даже по относительно гладкой поверхности. Такая подвижность — следствие своеобразного устройства предплечья. Особая кожистая складка, выступающая вдоль предплечья, позволяет зверьку при ходьбе использовать крыло как нормальную конечность четвероногого наземного животного. Активность сразу после наступления сумерек.
Распространена в Новой Зеландии и на о-ве Стюарт.
СЕМЕЙСТВО БУЛЬДОГОВЫХ ЛЕТУЧИХ МЫШЕЙ
К FAMILIA MOLOSSIDAE GILL, 1872
Размеры от мелких до крупных. Длина тела 40—135 мм. Длина хвоста 14—80 мм. Длина предплечья 27—85 мм. Морда широкая, тупая, обычно покрытая волосами со своеобразными ложкообразными расширениями на вершинах. Около шеи имеются крупные складки кожи. Под каждым крылом у обоих полов расположено по большому железистому карману. Часто имеются горловые мешки. Глаза относительно маленькие. Ноздри обычно открываются на подушке: верхняя поверхность их часто несет тонкие роговые выросты. Носовой листок отсутствует. Губы очень крупные, причем верхняя губа часто собрана в крупные вертикальные складки. Уши у большинства видов толстые, очень широкие, хотя и относительно невысокие. Форма их разнообразна. Часто соединены между собой кожной перепонкой, проходящей по верху головы. Козелок очень небольшой, широкий. Крылья узкие, летательная перепонка толстая. Межбедренная перепонка узкая. Толстый мясистый хвост почти наполовину своей длины выдается за край межбедренной перепонки.
Задние конечности относительно короткие и сильные. Стопа укороченная, широкая. Второй палец передней конечности имеет хорошо развитый метакарпальный элемент и всего лишь одну рудиментарную фалангу. Туловище покрыто укороченными мягкими волосами.
Покрывающие тело волосы у представителей рода Cheiromeles настолько коротки, что летучие.мыши кажутся почти голыми. У некоторых видов бульдоговых летучих мышей на вершине головы расположен своеобразный гребень из удлиненных, вертикально стоящих волос. Волосяной покров j обычно бурый, грязно-желтый, серый или черный. На наружном пальце | задних конечностей имеются искривленные щетинки с ложкообразными расширенными вершинами. Эти щетинки служат для разглаживания и чистки меха. В черепе нет посторбитальных отростков. Предчелюстные кости
имеют небные отростки или лишены их. Зубная формула варьирует от 1113 1123
i — с — р — т— = 26 до i — с — р — т— = 32. Зубы примитив-1123 3123
но-насекомоядного типа. Плечевая кость имеет крупное дополнительное сочленение с лопаткой. Эта суставная поверхность почти равна по площади основной сочленовной поверхности. Малая берцовая кость развита довольно хорошо, составляя по диаметру примерно половину диаметра большой берцовой. Локтевая кость составляет примерно половину длины лучевой кости. Седьмой шейный позвонок срастается с первым грудным. Плечевой и тазовый пояса нормальные.
По сравнению с другими летучими мышами, бульдоговые летучие мыши имеют наиболее специализированный летательный аппарат: двойное сочленение плечевой кости с лопаткой у них развито лучше, чем у других летучих мышей, крыло относительно уже и острее. Характерные для них мускулистый хвост и костные шпоры на задних конечностях со своеобразными мышцами образуют наиболее совершенный среди рукокрылых тормозной аппарат.
Обитатели тропических и субтропических областей дневки устраивают в пещерах, расщелинах скал, зданиях, дуплах деревьев, среди листьев пальм. Отмечены миграции. Полет легкий и быстрый, со скоростью до 60, иногда до 95 км/ч. Часто летают группами. В северных частях ареала в настоящую спячку не впадают. Спаривание и оплодотворение происходят весной. Беременность примерно 90 дней. Самка приносит одного детеныша раз в год. Не способных еще к полету детенышей самки не носят во время кормовых полетов. Кормят молоком не обязательно только своего детеныша. В месячном возрасте становятся способными к самостоятельному полету и добыванию пищи. Большинство представителей семейства ведут колониальный и лишь некоторые виды одиночный образ жизни.
Распространены в Старом Свете от Южной Европы и Южной Азии к югу через всю Африку и Малую Азию на восток до Австралии и о-вов Фиджи; в Новом Свете от юго-западной Канады на юг через Вест-Индию и Центральную Америку до Южной Америки на юг до Чили и Аргентины.
В семействе 7 родов (119 видов):
эомопс — Eomops Thomas, 1905 (3 вида);
южноамериканских бульдоговых летучих мышей — Molossops Peters, 1865 (7 видов);
голокожих бульдоговых летучих мышей — Cheiromeles Horsfield, 1824 (2 вида);
ксифониктерис — Xiphonycteris Dollman, 1911 (1 вид);
складчатогубов — Tadarida Rafinesque, 1814 (81 видов);
бархатистых бульдоговых летучих мышей — Molossus Ё. Geoffroy, 1805 (24 вида);
плоскоголовое Нового Света — Neoplatymops Peterson, 1965 (1 вид).
Род эомопс
Genus Eomops Thomas, 1905
Размеры мелкие. Длина тела около 60 мм. Длина хвоста примерно 25 мм. Длина предплечья 35 мм. Уши относительно короткие, с округленными вершинами. Короткий козелок туповершинный. Верхняя челюсть, по сравнению с нижней, выдается довольно далеко вперед. Конец носа отделен от верхней губы. Ноздри открываются латерально. Волосяной покров на спинной стороне темно-бурый, на брюшной красновато-желтый или белый. Число зубов 26.
Экология не изучена.
Распространены в западной части Центральной Африки.
В роде 3 вида:
эомопс Уайтлея — Е. whitleyi Scharff, 1900 (Западная Африка); белый эомопс — Е. albatus Thomas, 1915 (Республика Заир); сенегальский эомопс — Е. senegalensis Oken, 1816 (Сенегал).
Род южноамериканских бульдоговых летучих мышей
Genus Molossops Peters, 1865
Размеры мелкие и средние. Длина тела 40—95 мм. Длина хвоста 14— 30 мм. Длина, предплечья 28—51 мм. Окраска волосяного покрова на спинной стороне желтовато-бурая, шоколадно-бурая, темно-бурая или черноватая, на брюшной обычно желтовато-белая, серая или пепельно-серая. Ряд о а л. .1113
видов имеет белое горло. Зубная формула: i -—- с — р — т~ =
= 26 — 28. Предчелюстные кости с небными отростками.
Экология не изучена. День проводят в дуплах деревьев, собираясь группами до 50—70 особей. Самцы и самки, по-видимому, держатся вместе в течение круглого года.
Распространение охватывает южную Мексику, Центральную Америку, о-в Тринидад и северную часть Южной Америки.
В роде 7 видов:
плоскорылая бульдоговая летучая мышь — М. planirostris Peters, 1865 (Панама, Бразилия, Колумбия, Гвиана);
мексиканская бульдоговая летучая мышь — М. malagai Villa, 1955 (южная Мексика и Гватемала);
бульдоговая летучая мышь Гринхалла — М. greenhalli Goodwin. 1958 (о-в Тринидад);
эквадорская бульдоговая летучая мышь — М. aequatorianus Cabrera, 1917 (Эквадор);
! бульдоговая летучая мышь Темминка — М. temmincki Burmeister, [ 1854 (Колумбия, Бразилия, Парагвай, Аргентина);
! перуанская бульдоговая летучая мышь — М. brachymeles Peters, 1865 [(Перу, Боливия, Парагвай, северная Аргентина, южная Бразилия, Гайана);
бульдоговая летучая мышь Миллера — М. tnilleri Osgood, 1914 (Перу).
Род голокожих бульдоговых летучих мышей
! Genus Cheiromeles Horsfield, 1824
[ Размеры крупные (наибольшие в семействе). Длина тела 115—135 мм. едина хвоста 50—65 мм. Длина предплечья 70—85 мм. Уши широко расставлены по бокам головы и не соединены между собой, а губы гладкие и не имеют складок. Это характерное отличие от других бульдоговых летучих мышей. Все тело-почти голое, покрытое лишь редкими, короткими и тонкими волосками. Довольно толстые грубые щетинковидные волосы имеются в районе горловой специфической кожной железы, расположенной в своеобразном кожном мешке. Эта железа продуцирует секрет с сильным запахом. Кожа на теле толстая, эластичная, почти черная у С. torquatus, темно-бурая у С. parvidens. В отличие от большинства других представителей семейства бульдоговых, крылья прикрепляются почти по средней линии спины. В подмышечной области находятся хорошо развитые кожные железы, которые тянутся почти вдоль всего бока тела. На задних конечностях первый палец противопоставляется другим и снабжен уплощенным ногтем вместо когтя. Вдоль боков тела располагаются кожные мешки: они открываются назад и имеют глубину 25—50 мм. В мешки помещаются сложенные крылья и в этом положении летучая мышь может легко передвигаться по твердому субстрату на всех четырех конечностях. Соски у самок располагаются около отверстий боковых мешков.
Экология не изучена.
Распространение охватывает Малайзию, о-ва Суматру, Яву, Сулавеси, Калимантан и Филиппинские.
В роде 2 вида:
голокожая летучая мышь — С. parvidens Miller et Hollister, 1921 (о-в Сулавеси);
ожерельевая летучая мышь — С. torquatus Horsfield, 1824 (Малайзия, о-ва Суматра, Ява, Калимантан, Сулавеси, Минданао в группе Филиппинских).
Род ксифониктерис
Genus Xiphonycteris Dollman, 1911
В роде единственный вид: ксифониктерис — X. spurrediDoWman, 1911.
Размеры мелкие. Длина тела около 56 мм. Длина хвоста 22—24 см. Длина предплечья около 27 мм. Крылья относительно невелики. Окраска волосяного покрова ржавчато-красная на спинной и грязно-белая на брюшной стороне.
Экология не изучена
Распространен на о-ве Фернандо-По, в Гане и Нигерии.
Род складчатогубов
Genus Tadarida Rafinesque, 1814
Размеры от мелких до средних. Длина тела 45—100 мм. Длина хвоста 20—60 мм. Длина предплечья 27—70 мм. Голова широкая и уплощенная (рис. 123). Верхняя губа имеет многочисленные свисающие складки и покрыта короткими щетинками. Ушные раковины с внутренней стороны имеют крупные кожные складки и, как правило, соединяются между собой над основанием носа тонкой перепонкой. Козелок широкий и короткий. Ноздри округлые. Крылья очень длинные, узкие, заостренные. Наружная сторона крайних пальцев задних конечностей усажена рядом желтых коротких волос. Окраска волосяного покрова красновато-бурая или почти черная. У некоторых видов от железистых полей позади ушей отходят пучки волос. Некоторые виды Старого Света имеют крупный гребень длинных прямых волос; он проходит по кожной складке, соединяющей уши. У самцов некоторых видов (в частности, у складчатогуба Мартиенссена) на горле есть специфическая железа в виде кожного мешка, причем, в отличие от голокожих бульдоговых летучих мышей, в нем нет удлиненных волос. Характерен удлиненный лицевой отдел черепа. Предглазничные отростки крупные. Скуловые дуги довольно широко расставлены. Сагиттальный гребень у боль-
„ - , .1 12 3
шинства видов не развит. Зубная формула: i ----с — р —т— =
2 — 3 1 2 3
= 30 — 32.
Обитатели тропических и субтропических областей. Экология изучена слабо. Многие виды держатся в горах, в скальных участках. День проводят в расщелинах скал, пещерах, дуплах деревьев. Селятся небольшими колониями по нескольку десятков особей. На охоту вылетают поздно, с наступлением темноты. Характерен быстрый и прямой полет. Некоторые виды, по-видимому, способны впадать в спячку. Наиболее охотно поедают жуков и молей. Размножение приходится на конец зимы — начало весны. Беременность примерно 11—12 недель. Самки, как правило, приносят одного детеныша в год. У большинства североамериканских видов детеныши появляются в июне — июле. Самки и самцы держатся вместе в течение всего
j
I г г
г
fl
Г
Д
Рис. 123. Египетский складчатогуб — Т. aegy-ptica и череп широкоухого складчатогуба— Т. teniotis
Распространены в тропических и субтропических областях Старого и Нового Света.
В роде 81 видов:
широкоухий складчатогуб — Т. teniotis Rafinesque, 1814 (Португалия, Франция, Италия, Сицилия, Югославия, Болгария, Турция, Греция, Швейцария; в СССР в отдельных пунктах Средней Азии и Кавказа; Марокко, Алжир, Ливан, Ирак, Израиль, Южный Китай, Корейский полуостров, п-ов Индокитай, о-в Тайвань, Япония);
египетский складчатогуб — Т. aegyptiaca Ё. Geoffroy, 1818 (АРЕ, Кения, Ангола, Замбия, ЮАР, Индия);
индийский складчатогуб — Т. tragata Dobson, 1874 (Индия);
африканский складчатогуб — Т. africana Dobson, 1876 (ЮАР);
складчатогуб Мастерсона — Т. mastersoni Roberts, 1946 (южная Родезия);
складчатогуб Сибра — Т. brunnea Seabra, 1900 (Ангола, Республика Заир);
складчатогуб Бокажа — Т. bocagei Seabra, 1900 (ЮАР, Юго-Западная Африка, Ангола);
складчатогуб Ансорга — Т. ansorgei Thomas, 1913 (Ангола, южная Родезия, Республика Заир, Танзания);
складчатогуб Беммелена — Т. bemmeleni Jentink, 1879 (Либерия);
суданскпй складчатогуб — Т. cisturus Thomas, 1903 (Судан);
мадагаскарский складчатогуб — Т. fulminans Thomas, 1903 (о-в Мадагаскар);
кенийский складчатогуб — Т. lobatus Thomas, 1891 (Кения);
охристый складчатогуб — Т. ochraceusi. Allen, 1917 (Республика Заир);
складчатогуб Тонга — Т. tongaensis Wettstein,1916 (Судан);
Т. latiei Taylor, 1934 (о-в Минданао в группе Филиппинских);
Т. plicata Buchannan, 1800 (Индия, о-в Цейлон, Тенассерим, Хайнань,
Малайзия, Суматра, Калимантан, Ява, вероятно, также Филиппинские острова, Северная Австралия);
карликовый складчатогуб — Т. pumila Cretzschmar, 1830 (Аравийский полуостров, Эфиопия, Судан, Уганда, Кения, Танзания, Ангола, Мозамбик, Ботсвана, ЮАР);
складчатогуб Спиллмана — Т. spillmanni Monard, 1933 (Ангола,Замбия);
малый складчатогуб — Т. limbata Peters, 1852 (ЮАР, Мозамбик, Ботсвана, .Малави, Ангола, Танзания, о-ва Пелиба и Занзибар, Республика Заир, Эфиопия);
длинногребешковый складчатогуб — Т. chapini J. Allen, 1917 (Юго-Западная Африка, Замбия, Ангола, Республика Заир);
украшенный складчатогуб — Т. bivittatus Heuglin, 1861 (Судан, Эфиопия);
Т. frater J. Allen, 1917 (Республика Заир);
гамбийский складчатогуб — Т. gambianus De Winton, 1901 (Гамбия);
Т. hindei Thomas, 1904 (Кения);
Т. leucogaster Grandidier, 1869 (о-в Мадагаскар);
пятнистобелый складчатогуб — Т. leucostigma G. Allen, 1918 (о-в Мадагаскар);
большой складчатогуб — Т. major Trouessart, 1897 (Танзания);
нигерийский складчатогуб — Т. nigeriae Thomas, 1913 (Нигерия);
алдабрский складчатогуб — Т. pusillus Miller, 1902 (о-в Алдабра);
красный складчатогуб — Т. russatus J. Allen, 1917 (Республика Заир);
складчатогуб Вебстера — Т. websteri Dollman, 1908 (Нигерия);
йохорский складчатогуб — Т. johorensis Dobson, 1873 (о-в Суматра);
лузонский складчатогуб — Т. luzonis Hollister, 1913 (о-в Лусон из группы Филиппинских);
складчатогуб Аба — Т. abac J. Allen, 1917 (Аба в Республике Заир);
черный складчатогуб — Т. nigri Hatt, 1928 (Судан);
ангольский складчатогуб — Т. condylura Smith 1833 (АРЕ, Ангола, Замбия, Малави, Мозамбик, Танзания, Судан, Республика Заир);
читауский складчатогуб — Т. chitauensis Hill, 1937 (Читау в Анголе);
белогрудый складчатогуб — Т. brachyptera Peters, 1852 (о-в Занзибар, Танзания);
Т. midas Sundevall, 1843 (Сенегал, Нигерия, Судан, Эфиопия, Аравийский полуостров, Малави, о-в Мадагаскар);
конголезский складчатогуб — Т. congicus J. Allen, 1917 (Республика Заир);
Т. demonstrator Thomas, 1903 (Судан);
леонский складчатогуб — Т. leonis Thomas, 1908 (Сьерра-Леоне);
складчатогуб Миари — Т. miarensis Grandidier, 1869 (Миари на о-ве Мадагаскар);
ниангарский складчатогуб — Т. niangarae J. Allen, 1917 (Ниангара в Республике Заир);
складчатогуб Руппелла — Т. riippellii Temminck, 1827 (АРЕ, Судан, Эфиопия);
камерунский складчатогуб — Т. thersites Thomas, 1903 (Камерун);
складчатогуб Тревора — Т. trevori J. Allen, 1917 (Республика Заир);
сулавесский складчатогуб — Т. sarasinorutn Meyer, 1899 (о-в Сулавеси);
Фараджский складчатогуб — Т. faradjius J. Allen, 1917 (Фарадже в Республике Заир);
складчатогуб пигмей — Т. nanulus J. Allen, 1917 (Республика Заир);
авакубийский складчатогуб — Т. occipitalis J. Allen, 1917 (Республика Заир);
складчатогуб Осборна — Т. osborni J. Allen, 1917 (Республика Заир);
Т. mops Blainville, 1840 (Малайзия, Суматра);
натальский складчатогуб — Т. acetabulosa Hermann, 1804 (ЮАР, о-в Мадагаскар);
белобрюхий складчатогуб — Т. albiventer Dobson, 1876 (о-в Мадагаскар);
щетинистый складчатогуб — Т. setiger Peters, 1881 (Кения);
складчатогуб Дориа — Т. doriae Anderson, 1907 (о-в Суматра);
складчатогуб Вроугтона — Т. wroughtoni Thomas, 1913 (Южная Индия);
складчатогуб Мартиенссена — Т. martiensseni Matschie, 1897 (ЮАР, Ангола, Танзания, Республика Заир);
складчатогуб Икара — Т. Icarus Chubb, 1917 (ЮАР);
яванский складчатогуб — Т. formosus Chasen, 1939 (о-в Ява);
папуасский складчатогуб — Т. papuensis Lawrence, 1948 (Новая Гвинея);
Т. secundus Hayman, 1952 (Новая Гвинея);
складчатогуб Макмиллана — Т. macmillani Thomas, 1906 (Эфиопия);
складчатогуб Хаагнера — Т. haagneri Roberts, 1917 (Юго-Западная Африка);
плоскоголовый складчатогуб — Т. petrophilus Roberts, 1917 (ЮАР, Юго-Западная Африка);
австралийский складчатогуб — Т. australis Gray, 1838 (Новый Южный Уэльс, Южная Австралия);
куборский складчатогуб — Т. kuboriensis McKean et Calaby, 1968 (Новая Гвинея) (возможно, является лишь подвидом австралийского складчатогуба);
норфолкский складчатогуб — Т. norfolkensis Gray, 1839 (о-в Норфолк, Новая Гвинея, Новый Южный Уэльс, Квинсленд);
складчатогуб Беккара — Т. beccarii Peters, 1881 (о-в Амбоина, Новая Гвинея);
бразильский складчатогуб — Т. brasiliensis I. Geoffroy St.-Hilaire, 1824 (юг США, Мексика, Центральная Америка, Большие и Малые Антильские острова, Южная Америка на юг до Аргентины);
мешотчатый складчатогуб — Т. femorosacca Merriam, 1899 (штаты Калифорния и Аризона в США, север Мексики);
широкохвостый складчатогуб — Т. laticaudata Geoffroy, 1805 (центральная Мексика, Бразилия, Парагвай);
юкатанский складчатогуб — Т. yucatanica Miller, 1902 (юг Мексики, Гватемала, Гондурас, Сальвадор);
Т. molossus Pallas, 1766 (юго-запад США, Мексика, Центральная Америка и Южная Америка с севера на юг до Перу, Парагвая, Бразилии);
колючкоухий складчатогуб — Т. aurispinosa Peale, 1848 (Бразилия);
венесуэльский складчатогуб — Т. europs Н. Allen, 1899 (Бразилия, Венесуэла);
стройный складчатогуб — Т. gracilis Wagner, 1843 (Бразилия);
Т. similis Sanborn, 1941 (Колумбия, Перу);
складчатогуб Калиновского — Т. kalinowskii Thomas, 1893 (Перу); кубинский складчатогуб — Т. minutus Miller, 1899 (о-в Куба).
Некоторые ученые выделяют из рода складчатогубов роды Eumops Шег, 1906; Promops Gervais, 1855; Otomops Thomas, 1913; Platymops Tho-3S, 1906; Mormopterus Peters, 1865, иногда и некоторые др. Вероятно, азанные роды следует рассматривать в качестве подродов рода Та-rida.
В СССР: широкоухий складчатогуб (окрестности Бухары, горы Баба-г на границе Узбекистана и Таджикистана, близ г. Ош в Киргизии, дома Сумбара, Шушинское ущелье в Нагорном Карабахе, у г. Кисловодска на юге Кабардино-Балкарии).
Род бархатистых бульдоговых летучих мышей
Genus Molossus Ё. Geoffroy, 1805
Размеры от мелких до крупных. Длина тела 40—130 мм. Длина хвоста 20—80 мм. Длина предплечья 33—80 мм. Уши относительно короткие, широкие, с тупыми или округлыми вершинами, обычно соединенные между собой на передней части головы. У самцов и самок большинства видов имеется специфическая кожная железа в виде горлового мешка, лучше развитая у самцов. У некоторых видов волосяной покров длинный, у других более короткий. Окраска его красновато-бурая, темно-шоколадно-бурая, темно-бурая, рыжевато-черная или черная, на брюшной стороне тела может быть светлее.
День проводят в дуплах деревьев, пещерах, расщелинах скал, зданиях. На дневки собираются небольшими группами или крупными колониями по нескольку сотен особей. Миграции не отмечены. На холодный период года могут впадать на короткое время в сон. Некоторые виды на охоту вылетают рано, еще при солнечном свете. Полет прямой и быстрый, иногда высоко в воздухе, иногда над самой землей. Питаются насекомыми. У рыжей буль-договой летучей мыши есть защечные мешки: во время охоты она наполняет их пойманными насекомыми, а возвращаясь к месту дневки, пережевывает и поедает добычу. Самка приносит летом обычно одного, иногда двух детенышей.
Распространение охватывает Мексику, юго-западую часть США, Центральную Америку, Антильские острова и Южную Америку.
В роде 24 вида:
рыжая бульдоговая летучая мышь — М. rufus Geoffroy, 1805 (южная Мексика, Центральная Америка, о-в Тринидад, Парагвай, Аргентина и от Гвианы и Венесуэлы до Перу);
А4. pretiosus Miller, 1913 (южная Мексика, Центральная Америка);
М. sinaloae J. Allen, 1906 (южная Мексика, Центральная Америка); бульдоговая летучая мышь Бонда — А4. bondae J. Allen, 1904 (Колумбия, Центральная Америка, на север до Гондураса);
большая бульдоговая летучая мышь — М. fortis Miller, 1913 (о-в Пуэрто-Рико);
ямайская бульдоговая мышь — А4. milleri Johnson, 1952 (о-в Ямайка);
А4. major Kerr, 1792 (южная Мексика, Центральная Америка, о-ва Гаити, Малые Антильские, Южная Америка от Гвианы до северной Аргентины и Парагвая);
бульдоговая летучая мышь Сан-Киттса — А4. debilis Miller, 1913 (Малые Антильские острова);
кубинская бульдоговая летучая мышь — А4. tropidorhimchiis Gray, 1839 (о-в Куба); У
тринидадская бульдоговая летучая мышь — М. trinitatus Goodwin, 1959 (о-в Тринидад);
А4. barnesi Thomas, 1905 (Гвиана, Бразилия);
центральноамериканская бульдоговая летучая мышь — А4. centralis Thomas, 1915 (южная Мексика, Гватемала, Гондурас);
бульдоговая летучая мышь Дависона — А4. davisoni Thomas, 1921 (Перу);
носатая бульдоговая летучая мышь — М. nasutus Spix, 1923 (Аргентина, Парагвай, Бразилия);
бразильская бульдговая летучая мышь — М. ратапе Miller, 1813 (Бразилия);
парагвайская бульдоговая летучая мышь — М. occultus Thomas, 1915 (Парагвай);
усатая бульдоговая летучая мышь — А4. perotis Schinz, 1821 (юго-восток США и север Мексики);
М. abrasus Temminck, 1827 (юг Мексики, Центральная Америка и
Южная Америка от Колумбии, Венесуэлы и Гвианы до Перу, Боливии и Бразилии);
гайанская бульдоговая летучая мышь — М. mauriis Thomas, 1901 (окр. г. Вальядолид в южной Мексике и Гайана);
бульдоговая летучая мышь Ундервуда — М. underwoodi Goodwin, 1940 (запад Мексики, Гватемала, Сальвадор, Гондурас);
М. glauciniis Wagner, 1843 (южная Мексика, Центральная Америка, Флорида, о-ва Куба и Ямайка, Колумбия, Эквадор, Бразилия);
М. bonariensis Peters, 1874 (Панама, о-в Тринидад, Бразилия, Боливия, Парагвай, Уругвай, Аргентина);
М. hansae Sanborn, 1932 (Бразилия);
амазонская бульдоговая летучая мышь — М. trumbulli Thomas, 1901 (район р. Амазонки от штата Пара в Бразилии до северного Перу).
Некоторые исследователи выделяют из рода Molossus роды Promops Gervais, 1855 и Eumops Miller, 1906.
Род плоскоголовое Нового Света
Genus Neoplatymops Peterson, 1965
В роде единственный вид: плоскоголов Нового Света — Р. mattogros-sensis Vieira.
Размеры небольшие. Длина тела 50—56 мм. Длина хвоста 20—23 мм. Длина предплечья 27—30 мм. Уши широко расставлены и не соединяются между собой. На дорзальной стороне предплечья имеются бородавчатовидные бугорки. Волосяной покров относительно короткий и редкий бурого цвета. Основания волос белые. Брюхо желтоватое, сероватое или беловатое. У обоих полов имеется горловая кожная специфическая железа, сильнее развитая v взрослых самцов. Зубная формула:
. 1 1 2 3 '
i — с — р — т -— = 30.
2 12 3
Экология не изучена.
Известен из трех областей Южной Америки: западной Бразилии к северу от Мату-Гросу, Гайаны и Венесуэлы.
ОТРЯД ПРИМАТОВ
ORDO PRIMATES LINNAEUS, 1758
Размеры изменчивы. Длина тела от 8,5—10—12 см (долгопяты, лемуры, тупайи) до 180 см (гориллы). Хвост может отсутствовать (человекообразные обезьяны, человек), быть слабо развитым (некоторые лемуры) или сильно развитым (многие обезьяны, тупайи). У ряда видов хвост хватательный (некоторые цепкохвостые обезьяны Нового Света). Большинство приматов имеет пятипалые стопоходящие конечности. Первый палец, как правило, противопоставляется остальным, образуя хватательные конечности. У большинства видов на пальцах есть ногти.
Волосяной покров у низших приматов густой и мягкий, хорошо делится на все категории. У большинства высших приматов он также густой и мягкий, но деление на категории выражено слабее. Самки тупай имеют от 1 до 3 пар сосков, лори — 2—3 пары, долгопятов — 2 пары, а самки всех других приматов — одну пару сосков' в области груди или подмышек.
Для черепа характерен крупный мозговой отдел. Глазные орбиты направлены больше вперед, чем в бока; зрение стереоскопическое. Зубная
О__2 0—1 2_4 2_3
формула: i с ——- р ——- т——- =18 — 38. Зубы гетеродонтные, дифиодонтные. Резцов может быть меньше или они совсем отсутствуют (верхние резцы некоторых лемуров), в других случаях они сильно увеличены в размерах (руконожки). Предкоренные сходны по форме с клыками или коренными зубами, обычно двувершинные. Коренные трех-, четырех-и пятивершинные, у примитивных форм брахподонтного и туберкул о-сек-ториального типов, а у более развитых форм — бунодонтного.
Печень лопастная. Есть желчный пузырь. Желудок у большинства видов простой. Слепая кишка развита. Матка низших приматов двурогая, у высших простая, плацента у лемуров эпителиохориальная, не отпадающая; у остальных приматов — гемохориальная, отпадающая, дискоидаль-ная. У большинстйа (за исключением долгопятов, шерстистых обезьян и людей) имеется os penis. Семенники расположены в мошонке или подкожно.
Ведут преимущественно древесный образ жизни (некоторые тупайи, кольцехвостый лемур и бабуины в основном наземные формы). Держатся небольшими, реже крупными группами. Активность, как правило, дневная и лишь у немногих представителей — ночная. Некоторые лемуры (крысиные и мышиные маки) впадают в летнюю спячку на жаркий, сухой сезон. По характеру питания имеют тенденцию к всеядности с различными отклонениями в сторону растительноядности или плотоядности в зависимости от вида, времени года и местообитания. Высшая нервная деятельность некоторых представителей достигает наиболее совершенного развития по сравнению с другими млекопитающими.
У большинства видов самка приносит по одному, в большей или меньшей степени беспомощному, детенышу. У низших приматов, по-видимому, размножение сезонное, некоторые виды имеют по два помета в год. У высших
приматов сезонности в размножении нет. Самки имеют по одному помету в год.
Распространение охватывает субтропические и тропические области Азии, Африки и Америки. Люди населяют весь Земной шар.
В отряде 2 подотряда:
низших приматов, или полуобезьян,— Prosimii liliger, 1811 (6 семейств, 22 рода и 53 вида);
высших приматов, или обезьян,-—Anthropoidea Mivart, 1864 (5 семейств, 31 род и 139 видов).
В подотряде полуобезьян 10 семейств:
тупай — Tupaiidae Mivart, 1868 (5 родов);
лемуров — Lemuridae Gray, 1821 (6 родов);
индри — Indridae Burnett, 1828 (3 рода);
руконожек — Daubentoniidae Gray, 1870 (1 род);
лори — Lorisidae Gregory, 1915 (6 родов);
долгопятов — Tarsiidae Gill, 1872 (1 род).
В подотряде обезьян следующие семейства:
цепкохвостых обезьян, или обезьян-капуцинов, — Cebidae Swainson, 1835 (11 родов);
игрунковых обезьян — Callithricidae Thomas, 1903 (4 рода);
низших узконосых обезьян, или мартышек,—Cercopithecidae Gray, 1821 (11 родов);
высших узконосых обезьян, или человекообразных обезьян, — Роп-gidae’ Elliot, 1913 (4 рода);
гоминид, или людей, — Hominidae Gray, 1825 (1 род).
Некоторые исследователи выделяют в пределах отряда приматов большее число подотрядов. Так, Нестурх (1960) насчитывает в отряде 4 подотряда: высших приматов Anthropoidae; долгопятов — Tarsioidea; лемуров, или полуобезьян, — Lemuroidea; и тупай — Tupaioidea. Ряд ученых, например Валлоа (Vallois, 1955), относят тупай к насекомоядным \ выделяя пх в особый подотряд Menotyphla Haeckel, 1866 вместе с прыгунчиками, а в пределах отряда приматов, помимо подотрядов Prosimii и Anthropoidea, обособляют долгопятов в особый подотряд Tarsioidea. Иногда отряд приматов разделяют на большее число семейств. Так, из семейства лори выделяют дополнительно семейство Galagidae, из семейства игрунковых обезьян — семейство Callimiconidae, из семейства мартышек — семейство Colobidae, из семейства человекообразных обезьян — семейство Hylobati-dae. Видимо, более правильно считать указанные семейства подсемействами.
Предки приматов — примитивные верхнемеловые насекомоядные. Центром возникновения приматов была, по-видимому, Азия, откуда приматы расселялись по другим материкам Старого Света и по Северной Америке. В меловом периоде древнейшие приматы разделились на 2 группы: протоле-муроидов (предков тупай и лемуров) и прототарзиоидов (предков долгопятов). Большинство исследователей предполагает, что эоценовые примитивные формы долгопятов стали исходными формами обезьян Е1ового и Старого Света. Е1екоторые ученые считают, что предками обезьян явились лемуры. В отложениях палеоцена обнаружены ископаемые остатки представителей тупай, лемуров и долгопятов. Исходными формами американских широконосых обезьян послужили, по-видимому, североамериканские долгопяты. Из Северной Америки в Южную предки обезьян проникли, очевидно, в палеоцене, когда между этими двумя материками существовал перешеек, затем разрушившийся и вновь образовавшийся к концу третичного периода. Низшие узконосые обезьяны Старого Света произошли, по-видимому, от азиатских долгопятов в начале или середине эоцена. Предками человеко-
1 Обсуждение вопроса о принадлежности тупай к насекомоядным см. в диагнозе отряда насекомоядных.
образных обезьян послужили нижнеолигоценовые проплиопитеки (Propli-opithecus Schlosser, 1911). В одну сторону от них пошла ветвь вымерших плиопитеков (Pliopithecus Gervais, 1849) и развившихся от них гиббонов, а в другую сторону — ветвь крупных ископаемых человекообразных обезьян (например, миоценовых представителей подсемейства Dryopithecinae Gregory and Hellman, 1939), в результате давших современных человекообразных обезьян и человека.
Подотряд низших приматов, или полуобезьян,
Subordo Prosimii liliger, 1811
Приматы мелких и средних размеров. Длина тела от 8,5 до 106 см. Хвост короткий, средней длины пли длинный; в последнем случае может вдвое превышать длину тела. Хвост не хватательный, густо покрыт волосами. Первые пальцы на конечностях не противопоставляются или противопоставляются другим пальцам. Пальцы у большинства видов вооружены когтями, иногда ногтями. Второй палец на передних и задних конечностях часто невелик по размерам, в большинстве случаев четвертый палец на обоих парах конечностей наиболее длинный.
Волосяной покров густой и мягкий с хорошо выраженными категориями волос. Самки имеют от одной до трех пар сосков.
Лицевой отдел черепа довольно сильно выдается вперед или развит слабо. Зубная формула: i - °-С~ 1~? с р ——2 ~..3_ щ JL = jg— 3g.
J -г г 1 — 2 — 3 0—1 0 — 2 — 3 3
Поверхность больших полушар пев мозга гладкая или с немногими извилинами и бороздами.
СЕМЕЙСТВО ТУПАЙ
FAMILIA TUPAIIDAE MIVART, 1868
Небольших размеров полуобезьяны. Длина тела 10—22 см. Длина хвоста 9—22 см. Примитивные приматы, имеющие ряд признаков, сходных с представителями отряда насекомоядных. Туловище удлиненное. Конечности короткие, пятипалые. Лапы не хватательные: большие пальцы не противопоставляются остальным. Третий палец самый длинный. Пальцы вооружены когтями, но у ископаемой тупайи — Anogale gobiensis Simpson, 1931 пальцы задних конечностей имели ногти. Примитивные кожные узоры
на ладонях и подошвах задних лап сходны с таковыми лемур и долгопятов. Голова относительно крупная, с удлиненной мордой, клинообразно заостренной к носу. Ушные раковины крупные, широкие в основании, округлые. Глаза большие, обращенные в стороны. Хвост длинный, пушистый, только у перьехвостой тупайи он голый и имеет на конце знамя из удлиненных
волос.
Волосяной покров умеренной длины, состоит из остевых и пуховых волос. Некоторые виды имеют продольные бледные полосы на плечах, у других имеются отметки на морде. У самок от 1 до 3 пар сосков.
Череп с расширенным и округлым мозговым отделом, не имеющим греб-
ней. По строению, особенно слуховой и глазнично-височной областей, он сходен с черепом лемуров. Костные слуховые барабаны вздуты. Заднеглазничные отростки сильно развиты и соединяются со скуловой костью. Име-
2 13 3
ется крылоклиновидный канал. Зубная формула: i— с — р — т— = 38. 3 13 3
Верхние резцы длинные, клыковидной формы. Верхние клыки небольшие и сходны с предкоренными зубами. Третий нижний резец маленький. Коренные зубы тритуберкулярные, с широкой жевательной поверхностью.
Позвонков шейных 7, грудных 13—14, поясничных 5—6, крестцовых 3, хвостовых 22—31.
Печень и легкие многодольчатые. Левая почка лежит каудальнее по отношению к правой, как у лемуров. Семенники располагаются в мошонке, которая находится рядом с половым членом или впереди от него. Матка двурогая. Плацента дискоидальная, гемохориальная, отпадающая, состоит из двух участков удлиненной формы.
Обитают, как правило, в лесных областях. Большинство ведет преимущественно древесный образ жизни, превосходно лазая по деревьям. Пищу разыскивают также и на земле. Активность у большинства дневная. Перье-хвостая тупайа, отличающаяся в основном ночной активностью, ведет и наземный образ жизни. Из органов чувств наиболее сильно развиты слух и зрение. Питаются в основном насекомыми и плодами, но употребляют и многие другие животные и растительные корма. Сезонность в размножении не отмечена. Беременность от 46 до 56 дней. Число детенышей обычно от 1—2 до 4.
Распространены в Юго-Восточной Азии от Индии через Бирму, Суматру, Калимантан и некоторые прилежащие области до Филиппин.
В семействе 5 родов (18 видов):
обыкновенных тупай — Tupaia Raffles, 1822 (13 видов);
индийских тупай, или анатан,—Anathema Lyon, 1913 (1 вид); гладкохвостых, или горных тупай, —Dendrogale Gray, 1848 (2 вида); филиппинских тупай^— Urogale Mearns, 1905 (1 вид);
перьехвостых тупай — Ptilocercus Gray, 1848 (1 вид).
Род обыкновенных тупай
Genus Tupaia Raffles, 1822
Длина тела примерно 15—22 см. Длина хвоста чуть меньше или чуть больше длины тела. Лицевой отдел головы удлинен (рис. 124). В отличие от представителей других родов семейства обыкновенные тупайи имеют более редкий волосяной покров. Окраска его на спинной стороне охристая, красноватая, оливковая или почти черная, на брюшной от беловатой или темно-желтой до темно-коричневой. Могут быть светлые полосы на плечах. Характерно, что нижняя доля ушной раковины меньше по размерам верхней. Безволосый участок у носа срезан прямо, тогда как у других тупай он слегка вытянут назад по средней линии. Хвост покрыт длинными волосами. Имеется горловая кожная специфическая железа, сходная с железами
Рис. 124. Обыкновенная тупайа — Т. gilis
некоторых насекомоядных и отличающаяся от желез других приматов. Диплоидное число хромосом у обыкновенной тупайи 60.
Обитатели лесов. Большинство видов ведет наземный образ жизни, лишь некоторые — древесный. В качестве убежищ строят гнезда среди корней деревьев на земле, в кучах валежника. Активность преимущественно дневная. Встречаются, как правило, поодиночке, иногда парами. Питаются в основном насекомыми, но также и другими мелкими животными и некоторыми растительными объектами (сочными плодами, семенами, листьями). Нуждаются в воде для питья и для купания. Беременность 46—50 дней. В помете обычно 2 детеныша.
Распространение охватывает Юго-Западный Китай, Сикким, Манипур, Ассам, Юго-Восточную Азию, Суматру, Яву, Калимантан и Филиппины.
В роде 13 видов:
обыкновенная тупайа — Т. glis Diard, 1820 (Юго-Западный Китай, Сикким, Манипур, Ассам, Бирма, Тенассерим, п-ов Индокитай, Таиланд, Малаккский полуостров, о-ва Суматра, Ява, Калимантан);
никобарская тупайа — Т. nicobarica Zelebor, 1869 (Никобарские острова в Бенгальском заливе);
малая тупайа — Т. minor Giinther, 1876 (Малаккский полуостров, .южный Таиланд, о-ва Суматра и Калимантан);
краснохвостая тупайа — Т. splendidula Gray, 1865 (о-в Калимантан);
кариматская тупайа — Т. carimatae Miller, 1906 (о-в Каримата у западного побрежья о-ва Калимантан);
тупайа Мюллера — Т. mulleri Kohlbrugge, 1895 (о-в Калимантан);
горная тупайа — Т. montana Thomas, 1892 (о-в Калимантан);
яванская тупайа— Т. javanica Horsfield, 1821 (о-ва Ява и Суматра);
стройная тупайа — Т. gracilis, Thomas, 1893 (о-в Калимантан);
украшенная тупайа — Т. picta Thomas, 1892 (о-в Калимантан); полосатая тупайа — Т. dorsalis Schlegel, 1857 (о-в Калимантан);
большая тупайа — Т. tana Raffles, 1821 (о-ва Суматра и Калимантан);
палаванская тупайа — Т. palawanensis Thomas, 1894 (Филиппинские острова).
Некоторые исследователи сокращают число видов рода до 10 (Андерсон и Джонс, 1967 и др.) или увеличивают до 32 (Бютнер-Януш — Buetner-Janusch, 1963). Иногда полосатую и большую тупай выделяют в отдельный род — больших тупай — Tana Lyon, 1913.
Род индийских тупай, или анатан,
Genus Anathana Lyon, 1913
В роде единственный вид: индийская тупайа, или анатана, — A. ellioti Waterhouse, 1850.
Длина тела 17—20 см. Длина хвоста 16—19 см. По внешнему виду сходна с обыкновенными тупайами, но в отличие от них ушные раковины более крупные и более густо покрыты волосами и верхние клыки напоминают резцы. Лицевой отдел головы укорочен. Окраска волосяного покрова на спине красноватая, иногда желтовато-бурая или красновато-бурая, у некоторых особей черноватая или оранжеватая, на брюхе обычно с желтыми или бурыми пятнами на грязно-желтом фоне. На плечах беловатые или кремовые полосы.
Обитатель лесов. Экология изучена слабо, но, по-видимому, напоминает обыкновенных тупай. Ведет древесный образ жизни, питается насекомыми, а также другими мелкими животными и плодами.
Распространена на п-ове Индостан.
Некоторые исследователи разделяют род на 3 вида.
Род гладкохвостых, или горных, тупай Genus Dendrogale Gray, 1848
Мелких размеров тупайи. Длина тела И—15 см. Длина хвоста 9— 14 см. Голова с относительно сильно вытянутой лицевой частью. Уши более крупные, чем у представителей родов тупай и анатан. Хвост, в отличие от других тупай, покрыт равномерными короткими волосами по всей длине. Когти на лапах короткие (северная тупайа) или длинные (южная тупайа). У северной тупайи окраска волосяного покрова на спинной стороне тела буроватая или черноватая с охристым или ржавым оттенком. Брюшная сторона темно-желтая. По бокам головы есть черные полосы, которые тянутся от основания вибрисс через глаза к уху, по бокам их окаймляют светлые полосы. Хвост сверху темный, темнеющий в дистальном направлении, снизу охристый с темной полоской. Южная тупайа на спинной стороне черноватая с охристым оттенком, причем доминируют темные тона, на брюшной и внутренних поверхностях конечностей — охристые. Полос на голове нет.
Обитатели густых джунглей как на равнинах, так и в горах на высоте от 900 до 1500 м над уровнем моря. Ведут преимущественно древесный образ жизни. Активность дневная. Питаются в основном насекомыми.
Распространены на п-ове Индокитай и о-ве Калимантан.
В роде 2 вида:
южная тупайа — D. melanura Thomas, 1892 (о-в Калимантан);
северная тупайа—D. murina Schlegel et Miiller, 1845 (п-ов Индокитай).
Род филиппинских тупай
Genus Ur о gale Mearns, 1905
В роде единственный вид: филиппинская тупайа — U. everetti Thomas, 1892.
Длина тела 17—22 см. Длина хвоста 11—17 см. Голова с вытянутой мордой. Хвост покрыт удлиненными, равными по длине волосами. Окраска волосяного покрова на спинной стороне черноватая, с бурым оттенком, на брюшной оранжевая или оранжево-красная. У некоторых особей на плечах оранжевые полосы. Вторая пара верхних резцов увеличена в размерах и напоминает клыки, третья пара сильно редуцирована. Диплоидное число хромосом 44.
Хорошо лазает по деревьям и быстро бегает по земле. Всеядная. Наибольшая активность утренняя. Беременность 54—56 дней. В помете 1—2 детеныша.
Распространена на о-ве Минданао в группе Филиппинских.
Род перьехвостых тупай
Genus Ptllocercus Gray, 1848
В роде единственный вид: перьехвостая тупайа — Р. lowi Gray, 1848 (рис. 125).
Размеры наименьшие в семействе. Длина тела 10—14 см. Длина хвоста 13—19 см. Голова с умеренно удлиненной мордой. Ушные раковины тонкие, крупные. Передние и задние конечности примерно одинаковые по длине. Кисть и стопа относительно более крупные, чем у других родов семейства. Пальцы передних и задних конечностей вооружены короткими острыми когтями. Хвост имеет в концевом отделе белую кисть удлиненных волос; основная часть хвоста покрыта кожными чешуйками, позади каждой из которых растут по три коротких волоска. Окраска кожи хвоста темная. Волосы его концевого отдела «расчесаны» в две стороны, создавая вид пера. Волосяной покров туловища густой и мягкий, на спинной стороне обычно
Рис. 125. Перъехвостая тупайа — Р. lowi
темно-серовато-бурый, на брюшной желтовато-серый. На плечах полос нет. По бокам морды проходит темная полоса. Верхние резцы очень крупные.
Обитатель лесных районов. Экология изучена слабо. Активность преимущественно ночная. Питается главным образом насекомыми и плодами. Превосходно лазает по деревьям. Хвост служит в качестве баланса; вероятно, имеет некоторое значение и как тактильный орган. По земле передвигается прыжками, при этом конец хвоста поднимает вверх.
Распространение охватывает Малаккский полуостров, Суматру, Калимантан и мелкие прилежащие острова.
СЕМЕЙСТВО ЛЕМУРОВ
FAMILIA LEMURIDAE GRAY, 1821
Мелкие и средней величины полуобезьяны. Длина тела 12—46 см. Длина хвоста 13—51 см. Тело вытянутое. Конечности средней длины. Задние конечности длиннее. Второй палец задних конечностей с когтем, с помощью которого животное расчесывает мех. Остальные пальцы имеют ногти. Четвертый палец наиболее длинный. Голова с расширенной, довольно крупной мозговой коробкой и относительно укороченным лицевым отделом. Верхняя губа голая, неподвижная. Уши короткие или средней длины, обычно покрытые густо волосами. У большинства глаза крупные, что, очевидно, связано с ночным образом жизни. Хвост обычно длинный, густо покрытый волосами. Подошвы кистей и стопы голые, со своеобразным кожным узором.
Волосяной покров на туловище густой и мягкий, окраска его черная, бурая, серая или рыжеватая. Хвост одноцветный, иногда с чередующимися светлыми и темными кольцами. У некоторых видов на голове и спине расположены продольные полосы. Самцы могут отличаться по окраске от са
А
(
Л
А
С
Р Г
Д
мок, особенно у черного лемура. У некоторых видов окраска полиморфная: серая или красная. Самки имеют одну или две пары сосков, причем одна обязательно в области груди. У серого лемура есть добавочная пара сосков, расположенная на плечах.
В черепе костные слуховые барабаны сильно вздуты. Зубная формула: О_2 1 3 3
i —-— с — р — т~ = 32 — 36. Верхних резцов нет или они имеют небольшие размеры. Нижние клыки сходны с резцами. Первый нижний предкоренной зуб имеет форму, сходную с клыком. Позвонков шейных 7, грудных и поясничных 18—22, крестцовых 2—4, хвостовых 20—27. Имеется os penis.
Матка двурогая. Плацента не отпадающая, эпителиохориальная, диффузная с крупным аллантоисом. Длина кишечника в 1,5 — 2,5 раза превышает длину тела. Печень и легкие, как и у тупай и долгопятов, многодольчатые. Хрящевые кольца трахеи сзади замкнуты. Кровеносные сосуды конечностей образуют «чудесную сеть». Головной мозг весьма примитивный, имеет небольшое число борозд и извилин, а у маленьких лемуров вообще гладкий. Мозжечок не покрывается большим мозгом. Обонятельные доли сильно развиты, мозг макросматического типа.
Обитатели лесов. В большинстве ведут древесный образ жизни (за исключением кольцехвостого лемура), но в поисках пищи часто спускаются на землю. У большинства ночная активность. Держатся поодиночке, парами, семейными группами или довольно многочисленными стадами — до 10—20 особей вместе. Питаются различными растительными объектами, насекомыми (карликовые лемуры) или всеядны. Убежищем служат главным образом дупла или устраивают себе гнезда из листьев и травы. Карликовые и мышиные лемуры на засушливый сезон впадают в спячку. Период размножения длится с августа по март или апрель. Беременность от 2 до 5 месяцев. В помете 2, редко 3, 4 детеныша. О детенышах заботятся оба роди
теля.
Распространение охватывает о-ва Мадагаскар и Коморские.
В семействе 6 родов (16 видов):
полумаки, или кротких лемуров, — Hapalemur I. Geoffroy, 1851 (2 вида);
маки, или обыкновенных лемуров, — Lemur Linnaeus, 1758 (6 видов);
тонкотелых маки, или ласковидных лемуров, — Lepilemur I. Geoffroy, 1851 (2 вида);
крысиных маки, или карликовых лемуров, — Cheirogaleus Е. Geoffroy, 1812 (3 вида);
мышиных маки, или мышиных лемуров,—Microcebus Ё. Geoffroy, 1828 (2 вида);
вильчатополосых маки, или вильчатополосых лемуров, — Phaner Gray, 1870 (1 вид).
Род полумаки, или кротких лемуров,
Genus Hapalemur I. Geoffroy, 1851
Размеры средние и крупные (наибольшие в семействе). Длина тела примерно 28—46 см. Длина хвоста в общем равна длине тела. Голова округлая (рис. 126). Морда укороченная. Низкие ушные раковины густо покрыты волосами. Конечности относительно короткие, с широкими кистями и стопами. Концевые фаланги пальцев с крупными подушками. Волосяной покров средней длины, мягкий. Окраска его буровато-серая, красновато-серая, серовато-зеленая или красновато-зеленая на спинной стороне и беловатая, грязно-желтая, серая или желтоватая на брюшной. Голова более темная. У самок две пары сосков, причем одна располагается почти на плечах, а другая на брюхе. На внутренней стороне запястья у серого лемура находит-
Рис. 126. Широконосый лемур — И. gri-seus. А — кисть; Б стопа
ся специфическая кожная железа. Все зубы, кроме заднекоренных, имеют острые режущие края. Диплоидное число хромосом у серого лемура 54—58.
Обитатели лесных районов, особенно там, где есть заросли бамбука. Держатся чаще всего семейными группами, иногда поодиночке. Активность ночная. Питаются главным образом бамбуком, но охотно поедают насекомых, особенно прямокрылых. У самок серого лемура в декабре—январе в гнезде рождается один детеныш. Беременность примерно 160 дней. Лактация около 6 месяцев. Полумаки хорошо уживаются в неволе, быстро приручаются. В неволе они доживали до 12 лет.
Распространены на Мадагаскаре.
В роде 2 вида:
серый лемур — И. griseus Link, 1795 (восток и запад Мадагаскара); широконосый лемур — Н. simas Gray, 1871 (окрестности оз. Алаотра). Как серый, так и широконосый лемуры малочисленны. Оба вида включены в «Красную книгу».
Род маки, или обыкновенных лемуров,
Genus Lemur Linnaeus, 1758
Размеры средние и крупные. Длина тела 30—45 см. Длина хвоста 40—51 см. Морда заостренная к носу. Волосяной покров мягкий, относительно длинный. Окраска его сильно варьирует: некоторые формы пятнистые, другие — одноцветные, рыжеватые, буроватые или черноватые. Самцы черного лемура обычно черные, а самки бурые. Катта буровато-серый сверху, светлый снизу, а на хвосте чередующиеся черные и белые кольца (рис. 127). У катта кисть и стопа длинные, гладкие снизу, большой палец значительно меньше других, в противоположность остальным представителям этого рода. На передних конечностях у катта находятся специфические кожные железы; секретом этих желез лемур смазывает себе хвост. Кроме
того, у него сильно развиты и анальные кожные железы. Верхние резцы маленькие. Диплоидное число хромосом у разных представителей рода изменчиво: 44, 46, 48, 52, 56, 58, 60.
Большинство обитает в лесных районах, ведет древесный образ жизни. Катта — наземная форма, обитает среди скал мало лесных сухих районов. Обычно держатся группами от 5 до 20 особей. Ведут, как правило, дневной образ жизни. Гнезда делает только вари, остальные находят себе убежище под камнями, в пещерах, кронах деревьев. Питаются как животной, так и растительной пищей. Предпочитают плоды и насекомых. При еде добычу держат в передних лапах. Беременность 4—-5 месяцев. Обычно в марте— апреле самка приносит одного-двух детенышей. Лактационный период около 5 месяцев. В неволе обыкновенные лемуры жили до 25 лет.
Распространены на Мадагаскаре.
В роде 6 видов:
катта — L. catta Linnaeus, 1758;
вари — L. variegaius Kerr, 1792;
черный лемур — L. macaco Linnaeus, 1766;
бурый лемур — L. fulvus Ё. Geoffroy, 1812;
монго — L. mongoz Linnaeus, 1766;
рыжебрюхий лемур — L. rubriventer I. Geoffroy, 1850.
Черный лемур и монго редки и внесены в «Красную книгу».
Род тонкотелых маки, или ласковидных лемуров,
Genus Lepiletnur I. Geoffroy, 1851
Размеры средние. Длина тела 31—35 см. Длина хвоста 25—30 см. Конечности укороченные (рис. 128.) Четвертый и пятый пальцы наиболее длинные. Ноготь имеется только на первом пальце, причем он большой и плоский. Голова конической формы, с короткой мордой. Ушные раковины крупные, округлые, кожистые, тонкие, безволосые. Хвост длинный, покрытый довольно длинными, густыми, равномерной длины волосами. На туловище волосяной покров средней длины, густой и мягкий. Окраска его довольно сильно варьирует, хотя в общем туловище имеет рыжеватый цвет, а голова —черноватый. Брюшная сторона белая или желтоватая. Некоторые особи сплошь серые, в то время как другие — бурые. Межчелюстная кость редуцирована. Верхние резцы очень малы или чаще редуцированы. Зубная формула:
• 0 1 3 3
i — с — р — tn— = 32.
2 13 3
Обитатели лесных районов. Активность ночная. Скрытны. На день прячутся в дуплах деревьев,
Рис. 128. Ласковидный лемур — L. muste- гнездах, которые делают из листь-4nus ев или Веточек. Часто встречаются
небольшими группами. Весьма подвижны, могут делать длинные прыжки с ветки на ветку. Питаются главным образом различными растительными объектами: листьями, проростками, плодами, но также и насекомыми. Самка, как правило, приносит одного детеныша. Период размножения, очевидно, приходится на позднюю половину дождливого сезона и начало сухого. Новорожденный покрыт мехом, глаза открытые. До годовалого возраста детеныш остается с матерью. Половозрелость наступает примерно в 18 месяцев.
Распространены на Мадагаскаре.
В роде 2 вида:
ласковидный лемур — L. mustelinus I. Geoffroy, 1851; краснохвостый лемур — L. ruficaudatus A.. Grandidier, 1867. Некоторые ученые объединяют оба вида в один вид L. mustelines. Оба вида редки и занесены в «Красную книгу».
Род крысиных маки, или карликовых лемуров, Genus Cheirogaleus Ё. Geoffroy, 1812
Размеры мелкие (как и у мышиных маки, наименьшие в семействе и средние). Длина тела 12—25 см. Длина хвоста примерно равна длине тела. Ушные раковины тонкокожистые, голые, за исключением волосатоухого
Рис. 129. Мышиный лемур — М. murinus
лемура, у которого уши имеют небольшие кисточки волос. Концевые фаланги пальцев расширенные. Волосяной покров очень густой, мягкий, средней длины. Спинная сторона буроватая, серая пли красновато-бурая, в то время как брюхо беловатое, часто с желтым оттенком. Характерно удлинение os calcaneus и os navicularae. Средние верхние резцы очень крупные. Диплоидное число хромосом у большого крысиного маки 66.
Обитатели лесных районов. Активность ночная. День проводят в гнездах шаровидной формы, которые делают из веточек и листьев на вершине или в дуплах высоких деревьев. К сухому сезону года запасают большое количество жира в проксимальной части хвоста и на сухой сезон могут впадать в спячку. Питаются преимущественно листьями и плодами, а также насекомыми и мелкими птицами. В помете 2—3 детенышей.
Распространены на Мадагаскаре.
В роде 3 вида:
крысиный маки — С. major Ё. Geoffroy, 1812;
толстохвостый лемур — С. medius Ё. Geoffroy, 1812;
волосатоухий лемур — С. trichotis Gunther, 1875.
Толстохвостый лемур редок, а волосатоухий лемур известен всего по трем экземплярам. Оба вида внесены в «Красную книгу».
Род мышиных лемуров
Genus Mycrocebus Ё. Geoffroy, 1828
Размеры небольшие. Длина тела 12—25 см. Длина хвоста 15—28 см. Одни из самых маленьких представителей отряда приматов. Вес примерно от 45 до 85 г. Голова округлая, морда укороченная. Уши крупные, перепончатые, не покрытые волосами (рис. 129). Задние конечности удлинены. Волосяной покров довольно короткий, густой и мягкий, на верхней I части тела сероватый, иногда с рыжеватым оттенком, на нижней — белый или желтовато-серый. Вдоль средней линии морды проходит белая полоска (иногда отсутствует). Диплоидное число хромосом у мышиного лемура 66.
Обитатели главным образом лесных районов, но встречаются и в дру-I гих биотопах. Ведут древесный образ жизни. Активность ночная. Перед
сухим сезоном в проксимальной части хвоста накапливается жир, и в сухой период года могут впадать в спячку. В качестве убежища используют чаще всего дупла, но строят также и гнезда из веточек и листьев в кронах деревьев. Мышиный лемур насекомояден, но поедает также плоды, листья и, вероятно, мед. В течение года два сезона размножения. Беременность 59—62 дня.
В помете от 1 до 4 детенышей. Длина тела новорожденных 3,7 — 5 см, вес — 2,7 — 4,3 г. В возрасте двух месяцев детеныши достигают размеров взрослых. Глаза открываются на 4-й день после рождения. В неволе приручается плохо. Продолжительность жизни в зоопарке до 7 лет.
Встречаются на Мадагаскаре.
В роде 2 вида:
мышиный лемур—М. murinus J. Miller, 1777;
мышиный лемур Кокуерела — М. coquereli Grandidier, 1867.
Численность мышиного лемура Кокуерела низка. Внесен в «Красную книгу»
Род вильчатополосых лемуров
Genus Pkaner Gray, 1870
В роде единственный вид: вильчатополосый лемур — Р. furcifer Blain-ville, 1839 (рис. 130).
Размеры средние. Длина тела 25—27 см. Длина хвоста 30—35 см. Уши большие, голые, закругленные. Концевые фаланги пальцев сильно расширены. Туловище покрыто густым, мягким волосяным покровом, общая окраска его красновато-серая или буровато-серая. Наибольшей яркости окраска
достигает на голове и на шее. На голове от глаз к затылку прбходят две черные полосы, которые в области затылка соединяются вместе и далее назад тянется единая черная полоса вдоль середины шеи и вдоль всей спины. Горло и брюхо бледно-рыжеватые или желтоватые. Кисти и стопы бурые, а хвост покрыт длинными густыми волосами темно-красновато-бурого цвета с черным или белым кончиком.
Обитатели лесных районов, а также ландшафтов типа саванн. Активность главным образом ночная. Днем находит убежища в дуплах деревьев или в гнездах, которые делает из веточек и сухих листьев. Обычно держатся по 2—3 вместе. Очень подвижный лемур, способный делать длинные прыжки. В спячку на время сухого сезона, по-видимому, не впадает. Питается преимущественно зелеными растениями, а также медом и, вероятно, насекомыми. В помете 2—3 детеныша.
Распространен по северному и западному побережью Мадагаскара. Вильчатополосый лемур встречается редко. Занесен в «Красную книгу».
СЕМЕЙСТВО ИНДРИ
FAMILIA INDRIDAE BURNETT, 1828
Мелких, средних и крупных размеров приматы. Длина тела от 30 до 106 см. Длина хвоста от 5 до 53 см. Внешний вид обезьяноподобный. Ко
нечности относительно удлиненные, причем задние примерно на У3 длиннее передних. Первый палец передних конечностей довольно короткий и противопоставляется остальным. На задних конечностях первый палец очень
крупный и сильно противопоставляется остальным четырем пальцам, сращенным в основании общим кожным покровом. Пальцы передних и задних конечностей снабжены ногтями. Вдоль наружных краев передних конечностей до боков груди тянется складка кожи, представляющая как бы рудимент летательной перепонки. Голова округлая, с укороченной лицевой частью. Морда не покрыта волосами. Вибриссы немногочисленны, небольших размеров. Глаза крупные. Ушные раковины хорошо развиты.
Волосяной покров средней длины, мягкий, густой. Окраска бурая, черная, красноватая, желтоватая или белая, одноцветная или из нескольких цветов. В коже расположены крупные потовые железы. У самок одна пара сосков в грудной области.
В черепе характерны расширенная и округлая мозговая часть и сильно укороченная лицевая. Симфизис в нижней челюсти длинный. Угловой J 2 12 3
отросток вытянутый. Зубная формула: i — с — р — т— = 30. Верх-2 0 2 3
ние резцы увеличены в размерах. Первый резец может быть крупнее или меньше второго. Резцы на нижней челюсти направлены вперед и лишь отчасти вверх. Первый нижний предкоренной зуб имеет форму клыка. Позвонков шейных 7, грудных 12, поясничных 8—9, крестцовых 3—4, хвостовых 11—30. Имеется os penis.
Сильно развиты слюнные железы. Желудок простой, но больших размеров. Кишечник длинный — в 9—14 раз превышающий длину тела. Слепая кишка большая, спирально закрученная.
Обитатели лесных районов. Ведут древесный образ жизни, хотя часто спускаются на землю. Способ передвижения по деревьям и по земле отличается от представителей семейства лемуров. При лазании по ветке индри медленно перехватывает ее поочередно передними и задними лапами. Вниз по стволу спускается хвостом вперед. По земле передвигаются короткими прыжками. Мохнатые индри имеют ночную активность, остальные представители семейства — дневную. В спячку не впадают. Держатся поодиночке, иногда парами или объединяются группами до 10 и более особей. Питаются различными растительными объектами: листьями, сочными плодами, орехами, корой, цветками. Пищу берут сразу ртом или придерживают передни-
Рис. 131. Мохнатый индри — L. laniger и его череп
ми конечностями. Размножение сезонное. Гон приходится, очевидно, на январь — февраль. Беременность от 2 до 5 месяцев. В помете один детеныш. Новорожденные имеют открытые глаза и несколько уже прорезавшихся зубов. Детеныш находится при матери в течение нескольких месяцев; сначала повисает на ее брюхе, а затем переносится самкой на спине. Половозрелость наступает примерно в 30 месяцев (у хохлатых индри). Короткохвостые индри способны издавать громкие крики, усиленные за счет специального гортанного мешка. Самцы могут сражаться между собой в период размножения.
Распространены на Мадагаскаре.
В семействе 3 рода (4 вида); мохнатых индри — Lichanotus liliger, 1811 (1 вид);
хохлатых индри — Propithecus Bennett, 1832 (2 вида);
короткохвостых индри — Indri Ё. Geoffroy et G. Cuvier, 1795 (1 вид).
Род мохнатых индри
Genus Lichanotus liliger, 1811
В роде единственный вид: мохнатый индри — L. laniger Gmelin, 1788 (рис. 131).
Наименьший представитель семейства. Длина тела 30—54 см. Длина хвоста 32—40 см. Хвост очень длинный, покрытый равномерными довольно длинными густыми волосами. Голова почти сферической формы, с сильно укороченной мордой. Глаза крупные. Относительно небольшие уши покрыты шерстью. Общая окраска волосяного покрова буро-серая. Конечности белые. Хвост красновато-оранжевый. Между верхними средними резцами имеется крупная диастема.
Обитатель лесов. Держатся чаще поодиночке, иногда по две или
три особи вместе. Большую часть времени проводит на деревьях, лишь иногда спускается на землю. Активность ночная. Днем прячется в дуплах деревьев или среди густо переплетающихся ветвей. Питается главным образом листьями, почками и корой деревьев. Беременность 4—5 месяцев. В помете один детеныш. При рождении имеет длину тела около 9 см.
Встречается в северо-западной части Мадагаскара.
Подвид мохнатого индри A. laniger occidentalis Lorenz, 1898 редок. Внесен в «Красную книгу».
Род хохлатых индри
Genus Propithecus Bennett, 1832
Наиболее крупные и средних размеров представители семейства. Длина тела 50—106 см. Длина хвоста 43—53 см. Задние конечности длинные и сильные (рис. 132). Лицевой отдел головы вытянут несколько больше, чем у мохнатых индри. Волосяной покров довольно длинный и мягкий, значительно более густой на спинной стороне, чем на брюшной. Окраска его сильно изменчива: от белой с желтоватыми оттенками до черной и серой или красновато-бурой. Безволосая часть морды черная. Уши относительно небольшие, скрытые в меху головы. Хвост относительно короче, чем у мохнатых индри, и значительно более длинный, чем у короткохвостых индри; покрыт довольно короткими, равномерной длины волосами. Диплоидное число хромосом у хохлатого индри 48.
Обитатели лесов. Встречаются группами от 6 до 10 особей. Большую часть времени проводят на деревьях. Способны делать длинные прыжки до 10—12 м. Спускаются также и на землю, где передвигаются прыжками на задних.лапах; при этом держат передние конечности поднятыми над головой и балансируют ими. Пищу с земли подбирают ртом, лишь изредка употребляют для этого передние конечности. Питаются преимущественно листьями, цветками, почками, иногда плодами. Активность дневная, в самое жаркое время дня отдыхает. Беременность 4—5 месяцев. Самки приносят единственного детеныша обычно в сентябре. Лактация 3—4 месяца. Поло-возрелость наступает в двухлетнем возрасте.
Распространены на Мадагаскаре.
В роде 2 вида:
белолобый индри — Р. diadema Bennett, 1832;
хохлатый индри — Р. verreauxi Grandidier, 1867.
Оба вида редки (особенно последний). Занесены в «Красную книгу».
Род короткохвостых индри
Genus Indri Ё. Geoffroy et G. Cuvier, 1795
- В роде единственный вид: короткохвостый индри — I. indri Gmelin, Г 1788 (рис. 133).
L Размеры средние. Длина тела 61—71 см. Длина хвоста 5—6 см. Голова I со слегка вытянутой мордой. Уши довольно крупные, покрытые волосами. F Волосяной покров на спине длинный, густой, шелковистый, на брюхе зна-I чительно короче. Окраска волосяного покрова различных оттенков серого, ! бурого и черного с большими вариациями у разных особей. Некоторые совсем черные, другие — почти белые. Голова, уши, спина и передние конечности обычно черные. Сзади на туловище обычно расположено треуголь-|! ной формы большое белое пятно, охватывающее и верхние части задних конечностей. Имеется гортанный мешок, который служит резонатором.
Обитает в дождевых лесах до 1800 м над уровнем моря. Живет обычно семейными группами по 3—4 особи. Ведет почти исключительно древесный образ жизни. Активность дневная. По земле передвигается прыжками на задних конечностях, причем туловище держит вертикально, а передние i лапы поднимает над головой. Питается листьями, плодами и цветками,
Рис. 132. Белолобый индри — Р. diadema. А — кисть; Б — стопа; В — череп
Рис. 133. Короткохвостый индри— /. indri
преимущественно рано утром и к вечеру. Беременность около двух месяцев. В помете один детеныш.
Распространен на восточном побережье Мадагаскара.
Численность мала. Занесен в «Красную книгу».
СЕМЕЙСТВО РУКОНОЖЕК
FAMILIA DAUBENTONIIDAE GRAY, 1870
В семействе единственный род: руконожек — Daubentonla Ё. Geoffroy, 1795 и единственный вид: руконожка, или ай-ай, — D. madagascariensis Gmelin, 1788 (рис. 134).
Размеры небольшие. Длина тела 36—44 см. Длина хвоста 50—60 см. Вес около 2 кг. Туловище удлиненное, стройное. Конечности умеренной длины, относительно тонкие. Первый палец передних конечностей подвижный, но не очень сильно противопоставлен остальным. Первый палец задних конечностей хорошо противопоставляется другим пальцам. Все пальцы имеют когтеобразные ногти, за исключением первого пальца задних ног, который несет плоский ноготь. Пальцы на конечностях, особенно передних, сильно удлинены. Голова округлая, с укороченным лицевым отделом. Глаза крупные. Ушные раковины очень большие, перепончатые, не покрытые волосами, подвижные. Зрачок на глазах вертикальный; имеется хорошо развитая мигательная перепонка. На морде волосы укороченные. Ноздри щелевидные. Лицо и губы относительно мало подвижные. Хвост длинный, покрытый удлиненными косматыми волосами.
Волосяной покров довольно грубый и не очень густой. Окраска его темно-бурая или черная сверху и снизу. Окологлазные пятна, щеки и горло желтовато-белые. У самок одна пара паховых сосков.
В черепе характерна округлая мозговая часть и сильно укороченная лицевая (см. рис. 134). Предчелюстные кости хорошо развиты. Лобные кости не срастаются между собой и не соприкасаются с верхнечелюстными костями. Имеется межтеменная кость. Угловые отростки нижней челюсти небольшие. Носовые кости средних размеров. Орбиты направлены больше вперед, чем у лемуров. Костные слуховые барабаны уплощенные. Зубная формула: i -J- с Р ~~ ~ или Клыки часто отсутствуют.
Позади резцов обширная диастема. Щечные зубы с уплощенными коронками. Резцы сходны в известной степени с резцами грызунов. Они большие, изогнутые, покрыты эмалью только на передней поверхности, имеют постоянный рост. Позвонков шейных 7, грудных 13, поясничных 6, крестцовых 2—4, хвостовых 18—24. Имеется os penis.
Слюнные железы крупные, мягкое небо имеет такие же валики, как у лемуров. Язык узкий, его свободная часть укороченная и толстая. Желудок простой. Хрящевые кольца трахеи с задней стороны незамкнутые. Левая почка располагается каудальнее правой. Матка двурогая.
Обитает в лесах, мангровых зарослях, бамбуковых чащах. В основном ведет древесный образ жизни. Держится поодиночке или парами. Активность ночная. Днем спит в дуплах или среди густых ветвей в кроне деревьев. Ночью очень активен и способен делать довольно длинные прыжки с ветки на ветку. Обычно молчалив, лишь иногда издает короткий крик. Питается в основном насекомыми и их личинками, иногда яйцами птиц и некоторыми растениями (например, сердцевиной бамбуковых ростков). Разыскивает личинок насекомых и самих насекомых на слух и вытаскивает их из щелей коры и древесины длинным средним пальцем, иногда резцами. Самка приносит единственного детеныша в феврале — марте в гнезде. Оно имеет диа
Рис. 134. Руконожка —D. madagascarien-sis. А — кисть; Б — стопа; В — череп
метр около 50 см, построено из листьев, с боковым входом. Новорожденный имеет около 16 сл/ в длину и покрыт коротким редким мехом. В неволе руконожки жили до 9 лет.
Ареал охватывает Мадагаскар.
Численность чрезвычайно мала. К 1969 г. оставалось, по-видимому, всего около 50 особей. Включен в «Красную книгу».
СЕМЕЙСТВО ЛОРИ
FAMILIA LORISIDAE GREGORY, 1915
Размеры небольшие. Длина тела 11—39 см. Хвоста нет или он составляет примерно треть, половину или чуть больше половины длины тела. Туловище удлиненное. Шея короткая, конечности средней длины или несколько укороченные. Передние и задние конечности одинаковой длины или задние развиты сильнее. Первый палец слабо или сильно противопоставляется остальным. Пальцы снабжены ногтями, за исключением второго пальца задних конечностей, который имеет когтеобразный ноготь. Кисть и стопа широкие. Голова округлая, с укороченным лицевым отделом. Глаза боль-шиё. Уши различных размеров от маленьких, скрытых в меху головы, до длинных, заостренных, перепончатых и очень подвижных. На морде имеются вибриссы, менее многочисленные по сравнению с лемурами. Волосяной покров относительно короткий, густой и мягкий. Окраска его обычно серая, бурая или золотистая, более светлая на брюшной стороне. Область вокруг глаз обычно темная. На голове имеются иногда темные полосы и светлые пятна. У самок 2 или 3 пары сосков. Половой деморфизм проявляется в том, что самки некоторых видов немного крупнее самцов или имеют с ними различную окраску.
В черепе мозговая коробка округлая, лицевой отдел укорочен. Имеется межглазничное сужение. Слезные кости редуцированы. Лобные кости соприкасаются с верхнечелюстными. Орбиты крупные. Костные слуховые j_______________________________________________2 1 3 3
барабаны хорошо развиты. Зубная формула: i —— с — р ~^т~ — = 34 — 36. У толстых лори крайние верхние резцы иногда отсутствуют. У большинства видов нижние резцы направлены вперед и слегка вверх. Верхние клыки длинные и острые, а нижние напоминают резцы. Коренные зубы квадритуберкулярные. Позвонков шейных 7, грудных 13—18, поясничных 6—9 (всего грудных и поясничных 19—24), крестцовых 2—9, хвостовых 5—25. У лори хвостовые позвонки срастаются между собой. Имеется os penis.
Желудок простой, округлый. Хрящевые кольца трахеи с задней сто-i роны не замкнуты. Матка двурогая. Плацента, как и у лемуров, за исклю-: чением галаго Демидова, у которого плацента эндотелиохориальная. Обо-! нательные доли мозга развиты не особенно сильно.
Обитатели тропических лесов, обычно густых, влажных или сухих. Поднимаются в горы до 2000 м над уровнем моря, иногда живут на более или менее открытых равнинах или в саваннах (галаго). Ведут в основном 1 древесный образ жизни, а галаго часто спускается и на землю. Активность I ночная; галаго бывают активны и в дневное время. День проводят в дуплах деревьев или на ветвях, в кроне. Спят, свернувшись клубком и поместив i голову между задними конечностями. Лори и потто передвигаются по де-I ревьям медленно, переставляя лапу за лапой. Галаго двигается прыжками. 1 По земле галаго передвигается прыжками на задних лапах. Для лори ха-! рактерна мощная хватка, так что они могут повиснуть на ветке, держась s только задними лапами. У них исключительно подвижные запястья и го-; леностопные сочленения, есть «чудесная сеть» в конечностях. Держатся : поодиночке, парами или небольшими группами (галаго). Для представителей \ семейства характерны довольно разнообразные звуки. По характеру пита
1 я
ния главным образом насекомоядные или хищники, хотя поедают также яйца птиц, различные растительные объекты: плоды, листья и т. п. При еде помогают себе передними конечностями.
Четкой сезонности в размножении не отмечено. Беременность от 6 недель до 6 месяцев. В помете 1—2 детеныша. Новорожденные покрыты волосами, глаза открыты. Лактационный период длится примерно 3,5 месяца. С матерью остаются в течение года. Половозрелость наступает примерно в 1,5 года.
Распространены в Индии, на Цейлоне, в Юго-Восточной Азии и Африке к югу от Сахары.
В семействе 6 родов (11 видов):
тонких лори — Loris Ё. Geoffroy, 1796 (1 вид);
толстых лори — Nycticebus, Ё. Geoffroy, 1812 (2 вида);
золотых потто, или медвежьих маки, — Arctocebus Gray, 1863 (1 вид);
потто — Perodicticus Bennett, 1831 (1 вид);
галаго — Galago Ё. Geoffroy, 1796 (4 вида);
иглокоготных галаго — Euoticus Gray, 1863 (2 вида).
Некоторые исследователи выделяют 2 последних рода в отдельное семейство Galagidae. По-видимому, более правильно считать их в подсемействе Galaginae. Иногда роды галаго и иглокоготных галаго соединяют в один род.
Род тонких лори
Genus Loris Ё. Geoffroy, 1796
В роде единственный вид: тонкий лори — L. tardigradus Linnaeus, 1758 (рис. 135).
Размеры небольшие и средние. Длина тела 17—25 см. Снаружи хвост не заметен. Вес достигает примерно 340 а. На передних и задних конечностях первые пальцы сильно противопоставляются остальным. Задние конечности примерно такой же длины, как передние. Голова округлая, со слабо вытянутой лицевой частью. Глаза крупные, круглые. Уши довольно боль-
Рис. 135. Тонкий лори — L. tardigradus
с Р п С т и н р
Рис. 136. Толстый лори — N. coucang
шие, тонкие, безволосые по краям. Волосяной покров густой и мягкий. Окраска его варьирует от желтовато-серого до темно-бурого на спинной стороне и от серебристо-серого до грязно-желтого на брюшной. Диплоидное число хромосом 62.
Обитатель лесных районов; в горы поднимается до 1800 м надуровнем моря. Активность ночная. День проводит на ветвях деревьев, ухватившись за ветку передними и задними лапами и свернувшись в шар. Ведет почти исключительно древесный образ жизни. Движения медленные. Передвигается по ветвям, перехватывая их поочередно передними и задними конечностями. Метит индивидуальный участок мочой. Питается главным образом насекомыми, ящерицами, в особенности гекко, древесными лягушками, птицами и их яйцами. Беременность примерно 6 месяцев. Самка приносит, по-видимому, два потомства в год. Число детенышей в помете 1—2. Они остаются с матерью по крайней мере в течение года. Хорошо уживается в неволе.
Ареал охватывает Южную Индию и Цейлон.
Род толстых лори
Genus Nycticebus Ё. Geoffroy, 1812
Мелких, средних и крупных размеров лори. Длина тела 18—38 см. Хвост снаружи почти не заметен. Вес 0,5—1,5 кг. Голова округлая, с сильно укороченной мордой (рис. 136). Первые пальцы на конечностях поставлены почти под прямым углом к остальным. Волосяной покров густой и мягкий. Окраска его на спинной стороне тела от светло-буровато-серой до красновато-бурой, на брюшной несколько светлее, от почти белой до грязно-желтой или сероватой. Вдоль шеи и спины по средней линии обычно проходит темная полоса. Посередине морды между темными окологлазничными кольцами располагается белая полоска. В черепе характерны увеличенный лицевой
отдел, небольшие орбиты и несколько уплощенный мозговой отдел. Диплоидное число хромосом как у толстого, так и у малого лори 50.
Обитают в лесных районах. Ведут древесный образ жизни, на землю спускаются* редко. Активность ночная. День проводят в дуплах или на ветвях деревьев, свернувшись в шар. Питаются главным образом насекомыми, а также мелкими млекопитающими, плодами, листьями, проростками. Беременность примерно 90 дней (по другим данным 174—180 дней). В помете обычно один детеныш.
Распространены от Ассама, Бирмы, Таиланда, п-овов Индокитай и Малакка до о-вов Суматра, Ява, Калимантан и Филиппинских.
В роде 2 вида:
толстый лори — N. coucang Boddaert, 1785 (Ассам, Читагонг, Бирма, Тенассерим, Таиланд, п-ова Индокитай, Малакка, о-ва Суматра, Ява, Калимантан и прилежащие до Филиппинских);
малый лори — N. pygmaeus Bonhote, 1907 (п-ов Индокитай).
Род золотых потто, или медвежьих маки,
Genus Arctocebus Gray, 1863
В роде единственный вид: золотой потто, или медвежий маки, — А. са-labarensis J. Smith, 1860.
Размеры средние. Длина тела от 23 до 30 см. Хвост снаружи едва заметен. Голова с относительно удлиненной, заостренной мордой. Глаза и уши крупные. Второй палец на передней конечности редуцирован почти полностью, так что снаружи от него остается лишь небольшой выступ. Волосяной покров довольно длинный, густой и мягкий. Окраска его золотистая, красновато-золотистая, желтовато-бурая на спинной стороне и светлая, почти белая на брюшной. Лицевая часть головы более темная, чем спина. Как и у предыдущего рода, мозговая коробка уплощена, орбиты небольшие.
Обитают в крупных лесных массивах. Экология изучена слабо. Питается, по-видимому, беспозвоночными и мелкими позвоночными животными, а также, вероятно, и растительными объектами.
Распространение охватывает западные районы Центральной Африки: Камерун, Нигерию на север до границы леса и на запад до р. Нигер.
Род потто
Genus Perodicticus Bennett, 1831
В роде единственный вид: обыкновенный потто — Р. potto Muller, 1766 (рис. 137).
Крупный лори — наибольший в семействе. Длина тела 32—39 см. Длина хвоста 5—10 см. Вес 1—1,5 кг. Глаза средних размеров, уши небольшие, кожистые. На передних конечностях второй палец сильно редуцирован, первый палец противопоставляется остальным, и кисть в высокой степени приспособлена к хватанию. На задних лапах второй палец слегка укорочен и имеет сильно изогнутый длинный и острый коготь. В отличие от золотого потто, имеет хвост умеренной длины, равномерно покрытый довольно густыми волосами. Волосяной покров средней длины, густой и мягкий. Окраска его на спине от буровато-серой до темно-красновато-бурой; на брюхе слегка более светлая. Характерны сильно увеличенные в размерах остистые отростки последних шейных и первых грудных позвонков. Своими вершинами они поднимаются непосредственно к коже, в которой в этих местах образовались ороговевшие бугорки. Диплоидное число хромосом 62.
Обитатель тропических лесов. Активность ночная. День проводит в дуплах деревьев и в развилках ветвей. Ведет древесный образ жизни. Пи-312
Рис. 137. Обыкновенный потто — Р. potto тается насекомыми, различными другими беспозвоночными, плодами, листьями. Поедает, вероятно, также яйца птиц, птенцов и ящериц. В течение года бывает, по-видимому, два сезона размножения. В помете, как правило, один детеныш.
Распространен в Западной, Центральной и Восточной Африке (Камерун, Нигерия, Гвинея, Республика Заир, Кения).
Род галаго
Genus Galago Ё. Geoffroy, 1796
Лори мелких и крупных размеров. Длина тела И—38 см. Длина хвоста 15—41 см. Голова округлая с укороченной мордой (рис. 138). Глаза большие, близко поставленные друг к другу. Уши крупные, перепончатые, подвижные, способные независимо друг от друга двигаться в разные стороны. Стопа сильно удлинена. Концевые фаланги пальцев с расширенными подушечками, имеющими снизу вид уплощенных дисков. Волосяной покров довольно длинный, густой и мягкий, слегка волнистый. Окраска его у большинства видов от серебристо-серой до бурой, брюшная сторона немного светлее спинной. У самок пара брюшных и пара грудных сосков. У сенегальского галаго диплоидное число хромосом 38, у толстохвостого — 62.
Обитатели лесных районов и участков, поросших кустарником в саваннах и степях. Ведут древесный образ жизни. День проводят в дуплах деревьев, среди густых ветвей, устроившись на развилке веток, иногда в старых гнездах птиц. Держатся поодиночке или парами. В ночное время очень активны; способны делать длинные прыжки с ветки на ветку. По земле передвигаются прыжками на задних конечностях, наподобие миниатюрных кенгуру. При этом они иногда делают прыжки до 3—5 м длиной. Питаются разнообразными объектами: насекомыми (преимущественно пря-
Рис. 138. Сенегальский галаго — G. sene-galensis
мокрылыми), мелкими птицами, их яйцами, сочными плодами, семенами и цветами. Беременность ПО— 120 дней. Самки приносят два помета с апреля по ноябрь; в каждом 1—2 (редко 3) детеныша. Перед рождением детенышей самка строит гнездо. Новорожденные сенегальского галаго весят около 15 а, и имеют длину тела примерно 6 см. В возрасте примерно двух недель детеныши способны самостоятельно двигаться и делать небольшие прыжки. Лактация длится примерно 3,5 месяца. Продолжительность жизни, вероятно, около 10 лет.
Ареал охватывает Африку к югу от Сахары.
В роде 4 вида:
толстохвостый галаго — G. crassicaudatus Ё. Geoffroy, 1812 (Южно-Африканская Республика, Мозамбик, южная Родезия, Замбия, Малави, Ангола, Танзания, Занзибар, Кения, Сомали);
сенегальский галаго — G. sen-egalensis Ё. Geoffroy, 1796 (Южно-Африканская Республика, Юго-Западная Африка, Ботсвана, южная Родезия, Замбия, Малави, Ангола, Мозамбик, Западная Африка от Республики Заир до Ганы и Сенегала, большая часть Восточной Африки до Судана и Сомали);
галаго Аллена — G. alien i
Waterhouse, 1837 (от р. Убанга и нижнего течения р. Нигер до Гвинейского залива; о-в Фернандо-По);
галаго Демидова — G. demidovi Fischer, 1808 (северо-восточная Ангола и тропическая Африка от Сенегала до Уганды и Танзании).
Некоторые исследователи включают в род галаго иглокоготных галаго и выделяют их в отдельное семейство Galagidae Anthony et Vallois, 1926. Видимо, более правильно считать галаго подсемейством семейства лори.
Род иглокоготных галаго
Genus Euoticus Gray, 1863
Крупные лори. Длина тела около 33 см. Длина хвоста примерно 25 см. Все пальцы, за исключением первого, на задних конечностях имеют заостренные и узкие ногти. Первый палец на задней конечности несет уплощенный ноготь. Голова уплощена. Уши расположены по бокам головы. Конечности укороченные, сильные. Характерен длинный хвост, покрытый равномерно по всей длине густыми волосами. Волосяной покров густой, мягкий, средней длины. Окраска его на спинной стороне тела оранжево-светло-коричневая или светло-коричнево-серая, на брюшной более светлая, часто сероватая.
Обитают в лесных районах. Экология изучена очень слабо. Ведут древесный образ жизни. Питаются, вероятно, насекомыми, плодами, птенцами. 314
Сезонности в размножении, по-видимому, нет, но в дождливый сезон не размножаются.
Распространены в западной экваториальной Африке, найдены также на о-ве Фернандо-По.
В роде 2 вида:
западный галаго — Е. elegantulus, Leconte, 1857 (от р. Санга, Убанга и Конго на запад до Камеруна и Габона);
восточный галаго — Е. innstos Schwarz, 1930 (Республика Заир).
Иглокоготных галаго и галаго иногда объединяют в один род.
СЕМЕЙСТВО ДОЛГОПЯТОВ FAMILIA TARSIIDAE Gill, 1872
В семействе единственный род: долгопятов — Genus Tarsius, Storr, 1780 (рис. 139).
Небольшие по размерам полуобезьяны. Длина тела 8,5—16 см. Длина хвоста 13— 27 см. Вес от 80 до 150 г. Голова округлая, со слабо развитым лицевым отделом. Глаза огромные, обращенные вперед сильнее, чем у лемуров. Зрачок способен сокращаться до узкого, горизонтального овала. Ноздри округлые, открываются на покрытую волосами подвижную верхнюю губу. Уши большие, безволосые. Лицевые мышцы хорошо развиты, и долгопяты могут гримасничать наподобие обезьян. Туловище худощавое, но не удлиненное. Шея короткая. Хвост длинный, покрытый редкими волосами, и лишь в концевой части имеет кисточку волос. Концы пальцев имеют расширения в виде подушечек (см. рис. 139). Волосяной покров шелковистый, слегка извитый, окраска его сероватая до красновато-бурой; с вентральной стороны более светлая. Самки имеют по паре подмышечных и паре паховых сосков.
Череп с расширенной округлой мозговой частью и укороченной лицевой (см. рис. 139). Лобные кости сочленяются со скуловыми и небными. Межчелюстные и носовые кости небольшие. Костные слуховые барабаны крупные. Зубная формула: 2 13 3
i — с — р — т — = 34. Верхние резцы крупные и острые. Нижние резцы направлены вверх, а не вперед. Верхние клыки относительно небольших размеров. Верхние коренные тритуберкулярные, нижние—
Рис. 139. Филиппинский долгопят — Т. syrichta. А — кисть: Б — стопа; В — череп долгопята-привидения — Tarsias sp.
туберкулосекториальные. Кости задней конечности удлинены, особенно os calcaneus и os naviculare. Дистальные отделы большой и малой берцовых
костей срастаются между собой. Позвонков шейных 7, грудных 13—14, поясничных 6—8, крестцовых 2—3, хвостовых 25—27. Os penis отсутствует.
Толстая кишка короткая и только с одним изгибом. Внутренняя подвздошная, бедренная и хвостовая артерии имеют «чудесную сеть». Полушария большого мозга без извилин или борозд. Мозжечок не полностью прикрыт полушариями большого мозга. Легкие многодольчатые (6 долей в правом и 4 в левом). Матка двурогая. Плацента дискоидальная, гемохориальная, отпадающая. Диплоидное число хромосом у западного долгопята 80..
Обитатели лесов. Ведут древесный образ жизни. Активность ночная. День проводят чаще всего в густых переплетениях ветвей, тесно прижавшись к вертикальной ветке, реже в дупле. Превосходно лазают по деревьям, способны делать прыжки до нескольких метров длиной. Хвост служит балансиром. По земле передвигаются прыжками, причем прыгают до 120—170 см в длину и до 160 см в высоту. Держатся поодиночке, парами или группами, иногда по 3—4 особи. Питаются в основном насекомыми, но поедают также мелких позвоночных и яйца птиц. При еде помогают себе передними лапами. Способны к размножению круглый год, но наиболее активный период гона в ноябре — феврале. Длительность беременности не установлена. В помете один детеныш. Новорожденный покрыт мехом, имеет открытые глаза и вскоре после рождения способен карабкаться по ветвям.
Распространены на о-вах Суматра, Банка, Биллитон, Каримато, На-туна, Калимантан, Саву, Селаяр, Сулавеси, Сангихе и Филиппинских (Минданао, Бохол, Лейте и Самар).
В роде 3 вида:
филиппинский долгопят—T.syrichta Linnaeus, 1758 (Филиппинские острова);
западный долгопят — Т. bancanus Horsfield, 1821 (о-ва Суматра, Калимантан и прилежащие острова);
долгопят-привидение, или восточный долгопят, — Т. spectrum Pallas, 1779 (о-в Сулавеси и соседние острова).
Подотряд высших приматов, или обезьян,
Subordo Anthropoidea Mivart, 1864
Мелкие, средние и крупные приматы (наиболее крупные в отряде). Длина тела от 15 до 180 см. Хвост отсутствует, развит слабо или сильно, достигая в последнем случае длины тела или превышая ее; может быть хватательным. У большинства видов первый палец противопоставляется остальным и все пальцы вооружены ногтями.
Волосяной покров густой и мягкий, обычно со слабо выраженными категориями волос. У самок одна пара функционирующих сосков. В черепе сильно развит мозговой отдел, лицевой относительно слабо выдается вперед. Зубная формула: i ~ с р 4—3 m~ 3- = 32 — 36.
Мозг с имеющими
2 ’ 1 r 2 — 3 2 — 3
крупными большими полушариями, иногда гладкими, но чаще многочисленные борозды и извилины.
СЕМЕЙСТВО ЦЕПКОХВОСТЫХ ОБЕЗЬЯН, ИЛИ КАПУЦИНОВ,
FAMILIA CEBIDAE SWAINSON, 1835
Высшие приматы мелких и средних размеров. Длина тела от 24 до 91 см. Хвост покрыт волосами, у всех представителей семейства длинный, за исключением уакари. Цепкий хвост имеют представители четырех родов:
коаты, паукообразные обезьяны, шерстистые обезьяны и ревуны. У этих обезьян нижняя часть хвоста у терминального конца не имеет волос, мягкая и обладает способностью к осязанию. На всех пальцах ногти уплощенные; у некоторых видов они узкие с килем. Первый палец передних конечностей не противопоставляется, а на задних противопоставляется остальным. Голова округлая, лицевая часть укороченная и уплощенная, иногда несколько вытянутая. Ноздри хорошо отделены друг от друга (за исключением мирикин, ревунов и паукообразных обезьян) и направлены лате-рально. Глаза большие с хорошо развитыми веками (у активных ночью мирикин глаза очень большие). Защечных мешков нет. Седалищных мозолей нет.
Волосяной покров довольно густой, разной длины. Он может быть шелковистый, мягкий или грубый. Окраска обычно одноцветная, черноватая или буроватая, иногда желтоватая или красноватая. У некоторых видов на голове или лице, а иногда и туловище находятся черные, белые или другого цвета отметки. Имеются короткие лицевые вибриссы. У некоторых видов удлиненные волосы образуют на лице своеобразную бороду. Иногда проявляется половой диморфизм в размерах: самки крупнее самцов (коаты) или самцы крупнее самок (ревуны).
В черепе мозговая часть округлена и расширена, лицевая укорочена и уплощена. Орбиты крупные, особенно у мирикин, и направлены вперед. Костные слуховые барбаны относительно невелики. Зубная формула у
2 13 3
большинства видов: I — с — р ~ 36. Коренные зубы обычно
квадритуберкулярные. Последние коренные, как правило, небольших размеров. Позвонков шейных 7, грудных 12—15, поясничных 5—8, крестцовых 2—5, хвостовых 17—34. Os penis у большинства представителей семейства имеется, но отсутствует у родов мирикин, коат, ателис и шерстистых обезьян.
Ведут древесный образ жизни. Приспособлены к дальним прыжкам и бегу по деревьям. Некоторые виды неторопливы в движениях. У большинства активность дневная, только у мирикин — ночная. Держатся обычно семейными группами, иногда собираясь до 50 особей вместе. Многие способны издавать громкие звуки. Питаются преимущественно растительными объектами: плодами, проростками и листьями (особенно ревуны), но поедают и яйца птиц, а также насекомых и других мелких животных. Передние конечности используют, чтобы добыть пищу и отправить ее в рот. Лицо цепкохвостых обезьян может выражать различные эмоции. Уши малоподвижные. Сезонности в размножении нет. Беременность примерно 180 дней. Самка, как правило, приносит одного детеныша, имеющего при рождении примерно 8% веса матери. Первое время детеныш висит, уцепившись на брюхе матери, а затем перемещается на ее спину. В неволе жили до 25 лет.
Распространены в Южной Америке к востоку от Анд примерно от 27° ю. ш., в Аргентине на север через Центральную Америку до в Мексике.
В семействе 11 родов (35 видов): мирикин, или ночных обезьян,—Aotes Humboldt, 1811 (1 прыгунов — Callicebus Thomas, 1903 (8 видов);
уакари, или какайо, — Cacajao Lesson, 1840 (3 вида);
саки — Pithecia Desmarest, 1804 (2 вида);
красноспинных саки — Chiropotes Lesson, 1840 (2 вида);
ревунов — Alouatta Lacepede, 1799 (6 видов); капуцинов — Cebus Erxleben, 1777 (4 вида); саймири—Saimiri Voigt, 1831 (2 вида);
коат — A teles Ё. Geoffroy, 1806 (4 вида);
паукообразных обезьян — Brachyteles Spix, 1823 (1 вид); шерстистых обезьян — Lagothrix Ё. Geoffroy, 1812 (2 вида).
23° с. ш.
вид);
Род мирикин, или ночных обезьян,
Genus Aotes Humboldt, 1811
В роде единственный вид: мирикина, или ночная обезьяна, — A. tri-virgatus, Humboldt, 1811.
Мелких размеров цепкохвостая обезьяна. Длина тела 24—37 см. Длина хвоста 31—40 см. Вес около 1 кг. Форма головы почти круглая, глаза очень крупные, с сильно выгнутой роговицей. Наружные носовые отверстия расположёны близко друг к другу. Хвост длинный, густо покрытый волосами, не хватательный. Уши относительно невелики и почти полностью скрыты в меху. В районе подбородка имеется специальный мешок, который резонирует голос. Кисти относительно короче и шире, чем у других цепкохвостых обезьян. Стопы длинные и узкие. Волосяной покров средней длины, плотный и мягкий. Окраска его изменчива у различных особей, но обычно серебристо-серая до темно-серой на спинной стороне и серая или буроватая на брюшной. В большинстве случаев по верху головы проходят три черные продольные полосы, отделенные друг от друга серыми полями. Резцы располагаются вертикально. Диплоидное число хромосом 54.
Обитатель различного типа лесов, поднимается в горы до 2100 м над уровнем моря. Активность ночная. День проводит в дуплах. Держится небольшими группами. Питается плодами, листьями, пауками, мелкими птицами и, вероятно, другими мелкими позвоночными. Способна издавать громкий крик.
Распространена от Никарагуа на юг до северо-восточной Аргентины и от Гвианы и Бразилии до Перу и Эквадора.
Род прыгунов
Genus Callicebus Thomas, 1903
Размеры от мелких до средних. Длина тела 24—61 см. Длина хвоста 26—55 см. Хвост не хватательный, густо покрыт волосами по всей длине; на конце особо удлиненные волосы (рис. 140). Голова небольшая, округлая, с уплощенным лицевым отделом. Уши почти полностью скрыты в меху. Волосяной покров длинный, мягкий и густой. Окраска его разнообразная: от красновато-серой или желтовато-серой до красновато-бурой или почти черной на спинной стороне и обычно более светлой на брюшной стороне. Нижняя челюсть в задней части очень высокая. Диплоидное число хромосом у красного прыгуна 46.
Обитают в лесах разнообразных типов. Активность дневная. Держатся парами или семейными группами. Большинство видов обладает громким голосом. Питаются плодами, проростками, листьями, насекомыми, яйцами птиц и, вероятно, мелкими птицами.
Распространены в лесах Южной Америки.
В роде 8 видов:
воротничковый прыгун — С. torquatus Hoffmannsegg, 1807 (бассейн Амазонки в Бразилии);
красный прыгун — С. cupreus Spix, 1823 (Бразилия, Эквадор, Перу);
бурый прыгун — С. brunneus Wagner, 1842 (Мату-Гросу в Бразилии, Боливия);
прыгун-молох — С. moloch Hoffmannsegg, 1807 (Парагвай, Перу, Бразилия);
пепельный прыгун — С. cinerascens Spix, 1823 (западная Бразилия, восточные районы Перу);
серый прыгун — С. melanochir Kuhl, 1820 (восточная Бразилия);
черноголовый прыгун — С. personatus Ё. Geoffroy, 1812 (восточная Бразилия);
Рис. 140. Красный прыгун — С. cupreus
чернолобый прыгун — С. nigrifrons Spix, 1823 (восточная Бразилия).
Систематика рода прыгунов окончательно не выяснена. По-видимому, некоторые из перечисленных видов имеют лишь подвидовой ранг.
Род уакари, или какайо,
Genus Cacajao Lesson, 1840
Средних размеров цепкохвостые обезьяны. Длина тела 51—57 см. Хвост довольно короткий, не превышает треть длины тела (рис. 141) Единственные из американских обезьян имеют короткий хвост. Голова довольно большая. Уши крупные, по форме сходны с ушами человека. Характерно, что лицо и верх головы почти полностью лишены волос, в то время как челюсти и горло довольно хорошо покрыты волосами. Иногда имеется небольшая борода. Окраска волосяного покрова различна. Лысый уакари имеет беловатый мех, за исключением лица, которое имеет красную окраску. У красного уакари волосяной покров красновато-бурый или бурый, причем кисти, стопы и уши бурые, а лицо и лоб — красные. Черноголовый уакари по спинной поверхности, бокам тела и верхней части хвоста бурый, а лицо,
Рис. 141. Красный уакари—С. rubicundus
плечи, конечности и нижняя поверхность хвоста — черные. Диплоидное число хромосом у красного уакари 46.
Обитатели лесов. Активны в дневное время. Держатся небольшими группами. Более молчаливые, чем все другие представители семейства. Питаются преимущественно плодами, но поедают, видимо, также листья, насекомых и мелких позвоночных.
Распространены в бассейнах рек Амазонки и Ориноко.
В роде 3 вида:
лысый уакари — С. calvus I. Geoffroy, 1847 (западная Бразилия);
красный уакари — С. rubicundus I. Geoffroy et Deville, 1848 (западная Бразилия и восточный Перу);
черноголовый уакари — С. melanocephalus Humboldt, 1811 (долина р. Рио-Негро).
Иногда в роде уакари выделяют и четвертый вид — С. roosevelti J. Allen, 1914.
Все три вида редки и внесены в «Красную книгу».
Род саки
Genus Pithecia Desmarest, 1804
От мелких до крупных размеров цепкохвостые обезьяны. Длина тела 30—70 см. Длина хвоста 25—54 см. Волосяной покров длинный, густой и мягкий. Волосы на голове обычно спадают от середины на стороны; иногда они растут радиально от центра. У саки-монаха волосяной хохол на голове частично прикрывает уши и лицо и не контрастирует по окраске со лбом и щеками. У взрослых самцов бледного саки хохол не закрывает лица, и голова имеет белую или желтую окраску. У взрослых самок на каждой стороне ноздрей по белой полоске. У саки-монаха волосяной покров верхней
части тела имеет сероватую окраску, а нижней варьирует от бледно-желтовато-бурой до рыжей. У бледного саки окраска тела однородно черная. Диплоидное число хромосом у бледного саки 46.
Ведут главным образом древесный образ жизни. Иногда спускаются на низкие кусты или на нижние ветви деревьев в поисках пищи. В случае опасности передвигаются по деревьям с большой быстротой. Способны бегать по горизонтальным ветвям на задних конечностях. Активность дневная. Часто собираются в группы до дюжины особей, но встречаются также и поодиночке. Питаются разнообразными плодами, медом, листьями, цветками, мелкими млекопитающими, как мышевидные грызуны или летучие мыши, и мелкими птицами.
Распространены в северной части Южной Америки.
В роде 2 вида:
саки-монах — Р. monachus Е. Geoffroy, 1812 (бассейн Амазонки от Пары до восточного Эквадора, Перу и Гвианы);
бледный саки — Р. pithecia Linnaeus, 1766 (от Рио-Негро на восток до Французской Гвианы).
Род красноспинных саки
Genus Chiropotes Lesson, 1840
Размеры от мелких до средних. Длина тела 36—52 см. Длина хвоста Зб—50 см. Характерно развитие большой бороды, хвост покрыт очень длинными и густыми волосами. Окраска несколько различная у разных видов: у белоносого саки волосяной покров на теле черный, а нос и верхняя губа покрыты растущими вниз грубыми желтовато-белыми волосами, черный саки имеет голову и бороду черные; плечи и спина, включая бока тела, от темно-красновато-коричневого до черно-бурого; хвост черный.
Обитатели густых, непроходимых лесов. Экология изучена слабо. Держатся поодиночке, парами или небольшими группами до 6 особей. Питаются плодами, листьями и, видимо, мелкими животными. Крик похож на громкий свист.
Ареал охватывает Бразилию к югу и северу от Амазонки, южную Венесуэлу и Гвиану.
В роде 2 вида:
черный саки — С. satanas Hoffmannsegg, 1807 (Бразилия к северу от Амазонки, южная Венесуэла и Гвиана);
белоносый саки — С. albinasus I. Geoffroy et Deville, 1848 (Бразилия к югу от Амазонки).
Иногда выделяют и третий вид: красноспинного саки — С. chiropotes Humboldt, 1811.
Белоносый саки редок и внесен в «Красную книгу».
Род ревунов
Genus Alouatta Lacepede, 1799
Средних размеров или крупные цепкохвостые обезьяны (наибольшие среди представителей семейства). Длина тела 56—91 см. Длина хвоста 58— 91 см. Вес от 7 до 9 кг. Хвост хватательный и обнажен по нижней стороне терминальной трети. Волосяной покров довольно короткий, у некоторых видов на голове и плечах сильно удлиненный, лицевая часть голая (рис. 142). Окраска его желтовато-бурая, красновато-бурая или черная, причем имеют место значительные возрастные и индивидуальные вариации. Лицевая часть черепа несколько вытянута. Затылочная кость поставлена вертикально. Щечные зубы крупные. Верхние резцы направлены наискось вперед, а
Рис. 142. Колумбийский ревун — A. palliata и череп рыжего ревуна
нижние — вертикально вверх. Клыки очень крупные, особенно у старых самцов. Диплоидное число хромосом 44 у рыжего и 52 — у черного ревунов.
Обитатели тропических лесов. Наибольшая активность в ранние утренние и поздние вечерние часы. Встречаются группами от 4—5 до 20—30, иногда до 40 особей, причем такие стаи имеют одного или двух предводителей из старых самцов. В группы объединяются обезьяны разных возрастов и полов. Обычно держатся на самых вершинах высоких деревьев. Характерны очень громким криком, некоторым образом напоминающим рев львов; в джунглях крик слышен километра за три. Питаются главным образом плодами и листьями, а также орехами, семенами, вероятно, птицами и их яйцами, ящерицами, мелкими млекопитающими, насекомыми. Беременность около 140 дней.
Распространены в Южной Америке от Веракрус в Мексике на юг до Гуаякиля в Эквадоре, на восток от Аид через Боливию и большую часть Бразилии до Парагвая, а также встречаются на о-ве Тринидад.
В роде 6 видов:
гватемальский ревун — A. villosa Gray, 1845 (Гватемала);
колумбийский ревун — A. palliata Gray, 1848 (Колумбия и Эквадор);
рыжий ревун — A. seniculus Linnaeus, 1766 (Венесуэла, Боливия, Колумбия, Эквадор, Перу, Гвиана, Бразилия);
бурый ревун — A. quariba Humboldt, 1812 (север Боливии, восточная Бразилия, северная Аргентина);
краснорукий ревун — A. belzebul Linnaeus, 1766 (Бразилия);
черный ревун — A. caraya Humboldt, 1812 (Боливия, Парагвай, Бразилия) ;
Некоторые исследователи сокращают число видов в роде ревунов до пяти.
Род капуцинов
Genus Cebus Erxleben, 1777
Размеры небольшие. Длина тела 30—38 см. Длина хвоста 38—50 см. Вес примерно от 2 до 4 кг. Хвост длинный, плохо приспособленный к хватанию (рис. 143). Лицо голое. Виды, представленные в роде, разделяются на 2 группы: к одной принадлежат обезьяны, у которых волосы на голове образуют два своеобразных «рога», или гребня, по бокам вершины головы. Эти обезьяны обычно имеют одноцветную серовато-бурую окраску меха. Обезьяны второй группы не имеют пучков волос на голове, у них белая окраска лицевой части головы, горла и груди и черная — остальных частей. Волосяной покров у большинства густой и мягкий. Резцы мощные. Мозг развит очень хорошо. Диплоидное число хромосом 54 (обыкновенный капуцин и капуцин-фавн). Обитатели лесов. Редко поднимаются в горы выше 1500 м над уровнем моря. Встречаются семейными группами, иногда собираются более крупными стаями до 40 особей. Обычно держатся по вершинам больших деревьев. Питаются главным образом плодами, а также проростками, листьями, насекомыми, яйцами птиц и птенцами, а также, вероятно, мелкими птицами и млекопитающими. Беременность примерно 6 месяцев. В неволе капуцины жили до 32 лет.
Распространены на обширной территории Южной Америки, но ареал прерывистый: на севере он достигает Коста-Рики, на юге — Парагвая, Атлантического океана у берегов Бразилии и Тихого океана у побережья Эквадора; встречаются также на о-ве Тринидад, однако отсутствуют на большей части территории северо-запада Южной Америки.
В роде 4 вида:
обыкновенный капуцин — С. capucinus Linnaeus, 1758 (Колумбия, Эквадор, Венесуэла);
Рис. 143. Белолобый капуцин — С. albifrons и череп капуцина
Рис. 144. Беличий саймири — С. sciureus и его череп
бурый капуцин — С. nigrivit tains Wagner, 1847 (Венесуэла, Колумбия, Гвиана, Бразилия);
белолобый капуцин — С. albifrons Humboldt, 1812 (Колумбия, Венесуэла, Эквадор, Перу, Бразилия);
капуцин-фавн — С. apella Linnaeus, 1758 (Бразилия, Колумбия, Венесуэла, Гвиана, Боливия, Парагвай, Аргентина).
Род саймири
Genus Saimiri Voight, 1831
Размеры небольшие. Длина тела 26— 36 см. Длина хвоста 35—42 см. Вес примерно 750—1100 г. Характерны крупные
глаза, посаженные близко друг к другу, и большие уши (рис. 144). Волосяной покров короткий, густой и мягкий, яркой окраски. Кожа в области передней части морды, губы и участок вокруг ноздрей почти лишены волос, черного цвета. Остальная лицевая часть
в области глаз, уши, горло и бока шеи белые. Вершина головы черная или сероватая. Спина, передние конечности и задние конечности красноватые или желтые. Брюшная поверхность белая или слегка охристая. Хвост такой же окраски, как туловище, за исключением терминальной части черного цвета. Диплоидное число хромосом у беличьего саймири 44.
Обитатели лесов с густым подростом, атакже вторичных лесов в районах, освоенных человеком, обычно вдоль водоемов. Активность дневная. До-
вольно часто спускаются в поисках пищи на землю. Встречаются группами от 10 до 100 особей. Питаются главным образом насекомыми, пауками, яйцами птиц, птенцами, сочными плодами, орехами. В неволе жили до 9 лет.
Распространены в Коста-Рике, Панаме и на юг примерно до 25° ю. ш.
В роде 2 вида:
рыжеспинный саймири — S. oerstedi Reinhardt, 1872 (Коста-Рика » Панама);
беличий саймири — S. sciureus Linnaeus, 1758 (от северных районов Южной Америки на юг до Перу, Боливии, Парагвая и Бразилии).
Иногда в роде саймири насчитывают не два, а четыре или пять видов: дополнительно S. boliviensis, S. madeirae, S. usta.
Род коат
Genus Ateles Ё. Geoffroy, 1806
Мелких и средних размеров цепкохвостые обезьяны. Длина тела 38— 63 см. Длина хвоста 50—89 см. Характерны очень длинные конечности и хвост. Хватательная способность хвоста развита в высокой степени. Волосяной покров у большинства видов грубый и редкий, у некоторых мягкий и довольно густой. Окраска его на спинной стороне желтовато-серая или темно-серая, красновато-бурая, темно-бурая, иногда почти черная, на боках золотисто-желтая или рыжая, на брюшной стороне светлее, чем на боках и спине, обычно беловатая или желтоватая. У большинства видов лицевая часть головы черная, с белыми окологлазничными кольцами, но у некоторых видов красноватая. Диплоидное число хромосом 34 (черная коата и коата }Коффруа).
Обитатели тропических лесов, поднимаются довольно высоко в горы. Активность дневная. Держатся небольшими группами до 15—25 особей вместе. Питаются главным образом сочными плодами и орехами. Беременность около 140 дней.
Ареал охватывает территорию от южной Мексики на юг до Центральной Боливии и Мату-Гросу в Бразилии.
В роде 4 вида: ,
коата Жоффруа — A. geoffroyi Kuhl, 1820 (от Колумбии на север до Панамы);
черная коата — A. paniscus Linnaeus, 1758 (Бразилия, Перу, Боливия);
светлолобая коата — A. belzebuth Ё. Geoffroy, 1806 (Венесуэла, Бразилия, Колумбия, Эквадор, Перу);
буроголовая коата — A. fusciceps Gray, 1866 (Эквадор, Колумбия).
Род паукообразных обезьян
Genus Brachytetes Spix, 1823
В роде единственный вид: паукообразная обезьяна — В. arachnoides Ё. Geoffroy, 1806 (рис. 145).
Размеры средние. Длина тела 61 см. Длина хвоста 67 см. Туловище довольно массивное. Конечности относительно длинные и тонкие. Длинный хвост хватательный, имеет на вентральной стороне терминальной части голую поверхность. Большой палец на передней конечности сильно редуцирован или его совсем нет. Ногти на пальцах острые и сжатые с боков. Волосяной покров средней длины, мягкий. Превалирующая окраска его на теле желтовато-серая или серовато-бурая, у некоторых особей красноватая. Голова черновато-бурая с примесью желтого или темно-серого; лоб и задняя часть шеи могут быть оранжево-рыжими, а вершина головы бурой. Голая часть морды обычно красного цвета.
Экология изучена слабо. Активность дневная. Держится группами. Питается главным образом плодами.
Рис. 145. Паукообразная обезьяна — В. arachnoides, вентральная поверхность конца хвоста, кисть (А) и стопа (Б)
Распространены в юго-восточной Бразилии от г. Салвадор до г. Сан-Пауло.
Численность чрезвычайно низка. Вид включен в «Красную книгу».
Род шерстистых обезьян
Genus Lagothrix Ё. Geoffroy, 1812
Средних размеров цепкохвостые обезьяны. Длина тела 50—68 см. Длина хвоста 60—72 см. Вес 5—6 кг. Голова относительно крупная, массивная, округлая (рис. 146). Туловище довольно тяжелое. Хвост длинный, хватательный, с обнаженной поверхностью на вентральной стороне концевой части Конечности относительно длинные и тонкие. Большой палец на передних конечностях развит хорошо. Пальцы относительно короткие, толстые, с длинными заостренными ногтями Волосяной покров короткий, густой, мягкий, с хорошо развитым пухом. Окраска его на спинной стороне серая, голубовато-серая, темно-бурая или черно-бурая, на брюшной слегка светлее. Обнаженная часть морды почти черпая. Диплоидное число хромосом у шерстистой обезьяны 62.
Обитают в различного типа лесах. Поднимаются в горы примерно до 2000 м над уровнем моря. Активность дневная Объединяются в стада до 12 и более особей. Часто держатся с другими обезьянами, например капуцинами. Довольно часто спускаются на землю в поисках пищи. Питаются в основном плодами. Беременность 4,5—5 месяцев.
Распространены в Колумбии, Эквадоре, Перу и Бразилии.
Рис. 146. Шерстистая обезьяна — L. lago-thricha
В роде 2 вида:
шерстистая обезьяна — L. lagothricha Humboldt, 1812 (Колумбия); желтохвостая обезьяна L. flavicauda Humboldt, 1812 (Бразилия^в верхней части бассейна Амазонки, Колумбия, Перу, Эквадор).
СЕМЕЙСТВО ИГРУНОК, ИЛИ МАРМОЗЕТОК,
FAMILIA CALLITHRICIDAE THOMAS, 1903
Самые маленькие представители высших приматов. Длина тела 15— 50 см. Длина хвоста 18—42 см. Вес от 70 до 1 кг. Самки несколько крупнее самцов. Телосложение довольно изящное, пропорциональное. Конечности относительно укороченные, причем передние слегка длиннее задних. Стопа короткая. Пальцы довольно короткие. Большой палец на передних конеч ностях не противопоставляется остальным. Все пальцы, за исключением большого пальца задних конечностей, который имеет уплощенный ноготь, вооружены серповидными когтями. Хвост длинный, не хватательный. Голова небольшая. Уши относительно крупные. Глаза небольшие, ноздри отстоят довольно далеко друг от друга и направлены в стороны. Волосяной покров мягкий и густой. На голове часто удлиненные волосы в виде шапки или в виде других образований. Иногда на голове и плечах грива. Лицевая часть головы голая или покрыта редкими волосами. Окраска волосяного покрова значительно варьирует от серой, черной до красновато-бурой или золотистой. У самок одна пара сосков.
Череп удлинен в лицевой части и закруглен в мозговой. Зубная форму-2 j з 2______________3
ла: i — с — р — т ——— = 32 — 36. Предкоренные зубы двувершинные. Верхние коренные триангулярные; последний (или два последних у рода мармозеток) коренной наименьший. Клыки хорошо развиты. Позвонков шейных 7, грудных 11—13, поясничных 6—8, крестцовых 2— 3, хвостовых 25—33. Таз узкий. Симфиз лобковых костей удлинен. Имеется os penis.
Большие полушария мозга относительно гладкие. Семенники в мошонке.
Очень подвижные и быстрые в движениях древесные обезьяны. Активность дневная, ночь проводят в основном в дуплах деревьев. Питаются главным образом насекомыми, но также поедают птиц, сочные плоды, семена. При еде помогают передними лапами. Держатся в основном семейными группами. Способны выражать свои эмоции движениями губ, век, ушей, и пучков волос на голове (если имеются). Издают разнообразные звуки. Беременность 140—150 дней. В помете от 1 до 3, обычно 2 детеныша. Продолжительность жизни 16 лет.
В семействе 4 рода (35 видов):
мармозеток — Callitnico Mirando Ribeiro, 1911 (1 вид);
игрунок — Callithrix Erxleben, 1777 (11 видов);
львиных игрунок — Leontideus Cabrera, 1956 (3 вида);
тамаринов—Sagainus Hoffmannsegg, 1807 (20 видов).
Распространены в Южной Америке к востоку от Анд и к северу от южной Бразилии, а также в Панаме.
Иногда семейство игрунок присоединяют в качестве подсемейства к семейству капуцинов.
Род мармозеток
Genus Callimico Mirando Ribeiro, 1911
В роде единственный вид: мармозетка — С. goeldii Thomas, 1904 (рис. 147).
Мелких размеров игрунковая обезьяна. Длина тела 18—21 см. Длина хвоста 25—32 см. Вес около 280 г. Волосяной покров довольно длинный, густой и мягкий. На верху головы, на шее и плечах небольшая грива. В задней части туловища растут удлиненные вол осы,спускающиеся на основание хвоста. Пучков волос на ушах нет. Окраска волосяного покрова буровато-черная, с желтоватыми отметинами на задней части хвоста. Иногда на голове и спине могут встречаться белые отметины. Диплоидное число хромосом 48.
Экология изучена крайне слабо. Держатся стаями по 20—30 особей в нижних и средних частях крон деревьев. Питаются, вероятно, сочными плодами, листьями, семенами, насекомыми и другими мелкими животными.
Распространены в бассейне Амазонки в западной Бразилии, в восточном Перу и северной Боливии. Некоторые исследователи относят род мармозеток к семейству капуцинов или выделяют в особое семейство Callimico-nidae.
Численность мала. Вид внесен в «Красную книгу».
Род игрунок
Genus Callithrix Erxleben, 1777
Размеры от мелких до средних. Длина тела 15—30 см. Длина хвоста 18—40 см. Вес от 230 до 450 г. Волосяной покров обычно густой и мягкий. У серебристой игрунки окраска его серебристо-белая, иногда с примесью желтовато-серого цвета, особенно на спине; хвост черный, резко контрастирующий с окраской туловища; лицо и уши лишены волос, красноватого цвета. Окраска волосяного покрова у остальных видов варьирует от серой, белой, темно-бурой до почти черной; голова может быть желтовато-бурая, серая, желтоватая, на брюшной стороне тела может проходить медиальная продольная черная полоса. У остальных представителей рода хвост в темных и светлых кольцах, чередующихся на всем его протяжении. Уши скры-
Рис. 147. Мармозетка — С. goeldii
ты в меху головы. Лицевая часть головы покрыта короткими волосами. У серебристой игрунки на голове и ушах нет удлиненных пучков волос. У большинства других видов на ушах имеются длинные пучки волос от серебристого и серого до черного цвета. Характерно, что нижние клыки игрунок имеют фор му резцов. Диплоидное число хромосом у обыкновенной и желто-ногсй игрунок 46.
Обитатели тропических и субтропических лесов. Держатся в различных частях крон деревьев, иногда спускаются на землю. Активность дневная. Встречаются группами от 3 до 12 особей. Питаются насекомыми, пауками, мелкими птицами и их яйцами и растениями, в частности плодами. Беременность 142—150 дней. В помете обычно 2 детеныша. Половозрелость у самок наступает в возрасте примерно 14 месяцев. Продолжительность жизни в естественных условиях около 10 лет, в неволе — до 16 лет.
Распространены в Бразилии, Перу и Эквадоре.
В роде 11 видов (иногда их насчитывают 8,9 или 10):
обыкновенная игрунка — С. jacchus Linnaeus, 1758 (северо-восток Бразилии);
желтоголовая игрунка — С. flaviceps, Thomas, 1903 (восток Бразилии);
чернокисточковая игрунка — С. penicillata Е. Geoffroy, 1812 (Бразилия);
белоухая игрунка — С. aurita Ё. Geoffroy, 1812 (восточная Бразилия);
белолицая игрунка — С. leucocephalai,. Geoffroy, 1812 (восточная Бразилия);
белоплечая игрунка — С. humeralifer Ё. Geoffroy, 1812 (Бразилия);
белошейная игрунка — С. albicollis Spix, 1823 (Бразилия);
длинноухая игрунка — С. santaremensis Matschie, 1893 (Бразилия)
желтоногая игрунка — С. chrysoleucos Wagner, 1842 (Бразилия);
серебристая игрунка — С. argentata Linnaeus, 1766 (Пара в Бразилии);
карликовая игрунка — С. pygmaea Spix, 1823 (Эквадор, Перу, Бразилия).
Род львиных игрунок
Genus Leontideus Cabrera, 1956
Рис 148 Золотистая игрунка — L. rosa.Ua
Мелкие и крупные игрунковые обезьяны. Длина тела 16,5—50 см. Длина хвоста 22,5—38,5 см. Характерна длинная, узкая кисть и соединенные кожей третий и четвертый пальцы и первые фаланги второго и третьего пальцев. Волосяной покров длинный, шелковистый. На голове и плечах удлиненные волосы образуют гриву, давшую этим обезьянам родовое название (рис. 148). Окраска волосяного покрова одноцветная, золотистая или желтая у золотистой игрунки; черная на большей части тела и золотистая на голове, крестце, верхней стороне хвоста, кистях и стопах у золотистоголовой игрунки; черная на теле с золотистым лбом, крестцом, основанием хвоста и ляжками и красными коленями ' у золотистозадой игрунки Нижние клыки длиннее резцов. Диплоидное число хромосом у золотистой игрунки 46.
Держатся небольшими группами. Питаются насекомыми и мелкими ящерицами, плодами, а также, вероятно, птичьими яйцами и мелкими птицами. Беременность 132—134 дня. Детенышей в помете обычно 1 или 2. Самец принимает участие в воспитании детенышей. В неволе жили до 15 лет.
Ареал охватывает восточную Бразилию.
В роде 3 вида (иногда их объединяют в 1 вид):
золотистая игрунка — L. го-salia Linnaeus, 1766;
золотистоголовая игрунка— L. chrysomelas Kuhl, 1820,
золотистозадая игрунка —
L. chrysopygus Mikan, 1820.
Львиные игрунки встречаются редко, и все три вида внесены в «Красную книгу».
Род тамаринов
Genus Saguinus Hoffmannsegg, 1807
Рис. 149 Сверху вниз: пегий тамарин — S bicolor, тамарин Жоффруа S. geoffroyi и императорский тамарин —S. imperator
Мелкие и средние представители семейства. Длина тела 18—31 см. Длина хвоста 20—42 см. Вес обычно 225—900 г. Характер расположения пучков вопос на голове и окраска меха сильно варьирует у различных видов (рис. 149). У эдипова тамарина и Тамарина Жоффруа удлиненные волосы на голове образуют медиальный гребень, причем у первого из них он не продолжается на затылок, а у второго заходит на затылок и верхнюю часть шеи в виде гривы. Эти два вида имеют слабо покрытый волосами лицевой отдел головы и большие голые уши. Три других вида (пегии тамарин, белоногий тамарин и тамарин Мартинса) не имеют волосяных пучков на голове, лица у них почти без волос и уши голые. Пегий тамарин имеет белую окраску на голове и передней части тела, в то
время как остальная часть его тела желтовато-бурая. У белоногого тамарина передние и задние конечности белые Тамарин Мартинса совсем не имеет белого цвета на теле. У обыкновенного и краснорукого тамаринов на морде нет белых волос, усов или пигментированных пятен, а лицевая часть морды черная. Конечности обыкновенного тамарина черные, а краснорукого тамарина—желтые или оранжевые. У остальных видов рода на морде имеются белые волосы или непигментированные участки кожи вокруг рта, в то время как остальная часть лица пигментирована и обычно покрыта волосами. Один из этих видов (императорский тама-рнн) обладает белыми усами, длина которых настолько велика, что они могут достигать плеч. Нижние клыки больше, чем резцы. Диплоидное число хромосом у эдипова, черноспинного и рыжего тамаринов 46.
Населяют леса Обычно держатся небольшими группами. Активность дневная. Краснорукие тамарины часто объединяются в одни стаи с капуцинами. Способны издавать разнообразные громкие крики. Питаются насекомыми, пауками, мелкими позвоночными, вероятно, яйцами птиц, а также плодами. Беременность около 5 месяцев. Самец помогает в воспитании детенышей. Половозрелость наступает в 15 месяцев. В неволе жили до 15 лет.
Ареал охватывает Панаму и Южную Америку, на юг до южного Перу, северной Боливии и южной Бразилии.
В роде 20 видов (некоторые исследователи насчитывают 18 или 21 вид); эдипов тамарин —S. oedipus Linnaeus, 1758 (север Колумбии);
тамарин Жоффруа S. geoffroyi Pucheran, 1845 (от северо-востока Колумбии на север до Панамы);
пегий тамарин—S. bicolor Spix, 1823 (север Бразилии);
тамарин Мартинса-—S. martinsi Thomas, 1912 (север Бразилии); обыкновенный тамарин—S. tamarin Link, 1795, (север Бразилии); краснорукий тамарин —S. midas Linnaeus, 1758 (бассейн Амазонки в Бразилии, Гвиана);
черноспинный тамарин —S. nigricollisSpix, 1823 (западная Бразилия);
тамарин Уэдделла — S. wedelli Deville, 1849 (восток Бразилии, север Боливии);
тамарин Девилла—S. devillei I. Geoffroy, 1848 (восток Перу);
буроголовый тамарин —S. fuscicollis Spix, 1823 (западная Бразилия и восточные районы Перу);
рыжий тамарин—S. illigeri Pucheran, 1845 (северо-восток Перу, восток Эквадора, Колумбия, Бразилия);
золотистый тамарин—S. tripartitus Milne-Edwards, 1878 (Перу, Эквадор);
усатый тамарин — S. mystax Spix, 1823 (запад Бразилии);
тамарин Леннберга — S. pluto Lonnberg, 1926 (запад Бразилии).
тамарин Рио Напо—S. graellsi Espada, 1870 (Эквадор, Перу).
красногривый тамарин —S. pileatus I. Geoffroy et Deville, 1848 (запад Бразилии);
краснобрюхий тамарин —S. labiatus Humboldt, 1812 (запад Бразилии, Перу);
императорский тамарин •—S. imperator Goeldi, 1907 (запад Бразилии);
белый тамарин —S. melanoleucus Mirando Ribeiro, 1912 (бассейн Амазонки в Бразилии);
белоногий тамарин—S. leucopus Gunther, 1876 (Колумбия).
СЕМЕЙСТВО НИЗШИХ УЗКОНОСЫХ ОБЕЗЬЯН, ИЛИ МАРТЫШЕК,
FAMILIA CERCOPITHECIDAE BLANFORD, 1888
Мелких и средних размеров высшие приматы. Длина тела от 32 до ПО см. Самцы обычно крупнее самок. Хвоста нет или он может достигать длины от 2 до 106 см. Телосложение различное от легкого изящного до довольно тяжелого. Передние конечности немного длиннее задних. Кисть короче стопы. Самый длинный палец третий, затем по длине идет четвертый, второй, пятый и первый. Большие пальцы передних и задних конечностей противопоставляются остальным (у толстотелых обезьян большой палец передней конечности отсутствует или очень мал), ногти уплощенные. Голова относительно небольшая или крупная, округлая или с удлиненной лицевой частью. Ноздри расположены близко друг к другу и направлены вперед ИЛИ ВНИЗ. Г fc
Волосяной покров обычно длинный, шелковистый, окраска его, как правило, буроватая, иногда черноватая, бело-желтоватая или зеленоватая. Часто удлиненные волосы образуют своеобразную шапку на голове или небольшую гриву или усы. Шерсть полностью покрывает все тело и хвост, за исключением лица, седалищной части, подошв и задних конечностей, иногда груди. Обнаженные лицевые и седалищные части кожи часто яркой окраски, обычно красноватой, а иногда голубоватой. Окраска может варьировать, становясь у самцов более яркой в сезон размножения. Густота волос значительно меньше, нежели у цепкохвостых обезьян. Самцы у ряда видов
окрашены более ярко, чем самки. Имеются седалищные мозоли, представляющие собой жировые подушки с утолщенной яркой голой кожей. У некоторых обезьян развиты защечные мешки (макаки, мангабеи, мартышки, хохлатые обезьяны, павианы и гелады). У самок одна пара сосков в области груди.
В черепе ростральный отдел обычно удлинен. Костные слуховые барабаны небольших размеров. Число зубов, как у человекообразных обезьян 2 12 3
и людей. Зубная формула: i’ — с — р — т— = 32. Коренные зубы четырехугольные в поперечнике и обычно четырехвершинные. Последний нижний коренной зуб пятивершинный. Четыре бугорка на нижних коренных зубах соединяются попарно высокими гребнями. Все эти зубы билофодонтного типа. Верхние предкоренные имеют по 3 корня, а нижние-—по 2. Позвонков шейных 7, грудных 12—13, поясничных 6—7 (общее число грудных и поясничных позвонков 18—20), крестцовых 2—4, хвостовых 5—27. Ключица хорошо развита. Грудная клетка узкая и килевидная. Грудина длинная и узкая, состоят из нескольких косточек. Имеется os penis (нет у хохлатых обезьян).
Головной мозг хорошо развит, с извилинами и бороздами. Обонятельный отдел его развит слабо (микросматический тип). Лицевая мускулатура хорошо развита. Желудок у тонкотелых обезьян сложный. Червеобразный отросток слепой кишки, как правило, отсутствует. В почке лишь один сосочек. Плацента у самок би- или монодискоидальная, гемохориальная, отпадающая.
Обитатели различных ландшафтов: джунглей, мангровых болот, открытых равнин, каменистых мест. Поднимаются в горы до 4 000 м над уровнем моря. Ведут наземный образ жизни (павианы), наземный и древесный (макаки) или древесный (все остальные представители семейства). Почти все виды семейства дневные животные. Обычно объединяются в семейные группы, но иногда встречаются стада до 50 или нескольких сотен особей. В период размножения самцы могут драться из-за самок. Ночь проводят на деревьях и скалах или в пещерах. Растительноядные или всеядные. Способны издавать разнообразные звуки, которые служат, в частности, средством общения между отдельными особями. Размножаются круглый год. У самок наблюдаются ежемесячные менструальные явления. Половозрелость наступает в возрасте 3—5 лет. Беременность 150—200 дней. В помете, как правило, один, редко два детеныша. Продолжительность жизни в неволе до 30—40 лет. За некоторыми представителями семейства охотятся из-за мяса или шкуры.
Распространение широкое: ареал охватывает Африку, Аравийский полуостров и Юго-Восточную Азию; в Европе встречаются только в Гибралтаре (возможно, они здесь были акклиматизированы).
В семействе 11 родов (61 вид):
макак — Масаса Lacepede, 1799 (12 видов);
хохлатых, или черных, павианов — Cynopithecus I. Geoffroy, 1835 (1 вид);
мангабеев — Cercocebus Ё. Geoffroy, 1812 (4 вида); j
павианов — Papio Erxleben, 1777 (7 видов);
гелад — Theropithecus I. Geoffroy, 1843 (1 вид);
мартышек — Cercopithecus Brunnich, 1772 (15 видов);
тонкотелых обезьян, или лангуров, — Pygathrix Ё. Geoffroy, 1812 (14 видов);
ринопитеков — Phinopithecus Milne-Edwards, 1872 (2 вида);
симиасов—Simias Miller, 1903 (1 вид);
носачей —Nasalis Ё. Geoffroy, 1812 (1 вид).
толстотелых обезьян, или колобусов, — Colobus liliger, 1811 (3 вида).
Систематика семейства имеет ряд спорных моментов. Так, ряд ученых выделяют роды тонкотелых и толстотелых обезьян, симиасов, ринопитеков и носачей в отдельное семейство Colobidae.
По-видимому, более правильно считать эти 5 родов принадлежащими подсемейству Colobinae в составе семейства Cercopithecidae. Некоторые исследователи предлагают более дробную систему родов или менее дробную, чем указанная выше.
Род макак
Genus Масаса Lacepede, 1799
Мелкие и средние представители семейства. Длина тела 38—76 см. Длина хвоста варьирует, у некоторых видов его нет совсем или может достигать 61 см. Вес до 13 кг. Телосложение плотное, конечности относительно короткие и сильные (рис. 150). Лицевой отдел головы умеренно удлиненный и округлый. Ноздри расположены не на конце морды и обращены вперед и вверх. Между пальцами имеются кожные перепонки, более короткие на передних конечностях, несколько более длинные на задних. Между вторым и третьим пальцами задних конечностей перепонка развита сильнее и достигает первого сочленения между фалангами. Первый палец передних конечностей большой и короткий. Волосяной покров обычно средней длины и густоты, но, например, у японского макака довольно длинный, густой и мягкий. Окраска меха обычно желтовато-бурая на спине и более светлая на брюхе, но некоторые виды целиком оливковые или почти черные. У некоторых видов на голове имеется шапка удлиненных волос. У вандеру длинные серые волосы обрамляют с боков лицевую часть морды. Длинные бакенбарды и бороду имеет японский макак. Лобные кости с толстыми валиками, ограничивающими глазницу сверху. Нижний третий коренной зуб крупнее, чем коренные, стоящие перед ними, и имеет пять бугров на своей жевательной поверхности. Клыки большие и острые. Диплоидное число хромосом 42.
Обитатели лесов, но часто встречаются также и на безлесых местах, в горах среди скал и камней, некоторые обитают в мангровых болотах. Ведут древесный и наземный образ жизни. Активность главным образом дневная. Питаются широким набором растительных объектов и различными животными, в том числе насекомыми и мелкими позвоночными. Способны выживать при низкой температуре и в Японии хорошо переносят довольно суровые зимы. Держатся группами до 20—25 особей разного возраста и пола. Хорошо плавают. Сезонности в размножении, по-видимому, нет. Беременность 5—7 месяцев. В помете обычно один детеныш, иногда два. Детеныш остается
Рис. 150. Львинохвостый макак — М. пе-mestrina и череп макака-резуса — М. mulatto
с матерью примерно в течение года. Половозрелость наступает около четырех лет. Длительность жизни в неволе примерно 30 лет. Широко используются как лабораторные животные. Макаки были первыми обезьянами, запущенными в ракете в стратосферу. В их крови впервые был показан резус-фактор. Яванский макак служил лабораторным животным для выработки вакцины против полиомиэлита.
Ареал охватывает Южную Азию, Индонезию и прилежащие острова на восток до Филиппин, Хайнань, Тайвань и Японию, Северную Африку; ма-гот обитает на Гибралтаре — это единственный вид, встречающийся в диком состоянии в Европе.
В роде 12 видов:
цейлонский макак — М. sinica Linnaeus, 1771 (Цейлон);
индийский макак — М. radiata Ё. Geoffroy, 1812 (п-ов Индостан); вандеру, или львинохвостый макак, — М. silenus Linnaeus, 1758
(п-ов Индостан);
свинохвостый макак — М. nemestrina Linnaeus, 1766. (Ассам, Бирма, Таиланд, Малаккский полуостров, о-ва Суматра и Калимантан);
яванский макак — М. irus Cuvier, 1818 (Бирма, Никобарские острова, п-ов Индокитай, Таиланд, Малаккский полуостров, о-ва Суматра, Калимантан и Ява);
макак-резус — М. mulatto Zimmermann, 1780 (восточный Афганистан, Кашмир, Пенджаб, Ассам, Бирма, северные районы п-ова Индостан, Таиланд, п-ов Индокитай, Сычуань, Юньнань, Хайнань, Тибет):
горный резус —М. assamensis M’Clelland, 1839 (Непал, Сикким, Бутан, Ассам, Северная Бирма, Юньнань, п-ов Индокитай);
тайванский резус — М. cyclopis Swinhoe, 1862 (о-в Тайвань);
медвежий макак — М. speciosa Cuvier, 1825 (Сычуань, Юньнань, Ассам, Бирма, п-ов Индокитай);
японский макак — М. fuscata Bly th, 1875 (Японские острова);
магот — М. sylvana Linnaeus, 1758, (Марокко, Алжир, Гибралтар); Целебесский макак — М. mounts Cuvier, 1823 (о-в Сулавеси).
Некоторые исследователи выделяют Целебесского макака в отдельный род Супотасаса Khajurian, 1953. Иногда в роде насчитывают не 12, а 7 или 8 видов.
Львинохвостый макак малочислен и внесен в «Красную книгу».
Род хохлатых, или черных, павианов Genus Cynopithecus I. Geoffroy, 1835
В роде единственный вид: хохлатый, или черный, павиан-—С. nigerDes-marest, 1822 (рис. 151).
Размеры мелкие и средние. Длина тела примерно 47—66 см. Длина хвоста 2—6 см. Занимает промежуточное положение между родами макак и павианов. Морда сильно вытянута, надглазничные валики очень крупные. Лицевая часть головы, стопы и кисти безволосые. Ноздри находятся не на конце морды, а отнесены довольно далеко назад. Волосяной покров плотный на туловище и более редкий на конечностях. Спинная часть тела темнобурая или черная, брюшная — несколько светлее, иногда приближается к серой. Молодые животные имеют бурую окраску. Седалищные мозоли развиты хорошо и имеют красноватый цвет. На голове удлиненные волосы образуют своеобразный гребень; когда животное раздражено, волосы на этом гребне поднимаются вертикально. Лицевой отдел черепа массивный и удлиненный. Диплоидное число хромосом 42. Экология изучена крайне слабо. Ведет в основном древесный образ жизни, но проводит много времени и на земле в поисках пищи. Питается, по-видимому, в основном плодами.
Распространен на о-ве Сулавеси.
Рис. 151. Хохлатый павиан — С. niger и его череп
Род мангабеев
Genus Cercocebus Ё. Geoffroy, 1835
Мелкие, средние и крупные представители семейства. Длина тела 38— 89 см. Длина хвоста 43—76 см. Характерна овально удлиненная голова с вытянутой лицевой частью (рис. 152). Между пальцами есть короткие перепонки: второй и третий пальцы соединены почти по всей своей длине между собой, а третий, четвертый и пятый — примерно до половины длины. Хвост длинный, слабохватательный. Окраска разнообразная. У гривистого манга-бея волосы мягкие и тускло-черные. На щеках волосы короткие, удлиненные волосы образуют хорошо выраженные брови и гребень на голове; кроме того, на плечах волосы удлинены в слабо выраженную гриву. Бородатый мангабей имеет короткий и блестящий черный мех, длинные сероватобурые бакенбарды частично закрывают ушные раковины. На голове волосы образуют высокий конический гребень; однако удлиненные волосы на надбровных дугах и грива на плечах отсутствуют. У губастого мангабея спинная сторона тела золотисто-бурая с черными пятнами или темно-сероватобурая с желтовато-золотистыми пятнами, брюшная поверхность охристооранжевая или беловатая, на голове небольшой хохол. У воротничкового мангабея спинная часть туловища темно-серая или серовато-бурая до темнобурой, а брюшная сероватая; обычно цветных пятен не имеет. У большинства особей этого вида на голове располагается пятно неправильной формы оливкового или черного цвета, черновато-бурое или буровато-красное, удлиненных волос на голове нет. Головное пятно окружено черной или белой по
лосой, которая продолжается назад в шейное пятно, которое в свою очередь, переходит в черную продольную линию, тянущуюся посредине спины и переходящую на хвост. На щеках обычно имеется борода. Защечные мешки сильно развиты. По крайней мере, у гривистого мангабея имеется горловсй мешок, резонирующий голос; он достигает небольших размеров у самок и очень крупный у самцов. Диплоидное число хромосом 42 (нет данных по бородатому мангабею).
Обитают в экваториальных лесах. Экология изучена слабо.
Ведут преимущественно древесный образ жизни. Питаются главным образсм побегами, листьями, а так
Рис. 152. Сверху вниз: бородатый маигабей — С. aterrimus, гривистый мангабей — С. albigena
же некоторыми животными. Держатся небольшими группами от 4 до 12 особей.
Распространены в Африке от Гвинеи на востоке до Кении на западе.
В роде 4 вида:
воротничковый мангабей — С. torquatus Kerr, 1792 (от нижнего течения Нигера на юг до Габона);
гривистый мангабей — С. albigenaGray, 1850 (Конго-Браззавиль, Республика Заир, Габон);
бородатый мангабей — С. aterrimus Oudemans, 1890 (Республика Заир);
чубастый мангабей — С. galeritus Peters, 1879 (Конго-Браззавиль, Республика Заир, Кения).
Один из подвидов чубастого мангабея — С. g. galer it us Peters, 1879 редок и внесен в «Красную книгу».
Род павианов
Genus Papio Erxleben, 1777
Размеры мелкие, средние и крупные (наибольшие в семействе). Длина тела от 50 до 110 см. Длина хвоста от 5 до 71 см. Вес взрослых самцов может достигать 54 кг. Сложение довольно тяжелое, конечности относительно укороченные (рис. 153). Кисти и стопы короткопалые, сильные. Голова внешне похожа на собачью за счет удлинения морды. Ноздри расположены терминально, не имеют формы щелей и срезаны вертикально. Хвост может быть очень коротким (мандрилы) или средней длины (павианы). Седалищные мозоли сильно развиты и могут быть яркого фиолетового цвета. Кожа около половых органов и анального отверстия красная. Волосяной покров довольно грубый. Лицевая часть морды и уши покрыты редкими волосами. У самцов вокруг шеи на плечах развивается довольно длинная грива. Окраска воло
сяного покрова пепелБно-серая, бурая, оливково-бурая, желтовато-бурая, желтая или зеленоватая. В верхней части морды мандрилов располагаются своебразные продольные валики. По средней линии проходит продольный валик красной окраски, а по бокам от него по 6 синих более мелких. Срединный валик продолжается вперед и окружает ноздри, которые тоже имеют ярко-красный цвет. Такая окраска свойственна главным образом взрослым самцам. Если животное возбуждено, то окраска становится более яркой. Череп с сильно развитым лицевым отделом. Клыки очень крупные. Диплоидное число хромосом 42.
Обитатели лесных и открытых каменистых и скальных районов. Ведут наземный образ жизни. Активность дневная. Держатся группами от нескольких до 25 и иногда даже 300 особей. Предводителями в стаде являются взрослые самцы. Взрослые самцы с необычайно сильно развитой мускулатурой, мощными зубами не боятся вступать в единоборство даже с леопардом. Питаются различными растительными объектами и животными (насекомыми, мелкими позвоночными). Сезонности в размножении, по-видимому, нет. Беременность 6—7 месяцев. В помете один, редко два детеныша. В неволе павианы жили до 45 лет, хотя в естественных условиях, по-видимому, они живут не дольше 20.
Ареал охватывает Африку.
В роде 7 видов:
гамадрил —- Р. hamadryas Linnaeus, 1758 (Аравийский полуостров, Эфиопия, Судан);
бабуин — Р. cynocephalus Linnaeus, 1766 (Мозамбик, Замбия, Малави, Ангола, Танзания, Кения, Республика Заир);
чакма — Р. ursinus Кегг, 1792 (Южно-Африканская Республика, Мозамбик, южная Родезия, Ботсвана, Юго-Западная Африка, Ангола, Ма-лавия, Танзания, Кения, Уганда, Судан, Аравийский полуостров, Эфиопия);
анубис — Р. anubis Fischer, 1829 (Кения, Республика Заир, Танзания);
гвинейский павиан — Р. papio Desmarets, 1820 (Гвинея, Камерун); мандрил — Р. sphinx Linnaeus, 1758 (Западная Африка);
дрил — Р. leucophaeus Cuvier, 1807 (Западная Африка).
Некоторые исследователи выделяют гамадрилов в отдельный род Comopithecus Allen, 1925 (Симпсон, 1945; Уолкер, 1964); мандрилов и
Рис. 153. Гамадрил — Р. hamadryas
дрилов в отдельный род — Mandrillus Ritgen, 1824 (Симпсон, 1945; Нестурх, 1960; Уолкер, 1964). Вероятно, правильнее рассматривать этих обезьян как отдельные подроды рода павианов.
Род гелад
Genus Theropithecus I. Geoffroy, 1843
В роде единственный вид: гелада — Т. gelada Riippell, 1835.
Размеры мелкие и средние. Длина тела 50—72 см. Длина хвоста 70—-80 см. Общий облик сходен с павианами. Есть и отличия. Ноздри располагаются не на конце, а по бокам морды, которая относительно укорочена (короче, чем мозговой отдел черепа). Хвост довольно длинный, толстый, на конце имеет небольшую кисть. Ниже орбит находятся глубокие впадины. Первый и второй пальцы передних конечностей почти одинаковы по длине. Конечности слабо приспособлены к схватыванию. Седалищные мозоли развиты сильно и имеют черную окраску. Волосяной покров густой и пушистый. Сильно развитые волосы по бокам морды образуют бакенбарды. На голове имеется срединный гребень из удлиненных волос. На плечах удлиненные волосы образуют гриву. Общая окраска волосяного покрова черно-бурая или черная, брюшная сторона светлее спинной, обычно сероватая или серовато-белая. На груди находится безволосый участок красного цвета. Диплоидное число хромосом 42.
Обитает в горах на скалистых участках или участках, покрытых кустарником, выше 1800 м над уровнем моря. Ведет в основном наземный образ жизни и лишь изредка вскарабкивается на деревья. Активность дневная. Ночью прячется в пещерах или в пустотах среди скал. Держится большими группами; обычно предводительствуют старые самцы. Питается широким набором растительных объектов (корни, луковицы, семена, сочные плоды, листья) и различными беспозвоночными и позвоночными животными. Иногда совершает набеги на поля. Детали размножения неизвестны.
Распространена в горах Эфиопии.
Род мартышек
Genus Cercopithecus Briinnich, 1772
Мелкие (наименьшие в семействе), средние и крупные представители семейства. Длина тела от 32 до 87 см. Длина хвоста от 36 до 87 см. Сложение изящное (рис. 154). Туловище удлинено. Голова округлая, лицевой отдел укорочен, нос не выступает, ноздри расположены близко друг к другу. Се-1 далищные мозоли небольших размеров и отделены друг от друга. Хвост обычно длинный. Кожные перепонки между пальцами если имеются, то только у их основания. Стопа удлинена, первый палец задней конечности сильно противопоставляется остальным. Защечные мешки очень большие. Волосяной покров на туловище обычно средней длины, довольно густой и мягкий. Окраска его весьма разнообразна и может варьировать от оливковой до настоящего зеленого цвета, может быть сероватой или почти черной. Брюшная поверхность белая или сероватая, розоватая или оранжевая, иногда с черными или желтыми пятнами. У некоторых видов белые или грязножелтые полосы на ляжках. Некоторые мартышки имеют белые или иного цвета бакенбарды, белые надбровные полосы, пучок волос на подбородке, белое, красное или голубое носовое пятно или желтое или серовато-желтое пятно неправильной формы на щеках, ограниченное снизу и сверху черными полосами. Кожа на отдельных участках лица может быть синей или фиолетовой. Диплоидное число хромосом 54, 58, 60, 62, 66, 68, 70, 72.
Населяют леса. Большинство видов ведет древесный образ жизни. Не-' которые, например мартышка-гусар, проводят много времени на земле.
Рис. 154. Мартышка Диана — С. diana
Держатся группами от 6 до 30, иногда до 200 особей. Обычно такие группы возглавляют взрослые самцы. Питаются в основном растительными объектами: плодами, листьями, а также и различными животными — насекомыми, птенцами и яйцами птиц, мелкими пресмыкающимися. Иногда совершают набеги на поля. Способны производить разнообразные крики. Вероятно, для большинства видов характерно отсутствие сезонности в размножении. Беременность примерно 7 месяцев. В помете один детеныш, иногда два. В неволе жили до 24 лет.
Распространены широко по Африке от Гамбии на западе до Эфиопии на востоке и Южно-Африканской Республики на юге.
В роде 15 видов:
зеленая мартышка — С. aethiopsLinnaeus, 1758 (Эфиопия, Судан, АРЕ, Кения, Мозамбик, Танзания, Уганда, Республика Заир, Южно-Африканская республика, Ботсвана, Ангола, Замбия);
бородатая мартышка — С. I'hoesti Sclater, 1899 (Республика ЗАИР, Камерун);
коронованная мартышка — С. mitis Wolf, 1822 (Южно-Африканская республика, южная Родезия, Мозамбик, Ангола, Малави, Замбия, Республика Заир, Танзания, Эфиопия, Сомали, Кения, Уганда);
мартышка-диана — С. diana Linnaeus, 1758 (Либерия, Гвинея, Республика Заир);
мартышка Бразза — С. neglectus Schlegel, 1876, (Конго-Браззавиль; Республика Заир, Судан);
мои а — С. топа Schreber, 1775 (Сенегал, Сьерра-Леоне, Гвинея, о-в Фернандо-По, Республика Заир, Габон, Берег Слоновой Кости);
чубатая мартышка — С. pogonias Bennett, 1883 (о-в Фернандо-По);
малая белоносая мартышка — С. petaurista Schreber, 1774 (Гвинея, Берег Слоновой Кости, Гана);
краснобрюхая мартышка — С. erythrogaster Gray, 1866 (Нигерия);
большая белоносая мартышка — С. nictitans Linnaeus, 1766 (Ангола, Замбия и далее на север через Кению, Уганду и Танзанию и тропики Западной Африки, по крайней мере до Либерии);
голуболицая мартышка — С. cephus Linnaeus, 1758 (Габон, Камерун, о-в Фернандо-По, Нигерия);
совинолицая мартышка — С. hamlyni Pocock, 1907 (Республика Заир),
Рис. 155. Гривистый тонкотел — Р. crista-tus и череп немейского тонкотела — Р. пе-maeus
крошечная мартышка — С. talupoin Schreber, 1774 (Ангола, Республика Заир, Камерун);
чернозеленая мартышка — С. nigroviridis Pocock, 1907 (Республика Заир);
мартышка-гусар — С. patas Schreber, 1774 (Сенегал, Республика Заир, Того, Нигерия, Камерун, Судан, Уганда, Кения).
Некоторые исследователи выделяют в качестве отдельных родов чернозеленую мартышку (Allenopythecus Lang, 1923), мартышку-гусара (Erythro-cebus Trouessart, 1897), крошечную мартышку (Miopithecus I. Geoffroy, 1842). По-видимому, правильнее их считать подродами в роде мартышек. Иногда в роде мартышек насчитывают не 15 видов, а 20 или 12. Таким образом, очевидно, что систематика мартышек нуждается в тщательной ревизии.
Род тонкотелых обезьян, или лангуров, Genus Pygathrlx Ё. Geoffroy, 1812
Размеры мелкие и средние. Длина тела 43—78 см. Длина хвоста 53— 106 см. Телосложение изящное. Голова округлая с укороченной лицевой частью. Конечности тонкие, удлиненные (рис. 155). Кисть и стопа также удлинены. Первый палец передней конечности короткий, остальные длинные и сильные. Защечных мешков нет. Имеется гортанный мешок. Область носа вперед не выступает. Окраска волосяного покрова может быть одноцветной, спинная сторона в основном буроватая, сероватая или черноватая, а брюшная несколько более светлая. У некоторых видов есть светлые отметины на голове. Некоторые имеют удлиненные волосы, расходящиеся по бокам головы в виде короны, у других удлиненные волосы образуют на голове гребень. Немейский тонкотел окрашен очень ярко: голова у него бурая с ярко-ко
ричневой полосой ниже ушей, лицо ярко-желтое, бакенбарды, туловище сероватое, более темное на спинной стороне, крестец и хвост белые.
. Число хромосом в диплоидном наборе у гульмана 50.
Обитают в основном в лесах, лишь иногда на открытых пространствах. Встречаются в горах до 4000 м над уровнем моря. Ведут преимущественно древесный образ жизни. Наибольшая активность ранним утром и поздним вечером. Обычно держатся высоко в кронах деревьев, но часто спускаются на землю на водопой, в поисках пищи или передвигаясь от одних деревьев к другим, причем по земле могут развивать скорость до 37 км/ч. Держатся группами до 40 особей, предводительствуемые взрослыми самцами. Питаются различными растительными объектами: плодами, листьями, цветками, в меньшей степени мелкими животными. В северных частях ареала, вероятно, имеет место сезонность в размножении. Беременность около 196 дней.
Распространены в Южной Азии от Цейлона и Индии на восток до Южного Китая, п-ова Индокитай и Индонезии.
В роде 14 видов:
гульман — Р. entellus Dufrense, 1797 (Цейлон, Индия, южный Тибет); белобородый тонкотел — Р. senex Erxleben, 1777 (Цейлон);
нилагерийский тонкотел, — Р. johni Fischer, 1829 (Южная Индия);
рыжий тонкотел — Р. elaphus Raffles, 1821 (Тенассерим, Таиланд, Малаккский полуостров, Суматра, Калимантан и прилежащие острова);
гривистый тонкотел — Р. cristatus Raffles, 1821 (Тенассерим, Таиланд, п-ова Индокитай, Малаккский, о-ва Суматра, Ява, Калимантан и мелкие со-.седние);
хохлатый тонкотел — Р. pileatus Blyth, 1843 (Ассам и Бирма);
очковый тонкотел — Р. obscurus Reid, 1837 (Тенассерим, Таиланд, Малаккский полуостров и мелкие прилегающие острова);
тонкотел Фейера — Р. phayrei Blyth, 1847 (Бирма, Тенассерим, Таиланд);
Тонкинский тонкотел — Р. francoisi Pousargues, 1898 (Южный Китай, 11-ов. Индокитай);
ментавайский тонкотел — Р. potenziani Bonaparte, 1856 (о-ва Ментавай у Суматры);
чубастый тонкотел — Р. aygula Linnaeus, 1758 (Ява, Суматра, Калимантан);
каштановый тонкотел — Р. rubicunda Miiller, 1838 (о-в Калимантан);
гололобый тонкотел —• Р. frontatus Muller, 1838 (о-в Калимантан);
немейский тонкотел — Р. nemaeus Linnaeus, 1771 (п-ов Индокитай, о-в Хайнань).
Некоторые исследователи включают в род тонкотелых обезьян лишь один вид немейский тонкотел, а все остальные относят к роду Presbytis Eschscholtz, 1821 (Симпсон, 1945). Правильнее объединить все эти виды в один род, подразделив их на 2 подрода. Несмотря на то, что название рода Pygathrix имеет приоритет (описан в 1812 г.), некоторые исследователи все виды объединяют под названием Semnopithecus, введенным Кювье в 1821 г. ..(Валуа, 1955; Нестурх, 1960).
Немейский тонкотел встречается очень редко, находится под угрозой уничтожения и включен в «Красную книгу».
Род ринопитеков
Genus Rhino pit hecus Milne-Edwards, 1872 c
Размеры мелкие и средние. Длина тела 52—83 см. Длина хвоста 51— J97 см. Лицевой отдел головы укорочен. Характерен короткий, вздернутый вверх нос. Защечных мешков нет. Волосяной покров обычно очень длинный — До.15—17 см и густой.. На голове может быть гребень из удлиненных
волос. Окраска меха изменчива. У рокселланова ринопитека спинная часть J светло-бурая или пепельно-серая с белой полосой по средней линии между I плечами; верх головы, затылок и плечи могут быть серовато-черные или лоб, бока головы, бока шеи и брюшная часть туловища золотистые, или бока ' головы и брюхо белые или серые. Передние конечности обычно желтоватые или беловатые, задние сероватые. Тонкинский ринопитек имеет лоб и щеки кремовые, бока шеи оранжевые, спину и конечности черные, на боках тела по желтовато-белой отметине и такого же цвета бока хвоста.
Обычно обитают в высокогорных лесах, но зимой могут спускаться довольно низко. Экология изучена слабо. Держатся, по-видимому, большими стадами, питаются преимущественно плодами, листьями, молодыми проростками бамбука.
Ареал охватывает Сычуань, Юньнань, Гуйчжоу, Бакбо на п-ове Индокитай.
В- роде 2 вида:
рокселланов ринопитек — /?. roxellanae Milne-Edwards, 1870 (Сычуань, Юньнань и Гуйчжоу в Китае, Ассам);
тонкинский ринопитек — /?. avunculus Dollman, 1912 (Бакбо на п-ове Индокитай).
Некоторые ученые выделяют из вида рокселланов ринопитек еще два вида: биэтовский ринопитек — R. bieti Milne-Edwards, 1897 и брелический ринопитек — R. brelichi Thomas, 1903. Правильнее рассматривать их в качестве подвидов R. roxellanae.
Рокселланов ринопитек немногочислен и внесен в «Красную книгу».
Род симиасов.
Genus Simlas Miller, 1903.
В роде единственный вид: одноцветный симиас—S. concolor Miller,, 1903.
Размеры мелкие. Длина тела 45—52 см. Длина хвоста 13—18 см. Общий облик сходен с ринопитеками. Нос тоже короткий и вздернутый. Характерен сильно укороченный хвост, голый на всем протяжении, за исключением конца, на котором имеется небольшая кисточка буроватых волос. Конечности относительно укороченные. Защечных мешков нет. Окраска волосяного покрова одноцветная, бурая, иногда плечи, затылок, спина и верхние части передних конечностей бывают несколько более светлыми. Волосы посредине головы растут назад, в то время как по бокам они образуют своеобразные пучки, спадающие на уши. Волосы выше середины щек растут вверх, а нижней половины щек — вниз.
Обитатель дождевых лесов. Экология изучена слабо. Питается в основном, по-видимому, растительными объектами, вероятно, использует в пищу • и некоторых водных животных.
Распространен на о-вах Пагай-Селантан, Сипора и Сиберут в группу о-вов Ментавай у западного побережья Суматры.
Везде малочисленен, включен в «Красную книгу». ">
Род носачей'
Gent*s Nasalis Ё. Geoffroy, 1812
В роде единственный вид: носач — N. larvatus Wurmb, 1781 (рис. 156). Размеры средние. Длина тела 66—76 см. Длина хвоста 60—76 см.. У взрослых, особенно старых, самцов нос хоботообразно удлинен, свисает над ртом и служит, цо-видимому, резонирующим органом. У самок и молодых животных он удлинен слабо. Крупные ноздри открываются вниз. Меховой покров относительно короткий, но довольно густой. Спинная сторона
Рис. 156. Носач — N. larvatus
туловища изменяется по окраске от бурой до светло-коричневато-рыжей. Брюшная поверхность значительно светлее — беловатого и сероватого цвета. Диплоидное число хромосом 48.
Экология изучена слабо. Обычно держится в лесах, поблизости от пресной воды, населяя мангровые болота, устья и берега рек. Ведет древесный образ жизни. Питается в основном листьями и плодами. Хорошо плавает и даже ныряет. Активность дневная, преимущественно утренняя. Живет небольшими группами.
Распространен на о-ве Калимантан.
Род толстотелых обезьян, или колобусов,
Genus Colobus liliger, 1811
Размеры мелкие и средние. Длина тела 50—70 см. Длина хвоста примерно 62—88 см. Сложение изящное, тело относительно тонкое (рис. 157). Большой палец передней конечности отсутствует или очень мал. Хвост обычно с большой кистью на конце. Окраска волосяного покрова сильно варьирует. Только один вид королевского толстотела можно разбить по признаку окраски на 4 группы. Представители одной группы имеют длинные бакенбарды, удлиненные волосы на верху головы, обычно слаборазвитые удлиненные волосы на горле, белые полосы над глазами, белые удлиненные волосы по бокам туловища и слабо развитый хвостовой пучок. Обезьяны второй группы не имеют белых отметин. У обезьян третьей группы отсутствуютзавитки удлиненных волос на голове, может быть хорошо развитая борода и белые отметины на теле, хвост полностью белый, концевая кисть развита слабо. В четвертой группе колобусы имеют удлиненные волосы на голове, направленные назад, хорошо развитую бороду, длинный и густой пучок волос на конце хвоста и белые полосы удлиненных волос по бокам тела и в области таза. Плечи у обезьян этой группы бурые. Красный толстотел имеет черное туловище и красную или коричнево-бурую голову, кисть и стопу. Удлиненные волосы по бокам туловища и хвостовой пучок отсутствуют. У молодых особей волосяной покров в большинстве случаев белый. Диплоидное число хромосом 44.
Обитатели густых лесов. Поднимаются в горы до 3000 м над уровнем моря. Ведут почти исключительно древесный образ жизни. Активность ночная. Передвигаются по деревьям очень быстро. Живут семейными группами, объединяясь иногда в стада до 20 особей, питаются в основном листьями и почками. Характерен громкий, тревожный крик. Служили объектом' охоты и подвергались значительному истреблению, так как мех шел на украшение женской одежды.
Распространены в Африке от Сенегала через Центральную Африку до Эфиопии и на юг до Анголы.
В роде 3 вида:
королевский толстотел — С. ро-lykomos Zimmermann, 1780 (Сьерра-Леоне, Гвинея, Берег Слоновой Кости, Либерия, Гана, Того, о-в Фернандо-По, Габон, Камерун, Эфиопия, Танзания, Судан, Кения, Конго-Браз-завиль; Республика Заир, Ангола, Малави);
красный толстотел — С. badius Kerr, 1792 (Сьерра-Леоне, Либерия, о-в Фернандо-По, Камерун, Сенегал, Гамбия, Гвинея, Гана, Того, Конго-Браззавиль; Республика Заир, Танзания, Кения, Уганда, о-в Занзибар);
зеленый толстотел— С. verus Van Beneden, 1838 (Западная Африка).
Некоторые исследователи выделяют из вида королевский толстотел еще один вид: гверецу — С. abyssini-
Рис. 157. Королевский толстотел — С. polykomos
cus Oken, 1816, хотя правильнее считать гверецу подвидом королевского толстотела.
Численность красного и зеленого толстотелое очень низка, и они включены в «Красную книгу».
СЕМЕЙСТВО ВЫСШИХ УЗКОНОСЫХ ОБЕЗЬЯН, ИЛИ ЧЕЛОВЕКООБРАЗНЫХ
ОБЕЗЬЯН, FAMILIA PONGIDAE ELLIOT, 1913
Мелких, средних и крупных размеров высшие приматы. Длина тела от 45 см у гиббонов до 180 см у горилл. Хвоста нет. Вес от 5 до 300 кг. Тело сложение довольно тяжелое, иногда массивное. Передние конечности длинные, задние короткие. Первый палец противспсставляется остальным, причем на передних конечностях он может быть слабо развит. На всех пальцах уплощенные ногти или (у гиббонов) ногти с небольшими килями (за исключением первого пальца). Небольшие седалищные мозоли развиты только у гиббонов. Защечных мешков нет. Лицевой отдел головы несколько выдается вперед. Голова округлая. Уши небольшие, у горилл и орангутанов рудиментарные, округлые. Глаза маленькие.
Волосяной покров грубый (у гиббонов более мягкий), короткий, иногда косматый. Окраска его черноватая, иногда серая (у старых самцов горилл),
красноватая (у орангутанов), серая или темно-желтая. У. некоторых гиббонов могут быть белые отметины на передних-конечностях и лице. Кожа более или менее пигментированная. У шимпанзе пигментация усиливается с возрастом. Лицевой отдел головы слабо покрыт волосами. Ладони кистей и подошвы стоп и уши без волос. У некоторых орангутанов развивается борода. Ладони и подошвы покрыты папиллярными узорами. Половой диморфизм проявляется хорошо. Самцы значительно крупнее самок, за исключением некоторых видов гиббонов. У сиаманга синдактилия развита сильнее, чем у самок. Горловой мешок, если он имеется, сильнее развит у самцов.
Длина черепа больше, чем его ширина. Орбиты направлены вперед; скуловые дуги сильно развиты. Гребни на черепе обычно крупные, особенно 2 12 3
у старых самцов. Зубная формула: i — с — р — т— = 32. Щечные зубы располагаются почти параллельно друг к другу. Клыки относительно крупные. Щечные зубы четырехугольные в поперечнике, большей частью с морщинками на жевательной поверхности. Позвонков шейных 7, грудных 12—13, поясничных 3—5, крестцовых 3—8, хвостовых 2—3. Грудина широкая. Позвоночный край лопатки увеличен. Таз удлинен. Имеется os penis.
Слепая кишка и червеобразный отросток имеются. Желудок простой. Головной мозг развит сильнее, чем у других обезьян. У крупных обезьян он достигает в объеме 600 см3, сложно устроен; поверхность его покрыта многочисленными извилинами и бороздами. В крови имеются все четыре группы крови, как и у человека. Плацента дискоидальная, отпадающая, гемохориальная. Диплоидное число хромосом у орангутанов, шимпанзе и гориллы 48, у сиамангов — 50, а у гиббонов — 44.
Обитатели тропических лесов. Ведут преимущественно древесный образ жизни. Гориллы полностью перешли к наземному существованию. Активность дневная. При передвижении по деревьям держатся в полувыпрям-ленном или почти выпрямленном положении. При движении по земле опираются на тыльную сторону средних или концевых фаланг пальцев передних конечностей, туловище при этом держат в наклонном положении. Исключительно на задних конечностях ходят редко, только гиббоны по земле и ветвям нередко передвигаются на задних конечностях. Питаются преимущественно растительными объектами, плодами, проростками, а также мелкими животными, птицами и их яйцами. Пищу отправляют в рот передними конечностями. На ночь устраивают своеобразные гнезда или платформы из сучьев и листьев на деревьях или на земле. Держатся небольшими группами от 2 до 5—15 особей. Между человекообразными обезьянами существует довольно сложная звуковая сигнализация. Крупные человекообразные обезьяны обладают высокоразвитой высшей нервной деятельностью. В опытах они способны употреблять палки в качестве орудий. У самск эстральный цикл составляет примерно 28 дней. Длительность беременности 200—-260 дшй. Сезонности в размножении не отмечено. Самка приносит обычно одного детеныша. Детеныши совершенно беспомощны в течение нескольких месяцев (до полугода). Лактация продолжается до года и дольше. Половозрелость наступает в возрасте от 5 до 10 лет. Продолжительность жизни в неволе до 46 лет. В результате преследования человеком численность крупных человекообразных обезьян сильно сократилась, и в настоящее время имеется примерно 2,5 тыс. орангутанов и около 10 тыс. горилл.
Распространены в Юго-Восточной 4зии и на прилегающих островах на запад до о-ва Калимантан, а также в Э.:вчториальной Африке.
В семействе 4 рода (10 видов):
гиббонов — Hylobites liliger, 1811 (7 видов);
орангутанов — Pongo Lacepede, 17Э9 (1 вид);
шимпанзе — Pan О ken, 1816 (1 вид);
горилл — Gorilla I. Geoffroy, 1852 (1 вид).
Некоторые ученые выделяют из рода гиббонов род сиамангов — Sym-
Рис. 158. Гиббон — Hylobates sp., кричащий обыкновенный сиаманг — Н. syndac-tylus (виден горловой мешок), череп белорукого гиббона — Н. lar
phalangus Gloger, 1841 и обособляют их в отдельное семейство Hylobatidae (Вебер, 1828; Валуа, 1955).
Род гиббонов
Genus Hylobates liliger, 1811
Размеры наименьшие в семействе. Длина тела 45—90 см. Обычный вес 5—8 до 13 кг. Телосложение довольно грациозное. Передние конечности очень сильно удлинены (рис. 158). У сиамангов на задних конечностях второй и третий пальцы сильно срастаются. Есть небольшие седалищные мозоли. Первый палец кисти довольно длинный. В запястье имеется центральная кость. Наружный нос хорошо развит. У сиамангов есть гортанный мешок, который снаружи покрыт безволосой кожей. Мешок представляет собой тонкостенное выпячивание слизистой оболочки гортани. Когда
Рис. 159. Белорукий гиббон — Н. lar
животное кричит, мешок сильно раздувается и значительно усиливает звук (рис. 158). Волосяной покров густой, окраска его сильно варьирует от черной или бурой до темно-желтой, почти кремовой или беловатой. У белорукого гиббона кисти и стопы белые и лицо окружено белыми волосами. У одноцветного гиббона волосы на верху головы стоят вертикально, образуя своеобразный гребень. Щечные зубы гиббонов четырехбугорчатые.
Обитатели густых тропических лесов — до примерно 2400 м над уровнем моря. Ведут древесный образ жизни, на землю спускаются редко. Питаются преимущественно растительными объектами (листьями, плодами), но поедают также и различных беспозвоночных и позвоночных животных (насекомых, пауков, птенцов и яйца птиц). По ветвям передвигаются с помощью брахиации (см. рис. 159). Держатся небольшими группами из 2—6 особей, как правило, представляющими отдельную семью. Беременность 200—212 дней. В помете обычно один детеныш. Половозрелость наступает в 6—10 лет. В неволе жили до 23-х лет.
Распространены в Ассаме, Бирме, Юньнани, на п-ове Индокитай, Хай-нани, Тенассериме, Таиланде, на Малаккском полуострове, о-вах Суматре, Ментавай, Яве и Калимантане.
В роде 7 видов:
лар, или белорукий гиббон, — Н. lar Linnaeus, 1771 (Малаккский по луостров, Таиланд, Тенассерим, Камбоджа, Суматра);
быстрый гиббон — Н. agilis Cuvier, 1821 (Южная Азия, Суматра);
губок — Н. hoolock Harlan, 1834 (Юньнань, Ассам, Бирма);
одноцветный гиббон — Н. concolor Harlan, 1826 (Хайнань, п-ов Индоки" тай, Таиланд);
серебристый гиббон— Н. moloch Audebert, 1797 (о-ва Ява, Калимантан), карликовый сиаманг — Н. klossi Miller, 1903 (о-ва Ментавай);
обыкновенный сиаманг — Н. syndactylus Raffles, 1821 (Малаккский полуостров, Суматра, Тенассерим).
Иногда обыкновенного сиаманга и карликового сиаманга выделяют в особый род Symphalangus Gloger, 1841. Некоторые исследователи насчитывают в роде не 7 , а 9 видов.
Н. klossi, Н. lar pileatus очень малочисленны и внесены в «Красную книгу».
Род орангутанов
Genus Pongo Lacepede, 1799
В роде единственный вид: орангутан Р. pygmaeus Linnaeus, 1760 (рис. 160).
Крупная обезьяна. Длина тела до 150 см. Вес взрослых самцов до 75— 100 кг. Передние конечности сильно удлинены, их размах может достигать 2,25 м. Сложение довольно неуклюжее, живот обычно большой. Первые пальцы на передних и задних конечностях развиты слабо, на стопе они лишены ногтя; остальные пальцы длинные и сильные. Голова округлая, лицевая часть широкая, несколько выдвинута вперед и имеет сферическую форму. У самцов на щеках находятся круглые, голые упругие наросты, образованные соединительной и жировой тканью (см. рис. 160). Седалищных мозолей обычно нет, встречаются лишь иногда небольших размеров. Самки значительно меньше, чем самцы. Волосяной покров довольно редкий, длинный, лохматый. Окраска его темно-рыжая или красновато-бурая. Череп довольно высокий, с вогнутым профилем. У самцов сильно развиты сагиттальный и лямбдоидальный гребни. Над глазницами имеются надбровные дуги. Жевательная поверхность щечных зубов покрыта очень мелкими бороздками. Легкие не подразделяются на доли. У самцов имеется огромный непарный гортанный мешок со многими ответвлениями. Головной мозг сравнительно большой, достигает 300—500 см? в объеме и похож на человеческий.
Обитатель леса. Ведет древесный образ жизни. Обычно встречается на равнинах, в болотистых лесах, но иногда поднимается в горы на высоту до 1500 м над уровнем моря. Активность дневная. Держится поодиночке, парами или небольшими группами, обычно семейными. Передвигается не спеша. При движении по земле опирается на нее наружным краем стопы. На ночь строит гнездо или платформу на деревьях. По-видимому, повторно эти гнезда не использует. Питается в основном плодами, особенно плодами дуриана, а также листьями, семенами, проростками, птенцами и яйцами птиц. Беременность 8—9 месяцев. В помете один детеныш. В неволе живет до 27 лет.
Распространен на о-вах Суматре и Калимантан. За последние 100 лет численность орангутанов резко упала и к настоящему времени на Суматре их сохранилось примерно 1 тыс. особей и на Калимантане около 5 тыс. Вид внесен в «Красную книгу».
Род шимпанзе
Genus Pan Oken, 1816
В роде единственный вид: шимпанзе Р. troglodytes Blumenbach, 1799 (рис. 161).
Небольшая человекообразная обезьяна. Длина тела 63—94 см. Вес самцов 56—80 кг. Когда животное стоит на задних конечностях, в высоту оно может достигать 170 см. Самки мельче самцов, но эти различия меньше, чем у орангутана или гориллы. Первый палец на передних конечностях небольшой, на задних •— крупный. Между пальцами задних конечностей развиты небольшие перепонки. Уши большие, нос маленький, верхняя губа высокая. Надбровные дуги хороши развиты. Лицо лишено волос, обычно черного цвета. Нос, уши, кисти и стопы розоватые. Волосяной покров на туловище черный, почти без подшерстка. Глазницы сильно сдвинуты вперед: над ними располагаются массивные надглазничные валики. Лоб довольно сильно ’покат. Щечные зубы имеют низкие бугорки с многочисленными бо-
Рис. 160. Орангутан — Р. pygmaeus
Рис. 161. Шимпанзе — Р. troglodytes и его череп
роздками. Клыки развиты относительно слабо. Горловые мешки средних размеров. В диплоидном наборе 48 хромосом.
Обитатель тропических лесов. Ведет как древесный, так и наземный образ жизни. При ходьбе по земле опирается на нее всей подошвой стопы и тыльной стороной пальцев передней конечности. Активность дневная. Держится обычно группами от 2 до 20 особей, возглавляемых взрослым самцом. На ночь располагаются в гнезде, которое строят на деревьях. Повторно это гнездо не используется. Слух и зрение — наиболее развитые органы чувств. Высшая нервная деятельность шимпанзе стоит на более высоком уровне, чем у других млекопитающих (кроме человека). Зарегистрировано по крайней мере 32 различных звука, издаваемых шимпанзе. Питается в основном растительными объектами: плодами, листьями, корнями, употребляют в пищу также и различных беспозвоночных и мелких позвоночных животных. Беременность 232—261 день. В помете обычно один, редко два детеныша. Детеныш остается с матерью самое меньшее 2 года. Половозрелости достигает примерно в 12-летнем возрасте. Продолжительность жизни, по-видимому, около 40 лет.
Распространен в Африке примерно от 14° с. ш. до 10° ю. ш., Численность невелика. Внесен в «Красную книгу».
Род горилл
Genus Gorilla I. Geoffroy, 1852
В роде единственный вид: горилла G. gorilla Savage et Wyman, 1847 (рис. 162).
Наиболее крупный представитель семейства человекообразных обезьян, приматов. Длина тела до 180 см. Вес самцов до 280 кг. Телосложение довольно неуклюжее. Туловище бочкообразное, с мощной трудней клеткой, толстым животом, короткой шеей, большой головой, длинными передними конечностями и укороченными сильными задними конечностями. Объем груди у самцов может достигать 125—175 см. Пальцы укороченные, на задних конечностях они соединены перепонками почти до ногтевых фаланг. Голова округлая, с относительно слабо выступающим лицевым отделом. Нос имеет хрящевой
скелет. По его спинке проходит продольная борозда. Верхняя губа короткая. Уши маленькие, прижатые к голове. Надбровные дуги^сильно развиты и нависают над глазами. Ноздри крупные. Волосяной покров грубый, средней длины, довольно редкий, окраска его черная, у старых самцов становится серебристо-серой. Лицо, уши, кисть и стопа голые. Грудь старых самцов лишена волос. Бороды нет. На верху головы имеется своеобразная подушка, образованная утолщением кожи из соединительной ткани. Эту подушку покрывают волосы. У горной гориллы на верху головы удлиненные волосы могут образовывать гребень. В черепе, особенно у старых самцов, очень развиты гребни — сагиттальный и лямбдоидальный. Надглазничные валики мощно развиты, есть лобные пазухи. Щечные зубы с крупными бугорками. Клыки большие. В кисти центральная кость запястья сращена с ладьевидной, как у шимпанзе и человека. Горловые мешки крупные. Абсолютный объем головного мозга больше, чем у других обезьян: 400— 600 см3, в одном случае даже 685 см3.
Обитатель равнинных и горных лесов. В горы поднимаются до 2300— 3500 м над уровнем моря. Ведут наземный образ жизни. При ходьбе по земле ступают на нее всей подошвой ступни и опираются, кроме того, тыльной
Рис. 162. Горилла — G. gorilla
поверхностью пальцев передних конечностей. Активность дневная. Каждый вечер строят гнезда, обычно на деревьях, реже — на земле. Держатся группами по 5—15 особей; иногда соединяются в стада до 40 осе бей вместе. Обычно предводителем является крупный самец. Питаются главным образом различными растительными объектами. Беременность примерно 9 месяцев. В помете один детеныш. Половозрелость, по-видимому, наступает в возрасте около 8 лет. В неволе жили до 34 лет; в естественных условиях, вероятно, живут до 50 лет.
Распространены в бассейне Конго и Западной Африке (береговая горилла ) и в горах восточной части Республики Заир и западной Уганды (горная горилла). Численность горилл за последние годы резко сократилась. Основным естественным врагом является леопард.
Численность горных горилл определяют в пределах 5—15 тыс. особей. Включены в «Красную книгу».
СЕМЕЙСТВО ЛЮДЕЙ
FAMILIA HOMINIDAE GRAY, 1825
В семействе единственный род: люди — Homo Linnaeus, 1758, с одним видом — человеком — Н. sapiens Linnaeus, 1758.
Средних размеров высшие приматы. Длина тела около 90 см. Высота человека обычно от 140 см до 190 см. Имеется половой диморфизм: женщины меньше мужчин на 9—12 см. Характерной особенностью является вертикальное положение тела и приспоссбление к передвижению только на нижних конечностях. Первый палец нижних конечностей в значительной степени утрачивает подвижность и не противопоставляется остальным. Длина нижних конечностей значительно превышает длину верхних. Большое значение для трудовых процессов имеет сильное развитие первого пальца верхних конечностей. Голова округлая, характеризуется сильным развитием мозговой части и слабо выступающей вперед лицевой. Настоящий волосяной покров на туловище утрачен. Пигментные клетки находятся в соединительной ткани кожи и определяют ее окраску, варьирующую от белой до черной. Кожа обладает сальными и потовыми железами. Имеется пара грудных сосков.
В черепе характерно сильное развитие мозговой коробки. Лицевой отдел помещается не впереди мозгового, а под ним. По объему мозговая часть черепа значительно превышает лицевую. Большое затылочное отверстие 2 12 3
направлено вертикально вниз. Зубная формула: i— с — р — т— = 32.
Клыки развиты слабо и по размерам почти не отличаются от резцов. Пред-коренные зубы, как правило, имеют по одному корню. Коренные имеют ! уплощенные бугорки на жевательной поверхности, причам верхние, как . правило, по 4 бугорка, а нижние по 5. Позвоночный столб имеет S-образ-; ное искривление, что связано с вертикальным положением человека при ходьбе. Позвонков шейных 7, грудных 12, поясничных 5, крестцовых 5, хвостовых 4. Крестцовые и хвостовые позвонки срастаются в сложные кости — крестец и кспчик. В скелете нижней конечности характерно сильное развитие бедренной кости, которая длиннее соответствующей кости верхней конечности (плечевой). Сильно удлинены кости предплюсны. Центральная кость отсутствует.
Слепая кишка имеет червеобразный отросток. Желудок простой. Характерно необычайно сильное развитие мозга, особенно больших полушарий, < имеющих на поверхности большое число борозд и извилин. Неопаллиум ! мозга развит особенно сильно. Плацента дискоидальная, отпадающая, гемохориальная. Диплоидное число хромосом 46—48. Беременность 243—298,
обычно около 280 дней. Как правило, рождается один ребенок, иногда два, три, редко четыре. Для человека свойственно наиболее длительное среди млекопитающих развитие ребенка.
Высшая нервная деятельность достигает у человека наивысшей среди млекопитающих степени развития.
Распространение всесветное. Во второй половине XX в. начато освоение космического пространства.
ОТРЯД НЕПОЛНОЗУБЫХ
ORDO EDENTATA GRAY, 1798
Размеры изменчивы: длина тела от 12 см у плащеносного броненосца до 120 см у гигантского муравьеда. Современные представители отряда ведут разнообразный образ жизни и сильно отличаются друг от друга по внешнему виду. Среди них есть древесные формы (ленивцы и муравьеды), наземные (муравьеды) и наземные хорошие копатели (броненосцы). Кожный покров может иметь волосы (муравьеды, ленивцы) или представлять собой костный панцирь, образующийся из кожных окостенений и покрытый сверху ороговевшим эпидермисом (броненосцы). Передние конечности двух-, трех-, четырех- или пятипалые, причем два или три пальца значительно превышают по величине остальные (если последние имеются). Пальцы вооружены сильными когтями. Задние конечности трех-, четырех- или пятипалые. Имеет место синдактилия. Конечности стопоходящие. Форма головы различная— от округлой до сильно вытянутой. Для ряда форм (муравьеды) характерен длинный язык. Хвост длинный (муравьеды) или короткий (ленивцы). Соски у самок располагаются в области груди, на брюхе, реже в пахах.
Череп укорочен (ленивцы) или сильно удлинен (муравьеды) в лицевом отделе. Предчелюстные кости развиты слабо или довольно хорошо. Скуловая дуга может быть полной, неполной или рудиментарной. Зубы могут полностью отсутствовать (муравьеды). У броненосцев и ленивцев нет резцов и клыков. Предкоренные и коренные зубы колышковидной формы с одним корнем. Зубы лишены эмали и обладают постоянным ростом в течение жизни животного. В шейном отделе позвоночника 7 позвонков у муравьедов и броненосцев и варьирует от 6 до 9 у ленивцев. Последние грудные и поясничные позвонки обладают дополнительными сочленовными отростками. Локтевая и лучевая кости разделены, а большая и малая берцовые могут быть разделены или сращены в проксимальных отделах, а иногда и в дистальных. Имеется ключица. Коракоидный отросток лопатки развит относительно сильнее, чем у других плацентарных млекопитающих.
Желудок простой (муравьеды, броненосцы) или сложный (ленивцы). Слепая кишка развита слабо или полностью отсутствует. Мозг макросмати-ческий. Извилины больших полушарий немногочисленны. Задняя полая вена двойная. В конечностях и хвосте имеется «чудесная сеть». Матка простая, плацента дискоидальная. Семенники расположены в брюшной полости.
Среди неполнозубых есть формы, живущие на земле и активные в течение дня и ночи (гигантский муравьед, броненосцы); держащиеся в основном на деревьях и обычно активные ночью (тамандуа, карликовый муравьед) и исключительно древесные виды, активные по ночам (ленивцы). Держатся поодиночке или парами и лишь иногда небольшими группами (броненосцы). Пищевой режим сильно варьирует у различных представителей. Муравьеды и броненосцы питаются в основном насекомыми, а ленивцы — растениями. В помете у муравьедов, ленивцев и некоторых броненосцев один детеныш, а
у ряда броненосцев от 2—4 до 12. Для броненосцев характерна задержка имплант; ции яйцеклетки и полиэмбриония1.
Распространены в Южной и Центральной Америке, Мексике, южных и юго-восточных штатах США.
В стпяле 3 семейства:
муравьедов — Myrmecophagidae Bonaparte, 1838 (3 рода);
ленивцев — Bradypodidae Bonaparte, 1831 (2 рода);
броненосцев — Dasypodidae Bonaparte, 1838 (9 родов);
В прошлом некоторые систематики резко отделяли неполнозубых от других млекопитающих, помещая их в отдельный подкласс или даже считали, что они произошли, независимо от других млекопитающих, от рептилий. В настоящее время нет сомнений, что неполнозубые принадлежат к подклассу Eutheria и развились в начале палеоцена из примитивных насекомоядных параллельно с некоторыми другими архаичными отрядами.
СЕМЕЙСТВО МУРАВЬЕДОВ
FAMILIA MYRMECOPHAGIDAE BONAPARTE, 1838
Размеры от небольших до наибольших в отряде. Длина тела 15—120 см. Вес от 350 а до 25 кг. Самцы крупнее самок. Тело покрыто густыми волосами. Голова удлиненная, часто с трубковидным лицевым отделом. Ротовая щель маленькая, узкая. Уши небольшие, округлые. Длинный хвост может быть хватательным (у родов тамандуа и карликовых муравьедов), и тогда он лишен волос на кончике. Передние лапы пятипалые, а задние четырех- или пятипалые. Когти на передних лапах длинные и острые. На задних лапах когти короткие. Третий палец вооружен особенно сильным когтем. Очень длинный язык служит ловчим органом: он смачивается липкой слюной, выделяемой слюнными железами (в частности .очень крупной подчелюстной железой). Волосяной покров плотный, короткий и мягкий или длинный и грубый. Окраска его на спинной стороне тела от серой до золотисто-бурой, часто с черноватыми полосами или с обширной черной зоной на туловище (тамандуа), на брюшной желтоватая или серовато-белая. Соски у самок расположены в грудной и брюшной областях.
Череп удлинен и выглядит хрупким, но на самом деле кости черепа толстые и обладают прочной структурой. Рострум может быть трубчатым, округлым сверху и уплощенным снизу. Предчелюстная кость небольшая. Скуловая дуга незамкнутая. Слезная кость хорошо развита. Крыловидные кости, примыкающие друг к другу по средней линии, значительно увеличивают протяженность твердого неба (гигантский муравьед и тамандуа). Ключица рудиментарна. Зубов нет.
Желудок простой, с тонкостенным кардиальным отделом и толстостенным пиш рическим. Матка простая. Плацента дискоидальная, гемохориальная. Темпера тура тела колеблется от 27,8 до 31,3° С у карликового муравьеда, от 33,7 до 35,7° С у тамандуа и от 32 до 35° у гигантского муравьеда.
Держатся главным образом в тропических лесах, но встречаются и в открытых районах типа саванн. Есть наземные и древесные формы. Активность в основном ночная. Встречаются поодиночке или парами, обычно самка и ее детеныш. Энтомофаги. Из органов чувств наиболее сильно развито обоняние. В настоящее время муравьеды довольно многочисленны.
Распространены от южной Мексики через Центральную Америку на юг до Парагвая в Южной Америке.
В семействе 3 рода (3 вида):
гигантских, или трехпалых, муравьедов — Myrmecophaga Linnaeus, 1758 (1 вид):
1 Полиэмбриония — развитие нескольких эмбрионов из одной яйцеклетки.
Рис. 163. Самка гигантского муравьеда— М. tridactyla с детенышем, череп
тамандуа, или четырехпалых муравьедов, — Tamandua Gray, 1825 V вид);
карликовых, или двупалых, муравьедов,— Cyclopes Gray, 1821 (1 вид).
Род гигантских, или трехпалых, муравьедов
Genus Myrmecophaga Linnaeus, 1758
В роде единственный вид: гигантский, или трехпалый, муравьед— М. tridactyla Linnaeus, 1758 (рис. 163).
Наиболее крупный в семействе муравьед. Длина тела 100—120 см. Длина хвоста 60—90 см. Вес 18—25 кг. Тело сжато с боков. Голова сильно вытянута в лицевом отделе, имеющем трубковидную форму. Передние конечности сильнее задних: первый палец очень маленький, высоко расположенный, второй и третий очень большие, с мощными когтями (на третьем пальце коготь может достигать 10 см в длину), четвертый палец небольшой, пятого нет совсем. На задних конечностях первый и пятый пальцы небольшие, три средних развитыхорошо.Окраска волосяного покрова серая, а диагональная полоса, проходящая по телу от груди к крестцу, черная с белым окаймлением. Волосы длинные (на голове короткие), густые, но грубые. Длинные волосы (до 40 см длиной) образуют на вентральной и дорзальной сторонах хвоста высокие гребни; животное накрывает себя хвостом во время отдыха. Вдоль середины спины также проходит гребень удлиненных волос. У самок 2 соска в грудной области. Язык может вытягиваться на 61 см, причем его диаметр в самом толстом месте составляет 10—15 мм.
Обитатель болотистых районов, влажных лесов и саванн. Ведет наземный образ жизни. Активность дневная. Не имеет постоянного убежища; все время проводит в поисках пищи. Кормом служат муравьи и термиты, убежища которых муравьед разрывает мощными когтями передних лап; может поедать и личинок насекомых. Охотно идет в воду и может переплывать широкие реки. В случае нападения врага защищается передними лапами, нанося опасные раны своими огромными когтя
ми; как правило, ягуары не рискуют нападать на взрослых животных. Беременность 190 дней. Самка рождает одного детеныша (по наблюдениям в Парагвае — весной) весом около 1700 г. Самка носит детеныша на спине. Молодой остается с матерью до ее следующей беременности. Продолжительность жизни в неволе до 14 лет.
Распространен в Центральной и Южной Америке от юга Гондураса до северной Аргентины.
Численность везде мала и продолжает снижаться. Включен в «Красную книгу».
Род тамандуа, или четырехпалых муравьедов,
Genus Tamandua Gray, 1825
В роде единственный вид: тамандуа, или четырехпалый муравьед — Т. tetradactyla Linnaeus, 1758 (рис. 164).
Средних размеров муравьед. Длина тела 54—58 см. Длина хвоста 54— 55 см. Вес 3—5 кг. Морда укороченная. Уши относительно крупные, широко расставленные. Глаза и рот маленькие. Хвост хватательный, его нижняя поверхность и весь концевой отдел (V3 длины) лишены волос. Конечности сильные и наименее специализированы по сравнению с другими представителями семейства. На передних лапах третий палец самый крупный, с когтем длиной около 5 см, второй и четвертый пальцы одинаковой длины, с небольшими когтями, первый палец очень мал, пятого нет совсем. На задних лапах пять пальцев, первый из них развит слабо. Тело покрыто густым, коротким и грубым волосяным покровом, окраска которого значительно варьирует; общая окраска рыжевато-коричневая (у отдельных особей кремовая, коричневая или черноватая). Обычно имеется черный участок между задними и передними конечностями, который продолжается вокруг шеи в виде ошейника. Кожные специфические железы продуцируют секрет с сильным запахом.
Рис. 164. Тамандуа — Т. tetradactyla и его череп
Рис. 165. Карликовый муравьед — С. didactylus и его череп
Обитатель тропических лесов, иногда саванн. Большую часть жизни проводит на деревьях. Ведет в основном ночной образ жизни, но бывает также активен рано утром или поздно вечером. Днем находит себе убежище в дуплах. Питается муравьями, термитами и пчелами. Пищу добывает на деревьях и на земле. Самка приносит одного детеныша, обычно весной.
Распространен от южной Мексики к югу до Боливии и Бразилии.
Род карликовых, или двупалых, муравьедов
Genus Cyclopes Gray, 1821
В роде единственный вид: карликовый, или двупалый, муравьед — С. didactylus Linnaeus, 1758 (рис. 165).
Размеры наименьшие в семействе. Длина тела 15—18 см. Длина хвоста — 17—20 см. Вес около 500 г. Хвост хватательный, обнаженный по нижней стороне. Морда несколько удлиненная, изогнутая концом вниз. Ротовое отверстие относительно крупное. Глаза и уши небольшие. Конечности сильные. На передних лапах второй и третий пальцы крупные, примерно одинаковых размеров, сращены друг с другом, когти на этих пальцах большие, острые, пятого пальца нет совсем. Задние конечности длинные, имеют пяточную подушку и особое «сочленение» в подошве, позволяющее трем
пальцам поворачиваться назад, под подошву: образуется хватательная лапа.
Волосяной покров короткий, густой и мягкий, окраска его от темножелтой до золотисто-желтой, более темной на спинной стороне тела.
Обитает в тропических лесах. Ведет древесный образ жизни. Активность ночная. День проводит свернувшись в шар в дупле дерева или в развилке сучьев. Питается термитами, в меньшей степени различными насекомыми.
Распространен от южной Мексики до Боливии и Бразилии.
СЕМЕЙСТВО ЛЕНИВЦЕВ
FAMILIA BRADYPODIDAE BONAPARTE, 1831
Средних размеров неполнозубые. Длина тела 50—64 см. Вес от 4 до 9 кг. Хвост очень короткий (около 7 см) или его нет совсем. Голова короткая и округлая. Ушные раковины маленькие. Глаза направлены вперед. Конечности длинные (передние длиннее задних). Кисть и стопа узкие и изогнутые. Пальцев 2 или 3; они срастаются между собой по всей длине. Когти на обеих конечностях раздельные, длинные, сильные и изогнутые. Конечности приспособлены поддерживать животное в висячем состоянии на ветке спиной вниз. Тело густо покрыто волосами с характерным направлением на туловище от брюха к спине. Морда беловатая или буроватая. Окраска волосяного покрова желтовато-белая или светло-бурая, может быть с зеленоватым оттенком из-за поселения зеленых водорослей в волосах (особенно в сезон дождей). Соски у самок располагаются в грудной области.
Череп характерен: укороченный лицевой отдел, неполная скуловая дуга, небольшие предчелюстные кости. Орбиты сближены и направлены вперед. Слезные кости малы. Крыловидные кости не соприкасаются между собой по средней линии. Нижняя челюсть с сильно развитым венечным отростком. Зубная формула: * с ~~ Р + т 4 5 ~ =18 — 20. Жевательная поверхность зубов чашковидная. Зубы не прекращают роста в течение всей жизни животного. В центре каждого зуба находится вазодентин, окруженный тонким слоем твердого дентина и толстым слоем цемента. Характерно изменчивое количество шейных позвонков: 6, 7, 8 или 9; шея настолько подвижная, что животное может поворачивать голову на 270°. Туловищно-поясничных позвонков 18—27. Акромиальный и коракоидный отростки лопатки срастаются между собой.
Желудок сложный. Слепой кишки нет. Матка простая, глобулярной формы. Плацента диффузная, дискоидальная, эндотелиохориальная.
Обитают в тропических лесах. Ведут древесный образ жизни. Активны в основном ночью. На землю спускаются крайне редко, однако умеют хорошо плавать. Проводят ряд лет на очень ограниченном участке. Держатся поодиночке или парами. Температура тела может значительно варьировать в зависимости от температуры окружающей среды (у двупалого ленивца от 24 до 35° С). По-видимому, в связи с этим их распространение ограничено экваториальной областью постоянных температур. Растительноядны. Слух развит слабо, обоняние — хорошо.
Беременность 4—6 месяцев. Самки рождают в сезон всего одного детеныша длиной около 20 см, которого затем носят на груди.
Распространены от Гондураса на юго-восток до северной Аргентины, Парагвая и Бразилии.
В семействе 2 рода:
трехпалых ленивцев — Bradypus Linnaeus, 1758 (3 вида);
двупалых ленивцев — Choloepus liliger, 1811 (2 вида).
Возможно, двупалых ленивцев следует выделить в особое семейство Choloepodidae (Хоффстеттер, Hoffstetter, 1969).
Род трехпалых ленивцев
Genus Bradypus Linnaeus, 1758
Длина тела 50—60 см. Длина хвоста 6—7 см. Вес 4—5 кг. На передних и задних конечностях по 3 сросшихся между собой пальца. Подошвы лап покрыты волосами. Общая окраска волосяного покрова серовато-коричневая, немного темнее на голове, на плечах обычно более светлая с коричневыми отметинами. В шерсти поселяются зеленые водоросли: Trichophilus welckeri и Cyanoderma bradypi. Шейных позвонков 8 или 9.
Обитают по окраинам лесов, в долинах рек. Держатся поодиночке. Редко спускаются на землю, по которой почти не в состоянии ходить. В некоторые годы в начале дождевого сезона отмечались миграции протяженностью в несколько километров. Питаются молодыми листьями, побегами, почками деревьев (преимущественно Cecropia peltata). Характерен крик, напоминающий «ай». Спаривание у одних видов происходит в течение всего года, у других в марте — апреле. Беременность 120—180 дней. Самка рождаетодно-го детеныша обычно в начале засушливого сезона.
Распространение совпадает с ареалом семейства.
В роде 3 вида:
трехпалый ленивец, или ай,— В. tridactylus Linnaeus, 1758 (ареал сов-
падает с ареалом семейства);
капюшонный ленивец— В. cuculliger Wagler, 1831 (Бразилия);
ошейниковый ленивец— В. torquatus liliger, 1811 (западная Бразилия, Боливия).
Ошейниковый ленивец редок и внесен в «Красную книгу».
Род двупалых ленивцев
Genus Choloepus liliger, 1811
Длина тела до 60— 64 см. Хвост рудиментарен. Вес около 9 кг. Окраска волосяного покрова серовато-коричневая, светлая на морде. В шерсти поселяются зеленые водоросли: Irichopilus sp. и Cyanoderma choloepi. На передних конечностях по два сросшихся между собой пальца (II и III), вооруженных крупными когтями длиной 75 мм (рис. 166). Задние конечности трехпалые: II, III и IV пальцы сращены между собой. Подошвы лап голые. В шейном отделе позвоночного столба 6, 7 илиъ8 позвонков.
Рис. 166. Двупалый ленивец — Ch. didactylus и его череп
Питание более разнообразное, чем у трехпалого ленивца: листья, нежные побеги и плоды различных видов деревьев (предпочитаютХ/юпДпз luted}. Это разнообразие обусловливает лучшую выживаемость двупалых ленивцев в неволе. Скорость передвижения двупалых ленивцев примерно в 3 раза выше, чем трехпалых. Беременность 5—6 месяцев. Самка приносит одного детеныша длиной около 25 см. В неволе жили до 23 лет.
Распространены в Центральной Америке и в северных и центральных районах Южной Америки.
В роде 2 вида:
ленивец Гоффмана — С. hoffmanni Peters, 1858 (от Никарагуа на юг до Перу и центральной Бразилии);
двупалый ленивец, или унау, — С. didactylus Linnaeus, 1758 (Венесуэла, Гвиана и северная Бразилия).
СЕМЕЙСТВО БРОНЕНОСЦЕВ
FAMILIA DASyPODIDAE BONAPARTE, 1838
Мелкие, средние и крупные неполнозубые. Длина тела от 12 до 100 см. Длина хвоста от 2,5 до 50 см. Вес до 55 кг.
Характерен наружный костный панцирь, образованный кожными окостенениями. Панцирь состоит из сплошных обручевидных полос, опоясывающих тело сверху и с боков, и щитков квадратной или полигональной формы. Хвост покрыт коническими щитками. Сплошные пояса и отдельные щитки соединены между собой соединительной тканью, обусловливающей подвижность всего панциря. Пояса и щитки снаружи покрыты слоем рога, образовавшегося из эпидермиса. На брюшной поверхности тела и внутренних частях конечностей кожа покрыта редкими волосами. У многих видов отдельные волосы растут между щитками на спине. Окраска чешуй от коричневой до розовой, а волос — от серовато-коричневой до белой. Соски у самок в грудной области, но иногда имеется и пара паховых сосков. Телосложение тяжелое. Форма морды изменчива — от короткой до вытянутой. Уши небольшие. Глаза маленькие, с толстыми веками. Конечности короткие. Передние лапы четырех- или пятипалые, задние пятипалые. Когти на пальцах передних конечностей мощные, длинные, изогнутые.
Череп уплощен в дорзо-вентральном направлении. Слезные кости маленькие. Предчелюстные кости небольшие. Скуловая дуга полная. Нижнечелюстные кости удлиненные. Зубы дифиодонтные, небольшие, колышкооб-7 9
разные, с постоянным ростом. Число зубов сильно варьирует от ——- до 25 — . На предчелюстных костях зубов нет (за исключением единственной пары у шестипоясного броненосца). В шейном отделе позвоночника по крайней мере 2 или 3 позвонка срастаются между собой. Большая и малая берцовая кости срастаются друг с другом в проксимальных и дистальных отделах. Акромиальный отросток сильно развит. Имеется ключица.
Подкожная мускулатура сильно развита. Желудок простой. Подчелюстная слюнная железа сильно развита. Плацента дискоидальная, гемохориальная.
Обитатели главным образом открытых пространств, но живут и в лесах. Ведут наземный образ жизни и превосходно копают землю. Активность ночная или дневная. Держатся поодиночке, парами или небольшими группами. Убежищем служат подземные норы. Могут довольно быстро бегать. Передвигаются, обычно опираясь кончиками когтей передней лапы и всей подошвой стопы задней. При опасности спасаются бегством или быстро зарываются в землю; некоторые виды сворачиваются в шар. Обоняние, зрение и слух хорошие. Температура тела колеблется от 32 до.36° С. Питание весьма разнообразное: пищей служат различные насекомые и другие беспозвоночные, 362'
мелкие позвоночные, некоторые растения и падаль. Беременность продолжительная вследствие задержки имплантации яйцеклетки после ее оплодотворения (латентный период). Самка обычно рождает несколько двоен, развивающихся из одной яйцеклетки. В помете детенышей бывает от 2—4 до 12, но иногда всего один. Детеныш рождается в гнезде в конце норы и остается с матерью в течение нескольких месяцев. Новорожденные имеют открытые глаза и покрыты мягкой кожей, которая вскоре становится жесткой. Через несколько часов после рождения способны ходить. Половозрелости самки достигают в 2 года. Ареал обширный: от юго-центральных и юго-восточных штатов США на юг через Мексику и Центральную Америку до Чили, Аргентины и Уругвая.
В семействе 9 родов (21 вид):
щетинистых броненосцев — Chaetophractus Fitzinger, 1871 (3 вида); шестипоясных броненосцев — Euphractus Wagler, 1830 (1 вид); карликовых броненосцев — Zaedyus Ameghino, 1889 (1 вид); гигантских броненосцев — Priodontes F. Cuvier, 1827 (1 вид);
голохвостых броненосцев — Cabassous McMurtrie, 1831 (5 видов); трехпоясных броненосцев— Tolypeutes liliger, 1811 (2 вида);
Девятипоясных броненосцев — Dasypus Linnaeus, 1758 (6 видов); плащеносных броненосцев — Chlamyphorus Harlan, 1825 (1 вид); щитоносных броненосцев — Burmeisteria Gray, 1865 (1 вид).
Род щетинистых броненосцев
Genus Chaetophractus Fitzinger, 1871
Мелких размеров броненосцы. Длина тела 22—28 см. Длина хвоста до 10 см. На передних и задних конечностях по 5 пальцев. Голова покрыта сверху щитками, имеющими зернистую поверхность (рис. 167). Спинной панцирь насчитывает примерно 18 поясов, из них 7—8 подвижно сочлененных. По сравнению с другими броненосцами, имеют более густой волосяной покров на брюхе и проксимальных отделах конечностей. Волосы выступают также из-под щитков на спине и боках тела. Окраска волос от беловатой до светло-коричневой.
Придерживаются открытых мест. Активность в основном ночная. Если
Рис. 167. Щетинистый броненосец — С. viltosus
волосатый броненосец не имеет возможности спастись от врага бегством или спрятаться в нору, он прижимается всем телом к земле так, что края панциря плотно касаются грунта, и становится недоступным для хищника. Питание разнообразное. Самка обычно приносит 2 помета в течение года. Беременность около двух месяцев. Детенышей в помете по большей части 2 (часто один самец и одна самка). Мясо используется местным населением в пищу.
Распространены в Боливии, Чили, Аргентине и Уругвае.
В роде 3 вида:
чилийский броненосец — С. nations Thomas, 1894 (Чили, Перу);
длинноволосый броненосец—С. vellerosus Gray, 1865 (Аргентина);
щетинистый броненосец — С. vitlosus Desmarest, 1804 (Аргентина).
Некоторые исследователи объединяют все три вида в один — С. villo-sus и включают его в род шестипоясных броненосцев.
Род шестипоясных броненосцев
Genus Euphractus Wagler, 1830
Вроде всего один вид: шестипоясной броненосец — Е. sexcinctusLinnaeus, 1758.
Средние по размерам броненосцы. Длина тела примерно 40—50 см. Длина хвоста 20—27 см. Вес около 5 кг. Передние и задние конечности пятипалые. Характерны заостренная и уплощенная голова и от 6 до 9 подвижных поясов в панцире. Головной щит состоит из сравнительно крупных пластин, расположенных довольно беспорядочно. На некоторых щитках у основания хвоста имеются отверстия, служащие, по-видимому, выводными протоками специфических желез, которые придают животному характерный запах. Окраска тела от сероватой до коричневатой.
Придерживается сухих саванн или сухих районов влажных саванн. Активность ночная.Питается преимущественно растениями (иногда культурными) и насекомыми. Беременность примерно 2 месяца. У самок в течение года обычно бывает два помета, в каждом по 2 детеныша.
Распространение охватывает большую часть Южной Америки к востоку от Анд.
Некоторые ученые включают в род шестипоясных броненосцев роды щетинистых и карликовых броненосцев.
Род карликовых броненосцев
Genus Zaedyus Ameghino, 1889
В роде единственный вид: карликовый броненосец — Z. pichi Desmarest, 1804 (рис. 168).
Размеры средние. Длина тела около 40 см. Длина хвоста примерно 15 см. Вес достигает 1—2 кг. Уши невелики. Передние и задние конечности пятипалые. Задние края спинных щитков густо окаймлены тонкими черноватыми волосами, перемежающимися с длинными желто-коричневыми и беловатыми щетинами. Нижняя поверхность тела покрыта редкими желтовато-белыми волосами. Окраска головного и спинного панцирей темно-коричневая с желтыми или беловатыми краями. Плечевые и тазовые щитки прикреплены к телу почти до основания конечностей.
Обитают в степи (пампе), предпочитают теплую, песчаную почву. Обычная защитная реакция-—плотно прижимается к земле. В ряде районов с холодной зимой, по-видимому, впадают в спячку. Питается насекомыми, червями, другими беспозвоночными и падалью. Местное население употребляет мясо карликового броненосца в пищу. В некоторых районах содержится для этой цели в неволе.
Распространен в Аргентине и Патагонии. Был ввезен в Чили.. Иногда этот вид включают в род шестипоясных броненосцев.
Рис. 168. Карликовый броненосец — Z. pichi
Род гигантских броненосцев
Genus Priodontes F. Cuvier, 1827
В роде единственный вид: гигантский броненосец — Р. giganteus Geoffroy. 1803 (рис. 169).
Размеры крупные, наибольшие в семействе. Длина тела до 100 см. Длина хвоста около 50 см. Вес до 55 кг. Форма головы овальная. Уши широкие. Конечности пятипалые. Третий палец и его коготь на передних лапах развиты особенно сильно. В спинном панцире 11—13 подвижных поясов. Щитки на хвосте плотно примыкают друг к другу и не образуют рядов. Волосы на теле редкие. Окраска тела темно-коричневая, исключая беловатые голову, хвост и полосу по нижнему краю спинного щита. В каждой половине челюсти обычно по 15—28 мелких зубов, выпадающих с возрастом.
Встречается обычно у воды. Ведет ночной образ жизни. Часто держится только на задних лапах и хвосте, не касаясь передними земли. Питается муравьями, термитами, другими насекомыми и их личинками, червями, пауками, змеями и падалью. В помете 1—2 детенышей.
Распространение охватывает восточные районы Южной Америки.
Численность гигантского броненосца невелика. Внесен в «Красную книгу».
Род голохвостых броненосцев
Genus Cabassous McMurtrie, 1831
Размеры от мелких до крупных. Длина тела от 30 до 70 см. Длина хвоста 10—18 см. Вес до 6 кг. Морда короткая и широкая. Уши широко расставлены. Конечности пятипалые. Третий палец и его коготь на передних лапах особенно крупные. Подвижные пояса в панцире спины наиболее многочисленны по сравнению с другими представителями семейства, их число достигает 10—13. Отсутствие щитков на хвосте является характерным признаком рода. Хвост защищен лишь небольшими, тонкими, редко расположенными пластинками. Плечевой и тазовый панцири прикреплены к телу почти до основания конечностей. Окраска спинной стороны от темно-бурой до почти черной. Боковые края панциря желтоватые. Брюшная сторона желтовато-се-9
рая. Число зубов обычно — = 34.
Обитатели открытых пространств и влажных лесов. Активность ночная. Норы часто устраивают в основании сооружений термитов. Норы до 5 м глубиной. В случае опасности спасаются бегством (на короткой дистанции),
Рис. 169. Гигантский броненосец — Т. giganteus и его череп
зарываются в землю, прячутся в нору, бросаются в воду или сворачиваются в шар. Питаются в основном термитами и муравьями.
Распространены в Центральной и Южной Америке к югу до северной Аргентины и Парагвая.
В роде 5 видов:
малый броненосец — С. hispidus Burmeister, 1854 (южная Бразилия);
голохвостый броненосец — С. loricatus Pelzen, 1883 (северная Аргентина и Мату-Гросу в Бразилии);
тату — С. tatouay Desmarest, 1819 (юг Бразилии, восток Парагвая и северо-восток Аргентины);
большой броненосец — С. unicinctus Linnaeus, 1758 (Гвиана и восточная Бразилия);
центральноамериканский броненосец — С. centralis Miller, 1899 (Гондурас, Коста-Рика и Панама).
Некоторые ученые рассматривают С. loricatus, С. tatouay и С. unicinctus в качестве подвидов С. hispidus.
Род трехпоясных броненосцев
Genus Tolypeutes liliger, 1811
Размеры мелкие. Длина тела примерно 35—45 см. Длина хвоста около 7—9 см. Голова узкая, с очень крепкими щитками. Уши короткие, мясистые, с широким основанием.В панцире тела три подвижно сочлененных пояса (иногда 2 или 4). Панцирь тяжелый, прочный, лишенный волос, от темно- до серо-бурого цвета. В отличие от большинства броненосцев боковые отделы Двух больших щитов трехпоясных броненосцев свободно налегают на тело и не срастаются с ним. Для трехпоясных броненосцев характерна способность сворачиваться в плотный шар и делаться таким образом недоступными для
хищников (рис. 170). Толстый хвост почти неподвижен. Второй, третий и четвертый пальцы задних конечностей срастаются между собой, их когти напоминают копыто, а первый и пятый пальцы слегка отделены от остальных и имеют нормальные когти. На передних конечностях 4 пальца (шаровидный броненосец) или 5 (трехпоясный броненосец).
Обитатели открытых травянистых и болотистых районов, саванн и влажных лесов. Основной пищей служат термиты и муравьи. Беременность 5—6 месяцев. Самка рождает в ноябре всего одного детеныша.
Распространены в восточных районах Южной Америки.
В роде 2 вида:
шаровидный броненосец — Т. matacus Desmarest, 1804 (Боливия,Мату-Гросу в Бразилии, Парагвай и Аргентина);
трехпоясный броненосец—Т. tricinctus Linnaeus, 1758 (северо-западная часть центральной Бразилии).
Трехпоясный броненосец, возможно, вымер. За последние 300 лет добыто 6 экземпляров этого вида. Включен в «Красную книгу».
Рис. 170. Трехпоясный броненосец — Т. tricinctus. Внизу он же, свернувшийся в шар
Рис. 171. Девятипоясный броненосец — D. novemcinctus и его череп
Род девятипоясных броненосцев
Genus Dasypus Linnaeus, 1758
Размеры небольшие. Длина тела 37—56 см. Длина хвоста 24—43 см. Вес взрослого до 4—8 кг.
Тело широкое и низкое, морда узкая, заостренная, конечности короткие (рис. 171). Уши длинные, стоят близко друг к другу. Число подвижно сочлененных поясов в панцире обычно 9 (может варьировать от 7 до 11). Панцирь тонкий и легкий. Волос почти нет на спинной части и они редки на брюшной. Окраска тела от темно-коричневой до желтовато-белой. Конечности относительно длинные. Передние конечности, как правило, четырехпалые, задние — пятипалые. Второй и третий пальцы на передних конечностях длиннее первого и четвертого, и когти на них достигают в длину 3,5 см. На задних конечностях третий палец наиболее длинный, все остальные имеют примерно одинаковые размеры. В каждой половине челюстей по 7—8 зубов. В диплоидном наборе хромосом у девятипоясного броненосца 60.
Обитатель густых кустарниковых зарослей. В горы поднимаются до 3000 м над уровнем моря. Активны главным образом ночью. Держатся группами, обитая по нескольку особей (обычно звери одного пола) в общей норе. Норы достигают 0,5—3,5 м глубины, слабо ветвятся. Питание разнообразное: в основном различные насекомые и их личинки, другие мелкие беспозвоночные, земноводные, пресмыкающиеся и растительные объекты (сочные плоды, семена, грибы и пр.). Спаривание приходится на июль — август. Имплантация яйцеклеток задерживается до ноября и роды происходят на 120 дней позже. В помете обычно 4 детеныша одного пола. Половозрелости достигают в возрасте одного года. Длительность жизни составляет примерно 4 года.
Ареал наиболее обширен по сравнению с другими броненосцами: от южных штатов США (Канзаса и Миссури) на юг через Мексику в Южную Америку до Гран-Чако в Аргентине. За последние 100 лет ареал рода постепенно расширился на север и восток. В настоящее время северная граница распространения совпадает с линией замерзания водоемов в зимний период.
В роде 6 видов:
короткохвостый броненосец — D. hybridus Desmarest, 1804 (юг Бразилии, Парагвай, Уругвай, Аргентина);
броненосец Капплера — D. kappleri Krauss, 1862 (Венесуэла, Гвиана, Бразилия, Эквадор, Перу, Боливия);
аргентинский броненосец — D. mazzai Yepes, 1933 (северо-восток Аргентины);
девятипоясный броненосец — D. novemcinctus Linnaeus, 1758 (южные штаты США, центральная Америка и Южная Америка к востоку от Анд и от Колумбии и Венесуэлы на севере до северной Аргентины и Уругвая на юге);
семипоясный броненосец — D. septemcinctus Linnaeus, 1758 (восточная Бразилия);
мохнатый броненосец — D. ptlosus Fitzinger, 1856 (Эквадор, Перу и Боливия).
Некоторые ученые насчитывают в роде не 6, а 4 вида.
Род плащеносных броненосцев
Genus Chlamyphorus Harlan, 1825
В роде один вид: плащеносный броненосец— С. truncatus Harlan, 1825 (рис. 172).
Размеры наименьшие среди броненосцев. Длина тела 12—15 см. Длина хвоста — 2,5—3,5 см. Вес примерно 90 г. Наружные ушные раковины представлены небольшими складками кожи. Глаза очень маленькие. Головной панцирь приклеплен на голове лишь к двум большим выступам над глазами. Туловищный панцирь соединен с телом лишь узкой полоской, проходящей вдоль середины спины. По всему остальному пространству панцирь свободно налегает на тело. Все пояса панциря на шее и туловище соединены между собой подвижно. Окраска панциря бледно-розовая. Кожа под панцирем.
Рис. 172. Плащеносный броненосец — С. truncatus
нижние части конечностей и брюшная сторона тела покрыты нежными, мягкими, бело-серыми волосами. Конечности короткие; передние сильнее задних. На передних и задних конечностях по пять пальцев. Лопатообразный хвост выступает на нижнем крае задней пластины и не может подниматься, а волочится по земле. Кости черепа очень тонкие. В каждой половине челюстей по 8 зубов.
Обитатель теплых сухих песчаных равнин, поросших колючими кустами и кактусами. Активность ночная. Малоподвижное, вялое животное, но прекрасный копатель. Питается в основном муравьями, а также червями, улитками, растениями, их корнями.
Распространение ограничено западными областями Аргентины. Хотя численность плащеносного броненосца довольно высока, но она сокращается из года в год. Вид включен в «Красную книгу».
Род щитоносных броненосцев
Genus Burmeisteria Gray, 1865
Единственный вид рода: щитоносный броненосец — В. retusa Burmeister, 1862.
Размеры мелкие. Длина тела 14—18 ши. Длина хвоста 2—3,5 ши. Наружные ушные раковины и глаза очень малы. На передних и задних лапах по 5 пальцев. Головной и спинной панцири прикреплены к голове и телу по всей внутренней поверхности. Задний щит, в отличие от цельного щита плащеносного броненосца, образован небольшими, раздельно расположенными пластинами. Тело снизу покрыто мягкими волосами.
Экология изучена слабо. Щитоносный броненосец не такой хороший копатель, как плащеносный броненосец. Питается насекомыми и их личинками.
Распространен от западной и центральной Боливии на юг до крайнего севера Аргентины.
Численность низкая, внесен в «Красную книгу».
ОТРЯД ЯЩЕРОВ
ORDO PHOLIDOTA WEBER, 1904
В отряде одно семейство: ящеров — Manidae Gray, 1821 и один род ящеров — Manis Linnaeus, 1758 (рис. 173).
Размеры довольно изменчивы. Длина тела 30—88 см. Длина хвоста 35—80 см. Самцы обычно крупнее самок. Вес от 4,5 до 27,2 кг. Отличаются от других млекопитающих черепицеобразно расположенными, подвижными роговыми чешуями, покрывающими тело животного. Форма чешуи разнообразна: треугольная, эллипсоидная, усеченная. Чешуи отсутствуют па боках морды, подбородке, горле, брюхе, внутренних поверхностях конечностей и у некоторых видов — на наружных отделах конечностей. Чешуи имеют эпидермальное происхождение. Они прикрепляются к коже передними отделами и свободно налегают друг на друга задними заостренными частями. В основании чешуй азиатских видов ящеров растут короткие щетинковидные волосы (по 3—4 у каждой чешуи). Окраска чешуй темно-коричневая, темно-оливково-коричневая, бледно-оливковая, желто-коричневая или жел-
Рис. 173. А — ящер (sp.); Б — белобрюхий ящер — М. tricuspis; В — череп яванского ящера — М. javanica
товатая. На местах, лишенных чешуй, растут редкие волосы от беловатого, бледно-коричневого до красновато-коричневого или черноватого цвета.
Пятипалые конечности вооружены крупными когтями, сильнее развитыми на передних лапах. Задние конечности длиннее передних. Ящеры часто передвигаются на задних конечностях, хвост волочится по земле. Когда зверь идет, опираясь на все четыре конечности, то на передних он подворачивает пальцы таким образом, что ступает на наружные поверхности когтей и пальцев. Задние лапы стопоходящие. Хвост в поперечнике выгнут сверху и уплощен снизу. Средний ряд дорзальных чешуй продолжается до самого кончика хвоста у азиатских видов и не доходит до конца хвоста у африканских.
Голова заостренная, с маленькими глазами, защищенными толстыми веками. Ушные раковины отсутствуют (у африканских видов) или вместо них имеются утолщенные валики (у азиатских видов). Ротовое отверстие мало и располагается вентрально. Характерен длинный, червеобразный, очень подвижный язык, способный вытягиваться до 25 см, диаметр его около 5 мм; служит для ловли насекомых. У самок по одной паре сосков, расположенных в грудной области. Некоторые виды имеют анальные железы, продуцирующие секрет с сильным запахом.
Череп удлиненный, конически заостренный кпереди и лишенный гребней. Скуловая дуга не замкнута (отсутствует скуловая кость). Нет слезной и межтеменной костей. Носовые кости крупные, расширенные в основании. Костное небо узкое и длинное. Крыловидные кости разделены и продолжаются назад до уровня костных слуховых барабанов. Нижнечелюстная кость в виде простой уплощенной тонкой пластины. Швы черепа зарастают у самок в более раннем возрасте, чем у самцов. Зубы полностью отсутствуют. Позвонков шейных 7, грудных и поясничных 19—22, крестцовых 3—5, хвостовых 26—49. Задний конец грудной кости вытянут у азиатских видов в лопатообразный, у африканских видов в раздвоенный отросток. У последних отростки грудины тянутся по бокам брюха далеко назад, почти достигая задних конечностей. Мышцы языка прикрепляются к грудине. Ключица и сочленовные отростки на поясничных позвонках отсутствуют.
Мозг макросматический. Желудок простой, с пилорической зоной, выстланной ороговевшим эпителием, несущей функцию механического измельчения пищи взамен отсутствующих зубов. Слепой кишки нет. Сильно развита предчелюстная слюнная железа. Матка двурогая: плацента неотпадающая, диффузная. В конечностях имеется «чудесная сеть» кровеносных сосудов. Мошонки нет, семенники находятся под кожей в паховой области. . Куперовых желез нет.
В диплоидном наборе у китайского ящера 42 хромосомы. Температура тела 32,6—33,6° С.
Обитают в лесах, густых кустарниках и открытых ландшафтах типа саванн. Ведут наземный (гигантский ящер) и древесный (длиннохвостый ящер) образ жизни. Активность ночная. Днем наземные виды прячутся в чужие или вырытые ими самими норы глубиной 3,5—6 м и имеющие в конце гнездовую камеру иногда до 2 м в диаметре. Древесные ящеры на день укрываются в дуплах. Держатся обычно поодиночке, иногда парами. Будучи застигнутыми хищниками вдалеке от убежища, сворачиваются в шар. Древесные виды прекрасно лазают и имеют хватательный хвост. Питаются в основном муравьями и термитами, а также другими насекомыми и их личинками. Добычу находят при помощи хорошо развитого обоняния. Слух и зрение плохие.
В помете один детеныш (редко 2). Новорожденный длиной около 30 см имеет мягкую чешую, отвердевающую лишь на второй день после рождения. Самка приносит детеныша в гнездовой камере норы в ноябре — декабре (белобрюхий ящер) или в январе — марте (китайский ящер). При передвижении матери детеныш удерживается на ее спине или хвосте при помощи своего цепкого хвоста.
Распространены в Африке к югу от пустыни Сахары (от Сенегала и Судана на юг до Южно-Африканской Республики) и на о-ве Фернандо-По и в Юго-восточной Азии — от Индии (на север до подножья Гималаев). Цейлона и юго-восточного Китая, Тайваня и Хайнаня на юг через Малаккский полуостров до о-вов Суматры, Явы, Бали, Калимантана, Палавана и некоторых мелких близлежащих островов.
В роде 7 видов:
гигантский ящер — Л4. gigantea liliger, 1815 (Нигерия);
степной ящер — М. temmincki Smuts, 1832 (Южно-Африканская Республика, Юго-Западная Африка, Ангола, Ботсвана, южная Родезия, Малави, Замбия, Танзания, Уганда, Кения, Судан);
белобрюхий ящер — М. tricuspis Rafinesque, 1820 (тропическая Африка от Анголы и Уганды до Либерии);
длиннохвостый ящер — М. longicaudata Brisson, 1762, (Сенегал, Гвинея, Гамбия);
китайский ящер — М. pentadactyla Linnaeus, 1758 (о-в Тайвань, Южный Китай, Бирма, п-ов Индокитай).
индийский ящер — М. crassicaudata Gray, 1827 (о-в Цейлон, Индия);
яванский ящер — М. javanica Desmarest, 1822 (Бирма, Тенассерим,
п-ов Индокитай, Таиланд, Малаккский полуостров, о-ва Суматра, Ява, Калимантан и многочисленные мелкие к востоку до Филиппинских).
Одно время ящеров объединяли вместе с трубкозубами и американскими неполнозубыми в один отряд. Затем было показано, что ряд общих признаков у этих форм объясняется сходным образом жизни, а не истинным родством. Ящеры, по-видимому, имели самостоятельное эволюционное развитие, хотя возможно, что ящеры и американские неполнозубые обладали общими предками, но, вероятно, еще до того, как неполнозубые отделились от насекомоядных предков.
ЛИТЕРАТУРА
В списке литературы приведены лишь основные, обзорного характера работы, касающиеся таксономии рассматриваемых в т. I отрядов млекопитающих. Как правило, статьи первоописаний новых видов, публикации, посвященные частным вопросам экологии, физиологии и биохимии млекопитающих, в список не включены.
Абеленцев В. И., Пидопличко И. Г., Попов Б. М. 1956. Фауна Украины, т. I, Ссавцц В. I. Изд-во АН УССР, Киев.
Адлерберг Г. П., Виноградов Б. С., Смирнов Н. А., Ф л е-р о в К- К.; 1935. Звери Арктики. Изд. Главсевморпути, Л.
Аргиропуло А. И. 1940. Мыши. Фауна СССР. Млекопитающие, т. III, вып. 5. Изд-во АН СССР, М.—Л.
Б а р а б а ш-Н икифоров И. И. 1957. Звери юго-восточной части черноземного центра. Воронежское книжное изд-во, Воронеж.
Бобринский Н. А., Кузнецов Б. А., Куз я кин А. П. 1944, 1965. Определитель млекопитающих СССР. «Просвещение», М.
Б ю ф ф о н. 1802—1804. Всеобщая и частная естественная история. Перевод И. Лепехина, СПб.
Верещагин Н. К. 1959. Млекопитающие Кавказа. Изд-во АН СССР, М.—Л.
Вин о'г радов Б. С. 1933- Грызуны. Определители по фауне СССР. Зоологический и-т АН СССР. 10. Млекопитающие.. АН СССР, Л.
Виноградов Б. С. 1937. Тушканчики. Фауна СССР, т. III, вып. 4, Изд-во АН СССР, М.—Л.
Виноградов Б. С. и Аргиропуло А. И. 1941. Определитель грызунов. Фауна СССР, сер. 29. Изд-во АН СССР, М.-—Л.
Виноградов Б. С. и Громов И. М. 1952. Грызуны фауны СССР. Изд-во АН СССР, М,— Л.
Виноградов Б. С., Павловский Е. Н., Флеров К- К. 1935. Звери Таджикистана, их жизнь и значение для человека. Изд-во АН СССР, М.—Л.
Воронцов Н. Н. 1958. Значение изучения хромосомных наборов для систематики млекопитающих. Бюлл. МОИП, т. 63, вып. 3.
Воронцов Н. Н., Раджабли С. И., Волобуев В. Т. 1969. Сравнительная кариология летучих мышей семейства Vespertilionidae (Chiroptera). Млекопитающие. Тезисы, Новосибирск.
Гептнер В. Г. 1937. Подсемейство песчанок. В кн.: Виноградов Б. С., Гептнер В. Г., Агриропуло А. И. Грызуны Средней Азии. Изд-во АН СССР, М.—Л.
Гептнер В. Г. 1956. Фауна позвоночных Бадхыза. Изд-во АН Туркм. ССР, у Я^)Я JT
Г е п т я е р В. Г., Ц а л к и н В. И. 1947. Олени СССР. Изд. МОИП. М.
ГептнерВ. Г., Наумов Н. П., ред. 1961—1967. Млекопитающие Советского Союза, т. I, 1961. Парнокопытные и непарнокопытные, т. II (ч. 1) 1967. Морские коровы и хищные. «Высшая школа», М.
Г м е л и н С. Г. 1771—1785. Путешествие по России для исследования трех царств естества, СПб.
Громов И. М., Бибиков Д. И., Кал абу хо в Н. И., Мейер М. Н. 1965- Наземные беличьи (Marmotinae). Фауна СССР. Млекопитающие, ,т. III, вып. 2. «Наука», М.—Л.
Громов И. М., Гуреев А. А., Новиков!’. А., Соколов И.И., Стрелков Н. П., Чапский К- К. 1963. Млекопитающие фауны СССР, ч. 1 и 2. Изд-во АН СССР, М.—Л.
Гуреев А. А. 1964. Зайцеобразные. Фауна СССР. Млекопитающие, т. III, вып. 10. «Наука», М.—Л.
Гюльденштедт И. А. 1787—1791. Reisen durch Russland und im Kauka-sischen Gebirge. Bd 1—2. St. Petersburg.
Д и н н и к Н. Я- 1910—1914. Звери Кавказа, ч. 1, 2. Записи Кавказского отдела императорского русского географического общества. Книжка 27, вып. 1 и 2, Тифлис.
Карелин Г. С. 1883. Путешествия Г. С. Карелина по Каспийскому морю. Тип. Имп. АН, СПб.
Кащенко Н. Ф. 1900. Определитель млекопитающих животных Томского края, Томск.
К а щ е н к о Н. Ф. 1905. Обзор млекопитающих Западной Сибири и Туркестана. Вып. 1 (Chiroptera—Insectivora), Томск.
Кузнецов Б. А. 1948. Фауна млекопитающих Казахстана. Изд-во МОИП, М.
Кузнецов Б. А. 1948. Звери Киргизии. Изд. МОИП, М.
Кузякин А. П. 1950. Летучие мыши. «Советская наука», М.
Лепехин И. И. 1771—1804. Дневные записки путешествия доктора и Академии наук адъюнкта Ивана Лепехина по разным провинциям Российского государства. т. I—IV. Имп. АН, СПб.
МиддендорфА. Ф. 1869. Путешествие на Север и Восток Сибири.
НестурхМ. Ф. 1960. Приматология и антропогенез. Медгиз, М.
Новиков Г. А. 1956. Хищные млекопитающие фауны СССР. Изд-во АН СССР, М—Л.
О г н е в С. И. 1913. Млекопитающие Московской губернии. Изд. Комис, для исследования фауны Московской губернии, состоящей при зоологическом отделении Имп. о-ва любителей естествознания, антропологии и этнографии, М.
О г н е в С. И. 1928—1950. Звери Восточной Европы и Северной Азии. С третьего тома «Звери СССР и прилежащих стран». Т. I—И, 1928—1931. Насекомоядные, летучие мыши. Госиздат, т. VII. 1935. Хищные, ластоногие. Биомедгиз, т. IV—VII, 1940—1950. Грызуны. Изд-во АН СССР.
П а л л а с П. С., 1778. Novae species Quadrupedum е Glirium Ordine Petropoli ed. 1831 Имп. Акад. наук.
П а л л а с П. С. 1811. Zoographia Rosso—asiatica Т. I—III. Petropoli Ex offi-cina caes. academiae scientiarum.
Попов В. A. 1960. Млекопитающие Волжско-Камского края. Изд-во АН СССР, Казань.
Сатунин К. А., 1895. Позвоночные Московской губернии. Вып. 1. Млекопитающие. Известия общества любителей естествознания, антропологии и этнографии, т. 36.
Сатунин К- А. 1915—1920. Млекопитающие Кавказского края, т. I. Рукокрылые, насекомоядные, хищные, 1915; т. II. Ластоногие, грызуны, 1920, Тифлис.
Сержанин И. Н. 1961. Млекопитающие Белоруссии, изд. 2-е. Изд-во АН БССР, Минск.
Симашко Ю. 1851. Описание всех зверей, водящихся в империи Российской. Русская фауна, ч. 2. Млекопитающие, СПб.
Слудский А. А. 1969. Млекопитающие Казахстана. Изд-во «Наука» Каз. ССР, Алма-Ата.
Смирнов Н. А. 1908. Очерк русских ластоногих. Записи Имп. АН, т. 23, № 4. Петербург.
Соколов И. И. 1959. Копытные звери (отряды Perissodactyla и Artyodacty-1а). Фауна СССР, Млекопитающие, т. I, вып. 3. Изд-во АН СССР, М.-—Л.
Строганове. У. 1948. Систематика кротовых Talpidae Труды Зоол. ин-та, т. VIII, вып. 2. Изд-во АН СССР, М.—Л.
Строганов С. У. 1957. Звери Сибири. Насекомоядные. Изд-во АН СССР, М.
Строганове. У. 1962. Звери Сибири. Хищные. Изд-во АН СССР, М.
Татаринов К. А. 1956. Ssipi захщних обласпй Укра'1ни. Киев.
Татаринов Л. П. 1969. Проблемы эволюции териодонтов. Докт. дис. Палеонтологический институт АН СССР. М.
Томилин А. Г. 1957. Китообразные. В кн.: «Звери СССР и прилежащих стран», т. IX. Изд-во АН СССР, М.
Топачевский В. А. 1969. Слепышовые. Фауна СССР. Млекопитающие, т. III, вып. 3. «Наука», Л.
Туркин Н. В., Сатунин К. А. 1902. Звери России. Cervidae, М.
Флеров К. К- 1952. Кабарги и олени. Фауна СССР. Млекопитающие, т. I, вып. 2. Изд-во АН СССР, М.—Л.
Ф л и н т В. Е., Ч у г у н о в IO. Д., С м и р и и В. М. 1965. Млекопитающие СССР. «Мысль», М.
Цалкин В. И. 1951. Горные бараны Европы и Азии. Изд-во МОИП, М.
ЭверсманнЭ. А. 1823. Путешествие из Оренбурга в Бухару. СПб.
ЭверсманнЭ. А. 1840—1866. Естественная история Оренбургского края, т. I—III. СПб.
Anderson S. and Jones К., Ed. 1967. Recent Mammals of the World. The Ronald Press Company, New York.
De Blainville 1816. Prodrome d’une nouvelle distribution systematique
du regne animale. Bull, de la Soc. Philom pour annee, 1816, p. 105. Journ. de Phvs.. t. LXIII, p. 244.
Blumenbach J. F. 1779. Handbuch der Naturgeschichte. Gottingen.
Bourliere F. 1955. In: Grasse P. P. (Ed.). Traite de Zoologie. Mammiferes. Vol. 17 (1 et 2 fasc). Mason et Cie. Paris.
В r i s s о n M. J. 1756. Regnum Animale in Classes IX. Distributum sive Synopsis Methodica. 4° Paris.
Broom R., 1932. The mammal-like reptiles of South Africa and the origin of Mammals. London: Withbery. XVI. p. 376.
Buffon G. L. L. etDaubentonL. J.M. 1753—1767. Histoire de Quad-rupedes. IV—XV. Paris.
Cabrera A. 1925. Genera mammalium. Insectivora, Galeopithecia. Museo National de Ciencias Naturales, Madrid.
Car Isson A. 1909. Die Macroscelididae und ihre Beziehungen zu den iibrigen Insectivoren. Zool. Jahrb., Abr. Syst. Okol. Geogr., Tiere 28, p. 349—400.
Ca.rlsson A. 1922. Ober die Tupaiidae und ihre Beziehungen zu den Insectivoren und den Prosimae. Acta Zool. 3, p. 227—270.
C h a s e n F. N. 1940. A handlist of Malaysian mammals. Bull. Raffles Mus. 15, p. 1—209.
Clark W. E., L. Gross. 1933. Brain of the Insectivora. Prov. Zool. Soc. London 1932, p. 975—1013.
С о r b e t G. B. and H a n к s J. 1968. A revision of the elephant-shrews family Macroscelididae. Bulletin of the British Museum (Natural History) v. 16 No 2, p. 47—112
Crompton A. W. 1958. The cranial morphology of a new genus and species of ictidosaurian. Proc. Zool. Soc. , London 130, p. 183—216.
Cuvier G. 1798. Tableau Elementaire de 1’Histoire Naturelle des Animaux.
Cuvier G. 1800. Lemons d’Anatomie Comparee. Paris.
Cuvier G. 1817. Le Regne Animal. Tome I—IV, 8°. Paris.
D a r 1 i n g t о п P. J., Jr. 1957. Zoogeography. The geographical Distribution of-Animals. John Wiley and Sons, Inc. New York.
Eller man J. R. and Morrison-Scott T. C. 1951. Checklist of Palearctic and Indian Mammals, 1758 to 1946. British Museum of Natural History.
El 1 e r m a n J. R., ,M о r r i s о n- S с о t t T. C. S. and Hayman R. W., 1953. Southern African Mammals, 1758—1951. A Reclassification, British Museum of Natural History.
Flower W. H. 1883. On the arrangement of the orders and families of existing mammalia. Proc. Zool. Soc. Apr. 17, p. 178—186.
Flower W.H. and Lydekker R. 1891. An introduction to the study of mammals living and extinct, 8°. London.
G e s n e r C. 1951. Conrad Gesneri Historiae Animalium Lib. J de Quadrupedibus viviparis. Folio, Tiguri.
Gill T. 1870. On the relations of the orders of mammals. Proc. Amer. Assos. Adv. Sci., 19th Meeting, p. 267—270.
Gill T. 1872. Arrangement of the families of mammals with analytical tables. Smithonian Miscellaneous Collections No 230.
Grasse P. P. (ed). 1955. Mammiferes. vol. 17 (1 et 2 Fasc). Traite de Zoologie, Masson et Cie. Paris.
Gregory W. K. 1910. The orders of mammals. Bull. American Mus. of Nat. History, v XX VII, p. 1—524.
G r z i m e k’s Tierleben. 1967. T. X, XI, XIII. Kindler Verlag. Miinchen.
Haeckel E. 1866. Generalle Morphologie der Organismen. Bd ii, 8°. Berlin.
Hall E. R. and Kelson K. R. 1959. The Mammals of North America. The Ronald Press Co.
Haltenorth T. 1958. Klassification der Saugetiere, 1 (Monotremata. Marsupialia), Handbuch der Zoologie, Band 8, Lieferung 16, S 1—40.
Heimde BalsacH. 1956a. Un Soricidae inedit et aberrant du Kasai exige le creation d’un Genre nouveau. (Mamm. Insectivores). Rev- zool Bot. Africaine 54, p. 137—146.
Heim de BalsacH. 1956b. Diagnoses de Crocidura inedites d’Afrique Occidentale. Mammalia, Paris 20, p. 131—139.
Heim de BalsacH. 1957. Insectivores Soricidae du Mont Cameroun. Zool. Jahrb (Syst) 85, p. 607—618.
Heim de BalsacH. 1958. La reserve naturelle intcgrale du Mont Nimba XIV. Mammiferes insectivores. Mem. Inst. Franc, Air. Noire 53 p. 301—337.
Heim ,de Balsac H., A e 1 1 e n V. 1958. Les Soricidae de basse Cote— de Ivoire. Rev. Suisse Zool, 65, p. 921—956.
I m a i z u m i Y. and О b a r a H. 1966. A natural history of mammals of the world. T. 1—Monotremata, Marsupialia. T. 2 Insectivora, Dermoptera. Tokyo
J о n e s F. W. 1923. The Mammals of South Australia. Part I. The Monotremes and the Carnivorous Marsupials. Government Printer, Adelaida.
Lacepede B. G. E. 1799. Tableau de Divisions, Sous—divisions, Orders et Genres des Mammiferes, 83. Paris.
Laurie E. M. O. and Hill J. E. 1954. List of Land Mammals of New Guinea, Celebes and Adjacent Islands, 1758—1952. British Museum of Natural History, 175 pp.
L i n n a e u s C., 1735. Systema Naturae, sive Regna tria Naturae systematice proposita per Classes, ordines, Genera et Species, Fol. Lugduni Batavorum.
Linnaeus C. 1758—1759 Sistema Naturae per Regna tria Naturae , secundum Classes, Ordines, Genera, Species, cum Characteribus, Differentiis, Synonymis, Locis. Editio Decima. Holmiae.
Matthew W. D. 1915. Climate and Evolution. Ann. New York Acad. Sci 24, 171—318.
M i 1 1 i e r G. and К e 1 1 о g R. 1955. List of North American recent mammals. Bull U. S. Matl. Mus. 205, p. 1—954.
R ay J. 1693. Synopsis Methodica Animalium Quadrupedum et Serpentini Generis, 8°. London.
R i d e W. D. L. 1964. A review of Australian fossil marsupials. J. Royal Soc. Western Australia 47 (4), p. 97—131.
Sanborn С. C. 1937. American bats of the subfamily Emballonurinae. Field Mus. Nat. Hist. Zool. Ser. 20. p. 321—354.
,S с о p о 1 i J. A. 1777. Introductio ad Historium Naturalem sistens Genera La-pidus, Plantarum et Animalium 8°. Prague.
S i m p s о n G. G. 1945. The principles of classification of mammals Bull. Am. Mus. of Nat. Hist., v. 85, p. 1—30.
T a t e G. H. H. 1933. A systematic revision of the marsupial genus Marmosa. Bull. Amer. Mus. N. H., New York, 66.
T a t e G. H. H. 1947. Results of Archbold expeditions. No 56. On the anatomy and classification of the Dasyuridae (Marsupialia). Bull. Am. Mus. Nat. Hist., 88 p. 97—156.
Taylor E. H. 1934. Philippine Land Mammals. Monogr. Bur. Sci. Manila 1—548.
T h e n i u s E. and Hofer H. 1960 Stammesgeschichte der Saugetier Springer —.Verlag. Berlin.
Wai ker E. P. et al. 1964. Mammals of the World. Johns Hopkins Press, Baltimore, 3 vols.
Watson D. M. S. 1953. The evolution of the mammalian ear. Evolution v. 7, p. 159—177.
Weber M. 1927—1928. Die Saugetiere. Gustav Fischer. Jena. Bd. I, II.
Wharton С. H. 1950. Notes on the life history of the flying lemur Cynocephalus volans. J. Mamm. 31, p. 269—278.
Указатель русских названий животных
Ай см. Ленивец трехпалый
Ай-ай 9, 83, 307
Анабис 256
Анамигдон 256, 278
Анатан см. Тупайа индийская
Анубис 338
Ацедорон 178, 189
— гривастый 189
— дневной 189
— макклота 189
---обыкновенный 189
— целебесский 189
Бабуин 290, 338
Бандикут 43, 72
— : Барроу 76
— большой 75, 76
— грубошерстный 73
— длинноногий 42
— длинноносый 72, 73
— длиннохвостый 74
— Керама 73, 75
— колючий 73, 74
— коротконосый 73, 75
— кроличий 73, 77
— — малый 77
— малый 75, 76
— мышевидный 73, 74
— • новогвинейский 73, 74
— папуасский 73, 74
— плоскоиглый 75
— • полосатый 73
— пустынный 73
— свиноногий 72, 73, 76
— тасманийский 73
— толстоголовый 75
Барсук сумчатый, или бандикут, 43, 72
Белка 79
Белозубка 148, 162
— Аагаарда 161
— Аллекса 158
— андаманская 161
— аорская 161
— асабская 159
— балийская 161
- — банкская 161
— Бартельса 161
— белобрюхая 158, 161
— белохвостая 158, 161
— Боливара 158
— большая 158, 161
— Блойета 160
— бурая 159
— Бутлера 158
Белозубка Бюттикофера 158
— Вебера 161
— Виммера 160
— водяная 149
— —- азиатская 148
— — гималайская 166
— — калимантанская 166
— — тибетская, или перепончатолапая, 148, 166, 167
— восточная 161
— выдровая 159
— габонская 158
— гамбийская 159
— гамугофская 158
— гара-мулатская 159
— гигантская 158, 162
— — африканская 162
— гладкая 160
— горная 148, 160
— — малая 165
— — ниерская 165
— Грассе 159
— Грея 161
— Гринвуда 160
— дайангская 161
— дарфурская 159
— Джексона 159
— домовая 162
— Дуцета 160
— дымчатая 159
— египетская 157
— — большая 158
— — малая 157
— Жоффруа 159
— занзибарская 160
— зунгерская 159
— инголдби 160
— итурийская 160
— килиманджарская 159
— калимантанская 161
— камерунская 159
— карликовая 111, 162, 164
— касайская 159
— катабатская 160
— каффская 160
— кенийская 158
— когтистая 148
— — Келаарта 148, 164
— — Пирсона 148, 164
— колючая 157
— коморская 158
— конголезская 148, 162
— короткохвостая 161
— кохская 161
— Клосса 161
хПолужирным шрифтом даются ссылки на рисунок.
Белозубка клюворылая 161
—красносерая 158
— кротовая 148, 166
— лагосская 159
— Ламотта 159
— лаулабская 160
— легкая 160
— либерийская 159
— лунная 159
— львиная 160
— мавританская 159
— Макартура 159
— малая 157, 158, 161
— малайская 161
— мапорская 161
— Мартиенссена 159
— Минданао 160
— Миндоро 161
— многозубая 148, 162, 163, 164
— мохнатая 161
— — мышевидная 164
— нигерийская 158
— ниерийская 160
— нижнеконголезская 158
- Никобарская 161
— нимбская 160
— обыкновенная 158, 161
— огненная 160
— острова Рождества 161
— палаванская 161
— пахучая 160
— пляшущая 159
— прибрежная 159
— пустынная 158
— Райнея 160
— рино-капская 160
— ровнохвостая 161
— рувензорийская 160
— Рузвельта 160
— рыжая 158
— — Сундевалла 159
— Сан-Томе 160
— светлая 160
— северонигерийская 159
— семиенская 158
— сенегальская 160
— серая 158
— Смита 158
— сомалийская 160
— средиземноморская 158
— странная 161
— стройная 159
— суданская 159
— сулавесская 160
— Сула 161
— тиоманская 161
— темная 159
— темнобурая 159
— Терезы 160
— угандская 159
— усамборская 160
— Фишера 159
— Флауера 157
— Хорсфилда 158
— цейлонская 148
— черная 158
— черноватая 160
— черномордая 161
— Швейтцера 160
— шелковистая 160
— Эйзентраута 160
Белозубка Ювената 160
— яванская большая 161
— — малая 161
Белозубка-броненоска 147, 148
— конголезская 168, 168
— угандская 168
Биндем 177, 178
Бобер 5, 13, 92, 93
Бонейя 178, 189
Брахифилла 225, 227
— малый 241
Броненосец 7—9, 23, 70, 355, 362, 365
— аргентинский 369
— большой 366
— гигантский 363, 365, 366
— голохвостый 363, 365, 366
— девятипоясный 363, 368
— длинноволосый 364
— Капплера 368
— карликовый 363, 364 , 365
— короткохвостый 369
—малый 366
— мохнатый 369
—- плащеносный 363, 369, 369
— семипоясный 369
— тэту 366
— трехпоясный 363, 367, 367
— центральноамериканский 366
— чилийский 364
— шаровидный 367
— шестипоясный 362, 363
— щетинистый 363, 363, 364
— щитоносный 363, 370
Бурозубка 148, 149
— азиатская 153,148
— Азуми 150
— альпийская 150
— арктическая 150
— баргузинская 150
— болотная 151
— большезубая 151
— бухарская 150
— Вагранта 151
— Верапаза 151
— водяная 149, 151
— Гаспе 151
— гватемальская 156
— гигантская
— Троубриджа 151
— Гудвина 156
— Гуерреро 156
— дальневосточная 150
— Джексона 151, 156
— длинномордая 151
— длиннохвостая 151
— — европейская 150
— — мексиканская 151
— дымчатая 151
— Заленского 152
— Зетека 156
— индийская 152
— иркутская 150
— кавказская 150
— калифорнийская 151
— карликовая 151
— когтистая 150
— короткохвостая 143, 155, 155
— — азиатская 155, 148
— — американская 148
— — болотная 155
— — обыкновенная 155
— короткоухая 148
Бурозубка короткоухая обыкновенная 155, 156
— — темная 156
— костариканская 156
— Крауфорда 157
— крошечная 150
— крупнозубая 150
— Лиелла 150
— Лове 153
— малая 150
— — кавказская 150
— мексиканская 156
— Мерриама'151
— Нельсона 156
— никарагуанская 156
— обыкновенная 149, 150
— орегонская 156
—• панамская 156 >
— плоскочерепная 150
— полосатоспииная 150
— прибыловская 151
— пустынная 157
— саикристобальская 151, 156
— Санта-Каталинская 151
— североамериканская 150
— сибирская 150
— сиккимская 152
— Склатера 151
— Соссюра 151
— средняя 150, 151
— стройная 156
— сумеречная 156
— украшенная 151
— Филлипса 157
— черноватая 156
— цейлонская 148, 153
— швейцарская 150
— широкомордая 156
— Эндерса 156
— юкатанская 156
•— японская 150
Бурозубка-пигмей 148, 152
Бык 7
Валлэби 107
— болотный 107, 108
— Ирмы 107
— обыкновенный 107, 108
— перчаточный 107, 108
— прыткий 108
— рыжесерый 107, 108
— чернополосый 108
Вампир 202, 228
— ложный 202, 227, ,234
-----австралийский 231,' 214, 215
— — азиатский 213
— — афроазиатский 212, 213
— — африканский 214, 214
— — длинноволосый 227, 234
— — желтокрылый 213, 215
— — индийский 214
— — малайский 214, 214
Вампир белокрылый 249
— мохноногий 249
— обыкновенный 248, 249
Вари 299, 299
Верблюд 5, 7
Вечерница 256
— азорская 266
— гигантская 255
— Жоффруа 266
— малайзийская 266
Вечерница малая 266
— рыжая 266, 266
— японская 266
Виверра 7, 8
Волк 119
— сумчатый, или тасманийский, 57, 68
Волосатохвост
— галапагосский 271
— красный 271
— малый 271
— мохнатый 271
— Пфейффера 271
— Семинола 271
— ямайский 271
Вомбат 43, 92
— длинношерстный 93, 94
— короткошерстный 93
— шерстоносый 93
— — Барнарда 93
Воронкоух 202
— багамский 251
— большой 251
— бразильский 251
— венесуэльский 251
— длинноногий 251
— доминиканский 251
— кубинский 251
— мексиканский 250, 251
— широколапый 251
— ямайский 251
Выдра 5, 133, 140
Выростогуб
— бабочковидный 270
— Гульда 270
— камерунский 270
— конголезский 270
— нигерийский 270
— новозеландский 270
— пятнистобелый 270
— серебристый 270
— смолистый 270
— уэльсский 270
— угандский 270
— южноавстралийский 270
Выхухоль 12, 120, 121—123
— пиренейская 121, 124
Галаго 309
— Аллена 314
— восточным 315
— Демидова 314
— западный 315
— иглокожий 310, 315
— сенегальский 314
— толстохвостый 314
Гамадрил 338, 338
Гверецу 345
Гелада 333, 339
Гепард 5
Гиббон 292, 345, 346, 347, 348
— белорукий 347, 348 , 348
— быстрый 348
— одноцветный 348
— серебристый 348
Гимнура 113, 114, 115
— малая 115, 117
— филиппинская 116
Гиппопотам 6, 7, ?
Гладконос австралийский 254, 256, 280
— — карликовый 279
— — малый 279
— американский 268
Гладконос арнхемский 279
— бледный 279
— большеухий 262
— волосатохвостый 256
— выростогубый 256
— Грея 268
— дисконогий 256, 267
— домовый 256
— — большой 269
— — желтый 269
— — малый 269
— желтый
— — малый 269
— Жоффруа 279, 280
— зеленый 270
— исполинский 269
— кенийский 263
— коричневый 269
— кубинский 268
— Купмана 279
— ламингтонский'256, 279
— мадагаскарский 270
— медноцветный 278
— Мейера 269
— наяритский 279
— нигерийский 268
— новогвинейский 279
— пустынный 256, 268
— Санборна 262
— светлокрылый 262
— севёроквинслендский 279
— серебристый 255, 256, 260
— стройный 269
— сумеречный 256
— темнокрылый 263
— тиморский 279
— украшенный 256, 277
— —• африканский 278
— — ганский 278
— — Гаррисона 278
— — дамарский 278
— — двуцветный 277
— — калимантанский 277
— — камерунский 278
— — карликовый 277
— — малый 278
— — Миллера 277
— — новогвинейский 277
— — папуасский 277
— — самарский 278
— — светлый 277
— — сент-аигнанский 277
— — Смита 278
---Уайтхэда 277
— — Хардвика 277
--- эфиопский 278
— — яванский 277
— Уолкера 279
— черный 270
— целебесский 270
— Шлиффена 268
— эфиопский 270
Гоминиды 290, 291
Горилла 345, 346, 349, 351, 352, 353
Грызуны 7,9—11, 13, 15, 21, 26, НО
Губок 348
Гульман 342
Даманы 10, 21, 110
Депаииктерис 204
— Изабеллы 209
Длиннокрыл 256
Длиннокрыл африканский 275
— большой 275
— восточный 275
— камерунский 275
— мадагаскарский 275
— малый 275
— новогвинейский 275
— Ньютона 275
— обыкновенный 274, 275
— рыжий 275
— серый 275
— Уиткампа 275
— Эшшольтца 275
Длиннонос Годмана 238
— инков 238
— малый 238
— мексиканский 226, 227, 237, 238
— средний 238
Долгопят 9, 290, 291, 292, 315
— западный 316
— филиппинский 315, 316
Долгопят-привидение 315, 316
Дрил 338
Дьявол тасманийский см. черт сумчатый
Дюгони 9
Ёж 5, 20, 112, 113, 114, 117, 132, 135, 136
— алжирский 118
— афганский 119, 120
— белобрюхий 118
— волосатый 113
— даурский 118, 119
— землеройковый 113, 114 117
— индийский 119
— лысый 114
— обыкновенный ИЗ, 118, 119
— темноиглый 119, 120
— ушастый 12, 114. 119, 120
— хайнаньский 116
— центральноафриканский 113, 114
— эфиопский 119
— южноафриканский 114, 118
— крысиный
— — большой 111, 113, 116
— — малый 115
Енот 9
Ехидна 34, 37
— австралийская 32, 36, 37
— тасманийская 37
Зайцеобразные 10, 15, 21, НО
Заяц 76
Звездорыл 121, 131, 132
Зебра 6
Землеройка 47, 77, 112, 121, 133, 138, 140, 148, 156, 165—167
— выдровая 111, 133, 140, 141
— -— карликовая 133
— — рувензорийская 142
— серая 148
Златокрот 112, 113
— африканский 143
— Винтона 143, 144
— готтентотский 144
— грубошерстный 144
— Гуннинга 144
— желтый 144
— исполинский 143, 144
— капский 143, 143
— конголезский 143
—• обыкновенный 142, 143
— пустынный 143, 144
Златокрот светлоносый 144
— Склатера 144
— Томаса 143
— тропический 144
— Фостера 144
— Ш'тульманна 143
Игрунка 328
— белолицая 330
— белоплечая 330
— белоухая 329
— • белошейная 330
— длинноухая 330
— желтоголовая 329
— желтоногая 330
— золотистая 330
— золотистоголовая 330
— золотистозадая 330
— карликовая 330
— львиная 328, 330
— обыкновенная 329
— серебристая 330
— чернокисточковая 329
Индри 291
.— белолобый 305, 306
— короткохвостый 304, 305, 307
— мохнатый 304, 305
— хохлатый 304, 305
Кабарга 7
Кагуан см. Шерстокрыл
Казиниктерис 179, 195
Какайо см. Уакари
Капибара 6
Капуцин 317, 326
— белолобый 323, 324
— бурый 324
— обыкновенный 323
Капуцин-фавн 324
Катта 299
Кенгуру 43, 69, 106
— гологрудый 96, 99
— Гудинафа 104
— Дельта 105
— Дориа 105, 105
— длинноухий 102
— древесный 96, 104
— — западный 105
— исполинский 94—96
— — горный 109
— Квинсленда 101
— кистехвостый 99, 100, 101
— когтехвостый 94, 96
— кольцехвостый 100, 101
— короткокоготный 102, 102
— короткомордый 96, 99, 100
— короткоухий 101
— короткохвостый 96, 106
— крысиный 96, 100
— — большой 96
— — кистехвостый 100
— — мускусный 94, 95
— — широкомордый 99
— кустарниковый 96, 104
— — горный 104
— — полосатый 104
— Лонгмана 101
— Лумхольтца 105
— луннокоготный 102
— Маклая 104
— Матши 105
— медвежий 105
Кенгуру мускусный'96,^97
— очковый 102, 103
— плоскокоготный 102
— полосатый 103
— пучкохвостый 102, 103
— рыжий 109
— серый 108, 109
— скалистый 94, 100, 104
— — карликовый 94,Е96, 103
— — кистехвостый 101, 101
— средний 96, 107, 108
— тасманийский 100
— Уилкинса 101
— Хагена 104
— широкоголовый 97, 98
Кит 5
— зубатый 10
— усатый 10
Китообразные 9, 13, 21, 22, 23, 26, 30, НО
Коала 44 , 79, 80 , 90 , 91, 93
Коата 316, 317, 325
— буроголовая 325
— Жоффруа 325
— светлолобая 325
— черная 325
Кожан 256, 261
— ангольский 262
— арабский 262
— аргентинский 262
— белокрылый 262
— Бланфорда 263 р
— Бобринского 261, 265
— большекрылый 263
— бурый
— — большой 262
— бразильский 262
— восточный 261
— ганский 262
— гарамбский 262
— гвинейский 262
— горный 263
— Грандидье 262
— двуцветный 261, 265
— длиннохвостый 262
— желтоватый 262
— Жоффруа 263
— западноафриканский 262
— зулусский 262
— калимантанский 263
— капский 262
— косолапый 261, 263
— • Линна 262
— Лове 263
— Ловена 262
— ложный 263
— мадагаскарский 262
— малайский 263
— малый 262
— Молони 263
— новогвинейский 263
— • Огнева 262
— панамский 262
— перуанский 262
— поздний 262
— Рендалла 262
— роговокожий 263
— северный 265
— серовато-желтый 261
— синдский 261
— суданский 262
— таиландский 262
Кожан Тиккела 263
— толстоухий 261
— Томеса 263
— Уалла 261
— угандский 262
— Уинтона 263
— фараджский 262
— Флоуэра 263
— Хумблота 262
— чернокрылый 262
— южноавстралийский 263
— южноафриканский 262
Кожан-привидение 262
Козел 7
Копытные 10, 13, 14, 20, 22, 30
Копьенос 227
— Беннетта 232
— бледный 233
— бразильский 236
— Грея 227
— длиннолистый 233
— западный 236
— обыкновенный 233
— панамский 236
— Петерса 227
— северный 233
— Томаса 227, 236
— тринидадский 232
— широколистый 233
— южный 233
Корвира 227, 241
Корова 194
— морская 9, 21
Кошка 7
— домашняя 19
Кролик 22
Крот 70, 71, 111, 112, 113, 120, 121, 126, 127,129—131, 138
— алтайский 126
— волосатохвостый 121, 129, 129
— восточноамериканский 121, 131
— горный 131
— длиннохвостый 128
— западноамериканский 121, 130, 131
— западнокитайский 121, 129
— землеройковый 121, 122
— - американский 121, 128
---Тру 127
— — японский 121, 127, 128, 129
— калифорнийский 130
— мексиканский 131
— могера 126
— обыкновенный 125, 126, 127, 142
— слепой 126, 127 •
— сумчатый 43, 70
— — большой 71
— — малый 71
— тихоокеанский 130
Крылан 177, 201
— австралийский 198
— ангольский 192, 193
— Анхиеты 179, 195
— аравийский 196
— Бланфорда 178, 182
— бурый 178, 181
— Бюттикофера 193
— Валберга 192
— Ветмора 182
— гамбийский 192
— голоспинный 177, 178, 190
— — малый 191
Крылан голоспинный сулавесский 191
— Добсона 193
— длиннохвостый 177, 179, 197, 199, 200
— длинноязыкий 177, 179, 198
— — африканский 179, 198
— — соломонов 199
• — — малый 197
— — малаккский 197
— — цветочный 179
— зеленый 190
— карликовый 178, 191
— коровьемордый 179, 194
— коротконосый 178, 179, 180
— — индийский 180
— — Лукаса 180
— — малайский 180
— — филиппинский 180
— — Хорсфилда 179, 180
— — цепколапый 180
— либерийский 194
— лузонский 197
— мадагаскарский 196
— молотоголовый 177, 179, 194, 195
— молуккский 190
— Неуманна 192
— ночной 178, 191
— ошейниковый 178, 184
— — малый 184
— — либерийский 184
— — Вроугтона 184
— — Сан-Томе 185
— папуасский 198
— пальмовый 178, 196
Перрона 190
— пещерный 177, 179, 197
— — большой 197
— плодоядный 197
— Пусаргуеза 192
— пятнокрылый 178, 182
— Розенберга 197
— соломенноцветный 179
— суданский 192
— сулавесский 177, 178, 188
— толстоухий 178, 191
— Темминка 178, 182
— темнобрюхий 179, 199
— — флоридский 199
— темный 178, 180
— тробрианский 191, 198
— трубконосый 177, 179, 200, 201
— — белобрюхий 200
— — большеголовый 200
— — большой 200, 201
— — карликовый 179, 201
— — малый 177, 200
— — новогвинейский 201
— — Томаса 201
• — — триопский 201
— — Робинсона 201
— Уоллэса 178, 190
— Фишера 178, 191
— Франкуета 193
— хальмахерский 190
— хищный 190
— Ценкера 194
— черношапочный 178, 181
— эполетовый 177, 178, 192—196
— — гамбийский 192
— — карликовый 177, 178, 193
• — — малый 192
— Ягора 178, 182
Крылан-гарпия 179, 201
Крыса 29, 62, 77, 79, 96, 178
— • сумчатая 57, 59
— —• большая 60
— — буробрюхая 61
— —• гребнехвостая 57, 61
— — двугребнехвостая 62
— — длиннохвостая 61
— — западная 6 1
— — Лорентца 60
— — малая 60, 60
— — новогвинейская 57, 60, 61
— — оранжевобрюхая 61
---полосатая 61
Ксифоииктерис 282, 284
Кузу лисий 80, 81
— собачий 80
Куокка см. кенгуру короткохвостый
Куница 7, 9, 79
— сумчатая 57, 65
— — карликовая 65, 66
— — крапчатая 65, 66
---полосатая 57, 66
— — пятнистохвостая 65, 66
---чернохвостая 65, 66
Кускус 43, 79, 80
— беличий 80, 85
— блестящий 90
— большой 83
— Виктории 89
— восточный 89
— Герберта 89
— длинношерстный 90
— карликовый 79, 80, 84
— кистехвостый 79, 80
— • кольцехвостый 80, 88
— — тасманийский 89
— —• карликовый 90
— красный 89
— лемуровидный 90
— летучий 91
— — гигантский 79, 80, 91
—--карликовый 79, 80, 87
— малый 83
— медвежий 83
— медноволосый 90
— наземный 89
— одноцветный 82
— пестрохвостый 83
— перьехвостый 80, 86
— полосатый 80, 83, 90
— пушистый 82, 83
— пятнистый 82, 83
— — Тейта 83
— скалистый 90
— собачий 90
— соневидный 80, 84
— тасманийский 84
— толстохвостый 85
— тонкохвостый 85
— Форбеса 89
— хвостатый 84
— хоботноголовый 42, 79, 80, 88
— целебесский 79, 83
— чешуехвостый 82
— • шелковистый 82
— Шлегера 90
Кутора 147, 148
— малая 153, 154
— обыкновенная 153, 154
Ламантин 7, 9
Ланцетонос 202
— большой 203, 203
— малый 203
— суматранский 203
Лар 348
Ласка 53
Ластоногие 10, 21, 22, 31,г 110
Ленивец 7—9, 174, 354, 355
'— Гоффмана 362
—• двупалый, или унау, 360 , 361, 362
— капюшонный 361
— ошейниковый 361
— трехпалый, или Ай, 360, 361
Лемур 79, 290, 291, 292, 299, 303, 315
— бурый 299
— вильчатополосый 302, 303
— волосатоухий 30 1
— карликовый 297
— кольцехвостый 290
— краснохвостый 300
— ласковидный 300
— мышиный 301, 302
— — Кокуерела 302
—• рыжебрюхий 299
— серый 297, 298
— толстохвостый 301
— черный 296, 299
— широконосый 298
Леопард 353
Летяга
— сумчатая 79, 80, 86, 91
— — гигантская 86
— — карликовая 86
— — средняя 86
Лиониктерис 227, 235
Лисица 5, 24, 26, 27, 79, 119, 124
— летучая 177, 186, 188, 189, 200
— — адмиралтейская 178
— — альдабрская^ 188
------- Ару 187
— — бавеанская 187
— — балутская 187
— — беловатая 187
— — большая 187
— — восточноавстралийская 178, 188
------- Вуудфорда 187 j
— — Гулда 185, 188
— — Жиллиарда 188
— — золотистая 1§7/
— — индийская 187
— — карликовая 187
— —• коморская 188
— —• Ливингстона 187
— — ломбокская 187
— — лузонская 187
— — маврикийская 187
— — малая 186
— — марианская 186
— — махагская 187
— — Мирпса 187
— — моротайская 187
— — Никобарская 187
— — новогвинейская 187
— — Остина 188
- — пембская 188
— — Райнера 187
— — родригесская 188
— — рыжая 188
— — Рюкю 186
— — сейшельская 188
— — седая 187
Лисица летучая серая 187
— — серебристая 187
— — сибутская 186
— — соломонова 187
— — сулавесская 187
— — табласская 188
— - — таиландская 187
— — Темминка 187
— — тонганская 186, 187
— — церамская 187
— — черноватая 188
— — черноспинная 187
— — япенская 187
Листонос абасский 221
— американский 202, 228,1231
— — рыжий 228
— банановый 227, 237
— барбадосский 236
— бахромчатогубый 233
— бахромчатый 227
— белый 227, 243
— бесхвостый 220, 224
— большеглазый 227
—. — малый 243
— большеухий 227, 230
— — волосатый 230
— — малый 226, 230
— большой 222
— бразильский 230
— бурый 230, 239
— Ватерхауза 230
— венесуэльский 243
— волосатый 224
— восточнобразильский 243
— гаитинский 241
— ганский 22 1
— гваделупский 244
— гвианский 244
— гигантский 221
— гималайский 222
— Годмана 227
— голоспинный 226, 228
— далекий 222
— двуцветный 222
— длиннолапый 227, 231
— длинномордый 235
— длинноносый 227, 237
— длиннохвостый 222
— длинноязыкий 227, 235
— Дориа 222
— доминиканский 240
— древесный 245
— — доминиканский7244
— — сект-люсийскиш 244
— • Дэви 229
— желтоплечий 227
— — большой 240
— Жоффруа 237
— золотистый 220, 221, 224
— индийский 222
— ипанемский 228, 245
— калимантанский 222
— калифорнийский 230
— камерунский 221
— карликовый 227, 240
— — обыкновенный 221
— Каррикера 231, 232
— колумбийский 243
— Коммерсона 221
— короткохвостый 227
— красный 222
— круглоухий 227, 231
Листонос круглоухий бразильский 231
— — малый 232
— кубинский 226, 236
— куракаосский 235
— Людовика 240
— лузонский 222
— Макконелла 243
— Маклеа 228
— малайзийский 222
— малый 222
— мексиканский 227, 228
— монтсерратский 244
— мохнатый 244
— мохноногий 240
— Миллера 239
— минданаоский 222
— Найсфора 230
— никарагуанский 232
— новогвинейский 222
— обыкновенный 220, 221, 223
— островной 236
— панамский 237
— папуасский 222
— пеленгский 222
— пепельный 222
— подбородколистый 226, 229
— Пратта 222
— пуэрториканский 236
— Райта 222
— Ридле я 222
— Робинсона 224
— Салвина 243
— Сандерсона 222
— Семона 222
— сент-винсентский 241
— сент-люсийский 236
— серпокрылый 244, 245
— — кубинский 244
— серый 244
— складчатомордый 225, 226, 228, 245, 246
— Соломоновых островов 222
— Соссюра 227, 239
— Старого Света 202, 216
— сулавесский 222
— темный 221
— Тиморский 222
— Торрейа 228
— трехцветный 240
— тринидадский 240
— увенчанный 222
— Ундервуда 227, 238
— Уолластона 222
— усатый 226, 228
— фигоядный 244, 245
— фруктоядный 227
— -— большой 244
— — карликовый 244
— — красный 228, 245
— -— ямайский 244
— хохлатый 222
— черноватый 228
— цветочный 225, 246
— — бурый 247
— — гаитинский 246
— — кубинский 246
— — пуэрториканский 246
— — светлый 228
— темножелтый 247
— • — ямайский 246
— цветкомордый 222, 223
— цейлонский 222
Листонос центральноамериканский 243
— широкомордый 237
— Шмидта 230
— шпороносный 222
— эль-гуамский 241
— эфиопский 221
— южноафриканский 221
— южноамериканский 244
— ямайский 236
Листонос-строитель 227, 241
Лихониктерис 227
— бурый 239
Лори 290', 291, 309
— малый 312
— толстый 309, 310, 311, 312
— тонкий 310
Лошадь 6, 7, 9
Люди 9, 333, 353
Магот 335
Маки 297
— вильчатополосый 297
— крысиный 290, 297, 301, 302
— мышиный 290, 297, 300
— тонкотелый 297
Макак 333
— вандеру 334, 335
— горный 335
— индийский 335
— медвежий 335
— свинохвостый 335
— тайванский 335
— цейлонский 335
— Целебесский 335
— яванский 335
— японский 334, 335
Макак-резус 334, 335
Мангобей 333
— бородатый 336, 337
— воротничковый 336, 337
— гривистый 336, 337
— чубастый 336, 337
Мангуста 66
Мандрил 337, 338
Мармозетка 327, 328, 329
Мартышка 327, 328, 329
— белоносая
— — большая 340
— — малая 340
— бородатая 340
— Бразза 340
— гол у бол и ца я 340
— зеленая 340
— - коронованная 340
— краснобрюхая 340
— крошечная 341
— мона 340
— совинолицая 340
— чернозеленая 341
— чубатая 340
Мартышка-гусар 341
Мартышка-диана 340
Медведь 7, 9, 20, 90
— бурый 5
— сумчатый 90
Меченос 226, 227, 231
Мешкокрыл 202, 204
— австралийский 208 •
— аденский 205
— африканский 204
— Беккара 205
— белокрылый 207
Мешкокрыл белый 204, 209 — бразильский 207 —• бурый 207 — венесуэльский 207 — Гамильтона 208 — Гильдегарда 208 — голобедрый 208 — голобрюхий 208 — двулинейный 206, 207 — длинноногий 208 — желтобрюхий 208 — Жоффруа 208 — Капплера 207 — колумбийский 209 — короткомордый 207 — мадагаскарский 205 — малый 207 — Максимилиана 207 — мексиканский 209 — могильный 204 — — новогвинейский 208 — новогвинейский 205 — обыкновенный 204 — Рафрайана 205 — сейшельский 205 — седой 207 — сенегальский 208 — собаковидный 207 — Старого Света 204, 205 — суданский 208 — Теобальда 208 — Темминка 208 — Троугтона 208 — филиппинский 208 — футлярохвостый 205 — хоботковый 204, 205, 206 — чернобородый 208 — черноватый 205 — черномордый 207 — юго-восточный 205 — южноафриканский 208 — южный 205 — Мирикина 317, 318 Многобугорчатые 18, 19, 20, 21 Многозубка — большая 163 — габонская 164 — Гранта 164 — Дейа 163 -— Джонстона 164 — домовая 163 — занзибарская 164 — камерунская 164 — карликовая 163 — кенийская 164 — лазающая 163 — лесная 163 — лусонская 163 — мадагаскарская 164 — малайская 163 — нигерийская 164 — обыкновенная 164 — палавапская 163 — руандская 164 — скрытная 163 — темполапая 163 — угандская 163 •— — Хозе 163 — Шаллера 163 Могера 126, 127 моиго 299 Морж 8, 9
Муравьед 7, 9, 354, 355
— гигантский, или трехпалый, 355, 356, 357
— карликовый, или двупалый, 355, 357, 359
Муравьед сумчатый 42, 43, 44, 69, 69
Мышехвост 202, 203
— большой 203
— индийский 203
— малый 203
— суматранский 203
Мышь 62, 74, ,79, 147, 178, 233
— домовая 18
— летучая 15, 24, 31, 209, 212, 213, 215, 218, 233, 244-, 255, 272
— — бульдоговая 202, 294
— — — амазонская 289
— — — бархатистая 282
— — — бразильская 288
— — — большая 288
— — — Бонда 288
— — — гайанская 289
— — — голокожая 282
— — Гринхалла 283
— — — Дависона 288
— — — кубинская 288
— — — мексиканская 283
__ — — Миллера 283
— — — носатая 288
— — — парагвайская 288
— — — перуанская 283
— — — плоскорылая 283
— — — рыжая 288
— — — Сан-Киттса 288
— — — Темминка 283
— — — тринидадская 288
— — — Ундервуда 289
— — — усатая 288
— — — центральноамериканская 288
— — — эквадорская 283
— — — южноамериканская 282
— — — ямайская 288
— — голокожая 284
— — дымчатая 202, 250
— — — восточная 252
— — — обыкновенная 252, 260
— — новозеландская 280
— — обыкновенная 202
— — ожерельевая 284
— — рыбоядная 202, 206
— — — мексиканская 210, 211
— - — южная 211
Мышь сумчатая, 57
— — Ару 64
— — Баркли 59
— — бархатистая 58
— — белоногая 63
— Беркли 63
— — Вильгельмины 57
— — гребнехвостая 57, 61
— — двугребнехвостая 57, 62
— — длиннохвостая 63
- - - Дэрби 63
— — желтая 58
— — западная 61
— — западноавстралийская 59
— — зернистоподошвенная 63
— — карликовая 58
— — Квинсленда 69
— — крапчатая 59
— — Лаусоиа 64
Мышь сумчатая малая 58
— — новогвинейская 59
— — песчаная 63
— — плоскоголовая 57, 59
— — полосатоголовая 63
— — пятнистоглазая 58
— — Свайнзона 58
— — северная 59 '
— — толстохвостая 59
— — Томаса 63
— — узколапая 57
— — — малая 63
— - — толстохвостая 63
— — — длиннохвостая 64
— — шелковистая 59
— — южная 59
Насекомоядные 9, 10, 14, 18. 20, 21, 110
Непарнокопытные 10, 21
Непарнопалые 110
Неполнозубые 21, 26, НО, 372
Носач 333, 343, 344
Носорог 7,9, 20, 21, 23
Ночница 255
— аризонская 258
— африканская 258
— Бартелса 258
— бахромчатая 258
— большая 255, 257, 259
— большеногая 258
— Брауна 258
— бурая
— — малая 258
— Велвитша 257
— водяная 257, 259
— Германа 258
— гималайская 257
— Гудота 258
— Давида 258
— длинноногая 259
— длиннопалая 257
— длинноухая 257, 259
— длиннохвостая 257, 259
— железистокрылая 259
— — ангольская 258
— — Лесуеура 258
— индианская 259
— • калимантанская 258
— калифорнийская 259
— Кина 258
— красная 259
— крупнопалая 258
— курносая 259
— малайзийская 258
— малая 257, 259
— малоногая 259
— Миллера 258
— минданаоская 258
— монтерейская 258
— Наттерера 257, 259
— остроухая 257, 259
— парагвайская 259
— пекинская 258
— пещерная 258
— плоскоголовая 259
— прудовая 258, 259
— Риккета 258, 259
— • рыбоядная 259
— рыжая 257
— самарская 258
— светлобурая 258
Ночница серая 258
— серебристая 259
— сиккимская 257
— Скотта 258
— стройная 257
— сычуанская 257
— танзанийская 258
— трехцветная 257, 259
— усатая 257, 259
— филиппинская 258
— Хасселта 258
— Херре 258
— Ходгсона 256
— Хорсфилда 258
— черная 259
— Чилоэ 259
— Шталькера 258
— юго-восточная 258
— южноафриканская 257
— Юма 258
— японская 257
Нетопырь 256
• — абасский 264
— ангольский 264
— - банановый 264
— большой 264
— восточноамериканский 264
— восточный 263, 264
— Дормера 264
— египетский 264
— западный 264
— индийский 263
— индонезийский 264
— камерунский 264
— карлик 263
— — индийский 263
— — камерунский 264
— кенийский 264
— кожановидный 261, 263, 264
— Коромандельский 263
— лесной 263, 265
— маврский 264
— мадейрский 264
— малый 264
— марасский 264
— • Минахасса 264
— Мюррея 264
— островной 264
— Пагенштехера 264
— папуасский 264
— Петерса 264
— пустынный 263
— Ридли 264
— Руп пела 264
• — рыжий 264
— серый 263
— солнечный 264
— средиземноморский 263, 265
— средний 264
— суматранский 264
— тасманийский 264
— тенассеримский 263
— толстопалый 264
— толстый 264
— тонкий 264
— цейлонский 263
— черный 264
— шоколадный 263
— Штампфла 264
— Эндо 263
— Юма 258
— яванский 264
Нетопырь японский 264
Нетопырь-красавец 264
Обезьяна 7, 9, 31, 290
— желтохвостая 327
— игрунковая 291
— ночная 318
— паукообразная 317, 325, 326
— толстотелая 332, 333
— тонкотелая 333, 342
— узконосая 291
— хохлатая 333
— цепкохвостая 291, 318, 320, 321
— шерстистая 317, 326, 327
Обезьяна-капуцин 291
Овца 7, 69
Однопроходные 9—11, 17-19, 20, 26
32, 40
Олень 7, 19
Ондатра 133, 139
Опоссум 7, 8, 18, 42, 43
— Аллена 51
— Алстона 50, 51
— амазонский 51
— андский 51
— Ацерамарка 51
— бархатистый 52
— белобрюхий 51
— Блазе 52
— боготский 51
— большеухий 52
— буробрюхий 5
— венесуэльский 51
— водяной 55, 56
— восточный 51
— высокогорный 52
— Германа 51
— голохвостый 46, 47, 48
— — перуанский 48
— грациозный 52
— густошерстный 47
— — полосатый 47
— Дамара 52
— длиннохвостый 47, 53
— длинношерстный 51
— домовый 48
— желтый 47
• — Кана 51
— карликовый 48
— колумбийский 48
— кольцеглазый 51
— короткохвостый 46, 48
— — полосатый 49
— красноголовый 46, 48
— красный 51
— лимский 50
• — малопалый 51
— Мапири 51
— Марайо 48
— Марика 51
— Мату-Гросу 52
— мексиканский 51
— многососковый 51
— Морейра 52
— Муйена 52
— мышевидный 46
— — белобрюхий 51
— — карликовый 50, 51
— —• короткохвостый 52
— — обыкновенный 52
— — перуанский 51
— — толстохвостый 52
Опоссум мышевидный ночной 51
— однополосый 48
— обыкновенный 46, 54
— пампасский 49
— Пары 51
— патагонский 46, 49
— пепельный 50
— пушистохвостый 46, 49
— Раппоза 51
— роатанский 51
— Рондона 52
— рыжий 47
— сан-луисский 52
— светлобрюхий 51
— северный 55
— серый 49
— Симмонса 51
— Столле 52
— стройный 51
— Тейта 52
— толстохвостый 47, 53, 54
— Томаса 51
— трехполосый 48
— трехцветный 48
— тринидадский 5 1
— Ундуави 5 1
— Формозы 52
— четырех глазый 46, 52,^53
— чилийский 52
— Чилоэ 46, 49
— щетиноплечий 51
— эквадорский 49
— Эрхардта 52
— южный 55
— Юнга 51
— Юнина 52
Опоссум-красавец 52
Орангутан 346, 349, 350
Осел 6
Павиан 333, 337
— гвинейский 338
— хохлатый 333, 335, 336
Парнокопытные 6, 9, 10, 21
Парнопалые 110
Платалина 227, 237
Плацентарные 10, 19, 21, 27, 40, 45
Плоскоголовы Нового Света 282, 289
Подковолист Блайнвилла 229
Подковонос 202
— абасский 218
— австралийский 219
— азиатский 2 17
— ангольский 218
— Андерсена 219
— • Андрейна 2 18
— аравийский 2 17
— бирманский 218
— блестящий 2 18
— большеухий 2 16, 219
— большой 215, 217
— борнейский 218
— Брокмана 218
— Бункера 219
— бурусский 2 19
— бухарский 2 17
— восточный 2 19
— — большой 217
— ганский 218
— Гильдербрандта 2 18
— дамарский 218
— Дарлинга 218
Подковонос Декена 218
— Дента 218
— длинноухий 218
— - Добсона 2 18
— Жоффруа 2 17
— индийский 217
— калимантанский 219
— карликовый 218
— каштановый 217
— кенийский 219
— Клосса 218
— конголезский 218
— королевский 218
— Крега 219
— кустарниковый_218
— Ландера 218
— ложный
— ломбокский 219
— лисий 218
— лузонский 219
— мадурский 218
— Маклауда 218
— малайзийский 218
— малайский 217
— малый 217
— - Мегели 217
— минданаоский 219
— мохнатый 219
— однорогий 217
— Пирсона 217
— Робинсона 218
— Ру 217
— рыжеватый 219
,— Свинни 218
’— сиантанский 218
— слабый 219
— средиземноморский 217
— Томаса 217
— трехлистный 217
— угандский 218
— филиппинский 218
— целебесский 219
— южноафриканский 218
— южный 217
— яванский 218
— японский 217
Полумаки 297
Полуобезьяны 9
Потору 98
— Гильберта 98
— трехпалый 98, 98, 98
Потто 310
— золотой 310, 312
— обыкновенный 312, 313
Приматы 7, 18, 20, 21, 22, 30, 110, 112, 169, 303, 327
Присосконог американский 250, 252
• — гондурасский 253
— мадагаскарский 253, 254
— трехцветный 253
Проехидна 37
— Бартона 37, 38
— Бруйпа 32, 38
— Б у бу 38
Прыгун 317, 318
— бурый 318
— воротничковый 318
— красный 319
— пепельный 318
• — серый 318
— черноголовый 318
— чериолобый 319
Прыгун-молох 318
Прыгунчик 10, 111— 113, 169, 171
— голохвостый 172
— длинноухий 169, 171
— катангский 171
— коротконосый 172
— короткоухий 169, 171
— кустарниковый 172
— лесной 169, 170, 171
— Матти 171
— пестроспинный 169
— Ревоала 172
— рыжий 172
- — североафриканский 172
— скалистый 172
— темный 172
— черный 171
— четырехпалый 170
— Эдварда 172
Птералопекс 178
— черный 189
Путорак 148
— пегий 165
Ревун 316, 317
— бурый 323
— гватемальский 322
— колумбийский 322
— краснорукий 323
— рыжий 322
— ' черный 323
Ринопитек 333
— биэтовский 343
— бреличевский 343
— рокселланов 343
— Тонкинский 343
Росомаха 9
Рукокрылые 8—10, 14, 21, 22, 30, 110, 176, 177
Руконожка, или ай-ай, 290, 291, 307, 308
Саймири 317, 324
— беличий 324, 325
— рыжеспинный 324
Саки 317
— белоносый 321
— бледный 321
— красноспинный 317, 321
— черный 321 (
Саки-монах 320, 321
Свинья 7, 9, 25
Сиаманг 346, 347
— карликовый 348
— обыкновенный 347, 348
Симиас 333
— одноцветный 343
Симметродонты 18, 19
Сирены 10, 22, 31, 95, НО
Складчатогуб 282
— Аба 286
— авакубийский 286
— австралийский 287
— алдарский 286
— ангольский 286
— Ансорга 285
— африканский 285
— Беккара 287
— белобрюхий 287
— белогрудый 286
— Беммелена 285
— Бокажа 285
Складчатогуб большой 286
— бразильский 287
— Вебстера 286
— венесуэльский 287
— Вроугтона 287
• — гамбийский 286
— длинногребешковый 286
— Дориа 287
— египетский 285
— Икара 287
— индийский 285
— йохорский 286
— Калиновского 287
— камерунский 286
— карликовый 286
— кенийский 285
— колючкоухий 287
— конголезский 286
— красный 286
— кубинский 287
— куборский 287
— леонский 286
— лузонский 286
— мадагаскарский 285
— Макмиллана 287
— малый 286
— Мартиенссена 287
— Мастерсона 285
— мешотчатый 287
— миары 286
— Натальский 287
— ниагарский 286
— нигерийский 286
— Норфолкский 287
— Осборна 286
— охристый 285
— папуасский 287
— пигмей 286
— плоскоголовый 287
— пятнистобелый 286
— Руппелла 286
— Сибра 285
— Спилмана 286
— стройный 287
— суданский 285
— сулавесский 286
— Тонга 285
— Тревора 286
— украшенный 286
— Фараджский 286
— Хаагнера 287
— черный 286
— читауский 286
— широкоухий 285, 287
— широкохвостый 287
— щетинистый 287
— юкатанский 287
— яванский 287
Складчатоморд малый 228, 246
— — бразильский 246
Склерониктерис 227, 238
Слепыш обыкновенный 13
Слон 5, 7—10, 18, 20, 25, 30, 95
Собака 7, 68, 119, 173, 178
— летучая 177
— — ангольская 177, 178, 184
— — аравийская 184
— — египетская 184
— кенийская 184
— — Ленолта 184
— — мадагаскарская 184
— — Смита 184
Собака летучая угандская 184
---Фернандо-По 184
--- цейлонская 184
— — Целебесская 184
— — цепкохвостая 183, 184
---Штреземанна 184
Собачка хоботковая 170
--- занзибарская 170
— — золотистая 170
— — Клауда 170
— — Петерса 170
— — пятнистая 170
— — Свиннертона 170
— — Штульманна 170
Соболь 13
Соня 59, 60
Стрелоух 256
— белобрюхий 267
Сумчатые 9—11, 18, 20, 30, 40, 41, 44
Сурок 19
Суслик 13
— крапчатый 13
Сферониктерис 225, 245
Тамандуа 355, 357, 358
Тамарин 328
— белоногий 331, 332
— белый 332
— буроголовый 332
— Девилла 332
— Жоффруа 331, 332
— золотистый 332
— императорский 331, 332
— краснобрюхий 332
— красногривый 332
— краснорукий 331, 332
— Леннберга 332
— Мартина 331
— обыкновенный 331, 332
— пегий 331, 332
— Рио Напо 332
— рыжий 331, 332
— усатый 332
— Уэделла 332
— черноспинный 331, 332
— Эдипов 331, 332
Тапир 9, 20, 25
— бразильский 6
Тенрек 11, 112, 113, 134, 136, 140
— большой, или ежовый, 133, 136, 137
— водяной
— — карликовый 141
— — рувензорийский 141
— Декари 139
— Диего-Суареза 139
— длинномордый 139
— длиннохвостый 132, 138, 139
---малый 139
— Добсона 139
— Дроухарда 138
— западный 139
— землеройковый 139
— земляной 133, 140
— Кована 138
— короткохвостый 138
— кротовидный 138
— крошечный 139
— Майора 139
— малый 132, 133, 137
— Мидонги 139
— Мильн-Эдвардса 138
— обыкновенный 132, 133, 134
Тенрек перепончатолапый, или болотный, 133, 139, 140
— полосатый 133, 135, 136
—• рисовый 133
— стройный 139
— Талазака 139
— Танала 139
— Томаса 139
— Фонтуанонта 133, 136
— черноголовый 136
— четырехпалый 138
Толстотел
— зеленый 345
— королевский 345
— красный 344, 345
Томопеас 256, 280
Тонкотел белобородый 342
— гололобый 342
— гривистый 341, 342
— Джона 342
— каштановый 342
— ментавайский 342
— немейский 341, 342
— очковый 342
— рыжий 342
— Тонкинский 342
— Фейера 342
— хохлатый 342
— чубастый 342
Трезубценос 221
— африканский 221, 224, 225
— обыкновенный 223
— моллукский 223
— эфиопский 223
— южноазиатский 223
— Тейта 221
Трилистонос 221
— африканский 225
— красный 225
— персидский 220, 225
— мадагаскарский 225
— Хумблота 225
Трубкозуб 9
Трубкозубые 21, ПО, 372
Трубконос 256
— большой 276
— малайзийский 276
— малый 276
— седой 276
— серый 276
— сиккимский 276
— флоресский 276
— Хуттона 276
— шерстокрылый 276
— яванский 276
— японский 276
Тупайа 10, 112, 169, 290, 291, 293
— большая 294
— горная 294
— гладкохвостая, или горная, 293
— индийская, или анатан, 293, 294
— кариматская 294
— краснохвостая 294
— малая 294
— Мюллера 294
— Никобарская 294
— обыкновенная 293, 294
— палаванская 294
— перьехвостая 293, 295, 296
— северная 295
— стройная 294
— украшенная 294
Тупайа филиппинская 293, 295
— южная 295
— яванская 294
Тушканчик 168
• — сумчатый 57, 64
• — — восточноавстралийский 64
— — центральноавстралийский 64
Уакари, или какайо, 317
— красный 319, 320
— лысый 319, 320
-— черноголовый 319, 320
Унау см. ленивец двупалый
Утконос 32, 33, 35, 38, 39, 39
Ушан 278
— аризонский 272, 273
— обыкновенный 272, 273
— пятнистый 256, 273
— Рафинеска 273
— Таунсенда 272, 273
Филандер 96, 106
— белогрудый 106
— Евгении 106
— краснобрюхий 106
— красноногий 106
• — красношеий 106
— новогвинейский 106
Хищные 10, 13, 14, 21, 30, 31, 110
Хоботные 10, 18, 21, 31, ПО
Хорь 124
Цоелопс 221
Ценолест, или крысовидный опоссум, 43, 77
— перуанский 78
— чилийский 78, 79
— эквадорский 78
Циттаропс 204, 209
Чакма 338
Человек 7, 23, 25, 352, 353
Черт сумчатый 57, 67
Шерстокрыл ПО, 173, 174
— малайский 174
•— филиппинский 174
Шимпанзе 346, 349, 351
Шиншилла 18
Широконос большой 242
— беловолосый 227, 242
— бразильский 242
— венесуэльский 242
— желтоухий
Широконос большой 242
— — гвианский 242
— — Добсона 242
— малый 242
— — перуанский 242
— — эквадорский 242
— Караккиоло 242
— колумбийский 242
— линейчатый 242
— украшенный 242
— Хеллера 242
— эквадорский 242
Широкоушка 256
— азиатская 272
— европейская 272, 272
Щелезуб 145
— гаитянский 146, 147
— кубинский 147
Щелеморд 202, 2 11
— авакубийский 212
— бледный 212
— большеухий 212
— большой 212
• — Бейтеса 212
— Винсона 212
— Вууда 212
— гамбийский 212
— египетский 211, 212
— желтый 212
— золотистый 212
— конголезский 212
• — мадагаскарский 212
— малый 2 12
— мохнатый 212
— сомалийский 212
— танзанийский 212
— эфиопский 212
— яванский 211, 212
Эомопс 282
— белый 283
— сенегальский 283
— Уайтлея 283
Ящер 8, 9, 369, 370, 371, 372
— белобрюхий 371
— гигантский 372
— длиннохвостый 372
— древесный 133, 372
— индийский 372
— китайский 372
— степной 371
— яванский 370, 371, 372
Указатель латинских названий животных
aagaardi см. Crocidura
abae см. Hipposideros, Rhinolophus, Та-darida
abaensis см. Vespertilio abbicauda см. Crocidura abramus cm. Vespertilio abrasus cm. Molossus abyssinicus cm. Coleura acciticus cm. Sorex Acerodon 178, 188 — celebensis 189 — humilis 189 — iubatus 189 — lucifer 189 — mack loti 189 acermarcae cm. Marmosa acetabulosa cm. Tadarida Acrobates 80, 87 — pygmaeus 87 — pulchellus 88 acrotis cm. Rhinolophus aculeatus cm. Tachyglossus acuminatus cm. Rhinolophus adersi cm. Rhynchocyon admiralitatum cm. Pteropus adversus cm. Myotis adusts cm. Monodelphis aegyptiaca cm. Tadarida aegyptiacus cm. Rousettus aello cm. Nyctimene Aepyprymnus 96, 100 — rufescens 100 aequalis cm. Aethalops aequatorialis cm. Glironia aequatorianus cm. Molossops aequicauda cm. Crocidura Aerhechinus 118 aero cm. Vespertilio aerosa cm. Kerivoula Aethalops 178, 190 — aequalis 190 — alecto 190 Aethechinus 114 aethiopica cm. Nycteris aethiopicus cm. Hemiechinus aethiops cm. Cercopithecus africana cm. Kerivoula afer cm. Triaenops affinis cm. Rhinolophus, Vespertilio afra cm. Coleura africana cm. Tadarida
africanus cm. Miniopterus, Myotis
agilis cm. Hylobates, Marmosa, Protem-nodon
agnella cm. Kerivoula agricolai cm. Marmosa alba cm. Ectophylla
albatus cm. Eomops
albertisi cm. Pseudocheirus albescens cm. Myotis albicollis cm. Callithrix albifrons cm. Cebus albigena cm. Cercocebus albigularis cm. Vespertilio albinasus cm. Chiropotes albiventer cm. Nyctimene, Tadarida albiventris cm. Erinaceus albofuscus cm. Nycticeius alboguttatus cm. Chalinolobus, Diclidu-rus
alcyone cm. Rhinolophus aldabrensis cm. Pteropus alecto cm. Aethalops, Cyttarops, Embal-lonura, Pteropus
alleni cm. Galago Allenopythecus 341 alienus cm. Vespertilio Alionycteris 178, 191 — paucidentata 191 algirus cm. Erinaceus al lex cm. Crocidura Allotheria 21 Alouatta 317, 321 — belzebul 323 — caraya 323 — palliata 322, 322 — quariba 323 — seniculus 322 — villosa 322 alpinus cm. Sorex alspanus cm. Sorex alstoni cm. Marmosa altaica cm. Talpa altarium cm. Myotis alticola cm. Cryptotis Amblysomus 143, 144 — fosteri 144 — gunningi 144 — hottentotus 144 — leucorhinus 144 — obtusirostris 144 — sclateri 144 — tropicalis 144 americana cm. Monodelphis Ametrida 228, 246 — centurio 246 — minor 246 Amorphochilus 252 — schnablii 252 Amphibies 9 amplexicaudatus cm. Rousettus anamalus cm. Neomys Anamigdon 256, 278 — solomonis 278
Anathana 293 , 294
— ellioti 294
anatinus cm. Ornithorhynchus
anchietai cm. Pierotes, Vespertilio andamensis cm. Crocidura anderseni см. Rhinolophus andersoni см. Rhynchonax andreinii cm. Rhinolophus
angeli cm. Sturnira
angolensis cm. Epomophorus, Rhinoli-phus, Rousettus
angulatus cm. Vespertilio
annectens cm. Stenoderma, Vespertilio — anomalis cm. Neonys
Anoura 227, 236
— brevirostrum 237
— caudifera 237
— cultrata 237
— geoffroyi 237
Anourosorex 148, 166
— squamipes 166
ansorgii cm. Tadarida
Antechinomys 57, 64
— laniger 64
— spenceri 64
Antechinus 57, 58
— apicalis 59
— bellus 58
— flavipes 58
— macdonnelensis 59
— maculatus 58
— melanurus 58
— mimulus 59
— minimus 58
— naso 59
— rosamondae 59
— swainsoni 58
— Wilhelmina 59
anthonyi cm. Scapanus
Anthops 223
antricola cm. Hipposideros
Anthropoidea 291, 316
Antrozous 256, 279
— dubiaguercus 279
— koopmani 279
— pallidus 279
anubis cm. Papio
anurus cm. Epomophorus aoris cm. Crocidura
Aotes 317, 318
— trivirgatus 318
apella cm. Cebus
aphylla cm. Phyllonycteris
apicalis cm. Antechinus aquaticus cm. Scalopus arabicus cm. Rouretius arachnoides см. Brachyteles araneus см. Sorex archeri см. Pseudocheirus archipelagus см. Cynopterus arcticus см. Sorex Arctocebus 310, 312 — calabarensis 312 arcuatus cm. Rhinolophus Ardops 245 arethusa cm. Crocidura arge cm. Nycteris
argentata cm. Callithrix, Chalinolobus, Kerivoula
argentatus cm. Myotis, Pteropus argynnis cm. Casi nycteris ridttla cm. Crocidura
ariel cm. Vespertilio Ariteus 245 armiger cm. Hipposideros arnhemensis cm. Nyctophilus arquatus cm. Pteropus Artibeus 227, 243 — cinereus 244 — concolor 244 — harti 244 — hirsutus 244 — jamaicensis 244 — lituratus 244 — nanus 244 — turpis 244 artinii см. Nycticeus Artiodactyla 21, 110 assamensis cm. Macaca Asellia 221, 223 — patrizii 223 — tri dens 223 Aselliscus 221, 223 •— stoliczkana 223 — tricuspidatus 223 asiatica cm. Chrysochloris asper см. Sorex Astrapotheria 20 Atelerix 114, 118 Ateles 317, 325 — belzebuth 325 — fusciceps 325 — geoffroyi 325 ‘
— paniscus 325 attenuate см. Crocidura atterrimus см. Cercocebus atrata см. Dorcopsis, Emballonura, Pte-ralopex
atrimaculatus cm. Phalanger aurata cm. Murina
aurantius cm. Melonycteris, Rhinonicte-ris
aurispinosa cm. Tadarida
aurita cm. Callithrix, Geogale, Lonchor-china, Myzopoda, Nycteris
auritus см. Chrotopterus, Hemiechinus, Plecotus
austini cm. Pteropus
australis cm. Dromiciops, Miniopte
Petaurus, Syconycteris, Tadaridarus austroriparius см. Myotis avakubia cm. Nycteris avunculus cm. Rhinopithecus axillaris cm. Rhinolophus ayguta cm. Pygathrix azoreum cm. Nyctalus
babu cm. Vespertilio badius cm. Coleura baileyi cm. Crocidura Balantiopteryx 207 Balionycteris 178, 182 — maculata 182 balstoni см. Murina baluensis cm. Crocidura balutus cm. Pteropus bancanus cm. Tarsius Barbastella 256, 271 — barbastellus 272 — leucomelas 272 barbastellus cm. Barbastella barnardi cm. Lasiorchinus barnesi cm. Molossus barrowensis cm. Thylacis
bartelsii см. Crocidura, Myotis batesi cm. Crocidura bartoni cm. Zaglossus beatrix cm. Chalinolobus beatus cm. Crocidura, Hipposideros bechsteini cm. Myotis beccarii cm. Crocidura, Emballonura, Tadarida
bedfordi cm. Protemnodon behni cm. Micronycteris Belluae 7 bellus cm. Antechinus belzebul см. Alouatta belzebuth cm. Ateles bemmeleni cm. Tadarida bendirii cm. Sorex bennettii cm. Mimon beta cm. Crocidura Bestiae -43 Bettongia 96, 99 — cuniculus 100 — gaimardi 100 — lesueur 99, 100
bicolor cm. Hipposideros, Kerivoula, Protemnodon, Saguinus, Vespertilio bidens см. Boneia, Corvira, Tonatia,
Vampyrops bieti cm. Rhinopithecus bifax см. Nyctophilus bilabiatum см. Pygoderma bilineata- cm. Saccopteryx billardieri cm. Protemnodon billardirre cm., Thylogale bilobatum cm. Uroderma Bimanes 9 bivittatus cm. Tadarida blainvillii cm. Mormoops blanfordi cm. Sphaerias, Vespertilio Blarina 148, 154 — brevicauda 155 — telmalestes 155 blarina см. Suncus Blarinella 148, 155 — quadraticauda 155 blasei см. Marmosa blasii см. Rhinolophus bloyetti см. Crocidura blythi cm. Dasycercus, Myotis bobrinskii cm. Vespertilio bocagei cm. Myotis, Tadarida bocharicus см. Rhinolophus bolivari см. Crocidura boliviensis см. Saimiri bombifrons см. Erophylla, Kerivoula bonariensis см. Molossus bondae cm. Molossus Boneia 178, 189 — bidens 189 borealis cm. Lasiurus borulensis cm. Rhinolophus bottegi cm. Crocidura bougainvillei cm. Perameles bovei cm. Crocidura boy di см. Crocidura brachycephala см. Myonycteris brachyotis см. Cynopterus, Lasiurus,
Micronycteris brachymeles см. Molossops Brachyphylla 227, 241 — cavernarum 241 — minor 241 — nana 241
Brachyphylla pumila 241 brachyptera см. Tadarida brachypterus cm. Vespertilio brachyrhynchus cm. Elephantulus Brachyteles 317, 325 — arachnoides 325, 326 brachyurus см. Setonyx Bradypodidae 356, 360 Bradypus 360 — cuculliger 361 — torquatus 361 — tridactylus 361 brasiliensis Tadarida, Tonatia, Vespertilio brelichi cm. Rhinopithecus brevicauda см. Blarina, Crocidura brevicaudata см. Microgale, Monodel-phis breviceps см. Petaurus brevimanus см. Natalus brevirostris cm. Saccopteryx brevirostrum см. Anoura brocki см. Vampyrops brockmani см. Rhinolophus breweri см. Parascalops brooksi см. Dyacopterus browni cm. Myotis bruchi cm. Marmosa bruijni cm. Zaglossus brunii cm. Thylogale brunnea cm. Crocidura, Tadarida brunneus cm. Callicebus, Vespertilio Bruta 7 bubuensis cm. Zaglossus buchariensis cm. Sorex bunkeri cm. Rhinolophus Burmeisteria 363, 370 — retusa 370 butteri cm. Crocidura bflttikoferi cm. Crocidura, Epomops byrnei cm. Dasyuroides Cabassous 363, 365 — centralis 366 — hispidus 366 — loricatus 366 — tatouay 366 — unicinctus 366 >
Cacajao 317, 319 — calvus 320 — melanocephalus 320 — rubicundus 320 — roosevelti 320 caeca см. Talpa caecutiens см. Sorex Caenolestes 78 — fuliginosus 78 Caenolestidae 43, 44, 77 cafer см. Suncus caffer см. Hipposideros calabarensis cm. Arctocebus calcaratus cm. Hipposideros Callicebus 317, 318 — brunneus 318 — cinerascens 318 — cupreus 318, 319 — melanochir 318 — moloch 318 — nigrifrons 319 — personatus 318 — torquatus 318 californicus см. Macrotis, Myotis caliginea cm. Crocidura
Callimico 328 — goeldii 328, 329 Callimiconidae 291 Callithricidae 291, 327 Callithrix 328 — albicollis 330 — argentata 330 — aurita 329 — chrysoleucos 330 — flaviceps 329 —• jacchus 329 — leucocephala 330 — penicillata 329 — pygmaea 330 — santaremensis 330 Caloprymnus 96, 99 — campestris 99 calura cm. Phascogale Caluromys 46, 47 — irrupta 47 — laniger 47 — philander 47 calvus см. Cacajao Callynicteris 197 campestris cm. Caloprymnus canescens cm. Marmosa, Murina, Pseudo-cheirus, Saccopteryx
caniceps см. Pteropus caninus cm. Trichosurus canguru cm. Protemnodon capaccinii cm. Myotis capensis cm. Rhinolophus, Vespertilio capucinus cm. Cebus caraccioloi см. Vampyrops caraya см. Alouatta carri см. Marmosa carrikeri cm. Tonatia carimatae cm. Tupaia caroli cm. Pseudocheirus Carnassiers 9 Carnivora 20, 21, 110 Carnivores 9 Carollia 227, 239 — castanea 240 — perspicillata 240 — tricolor 240
cartilagonodus cm. Otopterus Casinycteris 179, 195 — argynnis 195 castanea см. Carollia catta cm. Lemur caucae cm. Marmosa caucasica см. Talpa caucasicus см. Sorex caudata cm. Crocidura, Eudromicio caudatus cm. Soriculus caudifera cm. Anoura caurinus cm. Notoryctes cavernarum cm. Brachyphylla Cebidae 291, 316 Cebus 317, 323 — albifrons 323, 324 — apella 324 — capucinus 323 — nigrivittatus 324 cealatus см. Cryptotis
celebensis cm. Acerodon, Harpyionycte-ris, Phalanger, Rhinolophus, Scotophilus
centralis см. Cabassous, Molossus Centronycteris 207 Centurio 228, 245
Centurio senex 226, 245 centurio cm. Ametrida cephalotes cm. Nyctimene cephus cm. Cercopithecus Cercaertus 80, 84 — concinnus 85 — nanus 85
Cercocebus 333 , 336
— albigena 337
— aterrimus 337
— galeritus 337
-----galeritus 337
— torquatus 337
Cercopithecidae 291, 332
Cercopithecus 333, 339 — aethiops 340 — cephus 340 — diana 340 — erythrogaster 340 — hamlyni 340 — mona 340 — mitis 340 — neglectus 340 — nictitans 340 — nigroviridis 341 — 1’hoesti 340 — patas 341 — petaurista 340 — pogonias 340 — talapoin 341 cervinus cm. Hipposideros
Cetacea 21, 110
Cetaces 9
Cete 5, 7 ceylonicus cm. Vespertilio Chaeropus 73, 76 — ecaudatus 76
Chaetophractus 363 — nations 364 — vellerosus 364 — villosus 363, 364 Chalinolobus 256 , 270 — alboguttatus 270 — argentata 270 — beatrix 270 — dwyeri 270 — egeria 270 — gouldi 270 — humeralis 270 — morio 270 — picatus 270 — poensis 270 — tuberculatus 270 — variegata 270 chapini cm. Tadarida Cheirogaleus 297 , 300 — major 301 — medius 301 — trichotis 301
Cheiromeles 281, 283
— parvidens 284
— torquatus 284 chiloensis cm. Myotis Chilonycteris 226, 228 — fuliginosa 228 — macleayii 228 — personatus 228 — rubiginosa 228 — torrei 228
Chimarrogale 148, 166
— phaeura 166
— platycephala 166
chiriquinus см. Vespertilio Chiroderma 227, 243 — doriae 243 — isthmicum 243 — jesupi 243 — salvini 243
— trinitatum 243 %
— villosum 243
Chironax 178, 181
— melanocephalus 181
Chironectes 47, 55
— minimus 55, 56 Chiropotes 317, 321 — albinasus 321 — chiropotes 321 — satanas 321 chiropotes cm. Chiropotes Chiroptera 21, 110 Chiropteres 9 chitauensis cm. Tadarida Chlamyphorus 363, 369 — truncatus 369 Chodsigoa 153
< Choeroniscus 227, 238 — godmani 238
• — inca 238
, — intermedius 238
— minor 238
Choeronycteris 227, 237 — mexicana 226, 227 Choloepodidae 360 Choloepus 360, 361 — didactylus 361, 362 — hoffmanni 362 Chrotopterus 227, 234 — auritus 234 Chrysochlorida 112 Chrysochloridae 112, 113, 142 Chrysochloris 143 — asiatica 143 — congicus 143 — stuhlmanni 143 — vermiculus 143 chrysoleucos см. Callithrix chrysomelas cm. Leonti deus chrysoproctus cm. Pteropus chrysopygus cm. Leonti deus. Rhyncho-cyon
Chrysospalax 143, 144
— trevelyani 144 — villosus 144 Cinderella cm. Crocidura cineraceus cm. Hipposideros cinerascens cm. Callicebus cinerea см. Marmosa cinereus cm. Artibeus, Lasiurus, Phasco-larctor, Sorex
cinnamomeus cm. Nycticeius circumdatus см. Vespertilio cirnei cm. Rhynchocyon cirrhosa см. Trachops Cistugo 259 clara cm. Echymipera claudi см. Rhynchocyon clinedophus cm. Monophyllus clivosus cm. Rhinolophus Cloeotis 221, 224 — percivali 224 clossi см. Rhinolophus coelophyllus cm. Rhinolophus Coelops 221, 223 — frithi 220, 224
Coelops hirsutus 224 — robinsoni 224 cognatus cm. Pteropus Coleura 204, 205 — afra 205 — gallarum 205 — seychellensis 205 Colobidae 291, Colobus 333, 344 — abyssinicus 345 — badius 345 — polykomos 345 — verus 345 commersoni cm. Hipposideros Comopithecus 338 comorensis cm. Pteropus concinna cm. Perardorcas concinnus cm. Cercadrtus concolor cm. Artibeus, Hylobates, Si-mi as
Condylarthra 20, 21
Condylura 121, 131 — cristata 131, 132 condylura cm. Tadarida congicus cm. Chrysochloris, Scutisorex, Tadarida
conspicillatus cm. Lagorchestes, Pteropus
congobelgica cm. Crocidura constaninae cm. Marmosa convoluter cm. Pseudocheirus coquereli cm. Mycrocebus cor cm. Megaderma corinnae cm. Pseudocheirus Cormura 207 •-
cornutus см. Rhinolophus coromandra cm. Vespertilio Corvira 227, 241 — bi dens 241
coucang cm. Nyctoicebus cowani cm. Microgale coronatus cm. Hipposideros coxi cm. Hipposideros crassa cm. Syconycteris crassicaudata см. Lutreolina, Manis, Sminthopsis
crassicaudatus cm. Galago crassipes cm. Microgale crawfordii cm. Notisorex creaghi cm. Rhinolophus crenata cm. Crocidura, Dobsonia crenulatum cm. Mimon Creodonta 20
cristata cm. Condylura cristatus cm. Pygathrix cristicauda cm. Dasycercus croassulus cm. Vespertilio Crocidura 148, 157 — aagaardi 161 — aequicauda 161 — albicauda 158 — allex 158 — andamensis 161 — aoris 161 — arethusa 158 — aridula 158 — attenuate 158 — baileyi 158 — baluensis 161 — bartelsii 161 — batesi 158 — beatus 160
Crocidura beccarii 161
— beta 158
— bioyeti 160
— bolivari 158
— bottegi 158
— bovei 158
— boydi 158
— brevicauda 161
— brunnea 161
— butleri 159
— bilttikoferi 158
— caliginea 159
— caudata 158
— Cinderella 159
— congobelgica 159
— crossei 159
— crenata 159
— cyanea 158
— darfurea 159
— dolichura 159
— doriae 161
— douceti 160
— dracula 158
— edwardsiana 161
— eisentrauti 160
— elongata 160
— ferruginea 159
— fischeri 159
— flavescens 158, 159
— flowed 157
— foetida 161
— foxi 159
— fuliginosa 161
— fulvastra 159
— fumosa 159
— fuscosa 159
— geoffroyi 159
— giffardi 160
— glebula 159
— goliath 160
— glassi 159
— gracilipes 159
— grandis 161
— gravida 161
— grayi 160
— greenwood! 160
— grassei 159
— halconus 160
— hildegardeae 159
— hindei 159
— hirta 158
— hispida 157
— hedenborgiana 159
— horsfieldi 158
— ingoldbyi 160
— jacksoni 159
— jouvenetae 160
— klossi 161
— lamottei 159
— langi 159
— latona 159
— lasiura 158
— lea 160
— lepidura 161
— leucodon 158
— levicula 160
— littoralis 159
— ludia 159
— luluae 159
— luna 159
— lusitania 159
— lutrella 159
Crocidura macarthuri 159
— malayana 161
— manni 159
— maporensis 161
— marita 159
— martiensseni 159
— maurisca 159
— maxi 161
— melanorhyncha 161
— mindorus 161
— minuta 161
— mi ya 157
— monax 159
— monticola 160
— muricauda 159
— negligens 161
— nicobarica 161
— nigripes 160
— nigrofusca 159
— nimbae 160
— niobe 159
— odorata 160
— olivieri 158
— orientalis 161
— oritis 160
— palawanensis 161
— paradoxura 161
— parvipes 160
— parvacauda 160
— pasha 160
— percivali 160
— pergrisea 158
— pilosa 158
— poensis 160
— polia 160
— raineyi 160
— religiosa 157
— rhoditis 160
— roosevelti 160
— russula 158
— sansibarica 160
— schweitzeri 160
— sericea 160
— smithi 158
— somalica 160
— suaveolens 157, 158
— sururae 160
— tenius 160
— thomensis 160
—- theresae 160
— tionis 161
— trichura 161
— ultima 160
— velutina 160
— viaria 160
— villosa 161
— vosmaeri 161
— vulcani 160
— weberi 161
— wimmeri 160
— xantippe 160
— zaphiri 160
— zimmeri 160
crossei см. Crocidura
crumeniferus cm. Hipposidercs
Cryptoch loris 143, 145
— wintoni 145
Cryptotis 148, 155
— alticola 156
— cealatus 156
— endersi 156
— eurirhynchis 156
Cryptotis fossor 156
— frontalis 156
— goodwini 156
— griseoventris 156
— guerrerensis 156
— jacksoni 156
— magna 156
— mayensis 156
— mera 156
— micrura 156
— nelsoni 156
— nigrescens 156
— obscura 156
— olivacea 156
— orphila 156
— parva 156
— pergacilis 156
— soricina 156
— tersus 156
— zeteki 156
crypturus cm. Epomophorus cyanea cm. Crocidura
Cyclopes 357, 359
— didactylus 359 cyclopis см. Macaca cyclops cm. Hipposideros cyclotis cm. Murina cylindricauda см. Sorex Cynocephalidae 173 Cynocephalus 173 — temminckii 174 — volans 174 cynocephalus см. Papio, Thylacinus Cynomacaca 335 Cynopithecus 333, 335 — niger 335, 336
Cynopterus 178, 179, 181, 184
— archipelagus 180
— brachyotis 180
— harpax 180
— horsfieldi 180
— sphinx 180
Cyttarops 204, 209
— alecto 209
cubanus cm. Monophyllus, Nycticeus, Solenodon
cuculliger см. Bradypus culex см. Vespertilio cultrata см. Anoura cuniculus см. Bettongia cupidus см. Hipposideros cupreus cm. Callicebus, Pseudocheirus cuprosa см. Kerivoula
Dactylopsila 80, 83
— palpator 83
— trivirgata 83
daedalus см. Nyctophilus dahli cm. Pseudocheirus daphaenodon cm. Sorex darfurea cm. Crocidura Dasogale 133, 136 — fontoynonti 136 Dasycercus 57, 61 — blythi 61 — cristicauda 61 dasycneme cm. Myotis dasymallus cm. Pteropus Dasypodidae 356, 362 Dasypus 363, 368 — hybridus 369 — kappleri 369
Dasypus mazzai 369 — novemcinctus 368, 369 — pilosus 369 — septemcinctus 369 Dasyuridae 43, 45, 56 Dasyurinus 66 Dasyuroidea 45 Dasyuroides 57, 62 — byrnei 62 Dasyurops 66 Dasyurus 57, 65 — geoffroyi 66 — hallucatus 66 — maculatus 65, 66 — guoll 65, 66 daubentoni cm. Myotis Daubentonia 307 — madagascariensis 307, 308 Daubentoniidae 291, 307 dauricus cm. Erinaceus davidi см. Myotis davisoni см. Molossus davyi см. Pteronotus dayi см. Suncus debilis см. Molossus decaryi cm. Microgale deckenii cm. Rhinolophus degener cm. Lichonycteris deltae cm. Dendrolagus demararae cm. Marmosa demidovi cm. Galago demissus cm. Vespertilio demonstrator cm. Tadarida Dendrogale 293, 295 — melanura 295 — murina 295 Dendrolagus 96, 104 — deltae 105 — dorianps 105 — goodfellowi 105 — lumholtzi 105 — matschiei 105 —- spadix 105 — ursinus 105 denti cm. Rhinolophus denticulus cm. Eudiscopus Depanycteris 204, 209 — Isabella 209 Dermoptera 21, 110, 173 Desmana 121, 122 — moschata 122 Desmanidae 113 Desmodontidae 202, 247 Desmodus, 249 — rotundus 248, 249 desserti cm. Vespertilio devillei cm. Saguinus diadema cm. Hipposideros, Propithecus Diaemus 249 — youngi 249 diana cm. Cercopithecus Diclidurus 204, 208 — albus 209 — ingenc 209 — virgo 209 didactylus см. Choelopus, Cyclopes Didelhes 9 Didelphidae 43, 44, 45, 46 Didelphodon 45 Didelphoidea 44, 45 Didelphis 47, 54 — marsupialis 55
Didelphis paraquayensis 55 Digitigrades 9 dimidiata cm. Minuania diminutus cm. Vespertilio dinops cm. Hipposideros Dinocerata 20 Diphylla 249 — ecaudata 249 Diplarthra 10 Diplomesodon 148, 165 — pulchellum 165 discolor cm. Phyllostomus disci fera cm. Thyroptera dispar cm. Sorex Distoechurus 86 — pennatus 86 Dobsonia 178, 190 crenulata 190 — exoleta 191 — inermis 190 — minor 191 — moluccensis 190 — peroni 190 — praedatrix 190 ’— remota 191 — viridis 190 dobsoni cm. Epomops, Microgale, Rhinolophus
Docodonta 19
dolichura cm. Crocidura domestica cm. Monodelphis domina. cm. Marmosa dominicensis cm. Natalus Dorcopsis 96, 104 — atrara 104 — hageni 104 — macleayi 104 — vanheurni 104 — veterum 104
doriae cm. Chiroderma, Crocidura, Hipposideros, Phascolosorex, Tadarida, Vespertilio
dorianus cm. Dendrolagus dormeri cm. Vespertilio dorsalis cm. Phascolosorex, Protemno-don, Tupaia, Vampyrops
douceti см. Crocidura dracula cm. Crocidura Dromiciops 46, 49 — australis 49 drouhardi cm. Microgale dryas cm. Marmosa Dryopithecinae 292 dubiaguercus cm. Antrozous dupreanum cm. Eidolon dwyeri cm. Chalinolobus Dyacopterus 178, 180, 181 — brooksi 181 — spadiceus 181 dyacorum cm. Hipposideros Dymecodon 121
ecaudatus cm. Chaeropus, Megaerops, Tenrec
Echinops 133, 137
— telfairi 137
Echinosorex 114, 115
— gymnurus 114, 115
Echymipera 73, 74
— clara 74
— kalubu 74
Echymipera rufescens 74 Ectophylla 227, 243 — alba 243 — macconelli 243 Edentata 20, 21, 110, 355 E dentes 9 edwardi cm. Elephantulus edwardsiana cm. Crocidura ega cm. Scleronycteris egeria cm. Chalinolobus Eidolon 179, 196 — dupreanum 196 — helvum 196 — sabaeum 196 eisentrauti cm. Crocidure elegantulus cm. Euoticus elegans cm. Nectogale Elephantulus 169, 171 — brachyrhynchus 172 — fuscipes 172 — edwardi 172 — intufi 172 — fuscipes 172 — myarus 172 — revoili 172 — rozeti 172 — rufescens 172 — rupestris 172 ellioti cm. Anathana elongate cm. Crocidura, Glossophaga elongatus cm. Phyllostomus eloquensis cm. Rhinolophus emarginatus cm. Nycticeus emarginatus cm. Myotis Emballonura 204, 205 — alecto 205 — atrata 205 — beccarii 205 — furax 205 —• monticola 205 — nigrescens 205 — raffrayana 205 Emballonuridae 202, 203 emi Пае см. Marmosa Enchisthenes 244 endersi cm. Cryptotus entellus cm. Pygathrix Eomops 282 — albatus 283 — senegalensis 283 — whitleyi 283 Eonycteris 179, 196, 197 — langicauda 197 — major 197 — robusta 197 — rosenbergi 197 — spelaea 197 Eoscalops 126 — longirostris 126 — micrura 126 Epomophorus 178, 192, 193 — angolensis 192 — anurus 192 — crypturus 192 — gambianus 192 — neumanni 192 — labiatus 192 — pousarguesi 192 — wahlbergi 192 Epomops 178, 192, 193 — biittikoferi 193 — dobsoni 193
Epomops franqueti 193
Eptesicus 265
eremiana cm. Perameles Eremitalpa 143, 145 — granti 145 erhardti cm. Marmosa Erinaceidae 112, 113 Erinaceomorpha 112 Erinaceus 114, 117 — albiventris 118 — algirus 118 — dauricus 118 — europaeus 118 — frontalis 118 eriophora cm. Kerivoula Erophylla 228, 247 — bombifrons 247 — sezekorni 247 Erythrocebus 341 erythrogaster cm. Cercopithecus eschscholtzii cm. Miniopterus etruscus cm. Suncus Euderma 256, 273
— maculatumI273 Eudiscopus 256, 267 — denti cuius 267 Eudromicia 80, 83 — caudata 84 — lepida 84 — macrura 84
Euinsectivora 112 eugenii cm. Thylogale Eumops 289 . Euoticus 310, 314 — elegantulus 315 — inustus 315 Euphractus 363 , 364 — sexci nctus 364 eurirhynchis cm. Cryptotis europaea cm. Talpa europaeus cm. Erinaceus europs cm. Tadarida Euroscaptor 126 — grandis 126 — klossi 126 — parvidens 126 euryale cm. Rhinolophus euryotis cm. Rhinolophus Eutheria 9, 10 , 21, 40, 109, 356 everetti cm. Urogale evotis cm. Myotis exoleta cm. Dobsonia falabae cm. Nycticeius falcatus cm. Stenoderma faradjius cm. Tadarida, Vespertilio fasciata cm. Perameles
fasciatus cm. Lagostrophus, Myrmeco-bius
femorosacca cm. Tadarida
Ferae 10
Feroculus 148, 164
— feroculus 164
feroculus cm. Feroculus ferruginea cm. Crocidura ferrumequinum cm. Rhinolophus fidelis cm. Vespertilio
fischeri cm. Crocidura, Haplonycteris flavescens cm. Crocidura, Stenoderma, Vespertilio
flavicauda cm. Lagothrix flaviceps cm. Callithrix flavipes cm. Antechinus
florium cm. Murina
floweri cm. Crocidura, Vespertilio fodiens cm. Neomys foetida cm. Crocidura forbesi cm. Pseudocheirus formosa cm. Marmosa
formosus cm. Myotis, Tadarida forsteri cm. Amblysomus fortidens cm. Myotis fortis cm. Molossus fontoynonti cm. Dasogale fossor cm. Cryptotis forsteri cm. Minuania foxi cm. Crocidura, Rhinolophus fraenata cm. Onychogalea franqueti cm. Epomops francoisi cm. Pygathrix
frater cm. Monophyllus, Myotis, Tadarida
frithi cm. Coelops
froggatti cm. Sminthopsis frons cm. Lavi a
frontalis cm. Cryptotis, Erinaceus frontatus cm. Pygathrix frosti cm. Neopterys fructivorous cm. Macroglossus fuliginosa cm. Crocidura
fulliginosus cm. Coenolestes/ Hipposideros
fulliginosa cm. Chilonycteris fulminans cm. Tadarida fulvastra cm. Crocidura fulvus cm. Lemur fumosa cm. Crocidura fumeus cm. Sorex fumigatus cm. Rhinolophus furax cm. Emballonura furcifer cm. Phaner furcula cm. Triaenops furinalis cm. Vespertilio Furipteridae 202, 251 Furipterus 252 — horrens 252 furvus cm. Nyctalus fuscata cm. Macaca, Marmosa fusciceps cm. Ateles fuscicollis cm. Saguinus fuscipes cm. Elephantulus fuscosa cm. Crocidura fuscus cm. Vespertilio fusicaudus cm. Scaptonyx
gaimardi cm. Bettongia Galago 310, 313 — alleni 314 — crassicaudatus 314 — demidovi 314 — senegalensis 314 Galemys 121, 124 — pyrenaicus 124
galeritus cm. Cercocebus, Hipposideros gallarum cm. Coleura
gambianus cm. Epomophorus, Tadarida gambiensis cm. Nycteris garambae cm. Vespertilio gaspensis cm. Sorex gamellus cm. Sorex ganovensium cm. Platalina gelada cm. Theropithecus
geoffroyi cm. Anoura, Ateles, Crocidura, Dasyurus, Nyctophilus, Saguinus
Geogale 140 — aurita 133, 140 germana cm. Marmosa gibbsii cm. Neurotrichus giffardi cm. Crocidura gigantea cm. Macropus, Manis . giganteus cm. Priodontes, Pteropus gigas cm. Hipposideros, Macroderma, Notisorex, Scotophilus
gilberti cm. Potorous gilliardi cm. Pteropus glaucinus cm. Molossus glassi cm. Crocidura glebula cm. Crocidura Glires 7, 9 Glironia 46, 49 — aequatorialis 49 — venusta 49 glis cm. Tupaia Glischropus 265 Glossophaga 227, 234 — elongata 235 — longirostris 235 — soricina 227, 235, 235 godmani cm. Choeroniscus goeldii cm. Callimico goliath см. Crocidura, Praesorex goodfellowi cm. Dendrolagus goodwini cm. Cryptotis
Gorilla 346
— gorilla 351, 352 gorilla cm. Gorilla goudotii cm. Myotis gouldi cm. Chalinolobus, Pteropus gracilipes cm. Crocidura
gracilis cm. Microgale, Nasillus, Tadarida, Tupaia, Rhogeesa
gracillimus cm. Sorex graellsi cm. Saguinus grandidens cm. Parascaptor grandidieri cm. Vespertilio grandis cm. Crocidura, Euroscaptor, Micropteropus, Nycteris
granti cm. Eremitalpa, Suncus granulipes cm. Sminthopsis grassei cm. Crocidura gravida cm. Crocidura grayi cm. Crocidura greeghalli cm. Molossops greenwoodi cm. Crocidura greyi cm. Nycticeus, Protemnodon grisea cm. Murina griseoventris cm. Cryptotis grisescens cm. Myotis griseus cm. Hapalemur, Pteropus guerrerensis cm. Cryptotis guineensis cm. Vespertilio gunni cm. Perameles gunningi cm. Amblysomus Gymnobelideus 80, 85 — leadbeateri 85 gymnotis cm. Phalanger gymnura cm. Saccopteryx gymnurus cm. Echinosorex
haagneri cm. Tadarida hageni cm. Dorcopsis hainanensis cm. Neohylomys haitiensis cm. Stenoderma halconus cm. Crocidura
halli cm. Lestodelphys hallucatus cm. Dasyurus hamadryas cm. Papio hamilton cm. Tephozous hamlyni cm. Cercopithecus hansae cm. Molossus Hapalemur 297 — griseus 298 — simus 298 Haplonycteris 178, 191 — fischeri 19 hardwickei cm. Kerivoula, Rhinopoma harpax cm. Cynopterus harpia cm. Harpiocephalus Harpiocephalus 256, 276 — harpia 276 Harpyionycteris 179, 201 — celebensis 201 — whiteheadi 201 harrisoni см. Kerivoula, Musonycteris harrisi cm. Sarcophilus harti см. Artibeus hasseltii см. Myotis hastatus cm. Phyllostomus heathi cm. Scotophilus hedenborgiana cm. Crocidura helios см. Vespertilio helleri cm. Vampyrops helvum cm. Eidolon Hemicentetes 133, 135 — nigriceps 136 — semispinosus 135 Hemiechinus 114, 119 — auritus 119 — aethiopicus 119 — megalotis 119 — micropus 119 — hypomelas 119 hemprichi cm. Otonycteris henseli см. Monodelphis herbertensis см. Pseudocheirus hermani см. Myotis herrei см. Myotis hesperia cm. Lonchophylla Hesperoptenus 265 hesperus см. Vespertilio hildebrandti cm. Rhinolophus hildegardeae cm. Crocidura, Tapho-rous
hindei cm. Crocidura, Tadarida Hipposideridae 202, 219 Hipposideros 221 — abae 221 — antricola 222 — armiger 221, 222 — beatus 221 — bicolor 222 — caffer 221 — calcaratus 222 — cervinus 222 — cineraceus 222 — commersoni 221 — coronatus 222 — coxi 222 — crumeniferus 222 — cupidus 222 — cyclops 221 — diadema 222 — dinops 222 — doriae 222 — dyacorum 222 — fuliginosus 221
Hipposideros galeritus 222
— gigas 221
— inexpectatus 222
— lankadiva 222
— larvatus 222
— longicauda 222
— megalotis 221
— muscinus 222
— nanus 221
— nequam 222
— obscurus 222
— ornatus 222
— papua 222
— pelingensis 222
pomona 222
— pratti 222
— pygmaeus 222
— ridleyi 222
— ruber 222
— sabanus 222
— sandersoni 222
— semoni 222
— speoris 222
— stenotis 222
— wollastoni 222
— wrighti 222
hipposideros cm. Rhinolophus hirmeralis cm. Nycticeius hirsute cm. Micronycteris hirsutus cm. Artibeus, Coelops, Lagor-chestes, Rhinolophus
hirta cm. Crocidura hirtipes cm. Sminthopsis hirundo cm. Nycticeius hispida cm. Crocidura, Nycteris hispidus cm. Cabassous Histiotis 265
hoffmanni cm. Choloepus Hominidae 291, 353 Homo 8, 353 — sapiens 353 hoolock cm. Hylobates horrens cm. Furipterus horsfieldi cm. Crocidura, Cynopterus horsfieldii cm. Myotis hosei cm. Suncus hosonoi см. Sorex
hottentotus cm. Amblysomus,"^ Vespertilio
hova cm. Oryzorictes hoyi Cm. Microsorex humbloti cm. Triaenops, Vespertilio humeralifer cm. Callithrix humeralis cm. Chalinolobus humilis cm. Acerodon huttoni cm. Murina hybridus cm. Dasypus Hycteridae 202 hydrodromus cm. Sorex Hylobates 346, 347 — agilis 348 — concolor 348 — hoolock 348 — klossi 348 — lar 347, 348 — moloch 348 — syndactylus 347, 348 Hylobatidae 291 Hylomys 114 — suillus 115 Hylonycteris 227, 238 — underwoodi 238
hypomelanus cm. Pteropus hypomelas cm. Hemiechinus Hypotheria 9 hypsibius cm. Soriculus Hypsignathus 179, 192, 194 — monstrosus 194, 195 Hypsiprymnodon 96 — moschatus 96, 97 Hyracoidea 21, 110 icarus cm. Tadarida Ichtyes 5
illigeri cm. Saguinus imbricatus cm. Vespertilio imogene cm. Nyctophilus imperator cm. Saguinus importunus cm. Rhinolophus inca cm. Choeroniscus, Lestores incana cm. Marmosa
indri cm. Indri
Indri 304, 305
— indri 305, 307
Indridae 291, 303 inermis cm. Dobsoni a
inexpectatus cm. Hipposideros infenitesimus cm. Suncus inflatus cm. Miniopterus.T Scalopus infusca cm. Saccopteryx infuscus cm. Vampyrops intufi cm. Elephantulus ingens cm. Diclidurus ingoldbyi cm. Crocidura ingrami cm. Planigale innesi cm. Vespertilio innoxius cm. Vespertilio inornata cm. Petrogale inops cm. Rhinolophus Insectivora 20, 110 Insectivores 111
intermedius cm. Choeroniscus inustus cm. Euoticus
invicta cm. Marmosa investigator cm. Nasillus io cm. Saccopteryx, Vespertilio irkutensis cm. Sorex
irma cm. Protemnodon irrupta cm. Caluromys irus cm. Macaca
isabella cm. Depanycteris isabellinus cm. Vespertilio isodon cm. Sorex isthmicum cm. Chiroderma
jacchus cm. Callithrix jackson cm. Sorex jacksoni cm. Crocidura, Cryptotis jagori cm. Ptenochirus jagorii cm. Kerivoula jamaicensis cm. Artibeus janetta cm. Marmosa japonensis cm. Vespertilio
javanica cm. Manis, Nycteris, Tupaia javanicus cm. Rhinolophus javanus cm. Vespertilio jeanni cm. Myotis jesupi cm. Chiroderma joffrei cm. Nyctahis johnstoni cm. Suncus johorensis cm. Tadarida jouvenetae cm. Crocidura jubatus cm. Acedoron juncensis cm. Sorex
juniensis см. Marmosa kaguyae см. Myotis Kalinovskii cm. Tadarida kalubu cm. Echymipera kappleri cm. Dasyrus, Saccopterys karimii cm. Marmosa keenii cm. Myotis Kerivoula 256, 277, 278 — aerosa 277 — africana 278 — agnella 277 — argentata 278 — bicolor 277
— bombifrons 277
— cuprosa 278
— eriophora 278
— hardwickei 277
— harrisoni 278
— jagorii 278
— lanosa 278
— myrella 227
— muscilla 278
— muscina 277
— papillosa 277
— papuensis 277
— pellucida 277
— phalaena 278
— picta 277
— pusilia 277
— rapax 277
— smithii 278
— whiteheadi 277 kempi cm. Rousettus keyensis cm. Rhinolophus kihly cm. Vespertilio kinneari cm. R hi nopoma klossi cm. Crocidura, Euroscaptor, Hy-lobates
koopmani см. Antrozous kuboriensis cm. Tadarida kura cm. Podihik
labialis cm. Noctilio
labiatus cm. Epomophorus, Saguinus
Laephotis 265
lagochilus cm. Macroglossus Lagomorpha 21, 110 Lagorchestes, 96, 102 — conspicillatus 103 — hirsutus 103 — leporides 103 Lagostrophus 96, 100 — fasciatus 103 lagothricha cm. Lagothrix Lagothrix 317, 326 — flavicauda 327
— lagothricha 327 , 327 lagotis cm. Thylacomys Lamingtona 256, 279 — lophorhina 279 lamottei cm. Crocidura, Micropota-mogale
landeri cm. Rhinolophus lanei cm. Tadarida langi cm. Crocidura
laniger cm. Antechinomys, Caluromys, Lichanotus
lankadiva cm. Hipposideros lanosa cm. Kerivoula lanosus cm. Rousettus lar cm. Hylobates
larapinta cm. Sminthopsis
larvatus cm. Hipposideros, Nasalis Lasionycteris 255, 260 — noctivagans 260 Lasiorchinus 93 — barnardi 93 — latifrons 93, 94 lasiura cm. Crocidura
Lasiurus 256, 270
— borealis 271
— brachyotis 271
— cinereus 271
— degelidus 271
— minor 271
— pfeifferi 271
— : seminolus 271
latifolius cm. Phyllostomus
latifrons cm. Lasiorchinus laticaudata см. Tadarida latimans cm. Spacanus latona см. Crocidura latouchei см. Mogera Lavia 213, 215 — frons 215 lea cm. Crocidura
leadbeateri см. Gymnobelideus leisleri cm. Nyctalus Lemur 8, 297, 299 — catta 299, 299 — fulvus 299 — macaco 299 — mongoz 299 — rubriventer 299 — variegatus 299 Lemuridae 291, 296 Lemuroidea 291
lemuroides cm. Pseudocheirus leonis cm. Tadarida
Leonti deus 328, 330
— chrysomelas 330
— chrysopygus 330
—- rosalia 330, 330
lenida см. Endromicia, Marmosa
lepidura cm. Crocidura
lepidus cm. Myotis, Rhinolophus Lepilemur 297, 300 — mustelinus 300, 300 — ruficadatus 300
leporides cm. Lagorchestes leporinus cm. Noctilio leptodon cm. Myonycteris Leptonycteris 227, 239 — nivalis 239
leptura см. Parascaptor, Saccopteryx leschenaulti cm. Rousettus
Lestodelphys 46, 49
— halli 49
Lestores 78
— inca 78
lesueur cm. Bettongia
lesueri cm. Myotis
leucastra cm. Marmosa
leucocephala cm. Callithrix leucodon cm. Crocidura
leucogaster cm. Nurina, Tadarida leucomelas cm. Barbastella leucophalus cm. Papio leucops cm. Soriculus leucoptera cm. Saccopteryx leucopterus см. Pteropus
leucopus cm. Saguinus, Sminthopsis leucorhinus cm. Amblysomus leucostigma cm. Tadarida
leucura см. Suncus, Thylacomys levicula cm. Crocidura Lichanotus 304 — laniger 304, 304 Lichonycteris 227, 239 — degener 239 — obscura 239 lilium cm. Sturnira lirr.ae cm. Marmosa limbata cm. Tadarida Limnogale 133, 139 — mergulus 139 lineatus cm. Vampyrops Lionycteris 227, 235 — spurelli 235 Lipotyphla 112 Lissonycteris 184 littoral.is cm. Crocidura lituratus cm. Artibeus livingstonii cm. Pteropus lixus cm. Suncus lobatus cm. Tadarida lombocensis cm. Pteropus Lonchophylla 227, 235 — hesperia 236 — mordax 236 — robusta 236 — thomasi 236 Lonchorchina 227, 231 — aurit.a 226, 231 longicauda cm. Eonycteris, Hipposi— deros, Peroryctes
longicaudata cm. Manis, Microgale, My-rexia, Sminthopsis
longmani cm. Petrogale longimanus cm. Taphorous longirostris cm. Eoscalops, Glosso-phaga, Sorex
lophorhina cm. Lamingtona lophurus cm. Vespertilio lorentzi cm. Phascogale loricatus cm. Cabassous Loris 310 — tardigradus 310, 310 Lorisidae 291, 309 loveni cm. Vespertilio lowei cm. Soriculus, Vespertilio lowi cm. Ptilocercus lucasi cm. Penthetor luciae cm. Monophyllus, Stenoderma lucifer cm. Acedoron lucifugus cm. Myotis luctus cm. Rhinolophus ludovici cm. Sturnira ludia cm. Crocidura luluae cm. Crocidura lumholtzi cm. Dendrolagus, Sminthopsis
luna cm. Crocidura lunata cm. Onychogalea lunaris cm. Suncus lusitania cm. Crocidura luteola cm. Nycteris lutrella cm. Crocidura Lutreolina 47, 53 — crassicaudata 53, 54 luzoniensis cm. Suncus luzonis cm. Tadarida lyilli cm. Sorex lylei cm. Pteropus lynni cm. Vespertilio lyra cm. Megaderma
Macaca 333, 334
— assamensis 335
— cyclopis 335
— irus 335
— fuscata 335
— maurus 335
— mulatta 334, 335
— nemestrina 334, 335
— radiata 335
— silenus 335
— sinica 335
335
335 Lemur
cm. Crocidura
— speciosa — sylvana macaco cm. macarthuri
macdonaldi cm. Notopteris macdonnelensis cm. Antechinus
maccogelli cm. Ectophylla macer cm. Natalus mackloti cm. Acerodon maclaudi cm. Rhinolophus macleayi cm. Dorcopsis macleayii cm. Chilonycteris macmillani cm. Tadarida macrodactylus cm. Myotis Macroderma 213, 214 — gigas 214 macrodon cm. Sorex Macroglossus 179, 197 — fructivorous 197 — lagochilus 197 — minimus 197
Macrophyllum 227, 231
— macrophyllum 231
macrophyllum cm. Macrophyllum
Macropodidae 43, 45, 94
Macropus 96, 108
— gigantea 108, 109
— robustus 109
— rufus 109
Macroscelides 169, 171
— proboscideus 171
Macroscelididae 113 macrotarsus cm. Myotis macrotis cm. Nycteris, Pteropus, Rhinolophus, Saccopteryx
Macrotus 227, 230
— californicus 230
— mexicanus 230
— waterhousi 230
macrotus cm. Vespertilio macrourus cm. Thylacis macrura cm. Eudromicia, Sminthopsis maculata cm. Balinonycteris maculatum cm. Eudefma
maculatus cm. Antechinus, Dasyurus, Phalanger
madagascariensis cm. Daubentonia, Nycteris, Rousettus, Suncus
madeirae cm. Saimiri
maderensis cm. Vespertilio
madurensis cm. Rhinolophus magna cm. Cryptotis, Sturnira mahaganus cm. Pteropus
major cm. Cheirogaleus, Eonycteris, Molossus, Natalus, Nycteris, Nyctimene, Phyllonycteris, Tadarida
majori cm. Microgale, Miniopterus malagai cm. Molossops
Malagidae 291
malayana cm. Crocidura
malayanus cm. Suncus, Rhinolophus
Mammalia 17 manavi см. Miniopterus Mandrillus 339 Manidae 370
Manis 370
— crassicaudata 372
— gigantea 372
— javanica 371, 372
- — longicaudata 372
— pentadactyla 372
— temmincki 372
— tricuspis 371, 372 manni cm. Crocidura maporensis cm. Crocidura mapiriensis см. Marmosa maraxina cm. Monodelphis marianus cm. Pteropus marica cm. Marmosa, Nycteris marita cm. Crocidura marmota cm. Marmosa Marmosa 46, 50 — acermarcae 51
— agilis 51
— agricolai 52
— alstoni 51
— blasei 52
— bruchi 52
— canescens 51
— carri 51
— caucae 51
— cinerea 50
— constaninae 51
— demararae 51
— domina 51
— dryas 52
— elegans 52
— emi Пае 51
— erhardti 52 — formosa 52 — fuscata 51 — germana 51 — incana 51 — invicta 51 — janetta 52 — juniensis 52 — karimii 52 — leucastra 51 — lepida 52 — limae 50 — mapiriensis 51
— marica 51
— marmota 52
— meridio.nalis 52
— mexicana 51
— microtarsus 51
— mitis 51
— moreirae 52
— murina 51
— noctivaga 51
— ocellata 51
— pallidior 52
— parvidens 52
— phaea 51
— pusilia 52
— quichua 51
— rapposa 51
— regina 51
— • rondoni 52
— ruatanica 51
— rubra 51
— scapulata 51
— slmonsi 51
Marmosa stollei 52
— tatei 52
— tyleriana 51
— unduaviensis 51
— velutina 52
— venusta 52
— yungasensis 51
marrensis см. Vespertilio
Marsupialia 10. 20, 21, 41 marsupialis cm. Didelphis Marsupiaux 9, 10 martiensseni cm. Crocidura, Tadarida martinsi см. Saguinus mastersoni cm. Tadarida matacus cm. Tolypeutes matroka cm. Vespertilio matschiei cm. Dendrolagus, Petrodro-mus
mattogrossensis cm. Neoplatymops maurisca cm. Crocidura mauritianus cm. Taphorous
maurus cm. Macaca, Molossus, Vespertilio
maxi cm. Crocidura
maximiliani см. Saccopteryx
mayensis cm. Cryptotes mazzai cm. Dasypus mearnsi cm. Pteropus medius cm. Cheirogaleus Megachiroptera 176 Megaderma 213 — cor 214, 214 — lyra 214 — spasma 214, 214 Megadermatidae 202, 212 Megaerops 178, 182 — ecaudatus 182 — wetmorei 182 Megaleia 109
Megaloglossus 179, 198 — woer manni 198, 198 megalophylla cm. Mormoops megalotis cm. Hemiechinus, Hipposideros, Micronycteris, Paracoelops megalura cm. Suncus megaphyllus cm. Rhinolophus mehelyi cm. Rhinolophus mel cm. Pygathrix melanocephalus cm. Cacajo, Chironax melanochir см. Callicebus melanoleucus cm. Saguinus melanopogon cm. Pteropus melanops cm. Melonycteris melanopterus см. Vespertilio melanorhyncha cm. Crocidura melanotus cm. Pteropus melanura cm. Dendrogale melas cm. Myoictus melckorum cm. Vespertilio mellissa cm. Vampyrops Melonycteris, 179, 199 — aurantius 199 — melanops 199 Menotyphla 10, 112 mera cm. Cryptotis mergulus cm. Limnogale meridional is cm. Marmosa merriami cm. Sorex Mesophylla 243 Metachirops 46, 52 — opposum 52 Metachirus 47, 53
Metachirus nudicaudatus 53
Metatheria 9, 10, 21, 40
mexicana cm. Choeronycteris, Cryptotis, Marmosa
mexicanus cm. Macrotus, Natalus meyeri cm. Rhogeesa miarensis cm. Tadarida Microchiroptera 176, 201 microdon cm. Nyctophilus Microgale 133, 138 — brevicaudata 138 — cowani 138 — crassipes 138 — decaryi 139 — dobsoni 139 — drouhardi 138 — gracilis 139 — longicaudata 138 — -longirostris 139 — majori 139 — occidentalis 139
- parvula 139
— principula 139
— prolixacaudata 139
— pusilia 139 — sorella 139 — taiva 139 — thomasi 139 — talazacki 139 Micronycteris 226, 229 — b'ehni 230 — brachyotis 230 — hirsuta 230 — megalotis 230 — minuta 230 — nicefori 230 — platiceps 230 — pusilia 230 — schmidtorum 230 — sylvestris 230 Microperoryctes 73, 74 — murina 74 microphyllum cm. Rhinopoma Micropotamogale 133, 141 — lamottei 142 — ruwenzorii 142 Micropteropus 178, 192 — grandis 193 — pussillus 193
micropus cm. Hemiechinus, Natalus Microsorex 148, 152 — hoyi 152 microtarsus cm. Marmosa, microtis cm. Nyctophilus
micrura cm. Cryptotis, Eoscalops, Talpa
midas cm. Saguinus, Tadarida milleri cm. Molossops, Molossus, Myotis, Sorex
Mimetillus 265
Mi mon 227, 232 — bennettii 232 — crenulatum 232 mimulus cm. Antechinus mimus cm. Pteropus, Vespertilio minahassae cm. Vespertilio mindorus cm. Crocidura minimus cm. Antechinus, Chironectes, Macroglossus
Miniopterus 256, 274
— africanus 275
Miniopterus australis 275 <
— eschscholtzii 275
— inflatus 275
— manavi 275
— majori 275
— newtoni 275
— ravus 275
— rufus 275
— schreibersi 274, 275
— tristis 275
— witkampi 275
minor cm. Ametrida, Brachyphylla, Choeroniscus, Dobsonia, Lasiurus; Tupaia
Minuania 46, 48
— dimidiata 49
— fosteri 49
— umbristriata 49
minura cm. Crocidura, Micronycteris, Tonatia
minutissimus cm. Sorex
minutus cm. Nyctimene, Tadarida, Sorex
Miopithecus 341
mitis cm. Cercopithecus, Marmosa
miya cm. Crocidura
Mogera 121, 126, 127
— latouchei 126
— robusta 126
— wogura 126
moloch cm. Callicebus, Hylobates moloneyi cm. Vespertilio
Molossidae 202, 281
Molossops 282, 283
— aequatorianus 283
— brachymeles 283
— greenhalli 283
— malagai 283
— milleri 283
— planirostris 283
— temmincki 283
Molossus 282, 288, 289
— abrasus 288
— barnesi 288
— bonariensis 289
— bondae 288
— centralis 288
— davisoni 288
— debilis 288
— fortis 288
— glaucinus 289
— hansae 289
— major 288
— maurus 289
— milleri 288
— nasutus 288
— occultus 288
— pamane 288
— perotis 288
— pretiosus 288
— rufus 288
— sinaloae 288 — trinitatus 28?
— tropidorhynchus 288
— trumbulli 289
— underwoodi 289 molossus cm. Tadarida moluccensis cm. Dobsonia) mona cm. Cercopithecus monachus cm. Pithecia
топах см. Crocidura mongoz см. Lemur monoceros см. Rhinolophus Monodelphes 9 Monodelphia 10 Monodelphis 46, 47 — adusta 48 — americana 48 — brevicaudata 48 — domestica 48 — henseli 48 — maraxina 48 — peruviana 48 — sorex 48 — scalops 48 — unistriata 48 Monodonta 19 Monophyllus 227, 236 clinedophus 236 — cubanus 226, 236 — frater 236 — luciae 236 — plethodon 236 — portoricensis 236 — red man i 236 Monotremata 10, 19, 21, 32 monstrosus cm. Hipsignathus montana cm. Tupaia montanus cm. Scalopus, Vespertilio monticola cm. Crocidura, Emballonura, ' Sminthopsis
montosus cm. Vespertilio montserratensis cm. Stenoderma mops cm. Tadarida mordax cm. Lonchophylla, Sturnira, Vespertilio
moreirae см. Marmosa morio cm. Chalinolobus, Suncus Morganucodon 19 Mormopterus 287 Mormoops 226, 229 — blainvillii 229 — megalophylla 229 moschata cm. Desmana moschatus cm. Hypsiprymnodon, Scaptochirus
mulatta cm. Macaca mulleri cm. Tupaia Multituberculata 19, 20 murrayi cm. Vespertilio Murexia 60 — longicaudata 61 — rothschildi 61 muricauda cm. Crocidura muricola cm. Myotis Murina 256, 275 — aurata 276, 276 — balstoni 276 — canescens 276 — cyclotis 276 — florium 276 — grisea 276 — huttoni 276 — leucogaster 276 — suilla 276 — tunebrosa 276 murina cm. Dendrogale, Marmosa, Mic-roperoryctes, Sminthopsis murino-flavus cm. Scotophilus murinus cm. Mycrocebus, Suncus, Vespertilio musciculus, cm. Vespertilio
muscilla cm. Kerivoula muscina cm. Kerivoula muscinus cm. Hipposideros Musonycteris 227, 237 — harrisoni 237 mustelinus cm. Lepilemur Mutica 7
myarus cm. Elephantulus Mycrocebus 297, 301 — coquereli 302 — murinus 301, 302 Myoictus 57, 66 •— melas 66
Myonycteris 178, 184
— brachycephala 185
— leptodon 184
— torquata 184
— wronghtoni 184 Myosorex 148, 164 Myotis 255, 256 — adversus 258 — africanus 258 — albescens 259 — altarium 257 — argentatus 259
• — austroriparius 258
— bartelsii 258
— bechsteini 257
— blythi 257
— bocagei 257
— browni 258
— californicus 259
— capaccinii 257
— chiloensis 259
— dasycneme 258
— daubentoni 257
— davidi 257
— emarginatus 257
— evotis 258
— formosus 257
— fort i dens 258
— frater 257
— goudotii 258
— grisescens 258
— hasseltii 258
— hermani 258
— herrei 258
— horsfieldii 258
— jeanni 258
— kaguyae 257
— keenii 258
— lepidus 258
— lesueuri 258
— lucifugus 258
— macrodactylus 258
— macrotarsus 258
— milleri 258
— muricola 257
— myotis 255, 257
— mystacinus 257
— nattereri 257
— nigricans 259
— occultus 258
— oreias 258
— patriciae 258
— pequinius 258
— planiceps 259
— ricketti 258
— ruber 259
— rufopictus 258
— scotti 258
— seabrai 258
Myotis sicarius 257
— siligorensis 257
— simus 259
— sodalis 259
— stalkeri 258
— subulatus 259
— thysanodes 258
— tricolor 257
— velifer 258
— venustus 258
— vivesi 259
— volans 259
— welwitschi' 257
— yumanensis 258 myrella cm. Kerivoula Myrmecobiidae 43, 44, 69 Myrmecobius 69 — fasciatus 69 Myrmecophaga 356, 357 •— tridactyla 357 Myrmecophagidae 356 Mystacina 280 — tuberculata 280 Mystacinidae 202, 280 mystacinus cm. Myotis mystax cm. Saguinus Myzopoda 253 — aurita 253, 254 Myzopodidae 202, 253
naias cm. Syconycteris
nana cm. Brachyphylla, Nycteris Nanonycteris 179, 192, 194 — veldkampii 194
nanulus cm. Tadarida, Vespertilio
nanus cm. Artibeus, Cercaertus, Hipposideros, Sorex, Vespertilio
Nasalis 333
— larvatus 343
Nasilio 172
Nasillus 121
— gracilis 121
— investigator 121
naso cm. Antechinus, Rhynchonycteris nasuta см. Perameles nasutus см. Molossus
Natalidae 202, 250
Natalus 250
— : brevimanus 251
— dominicensis 251
• — macer 251
— major 251
— mexicanus 250
— micropus 251
— primus 251
— stramineus 251
— tumidifrons 251
— tumidirostris 251
nathusii cm. Vespertilio nations cm. Chaetophractus nattereri cm. Myotis Nectogale 148, 166 — elegans 166
neglectus cm. Cercopithecus negligens см. Crocidura negrina cm. Crocidura nelsoni cm. Cryptotis nemaeus cm. Pygathrix nemestrina cm. Macaca Neophascogale 60 neohibernicus см. Pteropus
Neohylomys 114, 116 — hainanensis 116 Neomys 148, 152 — anomalus 153 — fodiens 153, 154 Neoplatymops 282, 289 — mattogrossensis 289 Neopteryx 178, 188 — frosti 188 Neotetracus 114, 117 — sinensis 117 nequam cm. Hipposideros nereis cm. Rhinolophus Nesonycteris 179, 199 — woodfordi 199 neumanni cm. Epomophorus Neurotrichus 121, 128 — gibbsii 128 newtoni cm. Miniopterus Niadius 180 niangarae cm. Tadarida nicaraguae cm. Tonatia nicefori cm. Micronycteris nichollsi cm. Stenoderma nicobarica cm. Crocidura, Tupaia nictitans cm. Cercopithecus nigaeriae см. Tadarida niger cm. Cynopithecus, Pteropus nigrescens cm. Cryptotis, Emballonura, Soriculus, Thoopterus
nigri cm. Tadarida nigricans cm. Myotis nigriceps cm. Hemicentetes nigricollis cm. Saguinus nigrifrons cm. Callicebus nigripes cm. Crocidura nigriseta cm. Petrodromys nigrita cm. Scotophilus nigritellus cm. Scotophilus nigrivittatus cm. Cebus nigrofusca cm. Crocidura nigroviridis cm. Cercopithecus nilssoni cm. Vespertilio nimbae cm. Crocidura / niobe cm. Crocidura nitela см. Sminthopsis nivalis см. Leptonycteris Noctilio 209 — labialis 210, 211 — leporinus 209, 210, 211 Noctilionidae 202, 209 noctivaga cm. Marmosa noctivagans cm. Lasionycteris noctula cm. Nyctalus norfolcensis cm. Petaurus, Tadarida Notiosorex 148, 156 — crawfordii 157 — gigas 157 —- phillipsii 157 notius cm. Vespertilio Notopteris 179, 199 — macdonaldi 199 Notoryctes 70 — caurinus 71 — typhlops 71 Notoryctidae 43, 44, 70 Notoungulata 20 novaeguineae cm. Planigale novemcinctus cm. Dasypus Nyctalus 256 , 265 •— azoreum 266 — furvus 266
Nyctalus joffrei 266
— lasiopterus 266
— leisleri 266
— noctula 266
— stenopterus 266
Nycteridae 202, 211
Nycteris 211
— aethiopica 212
— arge 212
— aurita 212
— avakubia 212
— gambiensia 212
— grandis 212
— hispida 212
— javanica 211, 212
— luteola 212
— macrotis 212
— madagascariensis 212
— major 212
— marica 212
— nana 212
— pallida 212
— parisii 212
— proxima 212
— revoillii 212
— thebaica 211, 212
— vinsonii 212
— wood! 212
Nycticeius 256 , 268
'— albofuscus 268
— artinii 268
— cinnamomeus 268
— cubanus 268
— emarginatus 268
— falabae 268
— greyi 268
— hirundo 268
—' humeralis 268
— pallidus 268
— sanborni 268
— schlieffeni 268
Nyctimene 179, 200
— aello 201
— albiventer 200
— cephalotes 200
— cyclotis 201
— major 200, 201
— minutus 200
— robinsoni 201
— tryoni 21
Nycticebus 310, 311
— coucang 312
— pygmaeus 312
Nyctophilus 256, 279
— arnhemensis 279
• — bifax 279
— daedalus 279
• — geoffroyi 279
— imogene 279
— microdon 279
— microtis 279
— timoriensis 279
— walkeri 279
nymphaea j cm. Vampyrops
obesolus cm. Thylacis
obscura cm. Cryptotis, Lichonycteris obscurus cm. Hipposideros, Pygathrix obtus асм. Phyllonycteris
ocellata cm. Marmosa
occidentalis cm. Microgale, Pseudochei
occipitalis cm. Tadarida occultidens cm. Suncus occultus cm. Molossus, Myotis ochraceus cm. Tadarida ocularis cm. Pteropus odorata cm. Crocidura oedipus cm. Saguinus oerstedi cm. Saimiri ognevi cm. Vespertilio olivacea cm. Cryptotis olivieri cm. Crocidura ollula cm. Suncus Onychogalea 96, 101 — fraenata 102, 102 — lunata 102 — unguifer 102 ophiodon cm. Scotonycteris opposum cm. Metachirops orarius cm. Scapanus oratus cm. Vampyrops oreias cm. Myotis oreopolus cm. Sorex orientalis cm. Crocidura, Phalanger, Talpa oritis cm. Crocidura oriundus cm. Suncus ornatus cm. Hipposideros,"Sorex Ornitodelphes 9 Ornithorhynchidae 35 Ornithorhynchus 38 — anatinus 38 orphila cm. Cryptotis Oryzorictes 133, 138 — hova 138 — talpoides 138 — tetradactylus 438 osborni cm. Tadarida Otopterus 178, 191 — cartilagonodus 191 Otonycteris 256 — hemprichi 267 oweni cm. Scapanulus
Pachydermes 9
pachyotis cm. Vespertilio pachypus cm. Vespertilio pacificus cm. Sorex pagenstecheri cm. Vespertilio palawanensis cm. Crocidura, Tupaia, Suncus
palliata cm. Alouatta pallida cm. Nycteris pallidior cm. Marmosa pallidus cm. Antrozous, Nycticeius palpator cm. Dactylopsila palustris cm. Sorex pamane cm. Molossus Pan 346, 349 — troglodytes 349, 351 paniscus cm. Ateles Pantotheria 10, 19—21 papillosa cm. Kerivoula Papio 333, 337 — anubis 338 — cynocephalus 338 — hamadryas 338, 338 — leucophaeus 338 — papio 338 — sphinx 338 — ursinus 338 papio cm. Papio papua cm. Hipposideros papuanus cm. Vespertilio
papuensis см. Kerivoula, Peroryctes, Tadarida
Paracoelops 221, 224
— megalotis 224
Paracrocidura 148, 162
— shoutedeni 162
paradoxolophus cm. Rhinomegalophus paradoxura cm. Crocidura paradoxus cm. Solenodon Paraechinus 119 Parantechinus 59 Paranyctimene 179, 201 — raptor 201
paraquayensis cm. Didelphis Parascalops 121, 129 — breweri 129
Parascaptor 121 — grandidens 127 — leptura 127 parisei cm. Nyctefis parma cm. Protemnodon, Thy logale parva cm. Cryptotis parvacauda cm. Crocidura
parvidens cm. Euroscaptor, Cheirome-les, Marmosa
parvipes cm. Crocidura
parvula cm. Microgale, Rhogeesa pasha cm. Crocidura patas cm. Cercopithecus patriciae cm. Myotis patrizii cm. Asellia paucidentata cm. Alionycteris
pearsoni cm. Rhinolophus, Solisorex Pecora 7
pelingensis cm. Hipposideros pellucida cm. Kerivoula pennatus cm. Distoechurus penicillata cm. Petrogale, Callithrix pentadactyla cm. Manis Penthetor 178, 180, 181 — lucasi 181 pequinius см. Myotis Perameles 73 — bougainvillei 73 — eremiana 73 — fasciata 73 — gunni 73 — nasuta 73
Peramelidae 43, 45, 72
Perardorcas 96, 103 — concinna 103 percivali cm. Cloeotis, Crocidura peregrinus cm. Pseudocheirus Perissodactyla 21, 110 perforatus cm. Taphorous pergacilis cm. Cryptotis pergrisea cm. Crocidura Perodicticus 310, 312 — potto 312. 313 peroni cm. Dobsonia Peronymus 207 Perop teryx 207 Peroryctes 72, 74 — longicauda 74 — papuensis 74 — raff ray anus 74 perotis см. Molossus persicus cm. Triaenops personatus cm. Callicebus, Chilonycteris,
Pteropus
perspicillata см. Carollia perspicillifer cm. Saccopteryx
peruviana cm. Monodelphis petaurista cm. Cercopithecus Petaurus 80, 86 — australis 86 — breviceps 86 — norfolcensis 86
petersi cm. Rhynchocyon, Vespertilio Peirodromus 169 — matschiei 171 — nigriseta 171 — robustus 171 — rovumae 171 — sultan 171 — tetradactylus 170 Petrogaie 96, 100 — brachyotis 101 — inornata 101 — longmani 101 — penicillata 101 — — penicillata 101 — wilkinsi 101 — xanthopus 101 petrophilus cm. Tadarida pfeifferi cm. Lasiurus phaea cm. Marmosa phaeura cm. Chimarrogale phalaena cm. Kerivoula Phal anger 80, 82 — atrimaculatus 83 — celebensis 79, 83 — gymnotis 82 — maculatus 82, 83 — orientalis 82 — — peninsulae 83 — vestitus 82 — ursinus 83
Phalangeridae 43, 45, 79 Phalangeroidea 44 Phaner 297 — furcifer 302 Phascogale 57, 59 — calura 60 — lorentzi 60 — tapotafa 60 Phascolarctos 80, 90 — cinereus 90, 91 Phascolomyidae 43, 92 Phascolosorex 57, 61 — doriae 61 — dorsalis 61 phasma cm. Vespertilio phayrei cm. Pygathrix philander cm. Coluromys philippinensis cm. Rhinolophus phillipsii cm. Notiosorex Pholidota 21, 110, 370 Ph у le tor 265 Phylloderma 227, 233 — septentrionales 233 — stenops 232 Phyllonycteris 228, 246 — aphylla 246 •— major 246 — obtusa 246 — poeyi 246 Phyllops 245
Phyllostomatidae 202, 225 Phyllostomus 227, 232 — discolor 233 — elongatus 233 — hastatus 233 — latifolius 233
phyllotus см. Plecotus
picatus см. Chalinolobus
pichi cm. Zaedyus
picta cm. Kerivoula, Tupaia
pileatus cm. Hylobates, Pygathrix, Saguinus
pilorostris cm. Urotrichus
pilosa cm. Crocidura pilosus cm. Dasypus Pinnipedia 21, 110 pipistrellus cm. Vespertilio Pithecia 317, 320 — monachus 321 — pithecia 321 Pizonyx 259
pithecia cm. Pithecia planiceps cm. Myotis Planigale 57, 59 — ingrami 59 — novaeguineae 59 — subtilissima 59 — tenuirostris 59 planirostris cm. Molossops Plantigrades 9 Platalina 227, 236 — genovensium 236 platiceps cm. Micronycteris Platymops 287 platycephala см. Chimarrogale platyops см. Potoroops, Vespertilio Plecotus 256, 272 — auritus 273 — phyllotus 273 — rafinesqui 273 — townsendii 273 Pierotes 179, 192, 195 — anchietai 195
plethodon см. Monophyllus Pliopithecus 292
plicata см. Saccopteryx, Tadarida pluto см. Saguinus
Podihik 148, 153
— kura 153
Podogymnura 114
— truei 116
poensis см. Chalinolobus, Crocidura poeyi см. Phyllonycteris pogonias см. Cercopithecus pohlei см. Pteropus polia см. Crocidura poliocephalus см. Pteropus polusus см. Surdisorex polykomos см. Coleura pomona см. Hipposideros
Pongidae 291, 345 '
Pongo 346, 349
— pygmaeus 349, 350
portoricensis см. Monophyllus Potamogale 133, 140 — velox 140, 141 Potamogalidae 133 Potamogalinae 134 Potoroops 96, 97 — platyops 97 Potorous 96, 97, 98 — gilbert! 98 — tridactilus 98 — — tridactylus 98 potto см. Perodicticus potenziani см. Pygathrix pousarguesi см. Epomophorus praedatrix см. Dobsonia
Praesorex 148, 162 — goliath 162 pratti см. Hipposideros prattorum см. Rhynchomeles preblei cm. Sorex Presbytis 342 pretiosus cm. Molossus preussi cm. Suncus pribilofensis cm. Sorex Primates 7, 20, 21, 110 primus cm. Natalus principula cm. Microgale Priodontes 363, 365 — giganteus 364, 365 Prionotemnus 108 Proboscidea 21, 110 proboscideus см. Macroscelides prolixacaudata см. Microgale Promops 287, 289 Propithecus 304' — aiadema 304 — verreauxi 305 Propliopithecus 292 Prosimii 291, 292 Protemnodon 96, 107 — agilis 108 — bedfordi 108 — bicolor 108 — billardieri 108 — canguru 108 — dorsalis 108 — eugenii 108 — greyi 108 — irma 108 — parma 108 Prototheria 9, 10 proxima см. Nycteris psammophila cm. Sminthopsis Pseudantechinus 59 Pseudocheirus 80, 88 — albertisi 90 — archeri 90 — canescens 90 — caroli 89 — corinnae 90 — convoluter 89 — cupreus 90 — dahli 90 — forbesi 89 — herbertensis 89 — lemuroides 90 — occidentalis 89 — peregrinus 89 — pygmaeus 90 — rubidus 89 — schlegeli 90 — victoriae 89 Ptenochirus 178, 182 — jagori 182 Pteralopex 178, 189 — atrata 189
Pteronotus 226, 228 — davyi 229 — suapurensis 229 Pteropidae 177 Pteropus 178, 185, 189 — admiralitatum 187 — aldabrensis 188 alecto 187 — argentatus 187 — arquatus 187
Pteropus austini 188
— balutus 187
— caniceps 187
— chrysoproctus 187
— cognatus 187
— comorensis 186
— conspicillatus 187
— dasymallus 186
— giganteus 187
— gilliardi 188
— gouldi 185, 188
— griseus 187
— hypomelanus 186
— leucopterus' 187
— livingstonii 188
— lombocensis 187
— lylei 187
— macrotis 187
— mahoganus 187
— marianus 186
— mearnsi 187
— melanopogon 187
— melanotus 187
— mimus 187
— neohibernicus 187
— niger 188
— ocularis 187
— personatus 187
— pohlei 187
— poliocephalus 188
— pumilus 187
— rayneri 187
— rodricensis 188
— rufus 188
— seychellensis 188
— speciosus 186
— subniger 188
— tablasi 188
— temmincki 187
— tonganus 187
— vampyrus 187
— voeltzkowi 188
— woodfordi 187
Ptilocercus 293
— lowi 295, 296
pulchellum cm. Diplomesodon
pulcher cm. Vespertilio pulchellus cm. Acrobates pulveratus cm. Vespertilio pumila cm. Brachyphylla, Saccopteryx,
Tadarida
pumilio cm. Rhynophylla
pumilus cm. Pteropus, Vespertilio
pusilia cm. Kerivoula, Marmosa, Microgale, Micronycteris, Vampyrops
pussillus cm. Micropteropus, Rhinolophus, Tadarida
Pygathrix 333, 341
— ayguta 342 cristatus 342
— entellus 342
— melaphus 342
— nemaeus 342
— obscurus 342
— phayrei 342
— pileatus 342
— potenziani 342
— rubicunda 342
— senex 342
pygmaea см. Callithrix
pygmaeus см. Hipposideros, Nycticebus, Pongo, Pseudocheirus
Pygoderma 228, 245
— bilabiatum 245 pyrenaicus cm. galemys Pyrotheria 20, 21
quadraticauda cm. Blarinella quariba cm. Alouatta quichua cm. Marmosa quoll cm. Dasyurus
rabhanurus cm. Rhyncholestes raddei cm. Sorex
radiata cm. Macaca
raffrayana cm. Emballonura raffrayanus cm. Peroryctes rafinesqui cm. Plecotus raineyi см. Crocidura rapax cm. Kerivoula rapposa cm. Marmosa raptor cm. Paranyctimene ravus cm. Miniopterus, Tomopeas rayneri cm. Pteropus recifinus cm. Vampyrops rectitragus cm. Vespertilio redmani см. Monophyllus refulgens см. Rhinolophus regina cm. Marmosa
religiosa см. Crocidura remota см. Dobsonia rendalli см. Vespertilio retusa см. Burmeisteria revoili cm. Elephantulus, Nycteris rex cm. Rhinolophus reymi cm. Suncus Rhinolophidae 202, 215 Rhinolophus 216 — abae 218 — acrotis 217
— acuminatus 218
— affinis 217
— alcyone 218
— anderseni 219
— andreinii 218
— angolensis 218 arcuatus 219
— axillaris 218
— blasii 217
— bocharicus 217
— boruensis 218
— brockmani 218
— bunkeri 2 19
— capensis 218
— celebensis 219
— clivosus 217
— clossi 218
— coelophyllus 218
— cornutus 2 17
— creaghi 219
— darlingi 218 — deckenii 218 — denti 218 — dobsoni 218 — eloguensis 218 — euryale 217 — euryotis 219 — ferrumequinum 217 — foxi 218 — fumigatus 218 — hildebrandti 218 — hipposideros 217
Rhinolophus hirsutus 219 — inops 219 — javanicus 218 — keyensis 219 — landeri 218 — lepidus 217 — luctus 217 — maclaudi 218 — macrotis 218 — madurensis 218 — malayanus 217 — megaphyllus 219 — mehelyi 217 — monoceros 217 — nereis 218 —• pearsoni 217 — philippinensis 218 — pusillus 218 refulgens 218 — rex 218 — robinsoni 218 — rouxi 217 — sedulus 219 — simplex 219 — simulator 218 — stheno 218 — subbadius 217 — subrufus 219 — swinnyi 218 — thomasi 217 — toxopeusi 219 — trifoliatus 217 — virgo 219
Rhinomegalophus 216, 219 — paradoxolophus 219 Rhinonicteris 221, 224 — aurantius 220, 224 Rhinopithecus 333, 342 — avunculus 343 — bieti 343 — brelichi 343 — roxellanae 343
Rhinopoma 202
• — hardwickei 203
— kinneari 203
— microphyllum 203
— sumatrae 203
Rhinopomatidae 202 Rhinopterus 265 rhoditis cm. Crocidura Rhogeessa 256, 269 — gracilis 269 — meyeri 269 — parvula 269 — velilla 269 Rhynchocyon 169 — adersi 170 — cirnei 170 —- chrysopygus 170 — claudi 170 — petersi 170 — stuhlmanni 170 — swynnertoni 170 Rhyncholestes 72, 78, 79 — raphanurus 79 Rhynchomeles 72, 75 — prattorum 75 Rhynchonax 121 — andersoni 121 Rhynchonycteris 204, 206 — naso 206
Rhynophylla 227, 240 — pumilio 240 ridleyi cm. Hipposideros, Vespertilio ricketti cm. Myotis robinsoni cm. Coelops, Nyctimene, Rhinolophus
robusta cm. Eonycteris, Mogera, Lonchophylla
robustula cm. Vespertilio
robustus cm. Macropus, Petrodromys,Scotophilus
Rodentia 10, 21, 110 rodricensis cm. Pteropus rohui cm. Vespertilio romana cm. Talpa rondoni cm. Marmosa roosevelti cm. Cacajao, Crocidura rosalia cm. Leontideus rosamondae cm. Antechinus rosenbergi cm. Eonycteris rothschildi cm. Murexia rotundus cm. Desmodus Rousettus 178, 183—185 — aegyptiacus 184 — amplexicaudatus 187 — angolensis 184 — arabicus 184 — celebensis 187 — crypticola 187 — kempi 184 1anosus 184 — leschenaulti 184 — madagascariensis 184 — smithii 187 — stresemanni 187 rouxi cm. Rhinolophus rovumae cm. Petrodromus roxellanae cm. Rhinopithecus rozeti cm. Elephantulus ruandae cm. Suncus ruatanica cm. Marmosa ruber cm. Myotis, Hipposideros rubicunda cm. Pygathrix rubicundus cm. Cacajao rubidus cm. Pseudocheirus rubiginosa cm. Chilonycteris rubra cm. Marmosa rubriventer cm. Lemur rufescens cm. Aepyprymnus, Echymipera, Elephantulus
ruficaudata cm. Lepilemur rufigenis cm. Sminthopsis ruforisea cm. Protemnodon rufopictus cm. Myotis rufum cm. Stenoderma rufus cm. Macropus, Miniopterus, Molossus, Pteropus, Triaenops rupestris см. Elephantulus rflppelli m. Tadarida, Vespertilio russatus cm. Tadarida russula cm. Crocidura rusticus cm. Vespertilio ruwenzorii cm. Micropotamogale
sabaeum см. Eidolon sabanus cm. Hipposideros Saccopteryx 204, 206 •— bilineata 207 — brevirostris 207 — canescens 207 — gymnura 207 — infusca 207
Saccopteryx io 207
— kappleri 207
— leptura 207
— leucoptera 207
— macrotis 207
— maximiliani 207
— perspicillifer 207
— plicata 207
— pumila 207
Saguinus 328, 331
• — bicolor 331, 332
— devillei 332
— fuscicollis- 332
— geoffroyi 231, 332
— graellsi 332
— illigeri 332
— imperator 331, 332
— labiatus 332
— leucopus 332
— martinsi 332
— melanoleucus 332
— midas 332
— mystax 332
— nigricollis 332
— .oedipus 332
— pileatus 332
— pluto 332
— tamarin 332
— tripartitus 332
— wedelli 332
Saimiri 317, 324
— boliviensis 325
— madeirae 325
— oerstedi 324
— sciureus 324, 325
— usta 325
salenskii cm. Soriculus salvini cm. Chiroderma sanborni cm. Nycticeius sandersoni cm. Hipposideros sansibarica cm. Crocidura santaremensis cm. Callithrix,-— sapiens cm. Homo /
sarasinorum cm. Tadarida Sarcophilus 57, 67 — harrisi 67
satanas cm. Chiropotes Satanellus 66 saussurei cm. Sorex savii cm. Vespertilio Scalophus 121, 131 — aquaticus 131 — inflatus 131 — montanus 131 scalops cm. Monodelphis Scapanulus 121, 129 — oweni 129 Scapanus 121, 130 — anthonyi 130 — latimans 130 — orarius 130 — townsendii 130 Scaptochirus 121, 126 — moschatus Scaptonyx 121, 128 — fusicaudus 128 scapulata cm. Marmosa Schoinobates 80, 91 — volans 91 schlegeli cm. Pseudocheirus schlieffeni cm. Nycticeius schmidtorum cm. Micronycteris
schnablii cm. Amorphochilus, schweit-zeri cm. Crocidura
sciureus cm. Saimiri
sclateri cm. Amblysomus, Sorex Scleronycteris 227, 238 — ega 238
scotti cm. Myotis
Scotonycteris 178, 192, 193
— ophiodon 194
— zenkeri 194
Scotophilus 256, 269
— celebensis 270
— gigas 269
— heathi 269
— murino-flavus 270
— nigrita 269
— nigritellus 270
— robustus 270
— temmincki 269
— viridis 270
Scutisorex 148, 167
— congicus 168 — somereni 168 seabrai см. Myotis secunadus см. Tadarida sedulus см. Rhinolophus seminolus см. Lasiurus seminudus см. Rousettus semispinosus cm. Hemicentetes semoni cm. Hipposideros senegalensis cm. Eomops, Taphorous senex cm. Centurio, Pygathrix seniculus cm. Alouatta septemcinctus cm. Dasypus septentrionales cm. Phylloderma sericea cm. Crocidura
serotinus cm. Vespertilio Setifer 133, 136 — setosus 136 setiger cm. Tadarida Setonyx 96, 106 — brachyurus 106 setosus cm. Tachyglossus, Setifer seychellensis cm. Coleura, Pteropus sexcinctus cm. Euphractus sezekorni cm. Erophylla shalleri cm. Suncus shinto cm. Sorex
shoutedeni cm. Paracrocidura sibiriensis cm. Sorex sicarius cm. Myotis silenus см. Macaca siligorensis cm. Myotis simplex cm. Rhinolophus Simia 8
Simias 333 , 343 — concolor 343 similis cm. Tadarida simonsi cm. Marmosa simulator cm. Rhinolophus simus cm. Hapalemur, Myotis sinaloae cm. Molossus sinensis cm. Neotetracus sinica cm. Macaca sinuosus cm. Sorex
Sirenia 21, 110 smithi cm. Crocidura smithii cm. Kerivoula Sminthopsis 57, 62 •— crassicaudata 63, 64 — froggatti 63 — granulipes 63
Sminthopsis hirtipes 63
— larapinta 63
— leucopus 63
— • longicaudata 63
— lumholtzi 63
— macrura 64
— monticola 64
— murina 63
— nitela 64
— psammophila 63
— rufigenis 64
— stalkeri 63
sodalis cm. Myotis
Solenodon 143, 145
— cubanus 147
— paradoxus 146, 147
Solenodontidae 112, 113 Sol isorex 148, 164 — pearsoni 164 solomonis cm. Anamigdon Somalia cm. Crocidura somereni cm. Scutisorex sorella cm. Microgale sorex cm. Monodelphis
Sorex 148
— alascanus 151
— alpinus 150
• — araneus 149, 150
— arcticus 150
— bendirii 151
— buchariensis 150
— caucasicus 150
— caecutiens 150
— cinereus 150
— cylindricauda 150
— daphaenodon 150
— dispar 151
— fumeus 151
— gaspensis 151
— gemellus 150
— gracillimus 150
— hosonoi 150
— hydrodromus 150
— • irkutensis 150
— isodon 150
— jacksoni 151
— juncensis 151
— longirostris 151
— lyilli 150
— macrodon 151
— merriami 151
— milleri 150
— minutissimus 150
— minutus 150
— nanus 151
— oreopolus 151
— ornatus 151
— pacificus 150
— palustris 149, 151
— preblei 150
— pribilofensis 151
— raddei 150
— saussurei 151
— sclateri 151
— sibiriensis 150
— sinuosus 151
— • shinto 150
— stizodon 151
— tenellus 151
— trigonirostris 151
Sorex trowbridgii 151
— unguiculatus 150
— vagrans 151
— veraepacis 151
— vir 150
— volnuchini 150
— willeti 151
soriceps см. Uropsilus
Soricidae 112, 113, 147
soricina см. Cryptotis, Glossophaga Soricomorpha 112 Soriculus 148, 152, 153 — caudatus 152 — hypsibius 152
— • leucops 152
— lowei 152
— nigrescens 152
— calenskii 152
spadiceus см. Dyacopterus spadix cm. Dendrolagus spasma cm. Megaderma speciosa см. Macaca speciosus cm. Pteropus spectrum cm. Tarsius, Vampyrum spelaea cm. Eonycteris spenserae cm. Tarsipes spenceri cm. Antechinomys speoris cm. Hipposideros Sphaerias 178, 182 — blanfordi 182 Sphaeronycteris 228, 245 — toxophyllum 245 sphinx cm. Cynopterus, Papio spillmanni cm. Tadarida splendidula cm. Tupaia
spurelli cm. Lionycteris, Xiphonycteris squamicaudata cm. Wyulda squamipes cm. Anourosorex stalkeri cm. Myotis, Sminthopsis stampflii cm. Vespertilio Stenoderma 228, 244, 245 — annectens 244 — falcatus 244 — flavescens 244 — haitiensis 244 — luciae 244 — montserratensis 244 — nichollsi 244 — rufum 245 stenops см. Phylloderma stenopterus см. Nyctalus stenotis см. Hipposideros stheno см. Rhinolophus stigmatica см. Thylogale stizodon cm. Sorex stoliczkana cm. Aselliscus stoliczkanus cm. Suncus stollei cm. Marmosa stramineus cm. Natalus
stuhlmanni cm. Chrysochloris, Rhynchocyon
Sturnira 227, 240
— angel i 240
— lilium 240
— ludovici 240
— magna 240
— mordax 240
— tildae 240
Sturnirops 240
Styloctenium 178 — wallacei 190 suapurensis см. Pteronotus
suaveolens см. Crocidura subbadius см. Rhinolophus subflavus cm. Vespertilio subniger cm. Pteropus subrufus cm. Rhinolophus subtilissima cm. Planigale subulatus cm. Myotis Sudani cm. Tadarida suilla cm. Murina suillus cm. Hylomys sultan cm. Petrodromus sumatrae cm. Rhinopoma suncoides cm. Suncus Suncus 148, 162 — blarina 163 — cafer 163 — dayi 163 — etruscus 163 — granti 164 — hosei 163 — imfinitesimus 164 — Johnstoni 164 — leucura 164 — lixus 163 — lunaris 164 — luzoniensis 163 — madagascariensis 164 — malayanus 163 — megalura 163 — morio 164 — murinus 163 — ollula 164 — occultidens 163 — oriundus 164 — palawanensis 163 — preussi 164 — reymi 164 — ruandae 164 — shalleri 163 — stoliczkanus 163 — suncoides 164 — varius 163 superans cm. Vespertilio Surdisorex 148, 165 — norae 165 — polulus 165 sururae cm. Crocidura Syconycteris 179, 198 — australis 198 — crassa 198 — naias 198 sylvana cm. Macaca sylvicola cm. Tonatia Sylvisorex 148, 164 sylvestris cm. Micronycteris Symmetrodonta 19 Symphalangus 346, 348 syndactylus cm. Hylobates syrichta cm. Tarsius swainsoni cm. Antechinus swinnyi cm. Rhinolophus swynnertoni cm. Rhynchocyon Tadarida africana 285 — albiventer 287 — ansorgei 285 — aurispinosa 287 — australis 287 — beccarii 287 — bemmeleni 285 — bivittatus 286 — bocagei 285 — brachyptera 286 — brasiliensis 287 — brunnea 285 — chapini 286 — chitauensis 286 — cisturus 285 — condylura 286 — congicus 286 — demonstrator 286 — doriae 287 — europs 287 — farad]ius 286 — femorosacca 287 — formosus 287 — frater 286 — fulminanus 285 — gambiaus 286 — gracilis 287 — haagneri 287 — hindei 286 — icarus 287 — johorensis 286 — kalinowskii 287 — kuboriensis 287 — lanei 285 •— laticaudata 287 — leonis 286 — leucogaster 286 — leucostigma 286 — limbata 286 — lobatus 285 — luzonis 286 — macmillani 287 — major 286 — martiensseni 287 — mastersoni 285 — miarensis 286 — midas 286 — minutus 287 — molossus 287 — mops 286 — nanulus 286 — niangarae 286 — nigeriae 286 — nigri 286 — norfolkensis 287 — occipitalis 286 — ochraceus 285 — osborni 285 — papuensis 287 — petrophilus 287 — plicata 286 — pumila 286
tablasi cm. Pteropus Tachyglossidae 35 Tachyglossus 36 — aculeatus 32, 34, 36, 37 — setosus 37 Tadarida 282, 284, 287 — abae 286 — acetabulosa 287 — aegyptiaca 285 — pusillus 286 — rtippellii 286 — russatus 286 — sarasinorum 286 — secundus 287 — setiger 287 — similis 287 — spillmanni 286 — teniotis 285 — thersites 286
Tadarida tongaensis 285
• — tragata 285
— trevori 286 —
— websteri 286
— wroughtoni 287
— yucatanica 287
Taeniodonta 21
taiva cm. Microgale
talapoin cm. Cercopithecus talazacki cm. Microgale Talpa 121, 125 — altaica 126 — caeca 126 — caucasica 126 — europaea 126 — micrura 126 — orientalis 126 — romana 126 — streeti 126 Talpidae 112, 113
talpoides cm. Oryzorictes, Urotrichus Tamandua 357 , 358 — tetradactyla 358 tamarin cm. Saguinus
Tana 294
tana cm. Tupaia
Taphozous 204, 207
— australis 208
— flaviventris 208
— hamiltoni 208
— hildegardeae 208
— kachhensis 208
— longimanus 208
— mauritianus 208
— melanopogon 208
— mixtus 208
— nudicluniatus 208
— nudiventris 208
— perforatus 208
— philippinensis 208
— saccolaimus 208
— senegalensis 208
— Sudani 208
— theobaldi 208
— troughtoni 208
tapotafa cm. Phascogale tardigradus cm. Loris Tarsiidae 291, 315 Tarsioidea 291
Tarsipes 80, 88
— spenserae 88
Tarsius 315
— bancanus 316
— spectrum 316
— syrichta 315, 316
tasmaniensis cm. Vespertilio
tatei cm. Marmosa
tatouay cm. Cabassous telfairi cm. Echinops telmalestes cm. Blarina temmincki cm. Manis, Molossops, Pte-
ropus, Scotophilus
temminckii cm. Cynocephalus tenellus cm. Sorex teniotis cm. Tadarida tenius cm. Crocidura
Tenrec 134
• — ecaudatus 134
Tenrecidae 112, 113, 132 tenuipinnis cm. Vespertilio tenuirostris cm. Planigale tenuis cm. Vespertilio
tersus cm. Cryptotis tetradactyla cm. Tamandua tetradactylus cm. Oryzorictes, Petrod-romus
thebaica cm. Nycteris
Therapsida 19 theresae cm. Crocidura Theria 21 Theriodontia 18 Theropithecus 333, 339 — gelada 339 thersites cm. Tadarida thetis cm. Thylogale thomasi cm. Lonchophylla, Microgale,
Rhinolophus thomensis cm. Crocidura Thoopterus 178, 180 — nigrescens 180 Thylacinus 57, 68 — cynocephalus 68 Thylacis 72, 75 — barrowensis 76 — macrourus 76 — obesolus 76
Thylacomys 72, 76 — lagotis 77 •— leucura 77 Thylogale 96, 106 — billardierri 106 — brunii 107 — eugenii 106 — parma 106 thetis 106 — stigmatica 106 thyone cm. Vampyrops Thyroptera 252 — discifera 253 — tricolor 253
Thyropteridae 202, 252 thysanodes cm. Myotis tickelli cm. Vespertilio tildae cm. Sturnira timoriensis cm. Nyctophilus tionis cm. Crocidura
Tolypeutes 363 , 366 — matacus 367 — tricinctus 367, 367 tomesi cm. Vespertilio Tomopeas 256, 280 — ravus 280 Tonatia 227, 231 — bidens 231 — brasiliensis 231 — carrikeri 232 — minuta 232 — nicaraguae 232 — sylvicola 232 tongaensis cm. Tadarida tonganus cm. Pteropus torrei cm. Chilonycteris torquata cm. Myonycteris torquatus cm. Bradypus, Callicebus, Cer-cocebus, Cheiromeles
townsendii cm. Plecotus, Scapanus toxopeusi cm. Rhinolophus toxophyllum cm. Sphaeronycteris Trachops 227, 233 — cirrhosa 233 tragata cm. Tadarida trevelyani cm. Chrysospalax trevori cm. Tadarida
Triaenops 221, 225 — afer 225 — furcula 225 —• humbloti 225 — persicus 220, 225 — rufus 225 Trichosurus 80 — caninus 81 — vulpecula 81 trichotis cm. Cheirogaleus trichura cm. Crocidura tricinctus cm. Tolypeutes tricolor cm. Carollia, Myotis, Thyrop-tera
Triconodonta 19 tricuspidatus cm. Aselliscus tricuspis cm. Manis tridactyla cm. Myrmecophaga tridactylus cm. Bradypus, Potorous tridens cm. Asellia trifoliatus cm. Rhinolophus trigonirostris cm. Sorex trinitatum cm. Chiroderma trinitatus cm. Molossus tripartitus cm. Saguinus tristis cm. Miniopterus trivirgata cm. Dactylopsila trivirgatus cm. Aotes troglodytes cm. Pan tropicalis cm. Amblysomus tropidorhynchus cm. Molossus trowbridgii cm. Sorex truei cm. Podogymnura trumbulli cm. Molossus tryoni cm. Nyctimene tuberculata cm. Mystacina tuberculatus cm. Chalinolobus Tubulidentata 110 Tubulidenata 21 tumidirostris cm. Natalus tunebrosa cm. Murina Tupaia 293 — carimatae 294 — dorsalis 294 — glis 294 — gracilis 294 — javanica 294 — minor 294 — montana 294 — mulleri 294 — nicobarica 294 — palawanensis 294 — picta 294 — splendidula 294 — tana 294 Tupaiidae 291, 292 Tupaioidea 291 turpis cm. Artibeus tyleriana cm. Marmosa tylopus cm. Vespertilio typhlops cm. Notoryctes Typhlosaurus 145 ugandae cm. Vespertilio ultima cm. Crocidura umbratus cm. Vampyrops umbristriata cm. Minuania underwoodi cm. Hylonycteris, Molossus unduaviensis cm. Marmosa Unguiculata 7 unguiculatus см. Sorex unguifer cm. Onychogalea Ungulata 7
unicinctus cm. Cabassous unistriata cm. Monodelphis Uroderma 227, 241 — bilobatum 241 Urogale 293, 295 — everetti 295 Uropsilus 121 — soriceps 121, 122 Urotrichus 121, 127 — pilirostris 127 — talpoides 127 ursinus cm. Dendrolagus, Papio, Phalan-ger, Vombatus
usta cm. Saimiri vagrans cm. Sorex Vampyressa 242 Vampyriscus 242 Vampyrodes 242 Vampyrops 227, 242 — bidens 242 — brocki 242 — caraccioloi 242 - dorsalis 242 — helleri 242 — infuscus 242 — lineatus 242 — major 242 — melissa 242 — nymphaea 242 — oratus 242 — pusilia 242 — recifinus 242 — thyone 242 — umbratus 242 — venilia 242 — vittatus 242 — zarhinus 242 Vampyrum 227, 234 — spectrum 227, 234 vampyrus cm. Pteropus vanheurni cm. Dorcopsis variegata cm. Chalinolobus variegatus cm. Lemur varius cm. Suncus velatus cm. Vespertilio veldkampii cm. Nanonycteris velifer cm. Myotis velilla cm. Rhogeesa vellerosus cm. Chaetophractus velox см. Potamogale velutina cm. Crocidura, Marmosa venusta cm. Glironia, Marmosa venustus cm. Myotis veraepasis cm. Sorex Verecundae 262 vermiculus cm. Chrysochloris verreauxi cm. Propithecus verus cm. Coleura Vespertilionidae 202, 254 Vespertilio 8, 256, 260 — abaensis 264 — a bra mus 263 — aero 264 — affinis 263 — albigularis 262 — alienus 263 — anchietai 264 — angulatus 264 — annectens 264 — ariel 264 — babu 263 — bicolor 262
Vespertilio blanfordi 263
— bobrinskii 261
— brachypterus 264
— brasiliensis 262
— brunneus 262
— capensis 262
— ceylonicus 263
— chiriquinus 262
— circumdatus 264
— coromandra 263
— crassulus 264
— culex 264
— demissus 262
-— deserti 263
— diminutus 262
— doriae 263
— dormer! 264
— endoi 263
— faradjius 262
— fidelis 262
— flavescens 262
— floweri 263
— furinalis 262
— fuscus 262
— garambae 262
— grandidieri 262
— guineensis 262
— helios 264
— hesperus 264
— hottentotus 262
— humbloti 262
— imbricatus 264
— innesi 262
— innoxius 262
— io 264
— isabellinus 261
— javanus 264
— japonensis 261
— kuhly 263
— lophurus 263
— loveni 262
— lowei 263
— lynni 262
— macrotus 263
— maderensis 264
— malayana 263
— marrensis 264
— matroka 262
— maurus 264
— melanopterus 262
— melckorum 262
— minahassae 264
— mimus 263
— moloneyi 263
— montosus 262
— montanus 263
— mordax 264
— murinus 261
— murrayi 264
— musciculus 264
— nanulus 264
— nanus 264
— nasutus 261
— nathusii 263
— nilssoni 261
— notius 263
— ognevi 262
— pachyotis 261
— pachypus 261, 263
— pagenstecheri 264
— papuanus 264
— petersi 264
Vespertilio phasma 262
— pipistrellus 263
— platyops 262
— pulcher 264
— pulveratus 263
— pumilus 263
— pusillus 262
— rectitragus 262
— rendalli 262
— ridleyi 264
— robustula 263
— rohui 263
— rusticus 264
— riippelli 264
— savii 261, 263
•— serotinus 262
— stampflii 264
— subflavus 264
— superans 261
— tasmaniensis 264
— tenuipinnis 262
— tenuis 264
— tickelli 263
— tomesi 263
— tylopus 264
— ugandae 262
— velatus 263
— walli 261
— wintoni 263
— zuluensis 262
vestitus cm. Phalanger veterum cm. Dorcopsis viaria cm. Crocidura victoriae cm. Pseudocheirus vilensis cm. Crocidura villosa cm. Alouatta, Crocidura villosum cm. Chiroderma
villosus cm. Chaetophractus, Chrysospa-lax
vinsoni cm. Nycteris vir cm. Sorex
virgo cm. Diclidurus, Rhinolophus viridis cm. Dobsonia, Scotophilus vittatus cm. Vampyrops vivesi cm. Myotis voeltzkowi cm. Pteropus
volans cm. Cynocephalus, Myotis, Schoino-bates
volnuchini cm. Sorex
Vombatidae 45
Vombatus 93
— ursinus 93
— — ursinus 93
vosmaeri cm. Crocidura vulcani cm. Crocidura vulpecula cm. Trichosurus
wahlbergi cm. Epomophorus walkeri cm. Nyctophilus Wallabia 108
wallacei cm. Styloctenium walli cm. Vespertilio waterhousi cm. Macrotus weberi cm. Crocidura websteri cm. Tadarida wedelli cm. Saguinus welwitshi cm. Myotis wetmorei cm. Megaerops whiteheadi cm. Harpyionycteris, Kerivoula
whitleyi cm. Eomops
Wilhelmina cm. Antechinus
wilkinsi см. Petrogale willeti см. Sorex
wimmeri cm. Crocidura wintoni cm. Cryptochloris witkampi cm. Miniopterus woermanni cm. Megaloglossus wogura cm. Mogera wollastoni cm. Hipposideros v'oodfordi cm. Nesonycteris, Pteropus woodi cm. Nycteris wrighti cm. Hipposideros
wroughtoni cm. Tadarida, Myonycteris
Wyulda 80, 81
— squamicaudata 81
xanthopus cm. Petrogale
xantippe cm. Crocidura Xyphonycteris 282, 284 — spurrelli 284
youngi cm. Diaemus yucatanica cm. Tadarida yumanensis cm. Myotis yungasensis cm. Marmosa Zaedyus 363, 364 — pichi 364, 365 Zaglossus 367 — bartoni 37, 38 — bruijni 38 — bubuensis 38 Zalambdodonta 112 zaphiri cm. Crocidura zarhinus cm. Vampyrops zenkeri cm. Scotonycteris zeteki cm. Cryptotis zimmeri cm. Crocidura zuluensis cm. Vespertilio
От автора.................................................................. 3
Введение .............................................. . . 5
Класс млекопитающих Classis Mammalia Linnaeus, 1758 . .17
Краткий морфологический обзор млекопитающих............................... 22
Подкласс яйцекладущих млекопитающих Subclassis Prototheria Gill, 1872 32
Отряд однопроходных Ordo Monotremata Bonaparte, 1838 ............... 32
Семейство ехидн Familia Tachyglossidae Gill, 1872 ............... 35
Род ехидн Genus Tachyglossus liliger, 1811.......................... 37
Род проехидн Genus Zaglossus Gill, 1877 ............................ 37
Семейство утконосов Familia Ornithorhynchidae Burnett, 1880 .... 38
Подкласс живородящих млекопитающих Subclassis theria Parker et has-well, 1879 ........................................................ 40
Инфракласс сумчатых Infraclassis Metatheria Huxley, 1880 ................. 41
Отряд сумчатых Ordo Marsupialia liliger, 1811 ....................... 41
Семейство опоссумов Familia Didelphidae Gray, 1821.................. 46
Род густошерстных опоссумов Genus Caluromys J. Allen, 1900 ... 47
Род голохвостых опоссумов Genus Monodelphis Burnett, 1830 ... 47
Род короткохвостых опоссумов Genus Minuania Cabrera, 1919 ... 48
Род опоссумов Чилоэ Genus Dromiciops Thomas, 1894 ................ 49
Род пушистохвостых опоссумов Genus Glironia Thomas, 1912 . . 49
Род патагонских опоссумов Genus Lestodelphys Tate, 1934 .... 49
Род мышевидных опоссумов Genus Marmosa Gray, 1821............. 50
Род четырехглазых опоссумов Genus Metachirops Matschie, 1916 . 52
Род длиннохвостых опоссумов Genus Matachirus Burmeister, 1854 . 53
Род толстохвостых опоссумов Genus Lutreolina Thomas, 1910 ... 53
Род обыкновенных опоссумов Genus Didelphis Linnaeus, 1758 ... 54
Род водяных опоссумов Genus Chironectes liliger, 1811............. 55
Семейство хищных сумчатых Familia Dasyuridae Waterhouse, 1838 . 56
Род сумчатых мышей Genus Antechinus McLeay, 1841.................. 58
Род плоскоголовых сумчатых мышей Genus Planigale Troughton, 1928 59
Род сумчатых крыс Genus Phascogale Temminck, - 1824 .... 59
Род новогвинейских сумчатых крыс Genus Murexia Tate et Archbold, 1937 .......................................................... 60
Род полосатых сумчатых крыс Genus Phascolosorex Matschie, 1916 61
Род гребнехвостых сумчатых мышей Genus Dasycercus Peters, 1875 61
Род двугребнехвостых сумчатых мышей Genus Dasyuroides Spencer, 1896 .............................................................. 62
Род узколапых сумчатых мышей Genus Sminthopsis Thomas, 1887 62
Род сумчатых тушканчиков Genus Antechinomys Krefft, 1867 ... 64
Род пятнистых сумчатых куниц Genus Dasyurus Geoffroy, 1796 . 65
Род полосатых сумчатых куниц Genus Myoictus Gray, 1858 ... 66
Род сумчатых чертей, или тасманийских дьяволов, Genus Sarcophilus Geoffroy et F. Cuvier, 1837....................................... 67
Род сумчатых, или тасманийских, волков Genus Thylacinus Temminck, 1827 ............................................................. 68
Семейство сумчатых муравьедов—Familia Myrmecobiidae Waterhouse, 1838 ...................................................... 69
Семейство сумчатых кротов Familia Notoryctidae Ogilby, 1892 ... 70
Семейство сумчатых барсуков, или бандикутов, Familia Peramelidae Waterhouse, 1838 ................................ . . . . 72
Род длинноносых бандикутов Genus Perameles Geoffroy, 1803 ... 73
Род новогвинейских бандикутов Genus Peroryctes Thomas, 1906 . 74
Род мышевидных бандикутов Genus Microperoryctes Stein, 1932 . . 74
Род колючих бандикутов Genus Echymipera Lesson, 1842 .... 74
Род бандикутов Керама Genus Rhynchomeles Thomas, 1920 . 75
Род коротконосых бандикутов Genus Thylacis liliger, 1811 . . 75
Род свиноногих бандикутов Genus Chaeropus Ogilby, 1838 . .. 76
Род кроличьих бандикутов Genus Thylacomys Owen, 1838 . . .. 76
Семейство ценолестов, или крысовидных опоссумов, Familia Caenole-stidae Trouessart, 1898 ....... . . . 77
Род эквадорских ценолестов Genus Caenolestes Thomas, 1895 . 78
Род перуанских ценолестов Genus Lestores Owen, 1934 .... 78
Род чилийских ценолестов Genus Rhyncholestes Osgood, 1924 . 79
Семейство лазящих сумчатых, или кускусов, Familia Phalangeridae Thomas, 1888 . .... .................. 79
Род кузу, или кистехвостых кускусов, Genus Trichosurus Lesson, 1828 80
Род чешуехвостых кускусов Genus Wyulda Alexander, 1919 81
Род кускусов Genus Phalanger Storr, 1780 ........................... 82
Род полосатых кускусов Genus Dactylopsila Gray, 1858 ... 83
Род карликовых кускусов Genus Eudromicia Mjoberg, 1916 ... 83
Род соневидных кускусов Genus Cercaertus Burmeister, 1837 . . 84
Род беличьих кускусов Genus Cymnobelideus McCoy, 1867 ... 85
Род сумчатых летяг Genus Petaurus Shaw et Nodder, 1791 .... 86
Род перьехвостых кускусов Genus Distoechurus Peters, 1874 ... 86
Род карликовых летучих кускусов Genus Acrobates Desmarest, 1817 87
Род хоботноголовых кускусов Genus Tarsipes Gervais et Verreaux, 1842 88
Род кольцехвостых кускусов Genus Pseudocheirus Ogilby, 1837 . 88
Род сумчатых медведей, или коала Genus Phascolarctos De Blainville, 1816............................................................... 90
Род гигантских летучих кускусов Genus Schoinobates Lesson, 1842 91
Семейство вомбатов Familia Phascolomyidae Goldfuse, 1820 ............. 92
Род короткошерстных, или голоносых, вомбатов Genus Vombatus Geoffroy St.-Hilaire, 1803 ......................................... 93
Род длинношерстных, или шерстоносых, вомбатов Genus Lasiorchinus Gray, 1863 ................................................. 93
Семейство кенгуру Familia Macropodidae Owen, 1839 .................... 94
Род мускусных кенгуру Genus Hypsiprymnodon Ramsay, 1876 . 96
Род широкоголовых кенгуру Genus Potoroops Matschie, 1916 . 97
Род потору Genus Potorous Desmarest, 1804 .......................... 98
Род гологрудых кенгуру Genus Caloprymnus Thomas, 1888 . 99
Род короткомордых кенгуру Genus Bettongia Gray, 1837 .... 99
Род больших крысиных кенгуру Genus Aepyprymnus Garrod, 1875 100
Род скалистых кенгуру Genus Petrogale Gray, 1837 ............. 100
Род когтехвостых кенгуру Genus Onychogalea Gray, 1841 .... 101
Род заячьих кенгуру Genus Lagorchestes Gould, 1841 ...........102
Род полосатых кенгуру Genus Lagostrophus Thomas, 1887 .... 103
Род карликовых скалистых кенгуру Genus Perardorcas Thomas, 1904 103
Род кустарниковых кенгуру Genus Dorcopsis Schlegel et Miiller, 1842 104
Род древесных кенгуру Genus Dendrolagus Miiller, 1839 .... 104
Род филандеров Genus Thylogale Gray, 1837 ............................ 106
• Род короткохвостых кенгуру, или куокка, Genus Setonyx Lesson, 1842 106
Род средних кенгуру, или валлэби, Genus Protemnodon Owen, 1873 107
Род исполинских кенгуру Genus Macropus Shaw et Nodder, 1790 . . 108
Инфракласс плацентарных Infraclassis Eutheria Gill, 1872 .............. 110
Отряд насекомоядных Ordo Insectivora Bowdich, 1821.............................Ill
Семейство ежей Familia Erinaceidae Bonaparte, 1838 .............. 113
Род гимнур Genus Echinosorex Blanville, 1838 ......................... 115
Род малых крысиных ежей, или малых гимнур, Genus Hylomys Miiller, 1839 ......................................................... 115
Род хайнаньских ежей Genus Neohylomys Shaw et Wong, 1959 ... 116
Род филиппинских гимнур Genus Podogymnura Mearns, 1905 ... 116
Род землеройковых ежей Genus Neotetracus Trouessart, 1909 ... 117
Род ежей Genus Erinaceus Linnaeus, 1758 ...................... 117
Род ушастых ежей Genus Hemiechinus Fitzinger, 1866 ... . 119
Семейство кротов Familia Talpidae Gray, 1821 ...................120
Род землеройковых кротов Genus Uropsilus Milne-Edwards, 1871 . 121
Род выхухолей Genus Desmana Giildenstaedt, 1777 ...... 122
Род пиренейских выхухолей Genus Galemys Каир, 1829 .... 124
Род кротов Genus Talpa Linnaeus, 1758 ................................ 125
Род японских землеройковых кротов Genus Urotrichus Temminck, 1842 127
Род длиннохвостых кротов Genus Scaptonyx Milne-Edwards, 1871 128
Род американских землеройковых кротов Genus Neurotrichus Gunther, 1880 . ........................................................... 128
Род западнокитайских кротов Genus Scapanulus Thomas, 1912 . . 129
Род волосатохвостых кротов Genus Parascalops True, 1894 . . . 129
Род западноамериканских кротов Genus Scapanus, Pomel, 1848 . 130
Род восточноамериканских кротов Genus Scalopus Е. Geoffroy St. Hilaire, 1803 ......................................................... 131
Род звездорылов Genus Condylura liliger, 1811..........................131
Семейство тенреков Familia Tenrecidae Gray, 1821.........................132
Род обыковенных тенреков Genus Tenrec Lacepede, 1799 ................. 134
Род полосатых тенреков Genus Hemicentetes Mivart, 1871 .... 135
Род больших, или ежовых, тенреков Genus Setifer Froriep, 1806 . . 136
Род тенреков Фонтуанонта Genus Dasogale Grandidier, 1928 . . . 136
Род малых тенреков Genus Echinops Martin, 1838 .............. 137
Род рисовых тенреков Genus Oryzorictes Grandidier, 1870 ... . 138
Род длиннохвостых тенреков Genus Microgale Thomas, 1882 . . . 138
Род перепончатолапых, или болотных, тенреков Genus Limnogale Major, 1896 ......................................................... 139
Род земляных тенреков Genus Geogale Milne-Edwards et Grandidier, 1872 ......................................................... 140
Род выдровых землероек Genus Potamogale Du Chaillu, 1860 . . 140
Род карликовых выдровых землероек Micropotamogale Heim de Balsac, 1954 ......................... ............................... 141
Семейство златокротов Familia Chrysochloridae Mivart, 1868. . . . 142
Род обыкновенных златокротов Genus Chrysochloris Laccpede, 1799 . 143
Род исполинских златокротов Genus Chrysospalax Gill, 1883 . . . 144
Род африканских златокротов Genus Amblysomus Pomel, 1848 . . 144
Род пустынных златокротов Genus Eremitalpa Roberts, 1924 .... 145
Род златокротов Винтона Genus Cryptochloris Shortridge et Carter, 1939 145
Семейство щелезубов Familia Solenodontidae Dobson, 1882 ................ 145
Семейство землероек Familia Soricidae Gray, 1821.........................147
Род бурозубок Genus Sorex Linnaeus, 1758 ...................... 149
Род бурозубок-пигмеев Genus Microsorex Coues, 1877 ................. 152
Род азиатских бурозубок Genus Soriculus Blyth, 1854 .......... 152
Род цейлонских бурозубок Genus Podihik Deraniyagale, 1958 . . . 153
Род кутор Genus Neomys Каир, 1829 .................................. 153
Род американских короткохвостых бурозубок Genus Blarina Gray, 1838 154
Род азиатских короткохвостых бурозубок Genus Blarinella Thomas, 1911 ............................................. 155
Род короткоухие бурозубки Genus Cryptotis Pomel, 1848 . . . 155
Род серых бурозубок Genus Notiosorex Coues, 1877 ............. 156
Род белозубок Genus Crocidura Wagler, 1832 .................. 157
Род конголезских белозубок Genus Paracrocidura Heim de Balsac, 1956 ............................................. 162
Род африканских гигантских белозубок Genus Praesorex Thomas, 1913................................................................ 162
Род многозубых белозубок Genus Suncus Ehrenberg, 1832 .... 162
Род когтистых белозубок Келаарта Genus Feroculus Kelaart, 1852 164
Род когтистых белозубок Пирсона Genus Solisorex Thomas, 1924 . . 164
Род горных белозубок Genus Surdisorex Thomas, 1906 ................. 165
Род путораков Genus Diplomesodon Brandt, 1852 .............. 165
Род кротовых белозубок Genus Anourosorex Milne-Edwards, 1870 . 166
Род азиатских водяных белозубок Genus Chimarrogale Anderson, 1877 166
Род тибетских водяных белозубок, или перепончатолапых белозубок, Genus Nectogale Milne-Edwards, 1870 ...................... 166
Род белозубок-броненосок Genus Scutisorex Thomas, 1913 .... 167
Семейство прыгунчиков Familia Macroscelididae Mivart, 1868 . . . 168
Род хоботковых собачек, или пестроспинных прыгунчиков, Genus Rhynchocyon Peters, 1847 ................................. 169
Род лесных прыгунчиков Genus Petrodromus Peters, 1846 .... 170
Род короткоухих прыгунчиков Genus Macroscelides A. Smith, 1829 171
Род длинноухих прыгунчиков Genus Elephantulus Thomas et Schwann, 1906 ................................................. 171
Отряд шерстокрылов, или кагуанов, Ordo Dermoptera liliger, 1811............ 173
Отряд рукокрылых Ordo Chiroptera Blumenbach, 1779.......................... 175
Подотряд крыланов Subordo Megachiroptera Dobson, 1875 .............. 177
Род коротконосых крыланов Genus Cynopterus F. Cuvier, 1825 . . 179
Род темных крыланов Genus Thoopterus Matschie, 1899 ............... 180
Род черношапочных крыланов Genus Chironax Andersen, 1912 . . 181
Род бурых крыланов Genus Dyacopterus Andersen, 1912 .... 181
Род коротконосых крыланов Лукаса Genus Penthetor Andersen, 1912 181
Род крыланов Бланфорда Genus Sphaerias Miller, 1906 ............... 182
Род крыланов Я гор a Genus Ptenochirus Peters, 1861........ 182
' Род крыланов Темминка Genus Megaerops Peters 1865 .... 182
Род пятнокрылых крыланов Genus Balionycteris Matschie, 1899 . . 182
Род летучие собаки, или ночные крыланы, Genus Rousettus Gray, 1821 183
Род ошейниковых крыланов Genus Myonycteris Matschie, 1899 . . 184
Род летучих лисиц Genus Pteropus Brisson, 1762 .................... 185
Род сулавесских крыланов Genus Neopteryx Hayman, 1946 . . . 188
Род ацеродон Genus Acerodon Jourdan, 1837 .................. 188
Род птералопекс Genus Pteralopex Thomas, 1888 ................. 189
Род бонейя Genus Boneia Jentink, 1879 ..................... 189
Род карликовых крыланов Genus Aethalops Thomas, 1923 . . . 190
Род крыланов Уоллэса Genus Styloctenium Matschie, 1899 . . . 190
Род голоспинных крыланов Genus Dobsonia Palmer, 1898 .... 190
Род крыланов Фишера Genus Haplonycteris Lawrence, 1939 . . . 191
Род ночных крыланов Genus Alionycteris Kock, 1969 .......... 191
Род толстоухих крыланов Genus Otopterus Kock, 1969 ............ 191
Род эполетовых крыланов Genus Epomophorus Bennett, 1836 .... 192
Род карликовых эполетовых крыланов Genus Micropteropus Matschie, 1899 ............ ....................... ... 192
Род биндемов Genus Epomops Gray, 1870 .............. . . 193
Род скотониктерис Genus Scotonycteris Matschie, 1894 ........ 193
Род коровьемордых крыланов Genus Nanonycteris Matschie, 1899 194
Род молотоголовых крыланов Genus Hypsignathus H. Allen, 1861 194
Род казиниктерис Genus Casinycteris Thomas, 1910................ 195
Род крыланов Анхиеты Genus Pierotes Andersen, 1910.............. 195
Род соломенноцветных, или пальмовых, крыланов Genus Eidolon Ra-finesque, 1815............................................ ... 196
Род пещерных крыланов Genus Eonycteris Dobson, 1873 ... 196
Род длинноязыких крыланов Genus Macroglossus F. Cuvier, 1824 197
Род цветочных длинноязыких крыланов Genus Syconycteris Matschie, 1899 .......................................................... 198
Род африканских длинноязыких крыланов Genus Megaloglossus Pagenstecher, 1885 ...............................................' 198
Род темнобрюхих крыланов Genus Melonycteris Dobson, 1877 . . 199
Род соломоновых длинноязыких крыланов Genus Nesonycteris Thomas, 1887 .......................................................... 199
Род длиннохвостых крыланов Genus Notopteris Gray, 1859 . . 199
Род трубконосых крыланов Genus Nyctimene Borkhausen, 1797 . . 200
Род карликовых трубконосых крыланов Genus Paranyctimene Tate, 1942 201
Род крыланов-гарпий — Genus Harpyionycteris Thomas, 1896 . . 201
Подотряд летучих мышей Subordo Microchiroptera Dobson, 1875 . . 201
Семейство ланцетоносов, или мышехвостов, Familia Rhinopomatidae Miller, 1907 ............................................................. 202
Семейство мешкокрылов Familia Emballonuridae Dobson, 1875 . . 203
Род мешкокрылов Старого Света Genus Emballonura Temminck, 1838 205
Род африканских мешкокрылов Genus Coleura Peters, 1867 ... 205
Род хоботковых мешкокрылов Genus Rhynchonycteris, 1867 .... 206
Род обыкновенных мешкокрылов Genus Saccopteryx liliger, 1811 206
Род могильных мешкокрылов Genus Taphozous Е. Geoffroy 1818 207
Род белых мешкокрылов Genus Diclidurus Wied-Neuwied, 1820 . 208
Род депаниктерис Genus Depanycteris Thomas, 1920 .......... 209
Род циттаропс Genus Cyttarops Thomas, 1913 ...... 209
Семейство рыбоядных летучих мышей Familia Noctilionidae Gray, 1821 209
Семейство щелемордов Familia Nycieridae Dobson, 1875 .... 211
Семейство ложных вампиров Familia Megadermatidae Gill, 1872 . 212
Род афроазиатских ложных вампиров Genus Megaderma Е. Geoffroy, 1810 ........................................................... 213
Род австралийских ложных вампиров Genus Macroderma Miller, 1906 214
Род желтокрылых ложных вампиров Genus Lavia Gray, 1838 . . 215
Семейство подковоносов Familia Rhinolphidae Bell, 1836 ... 215
Род подковоносов Genus Rhinolophus Lacepede, 1799 .... 216
Род большеухих подковоносов Genus Rhinomegalophus Bourret, 1951 219
Семейство ложных подковоносов, или листоносов Старого Света, Familia Hipposideridae Miller, 1907 ..................................... 219
Род обыкновенных листоносов Genus Hipposideros Gray, 1831 . . 221
Род трезубценосов Genus Asellia Gray, 1838 ................ 223
Род трезубценосов Тейта Genus Aselliscus Tate, 1941 ............. 223
Род цоелопс Genus Coelops Blyth, 1848 ........................ 223
Род африканских трезубценосов Genus Cloeotis Thomas, 1900 . . 224
Род воронкоухих листоносов Genus Paracoelops Dorst, 1947 . . 224
Род золотистых листоносов Genus Rhinonicteris Gray, 1847 . . 224
Род трилистоносов Genus Triaenops Dobson, 1871 .............. 225
Семейство американских листоносов Familia Phyllostomatidae Waterhouse, 1839 ......................................................... 225
Род усатых листоносов Genus Chilonycteris Gray, 1839 .... 228
Род голоспинных листоносов Genus Pteronotus Gray, 1838 . . . 228
Род'подбородколистых листоносов Genus Mormoops Leach, 1821 . 229
• Род малых большеухих листоносов Genus Micronycteris Gray, 1866 229
Род большеухих листоносов Genus Macrotus Gray, 1843 .... 230
Род меченосов Genus Lonchorhina Tomes, 1863 ...................... 231
Род длиннолапых листоносов Genus Macrophyllum Gray, 1839 . . 231
Род круглоухих листоносов Genus Tonatia Gray, 1827 .... 231
Род копьеносов Грея Genus Mimon Gray, 1847 ....................... 232
Род копьеносов Genus Phyllostomus Lacepede, 1799 232
Род копьеносов Петерса Genus Phylloderma Peters, 1865 .... 233
Род бахромчатогубых листоносов Genus Trachops Gray, 1847 . . . 233
Род длинноволосых ложных вампиров Genus Chrotopterus Peters, 1865 234
Род ложных вампиров Genus Vampyrum Rafinesque, 1815 . . 234
Род длинноязыких листоносов Genus Glossophaga Е. Geoffroy, 1818 234
Род лиониктерис Genus Lionycteris Thomas, 1913.................... 235
Род копьеносов Томаса Genus Lonchophylla Thomas, 1903 .... 235
Род платалина Genus Platalina Thomas, 1928 236
Род монофиллус Genus Monophyllus Leach, 1821 236
Род длинноносых листоносов Genus Anoura Gray, 1838 .... 236
Род банановых листоносов Genus Musonycteris Schaldach et Mczanghlin, 1960 ................................................... 237
Род мексиканских длинноносое Genus Choeronycteris Tschudi, 1844 237
Род длинноносое Годмана Genus Choeroniscus Thomas, 1928 . . . 238
Род склерониктерис Genus Scleronycteris Thomas, 1912 .... 238
Род листоносов Ундервуда Genus Hylonycteris Thomas, 1903 . . 238
Род листоносов Соссюра Genus Leptonycteris Lydekker, 1891 . . . 239
Род лихониктерис Genus Lichonycteris Thomas, 1895 .......... 239
Род короткохвостых листоносов Genus Carollia Gray, 1838 . . . 239
Род карликовых листоносов Genus Rhynophylla Peters, 1865 . . 240
Род желтоплечнх листоносов Genus Sturnira Gray, 1842 .... 240
Род корвира Genus Corvira Thomas, 1915 .................. 241
Род брахифилла Genus Brachyphylla Gray, 1834 .......... 241
Род листоносов-строителей Genus Uroderma Peters, 1865 . . . 241
Род белополосых широконосов Genus Vampyrops Peters, 1865 . . 242
Род большеглазых листоносов Genus Chiroderma Peters, 1860 . . 243
Род белых листоносов Genus Ectophylla H. Allen, 1892 .... 243
Род фруктоядных листоносов Genus Artibeus Leach, 1821 . . . 243
Род красных фруктоядных листоносов Genus Stenoderma Ё. Geoffroy, 1818 ................................................ 244
Род ипанемских листоносов Genus Pygoderma Peters, 1863 . . . 245
Род складчатомордых листоносов Genus Centurio Gray, 1842 . . . 245
Род сфаерониктерис Genus Sphaeronycteris Peters, 1882 ... 245
Род малых складчатомордов Genus Ametrida Gray, 1847 .... 246
Род светлых цветочных листоносов Genus Phyllonycteris Gundlach, 1865 246
Род цветочных листоносов Genus Erophylla Miller, 1906 .... 247
Семейство вампиров Familia Desmodontidae Gill, 1866 .... 247
Род обыкновенных вампиров Genus Desmodus Wied-Neuwied, 1824 249
Род белокрылых вампиров Genus Diaemus Miller, 1906 .... 249
Род мохноногих вампиров Genus Diphylla Spix, 1823 ................ 249
Семейство воронкоухов Familia Natalidae Miller, 1899 .......... 250
Семейство дымчатых летучих мышей Familia Furipteridae Miller, 1907 251
Род восточных дымчатых летучих мышей Genus Furipterus Bonaparte, 1837 ................................................ 252
Род обыкновенных дымчатых летучих мышей Genus Amorpliochilus Peters, 1877 ........................................ 252
Семейство американских присосконогов Familia Thyropteridae Miller, 1907 ................................................ 252
Семейство мадагаскарских присосконогов Familia Myzopodidae Thomas, 1904 253
Семейство обыкновенных летучих мышей Familia Vespertilionidae Gray, 1821 254
Род ночниц Genus Myotis Каир, 1829 ... ............. 256
Род серебристых гладконосов Genus Lasionycteris Peters, 1865 . . 260
Род кожанов и нетопырей Genus Vespertilio Linnaeus, 1758 . . . 260
Род вечерниц Genus Nyctalus Bowdich, 1825 ........................ 265
Род дисконогих гладконосов Genus Eudiscopus Conisbee, 1953 . 267
Род стрелоухов Genus Otonycteris Peters, 1859 . . .... 267
Род сумеречных гладконосов Genus Nycticeius Rafinesque, 1819 . 268
Род желтых гладконосов Genus R/iogees H. Allen, 1866 .... 269
Род домовых гладконосов Genus Scotophilus Leach, 1821 .... 269
Род выростогубых гладконосов Genus Chalinolobus Peters, 1866 . . 270
Род волосатохвостых гладконосов Genus Lasiurus Gray, 1831 . . 270
Род широкоушек Genus Barbastella Gray, 1821 ................. 271
Род ушанов Genus Plecotus Geoffroy, 1818 ................... 272
Род пятнистых ушанов Genus Euderma H. Allen, 1892 ... 273
Род длиннокрылов Genus Miniopterus Bonaparte, 1837 . . 274
Род трубконосов Genus Murina Gray, 1842 ................... 275
Род шерстокрылых трубконосов Genus Harpiocephalus Gray, 1842 276
Род украшенных гладконосов Genus Kerivoula Gray, 1842 . . . 277
Род анамигдон Genus Anamigdon Troughton, 1929 ............ 278
Род пустынных гладконосов Genus Antrozous H. Allen, 1862 . . 278
Род австралийских гладконосов Genus Nyctophilus Leach, 1822 . . 279
Род ламингтонских гладконосов Genus Lamingtona McKean et Calaby, 1968 ............................................ 279
Род томопеас Genus Tomopeas Miller, 1900 ...... 280
Семейство новозеландских летучих мышей Familia Mystacinidae Simpson, 1945 ............................................ 280
Семейство бульдоговых летучих мышей Familia Molossidae Gill, 1872 281
Род эомопс Genus Eomops Thomas, 1905 ...................... 282
Род южноамериканских бульдоговых летучих мышей Genus Molossops Peters, 1865 ........................................ 283
Род голокожих бульдоговых летучих мышей Genus Cheiromeles Horsfield, 1824 ........................................ 283
Род ксифониктерис Genus Xiphonycteris Dollman, 1911 .... 284
Род складчатогубов Genus Tadarida Rafinesque, 1814 .... 284
Род бархатистых бульдоговых летучих мышей Genus Molossus E. Geoffroy, 1805 ........................................ 288
Род плоскоголовое Нового Света Genus Neoplatymops Peterson, 1965 289
Отряд приматов Ordo Primates Linnaeus, 1758 .... . . , . . 290
Подотряд низших приматов, или полуобезьян, Subordo Prosimii liliger, 1811 292
Семейство тупай Familia Tupaiidae Mivart, 1868 . ................. 292
Род обыкновенных тупай Genus Tupaia Raffles, 1822 ................ 293
Род индийских тупай, или анатан, Genus Anathana Lyon, 1913 . . 294
Род гладкохвостых, или горных, тупай Genus Dendrogale Gray, 1848 295
Род филиппинских тупай Genus Urogale Mearns, 1905 . ... 295
Род перьехвостых тупай Genus Ptilocercus Gray, 1848 .... 295
Семейство лемуров Familia Lemuridae Gray, 1821 ................. 296
Род полумаки, или кротких лемуров, Genus Hapalemur I. Geoffroy, 1851 .............................................. 297
Род маки, или обыкновенных лемуров, Genus Lemur Linnaeus, 1758 298
Род тонкотелых маки, или ласковидных лемуров, Genus Lepilemur
I. Geoffroy, 1851 ....................................... 300
Род крысиных маки, или карликовых лемуров, Genus Cheirogaleus Ё. Geoffroy, 1812 ............................... . . 300
Род мышиных лемуров Genus Microcebus Ё. Geoffroy, 1828 . . 301
Род вильчатополосых лемуров Genus Phaner Gray, 1870 .... 302
Семейство индри Familia Indridae Burnett, 1828 ................. 303
Род мохнатых индри Genus Lichanotus liliger, 1811................... 304
Род хохлатых индри Genus Propithecus Bennett, 1832 .... 305
Род короткохвостых индри Genus Indri Ё. Geoffroy et G. Cuvier, 1795 305
Семейство руконожек Familia Daubentoniidae Gray, 1870 .... 307
Семейство лори Familia Lorisidae Gregory, 1915........................ 309
Род тонких лори Genus Loris Ё. Geoffroy, 1796 ...................... 310
Род толстых лори Genus Nycticebus Ё. Geoffroy, 1812 .... 311
Род золотых потто, или медвежьих маки, Genus Arctocebus Gray, 1863 312
Род потто Genus Perodicticus Bennett, 1831.......................... 312
Род галаго Genus Galago Ё. Geoffroy, 1796 .......................... 313
Род иглокоготных галаго Genus Euoticus Gray, 1863 .................. 314
Семейство долгопятов Familia Tarsiidae Gill, 1872 .............. 315
Подотряд высших приматов, или обезьян, Subordo Anthropoidea Mivart, 1864 316
Семейство цепкохвостых обезьян, или капуцинов, Familia Cebidae Swain-son, 1835 .............................................. . 316
Род мирикин, или ночных обезьян, Genus Aotes Humboldt, 1811 . . . 318
Род прыгунов Genus Callicebus Thomas, 1903 .................... 318
Род уакари, или какайо, Genus Cacajao Lesson, 1840 ................. 319
Род саки Genus Pithecia Desmarest, 1804 ............. .... 320
Род красноспинных саки Genus Chiropotes Lesson, 1840 .... 321
Род ревунов Genus Alouatta Lacepede, 1799 .................... 321
Род капуцинов Genus Cebus Erxleben, 1777 .................... 323
Род саймири Genus Saimiri Voight, 1831 ............ . . 324
Род коат Genus Ateles Ё. Geoffroy, 1806 ............ ... 325
Род паукообразных обезьян Genus Brachyteles Spix, 1823 .... 325
Род шерстистых обезьян Genus Lagothrix E. Geoffroy, 1812 . . . 326
Семейство игрунок, или мармозеток, Familia Callithricidae Thomas, 1903 .................................................. 327
Род мармозеток Genus Callimico Mirando Ribeiro, 1911................ 328
Род игрунок Genus Callithrix Erxleben, 1777 ........................ 328
Род львиных игрунок Genus Leontideus Cabrera, 1956 .... 330
Род тамаринов Genus Saguinus Hoffmannsegg, 1807 ........... 331
Семейство низших узконосых обезьян, или мартышек, Familia Cercopi-thecidae Blanford, 1888 ... ... ............. 332
Род макак Genus Масаса Lacepede, 1799 ... ........... 334
Род хохлатых,или черных, павианов Genus Cynopithecus I.Geoffroy, 1835 335
Род мангабеев Genus Cercocebus Ё. Geoffroy, 1835 .............. 336
Род павианов Genus Papio Miiller, 1777 . . ... 337
Род гелад Genus Theropithecus I. Geoffroy, 1843..................... 339
Род мартышек Genus Cercopithecus Briinnich, 1772 . 339
Род тонкотелых обезьян, или лангуров, Genus Pygathrix Ё. Geoffroy, 1812 .............................................. 341
Род ринопитеков Genus Rhinopithecus Milne-Edwards, 1872 . . . 342
Род симиасов Genus Simias Miller, 1903 ............................. 343
Род носачей Genus Nasalis Ё. Geoffroy, 1812......................... 343
Род толстотелых обезьян, или колобусов, Genus Colobus liliger, 1811 344
Семейство высших узконосых обезьян, или человекообразных обезьян, Familia Pongidae Elliot, 1913 ... ................... 345
Род гиббонов Genus Hylobates liliger, 1811 ................ 347
Род орангутанов Genus Pongo Lacepede, 1799 .......... 349
Род шимпанзе Genus Pan Oken, 1816 ........................ 349
Род горилл Genus Gorilla I. Geoffroy, 1852 ..... 351
Семейство людей Familia Hominidae Gray, 1825 ...................... 353
Отряд неполнозубых Ordo Edentata Gray, 1798 ....................... 355
Семейство муравьедов Familia Myrmecophagidae Bonaparte, 1838 . . 356
Род гигантских, или трехпалых, муравьедов Genus Myrmecophaga Linnaeus, 1758 ....................................... 357
Род тамандуа, или четырехпалых муравьедов, Genus Tamandua Gray, 1825 ............................................................ 358
Род карликовых, или двупалых, муравьедов Genus Cyclopes Gray, 1821 ............................................................ 359
Семейство ленивцев Familia Bradypodidae oBnaparte, 1831 .... 360
Род трехпалых ленивцев Genus Bradypus Linnaeus, 1758 .... 361
Род двупалых ленивцев Genus Choloepus liliger, 1811 ... 361
Семейство броненосцев Familia Dasypodidae Bonaparte, 1838 . . 362
Род щетинистых броненосцев Genus Chaetophractus Fitzinger, 1871 363
Род шестипоясных броненосцев Genus Euphractus Wagler, 1830 . 363
Род карликовых броненосцев Genus Zaedyus Ameghino, 1889 . . 364
Род гигантских броненосцев Genus Priodontes F. Cuvier, 1827 . . 365
Род голохвостых броненосцев Genus Cabassous McMurtrie, 1831 . 365
Род трехпоясных броненосцев Genus Tolypeutes liliger, 1811 . . 366
Род девятипоясных броненосцев Genus Dasypus Linnaeus, 1758 . 368
Род плащеносных броненосцев Genus Chlamyphorus Harlan, 1825 369
Род щитоносных броненосцев Genus Burmeisteria Gray, 1865 . . 370
Отряд ящеров Ordo Pholidota Weber, 1904 ........................... 370
Литература...................................................... . . 374
Предметный указатель............................................... 378
ВЛАДИМИР ЕВГЕНЬЕВИЧ СОКОЛОВ
СИСТЕМАТИКА МЛЕКОПИТАЮЩИХ
Редактор Л. П. Петровская Художник Л. В. Алексеев Художественный редактор Т. А. Коленкова Технический редактор Г. Г. Киселева Корректор С. К. Марченко
Сдано в набор 8/VIII—72 г. Поди, к печати 16/1—73 г. Формат 70Х108*/1с- Объем 27 печ. л. Усл. п. л. 37,80. Уч.-изд. л. 38,40. Изд. № Е—212. Тираж 20 000 экз. Зак. 565. Цена I р. 69 к.
План выпуска литературы издательства «Высшая школа» (вузы и техникумы) на 1973 г. Позиция № 84.
Москва, К-51, Неглииная ул., д. 29/14 Издательство «Высшая школа»
Ярославский полиграфкомбинат «Союзполиграфпрома» прн Государственном комитете Совета Министров СССР по делам издательств, полиграфии и книжной торговли. Ярославль, ул. Свободы, 97.