/























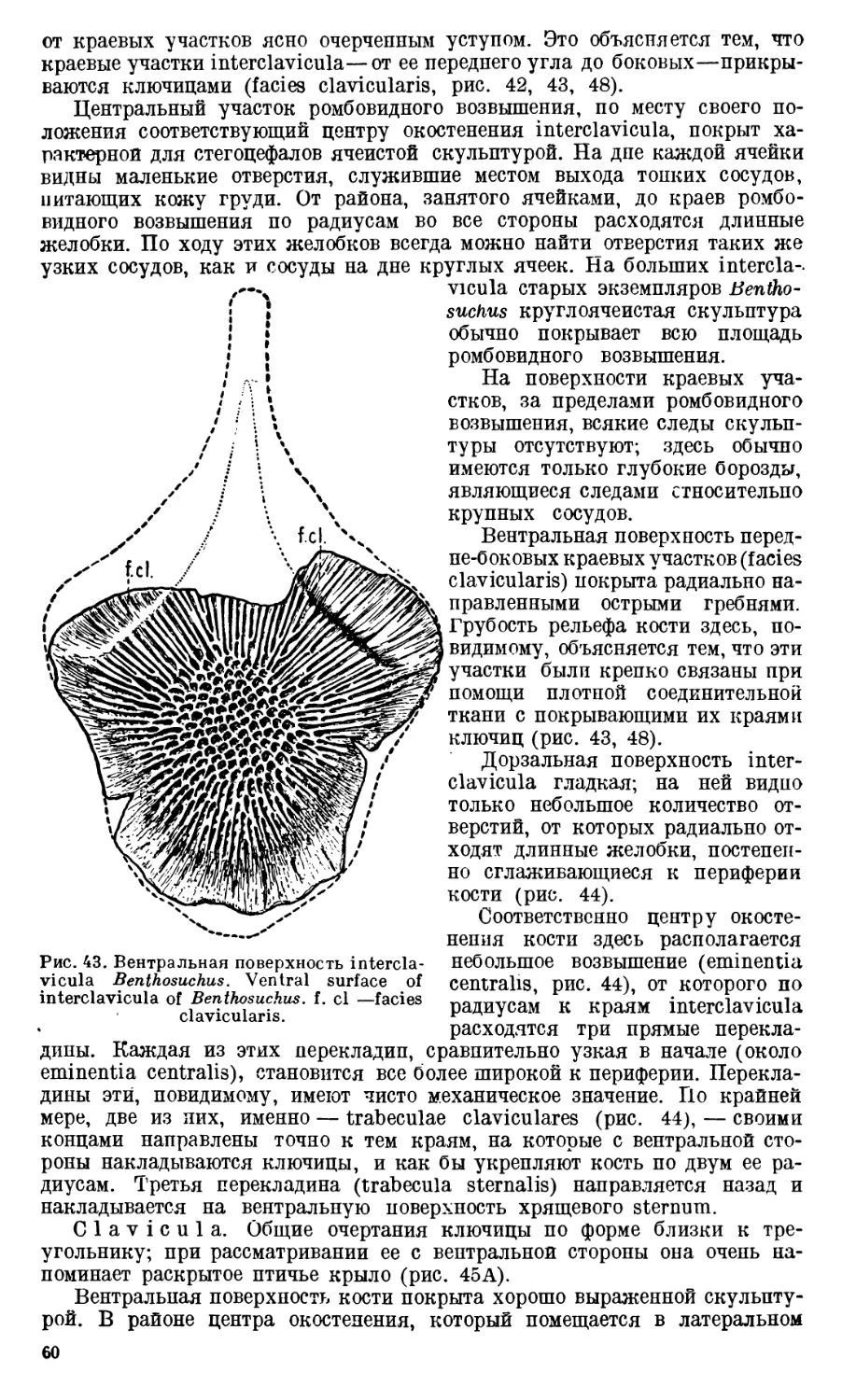
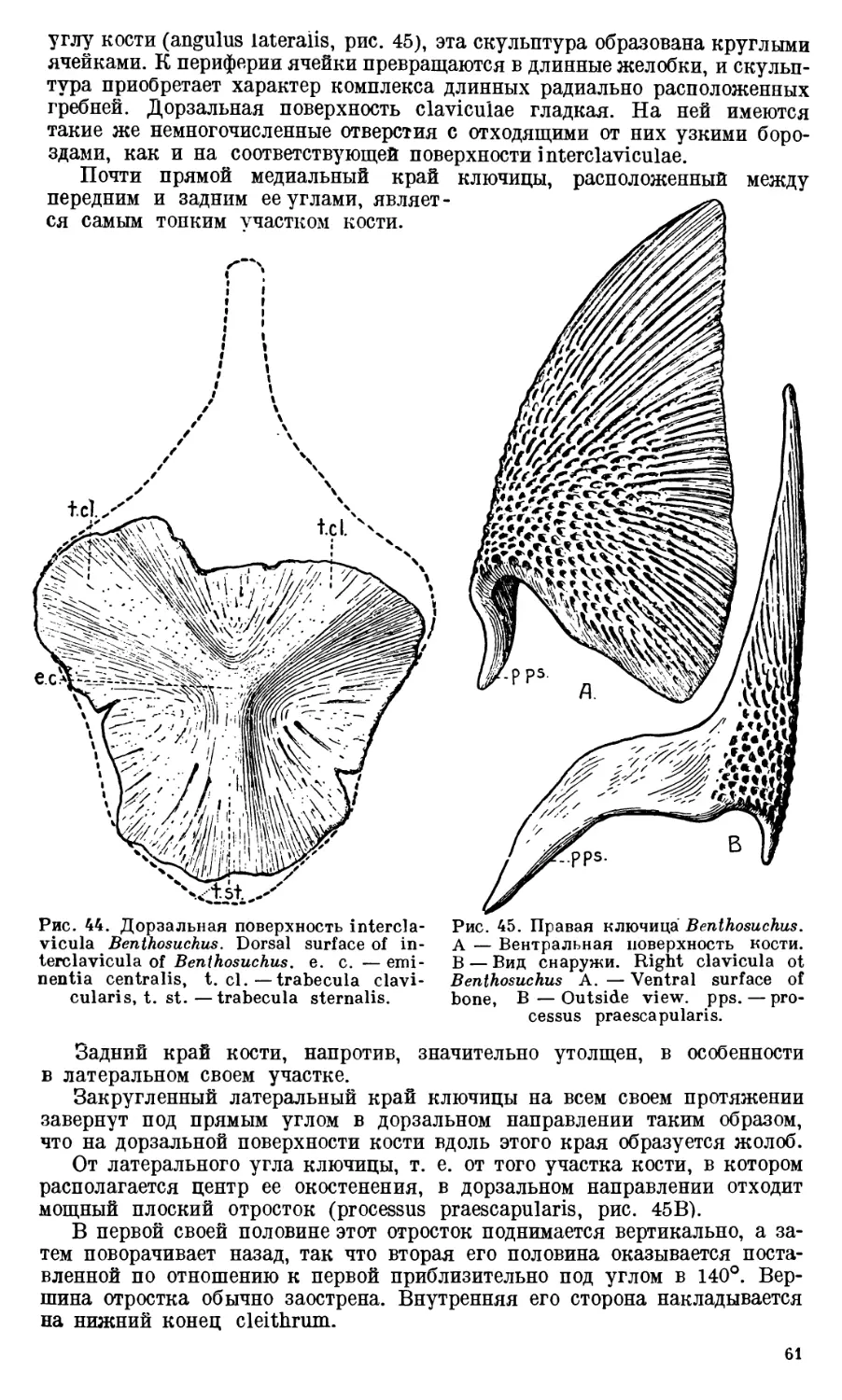
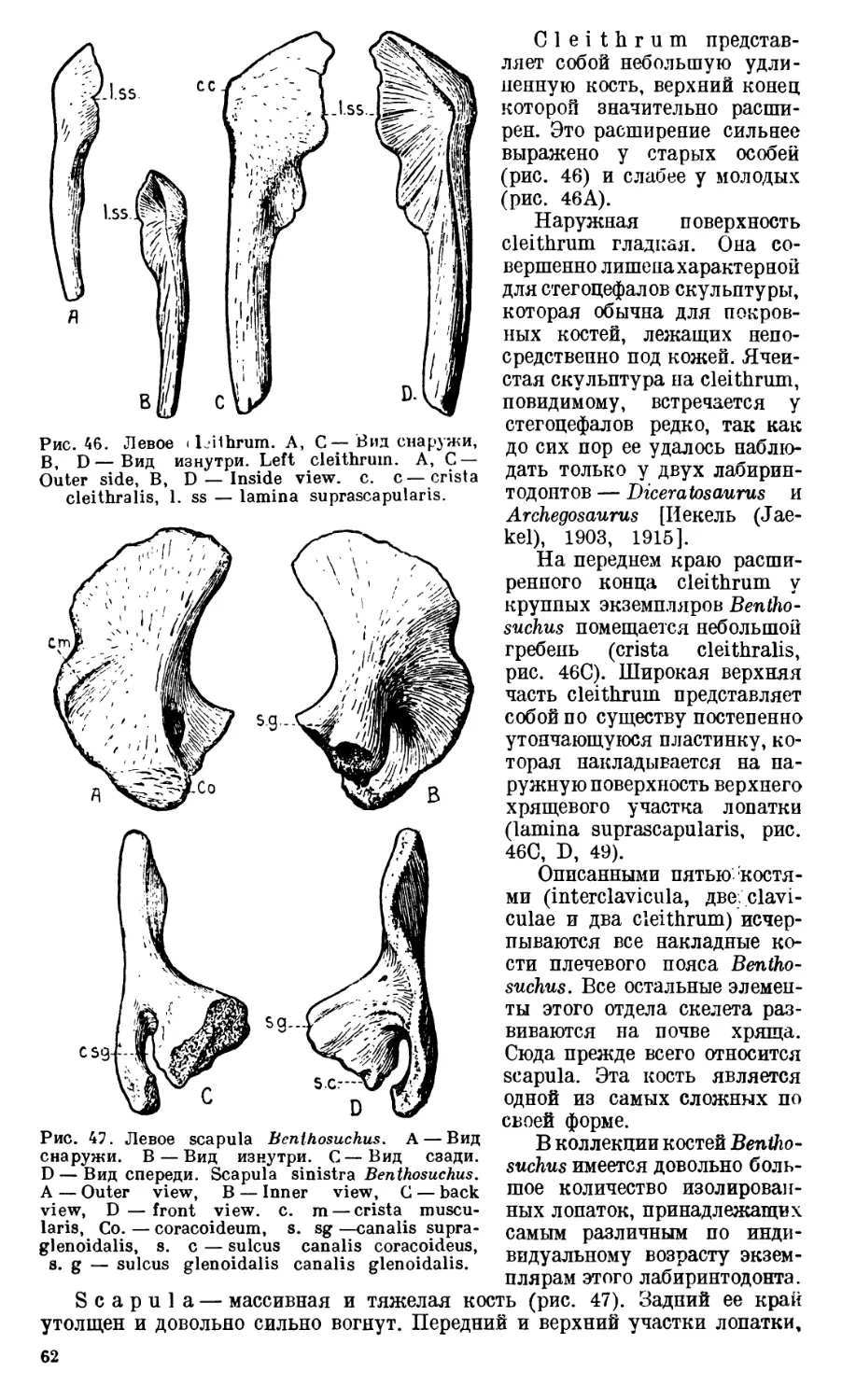




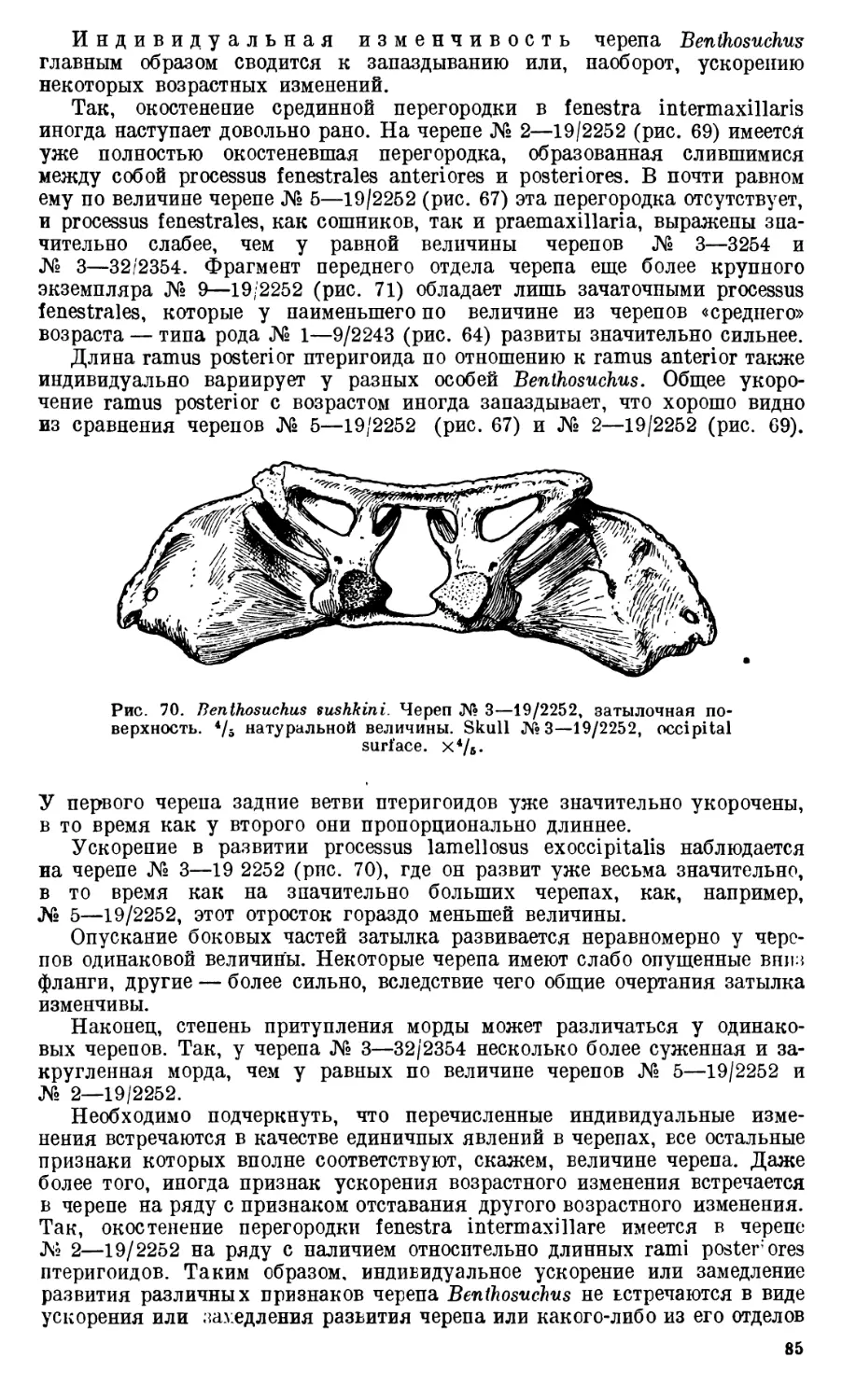
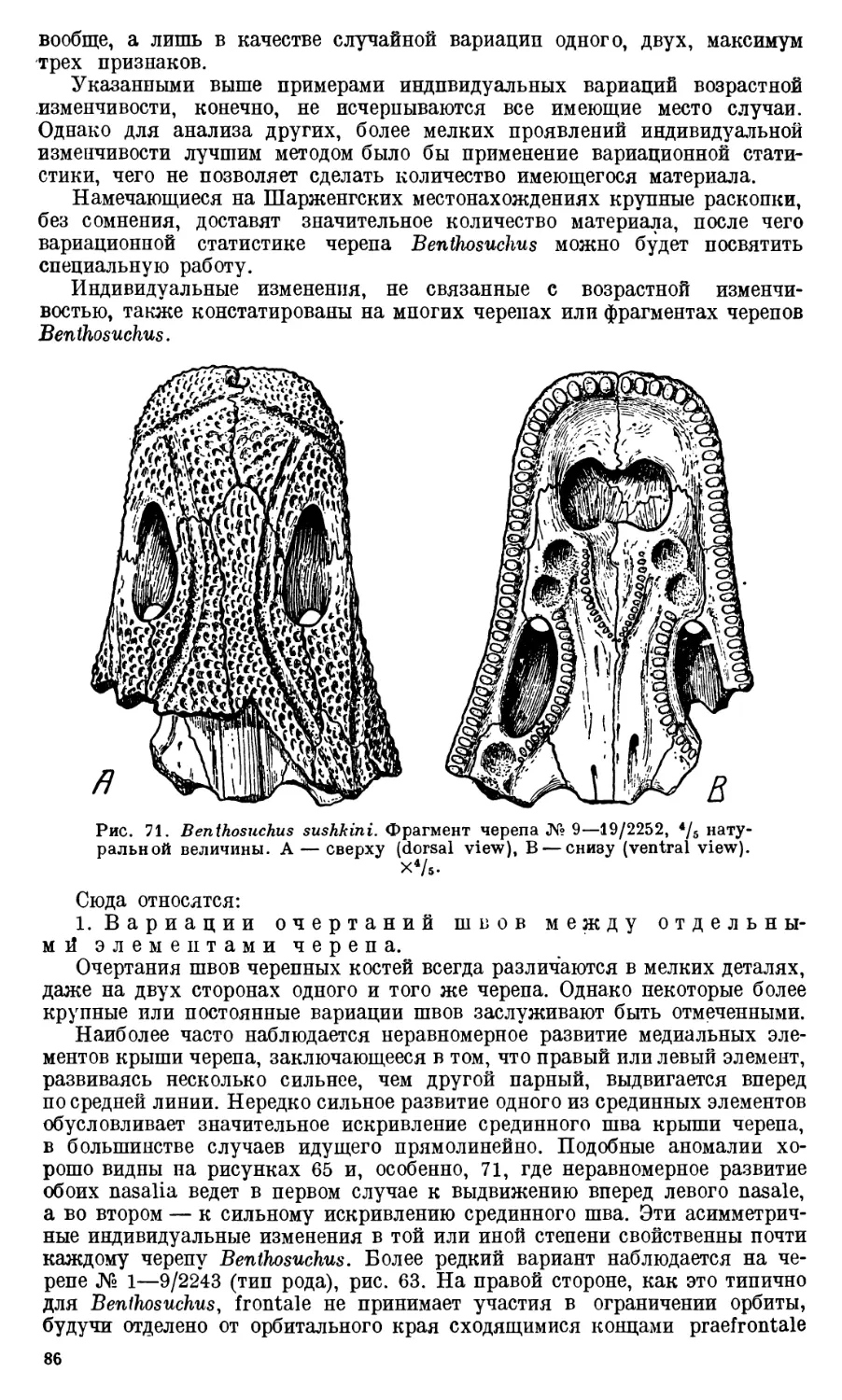








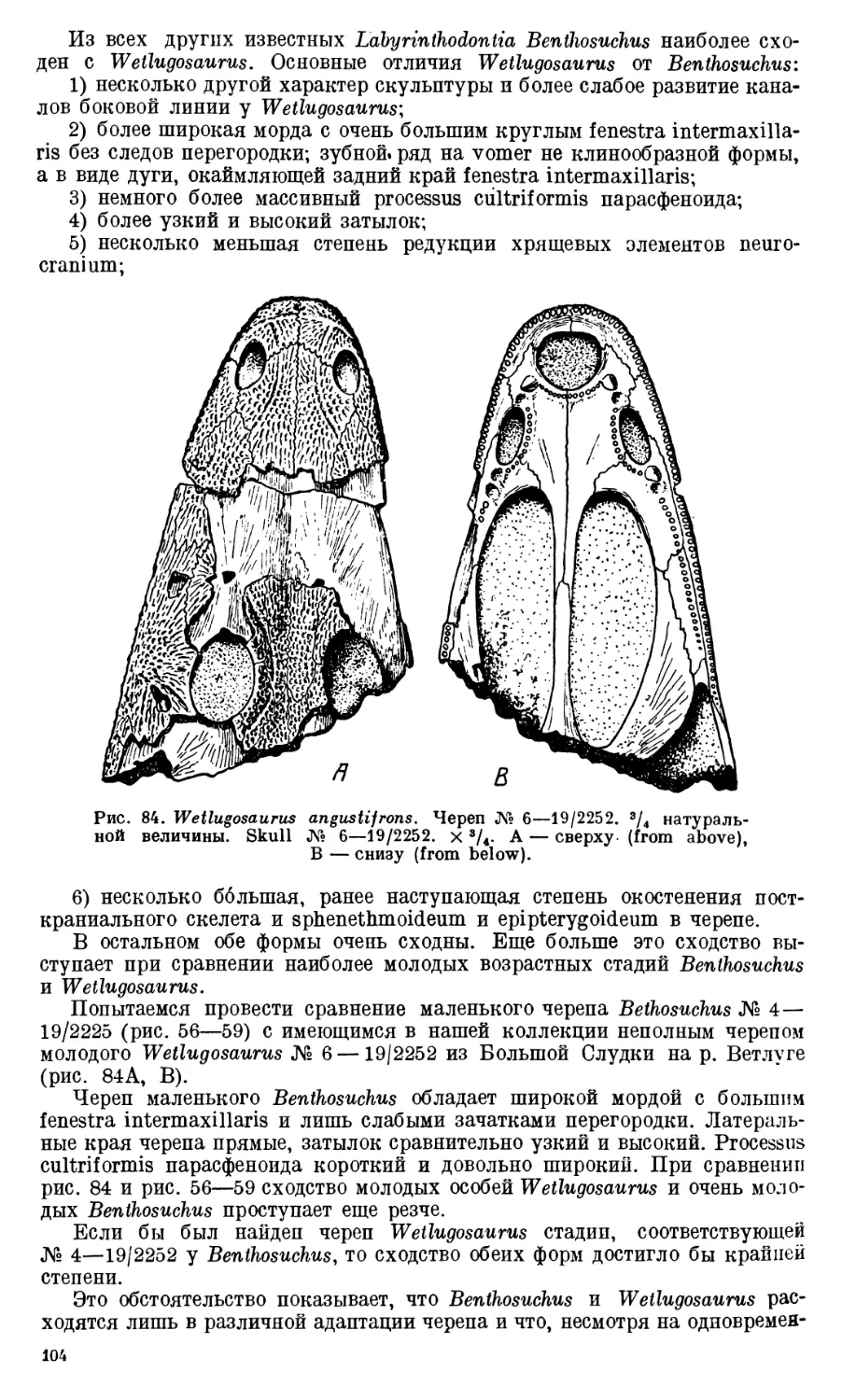
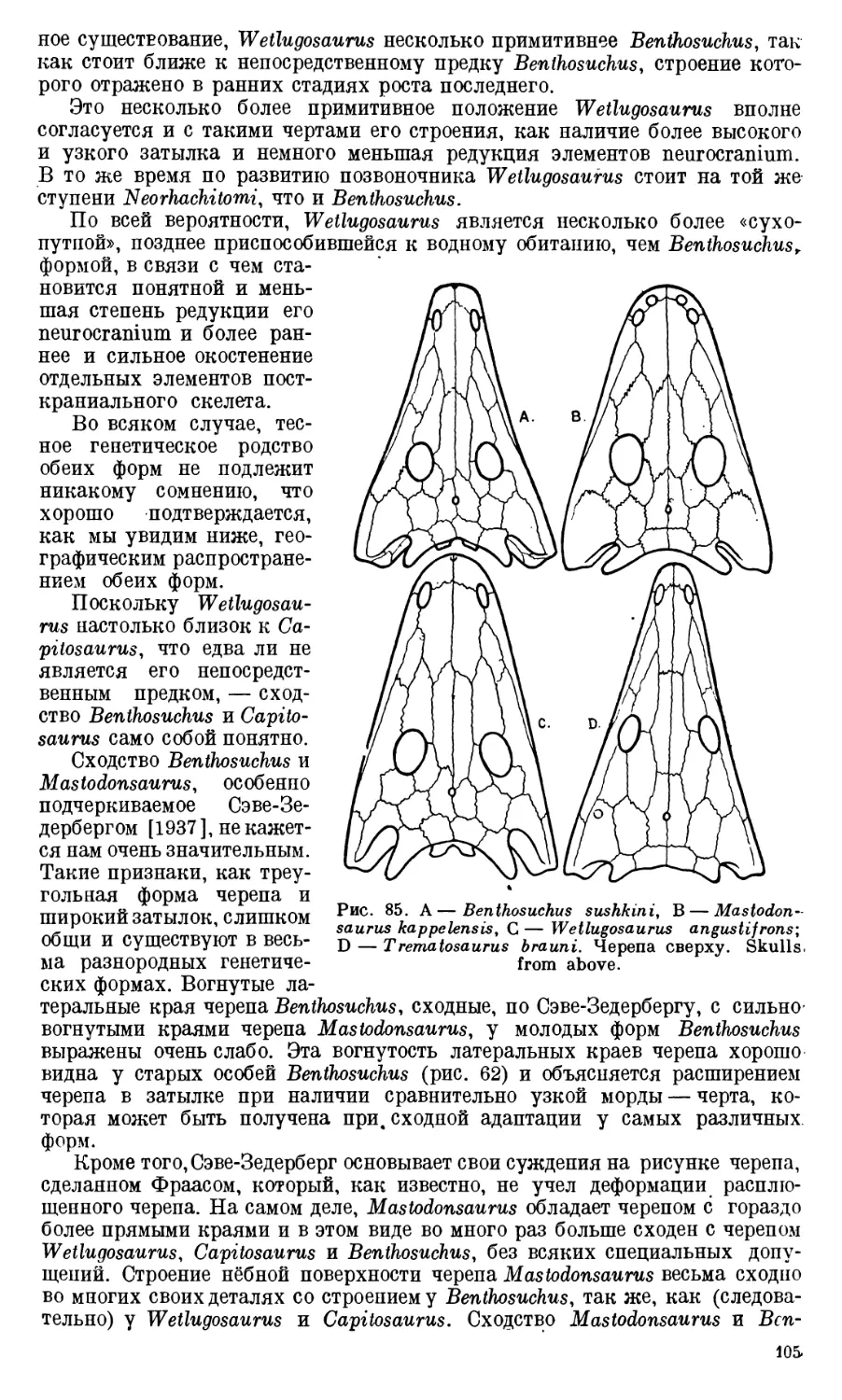
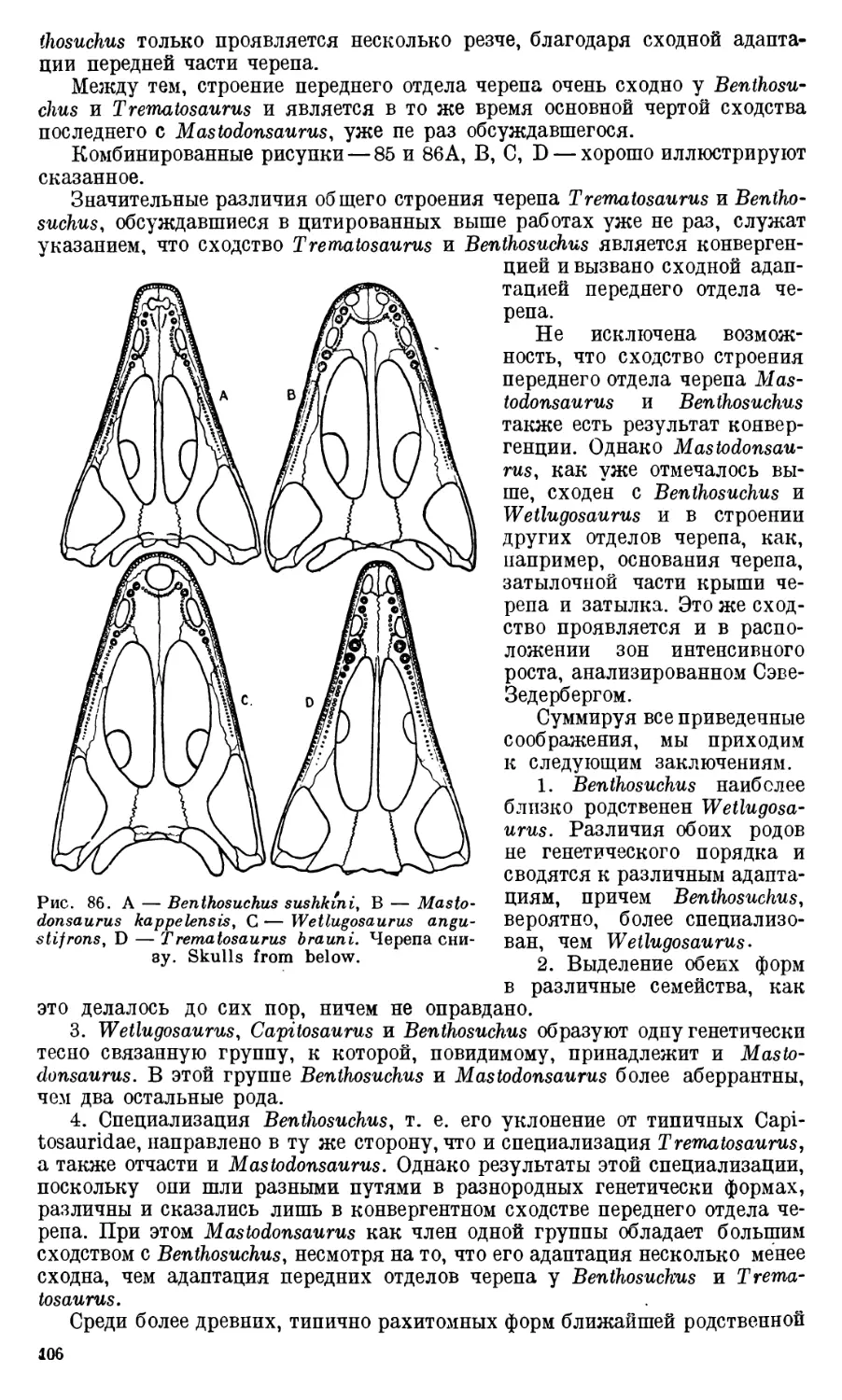


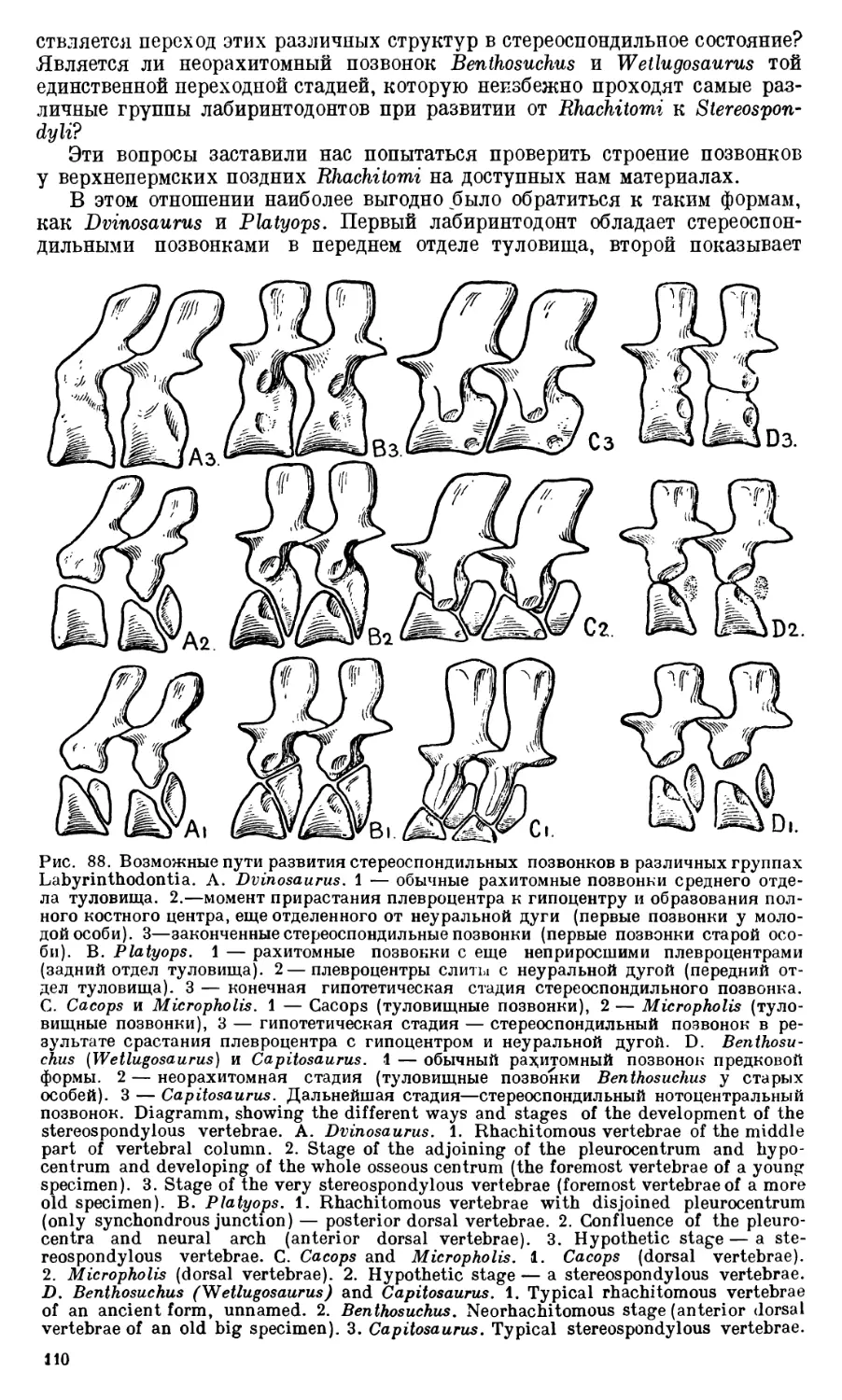












































Text
АКАДЕМИЯ НАУК
СОЮЗА СОВЕТСКИХ СОЦИАЛИСТИЧЕСКИХ РЕСПУБЛИК
ТРУДЫ
ПАЛЕОНТОЛОГИЧЕСКОГО
ИНСТИТУТА
ТОМ X
ВЫПУСК 1
А. И. БЫСТРОВ и И. А. ЕФРЕМОВ
BENTHOSUCHUS SUSHKINI EFR.— ЛАБИРИНТОДОНТ
113 ЭОТРИАСА РЕКИ ШАРЖЕНГИ
А. Р. BYSTROW and J. A. EFREMOV
BENTHOSUCHUS SUSHKINI EFR. —A LABVRINTHODONT
FROM THE EOTRIASSIC OF SHARJENGA RIVER
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
СОДЕРЖАНИЕ
5
7
12
Введение. ,.......................
Список исследованного материала .
Часть I
Анатомия скелета..................
Часть II
Introductory ................... . 12С
List of materials investiged . 121
Part I
Skeleton anatomy .......................121
Part II
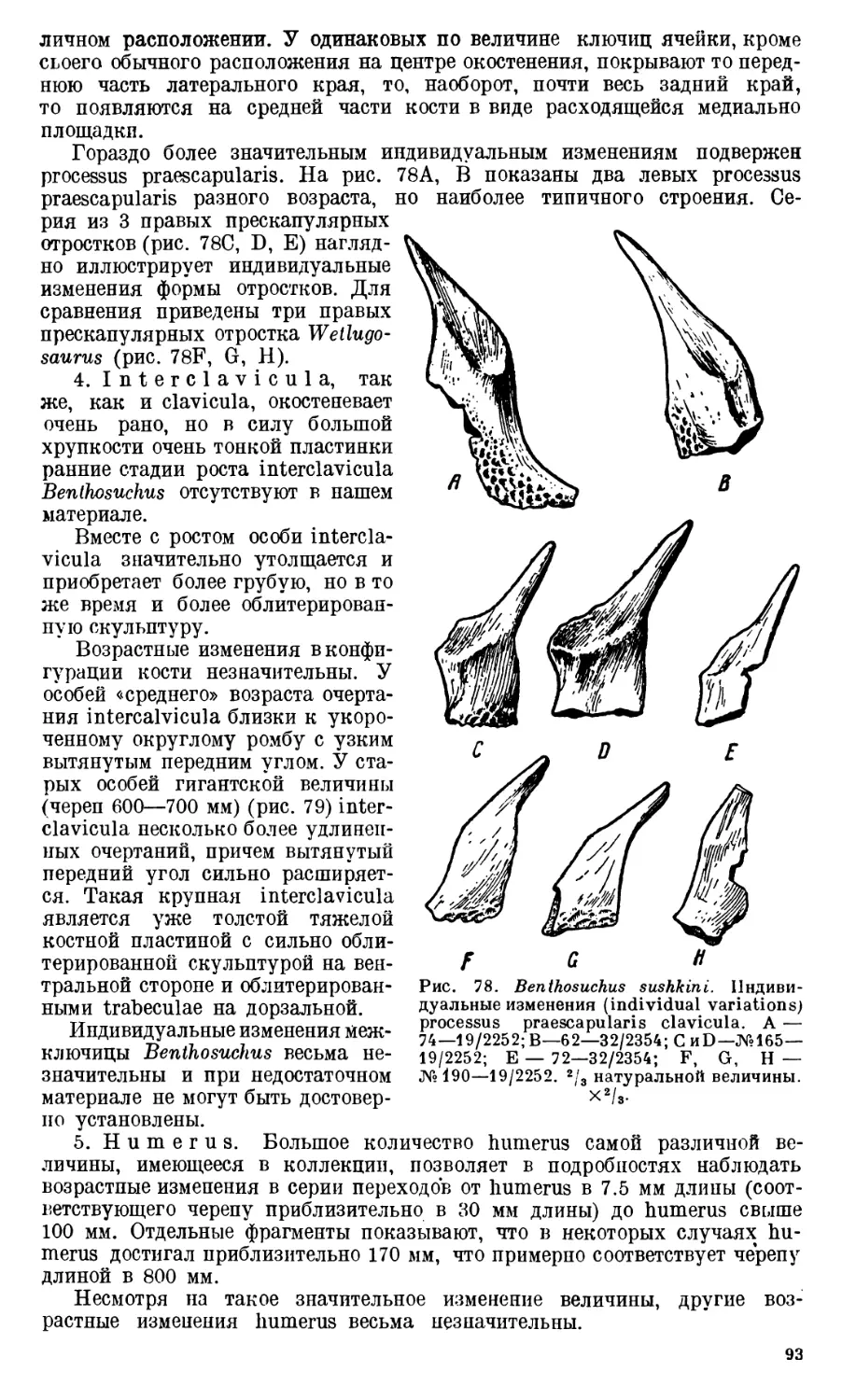
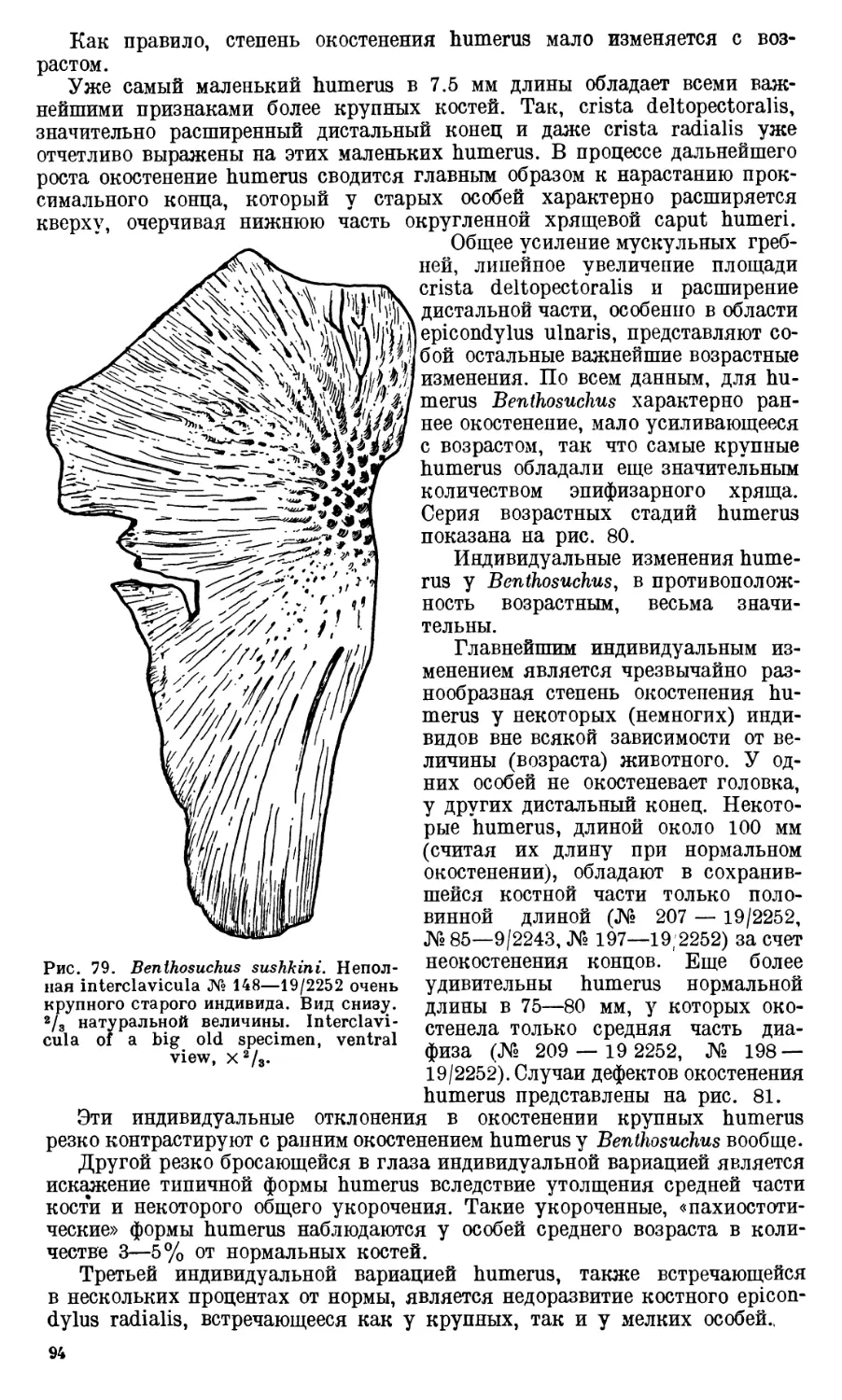
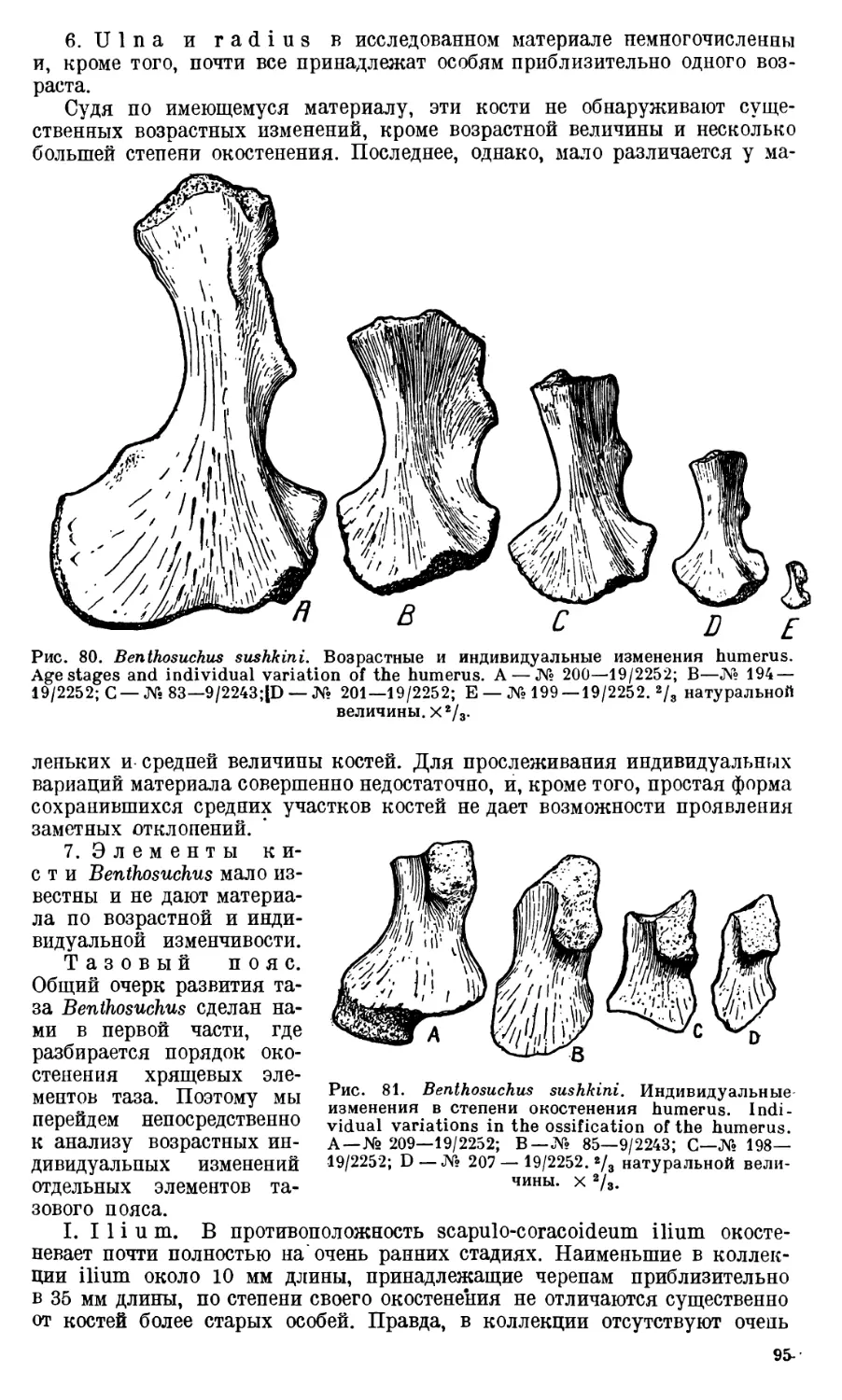
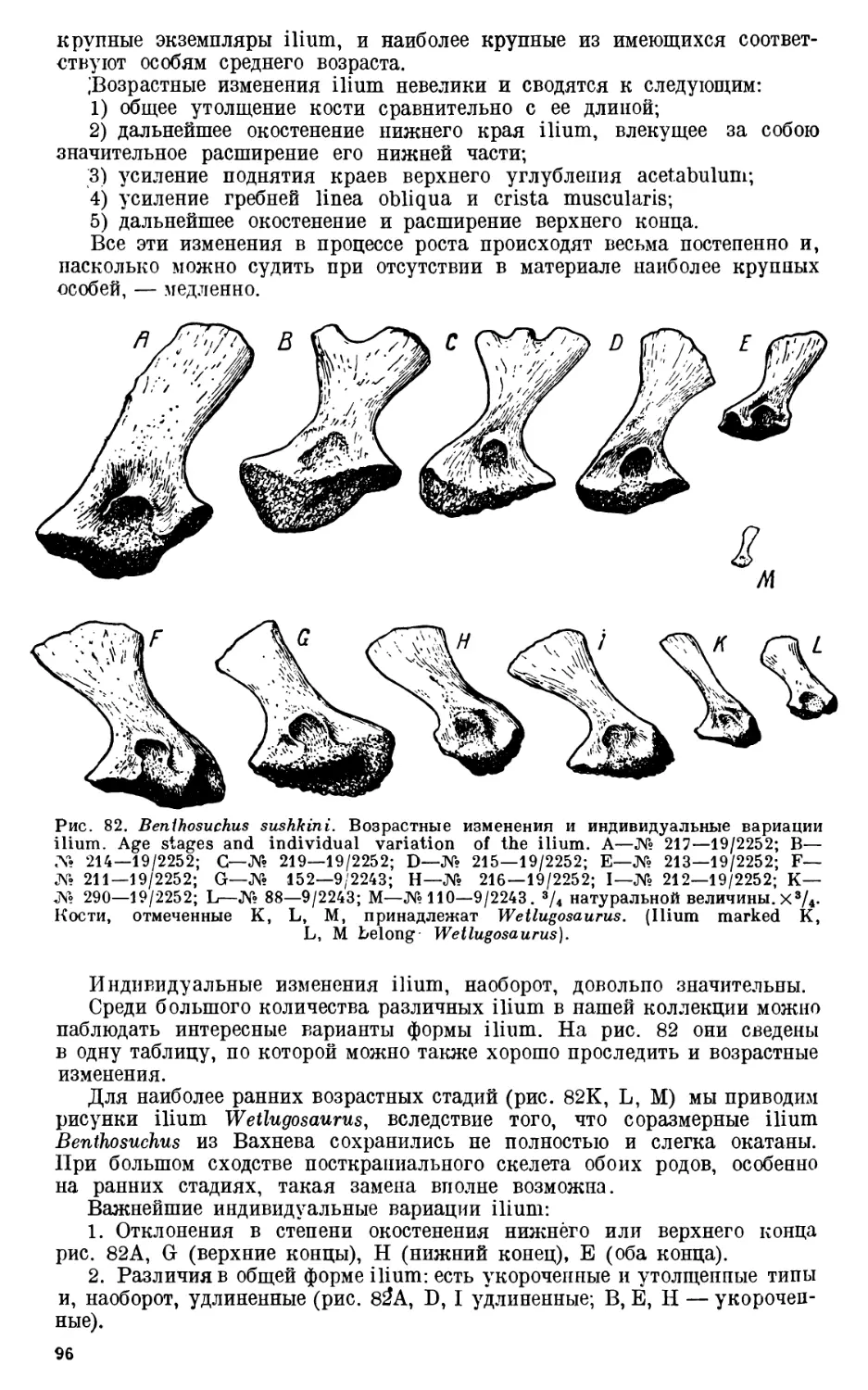
Глава I. Возрастные и индиви-
дуальные изменения скелета
Benthosuchus.....................72
Глава II. Систематическое поло-
жение и филогенетическое зна-
чение Benthosuchus sushkini
Efr.............................102
Глава III. Некоторые соображе-
ния о происхождении ^стерео-
спондильных позвонков у выс-
ших Labyrinthodontia...........108
. Глава IV. Условия захоронения,
распространение и фаунисти-
ческие связи Benthosuchus . 112
Chapter I. Age and individual
changes of the skeleton of
Benthosuchus .... 132
Chapter II. Systematic position ♦
and phylogenetic Significance
of Benthosuchus sushkini Efr. 139
Chapter III. Some considerations
on the Origin of Stereospondy-
lous Vertebrae in higher La-
byrinthodontia ..................143
Chapter IV. Conditions of Burial,
Spreading and faunistical Data
of Benthosuchus..................147
ACADEMIE DES SCIENCES
DE L’UNION DES REPUBLIQUES SOCIALISTES SOVIETIQJJES
TRAVAUX
DE L’INSTITUT PALEONTOLOGIQUE
ТОМЕ X
LTVRAISON 1
A. P. BYSTROW and J. A. EFREMOV
BENTHOSUCHUS SUSHKINI EFR.A LABYRINTHODONT
FROM THE EOTRIASSIC OF SHARZHENGA RIVER
EDITION DE L’ACADEMIE DES SCIENCES DE LU RSS
MOSCOU 1940 LENINGRAD
АКАДЕМИЯ НАУК
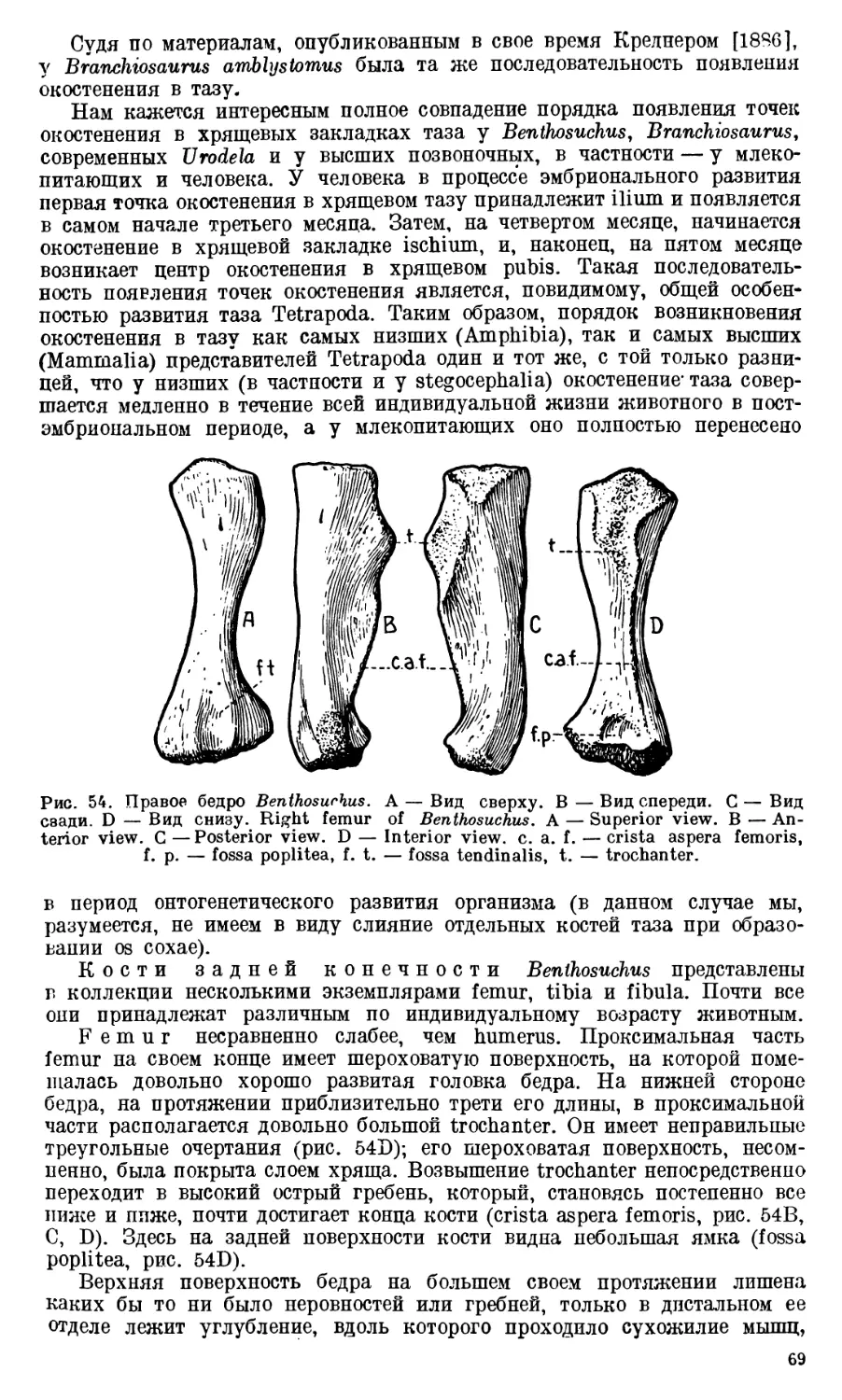
СОЮЗА СОВЕТСКИХ СОЦИАЛИСТИЧЕСКИХ РЕСПУБЛИК
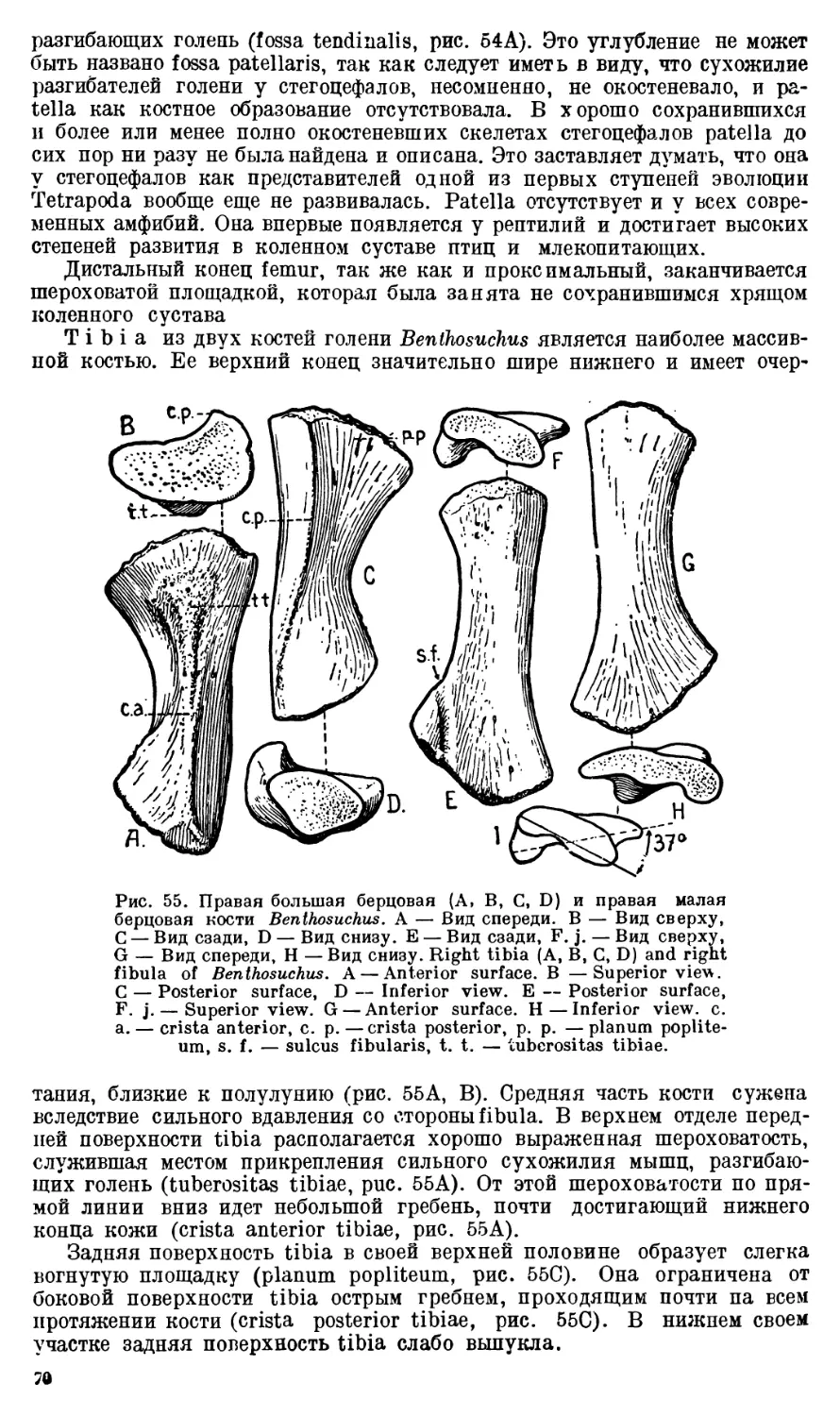
ТРУДЫ
ПАЛЕОНТОЛОГИЧЕСКОГО ИНСТИТУТА
ТОМ X
ВЫПУСК 1
А. П. БЫСТРОВ и И. А. ЕФРЕМОВ
BENTHOSUCHUS SUSHKINI EFR.- ЛАБИРИНТОДОНТ
ИЗ ЭОТРИАСА РЕКИ ШАРЖЕНГИ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК
МОСКВА 1940 ЛЕНИНГРАД
ОТВЕТСТВЕННЫЙ РЕДАКТОР
директор Палеонтологического института акад. Л. Л. Борисяк
Технический редактор А. П. Д р о н о в
Корректор И. Л. Го л о в ч и н е р
Слано в набор 11/VI 1939 г. Подписано к печати 110/IV* 1940г. Формат TOXlOSVie- Бум. л. 4’/*. Объем 94/а п. л.
В 1 п. л. 58000 печ. зн. Уч.-авт. 13,7 л. Тираж 850 экз. Уполн. Главлита № А-21572.
РИСО № 833. АНИ № 1371. Заказ № 2691.
1-я Образцовая тип. Огиза РСФСР треста .Полпграфкнига*. Москва, Валовая, 28.
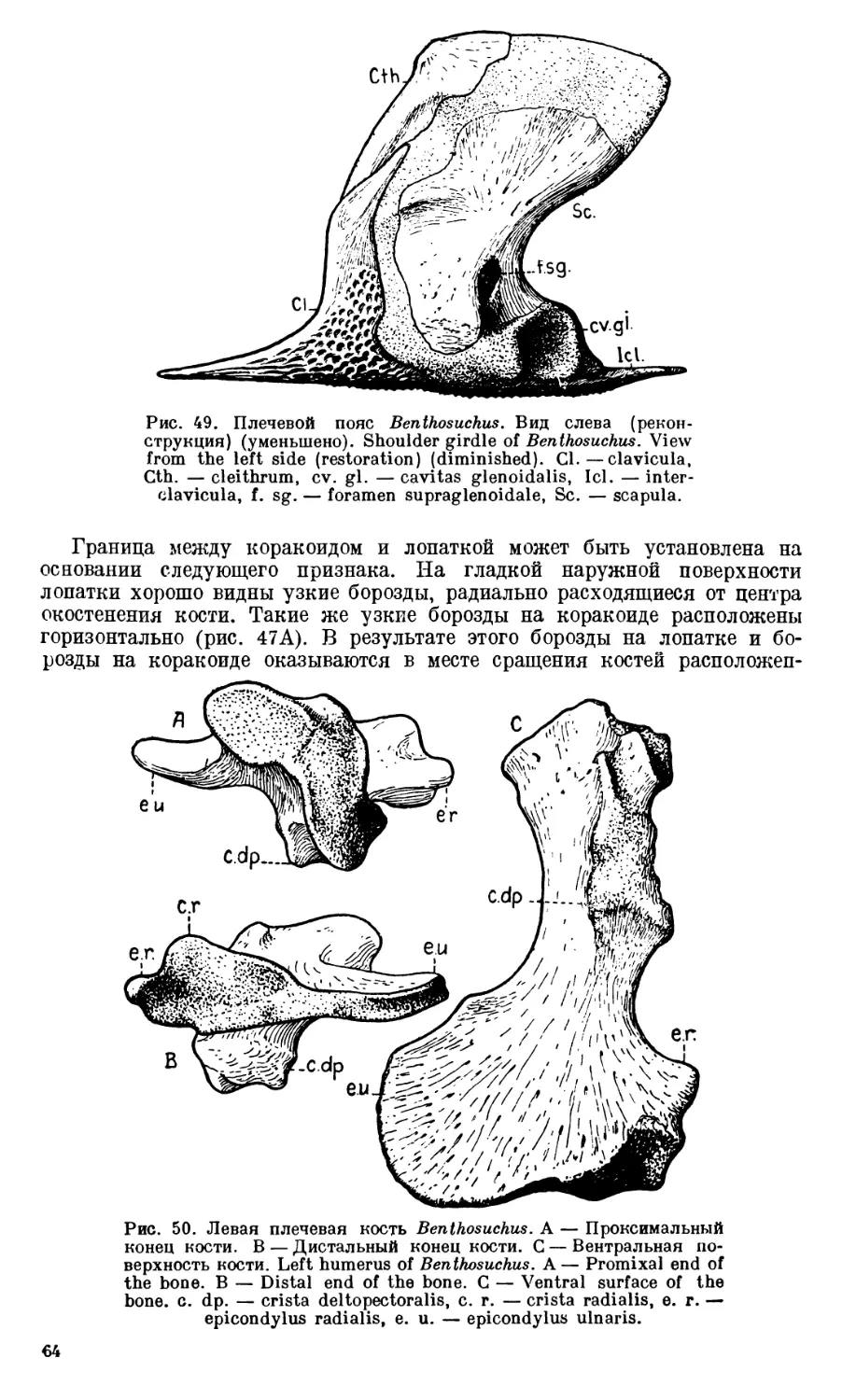
ВВЕДЕНИЕ
В 1927 г. в прослое серых песков и конгломератовидных песчаников,
залегающем среди толщи эотриасовых пестрых мергелей и глин по берегам
р. Шарженги (левого притока р. Юг), И. А. Ефремов открыл местонахожде-
ния, богатые остатками лабиринтодонтов. В последующие 1928—1929 гг. на
этих местонахождениях производились раскопки, доставившие большой
палеонтологический материал.
По полному черепу, найденному в 1927 г. И. А. Ефремовым [19291,
установлен новый род Benthosaurus sushkini, впоследствии [1936] переиме-
нованный в Benthosuchus. Предварительное изучение всего собранного ма-
териала показало, что в шарженгских местонахождениях представлен по
существу только один этот род лабиринтодонтов, так как среди нескольких
сот отдельных костей и фрагментов, принадлежащих Benthosuchus, найден
всего один обломок черепа, относящийся к другой, близко родственной фор-
ме. Таким образом, в Benthosuchus мы имеем лабиринтодонта, охарактери-
зованного девятью полными черепами (принадлежащими экземплярам
различного индивидуального возраста), большим количеством отдельных
черепных фрагментов и многочисленными разрозненными костями пост-
крапиального скелета. Все кости шарженгских местонахождений превосход-
ной сохранности и заключены в сравнительно рыхлых песчаниках, что поз-
волило отпрепаровать весь материал с исключительной полнотой и выяс-
нить такие детали в строении скелета, какие обычно оказываются или со-
вершенно недоступными для исследования, или обнаруживаются только
в случайно разбитых черепах.
Большое количество отдельных фрагментов позволяло не останавли-
ваться перед их разрушением для более подробного изучения. Большим
недостатком материала шарженгских местонахождений является отсут-
ствие полного скелета хотя бы одного экземпляра этого лабиринтодонта.
Процессы захоронения остатков Benthosuchus протекали таким образом,
что в костеносном пласту оказались беспорядочно перемешанными отдель-
ные разрозненные кости особей самого различного возраста. Поэтому точ-
ную реконструкцию всего скелета следует признать невозможной, так как
трудно составить себе точное представление о пропорциональных отноше-
ниях отдельных элементов скелета в целом. В качестве сравнительного ма-
териала при изучении Benthosuchus были использованы остатки другого,
очень близко родственного рода Wetlugosaurus, который распространен
в таких же конгломератовидных песчаниках эотриаса в бассейнах рр. Вет-
луги и Верхней Волги. Разрозненные остатки Wetlugosaurus имеются
в коллекциях И. А. Ефремова из дер. Большой Слудки и с. Зубовского на
р. Ветлуге и хранятся так же, как и коллекция шарженгских Benthosuchus,
в Палеонтологическом институте Академии Наук СССР.
Кроме того, для сравнения использованы любезно предоставленные
нам проф. А. Н. Рябининым оригиналы его работы о Wetlugosaurus апди-
stifrons из Большой Слудки на р. Ветлуге. В последней коллекции имеется
часть передней половины скелета Wetlugosaurus с позвоночником, ребрами,
5
плечевым поясом и затылочным отделов! черепа in situ, дающая полное пред-
ставление о пропорциях этих частей скелета.
Задачей настоящей работы является подробное описание Benthosuchus
как примера поздних лабиринтодонтов. В настоящее время большинство
известных Labyrinthodontia изучено совершенно недостаточно. Это объяс-
няется в огромном большинстве случаев неполной препаровкой изученного
материала или плохой его сохранностью. Наиболее полно известен Eryops,
которому посвящено несколько специально морфологических работ [Кейз
(Case), 1911; Хюне (V. Huene), 1912; Брум (Broom), 1913; Виллистон
(W. Williston), 1916; Демпстер (Dempster), 1935]. Однако и этот лабиринто-
донт еще нуждается в более детальной анатомической обработке.
В последнее время Сэве-Зедерберг (Save-Soderbergh, 1936) вновь пере-
работал материал, описанный в свое время Виманом (Wiman, 1914, 1916,
1917), и дал подробное описание черепов Aphaneramma, Lyrocephalus, Pla-
ty steда, сделав при этом попытку восстановить не сохранившиеся хряще-
вые части, сосуды и нервы головы этих форм.
Работа Сэве-Зедерберга дает наиболее полное представление о морфо-
логических особенностях черепа лабиринтодонтов. Однако плохая сохран-
ность материала не позволила Сэве-Зедербергу выяснить еще многие
интересные анатомические детали в строении черепа стегоцефалов. Что ка-
сается посткраниального скелета, то его в распоряжении Сэве-Зедерберга
совсем не было.
Приступая к совместной работе, оба автора разделили подготовку ма-
териала в целях ускорения обработки. Так, весь большой остеологический
материал был в деталях отпрепарован И. А. Ефремовым с выявлением всех
подробностей взаимоотношений отдельных элементов. Все черновые зари-
совки сделаны А. П. Быстровым, после чего оба автора приступили к со-
вместному изучению полученного фактического материала и его морфологи-
ческой трактовке. При этом мы встретились с необходимостью ввести не-
которое количество новых латинских наименований-для ряда специфических
деталей строения скелета Benthosuchus. В большинстве случаев мы сохра-
няли общепринятую латинскую номенклатуру и изменяли ее только тогда,
когда она представлялась нам неудачной.
Работа разделена на две части. В первой дается анатомическое описание
скелета Benthosuchus вообще, в средней норме различных индивидуальных
и возрастных особенностей.
Вторая часть заключает в себе описание условий захоронения остатков
Benthosuchus, возрастной и индивидуальной изменчивости элементов скелета,
общие выводы относительно эволюционной стадии и родственных связей
Benthosuchus, а также трактовку развития позвоночного столба у типичных
laby-rin thodon tia.
Составление первой части было выполнено А. П. Быстровым, второй —
И. А. Ефремовым. Все рисунки к обеим частям работы сделаны А. II. Бы-
стровым.
Палеонтологический институт
Академии Наук СССР
Москва.
6
СПИСОК ИЗУЧЕННЫХ МАТЕРИАЛОВ
(List of materials investigated)
Список дан по каталогам Палеонтологического института АН СССР
в хронологическом порядке поступления коллекций за три года сборов
с 1927 по 1929 г. Коллекции № 9/2253 и № 19/2252 разделены нами на два
отдела каждая. В первый отдел обеих коллекций включены остатки Bentho-
suchus с р. Шарженги, второй отдел выборочно составлен из сборов остат-
ков Wetlugosaurus на р. Ветлуге. Такое деление удобнее для восприятия,
каталоги же были составлены в большинстве случаев в анатомическом по-
рядке следования скелетных частей от черепа до хвоста. Остатки других
представителей эотриасовой фауны — рептилий и рыб — из списка исклю-
чены.
Для показа распределения материала по различным раскопкам в катало-
гах проставлены знаки Р с соответствующей цифрой. Эти знаки для шар-
женгских местонахождений ус. Вахнева означают: Рх—раскопка 1-я па
правом берегу р. Шарженги у мельницы, против села Вахнева; Р2 — там
же, немного выше по течению; Р3 — раскопка 3-ья на правом берегу р. Шар-
женги, сразу ниже устья рч. Медвежьей; Р4 — раскопка 4-я, левый берег
рч. Медвежьей, в 150 м выше устья; Р5— раскопка 5-я, правый берег рч. Мед-
вежьей, в 100 м выше ее устья. Для сборов всех трех лет обозначения раско-
пок одинаковы, так как работа производилась в одних и тех же точках.
Для р. Ветлуги обозначения читаются так: Рх — раскопка 1-я, правый
берег р. Ветлуги, выше дер. Большая Слудка, в 80 м выше оврага; Р2 — рас-
копка 2-я там же, непосредственно на мысу оврага; Р3 — правый берег
р. Ветлуги, у с. Зубовского, под церковью.
Нужно отметить, что по окончании изучения материала трактовка рас-
положения объектов на правой или левой стороне скелета в отдельных слу-
чаях изменилась. Однако мы даем в списке эти объекты так, как они запи-
саны в каталогах, — по первичным определениям, — чтобы избежать рас-
хождения с каталогами, которые остались пока неизмененными, полагая,
что в данном случае это не имеет существенного значения.
№ 9/2243 ПИН. Коллекция И. А. Ефремова 1927 г. Волго-Двинское
междуречье рр. Шарженга и Ветлуга
1. Benthosuchus sushkini Efr., р. Шарженга, с. Вахнево.
1. Cranium (генотип). Рх. 2. Cranium, pars medialis. Рх. 3. Cranium,
pars narialis. Px. 4. Cranium, pars narialis. P3. 5. Dermocranium, pars tem-
poralis dext. очень крупного черепа. P3. 6. Basis cranii. P3. 7. Stapes огром-
ной величины. P3. 8, Cranium, pars infraorbitalis. P3. 9. Cranium, 2 облом-
ка. P3. 10. Endocranium, pars pterygo-parasphenoidalis. P3. 11. Cranium,
pars praemaxillaris dext. Px. 12. Quadratojugale sin., крупной величины.
Pv 13. Endocranium, pars palato-transversalis, огромной величины. Px.
14. Quadratojugale dext. Px. 15. Corpus paras phen oidei, гигантской величины.
Px. 16. Cranium, pars articularis mandibulae dext. Pv 17. Pterygoideum
doxt. pars basalis. Px. 18. Corpus parasphenoidei, очень крупный экземпляр.
7
Рх. 19. Corpus parasphenoidei, неполный. Pv 20. Endocranium, pars vome-
ralis. Pj. 21. Corpus parasphenoidei. Px. 22. Pterygoideum, pars basalis-
Pv 23. Cranium, pars praemaxillaris. P3. 24. Quadratojugale dextrum, не-
большой величины. P3. 25. Cranium, pars praemaxillaris. P3. 27. Фрагменты
dermocranium. P3.28. Фрагменты endocranium.P3.32. Mandibula, pars anterior,
огромной величины, обнажение у мельницы. 33. Mandibula, неполная. Pv 34.
Mandibula sin., pars articularis. Px. 35. Mandibula, обломок. Px. 36. Mandi-
bula sin., неполная, окатанная. P2. 37—41. Обломкиmandibulae, 5 экз. P3.46.
Dentes, 5 экземпляров. P3. 47. Mandibula, неполная. Оползень у дер. Заха-
рово. 48. Arcus neuralis vertebrae. Р3.49. Hypocentrum (basiventrale), огром-
ной величины. Px. 50. Hypocentrum. P3. 51. Arcus neuralis vertebrae. P3. 52.
Arcus neuralis vertebrae. P3.53. Costa. Px. 54. Costa неполное. Px. 55—59. Cos-
tae pars inferior. 5 экз. Px. 60—64. Capitulum costae, 6 экз. разной величины.
Px, P3. 65. Costa, parsdistalis. P3. 66. Scapula, pars superior. Px. 67. Scapula,
неполная, оползень у дер. Захарово. 68—69. Interclavicula. 2 экз. непол-
ные. Р2. 73—74. Processus praescapulares claviculae. 2 экз. Р3. 75. Clavi-
cula, pars distalis. P3. 76—78. Cleithra, 2 неполных экз., один крупный.
Р3, Рх. 79. Clavicula sin. P3. 80. Фрагмент claviculae. P3. 81—86. Humerus
dext. etsin. 6 экз. P3. 91—93. Ilium dext. et sin. 3 экз. P3. Px. 94—98. Femur
dext. etsin. неполные, 5 экз., проксимальные и дистальпые концы. Р3. 99—
103. Radius, ulna, tibia et fibula dext. et sin., 5 экз. P3. 104—106. Элементы
acropodium, 3 экз. P3.107. Tibia, оползень у дер. Захарово. 121. Processus
cultriformis parasphenoidei, pars basalis, гигантской величины. Px. 134.
Cranium, pars narialis. Px. 136. Endocranium, pars praemaxillaris. P3. 137.
Endocranium, обломок P3. 138. Endocranium, condylus mandibularis sin.
P3. 139. Corpus parasphenoidei. P3. 140. Cranium, pars praemaxillaris dext.
P,. 141. Endocranium, фрагмент. P3. 142. Dens. P3. 143. Endocranium, обло-
мок. P3. 144. Mandibula, обломки, 3 экз. P3. 145. Interclavicula, обломок
от особи огромной величины. Р3. 146. Clavicula, неполная. Р3.147—-148. Cos-
tae, неполные, 2 экз. Р3. 149. Cleithrum, обломок. Р3. 150. Capitulum costae,
3 экз. Р3. 151. Ilium, pars acetabularis. Р3. 152. Ilium sin. P3.
2. Wetlugosaurus angustifrons Riab., p. Ветлуга, дер. Б. Слудка и с. Зубовское
26. Cranium, pars praeorbitalis. Рх. 30. Dermocranium, фрагмент. Р3.
31. Quadratojugale dext. крупной величины, неполное, с. Зубовское, в осы-
пи. 42. Mandibula sin., неполная. Мыс за оврагом, в осыпи. 43. Mandibula,
очень мелкий экз. Рх. 44. Mandibulasin., pars symphysalis, небольшой экз. Рх.
45. Dentes, 4 экз., с. Зубовское ид. Б.‘Слудка. 70—71. Intercl aviculae, непол-
ные, разбитые. Р3. 72. Interclavicula, неполная. Р3. 87. Humerus Рх. 88—89.
Ilium detx. etsin., небольшой величины, за оврагом, в осыпи. 90. Ischium, там
же. 108. Radius, там же. 109—111. Ilium dext. et sin., 3 экз. очень маленькой
величины, там же. 112—115. Femur dext. et sin. 4 экз. очень маленькой ве-
личины, там же. 116—120. Radius, ulna, tibia dext. et sin. 5 экз. очень ма-
ленькой величины, дер. Б. Служка, за оврагом, в осыпи, и Рх.
№ 19/2252 ПИН. Коллекция Шарженгской Палеонтологической экспедиции
Геологического музея Академии Наук 1928 г. рр. Шарженга (Вахнево) и Ветлуга
(Б. Слудка и Зубовское)
Benthosuchus sushkini Efr., р. Шарженга» с. Вахнево
1. Cranium, полный. Р3. 2. Крупный, почти полный cranium (разломан
по средней линии с утерей мелких частей). Р3. 3. Почти полный cranium
(разломан с утерей средней части). Р3. 4. Полный cranium очень маленькой
особи. Р3. 5. Полный cranium крупной величины. Р5. 8. Cranium, pars tem-
poralis sin. P3. 9. Cranium, pars narialis. P3. 10. Cranium, pars narialis,
сохранились хватательные зубы и vomera. Р3. 11. Cranium, pars narialis
et maxillo-jugalis sin. P6. 12. Basis cranii (дно мозговой коробки). Сохрани-
лись обе exoccipitalia. Крупная форма, с. Вахнево, ниже обнажения
у мельницы. 13. Cranium, pars praemaxillaris. Ръ Р2. 14. Cranium, pars
narialis, P5. 15. Cranium, pars narialis, из осыпи под- дер. Б. Слудкой.
8
16. Cranium, pars postchoanalis dext. Очень крупный череп. P3. 17. Фраг-
мент basis cranii с дном правой cavum tympani. Pv 18. Basis cranii, pars
vestibularis sin. P3. 19. Фрагмент cavum tympani sin. Pj. 20. Exoccipitale
dext. Ясно различимы швы, гребни сохранены. Pv 21. Фрагмент basis
cranii (часть левого птеригоида у сочленения с парасфеноидом). Р3. 22.
Tabulate sin. Имеется часть восходящих отростков exoccipitale. Р3. 23.
Dermocranium, pars dext. Р3. 24. Cranium, pars orbito-nasalis sin. P3. 25. Cra-
nium, pars maxillo-pterygoidea dext. Pj. 26. Cranium, pars temporalis sin.
27. Quadratojugale dext. P3. 28. Endocranium, pars vomeralis. Px. 33. Cra-
nium, pars postero-lateralis sin. Px. 34. Quadratojugalia, 5 экз. Pn P3. 35.
Обломки endocranium (basis cranii), 22 экз. Px, P3. 36. Обломки dermocra-
nium, 48 экз. Pn P3. 39. Cranium, pars pterygo-palatinalis. Pv 40. Cranium,
pars vomeralis. P3. 41. Mandibulasin., очень крупного индивида. P3.42. Man-
dibula dext. без артикулярного конца. P3. 43. Mandibulasin., неполная. P3.
44. Mandibula dext., неполная. P3. 45. Mandibula dext. без симфизах. P.
49. Mandibula sin., дистальная половина. P3. 50. Mandibula sin., pars arti-
cularis. Ps. 51—52. Mandibula dext., pars articularis, 2 экз. Px, P3. 53. Man-
dibula sin., pars articularis. P3. 55. Mandibula dext., pars distalis, крупный
экземпляр. Px. 56. Mandibulasin., parssymphysalis. Px- 57. Фрагмент mandi-
bulae. Px. 58. Фрагмент mandibulae, средняя часть. P3. 59. Mandibula
sin., parssymphysalis. Рх. 60. Mandibuladext., pars articularis. Px. 62. Фраг-
мент mandibulae. P3. 63. Mandibulasin., pars articularis. Px. 64. Mandibula,
pars distalis, небольшой экземпляр. P3. 65. Фрагмент madibulae. Px. 66. Man-
dibula angulare. P3. 67. Mandibula dext., без середины, зубы хорошо сохра-
нились. Р4. 68. Фрагмент mandibulae, средняя часть с зубами. Р5. 70. Об-
ломки mandibulae, 15 экз. Рх—Р3. 71—80. Hypocentra (basiventralia) раз-
ной величины, 10 экз. Р3.81. Arcus neuralis vertebrae, неполная (spina dor-
salis), очень крупной величины. Р3, Pj. 82. Arcus neuralis vertebrae thora-
calis, неполная. Pj. 83. Arcus neuralis, фрагмент. P3. 84. Фрагмент arcus
neuralis vertebrae. P3. 85. Фрагмент arcus neuralis vertebrae. Px. 86. Vertebra,
фрагмент. P3. 87. Фрагмент arcus neuralis vertebrae, небольшой экземпляр.
P3. 88—89. Hypopentra (basiventralia) очень хорошей сохранности. 2 экз.
Px, Р3. 101—108. Costae thoracales et cervicales. 8 экз. Px. 109—120. Costae
pars proximalis, 12 экз. Px. 121. Capitulum costae, 11 экз. Px. 122. Costa. P3.
123—124. Costae thoracales, 2 экз. P3. 125—130. Capitulum costae, 6 экз.
P3. 131. Costa, pars proximalis. P3. 132. Capitulum costae, 3 экз. P3. 133.
Costae, фрагменты, 6 экз. P3. 134—135. Scapulae sin., 2 экз. Px, P3. 138.
Scapulae pars distalis, 2 экз. Px, P3. 139—146. Claviculae dext. et sin., пол-
ные и неполные, 7 экз. Рх. 147—149. Interclaviculae, почти полные, 3 экз.,
одна—огромной величины. Pj. 150—151. 2 фрагмента interclaviculae. Рх.
152—154. Обломки interclaviculae, 11 экз. Рх. 155—156. Claviculae dext.,
2 экз., одна—огромной величины. Р3. 157—160. Claviculae sin., 4 экз.,
из них 2 на породе. Р3. 161—162. 2 почти полные interclaviculae. Р3. 163.
Неполная interclavicula, очень массивная. Р3. 164. Interclavicula-f-clavicvla
в породе, неполная. Р3. 165. Processus praescapularis claviculae, 5 экз.
Р3.193. Cleithrum. Р3. 194—197. Humerus dext. et sin., 4 экз. Рг. 198. Hume-
rus без эпифизов. Р. 199—202. Humerus dext. et sin., 4экз., один очень круп-
ной старой особи. Pj, Р3. 203. Pars distalis humeri. Р3. 204. Humerus, pars
proximalis. P3. 205. Часть проксимального эпифиза humerus. P3. 206. Hume-
rus «slender form». P4. 207. Humerus, грубой формы. Рх. 211. Ilium dext., не-
большой экземпляр. P3. 212—216. Iliumdext. et sin., 5 экз. P3, Px 217—218.
Ilium sin., 2 экз. Px. 219—221. Ilium dext., pars acetabularis, 3 экз.
Px. 232. Ischium. Px. 233. Femur. P^. 234—235. Femur, pars distalis. 2 экз. Pj.
236—238. Femur, pars proximalis, 2 экз. Px. 239. Femur, неполный.
Pi. 240. Femur, pars proximalis, P3. 241—242. Femur dext. etsin., pars di-
stalis, 2 экз. P3. 243—246. Femur, 4 экз. P3. 268—288. Radius, ulna, tibia
et fibula dext. et sin., 21 экз. Px, P3. 301—308. Неполные и полные элементы
acropodium, 8 экз. Pl Рз-
9
Wetlugosaurus angustifrons Riab.,
p. Ветлуга, дер. Б. Слудка и с. Зубовское
6—7. Cranium, неполный, 2 экз. Р2. 29. Cranium; pterygoideum sin. особи
гигантской величины. Р2. 30. Cranium; quadratojugale sin. особи очень боль-
шой величины. Р2. 31. Cranium; jugale sin. особи очень большой величины.
Р2. 32. Dermocranium, pars temporalis sin. особи огромной величины. Р2.
37. Фрагменты dermocranium. Р2, Рх. 38. Фрагменты endocranium. Р2, Р1#
46—48. Mandibulae dext., 3 экз., — одна из них очень крупной особи, —
неполные. Р2. 54. Mandibula dext., pars articularis. P2. 61. Mandibula sin.,
pars proximalis. P2. 69. Mandibula sin., pars distalis. Форма, отличная от
остальных. Р2. 90-^-92. Hypocentra (basiventralia), 2 экз., очень небольшие.
Р2. 93. Costa очень крупной величины. Р3. 94—99. Фрагменты costae,
6 экз. Р2. 100. Capitulum costae, 5 экз.Р2.136—137. Scapulae sin., 2 экз. не-
большой величины. Р2. 166. Clavicula sin., небольшой экземпляр Р2. 167.
Claviculasin.,неполная. Р2. 168. Обломок claviculae dext., очень крупный.
Р2. 169. Clavicula, pars proximalis. Р2. 170. Clavicula sin., неполная, неболь-
шой экземпляр. Р2. 171. Clavicula sin. То же. Р2. 172. Clavicula dext.,
неполная, большой величины. Р2. 173. Clavicula. Р2. 174. Clavicula, очень
неполная, небольшой величины. Р2. 175. Clavicula sin. То же. Р2. 176.
ломки claviculae, 2 экз. Р2. 177. Interclavicula, очень небольшая. Р2. 178.
Interclavicula, неполная, небольшая. Р2. 179. Interclavicula, небольшая.
180. Interclavicula, небольшая, неполная. Р2.181. Interclavicula, небольшая.
Р2.182. Interclavicula, окатанная, крупная. Р2. 183. Interclavicula крупной
величины, сильно окатанная. Р2. 184. Interclavicula, неполная, не-
большой экземпляр. Р2. 185. Отпечаток interclaviculae. Р2. 186. Inter-
clavicula, неполная, на породе. Р2. 187. Отпечаток и часть interclaviculae.
Р2. 188. Фрагменты interclaviculae большой величины,? экз. Р2. 189. Обломки
interclaviculae. Р2. 190. Processus praescapularis claviculae, 3 экз. Р2. 191.
Cleithrum, неполный, окатанный. Р2.192. Cleithrum, неполный. Р2. 208. Hume-
rus, небольшой величины, с хорошими эпифизами. Р2. 209. Humerus, без
эпифизов. Р2. 210. Humerus, очень небольшой, хорошо окостеневший. Р2.
222—224. Ilium, 3 экз. Р2. 225. Ilium, неполный, небольшой. Р2. 226. Ilium,
pars acetabularis. Р2. 227. Ilium, небольшой экземпляр. Р2. 228—229. Ilium,
2 экз. Р2. 230. Ilium, небольшой. Р2. 231. Ilium, pars acetabularis. Р2. 247—
251. Femur, 5 экз. 252. Femur, диафиз. Р2. 253.. Femur с неокостеневшими
эпифизами. Р2. 254. Femur. Р2. 255—256. Femur небольшой величины. Р2.
257—261. Femur, pars proximalis, 5 экз. Р2. 262—266. Femur, pars dista-
lis, 5 экз. P2. 267. Femur, неполный. P2. 289—300. Кости конечностей (ra-
dius, ulna, tibia, fibula), 12 экз. P2. 309—315. Фрагменты элементов acro-
podium, 12 экз. P2. 316. 3 phalanx in situ, небольшой величины. P2. 342.
Фрагмент mandibulae. P2. 343. Costa, pars distalis. P2. 344. Dens. P2.
№ 32/2354 ПИН. Коллекция Кузьмина 'Ф. M. 1929 г. р. Шарженга, с. Вахнево
Benthosuchus sushkini Efr.
1—3. Cranium, полные 3 экз. Р3. 4. Dermocranium, pars orbitalis sin.
P3. 5. Cranium, pars praeorbitalis sin. P3. 6. Cranium, pars pterygo-quadrato-
jugalis sin. P3. 7. Cranium, pars transverso-maxillo-jugalis dext., очень круп-
ный экземпляр P3. 8. Cranium, pars jugo-transversabis sin. гигантского эк-
земпляра. P3. 9. Endocranium, pars vomeralis sin. гигантского черепа. P3.
10. Dermocranium, pars temporalis sin. P3. 11. Endocranium, pterygoideum
dext. без задней части ramus quadrat!. P3. 12. Endocranium, exoccipitale
sin. P3. 13. Cranium, pars orbitalis dext. P3.14 Quadratum-|-ramus quadrati
pterygoidei. P3. 15. Ectopterygoideum (transversum), 2 крупных фрагмента.
P3.16. Squamosadext. etsin. P3.17. Dermocranium, pars oticalis, 2 экз. P3. 18.
Dermocranium jugale огромной величины. P3. 19. Rami quadrati ptery-
goidei, 2 экз. P3. 20. Cranium, pars praemaxillaris. P3. 21. Dermocranium, pars
tabularis, очень крупный экземпляр. P3. 22. Quadratojugalia, 5 экз. P3.
23. Dermocranium, pars quadrato-temporalis, огромный экземпляр. P3.
io
24. Обломки dermocranium. Р3. 25. Обломки endocranium. Р3. 27—28. Man-
dibula sin., pars angulo-complementaris, 2 экз. P3. 29. Mandibula dext.,
pars symphysalis. P3. 30. Mandibula dext., pars articularis, огромный
индивид. P3. 31. Angulare sin. P3. 32. Mandibula dext., pars articularis.
P3. 33. Mandibula sin., pars distalis. P3. 34. Mandibula dext., pars
proximalis. P3. 35. Mandibula sin., pars distalis, очень маленький инди-
вид. P3. 3G. Mandibula sin., pars anterior. P3. 37. Mandibula sin., pars arti-
cularis. P3. 38. Mandibula dext., pars posterior. P3. 39. Mandibula sin., pars
posterior. P3. 40 Mandibula sin., pars anterior. P3. 41. Mandibula dext., pars
articularis, крупный индивид. P3. 42. Mandibula sin., pars articularis, неболь-
шой экземпляр. P3. 43. Angulare sin. P3. 44. Mandibula sin., pars symphysalis.
P3. 44a. Обломки mandibulae, 19 экз. P3. 45—46.2 фрагмента arcus neuralis ver-
tebrae, очень крупные экземпляры. P3. 47. Arcus neuralis vertebrae. P3. 48.
Arcus neuralis vertebrae. P3. 49—52. Hypocentra (basiventralia), 4 экз. P3. 53.
Часть hypocentrum огромной величины. P3. 54. Costa thoracalis. P3. 55.
Costa lumbalis. P3. 56. Costa thoracalis. P3. 57. Costa thoracalis, pars proxima-
lis. P3. 58. Capitulum costae огромной величины. 59. Обломки costae. P3.
60. Costa thoracalis, pars distalis. P3. 61. Обломки costae, 10 экз. P. 62.
Processus praescapularis claviculae. P3. 63—67. Claviculae dext., 5 экз. P3.
68. Clavicula sin., неполная. P3. 69. Clavicula dext., гигантской величины.
P3. 70. Clavicula sin., небольшая. P3. 71. Clavicula sin., неполная, окатанная.
P3. 72. Processus praescapularis claviculae, 2 экз. P3. 73. Обломок claviculae
sin. P3. 74. Interclavicula, неполная. P3. 75. 2 окатанных обломка clavi-
culae. P2. 76. Обломки interclaviculae, 4 экз. P2. 78. Phalanx. P2. 81—82.
Scapulae dext. et sin. P3. 83. Cleithrum, неполный, небольшой экземпляр. P3.
84. Humerus dext. P3. 85 Humerus, pars proximalis, очень крупный экзем-
пляр. P3. 86. Humerus, pars proximalis, небольшой экземпляр. P3. 87. Ulna.
P3. 88. Fibula dext., pars proximalis, круппый экземпляр. P3. 89.Ulna. Ps.
90. Radius, неполные 3 экз. P3. 91. Ilium sin. P3. 92. Ilium dext. P3. 93.
Ischium. P3. 94. Femur sin. крупной величины. P3. 95. Femur, диафиз. P3.
96. Capitulum femoris P3. 97. Дистальный эпифиз femur sin. крупной вели-
чины. P3. 98—99. Tibia sin., 2 экз. P3. 100. Fibula. P3. 102. Обломки костей
конечностей. P3. 103. Dermocranium, part occipitalis. P3. 104. Dermocra-
nium, pars maxillaris. P5. 105. Jugale очень крупного индивида. P5. 106.
Endocranium, palatinum sin. P5. 107. Tabulate огромного черепа. Ps. 108.
Mandibula dext., pars articularis. P5. 109. Tabulate огромного черепа. P5.
110. Arcus neuralis vertebrae. P5. 111. Costa sacralis. P5. 112. Costa sacralis
крупной величины P5. 113—114. Costae sacrales, 2 небольших экз. P5.
115—117. Costae, разной величины, неполные, 19 экз. Ps. 118. Costa,
pars medialis с мощным proc, uncinatus. P5.119. Cleithrum средней величины,
P5. 120. Cleithrum, pars proximalis. P5. 121. Clavicula sin. P5. 122. Pro-
cessus praescapularis claviculae. P5. 123. Interclavicula, неполная. P3.
124. Humerus с окатанными эпифизами. P3. 129. Cranium, pars lateralis
на породе. P3.130. Обломки dermocranium крупной величины. P3.
№ 33/2355 ПИН. Коллекция Кузьмина Ф. М. 1929 г.
Benthosuchus sushkini Efr., р. Юг, дер. Кудрино
1. Pterygoideum, pars basalis. 2. Cranium, pars jugo-transversalis s’n.
3. Basis cranii, pars lateralis. 4. Mandibula dext., pars anterior. 5. Mandi-
bula, pars angularis, очень крупный индивид. 6. Mandibula sin., pars ante-
rior, мелкий экземпляр. 7. Palatinum маленького черепа. 8—10. Hypocen-
tra (basiventralia), 3 экз. 11. Costa sacralis, неполное. 12. Costa sacralis,
гигантской величины. 13. Costae обломки, 4 экз. 14. Interclavicula, неполная.
15. Interclavicula, обломок. 16. Clavicula sin., крупной величины с особой
скульптурой. 17—18. Claviculae dext. et sin., 2 экз. 19—20. Humerus sin.,
крупной величины и dext., небольшой.21. Ilium, pars acetabularis. 22. Ilium
dext. 23. Femur, без проксимального эпифиза, крупной величины.
11
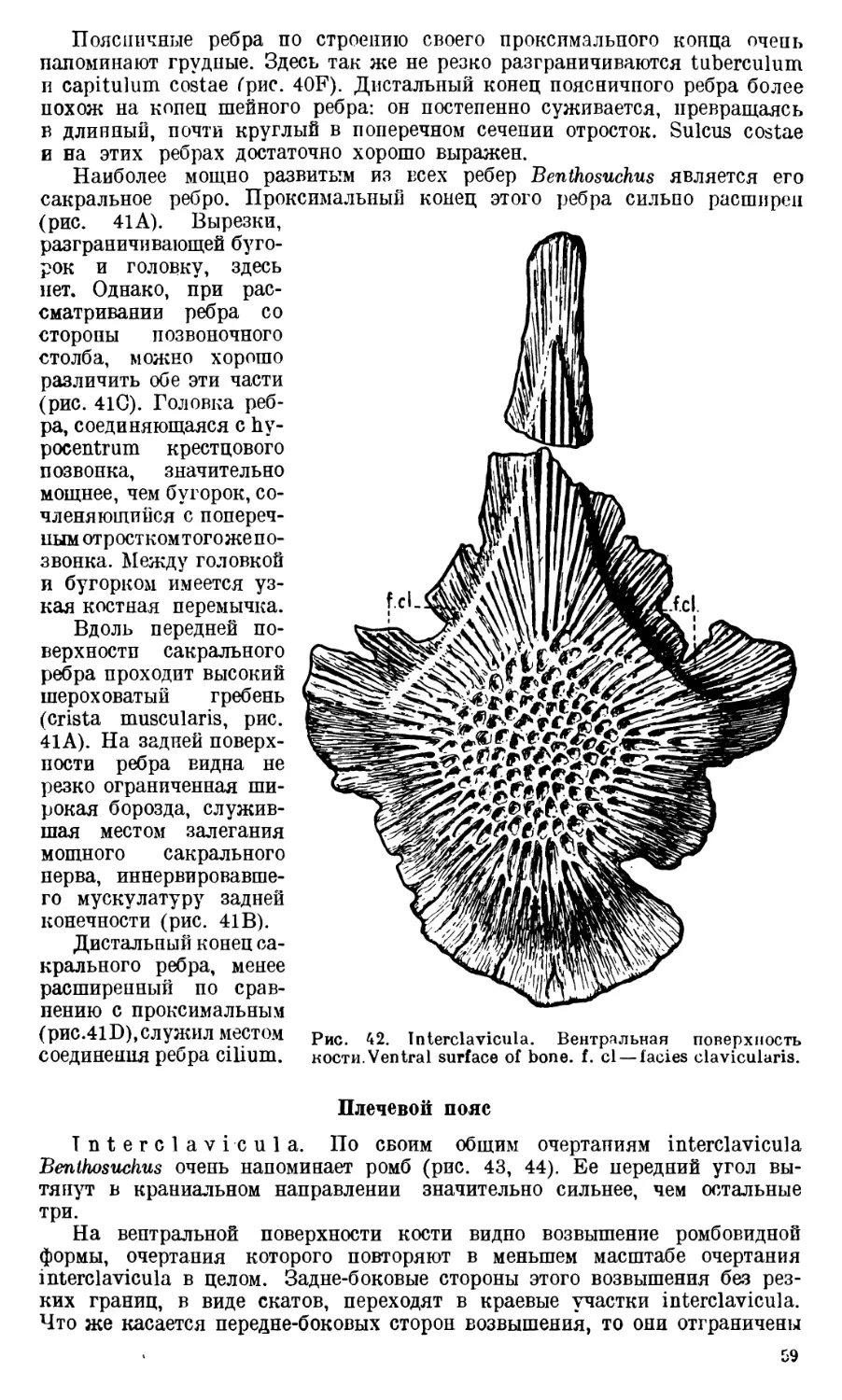
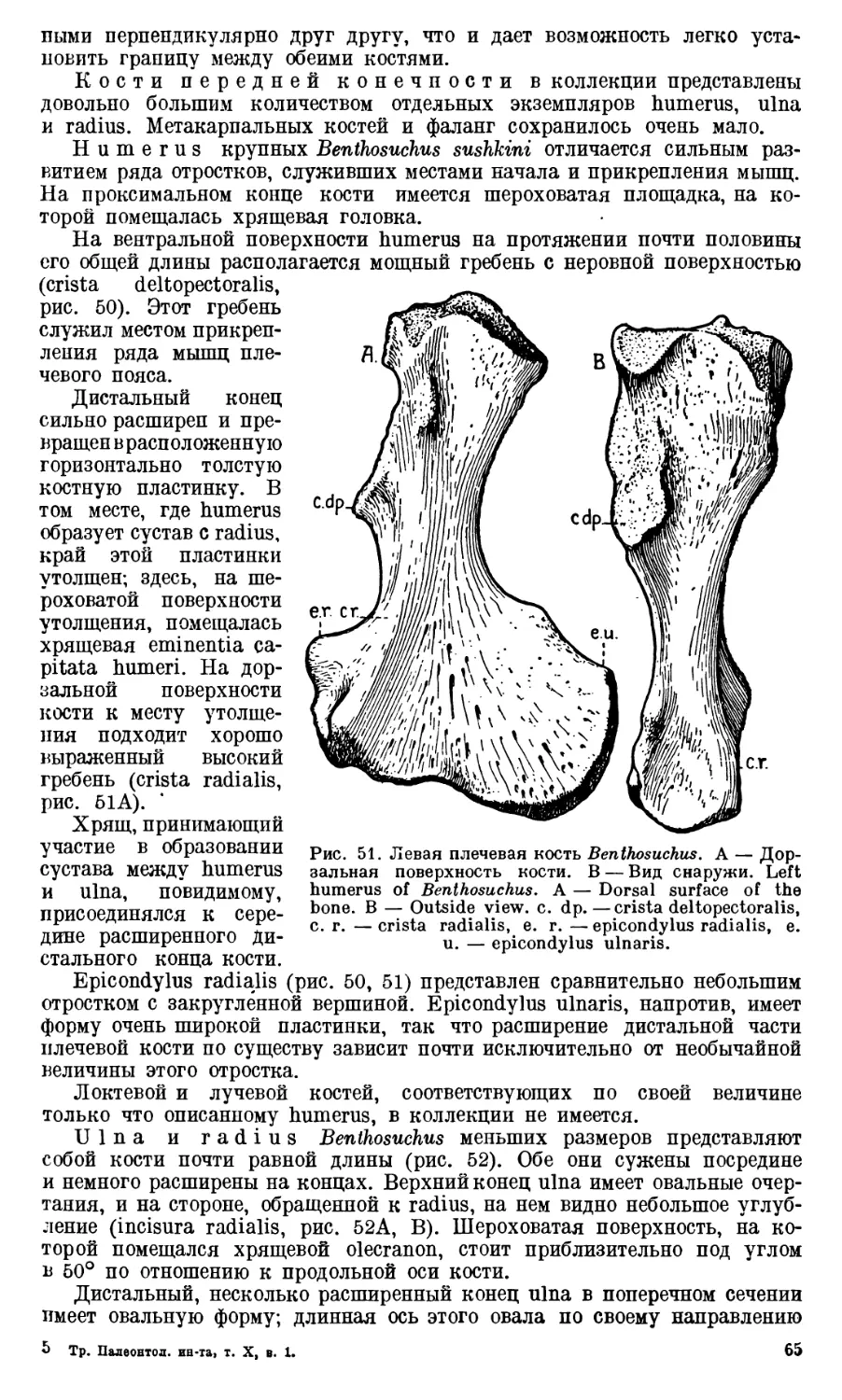
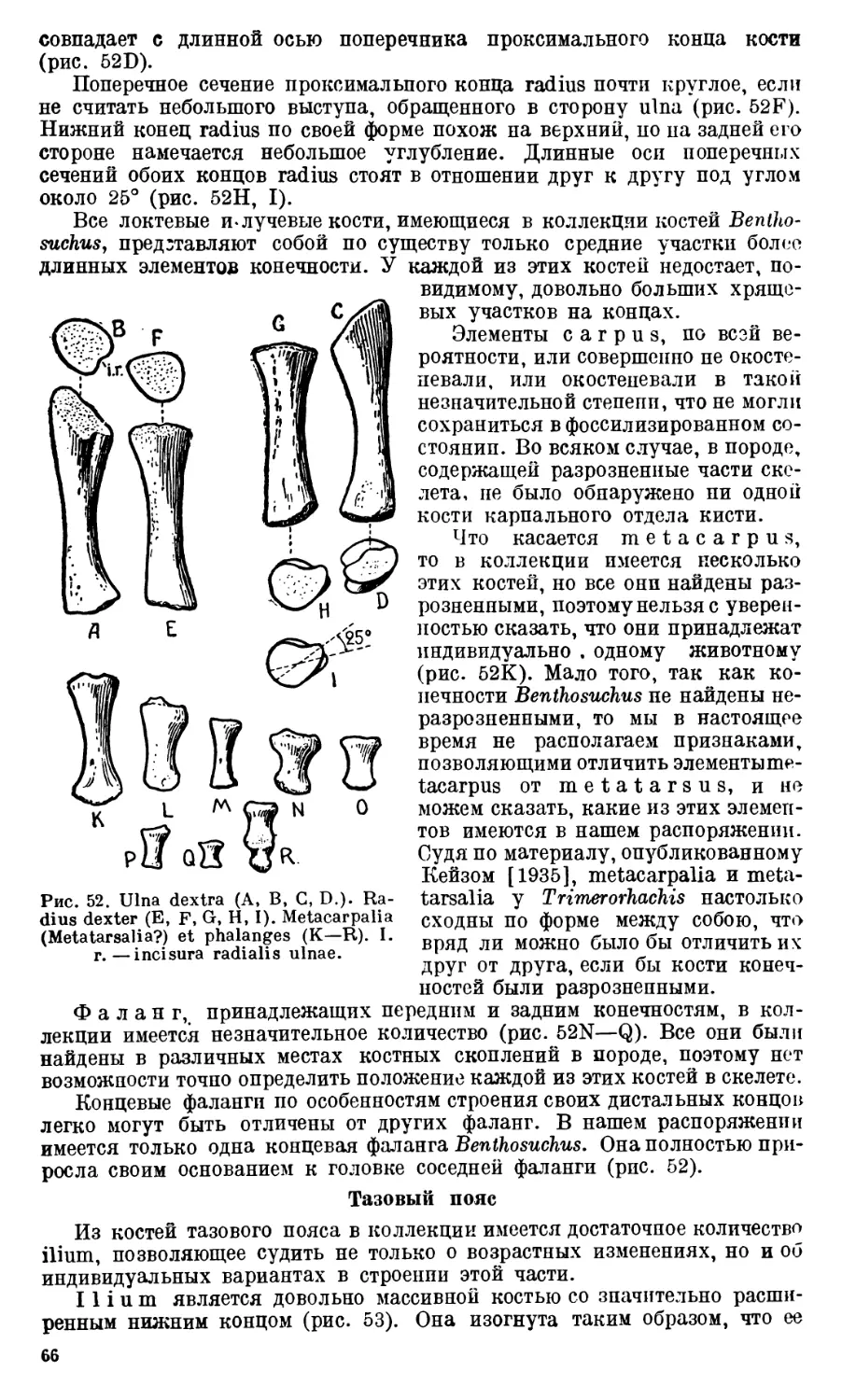
ЧАСТЬ I. АНАТОМИЯ СКЕЛЕТА
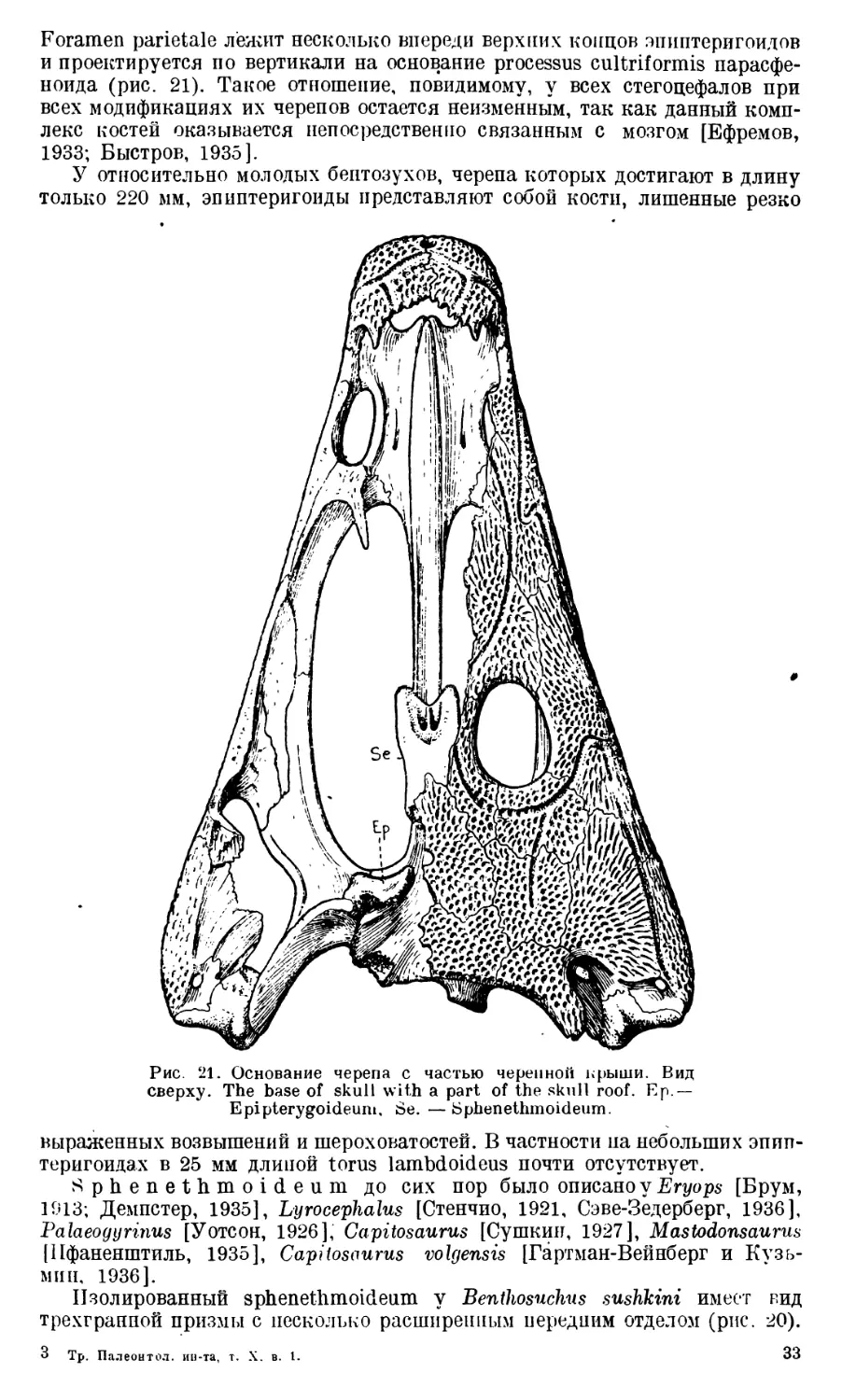
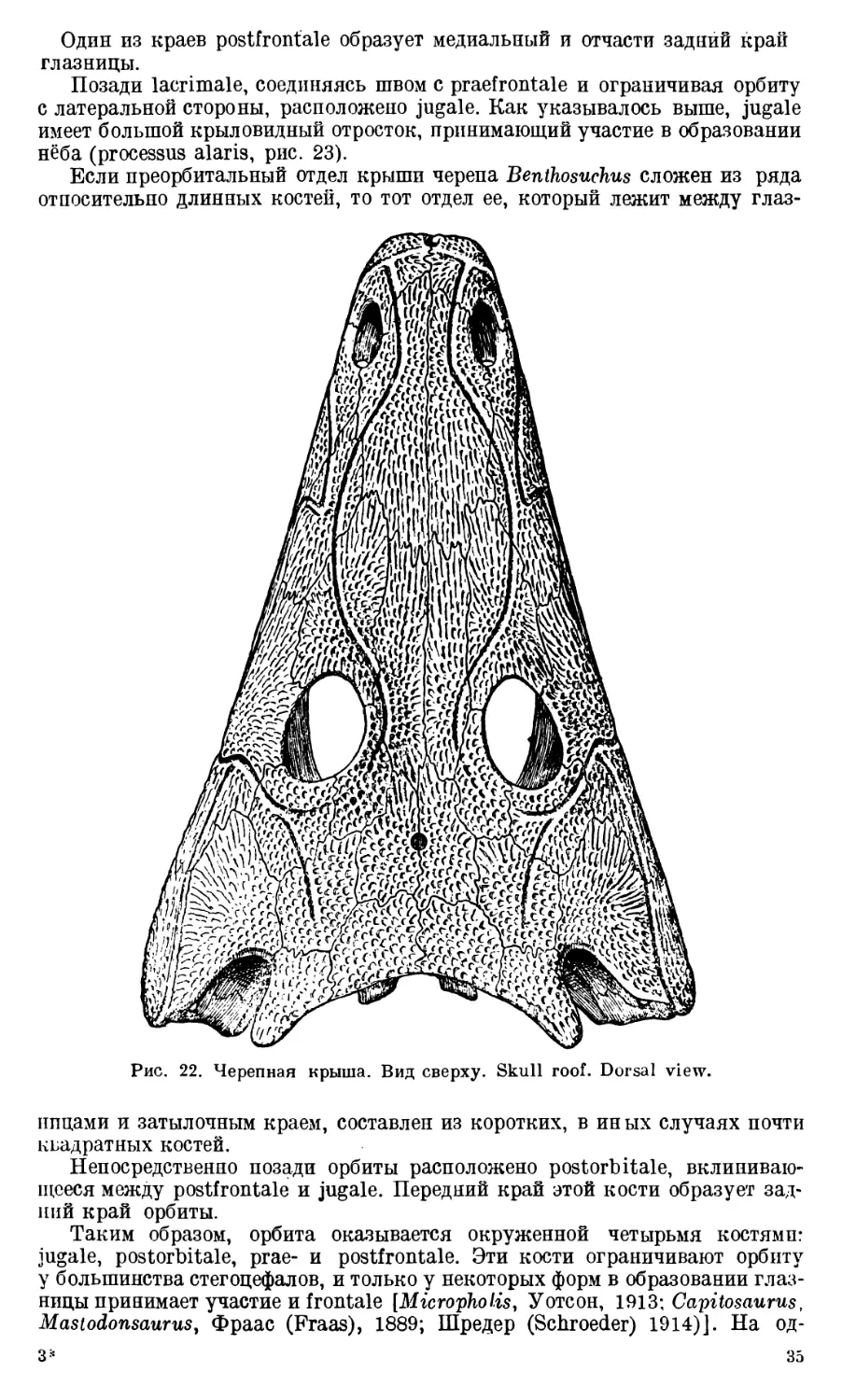
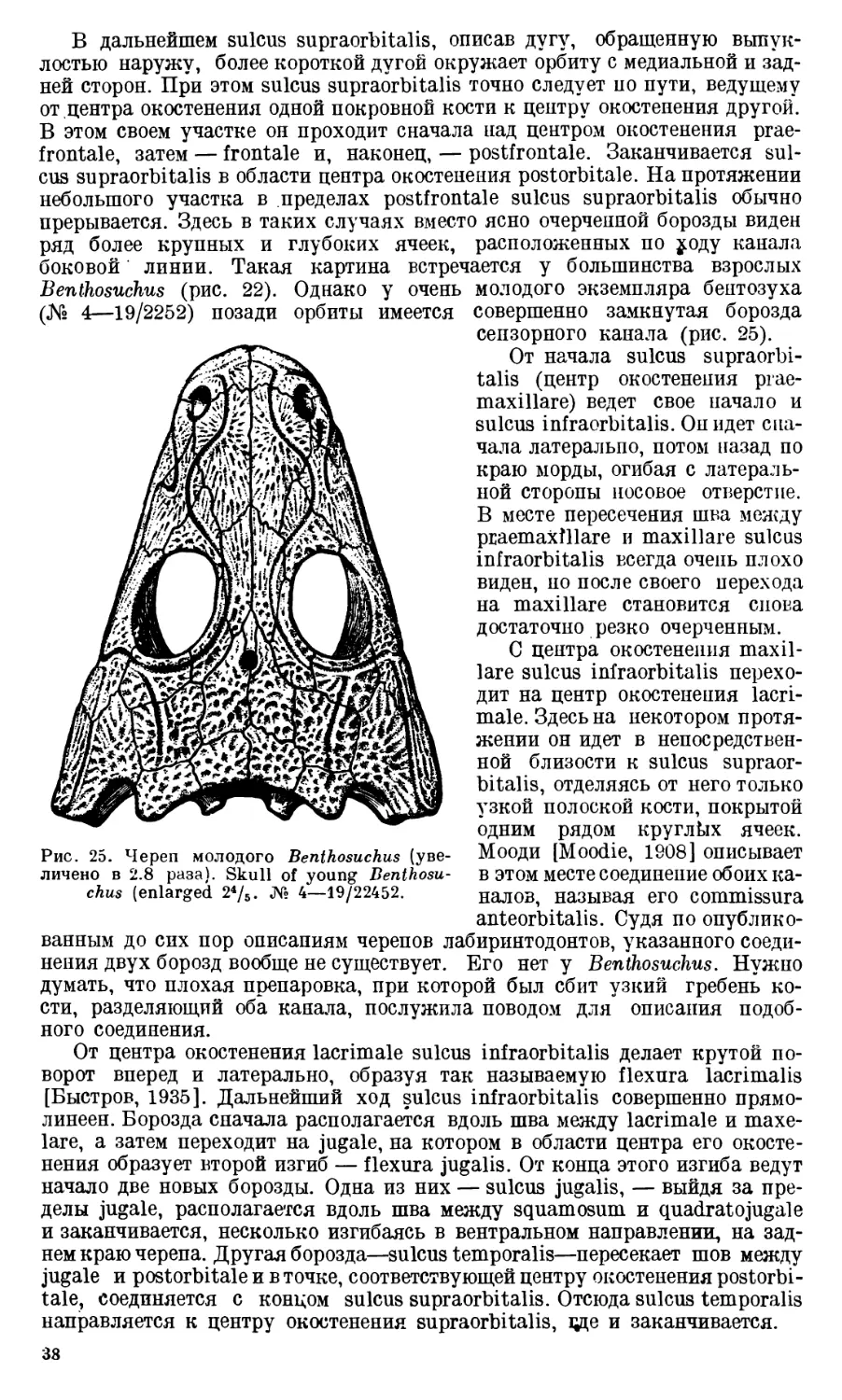
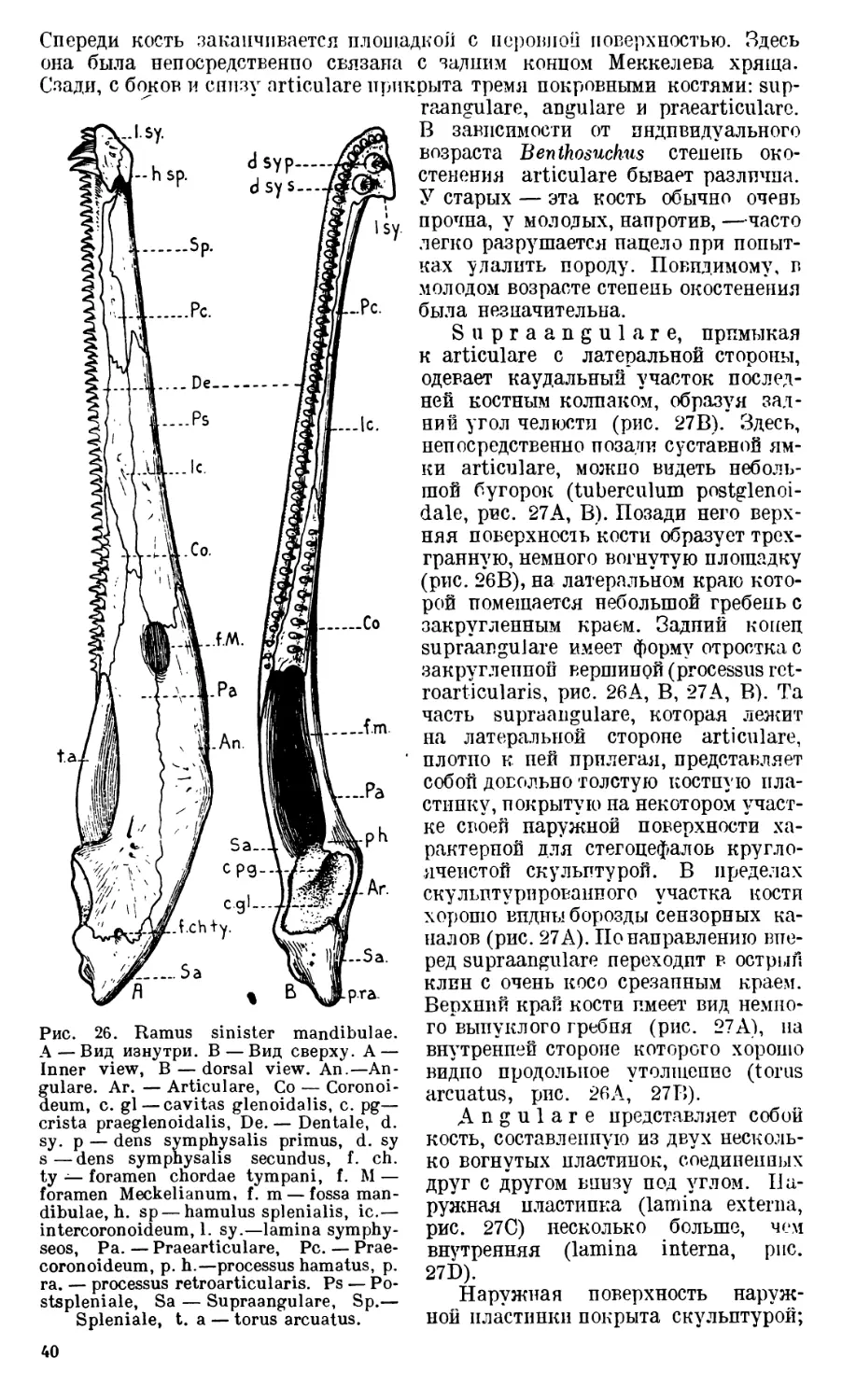
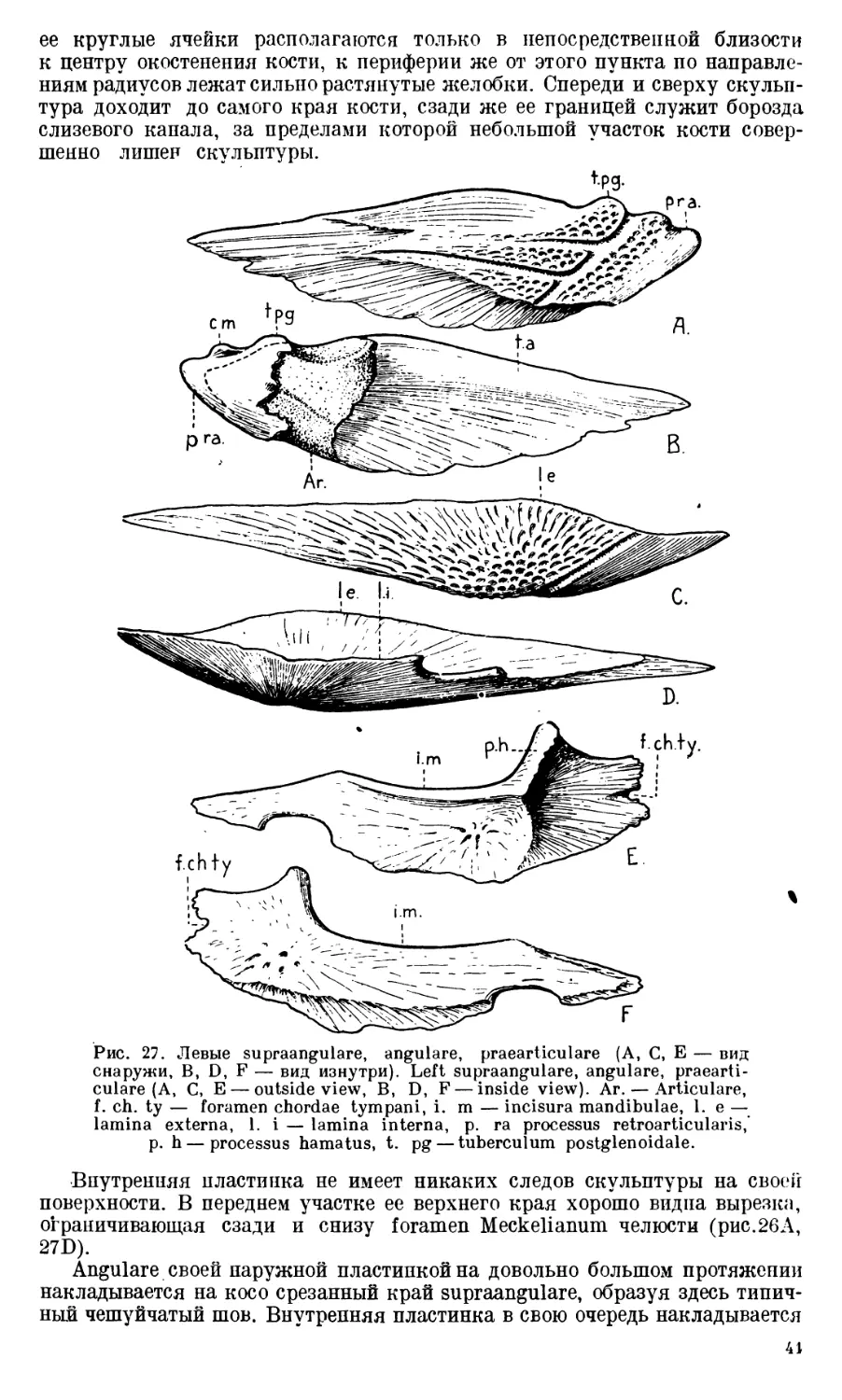
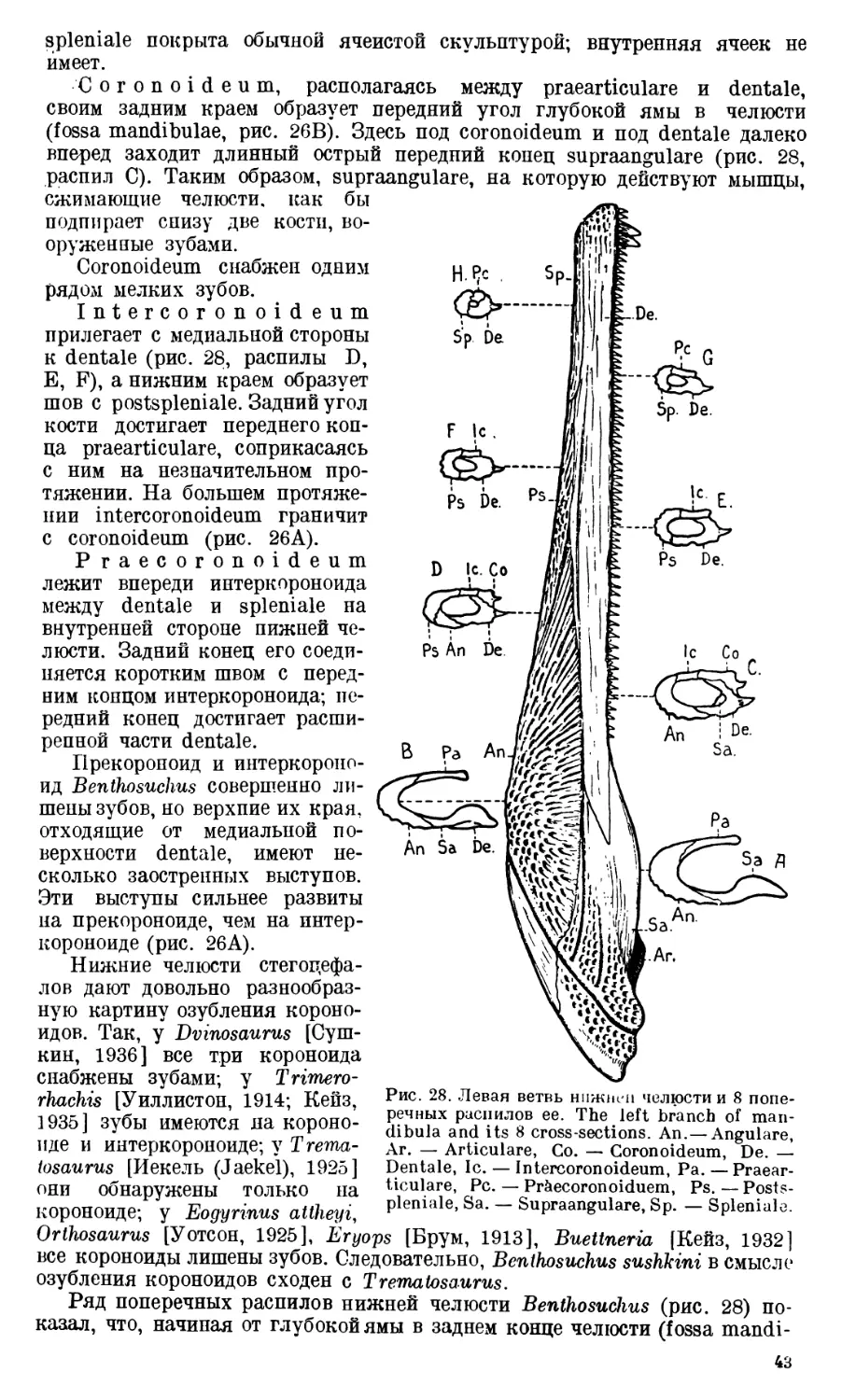
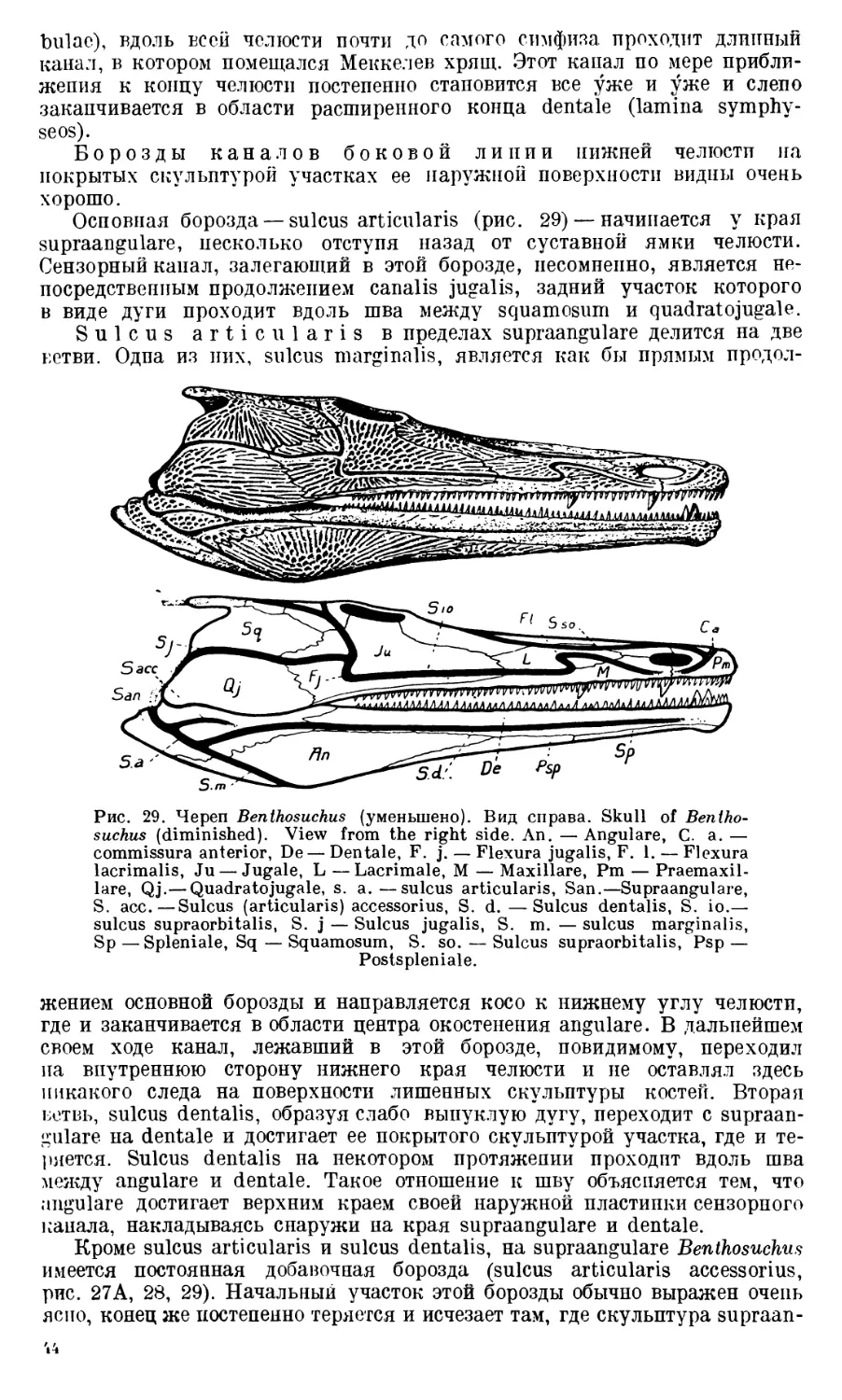
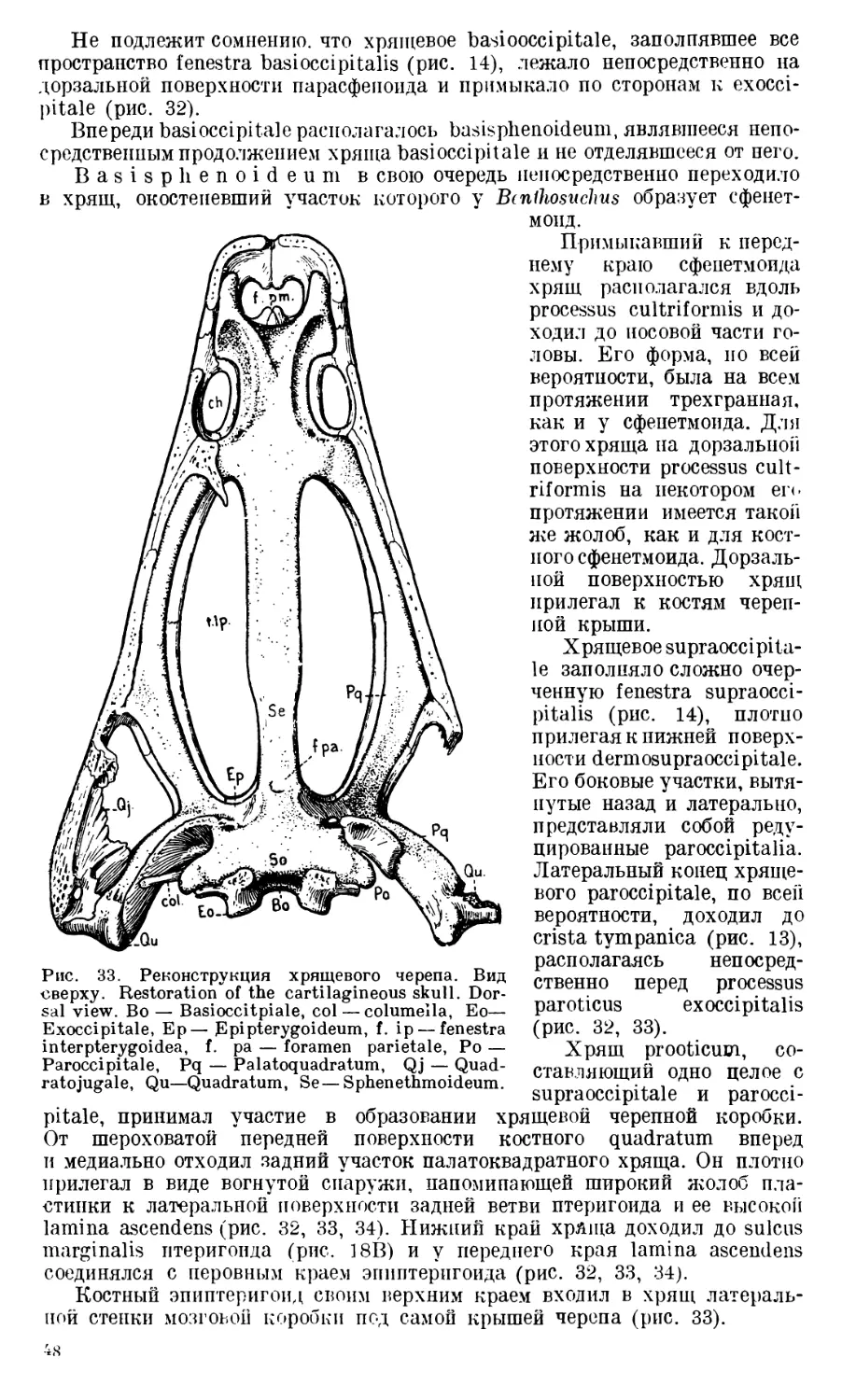
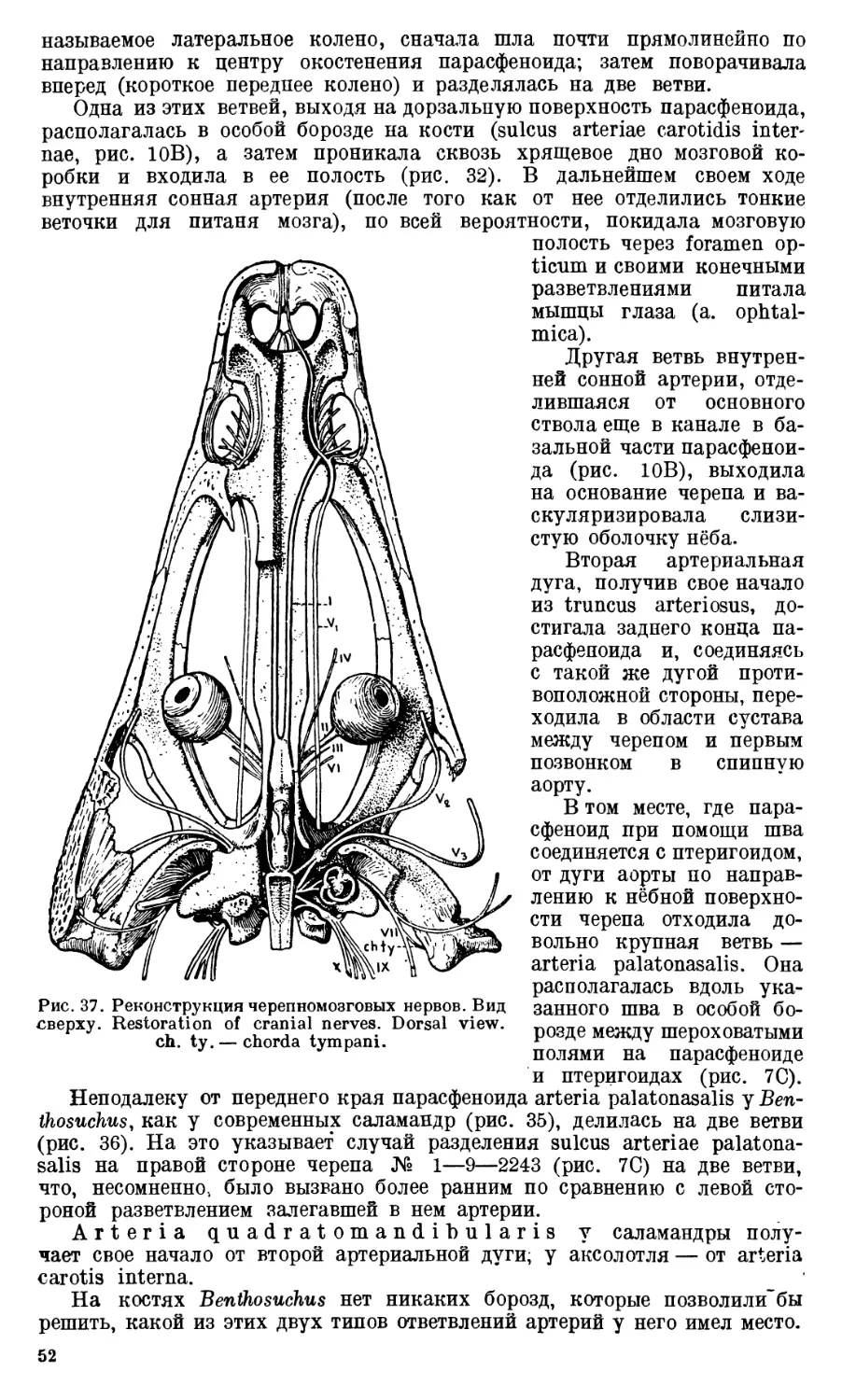
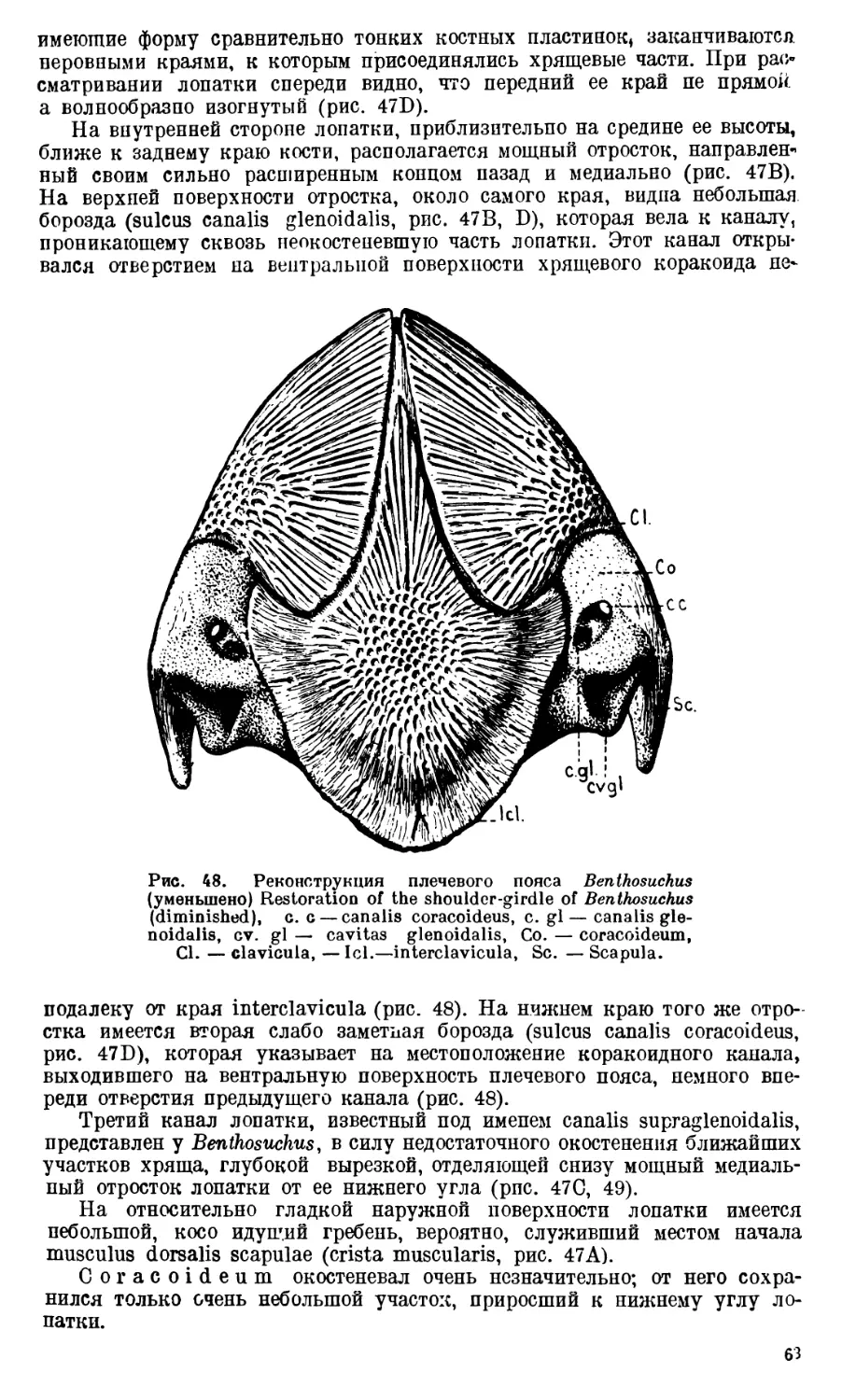
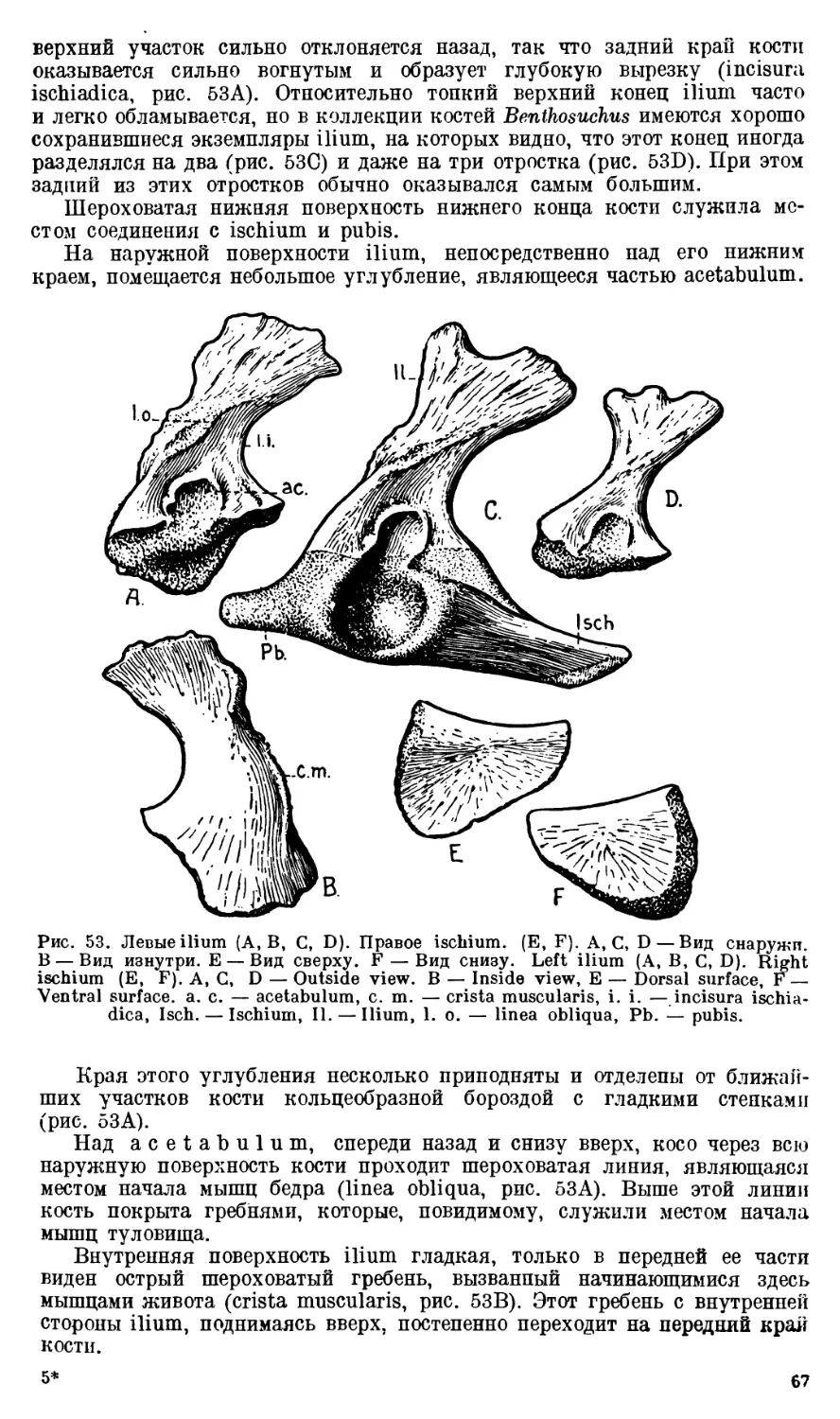
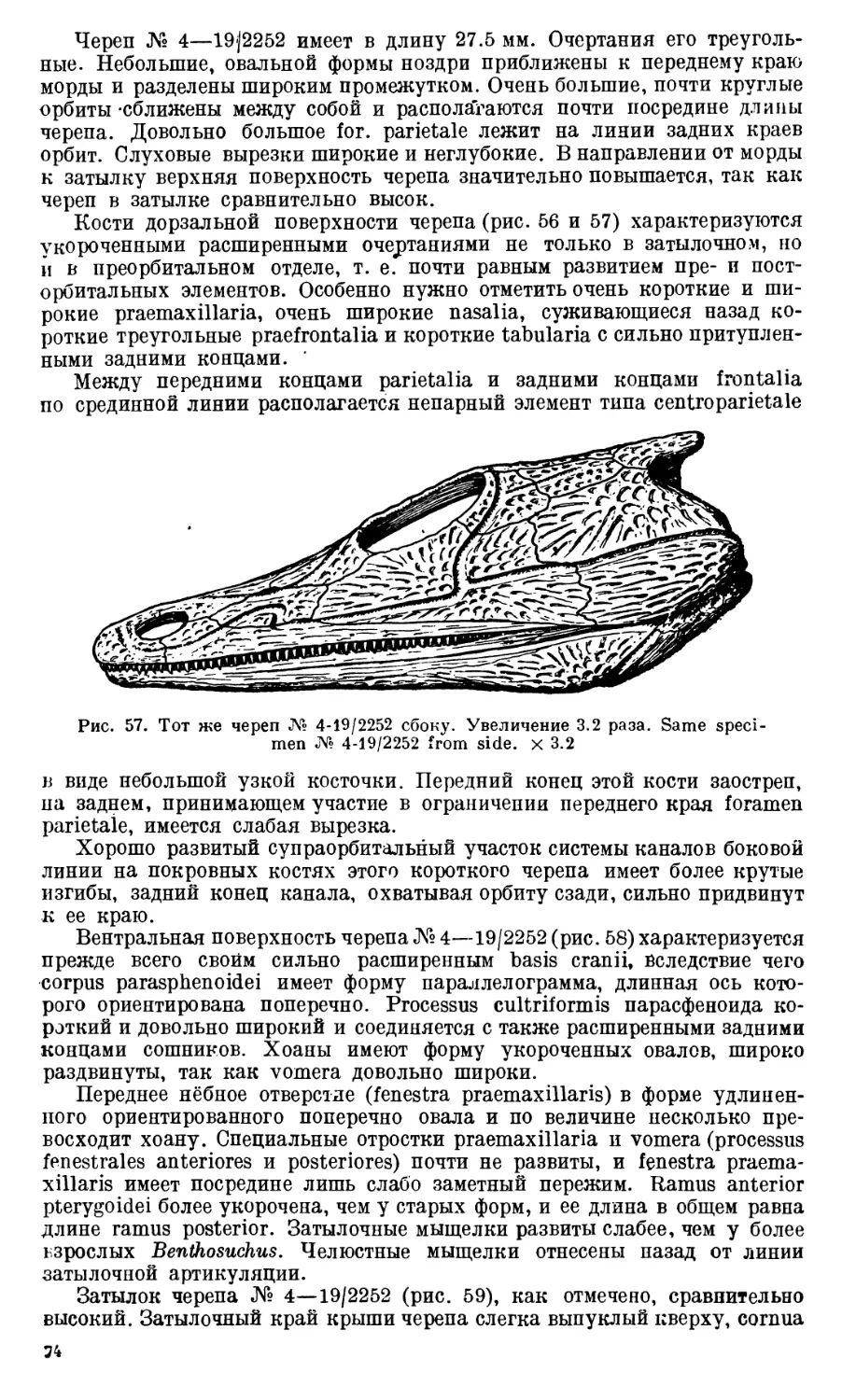
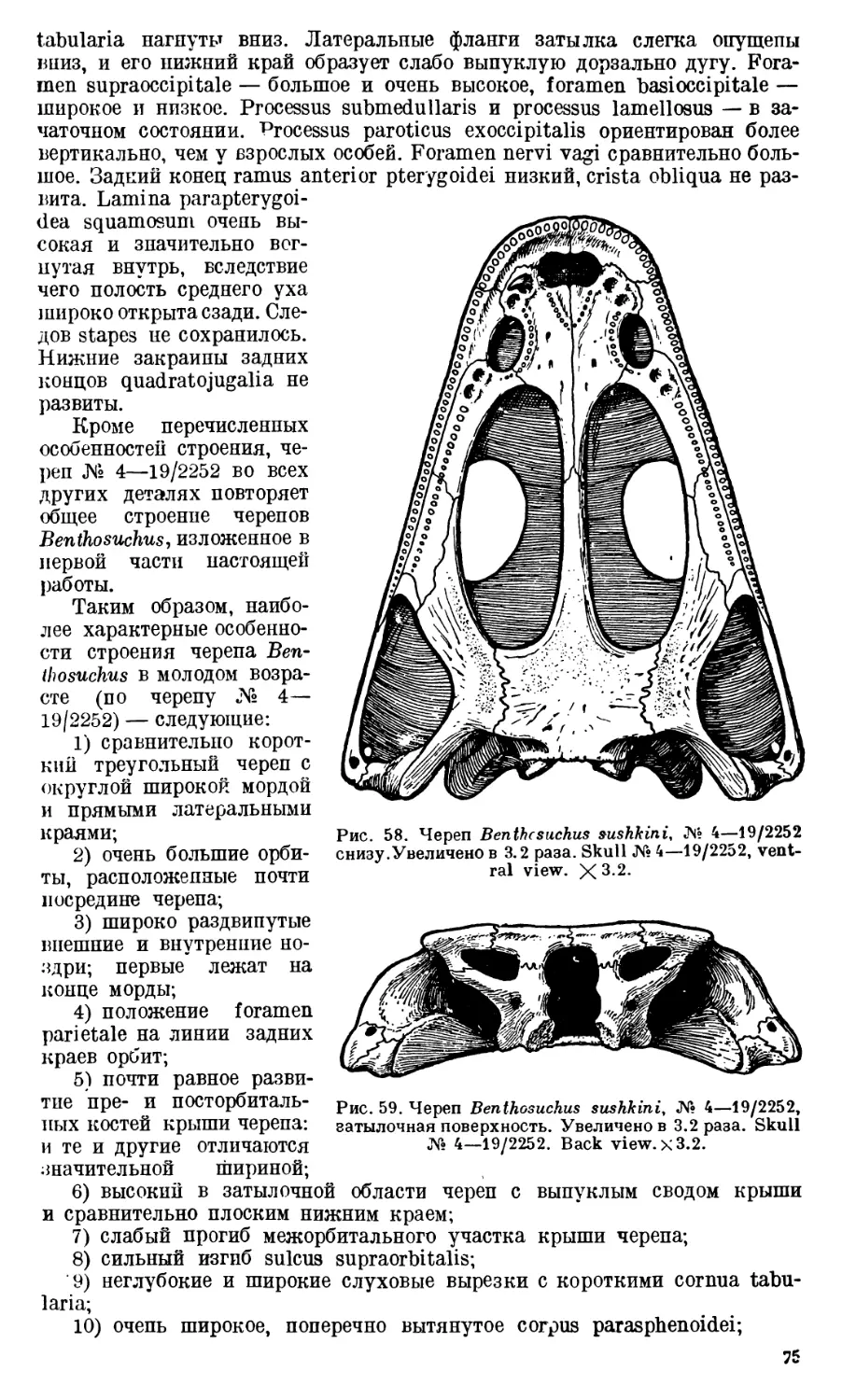
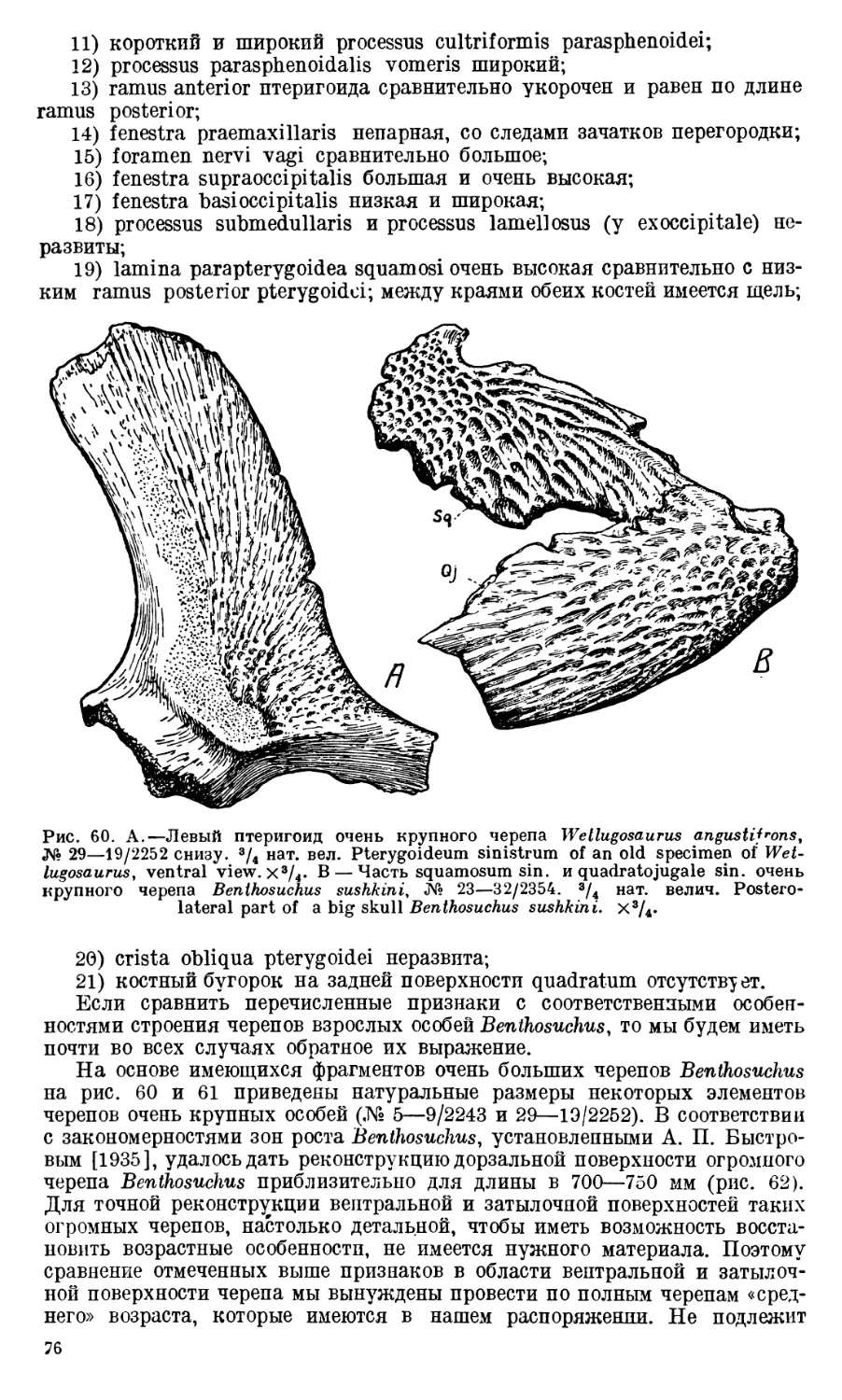
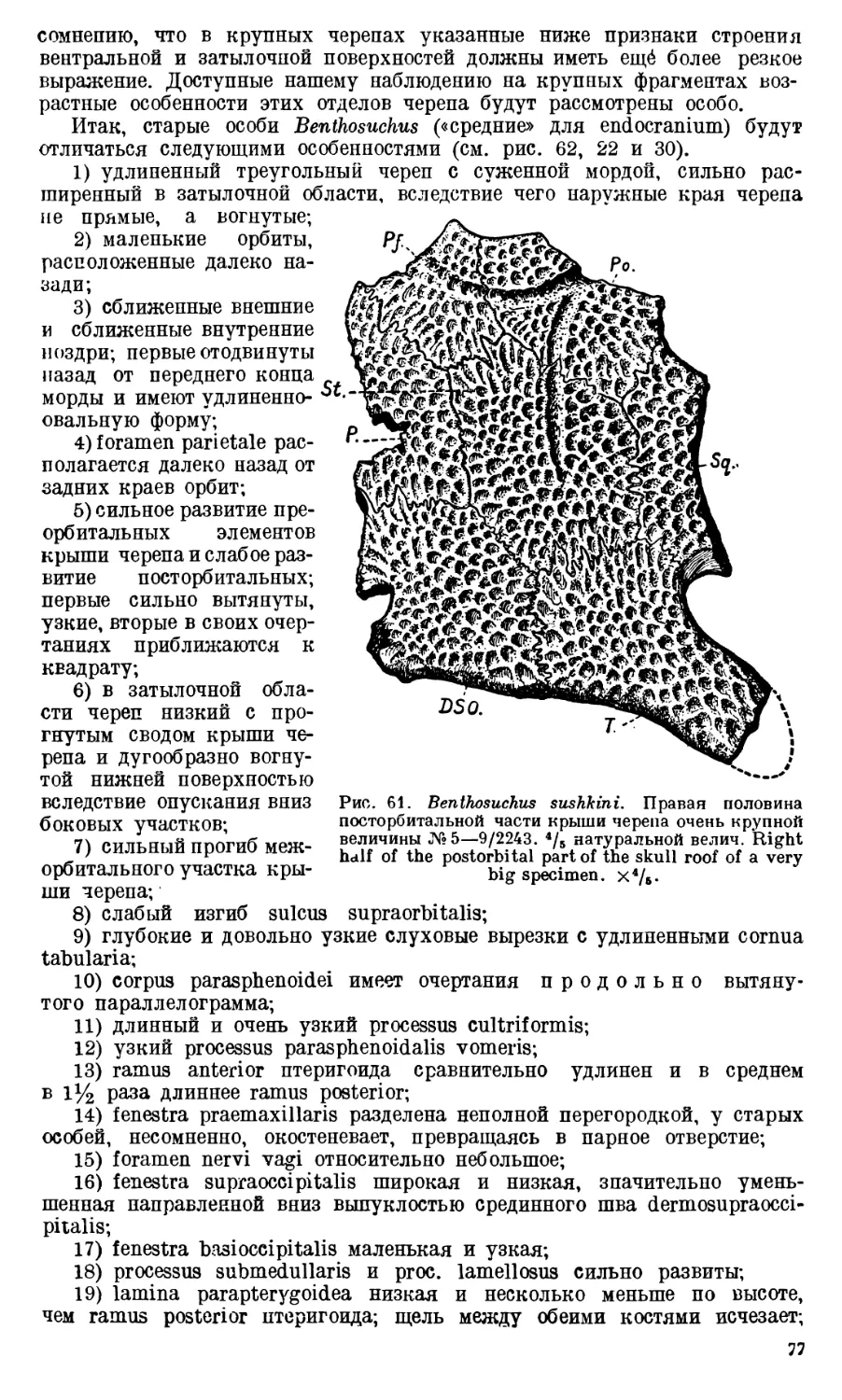
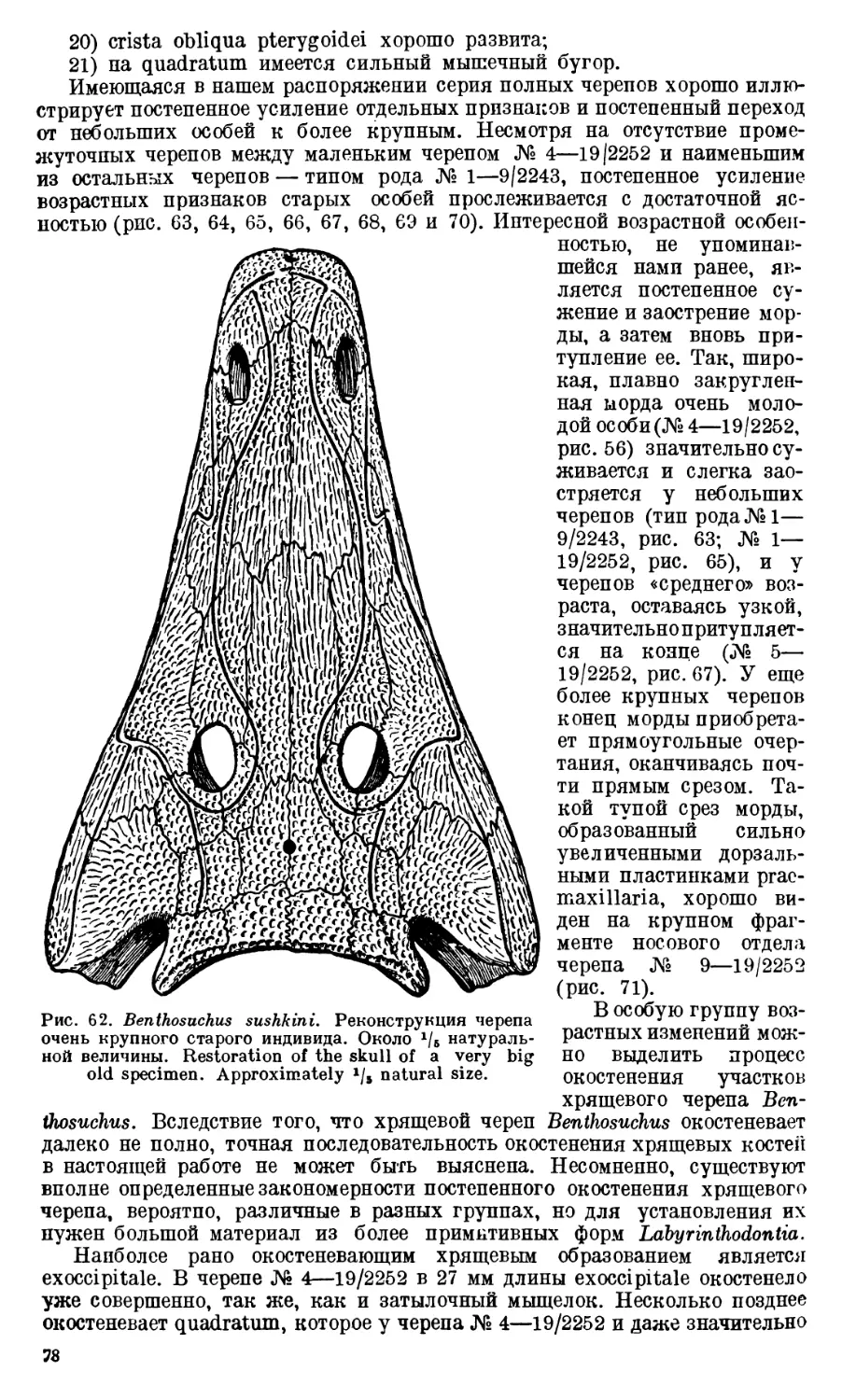
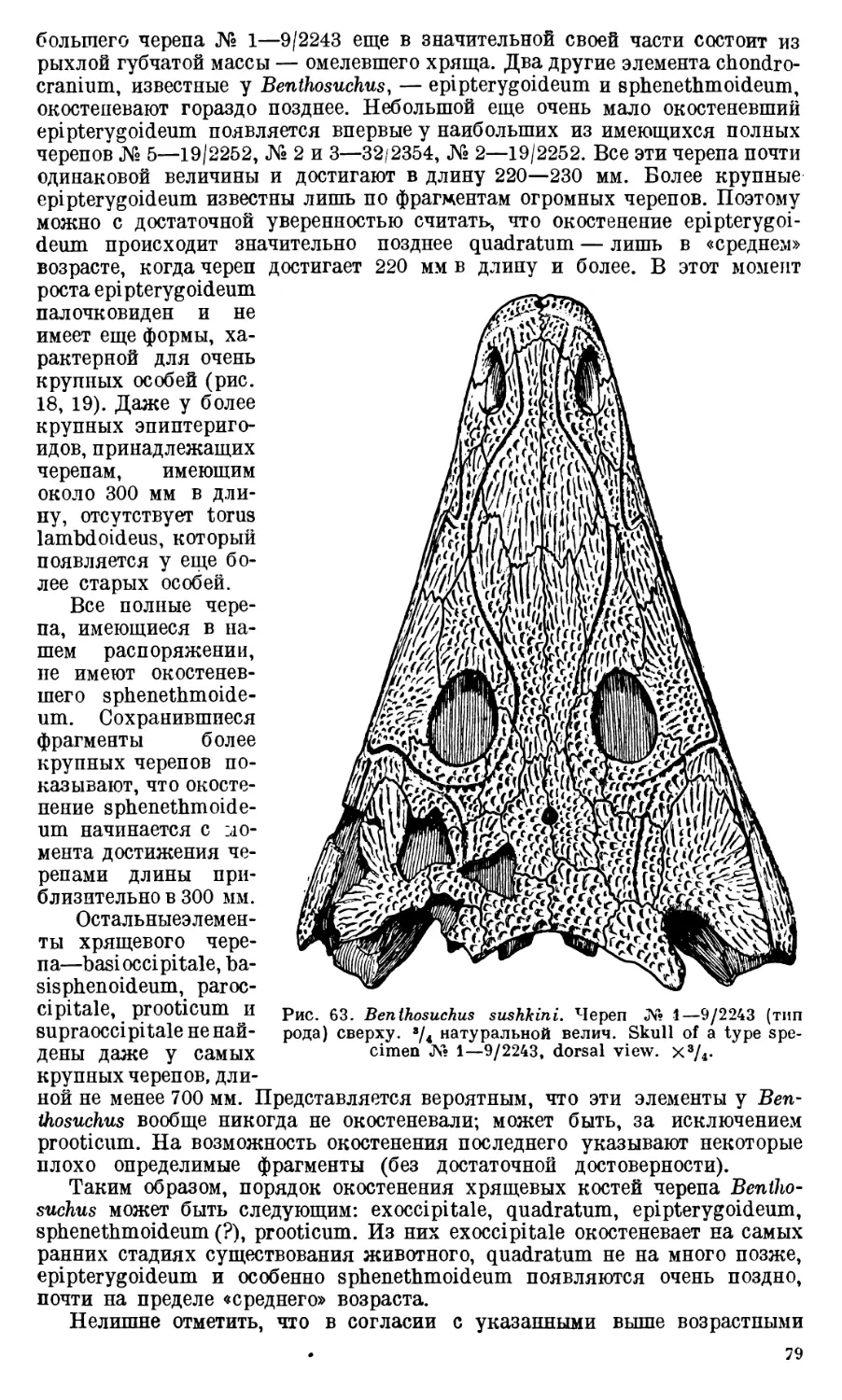
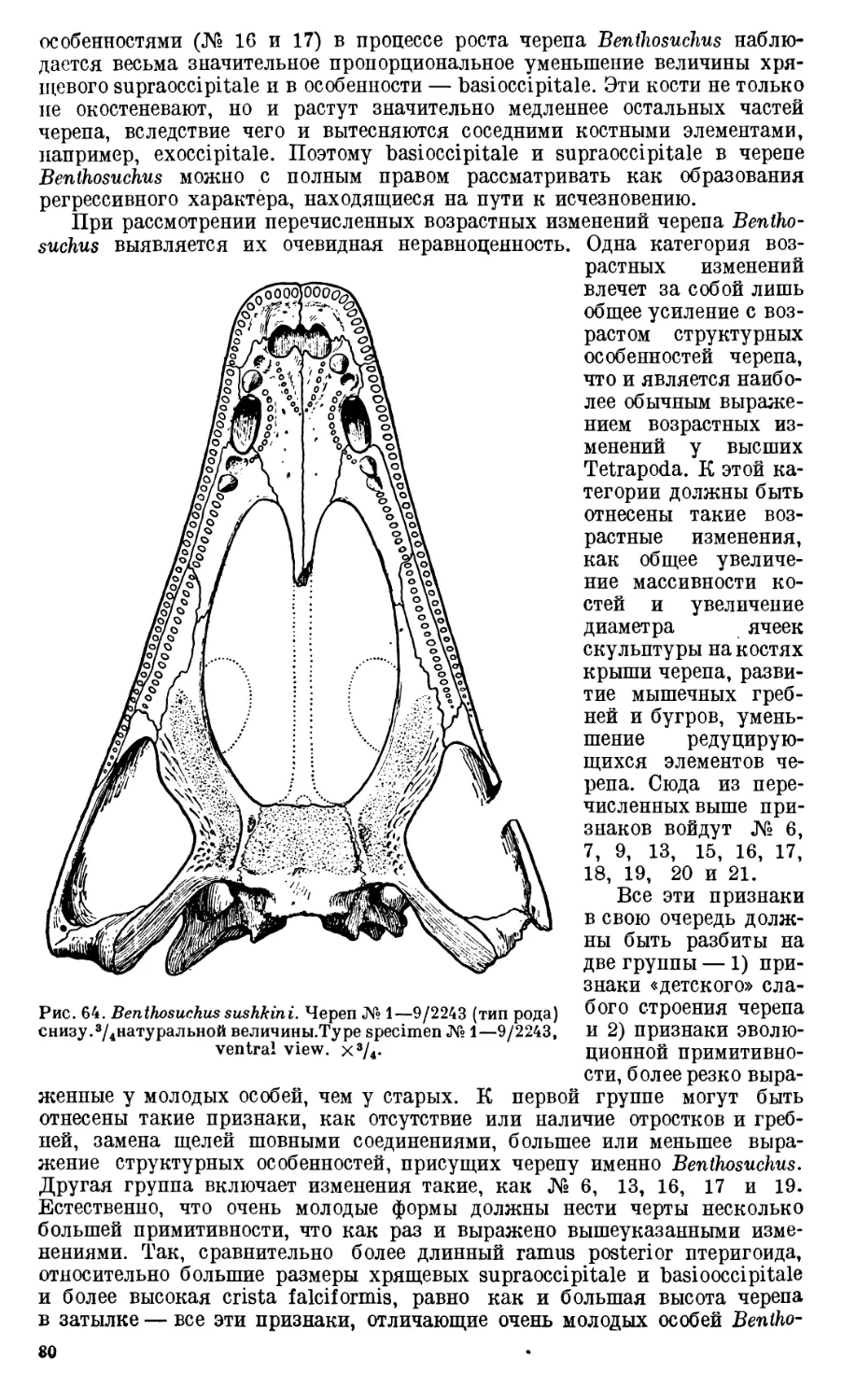
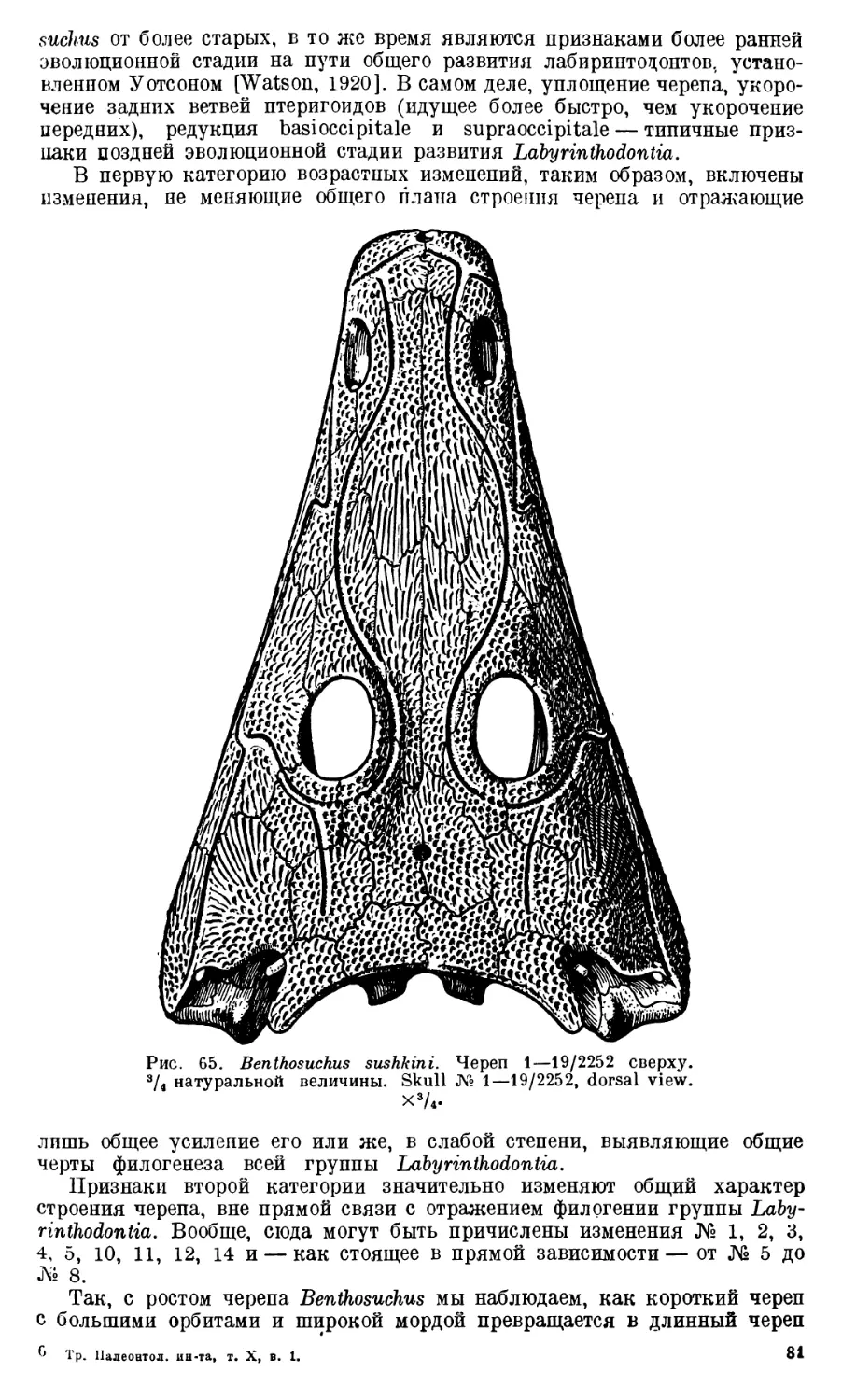
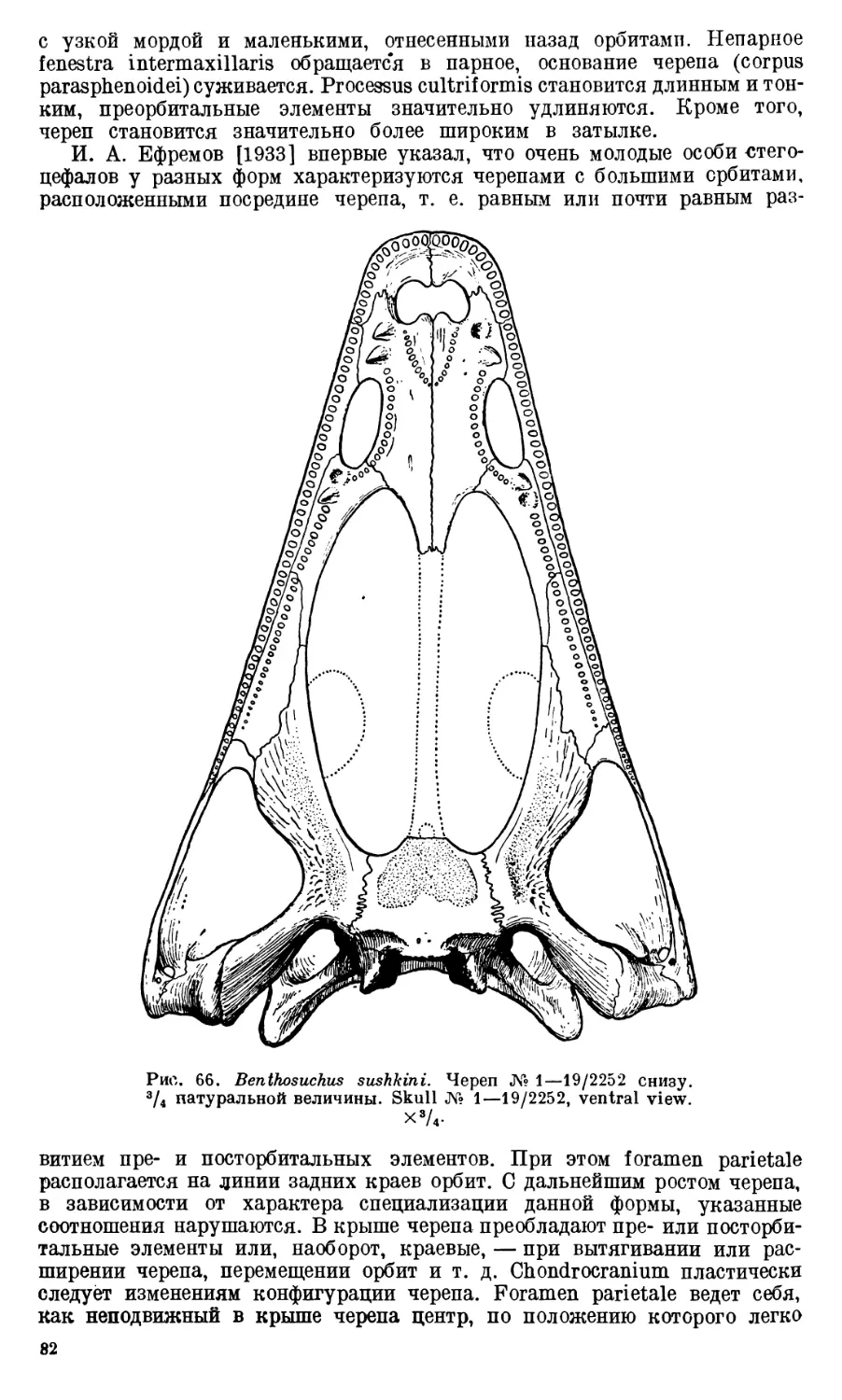
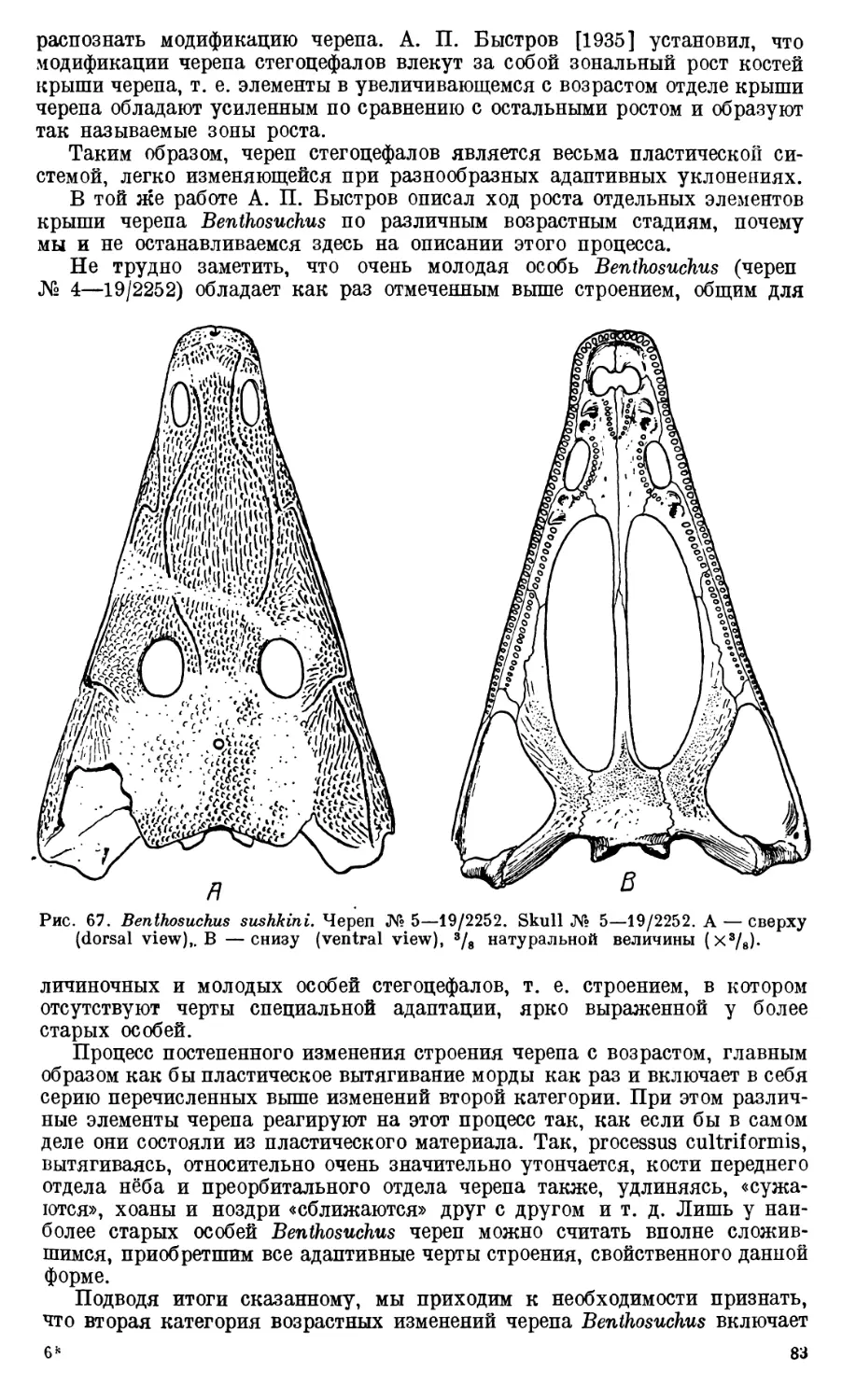
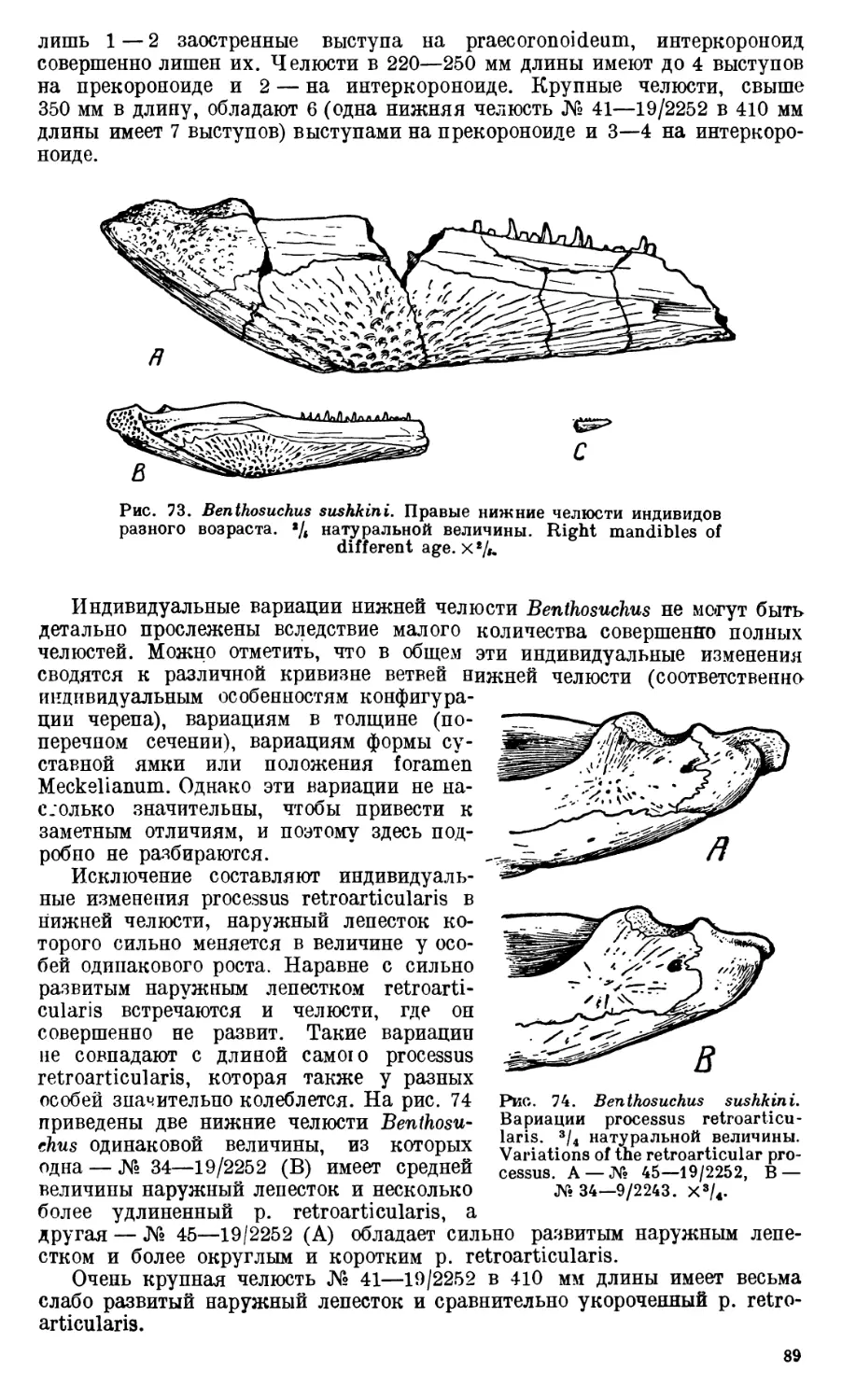
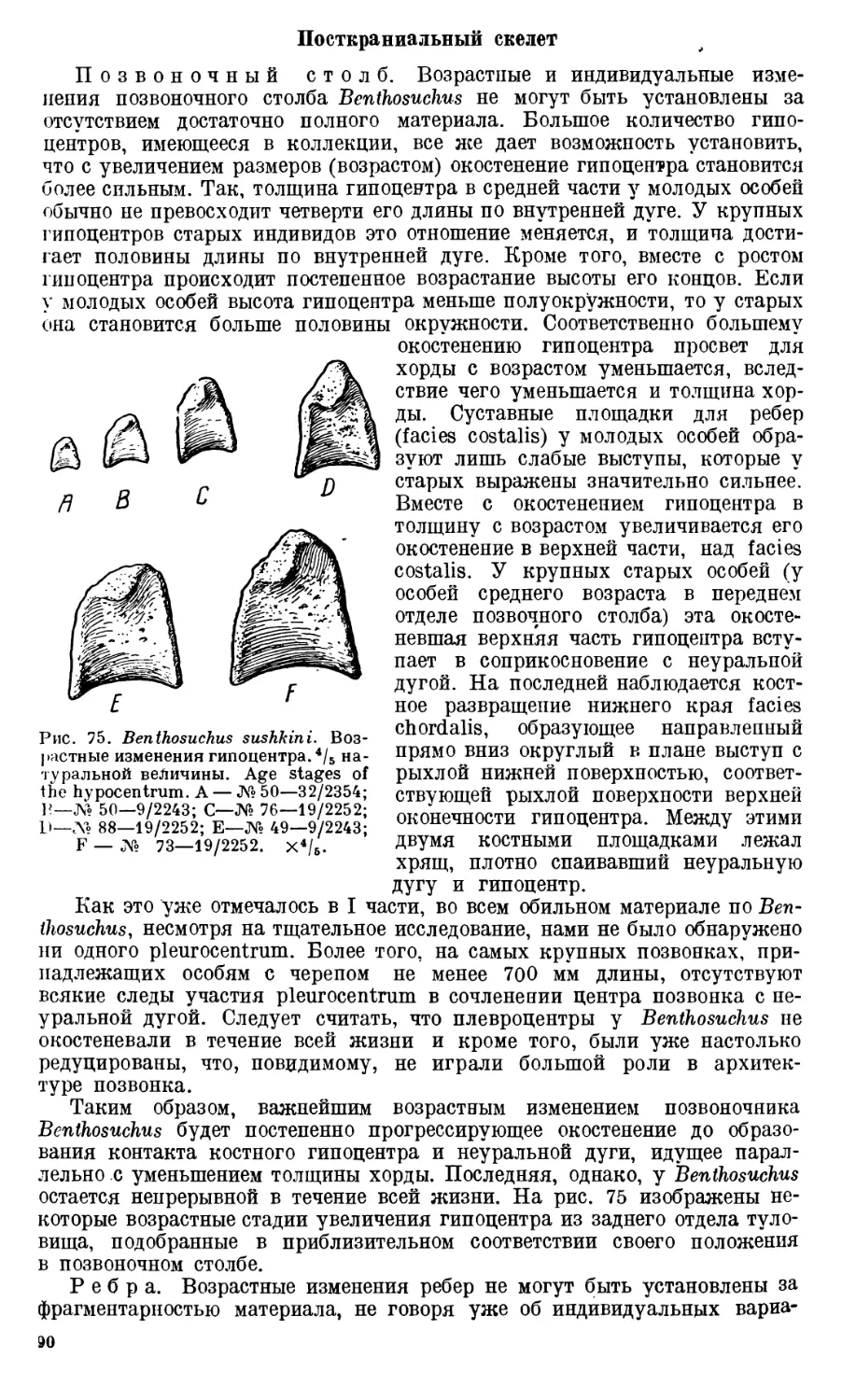
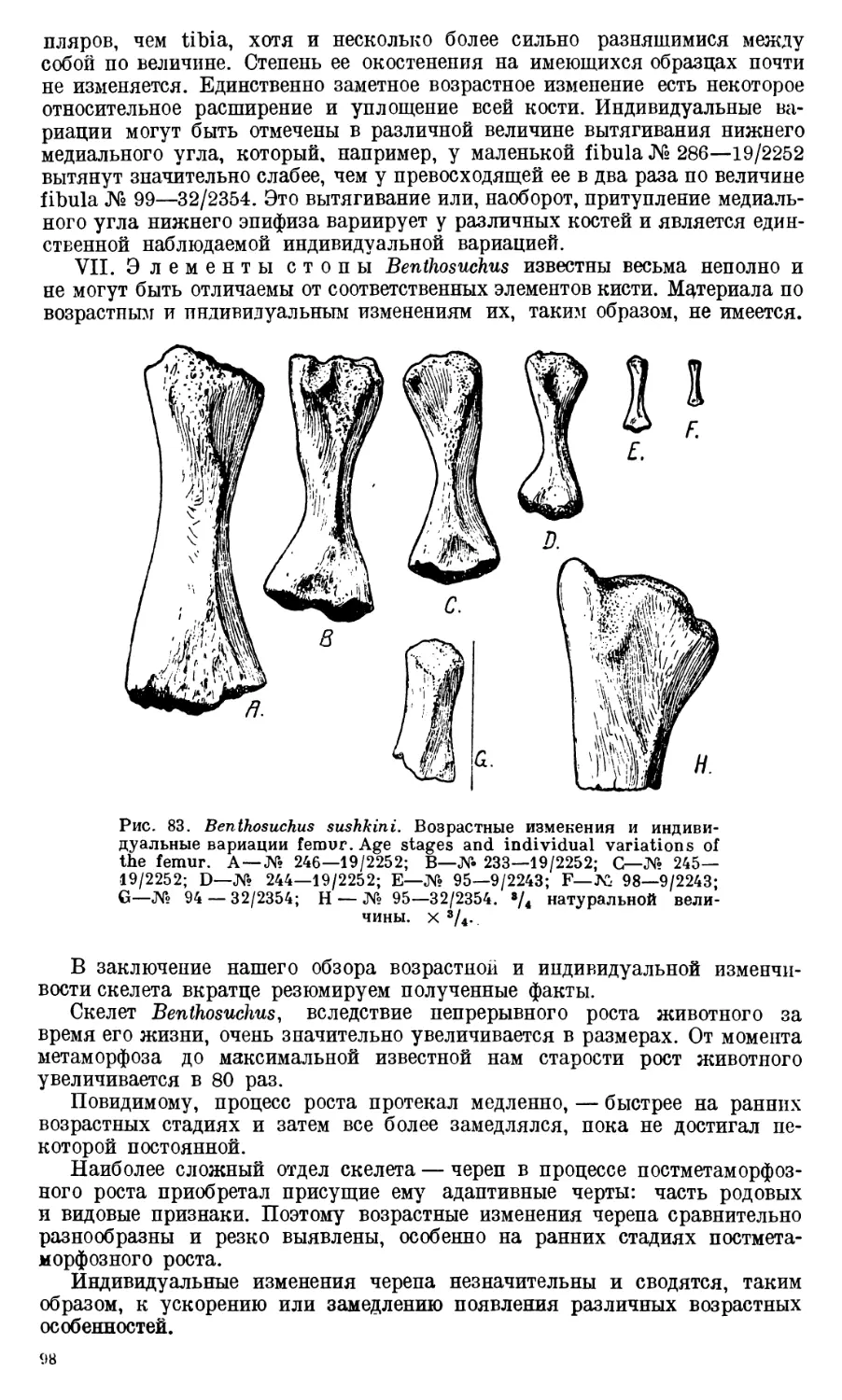
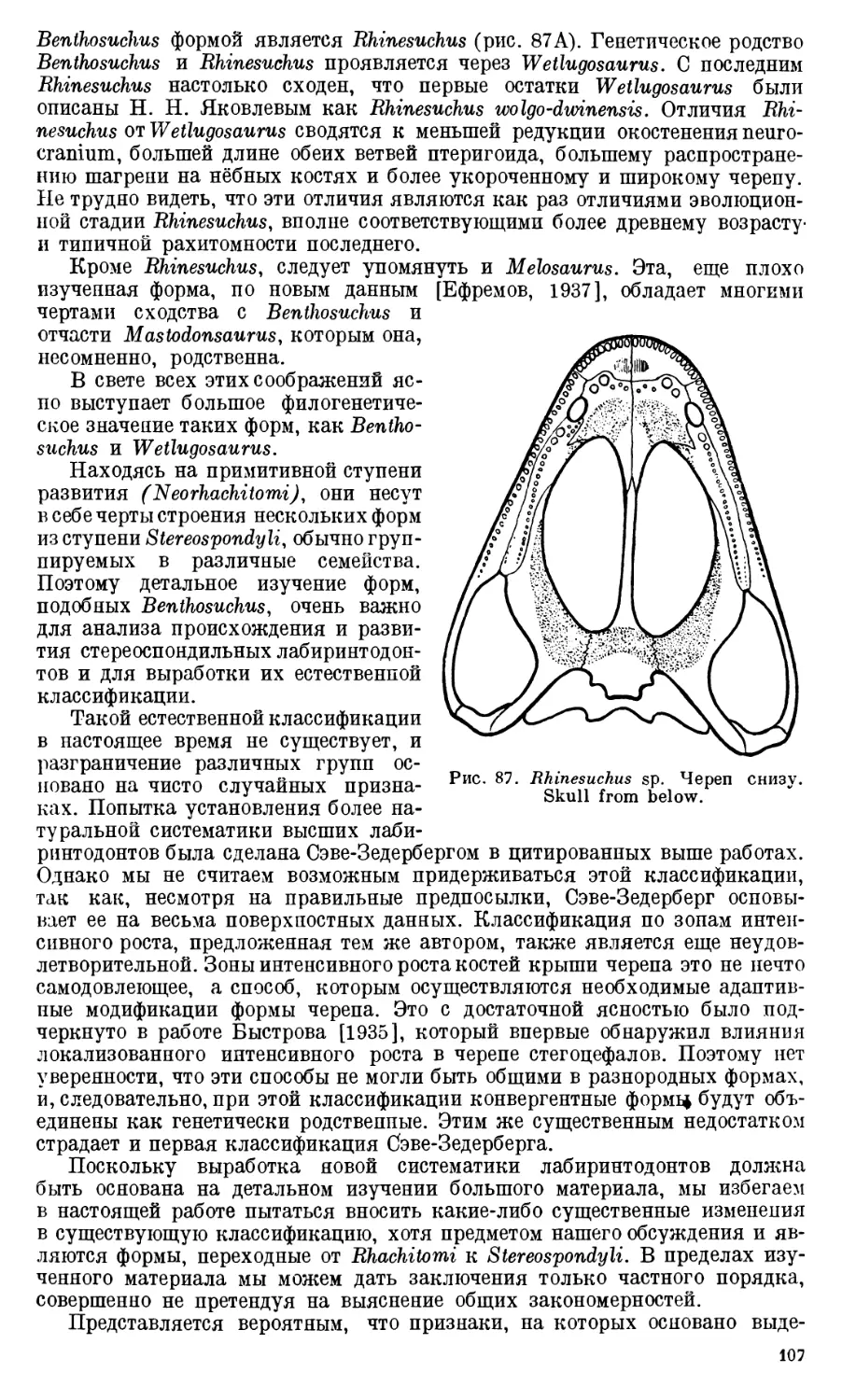
Череп Benthosuchus sushkini (табл. I, II) имеет очертания равнобедрен-
ного треугольника, стороны которого поставлены по отношению друг к другу
под углом приблизительно в 30°. Передний конец черепа слегка притуп-
лен. Носовые отверстия имеют овальную форму; ширина их в два раза мень-
ше длины.
Орбиты расположены на границе между средней и задней третями кры-
ши черепа. Они имеют овальную форму; длина их приблизительно в пол-
тора раза больше ширины.
Черепная крыша между орбитами вдавлена (impressio interorbitalis).
Круглое foramen parietale расположено несколько отступя назад от линии,
соединяющей задние края орбит.
Затылочный край черепа вогнут. Рога tabulare развиты хорошо и огра-
ничивают с медиальных сторон глубокие слуховые вырезки.
Девять целых черепов Benthosuchus, имеющиеся в коллекции Палеозооло-
гического института Академии Наук СССР, принадлежат разным по инди-
видуальному возрасту экземплярам. Длина черепов колеблется от 27 до
229 мм. Отдельные фрагменты покровных костей черепа и куски нижних
челюстей указывают, что длина черепа этого лабиринтодонта достигала
700—750 мм.
Изучение покровных костей черепа Benthosuchus выяснило, что в про-
цессе индивидуального роста его череп значительно удлинялся, приобре-
тая резко выраженную клинообразную форму. При этом оказалось, что
наиболее интенсивно растут кости, расположенные между носовыми отвер-
стиями и орбитами. Таким образом удалось установить в указанном участке
черепа так называемую «зону интенсивного роста» [Быстров, 1935]. Нали-
чие этой зоны является причиной возрастных изменений в пропорциях че-
репа. Череп Benthosuchus несколько более короткий в молодости, постепенно
становится длиннее у старых экземпляров.
Кости черепа Benthosuchus sushkini
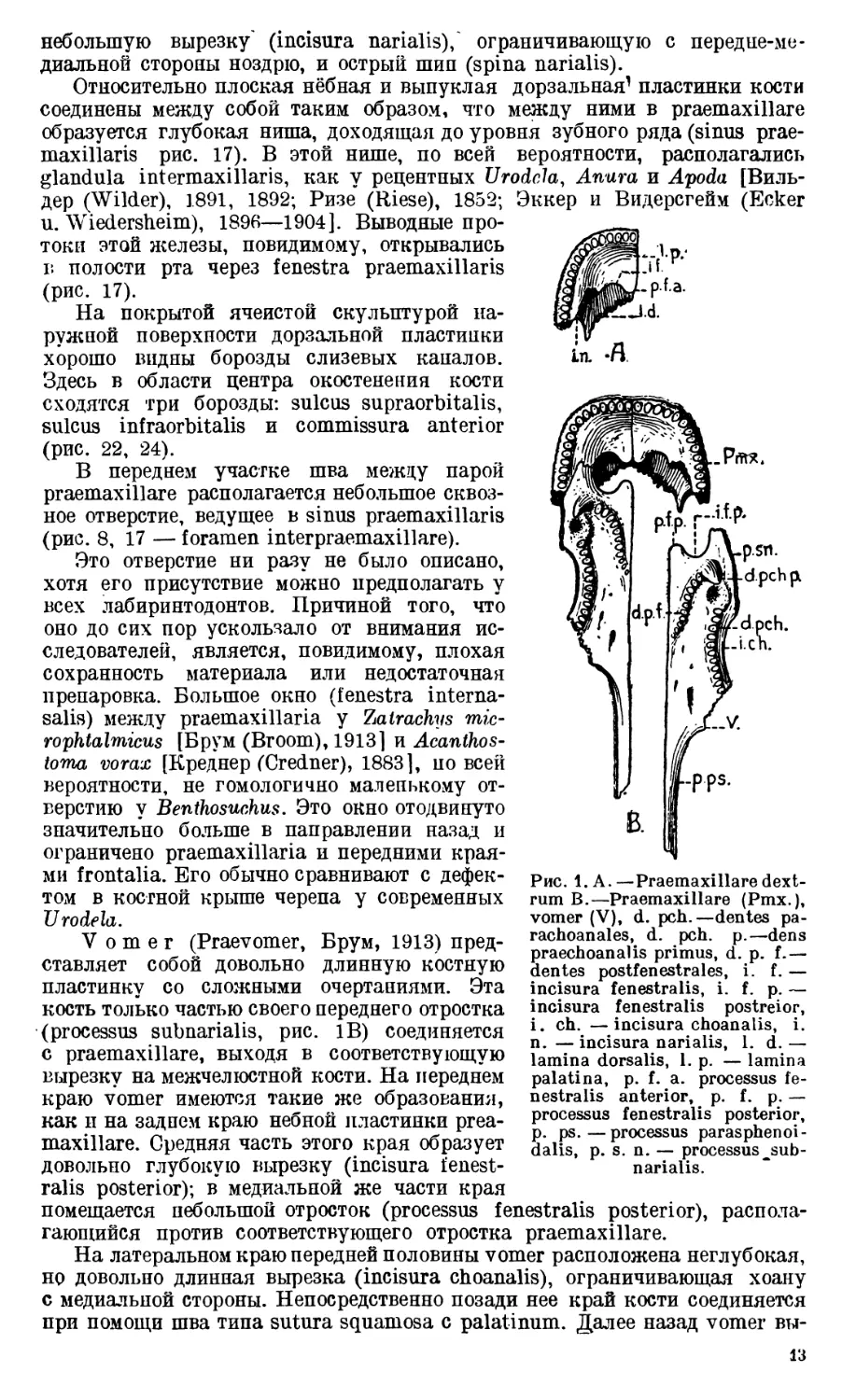
Praemaxillare, образующее несколько притупленный конец
морды, представляет собою сравнительно небольшую и короткую кость
и, может рассматриваться как две пластинки, соединенные друг с другом на
протяжении всего закругленного зубного края.
Небная пластинка (lamina palatina, рис. 1А) имеет форму плоского от-
ростка, образующего самый передний отдел костного нёба. Ее задний край
в своей медиальной части несет небольшой отросток с закругленной верши-
ной (processus fenestralis anterior). Латеральнее на том же краю расположе-
на довольно глубокая вырезка (incisura fenestralis); еще латеральнее имеется
небольшое углубление с неровными краями, куда входит vomer.
Дорзальная пластинка (lamina dorsalis, рис. 1А, 8) выпуклая. Ее наруж;
ная поверхность покрыта обычной для лабиринтодонтов ячеистой скульп-
турой. Задний край дорзальной пластинки в латеральной своей части имеет
<2
Эккер и Видерсгейм (Ecker
небольшую вырезку' (incisura narialis),' ограничивающую с передне-ме-
диальной стороны ноздрю, и острый шип (spina narialis).
Относительно плоская нёбная и выпуклая дорзальная1 пластинки кости
соединены между собой таким образом, что между ними в praemaxillare
образуется глубокая ниша, доходящая до уровня зубного ряда (sinus prae-
maxillaris рис. 17). В этой нише, по всей вероятности, располагались
glandula intermaxillaris, как у рецептных Urodela, Anura и Apoda [Виль-
дер (Wilder), 1891, 1892; Ризе (Riese), 1852;
u. Wiedersheim), 1896—1904]. Выводные про-
токи этой железы, повидимому, открывались
в полости рта через fenestra praemaxillaris
(рис. 17).
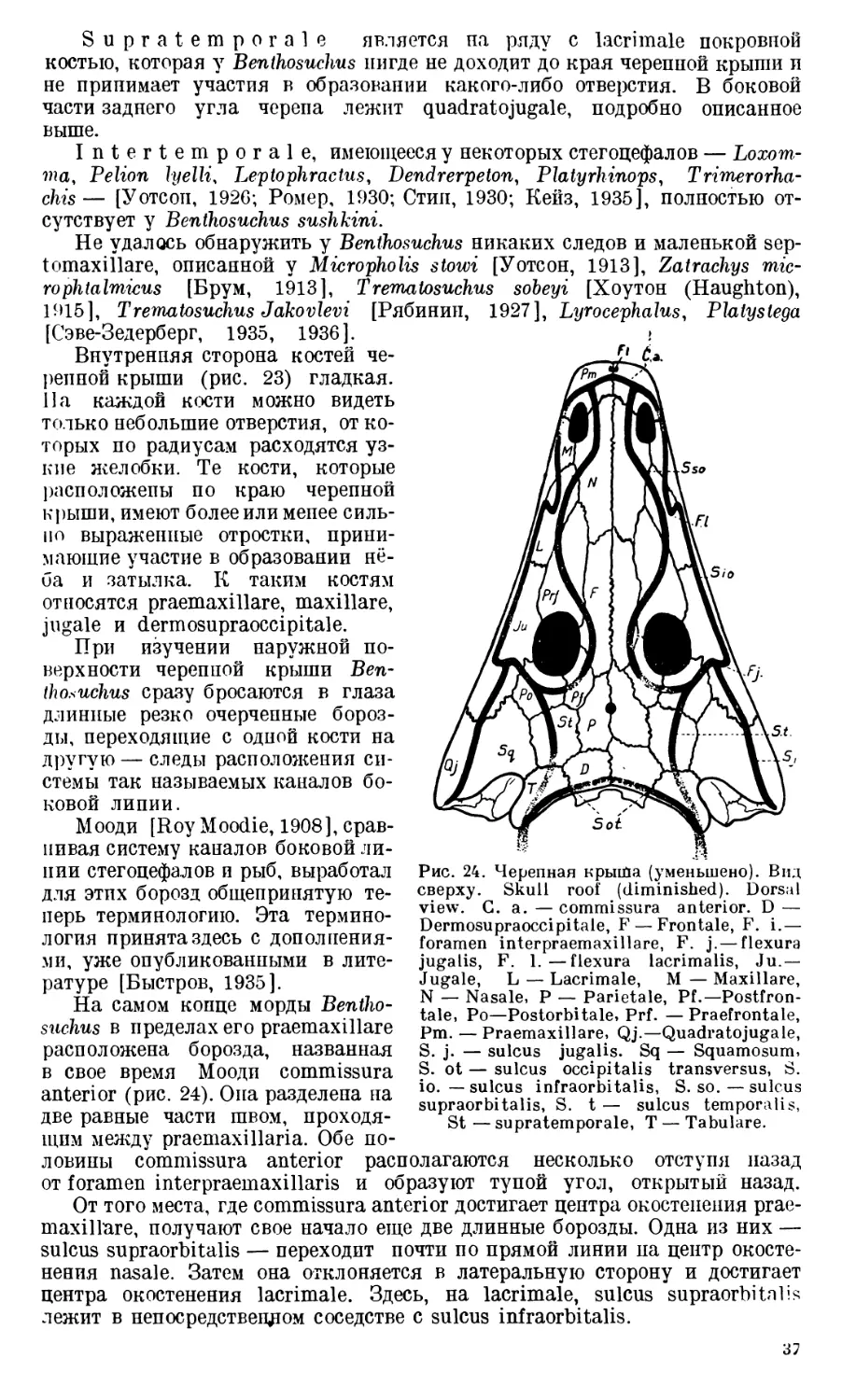
На покрытой ячеистой скульптурой на-
ружной поверхности дорзальной пластинки
хорошо видны борозды слизевых каналов.
Здесь в области центра окостенения кости
сходятся три борозды: sulcus supraorbitalis,
sulcus infraorbitalis и commissura anterior
(рис. 22, 24).
В переднем участке шва между парой
praemaxillare располагается небольшое сквоз-
ное отверстие, ведущее в sinus praemaxillaris
(рис. 8, 17 — foramen interpraemaxillare).
Это отверстие ни разу не было описано,
хотя его присутствие можно предполагать у
всех лабиринтодонтов. Причиной того, что
оно до сих пор ускользало от внимания ис-
следователей, является, повидимому, плохая
сохранность материала или недостаточная
препаровка. Большое окно (fenestra interna-
salis) между praemaxillaria у Pjatrachys mic-
rophtalmicus [Брум (Broom), 1913] и Acanthos-
toma vorax [Креднер (Credner), 1883], но всей
вероятности, не гомологично маленькому от-
верстию у Benthosuchus. Это окно отодвинуто
значительно больше в направлении назад и
ограничено praemaxillaria и передними края-
ми frontalia. Его обычно сравнивают с дефек-
том в костной крыше черепа у современных
Urodela.
Vomer (Praevomer, Брум, 1913) пред-
ставляет собой довольно длинную костную
пластинку со сложными очертаниями. Эта
кость только частью своего переднего отростка
(processus subnarialis, рис. 1В) соединяется
с praemaxillare, выходя в соответствующую
вырезку на межчелюстной кости. На переднем
краю vomer имеются такие же образования,
как и на заднем краю небной пластинки ргеа-
maxillare. Средняя часть этого края образует
довольно глубокую вырезку (incisura fenest-
ralis posterior); в медиальной же части края
помещается небольшой отросток (processus fenestralis posterior), распола-
гающийся против соответствующего отростка praemaxillare.
На латеральном краю передней половины vomer расположена неглубокая,
но довольно длинная вырезка (incisura choanalis), ограничивающая хоану
с медиальной стороны. Непосредственно позади нее край кости соединяется
при помощи шва типа sutura squamosa с palatinum. Далее назад vomer вы-
гз
Рис. 1.А.—Praemaxillare dext-
rum В.—Praemaxillare (Pmx.),
vomer (V), d. pch.—dentes pa-
rachoanales, d. pch. p.—dens
praechoanalis primus, d. p. f.—
dentes postfen estrales, i. f. —
incisura fenestralis, i. f. p. —
incisura fenestralis postreior,
i. ch. —incisura choanalis, i.
n. — incisura narialis, 1. d. —
lamina dorsalis, 1. p. — lamina
palatina, p. f. a. processus fe-
nestralis anterior, p. f. p. —
processus fenestralis posterior,
p. ps.—processus parasphenoi-
dalis, p. s. n. — processus^sub-
narialis.
тянут в длинный и узкий отросток (processus parasphenoidalis). Медиальный
край vomer прямой. Нёбная поверхность vomer снабжена двумя рядами
маленьких конических зубов и большими клыками. Обычно каждый vomer
несет по одному клыку, однако в коллекции имеются экземпляры, у кото-
рых сошники имеют по два клыка, расположенных один позади другого.
В тех случаях, когда сохраняется только один клык, он одинаково часто ока-
зывается как передним (первым), так и задним (вторым). Первый клык (dens
praechoanalis primus, рис. 1В) располагается непосредственно позади inci-
sura fenestralis posterior. Это — большой конусообразный зуб, направлен-
ный своей вершиной несколько назад. Второй клык (dens praechoanalis
secundus) располагается позади первого. В тех случаях, когда один из
клыков отсутствует, на его месте образуется небольшая ямка (fovea dentis
praechoanalis); она сравнительно неглубока, окружена небольшим кольце-
вым валиком, имеет шероховатое дно. Медиально и позади первого пред-
хоапного клыка тянется ряд маленьких зубов (dentes postfenestrales). Бли-
жайшие к клыку зубы этого ряда обычно несколько больше тех, которые
помещаются у самого края кости. Два ряда постфенестральных зубов па
двух сложенных вместе сошниках располагаются под углом приблизительно
в 60° друг к другу.
Позади второго постхоанального клыка, или его ямки в случае его от-
сутствия, помещается второй ряд мелких зубов, образующий слабо вогну-
тую в медиальном направлении дугу. Эти зубы (dentes parachoanales), как
и постфенестральный ряд, сидят вдоль небольшого костного валика, рас-
положенного параллельно краю хоанной вырезкн сошника.
Лишенная зубов нёбная поверхность vomer гладкая; на ней видны одно
или два довольно больших отверстия, от которых в каудальном направле-
нии тянутся длинные желобки. Отверстия эти проникают сквозь костную
пластинку vomer и видны на его дорзальной поверхности (рис. 1В, 8, 17 —
foramen nervi palatini). Через vomer, повидимому, проходила одна из ветвей
лицевого нерва (ramus palatinus nervi facialis). Сэве-Зедерберг [Save-So-
derbergh, 1936] описал подобное отверстие у Lyrocephalus euri, но в этом
случае нерв проникал в соответствующее отверстие с дорзальной, а не
с вентральной стороны vomer. Дальнейшего хода, начинающегося от этого
отверстия канала, Сэве-Зедерберг не проследил; таким образом, осталось
невыясненным, где у Lyrocephalus нёбпая ветвь nervi facialis выходила из
сошника.
На дорзальной поверхности vomer вдоль его хоанной вырезки хорошо
видно резко очерченное дугообразное утолщение кости (torus parachoana-
lis. рис. 8). Это утолщение, повидимому, имеет чисто механическое значе-
ние, укрепляя край хоаны, так как со стороны нёба здесь располагается ряд
парахоанальных зубов. Другой особенностью дорзальной поверхности сош-
ника является хорошо выраженный жолоб, идущий вдоль всего медиаль-
ного края кости. На двух сложенных вместе сошниках этот жолоб служит
местом, куда ложится передний конец processus cultriformis парасфеноида
(sulcus parasphenoidalis, рис. 8, 21).
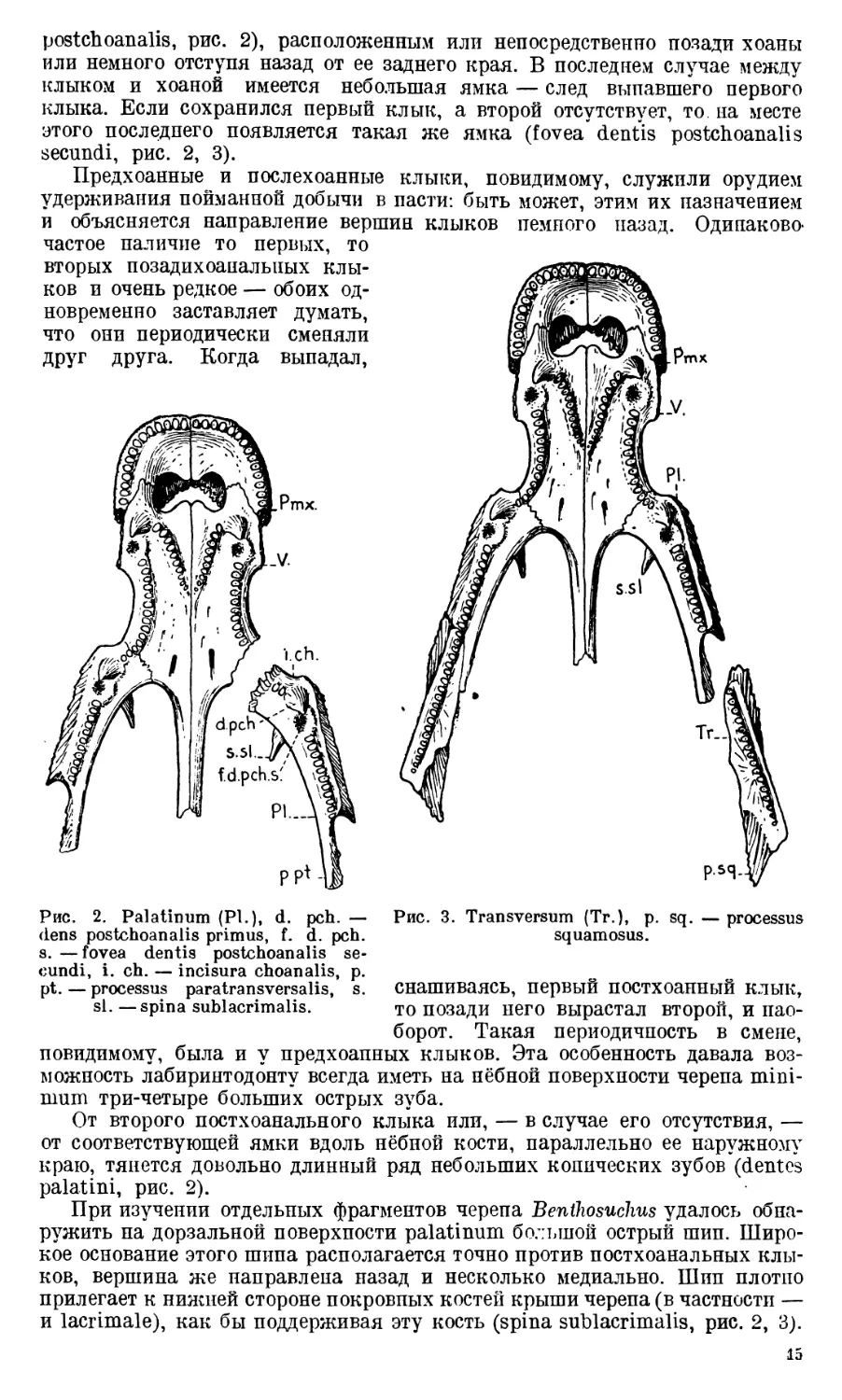
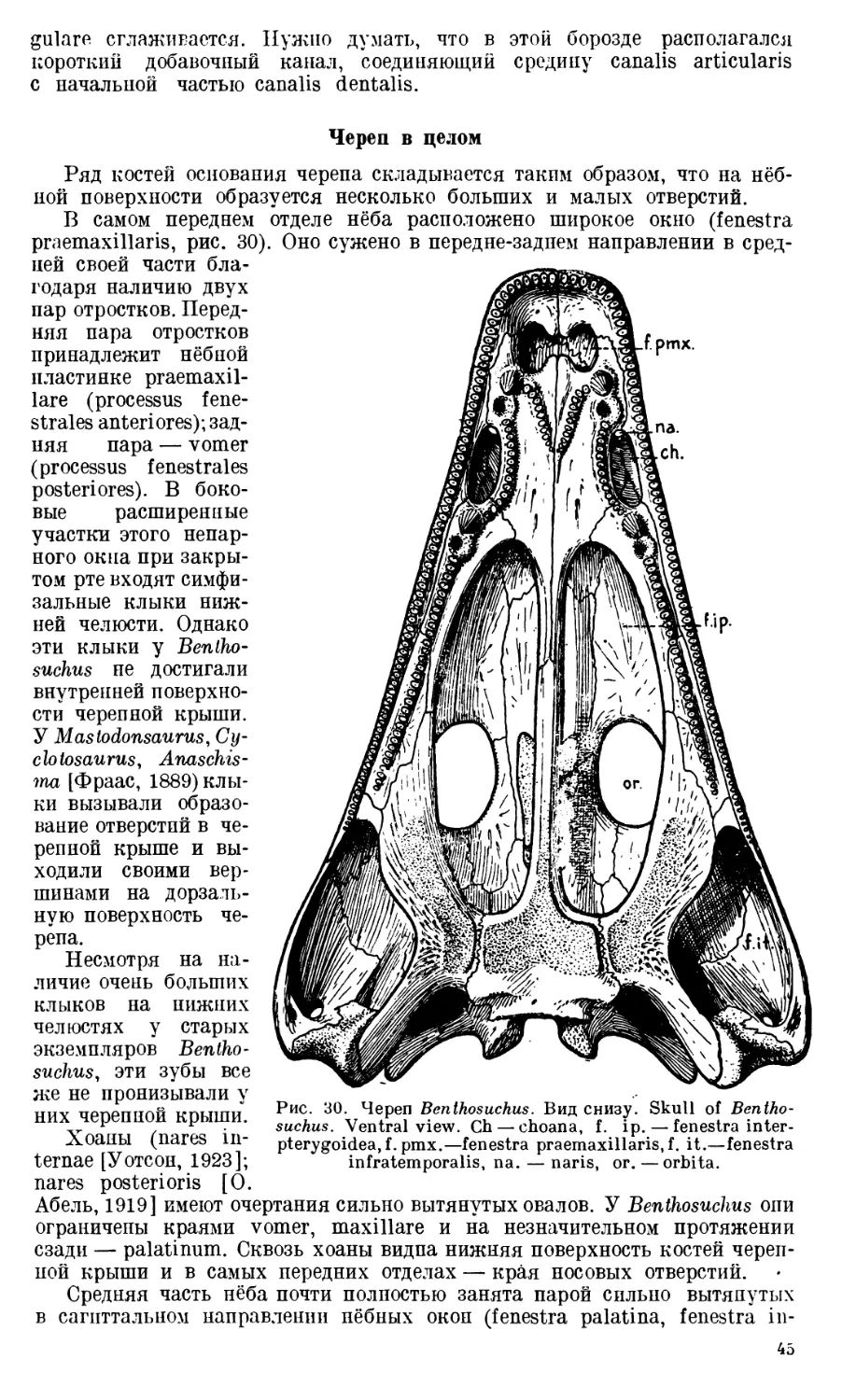
Palatinum присоединяется к латеральной стороне vomer непо-
средственно позади его хоапной вырезки. Передний край этой кости при-
нимает участие в образовании хоаны, ограничивая ее сзади (incisura choa-
nalis palatini, рис. 2). Palatinum соединяется с сошником при помощи
косо срезанного края, образуя своего рода sutura squamosa, причем pala-
tinum накладывается сверху на край vomer. Вдоль всего латерального края
нёбной кости располагается длинный гребень, обращенный своим острым
краем дорзально. Сзади palatinum переходит в узкий отросток (processus
paratransversalis, рис. 2), который своей латеральной половиной наклады-
вается в виде чешуи на помещающееся рядом transversum.
Передняя часть нёбной поверхности palatinum снабжена таким же боль-
шим клыком, как и сошник. Очень редко здесь наблюдаются два клыка, си-
дящие один за другим. Обычно же мы имеем дело с одним клыком (dens
14
postchoanalis, рис. 2), расположенным или непосредственно позади хоаны
или немного отступя назад от ее заднего края. В последнем случае между
клыком и хоаной имеется небольшая ямка — след выпавшего первого
клыка. Если сохранился первый клык, а второй отсутствует, то. на месте
этого последнего появляется такая же ямка (fovea dentis postchoanalis
secundi, рис. 2, 3).
Предхоанные и послехоанные клыки, повидимому, служили орудием
удерживания пойманной добычи в пасти: быть может, этим их назначением
и объясняется направление вершин клыков немного назад. Одинаково-
частое наличие то первых, то
вторых позадихоапальпых клы-
ков и очень редкое — обоих од-
новременно заставляет думать,
что они периодически сменяли
друг друга. Когда выпадал,
Рис. 2. Palatinum (Pl.), d. pch. — Рис. 3. Transversum (Tr.), p. sq. — processus
dens postchoanalis primus, f. d. pch. squamosus.
s. —fovea dentis postchoanalis se-
cundi, i. ch. — incisura choanalis, p.
pt. — processus paratransversalis, s. снашиваясь, первый постхоанный клык,
si. — spina sublacrimalis. то позади него вырастал второй, и нао-
борот. Такая периодичность в смене,
повидимому, была и у предхоапных клыков. Эта особенность давала воз-
можность лабиринтодонту всегда иметь на нёбной поверхности черепа mini-
mum три-четыре больших острых зуба.
От второго постхоанального клыка или, — в случае его отсутствия, —
от соответствующей ямки вдоль нёбной кости, параллельно ее наружному
краю, тянется довольно длинный ряд небольших конических зубов (dentes
palatini, рис. 2).
При изучении отдельных фрагментов черепа Benthosuchus удалось обна-
ружить па дорзальной поверхности palatinum большой острый шип. Широ-
кое основание этого шипа располагается точно против постхоанальных клы-
ков, вершина же направлена назад и несколько медиально. Шип плотно
прилегает к нижней стороне покровных костей крыши черепа (в частности —
и lacrimale), как бы поддерживая эту кость (spina sublacrimalis, рис. 2, 3).
15
Это образование, повидимому, имеет чисто механическое значение: шип,
упираясь в свод черепа, образованный плотно сомкнутыми покровными ко-
стями, создает прочную основу для обоих постхоапальных клыков. Шнп
не только дает опору для клыков, до и в значительной мере укрепляет пе-
редний конец palatinum, который в свою очередь служит опорой для зад-
него края vomer. Как указывалось выше, между vomer и palatinum имеется
чешуйчатый шов, в котором край palatinum накладывается сверху на край
vomer (рис. 2).
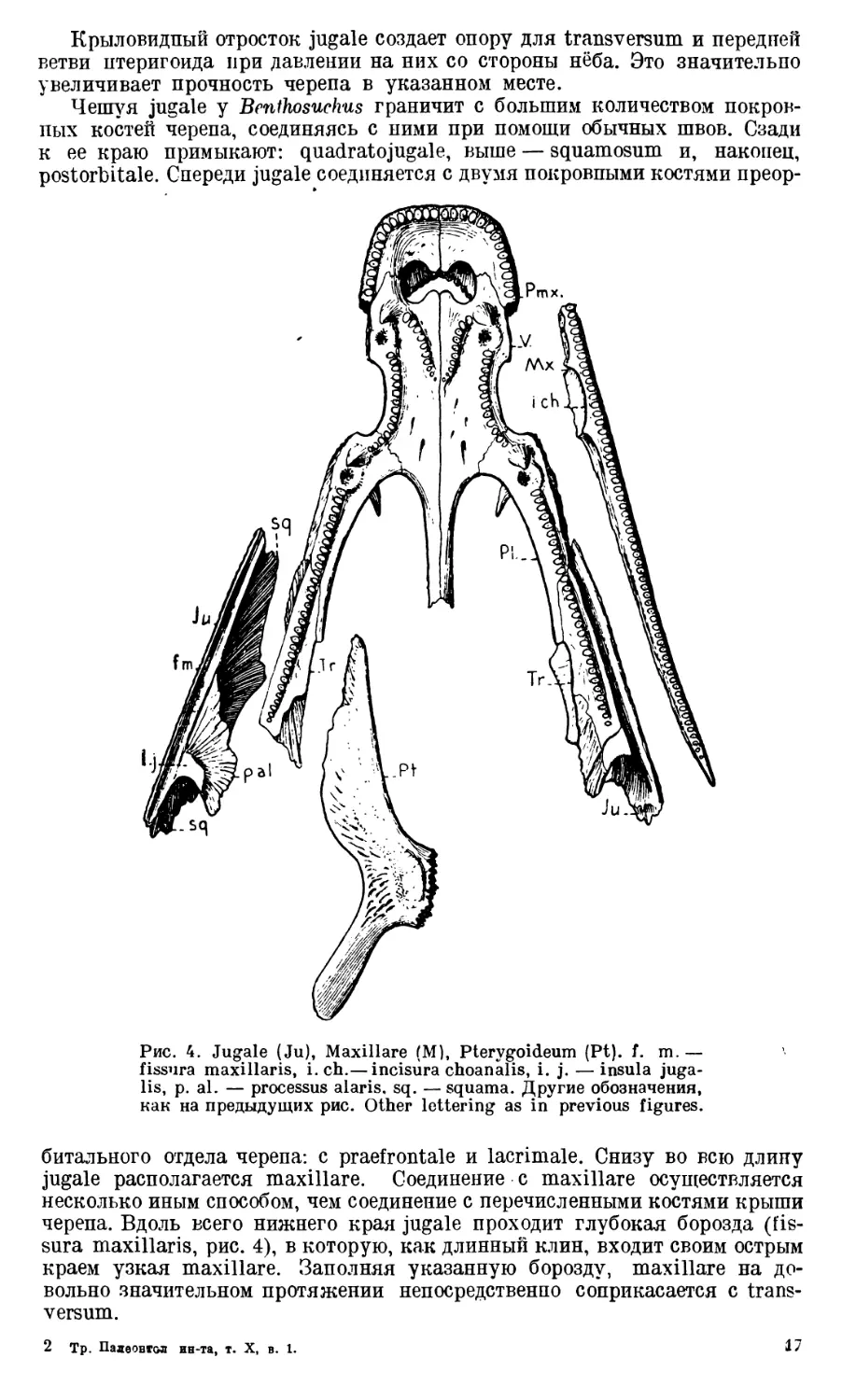
Transversum (Ectopterygoideum) представляет собой узкую и
длинную кость, несущую на своей вентральной поверхности один ряд зубов
(dentes transversi, рис. 3), расположенный параллельно его латеральному
краю. Передний угол transversum косо срезан и, входя в соответствующую
вырезку на palatinum, накладывается сверху на эту кость.
Выше уже упоминалось , что processus paratransversalis palatini своим
латеральным краем ложится на transversum с дорзальной его стороны. Таким
образом, соединение palatinum и transversum осуществляется при помощи
очень сложно построенного шва, представляющего собой комбинацию двух
sutura squamosa, что, несомненно, обусловливает большую прочность скреп-
ления обеих костей.
Задне-медиальный угол transversum вытянут в довольпо длинный пло-
ский отросток, который накладывается сверху на конец передней ветви
птеригоида (processus squamosus, рис. 3). С птеригоидом же граничит и почти
прямой медиальный край кости. На протяжении этого края transversum
соединяется с птеригоидом при помощи чешуйчатого шва; при этом передняя
ветвь птеригоида накладывается на transversum с дорзальной стороны
(рис. 4). В результате этого между указанными костями возникает доста-
точно прочное и сложно построенное соединение, являющееся комбинацией
двух sutura squamosa.
Задний край и латеральные края transversum граничат с jugale.
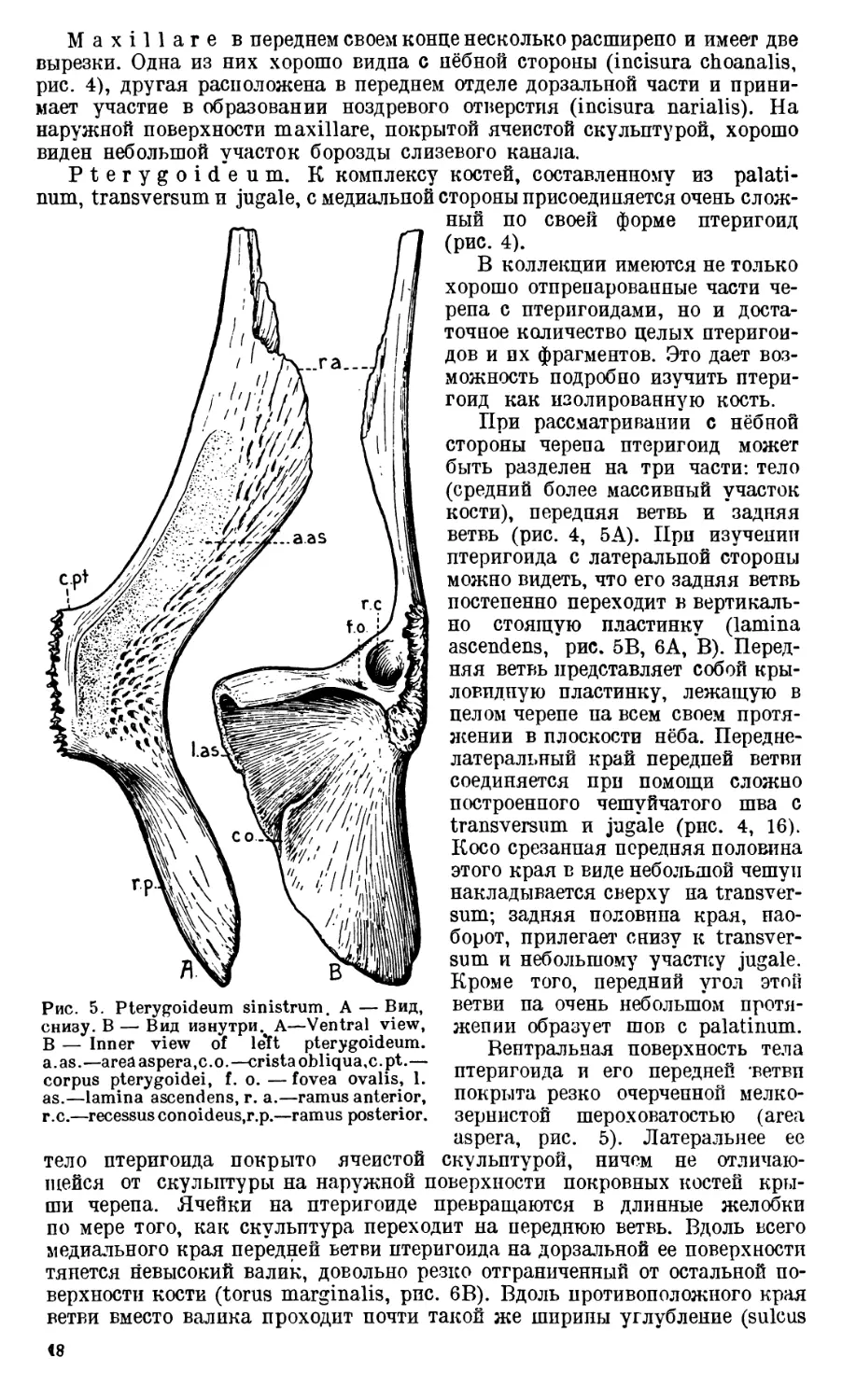
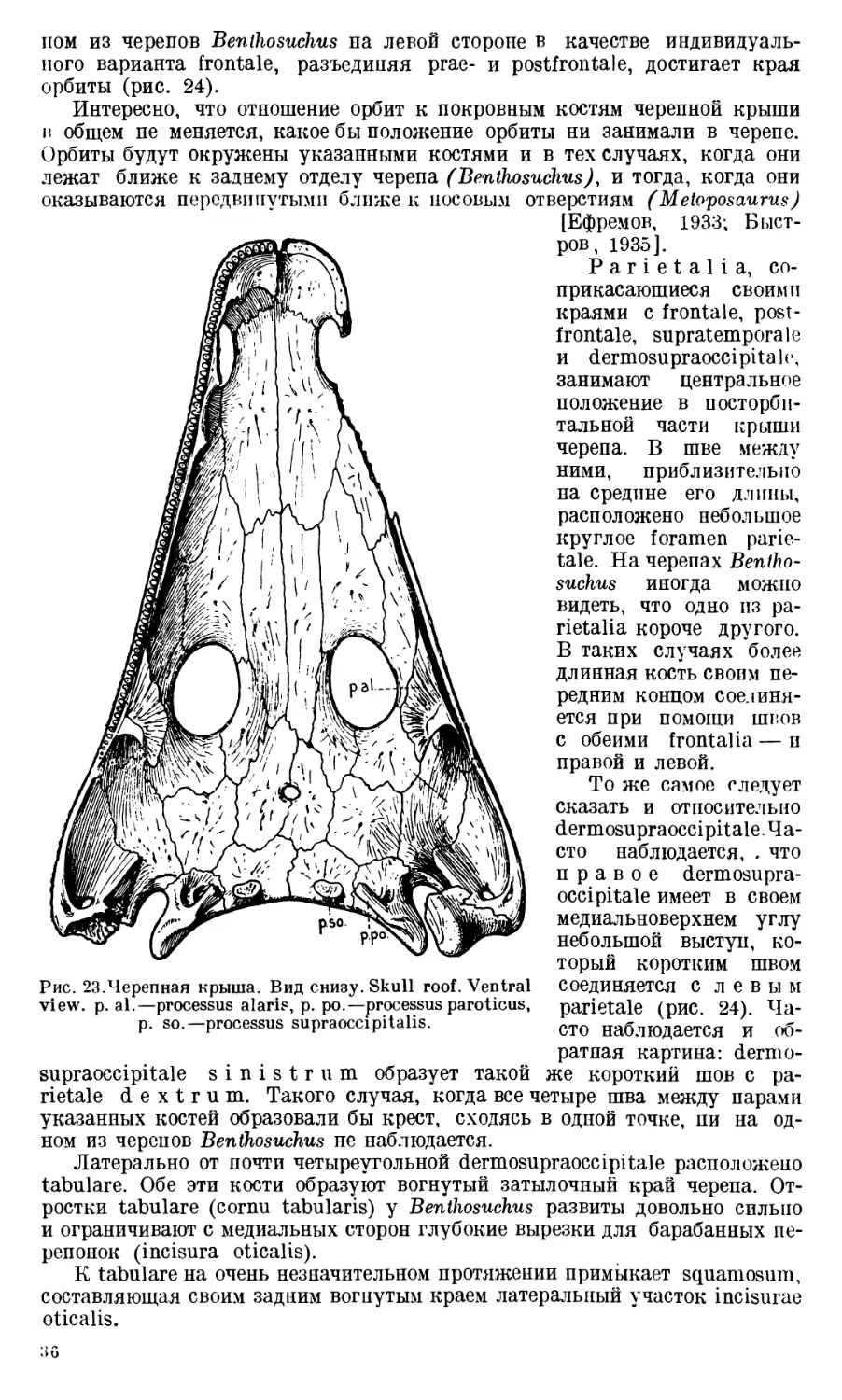
J u g а 1 е может рассматриваться как кость, состоящая из двух частей:
чешуи и крыловидного отростка (squama и processus alaris jugale, рис. 4).
Чешуя, покрытая с наружной стороны ячеистой скульптурой, на кото-
рой хорошо видны борозды слизевых каналов, принимает участие в обра-
зовании черепной крыши, а крыловидный отросток, отходящий от ниж-
него края чешуи в медиальном направлении, соединяется с костями нёба.
Впервые нёбный отросток jugale (processus alaris) был описан у Pelion
lyelli и у Macrerpeton huxleyi Ромером, [Romer, 1930]. Затем он был отмечен
без подробного описания у Benthosuchus (Benthosaurus) в работе о покровных
костях черепа стегоцефалов [Быстров, 1935]. Позднее Сэве-Зедерберг
[1936] подробно изучил с дорзальной стороны его форму у Lyrocephalus, Pla-
tystega и Aphaneramma.
Большая часть крыловидного отростка jugale у Benthosuchus, так же
как и у Lyrocephalus, Platystega и Aphaneramma, ложится на дорзальную
поверхность transversum, а меньшая — па переднюю ветвь птеригоида. При
этом оказывается, что небольшой участок вентральной поверхности отростка
остается не прикрытым указанными костями и хорошо виден со стороны
нёбной поверхности черепа в виде свободного островка кости (insula ju-
&alis, рис. 4).
Плохая сохранность материала, которым располагал Сэве-Зедерберг
[1936], не позволила ему обнаружить этот островок на нёбной поверхности
черепов Lyrocephalus, Platystega и Aphaneramma.
У Pelion, Macrerpeton, Benthosuchus, Lyrocephalus, Platystega n Apha-
neramma крыловидный отросток jugale ограничивает спереди височное от-
верстие (fenestra infratemporalis, рис. 16). По всей вероятности, те же отно-
шения были не только у форм, изученных Сэве-Зедербергом и Ромером, по и у
большинства, если пе у всех, стегоцефалов вообще. Что касается insula
jugalis, то, повидимому, только плохая препаровка не позволила обнару-
жить ее у большинства форм.
16
Крыловидный отросток jugale создает опору для transversum и передней
ветви птеригоида при давлении на них со стороны нёба. Это значительно
увеличивает прочность черепа в указанном месте.
Чешуя jugale у Benthosuchus граничит с большим количеством покров-
пых костей черепа, соединяясь с ними при помощи обычных швов. Сзади
к ее краю примыкают: quadratojugale, выше — squamosum и, наконец,
postorbitale. Спереди jugale соединяется с двумя покровными костями преор-
Рис. 4. Jugale (Ju), Maxillare (М), Pterygoideum (Pt), f. m. —
fissura maxillaris, i. ch.— incisura choanalis, i. j. —insula juga-
lis, p. al. — processus alaris. sq. — squama. Другие обозначения,
как на предыдущих рис. Other lettering as in previous figures.
битального отдела черепа: c praefrontale и lacrimale. Снизу во всю длину
jugale располагается maxillare. Соединение с maxillare осуществляется
несколько иным способом, чем соединение с перечисленными костями крыши
черепа. Вдоль всего нижнего края jugale проходит глубокая борозда (fis-
sura maxillaris, рис. 4), в которую, как длинный клин, входит своим острым
краем узкая maxillare. Заполняя указанную борозду, maxillare на до-
вольно значительном протяжении непосредственно соприкасается с trans-
versum.
2 Тр. Палеовтол ин-та, т. X, в. 1. ^7
Maxillare в переднем своем конце несколько расширено и имеет две
вырезки. Одна из них хорошо видна с нёбной стороны (incisura choanalis,
рис. 4), другая расположена в переднем отделе дорзальной части и прини-
мает участие в образовании ноздревого отверстия (incisura narialis). На
наружной поверхности maxillare, покрытой ячеистой скульптурой, хорошо
виден небольшой участок борозды слизевого канала.
Pterygoideum. К комплексу костей, составленному из palati-
num, transversum и jugale, с медиальной стороны присоединяется очень слож-
Рис. 5. Pterygoideum sinistrum. А — Вид,
снизу. В — Вид изнутри. А—Ventral view,
В — Inner view of left pterygoideum.
a.as.—areaaspera,c.o.—crista obliqua,c. pt.—
corpus pterygoidei, f. o. — fovea ovalis, 1.
as.—lamina ascendens, r. a.—ramus anterior,
r.c.—recessus conoideus,r.p.—ramus posterior.
тело птеригоида покрыто ячеистой
ный по своей форме птеригоид
(рис. 4).
В коллекции имеются не только
хорошо отпрепарованные части че-
репа с птеригоидами, но и доста-
точное количество целых птеригои-
дов и их фрагментов. Это дает воз-
можность подробно изучить птери-
гоид как изолированную кость.
При рассматривании с нёбной
стороны черепа птеригоид может
быть разделен на три части: тело
(средний более массивный участок
кости), передняя ветвь и задняя
ветвь (рис. 4, 5А). При изучении
птеригоида с латеральной стороны
можно видеть, что его задняя ветвь
постепенно переходит в вертикаль-
но стоящую пластинку (lamina
ascendens, рис. 5В, 6А, В). Перед-
няя ветвь представляет собой кры-
ловидную пластинку, лежащую в
целом черепе па всем своем протя-
жении в плоскости нёба. Передне-
латеральный край передней ветви
соединяется при помощи сложно
построенного чешуйчатого шва с
transversum и jugale (рис. 4, 16).
Косо срезанная передняя половина
этого края в виде небольшой чешуи
накладывается сверху на transver-
sum; задняя половина края, нао-
борот, прилегает снизу к transver-
sum и небольшому участку jugale.
Кроме того, передний угол этой
ветви па очень небольшом протя-
жении образует шов с palatinum.
Вентральная поверхность тела
птеригоида и его передней 'ветви
покрыта резко очерченной мелко-
зернистой шероховатостью (area
aspera, рис. 5). Латеральнее ее
скульптурой, ничем не отличаю-
щейся от скульптуры на наружной поверхности покровных костей кры-
ши черепа. Ячейки на птеригоиде превращаются в длинные желобки
по мере того, как скульптура переходит на переднюю ветвь. Вдоль всего
медиального края передней ветви птеригоида на дорзальной ее поверхности
тянется невысокий валик, довольно резко отграниченный от остальной по-
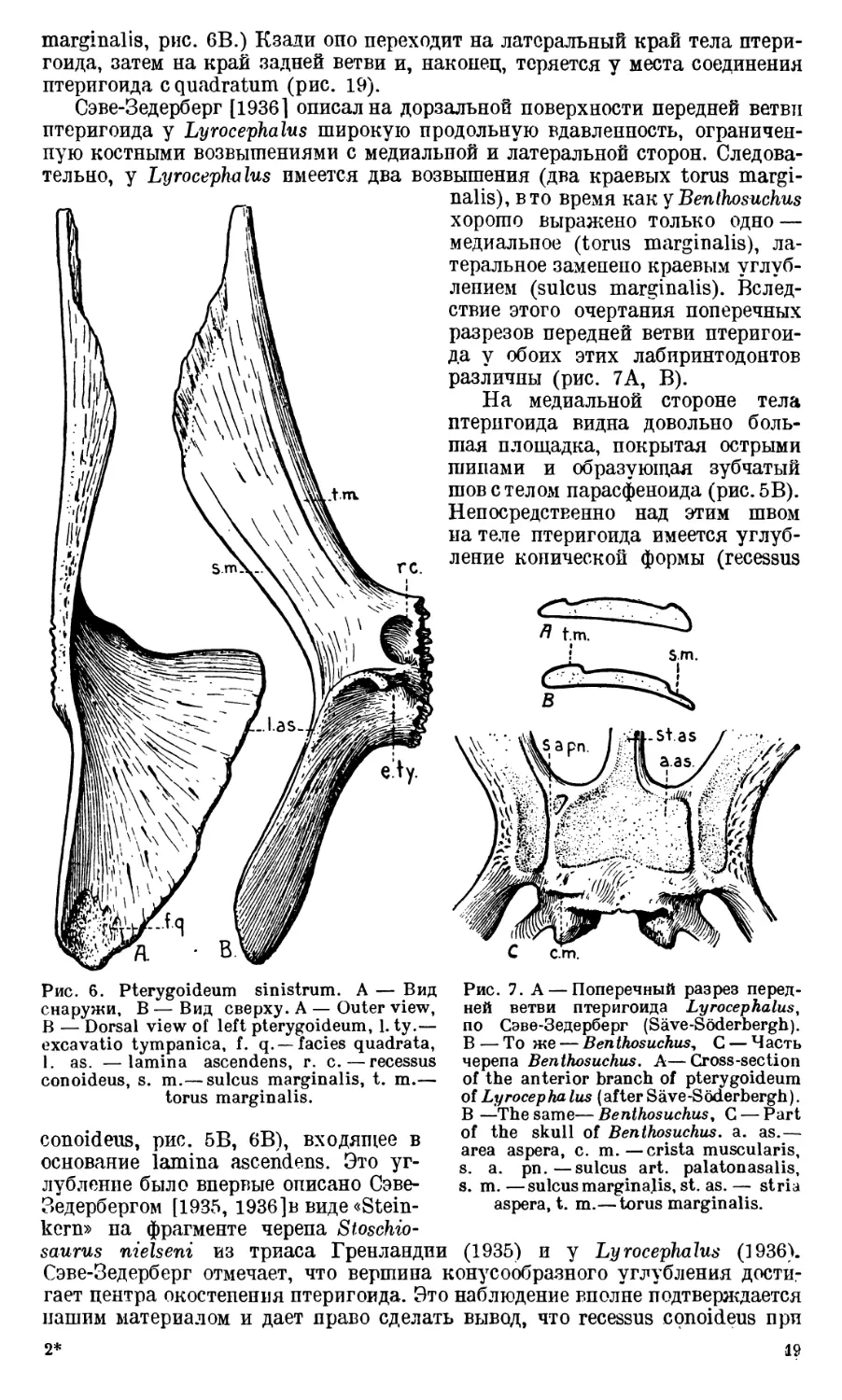
верхности кости (torus marginalis, рис. 6В). Вдоль противоположного края
ветви вместо валика проходит почти такой же ширины углубление (sulcus
<8
marginalis, рис. 6B.) Кзади оно переходит на латеральный край тела птери-
гоида, затем на край задней ветви и, наконец, теряется у места соединения
птеригоида с quadratum (рис. 19).
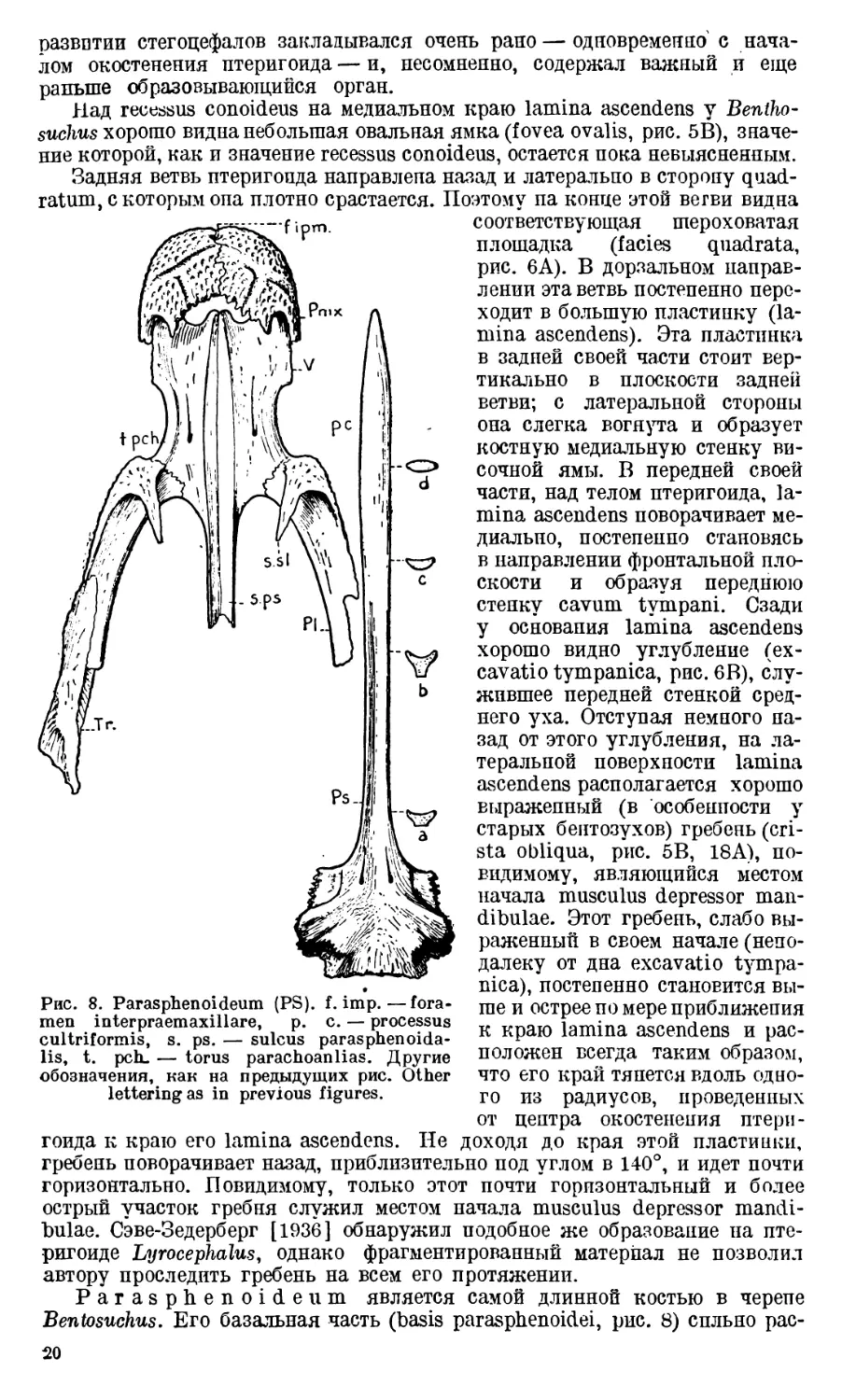
Сэве-Зедерберг [1936] описал на дорзальной поверхности передней ветви
птеригоида у Lyrocephalus широкую продольную вдавленпость, ограничен-
ную костными возвышениями с медиальной и латеральной сторон. Следова-
тельно, у Lyrocephalus имеется два возвышения (два краевых torus margi-
Рис. 6. Pterygoideum sinistrum. А — Вид
снаружи, В — Вид сверху. А — Outer view,
В — Dorsal view of left pterygoideum, 1. ty.—
excavatio tympanica, f. q. —facies quadrata,
1. as. —lamina ascendens, r. c.—recessus
conoideus, s. m.— sulcus marginalis, t. m.—
torus marginalis.
conoideus, рис. 5B, 6B), входящее в
основание lamina ascendens. Это уг-
лубление было впервые описано Сэве-
Зедербергом [1935, 1936]в виде «Stein-
kern» па фрагменте черепа Stoschio-
nalis), в то время как у Benthosuchus
хорошо выражено только одно —
медиальное (torus marginalis), ла-
теральное заменено краевым углуб-
лением (sulcus marginalis). Вслед-
ствие этого очертания поперечных
разрезов передней ветви птеригои-
да у обоих этих лабиринтодонтов
различны (рис. 7А, В).
На медиальной стороне тела
птеригоида видна довольно боль-
шая площадка, покрытая острыми
шипами и образующая зубчатый
шов с телом парасфеноида (рис.5В).
Непосредственно над этим швом
на теле птеригоида имеется углуб-
ление конической формы (recessus
Рис. 7. А — Поперечный разрез перед-
ней ветви птеригоида Lyrocephalus,
по Сэве-Зедерберг (Save-Soderbergh).
В — То же — Benthosuchus, С — Часть
черепа Benthosuchus. А—Cross-section
of the anterior branch of pterygoideum
of Lyrocephalus (after Save-Soderbergh).
В—The same—Benthosuchus, C — Part
of the skull of Benthosuchus. a. as.—
area aspera, c. m.—crista muscularis,
s. a. pn.—sulcus art. palatonasalis,
s. m. —sulcusmarginalis, st. as. — stria
aspera, t. m.—torus marginalis.
saurus nielseni из триаса Гренландии (1935) и у Lyrocephalus (1936).
Сэве-Зедерберг отмечает, что вершина конусообразного углубления дости-
гает центра окостенения птеригоида. Это наблюдение вполне подтверждается
нашим материалом и дает право сделать вывод, что recessus conoideus при
2*
19
развитии стегоцефалов закладывался очень рано — одновременно с нача-
лом окостенения птеригоида — и, несомненно, содержал важный и еще
раньше образовывающийся орган.
Над recessus conoideus на медиальном краю lamina ascendens у Bentho-
suchus хорошо видна небольшая овальная ямка (fovea ovalis, рис. 5 В), значе-
ние которой, как и значение recessus conoideus, остается пока невыясненным.
Задняя ветвь птеригоида направлена назад и латеральпо в сторону quad-
ratum, с которым опа плотно срастается. Поэтому па конце этой вегви видна
соответствующая шероховатая
площадка (facies qnadrata,
рис. 6A). В дорзальном направ-
лении эта ветвь постепенно пере-
ходит в большую пластинку (la-
mina ascendens). Эта пластинка
в задней своей части стоит вер-
тикально в плоскости задней
ветви; с латеральной стороны
она слегка вогнута и образует
костную медиальную стенку ви-
сочной ямы. В передней своей
части, над телом птеригоида, la-
mina ascendens поворачивает ме-
диально, постепенно становясь
в направлении фронтальной пло-
скости и образуя переднюю
стенку cavum tympani. Сзади
у основания lamina ascendens
хорошо видно углубление (ех-
cavatio tympanica, рис. 6В), слу-
жившее передней стенкой сред-
него уха. Отступая немного на-
зад от этого углубления, на ла-
теральной поверхности lamina
ascendens располагается хорошо
выраженный (в особенности у
старых бентозухов) гребень (cri-
sta obliqua, рис. 5В, 18А), по-
видимому, являющийся местом
начала musculus depressor man-
dibulae. Этот гребень, слабо вы-
раженный в своем начале (непо-
далеку от дна excavatio tympa-
nica), постепенно становится вы-
ше и острее по мере приближения
к краю lamina ascendens и рас-
положен всегда таким образом,
что его край тянется вдоль одно-
го из радиусов, проведенных
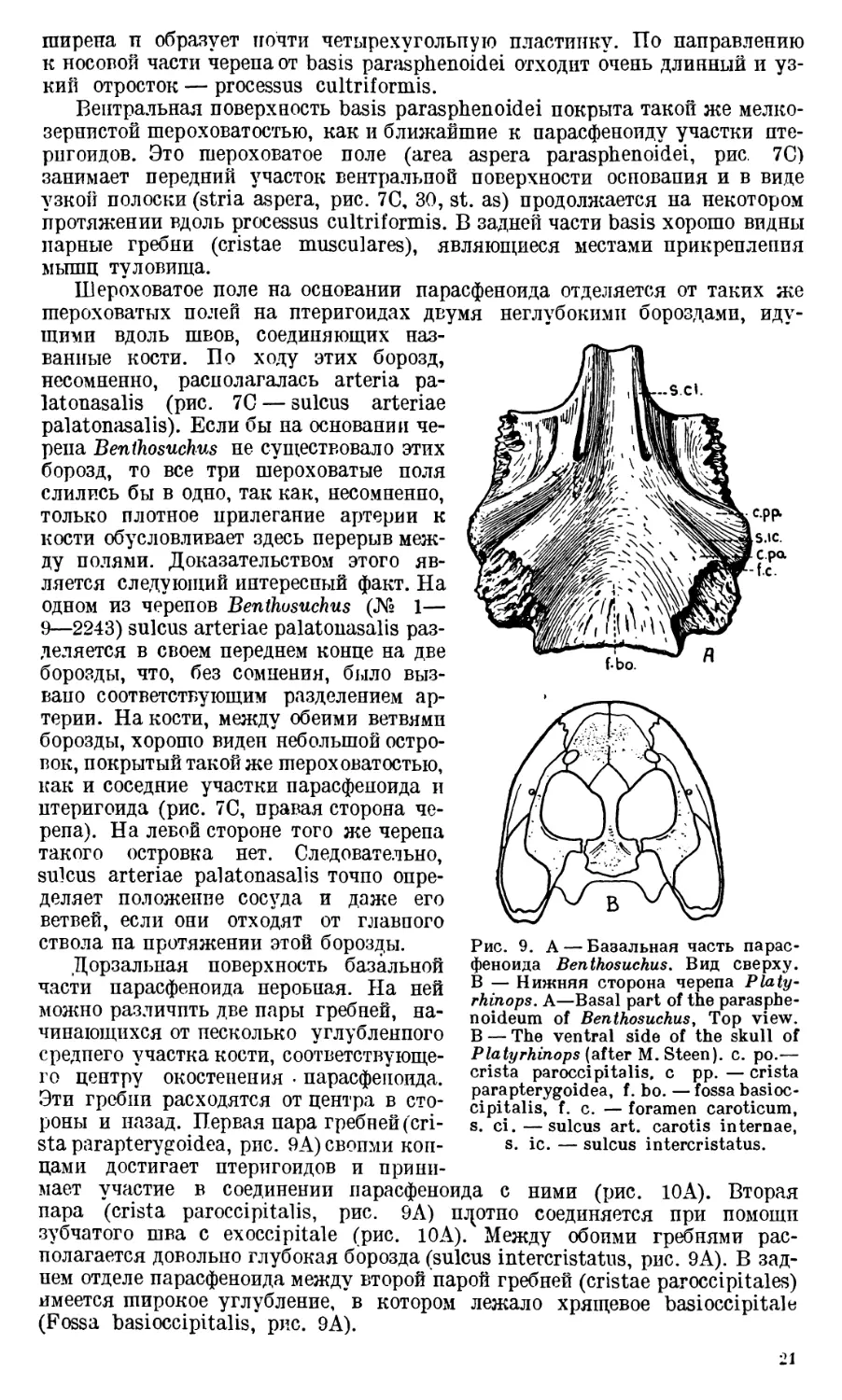
Рис. 8. Paras phen oi deum (PS), f. imp.—fora-
men interpraemaxillare, p. c. — processus
cultriformis, s. ps. — sulcus parasphenoida-
lis, t. pch. — torus parachoanlias. Другие
обозначения, как на предыдущих рис. Other
letteringas in previous figures.
от центра окостенения птери-
гоида к краю его lamina ascendens. Не доходя до края этой пластинки,
гребень поворачивает назад, приблизительно под углом в 140°, и идет почти
горизонтально. Повидимому, только этот почти горизонтальный и более
острый участок гребня служил местом начала musculus depressor mandi-
bulae. Сэве-Зедерберг [1936] обнаружил подобное же образование на пте-
ригоиде Lyrocephalus, однако фрагментированный материал не позволил
автору проследить гребень на всем его протяжении.
Parasphenoideum является самой длинной костью в черепе
Bentosuchus. Его базальная часть (basis parasphenoidei, рис. 8) сильно рас-
20
неглубокими оороздами, иду-
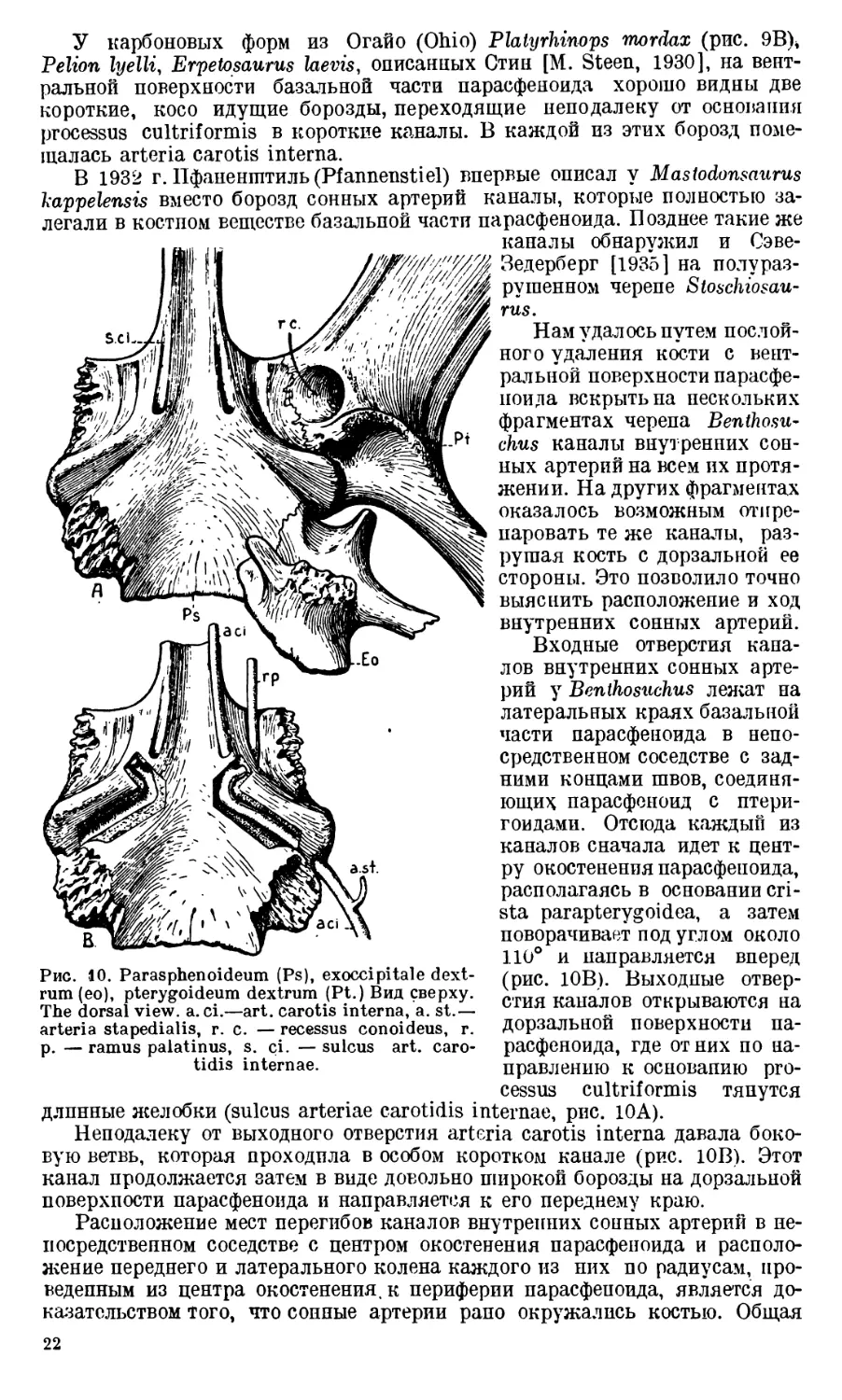
Рис. 9. А — Базальная часть парас-
феноида Benthosuchus. Вид сверху.
В — Нижняя сторона черепа Platy-
rhinops. А—Basal part of the paras phe-
noideum of Benthosuchus, Top view.
В — The ventral side of the skull of
Platyrhinops (after M. Steen), c. po.—
crista paroccipitalis, c pp. — crista
parapterygoidea, f. bo. — fossa basioc-
cipitalis, f. c. — foramen caroticum,
s. ci.—sulcus art. carotis internae,
s. ic. — sulcus intercristatus.
ширепа п образует почти четырехугольпую пластинку. По направлению
к носовой части черепа от basis parasphenoidei отходит очень длинный и уз-
кий отросток — processus cultriformis.
Вентральная поверхность basis parasphenoidei покрыта такой же мелко-
зернистой шероховатостью, как и ближайшие к парасфеноиду участки пте-
рпгоидов. Это шероховатое поле (area aspera parasphenoidei, рис. 7С)
занимает передний участок вентральной поверхности осповапия и в виде
узкой полоски (stria aspera, рис. 7С, 30, st. as) продолжается на некотором
протяжении вдоль processus cultriformis. В задней части basis хорошо видны
парные гребни (cristae musculares), являющиеся местами прикрепления
мышц туловища.
Шероховатое поле па основании парасфеноида отделяется от таких же
шероховатых полей на птеригоидах дв^
щими вдоль швов, соединяющих наз-
ванные кости. По ходу этих борозд,
несомненно, располагалась arteria ра-
latonasalis (рис. 70 — sulcus arteriae
palatonasalis). Если бы на основании че-
репа Benthosuchus не существовало этих
борозд, то все три шероховатые поля
слились бы в одно, так как, несомненно,
только плотное прилегание артерии к
кости обусловливает здесь перерыв меж-
ду полями. Доказательством этого яв-
ляется следующий интересный факт. На
одном из черепов Benthosuchus (№ 1—
9—2243) sulcus arteriae palatonasalis раз-
деляется в своем переднем конце на две
борозды, что, без сомнения, было выз-
вано соответствующим разделением ар-
терии. На кости, между обеими ветвями
борозды, хорошо виден небольшой остро-
вок, покрытый такой же шероховатостью,
как и соседние участки парасфеноида и
птеригоида (рис. 7С, правая сторона че-
репа). На левой стороне того же черепа
такого островка нет. Следовательно,
sulcus arteriae palatonasalis точпо опре-
деляет положение сосуда и даже его
ветвей, если они отходят от главного
ствола па протяжении этой борозды.
Дорзальная поверхность базальной
части парасфеноида неровная. На ней
можно различить две пары гребней, на-
чинающихся от несколько углубленного
среднего участка кости, соответствующе-
го центру окостенения • парасфеноида.
Эти гребни расходятся от центра в сто-
роны и назад. Первая пара гребней (cri-
sta parapterygoidea, рис. 9А) своими кон-
цами достигает птеригоидов и прини-
мает участие в соединении парасфеноида с ними (рис. 10А). Вторая
пара (crista paroccipitalis, рис. 9А) плотно соединяется при помощи
зубчатого шва с exoccipitale (рис. 10А). Между обоими гребнями рас-
полагается довольно глубокая борозда (sulcus intercristatus, рис. 9А). В зад-
нем отделе парасфеноида между второй парой гребней (cristae paroccipitales)
имеется широкое углубление, в котором лежало хрящевое basioccipitale
(Fossa basioccipitalis, рис. 9А).
21
Рис. 40. Paras phen oideum (Ps), exoccipitale dext-
rum(eo), pterygoideum dextrum (Pt.) Вид сверху.
The dorsal view. a. ci.—art. carotis interna, a. st.—
arteria stapedialis, r. c. — recessus conoideus, r.
p. — ramus palatinus, s. ci. — sulcus art. caro-
tidis internae.
У карбоновых форм из Огайо (Ohio) Platyrhinops mordax (рис. 9В\
Pelion lyelli, Erpetosaurus laevis, описанных Стин [M. Steen, 1930], на вент-
ральной поверхности базальной части парасфеноида хорошо видны две
короткие, косо идущие борозды, переходящие неподалеку от основания
processus cultriformis в короткие каналы. В каждой из этих борозд поме-
щалась arteria carotis interna.
В 1932 г. Пфаненштиль(Р1аппепз11е1) впервые описал у Mastodonsaurus
ka/ppelensis вместо борозд сонных артерий каналы, которые полностью за-
легали в костном веществе базальной части парасфеноида. Позднее такие же
каналы обнаружил и Сэве-
Зедерберг [1935] на полураз-
рушенном черепе Stoschiosau-
rus.
Нам удалось путем послой-
ного удаления кости с вент-
ральной поверхности парасфе-
ноида вскрыть па нескольких
фрагментах черепа Bentho.su-
chus каналы внутренних сон-
ных артерий на всем их протя-
жении. На других фрагментах
оказалось возможным отире-
паровать те же каналы, раз-
рушая кость с дорзальной ее
стороны. Это позволило точно
выяснить расположение и ход
внутренних сонных артерий.
Входные отверстия кана-
лов внутренних сонных арте-
рий у Benthosuchus лежат на
латеральных краях базальной
части парасфеноида в непо-
средственном соседстве с зад-
ними концами швов, соединя-
ющих парасфеноид с птери-
гоидами. Отсюда каждый из
каналов сначала идет к цент-
ру окостенения парасфеноида,
располагаясь в основании cri-
sta parapterygoidea, а затем
поворачивает под углом около
110° и направляется вперед
(рис. 10В). Выходные отвер-
стия каналов открываются на
дорзальной поверхности па-
расфеноида, где от них по на-
правлению к основанию pro-
cessus cultriformis тянутся
длинные желобки (sulcus arteriae carotidis internae, рис. 10A).
Неподалеку от выходного отверстия arteria carotis interna давала боко-
вую ветвь, которая проходила в особом коротком канале (рис. 10В). Этот
канал продолжается затем в виде довольно широкой борозды на дорзальной
поверхности парасфеноида и направляется к его переднему краю.
Расположение мест перегибов каналов внутренних сонных артерий в не-
посредственном соседстве с центром окостенения парасфеноида и располо-
жение переднего и латерального колена каждого из них по радиусам, про-
веденным из центра окостенения, к периферии парасфеноида, является до-
казательством того, что сонные артерии рано окружались костью. Общая
22
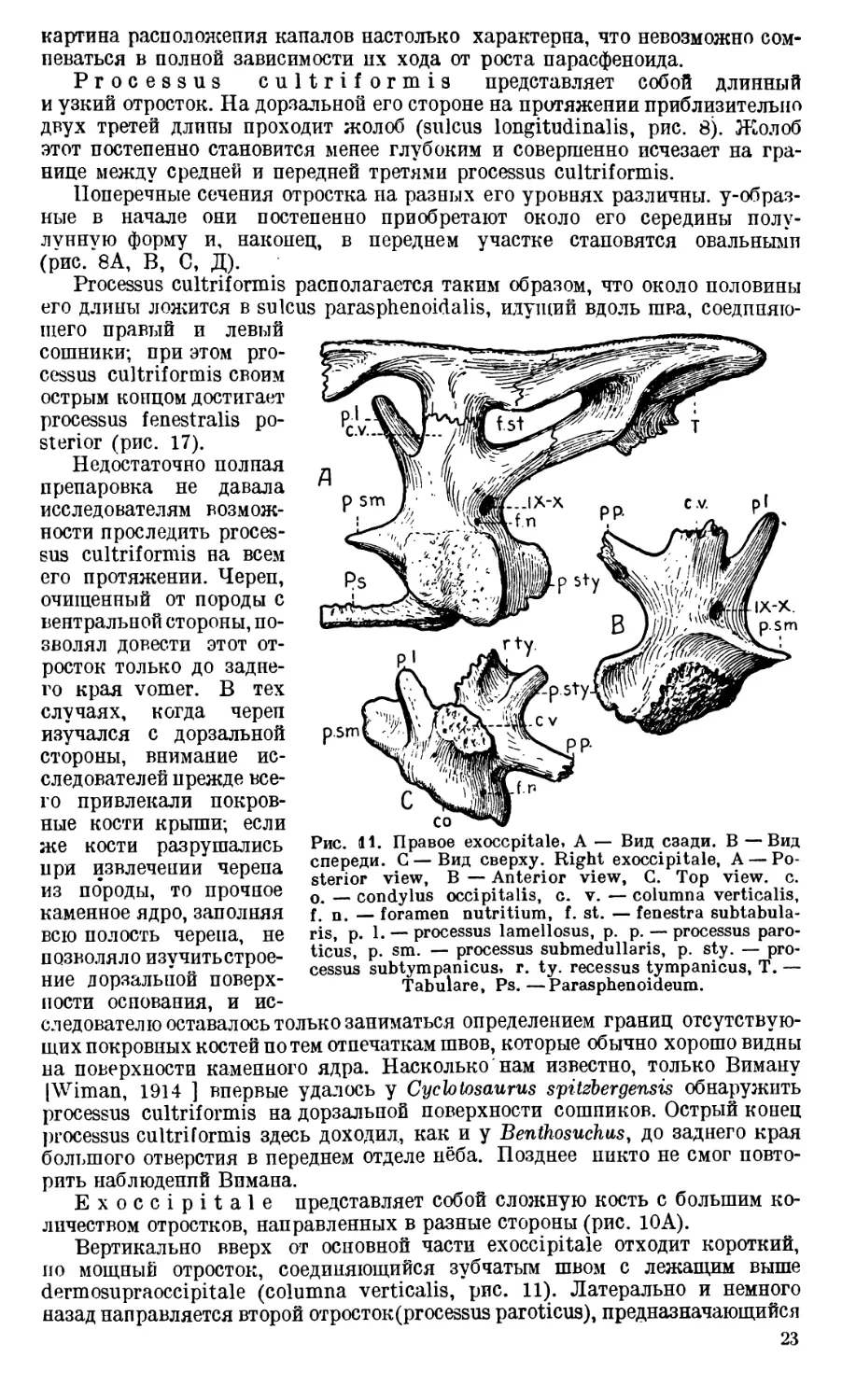
Рис. tl. Правое exoccpitale, А — Вид сзади. В —Вид
спереди. С — Вид сверху. Right exoccipitale, А — Po-
sterior view, В — Anterior view, G. Top view. c.
o. — condylus occipitalis, c. v. — columna verticalis,
f. n. — foramen nutritium, f. st. — fenestra subtabula-
ris, p. 1. — processus lameliosus, p. p. — processus paro-
ticus, p. sm. — processus submedullaris, p. sty. — pro-
cessus subtympanicus, r. ty. recessus tympanicus, T. —
Tabulare, Ps.—Paras phen oideum.
картина расположения каналов настолько характерна, что невозможно сом-
неваться в полной зависимости их хода от роста парасфеноида.
Processus cultriformis представляет собой длинный
и узкий отросток. На дорзальной его стороне на протяжении приблизительно
двух третей длины проходит жолоб (sulcus longitudinalis, рис. 8). Жолоб
этот постепенно становится менее глубоким и совершенно исчезает на гра-
нице между средней и передней третями processus cultriformis.
Поперечные сечения отростка па разных его уровнях различны, у-образ-
ные в начале они постепенно приобретают около его середины полу-
лунную форму и, наконец, в переднем участке становятся овальными
(рис. 8А, В, С, Д).
Processus cultriformis располагается таким образом, что около половины
его длины ложится в sulcus parasphenoidalis, идущий вдоль шва, соединяю-
щего правый и левый
сошники; при этом pro-
cessus cultriformis своим
острым концом достигает
processus fenestralis po-
sterior (рис. 17).
Недостаточно полная
препаровка не давала
исследователям возмож-
ности проследить proces-
sus cultriformis на всем
его протяжении. Череп,
очищенный от породы с
вентральной стороны, по-
зволял довести этот от-
росток только до задне-
го края vomer. В тех
случаях, когда череп
изучался с дорзальной
стороны, внимание ис-
следователей прежде все-
го привлекали покров-
ные кости крыши; если
же кости разрушались
при извлечении черепа
из породы, то прочное
каменное ядро, заполняя
всю полость черепа, не
позволяло изучить строе-
ние дорзальной поверх-
ности основания, и ис-
следователю оставалось только заниматься определением границ отсутствую-
щих покровных костей по тем отпечаткам швов, которые обычно хорошо видны
на поверхности каменного ядра. Насколько нам известно, только Виману
|Wiman, 1914 ] впервые удалось у Cyclotosaurus spitzbergensis обнаружить
processus cultriformis на дорзальной поверхности сошников. Острый конец
processus cultriformis здесь доходил, как и у Benthosuchus, до заднего края
большого отверстия в переднем отделе нёба. Позднее никто не смог повто-
рить наблюдений Вимана.
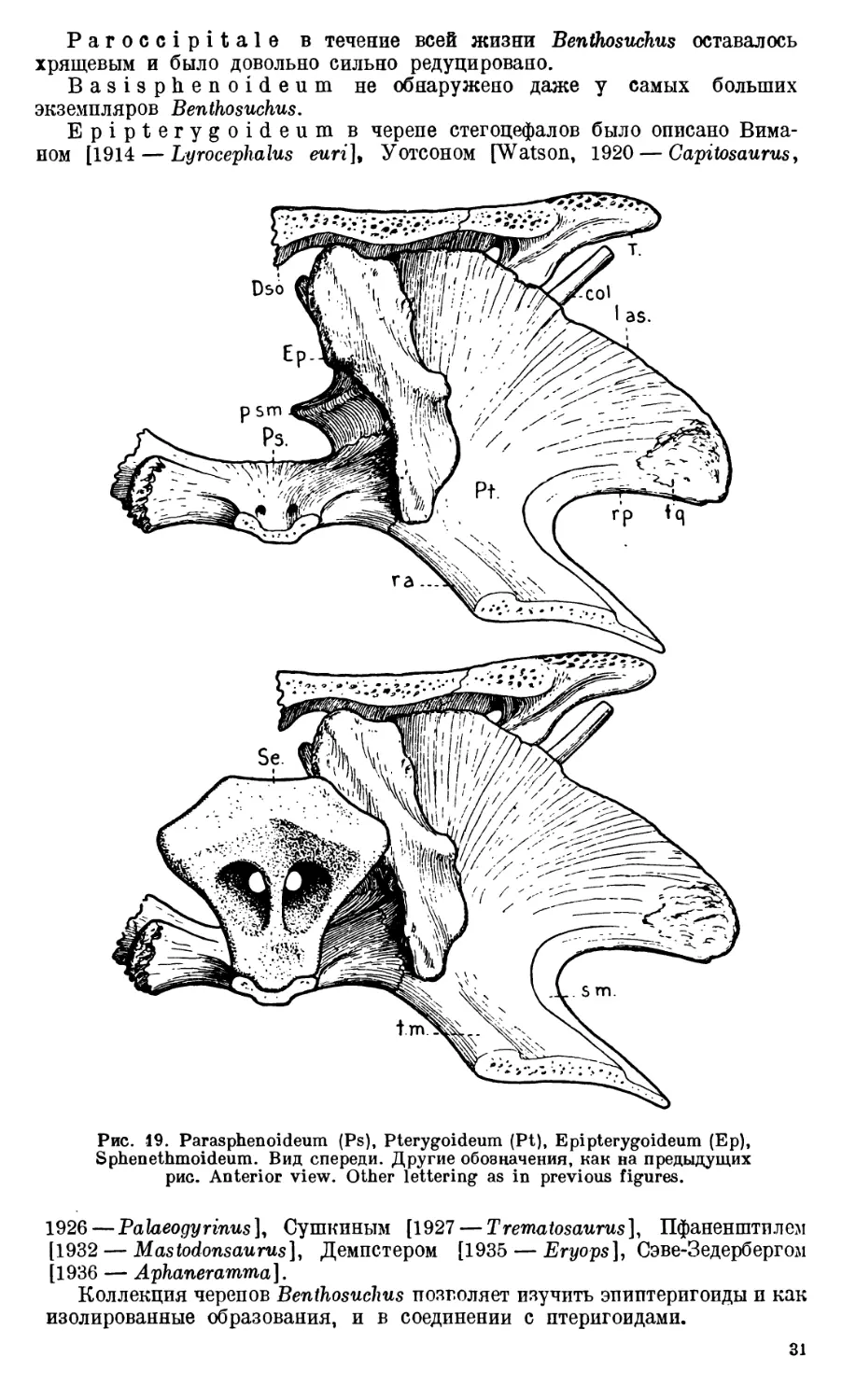
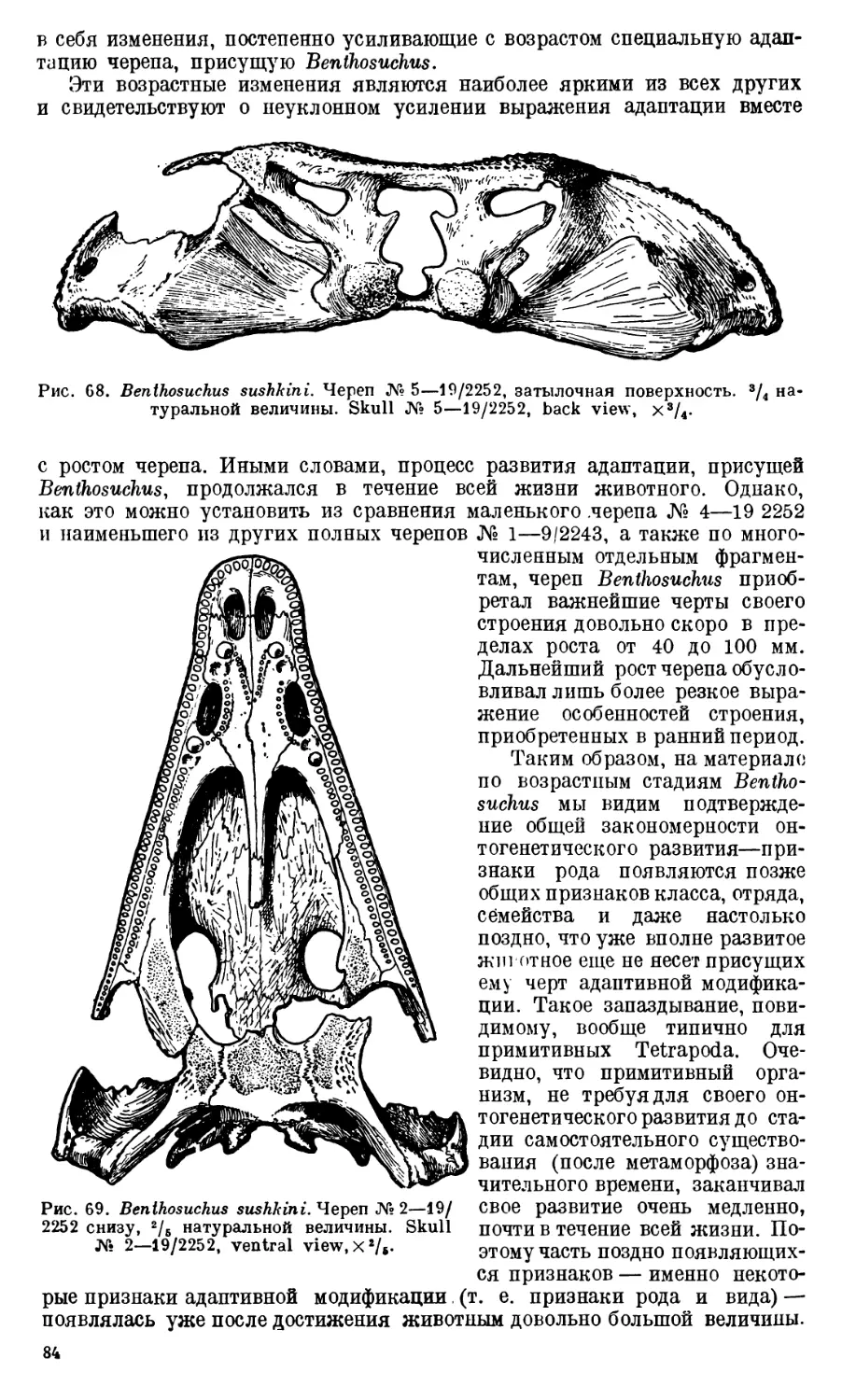
Exoccipitale представляет собой сложную кость с большим ко-
личеством отростков, направленных в разные стороны (рис. 10А).
Вертикально вверх от основной части exoccipitale отходит короткий,
по мощный отросток, соединяющийся зубчатым швом с лежащим выше
dermosupraoccipitale (columna verticalis, рис. 11). Латерально и немного
назад направляется второй отросток (processus paroticus), предназначающийся
23
для соединения с tabulare. От основания columna verticalis вверх, вперед
и медиально отходит сравнительно тонкий отросток, имеющий вид пластин-
ки— processus lamellosus. Он обычно мало доступен для исследования, так
как на черепах, содержащих породу, уходит в глубь каменной массы и по
большей части не отмечается при описаниях черепов лабиринтодонтов.
От основания exoccipitale отходят также три отростка. Один из них,
самый мощный, является суставным — condylus occipitalis. Неровная пло-
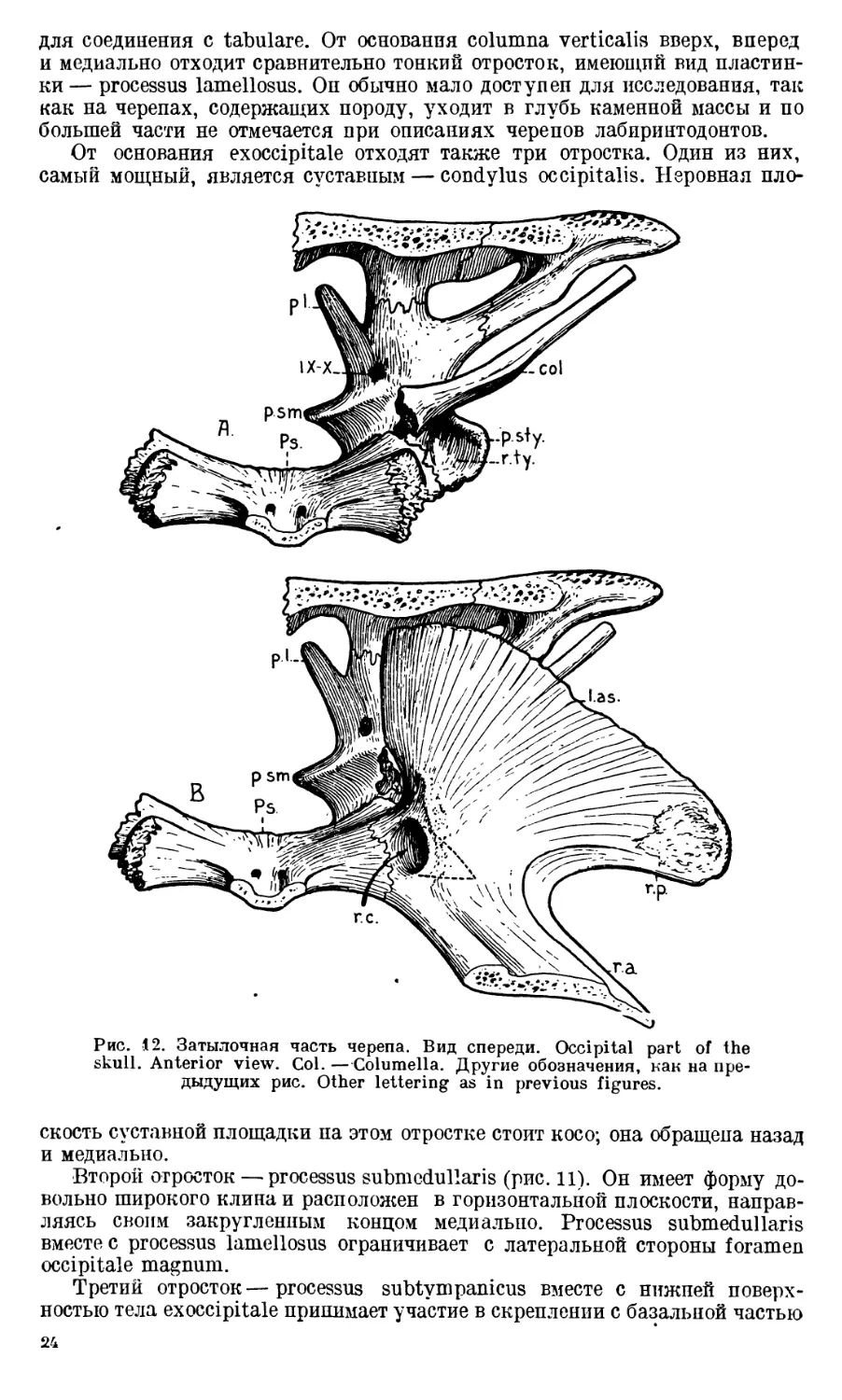
Рис. 12. Затылочная часть черепа. Вид спереди. Occipital part of the
skull. Anterior view. Col. — Columella. Другие обозначения, как на пре-
дыдущих рис. Other lettering as in previous figures.
скость суставной площадки па этом отростке стоит косо; она обращена назад
и медиально.
Второй отросток — processus submedullaris (рис. 11). Он имеет форму до-
вольно широкого клина и расположен в горизонтальной плоскости, направ-
ляясь своим закругленным концом медиально. Processus submedullaris
вместе с processus lamellosus ограничивает с латеральной стороны foramen
occipitale magnum.
Третий отросток—processus subtympanicus вместе с нижней поверх-
ностью тела exoccipitale принимает участие в скреплении с базальной частью
24
парасфеноида и прочно соединяется с концом crista paroccipitalis (рис. 10А).
На верхней поверхности отростка — большая впадина в виде короткого
жолоба с гладкими стенками. Она, несомненно, имела прямое отношение
к барабанной полости среднего уха, так как непосредственно над нею распо-
лагается basis columellae (рис. 12А). Поэтому наиболее подходящим для
этой впадины следует признать название recessus tympanicus (рис. 11).
Processus subtympanicus с его recessus tympanicus не доходит до птеригоида;
между птеригоидом и этим отростком имеется щель.
Тело exoccipitale пронизано довольно широким каналом, служащим
местом выхода черепномозговых нервов (рис. 11). Обычно этот канал при-
нято называть canalis nervi vagi; однако, несомненно, сквозь него, кроме
nervus vagus (X), проходил и IX черепномозговой нерв [Хюне (V. Huene),
1912].
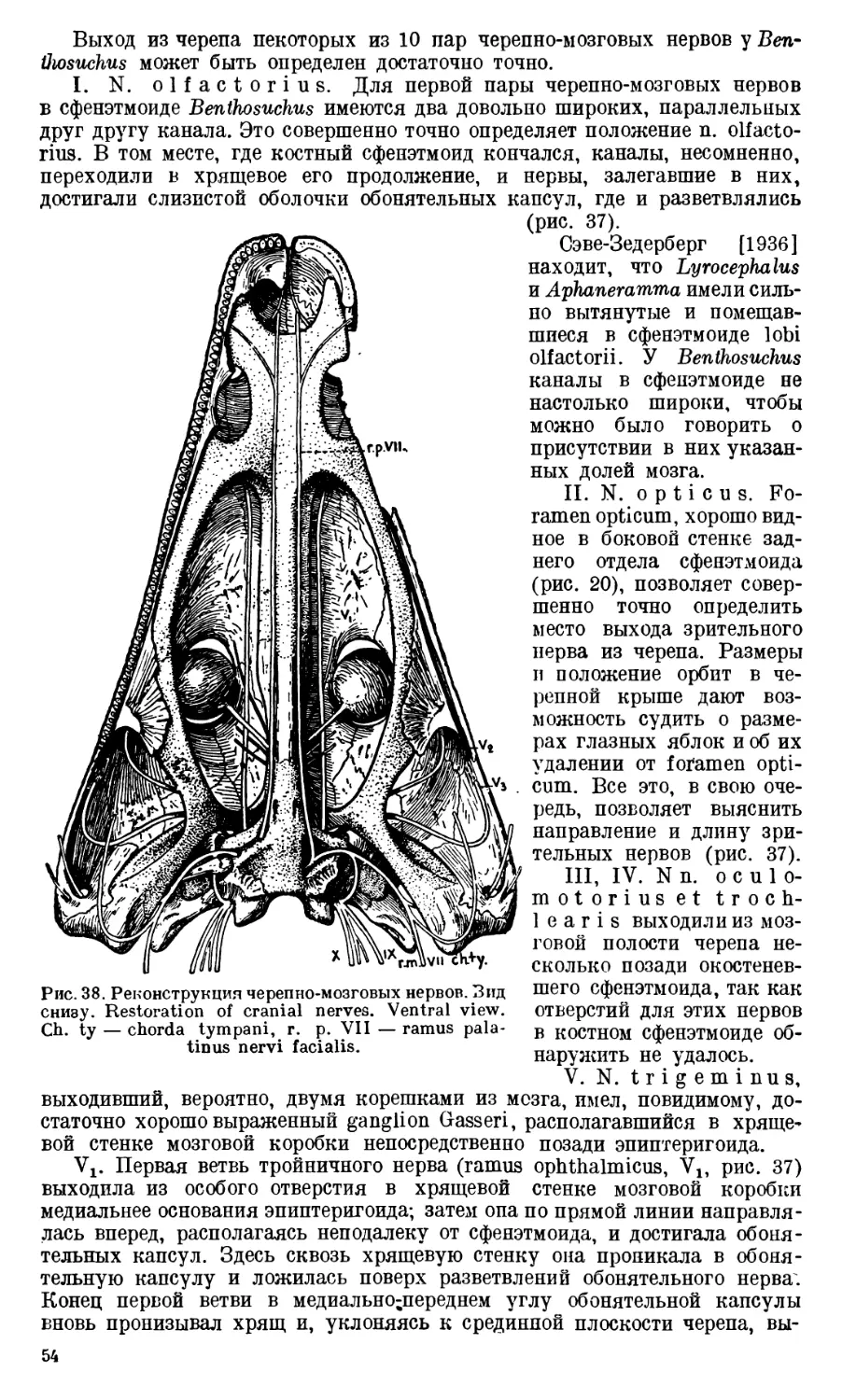
Кроме канала для IX и X пар черепномозговых нервов, в exoccipitale
Benthosuchus обычно наблюдается еще одно отверстие (рис. 11). Оно мень-
ше, чем canalis nervi vagi, и расположено сзади и медиально от него.
П. П. Сушкин [1927] считал подобное отверстие у Dvinosaurus местом
выхода nervus hypoglossus. Такого же взгляда держится и Сэве-Зедерберг
[1936] в отношении Lyrocephalus и Aphaneramma. Под именем foramen
nervi hypoglossi указанное добавочное маленькое отверстие было описано
и Ефремовым [1929] на первом экземпляре Benthosuchus sushkini. Однако
после изучения всего материала по Benthosuchus оказалось, что таких отвер-
стий на его exoccipitale может быть два и даже три; при этом часто на пра-
вом и левом exoccipitale одного и того же черепа число отверстий различно.
Кроме того, они сильно меняют свое положение, в то время как общее отвер-
стие для IX и X нервов всегда занимает строго определенное положение.
Такая неустойчивость в местоположении и такое непостоянство в числе
заставляет думать, что эти отверстия не служили местом выхода каких бы
то ни было черепномозговых нервов. Во всяком случае, nervus hypoglossus
через них не проходил; местом его выхода у лабиринтодонтов, как и у всех
амфибий, является промежуток между exoccipitale и первым позвонком.
Следует полностью присоединиться к мнению Демпстера [Dempster, 1935],
который считает небольшие отверстия позади канала vagus-группы пред-
назначающимися для небольших vasa nutritia.
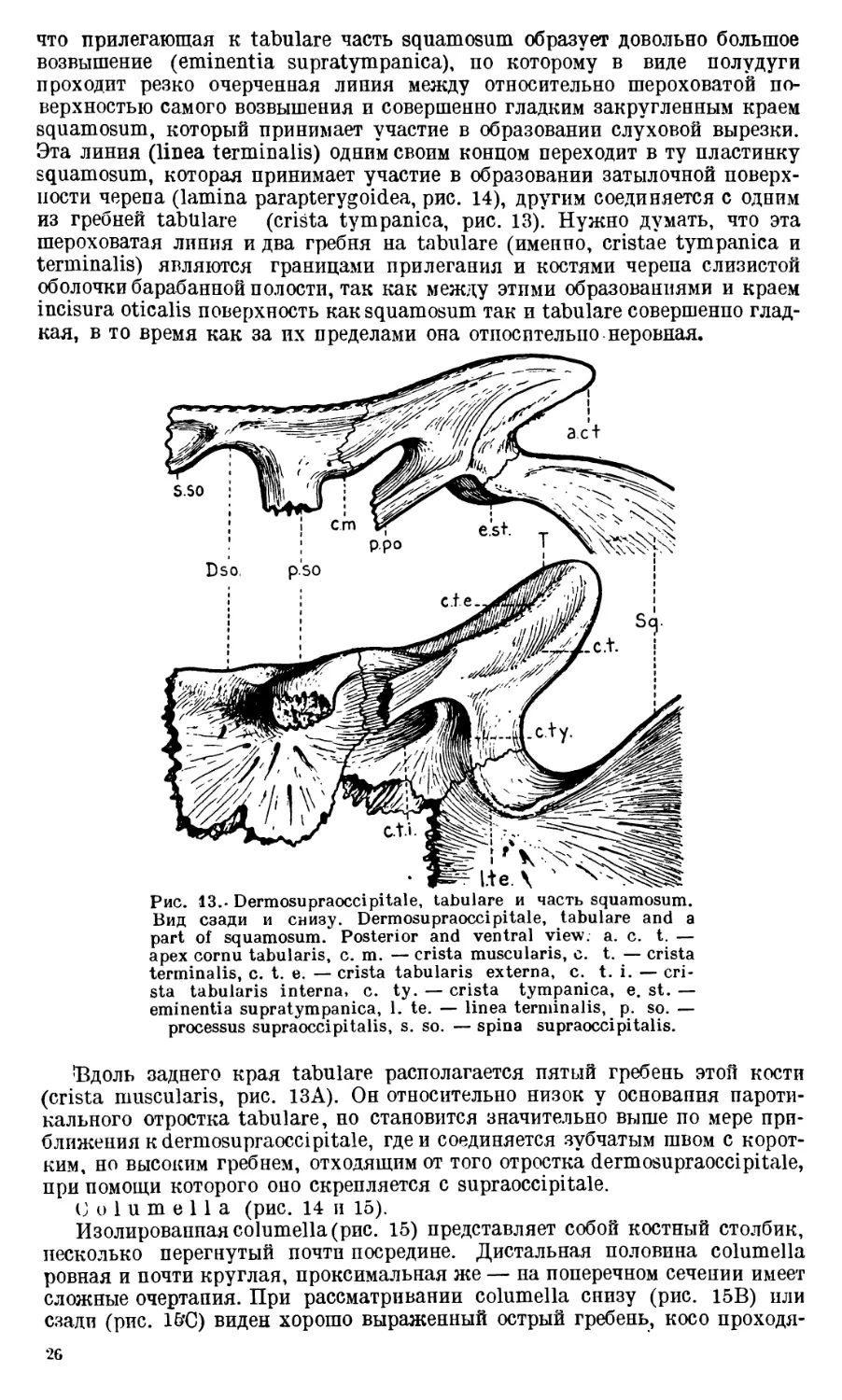
Dermosupraoccipitale соединяется при помощи короткого
отростка с columna verticalis exoccipitalis. Этот отросток (processus supraoc-
cipitalis, рис. 13) расположен на заднем краю почти четыреугольной пла-
стинки кости, лежащей в заднем отделе черепной крыши. В медиальном
участке заднего края dermosupraoccipitale имеется небольшой шип (spina
supraoccipitalis, рис. 13), на задней стороне которого у старых Benthosuchus
хорошо видно углубление (рис. 13А).
Tabular е, занимающее латеральный угол затылочного отдела череп-
ной крыши, является довольно сложной костью. На хорошо отпрепаровап-
ных фрагментах черепа Benthosuchus на нижней поверхности tabulare
можно видеть большой косо поставленный отросток (processus paroticus,
рис. 13), который соединяется своей вершиной с одноименным отростком
exoccipitale. Этот отросток служит как бы центром, от которого в разные
стороны расходится ряд более или менее резко выраженных гребней. Прежде
всего бросается в глаза высокий гребень, идущий от отростка tabulare
вдоль cornu tabularis.
Этот гребень заканчивается на оамом конце рога (apex cornu) — crista
tabularis externa (рис. 13). В противоположную сторону отходит более ко-
роткий и менее высокий гребень (crista tabularis interna). На передне-ла-
теральной стороне tabulare между указанными гребнями располагаются еще
два гребня. Один из них (crista terminalis), получив начало от косого от-
ростка (processus paroticus), направляется косо назад к краю рога tabulare
и теряется, не доходя до его вершины. Другой (crista tympanica), развитый
сильнее, кончается у шва, соединяющего tabulare и squamosum. Интересно,
25
что прилегающая к tabulare часть squamosum образует довольно большое
возвышение (eminentia supratympanica), по которому в виде полудуги
проходит резко очерченная линия между относительно шероховатой по-
верхностью самого возвышения и совершенно гладким закругленным краем
squamosum, который принимает участие в образовании слуховой вырезки.
Эта линия (linea terminalis) одним своим концом переходит в ту пластинку
squamosum, которая принимает участие в образовании затылочной поверх-
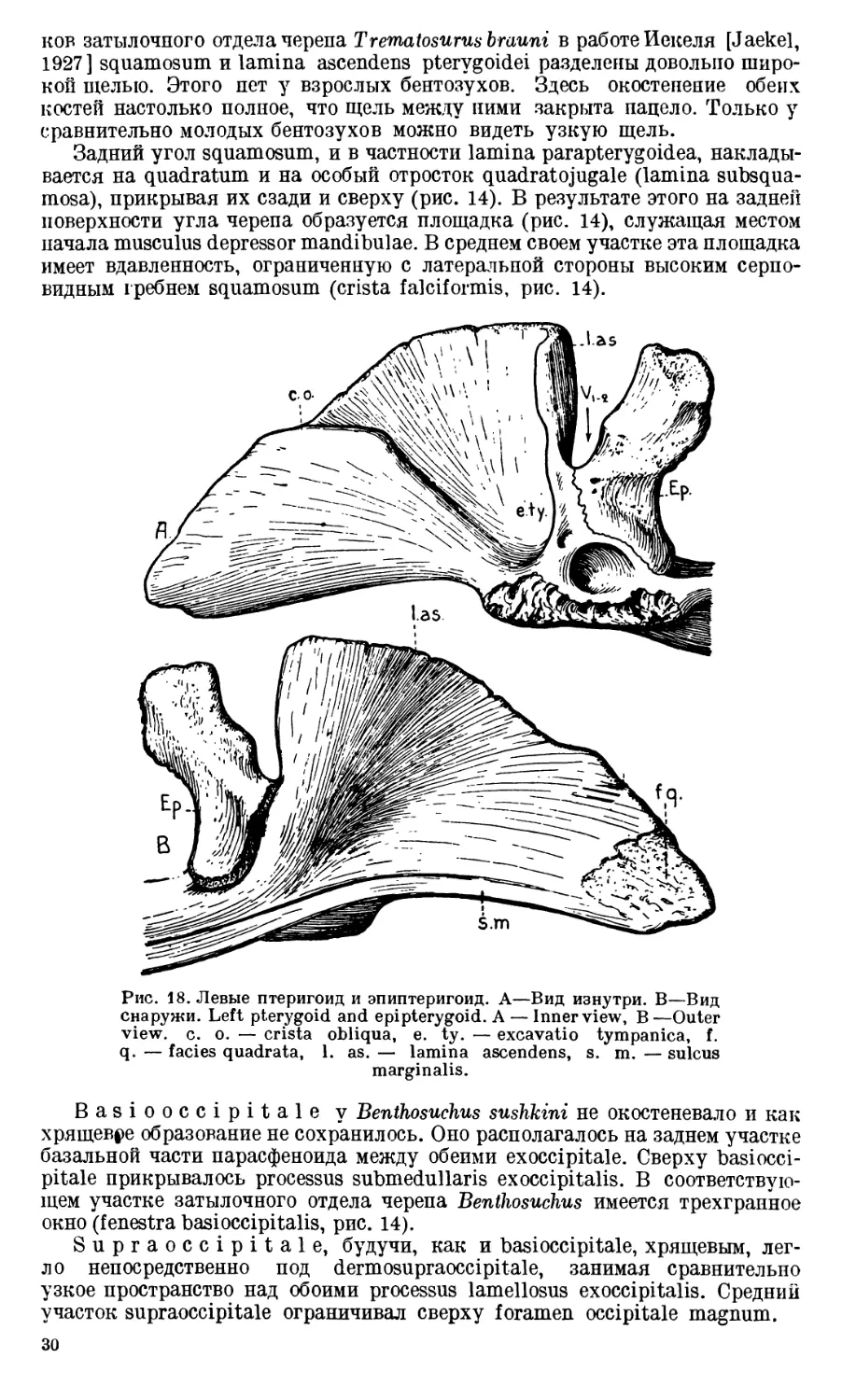
ности черепа (lamina parapterygoidea, рис. 14), другим соединяется с одним
из гребней tabulare (crista tympanica, рис. 13). Нужно думать, что эта
шероховатая линия и два гребня на tabulare (именно, cristae tympanica и
terminalis) являются границами прилегания и костями черепа слизистой
оболочки барабанной полости, так как между этими образованиями и краем
incisura oticalis поверхность как squamosum так и tabulare совершенно глад-
кая, в то время как за их пределами она относительно неровная.
Рис. 13.» Dermosupraoccipitale, tabulare и часть squamosum.
Вид сзади и снизу. Dermosupraoccipitale, tabulare and a
part of squamosum. Posterior and ventral view. a. c. t. —
apex cornu tabularis, c. m. — crista muscularis, c. t. — crista
terminalis, c. t. e. — crista tabularis externa, c. t. i. — cri-
sta tabularis interna, c. ty. — crista tympanica, e. st. —
eminentia supratympanica, 1. te. — linea terminalis, p. so. —
processus supraoccipitalis, s. so. — spina supraoccipitalis.
’Вдоль заднего края tabulare располагается пятый гребень этой кости
(crista muscularis, рис. 13А). Он относительно низок у основания пароти-
кального отростка tabulare, но становится значительно выше по мере при-
ближения к dermosupraocci pitale, где и соединяется зубчатым швом с корот-
ким, но высоким гребнем, отходящим от того отростка dermosupraoccipitale,
при помощи которого оно скрепляется с supraoccipitale.
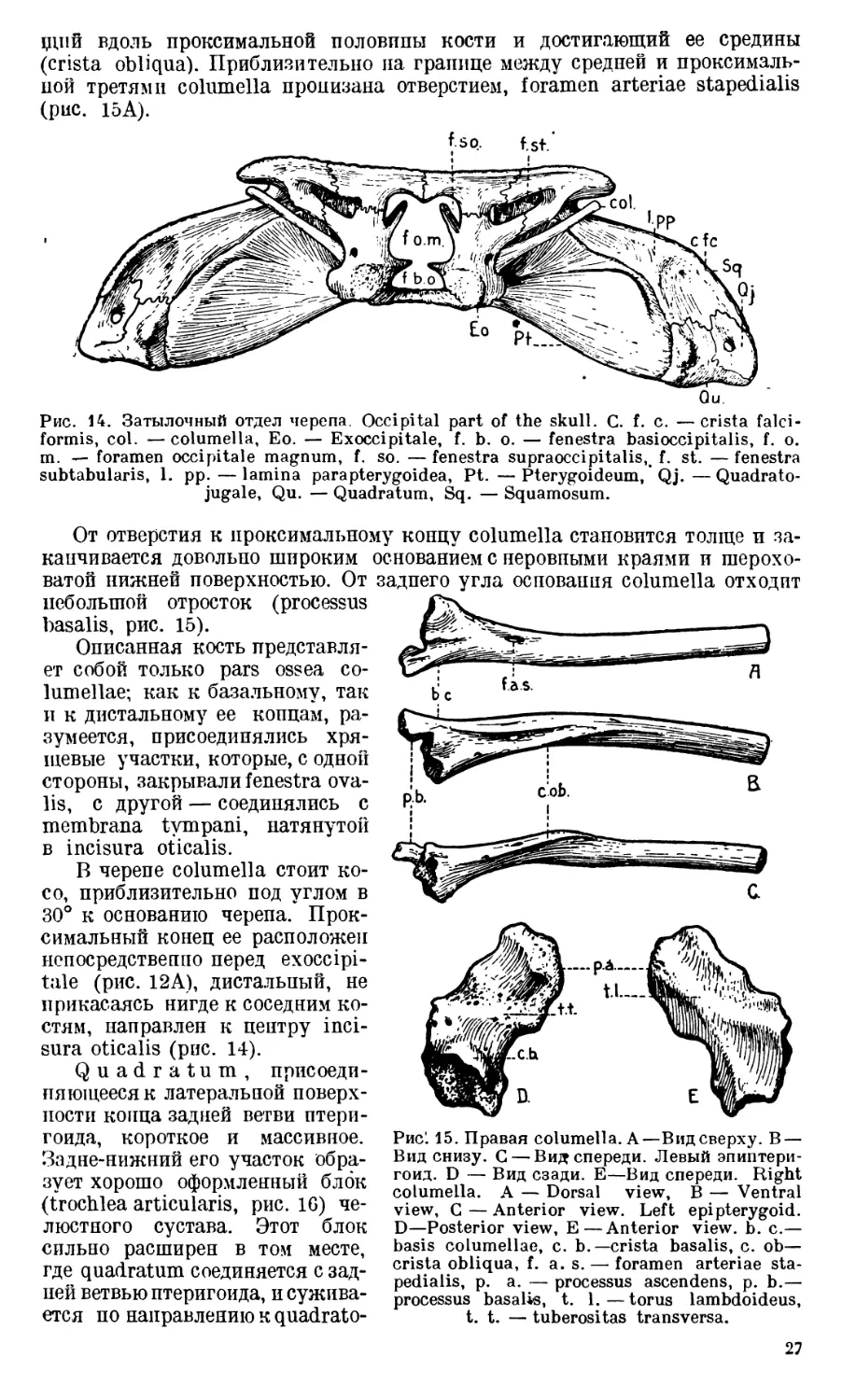
Columella (рис. 14 и 15).
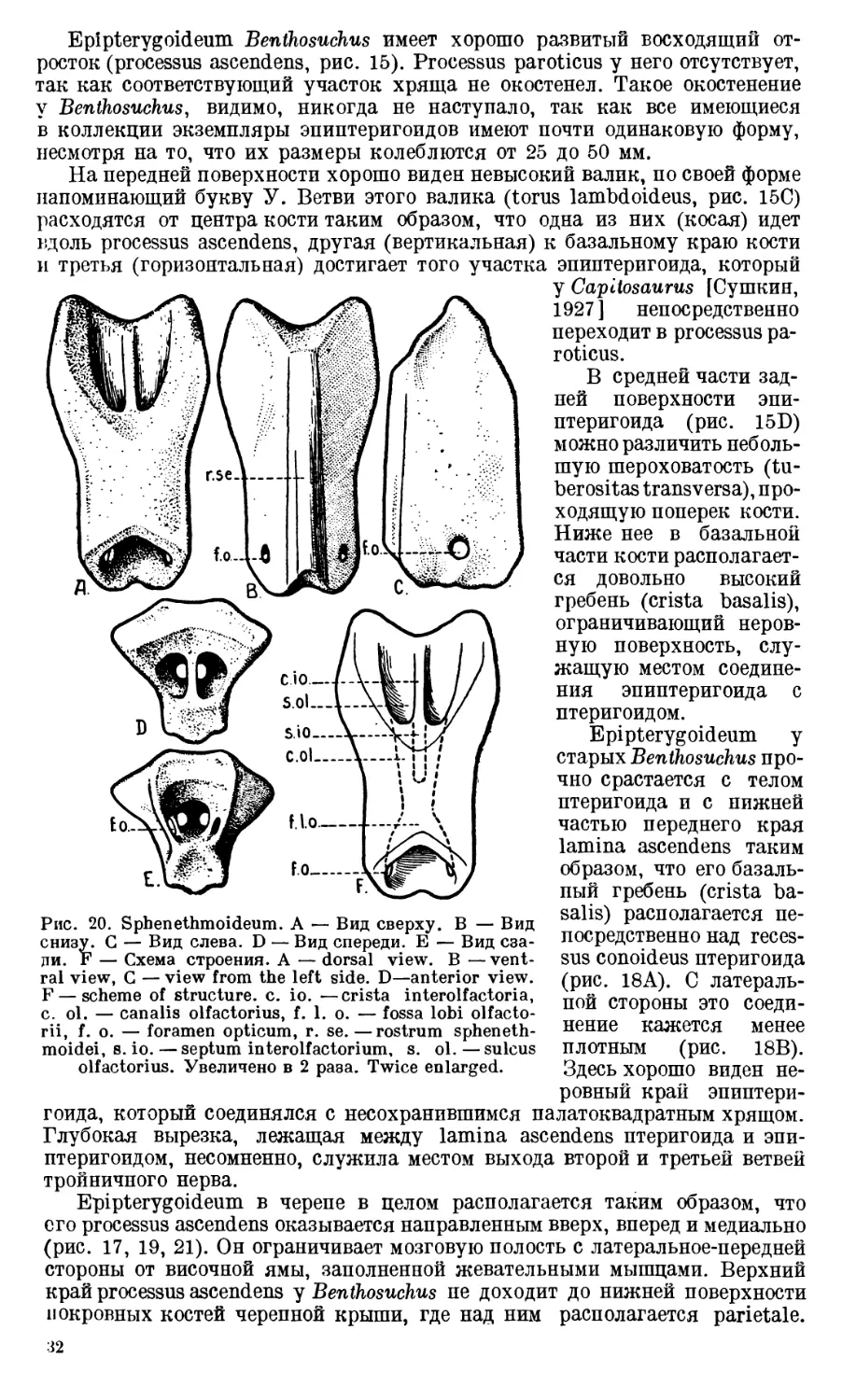
Изолированная columella (рис. 15) представляет собой костный столбик,
несколько перегнутый почти посредине. Дистальная половина columella
ровная и почти круглая, проксимальная же — на поперечном сечении имеет
сложные очертания. При рассматривании columella снизу (рис. 15В) или
сзади (рис. 15'С) виден хорошо выраженный острый гребень, косо проходя-
26
щпй вдоль проксимальной половины кости и достигающий ее средины
(crista obliqua). Приблизительно на границе между средней и проксималь-
ной третями columella пронизана отверстием, foramen arteriae stapedialis
(рис. 15А).
fso. f.st
Qu.
Рис. 14. Затылочный отдел черепа. Occipital part of the skull. C. f. c. —crista falci-
formis, col. — columella, Eo. — Exoccipitale, f. b. o. — fenestra basioccipitalis, f. o.
m. — foramen occipitale magnum, f. so. — fenestra supraoccipitalis,. f. st. — fenestra
subtabularis, 1. pp.—lamina parapterygoidea, Pt. —Pterygoideum, Qj.—Quadrato-
jugale, Qu. — Quadratum, Sq. — Squamosum.
От отверстия к проксимальному концу columella становится толще и за-
канчивается довольно широким основанием с неровными краями и шерохо-
ватой нижней поверхностью. От заднего угла основания columella отходит
небольшой отросток (processus
basalis, рис. 15).
Описанная кость представля-
ет собой только pars ossea со-
lumellae; как к базальному, так
и к дистальному ее концам, ра-
зумеется, присоединялись хря-
щевые участки, которые, с одной
стороны, закрывали fenestra ova-
lis, с другой — соединялись с
membrana tympani, натянутой
в incisura oticalis.
В черепе columella стоит ко-
со, приблизительно под углом в
30° к основанию черепа. Прок-
симальный конец ее расположен
непосредственно перед exoccipi-
tale (рис. 12А), дистальный, не
прикасаясь нигде к соседним ко-
стям, направлен к центру inci-
sura oticalis (рис. 14).
Quadratum, присоеди-
няющееся к латеральной поверх-
ности конца задней ветви птери-
Рис1 15. Правая columella. А—Вид сверху. В —
Вид снизу. С — Вид спереди. Левый эпиптери-
гоид. D — Вид сзади. Е—Вид спереди. Right
columella. А — Dorsal view, В — Ventral
view, G — Anterior view. Left epi pterygoid.
D—Posterior view, E—Anterior view. b. c.—
basis columellae, c. b.—crista basalis, c. ob—
crista obliqua, f. a. s. — foramen arteriae sta-
pedialis, p. a. — processus ascendens, p. b.—
processus basalis, t. 1.—torus lambdoideus,
t. t. — tuberositas transversa.
гоида, короткое и массивное.
Задне-нижний его участок обра-
зует хорошо оформленный блок
(trochlea articularis, рис. 1G) че-
люстного сустава. Этот блок
сильно расширен в том месте,
где quadratum соединяется с зад-
ней ветвью птеригоида, и сужива-
ется по направлению к quadrato-
27
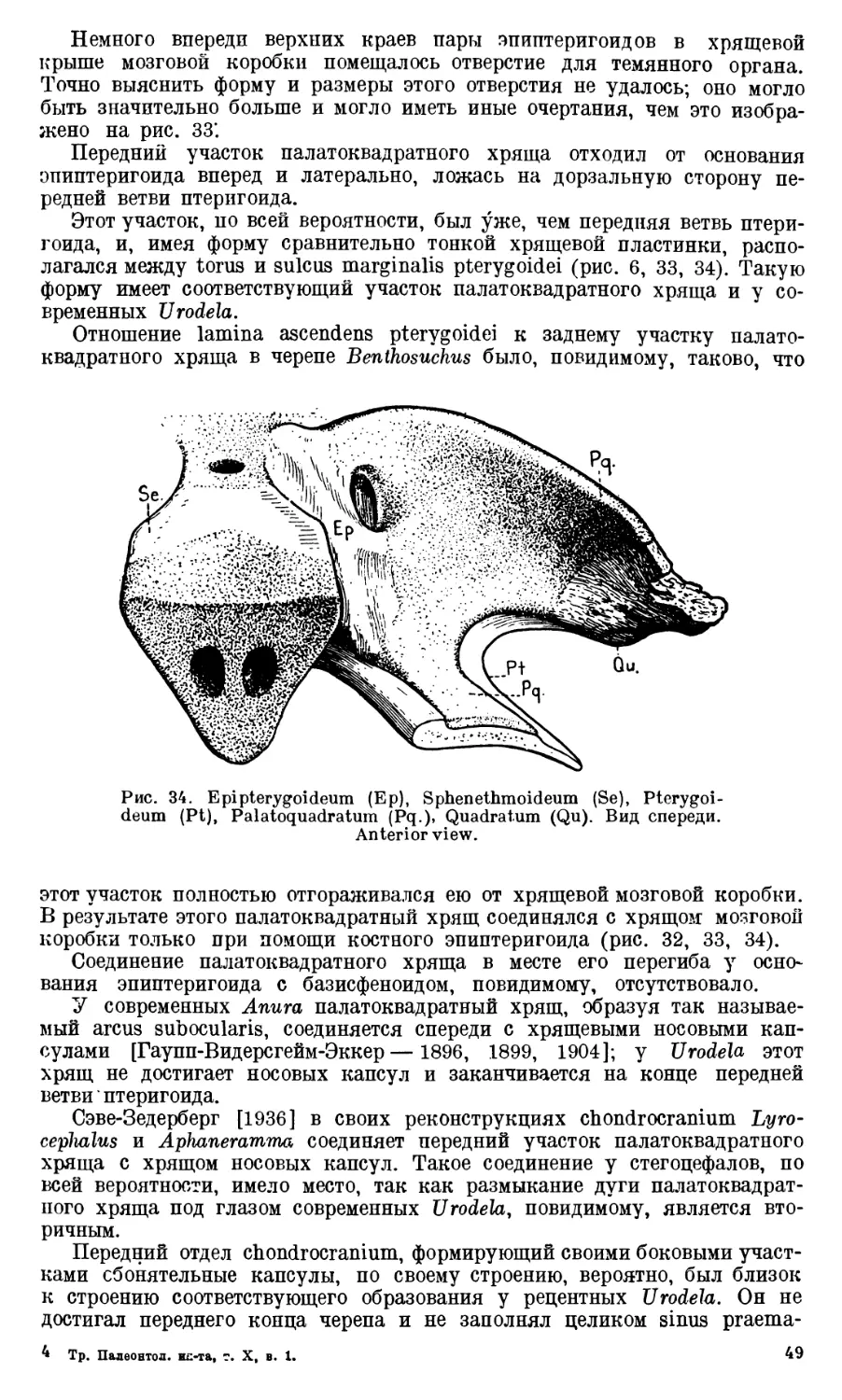
jugale. В задпем-медиальном участке quadratum, над его блоком, распола-
гается небольшой бугорок (tuberculum supratrochleare, рис. 17), видимо,
служивший местом прикрепления связок челюстного сустава и отчасти на-
чалом musculus depressor mandibulae. Латеральным краем quadratum со-
единяется с quadratojugale, которое особым выступом входит в соответствую-
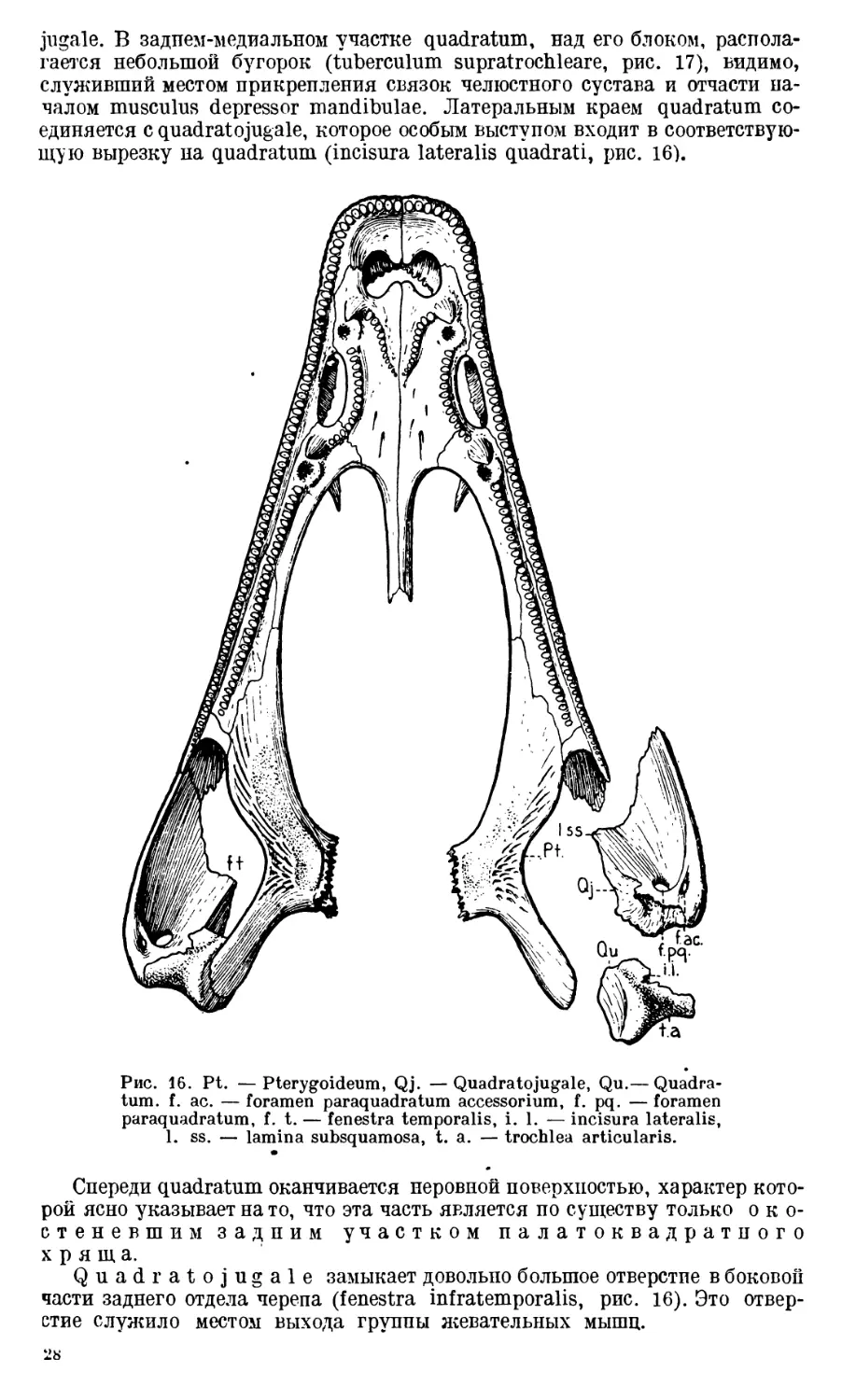
щую вырезку на quadratum (incisura lateralis quadrati, рис. 16).
Рис. 16. Pt. —Pterygoideum, Qj. —Quadratojugale, Qu.— Quadra-
tum. f. ac. — foramen paraquadra turn accessor ium, f. pq. — foramen
paraquadratum, f. t. — fenestra temporalis, i. 1. — incisura lateralis,
1. ss. — lamina subsquamosa, t. a. — trochlea articularis.
•
Спереди quadratum оканчивается неровной поверхностью, характер кото-
рой ясно указывает на то, что эта часть является по существу только око-
стеневшим задним участком палатоквадратного
хряща.
Quadratojugale замыкает довольно большое отверстие в боковой
части заднего отдела черепа (fenestra infratemporalis, рис. 16). Это отвер-
стие служило местом выхода группы жевательных мышц.
28
г По своей форме quadratojugale напоминает птичье крыло (рис. 16, 17).
Его вогнутая внутренняя поверхность гладкая, наружная — покрыта хо-
рошо выраженной скульптурой. Медиальная часть кости в виде чешуи за-
ходит под squamosum, образуя острый выступ (lamina subsquamosa, рис. 16,
17). В заднем своем участке кость пронизана большим овальной формы от-
верстием. В палеонтологической литературе это отверстие у лабиринтодон-
тов отмечалось много раз, однако без указаний на его назначение. Оно
обычно описывается чисто топографически и называется foramen quadrati,
хотя никогда не пронизы-
вает quadratum. Более
удачно было бы название
foramen paraquadratum. У
Benthosuchus foramen para-
quadratum (рис. 16, 17)
лежит полностью в преде-
лах quadratojugale. Однако
такое положение отверстия,
видимо, является типичным
не для всех стегоцефалов.
Сэве-Зедерберг [1935] у
Lyrocephalus kochi и Кейз
[Case, 1935] у Trimerorha-
chis наблюдали отверстие
па шве между quadratum
и quadratojugale.
Кроме большого отвер-
стия (foramen paraquadra-
tum), у Benthosuchus уда-
лось обнаружить маленькое
добавочное неподалеку от
первого и хорошо заметное
на quadratojugale с вент-
ральной его стороны. Это
отверстие (foramen paraqu-
adratum accessorium, рис.
16) до сих пор никем не
описывалось, но, судя по
многочисленным фрагмен-
там соответствующих уча-
стков черепа, у Benthosu-
chus sushkini является по-
стоянным образованием.
Squamosum — круп-
ная кость. Его большая
часть входит в состав тем-
порального отдела череп-
ной крыши, и только срав-
нительно небольшая плас-
тинка (lamina paraptery-
goidea, рис. 14) видна со
стороны затылочного отде-
ла черепа. Верхний угол
Рис. 17. Ео. — Exoccipitale, Ер. — Epipterygoi-
deum, Pt. — Pterygoideum, Se. — sphenethmoide-
um, f. ipm. — foramen interpraemaxillare, f. pm.—
fenestra praemaxillaris, f. pq. — foramen paraquadra-
tum, 1. ss. — lamina subsquamosa, p. c. —.processus
cultriformis, s. pmx.—sinus praemaxillaris,rt. pch.—
torus parachoanalis, t. st. — tuberculum supratroch-
leare.
squamosum соединяется c
tabulare (pnc. 13) и ограничивает спереди incisura oticalis. Co стороны
затылочного отдела черепа squamosum (его lamina parapterygoidea) сое-
диняется с верхним краем задней ветви птеригоида (точнее, с его lamina
ascendens). Обе костив месте соприкосновения не образуют настоящего шва,
но все же достаточно плотно прилегают друг к другу. На одном из рисун-
29
ков затылочного отдела черепа Trematosurusbrauni в работе Иекеля [Jaekel,
1927] squamosum и lamina ascendens pterygoidei разделены довольно широ-
кой щелью. Этого пет у взрослых бентозухов. Здесь окостенение обеих
костей настолько полное, что щель между ними закрыта нацело. Только у
сравнительно молодых бентозухов можно видеть узкую щель.
Задний угол squamosum, и в частности lamina parapterygoidea, наклады-
вается на quadratum и на особый отросток quadratojugale (lamina subsqua-
mosa), прикрывая их сзади и сверху (рис. 14). В результате этого на задней
поверхности угла черепа образуется площадка (рис. 14), служащая местом
начала musculus depressor mandibulae. В среднем своем участке эта площадка
имеет вдавленность, ограниченную с латеральной стороны высоким серпо-
видным гребнем squamosum (crista falciformis, рис. 14).
Рис. 18. Левые птеригоид и эпиптеригоид. А—Вид изнутри. В—Вид
снаружи. Left pterygoid and epi pterygoid. A — Inner view, В—Outer
view. c. o. — crista obliqua, e. ty. — excavatio tympanica, f.
q. — facies quadrata, 1. as. — lamina ascendens, s. m. — sulcus
marginalis.
Basiooccipitale у Benthosuchus sushkini не окостеневало и как
хрящевое образование не сохранилось. Оно располагалось на заднем участке
базальной части парасфеноида между обеими exoccipitale. Сверху basiocci-
pitale прикрывалось processus submedullaris exoccipitalis. В соответствую-
щем участке затылочного отдела черепа Benthosuchus имеется трехгранное
окно (fenestra basioccipitalis, рис. 14).
Supraoccipitale, будучи, как и basioccipitale, хрящевым, лег-
ло непосредственно под dermosupraoccipitale, занимая сравнительно
узкое пространство над обоими processus lamellosus exoccipitalis. Средний
участок supraoccipitale ограничивал сверху foramen occipitale magnum.
зо
Paroccipitale в течение всей жизни Benthosuchus оставалось
хрящевым и было довольно сильно редуцировано.
Basisphenoideum не обнаружено даже у самых больших
экземпляров Benthosuchus.
Epipterygoideum в черепе стегоцефалов было описано Бима-
ном [1914 — Lyrocephalus еигг], Уотсоном [Watson, 1920 — Capitosaurus,
Рис. 19. Parasphenoideum (Ps), Pterygoideum (Pt), Epipterygoideum (Ep),
Sphenethmoideum. Вид спереди. Другие обозначения, как на предыдущих
рис. Anterior view. Other lettering as in previous figures.
1926 — Palaeogyrinus], Сушкиным [1927 — Trematosaurus], Пфаненштилем
[1932 — Mastodonsaurus], Демпстером [1935 — Eryops], Сэве-Зедербергом
[1936 — Aphaneramma].
Коллекция черепов Benthosuchus позволяет изучить эпиптеригоиды и как
изолированные образования, и в соединении с птеригоидами.
31
Epipterygoideum Benthosuchus имеет хорошо развитый восходящий от-
росток (processus ascendens, рис. 15). Processus paroticus у него отсутствует,
так как соответствующий участок хряща не окостенел. Такое окостенение
у Benthosuchus, видимо, никогда не наступало, так как все имеющиеся
в коллекции экземпляры эпиптеригоидов имеют почти одинаковую форму,
несмотря на то, что их размеры колеблются от 25 до 50 мм.
На передней поверхности хорошо виден невысокий валик, по своей форме
напоминающий букву У. Ветви этого валика (torus lambdoideus, рис. 15С)
расходятся от центра кости таким образом, что одна из них (косая) идет
вдоль processus ascendens, другая (вертикальная) к базальному краю кости
и третья (горизонтальная) достигает того участка эпиптеригоида, который
у Capitosaurus [Сушкин,
1927 ] непосредственно
переходит в processus pa-
roticus.
В средней части зад-
ней поверхности эпи-
птеригоида (рис. 15D)
можно различить неболь-
шую шероховатость (tu-
berositas trans versa), про-
ходящую поперек кости.
Ниже нее в базальной
части кости располагает-
ся довольно высокий
гребень (crista basalis),
ограничивающий неров-
ную поверхность, слу-
жащую местом соедине-
ния эпиптеригоида с
птеригоидом.
Epipterygoideum у
старых Benthosuchus про-
чно срастается с телом
птеригоида и с нижней
частью переднего края
lamina ascendens таким
образом, что его базаль-
ный гребень (crista ba-
salis) располагается не-
посредственно над reces-
sus conoideus птеригоида
(рис. 18А). С латераль-
ной стороны это соеди-
нение кажется менее
плотным (рис. 18В).
Здесь хорошо виден не-
Рис. 20. Sphenethmoideum. А — Вид сверху. В — Вид
снизу. С — Вид слева. D — Вид спереди. Е — Вид сза-
ди. F — Схема строения. А — dorsal view. В —vent-
ral view, С — view from the left side. D—anterior view.
F — scheme of structure, c. io. —crista interolfactoria,
c. ol. — canalis olfactorius, f. 1. o. — fossa lobi olfacto-
rii, f. o. — foramen opticum, r. se.—rostrum spheneth-
moidei, s. io. —septum interolfactorium, s. ol. —sulcus
olfactorius. Увеличено в 2 раза. Twice enlarged.
ровный край эпиптери-
гоида, который соединялся с несохранившимся палатоквадратным хрящом.
Глубокая вырезка, лежащая между lamina ascendens птеригоида и эпи-
птеригоидом, несомненно, служила местом выхода второй и третьей ветвей
тройничного нерва.
Epipterygoideum в черепе в целом располагается таким образом, что
его processus ascendens оказывается направленным вверх, вперед и медиально
(рис. 17, 19, 21). Он ограничивает мозговую полость с латеральное-передней
стороны от височной ямы, заполненной жевательными мышцами. Верхний
край processus ascendens у Benthosuchus не доходит до нижней поверхности
покровных костей черепной крыши, где над ним располагается parietale.
32
Foramen parietale лежит несколько впереди верхних концов эпиптеригоидов
и проектируется по вертикали на основание processus cultriformis парасфе-
ноида (рис. 21). Такое отношение, повидимому, у всех стегоцефалов при
всех модификациях их черепов остается неизменным, так как данный комп-
лекс костей оказывается непосредственно связанным с мозгом [Ефремов,
1933; Быстров, 1935].
У относительно молодых бентозухов, черепа которых достигают в длину
только 220 мм, эпиптеригоиды представляют собой кости, лишенные резко
Рис. 21. Основание черепа с частью черепной крыши. Вид
сверху. The base of skull with a part of the skull roof. Ep.—
Epipterygoideum, Se. — Sphenethmoideum.
выраженных возвышений и шероховатостей. В частности на небольших эпип-
теригоидах в 25 мм длиной torus lambdoideus почти отсутствует.
Sphenethmoideum до сих пор было описано у Eryops [Брум,
1913; Демпстер, 1935], Lyrocephalus [Стенчио, 1921, Сэве-Зедерберг, 1936],
Palaeogyrinus [Уотсон, 1926], Capitosaurus [Сушкин, 1927], Mastodonsaurus
[Пфаненштиль, 1935], Capitosaurus volgensis [Гартман-Вейнберг и Кузь-
мин, 1936].
Изолированный sphenethmoideum у Benthosuchus sushkini имеет вид
трехгранной призмы с несколько расширенным передним отделом (рис. 20).
3 Тр. Палеонтол. ин-та, т. X. в. 1.
33
Верхняя поверхность его совершенно гладкая и плотно прилегает к ниж-
ней стороне покровных костей крыши черепа (рис. 21). У Benthosuchus на
сфенетмоиде лежат передние участки parietalia и задние половины frontalia.
В переднем отделе сфенетмоида при рассматривании его сверху видна
глубокая треугольная вырезка, образовавшаяся в силу неполной оссифи-
кации хряща в этом участке; на дне ее проходят два продольных желоба,
разделенных небольшим гребнем (рис. 20А).
Боковые стороны сфенетмоида гладкие. Они сходятся к нижнему ребру
кости rostrum sphenethmoidei (рис. 20В), приблизительно под углом в 80°.
В заднем отделе боковых поверхностей видпы отверстия для зрительного
нерва (рис. 20В, С).
Препаровка сфенетмоида сзади позволила обнаружить в нем глубокую
яму, в которой помещались обонятельные доли мозга (fossa lobi olfactorii,
рис. 20 Е, F). В относительно тонких боковых стенках этого углубления рас-
положены foramina optica. Яма для обонятельной доли мозга спереди посте-
пенно переходит в два канала обонятельных нервов (canafis olfactorium,
рис. 20D, E,F). Каналы отделены друг от друга костной перегородкой (septum
interolfactorium, рис. 20). В переднем отделе сфенетмоида, там, где сверху
имеется дефект в его костном веществе, каналы обонятельных нервов пере-
ходят в борозды, а перегородка между ними превращается в невысокий гре-
бень (sulci olfactorii, crista interolfactoria, рис. 20A).
Демпстер [1935] при изучении сильно окостеневшего сфенетмоида у
Eryops megacephalus обнаружил в нем, кроме canalis olfactorius, еще капал
для nervus. vomeronasalis: он располагался медиальнее канала обонятель-
ного нерва и открывался отверстием, лежащим впереди выхода nervus
olfactorius. Никаких следов подобного канала в сфенетмоиде Benthosuchus
обнаружить не удалось.
Нижнее ребро сфенетмоида (rostrum sphenethmoidei), направленное в сто-
рону нёба, входило в продольную борозду (sulcus longitudinalis) на дор-
зальной поверхности processus cultriformis парасфеноида (рис. 17, 19, 21).
Крыша черепа
Покровные кости крыши черепа у Benthosuchus на своей наружной по-
верхности, как и у всех лабиринтодонтов, покрыты своеобразной ячеистой
скульптурой.
Участки скульптуры каждой кости, в районе ее центра окостенения,
всегда состоят из относительно круглых ячеек. На периферии же'болыпинства
костей скульптура представляет собою сильно растянутые желобки, разгра-
ниченные радиально направленными гребнями. Центром всех линий, по. ко-
торым расходятся гребни скульптуры каждой покровной кости, всегда
является центр ее окостенения [Быстров, 1935].
В переднем участке шва между praemaxillaria располагается небольшое
сквозное отверстие, ведущее в так называемый sinus praemaxillaris. Задний
край дорзальной пластинки каждой praemaxillare соединяется при помощи
зубчатого шва с nasale, а латеральный угол кости на очень небольшом про-
тяжении имеет контакт с передним концом maxillare. Praemaxillare, na-
sale и maxillare ограничивают носовое отверстие овальной формы. Сквозь
него видно костное нёбо, в частности — часть vomer, отделенного швом
от praemaxillare и maxillare (рис. 22).
Между nasale и maxillare острым углом входит длинное и узкое lacri-
male, не принимающее участия в образовании носового отверстия.
К заднему краю nasale примыкает длинное frontale, задний конец кото-
рого лежит на уровне орбиты, но участия в образовании ее края у большин-
ства экземпляров Benthosuchus не принимает.
Латеральнее frontale, входя передним участком в виде острого клина
в промежуток между nasale и lacrimale, лежит praefrontale. Задний крап
этой кости ограничивает спереди орбиту и обычно на очень незначительном
протяжении соединяется швом с небольшим postfrontale.
34
Один из краев postfrontale образует медиальный и отчасти задний край
глазницы.
Позади lacrimale, соединяясь швом с praefrontale и ограничивая орбиту
с латеральной стороны, расположено jugale. Как указывалось выше, jugale
имеет большой крыловидный отросток, принимающий участие в образовании
нёба (processus alaris, рис. 23).
Если преорбитальный отдел крыши черепа Benthosuchus сложен из ряда
относительно длинных костей, то тот отдел ее, который лежит между глаз-
ницами и затылочным краем, составлен из коротких, в иных случаях почти
квадратных костей.
Непосредственно позади орбиты расположено postorbitale, вклиниваю-
щееся между postfrontale и jugale. Передний край этой кости образует зад-
ний край орбиты.
Таким образом, орбита оказывается окруженной четырьмя костями:
jugale, postorbitale, ргае- и postfrontale. Эти кости ограничивают орбиту
у большинства стегоцефалов, и только у некоторых форм в образовании глаз-
ницы принимает участие и frontale [Micropho Us, Уотсон, 1913; Capitosaurus,
Mastodonsaurus. Фраас (Fraas), 1889; Шредер (Schroeder) 1914)]. На од-
з*
35
пом из черепов Benthosuchus па левой стороне в качестве индивидуаль-
ного варианта frontale, разъединяя ргае- и postfrontalе, достигает края
орбиты (рис. 24).
Интересно, что отношение орбит к покровным костям черепной крыши
в общем не меняется, какое бы положение орбиты ни занимали в черепе.
Орбиты будут окружены указанными костями и в тех случаях, когда они
лежат ближе к заднему отделу черепа (Benthosuchus), и тогда, когда они
отверстиям (Metoposaurus)
оказываются передвинутыми ближе к носовым
Рис. 23.Черепная крыша. Вид снизу. Skull roof. Ventral
view. p. al.—processus alaris, p. po.—processus paroticus,
p. so.—processus supraoccipitalis.
[Ефремов, 1933; Быст-
ров, 1935].
Р а г i е t а 1 i а, со-
прикасающиеся своим и
краями с frontale, post-
frontale, supratempora 1 е
и dermosupraoccipitale,
занимают центральное
положение в пост орби-
тальной части крыши
черепа. В шве между
ними, приблизительно
на средине его длины,
расположено небольшое
круглое foramen parie-
tale. На черепах Benlho-
suchus иногда можно
видеть, что одно из ра-
rietalia короче другого.
В таких случаях более
длинная кость своим пе-
редним концом соединя-
ется при помощи швов
с обеими frontalia — и
правой и левой.
То же самое следует
сказать и относительно
dermosupraoccipitale. Ча-
сто наблюдается, . что
правое dermosupra-
occipitale имеет в своем
медиальноверхнем углу
небольшой выступ, ко-
торый коротким швом
соединяется с левым
parietale (рис. 24). Ча-
сто наблюдается и об-
ратная картина: dermo-
supraoccipitale s i n i s t г u m образует такой же короткий шов с pa-
rietale d е х t г u m. Такого случая, когда все четыре шва между парами
указанных костей образовали бы крест, сходясь в одной точке, пи на од-
ном из черепов Benthosuchus не наблюдается.
Латерально от почти четыреугольной dermosupraoccipitale расположено
tabulare. Обе эти кости образуют вогнутый затылочный край черепа. От-
ростки tabulare (cornu tabularis) у Benthosuchus развиты довольно сильно
и ограничивают с медиальных сторон глубокие вырезки для барабанных пе-
репонок (incisura oticalis).
К tabulare на очень незначительном протяжении примыкает squamosum,
составляющая своим задним вогнутым краем латеральный участок incisurae
oticalis.
36
, 1927], Lyrocephalus, Platystega
Рис. 24. Черепная крьпПа (уменьшено). Вид
сверху. Skull roof (diminished). Dorsal
view. G. a. — commissura anterior. D —
Dermosupraoccipitale, F — Frontale, F. i.—
foramen interpraemaxillare, F. j. — flexura
jugalis, F. 1. —flexura lacrimalis, Ju.—
Jugale, L — Lacrimale, M — Maxillare,
N — Nasale, P — Parietale, Pf.—Postfron-
tale, Po—Postorbitale, Prf. — Praefrontale,
Pm. — Praemaxillare, Qj.—Quadratojugale,
S. j. — sulcus jugalis. Sq — Squamosum,
S. ot — sulcus occipitalis transversus, S.
io. — sulcus infraorbitalis, S. so.—sulcus
supraorbitalis, S. t — sulcus temporalis,
St — supratemporale, T — Tabulare.
Supra temporale является па ряду с lacrimale покровной
костью, которая у Benthosuchus нигде не доходит до края черепной крыши и
не принимает участия в образовании какого-либо отверстия. В боковой
части заднего угла черепа лежит quadratojugale, подробно описанное
выше.
Intertemporal е, имеющееся у некоторых стегоцефалов — Loxom-
та, Pelion lyelli, Leptophractus, Dendrerpeton, Platyrhinops, Trimerorha-
chis— [Уотсон, 1926; Ромер, 1930; Стип, 1930; Кейз, 1935], полностью от-
сутствует у Benthosuchus sushkini.
Не удалось обнаружить у Benthosuchus никаких следов и маленькой sep-
tomaxillare, описанной у Micropholis stowi [Уотсон, 1913], Zatrachys mic-
rophtalmicus [Брум, 1913], T rematosuchus sobeyi [Хоутон (Haughton),
1915], Trematosuchus Jakovievi [Pj
[Сэве-Зедерберг, 1935, 1936].
Внутренняя сторона костей че-
репной крыши (рис. 23) гладкая.
11а каждой кости можно видеть
только небольшие отверстия, от ко-
торых по радиусам расходятся уз-
кие желобки. Те кости, которые
расположены по краю черепной
крыши, имеют более или менее силь-
но выраженные отростки, прини-
мающие участие в образовании нё-
ба и затылка. К таким костям
относятся praemaxillare, maxillare,
jugale и dermosupraoccipitale.
При изучении наружной по-
верхности черепной крыши Ben-
thosuchus сразу бросаются в глаза
длинные резко очерченные бороз-
ды, переходящие с одной кости на
другую — следы расположения си-
стемы так называемых каналов бо-
ковой линии.
Мооди [Roy Moodie, 1908], срав-
нивая систему каналов боковой ли-
нии стегоцефалов и рыб, выработал
для этих борозд общепринятую те-
перь терминологию. Эта термино-
логия принята здесь с дополнения-
ми, уже опубликованными в лите-
ратуре [Быстров, 1935].
На самом конце морды Bentho-
suchus в пределах его praemaxillare
расположена борозда, названная
в свое время Мооди commissura
anterior (рис. 24). Опа разделена на
две равные части швом, проходя-
щим между praemaxillaria. Обе по-
ловины commissura anterior располагаются несколько отступя назад
от foramen interpraemaxillaris и образуют тупой угол, открытый назад.
От того места, где commissura anterior достигает центра окостенения prae-
maxillare, получают свое начало еще две длинные борозды. Одна из них —
sulcus supraorbitalis — переходит почти по прямой линии на центр окосте-
нения nasale. Затем она отклоняется в латеральную сторону и достигает
центра окостенения lacrimale. Здесь, на lacrimale, sulcus supraorbitalis
лежит в непосредствепдом соседстве с sulcus infraorbitalis.
37
В дальнейшем sulcus supraorbitalis, описав дугу, обращенную выпук-
лостью наружу, более короткой дугой окружает орбиту с медиальной и зад-
ней сторон. При этом sulcus supraorbitalis точно следует но пути, ведущему
от центра окостенения одной покровной кости к центру окостенения другой.
В этом своем участке он проходит сначала над центром окостенения ргае-
frontale, затем — frontale и, наконец, — postfrontale. Заканчивается sul-
cus supraorbitalis в области центра окостенения postorbitale. На протяжении
небольшого участка в пределах postfrontale sulcus supraorbitalis обычно
прерывается. Здесь в таких случаях вместо ясно очерченной борозды виден
ряд более крупных и глубоких ячеек, расположенных по $оду канала
боковой линии. Такая картина встречается у большинства взрослых
Benthosuchus (рис. 22). Однако у очень
(№ 4—19/2252) позади орбиты имеется
Рис. 25. Череп молодого Benthosuchus (уве-
личено в 2.8 раза). Skull of young Benthosu-
chus (enlarged 24/5. № 4—19/22452.
молодого экземпляра бентозуха
совершенно замкнутая борозда
сепзорпого канала (рис. 25).
От начала sulcus supraorbi-
talis (центр окостенения ргае-
maxillare) ведет свое начало и
sulcus infraorbitalis. Он идет сна-
чала латеральпо, потом назад по
краю морды, огибая с латераль-
ной стороны носовое отверстие.
В месте пересечения шва между
praemaxfllare и maxillare sulcus
infraorbitalis всегда очень плохо
виден, но после своего перехода
на maxillare становится снова
достаточно резко очерченным.
С центра окостенения maxil-
lare sulcus infraorbitalis перехо-
дит на центр окостенения lacri-
male. Здесь на некотором протя-
жении он идет в непосредствен-
ной близости к sulcus supraor-
bitalis, отделяясь от него только
узкой полоской кости, покрытой
одним рядом круглкх ячеек.
Мооди [Moodie, 1908] описывает
в этом месте соединение обоих ка-
налов, называя его commissura
anteorbitalis. Судя по опублико-
ванным до сих пор описаниям черепов лабиринтодонтов, указанного соеди-
нения двух борозд вообще не существует. Его нет у Benthosuchus. Нужно
думать, что плохая препаровка, при которой был сбит узкий гребень ко-
сти, разделяющий оба канала, послужила поводом для описания подоб-
ного соединения.
От центра окостенения lacrimale sulcus infraorbitalis делает крутой по-
ворот вперед и латерально, образуя так называемую flexura lacrimalis
[Быстров, 1935]. Дальнейший ход sulcus infraorbitalis совершенно прямо-
линеен. Борозда сначала располагается вдоль шва между lacrimale и шахе-
lare, а затем переходит на jugale, на котором в области центра его окосте-
нения образует второй изгиб — flexura jugalis. От конца этого изгиба ведут
начало две новых борозды. Одна из них — sulcus jugalis, — выйдя за пре-
делы jugale, располагается вдоль шва между squamosum и quadratojugale
и заканчивается, несколько изгибаясь в вентральном направлении, на зад-
нем краю черепа. Другая борозда—sulcus temporalis—пересекает шов между
jugale и postorbitale и в точке, соответствующей центру окостенения postorbi-
tale, соединяется с концом sulcus supraorbitalis. Отсюда sulcus temporalis
направляется к центру окостенения supraorbitalis, оде и заканчивается.
38
Вдоль затылочного края черепа от вершины рога tabulare одной стороны
до такого же образования на другой стороне располагается слабо выражен-
ная борозда (sulcus occipitalis transversus), которая у Benthosuchus обычно
представлена'в виде ряда несколько более крупных ячеек, и только у очень
больших экземпляров она выглядит как достаточно резко очерченный sul-
cus, в особенности на tabulare.
В настоящее время трудно объяснить, почему sulcus temporalis у Bentho-
suchus, как и у многих других форм, доходит только до точки окостене-
ния supratemporale, а не продолжается до вершины рога tabulare, чтобы
соединиться здесь с концом sulcus occipitalis transversus. Однако не подле-
жит сомнению, что с a n а 1 i s temporalis, не оставлявший следа в виде
борозды на скульптуре покровных костей, все же соединялся с с a n а 1 i s
occipitalis transversus (рис. 24). На это указывают те случаи, когда sulcus
temporalis у некоторых форм стегоцефалов достигает шва между supratem-
porale и tabulare [Aphaneramma rostratum\ Виман, 1914] или, пересекая
этот шов, теряется среди ячеистой скульптуры tabulare [Lyrocephalus eun,
Lonchorhynchus oebergi, Виман, 1914—1917; Trewatosaurus brauni: Быстров,
1935].
Изучение хода каналов боковой линии на черепах Benthosuchus показало,
что их борозды располагаются таким образом, что последовательно пересека-
ют под разными углами швы, разделяющие кости. Только очень редко бороз-
да оказывается лежащей вдоль того или иного шва. В таких случаях дан-
ный шов всегда бывает чешуйчатым, и ход борозды вдоль него объясняется
исключительно тем, что одна из костей заходит под другую. По существу
борозда располагается на одной кости, но оказывается в непосредственной!
соседстве со швом только потому, что соседняя кость накладывается на
кость, связанную с каналом, и своим краем достигает борозды. Хорошим
примером таких отношений может служить задний участок sulcus jugalis
в том месте, где он проходит вдоль шва между squamosum и quadratnju-
gale. Югальный канал здесь, несомненно, связан с цептрсм окостенения
quadratojugale, лежащим в заднем углу кости. Squamosum, накладываясь
сверху на большую часть quadratojugale, только доходит до канала своим
латеральным краем. В редких случаях [Trematosaurus brauni, Бурмейстер
(Burmeister), 1849] латеральный край squamosum в процессе роста как бы
пододвигается под задний участок canalis jugalis, в результате чего канал
оказывается на некотором протяжении лежащим на squamosum. Эта особен-
ность в ходе борозды канала боковой линии должна рассматриваться как
вторичное явление.
Капал боковой линии не может располагаться и никогда не располагается
вдоль типичного зубчатого шва. Это объясняется тем, что у личинок стегоце-
фалов сензорный канал так или иначе был связан с первичными закладками
покровных костей. Эти закладки у взрослых по месту своего положения
соответствуют центрам окостенения костей черепной крыши. Ранняя связь
каналов с центрами окостенения является причиной, с одной стороны,
того, что у взрослого стегоцефала, какую бы форму его череп ни имел,
борозды сензорных каналов оказываются идущими от одного центра окосте-
нения до другого, с другой стороны, совершенно исключается возможность
расположения борозд вдоль зубчатых швов между двумя, соседними ко-
стями.
Нижняя челюсть
Mandibula Benthosuchus sushkini составлена из девяти покровных
костей, расположенных вокруг длинного Cartilago Meckeli, задний конец
которого, окостеневая, образует десятую кость — articulare.
А г t i с u 1 а г е. На целой нижней челюсти обычно видна только верх-
няя поверхность этой кости. Здесь она вогнута (cavitas glenoidalis), так как
образует сустав с блоком quadratum (рис. 26В). Передний край суставной
ямки поднимается в виде небольшого гребня (crista praeglenoidalis, рпс. 26В).
39
Спереди кость заканчивается площадкой с неровной поверхностью. Здесь
она была непосредственно связана с задним концом Меккелева хряща.
Сзади, с боков и снизу articulare прикрыта тремя покровными костями: sup-
гие. 26. Ramus sinister mandibulae.
А — Вид изнутри. В — Вид сверху. А —
Inner view, В — dorsal view. An.—An-
gulare. Ar. — Articulare, Co — Coronoi-
deum, c. gl — cavitas glenoidalis, c. pg—
crista praeglenoidalis, De. — Dentale, d.
sy. p — dens symphysalis primus, d. sy
s—dens symphysalis secundus, f. ch.
ty — foramen chordae tympani, f. M —
foramen Meckelianum, f. m — fossa man-
dibulae, h. sp — hamulus splenialis, ic.—
intercoronoideum, 1. sy.—lamina symphy-
seos, Pa. — Praearticulare, Pc. — Prae-
coronoideum, p. h.—processus hamatus, p.
ra. — processus retroarticularis. Ps — Po-
stspleniale, Sa — Supraangulare, Sp.—
Spleniale, t. a — torus arcuatus.
raangulare, angulare и praearticularc.
В зависимости от индивидуального
возраста Benthosuchus степень око-
стенения articulare бывает различна.
У старых — эта кость обычно очень
прочна, у молодых, напротив, —часто
легко разрушается нацело при попыт-
ках удалить породу. Повидимому, в
молодом возрасте степень окостенения
была незначительна.
Supraangulare, примыкая
к articulare с латеральной стороны,
одевает каудальный участок послед-
ней костным колпаком, образуя зад-
ний угол челюсти (рис. 27В). Здесь,
непосредственно позади суставной ям-
ки articulare, можно видеть неболь-
шой бугорок (tuberculum postglenoi-
dale, рис. 27А, В). Позади него верх-
няя поверхность кости образует трех-
гранную, немного вогнутую площадку
(рис. 26В), на латеральном краю кото-
рой помещается небольшой гребень с
закругленным краем. Задний конец
supraangulare имеет форму отростка с
закругленной вершиной (processus rct-
roarticularis, рис. 26А, В, 27А, В). Та
часть supraangulare, которая лежит
на латеральной стороне articulare,
плотпо к пей прилегая, представляет
собой довольно толстую костную пла-
стинку, покрытую на некотором участ-
ке своей наружной поверхности ха-
рактерной для стегоцефалов кругло-
ячеистой скульптурой. В пределах
скульптурнрованного участка кости
хорошо видны борозды сензорных ка-
налов (рис. 27 А). По направлению впе-
ред supraangulare переходит в острый
клин с очень косо срезанным краем.
Верхний край кости имеет вид немно-
го выпуклого гребня (рис. 27А), на
внутренней стороне которого хорошо
видно продольное утолщение (torus
arcuatus, рпс. 26А, 27В).
Angulare представляет собой
кость, составленную из двух несколь-
ко вогнутых пластинок, соединенных
друг с другом внизу под углом. На-
ружная пластинка (lamina externa,
рис. 27С) несколько больше, чем
внутренняя (lamina interna, рис.
27D).
Наружная поверхность наруж-
ной пластинки покрыта скульптурой;
40
ее круглые ячейки располагаются только в непосредственной близости
к центру окостенения кости, к периферии же от этого пункта по направле-
ниям радиусов лежат сильно растянутые желобки. Спереди и сверху скульп-
тура доходит до самого края кости, сзади же ее границей служит борозда
слизевого капала, за пределами которой небольшой участок кости совер-
шенно лишен скульптуры.
Рис. 27. Левые supraangulare, angulare, praearticulare (А, С, Е — вид
снаружи, В, D, F — вид изнутри). Left supraangulare, angulare, praearti-
culare (А, С, Е — outside view, В, D, F — inside view). Ar. — Articulare,
f. ch. ty — foramen chordae tympani, i. m — incisura mandibulae, 1. e —
lamina externa, 1. i — lamina interna, p. ra processus retroarticularis,’
p. h — processus hamatus, t. pg — tuberculum postglenoidale.
Внутренняя пластинка не имеет никаких следов скульптуры на своей
поверхности. В переднем участке ее верхнего края хорошо видна вырезка,
ограничивающая сзади и снизу foramen Meckelianum челюсти (рис.26А,
27D).
Angulare своей наружной пластинкой на довольно большом протяжении
накладывается на косо срезанный край supraangulare, образуя здесь типич-
ный чешуйчатый шов. Внутренняя пластинка в свою очередь накладывается
41
на нижний край praearticulare, по на меньшем протяжении и мепыпей пло-
щади. Таким образом, angulare, образуя нижний угол челюсти, захваты-
вает своими верхними краями снаружи и извнутри две расположенные над
нею кости.
Praearticulare (goniale) задним своим концом лежит на articula-
ге (рис. 26А) с внутренней стороны челюсти. Этот конец соединяется швом
с небольшим участком supraangulare, прикрывающим задний конец arti-
culare и заходящим на медиальную сторону челюсти. В непосредственном
соседстве со швом на краю praearticulare имеется небольшое отверстие, слу-
жившее местом входа в челюсть chorda tympani (рис. 27Е, G). На нижнем
краю дистального конца praearticulare расположена неглубокая вырезка,
принимающая участие в образовании foramen Meckelianum. Верхний край
кости несколько утолщен и образует вырезку нижней челюсти (incisura
mandibulae. рис. 26А, 27Е, G). При рассматривании нижней челюсти с внут-
ренней стороны хорошо видно, что этот край значительно ниже наружного,
образованного supraangulare.
На той стороне praearticulare, которая обращена в сторону articulare,
располагается плоский отросток (processus hamatus, рис. 26В, 27Е), прикры-
вающий верхнюю часть передней поверхности articulare. Таким образом,
articulare оказывается охваченной в заднем углу челюсти тремя покровными
костями: angulare прилегает к нему снизу, supraangulare прикрывает сна-
ружи, сверху и извпутри, praearticulare накладывается извнутри и отчасти
спереди. Свободными поверхностями у articulare оказываются только
верхняя, образующая суставную ямку, и передняя, непосредственно пере-
ходившая в cartilago Meckeli.
Dentale является самой длинной костью нижней челюсти. Опа
накладывается своим острым задним концом поверх supraangulare; отно-
шение же ее к angulare оказывается обратным; наружная пластинка этой!
кости ложится снаружи на нижний край dentale (рис. 28, распилы В, Е, D).
Вдоль всего свободного верхнего края dentale в один ряд располагаются
мелкие зубы, не отличающиеся от зубов maxillare и'ргаетахШаге. Передний
конец dentale несколько расширен и образует пластинку (lamina symphyseos,
рис. 26). На ее верхней поверхности расположены клыки (dentes
symphysales, рис. 26, 28). Отосительпо этих клыков следует сказать то же,
что говорилось выше о пре- и постхоанальных клыках.
Симфизальные клыки, —так же, как и клыки на нёбных поверхностях
vomer и palatinum,— встречаются в виде одного-двух острых, слегка загну-
тых назад зубов. Наличие у бентозуха одновременно двух клыков—явление
^вольно редкое. Обычно имеется один клык; при этом одинаково часто
приходилось обнаруживать как первый клык (dens symphysalis primus),
так и второй (d. s. secundus). При отсутствии одного из клыков на его месте
оказывается такая же ямка, какая была описана на vomer и palatinum. Все
это заставляет предполагать в смене симфизальных клыков такую же пе-
риодичность, как и в смене пре- и постхоанальных. Наружная поверхность
переднего конца dentale скульптурирована; кзади скульптура постепенно
сглаживается.
Postpleniale, расположенпое па нижнем краю приблизительно
посредине длины нижней челюсти, своим задним краем граничит с an-
gulare и praearticulare (рис. 26А). Вклиниваясь между передними кон-
цами, postpleniale вместе с ними принимает участие в образовании боль-
шого овальной формы foramen Meckelianum. Снаружи postspleniale на
некотором протяжении образует шов с dentale (рис. 28, распилы Е, G).
Нижне-наружная поверхность кости покрыта ячеистой скульптурой, тогда
как внутренняя — совершенно гладкая.
S р 1 е n i а 1 е занимает самый передний отдел челюсти снизу. За-
гнутый в виде крючка передний конец кости (hamulus splenialis, рис. 26А)
лежит непосредственно под расширенным концом dentale и принимает
участие в соединении обеих ветвей нижней челюсти. Нижняя сторона
42
spleniale покрыта обычной ячеистой скульптурой; внутренняя ячеек не
имеет.
С о г о п о i d е u т, располагаясь между praearticulare и dentale,
своим задним краем образует передний угол глубокой ямы в челюсти
(fossa mandibulae, рис. 26В). Здесь под coronoideum и под dentale далеко
вперед заходит длинный острый передний конец supraangulare (рис. 28
распил С). Таким образом, supraangulare, на которую действуют мышцы
сжимающие челюсти, как бы
подпирает снизу две кости, во-
оруженные зубами.
Coronoideum снабжен одним
рядом мелких зубов.
Intercoronoideum
прилегает с медиальной стороны
к dentale (рис. 28, распилы D,
Е, F), а нижним краем образует
шов с postspleniale. Задний угол
кости достигает переднего кон-
ца praearticulare, соприкасаясь
с ним на незначительном про-
тяжении. На большем протяже-
нии intercoronoideum граничит
с coronoideum (рис. 26А).
Praecoronoideum
лежит впереди интеркороноида
между dentale и spleniale на
внутренней стороне нижней че-
люсти. Задний конец его соеди-
няется коротким швом с перед-
ним концом интеркороноида; пе-
редний конец достигает расши-
ренной части dentale.
Прекороноид и интеркоропо-
ид Benthosuchus совершенно ли-
шены зубов, но верхние их края,
отходящие от медиальной по-
верхности dentale, имеют не-
сколько заостренных выступов.
Эти выступы сильнее развиты
на прекороноиде, чем на интер-
короноиде (рис. 26А).
Нижние челюсти стегоцефа-
лов дают довольно разнообраз-
ную картину озубления короно-
идов. Так, у Dvinosaurus [Суш-
кин, 1936] все три короноида
снабжены зубами; у Trimero-
rhachis [Уиллистон, 1914; Кейз,
1935] зубы имеются на короно-
пде и интеркороноиде; у Trema-
tosaurus [Некель (Jaeke'l), 1925]
они обнаружены только на
короноиде; у Eogyrinus attheyi,
Рис. 28. Левая ветвь нижней челюсти и 8 попе-
речных распилов ее. The left branch of man-
dibula and its 8 cross-sections. An.—Angulare,
Ar. — Articulare, Co. — Coronoideum, De. —
Dentale, Ic. — Intercoronoideum, Pa. — Praear-
ticulare, Pc. — Pr&ecoronoiduem, Ps. — Posts-
pleniale, Sa. — Supraangulare, Sp. — Spleniale.
Orthosaurus [Уотсон, 1925], Eryops [Брум, 1913], Buettneria [Кейз, 1932]
все короноиды лишены зубов. Следовательно, Benthosuchus sushkini в смысле
озубления короноидов сходен с Trematosaurus.
Ряд поперечных распилов нижней челюсти Benthosuchus (рис. 28) по-
казал, что, начиная от глубокой ямы в заднем конце челюсти (fossa mandi-
43
bulae), вдоль всей челюсти почти до самого симфиза проходит длинный
канал, в котором помещался Меккелев хрящ. Этот капал по мере прибли-
жения к концу челюсти постепенно становится все уже и уже и слепо
закапчивается в области расширенного конца dentale (lamina symphy-
seos).
Борозды каналов боковой линии нижней челюсти на
покрытых скульптурой участках ее наружной поверхности видны очень
хорошо.
Основная борозда — sulcus articularis (рис. 29) — начинается у края
supraangulare, несколько отступя назад от суставной ямки челюсти.
Сензорный капал, залегающий в этой борозде, несомненно, является не-
посредственным продолжением canalis jugalis, задний участок которого
в виде дуги проходит вдоль шва между squamosum и quadratojugale.
Sulcus articularis в пределах supraangulare делится на две
ветви. Одпа из них, sulcus marginalis, является как бы прямым продол-
Рис. 29. Череп Benthosuchus (уменьшено). Вид справа. Skull of Bentho-
suchus (diminished). View from the right side. An. —Angulare, C. a. —
commissura anterior, De — Dentale, F. j. — Flexura jugalis, F. 1. — Flexura
lacrimalis, Ju — Jugale, L —Lacrimale, M — Maxillare, Pm — Praemaxil-
lare, Qj.— Quadratojugale, s. a. —sulcus articularis, San.—Supraangulare,
S. acc.—Sulcus (articularis) accessorius, S. d. —Sulcus dentalis, S. io.—
sulcus supraorbitalis, S. j — Sulcus jugalis, S. m. — sulcus marginalis,
Sp —Spleniale, Sq — Squamosum, S. so. — Sulcus supraorbitalis, Psp —
Postspleniale.
жением основной борозды и направляется косо к нижнему углу челюсти,
где и заканчивается в области центра окостенения angulare. В дальнейшем
своем ходе канал, лежавший в этой борозде, повидимому, переходил
на внутреннюю сторону нижнего края челюсти и не оставлял здесь
никакого следа на поверхности лишенных скульптуры костей. Вторая
ветвь, sulcus dentalis, образуя слабо выпуклую дугу, переходит с supraan-
gulare на dentale и достигает ее покрытого скульптурой участка, где и те-
ряется. Sulcus dentalis па некотором протяжении проходпт вдоль шва
между angulare и dentale. Такое отношение к шву объясняется тем, что
angulare достигает верхним краем своей наружной пластинки сензорпого
канала, накладываясь снаружи на края supraangulare и dentale.
Кроме sulcus articularis и sulcus dentalis, на supraangulare Benthosuchus
имеется постоянная добавочная борозда (sulcus articularis accessorius,
рис. 27А, 28, 29). Начальный участок этой борозды обычно выражен очень
ясно, конец же постепенно теряется и исчезает там, где скульптура supraan-
'i4
gulare сглаживается. Нужно думать, что в этой борозде располагался
короткий добавочный канал, соединяющий средину canalis articularis
с начальной частью canalis dentalis.
Череп в целом
Ряд костей основания черепа складывается такпм образом, что на нёб-
ной поверхности образуется несколько больших и малых отверстий.
В самом переднем отделе нёба расположено широкое окно (fenestra
praemaxillaris, рис. 30). Оно сужено в передне-заднем направлении в сред-
ней своей части бла-
годаря наличию двух
пар отростков. Перед-
няя пара отростков
принадлежит нёбной
пластинке praemaxil-
lare (processus fene-
strates ant eri ores) зад-
няя пара — vomer
(processus fenestrates
posteriores). В боко-
вые расширенные
участки этого непар-
ного окна при закры-
том рте входят симфи-
зальные клыки ниж-
ней челюсти. Однако
эти клыки у Bentho-
suchus не достигали
внутренней поверхно-
сти черепной крыши.
У Mastodonsaurus, Cy-
clo tosaurus, Anaschis-
пга [Фраас, 1889) клы-
ки вызывали образо-
вание отверстий в че-
репной крыше и вы-
ходили своими вер-
шинами на дорзаль-
ную поверхность че-
репа.
Несмотря на на-
личие очень больших
клыков на нижних
челюстях у старых
экземпляров Ben tho -
suchus, эти зубы все
же не пронизывали у
них черепной крыши.
Хоаны (nares in-
ternae [Уотсон, 1923];
nares posterioris [О.
Рис. 30. Череп Benthosuchus. Вид снизу. Skull of Bentho-
suchus. Ventral view. Ch — choana, f. ip. — fenestra inter-
pterygoidea, f. pmx.—fenestra praemaxillaris, f. it.—fenestra
infratemporalis, na. — naris, or. — orbita.
Абель, 1919] имеют очертания сильно вытянутых овалов. У Benthosuchus опи
ограничены краями vomer, maxillare и на незначительном протяжении
сзади — palatinum. Сквозь хоаны видпа нижняя поверхность костей череп-
ной крыши и в самых передних отделах — края носовых отверстий.
Средняя часть нёба почти полностью занята парой сильно вытянутых
в сагиттальном направлении нёбных окоп (fenestra palatina, fenestra in-
45
Рис. 31. Поперечные разрезы зубов Bentho-
suchus. А — «клык» старого Benthosuchus,
В — Малый зуб старого Benthosuchus. С —
«клык» молодого Benthosuchus. D — Малый
зуб того же экземпляра (увеличено в 6 раз).
Cross-section of the teeth of Benthosuchus.
A — «tusk» of an old Benthosuchus.
В — small tooth of an old Benthosuchus.
C — «tusk» of a young Benthosuchus. D —
small tooth of the same specimen (\6).
terpterygoidea, рис. 30; f. mediopalatinalis). Длина их у Benthosuchus
равна половине общей длины черепа. Каждое окно ограничепо с медиаль-
ной стороны vomer и processus cultriformis, с латеральной — palatinum
и передней ветвью птеригоида. Через эти окна видна большая часть орбит,
только небольшие участки их прикрываются птеригоидами.
В области задних углов черепа располагаются довольно сложные по
своим очертаниям височные окна (fenestra infratemporalis, рис. 30; fenestra
basitemporalis [О. Абель, 1919]: f. adductoria [Сэве-Зедерберг, 1935]). Каждое
из этих окон с медиальной стороны ограничено двумя ветвями птеригоида,
сзади — quadratum, с латеральной стороны — quadratojugale, спереди —
insula jugalis (processus alaris jugalis). Височные окна служили местом
выхода из височной ямы черепа группы мышц, сжимающих челюсти.
Нижнечелюстная яма (fossa mandibulae, рис. 26) по своей длине равна
длине височного окна и располагается точно под ним. Fossa mandibulae
служила местом прикрепления жевательных мышц, поэтому она в черепе
вместе с височной ямой образует большое вместилище для мощного мышеч-
ного комплекса.
В затылочном отделе черепа, кроме описанных выше foramen occipitale
magnum, fenestrae basi и supraoccipitales, в латерально-верхнем углу
находится еще небольшое отвер-
стие — fenestra subtabularis (рис.
11, 14) (fenestra posttemporalis [О.
Абель, 1919]), ограниченное тремя
костями: tabulare, dermosupraocci-
pitale и exoccipitale. По принято-
му в настоящее время мнению, fe-
nestra subtabularis являлась одним
из мест прикрепления к черепу
мышц туловища.
При рассматривании черепа со
стороны его затылочного отдела,
видно, что боковые его участки,
сочленяющиеся с нижней челюстью,
опущены вниз, в результате чего
задний отдел основания черепа ока-
зывается вогнутым (рис. 14).
Columella, направленная
своим верхним концом к центру
слуховой вырезки, стоит по отно-
шению к плоскости базальной ча-
сти парасфеноида приблизительно
под углом в 30°.
Структура зубов
Зубы Benthosuchus типичны для
лабиринтодонта. По своему строе-
нию они очень близки к зубам
Mastodonsaurus giganteus [Фраас,
1889—1890],Buettneria [Кейз, 1932]
и значительно отличаются по своей
сложности от зубов Branchiosaurus,
Pelosaurus [Креднер, 1891, 1893],
Archegosaurus [Мейер (V. Meyer),
1856—1858], Тrimerorhachis [Кейз,
1935] и Dvinosaurus [Сушкин,
1936, Быстров, 1938].
Зубы Benthosuchus. расположенные па maxillare и dentale, имеют ко-
нусообразную форму, но не такую правильную, как у Branchiosaurus,
46
Pelosaurus, Acanthostoma и Archegosaurus [Кредпер, 1891]. Они, во-первых,
довольно сильно изогнуты, во-вторых, имеют на поперечном сечении не
круглые, а овальные очертания (рис. 31В, D). Зубы скреплены с костью
таким образом, что длинная ось их овального основания всегда стоит
перпендикулярно к наружному краю кости. В ту же сторону обращена
и выпуклая поверхность изогнутого по вертикальной оси зуба.
Фраас [1889—1890] при изучении зубов Mastodonsaurus giganteus уста-
новил, что структура зуба постепенно усложняется по направлению от
его вершины к осно-
ванию. Такое же стро-
ение имеют и зубы
Benthosuchus.
Кроме того, изу-
чение большого коли-
чества зубов Bentho-
suchus позволил о уста-
новить, что структу-
ра зуба оказывается
различной в зависи-
мости не только от
уровня, на котором
сделан распил зуба,
но и от размеров са-
мого зуба. Складча-
тость дентина на од-
ном и том же уровне
в общем менее слож-
на у маленьких зу-
бов по сравнению с
клыками (рис. 31А-В,
С-D). Но клыки моло-
дых Ben thosuchus в не-
которых случаях име-
ют складчатость еще
менее сложную, чем
маленькие зубы ста-
рых экземпляров (рис.
31 B-D).
Реконструкция хря-
щевого черепа
Chondrocra-
nium у Benthosuchus
окостенел в значи-
тельно меньшей сте-
пени, чем у Eryops
[Демпстер, 1935], Ly-
rocepha lus, A phane-
ra mma [ Сэве-Зедер-
берг, 1936] и неко-
Рис. 32. Реконструкция хрящевого черепа. Вид снизу.
Restoration of the cartilagineous skull. Ventral view. Bo.—
Basioccipitale, Bs. — Basisphenoidei, f. c. — foramen ca-
roticum, Eo. — Exoccipitale, Ep. — Epipterygoideum, f.
o.—foramen opticum, Po.—Paroccipitale, Pq.— Palato-
quadratum, Qu. —Quadratum, Se.—Sphenethmoideum,
T. — Tabulare.
торых других форм.
У Benthosuchus окостенели и сохранились только части chondrocranium,
известные как костные exoccipitale, quadratum, epipterygoideum и sphe-
nethmoideum.
Что касается других участков chondrocranium, то они со значительной
долей вероятности могут быть восстановлены по некоторым следам на
поверхности покровных костей.
47
Рис. 33. Реконструкция хрящевого черепа. Вид
сверху. Restoration of the cartilagineous skull. Dor-
sal view. Bo — Basioccitpiale, col — columella, Eo—
Exoccipitale, Ep— Epipterygoideum, f. ip — fenestra
interpterygoidea, f. pa — foramen parietale, Po —
Paroccipitale, Pq — Palatoquadratum, Qj — Quad-
ratojugale, Qu—Quadratum, Se—Sphenethmoideum.
Не подлежит сомнению, что хрящевое basiooccipitale, заполнявшее все
пространство fenestra basioccipitalis (рис. 14), лежало непосредственно на
дорзальной поверхности парасфеноида и примыкало по сторонам к exocci-
pitale (рис. 32).
Впереди basioccipitale располагалось basisphenoideum, являвшееся непо-
средственным продолжением хряща basioccipitale и не отделявшееся от него.
Basisphenoideum в свою очередь непосредственно переходило
в хрящ, окостеневший участок которого у Benthosuchus образует сфенет-
моид.
Примыкавший к перед-
нему краю сфепетмоида
хрящ располагался вдоль
processus cultriformis и до-
ходил до носовой части го-
ловы. Его форма, по всей
вероятности, была на всем
п ротяжении трехгранная,
как и у сфепетмоида. Для
этого хряща па дорзальной
поверхности processus cult-
riformis на некотором его
протяжении имеется такой
же жолоб, как и для кост-
ного сфепетмоида. Дорзаль-
ной поверхностью хрящ
прилегал к костям череп-
ной крыши.
Хрящевое supraoccipita-
1е заполняло сложно очер-
ченную fenestra supraocci-
pitalis (рис. 14), плотно
прилегая к пижней поверх-
ности dermosupraoccipitale.
Его боковые участки, вытя-
нутые назад и латерально,
представляли собой реду-
цированные paroccipitalia.
Латеральный конец хряще-
вого paroccipitale, по всей
вероятности, доходил до
crista tympanica (рис. 13),
расп ол агаясь неп осред-
ственно перед processus
paroticus exoccipitalis
(рис. 32, 33).
Хрящ prooticum, со-
ставляющий одно целое с
supraoccipitale и parocci-
pitale, принимал участие в образовании хрящевой черепной коробки.
От шероховатой передней поверхности костного quadratum вперед
и медиально отходил задний участок палатоквадратного хряща. Он плотно
прилегал в виде вогнутой снаружи, напоминающей широкий жолоб пла-
стинки к латеральной поверхности задней ветви птеригоида и ее высокой
lamina ascendens (рис. 32, 33, 34). Нижний край хряща доходил до sulcus
marginalis птеригоида (рис. 18В) и у переднего края lamina ascendens
соединялся с неровным краем эпиптеригоида (рис. 32, 33, 34).
Костный эпиптеригоид своим верхним краем входил в хрящ латераль-
ной стенки мозговой коробки под самой крышей черепа (рис. 33).
Немного впереди верхних краев пары эпиптеригоидов в хрящевой
крыше мозговой коробки помещалось отверстие для темянного органа.
Точно выяснить форму и размеры этого отверстия не удалось; оно могло
быть значительно больше и могло иметь иные очертания, чем это изобра-
жено на рис. 33*.
Передний участок палатоквадратного хряща отходил от основания
эпиптеригоида вперед и латерально, ложась на дорзальную сторону пе-
редней ветви птеригоида.
Этот участок, по всей вероятности, был уже, чем передняя ветвь птери-
гоида, и, имея форму сравнительно тонкой хрящевой пластинки, распо-
лагался между torus и sulcus marginalis pterygoidei (рис. 6, 33, 34). Такую
форму имеет соответствующий участок палатоквадратного хряща и у со-
временных Urodela.
Отношение lamina ascendens pterygoidei к заднему участку палато-
квадратного хряща в черепе Benthosuchus было, повидимому, таково, что
Рис. 34. Epipterygoideum (Ер), Sphenethmoideum (Se), Pterygoi-
deum (Pt), Palatoquadratum (Pq.), Quadratum (Qu). Вид спереди.
Anterior view.
этот участок полностью отгораживался ею от хрящевой мозговой коробки.
В результате этого палатоквадратный хрящ соединялся с хрящом мозговой
коробки только при помощи костного эпиптеригоида (рис. 32, 33, 34).
Соединение палатоквадратного хряща в месте его перегиба у осно-
вания эпиптеригоида с базисфеноидом, повидимому, отсутствовало.
У современных Anura палатоквадратный хрящ, образуя так называе-
мый arcus subocularis, соединяется спереди с хрящевыми носовыми кап-
сулами [Гаупп-Видерсгейм-Эккер— 1896, 1899, 1904]; у Urodela этот
хрящ не достигает носовых капсул и заканчивается на конце передней
ветви * птеригоида.
Сэве-Зедерберг [1936] в своих реконструкциях chondrocranium Lyro-
cephalus и Aphaneramma соединяет передний участок палатоквадратного
хряща с хрящом носовых капсул. Такое соединение у стегоцефалов, по
всей вероятности, имело место, так как размыкание дуги палатоквадрат-
пого хряща под глазом современных Urodela, повидимому, является вто-
ричным.
Передний отдел chondrocranium, формирующий своими боковыми участ-
ками обонятельные капсулы, по своему строению, вероятно, был близок
к строению соответствующего образования у рецептных Urodela. Он не
достигал переднего конца черепа и не заполнял целиком sinus praema-
4 Тр. Палеонтол. ш>та, т. X, в. 1.
49
xillaris, оставляя незакрытой fenestra praemaxillaris. Здесь над ним, в si-
nus praemaxillaris, лежала glandula praemaxillaris (рис. 33).
Боковые участки этого отдела chondrocranium, представляя собой
относительно тонкие хрящевые пластинки, составляли дно обонятельных
капсул и примыкали к медиальному краю torus parachoanalis (рис. 8).
Реконструкция кровеносных сосудов
Сэве-Зедерберг [1936] в качестве исходных пунктов для своей рекон-
струкции сосудов головы Lyrocephalus пользовался данными Гауппа-
Видерсгейма-Эккера [1896, 1899, 1904] в их «Анатомии лягушки». Повп-
Рис. 35. Сосудистая система саламандры.
Вид снизу. Vesselar system of a Salamandra.
Ventral view. Ao — aorta, ce. art. carotis
externa, ca—vena cardinalis anterior, cir—
art. carotis interna, cp — vena cardinalis
posterior, dC —ductus Cuvieri, gc —glomus
caroticum, mm.— art. mandibularis media-
ns, p.—art. pulmonalis, pn — art. palato-
nasalis, qm — art. quadratomandibularis, r.
m — ramus muscularis, r. oe—ramus oeso-
phageus, s — art. subcl avia, ta — truncus
arteriosus, th — glandula thyreoidea.
димому, правильнее будет рассмат-
ривать представителей современ-
ных Anura как очень своеобразно
и высоко специализированных по-
звоночных, ушедших далеко в сто-
рону от основного ствола амфибий.
Поэтому, если сосудистая система
головы стегоцефалов в какой-то
иной степени была сходна с сосу-
дистой системой соответствующей
области у рецентных амфибий, то
этого сходства следует ожидать
скорее между стегоцефалами и
Urodela, чем между стегоцефалами
и Anura. Большая примитивность
сосудистой системы хвостатых ам-
фибий видна хотя бы в том, что
у них сохраняются в течение всей
жизни животного три-четыре ар-
териальных дуги, из которых у
взрослых Anura остается по суще-
ству только одна.
Изучение борозд на кератобран-
хиальных костях и отверстий в че-
репе, оставленных сосудами, у
•неотенической формы стегоцефа-
лов — Dvinosaurus позволило уста-
новить (рис. 35) у него наличие
четырех жаберных артериальных
дуг [Быстров, 1938]. Это указывает
на то, что тип строения артериаль-
ной системы у двинозавра был
вполне тождествен с расположе-
нием артерий головы и жаберного
аппарата у рецентных неотеяиче-
ских форм и личинок Urodela. Та-
ким образом, двинозавр, являясь
неотенической формой стегоцефа-
лов, показал, что стегоцефалы в
личиночной стадии своего развития
обладали четырьмя артериальными
дугами. Весьма вероятно, что эти
дуги после атрофии жабер во вре-
мя метаморфоза сохранялись в
как это па-
том же числе в течение всей жизни стегоцефалов,
блюдается, например, у современных саламандр.
50
Если все эти соображения считать правильными, то следует допустить,
что от конца trunchus arteriosus сердца Benthosuchus вправо и влево отхо-
дили четыре артериальных дуги (рис. 36).
На первой из них располагалась так называемая Carotidendruse (glo-
mus caroticum). Это образование возникает у современных амфибий после
того, как они пройдут все стадии метаморфоза.
Изучение борозд на первой кератобранхиальной кости дышащего жаб-
рами двинозавра дало возможность обнаружить две идущие параллельно
друг другу глубокие борозды [Быстров, 1938]. Не подлежит никакому
сомнению, что два сосуда, залегавшие
между собой рядом тонких ана-
стомозов. Эти анастомозы у совре-
менных амфибий после метаморфо-
за дают начало Carotidendriise
(рис. 5).
Полное сходство сосудов жа-
берного аппарата у двинозавра и
у личинок рецептных Urodela поз-
воляет считать, что артериальная
система головы стегоцефалов в
постметаморфозном периоде была
сходна с сосудистой системой со-
временных взрослых хвостатых
амфибий. Следовательно, имеются
все основания допускать сущест-
вование Carotidendruse и у Bentho-
suchus.
Carotidendriise является тем
пунктом, от которого получают свое
начало arteriae carotides externa и
interna. Arteria carotis externa на-
правляется вперед, к мышцам подъ-
язычного аппарата и дна рта.
Выяснить подробности в ходе
этой артерии у Benthosuchus не
представляется возможным, так
как в нашем распоряжении нет ни-
каких остатков хрящевых элемен-
тов подъязычного аппарата, па
которых могли бы остаться какие-
нибудь следы этой артерии. Од-
нако отрицать ее существование
нет оснований.
Arteria carotis in-
terna, образуя большую часть
первой артериальной дуги у Bentho-
suchus, вероятно, соединялась
при помощи короткого ductus са-
roticus со второй артериальной ду-
гой (аорта). Это соединение, как
известно, существует у Proteus,
личинки саламандры, но отсутствует
1881].
в этих бороздах, были связаны
Рис. 36. Реконструкция сосудов Benthosu-
chus (уменьшено). Вид снизу. Restoration
of vessels of Benthosuchus (diminished). Ven-
tral view. m. 1 — art. mandibularis lateralis,
r. j. — ramus jugalis. Другие обозначения,
как на рис. 35, 36. Other lettering as on
. the fig. 35, 36.
Siredon, Necturus, Siren, Triton, у
у взрослой саламандры [Боас (Boas)
Проходя неподалеку от columella, arteria carotis interna давала тон-
кую a. stapedialis, которая проходила сквозь отверстие в костной части
columella (рис. 10, 37). Затем внутренняя сонная артерия входила в спе-
циальный канал в базальной части парасфеноида, описанный выше под
Ихменем canalis caroticus (рис. 10). В этом канале артерия, образуя так
51
Рис. 37. Реконструкция черепномозговых нервов. Вид
сверху. Restoration of cranial nerves. Dorsal view,
ch. ty. — chorda tympani.
называемое латеральное колено, сначала шла почти прямолинейно по
направлению к центру окостенения парасфеноида; затем поворачивала
вперед (короткое переднее колено) и разделялась на две ветви.
Одна из этих ветвей, выходя на дорзальную поверхность парасфеноида,
располагалась в особой борозде на кости (sulcus arteriae carotidis inter'
пае, рис. 10В), а затем проникала сквозь хрящевое дно мозговой ко-
робки и входила в ее полость (рис. 32). В дальнейшем своем ходе
внутренняя сонная артерия (после того как от нее отделились тонкие
веточки для питаня мозга), по всей вероятности, покидала мозговую
полость через foramen ор-
ticum и своими конечными
разветвлениями питала
мышцы глаза (a. ophtal-
mica).
Другая ветвь внутрен-
ней сонной артерии, отде-
лившаяся от основного
ствола еще в канале в ба-
зальной части парасфенои-
да (рис. 10В), выходила
на основание черепа и ва-
скуляризировала слизи-
стую оболочку нёба.
Вторая артериальная
дуга, получив свое начало
из truncus arteriosus, до-
стигала заднего конца па-
расфеноида и, соединяясь
с такой же дугой проти-
воположной стороны, пере-
ходила в области сустава
между черепом и первым
позвонком в спипную
аорту.
В том месте, где пара-
сфеноид при помощи шва
соединяется с птеригоидом,
от дуги аорты по направ-
лению к нёбной поверхно-
сти черепа отходила до-
вольно крупная ветвь —
arteria palatonasalis. Она
располагалась вдоль ука-
занного шва в особой бо-
розде между шероховатыми
полями на парасфеноиде
и птеригоидах (рис. 7С).
Неподалеку от переднего края парасфеноида arteria palatonasalis у Ben-
thosuchus, как у современных саламандр (рис. 35), делилась на две ветви
(рис. 36). На это указывает случай разделения sulcus arteriae palatona-
salis на правой стороне черепа № 1—9—2243 (рис. 70) на две ветви,
что, несомненно, было вызвано более ранним по сравнению с левой сто-
роной разветвлением залегавшей в нем артерии.
Arteria quadratomandibularis у саламандры полу-
чает свое начало от второй артериальной дуги, у аксолотля — от arteria
carotis interna.
На костях Benthosuchus нет никаких борозд, которые позволили бы
решить, какой из этих двух типов ответвлений артерий у него имел место.
52
Но откуда бы ни получала свое начало arteria quadratomandibularis, она
у Benthosuchus, повидимому, делилась, как у рецентных Urodela, позади
челюстного сустава на две ветви.
Одна из этих ветвей (arteria mandibularis, рис. 36) проходила вдоль
медиального края соответствующей ветви нижней челюсти; другая своим
концом проникала в foramen paraquadratum, где и делилась на две артерии
меньшего диаметра. Одна из этих артерий (ramus jugalis) направлялась
в мышцы, сжимающие челюсти; другая (arteria mandibularis lateralis)
переходила на латеральную поверхность нижней челюсти, пройдя в углу
черепа сквозь foramen paraquadratum accessorium.
Третья и четвертая артериальные дуги, располагаясь каудальнее пер-
вых двух, не имели прямого отношения к костям черепа, поэтому не могли
оставить на нем никаких следов.
Наличие в черепе Benthosuchus канала для внутренней сонной артерии,
отверстия в columella для arteria stapedialis, борозды для arteria palato-
nasalis и отверстий для ramus jugalis и arteria mandibularis lateralis —
является, по нашему мнению, достаточным доказательством того, что сосу-
дистая система головы Benthosuchus была построена по типу сосудов го-
ловы Urodela. Поэтому нужно думать, что изображенная на рис. 36
реконструкция сосудов более или менее близка к истине.
Реконструкция черепно-мозговых нервов
В виду того, что большая часть мозговой коробки у Benthosuchus в
течение всей жизни оставалась хрящевой, точно представить себе форму
его мозга трудно. Этому мешает еще то, что, повидимому, у него, так же,
как и у рецентных Urodela, мозг не полностью занимал всю мозговую
полость черепа и между ее стенками и мозгом, вероятно, было довольно
большое пространство.
Однако мозг Benthosuchus, по всей вероятности, был больше похож
па мозг современных хвостатых амфибий, чем на мозг Anura, как изобра-
жает его в своих реконструкциях Сэве-Зедерберг [1936]. В частности,
нужно думать, что lobus opticus, хорошо развитый у Anura, но отсут-
ствующий у Urodela, отсутствовал и у стегоцефалов. Демпстер [1935] не
обнаружил никаких следов lobus opticus при изучении мозговой полости
черепа Eryops megacephalus.
На изображенной на рис. 38 реконструкции черепно-мозговых нервов
Benthosuchus дан мозг, очень близкий по своей форме к мозгу Urodela.
Одной из несомненных его особенностей была относительно большая длина
его но сравнению с длиной мозга современных хвостатых амфибий. Это
объясняется тем, что хвостатые амфибии имеют короткую и широкую
голову, тогда как Benthosuchus обладал длинным клинообразным черепом.
А это, несомненно, должно было отразиться на сагиттальных размерах
его мозга. О длине мозга Benthosuchus можно судить по величине расстояния
от сфенэтмоида до exoccipitale (рис. 17, 37).
Положение сфенэтмоида у Benthosuchus выяснено совершенно точно.
Это, в свою очередь, позволяет определить положение переднего отдела
мозга, так как для его lobus olfactorius в заднем участке сфенэтмоида
имеется соответствующее углубление. Задний конец головного мозга (гра-
ница между myelencephalon и medulla spinalis), несомненно, располо-
гался на уровне краев foramen occipitale magnum.
Ширина мозга Benthosuchus не должна быть большой. На это указы-
вают небольшие поперечные размеры fossa olfactoria sphenethmoidei и
близко пододвинутые друг к другу эпиптеригоиды (рис. 17). Это опре-
деляет небольшую ширину telencephalon и отчасти diencephalon. Что же
касается mesencephalon, metencephalon и myelencephalon, то эти отделы
мозга у хвостатых амфибий обычно не превосходят ширины telencephalon.
Такие же отношения можно предполагать и у Benthosuchus.
53
Выход из черепа некоторых из 10 пар черепно-мозговых нервов у Ben-
thosuchus может быть определен достаточно точно.
I. N. olfactorius. Для первой пары черепно-мозговых нервов
в сфенэтмоиде Benthosuchus имеются два довольно широких, параллельных
друг другу канала. Это совершенно точно определяет положение n. olfacto-
rius. В том месте, где костный сфенэтмоид кончался, каналы, несомненно,
переходили в хрящевое его продолжение, и нервы, залегавшие в них,
достигали слизистой оболочки обонятельных капсул, где и разветвлялись
Рис. 38. Реконструкция черепно-мозговых нервов. Вид
снизу. Restoration of cranial nerves. Ventral view.
Ch. ty — chorda tympani, r. p. Vll — ramus pala-
tinus nervi facialis.
(рис. 37).
Сэве-Зедерберг [1936]
находит, что Lyrocephalus
и Aphaneramma имели силь-
но вытянутые и помещав-
шиеся в сфенэтмоиде lobi
olfactorii. У Benthosuchus
каналы в сфенэтмоиде не
настолько широки, чтобы
можно было говорить о
присутствии в них указан-
ных долей мозга.
II. N. opticus. Fo-
ramen opticum, хорошо вид-
ное в боковой стенке зад-
него отдела сфенэтмоида
(рис. 20), позволяет совер-
шенно точно определить
место выхода зрительного
нерва из черепа. Размеры
п положение орбит в че-
репной крыше дают воз-
можность судить о разме-
рах глазных яблок и об их
удалении от foramen opti-
. cum. Все это, в свою оче-
редь, позволяет выяснить
направление и длину зри-
тельных нервов (рис. 37).
Ill, IV. N п. о с u lo-
rn о t о г i u s е t troch-
lea г i s выходили из моз-
говой полости черепа не-
сколько позади окостенев-
шего сфенэтмоида, так как
отверстий для этих нервов
в костном сфенэтмоиде об-
наружить не удалось.
V. N. trigeminus,
выходивший, вероятно, двумя корешками из мозга, имел, повидимому, до-
статочно хорошо выраженный ganglion Gasseri, располагавшийся в хряще-
вой стенке мозговой коробки непосредственно позади эпиптеригоида.
Vv Первая ветвь тройничного нерва (ramus ophthalmicus, V\, рис. 37)
выходила из особого отверстия в хрящевой стенке мозговой коробки
медиальнее основания эпиптеригоида; затем опа по прямой линии направля-
лась вперед, располагаясь неподалеку от сфенэтмоида, и достигала обоня-
тельных капсул. Здесь сквозь хрящевую стенку она проникала в обоня-
тельную капсулу и ложилась поверх разветвлений обонятельного нерва;
Конец первой ветви в медиально;переднем углу обонятельной капсулы
вновь пронизывал хрящ и, уклоняясь к срединной плоскости черепа, вы-
54
ходил через foramen interpraemaxillare (рис. 17, 37) на кожу переднего
отдела морды.
V2 . Вторая ветвь тройничного нерва (ramus maxillaris V2, рис. 37) вы-
ходила из черепа позади эпиптеригоида и направлялась на дорзальную
поверхность палатоквадратпого хряща.
V3. Третья ветвь (ramus mandibularis V3, рис. 38) выходила из мозго-
вой полости вместе со второй ветвью. В дальнейшем своем ходе она откло-
нялась несколько назад и иннервировала короткими ветвями группу мыпщ,
сжимающих челюсти. Затем она вместе с этими мышцами выходила через
fenestra temporalis и проникала в fossa mandibulae. В нижней челюсти
третья ветвь, по всей вероятности, как и у современных Urodela, распо-
лагалась вдоль cartilago Meckeli, причем часть ее ветвей выходила сквозь
foramen Meckelianum ко дну рта.
VI. N. abducens. В черепе Benthosuchus специального отверстия
для этого нерва пе найдено. Он, вероятно, выходил из мозговой полости
позади окостеневшего сфенэтмоида в виде самостоятельного нерва или
вместе с первой ветвью тройничного нерва.
VII. N. facialis. Судя по ходу n. facialis у рецептных Urodela,
этот нерв у Benthosuchus, после выхода из мозга, располагался в хряще
мозговой коробки вентральнее тройничного нерва. Здесь, не выходя из
хряща, он делился на две ветви.
Первая из этих ветвей (ramus palatinus nervi facialis, рис. 38) распо-
лагалась вдоль processus cultriformis и своим передним концом входила
в специальное отверстие на вентральной поверхности vomer.
Вторая ветвь n. facialis направлялась к латеральному углу черепа.
Здесь она делилась на несколько ветвей. Одна из этих ветвей (ramus mus-
cularis, рис. 38) иннервировала m. depressor mandibulae; вторая — пред-
ставляла собой тонкую chorda tympani, которая входила на медиальной
поверхности нижней челюсти в предназначенное для нее особое отверстие
(рис. 26); третья, по всей вероятности, сопровождая ramus jugalis и arteria
mandibularis lateralis, проходила сквозь большое foramen paraquadratum
и маленькое foramen paraquadratum accessorium.
Кроме того, вторая ветвь n. facialis, несомненно, имела ramus anasto-
moticus, соединяющий ее с n. glossopharyngeus.
VIII. N. acusticus. Так как боковые стенки мозговой полости
у Benthosuchus не окостенели, то ход слухового нерва может быть опре-
делен только по положению слухового аппарата, который, в свою очередь,
может быть локализован в черепе по положению базального конца colu-
mella. Перепончатый лабиринт у Benthosuchus был со всех сторон окружен
хрящевыми образованиями, поэтому нам не удалось обнаружить его от-
печатков. На реконструкции (рис. 37) он изображен по аналогии с лаби-
ринтом амфибий вообще.
IX, X. N n. glossopharyngeus et vagus. Оба эти нерва
у Benthosuchus, вероятно, имели общий ганглий и выходили через большое
отверстие в exoccipitale общим стволом, но тотчас же разделялись на не-
сколько ветвей (рис. 37, 38).
XII. N. hypoglossus. Сэве-Зедерберг [1936] в своих рекон-
струкциях мозга и нервов Lyrocephalus и Aphaneramma изображает n. hypo-
glossus и считает, что он проходил сквозь особое отверстие в exoccipitale.
Небольшие отверстия позади отверстия так называемой vagus-группы,
как указывалось выше, имеются и в exoccipitale Benthosuchus. Однако
сквозь них, по всей вероятности, проходили небольшие сосуды (vasa nut-
ritia), а не нервы [Демпстер, 1935].
N. hypoglossus, иннервировавший мышцы подъязычного аппарата,
судя по размерам черепа Benthosuchus, должен был быть достаточно мощным
первом. Поэтому те очень небольшие отверстия, которые имеются в не-
постоянном числе (одно-два-три) па каждой exoccipitale, не могли служить
местом выхода XII черепно-мозгового нерва.
55
N. hypoglossus, по нашему мнению, выходил из мозга, располагаясь
между exoccipitale и неуральной дугой первого позвонка.
Позвоночный столб
Дозвоночный столб Benthosuchus sushkini не сохранился в таком виде,
чтобы можно было составить себе точное представление о его длине и ко-
личестве позвонков. Однако следует думать, что позвоночный столб Bentho-
suchus не был коротким.
В коллекции Палеонтологического института имеется достаточное
количество отдельных элементов позвонков. Эти элементы дают полную
возможность выяснить строение позвоночника.
Рис. 39. Позвонки Benthosuchus, Vertebrae of Benthosuchus.
A. В. G—Arcus neurales, D. E. F.—Hypocentrum, с. c—
capitulum costae, f. c — facies costalis, f. ch — facies
chordalis, f. n.— foramen neurale, he—hypocentruni, pt —
processuj transversus, p. s. — processus spinosus, t. c —
tuberculum costae, z. a — zygapophysis anterior, z. p —
zygapophysis posterior.
Неуральные дуги (Arcus neurales, basidorsale) каждого позвонка пред-
ставляют собой две изогнутые пластинки, сросшиеся друг с другом своими
верхними концами (рис. 39А, В, С). Непосредственно под местом их соеди-
нения расположено небольшое овальной формы, не замкнутое снизу,
отверстие, в котором помещался спинной мозг. На дорзальной части неу-
ральной дуги располагается довольно мощный и относительно короткий ос-
тистый отросток, на котором видны небольшие гребни, служащие местом на-
чала и прикрепления мышц и связочного аппарата позвоночного столба. Над
верхним краем foramen neurale от основания processus spinosus вперед
и назад отходят по две пары суставных отростков. Передние отростки
развиты несколько сильнее, чем задние (рис. 39С). Их суставные поверх-
ности обращены своими плоскостями вверх и немного медиально. Сустав-
56
ные поверхности задних отростков направлены в противоположную сто-
рону — вниз и латерально.
Вниз и в стороны неуральная дуга постепенно переходит в хорошо
развитые поперечные отростки. Эти отростки заканчиваются шерохо-
ватыми площадками, которые были покрыты хрящом и принимали участие
в образовании одного из суставов, соединяющих отдельные ребра с позво-
ночником.
Внутренние стороны неуральной дуги ниже отверстия для спинного
мозга так же шероховаты. Этими своими участками дуга накладывалась
на очень мощно развитую chorda dorsalis, проходившую вдоль всего позво-
ночного столба (facies chordalis, рис. 39А, В).
К нижней стороне хорды соответственно каждой неуральной дуге при-
легал базивентральный элемент hypocentrum.
Hypocentrum у Benthosuchus имеет форму полулунной костной
пластинки (рис. 39). Дорзальная, вогнутая сторона его шероховата и слегка
выпукла в передне-заднем направлении. Вентральная поверхность hypo-
centrum, напротив, в том же передне-заднем направлении вогнута (рис. 39F)
и выпукла — справа налево. На этой поверхности удается различить две
небольшие неглубокие ямки и слабо выраженные сагиттальные гребни,
повидимому, — следы прикреплявшихся здесь связок.
При рассматривании hypocentrum сбоку (рис. 39F), оно кажется костью
почти треугольной формы. В таком положении хорошо видна вогнутость
его нижней поверхности и небольшая площадка несколько позади верх-
него угла кости. Эта площадка (facies costalis, рис. 39Е, F), так же, как
и площадка на конце поперечного отростка дуги, служила местом соеди-
нения ребра с позвоночником.
Строение описанных двух элементов позвонка, несомненно, говорит
о том, что Benthosuchus принадлежит к рахитомным стегоцефалам. Однако
в каждом его позвонке недостает одного, постоянного у всех рахитомов,
костного элемента — pleurocentrum.
Среди многочисленных разрозненных остатков скелетов Benthosuchus
было собрано достаточное количество неуральных дуг и hypocentrum,
дающих представление не только о строении, но и о возрастных особен-
ностях этих элементов позвонка. Однако при этом не было обнаружено
ни одного pleurocentrum. Отсутствие pleurocentrum в коллекции дает
право предположить, что они у Benthosuchus даже и в тех случаях, когда
длина его черепа достигала 700—750 мм, оставались хрящевыми.
Ребра
Большое количество отдельных ребер, найденных вместе с другими
костями Benthosuchus, позволяет достаточно ясно представить себе их
строение.
Так называемые «шейные» ребра, соединенные с передним отделом
позвоночного столба (рис. 39, 40, А,В), у Benthosuchus являются сравнительно
небольшими косточками. Их проксимальный отдел разделяется глубокой
вырезкой на два довольно длинных отростка. Один из этих отростков,
являющийся головкой ребра, на своем конце несколько расширен (рис. 40А).
Этим отростком ребро соединяется с соответствующей площадкой на hypo-
centrum. Другой отросток ребра (tuberculum) такого расширения обычно
не имеет (рис. 40А, В).
Передняя поверхность первых ребер немного выпукла. Эта выпуклая
сторона кости обращена краниально, что дает возможность легко отличить
правое ребро от левого, если принять во внимание некоторые различия
в строении головки и бугорка ребра.
На немного вогнутой задне-нижней поверхности ребра можно разли-
чить слабо выраженную борозду (sulcus costae, рис. 40А), в которой рас-
полагался межреберный нерв.
57
У ребер, расположенных в том же начальном отделе скелета, но не-
сколько более каудально, головка становится более мощной, а бугорок
более узким и длинным (рис. 40В). Кроме того, у этих ребер вдоль борозды
(sulcus costae) на задней поверхности появляется хорошо выраженный
гребень (crista costae, рис. 40В, Ъ), что делает sulcus costae более глубоким.
Появление crista costae и яснее выраженного sulcus costae, несомненно,
свидетельствует о том, что вдоль этих ребер проходили более мощные
нервы, предназначавшиеся для иннервации мышц плечевого пояса и перед-
них конечностей.
Дистальный конец первых ребер
обычно узок, и ребро в целом имеет
клиновидную форму.
Ребра грудного отдела скелета
значительно длиннее «шейных»
(рис. 400). Их бугорок и головка
Рис. 40. Ребра Benthosuchus. Ribs of Bentho-
suchus. A — Costa cervicalis sinistra,
В — costa cervicalis dextra. C, D.—Costae
thoracales sinistrae, E — Costa thoracalis
dextra, c — capitulum, cc.—crista costae,
s. c — sulcus costal, t — tuberculum, p. u—
processus uncinatus (a — вид спереди,
b — вид сзади), (a — anterior view, b —
posterior view).
неразличимы, и вырезка на рас-
ширенном проксимальном конце
отсутствует. Кранио-дорзальная
поверхность этих ребер выпукла.
На противоположной, вогнутой,
стороне ребра обычно можно
Рис. 41. Крестцовое ребро Benthosuchus.
A — Вид спереди. В — Вид сзади.
Sacral rib of Benthosuchus. A—Anterior
view, В — Posterior view. с. c — capitu-
lum costae, c. m — crista muscularis,
s. c.—sulcus costal, t. c.—tuberculum
costae.
видеть достаточно хорошо выраженные sulcus и crista costae. На неко-
торых грудных ребрах имеется processus uncinatus (рис. 41D). У крупных
экземпляров Benthosuchus он достигает довольно больших размеров,
напоминая такие же образования на ребрах Eryops (рис. 40Е) [Olson,
1936].
Дистальные концы грудных ребер постепенно расширяются; они соеди-
нялись, повидимому, с широкими и хорошо развитыми хрящами.
58
Поясничные ребра по строению своего проксимального конца очепь
напоминают грудные. Здесь так же не резко разграничиваются tuberculum
и capitulum costae Грис. 40F). Дистальный конец поясничного ребра более
похож на копец шейного ребра: он постепенно суживается, превращаясь
в длинный, почти круглый в поперечном сечении отросток. Sulcus costae
и на этих ребрах достаточно хорошо выражен.
Наиболее мощно развитым из всех ребер Benthosuchus является его
сакральное ребро. Проксимальный конец этого ребра сильно расширен
(рис. 41А). Вырезки,
разграничивающей буго-
рок и головку, здесь
пет. Однако, при рас-
сматривании ребра со
стороны позвоночного
столба, можно хорошо
различить обе эти части
(рис. 41G). Головка реб-
ра, соединяющаяся с hy-
pocentrum крестцового
позвонка, значительно
мощнее, чем бугорок, со-
членяющийся с попереч-
ным отросткомтогожепо-
звонка. Между головкой
и бугорком имеется уз-
кая костная перемычка.
Вдоль передней по-
верхности сакрального
ребра проходит высокий
шерох оватый гребень
(crista muscularis, рис.
41А). На задней поверх-
ности ребра видна не
резко ограниченная ши-
рокая борозда, служив-
шая местом залегания
мощного сакрального
нерва, иннервировавше-
го мускулатуру задней
конечности (рис. 41В).
Дистальный конец са-
крального ребра, менее
расширенный по срав-
нению с проксимальным
Рис. 42. Interclavicula. Вентральная поверхность
кости.Ventral surface of bone. f. cl — facies clavicularis.
(рис.41D),служил местом
соединения ребра cilium.
Плечевой пояс
Interclavicula. По своим общим очертаниям interclavicula
Benthosuchus очень напоминает ромб (рис. 43, 44). Ее передний угол вы-
тянут в краниальном направлении значительно сильнее, чем остальные
три.
На вентральной поверхности кости видно возвышение ромбовидной
формы, очертания которого повторяют в меньшем масштабе очертания
interclavicula в целом. Задне-боковые стороны этого возвышения без рез-
ких границ, в виде скатов, переходят в краевые участки interclavicula.
Что же касается передне-боковых сторон возвышения, то они отграничены
59
от краевых участков ясно очерченным уступом. Это объясняется тем, что
краевые участки interclavicula—от ее переднего угла до боковых—прикры-
ваются ключицами (facies clavicularis, рис. 42, 43, 48).
Центральный участок ромбовидного возвышения, по месту своего по-
ложения соответствующий центру окостенения interclavicula, покрыт ха-
рактерной для стегоцефалов ячеистой скульптурой. На дне каждой ячейки
видны маленькие отверстия, служившие местом выхода топких сосудов,
питающих кожу груди. От района, занятого ячейками, до краев ромбо-
видного возвышения по радиусам во все стороны расходятся длинные
желобки. По ходу этих желобков всегда можно найти отверстия таких же
узких сосудов, как и сосуды на дне круглых ячеек. На больших intercla-.
vicula старых экземпляров Bentho-
• 5 suchus круглоячеистая скульптура
обычно покрывает всю площадь
р омб овид ног о возвышения.
На поверхности краевых уча-
стков, за пределами ромбовидного
возвышения, всякие следы скульп-
туры отсутствуют; здесь обычно
имеются только глубокие борозды,
являющиеся следами стносительпо
крупных сосудов.
Вентральная поверхность перед-
не-боковых краевых участков (facies
clavicularis) покрыта радиально на-
правленными острыми гребнями.
Грубость рельефа кости здесь, по-
видимому, объясняется тем, что эти
участки были крепко связаны при
помощи плотной соединительной
ткани с покрывающими их краями
ключиц (рис. 43, 48).
Дорзальная поверхность inter-
clavicula гладкая; на ней видно
только небольшое количество от-
верстий, от которых радиально от-
ходят длинные желобки, постепен-
но сглаживающиеся к периферии
кости (рис. 44).
Соответственно центру окосте-
нения кости здесь располагается
небольшое возвышение (eminentia
centralis, рис. 44), от которого по
радиусам к краям interclavicula
расходятся три прямые перекла-
дины. Каждая из этих перекладин, сравнительно узкая в начале (около
eminentia centralis), становится все более широкой к периферии. Перекла-
дины эти, повидимому, имеют чисто механическое значение. Г1о крайней
мере, две из них, именно — trabeculae claviculares (рис. 44), — своими
концами направлены точно к тем краям, на которые с вентральной сто-
роны накладываются ключицы, и как бы укрепляют кость по двум ее ра-
диусам. Третья перекладина (trabecula sternalis) направляется назад и
накладывается на вентральную поверхность хрящевого sternum.
Clavicula. Общие очертания ключицы по форме близки к тре-
угольнику; при рассматривании ее с вентральной стороны она очень на-
поминает раскрытое птичье крыло (рис. 45А).
Вентральная поверхность кости покрыта хорошо выраженной скульпту-
рой. В районе центра окостенения, который помещается в латеральном
f.cl.
f.cl.
Рис. 43. Вентральная поверхность intercla-
vicula Benthosuchus. Ventral surface of
interclavicula of Benthosuchus. f. cl —facies
clavicularis.
60
углу кости (angulus lateralis, рис. 45), эта скульптура образована круглыми
ячейками. К периферии ячейки превращаются в длинные желобки, и скульп-
тура приобретает характер комплекса длинных радиально расположенных
гребней. Дорзальная поверхность claviculae гладкая. На ней имеются
такие же немногочисленные отверстия с отходящими от них узкими боро-
здами, как и на соответствующей поверхности interclaviculae.
Почти прямой медиальный край ключицы, расположенный между
передним и задним ее углами, являет
ся самым топким участком кости.
Рис. 45. Правая ключица Benthosuchus.
A — Вентральная поверхность кости.
В — Вид снаружи. Right clavicula ot
Benthosuchus A. — Ventral surface of
bone, В — Outside view. pps. — pro-
cessus praescapularis.
Рис. 44. Дорзальная поверхность intercla-
vicula Benthosuchus. Dorsal surface of in-
terclavicula of Benthosuchus. e. c. — emi-
nentia centralis, t. cl.—trabecula clavi-
cularis, t. st.—trabecula sternalis.
Задний край кости, напротив, значительно утолщен, в особенности
в латеральном своем участке.
Закругленный латеральный край ключицы на всем своем протяжении
завернут под прямым углом в дорзальном направлении таким образом,
что на дорзальной поверхности кости вдоль этого края образуется жолоб.
От латерального угла ключицы, т. е. от того участка кости, в котором
располагается центр ее окостенения, в дорзальном направлении отходит
мощный плоский отросток (processus praescapularis, рис. 45В).
В первой своей половине этот отросток поднимается вертикально, а за-
тем поворачивает назад, так что вторая его половина оказывается поста-
вленной по отношению к первой приблизительно под углом в 140°. Вер-
шина отростка обычно заострена. Внутренняя его сторона накладывается
на нижний конец cleithrum.
61
Рис. 46. Левое < kiihrum. А, С — Вид снаружи,
В, D—Вид изнутри. Left cleithrum. А, С —
Outer side, В, D — Inside view. с. с—crista
cleithralis, 1. ss — lamina suprascapularis.
Рис. 47. Левое scapula Benthosuchus. A — Вид
снаружи. В — Вид изнутри. С—Вид сзади.
D — Вид спереди. Scapula sinistra Benthosuchus.
А — Outer view, В — Inner view, C — back
view, D — front view. c. m — crista muscu-
laris, Co. — coracoideum, s. sg —canalis supra-
glenoidalis, s. c — sulcus canalis coracoideus,
s. g — sulcus glenoidalis canalis glenoidalis.
Cleithrum представ-
ляет собой небольшую удли-
ненную кость, верхний конец
которой значительно расши-
рен. Это расширение сильнее
выражено у старых особей
(рис. 46) и слабее у молодых
(рис. 46А).
Наружная поверхность
cleithrum гладкая. Она со-
вершенно лишена характерной
для стегоцефалов скульптуры,
которая обычна для покров-
ных костей, лежащих непо-
средственно под кожей. Ячеи-
стая скульптура на cleithrum,
повидимому, встречается у
стегоцефалов редко, так как
до сих пор ее удалось наблю-
дать только у двух лабирин-
тодонтов — Diceratosaurus и
Archegosaurus [Некель (Jae-
kel), 1903, 1915].
На переднем краю расши-
ренного конца cleithrum у
крупных экземпляров Bentho-
suchus помещается небольшой
гребень (crista cleithralis,
рис. 46С). Широкая верхняя
часть cleithrum представляет
собой по существу постепенно
утончающуюся пластинку, ко-
торая накладывается на на-
ружную поверхность верхнего
хрящевого участка лопатки
(lamina suprascapularis, рис.
46С, D, 49).
Описанными пятью костя-
ми (interclavicula, две/clavi-
culae и два cleithrum) исчер-
пываются все накладные ко-
сти плечевого пояса Bentho-
suchus. Все остальные элемен-
ты этого отдела скелета раз-
виваются на почве хряща.
Сюда прежде всего относится
scapula. Эта кость является
одной из самых сложных по
своей форме.
В коллекции костей Bentho-
suchus имеется довольно боль-
шое количество изолирован-
ных лопаток, принадлежащих
самым различным по инди-
видуальному возрасту экзем-
плярам этого лабиринтодонта.
Scapula — массивная и тяжелая кость (рис. 47). Задний ее край
утолщен и довольно сильно вогнут. Передний и верхний участки лопатки.
62
имеющие форму сравнительно тонких костных пластинок^ заканчиваются
неровными краями, к которым присоединялись хрящевые части. При рас-
сматривании лопатки спереди видно, что передний ее край не прямой
а волнообразно изогнутый (рис. 47D).
На внутренней стороне лопатки, приблизительно на средине ее высоты,
ближе к заднему краю кости, располагается мощный отросток, направлен*
ный своим сильно расширенным концом назад и медиально (рис. 47В).
На верхней поверхности отростка, около самого края, видпа небольшая
борозда (sulcus canalis glenoidalis, рис. 47В, D), которая вела к каналу,
проникающему сквозь неокостеневшую часть лопатки. Этот канал откры-
вался отверстием на вентральной поверхности хрящевого коракоида не-
Рис. 48. Реконструкция плечевого пояса Benthosuchus
(уменьшено) Restoration of the shoulder-girdle of Benthosuchus
(diminished), с. c — canalis coracoideus, c. gl— canalis gle-
noidalis, cv. gl — cavitas glenoidalis, Co. — coracoideum,
Cl. — clavicula, — Icl.—interclavicula, Sc. — Scapula.
подалеку от края interclavicula (рис. 48). На нижнем краю того же отро-
стка имеется вторая слабо заметная борозда (sulcus canalis coracoideus,
рис. 47D), которая указывает на местоположение коракоидного канала,
выходившего на вентральную поверхность плечевого пояса, немного впе-
реди отверстия предыдущего канала (рис. 48).
Третий канал лопатки, известный под имепем canalis supraglenoidalis,
представлен у Benthosuchus, в силу недостаточного окостенения ближайших
участков хряща, глубокой вырезкой, отделяющей снизу мощный медиаль-
ный отросток лопатки от ее нижнего угла (рис. 47С, 49).
На относительно гладкой наружной поверхности лопатки имеется
небольшой, косо идущий гребень, вероятно, служивший местом начала
musculus dorsalis scapulae (crista muscularis, рис. 47A).
Coracoideum окостеневал очень незначительно; от него сохра-
нился только очень небольшой участок, приросший к нижнему углу ло-
патки.
63
Рис. 49. Плечевой пояс Benthosuchus. Вид слева (рекон-
струкция) (уменьшено). Shoulder girdle of Benthosuchus. View
from the left side (restoration) (diminished). Cl.—clavicula,
Cth. — cleithrum, cv. gl. — cavitas glenoidalis, I cl. — inter-
clavicula, f. sg. — foramen supraglenoidale, Sc. — scapula.
Граница между коракоидом и лопаткой может быть установлена на
основании следующего признака. На гладкой наружной поверхности
лопатки хорошо видны узкие борозды, радиально расходящиеся от центра
окостенения кости. Такие же узкие борозды на коракоиде расположены
горизонтально (рис. 47А). В результате этого борозды на лопатке и бо-
розды на коракоиде оказываются в месте сращения костей расположеп-
Рис. 50. Левая плечевая кость Benthosuchus. А — Проксимальный
конец кости. В — Дистальный конец кости. С — Вентральная по-
верхность кости. Left humerus of Benthosuchus. A — Promixal end of
the bone. В — Distal end of the bone. C — Ventral surface of the
bone. c. dp. — crista deltopectoralis, c. r. — crista radialis, e. r. —
epicondylus radialis, e. u. — epicondylus ulnaris.
64
пыми перпендикулярно друг другу, что и дает возможность легко уста-
новить границу между обеими костями.
Кости передней конечности в коллекции представлены
довольно большим количеством отдельных экземпляров humerus, ulna
и radius. Метакарпальных костей и фаланг сохранилось очень мало.
Humerus крупных Benthosuchus sushkini отличается сильным раз-
витием ряда отростков, служивших местами начала и прикрепления мышц.
На проксимальном конце кости имеется шероховатая площадка, на ко-
торой помещалась хрящевая головка.
На вентральной поверхности humerus на протяжении почти половины
его общей длины располагается мощный гребень с
(crista deltopectoralis,
рис. 50). Этот гребень
служил местом прикреп-
ления ряда мышц пле-
чевого пояса.
Дистальный конец
сильно расширен и пре-
вращен в расположенную
горизонтально толстую
костную пластинку. В
том месте, где humerus
образует сустав с radius,
край этой пластинки
утолщен; здесь, на ше-
роховатой поверхности
утолщения, помещалась
хрящевая eminentia са-
pitata humeri. На дор-
зальной поверхности
кости к месту утолще-
ния подходит хорошо
выраженный высокий
гребень (crista radialis,
рис. 51А).
Хрящ, принимающий
неровной поверхностью
Рис. 51. Левая плечевая кость Benthosuchus. А — Дор-
зальная поверхность кости. В — Вид снаружи. Left
humerus of Benthosuchus. A — Dorsal surface of the
bone. В — Outside view. c. dp.—crista deltopectoralis,
c. r. — crista radialis, e. r. — epicondylus radialis, e.
u. — epicondylus ulnaris.
участие в образовании
сустава между humerus
и ulna, повидимому,
присоединялся к сере-
дине расширенного Ди-
стального конца кости.
Epicondylus radialis (рис. 50, 51) представлен сравнительно небольшим
отростком с закругленной вершиной. Epicondylus ulnaris, напротив, имеет
форму очень широкой пластинки, так что расширение дистальной части
плечевой кости по существу зависит почти исключительно от необычайной
величины этого отростка.
Локтевой и лучевой костей, соответствующих по своей величине
только что описанному humerus, в коллекции не имеется.
Ulna и radius Benthosuchus меньших размеров представляют
собой кости почти равной длины (рис. 52). Обе они сужены посредине
и немного расширены на концах. Верхний конец ulna имеет овальные очер-
тания, и на стороне, обращенной к radius, на нем видно небольшое углуб-
ление (incisura radialis, рис. 52А, В). Шероховатая поверхность, на ко-
торой помещался хрящевой olecranon, стоит приблизительно под углом
в 50° по отношению к продольной оси кости.
Дистальный, несколько расширенный конец ulna в поперечном сечении
имеет овальную форму; длинная ось этого овала по своему направлению
5 Тр. Палеонтод. ин-та> т. X, в. 1.
65
длинных элементов конечности.
Рис. 52. Ulna dextra (А, В, С, D.). Ra-
dius dexter (Е, F, Gr, Н, I). Metacarpalia
(Metatarsalia?) et phalanges (К—R). I.
r.—incisura radialis ulnae.
совпадает с длинной осью поперечника проксимального конца кости
(рис. 52D).
Поперечное сечение проксимального конца radius почти круглое, если
не считать небольшого выступа, обращенного в сторону ulna (рис. 52F).
Нижний конец radius по своей форме похож на верхний, но на задней его
стороне намечается небольшое углубление. Длинные оси поперечных
сечений обоих концов radius стоят в отношении друг к другу под углом
около 25° (рис. 52Н, I).
Все локтевые и» лучевые кости, имеющиеся в коллекции костей Bentho-
suchus, представляют собой по существу только средние участки более
каждой из этих костей недостает, по-
видимому, довольно больших хряще-
вых участков на концах.
Элементы carpus, по вези ве-
роятности, или совершенно не окосте-
невали, или окостеневали в такой
незначительной степени, что не могли
сохраниться в фоссилизированном со-
стоянии. Во всяком случае, в породе,
содержащей разрозненные части ске-
лета, не было обнаружено пи одной
кости карпального отдела кисти.
Что касается metacarpus,
то в коллекции имеется несколько
этих костей, но все они найдены раз-
розненными, поэтому нельзя с уверен-
ностью сказать, что они принадлежат
индивидуально . одному животному
(рис. 52К). Мало того, так как ко-
нечности Benthosuchus не найдены не-
разрозненными, то мы в настоящее
время не располагаем признаками,
позволяющими отличить элементыте-
tacarpus от metatarsus, и не
можем сказать, какие из этих элемен-
тов имеются в нашем распоряжении.
Судя по материалу, опубликованному
Кейзом [1935], metacarpalia и meta-
tarsalia у Тrimerorhachis настолько
сходны по форме между собою, что
вряд ли можно было бы отличить их
друг от друга, если бы кости конеч-
ностей были разрозненными.
Фаланг, принадлежащих передним и задним конечностям, в кол-
лекции имеется незначительное количество (рис. 52N—Q). Все они были
найдены в различных местах костных скоплений в породе, поэтому нет
возможности точно определить положение каждой из этих костей в скелете.
Концевые фаланги по особенностям строения своих дистальных концов
легко могут быть отличены от других фаланг. В нашем распоряжении
имеется только одна концевая фаланга Benthosuchus. Она полностью при-
росла своим основанием к головке соседней фаланги (рис. 52).
Тазовый пояс
Из костей тазового пояса в коллекции имеется достаточное количество
ilium, позволяющее судить не только о возрастных изменениях, но и об
индивидуальных вариантах в строении этой части.
Ilium является довольно массивной костью со значительно расши-
ренным нижним концом (рис. 53). Она изогнута таким образом, что ее
бб
верхний участок сильно отклоняется назад, так что задний край кости
оказывается сильно вогнутым и образует глубокую вырезку (incisura
ischiadica, рис. 53А). Относительно топкий верхний конец ilium часто
и легко обламывается, но в коллекции костей Benthosuchus имеются хорошо
сохранившиеся экземпляры ilium, на которых видно, что этот конец иногда
разделялся на два (рис. 53С) и даже на три отростка (рис. 53D). При этом
задний из этих отростков обычно оказывался самым большим.
Шероховатая нижняя поверхность нижнего конца кости служила ме-
стом соединения с ischium и pubis.
На наружной поверхности ilium, непосредственно над его нижним
краем, помещается небольшое углубление, являющееся частью acetabulum.
Рис. 53. Левые ilium (А, В, С, D). Правое ischium. (Е, F). А, С, D —Вид снаружи.
В — Вид изнутри. Е — Вид сверху. F — Вид снизу. Left ilium (А, В, С, D). Right
ischium (Е, F). А, С, D — Outside view. В — Inside view, Е — Dorsal surface, F —
Ventral surface, a. c. — acetabulum, c. m. — crista muscularis, i. i. —incisura ischia-
dica, Isch. — Ischium, II. — Ilium, 1. o. — linea obliqua, Pb. — pubis.
Края этого углубления несколько приподняты и отделены от ближай-
ших участков кости кольцеобразной бороздой с гладкими стенками
(рис. 53А).
Над acetabulum, спереди назад и снизу вверх, косо через всю
наружную поверхность кости проходит шероховатая линия, являющаяся
местом начала мышц бедра (linea obliqua, рис. 53А). Выше этой линии
кость покрыта гребнями, которые, повидимому, служили местом начала
мышц туловища.
Внутренняя поверхность ilium гладкая, только в передней ее части
виден острый шероховатый гребень, вызванный начинающимися здесь
мышцами живота (crista muscularis, рис. 53В). Этот гребень с внутренней
стороны ilium, поднимаясь вверх, постепенно переходит на передний край
кости.
5*
67
Ischium — небольшая, клиновидной формы кость. Она лишена
каких бы то ни было гребней и отростков; обе ее поверхности исчерчены
слабо выраженными бороздами, расходящимися по радиусам от центра
окостенения (рис. 53Е. F).
На переднем краю ischium нет никаких следов ямки, которая бы явля-
лась частью acetabulum. Это указывает на неполное окостенение ischium.
Его передний отдел, повидимому, в течение очень продолжительного вре-
мени оставался хрящевым, и эта неокостеневшая часть принимала участие
в образовании задней стенки acetabulum (рис. 53С).
В собрании костей Benthosuchus совершенно отсутствует pubis.
Этот факт позволяет думать, что pubis у Benthosuchus окостеневал очень
поздно.
В том месте, где сходились целиком хрящевое pubis, хрящевая передняя
часть ischium и полностью окостеневший нижний конец ilium, на наруж-
ной поверхности таза располагалось довольно большое acetabulum (рис. 53С).
Небольшая часть acetabulum видна на ilium. Однако acetabulum в целом
следует представлять себе как углубление довольно больших размеров.
Хрящевое pubis и хрящевая часть ischium, несомненно, принимали боль-
шее участие в его образовании, чем ilium.
При препаровке породы, содержащей кости Benthosuchus, было найдено
довольно большое количество ilium самых разнообразных размеров и не-
сколько экземпляров ischium почти одинаковой величины. Что же ка-
сается pubis, то эта кость совершенно не была обнаружена. Такой состав
коллекции костей таза позволяет представить себе ту последователь-
ность, в какой наступало появление окостенения в его хрящевых частях.
Не подлежит сомнению, что все приблизительно одинаковые по своим
размерам ischium вместе с самыми большими экземплярами ilium образо-
вывали тазы взрослых Benthosuchus, у которых, однако, pubis все же оста-
вался еще хрящевым. Мы имеем только ilium более молодых экземпляров
Benthosuchus; соответствующих им по размерам ischium не найдено. Следо-
вательно, тазы этих молодых экземпляров Benthosuchus имели только
одну кость — ilium; ischium и pubis были хрящевыми. Если принять во
внимание еще более ранние стадии индивидуального развития Bentho-
suchus, когда таз полностью состоит из хряща, то можно говорить о трех
основных этапах возрастных изменений таза.
Первым этапом, несомненно, являлось образование полностью хряще-
вого таза.
Второй и довольно рано наступающий этап начинался с появления
окостенения в ilium. В течение, повидимому, довольно продолжительного
времени ilium, при постепенном увеличении степени своего окостенения
и своих общих размеров, оставался единственной костью таза.
В третьем этапе наступало окостенение ischium. В этот момент таз
содержал уже две кости; хрящевым оставалось только pubis.
Четвертого этапа, когда и pubis подвергалось бы окостененению, у Bentho-
suchus мы не можем констатировать. Однако не исключена возможность,
что у очень старых экземпляров полностью окостеневали все три части таза.
В коллекции имеются фрагменты нижних челюстей, принадлежащие экзем-
плярам Benthosuchus, череп которых достигал 700—750 мм в длину. Однако
несомненно, что ни одна из имеющихся в коллекции самых больших ilium
таким крупным стегоцефалам не принадлежала. Следовательно, нужно
признать, что тазовые кости этих индивидуально старых форм нам неизве-
стны. Судя по общим пропорциям скелета Benthosuchus, ilium наиболее
крупных экземпляров должно быть было в два или три раза больше самого
большого из имеющихся в коллекции ilium.
Трудно допустить, чтобы при таких больших размерах таза pubis все
еще оставалось хрящевым. Поэтому, вероятно, не следует исключать
у Benthosuchus того четвертого и последнего этапа развития таза, когда
все три его элемента полностью окостеневали.
158
Судя по материалам, опубликованным в свое время Крелпером [1886],
у Branchiosaurus amblystomus была та же последовательность появления
окостенения в тазу.
Нам кажется интересным полное совпадение порядка появления точек
окостенения в хрящевых закладках таза у Benthosuchus, Branchiosaurus,
современных Urodela и у высших позвоночных, в частности — у млеко-
питающих и человека. У человека в процессе эмбрионального развития
первая точка окостенения в хрящевом тазу принадлежит ilium и появляется
в самом начале третьего месяца. Затем, на четвертом месяце, начинается
окостенение в хрящевой закладке ischium, и, наконец, на пятом месяце
возникает центр окостенения в хрящевом pubis. Такая последователь-
ность появления точек окостенения является, повидимому, общей особен-
ностью развития таза Tetrapoda. Таким образом, порядок возникновения
окостенения в тазу как самых низших (Amphibia), так и самых высших
(Mammalia) представителей Tetrapoda один и тот же, с той только разни-
цей, что у низших (в частности и у stegocephalia) окостенение’ таза совер-
шается медленно в течение всей индивидуальной жизни животного в пост-
эмбриональном периоде, а у млекопитающих оно полностью перенесено
Рис. 54. Правое бедро Benthosuchus. А — Вид сверху. В — Вид спереди. С — Вид
сзади. D — Вид снизу. Right femur of Benthosuchus. A — Superior view. В — An-
terior view. C —Posterior view. D — Interior view. c. a. f. — crista aspera femoris,
f. p. — fossa poplitea, f. t. — fossa tendinalis, t. — trochanter.
в период онтогенетического развития организма (в данном случае мы,
разумеется, не имеем в виду слияние отдельных костей таза при образо-
вании os coxae).
Кости задней конечности Benthosuchus представлены
в коллекции несколькими экземплярами femur, tibia и fibula. Почти все
они принадлежат различным по индивидуальному возрасту животным.
Femur несравненно слабее, чем humerus. Проксимальная часть
femur на своем конце имеет шероховатую поверхность, на которой поме-
талась довольно хорошо развитая головка бедра. На нижней стороне
бедра, на протяжении приблизительно трети его длины, в проксимальной
части располагается довольно большой trochanter. Он имеет неправильные
треугольные очертания (рис. 54D); его шероховатая поверхность, несом-
ненно, была покрыта слоем хряща. Возвышение trochanter непосредственно
переходит в высокий острый гребень, который, становясь постепенно все
ниже и пиже, почти достигает конца кости (crista aspera femoris, рис. 54В,
С, В). Здесь на задней поверхности кости видна небольшая ямка (fossa
poplitea, рис. 54D).
Верхняя поверхность бедра на большем своем протяжении лишена
каких бы то ни было неровностей или гребней, только в дистальном ее
отделе лежит углубление, вдоль которого проходило сухожилие мыпщ,
69
разгибающих голень (fossa tendinalis, рис. 54А). Это углубление не может
быть названо fossa patellaris, так как следует иметь в виду, что сухожилие
разгибателей голени у стегоцефалов, несомненно, не окостеневало, и pa-
tella как костное образование отсутствовала. В хорошо сохранившихся
и более или менее полно окостеневших скелетах стегоцефалов patella до
сих пор ни разу не была найдена и описана. Это заставляет думать, что она
у стегоцефалов как представителей одной из первых ступеней эволюции
Tetrapoda вообще еще не развивалась. Patella отсутствует и у всех совре-
менных амфибий. Она впервые появляется у рептилий и достигает высоких
степеней развития в коленном суставе птиц и млекопитающих.
Дистальный конец femur, так же как и проксимальный, заканчивается
шероховатой площадкой, которая была занята не сохранившимся хрящом
коленного сустава
Tibia из двух костей голени Benthosuchus является наиболее массив-
ной костью. Ее верхний конец значительно шире нижнего и имеет очер-
Рис. 55. Правая большая берцовая (А, В, С, D) и правая малая
берцовая кости Benthosuchus. А — Вид спереди. В — Вид сверху,
С — Вид сзади, D — Вид снизу. Е — Вид сзади, F. j. — Вид сверху,
G — Вид спереди, Н —Вид снизу. Right tibia (А, В, С, D) and right
fibula of Benthosuchus. A — Anterior surface. В —Superior view.
C — Posterior surface, D — Inferior view. E — Posterior surface,
F. j. — Superior view. G — Anterior surface. H — Inferior view. c.
a. — crista anterior, c. p.—crista posterior, p. p. —planum poplite-
um, s. f. — sulcus fibularis, t. t. — tuberositas tibiae.
тания, близкие к полулунию (рис. 55А, В). Средняя часть кости сужена
вследствие сильного вдавления со стороны fibula. В верхнем отделе перед-
ней поверхности tibia располагается хорошо выраженная шероховатость,
служившая местом прикрепления сильного сухожилия мышц, разгибаю-
щих голень (tuberositas tibiae, рис. 55А). От этой шероховатости по пря-
мой линии вниз идет небольшой гребень, почти достигающий нижнего
конца кожи (crista anterior tibiae, рис. 55А).
Задняя поверхность tibia в своей верхней половине образует слегка
вогнутую площадку (planum popliteum, рис. 55С). Она ограничена от
боковой поверхности tibia острым гребнем, проходящим почти па всем
протяжении кости (crista posterior tibiae, рис. 55С). В нижнем своем
участке задняя поверхность tibia слабо выпукла.
70
Дистальный конец tibia на поперечном сечении имеет очертания тре-
угольника с закругленными углами (рис. 55D).
Fibula, равная по длине tibia, сильнее расширена в нижней своей
части, чем в верхней.
Очертания поперечного сечения ее верхнего конца близки к сильно
вытянутому овалу (рис. 55F). Задняя поверхность кости вогнута в верти-
кальном и поперечном направлениях. На ней нет таких гребней и шерохо-
ватостей, которые сразу бросаются в глаза на соответствующей поверх-
ности tibia.
В нижнем отделе задней поверхности кости, неподалеку от медиаль-
ного угла, на всех экземплярах fibula Benthosuchus можно видеть резко очер-
ченный жолоб, проходящий вертикально вниз (sulcus fibularis, рис. 55Е).
Передняя поверхность fibula слабо выпуклая и совершенно гладкая
(рис. 55).
Нижний конец кости на поперечном сечении вытянут значительно
сильнее, чем верхний (рис. 55Н).
Fibula немного скручена по своей продольной оси, так что длинные
оси поперечных сечений обоих ее концов перекрещиваются друг с другом
приблизительно под углом в 37° (рис. 551).
Каких бы то ни было элементов tarsus при препаровке пород,
содержащих разрозненные скелеты Benthosuchus, обнаружить не удалось.
Tarsus, повидимому, в течение большей части жизни Benthosuchus оставался
хрящевым.
Metatarsalis и phalanges, несомненно, окостеневали, но
среди пятнадцати экземпляров костей стопы и кисти, которые имеются
в коллекции, трудно различить кости, относящиеся к задним и передним
конечностям.
П. ОБЩАЯ ЧАСТЬ (GENERAL PART)
ГЛАВА I
ВОЗРАСТНЫЕ ИЗМЕНЕНИЯ И ИНДИВИДУАЛЬНЫЕ ВАРИАЦИИ СКЕЛЕТА
BENTHOSUCHUS
Обильный материал по Benthosuchus, принадлежащий особям самого
различного возраста, дает возможность подойти к оценке возрастной и ин-
дивидуальной изменчивости элементов скелета. Очевидно, что такая оценка
может быть сделана только в самых общих чертах и, охватывая наиболее
заметные изменения, тем самым наметить путь для дальнейшего изучения
вариаций скелета.
Выше отмечалось отсутствие взаимосвязанных элементов скелета в Шар-
женгских местонахождениях, в костеносных песчаниках которых беспо-
рядочно перемешаны самые различные части скелета крупных и мелких
особей. Однако во всей массе остатков Benthosuchus в местонахождениях
р. Шарженги найден всего один фрагмент, принадлежащей другому роду
лабиринтодонтов. Поэтому при большом количестве костей одного вида
В. sushkini, несмотря на отсутствие полных скелетов, возможно прямое
сравнение между собой одноименных скелетных элементов. Различия
в размерах одноименных костей мы принимаем за возрастные изменения
скелета Benthosuchus. Некоторые морфологические особенности равных
по своим размерам одноименных костей мы относим к группе индивидуаль-
ных вариаций.
Имеющийся в нашем распоряжении материал охватывает только рост
Benthosuchus sushkini после метаморфоза. Поэтому анализ изменений ске-
лета Benthosuchus — от личинки до дышащего легкими животного — не
может быть проведен.
Кроме того, в нашем материале далеко пе все части скелета предста-
влены с одинаковой полнотой. Так, возрастные и индивидуальные изме-
нения позвонков, ребер, ischium, tibia, fibula и acropodium не могут быть
прослежены в виду малого числа найденных костей. Ulna и radius, най-
денные в довольно большом числе, не выказывают, повидимому, никаких
возрастных особенностей, кроме прямого увеличения размеров, оставаясь
во всех своих частях вполне пропорциональными.
Рассмотрение изменчивости скелета у Benthosuchus, естественно, должно
быть направлено по двум основным группам изменений — возрастным
и индивидуальным. Несомненно, что в группу индивидуальных изменений
могли быть включены и изменения патологического или дегенеративного
характера, выделить которые в самостоятельную группу с уверенностью
в настоящее время невозможно.
Наиболее важным признаком возрастных изменений, общим для всех
отделов скелета, является величина.
В коллекции Палеонтологического института наименьшим из всех
известных черепов Benthosuchus является череп № 4-19(2252 в 27.5 мм
длины. Так как этот череп принадлежит уже вполне сформировавшемуся
72
. Указанное обстоятельство не дает возмож-
Рис. 56. Череп очень молодого Benthosuchus sushkini
№ 4—19/2252, сверху. Увеличено в 3.2 раза. Bentho-
suchus sushkini. bkull of a very young specimen,
№ 4—19/2252. Dorsal view. x3.2.
и дышавшему легкими животному, то черепа личинок Benthosuchus. не-
посредственно перед метаморфозом или в момент его, должны были быть
еще меньшей величины, по всей вероятности, пе на много превышавшей
10 мм. Обломки крупных черепов и элементы посткраниального скелета,
имеющиеся в коллекции, показывают, что наиболее крупные и старые
особи Benthosuchus обладали черепами, длиной не менее 700—800 мм.
Таким образом, в течение своей жизни уже вполне сформировавшееся
и прошедшее метаморфоз животное увеличивалось в росте примерно
в 70 раз.
Этот факт имеет важное значение, так как, с одной стороны, указы-
вает на длительный и, по всей вероятности, медленный рост животного
в течение всей жизни (т. е. на его значительную долговечность), а с дру-
гой — снижает до нуля значение величины животного как систематиче-
ского признака, если этот признак берется без учета других особенностей.
К сожалению, наиболее крупные черепа Benthosuchus известны лишь
по обломкам, что вполне согласуется с условиями захоронения, так как для
передвижения таких больших черепов * целиком требуется значительно
большая сила потока. Кроме того, несомненно, что наиболее крупные,
доживавшие до глубокой старости особи, были более редки, чем особи
средней величины и возрас
ности детально проанали-
зировать возрастные изме-
нения по наиболее край-
ним выражениям, поэтому
большинство возрастных
особенностей строения че-
репа даны для диапазона
27.5—350 мм, т. е. от наи-
меньшей известной нам ве-
личины до «средней». Ска-
занное относится и к эле-
ментам посткраниального
скелета, хотя некоторые
из последних известны
полностью или почти пол-
ностью для очень крупных
особей.
И зм енение величины
животного сопровождалось,
кроме того, изменениями
некоторых морфологиче-
ских деталей строения ске-
лета. Эти изменения луч-
ше всего прослеживаются
по наиболее сложному от-
делу скелета — черепу.
Возрастные изменения
черепа Benthosuchus
Сравнение самого ма-
ленького из имеющихся в
коллекции черепов Ben-
thosuchus — № 4—19/2252
изображениями среднего типа черепа Benthosuchus в первой части этой
работы (рис. 14, 22 и 30), так же как и с другими черепами, изображен-
ными в дальнейшем, выявляет значительные отличия как в общей конфи-
гурации черепа, так и в некоторых деталях строения.
(рис. 56, 57, 58 и 59) с соответственными
73
Череп № 4—19/2252 имеет в длину 27.5 мм. Очертания его треуголь-
ные. Небольшие, овальной формы ноздри приближены к переднему краю
морды и разделены широким промежутком. Очень большие, почти круглые
орбиты -сближены между собой и располагаются почти посредине длины
черепа. Довольно большое for. parietale лежит на линии задних краев
орбит. Слуховые вырезки широкие и неглубокие. В направлении от морды
к затылку верхняя поверхность черепа значительно повышается, так как
череп в затылке сравнительно высок.
Кости дорзальной поверхности черепа (рис. 56 и 57) характеризуются
укороченными расширенными очертаниями не только в затылочном, но
и в преорбитальном отделе, т. е. почти равным развитием пре- и пост-
орбитальных элементов. Особенно нужно отметить очень короткие и ши-
рокие praemaxillaria, очень широкие nasalia, суживающиеся назад ко-
роткие треугольные praefrontalia и короткие tabularia с сильно притуплен-
ными задними концами.
Между передними концами parietalia и задними концами frontalia
по срединной линии располагается непарный элемент типа centroparietale
Рис. 57. Тот же череп № 4-19/2252 сбоку. Увеличение 3.2 раза. Same speci-
men № 4-19/2252 from side, x 3.2
в виде небольшой узкой косточки. Передний конец этой кости заострен,
на заднем, принимающем участие в ограничении переднего края foramen
parietale, имеется слабая вырезка.
Хорошо развитый супраорбитальный участок системы каналов боковой
линии на покровных костях этого короткого черепа имеет более крутые
изгибы, задний конец канала, охватывая орбиту сзади, сильно придвинут
к ее краю.
Вентральная поверхность черепа № 4—19/2252 (рис. 58) характеризуется
прежде всего своим сильно расширенным basis cranii, вследствие чего
corpus parasphenoidei имеет форму параллелограмма, длинная ось кото-
рого ориентирована поперечно. Processus cultriformis парасфеноида ко-
роткий и довольно широкий и соединяется с также расширенными задними
концами сошников. Хоаны имеют форму укороченных овалов, широко
раздвинуты, так как vomera довольно широки.
Переднее нёбное отверстие (fenestra praemaxillaris) в форме удлинен-
ного ориентированного поперечно овала и по величине несколько пре-
восходит хоану. Специальные отростки praemaxillaria и vomera (processus
fenestrales anteriores и posteriores) почти не развиты, и fenestra praema-
xillaris имеет посредине лишь слабо заметный пережим. Ramus anterior
pterygoidei более укорочена, чем у старых форм, и ее длина в общем равна
длине ramus posterior. Затылочные мыщелки развиты слабее, чем у более
взрослых Benthosuchus. Челюстные мыщелки отнесены назад от линии
затылочной артикуляции.
Затылок черепа № 4—19/2252 (рис. 59), как отмечено, сравнительно
высокий. Затылочный край крыши черепа слегка выпуклый кверху, cornua
74
tabularia нагнуты вниз. Латеральные фланги затылка слегка опущепы
вниз, и его нижний край образует слабо выпуклую дорзально дугу. Fora-
men supraoccipitale — большое и очень высокое, foramen basioccipitale —
широкое и низкое. Processus submedullaris и processus lamellosus — в за-
чаточном состоянии. Processus paroticus exoccipitalis ориентирован более
вертикально, чем у взрослых особей. Foramen nervi vagi сравнительно боль-
шое. Задний конец ramus anterior pterygoidei низкий, crista obliqua не раз-
вита. Lamina parapterygoi-
dea squamosum очень вы-
сокая и значительно вог-
нутая внутрь, вследствие
чего полость среднего уха
широко открыта сзади. Сле-
дов stapes не сохранилось.
Нижние закраины задних
концов quadratojugalia не
развиты.
Кроме перечисленных
особенностей строения, че-
реп № 4—19/2252 во всех
других деталях повторяет
общее строение черепов
Benthosuchus, изложенное в
первой части настоящей
работы.
Таким образом, наибо-
лее характерные особенно-
сти строения черепа Ben-
thosuchus в молодом возра-
сте (по черепу «N® 4—
19/2252) — следующие:
1) сравнительно корот-
кий треугольный череп с
округлой широкой мордой
и прямыми латеральными
Рис. 58. Череп Benthcsuchus sushkini, № 4—19/2252
снизу.Увеличено в 3.2 раза. Skull № 4—19/2252, vent-
ral view. X 3.2.
краями;
2) очень большие орби-
ты, расположенные почти
посредине черепа;
3) широко раздвинутые
внешние и внутренние но-
здри; первые лежат на
конце морды;
4) положение foramen
pari etale на линии задних
краев орбит;
5) почти равное разви-
тие пре- и посторбиталь-
пых костей крыши черепа:
и те и другие отличаются
значительной шириной;
Рис. 59. Череп Benthosuchus sushkini, № 4—19/2252,
затылочная поверхность. Увеличено в 3.2 раза. Skull
№ 4—19/2252. Back view, х 3.2.
6) высокий в затылочной области череп с выпуклым сводом крыши
и сравнительно плоским нижним краем;
7) слабый прогиб межорбитального участка крыши черепа;
8) сильный изгиб sulcus supraorbitalis;
9) неглубокие и широкие слуховые вырезки с короткими cornua tabu-
laria;
10) очень широкое, поперечно вытянутое corpus parasphenoidei;
75
11) короткий и широкий processus cultriformis parasphenoidei;
12) processus parasphenoidalis vomeris широкий;
13) ramus anterior птеригоида сравнительно укорочен и равен по длине
ramus posterior;
14) fenestra praemaxillaris непарная, co следами зачатков перегородки;
15) foramen nervi vagi сравнительно большое;
16) fenestra supraoccipitalis большая и очень высокая;
17) fenestra basioccipitalis низкая и широкая;
18) processus submedullaris и processus lamellosus (у exoccipitale) не-
развиты;
19) lamina parapterygoidea squamosi очень высокая сравнительно с низ-
ким ramus posterior pterygoidci; между краями обеих костей имеется щель;
Рис. 60. А.—Левый птеригоид очень крупного черепа Wetlugosaurus angusti^ons,
№ 29—19/2252 снизу. 3/4 нат. вел. Pterygoideum sinistrum of an old specimen of Wet-
lugosaurus, ventral view. x3/4. В — Часть squamosum sin. и quadratojugale sin. очень
крупного черепа Benthosuchus sushkini, № 23—32/2354. 3/4 нат. велич. Postero-
lateral part of a big skull Benthosuchus sushkini. X3/4.
20) crista obliqua pterygoidei неразвита;
21) костный бугорок на задней поверхности quadratum отсутствует.
Если сравнить перечисленные признаки с соответственными особен-
ностями строения черепов взрослых особей Benthosuchus, то мы будем иметь
почти во всех случаях обратное их выражение.
На основе имеющихся фрагментов очень больших черепов Benthosuchus
на рис. 60 и 61 приведены натуральные размеры некоторых элементов
черепов очень крупных особей (№ 5—9/2243 и 29—19/2252). В соответствии
с закономерностями зон роста Benthosuchus, установленными А. П. Быстро-
вым [1935], удалось дать реконструкцию дорзальной поверхности огромного
черепа Benthosuchus приблизительно для длины в 700—750 мм (рис. 62).
Для точной реконструкции вентральной и затылочной поверхностей таких
огромных черепов, настолько детальной, чтобы иметь возможность восста-
новить возрастные особенности, не имеется нужного материала. Поэтому
сравнение отмеченных выше признаков в области вентральной и затылоч-
ной поверхности черепа мы вынуждены провести по полным черепам «сред-
него» возраста, которые имеются в нашем распоряжении. Не подлежит
76
Рис. 61. Benthosuchus sushkini. Правая половина
посторбитальной части крыши черепа очень крупной
величины №5—9/2243. 4/5 натуральной велич. Right
half of the postorbital part of the skull roof of a very
big specimen. x4/6.
supraorbitalis;
сомнению, что в крупных черепах указанные ниже признаки строения
вентральной и затылочной поверхностей должны иметь ещё более резкое
выражение. Доступные нашему наблюдению на крупных фрагментах воз-
растные особенности этих отделов черепа будут рассмотрены особо.
Итак, старые особи Benthosuchus («средние» для endocranium) будут
отличаться следующими особенностями (см. рис. 62, 22 и 30).
1) удлиненный треугольный череп с суженной мордой, сильно рас-
ширенный в затылочной области, вследствие чего наружные края черепа
не прямые, а вогнутые;
2) маленькие орбиты,
расположенные далеко на-
зади;
3) сближенные внешние
и сближенные внутренние
ноздри; первые отодвинуты
назад от переднего конца
морды и имеют удлиненно-
овальную форму;
4) foramen parietale рас-
полагается далеко назад от
задних краев орбит;
5) сильное развитие пре-
орб италь ных эл ем ент ов
крыши черепа и слабое раз-
витие посторбитальных;
первые сильно вытянуты,
узкие, вторые в своих очер-
таниях приближаются к
квадрату;
6) в затылочной обла-
сти череп низкий с про-
гнутым сводом крыши че-
репа и дугообразно вогну-
той нижней поверхностью
вследствие опускания вниз
боковых участков;
7) сильный прогиб меж-
орбитального участка кры-
ши черепа;
8) слабый изгиб sulcu
9) глубокие и довольно узкие слуховые вырезки с удлиненными cornua
tabularia;
10) corpus parasphenoidei имеет очертания продольно вытяну-
того параллелограмма;
11) длинный и очень узкий processus cultriformis;
12) узкий processus parasphenoidalis vomeris;
13) ramus anterior птеригоида сравнительно удлинен и в среднем
в 1^2 раза длиннее ramus posterior;
14) fenestra praemaxillaris разделена неполной перегородкой, у старых
особей, несомненно, окостеневает, превращаясь в парное отверстие;
15) foramen nervi vagi относительно небольшое;
16) fenestra supraoccipitalis широкая и низкая, значительно умень-
шенная направленной вниз выпуклостью срединного шва dermosupraocci-
pitalis;
17) fenestra basi occipitalis маленькая и узкая;
18) processus submedullaris и proc, lamellosus сильно развиты;
19) lamina parapterygoidea низкая и несколько меньше по высоте,
чем ramus posterior птеригоида; щель между обеими костями исчезает;
77
20) crista obliqua pterygoidei хорошо развита;
21) па quadratum имеется сильный мышечный бугор.
Имеющаяся в нашем распоряжении серия полных черепов хорошо иллю-
стрирует постепенное усиление отдельных признаков и постепенный переход
от небольших особей к более крупным. Несмотря на отсутствие проме-
жуточных черепов между маленьким черепом № 4—19)2252 и наименьшим
из остальных черепов — типом рода № 1—9/2243, постепенное усиление
возрастных признаков старых особей прослеживается с достаточной яс-
ностью (рис. 63, 64, 65, 66, 67, 68, 69 и 70). Интересной возрастной особен-
Рис. 62. Benthosuchus sushkini. Реконструкция черепа
очень крупного старого индивида. Около х/5 натураль-
ной величины. Restoration of the skull of a very big
old specimen. Approximately x/5 natural size.
thosuchus. Вследствие того, что хрящевой череп
ностью, не упоминав-
шейся памп ранее, яв-
ляется постепенное су-
жение и заострение мор-
ды, а затем вновь при-
тупление ее. Так, широ-
кая, плавно закруглен-
ная морда очень моло-
дой особи (№ 4—19/2252,
рис. 56) значительно су-
живается и слегка зао-
стряется у небольших
черепов (тип рода№1—
9/2243, рис. 63; № 1—
19/2252, рис. 65), и у
черепов «среднего» воз-
раста, оставаясь узкой,
значительно притупляет-
ся на конце (№ 5—
19/2252, рис. 67). У еще
более крупных черепов
конец морды приобрета-
ет прямоугольные очер-
тания, оканчиваясь поч-
ти прямым срезом. Та-
кой тупой срез морды,
образованный сильно
увел иченными дорзаль-
ными пластинками ргас-
maxillaria, хорошо ви-
ден на крупном фраг-
менте носового отдела
черепа № 9—19/2252
(рис. 71).
В особую группу воз-
растных изменений мож-
но выделить процесс
окостенения участков
хрящевого черепа Веп-
Benthosuchus окостеневает
далеко не полно, точная последовательность окостенения хрящевых костей
в настоящей работе не может быть выяснена. Несомненно, существуют
вполне определенные закономерности постепенного окостенения хрящевого
черепа, вероятно, различные в разных группах, но для установления их
нужен большой материал из более примитивных форм Labyrinthodontia.
Наиболее рано окостеневающим хрящевым образованием является
exoccipitale. В черепе № 4—19/2252 в 27 мм длины exoccipitale окостенело
уже совершенно, так же, как и затылочный мыщелок. Несколько позднее
окостеневает quadratum, которое у черепа № 4—19/2252 и даже значительно
73
большего черепа № 1—9/2243 еще в значительной своей части состоит из
рыхлой губчатой массы — омелевшего хряща. Два другие элемента chondro-
cranium, известные у Benthosuchus, — epipterygoideum и в phene thmoideum,
окостеневают гораздо позднее. Небольшой еще очень мало окостеневший
epipterygoideum появляется впервые у наибольших из имеющихся полных
черепов № 5—19/2252, № 2 и 3—32/2354, № 2—19/2252. Все эти черепа почти
одинаковой величины и достигают в длину 220—230 мм. Более крупные
epipterygoideum известны лишь по фрагментам огромных черепов. Поэтому
можно с достаточной уверенностью считать, что окостенение epipterygoi-
deum происходит значительно позднее quadratum — лишь в «среднем»
возрасте, когда череп достигает 220 мм в длину и более. В этот момент
роста epipterygoideum
палочковиден и не
имеет еще формы, ха-
рактерной для очень
крупных особей (рис.
18, 19). Даже у более
крупных эпиптериго-
идов, принадлежащих
черепам, имеющим
около 300 мм в дли-
ну, отсутствует torus
lambdoideus, который
появляется у еще бо-
лее старых особей.
Все полные чере-
па, имеющиеся в на-
шем распоряжении,
пе имеют окостенев-
шего sphenethmoide-
um. Сохранившиеся
фрагменты более
крупных черепов по-
казывают, что окосте-
нение sphenethmoide-
um начинается с мо-
мента достижения че-
репами длины при-
близительно в 300 мм.
Остальныеэлемен-
ты хрящевого чере-
па—basiocci pitale, ba-
sisphenoideum, paroc-
cipitale, prOOtiCUm И Рис. 63. Benthosuchus sushkini. Череп Л? 1—9/2243 (тип
SUpraoccipitale не пай- рода) сверху. а/4 натуральной велич. Skull of a type spe-
дены даже У самых cimen № 1—9/2243, dorsal view. х3/4.
крупных черепов, дли-
ной не менее 700 мм. Представляется вероятным, что эти элементы у Ben-
thosuchus вообще никогда не окостеневали; может быть, за исключением
prooticum. На возможность окостенения последнего указывают некоторые
плохо определимые фрагменты (без достаточной достоверности).
Таким образом, порядок окостенения хрящевых костей черепа Bentho-
suchus может быть следующим: exoccipitale, quadratum, epipterygoideum,
sphenethmoideum (?), prooticum. Из них exoccipitale окостеневает на самых
ранних стадиях существования животного, quadratum не на много позже,
epipterygoideum и особенно sphenethmoideum появляются очень поздно,
почти на пределе «среднего» возраста.
Нелишне отметить, что в согласии с указанными выше возрастными
79
особенностями (№ 16 и 17) в процессе роста черепа Benthosuchus наблю-
дается весьма значительное пропорциональное уменьшение величины хря-
щевого supraoccipitale и в особенности — basioccipitale. Эти кости не только
не окостеневают, но и растут значительно медленнее остальных частей
черепа, вследствие чего и вытесняются соседними костными элементами,
например, exoccipitale. Поэтому basioccipitale и supraoccipitale в черепе
Benthosuchus можно с полным правом рассматривать как образования
регрессивного характера, находящиеся на пути к исчезновению.
При рассмотрении перечисленных возрастных изменений черепа Bentho-
suchus выявляется их очевидная неравноценность. Одна категория воз-
растных изменений
влечет за собой лишь
общее усиление с воз-
растом структурных
особенностей черепа,
что и является наибо-
лее обычным выраже-
нием возрастных из-
менений у высших
Tetrapoda. К этой ка-
тегории должны быть
отнесены такие воз-
растные изменения,
как общее увеличе-
ние массивности ко-
стей и увеличение
диаметра ячеек
скульптуры пакостях
крыши черепа, разви-
тие мышечных греб-
ней и бугров, умень-
шение редуцирую-
щихся элементов че-
репа. Сюда из пере-
численных выше при-
знаков войдут № 6,
7, 9, 13, 15, 16, 17,
18, 19, 20 и 21.
Все эти признаки
в свою очередь долж-
ны быть разбиты на
две группы — 1) при-
знаки «детского» сла-
Рис. 64. Benthosuchus sushkini. Череп № 1—9/2243 (тип рода) бОГО Строения черепа
снизу.3/4натуральной величины.Туре specimen № 1—9/2243, И 2) признаки ЭВОЛЮ-
ventral view. х3/4. ЦИОННОЙ ПРИМИТИВНО-
СТИ, более резко выра-
женные у молодых особей, чем у старых. К первой группе могут быть
отнесены такие признаки, как отсутствие или наличие отростков и греб-
ней, замена щелей шовными соединениями, большее или меньшее выра-
жение структурных особенностей, присущих черепу именно Benthosuchus.
Другая группа включает изменения такие, как № 6, 13, 16, 17 и 19.
Естественно, что очень молодые формы должны нести черты несколько
большей примитивности, что как раз и выражено вышеуказанными изме-
нениями. Так, сравнительно более длинный ramus posterior птеригоида,
относительно большие размеры хрящевых supraoccipitale и basiooccipitale
и более высокая crista falciformis, равно как и большая высота черепа
в затылке — все эти признаки, отличающие очень молодых особей Bentho-
80
suchus от более старых, в то же время являются признаками более ранней
эволюционной стадии на пути общего развития лабиринтодонтов, устано-
вленном Уотсоном [Watson, 1920]. В самом деле, уплощение черепа, укоро-
чение задних ветвей птеригоидов (идущее более быстро, чем укорочение
передних), редукция basioccipitale и supraoccipitale — типичные приз-
наки поздней эволюционной стадии развития Labyrinthodontia.
В первую категорию возрастных изменений, таким образом, включены
изменения, не меняющие общего плана строения черепа и отражающие
Рис. 65. Benthosuchus sushkini. Череп 1—19/2252 сверху.
3/4 натуральной величины. Skull № 1—19/2252, dorsal view.
X3/v
лишь общее усиление его или же, в слабой степени, выявляющие общие
черты филогенеза всей группы Labyrinthodontia.
Признаки второй категории значительно изменяют общий характер
строения черепа, вне прямой связи с отражением филогении группы Laby-
rinthodontia. Вообще, сюда могут быть причислены изменения № 1, 2, 3,
4, 5, 10, 11, 12, 14 и — как стоящее в прямой зависимости— от № 5 до
№ 8.
Так, с ростом черепа Benthosuchus мы наблюдаем, как короткий череп
с большими орбитами и широкой мордой превращается в длинный череп
6 Тр. Палеонтол. ин-та, т. X, в. 1. 81
с узкой мордой и маленькими, отнесенными назад орбитами. Непарное
fenestra intermaxillaris обращается в парное, основание черепа (corpus
parasphenoidei) суживается. Processus cultriformis становится длинным и тон-
ким, преорбитальные элементы значительно удлиняются. Кроме того,
череп становится значительно более широким в затылке.
И. А. Ефремов [1933] впервые указал, что очень молодые особи стего-
цефалов у разных форм характеризуются черепами с большими орбитами,
расположенными посредине черепа, т. е. равным или почти равным раз-
Рис. 66. Benthosuchus sushkini. Череп № 1—19/2252 снизу.
3/4 натуральной величины. Skull № 1—19/2252, ventral view.
х3/4-
витием пре- и посторбитальных элементов. При этом foramen parietale
располагается на линии задних краев орбит. С дальнейшим ростом черепа,
в зависимости от характера специализации данной формы, указанные
соотношения нарушаются. В крыше черепа преобладают пре- или посторби-
тальные элементы или, наоборот, краевые, — при вытягивании или рас-
ширении черепа, перемещении орбит и т. д. Chondrocranium пластически
следует изменениям конфигурации черепа. Foramen parietale ведет себя,
как неподвижный в крыше черепа центр, по положению которого легко
82
распознать модификацию черепа. А. П. Быстров [1935] установил, что
модификации черепа стегоцефалов влекут за собой зональный рост костей
крыши черепа, т. е. элементы в увеличивающемся с возрастом отделе крыши
черепа обладают усиленным по сравнению с остальными ростом и образуют
так называемые зоны роста.
Таким образом, череп стегоцефалов является весьма пластической си-
стемой, легко изменяющейся при разнообразных адаптивных уклонениях.
В той же работе А. П. Быстров описал ход роста отдельных элементов
крыши черепа Benthosuchus по различным возрастным стадиям, почему
мы и не останавливаемся здесь на описании этого процесса.
Не трудно заметить, что очень молодая особь Benthosuchus (череп
№ 4—19/2252) обладает как раз отмеченным выше строением, общим для
Рис. 67. Benthosuchus sushkini. Череп № 5—19/2252. Skull № 5—19/2252. А — сверху
(dorsal view),. В —снизу (ventral view), 3/8 натуральной величины (х3/8).
личиночных и молодых особей стегоцефалов, т. е. строением, в котором
отсутствуют черты специальной адаптации, ярко выраженной у более
старых особей.
Процесс постепенного изменения строения черепа с возрастом, главным
образом как бы пластическое вытягивание морды как раз и включает в себя
серию перечисленных выше изменений второй категории. При этом различ-
ные элементы черепа реагируют на этот процесс так, как если бы в самом
деле они состояли из пластического материала. Так, processus cultriformis,
вытягиваясь, относительно очень значительно утончается, кости переднего
отдела нёба и преорбитального отдела черепа также, удлиняясь, «сужа-
ются», хоаны и ноздри «сближаются» друг с другом и т. д. Лишь у наи-
более старых особей Benthosuchus череп можно считать вполне сложив-
шимся, приобретшим все адаптивные черты строения, свойственного данной
форме.
Подводя итоги сказанному, мы приходим к необходимости признать,
что вторая категория возрастных изменений черепа Benthosuchus включает
6*
83
в себя изменения, постепенно усиливающие с возрастом специальную адап-
тацию черепа, присущую Benthosuchus.
Эти возрастные изменения являются наиболее яркими из всех других
и свидетельствуют о неуклонном усилении выражения адаптации вместе
Рис. 68. Benthosuchus sushkini. Череп № 5—19/2252, затылочная поверхность. 3/4 на-
туральной величины. Skull № 5—19/2252, back view, х3/4.
с ростом черепа. Иными словами, процесс развития адаптации, присущей
Benthosuchus, продолжался в течение всей жизни животного. Однако,
как это можно установить из сравнения маленького черепа № 4—19 2252
и наименьшего из других полных черепов № 1—9/2243, а также по много-
Рис. 69. Benthosuchus sushkini. Череп № 2—19/
2252 снизу, 2/6 натуральной величины. Skull
№ 2—19/2252, ventral view, Х2/6.
численным отдельным фрагмен-
там, череп Benthosuchus приоб-
ретал важнейшие черты своего
строения довольно скоро в пре-
делах роста от 40 до 100 мм.
Дальнейший рост черепа обусло-
вливал лишь более резкое выра-
жение особенностей строения,
приобретенных в ранний период.
Таким образом, на материале
по возрастным стадиям Bentho-
suchus мы видим подтвержде-
ние общей закономерности он-
тогенетического развития—при-
знаки рода появляются позже
общих признаков класса, отряда,
семейства и даже настолько
поздно, что уже вполне развитое
жпготное еще не несет присущих
ему черт адаптивной модифика-
ции. Такое запаздывание, пови-
димому, вообще типично для
примитивных Tetrapoda. Оче-
видно, что примитивный орга-
низм, не требуя для своего он-
тогенетического развития до ста-
дии самостоятельного существо-
вания (после метаморфоза) зна-
чительного времени, заканчивал
свое развитие очень медленно,
почти в течение всей жизни. По-
этому часть поздно появляющих-
ся признаков — именно некото-
рые признаки адаптивной модификации (т. е. признаки рода и вида) —
появлялась уже после достижения животным довольно большой величины.
84
Индивидуальная изменчивость черепа Benthosuchus
главным образом сводится к запаздыванию или, наоборот, ускорению
некоторых возрастных изменений.
Так, окостенение срединной перегородки в fenestra intermaxillaris
иногда наступает довольно рано. На черепе № 2—19/2252 (рис. 69) имеется
уже полностью окостеневшая перегородка, образованная слившимися
между собой processus fenestrales anteriores и posteriores. В почти равном
ему по величине черепе № 5—19/2252 (рис. 67) эта перегородка отсутствует,
и processus fenestrales, как сошников, так и praemaxillaria, выражены зна-
чительно слабее, чем у равной величины черепов № 3—3254 и
Кг 3—32/2354. Фрагмент переднего отдела черепа еще более крупного
экземпляра № 9—19/2252 (рис. 71) обладает лишь зачаточными processus
fenestrales, которые у наименьшего по величине из черепов «среднего»
возраста — типа рода № 1—9/2243 (рис. 64) развиты значительно сильнее.
Длина ramus posterior птеригоида по отношению к ramus anterior также
индивидуально вариирует у разных особей Benthosuchus. Общее укоро-
чение ramus posterior с возрастом иногда запаздывает, что хорошо видно
из сравнения черепов № 5—19/2252 (рис. 67) и № 2—19/2252 (рис. 69).
Рис. 70. Benthosuchus sushkini. Череп № 3—19/2252, затылочная по-
верхность. 4/5 натуральной величины. Skull №3—19/2252, occipital
surface. х4/б.
У первого черепа задние ветви птеригоидов уже значительно укорочены,
в то время как у второго они пропорционально длиннее.
Ускорение в развитии processus lamellosus exoccipitalis наблюдается
на черепе № 3—19 2252 (рис. 70), где он развит уже весьма значительно,
в то время как на значительно больших черепах, как, например,
№ 5—19/2252, этот отросток гораздо меньшей величины.
Опускание боковых частей затылка развивается неравномерно у чере-
пов одинаковой величины. Некоторые черепа имеют слабо опущенные вниз
фланги, другие — более сильно, вследствие чего общие очертания затылка
изменчивы.
Наконец, степень притупления морды может различаться у одинако-
вых черепов. Так, у черепа № 3—32/2354 несколько более суженная и за-
кругленная морда, чем у равных по величине черепов № 5—19/2252 и
№ 2—19/2252.
Необходимо подчеркнуть, что перечисленные индивидуальные изме-
нения встречаются в качестве единичных явлений в черепах, все остальные
признаки которых вполне соответствуют, скажем, величине черепа. Даже
более того, иногда признак ускорения возрастного изменения встречается
в черепе на ряду с признаком отставания другого возрастного изменения.
Так, окостенение перегородки fenestra intermaxillare имеется в черепе
№ 2—19/2252 на ряду с наличием относительно длинных rami poster’ores
птеригоидов. Таким образом, индивидуальное ускорение или замедление
развития различных признаков черепа Benthosuchus не встречаются в виде
ускорения или замедления развития черепа или какого-либо из его отделов
85
вообще, а лишь в качестве случайной вариации одного, двух, максимум
трех признаков.
Указанными выше примерами индивидуальных вариаций возрастной
изменчивости, конечно, не исчерпываются все имеющие место случаи.
Однако для анализа других, более мелких проявлений индивидуальной
изменчивости лучшим методом было бы применение вариационной стати-
стики, чего не позволяет сделать количество имеющегося материала.
Намечающиеся на Шарженгских местонахождениях крупные раскопки,
без сомнения, доставят значительное количество материала, после чего
вариационной статистике черепа Benthosuchus можно будет посвятить
специальную работу.
Индивидуальные изменения, не связанные с возрастной изменчи-
востью, также констатированы на многих черепах или фрагментах черепов
Benthosuchus.
Рис. 71. Benthosuchus sushkini. Фрагмент черепа № 9—19/2252, 4/5 нату-
ральной величины. А — сверху (dorsal view), В — снизу (ventral view).
Х4/б-
Сюда относятся:
1. Вариации очертаний швов между отдельны-
мй элементами черепа.
Очертания швов черепных костей всегда различаются в мелких деталях,
даже на двух сторонах одного и того же черепа. Однако некоторые более
крупные или постоянные вариации швов заслуживают быть отмеченными.
Наиболее часто наблюдается неравномерное развитие медиальных эле-
ментов крыши черепа, заключающееся в том, что правый или левый элемент,
развиваясь несколько сильнее, чем другой парный, выдвигается вперед
по средней линии. Нередко сильное развитие одного из срединных элементов
обусловливает значительное искривление срединного шва крыши черепа,
в большинстве случаев идущего прямолинейно. Подобные аномалии хо-
рошо видны на рисунках 65 и, особенно, 71, где неравномерное развитие
обоих nasalia ведет в первом случае к выдвижению вперед левого nasale,
а во втором — к сильному искривлению срединного шва. Эти асимметрич-
ные индивидуальные изменения в той или иной степени свойственны почти
каждому черепу Benthosuchus. Более редкий вариант наблюдается на че-
репе № 1—9/2243 (тип рода), рис. 63. На правой стороне, как это типично
для Benthosuchus. frontale не принимает участия в ограничении орбиты,
будучи отделено от орбитального края сходящимися концами praefrontale
86
и postfrontale. На левой стороне довольно значительный участок frontale
вклинивается между praefrontale и postfrontale и образует часть орбиталь-
ного края.
На вентральной поверхности наиболее часто отмечаются индивидуаль-
ные изменения в очертании швов corpus parasphenoidei с птеригоидами.
Здесь нередко corpus parasphenoidei развивается асимметрично, т. е одна
его половина больше другой. В этом случае искривления швов с обоими
птеригоидами значительно изменяют очертания corpus parasphenoidei.
Более ярко подобная вариация выражена на черепе № 2—19/2252 (рис. 69),
где правильная форма corpus parasphenoidei совершенно изменена.
2. Вариации общей формы черепа.
На некоторых черепах наблюдаются небольшие отклонения от обычно
присущей черепу Benthosuchus формы—встречаются сравнительно узкие
п сравнительно широкие черепа. Так, череп № 3—32/2354 является наи-
более узким из всех остальных черепов и ограничен прямыми латераль-
ными краями, в то время как другие черепа одинаковой с ним величины —
№ 2—32/2354, № 5—19/2252, № 2—19/2252 — уже обладают некоторым
прогибом латеральных краев черепа внутрь в связи с более резким пере-
ходом от черепа к вытянутой морде. В соответствии со своим узким габи-
тусом череп № 3—32/2354 обладает наиболее узким fenestra interptery-
goidea.
Пропорционально большей шириной обладает небольшой череп
№ 3—19/2252, который наиболее отчетливо контрастирует с узким чере-
пом № 3—32/2354.
Подобные изменения в общей конфигурации черепа, при наблюдении
на целой серии черепов, разумеется, весьма незначительны, но в крайних
своих выражениях могут представить заметные отличия.
3. Вариации в относительной величине нозд-
рей, орбит, хоан и глубине слуховых вырезок.
Череп № 1—19/2252 (рис. 65 и 66) обладает относительно большими
хоанами и орбитами, чем другие черепа Benthosuchus. Небольшой череп
№ 3—19/2252 отличается пропорционально наименьшими орбитами. Ноздри
черепа № 1—19/2252 пропорционально меньше ноздрей, например, черепа
№ 1—9/2243 (рис. 63). Отклонения эти, перечисленные с поправкой на раз-
личный возраст особи, не превышают 2 мм в максимальном диаметре, но
гее же хорошо заметны.
Различная глубина слуховых вырезок определяется различной длиной
cornu tabularia, наибольшая длина которых, относительно превышающая
среднюю длину cornu tabulare у Benthosuchus на 0.25, наблюдается на
черепе № 1—32/2354. Отклонения, наблюдаемые на других черепах, зна-
чительно менее.
4. Вариации хода каналов боковой линии в за-
метном выражении наблюдались только на черепе № 5—19/2252, где уста-
новлено отклонение заднего конца sulcus jugalis кверху от sutura squamo
quadrato jugalis.
5. Индивидуальные вариации в расположении
постфенестрального зубного ряда на сошниках
Benthosuchus заключаются в различной длине последнего. Обычно конец
клинообразного ряда этих зубов доходит почти до конца 1-й трети длины
хоан, по иногда не достигает даже их переднего края. Последнее наблю-
дается, например, на черепе № 5—19/2252 (рис. 67В) ср. с черепом № 2—
19 2252 (рис. 69).
6. Индивидуальные колебания в массивности
затылочных элементов черепа. Для одинаковых по ве-
личине черепов Benthosuchus характерно некоторое различие в массивности
строения затылка. Кости, образующие затылок, могут различаться по
своей толщине и толщине своих отростков почти на 73. Таким массивным
затылком обладает, например, череп № 5—19/2252 (рис. 67) в противо-
87
Рис. 72. Вариации fo-
ramen nervi vagi у
Benthosuchus sushkini
no exoccipitalia очень
крупных черепов. A—
нормальное положе-
ние, В, С, D — от-
клонения. А — колл.
№ 32/2354, фрагмент
без №; В —№12—19/
2252; С — фрагмент
без №, колл. №
19/2252; Ь — № 12—
32/2354. 3/4 натураль-
ной велич. Variations
of the foramen nervi
vagi. A — normal, B,
C, D — aberrations.
положность затылкам одинаковых по величине черепов № 2—19/2252
и № 2—32/2354.
7. Индивидуальные колебания в ходе нервов
могут быть прослежены лишь для группы glossopharyngeus-vagus, обла-
дающей точно фиксированным в хорошо окостеневшем exoccipitale местом
выхода. Отверстие и. glossopharyngeus-vagus у основания processus paro-
ticus часто изменяет свое положение, перемещается
несколько ниже или назад к condylus occipitalis.
Иногда одно большое отверстие распадается на два
или три небольших, нередко трудно отличимых от
for. nutritium отверстий. Для иллюстрации подобных
изменений приведем серию отдельных фрагментов
exoccipitalia, принадлежащих черепам больших осо-
бей Benthosuchus: № 12—19/2252, 12—32/2354 и два
фрагмента без № коллекции 19/2252. На рис. 72А
показано типичное расположение for. nervi vagi и
в В, С, D — его вариации.
8. В заключение отметим, что относительная ве-
личина и форма покрытого шагренью участка на
corpus parasphenoidei (area aspera parasphenoidei) под-
вержены значительным индивидуальным колебаниям
(рис. 63—69).
Возрастные изменения нижней
челюсти сводятся к следующим основным изме-
нениям.
1. Степень окостенения articulare. Эта кость, так
же, как и quadratum, более или менее полно око-
стеневает лишь в челюстях особей «среднего» возра-
ста, приблизительно с момента достижения челю-
стью длины около 200 мм.
Особи меньшей величины обладают articulare,
состоящими из очень рыхлой губчатой костной массы
или даже из только слегка объизвествленного хряща.
Такие кости в фоссильном материале на 80—90%
состоят из чистой породы, передающей лишь внеш-
нюю форму кости. По мере увеличения челюсти мы
можем проследить постепенное увеличение относи-
тельных размеров articulare за счет продвижения впе-
ред переднего конца кости, т. е. дальнейшего распро-
странения оссификации меккелеева хряща.
2. Увеличение относительной высоты нижней че-
люсти, особенно резко сказывающееся в задней ее
половине. Челюсти крупных особей обладают высо-
той относительно большей, чем челюсти молодых.
Возрастание высоты челюсти по мере ее роста в сред-
нем равно отношению 1.5 к 2, где 1.5 будет абсолют-
ная длина нижней челюсти, а 2 — ее абсолютная
высота от верхнего края crista muscularis supraangu-
laris до нижнего угла angulare. На рис. 73А, В, С
изображены три пижних челюсти особей различного
возраста из коллекции № 19/2252.
3. Увеличение относительной ширины симфизного
отдела по мере роста челюсти.
4. Уменьшение относительной величины foramen
Meckelianum (posterius).
5. Увеличение числа заостренных выступов на
прекороноиде и интеркороноиде. На небольших че-
люстях, длиной около 120—150 мм, наблюдаются
88
лишь 1—2 заостренные выступа на praecoronoideum, интеркороноид
совершенно лишен их. Челюсти в 220—250 мм длины имеют до 4 выступов
на прекороноиде и 2 — на интеркороноиде. Крупные челюсти, свыше
350 мм в длину, обладают 6 (одна нижняя челюсть № 41—19/2252 в 410 мм
длины имеет 7 выступов) выступами на прекороноиде и 3—4 на интеркоро-
ноиде.
Рис. 73. Benthosuchus sushkini. Правые нижние челюсти индивидов
разного возраста. «Д натуральной величины. Right mandibles of
different age. х8Д.
Индивидуальные вариации нижней челюсти Benthosuchus не метут быть
детально прослежены вследствие малого количества совершенно полных
челюстей. Можно отметить, что в общем эти индивидуальные изменения
сводятся к различной кривизне ветвей нижней челюсти (соответственно
индивидуальным особенностям конфигура-
ции черепа), вариациям в толщине (по-
перечном сечении), вариациям формы су-
ставной ямки или положения foramen
Meckelianum. Однако эти вариации не на-
столько значительны, чтобы привести к
заметным отличиям, и поэтому здесь под-
робно не разбираются.
Исключение составляют индивидуаль-
ные изменения processus retroarticularis в
нижней челюсти, наружный лепесток ко-
торого сильно меняется в величине у осо-
бей одинакового роста. Наравне с сильно
развитым наружным лепестком retroarti-
cularis встречаются и челюсти, где он
совершенно не развит. Такие вариации
не совпадают с длиной самою processus
retroarticularis, которая также у разных
особей значительно колеблется. На рис. 74
приведены две нижние челюсти Benthosu-
chus одинаковой величины, из которых
одна — № 34—19/2252 (В) имеет средней
величины наружный лепесток и несколько
более удлиненный р. retroarticularis, а
Рис. 74. Benthosuchus sushkini.
Вариации processus retroarticu-
laris. 3/4 натуральной величины.
Variations of the retroarticular pro-
cessus. A — № 45—19/2252, В —
№ 34—9/2243. X3/4.
другая — № 45—19/2252 (А) обладает сильно развитым наружным лепе-
стком и более округлым и коротким р. retroarticularis.
Очень крупная челюсть № 41—19/2252 в 410 мм длины имеет весьма
слабо развитый наружный лепесток и сравнительно укороченный р. retro-
articularis.
89
Рис. 75. Benthosuchus sushkini. Воз-
растные изменения гипоцентра. 4/5 на-
туральной величины. Age stages of
the hypocentrum. A — № 50—32/2354;
B—№ 50—9/2243; C—№ 76—19/2252;
D—A? 88—19/2252; E—№ 49—9/2243;
F — № 73—19/2252. x4/6.
Посткраниальный скелет
Позвоночный столб. Возрастные и индивидуальные изме-
нения позвоночного столба Benthosuchus не могут быть установлены за
отсутствием достаточно полного материала. Большое количество гипо-
центров, имеющееся в коллекции, все же дает возможность установить,
что с увеличением размеров (возрастом) окостенение гипоцентра становится
более сильным. Так, толщина гипоцентра в средней части у молодых особей
обычно не превосходит четверти его длины по внутренней дуге. У крупных
гипоцентров старых индивидов это отношение меняется, и толщина дости-
гает половины длины по внутренней дуге. Кроме того, вместе с ростом
гипоцентра происходит постепенное возрастание высоты его концов. Если
у молодых особей высота гипоцентра меньше полуокружности, то у старых
она становится больше половины окружности. Соответственно большему
окостенению гипоцентра просвет для
хорды с возрастом уменьшается, вслед-
ствие чего уменьшается и толщина хор-
ды. Суставные площадки для ребер
(facies costalis) у молодых особей обра-
зуют лишь слабые выступы, которые у
старых выражены значительно сильнее.
Вместе с окостенением гипоцентра в
толщину с возрастом увеличивается его
окостенение в верхней части, над facies
costalis. У крупных старых особей (у
особей среднего возраста в переднем
отделе позвочного столба) эта окосте-
невшая верхняя часть гипоцентра всту-
пает в соприкосновение с неуральной
дугой. На последней наблюдается кост-
ное развращение нижнего края facies
chordalis, образующее направленный
прямо вниз округлый в плане выступ с
рыхлой нижней поверхностью, соответ-
ствующей рыхлой поверхности верхней
оконечности гипоцентра. Между этими
двумя костными площадками лежал
хрящ, плотно спаивавший неуральную
дугу и гипоцентр.
Как это уже отмечалось в I части, во всем обильном материале по Веп-
thosuchus, несмотря на тщательное исследование, нами не было обнаружено
ни одного pleurocentrum. Более того, на самых крупных позвонках, при-
надлежащих особям с черепом не менее 700 мм длины, отсутствуют
всякие следы участия pleurocentrum в сочленении центра позвонка с не-
уральной дугой. Следует считать, что плевроцентры у Benthosuchus не
окостеневали в течение всей жизни и кроме того, были уже настолько
редуцированы, что, невидимому, не играли большой роли в архитек-
туре позвонка.
Таким образом, важнейшим возрастным изменением позвоночника
Benthosuchus будет постепенно прогрессирующее окостенение до образо-
вания контакта костного гипоцентра и неуральной дуги, идущее парал-
лельно с уменьшением толщины хорды. Последняя, однако, у Benthosuchus
остается непрерывной в течение всей жизни. На рис. 75 изображены не-
которые возрастные стадии увеличения гипоцентра из заднего отдела туло-
вища, подобранные в приблизительном соответствии своего положения
в позвоночном столбе.
Ребра. Возрастные изменения ребер не могут быть установлены за
фрагментарностью материала, не говоря уже об индивидуальных вариа-
90
циях. Можно отметить, что грудные ребра старых особей отличаются го-
раздо более сильным развитием processus uncinatus, чем соответственные
ребра молодых индивидов.
Плечевой пояс. 1. Scapul о-с oracoideum (рис. 76).
По особенностям захоронения материала лопатка в коллекции остатков
Benthosuchus представлена сравнительно малым количеством материала,
что затрудняет наблюдение возрастных изменений.
Главное возрастное изменение scapulo-coracoideum — постепенное око-
стенение его нижнего отдела. Раньше всего окостеневают средняя часть
самой scapula и ее мощный медиальный отросток, вследствие чего canalis
supraglenoidalis появляется на возрастных стадиях с черепом не более
60—70 мм в длину. С дальнейшим
ростом скапулокоракоида (рис. 76)
происходит постепенное расшире-
ние ее верхней части за счет око-
стенения и разрастания ее перед-
него края, и лопатка с возрастом
расширяется. Параллельно этому
расширению идет окостенение на-
ружного нижнего угла лопатки.
Этот угол у молодых особей
(рис. 76D, Е) значительно короче
медиального отростка. На стадии
с черепом около 120 мм в длину
наружный пижний угол scapula
становится по длине равным ме-
диальному отростку (рис. 76С). У
особей «среднего» возраста (длина
черепа 120—250 мм) пижний угол
лопатки разрастается еще более за
счет частичного окостенения сога-
coideum (рис. 76А, В) и становится
длиннее медиального отростка.
Достаточно полных фрагментов
scapulo-coracoideum старых круп-
ных особей в коллекции Палеон-
тологического института не имеет-
ся, поэтому дальнейший ход око-
стенения остается неясным. Во вся-
ком случае, можно с уверенностью
говорить, что окостенение корако-
ида начинается только в «среднем»
возрасте, а окостенение прокора-
коида наступает еще позже. Око-
стеневают ли оба названные элементы полностью, сказать пока нельзя,
но вероятно, что огромные старые особи Benthosuchus должны иметь до-
вольно полно окостеневший scapulo-coracoideum. В равной степени на
всех имеющихся костях не окостеневает и самый верхний конец лопатки.
Среди остатков W etlugosaurus в коллекции Центрального геологического
музея в Ленинграде имеется скапулокоракоид, послуживший А. Н. Ря-
бинину основой для описания этого элемента у W etlugosaurus. Он при-
надлежит довольно крупному черепу (около 300 мм в длину) и почти пол-
ностью окостенел. Равные или почти равные остатки scapulo-coracoideum
Benthosuchus показывают гораздо более слабое окостенение. Поэтому нужно
думать, что scapulo-coracoideum у Benthosuchus окостеневало значительно
позже, чем у Wetlugosaurus.
Индивидуальные вариации scapulo-coracoideum замечены нами только
в различном характере окостенения. У некоторых особей быстрее окосте-
Рис. 76. Benthosuchus sushkini. Возраст-
ные и индивидуальные изменения scapu-
la. 4/5 натуральной велич. А — №134—
19/2252; В—№ 135—19/2252; С —№ 82—
32/2354; D — № 67—19/2252; Е —№ 196—
19/2252. Age stages and individual varia-
tion of the scapula. x4/6-
91
ным возрастным изменением. Кроме общего
и развития более грубой, но в то же время и
Рис. 77. Benthosuchus sushkini. Возрастные измене-
ния ключицы. Age stages of the clavicula. A — № 66—
32/2354; В — № 139—19/2252; C — № 69—32/2354;
x2/5.
невает верхняя часть переднего края, у других, наоборот — нижняя.
Таким образом, некоторые scapula расширены вверху (рис. 76С), некоторые
более широки в нижней части и сужены кверху.
2. Cleithrum. Возрастные изменения cleithrum сводятся к по-
степенному развитию crista cleithralis и постепенному расширению lamina
suprascapularis (рис. 46А, В). Поэтому с возрастом верхняя часть cleithrum
расширяется и приобретает у крупных особей дополнительный выступ
внизу lamina suprascapularis, отсутствующий у молодых индивидов.
3. Clavicula. Ключица Benthosuchus отличается весьма характер-
значительного утолщения
менее резкой скульптуры,
ключица с возрастом пре-
терпевает заметное измене-
ние в очертаниях (рис. 77
А, В, С, D). Строение клю-
чиц у наиболее молодых
форм нам неизвестно, но
нужно думать, что ключи-
ца вполне окостеневала на
очень ранних стадиях, так
как самые маленькие из
известных нам ключиц не
имеют по сравнению с са-
мыми старыми никаких де-
фектов окостенения.
Имеющиеся в нашем
материале маленькие клю-
чицы принадлежат особям
с черепами около 80 мм
длины (напр., № 66 —
32/2354, №146—9/2243). Эти
ключицы очень характерны
своими очертаниями узкого
овала, их вентральная пла-
стинка плавно суживается, закругляясь в направлении не только
вперед, но и назад. Processus praescapularis очень узкий. Ключицы
более взрослых особей (череп около 120—170 мм длины) (рис. 77В)
отличаются узкими прямоугольными очертаниями и почти прямыми ме-
диальным и латеральным краями. Задний край срезан почти без всякого
сужения волнистой линией, к переднему краю вентральная пластинка
постепенно суживается. Еще более крупные ключицы, принадлежащие
скелетам с черепами около 200—230 мм длины, обнаруживают, расширение
заднего отдела вентральной пластинки и выпуклость латерального края.
У ключиц еще более крупных особей (череп около 300 мм длины) (рис. 770)
вентральная пластинка, сильно расширяясь по заднему краю, приобретает
треугольную форму со сходящимися впереди краями. Наконец, ключицы
огромных особей (с черепами около 500—600 мм в длину) № 69—32/2354,
139—19/2252 показывают очень сильное расширение вентральной пластинки,
которая получает очертания тупого треугольника.
Параллельно этим изменениям вентральной пластинки processus praesca-
pularis значительно расширяется в своей средней части и утолщается путем
образования сильного костного валикообразного возвышения ha внутренней
стороне.
Индивидуальные изменения clavicula также своеобразны. Вентральная
пластинка индивидуально изменчива только в ускорении или запаздывании
появления возрастных признаков и в характере скульптуры. Последние
вариации заключаются в различном числе анастомозов, появляющихся
между длинными радиальными лучами скульптуры (т. е. ячеек), и их раз-
92
наиболее типичного строения. Се-
Рис. 78. Benthosuchus sushkini. Индиви-
дуальные изменения (individual variations)
processus praescapularis clavicula. A —
74—19/2252; B—62—32/2354; C hD—№165—
19/2252; E — 72—32/2354; F, G, H —
№ 190—19/2252. 2/3 натуральной величины.
личном расположении. У одинаковых по величине ключиц ячейки, кроме
своего обычного расположения на центре окостенения, покрывают то перед-
нюю часть латерального края, то, наоборот, почти весь задний край,
то появляются на средней части кости в виде расходящейся медиально
площадки.
Гораздо более значительным индивидуальным изменениям подвержен
processus praescapularis. На рис. 78А, В показаны два левых processus
praescapularis разного возраста, но
рия из 3 правых прескапулярных
отростков (рис. 78С, D, Е) нагляд-
но иллюстрирует индивидуальные
изменения формы отростков. Для
сравнения приведены три правых
прескапулярных отростка Wetlugo-
saurus (рис. 78F, G, Н).
4. Interclavicula, так
же, как и clavicula, окостеневает
очень рано, но в силу большой
хрупкости очень тонкой пластинки
ранние стадии роста interclavicula
Benthosuchus отсутствуют в нашем
материале.
Вместе с ростом особи intercla-
vicula значительно утолщается и
приобретает более грубую, но в то
же время и более облитерирован-
ную скульптуру.
Возрастные изменения в конфи-
гурации кости незначительны. У
особей «среднего» возраста очерта-
ния intercalvicula близки к укоро-
ченному округлому ромбу с узким
вытянутым передним углом. У ста-
рых особей гигантской величины
(череп 600—700 мм) (рис. 79) inter-
clavicula несколько более удлинен-
ных очертаний, причем вытянутый
передний угол сильно расширяет-
ся. Такая крупная interclavicula
является уже толстой тяжелой
костной пластиной с сильно обли-
терированной скульптурой на вен-
тральной стороне и облитерирован-
ными trabeculae на дорзальной.
Индивидуальные изменения меж-
ключицы Benthosuchus весьма не-
значительны и при недостаточном
материале не могут быть достовер-
но установлены.
5. Humerus. Большое количество humerus самой различной ве-
личины, имеющееся в коллекции, позволяет в подробностях наблюдать
возрастные изменения в серии переходов от humerus в 7.5 мм длины (соот-
ветствующего черепу приблизительно в 30 мм длины) до humerus свыше
100 мм. Отдельные фрагменты показывают, что в некоторых случаях hu-
merus достигал приблизительно 170 мм, что примерно соответствует черепу
длиной в 800 мм.
Несмотря на такое значительное изменение величины, другие воз-
растные изменения humerus весьма незначительны.
93
Рис. 79. Benthosuchus sushkini. Непол-
ная interclavicula № 148—19/2252 очень
крупного старого индивида. Вид снизу.
2/3 натуральной величины. Interclavi-
cula of a big old specimen, ventral
view, x 2/3.
Как правило, степень окостенения humerus мало изменяется с воз-
растом.
Уже самый маленький humerus в 7.5 мм длины обладает всеми важ-
нейшими признаками более крупных костей. Так, crista deltopectoralis,
значительно расширенный дистальный конец и даже crista radialis уже
отчетливо выражены на этих маленьких humerus. В процессе дальнейшего
роста окостенение humerus сводится главным образом к нарастанию прок-
симального конца, который у старых особей характерно расширяется
кверху, очерчивая нижнюю часть округленной хрящевой caput humeri.
Общее усиление мускульных греб-
ней, линейное увеличение площади
crista deltopectoralis и расширение
дистальной части, особенно в области
epicondylus ulnaris, представляют со-
бой остальные важнейшие возрастные
изменения. По всем данным, для hu-
merus Benthosuchus характерно ран-
нее окостенение, мало усиливающееся
с возрастом, так что самые крупные
humerus обладали еще значительным
количеством эпифизарного хряща.
Серия возрастных стадий humerus
показана на рис. 80.
Индивидуальные изменения hume-
rus у Benthosuchus, в противополож-
ность возрастным, весьма значи-
тельны.
Главнейшим индивидуальным из-
менением является чрезвычайно раз-
нообразная степень окостенения hu-
merus у некоторых (немногих) инди-
видов вне всякой зависимости от ве-
личины (возраста) животного. У од-
них особей не окостеневает головка,
у других дистальный конец. Некото-
рые humerus, длиной около 100 мм
(считая их длину при нормальном
окостенении), обладают в сохранив-
шейся костной части только поло-
винной длиной (№ 207 — 19/2252,
№ 85—9/2243, № 197—19,2252) за счет
неокостенения концов. Еще более
удивительны humerus нормальной
длины в 75—80 мм, у которых око-
стенела только средняя часть диа-
физа (№ 209 — 19 2252, № 198 —
19/2252). Случаи дефектов окостенения
humerus представлены на рис. 81.
Эти индивидуальные отклонения в окостенении крупных humerus
резко контрастируют с ранним окостенением humerus у Benthosuchus вообще.
Другой резко бросающейся в глаза индивидуальной вариацией является
искажение типичной формы humerus вследствие утолщения средней части
кости и некоторого общего укорочения. Такие укороченные, «пахиостоти-
ческие» формы humerus наблюдаются у особей среднего возраста в коли-
честве 3—5% от нормальных костей.
Третьей индивидуальной вариацией humerus, также встречающейся
в нескольких процентах от нормы, является недоразвитие костного epicon-
dylus radialis, встречающееся как у крупных, так и у мелких особей..
94
6. Ulna и radius в исследованном материале немногочисленны
и, кроме того, почти все принадлежат особям приблизительно одного воз-
раста.
Судя по имеющемуся материалу, эти кости не обнаруживают суще-
ственных возрастных изменений, кроме возрастной величины и несколько
большей степени окостенения. Последнее, однако, мало различается у ма-
Рис. 80. Benthosuchus sushkini. Возрастные и индивидуальные изменения humerus.
Age stages and individual variation of the humerus. A — № 200—19/2252; В—№ 194 —
19/2252; C— № 83—9/2243;|D — № 201—19/2252; E — № 199 — 19/2252. 2/3 натуральной
величины. x2/3.
леньких и средней величины костей. Для прослеживания индивидуальных
вариаций материала совершенно недостаточно, и, кроме того, простая форма
сохранившихся средних участков костей не дает возможности проявления
заметных отклонений.
7. Э л е м е н т ы ки-
сти Benthosuchus мало из-
вестны и не дают материа-
ла по возрастной и инди-
видуальной изменчивости.
Тазовый пояс.
Общий очерк развития та-
за Benthosuchus сделан на-
ми в первой части, где
разбирается порядок око-
Рис. 81. Benthosuchus sushkini. Индивидуальные
изменения в степени окостенения humerus. Indi-
vidual variations in the ossification of the humerus.
A—№ 209—19/2252; B—№ 85—9/2243; C—№ 198—
19/2252; D —№ 207 — 19/2252. 2/3 натуральной вели-
чины. X 2/3.
стенения хрящевых эле-
ментов таза. Поэтому мы
перейдем непосредственно
к анализу возрастных ин-
дивидуальных изменений
отдельных элементов та-
зового пояса.
I. 11 i u m. В противоположность scapulo-coracoideum ilium окосте-
невает почти полностью на очень ранних стадиях. Наименьшие в коллек-
ции ilium около 10 мм длины, принадлежащие черепам приблизительно
в 35 мм длины, по степени своего окостенения не отличаются существенно
от костей более старых особей. Правда, в коллекции отсутствуют очень
95-'
крупные экземпляры ilium, и наиболее крупные из имеющихся соответ-
ствуют особям среднего возраста.
возрастные изменения ilium невелики и сводятся к следующим:
1) общее утолщение кости сравнительно с ее длиной;
2) дальнейшее окостенение нижнего края ilium, влекущее за собою
значительное расширение его нижней части;
3) усиление поднятия краев верхнего углубления acetabulum;
4) усиление гребней linea obliqua и crista muscularis;
5) дальнейшее окостенение и расширение верхнего конца.
Все эти изменения в процессе роста происходят весьма постепенно и,
насколько можно судить при отсутствии в материале наиболее крупных
особей, — медленно.
Рис. 82. Benthosuchus sushkini. Возрастные изменения и индивидуальные вариации
ilium. Age stages and individual variation of the ilium. A—№ 217—19/2252; В—
№ 214—19/2252; G—№ 219—19/2252; D—№ 215—19/2252; E—№ 213—19/2252; F—
№ 211—19/2252; G—№ 152—9/2243; H— № 216—19/2252; I—№ 212—19/2252; K—
№ 290—19/2252; L—№ 88—9/2243; M—№ 110—9/2243. 3/4 натуральной величины. X3/4.
Кости, отмеченные К, L, М, принадлежат Wetlugosaurus. (Ilium marked К,
L, M belong Wetlugosaurus).
Индивидуальные изменения ilium, наоборот, довольно значительны.
Среди большого количества различных ilium в нашей коллекции можно
наблюдать интересные варианты формы ilium. На рис. 82 они сведены
в одну таблицу, по которой можно также хорошо проследить и возрастные
изменения.
Для наиболее ранних возрастных стадий (рис. 82К, L, М) мы приводим
рисунки ilium Wetlugosaurus, вследствие того, что соразмерные ilium
Benthosuchus из Вахнева сохранились не полностью и слегка окатаны.
При большом сходстве посткраниального скелета обоих родов, особенно
на ранних стадиях, такая замена вполне возможна.
Важнейшие индивидуальные вариации ilium:
1. Отклонения в степени окостенения нижнего или верхнего конца
рис. 82А, G (верхние концы), Н (нижний конец), Е (оба конца).
2. Различия в общей форме ilium: есть укороченные и утолщенные типы
и, наоборот, удлиненные (рис. 8^А, D, I удлиненные; В, Е, Н — укорочен-
ные).
96
3. Различие в форме верхнего конца ilium. Обычно простой неразде-
ленный верхний конец ilium разделяется на два, иногда три отростка
(рис. 82В, С). Эти различия, повидимому, определяются совпадением боль-
шого индивидуального окостенения верхнего конца ilium с укороченным
типом, т. е. изменений 1 и 2 групп. При этом окостеневает почти самая вер-
шина ilium, у нормальных или, тем более, у удлиненных костей остающаяся
хрящевой.
II. Ischium, найденный в малом числе экземпляров одинаковой
величины, не дает возможности оценки его возрастной или индивидуальной
изменчивости.
III. Pubis, повидимому, не окостеневавший или окостеневавший
очень поздно, в имеющемся материале отсутствует.
IV. Femur. В коллекции имеются многочисленные экземпляры
femur, большинство которых принадлежит особям «среднего» возраста,
что типично и для всего материала. Однако, кроме этих средней величины
костей, имеются полные femur очень небольшой величины и, с другой сто-
роны, фрагменты, принадлежащие особям очень крупной величины. Таким
образом, мы можем проследить возрастные изменения femur от длины
в 9 мм до, приблизительно, 150 мм, т. е. от особей с черепом около 30 мм
длины до особей с черепами свыше 500—6£0 мм.
Возрастные изменения незначительны и сводятся к постепенно увели-
чивающемуся окостенению концов (в связи с чем пропорциональная ширина
эпифизов возрастает),, к увеличению высоты crista aspera и trochanter и
к увеличению глубины fossa tendinalis и fossa poplitea.
Все перечисленные признаки femur выражены уже на самых маленьких
костях; несмотря на очень значительное увеличение размеров кости, сте-
пень ее окостенения увеличивается очень мало. Это малое увеличение сте-
пени окостенения особенно хорошо заметно для диапазона 15—150 мм,
так как еще меньшие кости в верхних своих концах окостеневают слабее.
Индивидуальные изменения femur Benthosuchus сходны с вариациями
humerus, но выражены менее сильно.
В коллекции имеются крупные femur, окостеневшие значительно слабее
меньших по величине костей или, наоборот, маленькие, окостеневшие
несколько больше нормального, рис. 83Аи D. Равным образом встречаются
femur, имеющие укороченный habitus с сравнительно более массивными
эпифизами.
На рис. 83 представлены возрастные стадии femur и некоторые индиви-
дуальные отклонения (крупный, слабо окостеневший femur А и маленький,
хорошо окостеневший (несколько более нормы) femur D, вместе с очень
плохо окостеневшим femur Н).
V. Tibia. Для детального разбора возрастных и индивидуальных
вариаций tibia Benthosuchus имеющийся в коллекции материал (14 костей)
недостаточен, так как, кроме небольшого числа костей, все они принадле-
жат к «среднему^ возрасту и дают малый диапазон величины. Общим воз-
растным изменением tibia является, кроме увеличения размера, относи-
тельное ее утолщение в диафизе, резкое усиление гребней и tuberositas
tibiae и общее расширение кости за счет развития медиального края. По-
следний заостряется, превращаясь в тонкую острую пластинку.
Индивидуальные вариации tibia заключаются в том, что одинаковые
по величине кости имеют заметно различную толщину и массивность. Так,
tibia № 285—19/2252 почти в полтора раза толще tibia № 97—32/2354, пре-
восходящей первую на 2 мм по длине. Некоторые небольшие кости имеют
сильно расширенную верхнюю часть, которая значительно превосходит
нормальную ширину конца tibia.
Ускорение или замедление возрастных изменений сказывается в не-
равномерном развитии гребней у одинаковых по величине костей. Гребни
могут быть равиты то сильнее, то слишком слабо.
VI. Fibula Benthosuchus представлена еще меньшим числом экзем-
7 Тр. Палеонтол. ин-та, т. X, в. 1. 97
пляров, чем tibia, хотя и несколько более сильно разнящимися между
собой по величине. Степень ее окостенения на имеющихся образцах почти
не изменяется. Единственно заметное возрастное изменение есть некоторое
относительное расширение и уплощение всей кости. Индивидуальные ва-
риации могут быть отмечены в различной величине вытягивания нижнего
медиального угла, который, например, у маленькой fibula № 286—19/2252
вытянут значительно слабее, чем у превосходящей ее в два раза по величине
fibula № 99—32/2354. Это вытягивание или, наоборот, притупление медиаль-
ного угла нижнего эпифиза вариирует у различных костей и является един-
ственной наблюдаемой индивидуальной вариацией.
VII. Элементы стопы Benthosuchus известны весьма неполно и
не могут быть отличаемы от соответственных элементов кисти. Материала по
возрастным и индивидуальным изменениям их, таким образом, не имеется.
Рис. 83. Benthosuchus sushkini. Возрастные изменения и индиви-
дуальные вариации femur. Age stages and individual variations of
the femur. A—№ 246—19/2252; B—№ 233—19/2252; G—№ 245—
19/2252; D—№ 244—19/2252; E—№ 95—9/2243; F—№ 98—9/2243;
G—№ 94 — 32/2354; H — № 95—32/2354. s/4 натуральной вели-
чины. X 3/4.
В заключение нашего обзора возрастной и индивидуальной изменчи-
вости скелета вкратце резюмируем полученные факты.
Скелет Benthosuchus, вследствие непрерывного роста животного за
время его жизни, очень значительно увеличивается в размерах. От момента
метаморфоза до максимальной известной нам старости рост животного
увеличивается в 80 раз.
Повидимому, процесс роста протекал медленно, — быстрее на ранних
возрастных стадиях и затем все более замедлялся, пока не достигал не-
которой постоянной.
Наиболее сложный отдел скелета — череп в процессе постметаморфоз-
ного роста приобретал присущие ему адаптивные черты: часть родовых
и видовые признаки. Поэтому возрастные изменения черепа сравнительно
разнообразны и резко выявлены, особенно на ранних стадиях постмета-
морфозного роста.
Индивидуальные изменения черепа незначительны и сводятся, таким
образом, к ускорению или замедлению появления различных возрастных
особенностей.
98
Однако многие вариации черепа настолько сильно выражены, что,
будучи взяты отдельно, вне сравнения с различными градациями таких же
вариантов у других черепов, могли бы быть приняты за видовые особенности.
Позвоночный столб Benthosuchus в процессе роста окостеневает у ста-
рых особей сильнее, чем у молодых и среднего возраста индивидов, при-
обретая тесную связь центра с неуральной дугой, правда, еще путем син-
хондроза.
Повидимому, и у многих других стегоцефалов индивидуальный возраст
играет значительную роль в степени окостенения позвоночного столба.
Плечевой пояс Benthosuchus в области scapula-coracoideum, несмотря
на свою массивность, окостеневает даже у довольно крупных особей срав-
нительно слабо. Зато его накладные кости—interclavicula, clavicula и clei-
thrum окостеневают на ранних стадиях и дают целую серию возрастных
изменений (главным образом, — ключицы). Так, ключицы из сравнительно
узких овальных образований становятся широкими треугольными пласти-
нами, в связи с чем относительная ширина вентральной поверхности пле-
чевого пояса с возрастом увеличивается.
Humerus Benthosuchus, равно, как и femur, окостеневает в процессе
роста незначительно и так же незначительно изменяется по возрастным
стадиям. Однако отростки и шероховатости, служащие местами начал
и прикреплений мышц, с возрастом значительно усиливаются.
Зато у некоторых особей обнаруживаются значительные индивидуаль-
ные вариации окостенения humerus (femur) или отклонения humerus (femur)
от типичной формы, образование массивных укороченных костей.
Таз Benthosuchus окостеневает неполно, и стадии его развития еще
очень мало известны. Ilium по характеру возрастных и индивидуальных
изменений сходен с humerus и femur.
Кости предплечья и голени еще мало охарактеризованы возрастным
материалом, но, повидимому, слабо изменяются с возрастом. Среди кол-
лекции tibia имеется несколько сильно утолщенных экземпляров костей.
Таким образом, при рассмотрении возрастных и индивидуальных ва-
риаций скелета Benthosuchus в целом выявляется, что отделы скелета, не-
сущие ярко выраженную специфическую адаптацию, в наибольшей степени
изменяются с возрастом и мало меняются индивидуально. Примитивно
построенные (слабо моделированные) отделы скелета, — как скелет конеч-
ностей, плохо развитый у большинства водных форм, — слабо изменяются
с возрастом, но зато в отдельных случаях подвержены значительным инди-
видуальным колебаниям. Эта закономерность понятна, так как у прими-
тивных Tetrapoda давно установлено, что некоторые признаки, так сказать,
окончательно определяющие данную форму (род, вид), развиваются в пост-
эмбриональный период, в процессе роста особи. Поэтому специфические
адаптивные черты, появляясь сравнительно поздно, и определяют значи-
тельную возрастную изменчивость тех отделов скелета, в которых выра-
жена данная адаптация.
Что касается посткраниального скелета, то у большинства стегоце-
фалов он находится на низком уровне развития и слабо различается даже
у разных семейств. У водных и «сухопутных» лабиринтодонтов основные
различия посткраниального скелета сводятся к большему окостенению
его у последних.
Такой несовершенный посткраниальный скелет, в противоположность
пластичному к различным модификациям черепу, естественно слабо варии-
рует с возрастом, так как не имеет большого количества специфических
признаков, определяющих данную форму, т. е. адаптацию.
Весьма интересно, что у Benthosuchus степень окостенения конечностей
и их поясов мало увеличивается с возрастом. Это обстоятельство указывает
на то, что у низших Tetrapoda степень окостенения отдельных элементов
скелета есть некоторая постоянная для данной группы или формы, полу-
ченная в результате эволюционного процесса.
7*
99
В коллекции Палеонтологического института, на ряду с остатками
Benthosuchus из местонахождений р. Шарженги, имеются в большом коли-
честве остатки Wetlugosaurus из синхроничных местонахождений р. Ветлуги.
Эти остатки в настоящей работе не описываются, но были нами изучены
для сравнения с Benthosuchus. Черепной материал представлен лишь одним
более или менее полным черепом, но имеется большое количество отдельных
фрагментов черепов и костей посткраниального скелета; вместе с остатками
очень небольших особей имеются фрагменты черепа и кости конечностей,
принадлежавшие черепам, имеющим свыше 600 мм длины.
Посткраниальный скелет Wetlugosaurus имеет небольшие отличия от
Benthosuchus, но возрастные и индивидуальные изменения скелета обеих
форм в точности совпадают. На ранних возрастных стадиях сходство весьма
значительно.
В последнее время из сходных и синхроничных вновь открытых место-
нахождений Тиманской области доставлены обломки костей конечностей
и фрагменты основания черепа, принадлежащие еще более огромным особям
Benthosuchus, чем все известные с р. Шарженги.
Захоронение вместе остатков самых различных по возрасту особей
присуще всем многочисленным местонахождениям эотриасовых лабиринто-
донтов и зависит, как мы увидим ниже, от условий образования место-
нахождений. Благодаря этой особенности нам удалось наметить явления
возрастной изменчивости у Benthosuchus.
Разумеется, приведенные данные далеки от полного представления
о росте и индивидуальной изменчивости Benthosuchus. Неравномерное
количество остатков разного возраста, отсутствие полных скелетов за-
ранее сужают возможности исследования. Одним из важнейших вопросов,
для решения которого мы не имеем нужного количества фактов, является
вопрос о продолжительности жизни Benthosuchus и, следовательно, скорости
его роста. Этот вопрос и не обсуждается в нашей работе, так как, кроме
аналогии с современными крупными низшими Tetrapoda, нет никакой
отправной точки для его разрешения.
В литературе по стегоцефалам известно очень мало работ с описанием
возрастных стадий отдельных форм.
Общеизвестные классические работы Германа Мейера [Hermann von
Meyer, 1857] об Archegosaurus и Креднера [Credner, 1892] о Branchiosaurus
основаны на очень большом материале, по количеству и полноте остатков
значительно превосходящем материал по Benthosuchus. Однако плохая
сохранность материала и плохая препаровка не позволили указанным
авторам детально выяснить возрастную и индивидуальную изменчивость
этих форм. Кроме того, в силу условий захоронения или особенностей
животных, для Archegosaurus и Branchiosaurus не было найдено таких
крупных особей, как у Benthosuchus.
Оставляя в стороне Branchiosaurus как неспециализованную мелкую
форму из другого отряда стегоцефалов, можно отметить, что возрастные
изменения Archegosaurus весьма сходны с таковыми у Benthosuchus: череп
приобретает законченные черты своей адаптации лишь у более крупных
особей; черепа очень молодых особей Archegosaurus обладают тем же ха-
рактером строения, как и маленький череп № 4—19/2252 у Benthosuchus.
Так, большие орбиты располагаются в середине треугольного укорочен-
ного черепа, foramen parietale сильно приближено к орбитам и т. д. Однако
Archegosaurus, повидимому, еще до окончания метаморфоза обладал зна-
чительно большей величиной, чем уже прошедшие метаморфоз особи Bentho-
suchus. Если величина личинки у стегоцефалов определяет до известной
степени конечную величину старой особи, то возможно, что старые особи
Archegosaurus достигали еще более гигантской величины, чем старые бенто-
зухи.
Некоторым подтверждением этого предположения служит находка
в Акбатыровском руднике обломка очень крупного черепа Platyops stucken-
100
bergi — близко родственной Archegosaurus формы. Этот череп в своем
полном виде должен был достигать приблизительно 1200 мм в длину.
Индивидуальные изменения скелета Archegosaurus Мейером не отме-
чены, но, судя по некоторым изображениям, они также имели место.
Обильный по числу особей материал остатков стегоцефалов из Нюршана
в Богемии, Линтона в Огайо США и South Joggins в Канаде не подвергался
исследованию с точки зрения возрастных или индивидуальных изменений,
да и пе может дать нужного количества фактов без детальной препаровки.
Позднейшая переработка материалов из двух последних местонахождений,
выполненная Ромером [Romer, 1930] и Стином [Steen, 1934], ничего суще-
ственного для познания возрастных и индивидуальных изменений не дала,
что объясняется отчасти близко сходными размерами найденных особей.
Кейз [Case, 1935] опубликовал описание глыбы породы, где были най-
дены 16 черепов Trimerorhachis вместе с частями посткраниального скелета.
К сожалению, работа Кейза, вследствие поверхностного исследования,
плохих изображений и недостаточно тщательной препаровки, не дает пол-
ного представления о Trimerorhachis вообще, не говоря уже о возрастных
или индивидуальных изменениях этого животного. Кроме того, судя по
таблице измерений и по снимкам, все черепа мало вариируют в величине
(80—116 мм длины), принадлежа близким по возрасту особям.
Серии найденных одновременно, в одном и том же месте черепов
Microphalis и Lydekkerine, описанные Бройли и Шредером [1937], также
принадлежат почти равным по величине особям и не показывают сколь-
ко-нибудь значительных возрастных изменений.
Особо нужно отметить работу Вагнера [Н. Wagner, 1935], специально
посвященную индивидуальному росту Trematosaurus brauni. Материал,
положенный в основу исследования, состоит из 62 interclavicula, 21 clavi-
cula, 130 черепных крыш (25 полных черепов), 57 фрагментов нёбной по-
верхности черепа и 39 нижних челюстей. Не пытаясь детально отпрепа-
ровать имеющийся большой материал, автор произвел большое количество
измерений отдельных элементов черепа, нижней челюсти и плечевого пояса
в различных направлениях от центров окостенения. Из этих измерений не
получилось никаких ясных выводов, кроме общего представления о про-
порциональных отношениях деталей различных черепов Trematosaurus,
Исходя из неправильного представления о наличии последовательных
зон «ритмического роста» («годовых колец») на различных костях Trema-
tosaurus и главным образом — на clavicula, Вагнер вычерчивает кривую
роста черепа и приходит к выводу, что череп Trematosaurus brauni к концу
первого года жизни достигал 72, а к концу 28-го — 402 мм длины. Автор
приводит соответственные длины черепа для каждого года жизни. Кроме
этих, весьма спекулятивных построений, Вагнер не сообщает никаких
сведений о возрастных и индивидуальных изменениях Trematosaurus.
Пользуясь измерениями и немногочисленными изображениями автора,
можно установить, что размеры Trematosaurus также весьма значительно
увеличивались с возрастом и череп его проделывал целую серию возра-
стных изменений, сходных с возрастными изменениями ‘ Benthosuchus.
Сходство это далеко не полное и объясняется различным происхождением
и различной адаптацией черепа Benthosuchus и Trematosaurus. Однако
процесс возрастного изменения черепа Trematosaurus в точности подобен
возрастным изменениям Benthosuchus. Отсутствие очень маленьких чере-
пов Trematosaurus не дает возможности подробно анализировать весь ход
роста черепа.
Индивидуальные изменения черепа Trematosaurus, невидимому, про-
явлены даже несколько резче, чем у Benthosuchus. Интересен отмеченный
автором факт сильного сужения («перешнуровывания») черепа позади
орбит на одном экземпляре Trematosaurus.
Подводя итоги оценке работы Вагнера, следует отметить, что она не
дает ясного представления об изменении скелета Trematosaurus с возрастом.
101
Кроме того, в руках автора был хотя и обильный, но разрозненный материал,
собранный из разных мест. Таким образом, индивидуальные различия скелета
Trematosaurus могли быть вызваны и различными условиями существо-
вания.
Из приведенного краткого обзора литературных данных видно, что
материал по Benthosuchus sushkini, несмотря на существенные недостатки,
впервые дает представление о характере возрастных и индивидуальных
изменений высших стегоцефалов.
Значительное увеличение размеров животного с возрастом сводит к нулю
возможность установления видов по величине найденного объекта. Зна-
чительные возрастные изменения черепа свидетельствуют, что и череп
и некоторые другие отделы скелета могут значительно различаться на
разных стадиях жизни животного. Различия эти в крайних выражениях
имеют значение родовых признаков.
Крайние индивидуальные вариации черепа также могут быть ошибочно
приняты за видовые отличия.
Индивидуальные вариации некоторых элементов посткраниального
скелета, возможно, вызванные патологическими дефектами, могут в не-
большом числе случаев значительно изменить общую форму или степень
окостенения данного элемента. При единичных находках такие откло-
нения, несомненно, будут возведены в правило и признань! за отличия
крупного порядка, получив значение родовых или видовых признаков.
Наиболее важный вывод из всего вышесказанного, это — необходимость
тщательного учета возрастных изменений и индивидуальных отклонений
как при описании новых находок, так и при пересмотре старых материалов.
Несомненно, что многие единичные, да и серийные находки, описанные
в качестве новых родов и видов, являются возрастными стадиями или инди-
видуальными вариациями. Особенное значение мы придаем возможности
ошибок при описании фрагментарного, неполноценного материала, на
котором, к сожалению, построено наибольшее количество новых родов
и видов, только загромождающих ясное представление о фауне и не дающих
ничего существенного для ее познания.
Отсутствие специфической адаптации на ранних стадиях роста черепа
Benthosuchus (так же, как, повидимому, и черепов других стегоцефалов)
дает возможность более достоверно анализировать происхождение и род-
ственные связи данной формы. Это мы и попытаемся сделать для Bentho-
suchus.
глава и
СИСТЕМАТИЧЕСКОЕ ПОЛОЖЕНИЕ И ФИЛОГЕНЕТИЧЕСКОЕ ЗНАЧЕНИЕ
BENTHOSUCHUS SUSHKINI
Ефремов [1929] в первом описании Benthosuchus (Benthosaurus) sushkini
отметил значительное сходство строения черепа Benthosuchus с черепами
Trematosaurus и Mastodonsaurus, с одной стороны, и Capitosaurus — с дру-
гой. Однако ’большая примитивность Benthosuchus, проявляющаяся как
в деталях строения черепа, так и в строении позвоночника, не позволила
Ефремову включить Benthosuchus в одно из перечисленных семейств. Строе-
ние позвоночника, промежуточное между рахитомным и стереоспондиль-
ным типом, было названо в той же работе прогрессивно рахитомным или
неорахитомным, и род Benthosuchus Ефремов считал заслуживающим выде-
ления в особое семейство.
В 1930 г. А. Н. Рябинин описал новый род Wetlugosaurus из синхронич-
ных отложений р. Ветлуги и установил, что степень развития позвоноч-
ника Wetlugosaurus вполне соответствует таковой у Benthosuchus. Wetlu-
gosaurus был включен Рябининым в семейство Rhinesuchidae.
Позднее [1932] Ефремов объединил оба рода в одно семейство Bentho-
suchidae (Benthosauridae), считая их близко родственными формами. Вуд-
102
ворд [A. S. Woodward, 1932] в английском издании «Основ палеонтологии»
Циттеля включил и Benthosuchus и Wetlugosaurus в сем. Rhinesuchidae.
Хюне [1932] описал из пестрого песчаника Германии новую, особую фор-
му лабиринтодонта Sclerothorax hypselonotus, также обладающую прогрес-
сивно рахитомным строением позвоночника. Таким образом, наличие этой
ступени в разных группах вполне подтвердилось.
Сэве-Зедерберг [Save-Soderbergh G., 1935] причислил Benthosuchus
к подсемейству Lyrocephalinae сем. Trematosauridae.
Гартман-Вейнберг и Кузьмин [1936], основываясь на работе Сэве-Зе-
дерберга, подвергли критике объединение Benthosuchus и Wetlugosaurus
в одно семейство Benthosuchidae, считая Benthosuchus типичным тремато-
завридом.
Сэве-Зедерберг [1937] впоследствии отказался от своих прежних за-
ключений и пришел к выводам, сделанным Ефремовым еще в 1929 г., т. е.
что Benthosuchus ближе всего родственен Wetlugosaurus, Capitosaurus и
Mastodonsaurus и должен быть выделен в особое семейство сверхсемейства
Capitosauroideae. Это семейство есть особая ветвь отряда Capitosauroidei,
которая, возможно, оканчивается в Mastodonsaurus.
Ефремов [1937, 1938] разобрал некоторые ошибки прежних авторов
и считает Benthosuchus и Wetlugosaurus представителями особой ступени
в эволюции высших Labyrinthodontia — группы Neorhachitomi, из которой
впоследствии развились типичные триасовые стереоспондильпые формы.
Не останавливаясь вновь на деталях строения Benthosuchus, чтобы не
повторять сказанного в первой части, вкратце резюмируем общую характе-
ристику этой формы.
Benthosuchus является лабиринтодонтом с особым прогрессивно рахитом-
ным строением позвоночника. Этот тип строения характеризуется редук-
цией костных плевроцентров при наличии еще слабо окостеневших гипо-
центров. Последние у старых особей вступают в контакт с неуральной дугой
путем синхондрозного соединения и образуют таким путем низшую стадию
типичного нотоцентрального стереоспондильного позвонка. Неорахитом-
ный тип строения позвонков представляет собой очень яркую промежуточ-
ную стадию между рахитомным и стереоспондильным типами, вполне
соответствующую прежним теоретическим построениям.
Череп Benthosuchus обладает некоторыми сравнительно примитивными
чертами (по сравнению с триасовыми стереоспондилами), как, например,
более длинные ramus anterior и ramus posterior pterygoidei, отнесенные назад
челюстные мыщелки и т. д. В то же время череп Benthosuchus являет высо-
кую степень редукции хрящевых элементов, не уступающую типичным
стереоспондилам триаса и даже превосходящую таковую, например у Capito-
saurus. Слабое развитие epipterygoideum у Benthosuchus является единствен-
ной чертой, свидетельствующей о большей примитивности его chondrocra-
nium сравнительно с триасовыми стереоспондилами.
Таким образом, Benthosuchus представляет собой интересную форму,
в которой примитивное строение позвоночника и некоторые черты строения
черепа объединяются с высокой степенью редукции хрящевых костей моз-
гового черепа.
Объяснение этой особенности Benthosuchus заключается в приспособлен-
ности к водному образу жизни. Так, нижнепермский Trimerorhachis, яв-
ляющийся сравнительно с другими современными ему лабиринтодонтами
типичной водной формой, обладает очень высокой степенью редукции кост-
ной мозговой коробки.* По общему характеру своего черепа Trimerorha-
chis гораздо более близок к последним верхнепермским формам, чем к своим
современникам.
Очевидно, что и Benthosuchus как водная форма, подобно Trimerorha-
chis, имеет высокую эволюционную ступень строения neurocranium, не
соответствующую ни геологическому возрасту, ни общему эволюционному
уровню этой формы.
103
Из всех других известных Labyrinthodontia Benthosuchus наиболее схо-
ден с Wetlugosaurus. Основные отличия Wetlugosaurus от Benthosuchus:
1) несколько другой характер скульптуры и более слабое развитие кана-
лов боковой линии у Wetlugosaurus;
2) более широкая морда с очень большим круглым fenestra intermaxilla-
ris без следов перегородки; зубной, ряд на vomer не клинообразной формы,
а в виде дуги, окаймляющей задний край fenestra intermaxillaris;
3) немного более массивный processus ciiltriformis парасфеноида;
4) более узкий и высокий затылок;
5) несколько меньшая степень редукции хрящевых элементов neuro-
cranium;
Рис. 84. Wetlugosaurus angustifrons. Череп № 6—19/2252. 3/4 натураль-
ной величины. Skull № 6—19/2252. х 3/4. А — сверху (from above),
В — снизу (from below).
6) несколько большая, ранее наступающая степень окостенения пост-
краниального скелета и sphenethmoideum и epipterygoideum в черепе.
В остальном обе формы очень сходны. Еще больше это сходство вы-
ступает при сравнении наиболее молодых возрастных стадий Benthosuchus
и Wetlugosaurus.
Попытаемся провести сравнение маленького черепа Bethosuchus № 4—
19/2225 (рис. 56—59) с имеющимся в нашей коллекции неполным черепом
молодого Wetlugosaurus № 6 —19/2252 из Большой Слудки на р. Ветлуге
(рис. 84А, В).
Череп маленького Benthosuchus обладает широкой мордой с большим
fenestra intermaxillaris и лишь слабыми зачатками перегородки. Латераль-
ные края черепа прямые, затылок сравнительно узкий и высокий. Processus
cultriformis парасфеноида короткий и довольно широкий. При сравнении
рис. 84 и рис. 56—59 сходство молодых особей Wetlugosaurus и очень моло-
дых Benthosuchus проступает еще резче.
Если бы был найден череп Wetlugosaurus стадии, соответствующей
№ 4—19/2252 у Benthosuchus, то сходство обеих форм достигло бы крайней
степени.
Это обстоятельство показывает, что Benthosuchus и Wetlugosaurus рас-
ходятся лишь в различной адаптации черепа и что, несмотря на одновремен-
на
ное существование, Wetlugosaurus несколько примитивнее Benthosuchus, так
как стоит ближе к непосредственному предку Benthosuchus, строение кото-
рого отражено в ранних стадиях роста последнего.
Это несколько более примитивное положение Wetlugosaurus вполне
согласуется и с такими чертами его строения, как наличие более высокого
и узкого затылка и немного меньшая редукция элементов neurocranium.
В то же время по развитию позвоночника Wetlugosaurus стоит на той же
ступени Neorhachitomi, что и Benthosuchus.
По всей вероятности, Wetlugosaurus является несколько более «сухо-
путной», позднее приспособившейся к водному обитанию, чем Benthosuchus,
формой, в связи с чем ста-
новится понятной и мень-
шая степень редукции его
neurocranium и более ран-
нее и сильное окостенение
отдельных элементов пост-
краниального скелета.
Во всяком случае, тес-
ное генетическое родство
обеих форм не подлежит
никакому сомнению, что
хорошо подтверждается,
как мы увидим ниже, гео-
графическим распростране-
нием обеих форм.
Поскольку Wetlugosau-
rus настолько близок к Ca-
pitosaurus, что едва ли не
является его непосредст-
венным предком, — сход-
ство Benthosuchus и Capito-
saurus само собой понятно.
Сходство Benthosuchus и
Mastodonsaurus, особенно
подчеркиваемое Сэве-Зе-
дербергом [1937], не кажет-
ся нам очень значительным.
Такие признаки, как треу-
гольная форма черепа и
широкий затылок, слишком
общи и существуют в весь-
ма разнородных генетиче-
Рис. 85. А—Benthosuchus sushkini, В — Mastodon-
saurus kappelensis, G — Wetlugosaurus angustifrons;
D — Trematosaurus brauni. Черепа сверху. Skulls,
from above.
ских формах. Вогнутые ла-
теральные края черепа Benthosuchus, сходные, по Сэве-Зедербергу, с сильно'
вогнутыми краями черепа Mastodonsaurus, у молодых форм Benthosuchus
выражены очень слабо. Эта вогнутость латеральных краев черепа хорошо
видна у старых особей Benthosuchus (рис. 62) и объясняется расширением
черепа в затылке при наличии сравнительно узкой морды — черта, ко-
торая может быть получена при. сходной адаптации у самых различных.
форм.
Кроме того, Сэве-Зедерберг основывает свои суждения на рисунке черепа,
сделанном Фраасом, который, как известно, не учел деформации расплю-
щенного черепа. На самом деле, Mastodonsaurus обладает черепом с гораздо
более прямыми краями и в этом виде во много раз больше сходен с черепом
Wetlugosaurus, Capitosaurus и Benthosuchus, без всяких специальных допу-
щений. Строение нёбной поверхности черепа Mastodonsaurus весьма сходно
во многих своих деталях со строением у Benthosuchus, так же, как (следова-
тельно) у Wetlugosaurus и Capitosaurus. Сходство Mastodonsaurus и Всп-
105.
thosuchus только проявляется несколько резче, благодаря сходной адапта-
ции передней части черепа.
Между тем, строение переднего отдела черепа очень сходно у Benthosu-
chus и Trematosaurus и является в то же время основной чертой сходства
последнего с Mastodonsaurus, уже пе раз обсуждавшегося.
Комбинированные рисунки — 85 и 86А, В, С, D — хорошо иллюстрируют
сказанное.
Рис. 86. А — Benthosuchus sushkini, В — Masto-
donsaurus kappelensis, G — Wetlugosaurus angu-
stifrons, D — Trematosaurus brauni. Черепа сни-
зу. Skulls from below.
Значительные различия общего строения черепа Trematosaurus и Bentho-
suchus, обсуждавшиеся в цитированных выше работах уже не раз, служат
указанием, что сходство Trematosaurus и Benthosuchus является конверген-
цией и вызвано сходной адап-
тацией переднего отдела че-
репа.
Не исключена возмож-
ность, что сходство строения
переднего отдела черепа Mas-
todonsaurus и Benthosuchus
также есть результат конвер-
генции. Однако Mastodonsau-
rus, как уже отмечалось вы-
ше, сходен с Benthosuchus и
Wetlugosaurus и в строении
других отделов черепа, как,
например, основания черепа,
затылочной части крыши че-
репа и затылка. Это же сход-
ство проявляется и в распо-
ложении зон интенсивного
роста, анализированном Сэве-
Зедербергом.
Суммируя все приведенные
соображения, мы приходим
к следующим заключениям.
1. Benthosuchus наиболее
близко родственен Wetlugosa-
urus. Различия обоих родов
не генетического порядка и
сводятся к различным адапта-
циям, причем Benthosuchus,
вероятно, более специализо-
ван, чем Wetlugosaurus-
2. Выделение обеих форм
в различные семейства, как
это делалось до сих пор, ничем не оправдано.
3. Wetlugosaurus, Capitosaurus и Benthosuchus образуют одну генетически
тесно связанную группу, к которой, повидимому, принадлежит и Masto-
donsaurus. В этой группе Benthosuchus и Mastodonsaurus более аберрантны,
чем два остальные рода.
4. Специализация Benthosuchus, т. е. его уклонение от типичных Capi-
tosauridae, направлено в ту же сторону, что и специализация Trematosaurus,
а также отчасти и Mastodonsaurus. Однако результаты этой специализации,
поскольку они шли разными путями в разнородных генетически формах,
различны и сказались лишь в конвергентном сходстве переднего отдела че-
репа. При этом Mastodonsaurus как член одной группы обладает большим
сходством с Benthosuchus, несмотря на то, что его адаптация несколько менее
сходна, чем адаптация передних отделов черепа у Benthosuchus и Trema-
tosaurus.
Среди более древних, типично рахитомных форм ближайшей родственной
106
Benthosuchus формой является Rhinesuchus (рис. 87А). Генетическое родство
Benthosuchus и Rhinesuchus проявляется через Wetlugosaurus. С последним
Rhinesuchus настолько сходен, что первые остатки Wetlugosaurus были
описаны Н. Н. Яковлевым как Rhinesuchus wolgo-dwinensis. Отличия Rhi-
nesuchus от Wetlugosaurus сводятся к меньшей редукции окостенения пепго-
cranium, большей длине обеих ветвей птеригоида, большему распростране-
нию шагрени на нёбных костях и более укороченному и широкому черепу.
Не трудно видеть, что эти отличия являются как раз отличиями эволюцион-
ной стадии Rhinesuchus, вполне соответствующими более древнему возрасту
и типичной рахитомности последнего.
Кроме Rhinesuchus, следует упомянуть и Melosaurus. Эта, еще плохо
изученная форма, по новым данным [Ефремов, 1937], обладает многими
чертами сходства с Benthosuchus и
отчасти Mastodonsaurus, которым она,
несомненно, родственна.
В свете всех этих соображений яс-
по выступает большое филогенетиче-
ское значение таких форм, как Bentho-
suchus и Wetlugosaurus.
Находясь на примитивной ступени
развития (Neorhachitomi), они несут
в себе черты строения нескольких форм
из ступени Stereospondyli, обычно груп-
пируемых в различные семейства.
Поэтому детальное изучение форм,
подобных Benthosuchus, очень важно
для анализа происхождения и разви-
тия стереоспондильных лабиринтодон-
тов и для выработки их естественной
классификации.
Такой естественной классификации
в настоящее время не существует, и
разграничение различных групп ос-
новано на ЧИСТО случайных призна- Рис- 87• R^MSuchus sp. Череп снизу,
т-г J /г F Skull from below.
ках. Попытка установления более на-
туральной систематики высших лаби-
ринтодонтов была сделана Сэве-Зедербергом в цитированных выше работах.
Однако мы не считаем возможным придерживаться этой классификации,
так как, несмотря на правильные предпосылки, Сэве-Зедерберг основы-
вает ее на весьма поверхностных данных. Классификация по зонам интен-
сивного роста, предложенная тем же автором, также является еще неудов-
летворительной. Зоны интенсивного роста костей крыши черепа это не нечто
самодовлеющее, а способ, которым осуществляются необходимые адаптив-
ные модификации формы черепа. Это с достаточной ясностью было под-
черкнуто в работе Быстрова [1935], который впервые обнаружил влияния
локализованного интенсивного роста в черепе стегоцефалов. Поэтому нет
уверенности, что эти способы не могли быть общими в разнородных формах,
и, следовательно, при этой классификации конвергентные формц будут объ-
единены как генетически родственные. Этим же существенным недостатком
страдает и первая классификация Сэве-Зедерберга.
Поскольку выработка новой систематики лабиринтодонтов должна
быть основана на детальном изучении большого материала, мы избегаем
в настоящей работе пытаться вносить какие-либо существенные изменения
в существующую классификацию, хотя предметом нашего обсуждения и яв-
ляются формы, переходные от Rhachitomi к Stereospondyli. В пределах изу-
ченного материала мы можем дать заключения только частного порядка,
совершенно не претендуя на выяснение общих закономерностей.
Представляется вероятным, что признаки, на которых основано выде-
107
ление семейств Capitosauridae и Mastodonsauridae несущественны н тре-
буют пересмотра.
Если считать, что генетическая классификация должна быть вертикаль-
ной, а не горизонтальной, то семейства Capitosauridae, Mastodonsauridae и
Benthosuchidae1 должны быть объединены в одно семейство. В это же семей-
ство должен быть включен и Rhinesuchus, несомненно, стоящий очень
близко к предкам Wetlugosaurus. Такое семейство, проходящее через все
ступени эволюционного развития высших Labyrinthodontia, лучше всего
назвать семейством Rhinesuchidae.
Если принимать признаки семейства более мелкого значения для горизон-
тальной классификации, то все перечисленные формы уложатся в три
семейства: Rhinesuchidae — Rhachitomi, Benthosuchidae — Neorhachitomi,
Capitosauridae — Stereospondyli. Стратиграфическое распределение во вре-
мени соответственно: верхняя пермь, эотриас, триас.
В заключение обзора родственных связей Benthosuchus следует отметить,
что в последнее время в эотриасовых отложениях Верхней Волги и Тимана,
идентичных эотриасовым отложениям с Benthosuchus и Wetlugosaurus,
найдены новые формы лабиринтодонтов, очень близкие к Benthosuchus и
Wetlugosaurus и также, повидимому, стоящие на ступени Neorhachitomi
[Ефремов, 1938]. Эти формы еще не достаточно изучены, но показывают
иные адаптационные уклонения, в частности еще более близкие к Trema-
tosaurus.
Не исключена возможность, что при дальнейшем исследовании фауны
неорахитомных лабиринтодонтов будут собраны факты, доказывающие
возникновение таких форм, как Trematosaurus из этой же группы. Наличие
многих черт сходства такой формы, как Thoosuchus [Ефремов, 1938] с Tre-
matosaurus, с одной стороны, и с Benthosuchus — с другой, служит указа-
нием на неестественность глубокого классификационного разделения тре-
матозаврид и капитозаврид, проводимого Сэве-Зедербергом. По всей вероят-
ности, в группе трематозаврид смешаны формы как более глубокого во
времени происхождения, так и сравнительно недавно возникшие из группы
Neorhachitomi.
глава ш
НЕКОТОРЫЕ СООБРАЖЕНИЯ О ПРОИСХОЖДЕНИИ СТЕРЕОСПОНДИЛЬНЫХ
ПОЗВОНКОВ У ВЫСШИХ LABYBINTHODONTIA
Строение позвоночника у Benthosuchus и других форм, стоящих на одной
с ним ступени (Wetlugosaurus, Sclero thorax), наиболее хорошо соответствует
обычному представлению о генезисе нотоцентрального стереоспондилового
позвонка лабиринтодонтов.
Постепенная редукция плевроцентров в типичном рахитомном позвонке,
идущая параллельно со сращением гипоцентра с неуральной дугой и увели-
чением гипоцентра до образования монолитного круглого центра, — вот
путь, установленный многочисленными исследованиями.
Находка Benthosuchus и других форм с пеорахитомным строением позво-
ночника дает нам наиболее яркую промежуточную стадию между рахитом-
ным и стереоспондиловым типами. Момент редукции плевроцентров сопро-
вождался *перенесением всей опоры неуральной дуги исключительно па
гипоцентр, синхондрозно контактирующий с неуральной дугой. Такой позво-
нок является, если можно так выразиться, стереоспондиловым позвонком
in statu nascendi, притом происходящим из типично рахитомного.
Геологический возраст Benthosuchus и Wetlugosaurus как нельзя лучше
согласуется с их неорахитомными позвонками. Все триасовые лабиринто-*
1 Поскольку семейство Benthosuchidae предложено Ефремовым в 1929 г., а объ-
единение Benthosuchus и Wetlugosaurus сделано им же в 1932 г., то приоритет на-
звания семейства должен быть за Ефремовым. Сем. Wetlugosauridae установлено
Сэве-Зедербергом в 1934 г.
108
донты стереоспондильны, все пермские рахитомны, и наличие переходной
ступени между обоими типами в самых древних слоях триаса еще более
соответствует общепринятому представлению о развитии стереоспондилии.
Нам, однако, известны случаи, когда появление стереоспондильных
позвонков возникает во времени раньше или, наоборот, запаздывает. Хо-
роший пример мы имеем в Sclerothorax, сохранявшем позвонки неорахитом-
ного типа в то время, когда все остальные лабиринтодонты были уже сте-
реоспондильны. Может быть также упомянуто появление стереоспондиль-
ных позвонков в переднем отделе позвоночника у пермского Dvinosaurus.
Кроме того, с появлением детальных палеонтологических описаний и на-
ходками новых материалов лучшей сохранности прежний взгляд на рахи-
томные позвонки как позвонки, построенные по одному типу у всех Rha-
chitomi, потерял свое значение.
Чем больше накапливается сведений по морфологии рахитомных лаби-
ринтодонтов, тем больше нам известно вариаций структуры рахитомного
позвонка.
В качестве примера укажем на недавно описанное [Стин, 1934, 1937]
строение позвонков у Dendrerpeton и Acanthostoma.
Оставляя в стороне эти древние формы, особенное строение позвонков
которых может быть вызвано различными уклонениями от эмболомерного
к рахитомному типу или наоборот, — возьмем типичных Rhachitomi.
Весьма интересно строение рахитомного позвонка у Cacops (рис. eeCi).
Эти позвонки обладают очень низкими гипоцентрами и очень большими пле-
вроцентрами. Последние образуют плоский диск, по всей своей передней по-
верхности контактирующий с широкой специальной фасеткой неуральной
дуги. Задняя поверхность плевроцентра сочленяется с особой передней
фасеткой на неуральной дуге следующего позвонка и со скошенной передней
поверхностью его гипоцентра. Контакт неуральной дуги и гипоцентра го-
раздо слабее и осуществляется путем небольшой нижней фасетки на конце
сильно опущенной вниз базальной части неуральной дуги. Благодаря этим
особенностям центр позвонка Cacops приобретает интересное особенное
строение. Его нижняя часть на высоту не более трети толщины хорды обра-
зована целиком гипоцентром. Верхняя большая часть центра, назади еще
сильно вдающаяся в центр следующего позвонка, образована неуральной
дугой и плевроцентром. Совершенно очевидно, что в таком рахитомном
позвонке плевроцентр играет очень большую роль и позвонок Cacops
ближе к эмболомерному типу строения позвоночника, чем обычный рахитом-
ный тип.
Другой пример — отроение значительно более поздней формы: Micro-
pholis stowi, недавно подробно изученной Бройли и Шредером [Broili
F. u. Schroder J., 1938] (рис. 88С2). Здесь гипоцентр имеет мощный контакт
с неуральной дугой по широкой скошенной площадке, так что неуральная
дуга опирается исключительно на гипоцентр. Плевроцентр очень сильно
развит и по всей своей передней поверхности контактирует с неуральной
дугой и всем задним краем гипоцентра. Таким образом, контакт предыду-
щего позвонка с последующим осуществляется только через плевроцентр,
образующий весь задний отдел центра позвонка. Иными словами, в Micro-
pholis (весьма поздней форме) мы имеем также несколько эмболомерный ха-
рактер строения позвонков.
Несомненно, дальнейшее изучение строения рахитомного позвоночника
у различных форм покажет еще более разнообразные структуры позвонков.
Однако обсуждение и разбор этих случаев не составляет задачи нашей
работы. На приведенных двух примерах мы показали, что различные типы
строения рахитомных позвонков действительно существуют, и притом
у весьма различных по возрасту форм, как нижнепермский Cacops и триа-
совый Mwropholis.
Если рахитомные позвонки обладают значительно большим, чем это
ранее принималось, структурным разнообразием, то каким путем осуще-
109
ствляется переход этих различных структур в стереоспондильпое состояние?
Является ли неорахитомный позвонок Benthosuchus и Wetlugosaurus той
единственной переходной стадией, которую неизбежно проходят самые раз-
личные группы лабиринтодонтов при развитии от Rhachitomi к Stereospon-
dyli?
Эти вопросы заставили нас попытаться проверить строение позвонков
у верхнепермских поздних Rhachitomi на доступных нам материалах.
В этом отношении наиболее выгодно было обратиться к таким формам,
как Dvinosaurus и Platyops. Первый лабиринтодонт обладает стереоспон-
дилъными позвонками в переднем отделе туловища, второй показывает
Рис. 88. Возможные пути развития стереоспондильных позвонков в различных группах
Labyrinthodontia. A. Dvinosaurus. 1 — обычные рахитомные позвонки среднего отде-
ла туловища. 2.—момент прирастания плевроцентра к гипоцентру и образования пол-
ного костного центра, еще отделенного от неуральной дуги (первые позвонки у моло-
дой особи). 3—законченные стереоспондильные позвонки (первые позвонки старой осо-
би). В. Platyops. 1 — рахитомные позвонки с еще неприросшими плевроцентрами
(задний отдел туловища). 2—плевроцентры слиты с неуральной дугой (передний от-
дел туловища). 3 — конечная гипотетическая стадия стереоспондильного позвонка.
С. Cacops и Micropholis. 1 — Gacops (туловищные позвонки), 2 — Micropholis (туло-
вищные позвонки), 3 — гипотетическая стадия — стереоспондильный позвонок в ре-
зультате срастания плевроцентра с гипоцентром и неуральной дугой. D. Benthosu-
chus (Wetlugosaurus) и Capitosaurus. 1 — обычный рах;итомный позвонок предковой
формы. 2 — неорахитомная стадия (туловищные позвонки Benthosuchus у старых
особей). 3 — Capitosaurus. Дальнейшая стадия—стереоспондильный нотоцентральный
позвонок. Diagramm, showing the different ways and stages of the development of the
stereospondylous vertebrae. A. Dvinosaurus. 1. Rhachitomous vertebrae of the middle
part of vertebral column. 2. Stage of the adjoining of the pleurocentrum and hypo-
centrum and developing of the whole osseous centrum (the foremost vertebrae of a young
specimen). 3. Stage of the very stereospondylous vertebrae (foremost vertebrae of a more
old specimen). B. Platyops. 1. Rhachitomous vertebrae with disjoined pleurocentrum
(only synchondrous junction) — posterior dorsal vertebrae. 2. Confluence of the pleuro-
centra and neural arch (anterior dorsal vertebrae). 3. Hypothetic stage — a ste-
reospondylous vertebrae. C. Cacops and Micropholis. 1. Cacops (dorsal vertebrae).
2. Micropholis (dorsal vertebrae). 2. Hypothetic stage — a stereospondylous vertebrae.
D. Benthosuchus (Wetlugosaurus) and Capitosaurus. 1. Typical rhachitomous vertebrae
of an ancient form, unnamed. 2. Benthosuchus. Neorhachitomous stage (anterior dorsal
vertebrae of an old big specimen). 3. Capitosaurus. Typical stereospondylous vertebrae.
110
высокую степень окостенения рахитомного позвоночного столба, близкую
к переходу к стереоспондильному типу строения.
Позвонки Dvinosaurus детально описаны и изображены Быстровым [1938],
из работы которого мы и берем их изображение (рис. 88А1? А2, А3).
Позвонки Platyops stuckenbergi (рис. 88Bj и В2) изучены по полным двум
позвонкам старой особи Platyops из Акбатыровского рудника, впервые
изображенным Ефремовым [1933], и по почти полному скелету № 160/1
ПИН, недавно найденному у г. Белебея.
Позвонки Dvinosaurus обладают рахитомным строением наиболее обыч-
ного типа. Неуральпая дуга не контактирует или слабо контактирует с
треугольными высокими гипоцентрами. Парные узкие плевроцентры также
не имеют контакта с неуральной дугой и располагаются в просвете, обра-
зующемся в верхней части центра благодаря заострению концов гипоцент-
ров.
Прослеживая строение позвонков переднего отдела туловища сзади
наперед, мы отчетливо видим, как плевроцептры сначала тесно соединяются
с гипоцентром анкилозным соединением, затем плотно прирастают и, нако-
нец, нацело сливаются с гипоцентром в одну кость с неразличимым соеди-
нением. Это приращение плевроцентра идет параллельно все большему
окостенению как гипоцентра, так и плевроцентра, совместно обра-
зующих полный стереоспондильный костный центр.
У старых экземпляров Dvinosaurus плевроцентры в трех первых позвон-
ках нацело срастаются с гипоцентрами и неуральной дугой, превращая
эти позвонки в стереоспойдильные. Таким образом, у Dvinosaurus стерео-
спондильные позвонки образуются путем прямого сращения
всех элементов позвонка, без редукции какого-
либо из этих элементов.
Другое соотношение элементов рахитомных позвонков мы наблюдаем
у Platyops (рис. 88В).
Здесь также рахитомные позвонки построены по обычному типу, но
плевроцентры в своей расширенной верхней части имеют соединение со спе-
циальными фасетками.на отростке неуральной дуги (рис. 88Bi). У ста-
рых особей в переднем отделе туловища плевроцентры наглухо прирастают
только к неуральной дуге, сливаясь в одну кость. В результате (рис. 88В2)
часть центра позвонка слита с неуральной дугой, другая часть существует
еще раздельно.
Несомненно, что на дальнейшем этапе развития позвонков у Platyops
плевроцентр не уничтожится, а так же, как и у Dvinosaurus, плотно при-
растет к гипоцентру. В результате этого возникает тот же стереоспондиль-
ный тип позвонка, но иным путем, чем у Dvinosaurus (рис. 88В3).
Указанные выше позвонки Micropholis (рис. 88С2) по степени окосте-
нения и развитию широкого контакта неуральной дуги и гипоцентра, не-
сомненно, близки к переходу в стереоспондильное состояние. Это вполне
согласуется с поздним геологическим возрастом формы (триас). В то же
время нет никаких данных предполагать скорую редукцию большого плев-
роцентра. Наиболее вероятно сращение плевроцентра с неуральной дугой
и гипоцентром одновременно, что составит третий путь получения стерео-
спондильного позвонка.
Наконец, еще одним путем развития стереоспондильного позвонка
является вышеописанный путь редукции плевроцентров и расширение
верхней части гипоцентра с последующим срастанием неуральной дуги
и гипоцентра. Переходную стадию на этом пути образуют неорахитомные
позвонки Benthosuchus и Wetlugosaurus (рис. 88D2).
Таким образом, нам кажется, что стереоспондильный тип строения по-
звонка мог образоваться из рахитомного четырьмя следующими путями:
1) полное и одновременное слияние хорошо развитых плевроцентра,
гипоцентра и неуральной дуги — путь, демонстрируемый позвоночником
Dvinosaurus (рис. 88А);
nt
2) слияние плевроцентра сначала с неуральной дугой и только после
этого — сращение с гипоцентром, — путь, прослеженный нами па примере
отроения позвоночника Platyops (рис. 88В);
3) слияние гипоцентра с неуральной дугой и более позднее присоедине-
ние плевроцентров к обоим этим сросшимся в одну кость элементам — путь,
повидимому, намечающийся у Mwropholis (рис. 88С);
4) редукция плевроцентра до полного его уничтожения, вместе с расши-
рением верхних концов костного гипоцентра, до полного сращения послед-
него с неуральной дугой — путь, показываемый неорахитомпыми позвон-
ками Benthosuchus и Wetlugosaurus (рис. 88D).
Приведенные нами факты, разумеется, не могут служить основанием для
сколько-нибудь значительных обобщений, и мы пока не можем строить какие-
либо теории происхождения позвонков у стегоцефалов, без проверки ука-
панных положений на большом и разнообразном материале.
Нашей задачей было обратить внимание исследователей на накопляю-
щийся в морфологии позвонков высших лабиринтодонтов материал, позво-
ляющий притти к следующим выводам:
1) рахитомные позвонки у различных форм лабиринтодонтов обладают
разнообразной структурой в соотношениях слагающих их элементов;
. 2) эти разнообразные структурные модификации могут переходить в сте-
реоспондильное состояние различными путями;
3) стереоспондильный позвонок, полученный в результате сращения
плевроцентров с гипоцентром или неуральной дугой, не является ното-
центральным в точном смысле этого слова;
4) настоящими нотоцентральными стереоспондильными позвонками яв-
ляются те, которые проходят неорахитомную стадию, сопровождающуюся
редукцией плевроцентров.
Если при проверке высказанные положения окажутся правильными, то
станет несомненным, что переход к стереоспондилии в различных генети-
ческих группах осуществлялся разными путями и часто разновременно.
В этом случае не исключена возможность, что нотоцентральная стереоспон-
дилия, полученная через неорахитомную стадию, является присущей только
одной группе близких между собой форм и главным образом — капитозав-
роидов. Поскольку в эту группу входят все наилучше изученные формы,
пользовавшиеся широким распространением и бывшие многочисленными,
то становится ясным, что наши первоначальные представления о рахитом-
ных и стереоспондильных позвонках сложились не случайно.
ГЛАВА IV
УСЛОВИЯ ЗАХОРОНЕНИЯ, РАСПРОСТРАНЕНИЕ И ФАУНИСТИЧЕСКИЕ СВЯЗИ
BENTHOSUCHUS
Возраст Benthosuchus sushkini определяется его нахождением в эотриа-
совой пестроцветной толще, выделенной Ефремовым в V фаунистическую
зону [Ефремов, 1937] как наиболее низкие горизонты триаса.
Местонахождения лабиринтодонтов эотриаса СССР известны довольно
давно и первоначально описаны Н. Н. Яковлевым [1916]. Однако отсут-
ствие полных нах од ок и малое количество данных привели к тому, что как
фауна эотриаса, так и условия образования местонахождений практически
оставались неизвестными.
Крупные находки и более полное освещение местонахождений были ре-
зультатом работ Ефремова [1927—1929], во время которых и была собрана
коллекция, положенная в основу настоящей работы.
Геологическими работами последнего времени выявлено очень широкое
распространение эотриасовой пестроцветной толщи. При этом все пункты
ее выходов содержат слои с костями лабиринтодонтов сем. Benthosuchidae.
Площадь распространения местонахождений эотриаса занимает огромное
112
поле в северо-восточном углу Русской платформы и простирается по долготе
приблизительно от 43 до 59° Гринича, а по широте — от 56 до 66° с. ш.
Разумеется, распространение эотриасовой толщи в пределах этого
поля не непрерывно как вследствие последующих геологических процес-
сов, так, возможно, и по условиям ее образования.
Но литологические особенности пород эотриасовой толщи, характер
напластования и условия залегания костеносных пластов, условия захоро-
нения, сохранность и состав фауны во всех отдельных пунктах выходов,
хотя бы и удаленных очень значительно друг от друга, удивительно сходны.
Это неоспоримо свидетельствует об идентичности внешних условий и про-
цессов образования пород на всей площади развития эотриасовой толщи.
Все разрезы местонахождений эотриасовой пестроцветной толщи одно-
типны, отличаясь деталями напластования и содержанием костей лабиринто-
донтов в географически различных пунктах.
Сама пестроцветная толща представляет собой довольно правильное чере-
дование пластов красных известковистых глин, мощности, колеблющейся
от 10 до 50 см, и голубовато-серых глинистых рыхлых песчаников, или пес-
чанистых глин, залегающих более тонкими слоями от 5 до 30 см мощности.
Это правильное чередование красных глин и голубовато-серых песча-
нистых пород придает разрезам толщи характерный полосатый, пестроцвет-
ный вид.
Во всех выходах толщи в этих пестроцветных породах не было найдено
никаких органических остатков.
Предположительная мощность пестроцветной толщи равна величине
приблизительно 80—100 м.
Примерно в средней части разреза пестроцветной толщи залегает слой
серого песка, неравномерно сцементированного известковистым цементом
в сравнительно мягкий песчаник, часто содержащий гальки красных из-
вестковистых глин. Цементация захватывает различные прослойки в слое
серого песка, по простиранию переходящие снова в серый песок, который
затем снова цементируется уже в других своих прослойках. Мощность слоя
серого песка в различных местонахождениях колеблется от 50 см до 3—3.5 м.
Этот серый песок залегает на размытой поверхности подстилающих пород
пестроцветной толщи и сверху прикрывается такими же породами. Никаких
существенных различий в породах кровли и почвы прослоя серого песка
не установлено. Напластование самого песчаного слоя крайне беспорядочно.
Резко выражена косая слоистость, колебания в мощности и частые изме-
нения по простиранию.
Слой серого песка внутри эотриасовых пестроцветных напластований
и является костеносным, содержащим остатки рыб, лабиринтодонтов, реп-
тилий, мелких пресноводного типа беспозвоночных (эстерий и антракозий)
и очень редко находимые остатки растений, типа каламитов.
Очень часто покрывающая костеносный серый песок верхняя часть пестро-
цветной толщи почти нацело смыта, и в этом случае костеносный слой
непосредственно прикрыт более поздними образованиями, главным образом—
четвертичными песками.
Указанная общая картина повторяется за небольшими изменениями во
всех местонахождениях.
По характеру костеносного пласта все местонахождения можно разделить
на две группы — южную и северную. Южная группа, куда входят место-
нахождения Верхней Волги (Плес, Семигорье, Решма), Ветлуги (Б. Слудка,
Зубовское), Вятки (р. Кобра, Терюхан) и др., характеризуется более тонким
(50 см) прослоем костеносного серого песка.
Песок цементирован в песчаник сравнительно постоянными по прости-
ранию тонкими пластами до 20 см мощностью. Этот песчаник залегает либо
в верхней, либо в нижней части слоя песка, мелкозернист и обладает при-
месью глинистого материала. Гальки известковистых глин сравнительно
редки. Остатки позвоночных, как и всюду в местонахождениях эотриаса,
3 Тр. Палеонтология, ин-та, т. X, в. 1
113
приурочены именно к участкам цементированного песка — песчанику. Хотя
сохранность костей в южной группе местонахождений и очень хорошая, но
поверхность костей носит явственные следы коррозии. Цельные черепа и
кости попадаются редко, зато костеносный пласт изобилует мелкими, сильно
окатанными костными обломками. Изредка встречаются целые комплексы
частей скелета (серии позвонков с ребрами, более или менее полные части
конечностей и т. п.).
Северная группа местонахождений — р. Шарженга (Вахнево и Захаро-
во), р. Анданга, р. Юг (Кудрино), р. Луза (Черный Бор, Скоба и др.)
и серия недавно открытых А. А. Малаховым в Тиманской области место-
нахождений (рр. Мезень, Выбор, Саусара, Цыльма) характеризуются очень
мощным прослоем серого песка, свыше 3 м мощности.
Цементация песка чрезвычайно беспорядочна и дает целую серию пес-
чаниковых небольших линз самой различной мощности и протяжения, вкли-
нивающихся и выклинивающихся по простиранию внутри слоя серого
песка. Песчаник более грубозернист, без заметной примеси глинистого ма-
териала и содержит большое количество гальки из красных известкови-
стых глин, иногда очень плохо окатанной.
В Тиманских местонахождениях к гальке из известковистых глин при-
мешивается. галька из изверженных и метаморфических пород. Окатанные
обломки костей встречаются реже, чем в южной группе, цельные кости
(совершенно исключительной сохранности, с гладкой, блестящей поверх-
ностью, совершенно не окатанные) и черепа попадаются чаще.
Следует считать, что’костеносный пласт северных местонахождений отла-
гался в условиях более сильного течения, чем в южных. Более грубозерни-
стый состав отложений, отсутствие глинистого материала и большая мощ-
ность песков и песчаников вполне соответствует наличию в костеносном пла-
сту крупных отделов скелета, например, черепов. Превосходная сохранность
костей свидетельствует о быстром их захоронении, т. е. весьма кратком
сроке пребывания вне консервационных условий.
В южной группе, в условиях более тихого течения отлагавших осадки
вод, естественно, преобладают более мелкие кости и главным образом — их
обломки. Обломки эти проходили до своего отложения значительный путь
и сильно окатывались. Более крупные, цельные кости и черепа также пока-
зывают больший срок нахождения вне консервационных условий, отра-
зившийся в худшей сравнительно с северной группой сохранности поверх-
ности костей.
Связанные комплексы частей скелета своим присутствием, повидимому,
обязаны плавающим трупам, которые даже тихое течение могло свободно
переносить.
Характер захоронения остатков животных в обеих группах местона-
хождений во всех существенных деталях одинаков. В костеносном пласту
беспорядочно разбросаны, — то группами, то поодиночке — отдельные
разрозненные элементы скелетов самой различной величины (возраста).
В групповых скоплениях смешаны кости самых различных отделов скелета,
принадлежащие различным по величине особям. Черепа часто перевернуты
нёбом кверху или занимают наклонное относительно плоскости пласта по-
ложение. Нижние челюсти, за чрезвычайно редкими исключениями, отде-
лены от черепов и разнесены так, что нет никакой возможности ассоции-
ровать их с тем или иным черепом по месту нахождения.
В южной группе намечается как бы послойное расположение костей,
так как они рассеяны внутри слоя серого песка приблизительно на одном
уровне. Этим объясняется и пластовое расположение прослоя песчаника,
так как цементация песка происходила в тех частях слоя, которые были обо-
гащены животными остатками.
В северной группе местонахождений кости залегают отнюдь не в виде слоя,
а на самых различных уровнях мощного костеносного пласта. В соответ-
ствии с этим мощность песчаниковых прослоев внутри серого песка очень
114
сильно меняется, слои нередко располагаются в несколько пропластков,
один над другим, и быстро выклиниваются по простиранию.
Этот характер распределения костей в южной и северной группах как
нельзя более соответствует указанным выше условиям их отложения.
В северной группе воды с быстрым течением, нагромождавшие мощные
массы песка, турбулентно перемешивали кости животных вместе с осадком.
Более тихое течение южной группы обусловило тонкое наслоение костей
па определенном уровне отложения, возможно, соответствующем фазе наи-
более сильного течения, способного переносить и крупные фракции, какими
являются в данном случае кости.
Во всех местонахождениях участки костеносного пласта, содержащие
кости, часто перемежаются пустыми, не содержащими костей, участками:
общая площадь пустых участков значительно более площади участков, со-
держащих кости.
Это обстоятельство и обусловило то, что полные остатки фауны*, захоро-
ненной в эотриасовых местонахождениях, были найдены только при пра-
вильных раскопках, вскрывших довольно большие участки костеносного
пласта.
Фауна эотриасовых местонахождений довольно однообразна. Наибольшее
количество остатков принадлежит неорахитомным лабиринтодонтам Вен--
thosuchus и Wetlugosaurus. Гораздо реже встречаются остатки рептилий —
динозавров типа Thecodontosaurus— в виде обломков небольших позвонков'
и трубчатых костей конечностей, точно не определимых. Кроме того, в се-
верной группе встречены фрагменты челюстей с зубами и обломки черепных’
костей, принадлежащие мелким рептилиям типа Eosuchia.
Остатки рыб попадаются весьма редко. Это обломки щитков двоякоды-1
шащих (Dipnoi) и разрозненные ганоидные чешуи Acrolepis. Отпечатки
стеблей каламитов и ядра антрокозий и эстерий встречаются так же редко,
как и остатки рыб, причем в большем количестве они найдены в южной
группе.
До сих пор сколько-нибудь значительным раскопкам подвергались толь-
ко два местонахождения из всех многочисленных местонахождений эотриа-
совой пестроцветной толщи — правый берег р. Шарженги у с. Вахнево
и правый берег р. Ветлуги у деревень Б. Слудка и Зубовское.
Эти раскопки доставили значительный материал, описанию которого
и посвящена настоящая работа.
Разумеется, произведенные раскопки едва затронули имеющиеся в этих
местонахождениях скопления костных остатков лабиринтодонтов. Площади
костеносных пластов в районе Вахнева измеряются десятками квадратных
километров, в районе Б. Слудки— квадратными километрами. Трудно себе,
представить то количество остатков, которое еще должно содержаться,
в этих местонахождениях.
Весьма интересно, что вся масса остатков лабиринтодонтов, полученная;
при раскопках на р. Шарженге у с. Вахнево, принадлежит исключительно
Benthosuchus. Только один фрагмент относится к другому близкому роду/
значительно меньшей величины—Thoosuchus. Весь материал из раскопок*
на р. Ветлуге у деревень Б. Слудка и Зубовское принадлежит исключительно
Wetlugosaurus, опять-таки за исключением одного фрагмента, принадлежа**
щего Thoosuchus.
Таким образом, в своем распространении эти два рода как бы исключают-
Друг друга. На севере обитал Benthosuchus, на юге— Wetlugosaurus, при--
чем численпость особей обоих родов была очень значительна.
Материалы, собранные из других не раскапывавшихся местонахождений, •
несмотря на свою отрывочность, показывают, что на самом деле распро-
странение обоих родов было более сложным.
Так, в наиболее северных местонахождениях Тиманского района найдены
Benthosuchus и Wetlugosaurus в пунктах, очень близких друг от друга.
В наиболее южных местонахождениях Верхней Волги к Wetlugosuchus
8* <15
примешивается еще новая форма, пока еще неизученная — Volgosuchus.
Остатки Thoosuchus здесь встречаются чаще.
Ефремов [1938] считает, что наиболее северные местонахождения эо-
триасовых лабиринтодонтов являются относительно более древними, а юж-
ные — более молодыми по геологическому возрасту.
Это обстоятельство сходится и с географическим распространением
эотриасовых неорахитомных лабиринтодонтов. Вначале оба рода — Ben-
thosuchus и Wetlugosaurus существуют, если не совместно, то, во всяком
случае, в сближенных ареалах. Затем, повидимому, вместе с расхождением
адаптации ареалы обоих родов расходятся. В еще более позднее время в са-
мых южных областях распространения эотриасовой фауны к Wetlugosau-
rus примешиваются новые формы, появившиеся в результате дальнейшей
адаптивной радиации эотриасовых лабиринтодонтов.
В этом случае ясна важность исследования и раскопок наиболее север-
ных и наиболее южных местонахождений, при которых могут быть обнару-
жены более ранние и более поздние стадии эволюционного развития как
Benthosuchus так и Wetlugosaurus.
Вообще, крупные раскопки, поставленные на эотриасовых местонахож-
дениях, безусловно, дадут богатейший материал по поздним лабиринто-
донтам, равного которому по сохранности неизвестно.
Подведем итоги данным о местонахождениях эотриасовых лабиринтодон-
тов.
В пределах всей огромной площади развития эотриасовой пестроцветной
толщи почти в каждом ее выходе на дневную поверхность присутствует
костеносный прослой серых песков и песчаников, содержащий остатки не-
орахитомных лабиринтодонтов Benthosuchus и Wetlugosaurus.
Уже обследованные местонахождения показывают распространение ко-
стеносного прослоя на десятки квадратных километров, и нет основания счи-
тать, что в других малоизвестных местах мы встретимся с другими услови-
ями его залегания.
Наличие костеносного прослоя почти во всех выходах пестроцветной
толщи, в самых удаленных друг от друга точках, неоспоримо указывает на
громадное распространение костеносных отложений, измеряющееся уже
тысячами квадратных километров.
Если из раскопок на рр. Шарженге и Ветлуге, общей площадью разра-
ботанного костеносного пласта не свыше 200 м2, нами собрано свыше полу-
тора тысяч отдельных костей, обломков и фрагментов, принадлежащих сот-
ням разных по возрасту особей, то какими цифрами должно измеряться
число особей, захороненных на всем протяжении пестроцветной толщи?
Даже если считать, что мы при раскопках попали на наиболее богатые участ-
ки костеносного пласта, то при самом осторожном подсчете становится ясным,
что в местонахождениях эотриасовой толщи захоронены сотни тысяч осо-
бей Benthosuchus. Wetlugosaurus и других, близких, но более редких форм.
Такое огромное поле смерти лабиринтодонтов представляет собой исклю-
чительно своеобразное явление и не может быть объяснено с точки зрения
обычных жизненных процессов.
В самом деле, наличие повсюду костеносного прослоя серого песка
обусловливает как бы перерыв в отложении пестроцветной толщи. Этот
перерыв не был долгим, так как песчаные отложения при значительной силе
водных потоков могли быть отложены в весьма короткий срок и образуют,
сравнительно со всей толщей, незначительный прослой. Равным образом
этот перерыв произошел не в силу резкого изменения физикогеографиче-
ской обстановки, так как после отложения слоя серого песка попрежнему
продолжалось напластование той же пестроцветной толщи, неотличимой от
нижних, подстилающих слоев.
И тем не менее, в момент образования слоя серого песка на колоссаль-
ной площади отложилось огромное количество остатков лабиринтодонтов
самого различного возраста, беспорядочно перемешанных, частью разло-
116
манных и окатанных. Исключительная сохранность костных остатков может
быть объяснена только чрезвычайно быстрым их захоронением. Кроме этого
слоя песка, нигде во всей толще не обнаружено никаких органических
остатков.
Следует считать, что момент отложения слоя серого песка совпадал
или почти совпадал с моментом массовой гибели лабиринтодонтов. Гиб-
ли все без исключения, как старые, так и молодые особи. Наибольшее
количество остатков принадлежит особям «среднего» возраста, что также
показывает наличие в местонахождениях не выборочного захоронения, а
всего состава фауны лабиринтодонтов.
Если учесть размеры площади, на которой происходила массовая гибель
животных, а также то, что одновременно гибли формы с разнородной адап-
тацией, становится ясным, что мы имеем дело с катастрофой огромного мас-
штаба.
Эта катастрофа, отраженная в комплексе местонахождений эотриаса
Русской платформы, по масштабу, а, главное, по быстроте своего развития
является одним из очень немногих в истории древнейших Tetrapoda случаев.
Еще более интересно то, что, невидимому, причина эотриасовой ката-
строфы лежит не в анастрофических изменениях физикогеографической
обстановки.
Каковы были условия физикогеографической обстановки, вызвавшие
отложение пестроцветной толщи, — остается пока неизвестным. Отсут-
ствие органических остатков в толще может свидетельствовать, с равной
вероятностью, как об отложении в пустынных условиях или условиях без-
жизненных водоемов, так и всего лишь об отсутствии условий захоронения
в пригодной для животной жизни обстановке.
Разрешение спорного вопроса о происхождении пестроцветной толщи
может произойти только после детальнейших геологических исследований,
в настоящее же время и в настоящей работе оно невозможно.
Что бы ни представляла собой физикогеографическая обстановка на
всей обширной площади отложения эотриасовой толщи, остается ясным,
что эти условия существовали до отложения костеносного слоя серого песка
и продолжали в неизменном виде существовать и после. Лишь на короткий
срок в момент отложения серого песка эти условия изменились также по
всей обширной площади, вероятно, не вполне одновременно.
По всем данным, слой серого песка отлагался в условиях текучих вод,
причем довольно быстрых, на что указывает беспорядочное напластование
песков и хаотическая разбросанность органических остатков.
Таким образом, очевидно, что если пестроцветная толща отлагалась
в пустынных условиях, то отложение костеносного песчаного слоя было
вызвано вторжением массы текучих вод, разлившихся по всей равнине.
Если же пестроцветная толща отлагалась в бассейне, то отложение серого
песка было вызвано резким усилением течений с обычным приносом более
крупного материала и, так же, как и в первом случае, размывом уже отложен-
ных напластований.
Следовательно, и в том и в другом случае мы имеем дело с быстрым за-
топлением огромной площади, нагромоздившим повсюду массы песка,
смешанного с остатками лабиринтодонтов, и закончившимся сравнительно
быстро, после чего вновь восстановились прежние условия.
Это затопление, повидимому, распространялось с севера на юг, постепен-
но уменьшаясь в интенсивности.
Такое огромное количество трупов лабиринтодонтов вряд ли было при-
несено откуда-то с севера. Против этого предположения свидетельствует
то, что имеющиеся формы занимают определенные ареалы распространения.
Трудно допустить также, что массовая гибель животных была вызвана самим
затоплением области. Такие, хорошо приспособленные к жизни в воде
формы, как Benthosuchus и Wetlugosaurus, естественно, не могли погибнуть
только при увеличении водной массы.
117
Ключ к разрешению вопроса, очевидно, лежит в том общем и внезапном
нарушении привычной среды обитания, которое было вызвано процессами,
обусловившими обширное затопление области.
Отсутствие сколько-нибудь значительного количества остатков рыб,
в сравнении с массой остатков лабиринтодонтов, очень резко бросается
в глаза, так как рыбы составляли основной, вернее, единственный род пи-
тания Benthosuchus и Wetlugosaurus. Не менее важно отсутствие растительных
остатков, которые, если бы они имелись в большом количестве, конечно,
сохранились бы, хотя бы в виде неясных отпечатков.
Таблица I. Benthosuchus sushkini, № 1—19
На осповании этих данных предположение о гибели животных вслед-
ствие недостатка пищи, вызванного наводнением или предшествовавшими
ему явлениями, кажется вероятным.
Отдаленно аналогичные явления мы можем наблюдать в настоящее
время в огромных разливах тропических рек в низменных областях (на-
пример, так называемые гапо в Южной Америке), когда сотни и тысячи
квадратных километров затопляются текучими водами. Такие наводнения
вызывают соответственную гибель и захоронение животных, однако в го-
раздо меньших, чем это наблюдается в нашей катастрофе, размерах.
Конечно, массовая гибель лабиринтодонтов в эпоху эотриасового навод-
нения, так же, как и самое наводнение, должна была происходить в спе-
цифических условиях, возможно, в настоящее время неповторимых, и по-
дыскание аналогии этого в современных обстановках всегда будет лишь
очень приблизительным.
Таким образом, огромное поле смерти эотриасовых лабиринтодонтов
образовалось путем быстрого отложения их остатков в песчаных массах
118
в момент кратковременного затопления всей области сравнительно быстрыми
текучими пресными водами. Этот момент затопления области совпал с мас-
совой гибелью очень многочисленных животных в количестве сотен тысяч
особей. Животные обитали, несомненно, в пределах области развития
пестроцветпой толщи, но, повидимому, в нормальных жизненных условиях
не попадали в захоронение. Затопление всей области и отложение серых
песков создали прекрасные условия для захоронения остатков животных
и быстрой их консервации.
Благодаря крупной катастрофе до нас дошли обильные остатки эотриа-
совой фауны лабиринтодонтов, которая, несмотря на свою многочислен-
ность, нигде, кроме этого песчаного слоя, не была найдена. Предковые
формы эотриасовых Benthosuchus и Wetlugosaurus в более древних слоях
па территории Русской платформы не были найдены.
Таблица II. Benthosuchus sushkini, Л? 1 — 9/2243.
Такое внезапное появление обильной фауны, не имеющей известных
«корней» в более древних слоях, очень характерно для большинства находок
древнейших позвоночных и лишний раз свидетельствует о том, какая малая
часть действительно существовавшего животного мира доходит до нас в про-
цессах захоронения, причем всегда в результате крупных, катастрофиче-
ских или другого характера явлений.
В согласии с более ранним появлением фауны на севере области эотриа-
совой пестроцветйой толщи стоит недавнее обнаружение фауны лабиринто-
донтов пеорахитомного типа в Восточной Гренландии. Среди этих форм
найден Wetlugosaurus и другой, внешне близкий к Benthosuchus род, оши-
бочно отождествленный Сэве-Зедеребергом с Lyrocephalus [Ефремов,
1938]. По всей вероятности, фауна эотриасовых лабиринтодонтов типа Ben-
thosuchus и Wetlugosaurus занимала обширную область распространения
на севере современной Европы и далее за полярный круг. -Более точные
данные о происхождении и распространении этой важной и весьма много-
численной фауны будут, несомненно, получены при более детальном иссле-
довании различных местонахождений эотриаса как СССР, так и других стран.
119
BENTHOSUCHUS SUSHKINI —A LABYRINTHODONT FROM
THE EOTRIASSIC OF SHARZHENGA RIVER
BY A. P. BYSTROW and J. A. EFREMOV
Introductory
In 1927 J. A. Efremov discovered, some rich localities of Labyrinthodonts
in a streak of grey sands and conglomerate-like sand-stones, bedding amidst
the series of Eotriassic variegated marls and clays along the banks of Sharzhenga
(a left tributary of the Yug). The next years — 1928, 1929 — in these locali-
ties were organized successful excavations.
• In 1929 J. A. Efremov described from there a complete skull of a new
genus — Benthosaurus sushkini, which name later on (1936) was changed
into «Benthosuchus». In the localities is represented practically only this one
genus of Labyrinthodonts: among several hundreds of isolated bones and frag-
ments of Benthosuchus there was found only one piece of another Labyrin-
thodont. Benthosuchus is represented by 9 complete skulls of different ages
and by a great number of separate skull-fragments and other bones. All the
bones are perfectly preserved and enclosed in rather loose sandstones which
enabled us to prepare all the material exceptionally completely and to elu-
cidate such details of the structure, which usually are either quite unacces-
sible for examination, or observed only on accidentally broken skulls.
An important defect of the material is the lack of complete skeletons. In
the bone-bearing layers separate bones of individuals of most different ages
are found confusedly mixed up. Therefore, an exact restoration of Benthosu-
chus is impossible, as it is difficult to get a right idea of the correlation of
separate elements of the skeleton.
For comparison, remains of the closely related genus Wetlugosaurus from
the Eotriassic of the Vetluga and the Upper course of the Volga were studied.
This material containing remains of Benthosuchus and Wetlugosaurus belongs
to the Palaeontological Institute of the Academy of Sciences of the USSR.
We have also studied for comparison the type-specimens of Wetlugosau-
rus angustifrons Riab. from the Vetluga, kindly placed at our disposal by
A. N. Riabinin.
The object of this work is to give a possibly complete description of Ben-
thosuchus, a representative of late Labyrinthodonts. For the time being, the
most part of the known Labyrinthodonts are mostly quite unsatisfactorily
studied, because of the insufficient preparation, or the poor preservation.
Better studied is Eryops described in several morphological papers (Case,
1911; V. Huene, 1912; Broom, 1913; W. Williston, 1916; Dempster, 1935).
However, this Labyrinthodont also still requires a more detailed anatomical
study.
Lately, G. Save-Soderbergh (1936) revised the material previously des-
cribed by Wiman (1914, 1916, 1917) and gave a detailed description of the
skulls of Aphaneramma Lyrocephalus and Platy stega and made an attempt
120
of restoration of the chondrocranium of the vessels and nerves of the head of
these forms.
This work of Save-Soderbergh gives a most complete idea of the skull
of Labyrinthodonts. However, the poor preservation has not allowed him
to elucidate still many an interesting detail of the structure of the skull of
Stegocephalians. The postcranial skeleton was altogether absent from the Save-
Soderbergh’s collection.
The authors of the present paper divided their work, so that the material
was minutely prepared by J. A. Efremov, while the preliminary drawings
were drafted by A. P. Bystrow. After this we studied together the obtained
material.
We were forced to introduce new Latin names for a number of details
of structure of the skeleton of Benthosuchus, but usually we retained the uni-
versally adopted Latin nomenclature, changing it only when it seemed to
be inadequate* The first part of the paper prepared by A. P. Bystrow is de-
dicated to the general anatomic description of the skeleton of Benthosuchus.
The second part, written by J. A. Efremov, comprises the description of the
conditions of embedding of the remains of Benthosuchus, of the individual
growth and variability of the elements of skeleton and the general conclu-
sions concerning the evolutionary stage and relationships of Benthosuchus,
as well as a treatise of the development of the vertebral column in typical
Labyrinthodonts. All the drawings are made by A. P. Bystrow.
List of materials investigated
This list is compiled according to the catalogues of the Palaeontological
Institute of the Academy of Sciences of the USSR, in chronological order
(1927—1929). The collections No. No. 9/2243 and 19/2252 are divided into
two sections each. Into the first section were included the remains of Bentho-
suchus from the Sharzhenga river; while the second section includes specimens
selected from the collections of remains of Wetlugosaurus, from Vetluga
river.
The specimens are listed mostly according to their anatomic position,
from skull to tail.
To show the distribution of material according to the excavations it came
from, the localities are indicated by an «Р» with an index. For the locality
on Sharzhenga at Vachnevo, these indexes are: Pj — excavation No. 1, right
bank of Sharzhenga at the watermill opposite to Vachnevo; P2 — excavation
No. 2, somewhat farther up-stream; P8 — excavation No. 3, right bank of Shar-
zhenga, just below the mouth of Medvejia; P4—excavation No. 4, left bank,
of the Medvejia, 150 m. up-stream from the mouth; P5 — excavation No. 5,
right bank of the Medvejia, 100 m. up-stream from the mouth.
For the locality on the Vetluga the indexes are as follows: Pi — excava-
tion No. 1, right bank of the Vetluga, up-stream of Bolshaia Sludka and 80 m.
up-stream the ravine; P2—excavation No. 2, same place on the bank of the
mouth of the ravine; »P3 — excavation No. 3, right bank of the Vetluga at
Zubovskoie, below the church.
After the study of the material the interpretation of some bones as right
or left ones has been changed. But to avoid the divergence with the catalogues,
which for the time being remain not corrected, we have here retained the pre-
liminary designations of the catalogues.
I. SKELETON ANATOMY
The skull of Benthosuchus sushkini (pl. I, II) has the outlines of an acute
triangle, the sides of which are set at an angle of about 30° to each other. The
anterior end of the skull is slightly blunted. The narial openings are of oval
outlines, being twice as long as wide.
121
The orbits lie on the verge between the medial and posterior of the cra-
nial roof. They are of oval outlines, their length being about 1.% times as
much as their width. The cranial roof is impressed at the place between the
orbits, forming impressio interorbitalis. The round foramen parietale is situa-
ted somewhat posteriorly of the conjectural line drawn along the posterior
borders of orbits.
The occipital border of the skull is concave. The horns of tabulare are
well defined, and bound from medial side the deep otic notches.
Bones of skull of benthosuchus sushkini
Praemaxillare, forming the somewhat blunted end of the snout
of Benthosuchus, is a comparatively small and short bone. It appears in the
form of two laminae, joined with one another along the whole rounded den-
tal margin.
The lamina palatina (fig. la) has the appearance of a flat process, forming
the foremost portion of the bony palate. The posterior margin of this lamina
bears in its medial part a small process with rounded apex — the processus
fenestralis anterior. Farther laterally, on the same margin, lies the rather deep
incisura fenestralis. Still more laterally is situated a small hollow with rag-
ged borders into which enters the vomer. The dorsal lamina of praemaxilla-
re — the lamina dorsalis — is convex (figs, la, 8) The posterior margin of
lamina dorsalis bears in its lateral portion the small incisura narialis, boun-
ding the nostril from antero-medial side, and a small sharp spine — the spina
narialis.
The rather flat lamina palatina and the convex lamina dorsalis are joined
with one another in such a way, that a deep recess — sinus praemaxillaris —
attaining the level of the teeth row is formed between them in the praemaxil-
lare (fig. 17).
In the anterior part of the suture between two praemaxillare, there is
a. small through aperture — the foramen interpraemaxillare — leading into
sinus praemaxillaris (figs. 8, 17).
Vomer (Praevomer, Broom, 1913). The vomer represents a rather
long bony lamina of multiplex outlines. This bone is joined with praemaxilla-
re only in a part of its anterior process — the processus subnarialis (fig. lb), —
embedding into corresponding incisure of intermaxillare. On the anterior
border of vomer are observed formations similar to those present on the poste-
rior margin of lamina palatina of praemaxillare. The border forms in its
middle part a rather deep notch — incisura fenestralis posterior; while in
its middle part it bears a small process — processus fenestralis posterior, —
situated just opposite to the corresponding process of praemaxillare.
On the lateral border of the anterior half of vomer is observed a shallow,
but rather long notch — the incisura choanalis bounding the choana from me-
dial side. Immediately behind the incisura choanalis, the border of the bone in
question joins palatinum by means of a suture of the type of sutura squamosa.
Farther backward, vomer extends into a long and narrow process — processus
parasphenoidalis. •
The medial margin of vomer is straight. Its palatal surface is provided
with two rows of small conical teeth and large tusks. Usually there is one
tusk on each vomer, though in our collections are available specimens, on
which vomers have by two tusks set one after another. In cases, when only
one of the two tusks is preserved, it may equally often be the anterior (first)
or the posterior (second) one. The first tusk — dens praechoanalis primus
(fig. lb) — is situated immediately behind the incisura fenestralis posterior.
Medially and posteriorly of this tooth extends a row of small dentes post-
fenestralis.
Posteriorly of the second postchoanal tusk, or in the case when it is not
preserved — of its pit, extends the second row of small teeth, forming a con-
cave arch, with its concavity directed medially.
122
On the dorsal surface of vomer of Benthosuchus, along its incisura choana-
lis, extends a very distinct thickness of the bone of arch-like outlines — the
torus parachoanalis (fig. 8).
Another peculiarity of structure of the dorsal surface of vomer is the pre-
sence of a markedly expressed groove, running along the whole length of the
medial margin of the bone. When two vomers are put together, these grooves —
sulcus parasphenoidalis — form the bed for the anterior end of processus cult-
riformis of parasphenoid (figs. 8, 21).
Pal a t i num. This bone adjoins the lateral side of vomer immediately
behind the incisura choanalis of the latter. The anterior margin of palatinum
takes part in formation of the choana, bounding it from posterior side (inci-
sura choanalis palatini, fig. 2).
Palatinum and vomer join one another by their obliquely cut margins, in
such a way that palatinum overlays the border of vomer, but not vice-versa.
Along the line of their joining they form a kind of sutura squamosa.
Along the whole length of the lateral margin of palatinum extends a long
crest, the sharp side of which is directed dorsally. Backwards palatinum passes
into a narrow process — processus paratransversalis (fig. 2), the lateral half
of which covers like a squama the neighbouring it transversum.
The anterior part of the palatal surface of the bone under description bears
a large tusk, similar to the tusk on vomer. In very rare cases we observe here
two tusks, set one after another. But such cases must be considered as an ex-
ception; usually here is present only one tusk — the dens postchoanalis
(fig- 2).
Examining different fragments of the skull of Benthosuchus, we have de-
tected on the dorsal surface of its palatinum the presence of a large, sharp
spine. The broad base of this spine lies exactly opposite to the postchoanal
tusks, its apex being directed backwards and somewhat medially. This spi-
ne — spina sublacrimalis — is closely adjacent to the lower surface of der-
mal bones of the cranial roof — namely, to the lacrimale, — and appears as
if supporting this latter bone (figs. 2, 3).
Transversum (Ectopterygoideum). This is a long narrow bone
bearing on its ventral surface a row of dentes transversales (fig. 3), which
are arranged parallel to its lateral margin. The anterior angle of transversum
is obliquely cut, and entering the corresponding notch (incisure) of palatinum
covers this latter bone. The postero-medial angle of transversum is stretched
into a rather long, flat process — processus squamosus — covering the end
of the anterior branch of pterygoid (fig. 3). The almost straight medial bor-
der of the bone under description also adjoins the pterygoideum. For the whole
length of this margin, transversum is joined with pterygoideum by means
of a scaly suture; while the anterior branch of the latter bone overlays trans-
versum from the dorsal side (fig. 4).
J u g a 1 e may be considered as a bone composed by two components:
squama and processus alaris jugalis (fig. 4).
In Benthosuchus, the greater part of this process overlays the dorsal sur-
face of transversum; while by it smaller portion processus alaris covers the
anterior branch of pterygoideum. An inconsiderable space on the ventral
surface of the process in question remains uncovered by the said bones and
appears when viewed from the side of the palatal surface of the skull, as a
small free islet of the bone (insula jugalis) (fig. 4).
Maxiliar e. The anterior end of this bone is somewhat dilated and
has two incisures. One of these incisures (notches) — the incisura choanalis —
is perfectly visible when yiewed from the palatal side (fig. 4); the other — in-
cisura narialis — is situated in the anterior region of the dorsal part of the
bone, and takes part in the formation of the narial opening.
Pterygoideum. To the complex of bones, composed by palati-
num, transversum and jugale, joins from, the lateral side one more bone of
very complicated outlines— viz. the pterygoideum (fig. 4).
123
When examining pterygoideum from the palatal side of the skull, one
may discern on this bone three parts, namely: its corpus (the middle, more mas-
sive portion of the bone), the anterior branch, and the posterior branch
(figs. 4, 5a). Examining pterygoideum from the lateral side we see that the
posterior branch of this bone gradually transforms into a vertically set lamina—
lamina ascendens (figs. 5b, 6a, b). Its anterior branch represents a wing-like la-
mina; in complete skulls, the whole of this branch lies in the plane of the
palate.
On the dorsal surface of the anterior branch of pterygoideum, along the
whole length of its medial border, extends the rather low torus marginalis,
sharply distinct from the remainder of the surface of the bone (fig. 6b). Along
the opposite border of this branch, instead of torus, extends the about equally
wide sulcus marginalis (fig. 6b.).
Examining the medial side of the corpus of pterygoideum, one can observe
here a rather large surface covered by sharp spines and forming with the body
of parasphenoid an indented suture (fig. 5b). Immediately under this place,
the corpus of pterygoideum bears a hollow of conical outlines — the recessus
conoideus (figs. 5b, 6b) — penetrating into the base of lamina ascendens.
In Benthosuchus, on the medial border of lamina ascendens, over the re-
cessus conoideus, is observed a very distinct small pit of oval outlines — the
fovea ovalis (fig. 5b).
The posterior branch of pterygoideum is directed backwards and laterally,
towards quadratum, with which it becomes fused. This is why on the end of
this branch we see a corresponding rugosity — facies quadrata (fig. 6a). Dor-
sally the branch in question gradually transforms into the great lamina ascen-
dens.
Behind the base of lamina ascendens lies a very distinct hollow — exca-
vatio tympanica (fig. 6b), which served as the anterior wall of tympanical
cavity. Somewhat posteriorly of this hollow, over the lateral surface of lami-
na ascendens extends the markedly expressed — (especially in the aged rep-
resentatives of Benthosuchus) — crista obliqua, from which, apparently,
derived the musculus depressos mandibulae (figs. 5b, 18a).
Parasphenoideum is the longest bone in the skull of Bentho-
suchus. Its basal part — basis parasphenoidei — is strongly dilated, forming
an almost rectangular lamina (fig 8). The dorsal surface of the basis parasphe-
noidei is rugous. One can perceive here the two pairs of ridges deriving from the
somewhat depressed middle portion of the bone, which corresponds to the
punctum ossificationis of parasphenoideum. These ridges are diverging from
the centre in sideward and backward directions. The first pair of these rid-
ges — crista parapterygoidea (fig. 9a) — by its ends reaches the pterygoids
and takes part in bridging these latter with the bone under description (fig. 10a).
The ridges of the second pair — of crista paroccipitalis (fig. 9a) — are tigh-
tly joined to exoccipitale by means of an indented suture (fig. 10a). They
are separated from one another by a rather deep furrow — sulcus intercri-
status (fig. 9a). In the posterior part of the parasphenoideum, between the
ridges of the second pair — the cristae paroccipitales — lies a wide hollow —
fossa basi occipitalis (fig. 9a) — in which the cartilagous basioccipitale was
embedded.
Gradually removing from the ventral surface of parasphenoideum thin la-
mellae of bony matter, we finally succeeded in exposing on some fragments
of the skull of Benthosuchus the canals of internal carotids all along. On some
other fragments, it was possible to expose these canals by means of destroying
the bone from its dorsal side. All this allowed us to precisely establish the
arrangement and course of the internal carotids.
The inlet openings of the canals of internal carotids are situated in Bentho-
suchus on the lateral borders of the basis parasphenoidei, in close neighbour-
hood with the posterior ends of the sutures joining parasphenoideum with
pterygoids.
124
From here either canal goes first to the ossification centre of parasphe-
noideum, passes near the base of crista parapterygoidea, and then, turning
at an angle of about 110°, extends forward (fig. 10b). The outlet openings of
the canals of internal carotids come out on the dorsal surface of parasphenoi-
deum, where they send off long grooves — the sulcus arteriae carotidis inter-
nae — directed towards the base of processus cultriformis (fig. 10a).
Not far from the outlet opening of the canal of internal carotid, the arteria
carotis interna sent off a lateral branch, which passed through a special short
canal (fig. 10b). This canal farther continued over the dorsal surface of pa-
rasphenoideum in the form of a rather broad furrow directed towards the ante-
rior border of the latter.
The processus cultriformis is a long and narrow bone. On the dorsal sur-
face of this process, for about 2/3 of its total length, extends a groove — sul-
cus longitudinalis (fig. 8), — which gradually decreasing in depth, finally
disappears on the verge between the middle and the anterior г/3 of the said
processus cultriformis.
In the skull as a whole, the processus cultriformis is situated so that for
about a half of its total length it is embedded in sulcus parasphenoidalis,
which runs along the suture joining the right and left vomers; and by its
sharp end processus cultriformis reaches the processus fenestralis posterior
(fig. 17).
Exoccipitale of Benthosuchus sushkini represents a very compli-
cated bone, provided with a great number of processes extending in different
directions (fig. 10a).
From the basal part of this bone ascends vertically a short but powerful
process — columna verticals — joining the dermosupraoccipitale, — which
is situated somewhat higher than the bone in question,—by means of an inden-
ted suture (fig. 11). Another process — processus paroticus, — intended for
joining with tabulare, extends laterally and somewhat backwards. From the
base of columna verticalis derives a comparatively thin process — processus
lamellosus — having the form of a lamina, and directed upward, forward
and medially.
The base of exoccipitale also sends off three processes. One of them — con-
dylus occipitalis — which is the thickest of the three, is articulated. The se-
cond process is known under the name of processus submedullaris (fig. 11).
It has the form of a rather broad wedge, lies in horizontal plane, being directed
by its rounded end medially.
The third process — processus subtympanicus — is a rather complicated
bony formation. Together with the lower surface of the corpus of exoccipitale
it takes part in bridging this latter bone with the base of parasphenoideum,
being at the same time closely adjacent to the end of crista paroccipitalis
(fig. 10a). On the upper surface of the process under description is clearly
seen a large hollow in the form of a short groove with smooth walls. This hol-
low undoubtedly had direct relation to the tympanical cavity, as the basis
columellae is situated immediately above it (fig. 12a). And therefore the name
«recessus tympanicus» must be acknowledged as the most suitable term for
this formation (fig. 11).
Dermosupraoccipitale. This bone is bridged by a short
process with the columna verticalis exoccipitalis. The said process — processus
supraoccipitalis (fig. 13) — is situated on the posterior border, of the almost
rectangular lamina of the bone lying in the posterior portion of cranial roof.
The medial part of the posterior border of dermosupraoccipitale bears a small
spine — spina supraoccipitalis (fig. 13), — on the posterior side of which,
in aged representatives of Benthosuchus, is observed a very distinct hollow
(fig. 13a).
Tabulare, occupying the lateral corner of the occipital region of
cranial roof, represents a rather complicated bone.
On the thoroughly prepared fragments of the skull of Benthosuchus, on
125
the lower surface of this bone may be observed a large, obliquely set process —
processus paroticus (fig. 13), — the apex of which is joined with the corre-
sponding process of exoccipitale. This process represents as if a centre, from
which a number of more or less markedly expressed ridges extend in different
directions. Of them, the high ridge extending from the process along the cornu
tabularis immediately draws one’s attention. This ridge — crista tabularis
externa (fig. 13) — terminates on the very end of apex cornu. In opposite
direction from the said process extends a shorter and low’er ridge — the cri-
sta tabularis interna.
On the antero-lateral surface of tabulate, between the above mentioned,
are situated two ridges more. One of them — crista terminalis — deriving
from the processus paroticus extends obliquely backwards to the border of
horn tabulate and disappears not reaching its apex. The other ridge — crista
tympanica — somewhat more strongly deleloped than the first one, termi-
nates at the suture joining tabulate with squamosum.
Along the posterior border of tabulate extends the fifth ridge of the bone —
the crista muscularis (fig. 13a).
Columella. An isolated specimen of columella (fig. 15) represents
a little bony column, slightly bent at about its middle. The distal half of colu-
mella is smooth and almost round, while the proximal one in cross-section
appears of complicated outlines. Examining columella from below (fig. 15b),
or from behind (fig. 15c), one can perceive a very distinct sharp ridge — crista
obliqua, — extending obliquely over the proximal half of the bone and reaching
its middle part. About the verge between the middle and proximal % °f the
bone, columella is pierced by an aperture — foramen arteriae stapedialis
(fig. 12a) — through which passed the arteria stapedialis.
Beginning from the level, at which foramen arteriae stapedialis is situa-
ted, towards the proximal end, columella is gradually thickening, and then
terminates in rather large base w’ith ragged margins and rugose lower sur-
face. From the posterior angle of the base of columella derives a small process —
the processus basalis (fig. 15).
Quadratum. This is a short and massive bone. In its postero-lower
part it forms the finely defined trochlea articularis (fig. 16), which served as
an articular surface in the mandibular articulation. This trochlea in Benfho-
suchus is strongly dilated at the point where quadratum adjoins the poste-
rior branch of pterygoideum, and is narrowing towards quadratojugale. In
the postero-medial portion of quadratum, above its trochlea lies a small tuber-
cle — the tuberculum supratrochleare (fig. 17).
Quadratojugale. By its form quadratojugale reminds one of
a bird’s wing (figs. 16, 17).
In its posterior portion this bone is pierced by a large aperture of oval
outlines. "
Besides the large foramen paraquadratum, we have detected in Benthosu-
chus the presence of another small extra aperture, lying not far from the
first one, and perfectly visible on the ventral side of quadratojugale. This
aperture — foramen paraquadratum accessorium (fig. 16) — never before
was mentioned in literature.
Squamosum. The upper angle of this bone adjoins tabulare (fig. 13),
and bounds from the anterior side the incisura oticalis. From the side of the
occipital regipn of the skull, squamosum (its lamina parapterygoidea) is
adjoined by the upper border of the posterior branch of pterygoid (more exactly—
by its lamina ascendens).
Epipterygoideum. This bone in Benthosuchus is provided with
a strongly developed ascending process — processus ascendens (fig. 15).
The processus paroticus is altogether lacking, because of the corresponding
region of the cartilage being not ossified.
Examining the anterior surface of epipterygoideum, one can easily detect
the presence here of a rather low torus, the outlines of which remind one of
126
the letter Y. The branches of this torus —torus lambdoideus (fig, 15c)— are
diverging from the centre of the bone in the following manner: one of them
(oblique) runs along the processus ascendens; the other (vertical) extends to-
wards the basal margin of the bone; while the third one (horizontal) reaches
by its end that part of epipterygoideum, which in Capitosaurus (Sushkin,
1927) directly passes into the processus phroticus.
In the middle part of the posterior surface of epipterygoideum (fig. i5d)
is conspicuous a small tuberosity — the tuberosita transvers — extending
across the bone. Somewhat lower, in the basal part of the bone, there is situa-
ted a rather high ridge — crista basalis — bounding the rugosity, which served
as the facet for joining of epipterygoideum with pterygoideum.
Sphenethmoideum. The isolated sphenethmoideum of Bentho-
suchus sushkini has the form of a three-sided pyramid, with somewhat dilated
anterior part (fig. 20). Its very smooth upper surface is closely adjacent to the
lower surface of dermal bones of the cranial roof.
Preparing the posterior side of sphenethmoideum, we have detected the
presence on it of a deep fossa — fossa lobi olfactorii (fig. 20e, g) — in which
lied the lobi olfactorii. In the rather thin lateral walls of this fossa are situa-
ted the foramina optica. Anteriorly the fossa lobi olfactorii is gradually passing
into the two canalis olfactorius (fig. 20a, e, g), which are separated from
one another by the boni septum interolfactorium (fig. 20).
Cranial Roof
In Benthosuchus, as well as in all the Labyrinthodonts, the dermal bones
of the cranial roof are covered by a peculiar pitted sculpture.
In each bone, the parts of the ornamented surface situated in the region
of the ossification centre, are always formed by relatively round pits. While
in the peripheral parts of most bones the sculpture is formed by rather greatly
stretched grooves separated by radial ridges. The ossification centre of the
bone represents at the same time the centre of the lines, along which are exten-
ding the ridges of the scuplture of every dermal bone (Bystrow, 1935).
The arrangement and mutual relation of dermal bones, forming the cra-
nial roof of Benthosuchus, are shown on figs. 22. 23 24 and 25.
The furrows of the lateral line system of the skull of Benthosuchus are
pretty well conspicuous (figs. 22, 29).Examination of their courses showed
that they are arranged in such a way that successively cross the points of
ossification of dermal bones. This phenomenon may be explained by the
fact that at the initial stages of development of the skull of Stegocephalians,
the lateral line system was connected with the primary elements of its de-
veloping cranial bones. This initial connection is retained unchanged during
the whole lifetime of the animal.
Mandible
Mandibula of Benthosuchus sushkini is composed by nine dermal bones
arranged around the long Cartilago Meckeli, the posterior end of which ossi-
fying forms the tenth bone of this complex, viz. the articulare.
Articular e. On complete mandibles only the upper surface of this
bone is visible. Here it appears concave, due to its being articulating with
trochlea of quadratum into a joint, forming for the latter the cavitas gle-
noidalis of corresponding shape (fig. 26b). The anterior border of artied-
lare rises in the form of a small ridge — crista praeglenoidalis (fig. 26b).
Anteriorly the bone terminates in a facet with rugose surface. It was here
directly joined with Cartilago Meckeli. From behind, from sides and from
below the articulare is covered by three dermal bones: supraangulare, an-
gulare and praearticulare.
Supraangulare. This bone, adjoining articulare from the lateral
side, covers the caudal portion of the latter as if by a bony cap, forming
127
the posterior angle of the jaw (fig. 27b). Here, immediately behind the
articular fossa of articulare is seen the small tuberculum postglenoidalis
(fig. 27a, b). Posteriorly of this tubercle, the upper surface of the bone forms
a three-sided, slightly concave facet, on the lateral margin of which extends
a fine ridge with rounded border.
The posterior end of supraangulare has the appearance of a process with
rounded apex — processus retroarticularis — which just forms the posterior
angle of the jaw (figs. 26a, b, 27a, b.).
The upper margin of the bone under description has the appearance of
a slightly convex ridge (fig. 27a), on the inner side of which is present a very
distinct longitudinal thickening — the torus arcuatus (figs. 26a, 27b).
Angulare is a bone composed by two slightly concave laminae,
which adjoin one another below at a certain angle. Of them the external lami-
na (fig. 27c) is of somewhat larger dimensions than the internal one (fig. 27d).
Praearticulare (Goniale). The posterior end of this bone is over-
lying articulare (fig. 26a) from the internal side of the jaw.
On the posterior border of praearticulare there is a small aperture, through
which chorda tympani entered the jaw (fig. 27e, g). On the lower border of
its distal end is seen a shallow incisure, which takes part in the formation
of foramen Meckelianum.
On the side of praearticulare directed towards articulare, is situated the
flat processus hamatus (figs. 26b, 27e), covering the upper part of anterior
surface of this latter bone.
D e n t a 1 e. This is the longest bone in the mandible of Benthosuchus.
Its sharp posterior end overlays supraangulare; its position with respect to
angulare being just reverse, i. e. the lamina externa of the latter bone over-
lays from the external side the lower border of dentale (figs. 28, cross-sections
b, e, d).
The anterior end of dentale is somewhat dilated, forming a lamina — la-
mina symphyseous (fig. 26).
Postspleniale is situated on the lower border, at about the middle
of the length of the mandible, and adjoins by its posterior margin the angu-
lare and praearticulare (fig. 26a). Being wedged between the anterior ends of
the said bones, the postspleniale, together with the latter takes part in the
formation of large, oval foramen Meckelianum.
Spleniale. This bone occupies the lower part of the foremost portion
of the jaw. The anterior end of spleniale, which is curved hook-like — (hamu-
lus splenialis, fig. 26a) — is lying just under the dilated end of dentale, and
takes part in bridging the two branches of the mandible.
Coronoideum. This bone is situated between praearticulare and
dentale. Its posterior border forms the anterior angle of the deep fossa mandi-
bulae (fig. 26b). Here, under the coronoideum and dentale (fig. 28, cross-
section c), the long sharp anterior end of supraangulare is extended far for-
ward.
The coronoideum of Benthosuchus bears a row of small teeth.
Intercoronoideum. This bone adjoins dentale from the medial
side (fig. 28, cross-sections d, e, f), and its lower border forms with postsple-
niale a suture. The posterior angle of intercoronoideum reaches the anterior
end of praearticulare, being adjoined with it for a short distance. For a more
considerable distance intercoronoideum is adjoined with coronoideum (fig. 26a).
Praecoronoideum. This bone lies anteriorly of intercoronoideum,
between dentale and spleniale, on the internal surface of the mandible. Its
posterior end is joined by a short suture with the anterior end of intercoronoi-
deum; while its anterior end reaches the dilated part of dentale.
A number of cross-sections of the mandible of Benthosuchus (fig. 28) sho-
wed that from the deep fossa mandibulae situated on the posterior end of the
jaw, along the whole length of the mandible and nearly reaching the symphy-
sis, extends a long canal in which the Cartilago Meckeli was embedded.
428
The canals of the lateral line system of the
mandible are pretty well conspicuous on the ornamented portions of its ex-
ternal surface.
The main furrow — sulcus articularis (fig. 29) — derives from the border
of supraangulare, somewhat posteriorly of the fossa mandibularis. The late-
ral line, which was embedded in this furrow, undoubtedly represents the di-
rect continuation of canalis jugalis, the posterior portion of which extends
arch-like along the suture between squamosum and quadratojugale. The
sulcus articularis befurcates in the limits of supraangulare; one of its arms
(branches) — sulcus marginalis — appears as if its direct continuation, and
is extending obliquely towards the lower angle of the jaw, where it terminates
in the region of the punctum ossificationis of angulare. The other branch —
sulcfts dentalis — forming a slightly convex arch passes over from supraan-
gulare onto dentale, and reaching the ornamented portion of the latter, di-
sappears.
Besides sulcus articularis and sulcus dentalis there is on the supraangu-
lare of Benthosuchus one more constant additional furrow — the sulcus arti-
cularis accessorius (figs. 27a, 28, 29).
Only those parts of chondrocranium of Benthosuchus, which are known
as the bony exoccipitale, quadratum epipterygoideum were ossificated and
preserved. As to its other parts, — they may be rather correctly restored by
their scars left on the surfaces of dermal bones. Such restorations are shown on
figures 32, 33 and 34.
Basing on the sufficiently thorough study of the anatomy of recent Urode-
la, we have made an attempt to restore the general picture of disposition of
nerves and vessels in the head of Benthosuchus. The presence on the skull of
Benthosuchus of a number of furrows and apertures for vessels and nerves
which are homologous with the corresponding formations in recent Urodela,
allows us to suppose that the vascular and nervous systems of Benthosuchus
by the arrangement of their nerves and vessels in general features were simi-
lar to those in the head of recent Urodela.
Vertebral column
The neural archs of each vertebra appear in the form of two curved lami-
nae, the upper ends of which are fused with one another (fig. 40a, b, c). Di-
rectly under the point of their fusion lies a small, open below foramen of oval
outlines in which was enclosed the medulla spinalis. The dorsal part of the
neural arch bears the rather powerful and comparatively short processus spi-
nosus.
Above the upper border of foramen neurale, from the base of processus
spinosus extend forward and backward by two pairs of articular processes.
The anterior processes directed forward — zygapophyses anteriores — being
somewhat more strongly developed than the posterior ones (fig. 40c). Forward
and sideward the neural arch gradually passes into well defined transverse
processes, which terminate in rugosities.
The internal sides of neural arch, downward of the foramen for medulla
spinalis are also rugose. These portions of the arcus-facies chordialis — over-
laid the strongly developed chorda dorsalis, which extended along the whole
length of the vertebral column (fig. 40a, b).
Hypocentrum of Benthosuchus has the form of a semilunar la-
mina (fig. 40 d). In lateral view it appears as an almost triangular bone (fig.
40f), and the concavity of its lower surface as well as a small facet situated
somewhat posteriorly of the upper angle of the bone are perfectly visible.
The character of structure of the two described elements of a vertebra, assur-
edly indicates that Benthosuchus refers to the rhachitomous Stegocepha-
9 Тр. Палеовтол. ин-та. т. X, в. 1
129
lians. Though in all the vertebrae of Benthosuchus lacks the bony element,
which is constantly present in the representatives of Rhachitomi — viz. the
pleurocentrum.
Costae
The so-called, «cervicals ribs», joined with the fore part of vertebral column,
in Benthosuchus represent comparatively small bones (figs. 40, 41a, b). Their
proximal part is dissected by a deep incisure into two rather long processes.
The anterior surface of the first ribs is somewhat convex. On their slightly
concave postero-lower surface one can trace a poorly defined furrow — the
sulcus costae (fig. 41a). Their distal end usually is narrow, so that the rib
as a whole appears to be cuneiform.
The ribs of the thoracal part of skeleton are considerably longer than*the
cervical ribs (fig. 41c). Their tubercle and head merge into one another, and
no incisure is observed on their dilated proximal end. Some of them are bearing
the processus uncinatus (fig. 42d), which in big-sized specimens attains
rather great dimensions, reminding one of the corresponding formation on
the ribs of Eryops (fig. 41e).
The costae lumbales by the character of structure of their proximal
end resemble very much the ribs of the thoracal part of skeleton. Here the
tuberculum and capitulum costae are also hardly perceivably separated from
one another (fig. 41f). Though the distal end of the vertebra lumbalis resemb-
les more the corresponding end of the cervicals ribs: it is also gradully narrow-
ing and passes into a long, nearly round in cross-section, process. The sacral
rib is the most powerfully developed bone of all the costae of Benthosuchus.
The proximal end of this rib is greatly dilated (fig. 42a). No incisure separa-
ting the tuberculum from capitulum costae is observed; though when examin-
ing the rib from the side of vertebral column, one can readily discern these
two parts of the bone (fig. 42c). Over the anterior surface of the sacral rib
extends the high scabrous crista muscularis (fig. 42a).
Shoulder-girdle
Interclavicula. The outlines of this bone in Benthosuchus are
nearly rhomboidal (figs. 44, 45). Its anterior angle is extended in the cranial
direction much more than the three remaining angles.
On the ventral surface of the bone under description there is observed
a rhomboidal elevation, the outlines of which repeat in smaller dimensions
the outlines of interclavicula as a whole. The postero-lateral slopes of this
elevation are hardly perceptibly passing into the marginal portions of the
bone. As to the antero-lateral sides of this elevation, they are separated
from the peripheral portions of the bone by a distinct shelf. This feature of
structure is caused by the peripheral portions of interclavicula being covered
by facies clavicularis for the space from the anterior and up to the lateral
angles of the bone (figs. 43, 44, 49.)
The central part of this rhomboidal elevation — the position of which
corresponds to the punctum ossificationis of interclavicula — is ornamented
by the typical for Stegocephalians pitted sculpture.
The dorsal surface of interclavicula is smooth. Here, correspondingly to
the ossification centre, is observed a small elevation — the eminentia cent-
ralis (fig. 45) — from which derive three straight beams, extending radially
towards the borders of the bone. Either of these three beams being compara-
tively narrow in its initial part (nearer to the eminentia centralis), gradually
widens toward the peripheral portions of the bone. These beams apparently
were of but a purely mechanical significance. At least two of them — namely,
the trabeculae claviculares (fig. 45.) — are directed with their ends just
towards those borders, which from the ventral side are overlaid by claviculae,
and as if consolidate the bone along its two radii. The third beam — trabe-
130
cula sternalis — directed, backwards, covered the ventral surface of the carti-
laginous sternum.
Clavicula. This bone is of about triangular outlines; when viewed
from the ventral side it resembles very much a stretched bird’s wing (fig. 46a).
From the lateral angle of clavicula, from that part of the bone in which
punctum ossificationis is situated, derives the powerful flat processus prae-
scapularis directed dorsally (fig. 46b). In its first half this process is ascending
vertically, then it turns backward, so that its second half is set at an angle
of about 140° to the first one. The apex of this process usually is sharpened.
By its internal side it covers the lower end of cleithrum.
Cleithrum of Benthosuchus represents a small, elongated bone,
with considerably dilated upper end. In large specimens of Benthosuchus,
the anterior border of this dilated end of cleithrum bears a small ridge — the
crista cleithralis (fig. 47c).
Scapula of Benthosuchus is a massive heavy bone (fig. 48). On the
lateral side of scapula, about at the middle of its height, nearer to the poste-
rior border of the bone, is observed a powerful process, directed by its greatly
dilated end medially and backwards (fig. 48b). On the upper surface of this
process, just along its margin, runs a fine furrow — the sulcus canalis gle-
noidalis (fig. 48b, d), leading to the canal which pierced the unossified porti-
on of the bone. On the lower border of the same process, there is another hard-
ly conspicuous furrow — the sulcus canalis coracoideus (fig. 48d) — which
denotes the position of the coracoid canal.
The third canal of scapula, known under the name of canalis supraglenoi-
dalis, appears in Benthosuchus — due to the insufficient degree of ossifica-
tion of the neighbouring portions of cartilago — in the form of a deep inci-
sure, separating below the powerful medial process of scapula from the lower
angle of the bone (figs. 48 c, 50).
Bones of fore limb. In the available collections the bones of
the fore limb of Benthosuchus are presented by a lot of isolated specimens
of humerus, ulna and radius. As to the metacarpal bones and phalanges, there
are present very few of them.
Humerus. Over the ventral surface of this bone for about half its length
extends a powerful crest — the crista deltopectoralis — with rugose surface
(fig. 51). The distal end of humerus is greatly dilated, and has assumed the
form of a thick bony lamina, lying in horizontal plane.
The epicondylus radialis (figs. 51, 52) appears as a process of comparati-
vely inconsiderable dimensions, with rounded apex. The epicondylus ulna-
ris, on the contrary, has the form of a very wide plate.
Ulna and Radius. The upper end of ulna is of oval outlines;
on the side directed to radius it bears a small depression — incisura radia-
lis (fig. 53a, b). The distal, somewhat dilated end of this bone is oval in cross-
section.
The proximal end of radius in cross-section is almost round. The form of
the lower end of this latter bone is similar to that of the upper one, only on
its posterior surface are observed marks of a small hollow.
Pelvic girdle
Ilium of Benthosuchus represents a rather massive bone with greatly
dilated lower end (fig. 54). It is bowed in such away that its upper part .is
shown to be strongly inclined backwards, — and consequently the posterior
border of the bone is strongly concave, forming the deep incisura ischiadica
(fig. 54a).
On the external surface of ilium immediately under its lower border,
there is a small hollow, which forms a part of acetabulum.
Above the acetabulum, over the whole external surface of the bone, runs
a scabrous line — linea obliqua — directed obliquely backwards and upwards,
which represents the point of originating of the muscles of femur (fig. 54a).
9* *31
Ischium of Benthosuchus is a small cuneiform bone. It is deprived
of any ridges or processes.
Pubis is altogether absent from the collections of bones of Bentho-
suchus.
Bones of hind limb. Femur of Benthosuchus is by far not
so powerful as its humerus. In the proximal part of the lower surface of this
bone, for about 1/3 of its length, extends the rather large trochanter. Eleva-
tion of the trochanter abruptly passes into a high sharp ridge — crista aspera
femoris (fig. 55b, c, d) — which lowering little by little, nearly reaches the end
of the bone.
Tibia is a massive bone. Its upper end is of about semilunar outlines,
being considerably wider than the lower one (fig. 56a, b). The middle part of
the bone is narrowed, being strongly impressed from the side of fibula. In
the upper part of the anterior surface of tibia there is observed a very distinct
rugosity — the tuberositas tibiae (fig. 56a). From this rugosity derives a small
ridge—crista anterior tibia — extending along a straight line downwards and
nearly reaching the lower end of the bone (fig. 56a).
The posterior surface of tibia, in its upper half forms a slightly concave
facet — planum popliteum (fig. 56c). This facet is separated from the lateral
surface of tibia by a sharp ridge — crista posterior tibiae, extending for about
the whole length of the bone (fig. 56c). The posterior surface of tibia in its
lower part is slightly convex.
The distal end of tibia in cross-section has the outlines of a triangle with
rounded apex (fig. 56d).
Fibula is as long as tibia; its upper part is more strongly dilated
than the lower one.
The upper end of this bone in cross-section has the outlines approaching
that of greatly elongated oval (fig. 56f). The posterior surface of fibula bears
neither ridges, nor rugosities, which immediately catch one’s eye on the corres-
ponding surface of tibia.
On all the specimens of fibula of Benthosuchus, in the lower part of the
posterior surface of the bone, not far from its medial angle, is observed a sharply
defined groove — th» sulcus fibularis — extending vertically downward
(fig. 56e).
The anterior surface of fibula is slightly convex and very smooth (fig. 56).
The lower end of the bone in cross-section appears of much more elongated
form than the upper one (fig. 56h).
The fibula of Benthosuchus is gently twisted about its longitudinal axis
in such a way that the long axes of the cros-sections of its two ends cross one
another at an angle of about 37° (fig. 56i).
As to the bones of feet — we have not succeeded in discovering any ele-
ments of tarsus, when preparing the rock enclosing disunited skeletons of
Benthosuchus. Apparently for a long period of the lifetime of Benthosuchus,
its tarsus remained in cartilagineous state.
The metatarsalia and phalanges of Benthosuchus evidently were ossified,
but among the fifteen available specimens of the bones of limbs it is very
difficult to discern those belonging to the fore limb from those belonging to
the hind one.
II. GENERAL PART
CHAPTER I
AGE and INDIVIDUAL CHANGES OF THE SKELETON OF BENTHOSUCHUS
The abundant material on Benthosuchus, belonging to individuals of the
most different age, makes possible an evaluation of the age and individual
variability of the elements of the skeleton. It is obvious that such an evalua-
tion may be made only in the most general features, comprising the most
132
marked changes; thus the way may be paved for a further study of the skele-
tal variations.
In the preceding pages the absence of interrelated elements of the skeleton
in the Sharzhenga occurrences has been more than once noted; in the fossili-
ferous sandstones of these occurrences the most different parts of the skeleton
of large and small individuals are mixed in disorder. However, in the great
mass of remains of Benthosuchus in the occurrences on the Sharzhenga river
only one fragment has been found belonging to another genus of Labyrintho-
donts. Therefore, the number of bones of one species B. sushkini being great,
it is possible to have a direct comparison of the homonymous skeletal elements
in spite of the lack of complete skeletons. The differences in the size of the
homonymous bones are assumed to be age changes of the separate elements
of the skeleton of Benthosuchus. Some morphological peculiarities of homony-
mous bones which are of equal size are referred to the group of individual
variations.
The material available comprises only the growth of Benthosuchus sush-
kini after metamorphosis. No analysis of the changes of the skeleton of Ben-
thosuchus, from a larva to an animal breathing with lungs can be, therefore,
carried out.
In addition, in the material at our disposal not all the parts of the skeleton
are represented with equal completeness. Thus, the age and individual changes
of the vertebrae, ribs, ischium, tibia, fibula and aeropodium cannot be traced
due to the small number of bones found. Ulna and radius, found in a rather
large number, seem not to reveal any age peculiarities besides a direct increa-
se in size, remaning quite proportional in all their parts.
The examination of the variability of the skeleton of Benthosuchus should
naturally follow two main groups of changes — age changes and individual
changes. There is no doubt that into the group of individual changes also chan-
ges of a pathological or degenerative nature may be included, which at pre-
sent cannot be separated out with assurance to an independent group.
The most important feature of age changes common to all the divisions
of the skeleton, is the size or some element of the skeleton, or other.
In the collection of the Palaeontological Institute the smallest of all the
skulls of Benthosuchus known is skull No. 4—19/2252, 27.5 mm. long. Since
this skull belongs to a fully formed animal, breathing with lungs, the skulls
of the larvae of Benthosuchus immediately before metamorphosis or at the
moment of it should have been still smaller in size, probably but little excee-
ding 10 mm. Fragments of large skulls and elements of postcranial skeleton
available in the collection show that the largest and oldest specimens of Ben-
thosuchus possessed skulls, the length of which was not less than 700—800 mm.
Thus, during their life the quite formed animals, which had passed through
metamorphosis, increased in size about 70 times.
This fact is of utmost importance since on the one hand it points out to
a prolonged and most probably slow growth of the animal during the whole
course of its life, i. e. to its considerable longevity, and on the other hand
reduces to zero the importance of the size of the animal as a systematic fea-
ture, if this feature is taken without a consideration of the other peculia-
rities.
The change in the size of the animal was moreover accompanied by changes
of some morphological details of the structure of the skeleton. These changes
are best of all traced in the most complicated division of the skeleton — in the
skull.
The most characteristic peculiarities of the structure of the skull of Ben-
thosuchus at the young age (according to skull No. 4—19/2252) will be as fol-
lows: —
1. A comparatively short triangular skull with a rounded wide snout and
straight lateral margins.
2. Very large orbits located almost in the middle of the skull.
133
3. External and internal nostrils placed wide apart. The former lie on
the end of the snout.
4. The position of foramen pari etale on the line of the posterior margins of
the orbits.
5. An almost equal development of pre- and post orbital elements of the
cranial roof. Both are distinguished by a considerable width.
6. Skull high in the occipital region with a convex arch of the cranial
roof and a comparatively flat lower margin.
7. A slight bend of the interorbital part of the cranial roof.
8. A strong curvature of sulcus supraorbitalis.
9. Shallow and wide otic notches with a short cornua tabularia.
10. A very wide transversally elongated corpus parasphenoidei.
11. A short and wide processus cultriformis parasphenoidei.
12. Processus parasphenoidalis vomeris wide.
13. Ramus anterior of the pterygoid comparatively shortened and equal
in length to ramus posterior.
14. Fenestra praemaxillaris unpaired, with traces of cartilaginous of in-
terspace.
15. Foramen nervi vagi comparatively large.
16. Fenestra supraoccipitalis large and very high.
17. Fenestra basioccipitalis low and wide.
18. Processus submedullaris and processus lamellosus exoccipitale unde-
veloped.
19. Lamina parapterygoidea squamosi high as compared with the low
ramus posterior pterygoidei. Between the two above bones is a slit.
20. Crista obliqua pterygoidei undeveloped.
21. The osseous tuberculum on the posterior surface of quadratum is absent.
If we compare the features enumerated with the corresponding peculiari-
ties of the structure of skulls of adult specimens of Benthosuchus, we shall have
nearly in all cases their reverse expression.
From the available fragments of very large skulls of Benthosuchus in
Figs. 60 and 61 the natural size is shown of some elements of the skull of very
large specimens (No. 5—9/2243 and 29—19/2252). In accordance with the laws
of growth of Benthosuchus, established by A. P. Bystrow in 1935 , we succeeded
in giving a reconstruction of the dorsal surface of a huge skull of Benthosu-
chus for about the length of 700—750 mm. (fig. 62). For a precise reconstru-
ction of the ventral and occipital surface of also huge skulls no sufficient
material is available for a detailed reconstruction of the age peculiarities.
Therefore we are obliged to make the comparison of the above noted features
in the region of the ventral and occipital surface of the skull using full skulls
of middle age which are at our disposal.
Thus, the old specimens of Benthosuchus («medium» for endocranium) will
be distinguished by the following peculiar features (see figs. 62, 22, 30):
1. An elongated triangular skull with a narrow snout, strongly widened
in the occipital region, due to which the exterior margins of the skull are not
straight but concave.
2. Small orbits located further posteriorly.
3. Externa] and internal nostrils lying closer apart. The former are located
backwards from the anterior end of the snout and have an elongated-oval
shape.
4. Foramen parietale located far backwards from the posterior margins
of the orbits.
5. A strong development of the preorbital elements of the cranial roof and
a slight development of the postorbital ones. The former are strongly elonga-
ted, narrow; the latter in their outlines approach a square.
6. In the occipital region the skull is low, with the arch of the cranial roof
bending downwards and the lower surface concave in an arch-like manner,
due to a downward bending of the lateral parts.
134
7. A strong downward bend of the interorbital part (area) of the cranial
roof.
8. A slight curvature of sulcus supraorbitalis.
9. Deep and rather narrow otic notches with elongated cornua tabularia.
10. Corpus parasphenoidei has the outlines of a longitudinally elongated
parallelogram.
11. Long and very narrow processus cultriformis.
12. Narrow processus parasphenoidalis vomeris.
13. Ramus anterior of the pterygoid comparatively elongated on the ave-
rage 1.У2 times the length of ramus posterior.
14. Fenestra praemaxillaris divided by an uncomplete septum, in old
specimens is undoubtedly ossified, passing to a paired aperture.
15. Foramen nervi vagi relatively small.
16. Fenestra supraoccipitalis wide and low, considerably diminished by
a downward convexity of the median suture dermosupraoccipitalia.
17. Fenestra basioccipitalis small and narrow.
18. Processus submedullaris and processus lamellosus strongly developed.
19. Lamina parapterygoidea low and somewhat lesser in height than ramus
posterior of the pterygoid. The slit between the two bones disappears.
20. Crista obliqua pterygoidei well developed.
21. On quadratum there is strong muscular tubercle.
A series of complete skulls which are at our disposal illustrate well the
gradual strengthening of separate features and the gradual transition from
small individuals to larger ones.
The order of ossification of the cartilaginous bones of the skull of Bentho-
suchus may be the following: exoccipitale, quadratum, epipterygoideum,
sphenethmoideum, prooticum. Of these, exoccipitale ossify at the earliest sta-
ges of the existence of the animal, and qudratum a little later; epipterygoideum
and particularly sphenethmoideum appear very late, almost at the boundary
of «middle» age.
It would not be superfluous to note that in accordance with the above
mentioned age peculiarities of Nos. 16 and 17, in the process of growth of the
skull of Benthosuchus a very considerable proportional diminution of the size
of the cartilaginous supraoccipitale and especially basioccipitale is observed.
These bones not only do not ossify but grow considerably more slowly than the
rest of the cranial parts, due to which they are displaced by the adjacent osse-
ous elements, as, for instance, exoccipitale. Therefore we are fully justified in
considering basioccipitale and supraoccipitale in the skull of Benthosuchus
as features of a relict nature which are on the way towards disappearing.
A detailed examination of the above enumerated age changes of the skull
of Benthosuchus shows obvious unequality of their significance. One cate-
gory of age changes involves only a general intensification with age of the
structural peculiarities of the skull, which is the most usual expression of the
age changes in the higher Tetrapoda. To this category such changes should be
referred as the general increase of the massive nature of the bones and the
increase of the diametres of the cells of sculpture on the bones of the cranial
roof, the development of muscular ridges and tubercles, the decrease of the
reduced elements of the skull. Of the above enumerated features belonging
here, are Nos. 6, 7, 9, 13, 15,-16, 17, 18, 19, 20 and 21.
All these features, in turn, should be subdivided into two groups: features
of an «infantile» weak structure of the skull, and features of evolutionary pri-
mitiveness pronounced more distinctly in young than in old specimens.
Referred to the first group may be such features as the absence, or presence
of processes and ridges, the replacement of slits by sutural connections, better
or slighter pronounced structural peculiarities, inherent to (characteristic of)
the skull of Benthosuchus, in particular. The other group includes such chan-
ges as Nos. 6, 13, 16, 17 and 19. It is natural that very young forms must have
features somewhat more primitive, which is expressed by the above changes.
135
Thus, a comparatively longer ramus posterior of the pterygoid, a relatively
larger size of the cartilaginous supraoccipitale and basioccipitale and a higher
crista falciformis, as well as the grater height of the skull in the occiput, all
of these features distinguishing very young specimens of Benthosuchus from
older ones, are at the same time features of a lower evolutionary stage in the
course of general development of Labyrinthodonts as established by Watson
(1920). Indeed, the flattening of the skull, the shortening of the posterior bran-
ches of pterygoids, proceeding at a more rapid rate than the shortening of the
anterior ones, the reduction of basiocipitale and supraoccipitale are typical
features of the high evolutionary stage of the development of Labyrintho-
donts.
Thus, the first category of age changes includes those not altering the gene-
ral plan of structure of the skull and reflecting only its general stoutness,
or, to a slight degree, showing the general features of the phylogenesis of
the whole group Labyrinthodonts.
The features of the second category considerably alter the general character
of structure of the skull, without any direct connection with the reflection of
the phylogeny of the group Labyrinthodonts. Referred here may be changes
Nos. 1, 2, 3, 4, 5, 10, 11, 12 14, and as being directly dependent on Nos.
5 to 8.
Thus, with the growth of the skull of Benthosuchus we observe how a short
skull with large orbits and a wide snout passes to a long skull with a narrow
snout and orbits receding backwards. An unpaired fenestra praemaxillaris
becomes paired, corpus parasphenoidei becomes narrower. Processus cultri-
formis becomes long and thin; the preorbital elements, considerably longer.
In addition, the skull becomes considerably broader in the occiput.
J. A. Efremov (1933) was the first to show that very young specimens of
Stegocephalians in different forms are characterized by skulls with large or-
bits located in the middle of the skull, i. e. by an equal or nearly equal develop-
ment of pre- and postorbital elements. Foramen parietale is situated on the
line of the posterior margins of the orbits. W ith a further growth of the skull,
the above proportions are broken, depending on the character of specialization
of a given form. In the cranial roof prevail pre- or postorbital elements, or,
on the contrary, marginal ones, when the skull is elongated or widened, the
orbits are displaced, etc. Foramen parietale behaves as a centre fixed in the
cranial roof, according to whose position it is easy to distinguish the modi-
fication of the skull.
It may be easily seen that a very young specimen of Benthosuchus (skull
No. 4—19/2252) has exactly the above described structure, common for the
larvaland young specimens of Stegocephalians, i. e. it shows the absence of
features of special adaptation strikingly pronounced in older specimens.
Summing up all the aforesaid, we are led to the necessity to recognise that
the second category of age changes of the skull of Benthosuchus comprises
changes which are increasing with age the special adaptation of the skull
inherent to Benthosuchus.
These age changes are more distinct than all the others and evidence a
steady intensification of the expression of adaptation with the growth of the
skull. In other words, the process of development of adaptation, inherent to
Benthosuchus, continued throughout the whole life of the animal. However,
as it may be established from a comparison of the small skull No. 4 —18/2252
and the smallest of the other full skulls — No. 1—9/2243, as well as from nu-
merous separate fragments, the skull of Benthosuchus acquired the most
important features of its structure rather quickly, within the range of growth
from 40 to 100 mm. The further growth of the skull caused only a sharper
development of the peculiarities of structure acquired during the early period.
Thus, on the material on the age stages of Benthosuchus we see a corrobo-
ration of the general regularity of onthogenetic development — the features
of the genus appear later than the general features of the class, order, family,
136
and even so late that a fully developed animal has as yet no inherent features
of adaptive modification. Such a retardation seems to be typical in general
for primitive Tetrapoda. Obviously, a primitive organism, requiring no
considerable period of time for its onthogenetic development to the stage of
independent existence (after metamorphosis), completes its development very
slowly, almost during the whole of its life. Therefore, a part of the late appearing
features, namely, some features of adaptive modification (i. e. features of genus
and species) appeared already subsequent to the animal having reached a rat-
her large size.
The individual variability of the skull of Benthosuchus
consists chiefly in the retardation, or, on the contrary, acceleration of some age
changes.
Individual changes, not connected with age variability, have been also
observed in many skulls or fragments of skulls of Benthosuchus.
To these belong the following: —
1. Variations in the outlines of sutures between the separate parts of the
skull.
2. Variations in the general shape of the skull.
3. Variations in the relative size of narial openings, orbits, choanas and
in the depth of otic notches have been also established in different skulls of
Benthosuchus sushkini.
4. Variations in the course of the canals of the lateral line, distinctly
pronounced, were observed only in skull No. 5—19/2252, where a deviation
(deflection) of the posterior end of sulcus jugalis upward of sutura squamoquad-
rato-jugalis has been noted.
5. Individual variations in the location of the postfenestral dental row on
vomers of Benthosuchus.
6. Individual variations in the massiveness of the occipital elements of
the skull.
7. Individual variations in the issues of the nerves may be traced only
for the group glossopharyngeus vagus, possessing a point of outlet precisely
fixed in the well ossified exoccipitale.
To illustrate similar changes may serve a series of separate fragment of
exoccipitalia belonging to skulls of large specimens of Benthosuchus, Nos. 12—
19/2252, 12—32/2354, and two fragments without number from the collection
19/2252. Fig. 72A shows a typical arrangement of for. nervi vagi; and В, C
and D — its variations.
8. It should be noted that the relative size and shape of the area on cor-
pus parasphenoidei (area aspera parasphenoidei), covered with chagrin, are
subject to considerably individual variations (see figs. 61—69).
In concluding our review of the age and individual variability * of the
skeleton, a brief summar of the facts obtained will be given.
The skeleton of Benthosuchus due to an incessant growth of the animal
during the whole time of its life increases in size very considerably. From
the moment of metamorphosis until the maximum old age known, the size
of the animal increases 80 times.
It seems that the process of growth of the animal was going on slowly,
more rapidly in the early stages of age and then increasingly slowly, until
a certain constant was reached.
The most complicated part of the skeleton — the skull, in the process
of post-metamorphic growth acquired the adaptive features inherent to it —
a part of the generic and the specific features. Therefore, the age changes
of the skull are comparatively diverse and distinctly pronounced, especially
in the early stages of post-metamorphic growth.
The individual changes of the skull are insignificant and consist mainly
in an earlier or later appearance of different age peculiarities.
However, many variations of the skull are so strongly pronounced that
when taken separately, without a comparison with the different gradations
137
•of similar variations in other skulls, may be taken for specific peculiarities
by investigators inclined to create a large number of separate systematical
units.
The vertebral column of Benthosuchus in the process of growth is ossified
more strongly in old specimens than in young and middle-aged indivi-
duals, acquiring a close connection of the centrum with the neural arch, it is,
however, true by way of synchondrosis.
It seems that also in many other Stegocephalians the individual age plays
a considerable part in the degree of ossification of the vertebral column.
The shoulder girdle of Benthosuchus in the region of scapula-coracoideum,
in spite of its massiveness, even in rather large specimens ossify rather slight-
ly. But its dermal bones — interclavicula, clavicula and cleithrum, ossify in
the early stages and give a whole series of age changes (chiefly of the clavicula).
Thus, the claviculae from comparatively narrow oval bones, become wide
triangular plates, due to which the relative width of the ventral surface of
the shoulder girdle increases with age. z
Humerus of Benthosuchus, as well as femur, ossify in the process of growth
to an insignificant degree and change according to the age stages also insig-
nificantly. However, the processes and rugosities serving as places of origin
and attachment of muscles considerably increase with age.
On the other hand, in some specimens considerable individual variations
in the ossification of humerus (femur), or deviations of humerus (femur) from
the typical shape are observed, as well as the formation of massive shorter
bones.
The pelvis of Benthosuchus is not completely ossified, and the stages of
its development are so far very little known. Ilium in the nature of age and
individual changes is similar to humerus and femur.
The bones of antibrachium and crus are as yet little characterized by age
material, but seem to be slightly changed with age. In the collections of
tibia there are several strongly thickened specimens of bones.
Thus, from a consideration of the age and individual variations of the ske-
leton of Benthosuchus as a whole, it is evident that the regions of the skele-
ton, having a distinctly pronounced specific adaptation, to the greatest degree
change with age and little vary individually. The primitively built, slightly
modelled parts of the skeleton, as the skeleton of limbs, poorly developed in
the majority of aquatic forms, are slightly changing with age, but in individual
cases are subject to considerable individual variations. This regularity is
comprehensible, since in primitive Tetrapoda it has been long since established
that some features which, so to say, ultimately determine the given form
(genus, especies) develop during the postembryonic period, in the process of
growth of the individual. Therefore, the specific adaptive features, appearing
comparatively late, determine the considerable variability with age of those
parts of the skeleton, in which the given adaptation is pronounced.
As to the postcranial skeleton, the latter in the majority of Stegocepha-
lians is on a certain low level of development and is hardly distinguishable
even in different-families. In aquatic and «land» Labyrinthodonts the main
differences of the postcranial skeleton consist in its greater ossification in
the latter.
Such an imperfect postcranial skeleton, in contradistinction to the skull
plastic to different modifications, naturally but slightly varies with age,
since it has no large number of specific features determining a given form,
i. e. the adaptation.
It is very interesting that in Benthosuchus the degree of ossification of the
bones of extremities and their girdles little increases with age. This fact indi-
cates that in the lower Tetrapoda the degree of ossification of the separate
elements of the skeleton is a certain constant for a given group or form, pro-
duced as a result of the evolutionary' process.
As mentioned above, in the collection of the Palaeontological Museum,
138
along with remains of Benthosuchus from the occurrences on the Sharzhenga
river, there are in abundance remains of Wetlugosaurus from the synchro-
nous deposits of the Vetluga river. These remains are not described in the
present paper, but they have been studied by the author for the sake of com-
parison with Benthosuchus. The cranial material is represented by only one
more or less complete skull, but there is a great number of separate fragments
of skulls and bones of the postcranial skeleton. This genus reveals the same
series of changes with age as Benthosuchus, since along with remains of very
small specimens there are fragments of skulls and bones of extremities belon-
ging to skulls over 800 mm. in length.
Lately, fragments of bones of extremities and fragments of the basis cra-
nii, belonging to still larger individuals of Benthosuchus than all those known
from the Sharzhenga river, have been obtained from similar and synchro-
nous, newly discovered occurrences of the Timan region.
Of course, the data given here are as yet very far from a clear idea of the
size and individual variability of Benthosuchus. The unequal quantity of
remains of different age, the absence of full skeletons, from the start restrict
the possibilities of study. One of the most important questions, for the solution
of which we have as yet no sufficient amount of facts at our disposal, is that
of the duration of life of Benthosuchus and, consequently, of the rate of its
growth. This question is not discussed in our paper, since there is no olher
starting point for its solution besides an analogy with the present large-sized
coldblooded Tetrapoda.
From the brief review of literary data presented it follows that the mate-
rial on Benthosuchus sushkini, regardless of all its essential defects, for the
first time gives an idea of the character of the age and individual changes of
higher Stegocephalians.
The considerable age changes of the skull are an evidence that the skull
and some other regions of the skeleton may considerably differ at the various
stages of life of the animal. These differences when pronounced to the extreme
degree have the value of genetic features.
Individual variations of the skull, when pronounced to the extreme degree,
also may be erroneously taken for specific distinctions.
Individual variations of some elements of the postcranial skeleton, possibly
caused by pathological defect, may in a few cases considerably alter the general
shape or degree of ossification of a given element. When the findings are but
isolated ones, such deflections .will be undoubtedly taken to be a rule and
thought to be distinctions of a big order, having assumed the value of gene-
tic or specific features.
The most important inference from all the aforesaid will be the necessity
of a careful consideration of the age changes and individual deflections, both
in describing new findings and in revising the old materials. There is no doubt
that many isolated, and even serial findings, described as new genera and
species, are age stages or individual variations. We ascribe particular im-
portance to the possibility of errors in the description of fragmentary imper-
fect material, on which, unfortunately the greatest number of new genera and
species are based, only overcharging the clear idea of the fauna, without cont-
ributing essentially to the knowledge of it.
The absence of specific adaptation at the early stages of growth of the
skull of Benthosuchus, as well as probably of other Stegocephalians, makes pos-
sible a more certain analysis of the origin and relationships of the given‘form.
This we shall try to do for Benthosuchus.
CHAPTER II
SYSTEMATIC POSITION and PHYLOGENETIC SIGNIFICANCE
OF BENTHOSUCHUS SUSHKINI
Benthosuchus is a Labyrinthodont with a special progressively rhachi*
tomous structure of the vertebral column. This type of structure is characte-
I
139
rized by a reduction of the osseous pleurocentra, slightly ossified hypocentra
existing. The latter in old individuals come in contact with the neural arch
by way of a synchondroseous connection and thus form a lower stage of a typi-
cal notocentral stereospondylous vertebra. The neorhachitomous type of
structure of vertebrae is a very striking intermediate stage between the rha-
chitomous and the stereospondylous type, wholly corresponding to the for-
mer theoretic conclusions.
The skull of Benthosuchus has some comparatively primitive features
(with respect to Triassic Stereospondyli), such as, e. g. longer ramus anterior
and ramus posterior pterygoidei, jaws articulation shifted backwards, etc.
At the same time the skull of Benthosuchus represents a higher dergee of re-
duction of cartilaginous elements, on the whole not at all inferior to the typi-
cal Stereospondyli of the Triassic and even exceeding that of e. g. Capitosau-
rus. The slight development of epipterygoideum in Benthosuchus is the only
feature suggesting the greater primitivity of its chondrocranium when com-
pared to the Triassic Stereospondyli.
Thus, Benthosuchus is an interesting form in which the primitive structure
of the vertebral column and some features of the structure of the skull are
joined with a high degree of reduction of cartilaginous bones of the brain case.
The explanation of this peculiarity of Benthosuchus lies in the adaptation
to an aquatic mode of life. Thus, the Lower Permian Trimerorhachis, which
is a typical aquatic form, has a very high degree of reduction of the osseous
brain case. The general character of the skull of Trimerorhachis is much nearer
to the latest Upper Permian forms than to that of its contemporaries.
Evidently, Benthosuchus as an aquatic form, like Trimerorhachis,
also has a high evolutionary degree of structure of neurocranium, corresponding
neither to the age nor to the general evolutionary level of this form.
Of all the other Labyrinthodonts known, Benthosuchus most closely re-
sembles Wetlugosaurus. The distinctions of Wetlugosaurus from Benthosuchus
are essentially as follows:
1. A somewhat different character of the sculpture and a slighter develop-
ment of the canals of the lateral line in Wetlugosaurus.
2. A wider snout with a very large round fenestra praemaxillaris without
any traces of a septum. The dental row on vomer not wedge-shaped, but in the
form of a flat arch, bordering the posterior margin of fenestra praemaxillaris.
3. A slightly more massive processus cultriformis of the parasphenoid.
4. A comparatively narrower and high occiput.
5. A somewhat lesser degree of the reduction of the cartilaginous elements
of neurocranium.
6. A somewhat greater, earlier coming degree of ossification of the post-
cranial skeleton and sphenethmoideum and epipterygoideum in the skull.
In the other features the two forms are extremely alike. A still greater sig-
nificance is acquired by this similarity when comparing the youngest age stages
of Benthosuchus and Wetlugosaurus.
We shall make an attempt to compare the small skull of Benthosuchus,
No. 4—19/2252 (figs. 56—59), with the incomplete skull of a young Wetlu-
gosaurus No. 6—19/2252 in our collection, from Bolshaya Sludka on the Vet-
luga river (fig. 84 A, B).
The skull of the little Benthosuchus has a wide snout with a large fenestra
intermaxillaris and only slight traces of osseous septum. The lateral edges of
the skull straight, occiput comparatively narrow and high. Processus cultri-
formis of the parasphenoid short and rather broad. A direct comparison of
fig. 85 and figs. 57—60 shows very clearly that the similarity of young indi-
viduals of Wetlugosaurus and very young Benthosuchus stands still better
revealed.
If a skull of Wetlugosaurus were found representing a stage corresponding to
No. 4—19/2252 in Benthosuchus, the resemblance of the two forms would
reach an extreme degree.
140
This fact shows that Benthosuchus and Wetlugosaurus diverge only in the
different adaptation of the skull and that in spite of their simultaneous exi-
stence, Wetlugosaurus is somewhat more primitive than Benthosuchus since it
stands nearer to the direct ancestor of Benthosuchus, the structure of which
is reflected in the early stages of the growth of the latter.
This somewhat more primitive position of Wetlugosaurus is quite in agree-
ment with such features of its structure as the presence of a higher and narro-
wer occiput, a slightly lesser reduction of the elements of neurocranium. At
the same time, in the development of the vertebral column Wetlugosaurus
stands on the same step of Neorhachitomi as Benthosuchus.
Most probably Wetlugosaurus is a somewhat more «land» form, which
adapted itself to the life in water later than Benthosuchus, in connection with
which both the lesser degree of reduction of its neurocranium and the earlier
and stronger ossification of separate elements of the postcranial skeleton become
comprehensible.
In any case, the close genetic relationship of the two forms is beyond any
doubt, which is well corroborated, as it will be shown later, by the geogra-
phical distribution of the two forms.
Insofar as Wetlugosaurus is so close to Capitosaurus as to be possibly its
direct ancestor, the resemblance of Benthosuchus and Capitosaurus is under-
standable in itself.
The resemblance of Benthosuchus and Mastodonsaurus, especially stressed
by Save-Soderbergh (1937), although actually existing, does not seem to us
to be very considerable. Such features as a triangular shape of the skull and
a wide occiput are too general and exist in forms highly different genetically.
The concave lateral edges of the skull of Benthosuchus, according to Save-
Soderbergh, similar to the strongly concave edges of the skull of Mastodon-
saurus, are very slightly pronounced in the young forms of Benthosuchus.
This concavity of the lateral edges of the skull is well seen in the old specimens
of Benthosuchus (fig. 62) and is accounted for by a broadening of the skull
in the occiput, the snout being comparatively narrow, — a feature which
may be produced with similar adaptation in the most different forms.
Moreover, Save-Soderbergh bases his views on the drawing of the skull
made by Fraas who, as it is known, did not take into consideration the defor-
mation of the flattened skull. In reality Mastodonsaurus has a-skull with much
straighter edges and in this aspect resembles much more closely the skull of
Wetlugosaurus, Capitosaurus and Benthosuchus, without any special assum-
ptions. The structure of the palatal surface of the skull of Mastodonsaurus in
many details closely resembles that of Benthosuchus, as well as, consequently,
of Wetlugosaurus and Capitosaurus. The likeness of Mastodonsaurus and
Benthosuchus is only somewhat more sharply pronounced due to the similar
adaptation of the anterior part of the skull.
The structure of the anterior part of the skull is much alike in Benthosuchus
and Trematosaurus and is at the same time a fundamental feature of the re-
semblance of the latter to Mastodonsaurus, which has been already more than
once discussed.
The combined drawings Nos. 85 and 86А, В, C, D will illustrate the above
statements.
The considerable differences of the general structure of the skull of Tre-
matosaurus and Benthosuchus, discussed in the above cited papers more than
once, is an evidence that the resemblance of Trematosaurus and Benthosuchus
is a convergence and has been caused by a similar adaptation of the anterior
part of the skull.
It is not impossible that the resemblance of the structure of the anterior
part of the skull of Mastodonsaurus and Benthosuchus is also the result of
convergence. However, Mastodonsaurus, as noted above, is similar to Bentho-
suchus and Wetlugosaurus also in the structure of other parts of the skull,
as, e. g., of the base of the skull, of the occipital part of the cranial roof and
141
in the occiput. The same likeness is manifested in the arrangement of the zones
of intense growth, analyzed by Save-Soderbergh.
Summing up the above considerations, we arrive at the following concbu-
sions: —
1. Benthosuchus is most closely related to Wetlugosaurus. The distinctions
of the two genera are not of a genetic order and are restricted to different adap-
tations, Benthosuchus being probably more specialized than Wetlugosa-
urus.
2. The separation of the two forms to different families, as it has been done
so far, is not justified.
3. Together with Wetlugosaurus and Capitosaurus, Benthosuchus consti-
tutes one group closely related genetically, to which probably belongs also
Mastodonsaurus. In this group Benthosuchus and Mastodonsaurus are more
aberrant than the two other genera.
4. The specialization of Benthosuchus, i. e. its deviation from the typical Ca-
pitosaurus, is directed towards the same side as the specialization of Tremato-
saurus, and partly also Mastodonsaurus. However, the results of this speciali-
zation, insofar as they proceeded along different ways in genetically different
forms, are different and have been reflected only in the convergent similarity
of the anterior part of the skull. Mastodonsaurus, as a member of the same
group, has a great resemblance to Benthosuchus in spite of the fact that its
adaptation is somewhat less alike than that of the anterior parts of the skull
in Benthosuchus and Trematosaurus.
Among the older, typically rhachitomous forms, the one most closely
related to Benthosuchus is Bhinesuchus (fig. 87A, B). The genetic relationship
of Benthosuchus and Bhinesuchus is manifested through Wetlugosaurus.
To the latter Bhinesuchus is so similar that the first remains of Wetlugosau-
rus were described by N. N. Yakovlev as Bhinesuchus wolgodwinensis. The
distinctions of Bhinesuchus from Wetlugosaurus ‘consist in a lesser reduction
of the osseous neurocranium, greater length of the two branches of the pterygo-
id, greater development of chagrin on the palatal bones, and shorter and wider
skull. It is easily seen that these distinctions are those of the evolutionary
stage of Bhinesuchus, fully corresponding to the older age and typical rhachi-
tomous of the latter.
In the light jof all these considerations, the great phylogenetic significance
of such forms as Benthosuchus and Wetlugosaurus is apparent.
Standing on a primitive stage of ’ development (Neorhachitomi), they
bear features of structure of several forms from the stage of Stereospondyli,
usually grouped in different families. Therefore a detailed study of forms
similar to Benthosuchus is very important for an analysis of the origin and
development of stereospondylous Labyrinthodonts and for the working out
of their natural classification.
No such natural classification exists at present, and the dividing lines bet-
ween the different groups are often based on casual features. An attempt to
establish a more natural systematics of the higher Labyrinthodonts was made
by Save-Soderbergh in the above cited works. However, we do not consider
it possible to adopt this classification since, in spite of correct surmises Save-
Soderbergh bases it on very superficial data. A classification according to
zones of intense growth, proposed by the same author, is not satisfactory
either. The zones of intense growth of the bones of the cranial roof are not so-
mething self-sufficient, but only a way (mode) in which the necessary adaptive
modifications of the skull are realized. This has been sufficiently clearly
demonstrated in the work of Bystrow (1935), who was the first to discover
the phenomena of localized intense growth in the skull of Stegocephalians.
Therefore there is no certainty that these ways could not be common
in diverse forms and, consequently, with this classification the convergent
forms would be united as genetically allied. The same essential defect is
present also in the first classification of Save-Soderbergh.
142
Since the development of a new systematica of Labyrinthodonts must
be based on a detailed study of an abundant material, in the present work
we avoid any attempt to introduce any essential changes into the existing
classification, although forms transitionary from Rhachitomi to Stereos-
pondyli are the subject of our discussion.
Within the boundaries of the material studied we may make conclu-
sions only of a particular order without, in any way, pretending to give
an elucidation of general regularities.
It seems probable that the features, upon which the separation out of the
families Capitosauridae and Mastodonsauridae is based, are not essential and
require a revision.
If we assume that a genetic classification must be a vertical and not a ho-
rizontal one, the families Capitosauridae, Mastodonsauridae and Benthosu-
chidae 1 must be united in one family. Included in the same family must be
also Rhinesuchus, which is unoubtedly very close to the ancestors of Wetlu-
gosaurus. Such a family, passing through all the stages of evolutionary de-
velopment of higher Labyrinthodonts, should be best of all named the family
Rhinesuchidae.
If we assume the features of a family of small significance for a horizontal
classification, all of the forms enumerated will be included in three families:
Rhinesuchidae — Rhachitomi, Benthosuchidae — Neorhachitomi, Capitosau-
ridae — Stereospondyli. The stratigraphical distribution in time is res-
pectively: Upper Permian, Eotriassic, Triassic.
In concluding the review of the generic relations of Benthosuchus it should
be noted that in the recent years in the Eotriassic deposits of the Upper Volga
and Timan, identical to the Eotriassic deposits with Benthosuchus and Wetlu-
gosaurus, new forms of Labyrinthodonts have been found, very close to Bentho-
suchus and Wetlugosaurus, which seem to have been also at the stage of
Neorhachitomi (Efremov, 1938). These forms have not as yet been sufficiently
studied, but they reveal different adaptation deviations, in particular still
nearer to Trematosaurus.
It isxnot impossible that in the course of a further study of the fauna of
neorhachitomous Labyrinthodonts, facts will be collected proving the appea-
rance of such forms as Trematosaurus from the same group. The existence of
many features of resemblance of such a form as Thoosuchus (Efremov, 1938)
to Trematosaurus, on the one hand, and to Benthosuchus, on the other, is an
indication of the unnaturality of a deep classificational separation of Trematosau-
ridae and Capitosauridae carried out by Save-Soderbergh. Most probably, in
the Trematosauridae group the genera are mixed both of a more ancient ori-
gin and comparatively recent ones from the Neorhachitomi group.
chapter nr
SOME CONSIDERATIONS ON THE ORIGIN OF STEREOSPONDYLOUS
VERTEBRAE IN HIGHER LABYRINTHODONTS
The structure of the vertebral column in Benthosuchus and other forms,
which are on the same stage with it (Wetlugosaurus, Sclerothorax), best of
all correspond to the usual idea as to the genesis of notocentral stereospondy-
lous vertebrae of Labyrinthodonts.
The gradual reduction of pleurocentra in a typical rhachitomous vertebra,
proceeding parallel to a coossification of the hypocentrum with the neural arch
and an increase of the hypocentrum to the formation of a monolithic round
centrum, — this is the path established by numerous investigations.
1 Since the family Benthosuchidae was proposed by Efremov in 1929, and the
uniting of Benthosuchus and Wetlugosaurus was made by him in 1932, the priority of the
name of the family must belong to Efremov. The family W etlugosauridae was founded
by Save-Soderbergh in 1934.
143
The finding of Benthosuchus and other forms with a neorhachitomous
structure of the vertebral column reveals in the most striking manner the in-
termediate stage between the rhachi tomous and stereospondylous types. The
moment of reduction of pleurocentra was accompanied by a shiftingjof the whole
support of the neural arch exclusively to the hypocentrum, synchondrously
contacting with the neural arch. Such a vertebra is, if it may be said so, a ste-
reospondylous vertebra «in statu nascendi», originating from a typical rhachi-
tomous one.
The geological age of Benthosuchus and Wetlugosaurus agrees extremely
well with their neorhachitomous vertebrae. All the Triassic Labyrinthodonts
are stereospondylous, and all the Permian—rhachitomous ones, and the
existence of an intermediate stage between the two types in the oldest layers
of the Triassic still closer correspond to the generally accepted idea of the
development of stereospondylia.
However, we know of cases when the appearance of stereospondylous ver-
tebrae takes place earlier in time or, on the contrary, which retained verte-
brae of the neorhachitomous type at the time when all the other Labyrintho-
donts were already stereospondylous. The appearance of stereospondylous
vertebrae in the anterior part of the vertebral column in the Permian Dvino-
saurus may be also mentioned.
Moreover, with the appearance of detailed paleontological descriptions
and findings of new better preserved materials, the former view considering the
rhachitomous vertebrae as built according to the same type in all Bhachitomi,
has lost its significance.
The more abundant the information gathered with time on the morphology
of rhachitomous Labyrinthodonts, the greater number of variations of the
structure of the rhachitomous vertebra become revealed to us.
As an example let us point to the structure of vertebrae of Dendrerpeton
and Acanthostoma recently described by Steen (1934, 1937).
Leaving aside these old forms, the peculiar structure of whose vertebrae
may have been produced by different deviations from the embolomerous to
the rhachitomous type, and vice versa, we shall take typical Bhachitomi.
Of great interest is the structure of the rhachitomous vertebra in Cacops
(fig. 88, Cx). These vertebrae have very low hypocentra and very large pleu-
rocentra. The latter form a flat disc which throughout the whole of its anterior
surface is in contact with a special wide facette of the neural arch. The poste-
rior surface of the pleurocentrum is articulated with a special anterior facette on
the neural arch of the next vertebra and with the bevelled anterior surface of
its hypocentrum. The contact of the neural arch and of the hypocentrum is much
weaker and is accomplished by means of a small lower facette at the end of
a strongly lowered basal part of the neural arch. Due to these peculiarities
the centrum of the vertebra of Cacops acquires an interesting peculiar stru-
cture. Its lower part to the height of no more than one third of the thickness
of the notochord is wholly formed by the hypocentrum.
The upper part of the centrum, posteriorly strongly projecting into the cent-
rum of the next vertebra, is formed by the neural arch and the pleurocentra. It
is quite obvious that in such a rhachitomous vertebra the pleurocentrum play
a very great part, and the vertebra of Cacops is nearer to the embolomerous
type of structure of the vertebral column than the usual rhachitomous type.
Another example is shown by the structure of a considerably later form —
Micropholis stowi, recently studied in detail by F. Broili and J. Schroder
(1938) (fig. 88, C2). Here the hypocentrum has astrong contact with the neural
arch along a wide bevelled area, in such a way that the neural arch rests so-
lely on the hypocentrum. The pleurocentrum is very strongly developed and
along the whole of its anterior surface is in contact with the neural arch and
the whole posterior margin of the hypocentrum. Thus, the contact of the prece-
ding vertebra with the next one is accomplished only through the pleurocentra,
forming the whole posterior part of the centrum of the vertebra. In other words,
144
in Micropholis, a very late form, we have also a somewhat embolomerous
nature of the structure of the vertebrae.
There is no doubt that a further study of the structure of the rhachitomous
vertebral column in different forms will show a still greater diversity of the
structure of vertebrae.
However, a discussion and consideration of these cases is beyond the scope
of the present work. In the two examples cited we have shown that the diffe-
rent types of structure of rhachitomous vertebrae actually exist, and in forma
highly different in age at that, such as the Lower Permian Cacops and the Trias-
sic Micropholis.
If rhachitomous vertebrae possess a greater structural diversity than it
has been formerly assumed, in what way was the transition of these different
structures to the stereospondylous state accomplished? Is the neorhachitomous
vertebra of Benthosuchus and Wetlugosaurus the only one transitionary stage,
which is inevitably passed by the most different groups of Labyrinthodonts
in the development from Rhachitomi to Stereospondyli?
These questions induced us to make an attempt to verify the structure of
the vertebrae in the Upper Permian late Rhachitomi on materials available
to us.
In this connection it is most useful to take such forms as Dvinosaurus and
Platyops. The former Labyrinthodont has stereospondylous vertebrae in the an-
terior part of the body; the latter shows a high degree of ossification of the
rhachitomous vertebral column, closely approaching a transition to the ste-
reospondylous type of structure.
The vertebrae of Dvinosaurus are described in detail and figured by Byst-
row (1938), from whose work we take their picture (fig. 88AX, A2, As).
The vertebrae of Platyops stuchenbergi (fig. 88Bj andB2) have been studied on
two full vertebrae of an old specimen of Platyops from the Akbatyrovo mine,
first figured by Efremov (1933) and on a nearly complete skeleton No. 160/1,
Paleontological Institute, recently found near the Belebey.
The vertebrae of Dvinosaurus possess a rhachitomous structure of the
most common type. The neural arch is not in contact or in a slight contact
with triangular high hypocentrum. Paired narrow pleurocentra have no con-
tact with the neural arch either and are located in a clearance produced in
the upper part of the centrum as a result of a tapering of the ends of the hypo-
centra.
In tracing the structure of the vertebrae of the anterior part of the trunk from
the posterior to the anterior we distinctly see how the pleurocentra are first
tightly united with the hypocentrum with an ankylose junction, then tightly
growing toit and, finally completely fused with the hypocentrum into one bone
with undistingishable junctions. This coalescence of the pleurocentrum proceeds
parallel to an increasingly greater ossification both of the hypocentrum and of
the pleurocentrum, together forming a complete stereospondylous osseous
centrum.
In old specimens of Dvinosaurus the pleurocentra in the three first ver-
tebrae are completely grown together with the hypocentra and neural arch,
converting these vertebrae into stereospondylous ones. Thus in Dvinosaurus
the stereospondylous vertebrae are formed by way of a direct growing together
of all the elements of the vertebra without reduction of any of these elements.
Another correlation of elements of rhachitomous vertebrae is observed in
Platyops (fig. 88B).
Here too the rhachitomous vertebrae are built according to the usual type,
but the pleurocentra in their dilated upper portion have a junction with spe-
cial facettes on the process of the neural arch (fig. 881^). In old specimens in
the anterior part of the trunk the pleurocentra are tightly grown only to the
neural arch, being fused into one bone. As a result of this (fig. 88B2) one part
of the centrum of the vertebra is united with the neural arch, while the other
part still exists separately.
10 Tp. Палеовтол. ин-та. т. X, в. I
145
There ig no doubt that in a further stage of development of vertebrae in
Platyops the pleurocentrum will hot be destroyed, but as in Dvinosaurus
will become tightly grown to the hypocentrum. As a result of this the same
stereospondylous type of vertebra is produced, but in a way different from
that in Dvinosaurus (fig. 88, B3).
The above mentioned vertebrae of Micropholis (fig. 88, C2) in the extent
of ossification and development of a wide contact of the neural arch and hy-
pocentrum are undoubtedly near to a transition to the stereospondylous state.
This is in perfect agreement with the late geological development of the form
(Triassic). At the same time there are no data whatever to permit a rapid redu-
ction of the great pleurocentrum to be surmised. The most probable will be a
fusion of the pleurocentrum with the neural arch and with the hypocentrum
simultaneously, which will be the third way of producing a stereospondylous
vertebra.
Finally, still another way of development of a stereospondylous vertebra is
the above described way of a reduction of the pleurocentra and a widening
of the upper portion of the hypocentrum with a subsequent fusion of the neural
arch and the hypocentrum. A transitionary stage on this way is presented
by the neorhachitomous vertebrae of Bentosuchus and Wetlugosaurus
(fig. 88, D2).
Thus it seems to us that the stereospondylous type of structure of the vertebra
could have been formed from the rhachitomous type in following four ways:
1. A complete and simultaneous fusion of well developed pleurocentra,
hypocentrum and neural arch,— a way demonstrated by the vertebral column
of Dvinosaurus (fig. 88, A).
2. A fusion of the pleurocentrum first with the neural arch and only sub-
sequent to this, a close coalescence with the hypocentrum, — a way traced
by the authors in the instance of structure of the vertebral column of Platyops
(fig. 88, B).
3. A fusion of the hypocentrum with the neural arch and a later union of
the pleurocentra with both these elements grown together into ond'bone, —
a way, probably shown in Micropholis (fig. 88, C).
4. A reduction of the pleurocentrum to its complete obliteration together
with a widening of the upper ends of the osseous hypocentre to a complete
contact of the latter with the neural arch, — a way shown by the neorhachi-
tomous vertebrae of Benthosuchus and Wetlugosaurus (fig. 88, D).
Certainly, the facts cited here cannot serve as a basis for any considerable
generalizations, and so far we cannot construct any theory of origin of the
vertebrae of Stegocephalia without a verification of the above statements on
a vast and diverse material.
Our task consisted in attracting the attention of the investigators to the
materia] being accumulated in the morphology of vertebrae of higher Labyrin-
thodonts. This material enables us to reach the following conclusions: —
1. The rhachitomous vertebrae in different forms of Labyrinthodonts
possess a various structure in the correlations of the elements composing them.
2. These various structural modifications can pass to the stereospondylous
state in different ways.
3. The stereospondylous vertebra produced as a result of a fusion (growing
together) of pleurocentra with the hypocentrum or the neural arch is not noto-
central in the strict sense of this word.
4. Real notocentral stereospondylous vertebrae are those, which pass through
the neorhachitomous stage, accompanied by a reduction of the pleurocentra.
If in verifying the above statements they prove to be correct, it will be
certain that the transition to the stereospondylia in different genetic groups
was accomplished in various ways and often at different times. In this case the
possibility is not excluded that the notocentral stereospondylia, obtained
through the neorhachitomous stage, is peculiar only to one group of closely
allied forms and most Capitosauridae. Since entering into this group are all
146
the best studied forms, which were widely spread and numerous, it becomes
clear that our former ideas of the rhachitomous and stereospondylous vertebrae
had not shaped themselves in a casual manner.
CHAPTER IV
CONDITIONS OF BURIAL, SPREADING and FAUNISTICAL DATA
OF BENTHOSUCHUS
The age of Benthosuchus sushkini is determined by its being found in the
Eotriassic variegated beds separated out by Efremov in the Vth faunistical zone
(Efremov, 1937), as the lowest horizons of the Triassic.
The localities of Labyrinthondonts from the Eotriassic of the U. S. S. R.
have been known for rather a long time and were described for the first time
by N. N. Yakovlev (1916). However, due to the absence of complete findings
and the small amount of data both the fauna of the Eotriassic and the con-
ditions of formation of the occurrence remained practically unknown.
jLarge findings and a fuller elucidation of the occurrences were the results
of the works of Efremov (1927—1929), when the collection, which was laid at
the base of the present work, was put together.
The geological works of the recent years have revealed a very wide sprea-
ding of the Eotriassic variegated beds. All the points of their exposures con-
tain layers with bones of Labyrinthodonts of the family Benthosuchidae.
The area of distribution of the localities of the Eotriassic occupies a vast
field in the North-Eastern corner of Russian Platform and extends in lon-
gitude from about 48° from Greenwich to 50°, and in latitude from 56°
N. to 66° N.
Certainly, the development of the Eotriassic beds within this field is dis-
continuous due both to the subsequent geological processes and, possibly, the
conditions of its formation.
'However, the lithological peculiarities of the rocks of the Eotriassic beds,
the nature of bedding of the fossiliferous beds, the conditions of burial, the
state of preservation and the composition of the fauna at all the individual
points of the exposures, even though very remote from each other, are stri-
kingly similar. This is an indisputable evidence in favour of the identity of
the external conditions and processes of the formation of rocks over the whole
area of the development of the Eotriassic beds.
The sections of all the localities of the Eotriassic variegated beds are all
of the same type, differing in details of the bedding and content of bones
of Labyrinthodonts at geographically different points.
It is very interesting that the whole mass of remains of Labyrinthodonts
found in the excavations on the Sharzhenga river near the village of Vakh-
nevo belongs exclusively to Benthosuchus. Only one fragment belongs to
another closely allied genus of a considerably lesser size — to Thoosuchus.
All the material from the excavations on the Vetluga river near the villages
of B. Sludka and Zutovskoye belongs solely to Wetlugosaurus, with the ex-
ception of one fragment, belonging to Thoosuchus.
Thus in their distribution these two genera seem to exclude each other.
In the North dwelt Benthosuchus, in the South — Wetlugosaurus, the number
of specimens of the two genera being very considerable.
The materials collected from the other localities dug up, in spite of their
fragmentary nature show that actually the spreading of the two genera was
more complicated.
Thus, in the northernmost localities of the Timan region Benthosuchus and
Wetlugosaurus have been found at points very near to each other. In the sou-
thernmost occurrences of the Upper Volga a new, so far not studied form,—
Volgosuchus — is added. Remains of Thoosuchus are encountered more fre-
quently.
to*
147
Efremov (1938) considers that the northernmost localities of Eotriassic
Labyrinthodonts are relatively older, and the southern ones younger in geo-
logical age.
This fact is in agreement also with the geographical spreading of the Eo-
triassic neorhachitomous Labyrinthodonts. At the beginning the two genera
Benthosuchus and Wetlugosaurus existed, if not jointly, in any case, in clo-
sely lying areas. Then it seems that with a divergence of adaptation the area
of the two genera diverge. At a still later time in the southernmost areas of
development of the Eotriassic fauna to Benthosuchus new forms are added,
that appeared as a result of a further adaptive radiation of the Eotriassic La-
byrinthodonts.
Let us sum up the data as to the localities of the Eotriassic Labyrinthodonts.
Within the vast area of development of the Eotriassic variegated beds ne-
arly at every point of their exposure there is an ossiferous layer of grey sands
and sandstones containing remains of rhachitomous Labyrinthodonts, —
Benthosuchus and Wetlugosaurus.
The occurrences exlpored show a spreading of the ossiferous layer over tens
of square kilometers, and there are no reasons to believe that at other little
known points we shall meet with different conditions of its occurrence.
The presence of the ossiferous layer in nearly all of the exposures of the va-
riegated beds, at points extremely remote from each other, proves with certainty
the extensive development of the ossiferous deposits amounting to thousands
of square kilometers.
If from the excavations of the Sharzhenga and the Vetluga rivers, with a
total area of the worked ossiferous layer not exceeding 200 square meters,
we have collected over 1500 separate bones and fragments, belonging to hund-
reds of specimens of various age, then what must be the number of specimens
buried throughout the whole of the variegated beds? Even assuming that when
digging we have struck the richest portions of the ossiferous layer, then, accor-
ding to the most reserved estimates, it becomes evident that in the occurrences
of the Eotriassic beds there are buried hundreds of thousands* of specimens
of Benthosuchus, Wetlugosaurus and other closely allied, but much rarer forms.
Such a vast field of death of Labyrinthodonts is an exceptionally peculiar
phenomenon and cannot be accounted for from the standpoint of usual vi-
tal processes.
Indeed, the presence everywhere of the ossiferous layer of grey sand causes
as if a break in the deposition of the variegated beds. This break was not of
long duration, as the arenaceous deposits, the power of the water streams be-
ing considerable, could have been deposited during a very short period and for-
med a layer insignificant when compared to the total thickness. Likewise,
this break did not occur due to a sharp change of the physico-geographical
environment, since subsequent to the deposition of the layer of grey sand the
bedding of the same variegated beds, undistinguishable from the lower under-
lying layers, continued.
Nevertheless, at the moment of the formation of the layer of grey sand,
over an enormous area a vast amount of remains of Labyrinthodonts of the most
different age was deposited, mixed in disorder, partly broken and rolled. The
exceptionally good preservation of the osseous remains may be explained only
by the exceedingly rapid rate of their burial. With the exception of this layer
of bed, no organic remains have been found anywhere throughout the whole
thickness.
It should be considered that the moment of deposition of the layer of grey
sand coincided or nearly coincided with the moment of a mass destruction
of Labyrinthodonts. All specimens without exception, both old and young
ones, were perishing. The greatest quantity of remains belongs to individuals
of «middle» age, which shows also the existence in the occurrence not of a «se-
lective» burial processes, but of those of the whole fauna of Labyrintho-
donts.
148
If we consider the size of the area within which a mass destruction of ani-
mals occurred, and also that forms with a various adaptation were perishing
simultaneously, it becomes evident that we have to deal with a large-scale ca-
tastroph.
This catastroph reflected in the complex of the occurrences of the Eotri-
assic of the Russian Platform, is one of very few cases in the history of the ol-
dest Tetrapoda in its scale and chiefly in the rapid rate of its deve-
lopment.
It is still more interesting that probably the cause of the Eotriassic catast-
roph lies not in the anastrophic changes of the physico-geographical environ-
ment.
What were the conditions of the physico-geographical environment that cau-
sed the deposition of the variegated beds — this question remains as yet un-
settled. The absence of organic remains in the beds may be an evidence with
equal probability both of the deposition under desert conditions or condi-
tions of lifeless basins, and only of the absence of conditions of burial in an en-
vironment suitable for animal life.
The settling of the disputable question as to the origin of the variegated
beds may be arrived at only after detailed geological investigations; at the pre-
sent time and in the present paper it is as yet impossible.
Whatever was the physico-geographical environment throughout the vast
area of deposition of the Eotriassic beds, it remains clear that these conditions
existed before the deposition of the ossiferous layer of grey sand and conti-
nued to exist afterwards in an unchanged form. Only for a short time at the
moment of deposition of grey sand these conditions changed also over the whole
vast area, probably not quite simultaneously.
It is easy to see that the layer of grey sand was deposited under conditions
of flowing waters, rather swift at that, which is indicated by the disorderly
bedding of sands and the chaotic distribution of organic remains.
Thus it is evident that if the variegated beds were deposited under desert
conditions, the deposition of the ossiferous sandy layer was caused by an in-
vasion of a mass of flowing waters, that flooded the whole plain.
If the variegated beds were deposited in the basin, the deposition of grey
sand was caused by a sharp increase of the strength of currents with a usual
importation of coarser material and similar to the former case by an erosion
of previously deposited strata.
Hence, in both cases we have to do with a rapid flooding of a vast area,
which piled up everywhere masses of sand, mixed with remains of Labyrin-
thodonts and ended comparatively quickly, subsequent to which former condi-
tions were reinstated.
This flooding was probably spreading from the North southward, gradually
decreasing in intensity.
Such a huge amount of corpses of Labyrinthodonts hardly could have been
brought from some place in the North. An evidence against this assumption is
that the existing forms occupy definite areals of spreading. It is also difficult
to presume that the mass destruction of animals was called forth by the flo-
oding of the region itself. Such forms, well adapted to the life in water, as Ben-
thosuchus and Wetlugosaurus, naturally could not have perished only with
an increase of the volume of water.
A clue to the solution of this question evidently lies in that general and
sudden disturbance of the habitual conditions of habitation, which was pro-
duced by the processes causing a vast flooding of the region.
The absence of any considerable amount of remains of fishes as compared
to the mass of remains of Labyrinthodonts, is striking, since the fishes consti-
tuted the chief, or strictly sole kind of food of Benthosuchus and Wetlugosaurus.
Of no less importance is the absence of vegetable remains which, if they had
existed in a large amount, would have been preserved even as indistinct
impressions.
<49
From these data the assumption as to the destruction of animals due to a
lack of food caused by a flood or phenomena preceding it, seems probable.
Remotely analogical phenomena may be observed at present in the great
overflows of tropical rivers in lowland areas (for instance the so-called «hapo»
in South America), when hundreds and thousands of square kilometers are flo-
oded by flowing waters. Such inundations cause a corresponding destruction
and burial of animals, however on a much lesser scale than it is observed in
our catastroph.
Certainly, the mass destruction of Labyrinthodonts at the epoch of the Eot-
riassic inundation, as well as the inundation itself, must have taken place un-
derspecific conditions, possibly not reproducible at present, and any analogy
found in the present environment will always be only a very approximate
one.
Thus, the huge field of death of Eotriassic Labyrinthodonts was formed by
way of a rapid deposition of their remains in sandy masses at the moment of
a short flooding of the whole region by comparatively rapidly flowing fresh
waters. This moment of flooding of the region coincided with a mass destruc-
tion of very numerous animals, their number amounting to hundreds of thou-
sands specimens. The animals undoubtedly lived within the area of develop-
ment of variegated beds, but as it seems, under normal living conditions they
did not get into the burial place. The flooding of the whole region and
the deposition of grey sands created excellent conditions for the burial of
remains of animals and their conservation.
Due to a great catastroph abundant remains of the Eotriassic fauna of La-
byrinthodonts have reached us; this fauna, in spite of its numerous represen-
tatives, has not been found anywhere besides this sandy layer. The ancestral
forms of Eotriassic Benthosuchus and Wetlugosaurus in older layers of the
territory of the Russian Platform have not been found.
Such a sudden appearance of an abundant fauna, having no known «roots»
in the older layers, is very characteristic of the majority of findings of the ol-
dest Tetrapoda and is an additional evidence in favour of the fact that only
a very small portion of the actually existing animal really reaches us in the
processes of burial, always as a result of great catastrophical phenomena or
of those of a different nature.
«50
ЦИТИРОВАННАЯ ЛИТЕРАТУРА
(Literature cited.)
Abel О. 1919. Die Stamme der Wirbeltiere.
Boas. 1881. Ueber den Conus arteriosus u. die Arterienbogen der Amphibia. Mor-
phologisches Jahrbuch, Bd. 7.
Broili F. u. Schroder J. 1938. Beobachtungen an Wirbeltieren der Karroo-
formation. Micropholis stowi u. Lydekkerina huxleyi. Sitzungsberichte d. Bayer.
Akademie d. Wissensch. Naturwiss. mathem. Abt. 1938.
Broom R. 1913. Studies on the Permian Temnospondylous Stegocephalians of North
America. Bull. Am. Mus. Nat. History, vol. 31.
Burmeister H. 1849. Die Labyrinthodonten aus dem bunten Sandstein von Bern-
burg. I Abt. Berlin.
Burmeister H. 1850. Die Labyrinthodonten aus dem Saarbriicker Steinkohlenge-
birge. II Abt. Berlin.
Bystrow A. P. 1935. Morphologische Untersuchungen der Deckknochen des Scha-
dels der Wirbeltiere. I Mitt. Schadel der Stegocephalen. Acta Zoologica. Bd. 16.
Bystrow. A. P. 1938. Dvinosaurus als neotenische Form der Stegocephalen (Acta
Zoologica).
Case E. C. 1911. Revision of the Amphibia a. Pisces of the Permian of North
America.
Case E. C. 1935. Description of a collection of associated skeletons of Trimero-
rhachis. Contributions from the Museum of Paleontology, University of Michigan,
vol. 4'.
Credner H. 1881—1892. Die Stegocephalen u. Saurier aus dem Rotliegenden des
Plauenschen Grund bei Dresden. I—X. Zeitschr. d. Deutsch. Geol. Gesellsch. Bl.
Bd. 33—45.
Dempster W. T. 1935. The Brain-Case a. Endocranial Case of Eryops. The
Journal of Comparative Neurology, vol. 62.
Efremov J. A. 1929. Benthosaurus sushkini, ein neuer Labyrinthodont aus den
permo-triassischen Ablagetungen des Scharschenga-Flusses. Bull. Acad. Sciences
U. R. S. S., No 8.
Ефремов И. A. 1929. Местонахождения стегоцефалов на северо-востоке Евро-
пейской части СССР. Докл. Акад. Наук СССР, № 1.
Ефремов и Кузьмин. 1932. Пермотриас северной части Русской платфор-
мы и его местонахождения лабиринтодонтов. Тр. Палеозоологического инсти-
тута Акад. Наук. СССР, т. I.
Efremov J. А. 1933. Ueber die Labyrinthodonten der UdSSR. II. Permische
Labyrinthodonten des friiheren Gouvern ements Wjatka. Travaux de 1’Institut
Paleozoologique de I’Academie des Sciences de 1’URSS, t. II.
Ефремов И. A. 1933. Два поля смерти минувших геологических эпох. «Природа».
Ефремов И. А. 1937. Стратиграфическое разделение континентальных перми
и триаса СССР по фауне древнейших наземных позвоночных (On the stratigra-
phic subdivision of the continental Permian a. Triassic of the USSR on the ba-
sis of the fauna of early Tetrapoda). Докл. Акад. Наук, т. XVI, № 2.
Ефремов. И. А. 1937. О лабиринтодонтах СССР. III. Melosaurus uralensis
Н. v. М. Тр. Палеонтологического института Акад. Наук СССР, т. VIII, вып. I.
Ефремов И. А. 1938. Эотриасовые лабиринтодонты Верхней Волги. В серии
«Предварительное описание новых форм пермской и триасовой фауны Tetra-
poda СССР», № I. Тр. Палеонтологического института Акад. Наук СССР.
Fraas Е. 1889—1890. Die Labyrinthodonten aus der Schwabischen Trias. Palaeonto-
graphica. Bd. 36.
Gaupp-Wiedersheim-Ecker. 1896—1904. Anatomie des Frosches.
Haughton S. H. 1925. Investigation in South-African Fossil Reptiles a. Amphi-
bia. — Annals of the South-African Museum, vol. 22.
Hartmann-Weinberg A. u. Kusmin Th. 1935. Untertriadische Stegoce-
phalien der Oka-Zna Antiklinale. II. Capitosaurus volgensis. Problems of Paleon-
tology, vol. I,
151
Hartmann-Weinberg A. u. Kusmin. Th. 1935. ^Untertriadische Ste-
gocephalien der UdSSR. Lyrocephalus acutirostris nov. sp. Ibidem.
v. H u en e F. 1912. Beitrage zur Kenntnis des Schadels von Eryops. Anatomischer
Anzeiger. Bd. 41.
v. H u e n e F. 1932. Ein neuartiger Stegocephalen-Fund aus dem oberhessischen Bunt-
sandstein. Palaeont. Zeitschr. Bd. 14, No. 3.
J a e к e 1 O. 1903. Ueber Ceraterpeton, Diceratosaurtus u. Diplocaulus. Neues Jahrbuch
fur Mineralogie, Geologie u. Palaontologie, Bd. 1.
J a e к e 1 O. 1915. Die Wirbeltierfunde aus dem Keuper von Halberstadt. Palaontolo-
gische Zeitschrift.
J a e к e 1 O. 1925. Das Mundskelett der Wirbeltiere. Morphologisches Jahrbuch. Bd. 55.
J a e к e 1 O. 1927. Der Kopf der Wirbeltiere. Ergebnisse der Anatomie u. Entwick-
lungsgesch. Bd. 27.
v. Meyer H. 1857. Reptilien aus der Steinkohlenformation Deutschlands. Palaeon-
tographica. Bd. VI.
M о о d i e R. L. 1908. The lateral line system in extinct of Amphibia. Journal of Mor-
phology. vol. 19.
Olson E. G. 1936. The dorsal axial musculature of certain primitive permian Tetra-
pods. Journal of Morphology, vol. 59.
Pfannenstiel M. 1932. Gehirnkapsel u. Gehirn fossiler Amphibien. Monogra-
phien zur Geologie u. Palaeontologie. Serie II, Heft 6.
Riabinin A. 1928—1929. A Labyrinthodont Stegocephalian Wetlugosaurus
angustijrons nov. gen., nov. sp. from the Lower Triassic of Vetluga-Land in Nor-
thern Russia. Annuaire de la Societe Paleontologique de Russie, t. 8.
R i e s e. 1892. Beitrag zur Anatomie des Tylototriton verrucosus. Zoologische Jahr-
biicher. Bd. 5.
Romer A. S., 1930. The Pennsylvanian Tetrapods of Linton, Ohio. Bull, of the Ame-
rican Museum of Nat. Hist., vol. 59.
Save-Soderbergh G. 1935. On the dermal bones of the Head in Labyrin-
thodont Stegocephalians a. primitive Reptilia. Meddelelser om Grenland. Bd. 98.
Save-Soderbergh G. 1936. On the Morphology of triassic Stegocephalians
from Spitsbergen, a. the Interpretation of the Endocranium in the Labyrintho-
dontia. Kungl. Svenska Vetenskapsakademiens Handlingar. Bd. 16.
Save-Soderbergh G. 1937. On the dermal skulls of Lyrocephalus, Aphaneramma
a. Benthosaurus, Labyrinthodonts from the Triassic of Spitsbergen a. N. Russia.
Bull. Geol. Inst. Upsala, vol. XXVI1.
Schroeder. 1912. Ein Stegocephalen-Schadel von Helgoland. Kgl. Preuss. Geolog.-
Landesanstalt Jahrbuch, Bd. 33.
Steen M. 1930. The british Museum Collection of Amphibia from the Middle Coal
Measures of Linton, Ohio. Proceedings of the Zoological Society of London.
Steen M. 1934. The amphibian fauna from the South Joggins, Nova Scotia. Proceed.
Zool. Soc. Lond.
Steen M. 1937. On Acanthostoma vorax Credner. Proceed. Zool. Soc. Lond.
Sushkin P. P. 1927. On the modification of the mandibular a. hyoid arches and
their relations to the brain-case in the early Tetrapoda. Palaeontologische Zeit-
schrift., Bd. 8.
Sushkin P. P. 1936. Notes on the pre-jurassic Tetrapoda from USSR. III. Dvinosau-
rus Amalitzki, a perennibranchiate Stegocephalian from the upper Permian from
North Dvina. Travaux de 1’Institut Paleozoologique de I’Academie des Sciences
de 1’USSR, t. V.
Wagner H. 1935. Das individuelle Wachstum von Trematosaurus Brauni u. der
Nachweis von Mastodonsaurus im Bernburger Buntsandstein. Inaugural-Disserta-
tion. Halle (Saale).
Watson D. M. S. 1913. Micropholis stowi Huxley, a Temnospondylous Amphibian
from South Africa. Geolog. Magazin. vol. 10.
Watson D. M. S. 1920. The Structure, Evolution a. Origin of the Amphibia.
Philosophical Transactions, vol. 209.
Watson D. M. S. 1921—1923. The carboniferous Amphibia of Scotland. Palaeon-
tologia Hungarica, vol. I.
W a t s о n D. M. S. 1926. The Evolution a. Origin of the Amphibia. Philosophical
Transactions, vol. 214.
Wilder. 1891. A Contribution to the Anatomy of Siren lacertina. Zoologisches Jahr-
buch, Bd. 4.
Wilder. 1892. Die Nasengegend von M enopoma alleghaniense u. Amphiuma tri-
dactilum. Ibidem, Bd. 5.
Williston S. W. 1916. Synopsis of the Americ. Permo-carbonif. Tetrapoda. Cont-
ribut. from Walker Mus., vol. I.
Wiman G. 1914. Ueber die Stegocephalien aus der Trias Spitzbergens. Bull, of the
Geolog. Inst, of Upsala, vol. 13.
Wiman G. 1916. Neue Stegocephalienfunde aus dem Posidonomyaschifer Spitzber-
gens. Ibidem, vol. 13.
Wiman G. 1917. Ueber die Stegocephalen Tertrema u. Lonchorhynchus. Ibidem,
vol. 14.
152