/










































































Similar




Text
ТРУДЫ ПАЛЕОНТОЛОГИЧЕСКОГО ИНСТИТУТА
ГОМ VII
выпуск 3
ИСКОПАЕМЫЕ НАСЕКОМЫЕ
ИЗДАТЕЛЬСТВО АКАДЕМИИ НАУК СССР
ACADEMIC DES SCIENCES
DE l'CNION DES BEPUBLIQUES SOV1ET1QUES SOCIAL1STES.
TRAVAUX
DE L’INSTITUT PALEONTOLOGIQUE
ТОМЕ VII
LIVRA1SON 3
FOSSILES 1NSECTES
EDITION DE L ACADEMIE DES SCIENCES DE L’URSS
ТРУДЫ ПАЛЕОНТОЛОГИЧЕСКОГО ИНСТИТУТА
ТОМ VII
выпуск з
ИСКОПАЕМЫЕ НАСЕКОМЫЕ
ИЗДАТЕЛЬСТВО АКАДЕМИИ ПАУК СССР
ОТВЕТСТВЕННЫЙ РЕДАКТОР
ДИРЕКТОР ПАЛЕОНТОЛОГИЧЕСКОГО ИН-ТА АКАД. А. А. БОРИСЯК
Редактор издательства Б, Б. Родендорф
Технический редактор В. М. Юрова Корректор Л. Г. Афанасьева
Сдано в набор 15/V 1938 г. Подписано к печати 3/XI 1938 г. Формат ТОХЮв1/^. Объем 5 п. л. Bin. л. 62.000 печ. зн. Уч.-авт. л. 6,62. Тираж 1000 экз. Уполн. Главлита № Б-50427. РИСО №636.
АНИ № 995. Заказ № 2414.
1-я Образцовая тип. Огиза РСФСР треста „Полиграфииига". Москва, Валовая, 28.
СОДЕРЖАНИЕ
A. B. Martynov.
(with 2 maps) . .
А. В. Мартынов.| Местонахождения ископаемых насекомых в пределах СССР (с 2 картами) ................................................ 7
Review of localities of Fossil Insects in USSR ........................................... 26
Б. Б. Родендорф. Двукрылые насекомые мезозоя Кара-тау. 1. Brachycera
и часть Nematocera (с 21 рис.).............................. 29
В. В. Rohdendorf. Mesozoische Dipteren aus Kara-tau. I. Brachyceren und einige Nematoceren (mit 21 Abb.)................................... 57
А. В. M a p т ы н о в. | Пермские насекомые Архангельской области (б. Северного края). Часть V. Сем. Eutygrammidae и его отношения (с описанием одного нового рода и семейства из Чекарды) (с 5 рис.) . . 0 . . 69
| А. В. Martynov. | Permian Fossil Insects from the Arkhangelsk district.
PartV. The Family Eutygrammidae and its relationships (with the description of a new Genus and Family from Chekarda) (with 5 figures) ... 79
ВЕРХНИЙ ПАЛЕОЗОЙ МЕЗОЗОЙ КАЙНОЗОЙ
Схема ]
СТРАТИГРАФИЧЕСКАЯ СХЕМА МЕСТОНАХОЖДЕНИЙ ИСКОПАЕМЫХ НАСЕ КОМЫХ В ПРЕДЕЛАХ СССР
Плиоцен Река Ваенга, С. Двина. 10. Село Новокиевское, Посьет.
Миоцен И. Чон-Туз, Киргизия. 1. У Ставрополя. 2. Апшеронский п-в(диатомовые слои). 3. Курганский р-н, Краснодарский кр. 4. Орметы в Грузии. 5. Закавказье (материал И. В. Палибина). 9. Река Кудя, приток Амагу, ДВК.
Олигоцен 6. Уфалей. 7. Ашутас у Черного Иртыша. 8. Мгач. Сахалин.
Эоцен —
Мел —
Юра | Мальм 14—17. Река Товега у Нерчинска, Турга, Кондуев-ская, устье р. Бирки.
Доггер 10—1.1. Кузбасс: Нарык-Чернева и Тарбаганский камень. 6—8. Кара-тау: Галкино, Карабас-тау, Михайловка. 9. Окрестности Иссык-куля.
Лиас верхний 13. Усть-балей на Ангаре.
нижний 12. Буроугольный район у деревень Сухомесово и Коркино, Челябинского р-на. 3. Шураб I—III. 4. Сулюкта. 5. Кизил-кия.
Триас 1. Кузбасс, Бабий камень. 2. Шураб, 40 км к югу от Шураба II.
Татарский ярус —
Пермь I Казанский ярус конхи-феро-вый го- ризонт 17. Река Перебор на Печоре. 22. Река Б. Тыхтаудер. Соколовой. 23. Б. Байкаимская Листвяжка. 15. Река Сардык, бассейн Вятки. 16. Богатый лог на Каме.
спири-феровый горизонт 12. Река Уяза, Чотмак-Томак (?). 1. Река Сояна. 2. Тихие Горы. 13. Овраг Тахташур (?) 14. Дер. Щуня, р. Вятка (?). 3. Каргала, близ Оренбурга* 4. Дер. Чекарда на р. Сылве. 5—7. Суксун, Седы, Матвеево. 8. Савиновский лог, Пермск. кр. 10. Чердынь на Язьве. 11. Ирень у дер. Колпаковой (?). / 9. Река Вильва, ст. Чусовая (?). 24. Владивосток (?).
Кунгурский ярус
Артинский ярус
Карбон I Верхний 17. Балахонская свита: 18. Близ с. Верхотомского. 19. Река Томь, близ дер. Заломной. 20. У дер. Балахонки, 21. Лев. бер. Уньги у дер. Че-ремичкиной.
Нижний —
АКАДЕМИЯ НАУК СССР
Тэзв ТРУДЫ ПАЛЕОНТОЛОГИЧЕСКОГО ИНСТИТУТА. ТОМ VII, вып. 3
| А. В. МАРТЫНОВ I
МЕСТОНАХОЖДЕНИЯ ИСКОПАЕМЫХ НАСЕКОМЫХ В ПРЕДЕЛАХ СССР
(с 2 картами)
| А. В. MARTYNOV |
REVIEW OF LOCALITIES OF FOSSIL INSECTS IN USSR
(with 2 maps)
В настоящее время нам стало известно уже довольно много (до 53) местонахождений, в которых обнаружены отпечатки насекомых (схема I). Правда, из доброй половины их мы знаем всего по 1—2 формам, тем не менее во многих из них при постановке соответствующих поисков и раскопок может быть добыто и еще некоторое количество экземпляров, которые уже лучше ознакомят нас с характером отдельных форм или даже фауны данного района и данного времени. Так как сведения об этих местонахождениях разбросаны в ряде специальных журналов и некоторые из них уже основательно забыты, я считаю небесполезным дать здесь краткую сводку их.
Наибольшее количество пунктов с находками насекомых нам известно, как и следовало ожидать, из пермских отложений. Ряд таких местонахождений мы имеем в области развития красноцветных пород к востоку от р. Камы и далее к югу до р. Урала; однако в большинстве из них найдено по 1—2 или 3 вида, и только из Чекарды, на р. Сылве, мы обладаем значительным количеством остатков насекомых. Все это озерные и речные отложения; в некоторых из них (Чекарда, Каргала) обнаружены и водные личинки. Несравненно богаче остатками насекомых местонахождения в области морских прибрежных и лагунных отложений, расположенных к западу от первых (схема II, стр. 11). Сюда относятся знаменитые теперь районы Тихих Гор на Каме и Ива-Горы, Шеймо-Горы и Летополы на р. Сояне, Архангельской области. В бассейне Вятки есть еще некоторые пункты таких отложений, но пока из них известно по одной форме.
Другим районом пермских и верхнекаменноугольных местонахождений является Кузнецкий бассейн. Пока мы знаем оттуда не много видов, но в будущем можно ожидать там ряда новых находок.
Из Донбасса, из Минусинского и Тунгусского бассейнов ископаемых насекомых нам пока не известно, но вряд ли можно сомневаться в том, что они там есть, особенно в области огромного Тунгусского бассейна. На востоке нам известен только один отпечаток насекомого из окр. Владивостока, и только.
Из приведенного ясно, что о пермской и карбоновой фауне Северной Азии мы практически еще ничего не знаем, почему новые поиски были бы особенно желательны в угленосных бассейнах Сибири.
Мезозойские местонахождения сосредоточены у нас в двух районах — в Средней Азии и в Забайкалье, плюс прибайкальское местонахождение Усть-Балей, на Ангаре. Особенно много дали и дадут в будущем среднеазиатские местонахождения лиасовых и среднеюрских насекомых. Несколько форм было недавно обнаружено в буровых колонках в Челябинском буроугольном районе. Очень немного остатков триасовых насекомых найдено в области Шураба, в Фергане, и в Кузбассе, причем в Кузбассе найдены еще и многоножки.
В Средней Азии остатки насекомых по большей части вполне удовлетворительны, а нередко и очень хороши. В виду того, что нахождений мезозойских насекомых крайне мало за границей, следует вообще особо отметить, что среднеазиатские местонахождения дают нам исключительно ценные материалы для суждения о мезозойской энтомофауне вообще. Все это отложения озер.
Далее мезозойские местонахождения мы находим в Забайкалье, но там сделаны лишь единичные находки, почему всякий новый материал оттуда весьма желателен, особенно в виду спорности вопросов стратиграфии мезозойских угленосных месторождений этой области.
Что касается третичных местонахождений и наших сведений о третичных насекомых, то здесь положение дела пока совершенно неудовлетворительное. Нам известно только два местонахождения, откуда добыто по нескольку видов, — это холмы Ашутаса, у Черного Иртыша, и отложения по речке Кудя, притоку Амагу, Дальневосточного края.
После работ, произведенных в Ашутасе в 1928 г. М. Ф. Не й б у р г, здесь никто не был, между тем остатки растений и насекомых здесь изумительно хороши. Растений здесь добыто много, — их собирание было специальной задачей М. Ф. Нейбург, — но сборы насекомых были более случайны и скудны, почему организация новых работ в этом районе необходима.
В отдаленном местонахождении на речках Куде и Амагу, Приморской области, немного работал в 1923 г. проф. Т. Д. К о к к е р е л ь, специально приехавший для этих работ из Колорадо. Он и обработал свои сборы, но наших работников после этого там не было, о чем нельзя не пожалеть, так как фауна Амагу, по всей видимости, должна быть обильной.
В прочих, очень немногих, третичных местонахождениях найдено по 1—2 отпечаткам, и только в 1936 г. обнаружено новое местонахождение у Иссык-куля (миоцен), где встречается много отпечатков; материал оттуда еще не изучен.
Как сказано выше, из отложений каменноугольного возраста в пределах СССР остатков насекомых мы еще не знаем, если не считать балахон-ской свиты Кузбасса, откуда описано несколько форм. Трудно думать, чтобы остатки насекомых действительно отсутствовали в Донбассе, Караганде, а также в пермо-карбоновых Минусинском и Тунгусском бассейнах. Наверно они там имеются, только пока их не заметили, не обнаружили. Можно ожидать нахождения остатков насекомых не только в верхнем, но также в среднем и нижнем карбоне.
Ископаемые насекомые, помимо того общего интереса, который имеет для нас ознакомление с историческим развитием этой колоссальной группы, все более и более приобретают определенное стратиграфическое* значение. При оценке значения насекомых для стратиграфии, для суждения о климатах прошлого для уяснения вопросов миграций и т. п. следует всегда иметь в виду, что насекомые — существа наземные или пресноводные, почему они могут говорить, как и наземные растения, только о жизни на континенте. Поэтому служить они могут не стратиграфии вообще, а лишь стратиграфии континентальных, а также прибрежных морских (и скорее лагунных) отложений, куда насекомые часто сносятся и отлагаются в больших количествах, как, например, песчанистые отложения по р0 Сояне, глинисто-сланцевые отложения Тихих Гор.
С какими главными группами имеет дело палеонтолог или геолог, изучающий такие отложения? Практически он имеет дело только с тремя группами: с наземными растениями, насекомыми и наземными позвоночными. Встречаются, конечно, и другие группы, но они имеют сравнительно ничтожное значение; это — раковины наземных и пресноводных моллюсков, остатки ракообразных и других членистоногих. Раковины наземных моллюсков встречаются редко, спорадично (мы имеем в виду, главным образом, палеозойские и мезозойские отложения), да изучение одних раковин не дает, конечно, таких ценных данных для палеонтолога, какие может дать изучение остатков тела таких сложно построенных организмов, как позвоночные и насекомые. Ракообразные встречаются в ископаемом состоянии редко, за исключением групп Conchostraca из Phyllo-poda и Ostracoda^ но зато от этих рачков сохраняются лишь их створки, что сильно умаляет значение этих находок. О других членистоногих — паукообразных и многоножках — говорить вовсе не приходится, так как их остатки — великая редкость. Такие находки,конечно, представляют большой интерес, но больше с зоологической точки зрения. У нас, из пределов СССР, не известно с определенностью пока ни одной формы наземных паукообразных, а из многоножек нам попались до сего времени остатки только двух форм: одной из триаса Кузбасса, другой из перми с р. Сояны (еще не описана). Не приходится вовсе говорить о червях, в ископаемом состоянии почти не встречающихся.
Таким образом, для суждения о стратиграфии континентальных отложений могут иметь значение действительно лишь три названные выше группы. Не подлежит сомнению огромное значение для суждения о стратиграфии отложений континентального типа остатков растений, встречающихся к тому же весьма часто. Значение остатков позвоночных в этом отношении более скромное, и лишь для четвертичных и верхнетретичных отложений их значение выступает часто на первый план.
Действительно ли показания, даваемые со стороны ископаемых остатков растений, столь безупречны и столь универсальны, чтобы геолог мог игнорировать данные, предоставляемые такой огромной группой, как насекомые? Увы, этого сказать никак нельзя. Ископаемые растения представлены, как правило, остатками вегетативных органов, которые нередко, особенно в случаях с палео- и мезозойскими отложениями, оказываются недостаточными как для диагностики, так и для точного суждения о возрасте. Для ботаников эти соображения, конечно, понятны, но, чтобы не быть совсем голословным, я сошлюсь на длительное смешивание некоторых палеозойских и мезозойских типов в Кузбассе, на неправильные определения Геером возраста растений ряда североазиатских третичных местонахождений и т. п.
При таких условиях игнорировать насекомых нельзя. Никогда нельзя забывать того факта, что по количеству видов и родов насекомые превышают все остальные группы животных, вместе взятые, и количество уже зарегистрированных в каталогах видов достигает почти миллиона. Изучение ископаемых фаун показало, что такой же доминирующей на суше группой был класс насекомых и раньше, в мезозое и верхнем палеозое.
Насекомые встречаются реже растений, но зато часто сразу в значительном количестве видов, так что в таких случаях мы строим наши выводы на основании не отдельных видов, а целых комплексов их, нередко очень больших.
Далее, если в палеозойских и мезозойских отложениях насекомые сохраняются обычно не целиком, а в виде крыльев, то следует иметь в виду, что крылья насекомых как раз представляют собой те органы, на которых по преимуществу построена современная систематика — систематика родов, семейств и пр. Для характеристики видов требуется сохранность также и других органов, главным образом связанных с полом, однако обычно в крыльях мы находим сравнительно достаточные данные для ха
рактеристики также и видов. Следовательно, палеоэнтомологи находятся в этом отношении в более счастливом положении, чем ботаники, которые обычно имеют дело только с вегетативными органами.
Мы должны отметить еще и то обстоятельство, что фауны насекомых сравнительно быстро меняют свой состав и облик, переходя от одного горизонта к другому, так что по нескольким, часто по 2—3 формам мы с определенностью можем сказать, например, относится ли данная толща к нижнему лиасу, к нижнему или верхнему горизонту казанского яруса и т. п.
Состав фауны дает нередко достаточно определенные указания для суждения об орографии, орошении и даже климате местности, в которой обитали ископаемые насекомые. Нахождение цельных, неразрушенных водных личинок или куколок исключает всякие сомнения в том, что данное местонахождение есть озерное отложение; таковы отложения под с. Галкиным в Кара-тау и ряд пермских местонахождений. По нахождениям некоторых форм, например веснянок (Plecoptera), мы заключаем, что в данной области были быстро текучие воды, следовательно ландшафт был гористый или холмистый.
Нахождение ряда влаголюбивых форм (например, мошек — разных Fiingivoroidea — в отложениях Кара-тау) говорит о наличии в данной местности сырых лесов и т. п. В случае с третичными насекомыми наши суждения могут быть особенно точны.
Жизнь большей части насекомых как современных, так и ископаемых тесно связана с миром растений и вместе с последними находится в большой зависимости от климата. Климатические перемены вызывают сильные изменения как во флоре, так ивфауне насекомых, особенно чисто наземных, обусловливая вымирание одних, сохранение и миграцию других групп, в зависимости, главным образом, от их экологии. Имея сборы насекомых из нескольких местонахождений соседних горизонтов и ярусов, мы уже можем намечать эти миграции; наше положение будет много лучше, когда прибавится ряд новых местонахождений данного периода.
Для понимания современного географического распространения и состава фауны крупных зоогеографических единиц изучение ископаемых третичных, мезозойских и даже пермских фаун имеет огромное значение, но мы не имеем здесь возможности останавливаться на этом вопросе. В виду чрезвычайного разнообразия насекомых и адаптации их к самым различным биотопам, к самым различным условиям существования, выяснение истории этого огромного класса должно дать нам (и уже дает!) особенно ценный материал для суждений о закономерностях эволюции животного мира в связи с меняющейся обстановкой, но ответ на все такие вопросы будет тем компетентнее, чем глубже исследователь войдет в изучение организации, биологии и миграций вымерших животных, в частности насекомых, а также в изучение, целых фаун.
Переходим к обзору местонахождений.
ПАЛЕОЗОЙ
Из Европейской части СССР (схемы II и III) нам известно в настоящее время 17 пунктов, в которых были добыты пермские ископаемые насекомые. Эти местонахождения далеко не равноценны в том смысле, что из одних мы знаем в настоящее время уже много, десятки и даже сотни видов, в то время как в других добыты единичные экземпляры, отдельные виды. Кроме того, местонахождения эти имеют различный возраст: одни из них принадлежат казанскому, другие — кунгурскому ярусу, а одна находка отмечена как артинская.
1. 1 От ложе ния по р. Сояне, притоку К у л о я, и по речкам Большой и Малой Летополе, впадаю-
1 Порядковый № местонахождения всюду далее приведен на прилагаемых •схемах — картах.
Схема II
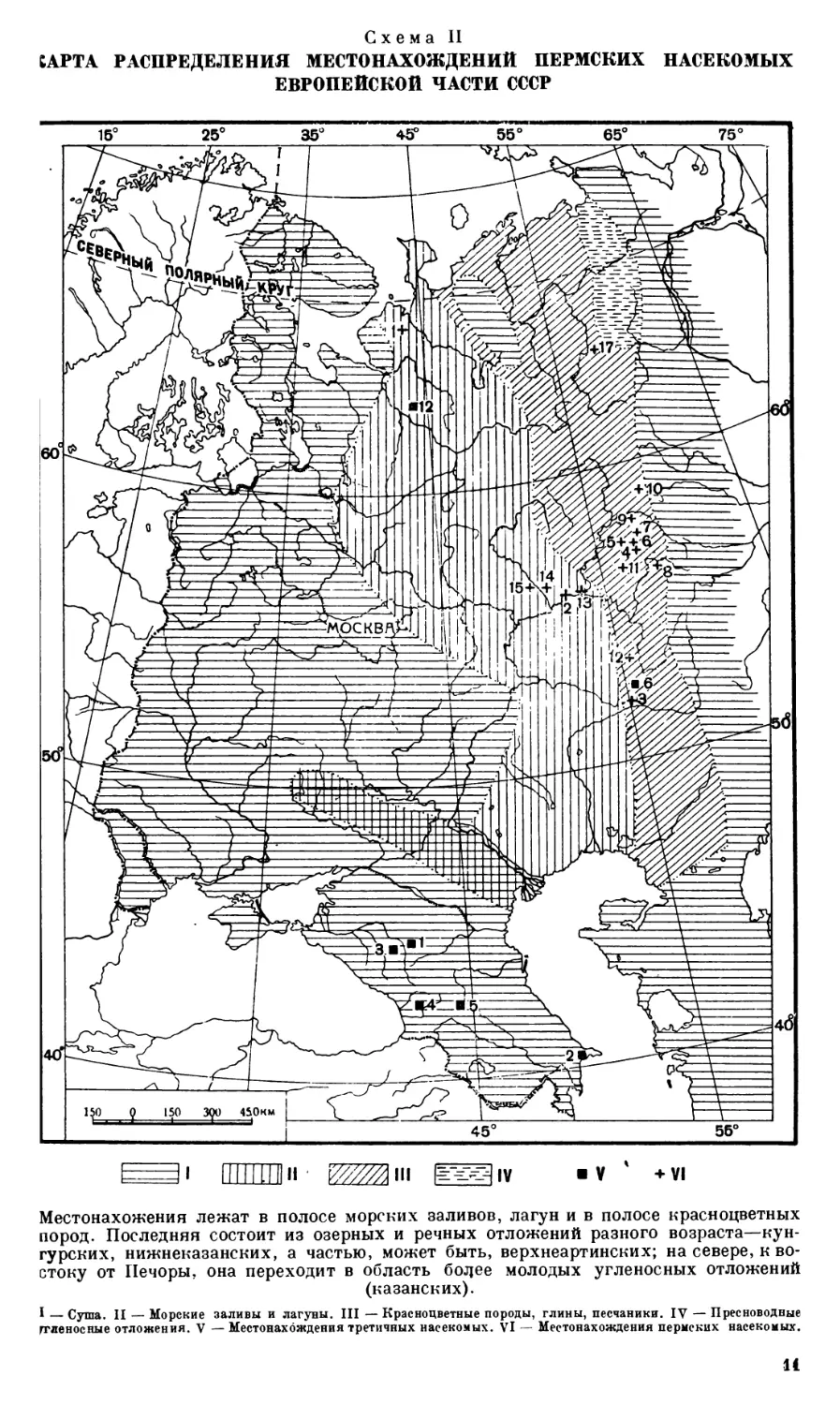
САРТА РАСПРЕДЕЛЕНИЯ МЕСТОНАХОЖДЕНИЙ ПЕРМСКИХ НАСЕКОМЫХ ЕВРОПЕЙСКОЙ ЧАСТИ СССР
15° 25° 35° 4ЬЮ 55° 65° 75°
ПТПТГЯн Mm g^iv "V ' +vi
Местонахожения лежат в полосе морских заливов, лагун и в полосе красноцветных пород. Последняя состоит из озерных и речных отложений разного возраста—кунгурских, нижнеказанских, а частью, может быть, верхнеартинских; на севере, к востоку от Печоры, она переходит в область бодее молодых угленосных отложений (казанских).
I — Суша. II — Морские заливы и лагуны. III — Красноцветные породы, глины, песчаники. IV — Пресноводные ггленосные отложения. V — Местонахождения третичных насекомых. VI — Местонахождения пермских насекомых.
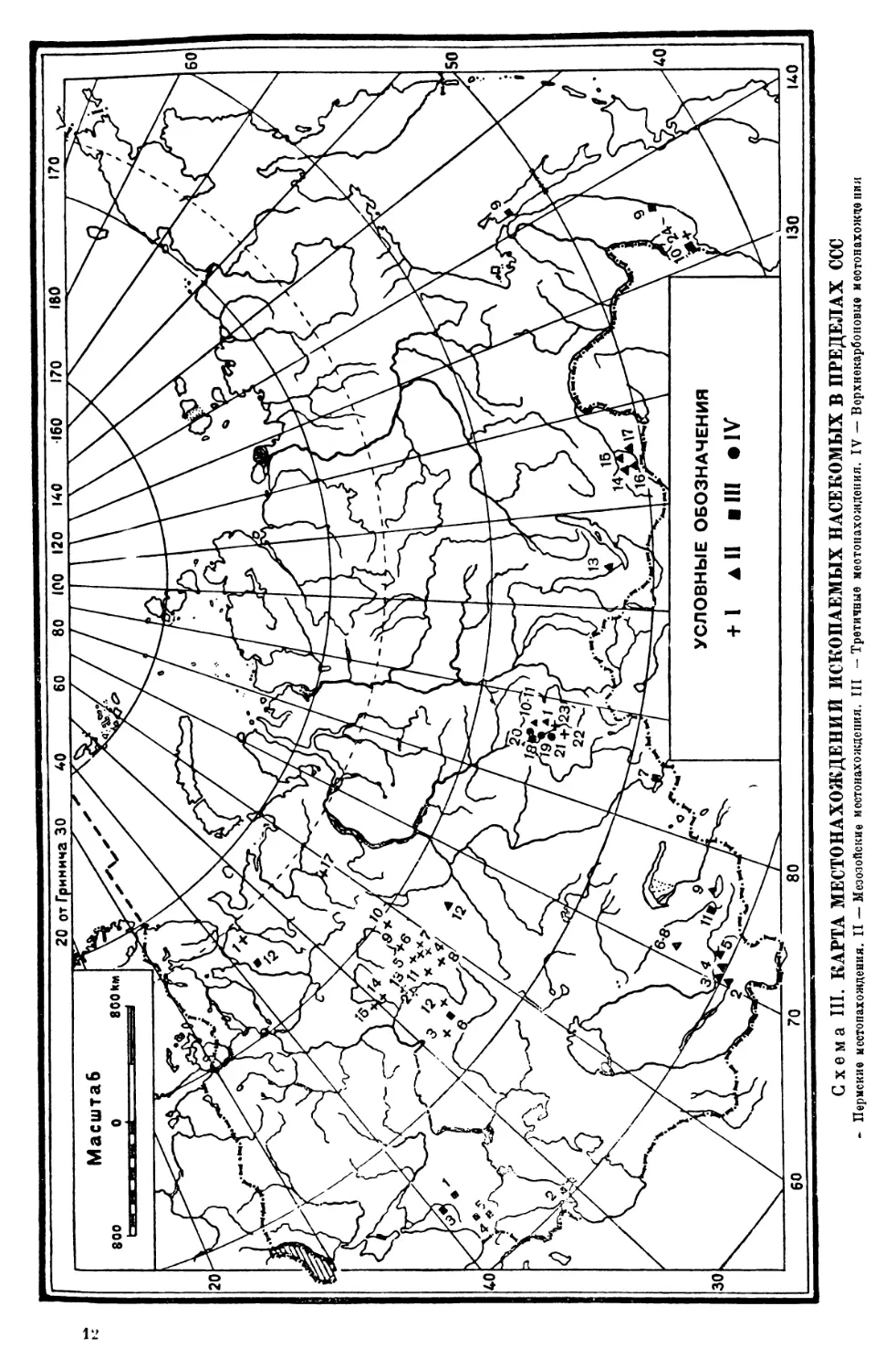
Схема III. КАРТА МЕСТОНАХОЖДЕНИИ ИСКОПАЕМЫХ НАСЕКОМЫХ В ПРЕДЕЛАХ ССС
- Пермские местонахождения. II — Мезозойские местонахождения. III —Третичные местонахождения. IV — Ворхнекарбоновые местонахождения
щ и м в С о я н у, Архангельской области; местности Ива-Гора, Шеймо-Гора, Летопола. Все эти три местности расположены по берегу р. Сояны на расстоянии 1.5—2 км; отложения представлены песчаниками, желтоватыми или желтоваторозоватыми, мощностью иногда до 2 м. Это местонахождение было открыто М. Б. Едемским в 1926 г., когда им было добыто в Ива-Горе и Шеймо-Горе небольшое количество насекомых, по которым было описано 18 видов. В дальнейшем там работали М. Б. Едемский, его сотрудник Бунде р с е н и затем два раза, в 1934 и 1935 гг., Я. Д. 3 е к к е л ь. Ими был собран огромный материал, содержащий около 5000 экземпляров насекомых. Кроме того, там же собрано и некоторое количество растений. Часть материала обработана и результаты опубликованы (Мартынов, 1928—1935), однако большая часть, главным образом сборы Я. Д. Зек-келя 1935 г., еще не изучены. До сего времени опубликованы результаты изучения первых сборов Едемского (1928), затем групп Palaeoptera, Mecoptera, Neuroptera, Coleoptera и Homoptera прежних сборов, до 1934 г. Уже в одних этих работах (1932—1935) описано до 75 новых видов и много новых родов и даже семейств. Между тем, главной группой на Сояне являются прямокрылые, которые еще не описаны. По приблизительным расчетам общее количество видов в собранных материалах должно достигать 170—200.
Когда будут обследованы все материалы этого местонахождения, оно даст нам исключительно полное знакомство с нижнеказанской энто-мофауной северных областей Европейской части СССР. Фауна эта была, видимо, богаче известной пермской фауны Эльмо в Канзасе. Как ясно уже из того, что опубликовано, фауна местонахождений р. Сояны очень своеобразна и очень походит, как и тихогорская (см. ниже), на вышеназванную фауну Канзаса.
Другую, несколько меньшую часть ее составляют группы сетчатокрылых (Neuroptera), часть Mecoptera^ Homoptera (сем. Prosbolidae и Scyti-nopteridae), отчасти пермские жуки (колеоптероиды), некоторые стрекозы (сем. Pholidop tilidae'), близко напоминающие и действительно родственные представителям тех же отрядов и семейств в фауне верхней перми Австралии. Совсем небольшую часть всего состава образуют формы, — сюда мы относим, главным образом, представителей Рrotorthoptera Salta tor ia, на которых можно смотреть, как на пережитки в иных условиях элементов верхнекарбоновой (стефанской) фауны западноевропейского субтропического типа.
Большая часть фауны, однако, явно родственна фауне Канзаса. Она состоит из Protohymenoptera, некоторых стрекоз, подсем. Typinae из Megan isoptera, комплекса семейств прямокрылых, объединяемых мною в группу Idelodea, отряда Protoperlaria, сем. Archescytinidae, псоцид (Permopsocidae'). Эта часть фауны резко отличается от верхнекарбоновой фауны европейского типа и от верхнекарбоновой фауны Северной Америки (Иллинойс), хотя корни значительной части ее, видимо, идут к к о р-н я м этой карбоновой фауны. Интересующая нас северорусская часть фауны отличается от канзасской по существу такими чертами, какими вообще отличается среди насекомых более молодая от более древней фауны того же состава. Это будут, во-первых, более крупные размеры северорусских представителей тех же или близких к канзасским родам. Рост размеров последовательных представителей эволюционирующих узких стволов является в разных группах насекомых совершенно определенной закономерностью (это отмечалось мною ранее, еще в 1923 г.), находящей, в частности, полное подтверждение в случае сравнения многих северорусских форм с канзасскими.
Далее следует отметить более высокую специализацию крыльев некоторых групп, например Protohymenoptera, отчасти некоторых Idelodea. Однако делать выводы о большей специализации северорусских форм по
сравнению с канзасскими весьма рискованно: наши формы не являются потомками канзасских, и у многих канзасских форм мы встречаемся с чертами большей специализации крыльев, ,например с чертами особого упрощения жилкования. Фауна отложений по р. Сояне очень сходна с фауной Тихих Гор как по своему составу, так и по наличию близких и даже некоторых тождественных видов, почему и возраст ее должен быть приблизительно тот же.
Возраст Тихих Гор (см. ниже) прежде считался конхиферовым, теперь же оценивается чаще как спириферовый, а некоторые (Ч е р д ы н-цев, Мазарович) считают его даже нижнеспириферовым. С последней оценкой мы согласиться не можем и считаем его верхнеспириферо-вым (верхняя половина этого горизонта). Таким же приблизительно должен быть и возраст фауны р. Сояны; возрастные различия здесь самые незначительные, трудно уловимые.
Состав фауны р. Сояны и сравнение ее с другими фаунами позволяют поставить вопрос и о физикогеографических и климатических условиях ее обитания. Остатки насекомых представлены почти исключительно крыльями, и очень редко встречаются и самые тела их. Насекомые разлагались, вероятно, частью здесь, частью в другом месте, крылья сносились течением и ветром и затем откладывались и засыпались песком в затишной мелководной бухте, связанной, однако, с морем, что доказывается нахождением здесь остатков Lingula. Хорошая сохранность крыльев, остатков растений и, наконец, сама порода — песчаник — говорят о том, что суша, с которой были снесены насекомые и части растений, была совсем недалеко.
При таких условиях в нашей бухте могла быть погребена большая часть жившей на этой части суши энтомофауны, кроме самых мелких представителей ее. Нельзя считать, конечно, что погребенная в песчанике фауна может дать нам точное представление об относительных частотах или количествах, в которых встречались на суше разные виды, роды и семейства. Конечно, в нашем кладбище эти отношения во многом извращены, но по отношению к летающим формам средних и крупных размеров они вряд ли сильно отличны от истинных отношений, а потому количественный состав разных групп ископаемой фауны, несомненно, дает нам право судить и о количественных отношениях в живой фауне, по крайней мере о летающих средних и крупных формах.
Обратимся теперь к общему составу материалов с р. Сояны. Они далеко еще не обработаны, и о сколько-нибудь точных относительных цифрах говорить не приходится; но все же совершенно ясно, что главную массу составляют цикадки (Homoptera, главным образом сем. Prosbolidae, затем Scytinopteridae и другие), разнообразные прямокрылые (Idelodea, Glosselytrodea, Caloneurodea и др.), Protoperlaria, скорпионницы (Me-coptera), псоциды. Гораздо меньше стрекоз, сетчатокрылых, прыгающих прямокрылых, жуков. Homoptera — несомненно, чисто наземная группа; к наземным же относятся псоциды, многие прямокрылые, скорпионницы, вероятно и часть сетчатокрылых. Группы с водными нимфами и личинками менее многочисленны; к ним относятся стрекозы и Meganisoptera, поденки, Protoperlaria, вероятно большинство Idelodea. Если судить по этой фауне, то можно думать, что озера и реки здесь были, но играли не очень большую роль; открытые пространства с цикадовыми были представлены хорошо. Климат, следовательно, был умеренно влажный. Что касается теплового режима, то, учитывая резкое отличие этой фауны от теплолюбивой фауны верхнего карбона Западной Европы, присутствие ряда элементов, близких и очень похожих на главные группы верхнепермской фауны Австралии (Homoptera, Mecoptera, Neuropterd), жившей в условиях умеренной и даже прохладной зоны, — мы и о фауне северной половины Европейской части СССР должны сказать, что она жила в умеренной зоне, хотя, несомненно, и более теплой, 14
чем та, в какой жила известная нам австралийская фауна верхней перми.
2. Местонахождение Тихие Горы на р. Каме. Местонахождение представлено известковистыми и глинистыми буроватыми, синеватыми и желтоватыми сланцами и песчаниками, а иногда и просто глинами, обнажающимися по крутому правому берегу Камы. Прослои с насекомыми покрываются слоем с Pseudomonotis garfortensis, а вместе с остатками насекомых и ниже попадаются Lingula orientalis и некоторые другие брахиоподы. Кое-где попадаются конкреции гипса. Остатки насекомых встречаются ниже слоя Pseudomonotis, но единичные остатки насекомых и растений изредка попадаются, как будто, и выше этого слоя, против пристани Тихих Гор. Слои с ископаемыми подстилаются немым желтовато-буроватым песчаником без ископаемых.
Первые насекомые были обнаружены здесь (1898 г.) немецким геологом Кокеном, бывшим здесь с экскурсией геологического конгресса из Москвы. Этот сбор в 6 видов был переслан австрийскому энтомологу и палеонтологу Г а н д л и р ш у, который и опубликовал результаты его изучения в 1904 г.
После этого долгое время здесь не находили никаких остатков насекомых. В 1926 г. это место посетил автор этой заметки и собрал здесь значительное количество ископаемых, среди которых оказалось 30 видов насекомых, в большинстве хорошо сохранившихся. Результаты изучения этой коллекции, а также первых сборов М. Б. Едемского нар. Со-яне были опубликованы в 1928 г. Затем это местонахождение вскоре же посетили Е. В. Пермякова и М. Д. Залесский; эти материалы были изучены М. Д. Залесским (1929, 1932), а частью Ю. М. Залесским (1931, 1934). Мартынов работал там еще дважды (1929, 1931) и собрал новые материалы; результаты опубликованы в 1931 и 1933 гг. В настоящее время описано из этого местонахождения около 80 видов, но осталось еще несколько неописанных форм.
Фауна Тихих Гор, как сказано выше, очень похожа на фауну с р. Со-яны, но немного беднее, особенно по числу видов скорпионниц (М ecoptera} и цикадок (Homoptera).
Возраст фауны Тихих Гор определялся прежде как конхиферовый. Я определял его раньше как нижнеконхиферовый или верхнеспириферо-вый, а в последнее время — как верхнеспириферовый. Геологи Ч е р-дынцев и Мазарович высказываются (конференция по пермским отложениям в конце февраля 1937 г.) в пользу нижнеспириферового возраста их.
Я должен сказать, что если мы согласимся с этим мнением, то мы неизбежно должны повысить соответственно возрасты Каргалы, отложений в бассейне р. Сылвы и других отложений с насекомыми как в СССР, так и в других странах, что наталкивается на некоторые неувязки и возражения. Поэтому, впредь до более несомненного и точного выяснения возраста отложений Тихих Гор и р. Сояны, я предпочитаю считать эти отложения принадлежащими верхней половине спириферового горизонта. Сходный состав фауны заставляет говорить и о сходном с соянским климате, может быть несколько более засушливом (присутствие .гипсов).
3. Карга л а, близ Оренбурга. Это местонахождение, в котором попадаются и насекомые, и позвоночные, было известно давно, но насекомых до недавнего времени было известно всего 5 видов, описанных Г андлиршем (1908). Позже оттуда было описано еще 4 вида М. Д. и Ю. М. Залесским и (1926, 1932), один вид Мартыновым (1932). За последние годы там работала Е. В. Пермякова. Собранный ею материал поступил в Палеонтологический институт и в настоящее время обработан. Он содержит еще не менее 20 новых видов и родов, так что общее число видов Каргалы повышается до 31.
Фауна Каргалы явно старше фауны насекомых Тихих Гор, но носит все же характер нижнего отдела казанского яруса. Если принимать, что фауны Тихих Гор и р. Сояны жили во вторую половину спириферового времени, то каргалинскую фауну следует считать происходящей из самых низов казанского яруса, частью из верхов кунгурского (николаевские отвалы, может быть и некоторые другие); если же тихогорскую фауну мы согласимся считать нижнеспириферовой, то фауну Каргалы нам пришлось бы считать кунгурской и даже глубококунгурской. Однако такой вывод неизбежно вызывает другие возражения, ибо если это так, то как же тогда оценивать ряд местонахождений Среднего Урала? Они обычно признаются кунгурскими, и, действительно, фауна их значительно архаичнее, старше каргалинской. Нам пришлось бы признать тогда эти местонахождения не кунгурскими, а артинскими, что натолкнулось бы на другие возражения. Фауна их, действительно, все же в большинстве моложе канзасской, происходящей из нижней перми. По этим основаниям мы пока воздерживаемся от всех этих перемещений и принимаем для Каргалы нижнеспириферовый возраст до верхнекунгурского (сюда относятся ископаемые из николаевских отвалов).
Фауна Каргалы сильно отличается от фауны Тихих Гор и р. Сояны отсутствием скорпионниц и цикадок и присутствием относительно большого числа водных фом — стрекоз, поденок, Protoperlaria, веснянок, из которых поденки и веснянки представлены нимфами. Такой характер фауны с определенностью говорит о том, что местность, в которой она обитала, содержала озера и реки и,видимо, хорошо орошалась. Ископаемые сохранились в осадках озера, а может быть и реки.
4. Побережье р. Сылвы, у дер. Чекарды и ниже. Песчанистые и глинистые сланцы. Первые насекомые отсюда были добыты Г. Т. Мауэром. Одно из них Мауэр переслал мне, и оно оказалось представителем особого семейства, родственного верхнекаменноугольному сем. Caloneuridae из Западной Европы (1930). Затем значительные сборы были произведены там Е. В. Пермяковой в 1931, 1935 и 1936 гг. и Г. Т. Мауэром. Эти материалы в настоящее время обрабатываются частью мною, частью Ю. Залесским (материалы Мауэра). Это по преимуществу озерные отложения, но по близости существовали, несомненно, и текучие воды. Это местонахождение интересно тем, что большая часть насекомых представлена здесь почти целыми экземплярами; к сожалению, жилкование крыльев во многих случаях не сохранилось вовсе. Фауна носит архаический характер и, несомненно, старше даже, чем фауна Каргалы, так что ее можно считать кунгурской. Преобладают веснянкообразные водные формы, имеются и скорпионницы, но цикадок {Homoptera) нет или почти нет. Элементы, близкие к северо-американским (Канзас), здесь имеются. Интересно нахождение одного представителя Palaeodictyoptera и нового рода стрекоз — Ditaxineurella gen. nov., довольно близкого к соответствующим родам из Канзаса. В коллекции, обработанной мною, было около 20—25 видов.
Из того же Кунгурского района за последние годы нам стали известны еще четыре местонахождения (5—8), из которых пока известно по одному-двум видам.
5. Окрестности дер. Су к с у н; сборы М. Н. Круглова. Я видел отсюда два отпечатка, один из которых относится к сем’. Idelidae. Возраст не выяснен; вероятно, верхнекунгурский.
6. Деревня Седы, километрах в 10 к юго-востоку от Суксуна. Около 25 отпечатков мелких насекомых здесь было добыто мною в 1931 г. Они оказались принадлежащими трипсам и отнесены мною к одному виду рода Permothrips Mart. (1936).
7. Окрестности с. Матвеева, у р. Барды, к северо-востоку от Кунгура. Несколько отпечатков отсюда было добыто Г. Т. М а у-э р о м, и один из них был переслан мне и вскоре описан (1930). Он 16
оказался прекрасно сохранившимся крылом нового крупного таракана— Kunguroblattina arcuata Mart. Порода — мелкозернистый темный песчанистый сланец, на котором остатки насекомых должны быть хороши. Здесь желательны новые поиски. Возраст, видимо, сходный с возрастом Чекарды.
8. Савиновский лог, по дороге от Красноуфимска к Артинскому Здесь Е. В. Пермяковой добыт один экземпляр крыла, принадлежащий к новому роду из группы Protohymenoptera (Scytohymen extremus Mar t.). Возраст этого местонахождения, видимо, такж^ кунгурский (верхнекунгурский).
9. Река Ви льва, левый берег, недалеко от ст. Чусовой. П. Шеиным (из геолого-разведочной партии Г. Н. Фредерикса) здесь был добыт один отпечаток крупного крыла, который был описан Г. М. Залесским как Vilvia densi-nervosa Z а 1. (прямокрылое). Возраст назван как артинский, но без каких-либо пояснений; точное определение возраста, конечно, дело будущего.
10. Чердынск. р., правый берег р. Язьвы у деревни того же имени. Прекрасный отпечаток большого крыла был добыт здесь в 1931 г. Н. А. 3 е н ч е н к о. Экземпляр попал к Ю. М. Залесскому и описан как прямокрылое Jasvia reticulata Ъ а 1. (1934). Возраст назван кунгурским.
11. Река Ирень, у дер. Колпаковой, Средний У рал. Из этого пункта уже давно описана Гандлиршем (1908) одна форма (Dyadentomum permense Н a n d 1.); новых материалов оттуда не было. Возраст не выяснен; скорее всего верхний кунгур (?).
12. Речка У я з а, приток Демы, дер Чотмак-То-м а к, Белебеевского района. Описан один экземпляр таракана из сем. Archimylacridae. М. Д. Залесский указывает конхиферовый горизонт, не давая каких-либо пояснений.
13. Овраг Тахташур близ речки Гулюшер-минки, впадающей в р. Иж, бассейн Камы. Отсюда также описана м. д. Залесским лишь одна форма —Pruvostites takhtachuren-sis М. Z а 1. из прямокрылых; возраст им называется «казанский».
14. Деревня Шуня, на р. Вятке. Отсюда известен фрагмент крыла неизвестного насекомого (прямокрылого), описанного еще Гандлиршем (1904). Новые поиски в этом районе не увенчались успехом.
15. Речка Сарды к, приток Буры, бассейн р. Вятки. М. Залесским описана отсюда одна форма таракана; возраст «пределен им как «верхнепермский».
16. Богатый лог на р. Каме. Добыт только один экземпляр жука (Elateridopsis permiensis Ъ а 1.), описанный М. Д. Залесским (1932); возраст указывается им «конхиферовый».
17. Речка Перебор, бассейн Печоры. М. Д. Залесский описал отсюда один вид Homoptera — Pereboria bella Z а 1. Возраст определялся как «верхнепермский».
Как видно из распространения местонахождений, большая часть их сконцентрирована к востоку от р. Камы и южнее, до Каргалы. Эти местонахождения, особенно расположенные в бассейне р. Сылвы, Вильвы, по речке Ирень и верхней Уфе (по дороге к Артинскому), находятся в области отложения красноцветных пород. Значительная часть их отложилась, видимо, в озерах; озерными же (и речными) являются и отложения Каргалы. В большей части этих местонахождений найдено пока по 1—2 ископаемым формам, но они найдены геологическими партиями скорее попутно, почему в дальнейшем можно ожидать нахождения в этих пунктах еще некоторых форм, но, вероятно, немногих. Несомненно, что в той же меридио-нально>~1гоя№е' краеноцветных пород (схема II) к западу от р. Камы и в басеейне-ф^ Уфй 4гожнЬ ожидать, что будет встречен еще ряд новых местонарс&вдедвж ископаемых насекомых и растений.
Возраст большинства этих местонахождений, рассмотренных в полосе красноцветных пород, согласно указывается как кунгурский, и только для р. Уязы (местонахождение 12) М. Д. Залесский указывает конхиферовый горизонт, а для местонахождения у ст. Чусовой (местонахождение 9) Ю. М. Залесский говорит об артинском возрасте. К артинскому ярусу Толстихина почему-то относит и Paleutky-gramma из Чекарды. На юге полоса этих местонахождений кончается Кар-галой. На севере полосу красноцветных пород А. Д. Архангельский1 проводит до Печоры; к востоку от этой реки появляются угленосные отложения, следовательно также пресноводные, но более молодые. Близ южной границы их, у речки Переборы, экспедицией А. А. Ч е р-н о в а добыт один экземпляр крыла Homoptera (Pereboria bella Z a 1.) казанского возраста. Архангельский, между прочим, высказывается против «обычного мнения», что красноцветные породы формировались в условиях пустынного жаркого климату. Для нас последнее мнение особенно неприемлемо: выше мы уже отмечали, что в области Чекарды, например, водные формы даже преобладали над наземными и формы, имевшие водных нимф, не отсутствовали и в других местонахождениях Среднего Урала. Я принимаю, что эти местонахождения располагались еще в умеренной, хотя бы и тепло-умеренной зоне.
Новые поиски насекомых, особенно в кунгурских и артинскпх отложениях, были бы очень желательны и для выяснения постепенной смены фауны и миграций. В кунгурских местонахождениях мы встречаемся с большим относительным количеством пережитков и дериватов верхне-каменноугольной фауны европейского типа и с несколько меньшим содержанием в них форм американского и австралийского типа. Как далеко вглубь идут такие формы? Каково их географическое распространение? Как объяснить постепенное исчезновение к югу большой группы цикадовых (Homoptera), столь богато представленной на севере? Ряд подобных вопросов может разрешиться лишь на базе новых находок в восточных и северных областях Европейской части СССР и в Северной Азии.
Каменноугольные насекомые из пределов Европейской части СССР не известны.
Азиатская часть СССР. В Северной Азии ископаемые верхнепалеозойские насекомые обнаружены пока в двух районах, в Кузнецком каменноугольном бассейне и в нижнепермских отложениях под Владивостоком. В Кузнецком бассейне насекомые были обнаружены в балахон-ской и кольчугинской свитах, в триасовом разрезе у Бабьего камня и, наконец, в отложениях юрского возраста.
Местонахождения насекомых, относящихся к балахонской свите, сконцентрированы в северной части бассейна, по правому берегу р. Томи, по речке Чесноковке и по левому берегу р. Северной Уньги.
18. Правый берег речки Чесноковки, выше с. Верхотомского (Александровский пласт). В этом пункте геолог М. Ф. Нейбург добыла в 1929 г. отпечатки крыльев двух замечательных форм, описанных мною в 1930 г., — Neuburgia altaica Mart, из Palaeodictyoptera и Narkemina angustata Mart, из прямокрылых (Protorthoptera). Первая форма особенно интересна в смысле определения возраста свиты. Из Palaeodictyoptera переходит из карбона в нижнюю пермь, как известно, одно сем. Spilapteridae. Из него нам известен один вид из Канзаса (нижняя пермь), один из Чекарды (Кунгур) и один, Neuburgia altaica, с речки Чесноковки, но последний стоит ближе к одной сте-фанской группе родов (из Комментри, Франция), чем к упомянутым ро-
1 Архангельский А. Д. Геологическое строение СССР. Западная часть, вып. 1, 1934. Отложения красноцветных пород к западу от Урала Архангельский обозначает «уфимской свитой». Насколько я понимаю, она носит смешанный характер и содержит осадки как нижней половины казанского яруса, так и части кунгурского яруса. Довольно сходное мнение высказывает теперь Тихвинская (1937).
дам из нижней перми. Narkemina относится к маленькому семейству, известному только из верхнекаменноугольных отложений Северной Америки. Уже одни эти данные склоняют к мысли, что слой с этими формами относится не к перми, а скорее к самым верхам карбона. Изучение растительных остатков привело М. Ф. Нейбург к тому же выводу.
19. Правый берег р. Томи в 3 км выше устья р. 3 а л о м н о й. Здесь также найдены М. Ф. Н е й б у р г два вида— Phyloblatta regularis Mart, и Tomioblatta furcatella Mart, (тараканы). Оба вида относятся к сем. Archimylacridae, известному по преимуществу из карбона, но также и из перми.
20. Лог Мосточек, впадающий в р. Б а л а х о н к у у дер. Б а л а х о н к и; свита Н г Здесь М. Ф. Нейбург нашла две формы насекомых: Oiratia calida Mart, из Protoperlaria (сем. Probnisidae) и Archeglyphis crassinervis Mart, из Homoptera, сем. Prosbolidae (цикадовые). Оба эти семейства известны из перми, но кузнецкие формы отличаются большой архаичностью, так что они могут происходить как из низов перми, так и из верхов карбона.
21. Левый берег Сев. Уньги под дер. Черемич-к и н о й. Здесь одна форма найдена М. Ф. Нейбург (1929) и пять— Е. Ф. Ч и р к о в о й. Первая была изучена мйою, описана под названием Ungoneurites paucinervis Mart. Крыло (заднее) обнаруживает черты сходства п родства ее сеем. Palaeocixiidae из верхнего карбона Комментри. Отпечатки других пяти форм были обработаны М. Д. Залесским (1930). Им были описаны два таракана — Phyloblatta sibirica Z а 1. и Caenoblatta angaridensis Z а 1.; оба они также относятся к сем. Archi-mylacridae и похожи на описанные мною. Следующая форма, Рагарапогра ungensis Z а 1., отнесена им к скорпионницам (М ecopter а), но, по моему мнению, она относится к тараканам, к сем. Archimylacridae. Следующие две формы, Tchirkovaea guttata Z а 1. и Philiosptilon maculosum Z а 1., очень оригинальны и отнесены автором к Megasecoptera, но на мой взгляд принадлежность их к этой группе не доказана. На основании нахождения предполагаемой панорпы М. Д. 3 а л е с с к и й отнес уньгинскую подсвиту к типичной перми, но, как мы сказали, это крыло принадлежит таракану из сем. Archimylacridae, вследствие чего этот аргумент в пользу типично пермского возраста соответствующего слоя отпадает. Эти пять насекомых говорят скорее о самых верхах карбона, переходных к перми.
Перечисленные четыре местонахождения знакомят нас с очень своеобразной фауной насекомых, напоминающей отчасти фауну верхнего карбона Северной Америки, но, конечно, этих остатков, к тому же частью плохо сохранившихся, недостаточно для составления сколько-нибудь ясного представления о фауне Кузнецкого района данного возраста (балахонская свита) и об ее отношениях; здесь нужны новые повторные исследования и новые сборы.
Остатки насекомых из к о л ь ч у г и н с к о й свиты добыты Т. П. Радченко в 1934 г. Они найдены в следующих пунктах.
22. Левый берег р. Б. Т ы х т ы в дер. Соколовой, крестьянская штольня: один экземпляр, отнесенный мною к роду Prosbole Handl. (Homoptera), известному только из верхней перми северорусских пермских местонахождении (Р. triangularis Mar t.).
23. Г о р е л ь н и к и кровли пласта надбайкаим-с к о г о, в устье л о г а Б о л ь ш а я Б а й к а и м с к а я Л и с т-в я ж к а. Отсюда происходят две формы—Dictyoprosbole membranosa М art. и Orthoprosbole congesta Mart. — обе из того же отряда Homoptera.
Все три формы говорят о верхней перми, вероятно, даже несколько более молодой, чем насекомоносные отложения Ива-Горы и Тихих Гор. Новые исследования в кольчугинской свите и новые сборы необходимы. 2=: 19
Из Минусинского и Тунгусского бассейнов насекомые не известны. С новым местонахождением (24) мы встречаемся на Дальнем Востоке. Здесь в 1932 г. П. В. Виттенбургом добыт один отпечаток крупного насекомого на куске бурого глинистого сланца в г. Владивостоке. Это насекомое было описано мною под именем Nemuropsis tenuis Mart.; оно относится к отряду Paraplecoptera (часть Prothorthoptera Handl.) и внешностью напоминает веснянок. Об отношениях его трудно сказать что-либо определенное, так как крылья не сохранились.
мезозойские местонахождения
В связи с большим развитием юрских отложений континентального типа в Средней Азии, Восточной, а отчасти и Западной Сибири, почти все местонахождения ископаемых насекомых происходят именно из юры. Триасовых местонахождений у нас за последнее время стало известно два, меловых же пока не известно.
Триас
1. Кузнецкий бассейн, район Бабьего камня. Остатки ископаемых членистоногих были отсюда добыты М. Ф. Ней-бург в 1935 г. Они были описаны Мартыновым (1936), который обнаружил среди них одного жука Ademosynoides asiaticus Mart., хорошо сохранившегося и близкого к подобным формам из верхнего триаса Австралии, затем одно прямокрылое, Tomia costalis Mart. Кроме того, отсюда же описана впервые в СССР одна многоножка, Tomiulus angulatus Mart., и очень неясные остатки какого-то ракообразного. Я отнес отложения Бабьего камня определенно к триасу. За триасовый возраст их высказались на основании изучения растений М. Ф. Н е й-бург и Т. Г. Радченко. В том же году Ю. М. Залесский опубликовал описание двух остатков насекомых из того же района, собранных В. Я. Яворским. Он высказался за лиасовый возраст их, но следует сказать, что первая из этих форм, Bittacomorpha javorskii Z а 1 ., сохранилась крайне недостаточно и принадлежность ее к Ortho-phlebiidae более чем сомнительна. Вторая форма, относимая им к Lemma to-phoridae, говорит скорее в пользу повышения возраста их. Таким образом, триасовый возраст этого местонахождения можно считать установленным; я бы принял его даже не за верхний, а за средний триас. В настоящее время триасовые отложения обнаружены не только в районе Бабьего камня, но и к юго-западу от него. Т. Радченко недавно доставил мне оттуда еще 4 отстатка насекомых, но, поскольку это лишь фрагменты брюшка, определить их сколько-нибудь близко невозможно. Очень желательны там новые поиски.
2. Шураб, Фергана, около 40 км к юго-западу от Шураба II. Местонахождение триасовых растений и насекомых здесь было обнаружено в 1933 г. геологом Кочневым. Насекомых здесь найдено до 8 видов, относящихся к жукам, 4 вида — к тараканам, прямокрылым и цикадовым (Homoptera'), но они еще не описаны мною. Жуки мелки и походят на жуков, известных из нижнего триаса Европы, но, с другой стороны, таракан близок к роду Hongaya Н а n d 1., известному из верхнего триаса Тонкина. По растениям возраст этих отложений М. И. Брик определяет как нижнетриасовый.
ЮРСКИЕ МЕСТОНАХОЖДЕНИЯ
3. Шураб, Фергана. В Шурабском каменноугольном местонахождении известны не один, а три пункта, где найдены насекомые, именно: Шураб I, Шураб II и Шураб III. Все они расположены на расстоянии нескольких километров друг от друга и обследованы, главным обра-20
зом, геологом Н.В. Шабаровым, открывшим все эти местонахождения насекомых.
Главную массу насекомых дал один пункт в области Шураба II; там большое количество ископаемых насекомых было собрано сначала Н. В. Шабаровым, а затем А. В. Мартыновым в 1933 г. В Шура-бе I несколько остатков насекомых было добыто Н. В. Шабаровым. Им и М. И. Брик в Шурабе собраны, кроме того, большие материалы по растениям. Насекомые Шураба II и I, кроме жуков, обработаны. Фауна оказалась весьма архаичной, и некоторые представители ее еще сильно напоминают верхнетриасовые формы Австралии. Возраст, однако, устанавливается мною не по этим (часть тараканов), а по другим группам, главным образом Mecoptera, которые в большинстве, несомненно, близки к нижнелиасовым видам Англии. Часть походит больше на известные верх-нелиасовые формы Западной Европы. Вывод определенно тот, что насе-комоносные отложения (слой от А до М приблизительно, большая часть происходит из слоя Н.) Шураба принадлежат нижнему лиасу. 3—4 экземпляра добыты в более высоких слоях Z и др., и эти слои, быть может, относятся уже к верхнему лиасу. СлойН, откуда добыта большая часть остатков, во всяком случае относится мною к нижнему лиасу. М. И. Брик этот слой склонна отнести к верхнему лиасу, с чем я на основании изучения насекомых согласиться не могу. В фауне Шураба хорошо представлены тараканы, жуки, затем скорпионницы (Mecoptera), прямокрылые и цикадовые; другие группы представлены бедно, а двукрылых вовсе не найдено. Крупные тараканы, как сказано, ближе всего стоят к верхнетриасовым тараканам Австралии.
В Шурабе III, отстоящем от Шураба II километров на 15, насекомые были открыты и собраны Н. В. Шабаровым в 1936 г. Они еще не обработаны мною. Состав видов несколько иной, но имеются очень сходные виды. Возраст, повидимому, тот же.
4. Сулюкта, в 40 км от Шураба. Немного остатков насекомых здесь было найдено М. И. Брик, но я их не видел.
5. К и з и л-к и я, Фергана; близ сел. Учкурган. Насекомые в этом местонахождении были обнаружены впервые инж. И. С. К а-м ы ш а н о м, который и собрал небольшое количество их. Позже они собирались М. И. Брик и Н. В. Шабаровым. Эти сборы были обработаны мною в 1934 г. (жуки еще не обработаны) и описаны вместе с насекомыми Шураба в общей работе. Фауна оказалась довольно сходной с шурабской, и возраст ее я определил тоже как нижнелиасовый.
6. Обнажения у ручья Бахай-бас-тау, вЗ км от с. Галкина, южные отроги хребта К а р а - т а у. Это известное местонахождение, откуда добыто огромное число рыб, насекомых и растений, было обнаружено Е. Ф. Гориздро-Куль-чицкой и В. В. Мухиным. Сборы там производились неоднократно. В 1924 г. там работали экспедиция Гориздро-Кульчиц-к о й и я; в 1925 г. я опять посетил это местонахождение и собрал богатую коллекцию. Затем там работали М. И. Брик (растения), В. Н. Принада. Материалы, в конце концов, оказались сконцентрированными в двух местах — в Музее местной природы в Ташкенте и в Палеонтологическом институте Академии Наук. Значительная часть материалов Палеонтологического института обработана и описана мною в 1925— 1930 гг. Материал оказался чрезвычайно интересным. Сохранность многих экземпляров очень хороша: во многихслучаях сохранилось строение лапок, волоски, окраска и т. п. Несколько экземпляров было описано позже американским профессором Коккерелем, осматривавшим сборы насекомых в Ташкенте, а одна крупная форма рода Kaligramina была описана Н. Ф. Безобразовой. Двукрылые в настоящее время изучаются Б. Б.Р одендорфом. Порода представлена известковистыми сланцами, легко раскалывающимися на тонкие плиты; твердые сланцы часто
переслаиваются более нежными, листоватыми сланцами, которые можно расслоить на листики, подобные бумаге (Галкино). В таких сланцах хорошо сохраняются не только рыбы, но и самые мелкие насекомые. Эти сланцы отложились, очевидно, в озере; среди остатков Diptera мы встретили здесь водных личинок и куколок комаров и др. водные формы. Местонахождение у Галкина занимает очень небольшую площадь, всего 8—10 м2. Там копали все, кто только заезжал сюда, а кроме того, более крупные плиты увозились крестьянами для полов и т. п. В результате это замечательное местонахождение было так выкопано и перекопано, что в настоящее время там уже трудно найти насекомых.
Следует отметить, что, помимо насекомых и огромного количества рыб, в Галкине и Карабасгтау найдены: отпечаток черепа мелкой рептилии, одна летающая рептилия (Pterosauria) и одна черепаха.
Об общем характере фауны мы скажем ниже.
7. Карабас-та у, берега ключа и речки Карабас-т а у в урочище того же имени, расположенном близ верховьев р. Бугунь, притока Боролдая. Урочище представляет собой возвышенность, глубоко прорезаемую мощным ключом Карабас-тау. Обрывистые, высокие берега этого ключа, по крайней мере на полкилометра в длину, сложены плитами твердых сланцев, подобных таким же сланцам Галкина. Остатки насекомых здесь впервые были обнаружены проф. В. В. Мухиным. Затем там работали экспедиция проф. Г о р и з д р о-К у л ь ч и ц к о й в 1924 г. и М. И. Бри к; я посетил это местонахождение дважды, в 1924 и 1925 гг. Наконец, в 1936 г. здесь и в Галкине производил наблюдения Р. Ф. Г е к к е р.
Как я мог убедиться во время своих работ, остатки и насекомых, и рыб здесь попадаются гораздо реже, чем в сланцах Галкина,иочень спорадично; нужно бывает просмотреть большое количество плит, прежде чем найти 2—3 остатка насекомых. Тем не менее и отсюда удалось добыть десятки экземпляров; эти сборы обрабатывались также мною.
Фауна Карабас-тау оказалась вполне сходной с фауной Галкина, почему можно обобщить замечания о той и другой. Больше всего встречается жуков — несколько десятков (вероятно, до сотни) видов, затем двукрылых, цикадовых, скорпионниц и тараканов. Эти насекомые образуют как бы основной фон, но, кроме них, добыто не мало остатков и других отрядов — прямокрылых (крупные кузнечики из нового сем. Aboilidae), фасмид, сетчатокрылых, перепончатокрылых (сидячебрюхие и паразитические группы), вымерших Paratrichoptera, уховерток, трипсов, псиллид, псоцид, стрекоз, поденок, палеонтинид (группа Homoptera^ которую раньше считали бабочками).
Это очень богатая фауна, которая содержит, несомненно, не одну сотню видов; пока изучена и описана лишь часть ее. По своему составу и облику она очень напоминает фауну Золенгофена, относимую к верхней юре, но было бы совсем неправильно приписывать ей тот же геологический возраст. Значительная часть ее видов, несомненно, ближе стоит к видам из верхнего лиаса Мекленбурга, и уже одно это обстоятельство не позволяет считать ее верхнеюрской. Сначала я считал ее верхнелиасо-вой, теперь отношу ее скорее к доггеру, к нижней его части. Многие элементы, например Palaeontinidae, появились раньше в Азии, и уже несколько позже в Западной Европе. В Карабас-тау найдены остатки одной рептилии.
8. Недалеко от Карабас-тау находится третье местонахождение той же фауны, именно ус. Михайловки (ур. Ак - Тас; Терс-Джейлау). Главные сборы насекомых там сделаны А. Н. Турутано-вой, изучившей там флору; несколько экземпляров было добыто Веде-н я п и н ы м. Эти сборы, хранящиеся в Палеонтологическом институте, пока еще как следует не обрабатывались. По всей видимости, здесь мы имеем продолжение фауны Карабас-тау.
9. Местонахождение у Иссык-куля. В окрестностях оз. Иссык-куль некоторое количество остатков насекомых, десятка два, было собрано Н. А. Куликом. Порода темная, грубая, несланцеватая и колющаяся неправильно. Отпечатки насекомых по большей части неполные, фрагментарные, хотя есть и хорошие экземпляры. Судя по тому, что я видел отсюда тех же рафидий (род Mesoraphidia Маг t.), какие мне известны из Галкина, я думаю, что это местонахождение имеет тот же или близкий возраст (нижний доггер— верхи лиаса).
10 и 11. Кузнецкий бассейн, левый приток р. Н а-р ык- Че р Н9 в о й и район Тарбаганского камня. Из этих двух мест М. Ф. Нейбург в 1929 г. добыты остатки четырех видов, которые оказались передним крылом нового представителя сем. Aboilidae, надкрыльем таракана из сем. Mesoblattinidae, элитрами жука и одним фрагментом крыла, по которому установите группу затруднительно. Эти формы говорят о юрском, вероятно среднеюрском возрасте этих отложений. К сожалению, остатки эти сохранились недостаточно.
12. Буровые колонки у поселков С у х о ме с обского и Коркинского, Челябинский буроугольный район. Насекомые в этих колонках были обнаружены в 1933 г. А. Н. Криштофовичеми описаны мною (1935). Они оказались следующими тремя формами: поденка — Mesephemeridae, gen? — ornata Mart.; экземпляр представляет собой отпечаток брюшка нимфы, почему и не мог быть определен точнее; сетчатокрылое — Archedilaropsis furcata Mart., из сем. Prohemerobiidae; жук — Mesosagrites multi-punctatus Mart., из сем. Chrysomelidae; жук этот, по всей видимости, родственен как Sagrini, так и Donaciini и, как указано в работе, вероятно, вел водный образ жизни. Наличие этой формы и нимфы поденки привело меня к заключению, что здесь мы имеем дело с отложениями крупного озера. Возраст, видимо, лиасовый.
13. Усть-Балей, на р. Ангаре. Это местонахождение Центральной Сибири известно уже давно. Насекомые отсюда в количестве до 19 видов были описаны в 1889 г. тремя авторами — Брауером, Редтенбахером и Гангльбауером и затем ревизиро-ваны Гандлиршем в его «Fossile Insekten» (1908). Здесь оказались: тараканы, 2 вида; жуки, 2 вида; веснянки, 3 вида, из которых две нимфы; стрекозы, 6 видов, из которых пять нимф; поденки, 2 вида нимф; одна па-норпа; две цикады (Palaeontinidae); два двукрылых и три формы невыясненного положения. В 1928 г. мною описана по сборам Ю. А. Жемчужникова еще одна форма Palaeontinidae — Pseudocossus Zemcuznikovi и показано, что как эта, так и два ранее известных вида относятся к Homoptera, а не Lepidoptera, как это полагал Гандлирш. Кроме того, в тех же сборах с Усть-Балея оказались личинки рода Ephemeropsis. Возраст отложений Усть-Балея мы относим к самым верхам лиаса. Насекомые в этом местонахождении встречаются редко, но сохранность их обычно очень хороша, так как порода очень мелкозерниста; это, несомненно, отложения крупного озера.
В Восточной Сибири насекомые констатированы, правда, в единичных формах, еще из следующих трех пунктов:
14. Р е к а Товега, близ Нерчинска. Описана по нимфе одна поденка — Ephemeropsis trisetalis Eichw. (1864). Возраст — мальм.
15. Бирка, Забайкалье. Гандлирш описал отсюда другой вид того же рода — Е. middendorfi Н a n d 1., однако Коккерель считает, что эта личинка относится к тому же виду trisetalis; возраст определяется как мальм.
16. Кондуевская на р. Турге, Забайкалье: поденка Ephemeropsis orientalis Е i с h w., которую Коккерель также справедливо считает за trisetalis. Т у р г а: отсюда Б р а у е р описал ту же Е. orientalis, но Гандлирш отделил эту форму в особый род
Phacelobranchus. Из всех этих местонахождений известны, как видим, лишь нимфы поденок; это говорит о том, что здесь существовали в верхнеюрское время крупные озера. Та же поденка Е. trisetalis в массах встречается и в Монголии, в налайхинской толще, недалеко от Улан-батора. и в формации Ondai Sair; из последнего местонахождения насекомые были описаны Коккерелем.
17. У устья р. Бирки, притока Турги. Оттуда я описываю одну крупную форму (Anaropsychops turgensis gen. s р. no v., in lit t.); сборы Толстихиной.
ТРЕТИЧНЫЕ МЕСТОНАХОЖДЕНИЯ
Нам известно в настоящее время до 10 пунктов, где встречаются третичные насекомые, но из них лишь три местонахождения дали десятки экземпляров и видов, в прочих же найдено по 1—3 экземпляра. При таком положении дела мы должны признать, что по третичным насекомым нашей страны мы обладаем пока ничтожными сведениями. Большинство местонахождений приходится на Азиатскую часть Союза.
1. С п а н и о д о н т о в ы е глины под Ворошиловском Кавк аз ким — миоцен. В этих отложениях М. В. Боярунасом добыты две формы прекрасной сохранности—муха Tubifera miocaenica S t а с k 1. и стрекоза Oplonaeschna (Epiaeschna) stauropolitana Mart.
2. Диатомовые миоценовые слои Апшеронско-г о п-ва. Отсюда был добыт В. Н. Вебером один отпечаток крыла стрекозы. Он былописан мною n^Heliaeschna (?) parallella Mart. Порода — песчаник, почему животное сохранилось недостаточно хорошо. Обе стрекозы относятся к родам, отсутствующим теперь в Европе и Северной Азии.
3. Курганский район, Краснодарского края. Отсюда В. В. Поповым был описан (1931) один муравей — Ропега (г) umbra Popov; порода — темный сланец. Возраст обозначен А. Н. Федоровым, добывшим этот отпечаток, как миоценовый.
4. Орметы, Горийского района, Грузия. Тем же В. В. Поповым описан отсюда (1932) другой муравей — Lasius martynovi Popov, добытый С. И. Ильиным. Возраст определяется им как нижнесарматский.
5. Закавказье; материал И. В. Палибина. Я видел отсюда отпечатки двух или трех видов жуков, оставшихся неописанными. Возраст указывается как «миоценовый».
6. У ф а л е й, Урал. Отсюда давно, в 1876 г., К а в а л л описал одну личинку—Т incites crystallin которой я не видал.
7. Высоты А ш у т а с, 90 км к северу от г. Зайсанска, на правом берегу Черного Иртыша; порода глинисто-известковистая или глинистая, реже песчанистая. Ашутас — замечательное местонахождение, в котором Н. А. Беляевым (1910) и особенно М. Ф. Нейбург (1927) собраны большие материалы превосходно сохранившихся растений и небольшие — по насекомым, в большинстве случаев также хорошей сохранности. Насекомые были обработаны большей частью мною (1930); здесь оказались следующие формы:
Термиты: два вида из родов Ulmeriella С о с к. и Diatermes Mart.
Стрекозы: 1 вид из рода Basiaeschna S е 1 у s, 1 — Projagoria Mart., 1 — Epacantha Mart., 3 — Necracantha Mart., 1 —Lithagrion S c u d d.
Ручейники: 1 вид Phryganea, недостаточно сохранившийся.
Чешуекрылые: 1 вид из нового рода Oligamatites Kusnezov.
Жуки: В В. Баровским отсюда определены два вида — Cala-thus sp. и Plateumaris sp.
Эта фауна родственна третичной фауне западной части Северной Америки, отчасти некоторым элементам современной фауны юго-восточной 24
Азии. Возраст отложений Ашутаса определяется как верхнеолигоце-новый.
М. Ф. Нейбург собирала в Ашутасе, главным образом, растения, насекомые же взяты попутно. Несомненно, при новых работах, новых поисках в этом районе можно найти еще много остатков насекомых, и следует признать крайне желательным новое посещение его.
8. С Сахалина, из окрестностей Мгача, Геер (Н е е г) еще в 1878 г. описал одного жука — Cistelites sachalinensis Heer, но с тех пор новых материалов оттуда не поступало. Возраст определяется Криштофовичем как олигоценовый.
9. Берега речки Кудя, притока Амагу, Приморской о б л., в 6 км от моря. Здесь добыт целый ряд растений, определенных А. Н. Криштофовичем, а также и насекомых, которые были изучены американским профессором Т. Д. Коккерелем. Последний сам посетил в 1933 г. это местонахождение и привез оттуда насекомых. Им описано оттуда до 23 видов.
Trichoptera (ручейники): сем. Phryganeidae, 1 вид; сем. Limnophilidae, род Indusia, 2 вида (домики); вид и род Lunnophiliis — 1 вид. Mecoptera (скорпионницы): род Dinopanorpa — 1 вид. Diptera: сем. Bibionidae, род Plecia — 6 видов. Hymenopteraz сем. Megachilidae, род Megachile— 1 вид; Coleoptera : сем. Carabidae, род Patrobus (?); сем. Curculionidae, род С1е-onus — 1 вид, Anthonomus — 1 вид; сем. Throscidae, род Throscus (?) — 1 вид; Homoptera'. сем. Cicadellidae, род Lavrushinia — 1 вид; сем. Cixi-idae, род Mundopodopsis — 1 вид; сем. Delphacidae^ род Amagu — 1 вид.
Возраст этой фауны им определяется как нижнемиоценовый или верх-неолигоценовый.
10. П о с ь е т, близ с. Новокиевского. Отсюда указан лишь крупный домик личинки Phryganea kryshtofovichi Cock; возраст, вероятно, верхнемиоценовый, а, может быть, плиоценовый.
11. Киргизия, бассейн р. Качку р, по р. Чон-туз; глины и сланцы. Это новое местонахождение ископаемых растений и насекомых было открыто в 1936 г. геологом А. Я. Петросянц. Насекомых было свыше 100 экземпляров, и они в настоящее время изучаются специалистами. По предварительным определениям здесь обнаружены следующие группы: стрекозы, которые представлены только нимфами (свыше 70 экз.), относящимися к 2 или 3 видам; сосущие: вида два цикадок, 1 вид тлей и 1—2 вида клопов; трипсы — 1 вид; жуки — 3 вида; муравьи — 3 вида; двукрылые — 19 видов; бабочки — 1 вид. Кроме насекомых, найдено до десятка отпечатков перьев птиц, по которым А. Я. Тугаринов определил пока три рода: утку, цаплю и, возможно, баклана.
Судя по большому числу остатков нимф стрекозы, а также перьев водных птиц, сланцы р. Чон-туза отложились в области мелководной части какого-то озера. Возраст их, по всей видимости, верхнемиоценовый.
Это местонахождение представляет очень большой интерес, и там нужны повторные исследования и сборы.
12. Отложения (темные сланцы) по р. Ваенге, притоку Северной Двины. Отсюда мне был прислан Я. Т. Богачевым на определение один отпечаток мелкого двукрылого. По определению Б. Б. Р о д е н д о р ф а, он оказался относящимся к роду Boletina (Ра-leoanaclinia), виды которого встречаются нередко и в настоящее время. Возраст этих отложений скорее всего верхнеплиоценовый.
Июль 1937 г.
SUMMARY
In this paper I give a review of the localities of fossil insects in USSR together with short characteristics of their entomofaunas. The greatest part of localities, not less than is known from the Permian deposits, to which one may also refer 4 points from the Uppermost Carboniferous series (Balachonian) of Kusnetzk basin.
Mesozoic insects are known from 13, Tertiary ones from 12 localities (see maps II a. III).
PALEOZOIC LOCALITIES
At present, nearly all Permian localities (map II) are known from the European part of USSR, and only 3—4 points with Permian insects — in Asia. European localities are arranged in two meridional streaks or zones, zone of the deposits of shallow marine bays and lagoons and, eastwards from it, — zone of so called «red coloured» deposits. This zone is composed usually of sandstones, sandy clays or slades, sedimentation of which took place in lakes or’variously formed under the forces of rivers and other waters, which flowed from the mountainous chain of the Permian Ural. In several (few) localities of this zone aquatic larvae are well preserved, what testifies, indeed, for the lacustrine nature of these deposits, — usually lakes, rarely slowly running waters. Brachiopoda or other marine animals are absent in such localities, whereas in localities, arranged in the first zone they are not unfrequent.
Richest localities are discovered in western «marine», partly brackish zone (map II). Arctic locality at the shores of the river Sojana, tributary of Kuloj, south-west from the peninsula Kanin, is especially rich of remains of insects and plants; it is represented by yellow, somewhat roseous sandstone about 1 m in thickness (points Iva-Gora, Sheimo-Gora, Letopola). Second rich locality is known as Tikhije Gory, — a village at the shores of Kama-river. It is composed by chalky sandy slates, partly by brownish clays; shells of various Brachiopoda are not unfrequent here. About 75 species of insects are described from this locality. Nearly 80 species are known also from Sojana-river, but when all materials from this last point would be examined, total number of species from should be, according to my opinion, not less than 150.
Only 1—3 species are known from each other locality of the «marine zone».
«Red coloured» zone contains more localities with insects but even «rich» ones are much poorer. From the old known locality at Kargala, 70 km from Orenburg, is known only 30 species, and I have seen about 30 species from the vicinities of Czekarda, on the shores of the river Sylva; however this locality is in all probability richer. From Kargala are described nymphs of Ephemeroptera and Plecoptera-, in Czekarda we also meet with aquatic nymphs of these orders and moreover I have seen an aquatic nymph from the order Parapiecopter a. Imagines of aquatic groups often occur in this locality.
From such facts one may deduce only the conclusion that the locality Czekarda also represents deposits of a lake.
AGE OF THE LOCALITIES
Age of the faunas from the Sojana-river, from Tikhije Gory and, probably, from other localities situated in the «marine» zone is mostly spiriferous (Lower Kazanian) and rather upper and middle spiriferous. Fauna of Czekarda is evidently older and its age may be estimated, as Kungurian. Greatest part of other points in the central part of «red-coloured» zone is, probably, also Kungurian (some authors attribute to one or to two of these points even Artinskian age, but this is in no way evident). Kargala 26
belongs to the Lowest Spiriferous and partly to the Upper Kungurian (Martynov, 1937).
The locality of fossil insects (only one species) near Petchora is younger, apparently Upper Kazanian.
CHARACTERS OF FAUNAS
As the author has indicated in his newer papers, nearly half of the whole entomofauna of Iva-Gora etc. and of Tikhije Gory is composed of families, genera and even some species, which are more or less nearly related to many genera of the rich Lower Permian fauna of Kansas, USA.
More than x/4 of the whole fauna reminds clearly of the Upper Permian fauna of Australia and is allied to it. The remaining part contains some near relatives of the Upper Carboniferous fauna of European type (Protortho-ptera Saltatoria) and some genera of indistinct systematical position. The same elements we meet with in the Kungurian or Lower Spiriferous faunas, above mentioned, but such groups, as Homoptera Prosbolidae and Scytino-pteridae (which are abundant at Sojana-river), in particular are here apparently absent. Mecoptera appear to become rare in southern localities, in Kargala in particular. Kansasian relatives are here well represented.
Such facts concur to the hypotesis, that Prosbolidae, Scytinopteridae and Mecoptera were adapted, probably, to more temperate areas. In the Permian of Australia the described genera and species of this groups lived, indeed, in the temperate and rather somewhat cold-temperate climate. Russian Permian fauna, as well as the fauna of Kansas, lived in the temperate zone, although evidently warmer zone. It is comprehensible, that the entomofauna of Sojana-river resembles Upper Permian fauna of Australia nearer than that of other localities.
Notes on the records in Kusnetzk basin Balachonian series has provided us but few (locality 12) insects one of which is a Palaeodictyopteron, one or two belong to Megasecoptera. This is a peculiar fauna, not resembling that from the above mentioned localities but unfortunately too poor. In the same basin there are two points with insects of the Upper Permian age (Koltchugian Series); all they (3) are Homoptera, related to those from Iva-Gora etc.
At last, one form was recorded from the Lower Permian of Wladivostok.
MESOZOIC LOCALITIES
They ёге known mainly from Central Asia (Turkestan) and from Siberia.
Main locality of middle Yurassic (Dogger) insects is situated in the mountainous chain Kara-tau (Galkino, Karabas-Tau, Michailovka); it is represented with litographic and «paper» shales, containing rich fauna of insects and rich flora. Many insects are already described, but many new forms remain yet undescribed. The presence of larvae and pupae of some «culicoid» forms proves, that these shales are deposited in bays of a lake or in several lakes.
Similar insects we meet with also near the lake Issyk-kul.
To south from these districts a rich locality of Lower Liassic insects is discovered in 1927 by N. V. S h a b а г о v. These insects are described by a Martynov (except Coleoptera). Similar insects of the same age are known also from Kisyl-kija (last from Shurab). Many elements of the fauna of these localities are evidently allied to Upper Triassic ones from Australia. Several Lower Liassic insects are described also from Chelia-binsk district from the villages Sukhomesovskoe and Korkinskoe.
The locality at Ustj-Balei, not very far from Irkutzk, is known long ago. It procured about two dozens of remains of insects. The rock is represented with yellowish or brownish very thiny sandstone, and remains of insects are here not unfrequently perfect. Age of this locality we estimate as Upper
most Liassic. Three genera of Paleontinidae are described from this locality, and even more archaic genera are known from Shurab and from the Triassic of Australia. This testifies for the hypothesis that this family has been developed earlier in the eastern lands and in Australia, and only at the end of the Liassic has invaded W. Europe.
4—5 points with fossil insects are known from Transbaicalia. Few species are known from them, but larvae of large Ephemeropsis trisetalis E i c h w. are recorded almost from each of them (as well as from the localities in Mongolia). Frequent occurence of the larvae of Ephemeroptera points to the presence in Uppermost Jurassic of Transbaicalia and Mongolia of numerens occasionally large lakes.
At present two triassic localities are known in the asiatic part of the USSR. One of them is situated south of Shurab and contains small Coleoptera, some Blattodea and other forms. These insects are examined, but not yet described by A. M a r t у n о v; their age is probably Lower or Middle Triassic.
Second locality we meet with in the Kuznetzk basin, in the district of Babij Kamenj. Few insects andMyriopods are described by Martynov, who indicated that the Ademosinoides asiaticus closely resembles Upper Tiassic species of Ademosynoides and Ademosyne from Australia.
Age of this locality is probably Upper or Middle Triassic. Nothing is known on cretaceus insects or localities of USSR.
TERTIARY LOCALITIES
Eocene insects unknown.
To the Upper Oligocene localities may be referred two asiatic ones, at the river Kudia, a tributary of Amagu, in the maritime province, and in the district Ashutas, near the river Black Irtysh, west Siberia. An amount of fossil insects mere recorded from Kudia-river and described by Prof. T. D. Cockerell, but he was here only three days, therefore many new materials may be collected here; unfortunately, the locality is too far for the scientists of Moscow or Leningrad. Cockerell described here various Diptera, Mecoptera, Hymenoptera, Trichoptera, Homoptera, Orthoptera.
Ashutas is also not easily accessible, but this is a remarkable locality. Sandy rock is composed of very thiny grains, therefore preservation of remains of insects and plants is usually fine.
Insects were examined by M a г t у n о v, who described 19 species mainly Odonata. Fauna proved to reveal affinities with the recent Indian and recent and miocene fauna of America (Odonata). Climate was warm enough, and the presence of two species of Termites concur such a conclusion. This locality appears to belong to the Uppermost Oligocene or to Lower Miocene.
Other localities are situated mainly in various parts of Caucasus (s. 1.), — Upper Miocene shales at Voroshilovsk Kavkazsky (two fine remains), Upper Miocene sandstones of Apsheron peninsule, Upper Miocene shales in Kuban district (auts.), Upper Miocene locality in W. Georgia with beetles. However from these points are recorded but 1—3 species; beetles from Georgia are not yet de-scribed.
A new and, apparently, rich locality of Uppermost Miocene insects is newly (1936) discovered by Dr. Petrosjan west from Issyk-kul. There are many Diptera (about 19 species), some Homoptera, Aphidae, Coleoptera, many specimens of Libellulidae, gen. sp. Description of these insects is not yet published. There are two more old and, perhaps, doubtful localities in Sakhalin and Ufa disctrict, from which two single insects were indicated by Heer and К a w a 11.
Dr. B. Rohdendorf has indicated (1937) a Boletina sp. from Arkhangelsk district; it testifies for very young age this locality, perhaps Upper Miocene. Thus, tertiary insects of USSR are yet very unsufficiently explored.
АКАДЕМИЯ НАУК СССР
1938. ТРУДЫ ПАЛЕОНТОЛОГИЧЕСКОГО ИНСТИТУТА, том VII, вып. 3
Б. Б. РОДЕНДОРФ ДВУКРЫЛЫЕ НАСЕКОМЫЕ МЕЗОЗОЯ КАРА-ТАУ. I. BRACH Y-СЕВА и ЧАСТЬ NEMATOCERA
(с 21 рис.)
В. В. ROHDENDORF
MESOZOISCHE DIPTEREN AUS KARA-TAU. I. BRACHYCEREN UND EINIGE NEMATOCEREN
(mit 21 Abb.)
Настоящая статья является первой в серии исследований, посвященных описанию остатков двукрылых насекомых из юрских сланцев хребта Кара-тау в Южном Казахстане. Возраст этих отложений соответствует нижнему доггеру (Мартынов, 1938). Основными материалами по Diptera Кара-тау является коллекция, собранная А. В. Мартыновы м в окрестностях с. Галкина в 1925 г., в количестве, превышающем 120 номеров. Большой интерес этой коллекции, имеющей почти уникальный характер, станет ясным, если принять во внимание то обстоятельство, что до сих пор известно лишь около 40 видов двукрылых из мезозоя, притом исключительно из юры. Кроме того, в это число входит целый ряд проблематичных видов (почти треть!), описанных по фрагментарным остаткам и не могущих быть определенными более точно. Остальные виды в своем большинстве принадлежат к подотряду Nematocera — длинноусым двукрылым, притом, главным образом, к группе Tipuloidea. Лишь один единственный известный представитель короткоусых — Brachycera — из юры отнесен Handlirsch (1906) к сем. Nemestrinidae\ это определение может возбуждать сомнения вследствие очень несовершенного описания и изображения остатков мухи.
Обращаясь теперь к насекомым из Кара-тау, можно отметить богатство видового состава, во много раз превышающее все ранее известное по другим юрским фаунам. Не касаясь подотряда длинноусых, обработка которого еще не закончена, достаточно сказать, что в юре Кара-тау найдено 8 видов короткоусых двукрылых (Brachycera), относящихся к 5 семействам. Остатки короткоусых, составляют лишь незначительное меньшинство всех материалов; несомненно, по окончании обработки всех длинноусых следует ожидать значительно большего разнообразия фауны этой группы.
В настоящей статье описываются из материалов ПИН почти все представители короткоусых и лишь часть длинноусых двукрылых из юры Кара-тау; часть, посвященная короткоусым, является более или менее законченной, почему и ставится в начале, не согласно с системой отряда.
Список описанных форм
Подотряд Brachycera
Сем. Stratlomyidae
Подсем.?
1. Archisargus pulcher gen. sp. nov.
Подсем. Palaeostratiomyiinae subfam. nov.
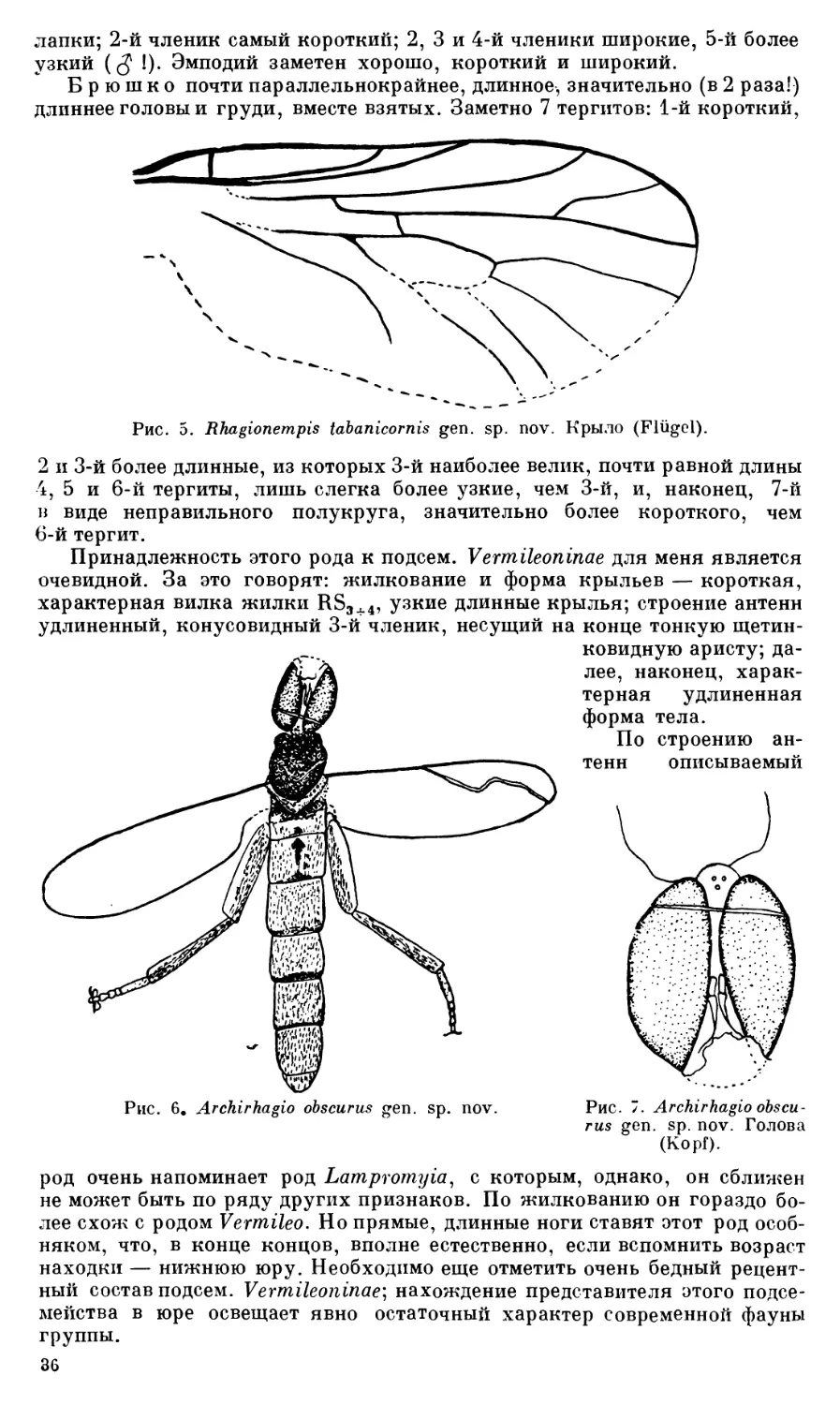
2. Palaeostratiomyia pygmaea gen. sp. nov. Сем. Rhagionidae
Подсем. RAiagionempidinhe subfam. nov.
3. Rhagionempis tabanicornis gen. sp. nov. Подсем. Vermileoninae
4. Archirhagio obscuvus gen. sp. nov. Подсем. Protorhagioninae subfam. nov.
5. Protorhagio capitatus gen. sp. nov.
Сем. Protocyrtidae fam. nov.
6. Protocyrtus jurassicus gen. sp. nov.
Сем. Omphalidae
7. Protomphale martynovi gen. sp. nov.
Сем. Archiphoridae fam. nov.
8. Archiphora ancestrix gen. sp. nov.
Подотряд Уematocera
Сем. Allactoiieuridae fain. nov.
9. Pleciojungivora latipennis gen. sp. nov.
10. Allactoneu rites jurassicus gen. sp. nov.
11. Pleciomima sepulta gen. sp. nov.
12. Fungivorites latimedius gen. sp. nov.
Подсем. Antiquamediinae subfam. nov.
13. Antiquamedia tenuipes gen. sp. nov. Подсем. Antejungivorinae subfam. nov.
14. Antejungivora prima gen. sp. nov. Сем. Penthetriidae
15. Mesoplecia jurassica gen. sp, nov.
ОПИСАНИЯ
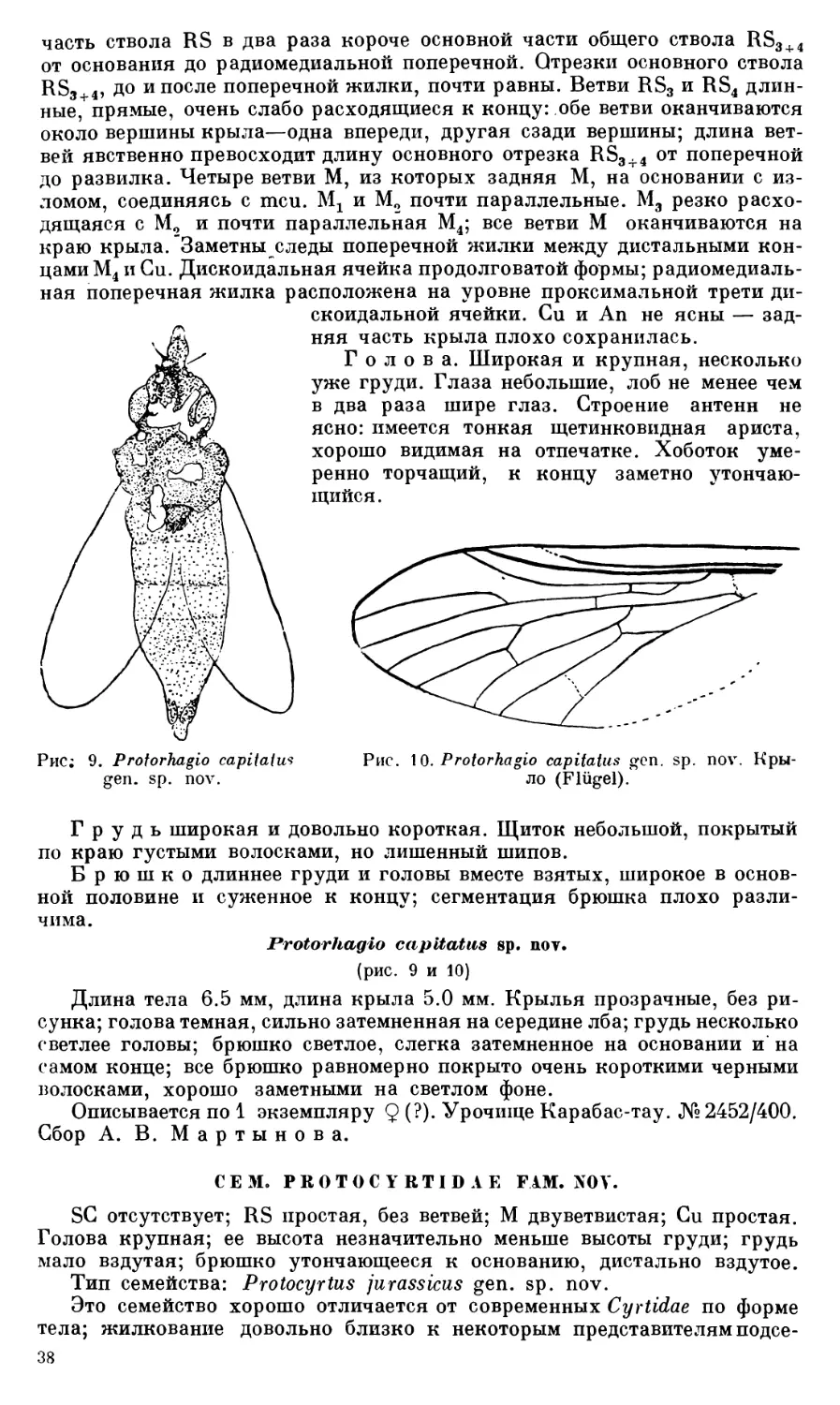
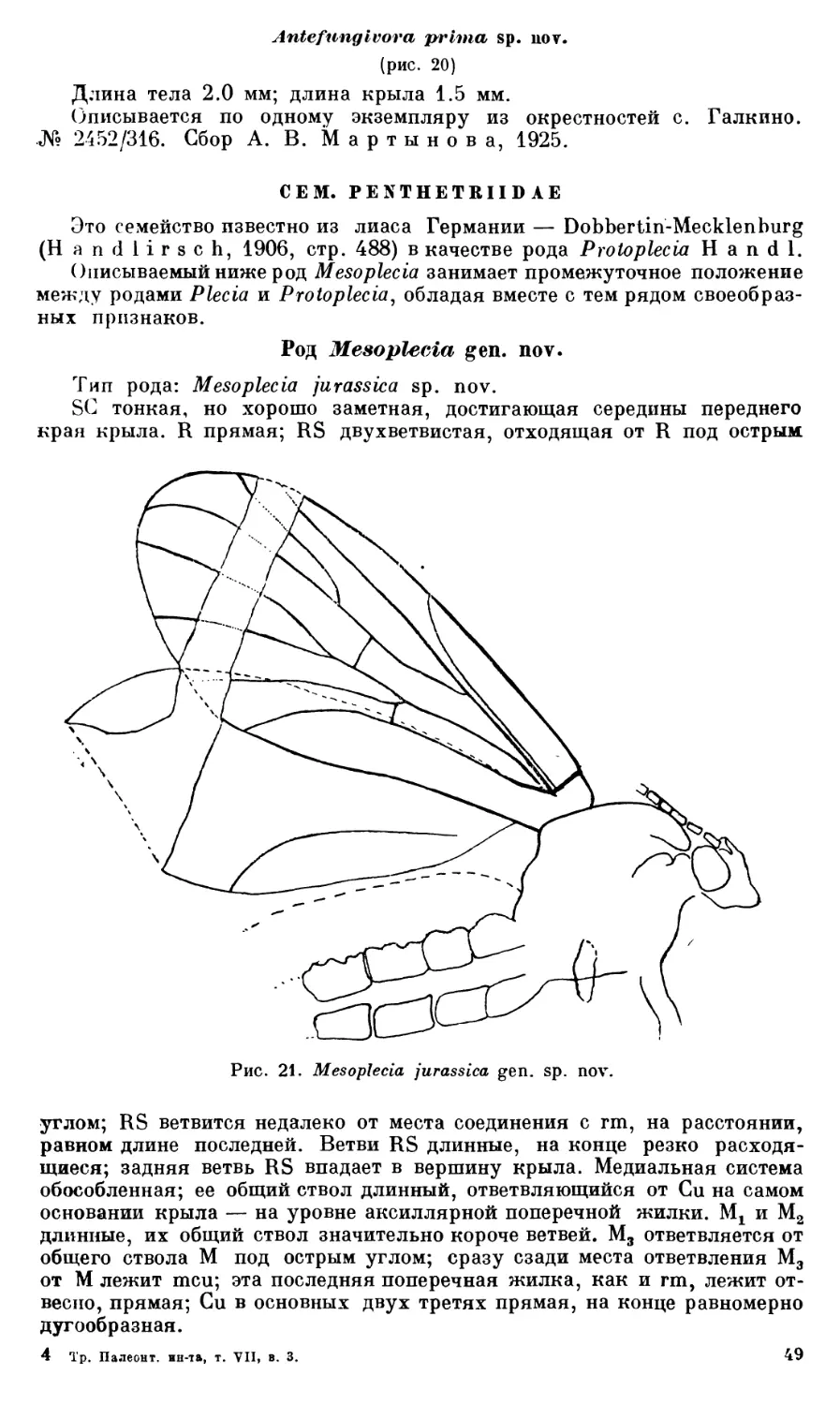
ПОДОТРЯД BRACHYCERA-КОРОТКОУСЫЕ
Описываемые ниже представители подотряда короткоусых двукрылых принадлежат к семействам, еще не известным для мезозоя.
СЕМ. S TRA TIOM Y1D АЕ
Род Archisargus gen. nov.
Тип рода: Archisargus pulcher sp. nov.
Крыл о. Форма крыла очень длинная и узкая, на основании сильно суженная; SC неясная; R длинная, конец ее значительно заходит за середину крыла; RS с 4—5 ветвями, из которых лишь RSX и RS2 и их общий ствол сильно склеротизированы. RS3 и RS5 более нежные, очень слабо заметные; жилка RS4 видна лишь на правом крыле — на левом не ясна; М и Си неясные; костальная жилка хорошо заметна на переднем крае, примерно до места впадения в нее жилки RS3.
Грудь небольшая, сверху квадратная, слегка вытянутая, с округленными углами; щиток короткий, почти прямоугольный, без шипов по углам; ноги тонкие, бедра 3 длинные, невздутые; голень 3 в два раза тоньше бедра.
Голова крупная; глаза овальные, на темени соприкасающиеся; глазки не ясны; передняя часть головы не сохранилась.
Брюшко длинное, повидимому, раза в два длиннее головы и грудиг вместе взятых: сохранились лишь четыре базальных тергита; 1 тергит заметно уже остальных, спереди суженный.
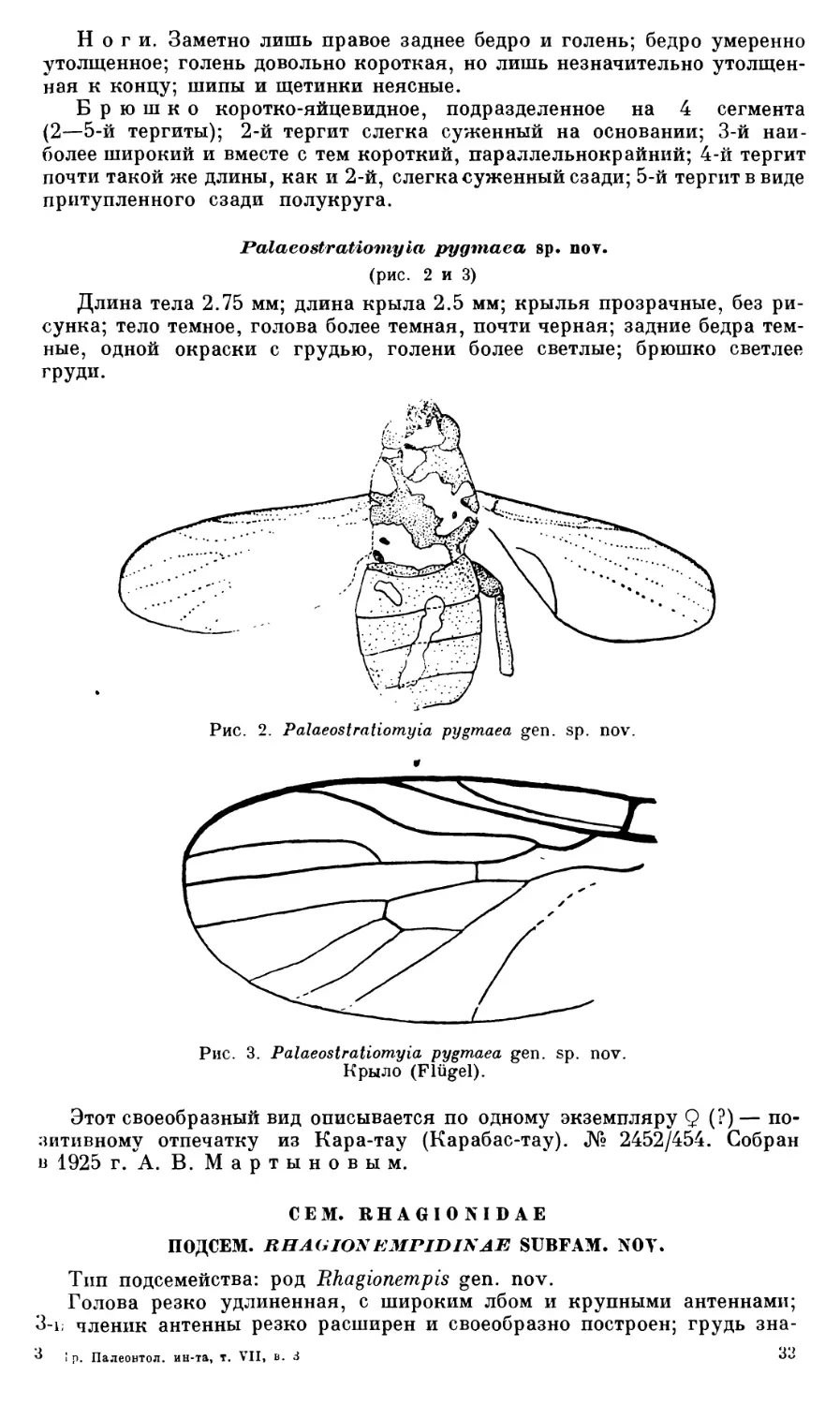
Archlsargus pulcher sp. nov.
(рис. 1)
Длина крыла 16.0 мм; длина остатков головы, груди и остатков брюшка 16.0 мм; тергиты брюшка с хорошо заметным рисунком: 1-й тергит с треугольным темным пятном на основании спинки, не доходящим до заднего края тергита; 2-й тергит с продолговатым, заостренным сзади пятном по средней линии спинки и каймой по заднему краю тергита, имеющей угловатый, направленный вперед выступ в середине; 3-й тергит без серединного переднего пятна, лишь с заднекрайней, спереди угловато выступающей полосой; 4-й тергит также окрашен.
Описывается по одному " $ из юры Кара-тау: урочище Качкар-Ата (колл. ПИН; сборщик неизвестен).
Описываемый несомненный представитель сем. Stratiomyidae с трудом может быть сближен с какими-либо рецентными группами семейства. Крайне вытянутая форма крыла, неясность субкостальной жилки, простое строение радиальной системы, повидимому, также отсутствие замкнутых ячеек, — все эти признаки резко отличают род Archisargus от известных представителей семейства Stratiomyidae.
ПОДСЕМ. PALAROSTRATIOMYIINAE SUBFAM. NOV.
Тип подсемейства: род Palaeostratiomyia gen. nov.
С по краю крыла достигает лишь его вершины, до конца жилки RS3; задний край крыла без жилки. RS ответвляется от R на самом основании крыла, лишь немного дистальнее плечевой поперечной жилки. Ветви RSX и RS2 слиты, не обособлены. Ветви RS3 и RS4 параллельны, образуя очень длинную и узкую вилку. Медиальные жилки образуют небольшую дискоидальную ячейку; жилка М4 отходит от ячейки на основании без излома; место соединения ее с Си не ясно.
Крылья притупленные, довольно широкие, значительно длиннее брюшка.
Тело короткое и широкое; голова незначительно уже груди, широкая, с умеренно выступающим лицом; лоб на темени шире глаза 'Q?); брюшк© коротко-яйцевидное, лишь с 4 видимыми сегментами — 2, 3, 4 и 5-м тер-гитами.
Род Palaeostratiomyia gen. nov., послуживший типом для установления особого подсемейства, обнаруживает целый ряд своеобразных черт, не позволяющих его поместить в рамки известных подсемейств Stratiomyidae. Наиболее характерно для него очень короткое 4-членистое брюшко и своеобразное, богатое жилкование, не обнаруживающее тенденции к сокращению и сдвигу жилок к переднему краю, что является правилом почти для всех известных современных групп семейства. Современные представители подсем. Beridinae, обладающие слабо редуцированным жилкованием, все же резко отличаются от описываемых Palaeostratiomyiinae иным жилкованием; наблюдается, хотя и слабый, сдвиг радиальной системы к переднему краю и редукция медиальных ветвей, в особенности строением, брюшка, которое является удлиненным и состоящим не менее чем из 7 хорошо заметных сегментов. Строение брюшка резко отличает описываемое подсемейство от Beridinae и с ясностью указывает на отсутствие прямых филогенетических связей между этими двумя группами. Наибольшее сходство по строению тела Palaeostratiomyiinae имеют со Stratiomyinae и Сlitellariinae, в особенности с последним, вследствие строения жилки М4, которая у Stratiomyinae обособлена от остальной медиальной сгстемы, неся на основании резкий излом. Все эти соображения позволяют нам в известной мере считать Palaeostratiomyiinae отчасти анцестральной группой или, во всяком случае, ставить ее в большой близости от исходного ствола подсемейств Сlitellariinae и Stratiomyinae. Плохая сохранность ископаемого, отсутствие данных о строении головы, именно антенн, не позволяет более точно выяснить генетические отношения.
Род Palaeostratiomyia gen» nov.
Тип рода: Palaeostratiomyiia pygmaea sp. nov.
Крыло. SC длинная, не особенно крепкая, достающая середины крыла. R крепкая, заходящая за середину крыла, прямая. Имеются трж ветви RS — RS1+2, RS3 и RS4; RS1+2 ответвляется от общего ствола RS примерно на уровне середины SC; RS1+2 резко волнообразно изогнутая; RS3 и RS4 очень характерной формы, длинные, почти точно параллельные, друг от друга ответвляющиеся на уровне конца дискоидальной ячейки, лишь едва далее конца R; основная часть ствола RS34-RS4, между его развилком и радиомедиальной жилкой, почти в 2 раза короче вилки; обе ветви вилки расположены близко друг от друга, образуя очень узкую ячейку. Поперечная гш короткая, расположенная ближе к основанию крыла, чем середина общего ствола RS3+RS4 и дискоидальной ячейки. Медиальные жилки в числе 4 ветвей, отходящих от дискоидальной ячейки; М4 прямая, составляющая продолжение общего ствола М, параллельная RS4; М2 и М3 короткие и слегка изогнутые, взаимно более или менее параллельные, но расположенные под углом к Мх; М2 ответвляется от заднего угла дискоидальной ячейки, слегка сходится с М3; жилки М2, М3 и М4 на концах нежные и слабые, плохо заметные; дискоидальная ячейка средней величины, вытянутая, неправильной пятиугольной формы; Си и Ап неясные: заметен лишь конец Си (?) в виде дуговидной жилки, впадающей в край крыла на значительном расстоянии от М4.
Голова широкая и короткая; лоб широкий, примерно в раза шире глаза; лицо умеренно выступающее; антенны и хоботок не сохранились.
Грудь массивная, плохо сохранившаяся; строение ее не ясно. •
Ноги. Заметно лишь правое заднее бедро и голень; бедро умеренно утолщенное; голень довольно короткая, но лишь незначительно утолщенная к концу; шипы и щетинки неясные.
Брюшко коротко-яйцевидное, подразделенное на 4 сегмента (2—5-й тергиты); 2-й тергит слегка суженный на основании; 3-й наиболее широкий и вместе с тем короткий, параллельнокрайний; 4-й тергит почти такой же длины, как и 2-й, слегка суженный сзади; 5-й тергит в виде притупленного сзади полукруга.
Palaeo stг atiomyia pygmaea sp. nov.
(рис. 2 и 3)
Длина тела 2.75 мм; длина крыла 2.5 мм; крылья прозрачные, без рисунка; тело темное, голова более темная, почти черная; задние бедра темные, одной окраски с грудью, голени более светлые; брюшко светлее
Рис. 2. Palaeostratiomyia pygmaea gen. sp. nov.
Рис. 3. Palaeostratiomyia pygmaea gen. sp. nov. Крыло (Flugel).
Этот своеобразный вид описывается по одному экземпляру Q (?) — по-зитивному отпечатку из Кара-тау (Карабас-тау). № 2452/454. Собран в 1925 г. А. В. Мартыновым.
СЕМ. RHAGIONIDAE
ПОДСЕМ. RHAGION EMPIDINAE SUBFAM. NOT.
Тип подсемейства: род Rhagionempis gen. nov.
Голова резко удлиненная, с широким лбом и крупными антеннами; 3-1; членик антенны резко расширен и своеобразно построен; грудь зна-
3 i р. Палеонтол. ин-та, т. VII, в. 3 33
чительно шире головы (при взгляде сверху); развилок RS3+RS4 короткий, его ветви резко расходятся в стороны; RS4 впадает в вершину крыла; имеются четыре ветви М. Жилка С доходит лишь до вершины крыла, до места впадения RS4.
Род Khagionempis gen. nov.
Тип рода: Rhagionempis tabanicornis sp. nov.
С доходит лишь до RS4; SC довольно крепкая, слабо изогнутая на конце; R прямая, слабо изогнутая, крепкая, соединяющаяся с С примерно на середине крыла; RS1+2 почти прямая, слегка дугообразная в последней четверти, ответвление этой жилки от R не ясно; общий ствол радиального сектора длинный — основание его не ясно; жилка RS ответвляется от RS1+2 значительно проксимальнее середины крыла; развилок RS3 + 4 короткий, менее отрезка RS3 + 4 от поперечной RM до развилка; жилка RS4 является прямым продолжением общего ствола RS3^.4, почти прямая; RS3 изогнутая; два последних отрезка С почти равной длины; имеются четыре ветви М — места их ответвления друг от друга не ясны; проксимальный угол дискоидальной ячейки острый, расположен почти на уровне ответвления RS3+4 от RS1+2. Крыло довольно широкое и притупленное иа вершине; проксимальная часть костального поля резко суженная.
Голова. Узкая, значительно уже груди, сильно вытянутая; глаза небольшие, в два раза уже лба (при взгляде сверху); глазки имеются — передний глазок значительно мельче задних. Лоб очень широкий и длинный; антенны хорошо развитые, длинные; 1 и 2-й членики антенн короткие, небольшие и закругленные; дистальная часть антенн состоит из крупного, угловатого 3-го членика, несущего хорошо выраженные бугорки, и концевой аристы, сильно заостренной и тонкой, состоящей из неясных двух члеников.
Тело довольно короткое и толстое; форма груди неясная, так как, повидпмому, у сохранившегося ископаемого она раздавлена; брюшко состоит, повидимому, из 5 довольно длинных сегментов.
Ноги почти не сохранились; заметны остатки задней (средней?) левой пары, в виде конца тонкого бедра и довольно тонкой голени; шипы и шпоры не ясны.
Взаимоотношения этого рода с другими близкими очень не ясны. Плохая сохранность задней части крыльев, к сожалению, не позволяет точно исследовать жилкование этой части крыла. Но характер R и число ветвей М достаточно ясны, и с определенностью можно утверждать о принадлежности этого рода к группе Eremochaeta. Дальнейшее уточнение положения этой формы — принадлежность ее к тому или иному семейству — наталкивается на большие трудности. Недостаточное знание строения тела — брюшка, груди и ног — и вместе с тем своеобразное устройство головы — ее необычайно удлиненная форма, — все это заставляет нас считать этот род представителем совсем особой группы, отличной от известных современных семейств. Наибольшее сходство по жилкованию описываемый род имеет с настоящими Rhagionidae, именно родом Spania. Все же другие особенности строения, упомянутые выше, не позволяют нам безоговорочно поместить этот род в семейство Rhagionidae^ и для него устанавливается особое подсемейство. Эта новая группа — Rhagionem-pidinae, провизорно названная подсемейством, обнаруживает ряд связен с другими семействами. Наиболее интересно строение антенн, которое живо напоминает строение их у представителей подсемейства Tabaninae. Из особенностей жилкования характерно окончание костальной жилки на вершине крыла; этот признак сближает группу с сем. Stratwmyidae. Наконец, необходимо отметить габитуальное сходство Rhagionempis с представителями сем. Empididae'. узкая голова и широкая грудь. Это сходство, правда, может быть лишь следствием сильного раздавливания этого ин-34
дивида при его гибели, на что указывают явно отодвинутые в стороны основания крыльев и неясные очертания груди. Кроме того, антенны могут также несколько напоминать некоторых Empididoe.
Rhaglonempis tabanicornis sp. nov.
(рис. 4 и 5)
Темно окрашенный вид, крылья прозрачные, без рисунка; антенны светло окрашенные, бледные; бедра и голени задней (средней?) пары ног светлые. Длина тела 3.75 мм, крыла — 3.5 мм.
Описывается по 1Q из юрских сланцев Карабас-тау. № 979/2465, Сбор А. В. Мартынова, 1925.
ПОДСЕМ. VHRMILEONINAK Род Archirhagio gen. nov.
Тип рода: Archirhagio obscuras sp. nov.
Крыло. С продолжается до середины заднего края крыла, до места соединения с ним жилки Си. SC длинная, достигающая до апикальной
трети крыла. R крепкая и длинная, почти прямая; сектор ответвляется
от него примерно на середине крыла. Имеются три ветви радиального сектора — RS1+2, RS3 и RS4. Ответвление передней ветви расположено примерно на середине, между основанием сектора и местом соединения с ним радиомедиальной поперечиной. Развилок жилки RS3+4 короткий — обе ее ветви RS3 и RS4 резко расходятся, так что RS впадает в передний край, a RS4 в задний край крыла; длина RS4 ясно короче общего ствола KS3 4, параллельна Мг Имеется хорошо выраженная дискоидальная ячейка. Четыре ветвиМ, из которых три передние отходят от дискоидаль-ной ячейки, а последняя, Мх, связанная поперечной шеи с кубитальной жилкой, имеет видоизмененное основание, превращенное в поперечную жилку. Си почти прямая и очень крепкая, особенно до соединения с жилкой meu; ее дистальная часть изогнутая и соединяющаяся с С недалеко от Ап; кубитальная ячейка открытая. Апх почти совсем прямая, довольно крепкая; Ап2 слабая и изогнутая.
Голова. Такой же ширины или шире груди. Глаза очень крупные, почти соприкасающиеся. Простые глазки в числе трех, очень мелкие, но ясные; глазковый треугольник не обособлен,
но теменная часть лба ясно выступает назад. Ан- рпс. 4. Rhagionempis ta-тенны короткие, довольно тонкие; различим ци- banicomis gen. sp. nov. линдрический 2-й членик, который в 2%—3 раза
длиннее своей толщины, и конусовидный 3-й членик, на конце несущий
тонкую, лишенную волосков и ясной членистости аристу. Антенны рас-
положены примерно на середине головы.
Грудь небольшой величины, сильно склеротизированная; щиток без шипов по краю.
Horn (задние) крепкие и довольно длинные. Задние бедра широкие, повидимому, сплющены с боков, более или менее параллельнокрайние. Голени задние длиннее бедер, на своем основании более чем в 2 раза тоньше последних, к концу слабо, но заметно утолщающиеся. Задние лапки лишь немного более 1/2 длины голеней. 1-й членик лапок равен половине длины
з*
35
лапки; 2-й членик самый короткий; 2, 3 и 4-й членики широкие, 5-й более узкий ((J !). Эмподий заметен хорошо, короткий и широкий.
Брюшко почти параллельнокрайнее, длинное, значительно (в 2 раза!) длиннее головы и груди, вместе взятых. Заметно 7 тергитов: 1-й короткий,
Рис. 5. Rhagionempis tabanicornis gen. sp. nov. Крыло (Flugel).
2 и 3-й более длинные, из которых 3-й наиболее велик, почти равной длины 4, 5 и 6-й тергиты, лишь слегка более узкие, чем 3-й, и, наконец, 7-й в виде неправильного полукруга, значительно более короткого, чем 6-й тергит.
Принадлежность этого рода к подсем. Vermileoninae для меня является очевидной. За это говорят: жилкование и форма крыльев — короткая, характерная вилка жилки RS3jl4, узкие длинные крылья; строение антенн удлиненный, конусовидный 3-й членик, несущий на конце тонкую щетин
Рис. 6. Archirhagio obscurus gen. sp. nov.
ковидную аристу; далее, наконец, характерная удлиненная форма тела.
По строению антенн описываемый
Рис. ". Archirhagio obscurus gen. sp. nov. Голова (Kopf).
род очень напоминает род Lampromyia, с которым, однако, он сближен не может быть по ряду других признаков. По жилкованию он гораздо более схож с родом Vermileo. Но прямые, длинные ноги ставят этот род особняком, что, в конце концов, вполне естественно, если вспомнить возраст находки — нижнюю юру. Необходимо еще отметить очень бедный рецент-ный состав подсем. V ermileoninae, нахождение представителя этого подсемейства в юре освещает явно остаточный характер современной фауны группы.
Archirhaglo obscuras sp. nov.
(рис. 6, 7 и 8)
Голова и грудь темно окрашенные; крылья прозрачные, с хорошо заметным, желто-коричневым жилкованием; антенны черные; задние ноги желтые; бёдра, кроме вентральной поверхности, покрыты короткими, тонкими, черными волосками, голени целиком покрыты волосками; 1-й членик лапок с редкими, плохо заметными волосками; все брюшко, так же как и голени и бедра, покрыто короткими волосками; 1-й тергит брюшка в основной половине темный, с так же окрашенным небольшим пятном на середине заднего края; 2-й тергит с продолговатым срединным пятном, постепенно суженным по направлению к заднему краю; задний край этого тергита на середине затемненный; темная кайма постепенно сходит
Рис. 8. Archirhagio obscurus gen. sp. nov. Крыло (Flugel).
на-нет к бокам; 3 и 4-й тергиты по заднему краю с темными, узкими перевязками, равномерной ширины; 5-й тергит с более узкой цдймой; 6 и 7-й тергиты одноцветные, светлые. Длина тела 21.5 мм, длина крыла 13 мм.
Описывается по 1 из юры окоестностей с. Галкино. № 2452/490.
Сбор А. В. Мар т ы н о в а.
ПОДСЕМ. VROTORHAGIONINAE SLBFAM. NOV.
Тип подсемейства: Protorhagio gen. nov.
Основной ствол RS очень короткий, более чем в 2 раза короче отрезка RS3 4 от основания до радиомедиальной поперечной; сумма этого отрезка и основного ствола RS примерно равна длине дискоидальной ячейки. Голова широкая. Тело довольно массивное и короткое; брюшко утончающееся лишь в апикальной трети. Строение ног не известно.
Описываемое новое подсемейство несомненно представляет своеобразную группу, отличную от всех известных представителей Rhagionidae. Своеобразное жилкование — характер основной части RS — резко отличает род Protorhagio от всех близких групп. Вместе с тем, плохая сохранность головы и ее придатков, задней части крыльев и ног не позволяет исследовать эти части тела, имеющие очень важное значение для выяснения генетических связей. Характер дистальных частей RS и общая форма тела приближают этот род к группе настоящих Rhagioninae (Chrysopi-lini и Rhagionini).
Род Protorhagio gen. nov.
Тип рода: Protorhagio capitatus sp. nova.
Крыло. Умеренно узкое, короче брюшка. Жилки крыла слабые. С заходит за вершину крыла; место окончания жилки не ясно. SC длинная, слабая на конце, почти прямая. R очень крепкая и длинная, дуговидно изогнутая. RS ответвляется почти на самом основании крыла, с тремя ветвями. RS1+2 дуговидно изогнутая; конец этой жилки заметно сближен к концом R, радиальная ячейка суженная к вершине. Основная
часть ствола RS в два раза короче основной части общего ствола RS3+4 от основания до радиомедиальной поперечной. Отрезки основного ствола RS3 + 4, до и после поперечной жилки, почти равны. Ветви RS3 и RS4 длинные, прямые, очень слабо расходящиеся к концу: обе ветви оканчиваются
около вершины крыла—одна впереди, другая сзади вершины; длина ветвей явственно превосходит длину основного отрезка RS3+4 от поперечной до развилка. Четыре ветви М, из которых задняя М, на основании с изломом, соединяясь с шеи. Мх и М2 почти параллельные. М3 резко расходящаяся с М2 и почти параллельная М4; все ветви М оканчиваются на краю крыла. Заметны следы поперечной жилки между дистальными концами М4 и Си. Дискоидальная ячейка продолговатой формы; радиомедиаль-
ная поперечная жилка расположена на уровне проксимальной трети дискоидальной ячейки. Си и Ап не ясны — зад-
Рис; 9. Protorhagio capitatus gen. sp. nov.
няя часть крыла плохо сохранилась.
Голова. Широкая и крупная, несколько уже груди. Глаза небольшие, лоб не менее чем в два раза шире глаз. Строение антенн не ясно: имеется тонкая щетинковидная ариста, хорошо видимая на отпечатке. Хоботок умеренно торчащий, к концу заметно утончающийся.
Рис. 10. Pro to r ha gio capitatus gen. sp. nov. Крыло (Flugel).
Грудь широкая и довольно короткая. Щиток небольшой, покрытый по краю густыми волосками, но лишенный шипов.
Брюшко длиннее груди и головы вместе взятых, широкое в основной половине и суженное к концу; сегментация брюшка плохо различима.
Protorhagio capitatus sp. nov.
(рис. 9 и 10)
Длина тела 6.5 мм, длина крыла 5.0 мм. Крылья прозрачные, без рисунка; голова темная, сильно затемненная на середине лба; грудь несколько светлее головы; брюшко светлое, слегка затемненное на основании и на самом конце; все брюшко равномерно покрыто очень короткими черными волосками, хорошо заметными на светлом фоне.
Описывается по 1 экземпляру 9 (?). Урочище Карабас-тау. № 2452/400. Сбор А. В. Мартынова.
СЕМ. PROTOCYRTIDAE FAM. NOV.
SC отсутствует; RS простая, без ветвей; М двуветвистая; Си простая. Голова крупная; ее высота незначительно меньше высоты груди; грудь мало вздутая; брюшко утончающееся к основанию, дистально вздутое.
Тип семейства: Protocyrtus jurassicus gen. sp. nov.
Это семейство хорошо отличается от современных Cyrtidae по форме тела; жилкование довольно близко к некоторым представителям подсе-38
мейства Cyrtinae — род Oncodes La t r. — и подсемейству Panopinae — роду Corononcodes S p e i s e г. Последнее подсемейство, повидпмому, наиболее близко к описываемому юрскому. Некоторые черты сходства — форма головы и ее положение — напоминают семейство Omphralidae.
Род Protocyrtus gen. nov.
Тип рода: Protocyrtus jurassicus sp. nov.
R равна 3/5 длины крыла, прямая; RS резко отходит от R, дугообраз-
ная; гт косо расположенная, являющаяся прямым продолжением основной части передней ветви М и дистальной части RS; М ветвится ближе середины крыла; ее основная часть короткая; ветви М слабо изогнутые
расходящиеся; mcu длинная, косо расположенная; Си равномерно изогнутая в виде дуги.
Голова. Глаза небольшие, занимающие меньше половины головы, голова округлая в профиль, слегка выступающая спереди; антенны расположены под глазами, небольшие; щеки очень широкие, вздутые сзади.
Грудь с небольшим вздутьем перед основанием крыльев, вытянутая, более низкая сзади.
Ноги короткие, заднее бедро
Рис. 1 'rotocyrtus jurassicus gen. sp. nov.
и голень параллельнокрайние,
почти равной толщины; голень несколько короче бедра; задняя лапка незначительно длиннее голени, заметно ее тоньше; 1 и 2-й членики лапки
самые крупные; 4-й членик наиболее короткий; коготки короткие. Б р ю ш к о. Сегменты брюшка и его придатка не ясны.
Protocyrtus jurassicus sp. nov.
(рис. 11)
Длина тела 5.0 мм, длина крыла 3.5 мм; крыло короче брюшка; гт равна основной части передней ветви М и вместе с тем отрезку М от ее ветвления до mcu; mcu длинная, раза в 2х/2 длиннее гт. Описывается по Q из окрестностей с. Михайловки. № 2231. Сбор Турутановой-Кетовой.
СЕМ. OMPHRALIDAE
Род Protomphrale gen. nov.
Тип рода: Protomphrale martynovi sp. nov.
Крыло. Значительно короче брюшка, SC короткая, повидимому, свободная; костальное поле широкое — R изогнута дугообразно; RS с 4 ветвями, равномерно дихотамически ответвляющимися от общего ствола; место ответвления RS от R не ясно; отрезки костальной жилки между ветвями R и RS почти равной длины, лишь отрезок между RS3 и RS4 значительно короче предыдущих; другие жилки крыла плохо сохранились — на отпечатке имеются лишь неясные следы. Голова небольшой величины; глаза очень малы, неправильно эллиптической формы; жужжальца короткие, но заметные. Голова выше глаз резко вздутая; антенны длинные, длиннее головы примерно в 1у2 раза; 2-й членик антенн короткий, короче своей высоты; 1-й членик в 2 раза длиннее 2-го, слегка вздутый; 3-й членик самый длинный, почти в 2 раза длиннее двух основных члеников, на конце ясно заостренный.
Грудь массивная, умеренно выпуклая.
Ноги короткие и тонкие; бедра наиболее толстые, короче более тонких голеней; задние лапки почти в 2 раза короче голени; средние лапки лишь слегка короче голени; задняя голень наиболее толста на ос-
Рис 12. Protomphrale marlynovi gen. sp. nov.
новании, постепенно утончаясь к концу; средняя голень параллельнокрайняя.
Брюшко длиннее головы и груди, вместе взятых, постепенно утончающееся к концу; хорошо заметны 8 сегментов и широкие межсегментные перепонки между 2 и 3, 3 и 4, 4 и 5, 5 и 6-м; последний сегмент брюшка закругленный.
Protomphrale martynovi sp. nov.
(рис. 12)
Длина тела 3.5 мм; длина крыла 1.75 мм; окраска
светлая.
Описываемый несомненный представитель сем. Ornphralidae заметно
отличается от всех известных современных родов присутствием хорошо развитых четырех ветвей RS, очень небольшими глазами и удлиненными антеннами.
Описывается по одному экземпляру из окрестностей с. Галкино. № 2452/366. Сбор А. В. Мартынова, 1925.
С YCLORRHAPHA
СЕМ. ARCHIPHORI DAE FAM. NOV.
Тип семейства: род Archiphora gen. nov
Голова небольшая, округлая; строение антенн не известно; глаза неправильной формы; хоботок, повидимому, умеренно удлиненный; щетинки на голове не ясны.
Грудь короткая, не резко вздутая, в профиль значительно превос ходящая по высоте голову; ноги слабые, не особенно длинные; крылья довольно узкие, с характерным жилкованием: SC отсутствует, R, RSt и RS.J сильно утолщены и сдвинуты к переднему краю; RS3+4 простая; медиальная система хорошо развита: имеется М2, расположенная в виде поперечной ветви между RS3+4 и М2; имеются, кроме того, задняя ветвь М4 в виде развилка с Си и поперечные: rm, tp и шеи; две жилки Ап.
Брюшко конической формы и состоит из 9 сегментов.
Описываемая группа мух, несомненно родственных современным Phoridae, обнаруживает ряд черт, заставляющих ставить ее в непосредственной близости от исходных форм Phoridae*, возможно даже, что Archipho-ridae явились непосредственными предками названного рецентного семейства.
Современные Phoridae очень резко обособлены от остальных семейств Diptera, хорошо характеризуясь крайней специализацией жилкования, сильным развитием ног и щетинок на теле. Лишь единственный австралийский род Sciadocera резко отличается от остальных по более примитивному жилкованию и, вероятно, должен быть выделен из сем. Phoridae в особую группу, лишь ошибочно сближаемую с этим семейством.
Сравнивая род Archiphora с Phoridae, необходимо отметить следующие особенности, которые заставляют считать этот род предком рецентной группы:
1. Присутствие ветвей М—Мп М4, далее поперечных жилок rm, tp, mcu и, наконец, Ап2._ Все эти жилки явно утрачены у современных Phoridae в процессе специализации крыла.
2. Короткие, слабо утолщенные ноги. Для Phoridae характерны крепкие, частью даже почти прыгательные ноги.
3. Отсутствие или незначительное развитие щетинок на теле. Неясность щетинок у Archiphora не доказывает, конечно, полного их отсутствия; возможно, что у этого рода имелись нежные, тонкие волоски, не сохранившиеся у ископаемого. У представителей рецентных Phoridae имеются крепкие и длинные щетинки, несомненно хорошо бы сохранившиеся в процессе фоссилизации. Считать присутствие щетинок на ногах за признак большей примитивности можно лишь в немногих, редких случаях.
4. Коническое брюшко, разделенное на 9 сегментов; каждый из сегментов брюшка незначительно отличается от соседних. Брюшко современных Phoridae состоит из меньшего количества сегментов, причем, как правило, более или менее видоизмененных.
Таким образом, можно считать, что отношения описываемой формы к современным представителям сем. Phoridae хотя и в малой степени, но в общем уже намечены. Гораздо сложнее вопрос о связях этого семейства с другими. Своеобразие жилкования очень затрудняет решение этого вопроса. Повидимому, наиболее близки исходные формы описываемой группы были к группе Empididae — на это указывает отчасти характер редукции медиальной системы. С другой стороны, по строению тела — головы и груди, а также и брюшка — эта группа несколько напоминает длинноусых, именно группу Fangivoroidea. Все эти последние заключения, разумеется, очень провизорны и нуждаются в дополнительных исследованиях на новом материале.
Род Archiphora gen. nov.
Тип рода: Archiphora ancestrix sp. nov.
Крыло. R равна лишь г/3 длины крыла, крепкая; RS1 + 2 очень крепкая, расщепленная на конце; RS3+4 ответвляется от общего ствола RS сразу после ответвления R, прямая, без ветвей, утолщенная на основании и тонкая на конце, неправильно изогнутая; поперечная rm косо лежащая, почти являющаяся продолжением основной части RS3+4; имеется крепкая поперечная жилка, лежащая между RS3 ,.4 и М1+2, повидимому, являющаяся передней ветвью М — Мг; М2 прямая, дистальнее Мх, почти параллельная RS3+4, без дистальных ветвей; имеется довольно тонкая задняя поперечная жилка (tp) — интермедиальная поперечная и более короткая, но толстая медиокубитальная (mcu); М3 отсутствует или слита со следующей; М4 имеется в виде короткой ветви Си — обе эти жилки, как и все последующие, очень нежные, плохо заметные; Апх на основании слита с Си, Ап2, сильно изогнутая, нежная, но ясная.
Ноги довольно короткие; передние и задние (?) бедра умеренно утолщенные; средние и передние голени в 1%—2 раза тоньше бедер, тонкие, почти равномерной толщины; лапки примерно равны голеням, пятичленистые:. 1-й членик передних лапок наиболее короткий; последовательность размеров члеников лапок для разных пар ног такова: передние лапки — 1, 5, 3, 4, 2; средние — 1, 3, 4, 2 (5-й не сохранился); задние — 2, 5, 4, 3, 1; щетинки на ногах не ясны.
Грудь выпуклая, щиток резко выдается, узкий, не выпуклый.
Голова без каких-либо особенностей: антенны и хоботок не сохранились; щетинки не ясны.
Брюшко очень широкое на основании, постепенно утончающееся к концу; 1 и 2-й сегменты слиты вместе, крупные; сегменты 3—9-й постепенно уменьшаются в величине; последний, 9-й сегмент несет пару цер-ков (9).
Archiphora ancestrix sp. iiov.
(рис. 13)
Темно окрашенный вид; длина тела 2.0 мм, длина крыла несколько менее.
Описывается по 1 2 из юрских сланцев окрестностей с. Галкино. № 2452/329. Сбор А. В. Мартынова, 1925.
Рис. 13. Archiphora ancestrix gen. sp. nov.
ПОДОТРЯД NEMATOCERA — ДЛИННОУСЫЕ
СЕМ. ALLACTONEURIDAE FAM. NOV.
Тип семейства: род Allactoneura de M e i j e г e, 1907.
SC хорошо развитая, примерно равная базальной трети крыла, впадающая в костальную жилку. RS простая, без ветвей, отходящая от R нормально, т. е. под острым углом; гт короткая, поперечно лежащая. Медиальная система хорошо развита; М3 длинная, слитая с Си; обе передние ветви М хорошо развиты. Антенны умеренной длины, тонкие, длиной равные голове и груди, вместе взятым. Тазики увеличенные, ноги тонкие, щетинки на голенях и бедрах умеренно развитые.
Это семейство описывается по неизвестному мне в натуре, рецентному роду Allactoneura de М., найденному на о. Яве, и шести ископаемым родам, найденным в юре Кара-тау. Взаимоотношения этого семейства ясны: по строению радиальной системы оно сближается с Diadocidiidae (имеющему лишь один известный, притом исключительно современный род), медиальные же жилки объединяют это семейство одновременно с Bolitophilidae и Fungivoridae.
Род Pleciofungivora gen. nov.
Тип рода: Pleciofungivora latipennis sp. nov.
Крыло. Форма довольно широкая; конец крыла закругленный; длина крыльев слегка превышает брюшко; SC длинная, более одной трети 42
«длины крыла, нежная, равномерно изогнутая вперед и впадающая в костальную жилку; R крепкая, почти прямая, впадающая в край крыла на уровне границы 2 и 3 трети крыла; RS крепкая, ответвляется от R значительно ближе к основанию крыла, чем место соединения SC с С; имеется более слабая передняя ветвь RS, ответвляющаяся несколько проксимальнее конца R и впадающая в С; RS после соединения с гт почти прямая
п лишь на конце, несколько проксимальнее уровня конца R, загнута назад; Мэ ответвляется от Си; гш короткая и прямая, расположенная значительно ближе к основанию крыла, чем к середине непарного отрезка М; развилок М расположен примерно на уровне места соединения R с С; передние ветви М почти параллельные, лишь едва сходящиеся к краю крыла; М3 почти прямая; Ап и Си не ясны.
Грудь выпуклая; щиток не выдается в виде угла; передняя часть груди лишь едва нависает над головой.
Голова в профиль значительно короче своей высоты, угловатая, с резким глазковым бугорком; глаза овальной формы, антенны не особенно удлиненные, тонкие; членики антенн неясные; хоботок плохо сохранился.
Ноги тонкие и длинные; тазики довольно короткие; бедра тонкие и длинные; передние голени длиннее и несколько тоньше бедер; задние голени равны по длине,
но несколько тоньше задних бедер;
передние и задние лапки плохо сохранились, равны по длине голеням.
Брюшко более чем в 1у2 раза длиннее головы и груди, вместе взятых; сегментация не ясна; конец брюшка заостренный.
Pleclofunglforci Icbtipennis sp. nov.
(рис. 14)
Длина тела 2,5 мм; длина крыла 20 мм. Заметна птеростигма в виде пятна, охватывающего конец R и переднюю ветвь RS.
Описывается по Q (?) из окрестностей с. Галкино. № 2452/332.
р А. В. М ар тын о ва, 1925.
Род Allactoneurites gen. nov.
Тип рода: Allactoneurites jurassicus sp. nov.
Крыло. Довольно длинное и узкое; SC нежная и тонкая, равномерно изогнутая к переднему краю, равная примерно 1/3 крыла, впадающая в передний край крыла; R более крепкая, очень слабо и равномерно дугообразно изогнутая, раза в два длиннее SC; RS отходит от R под острым углом несколько ближе к основанию крыла, чем место соединения SC с С; имеется косая передняя ветвь RS, впадающая в С; конец RS впадает в край крыла ровно на его вершине; передняя часть М жилки с хорошо
выраженной вилкой Мг +М2, ее общий ствол заметно длиннее вилки; RM короткая, расположенная близко от места ответвления RS от R; обе ветви М — Мх и М2 почти параллельные, очень слабо расходящиеся; М3 (или Си+М3) длинная, равномерно изогнутая; место соединения ее с М лежит очень близко к основанию крыла, на материале не ясно.
Тело очень плохой сохранности — детали его строения совсем не ясны.
Рис. 15. Allactoneurites jurassicus gen. sp. nov.
AUactoneiirttes j liras-sicus sp. nov.
(рис. 15)
Длина тела 2.5 длина крыла 3.5
Описывается по ному экземпляру
окрестностей с. Галкино. № 2452/356. Сбор А. В. Мартынова, 1925.
mm; mm.
од-из
Описываемый род наиболее близок к роду Pleciofungivora, отличаясь от него иными пропорциями жилок — более длинной RS и иной, более узкой формой крыла.
Род Pleciomima gen. nov.
Тип рода: Pleciomima sepulta sp. nov.
Крыло. Форма умеренно вытянутая, овальная; SC нежная и тонкая, впадающая в край крыла; R почти прямая, довольно толстая и крепкая; RS простая, отходящая от R под острым углом, впадающая в край крыла на его вершине; RM расположена недалеко от места ответвления RS, косая; М обособленная; непарный ствол очень короткий, впадающий в Си; непарный ствол передних ветвей М длинный, примерно равный ветвям Мг и М2; последние почти параллельные; М3 длинная, слегка изогнутая в дистальной половине; Си прямая в базальных двух третях, на конце резко дугообразно изогнутая .
Грудь. Сильно выпуклая, нависающая над головой; щиток округлый, умеренно выдающийся.
Голова в профиль неправильно округлая, с сильно выступающим вперед лбом, на котором расположены антенны; антенны умеренной длины, несколько короче или равны голове и груди, вместе взятым; антенны 12-члениковые; базальные членики не увеличены; антенны слегка зазубренные — членики не цилиндрические, несколько утонченные на основании; глаз овальный; его высота значительно превосходит его длину; глазковый бугорок не ясен.
Ноги крепкие, не особенно удлиненные; тазики длинные, сильно развитые; бедра слегка вздутые в середине, утончающиеся к обоим концам; задние голени равны, средние голени слегка длиннее бедер; лапки пятичленистые; границы члеников не ясные; лапки лишь слегка длиннее голени, передние ноги не сохранились.
Брюшко. Хорошо выражены 6 сегментов брюшка и генитальные концевые придатки; границы сегментов резкие.
Pleclomlnia sepulta sp. nov.
(рис. 16 и 17)
Длина тела 4 мм; длина крыла 2.5 мм.
Описывается по 1 $ из юры окрестностей с. Галкино. № 2452/323.
Сбор А. В. Мартынова, 1925.
Описываемый род по жилкованию приближается к роду Pleciofungivora, отличаясь от него хорошо развитой Си и крупными, массивными
Рис. 17. Pleciomima sepulia gen. sp. nov. Крыло (Flugel).
тазиками. Отличия в строении тела, кроме указанных выше, как, например, резкая сегментация брюшка, вероятно, являются следствием особенностей захоронения материала.
Род Fungivorite# gen. nov.
Тип рода: Fungivorites latimedius sp. nov.
Крыло. Форма крыла обычная, умеренно удлиненная, широкая; SC нежная, длинная; дистальный конец ее не сохранился; R почти прямая, крепкая; RS крепкая, волнообразно изогнутая; ее дистальная четверть резко загибается назад: эта жилка в большей своей части параллельна краю крыла; другие ветви RS отсутствуют; жилка гт прямая, расположенная близ места развилка R, лишь в 2 раза короче первого отрезка RS; М в основной части крыла слита с Си; жилка гт делит основную часть М на две почти равные половины; развилок М расположен почти на середине крыла или слегка проксимальнее; ствол М примерно в 3 раза короче ветвей М; ветви М очень резко расходятся в стороны; ячейка М,
Рис. 18. Fungivorites latimedius gen. sp. nov.
очень широкая, почти в 2 раза шире RS; изгиб ветвей неправильный, угловатый: к краю крыла обе ветви очень слабо расходятся, почти параллельные; Си без ветвей, простая, не ветвящаяся, почти прямая, расположенная близко от М2 — ячейка М2 очень узкая, особенно резко суженная на границе своих двух последних третей; части крыла сзади Си плохо сохранились, жилкование не ясно.
Голова. Плохо сохранилась — придатки ее не ясны.
Грудь небольшой величины, умеренно выпуклая, с резко выступающим щитком; жужжальца не ясны.
Ноги. Плохо сохранились; тазики, повидимому, длинные; бедра короткие и резко вздутые; голени цилиндрические; лапки не сохранились.
Брюшко. Крупное, в профиль яйцевидное, на конце закругленное; заметно 7 тергитов и 5 стернитов разной величины, с широкой плевральной перепонкой между ними; придатки брюшка не ясны.
Fungi го rites latimedius sp. nov.
(рис. 18)
Длина тела 3.5 мм; длина крыла 3 мм.
Описывается по 1 9 из юры окрестностей с. Галкино. ЛГ? 2452/548.
. ПОДСЕМ. ANTIQUAMEDIINAE SPBFAM. NOV.
Тип подсемейства: род Antiquamedia gen. nov.
SC короткая, загнутая назад и впадающая в R; длина ее менее х/4 длины крыла. RS простая, без ветвей, отходящая под острым углом почти на самом основании крыла от R. Поперечная жилка гт неясная, повидимому, отсутствующая. Медиальная система хорошо развита: общий ствол отходит от Си примерно на уровне середины SC; общий ствол двух передних ветвей М отходит от М3 на уровне конца SC, по длине равен своим дистальным ветвям — Мх и М2. Си и Ап хорошо развиты. Тазики, повидимому, короткие. Щетинки на голенях и бедрах не ясны.
Эта группа описывается по единственному роду из юры Кара-тау. Признаки подсемейства, с одной стороны, сближают его с сем. Fungivo-ridae — короткая SC и хорошо развитые Си и An. С другой стороны, строение медиальной системы и отсутствие гт ставят это подсемейство7 особняком. »
Род Antiquamedia gen. nov.
Тип рода: Antiquamedia tenuipes sp. nov.
Крыло. Форма обычная, умеренно вытянутая; SC очень короткая, менее х/4 длины крыла, на конце загнутая назад и впадающая в R; R длин
ная, равномерно изо-гнутац; RS простая, отходящая от R на уровне аксиллярной поперечной жилки, т. е. почти на' самом основании крыла; RS впадает в край крыла перед его вершиной; гт отсутствует; М имеет короткий общий ствол, отходящий от Си на уровне середины SC; вилка М (Mj-J-Mg) короткая, равная по длине переднему стволу М; М3 очень
Рис. 19. Antiquamedia tenuipes gen. sp. nov.
длинная, дугообразная;
Си дугообразная в конечной половине, с легким изломом; Ап длинная, хорошо развитая, брюшко короче крыльев; грудь выпуклая; голова крупная; хоботок длинный, равный по длине высоте головы; усики не
особенно тонкие, длинные, равные по длине голове и груди, вместе взятым; ноги тонкие и длинные, голени и бедра не утолщенные.
Antiquamedia tenuipes sp. nov.
(рис. 19)
Длина тела 2.75 мм; длина крыла 2.0 мм.
Описывается по одному экземпляру из юры окрестностей с. Михайловки. № 2335 75. Сбор Турутановой-Кетовой.
ПОДСЕМ. ANTEFUNGIVORINAE SUBFAM. NOV.
Тип. подсемейства: род Antefungivora gen. nov.
SC хдрошо развитая, не особенно короткая, несколько более х/4 крыла, впадающая в костальную жилку. RS простая, без ветвей, отходящая от R под острым углом, примерно на уровне места впадения в С жилки SC; гт короткая, поперечно лежащая. Медиальная система хорошо развита; М8 очень короткая, в виде короткой ветви общей жилки Си; обе передние
ветви М хорошо развиты, длинные, длиннее их ствола. Крылья широкие, более или менее треугольные. Антенны длинные и тонкие, равные по длине голове и груди, вместе взятым. Тазики крупные и массивные. Ноги тонкие. Щетинки не ясны.
Это подсемейство описывается по единственному роду из юры Кара-тау. Взаимоотношения его с другими подсемействами более или менее ясны: положение и строение ветвей М объединяют его с сем. Fungivori-dae. С другой стороны, ответвление RS сближает его с подсем. Allacto-neurinae и Antiquamediinae.
Род Antefungivora gen. nov.
Рис. 20. Antefungivora prima gen. sp. nov.
Тип рода: Antejungivora prima sp. nov.
Крыло. SC не особенно короткая, на конце дугообразно изогнутая, впадающая в край крыла; R слабо, незаметно волнистая, оканчивающаяся на границе второй и третьей трети крыла; RS сильно волнообразно изогнутая, впадающая в край крыла за его вершиной; таким образом, ячейка R резко расширенная на краю крыла, a RS заметно суженная на краю; жилка ни очень короткая, поперечно лежащая; общий ствол М короткий, не менее чем в 2 раза короче передней ветви; обе ветви М равномерно расходящиеся в стороны, передняя прямая, задняя изогнутая в осно вной поло вине; Си на основании очень сближена с В, слитая с М3 на большом протяжении; Си в своей дистальной трети ветвится; развилок Си4-М3 расположен на одном уровне с развилком М; Ап длинная, на кон
це резко загнутая назад, доходящая почти до середины крыла.
Голова. Глаза небольшие, овальные; передний край головы, под основанием антенн, со следами выступа, повидимому, бугорком с простыми глазками; антенны плохо сохранились; передний край головы выступающий в виде уголка.
Грудь довольно выпуклая, слегка нависающая над головой; хорошо видны жужжальца шаровидной формы.
Ноги плохо сохранились, тонкие, длинные; бедра и голени, повидимому, не вздутые.
Брюшко плохо сохранилось.
Antefunglvora prlma, sp. nov.
(рис. 20)
Длина тела 2.0 мм; длина крыла 1.5 мм.
Описывается по одному экземпляру из окрестностей с. Галкино. № 2452/316. Сбор А. В. Мартынова, 1925.
СЕМ. PENTHETKIIDAE
Это семейство известно из лиаса Германии — Dobbertin-Mecklenburg (Н a n d lir sc h, 1906, стр. 488) в качестве рода Proloplecia Handl.
Описываемый ниже род Mesoplecia занимает промежуточное положение между родами Plecia и Protoplecia, обладая вместе с тем рядом своеобразных признаков.
Род Mesoplecia gen. nov.
Тип рода: Mesoplecia jurassica sp. nov.
SC тонкая, но хорошо заметная, достигающая середины переднего края крыла. R прямая; RS двухветвистая, отходящая от R под острым
Рис. 21. Mesoplecia jurassica gen. sp. nov.
углом; RS ветвится недалеко от места соединения с гт, на расстоянии, равном длине последней. Ветви RS длинные, на конце резко расходящиеся; задняя ветвь RS впадает в вершину крыла. Медиальная система обособленная; ее общий ствол длинный, ответвляющийся от Си на самом основании крыла — на уровне аксиллярной поперечной жилки. Мх и М2 длинные, их общий ствол значительно короче ветвей. М3 ответвляется от общего ствола М под острым углом; сразу сзади места ответвления М3 от М лежит mcu; эта последняя поперечная жилка, как и гт, лежит отвесно, прямая; Си в основных двух третях прямая, на конце равномерно дугообразная.
4 Тр. Палеонт. ин-та, т. VII, в. 3. 49
Голова. Глаза овальные. Передний край головы снизу выступающий вперед. Антенны длиннее головы; их членики цилиндрические, в два раза длиннее своей толщины каждый.
Грудь умеренно вздутая, массивная.
Брюшко и ноги плохо сохранились.
Mesoplecla jurassica sp. nov.
(рис. 21)
Длина тела 7.5 мм; длина крыла 7.0 мм. Тело темное; крылья слегка, пигментированные, коричневые.
Описывается по одному экземпляру из окрестностей с. Михайловки. № 2231. Сбор Турутановой- Кетовой.
Описываемый род наиболее близок к рецентному роду Plecia, известному начиная с олигоцена. Отличается от указанного рода иным строением медиальной системы, именно ответвлением под острым углом жилки М3. Повидимому, также характерны для рода Mesoplecia* более длинные антенны.
ХАРАКТЕР ФАУНЫ DIPTERA КАРА-ТАУ
Проводя анализ описанной фауны двукрылых мезозоя Кара-тау, в первую очередь следует остановиться на представителях подотряда короткоусых —Brachycera.
' Первое, что бросается в глаза при рассмотрении списка Brachycera, — это явное преобладание представителей серии Eremochaeta — видов семейств Stratiomyidae, Bhagionidae и Protocyrtidae. Сем. Omphralidae, как и Archipho г idae, относится к совсем иным сериям Brachycera. Характерно отсутствие серий Microphona (сем. Empididae и Bolichopodidae), развитых в настоящее время во всех ландшафтных зонах. Особенно же важно полное отсутствие представителей всего подотряда Cyclorrhapha, за исключением сем. Archiphoridae, о чем речь будет несколько далее. Подобный характер юрской фауны очень интересен, так как он освещает происхождение ряда семейств Brachycera и, вместе с тем, объясняет генезис современной фауны.
Группа Eremochaeta может быть охарактеризована как заключающая в себе морфологически наиболее слабо специализированные формы из всего подотряда Brachycera. Основные черты слабой морфологической специализации группы таковы:
1. Как правило, многочленистые антенны, состоящие из сложно построенного дистального отдела — «3-го» членика; число члеников антенн иногда может превышать 20 (Erinnidae).
2. Богатое жилкование; частое присутствие 4 ветвей М, 3 ветвей RS и других жилок; прямые жилки, впадающие в край крыла; присутствие в некоторых случаях богатой сети мелких ячеек, несомненно первичного происхождения (Nemestrinidae).
3. Отсутствие или слабое развитие щетинок на теле, присутствующих в некоторых случаях лишь на голенях ног.
Современный состав фауны Eremochajeta своеобразен; всего известно 10 семейств, из которых лишь 5 обладают многочисленными родами и видами, будучи широко распространены во всех зоогеографических областях. Остальные 5 семейств очень невелики по объему и имеют частью реликтовый или остаточный характер. Часть из них обычно считается лишь подсемействами других крупных семейств группы; это вызывается не отсутствием морфологических гиатусов между ними, а лишь незначительным их объемом и так называемой «боязнью» мелких систематических категорий — семейств. Таковы, например, Erinnidae, Solvidae, Соепо-myiidae, Chiromyzidae— эти семейства произвольно включаются в Stratiomyidae и Bhagionidaen качестве крайних, так называемых «переходных»' 50
подсемейств. Явно реликтовая или рестантная природа семейств Рап-tophthalmidae, Erinnidae, Sohidae и Coenomyiidae вполне точно доказывается не только морфологическими их особенностями, но также и их географическим распространением: каждое из этих семейств очень широко распространено почти во всех зоогеографических областях. Сем. Chiromyzidae — несомненно, высоко специализированная группа, распространенная лишь в Австралии и Южной Америке и, вероятно, являющаяся производной от форм, близких к современным Stratiomyidae— Beridinae.
Возвращаясь к распространению «главных» современных семейств серии — Stratiomyidae, Bhagionidae, Tabanidae, Cyrtidae и Nemestrinidae, следует отметить, что лишь два семейства — Stratiomyidae и Tabanidae — во много раз превосходят остальные по количеству родов и видов, составляя один из характерных элементов фауны Diptera любого района той или иной зоогеографической области. При этом оба названных семейства крайне различны по своим особенностям. Tabanidae — одна из специализированных групп из всей серии Eremochaeta, несмотря на свой крупный объем (это семейство наиболее богато видами из всех других в серии), обладает очень мало изменчивой морфологией: очень константно богатое, мало специализированное жилкование крыльев, очень постоянное строение тела — широкое многочленистое брюшко, широкая голова с очень крупными глазами. Морфологическая специализация в этой группе есть следствие своеобразной биологии этих мух-кровососов. Таковы крупные глаза, строение ротовых органов, возможно редукция простых глазков. Строение антенн мало изменчиво в этом семействе. Вторичные подразделения, подсемейства, имеются в очень ограниченном количестве, притом их характер скорей приближается к трибальному, чем к подсемейственному, являясь сугубо подчиненным относительно семейственных признаков.
Второе семейство — Stratiomyidae — по количеству видов несколько беднее предыдущего, но зато в своем составе имеет целый ряд вторичных категорий — подсемейств и триб, резко отличных друг от друга. Особенности строения тела очень разнообразны в пределах различных подсемейств. На ряду с очень мало специализированными чертами строения, как, например, многочленистые антенны, многочленистое брюшко, 4 ветви М, удлиненное тело (что наблюдается, например, в подсемействе Beridinae), в этом же семействе имеет место и довольно высокая специализация, выражающаяся в образовании почти настоящих трехчленистых антенн, резком укорочении брюшка и редукции ветвей М (таковы, например, представители Stratiomyinae и Clitellariinae). Специализация жилкования особенно характерна для Stratiomyidae и выражается всегда в редукции С и более или менее развитом сдвиге всех жилок к переднему краю.
Остальные семейства серии Eremochaeta — Bhagionidae, Nemestrinidae и Cyrtidae — довольно бедны видами; Cyrtidae крайне специализированная группа, обладающая своеобразным жилкованием, редуцированными антеннами и рядом других признаков; личинки их ведут паразитный образ жизни. Nemestrinidae очень своеобразны, сохраняя в жилковании крыльев характерные архаические черты в виде многочисленных поперечных жилок. На ряду с примитивным жилкованием у этого семейства наблюдается специализация в виде укороченных антенн и брюшка.
Сем. Bhagionidae наименее специализировано по сравнению с двумя предыдущими: жилкование очень полное, почти без следов каких-либо редукций или перемещений жилок; тело удлиненное, брюшко часто цилиндрическое, многочленистое. Морфологическая специализация в сем. Bhagionidae выражается, главным образом, в укорочении и видоизменении антенн, превращающихся в трехчленистый орган, несущий тонкую щетинку — видоизмененный дистальный отдел антенн. В сем. Rhagionidae различается несколько вторичных подразделений — подсемейств, из ко-4* 51
торых лишь некоторые (V ermileoninae) действительно отдалены от других (Chrysopilini, Rhagionini, Bicalcarini), имеющих лишь явно трибальный характер.
В свете данного краткого очерка серии Eremochaeta анализ описанных юрских представителей Stratiomyidae, Rhagionidae и Protocyrtidae дает важные результаты. В первую очередь характерен общий состав фауны: присутствуют представители семейств Stratiomyidae и Rhagionidae и один вид из сем. Protocyrtidae, относительно близкого к рецентным Cyrtidae. При этом все найденные виды являются представителями особых подсемейств. Так, сем. Rhagionidae представлено тремя подсемействами: Rha-gionempidinae, Protorhagioninae и Vermileoninae. Два первых — ископаемые группы, а последнее имеет 2—3 рецентных рода с немногими редкими видами, нося явно рестантный характер. Описанные Stratiomyidae очень своеобразны, так же как и Rhagionidae, являясь представителями особых подсемейств. Protocyrtidae, в лице рода Protocyrtus, — высоко специализированная группа, довольно удаленная от рецентных Cyrtidae. Указанные подсемейства Rhagionidae носят отчасти характер исходных групп некоторых рецентных семейств. Таковы Rhagionempidinae, которые обладают чертами сходства с Tabanidae, ь одной стороны, и Empididae — с другой. Подсемейство Protorhagioninae является, повидимому, исходной группой современных Rhagioninae (Rhagionini+Chrysopilini). Представитель Vermileoninae — род Archirhagio — не отличается от рецентных какой-либо меньшей специализацией, являясь типичным для подсемейства и, вместе с тем, явно параллельным отделом, не имеющим прямого генетического отношения к современным формам. Два описанных рода Stratiomyidae очень далеки друг от друга, также при этом почти не обнаруживая какой-либо генетической близости к современным. Подсемейство Palaeostra-tiomyiinae очень любопытно своим мозаичным характером: на ряду с мало специализированным жилкованием наблюдается крайняя специализация тела — брюшко очень укорочено, состоя лишь из 4 видимых сегментов. Второй представитель Stratiomyidae — род Archisargus — также своеобразен, но вследствие плохой сохранности не может быть достаточно изучен для вынесения окончательного решения о его положении.
Необходимо отметить отрицательные черты характеристики фауны Eremochaeta юры Кара-тау, а именно: важно полное отсутствие представителей сем. Tabanidae, занимающего в современной фауне первое место.
В результате обзора всех данных о Eremochaeta можно вывести следующее заключение. Фауна этой группы семейств в юрское время в пределах Средней Азии слагалась в первую очередь из различных подсемейств Rhagionidae и Stratiomyidae, в своем большинстве отличных от рецентных; только одно современное подсемейство Vermileoninae уже было представлено в фауне. Лишь подсемейства Rhagionidae имеют характер исходных форм современных групп; оба же представителя Stratiomyidae очень своеобразны и специализированы, явно не имея прямых генетических связей с рецентнымп. Нахождение специализированного семейства Protocyrtidae, на ряду с присутствием уже указанных выше Stratiomyidae, указывает на древность и долгую предшествующую историю этих групп. Наибольшего расцвета в то время достигали, видимо, семейства Rhagionidae и Stratiomyidae, первое из которых сейчас явно имеет характер реликта. С другой стороны, характерный член рецентной фауны Eremochaeta — Tabanidae — вовсе отсутствует в мезозое Кара-тау.
Нахождение представителя семейства Omphralidae в юре Кара-тау представляет очень большой интерес. Известны лишь рецентные представители этого семейства, притом очень немногочисленные и носящие явно остаточный характер. Описанный юрский представитель семейства носит черты меньшей специализации жилкования; остальные признаки строения тела не выходят из рамок диагноза современных Omphralidae. Самый факт одновременного нахождения Omphralidae в юре вместе с представителями 52
Eremochaeta и при отсутствии других групп Brachycera очень интересен и доказывает большую древность этого семейства, до последнего времени сближаемого с сем. Bombyliidae.
Переходя к рассмотрению взаимоотношений семейства Archiphoridae, надо отметить неожиданность этой находки. Рецентное семейство Phort-dae, несомненный потомок Archiphoridae, является представителем третьего подотряда Diptera — круглошовных мух — Cyclorrhapha. Несмотря на известную отдаленность Archiphoridae от названного рецентного семейства, оно все же, повидимому, уже носило черты этого третьего подотряда — сдвиги жилок к переднему краю и обедненное жилкование. Если это предположение подтвердится, то можно будет говорить о появлении круглошовных двукрылых уже с юры.
Вторая половина статьи посвящена описанию 7 видов длинноусых двукрылых, что является лишь небольшой частью всей коллекции. Но и среди этих первых материалов оказались многочисленные новые формы, относящиеся не только к родовым, но и к более высшим категориям. Подробный анализ этой группы в настоящее время проводить еще преждевременно. Далее, лишь кратко рассматривается система серии Fungivo-roidea; провизорно принимается деление группы на семейства по Hen-del (1928), с некоторыми изменениями. Схема Н е n d е 1 наиболее последовательна; она выделяет ряд мелких семейств явно остаточного или реликтового характера, которые до сих пор считались лишь за подсемейства Fungiroridae.
Главнейшим недостатком системы Н е n d е 1 является недостаточность морфологической характеристики семейств., что привело в ряде случаев к чисто формальному выделению некоторых групп. Наиболее важно то обстоятельство, что Н е n d е 1 правильно базировался на жилковании, которое у этих насекомых отражает в большей степени, чем какой-либо иной орган тела, действительные филогенетические взаимоотношения. Это последнее и подтвердилось теперь, после исследования ископаемых юрских форм.
Современная фауна серии семейств длинноусых двукрылых Fungi-voroidea слагается из 10 семейств: Allactoneuridae. Diadocidiidae, Ditornyc-idae, Bolitophilidae, Mdcroceratidae, Ceroplatidae, Fungiroridae, Lycoriidae, Lestremiidae и Itonididae. Эта естественная группировка названных семейств хорошо характеризуется целым рядом морфологических признаков: строением головы, антенн, жилкованием крыльев — сильная редукция ветвей RS, строением грудного отдела и брюшка — укорочение и сильное вздутие, гипертрофия груди. Наблюдаются связи отдельных семейств этой группы с другими длинноусыми двукрылыми. Так, через сем. Lycoriidae намечается связь с группой Bibionoidea, именно с сем. Corynosce-lididae. С другой стороны, сем. Ditoinyiidae и в меньшей мере Масгосега-tidae с ясностью сближаются с группой Phryneodea через переходное семейство Mycetobiidae.
Описываемые выше юрские подсемейства этой группы двукрылых занимают промежуточные положения между различными рецентными семействами и в известной степени могут считаться их исходными формами.
Сем. Allactoneuridae занимает отчасти промежуточное место между семействами Bolitophilidae, Diadocidiidae и Fungiroridae. Некоторые роды этого семейства сближаются с Bolitophilidae — poft'Pleciominur, все остальные, благодаря редукции кубитальной системы, могут быть сближаемы с Fungiroridae. Характер радиальной системы, ее редукция, сближает сем. Allactoneuridae с Diadocidiidae.
Подсем. Antefungivorinae имеет более определенное положение, являясь связующим звеном между Diadocidiidae, Fungiroridae и основным подсемейством.
С первым семейством его сближает простое строение радиальной системы и хорошо развитая SC; с Fungiroridae, наоборот, это семейство очень
сходно по строению медиальной системы — далекое отхождение и слияние с Си третьей ветви М. Подсем. Antefungivorinae характерно еще своеобразной формой крыла и резким изгибом RS, чего мы не видим ни у каких современных форм.
Подсем. Antiquamediinae, пожалуй, наиболее своеобразно из всех вышеописанных групп; взаимоотношения его с другими семействами группы еще очень неясны. С одной стороны, строение радиальной системы так же просто, как у Diadocidiidae, Allactoneurinae и Antefungivorinae, — RS без ветвей, правильно ответвляющаяся от R. С другой стороны, медиальная система обнаруживает очень примитивные черты строения — существует общий ствол М в базальной части крыла, соединенный с Си. Характерно, что поперечная гш, признак, очень стойкий во всей группе этих семейств, у Antiquamediinae отсутствует вовсе. Наиболее близко это семейство, повидимому, к настоящим Allactoneurinae, в качестве подсемей
ства которого оно и ставится.
Перечисленные выше 10 современных семейств группы Fungiro го idea очень неравноценны по их объему. В настоящее время нет монографий, охватывающих всесветную фауну этих насекомых; поэтому для иллюстрации картины рецентной фауны Fungivoroidea будет наиболее уместно взять близкую нам область, именно Палеарктику, которая к тому же срав
нительно хорошо изучена.
Семейство
Источник
Число палеарктических родов видов
Allactoneuridae .... в палеарктической области не представлено
Diadocidiidae (no Landrock, 1927) 1 1
Ditomyiidae ... • . . » » » 2 3
Bolilophilidae » » » 1 15
Macroceratidae » » » 1 42
Ceroplatidae » » » 7 60
Fungivoridae » » » 47 630
Lycoriidae ....... (Lengersdorf, 1930) 22 223
Itonidida? (Kieffer, 1912) 185 1089
Lesteremiidae . . . • • » » 23 95
Как мы видим, лишь три семейства — Fungivoridae, Itonididae и Lycoriidae — очень разнообразны и наиболее богаты видами (на их долю падает более 4/5 всего количества родов и видов этой группы). Остальные семейства сравнительно невелики по объему. Особенно бедны видами первые три семейства списка: можно с несомненностью утверждать их древний, рестантный характер. В особенности это справедливо в отношении семейств Diadocidiidae, Ditomyiidae и Bolilophilidae, насчитывающих все вместе взятые всего лишь четыре рода с 20 видами.
Древний, рестантный характер этих семейств с очевидностью выясняется их сравнительно богатым жилкованием, особенностями строения головы и близкими отношениями к описываемым новым юрским семействам группы, связывающими их с другими, повидимому, более молодыми,— Fungivoridae, Lycoriidae и Itonididae.
ПРЕДВАРИТЕЛЬНЫЕ СООБРАЖЕНИЯ О ВОЗМОЖНОЙ ЭКОЛОГИИ ОПИСАННЫХ ВИДОВ ДВУКРЫЛЫХ ИЗ ЮРЫ КАРА-ТАУ
В настоящей статье еще преждевременно делать какие-либо заключения о характере ландшафта или климатических условий, в которых жили двукрылые доггера Кара-тау, — вся главная масса длинноусых Diptera еще остается необработанной. Разбирая весь список описанных семейств, в первую очередь группы Brachycera, сразу же сталкиваешься с целым рядом новых групп, судить о биологии представителей которых очень затруднительно. Таковы, например, Archiphoridae, Protocyrtidae и большинство длинноусых. Все представители рецептных семейств — Stratio-
myidae, Bhagionidae, Omphralidae и Penthelriidae —также крайне своеобразны, и суждение об их экологии должно быть составлено с большой осторожностью. Простой перенос на юрских мух тех биологических особенностей, которые характерны для рецентных представителей тех же семейств, не может иметь места. Лишь те немногие виды, которые отнесены не только к современным семействам, но также и к более низким категориям, подсемействам, могут быть сравнимы с рецентными. Таковых же видов очень немного: представитель Bhagionidae — Archirhagio obscuras, Omphralidae — Protomphrale martynovi, Pentheriidae — Mesoplecia jurassica. Биология близких рецентных форм своеобразна: Omphralidae и Vermileoninae— обитатели сухих субтропических областей, группа Pentheriidae наиболее богато развита во влажных тропиках. Личинки Vermileoninae — хищники, Omphralidae — сапрофаги, питающиеся животными веществами. Личинки Penthetriidae — все растительноядные сапрофаги.
Остальные ископаемые двукрылые остаются с биологической точки зрения загадочными формами. Большое семейство Stratiomyidae заключает в себе виды с самой разнообразной биологией; большинство из них растительноядные сапрофаги, иногда связанные с водной средой, часто известных видов — хищники. Личинки Bhagionidae также хищники. Что касается географического распределения этих двух семейств, то оно очень широко, захватывая как умеренные, так и тропические зоны. Оба названных семейства так же хорошо представлены и в пустынных районах, как и во влажных умеренных и тропических. Сем. Cyrtidae, близкое к описанным ископаемым Protocyrtidae, обнаруживает ясное тяготение к сухим субтропическим районам, являясь при этом группой высоко специализированных паразитов-пауков.
Переходя теперь к длинноусым двукрылым, именно к комплексу семейств Fungivoroidea, следует отметить большую трудность суждения о возможном образе жизни этих насекомых. Ближайшие рецентные группы то являются растительноядными сапрофагами (Lycoriidae, Piadocidiidae), М acroceratidae и др.), то живут в различных грибах (Fungivoroidae и др.) или являются галлообразователями на различных растениях (Itonididae). Географическое распространение их очень широко — охватывает почти все зоогеографические области. Наиболее богато представлены эти семейства во влажных ландшафтах, хотя галлообразователи, например Itonididae, также многочисленны и в пустынных, и полупустынных районах. Наиболее правильно, вероятно, будет считать всех описанных юрских Fungivoroidea растительноядными сапрофагами, тяготеющими к влажным ландшафтам. Itonididae — несомненно, наиболее молодая группа, еще не появившаяся в юре (об этом говорит тесная связь Itonididae с цветковыми растениями).
РЕЗЮМЕ
1. Описываются 15 видов двукрылых насекомых из юры Кара-тау г относящихся к 15 новым родам и к 9 семействам, из которых 5 оказались новыми.
2. Некоторые из описанных групп обладают ясным «предковым» характером, позволяющим выяснить филогению современных групп. Таковы Protomphrale martynovi, Archiphora ancestrix, сем. Allactoneuridae, Protorhagio capitatus и Rhagionempis tabanicornis, являющиеся предками ряда рецентных семейств и родов.
3. На ряду с указанными «предковыми» формами, обнаружен целый ряд своеобразных, частью специализированных видов, несомненно не стоящих в связи с какими-либо современными двукрылыми. Таковы Archisa г-gus pulcher, Palaeostratiomyia pygmaea, Protocyrtus jurassicus.
к. Проведен обзор описанной юрской фауны Diptera Brachycera и выяснены различия с современной. Основной группой, составляющей фон всей фауны, является серия семейств Eremochaeta — Stratiomyidae и Rha-gionidae. Отмечается высокая специализация ряда форм. Слабо специализированные, примитивные формы немногочисленны.
5. Описывается семейство Archiphoridae по новому роду Archiphora, который характеризуется рядом примитивных признаков и ясно связывает современное, высоцо специализированное сем. Phoridae с другими (Empididae), являясь его предковой группой. Нахождение сем. Archiphoridae в юре очень интересно вследствие принадлежности сем. Phoridae к подотряду круглошовных мух. Очень вероятно, что и описанный предок этого семейства также принадлежал к подотряду Cyclorrhapha, до сих пор известному лишь начиная с олигоцена.
6. Характеризовать описанную фауну двукрылых с экологической точки зрения очень затруднительно вследствие резкого ее отличия от современной. По присутствию представителей групп Vermileoninae, Omphralidae и Penthetriidae, а также по косвенным соображениям о биологии длинноусых серии Fungivoroidea можно предполагать, что большинство описанных юрских Diptera являлись растительноядными видами, повидимому, тяготеющими к влажным ландшафтам.
7. Описываемая юрская фауна Diptera Brachycera не может быть сравниваема с какими-либо другими мезозойскими фаунами, так как до сих пор известен лишь один представитель Brachycera из лиаса Германии (Nemestrinidael).
ЛИТЕРА ТУРА
Н andlirsch, 1906. Die fossilen Insecten und die Phylogenie der rezenten For-men. Leipzig.
H e n d e 1. 1928. Zweifliigler oder Diptera-, Allgemeiner Teil; in: Dahl — Die Tierwelt Deutschlands usw. II Teil. Jena.
Kieffer, 1912. Cecidomyiidae\ in: Wytsman — Genera Insectorum, fasc. 152. Bruxelles.
Landrock, 1927. Fungiroridae-, in: Lindner— Die Fliegen d. Palaarktischen Region.
Lengersdorf, 1930. Lycoriidae: in: Lindner — Die Fliegen d. Palaarktischen Region.
M а рты н о в, 1938, Местонахождения ископаемых насекомых в пределах СССР/ Тр, ПИН, VII, 3,
Meijere de, 1907. Tijdschrift voor Entomologie, L. pp. 196—264.
ZUSAMMENFASSUNG
NEUBESCHKEIBUNGEN
FAM. STRATIOMYIDAE
SUBFAM./
G. Archisargu» gen. nov.
. Archisa rgus rille he r sp. nov.
Typus generis: Archisargus pulchersp. nov.
Flugel. Sehr lang und schmal, an der Basis stark verschmalert; SC undeutlich; R lang, viel langer als die Halfte des Flugels; RS mit 4—5 Asten; RSt und RS2 stark sklerotisiert; US3 und RS4 viel feiner, nur we-nig deutlich; Ader RS4 nur auf dem linken Flugel bemerkbar, am rechten fehlend; M und Cu undeutlich; C bis zum Endeder RS3 reichend. —Thorax mittelgross, quadratisch, etwas verlangert, mit abgerundeten Ecken; Schil-dchen kurz, fast rechteckig, am Rande unbeborstet; Beine diinn, Schenkel lang, nicht verdickt; Schienen 3, zweimal diinner als die Schenkel. — Kopf gross; Augen oval, am Scheitel sich beriihrend; Ocelli undeutlich; Vorderteil des Kopfes undeutlich. — Abdomen lang, wahrscheinlich fast zweimal so lang als der Kopf und Thorax zusammen; nur 4 vordere Tergite vorhanden; Abdomen fast parallelseitig: 1 Tergit deutlich schmaler als die anderen, vorne stark verschmalert.
Archtsargus pulcher sp. nov.
Fliigellange 16.0 mm; Lange des Thorax und des Kofprestes-}-Abdomen 16.0 mm; Abdominaltergite mit gut entwickelter Zeichnung: 1. Tergit mit dreieckigem dunklem Mittelflecke; 2. Tergit mit verlangertem, hinten spitzem Sagittalflecke und dunkler Hinterrandbinde; 3. und 4. Tergit nur mit dunk-ler Hinterrandbinde; Mittelflecke fehlend.
1 (J—Lok. Katschkar-Ata; Kara-tau-Gebirge (unter. Dogger).
Diese interessante Fliege ist ohne Zweifel eine echte Stratiomyidae. Wegen des undeutlichen Fliigelgeaders und Kopfbaues ist die systematische Stellung der Gattung Archisargus mir unklar. Stark verlangerte Flugel, einfach gebaute R und RS u. undeutliche SC ist von alien anderen Stratiomyidae sehr verschieden.
SUBFAM. PALAEOSTRATIOMYIIN AE SUBFAM. NOV.
Typus subfam: g. Palaeostr a tiomyia gen. nov.
C am Rande des Flugels nur bis zum Ende der RS3 reichend. RS von R an der Basis des Flugels abgetrennt. RS4 und RS2 nicht voneinander getrennl, ganz verbunden. RS3 und RS4 parallel, eine sehr lange und schmale Gabel bildend. Medialadern mit mittelgrosser Discoidalzelle: M4 vom Hinterrande der Zelle gehend. Flugel abgestumpft, ziemlich breit, viel linger als das 57
Abdomen. Кбгрег kurz und breit; Kopf nur wenig schmaler als der Thorax, breit, mit ziemlich schwach vorragendem Gesicht; Stirn am Scheitel breiter als das Auge (9). Abdomen kurz eiformig, nur mit 4 Segmenten.
Diese Gruppe ist durch ihre charakteristischen Fliigelgeader und stark spezialisierten Korperbau von alien anderen bekannten Stratiomyiden weit entfernt. Nur das viergliederige Abdomen nahert etwas diese Subfa-milie den Clitellariinae und Stratiomyinae. ]
G. Palaeostratiomyia gen. nov.
Typus generis: Palaeostratiomyia pigmaea sp. nov.
F 1 u g e 1. SC lang, nicht besonders stark, nur bis zur Mitte des Flugels reichend. R lang und stark, fast gerade. 3 Aste der RS vorhanden: RS1+2, RS3 und RS4; RS1+2 stark wellenformig gebogen; RS3 und RS4 lang und parallel; Basalteil des gemeinsamen Stammes RS3 und RS4, zwischen der Basis und rm — Ader, fast zweimal so lang als die Gabel; rm diesseits der Mitle der Diskoidalzelle liegend. 4 Medialadern; M4 von der Diskoidalzelle gehend; Mj mit RS4 parallel; M2 und M3 kurz, etwas nach hinten gebogen, einander parallel; M4 mit M3 etwas konvergierend. Enden der M2, M3 und M4 sehr fein, 4^ undeutlich Diskoidalzelle mittelgross, unregelmassig fiinfeckig; Cu und An undeutlich.
Kopf breit und kurz; Stirn breit, ca 1so breit als das Auge; Gesicht nur etwas vorstehend; Antennen und Russel undeutlich.
Thorax massiv, schlecht erhalten.
В e i n e. Hinterschenkel nu> wenig verdickt; Hinterschienen ziemlich kurz, am Ende etwas verdickt; Dome und Borsten undeutlich.
Abdomen kurz eiformig, aus 4 (2.—5. Tergite) Segmenten bestehend; 2. Tergit an der Basis etwas verschmalert; 3. Tergit am breitesten, kurz und parallelrandig; 4. und 2. Tergit gleich lang, hinten etwas verschmalert; 5. Tergit halbkreisformig.
Palaeostratiomyia pygmaea sp. nov.
Korperlange 2.75 mm; Fliigellange 2.5 mm; Flugel glashell, ungezeichnet; Korper dunkel, Kopf dunkler, fast schwarz. Hinterschenkel und Thorax dun-kel, Schienen heller; Abdomen heller als der Thorax.
1 p (?) aus Lok. Karabas-tau; Kara-tau-Gebirge (unt. Dogger). №2452/454. A. B. Martynov, 1925.
FAM. RHAGIONIDAE
SUBFAM. RHAGIONEMPIDINAE SUBFAM. NOV.
Typus subfam.: Rhagionempis gen. nov.
Kopf stark verlangert, mit breiter Stirn und grossen Antennen; 3. Anten-nenglied stark erweitert und charakteristisch gebaut; Thorax viel breiter als der Kopf; Gabel derRS3+RS4 kurz und stark divergierend; RS4 in die Fliigelspitze miindend; 4 Aste der Medialader; C nur bis zum Ende der RS4 reichend.
G. Rhagionetnpis gen. nov.
Typus generis: Rhagionempis tabanicornis sp. nov.
G ziemlich stark, am Ende schwach gebogen; R gerade und stark, etwas gebogen, bis zur Mitte des Flugels reichend; RS1+2fast gerade, etwas bogenformig im distalen Viertel; der gemeinsame Stamm der RS-Ader lang, an der Basis undeutlich; RS3+4 aus RS1+2 weit proximalwarts von der Mitte abzweigend; Gabel der RS3+4 kurz; RS3 gebogen, RS4 fast gerade; die zwei letzten Abschnitte der C fast gleichlang; 4 Medialadern; Proximalecke der Diskoidalzelle spitz, auf dem Niveau der Basis der RS3+4 liegend. Flugel 58
ziemlich breiL und am Ende abgestumpft; Proximalteil des Kostalfeldes stark verengl.
Kopf schmal, viel schmaler als der Thorax, stark verlangert; Augen ziemlich klein, zweimal schmaler als die Stirn, Ocelli deutlich: vorderer Ocellus viel kleiner als die hinteren. Stirn sehr breit und lang; Antennen gut entwi-ckelt und lang; 1. und 2. Antennenglied kurz und rundlich; Distalteil der Antennen aus grossem winkeligem 3. Antennengliede bestehend; 3. died •am Ende mit einer spitzen und diinnen zweigliederigen Arista.
К or per ziemlich kurz und dick; Form des Thorax undeutlich; Abdomen aus 5 ziemlich langen Segmenten bestehend.
В e i n e undeutlich; hintere (oder mittlere?) Beine schlank; Dome und Borsten undeutlich.
Diese Gattung ist von alien anderen Rhagioniden durch reduzierte C, charakteristiscjien Antennenbau und Kopf stark entfernt und bildet eine be-sondere Subfamilie. Grosse, stark entwickelte Antennen nahern diese Sub-familie deutlich den Tabaninae, verkiirzte C — den Stratiomyidae, Den Empididen ist diese Gattung wahrscheinlich nur habituell ahnlich.
Khagtonempts tabanicornis sp. nov.
Dunkel gefarbte Art; Flugel glashell, ungezeichnet; Antennen hell gefarbt; Schenkel und Schienen des hinteren (mittleren?) Beinpaares hell; Korperlange 3.75 mm; Fliigellange 3.5 mm.
1 p Lok. Karabas-tau, Kara-tau-Gebirge (unt. Dogger). № 979/2465. Turutanov a-K e t о v a, 1930.
SUBFAM. VERMILEONINAE
G. Archirhagio gen. nov.
Typus generis: Archirhagio obscuras sp. nov.
Flugel G bis zur Mitte des Flugelhinterrandes zum Ende des Cu reichend; SC lang, bis zum Apikaldrittel der Flugel reichend. R stark und lang, fast gerade; RS von R in der Fliigelmitte abzweigend, in drei Aste gespaltet: RSr+2, RS3 und RS4. Gabel von RS3+4 kurz: beide Aste stark divergierend und in den Vorder- und Hinterrand einmiindend. Diskoidalzelle gut entwi-ckelt. 4 Medialader; M4 der RS4 parallel; M4 an der Basis abgebrochen, von der Form einer Querader. Cu fast gerade, sehr stark; Cubitalzelle offen. Anj fast gerade, ziemlich stark. An2 schwach und gebogen.
Kopf mit Thorax gleichbreit. Augen sehr gross, fast zusammenstoBend. 3 Ocelli, sehr klein, aber deutlich, Ocellardreieck nicht entwickelt, aber der Scheitel deutlich nach hinten vorragend. Antennen kurz und diinn; 2. Antennenglied zylindrisch, 2 x/2—3-mal so lang als dick; 3. Glied konisch, am Ende mit diinner, nackter, ungegliederter Arista.
Thorax mittelgross, stark sklerotisiert; Schildchen am Rande ohne Dome.
Beine. Hinteres Paar stark und ziemlich lang. Hinterschenkel breit und parallelseitig, wahrscheinlich stark kompressiert. Hinterschienen langer und fast zweimal schmaler als das Schenkel, am Ende schwach, aber deutlich verdickt. Hintertarsen nur wenig langer als die Halfte der Schienen; 1. Tarsenglied die Halfte der Tarsen einnehmend; 2. Glied am kiirzesten; 2., 3. und 4. Tarsenglied breit, 5. Glied schmaler; Klauen langer als das 5. Glied, diinn und schwach gebogen ((J). Empodium gut entwickelt, kurz und breit.
Abdomen fast parallelseitig, 2-mal so lang als der Kopf und Thorax zusammen und aus 7 Segmenten bestehend: 1. Tergit kurz, 2. und 3. langer, 4., 5. und 6. Tergit nur etwas kiirzer als das 3., 7. Tergit viel kiirzer als das 6., halbkreisformig.
Diese Gattung ist von alien bekannten Vermileoninen leicht kenntlich. Das Fliigelgeader ahnelt etwas Vermileo.
Archirhagio obscuras sp. nov.
Kopf und Thorax dunkel gefarbt; Flugel glashell, mit gut entwicke-Item, gelb gefarbtem Geader; Antennen schwarz: Hinterbeine gelb; Dorsalseite des Schenkels und ganze Schiene kurz und fein schwarz behaart; 1. Tarsen-glied undeutlich behaart. Abdomen, wie die Beine, kurz und fein behaart; 1. Tergit in der Basalhalfte dunkel, mit ebenso gefarbtem Fleck in der Mitte des Hinterrandes; 2. Tergit mit verlangertem, nach hinten allmahlich ver-schmalertem Mittelfleck; Hinterrand des 2., 3., 4. und 5. Tergites einfarbig hell. Korperlange 21.5 mm. Fliigellange 13.0 mm.
1 (J aus Lok. Galkino; Kara-tau Gebirge (unt. Dogger). № 2452/490. А. В. M a r t у n о v, 1925.
SUBFAM. PROTORHAGIONINAE SUBFAM. NOV.
Typus subfam.: G. Protorhagio gen. nov.
Der gemeinsame Stamm der RS sehr kurz, mehr als 2-mal kiirzer als der Abschnitt der RS3+4 von der Basis bis zur rm-Ader. Kopf breit. Korper ziem-lich massiv und kurz. Abdomen im Apikaldrittel verschmalert. Bau der Beine unbekannt.
Das Fliigelgeader und der Basalteil der RS, laBt diese Gruppe von alien anderen Rhagioniden leicht unterscheiden. Distales Geader den rezenten Rhagioninen (Rhagionini und Chrysopilini) ahnlich.
G. Protorhagio gen. nov.
Typus generis: Protorhagio capitatus sp. nov.
F 1 ii g e 1. Ziemlich schmal, kiirzer als das Abdomen. Geader schwach. C die Fliigelspitze iibergehend, Ende der Cundeutlich. SC lang und fast ge-rade, am Ende schwach. R sehr stark und lang, bogenformig. RS von R fast an der Basis abzweigend, mit 3 Asten. RS1 + 2 bogenformig, zum Ende deutlich mit R konvergierend. Gemeinsamer Stamm der RS zweimal kiir-zer als der 1. Abschnitt der RS3+4 (von der Basis bis zur rm). Abschnilte der RS3+4 diesseits und jenseits der rm gleich lang; RS3 und RS4 lang und gerade, nur sehr schwach divergierend: RS3 vor, RS4 hinter der Fliigelspitze miindend. 4 Medialaste; M4 an der Basis queraderformig, mit Cu durch mcu vprbunden; M4 und M2 fast parallel, M4 mit M2 stark divergierend und M4 parallel; alle M-Aste am Fliigelrand endend. Zwischen den distalen Teilen der M4 und Cu spurenformige Querader deutlich vorhanden. Diskoidalzelle verlangert; rm-Querader auf dem Niveau des Proximaldrittels der Disco-idalzelle liegend. Cu und An undeutlich. Kopf breit und gross, etwas schmaler als der Thorax. Augen klein, Stirn mindestens 2-mal so breit als das Auge. Antennenbau schlecht wahrnehmbar: Arista diinn und lang, borstenformig. Riissel nur wenig vorstehend, am Ende verdiinnt.
Thorax breit und ziemlich kurz. Schildchen mittelgross, am Rande dicht behaart, aber unbedornt.
Abdomen langer als der Thorax und Kopf suzammen, in der Basalhalfte breit, am Ende verschmalert; Abdominalsegmentierung undeutlich.
Protorhagio capitatus sp. nov.
Korperlange 6.5 mm; Fliigellange 5.0 mm. Fliigel glashell, ungezeichnet, Kopf dunkel, in der Mitte der Stirn stark verdunkelt; Thorax etwas heller als der Kopf; Abdomen noch heller, an der Basis und am Ende etwas dunk-ler; Abdomen fast ganz regelmassig sehr kurz, aber deutlich schwarz behaart.
1 (?) aus Lok. Karabas-tau, Kara-tau-Gebirge (unt. Dogger).
№ 2452/400. A. B. Martynov, 1925.
FAM. PROTOCYRTIDAE FAM. NOV.
Typus familiae: g. Protocyrtus gen. nov.
SC ganz fehlend. Rs einfach, ohne Aste. 2. Medialader. Cu einfach. Kopf gross, nur etwas niedriger als der Thorax. Thorax schwach gewdlbt. Abdomen an der Basis verschmiHert, distalwarts verdickt.
Diese neue Familie ist durch ihren anders gebauten Korper von rezenten Cyrtidae leicht kenntlich. Fliigelgeader einigen Cyrtinae — g. Oncodes L a t r. und Panopinae — g. Corononcodes S p e i s e r ahnlich. Letzte Subfamilie. Panopinae^ ist wahrscheinlich den Protocyrtidae am nachsten.
G. Protocyrtus gen. nov.
Typus generis: Protocyrtus jurassicus sp. nov.
R 3/5 der Fliigellange einnehmend, gerade, RS stark von R abzweigt. bogenformig; rm schief liegend, mit dem Basalteil von M und Distalteil von RS eine gerade Linie bildend; M in der Fliigelmitte gegabelt, mit kur-zem Basalteil; Aste der M schwach gebogen, stark divergierend; mcu lang, schief; Cu regelmasssig bogenformig.
Kopf. Augen klein; Kopf im Profit rundlich, vorne etwas gewolbt; Antennen unter den Augen liegend, undeutlich; Backen sehr breit, hinten gewolbt. Thorax gross undmassiv, vor den Fliigelwurzeln etwas gewdlbt; Thorax verlangert, hinten niedriger.
В e i n e kurz; Hinterschenkel und Hinterschiene parallelseitig fastgleich breit: Schienen etwas kiirzer als die Schenkel; Hintertarsen nur etwas langer als die Schienen, deutlich diinner; 1. und 2. Tarsenglied am langsten, 4. Glied am kurzesten. Klauen kurz.
Abdomen. Segmentierung undeutlich.
Protocyrtus jurassicus spz nov.
Korperlange 5.0 mm; Fliigellange 3.5 mm; Fliigel kiirzer als das Abdomen; rm dem Basalteil der Vorderaste der M und dem Abschnitt der M zwischen der Gabel und mcu gleichlang; mcu lang, ca 21/4-mal so lang als die rm.
1 (J aus unterem Dogger des Kara-tau-Gebirges — Lok. Michailovka. №2231; Turutanova-Ketova.
FAM. OMPHRALIDAE
G. Protomphrale gen. nov.
Typus generis: Protomphrale martynovi sp. nov.
F 1 ii g e 1 viel kiirzer als das Abdomen. SC kurz, am Ende zuweileh frei; Costalzelle breit, R bogenformig; RS charakteristisch dichotomisch in 4 Aste verzweigt; Stelle der Abzweigung der RS undeutlich; Costalabschnitte zwischen den R und RS fast gleichlang, nur der letzte Abschnitt zwischen RS3 und RS4 kiirzer als der vorletzte; Medialader und andere Adern undeutlich.
Kopf klein, viel kiirzer und niedriger als der Thorax; Augen sehr klein, unregelmassig eiformig; Scheitel stark gewolbt; Antennen lang, ca l^-mal so lang wie der Kopf; 1. Antennenglied 2-mal so lang wie das 2., letztes am kiirzesten, kiirzer als breits 3. Glied langer, fast 2-mal so lang wie die ersten Glieder suzammen, zylindrisch, am Ende spitz.
Thorax massiv, nur wenig konvex; Halteren kurz. aber deutlich.
В e i n e kurz und diinn, Schenkel am dicksten, kiirzer als die diinneren Schienen; Hintertarsen fast 2-mal kiirzer als die Schienen; Mitteltarsen nur etwas kiirzer als die Schienen; Hinterschienen an der Basis am dicksten, zum Ende allmahlich verdiinnt; Mittelschienen parallelseitig.
Abdomen etwas langer als der Thorax und Kopf zusammen, nach dem Ende allmahlich verschmalert und aus 8 deutlichen Segmenten beste-hend; letztes Segment abgerundet.
Protomphrale martynovi sp. nov.
Korperlange 3.5 mm; Fliigellange 1.75 mm, Kbrperfarbung hell. — 1 Exemplar aus Lok. Galkino (unt. . Dogger d. Kara-tau). № 2452/366. A. B. Martynov, 1925.
Diese neue Omphralidengattung ist von alien anderen Omphralidae durch Vorhandensein der vier RS-Asten leicht zu unterscheiden.
FAM. ARCHIPHORIDAE FAM. NOV.
Typus familiae: g. Archiphora gen. nov.
Kopf klein, rundlich; Augen unregelmassig oval; Antennen, Russel undeutlich, Kopfbeborstung wahrscheinlich fehlend.
Thorax kurz, nicht besonders stark gewdlbt, im Profil viel holier als der Kopf. Beine diinn, nicht besonders lang, unbeborstet. Fliigel ziemlich schmal, mit charakteristischem Geader: SC fehlend; R, RSX und RS2 stark und dick, dem Vorderrande stark genahert. RS3+4 einfach; Mx quer liegend; von der Form einer Querader, zwischen der RS3+4und M2; M4 vorhandenund mit Cu verbunden eine kurze Gabel bildend; zm, tp und mcu vorhanden: zwei An-Ader.
Abdomen konisch und aus 9 Segmenten bestehend.
Diese Familie ist sehr interessant und zeigt eine ausgesprochene Ver-wandtschaft mit rezenten Phoriden.
G. Arcltiph&ra gen. nov.
Typus generis: Archiphora ancestrix sp. nov.
F 1 ii g e 1. R nur x/3 der Fliigellange einnehmend, stark; RS1+2 sehr stark, am Ende gegabelt; RS3+4aus gemeinsamem Stamm von RS neben der Basis abzweigend, gerade und ohne Aste, an der Basis verdickt und nach dem Ende verdiinnt, unregelmassig gebogen; rm schief, fast in gerader Liniemit RS3+4 liegend; Mx stark, queraderformig, zwischen der M2 und RS3+4 liegend; M2 gerade, RS3+4 parallel, intermediate Querader tp nicht besonders kurz und ziemlich diinn; mcu kiirzer und dicker; M3 fehlend (oder mit M4 verschmol-zen?); M4 als ein kurzer Ast der Cu vorhanden; Anx an der Basis mit Cu ver-schmolzen, An2 stark gebogen; M4, Cu, Anx und An2 sehr fein und diinn.
Beine ziemlich kurz; vordere und hintere (?) Schenkel nur wenig verdickt; mittlere und vordere Schienen l1^ — 2-mal diinner als die Schenkel, fast parallelseitig; Tarsen den Schienen fast gleichlang, fiinfgliederig; 1. Vordertarsenglied am kiirzesten, 2. — am langsten (Verhaltnis der Glie-der=2>*4>>3>>5>>l; Verhaltnis der Mitteltarsenglieder=2>4>3>1, der Hintertarsenglieder — 1>3>4>5>*2); Borsten an den Beinen undeutlich. Thorax stark konvex, Schildchen vorragend, schmal, aber stark kon-vex.
Kopf schlecht erhalten.
Abdomen sehr breit an der Basis, zum Ende allmahlich verschmalert; 1. und 2. Tergit miteinander verschmolzen, gross; 3. bis 9. Tergit schmaler, distalwarts verringert; 9. Tergit mit einem Paar von Cerci (^).
\ Archiphora^ancestrin nov.
Dunkel gefarbte Art; Korperlange 2.0 mm; Fliigellange etwas kleiner. 1 Q aus Lok. Galkino, Kara-tau-Gebirge (unt. Dogger). № 2452/329.
А. В. M a r t у n о v, 1925. f
FAM. ALLACTONEURIDAE FAM. NOV.
Typus familiae: g. Allactoneura M e i j e г e, 1907.
SC gut entwickelt, ca ^der Fliigellange einnehmend, in Cmiindend; RS einfach, ohne Aste, von R normal abzweigend; rm kurz, quer liegend. Medial-62
adern gut entwickelt; M3 lang, von Cu abzweigend, beide vordere Medial-aste gut entwickelt. Antennen nicht besonders lang, diinn, dem Thorax und Kopf zusammen gleich. Coxen gross; Beine diinn; Abdominalbeborstung nur wenig entwickelt.
Diese Familie ist auf einer rezenten javanischen Gattung und vier juras-sischen Gattungen aus Kara-tau begriindet. Durch den Bau der R,RS nahert diese Gruppe den Diadocidiidae, durch Medialader — den Bolitophilidae und Fungivoridae.
G. Pleciofungivora gen. nov.
Typus generis: Pleciofungivora latipennis sp. nov.
F 1 ii g e 1 ziemlich breit, am Ende abgerundet; Flugel etwas langer als das Abdomen; SC lang, fein, regelmassig bogenformig und in die Costalader ein-miindend; R stark, fast gerade; RS stark, naher zur Fliigelbasis als Niveau dem Ende der SC aus R abzweigt, mit einem Vorderaste; RS distalwarts von rm fast gerade und nur am Ende etwas nach hinten gebogen, — M3 von +2 deutlich abgetrennt; rm kurz und gerade, naher der Fliigelbasis als der Mitte des gemeinsamen Stammes von M1+2; Mx und M2 fast parallel, nur etwas konvergierend; Cu und M3 verschmolzen; An undeutlich.
Thorax gewolbt; Schildchen nicht vorragend; Vorderteil des Thorax nur etwas iiber dem Kopf vorstehend.
Kopf im Profil viel kiirzer als hoch, winkelig, mit starkem Ocellenhocker; Augen oval; Antennen nicht besonders lang, diinn: Zahl der Antennenglieder unklar. Riissel undeutlich.
Beine diinn und lang; Coxen ziemlich kurz; Schenkel diinn und lang; Vorderschienen langer und etwas diinner als die Schenkel, Hinterschienen so lang, aber etwas diinner als die Hinterschenkel. Tarsen den Schienen gleich lang, sehr schlecht erhalten.
Abdomen mehr als l^-mal so lang als der Kopf und Thorax zusammen, am Ende spitz; Abdominalsegmentierung undeutlich.
Pleciofungivora latipennis sp. nov.
Korperlange 2.5 mm; Fliigellange 2.0 mm.
1 Q (?) aus Galkino, Kara-tau-Gebirge (unt. Dogger). № 2452/323. А. В. M a r t у n о v, 1925.
G. Allactoneurites gen. nov.
Typus generis: Allactoneurites jurassicus sp. nov.
F 1 ii g e 1 lang und schmal; SC ca 1/3 der Fliigellange einnehmend, in C mii-ndend, fein und diinn, regelmassig gebogen; R starker, sehr schwach bogenformig, 2-mal so lang wie die SC; RS aus R spitzeckig abzweigend, in die Fliigelspitze einmiindend, mit einem Vorderaste; gemeinsamer Stamm von M1+2 deutlich langer als die Gabel; rm kurz neben der Gabelung der R und RS liegend; Mx und M2 fast parallel, sehr schwach divergierend; M3 (oder Cu4-M3?) lang, regelmassig gebogen, an der Basis der Fliigel von M abzweigend. Korper sehr schlecht erhalten.
Allactoneurites jurassicus ep. nov.
Korperlange 2.5 mm; Fliigellange 3.5 mm.
1 Exemplar aus Galkino, Kara-tau-Gebirge (unt. Dogger). № 2452/356. А. В. M a r t у n о v, 1925.
Diese neue Gattung ist Pleciofungivora am nachslen und durch langere RS und Fliigel leicht kenntlich.
G. Pleciomima gen. nov.
Typus generis: Pleciomima sepulta sp. nov.
F 1 ii g e 1 etwas verlangert, oval; SC feinund diinn, in C miindend; R fast gerade, ziemlich dick und stark; RS einfach, aus R spitzeckig abzweigend, in die Fliigelspitze miindend; rm schief, neben der Abzweigung von RS liegend; gemeinsamer Stamm von M sehr kurz, aus Cu abzweigend; gemein-samer Stamm der vorderen Aste der MiMjUnd M2) lang, der oder M2fast gleichlang; Mj und M2 fast parallel; M3 lang, in distaler Halfte etwas gebo-gen; Cu in basalen 2/3 gerade, am Ende stark bogenformig.
Thorax stark konvex, iiber dem Kopf vorragend. Schildchen rundlich, nur wenig vorstehend.
Kopf im Profil unregelmassig abgerundet mit stark vorragender Stirn, welche die nicht besonders lange Antennen tragt. Antennen dem Kopf und Thorax zusammen etwa gleichlang; Antennen etwas gezahnt, 12-gliederig; Augen oval, viel hoher als lang; Ocellenhocker undeutlich.
Beine stark, nicht besonders lang; Coxen lang, gutentwickelt; Schenkel in der Mitte etwas verdickt, zu beiden Enden verdiinnt; Hinterschienen so lang, Mittelschienen etwas langer als die Schenkel; Tarsen fiinfgliederig, mit schwach erhaltenen Gliedergrenzen; Tarsen nur etwas langer als die Schienen; Vorderbeine ganz undeutlich.
Abdomen aus 6 gut abgegrenzten Segmenten undGenitalien bestehend.
Pleciomima sepulta sp. nov.
Korperlange 4 mm; Fliigellange 2.5 mm.
1 aus Galkino, Kara-tau-Gebirge (unt. Dogger). № 2452/323. A. B. Martynov. 1925.
Diese neue Gattung ahnelt durch den Fliigelgeader Pleciofungivora, ist aber wegen der gut entwickelten Cu und starker Coxal leicht kenntlich.
G. Fungivorites gen. nov.
Typug generis: Funqivorites latimedius sp. nov.
F 1 ii g e 1 von gewohnlicher Form, nur wenig verlangert, breit; SC fein und lang, am Ende undeutlich; R fast gerade, stark; RS stark, im distalenVier-tel deutlich nach hinten gebogen; diese Ader dem Fliigelrande parallel; rm gerade und kurz; basaler Abschnitt der RS zweimal so lang als die rm; M im Basalteile des Fliigels mit Cu verschmolzen; rm fast in der Mitte des unpaarigen Teiles der M liegend; M£ und M2 an der Basis stark divergierend, ungerelmassig gebogen; diese Adern distalwarts fast gerade (Cu4-M3?); Flii-gelteile hinter der Cu schlecht erhalten, Geader undeutlich.
Beine schlecht erhalten; Coxen wahrscheinlich lang; Schenkel kurz und stark verdickt; Schienen zylindrisch; Tarsen undeutlich.
Abdomen gross, im Profil oval, am Ende abgerundet; 7 Tergite und 5 Sternite deutlich, mil breiter pleuraler Membran; Cerci undeutlich.
Fung Ivor ites latimedius sp. nov.
Korperlange 3.5 mm; Fliigellange 3.0 mm.
1 Q aus Lok. Galkino. Kara-tau-Gebirge (unt. Dogger). № 2452/548. A. B. Martynov. 1925.
SUBFAM. ANTIQUAMEDIINAE SUBFAM. NOV.
Typus subfamiliae: g. Antiquamedia gen. nov.
SC kurz, nach hinten gerichtet, und in R miindend. RS einfach, ohne Aste, aus R spitzwinkelig abzweigend; rm undeutlich, wahrscheinlich ganz fehlend. M gut enlwickelt: der gemeinsaine Stamm der Mx und M2 auf dem 64
JNiveau des Endes von SC aus M3 abzweigend; distalen Asten (Mx, M2) fast gleichlang. M3, Cu und An gut entwickelt. Coxae wahrscheinlich kurz und schwach. Borsten an den Beinen undeutlich.
Diese Subfamilie ist durch charakteristisch entwickelte M und undeut-liche rm leicht kenntlich. Durch SC und starke Cu und An ist sie den Fun-givoridae. durch die Gabel RS—R mit Allqctoneurinae verwandt.
G; Antiquamedia gen. nov.
Typus generis: Antiquamedia tenuipes sp. nov.
IF 1 u g e 1 nur etwas verlangert; SC sehr kurz, deutlich kiirzer als x/4 der Fliigellange, am Ende nach hinten gebogen und in R miindend; R lang, regelmassig gebogen; RS einfach, aus R fast an der Fliigelbasis abzweigend; vor der Fliigelspitze in C miindend; M3 sehr lang, bogenformig; Cu gebogen; in der Distalhalfte etwas gebogen; An lang und gut entwickelt. Abdomen kiirzer als der Fliigel.
Thorax konvex. Kopf gross; Russel der Kopfhohe gleichlang; Antennen nicht besonders diinn, dem Kopf und Thorax zusammen gleichlang.
Beine diinn und lang. Schienen und Schenkel nicht verdickt.
Antiquamedia tenuipes sp. nov.
Korperlange 2.75 mm; Fliigellange 2.0 mm.
1 Exemplar aus Lok. Michajlovka, Kara-tau-Gebirge (unt. Dogger). -№ 2335/75. Turutanova-Ke tova.
SUBFAM. ANTEFUNGIVORINAE SUBFAM. NOV.
Typus subfamiliae: G. Antehtngivora gen. nov.
SC gut entwickelt, nicht besonders kurz, ca x/4der Fliigellange einnehmend, in C miindend. RS einfach, aus R spitzwinkelig abzweigend; rm kurz, senkrecht liegend. Medialadern gut entwickelt; M3 sehr kurz, aus distalem Teile von Cu abzweigend, und M2 lang und gut entwickelt. Fliigel breit, Hlreieckig. Antennen lang und diinn, dem Thorax und Kopf zusammen gleichlang, Coxae gross und massiv; andere Beinenteile diinn. Borsten undeutlich.
< Die Subfamilie Antefungivorinae ist durch den Bau der M den Fungiroridae deutlich genahert. Anderes Merkmal die Gabel der RS — R weist deutlich auf Ailactoneurinae und Antiquamedijnae hin.
G. Antefungivora gen. nov.
Typus generis: Antefungivora prima sp. nov.
iF 1 ii g e 1. SC am Ende bogenformig, in C miindend; R schwach, aber deutlich wellenformig, RS starker wellenformig gebogen, hinter der Fliigelspitze in den Fliigelrand einmiindend; Zelle R im Distalteile stark erweitert, der gemeinsame Stamm von M kurz, ca 2-mal kiirzer als die Mx; Mx und M2 regelmassig divergierend; Mx gerade, M2 in der Basalhalfte gebogen. Cu an der Basis der M stark genahert und mit M3 bis zum distalen Drittel ver-schmolzen; An lang, am Ende stark nach hinten gebogen.
Kopf. Augen mittelgross, oval; Vorderrand des Kopfes mit undeutli-chen Ocellenhocker; Antennen schlecht erhalten.
Thorax ziemlich konvex, etwas iiber den Augen vorragend, Halte-ren gut bemerkbar, kugelig.
Beine schwach erhalten, diinn und lang; Schenkel und Schienen wahrscheinlich nicht verdickt.
Abdomen schlecht erhalten.
э Tp. Палеоыг. ин-та, т. VI, в. 3. 65
Antefungivora prima sp. nov.
Korperlange 2.0 mm; Fliigellange 1.5 mm.
1 Exemplar aus Lok. Galkino, Kara-tau-Gebirge (unt. Dogger). № 2452/332. A. B. Martynov, 1925.
FAM. PENTHETRIIDAE
G. Mesoplecia gen. nov.
Typus generis: Mesoplecia jurassica sp. nov.
SC diinn, aber deutlich, bis zur Mitte des Vorderrandes gehend. R gerade; RS zweiastig, aus R spitzeckig abzweigend; RS neben der rm gegabelt; vorderer Ast der RS weit vor der Fliigelspitze endend; hinterer Ast viel langer als der vordere, am Ende nach hinten gebogen und in die Fliigelspitze miindend; rm lang, senkrecht. Der gemeinsame Stamm der M an der Fliigelbasis aus Cu abzweigend, lang; Mx und M2 lang; ihr gemeinsamer Stamm viel kiirzer als die Aste; M3 aus dem gemeinsamen Stamm spitzwinkelig abzweigend; mcu — Querader nur etwa jenseits der Abzweigung der M3 liegend. Cu gerade und nur im Distaldrittel nach hinten gebogen.
К о p f. Augen oval. Vorderrand des Kopfes hinten stark vorstehend. Antennen langer als der Kopf; Antennenglieder zylindrisch, zweimal so lang als dick.
Thorax massiv, aber nur etwas gewolbt.
Abdomen und Beine schlecht erhalten.
Mesoplecia jurassica sp. nov.
Korperlange 7.5 mm; Fliigellange 7.0 mm; Korper dunkel gefarbf; Fliigel etwas pigmentiert, braunlich.
1 Exemplar aus Lok. Mighajlovka, Kara-tau-Gebirge; (unt. Dogger). №2231. Turutanova-Ketova.
Diese neue Gattung ist Plecia am nachsten und durch spitzwinkelige Abzweigung von M3 und lange Antennen leicht kenntlich.
ZUSAMMENFASSUNG
1. Es werden 15 Dipterenarten aus dem Jura von Kara-tau beschrieben, die zu 15 neuen Gattungen und zu 9 Familien gehoren, von denen 5 sich als neu erwiesen.
2. Einige von den beschriebenen Gruppen zeigen ein deutliches «Vorfah-ren»-Geprage, was uns die Aufklarung der Phylogenie rezenter Gruppen ermoglicht. Hierher gehoren: Protomphrale martynovi, Archiphora ance-strix. Fam. Allactoneuridae, weiter Protorhagio capitatus, Rhagionempis tabanicornis, welche Vorfahren einer Reihe von rezenten Familien und Gattungen darstellen. *
3. Neben den angegebenen «Vorfahren»-Formen wurde eine ganze Reihe eigenartiger, teils spezialisierter Arten entdeckt, die zweifellos keinerlei Zusammenhang mit irgendwelchen rezenten Dipteren haben. Hierher gehoren: Archisargas pulcher^ Palaeostratiomyia pygmaea, Protocyrtus juras-sicus.
4. Der Verfasser gibt eine Ubersicht der Beschreibungen jurassischer Diptera Brachycera und stellt die Unterschiede zwischen dieser Fauna und der gegenwartigen fest. Als Hauptgruppe, welche die Grundlage der ganzen Fauna bildet, ist die Familien-Serie Eremochaeta — Stratiomyidae und Bhagionidae anzusehen. Die hohe Spezialisation einer Reihe Formen wird konstatiert. Wenig spezialisierte, primitive Formen sind nicht zahlreich.
5. Die Familie Archiphoridae wird auf Grund der neuen Gattung Archiphora aufgestellt, die sich durch eine Reihe primitiver Merkmale auszei-chnet, ein deutliches Bindeglied zwischen der rezenten hoch spezialisierten Familie Phoridae und anderen (Empididae) bildet, indem sie deren Vorfah-ren-Gruppe reprasentiert. Das Auftreten der Fam. Archiphoridae in der Jura ist sehr bemerkenswert, da die Fam. Phoridae zur Unterordnung der Diptera Cyclorrhapha gehort. Es ist sehr wahrscheinlich, dass auch der beschriebene Vorlaufer dieser Familie ebenfalls zur Unterordnung Cyclorrhapha ge-hort, die bis jetzt nur vom Oligocan an bekannt war.
6. Es ist sehr schwer, die beschriebene Dipterenfauna vom okologischen Standpunkt zu charakterisieren, da sie von der gegenwartigen Fauna scharf abweicht. Auf Grund des Vorhandenseins der Vertreter solcher Gruppen wie Vermileoninae, Omphralidae und Penthetriidae, sowie auf Grund indirekter Erwagungen uber die Biologie der Nematocerenserie Fungivoroidea kann angenommen werden, dass die Mehrzahl der beschriebenen Juradipteren aus pflanzenfressenden Arten bestand, die offenbar zu feuchten Landschaf-ten hinneigteh.
7. Die geschilderte jurassische Fauna der Diptera Brachycera kann mit keinerlei anderen mesozoischen Dipterenfaunen verglichen werden, da bis jetzt nur ein einziger Brachycerenvertreter aus dem Lias Deutschlands bekannt ist (Nemestrinidae?).
5*
, €7
АКАДЕМИЯ НАУК СССР
1938. ТРУДЫ ПАЛЕОНТОЛОГИЧЕСКОГО ИНСТИТУТА, том VII, вып. 3
А. В. МАРТЫНОВ
ПЕРМСКИЕ НАСЕКОМЫЕ АРХАНГЕЛЬСКОЙ ОБЛАСТИ (Б. СЕВЕРНОГО КРАЯ). ЧАСТЬ V. —СЕМ. EVTHYGRAMMIDAE И ЕГО ОТНОШЕНИЯ (С ОПИСАНИЕМ ОДНОГО НОВОГО РОДА И СЕМЕЙСТВА ИЗ ЧЕКАРДЫ)
(с 5 рис.)
А. В. MARTYNOV
PERMIAN FOSSIL INSECTS FROM THE ARKHANGELSK DISTRICT. PART V.—THE FAMILY EUTHYGRAMM1DAE AND ITS RELATIONSHIPS (WITH THE DESCRIPTION OF A NEW GENUS AND FAMILY FROM CHEKARDA)
(with 5 figures)
Семейство Euthygrammidae Mart, с родом Euthygramma Mart, из Ива-Горы (Eu. parallellum Mar t.) было описано мною в 1928 г., и тогда же я определенно указал, что это семейство родственно каменноугольному семейству Caloneuridae Handl., но по крыльям являет высшую ступень его специализации.
Вскоре мною был описан (1930) другой вид рода Euthygramma, Ей. curvatum из Тихих Гор, мало отличающийся от первого. Было отмечено, что жилкование крыльев Euthygrammidae и Caloneuridae в некоторых чертах еще не выяснено.
В том же 1930 г. описан новый род Р aleu thy gramma, с видом Р. tenuicor-nis Mart., происходящий из окрестности дер. Чекарды на р. Сылве (кунгурский ярус). Передние крылья этого рода несколько примитивнее в том отношении, что RS образует здесь небольшой развилок. У описанного экземпляра сохранились голова, грудь, части ног, и видно, что усики были тонкие, с вздутым основным члеником, переднегрудь небольшая, ноги довольно длинные, бегательные или ходильньр. Как отмечено на рис. 7 (стр. 46 упомянутой работы), М занимает на крыле вогнутое положение, подобно RS.
Ю. М. Залесский описал zb 1933 г. третий род семейства — Vilvia Z а 1. (У. densinervosa Z а 1.) — с р. Вильвы, близ ст. Чусовой. У этой формы RS давал Htf одну, а две ветви сзади; по этому признаку этот род занимает промежуточное положение между Caloneuridae и Euthygramma, однако анальная область была у него так же вытянута, как, напрпйер, у Р aleu thy gramma.
Вот все, что описано до сих пор из этого семейства. В новых материалах с р. Сояны я обнаружил около двадцати пяти отпечатков передних крыльев вида, который я склонен считать за Ей. parallellum, несмотря на не
которые мелкие отличия. Кроме того, оттуда же имеется один экземпляр другого вида и один экземпляр, относимый мною к новому роду.
Интерпретация жилкования дается мною несколько иная, чем раньше; основания ее будут рассмотрены ниже.
Октябрь 1937 г.
СЕМ. EUTHYGRAMMIDAE MART.
Martynov, 1928. Trav. Mus. Geol. Acad. Sci. USSR, IV
Передние и задние крылья вытянутые; прекостального поля нет; продольные жилки прямые и идут наружу почти параллельно друг другу; SC длинная; RS отходит от R около середины крыла, обычно простой, реже дает 1—2 ветви в дистальной части. М рано, еще в основной части крыла, делится на две ветви, из которых передняя, или МА, обычно дает довольно короткий развилок, а задняя, или МР, простая. Кубитус весьма рано, почти в основании крыла, делится на две простые прямые ветви, обычно сближенные, но иногда, наоборот, СиА сближен с MP, а СпР протекает на том же расстоянии от СиА, как и другие жилки. Анальных жилок две; обе они длинны и идут почти параллельно наружу.
В отличие от обычного, МА вогнутая, а МР — выпуклая жилка; СиА, как и СиР, вогнутые, анальные жилки выпуклые. Продольные жилки связаны довольно правильными густыми рядами поперечных.
Задние крылья сходны с передними по жилкованию, размерам и форме: анальная область, следовательно, очень слабо расширена. Голова небольшая: основной членик усиков толстый, вздутый, сами усики тонкие, длинные. Переднегрудь небольшая, средне- и заднегрудь значительно больше и шире. Ноги тонкие, сравнительно длинные; задние ноги немногим длиннее прочих, не прыгательные.
К этфму семейству относятся три рода: Euthygramina Mart. ср. Сояны и из Тихих Гор, Paleuthy gramma Mart. ср. Сылвы, у Чекарды и Vilvia Z а 1. ср. Чусовой х.
Мы дали новое, более подробное описание сем. Euthygrammidae потому, что, во-первых, в прежнем (1928) определении рассматривались только крылья и, во-вторых, теперь я даю иную интерпретацию жилкования, отличную от прежней. Прежде я считал медиану простой, дающей лишь концевой развилок, теперь я должен считать ее двойной, состоящей пз длинных МА и МР. Соответственно изменяются обозначения следующих жилок, которые будут, следовательно, СиА, СиР, Ах и А2.
Сем. Euthygrammidae близко к каменноугольному сем. Caloneuridae, но последнее отличается более сложным строением RS, а в связи с этим и направлением ветвей кубитуса, анальных, а отчасти и медианы, которые направлены косо назад и наружу. В филогенезе Euthygrammidae, несомненно, имело место сжатие RS и выпадение его ветвей, а также сжатие кубитальной и медиальной областей и вытягивание анальной. Некоторым подтверждением этого вывода является 1) то, что более древний (кунгурский) род Ра leu thy gramma имел небольшой развилок на RS, а ветви кубитуса были немного более косы, чем у Euthy gramma, и 2) то, что у еще более древнего рода Vilvia RS давал сзади не одну, а даже две ветви.
Род Euthygram/ma Mart.
Martynov, 1928. Ann. Mus. Geol. Acad. Sci. USSR, T. IV, p. 50
RS отходит от R против середины крыла, простой; МА кончается развилком, длина которого лишь немного больше половины длины RS; СиР
1 Отложения под дер. Чекардой мы считаем кунгурскими. Отложения по р. Чусовой 10. Залесский называет артинскими, но для меня возраст их не совсем ясен, так как каких-либо доказательств или ссылки у автора нет; возможно, что они также относятся к кунгуру.
Отложения по р. Сояне синхроничны тихогорским, и мы считаем их принадлежащими спириферовому горизонту (скорее второй его половине).
сближен с CuA, и лишь в основной части эти жилки слегка расходятся; концы ветвей кубитуса лишь немного не доходят до конца крыла; Ах и А2 в основной части сближаются и даже соединяются друг с другом, а затем в самом основании опять расходятся.
Euthygramma parallelism Mart.
(рис. la, 1b и 2)
Martynov, 1928.fAnn. Mus. Geol. Acad. Sci. USSR, T. IV, p. 50, Pl. II, 5, XI, 1
Этот вид был описан и изображен мною по экземпляру, у которого жилкование в апикальной части сохранилось недостаточно. Мы даем теперь новое описание по экземплярам, жилкование которых сохранилось лучше; интерпретация жилкования дается теперь иная.
Рис. 1а. Переднее крыло Euthygramma parallellum Mart, (fore wing).
Передние крылья вытянутые, суживающиеся к основанию, SC тонкая, в дистальной части сближающаяся с передним краем и, наконец, исчезающая. RS отходит от R посередине длины крыла; М делится обычно в конце основной четверти крыла (рис. 1 и 2) или несколько раньше; МА образует развилок, который лишь немного длиннее половины RS; передняя ветвь его параллельна RS, задняя на конце загибается более или менее вперед. Обе ветви кубитуса сближены, но в основной четверти расходятся с тем,
Рис. 1b. Основная часть крыла (basal portion of fore wing).
чтобы в самом основании соединиться в очень короткий общий ствол. At проходит почти параллельно СиР, а на конце слегка загибается вперед; А2 кончается значительно раньше Ах; в основной части обе жилки сближаются или даже касаются друг друга, а в самом основанииг в передних крыльях опять расходятся (рис. 1).
Поперечные жилки расположены правильно и довольно часто. Длина передних крыльев вариирует от 18 до почти 25 мм, но обычная длина их 21— 24 мм.
Сюда относится до 25 экземпляров, в большинстве из Летополы и Ива-Горы.
Экземпляры пз Летополы: № 451/117, 447/117, 390/117, 380/117 402/117, 317/117, 432/117, 510/117, 385/117 (Я. Д. Зеккель).
Экземпляры из Ива-Горы: №*315/2334, 316/2334, 336/2334, 357/ 2334, 283/2334, 160/94,191/94, 101/94,603/94, 182/94, 625/94 (М. А. Е дем с к и й).
Экземпляры из Шеймо-Горы: № 510/117, 511/117 (Зеккель). 100/94, 169/94 (Е д е м с к и й).
Почти все экземпляры представляют собой отпечатки передних
крыльев; в двух случаях видны сверху два передних крыла и остатки
груди между ними, но сохранность этих отпечатков не очень хорошая.
Размеры, как указано, сильно вариируют, жилкование также подвержено некоторой вариации: развилок МА то длиннее половины RS, то равен ей; М делится то в концевой части первой четверти, то раньше, почти как на рис. 1, табл. XI работы 1928 г. СпА по большей части расположен так, как это показано у нас на рис. 1, но иногда он кажется проходящим ближе
к MP, как на рис. 1, табл. XI работы 1928 г. Задняя ветвь развилка медианы загибается вперед то резко, то более слабо.
Цепь переходов, связывающая форму, изображенную нами на рис. 1 и 2, с описанной в 1928 г., склоняет нас к заключению, что все это — один, сильно вариирующий и очень обычный вид. Так как у экземпляра, описанного нами в 1928 г., жилки в апикальной части не яв-
Рис. 2. Основная часть переднего крыла другого экземпляра Euthygramma parallellum Mart. (basal portion of fore wing of another specimen).
ственны, то я не думаю, чтобы он составлял исключение; наверно, и тут МА2 загибается вперед.
В 1930 г. мы описали из Тихих Гор часть крыла Euthygramma сиг-vaturn Mart, с жилкованием, очень похожим на жилкование многих наших экземпляров с р. Сояны. Тихогорский экземпляр отличается лишь немного более густым расположением поперечных жилок и некоторым отдалением RS от R. Если бы такой экземпляр нам попался с р. Сояны, мы зачислили бы его в Ей. parallellum; очень возможно, что это тот же вид, а если и особый, то очень близкий к parallellum. Я считаю теперь медиану Euthygrammidae состоящей из двух главных ветвей, МА и МР, так как всюду они соединяются в основной четверти нормальным порядком. Равным образом кубитус, как это ясно на рис. 1, тоже состоит из двух главных ветвей, СпА и СпР, которые в дальнейшем сближаются между собой. Как сказано выше, МР выпуклая, СпА и СпР вогнутые. Это противоречит обычному представлению, по которому СпА всегда выпуклый, а МР вогнутая. Однако это представление вполне правильно лишь в отношении РРаео-ptera, у Aeoptera же дело обстоит нередко совершенно иначе. Так, у Tct-tigoniodea и Haglodea СпА вогнутый, а СпР выпуклый; сходное положение имеем и у Gryllodea ((J). Выпуклый сильный СпР мы находим также у Glosselytrodea.
Правильность чередования выпуклостей п вогнутостей установилась только у Palaeoptera, у которых крылья рано сделались перепончатыми. Надкрылья не приобрели гофрированности, а выпуклости и вогнутости у них образуются под влиянием других причин, чего я кратко коснулся в своей статье в 1930 г.
Задние крылья должны быть сходны с передними, но анальная область, наверно, была расширена, хотя и очень слабо. Удовлетворительных остат-
ков тела, ног и головы у Ей, parallelism не сохранилось; о них мы можем судить по остаткам их у Paleuthygramma tenuicornis Mart., описанных и изображенных нами в статье 1930 г. Усики у этой формы были тонкие, длинные, с сильно вздутыми основными члениками; переднегрудь была небольшая, значительно уже средне- и заднегруди; ноги довольно тонкие и длинные, задние почти таких же размеров, как и средние и передние.
Euthygranima aberrans sp. nov.
? (рис. 3)
№ 313/2334. Ива-Гора. Е д е м с к и й, 1929.
Переднее крыло имеет ту же форму, как у Ей. parallellum, но немного меньше; длина тела его 17.5 мм.
Субкоста, радиус, анальные жилки и полоса из двух кубитальных жилок расположены, как у предыдущего вида, но существенные отличия имеются в строении RS и М. RS отходит также против середины крыла, но дает с задней стороны две ветви, из которых задняя к концу загибается вперед. Медиана представлена одной простой жилкой.
С первого взгляда кажется, что особенности RS и М здесь настолько своеобразны, что в описанной форме мы должны видеть, по крайней мере,
Рис. 3.. Переднее крыло Euthygrarnma aberrans sp. nov. (fore wing).
особый род; однако задняя ветвь RS так сходна с задней ветвью МА у Eulhy-grammd parallellum, что ее принадлежность к передней медиане (МА) мне представляется несомненной. На мой взгляд и передняя ветвь развилка МА не исчезла, а представлена средней веточкой RS. Все это приводит нас к заключению, что МА объединилась с RS, в результате чего и получилось описанное выше уродливое расположение. Мы не знаем, устойчиво оно, или нет; в последнем случае описанная форма должна представлять собою лишь аберрацию Ей. para Helium.
Род Vilviopsis gen. nov. ♦
Сектор радиуса отходит от R значительно раньше середины крыла и на конце дает сзади одну или две ветви; медиана делится, как обычно, рано, причем МА образует в дистальной части крыла развилок, а МР остается простой. СиА сближен с МР на всем протяжении, а СиР проходит на некотором расстоянии от него; А] иА2 — обычные.
Сюда относится пока один вид — Vilviopsis extensa sp. nov. из Шеймо-Горы.
Vilviopsis extensa sp. nov.
(pnc. 4)
№ 498/117. Шеймо-Гора, на p. Сояне. 3 e к к e л ь, 1935.
Передние крылья сильно вытянутые, узкие. RS отходит от R в начале второй четверти крыла и дает в дистальной части две ветви, идущие наружу. МА образует развилок, длина которого^равна половине длины RS; МР простая и сближена с СиА, образуя вместе двойную полосу, на конце слегка загибающуюся назад и кончающуюся перед апикальной частью крыла, на уровне начала первой ветви RS. СиА не сближен с СиР и проходит
параллельно ему; АхиА2 кончаются по заднему краю раньше, чем у предыдущего рода. Поперечные жилки располагаются правильными рядами; в узкой полосе МР—CuA они присутствуют.
Крупная форма: длина переднего крыла 28 мм. Описанная форма очень напоминает по своему жилкованию род Vilvia G. Z а 1. (V. densinervosa Z а 1.), но у последней формы МА, по изображению Залесского, простая. Кроме того, ветви RS отходят от R под большим углом, а поперечные жилки расположены значительно гуще; размеры сходные.
Крыло Vilvia densinervosa сохранилось не полностью, как и крыло нашей' формы; но все же сохранившиеся части позволяют заключать, что они относятся к двум близким родам; считать их одним родом, однако, уже представляется невозможным, раз МА в одном случае простая, в другом с развилком.
Рис. 4. Переднее крыло Vilviopsis extensa sp. nov. (fore wing).
Vilvia densinervosa найдена в отложениях по р. Вильве близ ст. Чусовой. Возраст ее обозначается Ю. Залесским как артинский, но у меня нет уверенности, что он действительно столь древний.
ОТНОШЕНИЯ СЕМ. Е U ТН Y G R АМ MI D А Е
Известное только из перми сем. Euthygrarnmidae близко стоит к сем. Caloneuridae Н a n d 1., известному из каменноугольных отложений Западной Европы (Commentry и Северная Франция). У Caloneuridae мы встречаемся с подобными же прямыми, неанастомозирующими продольными жилками и с подобными же густыми рядами поперечных, но RS давал всего 4—7 ветвей, в связи с чем ветви медианы и кубитуса шли более косо назад. Медиана также состояла из двух главных ветвей, из которых передняя (МА) в роде Caloneura Br’ongn. давала развилок. CuA проходит то довольно близко к MP (Caloneura В г о n g п..Соп/usio Hand 1.), то* на обычном расстоянии. Если судить по рисункам Гандлирша для Homaloptila similis Meunier (1919, фиг. 42 и 43), то здесь в задних крыльях М состояла из двух ветвей, а в передних крыльях МР, повидимому, даже сливалась с CuA, образуя одну более толстую жилку. Все эти картины очень напоминают таковые у Euthygrarnmidae. Си состоял из подобных же CuA и СиР, но анальных жилок у некоторых форм было не две, а три, т. е. А2, вероятно, давала сзади одну ветвь.
Некоторое вытягивание кубитальных и анальных жилок мы замечаем уже у Confusio Handl., ay Pruvostiella Handl. (Euthyneura P r u-v о s t) оно становится еще яснее.
У Euthygrarnmidae этот процесс продвинулся еще дальше, что повлекло за собою полное исчезновение ветвей RS у Euthygramma. Euthygram-midae являют собою, таким образом, дальнейший этап специализации жилкования, дошедший до своего предела в верхнепермском роде Euthygramma.
У Caloneuridae задние крылья также мало отличались от передних, как у Euthygrarnmidae. Ноги были относительно довольно тонкие и длинные, задние немного длиннее и крепче средних, усики длинные, с утолщенным основанием. Повидимому, сходны были и некоторые другие семейства.
Итак, из предыдущего, я думаю, совершенно ясно, что сем. Euthygrammidae близко родственно каменноугольному сем. Caloneuridae, и можно удивляться тому, что Гандлирш, выделяя в отряде Protorthoptera группу Caloneurodea (1930, стр. 688), вовсе не упомянул о сем. Euthygrammidae, а включил в нее семейства Emphylopteridae Р г п v о s t, Stenaro-ceridae H a n d 1. и, предположительно, Laspeyresiellidpe Schlecht.
СОСТАВ ОТРЯДА CALONEURODEA
Мы выделили в другой работе (1938) группу Caloneurodea в особый отряд, но состав его принимаем несколько иным, чем у Гандлирша. В отряд этот входит несколько семейств из карбона и перми. Из каменноугольных семейств, кроме центрального сем. Caloneuridae, сюда входят следующие семейства:
1. Protocollariidae Handl. На сходство образующего это семейство рода Protocollaria (Р. ingens В г о n g п., Комментри) с родом Caloneura обратил внимание Л я м е е р еще в 1917 г. и на основании нового отпечатка этой формы привел некоторые дополнительные данные об антеннах, груди и крыльях, которые также свидетельствуют в пользу известной близости этого рода к Caloneuridae.
2. Сем. -Stenaroceridae Hand]. Сюда относится род Stenarocera В г о n g п. с двумя видами из Комментри. Насколько можно судить по рисунку S. pachytyloides В г о n g п., в характере жилкования этой формы, а также в очертаниях небольшой переднегруди и передней ноги есть определенное сходство с Caloneuridae, почему мы также склонны включить это семейство сюда же. R отличается богатством своих косых ветвей, if ветви эти связаны такими же тесными рядами поперечных жилок, как у Caloneuridae и Euthygrammidae.
Можно ли отнести к тому же отряду сем. Laspeyresiellidae Schlecht.? Мне не совсем ясно жилкование крыльев у относящегося сюда вида рода Ldspeyresiella Schlecht., так как оно сохранилось недостаточно.
Наконец, что касается сем. Emphylopteridae Р г u v о s t, которое этот автор включил - было в отряд Hapalopteroidea, а Гандлирш перенес в свои Protorthoptera, то я не стану отрицать того, что в жилковании переднего крыла Emphyloptera pecrivaini Р г u v о s t (Северная Франция) есть некоторое сходство с Caloneuridae, тем не менее все же отцошения здесь более отдаленные. Жилкование переднего крыла Emphyloptera еще весьма примитивно, по крайней мере в том отношении, что анальные жилки, ку-битус и медиана здесь еще совершенно не вытянуты в длину.
Итак, в каменноугольное время отряд Caloneurodea был представлен довольно богато и включал в себя разнообразные семейства. Все они обитали в странах Западной и Центральной Европы и к концу карбона, видимо, вымерли. В перми, а именно в Канзасе и в северной Европейской части СССР, т. е. уже в умеренной зоне, мы встречаем другие семейства.
Основным семейством является сем. Euthygrammidae из отложений кунгурского и казанского ярусов Европейской части СССР. Это семейство явно родственно Caloneuridae, но обнаруживает черту более высокой специализации жилкования крыльев.
Кроме этого семейства, мы причисляем сюда еще два семейства — Permobiellidae (Till.) Mart., из отложений Канзаса, и Synomalopti-lidae, fam. nov., из отложений Чекарды.
Род Permobiella с видом Р. perspicua Till, был описан Т и л л ь я р-д о м в 1937 г. и ошибочно отнесен к сем. Permoberothidae Т i 1 1. из Neu-roptera; однако уже весь habitus жилкования говорит против принадлежности крыл$ Permobiella к Neuroptera, а ряды поперечных жилок, связывающие продольные, указуют в сторону Caloneuridae и Euthygrammidae. Дальнейшее изучение жилкования Permobiella perspicua Till, убеждает нас, что оно построено, в сущности, по тому же плану, как у Caloneuri-
dae. RS также отходит перед серединой крыла и образует в дистальной части всего 3 ветви. М описывается Тилльярдом как вогнутая жилкаг образующая затем развилок. Эта жилка, очевидно, вполне сходна и вполне отвечает МА Euthygrammidae', в основании она таким же образом соединяется со следующей жилкой, которую Тилльярд называет СиА, но которая представляет собою МР. Истинный СиА (СиР Тилльярда) сближен с МР, как у Paleuthygramma, только идет не наружу, а косо назад. Далее следуют СиР, Ах и А2. Особенностями Permobiella являются укороченная SC, длинный, загибающийся назад R и присутствие очень коротких вставочных жилок между концами продольных жилок; анальные жилки и ветви кубитуса еще вовсе не вытянуты наружу.
Напоминая своим жилкованием такие формы, как Paleuthygramma, род Permobiella все же не может быть включен ни в Euthygrammidae, ни в Caloneuridae, а должен считаться в недрах Caloneurodea представителем особого семейства — Permobiellidae.
В особое семейство я выделяю также одну форму из кунгурских отложений по р. Сылве. Даю здесь краткое описание этой формы.
СЕМ. SYNOMALOP^LIDAE FAM. NOV.
В передних крыльях RS отходит от R рано и дает несколько ветвей сзади; М состоит из двух длинных простых ветвей, соединяющихся лишь в самом основании крыла; МА лежит свободно, МР в большой средней части слита с СиА и отделяется от него лишь в концевой и в основной частях. СиР, Ах и А2 умеренно вытянутые, простые.
Род Synomaloptila gen. nov.
SC постепенно приближается к переднему краю и в апикальной части сливается с ним. RS образует в дистальной половине три ветви сзади. МА слегка изогнута против начала свободной концевой части МР и идет почти прямо наружу; МР слита в большей части с прямым СиА, но отходит от него в дистальной части, образуя как бы развилок его; в основной части ствол MP-j-CuA также разделяется, и МР направляется несколько вперед, а СиА идет внутрь, на соединение с СцР. Общий ствол МР + СиА^ как и концевое продолжение СиА, имеет вид прямой выпуклой жилки. СиР параллельно СиА, вогнутый; Ах и А2 прямые, параллельны СиР. Между продольными расположены обычные ряды поперечных жилок. В задних крыльях RS образует сзади не 3, а 4 ветви; объединение в основании МА и МР яснее; направление кубитуса и анальных сходно (но сохранность этих жилок недостаточна).
Переднегрудь небольшая, значительно уже среднегруди. Тип рода — Synomaloptila longipennis Mart., из отложений Чекарды.
Synomaloptila longipennis sp. nov.
(рис. 5)
№24/118. Сланцы по берегу р. Сылвы у дер. Чекарды; сборы Е. В. П е р-м я к о в о й, 1936.
Экземпляр представляет собой довольно хороший отпечаток, на котором видимы размытые очертания груди и три крыла: переднее и заднее на левой стороне и переднее и следы заднего — на правой. Передние крылья длинные и довольно узкие; длина их — 27.5 мм. SC прямая и в дистальной трети сливается с передним краем. Ветви RS не пряЛы, а слегка изогнуты, так что в конце направляются почти прямо наружу. МА тонкая. В передних крильях МР как бы внезапно отходит вперед от прямого кубитуса (СиА); в основании расхождение МР п СиА также ясно.
В задних крыльях RS образует 4 ветви. МА имеет сходный вид, как в передних крыльях, и в основании соединяется с МР, но далее жилкование становится очень неясным. Анальная область, видимо, едва цли вовсе не расширена, как у Caloneuridae и Euthygrarnmidae.
Описанный род по форме крыльев и строению их жилкования очень напоминает Euthygrarnmidae и Caloneuridae, но в то же время резко'хкгди-чается от этих семейств полным слиянием МР и CuA в средних частях их.
В этой особенности на крыло Protocollaria ingeus В г о n g п. из отложений Коммен-три, в особенности рисунок, данный самим Броньяром (1893, Pl. XL, Fig. 6). В других отношениях крыло Protocollaria сильно отличается от нашего; в нем еще сохраняется примитивная сетка, Аа и А2 образуют каждая по нескольку ветвей, часто вновь бифурциру ющих, СиР также дает ветви. Все это говорит еще о большей архаичности жилкования этогб крыла, и тем более неожиданным представляется очень большое сходство с Synomaloptila в отношениях МР с CuA. Имеем ли мы здесь дело с результатом некоторого конвергентного процесса развития или же эта особенность говорит о каких-то генетических, родственных отношениях нашего
нашу форму странным образом очень походит
пермского рода С ка- Рис. 5. Отпечаток Synomaloptila longipennis sp. nov. менноугольным ро-
дом Protocollaria? На это пока очень трудно ответить, тем^более что крыло Protocollaria сохранилось недостаточно. Во всяком случае действительное родство Synomaloptila с Protocollaria мне не кажется невероятным, и если это так, то интересно, что пермские роды, связанные с различными семействами карбона, в конце концов, пойдя по общему пути редукции лишних(с механической точки зрения!) жилок и выпрямления продольных, дали очень сходные с внешней стороны формы.
Итак, пермские роды Caloneurodea? несмотря на их взаимное сходство, не образуют одного цельного комплекса, а родственны разным каменноугольным семействам и составляют при этом особые семейства. Это показывает, что в них вряд ли можно видеть непосредственных потомков известных нам верхнекаменноугольных родов; вероятнее тот вывод, что
пермские семейства отделились от общих карбоновыми предков ранее — в средне- или нижнекаменноугольное время. Оставшиеся в тропической зоне роды быстро расцвели, но к перми вымерли; попавшие в болео северные области предки пермских семейств удержались дольше, почти до конца перми, но в триас, невидимому, они все же не перешли.
ОТНОШЕНИЯ CALONEURODEA
Хотя передние крылья по своему жилкованию иногда, может бытьг и походят на надкрылья некоторых современных саранчовых, тем не менее сходство это чисто внешнего, поверхностного характера, и о действительном родстве Caloneurodea с саранчовыми, да и вообще с Orthoptera Saltatoria, говорить не приходится. Caloneurodea, конечно, представляют собою группу ортоптероидов, но группу глубоко своеобразную и лишь довольно отдаленно родственную с Saltatoria п Phasmatodea. Чрезвычайно своеобразной особенностью ее является почти гомономное состояние крыльев их, отсутствие (первичное!) веера на задних крыльях, а затем и само жилкование их.
В палеозое были и некоторые другие группы с почти гомономными крыльями, например, Hypoperlidae, но ни одна из них не дожила даже до мезозоя.
ЛИТЕРАТУРА
Brongni art Ch., 1893. Recherches pour servir a 1’histoire des Insectes Fossiles des temps prinraires. S.-Etienne.
H andlirsch A., 1908. Die Fossilen Insecten etc..
Handlirsch. A., 1919. Revision der palaeozoischen Insecten. — Denkschrift. Acad. Wiss, Wien. Math. Naturw. KI., 96. Band.
H andli rsch A., 1911. New paleozoic Insects from the vicinity of Mazon Creek, YIl.-Amer. Journ. Sci., 31.
H andlirsch A., 1930. Insecta, in Handbuch der Zoolqgie, von W. Kukenthal u. Th. Krumbach, IV. B., 8. Lief.
L a m e ё r e A., 1917. Revision sommaire des Insectes Fossiles du 81ёрЬатеп de Commentry. Bull. Mus. Nat. d’Histoire Naturelle, № 3, Paris.
M artynov A., 1928. Permian Fossil Insects of N. E. Europe. Trav. Mus. вёо1од. Acad. Sci. URSS. T. IV.
M а ртыно в A., 1930. О двух новых прямокрылых насекомых из пермских отложений Кунгурск. у. Пермского края. Ежег. Гос. палеонт. о-ва.
Мартынов А., 1938. Очерки геологической истории и эволюции насекомых. Часть I. Palaeoptera, и Neo pie га Polyneoptera. Тр. ПИН, т. VII, вып. 4.
Р г u v о s t Pierre,k 1919. Faune Continentale du terrain Houiller du Nord de la France. Мёт pour servir a 1’expl. de la Carte Geolog. de la France. Paris.
Zalessky G., 1933. Sur de nouveaux insectes permiens. —Ann. Soc. вёо1. du Nord. T. LVII, p. 137.
SUMMARY
The fam. Euthygrammidae Mart, is described in 1928. At presents it contains three genera — Euthygramma Mart., with two species, Eu. parallellum and Eu. curvatum Mar t., Paleuthygramma Mar t., with P. tenuicornis Mart, and Vilvia G. Z a 1., with V. densinervosa Z a 1. This family is evidently allied to Caloneuridae, but the morphology of its wing-venation till now remains somewhat obscure.
New records at Sojana-river produced us many additional materials, and at present we may give a more adequate description of the wings of this family and elucidate somewhat better its relationship.
\ FAM. EUTHYGRAMMIDAE MART.
Wings elongated, with straight longitudinal veins, running outwards; precostal area absent; RS deriving from R at the middle of wing, usually imple, but somewhere giving 1—2 branches; M early dividing into two main branches, MA and MP; MA concave and usually forming a fork, MP convex, but simple. Cu also divided in the basal part of wing into two main branches, which are usually approximated to each other, but sometimes CuA is approximated to MP and CuP runs like other veins. Two long anal veins, running outwards. Longitudinal veins connected by series of dense cross-veins. Posterior wings similar to the anterior ones, anal region being not or scarcely dilated. Head rather small, antennae long, slender, with basal joint robust. Prothorax rather small and short, meso- and metothorax longer; legs long, ambulatory, posterior ones only slightly longer, than the other.
Earlier (1928, 1931) I considered Mediana to be onebranched, forming but a fork, but now the examination of several good remains of wings showed, that M is composed of two main branches,‘tMA and MP, but MA is concave and MP convex, Cu is also clearly composed of two long branches, which in Euthygramma are running near to each other and rather concave; behind CuP there are only two anal veins, Ax and A2; they are convex and long, running outwards.
Gen. EiithygrammaMart.
RS deriving at the middle of wing, simple; MA ending with a fork; CuP approximated to CuA, but in the basal part they are somewhat diverging from each other; Ax and A2 are approximated near the base of wing.
Euthygramma parallellum Mart.
(Fig. la, 2)
About 25 remains of anterior wings have been examined; they are recorded at Letopola, Iva-Gora and Sheimo-Gora, along the river Sojana.
Length is varying from 18 up to 25 mm, usual length being 21—24 mm. Venation is also varying: M sometimes divides at the base of wing, but sometimes more late, as in fig. 1; Cu divides earlier than M; MA2 at its end is somewhat curved forwards.
Eu. curvatum from Tikhije Gory is a closely allied species.
Euthygramma aberrans sp. nov.
(Fig- 3)
One specimen from Iva-Gora.
Anterior wing is of same shape (fig. 3), as in Eu. parallellum, but smaller, its length being 17.5. Venation is somewhat abnormous; RS is not simple, but forms two additional branches'from'behind; M simple; branches of Cu, Ax and A2, as in the foregoing species.
It is clear, that MA is detached from MP and more or less united with RS, posterior and median branches of the latter belong probably not to
RS, but to MA; branches of Cu and two analia are running, as in the foregoing species.
Gen. Vilviopsis gen. ndv.
RS deriving from R before the middle of wing and forming 2 branches from behind; M dividing earlier than RS, MA forming a fork. MP approximated to CuA, CuP running not very near to CuA; analia, as usual.
Vilviopsis extensa sp. nov.
(Fig. 4)
One specimen from Sheimo-Gora, on Sojana-river. Anterior wings long and comparatively narrow; RS forming three branches altogether; fork of MA reaching half of the length of RS, MP simple, running near to CuA. Length of wing — 28 mm.
This species reminds of the gen. Vilvia Z a 1., but in the latter MA is simple, according to Zalessky; moreover the branching of RS is somewhat different.
RELATIONSHIPS OF THE FAM. EUTHYGBAMMID AE
Euthygrarnmidae are evidently allied to the Uppercarboniferous family Caloneuridae, but in their wing-venation are more specialized. Some stretching of the cubital and anal veins is perceptible in the gen. Confusio Handl, and Pruvostiella Handl. In Euthygrarnmidae such a stretching was much stronger arid caused an elimination of the branches of RS. Hind wings in Caloneuridae, Euthygrarnmidae and, probably, in other families were nearly alike to the anterior ones.
The author concurs with Handlirsch in the necessity of separation of the fam. Caloneuridae with allied families in a separate group, but believes this group to represent a distinct order Caloneurodea. He includes in it a series of families, as follows: Carboniferous families: Caloneuridae, Proto-collariidae, Stenaroceridae and Emphylopteridae-, Permian families: Euthy-grammidae, Permobiellidae (Til 1.) and Synomaloptilidae Mart., fam. nov. The author believes the gen. Permobiella Till, is belonging not to Neuroptera, but to the Caloneurodea, and .must be separated into a distinct family. The third Permian family of Caloneurodea — Synomaloptilidae fam. nov., is represented with the gen. Synomaloptila gen. nov. from the deposits at Chekarda, on the shores of the river Sylva.
Gen. Synomaloptila gen. nov.
SC uniting with C in the apical portion of wing; RS forming 3—4 branches from behind; MA a little curved; MP is united with CuA, but in its distal part separates, forming as a form of CuA; in the basal part the stem MP+CuA also divides into MP and CuA, the latter soon uniting with CuP; Aj and A2 straight, parallel to CuP. Cross veirts numerous, regularly arranged.
Synomaloptila longipennis sp. nov.
(Fig. 5)
A good reprint of the thorax and of wings. Anterior wings rather long and narrow; length 27.5 mm. Branches of RS somewhat curved outwards; MA slender, MP forming with the distal part of CuA as a broad fork.
Anal region in the hind-wings not dilated. The gen. Synomaloptila resembles Euthygrarnmidae and Caloneuridae, but differs in the union of MP and CuA, in what a character it evidently reminds of the gen. Protocollaria. Perhaps it is allied more to this genus. Permian families have been originated from some common ancestors with the uppercarboniferous families, probably in the Middle- or Lower Carboniferous. Caloneurodea seem to be distantly allied to Phasmatodea and to Saltatoria^ " * Г / ’ ~ ।
СПИСОК ОПЕЧАТОК
Стр. Строка Напечатано Следует читать
6 5 сн. в схеме 27 18 св. 28 16 сн. 51 20 св. 56 10 сн. 57 18 сн. 77 34 св. 17. Балахонская Балахонская свита: свита: Kansasian Kansan Kavkazsky Kavkazskij постоянное постоянно II Teil 11. Teil /’Kofprestes Kopfrestes Synomaloptila Protocollaria
Tdvum ПИН. т. VII. вып. 3