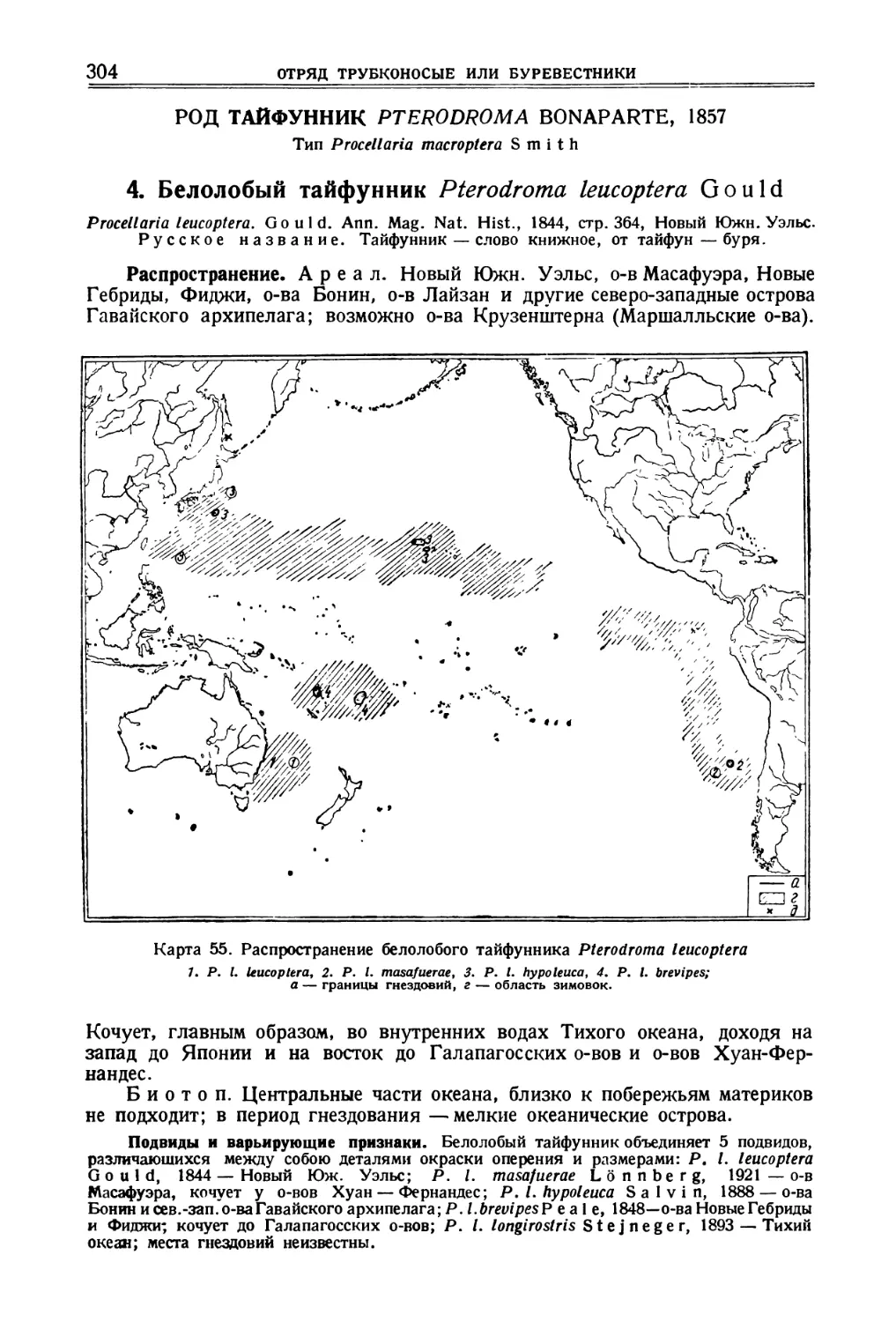
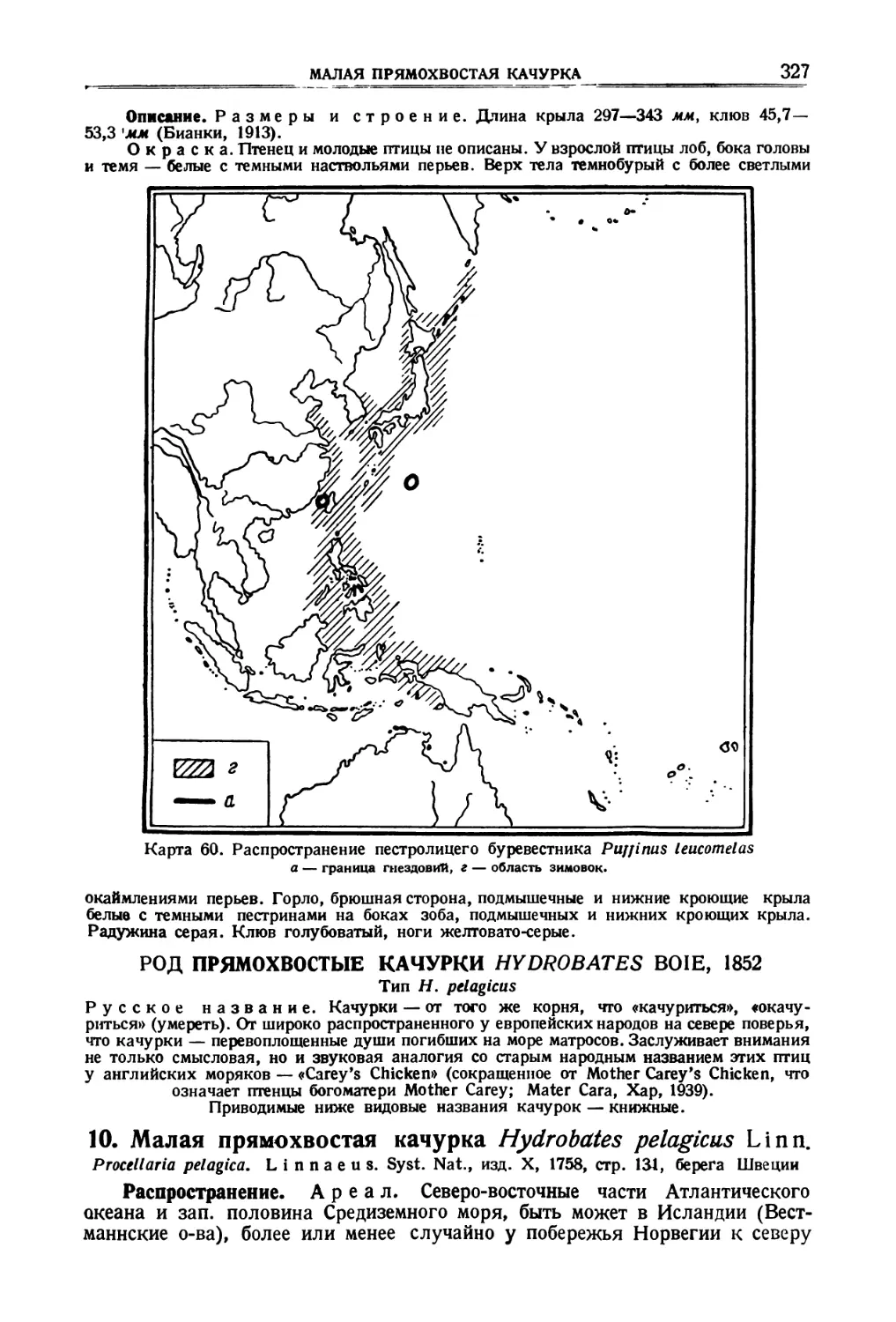
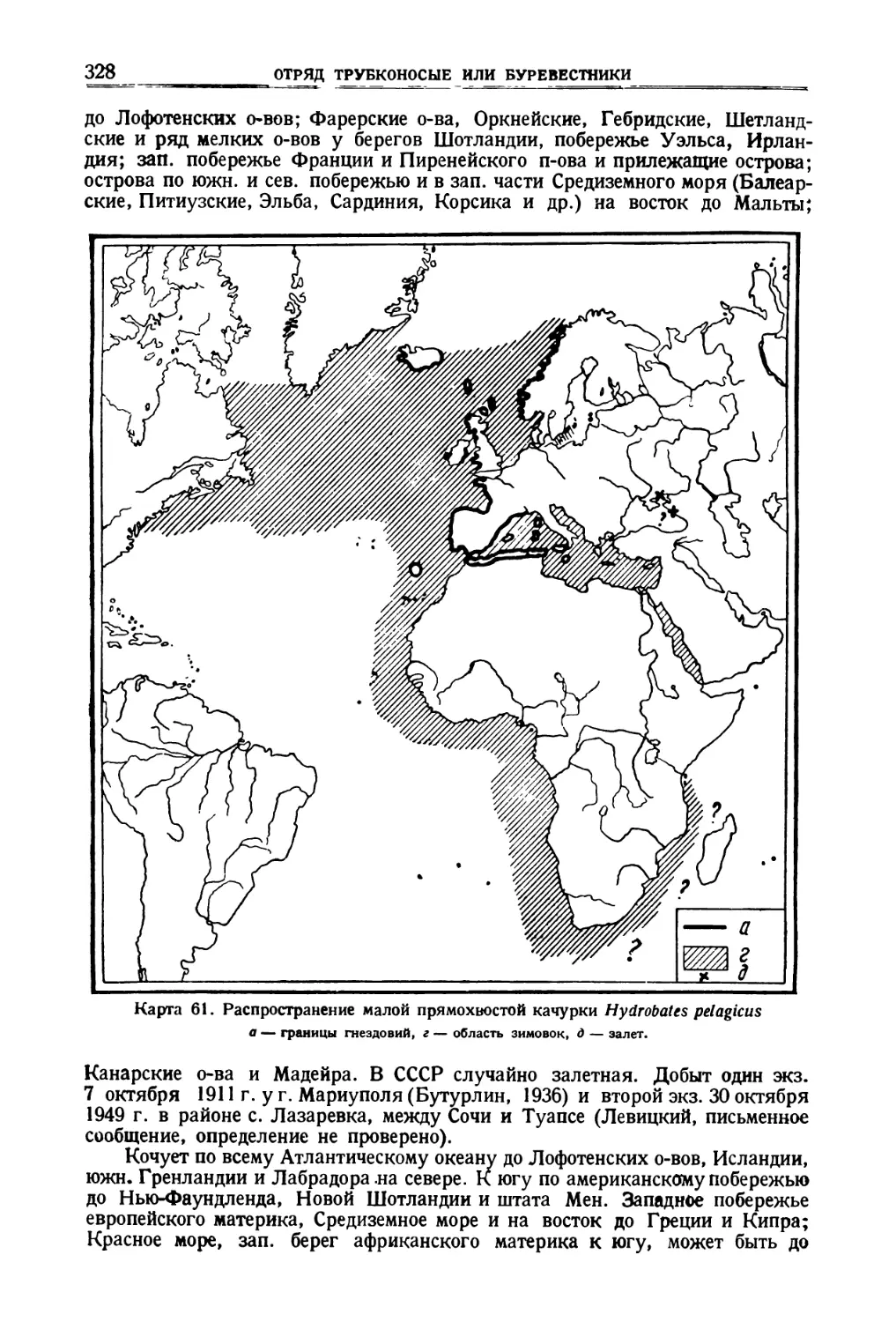
/


















































































![[Сенегальский рябок]](https://djvu.online/jpg/d/e/z/dezr5znAJihgB/083.webp)
















































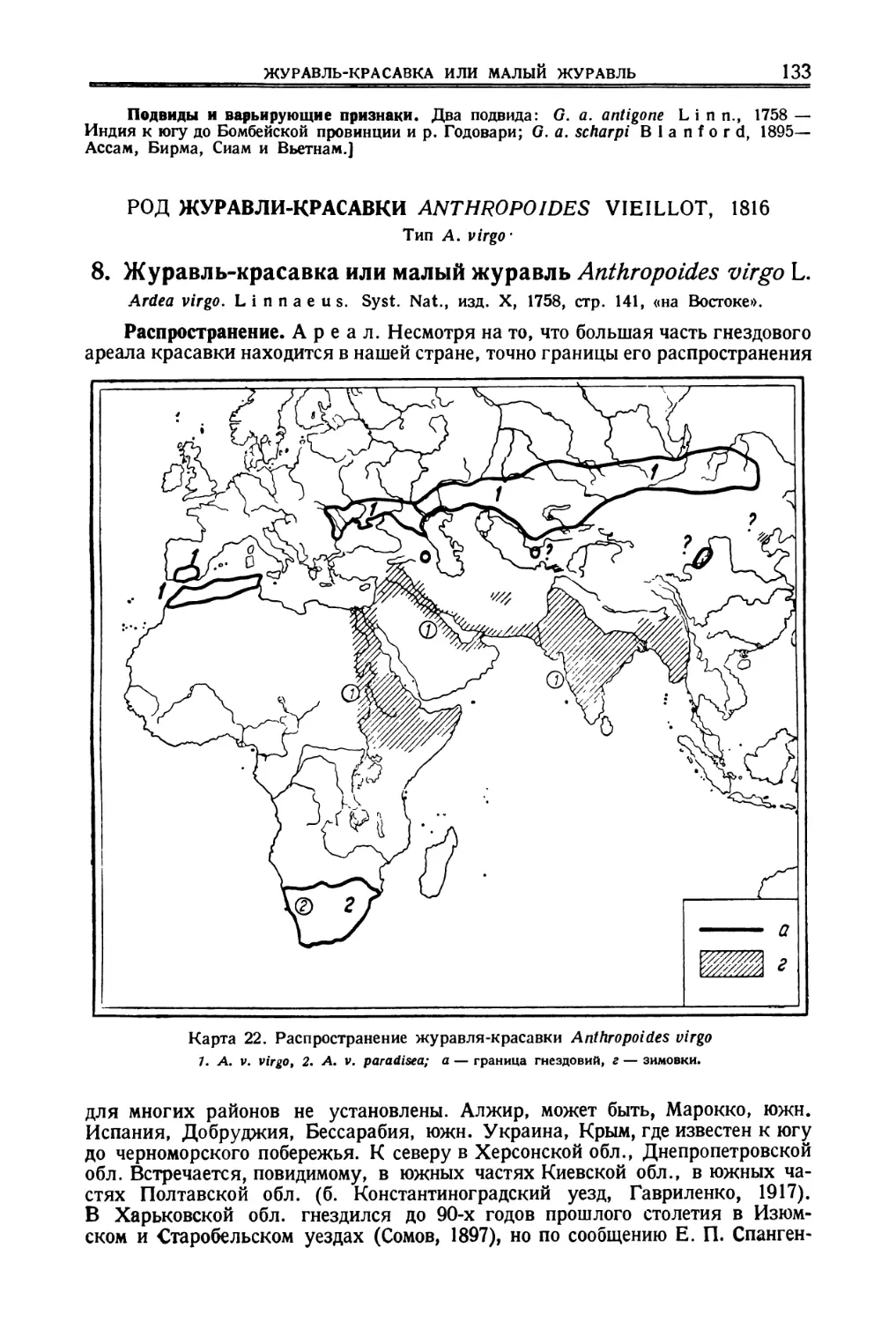
![[Индийский журавль]](https://djvu.online/jpg/d/e/z/dezr5znAJihgB/132.webp)
















































































![[Алеутский пыжик]](https://djvu.online/jpg/d/e/z/dezr5znAJihgB/214.webp)





















































































































































































































































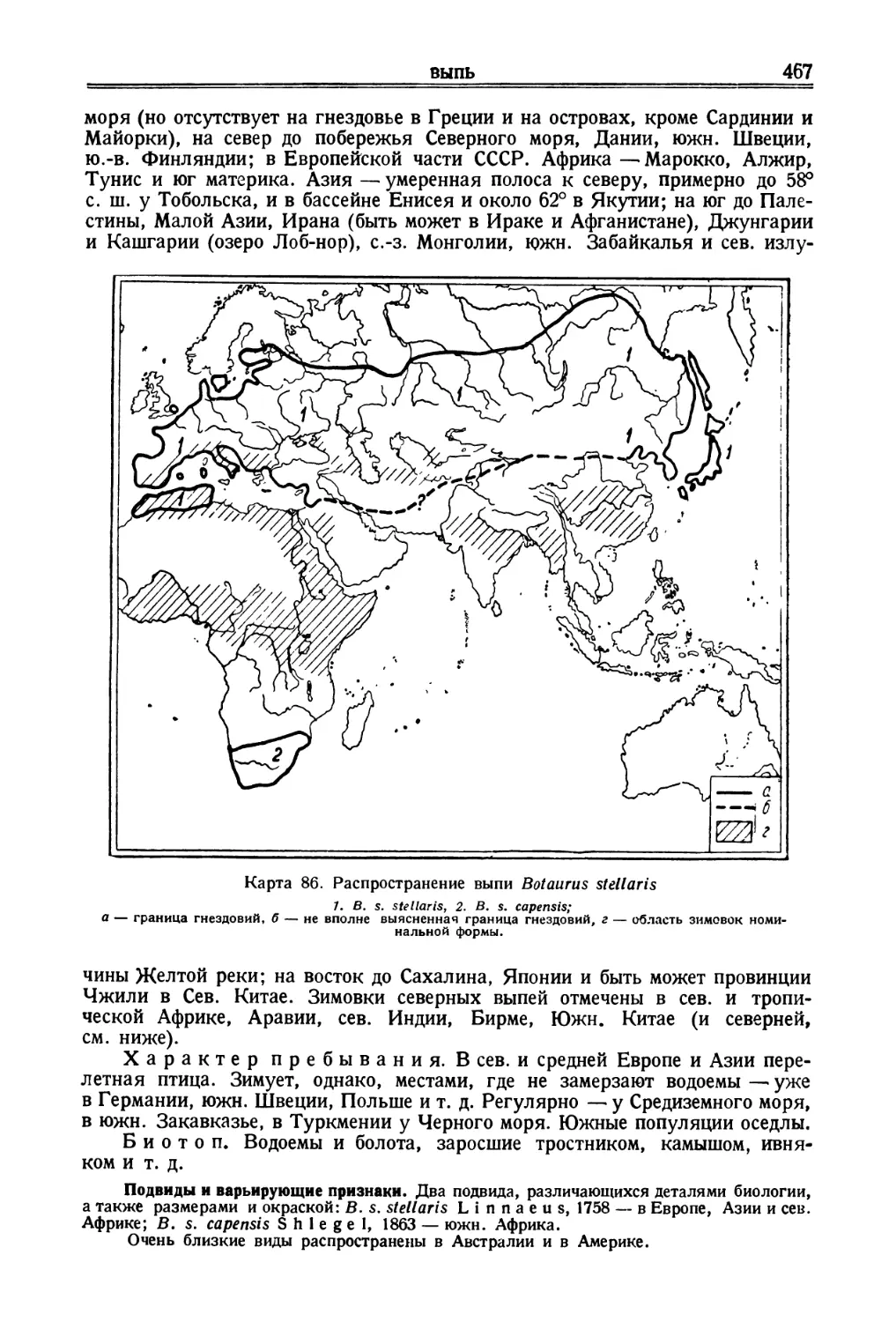






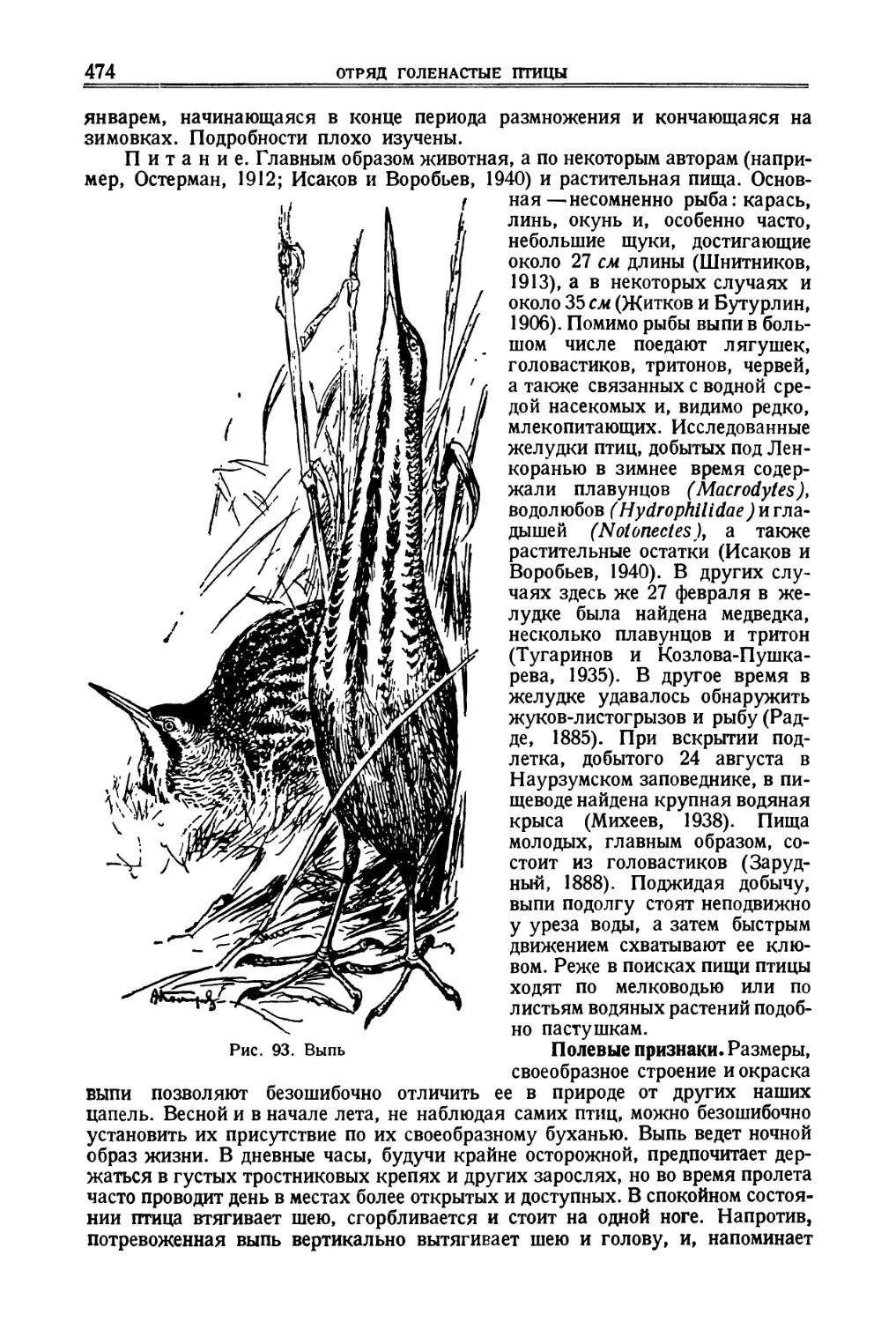













Author: Дементьев Г.П. Спангенберг Е.П. Гладков Н.А. Птушенко Е.С. Судиловская А.М.
Tags: птицы фауна ссср птицы советского союза
Year: 1951




Text
Г. П. ДЕМЕНТЬЕВ, Р. Н. МЕКЛЕНБУРЦЕВ,
А. М. СУДИЛОВСКАЯ, Е. П. СПАНГЕНБЕРГ
ПТИЦЫ
СОВЕТСКОГО
СОЮЗА
Том II
Под общей редакцией
Г. П. Дементьева и Н. А Гладкова
ГОСУДАРСТВЕННОЕ ИЗДАТЕЛЬСТВО
„СОВЕТСКАЯ НАУКА"
Москва—1951
ОТРЯД ГОЛУБИ
COLUMBAE или COLUMD1FORMES
Характеристика отряда
Морфология. Внешние признаки. Большинство голубей — средней
величины, немногие (венценосные голуби) — крупные птицы, а горлицы родов
Oxypelia и Geopelia не превосходят дроздов. Клюв обычно относительно тонкий,
утолщенный в конечной части. Ноздри узкие, щелевидные; у большинства
видов они открываются в восковице. Граница оперения лба выдается вперед.
Голова небольшая. Шея короткая, тонкая у головы и сильно расширяющаяся
(большой зоб) у груди. Туловище короткое и широкое, с сильно развитой
грудью. Крылья обычно длинные и острые, с вершиной, образованной вторым
или третьим маховым (реже третьим или четвертым). Хвост чаще короче крыла,
иногда равен крылу по длине, слегка или значительно закругленный. Цевка
короткая, голая или только слегка оперенная у межплюсневого сустава;
спереди цевка покрыта щитками, сзади и по бокам сетчатая (у венценосных
голубей сетчатая со всех сторон). Пальцы длинные, с короткими, но сильными
когтями. Задний (первый) палец хорошо развит и находится на одном уровне
с передними. Оперение густое, сравнительно жесткое и плотное. Окраска раз-
нообразна, иногда очень яркая. Возрастные изменения несложны.
Птенцы вылупляются слепыми, покрытыми редким волосовидным пухом.
Пуховой наряд сменяется ювенальным оперением, которое в первую же осень
или зиму заменяется в результате полной линьки окончательным нарядом.
Ювенальное оперение незначительно отличается от окончательного: оно более
тусклое и без металлических отливов. Половой диморфизм большей частью
выражен в размерах — самцы крупнее самок. У некоторых видов самец
окрашен ярче. Первостепенных маховых 10 или 11; рулевых обычно 12—14,
реже 16—18 и даже 20.
Анатомические признаки. Череп схизогнатический и схи-
зоринальный. Носовые перегородки и базиптеригоидные отростки имеются,
но сошник отсутствует (исключение составляет Didunculus). Шейных позвон-
ков 14—16. От 3 до 5 грудных позвонков сливаются и образуют os dorsale.
Грудина с двумя парами вырезок. Пищевод имеет два больших боковых выро-
ОТРЯД ГОЛУБИ
ста, образующих хорошо развитый зоб. Стенки зоба во время выкармливания
птенцов сильно утолщаются, эпителий претерпевает жировое перерождение
и выделяет особый продукт (так называемое «голубиное молоко»), служащий
для кормления птенцов в первые дни их жизни. Мускульный желудок у зер-
ноядных видов весьма массивен, с сильно развитой мускулатурой. У плодояд-
ных видов стенки желудка слабые. Кишечник у зерноядных длинный, а у пло-
доядных короткий и широкий. Слепые кишки отсутствуют или рудиментарны.
Желчный пузырь у большинства видов отсутствует. Три последних кольца
трахеи, участвующие в образовании нижней гортани, очень тонкие и иногда
вовсе исчезают на дорсальной стороне. На вентральной стороне эти кольца
соединены костным или хрящевым мостиком. Перья без побочных стволов.
Стволы контурных перьев плоские и широкие, резко суживающиеся к вер-
шине. Пуха нет, но значительно развиты пуховые основания перьев. Между
контурными перьями расположены местами тонкие волосовидные перышки.
Кожа тонкая, перья в ней сидят слабо. Некоторые перья продуцируют поро-
шок, покрывающий легким налетом все оперение. Крыло, за некоторыми
исключениями, диастаксическое.
Образ жизни. Все виды ведут строго дневной образ жизни. Большинство —
оседлые птицы; лишь немногие в умеренных широтах совершают перелеты.
Голуби связаны главным образом с лесными местообитаниями; меньшее число
видов живет в скалах, обрывах, человеческих сооружениях. Пищу обычно
собирают на земле. В связи с этим голуби хорошо ходят (но бегают плохо).
Взлетают свободно, без разбега, сильно хлопая крыльями. Полет легкий,
сильный и быстрый, с частыми взмахами крыльев. При необходимости голуби
могут на лету делать быстрые и резкие повороты и способны к значительному
ускорению полета. Большинство видов гнездится группами и даже в гнездовое
время летает на кормежку стаями. В негнездовое время образуют стаи, иногда
громадных размеров.
Пища в огромном большинстве случаев растительная, чаще всего — семена
разных травянистых растений. Некоторые виды голубей специализировались
на питании плодами. Животная пища — мелкие беспозвоночные — служит
незначительным добавлением к растительной. Голуби не могут обходиться без
воды и иногда летают на водопой на большие расстояния. При питье погружают
клюв почти до ноздрей в воду и втягивают ее, не поднимая головы при глотках,
как это делают куриные.
Гнезда строятся из небольшого количества сухих веточек, травинок или
корешков, с неглубоким и плоским лотком, обычно без всякой выстилки.
Нередко яйца бывают видны сквозь дно гнезда. Располагаются гнезда чаще
всего на деревьях, реже в дуплах, на скалах или в постройках; еще реже
прямо на земле. Число яиц в кладке обычно два, у некоторых видов три, или
одно. Скорлупа белая. В течение года обычно бывает две или более кладок;
даже перелетные виды гнездятся дважды, а оседлые иногда 4—5 раз. Эмбрио-
нальное развитие быстрое — от 14 до 30 дней (у разных видов).
Птенцы развиваются по птенцовому типу. У видов средней величины они
вылетают из гнезд в возрасте около месяца. В известной мере продолжитель-
ность постэмбрионального развития зависит от особенностей гнездования:
например, у гнездящегося открыто вяхиря птенцы покидают гнездо в воз-
расте около 20 дней, а у гнездящегося в дуплах, несколько меньшего по раз-
мерам, клинтуха — в возрасте 25 дней.
Систематика и географическое распространение. Отряд содержит два
семейства, одно из которых вымерло уже в XVI веке. Это дронты Raphidae
с 2 родами Raphus B вида) и Pezophaps A вид). Все они были крупными, утра-
тившими способность к полету птицами; распространение их ограничивалось
островами св. Маврикия, Бурбон и Родригец. Второе семейство Columbidae
состоит из 292 видов. Из них в фауне СССР представлено 11 или 3,77%
ХАРАКТЕРИСТИКА ОТРЯДА
(в том числе 6 видов рода Columba, 4 вида рода Streptopelia и 1 вид рода
Oenopopelia).
Ископаемые остатки голубей немногочисленны и не позволяют восстано-
вить хотя бы приблизительно историю отряда. В известной мере за центр
распространения отряда может быть принят Индо-австралийский архипелаг
и Австралия. Наиболее древний ископаемый представитель отряда Gerandia
calcaria известен из верхнего олигоцена (Франция). В верхнем плиоцене появ-
ляются современные роды, например, Columba (С. sicula в Сев. Америке).
Из плейстоцена известны как вымершие роды (Progoura), так и современные
(Columba, Leucosarcia). Кроме того, из плейстоцена известны и современные
виды (клинтух, вяхирь, сизак, обыкновенная горлица). В историческое время,
как указывалось, было уничтожено семейство дронтов. Надо еще упомянуть
об истреблении совсем недавно в Сев. Америке странствующего голубя Ecto-
pistes migratorius (последний экземпляр этого вида дожил в неволе до
1914 года).
Голуби в настоящее время распространены почти по всему свету, за исклю-
чением полярных стран — Арктики и Антарктики.
Распространение голубей в СССР таково. Лесной зоне свойственно не-
сколько видов, причем 3 из них — вяхирь, клинтух и обыкновенная горлица —
населяют преимущественно лиственные леса в Европейской части, Союза и
на западе Азиатской части. В зоне открытых пространств из этих видов распро-
странена только горлица, для которой достаточно и скудной кустарниковой
растительности. Упомянутые виды только незначительно проникают в область
хвойных лесов, в частности в горах: в последних все 3 встречаются в поясах
хвойного и лиственного леса. Леса таежного типа населены в сущности одним
характерным видом — большой горлицей, причем она свойственна и южной
половине тайги. В горную зону эта горлица проникает на западе до прилегаю-
щих к Аму-Дарье окраин Памиро-Алая, населяя пояс хвойного леса.
В зоне открытых пространств (на восток до Енисея) широко распростра-
нен сизак, но еще больше обычен он в горах (все пояса до нижней окраины
альпийского); характерен он и для культурного, антропического ландшафта.
Для среднеазиатской части зоны открытых пространств весьма характерен
бурый голубь. Сюда же могут быть причислены малая и кольчатая горлицы,
в нашей стране приуроченные к оазисам низменностей Средней Азии.
Горная зона имеет и своих специфических представителей: скалистого
и белогрудого голубей. Первый широко населяет все горы юга Сибири и
проникает в Среднюю Азию до западных окраин Памиро-Алая, второй известен
только по единичным находкам на высоких горах Средней Азии.
Наконец, китайская короткохвостая горлица в фауне СССР встречается
случайно и вряд ли может быть приурочена к какой-либо зоне. Все находки
этой птицы относятся к области смешанных лесов Дальнего Востока. Таким
образом, в пределах СССР Средняя Азия наиболее богата голубями.
Практическое значение. Хотя мясо голубей отличается хорошими качест-
вами, организованного охотничьего промысла на них нет. Крупные виды служат
предметом спортивной охоты. По характеру питания голуби, кормясь зернами
злаковых культур, могут местами приносить известный ущерб сельскому хозяй-
ству. Однако при оценке этого вреда надо учитывать то, что голуби кормятся
на убранных полях выпавшими из колосьев зернами, а также и то, что наряду
с семенами культурных растений голуби поедают в большом количестве семена
сорняков, способствуя этим очистке полей. Наконец, голуби не так многочис-
ленны, чтобы приносимый ими вред мог принять где-либо массовый, реаль-
ный, характер.
Нельзя не отметить значения голубей в качестве домашней птицы. Помимо
множества декоративных пород, имеются и мясные и почтовые (последние
с успехом применяются при некоторых обстоятельствах и в настоящее время).
ОТРЯД ГОЛУБИ
Кроме того, домашние голуби — один из лучших видов подопытных птиц
для научных исследований по физиологии, эмбриологии и т. д. Большой инте-
рес представляют отечественные породы домашних голубей (русские чистые,
чеграши, турманы и другие; среднеазиатские бухарские).
Таблица для определения видов голубей
гB). Общая окраска сизая или белая. На шее нет черных пестрин и черной серпо-
образной полосы. Концы рулевых черные 3
2A). В общей окраске преобладает бурый, рыжеватый или охристый цвет. У взрослых
птиц на шее черные пестрины или черная серпообразная полоса. Концы рулевых
белые 13
3D). На боках шеи с каждой стороны по белому или охристому пятну. Первостепенные
маховые с белым краем наружных опахал. На кистевом сгибе крыла белое пятно
вяхирь Columba palumbus
4C). Бока шеи без белых или охристых пятен. На наружных опахалах первостепен-
ных маховых и на кистевом сгибе крыла белого цвета нет 5
5F). Голова черноватая, резко отграниченная от белой шеи. Спина, шея и зоб белые;
у старых птиц шея без металлического блеска
белогрудый голубь Columba leuconota
6E). Голова сизая, не отделяющаяся по окраске от шеи. Спина, шея и зоб сизые; у ста-
рых птиц шея и зоб с металлическим блеском 7
7(8). Клюв черный. Поперек сложенного крыла идут 2 явственных черных полосы . . .
9
8G). Клюв светлый. Поперек сложенного крыла идут неполные ряды темных пятен
9A0). Поперек хвоста явственная широкая белая полоса
скалистый голубь Columba rupestris
10(9). Хвост сизый без белой поперечной полосы посередине
сизак Columba livia
11A2). Верх головы чисто сизый. Поясница сизая, немного светлее спины или одного
с ней цвета клинтух Columba oenas
12A1). Верх головы с розоватым налетом. Поясница белая или светлосизая в кон-
трасте с окраской спины бурый голубь Columba eversmanni
13A4). Окраска спины, плечевых, кроющих крыла пестрая. На боках шеи по 2—3 ряда
мелких полу лунных пятнышек голубовато-белого или голубовато-серого цвета. Задняя
сторона шеи без черной серповидной полосы 15
14A3). Окраска спины, плечевых, кроющих крыла без пестрин. На боках шеи нет
голубоватых пятнышек 17
15A6). Зоб и грудь розоваты. Пятнышки на боках шеи светлые, почти беловатые . . .
обыкновенная горлица Streptopelia turtur
16A5). Зоб и грудь буроваты. Пятнышки на боках шеи темные, сероватые
большая горлица Streptopelia orientalis
17A8). Первое маховое короче третьего. Верхние кроющие первостепенных маховых
серые 19
18A7). Первое маховое длиннее третьего. Верхние кроющие первостепенных маховых
коричневые китайская горлица Oenopopelia tranquebarica
19B0). Крыло не менее 165 мм. На задней стороне шеи большое черное полулунное
пятно. На боках шеи нет черных пестрин
кольчатаягорлица Streptopelia decaocto
20A9). Крыло не более 155 мм. На задней стороне шеи нет черного полулунного пятна.
На боках шеи мелкие черные пестрины
малая горлица Streptopelia senegalensis
РОД ГОЛУБИ COLUMBA LINNAEUS, 1758
Тип С. oenas
1. Сизый голубь, сизяк, сизак Columba livia Gm.
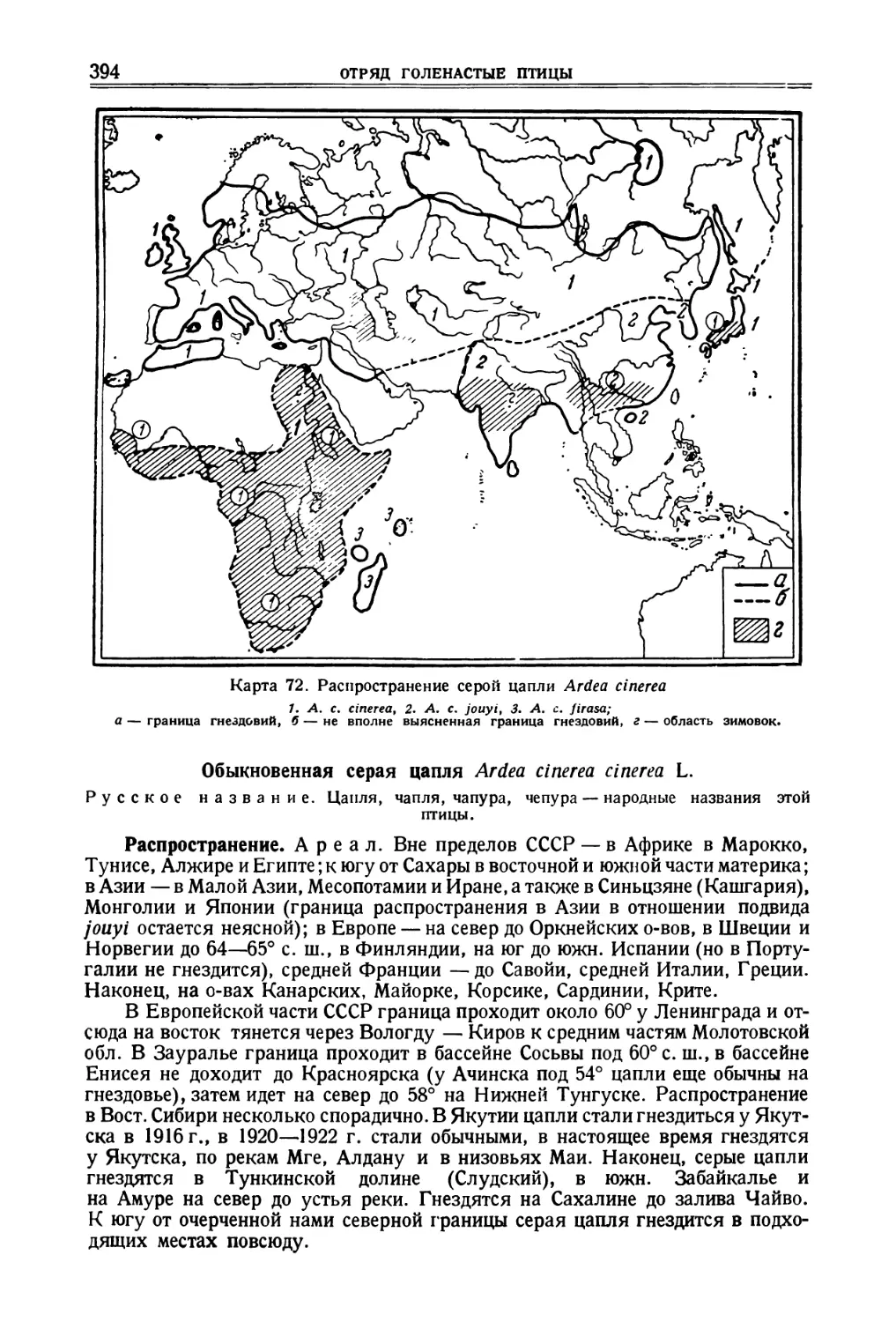
Columba livia. G me 1 i n. Syst. Nat. 1789, стр. 769, южн. Европа.
Распространение. Ареал. Широко распространен в Европе, сев. Аф-
рике и Азии. На севере доходит до Шотландии, Фарерских о-вов, о-ва Чен-
нель. Указывался для южн. Норвегии, но в новых сводках для этой области
не приводится. На материке населяет Южн. Францию, Швейцарию, сев.
СИЗЫЙ ГОЛУБЬ
части Балканского п-ова, Крым, низовья Дона, Волгу до Камы, Уральские
горы до Колвы. На восток распространяется до Чумыша и верховьев Енисея.
На юго-восток до Ассама. На юге ареал включает сухие части Бирмы, всю
Индию, Цейлон, Аравию, Нубию, Сахару и Сенегал, На западе ряд о-вов Атлан-
тического океана — Азорские, Канарские и Мадейру. В полудомашнем со-
стоянии идет вслед за человеком на север гораздо выше указанной линии и
поднимается до Балтийского моря, Мурмана, Печоры F6°), Тобольска, 59°30'
на Енисее. Кроме того, населяет Японию и обширные участки Китая.
Биотоп. В диком состоянии сизый голубь придерживается скалистых
или обрывистых берегов рек. Очень охотно селится на скалах, в горных ущельях.
Полудомашние голуби живут в постройках людей или на скалах вблизи
поселений.
Карта 1. Распространение сизого голубя Columba livia
7. С. /. livia, 2. С. /. atlantis, 3. С. I. canariensis, 4. С. I. gaddi, 5. С. /. palaestmae, 6. С. /. schimperi,
7. С. L targia, 8. С. 1. dakhlae, 9. С. I. butleri, 10. С. L gymnocyclus, 11. С. /. Uvidior, 12. С. /. neglecta,
13. С. I. intermedia, 14. С. I. nigricans;
a — граница области распространения, б — не вполне выясненная граница распространения, в — северная
граница распространения и отдельные участки, населенные полудомашними голубями.
Подвиды и варьирующие признаки. C.I. livia Gm., 1789. — Вся европейская и
сибирская часть ареала; в юго-зап. Азии до Мугоджар (может быть Усть-Урта) и
Сев. Кавказа; одни авторы включают целиком Малую Азию, другие только ее западную
часть и то под вопросом, третьи исключают весь полуостров из ареала типичной формы;
сев .-зап. Африка; С. /. atlantis Ban ner man — Мадейра, о-ва Азорские и о-ва
Зеленого мыса; С. /. canariensis Bannerman, 1914 — Канарские о-ва (некоторые
авторы объединяют последние подвиды); С. /. gaddi Zarudnyet L о u d о n, 1906 —
юго-зап. Иран, Ирак, Сирия, Месопотамия, сев.-зап. Египет (Сулум, Мирза-матру),
Киренаика, Крит, Палестина (за исключением низменности у Мертвого моря); по некоторым
авторам и Малая Азия; С/, palaesti пае Ze d I i t z, 1912 — южн. Палестина, Аравия, Синай;
C.I. schimperi Bonaparte, 1854 — дельта Нила и его долина вверх до Ассуана и
Вахи Хальфа; С. /. targia G е у г, 1916 — Центральная Сахара; С. /. dakhlae M e i n e г-
tzhagen, 1928 — оазисы Дакла и Та рта в Ливийской пустыни ;С. /. butleri M e i n e г-
t z h a ge n, 1921 — африканское побережье Красного моря; С. /. gymnocyclus G. R. Gray,
1856 — Сенегал; С. /. Uvidior Bates, 1932 — Гамбага, северная часть Золотого
берега; С. /. neglecta И п те, 1873 — Средняя Азия, Афганистан (в сев.-зап., по Мей-
нертцхагену, С. /. gaddi), Белуджистан, Иран (кроме юго-зап. части), Закавказье и южн.
склоны Кавказа, Пенджаб и Синд; С. /. intermedia Strickland, 1844 — Индия
(кроме Синда и Пенджаба), Цейлон, сухие части Бирмы, на восток до Аннама по одним дан-
ным и до Ассама — по другим; С. /. nigricans В u t и г 1 i n, 1908 — верховья р. Шара-мурена,
окрестности Тунцай-инцза (ю.-з. Манчжурии). Известен только в одном экземпляре. Был
описан в качестве особого вида.
Подвиды различаются интенсивностью общей окраски, цветом и шириной светлой
полосы на крестце, а также размерами. Экологические различия подвидов выражены в том,
8 ОТРЯД ГОЛУБИ
что одни держатся, главным образом, на скалах морских и речных берегов; другие на всякого
рода обрывах среди песчаных пустынь; третьи обитают преимущественно на невысоких,
скалистых знойных горах среди песчаных пустынь; наконец, четвертые держатся на скалах
высочайших горных массивов.
Обыкновенный сизый голубь Columba livia livia Gmel.
Синонимы. Columba livia 8. rustica. Богданов. Тр.СПБО. Естеств. 1881,
стр. 98 — Columba livia infuscata. Круликовский. Тр. Украинск. общ. люб. естеств.,
XXXV, 1915 A916), ст. 8, Сарапул.
Русское название. Сизак, сизарь — народное. Сизый голубь — книжное.
Голубь — старинное русское название разных видов отряда, несомненно родственное
«Columba».
Распространение. Ареал. В СССР сизый голубь в диком состоянии
занимает сравнительно небольшую территорию. Он встречается в Крыму (Пуза-
нов, 1933), по обрывистым берегам Азовского моря, быть может на Сиваше.
В незначительном количестве в низовьях Днепра (Вальх, 1911) и по правому
берегу Дона {Алфераки, 1910). Затем населяет северное Предкавказье и север-
ные склоны Кавказского хребта (Динник, 1886). На Волге, по старым данным,
он поднимается до устьев Камы, а по новейшим — признается возможным
гнездование на Волге, Оке или Суре и считается несомненным на Свияге (Пуза-
нов, 1942). Между Волгой и Уралом немногочисленен, отмечен в песках левого
берега Волги (Орлов и Фенюк, 1927) и в Губерлинских горах (Зарудный, 1888).
В бассейне Урала обнаружен в области верхнего течения этой реки (Снигирев-
ский, 1929). Редок в области верховьев Тобола (Михеев, 1937), гнездится в
Мугоджарах (Зарудный, 1888) и у верховьев Иргиза (Сушкин, 1908). В неболь-
шом числе найден на северном берегу Аральского моря (Бостанжогло, 1911).
В Тарбагатае сизый голубь гнездится довольно значительными колониями.
Впрочем в этих горах, так же как и во всей области между низовьями Урала,
северным берегом Арала, верховьями Тобола и Тарбагатаем попадается уже
много особей как переходного характера к туркестанскому голубю, так и
неотличимых от него. Алтай населяет типичная форма (Сушкин, 1938), рас-
пространенная небольшими колониями и в области верхнего течения Енисея
(Сушкин, 1914).
Полудомашний сизак идет к северу гораздо дальше, чем дикий. Он обычен
на гнездовье в поселениях Ленинградской, Калининской и Горьковской обла-
стей. Местами выходит за пределы этой границы, образуя колонии, например,
в Мурманске и низовьях Оби. Оторванный от Европы и зап. Азии участок охва-
тывает незначительное пространство Дальнего Востока (басе. р. Уссури).
В Японии полудомашний голубь обычен.
Характер пребывания. Дикий голубь является почти осед-
лой птицей, которой свойственны лишь не регулярные, но иногда очень зна-
чительные кочевки.
Биотоп. Избегает лесов и больших ровных площадей. Излюбленные
места гнездовий — всякого рода обрывы. Так сизый голубь с большой охотой
поселяется на высоких известковых скалах берегов Крыма, на меловых бере-
гах Волги и Дона, также на утесах Мугоджарских гор и в настоящей горной
обстановке на Алтае.
Численность. В настоящее время сизаки не образуют больших
колоний ни в одном из перечисленных мест. В европейской части ареала
они гнездятся большей частью одиночными парами, изредка образуя
небольшие колонии по несколько пар. Так же немногочисленны эти голуби
в западной половине азиатской части ареала, в восточной же половине они
гораздо многочисленнее. Например в Тарбагатае уже попадаются колонии,
содержащие до нескольких десятков пар. Десятками пар исчисляются коло-
нии на Алтае и Енисее. Около шестидесяти лет назад сизые голуби были го-
СИЗЫЙ ГОЛУБЬ
раздо многочисленнее; у Чкалова встречались кочующие стаи, численность
которых определялась в 50 тысяч экземпляров.
Экология. Размножение. Данные по гнездованию сизого голубя
в пределах Союза ничтожны и даже по полудомашним голубям, несмотря на
всю легкость наблюдений, отрывочны. Весеннее воркование и спаривание
начинается у полудомашних птиц необычайно рано — у Чкалова уже в сере-
дине февраля (Зарудный, 1888). У совершенно диких голубей брачный период
начинается позднее, и спаривание приходится на конец марта — начало ап-
реля, яйца откладываются к концу апреля. Образованию пары предшествует
энергичное воркование самца. Воркующий сизак низко наклоняет переднюю
часть туловища и кружится на одном месте то в одну, то в другую сторону,
полураспустив хвост. Заканчивая трель, он поворачивается к голубке и резко
поднимает вверх переднюю половину тела. Голуби часто «целуются» клювами,
перебирают друг у друга перья на головах и т. п. Эти же повадки предше-
ствуют совокуплению, после которого самец взлетает, поднимается, описывая
полукруг, на известную высоту и спускается вниз планирующим полетом, дер-
жа крылья над спиной под углом.
Гнездо располагается где-либо в трещине обрыва, нише скалы, на кар-
низе или выступающем камне и т. п. По данным западноевропейских орнито-
логов, самец отыскивает место для гнезда и носит материал, а самка лишь
укладывает его. Гнездо складывается из сравнительно грубых веточек и кореш-
ков и бывает весьма неискусным, представляя собой просто плоскую кучку
с очень неглубоким лотком. Гнездовый участок каждой пары невелик, так
как голуби селятся часто колониями, располагая гнезда близко друг к другу.
Однако каждый самец строго следит за тем, чтобы соседи не переходили уста-
новленных пределов, которые в больших и тесных колониях иногда сужива-
ются до двух — трех десятков сантиметров.
Кладка из 2 яиц, чисто белого цвета; размеры: G7) 36,4—43,3 х 27 —
31,5, в среднем 39,33 х 29,22 мм (Хартерт, 1915). Насиживают оба родителя
с первого яйца. Самка сидит на гнезде больше, чем самец, который проводит
в гнезде время с 10 утра до 2 часов дня. На восьмой день насиживания у старых
голубей начинается утолщение стенок зоба. Насиживание продолжается
17 дней. К моменту вылупления птенцов зоб уже выделяет белое кашицеобраз-
ное вещество, служащее пищей маленьким птенцам. Последние получают от
родителей корм, погружая в широко открытые рты старых птиц клюв и при-
нимая в них отрыгиваемое старыми птицами содержимое зоба. Через несколько
дней птенцы начинают получать размягченные в зобах старых птиц зерна, но
выделение кашицеобразного вещества заканчивается у домашнего голубя на
18-й день по вылуплении птенцов, а у дикого еще позднее.
Гнездовый период продолжается 30—35 дней. Даже покинувшие гнезда
птенцы получают корм от родителей, выпрашивая его с писком и своеобразным
подрагиванием крыльев. Поэтому у совершенно оперившихся молодых перья
у основания клюва — на лбу, по бокам и на подбородке — вырастают позднее
всех других, а клюв кажется более длинным, чем у взрослых птиц. Через
несколько дней после вылета родители отгоняют птенцов и приступают ко
второй кладке. Даже в самых северных участках ареала — в Шотландии
и на Фарерских о-вах, сизаки несутся не менее двух раз в лето. В средиземно-
морских странах они успевают высиживать яйца до трех раз. Летные молодые
первого вывода наблюдались здесь в конце мая.
Для наших пределов данные по размножению отсутствуют, если не счи-
тать указаний для Чкаловской обл. на то, что живущие там полудомашние
голуби имеют будто бы до четырех кладок в лето (Зарудный, 1888). Ставшие
самостоятельными птенцы образуют стаи, которые постепенно увеличиваются
за счет присоединения подрастающих молодых. После того, как выкормлены
последние молодые, старые присоединяются к стаям, и птицы начинают коче-
10
ОТРЯД ГОЛУБИ
вать у места своего гнездования, отлетая иногда не надолго на более далекие
расстояния.
Линька. Сизый голубь, как и другие виды голубей, имеет одну пол-
ную линьку в году. Смена оперения начинается в мае выпадением десятого
первостепенного махового и идет к переднему краю крыла. У некоторых особей
начало линьки приходится на апрель, так как в мае встречаются птицы с двумя
и даже тремя сменившимися маховыми. В июне попадаются особи, у которых
сменилось от двух до пяти маховых, в июле — от двух до семи, в августе — от
пяти до восьми. Сентябрьские материалы представлены одной птицей, имеющей
восемь новых маховых. В ноябре смена первостепенных маховых у всех ста-
рых птиц заканчивается (Ларионов и Щукина, 1948). Таким же порядком
идет смена маховых у домашних го-
лубей. Второстепенные маховые ли-
няют от двух центров — с первого и
десятого перьев. Линька идет навстре-
чу, центростремительно и у старых
заканчивается вместе с концом линьки
остального пера. У молодых иногда
успевают перелинять все второстепен-
ные маховые, и перья от четвертого
до восьмого остаются не сменившими-
ся. Линька мелкого пера начинается
в конце августа, интенсивно она идет
в течение сентября и октября.
Начало выпадения мелкого пера
совпадает со сменой шестого махово-
го, а конец — со сменой второго —
первого маховых. Прежде всего выпа-
дают перья спины и груди, но линька
очень быстро распространяется и на
остальные участки оперения. По всей
вероятности линька диких голубей про-
текает сходным образом.
У молодых смена нарядов идет
следующим порядком: птенцовый пух
вытесняется ювенальными перьями,
которые напоминают по цвету оперение старых птиц, но цвета более тусклые,
оперение рыхлее и с буроватым налетом. Это оперение в первую же осень
сменяется окончательным взрослым нарядом. Запоздалые птенцы заканчивают
линьку зимой. Последними отрастают как у молодых, так и у старых птиц,
своеобразные, продуцирующие тонкий порошок, перья. Они располагаются
узкой полосой от подмышки до хвоста, проходя по бедру. Этот ряд перьев
пробивается, когда линька уже закончена и отрастает только в январе —фев-
рале. Обыкновенное мелкое оперение также способно продуцировать порошок.
Во всех случаях он получается путем обламывания кончиков тонких бородок.
Порошок равномерно рассеивается по оперению и придает ему мягкость оттен-
ков, свойственную всем голубям.
Питание. Основную пищу сизаков составляют семена самых разно-
образных растений; из диких он предпочитает бобовые, из культурных —зла-
ки, преимущественно пшеницу. Зерна подбирает с земли, но не выбивает их
из растущих колосьев. Иногда, особенно после того, как землю засыплет
снегом, сизаки садятся на оставшиеся в полях копны и выклевывают зерна из
колосьев. Однако этот вред крайне незначителен потому, что, во-первых, голуби
в общем немногочисленны, во-вторых, они откочевывают при глубоких снего-
падах, в-третьих — зерно из колосьев, оставшихся в поле на зиму, все равно
Рис. 1. Сизый голубь
СИЗЫЙ ГОЛУБЬ 11
высыпается под влиянием атмосферных факторов и растаскивается грызунами.
Уничтожением семян сорной растительности голуби приносят определенную
пользу, способствуя очистке полей. Как и все зерноядные птицы, сизаки за-
глатывают крупные песчинки и мелкие камешки, которых в желудке одной
особи бывает до десятка. Кормятся они преимущественно в утренние и вечер-
ние часы, отдыхая где-либо в жаркую пору дня.
Полевые признаки. В полевых условиях сизак легко отличим от других ви-
дов наших голубей издали заметной на лету белой поясницей (при взгляде сверху
или сзади). При взгляде сбоку или снизу более заметен белый подбой крыла,
который находится в контрасте со сравнительно темно окрашенными грудью
и брюхом, чего нет у других видов. На самых близких расстояниях сидящего
сизака можно узнать по двум резко обозначенным полосам, которые идут
поперек крыла. Сизак — «общественная» птица и даже в период гнездования
редко приходится отмечать одиночных птиц. Там, где эту птицу не преследуют,
она становится очень смелой, доказательством чему могут служить полудо-
машние голуби, разгуливающие по улицам больших городов. При преследова-
нии сизак становится гораздо осторожнее и поднимается в воздух, заметив
опасность еще на большом расстоянии. Лесов сизак избегает и редко садится
на деревья. Для отдыха и ночлега стая выбирает какой-нибудь высокий утес,
обрыв, большую постройку и лишь в редких случаях отдельное дерево или
небольшую рощу, но только не глубокие участки обширных лесов. К другим
видам голубей сизаки присоединяются не особенно охотно, но иногда летают
кормиться вместе с клинтухами. Голос сизого голубя хорошо отличен от
монотонного воркования лесных голубей и сходен с воркованием домашних.
Описание. Размеры и строение. Длина крыла самцов C9) 204—250, самок
B3) 203—229, в среднем соответственно 221,15 и 214, 78 мм. Длина хвоста — около 100—
130 мм, клюва —около 17—22 мм, цевки —около 27—32 мм.
Окраска. Оба пола по окраске сходны. Общая окраска взрослых птиц сизая,
более темная на нижней стороне и надхвостье. Мантия несколько свет лее. Попе рек поясницы
проходит белая полоса около 5—б см ширины. Подбой крыльев и подмышечные также белые.
Рулевые и маховые темносизые — темнее надхвостья. По концам рулевых проходит черная
вершинная полоса, но самые кончики рулевых сизые. Внешнее опахало крайнего рулевого
почти целиком белое. Внутренние опахала маховых у основания беловатые. Второстепен-
ные маховые и их большие кроющие имеют черные сквозные пятна, сливающиеся в две
резко выраженных черных поперечных полосы на крыле. Шея с ярким металлическим зеле-
новатым отливом, зоб с меднокрасным (у самки эти отливы менее ярки). Окраска молодых
отличается наличием буроватого налета на мантии и передней части нижней стороны; метал-
лических отливов нет; некоторые перья с узкими беловатыми краями; белизна на спине
часто с примесью сизого. Глаз оранжевый, то с желтой, то с красной примесью. Ноги мали-
ново-красные, у молодых бледнее. Клюв черный с белой восковицей.
Туркестанский сизый голубь Columba livta neglecta Hume
Columba neglecta. Hume. Lahore ta Yarkand, 1873, стр. 272, Ладак.
Синоним. Columba livia korejewi. Sarudny und L о u d о п. Orn. Monatsber.
1906, стр. 134, Семиречье.
Распространение. Ареал. Туркестанский сизак населяет в пределах
СССР республики Закавказья и Средней Азии. Северная граница вырисовы-
вается неясно. По мнению некоторых (Бутурлин, 1934), все Закавказье и южный
склон Кавказа считаются заселенными этой формой, по другим (Хартерт, 1915)—
Передняя Азия вплоть до Ирана включается в ареал типичного подвида. Восточ-
ный берег Каспийского моря то признается заселенным диким голубем типич-
ной формы (Зарудный, 1918), то туркестанским голубем (Бутурлин, 1934).
Для северного берега Аральского моря приводится туркестанская форма (Бо-
станжогло, 1911). Для Темира и Мугоджар также приводится туркестанская
форма и переходы к типичной (Сушкин, 1908). На Усть-Урте кое-где попадается
небольшими колониями, как выяснено в последнее время С. /. neglecta. В Кара-
Кумах найдена эта же форма (Дементьев, 1950). На Аму-Дарье сизаки обычны.
12 ОТРЯД ГОЛУБИ
В Кызыл-Кумах не могут считаться редкими (Зарудный, 1914). В долине
Зеравшана сизак широко распространен как в равнинной, так и в горной
части (Даль, 1936). В Фергане это одна из самых обычных птиц (Кашкаров,
1927, 1934, Мальчевский, 1940). В среднем и нижнем течении Сыр-Дарьи
довольно редок (Спангенберг, 1936), но в бассейне Арыси многочисленен.
В горных системах Средней Азии сизак обычен повсюду: так, он широко насе-
ляет Копет-Даг, Памиро-Алай, за исключением Памирского нагорья (Иванов,
1940). Обычен в хребтах Тянь-Шаня—западного (Кашкаров, 1925, 1927;
Шапошников, 1931) и восточного (Зарудный, 1906, Кашкаров, 1937). Для
Тарбагатая вместе с типичной формой и одичалыми голубями приводится и
этот подвид (Селевин, 1929), так же, как и для Курчумских гор у озера Зайсан
(Поляков, 1912). В пределах всех этих территорий сизак является оседлой
птицей, хотя незначительные передвижения он кое-где и предпринимает.
Биотоп. Населяет различные вертикальные пояса, начиная от берегов
Каспийского моря, лежащих ниже уровня океана, и кончая хребтами Алая и
Памира, где он встречается до высот в 2500, а местами даже и 3000 м. По бере-
гам Каспийского и Аральского морей держится на обрывах, сложенных извест-
няками или песчаниками. В пустынях селится в стенах колодцев и по останце-
вым скалам. В долинах рек сизак населяет лессовые обрывы, в полосе предгорий
он селится на конгломератовых обрывах, в горах — на скалах, сложенных раз-
нообразными горными породами. Наконец, селится на развалинах и больших
каменных постройках. Во всех этих случаях требуется лишь наличие в обры-
вах или скалах каких-нибудь углублений, пещерок или трещин, где было бы
возможно соорудить гнездо. Кроме того, невдалеке должен находиться водоем,
предпочтительно проточный.
Численность. Количество сизаков сократилось за последние деся-
тилетия очень заметно. Еще два —три десятка лет назад в каждом городе
Средней Азии можно было насчитывать тысячи и десятки тысяч оседлых си-
зых голубей. В настоящее время в городах и кишлаках голуби не встречаются
совершенно. Чрезвычайно сильно сократилась численность этих птиц и вне
поселений. Тысячные стада голубей в настоящее время если и можно встре-
тить, то лишь в самых глухих углах Средней Азии. Даже сотенные стаи попа-
даются теперь сравнительно редко и лишь вдали от городов и крупных селений.
Причины остаются не вполне ясными.
Экология. Размножение. Зимние стаи сизаков содержат значи-
тельное количество неразошедшихся пар, но, наряду с этим, молодые прошед-
шего лета оказываются холостыми. Брачный период у этих холостых птиц
начинается позднее, чем у старых, так как некоторое время требуется для
образования пары. Как и у номинального подвида, самец воркует, кружась
около голубки. Затем следует поглаживание клювом головы, так называемые
«поцелуи», в которых может быть следует усматривать рефлекс кормления
молодых, и, наконец, образование пары.
Гнездо, как у всех голубей, делается очень рыхло и представляет собой
небольшую, плоскую кучку сухих стебельков, трав и веточек. Лоточек весьма
неглубокий. Гнездо всегда сильно загрязнено экскрементами птенцов, запол-
няющими все пустоты между основным материалом. Оно помещается в щели
обрыва, на выступе, в углублении, оставшемся после выпавшего камня,
в норе сизоворонки и т. п. местах. В пустынных местностях широко исполь-
зуются саксауловые крепления колодцев (Кара-Кумы, Бадхыз и Карабиль,
Кызыл-Кумы). Гнездовый участок иногда ограничивается несколькими
десятками квадратных сантиметров в непосредственной близости от гнезда.
При колониальном гнездовании голубей гнезда располагаются иногда почти
рядом друг с другом.
Кладка из 2 яиц, которые насиживаются обоими родителями. Насижива-
ние продолжается селшадцать дней. Птенцы остаются в гнезде, повидимому,
СИЗЫЙ ГОЛУБЬ 13
около месяца. Их довольно сильно беспокоят различные паразиты, как напри-
мер клещи Ornithodorus, в изобилии живущие в щелях лессовых обрывов и
скал. Сведения о сроках гнездования туркестанского сизака в литературе
почти отсутствуют. Начало постройки гнезда наблюдалось на р. Или уже
в середине марта (Шестоперов, 1926). Кладка из 2 яиц была обнаружена в гнезде
в начале июня (Спангенберг, 1936). Подлетки в Гязь-Гедыке в ю.-в. Туркме-
нии — 20 апреля; в конце апреля в Карабиле у самцов семенники достигли пол-
ного развития (Дементьев, Рустамов, Спангенберг, 1947). 22 июня яйца были
найдены на юге Закаспия (Зарудный, 1896), что указывает на возможность
третьей (I) кладки. Молодые добывались в различные сроки: 3 и 16 июня
у Зайсана (Поляков, 1912), 17 мая у Ташкента, 9 и 30 июня в Нура-тау и
Кызыл-Кумах и 17 сентября под Ташкентом (коллекционный материал). Из
этих данных можно видеть, что откладка яиц растягивается на все лето, и
голуби имеют по несколько кладок. Если считать, что насиживание и выкарм-
ливание птенцов одной кладки занимает около IV2 месяцев, то пара голубей
выводит птенцов не менее трех раз в лето, а загнездившиеся особенно рано
пары может быть успевают вывести и четвертых птенцов.
Из-за разновременности образования пар получается значительная нерав-
номерность в сроках откладки яиц и появления молодых. Поэтому различные
гнезда содержат в одно время либо яйца, либо птенцов различного возраста.
Молодые первых кладок образуют самостоятельные стаи, которые все лето
держатся возле колонии, и к которым постоянно присоединяются старые
птицы, выводящие птенцов. Осенью покончившие с гнездованием старые птицы
окончательно присоединяются к этим стаям.
Линька. Линька старых птиц начинается уже в середине мая, но она
охватывает в это время далеко не всех особей, и еще в июне встречаются ста-
рые экземпляры, не начавшие линять. Смена перьев начинается с крыла, в кото-
ром первым выпадает десятое первостепенное маховое. Затем линька распро-
страняется постепенно к концу крыла. Она идет медленно, и в течение июня
попадаются птицы, едва начавшие линьку, наряду с такими, которые меняют
седьмое, шестое или пятое из первостепенных маховых. В июле попадаются
голуби с семью, восемью, девятью старыми первостепенными маховыми, но
есть и такие, у которых остается лишь по три, четыре не сменившихся самых
длинных пера в крыле. К концу августа — началу сентября начинают встре-
чаться особи, заканчивающие линьку. Мелкое перо линяет очень быстро —
в срок около полумесяца или немногим больше. Вместе с мелким пером идет
смена рулевых, которые в противоположность маховым сменяются так же
быстро, как мелкое перо. Повидимому, некоторые экземпляры заканчивают
линьку к середине сентября, а к концу этого месяца большинство старых
птиц уже сменяет перо. Самые ранние птенцы начинают смену ювенального
пера уже в начале мая, «поздние» — в октябре. Туркестанский сизак имеет
в году одну полную линьку.
Возрастные изменения несложны, как у всех голубей. Редкий светложел-
тый пух птенцов сменяется рыхлым ювенальным пером, которое носится только
до первой осени и заменяется окончательным нарядом.
Питание. Насколько можно судить, голуби любят пшеницу больше
всякого другого корма. Поля ячменя и риса посещаются ими с гораздо мень-
шей охотой. Однажды в зобу голубя были найдены косточки дикой вишни
Prunus Machaleb (Иванов, 1940). На полях голуби собирают только опавшие
зерна и не повреждают колосьев, стоящих на корню. Кормятся они недолго
и, наполнив как следует зобы, летят на водопой. Последним служит участок
открытого берега полого опускающейся к воде песчаной или галечной отмели.
Пьют помногу, причем напиваются очень быстро. Если их не тревожат на ме-
стах водопоя, они подолгу остаются у воды и где-нибудь поблизости садятся
на дневной отдых. Если же место водопоя не безопасно, голуби быстро сни-
14
14 ОТРЯД ГОЛУБИ
маются и летят в ближайшие скалы или обрывы, где и рассаживаются, стараясь
обязательно скрыться в тень. Иногда садятся на сухие вершины старых боль-
ших деревьев. Особенно любят выступающие вершины или сухие боковые
ветки орехов, чинар и развесистых белых тополей, но в чащу кроны никогда
не садятся. Часто местом отдыха служит какая-нибудь постройка или разва-
лина, даже колодец, если вблизи нет более подходящих мест.
С другими видами птиц голуби если и соединяются, то обычно на кормежке
и в маленькие стаи. Большей частью рядом с голубями кормятся стайки галок
или грачей, иногда серых ворон, скворцов. Из других видов голубей сизаки
соединяются с вяхирем, клинтухом, скалистым голубем и особенно с бурым,
с которым часто даже гнездятся в самом ближайшем соседстве.
Описание. Размеры и строение. Крыло самцов A2) 214—239, самокA6)
216—231, в среднем 230 и 222 мм.
Окраска. По окраске сходны с типичным подвидом, с той лишь разницей, что
у туркестанского сизака общий тон бледнее, на спине обычно с большей или меньшей при-
месью голубовато-серого цвета. Ширина белой перевязи спины меньше — от 2 до А см.
2. Скалистый голубь Columba rupestris Pall.
Columba oenas rupestris. Pallas Zoographia Rosso-Asiatica, 1811, стр. 560, Даурия.
Распространение. Ареал. Южн. Сибирь, Сев. Китай и Централь-
ная Азия; северная граница в Казахском нагорье несколько юго-восточнее
истоков Ишима и идет к Иртышу, который и пересекает у Семипалатинска;
далее проходит северными подножиями Алтая, пересекает верховья Енисея,
охватывает с севера озе-
ро Байкал, примерно
следуя течению Лены,
доходит на ней до60° с. ш.
и, захватив Забайкалье
и бассейн Амура, выхо-
дит к берегу Великого
океана. Затем она свора-
чивает на юг и тянется
берегом материка до юга
Кореи; обогнув этот по-
луостров, граница пово-
рачивает к западу, выхо-
дит на материк между
устьями рек Желтой и
Голубой, повидимому
огибает с севера боль-
шую излучину Желтой
реки, захватывая горы
Алашань, и спускается
к югу до Голубой реки.
Здесь граница снова по-
ворачивает к западу, пе-
ресекает Меконг и Сал уэн
и тянется южными подножиями Гималаев до Гильгита; далее захватывает
Ладак, зап. Памир и предгорьями среднеазиатских хребтов доходит до Джун-
гарского Ала-тау. Затем, широко обогнув с востока озеро Балхаш, идет юж-
ными отрогами Казахского нагорья до соединения с северной границей в
указанном выше пункте, несколько юго-восточнее истока Ишима. Большие пло-
щади песчаных пустынь, располагающиеся внутри этого обширного простран-
ства, не заселены скалистым голубем и представляют собой пробелы в ареале.
Карта 2. Распространение скалистого голубя Columba ru-
pestris
7. С. г. rupestris, 2. С. г. turcestanica;
а — граница области распространения, б — не вполне выясненная
граница распространения, в — изолированные нахождения особей.
СКАЛИСТЫЙ ГОЛУБЬ 15
Характер пребывания. Голубь — повсюду оседлая птица и
зимует даже в Сибири. Однако местами он откочевывает с высоких гор в более
низкие области и кое-где предпринимает небольшие перелеты.
Биотоп. Селится, если не исключительно, то в огромном большинстве
случаев, в скалах. При этом проявляет некоторую привязанность к сравнительно
прохладному климатическому режиму, на юге ареала держится только в высо-
ких горах. На севере спускается и в обширные равнины, но селится здесь или
на скалистых берегах озер и рек, или следует за человеком, населяя постройки.
В этом случае голубь предпочитает крупные здания каменной кладки и, в силу
этой причины, встречается по большей части в городах, нисколько не пугаясь
движения и шума.
Подвиды и варьирующие признаки. С. г. rupestris Pallas, 1811 — восточная часть
ареала до верховьев Лены, Танну-олы и Урянхайского края на севере и верховьев рек Жел-
той, Голубой и Меконга на юге; С. г. turkestanica В u t и г 1 i n, 1908 — западная часть
ареала до указанной выше границы. Эта граница носит до известной степени условный харак-
тер. Изменчивость скалистого голубя выражена в постепенном посветлении окраски с востока
на запад. Наиболее светлые особи свойственны Средней Азии и Кашгарии, наиболее тем-
ные — Дальнему Востоку. Промежуточные особи были выделены Бутурлиным в качестве
особой расы С. г. intercedens (занимающей широкую полосу между двумя другими). Однако
позднейшие авторы (Козлова, 1930) не признают самостоятельности этого подвида и считают,
что он был описан по темным экземплярам западной формы и светлым — восточной. Эколо-
гические различия между подвидами выражены в том, что восточный подвид обитает на
меньших высотах, иногда даже спускаясь в равнины. Кроме того, эта форма часто встре-
чается в городах, где ведет жизнь, сходную с полудомашними сизаками Европы и Азии.
Западный подвид — обитатель высоких гор и на крайнем западе ареала никогда не
селится на постройках.
Северный скалистый голубь Columba rupestris rupestris Pall.
Распространение. Ареал. В Сибири обычен в Уссурийском крае (Шуль-
пин, 1936, Спангенберг, 1940) и в бассейне Амура (Радде, 1863, Тачановский,
1893). В Забайкалье обыкновенен во всех городах и селениях (Штегман, 1927);
на островах и берегах Байкала не представляет редкости (Штегман, 1936).
В бассейне Лены —в Якутске (Иванов, 1929). К западу от Байкала населяет
Минусинский район и Танну-туву (Сушкин, 1914).
Биотоп. Чаще всего поселяется в скалах, хотя бы и незначительных.
Обыкновенно голуби селятся вблизи от воды, будет ли это поверхность боль-
шого озера, широкая река или горный ручей. В Восточной Сибири постоянно
наблюдаются в человеческих постройках, заменяя отсутствующих там сиза-
ков. Как кажется, предпочитают каменные постройки, хотя кое-где живут
и в деревянных. В Монголии очень обычны в буддийских монастырях и нередки
в глинистых обрывах глубоких оврагов. На деревьях держатся редко и отме-
чены только на громадных ильмах и по садам оазиса Дынь-юань-инь и в Ала-
шанском хребте. Типичная форма скалистого голубя — вполне оседлая птица,
разве только откочевывающая к югу у северной границы ареала. Правильные
перелеты для этого вида на территории Вост. Сибири не отмечены, но у Пекина
ежегодные перелеты указываются в начале весны и конце осени.
Численность. Во многих местах своего ареала описываемый под-
вид далеко не представляет редкости. Особенно многочисленны голуби в мон-
гольских монастырях, где люди их совершенно не тревожат. В таких городах,
как, например, Якутск, где птиц мало беспокоят, они также сравнительно мно-
гочисленны и встречаются если не тысячами, то во всяком случае сотнями.
Экология. Размножение. Сведения до крайности скудны и поз-
воляют лишь заключить, что брачный период у этой птицы начинается очень
рано. Для Приморья существует указание на откладку яиц в феврале. Впрочем
непосредственное наблюдение здесь не имело места, а уже 26 апреля были
встречены хорошо летающие молодые (Шульпин, 1936). Пожалуй, правильнее
16
ОТРЯД ГОЛУБИ
будет относить в этом случае начало гнездования к первой декада марта.Непо-
средственными наблюдениями в Урге начало брачного периода зарегистриро-
вано во второй декаде марта (началось усиленное воркование и токовый полет
самцов, Козлова, 1930). По обобщенным данным для Вост. Сибири в целом,
начало гнездования относится к марту (ст. ст., Тачановский, 1893).
Гнезда располагаются в расщелинах скал. В поселениях гнезда строятся
под крышами, в различных удобных местах чердаков, карнизов и т. п. Дан-
ные о количестве кладок отсутствуют. Также не описывается для пределов Сою-
за устройство гнезд, число яиц,
выкармливание птенцов, но во
всем этом несомненно нет раз-
личий с другими близкими ви-
дами голубей. В гнездовое время
этот голубь живет колониями,
то небольшими, по 2—4 пары,
то гораздо более многочислен-
ными — до нескольких десятков
пар.
Линька. Как и у других
видов голубей, одна, полная
годовая, начинается выпадением
десятого первостепенного махо-
вого. Начало линьки в большин-
стве случаев приходится, видимо,
на май, но затягивается по край-
ней мере на месяц, так как птицы
с восемью и семью старыми махо-
выми встречаются в течение всего
июня, и даже в первой половине
июля имеется незначительное ко-
личество птиц с семью старыми
маховыми. Большинство же имеет
в это время по четыре старых
маховых, но начавшие рано
линять экземпляры сохраняют
только по три и реже по два ма-
ховых. В сентябре линька закан-
чивается, но некоторые птицы линяют до ноября. Рулевые начинают линять при
выпадении второго-первого маховых. Их смена идет быстро, и к концу линьки
маховых все рулевые обновляются. Мелкое перо сменяется в это же время.
У некоторых особей, сменяющих третье маховое, заметно разрежено мелкое
перо головы и шеи. У птиц с одним старым маховым мелкое перо в большинстве
случаев уже все новое.
Питание. Кормовой режим изучен крайне недостаточно. Известно
лишь, что скалистый голубь любит кормиться на полях, выгонах и т. п. местах,
не отличаясь, таким образом, в выборе кормовых мест от других видов голу-
бей. Несомненно не отличается в выборе объектов питания и заглатывает раз-
личные семена диких и культурных растений.
Полевые признаки. Общий облик скалистого голубя напоминает сизака.
Однако от последнего его очень легко отличить даже издали по наличию на
хвосте широкой белой перевязки, особенно хорошо заметной при взлете птицы.
Впрочем, в области, занятой типичной формой скалистого голубя, сизак не
встречается, если не считать небольшого района в истоках Енисея. От боль-
шой горлицы, которая встречается повсюду в области распространения описы-
ваемой формы, скалистый голубь отличается чисто сизым цветом и крупной
Рис. 2. Скалистый голубь
СКАЛИСТЫЙ ГОЛУБЬ 17
величиной. Полет его очень похож на полет сизака, но, пожалуй, несколько
быстрее, чем у последнего.
Описание. Размеры и строение. Длина крыла самцов A1) 214—230,6,
самок A0) 216—238, в среднем 221 и 232,7 мм.
Окраска. У взрослых самцов и самок общая окраска сизая, несколько темнеющая
на голове и шее. Последняя с зеленым и красным металлическим блеском; сизый цвет мантии
и особенно брюха часто бывает несколько светлее, чем на других частях тела. Задняя часть
спины чисто белая; надхвостье аспидное или свинцово-серое; на верхних кроющих крыла
и внутренних из второстепенных маховых проходят сквозные черноватые или темноаспидные
пятна, образующие поперек крыла две полосы; первостепенные маховые буровато-сизые
с светлосерыми внутренними опахалами; второстепенные—темносерые, с черноватыми
концами; рулевые имеют темносизый основной цвет; поперек всего хвоста проходит широкая
предвершинная полоса белого цвета; на средней паре она серовато-белая, а на крайней паре
и внешнее опахало, за исключением конца, белое. Молодые без блеска на шее; зоб коричне-
ватый, как и верхние кроющие крыла, на последних имеются серо-беловатые каемки. Клюв
черный, ноги красные, радужина оранжевая или красноватая.
Южный скалистый голубь Columba rupestris turkestanica But.
Columba rupestris turkestanica. Buturlin. Orn. Monatsb., 1908, стр. 49, nom. emend.
Синонимы. Columba rupestris p alii da. Rothschild und H а г t e r t. Ornith.
Monatsber. 1893, стр. 41, Алтай, nom. praeoccupatum.—Columba rupestris intercedens.
Бутурлин. Изв. О.Л.Е.А. и Э. A), 1913, стр. 10, верхняя Лена.
Распространение. Ареал. Западная половина ареала вида, примерно
от линии, тянущейся от верховьев Лены, Урянхайского края и Танну-олы
к верховьям рек Желтой, Голубой и Меконга. В пределах Союза населяет
Алтай (Сушкин, 1938). При этом в юго-вост. Алтае, в долине р. Чулышман
он спускается до Кату-ярыка (Фолитарек и Дементьев, 1938). В сев.-зап. Ал-
тае идет по Чарышу до ст. Чарышской и по Аную до Солонешного (Залесский,
1939). В зап. части Алтая спускается до Усть-Каменогорска (Штегман, 1926),
Колыванского хребта и Мохнатых сопок (Селевин, 1928) и Семипалатинска
(Хахлов, 1928). Отмечен на Иртыше между Шульбинским и Глубоким (Поля-
ков, 1912—1914). По левобережью Иртыша известен из гор Кок-тау в Калбин-
ском Алтае (Селевин, 1929). Многочисленен в Коконе, но редок в Чивиз-тау
и случаен в Семей-тау (Залесские, 1931). Отмечен в мае, скорее всего на гнез-
довье, у Лепсы и г. Аркат (Финш, 1879). В небольшом количестве гнездится
в долине Тышкана в Д^кунгарском Ала-тау (Зарудный и Кореев, 1907). Заре-
гистрирован в Тарбагатае (Поляков, 1915, Селевин, 1929). В долине р. Или
гнездится в Кульдже, по р. Текес в Бик-тау (Шестоперов, 1929). В горах Кир-
гизии обычен по р. Чу, на Сусамыре, Алабуге, Нарыне (Шнитников, 1930),
Сары-джасе (Иоганзен, 1902), Куэлю, Торпу (Кашкаров, 1937). Далее к западу
найден в хребте Киргизском (Спангенберг и Дементьев, 1935), верховьях
Ангрена (Зарудный, 1915), в хребтах Туркестанском, Зеравшанском (Даль,
1936), Алайском (Штольцман, 1897). В Гиссарском хребте отсутствует в запад-
ных отрогах, обычен в восточной его части (Иванов, 1940). Наконец,
обыкновенен на восточном Памире (Мекленбурцев, 1945) и прилежащих частях
западного (Мекленбурцев, 1936). В Туркмении отсутствует.
Характер пребывания. Настоящие перелеты для скалистого
голубя не описываются. На Алтае он встречается в течение круглого года,
лишь приближаясь зимой к поселениям. В высокогорьях Средней Азии (Па-
мир) на зиму остается в очень небольшом количестве, причем держится у посе-
лений. В равнинах и предгорьях никогда не наблюдался, что заставляет пред-
полагать откочевку в несколько более низкие области гор, а может быть и
отлет через горы на юг.На плоскогорьях Памира появление отмечено весной —^
23 марта еще при сильных морозах и сравнительно глубоком снеге. В конце
марта и начале апреля голуби были уже обычны (Мекленбурцев, 1945).
Биотоп. Западная форма скалистого голубя еще более, чем восточная,
связана с горами и скалами. На крайнем западе ареала птица только в виде
2 Птицы СССР, т. II
18 ОТРЯД ГОЛУБИ
исключения попадается у поселений. Впрочем и на востоке ареала — в Алтае —
голуби только осенью и зимой посещают деревни, но в них не гнездятся. Лишь
в Кульдже голуби живут, хотя и в небольшом количестве, как полудомашние
птицы. Избегает этот голубь и гор, покрытых лесами. Типичное местообита-
ние — скалы безлесных гор, одетые травянистой растительностью. Характер
скал не имеет особого значения и, например, на Алтае голуби одинаково охотно
селятся в обрывистых каменных буграх степи и пустыни у Колыванского озера,
в грядах красного гранита в Хара-гоби, в высочайших обрывах у Чулышмана
и в разрезах древней морены у Кор-кечу. В горах Средней Азии заселяют
исключительно скалы. Чем дальше к западу, тем выше поднимается нижняя
граница вертикального распространения каменного голубя. В Чингиз-тау
и Тарбагатае он селится чуть ли не у подножий гор, а на Иртыше и Зайсане —
на скалах у самой воды. В Джунгарском Ала-тау также встречается совсем
невысоко — например у Лепсы. Между Терскей и Кунгей Ала-тау — в Буам-
ском ущелье опускается до уровня, примерно, тысячи метров. В Киргизском
хребте был найден только выше 2000 метров, на Зеравшане около 1700 (Артуч,
Даль, 1936). Наконец, в бассейне Аму-Дарьи, на р. Гунт и Шах-дара попадается
от высот примерно 3000 м. Верхняя граница на Алтае и в хребтах Средней
Азии у снеговой линии, а местами даже и выше. В центральном Тянь-Шане
найден на р. Торпу 2880 м и Куэлю 3350 м, но это, конечно, не предельные
высоты. Особенно высоко поднимается на Памире, где неоднократно наблю-
дался на высотах свыше 4000 м.
Численность. Во многих местах скалистый голубь обычная, а
местами сравнительно многочисленная птица, образующая стаи до несколь-
ких десятков или даже сотен особей. Вследствие трудной доступности биото-
пов и местообитаний, скалистый голубь подвергается уничтожению меньше
других видов голубей, особенно сизого голубя.
Экология. Размножение. Скалистый голубь приступает к размно-
жению очень рано. На Памире воркование самцов начинается в конце апреля
при морозах и снеге. В середине мая голуби часто попадаются парами. Приб-
лизительно в это время начинается витье гнезд и откладывание яиц. Несом-
ненно, что в местах с менее суровым климатом размножение начинается зна-
чительно раньше. Даже на Алтае известны случаи добывания летных молодых
уже 23 мая, что указывает на откладку яиц в самом начале апреля или конце
марта; на Алтае сроки вывода птенцов, повидимому, чрезвычайно растянуты,
и последние молодые вылетают из гнезд в начале августа (Сушкин, 1938).
Однако скорее можно допустить, что здесь имеют место нормальные повторные
циклы. На это указывает наличие желтков у самки, добытой в Алтайском
заповеднике 23 июня (Фолитарек и Дементьев, 1938)- В центральном Тянь-
Шане на р. Сары-джас и юго-зап. Алтайской системе — Курчумском хребте —
молодые были добыты 9 июля (Иоганзен, 1907) и 2—3 июля (Поляков, 1912).
Обе находки слишком ранни для второго вывода и их приходится отнести
к первому. Недостаток данных не позволяет все же высказаться с полной уве-
ренностью, но для Тянь-Шаня и Памиро-Алая вполне можно допустить два
вывода в лето. Гнездятся скалистые голуби колониями, иногда небольшими,
иногда сравнительно многочисленными — до нескольких десятков пар. На
Памире в подобных колониях гнезда не бывают так тесно расположены, как,
например, у сизака, и отдельные пары голубей занимают удобные места
в нескольких десятках метров друг от друга. Однако, члены подобной колонии
вылетают на кормежку все в одно место и держатся там общей стаей. По
окончании брачного периода голуби кочуют выводками, которые на Алтае
в сентябре собираются во все более крупные стаи и посещают поля,
гумна и деревни, иногда присоединяясь к сизакам. На Памире большие
стаи были отмечены в начале ноября, в среднем течении р. Гунт (Меклен-
бурцев).
БЕЛОГРУДЫЙ ИЛИ БЕЛОСПИННЫЙ ГОЛУБЬ
19
Линька. Общая схема — как у других видов голубей. Продолжитель-
ность процесса линяния — около четырех месяцев. Линька начинается в мае —
июне выпадением десятого первостепенного махового. Июньский экземпляр
с Алтая имеет 4 старых пера. Экземпляры с Марка-куля, добытые в начале
июня, обладают четырьмя — пятью старыми маховыми. Птица из Джунгар-
ского Ала-тау, добытая в этом же месяце, содержит в крыле пять старых перьев,
из Киргизского хребта — шесть, а из Туркестанского — шесть — семь.
Может быть алтайские голуби начинают линьку раньше туркестанских, так
как многие особи заканчивают ее уже к началу августа, хотя другие не выли-
нивают в это время и наполовину. Июльские птицы с Марка-куля имеют по
три — четыре старых пера, с Киргизского хребта —шесть, а из Туркестан-
ского — четыре. Наконец августовские экземпляры из Джунгарского Ала-тау
и Памира несут по три старых пера. Эти отрывочные данные как будто
показывают более позднее начало линьки у среднеазиатских голубей в сравне-
нии с алтайскими. Однако материал слишком незначителен, и большие серии
могут показать совпадение в сроках хода смены пера. Рулевые и мелкое перо,
повидимому, линяют вместе со сменой первого — третьего маховых, которые
выпадают последними в августе — сентябре.
Питание. Состав пищи туркестанского скалистого голубя не изу-
чался. Для Алтая имеется указание на питание культурными злаками (Суш-
кин). На Памире эти голуби обычно кормятся в долинах рек, где наиболее
распространенным растением является осока, зерна которой, видимо, и соби-
раются голубями (Мекленбурцев).
Описание. Размеры и строение. Размах крыльев самца A) 667, самки A)
666 мм. Длина крыла самцов A3) 213—236, самок (И) 215—235, в среднем 225 и 224 мм.
Окраска. Оба пола окрашены одинаково. Общая окраска у взрослых птиц сизая,
обычно несколько более светлая, чем у типичной формы. Особенно светло окрашена ниж-
няя сторона.
3. Белогрудый или
белоспинный голубь
Vig.
Colutnba leuconota
Columba leuconota. Vigors. Proceed. Committe Zoolog. Soc. London I. 1831, стр. 23,
Гималаи к западу от Сикима.
Русское название. Книжное.
Распространение. Ареал. Западные части Памиро-Алая, вост. Гиндукуш,
Кашмир, Гималаи, область верховий Салуэна, Меконга, Голубой и Желтой рек.
Биотоп, Высокие горы от 2700
до 5000 м. Гнездится только на ска-
лах, повидимому, выбирая высокие и
труднодоступные места. Вне гнездового
периода совершает вертикальные миг-
рации, не покидая, впрочем, горных
районов.
Подвиды и варьирующие признаки.
С. /. leuconota Vig., 183U — заЬ. часть
ареала до Сикима на востоке; С. /. gradaria.
Н а г t е г t, 1916— вост. часть ареала —
Ганьсу, Сычуань, Тибет. Подвиды отличаются
окраской спины, поясницы и надхвостья,
а также размерами. Экологические различия
между подвидами не выяснены. Возможно,
что западный подвид не поднимается так вы-
соко в горах, как восточный. Возможно так-
же, что наиболее северо-западная популяция
является перелетной, в то время как осталь-
ные оседлы или предпринимают только
вертикальные кочевки.
Карта 3. Распространение белогрудого го-
лубя Columba leuconota
1. С. 2. leuconota, 2. С. I. gradaria;
а — граница области распространения, б — на
вполне выясненная граница.
20
ОТРЯД ГОЛУБИ
Западный белогрудый голубь Columba leuconota leuconota V i g.
Распространение. Ареал. Первый раз эта птица была указана для Тур-
кестана и вместе с тем вообще для России по сборам Барея с перевалов Тенгиз*
бай и Кара-казык, зап. часть Алайского хребта (Штольцман, 1897). Затем
белогрудый голубь был найден на р. Кшемыш в вост. части Туркестанского
хребта (Зарудный, 1910). Следующее указание относится к долине Пянджа
между г. Хорогом и кишлаком Ишкашим (Зарудный, 1915). В последние годы
стайка из трех экземпляров наблюдалась в Заилийском Ала-тау, невдалеке
от г. Алма-Ата (Штегман,
1948).
Биотоп. Немногочис-
ленность находок не дает воз-
можности судить о приурочен*
ности этого вида к каким-либо
определенным ландшафтам.
Можно заключить лишь, что
эта птица держится в очень
высоких горах —у таких пере-
валов, как Тенгиз-бай (около
3300 м) или Кара-казык (око-
ло 4750 м). В долине Пянджа
и у перевала Кум-бэль голуби
отмечались во всяком случае
выше 2500 м. Несомненно вид
этот приурочен к скалам, как
сизый и скалистый голуби, но
в отличие от первого селится
в самых высоких областях гор.
Численность. В на-
ших пределах белогрудый го-
лубь весьма немногочисленная
птица (граница ареала). Боль-
шей частью он наблюдается
одиночными особями или парами. Самая крупная колония, найденная у пере-
вала Кум-бэль, состояла не более, чем из пяти пар. В Гималаях местами
образует большие гнездовые колонии, а вне периода размножения—боль-
шие стаи (Бекер, 1928).
Экология. Размножение белогрудого голубя остается до настоя-
щего времени для пределов Средней Азии совершенно неизвестным, как и все
другие стороны его экологии. В Гималаях гнездится колониями на высотах
примерно в 3000—3600 м, иногда ниже или выше. Гнезда расположены на ска-
лах, в щелях и пещерках, реже на карнизах; как у других голубей, они
построены из веточек с примесью травы, перьев и т. п. Гнезда используются
много лет, поэтому очень грязны и изобилуют паразитами; птицы в брачный
сезон ограничиваются ремонтом старых построек. Яиц в кладке 2 (в виде
исключения 3). Размеры B6) 34,6—42,8 х 26,3—31,2, в среднем 38,2—28,0 мм
(Бекер, 1928).
Линька. Единственный осмотренный линный экземпляр (взрослая
самка, добыта 21.VI. 1893, перевал Кара-казык) имеет по 8 старых маховых,
девятые в виде пеньков, десятые до конца не доросли. Все остальное перо
старое. Таким образом, линька этого экземпляра началась в конце мая или
первой трети июня.
Некоторые из представителей восточной формы (всего осмотрено 27 шку-
рок в Зоол. институте Акад. наук и музее Моск. университета), добытые в
Рис. 3. Белогрудый голубь
БУРЫЙ ГОЛУБЬ
верховьях рек Желтой и Голубой в конце июня, имели большей частью по б ста-
рых маховых. Иногда линька начиналась позднее, и к этому времени птицы
сохраняли по 8 старых маховых. Добытые в декабре в стране нам птицы закан-
чивают линьку, имея сменившиеся маховые, сменяющиеся рулевые и свежее
мелкое перо.
Питание, Вопросы питания этого вида в условиях Средней Азии со-
вершенно не изучены. В Гималаях в качестве кормов указываются ягоды, поч-
ки, семена различных растений.
Полевые признаки. Белогрудый голубь похож (Бекер, 1928) на сизака,
но очень легко отличается от него и ют всех других наших голубей белым цве-
том всей нижней и верхней стороны тела, при более темных крыльях, очень
темной голове и хвосте. При этом на последнем вырисовывается широкая
белая полоса, похожая на таковую у скалистого голубя, но не прямая, а имею-
щая вид тупого угла, направленного вершиной к основанию хвоста.
Описание. Размеры и строение. Единственная добытая в СССР самка имеет
крыло 229 мм. По литературным данным крыло у обоих полов от 215 до 245 мм
(Бутурлин, 1934).
Окраска. Голова и прилегающие части шеи—аспидно-серые. Передняя часть
спины, плечевые и самые внутренние из малых верхних кроющих крыла — буровато-
серые; кольцо вокруг шеи, зад спины и вся нижняя сторона — чисто белые; бока тела, под-
хвостье и нижние кроющие крыла—светло-серые; поясница, надхвостье и верхние
кроющие хвоста черновато-аспидного цвета; маховые серые; концы первостепенных и
каемки их опахал — бурые, верхние кроющие крыла — серые; на самых больших — бурые
поперечные пятна, образующие поперек крыла полосу. Такие же пятна имеются на концах
второстепенных маховых, рулевые — аспидно-черноватые; на каждом пере имеется широкая
белая предвершинная полоса, которая на крайних рулевых расположена почти у вершины,
а на средних — отступя от вершины наЗ—А см. Благодаря этому белая хвостовая полоса
имеет вид тупого угла вершина которого направлена к основанию хвоста. Клюв черный.
Глаз золотисто-желтый. Ноги ярко красные. Молодые с охристыми каемками перьев
верха и охристым налетом низа.
4. Бурый голубь Columba eversmanni В p.
Columba eversmanni. Bonaparte. Compt. Rend. Acad. Paris, 63, 1856, стр. 838 «Зап.
и центр. Азия»
Синоним. Columba fusca. Pallas. Zoographia Rosso-Asiatica. I, 1811, стр. 567,
nom. praeoccupatum.
Распространение. Ареал. От юго-вост. угла Каспийского моря по север-
ным предгорьям Копет-Дага, в бассейнах Теджена, Кушки, Мургаба, долине
Аму-Дарьи вниз до Аральского моря (Зарудный, 1896), Урги, Судочьего озера
и обрывов Усть-Урта (Гладков, 1835); вверх по Аму-Дарье поднимается при-
мерно до Вахша, а может быть и еще выше. В низовьях Аму-Дарьи гнездится
в Айбугирской котловине (Молчанов, 1913). Восточнее Аму-Дарьи известен
из Кызыл-Кумов (Спангенберг, 1926), из долины Зеравшана (Лоудон, 1909,
Даль, 1936) и всех правых притоков Аму-Дарьи — Сурхана, Кафирнигана,
Вахша, Ях-су (Иванов, 1940). В системе Сыр-Дарьи распространен, как в
нижнем и среднем течении (Спангенберг, 1934), так и в более высоких участ-
ках долины —-у Чиназа (Плеско, 1888) и в Фергане (Кашкаров, 1934, Маль-
чевский, 1940). По правым притокам Сыр-Дарьи — Чирчику, Келесу, Арыси —
бурый голубь гнездится повсюду (Северцов, Лоудон, 1909, Сатаева, 1937),
Еще далее к востоку бурый голубь встречен в юго-вост. части хребта Кара-
тау (Колесников, 1934), на р. Чу (Гладков, 1932, Селевин, 1935). По долинам
впадающих в Балхаш рек бурый голубь встречен на Или (Шестоперов, 1929),
на Лепсе (Финш, 1876). В этой же области найден у подножий Джунгарского
Ала-тау в Саркане (Селевин, 1935). Самое восточное из гнездовых нахождений
в СССР относится к долине Черного Иртыша (Поляков, 1912). Существует
указание на наличие бурого голубя на Алтае (Иоганзен, 1898), но тщатель-
22
ОТРЯД ГОЛУБИ
ными последующими исследованиями оно не подтвердилось. За пределами
СССР бурый голубь населяет Кашгарию, Кашмир, Ладак, сев. Афгани-
стан и сев.-вост. Иран. Зимует от северного подножья Коиет-Дага,
долины Мургаба и Афганистана. Но основные места зимовок охватывают
Синд, южн. Пенджаб, соединенные провинции и Бихар. Известны слу-
чайные залеты до Красноярска на Енисее, Чкалова, Бугуруслана и даже
среднего течения Суры и гор Бей-шань к востоку от озера Лоб-нор. Вид моно-
типический.
Характер пребывания. Бурый голубь почти на всей части
своего ареала, входящей в пределы СССР, — перелетная птица, появляющаяся
сравнительно поздно. Од-
нако, имеющиеся в литера-
туре данные о сроках про-
летов настолько отрывочны,
что не дают возможности
судить о последовательно-
сти этого явления. Появле-
ние в Туркмении отмечается
в конце марта —первой по-
ловине апреля (Дементьев,
1950), отлет там происхо-
дит, повидимому, в сентяб-
ре. В первой трети апреля
— 5 числа —голуби были
отмечены у Ташкента (Се-
верцов), 5—-8—у Кермине
на Зеравшане (Лоудон,
1909), а 7 — в Дуруне (Рад-
де, 1889). Во второй трети
апреля — 13 — их появле-
ние было отмечено у ст.
Вревской на р. Чирчик,
17 — у Алма-Ата (Зарудный
и Кореев, 1906) и 18 —у
Чимкента (Северцов). На-
конец, в последней трети апреля—21 —их встречали около Джаркента
(Зарудный и Кореев, 1906), 22 — под Туркестаном (Северцов), 24—на озере
Балты-Куль у Ташкента, в конце апреля — в низовьях Сыр-Дарьи (Спан-
генберг, 1936) и 28 числа на Памире (Мекленбурцев, 1945). В Кульдже
эти птицы появились только 3 мая, но были уже обычны (Шестоперов, 1929).
В этом же месяце —12 числа бурый голубь был добыт у Чкалова (Зарудный,
1888), а 20—24 отмечен в Кызыл-Кумах (Зарудный, 1914). Зимовки бурого
голубя лежат почти полностью за пределами Союза. Лишь на крайнем юго-
зап. Средней Азии —у северных подножий Копет-Дага и в бассейнах Мургаба
и Теджена —эта птица зимует. Однако, основная область зимовок лежит
значительно южнее и охватывает Белуджистан, Синд и Пенджаб. Менее регу-
лярно посещает бурый голубь зимой соединенные провинции Индии — Уд и
Бихар. На зимовках в Индии наблюдается с октября — ноября по март.
Пролет бурых голубей идет главным образом равнинами; в больших.горах
они встречаются редко и только одиночками или маленькими группами.
Биотоп. Главным образом низменности. Пожалуй, с наибольшей охо-
той бурый голубь поселяется на высоких лессовых обрывах, образующихся
по берегам рек Тянь-Шаня и Памиро-Алая, по выходе их из предгорий в рав-
нины. В этих обрывах голуби образуют колонии, никогда не бывающие такими
многочисленными, как у сизого голубя. Помимо обрывов, голуби любят селить-
Карта 4. Распространение бурого голубя Columba
eversmanni
а — граница гнездовий, б — не вполне выясненная граница
гнездовий, в — изолированные нахождения особей, г — зимовки.
БУРЫЙ ГОЛУБЬ 23
ся в древесных насаждениях, предпочитая старые, малопосещаемые сады из
карагачей или платанов. Голуби охотно заселяют рощи старых развесистых,
дуплистых тополей в поймах больших рек, например Мургаба, Аму- и Сыр-
Дарьи и Или. Поднимаются довольно высоко в горы, например, в северной
Фергане у Арсланбоба. В горных лесах всякого иного характера, даже обра-
зованных такими породами лиственных деревьев, как орех, поселяются с го-
раздо меньшей охотой. В яблоневых лесах, так же как и в можжевеловых,
селится очень редко и в последних отмечался лишь в горах Копет-Дага. Непре-
менным условием возможности поселения голубей служит наличие воды—
большей частью речки или ручья.
Численность. Бурые голуби никогда не образуют таких больших
скоплений, как сизаки. Вряд ли можно встретить стаю, превышающую пол-
торы — две сотни экземпляров. В настоящее время осенью еще можно встре-
тить стаи, насчитывающие по 2—3 десятка птиц. Весной большей частью попа-
даются общества, объединяющие не более десятка. Количество пар в колонии
исчисляется двумя —тремя десятками. Чаще же всего встречаются одиночные
пары, или немногочисленные колонии по 4—5 пар.
Экология. Размножение. Прилетающие с зимовок стайки голубей
содержат в своем составе уже готовые к гнездованию пары старых птиц, но
по крайней мере часть прошлогодних молодых прилетает холостыми и присту-
пает к спариванию сейчас же по прилете. Самцы начинают ворковать, едва
успев появиться у своих гнездовых биотопов. Если местом гнездования
служит обрыв, то голубь садится на какой-нибудь наиболее выделяющийся
бугорок, нависший над самым обрывом. Если же гнездо будет строиться
в дупле, то токующий самец скрывается где-либо в гущине кроны, сидя
на толстой ветви. Где бы ни располагался воркующий голубь, время от
времени он взлетает и, сделав небольшой круг, возвращается на прежнее
место.
После образования пары гнездо строится очень быстро. Оно представляет
собой обычную для голубей рыхлую просвечивающую постройку и в гро-
мадном большинстве случаев располагается где-либо в закрытом месте: дупле
дерева, в расщелине лессового обрыва или в выкопанной в нем норе сизово-
ронки. Последнюю голуби занимают очень охотно, тем более, что приступают
к гнездованию несколько раньше, чем исконные хозяева нор. Гнездятся
голуби и в развалинах крупных построек, строя здесь гнездо где-либо в глу-
бокой щели, на месте выпавшего кирпича и т. п. Открытые гнезда делают
редко и располагают их, по большей части, высоко на деревьях.
Дупла, в которых закладываются гнезда, также находятся обычно высоко
над землей в главном стволе дерева или в одной из крупных ветвей. Вход в дуп-
ло бывает иногда очень широким, а само дупло коротким и открытым, так что
яйца лежат на виду.
Яиц в кладке, как и у всех голубей, — два, цвет их чисто белый. Размеры
(9) 33,2—38,1 х 25,7—27,9 мм (Спангенберг, 1936). Наиболее ранние на-
ходки яиц относятся к окрестностям Джаркента, где валовая кладка при-
ходится на конец апреля (Зарудный и Кореев, 1906). Впрочем здесь же находки
свежих яиц известны от 20 и 26 мая и 18 июня (Шестоперов, 1929). Такие же
сроки приведены для Ташкента (Сатаева, 1937). В долине Сыр-Дарьи ниже
Чиили яйца находили от середины мая до середины июня (и даже позднее).
Массовое гнездование здесь приходится на конец мая — первую половину
июня (Спангенберг, 1936). В долине р. Чу кладки были обнаружены в начале
июня и конце июля (Гладков, 1932). По мнению Северцова, под Ташкентом
в середине июля вылетают из гнезд птенцы второй кладки. В Туркмении у
Карабента яйца были найдены, примерно, 10 мая, а у Сулюкли — 21 мая
(Зарудный, 1896). Однако, учитывая ранее приведенные сроки, эти находки
нельзя считать за начало откладки яиц.
24 ОТРЯД ГОЛУБИ
Насиживают оба родителя попеременно. Немного подросшие птенцы были
обнаружены в середине мая (Сатаева, 1937), а летные —в последних числах
этого месяца (Северцов). Молодые, едва покинувшие гнездо, попадаются еще
в начале сентября. Это несомненно указывает на наличие третьей кладки,
как и яйца, обнаруженные в конце июля. Сроки пребывания молодых в гнезде
неизвестны. Молодые первых выводов держатся по несколько дней с родите-
лями, а затем соединяются в особые стаи.
Линька. Старые птицы начинают линять в июле, т. е. позднее, чем
другие виды голубей. Смена оперения начинается с десятого махового и идет
к концу крыла. В третьей декаде июля попадаются особи, у которых осталось
по 8 или даже по 7 старых первостепенных маховых (возможно, что в этом слу-
чае начало линьки падает на июнь) и уже отросло по два новых пера. В первой
декаде августа встречаются птицы с пятью старыми первостепенными махо-
выми. В конце августа — начале сентября начинается интенсивная смена всего
мелкого оперения. Вместе с ним сменяются рулевые, начиная от крайних к сере-
дине хвоста. К концу первой декады сентября у многих старых голубей линька
уже заканчивается. Однако некоторые особи очень запаздывают и еще в конце
сентября встречаются старые птицы в изношенном пере. Птенцы самых позд-
них выводков отлетают не перелинявшими. К концу лета оперение бурых
голубей сильно выцветает. От этого мелкое оперение заметно бледнеет, а
маховые буреют. Таким образом старое перо отличается у линяющих птиц
от нового.
Возрастные изменения бурых голубей аналогичны изменениям других
видов голубей. Птенцы одеты редким пухом, который сменяется рыхлым юве-
нальным пером. Последнее в первую же осень или зиму заменяется окончатель-
ным нарядом.
Питание. Подобно другим голубям, бурый кормится всегда на земле.
Его пищей служат, главным образом, семена бобовых и злаковых растений.
Из первых он охотно клюет семена люцерны, из вторых — предпочитает пше-
ницу, так как ее зерна лишены колючих остей. Весной, посещая только что
засеянные поля, голуби могут причинять им некоторый вред. Однако в это
время года птицы никогда не бывают многочисленными, что сильно сни-
жает их вредную деятельность. Осенью голуби довольно охотно посещают поля
сафлора, заглатывая семена этого растения вместе с их твердыми оболочками.
Иногда в желудках птиц обнаруживаются мелкие раковинки улиток или клу-
беньки растений.
Кормятся бурые голуби, подобно сизым, утром и вечером, летая иногда
за кормом за пять — десять километров. После кормления летят на водопой
и рассаживаются на отдых. Отдыхать предпочитают на деревьях, садясь, в
противоположность сизакам, не открыто, а в самой гущине кроны. Сидят очень
спокойно и настолько малр заметны среди густых листьев, что и зная, на кото-
ром дереве сидит голубь, его почти невозможно увидеть, даже если подойти
вплотную к дереву.
Полевые признаки. Бурый голубь легче всего может быть смешан с сиза-
ком благодаря наличию у обоих белой поясницы, издали бросающейся в глаза.
Этот же признак отличает его от клинтуха. От сизака бурый голубь отличается
меньшим ростом и не такой резкой разницей в окраске между подкрыльями
и нижней стороной тела. На лету он кажется более короткохвостым, чем дру-
гие виды голубей. Кроме всего этого, бурый голубь, подобно клинтуху, иногда
издает при взлете своеобразный свист крыльями. Ненапуганные охотой
голуби безбоязненно подпускают к себе на самое близкое расстояние, как во
время кормежки, так и во время отдыха. К напуганным же птицам невозможно
приблизиться даже на сотню метров, и не только стаи, но и одиночки никогда
не налетают на выстрел. С другими птицами бурые голуби не образуют общих
стай и даже с другими видами голубей обычно не держатся вместе. Чаще всего
КЛИНТУХ
25
их можно видеть вместе с сизыми голубями, отдыхающими после кормежки
на каком-нибудь обрыве. Иногда они вместе кормятся на полях.
В других случаях бурые голуби объединяются с большими горлицами,
выбирая для дневного отдыха
одну и ту же группу деревьев.
Однако оба вида держатся и в
этих случаях совершенно само-
стоятельно, а спугнутые разле-
таются в разные стороны. Голос
бурого голубя приходится слы-
шать только во время гнездова-
ния. Он весьма напоминает вор-
кование клинтуха и представляет
собой такие же глухие «уу-уу-
уу». Воркующий самец не кру-
жится вокруг голубки, как про-
делывает это сизак, а сидит спо-
койно на избранном им месте.
Голос голубя разносится далеко
и звучит так глухо, что найти
его по звуку обычно не сразу
удается.
Описание. Размеры и стро-
ение. Крыло самцов A4) 195—208,
самок A0) 190—201, в среднем, соот-
ветственно, 201 и 197 мм.
О к р а с к а. Основной цвет
серо-или пепельно-сизый; подхвостье
и нижние кроющие хвоста значи-
тельно темнее — темносизые; мантия
и зерхние кроющие крыла с сильным буроватым налетом; спина и поясница гораздо
светлее — от светло-сизой до почти белой окраски; надхвостье очень темное — аспидно-
сизое. Верх головы и грудь с сильным винно-розовым налетом; бока и задняя часть
шеи с зеленым металлическим блеском, переходящим при некоторых положениях в пур-
пуровый; на верхних кроющих крыла черноватые пятна, образующие на крыле две
неявственные прерывистые полосы: нижние кроющие крыла и подмышечные — белые
или беловато-сизые; первостепенные маховые — черновато-бурые с светло-серой основ-
ной половиной внутреннего опахала; второстепенные — светло-серые с черноватыми вер-
шинами; рулевые — сизые с черноватой широкой вершинной полосой, от которой сизой
полоской отделена более узкая часть, лежащая ближе к основанию пера. Глаз желтый.
Клюв у основания черноватый или аспидный, его конечная половина — зеленовато-жел-
тая. Ноги — от бледно-малиновых до желтовато-розовых.
Молодые в первом пере окрашены значительно тусклее старых. Перья зоба и груди
с неясными буроватыми каемками. Блеска на шее нет.
5. Клинтух Columba oenas L.
Columba oenas. Linnaeus. Syst. Nat., изд. X, 1758, стр. 162 , Швеция.
Русское название. Народное, применяется к этому виду по крайней мере
с XVIII в. (Левшин, 1774) в написании «клиндух». Другое народное имя — «лесовой голубь».
Распространение. Ареал. Более или менее регулярно гнездится в зна-
чительной части Европы, на запад до берегов Пиренейского п-ова и Франции.
Британские о-ва граница пересекает таким образом, что Ирландия, Шотлан-
дия и часть Великобритании к северу от Нортумберлэнда остаются вне гнездо-
вого ареала. На Скандинавском п-ове клинтух встречается только в южных
частях Норвегии и Швеции, а в Финляндии найден лишь на юго-западе. В СССР
северная граница ареала проходит, примерно, у Ленинграда, оттуда тянется
к истокам Печоры и среднему течению Камы, пересекает Уральский хребет
у Свердловска, в бассейне Ишима и Тобола, в низовьях Иртыша у Тары.
Рис. 4. Бурый голубь
26
ОТРЯД ГОЛУБИ
Далее граница, примерно, идет от Колывани до Барнаула, снова пересекает
Иртыш у Семипалатинска, Ишим и Тобол у верховий, захватывает нижнее
течение Илека, среднее течение Урала и выходит на Волгу у Саратова. Пустын-
ные области между низовьями Волги, Доном и Тереком не входят в гнездовой,
ареал клинтуха. Каспийское море граница пересекает, примерно, от низовий
Терека к Чикишляру, после чего тянется к востоку южными предгорьями
Копет-Дага и, обогнув Астрабадские горы, поворачивает на запад. Затем она
захватывает Эльбрус (Мазандеран) и Загрош, Сирию и Палестину, Малую
Азию, Болгарию, Македонию, Италию и всю Малую Африку, т. е. Тунис,
Алжир и Марокко.
Большой изолированный участок ареала лежит к востоку от очерченной
территории. Он включает весь Тянь-Шань (быть может и Каратау) и всю до-
Карта 5. Распространение клинтуха Columba oenas
7. С. о. oenas, 2. С. о. hyrcana, 3. С. о. yarkandensis;
а — граница гнездовий, г — места зимовок перелетных форм.
лину Тарима до Лоб-нора. ВПамиро-Алайскойсистеме,в бассейне Аму-Дарьи,
имеются лишь отдельные разрозненные находки. В Туркмении гнездовые
находки относятся лишь к бассейну Мургаба и Аму-Дарьи.
Область зимовок клинтуха незначительно выходит за пределы гнездового
ареала. Регулярные зимовки захватывают Францию, средиземноморские стра-
ны Европы, Переднюю Азию и Кашгарию. В Африке зимует главным обра-
зом в Алжире, Тунисе и Марокко. В Египте попадается реже. Отдельные
особи не регулярно перезимовывают в Англии, Германии и в СССР даже
до Калуги.
Биотоп. Клинтух — обитатель лесов, главным образом, лиственных,
с большими старыми дуплистыми деревьями. В редких случаях Селится на вы-
соких обрывах.
Подвиды и варьирующие признаки. С. о, oenas Linnaeus, 1758— весь ареал вида,
за исключением участков к востоку и юго-востоку от Кавказа; С. о. hyrcana Zarudny
et Bilkewitsch, 1913 — горы Эльбрус, Загрош и Месопотамия; Co. yarkandensis
В u t u г 1 i n, 1908—среднее течение Аму-Дарьи, Зеравшан, Кашка-Дарья, Тянь-Шань,
Кашгария. Изменчивость у клинтуха выражается в более темном или светлом оттенке
основной окраски и, в меньшей степени, в размерах. Экологические различия между
клинтух 27
подвидами сводятся к тому, что одни из них свойственны прохладным и влажным лист-
венным лесам Европы или гор Азии, а другие обитают в сухом и жарком климате азиат-
ских пустынь. Некоторым подвидам свойственны перелеты, другие же являются оседлыми
или только незначительно откочевывают зимой.
Европейский клинтух Columba oenas oenas L.
Распространение. Ареал. Населяет большую часть европейской тер-
ритории СССР. В Подолии этот голубь местами нередок на гнездовье (Портенко,
1928, Герхнер, 1928), на Волыни он так же обычен (Шарлемань и Портенко,
1926). В бассейне верхнего Днепра это гнездящаяся птица. Далее к северу
клинтух гнездится в области верховий Зап. Двины, в Калининградской обла-
сти, в Прибалтике и вокруг озер Псковского и Ильмень, где он, однако, редок.
Еще реже встречается этот голубь в Ленинградской обл. (Бианки, 1901),
не встречаясь к северу от Ладожского озера. В Горьковском крае, через кото-
рый проходит северная граница распространения, он очень редок (Серебров-
ский, 1917, Пузанов, 1942). В бассейне Оки гнездится. В междуречье Оки и
Волги редок (Лоренц, 1892), как и в области верхнего течения Волги (Третья-
ков, 1940). В Поволжье обычен, но в низовьях Волги не гнездится (Воробьев,
1936), как и в низовьях Дона и в Присивашье (Воронцов, 1937). Немногочис-
ленна эта птица и в области среднего течения Дона — на р. Хопер (Измайлов,
1940), однако далее, вверх по левобережью Дона, этот голубь обычен (Резцов,
1910). В области среднего течения Днепра (низовья Десны) клинтух обычен
(Шарлемань, 1936), в нижнем течении спорадичен, на Днестре многочисленен
(Вальх, 1911). Вообще на Украине гнездится повсюду, местами даже в боль-
шом количестве, однако в степной части бывает только на пролетах (Шарле-
мань, 1938). В Крыму клинтух гнездится в небольшом количестве в степной
части, но в горах более обычен. На Кавказе, как и в Закавказье, клинтух —
обыкновенная гнездящаяся птица во многих местах. В долине р. Урала клин-
тух гнездится только в среднем и верхнем течении, по нижнему течению Илека
и изредка попадается отдельными парами в верховьях этой реки (Зарудный,
1888). В Башкирии он более обычен. В Уральском хребте гнездится в южной
половине (Теплоухов, 1921, Снигиревский, 1929), доходя к северу до 58° с. ш.
(Куклин, 1936). В сибирской части ареала клинтух гнездится в низовьях
Тобола (Тарунин, 1928) и Ишима, в среднем течении этих рек и на Иртыше
между Тарой и Семипалатинском (Иоганзен, 1907, Залесские, 1931). Впро-
чем, в районе этого города клинтух не гнездится. На правой стороне Оби най-
ден в Кузнецкой степи и Салаирском кряже (Хахлов, 1937). Залет клинтуха
был отмечен у Красноярска (Сибом, 1878).
Область зимовок клинтуха начинается от средней Франции и захватывает
ее юг, весь Пиренейский п-ов, Италию и Балканский п-ов. В СССР этот
голубь в небольшом количестве зимует по берегам Черного и Азовского морей,
на Кавказе и, особенно, в Закавказье. На юге область зимовок захватывает
Иран, Месопотамию, Сирию, Палестину и Малую Африку. В Египте и Синае
зимовки отмечались как редкое явление. Таким образом, область зимовок
сильно налегает своей северной окраиной на южную окраину гнездового
ареала, что обусловливает незначительную длину пролетных путей.
Характер пребывания. В связи с близостью мест зимовок
клинтух прилетает рано. Например, в Подолии первое появление отмечается
в 20-х числах февраля (Герхнер, 1928). В районе Черкасс в теплую
зиму 1938 г. клинтухи также были обычны в конце этого месяца, хотя
в нормальные годы они прилетают в эту область в начале марта (Орлов, 1947).
В Калининградской обл, появляются в конце февраля или в начале
марта (Тишлер, 1914). В это же время появляются они под Киевом (Шарле-
мань, 1913) и у Чернигова (Емельяненко, 1913). 11—13 марта прилет отмечен
в Полесье (Шнитников, 1913). У Тамбова и Калуги их появление приходится
28 ОТРЯД ГОЛУБИ
на середину марта (Резцов, 1910, Генглер, 1909, Филатов, 1915), как
и у Чкалова (Зарудный, 1888). У Пскова прилет клинтухов относится к послед-
ним числам этого месяца (Зарудный, 1910). Данные за ряд лет, собранные в
средних частях западной области, показывают, что клинтухи появляются здесь
между 22 марта и 15 апреля (Меландер, 1934). 8 апреля клинтухи замечены
в Горьковской обл. (Серебровский, 1918), а 14—в б. Бугурусланском
уезде (Карамзин, 1913). В середине апреля они прилетают в Калининскую
обл. (Третьяков, 1940) и район Казани (Богданов, 1871), а к его. концу до-
стигают северной границы своего ареала, долетая до Лахты (Лауниц, 1913),
Ярославля. Очень поздняя дата — 16 мая, указывающая на длительный про-
лет, приводится для дельты Волги (Воробьев, 1940).
Для зап. Азии имеется мало данных. У Актюбинска появление клинтухов
отмечено 2 мая (Сушкин, 1908). В Кызыл-Кумах отмечены в марте (Богданов,
1882), а на Сыр-Дарье в конце апреля (Спангенбсрг, 1936). В устьях Урала они
появляются уже в конце марта (Бостанжогло, 1911). У Тобольска указывается
очень позднее появление —Змая(Ланин, 1907). Весенний пролет, таким обра-
зом, значительно растянут. Например, у Чкалова первые пролетные птицы
показываются в середине марта, а валовой лет идет в начале апреля (Заруд-
ный, 1888). Под Псковом первое появление приурочено ко второй декаде ап-
реля, а конец весеннего пролета падает на первую — вторую декаду мая
(Зарудный, 1910). В низовьях Урала пролет длится около полутора месяцев
(Бостанжогло, 1911).
Отлет у северной границы ареала начинается в сентябре, но даже у Ка-
зани, где главный лет приходится на конец этого месяца, отдельные особи
задерживаются иногда до конца октября (Богданов, 1871), а у Пскова даже
до первой декады ноября (Зарудный, 1910). Поздний отлет указан для Кали-
нинградской обл., где пролетные клинтухи появляются в октябре, наиболь-
шей силы пролет достигает к концу этого месяца и кончается в середине ноября;
отдельные отсталые особи иногда даже зимуют в этой области. Под Минском
последний клинтух был отмечен 29 ноября (Шнитников, 1913). У Калуги та-
кое же наблюдение было сделано двумя месяцами ранее — 28 сентября, при-
чем уменьшение численности голубей началось уже в конце августа (Филатов,
1915). В Калининской обл. отлет приходится на конец сентября (Третья-
ков, 1940). В эти же дни B8 сентября) пролет громадных стай наблюдался
в Поволжье (Житков и Бутурлин, 1906).
Под Чкаловом отлет начинается с 20-х чисел сентября, валовой
лет приходится на две первых трети октября, в теплые осени клинтухи попа-
даются до конца ноября (Зарудный, 1888 и 1897). В области между Мугоджа-
рами и низовьями р. Урал осенний пролет приходится на вторую половину
сентября, октябрь и первую половину ноября (Сушкин, 1908). В устьях Урала
интенсивный лет падает на вторую половину сентября, но некоторые особи
остаются там почти до середины ноября — до сильных морозов (Бостанжогло,
1911). В дельте Волги клинтух встречается на пролетах видимо реже, чем
в дельте Урала, и отмечен 2 ноября (Воробьев, 1936). На Кавказе клинтухи
летят в октябре. В Зап. Закавказье начало пролета отмечено 10, а конец —
20 октября (Вильконский, 1897). В Приазовье клинтухи отмечались в сентябре
(ст. ст.). Наконец в Подолии этот голубь летит в октябре (Шарлемань, 1936).
Биотоп. Клинтух — исключительно лесной обитатель, однако он
мирится с лесами довольно различного характера, требуя только наличия
крупных старых дуплистых деревьев. У северных границ ареала, например
в Горьковской обл., он селится в смешанных лесах, в сосновых борах,
в еловой рамени с обильным, разнообразным подлеском, в березовых насаж-
дениях. В долине Волги населяет пойменные дубово-осокоревые леса, а в пой-
мах Суры и Алатыря попадается в смешанных лесах. Под Москвой клинтух
найден в сосново-еловой и лиственно-еловой субори и в смешанных лесах.
клинтух 29
В Калининградской обл., в Белоруссии и на Украине гнездится в ста-
рых дубовых лесах. Кроме того, в Белоруссии гнездование обнаружено в оси-
новых и сосновых лесах. У Тобольска и Кокчетава местообитания этого голу-
бя — сосновые леса, и на южном Урале (у Миасского озера) — рощи старых
берез. Наконец, на Кавказе клинтух — обитатель буковых и других листвен-
ных лесов. Клинтух гнездится в дуплах, и приведение лесов в культурный
вид, когда уничтожаются дуплистые деревья, катастрофически сказывается
на численности этой птицы. Только в некоторых местах Зап. Европы, где со-
вершенно уничтожены дуплистые деревья, но вблизи имеются обрывы, клин-
тухи гнездятся в норах и трещинах, подобно сизым голубям. В больших лес-
ных массивах клинтух обычно не встречается. Он предпочитает небольшие
рощи, опушки лесов или леса, чередующиеся с открытыми местами. Наличие
лесных полян или речных лугов для него необходимо, так как на такие места
клинтухи летают кормиться.
Численность. Своеобразные местообитания клинтуха, с непре-
менным условием наличия дупел для гнезд, определяют возможность суще-
ствования и численность птицы. В настоящее время этот голубь повсюду срав-
нительно редок, и наблюдать в один день во время пролета около 30 тысяч
особей, как это мог под Чкаловом в 80-х годах прошлого века Зарудный, вещь
невозможная. В наших пределах клинтухи обычно встречаются стаями не боль-
ше 30—40 особей, даже в области зимовок в Закавказье. Лишь в западных
частях Украины, во время осеннего пролета, еще попадаются стаи, содержащие
до нескольких сот особей.
Экология. Размножение. Цикл размножения ранний. Воркование
начинается немедленно по прилете. Воркующий самец устраивается где-либо
на толстой ветви вблизи от будущего гнезда. Гнезда, как правило, в дуплах.
Очень часто клинтух занимает дупла, выдолбленные черным дятлом. Гнездо-
вой участок пары клинтухов больше, чем у других наших голубей, но это объяс-
няется не неуживчивостью птицы, а редкостью дуплистых деревьев. В тех
случаях, когда дупла многочисленны, клинтухи образуют своеобразные коло-
нии. Для Зап. Европы существуют наблюдения, что при повторных вывод-
ках в одно лето пара занимает каждый раз новое дупло. Может быть, это объяс-
няется тем, что птиц сильно тревожили. В Финляндии сделаны обратные,
наблюдения, показывающие, что пара занимает из года в год одно и то же
гнездо. Гнездовая выстилка, как у всех голубей, скудна и состоит из корешков,,
веточек, листьев и мха. Гнездо имеет наружный диаметр 18—29 см, внутрен-
ний 10—14, высота гнезда от 12 до 19 см, а глубина лотка от 2 до 7 см. Ко
времени вылета птенцов стенки гнезда сильно утолщаются и уплотняются
экскрементами молодых.
Кладка из 2 яиц. Размеры: E) 36—37 х 26—28, среднее 27 X 36,4 мм
(Гебель, 1879). Начало кладки приходится на апрель и май. В Белоруссии
свежие яйца были найдены 23—30 апреля и 13 мая (Шнитников, 1913).
В Калининградской обл. находки яиц известны 23 мая (Тишлер, 1914).
Под Псковом гнездо с двумя сильно насиженными яйцами было обнаружено
10 мая (Зарудный, 1910). У Жиздры 31 мая были найдены уже оперившиеся
молодые (Филатов, 1915). Для Поволжья вылет первых молодых приходится
на конец июня. В Подолии клинтухи воркуют до конца июля и даже до начала
августа, что указывает на вероятность повторных циклов размножения. На
Волыни полуоперенные птенцы были встречены в середине мая. Однако
в Витебской обл. была найдена кладка 18 июля, но в эти же дни уже имелись
летные молодые (Федюшин, 1926). Это подтверждает наличие двух кладок.
На двукратное гнездование клинтуха в течение лета имеются указания для
Горьковской обл. (Серебровский, 1925). В Крыму гнездо с очень молодым
птенцом было найдено 16 августа (Никольский, 1892), что может указывать
только на вторую, если не на третью кладку.
30 ОТРЯД ГОЛУБИ
Насиживание длится, по одним данным, 16—18 дней (Уайзерби, 1940),
по другим, 20—21 день. Молодые проводят в гнезде 25—28 дней. Таким образом,
для одного вывода гнездо бывает занято 41—49 дней. Насиживают самец и
самка, но самка в несколько большей степени, чем самец. Последний часто и
подолгу воркует у гнезда, которое, однако, найти нелегко, так как при тревоге
самец слетает с гнезда и скрывается, ловко лавируя между деревьями. Выкарм-
ливают птенцов оба родителя, первое время кормом молодых служит выделение
стенок зоба, а затем различные зерна, приносимые в зобах. Птенцы, как ука-
зано выше, покидают гнезда в возрасте около месяца. После того, как молодые
приобретут полную самостоятельность, старые птицы приступают к последую-
щей кладке. Закончив выкармливание птенцов последней кладки, старые птицы
присоединяются к стаям молодых, более ранних выводков или образуют осо-
бые стаи, вместе с молодыми последнего выводка.
Линька. Общая схема — как у других голубей. Одна полная годовая
линька. Начинается она выпадением 10-го махового, что обычно происходит
в мае. В июне часто встречаются особи с 9, 8 или 7 старыми маховыми. В июле
этих перьев остается от 7 до 4, а в августе — от б до 4. Сентябрьские птицы имеют
2—3 старых маховых, и в октябре большинство птиц заканчивает линьку.
Рулевые быстро сменяются, вместе с 3-м, 2-м и 1-м маховыми. Их смена идет
сразу от центра и от края. В это же время сменяется мелкое оперение, хотя
перья головы иногда начинают постепенно редеть вместе с выпадением 7-го и
8-го маховых. Наиболее интенсивная смена мелкого пера идет в то время, когда
в крыльях остается по два старых, маховых. Примерно таков же общий ход
линьки молодых, только он иногда значительно смещается во времени, и линька
заканчивается в мае. Последовательность смены нарядов сводится у клинтуха
к тому, что птенцовый редкий пух сменяется ювенальным оперением, которое
покрывает птенца уже на 24—26-й день. В этом наряде молодые клинтухи
остаются некоторое время по вылете из гнезд и в большинстве случаев сменяют
его на взрослый наряд в первую же осень.
Питание. Клинтух — исключительно растительноядная птица. Основ-
ную пищу его составляют разнообразные зерна и семена культурных и диких
растений. Летом клинтух любит кормиться на лугах и лесных полянах, и тогда
в его питании преобладают дикие травы. С конца июля и осенью во время пере-
летов стаи клинтухов жируют, главным образом, на скошенных хлебных
полях. Корм эти голуби обычно собирают с земли, не выклевывая зерен из
стоящих на корню колосьев. Впрочем, если хлеб собран в копны, то это уже
защищает зерно, так как клинтухи предпочитают собирать те зерна, которые
выпали на землю. Зимой по возможности придерживаются полей, но при выпа-
дении снега вынуждены разыскивать проталины. Кроме семян, клинтух клюет
жолуди, буковые орешки и даже ягоды, например, чернику. Крайне редко
эти голуби заглатывают улиток, может быть, в качестве жерновков. Мелкие
камешки обычно присутствуют в желудках клинтухов.
Полевые признаки. В природе клинтуха скорее всего можно смешать с си-
зым голубем. На лету, если смотреть сверху, клинтух отличается от сизака
более светлыми крыльями и одноцветной спиной. Снизу крылья клинтуха
заметно темнее окрашены и почти не отделяются по цвету от груди и брюха,
которые у него светлее, чем у сизака. У последнего низ тела резко контрасти-
рует с подкрыльями. Взлетая, клинтух издает своеобразный сильный свист
крыльями. По этому звуку его легко отличить от всякого другого из наших
голубей, кроме бурого. От последнего клинтух лучше всего отличается одно-
цветной спиной. Клинтух осторожная птица. В период размножения клинтухи
держатся скрытно, прячась в густых кронах деревьев близ гнезда. Даже
воркующего самца бывает трудно обнаружить, тем более, что он немедленно
прекращает свою «песню», заметив что-либо подозрительное. Кормятся насижи-
вающие голуби тут же, невдалеке от гнезда, на одной из ближайших лужаек или
КЛИНТУХ
31
просто под деревьями. Вылеты на кормежку начинаются позднее, когда со-
бьются в стаи молодые первого поколения. Кормятся эти стаи утром и вечером,
наевшись, летят на водопой, а жаркое время дня и ночь проводят в лесах.
Пролетные стаи клинтухов держатся еще осторожнее, чем гнездящиеся. Обычно
они садятся кормиться лишь в таких
местах, где к ним невозможно подойти
близко. Ночуют в лесах или хотя бы
на отдельно стоящих деревьях, выби-
рая наиболее высокие и развесистые.
Голос клинтуха чаще всего приходится
слышать во время гнездования, и тогда
воркование этого голубя звучит как
монотонное, повторяемое до 10 и даже
15 раз подряд «ху-хуу...»
Описание. Размеры и строе-
ние. Крыло самцов A9) 196—228 мм, са-
мок A0) 200—215, в среднем, соответ-
ственно, 212 и 208 мм.
Окраска. Взрослые самцы. Общая
ркраска серо-сизая; мантия часто более
темная, иногда даже (особенно в свежем
пере) с легкой буроватой примесью; брюш-
ная сторона более светлая, иногда пепельно-
сизая; такого же цвета часть больших
кроющих крыла и наружные опахала вто-
ростепенных маховых, так что проксималь-
ная половина развернутого крыла значи-
тельно светлее всей остальной верхней сто-
роны; на внутренних второстепенных махо-
вых и их больших кроющих—округлые чер-
новатые пятна, сливающиеся на сложенном
крыле в две неясные, разорванные полосы;
зоб с буровато-розовым налетом; бонд и зад-
няя часть шеи с ярким металлическим блес-
ком — зеленым на боках и пурпуровым на
зашейке; подкрылья — пепельно-сизые, иногда несколько светлее остальной нижней стороны,
иногда почти одного с ней цвета, маховые в свежем пере — аспидно-черноватые, в изношен-
ном—сильно буреют; рулевые — сизые с двумя черноватыми полосами, разделенными
нерезкой узкой светлосерой полоской, на наружных опахалах крайней пары — светлосизое,
почти беловатое, неявственное пятно. Ноги бледнокрасные. клюв красный в основной
половине и желтый —в конечной; крышечки ноздрей белые. Радужина темнокоричневая.
Самка окрашена несколько тусклее. Блеск на перьях шеи менее развит. Молодые еще
более тусклы, с большим развитием бурой примеси на спине и почти полным отсутствием
блеска на шее.
Туркестанский клинтух Columba oenas yarkandensis But.
Columba oenas yarkandensis. Бутурлин. Ежег. Зоол. музея Акад. наук, XIII, 1903,
стр. 325, Кашгария.
Синоним. Columba oenas tianshanicus. Бутурлин. Наша охота, 1916 (IX),
стр. 9, Нарын.
Распространение. Ареал. Туркестанский клинтух найден на гнездовье
в пределах Союза на крайне незначительной территории. В Тянь-Шане на реках
Кунгес и Нарын (Бутурлин, 1916), перевал Кызыл-кия к востоку от озера
Иссык-куль (Кашкаров, 1937), из бассейна р. Чирчик (Зарудный, 1910). Кроме
того, он найден и в Ферганской части Тянь-Шаня — у хр. Чаткальского (Каш-
каров, 1927) и Ферганского (Кашкаров, 1934). Указан клинтух и для Кара-тау
(Шапошников, 1931), впрочем, здесь, при отсутствии настоящих лесов с боль-
шими дуплистыми деревьями, трудно допустить гнездование. В Гиссаре из-
вестны только три летние находки, в конце мая и июле (Иванов, 1940) неопре-
деленного значения. В равнинах Средней Азии клинтух: найден по р. Зерав-
Рис. 5. Клинтух
32 ОТРЯД ГОЛУБИ
шан и Кашка-Дарья (Мекленбурцев, 1940). Кроме того, гнездится по Аму-
Дарье между Карабекаулом и Саятом. Наконец, гнездится в очень небольшом
числе у Ташкента. Область зимовок выяснена крайне схематично. Нерегуляр-
ное зимование отмечено уже у Ташкента, клинтухи наблюдались зимой —
14 декабря —в Гиссаре. Северцовым доставлены зимние экземпляры из Фер-
ганы от 30 декабря и 1 января.
В Кашгарии зимует регулярно, и возможно, что зимой к местным птицам
там присоединяются прилетные из вост. Тянь-Шаня. Может быть, изредка
зимует в с.-з. Афганистане.
Характер пребывания. Гнездящаяся пролетная птица. Весен-
ний пролет мало изучен. В Туркмении появление клинтухов отмечалось
в марте. В Гиссаре добыт Ивановым 24 марта и 1 апреля. Под Ташкентом
наблюдался 24 февраля, 20 марта и 9 апреля (Сатаева, 1937). У Наман-
гана клинтухи были встречены 16 февраля и 1 марта, а у ст. Урсатьевской —
19 февраля (Мекленбурцев). У Балыкчи (Фергана) экземпляр добыт Северцо-
вым 27 апреля. Данных относительно осеннего пролета в литературе нет. Семь
особей было отмечено Мекленбурцевым 16 ноября у ст. Мирзачуль и 3 особи —
в начале ноября у кишл. Беговат на Сыр-Дарье.
Биотоп. Туркестанский клинтух требует наличия крупных, дупли-
стых деревьев. Наиболее обычен он в Тянь-Шане в лиственных лесах, состоя-
щих из ореха с примесью клена, ясеня, яблони, боярышника, кое-где тополей.
Подобные леса разбросаны, по склонам гор, большей частью, сравнительно
мягким, но иногда изобилующим выходами скал. В вост. Тянь-Шане
клинтух найден в еловых лесах, состоящих из огромных деревьев с незначи-
тельным подлеском. В долине Зеравшана у Бухары он довольствуется обшир-
ными старыми садами, главной породой в которых служит урюк, к которому
примешано немного декоративных пород. В долине Кашка-Дарьи и Аму-Дарьи
найден в тутовых рощах, состоящих из крупных, старых и, благодаря ежегод-
ной рубке ветвей, чрезвычайно уродливых деревьев. Наконец, он доволь-
ствуется и лессовыми обрывами, гнездясь в этих условиях у Ташкента по
отвесным, высоким берегам р. Кара-камыш.
Численность. Туркестанская форма клинтуха не может считаться
многочисленной. Даже во время пролета (Кашка-Дарья) наблюдались скопле-
ния лишь до нескольких десятков особей. В противоположность другим голу-
бям численность клинтухов в Туркестане нельзя считать уменьшающейся за
последние годы.
Экология. Размножение не изучено. Воркование самцов под
Ташкентом начинается уже в конце февраля. Совершенно взрослые молодые,
уже оставленные родителями, наблюдались здесь же 7 июля. В этом же месяце—
4—б числа — были найдены под Самаркандом кладки слегка насиженных яиц,
относящихся по крайней мере ко второму выводку. Обе эти кладки были в гнез-
дах сорбк (Спангенберг). Самец, добытый под Ташкентом 12 июля, имел сильно
развитые семенники; он добыт в гнезде. Благодаря раннему началу гнездова-
ния и длинному лету туркестанский клинтух несомненно успевает высидеть
три а, может быть, и четыре кладки за лето. Гнезда, яйца и нелетные птенцы
не описаны, но несомненно здесь нет особой разницы с типичным под-
видом.
Линька. Крайне отрывочные данные позволяют заключить, что общий
ход линьки сходен с таковым у типичной формы. Начало выпадения маховых
приходится, видимо, на апрель или даже март, так как среди птиц, добытых
в мае, уже имеются экземпляры с 8, 7 и 6 старыми первостепенными маховыми.
Один из июньских экземпляров имеет шесть старых маховых, а июльский
только три. По одному старому (или отрастающему новому) маховому имеют
экземпляры от октября — декабря. Наряду с этим встречаются птицы, конча-
ющие линьку в январе — апреле. Повидимому, нормальная линька старых
ВЯХИРЬ ИЛИ ВИТЮТЕНЬ 33
птиц начинается месяцем раньше, чем у типичной формы, и имеет, соответ-
ственно, более раннее окончание.
Питание. В литературе нет никаких сведений по питанию туркестан-
ского клинтуха. По указанию Иванова A940) в зобу клинтуха были обнару-
жены зерна бобовых, кукурузы и шкурки личинок. При случае он кормится
на старых хлебных полях, но это сравнительно редкое явление, так как в обла-
сти горных лесов посевов немного.
Полевые признаки. В природе туркестанский клинтух может быть спутан
с сизым или бурым голубем. От обоих отличим уже издали по темной пояснице
и светлой верхней стороне крыльев. Кроме того, отличается от сизака более
светлой нижней стороной тела и более темными подкрыльями. При взлете
издает резкий свист крыльями, в чем проявляется его отличие от сизого
голубя и сходство с бурым.
Даже в тех местах, где его мало преследуют, клинтух бывает очень осторо-
жен. На водопой прилетает на открытые берега ручьев или речек, но и здесь
никогда долго не задерживается, а, напившись, немедленно улетает. Жаркую
пору дня наевшиеся голуби проводят на деревьях, причем стараются выбирать
отдельно стоящие, большие деревья, с которых слетают, едва заметив опасность.
После вечерней кормежки прилетают к местам ночлега очень поздно, почти
в темноте, быстро рассаживаются по кронам среди густой листвы и сидят не-
подвижно. Осторожность сказывается и в том, что над подстреленным клинту-
хом остальные птицы стаи не вьются, так неосмотрительно налетая вновь на
выстрел, как это делает сизак. Голос тянынанского клинтуха вполне сходен
с голосом европейского и звучит как многократно повторяемое «уу-уу-уу»
или «хуу-хуу-хуу».
Описание.Размеры и строение. Длина крыла самцов A8) 207—230, самок
(9) 205—221, в среднем, соответственно 220 и 214 мм.
Окраска. Сходна с номинальной формой, но все цвета несколько светлее. Брюш-
ная сторона не серо-, а пепельно-сизая, мантия почти, а иногда и совершенно, без бурой»
примеси.
6. Вяхирь или витютень Columba palumbus Linn.
Columba palumbus. Linnaeus. Syst. Nat., изд. X, 1758, стр. 163. Швеция.
Русское название. Народное, встречается по крайней мере с XVIII в. (Лев*
шин). Старая транскрипция ¦ветитень», «ветитен». Этимология неясна (быть может, связана
с украинским «трепетен»).
Распространение. Ареал. Европа — до 65° с. ш. в Норвегии, 64° в Шве-
ции и Финляндии, Архангельска и верховьев Камы в СССР. О-ва Атлантиче-
ского океана (Азорские, Мадейра) и Средиземного моря, Ирландия и Велико-
британия. В Африке — Марокко, Алжир и Тунис. В Азии к востоку от южной
половины Уральского хребта до среднего течения Ишима. Второй азиатский
выступ ареала захватывает Малую Азию, Месопотамию, Сев. Иран, Афгани-
стан, Памиро-Алай, Тянь-Шань и Гималаи. Зимует, начиная уже от южн.
Великобритании и Франции. На юге Европы и о-вах Средиземного моря зимой
обычен, реже — в Крыму, на Кавказе и на юге Туркмении (до среднего тече-
ния Аму-Дарьи). В сев.-зап. Индии граница ареала смещается до Пенджаба
и Синда, в Афганистане — до Кветты. Зимует в горах Ирана, в Месопотамии.
Особенно обычен в Палестине, где не гнездится. В Алжире» Марокко и Тунисе
к местным особям присоединяются прилетевшие из Европы. Таким образом, по
характеру пребывания вяхирь — перелетная птица, но на юге гнездового
ареала постоянно зимует.
Биотоп. Вяхирь — чисто лесная птица и всюду придерживается дре-
весной растительности или, в крайнем случае, крупных кустарников. В этих
условиях встречается как по окраинам, так и в глубине лесных массивов.
3 Птицы СССР, т. II
34
ОТРЯД ГОЛУБИ
Подвиды и варьирующие признаки. С. p. azorica Н а г t e r t, 1905, — Азорские
о-ва, за исключением Флорес и Корво; С. p. maderensis T s с h u s i, 1904 —о-в Ма-
дейра; С. p.exceisa Bonaparte, 1856 — горно-лесные области Марокко, Алжира и
Туниса; С. p. palumbus Linnaeus, 1758 — весь ареал вида, за исключением указанных
е-ов Атлантического океана, сев. Африки и азиатской части ареала от Ирана до Копет-
Карта 6. Распространение вяхиря Columba palumbus
1. С. p. palumbus, 2. С. p. azorica, 3. С. p. maderensis, 4. С. p. exceisa, 5. С. p. casiotis;
а — граница области распространения, г — зимовки перелетных форм, д — изолированные нахождения
вяхиря.
Дага; С. p. iranica Z a r u d n у, 1910 — Иран, в СССР — Копет-Даг; С. p. casiotis Bonapar-
te, 1854 —горы Памиро-Алайской, Тяньшанской, Каракорумской, Гиндукушской и
Гималайской систем, на запад до Кугитанга и Аму-Дарьи. Внутривидовая изменчивость
вяхиря выражена в большей или меньшей насыщенности окраски, в различном развитии
темного цвета на рулевых, в окраске боковых пятен шеи и в вариациях общих размеров.
Некоторые формы свойственны главным образом лиственным лесам, другие — хвойным.
Одни подвиды совершают довольно значительные перелеты, в то время как другие оседлы
или почти оседлы.
Европейский вяхирь Columba palumbus palumbus L.
Распространение. Ареал. На гнездовье встречается во многих местах
европейской части СССР. В лесах Подолии это нередкая птица (Портенко,
1928). Довольно обычен он на Волыни (Шарлемань и Портенко, 1926) и в других
частях Украины, но редок в Белоруссии (Шнитников, 1913) и Прибалтийских
республиках. Однако в Новгородской обл. вяхирь как гнездящаяся птица
обыкновенен, также и в Ленинградской (Бианки, 1910—1914). Далее к северу
он становится реже, но гнездится вплоть до южной оконечности Белого моря
и верховьев Камы. В Горьковской обл. —обычная гнездящаяся птица
(Пузанов, 1942). В Пре/гуралье гнездится до 60° с. ш. (Куклин, 1936). В Зап.
Сибири населяет незначительный участок до района Тюмени к северу, среднего
течения Ишима—к востоку и верховьев Тобола—к югу (Гептнер, 1925; Залес-
ские,.192б). ВЧкаловской обл. обыкновенен к северу от р. Урала (Зарудный,
1888), но в ее нижнем течении не гнездится (Бостанжогло, 1911). В пустынной
местности между нижним течением Волги, Доном и Предкавказьем на гнездовье
не попадается. В Приазовье гнездится (Алфераки, 1910). В бассейне Дона указан
для Хоперского заповедника (Измайлов, 1940) и Воронежской обл. (Север-
ВЯХИРЬ ИЛИ ВИТЮТЕНЬ 35
цов, 1855; Огнев, 1924), в Поволжье для бассейнов Суры и Алатыря (Житков и
Бутурлин, 1906) по водоразделу Волги и Дона (р. Иловля) к югу почти до
Сталинграда (Гладков) и по всему течению Волги от Камы до устьев (Богда-
нов, 1871). В центральных частях РСФСР вяхирь гнездится повсюду, где есть
подходящие условия. На Кавказе — это также повсеместно гнездящаяся птица.
То же самое в Крыму (Пузанов, 1933). Залеты типичной формы известны
с верховьев Печоры на Усть-Уньи (Самородов, по Дементьеву, 1941) и с Енисея
у Красноярска (Зибом, 1878); впрочем последний случай вероятнее отно-
сится к туркестанскому вяхирю.
Характер пребывания. Везде в пределах очерченного выше
пространства вяхирь является перелетной птицей, но в Крыму и на Кавказе
кроме того и зимует. Он рано прилетает, а отлетает, напротив, очень поздно.
Появление весной у южных и западных окраин Союза отмечается уже в на-
чале марта. У Бартенштейна (Калининградскаяобл.), первое появление за ряд
лет отмечено в марте (Тишлер, 1914). 16 и 21 марта вяхири были встречены
у Чкалова (Зарудный, 1897). 30 марта вяхирь наблюдался в Прибалтике у Тарту
(Герме, 1913) и 3 апреля на водоразделе р. Аа и Буртнекского озера (Лоудон,
1913). Под Гурьевом пролет был указан с 5 апреля (Бостанжогло, 1911).
У окрестностей Смоленска вяхиря отмечали 7 апреля (Гржибовский, 1912).
В Смоленской обл. —8 апреля (Меландер, 1913). В Новгородской обл.
около середины этого месяца так же, как и в Псковской обл. У Казани вяхирь
появляется также около середины апреля. Еще позднее его прилет приводится
для Молотовской обл., окрестностей Москвы и Калинина. У Тюмени появление
отмечено наиболее поздно — в конце апреля.
Отлетает вяхирь не ранее сентября. Под Чкаловом первые пролетные стаи
показываются в начале этого месяца. В эти же дни отлет начинается у Пскова.
В Калининградской обл. —на Куришской косе —осенний пролет в 1905 г.
кончился 21 сентября, в 1906 г. — 11 сентября и в 1910 г. —20 сентября
(Тишлер, 1914). В другие годы количество голубей на Куриш-Гаффе особенно
многочисленно около 1 октября. В прибалтийских республиках вяхирей нахо-
дят до середины сентября. В это же время пролетные вяхири несколько раз отме-
чались на Сыр-Дарье. В Поволжье отлет начинается в середине сентября и за-
канчивается в середине октября. Под Бугурусланом большие пролетные стаи
были отмечены 21 сентября (Исполатов, 1911). В Калининской обл. пролет
зарегистрирован позднее — в последней трети сентября, в Новгородской—
даже в конце этого месяца. Для Московской обл. имеются указания на еще
более поздние даты отлета, начало которого приходится на первую декаду октя-
бря, а конец — на середину этого месяца (Лоренц,. 1892). В дельте Волги пролет
приходится на первую половину октября (Воробьев, 1936). Наконец, в Крыму
вяхири отлетают в начале ноября (Никольский, 1892). Исключительные случаи
зимовок известны для окрестностей Козельска D. I, Генглер, 1909). Чаще
попадается зимой в Калининградской обл. (Росситен — 29 января, 31 января;
Тишлер 1914). Отдельные особи зимуют в Крыму и на Кавказе.
Биотоп. Разнообразные леса. У северной границы вяхирь гнездится
в хвойных лесах, ельниках различного типа. Далее к югу населяет смешанные
леса, хотя и здесь не чуждается участков чистого елового леса, а кое-где даже
предпочитает хвойные леса другим, как, например, в Прибалтике, Псковской
и Горьковской областях. В области широколиственных лесов селится в дубовых
или смешанных лесах самого разнообразного характера. В степной полосе
населяет речные уремы с их разнообразной древесной растительностью. На
Кавказе — в лиственных лесах, главным образом буковых, поднимаясь до их
верхней границы. В бассейне Иртыша попадается в березовых колках и при-
брежных лесах Тобола и Ишима. В подобных же условиях селится в долине
Урала. Чистых сосновых насаждений вяхирь не любит, но местами гнездится
в них (окрестности Камышина).
36 ОТРЯД ГОЛУБИ
Численность. Количество вяхирей, в связи с вырубкой лесов, с по-
степенным освоением новых лесных массивов, а может быть и с отстрелом, со-
кращается довольно заметно. Во всяком случае, стаи в несколько сот экземпля-
ров, какие встречал Богданов в Поволжье, позднейшими авторами не упоми-
наются. Лишь Сушкин A908) встречал на Эмбе скопища до 800 экземпляров.
Вяхирь, как и все голуби, общественная птица и встречается парами только
во время гнездования. Количество особей в стаях весьма разнообразно и ко-
леблется от нескольких штук до нескольких десятков. Особенно крупные стаи
образуются во время осеннего пролета. Еще более крупные скопления образуют
вяхири на зимовках и, например, в Палестине стаи бывают иногда очень много-
численными.
Экология. Размножение. Стаи вяхирей, возвращающихся весной
к местам гнездовий, состоят, наряду с образовавшимися парами, из значительного
количества молодых птиц. «Ток» начинается вскоре после того, как оседают
на летний период прилетевшие птицы. Холостые самцы воркуют на высоких
деревьях. Наиболее энергичное воркование происходит утром. Период разбивки
на пары —с прилета, примерно, до 15—20 апреля. К этому сроку образуется
большинство пар, и вяхири принимаются за постройку гнезд. Гнездо строится
в течение нескольких дней. Местом для него служит основание толстой горизон-
тальной ветви, сплетение нескольких веток или чье-либо старое гнездо, большей
частью белки, вороны, сороки, мелких соколов и т. п. В дуплах вяхирь не гнез-
дится. Постройка большей части гнезд бывает закончена к концу апреля,
и пара приступает к откладке яиц. Брачные игры вяхирей выражаются в том,
что самец время от времени косо взлетает на 1У2—2 десятка метров, сильно
хлопая при этом крыльями. Достигнув наивысшей точки полета, он скользит
вниз на распущенных крыльях и, описав широкий круг, возвращается к самке.
Перед взлетом и после посадки птица обычно воркует.
Гнездо вяхиря представляет собой тонкую, рыхлую, обычно просвечиваю-
щую постройку с очень плоским лоточком. Строительным материалом для гнезда
служат тонкие веточки тех древесных пород, которые растут около него. Иногда
лоток образован более тонким материалом. Гнездо располагается на самой
различной высоте от I1/»—2 м до тонких вершин высоких деревьев, иногда
довольно далеко от ствола, иногда вплотную к нему. Размеры гнезда от 30 до
40 см по наружному диаметру йот 11 до 14—по внутреннему. Глубина лотка
3—5 см, а общая высота гнезда от 7 до 14. Паразитов в гнездах не обнаружено.
Кладка из 2 яиц; размеры: F5K7—44 х 26,5—34,5, в среднем 41 х 30,4 мм
(Гебель, 1879). Ранние сроки откладки отмечены для Калининградской обл.,
где слабо насиженные яйца найдены уже 22—27 апреля (Тишлер, 1914).
8 Прибалтике кладки встречены 3 мая (Руссов, 1880), а у Пскова сильно наси-
женные яйца были найдены 5 мая (Зарудный, 1910). Однако это — случай осо-
бенно раннего гнездования, а большая часть яиц откладывается у Пскова
между Ю и 15 мая (Зарудный, 1910). На такие же сроки откладывания яиц
указывает нахождение в Минской обл. хорошо летающих молодых от начала
до середины июня (Федюшин, 1912). В Чкаловской обл. начало спаривания отме-
чалось приблизительно в середине апреля, а полувзрослый птенец был найден
9 июня (Зарудный» 1888). На откладывание яиц в конце апреля в Новгородской
обл. указывает появление молодых в середине июня (Бианки, 1910). В По-
долии молодые были отмечены 2 июля (Портенко, 1929). Это позволяет допу-
стить, что кладка началась около середины мая. Аналогичные сроки приведены
для окрестностей Бугуруслана, где насиженные яйца отмечены 28 мая (Испо-
латов, 1911). В дельте Волги гнезда встречаются с конца апреля (Бостанжогло,
1911). В Крыму молодой наблюдался 10 июля (Никольский, 1892), что указы-
вает на начало гнездования 15—20 мая. Очень поздняя дата находки молодых—
21 июля —приводится для Минской обл. (Шнитников, 1913). В этом случае
птицы должны были отложить яйца приблизительно в начале июня. Вероятно,
ВЯХИРЬ ИЛИ ВИТЮТЕНЬ 37
в данном случае имеет место вторая кладка. Второе яйцо откладывается дня
через 2—3 после откладки первого.
Насиживание происходит после откладки первого яйца, поэтому птенцы
разновозрастны. Насиживание продолжается 17—18 дней, при участии обоих
родителей, но самка проводит в гнезде гораздо больше времени, чем самец.
Птенцов кормят оба родителя, сначала творожистым выделением стенок зоба,
затем принесенной и размягченной в зобе пищей. Птенцы проводят в гнезде
30—38 дней (Нордберг, 1936). Впрочем, по некоторым данным этот срок огра-
ничивается 20—21 днями (Шульпин, 1940). Первое время по вылете молодые
держатся вместе с родителями, которые еще несколько дней продолжают кор-
мить их размягченными в зобе зернами. Вскоре молодые отбиваются от старых,
образуют стаи, к которым, может быть, присоединяются холостые старые птицы,
и бродят по окрестным лесам. Старики приступают к вторичной кладке. У се-
верной границы ареала вяхирь выводит птенцов только один раз в лето. Далее
на юг, во всяком случае от Новгорода, некоторые пары выводят дважды в лето.
Молодые последнего вывода и закончившие размножение старые птицы посте-
пенно присоединяются к стаям, состоящим из молрдых первых выводов.
Осенние стаи вяхирей широко кочуют, постепенно отлетая к местам своих,
зимовок.
Линька. Вяхирь, как и другие голуби, имеет в году одну полную линьку.
Сроки ее не совпадают у разных особей, повидимому, в связи с размножением.
Оперение начинает сменяться с 10-го махового, и смена перьев идет к переднему
краю крыла. Когда линька доходит до 3-п>—*-го и 1-го маховых, начинается
смена рулевых и мелкого оперения. Начало линьки (выпадение 10-го махо-
вого) приходится у большинства особей на май. Линька идет сравнительно мед-
ленно и в июле сменяются 6-е— 5-е—4-е маховые. Мелкое перо, вместе со 2-м—
3-м маховыми и рулевыми меняется в октябре, а в ноябре линька заканчи-
вается. Молодые линяют не одновременно. Птенцы последних выводков иногда
не успевают перелинять в течение зимы и заканчивают линьку весною, имея еще
в апреле по 3—2 старых маховых, все рулевые и мелкое перо. Последова-
тельность смены нарядов несложна. Пух птенцов сменяется рыхлым юноше-
ским оперением, которое в первую же осень заменяется окончательным нарядом.
Питание. Пища вяхиря состоит, главным образом, из растительных
объектов. Животные компоненты встречаются в исключительных случаях.
Основой питания служат зерна растений. Из них чаще других встречаются се-
мена культурных и диких злаков и бобовых. В тех местах своего ареала, где
господствует хвойная растительность, вяхирь поедает очень много семян этих
деревьев, главным образом, ели. Далее к югу, где хвойные породы вытесняются
дубом, вяхирь поедает много желудей, а еще далее к югу, где в лесах господ-
ствует бук, буковые орешки. Наконец, вяхирь ест и кое-какие ягоды — земля-
нику, чернику, голубику, бруснику и даже калину. Зоб вяхиря очень объемист
и вмещает до 7 штук жолудей или около блюдца пшеницы. В зобах молодых на-
ходили мелкие зерна бобовых (мышиный горошек).
Кормится вяхирь главным образом на земле, собирая выпавшие из расте-
ний зерна. Высоких трав вяхирь не трогает, но низкие растения, может быть,
сщипывает на корню. Ягоды, жолуди и буковые орешки склевывает и с ветвей
или кустов, а не только собирает на земле.
Полевые признаки. В природе вяхирь легко отличим от других наших голу-
бей своим крупным ростом. На близких расстояниях характерным признаком
служат белые пятна на боках шеи. В полете его сразу выдают относительно более
короткие, чем у других голубей, крылья и более длинный хвост. На расстояниях
до сотни метров при взгляде сбоку или сверху безошибочно определим по нали-
чию больших белых, пятен на крыльях. Вяхирь осторожная птица. В лесу он
держится скрытно, выбирая деревья с густой хвоей или листвой. В осеннем
пролете на Украине и в Чкаловской обл* он предпочитает дубовые рощи
38
ОТРЯД ГОЛУБИ
так как эти деревья сохраняют листья дольше других, давая поэтому надежное
укрытие.
На кормежку вылетает утром и кормится на полях, пойменных лугах, лес-
ных лужайках, и опушках и т. п. местах. Иногда посещает солонцы или соле-
ные источники, как напри-
мер, на Кавказе (Аверин и
Насимович, 1938), видимо
с целью заглатывания соли
или соленой воды. Голос
вяхиря бывает слышен
только в период размноже-
ния и представляет собой
воркование, которое иногда
сравнивается со стоном и пе-
редается звуками «куу-у-у,
куу-рруу» или «гурр-гру-
гурр-гурр».
Описание. Размеры и
строение. Длина самца A)
450, самок B) 422—436 мм. Вес
самцов C) 530—625 г, в среднем
588 г. Крыло самцов B0) 237—
262, самок A3) 225—257, в сред-
нем, соответственно, 248,8 и
243,7 мм.
Окраска. Взрослые
самцы и самки на спинной сто-
роне окрашены в сизый цвет,
на передней части спины, плече-
вых и кроющих крыла — с при-
месью буроватого; задняя часть
спины, надхвостье и верхние
кроющие хвоста — чистого си-
зого цвета, более темного на
спине и слегка светлеющего на
надхвостье. Наружные кроющие второстепенных маховых имеют широкие белые каймы
на наружных опахалах. Эти каймы, сливаясь, образуют на крыле белое пятно; махо-
вые и кроющие первостепенных — черновато-аспидные с узкими белыми каймами на
наружных опахалах первостепенных; подбой крыла беловатый с легким сизым оттенком;
рулевые — аспидно-серые, светлеющие к вершине до сизого (с нижней стороны до бело-
ватого) цвета; широкая вершинная полоса аспидная. Зоб и грудь — сизовато винно-крас-
ные; бока, брюхо, подхвостье и нижние кроющие хвоста — светлого сизого цвета; шея
и голова — темносизые; на боках шеи — большое белое ожерелье; бока и задняя сто-
рона шеи — с металлическим зеленым и розовато-красным блеском. Клюв у основания
красный. Вершина — желтая. Ноги — малиново-красные. Радужина бледножелтая. Моло-
дые сходны со старыми, но спина более буровата, белые шейные пятна отсутствуют, розо-
ватый цвет зоба и груди заменен грязно-коричневым.
Рис. 6. Вяхирь
Иранский вяхирь Columba palumbus iranica Zar.
Palumbus palumbus iranicus. 3 a p у д н ы й. «Наша Охота», 1913, стр. 116, Копет-Даг.
Распространение. Ареал. Иран и южн. Туркмения — система Копет-
Дага от западных его частей (бассейн Чандыра и Сумбара) до центральных;
восточная граница не вполне ясна, так как в Кугитанге гнездится туркестан-
ский подвид casiotis.
Характер пребывания. Гнездится, на зиму улетает, по край-
ней мере из Туркмении. Прилет в разные числа марта (по Шестоперову, 1937,
еще в начале апреля).
Биотоп. Гнездовый — высокие ступени гор: в центральном Копет-Даге
в поясе арчи, в зап. Копет-Даге и в Иране — в лиственных лесах.
ВЯХИРЬ ИЛИ ВИТЮТЕНЬ 39
Численность. Невысокая, чаще встречается в зап. Копет-Даге, где
лучше сохранились леса. Так как в центральном Копет-Даге арчевников мало
и они расположены отдельными изолированными участками, то распростране-
ние вяхиря здесь спорадично; отсутствие арчи, повидимому, ограничивает
распространение вяхиря в вост. Копет-Даге.
Экология. Размножение. Начало кладки — в конце мая. В Сулюкли
в 2 гнездах 25 мая найдено по одному яйцу (Шестоперов, 1928), 27 мая гнездо
с одним яйцом найдено в горах Асильма (Зарудный, 1898). Гнездо —как всюду,
небрежная, просвечивающая снизу постройка — на высоких деревьях, осо-
бенно предпочитается арча. Кладка из 2 яиц (Александров). Насиженные
яйца найдены 8 августа, что указывает на 2, быть может 3 кладки в году (см.
также данные о линьке).
Линька. Смена нарядов происходит, как у номинальной формы. Осен-
няя линька молодых растянута, что указывает на вероятность двух выводов за
лето; так, у одной молодой птицы 17 сентября смена гнездового наряда еще не
началась, а у другой 22 сентября сменилось несколько задних первостепенных
маховых, у третьей 10 октября —пять маховых. Взрослые самцы начинают
смену маховых (как у других голубей, с заднего края партии) в начале июля;
в конце июля смена первостепенных у некоторых закончена наполовину, но
птицы в таком же состоянии линьки добыты и 27 августа, повидимому, от вто-
рой кладки.
Питание. Осенью вяхири кормятся в предгорьях и у горных поселков
по озимым и на местах обмолота: у одного голубя (Иол-дере, 27 мая) в зобе
найдено 100 г пшеницы.
Описание. Размеры и строение. Крыло самцов A5) 240—260, самок B)
250—252, в среднем, соответственно, 258 и 251 мм.
Окраска. От европейского вяхиря отличается бледной общей окраской, близкой
к окраске туркестанских птиц; но ошейник широкий, как у европейских.
Туркестанский вяхирь Columba palumbus casiotis В p.
Palumbus casiotis. Bonaparte. Consp. Gen., II, 1854, стр. 42, Гималаи.
Распространение. Ареал. Гнездится в пределах Союза в горных лесах
Средней Азии. Здесь он найден в западных отрогах Гиссарского хребта (Иванов,
1940), в хребтах Петра Первого, Зеравшанском (Даль, 1936), Туркестанском
(Плеске, 1888), Алайском и всей системе Тянь-Шаня, на запад до Кугитанга
(Дементьев, 1950). Единичные нахождения известны с Алтая и верховий Иргиза
(Сушкин, 1908, 1938). В очерченных таким образом пределах вяхирь встре-
чается летом и является перелетной птицей. Область зимовок этой формы начи-
нается от среднего течения Аму-Дарьи (Зарудный, 1918) и вост. Ирана (где
встречается нерегулярно) и захватывает Афганистан, Пенджаб, низовья Инда
и подножья Гималаев до Уда и Непала (Рунвал, 1941). Возможно, что отдель-
ные особи перезимовывают даже в бассейне Сыр-Дарьи, откуда известны зимние
экземпляры (Канджигалы в долине Ангрена). На пролетах вяхири попадаются
и в долинах. Однако такие случаи редки и основной пролет, повидимому,
идет по горам.
Даты. Известны наблюдения у Ташкента 20 марта (Сатаева, 1937) и
24 апреля (Зарудный, 1923) и у ст. Ломакино 26 числа этого месяца. 12 апреля
вяхирь был встречен в долине Варзоба, а в середине этого месяца — на Искан-
дер-куле (Иванов).
Биотоп. Подобно типичному подвиду, туркестанский вяхирь — чисто
лесная птица, приуроченная в Средней Азии к различным типам горных лесов.
Наиболее обычны эти голуби в арчевых лесах Гиссаро-Алая и зап. Тянь-Шаня.
Охотно селятся они в искусственных лиственных насаждениях Зеравшанского
хребта, состоящих из ореха, акации, айланта. Так же охотно поселяются и
40 ОТРЯД ГОЛУБИ
в естественных ореховых лесах Ферганы. В вост. Тянь-Шане вяхирь — птица
ельников и отчасти яблочников. Повидимому, он предпочитает хвойные леса,
обращая мало внимания на прочие условия, так как легко уживается в еловом
поясе Тянь-Шаня с его прохладным и дождливым летом и столь же легко
переносит сухость и зной Гиссарских арчевников.
Численность. Наиболее многочисленны вяхири в Гиссарском, Фер-
ганском и Туркестанском хребтах. Однако стаи даже в сотню особей для этих
областей не отмечались. Повидимому, эта форма, в отличие от типичной, не
образует особенно больших скоплений даже на зимовках.
Экология. Размножение. Видимо, нет различия с типичным под-
видом, и разбивка на пары, отношение к гнездовому участку и брачные «игры»
у обоих подвидов одинаковы. Календарные сроки начала устройства гнезд,
откладки яиц и т. п. очень слабо изучены. Находки гнезд с полувзрослыми
птенцами — в начале июня (Колесников) и со взрослыми —в конце июня (Даль,
1936) указывают на начало откладки яиц в первых числах мая. На гораздо более
позднее начало гнездовья указывают гнезда с яйцами, найденные в середине
июня (Плеске, 1888), и на еще более позднее —оживленное воркование самцов
в середине июля. Вероятно, это вторые кладки, но возможно эти более поздние
сроки находят объяснение в том, что в горах весна начинается поздно, и снег
сходит здесь лишь к началу мая. Способ постройки гнезда, располагающегося
на дереве или крупном кусте, насиживание и уход за птенцами—аналогичны
таковым у европейского вяхиря. Число кладок в лето неизвестно, но их во
всяком случае не менее двух; почти наверное можно думать, что отдельные
пары гнездятся и три раза в лето.
Линька. Характер и ход линьки туркестанского вяхиря совершенно
сходны с тем, что было описано для типичного. Незначительный изученный
материал показывает, что у описываемого подвида маховые начинают выпадать
несколько позднее, и линька отстает в среднем на одно маховое. Особи с 9 ста-
рыми первостепенными маховыми известны от второй половины мая. Через
месяц многие птицы имеют по 7 и 8 старых маховых, а в первой половине
июля — от 5 до 8. У августовских птиц обнаружено от 4 до 7 маховых, а у
октябрьских — от 3 до 7 (возможно, что в последнем случае имеет место запо-
здалая линька, или линяет молодая птица). Относительно линьки мелкого пера
данные еще более незначительны. Известна добытая 18 августа птица, меняю-
щая крупное и мелкое перо (Зарудный, 1926). У экземпляров от 23 сентября
из Арчата, от 7 декабря из Канджигалы и 23 февраля из Багира вся
голова покрыта пробивающимися перьями. У второго из этих экземпляров
сменяются 4 средних рулевых, а у третьего—2. Видимо, линька проходит
так же быстро, как и у других голубей. Возрастные изменения и смена
нарядов туркестанского вяхиря ни в чем не отличаются от типичной
формы.
Питание. Отрывочные данные по питанию показывают, что турке-
станский вяхирь клюет много ягод жимолости, дикой вишни, барбариса, чер-
ной смородины (Иванов). Кроме того, он поедает плоды шелковицы и шиповника.
В те времена года, когда ягоды отсутствуют, его основным кормом должны
быть семена различных травянистых растений, так как в области распро-
странения этого подвида совершенно нет дуба и бука, плоды которых во
множестве поедаются этим голубем в Европе. Почти нет в горах Средней
Азии и хвойных пород, семена которых могли бы быть кормом для
вяхиря.
Описание. Р а з ме р ы и ст р о е н и е. Крыло самцов B3) 238—269, самок A0)
233—266, в среднем, соответственно, 256 и 252 мм, т. е. немного крупней, чем у евро-
пейского подвида.
Окраска. Общий тон спинной стороны бледный, ошейник узкий и не белый, а охри-
стый; нет белой каймы на наружном опахале первого первостепенного махового.
ГОРЛИЦА
41
РОД ГОРЛИЦЫ STREPTOPELIA BONAPARTE, 1854
Тип. S. risoria
7. Горлица Streptopelia turtur L.
Columba turtur. Linnaeus. Syst. Nat., изд. XI, 1758, стр. 164, Англия.
Русское название. Народное, относится, в сущности, ко всем видам мелких
голубей.
Распространение. Ареал. Большая часть Европейского материка, за
исключением средней и северной Англии и Шотландии, Скандинавского п-ова,
северной половины Эстонии, области к северу от бассейна верхней Волги,
60° между Волгой и Камой, верховьев последней, 58° с. ш. на Урале. В Азии
она занимает юго-западную часть советской территории до среднего течения
Тобола и Ишима, Омска, Семипалатинска, Зайсанской низменности, Черного
Карта 7. Распространение обыкновенной горлицы Streptopelia turtur
7. S. U turtur, 2. 5. t. arenicola, 3. S. t. Ш, 4, S. U hog gar a, 5. S. t. isabellina;
a — граница области распространения, б — не вполне выясненная граница рас-
пространения, • — изолированные нахождения горлицы, г — зимовки пере-
летных форм.
Иртыша, Джунгарского Ала-тау и южных склонов Кок-шаал-тау. В Кашгарии
гнездится до оазисов в Хами и Са-чжеу. Населяет самую северную окраину
Индии, сев. Афганистан, весь Иран, Месопотамию и Палестину. В Африке
встречается в долине Нила до Нубии, в горах и оазисах Сахары. Западная
граница захватывает о-ва Канарские и Мадейру, Пиренейский п-ов, Францию
и выходит к середине западного берега Великобритании. Основная область
зимовок охватывает южные части Сахары, Сенегал и Судан.
Б и о т о п. На гнездовье леса, рощи, сады или хотя бы невысокие кустар-
ники; встречаются при наличии последних даже в пустынях, но везде невда-
леке от воды. Хвойных лесов избегает, но легко мирится с самыми разнообраз-
ными лиственными насаждениями.
42 . ОТРЯД ГОЛУБИ
Подвиды и варьирующие признаки. 5. /. turtur Linnaeus, 1758.—Вся евро-
пейская часть ареала; в азиатской части этот подвид населяет Малую Азию, Армению,
низовья Урала, долину Эмбы до Кок-джиды, Тургая — до низовьев, Тобола и Ишима до
истоков; на Иртыше — у Омска, но у Семипалатинска уже следующий подвид; 5. /. areni-
cola H a r t e r t, 1894 — вся азиатская часть ареала от указанной южной границы преды-
дущего подвида; в Африке населяет побережье Средиземного моря и север Сахары
(вероятно сюда же относятся горлицы с Балеарских о-вов); S. t. hoggara G е у г,
1911 — Центр. Сахара (Ахаггара, Тассили, Аир); 5. t. i sab el И па Bonaparte, 1856 —
оазисы Ливийской пустыни, Египет, Нубия, Судан. Изменчивость выражается в большей или
меньшей интенсивности окраски разных каемок плечевых и верхних кроющих крыла и в
меньшей степени остального оперения; кроме того, варьируют размеры. Экологические
различия между подвидами заключаются в том, что одни связаны со сравнительно высоко-
ствольной и влаголюбивой лиственной растительностью умеренного климата, другие подвиды
обитают в условиях субтропических пустынь или пустынь юга умеренной полосы и связаны
здесь с ксерофитными кустарниками вблизи от оазисов, искусственными древесными наса-
ждениями и с пойменной растительностью рек.
Европейская горлица Streptopelia turtur turtur L.
Распространение. Ареал. В пределах СССР горлица встречается, начи-
ная от западной государственной границы. В Подолии обычна (Портенко, 1928,
Герхнер, 1928), как и на Волыни (Шарлемань и Портенко, 1926). Гнездится
в среднем течении Днепра (Шарлемань, 1936) и в верхних частях бассейна этой
реки (Шнитников, 1913, Федюшин, 1926). В Калининградской обл. нередка
на большей части ее территории (Тишлер, 1914). В Прибалтийских республиках
гнездится реже (Руссов, 1880) и редка в области Псковского озера и реки Ве-
ликой (Зарудный, 1910). Под Ленинградом —только залеты (Бианки, 1916),
но в Калининской обл. гнездится, хотя и редко (Третьяков, 1940). У Москвы
нередка на гнездовье (Лоренц, 1892—1894), а у Калуги даже обычна (Филатов,
1915). В области верховьев Днепра местами редка (Огнев, 1904). В Карачев-
ском районе очень обыкновенна (Суслова, 1937), как и в Полтавщине (Гаври-
ленко, 1929). В низовьях Днепра — многочисленная гнездящаяся птица
(Вальх, 1918). В Крыму гнездится везде (Никольский, 1892, Пузанов, 1933),
как и на восточном берегу Азовского моря (Алфераки, 1910). В бассейне Дона
указывается как гнездящаяся для р. Хопер (Измайлов, 1940) и Воронежской
обл. (Огнев, 1924), для р. Цны (Резцов, 1910) и для истоков Дона (Сушкин,
1892). В бассейне Волги обычна по течению Оки (Хомяков, 1900), на самой
Волге у Костромы (Кирпичников, 1915), по всему Горьковскому краю (Ворон-
цов, 1937, Пузанов и др., 1942). Далее вниз по Волге встречается в поймах
Алатыря и Суры (Житков и Бутурлин, 1906) и вплоть до Каспийского моря.
Предкавказье повсюду населено этой птицей, как Кавказский хребет и Закав-
казье. В междуречье Волги и Урала гнездится везде в подходящих местах
(Бостанжогло, 911, Волчанецкий, 1937). На р. Урал очень обычна (Зарудный,
1888 и 1897). К югу и юго-востоку от этой реки гнездится на Эмбе до Кок-
джиды, в Мугоджарах, до Иргиза в низовьев Тургая (Сушкин, 1908), в Наур-
зуме (Деливрон, 1937). В системе Уральского хребта весьма обыкновенна в
Ильменских горах (Теплоухов, 1921, Снигиревский, 1929) и Общем Сырте, но
далее к северу по Уральскому хребту становится реже (Снигиревский, 1929),
хотя доходит до 58° с. ш. (Куклин, 1936). В пределах юго-зап. Сибири найдена
в Барабинской степи и у Омска (Коте, 1910), в бассейне Ишима (Курганский
район) и Тобола (Залесские, 1931). На пролете отмечена на вост. берегу Каспия
в Туркмении между Кара-Богазом и Гассан-кули (Дементьев, 1950). Места
зимовок горлицы расположены на юге Сахары, в Сенегале и Судане. Таким об-
разом, ее пролетные пути достаточно длинны и отчасти поэтому горлица поздно
появляется у своих гнездовых мест.
Характер пребывания. Горлица —перелетная птица. Несмотря
на обилие данных по появлению горлиц весной, планомерную картину их про-
движения по территории Союза все же можно дать только схематично, так как
эти сведения носят несколько сбивчивый характер.
ГОРЛИЦА 43
Даты. На крайнем юге Европейской части СССР — в Закавказье—
появление этих птиц отмечалось 20 апреля (Домбровский, 1913), в Крыму — 25,
а по средним данным за двенадцать лет — 29 числа этого месяца (Никольский,
1892). На юго-западном Кавказе горлицы отмечались в середине апреля, а на
северо-западном — несколькими днями позднее — в середине последней трети.
Весенний пролет на ю.-в. Каспии в Туркмении продолжается еще в конце
апреля — начале мая. Значительно раньше — 30 марта (что не может не вы-
звать сомнения) — первое появление горлиц отмечено на р. Хопер (Измайлов,
1940). В те же сроки, как на Кавказе, около середины апреля—горлицы по-
являются в бассейне Оки (Хомяков, 1900), 15 апреля у Лепеля (Федюшин, 1926),
во второй половине этого месяца у Сары-Чеганака, а около 25 числа у устья
Урала (Бостанжогло, 1911). 20 апреля горлицы наблюдались в Смоленской
обл. (Гржибовский, 1912), а в конце этого месяца в бассейнах Суры и Ала-
тыря, под Москвой, у Киева, Минска, окрестностей Бугуруслана. В последней
же трети апреля горлицы прилетают, по наблюдениям за ряд лет, в Калинин-
градскую обл. 1 мая они были встречены у Тамбова (Резцов, 1910). В конце
первой декады мая их появление отмечалось вблизи от северной границы ареа-
ла — у Казани, в это же время у Чкалова. У Чернигова они были замечены
2 мая (Емельяненко, 1913), что согласуется с приведенными выше данными для
Киева. Во второй трети мая прилет горлиц наблюдался у Горького, Калуги и
Омска. Валовой лет приходится на пятый — шестой день после первого появле-
ния, о чем можно судить по тому, что в устьях Урала он падает на первую треть
мая, а у Чернигова на 8 число этого месяца (Емельяненко, 1913). Особняком
стоят указания на появление горлиц 16 марта у Николаева (Токарь, 191&)
и 7 апреля у Гатчины (Диц, 1913). Здесь возможно предположение о какой-
нибудь путанице или неверном определении. Осенний отлет начинается рано
и по некоторым данным первыми начинают откочевку молодые. Например они
появляются у Гурьева в начаде второй трети августа. На восточном берегу Кас-
пийского моря появление пролетных горлиц приходится на вторую треть ав-
густа. В середине этого месяца горлицы исчезают из окр. Карачева (Суслова,
1927) и Калужских лесов (Филатов, 1915). В начале августа отлет начинается
в таких различных местах как Приазовье и Калининградская обл., где наиболее
интенсивный лет приходится на август — сентябрь, причем последние наблю-
дения относятся к 18 и 29 сентября (Тишлер, 1914). На юге Каспийского моря
пролетные горлицы показываются во второй половине августа. Интенсивный
пролет идет в сентябре, и последние горлицы отмечались 2 октября (Воробьев,
1940). В первой трети сентября начинается пролет в Крыму, во второй трети
последние горлицы отмечались у Тамбова, в бассейнах Алатыря и Суры, у Бу-
гуруслана и в верховьях Тобола. Последняя треть сентября знаменует исчез-
новение горлиц у Сызрани B4 числа, Богданов, 1871), у Чкалова, верховьев
Иргиза, бассейна Оки. На крайнем юге Европейской части Союза, как, напри-
мер, на ю.-з. Кавказе, пролет кончается в начале октября (Домбровский,
1913). Некоторые птицы задерживаются весьма надолго и попадаются даже на
севере Каспия в середине ноября (Бостанжогло, 1911). Продолжительность
осеннего пролета больше, чем весеннего, и он занимает обычно около месяца,
но иногда затягивается на значительно большее время. Особняком стоят до-
вольно частые случаи осенних залетов горлиц далеко к северу от своего гнездо-
вого ареала. Так, один экземпляр был добыт 30 сентября под Павловском.
Чаще горлицы отмечались в Финляндии. В Лапландии известны залеты до
Варсе и Варде. Случаи зимовок горлицы в СССР крайне редки. 2 экземпляра
необычайно зажиревших птиц, из которых одна молодая, были добыты в Бай-
рам-али 25. XII. 1912 г.
Биотоп. Горлица — прежде всего птица лиственных лесов. Однако,
в некоторых случаях она живет и в хвойных лесах, особенно у северной границы
ареала, а в Калининградской, Горьковской и Московской обл. предпочитает
44 ОТРЯД ГОЛУБИ
хвойные леса. В Горьковской обл. горлица поселяется главным образом в сос-
ново-ягодниковых борах и сосновых травянистых борах с подлеском из раз-
личных кустарников (можжевельник, крушина, рябина, ракиты, осины и т. д.).
Однако и здесь горлицы охотно гнездятся в уремах Ветлужской поймы. Расти-
тельность ее складывается, главным образом, из лиственных пород с под-
леском из дубняка, лещины, ели, можжевельника, шиповника, малины, смо-
родины и т. п. Больших боров северной части этой области горлицы избегают.
В Белоруссии она предпочитает лиственные и смешанные насаждения,
причем не гнездится в глубине леса, а выбирает обособленные рощи и парки.
Охотно селится горлица здесь и в мелколесье и кустарниковых зарослях с от-
дельными деревьями. В Подолии гнездится в пойменных зарослях чаще, чем
в больших участках лесов. На Украине населяет дубовые леса с сопутствующими
дубу ясенем, липой, кленом, лещиной, с богатым подседом из разнообразных
трав. На крайнем западе Украины, где дуб местами уступает свое место грабу
или буку, горлица придерживается этих насаждений.
Во многих местах средней полосы СССР, например, в бассейнах Суры и
Алатыря, горлицы с наибольшей охотой поселяются в пойменных зарослях
(уремах), состоящих здесь из дубняка, черемухи и татарского клена. В Чкалов-
ской обл. горлицы гнездятся всюду, где есть леса и кустарники, однако осо-
бенно многочисленны они здесь в долинах рек, где развиваются заросли ольхи
и ивняка, особенно густые на болотистых местах. Далее к югу и юго-востоку—
в степях сев. Казахстана — горлицы населяют березовые колки и сосновые
боры, располагающиеся на водораздельных пространствах. Иногда селятся
здесь в зарослях высоких трав у казахских зимовок и на развалинах последних.
На Кавказе горлицы поселяются в разнообразных лесах. Начиная с уремы
Кубани, представляющей собой заросли невысокого ивняка, они распростра-
няются в дубовые или буковые леса горных склонов, где к основным древесным
породам примешаны: граб, вяз, ясень, липа, местами дзелква и самшит и масса
кустарников. Кое-где на Кавказе еще сохранились остатки брошенных когда-то
жителями садов. Здесь древесная растительность представлена фруктовыми
деревьями: алычей, орехом, сливой, черешней, яблоками, грушами и т. д.
В Закавказье — в низменностях Риона и Талыша, где леса располагаются на
болотах они состоят преимущественно из ольхи. Все эти насаждения горлицы
населяют с охотой, но хвойной растительности верхнего пояса гор избегают,
хотя в Армении иногда гнездятся в можжевельниках. В Крыму местообитанием
этих птиц служат леса, подобно кавказским, состоящие из дуба или бука, но
с обильной примесью хвойных пород.
В степной части ареала горлица повсюду довольно охотно поселяется
в садах из фруктовых и декоративных деревьев и в полезащитных полосах леса,
которые в последние 10—15 лет насаждаются в засушливых участках Украины,
Предкавказья и Поволжья.
В каких бы условиях ни селились горлицы, они никогда не удаляются от
воды, хотя бы это был незначительный ручей, или даже колодец, из которого
поят скот, как это бывает в сев. Казахстане.
Численность. Больших скоплений горлицы в пределах Союза не
образуют. На гнездовье они держатся парами, никогда не объединяясь в коло-
нии. Выросшие птенцы собираются в небольшие стайки — по 15—20 шт. К осени
горлицы образуют несколько более многочисленные стаи, которые тем не менее
редко достигают сотни. Правда, во время пролета на местах ночлега иногда
собираются и более многочисленные общества, но они крайне нестойки, и утром
птицы разлетаются стайками особей по тридцать. Крупные стаи образуются
из более мелких по мере приближения к области зимовок. В Палестине и Си-
рии эти стаи достигают до нескольких сот экземпляров. Так же многочисленны
бывают они в сев. Африке. На весеннем пролете бесчисленные скопления
горлиц отмечали в Месопотамии и внутренней Сирии. Однако приближаясь
горлица 45
к границам СССР, эти скопления постепенно разбиваются, и у нас горлицы
летят уже сравнительно небольшими стайками.
Экология. Размножение. Прилетные стаи горлиц содержат самцов
и самок, причем часть птиц уже образует пары. Этим объясняются случаи, когда
гнездование следует непосредственно за прилетом. Другая часть горлиц при-
летает еще не разбившись на пары, и сейчас же после прилета начинается вор-
кование самцов. На р. Хопер оно отмочено 30 марта (Измайлов, 1940; этот
срок кажется, однако, слишком ранним), под Тамбовом 13 мая (Резцов, 1910),
а в Брянской обл. 8 мая (Суслова, 1937). Воркуют самцы, сидя где-либо на дереве,
время от времени взлетая вверх и вновь спускаясь на свое место. Воркование
продолжается и после того, как горлицы образуют пары. В северных частях
ареала, например, у Калуги, конец воркования отмечался в середине июля
(Филатов, 1915), а у южных окраин СССР —в Крыму —в первой трети августа
(Никольский, 1892).
Гнездо располагается на различной высоте, иногда почти у поверхности
земли, в основании какого-нибудь куста. В других случаях оно закладывается
на высоте более, чем десять метров. Чаще всего гнезда вьются на высоте от 3
до 6 метров. В виде исключения горлицы гнездятся и на земле. Гнездо свито
небрежно, и сквозь него почти всегда видны яйца. Последних, как и вообще
у голубей, в кладке 2. Цвет яиц белый. Размеры; F9) 28,5—33 х 22—24,5,
в среднем 30 х 22,9 мм (Гебель, 1879).
Откладка яиц происходит в разные сроки, так как одня пары уже начинают
нестись, в то время как другие еще не образовались. Наиболее ранние даты
находок яиц относятся к Закавказью, где 5 мая яйца былн найдены у Еревана
(Соснин и Ляйстер, 1942). В середине мая — 17 числа — свежие кладки при-
водятся уже для Белоруссии. Здесь же они были обнаружены 10 июля, что ука-
зывает на вторую кладку (Шнитников, 1913). У Воронежа гнездо с 2 яйцами
было найдено 18 мая (Резцов, 1910). В Подолии известны свежие яйца от 24 мая
(Герхнер, 1928) и 2 июня (Портенко, 1928). В Чкаловской обл. валовая кладка
яиц приходится на первую и вторую трети июня, но свежие яйца находят там
до середины июля (Зарудный, 1888, 1897). В Сочинском округе гнездо с неопе-
ренным птенцом было отмечено 10 июня (Кудашев, 1917). 15 июня были найдены
слегка насиженные яйца у Бугуруслана (Исполатов, 1911). В Хоперском запо-
веднике насиженные яйца найдены 17 и 23 июня (Измайлов, 1940). Эти кладки,
если даже они являются запоздалыми, указывают на то, что наблюдение, сде-
ланное 30 марта над воркующими горлицами, может быть ошибочно или отно-
сится к другому виду. Почти зрелые яйца имела в яйцеводе самка, добытая
в Армении 28 июня. Гнездо со слабо насиженными яйцами найдено 30 июня
в Псковской обл. (Зарудный, 1910). 6 июля в Закавказье обнаружено гнездо
с слабо насиженными яйцами (Нестеров, 1911). Поздние кладки были найдены
в Барабинской степи 22 июля (Коте, 1910) и поймах Суры и Алатыря в первой
трети августа (Житков и Бутурлин, 1906). К июлю приурочивается вылет птен-
цов из гнезд у Казани. Поздняя дата B9 августа) добычи молодого с пухом от-
мечается для Горьковской обл. (Серебровский, 1908). 11 августа наблюдались
молодые с неотросшими маховыми в Закавказье (Ляйстер, 1942). Все эти дан-
ные позволяют заключить, что горлицы имеют не менее двух кладок в
лето даже у северной границы ареала. Очень поздние кладки, найден-
ные в августе в бассейне Суры, может быть принадлежат уже третьему
выводку.
В насиживании яиц принимают участие оба пола. Оно продолжается 13—
14 дней (Уайзерби). Птенцов кормят оба родителя обычным для голубей спосо-
бом, т. е. отрыгивая содержимое своих зобов в клювы птенцов. Последние оста-
ются в гнездах около 18 дней, а еще через три дня начинают как следует ле-
тать. Вылетевшие молодые вскоре собираются в самостоятельные стайки по
10—15 особей. Возможно, что эти стайки постепенно пускаются в отлет, так
46
ОТРЯД ГОЛУБИ
как на восточном берегу Каспийского моря, где гнездящихся горлиц нет, уже
во второй трети августа появляются молодые птицы.
Линька. Вопросы линьки горлиц в литературе освещены очень слабо.
Взрослые птицы улетают в старом оперении и возвращаются в свежем. Таким
образом, их линька происходит на зимовках. Молодые первых выводков ли-
няют до отлета, надевая окончательный наряд взрослых птиц. Видимо к таким
молодым и относится указание Богданова A871) на линьку горлиц в конце
июня ст. ст. Молодые последнего вывода улетают в гнездовом пере и возвраща-
ются перелинявшими во взрослый наряд. Добытые 25.XII. 1912 г. у Байрам-али
взрослая и молодая горлицы находятся в стадии линьки. У старой птицы смени-
лись 10-е и 9-е маховые и неболь-
шое количество мелких перьев
боков. У молодого экземпляра
сменилось только 10-е маховое
каждого крыла. Мелкое перо еще
не имеет следов линьки.
Питание. Кормом горлиц
служат семена разнообразных
растений. В первую половину
лета они кормятся, главным об-
разом, дикой растительностью,
разыскивая ее зерна по лугам,
выгонам, берегам рек и т. п.
местам. После созревания хлебов
и их уборки перебираются на
поля, предпочитая пшеницу, про-
со или гречиху. В тех случаях,
когда имеются масличные куль-
туры, горлицы отдают им пред-
почтение. Животный корм зани-
мает в питании этих птиц очень
немного места и состоит главным
образом из мелких раковин, кото-
рые может быть заглатываются
вместо камешков. Кормятся гор-
лицы обычно утром и как только начинает пригревать солнце, летят на водо-
пой, а затем прячутся в деревья.
Полевые признаки. Общий склад горлицы напоминает (формой головы,
короткой шеей и ногами и длинным туловищем) склад домашнего или дикого
голубя. Однако она гораздо тоньше, стройнее и изящнее голубей. Кроме того,
отличается от них более мелкими размерами и пестрой, коричневатой окраской
верха. В полете горлиц отличает темный подбор крыльев, которые гораздо
темнее нижней стороны тела, и сильно развитая белизна на концах рулевых. За
исключением брачного периода, когда горлиц выдает воркование, это тихие и
малозаметные птицы. Большую часть суток они проводят в ветвях деревьев
и слетают на землю только для кормежки и водопоя. Напившись, любят про-
вести некоторое время у берега за чисткой и разборкой перьев. Несмотря на
свою скрытность, горлица не может считаться очень осторожной птицей. Она
быстро привыкает к людям, и там, где ее не преследуют, безбоязненно посе-
ляется у самых деревень и кормится по их окраинам вместе с полудикими
голубями.
Голос горлицы слышен только в брачное время. Это — воркование, очень
легко отличимое от голосов других видов голубей Европы. Оно легко запоминает-
ся и довольно точно передается звуками — «турр—турр» или «курр—курр».
Этот слог обычно повторяется три — четыре раза, после чего следует короткая
Рис. 7. Обыкновенная горлица
горлица 47
пауза и новое повторение. Иногда воркование прерывается более длинными
паузами.
Описание. Размеры и строение. Длина самцов (8) 288—310, самок (9) 279 —
310 мм, в среднем 296,5 и 299 мм. Размах самцов F) 530—593, самок (9) 505—54О,в среднем
548 и 520,5 мм. Крыло самцов D8I65—184, самок B7I63—178, в среднем 174,7 и1б9,4 мм.
Окраска. Окраска горлиц состоит из сочетания неярких цветов: голубовато-серого,
рыжевато-коричневого, бурого и черного; у старых птиц голова и шея сверху окрашены
в голубовато-серый цвет, бледнеющий ко лбу; спина светлобурая, к пояснице и подхвостью
этот цвет все более вытесняется темносерым. У некоторых особей надхвостье совершенно
теряет буроватые участки, которые дольше всего удерживаются по средней линии, и делается
чисто темносерым. Плечевые и верхние кроющие крыла (кроме наружных) темно бурого,
иногда даже черного цвета с широкими рыжевато-коричневыми краями перьев. Кроющие
первостепенных маховых одноцветные — темносерые. Зоб и грудь — сизовато-розовые, к под-
бородку этот цвет сильно светлеет, к брюшку и подхвостью становится совсем белым. Бока,
подмышечные и подбой крыльев — голубовато-серые. Маховые — буровато-черные, в свежем
пере с узкими беловато-коричневыми каймами на внешних опахалах (на которых у второ-
степенных маховых имеется еще и серый налет. Рулевые, кроме средней пары, аспидно-
черные, на концах белые, на крайних рулевых и наружное опахало белое. Средняя пара,
как и верхние кроющие хвоста, бурые. На боках шеи несколько рядов черных перышек
с очень светлыми серовато-голубыми вершинными каймами, образующими ряд полулун-
ных пятен; голая кожа вокруг глаз малиново-красного цвета; ноги красные, глаз оран-
жево-красный или оранжево-бурый; клюв черноватый.
Молодые сверху окрашены однообразнее; все серые тона заменены коричневатыми;
темные центры плечевых и кроющих крыла светлее,чем у старых, коричневато-буры;на ма-
ховых сильный ржаво-коричневый налет. Зоб и шея без розового оттенка, сероватые с узкими
коричневато-охристыми краями перьев; горло того же цвета, но светлее. Остальной низ —
белый; на боках шеи нет темных и голубоватых пятен.
Азиатская горлица Streptopelia turtur arenicola Hart.
Turtur turtur arenicola. H a r t e r t. Nov. zoological, 1894, стр.42, Фао, Персидский
залив.
Синоним. Turtur communis var. gregorjewi. Z a r u d n у und L о u d о n. Orn. Monatsber.
1902, стр. 149, Гильменд, Белуджистан.
Распространение. Ареал. В СССР — к востоку от Каспийского моря.
Северная граница ареала совпадает с южной границей ареала номинального
подвида. По Эмбе поднимается к северу до Кок-джиды, по Иргизу—до одноимен-
ного города и по Тургаю захватывает только низовья (Мугоджары населяет
европейская форма, Сушкин, 1908). На Иртыше наиболее северной находкой
служат окрестности Семипалатинска (Хахлов, 1923). Южнее добыта в При-
алайской степени у пос. Новенского, а может быть и у Усть-Каменогорска
(Селевин, 1935). Выше Усть-Каменогорска по Иртышу гнездится в большом
количестве между протоком НДутуй и устьем Кальджира (Поляков, 1912—
1914). Азиатская горлица гнездится в Зайсанской котловине и Тарбагатае
с предгорьями (Поляков, 1915), так же, как и в долине р. Или (Зарудный, 1906,
Шестоперов, 1929). В центральном Казахстане распространена в долине Сары-
Су (Афанасьев и Слудский, 1947). К северу от Балхаша идет по возвышенно-
стям до станции Жарык, но у гор Кызыл-рая и у Каркалинска отсутствует
(Долгушин, 1947). На р. Чу прослежена, по всей равнинной части (Гладков,
1932). В Кара-тау гнездится в поселках (Шапошников, 1931). В долине нижней
Сыр-Дарьи вполне обычна (Спангенберг, 1936), как и в долинах ее крупных при-
токов, например, Чирчика (Сатаева, 1937, Гладков, 1938). Вверх по Сыр-Дарье
поднимается до Ферганы (Штольцман, 1897, Кашкаров, 1927, 1934, Мальчев-
ский, 1940), идет и еще выше в горы по Нарыну, поднимаясь до озера
Коккуль, пер. Эмель и может быть долины Чаманды (Шнитников, 1930). Широко
распространена в Кызыл-Кумах (Зарудный, 1914). По южной окраине этой
пустыни — в горах Нура-тау — не особенно многочисленна (Мекленбурцев,
1934). Встречена в долине Зеравшана от 200 до 1300—1400 м (Каррузерс, 1910).
В западных отрогах Гиссарского хребта и по долинам текущих с него рек попа-
дается всюду (Иванов, 1940). По Вахшу (Сурхоб) доходит до Богорака (Заруд-
48 ОТРЯД ГОЛУБИ
ный, 1926), а в бассейне Пянджа гнездится, начиная с высоты около 2500 м
(киш. Рош-кала на Шах-Даре). Далее, вниз по долинам Аму-Дарьи, ука-
зана вплоть до Аральского моря. К западу от Аму-Дарьи встречается по Мур-
габу и Теджену, широко распространена в Кара-Кумах в особенности на зап.
Узбое (Дементьев, 1950 и др.). Распространена по северным подножьям и скло-
нам Копет-Дага. Область зимовок азиатской горлицы лежит в тех же местах
южн. Сахары и северных Сенегала и Судана, как и типичного подвида.
Характер пребывания. В среднеазиатской части ареала это
перелетная птица, появляющаяся поздно весной.
Даты пролета азиатской горлицы носят пестрый характер. Во второй поло-
вине апреля горлицы были отмечены в долине р. Сыр-Дарьи и ее среднем тече-
нии (Спангенберг, 1936) и у Ташкента (Сатаева, 1937). В последней трети этого
месяца появление зарегистрировано между Мервом и Аму-Дарьей, на р. Душак
и Теджен, в долине Арыси. 26 числа в Таджикистане у Гиссара (Иванов, 1940).
В первых числах мая C и 7) горлицы достигают Джаркента (Зарудный, 1906),
но у Чарджоу были отмечены только шестого, а у Коканда —десятого (Штольц-
ман, 1897). В общем, в Туркмении прилет и пролет относится к концу апреля—
началу мая (Дементьев, 1950). В середине мая они были встречены у Казалинска
и Аральского моря (Никольский, 1892). 15 мая первое появление наблюдалось
в низовьях Иргиза (Сушкин, 1908). Существуют указания на более раннее по-
явление горлиц под Ташкентом — 23 и 27 марта и 3 и 5 апреля, приведенные
Сатаевой A937) по данным 1928 года. Однако они настолько не согласуются
с остальными литературными данными и имеющимися материалами, что их
невозможно принимать во внимание. Отлет горлиц начинается рано, но тянется
продолжительное время. В Кара-Кумах в Туркмении в саксаульниках у Ре-
петека местные горлицы исчезли уже в начале июля, а в августе появились
птицы, повидимому, с севера (Снигиревский, 1928). На крайнем юге, у Кушки,
осенний пролет 26 сентября. В общем же отлет и пролет отмечается осенью,
в Туркмении—в сентябре (Дементьев, 1950). Уже 12 августа их появление отме-
чено на побережье Каспийского моря у залива Михайловского (Зарудный, 1896).
В конце этого месяца они обычны на пролете у Джаркента (Зарудный, 1906),
у Ташкента • В начале сентября F-го) были замечены на озере Яшиль-куль на
Памире (Мекленбурцев, 1935). В эти же дни под Ташкентом число горлиц убы-
вает. В начале второй трети сентября горлицы наиболее многочисленны у Джар-
кента, но в середине месяца их количество здесь несколько убывает, а к последней
трети они становятся редкими (Зарудный, 1906). У Ташкента в это время от-
мечаютея только одиночные птицы, но порядочные стаи были встречены на
Кашка-Дарье у Чиракчей. Однако к концу месяца горлиц стало здесь меньше,
и они попадались, главным образом, одиночками. Наиболее поздние даты на-
ходок относятся к Памиру, где горлицы были добыты 27 сентября на Ранг-куле
и 3 октября на Кара-куле (Тугаринов, 1930), к долине Сыр-Дарьи, где они от-
мечались в конце октября и даже начале ноября (Спангенберг, 1936). Так же
поздно — 5 октября горлицы наблюдались у Чимкента и 4-го у Ташкента
(Сатаева, 1937).
Биотоп. Азиатская горлица довольно широко приспосабливается к раз-
личной древесной растительности в том случае, если вблизи имеется вода. Гнез-
дится у колодцев пустыни, на кустах саксаула или других песчаных растений.
В низовьях рек довольствуется зарослями тамариска или разнолистного тополя.
Иногда гнездится даже в камышах. В долинах среднего течения рек горлицы
многочисленны в тугаях, состоящих из ив, джиды, облепихи, кое-каких ши-
повников и жимолостей. В культурной полосе постоянно селится в садах киш-
лаков и городов, не исключая таких крупных, как Ташкент. В последнем случае
горлицы гнездятся исключительно на высоких деревьях акаций, тополей, ка-
рагача и т. п. По долинам рек поднимается в горы, следуя за лиственной расти-
тельностью, примерно до высоты 1000 м в зап. Тянь-Шане, 1400—1600 м в Гис-
горлица 49
сарском хребте и около 2500 м в горных долинах Пянджа и Шах-Дары;
в Копет-Даге до арчевников, на высотах около 2000 м (Чаек, Хейрабад). Всюду
в горных долинах горлицы держатся в пойменных лесах из ив, тополя, клена
с подседом из облепихи и густых травянистых зарослей.
Численность. Количество азиатских горлиц уменьшается не так
быстро, нежели сизых или даже бурых голубей и малых горлиц: горлицы
гнездятся одиночными парами в густых кронах деревьев и держатся у гнезда
более скрытно, чем названные виды. Кроме того, они в меньшей степени пре-
следуются охотниками. Благодаря всем этим причинам азиатские горлицы еще
очень обычны и даже многочисленны во многих местах. Наибольшая плот-
ность их приурочена к полосе предгорных кишлаков и долин ручьев, сбегаю-
щих с гор и поросших густыми зарослями ивняка и облепихи. В предгорьях
Тянь-Шаня в некоторых мало посещаемых местах или глухих кишлаках гор-
лицы настолько обычны, что в период гнездования можно сразу видеть несколь-
ко воркующих самцов, а гнезда располагаются на расстоянии 100—150 шагов
одно от другого. Однако больших стай горлицы не образуют даже осенью.
На кормежках в этот сезон они держатся одиночками или самыми малень-
кими обществами и лишь на дневной отдых или ночлег собираются по несколько
десятков или даже до сотен особей вместе. Сборища эти очень неустойчивы;
покидая избранное место, горлицы разлетаются мелкими стайками. На весен-
нем же пролете горлицы еще малочисленнее и не образуют скоплений даже
на отдыхе.
Экология. Размножение. Повидимому, часть горлиц прилетает уже
разбившись на пары. С другой стороны большое число птиц прилетает холо-
стыми. Немедленно после появления начинается воркование горлиц. Воркую-
щие самцы держатся у места будущего гнезда. Воркование слышится в течение
всего дня, но наиболее усиленно утром и вечером.
Токующий самец воркует, сидя совершенно спокойно, но часто прерывает
воркование и косо взлетает на высоту 2 — 3 десятков метров, сильно хлопая
крыльями. Достигнув предельной высоты, он широко раскрывает крылья и летит
вниз, не шевеля ими, описав дугу, возвращается к избранному месту. Самка
скрывается где-либо поблизости в чаще деревьев или кустов. Время от времени
она взлетает, чтобы отлететь несколько десятков шагов и снова сесть в чащу.
Самец, увидя взлетевшую самку, немедленно гонится за ней с характерным
тихим причмокиванием. Стоит только парочке сесть, как самец сейчас же начи-
нает особым образом ворковать, ритмично и сильно выпрямляясь на вытяну-
тых ногах. Такого рода воркование самец часто издает и на земле у водопоя,
куда после образования пары птицы обычно летают вместе. Иногда после подоб-
ного воркования следует спаривание.
Гнездо помещается где-нибудь невдалеке. Если птицы не напуганы и живут
в безлюдной местности, гнездо часто расположено на невысоком кусте, который
иногда бывает ниже роста человека. В других случаях место для гнезда выби-
рается очень высоко, на 12—15 метров над землей — в кроне большого дерева.
Гнездо обычного для голубей строения бывает иногда настолько неплотно, что
яйца хорошо видны снизу. Гнездовый материал состоит из тонких сухих пру-
тиков, стеблей трав и т. п. Лоточек часто бывает очень плоский, без всяких
бортов. Иногда горлицы занимают гнездо грача или сороки. Гнездовый участок
занимает территорию около 200—300 м2 и, как кажется, самец прогоняет зале-
тевших туда других горлиц. Впрочем, иногда две пары гнездятся на двух рядом
стоящих деревьях и даже на одном дереве.
Кладка состоит из двух яиц, слегка удлиненных, с почти одинаковыми кон-
цами. Размеры (8) 30,7—35,0 х 22,8 —25, в среднем 31,8 х 23,7 мм (Заруд-
ный, 1896). Наиболее ранние кладки в пределах Союза отмечены для Туркме-
нии и были найдены у Уч-аджи 8 апреля (Зарудный, 1896). В конце этого месяца
и начале мая свежие яйца были обнаружены во многих гнездах в Кызыл-Кумах
4 Птжцы СССР, т. II
50 ОТРЯД ГОЛУБИ
(Зарудный, 1915). В середине мая гнезда со свежими яйцами были найдены в до-
лине р. Или (Шестоперов, 1929). В самом конце июня и начале июля свежие
кладки зарегистрированы у Ташкента (Сатаева, 1937) и Куляба (Иванов 1940).
8—10 июля яйца были отмечены в ур. Чаманды в области верхнего течения
р. Нарын (Шнитников, 1930), а в середине июля на Сыр-Дарье между Чиили
и Казалинском (Спангенберг, 1936). Наконец под Ташкентом самка с крупными
яйцами в яйцеводе добыта в начале августа, а летные птенцы со следами пуха —
в самом конце сентября — начале октября, что указывает на кладку яиц во
второй половине августа. Наибольшее количество упоминаемых в литературе
кладок приходится на конец апреля и начало мая. Затем количество их посте-
пенно падает и к осени становится весьма незначительным. Некоторые пары,
видимо, гнездятся до трех раз в лето, в то время как другие успевают спра-
виться только с двумя выводками птенцов.
Насиживают и кормят птенцов оба родителя. Сроки насиживания и пребы-
вания птенцов в гнезде видимо сходны с таковыми у типичного подвида. Птен-
цы, достигшие несколько больше половины роста взрослых, часто выбираются
из гнезда и сидят рядом с ними на ветвях. Иногда случается, что они падают на
землю и, будучи еще совсем беспомощными, обычно погибают. Летные молодые
начинают встречаться в Туркмении уже в конце мая. Последние птенцы, недавно
покинувшие гнездо, попадаются до конца сентября. Птенцы первых выводков,
покинув родителей, бродят самостоятельно, большей частью парами или оди-
ночно, лишь редко объединяясь в небольшие общества. К концу лета к этим
бродячим птенцам все больше присоединяется старых особей, то закончивших
выведение птенцов, то потерявших одного из членов пары.
Линька. Часть молодых — именно те, которые вылупились из первых
кладок, —линяет уже в августе и к концу лета успевает надеть оперение взрос-
лых птиц. Более поздние птенцы линяют во время пролета и птенцы последних
выводков сменяют оперение во время зимовок. Видимо в это же время линяют и
старые птицы. Во всяком случае среди экземпляров, добытых летом и осенью
в Средней Азии, нет таких, которые бы линяли. Улетают горлицы в сильно по-»
трег^анном оперении, но возвращаются в совершенно свежем. В литературе опи-
сывается только один случай линьки горлицы летом, когда несколько птиц
перелиняли в условиях неволи (Снигиревский, 1927). При этом их оперение
приняло цвета, характерные для типичного подвида. Однако эти птицы были
чрезвычайно зажиревшими. Повидимому ненормальные условия вызвали не-
своевременную линьку. Кроме того, пропитывание оперения жиром придало
ему темную окраску, напоминающую окраску типичной формы. Смена нарядов
азиатской горлицы идет общим для всех наших голубей порядком, т. е. эмб-
риональный покров из редкого желтоватого пуха сменяется ювенальным
рыхлым оперением, а последнее — взрослым нарядом, надеваемым не позже
первой зимы жизни.
Питание. Точные данные по питанию горлиц в Средней Азии весьма
невелики. По аналогии с европейским подвидом их пища должна состоять из
разнообразных семян и зерен. Немногочисленные вскрытые зобы содержали
большей частью зерна пшеницы, различных сорняков и сафлора. Поля, засе-
янные последним, горлицы посещают с особенной охотой. Кормятся, подобно
другим голубям, только на земле и не трогают неповрежденных растений. Из-
вестны случаи поедания ягод шелковицы.
Полевые признаки и повадки. Подобно типичной форме, азиатская гор-
лица мало заметная птица. Будучи сравнительно обыкновенной, даже в боль-
ших городах, она редко попадается на глаза, так как держится в листве
больших деревьев и не слетает на землю даже кормиться, улетая для этого
из города. В кишлаках держит себя не так скрытно, и иногда опускается
на землю даже во дворах и на улицах. Однако на кормежку и водопой гор-
лицы и здесь летают на соседние поля и речки. Кормятся утром, иногда
БОЛЬШАЯ ИЛИ СТЕПНАЯ ГОРЛИЦА 51
появляясь на полях еще до восхода солнца. Наевшись, летят на водопой,
но никогда не собираются для этого стаями, а держатся не более, чем по
две пары вместе. Местами водопоя служат галечниковые или песчаные отмели.
Здесь горлицы любят отдохнуть, почиститься, поклевать песчинки и камешки,
но долго на местах водопоя не задерживаются. Старые птицы летят к гнез-
дам, а молодые садятся в кроны густых деревьев, где пережидают всю жар-
кую пору дня. Вечером они вновь летят на поля кормиться. Ночуют на тех же
деревьях, где отдыхают днем. Сидящих в кроне густого дерева горлиц очень*
трудно заметить, даже если при подходе не спускать глаз с вершины, на
которую село несколько птиц. Вылетают они из листвы стремительно, с особым
шумом крыльев и быстро разлетаются в разные стороны. Точно так же реаги-
руют горлицы на появление вблизи хищной птицы.
С другими видами голубей горлицы образуют смешанные стаи только в ме-
стах дневного отдыха или ночлега. Здесь их можно встретить вместе с бурыми
голубями или большими горлицами. На местах кормежки они держатся иногда
в самом близком соседстве с сизаками. Все эти стаи очень нестойки. Лишь иногда
одиночные особи азиатских горлиц упорно держатся в больших стайках. Голос
азиатской горлицы не отличим от голоса европейского подвида. Он слышен так-
же только в брачное время и может быть передан теми же слогами «курр-куррк
Гоняясь за самкой, самец издает тихое причмокивание, слышное на близком
расстоянии.
Описание. Размеры и строение. Длина самцов E) 280—305, в среднем
297,4 мм; размах E) 490—592, в среднем 541,4 мм. Крыло самцов C4) 161—179, самок C0)
161—175, в среднем, соответственно, 170 и 165,5 мм.
Окраска. По расположению цветов совершенно сходна с окраской европейской
горлицы. Однако все рыжевато-коричневые цвета у азиатского подвида по большей части
светлее. У большинства особей каемки перьев плечевых и верхних кроющих крыла не рыже-
вато-коричневые, а охристо-коричневые; на пояснице и надхвостье темносерый цвет заме-
нен более или менее чисто серым; зоб и грудь охристо-серовато-розовые; спина более
рыжеватая. Молодые также более светлого основного цвета, иногда почти соловые сверху
и беловатые снизу.
8. Большая или степная горлица Streptopelia orientalis Lath.
Columba orientalis. Latham. Ind. Ornith. II, 1790, стр. 606, Китай.
Русское название Книжное.
Распространение. Ареал. В качестве гнездящейся птицы населяет
восточные части азиатского материка, охватывая территорию, ограниченную на
севере южными предгорьями Верхоянского хребта, районом Якутска, Прибай-
кальем, междуречьем Ангары и Енисея, районом Новосибирска, Нарымом,
устьями Ишима и Тобола и районом Тобольска и Тюмени. Далее граница пово-
рачивает на юг, охватывая бассейны Тобола и Ишима и горы Казахстана. Затем
она делает петлю, исключающую Прибалхашье и Алакульскую равнину, и
поворачивает к юго-западу, захватывая Джунгарский Ала-тау и всю систему
Тянь-Шаня и Памиро-Алая. Отсюда граница ареала идет к юго-востоку, вклю-
чает зап. Памир, сев.-вост. угол Афганистана, Кашмир, Ладак и Гималаи.
Далее ареал большой горлицы захватывает центральные провинции Индии, Би-
хар, Ориссу, Хайдерабад (до 15° с. ш.), Бенгалию, Ассам и Бирму до Тенас-
серима. Затем он включает о-ва Хайнан, Формозу и Риу-Киу, Японские и
Сахалин. Внутри этого пространства распространение большой горлицы обра-
зует большие пробелы. Она не гнездится во всех пустынях внутренней Азии»
в степях Монголии, на плоскогорьях Тибета и Памира. Область зимовок боль-
шой горлицы начинается от южных подножий Гималаев и Южн. Китая.
В небольшом количестве зимует на крайнем юге Ирана (Бампур?). В Индии
на зимовках была найдена вплоть до Цейлона. Далее к востоку область зимовок
распространяется на Индо-Китай, о-ва Хайнан, Формозу и Японские.
52
ОТРЯД ГОЛУБИ
Биотоп. Леса самого разнообразного характера. В сев. участках аре-
ала — в равнинной тайге, поднимаясь в горы в лесную зону. На западе и
юге ареала большая горлица живет в горах и приурочена здесь главным обра-
зом к хвойной растительности, реже к лиственным лесам (впрочем, местами пре-
имущественно в лиственных насаждениях).
Карта 8. Распространение большой горлицы Streptopelia oriental is
7. S. о. orientalis, 2. S. о. теепа, 3. S. o. stimpsoni, 4. S. o. orii, 5. S. o. agricola;
a — граница области распространения, в — изолированные нахождения большой горлицы, г — места
зимовки перелетных форм.
Подвиды и варьирующие признаки. S. о. orientalis Latham, 1790 — восточная часть
ареала от Сиккима на юге и бассейна Енисея на севере (за исключением о-вов Риу-Киу и
Формозы); 5. о. теепа S у k e s, 1832 — западная часть ареала от границы, указанной для
предыдущего подвида; S. о. stimpsoni Ste jneger, 1887 — о-ва Риу-Киу; S. о. orii
Yamashina — о-в Формоза; 5. о. agricola T i с к е 1 1, 1832 — с.-в. Индия (Бихар,
Орисса, Бенгалия), Ассам, Бирма до Тенассерима. Подвиды большой горлицы в одних слу-
чаях различаются развитием на подхвостье и рулевых белого или сизого цвета, в других —
развитием винных тонов на нижней стороне и красноватых — на верхней и, в-третьих,
колебаниями общих размеров.
Экологические особенности подвидов выражаются в том, что местообитанием одних
служит высокоствольная тайга или лиственные древесно-кустарниковые заросли. Другие
приурочены к лесам, состоящим из древовидного можжевельника. Индийские подвиды гнез-
дятся в тропических лесах из разнообразных деревьев. Северные формы — типичные пере-
летные птицы, а южные — вполне оседлы.
Таежная большая горлица Streptopelia oriental is orientalis Lath.
Синоним. Columba rupicola. Pallas. Zoographia Russo-Asiatica, 1811, стр. 566, Лена.
Распространение. Ареал. В СССР южные области Восточной и средней Си-
бири. На Сахалине большая горлица местами гнездится в значительном количе-
стве (Никольский, 1889). На материке Азии в пределах Союза она обычна в Уссу-
рийском крае. В бассейне Амура отмечена у рек Хунгари и Горюн (Воробьев,
1947). На Лене известны у Якутска (Воробьева, 1931) и до 64° с. ш., по Алдану
прослежена до 63°30' (Иванов, 1929). От устья этой реки к югу гнездится нор-
мально, как и в районах ее верхнего течения; так же гнездится в области верх-
БОЛЬШАЯ ИЛИ СТЕПНАЯ ГОРЛИЦА S3
него течения Олекмы и Май (Тугаринов, Иванов, Смирнов, 1934). В Забайкалье
часто встречается в лесной области, на Байкале очень обыкновенная птица
(Штегман, 1928, 1936). К Западу от этого озера гнездится по р. Уса, в долине
Енисея, у Минусинска, в Мирском хребте (Сушкин, 1914, Залесский, 1921).
Обычна в р-не Канска и окрестностях Иланской (Залесский, 1921). В вост.
Алтае (Яйлю, Кезебень) встречаются особи, переходные к западному подвиду.
Область зимовок — юг Китая от долины Голубой реки, Сиамский п-ов от
Сычауни и Бирмы и вост. Индия до соединенных провинций и Декана.
Характер пребывания. Отдаленность зимовок вызывает позд-
ние сроки пролета и прилета. Наиболее ранние особи отмечены на край-
нем юге Приморья—на берегу Амурского залива —31 марта (Медведев,
1913). 10 апреля горлицы были замечены у истоков Сунгари, 11 —у поселка
Нововладимировка, а 14—у ст. Бейцуха (Шульпин, 1936). Следующая дата
относится к Олекминску, где эти птицы наблюдались месяцем позднее —
12 и 14 мая (Воробьев, 1931). 18 мая первое появление горлиц было отме-
чено у Якутска (Воробьева, 1931) и 20 — около Урги (Козлова, 1930). Пови-
димому, последнее наблюдение нельзя относить к началу пролета. Точно так
же, по всей вероятности, не первые появившиеся птицы были отмечены 3 июня
у Николаевска и на западных склонах Станового хребта (Иванов, 1929). У запад-
ных окраин своего ареала — в Минусинском крае — прилет отмечен очень
поздно — 5 июня (Сушкин, 1914). Это позволяет предположить, что пролет
горлиц идет в Вост. Сибири с востока на запад и тянется очень продолжи-
тельное время.
Сведения по осеннему отлету еще более отрывочны, нежели данные по при-
лету горлиц весной и сводятся к указаниям на встречи с этой горлицей 5 сен-
тября у р. Халхин-гол (Тугаринов, 1928),24—у сопки Рябоконь (Шульпин, 1936)
и 28 в ю.-в. Хангае (Козлова, 1930). 14 октября пролетные горлицы наблю-
дались в Кентее (Козлова, 1930), и в этот же день они появились у устья
Ян-цзе (Воган, 1913). Самые поздние даты, в которые горлицы были отмечены
на территории СССР, относятся к о-ву Аскольд, где они наблюдались до ноября
(Воробьев, 1937), и Седанке, где экземпляры зарегистрированы 29 декабря
(Шульпин, 1936). Последняя дата может быть относится к зазимовавшей птице.
Зимовки этой горлицы указаны для Уссури (Маак, 1861) и Забайкалья
(Тачановский, 1873). Несомненно, что это явление имеет характер исключения.
Иногда большие горлицы залетают очень далеко к западу от границы за-
нятой ими области. Несколько особей было добыто в различных местах Скан-
динавии (Финмаркен, Херьедаль и Питеа). Находки известны из Дании, с вос-
точного берега Великобритании (Скарборо) и одна из Италии.
Биотоп. Селится большая горлица в разнообразных местах. На крайнем
востоке — на о-ве Сахалине — приурочена к лиственным лесам, состоящим
из ив с примесью берез, ольхи, тополей, дальневосточного береста и манчжур-
ского ясеня. В Приморье горлица любит уремные леса, характеризующиеся
богатым развитием манчжурской флоры с такими представителями, как амур-
ская липа, манчжурский ясень и дуб, мелколистный клен, амурская сирень,
бархат. На склонах гор Сихотэ-Алиня горлица обитает в мелком кустарном
лесу из дуба, вяза и ряда других кустарников. Селится она и в хвойных лесах,
представленных саянской елью, белокорой пихтой и корейским кедром. При
всем этом для горлицы необходимо сочетание густых зарослей с полянами,
лишенными деревьев и поросшими травянистой растительностью.
На Амуре горлицы живут в ивняках островов. На Байкале они наиболее
многочисленны у берегов озера и в глубоких долинах, где произрастают лист-
венничные леса с примесью елей, пихт и кое-каких лиственных деревьев,
например, березы и благовонного тополя. Единичные пары горлиц поднимаются
в Прибайкалье высоко в горы до границы древесной растительности. У берегов
Енисея горлицы гнездятся и в лесостепной зоне, и в равнинной тайге, и по
54
ОТРЯД ГОЛУБИ
окраинам таежных массивов, где к хвойным породам примешивается береза,
и в глубине обширных таежных площадей.
Численность. В литературе существуют указания лишь на то, что
вылетевшие в половине июня птенцы образуют группы в 6—7 штук. В сентябре
горлицы кочуют по полям стаями по 12—15 особей.
Экология. Размножение. Брачное воркование самцов начинает раз-
даваться немедленно после прилета и сейчас же начинается гнездостроение и
откладывание яиц. Гнезда расположены на деревьях, кустах (иногда над водой),
в виде исключения на земле. Бывали случаи нахождения гнезд на кучах хво-
роста. Постройка гнезда крайне примитивная, типичная для голубей. В кладке
2 яйца белого цвета. Размеры (8) 22,6—26x32—36,6, в среднем 24,5x34,0 мм
(Спангенберг, 1940). Свежие яйца были найдены в Приморье уже 27 апреля
(Шульпин, 1936) и попада-
лись в самые различные
числа мая и июня; поздняя
свежая кладка найдена 30
июня (Спангенберг, 1940).
На о-ве Аскольд 10 мая была
обнаружена кладка из на-
сиженных яиц (Воробьев,
1937). Прозревшие птенцы
были найдены в Приморской
области 11 мая, в пуху —12
июня (Шульпин, 1936). У р.
Имана птенцы горлицы были
отмечены 28 мая (Спанген-
берг, 1940). В бассейне Ени-
сея вылетевшие из гнезд
молодые наблюдались в се-
редине июля и в то же
время начали встречаться
яйца второй кладки (Суш-
кин, 1914). Существуют
указания на то, что выле-
тевших из гнед молодых сопровождает некоторое время самец, а самка присту-
пает к последующей кладке. О сроках насиживания и пребывания птенцов
в гнезде указаний нет.
Линька. В литературе существуют указания на то, что линька взрос-
лых птиц в бассейне Енисея начинается в конце июня и кончается в августе
(Сушкин, 1914). Сходные данные указываются для Приморья, где взрослые вы-
линивают к концу августа (Маак, 1861). Однако, в коллекциях нигде нет линяю-
щих взрослых птиц, и все эти сведения относятся к молодым, первые выводки
которых действительно линяют летом, или к ненормальным случаям линьки
старых. Вообще же старые птицы вместе с молодыми последних выводков ли-
няют на зимовках, как и обыкновенная горлица.
Питание. Сведения по питанию незначительны. На Сахалине в зобах
горлиц находили, главным образом, корневища диких трав. Горлицам припи-
сывается здесь порча овощей, особенно огурцов и капусты. В Приморье в зо-
бах этих птиц находили семена ячменя, чумизы, песок и немного улиток. В неко-
торых случаях горлицы кормятся животной пищей, так как у экземпляров,
добытых на отмелях рек, в зобах обнаруживалось множество ручейников.
Обычными местами кормежек служат, помимо отмелей рек, полянки, тропы,
поля, огороды.
Полевые признаки. Большая горлица имеет общий склад более массивный»
чем обыкновенная, и поэтому больше напоминает голубя. В пределах своего
Рис. 8. Большая горлица
БОЛЬШАЯ ИЛИ СТЕПНАЯ ГОРЛИЦА 55
ареала она может быть спутана только со скалистым голубем, но место обита-
ния легко позволяет различить эти два вида. По внешним признакам горлица
отличается от голубя отсутствием сизого цвета на спине и зобе и темным под-
боем крыльев. В очень редких случаях большая горлица может быть спутана
с кольчатой и короткохвостой горлицами. От первой ее отличают длинные,
острые крылья и пестрая окраска спины. От второй — этот же последний
признак и более крупная величина.
Описание. Размеры и строение. Длина крыла самцов A1) 184—198, самок
A3) 177—193, в среднем 192 и 193 мм. Длина самки A) 342, размах A) 593 мм.
Окраска. У таежной горлицы голова темносерая с легким буроватым оттенком;
задняя часть шеи — винно-красноватая, по бокам основания шеи имеется по пятну из дву-
цветных перьев — черных с серебристо-серыми вершинками; верхняя часть спины, плече-
вые, верхние кроющие крыла — бурые с рыжими краями перьев; задняя часть спины, пояс-
ница и самые внешние кроющие крыла — аспидносерые; горло — беловатое; зоб, грудь и
передняя часть брюха — розовато-рыжие с охристым оттенком; все остальное брюхо и под-
хвостье— бледносизые; маховые — аспидные, с узкими бледно-коричневыми каемками,
которые на второстепенных становятся немного пошире, подбой крыла — серый, рулевые —
темносерые, почти черные с широкими темное изыми концами, более узкими на средней паре.
Глаз оранжевый или светло-красный; голая кожа на веках — тёмнокрасная, клюв — буро-
ватый или сероватый, ноги—красные. У молодых верх головы и шея коричневые. Каймы
на перьях верха спины, плечевых и кроющих крыла — бледноржавые и уже, чем у старых.
Низ бледнее и более коричневатый. Перья зоба с бледными ржавчатыми каемками.
Туркестанская большая горлица Streptopelia orientalis meena Sykes
Columba meena. Sykes. Proc. Comm. Zool. Soc. London. 11, 1832, 149, 1832, Декан.
Синоним. Columba ferrago. Eversmann. Addenda ad eel. Pallasli, стр. 17,
Тарбагатай.
Распространение. Ареал. В пределах Союза —юго-запад Сибири и вос-
ток Средней Азии. Крайние северо-восточные точки гнездования туркестан-
ской горлицы лежат у р-на Ачинска и реки Четь (Иоганзен, 1916). Самые юго-
восточные нахождения в бассейне Енисея лежат поблизости от границ СССР
в кр. Танну-ола, у озера Усуа-нор и на р. Тесь (Тугаринов, 1916). К западу от
линии, соединяющей северо-восточные и юго-восточные места находок, горлица
отмечена на гнездовье у Мариинска и Томска (Залесский, 1915, 1921). Найдена
в Кузнецкой степи (Хахлов, 1937). Часто встречается на гнездовье в Салаирских
горах (Иоганзен, 1935, Хахлов, 1937). К северу поднимается до Нарыма (Иоган-
зен, 1907). Гнездится у Новосибирска. В большом числе гнездится вокруг Бар-
наула. Широко распространена на гнездовье по системе Алтая, где найдена в за-
падной и северо-западной частях, в южн. Алтае обычна (Сушкин, 1938). В зап.
отрогах Алтайской системы —по Большой Убе, в Калбинском Алтае гнездится
(Селевин, 1929, 1935). Встречается повсюду в предалтайских степях (Штегман,
1926). Обычна в Барабинской степи и по берегам озера Чаны (Лавров, 1927).
В долине Иртыша изредка гнездится по реке Тара в Тюменской обл. (Ушаков,
1913). У Семипалатинска многочисленна на гнездовье (Хахлов, 1928). Еще
дальше вверх по Иртышу найдена, как обычная птица по берегам Марка-куля
(Поляков, 1912—1914). Будто бы гнездится у озера Ала-куль (Финш, 1879).
В сев.-зап. углу ареала гнездится в Курганской обл. (Иоганзен, 1898, Сниги-
ревский, 1929) и доходит до Ильменского заповедника в Зауралье (экземпляры
с р. Синары, Миаса, Тургояка и Ильменского озера). Обычна на гнездовье во
всех лесных массивах Наурзумского заповедника (Михеев, 1937) и весьма
обычна в Кокчетавских горах (Штегман, 1924). В Казахском нагорье многочис-
ленна на гнездовье в горах Баян-аула, Каркалинска, Кызыл-рая. В сев.
Прибалхашье так же многочисленна у Токрау (Долгушин, 1947). Гнездится
в Тарбагатае и его предгорьях (Поляков, 1915). Населяет Джунгарский Ала-
тау. В долине р. Или не гнездится, но в предгорьях ограничивающих ее хребтов
имеется. В Терскей Ала-тау обнаружена на гнездовье во многих местах (Шаров»
56 ОТРЯД ГОЛУБИ
1908, Кашкаров, 1937). В центральном Тянь-Шане встречена в долине Чаманды
(Шнитников, 1930).
В Киргизском хребте также принадлежит к гнездящимся птицам (Спан-
генберг, 1935). Зарегистрирована в Таласском Ала-тау (Кашкаров, 1925). Изо-
билует в Чаткальском (Кашкаров, 1927) и Ферганском хребтах (Кашкаров,
1934). В Алайском хребте большая горлица также гнездится (Штольцман, 1897*
Мальчевский, 1940). Обычна на гнездовье в Туркестанском, Зеравшанском и
Гиссарском хребтах (Иванов, 1940, Даль, 1936). На зап. Памире встречается
несколько реже, но все же гнездится (Зарудный, 1926, Мекленбурцев, 1945).
Крайней западной точкой гнездования служит хребет Кугитанг (Шестоперов,
1937), пролетает у Кушки (Дементьев, 1950); залеты западной формы большой
горлицы отмечены до Чкалова, различных мест Туркмении, вплоть до о-ва
Челекен на Каспийском море (Левчук, 1906) и даже Крыма (Молчанов, 1906).
Область зимовок туркестанской большой горлицы начинается уже от южн.
Ирана. Однако здесь она зимует не регулярно. Основные зимовки находятся
в Индии, начиная от южных подножий Гималаев. К востоку идет до Бихара.
Случайно встречается зимой на Цейлоне.
Характер пребывания. Перелетная птица. Весенний пролет начи-
нается поздно—не ранее второй половины весны, когда начинает распускаться
самая поздняя растительность, и ночные температуры не опускаются до нуля.
Вместе с тем данные по пролету настолько отрывочны, что не позволяют устано-
вить общий ход поступательного движения горлиц, а лишь отмечают его отдель-
ные моменты. Наиболее ранние даты, в которые были отмечены туркестанские
большие горлицы в СССР — это 22 апреля, когда птица наблюдалась у Джу-
лека (Спангенберг, 1936) и Ташкента, т.е. уже очень далеко от южных границ
средне-азиатской части ареала (под Джулеком горлицы держались до 5 мая).
Следующая дата относится к Джунгарскому Ала-тау, где горлицы были отме-
чены 25 апреля (Кузьмина, 1945). Другие наблюдения у Репетека (Зарудный,
1896) и Пржевальска. В обоих этих местах горлицы наблюдались 1 мая; 4 мая
одиночная особь наблюдалась на Памире в верховьях р. Аличур (Мекленбур-
цев, 1946); 6 мая горлицы уже достигли Джаркента (Шестоперов, 1926). 8-го
они были отмечены у Сергиополя, а 10— у озера Ала-куль, где они будто бы
гнездились в норах глинистых обрывов (Финш, 1879). Это указание кажется
сомнительным и скорее относится к пролетным птицам.
Далее горлицы были отмечены в садах гор Нура-тау, где они наблюдались
10 и 15 мая (Мекленбурцев, 1934). 13 числа этого месяца их наблюдали в Куз-
нецкой степи, т. е. у северных границ ареала. 14 мая несомненно пролетные
птицы отмечены у Чиназа, где большая горлица не гнездится. 16 и 18 мая наблю-
дались на р. Лепсе и Джунгарском Ала-тау (Финш, 1879) и 18 мая в сев.-зап.
Кызыл-Кумах, где могли быть только пролетные птицы (Спангенберг, 1936).
Экземпляры от 15 мая имеются из Алтайского государственного заповедника,
а 20 мая один экземпляр был добыт у поселка Лебяжье (Сушкин, 1938).
7 июня залетная особь была добыта у Чкалова (Зарудный, 1888).
Весенний пролет горлиц тянется долго. На это указывают наблюдения под
Джаркентом, где первое появление зарегистрировано 6 мая и где птицы держа-
лись весь этот месяц, а последние наблюдения были сделаны только 13 июня
(Шестоперов, 1926). Возможно, что горлицы, наблюдавшиеся стайкой 22 июня
в Курчумских горах (Поляков, 1912—1914), еще не успели разбиться попарно
и представляли собой запоздалую пролетную стайку.
По сведениям еще более неполным, чем о весеннем пролете, осенний про-
лет начинается сравнительно рано и тянется долго. Наиболее ранние наблю-
дения относятся к высокогорью Памира, где горлица не гнездится, и где она
была отмечена 22 августа; 26 августа первое появление одиночных горлиц отме-
чено в долине р. Чирчик у города того же имени. 3—6 августа здесь наблюда-
лись уже небольшие пролетные стайки. Пятью днями позднее они были отме-
БОЛЬШАЯ ИЛИ СТЕПНАЯ ГОРЛИЦА 57
чены в низовьях этой реки. 12 сентября добыта под Чкаловом залетная птица
(Зарудный, 1888). 15 сентября отмечено первое появление под Чимкентом.
16 сентября добыты пролетные из южн. Кызыл-кумов. У Кушки пролетные до-
быты 11—17 сентября (Дементьев). 22 сентября была добыта под Самаркандом
молодая пролетная птица (Плеске, 1888). Этим же днем датируется наблюдение
довольно большого количества горлиц в среднем течении Кашка-Дарьи, где
они регулярно отмечались до 26 сентября. Для Чимкента наиболее поздней
датой является 27 сентября, для Ташкента 7 октября, но в Джулеке З.Х появи-
лись первые стаи (Грачев, 1939). Очень позднее для Средней Азии наблюде-
ние — 6 ноября —указывается для р. Кашка-су в р-не г. Пржевальска (Шалов,
1908). Еще позднее —между 26 ноября и 2 декабря добыта у Джамбула
(Северцов, 1873 и 1947). Таким образом, если не принимать во внимание этих
исключений, пролет больших горлиц следует признать уже закончившимся
к середине октября, так как даже на юге Средней Азии—в Гиссарском хребте —
последние наблюдения были сделаны в первых числах октября (Иванов, 1940).
Залетный экземпляр из Крыма был добыт 23 октября.
Биотоп. Лесная птица, придерживается на гнездовье древесной рас-
тительности разнообразного характера. На крайнем юге ареала в СССР — на
зап. Памире —туркестанская горлица гнездится по долине реки Шах-
Дарьи в пойменных лесах, состоящих почти целиком из высоких деревьев, ивы
с примесью тополя на более сухих местах. В Гиссарском хребте поселяется в ар-
чевниках, так же в Туркестанском хребте и зап. Тянь-Шане. Горлицы также
гнездятся в лиственных лесах, приуроченных к более низким высотам, нежели
арчевники. В большом количестве населяют лиственные леса сев. Ферганы*
состоящие из грецкого ореха, клена, яблони, кое-где ясени и тополя. В вост.
Тянь-Шане горлицы обычны в елово-пихтовых лесах, расположенных то по
крутым склонам гор, то по днищам долин. В Джунгарском Ала-тау места оби-
тания горлицы имеют такой же характер.
В Алакульской равнине гнездится в норах глинистых обрывов (Финш,
1879). Эти сведения не могут не вызвать сомнения, тем более, что дата наблю-
дения A0 мая)—ранняя для гнездовья. На берегах Марка-куля —в зарослях
лиственниц. В Казахском нагорье поселяется в сосновых борах с примесью
березы, а по небольшим горам левобережья Иртыша (Альджан и Байджан)
в долинках с уремой из осин, ив, черемухи, жимолости. В Калбинском Алтае
населяет тополевые леса с кустарным подседом. Уремы многочисленных рек—
Тобола, Ишима, Иртыша, Оби и всех их притоков, поросших то тальником, то
тополями или березой, повсюду населены туркестанской горлинкой. Селится
она и по водораздельным пространствам в сосновых борах или осиново-березо-
вых колках. У Томска поселяется в кедровых борах. По наблюдениям Суш-
кина туркестанская большая горлица на Алтае не заходит вглубь лесов, а дер-
жится по опушкам, редколесью, лиственным перелескам. В степи селится в свет-
лых борах и иногда довольствуется даже кустарниками. Таким образом, в юго-
зап. Сибири и сев. Казахстане горлица распространена в низменностях, на
Алтае она поднимается на высоту до 1500 м. Еще выше поднимается в Средней
Азии, где нижняя граница ее распространения лежит уже не в равнинах, а
поднимается в Киргизском хребте и Таласском Ала-тау до 1000 м, в Ферган-
ском хребте—до 1500л?. Верхняя граница исчисляется в Киргизском хребте вы-
сотой в 2000 м, в Зеравшанском — в 3250 м и на зап. Памире — в долине Шах-
Дарьи — высотой около 3000 м.
Численность. Стаи больших туркестанских горлиц не бывают осо-
бенно многочисленными, достигая не более, чем нескольких десятков особей.
В то же время эта горлица не может считаться малочисленной, и в некоторых
местах гнездящиеся пары располагаются на расстоянии 150—200 шагов друг
от друга. Деятельность человека не снижает заметно численности этой птицы.
После вырубки лесов она довольствуется подросшей там через некоторое время
58 ОТРЯД ГОЛУБИ
свежей порослью и только распашка лесных площадей заставляет ее бросить
привычные места. Охота на эту горлицу ведется только случайно, так как она
селится вдали от людей и держится скрытно.
Экология. Размножение. Брачные игры, предшествующие образо-
ванию пар, сходны с таковыми у обычной горлицы, с той лишь разницей, что
воркование описываемой птицы значительно более низкого тона и разносится
дальше. Яйца попадаются в гнездах даже на широте Барнаула уже около
середины мая (указанием на это служит находка подросших птенцов в конце
июня, Штегман, 1926). Около середины июня кладки отмечены для Томска и
стариц Ишима (Штегман, 1924), Заамина в Туркестанском хребте (Плеске, 1888)
и Джунгарского Ала-тау (Зарудный, 1914). 24 июня найдены у Томска (Залес-
ские, 1915), Баян-аула в сев. Прибалхашье (Долгушин, 1948). В дальних
окрестностях Джаркента 30 июня были обнаружены гнезда со свежими яйцами
(Шестоперов, 1926). 10 июля у Исфары найдена кладка свежих яиц одновременно
с сильно насиженными (Иванов, 1940). Наиболее поздние кладки известны из
хр. Киргизского, они датируются концом июля (Спангенберг, 1935). Видимо
существуют еще более поздние кладки, так как вылетевшие из гнезд молодые
еще без следа линьки попадались 6 октября. Такая разница в сроках несом-
ненно указывает на наличие у горлиц двух кладок в течение лета. Может быть
некоторые пары, начавшие гнездование особенно рано и не делающие больших
перерывов между кладками, успевают вывести птенцов и три раза.
Гнездо большой горлицы помещается на самых разнообразных деревьях.
Его можно найти высоко на ели, пихте или орехе, можно обнаружить на дереве
средней величины, вроде яблони или алычи, и, наконец, некоторые гнезда распо-
лагаются совсем низко — на кустах ивы, боярышника и даже шиповника.
В виде исключения гнезда строятся и на земле. Постройка гнезда такая же рых-
лая, иногда совершенно просвечивающая, как у обыкновенной горлицы, но
несколько крупнее (диаметр до 20 см). Материалом для гнезда служат сухие
веточки, корешки или травинки, собранные в непосредственной близости.
Как и у всех голубей, в кладке 2 яйца, несколько более крупных, нежели яйца
обыкновенной горлицы; размеры33—35 х24—25мм (Плеске, 1888). Сроки наси-
живания яиц, так же как и сроки пребывания птенцов в гнездах, для туркестан-
ской горлицы в литературе не указываются. Видимо насиживание длится около
20 дней, и несколько больший срок требуется для того, чтобы птенцы выросли.
Линька. Сведения о линьке большой туркестанской горлицы весьма
отрывочны. Залесские указывают, что ими был добыт 14 июня взрослый экземп-
ляр в свежем оперении. Поляков дает несколько противоречащие этому ука-
зания, сообщая о двух взрослых птицах, у которых линяло все мелкое оперение,
рулевые и внутренние маховые и которые были добыты 22 июня. Третий экзем-
пляр в таком же состоянии был добыт 30 июня. Все экземпляры добытые Поля-
ковым, — молодые птицы, перелинивающие во взрослый наряд. Экземпляр,
упоминаемый Залесскими, мог и не линять, а сохранить до июня свежее перо,
полученное зимой. Таким образом, фактические указания на линьку старых
птиц летом отсутствуют. В материале зоологических музеев Академии наук,
Московского университета и Среднеазиатского университета нет старых птиц,
линяющих летом. Вместе с тем осенние птицы находятся в поношенном опере-
нии, а весенние экземпляры в самом свежем пере. Все эти данные говорят о том,
что взрослые туркестанские горлицы перелинивают на зимовках.
Питание. Подобно другим голубям, туркестанская горлица кормится
большей частью на поверхности земли. Она предпочитает садиться на открытые
места, хотя не избегает и густых травянистых зарослей и иногда садится на кор-
межку на несжатые поля. В зобах горлиц, добытых в сосновых лесах, были
обнаружены, главным образом, сосновые семена. Повидимому птицы собирают
выпавшие из шишек семечки, так как они вряд ли могут доставать их из-под
чешуек своим слабым клювом. В Средней Азии во время весеннего пролета гор-
КОЛЬЧАТАЯ ГОРЛИЦА 59
лицы часто кормятся на засеянных пшеницей полях, собирая оставшиеся на по-
верхности зерна. Осенью они так же посещают поля, где собирают опавшие
зерна пшеницы и других злаков. Данные по питанию горлиц летом в горных
лесах, где они гнездятся, в литературе отсутствуют. Здесь этих птиц довольно
часто приходится видеть на деревьях Prunus Mackaleb или у кустиков Prunus
prostrata, клюющими ягоды этих растений. В зобах горлиц, добытых осенью
на пшеничных полях, кроме пшеницы найдено не малое количество зерен раз-
личных сорняков и в редких случаях улитки.
Полевые признаки и повадки. В природных условиях туркестанскую гор-
лицу можно встретить в одних и тех же местах с вяхирем, клинтухом, реже
с бурым голубем и сизаком и еще реже со скалистым голубем. Из горлиц она
встречается в соседстве, иногда даже в общих стайках, с обыкновенной горли-
цей. Природная осторожность туркестанской горлицы развита гораздо сильнее
нежели у других горлиц, встречающихся на одной с ней территории. Кормя-
щиеся на земле небольшие стайки осенне-пролетных птиц не позволяют подойти
к ним ближе сотни метров. Сидящие на отдыхе в густых вершинах деревьев
птицы иногда позволяют подойти под дерево, но стремительно срываются и уле-
тают; часто их совсем не удается заметить, даже вблизи, так как птицы сидят
не шевелясь. Во время осеннего пролета еще более осторожны. Вместе с тем про-
летные горлицы весьма привязаны к определенным местам— к отдельным груп-
пам деревьев и даже к одиночным деревьям, где они отдыхают днем и проводят
ночь. Они не бросают этих мест даже при преследовании. На дневной отдых
и особенно на ночлег в этих излюбленных местах иногда собирается до несколь-
ких десятков и даже сотен особей, но эти сборища крайне неустойчивы и разле-
таются во все стороны маленькими стайками. Большую часть суток пролетные
горлицы проводят, сидя в гуще древесных ветвей, уже одетых листвой, если дело
происходит весной, или еще не потерявших ее в осеннюю пору года. На кор-
межку вылетают ранним утром и возвращаются на отдых, начиная часов с 11.
Всю жаркую пору дня пережидают в тени дерева и вечером кормятся вторично.
На ночлег прилетают поздно, а если заметят опасность, то при густых сумерках.
Более подробные данные по экологии туркестанской горлицы нет возмож-
ности привести, так как она до настоящего времени остается еще недостаточно
изученной птицей. Голос этой формы похож на голос типичного подвида и обыч-
но передается слогами «ку-ку-ку» или—«ху-ху-ху». Первый слог произносится
птицей коротко — второй и следующие более протяжно. После нескольких
повторений следует пауза, иногда очень продолжительная. Воркование турке-
станской горлицы слышится только в брачное время.
Описание. Размеры и строение. Длина крыла самцов C5) 181—202, самок
<24) 180—198, в среднем 193 и 185 мм.
Окраска. Окрашена сходно с типичной формой. Отличия в том, что у туркестан-
ского подвида задняя часть головы, шеи и передняя часть спины окрашены в серо-коричне-
вый цвет, иногда с розовато-винным налетом; мантия, плечевые и верхние кроющие крыла —
бурые с ржавчато-охристыми каймами перьев; горло и подбородок — сливочно-беловатые;
особенно отличается окраска низа, так как брюшко и подхвостье не сизые, а чисто белые;
так же белые и те части рулевых (концы и внешнее опахало крайней пары), которые у типич-
ной формы сизые. Зоб и грудь иногда со слабым охристым налетом. Остальные части по
окраске не отличаются от типичной формы.
9. Кольчатая горлица Streptopelia decaocto Friv.
Columba risoria var. decaocto. Frivalszky. Tarsasag evkonyvei, 1834—1836, 111C),
стр. 183, Турция.
Русское название. Книжное.
Распространение. Ареал. В Европе кольчатая горлица распространена
до р-на г. Ужгорода, откуда граница идет к сев.-вост. углу Адриатического моря.
К юго-востоку от этой линии она населяет долины Тиссы и Дуная и почти
весь Балканский п-ов, кроме его самой южной части. В Азии область ее
60
ОТРЯД ГОЛУБИ
распространения охватывает южную половину Малой Азии, Сирию, Палестину,
Месопотамию, южные половины Ирана и Афганистана. Затем эта птица
населяет всю Индию, Бирму и Индо-Китай без Малакки. В Китае встречается по
всей его территории, кроме высокогорных областей, причем не идет к северу от
Пекина, Баоту и Лобнора. Из Кашгарии проникает в СССР — в долину р. Или.
К востоку от Китая населяет Корею и небольшой участок Японии в окрестно-
стях Токио, куда, повидимому, была завезена человеком.
Характер пребывания. Кольчатая горлица в Европе и Передней
Азии — оседлая птица, но в Индии отмечаются довольно значительные сезон-
ные перелеты. Кроме того, во многих местах ареала ей свойственны миграции,
имеющие характер незначительных выселений за пределы области распростра-
нения и ведущие к постепенному заселению новых участков в виде небольших
выступов основного ареала.
Карта 9. Распространение кольчатой горлицы Streptopelia decaocto
1. S. d. decaocto, 2. 5. d. xanthocyclus, 3. S. d. stoliczkae;
a — граница области распространения, б — не вполне выясненные границы распространения, в —
изолированные нахождения кольчатой горлицы.
Биотоп. Кольчатая горлица везде тесно связана с человеком, и поэтому
заселенные ею биотопы изменяются соответственно с особенностями культуры
и архитектуры различных населенных ею стран. В Европе она живет как в боль-
ших городах (Будапешт, Белград, Константинополь), так и в небольших посел-
ках. В Передней Азии населяет поселения, то лишенные зелени, то окруженные
большими садами. В Индии и Индо-Китае приурочена к легким постройкам
с вечнозеленой растительностью кругом. На севере Китая живет в оазисах среди
суровых пустынь. Наконец, у Токио обитает у маленьких крестьянских по-
строек среди садов из старых деревьев, окружающих дворы.
Подвиды и варьирующие признаки. Большая индивидуальная изменчивость кольчатой
горлицы служит причиной того, что у различных авторов нет единогласия ни в количестве
принимаемых подвидов, ни в проведении границ между ними. В монографических обработках
Рунвала и Питерса принимаются три подвида в следующих границах: 5. d. decaocto F г i-
v a 1 s z k у, 1838 — вся европейская часть ареала, Передняя Азия, Индия к северу до
Гималаев, зап. Ассам, зап. и Сев. Китай, Япония, так же и вост. Китай; 5. d. xantho-
cyclus Newman, 1906 — Бирма, вост. Ассам, Юннань, Кохинхина (Питере включает
вост. Китай ниже долины Янцзе и Фудзяня); S. d. stoliczcae Hume, 1874—Кашга-
рия, Алашань, Ганьсу. Наиболее типичны, т. е. крупны и бледны, кашгарские птицы.
Экземпляры из Алашаня и Ганьсу в общем мельче и темнее, будучи, таким образом, до
некоторой степени переходными. Некоторые особи настолько мелки и темно окрашены,
что более сходны с представителями типичной формы, а не с S. й. stoliczkae. Кроме указанных
КОЛЬЧАТАЯ ГОРЛИЦА 61
подвидов Бутурлиным A934) была описана 5. d. koreensis с ареалом, охватывающим вост.
Китай, Корею и Японию. Однако позднейшие авторы не признают этой формы. Подвиды
кольчатой горлицы отличаются друг от друга темнотой или бледностью основного цвета,
общими размерами и окраской голого кольца вокруг глаза.
Экологические различия между ними выражены в том, что одни держатся у человече-
ских поселений умеренного климата Европы. Другие популяции этого подвида населяют
поселения и оазисы Передней Азии, субтропики Китая и настоящие тропики Индии. Один
из подвидов обитает в обстановке тропиков Индо-Китая и один свойственен исключительно
пустыням внутренней Азии.
Западная кольчатая горлица Streptopella decaocto decaocto F г i v.
Распространение. Ареал. В пределах СССР ареал типичной формы
кольчатой горлицы заходит двумя чрезвычайно небольшими выступами.
Первый из этих выступов является крайним сев.-зап. концом ареала этой птицы
и захватывает незначительный участок Закарпатской Украины. Появление
здесь горлиц относится к 1944 г., когда в Ужгороде было отмечено сразу три
пары. Начиная с этого времени, количество горлиц здесь непрерывно увели-
чивается. 17 июля 1946 г. пара была найдена в Берегово (Страутман, 1947).
В 1947 г. эти птицы встречались в Севлюже и у станции Чоп (Страутман).
Второй участок Союза, где встречается типичная форма кольчатой горлицы,
так же очень невелик и захватывает самый южный выступ нашей территории
на р. Мургаб у границы с Афганистаном. Первое нахождение здесь горлиц
относится к 1942 г., когда они были обнаружены Гептнером в поселке Мор-
гуновском у Кушки. Далее горлицы были добыты в этом же месте 26 апреля
1946 г. Затем экземпляры от 27 и 28 апреля и 2 мая доставлены из аула Байрач
у Тахта-базара. Вообще, кольчатая горлица обыкновенна между Кушкой
и Тахта-базаром (Дементьев, Рустамов, Спангенберг, 1946). Наконец, залет
этой же формы отмечен один раз в низовьях р. Амура уМариинска, где она
была добыта 5 сентября 1855 г. Максимовичем (Тачановский, 1893).
Б и о т о л. В Закарпатской Украине горлицы держатся у городов по
небольшим садам из дуба, бука и «смереки». В Туркмении приурочены к куль-
турному ландшафту и держатся как в русском поселке Моргуновском, так и
в туркменском ауле Байрач.
Численность. В наших пределах кольчатая горлица немного-
численна. На Украине встречаются одиночные пары. В Туркмении много-
численнее и местами становится даже обыкновенной птицей.
Экология. Размножение. Данных по размножению кольчатой гор-
лицы в пределах СССР в литературе почти не имеется. В Туркмении у аула
Байрач в конце апреля у самцов были сильно развитые семенники и птицы
токовали (Дементьев). На Балканском п-ове подготовка к гнездованию
начинается уже в середине января, когда у Адрианополя наблюдались бурно
воркующие самцы. В Алексинасе в апреле горлицы насиживали яйца. Наконец,
в Ускюбе 22 октября еще отмечались не вполне хорошо летающие молодые
(Генглер, 1920). В Сеистане кольчатая горлица гнездится на невысоких
деревьях и реже на кустах.
Гнезда обычно располагаются открыто, чаще всего на высоте 2—3 м от
земли и представляют собой чрезвычайно легкую постройку, сложенную из
тонких, крепких, ветвистых прутиков, которых обычно бывает не больше
7—10 штук. Гнездо настолько прозрачно, что яйца всегда видны снизу. Одно из
измеренных гнезд имело следующие размеры: ширина — 145 мм, диаметр
лотка — 75 мм, высота в центре — 30 мм.
В мае и начале июня (ст. ст.) в гнездах были свежие и насиженные яйца
и птенцы до двухнедельного возраста. Кладки содержали по два яйца (в ред-
ких случаях одно) сливочно-белого цвета. Размеры яиц: G) 30—32,1 х 22,7—
24,3, в среднем 31 х 23,9 мм. В некоторых случаях около основного гнезда
было еще по несколько гнезд, свитых также рыхло и выстланных свежими
листьями джиды. В таких гнездах часто отдыхали самцы в то время, когда
62
ОТРЯД ГОЛУБИ
самки сидели на яйцах (Зарудный, 1900). Вполне развитые молодые в южной
Туркмении встречены 13—18 августа (Дементьев, 1950).
Линька. Материалы по линьке настолько незначительны, что не дают
возможности установить ее ход. Два линяющих экземпляра из Сеистана, добы-
тые между 2 и 13 июня, имеют рулевые, еще не начавшие линьку. Мелкое
перо у одного из этих экземпляров линяет на голове и шее. У этой птицы сме-
няется восьмое маховое, доросшее только до половины длины. Второй экзем-
пляр имеет отрастающее девятое маховое
и не затронутое линькой остальное опере-
ние. В Туркмении 30 июля у добытых
птиц сменились 7—10 маховых, сменяется
мелкое перо; начало линьки маховых —
в конце июня; у самца, добытого 26 ав-
густа, сменилась половина первостепен-
ных маховых (Дементьев, 1950).
Питание. В нашей литературе
никаких данных по питанию кольчатой
горлицы нет. Повидимому, она сходна
в этом отношении более всего с малой
горлицей.
Полевые признаки. Кольчатая гор-
лица —птица обычного голубиного склада.
Легко отличается от обыкновенной гор-
лицы короткими, тупыми крыльями, длин-
ным хвостом и бросающейся в глаза черной
полосой на задней стороне шеи. Кроме
того, хорошо заметна одноцветная, не
пестрая окраска верха. От голубой гор-
лицы кольчатая горлица отличается
отсутствием в окраске сизого цвета.
Описание. Размеры и строение. Длина крыла самцов F) 164—173 мм,
самок G) 160—174, в среднем 167,8 и 167,6 мм.
Окраска. Верх головы с прилегающей частью шеи — серые с винно-розовым нале-
том; поперек задней стороны шеи проходит черная серпообразная полоса, отороченная
спереди и сзади узкими беловатыми каемками; верхняя сторона коричнево- или землисто-
буланая, на надхвостье эта полоса часто бывает с серой примесью; горло беловатое; ниж-
няя сторона головы и шеи — бледные, серовато-розовые; зоб и грудь такого же цвета;
брюхо и бока—голубовато-серые, как и подмышечные; верхние кроющие крыла одного
цвета со спиной, кроме кроющих первостепенных маховых, которые голубовато-серые;
подбой крыла бледнее—беловато-серый; второстепенные маховые серые; — первостепен-
ные — буроваты с бледноохристыми концами и краями внешних опахал; рулевые, кроме
средних — серые, с беловато-серыми концами; этот беловато-серый цвет на крайней паре
становится чисто белым, а на соседней с центральной парой — серым; средняя пара — цвета
спины. Глаз, края век и ноги — красные. Клюв черный. Молодые более тусклые, без винно-
розового цвета на зобе и груди. Черный полуошейник почти не выражен. Кроющие пер-
востепенных маховых — коричневатые с более светлыми, тусклыми каемками.
Центральноазиатская кольчатая горлица Streptopelia decaocto stoliczkae Hume
Turtur stoliczkae. Hume. Stray Feathers 11. 1874, стр. 519, Кашгар.
Распространение. Ареал. В пределах Союза встречается на незна-
чительной территории, прилегающей к равнинной части Синь-цзяня. Эта
птица обнаружена вблизи от границы в г. Джаркенте и поселке Борохуд-
зир (Зарудный, 1906, Шестоперов, 1929). Крайними восточными местами ее
нахождений служат поселки: Карабулак (ШнЯгников, 1925) и Саркан (Селе-
вин, 1935). К западу она доходила до Алма-Ата (Шнитников, 1948) и Фрунзе
(Кашкаров, 1937). В 1926 г. единичная особь наблюдалась Мекленбурцевым
в Чимкенте. 1918 г. добыта им же в Ташкенте, но эта птица была с сильно
Рис. 9. Кольчатая горлица
КОЛЬЧАТАЯ ГОРЛИЦА 63
потрепанными и попачканными крыльями и хвостом и, видимо, вылетала из
клетки. Один экземпляр из Ташкента, на этикетке которого нет ни даты,
ни фамилии сборщика, находится в коллекции Зоологического музея Москов-
ского Государственного университета. По всей вероятности, входящий в наши
пределы участок ареала периодически сильно колеблется, и в годы наиболь-
шего увеличения численности горлиц отдельные особи может быть дости-
гают Чимкента и Ташкента. В настоящее время этот вид исчез или почти
исчез из Семиречья (Шнитников, 1948).
Биотоп. Азиатская кольчатая горлица так же тесно связана с куль-
турным ландшафтом, как и типичная форма. Живет в городах даже таких
сравнительно многолюдных и крупных, как Фрунзе и Алма-Ата. Здесь она
селится и по окраинам, и в центральных частях, и на шумных базарах. В посел-
ках живет лишь в случае, если они бывают достаточно велики и люди не поки-
дают их в течение всего года. Для временных поселков типа казахских
зимовок или летовок эта горлица никем не указывается. Любит она нали-
чие садов.
Численность. В пределах Союза кольчатая горлица не может
считаться многочисленной, так как встречается большей частью отдельными
парами. Впрочем, в годы сильного увеличения численности пары образуют
очень плотное население и гнездятся повсюду.
Экология. Размножение. Приступает к размножению рано.
Ее воркование начинается уже в конце февраля. Гнезда охотно закладываются
в развилинах таловых деревьев. Иногда местом для гнезда служит какой-
нибудь карниз или застреха под нависающей крышей. Гнезда обычной для
голубей формы часто устраиваются на остатках гнезд прошлого года.
Одно из измеренных гнезд имело следующие размеры: наружный диа-
метр 185—135 мм, диаметр лотка 105—75 мм, высота 105 мм (Шестоперов,
1929).
Насиживание начинается после откладки первого яйца; большей частью
в кладке два и реже одно яйцо. Размеры яиц F) 29,8-^33,7x23,5—25,8,
в среднем 31,0x24,4 мм. Насиженные яйца в гнездах были найдены уже
9 апреля, в этот же день был встречен вылупившийся птенец. 18 апреля
найдены и свежие яйца. Следующие находки свежих яиц относятся уже
к 9 и 27 мая. Из всех этих данных следует, что описываемый вид имеет не
менее 2-х кладок в лето (Шестоперов, 1929).
Линька. Бывает одна полная линька в году. Ее началом служит
выпадение десятого первостепенного махового. Далее постепенно сменяются
перья по направлению к концу крыла. Рулевые выпадают и быстро отрастают
вновь, причем начало их смены не приурочено к линьке трех крайних наибо-
лее длинных маховых, как это свойственно остальным голубям нашей фауны,
а может произойти с выпадением четвертого, пятого и даже шестого махового.
Смена мелкого пера часто не совпадает со сменой рулевых и начинается при
выпадении то седьмого или шестого, то пятого или четвертого маховых. У неко-
торых птиц к выпадению первого махового рулевые и мелкое перо уже сме-
нились. Календарные сроки линьки весьма непостоянны, и экземпляры с
одним из отсутствующих с десятого по седьмое маховых встречаются как
весной, так и осенью. В различные месяцы с февраля по декабрь можно встре-
тить птиц, линька которых заканчивается, и у которых осталось по одному-
два старых маховых, иногда при обновленных уже рулевых и мелком
пере.
Описание. Размеры и строение. Длина крыла самцов A5) 175—191 мм,
самок A8) 169—188, в среднем 179,5 и 178,6 мм.
Окраска. Имеет большое сходство с окраской типичной формы, но более бледная,
голова и шея винно-розовые с светло-серым налетом; верхняя сторона, включая кроющие
крыла (кроме кроющих первостепенных маховых)—как у типичной, но светлее; зоб, грудь
и брюхо — светловинно-розовые, на брюхе с сильной светлосерой примесью.
64
ОТРЯД ГОЛУБИ
10. Малая или египетская горлица Streptopelia senegalensis
L.
Columba senegalensis Linnaeus. Syst. Nat., изд. XII, 1766, стр. 283, Сенегал.
Русское название. Книжное.
Распространение. Ареал. Вся Африка, в Азии —Аравия, Месопотамия,
Сирия, Палестина, Малая Азия; в сев. Иране граница неясна; далее к востоку
населяет весь Афганистан, Индию, западные части Кашгарии и Советскую Сред-
нюю Азию до р. Лепсы на востоке, Аральска на севере и Ашхабада на западе.
Биотоп. Малая горлица везде связана с человеком — большей частью
с постройками. В сев. Африке обитает в садах и рощах.
Карта 10. Распространение малой горлицы Streptopelia senegalensis
7. S. s. senegalensis, 2. S. s. aequatorialis, 3. S. s. thomae, 4. 5. s. dakhlae,
5. S. s. phoenicophila, 6. S. s. aegiptiaca. 7. S. s. sudanensis, ?, 5. 5. sokotrae,
9. S. s. cambayensis, 10. S. s. ermanni;
a — граница области распространения, б — не вполне выясненная граница, в —
изолированные нахождения.
Подвиды и варьирующие признаки. 5. s. senegalensis Linnaeus, 1766 — Сенегал,
Гвинея, Золотой берег до Нигера. По некоторым данным (Бутурлин, 1934, Хартерт, 1932)
Аравия, Палестина, Малая Азия; 5. s. aequatorialis, E r 1 a n g е г, 1905 — южн. Нигерия,
Дарфул, Нардофан, Эритрея; на юг до Капа; по некоторым авторам (Питере, 1931) — Ара-
вия, Палестина, Сирия, Малая Азия; S. s. thomae Bannermann, 1931 — о-ов Томе;
5. s. dakhlae Meinertzhagen, 1928 — оазис Дакла; S. s. phoenicophila Н а г t e г t,
1916 — сев.-зап. Африка к югу от гор Атласа (оазисы Алжира и Туниса) на восток до Три-
поли; S. s. aegyptiaca Latham, 1790 — Египет к югу до Вади-хальфа; S. s. sudanensis
Sclater et Praed., 1920 — долина Нила от Вади-хальфа до Хартума; 5. s. sokotrae
О. G г a n t, 1914—о-ов Сокотра; 5. s.cambayensisGmel., 1789 — Индия к северу от Гима-
лаев, к западу до вост. Ирана и Белуджистана (Питере, 1931), а по другим авторам даже
до Месопотамии (Молиньюкс); 5. s. ermanni В о n a p., 1856 — Афганистан, Средняя Азия.
Отличия между подвидами выражены в разной окраске спины и надхвостья, в интен-
сивности общей окраски и в величине. Экологически подвиды отличаются тем, что одни дер-
жатся и гнездятся в кронах пальм и по окраинам рощ, а другие целиком связаны с построй-
ками человека.
МАЛАЯ ИЛИ ЕГИПЕТСКАЯ ГОРЛИЦА 65
Туркестанская малая горлица Streptopelia senegalensis ermanni Bon.
Turtur ermanni. Bonaparte. Compt. Rend. 43. 1856, стр. 942, Бухара.
Синоним. Peristera cambayensis centralasiae. Зарудный. Изд. Турк. О. ИРГО, XIII,
1917, стр. 99, Ташкент.
Распространение. Ареал. Самым западным нахождением в пределах
СССР является станция Безмеин, расположенная в 40 км к западу от Ашха-
бада. В Ашхабаде и Мары она появилась и стала вполне обычной, повидимому,
в 1904—1909 годах (Зарудный, 1910). В 1886 г. уМургаба птица еще не встре-
чалась (Зарудный, 1896). ВИолатани была добыта в 1891г. (Штольцман, 1892),
а в Репетеке наблюдалась в 1911 г. (Лоудон, 1913). В долине Аму-Дарьи осо-
бенно многочисленна в области среднего течения. Вниз по этой реке спускается
до Ходжейли и Нукуса (Гладков, 1932). Вверх по Пянджу поднимается до Писта-
мазара и Исла (Иванов, 1940). Указывается и для Памира, но экземпляры
оттуда в литературе не приводятся и в коллекциях также отсутствуют.
В сев. и зап. Таджикистане широко распространена (Иванов, 1940). В долине
Зеравшана многочисленна в Бухаре (Зарудный, 1896) и ряде других мест.
Поднимается по этой реке не выше кишлаков Пенджикент и Урмитан
(Даль, 1936; Иванов, 1940). К северу от Зеравшанской долины очень обычна
в г. Нура-ата и по крупным кишлакам северных подножий Нуританского хребта
(Мекленбурцев, 1934). В поселениях Кызыл-Кумов не найдена. В долине
Сыр-Дарьи это обыкновенная птица у Чиназа (Плеске, 1888). Далее вниз —
между Чиили и поселком Александровским (между г. Кзыл-Орда и ст. Кара-
узяк) многочисленна во всех подходящих местах (Спангенберг, 1936). В Фер-
гане обитает в низменной части — городах Коканд, Фергана (Штольцман,
1857), Андижан (Кашкаров, 1927), Балыкчи (сборы Северцова) и во многих
поселениях (Мальчевский, 1940). В горы, окружающие Фергану, идет невы-
соко, достигая, например, курорта Арслан-боб (Кашкаров, 1934). В долине
р. Чирчик обычна повсюду, включая г. Ташкент (Сатаева, 1937). Вверх по
Чирчику поднимается до впадения р. Угам. В долине Арыси —многочисленная
птица, может быть прослежена почти до самых истоков. Далее на восток най-
дена в поселках, разбросанных вдоль северных подножий Киргизского хребта
(Дементьев и Спангенберг, 1935). Еще далее к востоку добыта в Алма-Ата
(Иоганзен, 1909), Кашка-су (Шалов, 1908) и Карабулаке (Иоганзен, 1909).
Это место — самая восточная точка ареала малой горлицы в СССР. За пре-
делами Союза отмечена еще далее к востоку — у г. Суйдина в Китае на р. Или
(Шестоперов, 1926). Существуют указания, что в настоящее время этот вид
горлицы сократил свой ареал в Казахстане и исчез из бассейна оз. Балхаш (быв-
шего Семиречья) (Шнитников, 1948), долины Сыр-Дарьи ниже поселка Алек-
сандровского и Аральска (Спангенберг, 1936). Вертикальные границы распро-
странения малой горлицы лежат в пределах от 50 м от уровня моря (Аральск)
до высоты 1000 м (Ош, Хумсан), 1300 м (Нанай), 1400 м (Гарм), 1470 м (Гульча),
1600 м (Арслан-боб). Залетные отмечены до высоты 3000 м (Искандер-куль).
Известен еще один залет 14 B6) мая 1884 г. в г. Чкалов (Зарудный, 1888).
Характер пребывания. За редкими исключениями малая гор-
лица является вполне оседлой птицей, и не только перелеты, но даже незна-
чительные сезонные кочевки для нее не указываются. Зимой количество этих
горлиц в населенных ими поселках не уменьшается и не увеличивается.
Биотоп. Малая горлица тесно связана с человеком и совершенно ire
встречается вдали от поселений людей. Наиболее охотно она селится в кишла-
ках и городах, состоящих из построек старинного типа, которые представ-
ляют собой небольшие глинобитные сооружения с плоскими крышами. Среди
построек европейской архитектуры эта горлица поселяется не так охотно.
Видимо, некоторую роль играет скученность строений, так как в узбекских
городах, где дома расположены очень тесно, горлицы наиболее многочисленны.
5 Птицы СССР, т. II
66 ОТРЯД ГОЛУБИ
Весьма обычны они в кишлаках, причем скорее всего могут быть найдены в их
центральных частях, где постройки наиболее скучены. В казахских аулах
старого типа, где отдельные зимовки были сравнительно далеко разбросаны
одна от другой, горлицы совсем не встречались.
Численность. Малая горлица до сих пор заслуживает названия
одной из самых многочисленных птиц Средней Азии. В крупных населенных
ею пунктах она скапливается в местах кормежки обществами до нескольких
десятков. Однако теперь уже не удается наблюдать сотенных стай горлиц,
собиравшихся ранее на базарах Самарканда, Бухары и Ташкента. В последние
2—3 десятилетия количество этих птиц то резко сокращается, то вновь повы-
шается. Причиной этого служит, повидимому, охота. В то же время, благо-
даря своей незначительной величине, малая горлица не является завидной
добычей. При достатке мясных продуктов она оставляется без внимания и
количество птиц быстро восстанавливается. В настоящее время в таких горо-
дах, как Ташкент, можно встретить (в тех местах города, где сохранились узбек-
ские постройки) на кормежке или отдыхе стаи этих птиц экземпляров по 10—15.
Чаще же они попадаются обществами, состоящими из 2—3 экземпляров.
Экология. Размножение. Будучи оседлой птицей и живя в теп-
лых областях СССР с ранней весны, малая горлица очень рано гнездится. Пер-
вое воркование отмечается для Ташкента в начале февраля, а для Сталина-
бада — в январе. В солнечные теплые дни марта и апреля воркование раздается
повсюду. Воркующий самец всегда выбирает какой-нибудь сильно возвышаю-
щийся предмет — телеграфный столб, сухую торчащую вершину дерева,
трубу, конек высокой крыши и т. п. Воркуя, он сидит на месте спокойно, сильно
раздувая зоб. Время от времени самец косо взлетает, сильно хлопая крыльями,
а затем, широко распустив их, скользит вниз и усаживается на прежнее или
соседнее место. Подобное поведение свойственно самцу и до образования пары,
и когда самка уже сядет на яйца. Пары образуются очень рано; возможно,
что старые птицы не расходятся в течение зимы. Спаривание горлиц удается
видеть уже около середины февраля. Перед спариванием самец воркует, при
этом он бежит за самкой, волоча хвост и быстро поднимая и опуская перед-
нюю половину туловища. В теплую погоду воркование и спаривание бывает
уже в декабре (Ашхабад, Дементьев, 1950).
Гнездо располагается большей частью на постройке — в щелях стен,
на карнизах, под крышами и т. п. Реже гнездятся на деревьях и сплетениях
виноградных лоз. В этих случаях всегда выбираются самые густые вершины
или сплетения. Гнездо обычного голубиного типа. Оно представляет собой
небольшую плоскую кучку веточек и травинок, лишенную всякой выстилки
и просвечивающую насквозь. Пары малых горлиц обычно селятся обособленно
друг от друга, занимая отдельное дерево или постройку. Впрочем, во время
увеличения численности возможны случаи и более тесного соседства. Самец
внимательно следит за своим участком и быстро прогоняет всякую другую
горлицу, случайно залетевшую на занятую парой крышу, дерево или двор.
Кладка обычно из 2 яиц, белого цвета. Размеры B) 25,4—28,6x19,2—
22,0 мм (Спангенберг, 1936). Свежие яйца попадаются в гнездах уже в конце
февраля, на что указывает спаривание, наблюдавшееся 14 числа этого месяца.
В Чарджоу яйца были отмечены 20 марта (Зарудный, 1896). Для Самарканда
известны свежие кладки, добытые 25 мая и 6 июня. В Термезе 9 июня было
найдено гнездо с одним яйцом (Иванов, 1940). В Кзыл-Орде яйца были найдены
29 и 30 апреля, а в других местах долины Сыр-Дарья, но не ниже названного
города и не выше Чиили, они попадались все лето и осень до заморозков,
которые губят поздние кладки (Спангенберг, 1936). В этих же местах летаю-
щие птенцы отмечались 23 апреля. Однако эта находка не может служить
точным указанием на время откладки яиц, так как сроки насиживания и
пребывания птенцов в гнезде в литературе по Средней Азии не приводятся.
МАЛАЯ ИЛИ ЕГИПЕТСКАЯ ГОРЛИЦА
67
Нет также указаний на смертность птенцов, если не считать сведений о том,
что они часто выпадают из гнезд и становятся добычей кошек (Кашкаров, 1932).
По вылете молодых парочка немедленно приступает к последующей кладке,
а подросшие птенцы присоединяются к своим сверстникам или ведут одиночный
образ жизни, бродя вблизи места своего вывода. В насиживании яиц и
выкармливании птенцов участвуют и самец и самка.
Линька. Вопросы линьки малой горлицы в литературе, касающейся
Союза, не освещены совершенно. Имеющиеся в музеях материалы не дают
возможности точно проследить ход этого процесса. Создается впечатление,
что линька взрослых горлиц не приурочена к осени и вообще не имеет таких
определенных календарных сроков, как у голубей рода Columba. Самый ход
линьки также не имеет такой строгой последовательности, как у последних.
Смена пера начинается вы-
падением десятого перво-
степенного махового и идет
к концу крыла. Рулевые
начинают сменяться при
различном числе старых ма-
ховых, как и мелкое перо.
Так, например, экземпляр
от 7 июля, самка, Керки:
имеет по 4 самых длинных
маховых, еще не сменив-
шихся. 3 средних рулевых
не доросли, одно отсут-
ствует совершенно. Мелкое
перо все в линьке. Птица
от 12 сентября, самка, Таш-
кент : в каждом крыле по 2
старых маховых; рулевые
все старые, кроме двух
средних, начавших пробиваться; мелкое перо все старое. Экземпляр от 23 ав-
густа, самец, Аулие-ата: имеет по 4 старых маховых; рулевые с правой сто-
роны : 1-е и 2-е — старые, 3-го— нет, 4-е и 5-е —новые. 6-е — отрастает; с левой
стороны: 1-е, 2-е, 3-е и 5-е —новые, 4-е —старое, 6-го — нет; мелкое перо про-
изводит впечатление свежего. Экземпляр от 7 сентября, самец, Ашхабад:
по 2 крайних маховых старые; рулевые все старые; мелкое перо все линяет.
Экземпляр с той же датой и из того же места, как предыдущий, самец: по 2
крайних маховых старые, но крайние рулевые сменились, мелкое перо старое,
и только на шее пробиваются пеньки. Птица от 29 декабря, самец, Балыкчи:
шесть маховых старые; все остальное перо еще не имеет следов линьки.
Экземпляр от 9 мая, самец, Бишкент: первые маховые старые, все остальное
оперение свежее. Реже всего линяющие птицы встречаются в январе — апреле,
но и в это время некоторые особи линяют.
Питание. Пищевой режим малой горлицы в литературе по Средней
Азии освещен далеко недостаточно и ограничивается указанием на нахождение
в зобах и желудках зерен ячменя, джугары, клевера, дыни, люцерны (Заруд-
ный, 1890). У девяти горлиц, добытых осенью 1940 г., в зобах было обнаружено
много маша (культурное растение из бобовых), пшеницы и ячменя. Кроме
того, там находилось значительное количество семян сорных растений и,
в меньшей степени, зерна огородных культур (дыня, помидоры). В виде исклю-
чения попадались раковинки улиток и остатки жуков. Кормятся эти горлицы
обычно на земле, бродя по улицам, базарам и дворам. В очень большом числе еще
два-три десятка лет назад собирал йсь для кормежки на мельницах, рисовых толче-
ях и зерновых базарах. Теперь в подобных условиях собираются стаями не свы-
Рис. 10. Малая горлица
68
ОТРЯД ГОЛУБИ
ше нескольких десятков. На огородах и пашнях кормятся гораздо реже и толь-
ко в тех случаях, если они расположены непосредственно у построек. Никаких
перелетов на близлежащие поля горлицы не предпринимают и поэтому не вре-
дят сельскохозяйственным культурам. Малая горлица—наиболее доверчивый
из видов голубей нашей фауны и даже при преследовании не избегает соседства
с людьми, становясь только немного более осторожной у самых опасных мест.
Полевые признаки. Малая горлица имеет общий склад, характерный для
голубей, с той лишь разницей, что ее туловище относительно короче, а хвост
длиннее, чем у других голубей и горлиц (кроме полуошейниковой). Благодаря
тому, что птица не боязлива, подпускает на самое близкое расстояние и сво-
бодно позволяет рассматривать себя, ее определение не представляет затруд-
Карта 11. Распространение короткохвостой горлицы Oenopopelia tranquebarica
1. О. t. tranquebarica, 2. О. f. murmensis, 3. О. t. hum His.
a — границы области распространения, б — не вполне выясненные границы, в — изолиро-
ванные нахождения.
нений. Она легко узнается по своему маленькому росту, окраске без пестрин
на спине и отсутствию черной полосы на задней стороне шеи и рядов светлых
и черных пятнышек по ее бокам. На лету малую горлицу сразу выдают ее
короткие, округлые крылья и длинный хвост. От похожей на нее по этим при-
знакам ошейниковой горлицы, малая горлица отличается (кроме меньшей
величины) более темной окраской.
Описание. Размеры и строение. Длина самцов D) 275—288 мм, в среднем
281.5 мм, самок E) 265—285, в среднем 276 мм. Размах самцов E) 410—430, в среднем
421.6 мм. Крыло самцов C4) 128—143,5, в среднем 134,8 мм, самок B0) 124—142, в сред-
нем 132,3 мм. Вес самцов E) 104—121, в среднем 110,4, самок E) 95—119, в среднем 108,8г.
Окраска. Оба пола окрашены одинаково. Верх и бока головы и шеи — сизовато
винно-розовые; наибольшей густоты этот цвет достигает на верхних частях, на боках он
бледнее, нижняя сторона шеи и зоб того же, но еще менее насыщенного цвета, подборо-
док и горло беловатые, на боках шеи по большому пятну. Каждое перо этого пятна имеет
раздвоенную вершинку ржавчато-винного цвета, из-под которого заметны черные основания
перьев, так что все пятно кажется пестрым; весь верх — спина, плечевые, поясница, над-
хвостье и верхние кроющие хвоста — однообразного землисто-бурого цвета; кроющие кры-
ла— голубовато-сизые, кроме кроющих первостепенных маховых, которые черны; брюхо и
подхвостье — белые, низ крыльев и подмышечные — серые; маховые буровато-черные в све-
КОРОТКОХВОСТАЯ ГОРЛИЦА
69
жемпере, сильно буреющие при выцветании; на внешних опахалах нескольких первостепен-
ных — узенькие, беловатые каемки; рулевые (кроме средних) — темносерые; на трех край-
них парах — широкие белые вершины, отделенные от серого основания черной полосой;
на втором и особенно первом рулевом эта полоса часто вытесняет основной серый цвет;
на третьем черная полоса менее развита и белая вершинная часть часто сильно сереет; чет-
вертая пара почти теряет белую вершину, сохраняя черную полосу; пятая имеет только
следы черной полосы и землисто-бурую примесь в окраске; средняя пара — одноцветная —
землисто-бурая, как спина, но с сероватым налетом. Глаз — белый, с темнобурым краем
радужины. Клюв — черный. Ноги малиновые или малиново-красные. Молодые окрашены
более тускло, без заметного винно-розового цвета головы, шеи и зоба. На шее нет рыже-
вато-винных перьев с черными основаниями. Перья мантии с узкими белесоватыми каемками.
РОД КОРОТКОХВОСТЫЕ ГОРЛИЦЫ OENOPOPELIA BLANFORD, 1898
11. Короткохвостая горлица Oenopopelia tranquebarica Herman
Columba tranquebarica. Herman. Observ. Zool. 1804, стр. 200. Индия.
Распространение. Ареал. Юго-восток Азии; ареал охватывает всю
Индию до Инда и Кашмира, Бирму, Индо-Китай (без Малакки). Населяет
о-ва Хайнан, Формозу и сев. Филиппины. В Китае занимает бассейны рек
Голубой и Желтой (кроме
северного выступа ее боль-
шой излучины). Известны
залеты в Афганистан (Дже-
лалабад), северную часть
Борнео, Японию (Иока-
гама), о-в Аскольд, Уссу-
рийский край, Забайкалье
и Манчжурию.
Биотоп. Указания
на необходимые для этого
вида условия существова-
ния довольно неопреде-
ленны. Часто короткохво-
стых горлиц наблюдали и
в добывали у поселков.
Гнезда большей частью об-
наруживают на деревьях в
садах и рощах у поселений.
Подвиды и варьирующие
признаки. О. t. tranquebarica
Herman, 1804 — Индия к западу до Пенджаба и Синда. Однажды добыта в Афгани-
стане (Джелалабад); к северу идет до Гильгита и Гималаев, на востоке — до Бенгалии,
Бихара и Зап. Непала; О. t. murmensis H a r t e r t, 1915 — Вост. Непал, Сикким и зап.
Ассам к северу до Брамапутры; О. t. humilis T e m m i n с k, 1824 — Бенгалия, вост.
Ассам, Бирма, Индо-Китай, бассейны рек Голубой и Желтой, о-ва Хайнан, Формоза и
северная часть Филиппинского архипелага. Подвиды отличаются друг от друга, главным
образом, окраской нижних кроющих крыла и подмышечных. Кроме того, есть различия
в размерах.
Рис. 11. Короткохвостая горлица
Китайская короткохвостая горлица Oenopopelia tranquebarica humilis
Те mm.
Columba humilis. Temminck. PI. col. 1824, табл. 259, Бенгалия.
Распространение. Ареал. В пределах СССР известны крайне немного-
численные случаи нахождений короткохвостой горлицы. Первая находка
относится к июню 1856 г., когда один экземпляр был добыт Валецким у Нер-
чинского завода. Второй раз эта горлица была добыта на о-ве Аскольд осенью
70 ОТРЯД ГОЛУБИ
1876 г. Янковским. В следующем году на этом же острове наблюдалась небольшая
стайка, и еще 9 особей отмечено здесь 1 октября того же года. Один экземпляр
доставлен известным путешественником по Уссурийскому краю Арсеньевым.
Этот экземпляр был добыт им 5 A8) октября 1908 г. на Уссурийском побережье
между устьями рек Касьга и Луговая. Наконец, взрослый самец добыт в сен-
тябре 1948 г. под Хабаровском К. А. Воробьевым (коллекция Зоологиче-
ского музея Московского университета).
Биотоп. Один из добытых у нас экземпляров держался в лесу. Второй,
видимо, невдалеке от поселка. За пределами Союза часто держится у посел-
ков, в садах.
Экология. В пределах СССР этот вид появляется только случайно, и
поэтому данные по его экологии приходится брать из источников, относящихся
к другим частям ареала. Во внутренней Монголии и Китае короткохвостая
горлица гнездится обычно на деревьях. Гнезда устраивает скрытно, но, будучи
пугливой, легко обнаруживает их, слетая при приближении человека.
Гнезда этого вида по своей внешности представляют собой обычную голу-
биную постройку. Яиц в кладке два. Их цвет бывает белым или кремовым
(цвета слоновой кости). Размеры B яйца) от 20,5 до 20,7 мм ширины и от 25,3
до 27,5 мм длины (Латуш). По сведениям других авторов размеры серии яиц
имели от 19,8 до 21,5 мм ширины и от 24,9 до 28,5 мм длины (Хартерт, 1915).
Гнездится в июне и июле. Осенью в северных областях ареала наблюдаются
большие стаи пролетных горлиц. Вместе с тем попадаются и одиночные птицы.
Линька. Повидимому, популяция, приуроченная к определенной области,
линяет одновременно. Из десяти горлиц, добытых в Цайдаме и сев. Ганьсу
в сентябре, нет ни одной не линяющей птицы. Большинство этих птиц имеет
по 3—4 еще не сменившихся маховых. Только по одной птице из этой серии
имеет по 6 и 5 старых первостепенных. Рулевые и мелкое перо не линяют ни
у одной.
Полевые признаки. Короткохвостая горлица имеет склад настоящего
голубя, но легко узнается по своему очень маленькому росту. Кроме того,
самцов этого вида сразу выдает своеобразный красно-коричневый цвет верх-
ней стороны. Голос этой горлицы — глубокое хрюкающее короткое «у».
Описание. Размеры и строение. Крыло самцов G) 132—137, в среднем
134,6 мм, самок F) 129—137, в среднем 132 мм.
Окраска. Короткохвостая горлица отличается от других видов голубей нали-
чием полового диморфизма в окраске. Голова взрослого самца и шея — пепельно-серые;
спина, плечевые, верхние кроющие крыла, зоб, грудь и брюхо — коричнево-винно-красные,
в свежем пере с сизоватым налетом; на нижних частях тела основной цвет бледнее, чем на
верхних; между серым цветом шеи и красноватым спины проходит черная полу лунная
полоса; низ спины, надхвостье и верхние кроющие хвоста — серые; подхвостье — бледно-
серое или беловатое; бока, нижние кроющие крыла и подмышечные — светлосерые; махо-
вые — темнокоричневые, рулевые — аспидно-черные с белыми концами у крайней пары; эти
концы по мере приближения к центру хвоста становятся все более серыми; средняя пара
рулевых — серовато-коричневая.
Взрослая самка отличается от самца тем, что коричнево-красный цвет верха заменен
у нее землисто-бурым; на груди выступает розовато-серый оттенок; серый цвет головы на
лбу светлеет, черное полукольцо шеи с серовато-белым узким окаймлением. Молодые птицы
похожи на самок, но черное полукольцо шеи развито слабее; перья груди, спины, верхние
кроющие крыла, плечевые и маховые — с бледноохристыми каемками. Глаз—бурый,
клюв— черноватый, ноги — буровато-красные.
ОТРЯД РЯБКИ
PTEROCLETES или PTEROCLETIFORMES
Характеристика отряда
Строение. Внешние признаки. Птицы средней величины,
с небольшого голубя. Средний вес около 300—500 г. Клюв короткий, с узкими
щелевидными ноздрями, прикрытыми кожистой складкой. Голова небольшая,
шея короткая, туловище плотное, с сильно развитыми грудными мыщцами.
Крылья длинные и острые, хвост ступенчатый и часто заостренный. Ноги
короткие, оперенные до пальцев. Задний палец слабо развит или отсут-
ствует.
Оперение плотное и густое, но сидит в коже слабо, как у голубей, особенно
на спинной стороне. Окраска характерного «пустынного» типа, с большой
примесью желтоватых, песчаных, охристых тонов. Возрастные изменения
окраски незначительны; самцы окрашены ярче самок. Маховые и рулевые
жесткие, число первостепенных маховых 10 (кроме того, скрытое под крою-
щими крыла «абортивное» маховое, в сущности первое); рулевых обычно
16, иногда 14.
Анатомические признаки. Череп схизогнатический и голо-
ринальный; сошника нет; рострум парасфеноида сильно вытянут, небные
кости разделены; имеются базиптеригоидные отростки. Скелет сильно пнев-
матизирован.
Зоб большой, но без характерных для голубей выступов; сильно разви-
тый мускульный желудок, длинные слепые кишки; большой желчный пузырь.
Нижняя гортань с симметричными стенотрахеальными мышцами и двумя
парами трахеобронхиальных мускулов; кольца трахеи хрящевые.
Перья со слабо развитым побочным стволом. Копчиковая железа голая
и рудиментарная. Пуховые части перьев отделяют «пудру». Пух расположен
и по аптериям. Крыло аквинтокубитального типа.
Экология. Рябки — дневные наземные птицы открытых сухих ландшаф-
тов, оседлые и полуоседлые, за исключением самых северных популяций
в Азии, которые перелетны.
72 отряд рябки
Один вид — обыкновенная саджа — совершает временами огромными
стаями за пределы гнездовой области вылеты, связанные с особо неблагопри-
ятной фенологической обстановкой.
Полет рябков очень быстрый и стремительный, напоминающий голубиный.
Они хорошо ходят и бегают мелкими шажками. Характерно, что рябки в
известной мере сохраняют «стайность» и в гнездовое время. Гнездовья часто
расположены неплотными, но обширными колониями. Перелеты на водопой
и т. п. также нередко совершаются стаями и в период размножения. В поведе-
нии очень развиты подражательные реакции, например, при взлете с гнезда
одной потревоженной чем-либо птицы, снимаются с гнезд и соседние.
В холодное время года рябки ведут стайный образ жизни.
Питаются рябки, главным образом, семенами различного рода пустынных
и степных растений, подбираемыми с земли. Повидимому, иногда они разы-
скивают пищу, разрывая по-куриному почву. Охотно купаются в пыли.
Рябки регулярно пьют воду и совершают довольно далекие перелеты
на водопой — к рекам, пустынным колодцам, выходам родников. Так как
старые птицы вынуждены поить нелетных птенцов, принося им воду в зобе,
то, естественно, они должны гнездиться не слишком далеко от водопоев. Во
всяком случае, в глубине безводных участков пустынь рябки не встречаются.
Молодые выкармливаются взвесью мелких семян в воде — из зоба роди-
телей.
Гнездятся рябки относительно поздно, гнезд, в сущности, не устраивают,
откладывая яйца в ямку на земле, иногда слегка выстланную сухими стеблями
и т. п. Кладки состоят из 2—4 пестрых яиц. Насиживание начинается
с первого яйца, при участии обоих полов. Срок насиживания около месяца.
Развитие птенцов —по выводковому, в отличие от голубей, типу. Постэмбрио-
нальное развитие быстрое.
Неясным представляется вопрос, имеют ли рябки в норме одну или две
кладки в году или у них часто бывают дополнительные кладки взамен уте-
рянных, что должно быть нередким при открытом гнездовании. Период раз-
множения рябков очень растянут, что приводит к большой разновозрастности
птенцов (в особенности при насиживании с откладки первого яйца), а послед-
няя может вызвать разновременность наступления половой зрелости различ-
ных возрастных групп и — как следствие этого — различия в начале периода
размножения у них.
Линька у большинства видов бывает раз в году, полная. У некоторых,
например, у белобрюхого рябка, имеется еще и частичная предбрачная линька
мелкого оперения. Вопрос о возрастных изменениях наряда рябков не вполне
ясен: после пухового наряда надевается первый птенцовый, быстро сменяемый
вторым птенцовым, но быть может только частично, а затем первым годовым,
в котором мелкое оперение как у взрослых, но маховые и рулевые остаются
от птенцового наряда. Только с первого годового наряда проявляется половой
диморфизм в окраске, вообще говоря, довольно резкий.
Схема развития окраски такова: оба пола окрашены в окончательном
оперении различно; возрастные изменения отражают различия между первым
и вторым перьевым (гнездовым) нарядом и последующими, не считая пухового
наряда; некоторые виды географически мономорфны, другие проявляют неко-
торые, обычно незначительные, вариации окраски в разных частях ареала;
группового диморфизма не имеется.
Систематика. Одно семейство Pterocletidae из двух родов: Pterocles с
широким распространением в южн. Европе, Африке, на Мадагаскаре, в Перед-
ней, Центральной и ю-з. Азии; Syrrhaptes—в Средней и Центральной Азии.
Второй род отличается от первого главным образом отсутствием заднего пальца
ноги и сращением передних пальцев. Число видов рода Pterocles — 14, Syr-
rhaptes —. 2.
ЧЕРНОБРЮХИЙ РЯБОК 73
Ископаемые рябки известны с олигоцена. Плейстоценовые остатки саджей
найдены в Венгрии и в Ордосе в ю.-в. Монголии.
В систематическом отношении рябки ближе всего стоят к голубям.
Географическое распространение. Рябки — обитатели открытых и сухих
ландшафтов Восточного полушария. Больше всего видов — в Африке (вероят-
ный центр распространения, связанный, судя по современным ареалам рябков
в Африке и Азии, с возникновением среднетретичных пустынь). Общее число
видов в фауне СССР — четыре, из них два монотипических. Процент числа
видов рябков в фауне СССР к общему числу видов в отряде —25%.
Распределение рябков по жизненным зонам в фауне СССР однообразно:
все виды и расы относятся к зоне открытых сухих ландшафтов, к подзоне
полупустынь и пустынь.
Как уже указывалось, по характеру пребывания эти птицы в пределах
СССР либо оседлы, либо кочуют, либо —у северных границ ареала—перелетны.
Практическое значение. При относительно ограниченном распространении
рябки имеют небольшое значение в качестве охотничьих птиц (мясо их вполне
доброкачественно). Местами (Туркмения) на осеннем пролете белобрюхие
рябки появляются к количестве, имеющем реально практическую ценность.
Бутурлин A934) указывал, что массовые гнездовья этих птиц в пустыне могут
иметь известное положительное значение в отношении внесения в почву высоко
эффективных удобрений.
Таблица для определения видов рябок
1 B) Заднего пальца нет; задняя сторона цевки оперена 3
2 A) Задний палец имеется (хотя слабо развит); цевка сзади голая 5
3 D) Первостепенные маховые серые, подмышечные белые или беловатые; концы зад-
них первостепенных маховых доходят до вершины 7-го махового
обыкновенная саджа Syrrhaptes paradoxus
4 C) Первостепенные маховые черноваты, подмышечные черно-бурые; концы задних
первостепенных маховых доходят до вершины 4-го махового
тибетская саджа Syrrhaptes tibetanus
5 F) Брюхо белое, без пестрин; крыло короче 220 мм
белобрюхий рябок Pterocles alchata
6 E) Брюхо не белое, черное или темнобурое; крыло длинней 220 мм
чернобрюхий рябок Pterocles orientalis
РОД РЯБКИ PTEROCLES TEMMINCK, 1815
Тип P. alchata
1. Чернобрюхий рябок Pterocles orientalis L.
Tetrao orientalis. Lin пае us. Syst. Nat. ed. X, v. 1, 1758, стр. 161, M. Азия.
Распространение. Ареал. Вост. Канарские о-ва; Пиренейский п-ов
и с.-з. Африка; Кипр, Палестина; М. Азия и Закавказье; южн. Заволжье;
Туркестан; зап. Синьцзянь; Иран и Афганистан. В холодное время года до
Месопотамии и с.-з. Индии.
Характер пребывания. Оседлый и кочующий, на севере
перелетный.
Биотоп. Пустыни и полупустыни, главным образом равнинные,
и предгорья.
Подвиды и варьирующие признаки. В западной части ареала — в Африке, Южн.
Европе, М. Азии и Закавказье, в Палестине — Р. о. orientalis Linn., 1758; на востоке — в
Туркестане, Иране, Афганистане — Р. о. arenarius Pall., 1775; различия незначительны
и сводятся к особенностям окраски самок — быть может эти формы не должны разделяться
и не представляют собой географические рассы в строгом смысле слова.
74
ОТРЯД РЯБКИ
Восточный чернобрюхий рябок Pterocles orientalis arenarius Pal.
Tetrao arenarius. Pallas. Novi Commentarii Acad. Caesar. Petropolitanae, XIX, 1775,
стр. 418, табл. VIII, степи Заволжья.
Синонимы. Pterocles orientalis var. magna Za roudnoi. Bull. Soc. Imp. Natu-
ralistes, Moscou, 1889 A890), стр. 811. Пендинский оазис и низовья Кушки,— Pterocles orien-
talis Kozlovae. Meinertzhagen. Bull. Brit. Orn. Club, L., V, 1934, стр. 59, Эмба.
Русское название. Чернобрюхий рябок — книжное. У русских охотников
в Туркмении — тураль. У народов востока —карабаур, карабаур-бульдерюк (-чернобрюхий
рябок), у туркмен еще багыртлак или кара-багыртлак.
Распространение. Ареал. Заволжье (Рын-пески), Кумо-Манычские степи
(Богданов, 1879), с.-з. Казахстан примерно до 48°30'—49°30' с. ш., восточней
до Зайсанской котловины и южных предгорий Алтая, в центральном Казах-
Карта 12. Распространение чернобрюхого рябка Pterocles orientalis
1. Р. о. orientalis, 2. Р. о. arenarius;
a — границы гнездовий, г — область зимовок.
стане около 47° с. ш., на юг до долины реки Кран в верховьях Черного Иртыша
(Сушкин, 1938) и с.-з. Синьцзяня (Уч-турфан, Урумчи, Аксу), Тянь-Шаня и
ю.-з. Таджикистана, Туркмении, Ирана (к югу до Загроша на западе, но там,
быть может, и номинальная раса; на востоке до Саргада), Афганистана и Белу-
джистана, быть может в Синде (где добыт 20 мая у Хайдерабада Бернсом,
Тайсхерт, 1923, но редок здесь и зимою). В холодное время года на юг до Месо-
потамии и с.-з. Индии (Пенджаб, Синд, Биканир в с.-з. Раджпутане; Куч;
редко до Катиавара и Бопала, т. е. лочти до тропиков; к востоку до Непала,
Лукноу, Аллахабада). Детали распространения в Туркестане таковы: на
р. Урал к северу до Бударминской станицы (Северцов, по Сушкину, 1908,
но повидимому только залеты), в верховьях Сагиза и у Улу-чима (Зарудный,
1897), на Иргизе от Кумсая между Карабутаком и Иргизом к югу, далее
в низовьях Тургая (Сушкин, 1908), обычен в песках Б. Барсуки; в Приараль-
ских Кара-Кумах, на о-вах Аральского моря (Барса-кельмес), далее на
восток в пустыне Бек пак дала (Селевин, 1935) и по реке Чу (Гладков и Грин-
берг, 1931; Долгушин, 1939), по Сары-Су и Каракенгиру до 47° (Слудский
и Афанасьев, 1947) в Муюн-Кумах, у южн. Балхаша (Или), в сыртах цен-
трального Тянь-Шаня (у Джаркента, в долине Музарта, у Койсара и т. д.,
даже у Хан-тенгри, Леннберг, 1905); на северо-востоке —в Зайсанской котло-
вине, в степях Тарбагатайской системы (Финш, 1879), в Барлыке (Хахлов,
1926) и в южных предгорьях Алтая у Теректы (Поляков и Велижанин, 1909;
ЧЕРНОБРЮХИЙ РЯБОК 75
быть может даже у Ульбинской, по Никольскому, 1882, что, повидимому,
сомнительно); далее к западу южная граница поднимается на Север и огибает
зап. Тянь-Шань по предгорьям Киргизского хребта и у Нура-тау; не встре-
чен в Фергане восточней Джелалабада и Намангана и на Памире, в Таджики-
стане гнездится в низинах северной и юго-западной части страны — на востоке
до Куляба, обычен в степях по верхней Аму-Дарье, в низовьях Кафирнигана
и Вахша, в долине Сурхана; в Гиссарской долине редок (Иванов, 1940); далее
к западу встречен в Туркмении от Кара-Богаза и Усть-Урта до приатрекских
пустынь и предгорий Копет-Дага, в предгорьях Кугитанга (Гуреев, 1937),
но в песчаной степи области предгорий Паропамиза в Бадхызе и Карабиле
повидимому не гнездится (хотя встречен весной 1946 г. у Навруз-абада на
Герри-руде, у Беюк-абайчешме в Гязь-гедыке, на Кушке за Кала-и-мором,
Дементьев, 1950).
Характер пребывания. На севере ареала чернобрюхий
рябок перелетный; на юге, начиная с Таджикистана и Туркмении — оседлый,
хотя, быть может, там часть популяций отлетает. В Кызыл-Кумах некоторые
особи уже зимуют в большем числе в относительно теплые года (Зарудный,
1915). В Таджикистане зимою в небольшом числе (Иванов, 1940, 1945). Зи-
мовки в Туркмении вполне регулярны—как на Аму-Дарье, Мургабе, Тед-
жене, так и в подгорной полосе Копет-Дага и у Каспийского побережья.
Однако и здесь замечаются местные передвижения, рябки в холодное время
года сосредоточиваются в большом числе по окраинам культурного ланд-
шафта. Залеты этого рябка указаны для Семипалатинска (Хахлов, 1928),
к низовьям Дона (Бостанжогло, 1911), к Астрахани, в низовьях Урала, к
Чкалову (Зарудный) и Орску (Назаров, 1886), будто бы поздней осенью «по
снегу»в Рязанской обл. (Хомяков, 1901), к Тобольску A0. IV. 1924г., Тарунин).
Даты. Прилет относительно ранний, отлет поздний и постепенный,
поздней, чем у белобрюхого рябка. Весенний прилет небольшими стайками.
На месопотамских зимовках отлет заканчивается к апрелю B7 апреля, р.
Карун, Тайсхерст, 1922). Пролет и прилет в Туркмении, повидимому, уже
в марте (8—9 марта, Радде и Вальтер, 1889; 12 марта — 1 апреля, низовья
Атрека, Мешеди-Мессерианская пустыня; 14 апреля у Чаача —стаи и пары,
Дементьев, 1950). Быть может к началу прилета относится наблюдение ряб-
ков 16 февраля на Вахше в Таджикистане (Иванов, 1945). На севере ареала
на Или прилет 23 марта, много стай в степях 30 марта (Шестопе-
ров, 1929), в низовьях Сыр-Дарьи прилет с 23 марта (Кара-узяк, Спан-
генберг, 1936) и до конца месяца, одиночками и небольшими стаями. Прилет
на Эмбу и Кокджиду—12 апреля, в низовьях Иргиза—13 апреля (Суш-
кин, 1908). Стаенье осенью, в августе, птицы, повидимому, после этого
медленно начинают подвигаться к югу, в сентябре —октябре заметный пролет
в зап. Туркмении через Узбой к Атреку. На Или рябки в стаях бывают в сентя-
бре —октябре, в ноябре их уже нет; последние рябки встречались на Сыр-Дарье
у Джулека в начале ноября (Спангенберг, 1936). На месопотамских зимовках
рябки появляются с середины ноября, в Индии в Пенджабе — в ноябре, редко
в последних числах октября, остаются они до марта (Уистлер, 1922).
Биотоп. В отличие от белобрюхого рябка, менее связан с пустынями,
охотно держится в пустынно-степном и степном ландшафте; отличие и в том,
что чернобрюхий рябок поднимается в предгорья, а в Тарбагатае и особенно
в центральном Тянь-Шане встречается, повидимому, и по горным степям.
Типичный биотоп — ровные или слегка холмистые площади с плотной гли-
нистой почвой, поросшие полынью и злаками; участки песков, перемежаю-
щиеся с глинистыми, более или менее густо покрытыми растительностью;
щебнистые участки предгорий (с.-з. Кара-тау, Спангенберг; долины рек Дюшам-
бе и Ях-су, Иванов); глинистые прибрежные участки Каспия с солянковой
растительностью; окраины культурного ландшафта. Так как чернобрюхий
76 ОТРЯД РЯБКИ
рябок, как и другие виды отряда, регулярно нуждается в воде, то распро-
странение его в той или иной мере связано с близостью речных долин, колод-
цами, выходами родников в предгорьях и т. п. На обширных безводных пло-
щадях центральных частей Кара-Кумов и Кызыл-Кумов этой птицы нет.
В вертикальном отношении по южным предгорьям Алтая отмечен до 400 м
(Сушкин, 1938), в Таджикистане до 800 мм (Каррузерс, 1910).
Численность. Как и другие рябки, распространен довольно
спорадично, но обычен в подходящих местах по Чу, в Кызыл-Кумах и Кара-
Кумах. Относительная численность в разных биотопах по среднему течению
Зеравшана такова: на 1 гектар в глинистой пустыне —0,33, в каменистой —
0,12, в ковыльной степи—0,01 особи (Даль, 1937). На зимовке в Месопотамии
многочисленен.
Экология. Размножение. Чернобрюхий рябок бывает в парах
уже с прилета, в марте. Однако период размножения начинается относительно
поздно, в середине апреля A4. IV. Чаача в южн. Туркмении—начало развития
половых органов у самцов. Тогда же происходит брачный полет. Самец с кри-
ком летает за самкой, когда последняя садится — ходит вокруг нее по земле
и т. п.). Полные кладки, повидимому, только в начале мая. Гнезд собственно
нет. Яйца откладываются на площадку на глинистой почве, обозначенную
по краям сухими былинками или стеблями, чаще в тени кустика полыни и
подобных растений (Туркмения, Сыр-Дарья).
Весь период кладки весьма растянут, это объясняется, вероятно, или тем,
что рябки нормально имеют две кладки в году, или тем, что часто бывают
дополнительные кладки взамен утраченных, или, наконец, тем, что молодые
птицы разного возраста приступают к размножению разновременно; воз-
можно, что действуют все эти причины. В Туркмении, например, полные
кладки найдены 24 мая (Чаача, Дементьев, 1950) и даже 17 мая (Карабент),
но, с другой стороны, из 11 добытых 20 июля самок у 5 в яйцеводе находились
еще яйца (Сары-язы, Зарудный, 1896). Насиженная кладка на Или найдена
3 июня (Шестоперов, 1929). В Кызыл-Кумах 14 июня —свежие кладки и
птенцы, 10 августа из 27 убитых самок у 20 кладка закончена давно, у 5 —
только что, у 2 в яйцеводах зрелые яйца (Зарудный, 1915). В низовьях
Сыр-Дарьи размножение начинается с первой трети мая, а кончается—в на-
чале августа; по опросным данным, поздние кладки еще до середины июля;
слабо насиженные кладки найдены 20 мая и 20 июня (Спангенберг, 1936).
В Таджикистане насиженная кладка найдена 15 июня, но 14 июня добыта
самка, не закончившая кладку (Сурхан, Иванов, 1940). В сев. Казахстане
в Таупе у самки 15 июня развитые яичники; кладки на Эмбе, конечно,
запоздалые, будто бы даже в середине сентября (Сушкин, 1908). В Афганистане
кладка в конце апреля —начале мая (Бекер, 1928).
Число яиц в кладке обычно 3, реже 2; число птенцов в выводке чаще 2
(Зарудный, 1896). Яйца вытянутой эллипсоидной формы, не заостряющиеся
на конце, блестящие; фон окраски скорлупы от светлосерого с слегка зеле-
новатым оттенком до светлооливкового; поверхностные пятна скорлупы серые
с коричневым налетом, внутренние слабо выражены, свинцово-серые. Раз-
меры: (8) 44,1—48,8x31,1—33,7 мм (Сыр-Дарья, Спангенберг, 1936); C)
41,6, 43,9, 44,6x30,7, 32,3, 32,4 мм (Или, Шестоперов, 1929).
Насиживанье, как у других рябков, с первого яйца (что повидимому
связано и с необходимостью защиты их от солнечных лучей), насиживают
оба родителя. Во второй части периода насиживания старые птицы сидят
крепко и отводят от гнезда. Срок насиживания — немного менее месяца (точно
не прослежен), птенцы появляются с середины июня (Кызыл-Кумы, Туркме-
ния) и позднее. Подлетки — в июле, хорошо летные молодые в начале августа.
От запоздалых (вторых) кладок выводки, разумеется, позднее. Отмечены
случаи «негнездования*, затухания начавшегося цикла размножения в случае
ЧЕРНОБРЮХИЙ РЯБОК
77
наступления особо неблагоприятных условий; у Кара-Богаза в 1939 г. при
необычайно засушливой весне у добытых самок установлена дегенерация
уже начавших развиваться яиц (Исаков).
Линька. Раз в году, полная. В соответствии с растянутостью периода
размножения, протекает у разных особей или групп особей (возрастных)
разновременно. Крупные перья (маховые) начинают сменяться у самцов с
последней трети мая, в середине июня смена маховых проходит на 40%, в
начале августа — на 70%; в октябре старые птицы в свежем пере. Общая
продолжительность процесса, таким образом, около 5 месяцев (у особи). У самок
линька наступает повидимому несколько позднее, чем у самцов. Последо-
Рис. 12. Чернобрюхий рябок (самец и самка)
вательность смены маховых — от заднего края партии к переднему, т. е.
от 10-го к 1-му (считая «видимые» первостепенные маховые). Ювенальный
наряд сменяется осенью лишь частично (мелкое перо, иногда, впрочем, не
полностью; у некоторых также 2—3 задних первостепенных маховых) — в
августе и поздней (в зависимости от даты вылупления). Последовательность
смены нарядов: пуховой — первый птенцовый — второй птенцовый (?) —
первый годовой (комбинированный, с птенцовыми маховыми или хотя бы
большинством из них) — второй годовой и т. д.
Питание. Пищу чернобрюхого рябка составляют семена и побеги
степных и пустынных растений — полыней, солянок, верблюжьей колючки
и т. д. и в небольшом числе — насекомые. Регулярно пьют даже солоноватую
воду, слетаясь на водопой по утрам, а также перед вечерним закатом. В начале
лета эти птицы летят на водопой парами или одиночками, позже стайками,
с характерным криком. Старые птицы поят птенцов, отрыгивая воду; в желудке
добытых на водопое находили до «чашки» воды (Назаров, 1886). Сообщения
разных авторов (в частности, Мид-Уолдо, 1922) о том, что рябки поят птенцов
из смоченных на водопое грудных перьев, не подтверждаются.
Полевые признаки. Типичный рябок—голова «голубиная», окраска«пустын-
ного» типа на спинной стороне, с черным, бросающимся в глаза на полете
брюхом. Хвост короткий, ноги относительно длинные. Хорошо передвигается
78 отряд рябки
по земле, держась при этом более или менее горизонтально. Охотно затаивается.
Крылья длинные и острые. Полет быстрый, но медленней, чем у саджей, голос
«тчурр-тчурр», резко отличный от других видов этого отряда, живущих в СССР.
Описание. Размеры и строение. Клюв «голубиного» типа, ноздри узкие
щелевидные, с сильно развитой кожистой лопастью (предохранение от песка и пыли). Крылья
длинные и острые, формула: 1>2>3 (не считая скрытого под кроющими «абортивного» махо-
вого, в сущности первого). Хвост короткий, из 14 перьев, средние рулевые удлинены незна-
чительно. Задний палец, как у других рябков рода Pterodes, имеется, но развит слабо и
высоко прикреплен. Цевка оперена спереди и с боков до пальцев, сзади обнажена. Длина
самок E) 350—330, в среднем 342 мм. Размах у самок E) 680—735, в среднем 712мм. Крыло
самцов: C8) 224—256, самок A9) 220—235, в среднем, соответственно, 237,5 и 226,2 мм.
Вес самцов 480, 507, 520, 550 г, самок 410, 422, 440, 465 г.
Окраска. Пуховой птенец из СССР не описан, по экземплярам из Африки — ржаво-
бурый с черными пестринами (Лош). Взрослый самец: голова и шея серые, вокруг шеи ржаво-
желтое «ожерелье*, спина, плечевые, кроющие крыла и хвоста серые — с большими желтыми
вершинными пятнами; большие кроющие крыла — охристые с ржаво-желтыми вершинами и
наружными опахалами; самые длинные кроющие хвоста с темносерыми и охристыми попе-
речными полосами; первостепенные маховые — темносерые с более темными стволами; второ-
степенные маховые — темносерые с белыми основаниями; кроющие кисти — серовато-белые;
рулевые — серые у вершины, с белой каймой на конце и с охристо-желтоватым поперечным
рисунком; средняя пара с широкой сероватой вершиной и с поперечным серым и охристым
рисунком; кроющие уха и уздечка бледносерые; подбородок и горло — каштаново-рыжие,
отделены от зоба черной поперечной полосой; зоб и грудь бледносерые, окаймленные чер-
ным; передняя часть боков и брюха — охристо-палевая, задняя часть—черная; оперение
подкрылий, подхвостья и цевок — белое, в подхвостье нерезкий темный поперечный рису-
нок. Самка на спинной стороне охристо-желтоватая с продольным на голове, поперечным
на спине, крыльях, хвосте рисунком черноватого цвета; маховые и большие кроющие
крыла — как у самцов; горло беловатое с черной поперечной полосой; зоб и грудь —
охристые с черноватыми пестринами; передняя часть брюха — беловатая, отделенная от
груди черной полосой; остальное брюхо и бока черные; подхвостье — беловатое с бурыми
основаниями перьев; подкрылья —белые. Молодые птицы в первом (втором?) птенцовом
наряде — как самки, но общий тон окраски более тусклый и маховые с широкими охри-
стыми каймами. Клюв серовато-бурый, пальцы серые, когти черные; радужина бурая.
Западный чернобрюхий рябок Pterocles orientalis orlentalis L.
Распространение. Ареал. Канарские о-ва (Фуэртевентура), централь-
ные и южные части Марокко, южн. Алжир, центр, и южн. Тунис, Три-
поли; Испания и Португалия; Кипр; М. Азия к востоку до турецко-иранской
границы (и быть может до Загроша?); к югу —до Палестины и Сирии,
на севере до Закавказья и в прилежащих частях Ирана (Котуро-Урмийский
район, Радугин, 1917). В Закавказье найден на Армянском плоскогорье, в
пустынных степях по Куре и Араксу, также в прикаспийских (куда, вероятно,
только залетает); наконец, на плоскогорье Карской обл. (Сатунин, 1907).
Залеты отмечены в Ленкорани и в долину Куры километрах в 40 ниже Тбилиси,
где возможно гнездился (Радде, 1885). Вопрос о гнездовании в Армении, пови-
димому, требует еще некоторого выяснения, оно вероятно для районов Ведин-
ского (у ст. Арарат) и Октемберянского (Даль); летние находки из советской
Армении, как и из Азербайджана (Шапоротов, Шуша), имеются; с несомнен-
ностью найден на гнездовье в нынешней Турецкой Армении (Бобринский, 1917).
Характер пребывания. Оседлая и кочующая птица. У озера
Урмия в сев. Иране оседла. В Армении отмечен пролет из турецких пределов на
север в сентябре (Даль), на Араксе — и зимние откочевки к югу (Беме, 1936).
Биотоп. Равнины и горные долины, плато, холмы предгорий —
пустынные и полупустынные, с щебнистым или глинистым грунтом и ксеро-
фильной растительностью, до зысот 1400 и даже 1875 м.
Численность. В Закавказье редок (граница ареала), несколько
чаще, повидимому, в холодное время года.
Экология. Размножение. На территории СССР экология почти
не изучена, повидимому, в основных чертах сходна с образом жизни Р. о. arena-
rius. В апреле птицы держатся еще в стаях (Бобринский, 1917), в парах по край-
БЕЛОБРЮХИЙ РЯБОК
79
ней мере до начала августа (Радде, 1885). Во второй половине июня у Игдыря
в гнезде — ямке без выстилки — 3 яйца (Бобринский). Линька. У взрослого
25 июля сменились 5 первостепенных маховых, удругого 13 августа—7 маховых.
Описание. Размеры и строение. Как у восточного подвида. Крыло самцов
CJ20—235, самок E) 220—235, в среднем 229 и 226,8 мм. Самцы окрашены как arenarius,
но у самок грудь ярче, не беловатая, а охристо-песочная.
Систематические замечания. Реальность различий восточных чернобрюхих рябков от
западных вызывает сомнения; вид, в сущности, монотипичен.
2. Белобрюхий рябок Pterocles alchata L.
Tetrao Alchata. Linnaeus. Syst. Nat., ed. XII, V. 1, 1766, стр. 276, Монпелье.
Распространение. Ареал. Португалия, Испания, южн. Франция
(Кро), сев. Африка от центр. Марокко до Киренаики и Сахары; Передняя
Азия от Анатолии и Закавказья до Закаспия, Туркестана, Ирана, Палестины,
Месопотамии на юго-западе и Белуджистана и сев.-зап. Индии на юго-востоке.
Карта 13. Распространение белобрюхого рябка Pterocles alchata
7. P. a. caudacntus, 2. P. a. alchata;
a — границы гнездовий, г — область зимовок.
Характер пребывания. На самом севере ареала белобрюхий
рябок — перелетная, на западе и юге — оседлая или кочующая птица.
Биотоп. Пустыни и полупустыни, глинистые, щебнистые или песчаные.
Подвиды и варьирующие признаки. В ю.-з. Европе — на Пиренейском п-ове и во
Франции — P. a. alchata Linn., 1766; в сев. Африке и Азии—P. a. caudacutus Gme lin,
1774. Морфологические различия — в окраске, восточный подвид бледней,в частности у сам-
цов светлее зоб, каймы верхних кроющих крыла беловатые, а не желтоватые.Экологических
различий не установлено.
Восточный белобрюхий рябок Pterocles alchata caudacutus G m.
Tetrao caudacutus. Gmelin. Reised. Russland 111, 1774, стр. 93, таб. 18, Гилян.
Синонимы. Tetrao chata. Pallas. Zoogr. Rosso-Asiat., 11, 1811, стр. 73, Киргиз-
ские степи. — Pterocles caspius. Menetries. Catal. Raisonne.., 1832, стр. 47, Баку. — Pteroc-
les sewerzowi. Богданов. Изв. Имп. Акад. наук, XXVII, 1881, стр.165, Арало-Каспий-
ские степи.
Русское название. Белобрюхий рябок — книжное название. У русских охот-
ников в Туркмении — туртушка (по голосу), у местного населения — ак-баур, кулан-
баур-бульдерюк, у туркмен еще киль-куйрюк (-острохвост).
Распространение. Ареал. Сев. Африка—Марокко, Алжир, южн.
Тунис, Триполи, Киренаика (быть может Синай), Палестина, Сирия,
80 ОТРЯД РЯБКИ
Малая Азия, Месопотамия; в СССР — в юж. Закавказье по Араксу (Богда-
нов, 1878), между Эчмиадзином и Джульфой в Армении (Краткий, 1894),
на западном берегу Каспия (Радде, 1885), но вероятно только залеты; Турке-
стан к северу до 47° с. ш. у Аральского моря (к западу от зал. Паскевича),
в бассейне Чу к востоку до гор Кан-тау или Хун-тау (Долгушин, 1939); на
Сары-Су и Каракенгире до 47°5Г (Слудский и Афанасьев, 1947); далее на юг
в Кызыл-Кумах, в равнинном Узбекистане; в Таджикистане в низменных
северных и юго-западных частях страны, на юго-западе по Кафирнигану,
в низовьях Вахша, у Джиликуля и Микоянабада, в предгорьях Кара-тау,
в степях от Кировабада от Термеза, на запад до Сурхана (Иванов, 1940); в
Туркмении повсеместно, но на гнездовье в холмистой песчаной степи Бад-
хыза и Карабиля; далее, вне пределов СССР, в Иране на юг до Сеистана,
Кугистана и Кермана (Зарудный, 1911); в Афганистане, Белуджистане и
с.-з. Индии в долине Пешавара. Зимой в с.-з. Индии к югу до Самбура, Джод-
пура, Биканира, Деза, на востоке в Пенджабе до Лудиана и Дели. Регулярные
зимовки от бассейна Гюргеня в сев. Иране до Ирака, Белуджистана и с.-з.
Индии. Залеты в сев. Казахстан (до Киш-копа и даже Чкалова, Зарудный,
1897); на Сев. Кавказ на Терек (Гмелин, 1789); к Ленкорани (Радде, 1885,
1889 и др.), в Армению (Соснин и Ляйстер, 1942); в Волжско-Уральскую степь
к Камыш-самарским озерам (Мейснер, Лавров, Киселев, 1907), в окрестности
Астрахани (Паллас, 1811; Хлебников, 1928), в Крым (Аверин, 1929), в
Днепропетровскую обл. (Нордман, 1840).
Характер пребывания. В северной части ареала — перелетная
птица, на юге, вероятно, оседлая (см. выше).
Даты. Пролет в Месопотамии весною отмечен в марте, осенью — в авгу-
сте — октябре. В Туркмении весенний пролет наблюдался с середины марта
и до начала апреля. На севере ареала на Сыр-Дарье пролет —с конца марта и
в начале апреля (начало его уДжулека—23 марта, валовой пролет—до 7 апреля,
большими стаями, Спангенберг, 1936). В конце марта —начале апреля пролет
отмечен и у Ленкорани (Радде, 1885, 1889). В вост. Иране пролет наблю-
дался и поздней; 15—17 апреля, в массе еще19—22 апреля (Зарудный, 1903).
В Туркестане весенний пролет идет с юго- и юго-запада и возможно связывается
с вегетацией пустынных растений (Каррузерс, 1910). Осенью, вероятно в
связи с иными кормовыми условиями, белобрюхие рябки собираются в громад-
ные стаи, самый отлет из Туркестана происходит поздно: на Сыр-Дарье послед-
ние особи встречаются еще в конце ноября B7 ноября, Спангенберг, 1936);
массовые скопления отмечены в с.-з. Узбекистане в долине р. Арысь в сен-
тябре — октябре (Мекленбурцев). В Туркмении этот осенний пролет бело-
брюхих рябков выражен особенно хорошо: птицы громадными стаями летят
от Мерва долиной Теджена к Копет-Дагу и затем поворачивают вдоль этого
хребта в западном направлении, долетая подгорными равнинами примерно
до меридиана Кызыл-арвата; отсюда рябки летят горными долинами (Ходжа-
калинской и др.). Через Атрек к Гюргенским зимовкам. В благоприятные
зимы рябки остаются в ю.-з. Туркмении и зимой встречаются у Чата (декабрь —
январь). Этот пролет проходит в последней трети октября — первой трети
ноября. Значительные пролеты идут по восточному берегу Аральского моря,
где эта птица не гнездится (Зарудный, 1916). Стаенье белобрюхих рябков
начинается сразу после окончания размножения в августе.
Биотоп. Полупустынные и пустынные пространства, в особенности
глинистые участки, перемежающиеся с песчаными; бугристая песчано-зла-
ковая степь; развеянные бугристые пески. Связь с песками более выражена,
чем у чернобрюхого рябка, который их в общем избегает. В отличие от послед-
него и саджей белобрюхий рябок не поднимается в горы и даже в предгорья
и не водится, по крайней мере в гнездовое время, в полынных степях. Рас-
пределение в ареале связано с относительной близостью водоемов — рек
БЕЛОБРЮХИЙ РЯБОК 81
(Сыр-Дарья в Кызыл-Кумах, Чу, Аму-Дарья), колодцев, родников, в особен-
ности в гнездовое время. На пролете и кочевках — и по окраинам культурного
ландшафта. Существенное значение из кормовых условий биотопа имеет
состояние растительного покрова, поэтому рябки не гнездятся в наиболее
«глухих» участках Кара-Кумов и Кызыл-Кумов, даже в холмистой песчаной
степи Теджено-Мургабского и Мургабо-Кушкинского междуречий в области
предгорий Паропамиза (где весной развивается пышный и сомкнутый, ни
в коей мере не пустынный растительный покров, который рано полностью
выгорает).
Численность. Биотопические требования белобрюхого рябка
делают распространение его спорадическим и быть может приводят к «коло-
ниальному» или «полуколониальному» расположению гнездовий. На севере
ареала у Аральского моря и Чу немногочисленен, обычен местами в Кара-
Кумах и Кызыл-Кумах, в низменных частях Узбекистана и Таджикистана,
в сев. Иране в бассейне Геррируда, в Месопотамии. На осеннем пролете —
в огромных стаях (например, по подсчету Лаптева, 1932, в подгорной полосе
Копет-Дага на пространстве в 1 км за 1 час пролетает до 650 птиц, а всего
по подгорной полосе не менее 780 000 особей). В последнее время отмечается
некоторое падение числа пролетных и кочующих осенью в Туркестане рябков
(в долине Арыси, Мекленбурцев; в Туркмении, Дементьев; впрочем в иные
годы рябков пролетает больше, в иные меньше, так что, вероятно, имеют место
и периодические колебания их численности).
Экология. Размножение. Половая зрелость у белобрюхих ряб-
ков наступает, повидимому, в первую весну после рождения в возрасте несколь-
ко менее года, но возможно в разные сроки, в зависимости от даты рождения.
Период размножения, как и у других рябков, весьма растянут. Это вероятно
связано или с наличием в норме двух кладок в году, или с дополнительными
кладками взамен погибших (гибель отложенных яиц у рябков при открытом
гнездовании на земле нередка), или отчасти с возрастным составом популя-
ции. Точных сведений о нормальных двух циклах в году нет. Стаи не раз-
биваются до конца апреля, когда начинается кладка (Аральское море, Бостан-
жогло, 1911, Туркмения, Зарудный, и др.; Сыр-Дарья, Спангенберг; Месо-
потамия, Тайсхерст, 1922). В первой половине мая —полные кладки. Гнезда
расположены прямо на земле, в небольшой ямке или на расчищенной пло-
щадке, иногда окружены стебельками, часто в тени осочки, саксаула и т. п.,
иногда замаскированные. Обычно они устраиваются на склонах бархана, даю-
щего известную тень. Подстилки в гнезде нет. Расположены гнезда колониями,
иногда на расстоянии 20 шагов одно от другого (Зарудный и др.), чаще менее
плотными. Поведение рябков носит известный отпечаток «стайности»: у них очень
развиты подражательные реакции, например, при взлете с гнезда одной птицы
поднимаются и другие, стаями птицы летают на водопой и т. д. (биологическое
значение этой «подражательности» поведения заключается, вероятно, в защите
от опасностей этих открыто гнездящихся птиц).
Яиц обычно 3, иногда 2, редко 4 (в Туркмении, Зарудный, 1896).
Окраска: бело-буроватый или желтовато-охристый фон, покрытый поверх-
ностными яркокоричневыми пятнами и внутренними расплывчатыми, светло-
свинцово-серыми. Размеры яиц в среднем 4бх 35 мм (Аралокаспийские степи,
Бостанжогло, 1911); A2) 44,4-^16,9x29,6—31,1 мм (Сыр-Дарья, Спанген-
берг, 1936); A7L0—45X30—32,5, в среднем44,03x31,И ш<;водномслучае —
четвертое яйцо в кладке—38x27 мм (Туркмения, Зарудный, 1896); 39—
49x28,5—30 мм, в среднем 44,6X30 мм (Месопотамия, Тайсхерст, 1922).
Яйца встречаются и значительно поздней, по указанным выше причинам
A5—18 мая в отрогах Кара-тау добыты самки с яйцами в яйцеводах; 23 мая
в гнезде в Кызыл-Кумах только одно свеже-отложенное яйцо, Спангенберг,
1936; 28 июля гнездо с яйцами в Кызыл-Кумах, Зарудный, 1915; 14 июля
6 Птицы СССР, т. II
82 отряд рябки
у самки яйцо в яйцеводе без известковой скорлупы, Туркмения, Зарудный,
1896; 28 июля в Кызыл-Кумах 3 гнезда с полными кладками, Зарудный, 1915).
Насиживание происходит с первого яйца, с участием обоих родителей.
Самец сидит по утрам, когда самка кормится, самка сидит в послеполуденные
часы (быть может самец насиживает ночью?). Продолжительность насижи-
вания, вероятно, около 4 недель. Тип развития, как у других рябков —вывод-
ковый. Первые подлетки отмечены во второй половине июля, 14—21 (Туркме-
ния, Зарудный, 1896); в Кызыл-Кумах — около середины июня птенцы в чет-
верть роста молодых были еще в гнездах, тогда же найдены и насиженные
яйца; летные молодые 21—23 июня отсутствуют, Тохта-тау в Кызыл-Кумах.
В Кызыл-Кумах подлетки и хорошо летающие молодые отмечены 9 августа
(Зарудный, 1915).
Линька. Белобрюхий рябок отличается от других наличием пред-
брачной линьки кроме послебрачной. Таким образом, две линьки в году:
послебрачная полная и предбрачная частичная, захватывающая мелкое
оперение, но не маховые и рулевые. Последовательность смены маховых от
10-го к 1-му (видимому). Предбрачная частичная линька происходит на
зимовке, птицы прилетают в Туркестан в свежем брачном наряде. После-
брачная линька зависит от окончания размножения: начинается в июне
или в начале июля и заканчивается в сентябре—октябре. Характерно,
что в результате послебрачной линьки наряд самцов носит «смешанный»
характер: голова окрашена как у самок, как и часть перьев спины и плече-
вых; остальное оперение типичное «самцовое». Последовательность смены
нарядов: пуховый наряд — первый птенцовый наряд — второй птенцовый
наряд (?) — первый предбрачный наряд — первый брачный наряд — после-
брачный наряд —брачный наряд и т. д. Хартерт A916) считает, что имеется
еще особый осенний наряд и что у белобрюхого рябка три линьки, но на нашем
материале подтверждений этому нет.
Питание. Пищу белобрюхого рябка составляют, таким образом,
как и у других рябков, семена растений, отчасти их побеги: полыни, солянок,
верблюжьей колючки и т. п. Меньшее значение имеют насекомые (Даль, 1937).
Питание сухими семенами связано со значительной потребностью в питье:
рябки даже в гнездовое время совершают регулярные перелеты на водопой,
при этом пьют не только пресную, но и солоноватую воду (Аральское море,
Бостанжогло, 1911 и др.). Перелеты на водопой рябки совершают утром, между
8 и 12 часами, и вечером, после 15 часов (Шестоперов, 1936; Молчанов, 1912
и др.). Особенно значительны утренние перелеты на водопой, вечерние менее
регулярны. Рябки поят нелетных и плохо летающих птенцов водой, которую
отрыгивают им из зоба (но не из смоченных водой перьев). Пьют рябки, загла-
тывая воду, запрокидывая голову, как куриные, и втягивая ее клювом, опустив
голову, как голуби.
Полевые признаки. От саджей отличается неудлиненными маховыми,
как и чернобрюхий рябок; от последнего меньшими размерами, удлиненными
и заостренными средними рулевыми и светлым (на полете) брюхом. Полет
очень быстрый. На земле держится горизонтально. Охотно «купается* в песке.
Крик «га-га» или «га-нг» издается на полете.
Описание. Размеры и строение. Как у других рябков, голова и клюв «го-
лубиного» типа. Ноздри узкие щелевидные, с кожистым клапаном. Крылья длинные и ост-
рые, формула их 1 > 2 > 3 > 4 (считая за первое видимое неприкрытое кроющими, а
не абортивное маховое). Хвост резко ступенчат, из 14 рулевых, средние рулевые заострены
и удлинены. Задний палец имеется, но очень короток и прикреплен выше передних; цевка
короткая, оперенная до пальцев с передней и боковых сторон (но не с задней). Длина сам-
цов D) 365—383, самок D) 340—365, в среднем 370,1 и 351,2 мм. Размах самцов D) 640—
655, самок D) 650—660, в среднем у обоих полов 647,5 мм. Вес самцов около 250, самок
около 225 г, наибольший у самцов 290, наименьший 230 г. Крыло самцов B5) 201—217, са-
мок A0) 196—216, в среднем 209 и 204,8 мм.
БЕЛОБРЮХИЙ РЯБОК
83
Окраска. Взрослый самец в брачном наряде оливково-серовато-бурый на спине,
плечах и малых кроющих крыла, с большими охристо-желтыми вершинными пятнами;
голова желтовато-серая с серым теменем и затылком; поясница и надхвостье с черным и
желтоватым поперечным рисунком; кроющие крыла белые с сероватыми основаниями
и каштаново-бурыми краями внешних опахал, окаймленными белым, задние большие крою-
щие с золотисто-желтыми пятнами; первостепенные маховые серые с темным стволом, зад-
ние с темной вершиной и беловатой каймой; наружное опахало первого (не абортивного)
махового темнобурое; второстепенные маховые белые с черной окаймленной белым вершин-
ной частью внутренних опахал и наружными опахалами такой же окраски; крайняя пара
рулевых и большая часть внешнего опахала — шиферно-серые с белой вершиной, остальные
рулевыг с более или менее развитым поперечным рисунком; средние рулевые с темной серо-
бурой удлиненной вершиной, темным стволом и желтоватым поперечным рисункам; за ухом
Рис. 13. Белобрюхий рябок (самец и самка).
проходит черная полоса, горло черное, передняя часть шеи и щеки желтые; зоб и грудь
розовато-рыжие, окаймленные черным; остальная брюшная сторона и оперение ног белые,
подхвостье с черноватым поперечным рисунком; подкрылья белые, кроме шиферно-бурого
наружного края. Послебрачный наряд отличается присутствием на спине и плечевых отдель-
ных перьев «самочьей» окраски, с черным и желтым поперечным рисунком; голова с охристым
и серовато-черным поперечным рисунком, щеки и передняя сторона шеи охристые с темными
пестринами, подбородок и горло белые.
Самка отличается тем, что вся спинная сторона у нее с поперечным охристым и черно-
ватым рисунком, на больших кроющих и некоторых плечевых — широкая серая предвершин-
ная полоса; кроющие крыла с чешуйчатым черно-белым рисунком; щеки желтые, горло бело-
ватое; остальная часть брюшной стороны окрашена, как у самца, но грудное пятно бледней,
над ним расположено сероватое поле. В послебрачном наряде самки с охристо-рыжеватыми
покрытыми черноватыми пестринками щеками и с темными пестринками на передней части
зоба, бока с охристым оттенком. Птенцовый наряд (первый? второй?) — как в зимнем пос-
лебрачном наряде, но бледней и без примеси серого цвета на спинной стороне.
Клюв темносерый, пальцы ног и кольцо вокруг глаз бледносерые, когти черноваты;
радужина бледнобурая. Пуховой птенец окрашен на спинной стороне в охристо-буроватый
цвет с беловатыми и черноватыми пестринками, на брюхе охристый (Месопотамия, Тайс-
херст, 1922; изображение —в Proceedings of the Soological Society of London, 1866, табл. 9;
из СССР не описан).
[Сенегальский рябок Pterocles senegallus L i n n a e u s, 1771
Pterodes senegallus Linnaeus, 1771, описанный из Алжира и распространенный
в сев. Африке и ю-з. Азии от Аравии, Палестины и Месопотамии до Афганистана, Белуджи-
стана и с.-з. Индии, внесен в нашу фауну Зарудным (Орнитологический вестник, 1910).
Пара отнесенных к этому виду птиц была добыта у аула Таклы на Аму-Дарье ниже Келифа
30. VI. 1889 г. Оба экземпляра были впоследствии утеряны. Самое определение было опу-
бликовано после утраты этих птиц и через 21 год после их добычи. При таких обстоятельствах
правильность определения может вызывать серьезные сомнения, и, так как никто и никогда
сенегальских рябков или даже похожих на них птиц вТуркестане не наблюдал и не добы-
вал, то от включения этого вида в фауну СССР приходится воздержаться.]
84
ОТРЯД РЯБКИ
РОД САДЖА SYRRHAPTES ILLIGER, 1811
3. Обыкновенная саджа Syrrhaptesparadoxus Pall.
Тип S. paradoxus
Tetrao paradoxus. Pallas. Reise d. verschied. Provinzen d. Russischen Reichs. Bd.
II, 1773, стр. 7J, пески Джидель-мамут, Киргизская степь.
Русское название. Этимология слова саджа неясна. Казахское название —
бульдрук или бульдерюк. В Поволжье и вост. России и Зап. Сибири — копытка.
Распространение. Ареал. Крайний ю.-в. Европейской России — в
низовьях Волги, примерно километрах в 100 на север от Астрахани (Яковлев,
1872), в Волжско-Уральской степи, в низовьях Урала и Эмбы (но факт гнездо-
вания требует подтверждения, как и неясным остается утверждение Мензбира,
1895, действительно ли увеличился в западном направлении ареал саджи
в России в XIX веке, или эти сведения основаны на временной задержке
и гнездовании саджи после
массовых залетов вблизи от
пределов гнездовой обла-
сти; регулярное гнездование
саджи в низовьях Волги и
Волжско-Уральских степях
на основании новых иссле-
дований кажется мало ве-
роятным); далее на востоке
— уже несомненно — в сев.
Казахстане примерно до
49° с. ш., в Каркаралинском
районе даже до 50° по лево-
бережью Иртыша (Селевин,
1929), на востоке у Семипа-
латинска и в Зайсанской
котловине с обращенными
к ней частями Алтая, в Тан-
ну-Тувинской обл. к югу
от гор Танну-ола, в Монго-
лии на север, вероятно до
ю.-з. Забайкалья (Тарей-
нор), на востоке — до зап.
Манчжурии (Буирнор, Ту-
гаринов, 1932); к югу до Усть-Урта (Богданов, 1882), низовьев Аму-Дарьи
(Куня-ургенч, Шестоперов, 1936, требуется подтверждение) и быть может Унгуза
(Зарудньгй, 1896), до Аральских Кара-Кумов, Джаны-Дарьи и северо-западных
предгорий Кара-тау (Спангенберг, 1941), Чу, Или, центр. Тянь-Шаня (достовер-*
ные находки на гнездовье обнаружены совсем недавно в Иссык-кульском на-
горье у Рыбачьего, Кызыл-омпола, на Семизбельском плато, на севере этого
озера до Чоктала, на юге—к востоку до Улахола, Д. Дементьев, 1943) и
вероятно до бассейна Тарима; в Монголии до ю.-в. Ордоса и с.-в. Ганьсу
(северные части хребта Рихтгофена).
В связи с периодическими выселениями саджей из пределов гнездовой
области отдельные пары и колонии саджей более или менее эпизодически
гнездятся за границами «нормального» ареала; так, при больших миграциях
в прошлом столетии и в начале текущего саджи встречались даже в зап. Европе
и Европейской России (например, в б. Минской губ., Шнитников, 1913;
в Крыму два вывода после миграции 1908 года, Разевиг, 1910; попытки гнездо-
вания в 1908 году отмечены в Коломенском уезде Московской губ. и т. д.);
Карта 14. Распространение саджей Syrrhaptes parado-
xus и 5. tibetanus
1. S. paradoxus, 2. S. tibetanus;
a — граница гнездовий.
ОБЫКНОВЕННАЯ САДЖА 85
более или менее регулярно в ю.-в. Забайкалье (по Далю, 1944, в 1929 г. были
пуховики, в 1930—1933 г. птиц не было вовсе), в Чуйской степи (Сушкин, 1938,
Щухов, 1946), в сев. Манчжурии (в 1923 и 1924 г., после массового налета
1922—1923 г., Лукашкин, 1931) и в Жехоле в Сев. Китае, на западе по среднему
течению Урала в 1888 и до 1891 года, в окрестностях Илецкой защиты и у
Актюбинска, в степях за Илеком у Буранной, у Тузкуля, у Биш-копа (Заруд-
ный, 1888, 1897; Сушкин, 1908); иногда по островам Аральского моря на
Куг-арал и Барсакельмес (Зарудный, 1915). В холодное время года птицы
частью остаются в гнездовой области (зимовки отмечены в области Араль-
ского моря и Сыр-Дарьи, Северцов и др., в центр. Тянь-Шане, в Синьцзяне,
в южн. Монголии), частью же двигаются на юг до Туркмении, равнинных
частей Таджикистана, быть может южнокаспийских провинций Ирана, залетая
в Закавказье, в области Хорасана и Паропамиза, на Алтай, в Приморье,
Сев. Китай, Корею и Манчжурию, изредка и далее (один случай в Индии
уДара, Бекер, 1928). В северных частях ареала повидимому регулярно не зимует.
Характер пребывания. См. выше. Миграции носят нере-
гулярный характер, будучи связаны с состоянием снегового покрова —
выпадение глубокого снега, а также образование наста лишают саджей воз-
можности кормиться. Именно с такими неблагоприятными условиями свя-
заны, повидимому, бывающие в иные годы массовые вылеты саджей из гнездо-
вой области в различных направлениях. К перенаселению, увеличению чис-
ленности саджей на местах гнездовья эти массовые кочевки не имеют отношения.
Даты. У северной границы распространения в с.-з. Казахстане прилет
наблюдался около середины апреля A4. IV. Иргиз, Сушкин, 1908). В Забай-
калье и Монголии появление весной в первой половине марта, при морозах
(Козлова, 1930), тогда же и в Туркестане (Сыр-Дарья, Спангенберг, 1936).
Осеннее движение в Монголии отмечено в октябре (Гобийский Алтай, Бианки,
1907). В Туркмении на зимовке и пролете саджи появляются обычно в конце
октября —начале ноября (Дементьев, 1950), иногда уже в сентябре (Ахал-
текинский оазис, Зарудный, 1896).
Залеты. Как уже отмечалось, саджа совершает непериодические
массовые вылеты из гнездовой области. Такие залеты в Европу отмечены в
1859, 1863, 1872, 1876, 1877, 1888, 1891; 1899, 1904, 1906, 1908, 1913, 1917,
1921, 1922; последний раз в 1944 году (Бежецкий район Калининской обл.
16 апреля, Даль, 1946). Некоторые миграции были грандиозны —в 1863 г.,
в 1888 г. (когда птицы достигли Архангельска, Финляндии, южн. и зап.
Норвегии, Швеции, Фарерских о-вов и Англии, также Франции, Италии,
Австро-Венгрии) и в 1908 году (до Англии, Франции и Италии на западе).
При этих массовых вылетах птицы, появляясь обычно весной в апреле, задер-
живались иногда до весны следующего года, а отдельные пары пытались
гнездиться. В таких случаях нерегулярное гнездование саджей имеет место
иногда в течение одного-двух лет после массового налета (Манчжурия, Лукаш-
кин, 1931 и приведенные выше данные). В 1908 году появление саджей было
отмечено на Европейской территории СССР в Саратовской обл. (с апреля
по июнь), Пензенской (апрель —май), Куйбышевской (апрель —май), Татар-
ской республике (май — июнь), Башкирии (апрель), Кировской (одиночка,
4 апреля), Горьковской (апрель—май), Ярославской (май), Калинин-
ской (апрель — май), Смоленской (апрель — май), Ленинградской, Москов-
ской (апрель — май), Калужской (май), Рязанской (апрель), Тамбов-
ской (апрель — май), Воронежской (апрель), Орловской (май), Курской
(апрель—май), Черниговской, Харьковской (апрель—май), Екатерино-
славской (май), Полтавской (май), Киевской (май), Волынской и Подольской
(май), в Крым (апрель — май и в июне, затем в августе, ноябре и декабре,
отдельные птицы до марта 1909), на Кавказе (апрель—май); в течение
весны и лета наблюдалось и незначительное обратное движение (Судилов-
86 ОТРЯД РЯБКИ
екая, 1933). Массовым миграциям саджи предшествуют—иногда задолго—
местные передвижения. Так, за несколько месяцев до миграции 1908 г., осенью
1907 г., саджи исчезли из Зайсанской котловины, где они обычно держатся
оседло; миграции 1863 г. предшествовали кочевки саджей в Киргизских сте-
пях и т. д. В восточном направлении миграции саджи также бывают, хотя
они и менее изучены, чем появление этой птицы в Европе. Массовый залет
в Минусинские степи был весной 1901 года (Сушкин, 1914), менее значительные
в 1903, 1904, 1905 г. В I860, 1897, 1904, 1912, 1913, 1916, 1922—1923 годах
были отмечены большие залеты в Сев. Китай и Манчжурию (Латуш, 1932 и др.).
В 1922—1923 г. залеты отмечались в области Иртыша (Селевин, 1927 и др.),
в 1907 и особенно в 1913 в Приморье (Арсеньев, 1923; Шульпин, 1936 и др.).
Наиболее значительные миграции происходили в 1860 г. в Сев. Китае, в 1912—
1913 г. в Манчжурии, в 1922—1923 г. там же. Миграции в западном и восточ-
ном направлении происходят, таким образом, несинхронно, и это говорит о
том, что они происходят в результате каких-то местных причин, и участвуют
в них только отдельные популяции.
Биотоп. Полупустыни и степи, ровные или всхолмленные, с глинистой
почвой, полынной и злаковой растительностью. Развеянных песков саджа
избегает и ее нет поэтому в Кара-Кумах и Кызыл-Кумах, в песках Тусум
в сев. Казахстане и т. п. В вертикальном отношении саджа не избегает и высо-
ких долин с подходящим ландшафтом — в с.-з. Монголии саджа найдена на
высотах 2400 м (Тошонгты, Олбо-нур, Сушкин, 1938), в Тянь-Шане в бас-
сейне Нарына до 3250 м (Шалов, 1908), нормально у Иссык-Куля от 1600
до 1900 м (Д. Дементьев, 1943), в Гобийском Алтае до 1850 м (Бианки, 1907),
в Синьцзяне около 1300 м (Ледлоу и Киннир, 1934).
Численность. Распространение несколько спорадическое; однако
гнездовые участки заняты плотно, птицы образуют колонии. На пролетах
и описанных выше залетах птицы образуют стаи самых разнообразных раз-
меров — от нескольких десятков до многих тысяч птиц.
Экология. Размножение. Как и другие рябки, моногамы. Когда
наступает половая зрелость, не выяснено. Цикл размножения относительно
поздний, его можно отнести приблизительно к середине апреля, когда в стаях
заметно распадение на пары. Самец в это время токует и кричит, бегая вокруг
самки по-голубиному, но не раздувая шеи и не кланяясь, иногда он летает,
делая вокруг самки круг; других самцов токующая птица отгоняет. Гнезда,
как и у других рябков, располагаются на земле; это —простая ямка, иногда
с немногими сухими стебельками, иногда среди кустика полыни или злака.
Расположены гнезда нередко вблизи одно от другого (на 4—6л<, Бианки, 1915),
как бы колониями. Кладка начинается с середины апреля (Забайкалье, Сыр-
Дарья, Монголия). Яйца можно найти и в середине и даже в конце июня (напри-
мер, в Аральских Кара-Кумах готовое к откладке яйцо у самки было обнаружено
еще 19 июня, малонасиженное яйцо —в конце этого месяца, Спангенберг, 1936).
Как и для других видов рябков, остается неясным, имеют ли место два
нормальных репродуктивных цикла или растянутость периода кладки проис-
ходит за счет дополнительных кладок взамен погибших по той или иной при-
чине, или младшие по срокам вывода прошлогодние птицы позже приступают
к размножению, или действуют все эти причины.
Кладка состоит из 3, иногда из 2 и, будто бы, даже из 4 яиц(Радде, 1863).
Яйца —в отличие от яиц других рябков —без блеска; основная окраска их
глинисто-серая, иногда со слабым желтоватым налетом; по этому фону раз-
бросаны поверхностные светлокоричневые пятна и внутренние расплывчатые
светлосерые. Размеры: E) 40,9—43,7x29,1—30,2 мм (Сыр-Дарья, Спанген-
берг, 1936); 44x35 мм (Арало-Каспийские степи, Бостанжогло, 1911); A00)
42,6x29,6 в среднем, максимум 46,5x29,9 и 42,5x32,4, минимум 39,2x28,5
и 40,3x27 мм (Уайзерби, 1940).
ОБЫКНОВЕННАЯ САДЖА 87
Насиживание начинается с откладки первого яйца. Днем насиживающие
птицы сидят неплотно и регулярно после восхода солнца летают на водопой.
Самки сидят утром и с 16—17 часов (Сушкин, 1908), самец также принимает
участие в насиживании (у него имеются наседные пятна; быть может, как другие
рябки, он сменяет самку по ночам, но это еще не выяснено). Продолжается наси-
живание около месяца. Недавно вылупившиеся птенцы в Туркестане обнару-
живаются со второй половины мая, в Забайкалье — примерно с половины
мая; в Монголии с конца мая попадаются подлетки; подлетки с остатками пуха
в Туркестане и Зайсанской котловине встречаются с последней трети июня.
Однако молодые — от вторых или запоздалых кладок — встречаются и
значительно позднее; 3 августа у Иссык-куля были обнаружены плохо летаю-
щие молодые, еще 10 августа—пуховик (Дементьев, 1943).
Линька. Полная годовая. Период линяния (у популяций) естественно
растянут в соответствии с размножением. Большинство особей линяют с конца
мая —начала июня, и в октябре надевают свежее оперение; продолжитель-
ность линяния, таким образом, около 4 месяцев. Порядок смены маховых от
10-го к 1-му (видимому, не абортивному), смена их медленная, к концу
июня обычно проходит на 40% (сменены 4 задних маховых). Рулевые сменяются
с конца июля —начала августа. Последовательность смены нарядов: пуховой
наряд — первый птенцовый наряд — второй птенцовый наряд — первый
годовой наряд (в нем иногда остается часть перьев от птенцового наряда,
часть передних маховых и больших кроющих) — второй годовой наряд и т. д.
Первый годовой комбинированный наряд надевается в возрасте около 2 меся-
цев, в июле — августе.
Питание. Пища — растительная, побеги и мелкие семена степных
и пустынных растений, которые саджа собирает, бегая по земле. Вероятно
с этим связано (при сильном нагреве почвы в весеннее и летнее время в гнез-
довой области саджи) наличие у нее толстой кожистой подушечки на лапах
и относительно толстая кожа на брюшной стороне (на спинной она тонкая,
как у голубей), это, вероятно, предохраняет саджу от перегрева теплом,
излучаемым раскаленной почвой. В Монголии в качестве основного корма
саджи указывается сульхир Agriophyllum gobicum, а также семена полыни и
бобовые (Пржевальский, 1876); в Забайкалье побеги Salicaria и злаков (Радде,
1863); в Туркестане — солянки, полыни, «черепашье просо» и т. п.; в Чуйской
степи и ю.-в. Алтае в зобу саджей найдены соцветия АШит tenuissimum,
прошлогодние семена Astragalus sp. и Elymus dasystachys (Шухов, 1926).
И в гнездовое время саджи обычно кормятся стайками или парами, слетая
с гнезда в жаркое время дня. Охотно роются в пыли шо-куриному». Как и
другие рябки, саджа пьет регулярно, летая на водопой стаями утром и вечером;
особенно значительны эти перелеты на водопой по утрам, около 9—10 часов.
Наличие водопоев определяет в известной мере распространение саджи, хотя,
при ее высоких летных качествах расстояние в несколько десятков километров
не имеет особого значения, за исключением однако времени вывода птенцов.
Полевые признаки. Характерная «пустынная» окраска делает птицу мало-
заметной на земле. На полете бросаются в глаза черное брюшко, заострен-
ные и удлиненные крылья и длинные средние рулевые. На земле держится
почти горизонтально, хорошо ходит; охотно «купается» в пыли. Взлет —с хло-
паньем крыльев, как у голубей, полет чрезвычайно быстрый. При приближе-
нии опасности часто затаивается или отводит от гнезда с сильно насиженными
яйцами. Голос «трю-трю» или <ггрюк-ти-трюю> на полете, «кек-кек» —на земле.
Описание. Размеры и строение. Ноги короткие, цевка оперена кругом,
пальцы широкие и оперенные, с толстыми подушечками на подошве, ерошенные до ногтевых
фаланг; заднего пальца нет. Голова небольшая «голубиного» типа, клюв короткий и сжа-
тый, с узкими щелевидными ноздрями; шея короткая. Крылья длинные и острые; самое
длинное маховое первое (не считая абортивного), вершина его вытянута и удлинена, особенно
у самцов. Формула крыла, следовательно 1>2>3... Хвост ступенчатый, из 16 рулевых,
88
ОТРЯД РЯБКИ
средняя пара рулевых удлинена и заострена. Длина самцов (8) 380—440, самок F) 355—389,
в среднем 413,25 и 373 мм. Размах самцов G) 675—711, самок F) 600—660, в среднем 693
я 634,16 мм. Хвост самцов A3) 160—235, самок (8) 163—205, в среднем 194,7 и 179, 75 мм.
Крыло самцов C0) 223—265, самок B5) 205—235, в среднем 239,4 и 222,84 мм.
Окраска. Пуховой наряд птенца охристого цвета с бледнобурым рисунком, на
голове черноватые пестрины; брюшная сторона—охристо-песочная. Птенцовый наряд блед-
ный охристо-песочный с бурыми поперечно вытянутыми на больших перьях пестринами;
горло и зоб — охристые; на брюхе — небольшое бурое пятно, на груди — темные пестрины;
маховые буро-серые с черноватым стволом и неправильным охристым рисунком и каймами у
вершин; рулевые с широким бурым поперечным рисунком; средняя пара рулевых удлинена
немного. Второй птенцовый наряд с несколько более насыщенным рисунком, при сходном
с первым птенцовым нарядом цветорасположении. В этом возрасте полового диморфизма
в окраске нет.
Рис. 14. Саджа (самец, самка и птенцы).
Взрослый самец на спинной стороне охристо-песочный с черноватым поперечным ри-
сунком на спине, плечевых, кроющих хвоста, рулевых и внутренних первостепенных маховых;
первостепенные маховые серые с черными стволами (а первое маховое и с черным наружным
опахалом), с охристой каймой; часть кроющих крыла с черными вершинными пятнами;
вокруг шеи желтое кольцо, охватывающее и кроющие уха; горло беловатое, общий тон брюш-
ной стороны песочнл-охристый, на груди с узким черноватым поперечным рисунком; задняя
часть груди и брюхо черно-бурые; подмышечные беловаты; все перья с прикрытыми
коричнево-бурыми основаниями.
Самка без поперечного рисунка на груди, с менее выраженным желтоватым пятном
на шее; темя в черноватых пестринах; больше развит черный рисунок на второстепенных
маховых и кроющих крыла, имеющийся — в отличие от самцов — и на средних перьях;
по бокам зоба и груди — черноватые пестрины; поперечный рисунок на спине и плечевых менее
правильный; средние рулевые менее удлинены и заострены. Клюв сероватый, радужина
бурая, когти черные или черноватые.
4. Тибетская или горная саджа Syrrhaptes tibetanus Gould
Syrrhaptes tibetanus. Gould. Proceed, of the Zoolog. Society of London, 1850, стр. 92,
Ладак.
Местное название. У памирских киргизов «уак», звукоподражательное (Меклен-
бурцев, 1936).
Распространение. Ареал. Центрально-азиатское нагорье — Тибет,
Ладак, Кара-корум, Памир. На север до Куку-нора, в южн. Нань-Шане и
до реки Мурус-су —на востоке (Пржевальский, 1876), до бассейна реки Кара-
каш (Юм, 1873) и Заалайского хребта —на западе. На Памире западная гра-
ница проходит примерно до линии пик Ленина — Таш-курган — Садым-Мац,
ТИБЕТСКАЯ ИЛИ ГОРНАЯ САДЖА 89
в глубоких долинах западного Памира отсутствует (Иванов, 1940). Южная
граница не вполне ясна, но птица встречена в Кашмире до Лагуля, в сев.
Сиккиме, в верховьях Сетледжа, а в Тибете —до Ялуня (Шефер, 1938).
Характер пребывания. Оседла, но совершает во второй поло-
вине зимы небольшие передвижения, что вызвано снегопадами. В это время
на Памире стайки тибетских саджей появляются на пастбищах архаров, где
последние в поисках корма разгребают снег (Мекленбурцев, 1946). При всех
обстоятельствах зимой саджа встречается на южных бесснежных или мало-
снежных склонах. На кочевках саджа выходит иногда несколько за пределы
гнездовой области (например, у Дараут-кургана на Алае, Головнин, 1901).
Однако эти кочевки нерегулярны и местами в Тибете птица, повидимому,
оседла. Вертикальный размах зимних миграций составляет примерно 650 м
(Бекер, 1928). Незначительный пролет на вост. Памире на запад, на боль-
шой высоте, наблюдался в начале ноября (Мекленбурцев, 1946).
Биотоп. Гнездовый — равнины у окраины снежных полей или речные
долины и приозерные котловины с галечником; щебнистая высокогорная полу-
пустыня (Памир); в Тибете — и по дюнам и пескам — в долинах. В гнездовое
время встречается примерно на высотах от 4400—5700 м (Тибет, Шефер, 1938);
гнездовые находки в Ладаке у озера Ко-морири на высоте 4375 м (Осместон,
1925) и—к югу от Пуга—5395 м (Мейнертцхаген, 1927); в Каракоруме
найдена на плато Депсанг и у Балти-Брангза на высотах около 5660—5947 м
(Ледлоу и Киннир, 1934); на Памире встречена в гнездовое время от 4897
(Ак-байтал) до 3615 м (Пост-Памирский).
Численность. Распространение несколько спорадично и птица
не может считаться частой (Тибет, Памир). С осени (начало августа на Памире,
Зарудный и Молчанов, 1915) попадается в стайках. Численность стай невелика,
от 5 до 30 особей, изредка и более, до 100 и даже выше (сев. Тибет, Пржеваль-
ский, 1876).
Экология. Размножение изучено слабо. Пары, повидимому, по-
стоянны; в Тибете саджи в парах встречены в последней трети марта, т. е. до
периода размножения (Шефер). По Бекеру A928), кладка состоит из 3 яиц,
которые помещаются в ямку; происходит она между маем и июнем, большин-
ство яиц сносится в апреле. Окраска яиц сходна с цветом их у обыкновенной
саджи, по светлому серовато-буроватому фону разбросаны бурые, охристо-
бурые или, реже, красновато-бурые пятна и крапины. Размеры яиц: 47,6—
50,6 х 31,8—35, но также 44-44,7 х 29—29,6 мм (Хартерт, 1915); C0)
44—54 х 29—34,8, в среднем 49,2 х 31,9 мм (Бекер). В насиживании, пови-
димому, как у других рябков, принимает участие и самец, так как у него
имеются наседные пятна. Два только что вылупившиеся затаившиеся пухо-
вика найдены в Ладаке у озера Ко-морири 18"июня (Осместон, 1925); однако
не вполне выросшие птенцы на Памире встречены еще в середине августа
(Северцов, 1879), что указывает либо на наличие дополнительных кладок,
откладываемых взамен утраченных, либо даже на наличие двух нормальных
выводков.
Линька. Полная годовая. Старые птицы начинают смену маховых
в начале июля, в начале августа линька маховых завершена примерно на 40%
(у самки с озера Кара-куль, Памир, 25 июля первые 4 маховых старые, 5-е выпало,
6-е — 10-е —новые). Начинается смена маховых с задних A0-го, 9-го и т. д.).
В октябре добываются птицы в свежем пере. Последовательность смены наря-
дов: пуховой наряд (вероятно, мезоптиль, но материала по этому вопросу
нет) — гнездовый наряд — послегнездовый наряд (комбинированный, с окон-
чательным рисунком мелкого оперения и неперелинявшими маховыми гнездо-
вого наряда, носится до следующей линьки в возрасте около одного года, по
Козловой, 1946) и т. д. В том, что маховые гнездового наряда сохраняются до
следующего года — отличие от обыкновенной саджи, у которой обычно хотя
90
ОТРЯД РЯБКИ
бы часть, а иногда и все маховые и большие кроющие крыла гнездового наряда
сменяются в первую осень жизни.
Питание. Пищу этих птиц составляет зелень и семена различных
растений, в сев. Тибете главным образом бобовых (Пржевальский). В зобу
у птиц, добытых в ноябре на Памире, были обнаружены почки и веточки Оху-
thropis immersa, Smelowskia calycina, Stellaria hemifusa (Мекленбурцев); у птиц,
добытых на гнездовье в Ладаке,— зелень, семена, надкрылья жука (Мейнертц-
хаген, 1927). В отличие от других рябков, и в частности обыкновенной саджи,
Рис. 15. Тибетская саджа (самец и самка).
повидимому, менее регулярно пьет воду. Так, например, на Памире зимою на
водопой эта птица не летает (Мекленбурцев). Однако у Куку-нора летавшие
днем около полудня на водопой стайки наблюдались в марте (Бианки, 1907);
лет стай на водопой в сумерках наблюдается на р. Каракаш (Юм, 1873).
Полевые признаки. Относительно крупный рябок с удлиненным и за-
остренным хвостом, с довольно широкими крыльями. Заметно больше обык-
новенной саджи и с менее быстрым полетом, напоминающим голубиный. Мало
осторожна (но зимой будто бы пуглива, Мекленбурцев, 1946). Вне гнездового
времени держится небольшими стайками. Голос «кага-кага-кага» (Пржеваль-
ский) или крякающий «уак-уак-кага» или «гок-гок» (Шефер, Бекер). Подается
голос только на лету, причем иногда слышен и ночью.
Описание. Размеры и строение. Оперение очень густое и плотное (высоко-
горная птица). Первого пальца нет, передние пальцы оперены до когтей и соединены в основ-
ной части толстой перепонкой (но не срослись до ногтевых фаланг, как, например, у обык-
новенной саджи); на нижней стороне пальцев — толстые подушечки; хвост из 16 рулевых,
средние рулевые удлинены и заострены. Формула крыла 1>2>3 , первое маховое без
заострения, характерного для обыкновенной саджи; вершина крыла относительно короткая,
внутренние первостепенные маховые удлинены (отношение вершины крыла к общей его длине
0,51, у настоящей саджи 0,61; задние маховые доходят до вершины 4-го махового, у обык-
новенной саджи — до 7-го махового, Козлова, 1946). Длина тела, размах крыльев, вес —
не описаны. Крыло самцов с Памира A0) 245—265, самок D) 245—255, в среднем, соответ-
ственно, 256,9 и 250 мм, хвост самцов (9) 196—230, самок E) 171—205, в среднем, соответстт
ТИБЕТСКАЯ ИЛИ ГОРНАЯ САДЖА 91
венно, 214,1 и 186,4 (Мекленбурцев и Дементьев). У тибетских птиц крыло 243—270 мм
(Шефер и де Шауензее, 1939). У птиц из разных мест крыло самцов 254—270, самок 248—
266, хвост, соответственно, 203—263 (?) и 203—216 мм (Бекер, 1928).
Окраска. Пуховой наряд (по Хартерту, 1915; у птиц из пределов СССР, насколько
знаем, неизвестен) сходен с нарядом соответствующего возраста обыкновенной саджи,
спинная сторона желтовато-охристая с черными и белыми пестринами. У взрослого самца
голова и шея — беловатые с черным поперечным рисунком, верхняя часть спины — охристая
с нерезкими тонкими сероватыми и поперечными полосками; остальная часть спины, плечевые,
кроющие крыла, второстепенные маховые — песочно-охристые с более или менее заметным
сероватым «мраморным» рисунком; на внутренних опахалах плечевых — черные пятна,
резко выступающие в обношенном пере; поясница и средние рулевые — беловато-охристые
с сероватыми «мраморными» пестринами, заостренные вершины средних рулевых — темно-
серы; большие кроющие крыла, передние второстепенные и первостепенные маховые —
темнобурые, на внутренних опахалах задних первостепенных маховых — светлое охристо-
сероватое поле; рулевые, кроме средней пары, песочно-рыжие с черным поперечным
рисунком и белой вершинной каймой; подбородок беловатый, вокруг шеи желтое «оже-
релье», захватывающее и оперение щек; зоб и грудь — беловатые с тонким поперечным
черновато-серым рисунком, нижняя часть груди — бледносерая; брюхо и бока, короткие
перья подхвостья — беловаты; длинные нижние кроющие хвоста — как боковые рулевые;
подкрылья серо-бурые с неясным мраморным серым рисунком, подмышечные и большие
нижние кроющие крыла — черно-бурые; оперение ног беловато. Развитие пестрин на
спинной стороне подвержено заметным индивидуальным вариациям.
Взрослая самка отличается от самца большим развитием поперечного рисунка, имею-
щегося и на нижней стороне груди, и на всем оперении спинной стороны; черноватые пест-
рины на плечевых более развиты, а желтоватый цвет на шее и на щеках — слабей. Клюв
серо-роговой, когти черно-роговые, радужина бурая. Молодые в гнездовом наряде сходны
с самками, но с еще более резким поперечным рисунком и с узкими светлыми каемками вну-
тренних опахал второстепенных маховых. В этом возрасте половой диморфизм в окраске
не проявляется.
Систематические замечания. При значительном морфологическом и экологическом
сходстве с обыкновенной саджей, тибетская саджа обладает некоторыми хорошо выражен-
ными особенностями (см. выше), на основании чего было предложено выделить этот вид
в монотипический род Tchangtangia Kozlova, 1917 (=Przewalskia К о z 1 о v a, 1946).
Нам кажется все же более оправданным считать тибетскую саджу за подрод рода саджей
Syrrhaptes.
ОТРЯД ТРЕХПЕРСТКИ
TURNICES или TURNICIFORMES
Характеристика отряда
Строение. Внешние признаки. Трехперстки — птицы некруп-
ных размеров, с перепела, которого весьма напоминают и по общему
внешнему виду. Размеры разных видов отряда в общем сходны. Вес трех-
персток около 35—50 г (Бутурлин, 1935). Клюв «куриного» типа. Задний
(первый) палец ноги у большинства видов отсутствует или, если имеется, то в
виде рудимента. Цевка неоперена и покрыта поперечными щитками. Оперение
довольно рыхлое, окрашено неярко, главным образом в охристые или рыжева-
тые тона с темными пестринами. Возрастные изменения окраски незначительны,
половой диморфизм выражается в большей яркости оперения самок (они при-
том несколько превосходят самцов и размерами). В проявлении полового
диморфизма, таким образом, отражаются особенности полового цикла трех-
персток. Крылья закругленные, состоят они из 10 первостепенных маховых;
хвост короткий и мягкий, рулевых 12.
Анатомические признаки. По строению нёба череп можно
отнести к форме, переходной между эгитогнатическим и схизогнатическим ти-
пами; сошник широкий и сзади раздвоенный, парасфеноид спереди вытянут,
небные кости с хорошо выраженными lamina interna. Базиптеригоидные отростки
начинаются на базисфеноиде, как у так называемых палеогнатических групп.
Ушной отдел квадратной кости с двумя сочленовыми буграми. Спинная кость
хорошо выражена, в образовании ее участвует до 9 позвонков, но задний
шейный позвонок с передним грудным не срастается. Грудина с одной лишь
парой вырезок на заднем крае. Вилочка с едва намеченным hypocleidium. Опор-
ный аппарат крыла вообще заметно слабей, чем у куриных. Зоба нет, передний
отдел пищевода сильно растяжим (резонатор, используемый самками при
«токовании»). Слепые кишки хорошо развиты и функционируют. Гортань
трахейно-бронхиального типа, трахейные кольца хрящевые. Перья с большим
побочным стволом. Копчиковая железа оперенная. Анатомические особенности
трехперсток указывают на их весьма обособленное положение в системе; внеш-
нее сходство с мелкими куриными представляет собой лишь конвергенция.
ХАРАКТЕРИСТИКА ОТРЯДА 93
Обрдз жизни. Дневные птицы, оседлые или полуоседлые, немногие, как
пятнистая трехперстка, у самой северной границы ареала перелетны. Пища
растительная, это преимущественно семена, нд также и мелкие беспозвоноч-
ные. Разыскивается корм на земле. На деревья трехперстки не садятся.
Размножение своеобразно. Пар не образуется, одна самка спаривается
с несколькими самцами, на последних ложится и вся «забота о потомстве».
Самка токует, бегая вокруг самца и издавая звуки, напоминающие ворко-
ванье; самец отвечает ей писком. Иногда самки дерутся из-за самцов. Кладка
у большинства видов состоит из 4 яиц (у рода Ortyxelus из 2 яиц), причем в
гнезде яйца располагаются, как и у куликов, весьма характерно, вершинами
к центру гнезда. Окраска яиц пестрая. Одна самка в сезоне делает, повиди-
мому, несколько кладок, каждая из которых насиживается разным самцом.
Сколько кладок в году насиживает самец —• одну или две — остается неясным.
Насиживание очень кратковременно, 12—13 дней, быстро протекает и разви-
тие птенцов, идущее по «выводковому» типу. В десятидневном возрасте птенцы
обычно летают, в возрасте 5—6 недель линяют, надевая оперение, сходное
в основных чертах с оперением старых птиц. В возрасте около месяца молодые
трехперстки становятся самостоятельными, а в тропиках, повидимому, присту-
пают к размножению, еще не достигнув 4 месяцев (Мерс, 1915). Гнезда распо-
лагаются на земле, делаются они из травы, устройство их несложно.
Систематика. Два семейства, резко различных: типичные трехперстки
Turnicidae и австралийские Pedionomidae.
У первых нога трехпалая; крыло квинтокубитального типа; имеется
только левая сонная артерия; яйца овальные, закругленные. Два рода Turnix
и Ortyxelus, в первом — 14 видов, во втором — 1.
У Pedionomidae имеется слабо развитый первый палец ноги, крылья аквин-
токубитальные, с резко укороченными тремя передними второстепенными
маховыми; сонные артерии парные; яйца грушевидной формы. Один род
Pedionomus с единственным видом.
Географическое распространение. Трехперстки населяют тропические и
субтропические части восточного полушария; два вида Turnix tanki и Г. sylva-
tica проникают в Палеарктику. Род Turnix имеет широкое распространение,
Ortyxelos встречается в южн. Судане, Pedionomus — в Австралии. Характерный
биотоп —травянистые степные и луговые пространства.
РОД ТРЕХПЕРСТКИ TURNIX BONNATERRE, 1790
Тип Т. sylvatica
1. Пятнистая трехперстка Turnix tanki Blyth
Turnix. tanki. Blyth. Journ. Asiatic Society of Bengal, v. 12, 1843, стр. 180, Бенгалия.
Распространение. Ареал. Южн. Азия — Индия от Пенджаба до
Ассама, к югу до Траванкора; Никобарские и Андаманские о-ва; Бирма;
Индо-Китай. Южн. и Вост. Китай, на север — до Манчжурии и Приморья.
Биотоп. Поля, луга, лесные опушки, кустарниковые заросли и т. д.
Характер пребывания. В большей части ареала оседла, на
севере (Чжили, вероятно в Шаньдуне, Манчжурии, Приморье) перелетна,
по крайней мере, частично.
Численность. На юге обычна; на севере относительно малочисленна
(возможно, здесь встречаются лишь перелетные популяции).
Подвиды и варьирующие признаки. В Индии на восток до Ассама—Т. /. tanki Blyth.,
1843; в остальной части ареала от Бирмы на восток—Т. t. blanfordii Blyth, 1863. Различия
этих подвидов заключаются в размерах (второй подвид крупней), и повидимому в экологи-
ческом отношении (строгая оседлость номинальной формы и частичная перелетность восточ-
ной; быть может в деталях цикла периодических явлений).
ОТРЯД ТРЕХПЕРСТКИ
Восточная или большая пятнистая трехперстка Turnix tanki blanfordii В1 у t h
Turnix Blanfordii. В 1 у t h. Journ. Asiatic Soc. of Bengal, XXXII, 1863, стр. 80, Тайет-
мио, Бирма.
Распространение. Ареал. Юго-вост. и вост. Азия: Бирма, Ассам
{к югу от Брамапутры), Тенассерим, Индо-Китай (Аннам, Кохинхина, Кам-
боджа, Лаос, Тонкий), Сиам, Шаньские государства, Китай — на гнездовье
в нем вероятно до долины Яньцзе-цзяня на юге (Латуш, 1932), на запад —
до среднего течения этой реки и Юннани, к северу до Манчжурии, Кореи и
южн. Приморья. На пролете и зимовках и на самом юго-востоке Китая,
в провинциях Фуцзянь и Гуандунь. В Приморье встречается в южной части
области; экземпляры добыты у озера Ханка, у залива Посьета, в окрестностях
Карта 15. Распространение пятнистой трехперстки Turnix tanki
7. Т. t. blanfordii. 2. Т. t. tanki;
a — граница гнездовий.
Владивостока, в низовьях реки Иман, в области Сихотэ-Алиня из долины
Сучана (Шульпин, 1936). Наблюдался у с. Маргаритовки на реке Псухун на
востоке. Северная граница проходит быть может у Хабаровска. На западе
добыта у Сретенска (вероятно залетный экземпляр), хотя граница ареала
в с.-з. Манчжурии подходит туда в сущности близко: добыта там не только
у Харбина, но и в окрестностях станции Джалантун в 115 км из. с.-з. от Цица-
кара, притом в гнездовое время (Мейзе, 1934). Наконец, молодая самка добыта
21.VIII.1941 г. на сухих лугах близ устья р. Кудалды, северо-восточное побе-
режье Байкала (Белышев, 1947).
Характер пребывания. В Приморье — перелетная птица, как
и в Сев. Китае (Чжили). Впрочем, для Порт-Артура приводится в качестве
редкой зимующей птицы (Курода, 1918), из южн. Манчжурии есть находки,
сделанные в начале февраля (Мейзе, 1934). Эти довольно противоречивые
сведения указывают, что часть популяций возможно зимует и на севере. В Сред-
нем Китае оседла, в Южном — область нормальных зимовок (Латуш, 1932).
ПЯТНИСТАЯ ТРЕХПЕРСТКА
95
Даты. Прилетает в Приморье одновременно с перепелом, сходные с кото-
рым биотопы занимает трехперстка (Медведев, 1909), около половины — конца
марта. Осенние экземпляры из Приморья известны от сентября и даже от 1.Х
(Шульпин). Весенняя миграция в Сев. Китае отмечается в мае, осенняя — в
Чжили с последней недели августа и до середины октября; появление в Южн.
Китае в Фуцзяне и Гуандуне отмечается в октябре—ноябре (Латуш,
1932).
Биотоп. В Приморье — поля, речные долины, холмы с густой травой
и кустарником, песчаные дюны с редкой растительностью (по реке Тумень-ула,
Шульпин). Влажных сырых участков с густой травой избегает. Встречается
и в горах (верховья реки Фаташи,
Шульпин).
Численность. При скры-
том образе жизни птицы трудно
поддается определению, тем бо-
лее, что в Приморье трехперстка
встречается у северной границы
ареала. У залива Посьета все же
повидимому нередка (Шульпин).
Экология. Размножение.
Сведения, относящиеся к нашей
стране, весьма отрывочны. Как
и другие трехперстки, у этой пти-
цы —полиандрия и агамия, самка
спаривается с несколькими сам-
цами; насиживание, забота о
выводке лежит только на самце
(но прямых наблюдений над этим
в СССР нет). В связи с этим, по-
видимому, самки численно пре-
обладают над самцами (Манчжу-
рия, Яковлев, 1929). Гнезда рас-
полагаются на земле, они пред-
ставляют собой стебли травы,
небрежно уложенные в ямке, под
прикрытием кустика травы (окрестности Харбина, Яковлев). Кладка состоит из
4 яиц; насиженная кладка у Владивостока найдена 19.VII. Яйца овально за-
остренные, по форме несколько шире чем у перепела; окраска их белая с бу-
ровато-серыми тонкими штрихами, густо расположенными и иногда закры-
вающими фон; кроме того, менее густо расположены серые, серо-голубова-
тые и черно-бурые пятнышки и пятна, обычно сосредоточенные на тупом
конце; размеры яиц Приморья 26 х 21, 25 х 21,5, 25,5x21,3, 25,5 х 21,7 мм
(Шульпин, 1930); для кладки из Харбина 25,8—27 х 19,6—20,1 мм (Яковлев,
1929); для яиц из Чженцзяня в Вост. Китае E4) 26 х 20,5 мм (Латуш, 1932);
для яиц разного происхождения A00) 25,5 х 20,8 мм (Бекер, 1928).
Насиживание начинается с откладки первого яйца, чем и объясняется то,
что в найденных яйцах зародыши разновозрастны (Харбин, Яковлев). Кладки
в Китае находили в июне —июле (Латуш, 1932; Кольтхофф, 1932). Старые
птицы у гнезд очень осторожны и исчезают при малейшей опасности. Нелетный
выводок из птенцов с остатками пуха найден у озера Ханка на п-ове Рябо-
конь 11.VII (Шульпин, 1930); 29.VII в Хасанском районе добыт подлеток
в гнездовом наряде (Горчаковская). Повидимому, в году бывает один цикл раз-
множения (Индо-Китай, Делакур, 1931). Продолжительность высиживания —
12 дней, через 10 дней после вылупления птенцы летают, в возрасте 5—6 недель
происходит ювенальная линька (Сет-Смис, 1907).
Рис. 16. Пятнистая трехперстка.
96 ___ ОТРЯД ТРЕХПЕРСТКИ
Питание. В желудках добытых в Приморье особей были обнаружены
мелкие насекомые, семена, песчинки. В желудке добытой близ Харбина
самки — две сухопутных улитки с их раковинами, у других — мелкие живот-
ные и растения (Яковлев).
Полевые признаки. Походит на перепела (закругленные крылья и корот-
кий хвост), но окраска однообразней, а полет слабей. Строго наземная, охотно
«купается» в песке. При опасности затаивается или, пролетев небольшое рас-
стояние в 50—60 шагов, убегает. Голос—писк (Пржевальский, 1876).
Описание. Размеры и строение. Крыло довольно длинное, но закруглен-
ное, из 10 первостепенных маховых. Формула 1>2>3>4.... Хвост мягкий, слабо развитый,
из 12 рулевых. Лапы трехпалые. Цевка неоперенная, покрытая спереди и сзади попереч-
ными пластинками. Самка заметно крупней самца. Длина самки 194, размах 340 мм (Тача-
новский,1893); длина молодого самца 160, размах 296 мм; вес 50,2 г. Крыло самцов 87,88,
самки 102 мм (Зоологический музей Московского университета); крыло самок B) 93, 103,
самца 88, птиц без определения пола 89,91 мм (Шульпин, 1930; экземпляры из Приморья
и Манчжурии); крыло самца 87, самки 96 мм (Приморье, Тачановский, 1891).
О краска. Пуховой птенец «на спинной стороне каштаново-бурый, вдоль темени
темнобурая полоса, от уздечки к глазам широкая белая полоска, на спине охристый и чер-
ный рисунок» (Хартерт, 1917). Взрослая самка — с черноватым теменем, каждое перо с
охристой каймой; иногда охристая полоска проходит вдоль темени; шея — ржавчато-ры-
жая; спина, плечевые и рулевые — серовато-бурые с тонким поперечным черноватым рисун-
ком и с разбросанными охристыми поперечными полосами; кроющие крыла — охристые с
глазчатыми черными пятнами; первостепенные маховые серо-бурые с охристыми краями
наружных опахал; рулевые цвета спины; горло охристое; щеки бурые с охристыми пестри-
нами; передняя сторона шеи и зоб — ржавчато-рыжие; остальная брюшная сторона —
охристая, темней в подхврстье и светлей на середине брюха и подкрыльях; по сторонам
зоба и груди и на боках—разбросанные черные круглые или полу лунные пятна. Повиди-
мому в году бывает 2 линьки, так как рыжий ошейник будто бы пропадает осенью после
периода размножения (Делакур, 1931).
Взрослый самец отличается от самки большей примесью охристых пестрин на спинной
стороне тела, меньшим развитием ржаво-рыжего цвета на нижней стороне шеи и на зобе
и отсутствием этого цвета в зашейке. Клюв желтый, буроватый у вершины; лапы желтые;
радужина бледнобурая или желтая. В гнездовом наряде вся окраска бледней, брюхо белова-
тое, охристые пестрины на спинной стороне сильно развиты, в особенности на кроющих
крыла (самец, 22.VII.1945 г., Чертова горка. Хасанский район, сб. Горчаковской).
ОТРЯД ЖУРАВЛИ
GRUES или GRUIFORMES
Характеристика отряда
Строение. Внешние признаки. Журавли — крупные птицы
с длинными ногами и шеей (только у трубачей шея умеренной длины). Голова
небольшая. Наиболее крупных размеров в нашей фауне достигают белые жура-
вли — высота их около 180 см, длина крыла — 600—690 лм, клюва 180— 200 лм,
вес около 10 кг, а вес манчжурского журавля достигает даже 15 кг. Самый
мелкий представитель журавлей у нас —журавль-красавка: вес его 2000—
2750 г. Самцы обычно одних размеров с самками, только у некоторых видов
незначительно крупнее их.
Оперение жесткое, плотно прилегающее к телу. Цевка и часть голени,
прилегающая к цевке, не оперены. Цевка покрыта спереди широкими крупными
поперечными щитками, сзади цевка — сетчатая. У многих видов семейства
журавлиных лицевые части головы, иногда темя и затылок частично не оперены,
а покрыты бородавчатой черной, красной или черной с красными пятнами кожей
с редкими волосовидными перьями (серый журавль, черный, даурский, стерх,
манчжурский и др.), у некоторых видов (журавль-красавка, четырехкрылый
журавль Anthropoides paradisea, трубачи и арама) голова вся оперена. На
голове и на теле у некоторых видов имеются удлиненные перья или хохлы,
последние образованы из длинных, жестких щетиноподобных перьев (венце-
носный журавль) или из перьев с несцепленными между собою бородками.
У журавля-красавки, четырехкрылого журавля и венценосного Balearica pavo-
nina перья зоба заострены и удлинены. У большинства журавлей третьесте-
пенные маховые длинные и обычно заходят за вершину крыла, реже они
бывают такой же длины как и крыло. У ряда видов (серый журавль,
канадский и др.) третьестепенные маховые рассучены, у журавля-красавки,
четырехкрылого журавля и венценосного они заострены и почти не рассу-
чены, а у венценосного журавля рассучены внутренние большие кроющие
крыла. Специальных украшающих перьев, развивающихся в брачный период,
у журавлей нет. Контурные перья имеют слабо развитый побочный ствол.
Пух располагается и по аптериям и по птерилиям.
7 Птицы СССР, т. II
98 ОТРЯД ЖУРАВЛИ
Окраска оперения за исключением тропических видов, скромная — серая
с черным, белая с черным, буровато-серая или буровато-черная. Большинство
тропических видов характеризуется наличием в окраске оперения блеска —
бронзового, пурпурового, фиолетового и зеленого. Полового диморфизма в
оперении нет. Возрастные отличия слабые, выражаются главным образом
в более сильном развитии буроватых тонов, отсутствии красного пятна на
темени у молодых. Окончательный взрослый наряд одевают поздно: например,
серый журавль —в возрасте 2*/2 лет, канадский — 3 лет, последний, по наб-
людениям Бента A926), приступает к размножению, не одев еще взрослого
наряда.
Крылья не длинные и широкие, вершина крыла образуется 2-м, 3-м и 4-м ма-
ховыми перьями, причем 3-е маховое перо часто очень незначительно длиннее 2-го
и 4-го, которые обычно равны между собою. У Gruidae крыло диастаксическое,
у Psophiidae — эутаксическое. Первостепенных маховых И, за исключением
трубачей арам и венценосного журавля, у которых их 10. Хвост короткий
и у Gruidae состоит из 12 рулевых перьев, у Psephiidae —из 10.
Клюв довольно длинный, особенно у трубачей, только у арам короткий.
Обычно он у видов, встречающихся в СССР (за исключением журавля-кра-
савки), прямой, цилиндрической формы, сильный и немного длинней головы;
у арам клюв по коньку надклювья сильно выпуклый. Ноздри сквозные удли-
ненные, расположены в основной половине клюва. Края клюва обычно гладкие,
только у некоторых видов журавлей в вершинной части слабо зазубрены
(стерх, манчжурский журавль, серый журавль и др.).
Ноги длинные, четырехпалые; задний палец D-й) поставлен несколько выше
остальных и не касается земли при хождении. Наружный и средний пальцы
соединены между собою небольшой перепонкой. Пальцы средней величины,
за исключением арам, у которых длинные. Когти тупые, широкие и короткие.
Анатомические признаки. Череп схизоринальный и схизо-
гнатический, а у трубачей —голоринальный. Базиптеригоидных отростков нет.
Затылочные фонтанелии, за исключением трубачей, имеются. Шейных позвон-
ков у настоящих журавлей 19—20, у арам — 17 и у трубачей 17—18. Несколько
грудных позвонков срастаются в спинную кость. Грудных ребер у Gruidae — 7
и у Psophiidae — 8. Грудина без вырезок на заднем крае. Трахея длинная,
часто образует извивы и петли, только у венценосного журавля трахея прямая.
У арам, четырехкрылого журавля и у самок некоторых видов журавлей
(Grus leucogeranus и Anthropoides virgo) петли образуются выше гру-
дины, у остальных трахея заходит в киль грудины и там образует петлю.
Нижняя гортань трахейно-бронхиального типа. Обводящая мышца имеется.
Две сонные артерии. Слепые кишки функционируют. Копчиковая железа
оперена.
Образ жизни. Дневные, наземные и отчасти древесные птицы. Большин-
ство наземных птиц связано с обширными открытыми болотами, расположен-
ными обычно среди разреженного леса, лесостепи, кустарника и степи (различ-
ные виды Gruidae), в тундру почти не заходят, но в горы при наличии подходя-
щих условий поднимаются высоко — до 2000 м. Меньшая часть (например,
род Anthropoides) — чисто степные и даже полупустынные птицы, селящиеся
от воды иногда в 2—3 км и прилетающие к воде только на водопой. Трубачи
и арамы обитают в обширных тропических болотистых лесах; они садятся
на деревья, хорошо передвигаются по земле, но плохо летают. Чисто наземные
виды (семейство настоящие журавли) никогда на деревья не садятся, хорошо
ходят по земле и хорошо летают, но полет медленный, с земли поднимаются
довольно легко, но с небольшого разбега, сделав 2—3 шага по земле.
Северные формы — перелетные, южные — оседлые. Зимовки гнездящихся
у нас видов журавлей расположены обычно далеко от гнездовий: в Китае, Индии,
Африке, а канадского журавля — в южных частях Сев. Америки. Северные
ХАРАКТЕРИСТИКА ОТРЯДА 99
области зимовок некоторых журавлей, например, даурского журавля, манч-
журского, канадского, серого—частично граничат с южными областями гнездо-
вого ареала. У журавля-красавки и черного журавля особенно сильно разоб-
щены область зимовок и гнездовая область. Трубачи и арамы оседлы. Прилетают
к нам рано, в особенности некоторые виды, например, манчжурский журавль
прилетает в Приморье, когда там стоят еще морозы и лежит снег. Летят обычно
стаями и, если птиц много, разбиваются на отдельные более мелкие стаи,
летящие на близком расстоянии друг от друга, но иногда летят и отдельными
особями, парами и небольшими группами. Во время пролета журавли летят,
выстроившись клином и меняясь но временам местами. При полете вытягивает
ноги и шею по прямой линии во всю длину.
По прилете и, главным образом, осенью перед отлетом держатся стаями,
совершая небольшие кочевки в поисках пищи. Не размножающиеся, холостые
птицы собираются в стаи и в продолжение всего гнездового сезона ведут коче-
вой образ жизни, и иногда во время кочевок встречаются далеко от своего
гнездового ареала, например, стерхи, черные журавли и др.
Пища преимущественно растительная и лишь в небольшом количестве
животная, но, повидимому, некоторые виды, например, стерх, манчжурский
журавль, поедают в большом количестве животные корма, а арамы питаются
преимущественно животной пищей, главным образом, улитками. В состав
растительной пищи входят: всходы и зерна хлебных злаков, почки, стебли,
ягоды, на севере — в особенности клюква, клубни и корни различных растений.
Из животных кормов — насекомые, преимущественно кобылки и водные насе-
комые, черви, моллюски, мелкие земноводные, рыба, иногда мелкие млекопи-
тающие и т. д. Корм добывают и собирают на земле при помощи клюва, только
арамы иногда придерживают пищу ногами, например, в момент освобождения
моллюска из раковины.
Моногамы. К размножению приступают на второй, быть может третий год
жизни. Гнездятся парами, на большом расстоянии пара от пары, только кра-
савки иногда устраивают гнезда близко друг от друга.
Настоящие журавли гнездятся на земле, арамы устраивают гнезда и на
земле и на деревьях и, наконец, трубачи гнездятся только на деревьях в гнез-
дах, устроенных на ветвях или в дуплах. Наши журавли, за исключением
красавки, гнездятся в обширных, труднопроходимых болотах, выбирая для
гнезд открытое, возвышенное, сухое место среди болота; если есть камыши и
тростники вокруг гнезда, то они обычно вытаптываются птицами. Гнездо
очень простое и обычно имеет вид набросанной кучи хвороста или тростника
и камыша, часто без всякой подстилки в лотке. Журавли-красавки гнезда со-
всем не строят и откладывают яйца чаще всего прямо на землю или на гальку,
иногда делая небольшое углубление в земле, выстилая его сухими травами или
огораживая кладку мелкими камешками. В кладке у настоящих журавлей
чаще 2 яйца, реже 1 или 3; у арам — 5—8 яиц и у трубачей — от 6 до 10 яиц.
Яйца пестро окрашены, за исключением яиц трубачей, которые белого цвета.
Окраска яиц сильно варьирует, даже у одного и того же вида. Для журавлей
очень характерны в весенний период брачные игры или как их обычно назы-
вают «журавлиные пляски». В местностях, где много гнездящихся пар журав-
лей, устраиваются общие «пляски», особенно многочисленные и азартные у жу-
равлей-красавок. Изредка «пляски» наблюдаются и осенью.
Выводковые птицы. Эмбриональное развитие медленное, но точно время
насиживания для большинства видов журавлей не выяснено. Серый журавль
высиживает 29—30 дней, журавль-красавка около 30 дней. Птенцы вылупляются
одетые пухом, у настоящих журавлей — охристо-рыжеватым, у арам — черным
и у трубачей — светло-коричневым. Птенцы, как только обсохнут после вылу-
пления, могут следовать за родителями, только у арам птенцы долгое время
остаются в гнезде и выкармливаются родителями.
100 ОТРЯД ЖУРАВЛИ
Смена нарядов у птенцов изучена слабо. У серого журавля два пуховых
наряда, для других видов количество пуховых нарядов не выяснено. Взрослые
журавли имеют 2 линьки в году: полная, послебрачная, начинается в конце лета
и заканчивается в начале зимы, и частичная —предбрачная —в конце зимы —в
начале весны. Линька маховых перьев (первостепенных и второстепенных) у на-
стоящих журавлей, за исключением журавля-красавки и венценосного, идет
одновременно, и птицы в течение нескольких дней C—5) теряют все маховые
и утрачивают на короткое время —дней на 12—15 —способность к полету.
У журавля-красавки и венценосного журавля линька маховых протекает по-
степенно. К отлету все крупное оперение у журавлей бывает уже сменено,
но среди мелкого пера имеются еще старые перья. Долинивают птицы в пути
и на зимовках.
Систематика. Отряд журавлей делится на 3 семейства: настоящие журавли
Gruidae, арамы Aramidae и трубачи Psophiidae.
Семейство трубачи или агамы Psophiidae. Размером с мелкого журавля
(журавля-красавку), с длинными ногами и средней длины шеей. Хвост короткий.
Шейных позвонков 17—18. Клюв очень короткий, слабый и сильно изогнут
по коньку надклювья. Верхний край глазницы образован из пяти отдельных
костей ossa supraobitalia. Грудных ребер —8.
Семейство трубачи состоит из одного рода трубачи Psophia с 3 видами, рас-
пространенными в Южн. Америке.
Семейство арамы или постушковые журавли Aramidae. Самые мелкие
птицы отряда журавлей несколько меньше журавля-красавки. Ноги длинные
с длинными пальцами. Шея умеренной длины. Шейных позвонков — 17. Клюв
очень длинный, прямой, только у вершины слегка изогнут вниз. Грудных
позвонков —7, Hypocleidium вилочки не развиты. Spina externa sterni неболь-
ших размеров. Трахея образует петлю выше грудины и в киль грудины не
входит.
Семейство арам содержит один род Aramus с одним видом Aramus scolo-
paceus, распространенным в Центр, и Южн. Америке, на п-ове Флорида
и на о-ве Куба.
Семейство настоящие журавли Gruinae. Самые крупные птицы отряда.
Шейных позвонков — 19—20. Грудина узкая и длинная, в особенности у пере-
летных видов, у них же и вилочка срастается с килем.
Семейство журавлей делится на 2 подсемейства: 1) венценосные журавли
Balearicinae и 2) настоящие журавли Gruinae. Настоящие журавли характери-
зуются отсутствием совершенно голых частей на голове (у некоторых видов
голова густо оперена, у других покрыта щетиноподобными волосовидными
перьями) и отсутствием хохла на затылке. У венценосных журавлей на голове
есть голые участки кожи и на затылке хохол. Трахея у настоящих журавлей
обычно очень длинная, делающая петлю в киле грудины, в связи с чем у них
развит громкий трубный голос. У венценосных журавлей трахея много
короче и прямая.
В подсемействе настоящие журавли Gruinae 3 рода: настоящие журавли
Grus с 10 видами, журавли-красавки Antrhopoides с 2 видами и африканские
журавли Bugeranus с 1 видом. Подсемейство настоящие журавли широко
распространено во всем свете, за исключением Южн. Америки. Подсемейство
венценосные журавли Balearicinae с 1 родом и 1 видом, венценосный журавль
Balearica pavonina, обитающим в южной и тропической Африке.
Ископаемые журавли известны уже с эоцена. В верхнем эоцене во внутрен-
ней Монголии был найден своеобразный вид Eogrus aeola, выделяемый в особое
семейство и обладающий чертами, переходными между журавлями, с одной
стороны, с кариамами и дрофами — с другой.
Из эоценовых отложений Англии, Франции, Италии и Сев. Америки
известны пять ископаемых родов: Geraaopsis (один вид), Palaeogrus (три вида),
ХАРАКТЕРИСТИКА ОТРЯДА 101
Paragrus (один вид), Geranoides (один вид) и Protogrus (два вида). Весьма инте-
ресна находка в верхнем олигоцене во Франции ископаемого вида Probalaerica
problematica, напоминающего живущих ныне э Африке венценосных журавлей.
Особый род Pliogrus (два вида) описан из плиоценовых отложений Герма-
нии и Пикерми.
Из нижнего плиоцена Ирана описан журавль Urmiornis maraghanus;
к этому же виду относятся, повидимому, находки ископаемых журавлеподоб-
ных птиц, сделанные в нижних меотических и сарматских отложениях у Ти-
располя.
В плейстоцене из известных бинагадинских Кировых отложений в Азер-
байджане описан стерх Leucogeranus bohatschevi (Серебровский, 1940). В этих же
отложениях найдены и остатки современного стерха, распространенного теперь
только на севере Сибири.
Ископаемые остатки представителей семейства Aramidae известны только
из Сев. Америки (роды Badistornis и Gnotornis в олигоцене и род Aramornis
в миоцене).
Географическое распространение. Журавли широко распространены по
всем странам света, за исключением крайнего севера и юга. Их можно назвать
материковыми птицами, на островах журавлей почти нет (они населяют только
некоторые острова северо-восточной части Сев. Америки, Кубу, Новую
Гвинею и Японию). Особенно широко распространено семейство настоящих
журавлей Gruidae, которых нет только в Центр, и Южн. Америке. Два
других семейства имеют ограниченное распространение: трубачи имеются
только в Южн. Америке, а арамы в Южн. и Центр. Америке, на п-ове Флорида
и на о-ве Куба. В фауне СССР представлено 2 рода журавлей и 7 видов, при-
надлежащие к семейству настоящих журавлей. Гнездящиеся в СССР виды
журавлей от общего числа видов отряда A8) составляют 38,9%. Из 7 гнездя-
щихся у нас видов журавлей 1 вид — стерх — гнездится только в СССР,
2 вида — черный и даурский журавли — почти не выходят за пределы нашей
страны и у 2 видов —журавля-красавки и манчжурского журавля —основной
гнездовый ареал лежит в нашей стране. Монотипических видов в фауне СССР—5.
Распределение журавлей на гнездовья по зонам в нашей стране следую-
щее (в числителе показано число видов, в знаменателе — число подвидов и
монотипических видов; характерные формы подчеркнуты):
I. Арктика (зона)
Grus grus, G. canadensis и G. leucogeranus 3
а. По бе ре ж ье (подзона) О
б. Тундра (подзона)
G. g. grus, G. g. lilfordiy G. canadensis и G. leucogeranus 3/2
в. Лесотундра G. g. grus, G.g. lilfordi, G. canadensis, G. leucogeranus 3/2
II. Лесная зона
G. grus, G. leucogeranus, G. canadensis 3
г. Тайга (подзона)
G. g. grus, G. g. lilfordi, G. canadensis и G. leucogeranus 3,2
д. Смешанный и лиственный лес (подзона)
G. g. grus, G. g. lilfordi, G. vipio, G. monachus 3/2
е. Л е состе п ь
G. g. grus, G. g. lilfordi, G. vipio, G. monachus 3/2
III. Зона открытых сухих пространств
G. grus и Anthropoides virgo 2
ж. Степь (подзона)
G. g. grus, G. g. lilfordi и Athr virgo (озерная сторона Зап. Сибири и Казахстана) 2/2
з. Полупустыни и пустыни (подзона)
Athr virgo, G. g. lilfordi 2/1
102 ОТРЯД ЖУРАВЛИ
IV. Горная зона
О. grus, A. virgo 2
и. Высокогорная борео-альпийская подзона 1/1
G. g. lilfordi
к. Горно-лесная подзона
G. g. lilfordi 1/1
л. Горно-степная и пустынная подзона
G. g. lilfordi и A. virgo 2/1
Хозяйственное значение. Промыслового значения не имеют. Иногда
приносят (серый журавль, журавль-красавка) вред сельскому хозяйству,
поедая и вытаптывая весною всходы зерновых культур, а осенью — созревшие
хлеба, однако вследствие малочисленности журавлей, особенно в местностях,
густо заселенных человеком, где хорошо развито хлебопашество, вред, прино-
симый ими, незначителен.
Таблица для определения видов журавлей
1 B) Преобладающая окраска оперения белая или, у молодых, охристо-рыжая . . 3
2 A) Преобладающая окраска оперения иная, белого цвета в оперении мало ... 5
3 D) Первостепенные маховые и верхние кроющие крыла черные. Остальное опере-
ние белое. Клюв и ноги красные стерх Grus leucogeranus
4 C) Второстепенные и третьестепенные маховые и верхняя часть шеи черная, осталь-
ное оперение белое. Клюв и ноги черные
манчжурский журавль Grus japonensis
5 F) На голове у взрослых птиц неоперенных участков кожи нет. По бокам головы
имеются пучки удлиненных белых или светлосерых (молодые птицы) перьев. Перья зоба
удлинены и заострены журавль-красавка Anthropoides virgo
6 E) На голове (взрослые) имеются неоперенные участки кожи, по бокам головы и
на зобе нет удлиненных перьев 7
7 (8) Третьестепенные маховые белые . ., . даурский журавль Grus vipio
8 G) Третьестепенные маховые серые или черные 9
9 A0) Оперение очень темное, почти черное, однообразное, только оперенная часть
головы и часть прилежащей к ней шеи — белые. . . черный журавль Grus monachus
10 (9) Окраска оперения тела и распределение белого цвета на шее и голове иное . . 11
И A2) Вся шея серая, третьестепенные маховые без черных концов. Первостепенные
маховые с бледножелтыми стволами перьев ...канадский журавль Grus canadensis
12 A1) На шее имеются белые участки перьев. Третьестепенные маховые с черными
концами. Первостепенные маховые с темными стержнями перьев
серыйжуравль Grus grus.
СЕМЕЙСТВО ЖУРАВЛЕЙ GRUIDAE
РОД ЖУРАВЛИ GRUS PALLAS, 1767.
Тип Ardea grus
Синонимы. Megalornis. Gray. 1841, тип M. grus.— Leucogeranus. Bonaparte.
1855, тип G. leucogeranus. — Pseudogeranus. S h a r p e, 1893, тип P. leucauchen. — Sarcogera-
nus Sharpe, 1893, тип. S. leucogeranus.
1. Серый журавль Grus grus L.
Ardea grus. Linnaeus. Syst. Nat., X, 1758, стр. 153, Швеция.
Синонимы. Grus vulgaris. Pallas. Zoogr. R-As., II, 1827, стр. 106. — Grus
communis. Bechstein. Gen. Nat. Deutsch., HI, 1793, стр. 60. — Gruscinerea. Meyer.
in Meyer und Wolf. Taschenb Deutsch., Vog., II, 1810, стр. 350.
Русское название народное несколько искаженное, звукоподражательное.
Распространение. Ареал. Сев.-вост. Германия, Скандинавия, Финляндия,
Кольский п-ов к северу до 68° с. ш., восточнее на север примерно до 66—
67° с. ш., нижнее течение Оби, Нижняя Тунгуска, Вилюй, Лена (до впаде-
ния Алдана), Индигирка (до устья Момы). На восток, может быть,—до Колымы,
СЕРЫЙ ЖУРАВЛЬ
103
Западная граница распространения идет от Прибалтики и Польши к югу до
средней Украины. Южная граница —до Киевской обл., центральной части
Днепропетровской обл.; нижней Волги (к югу до 47° с, ш.), среднего течения
Урала и Эмбы, сев.-вост. побережья Арала, низовьев Сыр-Дарьи, Сары-Су,
Чу, Или; Иссык-куль, Зайсан, Алтай, Забайкалье, Даурия. Изолированные
колонии на Сиваше, в Армении, в юго-зап. Закавказье, в Малой Азии, на
Балканах, в Венгрии, в сев. Италии, южн. Испании (Андалузия), в Тунисе.
Раньше гнездился в Австрии.
Карта 16. Распространение серого журавля Grus grus
1. Q. g. grus, 2. G. g. lilfordi;
a — граница гнездовий, г — область зимовок.
Зимует в сев. Африке, Месопотамии, Иране, сев. Индии и Китае. В неболь-
шом количестве остается на зиму на юге Европы и в Закавказье,
Характер пребывания. Гнездящаяся перелетная, местами осед-
лая птица. Прилетает в различные места, расположенные на одной и той же
широте, неодновременно, что, повидимому, зависит от близости и дальности
расположения зимовок отдельных популяций и от направления пролета.
Биотоп. Разнообразен, но всегда связан с глухими, обширными, топ-
кими болотами, озерами и пойменными лугами. Если такие места имеются
в тайге, лесостепи, степи, полупустыне или в горах, то серый журавль там
гнездится. В тундру не заходит и только, как исключение, иногда гнездится
в южной ее окраине.
Численность. Всюду обыкновенен, но немногочисленен.
Подвиды и варьирующие признаки. Два подвида, слабо различающихся морфологи-
чески и экологически: G. g. grus Linn., 1758 — Европа на восток до Печоры и Волги и сев.
Африка и G. g. lilfordi S h а г р е, 1894 — Малая Азия, Закавказье, Казахстан и Эап. и Сред-
няя Сибирь.
104 ОТРЯД ЖУРАВЛИ
Систематические замечания. Слабо различающиеся подвиды, быть может и не сущест-
вующие реально, географическую границу между ними провести трудно. В коллекциях
журавли представлены всегда довольно бедно, а возрастные различия у них очень велики.
Из всех приводимых отличий между подвидами наиболее стойким, повидимому, является
различие в общем сером тоне оперения — серый и светлосерый. Но чистый серый тон встре-
чается только весною, летом серый цвет обычно приобретает буроватый оттенок различной
интенсивности у разных особей, затрудняющий сравнение; все же птица из Сибири и восточ-
ных областей Европейской части Союза в среднем светлей. Величина красного пятна — ско-
рее признак возрастной и до некоторой степени половой. Различия в развитии черного цвета
на удлиненных третьестепенных маховых и окраску самих перьев я считаю также скорее
возрастным признаком, так как у всех неполовозрелых птиц черный цвет развит меньше,
чем у старых.
Хозяйственное значение и вред. Мясо журавлей вполне
съедобно, но промыслового значения журавли не имеют вследствие их мало-
численности и трудности охоты на них.
Посещая хлебные поля, журавли местами приносят известный вред, поедая
и вытаптывая весною всходы хлебов, а в конце лета и осенью — созреваю-
щие зерна хлебов. Но количество журавлей, как уже указано, невелико, в
особенности в местностях, густо населенных человеком, где хорошо развито
сельское хозяйство.
Журавли хорошо переносят неволю и быстро становятся ручными.
Западный серый журавль Grus grus grus L.
Распространение. Ареал. Сев.-вост. Германия, Скандинавия, Финляндия
к северу до 68° с. ш., Кольский п-ов к северу до бассейна р. Имандры и Пазы
(Гебель, 1902), Соловецкие о-ва (Поляков, 1929), Архангельская обл. к северу до
лесотундры (заходит в юго-западную часть Тиманской тундры по р. Пеше, Семе-
нов, 1939),средняя и нижняя Печора к северу от Усть-Усы F6° с. ш.),Усть-Цыль-
мы и быть может до полярного круга (со слов местных охотников, Дмохов-
ский, 1933). Далее на восток обитает, повидимому, другая форма, во всяком
случае на Оби — уже lilfordi, но весьма возможно, что птицы европейской
части Молотовской обл. также принадлежат к lilfordi. К югу доходит (запад-
ная граница указана в распространении вида) до Волыни, Подолии, Киевской,
Полтавской областей и до центральных частей Днепропетровской (в Павлодаре
и в заповеднике Конча-Заспа, в нижнем течении Днепра обычен, Шарлемань,
1930; но до Запорожья не доходит, Вальх, 1897). Повидимому, изолирован-
ная колония имеется в Сивашье и Присивашье (Воронцов, 1937). В Крыму
не гнездится (Пузанов, 1933, ошибочно считает его редко гнездящейся пти-
цей Крыма, ибо никто из исследователей Крыма его там на гнездовье не встре-
чал). На Дону доходит к югу приблизительно до 48°5'с. ш., на Волге —до 47°
(до Астрахани не доходит километров 150, Бостанжогло, 1911). В Заволжье
уже другая форма, граница между подвидами точно не выяснена, может быть,
проходит и несколько западнее. К северу идет по среднему Поволжью, воз-
можно захватывая и бассейн Суры. Изолированная колония есть на Балканах
(Сербия, Босния, Албания, Македония), в Добрудже, в низовьях Дуная и в
Венгрии, в сев. Италии (Венеция), в южн. Испании (Андалузия) и в сев.
Африке (Тунисе).
Зимует в сев.-вост. Африке от Марокко до Египта и к югу до Судана
и Абиссинии; в Месопотамии и в Загроше в Иране. Отдельные особи оста-
ются на зиму в Закавказье —в Муганской степи (Радде, 1885), у Ленкорани
(Спангенберг наблюдал в течение января — февраля пару журавлей в с.
Вель у иранской границы; ночевать прилетали на одно и то же дерево, стоя-
щее среди болот рисовых полей), на нижнем течении р. Храмы, на высоте более
300 м (Цветков, 1901), возможно изредка зимуют в Крыму и в небольшом
количестве в южн. Европе.
СЕРЫЙ ЖУРАВЛЬ 105
Характер пребывания. Журавль — гнездящаяся, перелетная
и отчасти зимующая птица. Птицы, зимующие в Закавказье, летят весною не на
север, а на юг в горы. К нам прилетают рано, двумя волнами — с юга и юго-
запада. У Ленкорани первые птицы появляются 16—19. III, валовый пролет
идет 24—26. III, по среднему течению Аракса —с 24.11 по 6. IV, в Армении
в районе Еревана и в Нахичевани —в среднем —26.III (Радде, 1885; Ляй-
стер и Соснин, 1942). В окрестностях Тбилиси первые птицы наблюдаются
между 12 и 15.III, валовой пролет — с 15—20.III (Радде, 1885). Пролет на
Кавказе тянется до конца апреля, но единичные пролетные птицы или пары
встречаются даже в мае (Беме, 1926, 21. V в Кизлярском районе). В южных
и юго-западных частях Украины первые пролетные журавли наблюдаются
поздней — со второй половины марта и пролет идет до середины апреля. В
Подолии (в Тростянцах, наблюдения за 16 лет) журавли в среднем прилетают
30—31.III, самый ранний прилет отмечен 24.III. 1906 г., самый поздний —
6. IV. 1899 и 1909 гг. (Бауман, 1924); в низовья Днепра (заповедник Конче-
Заспа) в среднем первые птицы прилетают 9. IV, самый ранний прилет —
25.III. 1925 г., самый поздний — 18.IV. 1928 г. (Шарлемань, 1930). В Павло-
даре пролет идет с середины марта и до середины апреля. В Ростове-на-Дону в
1906 г. первые журавли появились 5.III (ранняя весна), затем, вследствие
наступившего похолодания, пролет прекратился до 16. III и с 20. III наблю-
дался массовый пролет, к 31.111 пролет заметно ослабел (Сарандинаки, 1909).
В Харьковскую обл. первые птицы прилетают в среднем 16—17. III, вало-
вой пролет идет в первых числах апреля A—2) и заканчивается в начале вто-
рой половины апреля (Сомов, 1897, наблюдения за 11 лет). В центральные
области первые птицы в среднем прилетают 10—12. IV. В Московскую обл.
(наблюдения за 21 год) в среднем прилетают 10. IV, самый ранний прилет отме-
чен 31.III. 1924 г., самый поздний —20. IV. 1920г. (Паровщиков, 1941). В Ива-
новскую обл. в среднем прилетают также 10. IV, валовой пролет идет 18—19. IV
(Птушенко, 1937). В Новгородскую обл. в среднем прилегают — 11. IV (Бианки,
1910), в Псковскую — 13—14. IV (Нестеров и Никандров, 1913), в южных
частях Кировской обл. первые птицы отмечены во второй половине апреля,
в окрестностях Чердыни — 6.V. 1827, на Обве —25. IV. 1873 г. (Резцов, 1904),
над Усть-Цыльмой пролетные птицы наблюдались 21.V и 25.V (Сибом, 1876).
В конце июля или в начале августа семьи соединяются в небольшие стаи
и начинают вести кочевой образ жизни. В это время они откочевывают далеко
от мест своих гнездовий, иногда даже передвигаются к северу; в горных рай-
онах спускаются с гор в более низкие местности. Кочующие группы местных
птиц постепенно объединяются в большие стаи, уходят все дальше от мест
гнездовий и, наконец, покидают их совсем. Обычно через несколько дней
после отлета местных птиц наблюдается пролет более северных птиц. Момент
отлета местных птиц и начало пролета более северных очень трудно различимы.
Осенью журавли пролетают медленно. Чаще делают остановки, иногда
довольно длительные — до заморозков, собираясь тогда в большом количестве
на болотах или даже на отмелях рек и на хлебных полях. В ясную погоду летят
высоко, так что иногда птиц не видно, а только слышно, в пасмурные дни летят
низко. Стаи достигают большей величины, чем весною —от нескольких десят-
ков до нескольких сотен; при этом они разбиваются на несколько отдельных
клиньев, которые летят на близком расстоянии друг от друга.
Из северных частей своего распространения серый журавль начинает
улетать во второй половине августа и отлет идет до конца сентября, иногда
задерживаясь до начала октября, и случайные одиночные, повидимому заблу-
дившиеся птицы, встречаются и в конце октября B3.Х, Кировская обл., Кру-
ликовский, 1913). В средней полосе отлет идет с первых чисел августа, но иногда
наблюдаются пролетные птицы даже в июле (Московская обл., Воскресенский
район, 14.VII встречена пролетная стая в 44 особи, Евтюхов, 1928; в 1894 г.
106 ОТРЯД ЖУРАВЛИ
с 22.VIII шел валовой пролет журавлей, Семенов, 1894 г.). Валовой пролет
идет в сентябре и к концу месяца заканчивается, одиночные стаи встречаются
и в первой половине октября. На Украине отлет местных птиц начинается со
второй декады августа A1—13.VIII) и идет до конца декады и быть может
до середины третьей. Наибольшей интенсивности пролет достигает в середине
сентября. Заканчивается пролет в конце сентября и в первых числах октября,
но одиночные стаи и птицы встречаются и позднее.
В Абиссинии первые журавли осенью появляются в начале сентября
G. IX), в Нубии уже с конца августа и в течение всего сентября идет прилет
серых журавлей на зимовки. Покидают зимовки рано — из Нубии улетают
уже в феврале, в Абиссинии последние птицы улетают в середине мая.
Пролет идет в разное время ночи и дня. При сильном похолодании стаи
поворачивают обратно к югу и пролет на несколько дней прекращается. Летят
обычно треугольником или, как принято называть, журавлиным «клином».
Величина стаи различна, особенно большой величины стаи достигают осенью.
Во время валового пролета стая летит за стаей с очень короткими промежутками
времени. Если стая очень велика, то журавли обычно разбиваются на несколько
групп, которые летят на близком расстоянии друг от друга. Обычно в
стае от 10 до 50 особей. Но иногда летят и стаями в несколько сот особей (до
400 и более), в таком случае они держатся не клином, а волнистой линией или
очень тупым углом, часто в несколько рядов. Летят на большой высоте, в осо-
бенности весной. Останавливаются редко в безлюдных местностях — на боло-
тах или хлебных полях. Прежде, чем спуститься на землю, стая делает неко-
торое время круги и несколько раз поднимается вверх и опускается вниз. Во
время пролета над большими болотами стаи снижаются, кружат в воздухе,
но обычно не останавливаются, а летят дальше. Горные хребты не облетают,
летят прямо через горы. На Кавказе летят через Главный Кавказский хребет,
поднимаясь на высоту до 4000 м, пролетая через Казбек и Эльбрус (Радде, 1885).
Биотоп. Крайне разнообразен, но всегда связан с водой — с обшир-
ными глухими топкими лугами, болотами, озерами, если такие места имеются
в тайге, лесостепи, степи, полупустыне или в горах, то серый журавль обычно
там гнездится, в тундру не заходит, только, как исключение, гнездится в южной
окраине тундры. Для северной полосы Европейской части СССР характерные
места гнездовья — большие моховые болота, поросшие редким низким сосня-
ком, с кочками клюквы, окруженные водой, сфагновые болота, или обширные
сырые, топкие травянистые луга с разбросанными небольшими редкими груп-
пами низкорослой ольхи. Для средней полосы — кочковатые болота, иногда
окруженные лесом, поросшие кустарником ольхи и ивы; болота в пойме рек,
поросшие тростником, с небольшими кочкообразными возвышениями над уров-
нем воды; осоковейниковые болота с крупными кочками, заболоченные луга с
низким редким кустарником, поросшие камышом и осокой, а на мшистых
местах богульником. На Украине — крупные болота старицы, травянистые
лиманы, расположенные поблизости ольховых рощ и лесов; ольховые мок-
рые срубы с густой болотистой травой; болотистые понижения среди холми-
стых увалов украинской степи, поросшие по краям орешником и молодым
дубняком.
Численность. Обычен, как может быть обычна крупная осто-
рожная птица, подвергающаяся во многих местах постоянным преследова-
ниям. Абсолютная общая численность поэтому невысока, в особенности на юге
и в средней полосе.
Экология. Размножение. По прилете на места гнездовья журавли
держатся некоторое время стаями, хотя, повидимому, пары у них постоянные.
Сейчас же по прилете на места гнездовий у журавлей начинаются брачные
игры — «пляски*. Пляски в местностях, где серых журавлей много, иногда
принимают общий характер и похожи на пляски журавлей-красавок, но ни*
СЕРЫЙ ЖУРАВЛЬ 107
когда сборища не бывают так многочисленны и так азартны как у последних.
Иногда наблюдаются пляски и осенью (Круликовский, 1913), когда журавли
перед отлетом собираются уже в стаи —• имеется, следовательно, осенний
«абортивный» цикл размножения. Молодые, прошлогодние птицы, к размноже-
нию в первое лето не приступают, кроме того остаются иногда холостыми и
некоторые вполне взрослые птицы. Не гнездящиеся держатся стаями F—8—10
штук) и все лето кочуют.
Моногамы. Самец и самка сильно привязаны друг к другу. Гнезда обычно
устраивают на большом расстоянии друг от друга — в 5—6 км, и только в ред-
ких случаях несколько пар гнездится ближе одна к другой, но и то в двух-
трех километрах. На севере часто рядом с гнездами журавлей устраиваются
гнезда гусей, казарок, луней и т. д., с которыми журавли мирно уживаются.
Для гнезда выбирают сухое место среди болота, заросшего камышом, тростни-
ком или кустарником, но вокруг гнезда всегда имеется открытое пространство.
Гнездо устраивается на земле, в сухом месте. Иногда это просто небольшое
углубление в почве, выстланное тонким слоем сухих стеблей, трав, тростника
или камыша, но чаще — куча небрежно набросанного хвороста, с лотком,
выстланным небольшим количеством сухой травы и камыша, или плотно утоп-
танная куча перегнившего тростника, почти совершенно плоская. Поперечник
гнезда 80 см, вышина 20—30 см (Харьковская обл., Сомов, 1897). Журавли
в течение нескольких лет гнездятся на одном и том же болоте, занимают одни
и те же гнезда, даже в тех случаях, если в предыдущем году гнезда были
разорены или яйца вынуты.
В кладке обычно два яйца, редко одно или три, удлиненной формы с хорошо
выраженным тупым и острым концами. Скорлупа мелкозернистая, плотная,
почти без блеска. Основной фон скорлупы варьирует от красновато-бурого до
зеленовато-бурого, по нему разбросаны буроватые и буровато-серые глубокие
пятна и крапины и буровато-рыжие более поверхностные пятна. Размеры:
B> 101 х 60,2 мм и 100 х 59,3 мм (Харьковская обл., Сомов, 1897);
A) 96,5 х 60 мм (Калининская обл., Иоганзен, 1930); D) от 101—93 х
X 62—60 мм, в среднем 96,75 х 61 мм (Минская обл., Шнитников, 1913).
Среднее G) 97,8 х 61 мм. Вес яйца 167,4 г (Калининская обл., Иоганзен,
1930), 200 г (Хейнрот, 1924).
Данных о времени начала кладки очень мало; в среднем можно считать, что
кладка яиц происходит в начале апреля и до середины мая. В северной и средней
полосе Европейской части СССР в среднем начинается с конца апреля и с пер-
вых чисел мая; на Украине — во второй половине апреля до начала мая (Харь-
ковская обл., Сомов, 1897). Кладки найдены: в Московской обл., в Воскресен-
ском районе,—2.V (Евтюхов, 1928), в Минской губ. —29. IV. 1905 (в кладке
было два насиженных яйца), 14.V. 1905 иб.У. I. 1903 г. (в каждом гнезде было
по одному уже наклюнутому яйцу; Шнитников, 1913), в Новгородской обл.
в середине мая (Бианки, 1910). Если птиц беспокоят, они приступают к раз-
множению с большим запозданием. В таких случаях время начала кладки
в одном и том же районе у отдельных пар сильно растягивается иногда
даже на месяц. Промежутки между откладкой каждого яйца — около
двух дней.
Насиживание начинается, повидимому, с откладки первого яйца (в найден-
ном гнезде в Московской обл. первый птенец вылупился на 2 дня раньше вто-
рого; Евтюхов, 1928). В период высиживания журавли ведут скрытный образ
жизни. Насиживает преимущественно самка, но самец все время находится
около гнезда, зорко караулит его, предупреждает самку громким криком об
епасности и на короткое время кормежек сменяет ее. Вспугнутая самка не прямо
слетает с гнезда, а сначала отбегает на некоторое расстояние, и только затем
взлетает. Поэтому разыскать оставленное гнездо очень трудно. Продолжитель-
ность насиживания около месяца B9—30 дней).
108 ОТРЯД ЖУРАВЛИ
Птенцы вылупляются в средней полосе Европейской части СССР, начиная
с середины мая и до конца июня (в гнезде, найденном в Московской обл., первый
птенец вылупился 22.V и второй 24.V, Евтюхов, 1928). В Минской обл. 14.V.
1905 г. и 5.VI. 1903 г. были найдены надклюнутые яйца (Шнитников, 1913).
В Воронежской губ. были встречены 3—4-дневные птенцы: один раз 26.V,
второй —22.VI (Огнев, 1924). К птенцам и самец и самка сильно привязаны
и в выхаживании птенцов участвуют обе птицы. Постэмбриональное развитие
проходит следующим образом (Хейнрот, 1924). Вес птенца по вылуплении
около 120 г. Первые Здня птенцы мало прибавляют в весе —всего на 10 г,
затем идет быстрое прибавление в весе и на 12-й день птенец весит — 560 г,
на 16-й —800 г, на 26-й — 1350 г, на 32-й — 1880 г, на 70-й —3500 г, на
170-й —4800 г, т. е. достигает веса взрослой птицы. Первые перья —махо-
вые — появляются на крыльях на 15—17-й день, на 30-й день весь птенец
покрыт еще пухом, перья имеются только на крыльях, на 40—43-й день
птенец одет пером, но кое-где еще имеется пух.
Как только птенцы окрепнут, родители уводят их от гнезда — в камыши,
в приречные заросли, в кустарники. Когда птенцы приобретут способность
к полету, родители вылетают с ними на кормежку на близко расположенные
хлебные поля или луга, а на отдых (на ночь и полдень) возвращаются обратно
в камыши. В начале июля в средней полосе Европейской части СССР птенцы
летают, и в конце июля семьи переходят к кочевому образу жизни. В начале
августа семьи собираются в стаи и готовятся к отлету.
Линька. У взрослых птиц одна линька в году — полная послебрач-
ная. Быть может имеется и частичная предбрачная линька. Линька начи-
нается после вывода птенцов — в июле и даже в конце июня — и заканчи-
вается в октябре. Первостепенные маховые и наружные второстепенные
выпадают одновременно в течение двух-трех дней (по наблюдениям птиц в
неволе). Птицы на это время теряют способность к полету и уходят в совер-
шенно недоступные болота, куда к ним трудно подойти. Линька внутренних
второстепенных маховых и кроющих крыла идет постепенно. К концу июля
маховые и рулевые обычно уже все свежие, но украшающих маховых перьев
еще нет. Мелкое перо на голове и шее к отлету все свежее, а остальное мелкое
перо в перемежку — и новое и старое. В таком оперении птицы от нас уле-
тают и заканчивают линьку мелкого пера на зимовке.
У молодых птиц первая частичная линька происходит зимой — в ноябре —
феврале,—а первая полная линька —в годовалом возрасте, во второй половине
июня или даже несколько раньше — в первой половине июня, заканчивается
она в августе —сентябре. Во вторую зиму, в ноябре. — феврале, и молодые
птицы надевают взрослый наряд. В этом наряде птицы, в возрасте двух лет,
уже приступают к размножению.
Питание. Преимущественно растительная пища — различные ягоды
(на севере весною, главным образом, клюква), семена различных растений,
всходы хлебов, молодые побеги трав, водные растения и их корни, хлебные
зерна, в особенности пшеница, горох и овес. В небольшом количестве в состав
пищи входят и животные корма — различные насекомые, в особенности жуки
и саранча, лягушки, змеи, мышевидные грызуны, моллюски, черви, яйца
птиц и иногда даже мелкие птицы (однажды найдена в желудке камышевка
Acrocephalus paludicola, Англия, Уайзерби, 1945). В июне, как только под-
растут молодые птицы, журавли семьями, а затем и стаями, каждый вечер и
утро с громкими криками летают на кормежку на ближайшие хлебные поля;
если поблизости хлебных полей нет, то на луга, в степи, даже на солончаковые
степи и пески, где ловят саранчу и других насекомых. Отдыхать возвраща-
ются на болота в камыши. Воду пьет журавль в большом количестве и,
если на местах кормежек нет воды, то летает несколько раз в сутки на
водопой.
СЕРЫЙ ЖУРАВЛЬ
109
Полевые признаки. Наземная, дневная птица, только на перелетах летит
и по ночам. Легко узнается благодаря своим крупным размерам, длинным
ногам и шее, небольшой относительно голове и умеренной длины клюву.
Общая окраска серая. В полете вытягивают шею и ноги почти по прямой линии,
и все время машут крыльями, только в редких
случаях, например, при снижении —планируют.
Полет относительно медленный, даже во время
перелета. Журавль ^осторожная и чуткая птица.
Стаи всегда выставляют одного или двух сторо-
жей, обычно из старых птиц, которые своим кри-
ком предупреждают остальных журавлей об
опасности. Даже кормящиеся пары, подозревая
опасность, начинают кормиться по очереди.
Журавль крикливая птица, обладающая
исключительно громким голосом, который слы-
Рис. 17. Серый журавль
шен более, чем на 2 км. Чаще всего это трубный, курлыкающий крик. Но
когда пары птиц на близком расстоянии друг от друга «переговариваются»,
слышны низкие курлыкающие звуки, а резких, громких тонов, столь харак-
терных при перекличках журавлей в стае или во время полета, слышать не
удается (Спангенберг). Птенцы и молодые птицы вывода этого года издают
тонкий протяжный писк, который слышен на большом расстоянии. Писк этот
сохраняется до зимовок, во всяком случае во время осеннего пролета он слы-
шен. Вообще звуки, издаваемые журавлями, разнообразны.
Описание. Размеры и строение. Клюв прямой, конической формы и — отно-
сительно размеров тела — короткий. Вес 3950—7000 г, минимальные цифры относятся к
самкам, максимальные — к самцам. Длина тела самца A) 1200 мм, самок B) 1048 и 1130 мм.
НО ОТРЯД ЖУРАВЛИ
Размах крыльев самца A) 2295 мм, самок B) 2095 мм и 2100 мм. Длина крыла самцов A7)
600—660 лги, самок A0) 570—540мм, среднее соответственно —641 и 595 мм. Длина цевки
самцов 228—260 мм, самок 220—238 мм, среднее соответственно — 250 мм и 228 мм. Длина
клюва самцов 103—120 мм, самок 100—110 мм, среднее соответственно — 110 и 102 мм.
Первостепенных маховых—И, самое длинное 3-е; 4-е и 2-е почти равны между
собою и незначительно короче 3-го. Формула крыла 3^2 = 4>1>5 .... Третьестепен-
ные маховые удлинены и сильно рассучены. Хвост короткий, рулевых перьев 12. Уздеч-
ка, лоб и передняя часть темени у взрослых птиц не оперены, покрыты черной кожей и
редкими черными волосовидными перьями. На задней части темени и на затылке— голое
округлое кирпично-красное пятно, покрытое бородавчатой кожей. У молодых птиц вся
голова оперена.
Окраска. В первом пуховом наряде спинная сторона тела рыжевато-охристая,
более интенсивно окрашена вдоль середины спины и на плечах и светлеющая на верхней
стороне головы и на внешней поверхности ног. Брюшная сторона тела и головы и бока тела
и головы — беловатые, с легким охристым налетом, усиливающимся на зобе. Клюв корот-
кий, прямой, светлобурый, чуть темнеющий к вершине. Ноги светлобурые с легким
оливковым оттенком.
Во втором пуховом наряде верх тела буровато-серый, светлеющий на голове и шее,
и с охристым налетом на спине, голове, шее и на наружной стороне ног. Нижняя сторона
тела грязнобелая.
Молодая птица в птенцовом наряде. Оперение головы светлокоричневое. Шея—одно-
цветная, серая с мелкой охристой крапчатостью, более сильно развитой на верхней стороне
шеи, у основания головы и на подбородке. Некоторые из третьестепенных маховых рассуче-
ны, но без черных вершинных пятен. Маховые уже,чем у взрослых, крыло более тупое. Фор-
мула крыла 3 = 4>2>1 = 5. Вершины рулевых перьев еще неправильной округлой формы,
а стержни пера в виде острия выдаются над внешним краем пера. В оперении верхней и ниж-
ней сторон тела сильно развит охристый оттенок, кроме того, верхняя сторона тела пятни-
ста, вследствие того, что некоторые перья спины двухцветны—основная половина пера
серая, а вершинная — светлокоричневая или охристая. Клюв светло роговой. Ноги темно-
бурые. Этот наряд одевается в возрасте приблизительно около 2,5 месяцев — в августе.
В первом зимнем наряде оперение головы — охристое с черными крапинами (среди охри-
стых перьев передней части головы уже появляются волосовидные черные перья, а на зад-
ней части головы и передней части шеи — нормальные черные перья). За глазом — белое пятно
с мелкими темными крапинками у заднего края глаз. Белое пятно за глазом тянется к верх-
ней стороне шеи, постепенно переходя в охристый, а затем серый тон. Подбородок и нижняя
сторона шеи — крапчатая буровато-черная с отдельными охристо-серыми перьями. Третьесте-
пенные украшающие перья частично хорошо развиты и с черными вершинными пятнами,
но сохранились еще третьестепенные и первого наряда. Клюв — буроватый, светлеющий к
вершине. Ноги буровато-черные. Птицы одевают этот наряд в возрасте приблизительно
6 месяцев, с ноября. Первый брачный наряд сходен со взрослым. Голова голая, но крас-
ного пятна на затылке еще нет. Белый цвет на боках головы и шеи — чистый. Среди мелкого
пера спины еще имеются охристые перья. Этот наряд одевается в возрасте приблизительно
13—14 месяцев — с июня — июля второго после рождения года.
Взрослая птица. Оперение серое, за исключением черных первостепенных махо-
вых, их кроющих, наружных опахал второстепенных маховых, вершин третьестепенных
маховых, крылышка и вершин рулевых. Задняя часть головы, подбородок, нижняя сторона
шеи и бока шеи приблизительно на 2/з длины шеи буровато-черные. От глаз по бокам головы,
заднему краю головы и по верхней стороне шеи идет белая полоса, постепенно переходящая
у основания шеи в серый цвет. Клюв — зеленовато-бурый, светлеющий к вершине. Ноги и
когти — черные. Радужина — буровато-красная. В летнем пере на верхней стороне тела —
легкий охристо-бурый налет. Самцы и самки окрашены одинаково.
Восточный серый журавль Grus grus lilfordi S h a r p e
Grus lilfordi. S h a r p e. Cat. Birds Brit. Mus., XXIII, 1894, стр. 252, Вост. Сибирь.
Распространение. Ареал. Северная граница проходит на Оби у Об-
дорска (Шостак, 1926), по рекам Таз и Елагуй (Скалов и Слудский, 1941), на
Енисее может быть до Туруханска или несколько южней, до Нижней Тунгуски,
Вилюя, Лены (до впадения в нее Алдана), Верхоянска, Индигирки, до впадения
в нее р. Момы, т. е. до 66° с. ш. (Михель, 1935). Как далеко идет на восток от
Индигирки — не выяснено (на Колыме гнездятся какие-то журавли, скорее
всего это серые, а не канадские журавли, но в коллекциях экземпляров оттуда
нет). В Анадырском крае не гнездятся. О гнездовании серого Журавля на Кам-
чатке, Коряцкой земле и на Охотском побережье сведений нет. В Приморье
и Приамурье не гнездятся.
СЕРЫЙ ЖУРАВЛЬ 111
Западная граница точно не выяснена. Южная идет но Волжско-Уральским
степям. К югу доходит до 47°5' с. ш., захватывая Самарские озера, Эльтон,
Новоузенские степи (Волчанецкий, 1937). По р. Урал — до ее среднего тече-
ния; реки Хобда, Илек и Эмба, но как далеко идет по Эмбе к югу — не вы-
яснено (Сушкин, 1938). Далее южная граница доходит до северо-восточного
побережья Аральского моря, дельты Сыр-Дарьи, Камышлы-баш, озера Тели-
куль, Сары-Су, Чу, Иссык-куля, Алма-Ата, Или; Зайсанская и Эмильская
котловины, предгорья Барлыка, Тарбагатай (Хахлов, 1928), Алтай, Саяны,
Танну-тувинская обл. (Сушкин, 1938), Хангай, Прибайкалье, Забайкалье,
Даурия — озеро Тарей-нор, реки Онон и Аргунь. Далее на восток о гнездо-
вании серого журавля нет достоверных сведений, какие-то пролетные жура-
вли отмечены под Марийнском (Шренк, 1860) и в долине Уссури (Маак,
1861). Изолированные колонии имеются в Закавказье и Малой Азии: в Джа-
вахетии, в Армении на озере Севан, истоках Куры, на озере Чалдыр, в долине
верхнего течения Аракса, в сев.-зап. Иране — в Котурско-Урмийском районе
и в сев.-вост. Турции на озере Аджин-туз-гол.
Зимует в Китае — в Чжили, в долине Янцзе, Фуцзяне, Гуандуне и
Юннане; в сев. Индии к югу до Бомбейской провинции и Деккана в Сей-
стане и Белуджистане.
Залеты отмечены далеко на север, например, на о-в Врангеля, на Ново-
Сибирские о-ва, в дельту Индигирки, на Сев. Урал под 68°30' с. ш. и
в 1908 г. (Молчанов, 1911) и дважды в 1942 г. на южный о-в Новой Земли
(Успенский).
Характер пребывания. Перелетная и гнездящаяся птица.
В Армению на озеро Севан прилетает на гнездовье в половине апреля, пролет
западного серого журавля идет несколько раньше. В низовьях Урала первые
птицы появляются в конце марта, под Чкаловом и на Эмбе пролет начинается
с первой половины апреля и идет до конца месяца и даже начала мая B.V),
валовой пролет под Чкаловом наблюдается во второй половине апреля (Суш-
кин, 1908). В Башкирию и в Бугурусланский район, Куйбышевской обл.,
первые прилетают в среднем в конце апреля (Карамзин, 1909), в Среднем
Поволжье пролет идет с 28. III и до середины апреля (Волчанецкий и Яльцев,
1934). В Среднюю Азию прилетают очень рано. В Туркмении первые пролетные
журавли отмечены в самом начале марта, валовой пролет идет в конце этого
месяца и в начале апреля (Дементьев, 1950). В Таджикистане с 1. IV до сере-
дины апреля (П.IV, Иванов, 1940), на нижней Сыр-Дарье в первых числах
апреля G. IV. 1928, Спангенберг и Фейгин, 1936). В Семиречье первые журавли
прилетают очень рано: на р. Каскелен 20. III, под Алма-Ата 30. III, валовой
пролет — с 8. IV; под Джаркентом — с 4^8. IV, на р. Песчаной —1—4. IV; к
10.V пролет в Семиречье уже заканчивается, летят лишь отдельные пары и
одиночные птицы (Кореев и Зарудный, 1906). В Илийском крае первые жура-
вли встречаются на пролете 28.111 (Шестоперов, 1929). На Иргизе с 12. IV по
8.V (Сушкин, 1908), в Наурзуме в начале второй половины апреля (Михеев,
1938). На озере Чаны 28. IV (Данилович, 1935), в окрестностях Омска—в
начале второй половины апреля и пролет идет до начала мая. На Алтае — во
второй половине апреля B0. IV —на Телецком озере, 25. IV —в урочище Беле,
Фолитарек и Дементьев, 1936; 9 и 17. IV в с. Новенское, Лавров, 1911). На
Елагуе первые птицы появляются в середине апреля A5. IV. 1932, Скалой и
Слудский, 1941), у Витима в среднем прилетают 8.V D года наблюдений, Ска-
л он), в Якутск в среднем — I.V (данные Охлопкова за 22 года и других лиц,
Скалой), вскоре за появлением первых птиц начинается валовой пролет, кото-
рый идет бурно и в короткие сроки (в 1921 г. в Якутске первые птицы появились
7.V, валовой пролет наблюдался 10.V). Самый ранний прилет отмечен 29. IV.
1902 г., самый поздний—6. V. 1918 г. и 1925 г. и 7.V. 1921 г. (Скалой), в
Олекму — в среднем 29. IV C года наблюдений), в Усть-Мае — в среднем
112 ОТРЯД ЖУРАВЛИ
7. IV B года наблюдений), в Абага —9.V. D года наблюдений), в Татта —
2.V D года наблюдений), в Усть-Алданск — 9.V D года наблюдений), в Ви-
люйск —4.V D года наблюдений), в Верхоянск —9.V B года наблюдений,
Скалой, Воробьева, 1931; Тугаринов, 1928). На Индигирку в устье Момы при-
летели — 10.V.1929 (Михель, 1935).
Большую разницу в сроках прилета не только в Зап. и Вост. Сибири,
но и в западных и восточных частях Средней Азии и Казахстана, повиди-
мому, можно объяснить лишь расположением зимовок отдельных популяций
серого журавля и дальностью пролетного пути.
Осенний пролет. В Якутии отлет начинается с конца августа или
начала сентября и очень быстро — через несколько дней — все птицы исче-
зают и пролет заканчивается. В Верхоянске (данные Скалона) первые стаи уле-
тали 6,1Х. 1935 г., в Олекминске — 6.IX. 1937 г. и 15.IX отлет уже окон-
чился. На Абага первые стаи улетали 25.VIII. 1928 г. и к MX пролет
закончился, в Чуранча — улетели 2. IX. 1929, в Татта —13. IX. 1929 г. и
28.VIII. 1930 г., в Мегежене —4.IX. 1930 г. и на Витиме —5.IX. С Алдана
улетели первые стаи—28.VIII (Воробьева, 1928), в окрестностях Якутска
отлет и пролет идут с конца августа и до середины сентября. С верховьев
Таза последние птицы улетают в конце сентября B9. IX. 1929 г. и Скалой
и Слудский, 1941). В Минусинском крае и на Алтае пролет идет с конца
августа B5—29.VIII, на р.Чулыме, Иоганзен, 1923) и до конца сентября и
даже до начала второй половины октября A2 и 13.Х. 1935 г. наблюдался
хорошо выраженный пролет, Алтайский заповедник, Фолитарек и Дементьев,
1935; 16.Х летели многочисленные стаи ус. Новенское Змеиногорского уезда,
Аверин и Лавров, 1911). В окрестностях Кузнецка пролет шел 25.VIII (Залес-
ский, 1930). На Алтае птицы во время своих предотлетных ночевок спускаются
с гор в низины, но во время пролета не огибают горные хребты, а летят низко
над ними на высоте около 2200 м. В Зайсанской котловине пролет идет с конца
августа B8. VIII) по октябрь (З.Х). Направление пролета—на юг (Суш-
кин, 1938). Киргизские степи местные птицы покидают в августе, но пролет
идет в сентябре, в Наурзуме пролет наблюдался с 20. IX, валовой —с 23. IX
и отдельные стаи до первых чисел октября (Михеев, 1938). В Семиречье и в
Илийском крае пролет идет с середины сентября и до середины октября. На
Сыр-Дарье первая стая в окрестностях Джулека появилась 7.VIII. 1938 г.
(Спангенберг и Фейгин, 1936). В Кызыл-Кумах пролет наблюдался во второй
половине сентября. В Туркмении начало пролета—в конце августа (одиночки
и ранее, например, 5.VIII в низовьях Теджена, Зарудный, 1896), валовой —
во второй половине сентября (Дементьев, 1950). В Таджикистане летят с сере-
дины августа до сентября (с 17.VIII—I.IX, Иванов, 1940; Федченко в 1870 г.
наблюдал там большой пролет в сентябре в направлении на юго-запад).
В Башкирии, в Бугурусланском районе Куйбышевской обл. и в б. Орен-
бургской губ. отлет и пролет начинается в последних числах августа или
в первых сентября и заканчивается во второй половине сентября и в октябре;
направление пролета — на юго-запад.
В дельте Волги и на морском побережье Каспия журавли появляются очень
поздно — в конце сентября и пролет идет весь октябрь, особенно интенсивно —
в конце первой половины октября A2.Х. 1930, Воробьев, 1936), направле-
ние пролета — юго-западное. Повидимому птицы из Башкирии летят на зимов-
ки по Кавказскому побережью Каспия и через Главный Кавказский хребет.
На Кавказе пролетные журавли встречаются очень поздно —с середины сентя-
бря до конца октября и начала ноября. С озера Севан улетают во второй
половине сентября (с 22. IX до конца месяца, Радде, 1885). Отлет журавлей
стоит в большой зависимости от наступления холодов и на юге страны;
где есть подходящие условия существования и корм, птицы задерживаются
долго.
СЕРЫЙ ЖУРАВЛЬ 113
Биотоп. Как указано уже — крайне разнообразен и всегда связан
с водой. В Башкирии вершины степных оврагов, поросших лесом и густою
уремою. В Зауралье — густые болотистые низины, в степной части Зап.
Сибири — болота и озера с многочисленными плесами, заросшие тростником.
В лесистой части Зап. Сибири — болота среди смешанного и хвойного
леса. В Прибайкалье и Забайкалье — большие заболоченные пространства
с небольшими группами ельника, или болотистые луга, иногда расположенные
среди большого высокоствольного леса, с редким кустарником и отдельно
стоящими деревьями, преимущественно березами. В горных районах, если
есть подходящие местности, поднимается высоко в горы — на Алтае идет до
альпийской зоны и даже гнездится в нагорной тундре (Сушкин, 1938), в Семи-
речье до 1900 м (Кореев и Зарудный, 1906). В Армении поднимается в горы
до 2290 м в бассейне озера Севан, в истоках р. Балык-чай (Даль); гнездится
на высоких мокрых лугах, в болотистых местностях и на высокогорных озе-
рах; в низинах на гнездовьях не встречается. В полупустынях —заболочен-
ные берега озер и рек, поросшие невысокой травой и редкой кустарниковой
растительностью, например, тамариском.
Численность. Повсюду обыкновенен, но не многочисленен.
Экология. Размножение. Отличий от западного серого журавля
в устройстве гнезд, в количестве яиц, в кладке и в насиживании яиц — нет.
Молодые птицы к размножению в первый год не приступают (добытая 22. V на
Алтае молодая самка имела неразвитые половые органы, Сушкин, 1938). Время
размножения: в Армении близ озера Гилли 26.VI найдена кладка в два сильно
насиженных яйца (Даль), на нижней Сыр-Дарье кладка начинается в конце
апреля, на озере Тели-куль 21.V. 1927 г. найдена кладка в 2 яйца (Спангенберг
и Фейгин, 1936), в верховьях Или —в конце первой половины апреля A1. IV
найдена кладка с двумя ненасиженными яйцами, а 13.IV—2 гнезда: в одном
было 3 яйца, в другом — 1, Шестоперов, 1929); в Барабинских степях кладка
начинается в первой половине мая и заканчивается в середине июня, в окре-
стностях Каинска 24.V найдено гнездо с одним насиженным яйцом, 26.VH
29.V—2 гнезда с ненасиженными яйцами и 1 гнездо с двумя яйцами — на
озере Сартлан 10 июня (Иоганзен, 1907, 1930); на Алтае — в Катон-Карагае —
ненасиженная кладка, состоящая из двух яиц, найдена 7.V. 1931 г.
(Спангенберг). Размеры: B) 103,5 х 63,0 мм (Армения, озеро Гилли, Даль);
F) 88,8—96,0 х 57—61 мм, среднее 93,05 х 59,5 мм (Барабинские степи,
Иоганзен, 1907, 1930); C) 98,5—102,8 х 60.5—62 мм, среднее 100,8 х 61,1**
(Или, Шестоперов, 1929); B) 83,2 х 53,8 и 81,0 х 54,2 мм (озеро Тели-куль;
Спангенберг и Фейгин, 1938); B) 88,1 х 61,4 и 88 х 60 мм (окрестности
Катон-Карагая, Спангенберг). Вес:F) 150—200 г, среднее 182,4 г (Барабинские
степи, Иоганзен, 1907, 1930); B) 191 и 192 г (окрестности Катон-Карагая,
Спангенберг); B) 151 и 155 г (озеро Тели-куль, Спангенберг). Яйца несколько
мельче яиц западного серого журавля, окраска такая же как у западного.
В Армении на озере Гилли первые птицы вылупляются в самом конце июня
и в начале июля (вес только что вылупившегося птенца — 100 г; Даль); в конце
июля в Зап. Сибири появляются первые летные птицы. На Алтае (Чуйская
степь) 31.VII наблюдались первые летающие птенцы, но нелетные птенцы
встречались еще позже в течение двух-трех недель (Сушкин, 1838). В
первой половине августа семьи начинают собираться в стаи.
Линька. Общая схема — как у западного журавля. У взрослых (Алтай)
линька начинается во второй половине июля и к половине августа маховые и
рулевые уже полностью сменены, но мелкое перо находится еще в линьке.
Питание. То же, что и у западного серого журавля, лишь с некоторыми
изменениями в связи с наличием других растительных кормов. Например,
кедровые орехи (у убитого в Алтайском заповеднике журавля желудок и пище-
вод были полны кедровыми орехами; Фолитарек и Дементьев, 1938).
8 Птипы СССР, т. II
114
ОТРЯД ЖУРАВЛИ
Описание. Размеры и строение. Чуть мельче западного серого журавля.
Длина тела самок B) 1030 и 1100 мм; размах крыла — самок B) 1920 и 1945 мм. Длина
крыла самцов D) 580 — 640 мм, самок D) 560 — 610 мм, среднее, соответственно,
600 и 582 мм. Формула крыла как у G. g. grus.
Красное пятно на темени и затылке несколько меньших размеров. У только что вывед-
шихся птенцов на I и II пальцах передней конечности имеются хорошо развитые когти (Даль).
Окраска. Пуховой птенец и молодые птицы неотличимы от западного журавля.
У взрослой птицы окраска оперения несколько светлее, а черный цвет на вершинах второ-
степенных маховых развит слабее, чем у западного серого журавля.
2. Канадский журавль Grus canadensis L.
Ardea canadensis. Linnaeus. Syst. Nat., изд. X, 1758, стр. 141, Гудзонов залив.
Распространение. Ареал. Северо-восток Азии (Анадыр), о-в св. Лаврентия,
Сев. Америка от Аляски, Меккензи, земли Франклина на севере до Баффино-
вой земли, Гудзонова залива и Онтарио на востоке и на юг до Сев. Кали-
форнии, Аризоны, Невады, Колорадо, Небраски, Висконсина, Мичигана,
Иллинойса, Охайо; Южн. Георгия, п-ов Флорида, возможно в Луизиане
и Алабаме, о-в Пинос и западная часть о-ва Куба. Зимует в южных частях
Сев. Америки: Калифорния, Аризона, Техас, Луизиана, в низовьях Мис-
сисипи и в Мексике.
Карта 17. Распространение канадского журавля Grus canadensis
7. G. с. canadensis. 2, G. с. tabia, 3. G. с. pratensis. 4. G. g. nesiofles;
a — граница гнездовий, г — зимовки, д — изолированные находки особей.
Характер пребывания. Гнездящаяся и перелетная птица; только
журавли, обитающие на о-ве Пинос и в западной части о-ва Куба,— оседлы.
Биотоп. Тундра, обширные болота, побережья озер, рек и морей, тра-
вянистые склоны холмов до 1000 м над уровнем моря.
Подвиды и варьирующие признаки. 4 подвида, различающихся морфологическими
особенностями и экологией. G. с. canadensis Linnaeus, 1758 — Анадыр, северные части
Сев. Америки; G. с. tabia Peters, 1925 — Британская Колумбия, Саскачеван на
восток до Манитоба и Онтарио к югу до Сев. Калифорнии, Висконсина, Мичигана и
Охайо. Зимует в Калифорнии, Мексике, Техасе и Луизиане; G. с. pratensis Meyer, 1794—
Южн. Георгия, п-ов Флорида и вероятно, Южн. Луизиана и Алабама; зимует на реках
Миссисипи и Сабин в Луизиане и в Техасе; G. с. nesiotes Bangs, 1905 — о-в Пинос,
западные части о-ва Куба.
Обыкновенный канадский журавль Grus canadensis canadensis L.
Синоним. Grus niediecki. Reichenow. Orn. Monatsber, 1906, стр. 190, Анадыр.
Распространение. А р е а л. У нас достоверно гнездовья доказаны только
в восточной части Анадырского края — по Анадырскому лиману, в устье и
нижнем течении Анадыря и впадающих в него здесь рек, также по южному
склону Анадырского хребта. Под Марковым, повидимому, уже не гнездится
КАНАДСКИЙ ЖУРАВЛЬ 115
(Портенко, 1939). Но летние встречи и находки известны и с северо-восточного
побережья Северного Ледовитого океана и с Чукотки. Так канадский журавль
наблюдался летом 1927 г. и в 1933 г. на о-ве Врангеля (Минеев, 1946),
18.VII. 1912 г. —ум. Большой Баранов, к востоку от устья Колымы,
19.VII. 1912 г. встречен у м. Балаган, к западу от Чаунской губы, и в
Чаунской губе (Тайлер и Бенгс, 1914). Добыт у м. Северный 21.V. 1915 г.
(Артоболевский, 1926). Большие стаи встречены 16.VI. 1879 г. в губе Колю-
чинской, а поздней у м. Питлекай (Пальмен, 1887). Добыт на Чукотском
носу, на Командорских о-вах —18 и 20.V. 1914 г. (о-в Беринга) и наблю.-
дался там весною 1929 г. (Иоганзен, 1934). Повидимому, к этому виду отно-
сятся данные Палласа о наблюдении серого журавля на Камчатке — в Олю-
торске и Нижне-Камчатске. Гнездится на о-ве св. Лаврентия, в арктических
и северных частях Сев. Америки: на Аляске, Меккензи от их северных границ
до южных; на земле Банкса, Франклина, на п-ове Боотия и Мельвилья, на
восток до Баффиновой земли и Гудзонова залива, к югу по берегу залива до
м. Эскимос. Зимует в Техасе и Калифорнии к югу до Нижней Кали-
форнии.
Характер пребывания. Гнездящаяся и перелетная птица.
В Анадырский край прилетает во второй половине мая. В 1931 г. в Анадырском
лимане весенний пролет шел с 16.V—1.VI (Белопольский, 1933). В Новома-
риинске прилет журавлей отмечен в мае, но иногда прилетает и раньше — в
конце апреля (Сокольников, Портенко, 1939). На о-ве Беринга, повидимому,
именно пролетные журавли наблюдались 18 и 20.V. 1914 г. В Сев. Америку —
в южные области своего гнездового ареала — прилетает в среднем в сере-
дине апреля, в северные области — в первой половине мйя (в Меккензи —
в начале мая, на Аляску и на землю Франклина —в середине мая; Бент, 1926).
В двадцатых числах августа журавли в Анадырском крае группируются в
стаи — молодые вместе со старыми, и в конце августа начинается отлет, про-
должающийся до середины сентября и даже конца этого месяца. В Анадырском
заливе в 1930 г. пролет журавлей шел с 29.VIII—13. IX (Белопольский, 1933),
в устье р. Танюрера последние птицы были встречены в 1931 г. 15. IX A4. IX
выпал там снег до 18 ел* глубины), у Новомариинска последние птицы наблюда-
лись 9. IX. 1932 г. (Портенко, 1939). В1869г. 28. IX встречен одиночный журавль,
повидимому, отсталый, так как температура воздуха доходила до — 8° и лежал
уже глубокий снеговой покров (Майдель, 1894). Журавли с Анадыря обычно
летят на юг и где-то, вероятно около Командорских о-вов поворачивают на
восток, но иногда некоторые стаи летят осенью на север—вдоль южного
берега Чукотки. В Сев. Америке улетают на юг в среднем в первой половине
сентября (в 1898 г. журавли улетели очень рано, последние птицы на Аляске
отмечены 4. IX, в 1911—4. IX последние птицы покинули Меккензи); из
южных областей своего распространения журавли улетают в конце сен-
тября, одиночные птицы задерживаются иногда и до середины октября.
Залеты канадского журавля в Сев. Америке отмечены далеко от мест гнез-
довий, главным образом в восточных штатах Сев. Америки и преимущественно
осенью (Бент, 1926).
Биотоп. В Анадырском крае —кустарниковая тундра, хорошо увлаж-
ненная (болота, озера и реки). Не избегает невысоких горных хребтов и холмов
высотою до 1000 м, покрытых кустарниковой и травянистой растительностью.
Численность. В Анадырском крае обыкновенен и местами много-
численен. В устьях р. Танюрер обитает свыше 10 пар; на каждые 3—4 км прихо-
дится пара журавлей. В приморской полосе края более редок (Портенко, 1939).
Экология. Размножение. Наблюдений над особенностями размно-
жения в Анадырском крае нет. Сведения, касающиеся Сев. Америки, при-
водятся Бентом A926). Моногамы. Сейчас же по прилете на места гнездовий
разбиваются на пары и вскоре начинаются журавлиные «пляски». У канад-
116
ОТРЯД ЖУРАВЛИ
ского журавля обычно «танцуют» две птицы, повидимому самец и самка.
Танцы начинаются с того, что они с громким криком слетаются вместе и, описы-
вая в воздухе круги, опускаются на землю, поворачиваются друг к другу,
делают низкие поклоны, затем, полураспустив крылья, подпрыгивают высоко
в воздух, приседают и вновь подпрыгивают, и так продолжается до тех пор,
пока птицы не утомятся. Танцы наблю-
даются не только в весенний период, но
и когда птицы сядут на яйца, в течение
всего июня, причем обычно танцуют у гнезда 2
птицы, повидимому самец и самка. Молодые
птицы к размножению приступают в первое
лето, не одев еще полный взрослый наряд.
Гнездо устраивают недалеко от воды, иногда
под прикрытием кустарника. Оно представляет
собой неглубокую ямку (в диаметре около
Рис. 18. Канадский журавль
60 см) в земле или в песке. Подстилкой служит толстый слой сухой травы
с небольшим количеством перьев.
В кладке 2 яйца, овальной и овально-удлиненной формы. Скорлупа
обычно слегка шероховатая, но иногда и гладкая с небольшим блеском или
даже совсем без блеска. Основной тон скорлупы от буровато-коричневого до
буровато-оливкового. Крапины и пятна расположены в большом количестве на
широком конце яйца, и различных оттенков коричневого цвета. Глубокие пятна
размытые, тусклокоричневые, поверхностные — более резкие, более темные
и блестящие. Размеры: B5) 77,4—101 х 44—60, в среднем 84,5 х 51,1 мм
(Северная Америка, Бент, 1926).
ДАУРСКИЙ ЖУРАВЛЬ 117
Птенцы вылупляются покрытыми густым пухом. До конца июля и даже
середины августа журавли держатся семьями. Во второй половине августа
начинают группироваться в стаи и кочевать.
Линька. Взрослые птицы имеют 2 линьки в году: полная, после-
брачная, которая идет с августа до декабря и неполная, предбрачная, —
в конце зимы и в начале весны, во время которой сменяется только мелкое
оперение (Бент, 1926). Взрослый наряд канадский журавль надевает в возрасте
2х/2 лет, т. е. после второй полной линьки.
П и т а н и е. По наблюдениям в Америке преимущественно растительная
пища: ягоды, зерна и трава. В очень небольшом количестве —различные насе-
комые (Бент, 1926).
Полевые признаки. От серого журавля отличается меньшими размерами
и однообразной буровато-серой окраской оперения; на полете шея и ноги
вытянуты во всю длину. Во время пролета держится, на большой высоте. В ку-
старниковых зарослях тундры почти незаметен, только голова и шея выде-
ляются над кустарником. Очень осторожен. При приближении опасности жу-
равли вначале отходят, затем с криком поднимаются в воздух и, сделав не-
сколько кругов, улетают.
Голос — похож на голос серого журавля (Портенко, 1939).
Описание. Размеры и строение. Меньше серого журавля. Длина крыла
у самцов C) 520—580 мм, среднее -— 550 мм. Первостепенных маховых 11, самое длинное
3-е; 6-е первостепенное маховое не достигает вершины крыла на НО—115 мм. Третьестепен-
ные маховые и плечевые слабо рассучены и удлинены на 50—60 мм от вершины крыла. Фор-
мула крыла 3>2 = 4>1>5>6. Хвост короткий и прямой. Рулевых 12. Лоб и передняя
часть головы, включая и глаза, не оперены, покрыты буровато-красной кожей с черными
волосовидными перьями. Оперение задней части головы треугольником заходит на неопе-
ренную область головы.
Окраска. Пуховик имеет темный охристо-рыжий пух. Затылок, полосы на верх-
ней стороне шеи, вдоль спины и на верхней стороне крыльев — каштановые. Бока тела темно-
охристые. Горло и нижняя часть тела грязноватая серовато-белая (Бент, 1926).
В гнездовом наряде лоб, уздечка и темя оперены. Голова и шея рыжевато-бурые с ко-
ричневыми пятнами. Спина светлокоричневая, плечевые и кроющие крыла — более тусклого
светлокоричневого тона.
В первом брачном наряде лоб, уздечка и темя голые. Третьестепенные и плечевые
удлинены и слабо рассучены. В оперении несколько больше серого тона, чем у взрос-
лых птиц.
В первом зимнем наряде в оперении много серого, только голова, шея, плечевые и
кроющие крыла еще коричневые и кое-где на теле встречаются коричневые перья.
Во втором брачном наряде оперение серое, только кроющие крыла еще коричневые и
кое-где на голове, шее и теле имеются коричневые перья.
Взрослые самцы и самки имеют оперение серое, с сизоватым оттенком, несколько свет-
лей на нижней стороне тела. Первостепенные маховые — буровато-черные с более светлыми
стержнями перьев. Радужина коричневато-бурая, клюв буровато-роговой. Ноги черные.
3. Даурский журавль Grits vipio Pall.
Grus vipio. Pallas. Zoogr. Rosso-As., II, 1811, стр. Ill, Нерчинск.
Синоним. Grus leucauchen. Temminck. PI. Col., 1839, стр. 559, Япония.
Распространение. Ареал. Даурские степи, реки Онон и Аргунь, озеро
Торей-нор, может быть заходит и несколько далее, до Троицкосавска и Сев.
Монголии (добыт на р. Чикой и на р. Бура — к югу от Троицкосавска, Молле-
сон, 1906); низовья р. Буре (Шульпин, 1936), возможно гнездится в юго-вост.
Монголии (встречался Пржевальскому, 1876; летом на озере Далай-нор и на
территории от озера до Калгана); Манчжурия (найдены яйца под Харбином;
Иоганзен, 1930; птица добыта в 115 км к северо-западу отЦицикар;Мейзе,
1931); юго-западная часть Приморья: Даубиха, озеро ланка, реки Сунгачи,
Лефу, Мо, Сиянхе и другие мелкие речки бассейна озера Ханки, Уссури до
нижнего течения.
U8
ОТРЯД ЖУРАВЛИ
Зимовки. Корея; юго-вост. Китай—нижнее течение Янцзе, Фуцзян,
Шанхай и Фучжоу; юго-зап. Япония: обычен зимой у Арасаки на Кю-сю, редок
в южной части Хондо в Ияширо. Возможно отдельные особи остаются на зиму
в южной части Приморья на Суйфуне (Пржевальский).
Характер пребывания. Гнездящаяся и перелетная птица.
В Приморье прилетает рано — во второй половине марта. В 1868 г. в верховья
Сунгачи первые стаи прилетели 27.11, когда снег еще лежал глубоким слоем
и проталин нигде не было, температура ночью достигала — 20°Ц. Валовой
пролет наблюдался в середине апреля, причем журавли летели небольшими
стаями от 3 до 11 штук. В 1869 г. первые прилетные стаи отмечены 15—16. III,
затем до 12. IV пролет прекратился; 12. IV была встречена стая в 13 штук,
летевшая на север (Пржеваль-
ский, 1870, 1876). В 1928 г.
25. IV в устье Лефу наблю-
далась прилетевшая стайка
в 5 штук (Шульпин, 1936).
В окрестностях Троицкосав-
ска на р. Чикой 26. IV. 1896 г.
шел пролет даурского жу-
равля, и один из стаи добыт
(Моллесон, 1897). На пролете
и в первое время по прилете
даурские журавли держатся
на открытых болотистых рав-
нинах. Отлет идет постепенно
в течение всего сентября и
октября, а на юге Приморья—
и до конца ноября. Первые
пролетные стаи даурских жу-
равлей, летевшие на юг, от-
мечены 4. IX. 1859 г. на р.
Сунгаче(Маак, 1861). В 1867 г.
пролет наблюдался 15. IX на р.
Суйфун (у Чахузу), 13 и 18.Х в небольшом количестве в окрестностях Нов-
городской гавани и 22, 23 и 28.XI несколько стай замечены в заливе
Посьет (Пржевальский).
Залеты. Неоднократно наблюдались залеты даурского журавля весною,
летом и осенью, часто среди стай других видов журавлей, преимущественно
серых, и иногда далеко от мест гнездовий и зимовок. Встречен летом на западном
берегу Байкала (Радде, 1863), наблюдался на осеннем пролете в долине р. Хара
(подножье Кентея; Козлова, 1930) в стаях серого журавля. Указания на более
далекие залеты очень сомнительны: в большинстве случаев журавлей наблю-
дали на большом расстоянии, даже в тех случаях, когда они добывались,
определение производили не специалисты, и добытые экземпляры не со-
хранились. Дальние залеты даурского журавля отмечены в Средней Азии —
добыто 2 экземпляра из стаи в 5 штук на Сыр-Дарье в окрестностях Кзыл-Орды
в середине апреля и один экземпляр на озере Камышлы-баши, 10.Х. 1913 г.
(Зарудный, 1910, 1916), в степях восточного побережья Каспия, на нижнем
Урале, к западу от Ростова-на-Дону. Добыты 1 экз. 16.V. 1876 г. в южных
частях Ленкоранского уезда и 1 экз. 5. III. 1892 г. в окрестностях ст. Кум-
баши (в обоих последних случаях журавли добыты из стаи серых журавлей;
Тизенгаузен, 1902; Сатунин, 1907); наблюдался у Махач-Кала. Определения
в этих случаях все же сомнительны и позднейшими исследователями пока
эти данные не подтвердились. Вопрос о том, какие журавли встречались на
Каспии, до сих пор неясен. ЭверсманнA866) называл этих журавлей даур-
Карта 18. Распространение даурского журавля Grus
vipio
а — границы гнездовий, б — не вполне выясненные границы
гнездовий, г — зимовки.
ДАУРСКИЙ ЖУРАВЛЬ 119
скими G. antigone и описывал их оперение как у Grus vipio. Добытый
16.V, 1876 г. экземпляр не сохранился—Тизенгаузен A902) определил его
как О. monachus, но опубликованное им описание убитой птицы вполне соответ-
ствует О. vipio и Сатунин A907) на этом основании в свою работу «Материалы
к познанию птиц Кавказского края» включил О. vipio, правда, под вопросом.
Радде A855) называет встреченных у Махач-Кала журавлей G. antigone, причем
описания птиц не дает, повидимому, как и Эверсманн, он подразумевает под
этим именем G. vipio, так как ссылается на Палласа, который под именем
G. antigone понимал G. vipio.
Биотоп. Узкие горные заболоченные долины, с протекающими по ним
реками.
Численность. Был обыкновенен и довольно многочисленен на
Аргуни, Ононе и Тарей-норе (Тачановский, 1875) во времена первых иссле-
дователей Вост. Сибири и Приморья, многочисленен и встречался чаще,
нежели другие виды журавлей в бассейне озера Ханка, по горным речкам
южной оконечности Уссурийского края и в особенности на Сунгачинских
болотах (Пржевальский, 1876). В настоящее время стал редок и местами исчез
совсем; так, на Тарей-норе в настоящее время может быть уже не гнездится,
но в Даурских степях по озерам в бассейне Онона и Борзи и других рек много-
численен (Долгушин, 1941). Редок у озера Ханка, не встречается совсем в горах
к западу от Ханка — на р. Даубихе и по рекам, впадающим в Японское море.
(Шулышн, 1936). На нижнем течении р. Уссури всегда был редок и наблю-
дался в меньшем количестве, чем у Аргуни и Ононе (Тачановский,
1875), далее к югу по долине этой реки, вероятно, совсем не гнез-
дился. Одновременно с уменьшением количества даурского журавля,
повидимому, также сократилась и его гнездовая область распрост-
ранения.
Экология. Размножение. Моногамы. Сейчас же по прилете разби-
ваются на пары, иногда прилетают в парах, но первое время держатся все же
стаями на открытых местах; только после таяния болот, т. е. в Южно-Уссурий-
ском крае приблизительно с середины апреля, стаи разбиваются, и пары зани-
мают определенные гнездовые участки, на которых остаются до осени. Для
гнездовых участков обычно выбираются топкие непроходимые болота среди
уединенных узких горных долин.
В период ухаживания и даже во время насиживания яиц журавли часто
устраивают пляски, похожие на пляски журавля-красавки (Пржевальский,
1870, 1876). Происходят они по утренним и вечерним зорям. Обычно несколько
пар журавлей, живущих по соседству, собираются на раз выбранное среди
болота сухое, ровное место, не имеющее кустов, оврагов и других заслонов,
могущих скрывать врагов. С криком прилетают журавли на такое место и обра-
зуют круг, внутри которого находится свободное место для «танцев». На это
место выходят одна или две птицы и начинают прыгать, кивать головою, при-
седать, подскакивать, при этом журавли полураспускают крылья и издают
трубные звуки. Остальные присутствующие смотрят на них, затем сменяют
уставших, которые в свою очередь становятся зрителями. Пляска продолжается
часа два, пока птицы с громким криком не начинают разлетаться по своим
гнездовым участкам (бассейн озера Ханка, Пржевальский, 1870). Независимо
от плясок, самец, бродя с самкой по болоту, часто приседает перед нею,
полураспускает крылья вроде того, как это делает журавль-красав-
ка. Кроме того, весною самцы на лету иногда как-то особенно машут
крыльями сходно с тем, как это делают кроншнепы в период спаривания
(Пржевальский).
Гнездо, как и другие журавли, устраивают на земле.
В кладке 2 яйца. Размеры: 103,0 х 62,0 и 96,0 х 61,8. Вес 22,0 и 18,0 г
(Харбин, Иоганзен, 1930). Яйца похожи на яйца серых журавлей, зеленовато-
120
ОТРЯД ЖУРАВЛИ
бурые с темными буровато-коричневыми пятнами и крапинами. Кладка най-
дена 11.V. 1914 г. в окрестностях Харбина. В верховьях Сунгачи (Южно-Уссу-
рийский край) уже в конце апреля 1868 г. журавли наблюдались чаще по-
одиночке и лишь изредка парами. Это дает возможность предположить, что
насиживание началось (Пржевальский). Длительность насиживания не выяс-
нена. Время кладки, повидимому, с конца апреля и по конец мая. В долине
р. Сиянхе (Южно-Уссурийский край) 1.VI. 1869 г. добыт птенец, выведшийся
дня 2—3 тому назад. Первые птенцы встречены в бассейне озера Ханка 30.V;
в долине р. Лефу родители с птенцами
наблюдались в середине июня 1869 г.
(Пржевальский, 1876).
Линька, Не изучена.
Питание. Также не изучено,
повидимому, как и у большинства
видов журавлей, составляет преиму-
щественно растительная и в неболь-
шой степени животная пища.
Полевые признаки. Несколько
крупней серого журавля. По окраске
похож на серого журавля, но по
повадкам, пляскам больше напоминает
журавля-красавку. Осторожен, но
в уединенных местностях довольно
смел и подпускает к себе шагов на
150, чего никогда не делает на от-
крытых Сунгачинских болотах (Прже-
вальский). При полете, как и другие
журавли, вытягивает ноги и шею по
прямой линии во всю длину. Голос
похож на голос серого журавля
(Шульпин, 1936).
Описание. Размеры истроение.
Длина крыла 560—600 мм. Первостепенных
маховых 11, самое длинное 3-е, 2-е и 4-е
чуть короче 3-го. Третьестепенные маховые
удлинены. Рулевых 12. Лоб и бока головы
не олерены и покрыты красной кожей с ред-
кими черными волосовидными перьями.
Окраска. Пуховик C-4-дневный)
буровато-рыжий сверху и светложелтый
снизу.
В птенцовом наряде голова оперена.
Спинная сторона тела — рыжевато-бурая,
голова и шея сверху несколько светлее
спины. Горло — охристо-беловатое, шея спереди грязно-охристо-серая. Низ тела — серый
с узкими — охристыми окаймлениями перьев.
В первом брачном наряде лицевые части головы не оперены, красного цвета с редкими
черными волосовидными перьями. Оперение серое, сходное с оперением взрослой птицы.
Темя и затылок охристо-рыжие.
У взрослой птицы (самка и самец) общая окраска оперения серо-сизая с более свет-
лыми серыми окаймлениями перьев. Голова, за исключением серо-сизых перьев кроющих,
ухо, задняя часть шеи, горло и передняя часть шеи, соприкасающиеся с горлом, белые.
Остальная часть передней стороны шеи, спина, брюшная сторона тела, плечевые, рулевые,
за исключением их черных вершин, верхние и нижние кроющие хвоста и нижние кроющие
крыла — серо-сизые. Первостепенные маховые, кроющие их, крылышко — буровато-черные.
Стержни первостепенных маховых—белые. Второстепенные маховые у основания —
светлосерые, а к вершине — темнобурые. Третьестепенные маховые — белые с легким
сероватым налетом. Верхние кроющие крыла — серые, несколько светлее общего
оперения тела. Радужина — оранжево-желтая, клюв — желтовато-зеленый. Ноги
красные.
Рис. 19. Даурский журавль
ЧЕРНЫЙ ЖУРАВЛЬ ИЛИ ЖУРАВЛЬ-МОНАХ
121
4. Черный журавль или журавль-монах Grus monachus
Те mm.
Grus monachus. Temminck. PI. Col., 1835, табл. 555, Хоккайдо.
Распространение. Ареал. Гнездовый ареал не выяснен. Область летних
находок довольно обширна и занимает ряд районов Вост. и Зап. Сибири,
доходя в последней на запад до Каинска. Достоверно установленных — за
одним исключением — мест гнездования неизвестно. Единственный раз гнездо
найдено в АО км к западу от г. Каинска (Залесский, 1921); в Томске было
приобретено яйцо черного журавля (яйцо было сравнено с яйцом черного
Карта 19. Распространение черного журавля Grus monachus
а — границы гнездовий, в — места летних находок, г — зимовки.
журавля, отложенным в неволе в Англии; Иоганзен, 1907). Были встречены
или добыты черные журавли в Зап. Сибири, по данным Иоганзена A907,1930) и
Залесского-A921), в гнездовое время в следующих местностях: 1. Добыт Аники-
ным в с. Спасском A5 км к югу от Томска) 14.VI. 1893 г. (первая находка в Зап.
Сибири); 2. Добыт в дер. Пападейкино (близ Томска) 16.V, 1918 г.; 3. Под-
стрелен в середине мая 1915 г. около Томска; 4. Три журавля наблюдались
(в стае серых) 2.V. 1920 г. в окрестностях Томска; 5. Добыт близ Томска в дер.
Петровой 18.V. 1917 г.; 6. Добыт близ Томска у с. Коченево; 7. Наблюдалась
пара у дер. Поповой близ Томска в конце апреля и в начале мая 1917 г.;
8. Встречен в стае серых журавлей 19.VI. 1920 г. на Енисее; 9. Добыт на р. Кеть,
правом притоке Оби, в 1921 г. По сообщению Крохалева, черный журавль
гнездится на озере Чанах, но может быть не каждое лето (Залесский, 1921).
Организованная в 1909 г. Томским университетом экспедиция не только не
нашла там гнездовий, но и не видала там черных журавлей.
В Вост. Сибири в Забайкалье и в особенности в Даурии черный журавль
встречался в гнездовое время в ббльшем количестве, чем в Зап. Сибири- На
южном берегу Тарей-нора встречены Штегманом A928) пары, повидимому, гнез-
дящихся птиц и одновременно большие стаи холостых; в большом количестве
122 ОТРЯД ЖУРАВЛИ
встречены Долгушиным A941) летом по степным озерам Даурии. Самка добыта
30.VI. 1925 г. на Тарей-норе (Зоологический институт Академии наук СССР).
Молодой самец добыт 18.VI. 1939 г. Поповым на озере Зун-Аралантуй Бор-
зинского района, Читинской обл. Гнездится, по Моллесону A896), в верховьях
рек Чикой и Иро, но позднейшими исследованиями эти данные не подтвер-
дились. Наблюдались на среднем течении Амура, несколько пар в—Буреинских
горах и на р. Селбачи (вне наших государственных границ, Радде, 1863).
Добыт в сев. Манчжурии в первых числах июля 1933 г. в окрестностях ст.
Хайлар.
Зимует в юго-вост. Китае на нижнем течении Янцзе; в юго-зап. Японии:
на Кю-сю в Ятусиро, Акине и Арасаки и в южной части Хондо в Ияширо и
Ямагучи (Ян, 1942). Редко и может быть не ежегодно зимуют в вост. Индии,
в сев. Качаре, Субансири, в сев. Лахимпуре, Манипуре, в нижней Бирме
(единичные экземпляры).
Характер пребывания. Гнездящаяся и перелетная птица. На
зимовки в Китай прилетает в ноябре и остается там до апреля; в юго-зап.
Японию прилетает в середине ноября и держится там до конца февраля или
начала марта. Зимующие в Японии журавли летят к нам в Приморье через
Цусиму и юго-вост. Корею, где пролетные птицы наблюдались в Фузане.
В Южно-Уссурийском крае черные журавли появляются в небольшом числе
стайками в 3—11 штук в конце второй половины апреля и пролет идет до сере-
дины мая. Так в верховья Сунгачи в 1869 г. первые птицы прилетели 23. IV и
затем пролетные стаи наблюдались 5 и 15.V (Пржевальский). Далее пролет
идет на северо-запад — на озеро Ханка и на среднее течение Амура. Пролет
птиц, зимующих в Китае, идет, повидимому, по морскому побережью Китая
(в Чжили пролетные птицы отмечены в апреле и октябре). Затем пролетные
журавли в большом числе наблюдались уже далеко от морского побережья
в Калгане и на всем протяжении от Калгана до озера Далай-нор. Первые птицы
отмечены там 27.111. 1871 г., валовый пролет шел в том же году в первой поло-
вине апреля ст. ст. (Пржевальский, 1876). Отсюда журавли, вероятно, летят
прямо на север по западной окраине Большого Хингана. На северном берегу
озера Тарей-нора прилетевшие журавли наблюдались в мае, на южном берегу
Байкала, на Ононе и на Аргуни появляются уже в середине мая. В Зап. Си-
бири встречается с первых чисел мая и даже однажды в 1917 г. в конце ап-
реля близ Томска (дер. Попова) наблюдалась пара журавлей (Иоганзен,
1930).
Из Зап. Сибири отлетает во второй половине августа и осенний пролет
идет до первых чисел сентября. На курорте Карачи (озеро Колосово, Малый
Яркуль, Термакуль) два черных журавля наблюдались 2—4. IX. 1927 г. и
4 журавля—8.IX (Рузский, 1940); в окрестностях Томска—7.IX. 1919 г.—
3 журавля (Залесский, 1921); 20.VIII и затем в начале сентября 1920 г. — на
Оби около с. У ртам б. Томского уезда (Иоганзен, 1930); на р. Чулыме близ
с. Зырянское, к востоку от Томска,—в конце августа; на левом притоке Чулыма
в б. Минусинском уезде добыт 15.Х. 1912 г. один экземпляр (Тугаринов, 1913).
Встречается на пролете весною и осенью по среднему-течению Чулыма (сообще-
ние Аникина, Иоганзена, 1923), редкая пролетная птица на Курчуме, встре-
чается на пролете в верховьях Нижней Тунгуски (один экземпляр добыт,
Ткаченко, 1937). Добыт на пролете в сентябре на северо-восточном берегу
Байкала в окрестностях с. Кудалдов (Шульпин, 1936). Наблюдался на пролете
на южном берегу Байкала, на Ононе и на Аргуни с конца августа и до середины
сентября и в августе — на северном берегу Тарей-нора.
Биотоп. Мало выяснен, повидимому, как у других журавлей.
Экология. Размножение. Не изучено. В Томске 26. V. 1896 г. куплено
яйцо черного журавля, найденное, повидимому, близ этого города (Иоганзен,
1907) и в 1908 или 1909 гг. в 40 км к западу от Каинска найдено гнездо с клад-
СТЕРХ ИЛИ БЕЛЫЙ ЖУРАВЛЬ
123
кой из двух яиц (Залесский, 1921). Размеры: 106,0 х 60,0 мм (Томск) и
103,0 х 62,0мм (Каинск). Вес: 19,5г, 22,5г. Окраска их зеленовато-коричне-
вато-бурая с темными круп-
ными пятнами и крапинами.
Линька и питание
не изучены.
Полевые признаки. Менее
осторожен, чем другие виды
журавлей (Пржевальский).
Размерами меньше, чем серый
журавль. Хорошо отличается
от серого журавля белой голо-
вой и шеей, темной окраской
оперения и тем, что не имеет
черных и белых полос вдоль
шеи.
Описание. Строение и
размеры. Длина крыла: 470—
530 мм (Козлова, 1935), 460—545 мм
(Бутурлин, 1935). Первостепенных
маховых 11. Самое длинное 3-е
первостепенное маховое, 2-е и 4-е
чуть короче 3-го. Рулевых — 12.
Передняя часть головы до глаз и
темя голые, покрытые волосовид-
ными перьями.
Окраска. Пуховик не
известен.
В гнездовом наряде опере-
ние темносерое с буроватым
оттенком. Лоб, бока головы и гор-
ло бледноохристые. Темя, заты-
лок рыжевато-охристые. Шея
серая с узкими охристыми окайм-
лениями перьев.
Взрослые птицы имеют опе-
рение темного аспидно-серого цве-
та. Верхняя сторона — с бурова-
тым налетом, нижняя — с сероватым. Бока головы, затылок, горло, шея — сзади почти
до передней части спины и спереди — на три четверти своей длины — белые. Махо-
вые и рулевые черно-бурые. Неоперенные части головы черные, за исключением
красного пятна на темени, частично заходящего на затылок.
Клюв зеленоватый, переходящий в желто-зеленый на вершине и красноватый
у основания. Ноги черно-бурые. Радужина желтовато-бурая.
5. Стерх или белый журавль Grus leucogeranus Pall.
Grus leucogeranus. Pallas. Reise Versch. Prov. Russ. Reichs., 2, 1773, стр. 714,
болота окр. Ишима, Иртыша и Оби.
Синоним. Ardea gigantea. G m e 1 i n. Reise d. Russl. 11,1774, стр. 189, табл. 21,
Астрахань.
Русское название. Белый журавль—книжное; стерх, стерех, стрех— казах-
ское, ак-турка — башкирское.
Распространение. Ареал. Разорванный — одна гнездовая колония на
северо-западе, другая на северо-востоке Сибири. В настоящее время в Зап. Сиби-
ри гнездование стерха доказано только на нижней Оби — в бассейнахКонды и
Сосьвы на север до Березова (Дерюгин, 1898) или может быть северней, даже до
64°30' (Тарунин, 1928), и на юг, повидимому до с. Самарское, несколько северней
впадения Иртыша в Обь (Наумов); весьма вероятно, что стерх гнездится и на
болотах, окаймляющих с севера Барабинские степи (Рузский, 1940). В Вост.
Сибири — в бассейне Индигирки от устья до впадения в нее р. Момы, на запад
Рис. 20. Черный журавль
124
ОТРЯД ЖУРАВЛИ
до р. Хромы (Михель, 1935), нижнего течения Яны и Святого носа (Бируляг
1907; Бунге, 1887); может быть заходит и восточнее Лены между 68° и 72° с. ш.
(Аргентов, 1861) и по Колыме, повидимому, доходит до устья (Аргентов, 1876),
сюда же, вероятно, относится сообщение местных жителей о гнездовании на
Колыме «аиста» (Тайер и Бенге,1914); в окрестностях Якутска на озере Табога
(Миддендорф, 1855; Иванов, 1929). В долине р. Вилюя гнездование установ-
лено Мааком A859) и на Витимском плоскогорье Поляковым A873).
Карта 20. Распространение стерха Grus leucogeranus
а — граница гнездовий, б — не вполне выясненные границы гнездовий, г — зимовки (стрелки показывают
направление пролета).
Летом стерхи — очевидно холостые — встречаются далеко от гнездовой
области. Эта область залетов состоит из двух участков. Первый — от низовьев
Волги и Урала и Средней Азии на юге до северного Приуралья (среднее течение
Камы под 58°6' с. ш., Чердынь) и нижнего течения Оби на севере и до Барабин-
ских степей на востоке. Второй — от Забайкалья, сев. Монголии и Приморья
до Якутска, Яны, Индигирки и Колымы. В средней Сибири ни летом, ни на
пролете не наблюдался.
Очень сомнительны указания Карелина на гнездование стерха в начале
второй половины прошлого столетия 'в низовьях Урала между Гурьевом и
Редутской. Позднее стерх в окрестностях Гурьева был встречен летом лишь
в 1907 г. Даты встреч в Средней Азии относятся к весне и осени, за исключе-
нием одной находки на нижней Сыр-Дарье в районе Казалинска у Бишарны,
сделанной в июле, и это была, конечно, холостая кочующая птица. Более ве-
роятно, что в 80-х годах прошлого столетия стерх гнездился в Казахстане на
озерах Джиты-куль, Айке, Чалкар-игыз-кара и в истоках Тобола, но к 1894 г.
озера эти уже высохли и стерхи там исчезли. В конце прошлого столетия гнез-
СТЕРХ ИЛИ БЕЛЫЙ ЖУРАВЛЬ 125
дился на озерах водораздела Иргиза и Тобола, по Тургаю (урочище Сарыкопа),
в бассейне Ори, на Чалкар-тенизе и может быть у Наурзума (Сушкин, 1908).
В настоящее время в этих местах не гнездится. Гнездился у Барнаула, в
Барабинской степи, на Чанах, на озере Сартлан. В 70—80-х годах прошлого
столетия в Челябинской обл., по словам,местных жителей, стерх не представ-
лял на гнездовье большой редкости, но к 1898 г. ни одного стерха там не было.
В конце 80-х годов прошлого столетия может быть гнездился на озере Север-
ное близ Тюмени (в течение трех лет там наблюдалась пара стерхов).
Гнездование стерха в юго-вост. Забайкалье (Моллесон, 1896, 1897) ив сев.-
вост. Монголии (Радде, 1863) очень сомнительно. В настоящее время никем
из зоологов не встречен там на гнездовье (в коллекциях Троицкосавского
музея имеется чучело с этикеткой «С. Монголия, Моллесон, 1906», и в Читинском
музее естьэкземплярстерха, добытый в окрестностях г.Читы, без даты (Штегман,
1928). Возможно, что стерх гнездился ранее на Аргуни и Ононе (Тачановский,
1893), но последующие исследователи Аргуни и Онона его там не отмечали.
В Приморском крае стерх никогда не гнездился, все указания о гнездо-
вании его там в 50—60 годах прошлого столетия относятся к манчжурскому
журавлю, о распространении которого в Приморском крае в те годы не знали.
Позднейшие исследователи Приморского края стерха на гнездовье не нашли,
гнездование же манчжурского журавля в Приморье подтвердилось неодно-
кратными наблюдениями.
Области зимовок стерха разобщены друг от друга и их, как и гнездовую
область, можно разделить на два отдельных участка —восточный и запад-
ный. Восточные зимовки расположены в юго-вост. Китае на нижнем течении
Янцзе. Западные —в Сев. Индии на восток до Бихара; в Иране —в Сеи-
стане, Хорасане, Мазандеране и Гиляне; может быть в Закавказье (в феврале
1925 г. четырех стерхов встретил в Ленкорани Е. П. Спангенберг, но по его
мнению это были явно пролетные птицы).
Характер пребывания. Гнездящаяся и перелетная птица.
Летит стерх небольшими стаями, парами и в одиночку, осенью стаи достигают
несколько больших размеров, в особенности на местах отдыха и кормежек.
Ранее будто бы наблюдался на пролете большими стаями — до 300 шт. (Заруд-
ный,1896; Бостанжогло, 1911). Летят то клином, то шеренгой, перестраиваясь
на лету. Первые птицы появляются на юге нашей страны рано. Так, в Ленко-
рани пролетные стерхи наблюдались во второй половине февраля, в дельте
Волги — в самом начале апреля. На Эмбе — в середине апреля (ст. ст.),
в окрестностях Чкалова — в середине и в конце первой половины апреля.
На берегу Аральского моря у Камышлы-Баша наблюдался 5. IV, 1858 г., около
Карамакчи — 16. IV. 1858 г. (Северцов, Бостанжогло, 1911), в Соло-тюбе —
14. IV. 1927 г. (Спангенберг и Фейгин, 1936), в конце марта 1891 г. (ст. ст.) —
на берегу Сары-Камыша, в конце апреля — около Казалинска (Зарудный,
1896, 1910, 1916).
В Приморском крае стерх на пролете встречается только в южной части
края. Пролет начинается в первой половине апреля. В 1868 г. первая птица
появилась в верховьях Сунгачи 4. IV, в 1869 г.—1. IV. Валовой пролет наблю-
дался в середине апреля, в начале второй половины апреля птицы встречались
уже реже и к концу этого месяца исчезли совсем (Пржевальский). В Якутск
первые стерхи прилетели 10. IV. 1843 г. (Миддендорф,1855), в Верхоянске
пролетная стая наблюдалась 12.V (Бунге, 1887), в устье р. Момы (приток
Индигирки) первые птицы замечены 18.V, 1929 г. (Михель 1935).
После вывода птенцов в конце июля и в начале августа стерхи собираются
в небольшие стаи, молодые вместе со старыми, и начинают вести кочевой образ
жизни. Первое время кочуют близ гнездовий, затем постепенно уходят все
дальше и дальше от них. Даты и места встреч кочующих птиц: начало августа
1884 г.—близ Челябинска и 7.VI 11. 1936 г. —на р. Тартас, приток Оми, Омско-
126 ОТРЯД ЖУРАВЛИ
го района (Рузский, 1940), на озере Таволжан —8—10.VIII. 1896 г. (Рузский,
1897), на озере Терма-куль —14.VIII. 1923 г. и близ курорта Карачи 27V.II.
1938 (Рузский, 1940), 14V.III. 1915 г., на озере Кара-су Павлодарского района
(Шухов, 1930), 7.VIII—по дороге из Борового в Азбай (Словцов, 1878),
6.VIII. 1936 г.— у Наурзума (Михеев, 1938), 12. VI11 — в долине Вилюя у исто-
ков р. Чиили 65° с. ш. (Маак, 1859), 28.VIII — Витимское плоскогорье (Поля-
ков, 1873).
Пролет начинается в сентябре и идет до середины октября. На Индигирке
15. IX. 1929 г. еще добыт молодой стерх (Михель, 1935). С нижней Оби (район Кон-
до-Сосьва) улетают в начале второй половины сентябряA9. IX. 1941г.,Раевский).
В Наурзуме пролет наблюдался со второй половины сентября и до конца этого
месяца. Обычно летели стаями от 7 до 25 шт., на водопой и кормежки останавли-
вались на озерах в особенности на Ак-Суата, собираясь там в довольно большие
стаи, в 40—60 особей (Михеев, 1938). У Мугоджар пролетные стерхи встречены
30.VIII. 1857 г., на озере Ходжа-куль близ северного края Б. Барсуков
17.Х. 1857 г. (Бостанжогло, 1911), на нижнем течении Ангрена —в конце
сентября (ст. ст.) и на ст. Арысь 6.Х.1908г. (Зарудный, 1916). В конце
сентября 1913 г. — близ Дзауджикау.
Кроме вышеперечисленных местностей на пролете стерхи встречены (даты
не указаны) на юго-западном берегу Каспия около Дербента и к югу от Апше-
ронского п-ова, в низовьях Урала и Волги, у Чердыни и на Каме
58°5' с. ш., на Или, по южному берегу Балхаша и по р. Кара-талу, по Иртышу
и Курчуму, в районе Троицкосавска на р. Чикой, в Даурии в окрестностях
Цурухайтул, на озере Тарей-нор и в Буреинских горах.
Летние кочевки. Часть птиц, молодых, еще не половозрелых, и взрослых,
почему-либо не приступивших к размножению, собираются в стаи, а иногда
парами или в одиночку ведут все лето кочевой образ жизни. В это время они
встречаются далеко от мест гнездовий. Повидимому, именно такие птицы наблю-
дались летом в низовьях Урала (Рузский, 1940), на нижней Сыр-Дарье в районе
Казалинска у Бишарты (июль ст. ст. 1858), на озере Джумарт-куль близ
Б. Барсуков в половине мая (ст. ст.), на озере Кургальджин — 13.V. 1946 г. (по
сообщению местных жителей встречается ежедневно, Деревягин, 1947), на
Чанах — в июле (Коте, 1910), у Каинска 30.V. 1909 г. (Иоганзен, 1907), в окре-
стностях Семипалатинска 19.V (Хахлов и Селевин, 1928), на Абакане близ
горы Изых в июне (ст. ст.) 1877 г., в Называемском районе Омского округа
летом 1927 г. и добыт у южной границы Омского округа между Черлаком и
Урлютюиом летом 1928 г. Очевидно бродячие птицы встречены летом в окрест-
ностях г. Уфы, на Каме под 58°5' с. ш. и под Чердынью.
Добыт один экземпляр на Алтае близ ст. Катон-карагая в декабре 1882 г.
(Никольский, 1882). Добыт 1 экземпляр близ Ростова-на-Дону весной. Встре-
чен в Туркмении (Дементьев, 1950). Отмечен и на Сахалине. Большая стая
встречена на Куку-норе 20.Х.1872 г.
Биотоп. Большие недоступные, топкие, но неглубокие болота, озера
с низкими берегами, поросшие травянистой растительностью или зарослями
камыша и тростника, расположенные среди леса.
Численность. Всюду редок и спорадичен-
Экология. Размножение. О размножении стерха нет почти никаких
данных. К нам стерхи обычно прилетают уже в парах. По прилете на места гнез-
довий часть птиц вскоре приступает к размножению, а остальные ведут кочевой
образ жизни. Стерх токует, полураспустив крылья, опустив голову и вытянув
шею и клюв параллельно земле, издавая мелодичные трубные звуки (Кондо-
Сосьвинский заповедник, Раевский). Гнездо помещают на возвышенных сухих
местах, среди глухих болот; оно представляет собой небрежно набросанную
кучу камыша и осоки. К гнезду самец и самка очень привязаны и по очереди
стерегут его.
СТЕРХ ИЛИ БЕЛЫЙ ЖУРАВЛЬ
127
Кладка состоит из 2 яиц, основной тон которых желтовато-зеленый с тем-
ными пятнами (Паллас, 1811) или темнозеленый (Бируля, 1907). Продолжитель-
ность эмбрионального развития и постэмбрионального роста птенцов неиз-
вестна. Как только птенцы подрастут, стерхи собираются в стаи и проводят
день в степи, на полях или у
озера (Назаров, 1887).
Л и н ь к а. Не изучена.
Питание. Изучено слабо.
Пищу составляют мелкая рыба
(Паллас, 1811), ящерицы, ля-
гушки, змеи, мелкие грызуны,
рыба и пр. (Эверсман, 1866).
Бекер A929) считает стерха бо-
лее растительноядной птицей, по
сравнению с другими видами
журавлей, питающейся пре-
имущественно водными растени-
ями, но возделанным полям он не
приносит вреда. Повидимому,
наблюдение Бекера ошибочно,
так как и строение самого клюва
(на вершинной части клюва
имеются зазубрины, служащие,
вероятно, для удержания живой,
легко выскальзывающей добычи)
говорят о том, что стерх питается
преимущественно животной пи-
щей.
Полевые признаки. Крупнее
других видов наших журавлей
(за исключением манчжурского)
и хорошо отличается от них
белой окраской оперения. От
манчжурского журавля отли-
чается более сильным и массив-
ным клювом, красной окраской
клюва, неоперенных частей го-
ловы и ног и белой окраской
шеи, а также другим располо-
жением черного цвета на крыле.
Осторожная птица. Благодаря
своему росту, стерхи издали могут видеть опасность, заметив ее, они
с криком взлетают и долго не возвращаются обратно. Крик стерха не-
сколько напоминает крик серого журавля, но мелодичней и громче.
Описание. Размеры и строение. Вес стерха около 10 кг. Длина крыла C)
640—690 мм, в среднем 666 мм. Первостепенных маховых И; 2-е, 3-е и 4-е первостепенные
маховые между собою равны и образуют вершину крыла. Первое первостепенное маховое
равно пятому, остальные 5 маховых значительно короче. Из них самое длинное, 6-е, не
достигает вершины крыла на 145—150 мм. Хвост короткий, прямой, рулевых— ^.Третье-
степенные маховые удлинены (на 50—60 мм от вершины крыла) и рассучены, но менее,
чем у серого журавля. Лицевая часть головы, бока головы, не оперены и покрыты редкими,
короткими, рыжими перьями. Края клюва в вершинной части надклювья и подклювья
зазубрены.
Окраска. Птенец не описан.
Молодая птица в птенцовом наряде. Голова вся густо оперена. Верх тела — ржаво-
коричневый. Низ и бока тела и нижние кроющие хвоста — белые с ржаво-коричневыми
пятнами. Голова, шея и верхняя часть спины — интенсивно ржаво-коричневые, только
Рис. 21. Стерх
128
ОТРЯД ЖУРАВЛИ
у самого основания перья аспидно-серые. Верхние плечевые перья — аспидно-серые с белыми
основаниями и с ржаво-коричневыми окаймлениями. Нижняя часть спины и верхние крою-
щие хвоста пестро окрашены, перья их — белые с ржаво-бурыми окаймлениями. Средние
рулевые аспидно-серые с ржаво-бурыми вершинами и белыми основаниями. Крайние руле-
вые в основной половине — белые, постепенно переходящие в вершинной половине в ас-
пидно-серый тон и заканчивающиеся ржаво-бурыми вершинами. Первостепенные маховые —
черные, второстепенные — коричневые с пепельным налетом. Большие верхние кроющие
крыла — белые с небольшими ржаво-коричневыми пятнами на концах; плечевые аспидно-
серые с ржаво-коричневыми каймами и белыми основаниями. Подмышечные — белые. На
зобу и на боках тела у основания хвоста долго, до 15. IX., сохраняется пух — серый, бе-
леющий к основанию и бурый на вершине. Основание клюва — темнобурое с красноватым
оттенком, вершина желтовато-белая. Ноги — темные с красноватым налетом. Махи еще
не достигают размеров взрослой птицы (Михель, 1935).
Взрослая птица (самец и самка). Оперение белое, за исключением черных перво-
степенных маховых, верхних кроющих их и крылышка. Стволы первостепенных маховых
на наружной стороне пера — черные, на внутренней—буроватые, второстепенных маховых—
буровато-белые. Ноги — красные. Когти — черные. Клюв весной и летом — буровато-крас-
ный, светлеющий к вершине, причем у самок несколько светлее, чему самцов. Радужина —
яркожелтая. Голые части головы — красные. В осеннем наряде перья затылка не чисто
белые, а с легким сероватым налетом.
6. Манчжурский журавль Grits japonensis Mull.
Ardea japonensis. M ti 1 1 e r. Natursyst. Suppl., 1776, стр. ПО., Япония.
Синоним. Antigone montignesia. Bonaparte. Compt. Rend. Acad. Paris, XXXVIII,
1854, стр. 661, Манчжурия.
Распространение. Ареал. Приморье (Ханкайская и Уссурийская низ-
менности), к северу достоверно доходит до устья Уссури, может быть несколько
и северней, до низовьев Амура, но до дельты его не доходит (в Мариинске нет,
Шульпин, 1936). «Белые журавли»
на нижнем Амуре у устья Горин и
на о-ве. Челмок наблюдались 18.VII
и 27. IX (Шренк, 1861) и в гнездовое
время на Амуре и Уссури (Ма-
ак, 1861). Эти белые журавли
были отнесены Шренком и Ма-
аком к G. leucogeranus. Поздней-
шими исследователями гнездование
G. leucogeranus в Приморье не под-
твердилось. Пржевальским A870)
был найден в Приморье новый
вид —«китайский журавль» G. mon-
tignesia (G. japonensis). Повидимо-
му, именно этот журавль и наблю-
дался Шренком и Мааком во время
их путешествия по Амуру и Уссури.
На запад по Амуру может быть до-
ходит до Зеи (Шульпин, 1935). За
пределами СССР —в равнинах сред-
ней и западной Манчжурии, на р.
Сунгари, и может быть в долине
р. Халхин-гол (наблюдалась стая с 1—9. IX. 1928), кроме того, в Японии во-
сточней Хоккайдо.
Зимует в юго-вост. Китае, на нижнем течении Янцзе; в Сев. и центр.
Корее, где обычен до Сеула, далее становится редким.
Характер пребывания. Гнездящаяся и пролетная птица в
северных областях своего распространения, оседлая — в южных. В При-
морье прилетает очень рано — со второй половины марта и пролет идет до
середины апреля. В 1868 г. в верховьях Сунгачи небольшие стаи наблюдались
Карта 21. Распространение манчжурского жу-
равля Grus japonensis
в — граница гнездовий, б — не вполне выясненные
границы гнездовий, г — зимовки.
МАНЧЖУРСКИЙ ЖУРАВЛЬ 129
24 и 26. III, когда снег лежал еще глубоким слоем и проталин на озерах не
было. В 1869 г. первые птицы на Сунгачи отмечены 16. III, наиболее сильный
пролет шел между 6. IV и серединой апреля, но их летело немного (Пржеваль-
ский, 1870). На Уссури и Суйфун первые птицы прилетели 13—15.III парами
и небольшими стайками (Дорриес).
Осенний отлет очень растянут, идет с сентября и даже иногда с августа
и до ноября, даже до декабря. На Амуре у устья Уссури последние журавли
наблюдались 14. IX. 1854 г. В 1858 г. пролет шел с середины сентября до 28. IX
(Маак, 1861). В другие годы на Уссури наблюдались в ноябре непрерывно
летевшие на юг стаи манчжурского журавля. Река в это время была уже по-
крыта плавающим льдом, последняя стая в 4 особи встречена в декабре на неза-
мерзающем берегу Уссури (Тачановский, 1875). На озере Ханка у устья р.
Лефу 8.VIII.1926 г. наблюдались журавли, летевшие на юг(Шульпин, 1936).
В устье Суйфуна пролет шел с 22—28.Х.1867г., последние птицы встречены
3.IX, летели парами и небольшими группами вЗ—4 особи, 18. IX. 1869 г. на-
блюдался пролет на р. Лэфунцы под Ворошиловом (Пржевальский), а также,
вероятно, вдоль западных отрогов Большого Хингана. Встреченные здесь Туга-
риновым A932) 1—9. IX. 1928 г. стаи были пролетные. В Китае на пролете
встречаются редко и в небольшом количестве, обычно осенью.
Залеты. Отмечены у Дарасуна, в Танну-Туве на озере Гаголь (летом), и
в восточной части Танну-Ола у перевала Халдак-ажик. Залетает и на о-в Кюсю.
Биотоп. Большие открытые, но не топкие болота, поросшие в неболь-
шом количестве вейником (Calamagrostis neglecta), и побережья рек с песча-
ными отмелями почти без зарослей тальника, камыша или тростника.
Численность. Во второй половине прошлого столетия был обыкно-
венен и по количеству не уступал даурскому журавлю на озере Ханка, на р. Сун-
гачи, Мо, Сиянхе и Лефу (Пржевальский, 1870). По Уссури всегда был редок,
за исключением устья (Тачановский, 1875), позднее местами совсем исчез (Поля-
ков, 1915). В настоящее время сильно уменьшился в количестве и в ряде районов
исчез совсем, что до известной степени объясняется, может быть, усиленной
распашкой земель, осушкой болот и заселением этих мест. На озере Ханка
только на восточной стороне его еще наблюдается в значительном количестве;
по p.p. Сиянхе и Мо совсем перестал гнездиться, исчез на большом протяже-
нии реки Лефу, и только в небольшом количестве остался в устье реки, ре-
док на Суйфуне. На болотах р. Тумень-Ула гнездился в начале настоящего
столетия, но теперь будто бы не гнездится (Шульпин, 1936). Наблюдался
какой-то «белый» журавль в прошлом столетии на южн. Сахалине (Николь-
ский), повидимому, это был манчжурский журавль, а не стерх, но в настоя-
щее время здесь совсем не встречается.
Экология. Размножение. Первое время по прилете на Сунгачи жу-
равли держатся на реке в одних и тех же местах парами, реже небольшими стай-
ками в 3—4 особи и только в десятых числах апреля перемещаются с реки на
большие болота, где собираются у талых мест. Вскоре каждая пара выбирает
себе гнездовый участок, в котором живет до отлета. В отличие от даурского
журавля гнездовые участки манчжурского журавля расположены не в узких до-
линах, а на открытых равнинах (Пржевальский, 1870). На южной окраине озера
Ханка гнездовые участки достигают больших размеров и каждая пара отделена
друг от друга значительным расстоянием в несколько километров. Пары пере-
кликаются между собою трубным курлыканием, которое особенно часто раз-
дается на заре — утренней и вечерней, и в тихую погоду бывает слышно на
3—4 км. (Шульпин, 1936). Весенних плясок и игр у них не наблюдается.
Гнездо, найденное Шульпиным A936) у устья Лефу, близ Алтыновки,
помещалось среди обширной болотистой низменности, поросшей редким полу-
тораметровым вейником. Грунт болота был слегка кочковатым, покрытым
мелкими, чуть зеленевшими травянистыми растениями, а кое-где и мхами. В
9 Птицы СССР, т. II
130 ОТРЯД ЖУРАВЛИ
этой поросли на небольшой, в 4 метра, плешинке помещалось гнездо. Плешинка
была создана искусственно: вейник выдернут, трава помята, место утоптано,
а вейник уложен в беспорядке плоским настилом и несколько утрамбован
ногаМи птиц. Это и есть гнездо, диаметром около 98 см, по существу без лотка,
совершенно плоское, но на месте лотка лежали в изобилии широкие листья
вейника и сухая мелкая трава.
В кладке 2, реже 1, яйцо (данные Дорриеса, 1888, о нахождении кладок
в 3—4 яйца, как и нахождение гнезд преимущественно на деревьях, реже,
на болотах основаны, вероятно, на смешании с кладкой и гнездами аиста).
Срок насиживания в природных условиях неизвестен, в неволе 30 дней. Время
кладки в южн. Приморье, повидимому, с конца апреля по конец мая.
В конце апреля 1868 г. журавли на Сунгачинских болотах наблюдались
Пржевальским A870) чаще поодиночке (повидимому, один из пары сидел на
яйцах). В гнезде, найденном Шульпиным A936) в устье Лефу 23.V. 1928 г.
было 2 яйца, сдвинутых боком друг к другу. В одном яйце оказался вполне
сформированный, покрытый пухом птенец, другое же яйцо было неоплодотво-
рено. В 1927 г. 27.V на южн. Ханке близ д. Сиваковки добыто Шульпиным
A936) 1 яйцо с вполне сформировавшимся птенцом. В середине июня 1869 г.
на нижней Лефу птенцы уже вывелись (Пржевальский). На Уссури, близ
м. Цифяку у «G. leucogeranus», 20.VI 1.1858 г. птенцы были уже больших разме-
ров, но еще не летные (Маак, 1861). К своим птенцам родители менее привя-
заны, чем даурский журавль и при приближении опасности родители с гром-
ким курлыканием пробегают несколько сот шагов, поднимаются и улетают.
Птенцы же прячутся в густой траве и найти их невозможно.
Не все прилетевшие журавли приступают к размножению, часть их, не-
смотря на то, что даже образуют пары, к размножению не приступают. В 1869 г.
в середине мая на Сунгачинских болотах журавли наблюдались Пржеваль-
ским A870) парами, но как будто не имели гнезд. В 1927 г. 25 и 29. IV под Ново-
русановкой встречались еще журавли группами по 5 штук (Шульпин, 1936).
Надо думать, это были стаи холостых особей, тем более, что по наблюдениям
в неволе манчжурский журавль к размножению приступает в возрасте около
5 лет(?).
Линька. Детали неизвестны. Общая схема — как у других видов жу-
равлей. У добытого 25 июля самца все оперение старое, сильно изношенное.
Питание. Плохо изучено. Растительная и животная пища.
Полевые признаки. Один из самых крупных видов журавлей, много больше
серого журавля и чуть больше стерха. Очень осторожен, даже при птенцах.
На открытом месте не подпускает к себе ближе 200 метров. При полете, как и
все журавли вытягивает ноги и шею во всю длину. Издали, благодаря белой
окраске, хорошо заметен и от стерха, с которым может быть только спутан,
отличается окраской клюва и ног, другим расположением черного цвета на
крыле и черной шеей.
Голос — звонкое, трубное курлыканье, похожее на курлыканье серого
журавля, но мелодичней и громче. Вблизи друг от друга птицы издают более
тихие звуки —«тюрр-тюрр» (Шульпин, 1936). При тревоге —особое громкое
курлыканье, отличное от обыкновенного.
Описание. Размеры и строение. Вес самца 9—10,5 кг (Пржевальский,
1870, Яковлев, 1929), даже 15 кг (Шульпин, 1936). Размах крыла самца — 2200 мм (Прже-
вальский, 1870). Длина крыла самцов BN50j<jk и птицы с неопределенным полом—670 мм
(коллекции Зоологического музея Московского университета). Первостепенных маховых —
11, самое длинное 3-е, 2-е и 4-е, равны между собою и на 0,5 мм короче 3-го, 5-е короче 4-го
на 0,7 мм. Формула крыла: 3>2 = 4>5>1>б, остальные четыре пера короткие, не дости-
гают вершины крыла на 130—135 мм. Третьестепенные маховые и плечевые рассучены и уд-
линены (третьестепенные выступают за вершину крыла на 99 мм). Лоб и темя не оперены.
Края клюва в вершинной его части приблизительно на 30 мм слегка зазубрены, причем на
нижней челюсти слабее.
МАНЧЖУРСКИЙ ЖУРАВЛЬ
131
Окраска. Пуховик буровато-рыжий, более темный на спине и более светлый на
брюхе.
В гнездовом наряде голова вся оперена, охристо-бурая. Шея охристо-ржавая. Бе-
лые перья тела и верхних кроющих крыла — с широкими охристо-рыжими окаймлениями.
Второстепенные маховые — бурые с белыми основаниями перьев.
Молодая птица в первом наряде. Оперение тела белое, но среди белых перьев верхних
кроющих крыла встречаются перья с охристо-рыжими окаймлениями. Первостепенные
маховые белые с темными вершинами. Второстепенные маховые тем но бурые с белыми
основаниями перьев. Лицевые части головы не оперены, но неоперенные участки меньших
размеров, чем у взрослой птицы. Оперенная часть головы и шея — охристо-ржавые с ред-
кими отдельными темносерыми перьями на шее.
Рис. 22. Манчжурский журавль
Взрослые самцы и самки имеют оперение белое, за исключением черных второсте-
пенных и третьестепенных маховых, подбородка, горла и шеи. На боках головы, начиная
от глаз, захватывая ухо и переходя на затылок и верхнюю часть шеи, идет белая полоса.
Основание второстепенных и третьестепенных маховых перьев белое, количество белого
цвета постепенно уменьшается по направлению к последнему третьестепенному перу. У пер-
вого второстепенного махового наружное опахало белое и белое наружное окаймление
черного внутреннего опахала. Лоб—черный с короткими, черными волосовидными перьями,
темя — красное с бугристыми выростами кожи, покрытое редкими черными волосовид-
ными перьями. Ноги — черные. Клюв в основной части — темнооливковый, почти чер-
ный, в вершинной — оливковый. Радужина — коричневая. Сезонных изменений у взро-
слых нет.
7. Австралийский журавль Gras rubicundas Perry
Агйеа rubicundus. Perry. Arcana, VI, 1810, стр. 220, Ботани-Бей.
Синоним. Grus australasiana. Gould. Pr. Zool. Soc. London, 1847, стр. 220,
Новый Южный Уэльс.
Распространение. Ареал. Австралия и Новая Гвинея; в СССР один экзем-
пляр австралийского >куравля добыт Павловским под Якутском в 80-х годах
прошлого столетия и хранится в Зоологическом Музее Московского универ-
ситета. Каким образом попал в Якутию австралийский журавль — до сих пор
132
ОТРЯД ЖУРАВЛИ
остается загадкой. На родине эти птицы почти оседлы, совершают лишь не-
большие перелеты в пределах гнездового ареала. Залететь в стае с другими
журавлями этот журавль вряд ли мог, тем более, что ни один из видов журав-
лей, населяющих Си-
бирь, не доходит на
своих зимовках до Но-
вой Гвинеи и Австралии.
Подвиды и варьирую-
щие признаки.2 подвида. G.
г. rubicundus Perry, 1810 —
вост. и южн. Австралия и
южн. Новая Гвинея; G. г.
argenteus Mathews, 1912 —
сев.-зап. Австралия.
[Индийский журавль Grus
antigone L.
Ardea antigone. Linnaeus.
Syst. Nat., 1, 1758, стр. 142,
Азия.
Синоним. Grus collaris.
Boddaert. Tabl. PI. Enl.,
1783, стр. 52, Индия.
Распространение.
A p e а л. Индия к югу
до Бомбейской провин-
ции и р. Го довари; Ассам,
Бирма, Сиам и Вьетнам.
Характер пре-
бывания. Внесен в
фауну СССР как случай-
но залетная птица, на
основании данных ста-
рых авторов (Нордман,
Карелин, Радде и др.),
но, повидимому, ошибоч-
но. Очень сомнительны
указания на залет индий-
ского журавля в устья
Урала, на восточный
берег Каспия, в Махач-
Кала и в Ростов-на-
Дону. К тому же птиц
только наблюдали, а не
добывали. Кроме того,
точно не известно, о каком
виде журавля идет речь —о G. antigone Linn, или о G. antigone Pal las
A811), который под этим именем приводил даурского журавля G. vipio; правда
залеты последнего на Каспийском море также очень сомнительны. Поздней-
шими исследователями пока эти наблюдения не подтвердились, тем более, что и
условия обитания для индийского журавля в прикаспийских районах непод-
ходящие: в Индии этот журавль населяет болотистые местности с густой
кустарниковой и травянистой растительностью. Надо заметить, что даже в том
случае, если определения старых авторов были правильны, встреченные птицы
по всей вероятности улетели из неволи (как это имело место в некоторых стра-
нах Зап. Европы).
Рис. 23. Австралийский журавль
ЖУРАВЛЬ-КРАСАВКА ИЛИ МАЛЫЙ ЖУРАВЛЬ
133
Подвиды и варьирующие признаки. Два подвида: G. a. antigone Linn., 1758 —
Индия к югу до Бомбейской провинции и р. Годовари; G. a. scharpi В 1 a n f о г d, 1895—
Ассам, Бирма, Сиам и Вьетнам.]
РОД ЖУРАВЛИ-КРАСАВКИ ANTHROPOIDES VIEILLOT, 1816
Тип A. virgo'
8. Журавль-красавка или малый журавль Anthropoides virgo L.
Ardea virgo. Linnaeus. Syst. Nat., изд. X, 1758, стр. 141, «на Востоке».
Распространение. Ареал. Несмотря на то, что большая часть гнездового
ареала красавки находится в нашей стране, точно границы его распространения
Карта 22. Распространение журавля-красавки Anthropoides virgo
1. A. v. virgo, 2. A. v. paradisea; a — граница гнездовий, г — зимовки.
для многих районов не установлены. Алжир, может быть, Марокко, южн.
Испания, Добруджия, Бессарабия, южн. Украина, Крым, где известен к югу
до черноморского побережья. К северу в Херсонской обл., Днепропетровской
обл. Встречается, повидимому, в южных частях Киевской обл., в южных ча-
стях Полтавской обл. (б. Константиноградский уезд, Гавриленко, 1917).
В Харьковской обл. гнездился до 90-х годов прошлого столетия в Изюм-
ском и Старобельском уездах (Сомов, 1897), но по сообщению Е. П. Спанген-
134 ОТРЯД ЖУРАВЛИ
берга в настоящее время гнездится в южных частях области, не доходя до
Харькова 50—60 км. Нет его в Подолии (Храневич, 1925—1926). Сев. Кавказ —
Манычские степи (Сарандинаки, 1909), Моздокские и Ногайские степи (Беме,
1926) и Закавказье в районе озера Севан (Ляйстер и Соснин, 1942). Гнездится
в Астраханском крае, но к югу не доходит до дельты Волги, северного побе-
режья Каспия и устья Урала. К северу идет по Волге до Сталинграда, может
быть даже несколько северней (Аргирополо, 1928). В волжско-уральских сте-
пях — к северу до 49° с. ш. (Бостанжогло, 1911), может быть до 50° с. ш. Сред-
нее течение Урала к северу до Уральска и нижнего течения Илека, но до
Чкалова, вероятно, не доходит (имеется единственное указание на гнездование
его в 1888 г. в Донгузской степи в 15 км от Чкалова, Зарудный, 1896), на Ир-
гизе, верховья Тобола, но до Кустаная не доходит, северней Кара-тургай не
идет (Сушкин, 1908), Кургальджинский район, Барабинские степи (может быть,
в настоящее время остался только в южных частях, Рузский, 1940), Каинск,
Барнаул, Бийск, Алтай, за исключением сев.-вост. и юго-вост. Алтая, южные
части Минусинского края, Прибайкалье и юго-зап. Забайкалье, Даурия,
(Штегман, 1928) до Амура и Приморского края не доходит.
Южная граница в Азии идет от нижнего течения Урала до Уйлы 48? с. ш.
(Бостанжогло, 1911), верхнего течения Эмбы, (Кок-джиды, Сушкин, 1908);
Мугоджары до северного побережья Аральского моря и долины реки Сыр-Дарьи,
Сары-су, озера Тели-куль (Спангенберг и Фейгин, 1936), Чу, Или, может быть
сев. Таджикистан, район Ленинабада (в Туркмении не гнездится), Зайсан,
Тарбагатай, Танну-тува, Монголия, исключая центральную и южную; может
быть гнездится в сев.-зап. Китае на Алашане.
Зимовки. Сев.-вост. Африка до Белого и Голубого Нила, Абиссиния,
Палестина, Месопотамия, берега Персидского залива, Сеистан, южные части
Белуджистана, Индия к югу приблизительно до 12° с. ш., на восток до Ассама
и Бирмы; может быть зимует в сев.-вост. Китае—северное Чжили.
Характер пребывания. Гнездящаяся и перелетная птица.
Летят, как и серые журавли, «клином»; причем, нередко одна сторона угла много
длиннее другой. На пролете летят очень высоко, так что их чаще не видно,
а только слышно. Весною летят небольшими стаями, иногда даже парами. При-
летают красавки несколько поздней серых журавлей (начало пролета у кра-
савок совпадает с валовым пролетом серых журавлей). Зимовки, расположенные
в Африке, покидают в феврале—марте. В Бессарабию прилетают во второй
половине марта и только в холодные весны — в апреле. На Украину (Днепро-
петровская обл.) — в конце марта и в начале апреля. На Кавказе пролет идет,
начиная с последней трети марта и до второй половины апреля.
В Туркмении на пролете редок, пролет наблюдался с конца марта до начала
апреля: на Теджене с 26.III по 3.IV. 1886 г., у Чемен-и-бида и Кушки —
4. IV. 1887 г. (Радде и Вальтер, 1889). Встречается на пролете в Кызыл-Кумах,
в особенности в восточной части.
В Таджикистане пролет наблюдается в первой половине апреля G—16. IV,
Гиссарская долина, Иванов, 1940). На нижней Сыр-Дарье (окр. ст. Кара-узяк)—
во второй трети апреля, у Аральского моря у озера Камышлы-баш — в конце
апреля. На Эмбе в районе Кок-джиды первые птицы появляются в конце апреля,
в низовьях Иргиза — во второй половине апреля. На Или пролет начинается
в середине первой половины апреля (9. IV. 1913 г.), интенсивный пролет идет
во второй половине апреля (с 17—25. IV. 1913 г.). Отдельные пролетные пары
встречаются до конца апреля и начала мая (Шестоперов, 1929). На Алтай
прилетают, начиная с середины апреля (в долину Чулышмана — в середине
апреля) и пролет заканчивается в первой половине мая A3.Vy Бийска про-
лет уже закончился, Сушкин, 1938). В окрестностях Троицкосавска красавки
прилетают в конце апреля C0. IV, Меллесон, 1897). В сев. Монголии пролет
идет с конца апреля по первые числа мая (у юго-зап. Кентея валовой пролет
ЖУРАВЛЬ-КРАСАВКА ИЛИ МАЛЫЙ ЖУРАВЛЬ 135
наблюдается 28. IV, на озере Орок-нор первые журавли появились в апреле,
Козлова, 1930). В вост. Монголии первые птицы прилетели 16. IV (Бианки,
1906).
В первых числах августа отдельные выводки начинают уже соединяться
в стаи, которые некоторое время остаются еще на местах гнездовья, проводя
день на лугах и в степях. Вскоре кочевки начинают принимать все больший и
больший размах и постепенно местные птицы улетают. Отлет начинается почти
незаметно. На место улетевших местных птиц появляются новые стаи из более
северных районов и таким образом пролет растягивается на продолжительное
время — на месяц. Во время осеннего пролета стаи красавок достигают боль-
ших размеров, на Или наблюдались стаи до 400 и более особей (Шестоперов,
1929). Из Дуарии (озеро Тарей-нор), большая часть птиц к 27—28.VIII уже
отлетела и последние птицы исчезли 12. IX (Радде, 1863). На Алтае отлет начи-
нается в первой половине августа A7.VIII у Бийска встречены пролетные стаи
красавок, Сушкин, 1938) и идет до середины сентября (в долине р. Чулышман
отлет начался в начале сентября, Фолитарек и Дементьев, 1938), в низовьях
Черного Иртыша 7. IX встречена довольно большая пролетная стая (Сушкин,
1938). На Или пролет красавок наблюдается в течение всего сентября, но пер-
вые прилетные северные стаи встречаются в самых первых числах августа
B.VIII, Шестоперов, 1929); в низовьях Иргиза отлет местных журавлей на-
чинается в конце августа. В устье Урала — в конце августа и идет до конца
сентября. В Туркмении — на Ахал-Текинской равнине пролет идет в середине
сентября (в 1884 г. 16. IX, Зарудный, 1896). На Украине (Днепропетровская
обл.) отлет отмечен с первых чисел сентября. В долине Нила на пролете на-
блюдаются с первых чисел сентября и до конца месяца.
Залеты. Неоднократно отмечены залеты красавки в различные районы
Зап. Европы включительно до Скандинавии и Оркнейских о-вов. У нас как
залетная птица отмечена для Подолии, южной части Черниговской обл., в Пол-
тавской обл., Смоленской обл., в северных частях Чкаловской обл., в окрест-
ностях Кустаная, в Красноярском р-не, в долине Лены на 61°20' с. ш., под
Верхоянском, в окрестностях Владивостока.
Биотоп. Полупустыни и степи: разнотравные, ковыльные, ковыльно-
типичковые, кустарниковые, каменистые, глинисто-солончаковые, солончако-
вые, иногда почти пустынные, с каменистой или щебенистой почвой, также степ-
ные долины рек. Не избегает степей с вкрапленными в них пашнями или не-
большими лесами и поселений человека в степи.
Численность. В Крыму и на южн. Украине обычен, более много-
численен на Волге, в особенности под Сталинградом и на Маныче, где, по
сообщению Е. П. Спангенберга, особенно густо населены степи с солончако-
выми участками, лежащие между балками (на 10 км2 приходится 10 гнезд,
расстояние между некоторыми 200—300 м), и в сев.-зап. Казахстане. Далее
на восток становится более редкой птицей. Редок близ северной границы рас-
пространения и на границе настоящих пустынь (в Армении).
Экология. Размножение. Моногамы. Пары, как и у других журавлей,
постоянные. Одни и те же гнездовые места занимаются в течение ряда лет. Сразу
по прилете на места гнездовья начинается ток, так называемые «пляски»,
на которые собирается большое число особей. Для «плясок» выбирается ровная
сухая площадка, и на ней на утренней и, в особенности, на вечерней заре соби-
раются все гнездящиеся поблизости птицы. Собравшись, журавли образуют
круг, иногда в два или три ряда, оставляя середину свободной. На середину
круга выходят несколько птиц и начинают подпрыгивать, распускать крылья,
вытягивать шею, наклоняться, приседать, распуская перья зоба и издавая при
этом трубные звуки. Утомившиеся птицы возвращаются на свои места в кругу
зрителей, а на их места, на середину круга, выходят новые и снова начинаются
танцы. Затем красавки поднимаются всей стаей, описывая круги в воздухе, и
136 ОТРЯД ЖУРАВЛИ
улетают. Остается невыясненным, принимают ли участие в «плясках» самцы
и самки или только самцы, а самки остаются зрителями.
Тока красавок наблюдаются изредка осенью или летом, но они гораздо мало-
численней, (осеннее токование наблюдалось 30.VIII и 8, IX. 1905 г. в Днепро-
петровской обл.; три птицы танцовали в кругу стаи, Боровиков, 1907; на Алтае,
на озере Теньга с 1—3.VIII наблюдалась стая в 20 штук, устраивавшая под
вечер пляски, Шульпин, 1928). У них, следовательно, имеется и абортивный
осенний цикл. Часть птиц остаются холостыми и, собираясь в стаи, ведут коче-
вой образ жизни в течение всего лета (в Абаканской степи встретилась 30.VI.
1920 г. стая штук в 30, Залесский, 1921; на озере Теньга, Алтай, 1—3.VIII —
стая в 20 штук, состоявшая из одних взрослых птиц, Шульпин, 1928).
Гнездо красавки делают на земле в сухом месте, в степи, на пашне, на га-
лечнике, но обычно вблизи воды: речки, озера или болота. Иногда устраи-
вают гнездо в непосредственной близости от жилья человека. Гнездо предста-
вляет собой неглубокую ямку, но часто красавка откладывает яйца прямо на
земле, не делая даже какого-либо углубления в почве, или среди сухого кон-
ского навоза. Подстилкой в гнезде служат сухие стебли трав и злаков, набро-
санные небрежно и в очень незначительном количестве, иногда подстилка со-
всем отсутствует. Пара от пары гнездится примерно на расстоянии 3—4 км.
В кладке обычно 2 яйца, реже 1 или 3. Яйца красавки похожи на яйца се-
рого журавля, но несколько меньших размеров с шероховатой скорлупой,
оливково-бурого основного тона с желтовато-бурыми поверхностными пятнами
и серовато-бурыми глубокими пятнами.
Размеры: B) 94,0 х 59,0мм и 92,0 х 59,0 мм (Барабинская степь, Иоган-
зен, 1907); B) 83,2 х 53,8 мм, 81,0 х 54,2 мм (Ак-Булак, Казахстан, Спан-
генберг), B) 83,2 х 53,8 мм и 81,0 х 54,2 мм (Тели-Куль, Спангенберг, 1936).
Период кладки у красавки растянут иногда до месяца. В одном и том же
месте можно встретить одновременно гнезда с кладками яиц и уже довольно
больших птенцов. Причины этого явления остаются неясными. В Крыму и на
Украине кладка яиц идет, повидимому, с конца или даже с середины апреля и
до середины мая (в Евпатории 23.V. 1901 г. найдена кладка, состоящая из 2 яиц
с мертвыми зародышами, Молчанов, 1906; в Днепропетровской обл. найдены
ненасиженные кладки 1.V.1906 г. и 11.V-1898 г. и слегка насиженные 2.V.
1899 г., Боровиков, 1907). На Маныче Спангенбергом в 1950 г. найдены кладки:
20.V и 24.V — с сильно насиженными яйцами, в которых пищали птенцы;
28.V — кладка с яйцами в половине периода насиживания и две других
кладки — едва насиженные; 25.V — скорлупа яиц, из которых только что
вывелись птенцы. В Ак-Булаке (Казахстан) кладка идет с середины мая и до
конца июля B2.V. 1931 г. найдена кладка с 2 едва насиженными яйцами
и две кладки 31.V и 2.VII сильно насиженные, Спангенберг), на озере Тели-
куль 22.V. 1927 г. найдена кладка с 2 сильно насиженными яйцами —одно
яйцо надклевано (Спангенберг, 1936). В Илийском крае кладка начинается
с первых чисел мая и до начала июня (у убитой 10. IV самки из пары, яичники
были еще мало развиты), 15.V найдена кладка из 2-х совершенно свежих яиц,
13.VI кладка с одним сильно насиженным яйцом (Шестоперов, 1929). В Бара-
бинской степи близ Каинска 11.V.1903 г. найдена ненасиженная кладка
(Иоганзен, 1907, 1930). На Алтае, (озеро Теньга) 4.VI найдено гнездо с сильно
насиженными яйцами (Сушкин, 1938). В Сев. Монголии на р. Толе первая
кладка добыта 27. V, в конце июня найдена последняя кладка (Козлова, 1930).
Насиживают оба родителя, но участие самца в насиживании весьма незна-
чительно, но он всегда находится вблизи, оберегая самку от опасности, и
иногда «развлекает» ее танцами. Заметив что-то подозрительное, самец преду-
преждает самку отрывочным криком, и самка заблаговременно, шагов за 100 от
человека, сходит с гнезда. Высиживающая самка очень осторожна, спугнутая с
гнезда, она обычно сразу поднимается в воздух и, отлетев на большое расстояние
ЖУРАВЛЬ-КРАСАВКА ИЛИ МАЛЫЙ ЖУРАВЛЬ
137
от гнезда, спускается на землю (Спангенберг). Иногда пригнувшись, прячась в ку-
стах отбегает от гнезда и только тогда взлетает. К гнезду долго не возвращается.
Срок высиживания точно не установлен, вероятно около месяца. Вылупив-
шийся птенец, как только обсохнет, способен двигаться за родителями. Вылуп-
ление начинается с конца мая или начала июня и идет до середины июля.
(В Бессарабии птенец в трехдневном возрасте добыт 10.VI.1899 г., Остерман,
1918). Пуховики (возраст не отмечен) встречены на Алтае (низовья Чулышмана)
8 и 23.VI (Сушкин, 1938), на Тарбагатае (Чиликтинская степь) только что вылу-
пившиеся пуховички найдены
26.V (Финш, 1879), 8.VI.1936 г.
добыт пуховик на Сары-су (кол-
лекция Зоологического музея
Московского университета).
В сев. Монголии в самом
конце июня одновременно встре-
чались кладки и подросшие птен-
цы. В июне и в начале июля птен-
цы начинают одевать перовой по-
кров. Так на Алтае на Онгудае
2.VII пойманы 2 птенца перели-
нивающие в перовой покров,
27 VII наУймоне наблюдались 2
птенца, вполне оперившиеся, но
еще не летающие (Кащенко,
1899), 27.VII встречены в сев.-
зап. Казахстане оперившиеся, но
еще не летающие птенцы (Суш-
кин, 1908). На Украине (Днепро-
петровская обл.) в начале июля
птенцы начинают уже летать. На
Алтае (Абакан) 3.VIII наблю-
дался плохо летающий выводок
(Сушкин, 1914). В сев. Казахста-
не в начале августа многие мо-
лодые журавли уже летают. При
выводке находятся оба родителя.
Летные молодые журавли первое время держатся при родителях, и семьи весь
день проводят в степях или на лугах, а вечером всей семьей летают кормиться
на ближайшие хлебные поля. Вскоре семьи начинают собираться в стаи. В
Днепропетровской обл. журавли начинают стадиться в середине июля, в те
же сроки в Илийском крае; на Алтае (Абакан) —в середине августа.
Линька. Изучена недостаточно и, повидимому, происходит главным
образом на зимовках. Смена первостепенных маховых идет от внутренних и
крайних маховых к средним, в последнюю очередь сменяется 3-е первостепен-
ное маховое, или 2-е. Линька первостепенных маховых начинается в конце
июля и идет по декабрь, захватывая 4—5 месяцев. Так, у экземпляра, добы-
того в сентябре на сев. Цайдаме, все первостепенные маховые свежие, исклю-
чая 2 и 4, которые еще старые; у экземпляра с сев. Тибета от октября старое
первостепенное маховое только 3-е; у 2-х экземпляров, от 9.Х 11 с р. Бугунь (при-
ток Сыр-Дарьи) в одном случае — 2-е первостепенное маховое старое, в дру-
гом 3-е. По данным Сушкина A938) линька первостепенных маховых на Алтае
начинается в конце июля. Мелкое перо, рулевые, второстепенные и третьесте-
пенные маховые, повидимому, линяют позднее, так как у всех указанных выше
экземпляров, несмотря на сильно изношенное оперение, следов линьки нет.
Весною прилетевшие птицы в свежем наряде, даже прилетевшие очень рано
Рис. 24. Журавль-красавка
138 ОТРЯД ЖУРАВЛИ
имеют уже свежее перо, так экземпляр, добытый в начале апреля на р. Бу-
гунь, —в свежем оперении. Общая схема линьки —как у серого журавля.
Питание, Изучено слабо. Преимущественно растительная пища и
в очень небольшом количестве животная, главным образом насекомые. Во вто-
рой половине лета и осенью корм обычно состоит из поспевающих различных
хлебных растений. Красавки выводком, а затем и стаями, летают кормиться
на близрасположенные хлебные поля, иногда принося значительный вред
урожаю. Причем, красавка, обрывая колосья, заглатывает их целиком, повер-
тывая колосья по ходу их усиков (Фолитарек и Дементьев, 1938).
Полевые признаки. В природе хорошо отличается от других журавлей
своими мелкими размерами и хорошо видными издали, своеобразными белыми
пучками перьев по бокам головы. При полете держится, как и другие журавли,
вытянув шею и ноги во всю длину. По звуку голос красавки похож на голос
других журавлей—это курлыканье, но звук его более высокий и мелодичный,
чем у серого журавля.
Описание. Размеры и строение. Вес самца B) 2383 и 2540 г, самки — 1985г
(коллекции Зоологического музея Московского университета), 2000—2750 г (Бутурлин,
1935). Длина тела самца B) 760—920 мм, самки A) 800 мм. Длина крыла самца G) 510—
540 мм, самок E) 460—510 мм, среднее, соответственно, 530 и 490 мм.
Оперение жесткое, плотно прилегающее к телу. Голова оперена. По бокам головы
имеются удлиненные, рассученные перья. Первостепенных маховых 11. Самое длинное
4-е первостепенное маховое длиннее 3-го махового на 7—10 мм. Формула крыла
4^3>2>1>5>6. Третьестепенные маховые сужены к вершине, крайние из них слабо рас-
сучены и на сложенном крыле отстоят на 130—200 мм от вершины крыла. Перья зоба удли-
нены и сужены к вершине. Хвост короткий, из 12 рулевых перьев. Края клюва гладкие.
Окраска. Первый пуховой наряд. Верхняя сторона тела буроватая, светлеющая
на боках, с темнобурой полосой вдоль спины и с темными продольными пятнами на плечах.
Голова и шея рыжевато-охристые. Низ тела — беловато-грязный с легким охристым нале-
том, более сильно развитым на подбородке, передней стороне шеи и на боках тела. Вто-
рой пуховой наряд неизвестен. В птенцовом наряде верх и бока головы — светлосерые,
с буроватым налетом, более темным на темени и почти черным на затылке. Под глазом—тем-
ное, шиферно-серое продолговатое пятно. Ушные пучки перьев — грязнобелые и слабо уд-
линены, в значительной степени слабее, чем у взрослых птиц. Шея — коричнево-бурая.
Черные перья зоба — короткие, чуть длиннее остальных. Первостепенные и второстепенные
маховые и концы третьестепенных — темнобурые, почти черные. Рулевые — темнобурые,
но несколько светлее маховых. Остальное оперение — буровато-серое, более светлое на
нижней стороне тела и со слабым пепельным налетом на кроющих крыла. Ноги — серо-
бурые. Клюв у основания — серовато-желтый, постепенно переходящий в желтоватый и
в вершинной трети — красновато-оранжевый цвет. Радужина — красно-бурая. Молодая
красавка на ногах кажется более высокой, чем взрослая. Взрослые самцы и самки. Общий
тон оперения сизовато-серый, за исключением блестящей черной шеи, боков головы, лба,
задней части головы и удлиненных перьев зоба. От глаз к шее по бокам головы идет узкая
белая полоса, заканчивающаяся за ухом пучком удлиненных, рассученных белых перьев.
Первостепенные и второстепенные маховые и крылышко — тускло-черные с сероватым нале-
том, кроме того—второстепенные маховые у основания темносерые. Третьестепенные маховые
темносерые, постепенно темнеющие к вершинам, которые почти черные. Ноги — черные.
Клюв в основной половине — черный с оливковым оттенком, в вершинной половине — крас-
новато-бурый, светлеющий к вершине. Радужина —красная или краснобурая. Возрастные
изменения, в частности особенности первого брачного наряда, не выяснены. Окончательный
наряд — повидимому первый зимний или послебрачный, надеваемый в возрасте около
1 7а лет. Сезонного диморфизма у взрослых не установлено.
ОТРЯД ДРОФЫ
OTIDES или OTIDIFORMES
Характеристика отряда
Строение. Внешние признаки. Дрофы — крупные и средних
размеров птицы. У обыкновенной дрофы вес самцов обычно 7,5—12, реже
14 кг, старые особи достигают иногда 16 и, как исключение, 21 кг. Вес самок
колеблется между 3,8—6,5 кг. Самый мелкий наш вид—стрепет—весит
около 0,9 кг. Оперение жесткое, плотное, гладко прилегающее к телу. На
голове и шее у некоторых видов имеются удлиненные рассученные перья,
образующие хохол и воротник. Самцы других видов в брачном наряде обла-
дают усами. Контурные перья имеют побочный ствол. Пух и пуховые части
побочного ствола обладают своеобразной нежнорозовой окраской. Пух покры-
вает только аптерии. Хвост из 16—20 перьев. Крылья диастатаксические,
хорошо развиты, слегка выпуклы; первостепенных маховых 10.
Окраска оперения разнообразна. У наших дроф она состоит из сочетания
белого, рыжего, серого и черного цветов. Тропические (африканские) виды
окрашены более ярко, иногда с металлическим блеском. На спинной стороне
тела наших и многих экзотических представителей развит то мелкий, то круп-
ный струйчатый рисунок. Благодаря этому рисунку, окраска птиц становится
разбитой, покровительственной и позволяет дрофам искусно прятаться среди
скудной степной и полупустынной растительности. Птенцы дроф имеют также
покровительственную окраску и покрыты серо-дымчатым или песчано-желтым
пухом с резким черным или темным рисунком на спинной стороне тела. Самцы
отличаются от самок у большинства видов только более крупными размерами;
а у некоторых и окраской оперения.
Телосложение дроф плотное; шея довольно длинная, но толстая, голова
относительно большая. Форма клюва у различных видов неодинакова: обычно
клюв сплющен сверху и у конца верхней челюсти несколько вздут. Ноги сред-
ней длины, сильные и толстые. Цевка покрыта шестиугольными щитками;
голень оперена только наполовину. Пальцев ноги —три, они снабжены корот-
кими сильными тупыми когтями. Нижняя сторона пальцев и «пятка» покрыты
мозолистой толстой кожей. Глаза дроф относительно крупны; радужина окра-
140 отряд дрофы
шена разнообразно —в желтый, темнобурый, светлобурый, иногда неодно-
типный цвет.
Анатомические признаки. Череп схизогнатический, голо-
ринальный; затылочные фонтанели и базиптеригоидные отростки отсутствуют;
ноздри сквозные, шейных позвонков 16—18; в грудине отсутствуют spinasterni,
задний край имеет две пары вырезок, как у куликов; пищеварительный тракт
характеризуется наличием длинных, функционирующих слепых кишок с округ-
лыми железками и с ворсинками в концевом отделе; пищевод образует рас-
ширение; у самцов некоторых видов хорошо развит глоточный мешок; коп-
чиковая железа отсутствует; есть рудимент penis; голосовые мышцы или
полностью отсутствуют или развиты слабо; имеются отделяющие «пудру»
перья.
Образ жизни. Дрофы — дневные птицы открытого ландшафта, избегающие
древесной растительности и даже высоких и густых кустарниковых зарослей.
Характерные биотопы этих птиц — степи и полупустыни жаркого и умеренного
пояса. Виды, населяющие тропики и прилежащие к ним части умеренного пояса,
оседлы, остальные — перелетны. Преимущественно растительноядные птицы.
В большом количестве поедают сочные побеги степных растений. В теплое время
в питании имеют значение также насекомые, особенно прямокрылые. Крупные
виды изредка ловят ящериц и мелких грызунов. В поисках пищи птицы обры-
вают побеги трав и склевывают насекомых и других беспозвоночных с земли и
растений, руководясь при этом хорошо развитым зрением. Большинство видов
дроф не образуют во время размножения настоящих пар и за потомством уха-
живает самка. Среди наших видов обыкновенная дрофа может быть названа
полигамом. Напротив, стрепет и вихляй на время откладывания яиц разбиваются
на пары. У стрепета брачная связь нарушается, когда птенцы достаточно подра-
стут, а самцы к этому времени образуют группы и стайки. Самец покидает
самку, как только последняя приступает к насиживанию. Осенью и зимой
большинство представителей собираются в крупные стаи, реже держатся семь-
ями, но иногда встречаются и одиночно.
Сроки размножения у птиц в умеренных широтах падают на весенние
месяцы. Прилетая на родину очень рано или ведя оседлый образ жизни, дрофы
начинают гнездиться, когда степь или полупустыня покрывается молодой,
весенней зеленью. С момента прилета и до начала размножения протекает зна-
чительный промежуток времени, в течение которого птицы продолжают дер-
жаться стаями. Гнездятся дрофы отдельными парами, обычно далеко (на 1 —
1,5 и больше км) друг от друга. Гнездо чрезвычайно простого устройства. Среди
скудной растительности птица выкапывает пологую ямку и прямо на землю
кладет яйца. Спариванию предшествует своеобразное токование, имеющее свои
особенности у отдельных видов. Самцы одних дроф распускают хвост и крылья,
раздувают шею и вступают в ожесточенные драки; другие распускают перья
воротника, третьи барабанят ногами на одном месте, подпрыгивают, издают
звуки, некоторые поднимаются в воздух и устраивают воздушные игры. Кладка
у крупных видов состоит из 1—3 яиц, редко, но бывает и до 6; у мелких видов
3—5 яиц и очень редко до 9 и даже больше. Яйца дроф обладают покровитель-
ственной окраской. Основной цвет их оливковый и глинисто-песчаный с раз-
мытыми крупными и мелкими пятнами. Эмбриональное развитие сравнительно
медленное у крупных видов, более быстрое — у мелких. Насиживание, повиди-
мому, начинается после снесения второго или третьего яйца; насиживает только
самка, у которой в этот период развивается большое наседное пятно. Постэм-
бриональный рост медленный, и молодые птицы до осени отличаются от взрос-
лых размерами и деталями окраски оперения.
Систематика. Отряд дроф состоит из одного семейства дрофиных Otididae,
распадающегося на несколько родов: Otis, Eupodotis и др. Общее число видов 23,
из которых в пределах СССР встречается 3 A3%). В систематическом отноше-
СТРЕПЕТ 141
нии дрофы близки с одной стороны к журавлям и пастушкам, с другой, по
крайней мере в ряде морфологических черт — к куликам.
Ископаемая дрофа Palaeotis weigelti найдена в среднем эоцене (Германия).
Дрофы, относящиеся к современному роду Otis, описаны из верхнего миоцена
(Otis affinis из Германии) и из верхнего олигоцена (О. agilis из Франции).
Из плейстоцена известны современные виды — дрофа О. tar da и стрепет
О. tetrax.
Географическое распространение. Большинство видов дроф распростра-
нено в Африке, некоторые — в умеренной полосе южн. Европы и Азии,
другие — в южн. Азии, один вид Eupodotis australis населяет Австралию.
Дрофы живут в сухих открытых ландшафтах — степях, полупустынях и пусты-
нях. В самых северных частях области распространения дрофы перелетны,
в остальных — оседлы.
В СССР дрофы встречаются в степях от Украины до Забайкалья и в пусты-
нях Средней Азии.
Практическое значение. Дрофы —ценная дичь. Их добывают ради мяса и
ради трудного увлекательного процесса охоты. Промысловое значение дроф
ограничено. Так как дрофы крайне осторожны, то добываются они в общей
сложности в небольшом количестве. Однако численность этих птиц в настоящее
время сильно сокращается, сокращаются также и ареалы отдельных видов.
Это связано в значительной мере с охотой на птиц в тяжелое для них зимнее
время.
Большое значение имеет также распашка целинных степей. Все дрофы
полезны тем, что поедают в летнее время насекомых — вредителей сельского
хозяйства.
Таблица для определения видов дроф
1 B) Птица средних размеров: крыло короче 300 мм, плюсна короче 80 мм . . .
стрепет — Otis tetrax
2 A) Крупная птица: крыло длинее 300 мм; плюсна длиннее 80 мм 3
3 D) Ширина клюва у переднего края ноздрей равна или несколько больше высоты
клюва у заднего края ноздрей; у взрослых на верхней стороне головы и по бокам шеи пучки
из рассученных, удлиненных перьев вихляй — Otis undulata
4 C) Ширина клюва у переднего края ноздрей меныце высоты клюва у заднего края
ноздрей; удлиненные пучки перьев по бокам шеи и на верхней части головы отсутствуют —
„ обыкновеннаядрофа — Otis tarda
1. Стрепет Otis tetrax L.
Otis tetrax. Linnaeus. Syst. Nat. изд. X, 1758, стр. 154. Франция.
Русское название. Стрепет—народное название, широко распространенное в России,
известное по крайней мере с XVII в.
Распространение. Ареал. Зап. Палеарктика, от сев.-зап. Африки (Ма-
рокко, Тунис), Пиренейского п-ова (Испания, Португалия), Франции (к северу
до Шампани) через Зап. Европу и СССР до Афганистана, Зайсанской котловины,
Кашгарии и районов Каинска.
Характер пребывания. Гнездящийся, перелетный в северных и
восточных частях ареала, в южных частях —оседлый.
Биотоп. Целинные степи, культурные участки по окраинам песчаных
пустынь.
Подвиды и варьирующие признаки. Два подвида, отличающиеся размерами и дета-
лями окраски: О. /. orientalis Hartert, 1916—восточные части ареала вида —от Синьц-
зяня и Афганистана до Балканского п-ова и южн. Италии; О. /. tetrax L., 1758 — Франция,
Испания, Португалия и сев.-зап. Африка.
142
ОТРЯД ДРОФЫ
Восточный стрепет Otis tetrax orientalis H a r t e r t
Otis tetrax orientalis. H а г t e г t. Nov. Zool. 1916, стр. 339, Сарепта.
Распространение. Ареал. Польша, долина Дуная в южн. Чехословакии
и Венгрии; Румыния, Югославия, Болгария, Греция, Македония, Италия
(Абруцио, Апулио), о-ва Сардиния и Сицилия, Кипр, Палестина, Сирия, СССР,
Афганистан, вост. Туркестан. Залеты в Англию, Ирландию, Бельгию,
Голландию, Германию, Данию, Скандинавию, Финляндию, в Египет и сев.-зап.
Китай.
В СССР из-за распашки целинных степей и других причин, распространение
стрепета стало спорадичным. Во многих местах стрепет отступил в конце прош-
Карта 23. Распространение стрепета Otis tetrax
1. О. t. orientalis, 2. О. t. tetrax;
a — границы гнездовий, б — не вполне выясненная граница гнездовий, г — места
зимовок (цифры э кружках обозначают номера зимующих подвидов).
лого столетия к югу от северных границ ареала. Повидимому гнездится в Кали-
нинградской обл. на западе. В Бессарабии стрепет гнездится в Бендерском
и Измаильском районах (Остерман, 1912), затем северная граница его распро-
странения идет через Подолию, проникая в самые южные части Киевщины (Хра-
невич, 1925—26). На Волынь стрепет, вероятно, только залетает. В Полтавщине
уже в 1917 г. совсем перестал гнездиться, появляясь здесь лишь как залетная
птица (Гавриленко, 1929). Далее на восток граница с юга огибает большую часть
Харьковской обл., где стрепет гнездится лишь в восточной половине области
от р. Оскола и по Донцу (Сомов, 1897). Приблизительно до 1927 г. стрепеты еще
гнездились в районе Купянска. Отсюда граница круто поднимается на северо-
восток, охватывая степные пространства Воронежской обл., и проникает
в Тамбовскую обл., где стрепет несомненно гнездился до 1928 г. Волгу гра-
ница пересекает где-то в районе Саратова, к югу от которого стрепет распро-
странен равномерно, а к северу если и гнездится в наше время, то редко и спо-
радично. Найден в летнее время в районе Куйбышевской обл. и в южной поло-
вине Татарской АССР (Першаков, 1929). Если стрепета продолжают гнездиться
здесь, то это следует рассматривать как обособленные от остального ареала
гнездования. От Саратова граница идет на восток, почти совпадая с северной
границей Западно-Казахстанской обл., и выходит к Чкалову, где стрепет в изо-
билии гнездился прежде и нормально гнездится в наше время южней этого города
СТРЕПЕТ 143
(Спангенберг). Через восточные части Чкаловской обл. граница проникает
в Челябинскую обл. Далее стрепет гнездится под Омском, быть может, про-
должает гнездиться в южной части Тарского р-на; в Барабинской степи, доходя
на севере, быть может, до Каинска.
Еще далее на восток гнездование стрепета известно в Барнаульском округе,
в районе Змеиногорска, откуда граница идет к югу предгорьями Алтая, дости-
гая Зайсанской котловины (Поляков, 1915, Хахлов, 1928), Тарбагатая,бассейна
Или и территорий, прилегающих к Алма-Ате. К югу и западу от указанной гра-
ницы гнездится в соответствующих биотопах до южных границы СССР, кроме
Таджикистана и горных районов Киргизии и Узбекистана.
В Азии стрепет довольно равномерно населяет степи зап. Сибири и север-
ных частей Казахстана. Напротив, в области Средне-Азиатских пустынь он
или полностью отсутствует или распространен крайне спорадично. На Сыр-
Дарье найден не севернее Чийли. В Туркмении гнездится по степеподобным
плато в Копет-даге, в частности — в бассейне Сумбара на западе, у Сулюкли
в центральной части хребта (Дементьев, 1950). Спорадическое распространение
стрепета в Средней Азии безусловно связано с отсутствием в ряде районов
подходящих для гнездовья биотопов.
В Европейской части СССР спорадичность распространения особенно резко
выражена близ северной границы ареала, но имеет место и на юге, в районе
Днепропетровска и Запорожья. Спорадически распространен стрепет и в За-
кавказье. Новейшие исследования показали, что в настоящее время стрепет
гнездится в немногих местах этого края. Обнаружен на гнездовье близ с. Каря-
гино в Мильско-Карабахской степи, в степи Турут в долине Саруджа и близ
Кюрдамира в Ширванской степи. Недавно, вероятно на гнездовье (в июне),
отмечен километрах в десяти на север от Евлаха (Спангенберг).
В Европейской части СССР залеты известны: в Черниговскую, Киевскую,
Пензенскую, Московскую, Тульскую, Рязанскую области, в Горьковский край,
в районы Пскова, Ленинграда, под Казань, в южные части Башкирии, а
в Азии — в окрестности Томска и район Троицкосавска.
Характер пребывания. В большей части области распростра-
нения —оседлая, но на северо-востоке СССР —главным образом, гнездящаяся
и перелетная птица.
Основная масса птиц перезимовывает за пределами СССР — в Египте,
Месопотамии, Сирии, Малой Азии, Иране и сев.-зап. Индии. Регулярные зи-
мовки однако существуют и в наших пределах —в Крыму (Пузанов, 1933)
и в вост. Закавказье, в Средней Азии. В Закавказье места зимовок стрепета
начинаются от Тбилиси в долине р. Куры, в средней части долины р. Храма
и северо-западных частях Иорского и Кахетинского плоскогорий; к востоку они
тянутся до побережья Каспия. На указанной территории стрепеты регулярно
зимуют на обоих берегах Куры, в области рек Иоры и Алазани, в Аджинаур-
ской степи, в сев.-вост. и юго-вост. Ширвани, в Мильской степи, на Мугани и,
наконец, близ устья Куры в Сальянах. В других частях Закавказья, в частности
между Тбилиси и районом Евлаха, в районе Нахичевани, в районах Дивичей
и Кубы и близ залива Кирова стрепеты зимуют в ограниченном числе и не
ежегодно (Верещагин, 1940). В небольшом числе стрепеты бывают зимой даже
в приморской части Астраханского края (Хлебников, 1928), а в теплые годы —
и в степях, расположенных к западу от дельты Волги. В Азиатской части Союза
значительные зимовки стрепета известны в Таджикистане: на Аму-Дарье между
Келифом и Кировабадом, в большом числе в окрестностях Чубека, а также в Гис*
сарской долине (Иванов, 1940). В Туркмении стрепет остается зимовать в огра-
ниченном количестве на Атреке к западу от Кызыл-Атрека (Дементьев, 1950),
тогда как в смежных частях Ирана, между Атреком и Гюргеном стрепеты зи-
муют во множестве (Исаков и Воробьев, 1940). Стрепеты встречаются зимой
в оазисе Тамды в пустыне Кызыл-Кум (Зарудный, 1915).
144 отряд дрофы
Даты. В вост. Закавказье зимовка стрепета продолжается около шести
месяцев — с первых чисел октября до начала апреля. «Формирование» зимовок
нормально происходит в октябре—ноябре, но зависит от погоды в Предкав-
казье. В суровые зимы, сопровождающиеся снежными заносами и длительным
снеговым покровом, зимующие в Закавказье стрепеты попадают в крайне
неблагоприятные условия. В зиму 1924—1925 гг. часть зимующих стрепетов
продвинулась южней Ленкорани, множество их погибло здесь от голода.
Появление стрепетов на зимовке в Таджикистане начинается с последней
трети сентября, с этого времени стрепеты держатся здесь всю зиму. Последние
особи отмечены весной в конце апреля и даже 18 мая (Иванов, 1940). В Турк-
мении стрепет как правило не зимует, отлетая в ноябре в сопредельные части
сев. Ирана. В наших пределах —на Атреке и в Бадхызе остаются одиночки
и редкие пары.
Весной стрепеты начинают двигаться к северу в самом начале марта, а по-
являются всюду, где гнездятся, не позднее первой трети мая; чем дальше на
север, тем позднее прилетают весной птицы. На Украине, в зависимости от
широты местности стрепеты появляются между концом марта и последней
третью апреля. В Подолии — в самом начале апреля; в Днепропетровской обл.—
в последней трети марта или около середины апреля; в Харьковской обл. —
в последней трети апреля.
С закавказских зимовок стрепет начинает двигаться к северу в самом на-
чале марта, в середине этого месяца отлет достигает наибольшей силы и к началу
апреля полностью заканчивается. На Сев. Кавказе у Дзауджикау стрепеты
летят в марте и апреле. В окрестностях Кизляра огромные стаи пролетных стре-
петов, двигавшихся в северо-восточном направлении, наблюдались 21 и 22 марта
(Беме, 1925). В степях у Черного Яра и Владимировки в конце марта и начале
апреля, пролетные стрепеты уже не представляют редкости (Спангенберг),
и сотенными стаями отмечены в середине апреля и в других частях Сталинград-
ской обл. Района Бугуруслана, в Куйбышевской обл., птицы достигают
22 апреля (Карамзин, 1913). Около середины или во второй половине апреля,
редко в начале мая, стрепеты появляются в Чкаловской обл. (Спангенберг)
и вообще в степях б. Оренбургского края (Зарудный, 1888, 1897).
В Азиатской части Союза в Туркмении весеннее движение идет в марте,
иногда с конца февраля (Дементьев, 1950). На Сыр-Дарье между Чйили и Тур-
кестаном первые птицы появляются во второй половине марта, но летят и
в апреле. Между концом марта и серединой апреля появляются стрепеты в Кы-
зыл-Кумах. 2 апреля пара стрепетов отмечена близ Гуляевки на реке Чу
(Долгушин, 1939). В северных частях Казахстана —у Кок-джиды —наиболее
ранние особи отмечались 12. IV, в низовьях Иргиза— 19 апреля, настоящий же
пролет начался с 28 апреля и продолжался около 10 дней (Сушкин, 1908).
В конце апреля стрепеты нормально появляются в районе Наурзумского за-
поведника, между 2 и 24 апреля в бассейне верхнего Иртыша (Поляков,
1915) и около 3 мая в окрестностях Омска (Словцов, 1878).
Отлет осенью растягивается на продолжительное время. Он начинается
образованием стай, которые широко кочуют по степным пространствам. Кочевки
эти происходят с различных чисел августа; в августе стрепеты часто появляются
там, где за последнее время они перестали гнездиться. Кочевая жизнь стрепетов
продолжается несколько больше месяца и переходит затем в настоящий пролет.
Из верховьев Тобола стрепета улетают в конце первой трети сентября; из
верховьев Иргиза отлет происходит в конце сентября — начале октября, но
в низовьях этой реки стрепеты наблюдались еще в конце октября (Сушкин,
1908). Валовой пролет стрепетов в Оренбургских степях происходит впервой
трети октября, пролетные одиночки и маленькие группы наблюдаются до
10-х чисел ноября, и наиболее поздняя, видимо запоздавшая птица, была отме-
чена 13 декабря (Зарудный, 1888).
СТРЕПЕТ 145
К концу сентября стрепеты, повидимому, достигают своих азиатских зимо-
вок—в Алайской долине пролетные птицы отмечались 21. IX (Иванов, 1940).
В Туркмении пролет идет с конца сентября и заканчивается только в ноябре
(Дементьев, 1950).
В Европейской части СССР осенний пролет тоже растянут. В Харьковской
обл. появление и пролет стрепетов иногда наблюдается со второй половины
сентября, обычно же с конца первой трети октября, продолжаясь около месяца;
одиночки встречаются и позднее. Поздние встречи D ноября) известны и в Пол-
тавщине (Гавриленко, 1917). В начале сентября крупными стаями, а иногда и
массами летят стрепеты в Днепропетровской обл.; в начале октября, иногда
позднее, отлетают из Подолии. В дельте Волги стрепеты бывают в конце
сентября и половине октября, в низовьях Урала редкие стаи появляются
в середине октября, а Сев. Кавказа достигают в октябре и ноябре,
На своих закавказских зимовках стрепеты появляются обычно в самом
начале октября. В отличие от весеннего пролета, осенний во многих районах
носит характер медленной откочевки к югу.
На пролетах стрепеты по возможности придерживаются соответствующих
биотопов — степей, реже полупустынь и явно избегают настоящих пустынных
пространств. В связи с этим стрепеты редко проникают в Аральские Кара-Кумы,
на Сыр-Дарью и в Кызыл-Кумы. В сев. Казахстане на осеннем пролете основ-
ная масса птиц летит в юго-западном направлении, огибает с севера Аральское
море и, повидимому, пересекает Усть-Урт, далее — двигаясь вдоль восточного
берега Каспия. Хорошо выраженные пролетные пути намечаются в Туркмении.
Небольшая часть стрепетов летит здесь осенью с севера на юг вдоль береговой
линии; основная же масса двигается вдоль Копет-Дага.
На Кавказ осенью стрепеты, зимующие в Куро-Араксинской низменности,
проникают тремя путями. Основная масса птиц летит вдоль побережья Каспия
от Махач-Кала до Дербента и далее западнее Баку через Кабристан. В ограни-
ченном числе летят стрепеты через пониженные участки Главного Кавказского
хребта, особенно часто в пределах 45°36'—40°30' в. д. Еще в меньшем числе
наблюдается пролет пткц вдоль восточного побережья Черного моря. Осенние
направления пролета совпадают с весенними.
В северных частях ареала перелетные стрепеты двигаются группами и
небольшими стайками, иногда стаи достигают 50—70 особей; размер стай по-
степенно возрастает по мере движения к югу; особенно велики они бывают непо-
далеку от мест зимовок. Через Дербент и Дивичи летят стаи в несколько сот
и даже тысяч особей (Верещагин, 1940). В Туркмении пролетные стаи стрепетов
различны, от 2—ЗдоЗО—40 птиц (Тишкин), большие скопления наблюдаются
только на крайнем юге-западе по Атреку (Дементьев, 1950). В степях Украины
и сев. Казахстана пролетные стрепеты редко поднимаются на значитель-
ную высоту, предпочитая лететь метров на 30 от земли. На Кавказе во время
массового осеннего лёта стаи летят на высоте 100—300 м. При перелетах через
Главный Кавказский хребет стрепеты иногда поднимаются в воздух до 3500 м.
Биотоп. Гнездовый — травянистые целинные степи или давно непахан-
ные земли, заросшие степной растительностью, реже поросшие злаками и эфе-
мерами песчаные участки или сухие луга. Гнездовый биотоп меняется в различ-
ных частях ареала. В Воронежской обл. стрепеты гнездятся в солончако-
вой степи, поросшей полынью и лебедой, и избегают степных пространств,
покрытых густой травянистой растительностью. В Харьковской обл. стрепет
предпочитает гнездиться в слегка волнистых ковыльных степях на перелогах.
Известны также случаи гнездования среди возделанных полей и обширных лу-
гов. В районе Херсона стрепеты заселяют ковыли и залежи; залежи и песчаную
степь птицы предпочитают на левобережье Еруслана.
В Азии стрепет также гнездится в различных условиях. В Ак-булакском
районе Чкаловской обл. стрепеты положительно избегают степей, покрытых
10 Птицы СССР, т. II
146 отряд дрофы
густым и высоким разнотравием; не гнездятся стрепеты и в ковылях, если они
густы, высоки и на большом пространстве сплошь покрывают степную почву.
Птицы предпочитают слегка холмистую степь с большим разнообразием стаций.
В других районах б. Оренбургского края стрепеты гнездятся в ров-
ных глинисто-солонцеватых местностях, поросших невысокой травой. В такой
же обстановке стрепеты гнездятся в нижнем течении Урала и между Сталин-
градом и Астраханью.
В районе Наурзумского заповедника птицы избегают типчаковой и солон-
чаковой степи и гнездятся в ковыльных луговых участках (Михеев). К югу
отсюда, там, где травянистые степи постепенно сменяются полупустыней, рас-
пространение стрепета на гнездовье тесно связано с ковылем. Иногда птицы
могут выводить птенцов на полынных и даже на песчаных участках, если
поблизости есть ковыльная степь.
Своеобразен гнездовый биотоп стрепета в области средней Сыр-Дарьи и
пустыни Кызыл-Кум. В одних случаях стрепеты гнездятся здесь в ровных
глинистых степях, поросших довольно густой, но невысокой травянистой расти-
тельностью с обилием верблюжьей колючки, в других — населяют слегка
всхолмленные, реже ровные песчаные территории, покрытые на короткое ве-
сеннее время эфемерами —мелкой зеленой травой и цветущими тюльпанами
и маками. В Туркмении стрепет гнездится на высокогорных степных участках
Копет-дага с зарослями полыни, эспарцета и т. д.
Кормовой биотоп вида более широк и разнообразен: стрепеты кормятся
не только там, где обычно гнездятся, но охотно посещают в поисках пищи луго-
вые участки, а во второй половине лета вылетают на пахотные земли и дер-
жатся на зеленях вплоть до момента отлета к югу. На пролете стрепет также
чаще всего встречается в культурном ландшафте.
Численность. Общая численность весьма ограничена. Во многих
районах с каждым годом стрепет становится все более и более редкой птицей.
Совершенно неправильное представление о численности создается при озна-
комлении со стрепетом на закавказских зимовках. Здесь стаи в 500—2000
особей совсем не редкость; в некоторых случаях размер стаи определяется даже
в 20.000 особей (Верещагин, 1940). Следует помнить, что на закавказских зимов-
ках стрепеты собираются со значительной части гнездовой территории. Если
же обратиться к численности стрепета в отдельных частях гнездовой области,
то можно отметить следующее. В самых южных частях Украины и степных
районах Крыма стрепет еще продолжает быть обыкновенной птицей. К северу
отсюда по всей Украине, а также в областях Воронежской, Тамбовской, Пен-
зенской и в среднем Поволжье, стрепет редок и спорадичен. В степях Пред-
кавказья, в степях по среднему и нижнему Дону и в Сталинградской обл.
стрепет и сейчас обыкновенен. В Закавказье и на большем протяжении Средней
Азии стрепет на гнездовье очень редок. Местами в степях сев. Казахстана и
Западной Сибири — обыкновенен, даже многочисленен. Численность стрепетов
изменяется по годам, снижаясь, по всей вероятности, после неблагоприятных
суровых зим (в области зимовок).
Экология. Размножение. Гнездится отдельными парами. В благо-
приятных условиях гнездовые участки отдельных пар невелики и соприка-
саются друг с другом, например, в районе Ак-Булака Чкаловской обл. гнезда
располагались метров на 250 одно от другого (Спангенберг). Гнездо простого
устройства. Оно представляет собою иногда плоскую ямку (Сомов, 1897),
иногда довольно глубокое округлое углубление в почве, хорошо скрытое
между ковыльных кочек или кустиков. Лоток гнезда и его боковые стенки вы-
стилаются тонким слоем сухих стебельков и прочей растительной ветошью;
среди гнездовой выстилки часто попадаются перья самки. Поперечный диаметр
осмотренных нами гнезд 17—18 см, глубина лотка около 9 см. Таким образом,
когда самка насиживает яйца, ее спина находится на уровне почвы, и белые
СТРЕПЕТ 147
части оперения скрыты боковыми стенками гнезда. Строит ли гнездо одна самка
или в постройке участвуют обе птицы гнездовой пары — остается невыяснен-
ным. Законченные кладки нормально содержат 3—4 яйца; в некоторых случаях,
однако, яиц в кладке бывает значительно больше. В литературе известны случаи
нахождения гнезд стрепета с 9 яйцами (Аксаков), гнездо с 9 яйцами найдено
также в Барабинской степи (Иоганзен, 1911), с 11 яйцами —на Сыр-Дарье.
Последняя кладка вероятно принадлежала одной самке, так как все яйца этого
гнезда имели ненормальную голубовато-зеленую окраску, с едва заметными
пятнами. Яйца овальные, несколько вздутые, с хорошо выраженными острыми
и тупыми концами. Скорлупа яиц гладкая и обладает маслянистым блеском.
Основной фон ее — оливковый, иногда несколько рыжеватый. Изредка попа-
даются яйца с голубовато-зеленым основным фоном. По основному фону раз-
бросаны размытые крупные и мелкие пятна светлого рыжевато-бурого цвета.
На некоторых яйцах пятна едва заметны. Размеры A7) 47,0 — 58,2 х 37,2 —39,4,
в среднем 49,98 х 38,25 мм.
Как в южных, так и в северных частях ареала размножение у стрепета
начинается в одни и те же или, во всяком случае, в близкие сроки. В южных
частях ареала, рано прилетая весной на родину, стрепеты довольно значитель-
ный срок ведут стайный образ жизни. За это время самцы надевают брачное
оперение. У стрепетов, принадлежащих к северным популяциям, брачное опе-
рение развивается во время перелета, птицы появляются на местах гнездовья
в полном брачном пере и вскоре приступают к размножению. Полный брачный
наряд одевается стрепетами не ранее конца апреля. Спариванию и гнездо-
строению предшествует своеобразное токование самцов. Самец выбирает среди
степи открытый клочок почвы, так называемый точек, откуда ему видны бли-
жайшие окрестности. Токующая птица веером распускает хвост, опускает
крылья, несколько приподнимает перья ошейника и в такой позе то медленно
расхаживает по точку, то подпрыгивает вверх. Время от времени птица подает
голос — своеобразное короткое потрескивание. В местах, где стрепеты редки,
ток протекает в одиночку; там же, где птиц много, стрепеты иногда токуют
метров на 100 друг от друга. Между самцами, токующими по соседству, слу-
чаются ожесточенные драки. Птицы токуют ранним утром, на вечерней заре,
а в разгар тока и ночью. Самка приближается к токующему самцу и спаривается
с ним близ точка. Безусловно ошибочны указания в литературе (Алфераки,
1910) о вылете многих (до 50) самцов стрепета на токовище. Токовища у стре-
петов нет; есть индивидуальный точек, на котором случаются драки двух сам-
цов. Самое раннее токование в южных частях Чкаловской обл. происходит
в самых последних числах апреля, чаще с начала мая; в районе Наурзумского
заповедника—в первых числах мая; разгар тока под Кизляром—между
10—20 мая (Беме, 1925). Токующих самцов удается слышать (в Чкаловской и
Сталинградской областях) обычно до середины июня, значительно реже — до
конца этого месяца.
Массовая яйцекладка падает на различные числа мая, значительно реже
яйцекладка происходит в конце апреля или в июне. На Сев. Кавказе, в степях
под Кизляром, законченные кладки со свежими яйцами собраны 14 и 27 мая и
2 и 19 июня. В Чкаловской обл. под Ак-Булаком кладка в 4 свежих яйца най-
дена 12 мая, другая, еще незаконченная в 2 яйца —22 мая. Несколько южнее,
в Актюбинской обл., законченная кладка в 3 яйца найдена 20 мая (Карам-
зин, 1917), а на Сыр-Дарье в районе станции Тюмень-Арык законченная кладка
в 11 яиц найдена 15 июня. Приблизительно в те же сроки гнездятся стрепеты и
у северного предела своего распространения — в Зап. Сибири. В Барабинской
степи кладка в 9 яиц найдена 19 мая (Иоганзен, 1911). Поздние кладки, ве-
роятно, являются следствием гибели первых кладок.
Насиживает яйца только самка; самец в это время держится поблизости от
гнезда. В воспитании птенцов, во всяком случае до поднятия их на крылья»
148 отряд дрофы
участвуют оба родителя. Указания в литературе на то, что самцы оставляют
насиживающих самок и образуют стайки, вероятно, ошибочны. В Чкаловской
обл. при выводках наблюдались обе взрослые птицы; группы бродячих птиц,
наблюдавшихся в июне и июле, повидимому, состояли из холостых особей. После
откладки второго яйца птица уже остается на гнезде большую часть дня и сидит
настолько крепко, что ее можно схватить руками. ПродолжителъиеАнасижива-
ния —20—21 день. Первые птенцы появляются не ранее конца мал; обычно же в
различные числа июня. В степях Чкаловской обл. совсем маленькие пуховички
стрепета встречены 16 июня, птенцы ростом с голубя — 11 июля. К началу
августа молодые стрепеты отлично летают и образуют к этому времени крупные
стаи. Самка отводит от выводка, подражая раненой птице, и подпускает чело-
века на самое близкое расстояние, самец же убегает в сторону и близко не под-
пускает. В августе молодые стрепеты ростом не отличаются от взрослых и весят
около 800 г.
Линька. В году бывает две линьки — полная осенью, между июлем и
октябрем, и частичная предбрачная, захватывающая мелкое оперение, между
мартом и маем. Последовательность смены нарядов: пуховой —птенцовый —i
первый зимний — первый брачный — второй зимний — второй брачный. Вто-
рые наряды окончательные. Полная осенняя линька взрослых начинается с се-
редины мая; ко времени отлета успевают совершенно смениться рулевые и
маховые; несколько недоросшие маховые в количестве двух-трех могут быть
найдены и у пролетных птиц.
Перья брачного наряда начинают развиваться в различные числа марта.
У самца, добытого 23 марта под Кизляром, на груди и шее пробивались брачные
перья (Беме, 1925). В последних числах апреля и в самом начале мая все самцы
одевают полный брачный наряд. Экземпляры, добытые в Харьковской обл,
в самом конце августа в значительной степени вылиняли. К концу первой трети
октября все самцы одеваются в свежее зимнее оперение.
Питание. Пища растительная и животная. Состав и процентное соот-
ношение кормов меняется в зависимости от времени года. В теплый период,
наряду с растительными кормами, стрепеты, по наблюдениям в Чкаловской обл.,
в изобилии поедают разнообразных насекомых: кобылок на степных дорогах,
различных саранчевых, мелких жучков и улиток. Здесь же осенью стрепеты
регулярно вылетают на озимые посевы и тогда их желудок оказывается напол-
ненным зеленью молодых всходов. В конце августа в Харьковской обл.
стрепеты поедают большое количество всевозможных кузнечиков, заглатывают
раковины и песчинки.
Во время осеннего пролета в Туркмении стрепеты кормятся зелеными
листьями полыни и жуками чернотелками. Несравненно лучше изучено питание
стрепета на зимовках в Закавказье. Основные корма его на зимовках в Куро-
Араксинской низменности — осенние и весенние эфемеры полупустыни. Они
составляют около 70—73 процентов поедаемой пищи и относятся примерно
к 15 видам. Кроме того, в небольшом числе стрепетами поедаются на зимовках
насекомые: жуки, мухи, прямокрылые. Из механических примесей в желудках
удается обнаруживать песок, аморфный кварц, глинистый сланец и кусочки
ф^фора (Верещагин, 1940). Этот же автор сообщает, что экскременты стрепета,
содранные в Мильской степи в мае, состояли почти целиком из плохо перева-
ренных ростков гадючьего лука.
Стрепеты начинают кормиться ранним утром, вторично жируют после
3—4 часов дня. В жаркое время летом птицы кормятся только в утренние и
эечернне часы, а днем отдыхают. Известно, что стрепеты посещают водопои,
но, вероятно, долгое время могут ограничиваться влагой, получаемой при пое-
дании сочной, молодой, иногда покрытой росой, зелени.
Полевые признаки. Стрепет—самая маленькая дрофа нашей фауны, раз-
меры позволяют безошибочно отличить его от других близких видов.
Рис. 25. Стрепет (самцы в брачном и в осеннем наряде)
Рис. 26. Самка стрепета с птенцом
150 ___ отряд дрофы
В период размножения стрепеты встречаются одиночками и парами, а ран-
ней весной, осенью и зимой — небольшими группами и крупными стаями. Во вре-
мя полета самцы издают крыльями своеобразный мелодичный свист. Стрепеты
осторожны до крайности. При первой тревоге они легко поднимаются на крылья
и перемещаются на большое расстояние. Летят птицы то низко над землей, то,
напротив, высоко поднимаются в воздух. На лету птица часто и глубоко машет
крыльями и обычно двигается со скоростью 65 км в час. При сильном разлете
во время дальних перелетов стрепеты летят со скоростью 75—80 км в час (Вере-
щагин, 1940). Когда стрепеты заняты поиском корма, они передвигаются медлен-
ными шагами, но способны и на быстрый бег, не уступающий в скорости бегу
человека. Стрепеты используют свою покровительственную окраску и прячутся
с замечательным искусством. Стрепет — молчаливая птица, изредка он издает
негромкое — «пуль-пуль-пуль» (Огнев и Воробьев, 1924). Во время токования
самец издает короткое потрескивание — сгрр», причем эти звуки следуют друг
за другом со значительными перерывами.
Описание. Размеры и строение. Вес взрослых самок, добытых в мае в Чка-
ловскойобл. 740—910 г; взрослых самцов и самок, добытых в середине августа — 630—
820 г (Спангенберг); добытых в октябре и ноябре в Туркмении — 525—600 г (Исаков и Во*
робьев, 1940). Крыло самцов A9) 250—275, самок B2) 247—271, в среднем 261,9 и 256,6мм.
Помимо размеров и окраски, половой диморфизм выражен также в форме и относительной
длине четвертого первостепенного махового пера. Это перо у взрослых самцов меньше тре-
тьего и пятого, а у самок (и молодых самцов) длиннее. Как внешнее, так и внутреннее опа-
хало четвертого махового имеет у взрослых самцов резко выраженное сужение; это, вероятно,
способствует своеобразному звенящему свисту, слышимому при полете самца. Хвост из 18
рулевых, закругленный.
Телосложением стрепет напоминает обыкновенную дрофу. Клюв небольшой и широ-
кий, голова большая; шея довольно длинная; крылья широкие, ноги сильные, трехпалые.
Окраска. Основной тон окраски пуховичков — грязнобелый, иногда бледножелтый;
по этому фону на спинной стороне разбросаны частые темнобурые пятна и полосы. Светло-
бурые полосы и пятна с более темными оторочками размещены также и на подбородке, горле
и шее. Окраска оперения взрослых — пестрая из комбинации белого, темнобурого, серого,
охристого, а весной у самцов и черного цветов.
Спинная сторона тела самца в брачном наряде—темная из сочетания бурого и охристого
цветов со сложным струйчатым рисунком. Струйчатый рисунок проникает и на бока груди.
Грудь, брюшко, подхвостье, конец крыла — белые. Горло и бока головы ниже глаза и в
области ушей — темносерые. Широкий пестрый ошейник имеет на задней стороне шеи удли-
ненные перья. На зобе и боках шеи он состоит из трех черных и двух белых полос.
Передние первостепенные маховые, за исключением светлых кончиков и оснований,—
бурые; внутренние первостепенные маховые — белые с бурыми предвершинными пятнами;
второстепенные маховые почти сплошь белые. Средние рулевые — пестрые, крайняя пара
рулей — почти белая. Радужина — бледножелтая. Клюв — буровато-серый. Ноги — желто-
вато- или зеленовато-серые. В конце лета брачный наряд самца сменяется на зимний. Темно-
серая окраска горла и ушной области, а также пестрый ошейник исчезают. Горло становится
беловато-охристым, зоб — пестрым.
Оперение самок близко к осенне-зимнему оперению самцов, отличаясь от него более
грубым рисунком спинной стороны и большим развитием на зобе, брюшке и боках тела полу-
лунных пятен. Молодые стрепеты окраской оперения похожи на взрослых самок, отличаясь
от них матовым и бледным рисунком спины.
2. Вихляй или джек Otis undulata Jacquin
Psophia undulata. Jacquin. Beytr. Gesch. Vog., 1784, стр. 24, Триполи.
Русское название. Вихляй (народное). Казахское — джек. У русских
в Туркмении — красавчик.
Распространение. Ареал. Канарские о-ва, сев. Африка (Алжир, Египет,
Сахара), Аравия, Сирия, Месопотамия, Армения, Иран, Афганистан, Белуджи-
стан, вост. Туркестан и на восток до средних частей Монголии. В СССР —
по югу страны от Закавказья через Туркмению и Казахстан до юго-вост. Алтая
и Тувинской обл.
Характер пребывания. В северных и восточных частях ареала —
гнездящаяся, перелетная птица, в средних и южных — напротив — оседла.
ВИХЛЯЙ ИЛИ ДЖЕК
151
Биотоп. Пустыни, полупустыни, реже — участки солончаковой почвы
среди степи.
Подвиды и варьирующие признаки. Вихляй образует три подвида, отличающиеся
окраской оперения и размерами. О. и. macqueenii Gray, 1784 — Египет к востоку от
Нила и далее через Сирию, Аравию, Иран и пустыни Средней Азии до Тувинской обл. и
Карта 24. Распространение вихляя Otis undid at a
1. Of u. macquenii, 2. O. n. undulata, 3. О. и. fuertaventurae.
a — границы гнездовий, г — места зимовок (цифры в кружках обозначают номера зимующих
подвидов).
центр. Монголии; О. и. undulata J а с q u i п, 1784—Африка, сев. Сахара, Алжир, центр,
и южн. Тунис, Триполи, Киренаика, Египет на восток до Нила; О. и. fuertaventurae
Rothsch. et Hart., 1894 — вост. Канарские о-ва (Фуртавентура и Ланцароте).
Восточный вихляй Otis undulata macqueenii Gray
Otis Macqueenii. J. E. G r a y. in H a r d w i с k e's. 111. Ind. Zool., 2, 1834, табл. 47,
Гималаи
Распространение. Ареал. Египет на восток от Нила; Синай, Палестина,
Сирия, вост. Аравия, Месопотамия, Армения, вост. Иран, Афганистан, южн.
Белуджистан, Синьцзянь. Взрослые были добыты и наблюдались во многих
пунктах зап. Монголии (Банников); вероятно гнездование в Ордосе (Биадки,
1915). В пределах СССР широко распространен в полупустынях и пустынях от
Закавказья до юго-вост. Алтая. Недавно обнаружены особи, гнездящиеся в Ту-
винской обл. к югу от Танну-Ола (Терновский). Детали распространения
у нас таковы: в Европейской части СССР вихляй гнездится только в низмен-
ной Армении, где обнаружен в Сардар-Абадской степи и в б. Сурмалинском
уезде в долине Аракса (Бобринский, 1915). В пограничных частях Турции взрос-
лые и пуховички вихляев добывались в окрестностях поста Булган, неподалеку
от г. Нахичевани (Беме, 1926). Далее на восток в Закавказье вихляй отсутствует
и встречается на гнездовье лишь на северном и восточном берегу Каспия. Север-
ная граница распространения начинается в области низовьев Урала, подни-
маясь приблизительно до 49° с. ш. и населенного пункта Калмыкове. Отсюда
она идет на восток, огибает с юга бассейн Илека, выходит к Кара-Бутаку и,
следуя по речке Теке, достигает лежащих на Сары-Тургае местностей Сары-
Копа и Майкарау, далее по речке Кангабулак (приток Сары-Тургая) граница
тянется к горам Арганты. От этих гор она идет сначала на восток, а затем на
северо-восток, достигая 51° с. ш. у Семипалатинска (Хахлов и Селевин, 1928);
отсюда продолжается в Восточно-Казахстанскую обл., где вихляй гнездится на
западном берегу озера Зайсан и добывался в окрестностях Зайсанска (Поляков,
1915). Гнездится также в Чуйской и Курайской степи на юго-вост. Алтае.
153 отряд дрофы
К жует указанной границы вихляй широко распространен в соответствую-
щих биотопах всего Казахстана: сев. Прибалхашье, район нижней Сыр-Дарьи,
в пустыне Кызыл-Кум и на восточном берегу Аральского моря.
На Аральском море вихляй видимо населяет некоторые крупные острова,
так как оказался довольно многочисленным на о-ве Барса-Кельмес. Гнез-
дится в Узбекистане, в юго-зап. Таджикистане и, быть может, в районе Ленина-
бада. Населяет всю равнинную Туркмению от Кара-Кумов и прикаспийских
пустынь до долины Атрека и подгорной полосы Копет-Дага и Бадхыза. К северу
через юго-зап. части Усть-Урта проникает по побережью Каспия до Бузачи и
Мангышлака.
Во внегнездовое время, реже летом, вихляи иногда залетают далеко за
пределы гнездовой области. Известны залеты в Чкаловскую обл., под Астра-
хань, Сарепту и Харьков; в район Минусинска; на Кавказе — под Дзауджи-
кау и в б. Кубанскую обл.; в Прибалтику; в Зап. Европу—в Англию, Бель-
гию, Голландию, Францию, Швейцарию, Италию, Чехословакию, Румынию,
Данию, Швецию и Финляндию.
Зимовки отмечены в южных частях Средней Азии, в Индии, Белуджистане,
южной части Ирана, в Палестине и Аравии.
Характер пребывания. Гнездящаяся перелетная и местами
немногочисленная зимующая птица СССР. У нас зимовки не носят массового
характера. Редко и, вероятно, случайно одиночные особи попадаются зимой
в пустыне Кызыл-Кум, в оазисе Тамды; видимо регулярно, но в ограничен-
ном числе, перезимовывают вихляи в Туркмении: в низовьях Атрека, в Чат-
ской долине, в предгорной полосе Копет-Дага, где наблюдался в декабре —
феврале 1914—42 г. (Дементьев, 1950). В наиболее холодные зимы отмечается
гибель птиц. Из других частей ареала, лежащего в наших пределах, вихляи
на зиму улетают.
Сроки прилета и отлета выяснены плохо. Для Закавказья, например, они
остаются неизвестными. В пределах Средней Азии вихляи повсеместно появ-
ляются ранней весной в марте и во всяком случае не позже первой половины
апреля. К этому времени почти весь снег сходит, но ночная температура часто
понижается до 10—15°. Однако, вследствие сравнительной редкости птиц, их
осторожности и умения прятаться, прилетевших не сразу удается заметить.
Во многих случаях эти особенности птиц искажают наше представление о сро-
ках их весеннего прилета.
Наиболее раннее появление птиц в Туркмении отмечено у Уч-Аджи уже
в феврале. Не исключена возможность, что наблюдавшиеся в этих случаях вих-
ляи принадлежали к зимующим, так как нормально прилет в Кара-Кумах,
у Репетека и у озера Делели происходит между 6 и 20 марта. Птицы, встре-
ченные в различные числа апреля — на Теджене у Карабента, у Безмеана, на
Атреке у Беумбаша и в ряде других мест,— скорее могут быть отнесены к местным
гнездящимся, нежели к пролетным. Под Ташкентом весной вихляи добывались
не раньше 31 марта (Зарудный, 1915). На Сыр-Дарье между Арысью и Джуса-
лами пролет регулярно происходит в первой половине марта; севернее этого
пункта и до устья реки обычно во второй половине марта. В самом конце марта,
чаще в первых числах апреля, вихляи появляются на острове Аральского моря—
Барса-кельмес. На северо-восточном берегу Аральского моря вероятно непер-
воприлетные особи отмечены лишь 26 и 18 апреля, а наиболее ранние — 4 апреля
(Бостанжогло, 1911). Несколько северо-восточнее, у Кок-джиды, вихляи при-
летают около середины апреля, а на нижнем Иргизе наиболее ранние в 1899 г.
отмечены 27 апреля (Сушкин, 1908). Рано прилетают вихляи и в более восточ-
ных частях ареала, так как уже 20 апреля в сев. Прибалхашье удается нахо-
дить насиженные яйца (Долгушин, 1947). Осенний отлет сравнительно ранний.
В конце августа вихляи держатся семьями или небольшими стайками, а около
середины сентября начинают подвигаться к югу. С этого момента интенсивный
ВИХЛЯЙ ИЛИ ДЖЕК 153
отлет идет в течение второй половины сентября, а в южных районах и большую
часть октября. За исключением мест зимовок, в ноябре вихляи повсеместно
отсутствуют. В районе Семипалатинска еще отмечался 13 сентября (Поляков,
1915); с широты Иргиза вихляй отлетает в конце первой декады сентября,
на Эмбе отмечен 28 сентября (Сушкин, 1908); в нижнем отделе Сыр-Дарьи —
в конце сентября и начале октября; в Кызыл-Кумах в конце первой трети сен-
тября, на Чу в конце сентября (Гладков и Гринберг, 1932); осеннее движение
в Таджикистане около озера Кара-Куль отмечено 17 сентября (Иванов, 1940).
С этими же сроками совпадают случайные залеты пролетных вихляев, как на-
пример, в сентябре под Дзауджикау и 14 сентября в Кубанскую обл.
В Туркмении осенний отлет отмечен у Теджена в сентябре, в конце октября —
в приморской полосе у Красноводска; между октябрем и началом ноября —
во всей Туркмении.
Пролетные вихляи наблюдаются в дневные часы; они летят низко над
землей маленькими группками, парами и одиночками, изредка присаживаясь
и отдыхая в открытых солончаковых степях. Весной явно пролетные самцы,
останавливаясь на отдых, часто начинают токовать.
Биотоп. Гнездовый. Птица глинисто-солончаковых полупустынь и
закрепленных песков. В окрестностях поста Булган близ г. Нахичевани вихляй
гнездится на обширных пологих плато, покрытых редкими кустиками тип-
чака — Fesiuca sp. В Туркмении населяет глинистые и песчаные пустыни и
полупустыни с солянковой и полынной растительностью с относительно ровным
рельефом и наличием такыров; держится также по окраинам культурного
ландшафта, но избегает сильно всхолмленных песков. В пустыне Кызыл-Кум
излюбленными местами гнездования служат ровные, слегка волнистые про-
странства, завеянные песками и заросшие пышными, но редкими и невысокими
кустарниками. Еще лучше, если такие участки чередуются с полынной солон-
чаковой степью и небольшими такырами. В сходной обстановке вихляи гнес-
дятся в пустынях, прилежащих к низовьям Сыр-Дарьи и Сары-Су, но отнюдь
не избегают гнездиться в глубоких обширных котловинах, покрытых редкими
кустиками, деревцами саксаула и заключенных в рамки высоких песков.
Изредка выводит птенцов в щебнистых пустынях и среди редких тамарисковых
порослей, поблизости от пресноводных водоемов и в предгорьях.
В области Эмбы и Иргиза вихляи избегают барханов, избирая слабо-
всхолмленные пески с кустарниками, глинисто-солончаковые пески с кустар-
никами, глинисто-солончаковые и щебнистые участки с пучками ковыля, та-
волги, саксаула или лугоподобные площади с тамариском. На Алтае местами
вихляи проникают в горы, встречаясь на высоте до 1800 м.
Численность. У нас обычная, но вообще немногочисленная пъица,
не часто попадающаяся на глаза. Редки вихляи близ северной и восточной гра-
ницы распространения (Акмолинск, степи севернее Мугоджар). В Туркмении,
напротив, вихляи сравнительно обычны. На протяжении 280 км, покрытых
на автомашине, отмечено 7 особей (Дементьев).
Экология. Размножение. Вихляи гнездятся отдельными парами.
Гнездо от гнезда удалено редко на 1—2 км, чаще на 5—10 км (Кызыл-
Кумы).
Настоящего гнезда вихляй не строит. Самка выкапывает едва заметную
ямку в глинистой, иногда несколько завеянной песком почве и кладет в нее
яйца. Гнездо располагается то совершенно открыто, на солнцепеке, то под сла-
бой тенью кустика или деревца саксаула. Кладка окружена валиком из мелкой
растительной ветоши, образующей род гнездовых стенок. Высота их 1,5—2 см;
поперечный диаметр всего гнезда 13—14 см. Стенки гнездовой ямки, вероятно,
образуются благодаря частым ветрам, наносящим растительную ветошь вокруг
насиживающей самки, без активного участия самой птицы. Нормальное число
яиц в кладке — 3, реже 2, очень редко — 1 (указывается и 4, Уайзерби, 1940).
154 отряд дрофы
Яйца неправильной овальной формы, сильно вздутые. Основной фон окраски
скорлупы глинистый или оливковый, по нему разбросаны небольшие поверх-
ностные темносерые и глубокие светлосерые пятна. Размеры: (9) 54,6—
6О,6Х44,&—45,6 мм, в среднем 58,6X45,3 мм.
Яйцекладке предшествует своеобразное токование. Самцы ходят непода-
леку от самок, вытянув шею и распуская воротник то с одной, то с другой сто-
роны (обращенной к самке), при этом иногда несколько открывают крылья, но
звуков не издают (Дементьев, 1945). По сведениям других авторов, во время
токования самцы распускают также веером хвост (Бобринский, 1915, Спанген-
берг). Между самцами во время тока бывают ожесточенные драки (у многих
самцов, добытых в период размножнения и позднее, не хватает многих перьев
хвоста). Самцы, добытые во время весеннего пролета, обладают сильно увели-
ченными семенниками; яичники самок, напротив, в это время остаются на самой
ранней стадии развития. В Армении самка с яйцом в яйцеводе добыта 13 мая
(Бобринский, 1915), вероятно, это была поздно загнездившаяся птица. В Турк-
мении севернее Гассан-Кули кладка в 3 свежих яйца найдена Воробьевым
10 апреля, другая, повидимому незаконченная, —у Кызыл-Атрека 12 мая (Де-
ментьев, 1945). В Кызыл-Кумах неподалеку от Букан-Тау гнездо с двумя яйцами
найдено 13 апреля (Богданов, 1882).
В пустынях у нижней Сыр-Дарьи кладка происходит в первой половине
апреля, т. к. около десятых чисел этого месяца на о-ве Барса-Кельмес,
лежащем много севернее, находили яйца. Гнезда, осмотренные на Сыр-Дарье
уст. Байгакум ив низовьях реки Сары-Су, содержали. 17 мая единственное
яйцо, наклюнутое птенцом, а 22 мая — 3 яйца в последней стадии насиживания
(Спангенберг). Самка, добытая 22 апреля на Эмбе у Как-джиды, имела фолли-
кулы около 23 мм в диаметре; у Как-сая 2 яйца найдены 22 мая (Сушкин,
1908). Сравнительно рано вихляи приступают к откладке яиц в более восточных
частях страны. Например, в сев. Прибалхашье 3 насиженных яйца найдены
уже 20 апреля (Долгушин, 1947). Много позднее (приблизительно на месяц)
по сравнению со степями Казахстана, лежащими на 48° с. ш., гнездятся вихляи
в Чуйской степи на юго-вост. Алтае.
Насиживает только самка, она же и воспитывает птенцов (Сушкин, 1908).
Самцов у гнезд видеть не удается; после окончания яйцекладки самцы держатся
в степи одиночками, иногда небольшими группками (Спангенберг). Промежу-
ток между кладками яиц — сутки. Насиживание, вероятно, начинается
после отложения всей кладки, т. к, выклюнувшиеся пуховички слабо отли-
чаются размерами друг от друга. Продолжительность насиживания остается
невыясненной.
Самка сидит на яйцах очень крепко и подпускает человека совсем близко.
Будучи спугнутой с гнезда, иногда прекращает насиживание.
Вылупление птенцов происходит в мае и июне. Возможно, что первые
птенцы появляются в конце апреля, а запоздавшие — в западных частях
ареала — в июле.
У ст. Каракала в Армении оперившийся и хорошо бегающий птенец пой-
ман 28 июля (Бобринский, 1915). В апреле—мае маленькие птенцы добыты
у Булгана неподалеку от Нахичевани. В пустынях Туркмении вывод птенцов
происходит между серединой мая и началом июня. Между Суллы-Кужумды и
Хал-ата в Кызыл-Кумах птенцы пойманы 5 июня (Богданов, 1882). В той же
пустыне выводки в 2—3 птенца удавалось находить между серединой мая и
серединой июня. В низовьях Сыр-Дарьи и Сары-Су наиболее ранний вывод
птенцов в 1927 г. отмечен 17 мая; в другом гнезде, судя по насиженности яиц,
птенцы должны были появиться 27—29 мая (Спангенберг, 1936); 25 июня
начинавший оперяться птенец, добытый Слудским на Сары-Су, был ростом
с грача; в десятых числах июля здесь попадаются вполне оперившиеся птенцы
ростом немного меньше матки.
ВИХЛЯЙ ИЛИ ДЖЕК 155
В пустынях севернее Аральского моря пуховой птенец найден в гнезде
степного орла 15 июня, а 23 июня на западном берегу озера Челкар-Тениз птенцы
обладали переходным нарядом от аухового в птенцовый (Сушкин, 1908). У Кар-
сакпая 10 июня добыт пуховичок. В сев. Прибалхашье птенцы восьми
выводков, найденных в середине июля, уже достигли роста с полматки (Долгу-
шин, 1947). Вероятно позднее происходит вывод птенцов у северной границы
распространения вида —на Алтае, где совершенно оперившийся птенец вихляя
добыт в Чуйской степи 31 июля (Сушкин, 1938). В течение большей части июля
молодые вихляи отличаются от взрослых несколько меньшими размерами
и характером оперения. Полного роста они достигают в Туркмении к началу
августа (Дементьев, 1950). Под Иргизом, т. с близ северной границы рас-
пространения, молодая особь, уже державшаяся самостоятельно и одетая
в полный осенний наряд, была добыта 13 августа.
Пуховички остаются в гнезде самое непродолжительное время и в первые
дни искусно прячутся среди скудной растительности гнездового участка, вы-
давая себя, однако, писком. При маленьких птенцах самка ведет себя крайне
смело и в некоторых случаях бросается на человека. Когда птенцы подрастут,
самка часто прибегает к хитрости — быстро отбегает от места, где залегли
пуховички, на одно мгновенье поднимается во весь рост, затем присаживается,
похлопывая крыльями, как бы подбирая под себя птенцов. Повторив такой
прием раз пять и каждый раз меняя место, птица неожиданно улетает.
Около середины августа вихляев удается встречать выводками, иногда
стайками из 7—9 особей. Самки и молодые в это время присоединяются к сам-
цам; часть птиц, однако, держится в это время одиночками. Некоторое время
вихляи ведут бродячий образ жизни, а затем начинают постепенно отлетать
к югу (Спангенберг).
Питание. Вихляй кормится как растительной, так и разнообразной
животной пищей. В Кызыл-Кумах поедает плоды каперцов (Молчанов, 1913).
В пограничных частях Ирана неподалеку от Нахичевани охотится за ящери-
цами и прямокрылыми (Беме, 1926). В полупустынях, расположенных севернее
и восточнее Аральского моря, ест молодые побеги полыни, луковицы лука и
чеснока, различных жучков, в частности чернотелок и навозников, посещает
иногда в поисках насекомых истлевшую и высохшую падаль, ловит порой яще-
риц (Сушкин, 1908). В жаркое время года большое значение в питании взрослых
и особенно молодых особей имеют кузнечики, саранча и кобылки. Пойманные
молодые птенцы при содержании в неволе предпочитают этот корм всей осталь-
ной предлагаемой им пище. Желудки пяти взрослых вихляев, добытых 26 июня,
30 июля и 25 августа в зап. Монголии, оказались заполненными только саран-
човыми. Повидимому, не пьет, довольствуясь сочными побегами растений. Это
подтверждается наблюдениями над содержащимися в неволе птенцами (Бо-
бринский).
Линька. Общая схема смены нарядов — как у других дроф: пуховой^
птенцовый—первый брачный—первый зимний—второй брачный—второй зим-
ний и т. д. Второй наряд — окончательный. Две линьки в году: полная между
июнем —октябрем (однако, по Уайзерби, 1940, будто бы между августом
и декабрем), и частичная предбрачная в феврале — марте. При частичной пред-
брачной линьке сменяется мелкое оперение, но сезонного диморфизма нет.
Линька первостепенных маховых у самцов и у самок начинается в конце гнез-
дового периода. Последовательность смены маховых —от заднего края партии
к переднему, последним линяет, следовательно, первое первостепенное маховое.
Небольшой изученный нами материал таков: у взрослых самцов, добытых
12 марта и 22 апреля, линяет мелкое перо на спине, крыльях, голове и шее
(март), перо на голове и шее (апрель). Это —частичная предбрачная линька.
Майские экземпляры не линяют. У июньских особей сменяются задние перво-
степенные маховые (у птицы, добытой 10 июня, шесть задних первостепенных
156
ОТРЯД ДРОФЫ
маховых свежие; у двух других птиц, добытых в середине июня, у одного
выпало пятое маховое, шестое—десятое свежие; у другого — свежие седьмое —
десятое маховые). В таком же состоянии была линька у самки, добытой на
р. Кальджир 8 июля (что, очевидно, связано с более поздним размножением
вихляев в вост. Казахстане, о чем упоминалось нами выше). В конце августа
у •врех, добытых в сев.-зап. Монголии экземпляров, линька маховых приближа-
лась к завершению (у одной птицы 25 августа осталось старое первое маховое,
у двух других первое маховое растет, остальные маховые свежие). На сере-
дину — конец августа падает и интенсивная смена мелкого пера. В юго-зап.
Туркмении, на Атреке, 17 и 28 сентября добыты самец и самка, полностью
закончившие осеннюю линьку.
Полевые признаки. По общему складу вихляй — типичная дрофа средней
величины. От прочих дроф отличается воротником из длинных перьев на шее,
видимыми на большом расстоянии.
Окраска вихляя покровительствен-
ная, «пыльная», поэтому заметить
птицу среди пустынной обстановки
очень трудно. Напротив, во время
полета она кажется пестрой благодаря
черной и белой полосам на крыльях.
Спугнутый вихляй сначала быстро
бежит, время от времени взмахивая
крыльями. Сделав большой полукруг
по степи, птица поднимается и улетает
очень далеко. В других случаях вих-
ляй с места поднимается на воздух и,
опустившись метрах в трехстах, убе-
гает в сторону. Как и другие дрофы,
вихляи очень осторожны.
Бегают вихляи замечательно бы-
стро. Скорость бега, судя по определе-
ниям при помощи автомобильного
спидометра, —35—40 км в час (Демен-
тьев, 1945). Летают вихляи превос-
ходно, сравнительно медленно (по величине птицы) и очень глубоко взмахи-
вая своими длинными крыльями. Несмотря на хорошие летательные способ-
ности, вихляи не поднимаются высоко в воздух и предпочитают летать низко
над землей. Маленькие птенцы издают тонкий писк. Голоса вполне оперившихся
птенцов и взрослых особей никто не слышал.
Описание. Р а з м е р ы и строение. Вес самцов 1150 г (Монголия), 1500, 2000 г
(Туркмения), по Бутурлину A935)—до 2380 г; вес двух самок (Монголия) 1100 и 1100 г. Фор-
мула крыла: 3 = 4^2>1>5>6. Вырезки на наружных опахалах2-го — б-го, на внутрен-
них опахалах 1-го — 5-го первостепенных маховых. Хвост закругленный, из 20 рулевых.
Крыло самцов и самок A5) 340—425, в среднем 388 мм.
Окраска. Пуховые птенцы на спинной стороне покрыты по основному грязно-
белому или светлоохристому тону рыжеватыми и буроватыми полосками и пятнами. Такой
же рисунок в виде удлиненных полосок хорошо заметен на передней-части шеи. Горло и
брюшко птенцов — грязнобелое. Птенцовый наряд надевается пуховичками после пухового.
Окраска головы, спины, крыльев и хвоста у таких птенцов с хорошо выраженным
рыжим основным тоном и слабой пятнистостью. Интересно, что и в первом зимнем наряде
(когда птица размером с грача) птенцы имеют черные пятна на боках шеи (будущий
воротник).
Оперение молодых вихляев в течение первого года жизни отличается от оперения ста-
рых наличием светлоохристых пятен на перьях спинной стороны тела и бледноохристыми
кончиками рулевых. Молодые птицы уже в первую осень приобретают черный воротник,
который, однако, развит слабо. У взрослых верхняя часть головы (лоб и темя) — пепельно-
рыжеватая (золотистая) с крайне мелкими темными пестринами. На лбу и темени имеется
гахол из длинных очень тонких перьев. Окраска перьев хохла с белыми основаниями. На
Рис. 27. Пуховые птенцы вихляя
ДРОФА
157
боках шеи позади ушной области развит черный цвет, переходящий на большой воротник
из длинных частично черных, частично белых и пестрых перьев.
Бока зоба, вся спина, верхняя часть крыльев и хвоста — песчано-рыжеватые и сплоить
испещрены мельчайшим струйчатым темным (почти черным) рисунком и более крупными
темнооурыми пятнами. Рулевые перья — поперечно-полосатые с белыми вершинами; перьо
тТгпгнныг nfaYnnrtr— из сочетания черного и белого цветов. Брюшная часть тела (горло и
Рис. 28. Вихляй (взрослая птица и подросший птенец)
брюшко)—белые; щеки, низ шеи и верхняя часть зоба — пепельно-рыжеватые с мелким рыже-
ватым крапом. Нижняя часть зоба — голубовато-серая. Некоторые экземпляры, добытые
весной, имеют грязновато-белую окраску горла и шеи,*переходящую в голубовато-серый цвет
на зобе. Подбой крыла белый, подхвостье белое со слабо развитым темным мраморным
рисунком.
Радужина желтовато-бура я. Клюв бурый. Ноги светлосерые.
3. Дрофа Otis tarda L.
Otis tarda. Linnaeus. Syst. Nat. изд. X, 1758, стр. 154, Польша.
Русское название. Дрофа, драхва, дудак — народное.
Распространение* Ареал. Зап. Европа, Малая Азия, Иран, сев. Африка,
южная и средняя полоса СССР, на восток до Тувинской обл., Монголии и Манч-
журии.
Характер пребывания. Гнездящаяся, перелетная в северных частях
ареала; в южных —оседлая птица.
Биотоп. Открытые пространства степной и лесостепной полосы, полу-
лустыни и окраины пустынь, местами проникает в высокогорные степи.
Подвиды и варьирующие признаки. Имеет два подвида, отличающиеся в основном
деталями окраски оперения. О. t. tarda L., 1758— западная часть ареала вида на восток до
южн. и зап. Алтая; О. t. dylowskii T а с z., 1874 —восточная часть ареала вида к за-
паду до сев.-вост. частей Алтая.
158
ОТРЯД ДРОФЫ
Европейская дрофа Otis tarda tarda L.
Синоним. Otis tarda korejewi. S а г u d n y. Orn. Monatsber 1905, стр. 163, Семи-
речье.
Распространение. Ареал. Зап. Европа— Португалия, южн. Испания,
Германия к северу до Саксонии и Бранденбурга; Польша, южные части Чехо-
словакии, Венгрия, Румыния, Болгария, Греция. В Африке — в сев. Марокко;
в Азии — в Малой Азии, сев. Сирии и Иране.
В СССР широко распространена по южной и средней полосе от государствен-
ной границы до юго-зап. Алтая. Северная граница не вполне ясна, так как ме-
стами она отступила к югу, местами же, где лес сменился открытыми простран-
ствами, подвинулась к северу. Гнездится в Бессарабии, в Подолии. В восточ-
ной Белоруссии гнездится южнее Гомеля, тогда как в более северных частях
Карта 25. Распространение дрофы Otis tarda
7. О. t. tarda, 2. О. t. dybcwskii;
а — граница гнездовий, б — не вполне выясненная граница гнездовий, г — места зимовок (цифры
в кружках обозначают номера зимующих подвидов).
страны известны только залеты. Далее граница пересекает Черниговскую обл.
и переходит в Брянскую обл., где дрофа гнездится в районах Карачевском и
Шаблынинском (Суслова, 1935). Отсюда, следуя на восток, граница пересекает
Новосильский район Тульской обл., Ефремовский район и выходит на самый
юг (Даньковский район) Рязанской обл. В смежных восточных областях дрофа
несомненно гнездится в Тамбовской обл., в южных частях Пензенской обл.,
но не гнездится в Ульяновской обл. (Волчанецкий, 1937). Ранее про-
никала далее к северу, достигая юга Горьковской обл., куда сейчас лишь
залетает (Серебровский, 1918). В Куйбышевской обл., вероятно, гнездится
спорадично и отсюда проникает в южные части Татарской республики, а зале-
тает несколько севернее Чистополя (Першаков, 1929). Вполне вероятно хотя бы
редкое гнездование в степях южн. Башкирии, так как дрофы продолжают по-
являться в районе Стерлитамака, а в соседней Челябинской обл. гнездова-
ние известно в районе Миаса (Снигиревский, 1929). В Зауралье северная гра-
ница проходит примерно по 54° с. ш. Дрофа в районе Омска, достигает озера
Чаиы и Барабинской степи, откуда граница идет через Барнаульский округ
к предгорьям юго-зап. Алтая.
К югу от указанной границы ареал дрофы простирается до южного государ-
ственного рубежа СССР, здесь территория заселена дрофой не сплошь, а от-
дельными пятнами. В европейской части спорадическое распространение ха-
рактерно для местностей близ северной границы ареала, более равномерно
дрофа встречается на юге. Дрофа населяет степи и культурные земли почти
всей Украины, Молдавии, Крыма, Поволжья и Сев. Кавказа. Напротив,
в Закавказье гнездование дрофы носит в настоящее время случайный характер.
дрофа 159
Вероятно, не гнездится теперь в Армении, но гнездится в некоторые годы в
Муганской и в Ширванской степях (Верещагин, 1940).
Спорадическое распространение характерно также для Азиатской части
Союза. Дрофа гнездится в Чкаловской обл. и в южных степях Зап. Сибири.
В Казахстане она широко заселяет степные северные и восточные территории,
но положительно избегает настоящих пустынь и отчасти полупустынь. Найдена
по северной окраине аральских Кара-Кумов со стороны урочища Кара-Кум и
в восточных частях со стороны Голодной степи. Широко гнездится в степях,
примыкающих к Тянь-Шаню, а также в бассейне Или в районе озера Ала Куль,
в предгорьях Тарбагатая и в Зайсанской котловине. В Киргизии найдена
в котловине озера Иссык-Куль. Безусловно гнездится в южн. Таджикистане
и по восточному склону Кара-Тау, вероятно по степным участкам Копет-Дага
и, возможно, в его предгорьях.
За пределы гнездового ареала дрофы нередко залетают не только в теплое
время года, но и зимою. В зап. Сибири случайные залеты известны в окрест-
ности Каинска. Иногда птицы залетают в Прибалтику, Московскую обл. и
под Ленинград. Последний залет под Москву дрофы — самки известен в 1947 г.,
а в Котельнический район Кировской обл. —в середине декабря 1947 г.
Залеты дроф отмечены и в зап. и южн. Европе (Англия, Франция, Бельгия,
Голландия, Швейцария, Германия, Югославия, Албания, Италия и на острова
Средиземного моря), в сев. Африке — в Алжир, в Азии — в Палестину, Иран,
Афганистан.
Часть наших дроф живет оседло, большинство же откочевывает к югу,
перезимовывая на юге Украины, в Закавказье и Средней Азии.
Характер пребывания. Гнездящаяся и перелетная птица в северных
частях ареала, живет оседло или, повидимому, замещается зимой северными
популяциями в южных частях страны; массами перезимовывает на южных
окраинах СССР.
В Бессарабии обычно держится оседло, редко отлетая при выпадении боль-
ших снегов. В Крыму живет оседло и массами перезимовывает в степях. Зимую-
щие в Крыму дрофы отлетают крайне редко, появляясь на южном побережье
близ Судака и Алушты. На Украине живут оседло и перезимовывают в Подолии,
в южных частях Херсонской обл., на юге Днепропетровской. Напротив, в се-
верных частях упомянутых областей, а также в Воронежской, Полтавской и
Харьковской дрофы остаются на зиму сравнительно редко, лишь в малоснежные
зимы. В Черниговщине дрофа уже не зимует, но будто бы живет оседло в Брян-
ской обл. (Суслова, 1935). Отдельные особи перезимовывают даже в южных
частях Пензенской обл. (Федорович, 1910). Между Черным и Каспийским
морями дрофа живет оседло и зимует в вост. Приазовье. По устным данным,
регулярно зимует юго-западнее дельты Волги, в местности Черные Земли.
Кое-где зимует дрофа на Сев. Кавказе: Дагестан, частично под Дзауджикау
изредка на высоте 2000 м над уровнем моря, в северо-западных частях (Аве-
рин и Насимович, 1938). Местом массовых зимовок является Закавказье.
В восточных его частях дрофы регулярно зимуют по увалам Иорского и Кахе-
тинского плоскогорий. Особенно велики зимовки в степях Шираки, Аджинаур,
реже в сев. Ширвании и Мильской степи. Более часто в Муганской, юго-восточ-
ной части Ширванской и в Сальянской степи, а также на островах и по бере-
гам залива Кызыл-Агач. На запад отсюда дрофы отмечались зимой под Тби-
лиси и, видимо, в большом числе зимуют в долине Аракса и Сардар-Абадской
степи.
В Азиатской части Союза основным местом зимовки дроф служит Туркме-
ния. Особенно велики зимние скопления по Атреку — от Шарлаука и Чата на
востоке (Дементьев, 1950).
Большие зимние скопления дроф известны, наконец, в Таджикистане
(Иванов, 1940). В других районах азиатской части страны дрофа зимует нере-
160 ОТРЯД ДРОФЫ
гулярно и в ограниченном количестве. Иногда дрофы зимуют по окраинам пу-
стыни Кызыл-Кум со стороны Голодной степи; отдельные особи в малоснежные
зимы остаются в Чкаловской обл. и во второй половине зимы отмечались под
Семипалатинском (Хахлов и Селевин, 1928).
Размещение дроф на зимовках находится в соответствии с особенностями
снегового покрова. Весеннее движение к северу раннее. Птицы начинают подви-
гаться к местам гнездовья с первыми проблесками весны. В Аскании Нова про-
летная стая отмечена 10 марта (Дергунов, 1924). В южную половину Украины
дрофы обычно прилетают в первой половине марта и с этого времени пролет
затягивается до конца первой трети апреля (в Подолии в конце марта —на-
чале апреля), на юге Днепропетровской обл. —первые —в начале марта,
разгар — в середине этого месяца и в первой трети апреля. Под Киевом стая
дроф наблюдалась в марте ст. ст. (Шарлемань, 1909). В Харьковской обл.
в течение 9 лет пролет происходил между 13 марта и 7 апреля (Сомов, 1897);
в Орловской —отмечался 27 апреля (Ефимов, 1906); в Воронежской за 5 лет
между 7 и 19 марта (Огнев и Воробьев, 1924). В вост. Закавказье зимующие
дрофы начинают двигаться к северу с первых чисел марта, в середине марта про-
ходит валовой пролет, и к концу марта — началу апреля степи уже освобо-
ждаются от зимующих стай. В марте большие стаи птиц наблюдаются в степях
под Кизляром; б апреля отмечались на сев.-зап. Кавказе (Аверин и Наси-
мович, 1938); в течение всего марта, иногда и первую половину апреля,
летят дрофы в дельте Волги; 22 апреля первые птицы появляются в районе
Бугуруслана в Куйбышевской обл. (Карамзин, 1913).
Несколько позднее происходит пролет в Азии. Зимняя жизнь дроф в Турк-
мении продолжается до конца марта, однако на Атреке в марте пролет уже хо-
рошо заметен. На зимовках в Таджикистане последние особи еще наблюдаются
между 10 и 15 мая (Иванов, 1940). Вследствие столь длительных задержек на
зимовках в Азии и на пролетах нередко удается встречать там дроф поздней
весной. Первые птицы, однако, в Азии у мест гнездовья появляются ранней
весной — в марте, самое позднее в апреле. Во второй половине марта и в первой
половине апреля пролет отмечался в пустыне Кызыл-Кум; в апреле —на Чу;
между 23 марта и 10 апреля — на нижней Сыр-Дарье, где вообще пролет выра-
жен слабо (Спангенберг, 1936); 1 и 3 апреля—между Камышлыбашем и
п-овом Утюбас (Богданов, 1882); в последних числах апреля—на северо-
восточном побережье Аральского моря. В степях сев. Казахстана и при-
легающих местах Чкаловской обл., где годами снега выпадает много и про-
талины появляются поздно, сроки сильно колеблются в зависимости от хода
весны. Прилет передовых птиц обычно совпадает с появлением проталин и
укладывается в промежуток времени между 1 и 15 апреля. У Темира передо-
вые отмечены 1 апреля; между Темиром и Кок-джидой интенсивный пролет шел
7 апреля; в низовьях Иргиза — 19 апреля (Сушкин, 1908); в Наурзумском за-
поведнике — 9 апреля (Михеев, 1938).
Осенний отлет почти повсеместно тянется дольше, чем весенний. Часть осо-
бей (быть может, молодые птицы) из северных частей ареала начинают медленно
подвигаться к югу уже в августе, но часть упорно держится на местах гнездовья
до выпадения прочного снегового покрова и наступления настоящей зимы. До
самой зимы остаются дрофы в районе Зайсана; 2 октября наблюдался пролет
в сев. Прибалхашье (Долгушин, 1947); на высоте 2500 м 11 августа взрос-
лая особь добыта в Джунгарском Ала-тау Банниковым; до самого снега дер-
жатся дрофы в Наурзумском заповеднике (Михеев, 1938) и в степях Чкаловской
обл. В верховьях Тобола в некоторые годы с ранним похолоданием дрофы
исчезают около конца первой трети октября; в верховьях Иргиза первые осен-
ние стаи отмечены 10 октября, но валовой пролет шел между 3 и 8 ноября и
почти закончился 9 ноября (Сушкин, 1908). В пустыне Кызыл-Кум на осеннем
пролете дрофы попадаются с середины октября и до середины ноября.
ДРОФА 161
На местах зимовок дрофы начинают появляться очень рано. В Таджики-
стане первые осенние птицы прилетают в самом конце августа; на туркменских
зимовках —в последних числах октября; пролет продолжается иногда весь
ноябрь.
Столь же сильно растянут пролет дроф в Европейской части Союза. Из
Тульской обл. отлет, вероятно последних птиц, наблюдается с наступлением
морозов; в Орловской дрофы наблюдались еще в начале октября. Отлет птиц из
Харьковской обл. носит характер затяжной откочевки, начинаясь около сере-
дины октября и заканчиваясь к декабрю.
В Приерусланских песках осенний отлет заметен с конца сентября и длится
до середины декабря; в дельте Волги, вероятно, первые осенние пролетные
птицы появляются в конце октября. Под Дзауджикау пролетная стая дроф
отмечена 10 ноября, но весь пролет на Сев. Кавказе, видимо, затягивается
на длительное время, так как под Кизляром дрофы интенсивно летят в начале
декабря.
Вдоль Черноморского побережья Кавказа дрофы летят в течение октября;
в конце этого месяца отмечались на альпийских лугах Главного хребта. На
весеннем пролете птицы встречаются одиночками, чаще парами и небольшими
группами, летящими то низко над землей, то поднимаясь на значительную вы-
соту. На осеннем пролете чаще удается встречать крупные табуны — до 30—40,
редко до 200 особей. В противоположность весеннему пролету, осенью дрофы
летят низко, поднимаясь на большую высоту как исключение. В большинстве
случаев при перелетах птицы придерживаются открытого ландшафта и летят
широким фронтом. Однако на Кавказе области пролета хорошо выражены. При
осеннем движении дрофы летят в основной массе по побережью Каспия от Махач-
Кала на Дербент и далее западнее Баку. Начиная от ст. Килязи к югу полоса
пролета сильно расширяется, захватывая пространства между Шемахой и
Баку, т. е. так называемый Кабристан. Вторая волна незначительных размеров
следует вдоль восточного побережья Черного моря, частично задерживаясь на
зиму по северному склону Колхиды. Незначительное число дроф, наконец, про-
никает к закавказским зимовкам через Главный Кавказский хребет в пределах
45°36'—46°30' в. д. Весенний путь совпадает с осенним. При перелетах дроф
через Главный Кавказский хребет птицы поднимаются на большую высоту.
В Туркмении дрофы на пролете двигаются вдоль подгорной полосы Копет-
Дага на запад (Дементьев).
Биотоп, Более разнообразен, нежели у стрепета и вихляя. Птицы
населяют открытые пространства, избегая лишь настоящей пустыни и участ-
ков, заросших довольно высокой кустарниковой растительностью. Отсут-
ствие дроф среди кустарниковой поросли объясняется их чрезвычайной осто-
рожностью.
В Бессарабии и почти по всей Украине дрофы гнездятся в озимых хлебах,
на залежах и перелогах, на сенокосных участках, а в сухие годы, вероятно
нередко, среди травы настоящих лугов. Много реже птицы устраивают гнезда
на целинных участках ковыльной степи, используемых для выпаса скота, так
как в таких случаях гнезда часто вытаптываются домашними животными.
Сходны условия гнездования и в более северных частях ареала, В Чернигов-
щине известны случаи гнездования птиц среди песчаных холмов, поросших
кустарниками. В Татарской республике дрофы более тяготеют к целине, гнез-
дясь на залежах, в открытой степи на участках, поросших полынью. В б. Орен-
бургском крае дрофы гнездятся то в сильно всхолмленных, то в ровных черно-
земных и глинистых степях, устраивая гнезда по склонам балок, где ковыль
выше и гуще. Изредка гнездо устраивается среди зарослей чилиги и бобовника.
В смежных частях сев. Казахстана дрофа характерна для злаковых степей и
обширных лугов, причем заболоченность почвы, по крайней мере до известной
степени, вовсе не является помехой. В некоторых случаях птицы выводят на
11 Птицы СССР, т. 11
162 отряд дрофы
песчаной почве, если среди песков есть луговые участки или клочки, густо
заросшие злаками. У северных берегов Каспия дрофа гнездится среди сплош-
ных полыней, вблизи от многочисленных соленых озер и шоров. На окраинах
пустыни Кызыл-Кум дрофы избирают для гнездования песчаные и глинистые
равнины, заросшие полынью, злаками и эфемерами; на реке Чу —полынную
степь; в сев. Прибалхашье — полупустыни с ковылем; в районе Зайсана —
заросли молочая и чия.
Кормятся дрофы поблизости от места гнездовья. Птицы, собравшиеся в стаи,
наиболее охотно держатся в совершенно открытой степи или на вершинах поло-
гих холмов, откуда издали легко обнаружить приближение опасности. Как
видно, дрофа не избегает культурного ландшафта и часто гнездится на обрабо-
танных землях поблизости от населенных пунктов. Однако, там, где целинные
площади имеют широкое распространение и редко посещаются стадами домаш-
них животных, дрофы гнездятся особенно охотно. В вертикальном направлении
дрофа распространена в качестве гнездящейся птицы до 1000 м, а в Средней
Азии — до 3000 м.
Численность. Обычна, становясь местами многочисленной в южных
степных районах Европейской части Союза ССР; более редка близ северной гра-
ницы распространения и в Поволжье, в области лесостепи. В Закавказье, на-
против, гнездится в самом ограниченном количестве (Верещагин, 1940).
В Азии обыкновенна в северных, восточных и юго-восточных частях ареала,
но безусловно редка в гнездовое время в окраинах пустыни Кызыл-Кум, в Турк-
мении и Таджикистане. Общая численность дроф в СССР еще довольно значи-
тельна. Распашка земель сказывается на численности и распространении дрофы
в значительно меньшей степени, чем на стрепете, но неумеренное преследование
постепенно вытесняет дроф из ряда населенных районов. Громадные скопления
дроф еще наблюдаются на зимовках в Закавказье и в юго-зап. Туркмении.
Экология. Размножение. Птицы всегда гнездятся обособленными
парами. В районах, где дроф много, они держатся весной небольшими табу-
нами, и токование происходит на одних и тех же участках. В степях Чкалов-
ской обл. у места тока в радиусе 1 км удавалось находить 3 гнезда дрофы.
Вследствие этого гнездование дрофы приобретает характер густо заселенного
«пятна». Между отдельными гнездовыми «пятнами» существуют большие (в 10—
15 км) интервалы (Спангенберг).
Настоящего гнезда не бывает. Непосредственно перед откладыванием пер-
вого яйца самка сначала ногами выкапывает в почве пологую конусообразную
ямку, затем клювом выдергивает оставшиеся стебельки степных растений и,
наконец, вращательным движением тела округляет ее. Иногда гнездовой под-
стилкой служат примятые стебли. Диаметр гнездовой ямки 23—27 см
(Спангенберг). Гнездо помещается или совершенно открыто среди низко-
рослой полыни, или под кустиком среди густого и высокого ковыля. Число яиц
в кладке — 2, несколько реже — 3, еще реже 1 яйцо. Кладки в 4, 5 и 6 яиц,
указываемые в литературе (Мензбир, 1900 и др.), если и бывают, то предста-
вляют чрезвычайную редкость. Большинство яиц имеет овальную вздутую
форму; реже встречаются яйца вытянутой формы; скорлупа яиц блестящая, но
блеск слабее, чем у яиц стрепета. Основной фон окраски скорлупы зеленовато-
бурый и оливковый, по нему равномерно разбросаны крупные и мелкие буро-
ватые или рыжевато-бурые пятна, очертания и границы которых не вполне
ясны. Размеры B1) 76,9—86,6 х 55,0—61,0, в среднем 79,16 х 58,5 мм.
Сроки размножения в различных частях ареала выяснены недостаточно.
Однако известно, что чем дальше на север, тем позднее дрофы приступают
к кладке. В одной местности кладка протекает у большинства птиц в близкие
сроки; немногие самки отстают от других недели на две, даже на три. Токова-
ние происходит приблизительно с середины апреля и в мае. Одиночки или
группы самцов избирают для токования ровную открытую площадку или глад-
ДРОФА 1_fi3
кую вершину пологого холма. Раскрыв веером и вертикально подняв хвост,
опустив до земли крылья, плечевые перья которых в момент наивысшего воз-
буждения направлены вверх и вперед, самцы медленно расхаживают поодаль
друг от друга. Самки подлетают к току маленькими группами или прибли-
жаются к нему одиночками пешком. С появлением самок между самцами иногда
происходят ожесточенные драки. Чаще токующих самцов удается наблюдать
в утренние часы, но токуют они и перед вечером, а изредка и днем. Начало
кладки на юге Европейской части Союза происходит несколько раньше, чем
в Азии. Два ненасиженных яйца у Павлодара были найдены 11 апреля (Вальх,
1899); в других районах Днепропетровской обл. кладка происходит в конце
апреля B0—27 числа), а в холодные, запоздавшие весны — в середине мая.
В Подолии 3 свежих яйца найдены 11 мая; в Николаевской обл. —в сере-
дине мая, на островах Черного моря, вероятно насиженные, даже 20 июня (Шар-
лемань и Шуммер, 1930). Самые ранние кладки в Харьковской обл. удается
находить в конце апреля. Однажды гнездо со слабо насиженными яйцами было
найдено 7 июля (Сомов, 1897) — вероятно, вынужденная кладка. Не ранее мая
начинается кладка у дроф в азиатской части страны. По данным Зарудного
A888) и Карамзина A917) в б. Оренбургских степях дрофы кладут яйца около
середины мая. Впрочем, массовая кладка происходит здесь несколько раньше—
между 1 и 12 мая. Однако запоздавшие кладки со слабо насиженными яйцами
в районе Ак-булака удается изредка находить до самых последних чисел этого
месяца. В первых числах мая, основываясь на нахождении пуховичков, начи-
нается кладка у дроф и в сев. Прибалхашье.
Насиживание яиц и воспитание птенцов лежит только на самках. Самцы
после окончания токования собираются в небольшие стаи и кочуют по степям.
Отдельными стайками держатся холостые или потерявшие кладки самки.
Промежутки между откладкой отдельных яиц точно не выяснены.
Вероятно, птицы приступают к насиживанию после откладки первого яйца,
так как неоднократно приходилось спугивать самок с незаконченных кла-
док, а яйца одной кладки бывают насижены неодинаково (Спангенберг).
Срок насиживания —около месяца B5—28 дней по Уайзерби, 1940).
Самое раннее появление птенцов, видимо, совпадает с началом второй трети
мая в Европейской части Союза и с началом июня— в Азиатской. Конкретные
данные чрезвычайно скудны. 21 июня маленькие пуховички найдены на острове
Черного моря (Шарлемань и Шуммер, 1930); около середины июня, изредка не-
сколько раньше, происходит вывод птенцов в Харьковской обл. (Сомов,
1897); в Чкаловской обл. —в первой половине июня (Спангенберг); в сев.
Прибалхашье 3 птенца в возрасте 5 дней найдены 8 июня (Долгушин, 1947);
5—6 дней в Кустанайской обл. (Наурзум) — 15 июня (Келейников); птенцы
«ростом с курицу» в Приерусланских песках отмечены в середине июля. Хорошо
летающие дрофы наблюдались: 26 июля в Воронежской обл. (Огнев и Во-
робьев, 1924), между 10 и 15 июля — в Николаевской (Браунер, 1894); в начале
августа — в Чкаловской.
В первые дни жизни дрофята крайне слабы и беспомощны, напоминают
в этом отношении пуховичков домашней индейки. Обсохнув и покинув гнездо,
они неуверенно передвигаются медленными шагами и в течение 3—4 дней дер-
жатся вблизи от гнезда (Спангенберг, 1946). При опасности птенцы не пытаются
убегать, но затаиваются. Однако, едва пройдет испуг, дрофенок встает во весь
рост, вытягивает шейку и громким писком призывает матку. Дрофа — старка,
применяя различные уловки, отвлекающие врага, всеми силами пытается
отвести человека от места, где затаились пуховички. Отбежав в сторону с при-
поднятым хвостом и опущенными крыльями, она затем принимает обычный вид,
и как бы спокойно «прогуливается» или перелетает с места на место. В лите-
ратуре есть указания (Мензбир, 1900), что в некоторых случаях самка смело
бросается на человека, завладевшего ее птенцом. Птенцы приобретают способ-
164 отряд дрофы
ность к полету, достигнув веса около 2 кг, в возрасте немного более месяца.
Сравнительно долгое время каждая семья держится особняком и лишь после
того, как птенцы вполне окрепнут и научатся избегать опасность (в конце июля),
соседние семьи соединяются в общие стаи. С августа начинается «общественная»
жизнь дроф, продолжающаяся до весны следующего года.
Питание. Пища дроф весьма разнообразна: и животная, и раститель-
ная. В большом количестве дрофы поедают молодые побеги трав, семена разно-
образных диких злаков и земледельческих культур. В равной степени дрофы
питаются жуками и различными саранчевыми. Несравненно реже дрофы про-
глатывают лягушек, ящериц и мелких «мышевидных» грызунов. Известны слу-
чаи проглатывания птенцов воробьиных птиц. Кормовой режим меняется в за-
висимости от времени года. В общих чертах, весной и осенью дрофы главным
образом питаются растительной пищей, летом — животной.
В Бессарабии предпочитают растительную пищу, но охотно поедают саран-
чевых и ящериц (Остерман, 1912). В Харьковской обл. питаются семенами раз-
личных злаков, листьями степных кустарников и трав, кузнечиками и другими
насекомыми (Сомов, 1897). В желудках дроф, добытых 25 мая в Воронежской
обл., обнаружены растительные остатки и жуки-копры (Огнев и Воробьев, 1924).
У птиц, добытых в Волжско-уральских степях, среди другой пищи в желудках
найдены листья кермека и множество жуков-чернотелок (Волчанецкий, 1937).
Желудки дроф, добытых в летнее время в Наурзумском заповеднике, оказались
переполненными кобылками (Михеев, 1938). В степях Чкаловской обл. и
в соседних частях Казахстана весной птицы поедают молодые побеги трав и
жуков, а осенью вылетают на пашни, где кормятся зернами (Зарудный, 1888).
Весной дрофы так много поедают дикого чеснока, что мясо их пропитывается
чесночным запахом (Сушкин, 1908). В Туркмении пасущиеся дрофы охотно
держатся на зеленях по Атреку и в участках с вегетирующей растительностью —
мятликом, осочкой и т. д. (Дементьев, 1950).
Молодые дрофы питаются преимущественно животной пищей. По наблю-
дениям в Чкаловской обл. гнезда дрофы в большинстве случаев устраи-
ваются поблизости степных муравейников. После вывода птенцов самка пере-
водит их сюда и, раскапывая муравейник ногами, облегчает птенцам доступ
к насекомым. Подросшие дрофята во множестве поедают кобылок, саранчу,
богомолов и в меньшем количестве нежные молодые побеги трав. По другим
источникам летом молодые питаются почти исключительно кобылками, иногда
поедают ящериц; у одной молодой птицы были обнаружены в желудке два пу-
ховичка черного жаворонка.
Нередко дрофы заглатывают мелкие камешки.
Дрофы нуждаются в воде и регулярно посещают водопои — степные
роднички, окраины болотин. Там, где пресной воды нет, дрофы могут пить не-
сколько солоноватую воду (Зарудный, 1888). Однако наличие пресной воды
имеет для дроф большое значение, и в некоторых случаях может отражаться на
их распространении.
Пасутся дрофы в утренние и вечерние часы, а в жару залегают среди куку-
рузы, бурьяна, проса, бобовника, где, открыв рот, остаются на солнцепеке, не
пытаясь скрыться в тени. После отдыха летят к водопоям.
Линька. Изучена недостаточно, последовательность смены нарядов —
как у вихляя и стрепета. Две линьки в году — полная осенняя послебрачная
(смена первостепенных маховых у птиц в СССР начинается в разные числа
июля) и частичная весенняя предбрачная, охватывающая мелкое оперение.
Полевые признаки. Очень крупные размеры,толстая шея, большая голова и
пестрое оперение из сочетания рыжего и белого цветов, позволяют безошибочно
отличить дрофу от всех других птиц СССР. Обычно дрофы держатся неболь-
шими или более крупными стаями, изредка одиночками. Во время кормежки
широко разбредаются по степи. Пасущаяся стая всегда ведет себя крайне осто-
ДРОФА
165
рожно и улетает при первой опасности. Потревоженные птицы перед взлетом
пробегают метров 30 и, поднявшись против ветра невысоко над землей, летят
в прямом направлении. В жаркую погоду, заметив врага, предпочитают затаи-
ваться. Прячутся птицы — благодаря своей покровительственной окраске —
замечательно искусно. Близко подпустив к себе человека, они часто подни-
маются вертикально — ссвечой». Всякий подъем в воздух сопряжен с тяжелыми
взмахами крыльев, но, поднявшисьна известную высоту, дрофа летит уже сравни-
тельно легко и очень быстро (более 50 км в час). Во время полета стая дроф не
образует определенной фигуры, птицы летят вразброд, причем отдельные особи
Рис. 29. Птенцы дрофы
на различной высоте. Взмахи крыльев дроф относительно медленные и очень
глубокие. Предпочитают лететь в полветра или по ветру. Во время пастьбы
передвигаются медленными шагами, но в случае опасности могут бегать быстро.
Дрофа —молчаливая птица. Голос самца и самки различен; резко отли-
чается он и у птенцов. Крик самца негромкий, короткий и гортанный, передать
его крайне трудно. Подзывая птенцов, самка издает своеобразные короткие
звуки «тук». Маленькие птенцы издают громкий, протяжный писк; несколько
подросшие — неприятный стон или звуки с трелями, как бы издаваемые
дудкой, которые свойственны только раннему возрасту дрофят.
Описание. Размеры и строение. Очень крупная птица. Размеры и вес подвержены
сильной изменчивости, зависящей от пола, возраста и степени упитанности. Летний вес сам-
цов в районе Чкалова колеблется между 7,2—11,2 кг; редкие старые самцы достигают 16,8 кг.
Вес самок колеблется между 4 и 8 кг. Самец, добытый под Ак-булаком в середине мая,
весил 18 кг (Спангенберг). В исключительных случаях самцы достигают 21 кг (Бутур-
лин, 1935).
Телосложение тяжелое и мощное; клюв короткий и широкий; голова боль-пая, округ-
лая; шея толстая, средней длины; ноги сильные, довольно длинные, трехпалые; крылья
широкие, округлые, умеренной длины. Формула крыла: 5>3 = 4>1>2>б Вырезки
на наружных опахалах 2-го — 7-го, на внутренних опахалах 1-го — 5-го первостепенных
маховых. Хвост закругленный, из 20 рулевых.
Окраска. Пуховой птенец пестрый, на спинной стороне — грязнобелый, несколько
охристый фон испещрен темнобурыми пятнами и полосками; от лба к затылку, над глазом
и ухом идут темные, буроватые полоски; поперечные полоски — на задней стороне шеи и
166
ОТРЯД ДРОФЫ
продольные, прерывчатые—на спине и крыльях; брюшная сторона — бледноохристая; на
горле и передней части шеи— пятно темнобурого цвета.
Длина крыла самцов G) 610—700, самок A5) 490—660, в среднем соответственно 665,7
и 544,6 мм.
Молодые особи в первую осень отличаются от взрослых подробностями окраски. Верх-
няя часть головы у них в охристых темных пестринах; вдоль темени хорошо заметна темная
полоса; перья шеи и зоба с рыжими каймами.
Годовалые и двухлетние молодые похожи на самок, отличаясь от них, главным обра-
зом, наличием охристых пестрин на темени. Взрослые птицы и молодые на втором году жиз-
Рис. 30. Дрофа (самец и самка)
ни имеют: глаз темнобурый, клюв темносерый с черной вершиной, ноги зеленовато-бурые.
Окраска оперения самца весной пестрая, из комбинации рыжего, белого, черного и
светлосерого ЦЕетов. Белый цвет распространен на всем брюшке, подхвостье и частично на
крыльях и крайних рулевых хвоста.
Вся спинная сторона от задней части шеи и до хвоста — рыжевато-охристая с черным
поперечным струйчатым рисунком.
Большая часть головы и часть шеи—светлая дымчатосерая. На зашейке, зобу и частично
на передней стороне шеи — рыжий ошейник с редкими черными пестринами. По сторонам
горла— пучки длинных нитевидных перьев — так называемые «усы». Первостепенные и вто-
ростепенные маховые темнобурые. К концу лета самец теряет рыжий ошейник и шея и зоб
становятся серыми; исчезают также «усы».
Взрослая самка по окраске похожа на самца в осеннем наряде, отличаясь от него мень-
шими размерами, тонкой шеей и вообще более стройным сложением.
Восточно-сибирская дрофа Otis tarda dybowskii T а с z.
Otis dybowskii. Т а с z a n о w s k i. I. f. О. 1874, стр. 331, Даурия.
Распространение. Ареал. Манчжурия; Монголия; в СССР от Приморья
до северо-восточных частей Алтая.
Детали распространения у нас таковы:
Гнездится в Минусинском крае, под Минусинском, в Абаканской и север-
ных частях Сагайской степи, в Чуйской и Курайской степях юго-вост.
ДРОФА 167
Алтая, в предгорьях Саян и за Саянами, в Тувинской авт. обл. На восток
отсюда в степях южн. Сибири до Байкала и Буреинской степи. Далее в За-
байкалье распространение вида спорадично. Дрофы населяют среднее течение
Верхней Ангары (к северо-востоку от Байкала), Баргузинскую степь, южную
часть Витимского нагорья, к северу от озер Иван и Арахлей, степи в районе
Читы, нерчинские степи, долины рек Онон и Аргунь. Еще восточнее дрофа
гнездится в Зазейской равнине между Благовещенском и Малым Хинганом
и в степях у озера Ханка.
Возможен залет бродячих особей в западном направлении. Зимует в сте-
пях к югу от Ордоса и среднего Китая, в Корее, изредка проникает в сев. Ин-
дию. В СССР в небольшом числе встречается зимой в южн. Приморье и
южн. Сибири.
Вне пределов Союза — в сев.-зап. и вост. Монголии и в Манчжурии; ука-
зание на гнездование в Центр. Гоби (Бианки, 1915) — сомнительно.
Характер пребывания. Гнездящаяся, перелетная и отчасти
зимующая птица Союза ССР. Вероятно в самом ограниченном числе зимует
в окрестностях Сидеми, перекочевывая сюда с долины Сунгари и среднего
Амура. Отдельные случаи зимовок известны в Абаканской степи и более
регулярное перезимовывание части особей в Тувинской обл. На остальной
территории Союза дрофа перелетна. Сроки весеннего перелета выяснены крайне
слабо. На реке Чикой в окрестностях Троицкосавска прилет дроф отмечен
2 марта (Моллесон, 1897). Под Нерчинском в различные годы первые дрофы
отмечались весной 12 и 25 марта (Пуляевский, 1937). В Ольгинском районе
Приморья самец добыт 20 марта (Шульпин, 1936) и, вероятно, уже местная
птица добыта у Ханки 24 апреля (Бутурлин, 1909).
О сроках осеннего отлета мы почти ничего не знаем. Вероятно птицы
задерживаются у гнездовий до выпадения снега.
Биотоп. В отличие от европейского подвида, восточно-сибирская дрофа
охотно населяет степные пространства горных территорий, поднимаясь по вер-
тикали до 2000 м (Сушкин, 1938). Видимо птице безразлично, будет ли это
ровная степь или холмистый открытый ландшафт, изрезанный горными доли-
нами и балками и языками тайги по речкам. В Тувинской обл. дрофы не избе-
гают близости деревьев и населяют сухие поречные луга близ лесных опушек и
даже луга, вдающиеся в урему (Сушкин, 1914). В Чуйской равнине дрофы при-
держиваются глинистых степей с богатым растительным покровом. Однако
дрофы избегают как узких речных долин, высокогорий, где степь заменяется
альпийскими лугами или тундрой, так и более низких территорий, где степь
приобретает пустынный характер. В Приморье, в районе с. Красного, дрофы
иногда гнездятся по полям, а вдоль Тумень-Улы — по песчаным дюнам (Шуль-
пин, 1936). Места кормежки вероятно совпадают с местами гнездовья. В боль-
шей части своего ареала восточно-сибирская дрофа обитает на целине и лишь
местами мирится с близостью человека, гнездясь на культурных землях.
Численность. Редка под Минусинском; очень обыкновенна в Абакан-
ской степи; нередка в Сагайской степи и многочисленна в Тувинской обл. Чис-
ленность птиц, однако, подвержена сильным колебаниям в различные годы. В об-
ласти юго-вост. Алтая — редка в Курайской степи и довольно многочисленна
в Чуйской. Стала очень редка за последнее время в южной половине Приморья.
Экология. Размножение. Сведения о размножении ограничи-
ваются датами нахождения и добычи птенцов. В Чуйской степи дрофята
ростом с тетерева добыты 3 июля (Сушкин, 1938); в Тувинской авт. обл.
13 августа птенцы дрофы достигли 8/4 роста взрослой самки (Сушкин, 1938);
в Минусинском крае птенцы, видимо, появляются около середины июля
(Сушкин, 1914). В окрестностях Турьего рога в Ханкайской низменности
в найденном 24 августа выводке дрофята достигли роста тетерева-косача
(Пржевальский, 1870).
168 отряд дрофы
Линька. У взрослых особей наступает с очень большими колебаниями
в сроках: одни оказываются значительно облинявшими в середине июня,
другие начинают линять на целый месяц позднее. Две взрослых самки, добы-
тые 25 и 19 июля, начали линять. Старые самцы к концу сентября вполне
обновляют перо; перья «усов» к этому времени достигают больших размеров,
чем у О. /. tarda в брачном наряде. Полного размера эти перья достигают будто
бы к середине зимы.
У молодых к осени все перья гнездового наряда сменяются. Махи и рули
сменяются приблизительно в годовом возрасте. У некоторых годовалых самцов
уже появляются брачные, украшающие перья, «усы» и «борода».
Питание. Питание вероятно имеет много общего с питанием обык-
новенной дрофы. В желудках молодых особей, добытых летом в Тувинской обл.,
обнаружены саранчевые (Нестеров, 1909).
Описание. Размеры и строение. Общие размеры, повидимому, меньше,
чем у обыкновенной дрофы, но отличий в длине крыла нет. Слабо и не всегда отличима
более низким клювом. Длина крыла: самцов — 585; самок — 470—490 мм (Бутур-
лин, 1935).
Окраска. Взрослый самец отличается от самца номинальной формы следующими
особенностями: мелкие верхние кроющие крыла на сгибе серые, а не пестрые. Пять (а быть
может и больше) пар крайних рулевых белые с темной предвершинной полосой. Рыжие
пятна вблизи вершины появляются на внутренних рулевых, начиная с 6-й, а иногда с 8-й
пары. Рисунок верха более грубый вследствие того, что темные поперечные полосы шире.
Длинные нитевидные перья растут не только на боках горла, но в виде «бороды» на всем
горле. Взрослые самки отличаются от взрослых самок номинальной формы (за исключе-
нием нитевидных перьев на горле) теми же признаками, что и самцы. Молодые птицы
обеих форм, вероятно, неотличимы.
ОТРЯД ЧИСТИКИ
ALCAE или ALCIFORMES
Характеристика отряда
Морфология. Внешние признаки. Птица средней или даже мел-
кой величины. Мелкие виды при длине около 20 и размахе около 35 см весят
около 100 г (конюги); крупные имеют несколько более 40 см в длину при раз-
махе примерно в 70 см и при весе около 700—800 г и даже более килограмма
(кайры). Водные (морские) птицы типичного «ныркового» сложения с плотным
веретенообразным туловищем, короткими четырехпалыми далеко отнесен-
ными назад ногами, пальцы которых соединены перепонками; клюв умеренной
длины или короткий, обычно сжатый с боков, разнообразной формы, у многих
видов в брачном наряде со своеобразными выростами, пластинками и другими
«украшениями». Ноздри щелевидные или округлые, защищенные от воды
кожистым клапаном. Оперение короткое и жесткое, расстояния между бород-
ками небольшие, аптерии узкие; число перьев очень велико: у толстоклювой
кайры Кафтановским подсчитано 13821 перо, у тонкоклювой —12903,
у гагарки—11576; у тупика — 9073, у североатлантического чистика—8677;
плотность оперения примерно в 2 раза больше, чем у чаек. Оперение плотно при-
легает к телу и окрашено неярко: или в темные бурые, или серые, или черно-
ватые цвета, или, если оперение двухцветное, то спинная сторона темная,
а брюшная — светлая. Резко выраженный сезонный и возрастной диморфизм,
но окраска самцов и самок одинаковая. Цевка неоперенная. Крылья узкие
и короткие, хвост короткий. Число первостепенных маховых — 11 (первое
рудиментарно); внутренние (задние) второстепенные маховые укорочены;
рулевых 6—8 пар (у одного вида даже 9).
Анатомические признаки. Череп схизогнатический. Бази-
птеригоидные отростки у взрослых отсутствуют. Клюв схизоринального типа,
ноздри сквозные. Нередко имеются затылочные фонтанели. В хорошо выражен-
ных надглазничных ямках помещаются значительно развитые носовые железы.
Шейных позвонков 15; сращения грудных позвонков в спинную кость не
имеется; спинные позвонки опистоцельного типа. Ребер 7—8 пар. Грудина
длинная, узкая, с одной парой вырезок. Ключица с hypocleidium — самый
170 ОТРЯД ЧИСТИКИ
короткий отдел крыла (предплечья). Скелет крыла уплощенный. Скелет птицы
слабо пневматичен (относительный вес его около 7—8%). Таз узкий. Обводя-
щая мышца отсутствует. Перья с побочным стволом. Птерилия и аптерии
покрыты пухом. У планктоноядных форм (роды Aethia и Plautus) имеется
особый горловый (подъязычный) мешок — резервуар для приносимой птен-
цам пищи (мелкие ракообразные). Язык мясистый, вытянутый, иногда с шипи-
ками в основной части. Слепые кишки рудиментарны. Железистый желудок
развит хорошо, мускульный слабо (белковое питание); кишечник относи-
тельно длинный.
Образ жизни. Водоплавающие птицы, главным образом дневные (но неко-
торые виды активны и ночью), морские; с сушей связаны только в период
гнездования, кормятся на море, где проводят и весь негнездовый период.
Пища у крупных видов — преимущественно мелкая пелагическая рыба,
у мелких —морские беспозвоночные (ракообразные, полихеты и т. д.). Добы-
вается пища путем ныряния и плавания под водой (при этом птицы, наподобие
пингвинов, пользуются и крыльями). Плавая под водой, чистики держат
крылья в полусогнутом положении, делая ими короткие взмахи вниз и назад
(до 15—20 взмахов в минуту, Кафтановский). Перемены направления движе-
ния под водой производятся при помощи лапы. Средняя продолжительность
пребывания под водой крупных видов 30—40 сек. и до 1 минуты.
Добыча захватывается клювом, имеющим весьма разнообразную форму:
у чистиков он почти прямой и заостренный, у гагарок — высокий и сжатый
с боков (быть может в связи с рытьем нор), у конюг нижняя челюсть имеет
форму ложки (питание планктоном). Крючок на конце клюва у тупиков и
гагарки, повидимому, облегчает им ловлю рыбы, так как эти птицы —в отличие
от кайр — приносят к гнезду по 3—4 рыбки в каждом углу рта, зажимая их
плотно между разрезом клюва и языком. Мышцы, закрывающие клюв, очень
развиты, в связи с чем в затылочной области (в особенности на ossupraocci pi-
tale, где прикрепляются височные мышцы и мускул, закрывающий клюв
m. depressor mandibulae) имеются сильно развитые гребни (однако у тупиков
в связи с особенностями строения их клюва эти гребни едва ли видны). Сильно
развиты и мускулы, поворачивающие голову, что отражается на большой
величине остистых отростков первых трех шейных позвонков.
О глоточных мешках, имеющихся у мелких видов, кормящихся планкти-
ческими ракообразными, уже упоминалось (Портенко, 1934, 1948).
Полет чистиковых напоминает в известной мере движение по воздуху
нырковых уток и поганок. Весовая нагрузка крыла очень значительна —
примерно в 3 раза больше, чем у чаек (напр., у тонкоклювой кайры 2,60 г/см2;
у толстоклювой кайры —2,48; у гагарки —2,27; у тупика —2,27; у серебристой
чайки—0,76; у большой морской чайки—0,70; у моевки—0,58; но у чернозобой
гагары—2,08;* у краснозобой гагары—1,%г/см2, по Кафтановскому) *. Высокая
весовая нагрузка требует большой частоты взмахов крыльев (по киносъемке
в заповеднике «Семь Островов» на вост. Мурмане у кайр не менее 8 пол-
ных взмахов в секунду). Скорость полета весьма велика. Рулем служит не
рудиментарный хвост, а большие перепончатые расправленные лапы. Поворот-
ливость слабая, планирование и скольжение, повидимому, вовсе отсутствуют.
Полетом чистиковые пользуются ограниченно, лишь для перемещения с мест
отдыха и гнездовий на место кормежки. Даже сезонные миграции совершаются,
повидимому, главным образом вплавь.
Взлет с ровной поверхности суши для чистиков по всей вероятности невоз-
можен — слетают они с обрывов или хотя бы с наклонной поверхности или
¦ У вымершей недавно (около середины прошлого столетия) исландской бескрылой
гагарки Pinguinus impennis при весе около 4 кг весовая нагрузка составляла примерно
4 г/см1 — полет был уже невозможным.
ХАРАКТЕРИСТИКА ОТРЯДА 171
с воды, но на последней —после долгого разбега A0—12 м), с частым хло-
паньем крыльев.
Эти особенности полета (и плавания) хорошо выражены в структуре
двигательного аппарата. Крыло острое, но небольшое и короткое (особенно
по сравнению с чайками, укорочены предплечье и маховые перья); прокси-
мальная часть крыла уплощена; лучевые кости смещены вперед и наверх и
расположены впереди от локтевых (это — приспособление к «полету» под
водой, уменьшение встречного сопротивления воды); грудная мускулатура
как опускающая, так и поднимающая крыло — m. pectoralis major, m. sup-
racoracoideus, т. latissimus dorsi, m. deltoideus —развита очень сильно в связи
с тем, что при полете и при плавании под водой подъем крыла происходит
активно; грудина очень длинная, ключица прочная и выгнутая вперед, ребра
и реберная мускулатура сильно развита (энергичные изменения объема брюш-
ного воздушного мешка, т. е. удельного веса).
Передвижение чистика по суше крайне затруднено, хотя способность
к ходьбе у разных видов различная. Пальцы ног, как правило, длинные;
цевка — короткая. Лучше всего ходят по земле тупики и, повидимому, конюги
(у которых и цевки относительно длинней). Эти виды могут даже бегать. При
ходьбе тупики, подобно большинству других видов птиц, опираются на пальцы.
Возможно, что хорошее развитие лап у тупиков связано с рытьем ими нор для
гнезд. Другие виды чистиковых ходят хуже и обычно опираются при ходьбе
на цевку. При хождении тело занимает вертикальное положение — у кайры,
гагарки; полугоризонтальное—у тупика, горизонтальное—у чистика.
Обмен веществ у чистиковых интенсивный, и скорость пищеварения весьма
велика. Относительные размеры сердца высокие (но меньше, чем у чаек);
это связано, вероятно, с климатом (правило Гессе): у тонкоклювой кайры сер-
дечный индекс (относительный вес сердца в %0) равен 9,69, у толстоклювой —
8,53, у гагарки—11,74, у североатлантического чистика—10,49 (Кафтановский).
Объем воздушных мешков значительный, относительная величина его (отно-
шение к общему объему или общему весу тела) почти в 2 раза выше, чем у чаек,
более или менее приближаясь к единице (отношение объема мешков к весу
тела у тонкоклювой кайры 1,1 : 1 и 1,2 : 1; у толстоклювой кайры—1,1 : 1
и 1,2 : 1; у гагарки — 1 : 1 и 1,1 :1,у тупика — 1,1 : 1 и 1,2 : 1; у североат-
лантического чистика—1 :1,3 и 1 :1,2; у большой морской чайки—1 : 2,7,
у серебристой чайки—1 : 1,9, у моевки—1 : 1,6 или 1 : 1,5, по Слепцову).
Особенно развиты брюшные мешки у кайр C00—400 см3, по Слепцову). Зна-
чительные размеры мешков служат несомненно приспособлением к нырянию.
Удельный вес этих птиц высок и характерен для ныряющих птиц (у тонкоклю-
вой кайры 0,74, у толстоклювой—0,70, у гагарки — 0,76, у тупика — 0,72, у
североатлантического чистика—0,75), причем в результате подвижности гру-
дины объем может изменяться в значительной степени в пределах до У8. Ныря-
ющий чистик уменьшает свой удельный вес, сокращая объем тела и повышая
давление внутри тела (но не выдыхая воздух). Относительный вес слабо пнев-
матизированного скелета невысок: 6,58% —у тонкоклювой кайры, 7,12% —
у толстоклювой, 7,31% — у гагарки, 8,03% — у североатлантического чистика
(по Кафтановскому).
Чистиковые —моногамы. Вопрос о постоянстве пар неясен, но в пользу
этого говорит крайний консерватизм гнездовых местообитаний (результаты,
в частности, кольцевания на «Семи Островах», вост. Мурман). Потенциальная
половозрелость наступает, вероятно, в годовалом возрасте, но реализация ее
(возможно по причине отсутствия партнера или места для гнездования) начи-
нается обычно в возрасте около 2 лет, по крайней мере у крупных видов. На
это указывает и имеющийся летом «резерв» холостых годовалых особей у гнездо-
вых колоний. Гнездовья — колониальные (так называемые «базары»), раз-
меры колоний весьма различны: иногда в колонии объединены десятки тысяч
172 отряд чистики
птиц, а иногда лишь несколько пар. Некоторые виды (тупики) как правило
гнездятся небольшими группами; у других колониальность выражена очень
резко (люрики, кайры). Гнезда эти птицы собственно и не строят. Одни виды
гнездятся открыто на карнизах береговых обрывистых скал или —• реже —
на плоских вершинах островов (кайры, гагарки); другие —в щелях между
камнями и в других естественных убежищах (чистики, конюги); третьи — в вы-
капываемых ими норах (тупики). Подстилки не бывает, или бывает только
незначительная подстилка из сухих травинок. Цикл размножения поздний,
так как связан с началом таяния льдов и снега умеет гнездовья. Сильно выра-
женную, несмотря на колониальность, привязанность к месту гнездования
обнаружили окольцованные экземпляры кайры, которые на Семи островах
добывались по прошествии многих лет на тех же карнизах, где вывелись.
У крупных видов кладка состоит только из одного яйца, у некоторых мел-
ких — из двух. Величина яиц весьма крупна (у толстоклювой кайры — около
11,2% у тонкоклювой— 11,2%, у тупика—11,1%—14,2% веса взрослой птицы,
по Кафтановскому и Горчаковской). Форма яиц варьирует от своеобразно
грушевидной у открыто гнездящихся на карнизах видов (это, вероятно, за-
трудняет скатывание яйца при ветре, толчках и т. п.) до обычной овальной
у гнездящихся в норах тупиков. Различна и пористость скорлупы, и ее толщина
значительна у открыто гнездящихся видов). В случае утраты яйца чисти-
ковые откладывают второе и даже третье — на этом основано промысло-
вое использование их яиц. Насиживают оба родителя, крупные виды (кайры,
гагарки) около 35 дней, мелкие (люрики) около 24—25 дней. У закрыто гнездя-
щихся видов насиживание (как и постэмбриональное развитие) весьма дли-
тельно : 27—28 дней — у атлантического чистика, 35 дней— у тупика. Различно,
в зависимости от открытого или закрытого гнездования, и развитие птенцов.
Впрочем, у всех видов развитие птенцов протекает по типу, промежуточному
между выводковым и птенцовым, так сказать толу выводковому». Но у открыто
гнездящихся видов опушение птенца своеобразно, жестко, довольно плотно
сцепленное и хорошо защищает птенца от холода и ветра (кайры, гагарки),
у закрыто гнездящихся в общем сходно с пухом у чаек — мягкое и ветви-
стое. Закрыто гнездящиеся птицы долго находятся в гнезде и спускаются на
воду вполне развитыми, с выросшими маховыми, в возрасте 35—38 дней (атлан-
тический чистик) или 36—40 дней (тупики). Птенцы открыто гнездящихся
видов спускаются на воду, далеко не достигнув полного роста, с недоразвитыми
маховыми —при планировании со скал их функционально заменяют большие
кроющие, отчасти лапки; возраст птенцов при спуске — около 25 дней (кайры)
и даже 19—20 дней (гагарки). У птенцов последней группы развивается свое-
образный наряд—мезоптил ь.
Линька у взрослых проходит два раза в год. Неполная предбрачная линька
бывает в конце зимы — весною, она охватывает мелкое оперение, частично
(голова и шея) или полностью, но не маховые и рулевые. Осенью имеет место
полная послебрачная линька, когда сменяется все оперение. Маховые выпа-
дают почти одновременно, и птица, держась на воде, проходит «нелетную»
стадию. У многих видов при предбрачной линьке развиваются украшающие
перья на голове и различного рода выросты и пластинки на клюве (особенно у
тупиков и конюг). Половой диморфизм выражается в несколько больших (но не-
значительно) размерах самцов. Молодые птицы по окраске сходны со взрослыми
в э«шем наряде. В первом брачном наряде имеются незначительные отли-
чия в окраске от старых птиц, и часто клюв еще не достигает полных размеров.
Изменчивость окраски может быть выражена формулой: половой моно-
морфизм, возрастной диморфизм, сезонный диморфизм, географический поли-
морфизм (последний выражен, впрочем, нерезко, и некоторые популяции разли-
чаются толы» размерами). У некоторых видов имеется и групповой диморфизм,
«очковая» вариация у тонкоклювой кайры, темнокрылая вариация—у чистика.
ХАРАКТЕРИСТИКА ОТРЯДА 173
При изолированности расположения гнездовых колоний и консерватив-
ности местообитаний у некоторых видов чистиков довольно значительна, так
сказать, «популяционная» изменчивость (в размерах, окраске), послужившая
основанием для выделения ряда подвидов, которые, однако, едва ли могут
считаться равнозначащими географическим формам в обычном понимании
этого термина. Подробнее об этом ниже, в описании отдельных видов.
Систематика. Чистики образуют единственное семейство Alcidae, распа-
дающееся на роды: Alca A вид), Plautus{\ вид), Uria B.вида), Cepphus B вида),
Brachyramphus B вида), Ptychoramphus A вид), Endomichura B вида), Synthli-
boramphus B вида), Aethia C вида), Cyclorrhynchus A вид), Cerorhinca A вид),
Fratercala B вида), Lunda A вид). Из этих 20 видов в фауне СССР представлены
18, причем 16 из них гнездятся, и все роды, кроме калифорнийских Endomi-
chura.
Один вид — бескрылая гагарка Pinguinus impennis—вымер около 1844 г.
(в Исландии). Ископаемые чистики известны с эоцена. Ряд современных видов
найден в плейстоцене.
В систематическом отношении чистики стоят ближе всего к чайкам и от-
части к куликам.
Географическое распространение. Чистики распространены в северных
широтах, в гнездовое время — по берегам материков и островов, в негнездовое
время — в море, преимущественно в прибрежных его частях, но иногда (в особен-
ности молодые особи) встречаются и в открытом море. Связь чистиков с сушей
ограничивается только потребностью в подходящих местах для гнездования.
Специфические условия последних определяют и неравномерное распростра-
нение чистиков. Что касается негнездового времени, то тут распространение
чистиков в значительной мере определяется гидрологическими условиями
(в частности ледовым режимом; поэтому, например, чистики не встречаются
зимой в Охотском море, но держатся у севера Новой Земли), а также распро-
странением планктических организмов.
Два вида чистиков — обыкновенный чистик и толстоклювая кайра —
имеют почти кругополярное распространение, с некоторым перерывом на мери-
дианах Северной Земли в Азии и в области моря Бофорта и к востоку от него
в Америке. Тонкоклювая кайра имеет типичный амфибореальный ареал.
Остальные виды характерны либо для севера Атлантического, либо для севера
Тихого океана (большинство). Для первого эндемичны гагарка, люрик (он,
впрочем, отмечен и на Северной Земле), обыкновенный тупик. В Атлантике
гагарка, тупик и тонкоклювая кайра образуют южную группу, бореальную,
а толстоклювая кайра и люрик — северную, арктическую. Чистик распадается
на две группы подвидов — арктическую (mandtii и близкие формы) и бореаль-
ную (grylle в узком смысле).
В Тихом океане имеется ряд северных видов, характерных для Берин-
гова моря. Это крошечная конюга, пыжик Brachyramphus brevirostris, чистики
подвидов columba и kajurka. Другая группа распространена и в Беринговом
море, и южнее — ипатка, топорбк, тупик-носорог, белобрюшка, большая
конюга, старик, наконец обе кайры —тонкоклювая и толстоклювая. Из этих
групп видов большинство распространено несколько шире у азиатских бере-
гов, чем у американских. Только тупик-носорог и топорбк имеют по обоим
побережьям Тихого океана примерно равное амфипацифическое распростра-
нение.
К южной группе тихоокеанских чистиков, отсутствующих в Беринговом
море или слегка в него проникающих, относятся малая конюга, хохлатый
старик, пыжик Brachyramphus marmoratus, очковый чистик, курильский чистик
Cepphus grylle snowi, алеутский пыжик Ptychoramphus aleuticus, калифорний-
ские пыжики Endomichura hypoleuca, E. craveri, наконец, эндемичная для Кали-
форнии раса кайры Uriaaalge calif or nica. Из этой группы к Азии тяготеют ма-
174 отряд чистики
лая конюга, хохлатый старик, очковый чистик, курильский чистик и подвид
пыжика. В. /л. perdix, к Америке — P. aleuticus, В. т. marmoratus, Endo-
michura и калифорнийская кайра. Таким образом, на Тихом океане фауны
чистиковых обоих побережий к югу от Берингова моря заметно различны,
тогда как в Атлантике эти фауны сходны и в Европе и в Америке. Характерно,
что типичные пацифические формы не проникают или почти не проникают
в Северный Ледовитый океан, занятый обедненной атлантической фауной
чистиковых.
Приведенный анализ показывает, что центром возникновения чистиков
следует считать северные части Тихого океана (в пользу этого говорят и палеон-
тологические данные, так как ископаемые чистики в эоцене известны с берегов
Пацифики, в Атлантике же только с миоцена). Можно далее предполагать,
что в Атлантику чистики, представленные там хорошо выделившимися энде-
миками, проникли уже с миоцена. Полярный же бассейн был заселен атлан-
тической группой чистиков сравнительно недавно, повидимому, после четвер-
тичного оледенения (Кафтановский).
Надо еще отметить, что современное распространение чистиков, как гово-
рилось, определяется наличием удобных мест для гнездования (скалистые бе-
рега) и гидрологическими условиями моря, главным образом ледовыми, а также
продуктивностью. Наибольшая численность чистиковых отмечена вблизи от
линии так называемого полярного фронта, что особенно хорошо выражено
в Атлантическом океане.
Практическое значение. Громадные гнездовые колонии чистиковых из-
давна использовались населением севера для сбора яиц и добычи самих птиц
на мясо. В результате нерационального промысла местами количество чистиков
в XIX —начале XX столетия сильно сократилось, а нелетающий крупный
вид — бескрылая гагарка — подвергся полному истреблению. Это вызвало
необходимость создания охраны чистиков. В СССР для этой цели созданы запо-
ведники в Лапландии на Семи островах и на Айновских о-вах, а также на
западном берегу Новой Земли. При правильном промысле значение чистиков
в качестве источника питания для местного населения Севера может быть
значительным (см. в частности данные у Насимовича, 1934, Портенко, 1931,
Горбунова, 1925).
Таблица для определения видов чистиковых
1 B). Высота клюва менее 25 мм; граница оперения надклювья лежит обычно впе-
реди угла рта 7
2 A). Высота клюва более 25 мм; граница расположения оперения надклювья иная . .
• 3
3 D). Грудь и брюхо темнобурые (взрослые) или беловатые с темными вершинами
перьев; на подклювье поперечных полос нет (они имеются на надклювье)
топорик Lund a cirrhata
4 C). Грудь и брюхо белые; поперечные борозды имеются не только на надклювье,
но и на подклювье 5
5 F). Горло серое, темя черно-бурое тупик Fratercula arctica
6 E). Горло черное, темя дымчато-серое ипатка Fratercula corniculata
7 (8). Клюв длинней 25 мм 9
8 G). Клюв короче 25 мм 19
9 A0). Ноги красные или оранжевые И
10 (9). Ноги черноватые, сероватые или желтоватые 13
11 A2). На крыле нет белого зеркальца; у глаза белое пятно; подмышечные темно-
буры очковый чистик Cepphus carbo
12 A1). На крыле белое зеркальце; у глаза белого пятна нет; подмышечные белые
или сероватые обыкновенный чистик Cepphusgrylle
13 A4). На вершинах второстепенных маховых белая полоска; подкрылья белые,
с большей или меньшей примесью серо-бурых пестрин 15
14 A3). Вершины второстепенных маховых без белой полосы; подкрылья темные . •
тупик-носорог Cerorhinca monocerata
ГАГАРКА 175
15 Х16). Клюв высокий, с круто загнутым коньком надклювья; у взрослых на надклю-
вье поперечная белая борозда и 1—2 темных поперечных вдавления; надклювье (от опере-
ния лба) короче 30 мм гагарка Alca tor da
16 A5). Клюв ниже, без поперечных борозд; надклювье слабо изогнуто, длина его
от лба более 30 мм 17
17 A8). Клюв короче и выше; основная часть надклювья у разреза рта вздута и от-
деляется светлой окраской от остального (черного) надклювья; темя аспидно-черное или
серовато-черное толстоклюваякайра Urialomvia
18 A7). Клюв длиннее и ниже; основная часть надклювья у рта не вздута и не отли-
чается по цвету от остальных его частей; темя кофейно-бурое
тонкоклювая кайра Uria aalge
19 B0). Передняя сторона цевки покрыта поперечными пластинками 21
20 A9). Цевка вся — и с передней стороны — сетчатая 25
21 B2). Клюв черный, подкрылья темные люрик Plautus alle
22 B1). Клюв светлый, подкрылья белые 23
23 B4). Клюв длинней 16 мм; в брачном наряде на темени узкий, длинный, обращен-
ный назад, хохол хохлатый старик Synthliboramphuswumizusume
24 B3). Клюв короче 16 мм; хохла нет старик Synlhliboramphus antiquus
25 B6). Ноздри круглые; оперение лба достигает их заднего края 27
26 B5). Ноздри узкие щелевидные; оперение лба до ноздрей не доходит 29
27 B8). Клюв короче 15 мм; боковые рулевые белые
серый пыжик Brachyramphus brevirostris
28 B7). Клюв длинней 15 мм; боковые рулевые темнобурые
пестрый пыжик Brachyramphus marmoratus
29 C0). Клюв относительно низкий, высота его менее */з длины от оперения лба;
красного цвета на клюве нет алеутский пыжик Ptychoramphusaleuticus
30 B9). Клюв относительно высокий, высота его больше */з длины от оперения лба;
на клюве обычно примесь красного 31
31 C2). Крыло длинней 120 мм 33
32 C1). Крыло короче 120 мм 35
33 C4). Брюшная сторона темная .... большая конюга Aethia cristatella
34 C3). Брюшная сторона (грудь, брюхо, подхвостье) белые
белобрюшка Cyclorrhynchus psittacula
35 C6). Крыло длинней 100 мм малаяконюга Aethia pygmaea
36 C5). Крыло короче 100 мм конюга-крошка AethiapusiНа
РОД ГАГАРКИ ALCA LINNAEUS, 1758
Тип Alca tor da
1. Гагарка Alca tor da L.
Alca torde. Linnaeus. Syst. Nat. ed. X, t. 1. 1758, о. Стуре Карлсб, южная часть
Балтийского моря.
Русское название. На русском севере народное название гагарка приме-
няется и к кайрам. У Озерецковского A801) эта птица называется тупик, у Палласа A811) —
чистик. Этимология слова гагарка, повидимому, звукоподражательная. В применении к
этому виду термин носит книжный характер и встречается первый раз у Мензбира A895).
Распространение. Ареал. Северные части Атлантического океана —от
южн. Гренландии (Упернивик около 74° с. ш., Годтхоб, Фискенес, следова-
тельно на западе; на восточном побережье отсутствует), Исландия, о-в Мед-
вежий; восточные части Сев. Америки (Нью-Брунсвик, залив св. Лаврентия,
Нью-Фаундленд, Лабрадор, еще недавно на берегах штата Мен); на Фарер-
ских о-вах, в Англии, Шотландии и Ирландии, на побережье Бретани во
Франции, на о-ве Гельголанд, в Скандинавии (в Норвегии до Ставангера, на
юге в Швеции от северных частей Ботнического залива до Кельскеррен, в южн.
Сермланде, около 58° 40' с. ш.; недавно еще в юго-восточных частях провинции
Эстерготланд, около 58° 15' с. ш.; отдельные колонии у Карлсе 57° 15'; в Дании
у Христиансе, 55° с. ш.) по берегам Финского залива в Финляндии; в СССР
на Ладожском озере, на Мурмане (до Семи островов на востоке) и на Белом
море. В зимнее время на море: от Лабрадора до о-ва Лонг в Америке,
в Европе — у западных берегов (Голландия, Франция, Португалия) и до Среди-
земного моря (Тунис, Италия), в Северном и Балтийском морях. Случайные
176
ОТРЯД ЧИСТИКИ
залеты отмечены в Америке до местностей к западу от озера Онтарио, до Сев.
Каролины, в Атлантике у Азорских и Канарских о-вов; на севере ксев.-вост.
Гренландии (Маллемукфьелдет, 81° 12'), на Шпицберген и Новую Землю.
Характер пребывания. В негнездовое время, как указывалось,
кочует в море как вблизи от гнездовой области, так и южней.
Биотоп. В период размножения морские побережья — материковые
и островные. В остальное время ведет пелагический образ жизни.
Карта 26. Распространение гагарки Alca torda
1. A. U torda, 2. A. t. islandica;
а — граница гнездовий, г — область кочевок и зимовок.
Подвиды и варьирующие признаки. Обособленные географически и экологически
популяции гагарок образуют местные группировки с незначительными морфологическими
особенностями (размеры, в частности, клюва и крыла; развитие бороздок рамфотеки надклю-
вья), которым некоторые авторы (Саломонсен, 1944) придают значение диагностических при-
знаков подвидов. С теоретической точки зрения это весьма спорно. Если все же принять
такую точку зрения, то можно считать, что гагарки распадаются на два подвида: A. t. islan-
dica В г u n n i с h, 1764 — распространенная в Исландии, на Фарерских о-вах, в Вели-
кобритании и Ирландии, в Бретани, на Гельголанде (мелкая форма); А. /. torda L., 1758 —
в Прибалтике, на Ладожском озере, у побережий Норвегии, на Мурмане, Белом море, в Грен-
ландии и Канаде (крупная форма).
Северная гагарка Alca torda torda L.
Синоним. Alca pica. Linnaeus. Syst. Nat. ed. XII, 1766, стр. 210, Норвегия.
Распространение. Ареал. Занимает северо-восточную и восточную часть
ареала вида. В Америке от залива св. Лаврентия и Ньюфаундленда, по берегам
Лабрадора, в зап. Гренландии (см. выше); в Норвегии кюгуотСтавангера, но
более или менее многочисленна к югу до Бергена; о-в Медвежий (Кениг
и ле-Руа, 1912, Бертрам и Лек, 1933), в Дании на архипелаге Христансе, в Шве-
ции на о-вах Карлсе и у южн. Сермланда и далее до северной части Бот-
нического залива; в Финляндии по северному побережью Финского залива,
до Аландских о-вов; в сев.-зап. части Ладожского озера (Хильден, 1921);
у Мурманского побережья и на Белом море — по западному берегу от Онеж-
ского залива до Кандалакши; на Соловецких о-вах.
В негнездовое время у южн. Балтики (в частности Эстонии) и берегов
Норвегии. Немногие остаются у мест гнездовья (зап. Мурман). Окольцованные
на Семи островах (вост. Мурман) молодые птицы добывались зимой в южн.
ГАГАРКА 177
Норвегии, в Швеции и в Померании на о-ве Узедом (Дементьев, 1947).
Залеты отмечены у Шпицбергена (Мате—Дюпра, 1913), у Новой Земли (о. Ба-
зарный близ Малых Кармакул, Горбунов, 1924), на Канарские о-ва и к
Марокко, на Мальту, в Адриатическое и Ионическое моря, в Чехословакию,
Центральную Германию, Швейцарию, Египет и на Азорские о-ва. Изве-
стны две летних находки в Эстонии: на о-ве Эзель (Саремаа) 8.VI 1.1925 г.
и на северном побережье 24.VI. 1936 г. (Кумари).
Характер пребывания. Как и другие чистики, кочующая
птица, зимой держащаяся в море. Даты. Прилет на Мурман отмечен в конце
апреля, в стаях, в зависимости от ледовых условий. Отлет взрослых происходит
в конце августа, молодых — в начале сентября (Кафтановский, Горчаковская).
Отдельные особи на Мурмане у мест гнездовья задерживаются до середины
сентября.
Биотоп. В гнездовое время — прибрежные скалы как на материке,
так и на островах (чаще). В негнездовое время — в море, в латеральной поло-
се, быть может и в открытом (встречена например в 150 км от Ньюфаунд-
ленда).
Численность. Спорадична, на юге ареала редка. У Мурмана (Семь
островов—Харлов, Кувшин) многочисленна, на Белом море редка. Резких
колебаний численности не установлено (низкая плодовитость).
Экология. Размножение. Изучена главным образом по наблюде-
ниям на Семи островах (вост. Мурман Кафтановский, Горчаковская, Карта-
шев). Занятие гнезд происходит во второй половине мая. Гнезда встречаются
поодиночке или небольшими группами B—7), среди колоний кайр и моевок,
чаще в нишах или полуоткрытых местах, реже открыто. Иногда в гнездах
бывает скудная подстилка из сухой травы. Гнездовый консерватизм резко
выражен, одно гнездо из года в год занимается теми же птицами. Массовое
спаривание происходит в конце мая. Половая зрелость наступает, повидимому,
в возрасте около двух лет. Численное соотношение полов 1 : 1 (точней 5 сам-
цов на 4 самки). Кладка состоит из одного яйца. Происходит она в последних
числах мая или в начале июня B6 мая — 4 июня), иногда около середины
июня; дней на десять позднее, чем у кайр (вероятно, в связи с более поздним
исчезновением снега в укрытых гнездах гагарок). Заслуживает внимания то
обстоятельство, что на юге (Англия) кладка у гагарок протекает почти на
месяц раньше, в первой трети мая. В Норвегии около середины мая и в начале
июня, вФинмаркене — в первой половине июня (Коллетт). Окраска яиц белая,
иногда чуть зеленоватая, с коричнево-черными пестринами; форма яйца вы-
тянуто-овальная, менее грушевидная, чем у кайр. Размеры: B4) 71--80 х
X 44—51, в среднем 75,7 х 48,7 мм, вес A7) 81—105 г, в среднем 93,1 (Каф-
тановский); F2) 60—84 х 44—59, в среднем 76,71 х 48,5 мм; вес F2) 80,0—
115,0 в среднем 94,2 г (Карташев); D9) 71—86 х 44—51, в среднем 76,3 х
X 48,2 мм, вес B2) 85,7—114,0, в среднем 97,97 г (Горчаковская). Вес скор-
лупы 9,5—13, обычно 11—12 г. Вторая кладка взамен утерянной бывает в том
случае, если яйцо утрачено в течение первой недели насиживания. Насижи-
вают оба родителя в течение 35—36 дней, начинают насиживать через день или
два после откладки яйца. Наседных пятен у птиц два, по бокам брюшной сто-
роны, развиваются они к началу июня, зарастают во второй половине августа,
число наседных пятен, следовательно, больше, чем яиц в кладке. Сидят гагарки
на гнезде неплотно, спугнутые птицы всегда слетают и на более или менее
короткое время покидают гнездо. Процесс вылупления медленный, около
2—3 суток. Первые птенцы появляются в начале июня, массовое вылупление
происходит в середине этого месяца. Первые два дня птенец беспомощен и
малоподвижен, терморегуляция у него устанавливается между 4-м и 6-м днями.
Вес только что вылупившегося птенца 53,9—74 г в среднем около 63 г. Эмбрио-
нальный пух сменяется мезоптилем на 15-й—16-й день. Спуск на воду проис-
12 Птппы СССР, т. ТТ
178
ОТРЯД ЧИСТИКИ
ходит, когда птенцам всего 19—20 дней, в мезоптиле, при среднем весе птенца
около 250 г, или немного более одной трети веса взрослой птицы. Слетающий
на воду птенец пользуется не маховыми, в это время еще недоросшими, но хо-
рошо развитыми большими кроющими, образующими несущую поверхность,
крыла (как у кайр).
Линька. Последовательность смены нарядов: эмбриональный пух —
мезоптиль —первый осенний (зимний) наряд—первый брачный наряд —зим-
ний (послебрачный) наряд — брачный наряд и т. д. В году проходят две
линьки: предбрачная неполная, весной, заканчивающаяся во второй половине
мая (начинается, повидимому, в
марте); при ней не линяют маховые
и рулевые. Послебрачная полная
линька начинается в конце августа
с мелкого оперения на голове и шее,
и переходит на плечевые и крою-
щие ног, затем на брюшную сторо-
ну. Смена крупного пера на нашем
материале не прослежена и прохо-
дит, следовательно, в море после
того, как гагарки покинут гнездо-
вые колонии, в октябре. Маховые
линяют все более или менее одно-
временно, и птица на время теряет
способность летать. Первая после-
брачная линька годовых птиц начи-
нается уже ранее, с конца июня
(это указывает на то, что они, по-
видимому, не размножаются). Пер-
вый осенний наряд, взамен мезоп-
тиля, развивается в июле—сентяб-
ре (Уайзерби, 1941).
Питание. Кормятся гагарки
на море преимущественно мелкой
пелагической рыбой, в частности
сельдью Clupea harengus, мойвой
Mallotus villosus, песчанкой Ammo-
dytes tobiamib, молодой треской, реже молодыми зубатками Anarrhynchus
и ракообразными. За кормом летает далеко —иногда на 18—20 км от бере-
гов (Кафтановский).
Полевые признаки. Высокий клюв с белой поперечной полосой; двуцвет-
ная окраска — темная, черноватая на спинной, белая — на брюшной стороне
тела. При плавании характерно положение хвоста, поднятого кверху. На по-
лете кажется массивной, голова при этом вытянута вперед, хвост и лапы назад;
полет прямой и быстрый, с частыми взмахами крыльев. Отлично плавает и
ныряет под воду. Осторожна и молчалива. Только в период спаривания или
при испуге издает своеобразное скрипение.
Описание. Размеры и строение. Клюв высокий, сжатый с боков, в брачном
наряде вздутый у основания за счет развития пластин; имеются поперечные борозды, из
которых лежащая у основания клюва — белая; число бороздок 2, редко 3, считая от упомя-
нутой выше белой; вершина клюва загнута крючкообразно; ноздри щелевидны. Цевка короче
среднего пальца (с когтем). Рулевых 12—14. Длина мурманских и беломорских самцов A0)
410—480, самок (8) 410—460, в среднем 445,9 и 432,5 мм. Размах самцов A1) 620—780,
самок (8) 630—760, в среднем 678,5 и 680 мм. Вес самцов A1) 662—800, самок (8) 631—810,
в среднем 727 и 717 г. Крыло самцов A1) 192—205, самок (8) 196—214, в среднем 200,2 и
204,2 мм. По Горчаковской, длина самцов C1) 390—470, в среднем 438,7, размах C0) 620—
780, в среднем 707,6; длина крыла B9) 180—220, в среднем 206,2 мм; вес C1) 524—842,
в среднем 721,6 г. По тому же автору, у самок длина B5) 400—436, в среднем 434,4; размах
Рис. 31. Гагарка
ЛЮРИК, МАЛАЯ ГАГАРКА
179
B5) 630—760, в среднем 706,0; крыло B3) 195—230, в среднем 210,9 мм, вес B5) 631—910,
в среднем 747,6 г.
Окраска. У пухового птенца голова, грудь, брюхо белые; спина—бурая; шея,
бока, оперение ног — бурые с беловатыми вершинами. Мезоптилытемнобурый на спинной,
белый на брюшной стороне тела. Брачный наряд взрослых самцов и самок таков: спинная
сторона, подбородок, горло — шиферно-бурые, вершины второстепенных маховых— белые, от
глаза к клюву проходит белая полоса; зоб, грудь, брюхо, бока, подкрылья, подхвостье —
белые. Клюв — черный с белой поперечной полосой в основной части; лапы — черные; раду-
жина— темнобурая. Зимний наряд без вздутия у основания надклювья и без белой полосы
перед глазом, с белым или пестрым подбородком и горлом. Молодые в первом осеннем (зимнем)
наряде с относительно низким и гладким, лишенным бороздок, надклювьем, с темной серо-
бурой перевязью на горле, со слабо развитыми белыми отметинами на вершинах второсте-
пенных маховых. В первом брачном наряде молодые похожи на взрослых птиц, но попереч-
ные борозды на надклювье, повидимому, менее развиты (отсутствуют или менее числом).
Систематические заметки. Саломонсен A944) высказал предположение, что гагарки
из Норвегии, Гренландии и северных частей СССР отличаются от балтийских большим чис-
лом поперечных борозд надклювья (обычно тремя, не считая белой). Изученный нами мате-
риал из районов вост. Мурмана и Белого моря показывает, что у 19 птиц имеются две таких
бороздки, у одной только три, у пяти — одна, у одной (вероятно, прошлогодней) — ни одной.
Таким образом, птицы из сев.-вост. Атлантики не отличимы по этому признаку от бал-
тийских, и предложенное Саломонсеном выделение «pica» в отношении, по крайней мере, мур-
манских и беломорских птиц никак не может быть принято. Есть ли различия в этих деталях
строения рамфотеки между гренландскими, американскими и норвежскими — с одной сто-
роны, и мурманскими и беломорскими — с другой, остается для нас неясным.
РОД ЛЮРИКИ PLAUTUS GUNNERUS, 1761
Тип Alca alle L.
2. Люрик, малая гагарка Plautus alle L.
Alca alle. Linnaeus. Syst. Nat. ed. X, 1758, стр. 131, Гренландия.
Русское название. Слово «люрик» встречается у Палласа A811), вероятно народ-
ное; этимология неясна. Местами «портушка» (Бутурлин, 1934). Малая гагарка — книжное.
Распространение. Ареал. Самый северный вид из атлантических чисти-
ков, населяющий высокие широты Атлантического океана: Гренландия и
прилежащие с запада острова, сев. Исландия; Ян-Майен; Шпицберген;
Карта 27. Распространение люрика Plautus alle
I. P. a. nlle, 2. P. a. polarh;
a — граница гнездовий, г — область кочсьок и зимовок.
180 отряд чистики
Новая Земля; архипелаг Франца-Иосифа; Северная Земля. В негнездовое
время — на море как близ гнездовой области, так и южней, к югу до Англии,
Фарерских о-вов, в Америке до Нью-Джерси и даже Флориды, Бермудских
о-вов и Кубы.
Характер пребывания. Как видно из предыдущего, вне пе-
риода размножения кочует в море в свободных от льда участках.
Биотоп. В гнездовое время.— в прибрежных скалах, в негнездовое —
в море, часто далеко от берегов.
Численность. Самый многочисленный вид отряда.
Подвиды и варьирующие признаки. На архипелаге Франца-Иосифа гнездится Р. я.
polaris Stenho use, 1930, отличающийся более крупной величиной; в остальной части
ареала—P. a. alle L. Вопрос о том, имеются ли здесь географические различия в широком по-
нимании слова или только популяционные особенности, как и для многих других форм
чистиковых, остается неясным.
Обыкновенный люрик Plautus alle alle L.
Распространение. Ареал. Сев. Гренландия — восточное побережье (Ли-
верпуль, около 71° с. ш.) и западное (Туле, на север до Эта, в небольшом числе
на юге до Упернивика и о-ва Диско), Баффинова Земля, Земля Эльсмира,
Земля Гриннелля, быть может на о-ве Парри (Шалов, 1907); сев. Ислан-
дия (Гримсе), Шпицберген, Ян-Майен, о-в Медвежий, Новая Земля (губа
Архангельская на западном берегу Северного о-ва, 76° с. ш., Горбунов,
1929, быть может и южнее; гнездование на востоке возможно, но не доказано,
Антипин, 1938); повидимому, у м. Челюскина (Амундсен, 1919). Люрик гнез-
дится на базарах Северной Земли (Лактионов, 1946), но к какому подвиду отно-
сятся эти птицы — неясно. Сведения о гнездовании на Мурмане (Териберка,
о-в Гусиный, по Никольскому, 1885)—невероятны.
В негнездовое время в море, свободном от льда, как к северу, так и к югу
от области гнездования. Зимует у северо-востока и юга Новой Земли, в Ба-
ренцовом и Карском морях, в сев. Атлантике, у Мурмана (Екатерининская
гавань, Оленья губа, Оленьи о-ва, Пала-губа, Семь Островов и т. д.), у
берегов Норвегии, у Исландии, Фарерских о-вов, Шпицбергена, Ян-Майена,
на юг до Шотландии. Залетает до Прибалтики, Франции, Испании, Италии,
о-ва Мальты, Азорских, Канарских о-вов и Мадейры. В Америке в негнездо-
вое время доходит нормально от южн. Гренландии до Ньюфаундленда и
северных штатов (о-в Лонг, Нью-Йорк), изредка до Нью-Джерси, Виргинии,
Южн. Каролины, даже Флориды и Бермудских о-вов. Иногда имеют место
массовые миграции люриков на юг, например в Англии в 1895, 1900, 1910,
1912 гг. (Уайзерби, 1941), в Америке —в 1891, 1932, 1936, 1937 гг. (Саломон-
сен, 1944). В летнее время люрики встречаются также далеко на севере в мо-
ре — между 81°17' и 84°48' с. ш. (Коллетт и Нансен, 1899; Портенко, 1946).
Зимние кочевки связаны, повидимому, с холодными морскими течениями и
их высоко продуктивной кормовой базой (Уинн-Эдвардс, 1935); по этим же при-
чинам люрики на зимовке не встречаются в центральных частях Атлантическо-
го океана.
Даты. Прилет к местам гнездовья на Северной Земле начинается в начале
мая (Лактионов, 1946). На Новой Земле прилет не прослежен, на северо-востоке
встречен в апреле (Антипин, 1933). На Ян-Майене появление у мест гнездовья
отмечено 21—23 апреля; гнездовые колонии были заняты 2 мая (Фишер и Пель-
цельн, 1886). На Шпицбергене появление отмечено в начале апреля D апреля,
Норденшельд, 1875; 9 апреля по Бунге, Бианки, 1902) и ранее—в марте A7 марта,
Бируля, 1910 и др.), даже 27 февраля (Мунстеръельм, 1910). Наконец, в Грен-
ландии прилет в Девисоновом проливе отмечен 20 мая, в Баффиновом заливе —
18 мая, в заливе Болдуина — 8 мая, на севере вСмис-зунде — 8 мая, отдельные
особи в Баффиновом заливе появляются уже в апреле (Бент, 1919). Осенью
ЛЮРИК, МАЛАЯ ГАГАРКА 181
отмечен на северо-востоке Новой Земли, близ которой птица, впрочем, и зи-
мует. Движение люриков отмечено в ноябре, у южной части острова наблю-
дался около середины октября, иногда ранее, около начала этого месяца. Отлет
из колоний у губы Архангельской на Новой Земле отмечается во второй поло-
вине августа (Горбунов, 1929), в конце августа—начале сентября птицы
широко кочуют по Баренцову морю. Гнездовые колонии на Шпицбергене поки-
даются в августе, отдельные особи встречаются до октября..На Ян-Майене
массовый отлет отмечается в августе. С гренландских гнездовий птицы исче-
зают в августе —сентябре (в сев.-вост. Гренландии под 75° с. ш. уже 2 августа,
Бент, 1919). Растянутые сроки миграций объясняются рядом причин: с одной
стороны, наличием запоздалых выводков — люрики откочевывают от мест
гнездовья сразу после спуска молодых на воду, с другой — зависимостью
миграций и распределения птиц от ледовых условий, различных в разные
годы. Средней Атлантики кочующие стаи люриков достигают обычно в ноябре
и держатся там до середины февраля (Уайзерби, 1941).
Биотоп. Гнездовый — морские каменистые побережья, особенно ка-
менистые россыпи как у самого берега, так и высоко над ним (до 500 и даже
1000 м). Зимний — у кромки льдов, близ гнездовой области на севере, юж-
ней — в море, нередко открытом, в области холодных течений, в частности
Гренландского и Лабрадорского.
Численность. Весьма высокая, за исключением южной границы
гнездовой области, например, в Архангельской губе на западе Новой Земли
или на о-ве Гримсе у сев. Исландии, где к западу от Басавика гнездится только
до 10 пар (Холмс и Киз, 1936). Обычные гнездовые колонии крайне много-
численны, равно как велик и состав кочующих зимних стай. При больших
миграциях на юг, указанных выше, отмечалась массовая гибель кочующих
птиц.
Экология* Размножение. Достаточных наблюдений, относящихся
к нашей территории, нет. Половая зрелость не изучена. Гнезда размещаются
в щелях между камнями, где люрики вырывают небольшие углубления в
грунте. Подстилки нет. Кладка происходит, повидимому, около середины и
во второй половине июня (Шпицберген); тогда же на Ян-Майене и на о-ве
Медвежьем, в Гренландии—с половины июня и в начале июля (Бент, 1919). Яйцо
в кладке одно, но изредка бывают и два; окраска яиц бледноголубоватая,
иногда с желтовато-бурыми крапинами на толстом конце. Размеры: A40) 45—
53 х 30—36,7, в среднем 48,09 X 33, 75 мм (Уайзерби, 1941); D4) 45—51 х
х 32—35,5, в среднем 48,2 х 33 мм (Бент, 1919). Насиживают оба родителя,
каждый из которых имеет по два наседных пятна; срок насиживанья —24
дня (Фабер). Оба родителя выкармливают молодых, которые покидают гнездо
в возрасте несколько меньше одного месяца. Все развитие птенца, включая
рост первостепенных маховых, кроме нескольких передних (Саломонсен,
1944), протекает в гнезде (на Новой Земле птенцы перед вылетом были найдены
в гнездах еще 16 августа, Горбунов, 1929), хотя может быть, что летатель-
ные возможности молодых в момент спуска их на воду развиты непол-
ностью.
Линька. Изучена недостаточно. Общая схема —обычная для чисти-
ковых: пуховой наряд —первый зимний —первый брачный —зимний брачный
и т. д. В году проходят две линьки — частичная (мелкое перо) предбрачная
весной или в конце зимовки, повидимому, в марте — апреле (майские птицы
уже в брачном пере) и полная послебрачная осенью, в августе —сентябре;
рано закончившие линьку особи добывались, повидимому, уже 12 сен-
тября (Горбунов, 1929), хотя обычно в начале сентября у взрослых птиц
часть оперения на горле и зобе еще не перелиняла, маховые вырос-
ли только наполовину B сентября, Баренцово море под 79°7' с. ш. и
42° 8' в. д.).
182
ОТРЯД ЧИСТИКИ
Питание. Планктические беспозвоночные, у Новой Земли почти
исключительно Calanus finmarchicus (Горбунов, 1929). Имеется горловой
мешок — резервуар для пищи, которую приносят птенцам (Фильден, по Яр-
релю, 1878).
Полевые признаки. Мелкие размеры, короткий и прямой темный клюв,
черная окраска спинной и белая — брюшной стороны. Полет быстрый, обычно
низко над водо1О, с частыми взмахами крыльев. Обычно летят стаями или
группами. При плаваньи под водой, как и другие чистики, пользуется и
крыльями и ногами. Хорошо ныряет,
и в среднем под водой может нахо-
диться около полуминуты C3 сек. по
761 наблюдению, но иногда до 63 сек.,
Экблоу, по Бенту, 1919), глубина
ныряния до 2г/4 м (Дьюар, по Уайзер-
би, 1941) У мест гнездовья очень
криклив, голос вроде «петт-тет-тет-
тет» или «перре-тетете-тетт-тетт». Мало
осторожен (за исключением тех мест,
где сильно подвергается преследова-
нию, например, у о-ва Гримсе, сев.
Исландия).
Описание. Размеры и строение.
Клюв короткий, толстый, со слегка выпук-
лым надклювьем; ноздри овальные с кожи-
стым утолщением вокруг, не прикрыты перь-
ями; в брачном периоде у основания над-
клювья развивается поперечное утолщение.
Цевка короче среднего пальца (с когтем).
Шесть пар рулевых. Вес самки A) 135 г.
Крыло самцов (81) 116—129, самок D9)
113—125, в среднем 122 и 119,9 мм; клюв
13—17 мм (Саломонсен, 1944, по особям из
Гренландии, Шпицбергена, Медвежьего о-ва,
Исландии, Ян-Майена и зимним из Швеции
и Норвегии). У птиц из Баренцова и Карского моря — самцов и самок — крыло 110, 116,
116, 119, 127,5 мм (Зоологический музей Московского университета).
Окраска. Пуховой наряд темно-черновато-бурый на спинной стороне, бледный
на брюшной, середина которой сероватая (Уайзерби, Хартерт и др.). Молодая птица в пер-
вом осеннем наряде по расположению цветов, как взрослая в брачном наряде (см. ниже):
спинная сторона матово-буровато-черная, подбородок и зоб — бурые, подкрылья — бледно-
бурые; под глазом белое пятно; грудь, брюхо, подхвостье — белые, оперение боков с чер-
новатыми внутренними опахалами; все перо рыхлее и мягче, клюв слабей, чем у взрослых,
и бурого цвета. Брачный наряд самца и самки: спинная сторона — черная, голова, горло,
передняя часть шеи — дымчато-бурые; плечевые с белыми каемками; вершины второсте-
пенных маховых белые; грудь, брюхо, подхвостье — белые, оперение боков — белое с чер-
новатыми внутренними опахалами; подкрылья — бурые с темными пятнами у сгиба крыла,
задние большие, нижние кроющие с белыми вершинами. Радужина — бурая, клюв — чер-
ный; ноги — свинцово-серые с черными плавательными перепонками и когтями. В зим-
нем наряде подбородок, горло, зоб, бока шеи, затылка, задняя часть щек — белые; по
бокам зоба — черные пестрины.
Рис. 32. Люрик
Большой люрик Plautus alle polaris Stenhouse
Alle alle polaris. Stenhouse. Scottish Naturalist, 1930, стр. 47, Земля Франца-Иосифа.
Распространение. Ареал. Гнездится на Земле Франца-Иосифа, область
зимовок не выяснена.
Характер пребывания. Откочевывает на море после периода
размножения.
Даты. Появление у островов отмечается уже в конце февраля B5—
27. II, Джексон, 1899) и в начале марта B. III; 6—9. III, Кларк, 1898; 18—20.111
ЛЮРИК, МАЛАЯ ГАГАРКА 183
на о-ве Гукера при тридцатиградусном морозе, Демме, 1934; 10.III, Гор-
бунов, 1929 и т. д.). Однако, занятие скал происходит только в июне. Гнездо-
вые колонии люрики покидают в конце августа —- начале сентября после
спуска птенцов на воду (Джексон, 1899; Кларк, 1899; Горбунов, 1929; Демме,
1934 г.; последний автор отметил отлет 12 сентября, некоторые птицы задер-
живаются до начала октября — 1.Х, по наблюдениям Кеттлица, Кларк,
1898), когда уже замерзают проливы. Направление миграций не выяснено.
Возможно к этой форме относятся люрики, встреченные в высоких широ-
тах полярными экспедициями «Седова», «Малыгина» и «Фрама», некото-
рые—на меридианах Новосибирских о-вов (81° 17' с. ш., 138° 7' в. д.;
81° 21'с. ш., 137° 37' в. д.; 81° 23' с. ш., 137° 20' в. д.; 81° ЗГ с. ш.,
121° в. д.; между 81° 34' и 84° 48' с. ш., Портенко, 1946; Коллетт и Нансен,
19С0).
Биотоп. Гнездовый — обрывы или осыпи как у воды, так и в несколь-
ких сотнях метров от берега (по Горбунову, такая удаленность гнездовий от
воды объясняется, вероятно, тем, что покидают гнезда молодые люрики лет-
ными). В негнездовое время — среди разводий и в море.
Численность. Весьма высокая, больших колоний на архипелаге
Франца-Иосифа отмечено не менее 35 (Горбунов, 1929). Колонии часто состоят
из нескольких тысяч пар (на скале Рубини в 1931 г. около 5 000 птиц; Демме,
1934).
Экология. Размножение. Цикл, сходный с таковым у номинальной
формы. Основные наблюдения сделаны Горбуновым A929) и Демме A934).
Первые признаки половой активности — воркующее токование самцов —•
отмечены еще 28 марта, самки в это время еще не активны. Гнездятся в щелях
обрывов, россыпях туфа, нередко в колониях с чистиками и кайрами и т. д.,
но в последнем случае образуют обособленные группы. Гнездо представляет
собою норку в глубокой щели или среди камней. Используется оно в течение
нескольких лет, расположение гнездовых колоний также весьма постоянно.
Кладка происходит около середины июня (ранние яйца найдены 9—16 июня).
Яйца в гнездах попадаются до середины августа (имеются, повидимому, вто-
ричные кладки взамен погибших). Окраска яиц голубоватая, изредка с корич-
невыми крапинами у толстого конца. Заслуживает внимания то обстоятельство,
что размеры яиц не превосходят размеров яиц номинального подвида: A8)
46,8—52,5 х 32,8 —36,8, в среднем 49,29 х 34,62 мм (Горбунов). Насижи-
вают оба родителя, самка слетает с гнезда кормиться по утрам и вечерам.
Продолжительность насиживания не определена. Оперяющиеся птенцы в гнез-
дах найдены в начале августа, у птенцов, добытых около середины августа
A3 августа), обнаруживались только остатки пуха. Вылет из гнезда молодых
начинается с конца августа B8 августа —3 сентября; Демме; 28—29 августа,
Горбунов).
Линька. Предбрачная весенняя линька заканчивается, повидимому,
в марте, хотя некоторые птицы запаздывают. Конец полной послебрачной
линьки отмечается с 30 августа, хотя линяние иногда затягивается и до сен-
тября (Горбунов).
Питание. Преимущественно рачки Calanus finmarchicus (Горбунов),
также Atylus carinatus, Paramphitoe hystrix, Anonyx nugax, Acanthostepheia
behringensis; какая-то мелкая рыбка (Демме).
Описание. Размеры и строение. Гнездящиеся на архипелаге Франца-
Иосифа люрики отличаются от номинальной формы более крупными размерами: крыло
130—136 мм, клюв обычно 17—17,5 мм (Стенхоуз, 1930).
Систематические замечания. При обособленности колоний люриков, при полном сход-
стве их экологии естественно возникает вопрос, не являются ли крупные размеры
люриков с земли Франца-Иосифа только популяционными особенностями, и можно ли
их расценивать как проявление географической изменчивости в строгом смысле
слова.
184
ОТРЯД ЧИСТИКИ
РОД КАЙРЫ URIA BRISSON, 1760
Тип Colymbus troille = Uria aalge
3. Тонкоклювая или длинноклювая кайра Uria aalge Pontop.
Colymbus aalge. Pontoppidan. Danske Atlas v. 1, 1763, стр. 621, табл. 26. Исландия.
Русское название. «Кара», «арра» — звукоподражательное, у русских на Тихом
Океане (Паллас, 1811). Тонкоклювая и длинноклювая кайра — книжные имена.
Распространение. Ареал амфибореальный. Острова и побережья Ат-
лантического и Тихого океанов. В Европе: в Англии, Шотландии, Ирландии
на Фарерских о-вах, в Исландии, на о-ве Гельголанд, в Бретани и по островам
Атлантического побережья Франции, у северного берега Испании на юг до
о-вов Берленгас, в Португалии к северу от Лиссабона (Тейт, 1924); в Бал-
тийском море (Стура-Карльсе; Гресхольм в архипелаге Христиансе, на шхерах
Бонден к югу от Умеа в Ботническом заливе), у побережья Норвегии, на
Карта 28. Распространение тонкоклювой кайры Uria aalge
1. U. a, inornata, 2. U. a. californica, 3. U. a. hyperborea, 4. U. a. spiloptera, 5. U. a. albionis, 6. U. а.
aalge, 7. U. a. intermedia;
а — граница гнездовий (цифры в кружках обозначают номера подвидов и их местонахождения на зимовках
и кочевках); г — область зимовок.
Мурманском побережье и на Новой Земле; на о-ве Медвежьем; в зап.
Гренландии у Сермилингуака в округе Суккертоппен; на Атлантическом побе-
режье Америки от южн. Лабрадора и Ньюфаундленда до залива св. Лав-
рентия. В Тихом океане в Азии от Камчатки, Командорских и Курильских
о-вов, в Приморье, возможно на Охотском море, на о-ве Иезо в Японии.
В Америке от Аляски (Нортон-зунд), о-вов Прибылова, св. Матвея и Але-
утских до Вашингтона; изолированная колония на о-ве Фаральонес у Кали-
форнии. В негнездовое время отмечена, кроме того, у берегов Бельгии, Голлан-
дии, Германии, в датских водах у берегов Швеции, в Азии до Хондо; в Аме-
рике на востоке до Мена и изредка до Массачусетса, а на западе — до южн.
Калифорнии (Санта Моника). Отмечались редкие залеты к Шпицбергену,
в центральную Европу—в Германию и Швейцарию; даже на Канарские
о-ва; более обычно в Средиземное море у Франции, Италии, Мальты, Ма-
рокко. Особенно значительные миграции у молодых птиц в первую осень —
зиму их жизни, причем птицы передвигаются в разных направлениях, даже
к северу, и уходят в открытое море (Штехов, 1938).
Биотоп. Гнездовый — прибрежные скалы (птичьи базары), иногда
плоские поверхности скалистых островов (о-в Лабрадор, о-в Тюлений у Саха-
лина); зимой —как прибрежные части моря, так и вдали от берегов.
Численность. В отличие от толстоклювой кайры, тонкоклювая
более многочисленна в южной части ареала, например, в Исландии — у юж-
ного и западного берега острова; на Новой Земле встречается в ничтожном
ТОНКОКЛЮВАЯ ИЛИ ДЛИННОКЛЮВАЯ КАЙРА 185
количестве (на базаре в Пуховом заливе около 100 пар, Портенко, 1931)»
очень мало ее в Гренландии. Неумеренное преследование, если принять во
внимание и низкую плодовитость птицы, вызвало значительное падение чис-
ленности кайры во второй половине XIX — начале XX века во многих мест-
ностях, в частности на южн. Лабрадоре, в заливе св. Лаврентия, в южн.
Норвегии; кайра исчезла у берегов Пикардии и Нормандии, Ориньи и Серк,
во Франции; на о-ве Олдерней, Норманские о-ва; в Швеции у Кальмара
и на Борнгольме. С другой стороны, охрана гнездовий способствовала
увеличению численности кайр местами на Балтийском море (колония на
о-ве Карльсес 1918 по 1943 г. возросла с 2500 до, примерно, 50 000 птиц, и от
нее возникли новые колонии на о. Христиансе с 1928 г. и на шхерах Бонден
к югу от Умеа, Саломонсен, 1944), в СССР в заповеднике Семь островов на
вост. Мурмане.
Подвиды и варьирующие признаки. Морфологически варьируют размеры, в частности
длина крыла и клюва, и оттенки окраски.Кроме того, для разных частей ареала характерна
определенная пропорция в популяциях цветовой вариации — так называемых «очковых»
кайр, птиц с белым пятном у глаза (Uria Ringwia В г u n n i с h, 1764; Uria lacrymans
Valenciennes, 1822; эта вариация многими авторами считалась за особый вид, в
частности, в последнее время Красовским, 1936, указавшим на интересные скелетные особен-
ности этих птиц; однако, по исследованиям Кафтановского, взгляд на видовую самостоя-
тельность очковых кайр не выдерживает критики).
Количество очковых кайр постепенно возрастает с юга на север (Саломонсен, 1935,
1944; СоузерниРив, 1941), равняясь нулю в Португалии, 0,1% во Франции, около 1% з южн.
Англии, 1—2% на Балтийском море, примерно 10—30%—в Шотландии и Исландии, 18%—
на Атлантическом побережье Америки, 31—34% — на Фарерских о-вах, 53% — на о-ве Мед-
вежьем, более 50% — в сев. Норвегии, на Новой Земле; 42% — на Семи островах, вост.
Мурман (притом более 40% гнездяшихся там пар — смешанные, одна из птиц — «нормаль-
ная», другая — «очковая», Кафтановский).
У тихоокеанских кайр очковая вариация отсутствует. Это, повидимому, указывает
на возникновение «очковой» кайры на севере Атлантического ареала вида, уже в послелед-
никовое время, когда область распространения вида на западе и востоке стала разобщенной
и ареал стал амфибореальным.
Экологически варьирует календарь периодических явлений, более ранний на юге
и у омываемых Гольфштремом берегов (так, в Англии начало кладки относится к последней
трети мая; в Исландии —к концу мая и началу июня; на вост. Мурмане— к этому же вре-
мени, на о-ве Медвежьем — к середине июня и т. д.). Описаны подвиды: U. a. albionis
W i t h e r by, 1923 — Ирландия; Англия к северу до о-ва Фарн, Нортумберленд, на во-
стоке и Эльза-крейч, Айр, на западе; Гельголанд, Бретано, сев. побережье Испании;
о-в Берленгас у Португалии; зимою умеет гнездовий, но преимущественно у берегов
Ирландии, Ламанша, зап. Франции и сев. Испании, также у юго-зап. Норвегии, Гол-
ландии, Дании; U. a. intermedia N i I s s о n, 1835 — Балтийское море — Стуре
Карльсе, Христиансе, шхеры Бонден в Ботническом заливе; зимой до Каттегата и
Зунда; U. a. aalge Pontippidan, 1763 — Исландия, Великобритания к северу от
Ст. Абба, Бервик, —на востоке и Ислей—на западе; Гебридские и Шетландские о-ва; южн.
и зап. Норвегия, на юг, примерно, до Бергена, на север—до Лофотенских о-вов;
зап. Гренландия (Сермилингуак); атлантическое побережье Америки от Ньюфаундленда
(кроме западного берега) и южн. Лабрадора до залива св. Лаврентия (Берд-рок и острова
у северного побережья); зимой у берегов Швеции, Дании, в Северном море, Голландии,
Бельгии, Англии, зап. Франции, до Испании и Средиземного моря (Марокко и т. д.); встре-
чены и в открытом море (Иесперсен, 1930, Уинн-Эдвардс, 1935); U. a. spiloptera S а 1 о-
m о n s e п, 1932 — Фарерские о-ва, быть может самый север Шотландии и Шетландские
о-ва, более или менее оседла; окольцованная птица, впрочем, добыта зимою у Бергена
в Норвегии (А-Ригги, 1931); U. a. hyperborea Salomonsen, 1932 — о-в Медвежий,
Норвегия к северу от Тромсе, Мурманское побережье, Новая Земля; в негнездовое время
у Норвегии, на Мурмане, в Белом море, на юг до Дании (Никобинг); U. a. inornata Salo-
monsen, 1932 — северный Тихий океан, в Америке у Аляски, по островам Берин-
гова моря (Прибыловы, св. Матвея, Алеутские), на юг до Вашингтона; в Азии от Кам-
чатки до Охотского моря, Командорских и Курильских о-вов, Сахалина, Иезо;
зимой у Алеутских о-вов, Командорских, до Хондо; U. a. californica Bryant, 1869 —
на о-в Фаральонес у Калифорнии, зимой к югу до южн. Калифорнии (Санта
Моника).
Тихоокеанские подвиды хорошо отделяются от атлантических, среди последних резко
обособлен южный подвид U. a. albionis; остальные формы разграничены нерезко и, вероятно,
частично должны быть отнесены к категории популяционной, а не географической измен-
чивости.
186 ОТРЯД ЧИСТИКИ
Северо-атлантическая тонкоклювая кайраUria aalge hyperborea Salomonsen
Uria aalge hyperborea. Salomonsen. The Jbis. 1932, стр. 130, о. Медвежий.
Распространение. Ареал. О-в Медвежий, Норвегия северней Тромсе
(особенно в фиордах Порсангер и Варангер), Мурман (на восток до Семи
островов), западное побережье Новой Земли Русанов, 1924, указывает на
распространение кайры по западном берегу Новой Земли между 72° 5' и 76° с. ш.).
В негнездовое время—у Кольского п-ова, в сев. (Финмаркен) и зап.
(Берген) Норвегии, к югу до Дании (Никобинг, Морс). Залет, повидимому
этого подвида, отмечен в Черном море у Балаклавы.
Характер пребывания. Зимой отчасти находятся в гнездовой
области (Мурман, сев. Норвегия), отчасти передвигается к югу (см. выше).
Холостые (неполовозрелые годовалые) особи наблюдаются и к северу от гнез-
довой области G7° 16' с. ш. и 60° 29' в. д., 5.VIII.1930, по Горбунову, 1932;
отмечены у Шпицбергена, Плеске, 1928).
Даты. У мест гнездовья на Семи островах, вост. Мурман, появляются
уже с середины февраля и в начале марта, гнездовые колонии занимаются
в последней трети апреля (Горчаковская, Кафтановский). Покидаются коло-
нии в конце августа, причем взрослые частично отлетают ранее, чем молодые,
а частично вместе с последними. Откочевка от этих островов происходит в конце
августа—сентябре.
Биотоп. Скалистые обрывы на островах и материковых побережьях,
с карнизами, совершенно открытыми, предпочтительно широкими @,5—Л м)
и горизонтальными, или с небольшими уступами; реже на узких карнизах,
в 12—15 см ширины (Кафтановский, 1941). В негнездовое время в при-
брежных частях морей, но иногда и далеко от берегов (повидимому, моло-
дые птицы, которые и летом следующего после рождения года только случайно
появляются у гнездовых колоний).
Результаты кольцевания. Окольцованные на Семи островах
кайры встречены на зимовке и кочевках в Норвегии (Сольсвик к западу от
Бергена, Лофотенские о-ва, Лоппа в Финмаркене), в Полярном районе
Мурманской обл.; одна окольцованная птица добыта на следующий год
5 июня в Кандалакшском заливе; самая южная—у Никобинга в Дании.
Семь птиц были вновь добыты у мест гнездования на второй и третий год
после кольцевания. Крайний консерватизм кайр в отношении мест гнездования
находится в соответствии с хорошо развитой ориентацией. В 1939 г. взятые
с гнездовий на о-ве Харлов 10 кайр были завезены в закрытом ящике на раз-
ные расстояния от 25 до 250 км (до Мурманска); семь птиц* вернулись обра-
тно, в том числе две выпущенные у Мурманска (Кафтановский).
Результаты кольцевания показывают, что часть мурманских кайр и зимует
близ Мурманского побережья, часть же движется на запад-юго-запад, зимуя
у норвежских берегов и достигая Балтийского моря.
Численность. На Новой Земле весьма ограничена (в Пуховом
заливе, примерно, 100 птиц из 60 000 кайр, Горбунов, 1925). У Мурмана за-
метно преобладает над толстоклювой кайрой. Результаты кольцевания и дру-
гие данные показывают, что среди причин гибели кайр в освоенных человеком
местностях служат попадание кайр в рыболовные сети, загрязнение вод мазу-
том и т. п. Из естественных причин гибели птенцов большое значение имеет
хищничество крупных чаек, в особенности в малых колониях (смертность
птенцов на больших базарах Семи островов — 10%, на малых — от 30% до
70%, Кафтановский). В зимнее время кайры гибнут при штормах.
Экология. Размножение. Половая зрелость наступает, повидимому,
в возрасте около 2 лет, годовалые птицы, во всяком случае, как правило, не раз-
множаются. Количественное соотношение полов: ПО самцов на 101 самку,
т. е. 10% избытка первых (Семь островов). Спаривание происходит с начала
мая; у птиц, утративших кладку, еще 20 июня. Общая продолжительность
ТОНКОКЛЮВАЯ ИЛИ ДЛИННОКЛЮ^АЯ КАЙРА 187
периода размножения около 3V2 месяцев (вост. Мурман). Гнездовые колонии
занимаются из года в год и расположены на каменистых карнизах, в виде
исключения на торфе, а иногда кладка помещается даже в гнезде чайки-моевки.
Птицы гнездятся плотными группами, карнизы занимаются сплошь. Ни на-
стоящих гнезд, ни какой-либо подстилки не бывает. Кладка начинается с конца
мая, в первой половине июня принимает массовый характер (вост. Мурман,
1937 г. —20—25. V; 1938 г.—28. V; 1939 г.—последние числа мая—первые чис-
ла июня). На отдельных базарах и карнизах разница в сроке кладки может
доходить до 20 дней, в зависимости от экспозиции скалы и таянья на ней
снега. Кладка состоит из одного яйца, характерной кубовидной формы, затруд-
няющей скатывание яйца с карниза. Окраска скорлупы весьма изменчива:
по белому, зеленоватому, темнозеленому или голубоватому фону разной вели-
чины, формы и оттенков черные или бурые пятна и черточки. Размеры яиц
очень велики; относительно веса самки вес свежего яйца составляет около
11,2%. Вес яиц: G7) 101—139, в среднем 112,9 г; размеры G7) 79—93 х 49—55,
в среднем 84,4 х 51,8 мм (т. е. крупней, чем у номинальной формы). Скорлупа
очень пористая и толстая, толщина ее неравномерна, самая большая — в сере-
дине узкой части яйца, т. е. в точке опоры его при лежании на скале; незна-
чительное утолщение имеется у острого конца, толстый конец яйца имеет
самую тонкую скорлупу (Кафтановский, 1941).
Насиживают оба родителя (у старых птиц — одно наседное пятно, вытя-
нутой формы). Срок насиживания — 35 дней. Проклевывание птенца продол-
жается от одних до четырех суток. Появление птенцов на Семи островах в
1932 г. началось 3 июля, в 1937 г. — 29 июня, в 1938 г. — 1 июля, в 1939 г. —
4 июля, в первой трети июля происходило массовое вылупление. Только что
вылупившиеся птенцы имеют длину тела: A1) ПО—152, в среднем 134,3 мм,
при весе в 66—79, в среднем 71,3 г.
Со второго дня жизни птенцы начинают ходить по карнизу, терморегуля-
ция устанавливается у них между четвертым и шестым днем жизни, маховые
показываются на третий — пятый, рулевые на двенадцатый — пятнадцатый
день после рождения. Перелиняв в мезоптиль и достигнув около г/А веса взрос-
лой птицы, птенец спускается на воду, планируя и пользуясь при этом боль-
шими кроющими крыла, образующими несущую поверхность (маховые в это
время еще не развиты). Возраст птенца при спуске на воду около 25 дней.
Гнездовый период в целом равен для отдельной особи, таким образом, 60 дням
C5 дней насиживания и 25 дней выкармливания в гнезде). Приведенные выше
сведения относятся к Семи островам (Кафтановский, Горчаковская).
Линька. Смена нарядов типичная для кайр: пуховой наряд — мезоп-
тиль— первый зимний — первый брачный — второй зимний (послебрачный) —
второй брачный и т. д. У взрослых две линьки в году: предбрачная, охваты-
вающая главным образом голову, горло, шею (у годовалых неразмножающихся
птиц она частично распространяется на оперение груди и мантии, Саломонсен,
1944) и сопровождающаяся некоторыми изменениями клюва — увеличением его
размеров и изменением окраски от аспидно-серого до густочерного (Вервей,
1923). Протекает предбрачная линька поздней, чем у южных подвидов, в част-
ности— номинательной формы: предбрачная линька на Мурмане начинается
вэ второй трети апреля и кончается в конце мая. В июне птицы не линяют,
ь июле после вылупления птенцов у старых птиц начинается послебрачная
линька, полная, причем сначала сменяется пух на шее, голове, плечах, затем
мелкое перо на брюшной стороне; во второй трети июля линька мелкого опе-
рения охватывает все птерилии. Птицы покидают гнездовья, не сменив еще
крупного оперения, последнее линяет в море на зимовках. Конец линяния
отмечен в ноябре (Саломонсен, 1944). Маховые выпадают почти одновременно,
как первостепенные, так и второстепенные, вместе с большими кроющими,
так что кайры теряют способность к полету.
188
ОТРЯД ЧИСТИКИ
Питание. Кормятся на море, добывая пищу как с поверхности воды,
так — и преимущественно — ныряя и плавая под водой при помощи крыльев
(максимальная продолжительность пребывания под водой указывается в 68 сек.,
максимальная глубина ныряния —8,5 м, Дьюар). Пищу составляет мелкая
рыба: песчанка, сельдь, мойва, мальки тресковых и зубатки; кроме того,
ракообразные. Со времени прилета птица кормится главным образом мелкими
ракообразными (каланус) и полихетами; в гнездовое время — преимущественно
рыбой. Птенцы выкармливаются встречающимися! в массовом количестве пела-
гическими видами рыб —
песчанкой, сельдью, мой-
вой. Средний вес приноса
птенцу — около 10 г пищи,
кормят птенцов старики
2—3 раза в день (Кафта-
новский, Горчаковская).
Полевые признаки.
Размеры с небольшую утку;
окраска двуцветная, темная
аспидно-черновато-бурая на
спинной и белая на брюш-
ной стороне. Клюв почти
прямой, относительно длин-
ный. Держатся стайно как
на берегу в колониях, так
и на воде. Полет быстрый,
прямой, с частыми сильны-
ми взмахами крыльев. Си-
дящая и ходящая кайра
держится обычно прямо,
опираясь на цевки; наси-
живающая птица — более
горизонтально. Голос —
хриплое протяжное «кааар-
ра» или «арра», с различ-
ными модуляциями, иногда
короткое «арр». Птенцы из-
дают тонкий писк «уип-уип-уип», подросшие — звонкий крик «филип-фи-
лип». В отношении человека кайры на гнездовьях держатся неосторожно.
Описание. Размеры и строение. Клюв вытянутый, примерно равный по
длине голове, слегка сжатый с боков, с узкими щелевидными ноздрями, оперение верхней
челюсти доходит или почти доходит до разреза рта; ноздри прикрыты оперением. Хвост
короткий из 12 рулевых. Цевка короче среднего пальца, без когтя. Длина мурманских
самцов E0) 420—480, самок B9) 396—470, в среднем 464,1 и 484,4 мм; размах самцов E0)
720—790, самок B9) 710—780, в среднем 732, 2 и 735,5 мм. Вес мурманских самцов E0)
825—1185, самок B9) 820—1112, в среднем 1028,3 и 978 г. Крыло мурманских самцов E0)
200—209, самок B9) 200—220, в среднем 208,3 и 209,1 мм. Крыло новоземельских самцов
B) 218—219 мм, самок D) 208—212, 214, 217 мм. Вес новоземельских самцов B) 965—985 г,
самок D) 945, 950, 1060, 1127. Половых различий в размерах, таким образом, повидимому,
нет, индивидуальные колебания значительны. Сильно варьирует форма хребта надклювья.
Клюв относительно не длинный: у мурманских птиц G9) 45—56, в среднем 49,8 (т. е. все же
несколько больший, чем у птиц из типичной местности подвида — о-ва Медвежьего, судя по
данным Саломонсена, 1944). Может быть, что мурманские популяции занимают в отноше-
нии пропорций клюва промежуточное между северным и номинальным подвидом положе-
ние. По литературным данным (Красовский, 1937), для новоземельских птиц указываются
следующие размеры: крыло 208,5—215, в среднем 210,3 («ненормальные» особи); 212—221,2,
в среднем 214,9 мм (очковые особи); клюв 43,5—48,7, в среднем 45,6 («нормальные») и
46,2—50,3, в среднем 47,94 мм (очковые).
Окраска. Для подвида характерна темная окраска спинной стороны, значитель-
ная испещренкость бурым цветом боков и подкрылий (средние и малые кроющие крыла);
Рис. 33. Тонкоклювая кайра
ТОНКОКЛЮВАЯ ИЛИ ДЛИННОКЛЮВАЯ КАЙРА 189
часто, как указано, встречается «очковая» вариация. Пуховой птенец на спинной стороне
буровато-серый с белыми кончиками пушинок, заметно проступающими в особенности на
голове (птенец толстоклювои кайры в этом возрасте заметно рыжеватей или охристый);
гпудь, брюшко — белые, бока, анальная область — темнобуро-серая. Пух очень жесткий,
плотно сцепленный в проксимальной части, — этим создается хорошая защита птенца от
ветра на открытых гнездовых скалах. Клюв и ноги — темносерые. Мезоптиль (в возрасте
около 20 дней): спинная сторона — черновато-бурая, того же цвета щеки и горло; под-
крылья, зоб, грудь и брюхо — белые; маховых и рулевых нет — первые функционально
замещены большими кроющими крыла — вторые, повидимому, непропорционально боль-
шими лапами.
В брачном наряде общая окраска спинной стороны — черновато-шиферно-буроватая;
голова, горло, подбородок—буроваты; у нередкой «очковой» вариации—белое кольцо вокруг
глаза и белая полоса за глазами; маховые и рулевые — серовато-бурые, основания внутрен-
них опахал первостепенных маховых светлее; на вершинах второстепенных маховых—белая
каемка; грудь и брюхо — белые, бока — белые с бурыми продольными отметинами; подмы-
шечные бурые с белыми пестринами, подкрылья — белые с бурыми пестринами на средних
и малых кроющих крыла. Радужина — темнобурая, клюв — черный, лапы и плавательные
перепонки — черные, светлей (буроватые или желтоватые) на передней стороне цевки и на-
ружной поверхности пальцев. В зимнем наряде вся брюшная сторона—белая, за глазом белое
пятно, доходящее до шеи; клюв—сероватый и менее вздутый. В первом зимнем наряде —
как взрослые, но клюв меньших размеров. В первом брачном наряде обычно на подбородке
и горле, также на зобе остаются несколько белых перьев, маховые сильно обношены. Самцы
и самки окрашены одинаково.
Тихоокеанская тонкоклювая кайра Uria aalge inornata Salomonsen
Uria aalge inornata. Sa lomonsen. The Ibis, 1932, стр. 128, о-в св. Матвея
Распространение. Ареал. От Коряцкой земли (Олюторский залив)
и вост. Камчатки до Сахалина, о-ва Тюленьего, Курильской гряды (от
Кунашири до Парамушира), Командорских о-вов, севера Приморья
(бухта Мосолова, залив де Кастри) и, возможно, Кореи (Handlist of the Japa-
nese Birds, 1932); в северной части о-в Иессо; на Охотском море вероятно гнез-
дится на п-ове Тайгонос (Дементьев, 1940); на о-вах Утичьем и Птичьем
в группе Шантарских (Дулькейт и Шульпин, 1937), встречена летом у Аяна
(Шульпин, 1936). В Сев. Америке —на о-вах Прибылова и св. Матвея, на
Алеутских о-вах, на о-ве Кадьяк и у побережья Аляски (на севере до о-ва
Форрестер и Санта-Лазариа), Британской Колумбии и северо-западных штатов
(Вашингтон, Орегон). В зимнее время в свободных от льда участках Тихого
океана вблизи от гнездовой области. Распространение к северу от Коряцкой
земли не вполне выяснено. Старые указания Нельсона A887) о том, будто
бы эта птица была весьма обильной у о-ва Геральд 12 августа 1881 г. и сооб-
щение того же автора о встрече этой кайры у о-ва Врангеля, очевидно,
основано на смешении с толстоклювым видом. Тонкоклювой кайры нет и на
южном побережье Чукотской земли, также на Анадыре (Портенко, 1939),
куда она только изредка залетает в лиман; впрочем, Белопольский A934)
считает, что она обычна в северных частях Анадырской губы и ссылается
на встречу молодых птиц между заливом Креста и Анадырским лиманом.
Бутурлин A934) упоминает о встрече этой кайры у Уэллена в 1925 г.
Характер пребывания. Как и другие подвиды, зимою поки-
дает гнездовые колонии и держится в прибрежных частях моря, на север до
Прибыловых о-вов, Камчатки (Бергман, 1935, редка), Командорских
о-вов (Стейнегер, 1885), регулярно у Курильских о-вов, по берегам
Японского моря и в Японии до Хондо; в Америке на юг до Алеутских о-вов
и, вероятно, до Вашингтона и Орегона, но по литературе трудно решить из-за
смешения большинством авторов с калифорнийским подвидом.
Даты. Появление у гнезд на Командорских о-вах происходит, пови-
димому, в мае (Иоганзен, 1934). На острове Тюленьем —во второй половине
мая. На Прибыловых островах появление отмечено в конце апреля — начале
мая, колонии оставляются птицами в конце августа. С о-ва Тюленьего отко-
чевка в сентябре (Гизенко).
190 отряд чистики
Численность. Заметно уступает в численности толстоклювой кайре
на Прибыловых и Командорских о-вах, обычна на Алеутских и побережье
Аляски.
Экология. В азиатских частях ареала изучена очень мало.
Размножение. Гнездится колониями, отдельно или вместе с толсто-
клювой кайрой. Колонии на обрывах берегов с карнизами, но иногда на плато-
образных вершинах островов (Тюлений). На Прибыловых о-вах кладка
происходит в начале июня. Размеры яйца (конечно, единственного в кладке):
A0) 84,2—89,5 х 52,2—54,5, в среднем 87,2 х 53,42 мм (Джоурден, по Хар-
терту, 1917); B10) в среднем 82,9 х 50,3 мм, вес в среднем 112 г (сообщение
Гизенко). Птенцы на о-ве Тюленьем обнаружены в первой трети июля
(первые вылупляются 2—3 июля, в массе 5—7 июля). Молодые покидают
колонии на Прибыловых о-вах в конце августа, на Курильских — около 20
августа.
Линька. Птицы в зимнем наряде добыты 28 октября (Кайбато, южн.
Сахалин, Мунстеръельм, 1920) и 15 апреля (бухта Ольги, Камчатка). В ав-
густе взрослые уже в брачном наряде, осенняя линька, повидимому, проходит
в сентябре — октябре. Весной в брачном наряде экземпляры добывались
и мае.
Питание. Как у других подвидов. Анализ содержимого кишечника
добытых на Прибыловых о-вах кайр: амфиподы, главным образом Pontogeneia
sp.; также Апопух nugax, Allorchestes ochotensis, Hyale sp., Jassa sp., Metopa
sp.; изоподы Idothea ochotensis; Nereidae; из рыб бычки Cottidae. Кроме того,
гидроиды, моллюски и растительные остатки.
Описание. Размеры и строение. Клюв выше и массивней, чем у атланти-
ческих подвидов; общая величина больше. Клюв A0) 43—47, в среднем 44,7 мм (Саломонсен,
1932); A0) 42,5—48,1, в среднем 44,9 мм (Зоологический музей Московского университета);
крыло A0) 210—224, в среднем 215,5 мм (Саломонсен); самцов E) 206—228, самок B) 209—
210, в среднем 217 и 209,5 мм\ птиц с неопределенным полом (9) 210—222, в среднем 217 мм;
в общем 206—228, в среднем 216 мм (Портенко, 1939); самцов и самок A0) 205—217, в сред-
нем 212 мм (Зоологический музей Московского университета). Калифорнийская кайра
мельче; крыло F) 195—210, в среднем 204,4 мм (Саломонсен, 1932).
Окраска. Относительно бледная, светлее, чем у североатлантических кайр; горло
бурое; нижние кроющие крыла белые, без пестрин.
4. Толстоклювая или короткоклювая кайра Uria lomvia L.
Alca lomvia. Linnaeus. Syst. Nat. ed. X, 1, 1758, стр. 130, Гренландия.
Русское название. Толстоклювая и короткоклювая кайра — книжные имена.
О слове кайра см. предыдущий вид.
Распространение. Ареал. Северный Ледовитый океан и прилежащие
части Тихого и Атлантического, Мурман, Исландия, в особенности ее северные
побережья; Ян-Майен, о-в Медвежий, Шпицберген, архипелаг Франца-Иосифа,
Новая Земля, Новосибирские о-ва, о-в Вилькицкого, Бенетта, о-в Генриетты,
Геральда и Врангеля, залив Фаддея, о-в Преображения в Хатангской губе,
Чукотский п-ов (о-в Колючин), Коряцкая земля, Камчатка, Охотское
побережье, вероятно, у Татарского пролива и Сахалина, на Курильской
гряде на юг до Матсува, Командорские о-ва. В Америке, в Гренландии,
Баффинова Земля, о-в Сев. Девон, Земля Эльсмира, Земля Гранта, на юге до
залива св. Лаврентия (Берд-рок), Ньюфаундленда, на севере до Гудсонова
пролива по Лабрадору; в Аляске на Прибыловых о-вах, о-ве св. Лаврентия
и от м. Лисберн до Алеутских о-вов и с-ва Кадьяк (вероятно, случайно),
на севере до м. Барроу. В негнездовое время передвигается на юг, достигая
южн. Гренландии, побережья Мена в Америке, реже Массачусетса и даже
Южн. Каролины; в Европе —у берегов Норвегии и Исландии, изредка до
Англии и южнее; в Тихом океане до Аляски и сев. Японии.
ТОЛСТОКЛЮВАЯ ИЛИ КОРОТКОКЛЮВАЯ КАЙРА
191
Характер пребывания. Кочующая птица.
Биотоп. Гнездовый — скалистые острова и материковые побережья.
Зимний — свободные ото льдов части океана, причем, повидимому, более
придерживается открытого моря, чем тонкоклювая кайра.
Подвиды и варьирующие признаки. Тихоокеанские кайры хорошо различаются от
атлантических размерами и окраской. Кроме того, описано несколько слабо отличимых
подвидов из северных частей Тихого океана и из Северного Ледовитого, которые все-таки,
повидимому, не являются географическими формами в точном смысле слова. Экологические
Карта 29. Распространение толстоклювой кайры Uria lomvia
1. О. I. lomvia, 2. U 1. arroides, 3. U. 1. eleonorae, 4. U. I. heckeri, 5. U. I. arra;
a — границы гнездовий, г — область кочевок и зимовок.
различия плохо выяснены и сводятся, повидимому, в основном к разнице в календаре перио-
дических явлений, где описаны такие формы. В сев. Атлантике, на Новой Земле, Шпицбергене,
о-ве Медвежьем, Ян-Майене, Исландии, в сев.-вост. Америке, в Гренландии, на Мурмане —
U. 1. lomvia L., 1758; на архипелаге Франца-Иосифа — U. I. arroides Portenko, 1937; у
восточных частей сев. Сибири от бухты Преображения до Новосибирских о-вов— О. I. eleo-
norae Portenko, 1937; на острове Врангеля и на северном берегу Чукотского п-ова —
U. I. heckeri Porte nko, 1944; в северных частях Тихого океана к югу от Берингова про-
лива — U. 1. arra Pallas, 1811.
Атлантическая толстоклювая кайра Uria lomvia lomvia L.
Русское название. На Новой Земле у промышленников — «гагарка».
Распространение. Ареал. Западная Гренландия к северу до 82° с. ш.,
на юг до 64—65°; на восточном берегу до земли Ливерпуль, 72° с. ш., (гнез-
дится на о-ве Раффлса, ум. Брагастер, Сконнинг, 1938, добытые Денмарксхавне,
77° с. ш., Манних, 1910); западная часть Баффиновой земли, о-в Сев. Девон,
Земля Эльсмира, земля Гранта; сев. части Сев. Америки —Лабрадор, Нью-
фаундленд, на север от Гудсонова пролива; Исландия (особенно северный
берег); Ян-Майен, о-в Медвежий, Шпицберген, Новая Земля от 70° 50' на запад-
ном берегу до Оранских о-вов, на востоке —ум. Желания, 76° 57' с. ш.,
на о-ве Гемскерка; Мурманское побережье до Семи островов (Харлов и Кувшин),
на востоке. В негнездовое время в Баренцовом море, у берегов Новой Земли
(Антипин, 1938), у Ян-Майена (Берд, 1935), в горле Белого моря, у Мурмана,
сев. Норвегии, Исландии, у южн. Гренландии, у американских "берегов
Атлантики до Мена и Массачусетса, изредка до Южн. Каролины. В зимы
с тяжелыми ледовыми условиями на севере птицы появляются и далее на юге —
массовое появление их в такие годы отмечено в Канаде (Флеминг, 1907),
в Финляндии (Мела, 1909). Редкие залеты отмечены в Англию к Фарерским
о-вам, в зап. Швеции, зап. Германии, Голландии и Франции. Редкие залеты
внутрь континента, в частности в ноябре 1902 г. в окрестности Можайска;
в том же году в б. Нижегородскую, Владимирскую и Костромскую губерниг,
повидимому, массовый залет (Лоренц, по Полякову, 1910); залет отмечен
в Калининскую обл. (Бианки, 1915).
192 отряд чистики
Результаты кольцевания. Взрослая кайра, окольцованная
на Шпицбергене, была добыта зимой у юго-вост. Гренландии (Тидеманн, 1943).
Окольцованные в губе Безымянной, Новая Земля, кайры впоследствии добы-
вались на гнездовье там же, а одна, отмеченная кольцом в августе 1934 г.,
была добыта 3 июля 1939 г. на мысе Желания. Кайра, окольцованная на о. Хар-
лове, добыта в декабре у Яжмы на юге Канинского п-ова. Несколько
птиц, окольцованных на острове Харлове, были впоследствии добыты там же
на гнездовье (Дементьев, 1947).
Характер пребывания. Кочующая птица, на зиму покидаю-
щая гнездовые колонии и держащаяся в эта время в открытом море или у бере-
гов; более пелагическая, чем тонкоклювая кайра, и менее проникающая
на кочевках к югу. Впрочем, кайра лишь редко (возможно это надо отнести
за счет недостаточности наблюдений) встречена в центральных частях Атлан-
тики (Уинн-Эдвардс, 1935—в 170 милях к востоку от о-ва Белль-иль,
и в 400 милях к востоку от мыса Фаруелл, южн. Гренландия). Изредка кайры
встречались еще далее от берегов: 24 марта под 55° с. ш. и 24° з. д. в 5000
милях от континента; 1 октября под 53° с. ш. и 27° з. д. в 7 000милях от земли
(Говард, 1920).
Даты. Появление у мест гнездовий, в зависимости от ледовых условий,
отмечается в разные числа мая, но также и в апреле (Шпицберген, Кениг и
ле-Руа, 1911); на Новой Земле в конце апреля, больше в мае (Красовский,
1937); 4 мая (Кривошея, 1884); 24 мая (Сконнинг, 1916); на северо-востоке
Новой Земли у колоний 5 апреля (Антипин, 1938); на Семи островах, Мурман,
в последней трети апреля — начале мая (Кафтановский и Горчаковская).
Гнездовые колонии покидаются в разные числа августа — начале сентября
(Ян-Майен, Берд, 1935, отлет молодых; на Новой Земле 14 августа, Красов-
ский; во второй половине августа, Билькевич, 1907; последние 12 сентября,
Горбунов, 1925; базар покинут 23 августа, Антипин, 1938; Шпицберген —
вторая половина августа, Бируля, 1910J Мунстеръельм, 1910; Мурман, Семь
островов — конец августа — начало сентября, Кафтановский и Горчаковская).
Биотоп. Гнездовый — скалистые побережья островов и материка,
зимой — прибрежная полоса моря, иногда и в открытых его частях (см.
выше).
Численность. Высокая, общее количество кайр на западном по-
бережье Новой Земли ориентировочно определялось в 4000000 особей (Пор-
тенко, 1931); в одной колонии на западном берегу в губе Пуховой 600000 птиц
(Горбунов, 1925); на о-ве Гемскерка на северо-востоке. Новой Земли 30000 штук
(Антипин, 1938). На юге ареала малочисленней, чем тонкоклювая кайра,
на Семи островах занимает мелкие, менее удобные и укрытые карнизы, нередко
покрытые торфом. На о-ве Медвежьем реже тонкоклювой (Бертрам и Лек,
1933). В Исландии встречается главным образом на севере острова. Периоди-
ческих колебаний численности не отмечено. Как и у других кайр, устано-
влены случаи массовой гибели зимующих птиц в частности, при резком падении
весенней температуры в начале текущего столетия на Новой Земле (Портенко,
1931).
Экология. Размн ожение. Гнездится колониально, по карнизам
скал. Резко выраженная консервативность мест гнездовья (Семь островов,
Новая Земля). Половая зрелость потенциально наступает в возрасте около
года, одновременно с первой линькой (Слепцов), но реализация полового
цикла обычно бывает на второе лето, в двухлетнем возрасте. Соотношение
полов близко к 1 : 1 (на 11 самцов 10 самок, Семь островов, по Кафтановскому);
на Новой Земле, по Красовскому, 1937, будто бы значительно преобладают
самцы, что ошибочно. Спаривание начинается после занятия гнездовий,
в конце апреля — первых числах мая (Семь островов, Кафтановский, Горчаков-
ская). Кладка происходит в мае с конца месяца и в первой половине июня:
ТОЛСТОКЛЮВАЯ ИЛИ КОРОТКОКЛЮВАЯ КАЙРА 193
на Семи островах в 1937 г. в начале мая, обычно же в последних его числах
и в первых числах июня, в 1939—1941 г. между 20 мая и 6 июня (Кафтановский
и Горчаковская); на Новой Земле в конце мая — начале июля; в губе Безымян-
ной 10—12 июня (Красовский, 1937); на юге, повидимому, позднее —в конце
июня, но возможно, что найденные кладки были вторичными, сделанными
взамен собранных промышленниками; на севере—от п-ова Адмирал-
тейства до Горбовых о-вов — кладка бывает во второй половине июня
(Горбунов, 1925, 1929 г.); на северо-востоке на о-ве Гемскерка начало кладки
относится к 3 июня, валовая —в середине июня (Антипин, 1938); на Шпиц-
бергене — в конце мая (Кениг и ле-Руа, 1911); в Исландии 14—21 мая (Фабер
и др.); в области залива св. Лаврентия будто бы в последней трети июня —
24 июня (валовая, Бент, 1919); на Ян-Майене свежие.яйца 3—7 июля (Скон-
нинг, 1933); в Гренландии на о-ве Саундерса 25 июня (Бент). В разных частях
ареала кладка проходит, таким образом, неодновременно, но это не находится
в прямой связи с широтой местности, а главным образом с гидрологическими
условиями близ колоний (ранние кладки в области влияния Гольфштрема,
поздние на сев.-вост. Новой Земли, на Ян-Майене, в Гренландии). Кладка состоит
из одного яйца, удлиненно-грушевидной формы, придающей яйцу некоторую
устойчивость на карнизе (но яйцо все же нередко скатывается, и широко рас-
пространенный в литературе взгляд, будто бы яйца кайр «вертятся на месте»
при толчках и сильном ветре, неверен, Кафтановский). Скорлупа очень
пористая и толстая (самое толстое место —на середине острой половины яйца
и близ толстой его вершины — около 0,34 мм; в тонких местах 0,22—0,26 мм,
Кафтановский, 1941). Окраска скорлупы разнообразна, от темнозеленой или
голубовато-зеленой до беловатой, с различной формы и размеров коричневыми
и черными пятнами и штрихами, чаще расположенными наиболее густо у ту-
пого конца. Размеры меньшие, чем у тонкоклювой кайры: G9) 69—87 х
Х'41—59, в среднем 77,8 X 50,3 мм; средний вес ПО г или 11,9% веса взрос-
лой птицы (Семь островов, Кафтановский); в среднем 85 х 53 мм (Новая Земля,
Горбунов, 1925); в среднем 80,15 х 51,43 мм (Безымянная губа, Новая Земля,
Красовский, 1937); A2) 77—87,1 х 47,7—52,8, в среднем 80,7 х 50,7 мм,
средний вес A5) 100 г (Ян-Майен, Сконнинг, 1933). В случае пропажи яйца
(промысел, деятельность хищников) кайра через 16—18 дней кладет второе,
при гибели второго может отложить и третье, и, быть может, даже четвертое
(Слепцов). Размеры второго яйца обычно немного меньше первого.
Насиживают оба родителя; у самца и у самки имеется по одному насед-
ному пятну, расположенному вдоль продольной оси тела. При насиживании
кайра прижимает яйцо к наседному пятну, закрывая его с боков
перьями, а снизу подсовывает лапы, —тем самым компенсируется отсут-
ствие подстилки в гнезде (на севере Новой Земли кайры кладут яйца,
когда карнизы прикрыты еще снегом, птица слегка при этом разгребает
снег, Антипин, 1938). Участие обоих родителей в насиживании равно-
мерное, птицы сменяются 4—6 раз в сутки (Семь островов, Кафтанов-
ский). Продолжительность насиживания 33—35 дней (Семь островов,
Кафтановский; по Красовскому, 1937, на Новой Земле 30—35 дней). Вылуп-
ление птенца протекает от 1 до 4 дней. Оно отмечено в начале июля
(Семь островов 29 июня—4 июля, массовое — 10 июля, Кафтановский и
Горчаковская; 12 июля, губа Безымянная, Новая Земля, Красовский, 1937;
6—7 июля, Шпицберген, Мунстеръельм, 1910; Ромер и Шаудинн, 1900).
Птенцы от второй (дополнительной, взамен утерянной) кладки на Новой Земле
обнаружены 2—5 августа (Успенский). Размеры только что вылупившихся
птенцов: вес D) в среднем 71,5 г (Горчаковская); длина G) 111—146, в среднем
126,4, вес 65,1 —77, в среднем 70,6 г (Кафтановский); для Новой Земли вес 47—
63, в среднем 54,7 г (Красовский, —данные кажутся преуменьшенными).
Спуск на воду — в мезоптиле, с неразвитыми маховыми: птенец планирует
13 Птицы СССР, т. II
194
ОТРЯД ЧИСТИКИ
при помощи кроющих крыла, образующих несущую поверхность последнего,
и лап. На Новой Земле иногда птенец спускается на воду в 19—20-дневном
возрасте (Красовский, Успенский), на Семи островах в возрасте 23—25 дней
(Кафтановский, Горчаковская, Карташев). Даты спуска на воду указываются
такие: на Новой Земле в начале августа —5—13 августа (Красовский, 1937),
в конце июля (Билькевич, 1904); 10—19 августа в Безымянной губе (Успен-
ский); на северо-востоке северного острова — с начала и до 23 августа
(Антипин, 1938); на Шпицбергене— 17 июля (Мунстеръельм, 1910) и 25 июля
(Бируля, 1910); на Семи островах пер-
вые 25 июля, в массе — б августа (Каф-
тановский). Период размножения про-
должается, таким образом, 56—60
дней: 33—35 дней насиживания и
23—25 дней пребывания в гнезде (Семь
островов).
Линька. Возрастные измене-
ния, как у тонкоклювой кайры: пу-
ховой наряд — мезоптиль — первый
зимний наряд — первый брачный на-
ряд — второй зимний наряд — второй
брачный наряд и т. д. В году бывает две
линьки: частичная предбрачная, охва-
тывающая, главным образом, мелкое
оперение головы и шеи, и полная, по-
слебрачная. Во время последней махо-
вые выпадают почти одновременно,
и кайра на время утрачивает возмож-
ность полета (Саломонсен, 1944). Ве-
сенняя линька заканчивается в конце
апреля — начале мая (Семь островов,
Новая Земля), т. е. позже, чем у тон-
коклювой кайры. Годовалые птицы
линяют поздней и у них предбрачная
линька, повидимому, в корреляции с
более поздним и, быть может, неполным
развитием половых желез, —затяги-
вается до начала июня, а, возможно,
и позднее (например 21 июня в Баренцовом море по 76° с. ш. и 36° в. д.,
по экземплярам Зоологического музея Московского университета; то же
наблюдалось на Семи островах). Около середины сентября мелкое опере-
ние кайр—зимнее A2—15 сентября, Горбунов, 1929).
Питание. Кайры, как другие чистики, кормятся в море. Способы
добывания пищи те же, что и у тонкоклювой кайры. В кормовом режиме на
Новой Земле (Красовский, 1937) главное место занимает рыба, в том числе
треска Gaduscallarias, сайка Boreogadussaida, мойва Mallotus villosus, песчанка
Ammodytes tobianus, наконец Aspidophoroides alrickii. Основной пищей птен-
цам служит молодь трески G7%), затем мойва A8%) и Aspidophoroides E%).
На Мурмане в гнездовый период кайры также кормятся и выкармливают птен-
цов преимущественно рыбой: песчанкой, сельдью, мойвой, мальками трески
и зубатки Gymnacanthus tricuspis; меньшее место занимают беспозвоночные
Mysis oculata, Rhoda raschii, Hippolyte и др. (Горчаковская, Кафтановский
и др.). Птенцы выкармливаются мелкими пелагическими рыбами — сельдью,
песчанкой, мойвой; частота кормления 2—3 раза в день. В разные годы тот
или иной вид рыбы преобладает в кормовом режиме, повидимому, в связи
с подходом косяков рыб к местам гнездования кайр. В желудках кайр, добы-
Рис. 34. Птенец толстоклювой кайры
ТОЛСТОКЛЮВАЯ ИЛИ КОРОТКОКЛЮВАЯ КАЙРА
195
тых в негнездовое время у Шпицбергена, найдены остатки ракообразных
Thysanoessa inermis (Euphausiidaceae) и Spirontocerus gaimardii (Хартли и
Фишер, 1936). В гнездовое время питание взрослых птиц и птенцов в общем
сходно — и те и другие в основном ихтиофаги. В негнездовое время кай-
ры, повидимому, кормятся главным образом мелкими ракообразными, в
частности, каланусами, а также пелагическими полихетами (Кафтанов-
ский).
Полевые признаки. Сходна с тонкоклювой кайрой, но клюв короче и
толще. Повадки также сходны. Летает обычно низко над водой, быстро и часто
взмахивая крыльями. Голос «арра», при
беспокойстве «арр» или «орр», птенец сви-
стит.
Описание. Размеры и строение.
Клюв относительно короткий и высокий, угол
нижней челюсти более выдается по сравнению с
тонкоклювой кайрой; оперение верхней челюсти
не доходит до разреза рта. Хвост закругленный,
из 12 рулевых. Формула крыла 2>3>4>5._ A-е
рудиментарно). Длина тела самцовA4) 400—460,
самок A0) 390—480, в среднем 432,8 и 427 мм; раз-
мах самцов A4) 680—800, самок A0) 670—800, в
среднем 729,3 и 731,5 мм. Вес самцов A4) 605—
1050, самок A0) 617—1000, в среднем 927,3 и
913,9 г (Мурман, Семь островов). Вес новоземель-
ских птиц: самцов B25) 781—1120, самок (99)
872—1100, в среднем 934,9 и 917,8 г (Красовский,
1937). Вес самцов B) 1380—1470, самок F) 1300—
1440, в среднем 1425 и 1345 г (Сконнинг, 1933).
Это необычайно высокие цифры. Крыло самцов и
самок D0) 203—220, в среднем 210,4 мм (Зооло-
гический музей Московского университета); самцов
A4) 210—225, самок A0) 205—221, в среднем 216,2
и 214,3 мм (Семь островов, Кафтановский); самцов
C6) 205,8—224,5, самок B8) 192,5—217,5, в сред-
нем 218 и 210,4 мм (Новая Земля, Красовский,
1937); самцов B) 211—220, самок F) 213—220, в
среднем215,5и217*л<(Ян-Майен, Сконнинг, 1933);
самцов и самок D8) 207—230(Исландия, Ян-Майен,
Шпицберген; Саломонсен, 1944). Длина клюва
самцов C6) 3—30,5, самок B8) 31,1—45,5, в сред-
нем 40,96 и 38,1 мм; высота клюва F4) 13,5—15Д в среднем 14 6 мм (Новая Земля
Красовский); длина клюва самцов и самок 32,2—48,5, высота 13,16 л* (Семь островов).
В размерах, повидимому, имеются популяционные различия, но не подвидовые.
Окраска. Пуховой наряд: спинная сторона бурая с охристым оттенком и с белыми
вершинами пушинок, заметно проступающими на голове и крыльях, менее развитыми на
спине; горло — буроватое, бока — серовато-бурые; грудь и брюхо — белые; пух — жесткий,
волосовидный, с плотно сцепленными бородками базальной части (защита от потери тепла при
открытом гнездовании в холодном климате); лапы очень большие, темносерые; клюв— тем-
ный, рогового серого цвета.
Мезоптиль: спинная сторона, подбородок и горло — шиферно-черноватые, подкрылья—
темные, черновато-бурые со светлыми пестринами; зоб, грудь, брюхо, бока —белые;
маховых и рулевых нет.
Взрослые в брачном наряде: общая окраска спинной стороны —шиферно-черноватая;
бока головы, подбородок, горло — шоколадно-бурые; маховые бурые со светлыми осно-
ваниями внутренних опахал первостепенных, на вершинах второстепенных — белая по-
лоса; рулевые — черновато-бурые; грудь и брюхо —белые, бока со слабо выраженными
сероватыми штрихами; подкрылья — белые с буроватыми пестринами у карпального края и
на вершинах нижних больших кроющих. Радужина —темнобурая, клюв —черный, края
основания надклювья —зеленоватые или беловатые; лапы — черные, желтоватые на перед-
ней стороне цевки и на наружной поверхности пальцев.
В зимнем наряде подбородок, щеки и горло белые, клюв серовато-роговой, менее взду-
тый, чем в брачном наряде. В первом зимнем наряде птенцы напоминают взрослую птицу,
но клюв их меньше. В первом брачном наряде часто остаются белые крапины на
подбородке и зобе, маховые буроваты, сильно обношены. Самцы и самки окрашены
сходно. г
Рис. 35. Толстоклювая кайра
196 ОТРЯД ЧИСТИКИ
Полярная толстоклювая кайра Uria lomvia arroides Portenko
Uria lomvia arroides. Portenko. Mitteilungen aus demZoologischen Museum in Berlin, Bd.
22/2, 1937, стр. 227, о-в Гукера.
Распространение. Ареал. Архипелаг Франца-Иосифа, а именно —
южные побережья входящих в него южных островов, не северней 81° с. ш.
(Горбунов, 1932).
Даты. Начало появления у островов относится к первой половине
марта (самое раннее 9 марта), на севере — позднее, в начале апреля; у колоний
первые птицы появляются в первой половине апреля, валовое появление
отмечается во второй половине апреля — первой половине мая (Горбунов,
1932 и др.). На скале Рубини, о-в Гукера, первое появление !0 апреля, вало-
вое в ночь на 11 апреля (Демме, 1934). Откочевка происходит после спуска
молодых на воду, в сентябре; отдельные особи задерживаются в зависимости от
ледовых условий до октября.
Численность. На архипелаге не менее 14 колоний, а вероятно и
более (Горбунов). С прилета птицы держатся стайками по 15—20 и более, до
100—200 особей (Демме).
Экология. Размножение. Кладка происходит во второй половине
июня; у добытой 19 июня на о-ве Гукера самки было обнаружено готовое к сносу
яйцо. 22 июня птицы сидели на яйцах (Демме). Вылупление происходит во
второй половине июля B8июля, о-в Гукера, Демме; 25 июля, Горбунов). Спуск
птенцов на воду относится примерно к середине августа. Гнездование колония-
ми обычного для вида характера. Размеры яиц B) 80—83,1 х 48 х 9—61,6 мм
(Горбунов).
Л и н ь к а. Во второй половине апреля A3—27 апреля) птицы добывались
в брачном наряде (Горбунов); осенняя линька взрослых, смена нарядов птен-
цов не прослежены.
Питание. В желудках кайр, добытых на о-ве Гукера, найдены отолиты,
мелкая рыба, черви, бокоплавы Paramphiloe hystrix, Atyluscarinatus, Acantho —
stepheia behringensis, раковины двустворчатых моллюсков, также перья и ка-
мешки (Демме).
Описание. Размеры и строение. Как у номинальной североатлантической
расы: крыло E) 291—222, в среднем 214 мм (Портенко, 1937; цифры, приводимые тем же
автором в 1944 г., очевидно неверны и пользоваться ими невозможно).
Окраска. Спинная сторона темная, черноватая, напоминает в этом отношении тихо-
океанский подвид; белая кайма на вершинах второстепенных маховых будто бы уже, чем
у номинальной расы.
И для этой формы систематическое положение (подвид ли?) остается неясным.
Сибирская толстоклювая кайра Uria lomvia eleonorae Portenko
Uria lomvia eleonorae. Portenko. Mitteilungen aus dem Zoolog. Museum in Berlin, Bd. 22/2,
1937, стр. 227, о-в Преображения.
Распространение. Ареал. Остров Преображения в Хатангском заливе;
Новосибирские о-ва — Бельковский и Котельный, быть может, Столбовой. Ве-
роятно к этой же форме относятся птицы из залива Фаддея, Таймыр, с о-вов
Вилькицкого, Генриетта и Беннетта, а также наблюдавшиеся на Северной
Земле (может быть, на залете, Лактионов, 1946; по Демме колонии кайр имеются
там на севере на м. Молотова и на востоке в фиорде Матусевича). Миграции
не выяснены и, вероятно, ограничиваются Восточно-Сибирским морем. Есть
указания на то, что эта птица была встречена в негнездовое время в Беринговом
проливе (Портенко, 1937). В негнездовое время встречена у о-ва Медвежьего
близ устья Колымы (Пальмен, 1887). Норвежская полярная экспедиция
в мае встретила несколько кайр в океане к востоку от Новосибирских о-вов
между 74°42' и 76°46' с. ш. и 153°30' и 166°22' в. д. (Сконнинг, 1928).
ТОЛСТОКЛЮВАЯ ИЛИ КОРОТКОКЛЮВАЯ КАЙРА 197
Описание. Размеры и строение. Клюв послабей и короче, чем у кайр из
области Берингова моря и с о-ва Врангеля: длина ото лба у самцовCK,43—3,51, у самокC)
3,22—3,35, в среднем 3,47 и 3,29 мм (Портенко, 1944). Крыло — как у атлантических форм,
короче, чем у тихоокеанских: длина его у самцов DJ15—223, у самок C) 215—217, в среднем
219 и 216 мм (Портенко, 1937).
Окраска. Спинная сторона очень бледная, серая с шиферным или голубоватым от-
тенком, кофейный тон нижней стороны шеи светлее, пестрины на боках серые, слабо вы-
раженные или отсутствуют; белые вершины второстепенных маховых широкие; малые ниж-
ние кроющие крыла без пестрин. Подвидовое значение этой формы недостаточно ясно.
Чукотская толстоклювая кайра Uria lomvia heckeri Portenko
Urialomviaheckeri. По рте н к о. Доклады Академии наук СССР EI944, стр. 238, м. Уэринга,
северо-восточный берег о-ва Врангеля.
Распространение. Ареал. О-в Врангеля (большая колония нам. Уэрин-
га, немного южнее этого мыса, на северо-восточном побережье острова; запад-
ное побережье у лагуны р. Гусиной, Минеев, 1936,1946); о-в Колючин у север-
ного побережья Чукотского п-ова. Возможно к этой форме относятся и кайры
с о-ва Геральда. Миграции не выяснены.
Даты. Появление у о-ва Врангеля отмечается в первой половине мая,
откочевка—к концу сентября (Портенко, 1937).
Экология. Сведений в литературе нет.
Описание. Размеры и строение. Длина крыла, как у берингова подвида
U. 1. агга, значительная: крыло самцов A9) 213—234, самок A9) 204—240, в среднем 222
и 221 мм (Портенко, 1944). Клюв,по тому же автору, крупнее: длина от лба у самцов B2)
34,8—40,0, у самок A9) 33,9—41,5мм, в среднем 37,9 и 36, 4.
Окраска. Сходная с таковой у северосибирской формы U. I. eleonnae.
Подвидовая самостоятельность требует подтверждения.
Тихоокеанская толстоклювая кайра Uria lomvia arra Pall.
Cepphus Arra. Pallas. Zoographia Rosso-Asiatica, t. II, 1811, стр. 347. Камчатка.
Распространение. Ареал. Северные части Тихого океана к югу от
Берингова пролива. В Азии от южного побережья Чукотского п-ова
F7° с. ш.), в Коряцкой земле, повидимому, в Олюторском заливе, на Охотском
побережье (Тайгонос, Матуга, Аллен, 1905, Дементьев, 1940); о-в Утичий и
Птичий, Шантары (Дулькейт и Шульпин, 1937), на Камчатке, в особенности
на восточном берегу, может быть и на западном; на Курильских о-вах
на юг до Матсува в средней части архипелага (Ямасина, 1931), на Командорах,
на Сахалине и в сев. Приморье (Шульпин, 1936). В Америке на Прибыловых
о-вах, о-ве св. Лаврентия, по Алеутским о-вам, на о-ве Кадьяк, на побе-
режье Аляски от м. Лисберн и пролива Коцебу. Залеты отмечены в приморскую
полосу Анадырского края (Портенко, 1939), зимою от Камчатки (Усть-Кам-
чатск, Бергман, 1935) и Прибыловых о-вов до Командор, вероятно, в Приморье
(Шульпин); также в сев. Японии (Хоккайдо, изредка Хондо), также
у Курильских о-вов, у Алеутских, у южн. Аляски. В северной части ареала
зимою редка. Часть птиц, повидимому, держится зимою в открытом море.
Даты. В Японии на зимовке часть особей остается до мая. Появление
на Прибыловых о-вах отмечается во второй половине апреля (Прибл и
Мк. Эти, 1923). В Авачинской губе появление отмечено в мае (Бергман). На
Командорах у берегов — в конце марта—начале апреля (Стейнегер, 1885,
Иоганзен, 1934; по последнему автору самое раннее 24 февраля). Различия
в датах объясняются, повидимому, нерегулярным характером зимних мигра-
ций, возможно особенностями ледовой обстановки. Осенью покидает места
гнездовья с конца августа и в сентябре (Аляска, Бент, 1919; Курильские о-ва,
Сноу, 1896; Командоры).
198 отряд чистики
Численность. Высокая; в южных частях ареала — Сахалин, Ку-
рильские о-ва —реже, чем тонкоклювая кайра; на Командорах и Прибыло-
вых о-вах чаще последней. На западном берегу Камчатки редка. Отмечена
массовая гибель при штормовой погоде (при залете на Анадыр, Портенко,
1939; у Прибыловых о-вов, Прибл и Мк. Эти, 1923).
Экология. Размножение. Цикл относительно поздний, кладка проис-
ходит в первой половине и около середины июня (на Прибыловых о-вах самая
ранняя—1 июня, но обычно позже; в середине шрня—на Ко-
мандорах. Срок растянут, так как имеются вторичные дополнитель-
ные и даже третьи кладки. Гнезда обычные, колониальные, на карнизах
прибрежных скал, иногда будто бы в щелях (Командоры, Иоганзен). Размеры
яйца G9) 73—91 х 45—55,5, в среднем 82 х 51,5 мм (Аляска, Бент); A5)
76—88 х 49—52,2, в среднем 81,72 х 50,83 мм (Тачановский, 1893); (8)
78,5—84 х 48—53, в среднем 80,81 х 50,4 мм (Командоры, Стейнегер, 1885).
На Командорах пуховики встречаются в начале августа, крупные птенцы —
во второй половине этого месяца, спуск на воду молодых, за которым следует
откочевка, происходит с первых чисел сентября (Иоганзен, 1934).
Линька. Не изучена. В брачном наряде взрослые встречены уже во
второй половине апреля B1 апреля, бухта Ольги, вост. Камчатка); около
середины августа A3—17 августа) осенняя линька еще не началась.
Питание. Как у других подвидов, главным образом мелкая рыба, от-
части беспозвоночные. Анализ желудков добытых у Прибыловых о-вов кайр:
рыба D8,75% встреч), ракообразные, среди них Spirontovaris polaris и т. д.
(Прибл и Мк. Эти).
Описание. Размеры и строение. Размеры крупней, чем у атлантических
форм,в частности номинальной. Крыло самцов и самок A2) 214—235, в среднем 221,7 мм
(Зоологический музей Московского университета); крыло самцов A9) 213—232, самок 209—
226, в среднем 224 и 220 мм (Портенко, 1944). Клюв относительно длинный, около 35—45 мм.
Длина самца A) 430, самки A) 434, размах самца A) 800, самки A) 792 мм (Тачановский).
Вес самки A)830 г. Окраска. Весьма варьирует, но в среднем темнее, чем у других
подвидов, шиферно-черно вата я на спинной стороне, с сильно развитыми пеСгринами боков
и малых кроющих крыла.
РОД ЧИСТИКИ CEPPHUS PALLAS, 1769
Тип Alia grylle
5. Чистик Cepphus grylle L.
Alea Grylle. Linnaeus. Syst. Nat. X, t. I, 1758, стр. 130, о-в Готланд, Швеция.
Русское название. Русское название «чистик» — народное, звукоподражатель-
ное. Соответствует северо-европейскому Teiste, tysti, teisti и не имеет никакого отноше-
ния к «чистоте». На севере также «свистун».
Распространение. Ареал. Северные части Атлантического и Тихого
океанов, Северный Ледовитый океан. В Балтийском море по берегам Ботниче-
ского залива от северной его оконечности до Блекинге в Швеции, на берегах
Финляндии, к югу до Эстонии; на Ладожском озере (Хильден, 1921), у берегов
Дании (Каттегат), по берегам Норвегии; на Мурмане, на востоке до Белого
моря; у Ирландии, Шотландии, на Шетландских, Гебридских и Оркнейских
о-вах, на о-ве Мен, в Англии (Йоркшир, Уэльс); на Фарерских о-вах: в Сев.
Америке от Лабрадора, Гудзонова пролива, Ньюфаундленда до Мена и —
на юге — о-вов Матиникус, Грет-Менен и Сил в Новой Шотландии; в зап.
Гренландии от мыса Фаруэлл до бухты Диско G0° с. ш.); в вост.
Гренландии до Скоресби-зундаидаже 80°40'с. ш.(Сконнинг); на Земле Эльсмира
до Земли Гранта; Баффинова Земля, п-ов Мельвилля и по арктическим остро-
вам до о-ва Мельвилля; в сев. Аляске (мыс Барроу); на о-ве Медвежьем,
Ян-Майене, Шпицбергене, архипелаге Франца-Иосифа, на Вайгаче, Новой
Земле, на о-ве Преображения у устья Хатанги, на Сев. Земле, Новосибир-
чистик
199
ских о-вах, о-ве Беннета, Врангеля (быть может Геральд); по побережью Чукот-
ской земли, на островах и побережьях Берингова моря, на Курильских и Коман-
дорских о-вах, в Аляске на Прибыловых и Алеутских о-вах, на побережье
от мыса Лисберн и далее на юг до вджн* Калифорнии (о-ва Санта Барбара,
Фаральонес).
Характер пребывания. В южной части ареала более или
менее оседлая птица, из северных частей откочевывает в зависимости от ледовых
условий. Зимою встречен на южн. Балтике, случайно проникая до Северного
моря; у берегов Норвегии, Англии, изредка Голландии, Бельгии, сев. Фран-
ции; в Америке более или менее регулярно до Массачусетса (изредка до Кон-
нектикута, Лонг-Айленда и Нью-Джерси, Бент, 1919); у Фарерских о-вов
и Исландии; у берегов западной Гренландии, даже на севере ареала; в Кар-
ском и Баренцевом море, у берегов Мурмана; в Беринговом море, на юг до
сев. Японии. Кочующие чистики встречены в самых высоких широтах
Карта 30. Распространение атлантического чистика Cepphus grylle
7. С. g. atlantis, 2. С. g. grylle, 3. С. g. tajani, 4. С. g. mandtii, 5. C. g. columba, 6. C. g. snowi, 7. C.g,
kajurka, 8. C. g. ultimus, 9. C. g. islandicus, 10. C. g. faeroensis;
a — граница гнездовий (цифры в кружках обозначают номера подвидов).
Северного Ледовитого океана (в частности, Нансеном встречены к северу от
Шпицбергена за 84° с.ш. на разводьях; экспедицией ледокольного судна «Седов»
одиночная птица найдена 10. VII. 1939 г. под 86°8'с. ш. и 74°54' в. д.; 16июля
под 85°34' с. ш. и 62° 14' в. д.; советской дрейфующей станцией «Северный
полюс» в 1937 г. между 88° и 89° с. ш., Портенко, 1946).
Биотоп. Гнездовый — скалистые побережья островов и материков.
В негнездовое время — в море, обычно неподалеку от берегов (самая отда-
ленная находка сделана в 170 м восточней о-ва Бель-иль, Канада, поУинн-
Эдварсу, 1935). Чистики зимуют даже в замерзающих морях, собираясь по
полыньям и разводям. Молодые неполовозрелые особи до гнездования дер-
жатся в открытом море и передвижения и кочевки их, вероятно, более значи-
тельны.
Подвиды и варьирующие признаки. Систематика чистиков разработана еще недо-
статочно. В пределах вида можно установить три группы: североатлантическую, полярную
и тихоокеанскую, различающиеся экологией (календарь периодических явлений, большая
или меньшая оседлость) и морфологией (размеры, число рулевых перьев, окраска). При
обособленном расположении гнездовий и при обычной консервативности гнездовых место-
обитаний отдельные популяции представляют заметные различия, послужившие для ряда
авторов основанием для выделения нескольких, характеризующихся слабо выраженными
особенностями, подвидов (Саломонсен, 1944; Портенко, 1937, 1944). Сомневаясь в возмож-
ности считать эти популяционные различия признаком настоящих подвидов (географических
форм) и, во всяком случае, оставляя этот вопрос открытым, приводим все же полный каталог
описанных форм.
а.Северная группа С. g. mandtii M a n d t, 1822 — высокие широты Север-
ного Ледовитого океана — вост. Гренландия, Ян-Майен, о-в Медвежий, Шпицберген,
архипелаг Франца-Иосифа, Вайгач, Новая Земля, Северная Земля, Новосибирские о-ва;
C.g tajani Portenko, 1944 — о-в Врангеля и, возможно, Геральда; С. g. ultimus S a 1 о-
200 отряд чисгики
monsen, 1944 — сев.-вост. Америка, зап. Гренландия, на юг до сев. Лабрадора (Рама)
и Гудзонова залива, вероятно у м. Барроу.
Переходом от северной группы к южной, атлантической, служит С. g. arcticus В г е h m,
1824 — из зап. Гренландии и среднего Лабрадора (Саломонсен, 1944).
б. Атлантическая группа С. g. islandicus Н о г г i n g, 1934 — оседлый в
Исландии; С. g. faeroeensis В г е h m, 1837 — оседлый на Фарерских о-вах; С. g. atlantis
Salomonsen, 1944— зап. Швеция, вост. Дания, Норвегия, Мурман, Белое
море, Англия, Шотландия, Ирландия, Сев. Америка от южн. Лабрадора до Мена;
С. g. grylle L., 1758—из Балтийского моря и Ладожского озера.
в. Тихоокеанская группа С. g. columba Pall, 1811 — Берингово море
и сев.-зап. Америка; С. g. snowi S t e i n e g е г, 1897 — Курильские о-ва; С. g. kajurka Por-
te n k о, 1937 — Командорские и быть может Алеутские о-ва.
Последняя группа многими авторами рассматривалась как особый вид, но при наличии
географического викариата, большого морфологического и экологического сходства; — для
этого оснований нет (Кафтановский, 1941; Саломонсен, 1944).
Североатлантический чистик Cepphus grylle atlantis Salomonsen
Cepphns grylle atlantis. Salomonsen. Goteborgs Kungl. Vetenskaps-och Vitterhets.
Samhalles Handlingar., 1944, стр. 77, Бргуслан, Швеция
Распространение. Ареал. Северные части Атлантического океана —
у берегов Англии и Шотландии (на западе на о-ве Мен, ст. Бис, Уигмаун, Эйр,
Арджилл и далее к северу, на востоке до сев.-зап. Кезнесс, на Гебридских, Орк-
нейских и Шетландских о-вах, Уайзерби, 1941), в Ирландии, преимущественно
на западе; в западной Швеции к югу до Ведере, в Голландии в Каттегате
Eб°28' с. ш.); в Дании по островам Каттегата; по побережьям Норвегии до
Варангера; в СССР на Мурмане (Айновы о-ва, Пала-губа, Сайда-губа и т. д.;
Семь островов), в горле Белого моря (о-в Сосновец, Мела, 1882) и у западного
Карельского берега, на Соловецких о-вах, в Онежском заливе (о-в Шужмуй);
в Сев. Америке от побережья Мена и Новой Шотландии (о-ва Мати-
нукус, Грет-Менен, Сил) до южн. Лабрадора. В негнездовое время — как
вблизи мест гнездовий в море (в СССР на Мурмане по крайней мере на западе),
так и несколько южнее, до атлантического побережья Франции (окольцован-
ная в Осло-фиорде птица добыта в Бретани, Сконнинг, 1937), западной Гер-
мании, случайно до Бельгии и Голландии; американские чистики на зимовке
регулярно доходят до берегов Массачусетса. В СССР известен случай залета
к Пскову и устья р. Великой в 240 км от берега моря (Зарудный, 1911); све-
дения, возможно, относятся к номинальному подвиду.
Даты. Появление на Семи островах отмечается с конца февраля,
также в марте и начале апреля; во второй половине апреля птицы
уже многочисленны, но гнездовий еще не занимают (Кафтановский, Горчаков-
ская). В середине августа птицы, во всяком случае взрослые, покидают гнездо-
вые колонии. В Кольском заливе птицы оседлы (Спасский, 1925), в Онежском
заливе встречены в декабре и в феврале (Поляков, 1929).
Биотоп. Гнездовый — расщелины или пещеры морских побережий,
редко в торфе. Зимою в прибрежных свободных ото льда участках моря.
Численность. Распространение спорадично.
Экология* Размножение (Кафтановский, Горчаковская, Дарская).
У гнезд на вост. Мурмане чистики появляются в начале июня, когда базары
очищаются от снега. Так как чистики гнездятся в норах, медленно оттаиваю-
щих, то цикл размножения у них поздний. Гнездование колониальное, но также
и отдельными парами, в зависимости от биотопических условий. Гнездо — в це-
лях или осыпях, среди камней, реже в торфе. На Семи островах они располо-
жены как близко от берега (в 30—40 м), так и довольно далеко B50—300 м).
В гнезде имеется своеобразная подстилка из мелких камешков, предохраняю-
щая кладку от сырости. Гнездовый консерватизм выражен сильно — чистики
занимают одно гнездо из года в год. Спаривание происходит, повидимому, во
второй половине мая — начале июня. Кладка состоит из 2 яиц, изредка из
чистик 201
1 и даже 3. Дополнительные вынужденные кладки имеют место. Откладка яиц
происходит в конце мая —• начале июня, валовая же обычно около середины
июня (Семь островов, 1937 —начало 26—27 мая; 1939 —с начала июня по
20 июня; 1940 —с 7 по 24 июня, 1941 —с 29—30 мая по 10 июня). Форма
скорлупы яйцевидно-эллиптическая, сходная с чаячьими яйцами. Окраска
белая или зеленоватая с крупными черноватыми крапинами, сгущающимися
на тупом конце. Скорлупа относительно тонкая. Размеры F8) 57—65 х 38—44,
в среднем 59,9 х 40,8 мм; вес A8) 48,1—57, в среднем 52,1 г. Вес скорлупы
3,1—4,6 г. Насиживают оба родителя 27—28 дней, после откладки последнего
яйца. Вес только что вылупившегося птенца A0) 28,8—40, в среднем 33,9 г;
перед вылетом около 460 г. Вылупление продолжается два или даже три дня.
Массовое появление птенцов обнаруживается в начале июля и до 20 июля.
У птенца контурное оперение развивается в гнезде, мезоптиля нет; вылетает
из гнезда молодой чистик с развитыми крыльями (но не достигнув полного ро-
ста, с несколько недоросшими по длине маховыми). Терморегуляция у птенцов
устанавливается на 4-й день по вылуплении. В гнезде птенец находится 35—
38 дней.
Линька. Смена нарядов: пуховой — первый зимний — первый брач-
ный — второй зимний — второй брачный и т. д. У взрослых имеются две
линьки в году: полная, послебрачная, и неполная, предбрачная, охватывающая—•
в отличие от кайр — все оперение, кроме маховых и рулевых. Весенняя пред-
брачная линька заканчивается, как правило, в начале июня. Начало ее отме-
чено уже в конце декабря или первых числах января (Саломонсен, 1944). Со вто-
рой половины июля начинается послебрачная линька — сначала на голове и
шее, потом на спине. Линька эта продолжается до ноября, в конце этого месяца
птицы уже в зимнем наряде. Смена маховых протекает одновременно.
Питание. Пищу составляет мелкая рыба — мальки трески, песчанка,
мойва, сельдь, минога, ракообразные, моллюски (мидия). Птенцы выкармлива-
ются главным образом рыбой (треска, пикша, песчанка, Pholis gunellus, Myo-
xocephalus scorpio), также креветками (Горчаковская, Кафтановский). Кормо-
выми станциями являются, главным образом, прибрежные участки моря.
Чистик добывает корм, ныряя и плавая под водою (от 30 до 75 сек., в среднем
около 45 сек., Уайзерби, 1941).
Полевые признаки. Некрупный чистик. В брачном пере черноватой окрас-
ки с белым зеркальцем на крыле, с красными ногами. В зимнем наряде и в гнез-
довом (молодые) — пестрый, черновато-бурый с белыми пестринами, почти
белый на брюшной стороне. При плавании держит шею несколько вытянутой
кверху. Хорошо передвигается по земле, опираясь на пальцы; при сидении и
стоянии опирается обычно на цевку. Полет быстрый, с частыми ударами кры-
льев. Голос —свист «и-и-и»; потревоженная на гнезде птица издает шипенье.
Описание. Размеры и строение. Клюв почти прямой, слабо сжатый с боков.
Ноздри узкие, щелевидные, прикрытые оперением. Рулевых обычно 12, изредка 13—14.
Цевка сжатая с боков, сетчатая, короче среднего пальца с когтем. Когти тонкие, острые и за-
гнутые. Длина самцов A0) 330—380, самок G) 325—360, в среднем 351,7 и 343,6 мм. Размах
самцов A0) 480—590, самок G) 470—590, в среднем 542,8 и 541,4 лмс. Вес самцов A0) 350—
471, самок G) 398—451, в среднем 417,6 и 421, 4 г. Крыло самцов A5) 163—176, самок A0)
165—176, в среднем 109,06 и 171,1 мм (Зоологический музей Московского университета).
Саломонсен A944) приводит несколько меньшие размеры: крыло самцов и самок E8) 155—
173, в среднем 162,7 мм. Длина клюва около 30—35 мм, высота 10—12 мм.
Окраска. Пуховой птенец черный, пух рыхлый, слабо сцепленный, на брюшной
стороне несколько гуще (сырые гнезда). Клюв черный, розоватый у основания над-
клювья; ноги черные, радужина коричневая. Гнездовый наряд (надевается в возрасте 30—
32 дней): спинная сторона черная с большей или меньшей примесью беловатых пестрин,
в особенности на шее, плечевых, белое зеркальце на крыле испещрено черноватым; щеки
пестрые, черноватые с белыми пестринами; брюшная сторона белая с более или менее разви-
тыми черноватыми каймами перьев, маховые и рулевые черные; ноги лиловато-красноватые,
клюв черный. Перо отличается рыхлостью (как бы переход к мезоптилю). Взрослые птицы
в брачном наряде: спинная сторона черноватая, с более или менее заметным зеленоватым
202
ОТРЯД ЧИСТИКИ
отливом в свежем пере; брюшная сторона черновато-бурая, маховые черные с белыми осно-
ваниями внутренних пар, рулевые черные; на крыле белое зеркальце,образованное средними
кроющими и вершинами больших кроющих; подмышечные и часть подкрылий белые. Клюв
черный, ноги яркокрасные, когти черные, радужина бурая.
Взрослые в зимнем наряде черные, с белыми каймами перьев на спинной стороне тела,
каемки эти особенно широки на голове, шее, плечевых и пояснице; на уздечке и кроющих уха
черное пятно; брюшная сторона белая. Подобный зимний наряд надевают и молодые птицы
Рис. 36. Атлантический чистик
в первую осень своей жизни (между сентябрем — декабрем по Уайзерби, 1941), но белые пе-
стрины и каймы перьев развиты меньше. Годовалые чистики в первом брачном наряде (пови-
димому неразмножающиеся)отличаются от старых птиц наличием пестрин на зеркальце кры-
ла и иногда небольшой примесью таких же пестрин на брюхе. Самцы и самки окрашены оди-
наково.
Балтийский чистик Cepphus grylle grylle L.
Распространение. Ареал. Балтийское море — в Швеции по берегам
Ботнического залива от северной оконечности на юг до Блекинге, на о-в Гол-
ланд, по восточному берегу Ботнического залива в Финляндии, на северном
берегу Финского залива и на Аландских о-вах. В СССР у берегов Эстонии
уПакерорта (м. Пакри), нерегулярно на о-в Рогоэ (Кумари) и в северо-западном
углу Ладожского озера (Хильден, 1921). Залеты к Ленинграду (Мензбир).
Биотоп. Шхеры и литоральная часть моря.
Характер пребывания. Оседлая птица. В негнездовое время
совершает только небольшие кочевки — в Швеции у Сконе, в южн. Дании
и до Балтийского побережья Германии.
Численность. Ограниченная, гнездовые колонии невелики. В Эсто-
нии на м. Пакри гнездится не более 100 пар (Кумари).
Экология. Размножение. В Эстонии гнезда расположены в тре-
щинах известковой стены Пакрисского глинта, в \Ъм над уровнем моря. Тре-
щины глубокие — от 0,5 до 3 м. Выстилки в гнезде нет. Кладка из 2 яиц про-
исходит в середине июня (Кумари). В Финляндии кладка имеет место во
чистик 203
второй половине мая — начале июня. Размеры яиц E0) 54,2—64,8 х 35,4—
41,6 мм (Линдфорс, 1931, но все ли эти измерения относятся к балтийской фор-
ме — неясно). Насиживают оба родителя, повидимому до 28 дней (Хортлинг,
1931; в этой работе срок насиживания указывается в 21 день, что очевидно
неверно).
Линька. Послебрачная линька бывает во второй половине августа и
продолжается до октября — ноября. Весенняя предбрачная уже с января,
обычно же в феврале. В марте птицы в брачном наряде (Саломонсен, 1944).
Цикл, таким образом, более ранний, чем у северной группы подвидов.
Описание. Размеры и строение. Крупней других атлантических подвидов
(что, по Саломонсену, 1944, объясняется при оседлом образе жизни птиц относительно более
низкой температурой воды). Крыло B5) 167—182, в среднем 174,1 мм; клюв массивный, длина
30—35, в среднем 32,9 мм, высота у основания 10,5—12, в среднем 11,5 мм (Саломонсен,
1944)
Окраска. Как у предыдущего подвида. Самостоятельность этого подвида, быть мо-
жет ледникового реликта — вызывает сомнения, возможно он идентичен с предыдущим.
Полярный чистик Cepphus grylle mandtii M a n d t
Uria Mandtii. Mandt. Observ. Hist. Natur. Itin Groenl. 1822, стр. 30, Шпицберген.
Распространение. Ареал. Северная часть области распространения
вида—вост. Гренландия (бухта Франца-Иосифа, Сабина, о-в Шеннон;
к югу до Скоресби-зунда, на север до 80° 10' с. ш. Лида у Маллемукфь-
еллен, Манних, 1910), Ян-Майен, о-в Медвежий, Шпицберген, архипелаг Фран-
ца-Иосифа, Новая Земля (на юге от Карских ворот и по западному берегу; на
бостонном берегу между м. Желания и о-вом Гемскерк, Антипин, 1938; по осталь-
ным частям восточного побережья гнездование не доказано); может быть на
о-ве Уединения (Мон, 1879) и на вост. Таймыре (Плеске, 1928), на о-ве Преобра-
жения против устья Хатанги, на архипелаге Северная Земля (Лактионов,
1946); на о-вах де Лонга (о-в Беннетта, Генриетта, Вилькицкого), на Новоси-
бирских о-вах (Столбовой, Бельковский, Котельный, Новая Сибирь), наконец
на северо-западном побережье Чукотского п-ова (м. Иинретлен, Пальмен,
1887; м. Кибера и Северный или Иркайпий, Тайер и Бенгс, 1914).
В негнездовое время держится в свободных от льдов участках Северного
Ледовитого океана среди полыней и разводий, откочевывая недалеко от обла-
сти гнездовий (но не встречается, повидимому, у архипелага Франца-Иосифа
и у Северной Земли). Встречен в разных местах Баренцова и Карского морей
в районе Шпицбергена, у вост. Гренландии (за 74° с. ш., Сконнинг, 1933), у Но-
вой Земли (Экланд, 1928; Антипин, 1938; по последнему автору даже у северо-
востока), изредка к северу от восточного побережья Сибири G февраля 1902 г.
между Ляховским о-вом и материком Плеске, 1928; 9 августа 1915 у м.
Большой Баранов, Рил и, 1918); на востоке в области Берингова пролива.
Отдельные особи проникают южнее —до Мурмана (Смирнов, 1901; Гебель,
1901; в Зоологическом музее Московского университета хранятся экземпляры,
добытые в марте (ст. ст.) 1909 у св. Носа, взрослая и молодая птицы), до Коряц-
кой Земли (бухта Корфа), до о-ва св. Лаврентия (Фридманн, 1932) и Нортон-
зунда, Аляска (Бент, 1919). Кочующие (холостые) птицы встречены в разное
время в самых высоких широтах: к северу от Шпицбергена за 84° с. ш. (Коллетт
и Нансен, 1901); 31 июля 1938 под 81°51' с. ш. и 136°58' в. д., 10 июля 1939
под 86°08' с. ш. и 70°54' в. д. и 16 июля под 85°34' с. ш. и 62°14' в. д.
экспедицией ледокольного судна «Седов»; между 88° и 89° с. ш. станцией «Се-
верный полюс» (Портенко, 1946).
Характер пребывания. Как и у других подвидов, полу-
оседлая или кочующая птица.
Даты. Появление у мест гнездовья отмечено на Земле Франца-Иосифа
уже в последней трети февраля, валовое —в первых двух третях марта (Демме,
204 отряд чистики
1934; Горбунов, 1932); в это же время отмечен и на Шпицбергене (Бируля, 1910;
Норденшельд, 1875), но на Северной Земле —о-в Седова, будто бы лишь в сере-
дине мая (Лактионов, 1946). Для севера Новой Земли валовое появление отме-
чено в апреле (Антипин, 1938). Осенней откочевке чистиков предшествует
постепенное оставление ими базаров. На Земле Франца-Иосифа чистики по-
кидают гнездовья в последних числах августа и в первой половине сентября
B9 августа — 12 сентября, Горбунов, 1932; Демме, 1934), окончательно отко-
чевывают в октябре. Отлет на Северной Земле происходит около середины нояб-
ря(?). Откочевка на Шпицбергене—в первой половине сентября C0 августа —
15 сентября, Мунстерьельм, 1910). На Новой Земле чистики покидают гнезда
в конце августа — начале сентября, задерживаясь у южного острова близ
Маточкина Шара до середины ноября A8 ноября, Сконнинг, 1907). Следует
при этом иметь в виду, что часть птиц, как указывалось, зимует вблизи от мест
гнездовья.
Биотоп —как и у других подвидов —гнездится в скалах и среди каме-
нистых россыпей, иногда довольно далеко от моря —до 600 м над уровнем моря
(Шпицберген), даже в 2 км от берега (там же, Бируля, 1910). В негнездовое
время у кромки льда по полыньям.
Численность. Размещение колоний спорадично.
Экология. Размножение. Гнездится колониально, как и другие
подвиды чистиков, обычно незначительными группами, но на севере ареала
местами образует большие скопления (около 1000 пар на скале Рубини на о-ве
Гукера, архипелаг Франца-Иосифа, Демме, 1934). В некоторые годы наблю-
дается негнездование — прилетевшие на места гнездовья чистики не размно-
жаются (в 1934 г. на Северной Земле, Лактионов, 1946). На негнездование ука-
зывает и встреча в летнее время птиц с неразвитыми половыми железами (Кол-
летт и Нансен, 1901). Половая зрелость наступает, повидимому, в возрасте
около двух лет. Спаривание и токование начинается с появления птиц у гнез-
довых колоний. При токовании самец подходит к самке, кричит «и-и-и», затем
обе птицы сходятся и вытягивают шею (Демме).
Гнезда занимаются много лет подряд, расположены они в труднодоступ-
ных расщелинах и между камнями. Кладка состоит из 2 яиц, обычной для вида
окраски. Размеры C5) 54,6—64,3 х 37,6— 41,6, в среднем 58,58 х 39,44 мм
(Хартерт, 1917). Время кладки на Шпицбергене 10 июня —5 июля (Кениг и
ле-Руа, 1911), на Земле Франца-Иосифа 20 июня — 10 июля (Горбунов, 1932),
на севере Новой Земли 17 июня (Антипин, 1938). По тому, что яйца были най-
дены на Земле Франца-Иосифа еще в первой половине сентября (Горбунов,
1932), можно заключить, что, повидимому, бывают и вторые, «вынужденные»,
кладки. Насиживают оба родителя (наседные пятна и у самцов, и у самок).
Старые птицы «отводят» от гнезд (Антипин, 1938). Вылупление происходит,
повидимому, в июле. Спуск птенцов на воду отмечен с 16 августа на о-ве Гукера
(Демме, 1934), с 11 августа до начала сентября на Земле Франца-Иосифа (Гор-
бунов, 1932), во второй половине августа —начале сентября на Новой Земле
(Горбунов, 1929), 5 сентября на о-ве Вайгач (Хейглин, 1874), уже 10 августа и
позже на Шпицбергене (Мунстеръельм, 1910); на северо-западном побережье Чу-
котского п-ова в первых числах сентября (Тайер и Бенгс, 1914).
Линька. В конце марта на Земле Франца-Иосифа встречаются пест-
рые чистики, еще не закончившие линьку B5 марта, Демме, 1934). С другой
стороны, в это же время некоторые птицы уже перелиняли B3 марта, 29 марта,
Горбунов, 1932). В апреле птицы — в брачном наряде. Линяние в зимний наряд
протекает, повидимому, в сентябре — октябре, так как в последнем месяце
добывались уже вполне перелинявшие птицы (бухта Корфа, Коряцкая Земля;
Земля Франца-Иосифа, Джексон, 1899, но этот автор добыл и 23 сентября птицу,
не закончившую линьку; на Земле Франца-Иосифа, по Горбунову, 1932,
21 сентября был добыт чистик в зимнем насяде, но еще сохранивший пеньки).
чистик 205
Питание. Как и у других подвидов, пищу составляет мелкая рыба
и беспозвоночные. Полярные чистики кормятся, держась стайками, у берегов
и разводий, причем весьма деятельны даже полярной ночью (Антипин, 1938).
Отмечено, что они проплывают под водой 30—50 (Демме, 1934) и даже 75—80 м
(Горбунов, 1932), находясь там 50—60 сек. В пищеварительном тракте добытых
на Земле Франца-Иосифа чистиков обнаружена мелкая рыба, полихеты, но
главным образом, бокоплавы Atylus carinatus, Paramphitoe hystrix, Acanthos-
tepheia behringensis, Anonyx nugax (Демме, 1934).
Описание. Размеры и строение. Принадлежит к северной группе подвидов,
различающихся от атлантических, кроме окраски, меньшей величиной — длиной крыла и
высотой клюва. Крыло самцов и самок B2) 154—170, в среднем 164,48, в том числе у самцов
154—170, у самок 156—168 мм (Зоологический музей Московского университета); уновозе-
мельских самцов A2I58—177, у самокD) 170—175, в среднем 168,16 и 172 мм (сборы Успен-
ского); у самцов и самок 159—175, в среднем 166,9 мм (Саломонсен, 1944),; D0) 150—174,
в среднем 166 мм (Портенко, 1944). Длина клюва около 30—34 мм, высота около 8—10 мм
(у атлантических, как указывалось, высота клюва около 10—\2мм). Вес самцов A1) 342—
424, самок D) 377,5—446, в среднем 385 и 402,1 г (по новоземельским особям в Зоологическом
музее Московского университета). Число рулевых 12, изредка 13—14.
Окраска. В брачном наряде характерна большая величина белого зеркальца на
крыле, белый цвет распространяется на основные части средних и даже малых кроющих
крыла, на внутренних опахалах маховых белого цвета больше и он доходит обычно до ствола
пера. В зимнем наряде заметно светлее других подвидов на спинной стороне, белые каемки
перьев шире; голова, плечевые, поясница с преобладанием белого цвета, иногда почти вовсе
белые; черноватое пятно на уздечке и кроющих уха отсутствует. Молодые птицы тоже свет-
лее атлантических и с белыми вершинами второстепенных маховых (хотя более пестрые на
спинной стороне, чем взрослые в зимнем наряде).
Таянов чистик Cepphus grylle tajani Portenko
Cepphusgrylle tajani. Портенко. Доклады Академии наук СССР E), 1944, стр. 239,
м. Пиллер, о-в Врангеля.
Распространение. Ареал. О-в Врангеля, где гнездится от мыса Фомы
к северу до реки Клар; особенно большие колонии, по много тысяч особей, на
скалах у мыса Гаваи (Минеев). Быть может на островах Геральд и Генриетта.
Характер пребывания. Гнездящаяся перелетная птица, появ-
ление в начале мая, иногда в конце апреля, откочевка к концу сентября (Ми-
неев, Портенко, 1937).
Описание. Чистики с о-ва Врангеля выделены в особый подвид потому, что они отли-
чаются от других полярных чистиков (группа «mandtii») насыщенным черноватым с интен-
сивным зеленым отливом цветом, а также более крупными размерами. Крыло самцов и самок
D1) 166—181, в среднем 172 мм (Портенко, 1944), у двух взрослых особей с о-ва Врангеля в
Зоологическом музее Московского университета крыло 162,5 и 168,5 мм.
Систематическое положение этой формы в качестве подвида остается сомнительным.
Весьма вероятно, что особенности гнездящихся на о-ве Врангеля чистиков должны рассмат-
риваться как проявление популяционной, а не географической в строгом смысле слова, из-
менчивости.
Берингов чистик, берингова каюрка Cepphus grylle columba Pall.
Cepphus Columba. Pallas. Zoographia Rosso-Asiatica t. 11, 1881, стр. 348, Камчатка.
Русское название. Местное название — извозчик, каюр, каювер (Крашенинни-
ков, 1755), свистун, каюрок, каюрка.
Распространение. Ареал. Северная часть Берингова моря —восточный и
южный берег Чукотского п-ова (на север до Уэллена, но указания на о-ва
Геральда и Врангеля ошибочны, Стейнегер, 1885), восточное побережье Камчатки
с прилегающими островами: на западном —между м. Лопатка и Камбальной
(Бергман, 1935); быть может в Анадырском лимане и на Сахалине; на о-ве Диомид
в Беринговом проливе; в Америке и сев. Аляске от м. Лисберн (?), но главным
образом на южном ее побережье и у Британской Колумбии (Ванкувер, о-ва
Королевы Шарлотты), у Вашингтона (залив Педжета), у Орегона и Калифор-
206
ОТРЯД ЧИСТИКИ
нии (Санта Барбара, Фаральонес); на севере по островам Берингова моря —
на Прибыловых (?), о-вах св. Лаврентия, св. Матвея, Холл. Возможно к этой
же форме относятся и птицы Алеутских о-вов. В негнездовое время — от Але-
утских о-вов и Прибыловых до южн. Калифорнии (о-в Сан-Клементе, Бент,
1919), у сев. и средних Курильских о-вов, Камчатки и сев. Японии.
Характер пребывания. Частично оседлый в незамерзающих
участках моря. Часть птиц откочевывает к югу (см. выше).
Даты. Сроки сезонных перемещений не выяснены. Появление у мест
гнездовья раннее, но когда именно —¦ неясно. В Аляске в конце марта и в
начале апреля (Бент). Осенью молодые постепенно покидают гнездовые коло-
нии и откочевывают (в районе Нортон-зунда держатся до ноября). На При-
Рис. 37. Берингов чистик
быловых о-вах на зимовке встречены с ноября — января и до апреля, Прибл.
и Мк. Эти, 1923).
Биотоп. Гнездовый — скалистые побережья островов и материка.
В негнездовое время в прибрежных частях моря.
Численность обыкновенная, но немногочисленная птица вост. Кам-
чатки и Алеутской гряды; распространение спорадично.
Экология. Размножение. Изучено недостаточно. Гнездится в ска-
лах между камнями, не образуя больших скоплений. Иногда на низких берегах
в непосредственной близости от воды (Камчатка). Кладка в Калифорнии про-
исходит в первой половине мая, на Алеутских о-вах будто бы во второй поло-
вине июня (Бент, 1919). Кладка из 2 яиц, зеленоватых или зеленовато-белых,
с темными черными поверхностями и сероватыми глубокими крапинами". Раз-
меры E1) 57—68,5 х 38,5—43,5, в среднем 60,5 х 21 мм (Бент, 1919). Наси-
живают оба родителя, в течение 21 дня. 2—3 августа на о-ве Топорков, вост.
Камчатка, молодые выросли «на половину» (Бергман, 1935).
Линька. Как у других подвидов. Предбрачная линька взрослых охва-
тывает все контурное оперение, но не маховые и рулевые, и тянется иногда
с февраля и даже января до мая и даже июня; полная послебрачная линька
проходит с середины августа и в сентябре (Бент, 1919).
Питание. Мелкая рыба, моллюски, ракообразные. У зимующих у
Прибыдовых о-вов чистиков в желудке найдены амфиподы Allarchestes malleo-
lus, изоподы ldothea achotensis; кроме того Lithodes brevipes, Hapalogaster greb-
nitzkii, Dermaturus mandti; наконец, мелкие рыбы (Прибл и Мк. Эти, 1923).
чистик 207
Описание. Размеры и строение. Как и последующие подвиды, отличаются
от атлантической и полярной группы более крупной величиной и 14 рулевыми. Крыло самцов
A0) 181—189, самок A2) 184—195, в среднем 185 и 184 мм (Портенко, 1937). Вес гнездовых
самцов с вост. Камчатки 410, 475, 505 (Зоологический музей Московского университета).
Окраска. Как у предыдущих, но подкрылья в брачном наряде дымчато-бурые,
лишь по середине белые; белое зеркальце на крыле разделено бурой перевязью. В зимнем
наряде белого «а подкрыльях больше, но все же карпа льный край дымчато-бурый; зеркаль-
це с теми же особенностями, как в летнем наряде.
Систематические замечания. По всей вероятности к этому подвиду относятся пред-
ложенные Сторером названия С. с. adianta — тип. местн. Ванкувер и С. с. eureka — с
Фараллонских о-вов (S t о г е г. The Condar, 1950, стр. 28). К сожалению, в нашем рас-
поряжении не было птиц из американской части ареала.
Курильский чистик, курильская каюрка Cepphus grylle snowi Stejneger.
Cepphus snowi. Stejneger. The Auk. 1897, стр. 201, Райкоке, Курильские о-ва.
Распространение. Ареал. Курильские о-ва от Урупа на юге до Пара-
мушира на севере. Встречен у северной части Хоккайдо, но гнездование там
сомнительно. Быть может к этой форме относятся наблюдения над каюрками у
Сахалина B3 июня у м. Крильон, Никольский, 1889) и над пролетом их в
Приморье бухта Патрокл (в сентябре, Шульпин, 1936). Отмечен у м. Лопатки
(Кларе, 1910).
Характер пребывания. Более или менее оседлый.
Биотоп. Как и у других подвидов.
Численность. Очень обычен (Бергман, 1935).
Экология. Сведения отсутствуют. Размер яйца 62,5 х 39,6 мм (Хартерт,
1917).
Описание. Размеры и строение. Хвост, как у других близких подвидов,
из 14 рулевых. Размеры относительно крупные: крыло 184—187лш (Хартерт) и даже 190мм
у самца и 192 мм у самки (Саломонсен, 1944). Окраска. Как у предыдущей формы, но
белое зеркальце на крыле еще менее развито и обычно разбито поперечными перевязями
на 3—5 поперечных полосы, у некоторых особей белого зеркальца нет вовсе (такие птицы,
по Ямасина, 1931, встречаются и даже преобладают на средних Курильских о-вах и в этом
отношении они как бы представляют переход к очковому чистику, гнездящемуся на
южн. Курилах — Кунашире и Итурупе).
Командорский чистик, командорская каюрка Cepphus grylle kaiurka Por-
te nko.
Cepphus kaiurka: Portenko. Mitteilungen aus d. Zoolog. Museum in Berlin, Bd. 22B) 1937,
стр. 228, о-в Медный.
Распространение. Ареал. Командорские о-ва (о-ва Беринга и Медный).
Характер пребывания. Более или менее оседлый, но зимою не
держится непосредственно близ берегов.
Даты. Появление у гнездовых колоний отмечается в конце марта A4 мар-
та, по Стейнегеру, 1885). Откочевка постепенная и происходит после спу-
ска птенцов на воду в августе и до октября (Хартерт, 1920, Иоганзен, 1934).
Биотоп. В гнездовое время, как у других подвидов, скалистые побе-
режья с расщелинами и т. п., в зимний период литоральная полоса
моря.
Численность. Обычная птица, однако, при узкой области распро-
странения подвида — популяции в общем немногочисленны.
Экология. Размножение. Гнездится отдельными парами и неболь-
шими группами, не образуя значительных колоний. Кладка состоит из 2 яиц
и происходит около середины июня; размеры яиц B) 58—41 и 58-40 мм. На-
сиживание длится около месяца, пуховики появляются около середины и в
конце июля. Так как в гнездах добывались и самцы (Житков и Штехер, 1915),
то, как и следовало ожидать, в насиживании участвуют оба родителя. Около
середины августа птенцы уже летные и спускаются на воду и затем птицы начи-
208
ОТРЯД ЧИСТИКИ
нают постепенно покидать гнездовья. Впрочем, часть птенцов еще в конце
августа находится в гнезде B8 августа, о-в Медный, от дополнительной
кладки?).
Линька. Предбрачная линька заканчивается в мае, послебрачная
происходит с конца августа и кончается обычно к концу сентября и даже
в октябре (Хартерт, 1920).
Описание. Размеры и строение. Хвост из 14 рулевых. Величина относи-
тельно мелкая: крыло обычно 168—170 (Иоганзен, 1934), 174—177 мм (Хартерт, 1920, но у
одной птицы с о-ва Медного 182 мм)\ у самцов F) 171—176, у самок CI68—172,в сред-
нем 172 и 170 мм (Портенко, 1937); крыло самцов 168, 169, самок 173,177 (Стейнегер,
1885). Длина самцов B) 338, 347, самок
B) 342, 355 мм (Стейнегер). Окраска.
Как у беринговой каюрки.
Систематические замечания. Так-
сономическое положение этой формы
в качестве подвида неясно. Скорее всего
мы имеем дело в этом случае с популя-
цпонными, а не географическими особен-
ностями. Для птиц американских побере-
жий также указываются небольшие раз-
меры (Риджвей, 1919) и весь вопрос ос-
тается таким образом открытым.
6. Очковый чистик Cepphus
carbo Pall.
Cepphus carbo. Pallas. Zoographia
Rosso-Asiatica, t. II, 1811, стр. 351, Ку-
рильские о-ва (ошибочно указаны Але-
утские).
Русское название. Очковый
чистик книжное. У местных жителей,
повидимому, каюрка (Стейнегер, 1885).
Распространение. Ареал.
Побережья и острова Охотского
моря — как на севере (п-ов Тай-
гонос, Пенжинская губа, Гижигин-
ская губа, Тауиская губа), так и
на юге (например на Шантарских
о-вах); Сахалин, Курильские о-ва
(южная группа—Кунашир и Иту-
руп, Ямасина, 1931); острова и по-
бережья Приморья; вероятно в Ко-
рее (A Hand-list of the Japanese
Birds, 1932); в Японии на Хоккай-
до и сев. Хондо. Зимою, повидимо-
му, и в гнездовой области (зимовки
вероятны в южн. Приморье,
Шульпин, 1936, и на севере Охот-
ского моря, Аллен, 1905) и не-
сколько южнее — в Японии у Иокогама и о-ва Цусима. Залеты весной к
о-ву Беринга 28 апреля, Стейнегер, 1885). Указания на встречи у Камчатки
Тачановский, 1893) и на Алеутских о-вах (ПаЛлас, 1811) ошибочны.
Характер пребывания. Более или менее оседлый вид, зимую-
щий преимущественно в пределах гнездовой области (см. выше).
Даты. Вне гнездовой области на юге добывался в апреле (Иокогама).
В середине апреля птица держится в стайках (небольших, 10—12 особей) у бере-
гов гнездовой области (южн. Приморье, залив Посьет, Шульпин, 1936). В сен-
Карта 31. Распространение очкового чистика
Cepphus carbo
а — граница гнездовий, г — область зимовок и кочевок.
чистик
209
тябре молодые птицы на Охотском побережье держатся еще у гнездовий (Шуль-
пин и Дулькейт, 1937).
Биотоп. Гнездовый — как у других чистиков, скалы морских побере-
жий и островов, после гнездового периода — прибрежные части морей.
Численность умеренная. Обычен на Охотском море и в Приморье,
но не образует там, как и в других местах, больших колоний. На Сахалине,
Курильских о-вах, в Японии — малочисленен.
Экология. Сведения недостаточны.
Размножение. Гнездится закрыто, в трещинах между камнями по
крутым берегам, отдельными парами или небольшими группами. Гнезда как
в скалах в 6—9 м над морем, так и
среди камней близ береговой линии.
Подстилки нет. Цикл очень поздний.
В июне птицы держатся в парах. Клад-
ка из одного яйца (в отличие от других
представителей этого рода), весьма
похожего на яйцо обыкновенного
чистика, но более крупного и с более
грубым рисунком. Скорлупа белая или
охристо-беловатая с поверхностными
бурыми и глубокими сероватыми кра-
пинами. Размеры 66,66. 3, 67,69 х 41.4,
44,45, 45. 3 мм (Тачановский, 1893);
(8) 62,2—66x40,8—45,1 мм (Джо-
урден, по Хартерту, 1917). Насижи-
вают оба родителя (Шульпин, 1936).
По тому же автору птенцы будто бы
спускаются на воду с неразвитыми
или недоразвитыми маховыми, в по-
следних числах июля и начале августа.
Линька. Смена нарядов такая
же как у обыкновенного чистика. Пол-
ная линька осенью, послебрачная;
весенняя предбрачная линька охва-
тывает все мелкое оперение, но не ма-
ховые и рулевые. В начале апреля
взрослые —в полном брачном наряде,
но у молодых годовалых линяние за-
паздывает —• еще 30 апреля в Приморье такие птицы добывались со значи-
тельной примесью перьев зимнего наряда. Смена первостепенных маховых
начинается в первых числах августа.
Питание. Пищу составляет мелкая рыба, длиной в 6—8 см (Шульпин,
1936), вероятно и беспозвоночные.
Полевые признаки. Крупный чистик однообразной темной буроватой
окраски с белым пятном у глаза, черным клювом, красными лапами. На
воде держит голову высоко и приподымает хвост. Кормясь, опускает голову
в воду «до глаз», высматривает добычу и ныряет. Голос довольно разнооб-
разен — общий для настоящих чистиков свист, позыв «цик-цик-цик», при
беспокойстве «те-те-те-ти-лилилили», вторая часть этого звука—трель; иногда
«тюрр-тюрр-тюрр» (Шульпин, 1936).
Описание. Размеры и строение. Самый крупный из собственно чистиков.
Хвост из 14 рулевых. Оперение основания клюва не прикрывает полностью ноздревой
борозды, передняя часть ноздри—узкой и щелевидной — выходит за границу оперения.
Клюв относительно массивный, по форме сходный с клювом настоящего чистика. Цевка
короче среднего пальца (с когтем). Длина самцов B) 380, 382, самок B) 365, 400; размах
14 Птицы СССР, т. II
Рис. 38. Очковый чистик
210
ОТРЯД ЧИСТИКИ
самцов B) 655, 690, самок B) 665, 680 мм. Вес самца A) 510, самок B) 415,545 г.
Крыло самцов D) 190, 192, 198, 200, самок C) 184; 191, 5; 193 мм.
Окраска. Пуховой наряд неизвестен. Взрослые самцы и самки в брачном наряде
шиферно-бурые, брюшная сторона несколько светлее спинной; перья у основания клюва,
вокруг глаза и за глазом («очки») белые. Радужина темнобурая, лапы красные, клюв черный,
когти черные. В зимнем наряде брюшная сторона белая с более или менее развитыми серова-
тыми пестринами на вершинах перьев. Молодые в первом зимнем (осеннем) наряде — как
взрослые в зимнем, но со слабо выраженным белым пятном у глаза и с менее ярким красным
цветом ног.
Систематические замечания. Ареал очкового чистика географически исключает гнездо-
вые области обыкновенного чистика. В частности, указания о совместном гнездовании очко-
вого чистика и С. g. snowi на Курильских о-вах по всем вероятиям ошибочны: очковый
чистик гнездится только на южных о-вах Итурупе и Кунашире, aC.g. snowi от Урупа до
Парамушира. Заслуживает внимания отсутствие белого зеркальца у snowi на средних Кури-
лах. Эта темнокрылая вариация («mrtzfeldi») встречается и среди других подвидов настоя-
щего чистика. Очковый и обыкновенный чистики таким образом очень близки между собой.
РОД ПЫЖИКИ BRACHYRAMPHUS BRANDT, 1837
Тип Colymbus marmoratus
7. Пестрый пыжик Brachyramphus marmoratus Gmel.
Colymbus marmoratus. Gmelin.Syst. Nat., 1789, Vs стр. 583, бухта принца Вильяма,
южн. Аляска.
Русское название. У Двигубского A831) пестрый пыжик. Пыжик — народное
название этой птицы на Камчатке.
Распространение. Ареал. Северные прибрежья Тихого океана. На
западе от Камчатки, Курильских о-вов и Сахалина до Японии и Приморья;
Карта 32. Распространение пестрого пыжика Brachyramphus marmoratus
1. В. т, perdix, 2. В. т. marmoratus;
а — сбласть гнездовья, 6 — область кочевок и зимовок, г — отдельное местонахождение.
на востоке от южн. Аляски и Алеутской гряды до с.-з. Вашингтона и Кали-
форнии (Бумбольдт).
Характер пребывания. Оседлый. Зимой, повидимому, часть
птиц откочевывает к югу и выходит несколько за пределы гнездовой области.
ПЕСТРЫЙ ПЫЖИК
211
Подвиды и варьирующие признаки. Два подвида, различающиеся окраской и разме-
рами клюва (быть может крыла): азиатский пестрый пыжик В. т. perdix Pallas, 1811 и
американский В. т. marmoratus Gme lin. Экологические различия не выяснены.
Азиатский пестрый пыжик Brachyramphus marmoratus perdix Pall.
Cepphus perdix. P a 11 a s. Zoographia Rosso-Asiatica t. II, 1811, стр. 351, Берингово море
Распространение. Ареал. Не вполне еще выяснен. Птица в летнее время
отмечена у восточного берега Камчатки (пролив Литке по Белопольскому и
Роговой, 1947; Авачинская губа, Петропавловск) и в Приморье (залив де Ка-
стри); восточное побережье Камчатки служит, повидимому, местом гнездовий,
хотя гнезд этой птицы там пока не находили. Отмечен у Южн. Сахалина (Си-
рароро, в июне, Никольский, 1889 и др.), добыт у Курильских о-вов (Сибом,
Рис. 39. Пёстрый пыжик
1884). Добывался и гнездится на Охотском море (Аян, Мензбир, 1900, Шуль-
пин 1936; Магадан; Тауйская губа, Паллас, 1811; в устьях Пенжины в июле,
Дементьев и Волков, 1948; у Шантарских о-вов на с-ве Утичьем и Птичьем,
Дулькейт и Шульпин 1937), у юго-вост. берегов Коряцкой Земли, в бухте Корфа.
Относительно Командорских о-вов сведения противоречивы: был добыт 11 мая
у о-ва Медного (Хартерт, 1920), где по Дыбовскому будто бы гнездится (Тача-
новский, 1893), но в последнее время на Командорских о-вах вовсе не встречен
(Иогансен, 1934). В негнездовое время отмечен у берегов Японии (Хоккайдо,
Хондо, Кюсю, Амами-Осима).
Характер пребывания. Повидимому, отчасти оседлая, отча-
сти кочующая вдоль морских берегов птица. Сроки передвижений не изу-
чены.
Численность. В августе обычен у вост. Камчатки (Бергман, 1935).
Размножение. Найденное Б. Дыбовским на о-ве Медном и приписы-
ваемое этой птице яйцо бледноохристое с шиферно-серыми крапинами и с по-
верхностными ржавчато-бурыми пятнами, сгущающимися у тупого конца.
Размеры 62,5 х 41,2 мм (Тачановский). Вполне развитые молодые добывались
у берегов Камчатки около середины августа A8 августа в Авачинской
губе, Бергман, 1935; там же «совсем молодой» 17 августа, Бутурлин,
1909).
Линька. Изученные экземпляры в зимнем наряде добыты между кон-
цом октября и концом февраля; в брачном наряде между концом апреля и
сентября. Сильно линяющая птица без первостепенных маховых добыта на
Охотском море 31 августа.
212 отряд чистики
Полевые признаки. Мелкий, очень подвижной чистик с характерной «мра-
морной» окраской.
Описание. Размеры и строение. Клюв оперен более, чем наполовину, с от-
крытой ноздрей; лапы короткие; в хвосте 14 рулевых. Длина самцов 278, 289, самки 292
(Тачановский, 1893), самки 295 (Бутурлин, 1911). Размах самцов 487, 495 (Тачановский),
самки 511 (Тачановский),520 (Бутурлин). Крыло самцов 138 и 140(Тачановский), 139 (Берг-
ман, 1935), 141,5 (Бутурлин, 1911), птицы неизвестного пола 143 мм (Бутурлин, 1915). Крыло
самца 147, самок B) 138 и 141, особей неизвестного пола A1) 135—155, в среднем A4) 142,1 мм
(Зоологический институт Академии наук СССР, Зоологический музей Московского универ-
ситета). Клюв самцов 18,19, самок 19, 20 (Тачановский), самки 20 (Бутурлин, 1911), особей
неизвестного пола 19,5; 19,9 и 20 (Зоологический музей Московского университета), 23,5 мм
(Бутурлин, 1915).
Окраска. Пуховой наряд неизвестен. Взрослые самцы и самки в брачном наряде:
спинная сторона черноватая с буровато-охристыми вершинами перьев шеи, спины, надхво-
стья; на голове узкие беловатые каемки; маховые черноватые, кроющие крыла черноваты
с сероватыми каймами; бока головы черновато-серые с беловатыми пестринами; рулевые чер-
ные, боковые пары с более или менее развитым белым мраморным рисунком; брюшная сто-
рона белая с темными серо-бурыми каймами перьев, подкрылья серо-бурые. Радужина темно-
бурая, клюв черный, ноги спереди темносерые с синеватым оттенком, сзади и на перепонках
черноватые, когти черные. В зимнем наряде спинная сторона темиосерая со светлыми основа-
ниями перьев, вокруг глаза обычно белое кольцо, имеется белый прерванный посередине
ошейник, на плечевых примесь белого, грудь и брюхо белые; поперек зоба серо-бурая пе-
ревязь: подкрылья сероватые. Особых выростов на клюве и украшающих перьев на голове
в брачном наряде нет.
Молодые по Хартерту, 1917, повидимому, с буроватыми каймами на брюшной стороне
и узкими беловатыми краями перьев на спинной; по Тачановскому, 1893, молодые в первом
брачном наряде сходны со взрослыми, но с более развитым охристым оттенком на спинной
стороне и с беловатыми, испещренными буроватыми пятнами, средними и большими нижними
кроющими крыла.
Американский пестрый пыжик Brachyrampfius marmoratus marmoratus
Gmel.
Распространение. Ареал. Острова и побережья американских частей
Тихого океана — от восточных о-вов Алеутской гряды (Уналашка), вдоль
южн. Аляски (о-в Кадьяк, залив принца Вильяма, Ситка), на юг до Британской
Колумбии (о-ва королевы Шарлотты, Ванкувер) и с.-з. Вашингтона (Педжет-
зунд). Есть указания на гнездовья и севернее (Ном) и южнее (бухта Гумбольдта
в сев. Калифорнии). Зимой от Британской Колумбии до Калифорнии (бухта
Сан-Франциско, Санта Круц, бухта Монтерей, Санта Барбара, Сан Диего).
На территории СССР пара наблюдалась и одна птица добыта на о-ве Идлидля
в Колючинской губе (Тайер и Бенгс, 1914).
Характер пребывания. Частично оседлая птица, часть осо-
бей откочевывает на юг (см. выше).
Биотоп. Морские побережья, зимой в бухтах и заливах.
Экология. Размножение неизвестно. Описано только 2 яйца —
одно, извлеченное из самки, добытой 23 мая 1897 у архипелага принца Уэль-
ского (Кентуэлл, 1898), другое, найденное в расщелине скалы к северу от
Нома 10 июня 1904 (Денхем, по Бенту, 1919). Окраска скорлупы желтоватая
с темными черновато-бурыми мелкими крапинами. Размер яйца из Нома
60,5 х 37,5 мм.
Линька. Как и у других чистиковых, проходит дважды в году — пред-
брачная между февралем, мартом и маем и полная послебрачная между сентя-
брем — октябрем и ноябрем (Бент).
Питание составляет мелкая рыба.
Описание. Размеры и строение. Как и у предыдущего, но клюв короче,
длина его менее 16—18,5 мм (Хартерт, 1917). Окраска. В брачном наряде общий тон
спинной стороны темней, пестрины не охристо-бурые, а коричнево-бурые; боковые рулевые
одноцветны, без светлых пестрин. В зимнем наряде без белого кольца вокруг глаза. И у этой
формы пуховой наряд остается пока неизвестным.
СЕРЫЙ ПЫЖИК
213
8. Серый пыжик Brachyramphus brevirostris Vig.
Uria brevirostris. Vigors. Zoolog. Journ. IV, 1828A829), стр. 357, ошибочно Сан-Блас
в Мексике, на самом деле северные части Тихого океана.
Синоним. Brachyramphus Kittlitzi. Brandt. Bull. Acad. Imp. Sc. St. Petersbourg,
11, 1837, стр. 346, Петропавловск на Камчатке.
Распространение. Ареал. Выяснен недостаточно. Несомненно гнездится
у залива Павлова в южн. Аляске, к северо-западу от Шумагинских о-вов
(Тайер, 1914), у Хилл-пойнт Уэльс на Аляске (Форд, 1936) и, повидимому, в
Гласиер-бай (Гриннелл, 1909). Встречен у м. Барроу, Алеутских о-вов
(Уналашка, Нельсон, 1887, Атка, Амчитка, Тернер, 1886). У азиатского берега
Карта 33. Распространение серого пыжика Brachyramphus brevirostris
а — область гнездовья, г — область кочевок и зимовок, д — залеты.
отмечен на Чукотском п-ове у Восточного мыса (Бейли, 1936), в бухте Провиде-
ния в заливе Креста 22 июня, 1879 под 67° с. ш. (Пальмен, 1887); между Берин-
говым проливом и м. Якан (Тайер и Бенгс, 1914) на Камчатке (Карагинская
бухта, Белопольский и Рогова, 1947); у Петропавловска (Брандт, 1837,
Тачановский, 1884, 1893, Оджильви-Грент, 1898), на Курильских о-вах (на
Парамушире по Ямасина, 1931). В негнездовое время у азиатских берегов до
Курильских о-вов и севера Японии.
Характер пребывания. От мест гнездовья в Аляске откоче-
вывает к азиатским берегам (см. выше). Сроки передвижений неизвестны.
Биотоп. Гнездовый — высокогорье на довольно большом расстоянии
от моря (Тайер и Бенгс, 1914).
Численность. Редок и спорадичен (в большом числе — крупные
стаи —наблюдались в сущности только в Гласиер-бай, Гриннелл, 1909).
Экология. Размножение. Гнездо у бухты Павлова, Аляска, нахо-
дилось в скалах, выше верхней границы леса, на лаве среди снега; никакой
постройки в сущности не было. 29 мая у самки найдено готовое к сносу яйцо,
214 отряд чистики
10 июня в гнезде найдено яйцо (Тайер и Бенгс, 1914). Окраска скорлупы буро-
вато-желтовато-зеленоватая с бурыми или черноватыми различной величины
крапинами. Размеры 57,6 х 35,6 и 62,1 х 36,8 мм (Бент, 1919, Хартерт, 1917).
Линька. У взрослых бывает две линьки в году: предбрачная, охваты-
вающая большую часть мелкого оперения, но не рулевые и маховые, весною,
и полная послебрачная осенью. Детали неизвестны.
Питание. В качестве пищи этой птицы указываются мелкие рако-
образные (Нельсон, 1887).
Полевые признаки. От пестрого пыжика отличается коротким клювом и
белыми боковыми рулевыми. Полет быстрей, чем у близких видов. Очень осто-
рожен. Вне гнездового времени отмечен в больших стаях (до 500 птиц в каждой,
Гриннелл, 1909).
Описание. Размеры и строение. Клюв короткий, неоперенная часть его
10—11 мм, длина ото лба менее 15 мм,; крыло 146—151 (Хартерт, 1917). Крыло самцов
B) 136—145, самок C) 137—145, птиц неизвестного пола B) 140—141 мм, в среднем G)
140,1 мм (измеренные нами особи). Длина хвоста 35—36 мм (Хартерт); крыло 144—149,
хвост 37 мм (Тачановский).
Окраска. Пуховой наряд не описан. Первый зимний наряд молодых птиц тоже не-
известен . Взрослые в брачном наряде свинцово-серые с ржаво-желтыми пестринами на шее,
спине, плечевых; кроющие крыла серо-буроватые, маховые темнобурые со светлыми краями
внутренних опахал;средние второстепенные маховые с белыми пятнами на вершине и на вну-
треннем опахале; рулевые белые, средние рулевые бурые, ближайшие к ним две пары с бу-
рой примесью на наружных опахалах; подкрылья темнобурые; брюшная сторона белая
с черно-бурыми пестринами, густо расположенными на горле, шее, зобе. Радужина бурая,
клюв черный, лапы бледносерые. В зимнем наряде на спинной стороне нет ржаво-желтых
пестрин, ошейник и уздечка белые, под глазом белое пятно; на второстепенных маховых белое
вершинное пятно; в остальном спинная сторона свинцово-серая; брюшная сторона белая,
по бокам зоба бурые пятна.
[Алеутский пыжик Ptychoramphus aleuticus Pall.
Uria aleutica. Pallas. Zoogr. Rosso-As. t. II, 1811, стр. 370, вост. часть Берингова моря
Распространение. Ареал. Тихоокеанские побережья Сев. Америки
от южн. Аляски (о-ва Форрестер, Эгг и Санак) и Алеутских о-вов (Атка)
до Нижней Калифорнии (Серрос, Сан-Бенито, Сан-Херонимо, Сан-Мартин,
Тодос-Сантос, Лос-Коронадос). В зимнее время там же, на север до залива
Педжет. Брандт указывает, что алеутский пыжик был добыт Вознесенским
на Курильских о-вах. Есть сообщения о встрече на Камчатке (по Лихтенштей-
ну, 1854, экземпляр будто был в Берлинском университетском музее). И то и
другое сведения сомнительны.
Характер пребывания. В зимнее время кочует в пределах гнез-
довой области.
Биотоп. В гнездовый период — побережья, в негнездовое время —
прибрежные части океана.
Численность. Колонии многочисленны, но расположены споради-
чески.
Экология. Размножение. Гнездовья колониальные и располага-
ются в норах и расщелинах между камнями. Свежие кладки из одного яйца гряз-
нобелого цвета попадаются с середины марта. Размеры F0) 44—51 х 31,5—•
37,44, в среднем 46,9 х 34,5 мм (Бент, 1919). Насиживают оба родителя. Пе-
риод размножения длится до ноября, возможно бывают две нормальных кладки
(Брайент, 1888). Насиживание —по одним данным (Эмерсон, 1904) 21 день,
по другим (Литтлджон, по Бенту) — целых 30 дней.
Линька. Полная годовая бывает в августе—сентябре; частичная,
повидимому, весною (Бент).
Питание. Активен, главным образом, ночью. Пища состоит преиму-
щественно из Copepoda и Amphipoda.
СТАРИК
215
Описание. Крыло самцов и самок (8) 127—137, в среднем 131,25 мм. Пуховой наряд
темнобурый, бледнее на зобе, груди, брюхе. Взрослая птица серовато-бурая на спинной сто-
роне, с сероватым зобом, беловатой грудью и брюхом; у глаза белое пятно. Зимний наряд
сходен с брачным. Молодая птица тускло-буроватая на спинной стороне,без пятна на уздечке.]
РОД СТАРИКИ SYNTHLIBORAMPHUS BRANDT, 1837
Tun Alca antiqua
9. Старик Synthliboramphus antiquus Gmelin
Alca antiqua. Gmelin. Syst. Nat. I B) 1789, стр. 554, Берингово море.
Синоним. Uria senicula. Pallas. Zoographia Rosso-Asiatica t. II, 1811, стр. 367,
Камчатка. — Brachyramphusbrachypterus. Brandt. Bull. Ac. Imp. Sc. St. Petersb.,11,
1837, стр. 346. Уналашка.
Русское название. Народное, на Камчатке известно по крайней мере с XVIII
века (Крашенинников, 1755); также старичок. Повидимому, от белых перьев на голове,
как бы седины.
Распространение. Ареал. Гнездится по островам и побережьям частей
Тихого океана — у американских берегов от южн. Аляски (о-ва Санта Лаза-
риа и Форрестер) до о-вов королевы Шарлотты и Британской Колумбии, на
Карта 34. Распространение старика Synthliboramphus antiquus
а — граница гнездовий, е — область кочевок и эимовок.
Алеутских о-вах и на Прибыловых; у азиатских берегов, быть может в бухте
Эрулен, в заливе Креста (Белопольский, 1934); на вост. Камчатке, на Саха-
лине, Курильских о-вах, Командорских (обычен на о-ве Медном; отмечен
Стейнегером, 1885, на Арьем камне у о-ва Беринга, где добывался и Сокольнико-
216
ОТРЯД ЧИСТИКИ
вым; однако,в позднейшее время на о-ве Беринга на гнездовье не найден,Иоган-
сен, 1934); быть может у Б. Шантар (Дулькейт и Шульпин, 1937), по берегам
Приморья и Кореи; на юге отмечен на гнездовье у берегов Шаньдуня, Сев.
Китай (Латуш, 1934). Для Пенжинской губы отмечен еще Палласом A811)
и недавно добыт в июне на р. Пенжине (Дементьев и Волков, 1948). Гнездится
на западе Охотского моря на Мальминских о-вах. В негнездовое время
встречается как в пределах гнездовой области (Прибыловы о-ва; Алеутские
о-ва; Командоры, Курильские о-ва, южн. Приморье), так и далее к югу —
в Азии до Японии, особенно у средних островов, далее у о-ва Квельпарт, по
побережьям вост. Китая (Фуцзянь) и у о-ва Формоза; в Америке от Ванкувера
до Калифорнии (бухта Сан-
Франциско, Монтерей, о-ва
Санта Барбара, Сан-Диего).
Залеты внутрь материка
отмечены в Сев. Америке
(Уисконсин, Торонто, озеро
Эри, Бент, 1919).
Даты. Появление на
Командорах отмечается в
начале мая. В стайках в
южн. Приморье в заливе
Посьет 29 апреля (Шуль-
пин, 1936). В половине мая
и на Аляске. На зимовках
в Китае еще в апреле. Осен-
нее движение происходит
поздно. Еще в середине
октября держится у восточ-
ных берегов Камчатки
(бухта Моржовая, 14 октя-
бря, Слепцов); во второй по-
ловине этого месяца у Ку-
рильских о-вов (Бергман,
1935). В южн. Приморье
в заливе Абрек зимует с сентября до марта (Тачановский, 1893). Появление
на зимовку в Японии отмечается в октябре. В Калифорнии появляется в кон-
це октября и остается там до конца апреля. Миграции происходят, повиди-
мому, весьма постепенно, в области теплых морских течений.
Биотоп. Гнездовый — прибрежные скалы. В холодное время года на
море в литоральной полосе, но иногда вдали (в 400—500 милях от берега, Бен-
дайр, 1896).
Численность. Обычная птица. Зимою держится в стаях по 20—30
штук (Япония, Ян, 1942).
Экология. Размножение. Гнездится колониально, в расщелинах и
пустотах между камнями как у берега, так и высоко над морем (о-в Медный,
Иоганзен, 1934). Яйца — на земле, без какой-либо подстилки. В кладке обычно
2 яйца (иногда и одно, Паллас, 1811), откладываемые с промежутком в 2—3 дня
(Литтлджон, по Бендайру, 1895). Кладка, повидимому, вскоре после появле-
ния у мест гнездовий, точные даты не установлены. На юге ареала в Америке
(о-в королевы Шарлотты) уже с 20 апреля; в южн. Аляске с 1 мая (Бент, 1919).
По опросным сведениям на Командорах период размножения в июне — июле
(Иоганзен). Скорлупа яиц гладкая, блестящая и мелкозернистая. Окраска
светлая, желтоватая или желтовато-бурая, с темными бурыми и красноватыми
крапинами. Размеры 60—62 х 37,6—38 (Тачановский, 1893); E1) 57,5—-64,3 х
X 35,7—40,3, в среднем 61,1 х 38,6 мм (Бент, 1919). Насиживают оба роди-
Рис. 40. Старик
ХОХЛАТЫЙ СТАРИК 217
теля, у которых имеется по одному наседному пятну. Продолжительность наси-
живания неизвестна. Только что вылупившиеся птенцы на о-ве Форрестер,
Аляска, 29 мая (Хиз, 1913). В Приморье 23 июня найдены, однако, насиженные
яйца (быть может от дополнительной кладки?). Вполне развитые молодые
добывались в Приморье в июле A2 июля у м. Мосолова, 21 июля на о-ве Путя-
тина и т. д.).
Линька. Последовательность смены нарядов как у других чистиков.
В году бывает две линьки: предбрачная весенняя, в результате которой раз-
виваются брачные украшения на голове и шее (такие птицы добывались уже
в марте, но известны особи с остатками зимнего пера, добытые в апреле), но круп-
ное оперение не сменяется; и полная послебрачная осенняя, начинающаяся
уже в июле и заканчивающаяся в октябре (взрослые в зимнем наряде добыва-
лись в середине и во второй половине октября на Камчатке и Курильских о-вах).
Питание. Мелкие беспозвоночные, в частности, гаммариды (Командор-
ские о-ва).
Полевые признаки. Мелкий чистик, резко двухцветный, с серой спинкой
и белой брюшной стороной, с характерными пучками белых перьев на голове
и белыми полосками на шее, контрастирующими с черной головой. Полет очень
быстрый. Молчалив, голос — свист. На воде держится обычно группами. Мало
осторожен. Активность — дневная.
Описание. Размеры и строение. Клюв короткий, довольно высокий. Цевка,
примерно, одинаковой длины со средним пальцем. Ноздри открытые. Рулевых 14. Длина
самца A) 250 мм (Зоологический музей Московского университета), 266 (Тачановский), 260
и 273 (Стейнегер, 1885), самки 260 (Стейнегер), размах самца 464 мм (Тачановский). Вес
самцовDI91, 200, 210, 210, самки A) 200 г. Крыло самцов 135—143, самок 140—145 мм (Та-
чановский), самцов 129, 138, 135, самки 130 мм (Стейнегер); самцов D) 139—143, самок E)
131—137,5, в среднем 140,7 и 133,7 мм (Зоологический музей Московского университета),
самцов (8) 137—144, самок D) 141 — 145, птиц неизвестного пола B9) 134—149, в среднем
у самцов (8) 139,5, самок D) 142,7, у обоих полов D1) 141,2 мм (Зоологический институт
Академии наук).
Окраска. У пухового птенца спинная сторона черная с серым оттенком, за ухом
белое пятно, брюшная сторона охристо-белая. Взрослые самцы и самки в брачном наряде:
лоб, темя, щеки, кроющие уха и подбородок, горло, передняя и задняя сторона шеи черные,
бока шеи белые; над глазами и на затылке узкие белые перышки; спина серая, кроющие крыла
серовато-бурые, маховые серо-бурые с беловатыми стволами и основными частями внутрен-
них опахал передних первостепенных, рулевые черные; бока зоба черные с тонкими белыми
перышками, бока тела серовато-черные, подкрылья, средняя часть зоба, грудь,брюхо ипод-
хвостье белые. Клюв беловатый с темной серой полосой по коньку, ноги светлосерые,спереди,
сзади и на перепонках черноватые, когти черные; радужина темнобурая. Самцы и самки
в зимнем наряде без белых перьев на голове, с серым подбородком и горлом, бока серые с бе-
лыми пестринами. Молодые в первом зимнем наряде, как взрослые, но темный цвет на брюш-
ной стороне менее развит, бока только с незначительной примесью сероватого, подбородок и
горло белые со слабо развитыми сероватыми отметинками на вершинах перьев, иногда почти
незаметными.
10. Хохлатый старик Synthliboramphus wumizusume Те mm.
Uria wumizusume. T e m m i n с k. Planches Coloriees, № 579, 1836, Япония (от япон-
ского umi—suzume, но это название по-настоящему относится к обыкновенному старику;
хохлатый старик Kammuri-umi-suzume, по Яну, 1942).
Распространение. Ареал. Гнездится в Японии на «Семи Островах», Мия-
кесима, Сикинесима и о-ве Панафидина (Ян, 1942, по Кобаяси). В негнездо-
вое время у Хондо, Хоккайдо, Кюсю, Кореи и Сахалина.
Характер пребывания. Вне гнездового времени кочует у по-
бережий, выходя за пределы гнездовой области.
Биотоп гнездовый — морские побережья, зимний — прибрежные части
моря.
Численность. Редок.
Экология. Изучена слабо. Кладка из 2 яиц желтовато-бурых или беловатых
с немногочисленными сероватыми и буроватыми крапинами; размеры меньше,
218
ОТРЯД ЧИСТИКИ
чем у обыкновенного старика: A0) 52—56,1 х 33,6—36, в среднем 53,37 х
X 35,05 мм (Хартерт, 1917).
Описание. Размеры и строение. Крыло 126—136, клюв 16,5—18 мм (Хар-
терт). Окраска. Пуховой наряд на спинной стороне буровато-серый с беловатыми пестринами
на спине, брюшная сторона — белая (Хартерт). Взрослые птицы в брачном наряде имеют на
лбу обращенный назад хохол, состоящий из 10—12 узких перьев длиною в 6—7 см; лоб,
подбородок, щеки и хохол — черного цвета; темя, до глаз, и затылок — белые, с черными
перышками посредине; на шее длинные
нитевидные белые перышки; осталь-
ная спинная сторона аспидно-серая,
маховые— черновато-серые с белова-
тыми основаниями внутренних опа-
хал; рулевые черновато-серые; бока зоба
черноватые; грудь, брюхо и подхвостье
белые. Неоперенные части, как у обыкно-
венного старика. В зимнем наряде нет
хохла, вся верхняя часть головы — ши-
ферно-черная (Хартерт). Молодые птицы,
повидимому, сходны со взрослыми в зим-
нем наряде, быть может побурее на спин-
ной стороне (Темминк и Шлегель, 1847).
Оба пола окрашены одинаково.
РОД КОНЮГА AETHIA
MERREM, 1788
Тип A. cristatella
11. Большая конюга Aethia
cristatella Pall.
Alca cristatella. Pallas. Spicilegia
Zoologica I pax. 5, 1769, стр. 18, табл.
3, 5, фиг. 7—9, Курильские о-ва.
Синонимы: Alca tetracula. Pallas.
Spic. Zool. If. 5, 1769, стр. 23, Камчатка,
по птице в зимнем наряде. — Uria dubia.
Pallas. Zoographia Rosso-Asiatica v.
II, 1811, стр. 371, Авача, по молодым
птицам.
Русское название. Конюга —
местное название, повидимому, звукопод-
ражательное (Стейнегер, 1885). Так же
таратора, турутур (Паллас, 1811). По
Кращенниникову A755) — черный ста-
рик. Большая конюга — книжное имя.
Распространение. Ареал.
Сходный с ареалом белобрюшки —
Чукотский п-ов (губа Преобра-
жения, бухта Провидения) и о-ва
Диомида в Беринговом проливе
на севере;* о-ва Прибыловы, о-в св. Лаврентия; Алеутские о-ва, о-в
Кадьяк; Курильские о-ва; на юг—до Сахалина. Возможно гнездится
на группе Шантарских о-вов (наблюдение Миддендорфа у южного берега Охот-
ского моря; Дулькейт и Шульпин, 1937) и на Камчатке (Дыбовский, Тачанов-
ский, 1893). Вопрос о гнездовании на Командорских о-вах неясен: в конце
прошлого века найдена там на гнездовье Стейнегером A885, добыта в мае —
июне), но позже там не встречена и лишь редко добывалась вне гнездового
времени (Иогансен, 1934). Зимою как в северных частях Берингова моря (При-
быловы о-ва, Прибл и Мк. Эти, 1923), так и южнее у Курильских о-вов (уже
с конца сентября), о-в Шумагина, у Сахалина в Татарском проливе (Шренк,
I860), изредка у Хоккайдо и Хондо. Весною отмечена в заливе Петра Великого
Карта 35. Распространение хохлатого старика
Syntliborhamphus wumizusume
а — границы гнездовий, г — область кочевок и зи-
мовок.
БОЛЬШАЯ КОНЮГА
219
(Шульпин, 1936). В мае — июне встречена в заливе Креста, Анадырский край,
и будто бы гнездится у м. Беринга, но это еще требует подтверждения (Бело-
польский, 1934), скорее всего сюда только редко залетает (Портенко, 1939).
Случайно появляется у о-вов Врангеля и Геральда (Нельсон, 1863) и у сев. по-
бережья Чукотского п-ова B6—28 сентября ум.Иксурин,ТайериБенгс, 1914),
на сев. побережье Аляски (м. Барроу, залив Коцебу, Бент, 1919). Однажды
добыта в Атлантическом океане к северу от Исландии A5 августа 1912 г. под
66° 40' с. ш. и 12° 55' з. д. Саломонсен, 1944).
Характер пребывания. В негнездовое время откочевывает от бе-
регов в открытое море, в прибрежной полосе держится реже, чем другие конюги.
Карта 36. Распространение большой конюги Aethia cristatella
а — границы гнездовий, г — область кочевок и зимовок.
У мест гнездовий на севере (Чукотский п-ов) держится еще во второй
половине августа (Портенко, 1934). Откочевка с о-вов Диомида происходит
в первой трети сентября (Бент, 1919). У Прибыловых о-вов появляется в большом
числе в апреле, раннее появление отмечено 21 —18 марта (Прибл и Мк. Эти, 1923).
Биотоп. Гнездовый — скалы у морских побережий или каменистые
россыпи: вне гнездового времени в свободных ото льда участках моря, обычно
вдали от берега (быть может в связи с уходом в глубины мелких ракообразных,
составляющих основную пищу птицы).
Численность. Распространение спорадично, менее обычна, чем мел-
кие виды конюг. Зимою обычна у Курильских о-вов (Ямасина, 1931), в громад-
ном количестве встречена в августе на о-ве Шумагина (Таунсенд, 1913). На зи-
мовках наблюдалась иногда высокая смертность этой птицы (Прибыловы о-ва;
у берегов Сахалина, Шренк, 1860; в Приморье) —повидимому, в результате
штормов. На зимовке держится группами, парами и одиночками.
Экология. Размножение. Гнездится колониями, среди скал и кам-
ней, в расщелинах. Кладка происходит, повидимому, в июне (ранние находки
220
ОТРЯД ЧИСТИКИ
на Прибыловых о-вах 16 июня, Прибл и Мк. Эти, 1923), и состоит из одного
яйца, овальной или овально-закругленной формы, с мелко-зернистой матовой
белой скорлупой. Размеры: C0) 50—60 х 32,5—42,5, в среднем 54,2 х 37,9 мм
(Бент), т. е. относительно очень крупные. Насиживают, повидимому, оба роди-
теля. Продолжительность насиживания и гнездового периода неизвестны.
Летные молодые отмечены 10—15 агуста (Нельсон, 1887).
Линька. Как и у других конюг, два раза в год —частичная предбрач-
ная и полная послебрачная. Первая охватывает мелкое оперение, главным
образом, на голове и шее; при ней сильно развиваются укращающие хохолки
на голове и пластинки на клюве. Точные сроки ее не установлены, но около
середины марта встречаются уже перелинявшие особи. Послебрачная полная
Рис. 41. Большая конюга
линька имеет место в августе —сентябре (Бент, 1919). У птиц, добытых ум. Про-
видения 14 августа, проходила энергичная смена мелкого оперения и пле-
чевых. При послебрачнои линьке рамфотека утрачивает брачные украшения
в конце августа — сентября.
Питание. Мелкие беспозвоночные, добываемые нырянием, в зимнее
время со значительных глубин (до 9 м, Прибл и Мк. Эти). Преимущественно
копеподы, немного схизопод (Чукотский п-ов, Портенко, 1934), также амфи-
поды, среди которых Parathemisto oblivia, реже Socarnea vahli (Прибыловы
о-ва, Прибл и Мк. Эти, 1923), быть может изредка головоногие (Командоры,
Стейнегер, 1885). Имеется горловой мешок—резервуар для пищи, приноси-
мой птенцам, объем этого мешка около 16 см3 (Портенко, 1934).
Полевые признаки. Размеры птицы относительно крупные. Окраска
оперения — однообразно серая, более темная на спинной стороне, имеется обра-
щенный вперед лобный хохол из черных перышек; клюв —красный, изогну-
тый; сильно развита нижняя челюсть и имеется «розетка» у основания рта,
полет прямой, низко над водою. Активность дневная. В брачное время птица
очень криклива. Мало осторожна.
Описание. Размеры и строение. Клюв довольно массивный, надклювье
загнуто книзу, подклювье вверх; в брачном наряде у основания клюва появляется вздутая
пластинка, выдающаяся углом впереди ноздрей, под ноздрями имеется небольшая узкая
пластинка у разреза клюва; небольшая поперечная пластинка — у лба; розетка у угла раз-
реза рта; две больших выступающих пластинки с правой и левой стороны основания под-
клювья; таким образом, не считая розетки, рамфотека надклювья состоит из 4 парных (две
пары) и 2 непарных частей (поперечная пластинка и собственно рамфотека), рамфотека под-
клювья состоит из 2 парных (одна пара) и одной непарной части (собственно рамфотека).
МАЛАЯ КОНЮГА
.221
Хохол, загибающийся вперед и вниз, состоит из 14—18 тонких перьев на лбу; на голове име-
ются тонкие белые перышки; значительно удлиненные за глазом. Цевка короче среднего
пальца (с когтем). Хвост состоит из 14 рулевых. Длина самцов и самок 240—273, размах
500—507 мм (Тачановский, 1893), длина самца 245, размах 495 мм (Бутурлин, 1911); длина
самцов 240, 260, 273, самок 235, 247, 249, 254, 265, 266, 270 мм (Стейнегер, 1885). Крыло
самцов и самок 125—140 мм (Тачановский), самцов 134, 134,134, самок 125, 125, 133, 133, 134,
135, 138 мм (Стейнегер); самцов 135, 137, 140, самок 133, 134, 138мм (Зоологический музей
Московского университета); в среднем крыло самцов F) 135,6, самок A0) 133,9 мм.
Окраска. Пуховой птенец серовато-бурый, несколько светлей на брюшной стороне
(Бент, 1919, Хартерт, 1917). Самцы и самки в брачном наряде наспинной стороне шиферно-
серые, черноватые, с белыми узкими перышками на темени и щеках, с длинными тонкими
перьями за глазами; маховые и рулевые бурые; брюшная сторона светло шиферно-серая.
Клюв яркокрасный с восково-желтой вершиной; ноги серые с темными суставами и чернова-
тыми перепонками и когтями; радужина голубовато-белая. В зимнем наряде спинная сторона
темнее, менее серая; хохолок на лбу короче; украшающие белые перья сохраняются в не-
большом числе только за глазом; клюв буроватый без украшающих пластинок и розетки.
Молодые в первом зимнем наряде — как взрослые, но без черного хохла на лбу и без укра-
шающих перьев.
12. Малая конюга Aethia pygmaea Gm.
Alca pygmaea. G me 1 i n. Sys. Nat. I pt. 2, 1789, стр. 555, о-ва Берингова моря.
Синоним. Alca kamtschatica. L е р е с h i n. Nova Acta Acad. Caes. Petropol, XII,
1801, стр. 369, табл. 8, Камчатка.—Uria mystacea. Pallas. Zoographia Rosso-Asiatica t. II,
1811, стр. 372, Курильские о-ва.
Русское название. У русских на Дальнем Востоке тараторка или турутура
(с курильского, Паллас, 1811 и др.), вероятно звукоподражательное. По Стейнегеру A885)
петушок, конюга, турутурт. У Двигубского A831) — пыжик ушатый. Малая конюга — книж-
ное имя.
Распространение. Ареал. Южные части Берингова моря —гнездится
на Командорских о-вах, в большом числе на о-ве Медном, реже на о-ве Беринга,
Карта 37. Распространение малой конюги Aethia pygmaea
а — границы гнездовий, г — область кочевок и зимовок.
222
ОТРЯД ЧИСТИКИ
на Курильских о-вах(Уруп, Райкоке, Симушир по Ямасина, 1931); на о-ве св.
Лаврентия (Фридманн, 1932), быть может на Алеутских о-вах, на восток доходит
до Уналашки; Сахалин (Кайбато, Мунстеръельм, 1920). Зимою отмечена до
Хондо и Сикоку (A Handlist of the Japanese Birds, 1932). Добыта в Авачинской
губе (Бергман, 1935).
Характер пребывания. Более или менее оседлая птица, зимою
держится в море у мест гнездовий или немного откочевывает к югу (см. выше).
Биотоп. Гнездовый — прибреж-
ные скалы, в негнездовое время в при-
брежных участках моря.
Численность. Спорадична.
Обычна на о-ве Медном. У Командор-
ских о-вов более многочисленна в зим-
нее время (Иоганзен).
Экология мало изучена. Размно-
жение. У гнезд с начала мая. Гнез-
дится в расщелинах скал или между
камнями. Кладка происходит в июне,
вероятно в начале месяца, и состоит из
одного овального матово-белого яйца.
Размеры 45,2 х 32 мм (Тачановский,
1893), 48 х 33,5 мм (Бент, 1919). Пухо-
вые птенцы встречены в конце июня, в
середине и даже в конце июля молодые
в развитом первоосеннем пере уже спу-
скаются на воду и держатся в море (Стей-
негер, 1885), старые птицы, впрочем, но-
чуют в скалах у гнездовий еще в августе
(о-в Медный, Житков и Штехер, 1915).
Линька. В году бывает две линь-
ки, неполная предбрачная и полная
послебрачная; последняя начинается в
июле и заканчивается в августе (Бент,
1919). Смена первостепенных маховых, повидимому, как и у других конюг—-
замедленная по сравнению с другими чистиковыми, не одновременная (у экзем-
пляров, добытых 21 июля, 4—5 задних первостепенных махов сменились,
линяли кроющие крыла, у экземпляров, добытых 31 августа, были старые
первые два первостепенных маховых, третье росло, остальные же новые,
проходила линька рулевых и интенсивная смена мелкого оперения). Пред-
брачная линька начинается в конце декабря—начале января (Стейнегер)
и в это время особенно развиваются украшающие перья на голове, имею-
щиеся, впрочем, и в зимнем наряде, но, повидимому, менее в нем вы-
раженные.
Питание. Главным образом, амфиподы, в частности гаммариды; в мень-
шей степени декаподы и гастроподы (Стейнегер, 1885).
Полевые признаки. Окраска довольно однообразная буровато-серая,
несколько более светлая на брюшной стороне: клюв у взрослых красный; на
голове имеются белые украшающие перья и черный хохолок над лбом. Держится
на воде обычно небольшими группами B—4 птицы). Кормится ныряя. Голос
не описан.
Описание. Размеры и строение. Клюв выпуклый по коньку надклювья,
короткий, с узкими щелевидными прикрытыми перепонкой ноздрями. На лбу имеется длин-
ный, свисающий вперед хохолок, состоящий из 10—12 черных перьев, кроме того имеются
белые узкие перышки разной длины; такие же белые перья есть обращенные назад и вниз на
уздечке и за глазами. Хвост из 14 рулевых. Средний палец (с когтем) длиннее цевки.
Рис. 42. Малая конюга
КОНЮГА-КРОШКА
223
Длина самцов (9) 181, 185, 190, 191, 192, 193, 195, 195, 195, самок 184, 188, 192, 203,211 мм
(Стейнегер, 1885), самца 200 мм (Зоологический музей Московского университета). Вес самца
A)98г. Крыло самцов B3) 101—118, самок (8) 104—119, в среднем соответственно 107,8
и 109,2 мм. У одной птицы неизвестного пола крыло 121 мм.
Окраска. Пуховой наряд темносерый, на брюшной стороне более светлый (Стей-
негер). Самцы и самки в брачном наряде: спинная сторона темно шиферно-серая, серая на
пояснице и в надхвостье; маховые и рулевые черновато-серые; на лбу белые украшающие
перья, такие же на уздечке и за глазом; горло и зоб темносерые, остальная брюшная сторона
буровато-серая с беловатой средней частью брюха и подхвостьем. Клюв яркокрасный с бело-
ватой вершиной, ноги бледно синевато-серые с черноватыми перепонками и когтями; радужина
белая. В зимнем наряде окраска такая же, но клюв менее яркий (в результате линяния рам-
фотеки, Стейнегер, 1885); менее развиты украшающие черные и белые перья на голове.
Молодые в первом зимнем наряде без украшающих перьев, с более темной черноватой4
спинной стороной, с более слабым и черным по окраске клювом; окраска брюшной стороны
довольно значительно варьирует от серой, белеющей только на задней части брюха, до пре-
имущественно беловатой, с бледносерым горлом, зобом и боками.
13. Конюга-крошка Aethia pusilla Pall.
Uria pusilla. Pallas. Zoographia Rosso-Asiatica t. II, 1811, стр. 373, Камчатка;
Русское название. Конюга, как указывалось выше, народное, повидимому,
звукоподражательное. Конюга-крошка — книжное. У Двигубского A831) — пыжик
маленький. Другое местное название у русских — как для других конюг: турутура или
курукура (с курильского Паллас, 1811).
Распространение. Ареал. Северные части Тихого океана. У азиатских
берегов на Чукотском п-ове (у северного берега, Пальмен, 1887; Сердце-Камень,
Нельсон, 1883; м. Иксурин, Тайер и Бенгс, 1914), на о-вах Диомид; у амери-
Карта 38. Распространение конюги-крошки Aethia pusilla
а _ границы гнездовий, г — область зимовок и кочевок.
канских берегов у м. Лисберн и залива Коцебу в Аляске, на Прибыловых о-вах,
о-вах св. Лаврентия, св. Матвея, на Алеутской гряде, быть может на о-ве Кадьяк.
В негнездовое время у берегов Камчатки, Коряцкой Земли, у Командорских
о-вов, у Курильских (главным образом у Урупа), Итурупа, Симусира в При-
224
ОТРЯД ЧИСТИКИ
морье (Тачановский, 1893), у Сахалина, в Японии (Хоккайдо, Хондо, Тане-
гасима); в Америке — изредка у Прибыловых о-вов, обычно у Алеутских
о-вов, в небольшом числе и южнее (Педжет-зунд, Вашингтон). Случайная
встреча зимою у залива Коцебу (Бент, 1919), залеты к м. Барроу и
в дельту р. Мекензи (Порсилд, 1943).
Характер пребывания. Откочевывает в море от мест гнез-
довья, немногие остаются в гнездовой области. Повидимому, в бурную и холод-
ную погоду приближается к берегам и в негнездовое время (Стейнегер, 1885,
связывает с этим появление конюги-крошки в конце декабря и начале января
у Командорских о-вов).
Д а ты. Появление весной на Прибыловых о-вах обычно в апреле C0 марта —
10 мая), откочевка оттуда главным образом в августе, начало ее уже в конце
июля, в октябре очень редка. У Коман-
дорских о-вов отмечена в декабре, весг
ною последние 25 марта (Иогансен, 1934);
случайно, повидимому, задержавшаяся
особь еще 10 мая(Хартерт, 1920). У Ку-
рильских о-вов близ Итурупа добыты
уже 15 августа (Стейнегер, 1898). У бе-
регов Камчатки и Коряцкой Земли до-
бывались в середине и во второй поло-
вине октября. В Приморье в бухте Аб-
рек добыта 10 декабря (Тачановский,
1893).
Биотоп. Гнездовый — скалы, в
негнездовое время на море.
Численность. На местах гнез-
довья (Прибыловы о-ва, о-ва Диомид)
крайне многочисленна. В области зимо-
вок у берегов довольно редка, так как
держится повидимому, в открытом море.
Экология. Размножение. По-
явление у гнезда в конца апреля.
Гнездится в скалах, между камнями и
в расщелинах, без всякой подстилки.
Половая зрелость, повидимому, только во втором брачном наряде, в возрасте
несколько менее двух лет (Бент, 1919). Кладка из одного яйца, в разные числа
июня (ранняя дата на Прибыловых о-вах 28 мая, Прибл и Мк. Эти). Яйца
белые, матовые, относительно очень больших размеров: E7) 33,5—43 х 27—
33,5, в среднем 39,5 х 28,5 мм (Бент, 1919). Так как ненасиженные яйца попа-
даются еще и в середине июля, то, повидимому, бывает дополнительная кладка
взамен утерянной. Вылупление в июле, в августе птенцы спускаются на воду,
в конце этого месяца имеют уже полный наряд из контурных перьев и начи-
нается откочевка от мест гнездовий как старых, так и молодых птиц.
Линька. Смена нарядов: пуховой — первый зимний — первый брач-
ный—второй зимний—второй брачный и т. д. Полная осенняя (послебрач-
№ая) линька вероятно в сентябре, в октябре птицы в свежем зимнем пере. Ча-
стичная предбрачная линька, охватывающая, мелкое оперение или
часть его, с конца декабря (Стейнегер). При этой линьке развиваются белые
украшающие перья лба, а также линяет рамфотека — у основания ее появ-
ляется бугорок, вершина клюва краснеет. Маховые осенью сменяются, повиди-
мому, постепенно от заднего края партии к переднему. В конце сентября
B4 сентября) в бухте Эмма на Чукотском п-ове добыты птицы, у которых
остались старыми 2—4 передних маховых. Детали линяния требуют
изучения.
Рис. 43. Конюга-крошка
БЕЛОБРЮШКА 225
Питание. Мелкие беспозвоночные, преимущественно амфиподы. На
Командорах зимою при вскрытии найдены Gammaridae, Palaemonidae, Amphi-
poda (Стейнегер, 1885). Кормится днем, на море, добывая пищу как с поверхно-
сти, так и ныряя. Пища птенцам приносится в полости рта и в довольно разви-
том горловом мешке (Портенко, 1934).
Полевые признаки. Самый мелкий из чистиковых вид, с однообразно тем-
ной спиной и более или менее пестрым низом; украшающие перья мало заметны.
Голос — тонкий свист или писк. Неосторожна.
Описание. Размеры и строение. Клюв короткий, с бугорком — в брачном
наряде — у основания надклювья; подклювье изогнуто; ноздри щелевидные, с вздутыми
краями, не прикрыты оперением. Цевка короче среднего пальца (с когтем). Хвост из 14 руле-
вых. Длина самцов 168, 170, 175, 184 (Стейнегер, 1885), длина самца 190, размах его 340,
длина самки 180, размах 345, длина молодой самки 174, размах ее 344 мм (Тачановекий,
1893); длина самца 175, самки 185 мм (Зоологический музей Московского университета).
Вес самки A) 93 г. Крыло самцов A0) 92—97, самок B3) 90—98, птиц неизвестного
пола A2) 92—98; в среднем у самцов A0) 94,9, самок B3) 94,3, обоих полов D5) 94,6 мм.
Окраска. Пуховой птенец однообразно дымчато-бурый, несколько светлей
на брюшной стороне. По мере роста, как и у других чистиков, окраска несколько свет-
леет. Взрослые самцы и самки в брачном наряде: спинная сторона черная, передние
плечевые светлосерые, маховые черные с бурыми внутренними опахалами; вершины
наружных опахал внутренних второстепенных маховых с белыми каемками; рулевые
черные; на лбу, уздечке, у разреза рта, за глазом — тонкие белые «украшающие»
перышки; щеки и горло темносерые; подмышечные и бока буроватосерые, подкрылья
белые; зоб, грудь и брюхо белые с буровато-серыми пестринами. Радужина белая;
клюв красный, черноватый у основания; ноги бледносерые с черными когтями и перепон-
ками. В зимнем наряде клюв черноватый, без бугорка у основания надклювья; спин-
ная сторона черная, передние плечевые серые, брюшная сторона белая, иногда
с темными серыми немногочисленными пестринами на груди и брюхе; щеки сероватые;
украшающие перья слабо развиты и отсутствуют за глазом. В первом зимнем наряде моло-
дые — без белых украшающих перьев и с более интенсивной черной окраской спинной сто-
роны. В первом брачном наряде птицы без бугорка у основания надклювья, белые украшаю-
щие перья на голове развиты еще слабо (Бент, 1919).
РОД БЕЛОБРЮШКИ CYCLORRHYNCHUS KAUP, 1820
Тип С. psittacula
14. Белобрюшка Cyclorrhynchus psittacula Pall.
Alca psittacula. Pallas. Spicilegia Zoologica, t. 1. fasc. 5,1769, стр. 13, табл. 2, табл. 5,
фиг. 4—б, Камчатка.
Русское название. Па л л ас A811) приводит в качестве местного названия
«морской камас». Белобрюшка, повидимому, тоже народное имя (Стейнегер, 1885).
Распространение. Ареал. Северные части Тихого океана. В Азии от
Чукотского п-ова (Сердце-Камень на северном побережье, Нельсон, 1883);
о-ва Диомид (Нельсон, 1887); бухта Провидения и бухта Преображения (Пор-
тенко, 1934); Командорские о-ва (о-ва Беринга и Медный), быть может Куриль-
ская гряда (добывался, гнездование не доказано, скорее нечастые залеты);
Охотское море (Аян, Мальминские о-ва, северное побережье). В Аме-
рике на Прибыловых о-вах, о-в св. Лаврентия, св. Матвея, по всей Алеутской
цепи, на о-ве Кадьяк. Палласом указывался для Камчатки, где позже никто
не встречал. Зимний ареал плохо выяснен (зимовки в открытом море?). Встре-
чен на Сахалине, быть может зимует в Приморье (Шульпин, 1936), отмечен
у Курильских о-вов и у Хондо. В Америке встречен у берегов Орегона и Ка-
лифорнии (бухта Сан-Франциско, Монтерей, Гумбольдта). Залет в Швецию
(декабрь I860, Саломонсен, 1940).
Характер пребывания. В негнездовое время откочевывает
в море или к югу. Впрочем, наблюдалась у Прибыловых о-вов (Прибл и Мк.
Эти, 1923).
Даты. Появление на Командорских о-вах с конца апреля, массовое
в начале мая, тогда же на Прибыловых о-вах. Откочевывает после спуска птен-
15 Птиды СССР, т. II
226
ОТРЯД ЧИСТИКИ
цов на воду в конце августа и в начале сентября. Встречи на Чукотском п-ове
относятся к концу августа. Появление на зимовке в Калифорнии указано
в декабре.
Биотоп. Гнездовый — прибрежные скалы, кормится на море; зимовки,
повидимому, преимущественно вдали от берегов.
Численность. На гнездовьях образует значительные скопления, но,
как у других чистиков, общее распространение спорадично. На Чукотском п-ове
гнездовые колонии малочисленны (Портенко).
Экология. Размножение. Белобрюшка гнездится колониями в ска-
лах, .иногда высоко над морем (на о-ве св. Матвея до 200 футов, Бент). Гнезда
Карта 39. Распространение белобрюшки Cyclorrhynchus psittacula
а — границы гнездовий, г — область кочевок и зимовок.
устраивает, главным образом, в щелях между камнями, но иногда и открытые
(Командоры, Иоганзен, 1934). Кладка в середине или во второй половине июня
(Командоры, Иоганзен; однако, Дыбовский нашел яйцо на о-ве Медном 10
июня, Тачановский, 1893); на Прибыловых о-вах 8 и 13 июня (Прибл иМк.
Эти, 1923). В кладке одно яйцо овальной формы, окраска скорлупы матово-бе-
лая или голубовато-белая. Размеры 50,7 х 37,8 мм (Тачановский), C3) 51,5 —
— 58 х 33—40, в среднем 54,3 х 37,3 мм (Бент, 1919). Насиживание около
30 дней, пуховые птенцы на Командорах в середине июля (Иоганзен; Хдртерт,
1920). В конце августа выводки спускаются на воду и птицы откочевывают от
мест гнездовий.
Линька. Не изучена, повидимому, как у конюг, в частности, у малой.
Смена маховых у взрослых происходит вскоре после спуска молодых на воду.
Весенняя предбрачная линька заканчивается зимою.
Питание птицы составляют мелкие беспозвоночные. В желудках,
добытых у Чукотского п-ова найдены Calanidae, Parathemisto, полихеты;
наибольшее значение в кормовом режиме имеют, повидимому, планктонные
копеподы. Птица заглатывает их в большом количестве, причем у нее как
ТУПИК-НОСОРОГ
227
у конюг, имеется горловой мешок, позволяющий выкармливать птенцов непе-
реваренной нежной пищей (Портенко, 1934).
Полевые признаки. Белобрюшка крупнее других конюг и резко двуцвет-
ная, с темной спинной и белой брюшной стороной. Своеобразный красный
вздутый клюв. Полет быстрый по прямой линии. Голос — резкий дребезжащий
свист.
Описание. Размеры и строение.
Своеобразный выпуклый как по коньку надклювья, так и по подклювью клюв; над-
клювье уплощенное и в брачном наряде имеет четыре «украшающие» пластинки: беловатую
мягкую пластинку на надклювье у разреза рта; пластинку под ноздрями раздельно с правой
и левой стороны клюва; узкую поперечную пластинку на основании надклювья; небольшие
пластинки под этой поперечной пластинкой у основания надклювья (Стейнегер, 1885). Ноздри
щелевидны, не прикрыты перьями. Рулевых 14. Размеры: длина самцов 234, 256, 264; самок
Рис. 44. Белобрюшка
244, 256, 264 (Стейнегер, 1885); длина самки 272, размах 525 мм (Тачановский, 1893);
крыло самцов (8) 149—157, самок F) 145—154 птиц неизвестного пола (б) 151—157»
в среднем у самцов (8) 152,7; самок F) 149,6, птиц обоего пола B0) 152,05 мм
(Зоологический институт Академии наук, Зоологический музей Московского университета).
Вес самца A) 240 г.
Окраска. Пуховой птенец на спинной стороне и горле серовато-бурый, у основания
пух светлее, грудь и брюхо серовато-белые (о-в Беринга, Хартерт, 1920). У взрослых самцов
и самок в брачном наряде спинная сторона черная, за глазом пучок из узких белых перьев;
маховые черновато-бурые, рулевые черные, подбородок, горло, зоб темносерые, подкрылья
темно бурые, бока темные, серовато-бурые, грудь, брюхо, подхвостье белые. Клюв ярко-
красный, буроватый у ноздрей, с беловатым основанием надклювья; ноги серовато-белые,
буроватые у суставов, перепонки черные, когти черные. Радужина белая. В зимнем наряде
без пластинок на клюве (см. выше), брюшная сторона тела белая. Молодые не описаны.
РОД ТУПИКИ-НОСОРОГИ CERORHINCA BONAPARTE, 1828
Тип С. monocerata
15. Тупик-носорог Cerorhinca monocerata Pall.
Alca monocerata. Pallas. Zoographia Rosso-Asiatica, t. II, 1811, стр. 362, мыс св.
Ильи, Аляска.
Синоним. Chimerina cornuta, Ercholtz. Zool. Atlas Heft 111, стр. 2, табл. XII, 1829,
о-в св. Павла. — Cerorhinca orientalis. Brandt. Bull. Ac. Imp. Sc. St. Petersbourg,
11, 1837, стр. 348, западное побережье Сев. Америки.
Русское название. У Двигубского A831)—«чистик однорогий». Книжное —
длинноклювый тупик. Другое книжное название — шишконосая гагарка (Дулькейт и Шуль-
пин, 1937).
Распространение. Ареал. Северные части Тихого океана у побережий
Азии и Америки. В СССР указывается для Камчатки (Тачановский» 1893, но
228
ОТРЯД ЧИСТИКИ
основания неясны), для Командорских о-вов (залеты, Стейнегер, 1887), Ку-
рильской гряды (Итуруп, Кунашир, Сикотан, Шимушир, Райкоке—Бергман,
1935, Ямасина,1931), Сахалина (Анива, Кайбато, Монерон) и Приморья (Шуль-
пин, 1936); гнездится на Б. Шантарах (Дулькейт и Шульпин, 1937). К югу
до Японии (Хоккайдо, залив Аомори в северной части Хондо) и Кореи. В Аме-
рике — на Алеутских о-вах (Атка, Агатту, Умнак), на побережье Аляски,
к югу от Ситка и далее до Вашингтона. Ранее гнездился на о-вах Фаральонес.
Зимует от Приморья до Японии (до Нагасаки) и Кореи, в Америке — от Ва-
шингтона (о-в Форрестер) до северных частей южн. Калифорнии (о-в св. Неро-
нима, о-ва Серрос).
Карта 40. Распространение тупика-носорога Cerorhinca monocerata
а — границы гнездовий, г — область кочевок и зимовок.
Характер пребывания. Тупик-носорог — кочующая птица.
Зимовки — у морских побережий в южных частях гнездовой области и
за ее пределами. В СССР в Приморье. В Америке у мест гнездовья появляется
в апреле — мае, осенью откочевывает оттуда в сентябре — октябре.
Биотоп. Гнездовый — морские побережья с мягкой удобной для рытья
нор почвой, вертикально до 400—500 м, нередко среди деревьев (о-в Форрестер,
Хиз, 1915). Вне гнездового времени в прибрежных частях океана.
Численность. В азиатской части ареала редок, в американской
местами многочисленен (колония примерно из 10 000 птиц на о-ве Дистрекшен,
близ Вашингтона, Даусон, 1908), но спорадичей. Численность в Америке за-
метно снизилась за последние десятилетия.
Экология. Размножение. Относящихся к нашей территории сведе-
ний мало. Гнездится колониями. По американским данным (Бент, 1919) птица
на гнездовье ведет ночной образ жизни, днем прячется в норах. Гнездовые норы
на Сахалине имеют 1—\г/2 м в глубину и заканчиваются расширением, в ко-
тором, обычно без всякой подстилки, лежит единственное в кладке яйцо.
Цвет скорлупы грязнобелый, иногда с бледными сероватыми, желтоватыми или
ТУПИК
229
буроватыми крапинами. Размеры C9N3,7—73,5 х 42,6—50, в среднем 68,5 х
X 46,2 л?И1(Бент); в среднем 68 х 48,9 мм (Сахалин, сообщение Гизенко). Наси-
живают оба родителя, около трех недель. Появление птенцов, главным образом,
в первой неделе июля, спуск их на воду в начале августа (в бухте Таба молодые
на воде 7—9 августа, Шульпин, 1936).
Линька. Как и у других чистиков, детали мало известны. Полная
линька — послебрачная, осенью. При ней теряются брачные украшения
клюва (в Амурском заливе особь в зимнем наряде добыта 13 ноября,
Бутурлин, 1915). Брачное оперение и украшение на клюве разви-
ваются при частичной линьке весною.
Питание. Кормится беспозвоночными
и мелкой рыбой, иногда далеко в море (до
60 миль от мест гнездовий, Хиз, 1915).
Полевые признаки. Плавающий тупик-
носорог сильно втягивает шею, осадка
в воде глубокая (по крыло). Общая окраска
спинной стороны темная, брюхо светлое,
своеобразный вырост на клюве, пучок белых
перьев над глазами и на щеках (в брачном
наряде). Молчалив.
Описание. Размеры и строение.
Рулевых 18. Крыло самца 190, самки 185 мм (Тача-
новский, 1893); крыло молодых, добытых в ав-
густе: 146, 147, 148, 150, 152, 154 мм (Шульпин,
1936); крыло самцов B) 172—177, птиц неиз-
вестного пола G) 167—193, в среднем (9) 181,9 мм
(Зоологический институт Академии наук).
Окраска. Пуховой птенец однооб-
разно серовато-бурый, светлее на брюхе (Риджвей,
1887). У самцов и самок в брачном наряде спинная
сторона серовато-черная; маховые черные с буры-
ми основаниями внутренних опахал, хвост чер-
ный; шея, зоб, грудь, бока, голени, подкрылья
буро-серые; брюхо и подхвостье белые. Над
глазом и от угла рта — пучки тонких удли-
ненных белых перьев. Клюв оранжевый, чер-
ный по коньку надклювья, с выдающимся на-
ростом (рогом) и желтыми складками по разрезу рта; ноги желтовато-белые с черными
плавательными перепонками; радужина охристо-желтая. Самцы и самки в зимнем наряде с
менее развитыми белыми пучками перьев на голове (?), нет вздутия у разреза рта и рога
(только небольшой выступ у основания надклювья), цвет шеи и зоба серый, серая окраска
распространяется и на грудь. У молодых в первом зимнем наряде спинная сторона буровато-
серая, шея, зоб, бока, грудь и туловище, подкрылья, голени серовато-бурые, грудь и
брюхо — белые; клюв без рога и вздутия у основания.
Рис. 45. Тупик-носорог
РОД ТУПИКИ FRATERCULA BRISSON, 1760
Тип F. arctica
16. Тупик Fratercula arctica L.
Alca arctica. Linnaeus. Syst. Nat. ed X, 1758, стр. 130, сев. Норвегия.
Русское название. Тупик — местное название на Кольском п-ове, упоми-
нается еще Озерецковским A804). Кроме того — топорик, топорок, турпанчик. Этимология,
повидимому, от формы клюва. Местами — попугайчик (Бутурлин, 1934).
Распространение. Ареал. Северные и средние части Атлантического
океана и прилежащие части Северного Ледовитого. На Западе — в Гренлан-
дии, по ее западному берегу к северу до Смис-зунда, у берегов Лабрадора от
Унгава и залива св. Лаврентия (о-ва Брион, Берд-рок, Антикости, Болд-айс-
ленд, Брадор) и Ньюфаундленда до штата Мен (Матиникус-роки Метиас-сил-
230
ОТРЯД ЧИСТИКИ
айсленд); на востоке — в Исландии, на о-ве Медвежьем, Ян-Майене, Шпицбер-
гене, Новой Земле, на Мурманском побережье, в Норвегии к югу до Ставан-
гера и Осло, в зап. Швеции (две колонии у берегов Богуслена), в Шот-
ландии, Ирландии и Англии (на восточном побережье последней видимо только
на о-ве Фарн, Нортумберленд и у Фламборо в Йоркшире), на Гебридских,
Оркнейских и Фарерских о-вах, на Норманских о-вах, по островам у берегов
Бретани во Франции.
До тридцатых годов прошлого столетия гнездился на о-ве Гельголанд,
повидимому, также ранее гнездился на о-вах Берленгас у Португалии. В негне-
Карта 41. Распространение тупика Fratercula arctica
7. F. a. arctica, 2. F. a. naumanni, 3. F. a. grabae;
a — границы гнездовий, г — область кочевок и зимовок.
здовое время как в пределах гнездовой области, так и южнее. В Америке —
у берегов Массачусетса и изредка Нью-Йорка (Лонг-айсленд) и Пенсильва-
нии (Делавар); в Европе — у берегов Англии, Франции (до Пиренеев), в запад-
ном Средиземноморье (Балеары, Марокко и т. д.) до Корсики, Сицилии,
Мальты, изредка — в южных частях Адриатического моря. Залеты к берегам
Германии, на Канарские и Азорские о-ва, Мадейру, однажды к о-ву Раффлса,
Ливерпуль, в Гренландии (Педерсен, 1930), ив море к северу от Шпицбергена
иод 83°1Г с. ш. A2 июля 1896, Коллетт и Нансен, 1900).
Характер пребывания. В холодное время года тупик откоче-
вывает от мест гнездовий в море, частью в пределах гнездовой области,
частью к югу от нее.
Биотоп. Гнездовый — морские побережья, преимущественно на остро-
вах. Зимний — свободные от льда прибрежные районы морей, хотя в Атлан-
тическом океане встречен и в 450 милях от берегов Ирландии (Саломонсен,
тупик 231
1944) и в 400 милях от м. Фаруэлл, южн. Гренландия; наконец, в 300 милях
от Фарерских о-вов (Уинн-Эдварде, 1935).
Численность. В результате преследования человеком численность
тупика за последние 150 лет заметно сократилась. Тупик, например, ранее
гнездился в заливе Фенди, Лабрадор и в других местах у берегов этого полу-
острова — теперь он там исчез. Нет его более и на о-ве Гельголанд. Стал реже
в южн. Норвегии и Швеции, исчез в Англии на берегах Кента и Суссекса и т. д.
Немногочисленен тупик и на далеком севере, в частности на о-ве Медвежьем,
Шпицбергене, Новой Земле.
Подвиды и варьирующие признаки. Варьируют размеры, постепенно возрастающие от
юга на север. Вопрос осложняется большой индивидуальной изменчивостью и наличием
«карликовых» вариаций также и на севере (Саломонсен, 1944). Экологические различия огра-
ничиваются, повидимому, в основном разницей в сроках периодических явлений. Обычно
различают следующие подвиды: мелкий южный F. a. grabae В г е h m, 1831, — распространен-
ный на Фарерских о-вах, в южн. Скандинавии (южнее Ставангера), в Англии, Шотландии,
Ирландии, на Норманских о-вах, у берегов Бретани. Южно-арктический подвид
средних размеров F. a. arctica L., 1758, — гнездящийся в Сев. Америке, на за-
падном побережье Гренландии на север до бухты Мельвилля, в Исландии, в Норвегии
к северу от Бергена (разрыв между гнездовыми колониями этого и южного под-
вида между Хузо к северу от Бергена и скалами Ротт у Ставангера составляет около 200 км,
Саломонсен, 1944), на Медвежьем о-ве, Ян-Майене, у Мурмана и на Новой Земле (на о-ве
Медвежьем, Ян-Майене и Новой Земле, повидимому, переходы к следующей форме; Хартерт,
1917, относит к последней тупиков с Ян-Майена, но Сконнинг, 1933, считает их за представи-
телей номинальной формы). Наконец, крупный высокоарктический подвид F. a. naumanni
Norton, 1901, — гнездящийся на Шпицбергене, в с.-з. Гренландии к северу от 76? с. ш.
В качестве диагностического признака последнего подвида, кроме большой величины
(крыло 24 самцов и 21 самки 175—194, в среднем 185 мм, ле-Руа, 1911; крыло 20 самцов и
самок 172—193 мм, Саломонсен, 1944), приводится еще больший изгиб конька надклювья.
Обыкновенный тупик Fratercula arctica arctica L.
Распространение. Ареал. Норвегия к северу от Бергена; Кольский
п-ов — у Варде, на Айновских о-вах, на о-ве Гусинец, на Семи островах (Хар-
лов, Большой и Малый Зеленец, Кувшин, Малый Лицкий); в Белом море отсут-
ствует; далее на Новой Земле в немногих пунктах западного берега — в губе
Пуховой, Безымянной, Грибовой, быть может на Норвежском базаре в 30 км
от Крестовой губы, на севере в губе Архангельской, в общем между 72°3(У
и 76° с. ш.; на о-ве Медвежьем, в Исландии, в зап. Гренландии и по восточ-
ному побережью Сев. Америки (см. выше, в очерке распространения вида).
Возможно к этой же форме относятся тупики с о-ва Ян-Майен (Сконнинг,
1933, Берд, 1935; но Хартерт, 1917, считает из-за F. a. riaumanni, а по Саломон-
сену, 1944, там встречается более крупная переходная к F. a. naumanni по-
пуляция).
В негнездовое время близ гнездовой области и южнее —у Фарерских
о-вов, у Норвегии и зап. Швеции, изредка в датских водах и у Германии;
в Америке до Массачусетса. О залетах в открытое море см. выше. На Мурмале
уже весной, в марте, встречен у Варангер-фиорда (Мейнертцхаген, 1938) и
у о-ва Кильдина (Формозов). Возможно, что часть указаний на зимние встречи
тупиков в средних частях Атлантики также относятся к этой форме. Залеты
отмечены иногда и вглубь материка: у Эммериха в Гессене, в Финской Лаплан-
дии (в Энонтекис и у Усть-иоки), к Чун-озеру в Лапландском заповеднике в де-
кабре 1936. Однажды отмечен у Кандалакши в Белом море.
Характер пребывания. В зимнее время откочевывает от гнез-
довий, держась в море. Появление у мест гнездовий связывается с оттаиванием
гнездовых нор. На Семи островах (Мурман) появление во второй половине
апреля и до первых чисел мая (Кафтановский, Горчаковская). Откочевка
после спуска птенцов на море, в середине и второй половине августа (по тем
же авторам). У гнездовий тупики держатся таким образом около 4V2 месяцев.
232 отряд чистики
Отдельные особи отмечены на вост. Мурмане в ноябре (Кафтановский,
1941). На западе Новой Земли на о-ве Топорков в бухте Грибовой тупики дер-
жатся до конца августа — первых чисел сентября.
Биотоп. Гнездовый — склоны или площадки, главным образом, на
островах с мягкой торфяной почвой, удобной для рытья нор. Наличие гнездо-
вий тупика нередко приводит при этом к сильному развитию травянистой
растительности (Айновыо-ва). В негнездовое время на свободных от льда участ-
ках моря, главным образом, вблизи берегов.
Численность. В СССР большая колония тупиков — около 20 000
пар населяет Айновы о-ва. На Семи островах самая большая колония распо-
ложена на о-ве Большой Зеленец, в ней более 500 пар (Кафтановский и Моде-
стов, 1941). На Новой Земле немногочисленен —около 70 пар в губах Безы-
мянной и Грибовой (Успенский). Птица, несомненно, заслуживает охраны.
Экология. Размножение. (Семь островов, по Кафтановскому, Гор-
чаковской, Карташеву). Возраст наступления половой зрелости неизвестен. Со-
отношение полов —100 самцов на 110 самок. Гнездится колониально в закрытых
местах (норы в торфе, редко в расщелинах между камнями). Длина норы дости-
гает 1—2 и даже 3 л*. В норе одна или несколько гнездовых камер; в последнем
случае они последовательно занимаются в разные годы. Роет норы сам тупик при
помощи лап и быть может клюва. Диаметр хода норы у ее наружного отверстия
25—30 см, далее уже. Форма нор различна, изредка ход прямой, чаще дуго-
образный. Старые норы представляют собой сложное переплетение ходов с не-
сколькими гнездовыми камерами. Подстилка скудная, из сухой травы, перьев,
иногда водорослей.
Период размножения, и в частности кладки, зависит от протаивания почвы
в норах и может следовательно значительно колебаться (на о-ве Большой
Зеленец разница между сроками кладки у тупиков на южном и северном берегу
составляет до 15 дней, Кафтановский). Кладка протекает с конца мая, но
в среднем около второй трети июня; конец кладки на Семи островах, в зависи-
мости от указанных выше особенностей, 20—25 июня. Как правило, в кладке
одно яйцо, два бывают только в виде редкого исключения.
Яйца округло-эллиптические, скорлупа белая, иногда с лиловатыми кра-
пинами в глубоких слоях. Скорлупа относительно тонкая (норник), размеры
ее E5) 60—68 х 39—48, в среднем 62 х 44,2 мм (Кафтановский); E1) 60—
—68 X 32—48 мм, в среднем 63,8 х 44,4 (Шиханов); B3) 58,5—68 X 41,5—
—46,8, в среднем 63,9 х 44,8 мм (Горчаковская). Вес свежего яйца E5) 65,1 г;
B3) 66,42 г (Кафтановский, Горчаковская). Относительная величина яйца
таким образом очень значительная —около 11,1%—14,2% веса самки (Гор-
чаковская). При утрате яйца откладывается новое, что также влияет на растя-
нутость периода размножения.
Насиживают оба родителя, у которых имеется по два наседных пятна.
Срок — 35 дней; однако, в гнезде яйцо может находиться и более продолжитель-
ное время, так как плотно насиживать яйцо тупик начинает только через 4—6
дней, после его откладки. Появление птенцов с конца июня и в начале июля,
от запоздалых кладок и в конце этого месяца. На Новой Земле, повидимому,
позднее — проклюнутые яйца найдены только 3 августа (Успенский). Размеры
только что вылупившегося (однодневного) птенца: длина G) 120—138, средняя
126,3 мм; вес G) 40—53,5, в среднем 46,4 г (Кафтановский, Горчаковская).
Постэмбриональное развитие медленное. Пеньки маховых показываются на
10—11 день после рождения, рулевых — на 17—18 день; терморегуляция уста-
навливается на 6—7 день (до этого времени температура тела птенца коле-
блется между 27 и 35° Ц). К 40 дням маховые у птенца вполне развиты,
в оперении только остатки пуха. Между 36 и 40 днями молодой тупик слетает
на воду. На Семи островах спуск птенцов на воду отмечен между 5 и 23 августа,
обычно около середины месяца. Перед слетом на воду вес молодой птицы
тупик 233
несколько падает (средний вес 6 особей 332,3 г или около 67% веса взрослой
птицы, тогда как сидящие в гнезде птенцы — по 4 особям весят в среднем
404,5 г или 81,62% взрослой птицы). Повидимому, родители перестают кормить
выросшего птенца в гнезде, чем стимулируют его слет на воду.
Линька. Последовательность смены нарядов —типичная для отряда.
В половине августа или в начале сентября птенцы — в первом зимнем наряде,
сменяющемся следующей весной первым брачным, осенью — послебрачным
(зимним), на следующий год весною — брачным и т. д. Пуховой наряд моло-
дых заменяется первым зимним, мезоптиля нет. У взрослых две линьки в году—
частичная предбрачная, охватывающая мелкое оперение, но у некоторых осо-
бей по тем или иным причинам полная, со сменой рулевых и маховых, и после-
брачная (Саломонсен, 1944). При этом линянии развиваются и украшающие
выросты на клюве. Весеннее линяние происходит поздно, в апреле, у добытых
в середине апреля на Семи островах тупиков линяет голова, в конце этого
месяца линька переходит на шею, в последней трети мая заканчивается (птицы
в полном брачном наряде добыты 22—27 мая). В середине апреля клюв еще без
брачных выростов, но надклювье уже перекрасилось в красный цвет; в начале
мая поперечных желтых полос и выростов рамфотека еще не имеет, но в конце
мая брачные украшения клюва уже развиты полностью.
Послебрачная линька в зимний наряд также протекает очень поздно,
начинаясь, повидимому, не ранее ноября, при этом клюв теряет брачные укра-
шения, отпадают лопасти голой кожи у глаз, а лапы из красноватых становятся
желтыми; у некоторых особей при послебрачной линьке маховые и рулевые
не линяют до следующей весны (см. выше), у других — послебрачная линька
полная. В этом отношении тупик отличается, повидимому, от всех других чи-
стиков, в частности, от близких тихоокеанских видов.
У особей, имеющих частичную послебрачную линьку, смена маховых и
рулевых затягивается до весенней предбрачной. Маховые при линянии выпадают
в очень сжатые сроки, и птица, повидимому, на время утрачивает способность
летать (Саломонсен, 1944).
Питание. Тупик, по преимуществу, ихтиофаг. На Мурмане он в гнез-
довый период кормится и выкармливает птенцов преимущественно песчанкой,
реже—мойвой и сельдями; второстепенное значение в кормовом режиме
имеют беспозвоночные, среди них каланусы (Кафтановский). Для других райо-
нов в качестве второстепенных кормов тупика указываются ракообразные,
пелагические моллюски, полихеты (Уайзерби, 1941, Бент, 1919 и др.). Кормится
тупик в дневные часы, ночью отдыхает в гнездовых колониях. К гнезду тупик
приносит, подобно гагарке, иногда сразу по 10—12 рыбок, плотно зажимая их
между языком и клювом, причем рыбы свешиваются по бокам клюва. Пресле-
дуя рыбу, ныряет и плавает под водой, пользуясь при этом и крыльями и но-
гами.
Полевые признаки. В брачном наряде безошибочно различается от других
атлантических чистиковых ярко окрашенным высоким, напоминающим маску,
клювом. Окраска оперения в общем двуцветная — черная на спинной и белая
на брюшной стороне. Ноги красноватые или желтые. Полет быстрый но непо-
воротливый, с частыми взмахами крыльев. Стоит, опираясь на пальцы (не на
цевки, как большинство прочих видов чистиковых), нередко вертикально.
Хорошо ходит. Молчалив, в гнездовое время издает ворчащие звуки «арр»
или при испуге — крик вроде саааэ или topp». Птенец в гнезде издает писк
вроде «пинь-пинь». Держатся обычно группами. Гнездится колониально.
Описание. Размеры и строение. Резкий сезонный и возрастной диморфизм
в строении рамфотеки. В брачном наряде у взрослых птиц клюв высокий, сильно уплощен-
ный, почти треугольной формы, с крючкообразно загнутой вершиной надклювья; в прокси-
мальной части надклювья и подклювья у основания широкая пластинка; у основания над-
клювья поперечная выпуклая роговая складка; в дистальной части надклювья три-четыре
234
ОТРЯД ЧИСТИКИ
Рис. 46. Тупик
углубленных поперечных борозды, на подклювье одна-две борозды; разрез рта окаймлен взду-
той мягкой кожей, у угла рта розетка; под глазом голое пятно, над глазом мягкий кожистый
выступ (лопасть). В зимнем наряде боковые пластинки у основания клюва, поперечная склад-
ка, кожистая лопасть над глазом отсутствует; мягкая кожа у разреза рта и розетка спадаются.
Ноздри узкие, щелевидные, почти у самого края надклювья, когти сильные и круто загну-
тые, особенно на внутреннем пальце (рытье нор). Хвост слегка закругленный, из 14рулевых.
Длина мурманских самцов B0) 300—362, самок A7) 303—345, в среднем 336,8 и 329,6 мм
(Горчаковская); самцов G) 285—345, самок D) 300—335, в среднем316,8и 315мм (Карташев)
Размах самцов A7) 560—650, самок A9) 500—630, в среднем 622,3 и 600,1 мм (Гоочаковская).
Вес самцов A8) 466—586, самок A9) 445—565 г, в среднем 510,5 и 498,1 г (Гор-
чаковская); самцов G) 475—525, самок D) 461—517, в среднем 504,9 и 489,5 г (Карта-
шев). Вес новоземельских самцов C) 460—579, самок (8) 420—572, в среднем 524,7 и 475,7 г
(Успенский). Крыло мурманских
птиц — самцов A9) 150—190, са-
мок A9) 160—185, в среднем 173,7
и 170,9 мм (Горчаковская); сам-
цов G) 160—179, самок D)
171—175, в среднем 173 и 173 мм
(Карташев).Крыло новоземельских
самцов F) 162—188, самок (8)
168—182, в среднем 175,6 и 175,6 мм.
Среди изученного материала встре-
чаются изредка взрослые птицы
необычайно малых размеров.
Так, кроме перечисленных
экземпляров, в Зоологическом музее
Московского университета имеется
самка с Новой Земли, доставлен-
ная Носиловым, с крылом в 151,5
мм, два самца из губы Грибовой
имеют крыло в 162 мм и т. д.
Окраска. Пуховой птенец
с длинным, особенно на спинной
стороне пухом темного буровато-
серого цвета, с белой средней частью брюха и груди; лицо голое, клюв черноватый,
лапы черные. От других чистиковых пух тупика отличается большой длиной (вероятно,
в связи с гнездованием в норах) и относительно редким расположением бороздок (Кафтанов-
ский).
У взрослых птиц, самцов и самок, в брачном наряде спинная сторона черная, с буро-
ватыми внутренними опахалами маховых, на зобе матово-черное ожерелье; бока головы и
подбородок серые, иногда с темными пестринами; бока тела бледносероватые; остальная
брюшная сторона белая. Радужина ореховобурая; дистальная часть клюва ярко карминово-
красная с желтоватым концом; бороздки и складка у основания клюва желтые, «пластинки»
у основания клюва шиферно-серые; разрез рта и розетка желтые;голая кожа у глаза голубо-
ватая; лапы оранжеоо-красные, когти черные. В зимнем наряде окраска щек черноватая,
на клюве в основной части нет украшающих выростов, основная часть клюва темней, лапы
желтые; на спинной стороне нередко сероватые каемки перьев.
Молодые в первом зимнем наряде — как взрослые, но клюв низкий и без поперечных
борозд. В первом брачном наряде надклювье также относительно низкое, и поперечные бо-
розды развиты слабо или отсутствуют.
[Полярный тупик Fratercula arctica naumanni Norton
Fratercula arctica naumanni. Norton. Proceed of the Portland Soc. Nat. Historyvi 11E),
1901, стр. 144, Шпицберген.
В литературе имеется ряд указаний о встречах в пределах СССР этого подвида, но все
эти определения требуют подтверждения, хотя нахождение этого подвида в негнездовое время
у нас вполне возможно. Птицы с Новой Земли по размерам, как указано выше, все же не от-
личимы от номинального подвида и, следовательно, меньше шпицбергенских; заслуживает
все же внимания очень поздний цикл периодических явлений у новоземельских тупиков.
Экология последних требует однако еще детального изучения].
17. Ипатка Fratercula cornicalata Naumann
Mormon corniculata. Naumann. Isis. 1821, т. 2, стол. 782, фиг. 3—4, Камчатка.
Русское название. Народное, у жителей Камчатки — ипатка (Крашенин-
ников, 1755, Паллас, 1811). Этимология неясна.
Распространение. Ареал. Северные части Тихого океана. В Азии оди-
ночная птица наблюдалась у о-ва Геральд (Нельсон, 1883), повидимому, у о-ва
ИПАТКА
235
Врангеля (Пальмен, 1887) далее—у берегов Чукотского п-ова (м. Сердце-Ка-
мень, м. Восточный, на запад до Колючина о-ва, гнездовые колонии, Тайер и
Бенгс, 1915; на юге в бухте Провидения, Портенко, 1934), в устьях Анадырского
лимана на о-ве Алюмка (Портенко, 1939); по вост. побережью Камчатки, на юге
Коряцкой Земли (п-ов Тайгонос, о-в Матуга ниже устья Авековой, Аллен,
1905), на Охотских побережьях у о-ва Шантар (Дулькейт и Шульпин, 1937),
на Сахалине (во всяком случае у Кайбато, вероятно в других местах, Мун-
стеръельм, 1920), повидимому, в самых северных частях Приморья (Шульпин,
1936), на Курильских о-вах, на Командорах. В Америке—на север до м.
Карта 42. Распространение ипатки Fratercula corniculata
а — границы гнездовий, г — область кочевок и зимовок.
Лисберн, залива Коцебу, о-вов Диомид и Прибыловых, о-ва св. Лаврентия и
св. Матвея, по Алеутской гряде, на юг до южн. Аляски (о-в Форрестер, Санта
Лазариа, пролив принца Вильяма).
Зимует изредка в северной части гнездовой области (Прибыловы о-ва,
Прибл и Мк. Э^ги, 1923), обычно у Командорских и Курильских, в небольшом
числе у Хондо; кроме того, на востоке у южн. Аляски и Британской Колум-
бии, у о-ва Королевы Шарлотты. -Залеты до южн. Калифорнии (Монтерей,
Бент, 1919).
Характер пребывания. См. выше. Весеннее появление у мест
гнездовья связывается, повидимому, с таянием прибрежных льдов. Появление
у Прибыловых о-вов в первой трети мая, откочевка оттуда в сентябре. У Ко-
мандор — в начале мая, исчезает с гнездовых колоний в сентябре (Иоганзен,
1934). На о-ве Алюмка, Анадырь, отмечен около середины июня (Портенко,
1939).
Биотоп. Гнездовый —скалистые побережья, зимний —.прибрежные уча-
стки моря.
236
ОТРЯД ЧИСТИКИ
Численность. В азиатской части ареала у Командор, на Кам-
чатке, Курильских о-вах — нередок, но уступает в численности хохлатому
топорику.
Экология. Размножение. Цикл поздний. Гнездится ипатка коло-
ниально в скалах, в отличие от тупика не в норах, а в расщелинах скал (Кам-
чатка, Командоры), впрочем на о-ве Кадьяк и о. Форрестер будто бы и в норах.
Гнезда используются по крайней мере несколько лет. Кладка из одного,
иногда будто бы из двух яиц (Портенко, 1939). Размеры яйца относительно
очень велики, форма его более продолговатая, чем у атлантического тупика,
скорлупа матовая, грубозернистая, белого цвета, иногда с бледными крапинами.
Размеры C) 65,8—76 х 46,8—51,2 лм< (Тачановский, 1893), C8) 58,74 х 41,50,
5 мм (Бент, 1919). Насиживают оба
родителя, срок неизвестен. В гнезде
иногда подстилка из сухой травы.
Спуск молодых на воду, повиди-
мому, в августе; около середины сен-
тября, у Командорских о-вов встре-
чаются вполне оперенные молодые. В
случае гибели яйца птица несется
повторно (Бент).
Линька. Сходна с линькой у
атлантического тупика, два раза
в году — предбрачная частичная,
охватывающая мелкое оперение и
рамфотеку, протекающая вероятно в
конце зимы — начале весны (точные
даты не установлены), и послебрач-
ная. В связи с поздним периодом
размножения послебрачная линька
также начинается поздно, как у
обыкновенного тупика: еще в начале
сентября добываются птицы в полном
брачном наряде, без заметной линьки
(Анадырский лиман). Послебрачная
линька — полная, охватывающая мелкое оперение, маховые, рулевые,
рамфотеку. Указание Бента A919) о том, что послебрачная линька протекает
в августе — сентябре на азиатском материале не подтверждается.
Питание. Основной корм составляет мелкая рыба. На Анадыре в желуд-
ках ипаток найдены остатки рыбы и какого-то головоногого (Портенко, 1939).
Для Прибыловых о-вов указываются изоподы. Кроме того, моллюски (Тер-
нер, 1886). Кормовые вылеты —в разные часы дня.
Полевые признаки. Типичный тупик с массивным, ярко окрашенным клю-
вом у взрослых. От встречающегося на Тихом океане топорика отличается
белой брюшной стороной и отсутствием длинных пучков перьев за глазом.
Заметно крупнее атлантического тупика, с которым в общем сходен по повад-
кам. Полет быстрый, с частыми ударами крыльев и оттянутыми назад ногами.
В негнездовое время молчалив, в брачное издает крик «орр-орр» (Стейнегер,
1885) или тихое ворчанье. Мало осторожен.
Описание. Размеры и строение.В общем сходен с атлантическим тупиком.
Клюв массивный, уплощенный, с характерными украшениями в брачном наряде: у основа-
ния надклювья сильное вздутие в виде поперечной пластинки, рамфотеки; по бокам основания
надклювья большие пластинки; в вершинной половине надклювья обычно три поперечных
вдавления в виде борозд; у заднего края разреза рта мягкая розетка. Над глазом вытянутый
кожный голый лоскут, под глазом голое пятно. Ноздри близко к разрезу рта, вытянутые,
щелевидные. Когти сильные и загнутые, в особенности на внутреннем пальце. Рулевых 14.
Длина самцов 341—357, самок 342—343 мм (Стейнегер, 1885). Крыло самцов 173—183,
Рис. 47. Ипатка
ТОПОРИК, ХОХЛАТЫЙ ТОПОРИК 237
самок 170—194, птиц неизвестного пола 177,5—187; в среднем у самцов F) 178,5, самок (9)
176,1, у взрослых обоего пола A9) 178,2 мм.
Окраска. Пуховой птенец на спинной стороне серовато-бурый с беловатым брюхом.
У самца и самки в брачном наряде спинная сторона черная, за исключением серовато-бурых
лба, темени, затылка, внутренние опахала маховых буроваты; уздечка, щеки, широкая поло-
са над глазом и за глазом белые, кроющие уха сероваты, подбородок буровато-серый, горло
и зоб буровато-черные; бока буро-серые, подкрылья серые, чернеющие к кистевому сгибу;
остальная брюшная сторона белая. Голые выступы кожи у глаза черноватые, веки красные;
клюв красный в вершинной, лимонно-желтый в основной части; розетка в углу красная;
радужина сероватая; лапы оранжево-красные, когти бурые.
В зимнем наряде клюв без украшающих пластинок и вздутия у основания, нет и ро-
зетки в углу рта; вершина клюва красноватая, голая кожа у разреза рта желтая, основание
клюва бурое; веки бурые; голых выступов кожи у глаз нет; лапы оранжевые или бледно-
красноватые; оперение, как в брачном наряде, но на лице белого нет, щеки, брови и уздечка
серовато-бурые.
Молодые в первом осеннем (зимнем) наряде, как взрослые в зимнем, но с бурым низким
клювом, без борозд на надклювье, щеки окрашены обычно темнее. В первом брачном наряде
ипатки с относительно слабым развитием украшений клюва, поперечные борозды только
намечены, как и вздутие у основания надклювья и розетка; щеки не белые, а сероватые.
Систематические замечания. Морфологические различия ипатки и атлантического ту-
пика так значительны, что несмотря на явную генеалогическую близость этих птиц и строгий
географический викариат, их можно считать только за особые виды, хотя и довольно близкие.
Между ними есть и биологические различия (расположение гнезд, голос, детали линяния).
Характерно, что большая величина ипатки по сравнению с атлантическим тупиком — типич-
ное для тихоокеанских форм кругополярных видов чистиковых явление (Саломонсен, 1944).
РОД ХОХЛАТЫЕ ТОПОРИКИ LUND A PALLAS, 1811
Тип L. cirrhata
18. Топорик, хохлатый топорик Lunda cirrhata Pall.
A.lca cirrbata. Pallas. Spicilegia. Zoologica 1 E), 1769, стр. 7, табл. 1, табл.5, фиг. 1—3,
море между Камчаткой и Америкой.
Синоним Stagmatorrhina lathami. Bonaparte. Proceed. Zool. Soc. London,
1851, стр. 202, табл. X—IV, Берингов пролив.
Русское название. Топорик, топорок, повидимому, местное, от формы клюва птицы.
Распространение. Ареал. Северные части Тихого океана и прилежащие
районы Сев. Ледовитого. В Азии от сев. побережья Чукотского п-ова
(несколько гнездящихся пар на о-ве Колючин, Тайер и Бене, 1914); встречен
у м. Восточного и Сердце-Камень (Нельсон, 1883), в Анадырском лимане на
о-ве Алюмка (Портенко, 1939), у Коряцкой Земли, на востоке в Олюторском за-
ливе, в частности у устьев Апуки и Пахачи; на юге на о-ве Матуга, и по вост.
побережью п-ова Тайгонос (Дементьев, 1940); далее к западу по Охотскому
побережью до Шантарских о-вов (Дулькейт и Шульпин, 1937), у берегов Кам-
чатки (на востоке от м. Лопатки до м. Начиканскогоио-ваКарагинского; на
зап. берегу между Камбальной и Озерной и т. д., Бергман 1935, Белопольский,
1947); на Курильских и Командорских о-вах (чаще на о-ве Беринга), на Саха-
лине и в Приморье (залив Петра Великого, Шульпин, 1936, Тачановский,
1893). В Америке от с.-з. Аляски (м. Лисберн), Прибыловых о-вов, о-ва св. Лав-
рентия и Алеутской гряды до Британской Колумбии (Ванкувер, Триангль,
о-ва Королевы Шарлотты, Тавернер, 1934), Вашингтона, Орегона, Калифорнии
(Фаральонес, Санта Барбара, изредка Сан-Николас, Бент, 1919). Зимою в море
в пределах гнездовой области (по крайней мере до Прибыловых о-вов, Прибл
и Мк. Эти, 1923), но преимущественно южнее, на широтах Командорских и
Курильских о-вов и у Приморья, достигая сев. Японии, где редок (Хоккайдо,
Хондо). Залетный был добыт в устьях р. Кеннебек в штате Мен (Одюбон); указа-
ния старых авторов на случайные встречи в Гренландии, повидимому, оши-
бочны.
Даты. Стаи у берегов вост. Камчатки в начале июня E июня), у мест
гнездовья появляются там же во второй неделе июня (Бергман, 1935). В конце
238
ОТРЯД ЧИСТИКИ
мая отмечено появление у Командорских о-вов (Иоганзен, 1934). На сев. побе-
режье Охотского моря у Гижиги появлялся в первой неделе мая (Аллен,
1905). На Курильских о-вах в начале мая (Стейнегер, 1898, однако, на Итурупе
в конце мая, Бергман, 1935). У гнездовий на Прибыловых о-вах птица встре-
чена около середины мая (ранние одиночки, быть может зимовавшие, уже
5 марта, Прибл и Мк. Эти). Осенняя откочевка после спуска на воду молодых.
Она отмечена на Прибыловых о-вах с конца августа, на Охотском побережье
еще в середине октября (Аллен, 1905), на Курильских о-вах около середины
сентября (Стейнегер); в сентябре, повидимому, и на Командорах (Иоганзен).
Биотоп. Гнездовый — острова и материковые побережья, скалы у края
обрывов или площадки, обычно с травянистой растительностью и торфом.
Карта 43. Распространение топорика Lunda cirrhata
а — границы гнездовий, г — область кочевок и зимовок.
Небольшие острова, повидимому, предпочитаются. Зимою в море, обычно
довольно далеко от берегов. У последних, по Стейнегеру, преимущественно
в штормовую погоду.
Численность. Высокая. Впрочем, на севере ареала топорик малочисле-
нен: гнездовая колония на о-ве Алюмка в Анадырском лимане из 50—100 особей
Щортенко, 1939), на о-ве Колючин —только немного пар (Тайер и Бенгс, 1914).
На Прибыловых о-вах весьма обычен. Крупные колонии местами на вост.
Камчатке (например на о-ве Топорков, Бергман, 1935), на Охотском море (п-ов
Тайгонос, о-в Матуга), на Курильских о-вах. На о-ве Топорков близ о-ва Беринга
на площади не более V2 км2 подсчитано 100 000 пар этой птицы (Иоганзен).
Экология. Размножение. Половая зрелость, повидимому, в воз-
расте около 2 лет, во всяком случае в годовалом возрасте некоторые
птицы не размножаются, а держатся в открытом море (Стейнегер, 1885).
Топорик, хохлатый топорик
239
Как и другие тупики, гнездится в вырываемых самой птицей норах в торфяной
почве, колониально. На упомянутом выше о-ве Топорков близ западного
берега о-ва Беринга на 1 м2 площади приходится 3 норы (Иоганзен, 1934).
К норам ведут тропинки, протоптанные птицами в траве. Глубина нор не менее
1 Mt нередко глубже, очень редко до 0,5 м (Бергман, 1935). Яйца откладываются
прямо на землю или на редкую подстилку из сухой травы и Перьев (Бент, 1919).
Цикл размножения поздний и растянутый, последнее обстоятельство объяс-
няется и наличием вторых (дополнительных) кладок в результате промыслового
сбора яиц. Кладки отмечены в конце июня и в начале июля (Командоры, Стей-
негер, 1885, Иоганзен, 1934; самые ранние, по первому автору 12 июня) и
в конце июня (ранние 24—27 июня, Прибыловы о-ва, Прибл и Мк. Эти, 1923)
во второй неделе июня
(Гижига, Аллен, 1905); око-
ло середины июня (Ку-
рильские о-ва, Стейнегер,
1898).
В кладке 1 яйцо, ред-
ко —два (Житков и Штехер,
1915). Форма яйца овально-
коническая, размеры отно-
сительно крупные, окраска
белая или белая с серова-
тыми пятнышками. Размеры
68—71 х 45—50 мм (Стей-
негер); 69,2—78,2 х 48—
51,8 мм (Тачановский); D3)
65,5—78 х 45—51,5 в сред-
нем 72 х 49,2 мм (Бент,
1919). Насиживают оба ро-
дителя, повидимому около
месяца (однако, по Эммер-
сону, 1904, только 21 день,
что сомнительно).
Птенцы в возрасте око-
ло 2 недель добывались на
вост. Камчатке во второй
неделе августа, в начале —
середине августа отмечены на Командорах. В случае утраты первого яйца
имеются повторные кладки —до 2 и даже 3. От таких поздних кладок пухо-
вики в гнездах находились на Командорах еще в конце сентября (Стейнегер,
1885). Утверждение о наличии в норе двух кладок в году явно ошибочно. Пти-
цы, выводящие птенцов из дополнительных поздних кладок, задерживаются
на местах гнездовья B октября, Прибыловы о-ва, Прибл и Мк. Эти). Гнез-
довый период нормально продолжается до начала сентября, когда молодые,
надев первый зимний наряд, слетают на море. Выкармливают молодых оба
родителя, летая за кормом в море.
Линька. Смена нарядов обычная для чистиков. Птицы в первый зимний
наряд линяют в августе — сентябре, в соответствии с возрастом. Предбрач-
ная линька взрослых неполная, в конце зимы (даты не установлены; линные
птицы добывались в феврале), захватывает контурное мелкое перо, но не махс-
вые и не рулевые; тогда же развиваются украшающие перья на голове и пла-
стинки у основания клюва. Послебрачная линька полная в августе — сентябре,
иногда затягиваясь до конца октября (Стейнегер, 1885). Маховые сменяются,
повидимому, почти одновременно; утрачиваются украшающие перья на головг
и пластинки на клюве.
Рис. 48. Топорик (взрослый и молодой)
240 Отряд чистики
Питание. В пищу употребляется мелкая рыба (сельдевые и др.);
кормится топорик в море, кормовые вылеты преимущественно по утрам. Добы-
вает пищу нырянием, захватывая ее клювом поперек продольной линии тела*
Кроме того, в качестве пищи отмечаются моллюски, морские ежи, губки (Бент,
1919).
Полевые признаки. Крупная величина, однообразно-бурая окраска, бе-
лые щеки, длинные белые пучки перьев по обеим сторонам затылка, высокий
красный клюв хорошо отличают топорика в брачном наряде от всех другихчисти-
ков. Полет быстрый, по прямой линии, довольно высоко над водою; взлет и
повороты с затруднением. Довольно хорошо ходит, стоит и сидит, преиму-
щественно выпрямившись почти вертикально. Держится обычно стаями и
группами. Молчаливая, как и другие тупики, птица. Старые птицы, потре-
воженные в гнезде, издают своеобразное ворчание.
Описание. Размеры и строение. Топорик — самый крупный из тупиков.
Клюв очень массивный, в брачном периоде надклювье с 3 редко 4 поперечными бороздами;
у основания клюва над ноздрями большой щиток, под ноздрями у разреза рта узкая попереч-
ная пластинка с каждой стороны; выступ в основной части надклювья по гребню, пластинка
на лбу, испещренная углубленными точками; в углу рта розетка, подклювье массивное, вы-
ступающее углом примерно на уровне одной четверти длины от основания, с чуть намечен-
ными поперечными углублениями; вершина клюва слегка крючкообразна. В зимнем наряде
основание клюва без украшающих пластинок, с мягким беловатым вздутием по разрезу рта.
Крылья небольшие, формула обычная: 2>3>4...„(перо рудиментарно).Хвост из 14 рулевых.
Средний палец длиннее цевки. Когти острые, загнутые.
Длина самцов 391, 397, самок 367, 378, 385 мм (Стейнегер, 1885), самцов 396—405,
самки 395, размах самцов 740—744, самки 712 .мл* (Тачановский, 1893). Вес самца A) 869 г.
Крыло самцов 187, 189, 198, самок 187, 191, 192, 192, 193 мм (Стейнегер), самцов G) 191,
191,5, 193, 194, 194, 198, 201, самок 190, 197,5, птиц неизвестного пола 184, 186, 192, 194,
194, 197, 198 мм (Зоологический музей Московского университета). В среднем крыло самцов
A0) 192,6, самок G) 191,8 мм.
Окраска. Пуховой птенец серо-бурый, светлей на брюхе (Хартерт, 1917, Бент,
1919). По мере роста птенца на пухе проступает белый цвет. У самца и самки в брачном на-
ряде спинная сторона черная,брюшная буроватая; по сгибу крыла — белые перышки; подбо-
родок, щеки — белые; длинные украшающие перья за глазом —соломенно-желтые. Клюв
красный у вершины, с зеленоватым основанием надклювья и подклювья; крайние части осно-
вания рамфотеки красные; вокруг глаза красное пятно (голое); ноги красные, когти черные.
Радужина белая, в зимнем наряде украшающие перья за глазом отсутствуют, щеки и подбо-
родок бурые, на брюхе нередко беловатые пестрины; украшающие пластинки у основания
клюва отсутствуют, и проксимальная часть рамфотеки бурая, с беловатым разрезом рта;
радужина голубоватая. Молодые в первом осеннем (зимнем) наряде — без беловатых укра-
шающих перьев за глазом, на спинной стороне черные, на брюшной серые, на подбородке,
зобе, боках, подкрыльях, с белыми или беловатыми грудью и брюхом; клюв низкий, бурова-
тый; радужина бурая. В первом брачном наряде украшающие пластинки клюва развиты
слабо (Стейиегер, 1885); украшающие перья головы короткие и буроватые.
ОТРЯД ГАГАРЫ
GAVIAE или GAVIIFORMES
(Colymbi или Colymbiformes у разных авторов)
Характеристика отряда
Морфология. Внешние признаки. Гагары — крупные птицы.
Размеры самой большой гагары — белоклювой — около 850 мм в длину,
более 1500 мм в размахе, при весе, превышающем 4V2 кг. У самого небольшого
вида — краснозобой гагары — длина около 550—650 мм, размах около 1000—
1100 мм, вес около 2 кг.
Форма тела гагар вальковатая, шея довольно длинная, ноги далеко отне-
сены назад, и голени почти полностью покрыты общим с туловищем кожным
покровом.
Крылья хорошо развиты, узкие и острые, хотя в общем недлинные,
рулевые короткие и жесткие, почти полностью прикрыты кроющими хвоста.
Первостепенных маховых 11 (из них первое рудиментарно), второстепенных
22—23, рулевых 8—10 пар; уздечка оперенная; оперение жесткое, густое и
плотно прилегающее к телу, перья короткие.
Самцы и самки окрашены одинаково: брюшная сторона туловища у них
белая, спинная серовато-бурая (молодые, взрослые в зимнем наряде), серая
или черная с белыми пестринами (брачный наряд). Пуховые наряды без пест-
рин или полос.
Клюв длинный, прямой, иногда слегка выпуклый по коньку надклювья,
с заостренной вершиной. Ноздри узкие, щелевидные, с кожным клапаном,
закрывающим доступ воды при нырянии. Цевки голые, довольно длинные,
уплощенные, покрытые спереди сетчатой кожей. Пальцев ног—4, первый
(задний) слабо развит и прикреплен выше остальных, передние пальцы длин-
ные, соединены перепонками до конца дистальных фаланг. Когти узкие.
Анатомические признаки. Череп схизогнатический, голо-
ринальный, отростков основной крыловидной кости нет, ноздри сквозные.
Так как носовые железы сильно развиты, то и надглазничные ямки очень ве-
лики. Височные впадины широки, что, как и большая длина заднего отростка
нижней челюсти, связано с сильным развитием жевательной мускулатуры
(захватывание относительно крупной рыбы). Шейных позвонков 14—15. Груд-
ные позвонки не сращены. Грудина узкая и вытянутая с одной парой
16 Птицы СССР, т. II
242 ОТРЯД ГАГАРЫ
вырезок. Нижние поперечные отростки передних грудных A—3-го) позвонков
очень длинные. До грудины доходят 8—9 пар ребер. Длина кисти примерно
равна длине предплечья. Кнемиальный отросток processus rotularis tibiae
очень длинный, больше, чем бедро; коленной чашки нет (рудимент последней
существует в виде сезамоидной кости в сухожилии выпрямителя голени).
Малая берцовая кость длинная, доходящая почти до интердорзального сочле-
нения. Характерные особенности имеются в устройстве сочленения бедра
с голенью, позволяющие сгибание в сторону, а также в сочленении цевки с пе-
редними пальцами, ограничивающие возможности движения пальцев и увели-
чивающие общую прочность конструкции ноги.
Таз — длинный и узкий, как у всех ныряющих видов. Скелет не пнев-
матичный.
Перья — с побочным стволом. Пух расположен и по птерилиям, и по апте-
риям. Дорзальная аптерия не распространяется на шею. Крыло аквинтокуби-
тального типа. Копчиковая железа оперена.
Мозг — окципито-темпорального типа. Глаза расположены строго лате-
рально, умеренных размеров. Уши маленькие, со слабо развитыми кроющими.
Слепые кишки короткие. Две глубоких сонных артерии. Сердечный индекс —
15,18 у чернозобой гагары и 12,14 у краснозобой гагары (Кафтановский).
Гортанные мышцы группы m. m. sterno tracheales симметричны. Обводящая
мышца имеется.
Образ жизни. Гагары — дневные водоплавающие птицы, в значительной
мере морские и только в период размножения живущие на пресных водоемах.
Лишь в гнездовый период гагарам приходится бывать на берегу, вне этого
времени они держатся на воде, где и отдыхают.
Гагары — перелетные птицы (за редкими исключениями, относящимися
к отдельным популяциям).
Пища гагар — почти исключительно рыба средней и мелкой величины.
Подчиненное значение в кормовом режиме имеют беспозвоночные — моллюски,
ракообразные, черви, насекомые, в особенности в питании птенцов. Известное
значение имеют быть может и растения (Хр. Л. Брэм, 1831). Без рыбы гагары
обходиться не могут и в случае гнездования на безрыбных или малорыбных
озерах летают кормиться на соседние водоемы (в приморских тундрах и
на море).
Специализация гагар в ихтиофагии достигла высокой степени. Способы
добычи у всех видов однообразны. Гагары высматривают рыбу, плавая по по-
верхности воды или ныряя. При нырянии гагара погружает голову и шею
вперед и вниз, после чего следует и погружение тела под более или менее острым
углом к поверхности воды. Перед погружением гагара прижимает к брюху
цевку и лапу с вытянутыми вперед пальцами, а затем поворачивает под прямым
углом голень так, что пальцы обращаются латерально; следующее движение —
быстрое выпрямление цевки и отведение ног с раздвинутыми пальцами кзади и
немного кверху. В момент этого толчка лапы гагары широко расставлены, а
после него сближаются, сжимая воду и отбрасывая ее назад, что создает значи-
тельную опору. Нырянию способствует и удаление птицей воздуха из воздуш-
ных мешков и из оперения, которое прижимается к телу. Под водою гагары
могут пробыть несколько десятков секунд (по некоторым данным, до 2 минут,
или более) и покрыть расстояние до четверти мили. Рыба умерщвляется сжи-
манием клюва, чему способствует сильное развитие жевательных мышц.
Мелкие рыбы заглатываются под водою, крупные — выносятся в клюве на по-
верхность и заглатываются, начиная с головы.
Охотятся гагары поодиночке, но на зимовке и пролете — нередко на не-
большом расстоянии одна от другой.
Летают гагары быстро, но — из-за небольшой величины крыльев и высокой
весовой нагрузки — не могут изменять скорости полета, даже при спуске,
ХАРАКТЕРИСТИКА ОТРЯДА 243
и с трудом взлетают (чернозобая и полярная гагары — со значительным раз-
бегом по воде, а с суши взлетать не могут).
На суше гагары беспомощны, ходят с трудом и на самое короткое рас-
стояние; с трудом, при помощи согнутой шеи, поддерживают равновесие в
стоячем положении, держась при этом вертикально. При передвижении по
земле обычно ползают, отталкиваясь ногами (например, добираясь до гнезда
или спускаясь с него на воду).
Гагары —моногамы. Пары, вероятно, постоянные. Потенциально полово-
зрелыми становятся, вероятно, в годовалом возрасте, но такие птицы часто
остаются холостыми. Гнезда устраиваются в кучах отмершей растительности
или просто в ямках на берегу пресных водоемов — озер, реже речных заводей.
Кладка из 2, реже 1 и еще реже из 3 яиц, продолговатой формы, оливково-бу-
рых с поверхностными черноватыми и глубокими сероватыми пестринами.
В случаях утраты кладки устраивается дополнительная. Это обстоятельство,
в связи с зависимостью срока кладки от таяния водоемов, объясняет растяну-
тость периода размножения. Насиживают оба родителя (по большинству ав-
торов — с первого яйца), в течение 25—29 дней. Развитие птенцов —выводко-
вого типа.
В отличие от поганок, пуховики окрашены на спинной стороне в однообраз-
ный буроватый цвет без полос или пятен. Пуховых нарядов — два. Оперяются
молодые гагары поздно, в возрасте около 40 дней. Второй пуховой наряд сме-
няется первым контурным зимним, последний весною — первым брачным,
слегка отличным от последующих брачных. Взрослые гагары линяют два раза
в году: при этом у одного вида полная линька бывает осенью, в зимний наряд,
она охватывает и рулевые и маховые (краснозобая гагара), а частичная, охва-
тывающая мелкое оперение, —весной, когда образуется брачный наряд,
У других видов также две линьки в году, но осенняя послебрачная частичная,
а весенняя предбрачная полная (чернозобая и полярная гагары). При сходстве
цикла периодических явлений у всех гагар, причины таких различий в течение
линяния неясны. Выпадение первостепенных маховых при смене их происходит
у гагар одновременно, и птицы теряют на это время возможность полета. Зим-
ний наряд носится около 3 месяцев, примерно с декабря по март. Половой
диморфизм выражается только в несколько больших размерах самца.
Изменчивость окраски гагар может быть изображена следующей схемой:
половой и групповой мономорфизм, возрастной и сезонный диморфизм, геогра-
фический полиморфизм. Впрочем, географические расы отличаются по окраске
незначительно (эти различия коррелируют с разницей в размерах).
Систематика. Гагары — небольшая группа, состоящая из единственного
семейства Gaviidae, с одним родом Gavia, распространенным в холодном и уме-
ренном климате северного полушария и состоящим только из трех видов.
Будучи близкими к отряду поганок, гагары отличаются от последних
рядом анатомических и экологических признаков, так что большинство совре-
менных авторов справедливо считают эти группы за отдельные отряды.
Ископаемые гагары описаны из верхнего эоцена Англии и из верхнего
олигоцена Франции; недатированные находки ископаемых гагар имеются
и в Северо-Американских Соединенных Штатах. Рецентные виды в ископаемом
состоянии известны из плейстоцена.
Обычно полагают, что гагары — примитивная и древняя группа, имеющая
эволюционные связи с меловыми зубатыми птицами-гесперопнисами.
Географическое распространение. Гнездовая область гагар — тундра, лесо-
тундра и лесная полоса Европы, Азии и Сев. Америки; на зимовке главным
образом по морям умеренной зоны. В СССР представлены все три вида гагар.
Распространение гагар на гнездовье по зонам в нашей стране таково
(в числителе показано число видов, в знаменателе — число подвидов и моно-
типических видов; характерные формы подчеркнуты):
244 ОТРЯД ГАГАРЫ
I. Арктика (зона) Gavia arctica, G. immer, G. stellata 3
а. побережье (подзона) G. i. adamsii, G. s. stellata, G. Q. pacitica */6
б. тундра (подзона) G. i. adamsii, G. s. stellata, G. s. squamata, G. a. arctica,
G. a. viridigularis 8/5
в. лесотундра (подзона) G. s. stellata, G. a. arctica, G. a. viridigularis */»
II. Лесная зона G. stellata, G. arctica 2
г. тайга (подзона) G. a. arctica, G. a. viridigularis, G. s. stellata */з
д. смешанный лес и лиственный лес (подзона) О
е. лесостепь (подзона) О
III. Зона открытых сухих пространств G. arctica 1
ж. степь (подзона) G. a. arctica
(озерная страна Зап. Сибири и Казахстана) Vi
з. полупустыни и пустыни (подзона) О
IV. Горная зона G. arctica I
и. высокогорная борео-альпийская (подзона) G. a. arctica, G.a. viridigularib (?)Vi
к.горнолесная подзона О
л. горностепнаяи пустынная подзона О
Практическое значение. Широко распространенный взгляд о вреде гагар
для рыболовства едва ли оправдывается реальным количеством поедаемой ими
рыбы, в особенности на фоне промыслового рыболовства и по сравнению
с масштабами последнего- Шкурки гагар представляют собою хороший «пти-
чий» мех и находят себе применение для разных поделок (шапки, воротники,
отделка платья и т. п.).
Таблица для определения видов гагар
1B).Голова и шея серые или черные, в первом случае на горле каштаново-рыжее пятно
(брачный наряд взрослых) 3
2A). Голова и шея иной окраски, горло белое (зимний наряд взрослых, гнездовый
наряд) 7
3D). На горле каштаново-рыжее пятно . . . краснозобая гагара Gavia stellata
4C). На горле каштаново-рыжего пятна нет, окраска горла черная с синеватым, фио-
летовым или зеленоватым отливом 5
5F). Темя и задняя сторона шеи серые или сероватые
чернозобая гагара Gavia arctica
6E). Темя и задняя сторона шеи черные полярная гагара Gavia immer
7(8). На спинной стороне тела явственные белые или беловатые крапины
краснозобая гагара Gavia stellata
8G). На спинной стороне тела белых пятен нет 9
9A0). Крупней, высота клюва у проксимального края ноздри больше 22 мм; подмы-
шечные перья бурые с белыми краями опахал . . . . полярная гагара Gavia immer
10(9). Меньше, высота клюва у проксимального края ноздри менее 20 мм; подмышеч-
ные белые или белые с узкой бурой наствольной полосой
чернозобаягагара Gavia arctica
РОД ГАГАРЫ GAVIA FORSTER, 1788
1. Краснозобая гагара Gavia stellata Pont op p.
Colymbus stellatus. Pontoppidan. Danske Atlas. I, 1763, стр. 621, Дания.
Русское название. Слово «гагара» — звукоподражательное, передает крик
птицы. Местами в Сибири «краснозобая гагара», иногда «малая гагара». В сев. России,
как и другие гагары, «норь».
Распространение. Ареал. Субарктический и бореальный. Северная по-
лоса Европы и Азии и Сев. Америки — до островов Ледовитого океана, на юг
КРАСНОЗОБАЯ ГАГАРА
245
примерно до 57—59° с. ш. (детали см. ниже), в негнездовое время до Китая,
Черного и Средиземного морей, Флориды, Калифорнии, Британской Колумбии.
Характер пребывания. Перелетная птица. Биотоп. Озера в тундре,
лесотундре и лесной полосе в гнездовое время. На перелетах и зимою — как
Карта 44. Распространение краснозобой гагары Gavia stellata
7. G. s. stellata, 2. О. s. squama ta;
a — границы гнездовий, г — область зимовок.
внутренние водоемы (озера и реки — в этом отличие от других видов гагар),
так и прибрежные части морей.
Подвиды и варьирующие признаки. Два подвида, различающихся слегка окраской:
G. s. squamata Portenko, 1939 — с Земли Франца-Иосифа и возможно Шпицбергена
и G. s. stellata Pontoppidan, 1763 — в остальной части ареала. Возможность признания
первой расы за полноценную, впрочем, вызывает сомнения (см. ниже систематические
замечания), и вид, быть может, правильней считать монотипическим.
Обыкновенная краснозобая гагара Gavia stellata stellata Pontopp.
Распространение. Ареал. Север Европы, Азии и Сев. Америки. В СССР
на севере от Мурмана и прилежащих островов (Айновы, Семь островов), Кол-
гуева, Вайгача, Новой Земли до Новосибирских о-вов и о-ва Врангеля; далее
к югу по тундре и лесотундре и северной части лесной полосы до Латвии (уЦенн-
гофф к юго-зап. от Риги, Брандт, 1940), вероятно до озера Ильмень и Ладожского;
далее к востоку южная граница остается неясной, так как поздне-пролет-
ные птицы могли быть приняты за гнездовых, а точных находок на гнездовье
нет; регулярное гнездование отмечено в бассейне Удоры, Вологодская обл.
под 62° с. iil (Корсаков, 1941); в области Урала гагара встречается примерно
около 67° с. ш. на Оби у Березова, но далее и южнее, в бассейне Нарыма и
Васьюгана, в бассейне Таза (Б. Елагуй под 63°, Скалой и Слудский, 1941),
на Енисее примерно до 59° с. ш.; затем — на Байкале; в Якутии примерно до
бассейна Вилюя, Якутска, восточней до Май и Удского района (Шульпин,
1936); далее граница проходит по Охотскому побережью, достигая на юге
нижнего течения Амура (озеро Кизи), Шантарских о-вов; наконец, гнездится!
на Камчатке и Командорских о-вах, на Сахалине и Курильских о-вах (Пара-
мушир и Шимушир).
Вне пределов СССР в Европе гнездится в сев. Скандинавии, на Гебрид-
ских, Шотландских и Оркнейских о-вах, в Шотландии и Ирландии, в Ислан-
дии, на о-ве Медвежьем (Бертрам и Лек, 1933), вероятно на Ян-майене (Берд,
1935). В Америке, в Гренландии до 82°27\ по островам Ледовитого океана, на
Алеутских о-вах, на континенте до средних частей Квебека, Кьюотина, Боль-
шого Невольничьего озера, о-вов Королевы Шарлотты и южн. Аляски, на Нью-
фаундленде. Вне гнездового периода на пролетах встречается в разных мест-.
246 ОТРЯД ГАГАРЫ
ностях Европы, Азии и Сев. Америки, избегая пустынных и горных пространств
центральной Азии и будучи связанной с водоемами — реками и морями. Регу-
лярные зимовки этого вида — у берегов Европы от Скандинавии до Велико-
британии, на Балтийском и Северном морях; в Азии —от средних Куриль-
ских о-вов до Японии (чаще на море, реже на ее внутренних водоемах, Ян,
1942) и вост. Китая (на юг до Гуаньдуня, Латуш, 1934); в Америке главным
образом у морских побережий, на юг до Флориды и Калифорнии, на север до
области Великих озер и Алеутских о-вов. Отдельные особи встречаются в Среди-
земноморье (Испания), в Азии до Формозы.
В СССР зимовки отмечены у Кольского п-ова (Оленья губа, декабрь)
и на Камчатке (Ст. Бергман, 1935), нерегулярно на озере Севан в Армении
(Шелковников, 1934), а также на Черном и Каспийском морях, быть может
в Средней Азии (пролетные особи встречены на Аму-Дарье, Сыр-Дарье, на
Аральском море).
Характер пребывания. Перелетная птица. Как у всех гагар,
весенний пролет и прилет поздние. Осенью птица также летит поздно. Отдель-
ные особи еще в последней трети апреля встречаются в Южн. Китае (Фучжоу,
Латуш, 1934), в Средней Азии также в половине апреля (Керки на Аму-Дарье,
Зарудный; на Каспии встречается по крайней мере еще в первой половине
апреля, даже на юге у Чикишляра; в это же время и на Черном море, Норд-
ман, 1840 и др.). В основном же пролет в средней полосе европейской России
протекает с конца апреля до конца мая (в Московской обл. у Богородска самая
поздняя дата 21 мая). Появление гагар на севере у мест гнездовья падает на
разные числа мая, быть может и в конце апреля (впрочем и это относится к про-
лету). На Мурмане ранние встречи 17—24 апреля (Гебель, 1901), но северней—
позже — в начале и даже середине июня (например, на Таймыре 5 июня,
Миддендорф, 1855,10—16 июня, Вальтер, 1902, Бируля, 1907; на Новосибирских
о-вах 16 июня, Бируля, и 21 июня, Бунге, 1887; на Новой Земле 20 июня,
Сконнинг, 1907; тогда же примерно и в Гренландии).
Даты прилета колеблются по годам в зависимости от таяния местных водое-
мов. Осеннее течение на местах гнездовья растянуто и, начинаясь со времени
взматерения ставших на крыло молодых, продолжается до замерзания водое-
мов, т. е. с последней трети августа и до начала октября (последние на вост.
Таймыре 24 октября, Толмачев и Тугаринов, 1934; на Новой Земле 3 октября
Сконнинг, 1916; в Гыданской тундре 26 октября, Наумов, 1931; на Анадыре
5 октября, Портенко, 1939 и др.), южнее —Камчатка, Командоры —гагары
задерживаются до ноября. В октябре — ноябре гагары появляются на зимов-
ках на Черном море, хотя в южн. Украине пролет отмечается и в декабре
(Умань, Гебель, 1879). Летят гагары небольшими стайками из нескольких
особей, парами или одиночками.
Биотоп. Гнездовый —озера в тундре, лесотундре и северной части лес-
ной полосы, с пологими покрытыми растительностью берегами. Вне гнездового
времени — морские побережья, но также и внутренние водоемы. В отличие
от других гагар, на пролете краснозобая гагара менее связана с морскими
побережьями и охотно придерживается текучих водоемов.
Численность. В гнездовой области —высокая. На зимовках в Черном
л Каспийком море — умеренная.
Экология. Размножение. Брачные игры на местах гнездовья в конце
.мая — начале июня, весьма характерные. В это время птицы крикливы (го-
лос — каркающие и стонущие звуки). Токуют иногда несколько гагар сразу.
Самец, а иногда и самка выскакивают из воды, держась вертикально и прижи-
мая клюв к груди, к нему присоединяется другая птица, оба начинают кричать,
затем вытягивают головы вертикально. Эти игры сменяются плаванием и пого-
ней одной птицы за другой с хлопаньем крыльев; при этом гагары издают кар-
кающие звуки, иногда высокий свист. Токование продолжается 2—3 часа,
КРАСН030БАЯ ГАГАРА 247
в разное время суток (Лапландия, Хортлинг и Бекер, 1932). На места
гнездовья гагары перемещаются по мере очищения водоема ото льда, а до этого
держатся парами вблизи водоемов например, в море у берегов островов и т. п.
Срок наступления половой зрелости не ясен. Пары, повидимому, постоянные,
так как птицы на пролете и с прилета держатся парами.
Гнезда бывают двух типов: или из водяных растений, но более простого
устройства, чем у других видов гагар, или просто ямка без подстилки, обычно
близко (в полуметре) от береговой линии или даже в прибрежной полосе
в воде. Размеры небольшие, в поперечнике от 20 до ЬОсм. Дно лотка, как у дру-
гих гагар, обычно бывает мокрым. В пределах своего гнездового участка гагары
гнездятся то на одном месте,, то на другом; присутствие в гнездовом участке
других особей того же вида не терпится, хотя на одном озере — в зависимости
от его величины — могут гнездиться и несколько пар (Лапландия, Больше-
земельская тундра, низовья Колымы и т. д.).
Кладка обычно в первой половине июня F—8 июня, Семь островов, Мур-
ман, Кафтановский; в первой половине июня, Новая Земля, Горбунов, 1929;
около 10 июня, низовья Колымы, Бутурлин; на крайнем юге ареала, в Шотлан-
дии, с конца мая и в начале июня, изредка ранее, например 9 мая, Уайзерби,
1940; на юге американского ареала в мае, повидимому, обычно в начале этого
месяца, Бент, 1919). При этом в пределах ареала в СССР кладка происходит
более или менее одновременно и в сжатые сроки. При утрате кладки бывает
вторичная (этим объясняются поздние находки свежих яиц примерно в пределах
месяца после нормального окончания кладки, например, в низовьях Енисея
слабо насиженные яйца найдены 20 июля, Тугаринов; на Камчатке свежее
яйцо 14 июля, Ст. Бергман, 1935; свежие яйца на зап. Мурмане до 1 ав-
густа, Гебель, 1903 и т. д.). Кладка обычно из 2, иногда из 1 яйца (быть может
изредка даже из 3, Хортлинг и Ст. Беккер, 1932). Промежуток между откладкой
яиц 24—48 часов (Мурман, Кафтановский). Яйца крупнозернистые, темного
оливково-зеленого цвета с неправильно разбросанными темнобурыми крапинами.
Размеры B5) 68—82 х 43—48, в среднем 73,8 х 44,8 мм; вес 73—89,7, в сред-
нем 82,7 г (Мурман, Кафтановский).
Насиживают оба родителя, в течение 25 дней (Кафтановский). Остается
неясным, начинается ли насиживание с откладки первого яйца (Уайзерби,
или после окончания кладки (Кафтановский, также Хортлинг и Бекер, 1932).
Во всяком случае, оба птенца иногда вылупляются в один и тот же день.
Птенцы могут плавать уже через 10—12 часов после вылупления. Вес одно-
дневных, по 5 экземпляров, 49—61,6, в среднем 54,8 г (Кафтановский).
Птенцовый период — до замены второго пухового наряда контурным опере-
нием — продолжается 40 дней, хотя молодые не достигают еще в это время
полных размеров взрослых (вес добытых в конце августа и начале сентября
в низовьях Колымы молодых особей 1281,2, 1332,5, 1537,5 г). Вполне оперив-
шиеся молодые — с середины августа (Мурман, низовья Оби, Новосибирские
о-ва, Колыма, Коряцкая Земля, Камчатка). Отмечена высокая смертность
птенцов (Мурман).
Л и н ь к а. Последовательность смены нарядов: первый пуховой —второй
пуховой — птенцовый (первый зимний) — первый брачный — зимний (после-
брачный) — брачный и т. д. Окончательный наряд — первый брачный, хотя
он обычно все же может быть отличен от последующих (см. ниже, описание
окраски). Две линьки в году: осенняя послебрачная полная и весенняя частич-
ная. При послебрачной осенней линьке все маховые выпадают одновременно
и птица таким образом временно утрачивает способность к полету. Послебрач-
ная линька — на зимовке, в конце сентября —начале октября у пролетных га-
гар линька только начинается, заканчивается, повидимому, в декабре. Весен-
няя предбрачная линька проходит в марте — апреле и оканчивается в начале
мая, охватывая лишь мелкое оперение — не рулевые и маховые. У молодых
248
ОТРЯД ГАГАРЫ
птиц в гнездовом наряде предбрачная линька начинается рано, иногда уже
в декабре, но затягивается, вероятно в связи с половым циклом, до весны.
Питание. Как и другие гагары, кормится главным образом рыбой
(на Мурмане песчанками, мойвой и т. д., Кафтановский), которую добывает
ныряя. Дополнительный корм — моллюски, ракообразные, икра, водные
насекомые (Кандалакша, Протопопов). Охотясь за рыбой, гагара погружает
голову в воду и быстро ныряет и плавает под водою. Мелкую рыбу и беспозво-
ночных заглатывает под водою, более крупную добычу выносит на поверх-
ность. Срок пребывания под водою гагары, по наблюдениям в Зап. Европе,
определяется в 67 и даже 90 секунд (Уайзерби, 1940). Весной гагара ест и водя-
ные растения (Хр. Брэм, 1831).
Рис. 49. Краснозобая гагара
Полевые признаки. Наименьшая из наших гагар — размерами с крупную
утку. От других гагар в брачном пере легко отличается сероватой, а не черной,
спиной без резких белых пестрин и заметно более слабым клювом, как бы
вздернутым. В зимнем наряде отличается, кроме меньшей величины, наличием
беловатых пестрин на спине. Полет — быстрый и прямой, птица довольно легко
взлетает даже с небольших водоемов, как бы прямо выскакивая из воды (дру-
гие гагары нуждаются обычно в разбеге). Ходит по земле с трудом, иногда
ползет, отпихиваясь лапами. В брачный период часто подает голос. Звуки разно-
образны : как бы плачущее «уаа-уаа», затем грубое каркающее «га-га-гара-га-га-
гара>, иногда —мяуканье, заканчивающееся подобием хохота. На пролете
обычно негромкое «га-га-га» (Шульпин, 1936). При нырянии гагара гребет
ногами, помогая продвижению и крыльями.
Описание. Размеры и строение. Тело, как у других гагар, вальковатое
с далеко отнесенными назад ногами, голень мало выдается и прикрыта почти на всемсвоем про-
тяжении общим с туловищем кожным покровом; вершины второстепенных маховых с выем-
кой; оперение плотное и жесткое; рулевые короткие, почти прикрытые кроющими хвоста,
в числе 10 пар; первостепенных маховых II, 1-е рудиментарно, формула крыла 2>3>4 ...
Клюв сильный, заостренный у вершины, сжатый с боков, с щелевидными, снабженными ко-
жистым клапаном, ноздрями; надклювье слегка направлено кверху; первый (задний) палец
ноги короткий, прикреплен выше остальных; передние три пальца длинные, соединены пере-
понками. Самая мелкая из гагар. Крыло самцов D2) 265—308, самок D0) 260—298, в среднем
282,8 и 274,6 мм. Длина самцов G) 640—680, самок (9) 560—640, в среднем 664,8 и 600 мм.
Размах крыльев самцов D) 1090—1140, самок (8) 980—1110, в среднем 1109,25 и 1066,7 мм.
Вес самцов D) 1400—2460, самок A1) 1250—2255, в среднем 1887,5 и 1652,4 г.
Окраска. Птенец в первом пуховом наряде на спинной стороне однообразно черно-
вато-бурый, на брюшной стороне несколько светлей, особенно на груди и брюхе, имеющих
КРАСН030БАЯ ГАГАРА 249
сероватый оттенок; клюв и лапы черноватые. Второй пуховой наряд светлей и серей, вся
брюшная сторона, за исключением бурой шеи и горла, приобретает светлый сероватый цвет.
Гнездовый наряд: темя, затылок, шея серые с беловатыми каемками перьев; спина и крылья
темнобурые, у края каждого опахала по супротивному охристо-белому пятну; бока головы
и шея с бурыми пестрина ми, только горло чистобелое; брюшная сторона тела белая; маховые
и рулевые темнобурые; радужина темнобурая, клюв серо-роговой, беловатый у основания
подклювья; ноги черноватые, белеющие на внутренней стороне пальцев, кроме наружного
и цевки. Взрослые самцы и самки в зимнем наряде похожи на гнездовых птиц, но верх головы
и шеи темней и бурей, с более развитыми светлыми каймами; спинная сторона также темней,
а пятнышки на ней белей и контрастней; на боках головы и передней части шеи нет темных
пестрин.
В брачном наряде голова и шея пепельно-серые с темными черноватыми наствольями
на лбу, темени, затылке, с продольным черным и белым рисунком на задней стороне шеи,
причем черный цвет тут с зеленоватым отливом; спинная сторона тела и верхние кроющие
крыла серо-бурые, иногда с большей или меньшей примесью мелких белых пестрин; на пе-
редней стороне шеи более или менее треугольное каштаново-рыжее пятно; брюшная сторона
белая, с бурыми пестринами по краям зоба и брюха и с черной поперечной полосой в под-
хвостье. Радужина красноватая или ореховая; клюв черный, сероватый по коньку; ноги зе-
леновато-черные, с беловатым или желтоватым оттенком на внутренней стороне цевки, на
перепонках и пальцах (кроме наружного). Первый брачный наряд годовалых птиц отличается
значительной примесью старых перьев гнездового наряда (птица на спинной стороне кажется
поэтому пестрей ) и слабым развитием каштаново-рыжего горлового пятна, состоящего иногда
только из немногих перьев и более бледного, охристо-рыжеватого, по цвету. Самцы и самки
окрашены одинаково, половой диморфизм — в общих размерах (см. выше).
Пестрая краснозобая гагара Gavia stellata squamata Portenko
Colymbus st el I at us squamatus. Портенко. Фауна Анадырского края, II. Тр. Института
полярн. земледелия, животноводства и промыслов, хозяйства, 1939, стр. 155, Земля
Франца-Иосифа.
Распространение. Ареал. Южная часть архипелага Франца-Иосифа
примерно до 81° с. ш. (о-ва Белля, Мабель, Ньютона, Скотт-Кельти, Огорд, па
Горбунову, 1932) и Шпицберген. Характер пребывания. Перелет-
ная птица. Весенний прилет во второй половине июня B0 июня, Пинегин, 1924,
18 июня, Горбунов, 1932). Биотоп, Озера (гнездовый). Числен-
ность. Низкая.
Экология. Размножение. Цикл поздний, свежие и слегка насижен-
ные яйца —в конце июня (о-в Скотт-Кельти 24 июня, Горбунов, 1932; по тому
же автору на о-ве Огорд слабо насиженные яйца 31 июля, но 29 июля найден
птенец; на Шпицбергене яйца 26 июня и 6—7 июля). Вылупление, повидимому,
от конца июля до конца августа (быть может в связи с таянием льда на озерах).
В кладке 2 яйца. Гнездо на о-ве Скотт-Кельти — на мхе Drepanocladus,
диаметр его 54, диаметр лотка 27 см (Горбунов). На Шпицбергене гнездо —
вытоптанная в грязи и иле ямка, по краям ее — мох. Оперившаяся молодая
птица добыта на Шпицбергене 25 августа. Брачные игры продолжаются с при-
лета до периода насиживания; при этом птицы иногда преследуют друг друга,
как бы ища при этом что-то в воде. Иногда две птицы плывут рядом, вытянув
шеи и издавая ворчащие звуки. Проплыв некоторое расстояние, самка ныряет
и показывается из воды вертикально с пригнутой шеей и клювом и т. д.
Линька. Вероятно, как у номинальной расы, но возможны различия
в сроках. Питание. Не изучено.
Описание. Размеры и строение. Как у номинальной формы. Окраска. Спин-
ная сторона светлей и серей, чем у номинальной формы, на ней имеются обычно хорошо раз-
витые узкие бледносерые вершинные каемки.
Систематические замечания. Вопрос о том, можно ли придавать краснозобым гагарам
с Земли Франца-Иосифа и Шпицбергена (вся популяция из нескольких десятков пар) значение
географической расы в строгом смысле слова, т. е. формы, отражающей явления географи-
ческого изоморфизма, — остается, конечно, весьма сомнительным. В основе возникновения
особенностей этих гагар, к тому же слабо выраженных (часть птиц со Шпицбергена не отли-
чимы по окраске от обычных материковых особей), лежит не географическая изменчивость,
как таковая (географо-экологический диморфизм), а размножение в пределах небольших
изолированных популяций (так сказать «семейное сходство»).
650
ОТРЯД ГАГАРЫ
2. Полярная гагара Gavia immer Brunn.
Cotymbus immer В r ii n n i с h. Ornithologia Borealis, 1764, стр. 38У Фарерские о-ва.
Распространение. Ареал. Северные части Сев Америки, Азии и Европы,
в зоне тундры и в лесной полосе. Характер пребывания. Перелет-
ная птица, гнездящаяся на материке и зимующая на море. Б и о т о п. В гнез-
Карта 45. Распространение полярной гагары Gavia immer
7. G. /. adamsii, 2. G. i. immer;
a — границы гнездовий, 6 — вероятная южная гранииа гнездовий, г — область зимовки, д — изолирован-
ное гнездовое местонахождение на Охотском море.
довое время —озера, в холодное время года —литоральная полоса северных
частей Атлантического и Тихого океанов.
Подвиды и варьирующие признаки. Две географических расы, различающиеся морфо-
логией (размеры; величина, окраска и форма клюва; детали окраски оперения) и экологией
(биотопическое распределение, календарь периодических явлений): G. i. immer В г и п-
n i с h, 1764 (вероятный синоним G. i. elasson Bishop, 1921, — описана из Сев. Дакоты и
будто бы распространена на западе от сев. Уисконсина до сев. Калифорнии и Британской
Колумбии) — в Исландии, Гренландии, в Сев. Америке, кроме крайнего северо-запада;
G. /. adamsii Gray, 1859 — на крайнем севере Европы и Азии и на северо-западе Аме-
рики от сев. Аляски до дельты реки Мекензи.
Белоносая или белоклювая полярная гагара Gavia immer adamsii Gray
Colymbus adamsii. G. R. G r a y. Proceedings of the Zoological Society of London, 1859,
стр. 167, Берингово море, Аляска.
Русское название. Книжное. У русского населения сев. Сибири «тундреная
гагара» (Енисей, Шмидт, 1872), «морская гагара» (Туруханск, Миддендорф, 1843, Анадырь,
Сокольников), «большая гагара» (Яна, Бируля, 1907).
Распространение. Ареал. Крайний север Европы и Азии — от Печенги
до Чукотского п-ова; арктическая Сев. Америка — сев.-зап. Аляска, далее
на восток по побережью до заливов Ливерпуль и Франклина, до озер в низовьях
реки Мекензи, а также, повидимому, на лежащих к северу отсюда островах.
Гнездовое распространение в СССР спорадично: установлено на Айновых
о-вах (Хейнясаари у Печенги), вблизи от границы неоднократно наблюдалась
летом в вост. Финляндии (Гебель, 1903), также у Сайды-губы (Федюшин, 1925),
5 или 6 июня 1927 на Соловецких о-вах (Поляков, 1929), несомненно гнездится
на о-ве Колгуеве, на Новой Земле — на южном острове, быть может на север-
ном, где наблюдалась; указывается для нижней Оби (Паллас, 1776, Бутурлин,
1901, Шостак, 1921) — как у Березова, так и ниже по течению, но гнездование
тут не доказано; возможно на Ямале (Коллетт и Нансен, 1900), летом встречена
в Гыданской тундре (Наумов, 1931); далее к востоку отмечена в Туруханском
крае и в низовьях Енисея и несомненно гнездится на Таймыре — как на по-
бережье, так и внутри полуострова; из бассейна Лены и Яны достоверных
ПОЛЯРНАЯ ГАГАРА 251
сведений нет (Маак, 1886, упоминает о встречах в бассейне Вилюя; в бассейне
Яны встречается будто бы у Омолоя, Бируля, 1907); в низовьях Индигирки
на юг, повидимому доУяндиной (Михель, 1935); несомненно встречается на са-
мом северо-востоке Сибири в бассейне Колымы, на юг по крайней мере до Сред-
неколымска (пуховой птенец от 26 июля 1905 в Зоологическом музее Москов-
ского университета; Бутурлин, 1906, Тайер и Бенгс, 1914); наконец, в Чукот-
ской земле и в бассейне Анадыря; найдена на сев. побережье Охотского моря —
в Магаданском музее имеется самец, добытый там в мае 1941; другая птица из
гнездящейся пары добыта в бухте Нагаева 20 июля 1946 (Васьковский).
Старые указания на гнездование на Канине не подтверждаются, так как позже
никто этой птицы там не наблюдал. Определенно отсутствует в Тиманской и
Большеземельской тундрах, но быть может встречается на Вайгаче (Хейглин,
1872). Однажды добыта на Б. Ляховском о-ве на Новосибирском архипелаге
A4 мая 1940 г. в Зоологическом музее Московского университета). В негнездо-
вое время встречена у берегов Лапландии (в частности на о-ве Кильдин, Мид-
дендорф, 1843) и Норвегии, а также в южных частях Берингова моря (у нас
на Камчатке в Авачинской губе еще Стеллером и т. д., на о-ве Беринга Стей-
негервм, 1887). Это —область морских, как и других гагар, и регулярных
зимовок. Отдельные особи встречаются и южней — у Курильских о-вов, в Япо-
нии и Китае, в Европе—в Англии. Задеты у нас в Смоленскую обл.;
в Польшу, к устью Эльбы, в верхнюю Австрию и Италию.
Даты. Прилет и пролет связаны, повидимому, с замерзанием водоемов.
Основные места зимовок — у берегов Норвегии, там белоклювые гагары
появляются осенью в октябре (не ранее конца сентября), отлет оттуда проис-
ходит ранней весной, хотя отдельные особи даже летуют (Коллетт, 1894).
Однако на Новой Земле отдельные особи держатся до начала октября (Горбу-
нов, 1929), на Анадыре они наблюдались до 28 сентября (Портенко, 1939).
На севере СССР птицы добывались со второй половины мая, начала июня
(ранняя находка на Новосибирских о-вах 14 мая; прилет на Анадырь 2 июля,
Белопольский, 1934).
Биотоп. Гнездовый — озера, расположенные в приморской части
тундр, но также и в горах (с.-в. Сибирь, Портенко, 1939). Зимовки —в при-
брежных частях моря.
Численность. Редкая птица, более или менее обыкновенная лишь
на с.-в. Сибири. На пролете небольшими стаями (Портенко, 1939).
Экология. Размножение. Изучено слабо. Время наступления поло-
вой зрелости и образования пар не известно. Кладки, повидимому, в первой
половине июня (хотя экспедиция «Вега» добыла у м. Питлекай на Чукот-
ском п-ове только одно яйцо в гнезде 10 июля,Пальмен). Пуховой птенец у Сред-
неколымска добыт 26 июля. Гнездо — куча травы на тундровых озерах (Аля-
ска, Бент, 1919). Число яиц в кладке 1—2, чаще 2. Окраска яиц—как у номи-
нальной расы, бурая с темнобурыми мелкими крапинами; размеры яйца с Чукот-
ского п-ова 94,9 х 55,6 мм (Пальмен, 1887); у яиц из Сев. Америки A7) в сред-
нем 89 х 56,9, крайние величины 80—95 х 53,5—66 мм (Бент). Распадение вы-
водков и начало кочевок на Новой Земле отмечено в сентябре (Билькевич, 1904).
Линька. Последовательность смены нарядов — как у других гагар.
Полная годовая линька взрослых — осенью, перед зимовкой; в первой поло-
вине ноября (вероятно и ранее) птицы в полном зимнем наряде. Частичная
предбрачная линька, не охватывающая маховых и рулевых, протекает на зи-
мовке. В мае взрослые птицы несут полный брачный наряд.
Питание. Главным образом рыба. Наблюдаются перемещения на
кормежку с тундровых озер на море (Анадырь, Портенко, 1939).
Полевые признаки. Самая крупная из наших гагар, с большим белым
клювом, конек надклювья несколько «вздернутый». Голос громче и грубее,
чем у других видов (Бируля, 1907),
252
ОТРЯД ГАГАРЫ
Описание. Размеры и строение. Величина значительная. Длина самки A)
868, размах ее 1575 мм, вес 4715 г. Крыло самцов и самок B2) 364—405, в том числе 364—405
у самцов и 365—396 мм у самок; в среднем 383,8 мм. Рулевых — 9 пар. В остальном строе-
ние, как у краснозобой гагары.
Окраска. Первый пуховой наряд (Среднеколымск) на спинной стороне, подбородке,
горле — однообразно серовато-бурый, на брюхе серый; клюв серо-роговой. Второй пуховой
наряд не описан. В гнездовом наряде спинная сторона серовато-бурая, с охристо-серова-
тыми краями опахал на шее, спине, кроющих крыла, с беловатыми краями рулевых; бровь,
щеки, брюшная сторона белые, бока — с примесью серовато-бурого; подмышечные белые
с широкой продольной буроватой полосой у взрослых птиц, самцов и самок, в брачном наряде
общая окраска спинной стороны, шеи и горла, маховых и рулевых черная; на голове и шее
Рис. 50. Полярная (белоклювая) гагара (взрослая и молодая птицы)
синеватый или сине-зеленый оттенок, на маховых и рулевых буроватый; на горле продольные
белые отметины; спина, кроющие крыла — с супротивными белыми пятнами; плечевые с та-
кими же крупными более или менее прямоугольными пятнами; надхвостье с мелкими белыми
парными пестринами; брюшная сторона, кроме горла и шеи, белая; на боках зоба темные
настволья, бока груди и брюха цвета спины, черные, с парными супротивными белыми пят-
нами; подкрылья белые с темными наствольями; подхвостье белое с черной поперечной поло-
сой. Радужина красновато-бурая, клюв желтовато-белый, цвета слоновой кости; ноги бурые
с желтоватым или зеленоватым оттенком наружной поверхности цевки и третьего и четвертого
пальцев. Взрослые птицы в зимнем наряде — как молодые, но без заметных светлых краев
перьев на спине и крыльях.
Темноклювая полярная гагара Gavia immer immer В г u n n.
Распространение. Ареал. Заменяет на гнездовье белоклювую гагару
на большей части территории Сев. Америки, в Гренландии и сев. Ислан-
дии. В СССР достоверно встречена на о-ве Беринга. В Зоологическом музее
Московского университета имеются два экземпляра — взрослый и молодой —
вероятно происходящие из «Северной России» (пометка на этикетках). В лите-
ратуре, наконец, имеется ряд указаний на залет этой птицы до м. Городец-
кого в Белом море (Гебель, 1901, 1903, Бутурлин, 1935), но они требуют под-
тверждения. Несомненно встречена в негнездовое время у Варангерфиорда
(Сконнинг, 1907). Залетает в Прибалтику (Рига, Шведер, 1894) и быть может
в Литву (Эйхвальд, 1830, Платер, 1852). Указания о залетах к Одессе и в Крым
к Балаклаве несомненно ошибочны. Вне СССР на Алеутских о-вах, в северных
частях Сев. Америки, к югу до Мена, Нью-Гемпшира, Вермонта, Масса-
чусетса, Нью-Йорка (горы Адирондак), с.-в. Индианы, с.-в. Иллинойса, сев.
Айовы и с.-в. Калифорнии; отсутствует по всей вероятности в области рас-
пространения белоносой гагары в Аляске и на сев. Канады. Кроме того, ветре-
ЧЕРНОЗОБАЯ ИЛИ ПОЛОСАТАЯ ГАГАРА
253
чается в Гренландии, на севере Упернивика и Скоресби-зунда G6° с. ш.)
на о-ве Медвежьем (Бертрам и Лек, 1933), на Ян-майене (Фишер и Пельцам,
1886, Берд, 1935), в сев. Исландии на Гримсе. Зимой главным образом у бере-
гов Америки на юг до Флориды, Луизианы и Техаса, на север до Северных
Штатов и области Великих озер; в меньшем числе у атлантических берегов
Зап. Европы от Северного моря; отдельные встречи в Средиземном море
(Сардиния, Алжир), у Мадейры и Азорских о-вов. Летние находки на Фарер-
ских о-вах и на Шпицбергене веро-
ятно относятся к холостым птицам.
X ар актер пребывания.
Перелетная птица. Биотоп. В гнез-
довое время—озера в тундрах и лес-
ной полосе, зимою главным образом
в литоральной полосе океанов. Ве-
сенний пролет в апреле —мае, отлет
в сентябре—октябре (Бент, 1919).
Первыми летят молодые птицы.
Экология. Размножение.
Пары повидимому постоянные. По-
ловая зрелость в возрасте около 2 лет.
Кладки в Сев. Америке с конца
апреля—начала мая, на севере,
повидимому, позднее в связи с тая-
нием водоемов. Гнезда —на озерах,
по многу лет в том же самом месте,
больших размеров (около 60 см в ди-
аметре), из сухих растений. Кладка
обычно из 2 яиц, по окраске сходных
с яйцами белоносой гагары. Разме-
ры яиц D1) 82—96,5 х 52—61, в
среднем 88,9 х 56,2 мм. В случае
гибели кладки может иметь место
дополнительная. Насиживание при
участии обоих родителей, в течение
29 дней. Выводки распадаются в
сентябре. Линька. Как у белоклю-
вой полярной гагары. Питание.
Главным образом рыба, которая добывается под водой нырянием. Известное
значение имеют быть может водяные растения (Хетч, 1892).
Полевые признаки* От белоклювой гагары отличается темным цветом клюва
и несколько иной его формой — конек надклювья слегка выпуклый, не вздер-
нутый.
Описание. Размеры и строение. Несколько меньше белоклювой гагары:
крыло самцов и самок E) 340—380, в среднем 363,6 мм. Рулевых обычно 10 пар. Окраска.
Клюв темный серо-роговой (у взрослых).В брачном наряде отлив на голове и шее зеленовато-
синий; белые полосы на шее уже, чему белоносой гагары. Молодые птицы по окраске не от-
личимы от молодых белоносых гагар.
3. Чернозобая или полосатая гагара Gavia arctica L.
Colymbus arcticus. Linnaeus. Syst. Nat. ed. X, 1758, стр. 135, тип. местн. Швеция.
Русское название. Гагара — звукоподражательное, народное. Полосатая
или чернозобая гагара — книжное (первое упоминание у Кесслера, 1847). Местами имеются
другие, связанные с голосом птицы, названия — куковик, коковик, погиб, погибка, также
ревуха (у поморов).
Распространение. Ареал. Арктическая, субарктическая и бореальная
зона Европы, Азии и Сев. Америки. В Европе на юг до средней полосы СССР,
Рис. 51. Полярная гагара (темноклювая)
254
ОТРЯД ГАГАРЫ
в Азии — до Балхаша, Иссык-куля, Алтая, Саян, с.-з. Монголии, Приамурья
и Сахалина, к северу до пределов континента и островов Ледовитого Океана;
в Америке в Гренландии, по островам Ледовитого океана и в северных частях
континента до Кьюотина, Мекензи (Большое Невольничье озеро), Британской
Колумбии и с.-з. Аляски. Зимою до средней Европы (нерегулярно до Среди-
земного моря), на Черном и Каспийском морях, до Японии; в Америке, главным
Карта 46. Распространение чернозобой гагары Gavia arctica
7. G. a. pacifica, 2t G. a. arctica, 3. G a. viridigularis;
a — граница гнездовий, б — вероятная граница областей распространения G. a. arctica и С a. viridigu-
laris в Восточной Сибири, г — область зимовок (стрелки показывают направление перелета гагар по данным
кольцевания в СССР), д — залет.
образом по тихоокеанским побережьям до юга Британской Колумбии и Кали-
форнии.
Характер пребывания. Перелетная птица. Б и о т о п. В гнез-
довое время — материковые водоемы (равнинные и горные, на юге ареала —
главным образом по горным озерам), на зимовке —прибрежная полоса морей.
Численность. Высокая; обыкновенная птица.
Подвиды и варьирующие признаки. Три географических расы, отличающиеся размерами
и окраской. При этом две — из восточного полушария — очень близки, третья — амери-
канская — более диференцирована. В Европе и Азии, на восток до бассейна Лены —
G. a. arctica Linnaeus, 1758; в с.-в. Азии G. a. viridigularis D w i g h t, 1918; в примор-
ской части с.-в. Сибири от бассейна Индигирки и Колымы и до Анадыря, в Сев. Америке
G. a. pacifica Lawrence, 1860.
Европейская чернозобая гагара Gavia arctica arctica L.
Синонимы. Colymbus macrorhynchos. С h r. L. В г е h m. Isis, 1830, стр. 982, с.-в.
Европа. — Colymbus balticus. Chr.L.Brehm. Lehrbuch Naturgesch. europ Vb'gel, 11,1824,
стр. 888, Приуральские озера, зимою на Балтийском море. — Colymbus rossicus. A n d г -
ze i о w s k i.'Bull. Soc. Imp. Naturalistes Moscou, 1839, стр. 20, Днепр.— Colymbus megar-
hynchos. Chr.LBrehm. Der vollst. Vogelfang, 1855, стр. 405, на пролете в Германии.—
Urinator arcticus susehkini. За рудный. Орнитологич. Вестник 1912, пролетные птицы
из Туркестана, тип. местность — устье Аму-Дарьи.
Распространение. Ареал. Сев. Европа и с.-з. Азия. На север до
Мурмана, Тиманской и Большеземельской тундры, о-вов Колгуева, Вай-
гача, южн. части Новой Земли (примерно до 75° с. ш.), низовьев Оби, быть
может южн. Ямала, в низовьях Енисея, на Таймыре (о-в Вера, около 76°, Мид-
дендорф, 1855; озеро Портнягино 74° с. ш., Торгашев, шхеры Минина), в ни-
зовьях Анабары и Оленека; к югу до сев. Шотландии E6°), Гебридских
и Оркнейских о-вов, в Скандинавии E8°), до Померании и Пруссии,
в Прибалтике и Литве, далее на восток в Европейской части СССР примерно
до 55°, в Белоруссии (Минское Полесье, Шнитников, 1913), Псковской, Смолен-
ЧЕРНОЗОБАЯ ИЛИ ПОЛОСАТАЯ ГАГАРА 255
ской, Московской, быть может Рязанской обл., в Горьковской обл., в Татарской
республике (до Спасского района), вероятно в сев. частях Куйбышевской обл.
(б. Бузулукский уезд, Карамзин, 1909), в Приуралье до 53°(Темясова на Сак-
маре, Зарудный, 1888), в сев.-зап. Казахстане до 49° с. ш. (озеро Чушкалы,
Сушкин, 1908), у озера Кургальджин E0°30'), и далее до Балхаша (т. е.
46°30'); гнездится на Ала-куле, Сассык-куле, Калган-куле (Иоганзен,
1909) и выводок найден на озере в низовье р. Тьюп, по с.-в. побережью
Иссык-куля и на озере Кокуй-куль на Карабулунском мысе (Д. П. Де-
ментьев); далее на востоке —до ю.-в. Алтая (плато Укек 49°30', Сушкин,
1938), Танну-ола (Тугаринов), в Саяне, Минусинском крае и наконец в с.-з.
Монголии (Косогол, Хангай) и у Байкала. Какой подвид гнездится на нижнем
Амуре и на Витимском плато — остается неясным. Внегнездового времени на
зимовке на Балтийском и Северном морях, быть может случайно — в Порту-
галии и Средиземноморье; в Закавказье, Армении (Севан); на Черном и Кас-
пийском морях, в Туркестане (южн. часть Аральского моря, низовья Сыр-
Дарьи, иногда на Аму-Дарье). Один случай залета в Индию (Амбалла, Пенд-
жаб, 23 февраля 1922, Бекер, 1929). Холостые неполовозрелые особи летуют на
Черном море. Отдельные особи зимуют по озерам внутренних частей европей-
ской России.
Характер пребывания. Перелетная птица, оседлая повиди-
мому на крайнем юге (Балхаш). В благоприятные по гидрологическим (ледо-
вым) условиям годы зимует далеко на севере, у берегов Финмаркена (в 1858—
1859 у Карлсботтена, Зоммерфельдт, 1867).
Даты. Весеннее и осеннее течение связано с замерзанием и таянием
водоемов в гнездовой области и проходит растянуто. Весенний пролет на юге
и в средней полосе проходит в апреле —мае (зимующие гагары на Балтике
в Куришском заливе летят с середины апреля до конца мая, Шюц, 1935, что
указывает на участие в нем разных популяций и подтверждается результа-
тами кольцевания). У мест гнездовья на севере—в тундре и лесотундре —
гагары появляются в конце мая и начале июня; на Мурмане, впрочем,
отмечались в конце апреля. Отлет с черноморских зимовок происходит с начала
апреля, но тянется и в мае, отдельные особи двигаются на юге и ранее
(в б. Харьковской губ. пролет отмечен уже во второй половине марта, Сомов,
1897). На Каспии птицы отлетают уже в конце марта (Челекен, Левчук, 1906);
пролет в Туркестане отмечен в начале апреля, но на Сыр-Дарье птицы
встречены и в начале мая (Спангенберг, 1936).
Отлет на севере в разные числа сентября, иногда даже в начале октября,
но одиночки (молодые) начинают лететь уже в марте. В средней полосе между
60°—50° с. ш. птицы задерживаются до ноября, одиночки даже до начала де-
кабря (8—15 декабря, р. Нача, Смоленская обл.; 16 декабря, р. Свапа, Курская
обл. — в Зоологич. музее Московского университета. У разводий птицы задер-
живаются и на севере до 1 ноября (на с.-з. Таймыре, Гизенко). На зимовках на
Балтике гагары появляются уже в сентябре, в Туркестане — с начала октября,
отдельные особи и ранее (в августе 1854 добыта Бэром у Новопетровска на
Мангышлаке).
Результаты кольцевания. Встречи окольцованных на Бал-
тийском море в Куришском заливе гагар показали, что туда на зимовки соби-
раются птицы разных популяций — на севере от Лапландии до крайнего
востока ареала подвида — Авамской тундры на Таймыре, Хатанги, Анабары
и Оленека. Восточные популяции проделывают таким образом путь на зимовку
более б 000 км длиной. Подвид мигрирует на зиму на запад и юго-запад, в от-
личие от восточносибирской расы, летящей на юго-восток и, вероятно, восток.
Миграции на севере своеобразны: из лесотундры и тундры птицы двигаются
на север, а затем поворачивают к берегам Скандинавии. По всем вероятиям
значительную часть пути гагары совершают вплавь. Популяции гагар из лесной
256 ОТРЯД ГАГАРЫ
полосы Европейской части СССР (южные) также движутся, по крайней мере
частично, на Балтийские зимовки; эти же птицы частично зимуют на Черном
море, достигая иногда Средиземного моря (находки в Греции).
Гагары из лесной и лесостепной полосы юго-западной Сибири в Европе
на зимовке не отмечены, они зимуют, повидимому, на Каспии и в Туркестане.
Для южной популяции отмечена перемена мест зимовки — окольцованные
на весеннем перелете на Балтийском море у Куришского залива гагары добыты
были в последующие годы на Черном море у берегов Крыма и Кавказа. Околь-
цованные гагары добывались через два, три, четыре и даже девять лет после
кольцевания (Дементьев, 1948). Кольцевание показало консервативность гнез-
довых местообитаний: самка, отмеченная кольцом 21 мая 1948 на озере Б. Сло-
бодском в Холмогорском районе Архангельской обл., была добыта там же
27 мая 1949 г.
Биотоп. Гнездовый — озера с достаточной глубиной, большие по пло-
щади, чем обитаемые краснозобой гагарой, и обычно богатые рыбой (впрочем,
и безрыбные, Сушкин, Формозов), реже речные затоны. На юге высоко в горах
(на Алтае в альпийской тундре до 2 300 м, Сушкин, 1938; в Саянах по крайней
мере до 2133 мм, Нестеров, 1909).
Водоемы — в разных ландшафтах, в тундре, лесотундре, лесу, существен-
ным представляется наличие более или менее богатой травянистой расти-
тельности у берегов. На пролете внутри материка — главным образом по реч-
ным долинам, с выходом затем на морские побережья.
Зимовки—главным образом морские (см. выше).
Численность. Распространение в связи с условиями гнездова-
ния, несколько спорадическое. В общем птица многочисленна и в пределах
ареала на каждом небольшом водоеме гнездится по паре, а на больших —
по нескольку пар. Колебаний численности не установлено. На пролете
весною — в парах и небольшими группами (на с.-з. Таймыре до 30—40 особей,
Гизенко; в Куришском заливе в апреле —мае стаи по 10—30 и даже 100 птиц,
Шюц, 1935). Зимовки также группами. Осенний пролет небольшими стаями.
Экология. Размножение. Срок наступления половой зрелости потен-
циально, вероятно, в возрасте около одного года. Имеется резерв холостых осо-
бей. Пары, повидимому, постоянны, так как птицы летят на миграциях нередко
парами и держатся в парах с прилета весной. Места гнездовий очень постоянны
(Поляков, 1908 и др.), но гнезда меняются. Брачные игры не изучены, в общем
сходны с таковыми у краснозобой гагары, по зорям в период спаривания птицы
много кричат. Брачный период с прилета на севере в тундре в конце мая, в сред-
ней полосе месяцем ранее. Гнезда — как у других гагар — куча отмершей
растительности у берега водоема или простая утоптанная площадка на берегу
озера, с мокрым дном лотка. Размер гнезда обычно около 50—60 см в диаметре
с лотком в 20—25 см при высоте около 10 см. В виде исключения гагара строит
и пловучее гнездо (из отмерших корней и сухих трав с выстилкой из свежей
осоки, поддерживаемое водяными растениями, с общим диаметром до 60 см
(низовья Печоры, Сибом и Броун, 1876).
Кладка в средней полосе в первой трети и середине апреля (Московская
обл. Поляков, 1908; Новгородская обл., Хлебников, 1888; Латвия, Бутурлин,
1902 и т. д.), на крайнем юге на Балхаше с конца апреля B8 апреля, Николь-
ский, 1887, но в горах, например на Алтае и в Саяне — поздней), в лесотундре
и тундре — в разные числа июня, чаще, повидимому, около середины этого
месяца. Таким образом, в датах размножения значительная зональная раз-
ница между отдельными популяциями, как и у других водных птиц, размно-
жение которых зависит от времени таяния льда на водоемах. Яйца отклады-
ваются с промежутком в 48 часов (Московскаяобл., Поляков, 1908). В кладке
1—2 яйца, изредка 3 (Красноярск). Окраска яиц оливково-бурая в разных
оттенках, с поверхностными черноватыми и глубокими серыми пятнами.
ЧЕРНОЗОБАЯ ИЛИ ПОЛОСАТАЯ ГАГАРА 257
Размеры A00) 75,7—95 х 45,5—56, в среднем 84,5 х 51,74 мм
(Хартерт, 1915).
В случае утери кладки, бывающей нередко, дополнительная вторая, также
из двух яиц (Поляков, 1908), этим объясняются поздние находки свежих кладок
на севере, приводящие иногда к гибели молодых A5 сентября на южн. Таймыре,
Скалой, 1938). Частая гибель кладок объясняется возможно тем, что взрослые
птицы летают иногда кормиться на соседние водоемы. Насиживают оба роди-
теля, сидят относительно некрепко. Повидимому, начало насиживания с от-
кладки первого яйца (западноевропейские данные, Уайзерби, 1940; требуют
впрочем подтверждения). Продолжительность насиживания около 25 дней
(Поляков, 1908; по западноевропейским данным 28—29 дней). Сильно наси-
женные яйца в Тиманской тундре — 1 июля, а 22 июля птенцы имеют вели-
чину примерно в Vs взрослых (Гладков). Эмбриональная смертность высокая,
обычно бывает только один птенец. Птенцовый период, до оперения птенцов,
продолжается около 40 дней; в разные числа августа, первой половине сентября,
в зависимости от широты, начинают встречаться летные молодые (запоздавшие,
от вторых кладок и позже).
Линька. Последовательность смены нарядов—как у других гагар:
первый пуховой наряд — второй пуховой наряд — птенцовый (первый зим-
ний) наряд — первый брачный наряд — зимний наряд — брачный наряд и т. д.
Окончательный наряд —первый брачный. Две линьки в году: послебрачная,
осенью после периода размножения (не охватывающая рулевых и маховых),
и полная весной —предбрачная. Схема линяния таким образом отлична от та-
ковой у краснозобой гагары. Линька на зимовке и поэтому даты и последова-
тельность смены участков оперения требуют еще изучения. Начало послебрач-
ной линьки — осенью, обычно в октябре, — начало линяния очевидно варьи-
рует в корреляции с разницей в сроке периода размножения в разных широтах,
начинается линяние со смены оперения головы и шеи. Уже в январе, быть
может в декабре, начинается предбрачная линька со смены перьев на спине
и плечах. Предбрачная линька затягивается иногда до апреля (Шюц, 1935),
хотя с другой стороны в это время встречаются гагары в полном брачном на-
ряде. Продолжительность осенней линьки около 2, весенней около 3 месяцев.
Питание. Как и другие гагары, чернозобая гагара кормится преиму-
щественно рыбой, которую добывает нырянием. Отмечено, что ныряющая
гагара может находиться под водой до 53,95 и даже 120 сек. (Уайзерби, 1940),
проплывая в последнем случае до г/4 мили. Двигаясь под водой, иногда дей-
ствует не только лапами, но и крыльями (Яковлев, 1941). За рыбою летает и
на соседние водоемы. Подчиненное значение в качестве корма для гагар имеют
беспозвоночные, в частности насекомые (личинки стрекоз, плавунцы и другие
водяные жуки, Горьковская обл., Формозов). Из других беспозвоночных ука-
зываются Crustacea, Annelida, Mollusca. Есть наблюдение над удачным нападе-
нием чернозобой гагары на пуховика хохлатого нырка (Штегман, 1929).
Полевые признаки. По окраске напоминает полярную гагару, отличаясь
от нее меньшей величиной и серой головой и шеей (в брачном наряде). От
краснозобой гагары отличается — в брачном наряде — черной с белыми пе-
стринами спинной стороной, а также несколько большей величиной. Голос
грубей и ниже, чем у краснозобой гагары. Как у последней, звуки разнообразны:
напоминающие стон или лай, карканье, хохот и т. п. В брачный период «кук-
кук-уик»; кроме того, при тревоге «га-га-раа», свист и т. д. Как и другие га-
гары, чернозобая гагара с трудом передвигается по земле (иногда даже будто бы
вовсе не может взлететь, Хлебников, 1888), хорошо летает, но не может варьиро-
вать скоростей и быстро менять направление, превосходно плавает и ныряет(уже
пуховым птенцом). В воде гагара обычно погружается глубоко, оставляя на по-
верхности иногда только шею и голову. На воде нередко и спит. При полете гагара
часто взмахивает крыльями и вытягивает голову и шею, а также лапы, как утка.
17 Птицы СССР, т. II
258
ОТРЯД ГАГАРЫ
Описание. Размеры и строение. Строение — как у других видов гагар.
Рулевых 8—9 пар. Длина самцов D) 700, 720, 720 и 744, самок C) 673, 680, 720, в среднем
721 и 691 мм. Размах самцов C) 1220, 1235, 1235, самок C) 1080, 1140, 1180, в среднем
1230 и 1133,3 мм. Вес самца A) 3310, самок C) 2037, 2354, 2471 г. Длина крыла самцов
C5) 290—338, самок C0) 273—300, в среднем 305,9 и 292 мм.
Окраска. Первый пуховой наряд серовато-бурый на спинной стороне, буроватый,
более светлый, на подбородке, горле, груди и брюхе, заметно светлей, чем у краснозобой га-
гары. Второй пуховой наряд заметно бледней, на спинной стороне буровато-серый с сере-
ватым теменем, с беловатыми зобом, грудью, брюхом (дефинитивный пух у чернозобой гагары
белый). Гнездовый наряд: общая окраска спинной стороны буровато-серая, с серой головой
и шеей, на спине и верхних кроющих плеча светлые серовато-белые каемки; маховые и руле-
Рис. 52. Чернозобая гагара
вые черновато-бурые; бока головы и вся брюшная сторона белые, с темными наствольями по
сторонам груди и зоба. Клюв беловатый у основания, свинцово-серый у вершины; пальцы
зеленоватые, радужина ореховая.
Взрослые самцы и самки в зимнем послебрачном наряде похожи на гнездовых птиц, но
общий тон спинной стороны темней, черновато-бурый, без светлых каемок перьев, с серой
головой и шеей; на подхвостье темнобурая поперечная полоса; рулевые с беловатыми верши-
нами. Брачный наряд самцов и самок: лоб шиферно-сероватый, темя и шея светлосерые;
спина, плечи, кроющие крыла, надхвостье черное с более или менее выраженным зеленоватым
блеском; на спине поперечно-вытянутые белые пятна, такие же крупные пятна на плечевых;
кроющие крыла с белыми вершинными пятнами; маховые и рулевые черные; подбородок и
горло черноватые с фиолетовым отливом и с рядом белых продольных пестрин на горле;
щеки бурые; бока зоба и груди с черными продольными полосами, бока тела черные; осталь-
ная брюшная сторона белая, в подхвостье черная поперечная полоса; подмышечные иногда
белые, иногда же белые с черными наствольями. Клюв свинцово-серый, светлеющий у осно-
вания нижней челюсти; цевки и внешний палец зеленовато-черные с буровато-беловатыми
перепонками и внутренними пальцами; радужина красновато-каштановая.
Восточносибирская чернозобая гагара Gavia arctica viridigalaris D w i g h t
Gavia viridigalaris. D w i g h t. The Auk. 1918, стр. 198, низовья Колымы
Распространение. Ареал. Северо-восточная часть области распростра-
нения вида на европейско-азиатском континенте, на запад от низовьев Лены,
на восток до Анадыря, Коряцкой Земли, Камчатки, возможно о-ва Беринга,
Алеутских о-вов, на юг до Алдана и Май, Охотского побережья, Сахалина.
Вероятно к этому же подвиду относятся гагары с Амура (найдена у Джалинды,
гнездится по озерам в нижнем течении, например Кизи) и быть может Витим-
ского плато (на Байкале 24 августа добыта птица этой расы; в Зоологическом
музее Московского университета). В негнездовое время —в Приморье, в Японии
(до Кюсю, Ян, 1942), на Курилах. Залетает в Китай (Фуцзянь, Латуш, 1934),
в с.-з. Америку, в Туркестан (Аральское море, Козлова, 1947) и быть может
ЧЕРНОЗОБАЯ ИЛИ ПОЛОСАТАЯ ГАГАРА 259
в Германию (Штреземанн, 1936, но сведения относятся, вероятно, к особям
из «переходных» популяций с.-з. Якутии). Зимовки расположены таким образом
на юг и юго-восток от гнездовой области, и миграционные пути различны от
путей номинальной расы, летящей на запад-юго-запад. Следует отметить, что
крайний юго-восток Сибири — между низовьями Колымы (Большой Баранов
мыс) и Чукотским п-овом — занят на гнездовье повидимому только белошейной
гагарой, а восточносибирская раса начинает встречаться на гнездовье только
в бассейне Анадыря, замещая белошейную гагару южней — в Коряцкой Земле
и на Камчатке.
Характер пребывания. Перелетная птица. Даты. Миграции
в те же сроки, как у европейской расы. Весной пролет и прилет в мае —
начале июня (Забайкалье, Якутск, Индигирка, Колыма, Анадырь). Запоз-
давшие особи на юге встречены в конце мая B7 мая, Ханка; 31 января,
бухта де Кастри, Шульпин, 1936). Осенние кочевки в августе, отлете северных
мест гнездовья в сентябре, последние особи задерживаются до начала
октября (Коряцкая Земля, Аллен, 1905; Анадырь, Портенко, 1939). На про-
лете гагары, повидимому, медленно двигаются вдоль морских побережий (на
Командорских о-вах и на Камчатке наблюдаются до ноября, Бианки, 1911).
Биотоп. Гнездовый — большие озера, пролетный — озера, реки, мор-
ские побережья, зимний — главным образом прибрежная полоса морей.
Численность. Высокая.
Экология. Размножение. Сроки размножения — как у номинального
подвида. Имеется резерв холостых особей (Портенко, 1939). Число яиц в кладке
два. Насиженные кладки на Б. Барановом мысе 12 декабря, пуховики у Чаун-
ской губы 30 июня; крупные пуховики на озере Красное, Анадырь, 26 августа
(вторая дополнительная кладка?); 30 сентября у Маркова вполне оперившийся
птенец (Портенко, 1939; летные, по тому же автору, вероятно в начале сентября);
оперившийся молодой на Гижиге 16 сентября (Аллен, 1905); вполне оперив-
шийся молодой у бухты Кит, южн. Приморье, уже на кочевке, 14 августа.
Выводки не разбиваются до отлета.
Линька и питание. Повидимому, йак у номинальной расы. Начало
линяния у взрослой в зимний наряд 23 сентября (Авача, Петропавловск).
Описание. Строение. Как у номинальной расы, но величина в среднем больше.
Крыло самцов A2) 305—330, самок A3) 298—328, в среднем 317,6 и 307,3 мм. Длина самцов
B) 750, 765; самок D) 695, 702, 709, 718 мм. Размах самцов B) 1265, 1270; самок C) 1135,
1176, 1210 мм. Вес самцов B) 3280, 3792,5; самок C) 2255, 2665, 3075 г. Окраска как у номи-
нальной расы, но горло не с фиолетовым, а с зеленым отливом (против света).
Белошейная или тихоокеанская гагара Gavia arctica pacifica Lawrence
Colymbus pacificus. Lawrence in Baird. Birds of N. America, 1860, стр. 889, Кали-
форния.
Распространение. Ареал. Сев. Америка, Арктика и Субарктика. От
Берингова моря (м. Барроу) до п-ова Мельвилля и о-ва Саутгемптон,
юга Баффиновой земли, Гудзонова залива, западных частей Унгава, о-ва Кери
в с.-з. Гренландии; на юг до фактории Иорк (центральный Кьюотин), юга Мекен-
зи, Британской Колумбии и п-ова Аляски и о-ва Кадьяк. Зимою от ю.-в. Аляски
и Британской Колумбии, Алеутских о-вов (где возможно гнездится, Турнер,
1886) до южн. Калифорнии. В восточном полушарии —в прибрежной полосе
северо-востока Сибири. По Бутурлину A906), в сев. Якутии белошейная гагара
распространена на 15—20 км от моря, тогда как (тот же автор и Михель, 1935)
в лесной полосе и лесотундре гнездится уже восточносибирский подвид. На
запад до Колымы и Индигирки, на восток до Анадыря (Танлео, озеро Красное,
устье Анадыря); на Чукотском п-ове повидимому только этот подвид; в Коряц-
кой Земле —только viridigularis (Дементьев, 1941). Быть может гнездится на
Курильских о-вах, где добыта в июне на Парамушире, в июле на Симушире
(Ямасина, 1931). Добыта летом на о-ве Врангеля (Портенко, 1937). В Азии зимует
260 ОТРЯД ГАГАРЫ
регулярно в Японии на юг до Кюсю (Ян, 1942) и быть может на Курильских
о-вах (Стейнегер, 1888); залетный экземпляр добыт 9 октября 1907 в Приморье
(Бутурлин, 1915), пролетный на Камчатке в Кроноцком заливе у устья
р. Поворотной —31 мая 1947. Таким образом, области распространения О. а.
pacifica и viridigulariS перекрываются, повидимому, только в бассейне Анадыря.
Характер пребывания. Перелетная птица. Даты. Прилет
на Индигирке у Русского Устья отмечен в начале июня (б июня, Михель,
1935), около этого же срока и на Анадыре (Портенко, 1932), несколько позже,
чем появление О. a. viridigularis. Осенний отлет с августа, но птицы задер-
живаются почти до ледостава и иногда запоздавшие молодые погибают
(от поздних кладок? Индигирка, Михель, 1935). Последние на Индигирке
наблюдались у устья 8 сентября и даже 23 сентября (Бурулчан, Михель),
на Колыме заметный пролет у устья в конце августа и начале сентября (Бутур-
лин). Осенний пролет у Пенжины 14 сентября (Портенко, 1939). На Камчатке
у Усть-Камчатска гагары держались в ноябре —декабре, до замерзания залива.
В Сев. Америке пролет на север отмечается в апреле —мае, прилет на север —
в конце мая—начале июня; осенью движение на юг с конца августа и
в сентябре, прилет на зимовки в Калифорнию в сентябре. Отлет на северо-восток
Сибири происходит иногда в смешанных стаях с viridigularis.
Биотоп. В сев.-вост. Азии—озера в Приморских тундрах, в Сев. Америке
и в лесной полосе (так как там викариата с другими подвидами чернозобой
гагары нет). Численность в Азии умеренная. На Индигирке на пролете
наблюдались стаи до 10 птиц, на зимовках в Америке встречаются стаи из
50 птиц и более (Хоуэлл, 1910).
Экология. Размножение. Относящиеся к Сибири сведения отрывочны.
Сроки те же, что и у других чернозобых гагар на севере. На Чукотском
п-ове у Питлекая полная кладка из 2 яиц найдена 9 июня в гнезде диаметром
24 см и высотой 5—6 см (Пильмен, 1887). На' Анадыре близ реки Туманной
гнездо, расположенное на морском берегу, представляло собой кучу отмершей
водной растительности с мокрым лотком; 10 июля в нем было одно яйцо, там же
добыта самка с готовым к сносу яйцом (Портенко, 1939). Пуховики на Инди-
гирке наблюдались 28 июля, неразбившийся выводок там же 4 августа и не-
летный птенец 20 августа (Михель). Птенцов чаще бывает, повидимому, один
(Михель, Бутурлин), что указывает на высокую смертность молодых. Выводки
перед отлетом перемещаются с замерзающих тундровых озер на реки. Раз-
меры яиц B) 73,2—77,8x44,6—48,3 мм (Анадырь, Портенко), B) 75,8—77,3X
Х47,9—9—48 мм (Питлекай, Пальмен); по американским данным D1) 68,5—
— 87x44—51, в среднем 75,7x47 мм (Бент, 1919). Линька. Как у других
подвидов, материалов из СССР нет. Питание. Главным образом рыба.
Полевые признаки. Отличается от других чернозобых гагар светлой
беловатой шеей и затылком; величина меньше, голос тоньше, вся птица
кажется легче и подвижней; охотно, как краснозобая гагара, держится
по небольшим водоемам (Портенко, 1939). Голос —как у других гагар. Отды-
хает и спит на воде, положив голову на спину (Михель, 1937).
Описание. Размеры и строение. Самая мелкая раса чернозобой гагары.
Длина крыла птиц из сев.-вост. Сибири у самцов A2) 285—312, у самок A2J71—297,
в среднем 299,5 и 282,8 мм. Клюв заметно слабее: длина его у самцов (И) 35 — 45, у самок
E) 34—42, в среднем 36,7 и 38 мм.
Окраска. Самцы и самки в брачном наряде отличаются от других рас вида охри-
сто-беловатым, не серым, затылком и задней стороной шеи; щеки и кроющие уха шиферно-
черноватые, а не бурые; отлив зоба— зеленоватый; черный цвет спинной стороны без
зеленоватого блеска. Прочие наряды по окраске повидимому не отличимы от других подвидов.
ОТРЯД ПОГАНКИ
COLYMBI или COLYMBIFORMES
(Podicipites или Podicipitiformes у разных авторов)
Характеристика отряда
Морфология. Внешние признаки. Птицы средней и мелкой
величины. Самый большой из встречающихся в СССР видов — чомга —- имеет
около 550—600 мм в длину, около 850 мм в размахе и вес около 1000—1200 г;
самый мелкий вид—малая поганка — имеет в длину около 250 мм, размах около
300 мм и вес около 200 г.
Форма тела, как у других нырковых птиц, вальковатая; шея длинная;
ноги сильно отнесены назад и голени почти полностью прикрыты общим с туло-
вищем кожным покровом; крылья короткие и острые; рулевые перья рудимен-
тарны. Первостепенных маховых 12. Оперение густое и рыхлое. Уздечка, в от-
личие от гагар, не оперена. Самцы и самки окрашены одинаково, причем за
некоторыми исключениями брюшная сторона белая. Сезонный диморфизм, как
правило, хорошо выражен. Пуховые птенцы на спинной стороне пестрые,
с продольным рисунком (кроме рода Aechmophorus). Клюв прямой или слегка
загнутый с более или менее заостренной вершиной. Цевки голые, резко упло-
щенные, покрыты спереди поперечными щитками. Пальцев ног 4, каждый из
них оторочен лопастью; когти плоские.
Анатомические признаки. Череп схизогнатический, голо-
риналькый, ноздри сквозные; носовые железы слабо развиты (пресноводные
птицы). Пневматичны лишь некоторые кости черепа. Базиптеригоидные отро-
стки отсутствуют. Шейных позвонков 17—21. Грудина относительно широкая
и короткая, с одной парой вырезок. 3—4 грудных позвонка сливаются в os
dorsale. Коленная чашечка хорошо развита. Акромиальный отросток processus
rotaloristibiae очень велик. Голень длинная; малая берцовая кость не доходит
до дистальной трети большой берцовой кости. Сочленение цевки и основных
фаланг второго-четвертого пальцев имеет особое устройство, обеспечивающее
вращение пальцев при подтягивании ноги. Нет обводящей мышцы. Имеется
только левая сонная артерия. Musculi sterno-tracheales асимметричны. Копчи-
ковая железа оперенная. Язык продолговатый, узкий. Зоба нет. Пилорический
262
ОТРЯД ПОГАНКИ
отдел желудка хорошо развит. Слепые кишки рудиментарны, но функциони-
руют. Перья с побочным стволом, развитым слабо. Крыло диастатаксического
типа. Мозг окципито-темпоральный, относительно небольшой.
Образ жизни. Поганки — дневные водоплавающие птицы, распространен-
ные на гнездовье по внутренним, преимущественно пресным, водоемам. На
берег они выходят нерегулярно.
Пища поганок — водные животные организмы: мелкая рыба, ракообраз-
ные, насекомые; крупные виды преимущественно, если не исключительно,
рыбоядны. Добывается пища под водой, способы добывания корма — как у
гагар. Подводное движение совершается при помощи ног, крылья при этом —
в отличие от чистиков и пингвинов — не исполь-
зуются и остаются плотно прижатыми к тулови-
щу. Добыча захватывается клювом. Под водою
поганки могут оставаться долго, до трех минут,
но обычно — 30—60 секунд. В желудке поганок
часто находятся перья, имеющие значение гастро-
литов и выщипываемые у себя птицею.
Большинство поганок быстро летает; неболь-
шая величина крыльев и высокая весовая нагруз-
ка вынуждают птицу делать частые и сильные
взмахи. Полет прямолинейный, взлет и повороты
совершаются с затруднениями, перед тем, как
взлететь, поганки делают большой разбег по
воде. Передвижение по суше очень неуклюжее
и плохое.
Поганки — моногамы. Гнезда строят на
воде, из кучи водной растительности, более или
менее пловучие. Нередко гнезда расположены
колониями и группами. Брачные «церемонии»
очень сложны. Яиц в кладке 3—8, форма их
продолговатая. Скорлупа без пестрин. В гнезде
яйца находятся в воде или во всяком случае в сильно влажной среде. Наси-
живают оба родителя, обычно с откладки первого яйца, в течение 20—25 дней.
Птенцы развиваются по выводковому типу; первые дни держатся преиму-
щественно под крыльями старой птицы. Пуховой наряд один.
Линяние у видов, распространенных в умеренных широтах северного
полушария, дважды в году; у них хорошо выражен брачный наряд. Тропи-
ческие и субтропические формы имеют одну линьку в году и сезонный димор-
физм у них, следовательно, не выражен. Не выражен сезонный диморфизм и
у тропических форм широко распространенных видов (чомга, черношейная
поганка), палеарктические подвиды которых имеют и брачный и зимний наряд.
При одной годовой линьке она, конечно, полная. При двух линьках — весною
сменяется мелкое оперение, а осенью все оперение целиком. Маховые перья
выпадают почти одновременно и птица лишается на некоторое время способ-
ности к полету. У молодых птиц пуховой наряд сменяется птенцовым, а затем,
в первую же осень жизни, зимним. Половой диморфизм выражен главным
образом в больших размерах самцов.
Систематика. Поганки представлены единственным семейством Colymbidae,
распадающимся на четыре рода: Colymbus с 13 видами, Aechmophorus с 2 видами,
Centropelma с одним видом и Podilymbus с 2 видами. Первый род имеет космопо-
литическое распространение, не будучи, однако, представлен в Антарктике и
отсутствуя почти полностью в Арктике (тундра). Остальные роды — американ-
ские.
Будучи близкими к гагарам, поганки отличаются от них рядом существен-
ных морфологических и экологических особенностей и представляют собой
Рис. 53. Головы черношейной
(низ) и красношейной (верх)
поганок
БОЛЬШАЯ ПОГАНКА ИЛИ ЧОМГА 263
хорошо очерченную группу, несомненно отдельный отряд. Ископаемые поганки,
относящиеся, повидимому, к современным родам, известны в Сев. Америке из
олигоцена или плиоцена, в Европе—из верхнего плиоцена. Современные виды
найдены в плейстоцене.
Географическое распространение. Распространение родов указано выше.
Наибольшее разнообразие форм поганок свойственно Америке. Заслуживает
внимания крайне узкий эндемизм трех видов: Colymbus taczanowskii встречается
только на озере Юнин в Перу, Podilymbus gigas — на озере Атитлан в Гвате-
мале, Cetropelma micropterum — на озере Титикака в Боливии (у последнего
вида летательный аппарат находится, по всей вероятности, на пути к редукции).
В умеренных широтах поганки обычно перелетны, в тропиках и субтро-
пиках — оседлы.
В фауне СССР представлены 5 видов поганок. Распространены они преиму-
щественно в умеренной полосе и на юге; только один вид — серощекая поган-
ка — на крайнем северо-западе страны и на северо-востоке проникает в аркти-
ческую зону.
Практическое значение невелико. Шкурки крупных видов могут быть
использованы для изготовления различных меховых предметов. Мясо неудов-
летворительного качества. Вред от поедания поганками рыбы, за редким исклю-
чением, едва ли реален.
Таблица для определения видов поганок
1B) Крыло короче 110 мм; щитки на задней стороне цевки преимущественно тре-
угольные малаяпоганка Colymbus ruticollis
2A) Крыло длиннее 110 мм; щитки на задней стороне цевки преимущественно прямо-
угольные см. 3
3D) Клюв длиннее 27 мм (обычно более 30 мм); крыло длиннее 147 мм; цевка длин-
нее 50 мм см. 5
4C) Клюв короче 27 мм (обычно менее 25 мм); крыло короче 147 мм (обычно менее
145 мм); цевка короче 50 мм см. 7
5F) Уздечка, полоса над глазом, наружная часть плечевых белые (у взрослых в брач-
ном наряде хорошо развитый хохол и воротник) чомга Colymbus cristatus
6E) Уздечка, полоса над глазом, наружная часть плечевых не белая (хохла и ворот-
ника в брачном наряде нет) серощекая поганка Colymbus grisegena
7(8) Клюв слегка загнут вниз; над глазом рыжая или беловатая бровь; небольшие
пятна на внутренних опахалах задних второстепенных маховых только на одном-двух
перьях или отсутствуют (у взрослых в брачном наряде нижняя сторона шеи рыжая)
красно шейная поганка Colymbus auritus
8G) Клюв прямой; рыжей или беловатой брови нет; на задних второстепенных маховых
большие белые пятна, обычно на 3—5 перьях (у взрослых в брачном наряде нижняя сторона
шеи черная) . .черношейная поганка Colymbus caspicus
РОД ПОГАНКИ COLYMBUS LINNAEUS, 1758
Тип С. cri status
1. Большая поганка или чомга Colymbus cristatus L.
Colymbus cristatus. Linnaeus. Syst. Nat. ed. X, 1758, стр. 135, Швеция.
Русское название. Поганка — народное (вероятно от плохого вкуса мяса).
Чомга — также народное, на востоке России и в Сибири, от татарского названия этой птицы.
Большая поганка — книжное. Местные народные имена — гагара, нырец или норец.
Распространение. Ареал. Европа, кроме крайнего севера — до Англии,
Ирландии, Швеции (до Вермланда и Даларне) и Финляндии, Ладожского и
Онежского озера, бассейна реки Вятки (р. Нарым, около 60° с. ш.), Молотова,
в Сибири до Тюмени, Барабинской степи, Каинска и Ачинска, южн. и зап.
Алтая, далее еще южнее — в южн. Приморье (Ханка), на юг до Среди-
земного моря, Закавказья, Малой Азии, Туркестана, Палестины, Месопо-
тамии, Кашмира и сев. Индии, Монголии и Манчжурии, Сев. Китая
064
ОТРЯД ПОГАНКИ
(Чжили); в Африке на севере (Марокко, Алжир, Египет), а затем, после пере-
рыва, к югу от Сахары; наконец, в Австралии, Тасмании и Новой Зеландии.
Ареал, таким образом, состоит из нескольких разобщенных частей.
Характер пребывания. На севере — в Европе и Азии — пере-
летная, на юге — оседлая птица. Зимовки северных популяций — в Средизем-
номорье, на Черном море, на Каспии (Азербайджан, Иран, юго-зап. Туркмения),
в Месопотамии, в Иране, у Персидского залива, в Белуджистане, Индии и
Бирме, в Южн. и в Вост. Китае, в Японии.
Карта 47. Распространение чомги Colymbus cristatus
7. С. с. cristatus, 2. С. с. infuscatus, J. С. с. christiani, 4, С. с. australis;
а — граница гнездовий, г — область зимовок (цифры в кружках обозначают номера зимующих подвидов).
Биотоп. Пресноводные, заросшие растительностью водоемы, — в гнез-
довое время! Зимою также и на море (см. выше). Вертикально в Азии по край-
ней мере до 2000, местами даже до 4000 м.
Подвиды и варьирующие признаки. Несколько подвидов, различающихся окраской
и экологией (характер пребывания — оседлость и перелетность; линяние — южноафрикан-
ский подвид не имеет особого зимнего наряда и т. д.). В сев. Африке, Европе и Азии —
С. с. cristatus L.; в южн. Африке от Сенегала и Абиссинии до Капской земли —С. c.infuscatus
Sa I v a d о г i, 1884; в юго-вост. Австралии (Куинсленд, Новый Южный Уэльс, Виктория,
Южн. Австралия) и Тасмании — С. с. christiani M a t h e w s, 1911; на Новой Зеландии —
С. с. australis Gould, 1844.
Обыкновенная чомга Colymbus cristatus cristatus L.
Синоним. Podiceps wiadhalmi. G о e be 1. Journ. f. Orn. 1870, стр. 712, Черное море.
Распространение. Ареал. В Европе от 60° с. ш. в Норвегии и провинции
Вермланд и Даларне в Швеции, 66° с. ш. в Финляндии, на юг до Пиренейского
п-ова, Средней Италии, Сардинии и Сицилии, Балкан (Албания и Болгария),
Кипра; в сев.-зап. Африке, в сев. Марокко и Алжире, в Египте; в Азии,
в Палестине, Месопотамии, Малой Азии, в Иране на западе в южно-каспийских
БОЛЬШАЯ ПОГАНКА ИЛИ ЧОМГА 265
провинциях, на востоке в Сеистане, в Кашмире, Сиккиме, Ассаме, к северу от
Брамапутры, в Ладаке, южн. Тибете, Монголии, Манчжурии (Сунгари,
Гиринская провинция) и Сев. Китае (Чжили).
В СССР от Ладожского и Онежского озер (южная часть озер); восточнее
граница, вероятно, проходит около 60° с. ш. (птица встречена в бассейне реки
Вятки и в южной части Пермского Урала в окрестностях Молотова, на реке
Обва); за Уралом до широты Тюмени, Тюкалинска, Ишима, Барабинской степи
(многочисленна на озере. Чаны), Красноярска; в Забайкалье (низовья реки
Орхон, Леннберг, 1909), в южн. Приморье (Ханка, быть может Дорицени,
Шульпин, 1936). На юг до Черного моря, Закавказья, Туркмении, Таджики-
стана, Балхаша, Ала-куля, Сассык-куля и Иссык-куля. В негнездовое время
у Средиземного моря (североевропейские птицы), у западного побережья
Каспия (Дагестан) и в Закавказье (где местами оседла), на южном побережье
Каспия, в Таджикистане, на озере Иссык-куль, в южн. Иране, в Белуджи-
стане и сев. Индии (Ассам, Бихар, Бенгалия), в Вост. Китае и Японии.
Характер пребывания. Перелетная птица, кроме самых
южных частей ареала (в СССР в Туркмении, Таджикистане и Закавказье —
в Армении).
Даты. Движение к северу с зимовок в зап. Закавказье в марте
(Вильконский, 1897), на Сыр-Дарье с конца марта и в апреле, в Туркмении со
второй половины марта и в начале апреля, запоздалые в мае, в Азербайджане
отмечен в конце апреля, но, вероятно, начинается ранее; в Южн. Китае—в марте.
В средней полосе пролет связывается со сроками таяния водоемов, птицы летят
парами и поодиночке, иногда в стаях, обычно утром и вечером, реже ночью. На
Украине прилет и пролет протекает с конца марта и в апреле, тогда же в По
лесье (Пинск). В центральной полосе, в Московской обл., в Псковской,
Новгородской и Ленинградской, в апреле, от середины этого месяца и до начала
мая. В это же время и в Поволжье, в Башкирии; северней и восточней, в области
среднего Урала в Молотовской обл. прилет отмечен в первой трети мая. В сев.
Казахстане прилет и пролет кончается в начале мая. На озере Чаны чомга по-
является в первой неделе мая, в это же примерно время и у Барнаула. В южн.
Приморье пролет отмечен в первой трети апреля, но тянется до конца этого
месяца.
Осенний пролет проходит стайками. В центральной полосе он падает на
разные числа октября, на Украине задерживается до начала ноября. Пролету
предшествует перемещение птиц на более крупные водоемы, последние поки-
даются только после замерзания. Прилет на китайские зимовки отмечается
в ноябре. На озеро Иссык-куль — в октябре, в Туркмении — с середины
октября до середины ноября, в Закавказье — в октябре — декабре. Отдель-
ные особи по незамерзающим рекам задерживаются на севере значительно
позднее, например, добыта 13. XII, 1929 г. под Льговом, Птушенко; такие же
случаи известны для Финляндии, Ленинградской обл., и Украины.
Биотоп. Гнездовый — по преимуществу стоячие и достаточно обшир-
ные водоемы, с развитой водной растительностью, хорошо прогреваемые солн-
цем и содержащие много рыбы; плесы с тихим течением. В негнездовое время и
открытые водоемы реки — в особенности их дельты, также морские побережья.
Изредка холостые особи встречаются на море и летом. Хотя чомги преимуще-
ственно держатся на равнинах, но не избегают и горных озер (например, на
Алтае до 1500 и даже 2000 м, Сушкин, 1938; Иссык-куль—около 1700 м;
в Ладаке и южн. Тибете более 4000 м).
Численность. Самый обычный вид поганок в нашей фауне.
Экология. Размножение. Гнездится обычно отдельными парами, но
на больших озерах образует крупные колонии (например на озере Чаны, по
Приуральским озерам, в сев. Казахстане). Гнездовые участки занимаются из года
в год, весьма постоянны. Строят гнезда оба родителя, после того, как на водоеме
266 ОТРЯД ПОГАНКИ
подрастет растительность (тростник и т. п.). Обычно гнездо пловучее, реже
опирающееся на дно. Построены они из отмершей растительности, главным
образом, куги, камыша, тростника. Гнездо полузатопленное и мокрое, сверху
более или менее ровное с небольшим вдавлением. Подводная часть гнезда имеет
температуру окружающей среды, но температура надводной верхней несколько
выше, до 7,5°С при отсутствии птицы (Никифоров). Как у других поганок, гнезда
расположены так, чтобы быть защищенными от действия волны и ветра и до-
ступными для подплывания птицы (протока или редина среди зарослей и т. п.).
Размеры гнезд: общая высота 45—-60, высота надводной части 3—4,5, общий
диаметр 42—65, внутренний диаметр 18—22 см (Наурзум, Никифоров).
Кладка относительно поздняя, и, несмотря на различия в географическом
положении местности, протекает в СССР в относительно сжатые сроки. Кладке
предшествуют весьма характерные и сложные брачные игры — птицы плывут
навстречу одна другой, разворачивают ошейник, двигают головой и затем как
бы становятся одна против другой в воде, выставив грудь и брюхо; иногда
самец «стоит» таким образом на воде над самкой. При этом чомги много кричат.
Спаривание происходит на особой площадке из водяной растительности,
устраиваемой птицами, причем самка предварительно принимает характерную
«пригласительную» позу, лежа в воде с вытянутой вперед головой и шеей так,
что на поверхности воды заметна только спинка. В Алжире кладка отмечена
в конце апреля (Тристрам, 1860). В Туркмении свежие кладки найдены в пер-
вой половине мая. На Или 16 мая найдена насиженная кладка, там же 29—30
мая и вылупление (Долгушин). Полные кладки на Сыр-Дарье в конце апреля —
начале мая (Спангенберг). В это же время кладки встречаются на Украине
(Сомов, 1897) и в Псковской обл. 2 мая два яйца; 15 мая полная кладка
(Зарудный, 1910) и в средней полосе. В литературе имеется ряд указаний на
поздние кладки, очевидно повторные, взамен утраченных (главным образом
расклеванных воронами и Камышевыми лунями).
Число яиц в кладке обычно 3—4, реже 5 и даже 6 или 7. Дополнитель-
ные кладки имеют и по 1—2 яйца. Повидимому, на юге кладки меньше, чем на
севере. Но для окончательного суждения материалов нет. Окраска яиц зелено-
ватс-белая, вскоре, по мере насиживания, загрязняющаяся растительностью
в гнезде до бурой или желтой. Размеры: ширина варьирует обыкновенно между
39,5 и 33,0, реже 40,6—32,5; длина 60,0—49,0, реже 62,5—47,8 мм (Бианки,
1911); F) 54,9—57,6x35,9—36,0 мм (Спангенберг). Промежуток между от-
кладкой яиц 48 часов (Англия, Уайзерби).
Насиживают оба родителя, с откладки первого яйца (разновозрастность
птенцов), в течение 28 B7—29?) дней. Вылупление птенцов падает на разные
числа июня (от нормальных кладок), в начале июля молодые становятся уже
на крыло. Только что вылупившиеся чомги хорошо плавают, но при нырянии
старая птица берет их под крыло. Оба родителя долго выкармливают молодых.
Выводки распадаются перед отлетом,когда молодые в возрасте около 272месяцев.
Линька. Последовательность смены нарядов: пуховой —гнездовый —
первый зимний (предбрачный) —первый брачный — зимний (послебрачный)—
брачный и т. д. У взрослых две линьки в году. Неполная предбрачная,
когда сменяются все перья, кроме первостепенных маховых и больших крою-
щих крыла. Полная линька в зимний наряд охватывает все оперение, причем
маховые сменяются быстро и почти одновременно. Первый зимний наряд
надевается между октябрем и декабрем, в зависимости от возраста птицы.
Предбрачная линька взрослых в феврале—марте, в конце марта—начале
апреля взрослые (но не годовалые) птицы в брачном наряде. Послебрачная
(осенняя) линька взрослых птиц — в зависимости от размножения (запазды-
вают птицы, имевшие вторые вынужденные кладки) — начинается со смены
мелкого оперения (но не у украшающих перьев на голове) во второй половине
августа, около второй половины августа вырастают новые маховые. У некоторых
БОЛЬШАЯ ПОГАНКА ИЛИ ЧОМГА
267
особей вся линька заканчивается во второй половине сентября, обычно же
в начале октября.
Питание. В отличие от других поганок, в кормовом режиме чомги
рыба занимает относительно большое, если не главное, место (в особенности
весною и осенью), и распределение чомги в известной мере связано с наличием
рыбы. Кроме рыбы — земноводные, насекомые, ракообразные, моллюски,
также растения. В желудке обычно находятся перья (аналог гастролитов).
Птенцы выкармливаются преимущественно насекомыми. Пища добывается
в воде при плавании и нырянии.
Рис. 54. Чомга
Полевые признаки. Самый крупный вид поганок, с резко двуцветной
окраской (белая брюшная сторона и темная спинная), с хорошо развитым
каштаново-рыжим ошейником и с длинными сушками» в брачном наряде.
Молодые с пестрой головой и шеей и белой «бровью». На полете хорошо заметны
белые второстепенные маховые, полет быстрый, с частыми взмахами крыльев.
При плавании характерна вытянутая кверху шея. Ныряя, может находиться
под водою до 50 сек. (Уайзерби, 1940). На суше передвигается с трудом, дер-
жится более или менее вертикально. В период размножения встречается парами,
реже образует гнездовые колонии; зимою нередко в стаях. Голос разнообраз-
ный, в брачном периоде громкий горловой крик «ккуа»или«круа»; кроме того
«кек-кек»; опознавательный позыв «кьюию» и т. д.
Описание. Размеры и строение. Клюв прямой, заостренный _у_ вершины.
Ноздри вытянутые и узкие. Полоса на уздечке неоперена. Формула крыла 3>2>4>5>6...,
1-е маховое рудиментарно; вырезка на внешних опахалах 3-го и 4-го и на внутренних опа-
халах 2-го и 3-го, иногда и 4-го первостепенных маховых. Хвост из коротких и мягких руди-
268
ОТРЯД ПОГАНКИ
ментарных (перьев как и у других видов поганок). Цевка плоская, сжатая с боков; передние
пальцы плоские и относительно широкие, отороченные каждый отдельной лопастью, лопасти
эти соединяются лишь у проксимальной фаланги; задний палец короткий и прикреплен
к цевке выше остальных. Когти плоские и ногтеобразные. Длина самцов A5) 550—610, самок
E) 540—570, в среднем 583,3 и 558 мм. Размах самцов A5) 835—900, самок E) 810—850,
в среднем 869 и 832 мм. Вес самцов E) 1100—1200, в среднем 1140, самки A) 1050 г. Крыло
самцов E9) 177—205, самок D1) 168—194, в среднем 190,6 и 183,02 мм.
Окраска. Пуховой птенец на спинной стороне — пестрый, голова и шея черные
с белым, у основания надклювья с каждой стороны по белому пятну; спина и бока бурова-
тые, на спине с продольными бело-охристыми пестринами, горло и зоб белые; на уздечке,
под глазами, на темени голые пятна; клюв беловатый с двумя черными поперечными полосами;
ноги зеленоватые; радужина белая. От других поганок в пуховом наряде чомга отличается
большим развитием светлых полос на спинной стороне и разделяющейся книзу на две поло-
сой на нижней стороне шеи (вилообразно). Пестрый пух на голове и шее сохраняется отно-
сительно долго, до поздней осени, так как развитие здесь контурного оперения запаздывает.
В птенцовом наряде верх головы черновато-бурый с белыми пестринами на лбу; бока
головы белые с темной продольной полосой за глазами и другой, тянущейся1 от основания
надклювья к затылку; спинная сторона темнобурая, брюшная белая с буроватым оттенком
на боках; первостепенные маховые серо-бурые с беловатыми основаниями и белыми пестри-
нами на внутренних перьях; второстепенные маховые белые с бурыми пятнами на наружных
опахалах и темными основаниями стволов перьев; внутренние (задние) второстепенные
маховые черноваты; хвост черноватый с беловатыми пестринами. В первом зимнем наряде
темя темное без белых пестрин, «уши» слабо развиты, бока головы белые без украшающих
удлиненных перьев. Клюв бледнобурый, радужина оранжево-желтая.
Самец и самка в брачном наряде на спинной стороне буровато-черные с несколько
более светлыми краями перьев; крыло у кистевого сгиба, второстепенные маховые, подбой
крыла, зоб, грудь, брюхо — белые, на боках ржавчато-рыжий оттенок; уздечка и перья
у глаз белые, лоб, темя, затылок, перья «ушей» — черные; щеки и удлиненные, образующие
¦воротник» перья ржавчато-красноватые, «край воротника* — черный; маховые и рулевые —
как у молодых. Клюв темнобурый, синеватый у вершины, красный по бокам надклювья
и у его основания; ноги зеленоватые с темными пятнами; когти черноватые; радужина красная
с оранжево-желтоватым кольцом у зрачка.
Зимний (послебрачиый) наряд взрослых сходен с брачным, но общий тон спинной
стороны бурее, «ушные» пучки короче и со значительной примесью белого темнобурые со
слабым развитием рыжеватого оттенка.
2. Серощекая поганка Colymbus grisegena В odd.
Colymbus grisegena. Boddaert. Tabl. PI. Enlum. 1783, стр. 55, Франция.
Русское название. Книжное. Народное — как у чомги.
Распространение. Ареал. Европа к востоку от Рейна на север до
Голландии, Швеции (до южн. Лапландии, Оунасиоки, 67°30' с. ш.), Финлян-
Карта 48. Распространение серощекой поганки Colymbus grisegena
1- С. g. grisegena, 2. С. #: holMllii;
а — граница гнездовий, г — область зимовок (цифры в кружках обозначают номера зимующих подвидов).
дии, Кольского полуострова (Имандра), Кандалакши, Архангельска, далее
к востоку примерно до широты Уфы, на Урале около 56° с. ш., в зап.
Сибири, быть может в Сургутском районе и несомненно в области слияния
Иртыша и Тобола, на восток до Барабинской и Кулундинской степи, к югу до
СЕРОЩЕКАЯ ПОГАНКА 269
Баварии, Чехословакии, Австрии, Югославии (Словения), Болгарии и Румы-
нии (Добруджа); Украины, Закавказья, Малой Азии и низовьев Сыр-Дарьи;
кроме того, в Семиречье (Ала-куль, Баскан, Иссык-куль, Балхаш), в вост.
Сибири от верхнего Енисея, Забайкалья, в Якутии до Вилюйска, устья Алдана,
далее — до низовьев Колымы, на Анадыре на Камчатке и в Приамурье к югу
до сев. Манчжурии. Возможно гнездится в Марокко. В Сев. Америке
от низовьев Юкона, сев. Мекензи, сев. Унгава до северных частей штата
Вашингтон и сев.-зап. частей Миннесота. Зимовки в Англии, в сев. Африке
(Алжир, Тунис, Триполи, Египет), в Средиземноморье, в Крыму, Закавказье,
в Туркмении, в Малой Азии, сев.-зап. Иране, а также в южн. Приморье
(залив Посьета, бухта де Кастри), в Китае и Японии; в Америке до
сев. Каролины, долины Огайо, южн. Колорадо, севера Калифорнии.
Характер пребывания — перелетная птица.
Биотоп, как и у других поганок, гнездовый — заросшие тростником
и камышом водоемы — озера, пруды, речные старицы и т. п. В негнездовое
время у морских побережий и в устьевых частях рек.
Подвиды и варьирующие признаки. Два подвида, различающиеся размерами и, пови-
димому, биологическими деталями: С. g. grisegena В о d d. — в западной части ареала, на вос-
ток до Зап. Сибири, низовьев Сыр-Дарьи, Малой Азии и Закавказья и С. g. holboellii R e i n-
h a r d t, 1853— из Семиречья, вост. Азии и Сев. Америки.
Западная серощекая поганка Colymbus grisegena grisegena В о d d.
Синонимы: Colymbus rubricoll is. G me 1 i n. Syst. Nat. 1, 1789, стр. 592, «Норвегия и
Дания, Каспийское море». — Colymbus cucullatus Pallas. Zoographia Rosso-Asiatica
t. II, 1811, стр. 355. «Озера России и Сибири»—Colymbus naevius. Pallas, цит. соч.,
стр. 356, Волга.
Распространение. Ареал. Европа и Азия. В Европе гнездится в Гол-
ландии, Дании и Германии, в Швеции (на север до Пите- и Луле-Лаппмарка,
на юг до Шонен, Текерн и Эланда), вероятно, в юго-вост. Норвегии (Осло, Сунде-
валль, 1885), в Финляндии, в СССР — в северных частях Прибалтики, в Лап-
ландии (Имандра), у Белого моря (Кандалакша), у Архангельска; восточнее
граница, повидимому, резко спускается к югу, в бассейн Камы (Татарская
республика, Башкирия, у Уфы), на Урале отмечена несколько севернее E6°с.ш.,
Гаккель, 1898; на р. Сосьва, Портенко, 1937); в Зап. Сибири в бассейне Оби,
вероятно до Нарыма и Сургута (Паллас, 1800), несомненно в области низовьев
Тобола; восточная граница определяется Кулундинской и Барабинской степью
(гнездится на озере Чаны). К югу доходит до сев. Австрии, Югославии и
Болгарии, Румынии (Добруджа), наконец в Малой Азии (Эрзерум, Дрессер,
1891); вероятно гнездование изолированной колонии в Марокко (Ирби, 1875;
Отс, 1901); южная граница в СССР захватывает плавни Днестра, Украину,
дельту Волги и низовья Урала, далее проходит севернее Мугоджар примерно
к устью Иргиза, доходя, повидимому, до севера б. Семипалатинской обл.;
кроме того, птица гнездится по озерам в низовьях Сыр-Дарьи и низовьях Аму-
Дарьи, на Кавказе и в Закавказье. Вопрос о гнездовании в Крыму неясен
(возможно на Сиваше). В негнездовое время встречается в Англии и Шотландии,
реже в Ирландии; основная область зимовок у Средиземного моря (сев.-зап. Аф-
рика, Балканский п-ов), быть может в Бессарабии, в Крыму, в Закавказье, на
южн. Каспии; в Малой Азии, сев.-зап. Иране; изредка и случайно в ни-
зовьях Сыр-Дарьи. Залеты на Шпицберген, в Исландию, в Египет; в Турке-
стане к Термезу на Аму-Дарье (Зарудный).
Характер пребывания. В большей части ареала перелетная
птица, однако оседла в Скандинавии к югу от полярного круга и в Закавказье
(Армении, Соснин и Ляйстер, 1942). Надо полагать, что популяции серо-
щеких поганок из сев.-зап. и центральной России улетают на зимовку на
запад к Британским о-вам, а восточные — от бассейна Волги до Зап,
270 отряд поганки
Сибири — на Каспий, так как на Черном море зимовка этих птиц огра-
ничена.
Даты. На зимовке в Англии находится нормально с октября по март
(Уайзерби, 1940), в области южн. Каспия появляется в октябре —ноябре и
держится до апреля. Летит одиночками или парами, реже стайками. Уже
в конце марта серощекие поганки начинают появляться на Украине и в Бес-
сарабии, но главный пролет (и прилет) приходится на первую половину апреля,
в конце апреля птицы появляются и в средней полосе Европейской России, и
в сев. Казахстане B1—24 апреля, Наурзумский заповедник), и в низовьях
Сыр-Дарьи (отдельные особи добыты уже 16 апреля у Джалагаша). Прилет,
таким образом, заметно более ранний, чем у восточного подвида в Сибири (исклю-
чая Приморье). Осеннее движение протекает преимущественно в октябре. На
севере ареала у Архангельска отмечен 11 октября (Вальнев, 1880), в этом же
месяце птицы покидают Прибалтику (Лоудон, 1909) и Московскую обл. На
Украине пролет отмечен в октябре —начале ноября. Из Чкаловской обл.,
где местные передвижения начинаются в начале ноября, последние
поганки улетают в начале ноября (Зарудный, 1897). На Сыр-Дарье у Чиили
встречены 23 октября (Спангенберг, 1936), в Туркмении на зап. Узбое и
у Аму-Дарьи — в первой половине октября (Дементьев, 1950).
Биотоп. Гнездовый — стоячие водоемы, озера, болота, пруды, с обиль-
ными зарослями тростника, камыша рогоза и других растений. В негнездовое
время у морских побережий и в устьях рек. Иногда довольно далеко от берега
в море, например в январе 1938 г. добыта в 80 км от Гассан-кули на юго-вост.
Каспии. Преимущественно на равницах, хотя в Закавказье отмечена на гнез-
довье и на горных озерах (Севан).
Численность. Распространение спорадично и неравномерно.
Многочисленна в низовьях Днестра, у нижней Волги и в сев.-зап. Казахстане.
Экология. Размножение. Пары, повидимому, постоянны, так как
птицы держатся парами и на пролетах. Брачные церемонии напоминают
таковые у чомги. Гнездовые участки занимаются с прилета и весьма постоянны,
относительно невелики, так что на одном водоеме нередко гнездится по несколь-
ко пар (Карелия, Гебель, 1879 и др.). Гнезда обычного для поганок типа, среди
разреженных тростников и других растений, несколько меньше, чем у чомги,
обычно прикреплены к стеблям отмерших растений или хотя бы расположены
среди них. Все гнездо пропитано водою, яйца при отсутствии птицы при-
крываются кучкой водяных растений. Размеры гнезд: B) общая высота 35—48,
высота надводной части 3—4, общий диаметр 40—45, диаметр лотка 16—20 см
(Наурзум, сев.-зап. Казахстан, Никифоров).
Кладка происходит в мае — начале июня, точных дат немного (в б. Уман-
ском уезде Киевской губ. 28 мая неполная, Гебель, 1871; тоже неполная в Лат-
вии 6 мая; полные кладки на Ладожском озере 3—15 июня, Пальмгрен, 1902;
насиженные яйца в Карелии 20 июня, Гебель, 1879; около середины июня под
Архангельском, Вальнев, 1880; во второй половине мая на Украине; однако
в Полтавской обл. полная уже 26 апреля, Гавриленко, 1929; сильно наси-
женные яйца на Сыр-Дарье 3 апреля, Спангенберг 1936; кладки на озере Севан
в Армении 16 апреля —16 августа, Соснин и Ляйстер, 1942). Число яиц в кладке
3—6, обычно 4—5, иногда 2. Окраска яиц белая с зеленоватым налетом. Раз-
меры: ширина 38,2—32,5, длина 58,2—47,5 мм (Бианки, 1911); A00) 46,7—
57x30—36,5, в среднем 50,65x33,99 мм (Хартерт, 1915).
Насиживают оба родителя, но преимущественно самка, по некоторым
данным с первого яйца (Уайзерби, 1940), по другим — со второй половины клад-
ки, после откладки третьего яйца (Мензбир, 1918); птенцы в выводках во вся-
ком случае разновозрастные. Срок насиживания не определен, в неволе, по
Хейнроту, 23 дня. Одна кладка, во всяком случае как норма (но бывают, быть
может, дополнительные, так как вылупление птенцов под Архангельском на-
СЕРОЩЕКАЯ ПОГАНКА
271
блюдалось Вальневым, 4 августа, 1880). Пуховики разного возраста на Ладож-
ском и Онежском озерах, на Украине и в низовьях Волги, в сев.-зап. Казахстане
встречены в июле, в Армении от 28 июня и в июле, Даль, 1940; в низовьях Сыр-
Дарьи 6 июня. В конце августа молодые оперены, но держатся вместе со ста-
рыми до отлета.
Линька изучена недостаточно. Смена нарядов обычная для отряда:
пуховой — птенцовый — первый зимний (предбрачный) — брачный — после-
брачный (зимний) и т. д. Птенцовый наряд надевается в августе и сменяется
зимним — только за счет линьки головы и шеи — в ноябре — декабре. Брач-
ный наряд образуется за счет частичной линьки (без крупного оперения, в част-
ности первостепенных, передних второстепенных маховых и части кроющих),
Рис. 55. Серощекая поганка (взрослая и молодая птица)
на зимовке (повидимому, в феврале — марте), так как в апреле прилетающие
в СССР серощекие поганки уже в брачном пере. Исключением служат годова-
лые птицы, у которых появление брачного наряда затягивается до мая (очевид-
но, в связи с запозданием всего цикла эти птицы и не успевают, как правило,
приступить к размножению). Послебрачная линька осенью полная, протекает
во второй половине августа —в сентябре и заканчивается в начале октября;
первостепенные маховые выпадают при этом почти одновременно.
Питание изучено недостаточно. В гнездовое время — мелкая рыба,
лягушки, беспозвоночные —моллюски и насекомые (стрекозы, полужестко-
крылые, жуки и личинки разных групп), растения (Myriophyllum, семена осоки,
Армения, Соснин и Ляйстер, 1942). Зимою мелкая рыба и ракообразные (Англия,
Уайзерби, 1940).
Полевые признаки. Размеры несколько меньшие, чем у чомги, но большие,
чем у других поганок фауны СССР (с небольшую утку). Клюв относительно
короткий и толстый по сравнению с чомгой, почти прямой, резко двуцветный,
с желтым основанием. В брачном наряде характерны черный верх головы с чер-
ными «ушными» пучками и хохлом довольно коротким и прямо срезанным,
в резком контрасте с черным верхом головы находятся серые щеки, подбородок
и рыжая шея. В зимнем наряде отличается от чомги отсутствием белой «брови».
Движения — как у чомги, но, повидимому, лучше передвигается по суше. Про-
плывает под водою до 50—60 м (восточный подвид, Шульпин, 1936). Голос «кек-
272 ОТРЯД ПОГАНКИ
кек>, в брачный период оба пола издают громкий своеобразный крик, напоминаю-
щий ржание жеребенка; крик этот переходит затем в ворчание. Кричат в период
размножения и ночью. Чрезвычайно осторожна. При опасности стремится
спрятаться среди водной растительности, избегая открытого зеркала водоемов.
Описание. Размеры и строение. Формула крыла 3>2>4>5> 6...
1-е маховое рудиментарное. Длина самцов F) 440—456, самок C) 405—435, в среднем 445,5
и 421,7 мм. Размах самцов E) 740—765, самок C) 720—735, в среднем 749 и 727 мм. Вес
самца A) 650, самок B) 527 и 550 г. Клюв около 35—45 мм. Крыло самцов B0) 162—173,
самок A0) 160—170, в среднем 168,2 и 164,1 мм.
Окраска. Пуховый птенец на спине, крыльях, шее, боках черновато-бурый, грудь
и брюхо серовато-белые; голова и шея черные с белым продольным рисунком; на передней
стороне шеи и на зобу охристый оттенок; на затылке белое пятно; белый рисунок на темени
менее развит, продольные полосы уже, чем у чомги; белое пятно на затылке едва намечено;
полосы на зобу соединяются в форме V, а не U, как у чомги; радужина желтоватая, клюв
желтый с черной предвершинной поперечной перевязью; лапы зеленовато-желтоватые.
Взрослые самцы в брачном наряде черные на спинной стороне, в свежем пере со свет-
лыми краями перьев; с удлиненными «ушными» пучками и хохлом на затылке, под глазом
и за ухом беловатая полоса; щеки, подбородок и горло серые; передняя и боковая сторона
шеи и зоб ржавчато-рыжие; грудь и брюхо белые с неясными сероватыми крапинами, бока
черноватые со светлыми основаниями перьев; малые кроющие у края крыла белые, осталь-
ные — как средние и большие — серовато-бурые; первостепенные маховые серовато-бурые
с черными стволами; второстепенные маховые белые или белые с темными стволами; рулевые
черные с примесью белого. В зимнем наряде нет украшающих перьев — «ушных» и на за-
тылке; темя серо-бурое, спинная сторона бледнее, чем в брачном наряде; у глаза сероватые
пестрины; шеки, горло, верхняя часть зоба белые; нижняя часть зоба и бока шеи бурые
с беловатыми пестринами и с рыжеватым оттенком; остальной низ белый с черноватыми пест-
ринами на боках.
Молодые — как старые в зимнем наряде, но более буроваты, без рыжеватого оттенка
на зобе, с двумя сероватыми полосами по бокам головы и шеи; на второстепенных маховых
бурые пятна у вершины. Клюв на большем протяжении черный, яркожелтый у разреза
и в основании (зимою и у молодых бледнее); лапы черноватые с желтоватым и зеленоватым
оттенком на внутренней стороне цевки и на верхней поверхности окаймляющих пальцы лопа-
стей; радужина темнобурая у старых птиц, желтоватая у молодых.
Восточная серощекая поганка Colymbus grisegena holboellii Reinhardt
Podiceps Holboelii. Reinhardt. Videnskab. Meddelelser, 1853, стр. 76, Гренландия.
Синоним. Podiceps griseigena bergmani. Lonnberg. Fauna och Flora, 1936,
стр. 37, Камчатка.
Распространение. Ареал. Вост. Сибирь, от Енисейска (Попгем, 1901)
и Туруханского края (Третьяков, 1869), что требует выяснения; несом-
ненно — от бассейна Лены (Вилюй), на Колыме до 69°30' с. ш. (Бутурлин,
1908), в бассейне Анадыря (в последнем не доходит на север до Анадырского
хребта и на восток до моря, Портенко, 1939), на Камчатке, на Охотском море
(Шантары, Шульпин и Дулькейт, 1937), в Приамурье к югу до озера Дорицени
на границе с Кореей (Шульпин, 1936), за пределами СССР в сев. Манч-
журии (Цурухайтуй); не найдена в нижнем течении Лены, на Яне и Индигирке.
Имеются две изолированные колонии — одна в Забайкалье на Еравнинских
озерах (Бакутин, 1936), другая в Семиречье (Балхаш, Ала-куль, Баскан,
Иссык-куль). Птицы, добытые в гнездовое время в низовьях Сыр-Дарьи, по
размерам (см. ниже) промежуточны между западными и восточными подвидами,
в Сев. Америке на севере, на восток до Унгавы и Гудсонова пролива, на
юг до южн. Лабрадора, юго-зап. Нового Брауншвейга, вероятно Квебека и
Онтарио, также западных частей штата Миннесота и Сев. Дакота, на западе
до сев.-зап. Монтана, Вашингтона и в сев.-зап. Аляске, к северу до Юкона
(Форти-Майл) и сев. Мекензи (Бент, 1919). Зимою в южн. Приморье (залив По-
сьет, де Кастри), у Сахалина, Курильских о-вов, у Хоккайдо (Ян, 1942), в не-
большом количестве в юго-вост. Китае (Фуцзянь, низовья Янцзе, Латуш, 1934).
В Америке, где повидимому частично зимуют и азиатские птицы, в области
Великих Озер (Онтарио, Эри, Мичиган), но главным образом на атлантическом и
тихоокеанском побережьях от Мена и Нью-Йорка до Южн. Каролины и от юга
СЕРОЩЕКАЯ ПОГАНКА 273
Британской Колумбии до Вашингтона (Педжет Зунд), до южн. Калифорнии
(Санта Барбара). Залеты в Гренландию, Исландию, Шотландию, Францию,
в Америке на Прибыловы о-ва и Алеуты, в штаты Георгию и Теннесси.
Характер пребывания. Перелетная птица.
Даты. Прилет на севере в бассейн Анадыря в последней трети мая B3—
24 мая по Сокольникову, Портенко, 1939), на юге в Приморье, в парах, очень
рано, уже в конце марта —первой половине апреля (Шульпин, 1936 и др.).
Появление на Камчатке около середины мая (Бианки, 1911). Пролет в Забай-
калье отмечен в середине мая (Радде, 1863, Маак, 1859). Даты осеннего пролета
и отлета плохо выяснены, но несомненно поздние, на Камчатке, повидимому,
в октябре (Бергман, 1935) и даже в ноябре (Бианки, 1911); в ноябре птицы добы-
ваются у Командорских о-вов (Хартерт, 1920). На Шантарах отлет в сере-
дине октября (Дулькейт и Шульпин, 1937). Весенний пролет в Америке отме-
чен в апреле и в начале мая, осенний — в конце сентября — октября.
Биотоп. Гнездовый — болота и озера с густыми зарослями, особенно
тростника; в негнездовое время — морские заливы, большие озера.
Экология. Размножение. Отношение численности полов 1 : 1
(Шульпин, 1936). В брачном периоде птицы часто подают голос «кек-кек-кек»
(Бергман, 1935) или стонущий «хха-хаааа-ха-хаа-ха» (Шульпин, 1936) как днем,
так и ночью. Брачные игры сходны с таковыми у чомги: самец и самка плавают,
обливаются, подымаются из воды передней частью туловища, как бы стоят
одна перед другой, сблизив клювы и распустив хохол, потом отплывают в сто-
рону (Шульпин). Гнезда — пловучие, у зарослей рогоза и тростника, из водных
растений (рогоз, осока, пузырчатка), диаметром около 40 см (Шульпин, 1936).
В Приморье —21 мая кладки, частью сильно насиженные. Яйца зеленовато-
голубоватые, запачканные по мере насиживания ржавчатым или рыжеватым.
В кладке 3—4, иногда 1 яйцо (в Америке, по Бенту, до 6 и даже 8). Разме-
ры A2) 47,8—53,5x33,5—36,8, в среднем 51,5x35,77 мм (Приморье, Шуль-
пин, 1936). По Тачановскому A4) 51—57,5x33—38,2, в среднем 53,9х
Х35,5 мм; у американских F0) 49—64,6x30—37,5, в среднем 53,7x34,5 мм
(Бент, 1919).
Насиживание с первого яйца, в течение 20—23 дней (Сев. Америка,
Бент, 1919). Вылупление в Приморье в десятых числах июня (Шульпин, 1936).
В выводке 2—3, иногда 1 птенец (Камчатка, Бергман, 1935). Неперелинявшие
в контурное перо молодые добывались и наблюдались: на Омолоне 11 августа,
у Ключевского на Камчатке 14 июля, у Еловки 17 июля (Бианки, 1911), на
Камчатке 17 июля (Бергман, 1935), на озере Эворон в низовьях Амура 21 июля
(Воробьев, 1935), в Приморье 22 июня (Шульпин, 1936). Вполне выросшие
молодые наблюдались на Камчатке 5 сентября (Бианки, 1911), причем выводки
еще не разбились; в низовьях Или в начале августа (Гладков). Имеется абортив-
ный осенний цикл, брачный крик в октябре (Шульпин, 1936, Приморье).
К сожалению, мы не располагаем достаточными сведениями по экологии
южной киргизско-казахстанской колонии серощеких поганок.
Линька. Первый зимний наряд надевается, повидимому, в ноябре —
декабре; взрослые птицы в зимнем наряде встречены в сентябре (Маак, 1861;
Шульпин, 1936), в брачном — в апреле и мае; начало линяния в зимний наряд
у взрослых с июля (Бергман, 1935; Шульпин, 1936).
Описание. Размеры и строение. Восточная серощекая поганка отличается
от поминального подвида большими размерами; крыло самцов A0) 183—197, самок E) 173—
187, в среднем 189,6 и 186,7 мм. По Портенко A939), крыло самцов и самок C7) 180—206,
в среднем 192 мм. Длина клюва около 40—55 мм. Окраска — как у номинальной формы.
Систематические замечания. Серощекие поганки с низовьев Сыр-Дарьи по размерам
промежуточны между западной и восточной расами: длина крыла самцов D) 168—178, самок
E) 155—169, в среднем 174,2 и 165,1 мм. Птицы с Балхаша несколько уступают в размерах
восточносибирским, но крупнее западных: крыло 176—182 мм. Американские птицы, пови-
димому, крупнее в среднем, чем азиатские, крупнее у них и яйца (см. выше).
18 Птицы СССР, т. II
274
ОТРЯД ПОГАНКИ
3. Рогатая или красношейная поганка Colymbus auritus L.
Colymbus auritus. Linnaeus. Syst. Nat. ed. X, 1758, стр. 135, Швеция.
Синонимы. Podiceps auritus var. Korejewi. Zarudny und Loudon. Orn.
Monatsber. 1902, стр. 186, Оренбург, — Colymbus cornutus. Gmelin. Syst. Nat. 1 B) 1789,
стр. 591, Сев. Америка.
Русское название. Книжное. Народное — поганка, нырец, гагара.
Распространение. Ареал. Европа, Азия и Сев. Америка, умеренная
полоса. В Европе в Исландии, на Фарерских о-вах (очень редко, Саломонсен,
1935), местами в сев. Шотландии, Норвегии (до Тромсе, около 70° с.ш., и Варан-
гер-фиорда), Швеции и Дании (острова, сев.-зап. часть Ютландского полу-
острова, у Копенгагена), в Финляндии примерно до 66° с.ш., в СССР от Прибал-
тики (Латвия, Эстония) и Литвы и Онежского озера до бассейна Вычегды и
Сыктывкара, верхней Печоры на севере Европейской России (сведения о встре-
чах в Лапландии, у Белого моря и Архангельска сомнительны), на юг до Бело-
руссии (Минская обл.), возможно до северных частей Черниговской обл., далее
Карта 49. Распространение красношейной поганки Colymbus auritus
а — границы гнездовий, г — область зимовок.
до Смоленской, Калининской, Ярославской областей, Татарской республики,
Воронежской обл., бассейна реки Алатырь, Куйбышевской обл. (Бугульма,
Бугуруслан, Бузулук), Башкирии (Белебей, Бирск, Уфа), Чкаловской обл. на
юге; в Азии в Зап. Сибири, на севере до Верхотурья и Березова F3° с.ш.),
на Енисее около 57°30' с.ш., на Лене в бассейне Вилюя и у Якутска,
в верхнем течении Нижней Тунгуски около 6Г20' с.ш. (Ткаченко, 1937),
далее в среднем течении Анадыря, вероятна, в Коряцкой Земле, на
Охотском побережье (в июне добыта у Удского, также у Аяна, Шульпин,
1936, в июле на Шантарских о-вах, Дулькейт и Шульпин, 1937), на Камчатке
и Сахалине (Чайво); к югу до северных частей Тургайской степи (севернее
Кустаная), по озерам с верхнего Казахстана (Кокчетав, Атбасар, Акмолинск),
в Кулундинской, Барабинской (Чаны) и Кузнецкой степи (в последней
вероятно, Хахлов, 1937); на Алтае (кроме северо-запада, но, в противополож-
ность утверждению Сушкина, 1938, также и на Телецком озере и на озере Джулу-
куль) и в Тарбагатае, в Монгольском Алтае (озеро Дпин-гол, верховья Кобдо),
в Минусинском районе, в бассейне р. Уса и на озере Урюг-нор; в Туркестане
на Балхаше, по озерам в низовьях Аягуза, у Лепсы, или, вероятно, на Иссык-
куле и Сон-куле, в вост. Тянь-Шане в бассейнах Текеса и Кунгеса; в летнее
время добыта в низовьях Аму-Дарьи; в области Байкала и Забайкалья на
гнездовье, повидимому, отсутствует, но встречена в бассейне Амура в устьях
Аргуни, у Кумары, Благовещенска, в среднем течении Зеи; наконец, повиди-
мому, в устьях Уссури.
В Сев. Америке на гнездовье от нижнего Юкона, сев. Мекензи, Кьюотина,
сев.-зап. Унгава, Квебека, юго.-зап.. НьюгБруневика, вост. Мена на севере и
РОГАТАЯ ИЛИ КРАСНОШЕЙНАЯ ПОГАНКА 275
до южн. Висконсина и сев. Небраска на юге. В негнездовое время в зап.
Европе от Англии до Средиземноморья, в сев. Африке, в Крыму, вероятно на
Черном, Каспийском и Аральском море, в долине верхней Аму-Дарьи (Заруд-
ный и Билькевич, 1918), на озере Иссык-куль, в Китае (Фуцзянь, низовья
Янцзе, Латуш, 1934), Японии; в Сев. Америке главным образом на морских
побережьях от Британской Колумбии и Нью-Йорка до Флориды, южн. Кали-
форнии и Луизианы. Случайные встречи в Алашане, на Прибыловых, Алеут-
ских и Командорских о-вах, в южн. Гренландии, на Ян-Майене, Азорах.
Характер пребывания. Перелетная птица. Летит одиноч-
ками, парами и небольшими группами (редко). Сезонные перемещения в той
или иной мере связаны с замерзанием и таянием водоемов. В Англии по край-
ней мере отчасти оседла.
Даты. Весенний пролет в Европейской части СССР в конце апреля —
начале мая, тогда же и в Туркестане. У Тюмени 19—25 апреля (вероятно этот
вид, Словцов, 1892). В области Алтая во второй половине мая(Сушкин, 1938).
В Коряцкой земле на Пенжине отмечена в последней трети мая, тогда же и
у Удского острога (Миддендорф, 1853); в конце мая, повидимому, появляется и
на Анадыре. В Америке и на побережьях океанов и внутри страны пролет
весною в апреле и начале мая, прилет в Аляску около середины мая (Бент,
1919). Отлет с зимовок во Франции в марте (Майо, 1936), вюго.-вост. Китае
в Фуцзяне в апреле (Мартене, 1910).
Осенний отлет и пролет поздний, движение медленное. В октябре и даже
начале ноября птица еще встречается в средней России E ноября добыта на
Тростенском озере в Рузском районе Московской обл., в начале ноября Северцо-
вым отмечен в Воронежской обл.; в первой трети ноября — в Харьковской
губ., Сомов, 1897). В Туркмении осенью появляется в октябре. Сведения по
Сибири очень скудны: в конце октября птица добывалась в сев. Казахстане
на Иргизе и Тургае, в первой половине октября на Камчатке, на Анадыре
добыта 27 сентября (Портенко, 1939). В Америке осенний пролет отмечен в сен-
тябре — октябре (Бент, 1919).
Биотоп. В период размножения — стоячие водоемы, преимущественно
небольшие, с хорошо развитой растительностью; иногда участки торфяных
болот и речные заводи. На пролете по рекам и морским берегам, где и зимует.
В южной части гнездового ареала в горах (на Алтае обычно от 1700 до 2300 м,
не ниже 950 м, Сушкин, 1938; в Монгольском Алтае на высоте 2230 м, озере
Даин-гол; в Тарбагатае в долине Чиликты, около 2000 м).
Численность. В общем, ограниченная, птица может считаться
редкою (хотя местами, например на горных озерах альпийской зоны Алтая, в
западной Сибири в бассейне Оби и нижнего Иртыша —обыкновенна). В Вост.
Сибири положительно малочисленна и спорадична, что подтверждается и ред-
костью на зимовках в вост. Азии.
Экология. Размножение изучено слабо. Гнездится отдельными
парами (на малых водоемах) или небольшими колониями (на крупных озерах).
Период размножения поздний, с июня, когда часто слышится крик птицы.
Гнезда обычного для поганок типа, из отмерших водных растений, сырые,
расположенные близ берегов, чаще — пловучие. Кладка — как у многих
водных птиц — растянута во времени и не только в зависимости от широты
местности, но также, повидимому, от фенологической и экологической обста-
новки на том или ином водоеме. Имеются и дополнительные кладки взамен утра-
ченных (в Норвегии, по Коллетту). В Прибалтике, как и в Зап. Европе,
полные кладки отмечены со второй половины мая, в Псковской обл. в конце
мая; в Сибири, пцвидимому, около половины июня. В Америке яйца находили
в апреле — июне.
Яйца более или менее эллиптически-овальной формы, не отличимые, в сущ-
ности, от яиц черношейной поганки, гладкие со слабым блеском, беловатого
276
ОТРЯД ПОГАНКИ
или чуть зеленоватого цвета, вскоре после начала насиживания сильно загряз-
ненные. Размеры: «ширина обыкновенно 31,0—29,1, реже 32,8—28,5; длина
47,0^41,3, реже 49,8-46,8 мм» (Бианки, 1913); A00) 39,7—49x28,4—33,4,
в среднем 44,5x30,73 мм (Хартерт, 1915). Число яиц в кладке 4—5, изредка
3 или 6 (и будто бы даже до 8, Финляндия, Сандман, 1892 и др.; в Америке до 9
и даже 10, Бент, 1919).
Насиживают, повидимому, оба родителя, но преимущественно самка,
после окончания или с середины кладки (птенцы все же разновозрастны).
Продолжительность насиживания, вероятно, немного более трех недель;
в Исландии 20—24 дня (Хантш, 1907, в Шотландии 22—24 дня, Незерсол-
Томпсон). Пуховички, по всей вероятности недавно вылупившиеся, наблюда-
лись во второй половине июня в севе-
ро-западных частях СССР (Прибалти-
ка, Псковская обл.) 24—27 июня у
Березова на Оби (Дерюгин, 1898).
Остальные приводимые в литературе
сведения трудно использовать из-за
неопределенности возраста птенцов.
В разные числа августа добываются
уже вполне оперившиеся молодые, в
сентябре выводки разбиваются. В вы-
водках на Алтае наблюдалось 2—3
птенца (Дементьев и Фолитарек, 1938).
Линька. Последовательность
смены нарядов: пуховой — птенцо-
вый — первый зимний (предбрач-
ный)—первый брачный—послебрач-
ный — брачный и т. д., как у других
поганок. Птенцовый наряд надевается
в зависимости от возраста птенца —
в конце августа (Сон-куль, Ростов-
Ярославский) и в разные числа сен-
тября и сменяется на зимний в октябре,
иногда и позже. У взрослых две линь-
ки —частичная, предбрачная, охваты-
вающая контурное оперение кроме крупных перьев, первостепенных и частью
второстепенных маховых и их кроющих, происходит в апреле, начинаясь на
зимовке или в период отлета с зимовок, в марте и затягиваясь иногда до
начала мая. Полная послебрачная линька, во время которой маховые сме-
няются почти одновременно и птица не может летать, начинается во второй
половине июля и в августе (причем начинается линяние на голове с выпадения
украшающих перьев и т. д.) и заканчивается обычно в октябре, затягиваясь
у некоторых особей до октября. Географические особенности и подробности
линяния (как и размножения) требуют еще изучения.
Питание не изучено. У птенца, добытого на Сахалине, в желудке
плавунцы, личинки, в желудке взрослой птицы — семена, трава, плавунцы,
и их личинки, гаммариды. В Сев. Америке в качестве кормов этой поганки
указывается преимущественно мелкая рыба, также беспозвоночные — гамма-
русы и креветки и т. д.; лягушки и головастики, ручейники, мухи, жуки;
наконец растения (Бент, 1919). В желудках, как и у других поганок, находятся
обычно перья.
Полевые признаки. Средней величины поганка, несколько меньшая, чем
чирок. В брачном наряде характерны ржавчато-рыжие «ушные» пучки перьев
и хохол на затылке, рыжая шея, зоб и бока; от малой поганки в этом наряде
отличается контрастной окраской — черной на спинной стороне и белой на
Рис. 56. Красношейная поганка
ЧЕРНОШЕЙНАЯ ИЛИ УШАСТАЯ ПОГАНКА 277
брюшной. От черношейной поганки различается присутствием ржавчато-
рыжего цвета на верхней стороне головы, не распространяющегося ниже
глаз (у черношейной поганки верхняя сторона головы черная и рыжеватые
перья у глаза и за глазом). Отлична и форма клюва —относительно прямого
и более крепкого (у черношейной поганки клюв несколько вздернут кверху и
послабее). На лету —при сравнении с черношейной поганкой —менее броса-
ется в глаза белый цвет на крыле. При опасности охотнее подымается на крыло,
чем прочие поганки. Полет прямой, быстрый, с частыми ударами крыльев;
плавает обычно неглубоко погружаясь в воду, с вытянутой шеей. Голос разно-
образный, в брачном периоде довольно низкая трель; кроме того «ею» или «кею».
По сравнению с другими поганками менее осторожна.
Описание. Размножение и строение. Форма клюва — см. выше. Ноздри
овальные, сквозные, близко от основания надклювья, расположены в округленных спереди
ноздревых ямках. Формула крыла 3^2>4>5>6..., первое маховое рудиментарно. Длина сам-
цов C) 330, 340, 356 мм; самок B) 320, 341 мм. Размах самцов C) 540, 550, 606 мм; самок
B) 590, 594 мм. Вес самцов B) 385, 500, самки A) 375 г. Крыло самцов B0) 132—144, самок
A4) 124—140, в среднем 138,65 и 136,35 мм. Длина клюва около 20—27 мм.
Окраска. Пуховой птенец на спинной стороне черноватый с белым продольным
рисунком; на голове одна белая продольная полоса идет от надклювья, будучи ограничена
снизу черной полосой, другая — ото лба к ушам; на темени голый участок кожи, за ним —
белое продолговатое пятно; на шее две белых продольных полоски, на спине четыре узких
продольных белых полосы; подбородок белый с черными пестринками; на горле черное пятно;
бока и голени серовато-черные с белыми лестринами; зоб, грудь и брюхо белые. У подросшего
птенца белые полосы на спине почти исчезают, остаются лишь неправильно расположенные
беловатые пестрины, черные пятна на подбородке бледнеют, на зобе выделяется черное
пятно. Радужина беловата, клюв бледно желтоватый, черноватый у основания надклюоья,
с черным пятном у вершины; ноги желтоватые, темные (черноватые) на внешней стороне
цевки и по краям оторачивающих пальцы лопастей.
У взрослых самцов и самок в брачном наряде лоб, темя, задняя сторона шеи, подбородок
и спинная сторона тела черные с более светлыми на спине и плечах серединами перьев;
удлиненные перья ошейника черные; уздечка ржавчато-рыжая; удлиненные перья над гла-
зом и на затылке рыжие; передняя сторона шеи, зоб и бока рыжие; подкрылья белые с при-
месью буроватого у края крыла; грудь, брюхо, бока белые; рулевые черные с бопьшей или
меньшей примесью каштаново-рыжего цвета, рудиментарной структуры; кроющие крыла
серовато-бурые; внутренние большие кроющие белые с черно-бурыми крапинами; первосте-
пенные маховые бурые с черными стволами и с более светлыми, почти белыми у основания,
внутренними опахалами, передние второстепенные маховые белые с большей или меньшей
примесью бурого, средние чисто белые, задние черные с белым основанием. В зимнем (после-
брачном) наряде нет «ушных» пучков и «ошейника», общая окраска спинной стороны бледнее и
серее, по сторонам лба по беловатому пятну; за глазом рыжеватые пестрины; брюшная сто-
рона белая, на нижней части зоба обычно буроватый оттенок; бока с сероватыми пестринами.
У молодых птиц в гнездовом (птенцовом) наряде окраска спинной стороны еще более
буровата; на боках головы и шеи нерезкие темные пестрины, за глазом беловатая или охри-
стая полоса; в остальном — как взрослые в зимнем наряде. В первом зимнем наряде молен
дые — как взрослые, но без рыжеватых пятен за глазом. Клюв серовато-роговой, с беловатой
вершиной и красноватым основанием; ноги с желтоватыми краями пальцев; наружная сто-
рона цевки и нижняя поверхность пальцев и когти черные. Радужина красная или оранжевая
с беловатым кольцом вокруг зрачка. У молодых окраска неоперенных частей бледнее, чем
у старых.
4. Черношейная или ушастая поганка Colymbus caspicus
Hablizl.
Columbus caspicus. Hablizl. Neue Nordische Beitrage, 1783, стр. 9, Энзели.
Синоним. Podiceps nigricollis. Chr. L. В г е h m. Handbuch. d. Naturgesch. Vogel
Deutsche, 1831, стр. 963, Германия.
Русское название. Книжное. Народные — как у других поганок (гагара,
поганка, нырец и т. п.).
Распространение. Ареал. Европа, Азия, Африка, Сев. Америка.
В Европе от Англии, Ирландии, Дании, Литвы, Белоруссии (Минская обл.),
областей Московской и Горьковской, Татарской и Башкирской республик,
южной части Молотовской обл. на севере до Пиренейского п-ова, Италии,
Балканского п-ова, Крыма и Кавказа на юге. В Азии ареал разомкнут —
278
ОТРЯД ПОГАНКИ
с одной стороны, птица встречается в Зап. Сибири от Тюмени, Ишима,
Омска на севере до Алтая, к югу до Малой Азии, Палестины, Ирана, Турке-
стана (в последнем лишь в низовьях Аму-Дарьи, Сыр-Дарьи и в Семи-
речье, далее до Зайсана и Марка-куля, в средней Сибири изредка наблю-
далась в Кузнецкой степи, восточнее, повидимому, не гнездится. Затем
встречена в вост. Азии, где, повидимому, гнездится в южн. Приморье,
Манчжурии и Сев. Китае (Жехол, Шаньси). Возможно в юго.-вост.
Аравии. В Африке на севере в Тунисе, Алжире, быть может в нижнем,
Египте (Файюм, Бенгази); затем уже к югу от Сахары, Абиссинии и южн*
Карта 50. Распространение черношейной поганки Colymbus caspicus
7. С. с. caspicus, 2. С. с. gurneyi, 3. С. с. californicus;
а — граница гнездовий, г — область зимовок.
Анголы до Капской провинции. В Сев. Америке на западе до средних
частей Британской Колумбии, Большого Невольничьего озера и Манитобы до
Южн. Калифорнии, северных частей штатов Аризона и Айова. Зимовки
в Англии, у Средиземного моря, в Закавказье, в Туркмении, на озере Иссык-
куль, в Корее, Китае, Японии. В Америке главным образом у Тихого
океана к западу от Скалистых гор, на востоке до Невады и Техаса, к северу
до Вашингтона, к югу до Гватемалы (Бент, 1919).
Биотоп. Как и у других поганок, пресноводные (иногда солоноватые)
водоемы с густой растительностью в гнездовое время, зимою и у морских побе-
режий.
Подвиды и варьирующие признаки. Три подвида, из которых американский отличается
от прочих окраской; между двумя другими разница в размерах и в некоторых эмологических
особенностях. В Европе, Азии и северной (палеаркт и ческой) Африке — С. с. caspicus H a b 1.
1783; в Африке к югу от Сахары—С* с. gurneyi Roberts, 1919 (у этого подвида нет, повиди-
мому, особого зимнего наряда, Эллиотт и Фуггл-Кешмен, 1948); в Сев, Америке —
С. с. californicus Heermann, 1854.
Обыкновенная черношейная или ушастая поганка Colymbus caspicus casptcus
Hablizl
Распространение. Ареал. Европа, сев, Африка и сев. Азия. В Европе
от Ирландии, Великобритании, сев.-заи. Германии, Дании, южн, Шве-
ции и Польши до Португалии, южн. Испании, Франции, Италии, Бал-
канского п-ова, Кипра. В Африке, в Алжире, Тунисе, Марокко, быть может
в нижнем Египте. В Азии вне пределов СССР — в Малой Азии, Пале-
ЧЕРНОШЕЙНАЯ ИЛИ УШАСТАЯ ПОГАНКА 279
стине, Месопотамии, вост. Иране (Сеистан), в сев. Белуджистане; вероятно,
в Манчжурии, Корее и Сев. Китае.
В СССР ареал состоит из двух частей. Большая расположена
в Европейской части Союза, в Зап. Сибири и в Туркестане. Здесь черно-
шейная поганка гнездится от Прибалтики (Лоудон, 1909; требуются новые
подтверждения) и Белоруссии (Минская обл.), в средней полосе на север
до Московской и Горьковской областей, Татарской и Башкирских республик,
южной части Молотовской обл.; в Зап. Сибири от Тюмени, Ишима, Омска
и Барабы (озеро Чаны) до западной окраины Алтая; на юг до зап.
Украины, Крыма, Закавказья, Туркестана. В последнем в области низовьев
Аму-Дарьи (несомненно в области Ташаузских озер), Сыр-Дарьи и на северо-
востоке (Баскан, Аксу, Балхаш, низовья Или), достигая Зайсана и Марка-
куля. В средней Сибири встречается, повидимому, только случайно (отмечена
в Кузнецкой степи, Хахлов, 1937; однажды под Красноярском в мае; добыта
9 мая 1935 на Телецком озере). В Забайкалье и сев.-зап. Монголии встречена
только на пролете. С другой стороны, по всей вероятности гнездится в южном
Приморье, где добывался у Сидеми, на озере Ханка и у залива Посьет. В не-
гнездовое время встречается в Англии, во Франции, в Средиземноморье, в Египте
и на Синайском п-ове; в Иране, Туркестане, сев. Индии, в Корее, Японии,
Китае (Фуцзянь, Гуаньдунь, Юннань, Латуш, 1934), в СССР в Крыму, Закав-
казье, юго.-зап. Туркмении, на озере Иссык-куль. Залеты в Кольский залив
A9 августа 1921) в Финляндию, к Пскову и Ленинграду, к Кирову (в ноябре),
в Джунгарию и вост. Няньшань (Бианки, 1913), на Мадейру, Канарские
и Азорские о-ва.
Характерпребывания. Перелетная птица, но на крайнем юге
ареала, повидимому, оседлая (озеро Севан в Армении, по Шелковникову, 1934).
Даты. На зимовках в Китае у Фучжоу с ноября по март (Латуш, 1934).
В области Средиземного моря держится до марта — начала апреля. В западном
Закавказье до конца марта (Вильконский, 1897). Пролет и прилет на Украине
главным образом в апреле, тогда же и в средней полосе (Белоруссия, Москов-
ская, Тульская, Горьковская области, Поволжье и Заволжье), затягиваясь
до начала мая. Прилет на Эмбе, Сыр-Дарье, в Приалтайских степях отмечен
в конце апреля — начале мая. Поздний пролет — в последней трети мая
отмечен в Забайкалье. Летят птицы одиночками, парами, небольшими стай-
ками как днем, так и ночью. Осеннее движение по всей вероятности связано
с заморозками и очень растянуто. Первые пролетные особи даже на юге, в ни-
зовьях Сыр-Дарьи, встречены во второй половине сентября, однако возможно,
что это местные передвижения, так как птицы встречаются позднее и в зна-
чительно более северных местностях. Такие же местные передвижения в сен-
тябре наблюдались и на Украине (Харьковская обл., Сомов, 1897). Настоящий
отлет в средней полосе (Калужская, Московская области; Татарская и Баш-
кирская республики) происходит в октябре, тогда же ив сев.-зап. Казахстане.
Осенний пролет по Сыр-Дарье тянется по крайней мере еще в октябре, в Турк-
мении в октябре —середине ноября, на зимовках у южн. Каспия птицы по-
являются в ноябре —декабре.
Биотоп. Гнездовый — разной величины водоемы, преимущественно
стоячие, но также и речные протоки и заводи; менее связана с тростниками,
чем другие поганки, не избегает и открытых пространств. Зимою — открытые
внутренние водоемы и морские побережья. В вертикальном отношении поды-
мается до 2000 м в Закавказье (озеро Севан), в Туркестане до 1700 м (зимует
на озере Иссык-куль), на Алтае до 1500 ж (Сушкин, 1938).
Численность. Относительно высокая, в средней полосе и на юге
обычная птица. Местами образует значительные гнездовые колонии (на озере
Алатай-куль близ Караузяка в низовьях Сыр-Дарьи более 150 пар, Спанген-
берг, 1936).
2S0 ОТРЯД ПОГАНКИ
Экология. Размножение. Пары, повидимому, постоянные (парт-
неры держатся вместе уже на пролете; возможно образование пар происходит
на зимовках). Гнезда обычно расположены колониями, близко одно от другого.
Постройка обычного для поганок типа, мокрая, из отмершей водной раститель-
ности, обычно пловучая. Форма гнезда то более или менее округлая, то
продолговатая; размеры округлых в диаметре 16—20 см, продолгова-
тых 16 — 19,5 см в длину и 13—15 см в ширину; лоток то плоский, то
довольно глубокий, до 3,5 см (Сыр-Дарья); общая высота гнезда 16—45,
высота надводной части 2,5—3,5, общий диаметр гнезда 22—30, диаметр
лотка 13—16 см (Наурзум, сев.-зап. Казахстан, Никифоров). Располо-
жены гнезда то среди зарослей водяной растительности, то на открытом
зеркале.
Время кладки не установлено точно. На Украине в Харьковской обл. в мае
(Сомов. 1897); в первых числах июня кладки находили в сев. Казахстане
(Сушкин, 1908 и др.); в низовьях Сыр-Дарьи в начале июня насиженные кладки
(Спангенберг, 1936). В Англии кладки отмечены в мае, в Испании уже с поло-
вины апреля (Уайзерби, 1940). Число яиц в кладке обычно 4, иногда 3 или
до 6. Яйца по окраске сходны с яйцами предыдущего вида, овальные или элли-
птически-овальные, беловато-зеленоватые, вскоре после начала насиживания
сильно запачканные гниющей в гнезде водной растительностью, иногда до
черного цвета (Спангенберг).
Размеры: ширина варьирует обыкновенно от 32,0 до 28,5, реже может
быть уже от 34,0, длина от 47,0 до 39,0, реже может быть от 48,0 мм (Биан-
ки, 1913); 39,0—45,1 х 27,3— 30,9 мм (Сыр-Дарья, Спангенберг). Бывают
дополнительные кладки взамен утраченных.
Насиживают оба родителя; по западноевропейским данным насиживание
начинается с первого яйца, продолжительность его 20—21 день. Насижива-
ние неплотное, в жаркую погоду птицы часто оставляют гнездо, прикрыв кладку
растениями. Летные взматеревшие молодые добывались в конце июля и,
главным образом, в августе.
Линька. В общем сходна с линянием рогатой поганки, но изучена недо-
статочно. В конце августа — начале сентября молодые птицы сменяют птен-
цовый наряд на первый зимний (частичная линька на голове и шее). Полная
осенняя линька взрослых начинается постепенно в июле, будучи заметной
уже в конце этого месяца и в начале августа (голова, шея, спина); заканчивается
она до отлета к концу сентября — началу октября. Смена маховых (одновре-
менная) отмечена в первой половине августа (Ак-суат, сев.-зап. Казахстан, Ники-
форов).
Имеются индивидуальные различия в сроках линяния, вероятно связан-
ные с возрастом или* половым циклом (запаздывают особи, имевшие, пови-
димому, вторые «дополнительные» кладки).
Частичная предбрачная линька на зимовке, в марте, начинаясь, впрочем,
позже у молодых прошлогодних особей C1 марта, Челекен — полный еще зим-
ний наряд). Добытые во второй половине апреля взрослые в полном брачном
наряде, в начале же апреля попадаются еще особи с остатками зимнего
наряда на голове и шее. Предбрачная линька зимующих в Англии птиц
отмечена в феврале —мае (Уайзерби, 1940 —последнее относится, вероятно,
к молодым птицам).
Питание изучено слабо, но в общем сходно с питанием других видов
поганок. Рыба занимает в кормовом режиме небольшое место и преимущественно
в холодное время года. В период размножения главная пища —водные насеко-
мые и моллюски, ракообразные (гаммарусы, ослики), реже головастики и ра-
стения. Молодые выкармливаются, главным образом, насекомыми. Как у дру-
гих поганок, в желудках находятся обычно перья. Добывается пища из воды
и с воды главным образом нырянием.
ЧЕРНОШЕЙНАЯ ИЛИ УШАСТАЯ ПОГАНКА
281
Полевые признаки. Общие размеры сходны в общем с таковыми у рога-
той поганки, но верхняя сторона головы черная, а пучки удлиненных золотисто-
бурых перьев расположены по бокам головы у глаз; горло черное (в брачном
наряде). На полете заметно больше белого на крыле. Надклювье слегка во-
гнуто и клюв кажется вздернутым. Осторожнее рогатой поганки. Голос разно-
образный обычно «пипп» или «уипп», в брачное время напоминающее трель
«питепитепите».
Описание. Размеры и строение. Клюв относительно короткий, слегка вогну-
тый у ноздрей, конец надклювья приподнят кверху. Формула крыла 2>3>4>5>6...; первое
маховое рудиментарно; вырезки на внешних и внутренних опахалах 2-го, 3-го и 4-го перво-
степенных маховых. Длина самцов G) 310—336, самок F) 315—-321, в среднем 325,1 и 318,6 мм.
Рис. 57. Черношейная поганка
Размах самцов G) 532—595, самок F) 510—564, в среднем 567,4 и 546 мм. Вес самцов B)
300, 350, самки A) 320 г. Клюв около 20—25 мм. Крыло самцов E2) 120—141, самок C1)
118—136, в среднем 131,6 и 127,3 мм.
Окраска. Пуховой птенец на спинной стороне черный с нерезкими белыми поло-
сами по сторонам темени и над глазом, едва заметными на спине; на щеках полосы выра-
жены заметнее, на шее нерезки, зоб обычно белый, средняя часть груди и брюхо белые,
бока черные с белыми пестринами. Клюв мясного цвета с двумя черными поперечными поло-
сами. Голое пятно на темени и уздечка красные.
Взрослые самцы и самки в брачном наряде: спинная сторона, включая удлиненный
хохол на темени, черная, первостепенные маховые бурые с черными стволами, задние —
с большей или меньшей примесью белого; второстепенные маховые белые, на внутренних
примесь черного цвета, самое внутреннее перо — черное; за глазом пучок тонких, длинных
золотистых и рыжих перьев; теки, горло, верхняя часть зоба — черные; нижняя часть
зоба черная с белыми пестринами; грудь и брюхо белые; бока ржавчато-рыжие; подхвостье
рыжее; подкрылья белые.
В зимнем наряде (послебрачном) удлиненных перьев на темени и за глазами нет, спинная
сторона буровато-черная, по сторонам затылка белое пятно, края поясницы и боковые руле-
вые белые; брюшная сторона белая с буроватым оттенком на зобе и с примесью темносерого
цвета на боках груди и тела. Молодые сходны со взрослыми в зимнем наряде, но с пестринами
на боках головы. В первом зимнем наряде — как взрослые. Клюв темносерый с красноватым
основанием и беловатой вершиной, ноги зеленовато-серые с розоватой примесью на отороч-
ках пальцев, нижняя поверхность пальцев и наружная поверхность цевок черноват j.
Радужина оранжево-красная, беловатая вокруг зрачка; у молодых радужина корич-
невая.
282
ОТРЯД ПОГАНКИ
5. Малая поганка Colymbus ruficollis Pall.
Colymbus ruficollis. Pallas. Vroeg's Catal. Coll. 1764, Adumbrat, стр. б, Голландия.
Русское название. Книжное.
Распространение. Ареал. Южная и средняя Европа, средняя и южная
Азия, Африка и Мадагаскар, Новая Гвинея, Австралия, Тасмания. Харак-
тер пребывания. В северных частях ареала перелетная, на юге оседлая
птица. Биотоп. Гнездовый — пресные водоемы, как стоячие, так и речные,
с хорошо развитой водной растительностью, с илистым грунтом. В негнездо-
вое время и на больших реках и на взморьях.
Карта 51. Распространение малой поганки Colymbus ruficollis
1. С. г. ruficollis, 2. С. г. caper sis, 3. С. г. poggei, 4. С. г. japonicus, 5. С. г. kunikyonis, 6. С. г. philippen-
sis, 7. С. г. vu капо гит, 8. С. г. tricolor, 9. С. г. novaehollandiae, 10. С. г. longirostrls, 11. С. г. leucosternos,
12. С. г. rufilavatus;
а — граница гнездовий (цифры в кружках соответствуют номерам подвидов и показывают их зимние место-
нахождения).
Подвиды и варьирующие признаки. Морфологически варьируют пропорции (клюв)
и окраска, экологические различия плохо выяснены (характер пребывания, особенности
цикла периодических явлений). Описаны подвиды: С. г. ruficollis Pa I I a-s — из Европы
и с.-з. Африки, повидимому и в Малой Азии; С.г. capensis Salvador i, 1884 —из Кавказа,
Туркестана, Ирана, Палестины, Аравии, южн. Тибета, Кашмира, Гималаев, Индии,
Цейлона, Бирмы, в Африке, в Египте и от Золотого берега и Абиссинии до Капской Земли;
С. г. rufilavatus D е 1 а с о и г, 1932 — на Мадагаскаре; С. г. poggei Reichenow, 1902 —
в Приморье, Вост. Китае до Хайнаня и ю.-в. Юннани; С. г. japonicus H а г t e r t, 1920 —
в Японии (от Хоккайдо до Кюсо и с о-вов Квельпарт); С. г. kunikyonis Kuroda,
1927 — с о-вов Бородино, Окиназа, средних Риу-киу; С. г. iraquensis Ticehurst, 1923 —
из Ирака и быть может Палестины; С. г. philippensis Bonnaterre, 1791 — с Филиппин-
ских о-ов, о-ва Формоза, Суматра и Борнео; С. г. vulcanorum R e n s с h, 1929— с Тимора,
Флорес, Ломбока и Явы; С. г. tricolor Gray, 1861 — с Целебеса, Молуккских о-вов, Новой
Гвинеи, архипелага Бисмарка и Соломоновых о-вов; С. г. novaehollandiae Stephens,
МАЛАЯ ПОГАНКА 283
1826— из Австралии и Тасмании, зимою в северных частях Новой Гвинеи и быть может на
о-ве Алор; С. г. longirostris May г, 1931—на о-ве Реннелл из группы Соломоновых и на
новой Каледонии; С. г. leucosternos М а у г, 1931 — с о-ва Дельфин у Эспириту-Санто, Ново-
гебридские о-ва. Систематическое положение некоторых из перечисленных форм (в частно-
сти iraquensis, kunikyonis) остается неясным
Европейская малая поганка Colymbus ruficollis ruficollis Pall.
Распространение. Ареал. Европа и передняя Азия, с,-з. Африка. В Ев-
ропе на Британских о-вах, в южн. Скандинавии (в Норвегии примерно
до Бергена, в Швеции на западном побережье) и далее к югу до Средиземного
моря (включая Балеары, Корсику, Сардинию, Сицилию и Кипр), в Азии, в Ма-
лой Азии, Сирии\ в сев. Африке от Марокко до Туниса (граница на во-
стоке неясна, в Египте уже следующая форма). В СССР на западе в Калинин-
градской обл., быть может в Латвии, далее в Литве от Вильнюса, в Мин-
ской, Смоленской обл., спорадично в Молдавии и на Украине (Волынь,
Киевская, Полтавская, Херсонская, Днепропетровская области и др.), на во-
сток до Воронежской обл. (Огнев и Воробьев 1923) и быть может низовьев
Дона (Алфераки, 1910). Залеты в южн. Карелию, Ленинградскую, Псков-
скую (июнь), Московскую, Горьковскую обл„, обычно в сентябре — ноябре
и даже декабре. Вне СССР — в Финляндию, сев. Норвегию, на Канар-
ские и Азорские о-ва, Мадейру.
Характер пребывания. Оседлая птица, по крайней мере в большей
части ареала, где только есть незамерзающие водоемы. В негнездовое время все
же наблюдаются откочевки и тогда птица появляется в Крыму, на Сев. Кавказе
и становится более многочисленной в Великобритании, у Средиземного моря.
Даты. На севере ареала появляется повидимому в связи с таянием водое-
мов, движение отмечается в апреле (в Англии с середины марта до начала мая,
Уайзерби, 1940). Осеннее движение с сентября — ноября (появление в Крыму),
в октябре (низовья Дона); в сев. Африке в октябре—декабре (Ирби,
1875); появление птиц с материка в Великобритании с сентября до ноября.
Биотоп. Гнездовый — мелкие водоемы, главным образом стоячие или
с медленным течением (Вальх, 1900), с илистым грунтом и с зарослями водя-
ных растений. Численность в СССР ограниченная, за исключением,
возможно, низовьев Днестра и Днепра.
Экология изучена слабо.
Размножение. Относящиеся к территории СССР сведения отрывочны.
Гнездится парами и колониями. Пловучие гнезда обычно далеко от берега,
на открытой воде. Размеры гнезда меньшие, чем у других поганок, построено
гнездо из отмершей водяной растительности (в Воронежской обл. из Sparga-
nium minimum, Огнев и Воробьев, 1923). В Киевской обл. неполная кладка
найдена 29 мая (Умань, Гебель, 1879), яйца в последней стадии насиживания
и птенцы в Воронежской обл. 31 июля (Огнев и Воробьев). В Пинском Полесье
гнездо с 6 насиженными яйцами и двумя пуховиками 22 июля (Шнитников,
1913). Число яиц в кладке 4—5, иногда 2—3 и до 7, 8, 9 и даже 10 (Уайзерби).
По западноевропейским данным, насиживают оба родителя, с первого яйца,
19—20 дней (Уайзерби, 1940). Яйца продолговатые или заостренно-овальные
беловатые, быстро загрязняющиеся при насиживании Размеры: «ширина 28,0—
23,0, длина 39,4—33 мм> (Бианки, 1913); A00) 32,8—43 х 23,7—28,3, в сред-
нем 37,8 х 26,2 мм (Англия, Уайзерби, 1940), т. е. величина яиц значительно
меньшая, чем у других поганок.
Линька. Смена нарядов — как у других видов отряда в нашей стране.
В начале октября молодые —еще в птенцовом наряде, в конце этого месяца
добыты молодые птицы в первом зимнем наряде (Козлова, 1947). По западно-
европейским данным, смена птенцового наряда на первый зимний протекает
между июлем и декабрем (Уайзерби — в зависимости от возраста). Полная
284
ОТРЯД ПОГАНКИ
Рис. 58. Малая поганка
послебрачная линька взрослых в августе — сентябре и до октября. Частичная
весенняя в Англии отмечена в феврале, у некоторых особей (молодых) она
затягивается даже до июня—июля, кончаясь нормально в апреле. В конце
апреля в Крыму добыта взрослая птица в полном брачном наряде.
Питание. Не изучено, по западноевропейским данным преимуще-
ственно насекомые, также моллюски, ракообразные, мелкая рыба и растения.
Перья в желудках малых поганок находятся относительно редко.
Полевые признаки. Самая мелкая из поганок. В брачном наряде нет ни
ушных пучков, ни хохла. Шея кажется относительно короткой. В брачном опе-
рении характерна темная черноватая общая окраска с ржавчатым подбородком и
горлом, с зеленовато-желтым
клювом. В зимнем наряде (и у
молодых) спинная сторона бу-
рая, брюшная белая. Охотнее,
чем другие виды поганок, по-
дымается на крыло, летит бы-
стрыми взмахами крыльев,
обычно низко над водою. Го-
лос —мелодическое «уит-уит»,
нередко сливающийся в трель;
особенно в период размноже-
ния.
Описание.Размеры и стро-
ение. Клюв короткий и прямой,
сжатый с боков, но довольно широ-
кий в основании; ноздри овальные,
примерно на середине надклювья,
в глубокой округлой ямке. Цевка
покрыта на заднем крае трехуголь-
ными щитками, придающими ей пилообразную поверхность. Формула крыла 3>4>2>5>6 ...
первое маховое рудиментарное. Вырезки на внешнем опахале 3-го 4-го и на внутренних опа-
халах 2-го и 3-го первостепенных маховых. Вес (по Бутурлину, 1935) 125—200 и до 250 г.
Клюв около 16—20 мм. Крыло самцов C) 95—102, самок C) 92—95, в среднем 99 и 93,6 мм.
Окраска. Пуховой птенец на спинной стороне черный с нерезкими рыжими про*
дольными пестринами и с шелкоьисто-серой передней частью темени, на подбородке — чер-
ном — две белых продольных полосы; середина груди и брюха белые, остальная брюшная
сторона черная; клюв красноватый, ноги серые с примесью желтовато-телесного цвета на
внутренней стороне цевки и на пальцах.
Взрослые самцы и самки в брачном наряде: спинная сторона черная, маховые бурые,
второстепенные с примесью белого на вершинах и на внутренних опахалах; примесь белого
цвета имеется и на рудиментарных рулевых; подбородок черный, горло, щеки, передняя
часть шеияркокоричнево-рыжие; подкрылья белые; остальная брюшная сторона черноватая
с большей или меньшей примесью белого.
Взрослые птицы в зимнем (послебрачном) наряде: на спинной стороне рыжевато-бурые,
более темные на середине спины и на плечах; подбородок, горло, середина груди и брюха —
белые; щеки, шея, середина зоба, бока груди и тела рыжеватые, задняя часть брюха бурая.
Молодые птицы— как взрослые в зимнем наряде, но спинная сторона бледнее, а по
бокам головы, на щеках, на шее — рыжие и черноватые пестрины, зоб менее рыжеват, бока
бурые. В первом зимнем наряде птицы также бледнее, чем старые на спинной стороне;
пестрил на боках головы и шеи у них нет, но рыжеватый оттенок на шее выражен резче,
чему старых. Клюв черный, беловато-желтый у вершины, зеленоватый по разрезу; голая
уздечка черновата; ноги серовато-зеленые, светлей на внутренней стороне цевки и на сред-
них пальцах (в особенности у молодых); радужина красновато-бурая у взрослых, бледно-
бурая у молодых.
Белокрылая малая поганка Colymbus ruficollis capensis Salvadori
Podiceps capensis. Salvadori. Ann. Museo Civico Genova B), 1884, стр. 252, Шоа.
Распространение. Ареал. Весьма своеобразен — Африка к югу от
Сахары и северо-восточная часть севера Африки; Передняя Азия от Турке-
стана и Ирана до Гималаев, Индии> Бирмы и Цейлона; Кавказ. В Африке —
МАЛАЯ ПОГАНКА 235
от Золотого берега и Абиссинии до Капской Земли и в Египте (дельта Нила,
Вади-натрун, Файюм, Дакла, по Мейнертцхагену, 1931). В Азии, в Аравии,
Иране от южнокаспи^ских провинций до Мекранского побережья в Белуд-
жистане и до Сеистана на востоке, в Индии от Кашмира до крайнего юга,
в Бирме от низовьев Иравадди и Салуэна, в западных частях Юннани и южн.
Тибете (повидимому до Лхассы), на севере в Синьцзяне на восток до Хами
и Лоб-нора. В СССР на Кавказе от бассейна Терека и Кубани до Армении,
в Туркестане гнездится в Туркмении по Атреку, Теджену, Мургабу, Аму-
Дарье (как в нижнем течении, так и в верхнем), по рекам Кызыл-Су, Вахшу,
вероятно на Кафирнигане и Сурхане; по Сыр-Дарье, в среднем и нижнем тече-
нии; на озере Тели-куль и в низовьях Сары-Су; в Ташкентском оазисе; нако-
нец, на северо-востоке у Джаркента, на Балхаше и в низовьях Или и на Иссык-
к\\ле. Залет в дельту Волги, возможно к низовьям Илека (Бутурлин, 1935).
Характер пребывания — в большой части ареала оседлая птица,
совершает кочевки только на севере.
Даты. В ноябре малые поганки появляются на ю.-в. Каспии (Гассан-
кули) и иногда в октябре — ноябре на внутренних водоемах Туркмении, где
не гнездится. На Или пролет отмечен в ноябре —декабре (часть из этих наблю-
дений относится, вероятно, не к пролетным, а к зимующим птицам). Массовый,
будто бы, пролет в Закавказье в ноябре (Вильконский, 1897), но птица поло-
жительно оседла в Армении по горным озерам (Соснин и Ляйстер, 1942). В Фер-
гане у Маргелана осеннее движение имело место в декабре — январе (Север-
цов). Пролетные стайки в вост. Иране наблюдались в начале ноября (За-
рудный, 1903). Весеннее движение птиц отмечено с марта до начала мая,
но не ясно, есть ли это настоящий пролет.
Биотоп. Как у номинальной формы; зимою охотно по быстрым рекам,
но также и на море (южн. Каспий), близ берегов и по стоячим водоемам. На
гнездовье и зимою отмечена не только на равнинах, но и в горах (Севан,
Иссык-куль).
Экология. Сведения отрывочны.
Размножение. Кладка в Закавказье повидимому в мае (Радде,
1889). В Туркмении около середины мая (Александров, низовья Атрека).
В конце мая кладки и в вост. Иране (Зарудный). Гнезда в Иране иногда
расположены плотными колониями, по 7—8 пар, устройство гнезд обычного
типа. Число яиц в кладке 4—5, редко 3 или 6 (Туркмения, Иран). Размеры
37,5—39,9 х 25,0—28,8 мм (Зарудный, 1903). В середине июня в Иране только
что вылупившиеся птенцы; птенцы 25 июня наблюдались в Армении (Соснин
и Ляйстер, 1942). Около середины июня пуховики были найдены в ю.-з. Турк-
мении; оперяющиеся обнаружены в середине июня (Дементьев, 1950). Подлетки
в Туркмении в Мервском оазисе найдены 27 июля (Зарудный, 1889). Заслужи-
вает внимания большая изменчивость сроков кладки в разных частях ареала:
в Кашмире кладка имеет место в мае, в южн. Индии в мае — июне, в сев.
Индии в августе —сентябре, а на Цейлоне имеются два половых цикла
в году: в декабре —январе и в июне (Бекер, 1929).
Линька. Взрослые птицы в зимнем наряде добывались в октябре
(с конца сентября), в брачном в марте, но главным образом в апреле. Зимний
наряд у молодых птиц развивается к началу октября (затягиваясь у некоторых
особей, по Козловой, 1947, до ноября).
Описание. Размеры и строение. Крыло самцов A0) 90,5—98,5, самок A0)
91—103, в среднем 93,2 и 94,9 мм\ у птиц неизвестного пола (б) 90—95; в общем у обоих полов
B6) 90—103, в среднем 94,39 мм. Вес самки 188 г; длина ее 253, размах 315 мм.
Окраска. В общем сходна с окраской номинальной формы, но белый цвет на крыле
развит больше — основания первостепенных маховых белые, внутренние их опахала и второ-
степенные маховые совсем белые или белые с бурой полоской на внешнем опахале. На крыле
имеется, таким образом, белое зеркальце. Черный цвет в брачном наряде менее распростра-
нен на горле и по сторонам головы, рыжий цвет доходит до глаз.
2Р6 ОТРЯД ПОГАНКИ
Китайская малая поганка Colymbus ruficollis poggei Reichenow
Columbus nigricans poggei. Reichenow. Journ. f. Orn. 1902, стр. 125, Чжили.
Распространение. Ареал. Китай от севера до крайнего юга (Хайнань),
Индо-Китай (Кохинхина, Тонкий, Аннам), северная граница остается неясной;
быть может эта форма гнездится в Корее и вост. Манчжурии (Мейзе,
1934). Малая поганка повидимому гнездится в южных частях Приморья (Шуль-
пин, 1936). Наконец быть может к этому подвиду, но может быть и к японскому,
относится встречающаяся на Курильских о-вах поганка (Уруп, Бианки,
1913; гнездится на Кунашири, Бергман, 1935). Характер пребыва-
ния. В большей части ареала оседла, за исключением вероятно севера Китая
(Латуш, 1934).
Биотоп. Гнездовый — стоячие водоемы с густой растительностью, на
юге и в горах (Аннам, Делакур, 1931). Зимою на прудах, озерах и реках.
Численность. На севере, повидимому, редка (в Южно-уссурийском крае
добыто, повидимому, только четыре экземпляра), в Вост. Китае и Индо-
Китае обычна. В Тонкине иногда зимою наблюдается до сотни птиц на одном
пруду (Делакур, 1931).
Экология. По СССР сведений нет.
Размножение. Кладка в Китае с конца мая, яйца находят до сере-
дины июля. В кладке 3—7 яиц в Китае (Латуш, 1934), 3—5 в Индо-Китае (Де-
лакур, 1931). Размеры C3) 33—39,3 х 25,5—26,5 мм, в среднем 37 х 26 мм
(Латуш).
Л и н ь к а. Не изучена. Добытые в марте и ноябре особи — в зимнем на-
ряде. В начале апреля в Южн. Китае (Фучжоу) добыта линяющая в брачный
наряд особь. Питание составляет мелкая рыба, насекомые и их личинки,
моллюски, ракообразные (Индо-Китай, Делакур).
Описание. Размеры и строение. Клюв слабее, чем у нормального подвида,
с более узким и низким надклювьем. Крыло самца A) 103, самок C) 94, 105, 111 мм
(Зоологический музей Московского университета); крыло самцов и самок (б) 100—ПО
(Аннам, Делакур); 97—113 мм (Хартерт, 1915).
Окраска. Черный цвет на горле менее распространен, рыжий цвет на щеках
доходит до глаза. Белого зеркальца на крыле нет.
Систематические замечания. Из Приморья нами изучен только один экземпляр —
самец в зимнем наряде, добытый на реке Тачингоуза в Судзухинском заповеднике
28 ноября 1945 (Л. О. Белопольский). Эта птица не отличима от китайских и по
сравнению с японским обладает более слабым надклювьем. Впрочем, возможность разде-
ления С. г, japonicus от С. г. poggei остается неясной без исследования большого допол-
нительного материала.
ОТРЯД ТРУБКОНОСЫЕ
ИЛИ БУРЕВЕСТНИКИ
TUBINARES или PROCELLARIFORMES
Характеристика отряда
Морфология. Внешниепризнаки. Птицы различных размеров —
от очень крупных до мелких- У наиболее крупного представителя отряда
нашей фауны — белоспинного альбатроса — длина тела достигает около 550 мм
и размах крыльев до 2200 мм. Некоторые тропические формы альбатросов
имеют размах крыльев до 3750 мм и даже 4250 мм, вес у них 7500—8000 г.
Длина крыла у самой мелкой птицы, встречающейся у нас, —прямохвостовой
качурки —от 114 до 127 мм, у сизой качурки до 160 мм, вес тела у сизой
качурки около 42 г.
Оперение густое, плотное, гладко прилегающее к телу, довольно мягкое
на брюшной стороне и более жесткое на спинной (только у качурок все опере-
ние мягкое). Контурные перья у всех трубконосых птиц, исключая род аль-
батросов Diomedea, имеют хорошо развитый побочный ствол. Спинная птери-
лия у мелких форм (качурок, океанников и др.) узкая и, начиная от лопатки
до начала области безымянной кости, разделена на две; у крупных форм (аль-
батросов, глупышей и Pelecanoides) — широкая, разделена на две, которые
сливаются с бедренной птерилией; у буревестников (Puffinus) — в виде одного
сплошного тяжа без ветвей. Брюшная птерилия идет двумя ветвями по бокам
живота; у мелких видов образуется между ними широкое голое пространство —
брюшная аптерия; у крупных форм — брюшная аптерия узкая. Пух есть у всех
трубконосых и покрывает тело и по птерилиям, и по аптериям. Копчиковая
железа оперена и имеет по несколько выводных отверстий (у некоторых ви-
дов — до 6 отверстий на каждой стороне).
Крыло аквинто-кубитального типа, т. е. 5-е второстепенное маховое от-
сутствует; длинное, острое и узкое; вершину крыла образуют обычно 2-е
или 3-е, или 1-е и 2-е первостепенные маховые перья, не считая первого,
зачаточного махового (только у хорошо ныряющих форм крыло более округ-
лое). Первостепенных маховых И, первое первостепенное маховое — зачаточ-
ное, короче кроющих кисти. Второстепенные маховые очень короткие, не-
288 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
много длиннее кроющих. Число второстепенных маховых велико и сильно
колеблется у разных видов —от 10 перьев у мелких форм до 37 у крупных.
Кроющие первостепенных маховых и кроющие кисти хорошо развиты и до-
стигают большой длины; кроющие второстепенных маховых—короткие.
Рулевых от 12 до 16, но у большинства видов — 12. Хвост разнообразной
формы и различных размеров, в особенности у качурок: у буревестников, тай-
фунников, глупышей, альбатросов хвост короткий или средних размеров и
округлый, у качурок — вилообразный, выемчатый, прямой, клиновидный
и т. д.
Тело валькообразной формы и у крупных видов массивное и довольно
короткое, у мелких —удлиненное и тонкое. Окраска оперения или однотон-
ная от темносерой с различными оттенками до чисто белой или серая различ-
ных тонов с примесью белого. Некоторые виды диморфны — имеют темную
и светлую вариации окраски оперения, связанные между собою рядом перехо-
дов. Иногда эти вариации бывают более или менее географически локализо-
ваны. Полового диморфизма ни в окраске оперения, ни в размерах нет, так
же как нет и сезонного.
Клюв средних размеров с хорошо развитым загнутым вниз крючком над-
клювья; подклювье слегка загнуто кверху. У большинства видов, например,
у глупышей, альбатросов и др., клюв сжатый с боков и довольно высокий.
Рамфотека сложная, состоит из обособленных роговых пластинок различной
формы, которые по месту своего положения на клюве носят различные на-
звания: покрывающие конек клюва—culminicornu, бока надклювья late-
ricornu, ноздри —rhinotheca, крючок и вершину надклювья —d etrum.
Ноздри не сквозные, открываются в особые трубочки, образованные разраще-
нием стенок носовой капсулы, расположены в основной части клюва. Иногда они
лежат по бокам конька надклювья и разделены между собою широким простран-
ством хребта конька (альбатросы), или лежат близко друг к другу по хребту
конька, или у некоторых видов сливаются в одну трубочку, разделенную
внутри перегородкой различной толщины. У глупышей, качурок и тайфунников
перегородка тонкая, у буревестников толстая. У глупышей, качурок и тайфун-
ников ноздри открываются вперед, у буревестников и у ныряющего буре-
вестника кверху.
Ноги умеренной длины или короткие. Передние 3 пальца соединены хо-
рошо развитыми плавательными перепонками со слегка выемчатыми наруж-
ными краями. Задний палец обычно имеется, прикреплен он высоко, но развит
слабо, у альбатросов (Diomedea) почти совсем прикрыт кожей, виден только
коготь, за исключением рода Phoebetia, у которых задний палец хорошо заме-
тен, а у ныряющего буревестника Pelecanoides совсем отсутствует. Когти у ка-
чурок, глупышей и буревестников изогнутые, заостренные и узкие, а у аль-
батросов тупые и уплощенные. Цевка обычно сжата с боков и с острым перед-
ним и задним краями, лишь у качурок округлая. У большинства трубконосых
цевка покрыта мелкими многогранными щитками и только у океанников
Oceanites покрыта спереди поперечными щитками, иногда сливающимися в одну
сплошную пластинку.
Анатомические признаки. У крупных видов большинство костей
пневматичны, у мелких форм пневматичны только некоторые кости черепа и не-
которые шейные позвонки. Череп схизогнатический и голоринальный, лишь
у альбатросов замечается некоторая тенденция к десмогнатизму. Надглазнич-
ные ямы хорошо выражены, но у разных представителей трубоконосых бы-
вают различной глубины и формы. Места прикрепления жевательных мышц
выражаются в форме глубоких ям у буревестников, тайфунников, глупышей;
не глубоких, но широких —у альбатросов; совсем слабо развиты у качурок,
океанников. Базиптеригоидные отростки у некоторых трубконосых птиц совсем
отсутствуют, например, у альбатросов, океанников, Pelagodroma; обычно
ХАРАКТЕРИСТИКА ОТРЯДА 289
в зачаточном состоянии — у качурок, Cymodroma, но у некоторых видов
хорошо развиты, например у Pelecanoides, Oestrelata, Daption, Priocella,
глупышей и тайфунников; особенно сильно варьируют в развитии базипте-
ригоиды у буревестников Puffinus — от едва заметного бугорка до хорошо
развитого отростка. Слезная кость имеется, но у некоторых она свободна и
даже имеет длинный отросток (буревестники Puffinus), у других, например
у глупышей, она срастается с лобными и носовыми костями. Шейных позвон-
ков — 15. Грудина широкая и довольно короткая, со слабо развитым задним
отделом xiphosternum, только у хорошо ныряющих форм, например, у Peleca-
noides, грудина узкая, удлиненной формы, с хорошо развитым xiphosternum.
Задний край или с одной срединной вырезкой (альбатросы) или с двумя па-
рами вырезок, причем у буревестников, тайфунников вырезки глубокие, у глу-
пышей крайняя пара, а иногда и обе пары неглубокие и выражены слабо,
у качурок задний край грудины — без всяких вырезок. Ключица обычно соеди-
няется с килем грудины при помощи связок и только у небольшого числа форм—
синостотически. Скелет передних конечностей характеризуется своей длиною,
в особенности у крупных форм. У альбатросов сильно удлинено плечо и обе
кости предплечья, кисть »хотя и длинная, но короче первых двух отделов.
У средних и мелких размеров птиц кости плеча и предплечья не так сильно
удлинены, как у альбатросов, и равны кисти. Все трубконосые имеют хорошо
развитый крючкообразный отросток processus supracondyloideus плеча, который
особенно больших размеров достигает у парящих форм. Кроме того, в локте-
вом сочленении только у трубконосых имеются добавочные одна или две
сесамоидные косточки, не развивающиеся у всех других птиц. У альбатросов,
буревестников, Majaqueus и Adamastor их две; у тайфунников, качурок —
одна. Эти сесамоидные косточки и крючкообразный отросток служат местом
прикрепления ряда мышц крыла и образуют прочную основу, помогающую
при парении держать крыло в раскрытом состоянии.
Таз узкий, вытянутый в длину, в особенности у хорошо ныряющих форм,
например у Pelecanoides. Бедро и цевка одинаковой длины или даже бедро
несколько короче цевки. Большая берцовая кость имеет сильно развитый
кнемиальный отросток, различной формы. Малая берцовая — слабая и корот-
кая. Коленная чашечка незначительных размеров и имеется не у всех форм.
Задний палец, у тех форм, у которых он имеется, состоит из одной фаланги.
Обоняние развито хорошо, обонятельные лопасти значительных размеров.
Стенки носовой раковины завернуты в спираль и имеют сложное строение.
Нижняя гортань переходного строения между бронхотрахеальным и брон-
хиальным типами. Трахея обычно короткая и прямая, только у альбатросов
образует петли, причем у черноногого альбатроса петли имеются лишь у сам-
цов; у некоторых видов (глупыши, гигантские глупыши и др.) нижний ко-
нец трахеи разделен внутри перегородкой, как у пингвинов.
Язык различных размеров: у альбатросов и у буревестников он зачаточ-
ный, у Macronectes очень большой величины и заполняет всю ротовую полость.
Между этими двумя крайностями существует целый ряд переходов. В ротовой
полости имеются особые приспособления для удержания или процежива-
ния пищи. Например, у видов, питающихся преимущественно рыбой, на небе
имеются острые роговые выступы, загнутые внутрь, помогающие задерживать
скользкую добычу, а у видов, питающихся планктоном, на верхней челюсти
образуются выросты, "похожие на роговые пластинки, при помощи которых
они могут процеживать воду. Железистый желудок — больших размеров,
мускульный —очень мал, но, как и пищевод, сильно растяжим. Слепые
кишки или совсем отсутствуют или развиты очень слабо. Желчный пузырь
имеется, сонных артерий две.
Образ жизни. Трубконосые —птицы открытых морских пространств, с су-
шей связаны только в период размножения. Превосходно летают и плавают,
19 Птиды СССР, т. II
290 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
немногие виды хорошо ныряют, например ныряющий буревестник Peleca-
noides. По земле передвигаются с трудом, опираясь на цевку и даже крыло
(альбатросы). Большую часть жизни проводят в воздухе или на воде; на воде
они обычно и спят, и отдыхают. На крыло поднимаются с воды довольно легко,
иногда с небольшого разбега, с земли — с трудом, подыскивая обычно для
взлета какое-либо возвышение. Полет мелких видов, например качурок,
характерен частыми взмахами крыльев, но большинство видов, и в особенности
крупные, — альбатросы — могут в продолжение многих часов парить в воз-
духе, обычно летая низко над водою, то вдоль волны, то наперерез волне,
часто повторяя в воздухе движение волны. Почти все трубконосые летают во
всякую погоду, но некоторые виды с очень узким крылом, не обладают большой
способностью к активному полету, как например альбатросы, в особенности
приспособлены к парящему полету в штормовую погоду. Большинство видов
активны и днем и ночью, а в период размножения это настоящие ночные птицы.
Скорее кочующие, чем настоящие перелетные птицы, но некоторые из них
совершают и правильные перелеты, например, тонкоклювый буревестник.
От мест гнездовий в ю.-в. Австралии и Тасмании он летит по западной части
Тихого океана к Берингову проливу и к о-вам Врангеля и Геральда, а
обратно летит на гнездовье, придерживаясь восточной части Тихого океана,
и совершает, таким образом, путь не менее, чем в 25 000 км. Во время миграций
и кочевок держатся чаще небольшими стаями, но иногда наблюдаются громад-
ные скопления, иногда же попадаются и одиночные экземпляры.
Моногамы. На места гнездовий на севере птицы прилетают в марте —
апреле, редко в феврале—мае; в субтропиках, тропиках и в южном полушарии
на местах гнездовий появляются в сентябре—октябре, иногда в августе.
Гнездятся колониями или самостоятельными, или совместно с другими
птицами, например чайками и т. д. Большинство трубконосых гнездится из
года в год на одних и тех же местах, но некоторые виды, например глупыши,
меняют, повидимому, свои гнездовья в разные годы. По прилете на места
гнездовий у многих видов, например у глупышей, альбатросов и у ряда дру-
гих, наблюдаются брачные игры. Колонии устраивают, обычно, на пустынных
скалистых, океанических или морских островах. Гнезда располагаются у неко-
торых видов у самой воды, у других видов — на большом расстоянии от бе-
рега, например у некоторых альбатросов. Гнездо строят или на поверхности
земли, причем очень небрежно и примитивно (некоторые виды откладывают
яйца прямо на землю без всякой подстилки), или устраивают в норах, щелях
скал, под камнями, под корнями деревьев и т. д. Обычно, после окончания
постройки гнезда у многих видов трубконосых, все птицы покидают гнездовья
и улетают на 20—30 дней в море. Возвращаются они все одновременно перед
кладкой. В кладке обычно 1 яйцо белого цвета, у некоторых видов с тем-
ными мелкими крапинами. Форма яйца удлиненная, оба конца его почти оди-
наково закруглены.
Откладка яиц у альбатросов, буревестников и некоторых других видов
происходит почти одновременно у всей колонии.
Насиживают обычно оба родителя. Эмбриональное развитие медленное;
насиживание продолжается у крупных видов, например у темноспинного
альбатроса, 63 дня, у мелких — прямохвостой качурки — 36—40 дней.
Птенцы вылупляются покрытые густым пухом, но слепые. Постэмбрио-
нальное развитие медленное, и птенцы большинства видов имеют два пуховых
наряда. В гнезде птенцы остаются долго, некоторые виды альбатросов Diomedea
immutabilis и Diomedea nigripes — до 6 месяцев, северная качурка Осеапо-
droma leucorrhoa и малый буревестник Puffinus puffinus—до 70 дней, се-
рый буревестник Puffinus griseus — до 5 месяцев и тонкоклювый буревестник
Puffinus tenuirostris — до 4 месяцев. Из гнезда выходят или перед спуском на
воду, или перед поднятием на крыло, уже одетые пером. Птенцов выкармли-
ХАРАКТЕРИСТИКА ОТРЯДА 291
вают оба родителя, причем первое время — отрыгиваемой маслянистой жид-
костью — «ворванью», затем уже различными беспозвоночными; крупные виды
кормят птенцов, главным образом, моллюсками и рыбой. При получении
пищи птенец вкладывает свой клюв поперек клюва родителя и тот выбрасы-
вает отрыжку в клюв птенца или, поздней, когда кормит уже мелкими живот-
ными, подталкивает пищу языком в клюв птенца. Процесс кормления проис-
ходит обычно один раз в сутки, ночью. Птенцы быстро жиреют и еще будучи
в пуху достигают величины взрослой птицы. Незадолго до выхода птенцов
из гнезда взрослые птицы перестают их кормить и улетают. Птенцы, побуждае-
мые голодом, через несколько дней покидают гнезда и перебираются на воду.
Обычно оставление птенцами гнезд происходит ночью, и у некоторых видов*
например у альбатросов, буревестников и ряда других, одновременно почти
у всей колонии. Первое время молодые птенцы плохо еще летают и большую
часть времени проводят на воде близ гнездовий. Только вполне окрепнув,
они покидают гнездовья и начинают кочевать вслед за улетевшими уже взрос-
лыми птицами.
Трубконосые птицы на гнездовьях очень беспомощны, от преследователя
стараются уйти в море. Единственный способ защиты у них —это выбрасы-
вание изо рта струи вонючей маслянистой жидкости. Таким образом защища-
ются не только взрослые птицы, но и птенцы.
Питание. Пища состоит главным образом из различных беспозвоноч-
ных, планктона, рыбы, кроме того, многие виды поедают отбросы с судов,
падаль и т. д. Некоторые виды, собираясь в громадные стаи, сопровождают
суда, в особенности китобойные, ожидая, когда будут выброшены остатки
промысла и т. д. Трубконосые очень прожорливы и переваривание пищи у них
происходит очень быстро. Часа через два из насытившейся и отдыхающей
на воде стаи, например глупышей, начинают уже отделяться единичные особи,
которые вновь приступают к поискам пищи. Пищу большинство видов добы-
вает с поверхности воды или не глубоко ныряя за ней, находясь обычно на воде,
и только Pelecanoides добывают пищу, глубоко ныряя и плавая под водой.
Голос. Во время кочевок и миграций эти молчаливые птицы поднимают
крик лишь в моменты драки на кормежках. Они очень крикливы и шумливы
на гнездовьях ночью, во время кормления птенцов. Голос крупных видов —
хриплый стонущий крик, мелкие виды издают щебечущие звуки.
Хозяйственное значение. Мясо трубконосых птиц, очищенное от жира,
который придает ему неприятный запах, довольно хорошего вкуса и упо-
требляется в пищу. Кроме того, в большом количестве на гнездовьях загото-
вляются яйца и добываются птенцы. Последние идут, главным образом, на вы-
работку жира. В СССР трубконосых так немного, что практического значения
они не имеют.
Систематика. Отряд трубконосых содержит одно семейство — буревест-
ники Procellariidae, разделяющееся на 3 подсемейства: собственно буревестники
Procellariinae, альбатросы Diomedeinae и ныряющие буревестники Pelecanoi-
dinae.
I. Подсемейство собственно буревестники Procellariinae. Средних и мел-
ких размеров птицы, только гигантский глупыш Macronectes giganteus достигает
крупных размеров. Наружные носовые трубочки слиты в одну, разделенную
внутри перегородкой различной толщины у разных видов. Плечо, предплечье
и кисть обычно одинаковой длины, и только у рода буревестников Puffinus
кисть короче плеча и предплечья, которые равны между собою. Второстепен-
ных маховых не более 25, а у мелких форм —качурок —только 13. На по-
лете мелкие формы совсем не парят, полет у них неровный, напоминающий
полет стрижей, ласточек; средней величины птицы—буревестники, глупыши
и др. — парят, но имеют и хорошо развитый активный полет. Задний палец
имеется. На земле передвигаются с трудом, опираясь на пальцы и цевку>
292 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
в особенности плохо ходят по земле некоторые мелкие формы. Ныряют плохо,
не глубоко и не с разлета, а мелкие формы совсем не ныряют. Наряд молодых
не отличим от взрослых. Гнездятся обычно колониями и чаще гнездо строят
в норах, щелях скал.
Всего в подсемействе 21 род с 85 видами, населяющими, главным образом,
Южное полушарие, кроме нескольких видов качурок и буревестников, обитаю-
щих в Северном полушарии, и живущего только в арктических морях глупыша.
II. Подсемейство альбатросы Diomedeinae.
Крупные птицы. Наружные носовые трубочки расположены по бокам
клюва, отделены друг от друга широким пространством хребта надклювья.
Плечо короче предплечья, кисть немного короче плеча и достигает 3/4 длины
предплечья. Второстепенных маховых не менее 30. Летают преимущественно
парящим полетом. Задний палец отсутствует, скрыт складкой кожи, иногда
виден только коготь. По земле передвигаются довольно хорошо. К нырянию
совершенно не способны. Гнездятся колониями. Гнезда строят открыто на по-
верхности земли. Наряд молодых отличается от взрослых, и полный взрослый
наряд молодые птицы одевают на 2—4-й год жизни.
В подсемействе насчитывается 2 рода с 14 видами, распространенными
на островах в субтропических и тропических океанах и Антарктике. У нас
встречается 3 вида, только на кочевках.
III. Подсемейство нырковые буревестники Pelecanoidinae.
Средних размеров птицы. Ноздревые трубочки очень короткие, распо-
ложены у самого основания клюва близ края оперения и открываются кверху.
Грудина узкая и удлиненная. Крылья короткие и довольно широкие. Летают
хуже других трубконосых птиц и большую часть времени проводят на воде,
а не в воздухе. Превосходно ныряют и плавают под водой.
Подсемейство содержит только 1 род с 5 видами, распространенными
в Южном полушарии и не встречающимися у нас даже случайно.
Географическое распространение. Один из наиболее широко распростра-
ненных отрядов, населяющий побережья почти всех частей света и большин-
ство океанических островов — от Арктики на севере до Антарктики на юге.
Особенно широко распространены во время своих кочевок, встречаясь во
всех океанах и во многих морях: Средиземном, Черном, Азовском, Красном,
Балтийском, Белом и др. Ряд видов, гнездясь в Южном полушарии, совер-
шает кочевки и миграции далеко на север.
Интересно свойственное трубконосым биполярное распространение неко-
торых близких родов и видов. Глупыш Fulmarus glacialis гнездится в Арктике,
доходя на север до Гренландии, Шпицбергена и Земли Франца-Иосифа,
а два близких к нему вида Priocella antarctica и Thalassoica antarctica гнездятся
по побережью Антарктического материка и на близлежащих островах. Близ-
кий к качуркам вид — океанник Oceanites oceanicus распространен по побе-
режью Антарктики и на прилегающих островах, северная качурка Oceanodroma
leucorrhoa и вилохвостая качурка Oceanodroma furcata — на Курильских, Ко-
мандорских и Алеутских о-вах и прямохвостая качурка Hydrobates peladicus
от Средиземного моря до Исландии и Норвегии F9° с. ш.).
Необходимо отметить наличие в Арктике и Антарктике преимущественно
монотипических родов (с единственным видом). Так, эндемичный род Арктики
глупыш Fulmarus имеет один вид glacialis. Из 4 эндемичных родов Антарктики
только один род океанник Oceanites имеет два очень близких вида oceanicus
и gracilis, остальные три — с одним видом: Pagodroma nivea, Priocella antarctica
и Thalassoica antarctica.
Всего в отряде насчитывается 24 рода и 94 вида (за исключением сом-
нительных). Большинство распространено в Южном полушарии, в тропиках
и субтропиках. В Арктике представлен один только вид трубконосых птиц,
в умеренной зоне Северного полушария —21 вид, в тропиках —23, в умерен-
ХАРАКТЕРИСТИКА ОТРЯДА 293
ной зоне Южного полушария — 63, в Антарктике — 4. В период кочевок
количество видов во всех зонах сильно меняется — увеличивается в числе.
Наиболее богат трубконосыми Тихий океан F8 видов); в Атлантическом
всего 40 гнездящихся видов. В фауне СССР встречается 13 видов, из них гнез-
дящихся 4, остальные 9 — случайно залетные и кочующие.
Надо еще отметить вымирание отдельных видов, например, Pterodroma
cahow, живший на Бермудских о-вах, Pterodroma caribbaea —на Ямайке,
Puffinus newelli — на Гавайских о-вах.
Палеонтологические данные. Всего описано 12 ископаемых видов:
7 из Европы, 1 из Африки, 4 из Сев. Америки. Из эоценовых отложений южн.
Нигерии по грудине описана гигантская птица Gigantornis eaglesomei, превы-
шающая по величине почти вдвое альбатроса. В верхнеолигоценовых отложе-
ниях Франции найдена птица Hydrornis natator. Из среднего миоцена Европы
описан Plotornis delfortii, напоминающий средних размеров альбатроса.
Современные роды найдены уже в миоценовых отложениях Сев. Аме-
рики—буревестники Puffinus (P. diatomicus, P. conradit P. inceptor); из
плиоценовых отложений Франции описан Puffinus arvernensis. Род Procellaria
описан из средне-миоценовых отложений Европы. Род альбатроса Diomedea
описан из плиоценовых отложений Англии —Diomedea anglica. Из плейсто-
ценовых отложений известны уже современные виды: Diomedea exulans, Dio-
medea chlororhyncha, Fulmarus glacialis, Puffinus puffinus, Puffinus griseus,
Puffinus kuhli и др.
Таблица для определения видов трубконосых
1B) Ноздревые трубочки расположены далеко друг от друга по обеим сторонам конька
надклювья. Задний палец отсутствует. Крыло длиннее 400 мм 3
2A) Ноздревые трубочки расположены рядом друг с другом на коньке надклювья.
Задний палец имеется. Крыло короче 400 мм 7
3D) Клюв светлый, массивный, длиннее 115 мм, у взрослых спина белая
белоспинный альбатрос Diomedea albatrus
4C) Клюв темный или с темной вершиной или основанием, менее массивный, короче
114 мм 5
5F) Оперение почти все бурое, белый цвет развит очень незначительно (кроющие хво-
ста, кольцо вокруг глаз) черноногий альбатрос Diomedea nigripes
6E) Оперение двухцветное: голова, низ тела и кроющие хвоста белые или белые с серо-
ватым налетом, остальное оперение бурое
темноспинный альбатрос Diomedea immutabilis
7(8) Клюв от вершины до переднего края ноздри не более 12 мм. Крыло не более
170 мм 9
8G) Клюв от вершины до переднего края ноздри не менее 17 мм. Крыло более
200 мм 15
9A0) Хвост прямой. Крыло менее 130 мм
прямохвостая качурка Hydrobates pelagicus
10( 9) Хвост вилообразно вырезан. Крыло более 135 мм И
11A2) Оперение серо-сизое сизая качурка Oceanodroma furcata
12A1) Оперение темнобурое, иногда с белыми верхними кроющими хвоста .... 13
13A4) На верхних кроющих хвоста и частично на нижних есть белый цвет ....
северная качурка Oceanodroma leucorrhoa
14A3) Белого цвета в оперении нет . .ма лая качурка Oceanodroma monorhis
15A6) Ноздри расположены в двух рядом лежащих, слитых у основания трубочках;
расстояние между трубочками не менее 1,5 лш. Клюв длинный, низкий; высота клюва рав-
няется приблизительно 0,3 длины клюва 19
16A5) Ноздри соединены в одну трубочку, разделенную внутри перегородкой, толщи-
ною в 0,5 мм. Клюв высокий и относительно короткий; высота клюва приблизительно рав-
няется 0,2 длины клюва 17
17A8) Крыло не более 240 мм. Киль вершины подклювья изогнут вниз
среднийтайфунник Pterodroma leucoptera
18A7) Крыло не менее 270 мм. Киль вершины подклювья направлен прямо кверху . . .
глупыш Fulmarus glacialis
19B0) Оперение сверху темнобурое, снизу белое или белое с буроватым налетом или
нестринами 23
20A9) Оперение все темнобурое 21
294
ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
21B2) Подмышечные светлосерые. Клюв короче 36 мм
тонкоклювый буревестник Puffinus tenuirostris
22B1) Подмышечные темнобурые. Клюв длиннее 37 мм
серый буревестник Puffinus griseus
23B4) Лоб, бока головы, темя, иногда и бока зоба с темными пестринами
пестролицый буревестник Puffinusleacomelas
24B3) На лбу, боках головы, темени пестрины нет, оперение однотонное
малый буревестник Puffinus puffinus
РОД АЛЬБАТРОСЫ DIOMEDEA LINNAEUS, 1758
Тип Diomedea exulans
1. Белоспинный альбатрос Diomedea albatrus Pall as
Diomedea albatrus. Pallas. Spicil. Zool. 1780, V, стр. 28, Камчатка и Курильские о-ва.
Русское название. Альбатрос — широко распространенное название, принятое на
всех европейских языках, происходит от арабского слова al-kadus,4ro означает бадья, повиди-
мому данное птице из-за ее манеры держаться на воде.
Распространение. Ареал. Северные субтропические и тропические об-
ласти Тихого океана, к югу приблизительно до 18° с. ш., к северу до Чукот-
ского моря. Гнездовье доказано только на о-ве Бонин B8° с. ш. и 140° в. д.)
Карта 52. Распространение белоспинного альбатроса Diomedea albatrus
а — границы гнездовий, г — область зимовок.
и о-ве Уэйк A9° с. ш. и 166° в. д.); в Смитсонианском институте имеется яйцо,
добытое на о-ве Уэйк (Брьюер и Риджвей, 1884).
Характер пребывания. Регулярно появляется летом в тихо-
океанских водах СССР. Обыкновенен на кочевках с марта по октябрь в запад-
ной части Тихого океана по побережью и у островов азиатского материка,
реже наблюдается в центральной и восточной части Тихого океана. По запад-
БЕЛОСПИННЫЙ АЛЬБАТРОС 295
ному побережью Сев. Америки встречается от Аляски до Сан-Диего (Калифор-
ния). Наблюдается чаще к северу от 50° с. ш.; от п-ова Кеней это обыкновенная
птица, кочующая летом у берегов Аляски в Беринговом море.
К северу на кочевках доходит до Чукотского моря. В сентябре 1939 г.
на траверсе м. Сердце-Камень в 60 км от берега М. М. Слепцовым встре-
чено 8 альбатросов, из них 2 добыто. Неоднократно наблюдался в Берин-
говом проливе в июле (Нельсон, 1886), у бухты Провидения 10 августа,
к северу от о-ва Лаврентия 18 сентября и между бухтой Провидения и о-вом
Лаврентия, 26 июля и 22 сентября близ о-ва Матвея и на Прибыловых о-вах
(Бент, 1922)» Обыкновенен летом на Алеутских о-вах от о-ва Атту до о-ва
Уналашки (Кларк, 1910), на Командорских о-вах, где известен со времен
Стеллера, и в июне встречен между островами Медным и Атту (Кларк, 1910).
Наблюдается у берегов Камчатки (первые указания имеются еще у Краше-
нинникова, 1755), причем отмечено, что по восточному побережью встре-
чается реже, чем по западному; на Охотском море; у берегов Сахалина (из-
вестен там со времен первых исследователей Дальнего Востока); на о-вах
Шантарских встречен 21 октября 1934, Дулькейт и Шульпин A937); в южн.
Приморье (Тачановский, 1893); у Курильских о-вов, по берегам Японии,
Кореи и Китая.
У берегов ю.-в. Азии, у о-ва Формоза, о-ва Хайнань и в северной части
Южно-Китайского моря белоспинный альбатрос встречается круглый год,
в особенно большом количестве —зимой. В меньшем числе встречается зимой
и северней — до южной окраины Алеутских о-вов, к востоку от Командорских
о-вов.
Время появления альбатросов у наших берегов точно не выяснено, но,
повидимому, тесно связано с очищением вод от льда. Так, у Командорских
о-вов первые птицы наблюдались в половине марта (Стейнегер, 1885), у о-ва
Атту — в конце марта 1881 г. (Турнер, 1886), у берегов Южн. Сахалина около
середины апреля (Супруненко, 1890). Причем, повидимому, обычно прилетают
отдельные особи или небольшие группы, в первую очередь появляются старые
птицы, а много позднее — молодые; иногда птицы появляются сразу в большом
количестве. Последнее явление может быть объяснено запоздалым освобожде-
нием воды от льда. Весьма вероятно, что такие ранние и дальние миграции
совершают не гнездящиеся почему-либо в данном году старые птицы и неполово-
зрелые молодые.
Биотоп. Открытые океанические воды, но посещает и открытые моря,
побережья, заливы и бухты. В период размножения — мелкие скалистые океа-
нические острова.
Численность. Всюду, исключая места гнездовья, немногочисленен,
но и не редок. Зимой в большом количестве наблюдается у юго-вост. бере-
гов Азии иу прилегающих островов, но по мере движения к северу, в особенности
у берегов и в морях, количество птиц резко убывает, у Алеутских о-вов зимой
встречаются только одиночные особи.
Летом в ю.-в. Азии наблюдается редко, но становится более обычным по
мере движения к северу и не только в открытой воде, но и по побережьям ы
в морях. В западной части Тихого океана довольно обыкновенен и многочие-
ленен. У западных берегов Сев. Америки только к северу от Ванкувера счи-
тается нередким видом, а от п-ова Кеней — обыкновенным.
Экология. Размножение. Сведений о размножении очень мало.
Моногамы, в период размножения образуют колонии. Повидимому, размно-
жаются не ежегодно, так как старые белые птицы появляются у Командорских
о-вов уже в половине марта (Стейнегер, 1885), и в большом количестве зимой,т. е.
в период размножения, наблюдаются у берегов ю.-в. Азии и в открытом океане.
Предположение, что белоспинный альбатрос размножается не ежегодно,
подтверждается и наблюдениями над размножением у других видов альба-
296 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
тросов. К размножению приступают, вероятно, не ранее 3-й зимы после вы-
лупления. Срок насиживания неизвестен. Гнездо устраивают на земле в виде
небольшого бугра, который делают, повидимому, сами птицы при помощи клюва
(Годман, 1910) из размягченной дождями почвы, перемешанной с раз-
личными травами, мхом и корнями. Одно и то же гнездо служит в течение
нескольких лет и ежегодно подновляется птицами, так что очень старые
гнезда приобретают большие размеры. Само гнездо представляет собой не-
большое углубление на вершине бугра, иногда совсем без подстилки, иногда
выстланное свежей и сухой травой (Бент, 1922).
Время кладки точно не установлено, но на о-ве Уэйк насиживающие птицы
найдены 20 декабря, на о-вах Бонин — в ноябре. Кладка состоит из одного
яйца. Яйцо овальной формы, почти одинаково закругленное на тупом и остром
концах. Скорлупа без блеска, шероховатая. Окраска яиц с о-вов Бонин —сли-
вочно-белая или грязнобелая с красновато-рыжими пятнами и крапинами
на тупом конце, иногда сливающимися на вершине тупого конца в «шапочку»
(Оте, 1901). Яйца с о-ва Уэйк белые или грязнобелые, без пятен (Брьюер
и Риджвей, 1884). Размеры яиц D5) 98,1—120,7 х 56—-86, в сред-
нем 108,8 х 70,3 мм (Бент, 1922). Высиживают, повидимому, самец и
самка, во всяком случае у гнезда в период насиживания наблюдаются обе
птицы.
Линька. Как и у других альбатросов, одна полная линька в году
после периода размножения. Последовательность смены нарядов и детали
линьки остаются неясными. Окончательный наряд надевается, повидимому,
в трехлетнем (или даже в четырехлетнем возрасте (Кассин, 1858; Годман,
1910). Вероятно у кочующих птиц линька бывает летом, так как у старой
самки, добытой 29 июня в проливе Кука, сменялись первостепенные махо-
вые, из которых некоторые только начали появляться. У другой старой
самки, добытой у о-ва Атту 15 августа, наружные первостепенные маховые еще
не вполне доросли, остальные маховые были уже свежие (Бианки, 1913).
Судя по самке, добытой 15 августа, можно предположить, что линька маховых
идет от внутренних маховых к крайним (наружным), а также, что период
линьки, повидимому, весьма продолжителен, и смена маховых занимает,
по крайней мере, три месяца (июнь, июль и август).
Питание. Не изучено. Вероятно, как и другие виды альбатросов,
белоспинные альбатросы питаются, главным образом, мелкими моллюсками,
обитающими в поверхностных слоях воды, кальмарами и другими беспозво-
ночными, рыбой и отбросами с судов, преимущественно китобойных. Пищу
берет, как и все альбатросы, только с поверхности воды, не ныряет за ней
и не схватывает добычу налету, а опускается для этого на воду. Пищу во время
кочевок добывает не только днем, но и ночью (Сленцов). Птенцов выкармли-
вают отрыгиваемой пищей.
Полевые признаки. Старые птицы хорошо отличаются от других видов
альбатросов, встречающихся в наших водах, белым оперением тела. От дру-
гих морских птиц белоспинные альбатросы легко отличимы своими боль-
шими размерами.
Хорошо летает и плавает, но не ныряет. Все время, за исключением пе-
риода гнездования, проводит в воздухе или на воде. Тип полета, как и
у других видов альбатросов, —парящий, длительный и быстрый. При полете
крылья с телом птицы образуют прямую линию, ноги вытянуты назад и сбли-
жены друг с другом. Взлетать с ровной поверхности земли не может и для
взлета должен взобраться на какое-либо возвышение или скалу и оттуда
броситься вниз. С воды взлетает без большого труда, но с разбегу —
птица бежит но воде, взмахивая раскрытыми крыльями и вытянув шею.
Деятельны, по крайней мере в период кочевок, не только днем, но и
ночью.
БЕЛОСПИННЫЙ АЛЬБАТРОС
297
Осторожней других видов альбатросов, редко приближаются к судам.
На кочевках обычно держатся одиночками, но при обилии пищи несколько
птиц (до 10, даже 15) собираются вместе.
Белоспинные альбатросы —молчаливые птицы, только на гнездовьях во
время кормежек птенцов слышны звуки, похожие на гоготание, а при ссорах
из-за добычи издают крики, похожие на рев ослов (Брьюер и Риджвей, 1884).
Описание. Размеры и строение. Длина крыла 560—530 мм B экз.), 575—
475 мм (9 экз., Бианки, 1913); длина клюва 139—136 мм B экз.); 157—119 мм (9 экз.,
Бианки).
Рис. 59. Белоспинный альбатрос
Оперение густое, плотно прилегающее к телу птицы. Крыло острое, первостепенных
маховых И, самое длинное 2-е, 1-е зачаточное, короче кроющего его пера; формула
2>3>4>5. Второстепенных маховых обычно 37, они очень короткие и едва выдаются
из-под кроющих их перьев. Плечевые перья сильно удлинены. Хвост короткий, несколько
округлен, рулевых 12.
Клюв сильный, немного длиннее головы, сжатый с боков и довольно высокий. Носовые
трубочки короткие, расположены у основания клюва по его сторонам. Заканчивается клюв
крючкообразными коготками надклювья и подклювья, образованными из самостоятельных
пластинок, причем коготок более сильно развит на надклювье.
Ноги короткие, цевка сжата с боков, сетчатая. Первый задний палец совершенно скрыт
складкой кожи.
Окраска. Пуховой птенец не описан.
В первом гнездовом годовом наряде оперение темнобурое с несколько более светлым
низом тела, с рыжеватым оттенком на верхней стороне тела и с черноватой головой и шеей.
Наружные опахала маховых чернобурые, внутренние несколько светлей. Стволы перво-
степенных маховых светложелтые. В обношенном пере окраска чуть бледней. Радужина
темнобурая. Клюв желтый, ноги бурые. У живых птиц клюв и ноги фиолетово-телесные,
коготок на клюве голубовато-белый (Стейнегер, 1898).
298 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Второй гнездовый наряд темнобурый с примесью белых перьев, развивающихся на перед-
ней части лба и под глазом. Брюшная сторона тела серая. Перья низа тела с белыми основа-
ниями; на груди и внутренних опахалах второстепенных маховых—с узкими белыми окайм-
лениями.
Третий гнездовый наряд темнобурый, с большим количеством белого цвета. Лоб, уз-
дечка, брови и щеки белые. Верх головы темнобурый. Перья верхней стороны тела темно-
бурые, с белыми выступающими основаниями, в особенности на нижней части спины и на
надхвостье, где образуют пестрины. Внутренние из второстепенных маховых и их кроющие —
белые. Брюшная сторона тела белая, с сероватым налетом на боках, шеи, груди, брю-
ха и подхвостья. Подмышечные и нижние кроющие крыла — белые с бурыми верши-
нами.
В четвертом гнездовом наряде верх головы, шеи и бока шеи пестро-темнобурые, испещ-
ренные золотисто-желтыми перьями. На спине много белого. Нет, впрочем, уверенности,
что за четвертый наряд принимались различными авторами крайние вариации окраски
третьего наряда. Промежуточные наряды между первым гнездовым и взрослым описаны
по Годману A910).
Во взрослом наряде оперение белое, за исключением темнобурых крыльев и хвоста.
Лоб, уздечка, щеки, спина, верхние кроющие хвоста и низ тела белые, с легким желтоватым
налетом на лбу, уздечке и щеках. Верх головы и шеи и бока шеи — светложелтые. Наружные
опахала первостепенных маховых и крайних второстепенных маховых черновато-бурые,
внутренние опахала — буроватые с серым налетом. Внутренние второстепенные маховые
в основной половине белые, в вершинной — бурые. Малые кроющие крыла и кроющие перво-
степенных маховых — бурые. Кроющие второстепенных маховых — белые. Плечевые белые,
с серовато-бурыми вершинами. Подмышечные и нижние кроющие крыла, кроме самых на-
ружных, образующих по краю крыла бурую полоску, белые. Рулевые черновато-бурые
с белыми основаниями. Радужина темнобурая. Клюв желтый (шкурки) или телесный с лило-
ватым оттенком (живые птицы); ноги бледнобурые (шкурки) или серовато-голубые (у живых
птиц, Бианки, 1913).
2. Темноспинный альбатрос Diomedea immutabilis Rothsch.
Diomedea immutabilis. Rothschild. Bull. Brit. Orn. Club, 1893, стр. 48, Лайзан.
Распространение. Ареал. Гнездится на Гавайских о-вах к северо-за-
паду от главной группы: о-ве Лайзан, о-ве Мидуэй, о-ве Френч-Фрейгет, о-ве
Некер, о-ве Бэрд, о-ве Лисянского; повидимому, на о-ве Нихау в центральной
части архипелага (Мунро, 1944). Гнездился ранее на о-ве Маркус, но был
истреблен в конце прошлого столетия японцами. Гнездование, отмеченное
близ нижней Калифорнии на о-ве Гвадалупе и о-ве Сан-Херонимо (Питере,
1931) очень сомнительно.
Характер пребывания. В СССР случайно залетная птица,
добыт один раз 16 августа в Камчатских водах (экземпляр в Зоологическом
институте Академии наук СССР). Кочевки происходят преимущественно
в центральной части Тихого океана на север нормально до 40° с. ш. Случайно
доходит к западу до Японии (добыт 30 сентября 1893 г. у Риу-Киу, о-в Миуко-
шима), к востоку —до нижней Калифорнии и к северу до Камчатки. К югу
граница кочевок не выяснена.
Биотоп. Как и у других альбатросов. Гнездовый биотоп — внутрен-
ние части мелких океанических островов, поросшие кустарниковой раститель-
ностью.
Численность. На некоторых островах гнездовые колонии темно-
спинных альбатросов весьма значительны. На островке Санд у Мидуэя гне-
здится до 20 000 птиц (Мунро, 1944). Неумеренное преследование вызвало
резкое падение численности альбатросов на о-ве Лайзан: в 1902 г. там было
не менее миллиона птиц (Фишер, 1904), в 1911 г. только 180 000 особей (Дилл
и Брайен, 1912).
Экология. Размножение. Моногамы, гнездятся колониями. На
места гнездовий прилетают в ноябре; прилет захватывает очень короткое
время — в течение двух-трех дней после прилета первых птиц вся гнездовая
колония уже собирается на острове. Сейчас же по прилете начинаются брачные
игры — «танцы», сопровождаемые разными звуками. На о-ве Лайзан излю-
ТЕМНОСПИННЫЙ АЛЬБАТРОС
299
бленным местом гнездования служит равнина внутри острова, поросшая
кустарником, окружающая лагуну (Фишер, 1904).
Гнездо устраивают или в виде неглубокой ямки в земле, окружен-
ного невысокой насыпью из песка, или в виде небольшого возвышения, которое
сооружают сами птицы при помощи клюва из земли, с углублением на вер-
шине его, служащим собственно гнездом.
В кладке одно яйцо. По окраске и форме яйца сильно варьируют — от
сильно удлиненных продолговатых до коротких и тупых. Размеры: C9) 99—
116 х 62—75, в среднем 109,2 х 69,4 мм (Бент, 1922).
Карта 53. Распространение темноспинного альбатроса Diomedea immutabilis
а — граница гнездовий, г — область зимовок.
Окраска яиц от грязнобелого основного тона с крупными и мелкими буро-
вато-рыжими пятнами, крапинами, расположенными, главным образом, на
обоих концах яйца, до буровато-желтого, без пятен и крапин.
Кладка происходит в конце ноября, птенцы вылупляются в феврале.
Период насиживания 63 дня (Мунро, 1944). Насиживают оба родителя. Пост-
эмбриональное развитие медленное, и весь гнездовый период тянется около
полугода. Выкармливают птенца и самец и самка полупереваренной пищей
путем отрыжки «ворванью». В возрасте около 4 месяцев птенцы достигают
размеров взрослой птицы, но еще не одеты пером. В августе, т. е. через 6 меся-
цев после вылупления, птенцы уже одеты в первый наряд и покидают гнездо-
вья. Улетают с гнездовий на кочевки вместе со взрослыми птицами. Таким
образом, темноспинные альбатросы на местах гнездовий проводят около
10 месяцев.
Линька. Не изучена. Характерны крайне слабые возрастные изме-
нения окраски (см. ниже).
Питание. На местах гнездовья пищу составляют преимущественно
головоногие моллюски (Ommastrephes oualaniensis, О. sloanei, Onychotenthis
300
ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
baenksi). Непереваримые части пищи выбрасываются в виде погадок (Фишер,
1904). На корхмежку в период выкармливания птенцов вылетают ночью, между
заходом и восходом солнца; после 9 часов утра птицы у мест гнездовий
редко наблюдаются в море. Возвратившись перед восходом солнца с моря,
взрослые птицы приступают к кормлению птенцов. По окончании кормле-
ния, около 9—10 часов утра, взрослые птицы и птенцы спят, сидя на земле,
засунув клюв и голову «под крыло».
Рис. 60. Темноспинный альбатрос
Полевые признаки. От других альбатросов отличается окраской опере-
ния, напоминающей в общем расцветку большой морской чайки Larus
marinas.
Молчаливые птицы, голос похож на гогот, кончающийся завыванием;
во время брачных игр издают свистящие и стонущие звуки: «ах-х-х-х» (Фи-
шер, 1904). У птенцов —высокий, нежный писк.
Описание. Размеры и строение. Размеры: длина крыла 483—470 мм, клюва
113—106,7 мм (Бианки, 1913).
Окраска. Пуховой птенец серовато-охристый, с более темным основанием пуха;
по мере роста пуха и обнашивания вершин пушинок птенец становится темнобурым. Раду-
жина бурая, клюв черновато-бурый (Ротшильд, 1900).
Гнездовый наряд похож на окончательный.
Взрослые самцы и самки окрашены следующим образом: голова, шея, поясница,
надхвостье, основания рулевых, подмышечные и вся брюшная сторона — белые. Остальное
оперение бурое, с легким сероватым налетом и с белыми окаймлениями перьев. Маховые
и рулевые несколько более темного бурого цвета с белыми основаниями перьев. Нижние
кроющие крыла, пестрые белые и бурые, с белыми вершинами. Радужина бурая. Клюв жел-
тый с более темной (зеленовато-серой) вершиной, ноги желтовато-серые.
ЧЕРНОНОГИЙ АЛЬБАТРОС
301
3. Черноногий альбатрос Diomedea nigripes Aud.
Diomedea mgn>sAudubon. Orn. Biogr. V, 1839, стр. 327, Тихий океан под 30°44'с. ш.
и 146° в. д.
Распространение. Ареал. Гнездится на о-вах Волкано, на о-вах Лай-
зан, Гаспар-Рико, Мидуэй, Френч-Фрейгет и на Маршальских о-вах (Бент, 1922).
Характерпребывания. В негнездовый период кочует в север-
ных и центральных частях Тихого океана, к югу до тропика Рака. В это время
регулярно появляется в советских водах Тихого океана. До последнего времени
северным пределом летних кочевок считались Алеутские о-ва, а на запад-
о-в Медный. Недавно отмечен в Охотском море (у северной окраины Сахалина),
Карта 54. Распространение черноногого альбатроса Diomedea nigripes
а — границы гнездовий, г — область зимовок.
по южному и восточному берегу Камчатки у м. Лопатка, у м. Шипунского,
в Олюторском заливе и в сентябре 1939 г. на траверсе Анадырского залива(Слеп-
цов). Таким образом, северная граница находится под 65° с. ш. Южней кочует
у берегов Курильских о-вов, где особенно часто встречается в четвертом про-
ливе (Слепцов), по восточным берегам Японии, наблюдался в северной части
Японского моря у о-ва Рисири (Слепцов) и у берегов Китая в Восточно- и
Южно-Китайском морях от провинции Чже-Цзяня до 20° с. ш. к югу. В откры-
тых водах Тихого океана встречается на кочевках к югу до о-ва Гонолулу
B2° с. ш.), о-ва Гвадалупы B9°с. ш.) и на восток до берегов Сев. Америки. У бе-
регов Сев. Америки наблюдается от Нижней Калифорнии C2°35' с. ш.) на се-
вер до о-ва Королевы Шарлотты (Барретт-Гамильтон, 1903) и может быть
несколько северней; кочует у Алеутских о-вов.
В юго-западных частях области кочевок у о-ва Формоза и в Южно-Китай-
ском море альбатрос встречается круглый год. В северных областях — с по-
ловины июля по октябрь. У Командорских о-вов встречается в августе, сен-
302 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
тябре и начале октября (две особи добыты 5 октября 1947 г. Слепцовым), в про-
ливе между о-вами Медным и Атту —в июне (Кларк, 1910), в Анадырском за-
ливе —в сентябре (Слепцов). У берегов Сев. Америки наблюдаются с февраля,
особенно многочисленен между маем и июнем (Иоком, 1947), отдельные особи
и поздней, у о-ва Уналашка встречен в октябре — ноябре (Барретт-Гамиль-
тон, 1903).
Биотоп. Открытые океанические воды, но не избегает и больших зали-
вов. Однако к побережьям близко не подходит. С сушей связан только в период
гнездования. Гнездовый биотоп — песчаные берега мелких океанических
островов.
Численность. В наших водах в наибольшем количестве наблюдается
у Командорских о-вов (к месту убоя кита слетаются 8—10 особей, Слепцов).
Нередок у берегов Японии и в большом количестве встречается в Восточно-
и Южно-Китайских морях, и у о-ва Формозы. В местах гнездовий у северо-
западных островов Гавайской группы, в негнездовое время наблюдается
в небольшом количестве. У берегов Сев. Америки обычен между 30—
50° с. ш.
Экология. Размножение. На места гнездовий появляются в сере-
дине октября — в начале ноября. Моногам. Гнездится колониями. Самые
крупные колонии находятся на о-ве Сульфур (группа о-вов Волкано), на о-ве
Лайзан и несколько меньшая на о-ве Френч-Фригейт. На остальных островах—
небольшие колонии. По прилете на места гнездовий начинаются брачные
игры, которые продолжаются в течение всего гнездового периода, а иногда
и после. Во время брачных игр две птицы становятся друг против друга,
раскланиваются, приподнимая перья на груди, трутся клювами. Затем расхо-
дятся и вновь приближаются друг к другу, повторяя сначала все проделанные
церемонии, и так происходит несколько раз. Затем одна из птиц, полураскрыв
крылья, запрокинув голову и вытянув клюв кверху, издает глухой, наподобие
стона, крик; при этом другая громко хлопает клювом. Иногда обе птицы
запрокидывают голову и кричат.
На о-ве Лайзан гнездятся по песчаным берегам острова, вглубь острова
не заходят (Фишер, 1904). Яйцо откладывают в ямку, сделанную в песке без
всякой подстилки.
В кладке одно яйцо грязнобелого цвета. Форма и скорлупа такая же, как
и у белоспинного альбатроса. Размеры: D5) 98,1—120,7 х 56—86, в среднем
108,8 х 70,3 мм (Бент, 1922). Самые ранние находки кладок —в сентябре,
но большинство в ноябре—начале декабря. Насиживают самец и самка.
Насиживание продолжается около шести недель (Ричарде, 1909).
На крыло птенцы становятся в возрасте около шести месяцев; с мест
гнездовий в июле отлетают старые птицы, а в августе — молодые (Мун-
ро, 1944).
Линька. Смена нарядов, как и у других альбатросов. Возрастные из-
менения незначительны.
Питание. На кочевках пища состоит из головоногих моллюсков,
главным образом, мелких, распространенных в поверхностных слоях воды,
и рыбы, в наших водах преимущественно морского ленька, морского ерша,
наваги и мойвы (Слепцов), и отбросов с судов (Барретт-Гамильтон, 1903;
Иоком, 1947). В районе китобойного промысла питаются выброшенными
внутренностями кита, к месту убоя слетается до 10 птиц (Слепцов). Пищу
не берет с лета, и как в тихую, так в бурную погоду обязательно опу-
скается на воду.
Полевые признаки. Летает и плавает так же хорошо и неутомимо, как и
другие виды альбатросов. По земле передвигается довольно неуклюже, но
без труда, опираясь на пальцы. Взлетать с ровной поверхности земли не может;
даже с воды, если птица сыта, взлетает тяжело. Обычно же взлет с воды до-
ЧЕРНОНОГИЙ АЛЬБАТРОС 303
вольно легкий (Барретт-Гамильтон, 1903). Держится одиночками юж неболь-
шими стаями. Менее осторожен, нежели белоспинный альбатрос, часто близко
подлетает к судам и сопровождает их. Стаи черноногих альбатросов наблю-
даются у места обильной пищи, например, к месту убоя кита слетаются 8—Ю
птиц (Слепцов). Активны, по крайней мере на кочевках, не только днем, но
и ночью (Слепцов).
Описание. Размеры и строение. Несколько меньше белоспинного альбатроса.
Длина тела 830—810 мм B самки); длина крыла 520—500 мм, среднее 510 мм (Зэкз.); размах
крыла 2000 мм A), длина клюва 103—100 мм C). Длина крыла 546—457 мм, длина клюва
109,5—95,3 мм C экз., Бианки, 1913).
Оперение густое, плотно прилегающее к телу. Крыло острое, первостепенных махо-
вых 11,1-е первостепенное маховое зачаточное, 2-е — самое длинное; самое короткое, не счи-
тая зачаточного, 11-е. Формула крыла: 2>3>4>5>6>7>8>9>10>ll.
Рис. 61. Черноногий альбатрос
Второстепенных маховых 37, они очень коротки. Плечевые перья сильно удлинены.
Хвост короткий, чуть округлый, рулевых 12.
Клюв сильный, умеренной длины, почти равен длине головы, сжатый с боков. Строение
клюва такое же, как и у белоспинного, но меньших размеров. Ноги короткие, сильные.
Цевка сжата с боков, сетчатая. Первый (задний) палец скрыт под кожей.
Окраска. Пуховой птенец не описан.
В гнездовом наряде спинная сторона тела темнобурая, более темная на голове. Бока
головы с серым налетом. Брюшная сторона темносерая, почти черная на зобе и серая на гор-
ле и подбородке. Крылья черновато-бурые с серым налетом на внутренних опахалах. Раду-
жина темнобурая. Клюв буровато-черный, темнеющий на вершине и у основания. Ноги
черные. После первой годовой линьки в оперении появляется белый цвет. Вокруг основания
клюва образуется узкое белое кольцо; под глазом белая полоса, идущая к уху. Среди верх-
них кроющих хвоста имеются отдельные белые перья с буроватыми вершинами, иногда
же они все бывают только белыми. Значительные индивидуальные вариации делают затруд-
нительным описание дальнейших возрастных изменений (Миллер, 1940). Весьма вероятно,
что после второй годовой линьки черноногие альбатросы надевают окончательный наряд.
Взрослые птицы. Спинная сторона тела темнобурая, верхние кроющие хвоста белые.
Брюшная сторона серая, светлеющая к середине брюха, нижние кроющие хвоста белые.
Вокруг основания клюва белое кольцо; лоб серовато-белый. Под глазом и за глазом
проходит белая полоса. Передняя часть щек и подбородок белые. Задняя часть щек, горло
и ушная область — серые. Шея и зоб — буровато-серые. Маховые, кроющие крыла и руле-
вые темнобурые, несколько темнее верха тела.
Радужина золотисто-бурая. Клюв темный, буровато-серый, основание клюва и вер-
шина — черные. Ноги черные. Половых различий в окраске нет.
304
ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
РОД ТАЙФУННИК PTERODROMA BONAPARTE, 1857
Тип Procellaria macroptera Smith
4. Белолобый тайфунник Pterodroma leucoptera Gould
Procellaria leucoptera. Gould. Ann. Mag. Nat. Hist., 1844, стр.364, Новый Южн. Уэльс.
Русское название. Тайфунник — слово книжное, от тайфун — буря.
Распространение. Ареал. Новый Южн. Уэльс, о-в Масафуэра, Новые
Гебриды, Фиджи, о-ва Бонин, о-в Лайзан и другие северо-западные острова
Гавайского архипелага; возможно о-ва Крузенштерна (Маршалльские о-ва).
Карта 55. Распространение белолобого тайфунника Pterodroma leucoptera
7. P. /. leucoptera, 2. P. I. masafuerae, 3. P. I. hypoleuca, 4. P. I. brevipes;
a — границы гнездовий, г — область зимовок.
Кочует, главным образом, во внутренних водах Тихого океана, доходя на
запад до Японии и на восток до Галапагосских о-вов и о-вов Хуан-Фер-
нандес.
Биотоп. Центральные части океана, близко к побережьям материков
не подходит; в период гнездования —мелкие океанические острова.
Подвиды и варьирующие признаки. Белолобый тайфунник объединяет 5 подвидов,
различающихся между собою деталями окраски оперения и размерами: P. I. leucoptera
Gould, 1844 — Новый Юж. Уэльс; Р. /. masafuerae Lonnberg, 1921 — о-в
Масафуэра, кочует у о-вов Хуан — Фернандес; P. I. hypoleuca S а 1 v i n, 1888 — о-ва
Бонин и сев.-зап."о-ваГавайского архипелага; P. LbrevipesP е а 1 е, 1848—о-ва Новые Гебриды
и Фиджи; кочует до Галапагосских о-вов; Р. /. longirostris Stejneger, 1893 — Тихий
океан; места гнездовий неизвестны.
БЕЛОЛОБЫЙ ТАЙФУННИК
305
Бонинский белолобый тайфунник Pterodroma leucoptera hypoleuca S a 1 v i n
Oestrelata hypoleuca. S a 1 v i n. The Ibis. 1888, стр. 359, о-ва Крузенштерна.
Распространение. Ареал. О-ва Бонин, Лайзан, Мидуэй, Лисянского
и другие, расположенные к северо-западу от Гавайского архипелага,
вероятно о-ва Крузенштерна (Маршалльские о-ва). В СССР залетная птица
добыта в августе 1909 г. на Сахалине (Бутурлин, 1936). На кочевках наблю-
далась у о-вов Риу-Киу, однажды — у о-ва Хондо.
Экология. Размножение. Плохо изучено. Гнездится колониями.
Моногамы. На гнездовья на о-ве Лайзан прилетают около середины августа
сразу тысячными стаями (Годман, 1910). Сейчас же по прилете приступают
Рис. 62. Белолобый тайфунник
к исправлению старых нор и постройке новых. Норы роются самими птицами
в песчаной почве по ночам, с наступлением утра птицы покидают острова
и улетают в море. Расположены норы на очень близком расстоянии друг от
друга, ход норы достигает около 2 м длины. Кладка на о-ве Лайзан начинается
в первых числах января. В кладке одно яйцо, белого цвета. Размеры: 47—50,1 мм
36,5—37,1 мм (Годман, 1910). Продолжительность насиживания и время вы-
лупления неизвестны. На о-ве Лайзан 18 мая были найдены уже полуоперив-
шиеся птенцы, вылезшие к этому времени из гнезд (Фишер, 1903), на о-вах
Бонин 16 июня еще встречались птенцы (Сибом, 1890), 2 июня вокруг о-ва
Лайзан в большом количестве еще держались птицы (Ротшильд, 1893). Таким
образом, на гнездовьях тайфунники проводят около 10 месяцев.
Линька. Не изучена. Одна полная годовая. 2 июня на о-ве Лайзан
добыты 4 линяющих птицы (Ротшильд, 1893).
Питание. Повидимому, основную пищу этой птицы составляют мел-
кие морские животные.
Полевые признаки. Мелкий буревестник общей сероватой окраски. В пе-
риод гнездования — ночная птица, в остальное время активен как днем, так
и ночью. Голос — пронзительный крик различных интонаций от низких
тонов до звонкого.
20 Птицы СССР. т. ТТ
306
ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Описание. Размеры и строение. Длина крыла 225—231 мм (Бианки, 1913).
Окраска. Пуховой птенец. Сверху светлосерый пух, снизу — чисто бе-
лый (Фишер, 1903).
Взрослые самцы и самки. Спинная сторона темносерая со светлосерыми окаймлениями
перьев, низ тела и основания рулевых белые. Верх головы черновато-бурый. Передняя
часть лба, уздечка, передняя часть щек и узкая полоса над глазами — белые. Маховые
черные с сероватыми внутренними опахалами. Второстепенные маховые серые с белыми
основаниями перьев. Верхние кроющие крыла с серым налетом и с белыми окаймлениями
на больших кроющих крыла. Подмышечные и нижние кроющие крыла белые с черным
ободком по краю. Рулевые черные с серым налетом.
Птицы в гнездовом наряде повидимому сходны со взрослыми.
Радужина темнобурая, клюв черный. Ноги голубовато-розовые, наружный палец и
первые фаланги среднего и внутреннего пальцев черные.
Примечание. На основании ошибочных данных Шлегеля (Mus. Pays-Bas VI,
ProcelL, стр. 13, 1863) в русской литературе приводится залетPterodroma breuipes Ре a le
на Камчатку.
РОД ГЛУПЫШ FULMARUS STEPHENS, 1826
Тип Fulmarus glacialis
5. Глупыш Fulmarus glacialis Linn.
Procellaria glacialis. Linnaeus. Fauna Suecica, 1761, стр. 51, Шпицберген.
Русское название. Глупыш — народное название", из-за доверчивости птицы. Упо-
минается еще Палласом A811).
Распространение. Ареал. От северо-восточных частей Сев. Америки и Грен-
ландии до Земли Франца-Иосифа и Новой Земли и от островов в Беринговом про-
Карта 56. Распространение глупыша Fulmarus glacialis
1* F- 8- glacialis, 2. F. g. rodgersii.
a — граница гнездовий, г — область зимовок.
ливе и море до Командорских, Курильских и Алеутских о-вов. На кочевках к се-
веру доходит до кромки льда, местамидо85°—86° с. ш. к югу—до 43°—32° с. ш.
Биотоп. Океанические и морские воды, в период размножения — ска-
листые, пустынные берега материков и островов.
Подвиды и варьирующие признаки. Объединяет два подвида, различающиеся между
собою окраской оперения^, g. glacialis Li n п., 1761 — сев. Атлантика и Северный Ледови-
тый океан, от северо-восточных областей Сев. Америки до Земли Франца-Иосифа и Новой
Земли; F. g. rodgersi С a s s i n, 1862 — северная Пацифика: острова в Беринговом проливе
и море, Командорские, Курильские и Алеутские.
Атлантический глупыш Fulmarus glacialis glacialis Linnaeus
Распространение. Ареал. Северо-восточные области Сев. Аме-
рики от о-ва Мельвиля и Земли принца Альберта на западе до о-ва Северный
Девон и проливов Джонса и Дависа на северо-востоке и Баффиновой Земли
глупыш 307
на востоке; Гренландия от 69° с. ш. на юге до 81° с. ш., на север по восточному
берегу и 76° с. ш. по западному берегу; Ян-Майен, Исландия, Фареры, Геб-
риды, Оркнейские о-ва, на юг до Шотландии и окружающих ее островов, о-в Мен,
западная и ю.-в. Ирландия, побережья Норвегии у 62°5'с. ш., о-ва Медвежий,
Шпицберген, Земля Франца-Иосифа до 81° с. ш., северный о-в Новой Земли
(залив Кривошеина на западном берегу северного о-ва Новой Земли, Русанов,
1524; северо-восточное побережье у м. Желания и несколько восточнее,
у залива Константинова, Антипин, 1938). Это, повидимому, самый восточный
пункт гнездования атлантического глупыша.
Характер пребывания. В СССР гнездится и кочует. Часть
птиц (холостые) и летом ведет кочевой образ жизни, встречаясь или вблизи
мест гнездовий или заходя летом далеко к северу. В Сев. Америке кочующие
птицы наблюдались 26 июня у Земли Гриннеля на 82°30' с. ш. (Филден, 1877),
в Гренландии летом птицы встречены на севере до 85° с. ш. (Бент, 1922); к се-
веру от Шпицбергена экспедиция Нансена 22 мая на 83°45' с. ш. наблюдала
первых глупышей; северней Земли Франца-Иосифа (экспедиция на ледоколь-
ном судне «Седов», 1937—1940) первый глупыш отмечен 31 мая на 85°29' с. ш.
и 77°29' в. д. и 9 июня —на 86°0,2' с.ш. и 71° 19' в. д. затем в течение июня —
июля наблюдались там несколько раз. В августе и 6 сентября встречены
на 8б°35' с. ш. и 44°48' в. д. и отмечены между 28 августа и 3 сентября на о-ве Уе-
динения на 71°ЗГ с. ш., 77°42' в. д. и около 86° с. ш. (Мон, 1879). В море Лапте-
вых наблюдался между 6 и 11 августа (Русская полярная экспедиция, Бируля,
1907), к северо-западу от Ново-Сибирских о-вов и к востоку от Северной
Земли (экспедиция на Фраме), первые глупыши наблюдались в 1894 г. 22—
26 июня, в июле и в августе, последние два месяца довольно часто, но в конце
августа совсем исчезли. В 1895 г. первые птицы отмечены 29 мая (экспедиция
на Фраме) на 82°26' с. ш. и в июле были там обычны, но не многочисленны,
последние птицы — 16 августа. В 1938 г. судном «Седов» встречены к северу
от Ново-Сибирских о-вов 3 июля на 81°2Г с. ш. и 137°37' в. д. и 15 сентября
на 83°26' с. ш. и 141°32' в. д. (Портенко, 1946). Восточнее атлантический
глупыш не наблюдался, здесь вероятно проходит восточный предел его коче-
вок. Осенние кочевки и отлет, повидимому, сильно зависят от времени насту-
пления холодов и образования ледяного покрова у берегов материков и остро-
вов. В среднем можно определить время отлета с середины сентября по сере-
дину октября. С о-ва Медвежий главная масса отлетает в середине сентября
(Кельхау, 1831). У берегов Шпицбергена последние птицы улетают в начале
2-й половины сентября A7 сентября, Плеске, 1928), но изредка наблюда-
ются еще и в конце сентября. От берегов Земли Франца Иосифа начинают от-
летать с первых чисел сентября A0 сентября 1894), и отлет идет до конца
месяца (Горбунов, 1932), но в некоторые годы они еще наблюдались в конце
октября B8 октября 1881, Плеске, 1928), северного о-ва Новой Земли
отлет начинается в конце сентября (Плеске, 1928) и продолжается до начала
2-й половины октября A8 октября наблюдался последний экземпляр, Ан-
типин, 1938), но в 1896 г. последняя пара отмечена 20 сентября (Бильке-
вич, 1904). У восточного берега Новой Земли и у Карских ворот кочующие
птицы встречались уже с 12 сентября и держались до 22 сентября (Гор-
бунов, 1929; Антипин, 1938).
Кочует у берегов Колгуева, Канина, в горле Белого моря и, повиди-
мому, в небольшом количестве доходит до Соловецких о-вов (добыт 18 марта
1935), по берегам Кольского п-ова, Норвегии, в Северном море и проникает
в юго-западные части Балтийского моря; у берегов Дании, Голландии, Англии
и Франции.
По восточному побережью американского материка доходит на кочевках
до Ньюфаундленда и Массачусетса, в открытых водах Атлантического оке-
ана—до 43° с. ш. (Бент, 1922).
308 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Биотоп. В период зимних кочевок — открытые воды океанов и морей,
в летнее время кроме того и прибрежные воды. Гнездовый биотоп — скали-
стые пустынные побережья материков и островов.
Численность. Обыкновенен на гнездовье по восточному побережью
Баффиновой Земли, многочисленен по западному побережью Гренландии,
в Исландии, на о-ве Ян-Майен, Медвежьем, на Шпицбергене. Обыкновенен
и местами многочисленен на Земле Франца-Иосифа.
На кочевках многочисленен у северо-восточных берегов Лабрадора, у Нью-
фаундленда, у берегов Гренландии, Исландии и Скандинавии, в особенности
у Лофотенских о-вов. Обыкновенен в Баренцовом море, в южных его частях
даже многочислененч Обычен у Новой Земли, но редок в восточной части Кар-
ского моря, у Ямала, Таймыра, Северной Земли, в море Лаптевых и у западных
берегов Ново-Сибирских о-вов, у последних обычно уже наблюдается более
или менее случайно.
Экология. Размножение. К местам гнездовий прилетают рано,
но время появления у мест гнездовий довольно сильно варьирует, повидимому,
в зависимости от освобождения прибрежных вод от льда и от дальности рас-
стояния кочевок. Так, у о-ва Ян-Майена и у берегов Исландии, вблизи кото-
рых они кочуют почти в течение круглого года, на гнездовья прилетают в се-
редине марта, в с. в. Гренландию под 79° с. ш. в середине апреля (Бент, 1922),
на о-в Медвежий прилетели в 1825 г., даже в середине февраля (Кейльхау,
1831). У берегов некоторых островов архипелага Шпицбергена птицы начи-
нают появляться с конца января (о-в Бунге, 22 января 1900 г., Бианки,
1913, о-в Амстердам 11 февраля 1899, Дитмар, 1901); между о-вом Амстер-
дам и северо-западной оконечностью Шпицбергена—20 февраля 1889
(Чапман, 1897); на о-ве Зап. Шпицберген прилет, отмеченный 4 марта 1873,
продолжался до начала апреля (Норденшельд, 1875). На Земле Франца-Ио-
сифа прилет глупышей наблюдался с 13 марта по 15 апреля (Горбунов, 1932),
на о-ве Гукера —во второй половине апреля (Демме, 1934), На северо-во-
сточное побережье Новой Земли первые птицы прилетают в начале апреля
B апреля на м. Желания, 6 апреля на о-в Гемскерк), массовое появление
наблюдается в конце апреля (Антипин, 1938). По прилете к местам гнездовий
глупыши не сразу появляются на суше, а некоторое время проводят в море
у самых берегов, иногда даже залетая на короткое время на берег.
Моногам. На места гнездовий прилетают уже в парах, и спаривание,
повидимому, происходит на кочевках, еще до прилета на места гнездовий.
Так, у добытых самок, прилетевших 20 апреля на о-в Гукера, яичники были
хорошо развиты и эмбрионы достигали 5 мм величины, но иногда пары обра-
зуются и после прилета, так на Новой Земле пары разбиваются только
в начале мая. По прилете на гнездовья у них начинаются брачные игры:
у о-ва Гукера по утрам в тихую погоду на воде наблюдалось, как глупыши
через каждые 2 секунды ритмически выпрыгивали из воды на 3/4 своего
туловища, затем расправляли крылья, кричали и бросались на воду, ударяя
по ней крыльями и передвигаясь несколько вперед, и после этого вся про-
цедура начиналась сначала (Демме, 1934).
Гнездятся обычно колониями, только изредка отдельными парами. Коли-
чество пар в колонии сильно колеблется и иногда колонии достигают громад-
ных размеров. Часто глупыши гнездятся в колониях совместно с другими пти-
цами: чистиками, кайрами, тупиками и различными видами чаек, в особен-
ности с моевкой. Колонии глупышей в течение нескольких лет занимают одни
и те же места, но иногда перемещаются с одного места на другое или даже с од-
ного острова на другой. Обычно гнездовая колония располагается у самой
воды на высоких, скалистых, голых и обрывистых берегах, реже — на берегу
у подножья скал, как исключение—в некотором отдалении от берега (о-в Мед-
вежий). Однако, высота скал, повидимому, большого значения не имеет.
глупыш 309
Так гнездовые колонии глупышей найдены в Гренландии на высоте 240 м
над уровнем моря на о-ве Медвежьем — у подножья скал, на Шпицбергене —
на 4500 м, на о-ве Западный Шпицберген —1200—1500 м, на Земле Франца-
Иосифа, на о-ве Гукера — на высоте 100—120 м над уровнем моря. Гнездо
устраивает открыто на плоском выступе скалы, на земле или даже на льду
(на о-ве Медвежьем найдены были яйца, отложенные на нестаявшем еще на
скале льду, Тобльзер, 1882; также на Новой Земле, Бианки, 1913). Устрой-
ство гнезда очень несложное — это неглубокая ямка в почве, или, если
грунт мягкий, то более глубокая, так что насиживающая птица до половины
скрывается в ней, выстлана она небольшим количеством травы. В некоторых
случаях ямка окружена мелкими камешками.
Время начала кладки для большинства мест гнездовий точно не устано-
влено. На о-ве Кильда кладка начинается около середины мая; в Исландии
кладка обычно происходит в середине ,мая, но иногда находили яйца уже
с середины апреля; на Фарерских о-вах, начиная с половины мая и поздней,
на о-ве Ян-Майене первые яйца найдены 17 апреля (Сконнинг, 1933); на
о-ве Медвежьем кладка идет с половины мая и до второй половины июня
в 1907 г. —в середине июня все яйца оказались ненасиженными, найден-
ные в 1908 г. в самом начале июля яйца были сильно насижены (Кениг,
1911), в некоторые годы сильно насиженные яйца встречали уже в конце июня
(Свенандер, 1900). На Шпицбергене кладка яиц идет с первых чисел мая до
середины июля. Первые кладки найдены в начале мая (Кениг, 1911), но 18—
19 июня находили только слегка насиженные яйца, и 14 июля отмечены совер-
шенно не насиженные яйца (Плеске, 1928), повидимому, кладки от 14 июля
случайные, сильно запоздавшие. О времени размножения на Земле Франца-
Иосифа нет никаких данных, исключая единственное упоминание о нахожде-
нии гнездовий и яиц на о-ве Мэбель 5 мая (Кларк, Плеске, 1928).
В кладке одно яйцо (но встречались изредка гнезда с 2 яйцами). Яйца
овальной формы, иногда более сильно заостренные на одном конце. Скор-
лупа пористая, без блеска, грязнобелого цвета с красновато-бурым крапом.
Яйца очень крупные. Размеры: E2) от 68,1—80 х 45,5—53,5 мм (о-в Мед-
вежий, Кениг, 1911); A3) 70—80 х 43,5—53 мм (Гренландия, Шалоу; 1895);
B9) 66—80 х 47—53 мм (Гебель). Среднее по 77 экз. 74 х 51 мм (Бент, 1922).
Вес 65,2—98,3 г (о-в Медвежий, Кениг, 1911); 76,5—93,6 г (Гебель); 88—
117 г (Хантцш, 1905).
Насиживают самец и самка, причем у обеих птиц образуются на брюхе
наседные пятна. Время насиживания точно не установлено: по одним источ-
никам 42—49 дней (Кениг, 1911), по другим 56—60 дней (Эванс, 1891). О времени
вылупления птенцов почти нет никаких сведений. В Исландии только что вылу-
пившиеся птенцы встречаются в начале июля, на о-ве Ян-Майене птенцы най-
дены 26 июня и также встречаются в гнездах в середине августа (Сконнинг, 1933),
на о-ве Медвежьем в первые дни июля найдены в гнездах птенцы различного
возраста (Свенандер, 1900) и 12 августа встречены там же нелетные птенцы (Цед-
лиц, 1911), на Шпицбергене 18и 19июня найдены птенцы в гнездах (Кениг, 1911).
Гнездовый период у птенцов продолжается 48—57 дней (Уайзерби, 1940).
На Фарерских о-вах птенцы становятся летными в середине августа, в Ислан-
дии—в начале сентября. О птенцах с о-ва Медвежий имеется ряд противоре-
чивых данных: 18—19 июля наблюдались уже летные (Мальмгрен, 1865)
с другой стороны, птенцы 12—19 августа были найдены еще в гнездах (Цедлиц,
1911; Свенандер, 1900). На Новой Земле в середине августа молодые уже пла-
вали, в начале сентября стали подниматься на крыло (Антипин, 1938). Вскоре
после того, как молодые птицы поднимутся на крыло, они вместе со взрослыми
покидают места гнездовий и начинают кочевать.
Линька. Раз в году, полная линька между июнем и ноябрем. Смена
первостепенных маховых идет от внутренних к крайним маховым, причем
310 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
часто неодинаковое количество маховых перьев выпадает на каждом крыле.
(У самки, добытой 8 сентября в Баренцовом море, на правом крыле старых
перьев было 6, а на левом — 7.) Линька идет очень бурно и одновременно на
каждом крыле может выпасть сразу несколько первостепенных перьев. Обычно
одновременно выпадают внутренние 6—7 перьев и птица продолжает летать
на 5—4 крайних маховых, в некоторых случаях остаются только — 3 маховых
(коллекция Зоологического музея Московского университета) или даже 2 край-
них первостепенных маховых (Уинн-Эдвардс, 1935), но это не лишает птицу
способности к полету. Рулевые перья сменяются очень быстро и одновременно
выпадают почти все (Кениг, 1911).
Время начала линьки мало изучено. Повидимому, начало линьки насту-
пает не одновременно у всех птиц. Часть птиц линяет очень рано, когда еще
гнездовый период у глупышей не закончен. Вероятно эти рано начинающие
линять птицы принадлежат к птицам, не приступавшим в этом году к размно-
жению. Так, у Шпицбергена в июле наблюдались птицы в разных стадиях
линьки: у некоторых линька маховых была в самом разгаре (Хейглин, 1871),
у других она к половине июля уже заканчивалась (у добытых 15 и 16 июля птиц
все маховые уже сменились и достигли нормальных размеров, но 2-е и 3-е пер-
востепенные маховые были еще кровянистые, Кениг, 1911). Из 3, добытых
в Баренцовом море птиц, у одной от 20 августа линьки нет, у 2, добытых
в августе и 8 сентября, шла линька маховых (не сменились в одном случае 4-е,
3-е, 2-е и 1-е первостепенные махи, в другом 5-е, 4-е, 3-е, 2-е и 1-е, остальных
маховых перьев нет), у добытого самца на о-ва Кильдин 10 сентября четыре
первых первостепенных маховых были старые, остальные свежие, но короткие,
и у птицы от 14 августа, добытой на о-ве Колгуев, имелось по 3 старых перво-
степенных маховых, остальные были свежие, но короткие. Также неравномерно,
повидимому, идет и смена мелкого пера: ряд птиц на Шпицбергене к половине
июля сменили почти все мелкое перо, только кое-где оставались в оперении
старые перья (Кениг, 1911). Рулевые линяют в августе; на о-ве Медвежьем
в августе встречалось много птиц с выпавшими рулевыми перьями (Свенан-
дер, 1900). Вероятно птицы, начинающие линять поздней — в августе, —
принадлежат к птицам, размножавшимся в этом году, и у них линька идет
до сентября и может быть даже поздней.
Питание. Основную пищу глупышей составляет рыба, икра рыб,
моллюски, ракообразные и другие беспозвоночные, падаль, внутренности
китов, различные жирные отбросы (во время китобойного промысла) и даже
помет китов и других крупных водных млекопитающих. Кроме
того, на гнездовьях поедают в небольшом количестве различные
растения.
Кормятся всегда в море, но не с полета, а спустившись на воду или на
трупы китов, моржей и т. д. За пищей птица не ныряет, а только опускает
голову в воду до глаз. Заглатывает добычу сразу, даже довольно значительных
размеров, с костями (Кениг, 1911, в желудках находил остатки костей кита
в 9 см длиною и в 3 см шириной), и только в том случае, когда куски очень ве-
лики, поедает их постепенно, расклевывая по частям. Во время кормежки часто
бывают драки. Глупыши весьма прожорливы и иногда наедаются до того,
что прежде, чем подняться в воздух, должны отрыгнуть часть добычи. Часто
большими стаями сопровождают китобойные судна, подбирая отбросы с них.
Хозяйственное значение. Мясо взрослых птиц и птенцов пригодно в пищу,
только очищенное от жира, который имеет неприятный запах. Птенцов соби-
рают, солят и в таком виде употребляют в пищу. В Исландии добывали
птенцов глупышей ежегодно до 30 000 (Фабер, 1822), также большой промысел
на них идет в Гренландии, на Фарерских и других островах. В Сев. Америке
добывают и тысячи взрослых птиц. Особенно в большом количестве собирают
яйца глупышей, которые очень вкусны. Кроме того, собирается пух птенцов
ГЛУПЫШ
311
и перо взрослых птиц и маслянистая жидкость из желудков птиц и птенцов,
которая идет для технических надобностей.
Полевые признаки. В полете так же неутомим, как и другие виды трубко-
носых птиц. Полет легкий и плавный. Летает во всякую погоду, обычно низко
над водой, следуя волнообразным изменениям поверхности моря. Часто парит
в воздухе, наклоняя свое тело то в одну, то в другую сторону и чередуя паре-
ние с быстрыми и короткими взмахами крыльев. Ноги при полете всегда вытя-
нуты назад. Хорошо плавает, перебирая по очереди ногами, и много времени
проводит на воде, кормясь и отдыхая. Спит на воде, спрятав голову под крыло
Рис. 63. Глупыш
или только клюв в перья спины, высоко подняв заднюю часть тела, а подбо-
родком упираясь на вздутый зоб.
На воде сидит высоко и с воды поднимается легко, нос небольшого раз-
бега. Обычно раненая птица старается спастись вплавь, помогая крыльями.
Держится как на воде, так и в воздухе стаями, иногда достигающими очень
больших размеров, только изредка — поодиночке. Любит купаться, и при этом
перья птицы так сильно намокают, что она часто не может подняться с воды.
По земле передвигается неуклюже, опираясь на цевку, но стоять может
и на пальцах. С земли на крыло поднимается с трудом, выбирая для этого
какое-либо возвышение — камень, скалу. Активны как днем, так и ночью.
В длинные зимние ночи севера наблюдались летающими над морем (Дитмар,
1901; Бианки, 1913). Летом в период размножения они в светлые ночи летают
на кормежку, а день проводят у гнезда.
Глупыши очень доверчивые птицы, человека не боятся как на гнез-
довьях, так и во время кочевок. Защищаются при помощи выбрасывания изо
312 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕРЕСТНИКИ
рта струи маслянистой прозрачной желтой жидкости «ворвани», которая обра-
зуется в желудке и скапливается там в большом количестве. При помощи дви-
жения головы и шеи глупыш отрыгивает ее из желудка в рот, а затем двойной
струей, достигающей 50 см длины, она выбрасывается наружу, причем,
выбрасывание наружу жидкости может повторяться несколько раз, но самое
большое количество жидкости выбрасывается в первый раз. У птенцов, достиг-
ших половинного размера взрослой птицы, жидкость уже имеется (Кениг, 1911).
Глупыши на гнездовьях и во время кормежек, когда собираются в стаи,
очень крикливы и шумливы. Голос у них низкий, грубый, слышный на боль-
шие расстояния и похож на клохтание кур или гоготанье гусей. Одиночные
птицы, спокойно сидящие на воде, иногда издают тихие кряхтящие звуки.
Описание. Размеры и строение. Длина тела у самцов и самок F) 450—480,
в среднем 468,5 мм. Длина крыла A7) 300—350, в среднем 332,5 мм. Размах крыла D)
1110—1140, в среднем 1125 мм. Разницы в величине между самцами и самками нет. Вес (б)
665—880, в среднем 764 г. Клюв умеренных размеров, сжатый с боков и с сильно выпуклым
и резко отграниченным крючкообразным коготком на верхней челюсти. Носовые трубки
лежат в основной трети клюва по срединной линии надклювья и соединены в одну общую
трубку с тонкой перегородкой внутри нее. Крылья широкие, длинные, с острой вершиной.
Первостепенных маховых И, из них самое длинное 1-е B-е). Хвост средних размеров,
закругленный, состоит из 14 рулевых. Ноги сильные, цевка короткая, несколько короче
пальцев, сильно сжата с боков, передние и задние края ее — острые, покрыта она
шестигранными пластинками. Пальцы тонкие, наружный палец почти равен среднему,
имеется задний палец. Три передних пальца соединены плавательной перепонкой. Когти
узкие, с острыми краями. Оперение густое, довольно жесткое, только на нижней стороне
тела несколько мягче.
Окраска. Глупыши имеют две резко различные вариации окраски: светлую и тем-
ную. Птицы промежуточного типа окраски встречаются редко. У птенцов уже в пуховом
наряде оперение различно и дает возможность определить, к какой вариации окраски будут
они принадлежать, став взрослыми. Гнездовые пары обычно состоят из одинаково окрашен-
ных птиц. В области распространения глупыша вариации окраски распределены неравно-
мерно: в ряде местностей встречаются птицы преимущественно только одной окраски, а птицы
другой вариации наблюдаются среди них лишь единичными экземплярами или даже совсем
не встречаются. В других местностях встречаются птицы и той и другой вариации окраски
в почти одинаковом соотношении. Это наблюдается как среди кочующих стай птиц, так и в
гнездовых колониях. Например, у северо-восточных берегов Сев. Америки и у западных
берегов Гренландии встречаются, главным образом, светлые птицы, только изредка, чаше
у берегов Лабрадора, наблюдаются темные птицы; в последние годы у северо-восточных
берегов Сев. Америки стали чаще встречаться и темные птицы (Тавернер, 1934). В централь-
ных областях Атлантического океана наблюдаются обычно светлые птицы, на Гельголанде —
преимущественно светлые птицы; из 17 добытых только 1 темная (Гетке, 1891). В Исландии
и темные, и светлые, но светлых больше, на о-ве Медвежьем гнездятся те и другие, в одних
колониях. На Ян-Майене гнездятся и светлые, и темные,то же на Шпицбергене; здесь наблю-
даются и переходные особи. На Земле Франца-Иосифа наблюдаются преимущественно тем-
ные птицы. У побережья Мурмана, Соловецких о-вов, в Баренцовом море, у Колгуева
и Новой Земли встречаются преимущественно светлые птицы (коллекции Зоологического
музея Московского университета и Зоологического ин-та Академии наук СССР).
В первом пуховом наряде у светлой вариации голова, шея и нижняя сторона тела
белые с бледносерым пятном по середине брюха. Верхняя сторона тела и крылья серые.
Темная вариация — дымчато-серая.
Во втором пуховом наряде у светлой вариации голова несколько темней, чем в первом
наряде, такого же цвета как и верхняя сторона тела. Птенцы темной вариации во втором
пуховом наряде не описаны.
Молодые птицы в гнездовом наряде похожи на взрослых.
Взрослые птицы светлой вариации. Голова, зашеек и брюшная сторона белые, иногда
с легким сероватогрязным налетом. Спинная сторона тела серая, иногда с буроватым
налетом. Перья спины с едва заметными более светлыми каймами. Маховые, верхние кро-
ющие крыла темносерые, причем первостепенные маховые более темные — с темносерым
наружным опахалом и серым внутренним, светлеющим к внутреннему краю. Подмышечные
и нижние кроющие крыла светлосерые, но изредка встречаются экземпляры и с белыми
нижними кроющими крыла и подмышечными, только нижние кроющие первостепенных
маховых всегда серые. Кольцо вокруг глаз и пятно за глазом темносерые. Рулевые одно-
цветно серые или темносерые.
Иногда встречаются экземпляры почти белые с едва заметным светлосерым тоном на
спине и темносерыми первостепенными маховыми и их кроющими; иногда встречаются экзем-
пляры ровного перламутрово-серого цвета с несколько более темной спиной и крыльями.
глупыш 313
Темная вариация: дымчато-серое оперение то с сероватым, то с буроватым оттенком.
Перья спины с чуть более светлыми каймами. Первостепенные маховые несколько темнее
общего оперения. Встречаются экземпляры с белыми каймами перьев верхней и нижней
стороны тела. Ноги различных оттенков от бледнооливкового до желтовато-телесного.
Радужина темнобурая, у чисто белых — желтая.
Окраска клюва сильно варьирует по сезонам года. Весною и летом клюв светлооливко-
вый с темными носовыми трубочками и краями щитков на клюве, причем чем больше серого
цвета в оперении (голова, зашеек, брюхо), тем темнее окрашен клюв (иногда в почти черный
тон, за исключением ноготка и трубочек). Осенью клюв темнею л ивковый, только ноготок
и иногда подклювье светлые.
Тихоокеанский глупыш Fulmarus glacialis rodgersii С a s s.
Fulmarus rodgersii. C a s s i n. Proc. Acad. Philad., 1862, стр. 326. Индийский океан ошибка
вместо Тихого океана.
Синоним. Fulmarus glacialis glupischa. Stejneger. The Auk., 1884, стр. 234,
Командорские о-ва.
Распространение. Ареал. Острова в Беринговом проливе и море:
о-в Лаврентия, о-ва Прибыловы, о-в Матвея, о-в Галл я и другие; возможно
Камчатка (Аверин, 1948, считает его гнездящимся, но гнезд не нашел; Бент,
1922, приводит с Камчатки 4 кладки яиц, собранные с 4 по 20 июня без точ-
ного указания места, но ни Стейнегер, 1885, ни Бергман, 1935, не приводят
глупыша в качестве гнездящейся птицы Камчатки); Сахалин, Курильские
и Командорские о-ва (гнездование на Командорских и Курильских о-вах из-
вестно еще со времени Крашенинникова, 1755, подтверждается и новейшими
исследованиями), Алеутские о-ва (Турнер, 1886) и ряд других авторов указы-
вают все о-ва Алеутской гряды; Бент, 1922, приводит гнездование только для
одного о-ва Семихи в группе Ближних о-вов Алеутской гряды.
Характер пребывания. Гнездящаяся и кочующая птица.
На кочевках к северу доходит до о-ва Врангеля и о-ва Геральда (Нельсон,
1883, 1887), к югу наблюдается в Беринговом проливе и море, вдоль берегов
Чукотки, восточного берега Камчатки и у прилежащих островов, в Охотском
море (в частности, по сообщению М. М. Слепцова, у о-ва Ионы и Шантар-
ских), у берегов Сахалина, к северу приблизительно до 56° с. ш. в проливе
Лаперуза, в северных частях Японского моря (Слепцов), встречен у берегов
Кореи, по восточному берегу Японии до Иокогамы (Кларк, 1910).
По американскому побережью кочует от южных частей Берингова моря
и Аляски к югу до северных областей Нижней Калифорнии. Кочевки у глу-
пышей отмечены в течение круглого года, так как вероятно молодые птицы
приступают к размножению не в первый год жизни и даже не все взрослые
птицы ежегодно размножаются. Эти холостые птицы и наблюдаются на кочев-
ках в разное время года, вероятно именно они наблюдаются далеко на севере.
Например, у о-ва Врангеля и о-ва Геральда встречены в августе (Нель-
сон, 1883, 1§87), у Чукотского п-ова у м. Сердце-Камень в первых числах
сентября, в проливе Синявского 11 сентября (Слепцов). Откочевки к югу
начинаются с первых чисел сентября, у берегов Японии близ Иокогамы
птицы наблюдались 11, 12 и 13 сентября (Кларк, 1910), у американских берегов
первые кочующие птицы встретились у о-ва Баранова 6 сентября, у берегов
Британской Колумбии с 30 октября по 10 ноября и у берегов Калифорнии
с половины октября. Область зимовок весною глупыши покидают в апреле;
последние птицы отмечены в Калифорнии у м. Пинос 15 апреля и в Сан-Диего
26 апреля. Только единичные случайные экземпляры остаются там летом,
так, например, у берегов Калифорнии наблюдались глупыши в июле —
августе (Бент, 1922). В северозападной части Японского моря глупыши
встречались уже в мае — июне (Слепцов).
Большинство птиц как гнездящихся, так, повидимому, и холостых не
откочевывает далеко от мест гнездовий, а лишь отлетает от берегов материков
и островов вглубь морей и океанов.
314 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Численность. Численность глупышей, повидимому, подвержена
сильным колебаниям. Например, у Алеутских о-вов он был очень многочисле-
нен в 80-х годах прошлого столетия, но с начала 900-х годов стал там попа-
даться крайне редко (Кларк, 1910).
Обычен у берегов Сахалина;но у северных частей его в Сахалинском заливе,
на Шантарских о-вах и в центральных частях Охотского моря глупышей зна-
чительно меньше, несколько больше их в районе о-ва Ионы. В северной части
Японского моря малочисленен, но уже у о-ва Ребун и о-ва Рисири и Лапе-
рузовом проливе его количество увеличивается. Многочисленен у Курильских
о-вов, Командор, в особенности у о-ва Беринга и у юго-восточных берегов
Камчатки, но по мере приближения к Берингову проливу численность глупы-
шей постепенно снижается; в Беринговом проливе глупышей больше в вос-
точной части, чем в западной (Слепцов).
Большие гнездовые колонии имеются на Курильских о-вах и на о-ве
Беринга.
Биотоп. Во время кочевок — моря и прибрежная зона океанов,
в период гнездования — скалистые пустынные морские побережья и океа-
нические острова.
Экология. Размножение. К местам гнездования глупыши при-
летают очень рано: у Командорских о-вов первые птицы появляются в конце
марта и в начале апреля (Иоганзен, 1934), но на самых гнездовьях появляется
несколько поздней — в середине апреля. Первое время по прилете на местах
гнездовья они не живут постоянно и к размножению не приступают, повиди-
мому, сначала на гнездовья прилетают самцы, так как из 20 птиц, убитых
23 апреля, была только одна самка (о-в Медный, Бианки, 1913).
Моногамы. Гнездятся колониями, иногда достигающими громадных раз-
меров, часто вместе с другими птицами, преимущественно, с различными
чистиковыми и некоторыми чайками. Колонии иногда перемещаются с одного
места на острове на другое или с одного острова на другой. Так, он был очень
многочисленен в 80-х годах прошлого столетия на некоторых о-вах Алеут-
ской гряды (о-ве Семихи, о-ве Уналашка и других, Турнер, 1885; Нельсон, 1887)
и затем в 900-х годах стал там крайне редок; в конце мая и в начале июня
1906, на о-ве Семихи наблюдались всего 3 птицы и очень мало на о-ве Уналашка
(Кларк, 1910).
Гнездовые колонии располагаются на обрывистых скалах различной
высоты, лежащих у самой воды. Например, на Командорских о-вах колонии
помещаются на высоте от 80—230 м над уровнем моря (Стейнегер, 1885),
на о-ве Матвея на высоте от б—85 м (Бент, 1922).
Брачные игры тихоокеанского глупыша отличаются от брачных игр
атлантического глупыша. Птицы закидывают головы кверху и медленно пока-
чивают ими из стороны в сторону и при этом издают каркающие звуки, широко
открыв друг перед другом клювы с ярко окрашенной ротовой полостью.
Время начала кладки точно не установлено, повидимому она происходит
в мае — июне, причем птицы не одновременно начинают откладку яиц, даже
в одних гнездовых колониях. На о-ве Медном 2 кладки добыты 14 мая (Бент,
1922) и 12 и 13 июля —слабо насиженные яйца (Стейнегер, 1885); в конце июня
и в июле на Командорских о-вах находили гнезда с яйцами (Иоганзен,
1934). На Камчатке с 4 по 20 июня добыты 4 кладки; на о-ве Семихи
(Алеутские о-ва) 20 июня и 1 июля взяты две кладки; на о-ве Матвея кладка
найдена 9 июля; на Прибыловых о-вах яйца находили с конца мая по конец
июня (Бент, 1922).
В кладке одно яйцо, похожее по форме и окраске на яйцо атлантического
глуныша.
Размеры: E) 48,5—51 х 68—75 мм (о-в Медный, Стейнегер, 1885),
A9) 68—77,5 х 48—52, в среднем 72,7 х 50 мм (Бент, 1922).
глупыш 315
Срок насиживания не выяснен, вероятно такой же, как и атлантического
глупыша. Птенцы вылупляются покрытые густым пухом. Время вылупления
неизвестно: на о-ве Мацува, Курильская гряда, 14 августа найдены в гнездах
птенцы различного возраста (Бергман, 1935), 22—25 августа на Курильских
же о-вах все птенцы были еще в гнездах (Стейнегер, 1898).
В гнезде птенцы проводят около б недель (Эллиот, 1882). На Коман-
дорских о-вах молодые птицы покидают гнезда в половине сентября (Иоганзен,
1934), но в 1908 г. 20—23 сентября на о-ве Медном половина птенцов сидела
еще в гнездах, повидимому из-за поздней весны (Бианки, 1913). Первое время
после спуска на воду молод ле птенцы не могут подняться с воды и проводят
все время на воде. В октябре они уже отлетают от своих гнездовий (Коман-
доры). В период размножения г~упыши ведут строго ночной образ жизни.
Линька. Линька негнездящихся птиц начинается раньше, чем у
атлантического глупыша, в мае —июле; добытый самец 21 июля в Олютор-
ском заливе имел только 5 первых старых первостепенных маховых, остальные
отсутствовали (коллекции Зоологического музея Московского университета).
Птицы, добытые в мае и июле на Курильских о-вах, находились в линьке,
(Сибом, 1885), и у птиц, добытых 15 июня на о-ве Беринга, сменились махи и
мелкое перо (Бианки, 1913). Но линька у размножающихся птиц вероятно
начинается поздней, чем у атлантического глупыша, так как у птиц, добытых
в августе и сентябре у Командор и в августе в заливе Корфа, линька
первостепенных маховых еще не начиналась и ыла только смена мелкого
пера. У птицы, добытой в декабре у о-ва Беринга, только что сменились
все маховые, и крайние еще не достигли нормальных размеров. Судя по
этим отрывочным данным, можно предположить, что линька маховых у взрос-
лых, гнездящихся птиц идет в октябре, ноябре и декабре и, вероятно, у разных
птиц в разное время.
Сменяются первостепенные маховые так же, как и у атлантического глу-
пыша от внутренних к наружным перьям.
Питание. Пищу этой птицы составляют мелкие кальмары, планк-
тонные рачки, навага, песчанка и другие рыбы и отбросы производства с
китобойных судов (Слепцов). Глупыши очень прожорливы, съедают иногда
мяса и граксы столько, что не могут взлететь в воздух и отплывают от судна.
Могут заглатывать очень большие куски мяса или жира, до 400—500 г. Перева-
ривание происходит быстро, так что через 1—2 часа из стаи сытых птиц отде-
ляются одиночные глупыши, которые снова принимаются за кормежку. На-
брасываясь на выброшенную с судов добычу, глупыши быстро расталкивают
ногами в стороны жирные куски граксы, при этом сами птицы немного подни-
маются над водой, что предохраняет их от засаливания пера. Во время кормежки
часто происходят драки между птицами и они тогда издают короткие глухие
звуки (Слепцов). Собираясь в тысячные стаи, сопровождают китобойные судна,
подбирая выбрасываемые с них отбросы, граксу, содержимое желудков китов
и т. д. В период размножения уходят за флотилиями судов не более 30—40 км
от берега, по окончании гнездового периода — до 180 км.
В поисках пищи глупыши низко летают над водою и в тихую, и в штормо-
вую погоду. Временами взлетают вверх до 20—30 м, затем по косой линии
планируют к поверхности воды и, пролетев 40—50 м над самой водой, взлетают
снова. Такой характер полета, повидимому, помогает им высматривать добычу
(Слепцов).
Описание. Размеры и строение. Длина тела у самцов и самок A0)
450—490, в среднем 473,3 мм. Длина крыла A9) 290—340, в среднем 320 мм.
Окраска. Так же, как и у атлантического глупыша имеются две вариации окраски:
темная и светлая. Распределение птиц той и другой вариации неравномерно. В общей форме
можно отметить, что на юге, примерно до Олюторского залива, преобладают темные, на
севере —светлые птицы.
316 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Пуховой наряд. Светлая вариация белая — темная вариация не описана. Гнездовий
наряд — сходен с окончательным.
Взрослые самцы и самки светлой вариации отличаются от атлантического глупыша
светлой вариации меньшим развитием серого или серовато-бурого налета на голове,
зашейке и брюхе, белыми нижними кроющими крыла и подмышечными. Светлый край внут-
реннего опахала первостепенных маховых белый или почти белый. Рулевые двухцветны:
основание серое, вершина или вершинная половина темносерая с бурым оттенком. Темная
вариация: все оперение дымчато-серое с сероватым налетом, темнее, чем у атлантиче-
ского глупыша темной вариации.
Клюв светлый, варьирует от желтовато-оранжевого до желтовато-лимонного с темными
краями трубочек, и иногда с темным пятном у основания трубочек. Радужина темнобурая.
Ноги светлые с различными оттенками: голубоватым, зеленоватым, телесным.
РОД БУРЕВЕСТНИКИ PUFF IN US BRISSON, 1760
Тип Procellaria puffinus
6. Серый буревестник Puffinus griseus Gm.
Procellaria grisea. G m e 1 i n. Syst. Nat. 1789, стр. 564, Южное полушарие между 35° и
50°ю.ш.
Синоним. Neonectris griseus missus. Mathews et Iredale. The Ibis, 1915, стр. 603,
Курильские о-ва.
Русское название. Буревестник — широко распространенное название, свя-
занное с тем, что птицы бывают особенно активны в ветренную погоду. Видовые названия —
книжные.
Распространение. Ареал. Гнездовий—в Южн. полушарии. Отдельные
колонии по берегам Новой Зеландии и на мелких островах у ее берегов;
о-в Чатам, о-в Стюарт, о-в Снарс, Орклендские о-ва, о-в Кемпбел, о-в Мак-
куари, о-в Норфольк, о-в Капити, о-в Стефенс, некоторые из островов группы
Боунти и другие; Южн. Америка: острова в Магеллановом проливе, м. Горн,
Фалькландские о-ва, отдельные колонии по западному берегу и прилежащим
островам Южн. Америки до сев. Чили. Вне гнездового времени на кочевках
встречается в Тихом и Атлантическом океанах. Область кочевок тянется к
югу приблизительно до 60° ю. ш. В Тихом океане наблюдается на кочевках
в западной части Тасманского моря, у берегов Южного Нового Уэльса. Далее
к северу не отмечена ни в центральных, ни в северных внетропических водах
Тихого океана, ни по побережью ю.-в. Азии и прилежащих к ней островов.
Начинает вновь встречаться буревестник у Курильских о-вов. Причем здесь
и северней отмечены лишь единичные встречи и находки (Кларк, 1910;
Аверин, 1948). В последнее время, начиная с 1939 г., серый буревестник
стал встречаться в морях Дальнего Востока в большом количестве; в июле
1939 г. наблюдался в Охотском море и в Сахалинском заливе, в августе
того же года —в Охотском море у первого Курильского пролива, летом и
в начале осени 1947 г. —у Командор, вдоль берегов Камчатки и Чукотки
до Анадырского залива (Слепцов). Добыт между Камчаткой и Командорами
4 октября 1947.
У тихоокеанских берегов Сев. Америки серый буревестник встречается
от Ситки и может быть Алеутских о-вов до нижней Калифорнии. У берегов
Южн. Америки кочует на север до Перу. В прибрежных атлантических водах
южноамериканского материка встречается у берегов Уругвая, и у берегов
Сев. Америки от Каролины до Лабрадора (м. Чидли) и о-ва Революшен
(Тавернер, 1934). Кочует у южной оконечности Гренландии у м. Фаруэлл,
у с.-в. и ю.-в. Исландии, у Фарерских о-вов, Оркнейских и Гебридских.
Отмечен у юго-западного побережья Норвегии. Неоднократно наблюдался
у о-ва Гельголанд, у берегов Голландии, Великобритании, Франции, Порту-
галии, па атлантическому побережью Африки от Золотого берега до м. Доброй
СЕРЫЙ БУРЕВЕСТНИК
317
Надежды. В центральных, тропических и в субтропических водах Атланти-
ческого океана и в Индийском океане не встречается. Имеется указание
лишь на единственную встречу 16 октября 1868 г. к югу от Мадагаскара
(Лейард, 1863).
Даты. Время встречи на кочевках захватывает почти все месяцы года
(нет наблюдений в феврале, марте и апреле), но чаще — наблюдается у нас
в летние и осенние месяцы с мая по октябрь и даже в ноябре. У тихоокеан-
ского побережья южн- Африки и у южной оконечности Нижней Калифорнии
встречаются птицы в наши зимние месяцы с декабря по май, т. е. период раз-
множения серого буревестника (повидимому, холостые).
Трудно определить, в каком направлении идут кочевки и миграции:
наблюдения над ними отрывочны и случайны. У нас встречены на Курильских
Карта 57. Распространение серого буревестника Puffinus griseus
а и б — границы гнездовий, г — область зимовок.
о-вах с мая по август, в Охотском море с июля — августа, по побережью
Камчатки и Чукотки с июля по октябрь, у тихоокеанского побережья Сев.
Америки, за исключением южной ее части, с мая по октябрь. У атлантического
побережья Сев. Америки в мае — июне и с августа по октябрь, у южной
оконечности Гренландии — в июле, у берегов Исландии в июле — августе,
у Фарерских о-вов — в августе, на о-ве Гельголанде — с мая по октябрь (всего
встречены 2 раза), в Голландии — в октябре, у берегов Англии в августе,
сентябре и октябре, в Португалии — осенью. У атлантического побережья
Африки — с мая по сентябрь, и у атлантических берегов Южн. Америки,
у Уругвая — в июне и в сентябре.
Численность. В большом количестве встречен Слепцовым в по-
следние годы в наших дальневосточных морях, обычен у берегов Калифор-
нии, у Ситки редок. Многочисленен у атлантических берегов Южн. Америки,
обычен у юго-зап. оконечности Африки, но северней устья Оранжевой реки ста-
новится редким. Также обычен в северной части Атлантического океана вдоль
северо-американского и европейского материков, но у Лабрадора, в Гренлан-
дии и Исландии редок.
Биотоп. Прибрежная полоса океанов и морей в период кочевок,
каменистые пустынные берега материков и островов в период размно-
жения.
318 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Экология. Размножение. Мало изучено. Гнездятся колониально.
На гнездовьях появляются, повидимому, в различное время, на разных остро-
вах с конца октября до начала ноября. Однако в одну колонию птицы при-
летают почти одновременно. Прилетевшие в колонию буревестники при*
ступают к рытью норы в торфяной почве острова (Оливер, 1930). Нора
бывает длиною в 90—120 см (о-в Чатам), идет в начале прямо, затем несколько
изгибается в стороны и на конце образует расширение. В этом расширении
и помещается собственно гнездо, устланное небольшим количеством сухих
листьев и травой. После того, как птица сделает гнездо, она улетает обратно
в море. Возвращаются птицы только через несколько дней и сейчас же все
Рис. 64. Серый буревестник
откладывают свое единственное яйцо, дата кладки в колонии совпадает из года
в год. На о-ве Стюарт, например, кладка яиц происходит 25 ноября (Оливер,
1930) и только как исключение яйца бывают отложены поздней. В общем
период размножения падает на ноябрь — декабрь.
Форма яиц варьирует от закругленно-овальной до удлиненно-овальной.
Скорлупа крупно-зернистая, белая. Размеры: C4) 58,5—81,7 х 42—53,4,
в среднем 74 х 48 мм (Бент, 1922). Вес яйца от 69,1 до 77,75 г (Рейхенов, 1906).
Высиживают оба родителя. Время насиживания точно не установлено,
приблизительно 30—35 дней. Так, на о-ве Стюарт молодые вылупляются
в конце декабря (Оливер, 1930), одетые густым длинным пухом. В гнезде
остаются долго, до 5 месяцев, пока не сменят пуховой наряд на перо, что
обычно происходит в конце апреля. К этому времени родители перестают
кормить молодых птиц и они, побуждаемые голодом, выходят из нор и пере-
бираются в море. В течение мая все птицы покидают гнездовья и начинают
вести кочевой образ жизни.
Линька. Полная годовая, детали неизвестны. По Уайзерби A940)
между февралем — сентябрем и даже октябрем, т. е. после окончания размно-
жения.
ТОНКОКЛЮВЫЙ БУРЕВЕСТНИК 319
Питание. Мелкая рыба и различные мелкие ракообразные, моллюски
и другие планктонные животные, которых они собирают с поверхности воды
или ныряя за ними (Оливер, 1930), но ныряют не глубоко и не надолго. Во
время кочевок обычно отыскивают пищу в открытом море. Серый буревестник
в дальневосточных морях часто следует за китами, главным образом серыми,
питаясь отрыгнутой ими пищей, состоящей из мелких рачков и кальмар,
и выбирая также из экскрементов китов мелких рачков. Кроме того, киты,
всплывая из глубины, взбалтывают воду, выбрасывая на поверхность ее мел-
ких рыб и ракообразных, которых тут же хватает буревестник. В наших
водах особенно часто они наблюдаются в местах скопления планктонных
рачков и косяков мойвы, к промысловым судам подлетают редко (Слепцов).
Полевые признаки. Размеры средние, меньше глупыша. Окраска одно-
образная бурая. Хорошо летают и плавают. Обычно летают низко над водой,
почти касаясь ее крыльями. В тихую погоду характерны быстрые взмахи
крыльев, в ветреную погоду буревестник после частых взмахов крыльями пере-
ходит на парящий полет над волнами. С воды поднимается легко. На воде сидят
компактной группой, иногда цепочкой, у Командор на кочевках наблюда-
лись Слепцовым стаи до 400—500 особей. По земле передвигается неуклюже
и с ровной поверхности земли подняться не могут, для этого им необходимо
какое-либо возвышение.
Активны в течение круглых суток. В период кочевок, в лунные ночи
в дальневосточных морях наблюдаются стаи кормящихся птиц (Слепцов).
В период гнездования — это чисто ночная птица. Выкармливание птенцов про-
исходит ночью, день взрослые птицы проводят на море, отдыхая и кормясь.
На кочевках довольно молчаливы, на гнездовьях ночью во время кормления
птенцов и взрослые птицы и птенцы издают неприятные громкие, напоминаю-
щие плач крики.
Промысловое значение. Промысловое значение имеют только на местах
гнездовий, где собираются яйца, птенцы, которых употребляют в пищу,
а также и взрослые птицы ради их мяса, а главное — перьев.
Описание. Размеры и строение. Длина тела A) 480 мм; длина крыла самцов
и самок B) 290—310 мм, размах 950 мм A). Длина клюва B) 40—42 мм. Первостепенных
маховых 11. Формула крыла: 1B)>2C). Рулевых 12, хвост округлый, умеренной длины.
Клюв сжат с боков, вершина надклювья образует крючок. Носовые трубочки короткие,
слиты в одну, разъединенную внутри на две толстой перегородкой; расположены по коньку
надклювья у основания клюва.
Цевка сжата с боков с острым передним краем и покрыта многогранными щитками.
Три передних пальца соединены мало вырезанной, но узкой перепонкой. Задний палец
очень небольших размеров и прикреплен выше других. Оперение густое и довольно жесткое,
в особенности на верхней стороне тела.
Окраска. Пуховой птенец темный аспидно-серый несколько беловатый снизу
(Оливер, 1930).
Молодые птицы окрашены, повидимому, сходно со взрослыми.
Взрослая птица. Верх тела темнобурый, низ буровато-серый. Подбородок, горло и
нижние кроющие крыла серые с более темными пестринами на последних. Подмышечные
темнобурые. Радужина темнобурая. Клюв черный. Внутренняя сторона цевки, средний и
внутренний пальцы голубоватые. Наружная сторона цевки и наружный палец темнобурые
или черные.
Систематические замечания. Некоторые авторы выделяют южно-американских птиц
в особый подвид P. g. chilensis Bonaparte, 1856, но у нас для решения этого спорного
вопроса нет материала и мы присоединяемся к мнению большинства, что P. griseus не
имеет подвидов.
7. Тонкоклювый буревестник Puffinus tenuirostris Те mm.
Procellaria tenuirostris. Temminck. PI. Col., livr. 99, 1835. табл. 587, моря к северу
от Японии.
Распространение. Ареал. Гнездится в юго-зап. областях Тихого океана:
берега ю.-в. Австралии, Тасмании, о-ва в Бассовом проливе (Филипп, Флин-
дерс, Кинг, Зеленые, Биг-Дог и др.). Сведения о гнездовании на о-ве лорда
320
ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Гау, получившие в литературе широкое распространение, ошибочны и основаны
на смешении с P. carneipes (Хиндвуд, 1940). Область кочевок—в Тихом
океане, на севере до Берингова моря. Вдоль азиатского побережья наблюдается
на пролете в весенние и летние месяцы и только как исключение осенью (един-
ственная находка у южн. Курильских о-вов 30 сентября (Кларк, 1910). Встре-
чен у о-ва Бонин и в Японии. Осенние кочевки на юг проходят, наоборот,
вдоль американских берегов. В СССР с Курильских о-вов известен еще со
времен Крашенинникова A755), причем здесь он чаще встречается у северных
о-вов, чем у южных. Регулярно встречается на Командорских о-вах, с апреля
Карта 58. Распространение тонкоклювого буревестника Puffinus tenuirostris
а — граница гнездовий, б — стрелками обозначено направление зимних кочевок.
по август (Иоганзен, 1934 и др.). На Камчатке добыт в бухте Ольга 14 июня
(Аверин, 1948) и в Авачинской губе 25 апреля (Бианки, 1913), в Анадырском
заливе в середине июня (Портенко, 1939), в бухте Провидения 10 июля
(Бианки, 1913), наблюдался к сев.-зап. от Берингова пролива (Нельсон, 1883)
и обычен летом у берегов о-ва Врангеля (Портенко, 1937).
По берегам сев. Аляски редок и может быть лишь случаен, встречен только
два раза летом в заливе Коцебу. Обычен у Алеутских о-вов с конца июля и до
сентября (Бент, 1922), у берегов южн. Аляски с июля по октябрь, у берегов
Британской Колумбии в конце октября, у Соединенных Штатов с августа по
ноябрь, в Калифорнии с середины октября до конца января, особенно в ноябре
и декабре (Бианки, 1913; Бент, 1922).
Таким образом, у берегов Сев. Америки наблюдается на пролете и кочев-
ках осенью и в первой половине зимы.
У берегов Цент, и Южн. Америки совсем не встречается, также его не
наблюдали в центральных областях северной части Тихого океана.
ТОНКОКЛЮВЫЙ БУРЕВЕСТНИК 321
Имеются сведения о встречах тонкоклювого буревестника в осеннее и
главным образом в зимнее время в юго-зап. Полинезии у о-вов Товари-
щества, Туамоту и Самоа до Новой Каледонии (Годман, 1910), наблюдался
у о-вов Кермадек у берегов Новой Зеландии, в особенности у северного ост-
рова. Однажды встречен между Новой Померанией и Новым Мекленбургом.
Биотоп. Морские и океанические пространства прибрежных вод; в пе-
риод размножения —скалистые пустынные берега материков и мелкие острова.
Численность. На весеннем пролете в западной части Тихого
океана малочисленен, у берегов о-ва Врангеля и у Алеутских о-вов летом
обычен, в большом количестве встречается на осенних кочевках вдоль
Калифорнийских берегов. Очень многочисленен у мест гнездовий в течение
сентября — мая.
Экология. Размножение. Моногамы. На местах гнездовий в Бас-
совом проливе появляются с первых чисел до половины сентября. Подроб-
ные сведения о гнездовом периоде собраны Монтгомери A892) и Годман
A910). Гнездятся колониями, достигающими громадных количеств—до
нескольких сот тысяч на каждом из отдельных мелких о-вов Бассова пролива.
Гнезда устраивают в норах, которые роют сами птицы сейчас же по прилете.
Рытье нор происходит по ночам, день птица проводит в море. На рытье норы
уходит времени от 10 до 40 дней. По окончании рытья норы, приблизительно
около 1 ноября, все птицы покидают сушу и держатся в море около 20 дней.
В среднем 20 ноября, но никогда не раньше 18 ноября, птицы появляются на
гнездовья десятками тысяч, почти все в один день. Единственное яйцо откла-
дывают почти все птицы в один и тот же день; на различных островах время
кладки лишь незначительно отличается. Несмотря на то, что вся поверхность
острова изрыта норами, расположенными на близком расстоянии друг от
друга, многим птицам нехватает нор, и они откладывают свое яйцо на поверх-
ности земли. Так, на Зеленом о-ве (Бассов пролив) только У* прилетевших
птиц могла отложить яйцо в норы, 3/4i отложили свое яйцо на поверхности
земли, причем отложенные яйца лежали так близко друг от друга, что трудно
было при ходьбе не наступить на них.
Яйцо овальной формы, белое. Скорлупа гладкая, но без блеска. Размеры:
D0) 63,5—77,2 х 41—51,5, в среднем 71 х 47 мм (Бент, 1922).
Насиживают оба родителя 55—56 дней. Птенцы вылупляются весом около
15,1 г, одетые густым пухом. Подросшие пуховые птенцы очень жирны и скоро
достигают размеров взрослой птицы, а по весу превосходят их. Приблизительно
через 2 месяца, во второй половине марта, они начинают одеваться в перо и в
конце апреля могут уже подниматься на крыло. Вполне оперившихся птенцов
родители оставляют на гнездовье; птенцы, побуждаемые голодом, выходягг
из нор и самостоятельно переселяются в море в конце апреля или в начале мая
всегда ночью и почти одновременно все молодые птицы, образуя громадные
скопления в Бассоном проливе. Но здесь они держатся короткое время и скоро
разбиваются на более мелкие стаи и покидают побережья гнездовий.
Линька. Плохо изучена. У экземпляра, добытого 25 июня на Аляске,
шла сильная линька мелкого пера, у добытого в июле у о-ва Кадьяк мелкое
перо было уже свежее, сменялись маховые и рулевые, из первостепенных
маховых 3 были еще очень короткие (Бианки, 1913). У экземпляров, добытых
в декабре, линька еще не окончилась, маховые и рулевые коротки, у птицы
от 2 января сменялось мелкое перо на спине (Козлова, 1947).
Повидимому линька взрослых гнездящихся птиц происходит летом,
после окончания гнездового сезона, во время миграции с июня по октябрь.
Линька молодых, в данном году остающихся холостыми, происходит во время
их зимних кочевок с октября по январь.
Питание. Слабо изучено. Креветки и мелкие морские животные.
У берегов Алеутских о-вов присутствие тонкоклювого буревестника зависит
21 Птицы СССР, т. II
322
ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
от количества китов, что связано с однородной пищей у тех и других (Бент,
1922) или, что также весьма вероятно, с отбросами самих китов и китобойных
промыслов. Кроме того, киты при всплывании на поверхность воды выносят
с глубин моря мелких морских животных, могущих служить буревестникам
пищей.
Полевые признаки. Некрупный темноокрашенный буревестник, по повад-
кам сходный с другими видами этих птиц. Во время миграций и кочевок дея-
тельны в течение почти круглых суток, в особенности в пасмурную погоду.
В период гнездования, в особенности во время высиживания яиц, ведут чисто
ночной образ жизни, слетая с яиц только ночью и ночью же возвращаясь
обратно на гнездовье. Во время выкармливания птенцов взрослые тицы
проводят день в море и только с наступлением вечера начинают появляться
над островом с гнездовой колонией. Прилет птиц на острова происходит вечером
Рис. 65. Тонкоклювый буревестник
в полном молчании и в течение очень короткого времени — в течение часа—
все птицы колонии оказываются уже на острове. В это время птенцы
и взрослые птицы начинают издавать гортанные, хрипящие, стонущие звуки
и весь остров как бы оживает. Все птенцы колонии одновременно получают
пищу, отрыгиваемую родителями в открытый рот птенца. Вскоре колония зати-
хает — молодые переваривают пищу, родители отдыхают частью в норах,
частью на поверхности земли вблизи нор. Ночью же, еще задолго до наступле-
ния утра, колония начинает оживать — взрослые птицы молча передвигаются
к скалистым берегам или к удобному месту для взлета, и в полном молчании
почти все одновременно покидают гнездовья до следующей ночи (Монтгомери,
1892). Наиболее обычный звук, издаваемый буревестником, «кук-кук-а-оо»
повторяется им несколько раз с различной интонацией (Сибом, 1890). На
гнездовьях он издает хриплые, гортанные звуки.
Хозяйственное значение. Яйца и птенцы употребляются в пищу, и в опре-
деленное время года ведется на них промысел. Ежегодно добывают на о-вах
Бассова пролива до 400.000 штук птенцов. Взрослые птицы также употребляют-
ся в пищу, но они менее вкусны; кроме того собирается и их перо.
Описание. Размеры и строение. Длина тела B) 370—400 мм. Длина
крыла D) 260—330 мм, в среднем 297 мм; 254—290 мм (Бианки, 1913), длина клюва D)
31—35 мм, средняя 32,5; 30,5—34 мм (Бианки, 1913).
Первостепенных маховых 11, второстепенных 19; 2-е (видимое) первостепенное маховое
равно первому или немного короче. Хвост округлый, за пределы вытянутых ног не
выдается. Рулевых 12. Плюсна сильно сжата с боков, с острыми передними и задними
краями, покрыта многогранными щитками. 3 пальца соединены плавательной перепонкой,
которая довольно узка и слабо вырезана между пальцами. Задний палец очень небольших
МАЛЫЙ БУРЕВЕСТНИК
323
размеров и прикреплен выше остальных. Оперение густое, довольно жесткое, в особенности
на верхней стороне тела.
Окраска. Пуховой наряд серовато-бурый, бока тела светлей (Козлова, 1947).
Молодые птицы похожи на взрослых, но чуть светлее и с более сильно развитым буро-
ватым оттенком в оперении.
Взрослые птицы похожи на серого буревестника, отличаясь главным образом светло-
серыми подмышечными. Оперение темнобурое несколько светлей с сероватым оттенком
на нижней стороне тела и с узкими светлобурыми вершинными каймами перьев. Голова и
шея почти черная. Подбородок и горло светлее нижней стороны тела. Внутренние опахала
маховых частично светлосерые. Нижние кроющие крыла серые со светлосерыми вершинами.
В свежем оперении перья со светлобурыми окаймлениями. Радужина бурая. Клюв темно-
бурый с почти черным коньком надклювья и носовыми трубками. Ноги темнобурые с более
светлыми внутренней стороной плюсны, средним и внутренним пальцами.
8. Малый буревестник Puffinus puffinus Brunn.
Procellaria puffinus. В г и n n i с h. Orn. Bor., 1764, стр. 29, Фарерские о-ва.
Распространение. Ареал. Северные внетропические области Атлан-
тического океана: Исландия, Фарерские о-ва, Гебридские, Оркнейские и
Шетландские, Великобри-
тания (западный берег Ан-
глии, Ирландия), Бермуд-
ские и Азорские о-ва (Пор-
то-Санто), о-в Мадейра, зап.
и южн. Франция и о-ва Сре-
диземного и Мраморного
морей. Кочует в течение зим-
него периода с сентября —
октября по март — апрель,
преимущественно в северо-
восточной области Атлан-
тического океана, главным
образом, у Великобрита-
нии, западного побережья
Франции и Пиренейского
п-ова. К северу доходит до
берегов Норвегии, к югу
до Канарских о-вов. К
северо-западу кочует у бе-
регов Исландии и Фарер,
и изредка наблюдается в
южн. Гренландии и Сев.
Америке, регулярно встре-
чается на кочевках в Южн.
Америке, у берегов Брази-
лии и Аргентины к югу до
м. св. Антония (южнее Бу-
энос-Айрес), в южных об-
ластях Атлантического оке-
ана до 33°28' ю. ш. и 45°42'
в. д. (Мерфи, 1936), в Сре-
диземном и Черном морях.
Биотоп. Океанические и морские воды; в период размножения
мелкие скалистые океанические и морские острова и побережье крупных
островов и материков.
Подвиды и варьирующие признаки. 3 подвида, различающихся между собою особен-
ностями окраски оперения. P. p. puffinus Brii nn, 1764 — Исландия, Фарерские, Оркней-
ские, Шетландские и Гебридские о-ва, зап. побережье Англии (С. Уэльс), Ирландия, берег
Карта 59. Распространение малого буревестника Puf-
finus puffinus
7. P. p. puffinus, 2. P. p. mauretanicus, 3. P. p. yelkouan;
a — граница гнездовий, г — область зимовок.
324 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ ^^
Бретани, Азорские о-ва (Порто-Санто), Мадейра, о-в Сальважес и Бермудские о-ва; Р. р.
mauretanicus Lо we, 1921 — Балеарские о-ва, о-в Риу (близ Марселя), Корсика, Сардиния.
Мальта и может быть гнездится у берегов Алжира и Туниса; P. p. yelkouan Acerbi,
1821 — о-ва Эгейского моря и Принцевы о-ва в Мраморном море.
Систематические замечания. В 1935 г. Мэтьюз присоединил к малому буревестнику
P'Puffinus в качестве подвидов 3 вида буревестников, распространенных в Тихом океане:
P. opisthomelas Coues, 1864 — Сан-Бенито, о-в Нативидад и о-в Гвадалупа у зап,
берегов Нижней Калифорнии, P. auricular is To w se n d, 1890 — о-в Кларион, о-в Сан-
ренедикто в группе о-вов Ревиллья-Гигедоу зап. берегов Мексики и P. newelli Hens-
chaw, 1900 — о-вЛанаи и о-в Молокан в Гавайском архипелаге.
Таким образом, согласно взглядам Мэтьюза, P. puffinus имеет 6 подвидов, распростра-
ненных в Атлантическом и Тихом океанах. Так как у нас нет коллекционного материала из
выше указанных областей и проверить высказанный Мэтьюзом взгляд не можем, то мы
придерживаемся при описании P. puffinus общепринятого разделения видов буревестни-
ков, т.е. не присоединяем к нему 3, выше перечисленные, вида буревестников.
Средиземноморский малый буревестник Puffinus puffinus yelkouan Acerbi
Procellaria yelkouan. Acerbi. Bibl. Ital., 1827, стр.297, Средиземное море
Распространение. Ареал. Гнездится на о-вах у зап. побережья Греции
и в Эгейском море (Цикладские, Спорадские о-ва), Принцевы о-ва в Мра-
морном море, может быть гнездится у берегов Египта.
В Черном море на кочевках доходит до Украины, Крыма, Кавказа и
Азовского моря, где наблюдался у берегов Керчи и добыт у Мариуполя,
но не проникает в Каспийское море (упоминания старых авторов вероятно
ошибочны). Кочует по всей восточной части Средиземного моря, в Босфоре,
Мраморном море, в Дарданеллах. Частично оседлый в сев.-вост. обл. Среди-
земного моря и в Эгейском море, где встречается круглый год. Точно время
появления в Черном море и у наших берегов не установлено, но птицы, пови-
димому холостые, наблюдаются уже в июле и августе. Покидают Черное море
в середине или в конце апреля.
Биотоп. В период гнездования мелкие скалистые морские острова,
на кочевках на море.
Численность. В Эгейском море обычен и многочисленен как на
гнездовье, так и на кочевках; у южного побережья Средиземного моря
наблюдается реже, преимущественно зимой. В Черном море обыкновенен на
кочевках, в Азовском море очень редок, скорее случаен.
Экология. Размножение. На гнездовьях появляются в апреле.
Гнездятся колониями. Моногамы. Гнездо устраивают на скалистых островах,
иногда густо поросших кустарником, в расщелинах скал или в норах, которые
роют сами птицы под кучами камней (Северные Спорады). Длина хода норы
сильно варьирует и в некоторых случаях достигает значительных размеров,
до Р/г и более метров. Обычно ход заканчивается расширением, в котором
и помещается собственно гнездо. Дно гнезда, часто выстилается небольшим
количеством сухой травы, иногда яйца откладываются в гнезда без всякой
подстилки. Время кладки точно не установлено, — на Цикладах и Сев. Спорадах
с 20—23 мая обычно встречаются уже сильно насиженные яйца и только изредка
ненасиженные. Малый буревестник, повидимому, приступает к размножению
в различные даты апреля — мая на отдельных островах.
В кладке одно яйцо, овальной формы, иногда оба конца яйца почти оди-
наково широкие. Скорлупа гладкая, белая, но без блеска. Размеры: A8) 50—
64x38,2—43,7 мм (Греция, Рейзер, 1905); E) 55—58x39^12,5 мм, 55—58 мм
(Спорады и Циклады, Гебель). Вес A8) 31,6—46,2 г (Рейзер, 1905); E)
36,8—42,3 г (Гебель). Время насиживания не выяснено (номинальный под-
вид насиживает 52—54 дня, Локлей, 1930). В насиживании принимают уча-
стие оба родителя. Птенец вылупляется, одетый густым пухом, не одновре-
менно, даже на близко расположенных друг к другу островах. Большинство
птенцов вылупляется в середине июня,
МАЛЫЙ БУРЕВЕСТНИК
325
Родители кормят птенцов один раз в сутки — ночью. В возрасте около
40 дней птенцы начинают одевать перо и на 62—63-й день жизни уже одеты пером
(номинальный подвид). К этому времени родители перестают кормить молодых
птиц и они под влиянием голода, приблизительно на 72-й день своей жизни,
самостоятельно, без помощи родителей, покидают норы и уходят в море- Уход
птенцов в море всегда происходит ночью. Первое время, спустившись на воду,
молодые больше плавают, нежели летают. Таким образом эмбриональное и
постэмбриональное развитие птенцов занимают три месяца.
Линька. Раз в году, полная. Последовательность смены нарядов:
первый пуховой — второй пуховой — первый гнездовый — второй гнездо-
вый и т. д. Окончательная окраска приобретается уже в гнездовом наряде.
Линька первостепенных маховых идет постепенно, выпадает не более
Рис. 66. Малый буревестник
2 перьев одновременно на каждом крыле, начиная от внутренних первостепенных
маховых к наружным (Майо, 1931). Начинается линька у взрослых, гнездя-
щихся в данном году птиц в августе, но у холостых птиц, вероятно, линька
начинается раньше — в конце июня — в июле. Добытая 5 июля у Босфора
птица меняла маховые и мелкое оперение.
Питание. Питаются мелкой рыбой, различными мелкими ракооб-
разными и моллюсками, кроме того, в период насиживания в их желудках
найдены также остатки растений и водорослей (Молчанов, 1906 г.). На
кочевках у Крымского побережья питаются, главным образом, анчоусом
Engraulis encrasicholus (в желудке малого буревестника находили до 43 штук
мелкого анчоуса, Козлова, 1947).
Полевые признаки. Буревестник некрупных размеров, с буроватой спиной
и белой брюшной стороной. Летает легко, быстро и неутомимо, как и другие
виды буревестников. Может парить, образуя широко открытыми крыльями
и телом одну плоскость и наклоняя ее то в одну, то в другую сторону. Парение
чередуется с быстрыми короткими взмахами крыльев. Летают обычно низко
над водою почти всегда по прямой линии, на большом расстоянии друг от друга
и только изредка держатся близко друг к другу (Нордман). Ныряет с разлета, но
не на большую глубину, под водой держится довольно долго и может проплыть
нод водой значительное расстояние, работая при этом ногами и крыльями.
С воды поднимается легко, но с некоторого разбега, широко раскрывая крылья.
По земле передвигается неуклюже, наступая на цевку и пальцы. На кочевках
активен днем и ночью, но в период высиживания активен только ночью.
326
ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Звуки, издаваемые малым буревестником, очень разнообразны —то
звонкие и очень громкие, то хриплые, как бы приглушенные, то похожие
на воркование или мяуканье. Особенно они крикливы на гнездовьях ночью
во время кормления птенцов (Локлей, 1930).
Описание. Размеры и строение. Средних размеров птицы. Длина крыла D)
215—235, в среднем 252 мм. Первостепенных маховых— 11, из них первое короче
второго. Формула крыла: 2 C)> 1 B)> 3 D) Второстепенных маховых — 22.
Рулевых—12, хвост — округлый, короткий, не выдается за пределы вытянутых ног.
Плюсна сжата с боков, покрыта щитками, 3 передних пальца соединены перепонкой,
4-й палец короткий и поставлен выше других. Оперение густое, жесткое.
Окраска. Описания птенца P. p. yelkouan нет, ниже приводится описание птенца
P. p. puffinus (Локлей, 1930). Первый пуховый наряд. Темнобурый, с сероватым оттенком
пух, более светлый на нижней стороне тела. По середине брюха от горла идет*узкая белая
полоса, расходящаяся на 2 полосы в задней части брюха.
Второй пуховой наряд одевается приблизительно на 16-й день после вылупления.
Похож этот наряд на первый, но темнее. Отдельные пушинки первого наряда долго
остаются прикрепленными на вершинах пушинок 2-го наряда и длина такой пушинки
достигает о см. Клюв черный, ноги телесного цвета, наружный палец черный, ногти
черные с белой вершиной. Молодая птица похожа на взрослую.
Взрослая птица. Спинная сторона тела аспид но-бурая, брюшная сторона и нижние
кроющие крыла белые. Подмышечные белые с буроватым вершинным пятном на перьях.
Нижние кроющие хвоста беловато-серые или беловато-бурые. Наружные опахала маховых
и вершины их буровато-черные; внутренние опахала маховых светлобурые. Радужина
буровато-черная. Клюв черный, беловатый на подклювье. Ноги грязнобелые, внешняя сто-
рона наружного пальца и когти черные.
9. Пестролицый буревестник Puffinus leucomelas Те mm.
Procellaria leucomelas. Temminck. PI. Col. 1835, livre 99, табл. 587, Япония
Распространение. Ареал. Гнездовья на о-ве Бонин и Пескадор.
У нас случайно залетная птица Курильских о-вов (Курода, 1934; Синрио,
1936). На кочевках встречается у берегов Японии от Хоккайдо до Риу-Киу,
Рис. 67. Пестролицый буревестник
у южных берегов Кореи, у берегов Китая от Шандуня до Фу-чжоу, и
у Филиппинских о-вов, у северных берегов Борнео и Целебеса, у Молукских
о-вов и у западной части Новой Гвинеи.
Биотоп. Моря и окраины океанов, открытых океанических вод избе-
гает. Гнездится на мелких островах.
Численность. У о-вов Хоккайдо и У ондо и в Корее редок, к югу от
Корейского пролива обыкновенен и, начиная от Кюсю, к югу многочисленен.
Экология* Не изучена.
Полевые признаки. Отличается более крупным клювом, пестро оперенной
головой и длинным хвостом. Повадки, как у других буревестников.
МАЛАЯ ПРЯМОХВОСТАЯ КАЧУРКА
327
Описание. Размеры и строение. Длина крыла 297—343 мм, клюв 45,7—
53,3 мм (Бианки, 1913).
Окраска. Птенец и молодые птицы не описаны. У взрослой птицы лоб, бока головы
и темя — белые с темными наствольями перьев. Верх тела темнобурый с более светлыми
Карта 60. Распространение пестролицего буревестника Pujjinus leucomelas
а — граница гнездовий, г — область зимовок.
окаймлениями перьев. Горло, брюшная сторона, подмышечные и нижние кроющие крыла
белые с темными пестринами на боках зоба, подмышечных и нижних кроющих крыла.
Радужина серая. Клюв голубоватый, ноги желтовато-серые.
РОД ПРЯМОХВОСТЫЕ КАЧУРКИ HYDROBATES BOIE, 1852
Тип Н. pelagicus
Русское название. Качурки — от того же корня, что «качуриться», «окачу-
риться» (умереть). От широко распространенного у европейских народов на севере поверья,
что качурки — перевоплощенные души погибших на море матросов. Заслуживает внимания
не только смысловая, но и звуковая аналогия со старым народным названием этих птиц
у английских моряков — «Carey's Chicken» (сокращенное от Mother Carey's Chicken, что
означает птенцы богоматери Mother Carey; Mater Сага, Хар, 1939).
Приводимые ниже видовые названия качурок — книжные.
10. Малая прямохвостая качурка Hydrobates pelagicus Linn.
Procellaria pelagica. Linnaeus. Syst. Nat., изд. X, 1758, стр. 131, берега Швеции
Распространение. Ареал. Северо-восточные части Атлантического
океана и зап. половина Средиземного моря, быть может в Исландии (Вест-
маннские о-ва), более или менее случайно у побережья Норвегии к северу
328
ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
до Лофотенских о-вов; Фарерские о-ва, Оркнейские, Гебридские, Шетланд-
ские и ряд мелких о-вов у берегов Шотландии, побережье Уэльса, Ирлан-
дия; зап. побережье Франции и Пиренейского п-ова и прилежащие острова;
острова по южн. и сев. побережью и в зап. части Средиземного моря (Балеар-
ские, Питиузские, Эльба, Сардиния, Корсика и др.) на восток до Мальты;
Карта 61. Распространение малой прямохвостой качурки Hydrobates pelagicus
а — границы гнездовий, г — область зимовок, д — залет.
Канарские о-ва и Мадейра. В СССР случайно залетная. Добыт один экз.
7 октября 1911 г. у г. Мариуполя (Бутурлин, 1936) и второй экз. 30 октября
1949 г. в районе с. Лазаревка, между Сочи и Туапсе (Левицкий, письменное
сообщение, определение не проверено).
Кочует по всему Атлантическому океану до Лофотенских о-вов, Исландии,
южн. Гренландии и Лабрадора .на севере, к югу по американскому побережью
до Нью-Фаундленда, Новой Шотландии и штата Мен. Западное побережье
европейского материка, Средиземное море и на восток до Греции и Кипра;
Красное море, зап. берег африканского материка к югу, может быть до
МАЛАЯ ПРЯМОХВОСТАЯ КАЧУРКА
329
м. Доброй Надежды. Указания на кочевки по восточному побережью афри-
канского материка к северу до о-ва Занзибар (Бент, 1922) очень сомни-
тельны. Обычно залетная и случайно кочующая птица Северного моря и
южных частей Балтийского. Неоднократно отмечены залеты глубоко внутрь
материка — до Берлина, Вены и Женевского озера.
Характер пребывания. Оседлая и кочующая птица (см. выше).
Биотоп. Открытые и прибрежные воды океанов и морей. В период
гнездования пустынные и скалистые побережья материков и крупных остро-
вов и мелкие прибрежные острова.
Численность. Обыкновенна на гнездовьях у зап. берегов Англии,
Франции и на о-вах Средиземного моря. У Пиренейского п-ова и у берегов
Исландии редка. На кочевках обы-
кновенна и многочисленна по аме-
риканскому побережью, у южн-
Гренландии, Исландии, у зап. евро-
пейских берегов и у зап. берегов
Африки к югу до Французского
Конго. Далее к югу редка, и у
Капской Земли, повидимому, слу-
чайна. В Средиземном море обычна
в западной части и редка в восточ-
ной, как редка и в Красном море.
Рис. 68. Малая прямохвостая качурка
Экология. Размножение,
На местах гнездовий в Англии по-
является в конце апреля — начале
мая. Моногамы. Гнездятся коло-
ниями в щелях между камнями,
в норах, вырытых обычно дру-
гими птицами (топориками, чисти-
ками) и зверями и редко самими
птицами. Качурками нора роется
в мягкой почве и имеет обычно ход
в 30 см длины, но иногда ход достигает и метра. На конце ход расширяется
и образует камеру, где собственно и помещается гнездо. Гнездо выстлано
небольшим количеством небрежно набросанной травы.
Кладка бывает раз в году: в Англии обычно в июне — июле (Уайзерби,
1940), во второй половине июля на Фарерских о-вах (Саломонсен,
1935); на о-ве Мальта в июне—июле, у берегов Алжира—уже с мая
(Лош, 1867).
В кладке одно яйцо овальной формы с почти одинаково округленными
концами. Скорлупа без блеска, белая, иногда с мелкими красноватыми кра-
пинами на тупом конце.
Размеры: A00) 25,3—30,7 х 19,1—22,4 мм, в среднем 27,97 х 21,19 мм
(Уайзерби, 1940).
Насиживание продолжается 36—40 дней, высиживают оба родителя.
Птенцы вылупляются одетые густым пухом. Постэмбриональное развитие мед-
денное и птенцы долго остаются в гнезде — около двух месяцев E6—64 дня).
Покидают гнездовья обычно в сентябре.
Линька. Плохо изучена. Сильно растянута и у разных экземпляров
начинается неодновременно, повидимому, в зависимости от окончания гнез-
дового периода. Основная масса птиц линяет, начиная с сентября до мая,
но некоторые особи заканчивают линьку уже в ноябре.
Питание. Питаются различными мелкими ракообразными, моллю-
сками, медузами и планктонными животными, живущими в поверхностных
слоях воды. Пищу берут, не садясь на воду. Птицы спускаются низко к воде,
330 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ __=__
часто при этом поднимают крылья, вытягивают вниз ноги, касаясь ими поверх-
ности воды, на мгновение останавливаются, схватывают добычу и затем летят
дальше.
Полевые признаки. Мелкая птица темнобурой окраски с прямо обрезан-
ным хвостом, с белым надхвостьем. Как и все качурки превосходно летает
и плавает. В отличие от буревестников хорошо передвигается по земле —
бегает и легко взлетает. При полете часто взмахивает крыльями и никогда
не летит по прямой линии. Полет всегда очень извилист, и они особенно низко
летают над водою в бурную погоду, следуя по направлению волны.
В период кочевок это дневные и ночные птицы (Уинн-Эдвардс, 1935),
в гнездовое время активны только ночами.
В период кочевок качурка молчалива, лишь изредка издает чирикаю-
щие звуки; на гнездовьях, в особенности ночью, издаваемые звуки разнооб-
разней, у сидящих на яйцах—похожи на урчание, во время полета —
жалобные, горловые ноты.
Описание. Размеры и строение. Длина крыла 114—117 мм, длина клюва
10—12 мм. Хвост прямой; задний палец имеется, ноздри расположены так же, как
у вилохвостой качурки.
Окраска. Первый пуховой наряд серовато-бурый, несколько более светлый на
нижней стороне тела. Вокруг глаз, на горле, подбородке и щеках голое пространство.
Второй пуховой наряд более темный, голых частей на голове нет. Горло, подбородок
и щеки покрыты коротким пухом, на всем остальном теле длинным.
Молодые птицы темнобурые, похожие на взрослых, но с белыми узкими окай-
млениями на лопаточках и верхних кроющих хвоста перьев.
Взрослая птица темнобурая, несколько с более светлым низо.м тела и почти белая
по бокам брюшка. Рулевые и первостепенные маховые буровато-черные с белыми основа-
ниями перьев. Верхние кроющие крыла и второстепенные маховые буровато-черные с бе-
лыми узкими, почти незаметными, каймами на больших кроющих крыла и второсте-
пенных маховых. Радужина ореховая. Клюв и ноги черные. В свежем пере имеется
дымчато-серый налет в оперении. Белые каймы на верхних кроющих крыла и второсте-
пенных маховых сильнее развиты.
Систематические замечания. В 1843 г. более темно окрашенные птицы с о-ва Мальта
выделены в особый подвид Н. p. melitensis, но в дальнейшем они были сведены в синоним
Н. pelagicus. В 1936 г. Хартерт и Штейнбахер вновь восстанавливают подвид melitensis на
основании найденной на Питиузских о-вах гнездовой колонии более темно окрашенных птиц.
К сожалению из-за отсутствия материала невозможно решить вопрос — являются ли темно
окрашенные птицы самостоятельным подвидом или мы здесь имеем дело только с ярко выра-
женной индивидуальной изменчивостью. Оставляем Н.р. melitensis в синонимах Н.pelagicus.
РОД ВИЛОХВОСТЫЕ КАЧУРКИ OCEANODROMA REICHENBACH, 1852
Тип О. f urc at a
11. Северная качурка Oceanodroma leucorrhoa Vieillot.
Procellaria leucorrhoa. Vieillot. Nouv. Diet. d'Hist. Nat., XXV, 1817, стр. 422, Пи-
кардия, Франция
Распространение. Ареал. Сев. части Тихого и Атлантического океанов.
Гнездится на Курильских, Командорских и Алеутских о-вах; по тихоокеан-
скому берегу Сев. Америки от южн. Аляски до Нижней Калифорнии; по атлан-
тическому побережью Сев. Америки от п-ова Кумберленда (Баффинова Земля)
до штата Мэн; в ю.-з. Гренландии, Исландии, на Фарерских о-вах, юго-зап. по-
бережье Ирландии (пров. Керри) и о-ва Кильда, Сев. Рона, Сула-сгейр, Эйлли-
мор, Фленнен (Уайзерби, 1940). На кочевках в Атлантике несколько переходит
на юг за экватор, в Тихом океане до Галапагосских о-вов и Гавайских (Мидуэй).
Характер пребывания— гнездящаяся и кочующая птица,
местами — оседлая. Кочует обычно в южном направлении.
Биотоп— океаны, в моря залетает лишь случайно. С сушей связана
только в период гнездования.
СЕВЕРНАЯ КАЧУРКА
331
Подвиды и варьирующие признаки. Три подвида, различающихся между собою, глав-
ным образом, размерами и формой вырезки хвоста. О. е. leucorrhoa Vieillot, 1817 — рас-
пространение дано ниже при описании подвида; О. е. bealiE merson, 1906 — гнездится от
южных частей Аляски (Ситка) до Орегона и Калифорнии; О. е. kaedingi Anthony,
1898 — гнездится в Нижней Калифорнии, на о-вах Гвадалупе и Ревилья-Гигедос, кочует
от Сан-Диего до южной оконечности Нижней Калифорнии и в море до 5°30' с. ш. и 102° з. д.
(Бент, 1922).
Северная большая качурка Oceanodroma leucorrhoa leucorrhoa Vieillot
Распространение. Ареал. Курильские о-ва (указание на гнездование
на Сахалине пока не подтвердилось), Командоры (о-в Медный, для о-ва Беринга
гнездовье не доказано); Алеутские о-ва, южн. побережье Аляски и прилежащие
острова, к югу, вероятно, доходит до Ситки; восточный берег Сев. Америки от
Карта 62. Распространение северной качурки Oceanodroma leucorrhoa
7. О. I. leucorrhoa, 2. О. L beati, 3. О. /. kaedingi;
а — граница гнездовий, г — область зимовок (цифры в кружках обозначают номера зимующих подвидов).
южн. побережья п-ова Кумберленд (Баффинова Земля, 66°—65° с. ш.) по зап. и
южн. берегу Лабрадора, Нью-Фаундленд, Новый Брауншвейг, Новая Шотлан-
дия и штат Мэн, к югу, вероятно до Портленда. Гренландия, юго-зап. побережье,
северный предел точно не установлен, может быть до бухты Диско, 70° с. ш.
(Кемлин, 1898); Исландия, Фарерские о-ва (только недавно обнаружено
гнездовье у Мичченеса), Гебридские о-ва, Оркнейские, ряд мелких островов
у западных берегов Великобритании (Кильда, Фленнен и др.) и юго-зап. по-
бережье Ирландии.
Характер пребывания. Гнездящаяся, кочующая, только по
зап. побережью Гренландии и в некоторых других районах оседла.
Кочевки совершает обычно в южном направлении. Птицы, гнездящиеся
в северных областях Атлантического океана, на кочевках северней м. Фаруэль
(Гренландия), Исландии и Фарерских о-вов не встречаются. К югу кочуют
почти до экватора (в Британском музее имеются птицы, добытые в открытом
океане под экватором, Сальвин, 1896), на юго-запад—до Малых Антильских
о-вов и Бразилии до м. Сан-Рок, 5° ю. ш. На юго-восток доходит до о-вов
332 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Зеленого Мыса и зап. берегов Африки до Сиерра Леоне и Либерии (Бент, 1922,
Годман, 1910).
Птицы, гнездящиеся в сев. частях Тихого океана, кочуют в южн. частях
Берингова моря вблизи Алеутских о-вов (Нельсон, 1887) и к югу до Гавайских
и Галапагосских о-вов, на юго-запад до сев. Японии — Хоккайдо, на юго-
восток до Нижней Калифорнии.
Время начала кочевок, так же как и конец их, трудно установить, —
часть птиц остается вблизи мест гнездовий на зиму, часть птиц и летом (хо-
лостые) ведут кочевой образ жизни. Повидимому, нормальные кочевки на-
чинаются в первых числах октября и продолжаются до конца мая.
Первые осенние кочевки отмечены: 1 сентября Массачусетс, 9 сентября —
Атлантический океан 28°3б' с. ш. и 31°45' з. д., последние весенние —21 мая
Квебек, 15 мая —Н. Шотландия, 1 мая —Бермудские о-ва, 4 мая Атланти-
ческий океан и т. д. (Бент, 1922).
Залеты вглубь материков и морей редки, но неоднократно отмечены, в
частности в Средиземном и Балтийском морях.
В пределах Европейской части Союза отмечено 3 случая залетов одиноч-
ных птиц: близ Елгавы (Митавы) 1 декабря 1828 г., у Выборга в 1886 г.
и в Риге 26 декабря 1902 г. (Мензбир, 1918).
Залеты на побережья стран, омывающихся океанами, повидимому, пас-
сивны и могут быть объяснены только действием бурь и сильных ветров, несу-
щих птиц к материкам.
Биотоп. Открытые океанические воды, на кочевках избегает открытых
морей, прибрежной полосы воды, побережий материков и островов, только
в период размножения связаны с сушей, с отдаленными от материков мелкими
океаническими островами или с островами, омываемыми по крайней мере хотя
бы с одной стороны океаном.
Численность. У нас на гнездовье малочисленна, и на о-ве Медном
встречается в очень небольшом количестве, много реже, чем сизая качурка;
на Курильских о-вах, к югу от о-ва Парамушир, становится обычной, но к югу
от о-ва Симушир опять уже редка (Кларк, 1910). Обычна и многочисленна на
гнездовье по островам, прилежащим к берегам Сев. Америки, в особенности
многочисленна на островах у южн. Аляски, где на некоторых островах гнез-
дится до 150 000 птиц (Бент, 1922).
На кочевках у нас также малочисленна и встречается реже, чем сизая
качурка, более многочисленна у берегов Сев. Америки.
Экология. Размножение. Время появления весною на места гнездо-
вий в СССР точно не установлено. На зап. Алеутские о-ва прилетают в мае
(Даль, 1874), в штат Мэн в конце мая или в начале июня (Бент, 1922).
Моногамы. Гнездятся колониями, часто совместно с О. furcata. Гнезда
устраивают обычно в норах, выкопанных самими птицами. В щелях скал,
повидимому, не гнездятся. Норы расположены чаще на равнинных частях
острова, иногда высоко над уровнем моря, на скалах, покрытых слоем земли,
на торфяных болотах или на взморьях, заросших густою травою, кустарником
или одиночными деревьями; на Алеутских о-вах на торфяных берегах (Даль,
1874); на Аляске и прилежащих островах—на болотистых равнинах, по-
росших травою, кустарником или деревьями (Бент, 1922).
Норы размещаются на близком расстоянии друг от друга, в особенности
в больших колониях и при малой площади удобной земли. Вход в норы полу-
круглой формы, скрыт от глаз и лежит или под корнями деревьев, или у ос-
нования кустарника, или в кусте травы (Аляска). Ход норы идет с легким
наклоном вглубь почвы на 5—12,5 см, и затем извилисто направляется па-
раллельно поверхности земли. Длина хода очень различна: ot 15 см до 150 см.
Конец хода расширяется в диаметре на 12—13 см и до 7,5 см высоты, образуя
камеру, где собственно и помещается гнездо. Дно гнезда чаще покрыто незна-
СЕВЕРНАЯ КАЧУРКА 333
читальным количеством сухой травы и листьев, иногда яйцо лежит прямо на
голой земле. Земля, выброшенная птицами из норы, лежит у отверстия норы.
В кладке одно яйцо овальной формы, почти одинаковой ширины на обоих
концах. Скорлупа белая или грязнобелая без блеска с мелкими красноватыми
или фиолетовыми крапинами на тупом конце. Размеры (91): 30,2—35,3 х 22,4—
25,4, в среднем 32,76 х 23,85 мм (Уайзерби, 1940); E5) 30,5—35 х 22—26,
в среднем 32,5 х 24 мм (Бент, 1922).
Кладка яиц, повидимому, происходит несколько поздней, чему O.furcata,
начинается в самом конце мая (Англия), в Исландии отмечена в июле
(Хантцш, 1905). Точно время гнездования на о-ве Медном и Курильских о-вах
не выяснено, но у добытой 26 июля птицы имелись в яичнике рубцы от снесен-
ного яйца в июне или в начале июля(Слепцов). На Алеутских о-вах ненасиженные
яйца находили с 10 июня до конца июля (Даль, 1874), у о-ва Ситка, в половине
июля собрано 96 яиц, которые почти все были ненасиженные или слабо наси-
женные; на о-вах штата Мэн первые ненасиженные яйца находили 19 июня,
но иногда время кладки наступает поздней и ненасиженные яйца находили
до 15 августа (Бент, 1922). В 1850 г. в просмотренных там 24 июня гнездах
найдено только одно яйцо (Брьюер, 1884).
Время насиживания точно не установлено, по одним данным 42—50 дней
(Уайзерби, 1940), по другим около 35 дней (Бент, 1922). Насиживают оба роди-
теля. Смена птиц на гнезде происходит ночью и, вероятно, дважды в течение
одной ночи. С наступлением сумерек, сидящая днем птица слетает с гнезда,
и ее заменяет другая, под утро происходит, повидимому, еще раз смена — отдох-
нувшая в течение ночи птица вновь сменяет сидящую на гнезде птицу. Наси-
живающая птица во время нахождения на гнезде не ест. Выведшийся птенец
одет густым пухом, но первые дни обогревается одним из родителей и только
после 3—4 дней оставляется родителями в гнезде один на целый день. Кормле-
ние птенцов происходит ночью, попеременно отрыгиваемой родителями масля-
нистой жидкостью (Бент, 1922).
Оперение птенца идет очень медленно и он долгое время, около 70 дней,
остается в гнезде пока не оденет полного перового наряда, но рост и размеры
птенца увеличиваются довольно быстро и птенец скоро достигает размеров
родителей. Перья вначале появляются на крыльях, хвосте, затем на спине и
груди и пух остается только на брюхе. К моменту вылета из гнезда весь оде-
вается пером. Вылет из гнезда происходит, повидимому, в сентябре или
октябре.
Линька. Бывает раз в году, полная, между августом —мартом. Под-
робности неизвестны.
Питание. Креветки и другие мелкие ракообразные, мелкие мол-
люски, плавающие на поверхности воды, погибшие планктонные животные,
образующие на поверхности воды скопления, похожие на маслянистую жид-
кость, отбросы с пароходов, остатки рыб, в особенности внутренности, выбра-
сываемые с рыболовных судов. В желудке добытых птиц находили только масля-
нистое вещество «ворвань», которая, повидимому, является результатом бы-
строго переваривания мелких, главным образом, планктонных организмов
(Бент, 1922). В период размножения, у добытых на гнездовье птиц в желудках
вместе с «ворванью» находили кусочки щавеля (Бианки,1913).
Пищу схватывают, обычно, скользя в воздухе низко у поверхности воды,
время от времени наклоняя голову к воде, но более крупные куски схватывает
лапами (Бент, 1922). Но иногда кормится, плавая по воде (Мерфи, 1936).
Полевые признаки. В общем, похожа на прямохвостую качурку, но отли-
чается от нее вильчатым хвостом; крылья также относительно длиннее.
Большую часть времени проводит в воздухе, спит и отдыхает на воде; только
в период гнездования связана с сушей. По земле передвигается быстро и легко
взлетает даже с ровной поверхности.
334
ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Преимущественно дневная птица, только в период размножения — во
время высиживания птенцов — ведет строго ночной образ жизни, но уже
с момента вылупления птенцов активны и днем и ночью. День они проводят
в море, а ночью возвращаются на гнездовья для кормления птенцов. Холостые
птицы и в летнее время — в период размножения — ведут дневной образ жизни
(Уинн-Эдвардс, 1935).
На гнездовьях днем молчаливые птицы, с наступлением сумерек издают
громкие чирикающие звуки, несколько напоминающие щебетание ласточки,
только ниже тоном, которые издают птицы вылезая из норок и налету.
Рис. 69. Северная качурка
Описание. Размеры и строение. Длина тела A) 230 мм, крыла C) 150—
160 мм, длина клюва C) 15—16 мм. Вес A) 41 г.
Крылья длинные, острые, самое длинное 2-е первостепенное маховое. Первостепен-
ных маховых— 11, второстепенных— 12. Формула крыла г2C)>3D)>1B)>4E) . . . Ру-
левых— 12, хвост вилообразный.
Ноги слабые. Плюсна округлая. Пальцы длинные, есть задний палец, но очень неболь-
ших размеров и расположен выше остальных. Когти слабйс, изогнутые. Клюв слабый, удли-
ненный, в вершинной части сжатый с боков, у основания округлый, крючкообразно изогну-
тый на конце верхней и нижней челюсти.
Оперение густое, довольно мягкое и плотно прилегающее к телу.
Окраска. Пуховой птенец. Буровато-коричневый у основания и дымчато-серый на
вершинах пух (Бент, 1922).
Молодая птица похожа на взрослую птицу, но верхние кроющие крыла и внутрен-
ние второстепенные маховые с более широкими, белыми окаймлениями.
Взрослая птица темнобурая с легким сероватым налетом на верхней стороне тела
и с рыжеватым на нижней. Лоб, щеки и горло несколько светлей и с более ясно выраженным
серым оттенком. Внутренние из второстепенных маховых имеют беловатые окаймления.
Первостепенные маховые буровато-черные с сероватым налетом на внутренних опахалах.
Большие и средние кроющие крыла бурые, малые—темнобурые, почти черные. Рулевые —
черные. Верхние короткие кроющие хвоста — бурые с белыми вершинами, длинные
кроющие хвоста — белые. Нижние кроющие хвоста темнобурые, светлеющие к бокам
хвоста и белые на боках хвоста.
Радужина темнобурая, клюв и ноги — черные.
МАЛАЯ ВИЛОХВОСТАЯ КАЧУРКА
335
12. Малая вилохвостая качурка Oceanodroma monorhis S winh.
Tallasidroma monorhis. Swinhoe. The Ibis, 1867, стр. 386, Амой, Южн. Китай
Распространение. Ареал. О-в Верховского, близ Владивостока (Фирсов,
1928), на 2 островках у берегов Кореи (Hand-list of the Jap. Birds, 1832),
может быть на некоторых о-вах Риу-Киу (Суайно, 1871), на островах к се-
веро-востоку от Формозы (Латуш, 1934), острова у Нижней Калифорнии —
Карта 63. Распространение малой вилохвостой качурки Oceanodroma monorhis
7. О. т. monorhis, 2. О. m. socorroensis.
а — граница гнездовий, г — область зимовок (цифры в кружках обозначают номера зимующих подвидов).
Сан-Бенито и Лос-Коронадос. Кочует у азиатских берегов от Владивостока
и северной оконечности Хондо до Фу-Чжоу (у последнего наблюдалась в июле,
Латуш, 1934) и Амоя, ю.-в. Китай; у американских берегов от Калифорнии
Рис. 70. Малая вилохвостая качурка
до Мексики и Галапогосских о-вов. Вероятно, область кочевок азиатских и
американских птиц соединяется в центральной части Тихого океана, но до сих
пор ее в центральных водах Тихого океана не встретили.
Подвиды и варьирующие признаки. Объединяет два подвида, различающихся, главным
образом, размерами и слабой окраской. О. /п. monorhis Swinhoe, 1867 — азиатская
часть ареала и О. т. socorroensis Townsend, 1890—гнездится в американской части
ареала. Реальность второго подвида сомнительна (Харгерт, 1916).
336 ОТРЯД ТРУВКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Азиатская малая вилохвостая качурка Oceaaodroma monorhis monorhis
Swi nhoe
Распространение. Ареал. См. выше, в очерке вида.
Характер пребывания. Гнездящаяся и кочующая птица. На
кочевках отмечена от Владивостока и сев. оконечности Хондо до Фу-Чжоу и
Амона ю.-в. Китай.
Биотоп. Океанические воды прибрежной полосы; в период гнездова-
ния — мелкие пустынные острова, расположенные близ материков и крупных
островов.
Численность. В СССР очень редка. В колонии на о-ве Верхов-
ском имелось 15 гнезд (Фирсов, 1928).
Экология азиатского подвида неизвестна.
Описание. Размеры и строение. Длина крыла 165—151 мм (Бианки, 1913).
Окраска. Пуховой наряд азиатского подвида не описан; у американского — первый
пуховой наряд серовато-бурый; второй наряд — темнобурый (Бент, 1922).
Взрослые птицы темнобурые. На голове, горле, шее, спине с сероватым оттенком.
Первостепенные маховые, наружные опахала второстепенных маховых и малые кроющие
крыла буровато-черные. Средние и большие кроющие крыла и внутренние опахала второсте-
пенных маховых светлобурые. Рулевые черные. Верхние кроющие хвоста буровато-черные,
иногда частично белые.
Радужина темнобурая. Клюв и ноги черные (Козлова, 1947). Молодые окрашены, как
и взрослые.
13. Сизая качурка Oceanodroma furcata Gmel.
Procellaria furcata. Gmelin. Syst. Nat., 1789, стр. 561, Берингово море
Синоним. Procellaria orientalis. Pallas. Zoogr. Rosso-As., 1811, стр. 314, Куриль-
ские о-ва. — Thalassidroma cinerea. Taczanowski Faune Orn., Sib., Or., II. 1891,
стр. 1059, errore.
Распространение. Ареал. Сев. часть Тихого океана. Гнездится на Ку-
рильских о-вах к югу до о-ва Симушир (Сибом, 1903, Стейнегер, 1898, Кларк,
1910), на Командорских о-вах (для о-ва Медного гнездование доказано, для о-ва
Беринга нет прямых указаний на нахождение там гнезд или яиц), вероятно, по
ю.-в. побережью Камчатки от м. Лопатки до м. Козлова (Слепцов встречал
в гнездовое время —в июле, и у добытой там самки имелись в яичнике рубцы
от снесенных в этом году яиц). Ранее также на основании сборов Блакистана
высказывалось предположение о гнездовании сизой качурки на Камчатке (Стей-
негер, 1898). На Прибыловых о-вах птицы наблюдаются в течение всего лета, но
гнездовья там не найдены (ПриблиМк. Эти 1923). Западное побережье Сев.
Америки от южн. Аляски и Алеутских о-вов до штата Вашингтона — Клаллам
(Бент, 1922).
Часть птиц в течение круглого года держится у мест гнездовий, например
у о-ва Медного (Иоганзен, 1924), большинство же откочевывает осенью к северу
и югу (Слепцов, Бианки, 1913). Кочуют в Охотском море до Аянаио-ва Ионы,
по вост. берегу Сахалина, вдоль Курильских о-вов, по юго-вост. и вост. берегу
Камчатки (м. Лопатки 6 июля 1939, Авачинская губа 25 сентября и б июня),
у Петропавловска, в Беринговом море, в Анадырском заливе (9 октября, кол-
лекция Зоологического музея Моск. университета), в бухте Провидения
(Нельсон, 1887), в Беринговом проливе и у о-ва Лаврентия. По американскому
побережью и прилежащим островам к северу доходит до залива Коцебу, к югу
до залива Гумбольдта (Сев. Калифорния), т. е. приблизительно до 38° с. ш.,
южнее встречается лишь случайно (Бент, 1922).
Характер пребывания. Осенние кочевки в Беринговом море
и проливе наблюдаются с июля по октябрь и иногда даже до середины ноября.
16 ноября — в Авачинской губе (Бианки, 1913), в ноябре — в устье р. Юкон
(Нельсон, 1887). При наступлении морозов и замерзании морей, они некоторое
СИЗАЯ КАЧУРКА
337
время держатся еще у полыней, затем откочевывают к югу. В южных широтах
проводят всю зиму, задерживаясь иногда там до конца апреля.
Холостые, негнездящиеся в данном году птицы, ведут в течение всего гнездо-
вого периода кочевой образ жизни, встречаясь далеко от гнездовий, например,
у Аяна, и о-ва Ионы (июнь, Слепцов).
Залеты. Лишь случайно залетают в глубь материков (добыта 2 августа
1823 г. на Камчатке на верхнем течении р. Камчатки, Киттлиц, 1858; на р. Та-
нана в глубь Аляски — в ноябре (Бент, 1922).
Биотоп. Как у других качурок.
Карта 64. Распространение сизой вилохвостой качурки Oceanodroma furcata
а — граница гнездовий, г — область зимовок.
Численность. У мест гнездовий летом обыкновенна и многочисленна,
осенью и зимой редка. В северных областях своих кочевок редка осенью и
совсем не встречается зимой, в южных — обычна на кочевках и даже много-
численна в течение сентября — апреля, в особенности у зап. берегов Сев.
Америки.
Экология. Размножение. Время появления в период размножения
у мест гнездовий птиц, кочующих в южных широтах, трудно установить,
т. к. часть птиц остается зимовать у мест гнездовий или поблизости. Пови-
димому, с южных широт они прикочевывают на гнездовья в различные числа
мая в зависимости от дальности кочевок.
Моногамы. Гнездятся небольшими колониями как вблизи берега, так и на
некотором расстоянии от него (до 2 км), причем характер местности, повиди-
мому, не имеет большого значения — гнезда встречаются в прибрежной полосе,
среди скал (о-в Медный), на холмах, покрытых травянистой растительностью,
и в лесной местности (Бианки, 1913). Гнездо устраивают или в норах, выкопан-
ных самими птицами в мягкой почве, или в расщелинах скал. Щели в скалах,
занимаемые для нор, достигают размеров от 30 см (о-вУналашка, Далл, 1874)
до 90 см (о-в Медный, Стейнегер, 1885).
22 Птицы СССР, т. II
338 ОТРЯД ТРУБКОНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Норы, выкопанные в почве, имеют длину от 0,5 до 1 м, в диаметре от 6—
5 до 9 см. Ход норы идет почти параллельно поверхности земли на глубине
10—20 см (Майлльард, 1898, о-в Баранова). Заканчивается ход небольшим
расширением, в котором и помещается собственно гнездо. Гнездо, расположенное
или в скалах или в земле, выстлано небольшим количеством сухой травы.
Одна и та же нора, повидимому, служит птицам в течение нескольких лет.
В кладке одно яйцо, удлиненной формы с почти одинаково закруглен-
ными концами. Скорлупа без блеска или чисто белая, или белая с мелкими
фиолетово-черными крапинами на более тупом конце яйца, причем крапины
покрывают или сплошь тупой конец яйца или образуют венчик на тупом
конце.
Размеры: 25 х 32,5 мм, 26 х 34 мм, 26,5 х 34 мм (о-в Медный, Стейне-
гер, 1885); 26,5 х 36,5 мм (о-в Медный, Гребницкий); D0) 32—37,5 х 23,5—
27,5, в среднем 33,9 х 25,7 мм (Бент, 1922).
Кладка яиц на Командорских, Алеутских о-вах и у западного берега Сев.
Америки начинается с июня и продолжается до конца июля. На о-ве Медном
в половине июля 1883 г. найдены яйца различной степени насиженности,
(Стейнегер, 1885), в июне 1886 на о-ве Медном добыто яйцо (Бианки, 1913) и
16 июля 1911 г. на о-ве Медном пойман самец в гнезде на яйцах (Бутур-
лин, 1913). На о-ве Уналашки найдены ненасиженные яйца 2 июня 1872
(Даль, 1874), близ о-ва Баранова в проливе Ситки во 2-й половине июля 1896
добыты уже сильно насиженные яйца (Майлльард, 1898).
Насиживают самец и самка. Срок насиживания неизвестен. Птенец вылу-
пляется одетый пухом с голым подбородком и шеей. Постэмбриональное раз-
витие, повидимому, медленное и птенец долгое время, пока не заменит пуховой
наряд перьями, остается в гнезде. Перья начинают расти, когда птенец дости-
гает величины почти взрослой птицы. Первыми появляются маховые и рулевые
перья, затем мелкое перо на спине, последним сменяется пух на нижней по-
верхности тела.
Время вылета птенцов не установлено.
Линька. Схема линьки — как у других качурок. Смена маховых начи-
нается с первых чисел августа и заканчивается к середине октября. В первую
очередь сменяются второстепенные маховые, а затем первостепенные. Линька
первостепенных маховых идет от внутренних маховых к наружным и, пови-
димому, очень быстро и неодновременно у всех птиц. Например, у 2 самцов,
добытых 13 августа 1947 г. близ о-ва Беринга линька первостепенных маховых на
различных стадиях. У одного самца 5 первых первостепенных маховых старые,
остальные — уже сменившиеся и 6-е и 7-е маховые еще не достигли нормальных
размеров. У другого самца только 3 первых первостепенных маховых старые,
остальные сменившиеся и только 4-е маховое не достигло нормальных размеров.
У добытого 21 сентября близ о-ва Беринга самца 5 наружных маховых старые,
6-го и 7-го совсем нет, остальные свежие. У другого самца от 22 сентября, о-в
Беринг, первые 5 старые, 6-е и 7-е маховые свежие и почти нормальных разме-
ров (коллекция Зоологического музея Московского университета).
Линька рулевых идет в сентябре и октябре, причем сменяются рулевые
от центра к крайним рулевым (у экз., добытого 13 октября в Жупановском
заливе, Кроноки), средние рулевые свежие, две следующие пары свежие, но
недоросшие до половины своих нормальных размеров и крайняя пара — ста-
рая, не сменившаяся (коллекция Зоологического музея Московского уни-
верситета).
Мелкое оперение, повидимому, сменяется зимой. У всех осенних птиц
сильно обношено и менее обношено у весенних птиц. С окончанием линьки
маховых и рулевых перьев, повидимому, совпадает время (октябрь), когда
большая масса качурок покидает места гнездовий, области северных осенних
кочевок и направляется кочевать в южные широты.
СИЗАЯ КАЧУРКА
339
Питание. Плохо изучено. Повидимому, основную пищу составляют
мелкие животные, главным образом, различные ракообразные, живущие в
поверхностных слоях воды, и куски жира (китового, тюленьего и др.), плаваю-
щие на воде. Отрыгиваемая пища состоит из остатков мойвы, рачков Rhoda
i пег mis и кусочков китового жира G птиц, Слепцов). Иногда можно увидеть
целые стаи качурок, следующих за обработанными тушами тюленей и склевы-
вающих на воде кусочки жира (Турнер, 1886).
Схватывает добычу налету, не садясь на воду. Увидев добычу, качурка
низко спускается к поверхности воды и на момент остается парящей непо-
движно, вытягивая при этом обе ноги вперед, растягивая плавательные пере-
понки, но, коснувшись ногами воды, сейчас же отталкивается от нее (Слепцов),
таким образом, получается полное впечатление, что качурка бегает по воде
с раскрытыми крыльями.
Рис. 71. Сизая вилохвостая качурка
Полевые признаки. От других встречающихся в СССР качурок отличается
светлосерой окраской и несколько большей величиной. Повадки сходны. Дер-
жатся обычно одиночками, лишь ночью на свет пароходов слетаются в стаи.
Превосходно летают и большую часть жизни проводят в полете, хуже плавают.
Не ныряют, даже с разлета. Отдыхают и спят на воде. С воды
взлетают легко. Полет быстрый, легкий с частыми взмахами крыльев,
редко направленный по прямой линии, обычно с неожиданными поворотами
то в одну, то в другую сторону, с взлетами кверху, спусками вниз и лишь на
короткое время парящий (Слепцов), напоминающий несколько полет ласточек
и стрижей. Обычно летает низко над водой, в тихую погоду почти касаясь
крыльями воды, в штормовую — полет волнообразный, соответственно движе-
ниям волны, всегда на одном и том же расстоянии от поверхности воды. Она то
поднимается вместе с волной, то, пересекая ее, скрывается за гребнем, то летит
вдоль волны (Бианки, 1913, Слепцов).
Ночью, налетев на свет пароходов, с писком носится вокруг мачт, совер-
шая круги в вертикальной и горизонтальной плоскостях (Слепцов), часто
залетая в каюты и вентиляторы. По земле ходит мелкими шагами, опираясь
на пальцы, часто перебирая ногами и подгибая на ходу когти, стараясь заце-
питься ими за неровности почвы. Тело держит горизонтально, при ходьбе
сильно сутулится, крылья скрещивает над хвостом. Садится на цевку, вставая,
вначале опирается на цевку и затем только становится на пальцы (Слепцов).
Пойманная качурка защищается лишь тем, что моментально отрыгивает
темную струю жирной жидкости «ворвани».
Голос. На гнездовьях слыхали нежное щебетание (Майлльард, 1898),
во время полета и пойманные птицы издают тонкий писк (Слепцов).
340 | ОТРЯД ТРУБКЕНОСЫЕ ИЛИ БУРЕВЕСТНИКИ
Описание. Размер ыистроение. Длина тела самцов и самок A0) 230—210 мм
в среднем 216,5 мм; длина крыла A3) 155—170 мм, в среднем 160 мм, длина клюва A2)
14—16 мм, в среднем 14,8 мм. Вес A8) от 38,5 г до 45 г в среднем 42,4.
Крылья длинные, острые, самое длинное 2-е первостепенное маховое. Внутренние
из первостепенных маховых, начиная с 7-го махового, короче второстепенных маховых.
Первостепенных— И, второстепенных— 12. Формула крыла: 2C)>3D)>1B)>4E)...
Рулевых — 12. Хвост вилообразный, крайние рулевые длиннее центральных на 25—
30 мм. Ноги слабые. Плюсна округлая, сетчатая. Пальцы длинные, имеется задний палец
небольших размеров и расположенный выше остальных пальцев. Когти слабые, узкие и
сильно изогнутые.
Клюв слабый, удлиненный в вершинной части, сжатый с боков, у основания округлый
yi крючкообразно изогнутый на вершине верхней и нижней челюсти. Носовые трубочки рас-
положены у основания клюва и слиты в одну, внутри разделенную перегородкой.
Оперение густое, довольно мягкое и плотно прилегающее к телу.
Окраска. Первый пуховой наряд — дымчато-серый, несколько более светлый
на нижней стороне тела и серовато-белый на брюшке. Клюв черный, ноги желтоватые с тем-
ными ребрами плюсны и фаланг пальцев. Подбородок, горло, бока головы книзу от глаз го-
лые, кожа желтая. Второй пуховой наряд — спинная сторона тела с буроватым оттен-
ком, брюшная—темносерого цвета. Брюхо белесовато-серое. Темный цвет на фалангах,
распространяется на плавательную перепонку (Бианки, 1913).
У молодой птицы окраска оперения сходна со взрослой, но с буроватым налетом,
развитым особенно сильно на голове. На некоторых перьях имеются узкие белые каймы.
Взрослая птица серовато-сизая. Лоб темный с буроватым налетом, распространяю-
щимся иногда на всю голову. Вокруг глаза и за глазом темное пятно. Подбородок и горло
беловатые. Внешние опахала первостепенных маховых темнобурые, внутренние — буровато-
серые. Второстепенные маховые серовато-бурые, белеющие на вершине внешнего опахала.
Средние и большие кроющие крыла и рулевые серо-сизые, только крайняя пара рулевых
с беловатыми внешними опахалами. Малые кроющие крыла бурые с беловатыми окаймле-
ниями. Нижние кроющие хвоста — бледносерые, иногда даже белые.
Клюв и ноги черные. Радужина темнобурая.
ОТРЯД ФЛАМИНГО
PHOENICOPTERI или PHOENICOPTERIFORMES
Характеристика отряда
Морфология. Внешние признаки. Крупные, очень высокие на
ногах птицы, с длинной шеей, маленькой головой и клювом своеобразного
строения. Оперение довольно мягкое и рыхлое, за исключением, конечно,
первостепенных маховых перьев. Пух покрывает аптерии и птерилии; на перьях
хорошо развиты побочные стволы. Неоперенными остаются: узкое кольцо
вокруг глаза, уздечка и кожа «подбородка». Крыло, по сравнению с размерами
птицы, очень короткое. Тип крыла диастатаксический. При сложенном крыле,
внутренние второстепенные маховые длиннее первостепенных. Число перво-
степенных маховых — 12. Хвост короткий, рулевых 12—16, средняя пара
рулевых самая длинная. В оперении обычно имеется большая или меньшая
примесь розового или красного цвета. Разницы в окраске оперения самцов
и самок нет. В оперении молодых (неполовозрелых) особей преобладает серый,
грязнобелый цвет с буроватым оттенком. Клюв фламинго массивный, несколько
превышает длину головы и в средней части круто (коленообразно) изогнут
вниз. Верхняя челюсть сравнительно узкая, невысокая и едва прикрывает
край нижней челюсти, которая, напротив, широка и глубока. Края верхней
и нижней челюстей несут мелкие роговые пластинки и зубчики, образующие
цедильный аппарат, как у гусиных.
Ноги фламинго очень длинные, причем цевка почти втрое длиннее голени.
Задний палец ноги слабо развит и помещается несколько выше передних или
(у некоторых видов) отсутствует. Передние пальцы ног соединены хорошо раз-
витой плавательной перепонкой. Когти невелики. Они илк чуть выдаются
за концы пальцев, или только прикрывают последнюю фалангу. Голень неопе-
рена более чем на половину, цевка и основная часть пальцев покрыты попереч-
ными пластинками. Глаза маленькие, радужина желтая или (у молодых) —
свинцово-серая.
Анатомические признаки. Череп десмогнатический и голо-
ринальный, ноздри сквозные; шейных позвонксде 18—19; имеется os dorsale,
342 отряд фламинго
которая образуется с участием последнего шейного позвонка; сонные артерии
парные» соединяющиеся в проксимальной части шеи; гортань бронхотрахеаль-
ного типа; шейные воздушные мешки разделяются на четыре камеры, от воз-
душных мешков отходят связанные с ними хорошо развитые подкожные по-
лости; слепые кишки больших размеров, функционирующие; имеется обводя-
щая мышца.
Образ жизни. Фламинго — дневные птицы. Населяют они мелководные
соленые озера и морские отмели. Строго придерживаются открытого ланд-
шафта. Оседлы; только особи одного вида — Ph. ruber, гнездящиеся в умерен-
ном поясе, где водоемы хотя бы на короткий срок покрываются ледяной кор-
кой, перелетны.
Основная пища — ракообразные и моллюски, а также некоторые водо-
росли и их семена. Пища добывается на мелководных участках. Она захва-
тывается клювом вместе с илом и процеживается через цедильный аппарат,
образуемый нижней и верхней челюстями.
Все фламинго моногамы. До гнездового периода фламинго ведут бродячий
образ жизни. Гнездятся колониями, которые в некоторых случаях могут
достигать очень больших размеров.
Гнезда строятся на соленом мелководье, но позднее, вследствие обмеления
могут оказаться на лишенных воды косах и островах. Они представляют собой
довольно высокую колонку из ила с примесью мелких раковин моллюсков,
с усеченным конусом, на вершине которого помещается лоток, лишенный
выстилки. Такое гнездо используется птицами только одно лето и каждый год
сооружается вновь. Фламинго постоянно гнездятся на одних и тех же мес-
тах. Отдельные колонии размещаются то близко одна от другой, то очень
далеко.
Кладка состоит из 1—2, реже 3 яиц. Срок насиживания несколько более
месяца. В насиживании принимают участие самец и самка. Фламинго — вывод-
ковые птицы. Клюв птенцов в первое время жизни, в отличие от взрослых,
прямой и лишь позднее, когда возраст птицы достигает двух недель, искри-
вляется.
Пуховых нарядов у птенцов два. Первый пуховой наряд — белый. Он
носится в течение первого месяца жизни, а затем сменяется вторым пуховым
нарядом серого цвета.
Вначале родители выкармливают птенцов отрыгиваемой пищей.
Продолжительность жизни фламинго не выяснена. В неволе они живут
недолго, вероятно потому, что не получают необходимых условий — постоян-
ного мягкого грунта в водоеме и полноценного корма.
Систематика. Отряд фламинго содержит одно семейство Phoenicopteridae
и делится на 3 рода Phoenicopterus, Phoeniconaias и Phoenicoparrus. В первом
один, во втором также один и в третьем два вида.
Достоверные древнейшие остатки представителей отряда известны из
эоцена в Англии — Elornis; из олигоцена и миоцена Европы описан Palaelodus,
из миоцена Сев. Америки Megapalaelodus. Остатки современного рода Phoeni-
copterus найдены в олигоцене в Европе, в плиоцене — в Америке. Заслуживает
внимания, что древние роды ископаемых фламинго были в известной мере пере-
ходными к настоящим голенастым и в частности имели более короткие ноги
и более прямой клюв, чем современные фламинго.
В СССР близкая к фламинго ископаемая форма Agnopterus turgaiensis
найдена в эоценовых отложениях в Актюбинской обл. близ озера Челкар-
тениз (Тугаринов, 1940).
Географическое распространение. Тропические и субтропические части
Азии, Африки, Америки, южн. Европа. В СССР — один вид Ph. ruber с огра-
ниченным распространением в Казахстане и в области Каспийского
моря.
ОБЫКНОВЕННЫЙ ФЛАМИНГО
343
РОД ФЛАМИНГО PHOENICOPTERUS LINNAEUS, 1758
Тип Ph. ruber
\. Обыкновенный фламинго Phoenicopterus ruber L.
Phoenicopterus ruber. Linnaeus. Syst. Nat., изд. X, 1758, стр. 139, Вест-Индия
Распространение. Ареал. Средиземное море, Передняя Азия на восток
до с.-з. Казахстана и Каспийского моря, Ирана (Персидский залив), с.-з.
Индии и Цейлона, южн. Африка, о-в Зеленого Мыса; средняя и Южн. Америка.
Карта 65. Распространение фламинго Phoenicopterus ruber roseus
1. Ph. r. roseus, 2. Ph. r. ruber, 3. Ph. r. chilensts.
a — граница распространения, г — места зимовок гнездящихся в СССР птиц.
Биотоп. Побережья морей и внутренних соленоводных водоемов.
Характер пребывания. Оседлая птица, совершающая перелеты
только на крайнем севере ареала, в частности в СССР.
Подвиды и варьирующие признаки. Три очень близких по внешности подвида, разли-
чающихся деталями окраски и экологии: Ph. r. roseus Pallas, 1811—в Вост. полушарии;
Ph. r. ruber Linn., 1758 — атлантическое побережье тропической и субтропической зоны
Америки, на севере в Юж. Каролине, на юге до Бразилии и Галапагосских о-вов; Ph. г.
chilensis Molina, 1776 — Перу и Уругвай на востоке до Огненной Земли.
Европейский фламинго или краснокрыл Phoenicopterus ruber roseus Pall.
Phoenicopterus roseus. Pallas. Zoographia Rosso-Asiatica. II, 1811, стр. 207, устья Волги.
Распространение. Ареал. Имеет «пятнистый» характер. Гнездовья
известны в южн. Испании и в низовьях Роны (ла-Камарг) во Франции — в
Европе; в Африке — в Тунисе, нижнем Египте, в южной части материка и на
о-вах Зеленого Мыса; в передней Азии —кроме СССР —у берегов Персид-
ского залива и в Индии, на юг до Цейлона, на восток до Ассама и вост. Бен-
галии.
В СССР фламинго гнездится преимущественно на вост. побережье Каспий-
ского моря в заливах Комсомолец (б. Мертвый Култук) и Кайдак; до самого
344 отряд фламинго
недавнего времени — по крайней мере до 1937—гнездовая колония фламинго
была в южной части Кара-Богазского залива, но теперь, повидимому, фламинго
здесь не гнездится (Исаков, 1947). Последнее место гнездовья —озеро Чел-
кар-тениз в Тургайской степи и озеро Денгиз, расположенное в 150кл< юго-
западней Акмолинска (Долгушин).
Большая колония фламинго в прошлом веке была у вост. берега Индер-
ского озера (Карелин, 1875), но была покинута из-за неумеренного преследо-
вания. Повидимому, за счет этой колонии образовались новые гнездовья по
соленым грязам (хакам) к вост. от озера Баскунчак (Генке, 1882), быть может,
на севере достигавшие Черного Яра (Яковлев, 1863—1864). Теперь там фла-
минго не гнездятся. Сведения о гнездованьи фламинго в Армении на озере Се-
ван (Сатунин, 1907) —неверны, фламинго гнездятся только к югу от границы
СССР на озере Урмия (Радугин, 1917). Ошибочно и предположение о возмож-
ном гнездовании фламинго в Ленкоранском районе (Бутурлин, 1935).
В прошлом особи, вероятно бродячие, отмечались на Эмбе, но позднее здесь
не обнаружены. В некоторые годы бродячие фламинго наблюдаются летом в
степях между сев. побережьем Аральского моря и Мугоджарами (Спан-
генберг).
Нередко фламинго залетают далеко от мест гнездования. Большой залет
на север в Зап. Европу был отмечен в 1935 г., на юг —в 1933 г. Большие
залеты к Рейну отмечены в 1728, 1811, 1876 и 1878 гг. в разное время года,
но чаще осенью. В некоторые годы залеты носят массовый характер.
Известны редкие случаи залетов в низовья Сыр-Дарьи, в вост. части пу-
стыни Кызыл-Кум, в Мервский оазис, в низовье Теджена, в Закавказье и на
Сев. Кавказ к Новороссийску, Краснодару, Дзауджикау, в Дагестану Абхазию,
на озеро Севан в Армении, на Крымский п-ов у Джанкоя. Несколько чаще фла-
минго отмечались в различных местах Украины: близ Осипенко, под Киевом,
в Подолии, в Харьковской обл., в районе Чернигова. Затем — на ю.-в. РСФСР
в Чкаловской, Куйбышевской, Пензенской, Воронежской обл., на севере даже
до Ленинграда. Значительно чаще залеты совершаются в вост. и сев.-вос. на-
правлениях от мест гнездовья. Многие авторы отмечают фламинго в сев.-вост.
частях Казахстана, в Зап. Сибири, на Алтае, в районах Канска, Ачинска,
Красноярска и Енисейска и до Байкала включительно.
Подобные залеты наблюдаются не регулярно, причиной этого является
засуха, резко понижающая уровень озер в гнездовой области, что обрекает
птиц на голодовку и вынуждает их переходить на кочевой образ жизни. Для
основной массы птиц, залетающих далеко за пределы гнездового ареала, мигра-
ция обычно кончается гибелью (Исаков и Формозов, 1946).
Гнездящиеся в СССР птицы зимуют в южной части Каспийского моря в
Азербайджане на мелководье залива Кирова, в Туркмении между Чикишляром
и Гассан-Кули;такжеи на озере Урмия1В сев. Иране. Сравнительно недавно фла-
минго зимовали в устье Балханского залива близ о-ва Даг-ада и у железнодорож-
ной станции Койлю. Зимовка прекратилась видимо вследствие обмеления моря.
Места зимовок для одних и тех же особей постоянны. Окольцованная в Кызыл-
Агачском заповеднике 18 февраля 1948 г. птица —добыта в сентябре 1948 г. в
устье Куры; другая, окольцованная 25марта 1937 г. в Гассан-Кули, ю.-з. Турк-
мении — добыта там же 14 февраля 1939 г. (Дементьев, 1945). Хотя места зи-
мовок постоянны, но в суровые зимы, напр., 1924—1925, 1944—1945, 1948—
1949, 1949—1950 гг. птицы из Кызыл-Агачского залива и Гассан-Кули отко-
чевывали на юг, в пределы Ирана; имела место и значительная смертность.
Характер пребывания. Гнездящаяся, перелетная и зимующая
птица СССР.
Весенний отлет зимующих фламинго ранний. С залива Кирова птицы начи-
нают отлетать в первой половине марта (Тугаринов и Козлова-Пушкарева,
1935). 13 марта множество перелетных фламинго наблюдалось у Ленкорани
ОБЫКНОВЕННЫЙ ФЛАМИНГО 345
(Радде, 1885). После отлета основной массы зимующих птиц остаются небольшие
группы, которые наблюдаются в районе зимовья в течение всего лета. Тоже
явление отмечено и на Туркменских зимовках. Зимующие стаи отлетают отсюда
во второй половине марта и начале апреля постепенно, передвижение начи-
нается уже в начале марта, но некоторое количество птиц остается на лето на
взморье у Гассан-Кули.
Можно предполагать, что птицы, зимующие на заливе Кирова, пересекают
Каспий и, двигаясь к северу вдоль его вост. побережья, достигают Мертвого
Култука и п-ова Мангышлак, где пролет фламинго наблюдался 1 апреля 1904 г.
(Смирнов, 1916). Отсюда, повидимому, часть птиц летит к западу и выходит
к устьям Урала. Во всяком случае Бостанжогло A911) отмечает, что движение
фламинго весной на сев. берегах Каспия идет прямо с востока от берегов Усть-
Урта, где они появляются в половине мая. В устьях Урала, по Карелину,
A875) фламинго появляется в конце апреля по ст. ст. Птицы, зимующие на
Туркменских берегах Каспия, по всей вероятности, тоже двигаются вдоль
восточного побережья моря до Мертвого Култука. По данным СушкинаA908),
линия пролета к местам гнездовья на озере Челкар-тениз минует берега Араль-
ского моря, пересекая Орско-Казалинский тракт между станциями Джелавли и
Терекли. Нам кажется, что в этой области пролет идет более широким фронтом,
чем предполагал Сушкин, захватывая северную часть Аральского моря и
низовья Сыр-Дарьи к югу приблизительно до широты ст. Джусалы. В севе-
ро-восточном углу Аральского моря (залив большой Сары-Чеганак) пролет
фламинго будто бы ясно выражен. В 1905 г. в конце апреля ст. ст. наблю-
дались огромные стаи птиц, летевших с юга и направлявшихся прямо на север
(Бостанжогло, 1911). В низовьях Сыр-Дарьи птицы пролетают весной еже-
годно, но в самом ограниченном количестве. В бывшей Тургайской обл. в
сильно запоздавшую весну 1898 г. пролет фламинго шел между 12 и 20 мая
(Сушкин, 1908).
Данные относительно осеннего отлета отличаются крайней скудностью.
В устьях Урала осенью птицы пролетают уже в середине сентября (Карелин,
1875). Бостанжогло A911) предполагал, что в конце октября фламинго поки-
дают Арало-каспийские степи. В устьях Атрека осенью птицы появляются в
середине октября. Пролет идет ночью и по утрам большими стаями по 200—
300 особей в каждой (Дементьев, 1945). Под Ленкоранью первые осенние фла-
минго прилетают в конце ноября и в самом начале декабря, пролет обычно
заканчивается к концу этого месяца. В декабре же довольно часто фламинго
наблюдаются и добываются близ Баку, где они останавливаются на отдых и
кормежку на соленых озерах.
Биотоп. Острова обширных и мелких соленых водоемов, часто недоступ-
ные для человека, вследствие мелководья и топкого, затягивающего илистого
дна; берега замкнутых или почти замкнутых мелководных лиманов на морских
островах и косах. Колонии фламинго в юго-восточной части залива Кара-Богаз
(восточный берег Каспийского моря) были осмотрены Исаковым. Одна
из них располагалась на о-ве Тараба, узкой лентой вдоль края воды у неболь-
шого лиманчика, почти полностью отделенного от залива. Отмель, занятая
гнездами, была сложена из влажного соленого ила, покрытого твердой коркой
соли и шапками осевшего гипса. Основание гнезд находилось среди мелкой воды
или на сырых участках, которые заливались во время нагонных ветров. Другая
колония у косы Сары-Тамчи занимала бессточную впадину в центре отмели.
Грунт этой впадины был постоянно влажным вследствие просачивания воды
из залива. Во влажных местах у водоемов, полностью или почти полностью
отделившихся от залива, располагались три покинутых колонии. Колониаль-
ное гнездовье фламинго в 1876 г., посещенное Генке A882) на одном из соленых
озер Заволжской степи, хотя и помещалось на сухой почве, но носило следы
воды, покрывавшей основание гнезд на 30 см.
346 ОТРЯД ФЛАМИНГО
На озере Челкар-тениз характерные конические гнезда представляли собой
издали подобие высокого кочкарника; между некоторыми гнездами, повиди-
мому, еще стояла вода (Сушкин, 1908). Таким образом все известные в пределах
Союза колонии фламинго помещались на соленых водоемах или среди крайне
мелкой воды, или на влажных участках, где, видимо, во время постройки гнезд
была вода.
Кормовыми стациями птиц также являются соленые водоемы. Фламинго
кормятся на мелководных участках моря, на соленых и сильно опресненных
озерах и лишь при крайне неблагоприятных условиях, как исключение, посе-
щают в поисках пищи настоящие пресноводные бассейны. Глубина воды кор-
мовых участков ограничена.
На морском побережье в районе Госсан-Кули птицы кормятся на мелко-
водье с илистым грунтом, относительно богатым беспозвоночными животными,
глубина этих мест около 20—25 см (Дементьев, 1945). На заливе Кирова зи-
мующие стаи избирают мелководные участки с твердым, сильно опесчаненным
или раковистым дном (Тугаринов и Козлова-Пушкарева, 1938). Глубина воды
кормовых участков обычно колеблется от 15 до 50 см, иногда бывает и больше.
В пределах Союза птицы повсеместно избегают близости человека.
Численность. У нас численность фламинго ограничена. Благодаря
тому, что основная масса наших птиц проводит зиму на узко локализованных
участках Каспия, есть возможность дать ориентировочные цифры учета. Так,
в заливе Кирова в зиму 1933—1934 гг. держалось приблизительно около 20 000
птиц (Тугаринов и Козлова-Пушкарева, 1938). Приблизительно 20 000 фла-
минго, по мнению Воробьева A940), зимовало в 1938 г. на восточном берегу Кас-
пия от Чикишляра до иранской границы. Меньшие цифры указываются Лапте-
вым, который считает, что здесь зимует около 10 000 фламинго. В 1942 г. по
подсчетам Дементьева A945) там же держалось 6 000—8 000 фламинго, но под-
счет производился довольно поздно (в марте), когда часть особей уже могла
откочевать от мест зимовок по направлению мест гнездования. Количество
птиц, проводящих зиму на сев. Иране, остается неизвестным, но повидимому
оно меньше, чем в наших пределах. Используя эти подсчеты, можно предполо-
жить, что в пределах Союза проводит теплое время года не более 50 000 птиц.
Основываясь на данных литературы, можно думать, что в прошлом фламинго
встречались у нас в несравненно большем количестве и их численность к настоя-
щему времени резко снизилась.
Экология. Размножение. Гнездится всегда колониями. Размеры
гнездовых поселений различны, но, как правило, состоят из большого числа
особей. Одна из колоний, осмотренных на заливе Кара-Богаз, состояла из
184 гнезд, другая не менее чем из 300 гнезд (Исаков, 1948). Колония, осмотрен-
ная в июле 1876 г. в Заволжской степи, состояла из 419 гнезд (Генке, 1882).
Не менее 200 гнезд также насчитывалось в разоренной колонии на Индерском
озере (Карелин, 1875). Гнезда фламинго —конические колонки с усеченной
вершиной и кашеобразным углублением-лотком наверху. 14 вынуждено
покинутых птицами гнезд, измеренных Исаковым на заливе Кара-Богаз, имели
следующие размеры: высота колебалась от 10 до 32 см, диаметр основания от
40 до 48 см, диаметр лотка — от 21 до 23 см. Гнезда, осмотренные в свое время
Генке A882) в заволжской степи, достигали 61 см высоты. Более высокие
гнезда бывают повидимому в тех случаях, когда они закладываются среди
мелкой воды, где птищам легче брать строительный материал.
Возможно даже, что цифры, которые приводит Генке, несколько приумень-
шены, т. к. он имел дело с колониями, уже брошенными птицами. Гнезда в них
могли несколько деформироваться, в то время как жилые гнезда птицы ремон-
тируют в течение всего периода высиживания. Строительным материалом
служит ил (Генке, 1882), соленый ил с примесью мелких створок — Cardium
(Исаков, 1948). На о-ве Тараба большинство гнезд имели в основании большие
ОБЫКНОВЕННЫЙ ФЛАМИНГО
347
шапки осевшего гипса. В промежутках между ними, там, где ил более влажен,
птицы брали его клювом и укладывали на гипсовое основание. Клювом же они
приминали материал, а иногда утрамбовывали его ногами. Рядом с гнездом
образовывались ямки, ил из которых был выцарапан клювом. Подстилки
в лотке не бывает, и яйца откладываются прямо в высохшее углубление (Иса-
ков, 1948). В колониях фламинго отдельные гнезда размещаются близко друг
от друга. Так, на о-ве Тараба 184 гнезда занимали полосу около 67 м в длину
и в среднем 5—6 м в ширину. Местами эта полоса то суживалась до 2 м, то,
что бывало реже, расширялась до 7 м.
Некоторые гнезда почти примыкали
друг к другу. В среднем одно гнездо
занимало 1,14 м2.
Фламинго не пользуется старыми
гнездами, а, гнездясь в следующем
году на тех же косах и островах,
строит новые гнезда. Новая колония
помещается поблизости от прошло-
годней.
Количество яиц в кладке колеб-
лется от 1 до 3 (Генке, 1882). Яйца
удлиненно-овальной формы с сильно
заостренным концом. Основной цвет
скорлупы оливково-зеленый, но по-
крыт слоем белой извести. Разме-
ры: A00) 77—103,5 X 47,7—60,1, в
среднем 88,82 х 54,58 (Уайзерби,
1939).
Сроки размножения у фламинго,
повидимому, сильно растянуты, при-
чем отдельные пары одной и той же
колонии приступают к гнездованию
не одновременно. На восточном берегу
Каспия (залив Кара-Богаз) откладка
яиц начинается с первой трети мая *.
6 мая в ныне не существующей ко-
лонии на о-ве Тараба были найдены
скорлупки разбитых яиц. Три самки,
добытые 8 и 12 мая, имели готовые
к откладке яйца в яйцепроводах,
а добытая 11 мая уже отложила яйца
(Исаков, 1948). В Заволжской степи,
в колонии, посещенной 21 июля,
птенцы выклюнулись лишь из поло-
вины яиц. В остальных гнездах были яйца в количестве от 1 до 3 (Генке,
1882). На озере Челкар-тениз, по предположению Сушкина A908), фла-
минго также поздно приступает к размножению. Самки, добытые им там
в середине июня, обладали слабо развитыми наседными пятнами (повидимому
насиживание только начиналось). Согласно полученным тем же автором устным
сведениям, в один год, когда Челкар-тениз сильно обмелел и наступившие
ранние морозы сковали жидкую грязь, добравшиеся до гнезд фламинго казахи
обнаружили, что во многих случаях птенцы еще не могли летать. В насижива-
нии яиц принимают участие повидимому оба родителя, при этом птицы сидят
Рис. 72. Фламинго
* В сроках размножения имеются и географические различия — в южной Испании
оно начинается только в конце мая (Уайзерби, 1939}.
348 отряд фламинго
на гнездах, поджав ноги. Промежутки между откладкой самкой отдельных
яиц остаются невыясненными. Продолжительность насиживания (по наблю-
дениям в Испании) 30—32 дня.
В первые же дни жизни птенцы настолько самостоятельны, что, будучи
потревоженными, покидают гнезда. До двухнедельного возраста у птенцов
клюв остается прямым. Нормально птенцы находятся в гнездах около 4 дней,
а затем бродят около гнезд, возвращаясь в них при появлении родителей с пи-
щей. Вначале родители кормят птенцов отрыгнутой ими пищей. В возрасте
двух недель птенцы сами начинают добывать пищу, при этом ведут себя, как
взрослые, т. et опускают в воду клюв и голову.
Питание. Недостаточно выяснено. Указание Генке A882) будто бы
фламинго вскармливают птенцов лягушатами, добывая их в пресноводных озе-
рах, сомнительно. По Мензбиру A918) взрослые фламинго питаются мелкими
морскими слизняками. Еще в начале прошлого столетия подмечено, что большое
значение в питании фламинго имеет одноклеточная водоросль (Aphanothece
sabina). Взвешенные в воде массы ее, обладающие красным цветом и издающие
сильное зловоние, набиваются волнами в тихие морские заливчики, где по-
едаются птицами (Максимович, 1896). Значение этого корма подтверждается и
последними исследованиями Исакова A948) на восточном берегу Каспия. 7 же-
лудков фламинго, добытых им в первой половине мая 1938 г., содержали эту во-
доросль в значительном количестве. Во всех упомянутых желудках найдены
также семена руппии (Ruppiamaritima), занимавшие по объему не менее поло-
вины всего содержимого желудка. Эта водоросль растет в солоноватых родниках
и ее семена после созревания смываются в морские заливы. Зимующие под
Ленкоранью фламинго питаются моллюсками, главным образом мелкими
Cardium (Радде, 1885, Сатунин, 1912). По Исакову A948), зимнее питание
птиц состоит, главным образом, из ракообразных и мелких моллюсков.
Совершенно исключительное значение в питании фламинго на заливе Кара-
Богаз и на озере Урмия в сев. Иране, по мнению Исакова, имел в прошлом
рачёк Artemia salina и его икра. Однако ныне, вследствие повышенной солено-
сти воды в Кара-Богазе, рачек этот исчез совершенно. Пища всегда добы-
вается из воды, причем птицы топчутся на мелководном маленьком участ-
ке, процеживая через клюв воду. Клюв при питании обращен назад к
ногам, вверх нижней челюстью. По Дементьеву A945), птицы двигаются
вперед, поворачивая голову боком и зачерпывая верхней челюстью ил. В
одних случаях (например, на заливе Кара-Богаз) птицы погружают в воду
только клюв, что вызвано, вероятно, большой соленостью воды, действую-
щей на слизистую оболочку глаза (Исаков, 1948). В других (на Ленко-
ранских зимовках, в районе станции Челкар в Казахстане и в ряде других
пунктов) птицы погружают в воду не только голову, но и часть шеи (Спан-
генберг). Кормятся птицы, главным образом, ночью, но нередко и при
дневном освещении. По Дементьеву A945), утром и после полудня.
В поисках пищи в районах гнездования птицы предпринимают еже-
дневные далекие перелеты, в районе озера Челкар-тениз и в заволжских
степях до 30—40 км (Генке, 1882, Сушкин, 1908), на заливе Кара-Богаз до
50—60 км. В последнем месте перелеты были хорошо выражены до второй
половины мая, затем стали менее интенсивны и после 5 июня совершенно
прекратились (Исаков, 1948). Вероятно подобные перемещения птиц вызваны
ограниченностью кормовых территорий.
Линька. Фламинго линяют дважды в году. Полная линька — после-
брачная — начинается в конце периода размножения в июле и продолжается
до ноября или декабря. Почти сразу после ее окончания проходит частичная
предбрачная линька, захватывающая мелкое оперение; она отмечена между
январем и маем.
Смена нарядов следующая: первый месяц жизни птенцов — белый пухо-
ОБЫКНОВЕННЫЙ ФЛАМИНГО 349
вой наряд, второй месяц жизни — серый пуховой наряд. После этого — гнез-
довый (первый) наряд, причем первые перья этого наряда появляются на пле-
чах и груди птенца уже на пятой неделе. Гнездовий наряд сменяется на местах
зимовок на окончательный. Во время послебрачной линьки у фламинго одно-
временно выпадают все маховые и рулевые перья — птица в это время полно-
стью теряет способность к полету. На линьку фламинго массами собираются
на недоступных водоемах, где держатся вместе с линяющими лебедями и дру-
гими водоплавающими. С давних пор местом массовой линьки у нас является
залив Мертвый Култук, где с половины июля фламинго собирались раньше в
таком несметном количестве, что берег на большом протяжении казался как бы
покрытым алым сукном (Мензбир, 1918). По сообщениям Долгушина фламинго
линяет в большом числе на озере Денгиз в Акмолинской области, где часто
ловится и убивается местным населением палками и веслами.
Полевые признаки. В природе фламинго безошибочно отличаются от всех
птиц нашей фауны очень длинной шеей, загнутым клювом, длинными ногами
и яркой окраской (взрослые особи). Обычно фламинго держатся стаями, реже
маленькими группами и, как исключение, одиночками. Они то медленно
бродят по мелководью, часто опуская клюв и голову в воду, то плавают, изо-
гнув шею, как лебедь-шипун. При взлете пробегают по неглубокой воде до 5—
6 м. Во время полета птицы выстраиваются косой линией, но нередко летят
беспорядочной стаей. Благодаря длинным вытянутым ногам, шее и коротким
крыльям, на лету фламинго имеет форму креста. Летят птицы сравнительно
быстро, как гуси, часто взмахивая короткими крыльями. Голос — протяжное,
негромкое и спокойное гоготание, без резких, как у гусей, нот (выкриков).
Во время кормежки и отдыха фламинго очень осторожная птица. Они не под-
пускают близко человека. В то же время они могут налететь на охотника на
самое близкое расстояние.
Описание. Размеры и строение. Крупная, высокая на ногах птица. Размеры
сильно колеблются в зависимости от пола и возраста. Вес взрослых самцов, добытых в мае,
3 900—4 400 г, самок 3 050—3 400 г. Вес молодых особей, добытых в мае и феврале, 1960—
2 400 г (Исаков, 1948); самки, добытые в конце марта, весили 2200 и 2400 г (Дементьев,
1950). Крыло 21 самцов и самок, включая и несколько неполовозрелых птиц, 435—490, в
среднем 459 мм. Клюв птенцов до двухнедельного возраста прямой.
Окраска. Пуховички одеты белым пухом, который в месячном возрасте сменяется
на серый.
Молодые (неполовозрелые) особи, в отличие от взрослых, окрашены в грязновато-
белое и серое оперение со слабым развитием розового налета на нижней части шеи, зобе и
крыльях. Ноги, клюв и глаза свинцово-серые. Взрослый наряд одевается птицами на тре-
тий год жизни.
Окраска оперения взрослых самцов и самок (сходна и в брачном и в послебрачном
нарядах) — белая с розоватым налетом, развитым наиболее сильно на шее, темени и перед-
ней части спины. Верхние и нижние кроющие крыла, подмышечные и внутренние второсте-
пенные — ярко розовые. Розовый налет распространяется также на рулевые. Первостепен-
ные и второстепенные маховые черные. Розовая окраска особенно интенсивна у старых
особей. Неоперенные части головы: уздечка, кольцо вокруг глаза— красные; клюв из ком-
бинации красного и черного цветов. Ноги розовые, радужина желтая.
ОТРЯД
ГОЛЕНАСТЫЕ ПТИЦЫ
GRESSORES или CICONIIFORMES
Характеристика отряда
Морфология. Внешние признаки. Подавляющее большинство
относящихся к данному отряду видов — птицы среднего и крупного размера.
Особенно велики виды, населяющие тропические страны (некоторые аисты и
цапли). В нашей фауне самые крупные голенастые: белый аист — вес до 4 кг,
черный аист около 3 кг, серая цапля вес до 2 кг, колпица вес более 1,5 кг.
Другие представители отряда:выпь, кваква, каравайка, малая белая цапля и
пр. имеют средние размеры. Вес выпи немного менее 1 кг, кваквы несколько
более 0,5 кг, каравайки около 0,75 кг. Самые мелкие представители голенастых
птиц — волчки — не крупнее водяной курочки-камышницы. Вес волчка не
более 150 г, еще мельче амурский волчок.
Перья у мелких видов относительно мягкие, у крупных — более жесткие,
плотно прилегающие к телу. Неоперенными всегда остаются цевка и обычно
нижняя часть голени. Неоперены полностью или частично уздечка и кольцо
вокруг глаза; у некоторых ибисов во взрослом состоянии неоперена голова
и шея. У марабу, питающихся главным образом падалью, голова и шея по-
крыты редким пухом. Форма кроющих перьев — широкая, округлая. У всех
представителей отряда, за исключением некоторых аистов, имеются побочные
стволы пера. У ряда видов цапель в гнездовый период на голове, спине и зобе
развиваются украшающие рассученные перья; некоторые ибисы имеют слегка
рассученные внутренние второстепенные маховые перья. Число первостепен-
ных маховых 10—12, рулевых 8—12. Крыло диастатаксическое, сравни-
тельно с величиной птицы, большое, широкое и тупое, хвост короткий, округ-
лый. У многих видов (цапли) имеется так называемый порошковый пух.
Окраска оперения разнообразна. Как правило, оба пола окрашены оди-
наково или близки по своей окраске. Исключение в нашей фауне предста-
вляют волчки, обладающие хорошо выраженным половым диморфизмом.
ХАРАКТЕРИСТИКА ОТРЯДА 351
У многих видов молодые птицы по окраске оперения и неоперенных частей тела
отличаются от взрослых, но сезонные различия в оперении чаще отсутствуют
или выражены (например, у ряда цапель) лишь особыми «украшающими»
перьями, которые по окончании брачного периода выпадают.
Голенастые — длинноногие и длинношейные птицы, за редкими исключе-
ниями они обладают длинным клювом, форма которого разнообразна. У боль-
шинства цапель и аистов клюв более или менее прямой и острый, у некоторых
аистов вершина клюва несколько изгибается вверх, напротив, у многих ибисов
клюв изогнут вниз. У колпиков клюв плоский, расширенный к концу подобно
веслу или лопатке, у китоглавов —массивный, челнообразной формы.
Ноги четырехпалые с длинными пальцами, причем задний палец поста-
влен на уровне передних или несколько выше их. Обыкновенно между средним
и наружным пальцем имеется зачаточная перепонка, реже перепонкой обтя-
нуты основания трех пальцев. У большинства видов когти длинные, слабо
изогнутые, у других более короткие и тупые. Несмотря на длину ног голена-
стые не могут бегать, ходят медленно и, спасаясь от опасности, прибегают
только к полету.
Анатомические признаки. Череп десмогнатический, голо
ринальный (кроме Plegadidae), без базиптеригоидных отростков, две сонных
артерии, обводящей мышцы нет.
Шея у всех представителей отряда длинная, шейных позвонков от 16 до
20. Скелет туловища сравнительно легкий и слабый; пневматичность скелета
хорошо развита; шейные воздушные мешки разделены на четыре-пять камер;
большая грудная мышца из двух слоев (кроме ибисов); слепые кишки рудимен-
тарны; копчиковая железа оперенная; у самцов имеется рудимент полового
члена.
Образ жизни. Одни (аисты) ведут строго дневной образ жизни, другие
(цапли, ибисы) сумеречные птицы, но многие бывают деятельными и днем, и в
ночное время, наконец ряд видов (в нашей фауне выпь, кваква) настоящие ноч-
ные птицы. Тесно связаны с водой или хотя бы с влажными, затопленными
пространствами, лишь некоторые населяют сухие травянистые территории.
Виды, населяющие тропический пояс и южные части умеренного пояса, где
водоемы не замерзают в течение круглого года или замерзают на короткое
время, — оседлы, виды из более северных частей ареала — перелетны.
По роду своего питания голенастые должны быть отнесены к группе полк-
фагов или пантофагов, т. е. кормящихся разнообразной животной пищей (ры-
бой, амфибиями, рептилиями, насекомыми, в небольшой мере грызунами,
величиной до суслика, и молодыми птицами). Много реже встречаются настоя-
щие рыбоядные виды — ихтиофаги и падалыцики — сапрофаги (марабу).
Поедание растительной пищи, за исключением немногих видов, —довольно
редкое явление и происходит, вероятно, более или менее случайно при загла-
тывании животных. В небольшом количестве растительная пища в виде зеленых
частей растения и семян поедается некоторыми ибисами (у нас каравайкой и
колпицей). Добыча проглатывается всегда целиком, расчленять ее на части
голенастые не могут (исключение составляют марабу, отрывающие куски от
падали). Тем не менее размеры добычи могут быть довольно крупными. Серые
цапли свободно проглатывают взрослых крапчатых сусликов, аисты — домаш-
них цыплят и диких утят величиной с голубя, выпи — крупных водяных
крыс, кваквы — крупных лягушек.
Пища добывается то в воде, то на земле, в сухих лугах, реже склевывается
с травы и с ветвей деревьев (мелкие цапли). Способы добывания пищи у различ-
ных представителей отряда неодинаковы. Ибисы нащупывают пищу клювом
в иле мелководных, часто совершенно мутных водоемов или ловят насекомых
на сухих участках. Другие руководствуются только зрением, причем аисты
в поисках пищи неспеша расхаживают по берегам разливов или лугам, а
352 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
цапли — часами неподвижно стоят в неглубокой воде или в камышах, выжи-
дая, когда добыча сама приблизится на нужное расстояние, и значительно реже
ведут себя более активно — как аисты. Быстрота движения, необходимая при
ловле в воде рыбы, достигается у цапель благодаря своеобразному строению
очень подвижной длинной шеи, которая в спокойном состоянии бывает втянута
в плечи, а при ударе стремительно выбрасывается вперед. Пищеварение у голе-
настых птиц происходит весьма энергично, потребность в пище весьма значи-
тельна.
Голенастые птицы — моногамы. После окончания периода размножения
одни виды держатся семьями, другие образуют крупные стаи, но некоторые
(например, выпи, волчки) во внегнездовое время живут одиночками. Соответ-
ственно этому перелеты также совершаются то семьями (осень), то стаями, то
отдельными особями. Молодые птицы очень часто летят отдельно от взрослых.
У многих видов весной особи одного пола прилетают раньше другого. Неко-
торые виды, рано вернувшись на родину, вскоре приступают к разхмножению,
другие, прилетев также рано, сравнительно долго ведут бродячий образ жизни,
как бы выжидая благоприятного времени. Наконец, третьи прилетают поздно
и поздно гнездятся. К последним относятся виды, которые питаются и вскар-
мливают птенцов главным образом насекомыми. У некоторых видов в сроках
начала гнездовья северных и южных популяций бывает разница в 2—2,5 ме-
сяца. У других размножение начинается в одни и те же или близкие сроки
в различных частях ареала. Немногие цапли (у нас выпи, волчки, реже другие),
ибисы (китайский ибисы) и в подавляющем большинстве случаев аисты гнез-
дятся отдельными парами. Значительно чаще голенастые птицы гнездятся
колониями, нередко состоящими из нескольких видов. Сюда относятся многие
цапли, колпики, каравайки.
Большинство голенастых строят гнезда в зарослях тростника, камыша,
затопленных полой водой кустарников лозы, ольхи или на более значительной
высоте — на деревьях. Некоторые виды (например, амурский волчок) устраи-
вают гнезда среди высокой и густой травы или мелких кустарниковых зарослей
сухих участков. Аисты гнездятся на деревьях и на сооружениях человека (на
крышах построек, на стогах соломы и сена, на выставленных на шестах коле-
сах и боронах). В горных районах гнезда аистов (черный аист) часто поме-
щаются на карнизах скал. В различных частях ареала один и тот же вид в зави-
симости от местных условий может гнездиться по-разному. Серые цапли гнез-
дятся то на вершинах старых деревьев, то на земле среди зарослей, черные
аисты — на деревьях и на скалах, молотоглавы — на кустах, деревьях и ска-
лах. Гнезда сооружаются самими птицами и у некоторых видов (аисты) исполь-
зуются в течение многих лет подряд, причем вследствие ежегодной их до-
стройки, достигают иногда огромных размеров. Другие виды или подправляют
прошлогодние гнезда, часто принадлежащие другим парам или близким по
размерам видам, или строят новые гнезда. Для всех цапель характерны просто
устроенные легкие гнезда, сохраняющие свою форму (перевернутый конус)
при гнездовании в самой разнообразной обстановке — на ветвях деревьев, на.
стеблях тростника и на земле. Строительный материал—стебли расходятся
радиально и, если гнездо находится на дереве, сквозь них с боков и снизу
обычно видны яйца. У молотоглавов гнездо представляет собой шарообразное
сооружение из ветвей, достигающее двух метров в диаметре; оно помещается
на кустах, деревьях и карнизах скал. Внутри гнездо разделено на три камеры
и имеет входное отверстие сбоку. В постройке гнезд голенастых участвуют
оба родителя, но, как правило, самцы доставляют материал, а самки размещают
его в гнезде.
Спаривание происходит на ветвях гнездового дерева, на тростнике у гнезда
или в самом гнезде. У аистов ему предшествует своеобразное токование, выра-
жающееся в закидывании головы назад и усиленном трещании клюва.
ХАРАКТЕРИСТИКА ОТРЯДА 353
Кладка у крупных цапель из 2—5, реже 6 яиц, у мелких цапель даже из
9 яиц, у молотоглавов из 3 и больше, у китоглавов из 2, у аистов из 4—5,
реже 6 яиц, у ибисов обычно из 3—4 яиц. Число яиц в кладках нередко варьи-
рует географически. Наибольшее число яиц в кладках бывает близ границы
распространения, что можно объяснить, видимо, более частой гибелью молод-
няка в менее благоприятной обстановке. Колебание числа яиц в кладке отме-
чено и в различные годы.
Эмбриональное развитие относительно быстрое у мелких видов и более
медленное у крупных, постэмбриональный рост обычно быстрый. Насиживание
начинается после откладки первого яйца, но у большинства видов до откладки
третьего яйца самка не сидит на гнезде прочно, подолгу выстаивая на гнезде
над отложенными яйцами. Вследствие этого у многих видов разница в размерах
птенцов одного гнезда незначительна, у других, напротив, выражена очень
сильно. При больших кладках 1—2 яйца оказываются неоплодотворенными.
Насиживание производится обеими птицами гнездовой пары, причем у одних
самцы принимают в этом деле большое участие, у других они сменяют самку
на короткое время. В связи с этим наседные пятна у различных групп бывают
развиты неодинаково. У цапель самок наседные пятна вообще развиты слабо,
у самцов их удается обнаружить лишь при самом тщательном осмотре. Инку-
бационный период зависит от величины птицы и продолжается от 17 до 32 дней.
У основной массы представителей птенцы вылупляются совершенно беспомощ-
ными, имеют только один пуховой наряд. Напротив, птенцы аистов зрячи
с первого дня жизни и по типу постэмбрионального развития стоят на грани
между птенцовыми и выводковыми птицами. Их пуховой наряд впоследствии
заменяется более длинным и густым вторым пуховым нарядом.
У всех голенастых вскармливание птенцов производится обоими родите-
лями. Пока птенцы малы, самка большую часть времени остается на гнезде
и лишь после того, как птенцы достаточно подрастут, вместе с самцом улетает
за пищей. Вначале птенцы вскармливаются мелкими животными (насекомыми,
их личинками, позвоночными). Позднее родители передают им более крупную
добычу. Птенцы отличаются исключительной прожорливостью и их вскармли-
вание требует от взрослых большого напряжения. Мелкие виды остаются в
гнездах сравнительно короткий промежуток времени и очень часто, еще не
умея летать, перебираются на ветви гнездового дерева или лазают по тростни-
кам. Другие — крупные цапли, аисты — остаются в гнездах очень долго и
покидают их, когда крылья достаточно окрепнут. Незадолго до вылета молодых,
старики вначале сокращают кормление, а затем вовсе прекращают приносить
в гнездо пищу и тогда голод вынуждает птенцов покинуть гнездо и присоеди-
ниться к взрослым. Молодые надевают окончательный наряд на следующую
весну или через год и к этому времени становятся способными к размножению.
Потенциальные возможности однако не всегда реализуются, и молодые птицы
вероятно часто составляют резерв холостых производителей, не участвуя в
размножении-
Птенцы голенастых птиц растут быстро, особенно в первый период до фор-
мирования крупного оперения. Птенцы серой цапли к двухнедельному воз-
расту увеличиваются в весе с 39 до 1875 г. Дальнейший прирост оказывается
незначительным. Уменьшение скорости роста особенно резко выражено в
18-, 20-дневном возрасте, когда происходит наиболее интенсивный рост крупных
перьев (маховые увеличиваются в длине на б—8 мм в день). Приблизительно
в двухмесячном возрасте птенцы достигают почти окончательных размеров.
Рост птенцов кваквы продолжается около 25 дней. За это время, считая с мо-
мента вылупления и до конца постэмбрионального развития, их вес возрастает
с 25 до 650 г. Постэмбриональный рост кваквы по сравнению с ростом серой
цапли протекает менее интенсивно. Периодичность роста и в данном случае
ясно выражена, так как с 20-дневного возраста прирост становится очень незна-
23 Птшцы СССР, т. II
354 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
чительным (наиболее интенсивный рост маховых перьев кваквы наблюдается
как раз с 21-го дня жизни). Быстро растут и птенцы белого аиста. Начальный
вес птенца 67,2 г; к 20 дням жизни увеличивается в 20 раз, а к 40 дням, когда
рост почти закончен, — в 40 раз, доходя до 3 кг, т. е. почти до размеров взрос-
лой птицы.
Вопрос о продолжительности жизни голенастых птиц остается до послед-
него времени не разрешенным. Большое число особей погибает уже в первом
году жизни. Из результатов кольцевания следует, что серая цапля живет до
16 лет, рыжая цапля до 13 лет, кваква 10 лет, черный аист 8 лет и каравайка
10 лет. Цифры эти, конечно, приблизительны.
В неволе известны случаи более длительного выживания некоторых пред-
ставителей голенастых, вероятно близких к потенциальным возможностям жиз-
ни того или иного вида. В частности известен случай выживания черного аиста
в течение 30 лет, а серой цапли в течение 20—22 и даже 60 лет (Шульпин, 1940).
Систематика. К отряду голенастых относятся пять семейств: ибисы Pie-
gadidae, аисты Ciconiidae, китоглавы Balaenicipitidae, теневые цапли или моло-
тоглавы Scopidae и цапли Ardeidae. Ископаемые голенастые с признаками,
общими с собственно голенастыми и фламинго, описаны были уже из меловых
отложений (сем. Scaniornithidae в Дании). Но принадлежность их к голена-
стым остается не вполне ясной. Ибисы появляются в миоценовых отложениях
в Европе, аисты — в нижнем олигоцене в сев. Африке, несколько позже также
в плигоцене, а затем в миоцене—плиоцене —в Европе. Отдельные находки
аистов в плиоцене известны из Индии, сомнительные в Сев. Америке. Досто-
верные находки ископаемых аистов в западном полушарии относятся только
к плейстоцену.
К ибисам принадлежат около 30 видов средних размеров птиц, широко
распространенных в жарком и умеренном поясах земного шара. Сюда относятся
роды: Threskiornis, Plegadis, Nipponia, Guara, Comatibis, Platalea, Ajaja и др.
Характерные особенности их: хизоринальный череп; шейных позвонков
17—18; задний край грудины с двумя парами вырезок; большая грудная мышца
из одного слоя; зоба нет, первостепенных маховых 11, рулевых 12; пух распро-
странен и по птерилиям и по аптериям; устройство трахей разнообразно,
у некоторых трахеи образуют извивы. По ряду признаков (наличие затылоч-
ных фонтанелей, строение processus angularis posterior и др.) ибисы прибли-
жаются к фламинго.
К аистам относятся 17 видов крупных и очень крупных птиц, распростра-
ненных, преимущественно, в тропиках и отчасти в умеренном поясе Старого
и Нового света. К ним принадлежат роды: Ciconia, Sphenorhynchus, Anastomus,
Dissoura, Leptoptilus, Ephippiorhynchus и др. Их особенности заключаются
в голоринальном черепе, лишенном затылочных фонтанелей, в наличии пяти пар
ребер, в слабом развитии заднего пальца и обводящей мышцы, в наличии двух
слоев большой грудной мышцы, в частности редукции голосовых связок и пере-
понок (почти немые), в сильном развитии носоглоточной системы воздушных
мешков, в отсутствии зоба, в групповом расположении желез в железистом
желудке, в рудиментах двух слепых кишек, в наличии рудимента penis. Пух
распределен, как по птерилиям, так и по аптериям; первостепенных маховых
и рулевых — 12; на первом пальце кисти имеется коготь.
К китоглавам относится один род (Balaeniceps) с одним видом В. rex,
населяющим верховья рек Конго и Нила. Китоглавы — крупные птицы,
обладающие челнообразным клювом. Их череп голоринальный; затылочных
фонтанелей нет; ноздри не сквозные; шейных позвонков 16—17; мускулы
гортани и обводящая мышца редуцированы; только одна (правая) сонная арте-
рия; отсутствует мускульный желудок; единственная рудиментарная слепая
кишка; рудимент penis отсутствует; первостепенных маховых 11, рулевых 12;
на спине развит короткий пух.
ХАРАКТЕРИСТИКА ОТРЯДА 355
Молотоглавы или теневые цапли — птицы средних размеров — предста-
влены одним родом Scopus с одним видом S. umbretta, населяющим Африку и
Мадагаскар. Череп голоринальный с затылочными фонтанелями; ноздри не
сквозные; 16 шейных позвонков; шейные воздушные мешки не делятся на по
лости; обводящая мышца отсутствует; на когте среднего пальца имеются зазуб-
рины; маховых — 10; рулевых — 12.
К цаплям относятся чрезвычайно разнообразные по величине птицы совсем
мелких, средних и крупных размеров. Общее число видов около 65. Они при-
надлежат к 11 родам, распространенным, главным образом, в тропическом
и умеренном поясах земного шара. Сюда относятся роды: Ardea, Egretta, Bu-
bulcus, Ardeola, Butorides, Nycticorax, Ixobrychus, Botaurus, а из экзотических
родов — Cochlearia, Tigrosoma и другие.
Череп цапель голоринальный, ноздри сквозные, с отверстиями в между-
глазничной перегородке, без затылочных фонтанелей, шейных позвонков 19—
20, пара вырезок на заднем крае грудины, без обводящей мышцы, с гортанью
трахео-бронхиального типа, с двумя сонными артериями (кроме Ixobrichus),
с неразделенными на полости шейными воздушными мешками, без зоба, с одной
рудиментарной слепой кишкой, без penis. Первостепенных маховых 11, за
исключением рода Cochlearia, имеющих 19 первостепенных маховых. Рулевых
8—12, побочные стволы перьев сильно развиты, пух распределен только по
аптериям, а в области груди и поясницы с наличием порошкового пуха. Копчи-
ковая железа рудиментарна и оголена, ни первом пальце кисти коготь отсут-
ствует, на крае когтя среднего пальца ноги имеются зазубрины.
Географическое распространение. Голенастые птицы распространены очень
широко, но наибольшее число видов приурочено к жаркому поясу. В пале-
арктической области и, в частности, в СССР в качестве гнездящихся или залет-
ных встречаются представители родов: Platalea, Plegadis, Threskiornis, Nippo-
nia, Ciconia, Ardea, Egretta, Bubulcus, Ardeola, Butorides, Nycticorax, Ixobry-
chus, Botourus. Общее количество видов, встречающихся в СССР, 21, что соста-
вляет 20,56% от общего числа видов этого отряда в мировой фауне. Из этого
количества к Plegadidae относятся 4 вида, к Ciconidae 2 вида и к Ardeidae
16 видов. Виды, принадлежащие к семействам Balaenicipitidae и Scopidae, пол-
ностью отсутствуют в нашей фауне.
Распределение голенастых птиц фауны СССР по зонам неравномерно.
В арктической зоне до лесотундры включительно голенастые птицы на гнездовье
отсутствуют. Бедно населена голенастыми также таежная полоса нашей страны,
куда более или менее нормально заходят в качестве гнездящихся представи-
телей лишь 2 вида —черный аист и выпь.
Далее к югу, в области лиственного леса, лесостепи, степи и пустыни,
при наличии соответствующих биотопов численность резко возрастает, стано-
вясь особенно большой в равнинных южных частях нашей страны. В горных
областях голенастых птиц встречается мало.
На большей части территории Союза голенастые птицы ведут правильный
перелетный образ жизни, прилетая весной к периоду размножения и улетая
на холодное время года. Только в самых южных местностях Европейской и
Азиатской частей нашей страны, где заморозки кратковременны и где сохра-
няются незамерзающие водоемы, некоторые виды зимуют или, еще реже, живут
оседло.
Практическое значение голенастых весьма ограничено. Промысловыми
видами, используемыми на мясо, принято считать всех ибисов и немногих
цапель (кваква, выпь). Другие цапли («белые цапли») для охотников-промысло-
виков раньше представляли большую ценность в отношении использования
украшающих (брачных) и прочих перьев, которые до империалистической
войны пользовались большим спросом за границей. В результате этого про-
мысла численность большой белой цапли повсеместно была сокращена до мини-
356 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
мума. В настоящее время промысел цапель ради перьев запрещен и численность
видов, находившихся под угрозой полного исчезновения, за последнее время
вновь восстановлена.
Некоторые голенастые птицы, питаясь рыбой, приносят известный вред
рыбному хозяйству. Однако сейчас хорошо известно, что наряду с вредом
голенастые приносят несоизмеримо большую пользу, уничтожая вредных для
сельского хозяйства насекомых. Об истреблении голенастых птиц, мотивируя
их вредной деятельностью для человека, не может быть и речи.
Таблица для определения видов голенастых птиц
1 B). Клюв длинный и плоский, расширяющийся к концу подобно округлой лопатке
колпица Platalea leucorodia
2 A). Клюв различной формы, но всегда сужающийся к концу 3
3 D). Клюв заметно загнут книзу 5
4 C). Клюв почти прямой 9
5 F). Окраска оперения каштанового цвета с металлическим блеском в летнее время
и каштаново-бурая с белыми пестрянками на шее осенью и зимой * .
каравайка Plegadis falcinellus
6 E). Окраска иная 7
7 (8). Оперение почти все белое, только концы третьестепенных маховых и плечевых
темные, у старых голова и шея покрыта голой черной кожей, у молодых голова и шея покрыты
черно-бурыми с белыми краями перьями
священный ибис Threskiornisaecthiopica
8 G). Оперение все бело-розовое или из сочетания бело-розового (снизу) и серого (верх-
няя сторона тела) цветов, кожа неоперенных частей головы оранжево-красная
красноногий ибис Nipponia nippon
9 A0).Ноги красные. Окраска оперения из комбинации белых и черных цветов . . 11
10 (9). Ноги и оперение иной окраски 13
11 A2). Оперение на спинной стороне без белого цвета
черный аист Ciconia nigra
12 A1). Оперение белое, за исключением концов крыльев
белый аист Ciconia ciconia
13 A4). Все оперение белое 15
14 A3). Окраска оперения иная 21
15 A6). Рассученные перья спины много длиннее хвоста, на затылке хохол отсут-
ствует или слабо выражен 17
16 A5). Рассученные перья спины короче — почти равны хвосту, хохол на затылке
хорошо развит 19
17 A8). Размеры крупные, крыло не короче 360 мм
большая белая цапля Egretta alba
18 A9). Размеры мелкие, крыло короче 340 мм
средняя белая цапля Egretta intermedia
19 B0). На затылке хохол из 2—3 длинных перьев, клюв черный
малаябелаяцапля Egretta garzetta
20 A9). Хохол из 10—30 длинных перьев, клюв желтый
желтоклювая чепура нужда Egretta eulophotes
21 B2). Размеры крупные, крыло не короче 430 мм, преобладающая окраска светло-
серая сераяцапля Adrea cinerea
22 B1). Размеры мельче, крыло не более 400 мм, окраска разнообразная 23
23 B4). Преобладающая окраска коричневая, каштановая или охристая с темными
пятнами, длина крыла от 300 до 400 мм 25
24 B3). Окраска иная, длина крыла меньше 300 мм 27
25 B6). Окраска оперения с преобладанием коричневых и каштановых тонов . . .
рыжая цапля Ardea purpurea
26 B5).Окраска оперения с преобладанием охриаых тонов с мелкими черными пят-
нами выпь Botaurus stellar is
27 B8). Размеры мелкие. Крыло не более 160 мм 37
28 B7). Размеры крупные, крыло не менее 180 мм 29
29 C0). Преобладающая окраска оперения белая, летом верх головы, пятно на зобе
и спине — винно-охристые, зимой пятно только на голове
египетскаяцапля Bubulcus ibis
30 B9). Окраска иная 31
31 C2). Верхняя часть головы и спина блестяще-черные (брачный наряд), бурые со
светлыми пятнами (молодые птицы), размеры крупные, крыло от 26 мм и крупнее ....
квакваNycticorax nycticorax
КОЛПИЦА
357
32 C1). Окраска иная, размеры мельче, длина крыла меньше 260 мм 33
33 C4). Окраска оперения с явным преобладанием темноэеленых тонов на спинной
стороне тела зеленаякваква Butorides javanica
34 C3). Преобладающая окраска из комбинации желтых, охристых и кирпичных тонов,
на затылке хохол из удлиненных перьев 35
35 C6). Окраска бледноохристая или охристо-желтая, в брачном оперении хохол из
10 белых перьев с серыми опахалами желтаяцапля Ardeola ralloides
36 C5). В окраске оперения присутствуют кофейный, ржаво-кирпичный и бурый
цвет, весной хохол из 4—6 перьев белокрылая цапля Ardeola bacchus
37 C8). Верх головы блестящего черного цвета, спина черная или коричневато-бурая
волчок Ixobrychus minutns
38 C7). На голове нет блестящего черного цвета, спина иной окраски 39
39 D0). Темя темнобурое, спина или темнокорнчневая (самцы) или с присутствием
округлых белых пятен (самки) амурскийволчок Ixobrychus eurythmus
40 C9). На основном фоне темени продольные белые черточки. Спина бурая
длинноносый волчок Ixobrychus sinensis
СЕМЕЙСТВО ИБИСЫ IBIDIDAE
РОД КОЛПИЦЫ PLATALEA LINNAEUS, 1758
Тип P. leucorodia
1. Колпица Platalea leucorodia L.
Platalea leucorodia. Linnaeus. Syst. Nat., изд. X, 1758, стр. 139, южн. Швеция
Распространение. Ареал. В Европе на юго-востоке от Немзидлерского
озера в Австрии, в Венгрии, Югославии, Албании, в долине Дуная в Болгарии и
Румынии, на юге СССР до Закавказья; кроме того две изолированные обла-
Карта 66. Распространение колпицы Platalea leucorodia
7. P. i. leucorodia, 2. P. I. archeri;
a — граница гнездовий, б — не вполне выясненная граница гнездовий, г — область зимовок (цифры в круж-
ках обозначают номера зимующих подвидов).
сти —в южн. Испании и Голландии. В 1928 г. колпица гнездилась в Дании, до
XVI—ХУНвека местами в Англии (Суффольк, Норфольк, Суссекс,Миддлсекс).
В Азии — в Малой Азии, Сирии, Мессопотамии, вост. Иране, Афганистане,
Белуджистане, в Индии на восток до вост. Бенгалии, на юг до Цейлона; в
358 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Средней Азии, Казахстане, Туве, южн. Забайкалье, Монголии, Манчжу-
рии, на восток до Приморья. В Африке в Египте, Сомали, на островах южной
части Красного моря.
Характер пребывания. В северных частях ареала в Зап.
Европе и в СССР — перелетная, южнее — оседлая птица. Перелетные колпицы
зимуют в тропической Африке к югу от Судана, в небольшом количестве в ниж-
нем Египте. Гнездящиеся в Голландии птицы летят на юго-запад через Францию
и Пиренейский п-ов, гнездящиеся в бассейне Дуная — на юг через Италию,
Балканский п-ов, сев. Африку (Тунис, нижний Египет). Отмечены случаи
залетов колпиц, главным образом негнездящихся, в разные страны — в Анг-
лию, Германию, Швецию, Норвегию, Финляндию, в северные части СССР, в
Польшу, на юг до Канарских и Азорских о-вов, Мадейры и о-вов Зеленого
Мыса.
Биотоп. Влажные открытые местности с сильным развитием камышей,
кустарников и другой растительности.
Подвиды и варьирующие признаки. Два подвида Р. /. archeri Neumann, 1928 —об-
ласть Красного моря и Сомали и Р. /. leucorodia L., 1758—в остальной части ареала. Морфо-
логические различия незначительны и выражаются в том, что первый подвид меньше по
размерам, чем второй. Имеются различия и в деталях биологии — сомалийская колпица
строго оседла, биотопическое распределение ее отличается некоторыми особенностями,
также и размножение.
Европейская колпица Platalea leucorodia leucorodia L.
Синоним. Platalea major, Temminsk etSchlegel. Fauna Japonica
Aves, 1849, стр. 119, Япония
Русское название. Колпица, колпик — народное в южных частях страны.
Также лопатень — по форме клюва.
Распространение. Ареал. Область распространения колпицы за преде-
лами Союза указана выше, в очерке распространения вида. В СССР колпица
гнездится в низовьях Днестра и низовьях Днепра. Ранее гнездилась в Пинских
болотах, но теперь повидимому там не встречается. На пролете встречается
в Крыму, в вост. Приазовье, залетает в Подолию, Полтавскую и Харьковскую
обл. Восточнее колпица гнездится на озерах у восточного побережья Авовского
моря, вплавнях Кубани иТерека, на Маныче, по озерам в районе Кизляраидалее
к северу по западному побережью Каспия. В устьях Волги обычная гнездящаяся
птица, доходящая на север до Сарпинских озер и залетающая на север до Сыз-
рани, Куйбышева. Залеты колпицы известны и севернее — в южных частях
Молотовской обл., к Архангельску, в прибалтийские республики. В Закав-
казье колпица гнездится по среднему и нижнему течению Куры (Сатунин,
1907), доходя до устья, но несомненно отсутствует на гнездовье между устьем
Куры и иранской границей. В Армении гнездится в низменностях (Ереванская
равнина и долина Аракса), в горах у озера Севан.
В Азиатской части Союза колпица на гнездовье распространена спора-
дично. На северо-западе северная граница гнездового ареала пересекает,
р. Хобду под 50° с. ш. (Зарудный, 1888), в Наурзумских степях птица гнездится
на озерах Джар-куль и Сары-муюн (Деливрон, 1938), гнездовые находки
известны из района Кургана. В Барабинской степи, вопреки утвер-
ждению Палласа A811), колпиц нет. Восточнее гнездится на Зайсане,
в низовьях Черного Иртыша и в Тувинской обл. Отсюда колпица залетает
в Минусинский район, на Алтай (Уймон, Усть-каменогорск, Селевин, 1928);
впрочем, в Минусинском районе найдена на гнездовье в устьях Абакана
(Сушкин, 1938). В Вост. Сибири колпица гнездится в даурских степях (Штег-
ман, 1929), по нижней Зее, в окрестностях Благовещенска (Шульпин, 1936),
встречена на Амуре выше устья Сунгари и безусловно гнездится у озера Ханка.
Наконец, колпица спорадически гнездится в Средней Азии.
колпица 359
Гнездящиеся в СССР колпицы, кроме самых западных (украинских) попу-
ляций, зимуют вюжн. Азии. Окольцованная в дельте Волги 28 июля 1938 г.
птица добыта 28 октября 1938 г. у Кольхапура в провинции Бомбей, Индия.
Восточносибирские колпицы на зимовке отмечены в вост. Китае (Гуандунь,
Фуцзян, низовья Янцзе, на пролете в Чжили, на залете в Японии и один раз
на о-ве Формоза, Латуш, 1934). В Китае зимуют и гнездящиеся в Монго-
лии колпицы (гнездовая колония найдена в частности на озере Орок-нор).
Характер пребывания. В СССР гнездящаяся, перелетная
птица. Указание на то, что будто бы колпица зимует под Ленкоранью (Лоудон,
1909) не подтверждается позднейшими исследователями. За пределами нормаль-
ного гнездового ареала во многих районах колпица является или только про-
летным или только залетным видом. Весенний прилет ранний, в марте — мае,
но затяжной.
На северном берегу Черного моряпервые колпицы появляются в самом конце
марта. По наблюдениям Исакова и Воробьева A940) под Ленкоранью сроки
прилета значительно более ранние. В 1932 г. первый экземпляр колпицы был
отмечен здесь 19 февраля, второй 27 февраля; настоящий же пролет наблюдался
в первых числах марта. Значительно позднее появляются весной колпицы в
дельте Волги, где прилет первых особей происходит между 10 и 20 апреля
(Дубинины, 1940). В устье Урала появление птиц падает на конец марта. В Турк-
мении весенний пролет начинается в середине марта, продолжается весь апрель
и большую часть мая (Дементьев, 1950). Первые пролетные птицы на п-ове
Мангишлак отмечены 18 мая (Смирнов, 1916). На нижней и средней Сыр-Дарье
первые колпицы показываются в различные годы между 15 и 25 марта. Массо-
вый пролет происходит во второй трети апреля и, постепенно сокращаясь,
длится во всяком случае до конца этого месяца. Во второй половине апреля
колпицы видимо нередки уже на северном берегу Аральского моря. В Приморье
прилет идет в начале апреля. О сроках осеннего отлета сведения отличаются
большой скудностью. В Зап. Европе колпицы улетают между концом августа
и началом октября, отдельные особи даже в ноябре. Отлет в дельте Волги
происходит между 5 и 30 сентября (Дубинины, 1940). Наиболее поздний
экземпляр наблюдался здесь 19 октября (Воробьев, 1936). Зарудный A888)
указывает, что уже в июле (по ст. ст.) стаи колпиц сбиваются в низовьях Илека
и по Уралу. Из устьев Урала отлет происходит,по Бостанжогло A911),в сен-
тябре (ст. ст.), а по Карелину,—около второй трети сентября. Одиночная птица
была добыта южнее Гурьева 23 ноября (Спангенберг). На Черноморском
побережье Кавказа пролетная колпица добыта 19 сентября (Сатунин,
1913), а в Закавказье у Хармандали появление первых пролетных стай отмечено
в последних числах августа. В конце сентября происходит отлет с нижней
Сыр-Дарьи и в сентябре-октябре из пределов Туркмении. На самом юге
СССР отдельные особи задерживаются до поздней осени. Во время пролета
птиц удается наблюдать маленькими группами и крупными стаями в дневные
часы. Иногда птицы летят низко, но иногда на большой высоте, выстраиваясь
в поперечную линию и часто описывая парящим полетом большие круги.
Биотоп. Глухие заросли камыша, тростника, ивы или разнолистного
тополя и джиды, затерянные среди сети мелководных или глубоких озер,
речных стариц, труднодоступных топей или разросшихся по речным и морским
островам. В дельте Волги колпицы гнездятся то в чистых Камышевых и трост-
никовых зарослях, то среди кустарниковой и древесной ивы, пятнами вкраплен-
ной среди камышей (Воробьев, 1936). Два типа гнездовых биотопов отмечены
также на Сыр-Дарье в районе Кзыл-Орды. Большинство птиц выводит здесь
среди камышей и тростников, реже среди низкорослых тугайных зарослей
по глухим островам реки (Спангенберг и Фейгин, 1936). В Приморье гнез-
довый биотоп — густые затопленные полой водой ивовые заросли (Шульпин,
1936). Кормовые стации птиц повсеместно более или менее однообразны. Ими
360 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
являются мелководные, обычно лишенные камыша, тростника и осоки участки
водоемов. Охотно посещают колпицы в поисках пищи затопленные рисовые
поля, пока они не слишком заросли надводной растительностью. Глубина кор-
мовых участков редко превышает 30 см, колеблясь между 10 и 20 см. Таким
образом, при максимальной глубине, вода лишь достигает брюха кормящейся
птицы. Грунт водоемов илистый, топкий, нередко глинистый и весьма редко
мелкозернистый — песчаный. Водоемов с крупнозернистым песчаным и камени-
стым дном птицы избегают. Колпицы предпочитают кормиться на слабо теку-
щей воде. В связи с этим кормящиеся стаи сосредотачиваются часто на взморье,
в озере при устье впадающих рек, ериков или небольшого арыка. Будучи осто-
рожной птицей, колпицы предпочитают открытые участки, где они уже издали
могут заметить приближение опасности. В первые дни паводков колпицы посе-
щают вновь затопленные полой водой мелкие и редкие кустарники тамариска,
изобилующие в это время разнообразной пищей. Места кормежек иногда рас-
полагаются в непосредственной близости от мест гнездовья, но изредка колпицы
летят за пищей за 10—15 км от места гнездовья. Вообще говоря, колпица при-
держивается равнин, но в Армении гнездится и в горах на озере Севан.
Численность. Редкая птица в низовьях рек, впадающих в Черное
море. Не редка, а местами и обыкновенна, на Сев. Кавказе, во всем Закав-
казье и в дельте Волги. В последнем месте в пределах Астраханского заповед-
ника к концу гнездового периода 1939 г. было насчитано 710 колпиц (Рома-
шева, 1938). В устьях Урала ныне численность колпиц весьма ограничена.
Повидимому немногочисленны птицы в настоящее время также в области Иргиз-
Тургая и на Хобдах. Еще более редка колпица в гнездовый период в Туркме-
нии, где лишь во время пролета наблюдаются ее значительные скопления.
Наконец, немногочисленна птица и в Приморье. В противоположность этому
на всей нижней и средней Сыр-Дарье и отчасти на восточном берегу Аральского
моря колпица должна быть отнесена к обыкновенным, а в некоторые годы поло-
жительно к многочисленным птицам. В связи с редкостью крупных гнездовых
колоний и с большими интервалами между ними общая численность птиц в
нашей стране должна быть признана ограниченной.
Экология. Размножение. Колпица колониально гнездящаяся птица.
Образует или самостоятельные поселения, что бывает чаще, или присоеди-
няется к колониям других болотных и водяных птиц. При этом, если числен-
ность гнездящихся колпиц невелика, их гнезда бывают разбросаны среди
гнезд других видов колоний. При большом числе колпиц они в смешанных
колониях образуют самостоятельные или несколько обособленные поселения.
В дельте Волги колпицы присоединяются к различным видам цапель, каравай-
кам, большим бакланам. Однако, в некоторые годы, например в 1935 г., боль-
шинство местных птиц образовали самостоятельную колонию (Дубинины,
1940). На Сыр-Дарье в районе Кзыл-Орды колпицы гнездятся,по нашим наблю-
дениям, то самостоятельными колониями, то образуют колонии с каравайками,
то присоединяются к крупным гнездовым поселениям различных цапель,
караваек, больших и малых бакланов. Совместно с цаплями гнездятся колпицы
и в южн. Приморье. Самостоятельные колонии колпиц известны в пределах
Туркмении, местами в Закавказье, в устье Урала, на Хобде.
В подавляющем большинстве случаев гнезда колпицы устраивают на
заломе прошлогоднего камыша и тростника. Реже гнезда закладываются на
кустарниках и деревьях ивы, на разнолистном тополе или джиде. В дельте
Волги при гнездовании на древесной растительности птицы определенно пред-
почитают строить гнезда на отдельно стоящих деревьях ивы на высоте 3—5 м
над землей (Дубинины, 1938). То же наблюдается и на Сыр-Дарье в районе
Кзыл-Орды. Гнезда на деревьях расположены на высоте от 2 до 5, а иногда
до 7 м (Спангенберг, 1936). В Приморье на средней Лефу колпицы гнездятся
исключительно среди густых затопленных ивняков (Шульпин, 1936).
КОЛПИЦА 361
Гнездо — большая куча Камышевых и тростниковых стеблей и листьев,
сложенная то среди густых, то среди редких надводных зарослей. Если гнезда
расположены на деревьях, материалом для них служат ветви с небольшой
примесью тростника и листьев в лотке. Чем ближе к центру колонии, тем
теснее расположены отдельные гнезда. В некоторых случаях группы гнезд
образуют сплошные островки и тогда нет возможности наметить границы между
отдельными гнездами. На одной куче насиживают 2, иногда 3 колпицы. Чаще
между отдельными гнездами бывают промежутки от 1 до 2 м. Отдельные гнезда,
расположенные среди густых зарослей, в некоторых случаях бывают соединены
многочисленными коридорами, идущими среди тростника и протоптанными
самими птицами. Такие колонии отмечены Бостанжогло в устьях Урала и на
Сыр-Дарье (Спангенберг, 1936). Чем дальше от центра, тем гнезда расположены
реже. По краям колонии иногда они строятся на чистых мелководных участках,
опираясь своим основанием на щетку подрастающей зеленой растительности.
Форма отдельных гнезд то округлая, то, как например в устьях Урала, в виде
равнобедренного треугольника, с длиною сторон около 80 см и с высотою от
40 до 60 см.
Гнездо строится обеими птицами гнездовой пары, причем материал для
него собирается в непосредственной близости от колонии. В глухих, трудно-
доступных уголках нашей страны, где птиц не тревожат (например, Сыр-
Дарья), колпицы гнездятся на одних и тех же местах по нескольку лет подряд.
В этом случае гнезда достигают особенно большой высоты и состоят из двух-
трех слоев. Количество яиц в кладках колеблется от 3 до 5, причем 5 яиц —
относительная редкость. Число яиц изменяется в различные годы. В одни годы
подавляющее большинство гнезд содержат 3 яйца, в другие 4. Яйца колпиц
отличаются от яиц караваек, цапель, бакланов пятнистой окраской. Обычно
они или вздуты и округлы, или сильно вытянуты. Скорлупа известково-белого
цвета с резкими красновато-бурыми мелкими пятнами, точками и черточками,
собранными на тупой половине яйца, изредка в виде венчика. Размеры A7)
65,1—73,0 х 43,2—49,0 мм, среднее 69,5 х 45,0 мм.
Сроки размножения не одновременны. В среднем колпицы на юге
гнездятся недели на две раньше, чем близ северной и северо-восточной
границы распространения *. Значительные колебания в начале размно-
жения наблюдаются в одних и тех же географических районах, пови-
димому отчасти в зависимости от уровня воды на местах гнездовья.
В новых колониях птицы приступают к гнездованию недели на две
позднее, чем птицы, занявшие прошлогодние гнездовья. Наблюдается
большая разница в сроках размножения и у отдельных пар, принадлежащих
к одной и той же колонии. Последнее связано с гибелью некоторых кладок,
быть может и с другими не вполне ясными пока причинами. В Туркмении яйце-
кладка происходит в начале мая (Дементьев, 1950). Повидимому в те же сроки
колпицы кладут яйца и в Закавказье, что, однако, нуждается в проверке,
так как в гнездах колонии, осмотренной Сатуниным 17 мая, было по 2—3 све-
жих яйца, и кладки, вероятно, еще не были закончены. В дельте Волги начало
кладки нормально происходит между 15 и 25 мая (Дубинины, 1940).
В некоторые годы, например, в 1931 и ранее. Законченные кладки по 4 яйца
были обнаружены уже 11 мая (Воробьев, 1936). По данным Бостанжогло,
A911) в устьях Урала во второй половине мая колпицы уже прочно сидели на
гнездах. На Сыр-Дарье в районе Кзыл-Орды сроки кладки подвержены чрез-
вычайно сильным колебаниям. Свежие законченные кладки были взяты здесь
28 мая, 10 и 14 июня, а в другой колонии обнаружены недостроенные гнезда,
* В Зап. Европе в Испании начинается в конце апреля, в Голландии в конце апреля —
начале мая (Уайзерби, 1939), на Цейлоне кладка отмечена в марте, в Сев. Индии между
сентябрем и ноябрем (Бекер, 1929).
362 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
свежие и насиженные яйца и наряду с этим полуоперившиеся птенцы 5 июня.
Яйца откладываются со значительными промежутками иногда, повидимому,
в несколько дней.
Насиживание продолжается 24—25 дней (Тугаринов, 1947); по другим
данным однако только 21 день (Уайзерби, 1939).
Оно начинается с первого яйца, но пока кладка не закончена птица сидит
на яйцах лишь в ночное время. Оба родителя принимают участие в насижива-
нии и вскармливают птенцов, причем последние при получении пищи засовы-
вают клюв в рот родителей. По наблюдениям в Туркмении число птенцов
обычно бывает на один меньше, чем число яиц в кладке. Появление первых
птенцов в Астраханском заповеднике происходит между 1 и 10 июня. В массе
птенцы появляются между 15 и 20 июня. Начало вылета птенцов из гнезд
между 30 июня и 10 июля, массовый вылет между 10 и 20 июля. Таким образом
гнездовый период с начала яйцекладки и до вылета птенцов из гнезд продол-
жается несколько больше двух месяцев.
При потере первой кладки размножение отдельных пар чрезвычайно
сильно запаздывает. Воробьев A936) упоминает о случае нахождения еще нелет-
ных птенцов 5 сентября.
В первой половине июня у Мары в гнездах были обнаружены птенцы,
а первые летные наблюдались здесь 4 июля (Зарудный, 1890). В Армении на
озере Гилли летные молодые отмечены 10 и 15 июня (Ляйстер и Соснин,
1942), т. е. на две недели раньше, чем в дельте Волги. На Хобде хорошо
летающие молодые отмечены Зарудным A888) в начале второй трети июля.
В южн. Приморье 2 июля большинство гнезд оказалось пустыми, в редких
обнаружены крупные, но еще плохо летающие молодые (Шульпин, 1936).
Еще нелетные птенцы колпиц группируются на более крупных гнездовых
постройках и вытоптанном заломе камыша. Позднее, с приобретением способ-
ностей к полету, они сбиваются в крупные стаи и вскоре начинают посещать
кормовые участки, а затем откочевывают к югу, скопляясь в устьях круп-
ных рек.
Линька. Последовательность смены нарядов: первый пуховой наряд —
второй пуховой наряд — гнездовый наряд — первый брачный наряд — пер-
вый послебрачный наряд — второй брачный и т. д. Когда птица приобретает
окончательный наряд — неясно (повидимому окончательный наряд первый
послебрачный и второй брачный). Молодые птицы линяют в первый брач-
ный наряд между январем и мартом. У взрослых предбрачная линька про-
исходит в эти же месяцы, а послебрачная начинается в конце периода
размножения и тянется до поздней осени. Предбрачная линька частич-
ная, захватывающая мелкое оперение; послебрачная — полная. Детали
линьки требуют выяснения. Указания западноевропейских авторов (Уай-
зерби, 1939) о том, что у колпиц обе годовых линьки полные—неве-
роятно.
Питание. В Астраханском заповеднике особенно охотно колпицами
поедаются личинки водяных насекомых, но не брезгуют они также лягушками,
моллюсками и мелкой рыбешкой (Дубинины, 1940). Приблизительно такой же
состав пищи известен на Сыр-Дарье в районе Кзыл-Орды. При вскрытии много-
численных желудков и пищеводов взрослых и молодых птиц в большом числе
найдены личинки коромыслов, остатки хитина водолюбов, плавунцов и дру-
гих водяных жуков, довольно часто остатки рыбы и много реже мелкие лягу-
шата. Во время лёта и передвижения пешей саранчи, когда эти насекомые во
множестве попадают в водоемы, колпицы стаями появляются на таких участках.
Далеко не последнее место занимают в питании колпиц, по нашим наблюде-
ниям, личинки комаров и водяные растения. Насекомые, прочие беспозвоночные
и головастики обнаружены также в желудке птицы, убитой в пределах Туркме-
нии (Дементьев, 1946). При добывании пищи, колпицы ведут себя своеобразно.
КОЛПИЦА
363
В тех случаях, когда стая кормится на замкнутом водоеме, птицы «косят*
своим длинным клювом воду и временами щелочат йл. При этом они не соблю-
дают строя и охотятся маленькими группами и одиночками. Напротив, в водое-
мах, обладающих хотя бы слабым течением, колпицы всегда выстраиваются
косой линией и подвигаются навстречу течению. Каждый шаг вперед сопрово-
ждается широким движением опущенного в воду клюва в одну сторону, а при
следующем шаге в другую. Иногда птица, нащупав уходящую добычу, быстро
отбегает в сторону, но затем воз-
вращается на свое место, восста-
навливая правильность шеренги.
Полевые признаки. Белая
окраска оперения, черные ноги
и желтоватый клюв легко позво-
ляют узнать колпицу в природе.
Только на большом расстоянии
ее можно смешать с большой бе-
лой цаплей. Отыскивающая пищу
колпица узнается сразу («косит»).
На полете колпица то быстро
машет крыльями, то держит их
неподвижно; шея вытянута впе-
ред и слегка изогнута книзу,
ноги вытянуты назад. Может
парить. Активна преимущественно
в сумерки. Предпочитает дер-
жаться небольшими группами или
крупными стаями. Менее осто-
рожна нежели крупные цапли.
Подойти к колпице из-за при-
крытия нетрудно, а к кормящей-
ся птице удается иногда подой-
ти и на открытом месте. Спугну-
тая стая отлетает на некоторое
расстояние и вновь приступает к
поискам пищи. Молчаливая пти-
ца. Старые издают у гнезд вор-
чащие звуки, птенцы пищат.
Описание. Размеры и стро-
ение. У взрослых на голове имеет-
ся хохол, свисающий на шею. Форму-
ла крыла (не считая рудиментарного
махового): 1>2>3>4... На внутренних
опахалах 2-го — 4-го и на наруж-
ных опахалах 1-го—3-го первостепенных
маховых — вырезки. Хвост прямой, из 12 рулевых. На уздечке, вокруг глаз, на подбородке
и на середине зоба — пятно голой кожи. Клюв длинный, плоский, расширенный на конце
с шероховатой поверхностью. Ноздри узкие, вытянутые вдоль клюва. Ноги длинные, перед-
ние пальцы соединены в основной части перепонкой. Длина самок C) 777, 800, 830 мм; раз-
мах крыльев самок C) 1258, 1400, 1450 мм. Крыло самцов и самок <12) 370—410, в сред-
нем 387,5 мм.
Окраска. Первый и второй пуховой наряды — белые, во втором наряде — с охри-
стым оттенком. Клюв и ноги розоватые. Гнездовый наряд: хохла нет, общая окраска опере-
ния белая, но стволы и вершины первостепенных маховых и их кроющих и стволы второ-
степенных—черноватые, клюв сероватый, радужина бурозато-серая. В первом брачном
наряде сохраняются темные вершины перьев крыла. Взрослые самцы и самки белые,
но у основания шеи на брюшной стороне имеется охристый оттенок, перья хохла
желтоваты; голая кожа на голове и шее — желтого цвета, за исключением уздечки,
которая черного цвета; клюв черный с желтой вершиной, ноги черные, радужина
красная.
Рис. 73. Колпица
364
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
РОД КАРАВАЙКИ PLEGADIS KAUP, 1829
Тип P. falcinellus
2. Каравайка Plegadis falcinellus L.
Tantalus falcinellus. Linnaeus. Syst. Nat., изд. XII, 1766, стр. 241, Австралия, Италия.
Русское название. Каравайка — народное, повидимому от казахского назва-
ния этой птицы — «карабай».
Распространение. Ареал. Америка — Флорида, Луизиана, Орегон,
Юта и до юга Техаса и южн. Мексики. Южн. Америка от Перу и Бразилии до
Аргентины. Африка — Тунис и тропические части материка, Мадагаскар.
Южн. Европа — в дельте Роны во Франции (гнездование, впрочем, требует
подтверждения, Майо, 1936), южн. Испания, сев.-зап. Италия, придунайские и
балканские страны (кроме Греции), юг Европейской части СССР и Кавказ.
Карта 67. Распространение каравайки Plegadis falcinellus
7. P. J. Jalcinellus, 2. P. /. peregrinus, 3. P. j. guarana;
a — граница гнездовий, б — не вполне выясненная граница гнездовий, г — область зимовок (цифры в
кружках обозначают номера зимующих подвидов).
Азия — Малая Азия, Сирия, Ирак(?), Иран, Средняя Азия, Афганистан, Индия,
Бирма, Цейлон, Филиппины, Ява, вероятно Целебес, Австралия.
Характер пребывания. На севере — перелетная, на юге —
оседлая птица. Европейские птицы зимуют в тропической и южн. Африке,
птицы из умеренных широт Азии — в Индии. В Индии зимуют и каравайки,
гнездящиеся в низовьях Волги.
Биотоп. Богатые растительностью местности в долинах рек, болоти-
стые побережья других внутренних водоемов.
Подвиды и варьирующие признаки. Три подвида, различающиеся величиной, а один —
американский — и окраской. Кроме того имеются различия и в биологии сезонном, разме-
щении, особенностях размножения и т. д. В Европе, в Африке, в Азии, кроме островов к во-
стоку и юго-западу от нее, в Америке — во Флориде, Луизиане, на о-вах Гаити и Куба Р. /.
falcinellus L.\ л п.— 1766; на Яве, Филиппинах и в Австралии P.f.peregrinus, Bona-
parte—1855; наконец, в Сев. Америке от Орегона и Юта до Техаса и Мексики и
в Южн. Америке Р. /. guarana Linn,— 1766.
Европейская каравайка Plegadis falcinellus falcinellus L.
Синоним. Numenius igneus S. Gmelin. Reise d. Russland, I, 1760, стр. 166,
Южн. Россия — Numenius viridis. S. G me 1 i n, о. с. стр. 167, Южн. Россия.
Распространение. Ареал. Общее распространение см. выше. Детали рас-
пространения в пределах СССР таковы: повидимому гнездится в Бессарабии.
В небольшом числе гнездится в устьях Днестра, на Днепре бывает редко,
КАРАВАЙКА 365
только бродячими стайками. В вост. Приазовье, в дельте Дона и в Крыму встре-
чается не каждый год во время пролета. На Черноморском побережье Кавказа
бывает в небольшом количестве только на весеннем и осеннем пролетах. Пови-
димому в ограниченном числе гнездится в плавнях больших рек Сев. Кавказа,
в частности на Кубани и Тереке. Залетает во время полетов к Дзауджикау
(Беме, 1926). Довольно широко в качестве гнездящейся птицы распространена
в Армении, не только в низменностях, ной у озера Севан (Ляйстери Соснин, 1942).
Широко гнездится в восточных частях низменного Закавказья—по среднему и
нижнему течению Куры, по озерам Муганской степи, но в противоположность
данным Сатунина A907) отнюдь не под Ленкоранью, куда лишь, по нашим на-
блюдениям, сваливает во второй половине лета. Большими колониями гнездятся
птицы в дельте Волги и поднимаются вверх по течению реки до Сталинграда.
В прошлом нормально гнездилась по труднодоступным островам в устьях Урала
и спорадично —в нижнем течении реки (Бостанжогло, 1911). Повидимому, гнез-
дится здесь и в настоящее время, но в самом ограниченном количестве. В Турк-
мении найдена на гнездовье в Мервском оазисе, в низовьях Мургаба, в низовьях
Теджена, в Пендинском оазисе, в верхнем течении Мургаба и, повидимому, гнез-
дится по среднему и нижнему течению Аму-Дарьи, хотя точных данных о гне-
здовании каравайки по Аму-Дарье нет. В остальных частях Туркмении встре-
чается лишь во время пролетов. Очевидно, гнездится кое-где в низменном Тад-
жикистане, где наблюдалась в гнездовое время и на пролетах.
На Сыр-Дарье каравайка нормально гнездится на озерах в районе г. Тур-
кестана и отсюда вниз по реке до устья, а также на озере Камышлы-баш.
По расспросным сведениям в некоторые годы небольшая колония караваек гнез-
дится по восточному побережью Аральского моря и, несомненно, в богатые
водой годы на озерах Тели-куль и в самых низовьях реки Сары-Су. Нормально
гнездится в низовьях Чу и по южному побережью Балхаша западнее Или. На север
отсюда каравайка гнездится в низовьях Иргиза, приблизительно от города
Иргиза, и в низовьях Тургая не севернее одноименного города (Сушкин, 1908).
На северо-восток не переходит за Тарбагатай. Случайно гнездилась прежде и
возможно изредка гнездится в настоящее время в урочище Бишкопа на Хобде.
Во времена исследований Зарудного A888) иногда гнездилась под Чкаловом,
обычно же (как и в низовье Илека) только залетает туда. Отдельные случаи за-
летов известны в Скандинавии и Финляндии, в Исландию на Фарерские о-ва,
в Англию, Голландию, Бельгию, Данию, Германию, в южн. и юго-вост. Европу,
в сев. Африку, на о-в Мадейра, в Азии —в юго-вост. Китай (Сычуань, Фучжоу).
В СССР в Прибалтику и в разные районы средней полосы — в Подолию, к Пав-
лограду, к Харькову, Воронежу, Сызрани и т. д., в Средней Азии на Памир
(Мекленбурцев, 1936) и на озере Ала-куль. Исключительно далекий залет к се-
веру был отмечен летом 1926 г. Стайка из четырех птиц была отмечена в Моло-
товской обл., где близ Охайска птицы продержались все лето; 23 августа из
этой стайки удалось добыть одну птицу (Ушков, 1927).
Зимнее время европейские каравайки (западнее Волги) проводят в основ-
ной массе в Африке, часть летит в Индию. Птицы с низовий Волги и восточнее
зимуют в Индии.
Результаты кольцевания караваек в дельте Волги, в Астраханском запо-
веднике, показывают постоянство мест гнездования этих птиц, а также выясняют
общее направление пролетов и расположение зимовок этих птиц; в частности
то, что волжские каравайки, в отличие от других европейских, летят на юго-
восток и зимуют в Индии. Птица, окольцованная 9 июля 1931 г., добыта в дельте
Волги 1 сентября 1934 г. Другая, окольцованная 9 августа 1927 г., добыта
2 октября 1927 г. в низовьях Волги, повидимому, на пролете. На пролете же
добыта 7 августа 1943 г., в низовьях Сыр-Дарьи окольцованная 1 августа 1941 г.
птица — быть может в этом случае птица и переменила место гнездования. На-
конец, три птицы добыты на зимовке в Индии: окольцованная 1 августа 1941 г.—
Збб ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
зимой 1945—1946 гг.у Симри в Бихаре;окольцованная 12августа 1941 г.—^ап-
реля 1942 г. у Малда в Бенгалии; окольцованная 9 июля 1931 — Деолали близ
Бомбея (точная дата осталась невыясненной).
Характер пребывания. В СССР — гнездящаяся перелетная
птица. В Зап. Европе весенний прилет караваек падает на конец апреля, осен-
ний происходит в августе—сентябре. В СССР валовой весенний прилет про-
исходит поздно, когда вполне установится теплая погода, но первые особи
появляются вскоре по вскрытии мелководных водоемов. Между появлением
передовых особей и валовым пролетом проходит 2—3 недели. В Крыму, где
каравайка встречается только на весеннем пролете, известен экземпляр, добытый
под Симферополем 1 мая. Эту дату приходится считать поздней, так как в При-
азовье пролет караваек начинается в конце марта. Рано появляются каравайки
также на Черноморском побережье Кавказа; впрочем, весенний пролет птиц
растягивается здесь между 17 марта и 10 апреля (Вилькснский, 1897). Появле-
ние караваек на Куре и в других частях вост. Закавказья происходит, в зави-
симости от хода весны, с конца апреля и до середины мая (Сатунин, 1910). В конце
первой трети апреля или в половине апреля прилетает каравайка на Сев.
Кавказ (Мензбир, 1918). 9 мая стайка была отмечена на взморье в районе Киз-
ляра (Беме, 1925). В дельте Волги по ряду авторов первые особи в различные
годы появляются между 5 и 10 апреля. В 1935 г. основная масса прилетела
20 апреля (Аскаров, 1938). В более ранние сроки —в самом начале апреля—
каравайка отмечена Карелиным в устьях Урала, но Бостанжогло наблюдал ее
там в первой половине мая. В вост. Иране пролет наблюдался с 20 по 23 апреля
(Зарудный, 1930). В Туркмении весенний пролет идет двумя путями. Часть
караваек летит вдоль Копет-Дага (Геок-тепе, Ашхабад, поворачивая, повиди-
мому, через Ходжа-калинскую долину на Сумбар и далее к Атреку). Другие
птицы летят вдоль Каспийского побережья. Весь пролет протекает с конца пер-
вой трети марта и до последней трети мая, т. е. длится около 50 дней. Это
объясняется тем, что пролетные птицы, позидимому, принадлежат к разным
популяциям с различным календарем периодических явлений. Численность
пролетных вТуркмении караваек весной не велика по сравнению с численностью
птиц, пролетающих здесь осенью (Дементьев, 1950). На Мангышлаке пролет-
ные каравайки отмечались в различные даты между 16 и 18 мая (Смирнов, 1916).
Столь же длителен весенний пролет караваек и на Сыр-Дарье. Первые особи
появляются на вскрывшихся водоемах в районе Кзыл-Орды в конце марта, ва-
ловой пролет происходит с середины и до конца апреля. Направление пролета
на Сыр-Дарье идет вниз по реке, т.е. с юго-востока на северо-запад. В районе
станций Сало-тюбе — Байгакум, небольшая часть птиц круто поворачивает на
северо-восток и восток, огибает северо-восточные отроги Кара-Тау и выходит
к низовьям рек Чу и Сары-Су. Значительный весенний пролет караваек идет
также вдоль восточного побережья Аральского моря (Спангенберг, 1936). В ни-
зовьях рек Иргиза и Тургая весной каравайки появляются очень поздно, видимо
задерживаясь в южных частях Казахстана. В 1898 г. у озера Джалангач первая
стайка в 6 особей отмечена 17 апреля. Под Чкаловом самый ранний срок появле-
ния караваек 29 апреля (Сушкин, 1908). Что касается Таджикистана, то один
экземпляр был добыт в окрестностях Гиссара 28 апреля, а заканчивается весен-
ний пролет, согласно Иванову A940), в середине мая. Каравайки сразу по под-
нятии птенцов на крылья сбиваются в крупные стаи и начинают перемещаться
в поисках угодий изобилующих пищей. Эти перемещения в некоторых случаях
настолько значительны, что при движении в южном направлении приобретают
характер осеннего отлета. В южных частях страны (Ленкорань, вост. Закавказье)
стаи задерживаются на длительное время и ведут здесь бродячую жизнь, переме-
щаясь позднее к югу. Осенний пролет караваек в Приазовье происходит во вто-
рой половине августа и в первой половине сентября. На Черноморском по-
бережье Кавказа осенью каравайки начинают лететь к югу 23 июля (Вильксн-
КАРАВАЙКА 367
ский, 1897), а заканчивается пролет полностью к 14 октября. В Ленкоранской
низменности с середины августа каравайки в большом числе появляются на
истылях и беджарах между Ленкоранью и иранской границей, где раньше они
отсутствовали и держатся здесь до конца этого месяца и значительную часть
сентября. Взрослые и молодые птицы, как правило, образуют самостоятельные
стаи, причем численность последних несравнимо больше первых. 13 сентября
наблюдался хорошо выраженный отлет птиц к югу, которыйдлился около Здней.
Из Астраханского заповедника нормально каравайки отлетают с 25 августа по
10 сентября (Дубинины, 1940). Массовый же отлет, прослеженный за 4 года
(с 1926 по 1929), совпадал с датами: 6 сентября, 7 сентября, 15 сентября и
17 сентября (Аскаров, 1938). Отдельные запоздавшие особи были отмечены здесь
до 20 октября (Воробьев, 1936). С 10 августа и приблизительно до поло-
вины сентября осенне-перелетные каравайки в свое время были отмечены Север-
цовым на нижнем течении Урала на широте Антоньевской станицы. С Сыр-
Дарьи (район Кзыл-Орды) каравайки отлетают, вероятно, в середине сентября
или во второй половине месяца (Спангенберг, 1936). В Туркмении осенний про-
лет птиц чрезвычайно обилен. Он протекает в сжатые сроки, начинаясь в августе
и заканчиваясь в сентябре (Дементьев, 1950). Осенний пролет местами бывает
очень трудно отличить от летних кормовых вылетов. Уже в июне в Мервском
оазисе были в свое время отмечены стаи караваек до 500 особей (Зарудный,
1896). В дельте Аму-Дарьи 25 июля наблюдались стаи птиц, летевшие в южном
направлении (Молчанов, 1912). Во время осеннего пролета изредка каравайки
залетают в мало подходящие для них местности. Известен случай залета к Дзау-
джикау. Во время сезонных перемещений птицы летят в дневные часы, в утрен-
ние и вечерние сумерки (возможно и ночью). Наиболее часто каравайки летят
на сравнительно небольшой высоте, выстроившись поперечной линией и много
реже углом. Пролетные стаи нередко содержат более 500 и даже более 1000
особей.
Биотоп. Заросли камыша, тростника или древесная растительность
среди глухих озер, речных стариц, труднодоступных топей, а также по речным
и морским островам. В низменной и горной Армении каравайки заселяют ка-
мышевые и тростниковые крепи озер. В таких же зарослях птицы гнездятся по
нижнему течению Куры и в плавнях Кубани. В Астраханском заповеднике
гнездовым биотопом каравайки служат довольно густые заросли кустарниковой
и древесной ивы (Salix alba и Salix triandra), граничащие с тростниковыми кре-
пями и разбросанные среди многочисленных ериков. Места эти затопляются
весенним разливом и вода, глубина которой колеблется от 55 см до 1 м, сохра-
няется здесь в течение всего гнездового времени. Повидимому, в тростниковых
крепях гнездились прежде каравайки на морских островах в устье Урала.
В Туркмении гнездовый биотоп — сочетание воды и древесной растительности
и особенно такие угодия, которые затопляются речными разливами (Дементьев,
1950). На Сыр-Дарье в районе Кзыл-Орды и Чийли каравайки гнездятся в двух
резко отличных стациях. Они заселяют камышевые и тростниковые заросли
среди сети проточных глубоководных озер или — реже — избирают тугайные
заросли по островам Сыр-Дарьи (Спангенберг, 1936).
Кормовые стации крайне разнообразны, но мелководье и более или менее
открытый ландшафт — обязательные условия при выборе мест массовой кор-
межки караваек. В Закавказье, по нашим наблюдениям, излюбленными кормо-
выми стациями являются затопленные мелкой водой рисовые поля, даже в ав-
густе, когда рис довольно высок, он все же не скрывает кормящихся птиц и
позволяет им следить за окружающим. Охотно посещают каравайки также
спущенные водохранилища — истыли, бродя по жидкой грязи у заполненных
водой понижений и ямок, береговые отмели высыхающих озер и мелководные
участки моря. Реже птицы кормятся по отмелям речек и протоков, в значитель-
ной мере скрытых береговой растительностью. В дельте Волги со времени при-
368 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
лета места кормежки находятся за границей Камышевых зарослей на полях и
ильменях. Позднее (с июля) каравайки начинают посещать морские заливы, где
глубина воды в среднем достигает 35—40 см. Дно этих мелководий топкое, или-
стое. Местами встречаются участки, покрытые водой, или выступающие из
воды грязевые островки, лишенные растительности. На Сыр-Дарье в районе
Кзыл-Орды каравайки кормятся на мелководных пресных и солоноватых озе-
рах, где вследствие подсыхания заросли камыша оказываются далеко отодви-
нутыми от уреза воды. В период паводков каравайки скопляются во множестве
в редких тамарисковых порослях, где наряду с мелкой водой часто встречаются
сырые участки выступившей из воды почвы. Во многих случаях кормовые ста-
ции отстоят на большое расстояние от места расположения колоний. В низовьях
Сыр-Дарьи, каравайки улетали за кормом более чем на 20 км от колонии, пре-
небрегая находившимися поблизости глубоководными озерами. Эти перелеты
удавалось наблюдать как днем, так и на утренних и вечерних зорях. Во время
вскармливания одна из птиц гнездовой пары на вечерней заре улетает на кор-
мовые озера и с первыми лучами солнца возвращается обратно с кормом (Спан-
генберг и Фейгин, 1936). Почти повсеместно каравайки гнездятся в угодьях,
труднодоступных для человека. Напротив, отыскивая пищу не избегают и даже
предпочитают культурные ландшафты.
Численность. Каравайка редка в низовьях рек, впадающих в Черное
море, немногочисленна на реках Сев. Кавказа и более обычна в зап.
Закавказье. Обычна в вост. Закавказье и в Туркмении. Многочисленна на ниж-
нем и среднем течении Сыр-Дарьи, на юго-западе Балхаша и в дельте Волги.
В последнем месте только в пределах Астраханского заповедника в 1934 г.
общее количество караваек исчислялось^ 8 780 особей, в 1935 г. —в 12 620
особей (Ромашова, 1938). В низовьях Урала ныне каравайка немногочисленный,
даже редкий вид. Немногочисленна она также в низовьях Иргиза и Тургая.
Несмотря на резко выраженное спорадическое распространение и большие рас-
стояния между отдельными колониями, общая численность караваек в нашей
стране довольно велика.
Экология. Размножение. Колониально-гнездящийся вид. Размеры
отдельных колоний от 3 до нескольких сотен, даже тысяч пар. Как правило,
каравайки образуют смешанные колонии, гнездясь совместно с различными
видами цапель, колпиками, большими и малыми бакланами. В непосредственной
близости от таких поселений часто устраиваются и пеликаны. В Армении кара-
вайки присоединяются то к колониям бакланов, то к колониям чепуры нужды,
желтой цапли и кваквы (Ляйстер и Соснин, 1942). В Астраханском заповед-
нике гнездится совместно с серыми, большими белыми цаплями, чепурой нуж-
дой, кваквами, желтыми цаплями, колпицами, большими бакланами и грачами
(Ромашова, 1940). На Кубани, поПтушенко, присоединяется к колониям цапель.
На Сыр-Дарье в районе Кзыл-Орды гнездится с кваквами и грачами, с раз-
личными видами цапель (серая цапля, кваква, реже чепура нужда и желтая
цапля) с малыми и большими бакланами. В тех же колониях иногда гнездятся
пеликаны (Спангенберг, 1936). Свои гнезда каравайки устраивают или на за-
ломе прошлогоднего камыша и тростника, или, что бывает реже, на древесной
растительности. В Астраханском заповеднике избирают для гнезд древесную и
кустарниковую иву. На Сыр-Дарье в районе Кзыл-Орды и Чийли одни колонии
размещаются на низкорослых деревцах джиды и разнолистного тополя, другие
в Камышевых и тростниковых крепях. Гнездование на деревьях отмечено также
в Туркмении. В остальных частях страны каравайки строят гнезда только на
заломе тростниковых и Камышевых зарослей. При гнездовании на древесной
растительности гнезда помещаются низко над землей или водой, в нижнем ярусе
дерева (Туркмения) на низкорослой ветле и кустарниковой иве (дельта Волги),
в среднем и верхнем ярусе низких, но густых тугайных зарослей (Сыр-Дарья).
В общем высота, на которой строится гнездо, редко превышает 5—7 м. Часть
КАРАВАЙКА 369
гнезд скрыта в тени листвы, другие расположены на солнцепеке. В дельте Волги
гнезда сооружаются из сухих сучков ивы толщиной от 0,3 до 2 см и длиной от
10 до 82 см. Среди основного строительного материала в небольшом количестве
примешиваются сухие стебли тростника. Лоток выстилается сухими листьями
камыша и ивы.
Гнезда строятся искусно и отличаются небольшими размерами. Толщина
стенок гнезда от 4 до 27 см, глубина лотка от 5 до 8 см9 поперечник 30
(Аскаров, 1938). По Воробьеву A936) гнездо делается, главным обра-
зом, из сухих листьев тростника с небольшим количеством сухих веток.
В тугаях на Баубен-арале (низовье Сыр-Дарьи) каравайки строят гнезда из
колючих ветвей джиды, перевитых с небольшим количеством стеблей камыша.
Лоток обильно выстилается нежными стебельками камыша и травы. Размеры
гнезд относительно малы. При гнездовании в камышах и тростниках
гнезда размещаются среди гнезд других птиц колоний или как бы окружают по
краю воды гнездовые островки бакланов, помещаясь от нескольких сантиметров
до 7з метра над водой. Основанием гнезд служит полусгнивший камыш, скреп-
ленный пометом. Внутренние стенки гнезд тщательно выкладываются высох-
шими зелеными листьями камыша, а лоток—прошлогодней растительной ве-
тошью. Диаметр гнезд 35—50 см (Сыр-Дарья, Спангенберг, 1936). Таким об-
разом гнездо каравайки по размеру и характеру постройки хорошо отличается
от гнезд других обитателей гнездовых колоний. Изредка птицы занимают гнезда
квакв и других цапель, но при этом перестраивают их. Материал для постройки
гнезда собирается в непосредственной близости от колонии, но иногда птица
летит за веткой или камышом за несколько километров. В доставке материала
и в постройке гнезда участвуют обе птицы гнездовой пары.
Нормальное количество яиц в кладках от 3 до 5. Обычно яиц бывает 4,
несколько реже 3, еще реже 5. Кладки в 2 яйца — редкий случай, вызванный
гибелью части кладки. Яйца правильной яйцевидной формы с хорошо выра-
женными тупым и острым концами. Окраска скорлупы яркая, зеленовато-
синяя или голубая, более стойкая в отношении выцветания, чем у цапель. Вес
яиц B0) колеблется от 31,4 до 38,9 г (Аскаров, 1938). Размеры: C2) 49,0—54,5 х
X 33,1—37,5 мм, среднее 52,0 х 35,9 мм (Сыр-Дарья, Спангенберг).
В СССР начало гнездования у караваек происходит в одни и те же или близ-
кие сроки в различных частях ареала. Однако вследствие того, что в больших
гнездовых колониях часть кладок гибнет и вновь возобновляется, получается
впечатление большой растянутости гнездового периода. В Астраханском заповед-
нике наиболее ранняя откладка яиц отмечена 28 апреля, массовая яйцекладка
с 10 мая. Вторичные кладки (при потере первых) происходят до 1 июля (Аскаров,
1938). В сравнительно редких случаях яйца удается находитьв гнездах до 20июля
(Воробьев, 1936). В первой трети мая происходит массовая яйцекладка на Сыр-
Дарье в районе Кзыл-Орды. Как и в дельте Волги, первые яйца откладываются
в самых последних числах апреля, у других пар яйцекладка сильно запазды-
вает. В 1925 г. 28 мая в большинстве осмотренных гнезд мною были обнаружены
слегка насиженные яйца. В Туркмении, повидимому, полные кладки в гнездах
во второй половине мая (Дементьев, 1950), в Армении 10 июня еще были в гнез-
дах сильно насиженные яйца (Ляйстер и Соснин, 1942). Спаривание происходит
сразу после того, как гнездо построено. Птицы спариваются на ветвях близ
гнезд (Астраханский заповедник), на кучах слежавшихся стеблей камыша или
на гнездах (Сыр-Дарья). Яйца откладываются каждый день. Насиживание начи-
нается с первого отложенного яйца, но пока кладка не закончена, птица сидит
мало, большую часть дня простаивая над отложенными яйцами.
В насиживании принимают участие оба родителя, продолжая одновременно
подправлять гнездо. На Сыр-дарьинских гнездовьях большую часть дня на
гнездах сидят самцы, ночью сидят самки. В дельте Волги, по наблюдениям
Аскарова A938), смена производится два раза в сутки. С 10 часов утра и до 3 ча-
24 Птицы СССР, т. II
370 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ _^^___=„
сов дня насиживают самцы, а с 8 часов вечера исключительно самки. Свободная
от насиживания птица гнездовой пары занята доставкой корма. Продолжи-
тельность насиживания по Аскарову A938) — 15—18 дней, 21 день по Хейн-
роту A924); последнее более вероятно. Между появлением первого и послед-
него птенца в отдельных гнездах иногда протекает 6 дней. Весь период насижи-
вания в колонии растянут на 2,5 месяца. Первые птенцы появляются в гнездах
25—30 мая, в массе же с 10 по 20 июня (Дубинины, 1940). В кормлении уча-
ствуют оба родителя, причем самец только приносит пищу и передает ее самке,
самка же кормит птенцов в один или два приема, через 15—20 минут. Изредка
приносит пищу и самка. Это бывает либо при утренней первой кормежке, либо
при последней — вечером. При кормежке клюв птенца заходит в глотку роди-
теля довольно глубоко, так что голова его погружается до самых глаз и таким
путем птенец достает из глотки родителя пищу. Продолжительность самого
процесса кормежки, при двух приемах пищи, 25 минут, при одном приеме —
10—15 минут. Со дня вылупления до 5-дневного возраста количество кормежек
в сутки колеблется от 8 до 12, от 5- до 10-дневного возраста — 6—10 кормежек,
от 10- до 15-дневного возраста — 5—8 кормежек и свыше 15-дневного возраста
до поднятия на крыло кормежка происходит от 5 до 7 раз в сутки. Летающих
птенцов, не способных еще добывать себе пищу самостоятельно, каравайки про-
должают кормить до 30-дневного возраста.
Вес птенцов каравайки увеличивается с 25—30 г при вылуплении до 500 г
к 30-дневному возрасту. Таким образом, молодые птицы ко времени поднятия
на крыло не достигают веса взрослых.
Отдельные особи поднимаются на крыло с 20 июня. Массовый подъем на
крыло начинается 1 июля и продолжается до 15 июля. В некоторых колониях
запоздавшие, нелетные птенцы обнаруживались 17—18 июля. Со дня вылуп-
ления до поднятия на крыло одного птенца проходит приблизительно 24 дня.
В этом возрасте птенец перелетает небольшое расстояние (шагов 10—15) с де-
рева на другое. К 27-му дню жизни птенцы летают более или менее свободно,
а на 30-й день вместе с родителями перелетают на места кормежки (Аскаров,
1938). На Сыр-Дарье насиживание яиц протекает со второй трети мая до сере-
дины июня. В начале второй половины июня появляются птенцы. При посеще-
нии одной из колоний 20 июля 1926 г. мы нашли всех птенцов летающими.
Целые сотни молодых поднимались в воздух, кружились над колонией, посте-
пенно рассаживаясь на островки. Вполне окрепшая молодь долгое время оста-
валась на местах вывода. С середины августа птицы смешанными стаями (ста-
рики с молодежью) начинают попадаться на озерах Сыр-Дарьи, подготавли-
ваясь к отлету. В Туркмении не крупные, но порядочно развившиеся птенцы
найдены Зарудным в начале июля. Птенцы становятся летными около половины
июля и даже в конце июня. В Армении на озере Гилли 10 июля обнаружены
птенцы различного возраста — одни только что вывелись из яиц, другие до-
стигли размера взрослой птицы. 15 июля отмечены летные молодые каравайки,
а 24 июля общества, состоящие из молодых и взрослых (Ляйстер и Соснин, 1942).
Питание. По Мензбиру A918) птицы добывают из грязи червяков, рач-
ков, слизняков, лягушат и пр. Желудок каравайки, добытой 24 августа в Лен-
коранской низменности, был наполнен сильно перетертыми хитиновыми остат-
ками. В основном они принадлежали мухам и недавно перелинявшим прямо-
крылым (Исаков и Воробьев, 1940). На Сыр-Дарье в районе Кзыл-Орды боль-
шое значение в питании взрослых птиц имеют личинки коромысла; в значитель-
ной степени ими вскармливаются также птенцы. Рты и пищеводы у многочислен-
ных птиц, добытых в период вскармливания птенцов, содержали эту пищу,
а молодые, будучи пойманными, выбрасывали целые комки непереварившихся
личинок. В желудках караваек неоднократно обнаруживались остатки мелких
лягушек, рыбы и крылья мелких водяных жуков (Спангенберг). В желудках
караваек, вскрытых в Астраханском заповеднике Аскаровым обнаружены
КАРАВАЙКА
371
сверчки, стрекозы (Libellula) и их личинки, личинки мух, бабочки львинки
(Stratiomyia), жуки листогрызы (Chrysomela), водолюбы (Hydrophilus), жуки
радужницы (Donacia), жужжелицы (Carabidae), долгоносики (Curculionidae),
плавунцы (Dytiscidae) и другие насекомые, тарантулы (Trachasa cinerea),
прудовики (Limnaea), катушки (Planorbis), мелкие лягушата и головастики,
(в частности, Rana ridibunda), мальки рыб (лещ, сазан), в небольшом числе —
корешки растений.
Состав пищи изменяется в зависимости от времени года. При массовом
появлении головастиков и мелких лягушат караваики особенно часто поедают
эту пищу, в периоды мас-
сового появления саранчи
караваики долгое время в
основном кормятся за счет
этих насекомых.
Линька. Как у дру-
гих голенастых две линьки
в году — неполная, пред-
брачная, и полная, после-
брачная. Полная после-
брачная происходит между
июнем и ноябрем, частич-
ная предбрачная между
январем и маем (Уайзерби,
1939). Окончательный на-
ряд надевается в резуль-
тате полной послебрачной
линьки в годовалом возра-
сте. Детали линьки не вы-
яснены.
Полевые признаки. Ка-
равайка величиной и сло-
жением напоминает боль-
шого кроншнепа, но даже
на большом расстоянии без-
ошибочно отличается от
него своей темной кашта-
новой окраской и корот-
кими тупыми крыльями (хо-
рошо видно на лету). Во
время полета вытягивает
тело, откидывает назад ноги и то часто машет крыльями, то держит их непо-
движно. Летящая стая выстраивается в ряд. Караваики держатся обычно
стайками, иногда крупными стаями, редко одиночками. Молчалива, редко
подает голос на лету или во время кормежки. Чаще голос караваики удается
слышать в момент взлета при испуге, при выстреле по летящей стае и на гнез-
довых колониях во время ссор. Голос — глухие и грубые звуки, напоминаю-
щие своеобразное гоготание.
Описание. Размеры и строение. Клюв у караваск длинный, изогнутый,
длиной около 100—140 мм, с узкими щелевидными ноздрями. Длина самцов A4) 608—
660, самок A2) 512—588, в среднем 635 и 553 мм. Размах крыльев самцов A4) 1000—1500,
самок A2) 870—968, в среднем 1014 и 917 мм. Вес самцов 557—768, самок 530—679,5 г (Аска-
ров, 1938). Длина крыла самцов E) 265—285, самок F) 292—305 мм, в среднем 291,3 и 201,3мм.
(Зоологический музейкМосковского университета). Форму па крыла (не считая рудиментарного
1-го махового) 2>3>1>4>5... Вырезки на внутренних опахалах 1 -го 2-го и на внешних опа-
халах 2-го и 4-го первостепенных маховых. Хвост прямоусеченный, из 12 рулевых. На уз-
дечке у глаз, между ветвями нижней челюсти, на подбородке — голое пятно. Нижняя поло-
Рис. 74. Каравайка
372
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
вина голени голая. Основания 3-го и 4-го пальцев соединены перепонкой. Цевка спереди
покрыта пластинками, сзади — чешуйками. Когти довольно длинные, тупые; коготь сред-
него пальца с двумя-тремя зазубринами.
Окраска. Пуховой птенец черноватый, с беловатыми отметинами на горле и те-
мени; клюв красный с черными поперечными полосами. Взрослые самцы и самки в брачном
наряде — красновато-бурые на голове, шее, передней части спины, малых кроющих крыла,
брюшной стороне; лоб с зеленоватым отливом; маховые и рулевые, большие кроющие крыла
темнозеленые с пурпуровым отливом. В послебрачном (зимнем) наряде каравайка отли-
чается темнобурой с беловатыми пестринами окраской головы, а также темнобурой окраской
спинной стороны. У молодых в гнездовом наряде голова н шея светлобуры, белый рисунок
менее развит. У взрослых клюв и ноги — бурые; у птиц в гнездовом наряде клюв розовато-
бурый с темными поперечными полосами.
РОД БЕЛЫЙ ИБИС THRESKIORNIS GRAY, 1842
Тип Tantalus aethiopicus
3. Священный ибис Threskiornis aethiopicus Lath.
Tantalus aethiopicus. Latham. Ind. Ornith., II, 1790, стр. 790, Абиссиния.
Русское название. Ибис — широко распространенное на европейских языках
от классического у талышинцев, по Сатунину A907) «Имама л и Кут», т. е. птица пророка Али.
Распространение. Ареал. Африка к югу от Сахары, южн. Ара-
вия до берегов Персидского залива, о-ва Альдабра, Мадагаскар. В
прошлом — Египет. В СССР
залетает.
Характер пребыв а*
н и я. На большей части ареа-
ла — оседлая птица.
Биотоп. Равнинные мест-
ности у берегов водоемов, озер,
рек и т. п.
Подвиды и варьирующие при-
знаки. Три подвида, отличающиеся де-
талями окраски; Thr. aeth. aethiopi-
cas—ареал вида кроме перечисленных
ниже частей; Thr. aeth. abbotti R i d g-
way, 1893—о-ва Альдабра; Thr. aeth.
bernieri t Bonaparte, 1855—Мада-
гаскар.
Материковый священный ибис
Thresktornis aethloptcas aethiopi-
cus Lath.
Распространение. Ареал.
См. выше. В пределы нашей стра-
ны известны залеты белого ибиса
в области Каспийского и Черного
морей. Паллас A811) наблюдал
их на Сарпинских озерах и сооб-
щал, что в его время священные ибисы иногда попадались на Черноморском
и Каспийском побережьях. Позднее Сатунин A907), обработавший дневник
Тиэенгаузена, нашел в нем заметку о двукратной добыче (в 1874 и 1875 гг.)
белого ибиса в районе Колданинского истыля, расположенного между Ленко-
ранью и Иранской границей. Описание размеров и окраски добытых (но не
сохраненных) птиц дало повод Сатунину. высказать предположение, что это
были священные ибисы.
В противоположность этому Бутурлин A935), основываясь на окраске
глаза и других признаках, считал, что это был индийский ибис Threskiornis
Карта 68. Распространение священного ибиса
Threskiornis aethiopicus
7. Т. a. aethiopicus, 2. Т. a. abbotti, 3. Т. a. bernieri;
•а — граница гнездовий, б — не вполне выясненная граница
гнездовий, д — место залета.
КРАСНОНОГИЙ ИБИС
373
melanocephalus Latham, 1790. 4 апреля 1944 г. в окрестностях Сангачалы
(около 40 км к югу от Баку) семь белых ибисов, принадлежащих по мнению
наблюдателя несомненно к Т. aethiopicus, отметил Бурчак-Абрамович A946).
Птиц удалось хорошо рассмотреть на близком расстоянии. Последнее наблю-
дение позволяет с уверенностью сказать, что в наши пределы залетает именно
священный ибис.
Характер пребы-
вания. В пределах Союза —
только залетная птица.
Биотоп. В районе Лен-
корани птицы наблюдались на
рисовых полях — беджарах.
Экология. Размноже-
ние. Гнезда располагаются
на деревьях, сделаны они из
ветвей, с легкой подстилкой
из травы, иногда из перьев.
Кладка состоит из 3—4 яиц,
овальной или слегка закруг-
ленной формы, беловатые или
бледнозеленоватые, иногда с
мелкими бурыми пестринами.
Размеры 57—65 х 39,5—45мм.
Питание. Черви, пауки,
улитки и другие беспозвоноч-
ные, также лягушки и мелкая
рыба (Хартерт, 1914).
Полевые признаки. Не-
сколько напоминает каравай-
ку, отличаясь от нее более крупным ростом, белой окраской оперения, чер-
ной голой головой и некоторыми другими признаками.
Описание. Размеры и строение. Довольно крупная птица, несколько усту-
пающая по размерам колпице. Длина крыла 360—400 мм (Тугаринов, 1947).
Окраска. Преобладающая окраска взрослых — белая с черными вершинами
первостепенных и наружных второстепенных маховых. Внутренние второстепенные махо-
вые длинные и несколько рассученные, при основании пепельно-серые, к концу становятся
почти черными, с фиолетово-пурпурным отливом. Свешиваясь, эти перья закрывают хвост.
Голова и шея взрослых птиц — голые, черного цвета. Клюв и ноги — черные, радужина
темнобурая с красноватым наружным кольцом.
У птиц не вполне взрослых голова и шея покрыты короткими черными и белыми перыш-
ками, а внутренние второстепенные развиты слабо.
Общая окраска молодых грязнобелая, внутренние из второстепенных маховых удли-
нены, но не рассучены, голова и шея покрыты беловатыми перышками, бока головы, темя
и шея черновато-бурая с белыми основаниями перьев. Их вершины грязносерого цвета.
Клюв толстый, короткий, слабо изогнут, мясного цвета.
РОД КРАСНОНОГИЙ ИБИС NIPPONIA REICHENBACH, 1852
Тип Ibis Nippon
4. Красноногий ибис Nipponia nippon Те mm.
Ibis nippon. Temminck. PI. Col. V, 1835, 551, Япония.
Распространение. Ареал. Япония—Хоккайдо, Хондо, Садо; Корея;
Китай —Манчжурия, Чжили, Шандунь, Шаньси, Анхуэй, Сычуань, Ганьсу;
в СССР — Приморье.
Детали распространения в пределах Союза таковы. Дыбовский нашел
ибиса гнездящимся на Уссури (Тачановский, 1875), МаакA801) встретил ибиса
Рис. 75. Священный ибис
374
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
на озере Ханка и добыл одного в низовьях р. Сунгача, где, как и на р. Лефу
и Мурень, эта птица, по Пржевальскому, гнездится A876). На весеннем пролете
и на кочевках ибис встречен на р. Мо и у Турьего Рога; в марте (ст. ст.) 1908г.
один экземпляр добыт у г. Имана (Шульпин, 1936), 5 июня 1910 г. добыт у
с. Троицкого в устьях р. Манчжурки (Бутурлин, 1917). На пролете наблюдался
также у Сидеми (Тачановский, 1893); на р. Янчихе по сведениям, полученным
Пржевальским; зимою 1912 г. добыт под Владивостоком. Самые западные на-
ходки, таковы 17 апреля 1858 г. три птицы, отмечены Радде к востоку от Бурей
у Касаткиной, а Пржевальским было получено перо ибиса, убитого на Зее
в 25 км северней Благовещенска. Повидимому, в настоящее время ибис в При-
морье стал реже, а местами перестал гнездиться. Так, по Шульпину A936),
с 1917 г. ибис перестал гнездиться у Новорусановки, где бродячие птицы
наблюдались, впрочем, в 1927 г. Не гнездится теперь ибис в среднем течении
Лефу и у озера Ханка. Почти еже-
годно небольшое число птиц бывает
на пролете в Сидеми, у залива По-
сьет; экземпляр добыт 5 апреля
1927 г. на п-ове Песчаном (Шуль-
пин, 1936), 20 мая 1938 г. бродячий
ибис наблюдался у с. Лукьяновки
на Имане (Спангенберг, 1940), а по
сообщению В. А. Лебедева 27 авгу-
ста 1949 г. близ Елабуги на Амуре
в 90 км ниже Хабаровска.
Характер пребыва-
ния. В Китае ибис оседлый, в При-
морье —перелетный. По всем веро-
ятиям гнездящиеся у нас ибисы
зимуют в Китае. Только раз, в де-
кабре 1912г., ибис был добыт под
Владивостоком. В Приморье ибис
прилетает ранней весной. В 1868 г.
замечен еще при морозах. В 1858 г.
в истоках Сунгучи первое появле-
ние ибисов отмечено 5 апреля. За-
тем до 9 апреля пролетные одиночки
и парочки наблюдались почти ежедневно. Во второй половине апреля числен-
ность пролетных птиц резко сократилась, а к концу этого месяца пролет
полностью закончился. В 1869 г. там же массовый пролет птиц к северу про-
исходил 25 марта, позднее пролетные особи наблюдались более или менее
постоянно до 11 апреля. В течение указанного времени температура менялась,
но обычно стояла морозная погода, не раз были метели (Пржевальский, 1870).
Сроки осеннего отлета остаются невыясненными. Во второй половине сен-
тября и в первой половине октября птицы еще держатся в местах гнез-
довья.
Биотоп. Небольшие рощи лиственных древесных пород, состоящие из
липы, ильма, клена, дуба с обильным подседом, разбросанные подобно остро-
вам на повышенных участках почвы среди вейниковых и тростниковых болот
или среди пойм рек, протекающих в болотистой местности. Кормовые стации—
болота различного характера, сырые, вследствие паводка, участки почвы, по-
росшие редким низкорослым (в метр высотой) кустарником, подсыхающие бе-
рега озер, песчаные косы рек. Ранней весной, пока болота покрыты льдом и
снегом, ибисы держатся по берегам рек вместе с серыми и белыми цаплями. Те
же стации посещаются ими в поисках пищи осенью, после вывода птенцов.
В наших пределах красноногие ибисы избегают близости человека, но в Китае,
Карта 69. Распространение красноногого ибиса
Nipponia nippon
а — граница гнездовий, б — не вполне выясненная гра-
ница гнездовий, г — места зимовок.
КРАСНОНОГИЙ ИБИС
375
например в Ганьсу, селятся местами в культурном ландшафте, даже среди насе-
ленных пунктов.
Численность. Уже во времена исследований Пржевальского чис-
ленность красноногих ибисов была ограничена, ныне это исчезающий вид,
крайне нуждающийся в строгих мерах охраны.
Экология. Размножение. В противоположность другим близким
видам, у нас не образует колоний, всегда гнездясь отдельными парами. Эта осо-
бенность связана повидимому с тем, что Приморье — северный предел области
распространения птицы. Южнее—в частности в Китае — красноногий ибис
гнездится колониями. По данным Пржевальского A870), все известные гнезда
помещались на больших деревьях — были устроены в развилке сучьев высоко
Рис. 76. Красноногий ибис
над землей — изредка, впрочем, сравнительно низко. Гнездо представ-
ляет собой небрежное сооружение из маленьких веточек и отличается
малыми размерами по сравнению с величиной птицы. Количество яиц в
кладках, повидимому, от 2 до 4. По Тугаринову A947), яйца голу-
бовато-зеленого цвета с шероховатой поверхностью. Размеры: 50—65 х
X 32—40 мм.
Сроки яйцекладки, промежутки между откладкой яиц самкой и сроки на-
сиживания остаются неизвестными. У самки, добытой Пржевальским 2 апреля,
яичник был хорошо развит — одно яйцо достигло размера в мелкий лесной орех.
Однако, до 18 апреля, во всяком случае часть особей, не приступала к размно-
жению, так как их наблюдали то парами, то стайками. Судя по немногим отры-
вочным данным, птенцы появляются в середине мая. По Пржевальскому A870)
14 мая в гнезде были два недавно вылупившихся птенца. Два других гнезда,
найденные в мае в устье Уссури, содержали 3 и 4 птенца. Лётные молодые впер-
вые отмечены на Лефу 16 июня.
Питание. Произведено лишь пять вскрытий желудков, из которых один
оказался пустым. В желудке птицы, добытой в конце марта, была обнаружена
довольно крупная целая улитка и остатки жуков; у двух экземпляров, добытых
2 и 3 апреля, в желудках обнаружены остатки жуков и улиток и косточки мел-
376 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
кой рыбы; в желудке птицы, добытой 26 апреля, найдены неопределенные ко-
сточки (не рыбы) (Пржевальский, 1870).
Линька. Единственные данные у Пржевальского A870): старый самец,
добытый 28 июля, находился в сильной линьке. Старые перья оказались сильно
изношенными — они выцвели и не обладали ярким цветом. Новые перья появи-
лись на спине, но были не вполне развиты. Осенняя линька в Китае происходит
в конце сентября. Схема линьки, повидимому, как у других ибисов.
Полевые признаки. Сложение и окраска позволяет безошибочно отличить
красноногого ибиса от всех других наших пернатых. Летает вытянув вперед
шею и клюв и откинув назад ноги. Голос похож на карканье ворона, но зна-
чительно громче последнего. Его можно передать как «ка-га, ка-га, ка-га»,
а иногда, как — «каг, каг, каг». Чаще крик ибиса удается слышать во время
полета, но кричит он и сидя. Нередко ибисы садятся на деревья и на тальник
(Шульпин, 1936). Крайне осторожная птица, добыть которую трудно не только
на местах кормежки, но и у гнезда с птенцами.
Описание. Размеры и строение. Длина крыла 375—410 мм (Тугаринов, 1947).
Клюв длинный, несколько изогнутый вниз. Вся передняя часть головы, включая область
ушных отверстий, лишена перьев. Сильные короткие ноги с пальцами, обтянутыми на длину
первой фаланги перепонками. На затылке большой хохол — грива из многочисленных узких
перьев, ниспадающих до основания шеи. Шея сравнительно короткая; крылья длинные;
хвост слабо округлый.
Окраска. Общая окраска оперения взрослых белая с нежно розоватым налетом,
развитым наиболее сильно на первостепенных маховых и их стержнях и рулевых. Во время
полета снизу птица кажется розовой. Неоперенная часть головы — красная, клюв — чер-
ный с красной вершиной, вокруг глаза кольцо желтого цвета, радужина красная; ноги
грязновато-красные, почти бурые.
Ибисы, наблюдаемые и добываемые в наших пределах, окрашены несколько иначе.
Задняя часть их головы, шея, мантия и зоб покрыта пепельно-серым оперением. А. Я. Туга-
ринов A947) предполагает, что такая окраска связана с загрязнением перьев на тех участках
тела, которых птица не в состоянии достать клювом во время чистки.
По другим авторам серая окраска — проявление диморфизма (Хартерт, 1914).
Молодая птица (взятая из гнезда на Хоккайдо) серовато-бе лая. Буро-серый цвет
распространен на спине, плечевых, кроющих крыла, шее и передней части спины. Части
первых маховых, выставленные из пеньков (около 8—11 см) буро-серые, у третьего пера серо
только внешнее опахало и конец внутреннего; у остальных больших маховых только рас-
плывчатые серые пятна ближе к концу пера; самые маховые уже бледно розовые. Уже в этом
возрасте перья затылка и задней части шеи (верхняя часть) удлинены и образуют подобие
хохла (Шулышн, 1936).
СЕМЕЙСТВО АИСТОВ CICONIIDAE
РОД АИСТОВ CICONIA BRISSON, 1760
Тип Ciconia ciconia
5. Черный аист Ciconia nigra L.
Ardea nigra. Linnaeus. Syst. Nat., изд. X, 1758, стр. 142, сев. Европа.
Русское название. Аист — народное, черный аист — книжное.
Распространение. Ареал. Пиренейский п-ов (Алемтехо в Португалии,
южн. Испания), сев. Германия, южн. Швеция, Польша, Венгрия, Чехослова-
кия, Югославия, Болгария, Румыния, Албания, в СССР в Прибалтике, восточ-
нее, примерно до 60° с. ш., у Ленинграда, Вологды и Кирова к северу; в Азии
в Малой Азии, Иране, Средней Азии, в Сибири, сев.-зап. Монголии и Китае (горы
сев. Чжили, вероятно до Сычуаня), на восток до Приморья и Сахалина. Северная
граница в Сибири на восточном склоне Урала на р. Лозьва доходит до 61° с.ш.,
затем спускается к югу до Нарымского района, к северу до Томска, проходит на
Енисее до устья Каменной Тунгуски. В Якутии граница подымается к северу —
птица гнездится в бассейне Вилюя, у Якутска и в бассейне Алдана (Иванов,
ЧЕРНЫЙ АИСТ 377
1929). Восточнее граница спускается к югу и вероятно доходит до устья Амура.
Гнездится в Приморье и на Сахалине. Есть указания на гнездование чер-
ного аиста в южн. Африке (Гроте, 1932), но это еще требует подтвер-
ждения.
К югу от указанной границы распространение черного аиста в Союзе не-
равномерно. Гнездясь в горных и лесных местностях, он отсутствует в степных
районах или бывает в них только на пролетах. К югу он проникает на гнездовье
до Подолии, несомненно есть в областях Киевской, западной части Харьковской,
Карта 70. Распространение черного аиста Ciconia nigra
й — граница гнездовий, б — не вполне выясненная граница гнездовий, г — область зимовок.
в Курской, Воронежской, северной части Рязанской, вероятно в Саратов-
ской обл., безусловно в Ульяновской, Куйбышевской и в Башкирии
(Сушкин, 1897).
Кроме того, гнездится вероятно в горах Крымского п-ова, широко насе-
ляет низменные и горные области Сев. Кавказа и Закавказья. В Азиатской
части Союза, вероятно, гнездится в Бадхызском заповеднике (область отрогов
Паропамиза), безусловно гнездится в горах Куги-танга, в Таджикистане,
в Киргизии, спорадично в Тарбагатае, в центральном, южном и в более низких
частых юго-вост. Алтая (Сушкин, 1938). На запад от Алтая птицы гнездятся
в окрестностях Барнаула, но далее в западносибирских степях на гнездовье
представляют большую редкость. В прошлом, пока не были истреблены леса,
аисты гнездились между Орском и Чкаловом и близ Актюбинска, между Верхне-
уральском и ст. Нижнеозерной (Зарудный, 1888).
378
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Таким образом, в южной части Союза остается огромное незаселенное чер-
ным аистом пространство. Оно тянется от Молдавской ССР и всего восточного
побережья Каспия через пустыни Туркмении, Узбекистана и Казахстана до
Тарбагатая и Алтая на восток и до Актюбинска и Орска на север.
Гнездящиеся в Европе черные аисты зимуют в Африке, главным образом
в южной и юго-восточной, реже в западной от Сенегамбии до Анголы. Аисты
из зап. Азии зимуют в северной и средней Индии, восточносибирские в Китае,
в низовьях Яньцзе, в Фучжоу, в юго-вост. Юннане. Северокитайские аисты
(Чжили) и западнокитайские (Сычуань) повидимому оседлы. Залеты черных
аистов известны в Англии, Франции, Норвегии, Финляндии, Бельгии, Голлан-
дии, Швейцарии с о-ва Мадейры и из Японии.
Характер пребывания. В СССР гнездящаяся перелетная
птица. У северной границы области распространения местами только залетает.
Даты. Прилет в Европу в среднем недели на две позднее, чем у белого аиста.
В Германии черный аист прилетает между концом марта — началом апреля,
улетает от середины августа и в сентябре. В СССР нередко передовые черные
аисты появляются весной еще тогда, когда снег не стаял, а водоемы не полностью
освободились ото льда. Валовой прилет начинается через неделю, иногда через
две после появления передовых птиц и тянется долго, заканчиваясь нормально
в конце мая, а в исключительных случаях даже в половине июня. Поздно летя-
щие птицы вероятно холостые или относятся к самым северным популяциям.
На юге прилет раньше. В Ленкорани первые аисты отмечались между 26 фев-
раля и 20 марта, 15 марта прилет аистов отмечен на Куре в б. Арешском уезде
(Сатунин, 1907). Валовой прилет птиц в Закавказье происходит позже в сере-
дине апреля, а под Тбилиси аисты отмечались только 27 апреля (Радде, 1885).
В апреле аисты прилетают и в местности, расположенные севернее. У Новорос-
сийска Птушенко наблюдал черного аиста 11 апреля, под Таганрогом валовой
пролет протекал 16 апреля. В окрестносях Павлограда в низовьях Днепра пролет
отмечался 11 апреля (Вальх, 1899). В Бессарабии аисты появляются в середине
апреля. В разные числа апреля происходит прилет черных аистов и во многие
другие местности Европейской части СССР; так, на юге Сталинградской обл.
аисты отмечались 23 апреля (Аргиропуло, 1928), в Башкирии —в последней
трети апреля (Сушкин, 1897), в Московской обл. 21 апреля (Поляков, 1910),
в середине этого месяца в южных частях Минской обл. (Шнитников, 1913);
наиболее ранняя дата в районе Пскова 25 апреля (Зарудный, 1910); в половине
апреля в Новгородской обл. (Бианки, 1910).
В Азиатской части Союза весенний прилет сильно растянут и местами про-
исходит очень поздно. Под Ашхабадом в 1923 г. черные аисты впервые наблю-
дались 21 апреля, в 1924 г. 7 апреля (Шестоперов, 1928). В более северных
местах аисты наблюдались на средней Эмбе в начале апреля и в районе Чка-
лова ежегодно между 24 апреля и 10 июня (Зарудный, 1888). Напротив, у Зай-
сана первые особи появляются очень рано, уже 26 марта, но массовое движение
происходит здесь в самом начале апреля (Хахлов, 1914). В конце апреля птицы
появляются в западной окраине Алтая, но в самом Алтае и в Монголии зна-
чительно позднее. В горах на высоте 2000 м над уровнем моря у озера Кара-гель
стайка впервые была отмечена лишь 5 июня (Сушкин, 1938). Даты прилета
в различные части Сибири таковы: в районе Томска близ с. Новинского отме-
чен 16 апреля, в окрестностях Нерчинска прилет происходит в последней
трети апреля и в самом начале мая, в Яблоновом хребте 4 пролетных аиста отме-
чены 2 мая (Дорогостайский, 1915), в Амгинском районе Якутии 1 мая (Скалой).
Рано — во второй половине марта — появляются черные аисты в пределах
Приморья.
Осенний отлет начинается сравнительно рано, местами при высоких тем-
пературах, иногда в самом начале августа и затягивается до глубокой осени.
Как и весной, однако, массовое движение происходит в более сжатые сроки. На
ЧЕРНЫЙ АИСТ 379
Амуре, несколько севернее устья Уссури небольшие пролетные стайки черных
аистов отмечались 13 сентября и в последний раз близ устьяСунгари 15 сентября
(Маак, 1859). У Томска один экземпляр был добыт 22 сентября (Иоганзен,
1914). На зап. Алтае птицы отлетают около половины сентября A3 сентября,
Сушкин, 1938). Под Чкаловом осенний отлет начинается с конца августа и
продолжается до последних чисел сентября, одиночки изредка задерживаются
до половины октября, но в устьях Урала черные аисты отмечались уже в пер-
вой трети августа. В пределах Средней Азии во время осеннего пролета чер-
ные аисты задерживаются на длительное время. Наиболее поздний экземпляр
в районе Ашхабада наблюдался в 1928 г. 25 декабря (Шестоперов, 1937). Такие
задержки одиночных экземпляров безусловно следует рассматривать, как исклю-
чение, но не как нормальные зимовки. Что касается Европейской части Союза,
то в окрестностях Ленинграда (у пристани Лигово) одна птица отмечена 16 ноября
(Орнитолог, вестн. 1913), т. е. при наступлении зимних холодов. Нормально
же отлет происходит раньше и в районе Новгорода совпадает с первыми числами
сентября. В Минской обл. отлет начинается во второй трети августа и длится
почти до конца месяца. Позднее 28 августа птицы здесь не наблюдались (Шнит-
ников, 1913). Начало отлета в Харьковской обл. (по Салову, 1897) —30 ав-
густа; в конце первой трети сентября птицы исчезают из Башкирии. В течение
сентября и октября регулярно пролетают черные аисты на Сев. Кавказе. По
Мензбиру A918) черные аисты частично перезимовывают в Закавказье, что опять-
таки следует рассматривать, как редкое исключение.
Во время пролетов черные аисты летят небольшими стайками или семьями,
реже их удается наблюдать одиночками. В некоторых случаях птицы образуют
крупные пролетные стаи до 50 особей в каждой. Такие общества наблюдались
под Чкаловом. Птицы летят то придерживаясь определенного направления, то
начинают описывать парящим полетом круги, поднимаясь все выше и выше
в воздух. Пролетных аистов обычно удается наблюдать в дневные часы, но
вероятно они летят и в ночное время, не выдавая, как другие птицы, криком
своего присутствия. Изредка отдельные особи этого вида присоединяются к про-
летным стаям белых аистов, но чаще явно избегают держаться близко от своих
более крупных белых сородичей.
Результаты кольцевания. Аисты кольцевались в западных
областях Союза. Окольцованные в Латвии, добыты в сентябре в Польше, в ноябре
на юго-западе Малой Азии, в марте в Египте. Окольцованные в Литве аисты
добывались в Венгрии и средней полосе Союза;черный аист, окольцованный
в Калининградской обл., добыт в октябре в Пелопонесе.
Биотоп. Гнездовый — в связи с широким распространением — разно-
образен. Обязательным условием для гнездования вида является сочета-
ние старых лесных массивов или хотя бы отдельно стоящих крупных деревьев
или скал, с наличием поблизости болот разнообразного характера, открытых
берегов рек и озер. При этих условиях черные аисты населяют как низменности,
так и горы. В Закавказье черные аисты в гнездовый период явно тяготеют к сы-
рым низменным лесам, но гнездятся также довольно высоко в горах, где плот-
ность их населения меньше. В районе Сочи птицы гнездятся в нижнем поясе
гор (Кудашев, 1916), в вост. Аджарии до 1800 м над уровнем моря (Сату-
нин, 1913), заселяя широколиственные леса, покрывающие горные склоны.
В горной Армении гнездовая область аиста поднимается более, чем до 1000 м
над уровнем моря. Редкие пары птиц гнездятся в буковых лесах по крутым,
сырым (северным) склонам гор Делижанского ущелья в районе г. Кировакана.
В центральном Закавказье (Исмаилинский район) аисты также гнездятся
довольно высоко в горных котловинах, изобилующих речками и ключами. На
западном побережье Каспия между устьями Куры и Иранской границей черные
аисты в ограниченном количестве гнездятся в глухих лесах по нижним склонам
Талыша. Основная масса птиц выводит здесь в культурном ландшафте Ленко-
380 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
раненой низменности. Небольшие рощи, массивы старого леса, пятнами раз-
бросанные среди рисовых полей, или отдельные великаны деревья, растущие
среди открытого ландшафта, служат здесь излюбленным местом гнездовия
этого вида.
На Сев. Кавказе гнездовый биотоп повидимому схож с таковым в централь-
ном и зап. Закавказье, т.е. представляет собой старые широколиственные лесаг
покрывающие склоны гор. Здесь, по данным Беме A926) аисты в период размно-
жения проникают в горы до 1200—1500 м над уровнем моря. В пределах Турк-
мении, Таджикистана и Киргизии по данным ряда авторов гнездовым биотопом
черных аистов являются настоящие горы. В хребре Куги-танг, в Туркмении,
аисты гнездятся еще в относительно невысоких безлесных предгориях; в южн.
Киргизии на берегах озера Сары-Чилек уже на высоте 1925 м над уровнем моря,
в скалах, поросших смешанным лесом со значительным преобладанием хвойных
пород. Еще выше (до высоты 2000—2200 м) гнездятся аисты на озере Искандер-
куль в Гиссарском хребте. На Алтае между 1750—1800 м по окраинам тайги
(Сушкин, 1938). На южном берегу озера Марка-куль гнездо было найдено
в культурном ландшафте на дереве среди разделанного поля (Поляков, 1913).
В приморье птицы гнездятся в разнообразных горных стациях. По Шульпину
A936), в горной тайге, в верховьях рек, придерживаясь мест, где имеются раз-
бросанные среди лесных массивов поляны или открытые болотины близ рек;
заселяют горное чернолесье и густые ельники.
Несколько иной характер гнездования в равнинной и более северной
полосе Союза. В равнинной части Башкирской АССР птицы гнездятся в лист-
венных лесах (уремах), разросшихся по низким берегам рек и затопляемых
в период половодья. В Тюменской обл. гнездовый биотоп —смешанные
леса по долинам рек. В Тарском округе Омской обл. — хвойные леса (Ушаков,
1913), в Минской обл. —глухие участки леса среди болот (Шнитников, 1913).
Места кормежек черного аиста разнообразны и изменяются в различных
частях Союза. В Закавказье излюбленными местами добычи пищи птиц служат
рисовые поля (биджеры), сырые луга и болота на берегах озер. Не избегают аисты
лесных водохранилищ — истылей и мелких речушек, берега которых скрыты
лесной чащей. В горных областях Средней Азии аисты кормятся по берегам
речек и сырым луговинам близ горных озер, в средней и северной полосе посе-
щают разнообразные болота, в Приморье — сырые луга, болота, берега озер и
речные косы. На Алтае открытые места с заболоченными лугами и водоемы со
стоячей или тихо текущей водой.
В ряде случаев гнездовый и кормовой биотопы отстоят друг от друга на
5—10 км. Вообще говоря, черные аисты гнездятся и кормятся в глухих, редко
посещаемых человеком угодьях. Исключением из этого правила является За-
кавказье, где черные аисты не только кормятся в культурном ландшафте, но
и гнездятся в непосредственной близости от жилья, изредка даже в самых Азер-
байджанских селениях.
Численность. Черные аисты встречаются довольно редко, но вслед-
ствие чрезвычайно широкого распространения общая их численность значи-
тельна. В наиболее благоприятных условиях в районе Минска три пары гнез-
дились на территории около 10 000 га (Шнитников, 1913). Для Башкирской
АССР Сушкин считает черного аиста обычным гнездящимся видом, для Алтая
в общем редким. В Приморье (долина среднего течения реки Имана) обычен.
В других частях страны черные аисты в общем редки. Большая плотность на-
блюдается лишь в вост. Закавказье между устьями Куры и Иранской границей.
Здесь в соответствующих биотопах в 1934 году гнездо от гнезда редко распо-
лагалось на расстоянии более 1 км.
Экология. Размножение. Половая зрелость наступает в трехлетнем
возрасте (Нитхаммер, 1938). Никогда не образует настоящих колоний. В Лен-
коранской низменности (иногда и в Черниговской обл.) гнездятся близко друг
ЧЕРНЫЙ АИСТ 38Г
от друга и два обитаемых гнезда помещаются на одном дереве. Не избегают
близости гнездования других колониальных пернатых и под Ленкоранью,
строят гнезда на окраинах многочисленных поселений различных цапель,
больших и малых бакланов, населяющих лесные массивы и небольшие рощи.
Гнезда помещаются на старых деревьях или, что наблюдается несравненно
реже, в нишах скал и высоких конгломератовых обрывов. Гнезда, построенные
в скалах, известны в предгориях Куги-танга (Туркмения) в Таджикистане и
в Киргизии. В Киргизии найдено гнездо, помещавшееся в углублении отвесной
скалы метров на 16 над водой озера Сары-Чилек. Вероятно в нишах скал гнез-
дятся черные аисты и в других местах гор Средней Азии, а также Кавказа.
На Алтае аисты гнездятся на скалах и на деревьях (Сушкин, 1938). В остальных
частях СССР аисты гнездятся на деревьях, сооружая свои гнезда то на вер-
шине высокого дерева (озеро Марка-куль и под Верхнеуральском), то, напротив,
относительно низко, среди разветвлений толстых ветвей близко от основного
ствола или на боковой ветви. Для гнезд избираются в Приморье лиственницы и
черные березы (Шульпин, 1936), в Минской обл. огромные ольхи с голыми
стволами (Шнитников, 1913), в Черниговской обл. столетние дубы (Емелья-
ненко, 1916), тополя на Алтае (Сушкин, 1938), а местами также сосны (Менз-
бир, 1918). Под Ленкоранью особенно охотно черные аисты избирают для гнезд
великаны дубы, устраивая гнезда на большой высоте в верхней трети дерева,
но гнездятся также на ольхах или на вершине старого сломанного тутовника.
В одних случаях гнездо скрыто густой листвой кроны, в других предоставлено
лучам солнца и видно издали. Гнезда используются птицами в течение несколь-
ких лет и ежегодно ремонтируются и достраиваются. В таких случаях они до-
стигают чрезвычайно больших размеров и особенно толщины. Поперечный диа-
метр гнезда бывает от 1 до 1,5 м, но у вновь построенных значительно меньше.
Основной строительный материал — сухие сучья столь значительных размеров
и толщины, что птица с трудом справляется с ними. Лоток гнезда обильно вы-
стилается сухой травой, листьями, стеблями камыша и шерстью животных.
Нередко в гнездах удается находить куски бумаги, тряпки, куски шкур и
даже кошму.
Количество яиц в кладках от 2 до 5, очень редко 6. Птенцов в гнездах всегда
бывает меньше, чем яиц в кладках. При кладке в 5 яиц часто 2 яйца оказываются
болтунами. Яйца правильной яйцевидной формы или несколько вздутые, с силь-
но закругленными концами. Окраска скорлупы белая, несколько сероватая.
К концу насиживания яйца сильно загрязняются. От яиц белого аиста безоши-
бочно отличаются меньшими размерами и цветом скорлупы на просвет. Послед-
няя яркозеленая в отличие от желтой у предыдущего вида. Размеры: A1)
61,2—67,1 х 46,1-49,0 мм, в среднем 64,0 х 47,0 мм.
В северных частях ареала гнездование происходит значительно позднее,
чем в южных. В одних и тех же местах отдельные пары гнездятся не одновре-
менно. Под Ленкоранью в начале апреля большинство местных птиц держатся
у гнезд. Одни из них заняты постройкой гнезд или их подправкой, другие (ран-
ние) приступают к яйцекладке. В это время неоднократно наблюдались самки,
стоявшие на гнездах. Подлетающих самцов они встречали характерным кива-
нием головы. 4 апреля впервые удалось наблюдать спаривание на вполне за-
конченном гнезде (данные Птушенко). 13 апреля под Ленкоранью в осмотренном
гнезде нами обнаружена кладка в 4 едва насиженных яйца. В 1929 г. 4 мая
в одном гнезде у Порт-Ильича оказалось 3 совсем маленьких птенца и 2 яйца
болтуна, а 18 мая в другом гнезде 2 насиженных яйца. С этого времени и до
середины июня осмотрено 11 гнезд, содержавших маленьких и вполне оперив-
шихся птенцов. В июле приходилось наблюдать летных молодых, державшихся
семьями на рисовых полях и болотах. Однако, Сатунин A913) пишет о гнезде,
осмотренном в Мингрелии (зап. Закавказье) 11 августа, в котором нахо-
дились еще аистята. В Минской обл. по наблюдениям Шнитникова A913) чер-
382 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
ные аисты начинают гнездиться около середины апреля *, и около конца мая
в гнездах бывают птенцы. Однодневные птенцы найдены 24 мая. В Беловеж-
ской пуще слегка насиженные яйца взяты 23 апреля.
В Бузулукском бору совсем маленькие пуховые птенцы (величиной с куро-
патку) были вынуты из гнезда 10 июня (Карамзин, 1901). В Башкирии птицы
приступают к постройке гнезда в первой половине мая. В районе Новгорода
оперившийся птенец — 10 июля (Бианки, 1910). Раньше приступают к размно-
жению черные аисты в горах Тянь-Шаня. В гнезде, найденном на озере Сары-
Чилек (Киргизия) 29 июня, обнаружен Кашкаровым A927) крупный оперив-
шийся птенец. На Алтае на низовье р. Боку-Мерина, впадающей в озеро Ачит-
Нут, в найденном 3 июля гнезде птенцы оказались сплошь одеты пухом, и вер-
шины маховых только что начали показываться. В низинах гор Тарбагатая
4 августа в гнезде обнаружены крупные, но едва летающие птенцы, а хорошо
летающие на м. Тююк на озере Зайсан 30 августа (Сушкин, 1938). Полные
кладки в 3—4 яйца в Приморье удалось найти около 18 апреля (Дорриес, 1888).
Таким образом, у черных аистов наблюдается некоторое изменение сроков
начала размножения как в широтном, так и в долготном направлении. Птицы
раньше гнездятся на юге, чем на севере, и ранее на западе, чем, например,
в Башкирии и на Алтае.
Спаривание происходит на гнезде, когда постройка закончена или подходит
к концу. Яйца откладываются через день. Насиживание начинается с первого
яйца, но пока кладка не закончена птицы сидят на гнезде короткое время, но
и не отлучаются от него. Большую часть дня одна из птиц гнездовой пары стоит
на гнезде, оберегая яйца от пернатых хищников. Насиживание продолжается,
по одним авторам —32—38, по другим —35—46 дней (Уайзерби, 1939). На-
сиживают оба родителя. Пока птенцы малы одна из птиц постоянно остается на
гнезде, другая занята добычей пищи. Когда птенцы подрастут, обе птицы одно-
временно покидают гнездо, причем только для отыскания пищи. Как только
птенцы достаточно окрепнут и приобретут способность к полету, они следуют
за обоими родителями на места кормежки. Вначале взрослые и молодые птицы
держатся семьями, а ближе к осени сбиваются в стаи. Сроки поднятия на крылья
молодых изменяются в зависимости от широты и долготы. В общем гнездовый
период от вылупления до подъема на крыло продолжается около 60 дней F3 по
Хейнроту, 69—71 по Зиверту). Под Ленкоранью летные птенцы отмечались во
второй половине июля. В Минской обл. во второй трети августа семьи черных
аистов уже начинают подвигаться к югу. В Приморье кочующие выводки в стаях
белых и серых цапель отмечены 13 сентября.
Линька слабо изучена. По данным Сушкина A897) черные аисты при-
летают к нам весной далеко не окончив линьки. У самца, добытого 9 мая, боль-
шая часть маховых и больших кроющих крыла, перьев подхвостья и верхней
части головы старые, обношенные и только что начинают сменяться. Очень зна-
чительная примесь старых перьев замечается в мелких кроющих крыла и рулях.
Птица, добытая 8июля, почти кончила линьку. Старые перья сохранились только
на лбу, темени и в незначительном количестве среди маховых и плечевых. Белые
перья брюха заметно обносились и приняли грязный оттенок. По Сомову A897),
в конце мая новые все большие маховые, за исключением первых двух на каж-
дом крыле. Линька повидимому раз в году, весной начинается уже в феврале
и кончается в мае—июне.
Питание. Данные по питанию скудны. Наблюдения в районе Ленко-
рани показали, что в наибольшем числе птицы поедают рыбу, реже насекомых,
лягушек и змей, а из них те виды, которые населяют местные болота и сырые
¦ Мензбир A918),заимствуя эти данные у Шнитникова, пишет:«Черные аисты гнездятся
в конце апреля, так что в начале мая уже можно бывает найти выклюнувшихся птенцов».
Повидимому, здесь допущена ошибка: не в конце апреля,а в начале апреля, так как черный
аист насиживает около месяца.
БЕЛЫЙ АИСТ
383
луга, где обычно кормятся птицы. Меньшее значение в питании имеют мыше-
видные грызуны и птенцы воробьиных птиц.
Полевые признаки. Крупные размеры, своеобразное сложение и окраска
оперения, ног и клюва позволяют легко отличить черного аиста в природе от
других птиц, населяющих нашу страну. От белого аиста он отличается пре-
обладанием черного цвета в оперении и несколько меньшими размерами.
При взлете птица пробегает некоторое расстояние по земле и уже затем
поднимается в воздух, вытягивает длинную шею, откидывает назад
длинные ноги и медленно и глубоко взмахивает большими крыльями.
Часто аисты парят, как грифы, поднимаясь на большую высоту над
лесом. От взрослых птиц из-
редка удается слышать негром-
кое трещание клювом. Эти звуки
происходят вследствие частых
ударов нижней челюсти о верх-
нюю. Кроме того, аисты издают
негромкие звуки вроде «че-ли»
или «чи-лин». Молодые пищат.
Черный аист чрезвычайно осто-
рожная птица. Там, где его тре-
вожат, он не подпускает близко
человека даже у гнезда с яйцами
и птенцами. Напротив, во мно-
гих местах Закавказья, где этих
птиц не трогают, они относитель-
но доверчивы.
Описание. Размеры и стро-
ение. Клюв длинный, прямой, сужи-
вающийся к вершине; ноздри малень-
кие, щелевидные. Формула крыла (не
считая абортивного первостепенного)
4^3^5>2>1>6... Наружные опахала
2-го и 4-го первостепенных — с вырез-
ками. Хвост слегка закругленный, из
12 рулевых. Ноги длинные, передние
пальцы соединены у основания перепонками, когти тупые и короткие. Длина крыла самцов
и самок (8) 520—580, в среднем539мм. Клюв около 150—190 мм, цевка около 175—200 мм. Вес
около 3000.
Окраска. Первый и второй пуховые наряды белые, с сероватым оттенком на спине;
клюв желтый, ноги бледнорозовые. Взрослые самцы и самки: спинная сторона черная, с зе-
леноватым и медно-красным металлическим отливом, буроватые на надхвостье и рулевых.
Брюшная сторона, кроме шеи, белая. Клюв, ноги, голое пятно на уздечке и у глаз ярко-
красные, когти черные, радужина бурая. После окончания периода размножения, осенью,
клюв и ноги приобретают буроватый оттенок. Птицы в гнездовом наряде отличаются от
взрослых меньшим развитием металлического отлива в окраске, бурой головой, шеей и
грудью и тем, что на этих партиях, а также на кроющих крыла, плечевых и в подхвостье
имеются бледноохристые отметинки у вершин перьев.
6. Белый аист Ciconia ciconia L.
Ardea ciconia. Linnaeus. Syst Nat., изд. X, 1753, стр. 142, Швеция
Распространение. Ареал. Европа — в вост. Франции (Эльзас, деп. Мозель,
местами в Вогезах и на Сомме), Португалии и Испании, Голландии, южн. Шве-
ции (Сконе, юг пров. Халланд), Германии, Швейцарии, Австрии, Венгрии,
Чехословакии, на Балканах, на восток до Прибалтики, Пскова, Ленинградской
обл. (Луга), Смоленска, Харькова, Брянска, Орла; сев. Африка —сев. Тунис,
Алжир, Марокко; в Азии —в Малой Азии, зап. Иране, сев. Палестине, Ираке;
кроме того, в Закавказье и Средней Азии; кроме того на Дальнем Востоке —
в Японии (Хоккайдо, Хондо, Сикоку, Цусима), Корее, Приморье. Зимовки
Рис. 77. Черный аист
384
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
западных птиц отюго-зап. Аравии до центральной и южн. Африки, восточных —
в Индии и в юговост. Азии. Залеты в Англию, Норвегию, Финляндию, Бельгию,
на Мадейру, и Канарские о-ва. Область распространения в Зап. Европе сокра-
тилась в настоящее время, в частности аист истреблен в зап. Франции, в Бель-
гии, в средней и южн. Греции и в некоторых других местах.
Карта 71. Распространение белого аиста Ciconia ciconia
7. С. с. ciconia, 2. С. с. asiatica, 3, С. с. boyciana;
а — граница гнездовий, б — не вполне выясненная граница гнездовий, г — область зимовок.
Характер пребывания. В большей части ареала аист перелет-
ная птица; в Японии — оседлая.
Биотоп. Открытые и полуоткрытые пространства с отдельными деревьями
или группами деревьев, с наличием водоемов и болот; охотно селится в куль-
турном ландшафте. Гнездится как на равнине, так и в горах.
Подвиды и варьирующие признаки. Три подвида, из которых два очень близки (турке-
станский аист отличается от европейского большим и слегка загнутым вверх клювом), а
третий — китайский аист — имеет существенные отличия в окраске, в форме и цвете клюва,
а также и в биологии С. с. ciconia Linn., 1756 — в Европе, сев. Африке, зап. Азии; С. с.
asiatica S e v., 1873 ^- в Средней Азии; С. с. bojciana S w i n h о е, 1873 — в вост. Азии.
Европейский белый аист Ciconia ciconia ciconia L.
Местные названия: аист — широко распространенное название; на Украине—
черногуз и лей лека; в Закавказье — лейляк; в Армении —тарех-спилан.
Распространение. Ареал. Общее распространение описано выше, детали
распространения в наших пределах таковы: восточная граница СССР идет от Но-
воржевского, Псковского, Перховского, Островского районов через Смоленскую
обл., район Калуги, Брянскую, Орловскую и Харьковскую обл., охватывает
с востока Днепропетровскую обл. и спускается к Мариуполю. В Крыму не гнез-
БЕЛЫЙ АИСТ 385
дится. В Закавказье распространение несколько спорадично. Аист отсутствует
в самых западных частях Закавказья, но в Армении гнездится как в низменных
районах, так и на возвышенных равнинах. Гнездится также на берегах озера Се-
ван на высоте 2000 м над уровнем моря. Далее гнездится в низменностях вдоль
Куры в районах Арешском, Джевашинском, Шушинском и Джебраильском.
В районе Баку аистов нет, но к северу отсюда, в Кубинском районе, они вновь
оказываются обыкновенными. К югу от Куры вдоль морского побережья обычны,
во всяком случае до широты Ленкорани, но южнее последние, вероятно, отсут-
ствуют или встречаются крайне редко. На Сев. Кавказе в качестве гнездящейся
птицы видимо представляют большую редкость. Лишь в 1929 г. гнездящаяся
пара найдена в ауле Зрых, в Дагестане. Отмечено, что европейский аист посте-
пенно расширяет свою гнездовую область, продвигаясь к северу и востоку. За
пределами гнездовой области аисты встречаются во время пролетов на Сев.
Кавказе — в районе Краснодара, под Дзауджикау, изредка на Черноморском
побережье Кавказа, в вост. Приазовье, под Таганрогом. Случаи залета известны
в Воронежскую обл., довольно часто в Калужскую, где он на гнездовье редок,
был убит у станции Голицино под Москвой, известен в Тульской, Рязанской обл.
и в районе Пензы. Шестоперов A937) говорит также о залетах западной формы
в низовья Атрека и юго-зап. Туркмению. Зимовки европейских белых аистов
расположены в центральной и южной Африке до Капской колонии.
Результаты кольцевания. Аист в СССР перелетная птица,
хотя отдельные особи отмечены зимой у мест гнездовья в Калининградской обл.
(Нитхаммер, 1938); имеется наблюдение над задержавшимся в Полесье до января
аисте (Шнитников, 1913). На свои африканские зимовки европейские аисты
летят двумя направлениями: на западе через Испанию и Марокко, на востоке
через Босфор, Малую Азию, Палестину и Египет. Пролет продолжается долго;
в южн. Африке аисты появляются только в ноябре и даже декабре и остаются
там повидимому до конца февраля. Неполовозрелые особи в южн. Африке
проводят лето.
На Украине встречены аисты, окольцованные в Калининградской обл.,
быть может неполовозрелые особи. В частности, птица, окольцованная в Хел-
лингштедте близ Шлезвига 28 июня 1933, добыта 1 июля 1935 ус. Гушар в Ост-
ровском районе Черниговской обл.; окольцованный у Браунсбурга 25июня
1935, добыт 12 июля 1935 у с. Вербки в Семеновском районе Харьковской обл.;
окольцованный во Фриденау близ Инстербурга 1 июня 1930, добыт в Сочи ¦ле-
том» 1937. Аист, окольцованный где-то в Германии кольцом Гельголандской
станции, добыт 14 августа 1934 на юге Украины. Наконец, птица, окольцо-
ванная в Латвии у Люцина 18 июля 1940, добыта 10 сентября 1940 в среднем
течении Днестра на границе Украины и Бессарабии.
Надо добавить, что окольцованные в Калининградской обл. аисты добы-
вались на зимовках в Капской Земле, и что общие результаты кольцевания
аистов в этой области показывают на чрезвычайный гнездовый консерватизм
этой птицы.
Характер пребывания. Гнездящаяся перелетная птица. Весен-
ний прилет ранний, его сроки и направление хорошо выяснены. Кайгородов
A911) установил, что в среднем весь весенний пролет птиц в Европейской части
Союза протекает в течение 17 дней — с 23 марта по 9 апреля. Однако, в литера-
туре есть указания о пролете птиц в более поздние сроки. За ряд лет птицы при-
летали в район Пскова между 14 и 20 апреля, а в 1898 г. впервые появился 4 мая
(Нестеров и Никандров, 1913). В Армению птицы прилетают между 13 и 25 марта
(Ляйстер и Соснин, 1942), в Абхазии, где аист вообще бывает редок во время
пролета, одиночная птица была отмечена на берегу Пицундского мыса 27 марта
(Сатунин, 1913), в вост. Закавказье в некоторые годы аисты появляются чрез-
вычайно рано. Так, в 1895 г. аист появился уже 24 февраля (Сатунин, 1907),
но нормальный прилет происходит между 8 и 16 марта. Сначала прилетают оди-
25 Птицы СССР, т. II
386 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
ночные птицы, спустя же некоторое время идет массовый прилет. В районе
Ленкорани Лоудон A913) встретил аистов 14 марта, Тугаринов и Козлова-
Пушкарева A935) —2 февраля, а Исаков и Воробьев A940) первую прилет-
ную стаю наблюдали у Кумбашей уже 30 января. Крайне ранний прилет птиц
дал повод считать район Ленкорани местом зимовок аистов. Весной аисты летят
чаще одиночками, иногда стаями, самцы появляются недели на две ранее самок.
Образование стай перед осенним отлетом отмечено Сомовым на Донце A897)
в июле по старому исчислению. В середине августа (н. с.) огромные стаи на-
блюдались над Мариуполем. Отдельные особи держались здесь до 16 сентября
(Боровиков, 1907). По наблюдению Федюшина A928) в вост. Белоруссии 29 июля
аисты кормились уже стаями, а 20 августа все исчезли. В Минской обл. отлет
начинается около средины августа и заканчивается около средины сентября.
В 1903 г. был известен случай задержки одиночного аиста у проруби до 12 ян-
варя (Шнитников, 1913). Из Бессарабии птицы отлетают, видимо, в конце
августа. В районе Херсона перед отлетом аисты собираются в громадные стаи
у степных прудов, отлетая в первой трети сентября. На Сев. Кавказе пролет на-
чинается в августе и продолжается до самого начала октября. У Дзауджикау
птицы отмечались в сентябре и начале октября, на Кубани в 1914 г. пролет шел
с 16 августа и до 15 сентября (Птушенко, 1915). В Армении пролет происходит
в конце августа, в Гельской котловине в самом конце этого месяца или в пер-
вых числах сентября. Перелет совершается в дневные часы на большой высоте,
причем птицы не образуют строя, а летят парящим полетом, описывая широкие
круги и постепенно подвигаясь в определенном направлении.
Биотоп — гнездовый. Населенные пункты, отдельные крупные деревья
или группы деревьев, разбросанные среди открытого равнинного или холми-
стого ландшафта, изобилующего болотами и стоячими водоемами. В Армении
гнездится на высоте 2000 м над уровнем моря (озеро Севан). Последнее объяс-
няется наличием здесь обширных, безлесных и заболоченных площадей («мо-
крые» горы), с которыми экологически связан белый аист. Кормовой биотоп —
сырые луга, болота южного типа; мелководные участки стоячих водоемов (озера,
рисовые поля, речные разливы), реже водоемы, обладающие слабым течением и
берега крупных рек. Изредка посещает сухие луга, целинные степи и возделан-
ные поля. Места кормежки располагаются в непосредственной близости от мест
гнездования. В редких случаях аисты вынуждены летать от гнезда за пищей на
3—5 км, а иногда при скоплении в определенных участках саранчи или сусли-
ков и на более далекое расстояние.
Местами, как например в вост. Закавказье и в ряде других пунктов,
белые аисты поселяются вдали от поселков и здесь ведут себя, как настоящие
дикие птицы.
Численность. У нас в общем нередкая птица. Особенно обыкновенен
аист в юго-западных и западных частях ареала и местами в Закавказье. По мере
движения на восток и север, ближе к границе распространения,численность птиц
резко снижается. В частности, белый аист относится к обычным гнездящимся пти-
цам в Бессарабии, Днепропетровской, Запорожской, Полтавской, Киевской обл.,
зап. части Харьковской, Черниговской, Минской и Могилевской. Напротив,
в восточной части Харьковской, Орловской, южной части Калужской, восточной
части Смоленской, южной части Новгородской, Псковской и Ленинградской обл.
аист во всяком случае немногочисленен, а местами положительно редок на гне-
здовье. В Закавказье плотность населения аиста неравномерна. Он редок под
Ленкоранью, в Закаталах, в долине Алазани, в горах Армении—на берегах озера
Севан, но весьма многочисленен в Ереванской равнине по Араксу, в Гельской
котловине в районе Кюрдамира и обыкновенен, хотя и распространен спора-
дично, в Кубинском районе.
Экология. Размножение. Половая зрелость в возрасте двух лет, но
не все птицы двухлетки уже размножаются. Пары постоянные. Как правило.
БЕЛЫЙ АИСТ 387
белый аист гнездится отдельными, не связанными друг с другом парами. При
обилии пищи гнезда располагаются близко друг от друга. В маленьких селе-
ниях бывает 2—3, а изредка и 5—б гнезд. На одной крыше в некоторых случаях
гнездятся две пары. Однако, местами, в отличие от черных аистов, белые обра-
зуют нечто вроде настоящих колоний. В западных частях Харьковской обл.
группа в 6 обитаемых гнезд помещалась на ольхах среди обширных лугов(Сомов,
1897). По словам Емельяненко A916), в Черниговской обл. неподалеку от
р. Десны десятки дубов и сосен были покрыты гнездами белых аистов, иногда
на одном дубе помещалось 3—4 гнезда. Однако, это указание сомнительно.
В Кубинском районе (вост. Закавказье) аисты также гнездятся группами по
5—б гнезд, устраивая их на высоких деревьях (Сатунин, 1907). Наконец, ко-
лониями гнездятся аисты в Джаваохетии (Чхикишвили, 1930) и в районе Кюр-
дамира, в Азербайджане (Спангенберг). Гнезда сооружаются на крышах по-
строек человека, на развалинах зданий, на специально выставленных шестах,
столбах, колесах и боронах, на скирдах соломы и сена и на деревьях. При этом
в различных частях страны аисты предпочитают то тот, то иной тип гнездо-
вания.
При гнездовании на деревьях гнездо располагается то относительно низко
(на 3—5 м), то, напротив, на самой вершине. Птицы явно предпочитают соору-
жать свои постройки на солнцепеке, и потому большинство гнезд располагается
или на вершине сломанного дерева, или на сухих его ветвях. Для постройки
гнезда используются различной толщины сучья, ветви, иногда в перемежку
с сеном и прелой соломой. Лоток выстилается толстым слоем мягкого мате-
риала — сеном, соломой, тряпками, войлоком, обрывками газет, перьями птиц
и шерстью животных — все это птицы в изобилии находят на ближайших свал-
ках. Гнезда аистов — крупных размеров. Особенно велики они, когда исполь-
зуются много лет подряд и ежегодно ремонтируются и достраиваются птицами.
Поперечник их редко бывает менее 1 м и достигает 1,5 м. Общая толщина
(высота) гнезда колеблется в еще больших пределах. Вновь выстроенные гнезда
имеют высоту 40—50 см> многолетние достигают 1,5 л*. Лоток гнезда никогда
не бывает глубоким, а к концу размножения становится почти плоским вслед-
ствие утаптывания боковых стенок тяжелыми птенцами. Постройка гнезд про-
изводится обеими птицами гнездовой пары. Самец, вернувшийся к гнезду пер-
вым, в течение дня несколько раз посещает гнездо и проводит в нем ночь.
Гнездо ремонтируется позднее уже обеими птицами. Если у гнезда появ-
ляются две самки одновременно, между ними наблюдаются ожесточен-
ные драки и гнездо занимает более сильная. В этом самец не принимает
участия.
Количество яиц в кладках колеблется от 2 до 6, чаще всего 4, в исключи-
тельных случаях 1 или 7. Размер кладки изменяется, вероятно, в зависимости
от обилия и недостатка пищи в отдельные годы. В Бессарабии кладки содержат
от 2 до 5 яиц, но птенцов обычно бывает 2 (Остерман, 1915). На Украине нор-
мальная кладка содержит 4—5 яиц; в Закавказье под Ленкоранью часто б яиц.
При кладке в 5 яиц — 1 яйцо всегда оказывается болтуном. Яйца белого аиста
изменчивы по величине и форме. В одних случаях они правильной яйцевидной
формы, в других сильно вытянуты или вздуты. Скорлупа их крупнозернистая,
обладающая слабым блеском. Окраска скорлупы белая, сильно загрязняющаяся
к концу насиживания. В отличие от яиц черного аиста, скорлупа белого аиста
на просвет всегда желтая или желтоватая, но не зеленая.
Размер яиц: A6) 71,0—78,2 х 47,9—57,0 мм, среднее 73,8 х 53,8 мм.
В широтном направлении наблюдается значительное изменение сроков раз-
множения. Впрочем, в одних и тех же местностях аисты приступают к гнездо-
ванию не одновременно. Рано прилетая в Закавказье, под Ленкоранью, в пс-
следней трети марта белые аисты уже приступают к поправке гнезд. Яйцекладка
происходит между 1 и 8—10 апреля. 1 мая в осмотренных гнездах обнаружены
388 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
сильно насиженные яйца (Спангенберг). Сатунин, однако A907), находил
в вост. Закавказье ненасиженные яйца 24 и 25 апреля. Вероятно эти гнезда при-
надлежали поздно загнездившимся птицам. По наблюдениям БраунераA912),
у Херсона аисты закончили кладку 18 апреля, у Мариуполя откладка яиц на-
чинается в середине или в последней трети апреля, в Бессарабии в самом конце
апреля, в Харьковской обл. 29 апреля в гнезде оказалось 3 свежих яйца (Сомов,
1897). Спаривание происходит на гнездах. Первые яйца кладки откладываются
через день, но последние вероятно с более длительными интервалами. Насижи-
вание начинается с откладки первого яйца, но пока кладка не закон-
чена птицы сидят на них мало. В насиживании принимают участие обе
птицы.
Продолжительность насиживания 33—34 дня с небольшими отклонениями.
В наиболее южных частях страны птенцы вылупляются в самом начале мая. Под
Ленкоранью —3 мая (Спангенберг), у Херсона вывод птенцов отмечен 18 мая
(Браунер, 1912), в Кубинской низменности 18 июня птенцы были вполне опе-
рены, а 12 июля поднялись на крылья (Сатунин, 1907), в Черниговской обл.
в середине или в последней трети июня аистята стоят на гнездах (Емельяненко,
1916). В половине июля или в конце этого месяца молодые аисты видимо по-
всеместно приобретают способность к полету и покидают гнезда. Старые
птицы прекращают кормить своих птенцов, как только последние могут летать.
В районе Брянска в 1935 г. птенцы вылупились 23—24 мая, а 12 июля остава-
лись еще в гнездах (Суслова, 1935). В вост. Белоруссии аистята покинули гнездо
30 июля. Пока птенцы малы одна из птиц гнездовой пары (вероятно самка)
постоянно остается на гнезде, проявляя большую к ним заботливость. В жар-
кие дни — обливает водой из клюва и загораживает от солнца. По мере под-
растания аистят обе птицы заняты отыскиванием пищи для прожорливого по-
томства.
Неоднократно наблюдалось, что взрослые аисты выбрасывают из гнезда
одного из птенцов. Такие случаи отмечены в Бессарабии, в Черниговской, Мин-
ской обл. и у Запорожья. Быть может эти явления связаны с трудностями вы-
кормить птенцов в годы неурожая кормов. Рост птенцов относительно медлен-
ный. Они проклевываются из яиц слепыми, прозревая через несколько часов.
Вес таких птенцов 70—75 г. На 16-й день после вылупления птенцы способны
становиться на ноги, а свободно стоят на гнездах на 25-й день жизни. Маховые
и плечевые перья пробиваются на 20-й—22-й день. Гнездо оставляется молодыми
птицами в возрасте 54—63 дней, а достигшие 70 дней молодые аисты стано-
вятся самостоятельными.
Линька. В году — одна линька, происходящая крайне медленно. Кон-
кретные данные отсутствуют.
Питание. Белые аисты кормятся разнообразной животной пищей. Они
заглатывают различных насекомых, моллюсков, рыбу, амфибий, рептилий,
птенцов и небольших взрослых птиц, а из млекопитающих — грызунов до
маленьких зайчат включительно. Есть указания в литературе о поеданий аистом
растительной пищи, но какой именно — неизвестно. В желудке аиста, убитого
в Закавказье, были обнаружены три маленькие черепахи (Clemmys caspica),
четыре ужа Natris natris и шесть лягушек (Сатунин, 1907). По данным
ОстерманаA915), в Бессарабии аисты едят мышей, птиц, ужей, ящериц, три-
тонов, лягушек, саранчу и прочих животных. На границе Украины основ-
ную пищу аистов составляют: лягушки, ящерицы, мелкая рыба и насе-
комые. Изредка аисты поедают птенцов воробьиных птиц и пуховичков
водяных курочек. В период снятия хлебов аисты регулярно посещают ско-
шенные участки посевов, добывая здесь полевок и других мелких грызунов.
В желудках 2 особей, убитых в середине июля, обнаружены хитиновые остатки
насекомых, остатки рыбы, лягушек и шерсть полевок; у птицы, добытой в мае,
найден крапчатый суслик и несколько ящериц. При вскрытии крупного, погиб-
БЕЛЫЙ АИСТ
389
шего в гнезде аистенка мною найдена застрявшая в кишечнике челюсть неболь-
шой щуки.
У Херсона, по наблюдениям Браунера A912), маленькие птенцы аистов
вначале вскармливались кузнечиками и ящерицами, позднее лягушками,
змеями и сусликами. При случае аисты не прочь поживиться не только пухо-
вичками диких водяных и болотных птиц, но и домашними утятами и цыпля-
тами. Такие случаи наблюдались в Минской обл., а также на Украине.
Непереваримые части пищи (шерсть, перья
и пр.) отрыгиваются в виде погадок.
Полевые признаки. Птица крупных раз-
меров, высокая на ногах, с длинной шеей и
клювом с белым оперением, исключая кон-
цы крыла. Все эти признаки позволяют
безошибочно отличить белого аиста от чер-
ного аиста, а красные клюв и ноги — от
журавлей и цапель. В природе наблюдать
аистов приходится при различных услови-
ях. Весной одиночки и пары, а летом семьи
медленно бродят по сырым лугам, резко
выделяясь своей белой окраской, и время
от времени склевывают пищу. В жаркую
пору дня аисты охотно отдыхают, стоя на
одной ноге, то на массивном гнезде, то на
крыше строения или стоге сена, то на вер-
шине сломанного дерева. Чтобы взлететь на
воздух с земли, птица предварительно про-
бегает несколько метров, тяжело взмахивая
своими большими крыльями. Поднявшись
на достаточную высоту аист летит сравни-
тельно легко, вытянув шею и ноги, то мед-
ленно, то глубоко взмахивая крыльями, то
держа их неподвижно.
Во время отлета стаями и в жаркие
летние дни аисты летают парящим полетом,
поднимаясь на большую высоту. Исключи-
тельно молчаливая птица, но безусловно
обладает голосом. Слышать его приходится
чрезвычайно редко и лишь на близком рас-
стоянии. Для «выражения чувств» он поль-
зуется своеобразным трещанием клюва.
Птица закидывает голову и шею назад и начинает щелкать половинками
клюва, то, напротив, опускает клюв до самой земли. Щелканье при этом
то усиливается, то ослабевает, то становится резким, то, напротив, при-
глушенным и медленным. Там, где белые аисты пользуются покровитель-
ством человека — они не пугливы. Там, где их беспокоят, они становятся недо-
верчивыми и не подпускают к себе близко.
Описание. Размеры и строение. Вес взрослой птицы 3,5—4 кг, длина крыла
F) 58,5—60,5 мм. Самец отличается от самки более крупными размерами.
Окра ска. Окраска оперения у обоих полов белая, только первостепенные маховые
и их кроющие, наружные из второстепенных, большие кроющие предплечья и задние или
длинные плечевые — блестяще-черные. У молодых аистов эти части оперения буроватые.
Клюв красный, голая кожа вокруг глаза и передняя часть кожи подбородка — черные,
ноги красные, радужина бурая.
Детали строения — как у черного аиста. Птенцы в первом и втором пуховом наряде
одеты светлобелым пухом. Их ноги желтовато-розоватого цвета, клюв черноватый. У моло-
дых птиц черный цвет оперения взрослых заменен буроватым.
Рис. 78. Белый аист
390 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Туркестанский белый аист Ciconia ciconia asiatica Sev.
Ciconia alba asiatica, Северцов. Вертик. и гориз. распр. Туркест. жив., 1873, ст. 145,
Туркестан.
Синоним: Ciconia alba var. major (azreth. n. sp.) Северцов. Верт. и гориз. распр.
Туркест. жив. 1873, стр. 68; nomen nudum — Ciconia alba, var. orientalis. Sewertzoff,
Stray Feathers, III, 1875, стр. 431, Туркестан.
Распространение. Ареал. В пределах Союза гнездовым ареал тур-
кестанского белого аиста охватывает Узбекистан, сев. и зап. Таджики-
стан. В южн. Таджикистане аисты гнездятся на Кафирнигане у Микоян-
абада. Спорадически аисты гнездятся и в Семиречье, а именно, у р. Сукулук
к юго-западу от Фрунзе и у с. Кутентай (около 75 км к северо-востоку от Алма-
Ата),кроме того еще к востоку от Алма-Ата (бывш. Восточноталгарская волость).
Вероятно гнездится также в Кашгарии. Западная граница распространения этой
формы проходит в Туркмении по Аму-Дарье. Птицы спорадично гнездятся в сред-
нем течении реки и, по Зарудному A896), в очень ограниченном числе в Чар-
джоу. В последнем месте аист видимо не гнездится регулярно, так как Гладков
A932, 1937) его там не нашел. В остальных местах Туркмении белые аисты встре-
чаются во время пролетов и случайных залетов. В качестве залетной птицы добы-
вался в районе Ленкорани, и единственный раз отмечен на нижней Сыр-Дарье.
Зимует в Индии и в ограниченном числе на Цейлоне, по Зарудному A931) и
в вост. Иране.
Характер пребывания. Гнездящаяся перелетная птица. Сроки
весеннего появления и осеннего отлета плохо выяснены. Есть указание на зи-
мовку отдельных особей на Сыр-Дарье. В Туркмении — в низовьях Атрека —
пролетают весной в первой половине марта (Дементьев, 1950). Вероятно про-
летные птицы наблюдались 25 апреля в бассейне Мургаба Вальтером; одиноч-
ная птица, летевшая на север, отмечена у Сары-Язы 22 апреля (Лоудон, 1913).
Под Ташкентом стая в 15 особей отмечена 18 апреля (Сатаева, 1937). Сроки ве-
сеннего прилета в других частях страны остаются невыясненными. Вероятно
недавно прилетевшие на родину птицы были добыты в 1913 г. у Сарда бы —
14 марта и у Ташкента — 15 апреля.
Осенний отлет под Ташкентом начинается в последней трети августа и
в начале сентября, основная масса отлетает в середине октября. В Туркмении
отлет происходит в октябре. Стая отлетающих аистов на Атреке близ поста
Чатлы наблюдалась 5 октября (Шестоперов, 1927), запоздавшие птицы отме-
чены на Атреке 22 декабря (Исаков и Воробьев, 1940), в Таджикистане в кон-
це августа (Иванов, 1940).
Биотоп. Обязательное условие гнездования — наличие культурного
ландшафта с разбросанными высокими деревьями или высокими строе-
ниями человека (мечети). Есть данные о гнездовании белых аистов на лёс-
совых обрывах но Зеравшану. Кормовой биотоп — болотистые открытые мест-
ности. Так же, как и европейский подвид, предпочитает селиться в непосред-
ственной близости селений или в последних.
Численность. Местами (Таджикистан) образует большую плотность
населения, но в целом численность туркестанских белых аистов в нашей стране
весьма ограничена.
Экология. Размножение. Гнездится то отдельными парами, то 2—
3 пары размещаются на одном и том же дереве. Иванов A940) около Микоян-
абада видел небольшую группу чинар, «густо населенную аистами и коршу-
нами». Нижние части гнезд аистов заселяются воробьями. Таким образом
туркестанский белый аист в некоторых случаях может образовывать нечто вроде
колониальных гнездовий. Гнезда сооружаются или на высоких деревьях, или
на куполообразных крышах высоких зданий. В Туркмении гнездо, найденное
близ Карши, помещалось на дереве (Дементьев, 1950), в Бухаре на вершине
БЕЛЫЙ АИСТ 391
куполообразной высокой постройки. Напротив, в районе Намангана в Ферган-
ской долине, насколько известно, аисты гнездятся на деревьях, в кишлаках или
поблизости их, устраивая свои гнезда на самых вершинах чинар. В Таджики-
стане аисты гнездятся на деревьях, так как, за исключением Гиссара,
там нет старых высоких построек (Иванов, 1940). Гнезда представляют собой
массивные сооружения из толстых сухих веток и сучьев, не уступающие по
величине гнездам европейской формы. Гнезда используются в течение ряда лет
и тогда, вследствие ежегодной достройки, достигают особенно большой высоты.
Среди перевитых ветвей в толще гнезд во множестве гнездятся воробьи. Как и
европейская форма, этот аист не боится горячего среднеазиатского солнца и
устраивает гнезда на самом солнцепеке. По данным Молчанова A933), число
яиц в кладке колеблется от 3 до 5. По внешнему виду яйца не отличаются от
яиц европейского аиста. Наши сведения о сроках размножения птиц скудны.
В гнездах, осмотренных Ивановым A940) на Кафирнигане у устья Ханаки, самки
продолжали насиживать до конца апреля. Зарудным 7 июня найдены довольно
крупные аистята. В районе Намангана 3 июня Кашкаров A927) видел в гнездах
крупных оперившихся птенцов, вполне оперившиеся птенцы (по 4 птенца
в гнезде) наблюдались мною в кишлаках у Намангана между 18 и 22 июня.
Питание. Питаются мелкими животными: лягушками, рыбками, мы-
шами, ящерицами, насекомыми; при случае уничтожают саранчу (Молчанов,
1933). Видимо большое значение в питании этой формы имеют рептилии и насе-
комые. Последних (вероятно, цикад) аисты ловят не только на земле, но также
на ветвях, стоя на своих гнездах.
Описание. Размеры и строение. По общему сложению, за исключением формы
клюва, не отличимы от европейского белого аиста. Клюв этого подвида как бы слегка пере-
гнут кверху и несколько крупнее предыдущей формы. Крыло самцов и самок (9) 550—640,
в среднем 589,5 мм
Китайский белый аист Ciconia ciconia boyciana S w i n h.
Ciconiaboyciana. S w i n h о e. Proc. Zool. Soc. London, 1873, ст. 513, Иокогама, Япония.
Распространение. Ареал. Область распространения ограничена. Птицы
гнездятся в Приамурье, в Приморье, относительно недавно D5—55 лет тому
назад) стал гнездиться в Японии (Хондо, Хоккайдо, Сикоку, Цусима), быть
может гнездится также в вост. Манчжурии и Сев. Корее. Зимовки птиц распо-
ложены в Корее, в Японии (Хондо) и в зап. Китае. Кроме того отмечен
в Бирме, Ассаме и в Манипуре. Детали распространения этой формы аиста
в наших пределах таковы: когда-то был он добыт на левом берегу Амура близ
озера Болен-оджал, где встречался ежегодно. В Британском музее хранится
экземпляр из устья Амура. Наблюдался на Амуре в окрестностях Благове-
щенска и близ устья Уссури. Пржевальский A870) наблюдал аистов в бас-
сейне Ханки и нашел около двух десятков гнезд (истоки Сунгачи 3 гнезда,
на средней Лефу около 10 гнезд и на реке Мо несколько гнезд). Кроме того,
он имел сведения о гнездовании аистов в Прохоровских сопках у Сиваковки
ив верхней Уссури. Относительно недавно Шульпин A936) видел гнездо на
р.Красной вблизи деревни Новорусановка и имел сведения о гнездовании птиц
по восточной стороне Сихотэ-Алиня, в долине реки Самарги и на реке
Ботчи, где была добыта молодая птица. Есть указание о залете этого аиста
в Якутию. Одна птица отмечена в 20-х числах мая 1935 г. на о-вах Лены против
устья р. Ботомы (Скалой).
Характер пребывания. Гнездящаяся перелетная птица. Ве-
роятно, в исключительных случаях остается на зиму в устьях Уссури, где дер-
жится зимою у полыней. Данные относительно сроков весеннего прилета чрез-
вычайно скудны. В 1868 г. аисты наблюдались Пржевальским A870) только
13 апреля, но в 1869 г. первая птица была отмечена уже 20 марта и позже па-
392 t отряд голенастые птицы
рами и одиночками и лишь 4 апреля стаей в 16 особей. Сроки осеннего отлета
неизвестны. Есть только указание Шренка A861), что они улетают с озера Од-
жал в августе.
Биотоп. Сочетание болот и небольших перелесков или групп отдельно
стоящих деревьев. В районе озера Ханка болота чередуются с лесистыми воз-
вышенными сопками, поросшими липой, ильмом, кленом, дубом и кустарни-
ками. Эти сопки окружены обширными пониженными пространствами, заня-
тыми вейниковыми болотами и представляющими собою как бы лесные оазисы.
По средней Лефу и Мо, согласно Пржевальскому, местом гнездовья птиц служат
сопки, поросшие лиственным лесом, среди болот и предгорья, подступающие
к Ханкайской низменности. Кормовые стации, судя по вышеприведенным дан-
ным, бывают расположены в непосредственной близости от мест гнездовья, но
в Японии аисты летают за пищей очень далеко —до 20 км. С вылетом птенцов
из гнезд аисты начинают встречаться только в болотах (Шульпин, 1936). В пре-
делах нашей страны явно избегают не только близости человеческого жилья,
но вообще культурного ландшафта. Напротив, в Японии птицы гнездятся в насе-
ленных пунктах, ранее гнездился в Токио, но сейчас исчез оттуда и гнездится
лишь в некоторых селениях зап. Японии (Ян, 1942).
Численность. Был немногочисленен во времена исследования Уссу-
рийского края Пржевальским. В настоящее время должен быть отнесен к исче-
зающим видам, нуждающимся в полной охране со стороны человека. Причины
сокращения численности птиц не вполне ясны. Пржевальский A870) упоминает
о ряде случаев разорения гнезд тибетскими медведями, которые, по наблюде-
ниям автора добираются даже до гнезд, устроенных на вершинах труднодо-
ступных деревьев. Добывало аистов и местное население. Молодых
птиц, содержимых в неволе, Пржевальский видел на верхней Уссури
и по Амуру. Мясо аистов употребляют шаманы в пищу, длинные же кости ног,
скупаются китайцами, которые изготовляют из них обеденные палочки. Одна
из причин сокращения численности — нередкие случаи гибели гнезд. Числен-
ность птиц сократилась и в Японии. Ныне здесь насчитывается около 70—80
пар (Ян, 1942).
Экология. Размножение. Птицы гнездятся отдельными парами.
Гнезда иногда располагаются близко друг от друга. Строятся гнезда всегда на
высоких деревьях —липах, дубах, ильмах (Пржевальский, 1870; Шульпин,
1936). В Японии на старых соснах. Высота гнезд над землей от 6 до
17 м. По Шульпину A936), гнездо помещается на толстых сучьях у самого
ствола близ вершины старого высокого дерева и представляет собой массивную
постройку. Наблюдения Пржевальского A870) показывают, что гнезда исполь-
зуются птицами несколько лет подряд и ежегодно достраиваются и ремонти-
руются.
Насколько позволяют судить скудные данные, количество яиц в кладках
колеблется от 2 до 4. При этом число яиц в кладках сокращается по мере дви-
жения с юга на север. В бассейне Ханки три взятые кладки содержали по 4 яйца
(Пржевальский, 1870). В устье Уссури Дыбовский никогда не находил в гнез-
дах более 3 яиц (Тачановский, 1875), еще севернее на полтора градуса у озера
Оджал, по устным данным, аисты откладывают 2 или 3 яйца (Шренк, 1861).
На юге число птенцов видимо бывает меньше, чем яиц в кладке. В одном случае
Пржевальский A870) отметил, что в гнезде было 3 птенца. Яйца белого цвета,
похожие на яйца европейской формы, но больших размеров. Размер яйца, по
Тачановскому, 58 х 77 мм. Сроки размножения отдельных пар в одних и тех же
местах не одновременны. Вскоре по прилете птицы приступают к починке ста-
рых и постройке новых гнезд. В 1869 г. починка гнезд отмечена с 13по 16 апреля,
а в 1868 г. уже 8 апреля. Одна пара 27 апреля имела в гнезде яйца, другая,
запоздавшая с прилетом, в это время только приступила к починке гнезда.
В первом 4 мая оказались 4 довольно сильно насиженных яйца. В 1869 г*
СЕРАЯ ЦАПЛЯ 393
29 апреля гнездо содержало 4 слабо насиженных яйца, другое тоже 4 яйца, но
насиженных сильно. На средней Лефу 18 июня во многих осмотренных
гнездах были крупные молодые, повидимому, близкие к вылету. Вылет
молодых из гнезд на р. Мо отмечен в средних числах июля (Пржеваль-
ский, 1870).
Питание. Почти нет данных. По устным сообщениям местного населе-
ния поедает мелкую рыбу (Шренк, 1861). Оставаясь зимовать в Приморье китай-
ские белые аисты держатся все это время у незамерзающих полыней, питаясь
мертвой рыбой, выбрасываемой водой на лед в большом количестве (Точанов-
ский, 1893).
Полевые признаки. Китайский белый аист очень похож на своего европей-
ского сородича, отличаясь от последнего черным клювом. В отличие от европей-
ского, он крайне осторожен и дик. Даже от гнезда с яйцами или птенцами птица
слетает шагов за 200, летает на кругах некоторое время вокруг человека, затем
поднимается все выше и выше в подоблака и кружит здесь, наблюдая врага
сверху. Щелкает клювом, как европейский аист, но много короче (Шульпин,
1936).
Описание. Размер ыистроен и е. Общим сложением схож с европейским белым
аистом. Крыло (8) 620—670 лип, клюв 200—260 мм. Самец отличается от самки большими
размерами (Хартерт, 1914).
Окраска. Оперение белое, за исключением маховых кроющих кисти, больших
кроющих второстепенных маховых и длинных из плечевых, которые черные. Наружные
опахала второстепенных с серым налетом. Клюв черный, ноги красные, глаз красный. У мо-
лодых черная окраска оперения заменена бурой, на плечевых перьях есть бурые пятна.
СЕМЕЙСТВО ЦАПЛЕВЫЕ ARDEIDAE
РОД ЦАПЛИ ARDEA LINNAEUS, 1758
Тип A. cinerea
7. Серая цапля Ardea cinerea L.
Ardea cinerea. Linnaeus. Syst. Nat., изд. X, 1758, стр. 143, Швеция
Распространение. Ареал. Палеарктика; Европа, Азия, сев. Африка; за
пределами Палеарктики — несколько спорадически в вост. и южн. Африке на
Мадагаскаре, в Азии до Индии, Цейлона и Бирмы.
Характер пребывания. В северных частях Европы и в Сибири—
перелетная птица. Но уже в Закавказье, Средней Азии, в средней Японии и
в Китае к югу от Желтой реки серая цапля живет оседло.
Биотоп. Сочетание разнообразных водоемов — стоячих или проточ-
ных, пресных или солоноватых — с наличием кустарниковой или древес-
ной растительности или с тростниками. Предпочитает равнины, но встре-
чается и в горах. На пролете в открытых ландшафтах, особенно по реч-
ыым разливам.
Подвиды и варьирующие признаки. Кроме особенностей биологии (часть цапель оседлы,
часть перелетны; имеются различия в размножении т. п.), южно- и восточноазиатские цапли
выделяются по более бледной окраске (у взрослых шея белая, кроющие крыла бледносерые)
по сравнению с западными; на Мадагаскаре и о-ве Альдабра, повидимому и на Коморских и
Амирантских о-вах оседло живет подвид серой цапли с относительно большими клювом и
ногами. Таким образом, имеются три подвида: А. с. cinerea L.— Европа, зап. Азия и Сибирь,
на восток до Японии, Африка; А. с. jouyi С1 а г к,1907—вост. Азия от Алашаня, Нань-Шаня,
вероятно Сычуаня до Приморских провинций Китая (впрочем, границы между этими под-
видами все еше спорны); А. с. /irasa Hart., 1917 — с указанным выше распространением
на Мадагаскаре и т. д.
394
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Карта 72. Распространение серой цапли Ardea cinerea
7. А. с. cinerea, 2. А. с. jouyi, 3. А. с. firasa;
а — граница гнездовий, б — не вполне выясненная граница гнездовий, г — область зимовок.
Обыкновенная серая цапля Ardea cinerea cinerea L.
Русское название. Цапля, чапля, чапура, чепура — народные названия этой
птицы.
Распространение. Ареал. Вне пределов СССР — в Африке в Марокко,
Тунисе, Алжире и Египте; к югу от Сахары в восточной и южной части материка;
в Азии — в Малой Азии, Месопотамии и Иране, а также в Синьцзяне (Кашгария),
Монголии и Японии (граница распространения в Азии в отношении подвида
jouyi остается неясной); в Европе — на север до Оркнейских о-вов, в Швеции и
Норвегии до 64—65° с. ш., в Финляндии, на юг до южн. Испании (но в Порту-
галии не гнездится), средней Франции — до Савойи, средней Италии, Греции.
Наконец, на о-вах Канарских, Майорке, Корсике, Сардинии, Крите.
В Европейской части СССР граница проходит около 60° у Ленинграда и от-
сюда на восток тянется через Вологду — Киров к средним частям Молотовской
обл. В Зауралье граница проходит в бассейне Сосьвы под 60° с. ш., в бассейне
Енисея не доходит до Красноярска (у Ачинска под 54° цапли еще обычны на
гнездовье), затем идет на север до 58° на Нижней Тунгуске. Распространение
в Вост. Сибири несколько спорадично. В Якутии цапли стали гнездиться у Якут-
ска в 1916 г., в 1920—1922 г. стали обычными, в настоящее время гнездятся
у Якутска, по рекам Мге, Алдану и в низовьях Май. Наконец, серые цапли
гнездятся в Тункинской долине (Слудский), в южн. Забайкалье и
на Амуре на север до устья реки. Гнездятся на Сахалине до залива Чайво.
К югу от очерченной нами северной границы серая цапля гнездится в подхо-
дящих местах повсюду.
СЕРАЯ ЦАПЛЯ 395
Часть европейских цапель зимует в южн. Европе и в Африке, но уже в Анг-
лии и Средиземноморье цапли оседлы. Цапли, гнездящиеся на крайнем юго-восто-
ке Европы и в зап. Азии, зимуют преимущественно у южн. Каспия; восточные от
средней Японии до Индии и Китая. В благоприятные годы отдельные особи
остаются на зиму на южном побережье Крыма (автор наблюдал их в конце
декабря — начале января). В Закавказье цапли оседлы, сюда же прилетают
зимовать птицы северных популяций.
В Армении цапля оседла, в основном держится в низменностях, но при
наличии подходящих угодий поднимается до высот более 2 000 А над уровнем
моря у озера Севан. Одиночки зимуют у Тбилиси (Сатунин, 1907) и в Кахетии
(Чхиквишвили, 1930). В небольшом числе зимует на озерах и разливах Куры
у Евлаха, под Ленкоранью и на Муганской степи, но в суровые зимы из района
Ленкорани улетает. Северней, на западном побережье Каспия, более или менее
регулярно зимует по озерам и взморью в районе Кизляра и у Аграханского
залива. На юго-восточном побережье Каспия зимует в области низовьев Атрека,
в районе Гассан-кули и Чикишляра, местами в вост. Туркмении (Теджен,
Мургаб). Одиночки зимуют в Таджикистане.
Характер пребывания. В СССР — гнездящаяся перелетная
птица, оседла только на юге (см. выше.)
Даты. Прилет повсеместно ранний, во многих южных районах совпадает
с появлением на поверхности зимнеспящих и других грызунов. Первые птицы
появляются в то время, когда водоемы скованы льдом и, за исключением от-
дельных проталин, лежит сплошной снег. Резкое весеннее похолодание иногда
вызывает гибель передовых особей или их откочевку к югу (Вальх, 1899).
В южных частях Херсонской обл. первые птицы появляются с середины
марта, в северных —между 20 марта и 3 апреля (Браунер, 1898). В последней
трети марта отмечались под Мариуполем, но валовой пролет идет здесь в пер-
вой трети апреля. Наиболее ранний прилет под Харьков отмечен 13 марта, чаще
15—21 марта, валовой —в конце марта или в первых числах апреля (Сомов,
1897). Под Киевом самая ранняя птица наблюдалась 23 марта (Шарлемань, 1913),
в районе Чернигова —25 марта (Емельяненко, 1913). В марте цапли прилетают
на Сев. Кавказ и между 15 и 25 марта регулярно появляются в дельте
Волги (Дубинины, 1940). В Закавказье наблюдения над сроками прилета от-
сутствуют, так как этому мешают оседлый образ жизни и зимовка части особей.
В средней и северной полосе Европейской части СССР прилет начинается
со второй половины марта и длится до середины мая. Под Минском появляются
около середины марта, под Смоленском (вероятно, не первые) 30 апреля (Гржи-
бовский, 1919), у Калуги в различные годы между 26 марта и 8 апреля (Фи-
латов, 1915), у Тамбова между 9 и 19 апреля (Резцов, 1910), в Куйбышевской
обл. наиболее раннее появление 28 марта, обычно же между 6 и 21 апреля
(Карамзин, 1901, 1913), под Уфой в начале апреля.
В районе Пскова прилет укладывается между 7 и 20 апреля, в Ярославской
обл. наиболее ранняя особь отмечена 7 апреля, весь же прилет укладывается
между этой датой и серединой апреля; в Горьковскойобл.он происходит с се-
редины апреля и до половины мая, в Кировской — в середине апреля, в районе
Молотова —в первой половине мая.
В Азиатской части Союза сроки пролета также изменяются в зависимости
от широты местности. В вост. Иране пролет наблюдался между 20 и 23 апреля
{Зарудный, 1903). В Таджикистане, вероятно пролетные, цапли отмечались
в Гиссарской долине до конца апреля. В Туркмении Шестоперов A937) считает,
что отдельные птицы прилетают с середины февраля, по данным Дементьева
A950), валовой прилет позднее —в различные даты марта и первой половине
апреля. На реке Зеравшан под Самаркандом в раннюю весну 1947 г. хорошо
выраженный пролет отмечен Богдановым между 25 февраля и 10 марта. На
Сыр-Дарью в район Кзыл-Орды цапли регулярно прилетают в самом начале вто-
396 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
рой трети марта. В это время ночная температура нередко падает до—14°, водо-
емы скованы льдом и птицы бывают вынуждены кормиться в степи грызунами.
Массовый пролет идет в конце марта и заканчивается в начале апреля. Отстало-
пролетные иногда летят до середины апреля. Под Джаркентом наиболее ранние
птицы отмечались весной 12 марта (Кореев и Зарудный, 1906), в окрестностях
Зайсана в 1913 г. впервые отмечена 29 марта, а массовый пролет с 6 апреля
(Хахлов, 1913). В устьях Урала цапли появляются во второй половине мая и
летят на восток до начала второй трети апреля. На Эмбе у Кок-джиды первая
цапля наблюдалась 13 апреля и примерно тогда же под Иргизом (Сушкин,
1908).
В районе Чкалова весной серые цапли появляются в последней трети марта,
как только образуются проталины. Пролет достигает наибольшей силы между
концом апреля и началом мая, оканчивается в конце этого месяца. В Сибири
цапли появляются поздней. У Бийска пролетная стайка была отмечена 15 мая
(Сушкин, 1938), у Омска добывалась 28 апреля (Иоганзен, 1907), на Чанах
наиболее ранняя—ЗОапреля(Данилович), в районе Нерчинска за ряд лет между
16 апреля и 3 мая (Пуляевский, 1937). В Даурии около 10 апреля, а в Яку-
тии между 24 апреля и 22 мая (Скалой).
На Амурском заливе цапля отмечалась уже 13 марта, а в другой год 8
марта. Валовой пролет наблюдался 23 марта (Медведев, 1913, 1914). В устье
Лефу первая одиночка, летевшая на север, отмечена 18 марта (Шульпин, 1936).
По наблюдениям Пржевальского A870), первые цапли появляются в Уссурий-
ском крае рано, когда реки еще скованы льдом и, за исключением редких про-
талин, снег сплошь покрывает землю. В 1868 г. в истоках Сунгачи первая цапля
появилась 22 марта, сильный пролет шел между 5 и 13 апреля и закончился
в самом начале мая; в 1869 первая появилась 23 марта; между 25 марта и концом
первой декады апреля шел валовой пролет; группа, летевшая к северу, отме-
чена в середине апреля.
Осенний отлет начинается рано — при теплой погоде и затягивается до
осенних заморозков. В Уссурийском крае он слабо выражен — птицы отле-
тают исподволь и незаметно. В устье р. Суйфун вероятно пролетные цапли на-
блюдались 23—27 сентября (Шульпин, 1936), на южном берегу бухты Экспеди-
ции — 12 и 14 октября и в последний раз уже после морозных дней 22 октября
(Пржевальский, 1870). В Якутском округе обычно исчезают между 3 и 24 сен-
тября (Скалой), а в 1926 г. полностью отлетели уже 13 сентября (Иванов, 1929).
Под Барнаулом отмечались до 4 сентября (Велижанин, 1938).
В районе Чкалова отлет начинается в конце августа, достигает наибольшей
силы в конце первой трети сентября или в половине этого месяца и заканчивает-
ся в конце первой трети октября. В устье Урала птицы, встречая видимо более
благоприятные условия питания, задерживаются, и наблюдались здесь во вто-
рой половине октября, а редкие одиночки отмечались и в самом начале ноября.
В области верхнего течения Иргиза осенний пролет происходит во второй поло-
вине сентября и в первой половине октября. В одну осень хорошо выраженный
пролет начался только 27 сентября и через неделю окончился (Сушкин, 1908).
На Сыр-Дарье между станциями Кзыл-Орда и Тюмень-арык серые цапли обыч-
ны еще на озерах в течение всего октября, но с конца первой трети ноября ста-
новятся здесь редким явлением. Единственный раз вероятно отсталая пролет-
ная особь наблюдалась мною близ ст. Кара-узяк 12 сентября. Под Джаркентом
наиболее поздние экземпляры отмечались 16 ноября (Зарудный и Кореев,
1906). В пределах Туркмении осенний пролет происходит в сентябре —октяб-
ре. В конце августа — начале сентября пролетные особи отмечались на Памире.
В Европейской части Союза на Урале (Молотовская обл.) отлет начинается
со второй половины августа и длится до конца первой трети сентября. В Ки-
ровской обычно цапли отлетают в начале сентября, наиболее поздняя особь
была добыта 18 сентября (Плесский, 1927), В Ярославской отлетают в начале
СЕРАЯ ЦАПЛЯ 397
сентября, но запоздавшие одиночки отмечались до 12 октября (Птушенко,
1927). Гнездящиеся на Рыбинском водохранилище птицы отлетают в сентябре.
Валовое движение птиц к югу под Псковом наблюдается в первой трети
сентября, и в начале октября цапли здесь отсутствуют. У Рязани наиболее позд-
ний экземпляр был добыт 3 октября (Хомяков, 1901), под Калугой отлет про-
исходит в первой трети сентября, в Тамбовской пролет начинается в послед-
ней трети августа, а последняя птица у Моршанска наблюдалась 21 сентября
(Резцов, 1910), из Куйбышевской отлетают в самом начале сентября, в Баш-
кирии бывает еще обыкновенна до 3 октября (Резцов, 1904). В Белоруссии хоро-
шо выраженный отлет к югу укладывается между 16 и 22 сентября (Федюшин,
1928). В районе Харькова в сухие годы отлет начинается в первой трети сентября
или в середине этого месяца, пролет северных особей идет в конце сентября
и в первой трети октября. Запоздавшие молодые отмечались здесь 3 ноября
(Сомов, 1897). Приблизительно в те же сроки птицы отлетают из более южных
частей Украины: в половине октября из-под Херсона, в самом конце октября и
в первых числах ноября из района Мариуполя и между 25 сентября и 5 октября
из дельты Волги (Дубинины, 1940). На Сев. Кавказе пролет идет в сентябре и
октябре, заметное увеличение численности птиц осенью на Ленкоранских
зимовках отмечено 6 сентября (Тугаринов и Козлова-Пушкарева, 1938), а на
восточном берегу Каспия — в низовьях Атрека во второй половине октября.
Во время пролетов серые цапли летят обычно на большой высоте как
ночью, так и в дневные часы. Осенью цапли предпочитают лететь после вечерней
зари и останавливаться на дневку ранним утром. Часто птиц удается наблюдать
выстроившимися в косую линию. Одиночки во время пролета встречаются срав-
нительно редко. Обычно совершают перелет семьями или стайками размножав-
шихся в одних и тех же гнездовых колониях птиц, еще с лета ведших стай-
ную бродячую жизнь. В некоторых случаях, например, на восточном берегу
Каспия, пролетные серые цапли образуют крупные стаи, достигающие 200—250
особей (Исаков и Воробьев, 1940).
Результаты кольцевания. Цапли, которых на гнездах птен-
цами кольцевали в Дарвинском заповеднике, были найдены на осеннем про-
лете в Новгородской, Горьковской, Вологодской, Ярославской обл. и в Карело-
Финской ССР. Одна на зимовке в южн. Египте. Откочевка из района заповед-
ника идет, таким образом, на запад. Это подтверждается и находками птиц,
окольцованных весной в Калининградской обл., — в Смоленской и в Киевской
обл., а окольцованных в Польше —в Невельском районе Калининской обл.,
в Белоруссии и т. д. Еще в конце августа молодые птицы держатся относительно
недалеко от мест гнездования, а потом — до начала настоящего пролета —
разлетаются в разных направлениях. Есть основания предполагать, что рассе-
ление молодых цапель, как это повидимому характерно для колониально-
гнездящихся птиц со спорадическим распространением, довольно значительно.
Так, птенец, окольцованный у Змиева Харьковской обл., через восемь лет был
добыт у Славянска в Сталинской обл.; другая птица, окольцованная на Куриш-
ской косе в Прибалтике, была в августе того же года в Полоцком округе в Бе-
лоруссии. В последнем случае вопрос, вероятно, идет о разлете в послегнездо-
вый период (Дементьев).
Биотоп. Разнообразен и изменяется в различных частях страны в за-
висимости от местных условий. В выборе гнездовой обстановки птица неприхот-
лива. Ей необходимо сочетание кормовых водоемов самого разнообразного ха-
рактера и мест для устройства гнезд в виде камышей, тростников, затопленных
кустарников или высокоствольной древесной растительности. Населяет степ-
ные, лесостепные и лесные площади, избегая сплошных лесных территорий и
поселяясь лишь там, где наряду с лесом есть обширные открытые участки.
В поисках пищи птицы посещают берега и мелководье озер, прудов и рек,
кормятся на участках сухих лугов, полей и в степи вдали от воды. За пищей
398 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
нередко предпринимают далекие (до 10 км) вылеты от гнездовья. Местами гнез-
дятся в глухих и малодоступных зарослях или, напротив, не избегают культур-
ного ландшафта и близости человека, образуя колонии на окраинах населенных
пунктов и в парках.
Численность. Неодинакова в различных частях ареала. Особенно
многочисленны цапли по рекам низменного Кавказа, в плавнях низовий дру-
гих рек, впадающих в Черное и Каспийское моря. Только в пределах Астрахан-
ского заповедника в 1934 г. насчитывалось около 18,5 тысяч птиц и в 1935 —
около 23,5 тысяч (Ромашова, 1938). В средней полосе Европейской части Союза
цапли становятся только обыкновенными, а ближе к северной границе ареала
довольно редкими птицами. За последние годы, однако, большое количество
гнездящихся серых цапель наблюдается на Рыбинском море в Дарвинском
заповеднике. В азиатской части страны —обыкновенна, но немногочисленна
в Туркмении и Таджикистане, что объясняется недостатком соответствую-
щих биотопов. Напротив, в большом числе встречается на средней и нижней
Сыр-Дарье, Чу, в низовьях Сары-Су, на южном берегу Балхаша и местами в
южных районах Уссурийского края. В более северных районах в общем редка
и распространена спорадично. За последнее десятилетие стала более или менее
обыкновенной в Якутии. Общая численность серой цапли в пределах СССР дол-
жна считаться весьма значительной.
Экология. Размножение. Половая зрелость у самок в годовалом
возрасте, у самцов, повидимому, только в возрасте двух лет, но не все молодые
птицы в этом возрасте размножаются (Нитхаммер, 1938). Моногамы. Пары по-
стоянны. Хотя для серой цапли и характерно колониальное гнездование, но
в некоторых частях ареала птицы поселяются только отдельными парами.
Во многих местах цапли образуют самостоятельные колонии, в других — гнез-
дятся совместно с мелкими цаплями, а также с бакланами, колпицами и кара-
вайками. Приступая к размножению ранее прочих цапель, часто вид этот яв-
ляется основателем колониальных поселений, в которых позднее начинают гнез-
диться кваквы, желтые цапли и т. д. Бывают и такие случаи, когда серые цапли
присоединяются к колониям мелких видов цапель, строя свои гнезда в то время,
когда прошлогодние гнезда еще не заселены ее коренными обитателями.
Колонии, состоящие только из серых цапель, характерны для средней по-
лосы и особенно для северных частей ареала. К колониям цапель здесь присое-
диняются отдельные пары хищников: черные коршуны, белохвост, сапсан, бало-
бан, пустельга, чеглок, иногда беркут, ушастая сова, серые вороны, очень часто
грачи.
Размеры отдельных колоний бывают чрезвычайно разнообразны. Особенно
велики колонии в южных частях ареала. Под Ленкоранью, по моим наблюде-
ниям, каждая колония состоит из нескольких сотен серых цапель и многих сотен
прочих видов. В Астраханском заповеднике в 1935 г. на 1 га гнездилось до 382,
а в 1936 г. до 530 особей. В прошлом крупные колонии встречались и в северных
частях ареала. В бывшем Козельском уезде в районе Калуги иногда в колониях
насчитывалось до 100 гнезд (Филатов, 1915), в Серебряном бору, на окраине
Москвы, в 1917 г. колония состояла из 200 особей (Головин, 1917). Ныне круп-
ные колонии серых цапель в средней полосе встречаются много реже. Обычно
колония состоит из 5, иногда 10—20 пар. Однако, на Рыбинском море в Дарвин-
ском государственном заповеднике цапли недавностали гнездиться крупными по-
селениями. В 1949 г. три колонии содержали: 300, 105 и 200 гнезд (Скокова). На
юге Украины колонии бывают больших размеров. Под Запорожьем часто в коло-
ниях насчитывается 20—30 пар, изредка пар 50 (Спангенберг). Серые цапли не-
прихотливы в отношении выбора места для устройства гнезд. Известны случаи,
когда в одних и тех же районах птицы устраивают гнезда: в тростниках и камы-
шах, на затопленных ивах и на высокоствольной древесной растительности. В
других частяхареала, напротив, птицы гнездятся только в определенной обета-
СЕРАЯ ЦАПЛЯ 399
новке. В Бессарабии серые цапли строят гнезда исключительно в камышах
больших прудов (Остерман, 1915), в Белоруссии, несмотря на обилие древесной
растительности, цапли иногда строят гнезда на земле или на куче прошлогод-
него камыша (Федюшин, 1928). В камышах и тростниках птицы гнездятся
также в районе Херсона (Браунер) и в районе Мариуполя (Боровиков, 1907).
В Башкирии гнездование цапель отмечено в камышах и на деревьях (Сушкин,
1897; Штехер, 1915). В вост. Приазовье часть цапель также строит гнезда
в камышах, но часть гнездится по садам, расположенным вдоль речек (Алфе-
раки, 1910), вероятно на деревьях. У Павлограда гнезда помещаются на де-
ревьях (Вальх, 1899), под Харьковом на высоких затопленных ольхах,
иногда в сосновых лесах (Сомов, 1897). Большинство колоний под Ленкоранью
размещается на вершинах крупных дубов, карагачей или на затопленных
ольхах, а в низовьях Куры некоторые колонии строятся в тростниковых крепях.
В Астраханском заповеднике цапли гнездятся изредка в тростниках, вероятно
на кустарниковой иве, но чаще на относительно высоких деревьях. В противо-
положность этому, в северных частях ареала серые цапли, повидимому, устраи-
вают свои колонии обычно на высокоствольной древесной растительности; на
соснах—по реке Кинель, в Московской, Калининской обл., на Рыбинском море;
на соснах, березах и осинах — в районе Тулы; на соснах—в Кировской обл.;
на громадных то живых, то засохших елях г под Псковом.
В азиатской части страны места гнездовья столь же разнообразны. В Турк-
мении цапли гнездятся в камышах и на деревьях (Шестоперов, 1937). У Кара-
бента колония размещалась на деревьях среди затопленной площади. Радде и
Вальтер осмотрели колонию примерно из 100 пар, причем на одном дереве
помещалось 3—5 гнезд. На нижней Сыр-Дарье размер колоний бывает от 5 до
200 гнезд. Обычно колонии размещаются в тростниках и камышах крупных
озер, иногда на краях островков относительно маленьких водоемов (как,
например, Чиилинка). Значительно реже смешанные и чистые поселения
цапель удается находить на низкорослых турангах среди тугайных зарослей,
покрывающих острова реки. На Чу цапли гнездятся в камышах (Долгушин,
1939). В устье Урала серые цапли гнездятся колониально среди чащ камыша,
иногда рядом с колониями колпиков, причем отдельные гнезда располагаются
на 5—8 шагов друг от друга. В районе Иргиза цапли гнездятся преимуществен-
но в камышах, на Эмбе изредка на одиночных деревьях джиды (Сушкин, 1908).
В камышах, будто бы вследствие отсутствия древесной растительности, строятся
гнезда цапель в дельте Черного Иртыша. В Уссурийском крае —в устье
р. Лефу — основная масса серых цапель гнездится смешанными колониями,
располагая гнезда на 10—20 м одно от другого среди зарослей затопленного
тальника. Колония, осмотренная Шульпиным, состояла приблизительно из
50—70 пар. Много реже одно — два гнезда помещались на наваленном трост-
нике; изредка встречались и одиночные гнезда среди тростниковых зарослей.
Несколько севернее (на р. Или) цапли гнездятся колониями на высоких ли-
ственницах среди болота, устраивая по А—5 гнезд на одном дереве (Шульпин,
1936). На древесной растительности чистыми колониями гнездятся цапли и
на Имане. Поселения размещаются здесь на группах деревьев среди сельско-
хозяйственных угодий и на окраинах лесных массивов по островам реки
(Спангенберг, 1940). Колониями (вероятно чистыми) гнездятся серые цапли
также в Якутском округе (Иванов, 1929), но в какой обстановке —остается не-
выясненным.
Гнезда цапель в одних случаях закладываются прямо на земле или при-
легают своим основанием к воде, в других — на вершинах самых крупных
деревьев. Под Псковом были известны гнезда, выстроенные на старых елях на
высоте от 16 до 26 м (Зарудный, 1910), а в районе Москвы — в вершинах сосен на
40 м от земли (Мензбир, 1918). При гнездовании колониями с другими видами
цапель, серые всегда стремятся гнездиться выше своих сородичей. Гнезда серых
400 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
цапель отличаются чрезвычайной легкостью, поэтому их выдерживают тонкие
древесные ветви и верхушки тростниковых зарослей.
Форма вновь выстроенных гнезд, как и у прочих цапель, — перевернутый
конус с просвечивающими стенками, через которые хорошо видны яйца. При
гнездовании на деревьях и кустарниках основным строительным материалом
служат тонкие сухие прутья и ветки. В небольшом количестве используются
также стебли тростника. Гнездясь в тростниках, цапли строят гнезда из стеб-
лей этих растений, иногда с сухой травой в лотке. На нижней Сыр-Дарье
цапли используют для постройки дивную колючку. Колючие ветки тонким
слоем укладываются на тростниковую основу. При этом гнездо приобретает
особую прочность и не сбрасывается с камыша при сильных весенних вет-
рах. Одно из гнезд, осмотренных в Уссурийском крае, было построено из гру-
бых сучковатых стеблей какого-то «барьяна», напоминающего сухую крапиву
(Шульпин, 1936). Размер гнезд весьма различен: поперечный диаметр у свеже-
выстроенных гнезд от 50 еле и до 110 см у гнезд, сильно растоптанных птен-
цами. Различна также и высота гнезда. Свежие гнезда — около 60 см высоты,
но позднее становятся более низкими и плоскими. В крупных гнездовых посе-
лениях гнезда, обычно построенные небрежно, бывают сравнительно малы и
с плоским лотком. Напротив, гнезда птиц, гнездящихся отдельными парами,
построены лучше. Иногда гнезда используются в течение нескольких лет и еже-
годно подправляются. Такие постройки помещаются как бы на высоком фунда-
менте и достигают более метра в поперечнике.
Гнездо выбирается самцом, в постройке гнезда участвуют обе птицы, при-
чем материал приносится частично издалека, частично собирается поблизости.
В конце гнездостроения самка подолгу остается в гнезде, защищая его от соседей
и укладывая ветки, принесенные самцом. Постройка гнезда продолжается и в
начале насиживания. Количество яиц колеблется от 3 до 7, обычно 4—5 в кладке.
На нижней Сыр-Дарье в 1927 г. и 1928 г. во всех осмотренных гнездах было лишь
по 2 яйца, в 1930 г. в гнездах тех же колоний 3—4 яйца, а в некоторых 5 яиц
(Спангенберг, 1936), На севере ареала на Рыбинском водохранилище не-
редки кладки по 6 яиц. В 1950 г. из 160 осмотренных гнезд—18 содержали
по 6 яиц, 128 по 5 и 14 по 4 яйца. Как исключение, два гнезда содержали
по 7 яиц (Скокова). Форма яиц изменчива. Одни имеют правильную яйце-
видную форму, другие одинаково заострены с обоих конусов. Скорлупа свеже-
снесенных яиц лишена блеска, насиженных — со слабым блеском. Окраска
яиц зеленовато-голубая, часто с белыми известковыми мазками. Размеры:
D3) 55,3—60,4 мм X 41,0—46,5 мм, среднее 59,8 X 43,7 мм.
Промежутки времени между откладкой отдельных яиц, как правило,
48 часов. Насиживание начинается после откладки первого яйца, птенцы в гнез-
де бывают различного возраста. В насиживании принимают участие обе птицы
гнездовой пары, но самец сидит на гнезде значительно меньше самки. Насижи-
вание продолжается 26—27 дней (Спангенберг).
Близко зимуя, рано прилетая на родину, а местами и ведя оседлый образ
жизни, серая цапля в массе ранее других наших цапель приступает к размно-
жению, причем в северных частях ареала гнездится значительно позднее, чем
в южных. Неодновременно и гнездование у различных пар даже в одних и тех
же колониях. Эти колебания в Астраханском заповеднике выражены примерно
в 35 днях (Дубинины, 1940), в районе Харькова—в 30 днях, в Дарви-
новском заповеднике 40—60 дней (Скокова). Особенно рано начинают
гнездиться цапли в Закавказье. Под Ленкоранью первые яйца в гнездах
откладываются в десятых числах марта, а в самом начале мая гнезда содер-
жат птенцов разнообразного возраста и совершенно насиженные яйца. На
Черноморском побережье гнездостроение отмечено 3 марта, 10 марта были от-
ложены первые яйца, а в половине мая наблюдались летные птенцы (Вильков-
ский, 1897). В дельте Волги начало откладки яиц происходит между 5 и 15
СЕРАЯ ЦАПЛЯ 401
апреля, появление первых птенцов между 1—10 мая, а массовое между 20—30
мая (Дубинины, 1940). На Украине (под Запорожьем) гнезда осмотренных коло-
ний в половине и в самом конце апреля содержали незаконченные кладки и пол-
ные — с яйцами в начальной стадии насиженности. В районе Харькова яйца
в ненасиженном состоянии удается находить с конца апреля и с этого времени
до последних чисел мая. В первой трети июля часть птенцов еще не способна
к полету, но часть уже покидает гнезда, и в первой половине этого месяца их
удается наблюдать в стаях со взрослыми; у Одессы 30 мая в гнездах обнару-
жены птенцы (Зябрев, 1940). В районе Минска свежие и слегка насиженные
яйца бывают в последней трети мая. Дата эта несколько поздняя и вероятно
часть цапель здесь гнездится раньше. Во всяком случае даже под Псковом 5 мая
в одном из гнезд обнаружено б слегка насиженных яиц, 26 июля птенцы были
почти оперены, а 16 августа порядочно летали (Зарудный, 1910). Будто бы зна-
чительно раньше происходит размножение птиц под Калугой, где над ними ве-
лись наблюдения в течение ряда лет. Вероятно в указанных датах допущена
ошибка. Автор (Филатов, 1915) отмечает, что в 1910 г. цапли наблюдались на
Оке 26 марта. Наступившее похолодание, вероятно, вызвало временную отко-
чевку птиц, и они вновь появились лишь 2 апреля. Далее автор указывает»
что 19 апреля птенцы достигли роста голубя, а 3 июля почти роста взрослых
особей. В Куйбышевской обл. летные молодые отмечены в конце июля (Карам-
зин, 1901), в районе Уфы — 25 июня насиженные яйца и птенцы разного возра-
ста (Штехер, 1915), а в долине р. Оки (Московская обл.) 19 июля —летные и
вполне оперившиеся птенцы (Воробьев, 1925). В Дарвиновском заповеднике
в середине августа часть птенцов еще не способна к полету (Скокова).
Под Ленкоранью птенцы вылетают из гнезд в возрасте 6—7 недель. Первые
летные птенцы появляются около середины мая, массовый вылет их в конце
мая и в начале июня; в дельте Волги ранние летные птенцы между 20—30 июня,
массовое появление их 15—25 июля (Дубинины, 1940). В Ульяновской обл. во
второй половине июля. В Дарвиновском заповеднике 8 июля, массовое —
15—20 июля (Скокова).
В азиатской части страны сроки гнездовья также изменяются в зависимо-
сти от широты местности. В Туркмении гнезда содержали свежие яйца в сере-
дине мая (Шестоперов, 1937); на нижней Сыр-Дарье, в районе Кзыл-Орды, гнез-
да, осмотренные 14 апреля, содержали незаконченные кладки в 1 —2 яйца, слабо
насиженные яйца найдены 3 и 10 мая, а гнезда с сильно насиженными яйцами —
во второй половине июня (Спангенберг, 1936). В устьях Урала около середины мая
в гнездах обнаружены сильно насиженные яйца; некоторые из них надклюнуты.
В Уссурийском крае в устьях р. Лефу в половине апреля, когда лоза стоит
еще голая, большинство гнезд бывает уже отстроено. Свежие яйца в гнездах
обнаружены в 20-х числах апреля, но, с* другой стороны, 2 мая оказалось мно-
го недостроенных гнезд, 21 мая вскрытые яйца имели эмбрионов размером в 10—
15 и 20 мм, а 29 мая наблюдалась цапля с длинной хворостиной в клюве. Раз-
гар насиживания приходится на конец мая, в это время гнезда содержали на-
сиженные яйца, но попадались также неполные кладки (Шульпин, 1936).
Большая растянутость периода размножения в одних и тех же колониях
объясняется, с одной стороны, неодновременным образованием каждой коло-
нии, которая постепенно возрастает за счет присоединений новых групп; с дру-
гой стороны, откладкой новых яиц взамен погибших, а также вероятно тем,
что молодые особи, размножающиеся впервые, запаздывают в отношении разви-
тия гонад. При нормальных условиях в течение лета птицы успевают воспи-
тать только один выводок.
Появление первых птенцов в гнездах в Туркмении происходит, вероятно,
около середины мая B2 июня в Мервском оазисе молодые еще плохо летали).
Также и на нижней Сыр-Дарье и в устьях Урала к концу июля вся молодь
покидает гнезда. В районе Чкалова летные птенцы отмечались около сере-
26 Птицы СССР, т. II
402 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
дины июля, реже в первой трети августа. В двух гнездах, осмотренных 24 мая
в устьях р. Лефу, оказались большие птенцы, покрытые пухом. При посеще-
нии этой колонии 5 июня в ней оставалось лишь небольшое количество лазаю-
щих птенцов, остальные уже откочевали (Шульпин, 1936).
Выведшиеся птенцы совершенно беспомощны, но зрячи, со щелевидными
глазами. Вес птенца несколько больше 40 г. В самом начале взрослые кормят
их, отрыгивая в рот пищу. Первые перья появляются в возрасте 7—9 дней,
поднимаются птенцы на ноги к 16-му дню.
Оперившиеся птенцы иногда вылезают из гнезд на ветви гнездового дерева,
часто сваливаются на землю и бродят под деревьями колонии. Под Ленкоранью
такие птенцы в большом числе становятся жертвами шакалов и Камышевых
котов. После вылета птенцы некоторое время держатся семьями, встречаются
и одиночки. Иногда птицы сбиваются в крупные стаи. Стаи в сотни штук отме-
чались в Уссурийском крае, в 30 особей в дельте Волги, до 200—250 птиц на
юго-восточном берегу Каспия. До отлета серые цапли широко кочуют по мест-
ности, посещая наиболее кормные водоемы, а затем начинают подвигаться к югу.
Линька. Раз в году, полная годовая, между июлем и ноябрем. По
Уайзерби A939), у птиц в гнездовом наряде имеется и частичная линька мелкого
пера в первую осень жизни, между сентябрем и декабрем. Схема возрастных
изменений: первый пуховой наряд — второй пуховой наряд — гнездовый на-
ряд _ первый годовой наряд (окончательный) и т. д. В Уссурийском крае серые
цапли начинают линять около 20 июля (Шульпин, 1936). Вероятно в близкие
сроки линька начинается и в других частях ареала. Линька идет медленно и
заканчивается на местах зимовок. В районе Харькова у гнездящихся старых
птиц смена маховых второго порядка будто бы происходит в последней трети
мая, а в конце июля попадаются вполне вылинявшие взрослые птицы
(Сомов, 1897), что весьма сомнительно.
Питание. Разнообразная животная пища. Состав пищи меняется по
сезонам и в различных географических частях ареала. В общей сложности цапли
поедают: водяных и наземных насекомых и их личинок, разнообразную некруп-
ную рыбу, лягушек и их головастиков, ящериц, змей, грызунов. В Бессарабии
известны случаи поедания мышей и сусликов. В пищеводе одной добытой птицы
одновременно обнаружено 3 суслика (Остерман, 1915). Лягушки, головастики,
жабы, ящерицы и мыши составляют, по Сомову, пищу цапель в Харьковской
области.
Под Запорожьем основная пища состоит из рыбы, лягушек, ящериц и на-
секомых. Ранней весной прилетевшие цапли регулярно вылетают в степь и
охотятся здесь за сусликами; молодые зверьки поедаются также и летом (Спан-
генберг). В Астраханском заповеднике серые цапли поедают лягушек, рыбу,
водяных крыс, мышей, а в период вскармливания птенцов — ужей, крупных
насекомых и их личинок (Дубинины, 1940). Под Ленкоранью в летнее время
основной пищей несомненно служит мелкая рыбешка. Обнаружены в желудках
добытых птиц также хитиновые остатки насекомых (цикады, коромыслы и ко-
былки), лягушки и один раз песчанка (Спангенберг).
В зимнее время у Ленкорани цапли поедают рыбу (кутум, лещ и другие кар-
повые, щучки), крыс, мышей, лягушек, водяных насекомых и, вероятно случайно,
заглатывают части растений. В годы изобилия полевок (Microtus socialis) цапли
охотятся за ними в степи у кустов ежевики. На восточном берегу Каспия в теплое
осеннее время большое значение в питании серых цапель имеют бычки (Gobi-
idac). Их цапли отыскивают среди камней на мелководных участках. Позднее,
когда бычки отходят от берегов, цапли питаются почти исключительно раками
(Potamobius leptodactylus и P. pachypus), выброшенными волнами на берег
(Исаков и Воробьев, 1940). В низовях Сыр-Дарьи основная пища —мелкая
рыба. Во время подсыхания водоемов цапли концентрируются около сохранив-
шихся ям, переполненных мелкими карпами. Мелкие карпы и щучки обнару-
СЕРАЯ ЦАПЛЯ
403
жены и у гнезд серых цапель. В годы, богатые саранчей, на некоторое время
цапли полностью переключаются на эту пищу, а при затоплении весенним па-
водком участков степи в изобилии ловят песчанок. Поедают птицы также ко-
ромысла и их личинок (Спангенберг). На Илеке цапли вылетают в степи, пое-
дая здесь сусликов. По данным Зарудного A888), в бывшем Оренбургском крае
цапли посещают места лова рыбы бреднем и, опускаясь на цевку, подбирают
мелкую рыбешку. В Уссурийском крае у гнезд цапель обнаружены бычки,
касатки, мелкие сомики от 15 до 18—
25 см длиной. В поисках пищи в рай-
Фне озера Ханка птицы посещали мел-
ководные протоки, окна в болотах,
грязевые и песчаные берега (Шуль-
пин, 1936).
Добывая пищу, серые цапли по-
долгу выстаивают в мелкой воде, тер-
пеливо выжидая, когда жертва при-
близится на близкое расстояние. Долго
простаивают цапли также у норок
сусликов и полевок. Реже цапли в
поисках пищи расхаживают по отме-
лям и сухим участкам.
Полевые признаки. Крупными раз-
мерами и светлой окраской оперения
безошибочно отличается в природе от
всех наших цапель. По сложению ти-
пичная цапля, с узкой головой, боль-
шим клювом, длинной шеей, длинными
ногами, большими широкими крыль-
ями и коротким хвостом. Общая окра-
ска спинной стороны серая, брюшная
сторона белая, маховые черные. Не
видя птицы, ее легко узнать по гром-
кому голосу, издаваемому во время
полета. Птица бодрствует в течение
всей ночи. Часто ее удается наблюдать
неподвижно стоящей на речных косах,
на чистых мелководных плесах озер,
на торчащих из воды карчах, а в гнез-
довое время и на деревьях. Крайне
осторожна, зорко следит за человеком
и далеко улетает, будучи потревожен-
ной. Вспугнутая, усиленно машет
крыльями. Ноги неуклюже болтаются
в воздухе, но взлет относительно лег-
кий. Поднявшись высоко, летит уже плавным полетом, глубоко втянув длинную
шею в плечи и откинув назад ноги. Во время полета группы цапель часто об-
разуют угол или линию. Голос громкий и резкий, режущий звук, вроде «крянк».
Подрастающие птенцы издают однообразные трескучие звуки «кунго-ка-ка-ка»
(Филатов, 1915). Эти звуки сопровождаются киванием опущенной вниз
головы.
Описание. Размеры и строение. Клюв длинный, прямой, сжатый с боков;
ноздри длинные, щелевидные. Ноги длинные, нижняя треть голени не оперена. Пальцы
длинные и тонкие, передний и четвертый соединены у основания перепонкой. Когти длин-
ные, острые, слегка изогнутые, внутренний край когтя на среднем пальце с зазубринами.
На груди, брюхе, пахах — большие пятна «порошкового пуха». Формула крыла (не считая
рудиментарного первого махового) :3=4>2>1>5>6. Вырезки на наружных опахалах 2-го—
Рис. 79. Серая цапля
404
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
5-го, внутренних опахалах 1-го — 3-го маховых. Хвост прямо обрезанный, из 12 рулевых.
Перья затылка удлинены и образуют хохол.
Вес добытых на каспийских зимовках самцов B2) 1350—1770 г, самок B1) 1100—
1470 г (Воробьев и Исаков, 1940). По другим литературным данным максимальный вес до
2 000 е. Крыло самцов B0) 430—490 мм, самок A4) 440—475 мм, среднее 472 и 458 мм.
Окраска. Первый и второй пуховой наряды — из беловатого на вершинах и серо-
ватого у основания пуха, удлиненного на темени; спина серовата, брюшко беловато.
Взрослые самцы и самки: спинная сторона и рулевые серые; лоб и середина темени
белые; над глазом идет широкая черная полоса, соединяющаяся на затылке; перья хохла
черные; лоб и середина темени белые; шея серовато-белая, от горла по брюшной ее стороне
тянутся три полосы мелких черных продольно вытянутых пятен; перья зоба удлинены;
брюшная сторона серая, белеющая по середине груди, брюха и в подхвостье; первостепенные
маховые черные. Клюв желтовато-бурый, ноги зеленовато-серые, голень желтовата; голое
пятно на уздечке желтовато, вокруг глаз зеленовато; радужина желтая. В гнездовом наряде
хохол и удлиненные перья на зобе развиты слабо, цвет этих перьев серый; шея почти вся
серая, спинная сторона серая с буроватым оттенком; брюшная сгорона белая с буровато-
черными удлиненными пятнами. Клюв и ноги буроваты, радужина беловатая, позднее при-
обретающая желтоватый оттенок.
8. Рыжая цапля Ardea purpurea L.
Ardea purpurea. Linnaeus. Syst. Nat., изд. XII, 1766, стр. 236, Франция.
Русское название. Цапля, чепура — народное. Рыжая цапля — книжное.
Распространение. Ареал. Европа—Франция, кроме северной и восточ-
ной; Пиренейский п-ов —Испания, Португалия; Голландия, Италия, Австрия
(Нейзидлерское озеро), Венгрия, Балканы (в Греции только в Фессалии),
Карта 73. Распространение рыжей цапли Ardea purpurea
1. A, p. purpurea, 2* А. р. manillensis, 3. А. р. madagascariensis;
а — граница гнездовий, б — не вполне выясненная граница гнездовий, г — область shmobok.
о-в Майорка; южная полоса Европейской части СССР, примерно от 52° с. ш. до
Крыма и Закавказья. Африка, кроме Сахары—Марокко, Алжир, Тунис и спора-
дически в тропических и южных частях континента; Мадагаскар. Азия —от
Малой Азии, Месопотамии, Средней Азии, Ирана» Индии, Бирмы, Цейлона
РЫЖАЯ ЦАПЛЯ 405
до Приморья, с.-в. Китая, Манчжурии, Японии, Риу-Киу, Формозы, Филип-
пинских о-вов, Целебеса, Б. Зондских о-вов. Область зимовок северных попу-
ляций — от южн. Европы, Средней Азии, в степных районах зап., вост. и
южн. Африки, в Китае от Сычуаня до Гуандуня.
Характер пребывания. См. выше.
Биотоп. В общем сходный с биотопом серой цапли — берега водо-
емов с разнообразной растительностью; однако —в отличие от серой цапли—
положительно избегает лесистых местностей, придерживаясь полуоткрытого
ландшафта.
Подвиды и варьирующие признаки. Различия — в размерах и окраске, а также
деталях биологии (характер пребывания, детали репродуктивного цикла — в общем однако
сходного и т. п.). Три подвида: на Мадагаскаре—Л. p. madagascariensis Oort, 1910;
в южн. Азии — Индии, Бирме, Цейлоне, Зондских и Филиппинских о-вах, на Риу-Киу,
в Китае, на север до Манчжурии и Приморья — А. р. manillensis Meyen, 1831; в осталь-
ной части ареала— А. p. purpurea L., 1766.
Западная рыжая цапля Ardea purpurea purpurea L.
Синоним: Ardta Caspia. Gmelin. Reise d. Russland, II, 1774, стр. 193, Астрахань. —
Ardea, Botaurus. Gmelin. Syst. Nat., изд. XIII, 1789, стр. 636, Италия и Астрахань,
partim.
Распространение* Ареал. Общий ареал см. выше, в очерке вида. Детали
распространения в СССР таковы: северная граница проходит по линии Киев,
Чернигов, Харьков, Воронеж, низовья Волги, Урал, Иргиз и Тургай. К югу
отсюда рыжая цапля гнездится у нас в подходящих местах. Повсе-
местно. Более подробное распространение у нас этого вида таково. Регу-
лярно гнездится в плавнях Днестра и на степных ставках в Бессарабии. Широко
распространена по Днепру от устья этой реки вверх, во всяком случае, до райо-
нов Киева и Чернигова. Столь же широко встречается на гнездовье в гирлах
Дона и вверх по течению этой реки (Алфераки, 1910). Населяет подходящие
угодья Полтавской, Харьковской (Сомов, 1897) и реже Воронежской обл.
(Огнев и Воробьев, 1924). Местами (Черная речка, Салгир, Тамак, Тархан-
Сунак) гнездится в Крыму (Никольский, 1892). В Закавказье населяет низмен-
ные территории Армении (Ляйстер и Соснин, 1942), широко распространена
в среднем и нижнем течении Куры и гнездится между устьями этой реки и
г. Ленкоранью. Напротив, в лесистых истылях южнее Ленкорани в гнездовый
период положительно отсутствует. На Сев. Кавказе обыкновенная гнездящаяся
птица Кубани, во всяком случае до Краснодара, встречается в низовьях Те-
река, в районе Кизляра и далее по западному побережью Каспия до дельты
Волги включительно. Обычна также в камышах Сарпинских озер. На Урале
распространение приурочено к низовьям реки, на Эмбе встречается до устьев
Аты-Джаксы и в области пресных озер на север от морского побережья, напри-
мер, в урочище Сары-куль. Далее рыжая цапля гнездится в низовьях Иргиза и
Тургая, не доходя по Тургаю до одноименного города (Сушкин, 1908). В обла-
сти Аральского моря рыжая цапля гнездится на озере Камышлы-баш, на озерах
северного берега и, вероятно, по всему восточному и южному побережью
Аральского моря, где развиты тростниковые заросли. На Сыр-Дарье гнездовая
область охватывает нижнее и среднее течение реки, а на востоке от неё озера
Тели-куль, низовья Сары-су и Чу (Гладков и Гринберг, 1932), не достигая
Балхаша. Вероятно, гнездится в низовьях Атрека в Туркмении, на Аму-Дарье
по всему течению вверх до Вахша, на Мургабе и на западном Узбое (Дементьев,
1950). Известны случаи залета в районы Минска, Тамбова, Чкалова, Канска
и Томска и в горы Армении на озеро Севан, лежащее на 2.000 м над уровнем
моря (Ляйстер и Соснин, 1942); в Зап. Европе в Англию, Германию (где не-
сколько раз гнездилась), Швейцарию, Бельгию, Данию, Норвегию и южн.
Швецию; в Африке на Канарские и Азорские о-ва, на Мадейру. Особи север-
406 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
ной популяции европейской рыжей цапли нормально зимуют в Африке, часть
птиц, быть может, зимует и в более северных частях ареала и, в частности, в пре-
делах нашей страны. Молодая птица, окольцованная в Астраханском заповед-
нике 27 июня 1939 г., в ноябре 1939 г. добыта в Африке на реке Мунго, фран-
цузский Камерун.
Характер пребывания. Для большей части нашей страны —
гнездящаяся, перелетная птица.
Отдельные особи зимуют под Ленкоранью, но в более холодные зимы от-
кочевывает южнее. Для многих частей Союза рыжая цапля является залетной
или только пролетной птицей. Прилет ранний для южных районов и относи-
тельно поздний для средней полосы. Сроки прилета в различные точки Союза
не вполне выяснены. В Бессарабии птицы появляются весной около середины
марта, у Херсона неделей позже, чем серая цапля, т. е. в последних
числах марта, а под Мариуполем прилет начинается около половины апреля
и заканчивается в конце первой трети мая; в более северных частях (Днепро-
петровская обл.) главная масса птиц летит в конце апреля. В Харьковской обл.
появляется позднее серой цапли. Первые птицы прилетают около половины
апреля A3 апреля), валовой пролет идет до последних чисел этого месяца
(Сомов, 1897). В окрестностях Киева в 1926 г. первоприлетная птица отмечена
26 апреля (Шарлемань, 1926), в Подолии обычно появляется в начале апреля
(Храневич, 1925—1926). Рано прилетают птицы и в Закавказье. На Черномор-
ском побережье первые особи появляются около половины марта, и с этого вре-
мени пролет длится до конца первой трети мая. Еще более ранний прилет в во-
сточных частях Закавказья и в частности под Ленкоранью, где отдельные
особи, как уже было отмечено выше, перезимовывают в теплые зимы.
В Астраханском заповеднике прилет происходит, по Воробьеву A936),
в апреле, но изредка и в конце марта; по Дубининым A940), появление первых
особей здесь бывает между 1 и 10 апреля; в Туркмении первые рыжие цапли от-
мечены в самом начале второй трети марта и пролет затягивается на длитель-
ное время. Птицы наблюдались на Мургабе у Байрам-али 11 марта, у Юссуф-,
Хана 26 марта, у Иолотани 20—22 марта, у Ейлантепе 24 марта, в Мервском
оазисе 15 марта, у Теджена в большом числе 6 апреля, у Чикишляра, вероятно,
поздняя, пролетная особь 16 мая (Дементьев, 1950). На Мангышлаке наиболее
ранняя отмечена 10 апреля, самая поздняя 28 апреля (Смирнов, 1916). В сред-
нем и нижнем течении Сыр-Дарьи первые птицы появляются в самом начале
апреля, иногда в первой половине этого месяца и крайне редко — в последних
числах марта (Спангенберг и Фейгин, 1930, 1936). Окрестностей г. Иргиза
птицы достигают только в самых последних числах апреля. Осенняя откочевка
птиц к югу начинается с первым похолоданием. В Подолии она происходит
в октябре. В районе Харькова одиночки и группы рыжих цапель начинают
лететь к югу с первых чисел сентября. Заметный пролет затягивается прибли-
зительно до половины этого месяца. В более поздние сроки отмечаются запоздав-
шие одиночки. Самый поздний экземпляр отмечен Сомовым A897) 2 октября.
Когда происходит отлет в Воронежской обл., точных данных нет, но птицы
отмечались здесь 13 и 14 сентября (Огнев и Воробьев, 1924). Под Мариуполем
в самом начале сентября численность рыжих цапель возрастает, вследствие
подлета северных особей, заметный же отлет начинается с середины этого ме-
сяца. Более поздно,между 13 октября и 13 ноября —отлетают цапли из Бес-
сарабии (Остерман, 1915). В Астраханском заповеднике отлетает, по данным
Дубининых A940), между 20 и 30 сентября, но одиночки задерживаются до по-
следней трети октября (Воробьев, 1936). Также и в устьях Урала цапли отле-
тают в последних числах сентября. Осенний отлет в районе Дзауджикау на-
блюдается в сентябре и октябре. На Сыр-Дарье между станциями Джусалы —
Чийли отлет начинается с конца сентября, иногда в середине октября, а в ред-
кие годы бывает.хорошо выражен в самом начале сентября (Спангенберг и Фей-
РЫЖАЯ ЦАПЛЯ 407
гин, 1930, 1936). В вост. Иране пролетные рыжие цапли отмечались уже
15 сентября (Зарудный, 1903).
Во время пролетов большинство птиц летит в ночное время, поднимаясь
из камышей с наступлением вечерних сумерек или незадолго до захода солнца.
Утром пролет заканчивается после зари. Сравнительно редко эти цапли летят
большими обществами, чаще по несколько птиц на большой высоте, выстроив-
шись углом или поперечной линией. Есть наблюдения о присоединении одино-
чек рыжей цапли к пролетным стаям квакв.
Биотоп. Рыжая цапля повсеместно предпочитает обширные открытые
пространства, занятые водоемами, в значительной степени заросшими камы-
шами, тростниками и реже низкорослым кустарником ивы. В местностях, где
открытый ландшафт нарушается наличием лесных массивов, птицы становятся
малочисленными или не встречаются вовсе. Кормовые стации вида более разно-
образны. На Сыр-Дарье часть птиц добывают пищу поблизости от гнезд, стоя
на краю зарослей у маленького плеса. Другие птицы, напротив, летают далеко
от гнезд, посещая обмелевшие озера, реже затопленные полой водой тамари-
сковые поросли и еще реже отмели Сыр-Дарьи. На вост. побережье Аральского
моря цапли кормятся на морских отмелях, покрытых редкими тростниками.
При охоте рыжая цапля глубоко (по самое брюхо) заходит в воду и в ожидании
добычи целыми часами выстаивает на одном месте, не меняя позы. В отличие
от цапли серой, повсеместно избегает близости человека, гнездясь в наиболее
глухих и труднодоступных участках. В поисках пищи, напротив, встречается
поблизости от рыбачьих селений и в сельскохозяйственных угодьях.
Численность. Обыкновенный гнездящийся вид южной безлесной
полосы Европейской и Азиатской частей Союза. В лесостепной полосе числен-
ность птиц резко снижается. Может быть отнесена к обыкновенным для Бесса-
рабии, нижнего течения рек, впадающих в Черное море, для дельты Волги и
вообще для сев. побережья Каспия, для вост. побережья Аральского моря,
Иргиза и Тургая, а в Закавказье для низких районов Армении, но сравнительно
редка в Ленкоранской низменности. Очень обыкновенна в вост. Приазовье и
положительно многочисленна в дельте Аму-Дарьи и в нижнем течении Сыр-
Дарьи в районе Кзыл-Орды. В общем рыжая цапля довольно многочисленный
вид нашей страны.
Экология. Размножение. Гнездится отдельными парами или обра-
зует небольшие колонии, то самостоятельные, то вместе с серыми и белыми
цаплями и колпиками. Колониальные гнездовья известны в районах Харькова,
Киева, в дельте Волги, в низовьях Иргиза, в нижнем и среднем течении Сыр-
Дарьи. Однако и здесь многие птицы выводят отдельными парами. Гнезда
устраиваются на торчащем из воды старом камышевом и тростниковом заломе
или на смятых кучах этих растений. Много реже птицы воздвигают свои построй-
ки на низкорослых кустиках колючки, растущих среди тростников, или на за-
топленных водой кустарниках ивы и ольхи. Есть указания в литературе о гнездо-
вании рыжих цапель маленькими колониями на деревьях, иногда по нескольку
гнезд на одном дереве (Мензбир, 1918). Такие случаи должны рассматриваться
как исключение и не характерны для нашей страны. В зависимости от харак-
тера гнездования, материалом для постройки гнезд служат стебли и листья тро-
стникового и камышевого валежника или при постройке гнезд на кустарни-
ках — тонкие сухие прутья этих растений с примесью Камышевых листьев
в лотке. На Сыр-Дарье помимо сухих Камышевых стеблей и листьев рыжие
цапли часто используют сухие ветки колючих кустарников. По форме и харак-
теру постройки гнезда схожи с гнездами других цапель. В период откладки
яиц их форма — перевернутый конус с глубоким лотком и с просвечивающими-
ся стенками. С появлением птенцов, гнезда утаптываются, теряют свою первона-
чальную форму и становятся почти плоскими. Размер гнезд сильно варьирует,
но в общем они невелики по сравнению с размерами птицы. Поперечный диаметр
408 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
их достигает 60, редко 80 см, высота гнезд в период яйцекладки 40—50 см.
Птицы видимо не используют старых построек и ежегодно строят новые гнезда.
В постройке гнезд участвуют обе птицы гнездовой пары. Количество яиц в пол-
ных кладках 3—5 (в Зап. Европе даже б, 7 и 8). В низовьях Иргиза у самки,
добытой из гнезда с 5 яйцами, в яйцеводе был обнаружен крупный желток
{Сушкин, 1908), 3—5 яиц в кладках известны для Харьковской обл., для сред-
него и нижнего течения Сыр-Дарьи и для вост. Закавказья. Кладки по 3 и 5 яиц
найдены также в Ленкоранской низменности близ Порт-Ильича. Многочис-
ленные наблюдения на Сыр-Дарье позволяют предполагать, что в одни годы,
видимо благоприятные для птиц, яиц в гнездах бывает 5, в другие годы, напро-
тив, гнезда содержат не более 3 яиц (Спангенберг). Яйца рыжей цапли сильно
варьируют по величине и форме. Большинство яиц имеют яйцевидную форму,
вытянутую, или вздутую. На некоторых яйцах слабо выражен тупой и острый
конус. Окраска скорлупы, как у серой цапли — зеленовато-голубая при сне-
сении, а позднее сильно выцветает и загрязняется экскрементами. Размеры их
несколько меньше, чем у серой цапли, и этим признаком на массовом материале
они хорошо отличаются от яиц предыдущего вида. Размеры яиц A7) 53,0—
—57,5 мм X 37,9—39,2 мм, в среднем 54,8 х 38,5 мм.
В северных частях ареала кладка происходит значительно позднее, чем на
юге. Неодновременность наблюдается и в одних и тех же районах у отдельных
пар. В то время, когда запоздавшие пары только начинают класть яйца, в гнез-
дах ранних бывают птенцы, покрытые перьями. В Бессарабии рыжие цапли
строят гнезда и кладут яйца с середины мая, а 3 июля в двух гнездах были
обнаружены по 3 пуховых птенца (Остерман, 1915). В середине мая или в два-
дцатых числах этого месяца происходит яйцекладка и в нижнем течении Днепра,
южнее Запорожья (Спангенберг). В вост. Закавказье (бывш. Арешский уезд)
у самки, добытой 7 апреля, яичник оказался слабо развит, 4 мая в гнездах было
3—5 яиц, частично уже насиженных (Сатунин, 1907). В Ленкоранской низмен-
ности в окрестностях Порт-Ильича два гнезда содержали 20 мая незаконченную
кладку в 3 яйца и довольно сильно засиженную в 5 яиц (Спангенберг). В Харь-
ковской обл. по 3—5 свежих яиц найдены в гнездах у Змиева 28 мая, летные
птенцы наблюдались в последних числах июля (Сомов, 1897). В Киевской обл.
в колонии рыжих цапель, осмотренной Гебелем 11 июня, только в 1 гнезде ока-
залось 2 яйца, а 23 июня в 6 гнездах по 2—4 яйца уже насиженных. В Астра-
ханском заповеднике начало откладки яиц происходит между 1—10 мая, появле-
ние первых птенцов — между 30 мая и 5 июня, массовое — между 15 и 25 июня,
начало вылета молодых из гнезд между 10—20 июля, массовый вылет между
25 июля и 5 августа (Дубинины, 1940). В те же или близкие сроки происходит
размножение в устьях Урала. В низовьях Иргиза еще незаконченная кладка
в 5 яиц найдена 5 июля (Сушкин, 1908). На Сыр-Дарье в ее нижнем и среднем
отделах законченные, едва насиженные кладки удается находить с десятых чисел
мая и с этого времени до середины июня. Во второй половине июня подавляющее
большинство гнезд содержит насиженные кладки и птенцов различного возраста.
Промежутки времени между откладкой отдельных яиц в среднем 3 дня
(Мейлан). Первые 3 яйца откладываются через день (Спангенберг).
Насиживание начинается после снесения первого яйца, что доказы-
вается как прямыми наблюдениями, так и различной величиной эмбрионов
в яйцах кладки. При этом цапля, отложив первое яйцо, сидит на нем прочно,
покидая гнездо лишь при крайней необходимости. В насиживании принимают
участие обе птицы гнездовой пары, но участие самца, по сравнению с самкой,
меньше. Точный срок насиживания не установлен, предположительно 24—
28 дней (Уайзерби, 1939). По наблюдениям Остермана A915) в Бессарабии наси-
живание продолжается около месяца. Пока птенцы малы, пищу приносит только
самец» позднее в выкармливании принимают участие оба родителя. Оперив-
шиеся птенцы, оставшиеся одни в гнезде, при близком появлении луня реаги-
РЫЖАЯ ЦАПЛЯ
409
руют своеобразными ударами клювом по направлению врага. Вылетают птен-
цы в возрасте около 6 недель, двухмесячные держатся самостоятельно.
Питание. Кормится рыбой, лягушками, ящерицами, ужами, водяными
и наземными насекомыми и их личинками, изредка мышами, полевками, кры-
сами. Основную пищу птицы во многих местах страны все же составляет рыба.
На Сыр-Дарье рыжие цапли редко вылетают за пределы тростниковых и ка-
мышевых зарослей и кормятся здесь, главным образом, мелкой рыбешкой.
Во время движения пешей и летной саранчи, массами появляющейся в камы-
шах, желудки и пищеводы птиц оказываются наполненными этой пищей (Спан-
Рис. 80. Рыжая цапля
генберг). Остерман A915) считает главной пищей рыбу, упоминает также насе-
комых, лягушек и ящериц. В желудке одной цапли, убитой в Харьковской
обл., был обнаружен уж, у другого экземпляра—8 ящериц (Lacerta),
12 чесночниц (Pelobates), шерсть небольшого грызуна и остатки жуков (Сомов,
1897). Кроме того, автор упоминает о питании птиц лягушками, рыбой и насе-
комыми. Добыча пищи производится изредка днем, много чаще на зорях.
Линька. Последовательность смены нарядов — как у серой цапли.
У взрослых полная годовая линька с июня по ноябрь. Молодые рыжие цапли,
вылетевшие из гнезда, начинают сменять мелкое перо в конце этого же года
(ноябрь) и заканчивают свою первую линьку в марте — апреле следующей
весны. При этом крупное перо (маховые, большие кроющие крыла, рулевые)
не сменяются. Затем линька идет, как у взрослых (полная годовая). Некоторые
авторы (Уайзерби, 1939) указывают на наличие у рыжих цапель частичной пред-
брачной линьки (мелкое перо), это требует подтверждения.
Полевые признаки. Рыжая цапля очень напоминает серую, от которой,
однако, уже издали отличается меньшим размером и темнорыжей или даже каш-
410 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
тановой окраской оперения. Она менее осторожна, чем серая цапля. Охотясь
за рыбой в камышах, близко подпускает к себе лодку и человека, а затем круто
поднимается тяжелыми взмахами вверх, поднявшись, летит, как серая цапля.
На открытых местах рыжая цапля ведет себя более строго и улетает, заметив
человека издали. Меньшая осторожность, быть может, связана с сумеречным
и даже ночным образом жизни, а также с окраской птицы. Среди побуревших
прошлогодних тростниковых и Камышевых зарослей она менее заметна, чем
серая цапля. Раненая птица, упав в заросли, вытянув вверх голову и шею,
принимает неподвижную позу, как это делают волчки и выпи, так же ведут себя
птенцы рыжих цапель на гнездах. В отличие от серой цапли рыжая редко са-
дится на деревья и предпочитает скрываться в зарослях. Голос этого вида имеет
много общего с голосом серой цапли, но менее громок. Птенцы издают частые
короткие звуки, передать которые словами чрезвычайно трудно.
Описание. Размеры и строение. Формула крыла, как у серой цапли, но чет-
вертое маховое обычно пропорционально короче, а второе несколько длинней. Иное, по
сравнению с серой цаплей, соотношение длины цевки и среднего пальца: у серой цапли
цевка длинней среднего пальца (длина первой около 135—165, второго, измеренного с ког-
тем, около 95—110 мм), у рыжей цапли средний палец длинней цевки и по абсолютным раз-
мерам больше среднего пальца серой цапли (длина цевки около 115—130, среднего пальца
125— \35 мм). Вес B) 1220—1225г, по Зарудному A888) даже до 1650 г. Крыло самцов и
самок B5) 340—390, в среднем 367,2 мм. Отдельно, у самцов A0) 370—395, самок (9) 340—
370 мм, в среднем 366 и 378 мм.
Окраска. Первый и второй пуховой наряды: на спинной стороне буровато-серые,
на темени беловатый хохол.Брюшная сторона беловата;у глаза,на горле и по сторонам щек,
шеи, на зобе голое пятно. Взрослые самцы и самки: на затылке и спине длинные украшаю-
щие перья, которые у самки развиты менее, чему самца. Окраска оперения верхней стороны
головы и шеи, полоса на боках шеи, ряд пятнышек на нижней стороне шеи, полосы на зобе
и груди и середина брюха черные; передняя часть боков головы, герло и отметины в под-
хвостье белые; задняя часть боков головы, низ и бока шеи, полосы на зобе налет на мелких
кроющих крыла рыжие; удлиненные плечевые перья и боковые части груди и брюха темно-
рыжие; спинная сторона тела, хвост, большая часть кроющих крыла, бока с подмышечными
аспидно-серые. Спинная сторона тела у молодых птиц в гнездовом наряде с широкими рыже-
ватыми краями перьев, брюшная грязно охристо-белая. Клюв желтовато-бурый, ноги темно-
вато-бурые, глаз желтый.
Восточная рыжая цапля Ardea purpurea manillensis М е у е n
Ardea purpurea var. manillensis. M e у e n. Acta Acad. Leop. Carol. XVI, 1832, Supl.,
стр. 102, Филиппины.
Синоним. Ardea purpurea ussuriana. Шульпин. Ежегодн. Зоол. муз. А. Н. XXVIII,
1927A928), стр. 399, Ханка.
Распространение. Ареал. Общий указан выше в очерке вида. На севере
гнездится в Манчжурии (Цицикар-Хдрбин) и в южн. частях Приморья. Детали
распространения у нас таковы: гнездовье найдено в устьях Лефу (Шульпин,
1936), добывалась на западном берегу озера Ханка близ с. Троицкого (Чер-
ский, 1915), где найдено гнездо. Отмечена в летнее время Шульпиным A936)
в заливе Посьет, на озере Дорицени и на весеннем пролете у озера Тальмы.
Молодая, возможно залетная птица, в Хабаровском музее, вероятно, добыта
в окрестностях города. В устьях Лефу, видимо, начала гнездиться сравнительно
недавно, так как не встречена Пржевальским.
Характер пребывания. Гнездится, отлетает с наступлением
осени. Весной появляется ранее серой цапли, но точнее сроки прилета не про-
слежены. Имеется единственное указание Шульпина A936) о наблюдении стаи
цапель в устье Лефу 21 апреля 1928 г. Семь особей, образовав угол, летели до-
вольно высоко, прямо в СЗС направлении. Пролетные отмечены также у озера
Тальмы 27 апреля 1926 г. Когда происходит осенний отлет, неизвестно. На-
блюдались на западном берегу озера Ханка 9 и 12 сентября (Черский, 1915);
в музее Московского университета есть экземпляры из южной половины При-
морья, добытые 30 сентября и 20 октября.
РЫЖАЯ ЦАПЛЯ 411
Биотоп. Гнездовый — безлесные низменности, занятые болотами и
озерами с обширными пространствами тростника, камыша и лишь местами сме-
няющиеся тальниковыми порослями. В такой обстановке среди затопленных
водой тростников и тальников Шульпин нашел этих цапель на гнездовье на
р. Лефу. Места кормежки расположены в тех же стациях и представляют собою
мелководные протоки, окна чистой воды среди мелких тростниковых болот,
грязевые и песчаные отмели. Избегает человека, гнездясь в глухих и трудно-
доступных крепях.
Численность. В устьях Лефу по Шульпину многочисленная гнездя-
щаяся птица. Общая же численность в Приморье вследствие крайне ограничен-
ной области распространения ничтожна.
Экология. Размножение. Гнездится самостоятельными колониями
или присоединяясь к гнездовым поселениям серых цапель, иногда — отдель-
ными парами. Колонии найдены Шульпиным A936) в устье Лефу, одиночное
гнездо — на озерке на западном берегу Ханки, где этот вид вообще редок. В са-
мостоятельных поселениях рыжих цапель число гнездящихся пар невелико,
обычно 4—6 гнезд. В одних случаях гнезда устраиваются на высоте 1,5—2 м
от поверхности воды и укрепляются на толстых не качающихся при ветре
ветвях спутанной лозы. Для постройки основания гнезда птицы используют
сухие прутья тала и выстилают лоток тростником. Все сооружение бывает
мало, рыхло, боковые стенки его просвечивают насквозь. В тростниках гнезда
строятся еще ниже. Одно из осмотренных Шульпиным гнезд помещалось на
40 см над водой. Для постройки используются стебли тростника, складываемые
птицами в плоские кучи на тростниковый залом, торчащий из воды. Лоток
в таких гнездах бывает едва заметен, поперечный диаметр 46 см, высота или
толщина его 25,5 см. Полные кладки (по Шульпину, 1936) нормально состоят
из 5 яиц, иногда яиц бывает меньше (например 4)*. Число птенцов видимо не
соответствует первоначальному числу яиц. Шульпин не находил в гнездах
более трех птенцов. В гнездах с двумя птенцами иногда встречался болтун.
Яйца восточной рыжей цапли имеют бледноголубую окраску, но более насы-
щенную, чем яйца серой цапли.
Размеры 19 яиц, собранных Шульпиным: 54,7—60,5 мм х 39,0—43,4 мм,
в среднем 56,66 х 41,5 мм. Размеры яиц индийских птиц A00) 50—66,3х
X 38,1—46,4, в среднем 54,6 х 39,7 мм (Бекер, 1929).
К размножению не все птицы приступают одновременно. В 1927 г.
24 мая гнездование только начиналось (в большинстве гнезд по 1 яйцу, в одном—
4 ненасиженных яйца). В 1928г., вероятно 21 мая, Шульпин уже взял несколько
законченных кладок по 5 яиц**. Насиживание производится после снесения пер-
вого яйца. В каждой законченной кладке, найденной Шульпиным, часть яиц (по-
следние в кладке) оказывались свежими, остальные от 1 до 3 с кровяными
сосудами и хорошо видными эмбрионами. При осмотре колонии 5—в августа
отмечены птенцы разнообразного возраста, но все уже оперившиеся. Самые
маленькие достигли величины амурской кваквы. Часть птенцов выбралась из
гнезд и сидела поблизости на ветвях лозы. Первые летные молодые отмечены
18 июня, массовый подъем молодняка на крыло происходит несколько позднее,
и в частности, позднее, чем серой цапли.
Птенцы рыжей цапли при приближении человека к гнезду вытягивают
кверху шеи и головы и застывают в таком состоянии, то-есть ведут себя так, как
птенцы выпи и волчков. В самый же критический момент они пытаются спастись
бегством. Более полных данных по биологии размножения восточной рыжей
цапли мы не имеем.
* В Индии кладка из 3—5 яиц.
¦¦В Индии рыжая цапля размножается главным образом между июлем и сентябрем,
на Цейлоне между январем и мартом.
412
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Линька. Не изучена.
Питание. Согласно Шульпину A936) основную пищу взрослых особей,
повидимому, составляют мелкие бычки. Молодые вскармливаются разнообраз-
ной рыбой, которая доставляется стариками в большом количестве. При
осмотре гнезд были найдены мелкие сомики, достигающие иногда 16—17 см
длиной, мелкие сазанчики, верхоглядки, косатки и др.
Описание. Размерыистроение. Вес двух самцов, добытых между 7 сентября
и 20 октября 1940 г., 1310, 1358 г. У двух самцов в Зоологическом музее Московского
университета длина крыла по 370 мм. Размеры, по Шульпину A927): крыло самцов
370—401, самок 360—388; клюв самцов 121—137,3, самок 117,6—119,5; цевка самцов
128—142, самок 125,3—129 мм. Окраска. Задняя часть спины и подхвостье в отличие
от западной формы аспидно-черные. Черный цвет на брюхе более развит.
РОД БЕЛЫЕ ЦАПЛИ EGRETTA FORSTER, 1817
Tun. Ardea Garzetta
9. Большая белая цапля Egretta alba L.
Ardea alba Linnaeus. Syst. Nat., изд. X, 1758, стр. 144, Швеция.
Русское название. Белая цапля — народное название, применяемое и на дру-
гих языках народов Союза (ак-кутан у казахов, ак-хокор у туркмен, ак-вах у азербайджан-
цев).
Распространение. Ареал. Юго-вост. Европа; Африка к югу от Са-
хары (от Сенегала и Судана до Капской Земли), Мадагаскар; Передняя,
Средняя и южная Азия — на север до Аральского моря, Зайсана, сев.-зап.
Карта 74. Распространение большой белой цапли Egretta alba
7. Б. a. alba, 2. Е. a. modesta, 3. Е. a. maoriana, 4. Е. a. melanorhynchos, 5. Е. a. egretta;
а — граница гнездовий, б — не вполне выясненная граница гнездовий, г — область зимовок.
Монголии, южн. Забайкалья и Амура; Индо-австралийский архипелаг, Австра-
лия и Новая Зеландия; Америка от южных штатов до Патагонии, Антиль-
ские о-ва.
Характер пребывания. У северной границы ареала пере-
летная, на большей части гнездового ареала — оседлая птица.
Биотоп. Как у рыжей цапли.
БОЛЬШАЯ БЕЛАЯ ЦАПЛЯ 413
Подвиды и варьирующие признаки. Морфологические различия — в размерах и опе-
рении, окраске неоперенных частей (клюв, ноги), биологические— в характере пребывания,
деталях периодических явлений, трофических связях.
Описаны подвиды: Е. a. alba Linn., 1758 — в Европе, Передней, Средней Азии, на
восток до Сев. Китая и сев. Японии, на юг до Афганистана и Кашгарии; границы в Сев.
Китае остаются неясными; зимовки в Средней и южн. Азии, в южн. Европе, в сев. Африке;
Е. a. modesta Gray, 1831 — в Азии к югу от предыдущей, от средней Японии, Кореи,
Китая, до Австралии; Е. a. maoriana IredaleetMathews, 1914 — на южн. острове
Новой Зеландии; Е. a. melanorhynehos Wagler, 1827— Африка и Мадагаскар; ?. а.
egretta G m e 1 i n, 1789 — американская часть ареала.
Северная большая белая цапля Egretta alba alba L.
Синоним. Ardea egrettoides. S. G. Gmelin. Reise durch Russia nd, II, 1774,
стр. 193, «Астрахань и Персия».
Распространение. Ареал. Гнездится в Европе —в Австрии (в частности
на Нейзидлерском озере), Венгрии, Югославии, Албании, Болгарии, Румынии,
в Советском Союзе. В 1863 г. гнездилась у Глогау в Германии. В Азии —в
Малой Азии, Месопотамии, Иране, Афганистане, в Кашгарии (Лоб-нор), в Сев.
Китае и сев. Японии, в СССР от Средней Азии до Приморья. Залеты отмечены
на Пиренейский п-ов (Испания), во Францию, Бельгию, Голландию, Англию,
Германию, Польшу, Чехословакию, Швецию, на Канарские и Азорские о-ва,
на Мальту и Майорку, в разные местности СССР.
В пределах нашей страны гнездится в Бессарабии, в низовьях Днепра,
Днестра й Дона. Вверх по Днепру в небольшом числе гнездится в окрестностях
ст- Янчикрак под Запорожьем (Спангенберг). Далее к северу гнездится в райо-
нах Днепропетровском, Уманском, Валковском.
Гнездится в подходящих местах Закавказья: в низменных частях Арме-
нии (Ляйстер и Соснин, 1940), в Джавахетии, где колония была найдена близ
с. Гореловки на высоте около 1.800 м над уровнем моря (Чхиквишвили, 1934).
Видимо не гнездится в восточных частях Азербайджана; под Ленкоранью
во всяком случае этой цапли нет. На Сев. Кавказе гнездится в плавнях Кубани
(Птушенко), Терека и идет отсюда по западному побережью Каспия далеко
к северу. Весьма многочисленна в дельте Волги, изредка гнездится на Сарпин-
ских озерах и в самых низовых частях Урала. Ранее гнездилась на нижней
Эмбе до Кок-джиды, но гнездится ли там в настоящее время — неизвестно.
Гнездится на нижнем течении Иргиза и Тургая, найдена в бывшем Атбасарском
уезде, у Семипалатинска, Каменогорска и Павлодара (Селевин, 1928). В обла-
сти нижней и средней Сыр-Дарьи на гнездовье распространена от устьев реки
и вверх до Чиназа, в низовьях Сары-су, Чу и на озерах Тели-куль (Спанген-
берг и Фейгин, 1936). В Семиречье встречается на гнездовье в устьях рек:
Лепсы, Аксы, Каратале и Или, на озерах Ала-куль, Сасык-куль (Антонов,
1927), а также под Джаркентом (Кореев и Зарудный, 1906), в Зайсанской кот-
ловине (Хахлов, 1928). В верхнем течении Сыр-Дарьи гнездится у Ферганы.
В пределах Туркмении ныне с достоверностью гнездится в области Ташаузских
озер и в долине Аму-Дарьи. Вверх по Аму-Дарье заходит в Таджикистан,
но достоверно гнездится здесь только в урочище Джили-куль на Вахше
(Иванов, 1940).
Далее на восток, в Сибири, белая цапля гнездится в южн. Забайкалье.
В Уссурийском крае распространена неравномерно и более или менее обычна
только в южной части страны. В конце прошлого столетия была довольно много-
численна в устье реки Лефу, где продолжает гнездиться и в наши дни. Видимо,
изредка гнездится на нижнем течении р. Суйфуна, в небольшом числе на озере
Дорицени в районе Посьета (Шульпин, 1936). По старым авторам в прошлом
была довольно многочисленна на озере Ханка и редка на побережье Японского
моря. Раньше доходила также на гнездовье до устьев Амура. В общем северная
граница гнездования белой цапли в Азии проходит около 48° 5' с. ш. на мериди-
414 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
ане Аральского моря, на озере Зайсан и на Кара-Иртыше, на озере Хара-усу
в с.-з. Монголии, в южн. Забайкалье и в бассейне Амура. Залеты отмечались
в Крыму, в Прибалтике, в районе Минска, Харькова, Воронежа, Камышина,
в Наурзумском заповеднике, в южной и восточной частях Барабы и в окрест-
ностях Томска.
Зимовки находятся в сев. Африке, в Месопотамии, й Белуджистане,
в сев. Индии, в Китае, в Кашгарии и Алашане. У нас перезимовывает близ
южных границ.
Птица, окольцованная в устье Днепра (Рыбачье, Николаевской обл.),
добыта в устье Дуная, в Румынии. Птицы, окольцованные в Венгрии, добыты
в Прибалтике и на Украине.
Характер пребывания. На большей части своего ареала в СССР
эта цапля гнездящаяся, перелетная птица. Лишь местами — на юге Средней
Азии и на Кавказе она проводит зиму, но в очень небольшом количестве и не-
регулярно.
В небольшом количестве белые цапли зимуют на реках, впадающих в Чер-
ное море со стороны Кавказа (Вильконский, 1897). В Армении —это редкая
зимующая птица низменных частей страны (Ляйстер и Соснин, 1942). Редкие
особи, но довольно регулярно перезимовывают на Куре в низменном Азербай-
джане. В некоторые годы безусловно зимует в районе Баку. В значительно боль-
шем числе и нормально птицы зимуют в районе Ленкорани между устьем Куры
и Иранской границей. Однако, и в последнем месте численность зимующих
больших белых цапель подвержена чрезвычайно сильным колебаниям. Послед-
нее, по мнению Исакова и Воробьева A940), связано главным образом с услови-
ями зимнего питания. В зиму 1930—31 г. здесь было отмечено только 2 экзем-
пляра цапель, в зиму 1931—32 г. они изобиловали в этой местности, встречаясь
стайками от 20 до 36 экземпляров, в зиму 1933—34 г. отмечались в течение всей
зимы, но лишь одиночными особями. В крайне суровую для этих мест зиму
1924—25 г., сопровождавшуюся метелями и замерзанием большинства водо-
емов, несколько цапель благополучно пережили неблагоприятные условия
зимовки между Ленкоранью и Астарой. В январе 1950 г. при температуре
22° зимующие экземпляры наблюдались автором под Кирдамиром. Указания
Беме A926) о зимних залетах больших белых цапель в январе и начале фев-
раля в район Дзауджикау безусловно следует отнести к случайным встречам
заблудившихся на осеннем пролете особям. В пределах Средней Азии белая
цапля в ограниченном числе перезимовывает на Атреке и в предгорной полосе
близ Ашхабада, где добывалась в декабре. В Таджикистане зимует в долине
верхней Аму-Дарьи, в низовьях р. Вахш была многочисленна в зиму 1939—
1940, и сравнительно редка в зиму 1940—1941 г. (Иванов, 1945). Зимует
также в долинах Чирчика, Ангрена (Зарудный, 1915) и в пределах Семиречья.
Большая белая цапля принадлежит к раннеприлетным птицам; однако
сроки прилета плохо выяснены и нуждаются в уточнении. На северном берегу
Черного моря нормально птицы появляются во второй половине марта, однако
до южной окраины бывшей Киевской губ. достигают лишь в конце апреля или
в первой трети следующего месяца (Мензбир, 1918). Под Ленкоранью, по моим
наблюдениям, численность цапель возрастает с конца февраля; интенсивный
пролет идет в начале марта и заканчивается к концу этого месяца. Отлет зимую-
щих особей с Черноморского побережья Кавказа наблюдается в половине апре-
ля (Вильковский, 1897). В дельте Волги, согласно Воробьеву A936) и Дуби-
ниным A940), появление первых птиц происходит между 10 и 20 марта. Сведе-
ния о прилёте белых цапель на восток от Волги несколько противоречивы. Так
по Северцову в 1858 г. белые цапли прилетели на Иргиз уже 27 марта, а в 1898 г.
на нижнем течении Иргиза первая особь была отмечена Сушкиным A908)
только 26 апреля. В 1861 г. в устье Урала первые прилетные наблюдались
Карелиным 20 апреля. Более поздний прилет в устье Урала объясняется, по
БОЛЬШАЯ БЕЛАЯ ЦАПЛЯ 415
мнению Мензбира A918), тем, что цапли прилетают сюда с низовьев Сыр-Дарьи.
В низовья Сыр-Дарьи (между ст. Тюмень-арык и дельтой) белая цапля при-
летает когда озера бывают покрыты толстым льдом и лишь местами появляются
полыньи. В 1937 г. первая особь отмечена здесь 15 марта, в 1928 г. —в послед-
них числах марта. Ночная температура в это время нередко падала до 10—12°.
К 1 апреля на всем нижнем течении реки белые цапли уже довольно обычны.
В Гиссарской долине пролетные были отмечены 15 апреля (Иванов, 1940).
Странным образом в вост. Иране Зарудный A903) наблюдал пролетных цапель
между 20 и 23 апреля — очень поздно. В юго-восточной части Каспия интен-
сивный пролет наблюдается в марте. На Мургабе, Теджене и на западном Узбое
весенний пролет также происходит в марте, начинаясь с первых чисел этого
месяца и продолжаясь до первых чисел апреля (Дементьев, 1950).
В районе Джаркента большая белая цапля наблюдалась уже с 13 марта,
у Семипалатинска с 25 и 30 марта, а на Иртыше, несколько севернее Усть-
каменогорска, с 10 марта (Селевин, 1928). Также и в Уссурийском крае,
где весна наступает сравнительно поздно, большая белая цапля принадлежит
к ранне прилетным видам. В истоках р. Сунгачи в 1868 г. первые птицы были от-
мечены 24 марта и затем с 31 марта и до половины апреля наблюдался наиболее
сильный пролет одиночек и небольших групп. Во второй половине апреля про-
лет сильно сократился и почти прекратился к концу этого месяца. В 1869 г.
там же до 1 апреля пролетных цапель было очень мало. Птицы летели одиночка-
ми, парами и маленькими стайками в 5—7 особей. В период пролета разливов
еще не было, лежал снег, то и дело были метели (Пржевальский, 1870).
Сроки осеннего отлета известны еще менее сроков весеннего появления.
Из более северных частей своего ареала цапли продвигаются к югу, как только
молодые птицы вполне научатся летать. На юге, в наиболее кормных местах,
они задерживаются до поздней осени. В низовьях Днепра между г. Запорожьем
и Каховкой хорошо выраженный отлет отмечен во второй половине сентября
и уже в октябре белые цапли здесь большая редкость. В дельте Волги осенний
отлет начинается с 25 сентября и тянется весь октябрь, запоздавшие особи на-
блюдались до 8 ноября. На разливах Терека, в Аграханском заливе и по
озерам в районе Кизляра стаи пролетных птиц отмечались в течение всего но-
ября и в последний раз 6 декабря при значительных ночных заморозках. За-
держки с отлетом отмечались и на Сев. Кавказе в районе Дзауджикау, где
в январе и даже 3 февраля наблюдались одиночки (Беме, 1926). На о-ве Сара
под Ленкоранью пролет отмечен в первой половине ноября (Тугаринов и
Козлова-Пушкарева, 1938). На Черноморском побережье Кавказа белые
цапли появляются уже в конце августа и остаются здесь до половины
апреля.
До похолодания держатся белые цапли и в Зап. Сибири. Под Семипала-
тинском они отмечались до начала последней трети сентября, а в окрестностях
Томска заблудившаяся птица была добыта в начале ноября. В устьях Урала
нормальный пролет видимо в конце сентября, но редкие особи отмечались
у Гурьева до конца октября. В конце сентября начинают отлетать также с ни-
зовьев Сыр-Дарьи и к двадцатым числам октября становятся здесь положитель-
но редкими, одиночки задерживаются на текучей воде до первых чисел ноября,
а на среднем течении этой реки до декабря (Спангенберг). 29 сентября птицы на-
блюдались на р. Чу (Гладков и Гринберг, 1932). В Семиречье у Джаркента наи-
более поздние особи были отмечены 26 ноября (Кореев и Зарудный, 1906).
Пролет в юго-восточном углу Каспия происходит в течение всего октября и
ноября. В это время цапли держатся большими стаями. Напротив, в более позд-
ние сроки (зимой) они встречаются здесь в заметно меньшем количестве (Иса-
ков и Воробьев, 1940). Быть может к запоздавшим пролетным птицам следует
отнести экземпляр, добытый под Ашхабадом 10 декабря 1888 г. В Уссурийском
крае — наиболее поздние особи наблюдались 6 и 9 октября на р. Янчихе
416 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
(Пржевальский, 1870). Шульпин A936) упоминает об экземплярах, добытых 25
октября у с. Троицкого на п-ове Янковского.
Биотоп. Сочетание открытых плесов, озер, ериков, обладающих
слабым течением, подсыхающих мелководных водоемов с обширными тро-
стниковыми и камышовыми зарослями. В период размножения явно предпочи-
тает безлесные или бедные древесной растительностью низменные площади.
Лишь местами, например в Джавахетии, гнездится в горном ландшафте на вы-
соте 1800 м над уровнем моря (Чхиквишвили, 1930). Кормовые биотопы более
разнообразны. Часть особей добывают себе пищу в гнездовом биотопе, но мно-
гие вылетают за его пределы. Кормящихся птиц удается встречать на залитых
водой полях риса, на сырых, болотистых и сухих лугах (Закавказье), на от-
крытых морских мелководьях (дельта Волги), по берегам открытых речушек,
подсыхающих озер и на затопленных кустарниках тамариска (Сыр-Дарья), а
изредка по сырым понижениям в степной обстановке (низовье Урала, Или).
Во многих случаях места кормежек бывают расположены до 10—15 км от мест
гнездовья, обычно значительно ближе. Почти повсеместно эта цапля избегает
гнездиться вблизи жилья человека, избирая для размножения самые глу-
хие и труднодоступные уголки Камышевых крепей. Напротив, в поисках пищи
цапли посещают культурный ландшафт, но ведут себя здесь очень осторожно.
Численность. В дореволюционное время численность птиц в нашей
стране, вследствие неумеренного промысла, была доведена до минимума.
В дельте Волги и в Казахстане белая цапля была почти истреблена. В наше
время численность птиц быстро восстанавливается. Последнее целиком связано
с полным запретом охоты на белых цапель и организацией специальных заповед-
ников (Астраханский заповедник). Однако и в настоящее время белая цапля во
многих частях Союза не может быть названа не только многочисленной, но и
обыкновенной птицей. В низовьях рек, впадающих в Черное море, она редка и
еще реже становится по мере удаления от морского побережья. Несколько чаще
вид этот встречается в устьях Дона и еще чаще в вост. Закавказье, по рекам
Сев. Кавказа и в районе Кизляра. В дельте Волги большая белая цапля —
обыкновенный гнездящийся вид, и численность ее продолжает возрастать. В
1934 г. в Астраханском заповеднике насчитывалось 180 молодых и взрослых
особей. В 1935 г. численность возросла до 1070 птиц. В Казахстане восста-
новление численности птиц происходит значительно медленнее. В наиболее
глухих уголках (Сыр-Дарья, Тели-куль, Балхаш) она стала более или менее
обыкновенна, но местами все же продолжает быть немногочисленной птицей.
Умеренная численность птиц отмечается также в пределах Туркмении. Еще
реже цапли встречаются в более восточных частях нашей страны и в частности
в Приморье. Общая численность больших белых цапель в Союзе и до сего вре-
мени должна быть признана ограниченной.
Экология. Размножение. Там, где численность этих цапель значи-
тельна, они охотно гнездятся колониями. Число гнезд в колонии колеблется
между 3—8, реже их бывает 10—30 и еще реже (видимо только в Астраханском
заповеднике) колонии достигают больших размеров (от 30 до 180 гнезд на 1 га,
Ромашова, 1940). В районах, где белая цапля редка, удается встречать отдельно
гнездящиеся пары, иногда 2—3 гнезда, устроенных поблизости друг от друга.
В немногих местах большие белые цапли присоединяются к колониям других
голенастых птиц. В Астраханском заповеднике гнездится самостоятельными
колониями, а также в колониях серых и рыжих цапель, устраивает гнезда среди
крупных поселений мелких цапель и караваек. На Сыр-Дарье изредка присоеди-
няется к маленьким поселениям колпиков и к отдельным гнездам серых пели-
канов. В данном случае, видимо, птицы руководятся не общественным инстинк-
том, а чрезвычайно благоприятными условиями гнездования. В подавляющем
большинстве случаев гнезда устраивают на заломе тростника и камыша. Однако,
еще Мензбир A918) упоминает о гнездовании большой белой цапли и на деревь-
БОЛЬШАЯ БЕЛАЯ ЦАПЛЯ 417
ях, что подтверждается и западноевропейскими авторами. В Астраханском за-
поведнике по мнению Воробьева этот вид, несмотря на обилие рощ и кустарни-
ковых зарослей, гнездится только в тростниковых крепях. Однако, Рома-
шова A940), работавшая в Астраханском заповеднике несколько позже,
когда птиц стало больше, отметила, что в выборе места для гнезда большая
белая цапля гораздо менее разборчива, чем серая. Ее гнездо можно найти и
на высоких, и на низких ветлах, и на кустарниковой иве, и на камыше, при-
чем гнезда расположены не густо по одному на дереве, на десятки метров одно
от другого. В весну 1949 г. на одном дереве мне удавалось находить до 4—5 гнезд.
На Сыр-Дарье гнездование на деревьях — исключительно редкое явление.
Одно гнездо было найдено на низкорослом кустике джиды, растущем на клочке
сухой почвы среди обширных тростниковых и Камышевых зарослей. Постройка
гнезд на затопленной лозе известна также в Уссурийском крае на реке Лефу.
Высота, на которой строится гнездо птицы, бывает различна. В тростниковых
зарослях гнездо помещается то близ самой воды, прилегая к последней осно-
ванием, то до 4,5 м от поверхности воды на прошлогодних заломленных
тростниковых стеблях. При гнездовании в тростниковых и Камышевых крепях
для постройки гнезд используются сухие стебли и листья этих растений. Из
этого же материала птицы строят гнезда при гнездовании на кустарниковой
иве. Для гнезд, сооруженных на ветлах и джиде, используются сухие прутья
и колючие ветви. Гнезда большой белой цапли имеют форму перевернутого
конуса с просвечивающими стенками. Размеры гнезд сильно варьируют,
в общем сходны с гнездами серой цапли.
В сооружении гнезда участвуют обе птицы гнездовой пары. Доставка гнез-
дового материала в значительной степени лежит на самце, а его размещение и
утаптывание — на самке. Количество яиц в кладке 3—4, редко 5. Форма яиц
изменчива. Иногда яйца бывают правильной яйцевидной формы, иногда вздуты
или вытянуты с одинаково округлыми полюсами. Окраска скорлупы, как у серой
цапли, —зеленовато-голубая. В период яйцекладки и в начале насиживания
скорлупа лишена блеска, но к концу насиживания становится более гладкой
и блестящей.
Размер яиц из дельты Волги: 59,0—64,0 х 41,0—43,0 мм, в среднем
61,3 х 42,2 мм.
Насиживание начинается с момента отложения первого яйца, но прочно
садится птица ближе к окончанию кладки (примерно, с третьего яйца). Проме-
жутки между откладкой отдельных яиц и срок насиживания у нас точно не
установлены; по венгерским данным, насиживают обе старые птицы в течение
25—26 дней. К размножению белые цапли приступают сравнительно рано, но
неодновременно. В колониях в одни и те же даты находят маленьких и вполне
выросших птенцов. Растянутость срока начала гнездовья приблизительно
можно определить в 25 дней. В Астраханском заповеднике начало откладки яиц
происходит между 15 и 25 апреля, вылупление из яиц наиболее ранних птен-
цов между 1—10 июня, а массовое появление птенцов между 15 и 25 июня. Наи-
более ранний вылет птенцов из гнезд совпадает с 5—15 июля, а массовый вылет
с 20—30 июля (Дубинины, 1940). На Сыр-Дарье в окрестностях станции Соло-
Тюбе уже 5 мая 1926 г. мною обнаружены три вполне оперившихся, крупных
птенца, которые спустя несколько дней приобрели способность к полету.
В Туркмении откладка яиц происходит в апреле. Хорошо летающие молодые
отмечены Зарудным A896) в Мары 23 июня 1886 г. В Уссурийском крае (в устье
р. Лефу) 13 июня, судя по поведению взрослых цапель, в гнездах колоний
были уже птенцы, а 1 июля при посещении колоний многие птенцы летали,
другие (нелетные) оставались на лозе возле гнезд (Пржева;аскнй, 1870).
Некоторые особи приступают к размножению крайне рано. Можно думать,
что при потере первой кладки белые цапли не приступают к кладке повторно,
во всяком случае данные о нахождении гнезд с яйцами в июле отсутствуют.
27 Птжць* СССР, т. II
418
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
О том же свидетельствует появление в разгар размножения бродячих стаек
взрослых особей, повидимому потерявших кладки.
Как только молодые особи приобретут способность к полету, в возрасте
примерно б недель, они образуют семейные стайки. Позднее семьи соединяются
в более крупные стаи, которые до отлета придерживаются болот и водоемов,
отличающихся наибольшей кормностью. Кочевая жизнь выводков в Европей-
ской части Союза начинается лишь в августе; Северцовым в устье Урала стаи
белых цапель наблюдались с 10 августа и в более поздние сроки. Стаи, состоя-
Рис. 81. Большая белая цапля
щие из взрослых и молодых птиц, отмечались на Сыр-Дарье уже с конца июля
и много чаще в июле. В Уссурийском крае (Гнилой Угол) Шульпин A936) на-
блюдал кормившуюся стаю белых цапель, до 40—50 особей, 12 августа. В дельте
Волги в половине августа белые цапли отмечались уже стаями более, чем в
200 экземпляров (Воробьев, 1936).
Линька. Как у других крупных видов цапель. Молодые птицы сменяют
мелкое оперение в результате частичной предбрачной линьки между августом
и ноябрем первого календарного года жизни. В дальнейшем полная годовая
линька между августом и ноябрем. Имеется повидимому и вторая линька, будто
бы полная (Уайзерби, 1939); во всяком случае, в зимнем наряде украшающие
перья (плечевые) менее развиты.
Питание. Как у рыжей цапли, но добыча однако крупней, рыба, водя-
ные насекомые и их личинки, кузнечики и саранча, мелкие млекопитающие,
иногда птенцы воробьиных птиц. На Сыр-Дарье (окрестности ст. Соло-Тюбе),
по нашим наблюдениям, в питании птенцов вероятно большое значение имеют
БОЛЬШАЯ БЕЛАЯ ЦАПЛЯ 419
насекомые. Птенцы, осмотренные 5 мая 1926 г., стараясь при приближении
человека освободиться от съеденной пищи, выбрасывали целые комки еще не-
переварившихся стрекоз. В том же районе в желудке птицы, добытой в конце
июля, оказалось много саранчи и остатки какой-то рыбы. У экземпляра, добы-
того в январе 1925 г., несколько южнее Ленкорани, в желудке найдены форель
и 4 полупереварившихся полевки. У добытой под Ленкоранью 1 марта 1931 г.
найдена общественная полевка (Исаков и Воробьев, 1940). Форель и 2 экземп-
ляра водяной куторы найдены в желудке птицы, добытой 3 февраля под Дзау-
джикау (Беме, 1926), и несколько полевок у цапли, убитой в ноябре под Кизля-
ром (Спангенберг). Питание млекопитающими и наземными насекомыми позво-
ляет белой цапле держаться вдали от водоемов и долго задерживаться у нас
осенью.
Полевые признаки. Снежно-белая окраска и крупные размеры позволяют
легко отличить этот вид от других цапель. Однако, издали, ее легко смешать
с малой белой цаплей. Весной и летом встречаются одиночками, осенью чаще
стайками, штук 8—12, изредка крупными стаями или соединенными обществами
с серой цаплей. При взлете из камыша птица, тяжело взмахивая крыльями, круто
поднимается вверх, причем ее длинные ноги болтаются, как плети. Поднявшись
на известную высоту, птица летит плавным величественным полетом, вытянув
назад длинные ноги, глубоко втянув шею и медленно взмахивая широкими
крыльями. При далеких перелетах цапли нередко образуют в воздухе косую
линию или угол. Исключительно осторожная и пугливая птица, что вероятно
вызвано постоянным преследованием со стороны человека. Для кормежки пред-
почитает опускаться на открытые участки, где издали может увидеть приближаю-
щегося врага. Потревоженные птицы далеко улетают от беспокойного места.
Во время полета крайне внимательна и, неожиданно увидав под собой охотника,
взмывает вверх. На деревья садится относительно редко и много реже, чем
серая цапля. Крайне молчалива. Голос — грубое трещание, Чаще его
удается слышать во время полета.
Описание. Размеры и строение.В брачном наряде на затылке небольшой хо-
хол; на нижней части шеи удлиненные рассученные перья, свисающие «гриьой», на плечах
два пучка длинных рассученных перьев, заходящих даже за вершину хвоста («эгретки»).
Формула крыла (не считая рудиментарного первого махового): 2>3>4>1>5>6. Вырезки на
внешних опахалах 2-го — 4-го и на внутренних опахалах 1-го — 3-го первостепенных махо-
вых. Цевка значительно длиннее среднего пальца (длина первой около 170—215, второго
около 110—125 мм). Длина крыла самцов и самок A4) 412—465, в среднем 437,79 мм: в том
числе у самцов 451 мм, у самок 429 мм.
Окраска. Первый и второй пуховой наряды — белые, на темени хохолок из удли-
ненного пуха. Взрослые самцы и самки в зимнем и брачном нарядах белые. Клюв в брачном
наряде черный с желтым основанием, в зимнем наряде желтый;голая уздечка и кольцо у глаз
зеленые или зеленовато-желтые; ноги черные, пальцы черные с зеленоватым оттенком;
радужина желтая. У молодых клюв желтый.
Южная белая цапля Egretta alba modesta Gray
Ardea modesta. Gray. Zool. Miscell., 1831, стр. 19, Индия
Распространение. Ареал. Индия, Цейлон, ю.-в. Азия, Индоавстра-
лийский архипелаг, Австралия; на север до Гималаев, низовьев Янцзе (быть
может Чжили), средней Японии. В последней гнездится на Хондо, Кюсю, Ама-
ми-осима.
В СССР неоднократно добывалась в южн. Приморье. В конце прошлого века
три особи были добыты: экземпляр без точных дат в 1875 г. у залива Абрека,
самка б августа там же и самка 15 апреля 1885.
В последние годы в южн. Приморье был добыт ряд южных белых цапель,
9 мая 1945 г. самка найдена в устье р. Тачингоуза в Судзухинском заповеднике
Л. О. Белопольским. Семь птиц были добыты между 17 августа и 19 сентября
1946 г. в Хасанском районе Н. Н. Горчаковской, самец добыт 12 апреля 1947 г,
420
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
в Судзухинском заповеднике Бромлеем. Птицы ежедневно наблюдались
маленькими группами и большими стаями по берегам внутренних водоемов и
на взморье. Присутствие в августе и мае небольших групп птиц, по всем ве-
роятиям выводков, позволяет предположить, что птица эта возможно и гнез-
дится у залива Посьета.
Характер пребывания. На самом севере ареала в Японии соб-
ственно, в Сев. Китае — перелетная, в других местах оседлая птица. На
пролете отмечена на о-вах Риу-Киу, в Китае — в Гуандуне и Фуцзяне, на
зимовках в Гуандуне и в низовьях Яньцзе (Латуш, 1934).
Карта 75. Распространение средней белой цапли Egretta intermedia
а — граница гнездовий, б — не вполне выясненная граница гнездовий.
Биотоп. Как и номинальный подвид, южная белая цапля придержи-
вается влажных, богатых прибрежной растительностью (камыш, кустарники)
местностей.
Численность. В Китае белая цапля в конце прошлого — начале
текущего столетия была почти истреблена. В последней четверти XIX века коло-
нии этой цапли размещались на больших деревьях у дворцов, ямыней, кладбищ
ц даже охранялись. В тридцатых годах текущего столетия—по тем или иным
причинам птиц здесь повидимому не стало и вопрос о распространении их в насто-
ящее время в Китае остается открытым. В пределах Союза редка и встречена,
как указано выше, только на крайнем юге Приморья. В Индии цапля гнездит-
ся небольшими колониями и повидимому не очень многочисленна (Бекер, 1929).
Экология* Размножение. По данным Латуша (L934) и Бекера A929),
белые цапли гнездятся колониями, иногда вместе с другими цаплями и бакла»
СРЕДНЯЯ БЕЛАЯ ЦАПЛЯ
421
нами» в камышах или на деревьях. В сев. Индии период размножения падает на
время между июлем и сентябрем, в южн. Индии и на Цейлоне между ноябрем
и мартом. Яйца по размерам меньше, чем у номинальной формы:D0L8,5 —
60,5 х 36,8—40,6 в среднем 54,0 х 38,6 мм.
Полевые признаки. В природе различается от номинального подвида
по меньшим размерам.
Описание. От северной большой белой цапли отличается меньшими размерами. Вес
(8) 792—1108 г, в среднем 908,3 г. Крыло самцов и самок (8) 343—385, в среднем 357,3 мм,
цевка 13,8—15,8, в среднем 14,98 мм, клюв 92—109, в среднем 96 мм. Все измерения отно-
сятся к особям, добытым в Приморье.
10. Средняя белая цапля Egretta intermedia Wagler
Ardea intermedia. Wagler. Isis, 1829, столб. 659, Ява.
Распространение. Ареал. Африка—тропические области на северо-
востоке, востоке и юге от Кордофана и Египетского Судана до Капской Земли;
южн. Азия от Индии и Цейлона на западе до Малайи, Индо-Китая, Южн. и
Центр. Китая, Японии (Хоккайдо, Хондо), Филиппинских и Зондских о-вов;
е-ва Буру, Церам, Австралия; два раза
добыта в южн. Приморье.
Характер пребывания.
На севере ареала в Японии и местами
в Китае — перелетная, в остальных
частях ареала — оседлая птица.
Биотоп. Как у малой белой
цапли.
Подвиды и варьирующие признаки.
Морфологические различия — в размерах,
пропорциях, окраске неоперенных частей
тела. Биологические различия не выяснены.
Три подвида: Е. /. brachyrhynchus В г е h m,
1858—в африканской части ареала; Е. i. plu-
mi/era Gould, 1847 — в Австралии, на
о-вах Буру, Церам, Новой Гвинее; Е. i. inter-
media Wagler, 1829 — в азиатской части
ареала.
Азиатская средняя белая цапля Egret-
ta intermedia intermedia Wagler
Распространение. А р е а л. См.
выше. В пределах Советского Союза
добыта два раза. Одна птица в брачном
наряде встречена 9 мая 1922 г. на с-ве
Попова близ Владивостока, другая —
взрослый самец—9 мая 1944 г. добыта
Белопольским на болотистом берегу
озера Зора в Судзухинском заповеднике (обе птицы хранятся в Зоологи-
ческом музее Московского университета).
Характер пребывания. В СССР повидимому залетная птица.
Экология. Размножение. В Японии (Ян, 1942) гнездится колони-
ально, обычно с другими видами цапель. Гнезда то на группах деревьев среди
рисовых полей, то в зарослях бамбука, нередко вблизи от человеческих поселе-
ний. Яиц в кладке 3—4; они похожи на яйца большой белой цапли, но меньше
по размерам: F0) 42,6—52,8x33,1—38,6, в среднем 47,6х 35,8 мм (Индия, Бе-
кер, 1929). Период размножения в сев. Индии от июля до сентября, в Мадрасе
Рис. 82. Средняя белая цапля
422
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
и южнее в декабре — январе, на Цейлоне от ноября до марта. В Китае птенцы
вылупляются в середине июня, в июле встречаются подлетки (Латуш, 1934).
Линька. В Китае линяние начинается около начала июля, линные
птицы встречаются и в сентябре (Латуш, 1934).
Полевые признаки. От других белых цапель отличается желтой окраской
клюва, в гнездовый период также черной окраской ног.
Описание. Размеры и строение. Размеры добытых в СССР особей: крыло 307
и 316 мм; клюв 82 и 96 мм; цевка 128 и 131 мм; по Хартерту A915) крыло 290—320 мм,
ло Бекеру A929) 304—333 мм. Эгретки на спине и зобе менее развиты, хохла на затылке
нет. Общая окраска, как у большой белой цапли, но неоперенная часть голени черная. Клюв
желтый с буроватой вершиной зимой, черный в период размножения.
11. Чепура-нужда или малая белая цапля Egretta garzetta L.
Ardea Garzetta. Linnaeus. Syst. Nat., изд. XII, 1766, стр. 137, Малальберго, с.-в.
Италия.
Русское название. Чепура, чепура-нужда — народное. Малая белая цапля —
книжное, в зоологической литературе.
Распространение. Ареал. Европа — Португалия, южн. Испания, южн.
Франция (дельта Роны), сев. Италия, Венгрия, Югославия, Болгария, Ал-
бания, Румыния, южные части СССР от Днестра до Кавказа; Азия —Сирия,
Карта 76. Распространение малой белой цапли Egretta garzetta
7. Е. g. garzetta, 2. Е. g. nigripes, 3. E. g. immaculata, 4. E. g. dimorpha;
a — граница гнездовий, г — не вполне выясненная граница гнездовий.
Малая Азия, Иран, Средняя Азия и далее на восток через Китай до Япо-
нии (Хоккайдо, Хондо), на юг до Индии и Цейлона, Индо-Китая; споради-
чески в Африке — гнездятся регулярное дельте Нила и на о-вах Зеленого
Мыса, нерегулярно на озере Виктория и в Капской Земле, на Мадагаскаре,
о-ве Альдабра; Индо-Австралийский архипелаг от Зондских о-вов до Новой
Гвинеи, Австралия.
Характер пребывания. Гнездящиеся в Европе птицы зимуют
в области Средиземного моря, в сев. Африке и юго-зап. Азии; перелетны птицы
в Средней Азии. Остальные популяции оседлы.
ЧЕПУРА-НУЖДА ИЛИ МАЛАЯ БЕЛАЯ ЦАПЛЯ 423
Биотоп. Прибрежные, заросшие камышом, кустарниковой и древес-
н:й растительностью, низменные местности.
Подвиды и варьирующие признаки. Несколько подвидов, различающихся деталями
биологии, размерами и окраской. На островах: Мадагаскаре, Альдабра, Ассемпшен рас-
пространены диморфные цапли — белые и серые. В остальной части ареала чепура-нужда
мономорфная (белая).
Описаны подвиды: Е. g. immaculata Gould, 1846— Австралия и Папуасия; Е. g.
nigripes T e m m i n с k, 1840 — Филиппины, Зондские о-ва, Целебес; Е. g. dimorpha
Grant et Ma ck wort h-P гае d—Мадагаскар; Е. g. assumptionis Gruntet Mack-
wort h-P r a e d — о-ва Альдабра и Ассемпшен; Е. g. garzetta L i n п., 1766— в остальной
части ареала в Европе, Африке и Азии.
Северная чепура-нужда Egretta garzetta garzetta L.
Синоним. Ardea nivea. S.G. G me 1 i n. Reise d. Russland 1,1770, стр. 164, южн. Россия.
Распространение. Ареал. Ареал вне СССР указан выше, в очерке рас-
пространения вида.
В пределах Союза гнездится: в Бессарабии, в низовьях Днестра и Днепра,
на север до Днепропетровского и Золотоношского районов, в устье Дона
и вообще в удобных местах Приазовья, в дельте Волги, в устье Урала, подни-
маясь вверх по течению, приблизительно, до ст. Кулагинской. На Сев. Кавказе
гнездится в устьях Кубани, в дельте Терека. В Закавказье отмечена на проле-
тах и летом в районе Батуми (вероятно гнездится), в Армении гнездится только
в Ереванской равнине, но также на высокогорном озере Севан (Ляйстер и Сос-
нин, 1942), широко распространена в вост. Закавказье, сосредоточиваясь основ-
ной массой в долине средней и нижней Куры и к югу от устья этой реки до Иран-
ской границы. Гнездится по нижнему и среднему течению Аму-Дарьи вверх до
Чарджоу, в Мервском оазисе, в низовьях Теджена (Дементьев, 1950); иногда,
но не регулярно в Кзылординском районе на Сыр-Дарье (Спангенберг, 1936).
Бывает на пролетах, а в некоторые годы залетает и летом на Крымский п-ов.
Во время пролетов добывалась под Дзауджикау (Беме, 1926). В гнездовый
период залетает в район Киева. Весной и осенью, а крайне редко и летом, за-
летает в Харьковскую обл., еще реже в Воронежскую, Полтавскую, Чернигов-
скую и, повидимому, случайно в Прибалтику, в весеннее время в район Пензы
и в район Чкалова, Кузнецка. Вне СССР залеты в Англию, Бельгию, Голлан-
дию, Францию, Германию, Польшу, на Азорские и Канарские о-ва и на
Мадейру (Латуш, 1934). Зимовки в сев. Африке, Закавказье, южн. Туркмении,
Месопотамии, Белуджистане и Южн. Китае.
Птица, окольцованная 10 августа 1935 в устье Днепра в Черноморском
заповеднике, добыта 20 сентября 1935 у Коринфа в Греции.
Характер пребывания. В СССР чепура-нужда, гнездящаяся
перелетная птица. Для северных районов очерченного распространения только
залетный, а в более южных частях местами только пролетный вид. В ограни-
ченном числе и далеко не ежегодно зимует в вост. Закавказье. Одиночные особи
наблюдались на лимане в Дивичах (Азербайджанская ССР) в феврале (ст. ст.
Станчинский, 1914). Нерегулярные зимовки под Ленкоранью отмечаются мно-
гими авторами, хотя в более холодные зимы A903, 1908, 1924—25 и 1931—32)
отсутствует там. Известны зимовки также в Туркмении, в низовьях Атрека.
Сроки весеннего появления и осеннего отлета известны для немногих ме-
стностей. Весной первые птицы появляются во второй половине марта, но про-
лет затягивается, и многие особи летят еще в апреле. 27 марта впервые отме-
чена стаей в низовьях Дона (Сарандинаки, 1908). На Черноморском побережье
Кавказа по наблюдениям Вильконского A897) будто бы пролетает весной
между 1 и 20 мая. Повидимому, эти данные ошибочны. Во второй половине мар-
та прилетает в район Агдаша. В середине марта наблюдается массовый прилет
в район Ленкорани, а с 10—20 чисел этого месяца одиночные особи появляются
424 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
уже на взморье Астраханского заповедника (Дубинины, 1940). Большинство же
птиц прилетают сюда несколько позднее —в апреле. В Туркмении весенний при»
лет и пролет начинается в середине марта и длится в течение всего апреля (Де-
ментьев, 1950). Наиболее раннее появление птиц отмечено здесь у Дорт-Кую
14 марта, а массовый пролет в Марыйском оазисе 15 марта. На Мангышлаке ве-
сеннепролетная особь отмечена 3 мая (Смирнов, 1916). Осенний пролет на-
чинается в последних числах августа и продолжается весь сентябрь. Отдельные,
явно запоздавшие особи, встречаются в наиболее северных частях распростра-
нения до последних чисел октября, а на южных окраинах в течение всего ноября
и первой половины декабря. На Черноморском побережье Кавказа осенний
пролет идет с 26 августа по 27 сентября (Вильконский, 1897, Сатунин,- 1913).
В дельте Волги длится весь сентябрь, последние же птицы наблюдались до 20
октября (Воробьев, 1936). На вост. берегу Каспия (в районе Гассан-Кули) про-
должается весь ноябрь и первую половину декабря. Под Гурьевом птицы летят
в конце августа. С другой стороны на Украине, в местах, где названный вид
не гнездится, в иные годы цапли наблюдаются в последних числах октября
(район Волчанский на р. Донце) — 28 октября и на р. Поле в окрестностях
г. Сум—28 октября (Сомов, 1897). Это повидимому заблудившиеся особи.
На Сиваше пролетная птица добыта 10 сентября (Воронцов, 1937), в Крыму на
Бирючьей о-ве в первой трети сентября. Залеты на север, в области, лежащие
за пределами гнездового ареала, происходят в течение всего теплого времени
года — весной, летом и осенью.
Биотоп. Гнездовый, "в Бессарабии — пойменные леса в долинах рек.
В низовьях Днепра и в Днепровском лимане гнездится в рощах ольхи, затоплен-
ных полой водой. Под Ленкоранью и на Куре, в пределах Азербайджана,
гнездится в рощах широколиственных пород, разбросанных пятнами
среди рисовых полей, на группах ольхи среди водохранилищ — истылей или
в более крупных массивах леса, поблизости от болот и полей риса (Спангенберг).
В сходных условиях гнездится в низменной Армении, но в горных частях стра-
ны (озеро Севан) строит гнезда в Камышевых крепях (Ляйстер и Соснин, 19*2).
В Астраханском заповеднике для гнездовья избираются рощи ветлы (Salix
alba) с примесью кустарниковой ивы, протянувшейся лентой по берегам ери-
ков или врезающихся в тростниковые крепи. В Туркмении птицы устраивают
гнезда в камышах в поймах рек. В таких же условиях чепура-нужда гнездится
на глубоководных озерах, образованных Сыр-Дарьей в районе Кзыл-Орды
(Спангенберг, 1936). Кормовые биотопы разнообразны. Птицы предпо-
читают мелководные, стоячие и проточные водоемы. Их удается встречать не
только по берегам озер, проток, открытых и лесных речек, но также в болотах,
сырых и сухих лугах. В Астраханском заповеднике эти цапли кормятся на мел-
кой воде взморья. В б. Екатеринославской губ. по вылете птенцов группы
птиц встречаются в сухой степи (Вальх, 1911). Во всем Закавказье наиболее
охотно кормится в залитых водой рисовых полях — биджарах. Чепура-нужда
не избегает близости человека. Гнездясь местами (район Ленкорани) у самых
селений, она охотно кормится среди стад пасущихся буйволов. Птиц нередко
удается видеть сидящими на спинах этих животных.
Численность. Повидимому немногочисленна в Бессарабии, Обычна*
но спорадична в низовьях Днепра, Днестра, Дона и вообще в Приазовье. В на-
чале двадцатого века была почти полностью уничтожена в дельте Волги, но
ныне в Астраханском заповеднике многочисленна; по приблизительным под-
счетам осенью 1935 г. общая численность этого вида достигала там 22 000 особей
(Ромашова, 1938). Довольно обычна в низменной Армении, но редка на горных
озерах страны. Обычна, а местами многочисленна в долине Куры и крайне мно-
гочисленна между устьем этой реки и к югу до Иранской границы. В пределах
Туркмении редка на гнездовье. Крайне редка на Сыр-Дарье ш>Д Кзыл-Ордой и
л а Аральском море. В двух последних местах гнездится далеко не каждый год.
ЧЕПУРА-НУЖДА ИЛИ МАЛАЯ БЕЛАЯ ЦАПЛЯ 425
В устьях Урала чепура-нужда стала очень редкой. Повидимому, численность
ее здесь, вследствие отступления береговой линии, уничтожения и гибели камы~
шевых зарослей, продолжает уменьшаться. В Китае, где до конца XIX века
цапли были многочисленны на гнездовье, они в результате преследования
скупщиками перьев — почти истреблены, в частности в Фуцзяне, в низовьях
Голубой реки и т. д. Сильное сокращение численности этого вида отмечается
в Индо-Китае (Делакур, 1931).
Экология. Размножение. За редкими исключениями колони-
альная птица. Как правило, присоединяется к колониям других болотных
птиц или образует самостоятельные поселения. В низовьях Днепра гнездится
с кваквами и желтыми цаплями, в Астраханском заповеднике с крупными и
мелкими цаплями и каравайками, в Азербайджане с теми же видами, с обоими
бакланами и египетской цаплей, в Армении с цаплями каравайками и колпи-
ками. На озерах Сыр-Дарьи пара этих цапель была обнаружена среди колоний
других цапель, бакланов, караваек, колпиков. Гнезда устраиваются то на боль-
ших деревьях — карагача, грецкого ореха, шелковицы, черной ольхи (Лен*
корань, Кура), на крупных ольхах (Днепровский лиман), на ивах (Астрахан-
ский заповедник), реже — в камышах и тростниках озер (горная Армения,
Сыр-Дарья). Высота гнезд от поверхности земли или воды различна. Под Лен*
коранью не ниже трех метров, чаще же значительно выше, но не на вершинах
деревьев; наибольшее число гнезд, по нашим наблюдениям, расположено в сред-
нем ярусе. В Астраханском заповеднике —на вершинах низкорослых ветл,
кустарниковых ив или во второй трети высоких деревьев на высоте 7—
10 м иногда и ниже.
При гнездовании в камышах и тростниках гнезда помещаются низко (от
0,5 до 1,5 м) над поверхностью воды. В тех случаях, когда птицы гнездятся на
деревьях, их гнезда укрепляются на горизонтальных ветвях иногда очень да-
леко от ствола. Под Ленкоранью большое число гнезд строится среди густой
сети перепутанных колючих лиан и как бы висят между ветвей двух рядом
растущих деревьев. Такие гнезда оказываются совершенно недоступными.
Для постройки гнезд используются тонкие, но довольно длинные сухие прутики.
В камышах и тростниках гнезда помещаются на заломе и воздвигаются из стеб-
лей этих растений. По форме и характеру постройки гнезда схожи с гнездами
других цапель (перевернутый конус с просвечивающими стенками). Размеры
гнезд не отличимы от гнезд египетских цапель и чуть меньше гнезд квакв. В Аст-
раханском заповеднике чепура-нужда часто занимает старые гнезда других
мелких цапель, но строит, при недостатке старых, и свои собственные. Материал
для гнезда собирается в непосредственной близости от колоний. Гнездо соору-
жается обеими птицами гнездовой пары, причем самец приносит материал,
а самка укладывает его в гнездо и охраняет постройку от других цапель, гнездя-
щихся по соседству. Гнезда выбираются самками. При занятии гнезд наблю-
даются ожесточенные драки между отдельными парами, изгоняются из гнезд
кваквы и каравайки (Ромашова, 1940).
Количество яиц в кладках колеблется от 3 до 6, но чаще их бывает 5*.
Яйца имеют вытянутую форму со слабо выраженным острым и тупым концами,
иногда оба конца острые, реже яйца имеют правильную яйцевидную форму.
Окраска скорлупы свежих яиц насыщенная зеленовато-голубая, которая,
однако, вскоре выцветает. Размер яиц из Ленкорани B0) 41,6 — 50,1 X 30,0 —
34,8 мм, среднее 46,5 х 32,6 мм. Промежутки между откладкой яиц 24 и 48 час
Сроки гнездования у различных пар сильно растянуты. В Астраханском,
заповеднике появление первых птиц в пределах колоний происходит в середине
апреля, массовое в конце апреля и начале мая, но численность колониально
гнездящихся птиц продолжает возрастать до самого конца мая. Наиболее рано
* То же число яиц, в среднем 3—5, отмечается в Южн. Китае и в Индии.
426
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
загнездовавшиеся пары начинают кладку между 20 и 30 апреля (Дубинины,
1940). С этого времени яйца удавалось находить в гнездах до 13 июля (Воробьев,
1936). Под Ленкоранью яйцекладка начинается, повидимому, несколько рань-
ше — в конце апреля. 1 мая уже была взята кладка в 5 сильно насиженных яиц.
Кладки некоторых пар сильно запаздывают, и насиженные яйца можно найти
до начала июля. Безусловно, столь сильное запоздание гнездования вызывается
потерей первых кладок.
Насиживание производится обеими птицами гнездовой пары, но самец сидит
на гнезде значительно меньше, чем самка. Срок насиживания около 25 дней.
В Астраханском заповеднике первые птенцы в гнездах чепуры-нужды появля-
Рис. 83. Малая белая цапля
ются с конца мая по десятые числа июня, первые летные — с 1 по 10 июля
(Дубинины, 1940). На Аму-Дарье около Чарджоу 20 июня отмечена уже летная
молодь (Зарудный, 1896). В те же сроки летающие птенцы наблюдались в За-
кавказье близ Агдаша*.
Птенцы вскармливаются обоими родителями. Как только молодежь опе-
рится и подрастет, она перебирается на ветви дерева и проводит там большую
часть дня. При появлении родителей, птенцы спешат к своему гнезду, где и
получают пищу. Летающие молодые до середины июля кормятся поблизости
колоний и возвращаются сюда на ночевку. В конце этого месяца молодые
птицы вообще прекращают посещать место гнездовья и стаями широко бродят
по кормовым участкам. Таким образом, колониальная жизнь продолжается
с первой половины апреля и до середины, реже до конца июля. Как исключение,
запоздавшие нелетные птенцы были окольцованы на одной из колоний Астра-
ханского заповедника 22 августа (Воробьев, 1936).
Линька. Частичная смена мелкого пера у молодых птиц между августом
и ноябрем. Взрослые птицы имеют полную линьку между июнем и ноябрем.
Частичная предбрачная линька (по Уайзерби, 1941—полная)—между январем
и апрелем.
Питание. Мелкая рыба, лягушки, разнообразные водяные насекомые
и их личинки, в большом числе кузнечики, саранча, сверчки, коромыслы и их
¦ В южн. Европе (Испания; ла-Камарг в устьях Роны) кладки между концом
апреля — началом мая; в Южн. Китае в начале мая; в Индии в июле, на крайнем ее юге-и
на Цейлоне — в марте.
ЖЕЛТОКЛЮВАЯ ЧЕПУРА-НУЖДА
427
личинки. Во второй половине лета, у убитых птиц особенно часто попадаются
наземные насекомые. У птиц, добытых в зимнее время в районе Кзыл-Агачского
заповедника, в желудках найдены почти исключительно завезенные сюда с
целью борьбы с малярией рыбки гамбузии, по несколько мальков кутума и по
одному плавунцу. В одном желудке обнаружено 90 рыбок, в другом 15 (Туга-
ринов и Козлова —Пушкарева, 1935).
Полевые признаки. От большой белой цапли малая цапля отличается
мелкими размерами и значительно меньшей осторожностью. На местах гнез-
довья чепура-нужда подвижная и довольно крикливая птица. Голос вроде
«-арк-арк-арк» или «орк-орк». В гнездовое время часто садится на ветви
деревьев. Ведет стайный образ жизни, встречаясь маленькими группами,
стайками, а иногда (осень) крупными стаями. Обычно ее видно медленно бро-
дящей по неглубокой воде, по лугу и склевывающей добычу. Иногда цапля
неподвижно и подолгу стоит на берегу водоема, подстерегая рыбу. В Закав-
казье часто садится на спины отдыхающих буйволов. При полете втя-
гивает шею и вытягивает назад ноги, взмахивает крыльями несколько
чаще, чем большая белая цапля, но летит так же медленно, как и
последняя.
Описание. Размеры и строение. Формула крыла (не считая рудиментарного
первого махового): 2=3^1 ^4>5>б..., вырезки на внутренних опахалах 1-го—2-го и на-
ружных 2-го—4-го первостепенных маховых. Крыло самцов и самок A2) 275—300, в сред-
нем 287 мм, у самцов среднее 286, у самок — 285лш. Клюв около 90, цевка 90—110 мм. Вес
около 500 г.
Самец в брачном наряде имеет на затылке хохол из двух или трех нерассученных
очень длинных (до 15—20 см) перьев. Подобные же рассученные перья имеются на груди
и плечах (эгретки), последние до 20—24 см длиной и обыкновенно немного заходят за
конец хвоста; у самки украшающие перья менее развиты. В конце лета украшающие перья
выпадают и возобновляются к весне следующего
года.
Окраска. Пуховой наряд — белый. Мо-
лодые птицы по окраске оперения схожи со взрос-
лыми, но лишены украшающих перьев. Нижняя
челюсть клюва желтого цвета. Общая окраска
оперения взрослых снежно-белая. Клюв черный,
только у самого основания желтый; голая кожа
лица голубоватая или зеленовато-желтая; уздечка
черная.
12. Желтоклювая чепура-нужда
Egretta eulophotes Swinhoe
Herodias eulophotes. Swinhoe. The Ibis, I860,
стр. 64, Южн. Китай.
Распространение. Ареал. Гнездится
на о-ве Цусима, о-ве Квельпарт, в Корее,
в Китае (Гуандунь, Цзяньсу, в конце про-
шлого столетия в Фуцзяне). Из пределов
Советского Союза известна только взрослая
самка в брачном наряде, добытая 28 июня
1915 у м. Олимпиада. На зимовках на Фор-
мозе, Филиппинах и Целебесе.
Описание. Размеры и строение.
На затылке хохол из удлиненных перьев, плече-
вые — с удлиненными «эгретками», слегка выходящими за конец хвоста; на зобе удлинен-
ные перья. Крыло добытого в Приморье экземпляра 244 мм. Длина крыла самцов 249—
280, самок 236—250 мм, клюва 75—79 мм, цевки 71—78 мм (Хартерт, 1916).
Окраска. Пуховые птенцы и птенцы в гнездовом наряде не описаны. У взрослых
птиц оперение белое; клюв желтый; голая кожа на уздечке голубая; цевка черная; пальцы
желтоватые.
Карта 77. Распространение желток л ю-
вой чепуры Egretta eulophotes
а — граница гнездовий, г — область зимо-
вок, д — место залета.
428
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Рис. 84. Желтоклювая чепура-нужда
РОД МАЛЫЕ ЧЕПУРЫ BUBULCUS BONAPARTE, 1857
Тип Ardea ibis
13. Египетская цапля Bubulcus ibis L.
Ardea Ibis. Linnaeus. Syst. Nat., изд. X, 1, 1758, стр. 144, Египет
Распространение. Ареал. Пиренейский п-ов, Африка, Мадагаскар,
Аравия, Сирия, сев. Иран, Закавказье, Индия и далее к востоку до низовьев
Голубой реки в Китае, Индо-Китае, южн. Японии, Филиппинских и Зонд-
ских о-вов.
Характер пребывания. В большей части ареала — оседлая
птица, близ северной границы распространения (в наших пределах, и в север-
ных частях Ирана) гнездящаяся перелетная.
Биотоп. Низменности, изобилующие болотами, особенно рисовыми
полями и водохранилищами с вкрапленными перелесками и рощами или окай-
мленные крупными массивами леса; обычна в культурном ландшафте.
Подвиды и варьирующие признаки. Два подвида, различающиеся окраской, пропор-
циями и биологией; В. ibis ibis Linnaeus, 1758 — западная часть ареала — Европа,
Африка, Иран, Аравия; В. i. coromandus Boddaert, 1783 — восточая часть ареала от
Индии на западе.
Западная египетская цапля Bubulcus ibis ibis L.
Распространение. Ареал. Южн. Португалия и южн. Испания, сев.
Африка — Марокко, Тунис, Египет, тропическая и субтропическая Африка
к югу от Сахары, Мадагаскар, ю.-з. Аравия, Палестина, к северу до дельты
Волги и Закавказья, к востоку до сев. Ирана и ю.-з. Туркмении. Залеты
в Италию, Францию, на Балканский п-ов, изредка на Британские о-ва и
в Данию, Венгрию, на Канарские о-ва и Мадейру.
ЕГИПЕТСКАЯ ЦАПЛЯ
429
В СССР распространение таково: гнездится в низменности между г. Лен-
коранью и устьем Куры и по низменному течению этой реки. Южнее в 1929 и
1933 г. встречена только холостыми особями, но за самое последнее время
стали гнездиться у Асмары. По Куре распространение спорадично. Обычно
между крупными гнездовыми колониями большие перерывы. Регулярные
гнездовья известны с давних пор в районе Агдаша. Несравненно реже гнез-
дится в Ереванской равнине по долине Аракса (Ляйстер и Соснин, 1942). При-
водится Шелковниковым A934) в качестве гнездящейся для горного озера
Гилли в Армении на высоте 1925 м над уровнем моря, однако, по мнению
Ляйстера и Соснина A942), это сомнительно. Вероятно она только залетает
Карта 78. Распространение египетской цапли Bubulcus ibis
7. В. /. ibis, 2. В. /. coromandus.
а — граница гнездовий, 6 — не вполне выясненная граница гнездовий, д — место залета.
туда, хотя возможно также гнездится нерегулярно. Недавно гнездование
установлено Поярковым в дельте Волги в Астраханском заповеднике,
а Дементьевым — на озере М. Делили в низовьях Атрека в ю.-з. Туркмении.
В пределах нашей страны наблюдаются изредка довольно далекие залеты
за пределы гнездового ареала. Два экземпляра в свое время были добыты близ
местечка Квирии в западной Грузии, одна особь добыта 13 августа в окрест-
ностях Гизель на Сев. Кавказе, добывались залетные особи также в дельте
Волги, около Махачкала, осенью 1927 г. две молодые птицы под Ашхабадом и
на п-ове Мангышлак. В середине прошлого столетия египетские цапли
залетали на Крымский п-ов, однако, позднее здесь не наблюдалась и лишь
весной 1934 г. залетный экземпляр был добыт на Сиваше. Зимует в Африке.
Характер пребывания, В пределах Союза гнездящаяся
птица, отлетающая с наступлением холодного времени года. Единственное
указание на встречу птицы под Ленкоранью в зимнее время имеется у Радде
A885), повидимому, это была случайно задержавшаяся, быть может, больная
птица. Даты весеннего прилета точно не установлены. По опросным сведениям
у Ленкорани появлякугся в конце марта и реже в самом начале апреля, однако,
мною наблюдались с 11 апреля и позднее. Осенний отлет, повидимому, ран-
ний. В октябре в Закавказье египетской цапли уже нет.
430 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Биотоп. Гнездовый — небольшие группы высокоствольной ветлы,
чаще перелески и рощи широколиственных пород деревьев (дуб, грецкий орех,
тутовник, карагач), обвитых колючими и другими лианами и разбросанных
среди рисовых полей, болот или водохранилищ — истылей. Нередко птицы
гнездятся среди крупных участков широколиственного леса, расположенных
поблизости от кормовых, открытых участков. Последними, главным образом,
являются рисовые поля (беджары) и в меньшей степени луга, болота, подсы-
хающие водохранилища, берега озер и речек, протекающих в открытой мест-
ности. В период размножения и в другое время птица не избегает близкого
соседства с человеком, гнездясь и кормясь у самых селений. В поисках пищи
охотно держится среди стад буйволов, часто отдыхая и кормясь на спинах
этих животных.
Численность. Гнездясь большими колониями, достигающими
несколько сотен пар, египетская цапля местами многочисленна. Однако,
вследствие узкого и спорадического распространения, в общей сложности
численность ее у нас весьма ограниченна.
Экология. Размножение. Колониальная птица, как правило,
гнездящаяся на деревьях. В районе Ленкорани изредка образует самостоя-
тельные колонии, но значительно чаще гнездится совместно с другими цап-
лями (малой белой, желтой, кваквой, серой, а также и с большими и малыми
бакланами). Смешанные колонии образуют и в других местах. По наблюдениям
Шелковникова близ Агдаша гнезда строятся довольно высоко (8—10 м) над
землей. В районе Ленкорани на различной высоте, но не ниже 3 м от земли
или воды, вершин деревьев однако не занимает. В смешанных колониях египет-
ская цапля устраивается в среднем и нижнем ярусе, где гнезда располагаются
бок-о-бок с гнездами малой белой и желтой цапель, тогда как основная масса
других видов в колонии гнездится выше.
Особенно густо заселенными оказываются деревья, стволы и ветви кото-
рых обвиты колючими лианами, что позволяет птицам укреплять гнезда не
только на ветвях, но и среди перевитых лиан. Количество гнезд этого вида
на одном дереве бывает свыше 50. Вследствие большого числа гнездящихся
птиц, трава под колониальными деревьями, стволы, ветви и листья оказываются
густо покрытыми белым пометом. Такие деревья редко выдерживают два гнез-
довых сезона и, как правило, вскоре гибнут.
Вновь построенное гнездо имеет характерную для цапель форму — пере-
вернутого конуса с радиально расходящимися из вершины веточками. Оно
настолько рыхло, что яйца видны с боков и снизу. Материалом служат тонкие
сухие веточки от 0,1 до 0,5 см толщиной. Обычно материал для гнезда соби-
рается птицами на земле в той же роще или вытаскивается из пустующих
гнезд и гнезд соседей, в результате чего среди птиц колонии происходят частые
ссоры. Многие пары занимают прошлогодние постройки того же вида или жел-
тых и малых белых цапель. В этих случаях гнезда приводятся в порядок и
несколько достраиваются. Размеры гнезд варьируют чрезвычайно сильно.
Свежевыстроенное гнездо бывает сравнительно мало (в диаметре), имеет боль-
шую высоту и глубокий лоток. С подрастанием птенцов гнездо сильно растап-
тывается, его стенки становятся пологими, поперечный диаметр очень велик,
а лоток мелок. Особенно крупных размеров бывают прошлогодние достроен-
ные гнезда. Размеры гнезд, промеренных под Ленкоранью, таковы: поперечный
диаметр вновь выстроенного гнезда с незаконченной кладкой или с кладкой
со свежими яйцами около 30 см, диаметр лотка около 17 см, глубина
лотка до 12 см. В районе Агдаша поперечный диаметр гнезд, повиди-
мому, с подрастающими птенцами бывает свыше 50 см (Мензбир, 1918).
Гнездо сооружается обеими птицами гнездовой пары, причем самец
приносит материал, а самка укладывает его и защищает от других
особей.
ЕГИПЕТСКАЯ ЦАПЛЯ 431
Полная кладка содержит от 4 до 9 яиц, причем число яиц в кладках сильно
колеблется в различные годы. Последнее вероятно связано с условиями пита-
ния птиц на зимовках. Более 200 гнезд, осмотренных под Ленкоранью в 1929 г.,
содержали полные кладки в 4—5 яиц, а в 1933 г. большинство от 5 до 7 яиц,
изредка же до 8—9 яиц. От 7 до 9 яиц содержали гнезда, осмотренные у Агдаша
в 1894 г. (Мензбир, 1918). В южных частях ареала, в частности, в Африке,
число яиц в кладках никогда не бывает столь большим — обычно 2—3 яйца.
Массовая откладка яиц под Ленкоранью происходит в различные числа пер-
вой половины мая. В 1929 г. 1 мая постройка многих гнезд заканчивалась,
но только 1 гнездо из осмотренных содержало первое яйцо; 10 мая в гнездах
обнаружено 1—2 яйца, а 15 мая законченные кладки. (Возможно в данном
году размножение несколько запоздало). У Агдаша откладка яиц начинается
во второй половине апреля, но в конце первой трети мая яйца оказались
частью свежие, частью насиженные, свежие и частично насиженные яйца
обнаружены в гнездах в том же районе Смирновым A915) 11 июня. Из Армении
известны кладки в 3 и в 6 яиц, взятые 3 мая (Ляйстер и Соснин, 1942). Сроки
начала кладки у различных самок сильно варьируют. Насиженные яйца
в гнездах удается встречать даже в первой половине июля. Яйца египетской
цапли формой и окраской хорошо отличаются от яиц других цапель. Их форма
яйцевидная, несколько вытянута, но с хорошо выраженными острым и тупым
концами. Окраска скорлупы бледноголубая (бледней, чем у всех других
цапель, имеющих голубую окраску яиц). Размер яиц из-под Ленкорани B1):
43,9—50,0X32,8 — 34,8 мм, среднее 47,4 х 33,8 мм. Промежуток времени
между откладкой отдельных яиц у самок не выяснен.
Насиживают оба родителя, продолжительность насиживания 21—24 дня
(Уайзерби, 1939). У самцов, добытых под Ленкоранью в гнездовый период,
наседные пятна отсутствовали, а у самок оказались развитыми слабо. Доставка
пищи, пока птенцы малы, лежит на самце, самка же остается на гнезде, но
позднее, когда птенцы подрастут, оба родителя принимают участие в добыва-
нии пищи. По наблюдениям Шелковникова (Мензбир, 1918) у Агдаша первые
птенцы вылупляются в половине мая, 3 июля некоторые птенцы уже летали.
Подросшие, но еще нелетные птенцы покидают гнезда и, цепляясь ногами,
клювами и шеями, лазают по ветвям деревьев и лианам. Особенно охотно они
занимают сухие ветви по окраине колонии, покрывая их сплошной массой,
откуда криками встречают старых птиц, прилетевших с пищей. Летные птенцы
совместно со стариками образуют стаи, размер которых возрастает с каждым
днем. Эти стаи первое время кочуют по рисовым полям и болотам близ колоний,
возвращаясь ночевать на гнездовые деревья. Затем птицы покидают колонию
и, продолжая кочевать, медленно подвигаются к югу. В разгар размножения
южней Ленкорани удается встречать холостых египетских цапель. Числен-
ность их весьма ограничена — в стайке бывает 4—6 особей.
Питание. Основная пища — безпозвоночные (кузнечики, саранче-
вые, стрекозы, слепни, водяные жуки и их личинки, пауки, сколопендры,
медведки, моллюски), в меньшей степени лягушки. В большом числе по-
едаются кузнечики, которые, видимо, предпочитаются (Исаков и Воробьев,
1940; Мензбир, 1918).
Линька. Смена мелкого оперения взрослых особей начинается у гнезд
в 10-х числах июля, в это же время птицы теряют украшающие перья. Крупное
перо сменяется на зимовках; к середине февраля, иногда и раньше, в Африке
у птиц появляются украшающие перья на голове. В это время вторично проис-
ходит смена мелкого пера. Прилетевшие к нам цапли бывают одеты в брачное
оперение. Частичная предбрачная линька между мартом и маем, полная
послебрачная между июлем и ноябрем. У молодых частичная линька в пер-
вую календарную осень жизни — мелкое оперение сменяется между сентяб-
рем — декабрем.
432
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Полевые признаки. В области своего распространения в гнездовый период
безошибочно отличается от всех цапель яркой окраской клюва, которая видна
издали. Хорошо отличается от сходных с ней цапель коротким клювом, круп-
ной короткой головой, короткой шеей и яркой окраской головы, спины и зоба
и главным образом яркой окраской клюва. В отличие от других цапель много
бродит в поисках пищи по лугам и болотам. Ее удается видеть группами или
большими (до 300 особей) стаями и редко в одиночку. При взлете напоминает
квакву, полет легкий, но медленный, высоко поднимается редко. Любит кор-
миться среди буйволов и отдыхать на спинах этих животных, когда они спо*
койно лежат в воде мелких водоемов. Менее пуглива, чем многие другие цапли.
Рис. 85. Египетская цапля
Молчаливая птица, ведущая более древесный образ жизни, нежели ее соро-
дичи. Лишь на колониальных гнездовьях удается слышать голоса взрослых
птиц, —глухие односложные звуки — «корр» или «курр» (Мензбир, 1918).
Голоса птенцов не поддаются передаче — звучат по-разному и меняются
по мере подрастания птенцов.
Описание. Размеры истроение. Цапля средней величины. По размерам не-
сколько уступает квакве и значительно крупнее желтой цапли, обладает относительно
коротким клювом, большой, не столь вытянутой, как у многих цапель, головой, короткой
шеей и довольно длинными, толстыми ногами. На голове, груди и спине имеются удлинен-
ные рассученные перья, которые осенью выпадают. Общий облик ее имеет много общего
с кваквой. Длина крыла 233—253 мм (Уайзерби, 1939).
Окраска. Птенцы в гнездах одеты белым пухом. Молодая птица вся чистобелая.
Клюв серовато-черный с желтой вершиной, ноги почти черные, исключая сочленения го-
лени с цевкой, которое несколько зеленоватое. Кожа вокруг глаз евинцово-серая, раду-
жина грязнобелая. В брачном наряде окраска самца и самки белая, исключая верхние части
головы, спины и зоба. Эти части в виде изолированных друг от друга участков, винно-охри-
стого цвета и покрыты рассученными удлиненными перьями. Особенно интенсивна окраска
на темени, менее интенсивна на спине и зобе. Клюв лимонно-желтый, светлеющлй к углу
ЖЕЛТАЯ ЦАПЛЯ 433
рта, уздечка желтая с зеленым оттенком, голое кольцо вокруг глаз яркокрасное. Цевка
тёмнокрасная, пальцы темнобурые, неоперенные части голени желтоватые. Радужина крас-
ная. Самка отличается от самца меньшим размером рассученных перьев на голове, спине
и груди. Зимнее оперение снежно-белое, но на голове сохраняется рыжее пятно. Ноги
зимой становятся темными, буроватыми.
Восточная египетская цапля Bubulcus ibis coromandus В о d d.
Caucroma coromanda. Boddaert. Tabl. PI. Enlum. 1783, стр. 54, Коромандель, Индия.
Распространение. Ареал. Населяет восточную часть ареала вида.
Гнездится в Индии и Индо-Китае к западу включительно до Сеистана,
в вост. Иране (Зарудный, 1903), к северу до средних частей Китая (Сычуань —
низовья Янцзе), в Японии к северу до Токио. К югу до Зондских о-вов. Зале-
тает в Корею и крайне редко в южные части Приморья.
В пределах Советского Союза добыт взрослый самец 7 июня 1884 г. в южн.
Приморье на реке Сунгаче (Тачановский, 1893) и другой взрослый самец,
хранящийся в коллекции Зоологического музея Московского университета,
добыт 19 мая 1946 г. в Судзухинском заповеднике в устье реки Тачингоуза
D3° с. ш.) Бромлеем.
Характер пребывания. Ближайшие гнездовья находятся на
юге Японии, где, как и в Китае, птица перелетна.
Полевые признаки. От западной египетской цапли уже издали отличается
яркой окраской и более широким ее распространением на голове, шее и зобе.
Описание. Размеры и строение. Несколько крупнее западной египетской
цапли. Цевка обыкновенно длиннее 8,25 мм и достигает 9,25 мм, крыло 24,0—26,5 мм,
клюв 56—60 мм (Бутурлин, 1935). Размеры экземпляра Московского университета таковы:
крыло 249 мму цевка 88 мм, клюв 58 мм. Украшающие перья спины выдаются за хвост
на 22 мм.
Окраска. Хорошо отличается от западного подвида тем, что вся голова (а не только
верхняя ее часть) и вся шея и зоб окрашены в ярко оранжево-золотистый цвет, причем ок-
раска головы непосредственно (без границ) переходит в окраску зоба. Окраска украшающих
перьев спины ярко золотисто-винного цвета. Цевка иногда с красным оттенком. Молодая
птица — белая, без украшающих перьев весною, но с оранжевым оттенком на голове.
РОД КОСМАТЫЕ ЦАПЛИ ARDEOLA BOIE, 1822
Тип A. rail о ides
14. Желтая цапля Ardeola ralloides Scop.
Ardea ralloides. S с о р о 1 i. Ann. I Hist. Nat., 1769, стр. 88, Крайна.
Синонимы. Ardea pumila L e p e с h i n. Novi Comment. Acad. Caes. Petropol.
XV, 1770, стр. 502 —южн. Россия; Ardea castanea. S. G. G me 1 i n. Novi Comment.
Acad. Caes. Petropol. XV, 1771, стр. 454 — низовья Дона; Ardea Comata Pallas.
Reise d. ferschied. Prov. d. Russ-Reichs, II, 1778, стр. 715 — Каспийские побережья.
Русское название. Желтая цапля — книжное.
Распространение. Ареал. Западная Палеарктика в Эфиопской области.
В Европе гнездится в южн. Португалии и южн. Исрании, в дельте Роны и
быть может на Корсике, во Франции, в с.-з. Италии, в Венгрии, Югославии,
Болгарии, Румынии и на юге Европейской части Советского Союза. В Африке
к северу от Сахары от Марокко до Туниса, южнее —по всему континенту;
Мадагаскар, Передняя Азия от Малой Азии, Сирии, Ирака до Ирана и Сред-
ней Азии.
Подробности распространения в СССР таковы: гнездится в Бессарабии
в плавнях Днестра. По Днепру от устья в качестве спорадически гнездящейся
птицы далеко проникает вверх по течению, достигая б. Уманского уезда.
Местный выводок обнаружен под г. Каневым. Гнездится в гористой части
Крыма по рекам, стекающим с северных склонов Крымских гор (р. Салгир),
28 Птицы СССР, т. II
434
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
тогда как в других степных частях (Аскания-Нова, Тархан-Сунак)
бывает на пролетах. Быть может изредка гнездится в самых южных частях
Харьковской обл. (Сомов, 1897). Неоднократно отмечалась в Приазовье,
повидимому, гнездится здесь. Несомненно гнездится в плавнях Кубани,
откуда в коллекции Птушенко есть яйца, и в низменной части Осетии
(Беме, 1926). Отмечена в районе Кизляра (повидимому гнездится). Наверное
гнездится у Батуми. В Армении гнездится как в низменных районах, так и
на горных озерах страны, поднимаясь до 2 000 м над уровнем моря (Ляйстер
и Соснин, 1942). К востоку отсюда гнездится во всех подходящих местах Закав-
казья, но особенно в долине
средней и нижней Куры и
между устьем этой реки и
иранской границей. На север-
ном берегу Каспия регулярно
гнездится в дельте Волги и
редко в устье Урала. В 70-х
годах прошлого столетия нор-
мально гнездилась на Сарпин-
ских озерах (Мензбир, 1918).
В небольшом числе и споради-
чески гнездится на больших
водоемах Туркмении —Мерв-
ском оазисе, в низовьях Тед-
жена, по болотам у Чарджоу
и Карахоуза и, вероятно, в
низовьях Атрека и на Сумбаре
(Шестоперов, 1937, Дементьев,
1950). На нижней Аму-Дарье
гнездится в подходящих мес-
тах до дельты, где найдена
Гладковым. В свое время была
найдена гнездящейся в долине
верхнего течения Аму-Дарьи
между Кировабадом и Кели-
фом (Зарудный и Билькевич,
1918), но, повидимому, очень
редка здесь, так какие найдена
позднейшими исследователя-
ми Таджикистана (Иванов,
1940). Выводит также в ни-
зовьях Зеравшана (Мензбир, 1918) и в редкие годы на Сыр-Дарье в районе
Кзыл-Орды (Спангенберг, 1936). Случаи залета известны далеко к северу
от гнездового ареала. Они происходят в летнее время, весной и осенью.
Характер пребывания. Оседлая и кочующая птица. Большин-
ство особей, гнездящихся в Европе, улетает в Африку, но часть зимует на
крайнем юге Европы. В гнездовой области в Европе желтая цапля проводит
время между апрелем и сентябрем, но молодые покидают места гнездовий
ранее и до начала перелета — откочевывают в разных направлениях. В это
время — уже в июле и позднее — они встречаются в Сев. Франции, Бельгии,
Голландии, Германии, Дании, Австрии, Швейцарии, Чехословакии, Польше,
на Британских о-вах и т. д. Залеты известны на Канарских, Азорских о-вах
и на Мадейре.
В СССР отмечалась в Киевской обл., Харьковской обл., в Поволжье у Са-
репты, в Чкаловской обл., 25 июля и 8 августа у Буранной и у Бишкопа.
В мае 1873 г. добыта в Молотовской обл. (Ушков, 1927).
Карта 79. Распространение желтой цапли Ardeola
ralloides
а — граница гнездовий.
ЖЕЛТАЯ ЦАПЛЯ 435
Прилет относительно ранний, но сильно растянутый. Сроки прилета
плохо выяснены. В вост. Иране несомненно пролетный экземпляр был добыт
17 апреля (Зарудный, 1903). У нас наиболее ранние особи появляются в сере-
дине марта и с этого времени пролет длится до половины мая. Хорошо выра-
женный, массовый прилет происходит в конце апреля. Так, на северном берегу
Черного моря и в устьях Дона эта цапля появляется с конца первой трети апреля
(Мензбир, 1918), вдоль Черноморского побережья Кавказа птицы летят с поло-
вины марта и до половины мая (Вильконский, 1897). Отмечена у черноморских
берегов 25 мая одиночка, а 26 мая стайка в 7 особей (Нестеров, 1911). В Арме-
нии добывалась 29 апреля, 2 мая и в более поздние сроки (Ляйстер и Соснин,
1942). В Азербайджане в район Агдаша желтые цапли прилетают в половине
апреля. Под Ленкоранью известны случаи добычи 30 марта, но нормально они
появляются на беджарах близ колониальных гнездовий между 15 и 20 апреля.
В дельте Волги прилет происходит с 10 по 20 апреля (Дубинины, 1940), а по
Воробьеву в конце апреля. В устье Урала прилетает, повидимому, в конце
апреля или в конце первой трети мая. Пролетная особь под Кизляром на взморье
впервые отмечена 18 марта, затем 10 мая (Беме, 1925). В Туркмении наиболее
ранний прилетный экземпляр отмечен 26 марта у озера Юссуф-Хана.
Осенний отлет также растянут на длительный срок. Первые особи, пови-
димому, начинают подвигаться к югу во второй половине августа, массовый
отлет длится в течение всего сентября, одиночки в более северных частях
ареала задерживаются до половины октября, а в южных до начала ноября.
В Крыму бывает на пролете во второй половине августа и в первой сентября.
В низовьях Днепра желтая цапля добывалась 23 сентября. Под Дзауджи-
кау ежегодно пролетает с сентября по ноябрь. В дельте Волги отлет проис-
ходит в течение всего сентября, одиночки же задерживаются до половины
октября. В половине сентября пролет наблюдался на Черноморском побережье
Кавказа. Начало осеннего движения к югу в Туркмении происходит прибли-
зительно со второй половины августа.
Биотоп. Эта цапля наиболее охотно избирает для гнездовья небольшие
группы ветлы, рощи широколиственных древесных пород, разбросанных то среди
рисовых полей, озер и болот, то среди ериков и камышевых зарослей. Местами
заселяет и более крупные массивы леса в тех случаях, когда они не слишком
удалены от мест кормежек. При бедности и отсутствии древесной раститель-
ности устраивается в зарослях камыша по краям озер и проток. Кормовыми
биотопами служат главным образом берега и мелководные места стоячих водо-
емов самого разнообразного характера: залитые рисовые поля, водохрани-
лища — истыли, подсыхающие озера, реже берега ериков, открытых речу-
шек — канав. Нередко цапельки кормятся также по болотам и сухим лугам.
В Закавказье желтая цапля — характерная птица культурного ландшафта.
Гнездится она и кормится в непосредственной близости от жилья человека.
Численность. Местами в Днепровских плавнях довольно много-
численная птица. Во множестве гнездится в Закавказье по Куре и особенно
под Ленкоранью. Большая численность в некоторые годы наблюдается в пре-
делах Астраханского заповедника, в другие, напротив, гнездится редкими
парами. На остальном протяжении нашей страны и особенно в азиатской части
Союза редкая птица со спорадичным распространением. Общая численность
вида в Союзе ничтожна.
Экология. Размножение. Гнездится колониями. Самостоятельные
поселения образуют редко (у нас, повидимому, не гнездится обособленно).
Обычно присоединяется к колониям других цапель, караваек и колпиков.
В этих случаях гнезда то разбросаны среди гнезд других видов, то собраны
в одном месте. Под Ленкоранью, несмотря на большую численность этого вида,
желтая цапля всегда гнездится совместно с другими колониальными птицами.
В Азиатской части Союза, где эта цапелька крайне малочисленна, помещает
436 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
свои гнезда в колониях других цапель, бакланов, караваек и колпиков. При
гнездовании на деревьях желтые цапельки строят свои гнезда на различных
древесных породах, то на низкорослых, то на высоких ивах, на тополе, дубе,
грецком орехе, карагаче, тутовнике. Под Ленкоранью особенно охотно и
густо птицы заселяют такие деревья, ветви и стволы которых перевиты колю-
чими и другими лианами. В смешанных гнездовых колониях у Ленкорани
желтые цапли преимущественно занимают нижний и отчасти средний ярус
деревьев, где их гнезда располагаются то отдельными поселениями, то вкрап-
лены среди гнезд чепуры-нужды и египетской цапли. Выше численность гнезд
резко снижается.
В верхнем ярусе деревьев гнезда не устраивают. Высота расположения
гнезд от поверхности земли или воды колеблется, примерно, от 2 до 15—20 м.
На горных озерах Армении, на Сыр-Дарье и, повидимому, в других местах
Азиатской части Союза, бедных древесной растительностью, желтая цапля
устраивает свои гнезда на заломе прошлогоднего камыша и тростника. При
таких условиях гнездования на Сыр-Дарье легкие постройки желтых цапель
помещались выше гнезд всех остальных птиц, населявших колонию. Высота
гнезд колеблется от 2 до 3 м от водной поверхности. На деревьях гнезда обычно
укрепляются на горизонтальных ветвях или в разветвлении вертикально
растущей ветви, причем часто очень далеко от основного ствола.
У Ленкорани большое количество гнезд помещается среди перепутанных
колючих лиан и как бы висят в воздухе между ветвями. Для постройки гнезд
на деревьях используются тонкие, сухие и прямые прутики. В противополож-
ность другим цаплям в гнездах желтой бывает много сухих прошлогодних
листьев. В камышах и тростниках гнезда строятся из тонких, сухих стеблей
с небольшой примесью листьев этих растений. В тех и других случаях мате-
риал добывается в непосредственной близости от колоний. Форма гнезда, как
и у большинства других цапель, — перевернутый конус с просвечивающими
стенками. Лоточек у многих гнезд оказывается скрепленным пометом птицы
и илом. Малые размеры гнезда желтой цапли хорошо отличают его от гнезд
всех других цапель колоний, а форма — от гнезд малых бакланов и караваек.
Поперечный диаметр гнезд под Ленкоранью в период яйцекладки колеблется
от 17 до 23 см, диаметр лотка от 9 до 12 см, глубина лотка от 8 до 11 см. С появ-
лением птенцов гнездо сильно растаптывается и теряет свою первоначальную
форму. Боковые стенки гнезда становятся более покатыми, поперечный диаметр
сильно увеличивается. В постройке гнезда участвуют обе птицы гнездовой
пары, причем на самце лежит обязанность доставлять материал, а на самке —
строить гнездо.
Количество яиц в кладке обычно 4—5. Кладки в 6 яиц встречаются редко,
как исключение (для ю. з. Европы отмечались и 7 яиц). Сроки откладки
яиц различны не только в разных географических широтах, но и среди отдель-
ных парочек одной колонии. Это происходит вследствие того, что колония
в связи с растянутым весенним прилетом заселяется не одновременно. В то
время, как рано прилетевшие пары уже насиживают яйца, запоздавшие только
приступают к постройке и ремонту гнезд. В большинстве гнезд, осмотренных
на озере Гилли (горная Армения) 10 июня, оказались яйца (некоторые све-
жие), но были и птенцы (Ляйстер и Соснин, 1942). Под Ленкоранью закончен-
ные гнезда, но еще без яиц, найдены 12 мая; 14 мая в некоторых гнездах было
по 1 яйцу, а 19 мая законченные кладки в 4—5 яиц. С этого времени и до 20 июня
многие гнезда содержат яйца в различной степени насиженности, а в конце
июня и самом начале июля сильно насиженные яйца (Спангенберг). В Астра-
ханском заповеднике начало откладки яиц происходит между 10 и 20 мая
(Дубинины), но гнезда, содержащие яйца, попадаются еще в половине августа
(Воробьев, 1936). Можно думать, что это повторные кладки, вызванные гибелью
первых гнезд. На Кубани в Шаптерской плавне яйца были найдены Птушенко
ЖЕЛТАЯ ЦАПЛЯ 437
13 июня. На Сыр-Дарье в районе Кзыл-Орды 26 мая — 2 гнезда оказались
вполне законченными, одно из них содержало первое снесенное яйцо, два
других гнезда находились в стадии постройки (Спангенберг).
Яйца желтой цапли хорошо отличаются от яиц других цапель, гнездя-
щихся колониально мелкими размерами. Их форма вытянутая, как у чепуры-
нужды, с одинаково острыми концами. Несколько реже встречаются яйца
правильной яйцевидной формы. Окраска скорлупы на свежих яйцах насы-
щенная зеленовато-голубая, но подверженная довольно сильному выцвета-
нию. Размеры: 37,0—40,8 мм х 27,0—30,0 мм, среднее 38,6x28,5 мм.
Некоторые яйца бывают несколько меньших размеров. Одно яйцо из дельты
Волги имело размер 36,9 х 27,2 мм (Воробьев, 1936). Кладка в 4 яйца откла-
дывается за 4—5 дней, в 5 яиц за 5—6 дней, однако точными данными о вре-
мени между откладкой отдельных яиц мы не располагаем.
Большую часть времени на яйцах сидит самка, но, повидимому, и самец
принимает некоторое участие в насиживании. Самки, добытые в колониальном
гнездовье у с. Джиль (близ Ленкорани), обладали едва заметными наседными
пятнами, у самцов пятна отсутствовали. Срок насиживания не прослежен.
Наиболее ранние птенцы появляются, повидимому, в различные числа пер-
вой трети июля. На озере Гилли (Армения) 10 июня в некоторых гнездах были
уже птенцы (Ляйстер и Соснин, 1942). Под Ленкоранью 11 июня были совсем
маленькие птенцы в возрасте 1—2 дней (Спангенберг). В дельте Волги, появ-
ление первых птенцов наблюдается между 5—15 июня, массовое между 20—
30 июня (Дубинины, 1940), довольно крупные птенцы найдены 15 июня (Во-
робьев, 1936). Вылет птенцов из гнезд происходит здесь с 10 по 20 июля.
У Агдаша в Азербайджане 3 июля птенцы уже перепархивали с ветки на ветку
(Мензбир, 1918). Наконец, в Туркмении у Иолотани 27 июня Зарудным наблю-
дались подлетки (Дементьев, 1950). Необходимо отметить, что эта дата крайне
ранняя. В вскармливании птенцов принимают участие оба родителя, при-
чем на самце эта обязанность лежит в течение всего гнездового вре-
мени, а на самке лишь после того, как птенцы достаточно подрастут и
оперятся.
Вполне оперившиеся, но еще нелетные птенцы лазают по ветвям дерева
близ гнезда, а позднее, когда научатся перепархивать, в большом числе груп-
пируются в отдельных участках колонии. Нередко птенцы срываются и падают
на землю и затем взбираются обратно на дерево, пользуясь ногами, клювом
и крыльями. Благодаря этой способности к лазанию, они реже птенцов круп-
ных видов цапель становятся жертвой четвероногих хищников. По мере приоб-
ретения птенцами способностей к полету, молодые и старые птицы постепенно
покидают колонию. В июле у Ленкорани уже часто удается встречать малень-
кие группы (от 4 до 10—12 особей) желтых цапель, которые бродят по рисовым
полям, обмелевшим истылям, лугам и болотам. Позднее (в августе) числен-
ность таких стаек сильно возрастает, но число особей в отдельных стайках
сохраняется прежним. Видимо, у желтой цапли отсутствует стремление обра-
зовывать такие крупные общества, какие свойственны, например, цапле
египетской.
Линька. Плохо выяснена. Общая схема —как у других цапель. Моло-
дые птицы сменяют мелкое оперение между июлем и декабрем. У взрос-
лых две линьки в году: предбрачная между январем и маем, будто бы полная,
послебрачная полная, между июлем и декабрем. Возрастные изменения незна-
чительны.
Питание. По питанию у нас почти нет данных. Пища состоит из куз-
нечиков, пауков, различных личинок и мелких лягушек (Сатунин, 1907).
У птиц, добытых нами под Ленкоранью в июле — августе, желудки оказались
наполненными остатками насекомых: кузнечиков, коромысел и, повиди-
мому, цикад.
438
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Полевые признаки. Безошибочно отличается от других цапель размерами
и общим сложением. Сидящую птицу легко отличить от других и по окраске
оперения. Напротив, во время полета желтая цапелька кажется белой и на
большом расстоянии может быть легко спутана с другими видами, например,
с египетской цаплей и даже с чепурой-нуждой. На лету отличается от них
Рис. 86. Желтая цапля (взрослая и молодая)
особенно широкими и тупыми крыльями. Видеть ее удается в одиночку, спо-
койно сидящей на берегу водоема, на поваленном с краю уреза воды сухом
камыше, на кочке или коряге, при этом птица втягивает шею, сжимается и
кажется значительно меньше, чем есть на самом деле. Реже желтые цапли
кормятся небольшими группами, но и в этих случаях держатся поодаль друг
от друга.
БЕЛОКРЫЛАЯ ЦАПЛЯ
439
Иногда утрами и на заходе солнца приходится наблюдать цапель, летящих
маленькими стайками. Полет их медленный, но удивительно легкий с ред-
кими взмахами широких крыльев. В одних случаях цапли летят низко беспо-
рядочной стайкой, в других —выстроившись углом или в косую линию. Такие
перелеты стайками затягиваются до поздних сумерек, вообще же желтую
цаплю надо считать дневной птицей. Несмотря на светлую окраску оперения,
делающую ее заметной, желтая цапелька отличается особенной непугли-
востью и близко подпускает к себе человека. Ее голос, что-то вроде грубого
«карр», удается слышать очень редко, чаще — при испуге и взлете.
Описание. Размеры и строение.В брачном наряде на голове длинный хохол,
перья зоба и плечевые удлинены и рассучены. Формула крыла (без первого рудиментарного
махового) 2^3:^1>4>5... На наружных опахалах 2-гои 4-го махового имеются неглубокие
вырезки. Крыло самцов и самок A0) 202—240, в среднем 217,7 мм (по Тугаринову, 1947,
минимальная длина крыла 195 мм). Клюв около 60—70 мм, цевка около 55—65 мм, средний
палец (без когтя) около 60—70 мм. Самка чуть мельче самца и украшающие перья хохла
и спины менее развиты.
Окраска. Пуховой птенец белый, с охристым оттенком на плечевых, на темени
пух удлинен и имеет буровато-желтоватый цвет (Уайзерби). Молодые птицы похожи на
взрослых в зимнем оперении, отличаясь от них серо-бурым продольным рисунком. На крою-
щих крыла, маховые — с темными стволами, вершинами и наружными опахалами.
В брачном оперении бока головы и шея взрослой птицы — желтые. Нижняя часть
шеи, зоб и грудь яркоохристые, горло белое. На затылке и задней части шеи по желтому
фону — тонкие продольные, почти черные полоски. Наиболее длинные лентовидные перья
хохла белые с черными краями. Спина и украшающие перья спины — винные, несколько
желтеющие на концах. Крылья, подхвостье, хвост и вся нижняя сторона тела от зоба и ниже
белые, местами со слабым желтоватым налетом. Голая кожа вокруг глаз зеленоватая, клюв
голубой с почти черной вершиной, ноги темного цвета, подошвы желтые, когти черные.
Радужина светложелтая. В зимнем наряде украшающие перья отсутствуют, голова и шея с
черновато-бурым продольным рисунком. Спина, плечевые и внутренние из второстепен-
ных маховых — землисто-бурые. Ноги зеле-
новатые. В первом брачном наряде стволы
первостепенных маховых бурые (у взрослых
белые).
15. Белокрылая цапля Ardeola
bacchus В p.
Buphus bacchus. В о n a p а г t e. Consp. Av.
11, 1855, стр. 127, Малайский п-ов.
Распространение. Ареал. Ю.-в.
Азия от Бирмы и Ассама по Малай-
скому архипелагу на юг до Борнео,
на восток до Индо-Китая; Андаман-
ские о-ва; в Китае на север до Чжи-
ли, где вероятно гнездится, отмечена
в провинциях Гуаньдунь, Цзяньси,
Фуцзян, Сычуань, Хубей и в ни-
зовьях Яньцзе. Залеты в Японию
(Хоккайдо, Хондо, Семь островов,
Риу-Киу). Отмечена в южн. Манч-
журии. В пределах Советского Сою-
за известны четыре находки. 27 мая
птица была убита в Сидеми (Тачанов-
ский, 1893), два экземпляра 1 ав-
густа 1911 добыты на болоте в уро-
чище Славянка, Амурский залив (Бу-
турлин, 1911), одна птица добыта 20
мая 1926 в устьях р. Лефу (Иоган-
Карта 80, Распространение белокрылой
цапли Ardeola bacchus
а — граница гнездовий, б — не вполне выяснен-
ная граница гнездовий, д — места залетов.
440
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
сен, 1928). Одна птица 12 ноября была добыта у Кяхты вблизи нашей
i раницы.
Характер пребывания. В низовьях Яньцзе перелетный,
в остальном Китае повидимому оседлый вид (Латуш, 1934). Оседлая в Индо-
Китае, где однако вне периода размножения совершает довольно значительные
перемещения (Делакур, 1931). Вопрос о характере пребывания этой птицы
в южн. Приморье не совсем ясен. Быть может птица все же гнездится изред-
ка в южн. Приморье, так как в урочище Славянка наблюдались птицы, держав-
шиеся парой, а Шульпину A936)
сообщали, будто бы цапля гнез-
дится в устьях Лефу.
Биотоп. В Китае, Ассаме
и Бирме преимущественно в куль-
турном ландшафте у стоячих во-
доемов — прудов, по рисовым по-
лям и т. д.
Численность. В южн.
Азии весьма многочисленна.
Экология. Размножение.
Гнездится колониями, иногда с
другими цаплями, в частности с
малыми белыми. Гнезда находили
на деревьях, на бамбуке и т. п.
Слегка насиженные кладки в Ки-
тае найдены в конце мая. Период
размножения в Ассаме с конца
июня до августа. Кладка из 3—5
яиц. Размеры A7) 37,5—42x29^
30, в среднем 39,5 х 29,5 мм (Ки-
тай, Латуш, 1934); E0) 34—39,2 х
26,7—30,1, в среднем 37,7 х 28,4 мм
(Ассам и Бирма, Бекер, 1929).
Питание. Питается глав-
ным образом различными моллю-
сками.
Полевые признаки. Мелкая
цапля, сложением похожая на жел-
тую цаплю, от которой хорошо
различается окраской (см. ниже).
От восточных волчков отличается характером полета, в отличие от них, бе-
локрылая цапля медленно машет крыльями и втягивает шею.
Описание. Размеры и строение. В брачном наряде на голове хохол, на
шее и плечах удлиненные перья. Крыло самки — 218, самца 240 мм (Китай, Латуш);
крыло ассамских и бирманских самцов и самок 195—238 мм (Бекер). Клюв около 55—70 мм.
Цевка около 55—65 мм.
Окраска. Молодые похожи на взрослых в зимнем оперении, отличаясь от них
более светлой окраской головы с черными продольными черточками и присутствием схожих
пестрин на передней части шеи. В брачном наряде у птицы широко распространена белая
окраска. Она распространяется на горло, нижнюю сторону тела, включая и нижние крою-
щие крыла, заднюю часть спины, подхвостье, хвост и крылья. Голова с затылочным хохлом
яркоржавого цвета, длинные украшающие перья шеи — каштановые, плечевые — черные
с серо-зеленоватым оттенком, с каждой стороны верхней части груди пятно из удлиненных
черных перьев, на внешних маховых серовато-бурый налет. Клюв — желтый, чернеющий
к вершине с голубоватым основанием верхней челюсти, оголенные лицевые части и основа-
ние нижней челюсти зеленовато-желтое; радужина желтая; ноги бледнооранжевые с розо-
ватой голенью и буровато-зелеными когтями.
В послебрачный период голова птицы и шея темнобурая, с бледными охристыми
продольными пестринами, с более темным теменем и более светлой передней стороной шеи.
Рис. 87. Белокрылая цапля
ЗЕЛЕНАЯ КВАКВА
441
Спина, плечевые и самые внутренние из второстепенных маховых темнобурые с серым
налетом. Остальное оперение белое с широкими бурыми продольными пестринами на
груди и с буровато-охристым налетом на верхних кроющих крыла и наружных
первостепенных маховых.
РОД ЗЕЛЕНЫЕ КВАКВЫ BUTORIDES BLYTH, 1852
Тип. Ardea javanica
16. Зеленая кваква Butorides striatus L.
Ardea striata. Linnaeus. Syst. Nat., изд. X, 1758, стр. 144. Суринам
Распространение. Ареал. Крайне обширен. Птицы населяют умерен-
ные, но, главным образом, субтропические и тропические страны Азии, Африки,
Америки и Австралии.
Характер пребывания. Оседлый вид в южных широтах и
перелетный в самых северных частях ареала. Правильные перелеты свой-
ственны птицам, населяющим Японию, Сев. Китай, Корею и СССР.
Биотоп. Придерживается зарослей по берегам внутренних водоемов.
Карта 81. Распространение зеленой кваквы Butorides striatus
7. В. s. amurensis, 2. В. s. striatus, 3. В. s. robinsoni, 4. В. s. cyanurus, 5. В. s. brevipes, 6. B. s. atrtccapil-
tus, 7. B. s. rutenbergi, 8. B. s. degens, 9. B. s. crowfordl, 10. B. s. albolimbatus, 11. В s. albidulus,
12. B. s. spodiogaster, 13. B. s. patens, 14. B. s. connectens, 15. B. s. javanicus, 16. B. s. moluccarum,
17. B. s. macrorhynchus, 1&. B. s. stagnatilis, 19. B. s,- patruelis;
a — граница гнездовий, б — не вполне выясненная граница гнездовий, г — область зимовок.
Подвиды и варьирующие признаки. Много подвидов. Одни из них отличаются слабо,
другие, напротив, хорошо различаются размерами, окраской оперения и деталями биологии.
В. s. patens G г i s с о m, 1929 — Панамский перешеек; В. s. striatus Linnaeus, 1758 —
тропическая Америка от вост. Панамы до южн. Бразилии; В. s.robinsoni Richmond, 1896—
о-в Маргарита у побережья Венецуэлы; В. s. cyanurus V i e i 1 1 о t, 1817 — южн. Брази-
лия, Парагвай, Уругвай и Аргентина к югу до провинции Тукумак и Буэнос-Айрес; В. s.
brevipes Hemprichet Ehrenberg,l 833—Сомали и берег Красного моря; В. s. atricapil-
lus A f z e 1 i u s, 1805—тропическая Африка; В. s. rutenbergi H a r 11 a u b, 1880 —Коморские
о-ва, Мадагаскар, Родригес и Реюньон; В. s. degens H a r t e r t, 1915 — Сейшельские о-ва;
В. s. crawfordi N i с о 1 1, 1906 — о-в Ассемпшен, о-ва Альбадра; В. s. albolimbatus R e i-
chenow, 1900 — о-ва Диего-га рсия (Индийский океан); В. s. albidulus Bangs, 1913—
Маледивские о-ва; В. s. spodiogaster S h а г р е, 1898 — Анаманские и Никобарские о-ва;
В. s. amurensis Schrenck, i860 — Япония, Корея, с.-в. Китай, о-в Бонин, среднее
течение Амура, Приморье; В. s. connectens Stresemann, 1930 — Южн. Китай от Яньцзе
к югу до сев. Сиама и Индо-Китая; В. s. javanicus Hersfield, 1821 — Индия;
Цейлон, Малайский п-ов, страны южн. Индо-Китая, Филиппинские о-ва, Зондские о-ва,
Целебес; В. s. moluccarum Н а г t е г t, 1915 — Буру, Церам, Амбойна, Оби (?); В. s. macro-
442 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
rhynctms Gould, 1848 — Новая Гвинея, Соломоновы о-ва, о-ва Ару, Кеи, Австралия,
Новый южный Уэльс и Квинсленд; В. s. stagnatilis Gould, 1847 — с.-з. и сев. Австра-
лия; В s. patrutlisP е а 1 е, 1848 — Таити и, вероятно, другие острова южных частей Тихого
океана.
Амурская зеленая кваква Butorides striatus amurensis Schrenck
Ardea virescens var. amurensis. Schrenck. Reis. Amur Lande 1, 2, I860, стр. 441. Амур-
ская обл.
Синоним. Butorides Schrenckii. Богданов. Consp. Av. Imp. Rossici., 1884, стр. 115,
Приамурье.
Распространение. Ареал. Населяет Японские о-ва от Хоккайдо
до Кюсю, Квельпарт, Корею, отмечена в Манчжурии, где вероятно гнездится,
и в Сев. Китае у Киау-Чоу и в Чжили. У нас широко распространена в подхо-
дящих биотопах всего Приморья, среднего и нижнего Амура, быть может
до устья этой реки. Подробности распространения в наших пределах этой цапли
таковы. Характерна во всем Приморье, к северу во всяком случае до устья
Уссури. Основная масса птиц здесь приурочена к низменностям широких
речных долин, но местами (например, по Иману) в небольшом числе проникает
в предгорья и в горные части хребта Сихотэ-Алиня (Спангенберг). Встре-
чается также на берегах Японского моря. На Амуре правильно и широко
распространена на западе от устья реки Зеи и Благовещенска и вниз
отсюда до устья реки Горина. Ниже устья этой реки по Амуру никем из
орнитологов не наблюдалась, но так как этот вид хорошо известен гилякам,
населяющим устье Амура (Шренк, 1861), возможно его следует признать
гнездящимся и здесь или хотя бы залетным. Вероятно на Сахалине. Места
зимовок — Формоза, Филиппинские, Зондские о-ва, Целебес, Натуна.
Характер пребывания. В СССР гнездящаяся птица, совер-
шающая правильные перелеты. Сведения о весеннем появлении до последнего
времени были весьма противоречивы. Пролет в Китае — в мае весной, в сен-
тябре осенью. Однако нашими исследованиями A940) в низовьях р. Имана
определенно установлено, что амурская зеленая кваква прилетает в первые
дни мая. Наиболее ранний экземпляр наблюдался и был добыт 2 мая, с этого
времени птица становится обыкновенной. Массовый осенний отлет происходит,
повидимому, в первой половине сентября, так как позднее 13 сентября она
уже не была встречена на р. Уссури (Маак, 1861). Однако, есть основание
утверждать, что некоторые особи задерживаются у нас осенью несколько
дольше. Это доказывается наличием одного экземпляра, добытого на о-ве Фуру-
гельм 26 сентября, и другого в устье р. Хунгари 2 октября. Согласно Шуль-
пину A936) пролет протекает совершенно незаметно, видимо, ночью.
Биотоп. Тенистые прибрежные поросли лозы, ольхи, черемухи
и других древесных пород, обрамляющие реки. Особенно охотно птицы
избирают береговые заросли речных затонов, сильно захламленных протоков
среди многочисленных островов, где в изобилии встречаются коряги, подмытые
водой деревья с обнажившейся корневой системой и занесенный сюда в поло-
водье, сплавляемый по реке лес. Указанных стаций зеленые кваквы придержи-
ваются с большим постоянством. Здесь же, сидя на обнаженных водой корнях,
на коряге или на узкой отмели под круто обрывающимся берегом в быстро-
текущей воде, добывает себе пищу. В период подрастания птенцов эти цапельки
изредка вылетают за пределы береговых зарослей, посещая канавы, берега
прудов среди поселков, расположенных в непосредственной близости от рек.
На болотах и стоячих водоемах вдали от рек положительно не встречается.
В подобных стациях цапли живут отдельными парами, а позднее семьями,
никогда не образуя в нашей стране колониальных гнездовий. В противополож-
ность этому в Японии амурская зеленая кваква гнездится не только отдельными
парочками, но и колониями от 3 до 10 пар в каждой (Ян, 1942). На реках,
ЗЕЛЕНАЯ КВАКВА 443
стекающих с хребта Сихотэ-Алинь, обитает в нижних отделах с широкой
речной долиной и почти не заходит в горы. В средних и южных частях страны
(например, Иман), нередко проникает в горные долины.
Численность. На Амуре — немногочисленна. К югу численность
заметно возрастает и на Уссури цапля встречается очень часто и тем чаще, чем
далее на юг.Чаще же всего встречается на Сунгаче. Редка по нижней Лефу в рай-
оне Спасск-Яковлевска. В низовьях Имана на протяжении одного километра
вдоль по реке гнездится от 1 до 3 пар. В общем довольно многочисленная птица
в свойственных ей стациях, которые заселяет довольно плотно.
Экология* Размножение. Гнезда всегда устраиваются на деревьях
(ива, яблоня, ольха), то нависших над водной поверхностью, то растущих
в стороне до 30—35 метров от берега. Гнездовые постройки располагаются
на различной высоте от земли или воды. Когда гнездо строится над водой, оно
может помещаться совсем низко (около 1,5—2 м над водой), чаще несколько
выше, изредка на высоте 10—12 м. В большинстве случаев гнезда бывают
труднодоступны. Они помещаются или на тонких, перекрещивающихся лозах,
не выдерживающих тяжести человека, или на конце ветви яблони, далеко от
ствола.
По форме и расположению материала гнезда схожи с гнездами других
цапель, гнездящихся на деревьях. Их форма — перевернутый конус, то с очень
крутыми, то напротив, с пологими стенками. Часть тонких веточек расхо-
дится радиально из вершины конуса, где скреплены небольшим количеством
глины или помета птицы. Постройка неплотная, сквозь стенки с боков и снизу
видны яйца. Размеры гнезд значительно варьируют. В одних случаях они
не крупнее гнезд горлинки (Streptopelia orientalis L a t h), в других значи-
тельно больше. Гнездо, найденное на Суйфуне, имело диаметр в 28 см, при
глубине лотка — 6 см (Шульпин, 1936). Гнезда, просмотренные нами на Имане,
оказались значительно меньше. Самое крупное из них имело поперечный
диаметр в 19 см. В маленьком лотке гнезда боковые яйца при большой кладке
лежали несколько выше центральных. Немногие известные у нас гнезда содер-
жали кладки от 3 до 5 яиц. Законченные же кладки, судя по вскрытиям гнездо-
вых птиц, повидимому, могут содержать 7 и даже 8 яиц.
Сроки размножения остаются недостаточно выяснены. На Имане наиболее
ранняя незаконченная кладка в 5 яиц найдена 23 мая. Учитывая, что птица
откладывает первые 3 яйца ежедневно, а остальные с большими промежут-
ками, можно предполагать, что кладка началась 16 мая. Гнезда со свежими
яйцами просмотрены нами и значительно позднее (до 11 июня). На Суйфуне
гнездо в 5 совершенно свежих яиц найдено Шульпиным 4 июня. Яйца имеют
скорлупу — бледноголубого, как у кваквы, цвета. Форма их сильно варьирует.
Одни из них правильной яйцевидной формы, другие сильно вытянуты с одина-
ково округлыми концами. Они мельче яиц кваквы, малой белой цапли, цапли
египетской, но несколько крупнее желтой цапли. Длина яиц с Имана и Суй-
фуна B6) 37,4—43,0 мм х 29,0—31,1 мм, среднее 40,9 х 30,4 мм.
Насиживание начинается после отложения первого яйца. На первых
отложенных яйцах птица сидит мало и величина эмбрионов и птенцов одного
гнезда отличается слабо. Участие полов в насиживании не известно. На гнез-
дах обнаруживались самки, самцы же держались поблизости. Сидящая на
гнезде самка подпускает человека совсем близко. Встав с яиц, вытянув шею
и клюв, она замирает в такой типичной для некоторых цапель позе и слетает
только, когда наблюдатель качнет ветви гнездового дерева. Спугнутая с гнезда,
сравнительно скоро возвращается к насиженным яйцам, но долго не подлетает
к гнезду, кладка которого еще не закончена. Сроки насиживания яиц не выяс-
нены. Повидимому, как и у большинства цапель, птенцы в короткое время
приобретают способность к лазанью и полету. На р. Мо летный молодой экзем-
пляр добыт Пржевальским 12 июля, а с половины июля молодые попадались
444
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
ему вообще нередко (Шульпин, 1936). Вылетевшие из гнезд птенцы вместе
со стариками еще долгое время продолжают придерживаться гнездовых участ-
ков. На Имане старая самка и три ее летных птенца были добыты вблизи
опустевшего гнезда в промежуток времени с 24 по 29 августа. Таким образом,
семья продержалась в гнездовом участке в сущности до времени отлета. Быть
может и самый отлет происходит семьями, а не стаями, и потому его трудно
бывает заметить. Основная забота по вскармливанию птенцов лежит на самце.
Рис. 88. Зеленая кваква (взрослая и молодая птицы)
8 конце июня самцов, летающих за пищей, удается часто видеть не только
с наступлением сумерек, но и в течение целого дня.
Линька. Вследствие недостатка экземпляров в коллекциях линька
изучена слабо. Она начинается в конце июня (Шульпин, 1936). Птицы, добы-
тые в конце июля и в течение августа (ст. ст.), оказались линными, а убитая
9 сентября носила уже осенний наряд, за исключением двух маховых, бывших
еще в пеньках (Маак, 1861).
Пятнистый птенцовый наряд у молодых особей почти полностью сохра-
няется до конца лета следующего года. Окончательный наряд птицы надевают
вероятно со второго года жизни. Самка, добытая на Имане 16 июня, лишь
мелкими деталями в окраске отличается от птенцов, поднявшихся на крылья
и добытых в августе.
Питание. Повидимому, зеленая кваква питается преимущественно
рыбой, хотя указываются и беспозвоночные, также лягушки и т. д. Во всяком
случае при вскрытии птиц обнаружена только мелкая рыба.
Полевые признаки. Чаще всего зеленую квакву удается наблюдать взле-
тающей с берега или невысоко летящей над рекой. Размеры ее во время полета
приближаются к размерам желтой цапли или несколько мельче. Летит довольно
кваква 445
быстро, часто взмахивает крыльями и не столь сильно втягивает шею. Сидя
на берегу и завидя приближающегося человека или лодку, она сначала зами-
рает, вытянув шею, а затем перемещается на короткое расстояние вдоль берега.
Цапелька вновь садится на отмель под крутой берег на торчащий из воды
корень, перевернутую лодку, на нижнюю или среднюю сухую ветвь расту-
щего на берегу дерева, но не садится на верхушки деревьев. При этом
птица редко поднимается выше растущих на берегах деревьев и, как пра-
вило, летит метров на 15—20 над водной поверхностью. Обычно не перебе-
гает по земле, и ее наверное можно поднять там, куда она села. Однако,
раненная в крыло, бегает с замечательным проворством. В отношении чело-
века зеленая кваква ведет себя различно, в зависимости от обстоятельств,
но вообще менее осторожна, чем крупные виды цапель. Голос птицы удается
слышать довольно часто во время спокойного полета или при взлете. По
Мензбиру A916) он подобен короткому глухому карканью, что безусловно
ошибочно. Шульпин A936) передает его звуком «цик-цик-цик», с чем также
трудно согласиться. Голос птицы высокий и резкий, он звучит как резкое
сгильк» или чаще «тиуу».
Описание. Размеры и строение. Мелкая цапля. Вес от 266,0—370,9 г.
Длина крыла самцов и самок A4) 196—213 мм, среднее 201,5 мм. Размерами и внешним сло-
жением напоминает желтую цаплю.
Окраска. Молодые первогодки в отличие от второгодков обладают рыжими краями
кроющих крыла и большим развитием рыжих продольных пестрин на темени.
Молодые птицы второго года жизни отличаются от взрослых пестрым рисунком брюшной
стороны тела, черно-бурой окраской спины, темени и затылка и крайне слабым развитием
на этих частях тела металлического отлива и отсутствием хохла.
Половой диморфизм отсутствует. Общая окраска спинной стороны взрослых особей
серо-буро-стального цвета с хорошо выраженным зеленоватым металлическим блеском.
Нижняя (брюшная) сторона серовато-бурая за исключением белого с темными пестринами
горла и беловатой шеей и грудью. Темя и затылок, с удлиненными перьями хохла — чер-
ные с ярким зеленоватым блеском. Позади глаза буровато-белая полоса, граничащая с почти
черной полосой, расположенной под глазом и идущей к уху. Окраска хвоста сверху близка
к окраске темени и затылка. Крыло того же цвета, как и спина, но вследствие охристых
и белых узких каемок перьев, кажется пестрым. Маховые черные с белыми вершинными
пятнами и краями. Клюв черный с зеленоватым основанием; ноги буро-оливковые; когти
рогового цвета; радужина желтая.
РОД КВАКВЫ NYCTICORAX FORSTER, 1817
Тип N. nycticorax
17. Кваква Nycticorax nycticorax L.
Ardea Nycticorax. Linnaeus. Syst. Nat., изд. X, 1758, стр. 142, южн. Европа.
Русское название. Кваква, квак — народное, звукоподражательное, от голоса
птицы. Известно, по крайней мере, с XVIII века.
Распространение. Ареал. Европа, Африка, южн. и Средняя Азия на
восток до Японии, к югу до Индии, Зондских о-вов и Целебеса; Сев., средняя
и Южн. Америка, Фальклендские и Гавайские о-ва.
Характер пребывания. В южных широтах оседла. В Зап.
Европе, в Европейской и Азиатской части СССР, в Японии и в северных частях
американского ареала — перелетная.
Биотоп. Болотистые, поросшие древесной растительностью местности
вблизи водоемов, реже — камышовые и тростниковые заросли. Вне гнездо-
вэго времени — по берегам озер, прудов, рек и на болотах.
Подвиды и варьирующие признаки. Три слабо различающихся морфологией и биоло-
гией подвида: N. п. nycticorax Linnaeus, I758—европейская, азиатская и африканская
часть видового ареала; N. п. hoactli G me I i n, 1789 — американская часть ареала от се-
вера Орегона, юга Уайоминга и Маннитобы, сев. Квебека и Новой Шотландии, в Канаде,
446
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
через центральную Америку до севера Южн. Америки — до вост. Аргентины; также
на Гавайских о-вах; N. п. cyanocephalus Molina, 1782 — Южн. Америка от юга Перу,
в Чили и зап. Аргентине до Огненной Земли, Фальклендские о-ва.
Карта 82. Распространение кваквы Nycticorax nycticorax
7. N. п. nycticorax, 2. ft. л. hoactli, 3. Я. п. cyanocephalus;
а — граница гнездовий, б — не вполне выясненная граница гнездовий, г — область зимовок европейских
особей.
Обыкновенная кваква Nycticorax nycticorax nycticorax L.
Синоним. Ardea kwakwa. S. G. Gmelin. Novi Comment. Acad. Caes. Petrop. XV,
1771, стр. 542 низовья Дона.
Распространение. Ареал. В Европе гнездится в южн. Португалии и
Испании, в южн. Франции (Домб, ла-Камарг, быть может в деп. Форез и
на о-ве Корсика), нерегулярно на сев.-зап. в Вандее и на озере Гран-лье к югу
от Нанта; в с.-з. Италии, Югославии, Албании, Болгарии, Румынии и на юге
Советского Союза. В последней четверти прошлого века гнездилась в Бельгии
(Тиссельт, 1885—1886 годы и быть может гнездится и теперь, Ферхеейен, 1948).
Несомненно гнездилась в Германии, в последнее время у Геттингена, в Силезии
у Коствица, у Кенигсварта в Лаузице, на озере Драузен, где в 1914 г. были
найдены четыре пары. Гнездится ли теперь — неизвестно. Возможно редкое
и спорадическое гнездование в Польше. В Азии гнездится от Малой Азии,
Сирии, Палестины, Месопотамии, Ирана и Средней Азии до южн. Манчжурии,
Китая (от Чжили до Юннаня, Гуандуня, Фуцзяня), Кореи и Японии (на север
до Хондо), на юг до Филиппинских, Зондских о-вов и Целебеса. В Африке
к северу от Сахары от Марокко до Туниса, также в тропических и субтропи-
ческих частях южной части материка, на Мадагаскаре.
Детали распространения в СССР таковы. Гнездится в Бессарабии, в ни-
зовьях Днепра, в Крыму — как в горной части, так и в степной. Обыкновенна
в вост. Приазовье, на Сев. Кавказе по Кубани и Тереку, во всем низменном
Закавказье, достигая в Армении озера Севан, т. е. примерно высоты 2000 м.
Регулярно гнездится в устьях Волги и Урала. К северу достигает районов
Орла, Воронежа, на Волге —местностей несколько севернее Камышина, на
Урале до ст. Буранной (Зарудный, 1888). Северная граница в Азиатской части
Союза доходит до низовьев Тургая, района Атбасара, далее — Или. К югу
отсюда широко распространена по Средней Азии. В Туркмении гнездится
в поймах Аму-Дарьи, Мургаба и Теджена. Довольно широко гнездится в низ-
менном Таджикистане, в частности у ст. Мельниково, в урочище Джили-куль
КВАКВА 447
на Вахше, в долине Аму-Дарьи и Пянджа от Пархара до Термеза, повидимому,
у Куляба в тугаях по Ях-су. В Таджикистане на гнездовье в горы повиди-
мому не заходит, но в послегнездовое время кочующие птицы появляются
в горах довольно высоко, например 19 и 28 августа встречены на озере Искан-
дер-куль (Иванов, 1940). Птицы наблюдались также в Фергане, преимуществ
венно около Андижана, близ станций Чиназ и Сыр-дарьинской, а к северу
нормально гнездится вниз по течению реки, во всяком случае до широты ст.
Джусалы; редко гнездятся на озере Камышлы-баш. На восток от Сыр-Дарьи
в годы, богатые водой, населяет озера Тели-куль и низовья Сары-су (Спанген-
берг, 1936). Гнездится на Лепсе, найдена в низовьях Чу (Гладков и Гринберг,
1932), по тугаям в дельте Или (Долгушин, 1939), но не приводится для более
восточных частей Семиречья (Кореев и Зарудный, 1906). К северу отсюда еще
нормально гнездится в камышах дельты Тургая (Сушкин, 1908) и найдена
в Адбасарском районе Акмолинской обл. Как исключительный случай гнездо-
вание кваквы было отмечено Зарудным A888) на реке Мечетке в Кара-агаче,
но вообще названный исследователь считал ее редкозалетной птицей для
всего бывшего Оренбургского края, что вероятней; случаи залетов известны
в Прибалтику, в район Пскова и в Башкирию, где один экземпляр был добыт
в начале мая в окрестностях озера Шунгек-куля, другой (дата неизвестна)
в окрестностях г. Уфы, наконец 8 апреля 1947 г. взрослый экземпляр добыт
Бромлеем в Судзухинском заповеднике в Приморье. В Зап. Европе ква-
ква залетает в Англию, Францию, Бельгию, Голландию, Германию, Данию,
Скандинавию, Финляндию, Швейцарию, Польшу даже на Фареры. В Африке
на Азорские, Канарские о-ва. Отмечен залет в Гренландию.
Характер пребывания. В СССР гнездящаяся перелетная
птица. Только в Закавказье в Шемахинском районе и под Ленкоранью
частично зимует. Зимовки здесь носят нерегулярный характер, хотя иногда
птицы зимуют здесь в большом числе. Молодые кваквы зимой неоднократно
отмечались в дельте Волги (Хлебников, 1928), но эта зимовка кончается гибелью
птиц от голода. Весенний прилет ранний, но довольно растянут — с последней
трети марта и до половины мая. Изредка пролет затягивается до последней
трети этого месяца (в пустыне Кызыл-Кум —Зарудный, 1915). В Бессарабии
появляются во второй половине марта, в конце того же месяца под Херсоном.
27 марта на Черноморском побережье Кавказа, в середине марта наблюдалась
в Сальянах (возможно зимующая особь). В конце марта и апреле на Сев. Кав-
казе; 21 и 26 марта, у озера Юссуф-Хана и близ Мары (Дементьев, 1950),
в середине марта по всей Туркмении, у Коканда и в конце этого месяца в Астра-
ханском заповеднике. Однако и здесь основная масса птиц появляется позд-
нее и пролет, как правило, затягивается до конца первой трети мая на Черно-
морском побережье Кавказа и до начала мая в Туркмении. В районе Мариуполя
ранние появляются в первой трети апреля, но основная масса около половины
этого месяца (Боровиков, 1907). В районе Харькова прилет начинается также
приблизительно с середины апреля, валовой происходит в конце апреля, пол-
ностью заканчивается в первой трети следующего месяца. Под Киевом кваквы
прилетают в конце первой трети мая, изредка A925) уже 28 апреля. Несколько
раньше прилетают птицы на Сыр-Дарью в район Кзыл-Орды, где первые
особи появляются в начале апреля. Основная же масса прилетает в половине
этого месяца. Устьев Урала птицы также достигают в конце апреля.
Осенний отлет имеет еще более затяжной характер. Уже в конце июля,
начале августа собравшиеся стайки кваквы предпринимают широкие кочевки
в поисках пищи. В конце августа начинается слабо выраженное движение
птиц к югу, которое постепенно усиливается, приобретая характер настоящего
отлета. В конце сентября во многих северных районах своего распростране-
ния численность кваквы резко сокращается, но на юге, в местах, обильных
пищей, они задерживаются чрезвычайно долго. Под Харьковом осенний отлет
448 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
начинается в некоторые годы с последних чисел августа, но чаще с первых
чисел сентября. В конце сентября пролет достигает наибольшей силы — птицы
летят стаями в сотни особей. Затем численность пролетных квакв убывает
и движение к югу совершенно заканчивается в самом конце сентября или
в первых числах октября. Одновременно начинается пролет в низовьях Днепра,
достигая наибольшей силы в середине сентября и заканчиваясь в конце первой
трети следующего месяца. В Бессарабии кваквы улетают в октябре (ст. ст.,
Вальх, 1912). Заметный осенний пролет в районе Ростова начинается в послед-
ней трети сентября. В низовьях Волги по Воробьеву A936) отлет происходит
в сентябре, запоздавшая молодь задерживается до половины ноября, а по
Хлебникову A928) иногда кваквы наблюдаются в приморской части Волги и
в зимнее время. В центральной части Сев. Кавказа по Беме A926) пролет
происходит в сентябре — октябре, но на Каспийском побережье в Кизлярском
районе кваквы задерживаются на весь ноябрь и первые числа декабря. Взрос-
лый самец добыт здесь мною в окрестностях с. Юрковки б декабря. На Черно-
морском побережье Кавказа осенний пролет происходит в самом начале сен-
тября. Данные о сроках отлета для Азиатской части Союза отрывочны. Заруд-
ный A915) отметил крайне раннее появление квакв в пустыне Кызыл-Кум.
2 и 3 августа птицы летели прямо на юг. На нижней Сыр-Дарье Северцов
слышал квакв 4 октября, но по нашим наблюдениям в середине октября на
Сыр-Дарье кваквы отсутствуют. В Туркмении осенние кочевки квакв начинаются
с конца июля — начала августа, а в сентябре и октябре идет осенний пролет.
Перелеты совершаются, главным образом, в ночное время, начинаясь
на вечерних зорях, несколько ослабевая в глухую полночь и заканчиваясь
на восходе солнца, но изредка происходят и при дневном освещении. Птицы
летят то крупными, то небольшими стаями, обычно на значительной высоте,
выстроившись поперечной линией и часто издавая свои характерные крики.
Нередко кваквы летят совместно с серыми цаплями. Европейские кваквы
зимуют в Африке к югу от Сахары. Появление в западноевропейских местах
гнездовья от конца марта и в апреле, обычно около середины последнего ме-
сяца. В конце июля — начале августа молодые птицы начинают кочевать, не
придерживаясь определенных направлений; в это время и отмечаются залеты
квакв. Отлет взрослых в сентябре — октябре. Годовалые птицы, неполово-
зрелые, остаются в большинстве в первое лето своей жизни на местах зимовок.
Результаты кольцевания. Кольцевание квакв указывает на
большое развитие у них гнездового консерватизма: птицы, окольцованные
птенцами в Астраханском заповеднике, добывались там и в последующие
годы, в некоторых случаях через четыре, пять, шесть и семь лет после кольце-
вания. Зимовки астраханских квакв обнаружены у озера Чад в Африке. На
пролете птицы добывались в Малой Азии у Анкары. Весьма интересны находки
на территории СССР квакв, окольцованных птенцами на озере Балатон в Вен-
грии. В первую же осень одна из них встречена на Маныче, другая — у Куй-
бышеву, прочие — через два года и восемь лет — осенью в низовьях Волги.
Последние находки указывают на вероятное гнездование квакв далеко от места
рождения. К этому надо добавить встречу летом выведшейся в дельте Волги
птицы и находку другой птицы того же происхождения весной в Каракалпа-
кии (Дементьев).
Биотоп. Гнездовый — небольшие обособленные группы деревьев, рощи,
древесные питомники, тугайные заросли или более крупные массивы леса,
то пятнами разбросанные среди болот, сети озер, по берегам и островкам рек
или поблизости от них, то расположенные далеко (до 5—7 км) от ближайших
кормовых стаций. При недостатке древесной растительности — камышовые и
тростниковые крепи по берегам рек, водохранилищ, озер или более обширных
водоемов. Кормовые биотопы — берега рек, прудов, озер, камыши, участки
открытой воды среди болот, но не настоящие, поросшие травой болота и не
кваква 449
сырые луга. Не избегает близости человека, гнездясь у селений и кормясь на
водоемах, постоянно посещаемых рыбаками.
Численность. В местах нормального обитания квакву удается
встречать чаще всех других цапель. Число особей в некоторых гнездовых
колониях превышает 2—2,5 тысячи. У нас становится более редкой птицей
близ северных и восточных границ своего распространения. Для Союза в целом
многочисленная птица.
Экология. Размножение. Половая зрелость в возрасте одного
год&, хотя, повидимому, не все такие птицы размножаются. Настоящая коло-
ниальная птица, отдельными парами не гнездится. Колонии размещаются
на деревьях, в камышовых и тростниковых крепях. Изредка образует само-
стоятельные поселения, но значительно чаще присоединяется к колониям
других болотных и водяных птиц или к колониям грачей. К самостоятельным
и смешанным колониям квакв при их гнездовании на деревьях очень часто
присоединяются отдельные пары черных аистов, черных коршунов, в дуплах
гнездятся скворцы, а на Сыр-Дарье пустующие гнезда, расположенные наибо-
лее низко, занимаются парами пустельги, ушастой совы и буланой совки.
Гнезда устраиваются на различных древесных породах. В районе Харькова
и в низовьях Днепра наиболее охотно кваквы гнездятся большими колониями
в ольховых рощах, особенно в тех случаях, когда они затоплены полой водой;
в дельте Волги на деревьях ивы; под Ленкоранью то на ольхе среди водохра-
нилищ — истлей, то в сухих широколиственных лесах на тутовнике, кара-
гаче, дубе, грецком орехе. В Азиатской части Союза кваквы предпочитают
устраивать гнезда на разнолистных тополях в тугайных зарослях. Так, на
Теджене у Карабента до 500 пар квакв гнездилось на разнолистных тополях,
причем на одном дереве размещалось до 20—30 гнезд (Дементьев, 1950). На
о-ве Баубен-арал (Сыр-Дарья в районе Кзыл-Орды) кваквы заселяли тугай-
ные заросли, разместившись на низкорослых деревьях разнолистного тополя
и джиды. В том же районе другая крупная колония устроилась в роще кара-
гача среди закрепленных бугристых песков (Спангенберг).
Гнезда помещаются на различной высоте, что в первую очередь связано
с высотой гнездовых деревьев и составом колониального поселения. В местах,
где кваквы гнездятся совместно с серыми цаплями на высоких ольхах, их
гнезда устраиваются на высоте в полдерева, тогда как вершины занимаются
гнездами более крупных их сородичей. В смешанных колониях, размещенных
на больших деревьях под Ленкоранью, основная масса гнезд квакв сосредото-
чивалась несколько выше среднего яруса деревьев на одной высоте с гнездами
малых бакланов. Ниже гнездилась чепура-нужда, египетская и желтая цапля,
тогда как вершины были заняты крупными гнездами серых цапель и больших
бакланов. В дельте Волги гнезда квакв также помещаются относительно
низко, обычно на той же высоте, где гнездится чепура-нужда и каравайка.
В одной из колоний на Сыр-Дарье, где кваквы гнездились в роще карагача,
их гнезда занимали верхний ярус деревьев, чуть ниже основной массы грачи-
ного поселения. В другой колонии в тугае постройки квакв густо покрывали
вершины низкорослых деревьев джиды и тополя. Часть гнезд была выстроена
на высоте не более двух метров. При гнездовании в камышовых и тростнико-
вых зарослях легкие постройки квакв, как правило, строятся выше гнезд
колпиков, больших бакланов, иногда караваек. При этих условиях гнезда
помещаются до 1,5 му редко до 2 м от поверхности воды. На деревьях гнезда
укрепляются в разветвлении горизонтальных и вертикальных веток, под
Ленкоранью часто среди перепутанных колючих лиан, в тростниках и камышах
на прошлогоднем заломе этих растений. Характер и форма гнезд типичны
для данной группы, то-есть перевернутый конус с просвечивающими стенками.
Вся постройка и особенно в тех случаях, когда занимаются и достраиваются
старые гнезда, крупнее и грубее, чем у других мелких цапель, гнездящихся
29 Птицы СССР, т. II
450 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
на деревьях* В крупных смешанных колониях кваквы часто занимают старые
гнезда других мелких цапель, перестраивая их по своему вкусу. Вследствие
этого гнезда кваквы весьма изменчивы в размерах и форме. В постройке гнезда
принимают участие обе птицы гнездовой пары. Самец приносит гнездовый
материал, а самка укладывает его и охраняет от других птиц. По наблюдениям
в Астраханском заповеднике иногда приносит материал и самка (Ромашова,
1940). Материал собирается в непосредственной близости колониального гнез-
довья, а часто вытаскивается из гнезд, расположенных по соседству.
Спаривание происходит в ночное время. В начале размножения стайная
жизнь квакв нарушается слабо. Они продолжают стаями летать на кормежку.
Количество яиц в отдельных кладках от 3 до 5, чаще 4. Размер кладки
колеблется в различные годы, и в отдельных частях ареала. Несколько сотен
гнезд, осмотренных мною под Ленкоранью в два весенних сезона, содержали
законченные кладки в 3 яйца и лишь одна кладка —4 яйца. Не более 4 яиц
содержали кладки на Сыр-Дарье в районе Кзыл-Орды. В Астраханском запо-
веднике, судя по коллекционным материалам, нередки кладки в 4 яйца. По
4 яйца в кладках отмечено также в Туркмении (Дементьев, 1950). В районе
Харькова кладки содержат обычно 4—5 яиц (Сомов, 1897). Яйца имеют пра-
вильную яйцевидную форму и лишь, как редкое уклонение, вытянутую,
с одинаково-острыми концами. Окраска скорлупы свежеснесенных яиц
зеленовато-голубая (бледнее, чем у большинства других цапель, имеющих
голубые яйца, но насыщеннее, чем у египетской цапли). Уже к концу насижи-
вания яйца квакв сильно выцветают. Размер яиц C8) 44,0—52,0 х 32,0-^36,1 мм,
среднее 47,5 х 34,3 мм. По Уайзерби A939) размеры яиц A00) 43,7—
56,5 х 31—38, в среднем 49,52 х 35,3 мм.
С момента прилета и до начала гнездования проходит около 2—3 недель.
Сроки размножения у отдельных пар растянуты на значительное время, что
происходит вследствие неединовременного заселения колоний; годовалые
птицы, по всем вероятиям, гнездятся позднее; гнездовое поселение продол-
жает еще пополняться, когда многие пары уже успели закончить кладки и
приступили к насиживанию. Изменение сроков наблюдается также в зависи-
мости от широты местности. Наиболее рано яйцекладка происходит в Закав-
казье. Под Ленкоранью, где кваквы в некоторые годы частично зимуют и
вообще появляются рано, 15 апреля в гнездовых колониях многие гнезда
уже бывают закончены, в некоторых отложены первые яйца. В Астрахан-
ском заповеднике по данным Ромашовой A940) кваквы впервые появились
на колониальном гнездовье 29 марта, но долгое время проводили здесь только
день, улетая на кормежку с наступлением вечера, массовое появление на коло-
нии совпало с последней декадой апреля. С этого времени птицы приступили
к размножению. По Воробьеву A936) и Дубининым A940) яйцекладка начи-
нается в 20 числах апреля. Под Харьковом «ненасиженные» полные кладки
в 4—5 яиц бывают в середине мая (Сомов, 1897), то-есть почти на месяц позд-
нее, чем под Ленкоранью. Сравнительно позднее начало гнездовья отмечено
в Азиатской части Союза. У Теджена 18 мая многие гнезда оказались еще
не вполне достроены, в некоторых 1—3, в немногих 4 яйца. На Сыр-Дарье
в районе Кзыл-Орды в конце апреля кваквы появляются на колониальных
гнездовьях, но не ранее 10—12 мая в гнездах удается находить яйца. Начи-
ная с указанных чисел под Ленкоранью, в низовьях Волги и на Сыр-Дарье
яйца квакв в гнездах можно находить в мае, июне и в начале июля. В Астра-
ханском заповеднике Воробьев A936) нашел кладку даже во второй половине
июля и справедливо считает ее повторной после потери первой.
Насиживание начинается с первого яйца. В нем участвуют оба родителя,
но самец сидит на гнезде меньше, чем самка. Продолжительность насижива-
ния 21—22 дня. К концу насиживания почти исчезает инстинкт самосохране-
ния. Спугнутая с яиц птица тут же садится на гнездо Вследствие растянутого
КВАКВА 451
срока яйцекладки птенцы появляются также в различные сроки. Первые
етгенцы в районе Ленкорани в возрасте от 1 до 3 дней обнаружены при посеще-
нии колонии 10 мая. В Сальянах 2 июля Сатунин A907) наблюдал птенцов,
которые уже покинули гнезда, но еще не умели летать. На озере Гилли (гор-
ная Армения) 10 июня осмотрены уже оперившиеся птенцы, а в другом гнездо-
вом сезоне 4 июня летные молодые (ЛяйстериСоснин, 1942). В Астраханском
заповеднике в первой половине мая — маленькие птенцы, вполне летные
в конце июня и, как исключение, сильно запоздавшие нелетные в 20-х числах
августа (Воробьев, 1936). По данным Дубининых A940) появление первых
птенцов здесь происходит между 5—15 мая, массовое появление между 10—
20 июня *. В Харьковской обл. по Сомову A897) самостоятельные птенцы
в первой трети июня. На Теджене летная молодь еще в начале июня. Таким
образом гнездовой период у кваквы начинается в середине апреля (в наиболее
южных широтах) и затягивается до конца второй трети августа. Пока птенцы
малы и беспомощны, одна из птиц гнездовой пары постоянно остается на гнезде.
Позднее оба родителя заняты добыванием пищи. Отлет за пищей и возвра-
щение основной массы квакв совершается в сумерки, когда птенцы подрастут
и в дневные часы.
Оперившиеся птенцы вскоре начинают лазать по гнездовым деревьям,
собираясь в большом количестве на крайних ветвях, где ожидают появления
родителей с пищей. При гнездовании в Камышевых и тростниковых зарослях,
молодь группируется на крупных покинутых гнездах больших бакланов
(Сыр-Дарья). Как только молодые начинают летать,из них и взрослых особей
образуются стайки, которые все шире кочуют в поисках корма.
Рост птенцов до вылета из гнезд продолжается около 35 дней. За этот
период, считая с момента вылупления и до конца постэмбрионального разви-
тия, их вес возрастает с 25 до 650 г (Хейнрот, 1926—1934). Наиболее интенсив-
ный рост маховых наблюдается у птенцов кваквы с 21 дня жизни, что совпа-
дает с падением скорости общего роста.
Линька. Схема линьки —как у других цапель. Молодые птицы в гнездо-
вом наряде сменяют мелкое оперение в первую осень жизни между августом
и декабрем. В дальнейшем имеются две линьки в году, по некоторым авторам
(Уайэерби, 1939; Нитхаммер, 1938) обе полные. Полная предбрачная линька
между январем — февралем и апрелем, полная послебрачная линька между
августом и ноябрем. Остается все же неясным, является ли предбрачная
линька полной или ограничивается мелким оперением. Полный наряд, не отли-
чимый по окраске от старых птиц, приобретается в годовалом возрасте: это
второй зимний (а затем — второй брачный) наряд. Смена брачного оперения
начинается с выпадения украшающих перьев головы и мелкого оперения.
Питание. Животная пища; в питании молодых и взрослых основное
значение имеет рыба и лягушки. Птенцы первое время вскармливаются отрыг-
нутой иолупереварившейся пищей. По Воробьеву A936) в дельте Волги птенцы
вскармливаются рыбой, лягушками и водяными крысами, а по Дубининым
A940) кроме того и личинками водолюбов и других крупных насекомых. В Бес-
сарабии кваквы поедают рыбу, водяных насекомых и семена различных водя-
ных растений (Остерман, 1912). По Сомову A897) в районе Харькова кваквы
ночью охотно посещают илистые берега водоемов, где поят скот и где есть
навоз. Здесь они отыскивают различных червей и личинок лошадиных оводов.
Зимнее питание квакв под Ленкоранью состоит из рыб (семейства карпо-
вых), водных насекомых Macrodytes, Colymbetes, и др. и каких-то растений
(Исаков и Воробьев, 1940).
Полевые признаки* В природе хорошо отличается от всех наших цапель раз-
мерами (несколько крупнее вороны), общим коренастым сложением, окраской
• в вост. Китае вылупление птенцов квакв около середины июня (Латуш, 1034).
452 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
оперения и большими яркокрасными у взрослых птиц глазами. По крику,
издаваемому во время ночных перелетов на кормежку и во время сезонных
перемещений, может быть спутана разве только с выпью. Вообще кваква
более криклива, чем другие цапли. Ее обычный голос звучит как «квау, квау»
или «квак, квак, квак», повторяющееся несколько раз со значительными интер-
валами. Наиболее часто голос кваквы удается слышать с наступлением суме-
рек высоко в воздухе, иногда при взлете. На колониальных гнездовьях взрос-
лые и молодые кваквы чрезвычайно крикливы, причем их голос передать
Крайне трудно.
Рис. S9. Кваква
Кваква —типичная ночная птица. Повышенная жизнедеятельность начи-
нается с наступлением сумерек и заканчивается незадолго до восхода солнца.
В дневные часы кваквы стайками спокойно сидят на деревьях или в камыше
с краю чистого плеса, прячась от лучей солнца. Иногда их удается видеть
летящими, днем, причем во время полета они сильно втягивают и без того
короткую шею и обычно выстраиваются в поперечную линию. Будучи потре-
воженными в камыше, взлетают в воздух довольно быстро, но, под-
нявшись на известную высоту, летят медленно, легко, плавно взмахивая
крыльями.
Кваквы не очень осторожны, близко подпускают к себе человека, неожи-
данно взлетая почти вертикально. При добывании пищи птицы или медленно
бродят по мелководью или чаще неподвижно стоят, всматриваясь в воду.
Более древесные птицы* чем большинство других цапель.
МАЛАЯ ВЫПЬ ИЛИ ВОЛЧОК 453
Описание. Размеры и строение. Сложение плотное, голова большая, клюв сжа-
тий с боков, высокий и относительно короткий, с выпуклым коньком дистальной части над-
клювья, большие глаза. В брачном наряде на голове хохол из двух-четырех узких и длинных
перьев. Формула крыла (не считая первого рудиментарного махового): 2=3^4>1>5>6...
Вырезки на внутренних опахалах 1-го и 2-го первостепенных маховых, на наружных
опахалах 2-го и4-го маховых. Вес самцов и самок(8M50—650, в среднем 590г (Воробьев и Иса-
ков, 1940), по Хейнроту до 750 г. Крыло самцов и самок B3) 281—312, в среднем 203,2 мм.
Клюв около 65—80, цевка около 65—75, средний палец около 75—80 мм (с когтем).
Окраска. Пуховой птенец бледнобурый, с удлиненными пушинками на темени
па спинной стороне, брюшная сторона беловата. В гнездовом наряде темя черно-буроватое,
на каждом пере белое настволье; спинная сторона буровато-черноватая, с широкими белыми
наствольями с беловатыми вершинами перьев на плечевых; брюшная сторона охристо-бело-
ватая с темнобурыми продольными пестринами; последние особенно развиты на груди, где
перья имеют и буроватые каемки; маховые темнобурые с беловатыми краями, рулевые серо-
вато-бурые с более светлыми вершинами. Клюв бледнобурый, ноги зеленовато-серые, ра-
дужина бурая. Первый зимний (предбрачный) наряд, как предыдущий, но светлый; рисунок
на голове охристо-буроватый, спина почти одноцветно бурая.
В первом брачном наряде (в этом возрасте, примерно годовом, некоторые особи квакв
уже размножаются) перья темени черноваты с беловатым стволом, спинная сторона бурая
с сероватым оттенком и слабым зеленоватым отливом и со светлыми каймами перьев; брюш-
ная сторона охристо-беловатая, с более или менее развитыми буро-серыми каймами.
Второй брачный наряд — окончательный. Верх головы, плечевые черные с синеватым
металлическим отливом. Длинные украшающие перья головы белые. Лоб, надбровная
полоса и брюшная сторона белые. Крылья, поясница, хвост серые. Маховые серые, темнее
кроющих крыла. Клюв черный, радужина красная, ноги в период спаривания розоватые,
в остальное время желтые. Уздечка в разгар периода размножения тёмнокрасная или черно-
ватая, затем бледнее до зеленоватой или желтоватой. Второй зимний наряд от брачного
отличается тем, что голова и плечевые с зеленоватым металлическим отливом. См. также
выше, об окраске неоперенных частей.
РОД ВОЛЧКИ IXOBRYCHUS BILLBERG, 1828
Тип At delta minuta
18. Малая выпь или волчок Ixobrychus minutus L.
Ardetta minuta. Linnaeus. Syst. Nat., изд. XII, 1, 1766, стр. 246, Швейцария.
Русское название. Волчок народное.
Распространение. Ареал. Средняя и южн. Европа, Зап. Сибирь, Перед-
няя, Средняя и ю.-з. Азия на востоке до с.-з. Индии, Кашмира, Непала; Африка,
Мадагаскар; Австралия, Новая Зеландия.
Характер пребывания. Оседлая в южных частях ареала,
в СССР и в Зап. Европе — перелетная птица. Европейские птицы зимуют
в Африке.
Биотоп. Гнездится в Камышевых, тростниковых, кустарниковых зарос-
лях и среди древесной растительности, разросшейся по берегам и внутри водое-
мов. Кормится на крупных и мелких водоемах разнообразного характера.
Подвиды и варьирующие признаки. Шесть подвидов, отличающихся окраской и де-
талями биологии /. m. minutus Linnaeus, 1766—Европа, Азия, Африка к северу от Сахары;
/. т. payesii H a r 11 a u b, 1858 — тропическая Африка от Сенегала и Судана к югу до Кап-
ской провинции; ю.-з. Аравия; /. т. podiceps Bonaparte, 1855—Мадагаскар; /. т. dubius
Ma thews, 1912 — ю.-з. Австралия; /. m. alisteri M a t h e w s, 1913 —вост. Австралия
от Квинсленда до Виктории; /. т. novaezelandiae Potts, 1971 — Новая Зеландия.
Северная малая выпь Ixobrychus minutus minutus L., 1766
Распространение. Ареал. Гнездится в Европе всюду, кроме Британ-
ских о-вов, Дании, Скандинавии, сев. части Советского Союза, Греции; на о-вах
Средиземного моря только на Сицилии и Майорке. В Азии — в Малой Азии,
Сирии, Палестине, Месопатамии, Иране, Афганистане, с.-з. Индии и Кашмире;
в Средней Азии и ю.-з. Сибири (подробности см. ниже). В Африке от Марокко
454
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
до Туниса. Область зимовок в Африке к югу до Капской Земли и отчасти
в южн. Азии. На зимовках находится с начала октября до конца апреля.
Случайные зимовки отмечены в Венгрии, Англии, Южн. Франции. Сев. гра-
ница в СССР пересекает средние части Эстонии, достигает Ленинграда (Туга-
ринов, 1947), откуда снижается к Новгороду (Бианки, 1910) и подходит к Весье-
гонску, где гнездящиеся птицы обнаружены недавно в Дарвинском заповед-
нике (Спангенберг и Олигер, 1940). Далее на восток граница идет через Горь-
ковскую обл., по средней Каме, через Кировскую обл. и с севера огибает
Молотов (Ушков, 1927). В Сибири граница, несколько снижаясь, проходит
Карта 83. Распространение волчка Ixobrychus mi nut us
7. Y. т. minutus, 2. V. т. payesii, 3. V. m. podiceps, 4. Y. m. dublas, 5. Y. m. alls ten, 6. Y. m. novaezt-
landiae;
a — граница гнездовий, б — область зимовок.
южнее Тюмени через Шадринск и Курган, идет к Барнаулу и к Бийску. Отсюда
вдоль зап. Алтая и Тарбагатая граница идет в Киргизию. К югу от очерченной
сев. границы малая выпь гнездится в подходящих местах до южн. пределов
Союза ССР.
От гнездового ареала изредка залетает далее к северу. Залеты известны
на Британские и Фарерские о-ва, в Исландию, Данию, Южн. Скандинавию,
в разные районы Европейской части СССР, на Азорские и Канарские о-ва,
на Мадейру и т. д.
В Зап. Европе и в СССР — гнездящаяся перелетная птица.
Характер пребывания. В СССР гнездящаяся, перелетная
€1тица. Единственное указание о зимовках вида в ю.-з. Туркмении у Чатлы
на Атреке (Житников, 1900) нуждается в подтверждении. Прилет в южн.
части страны относительно ранний, более поздний в средней полосе и близ
северной границы ареала. Наиболее рано малые выпи появляются весной
в Туркмении. Первые птицы отмечены здесь Шестоперовым A037) во второй
МАЛАЯ ВЫПЬ ИЛИ ВОЛЧОК 455
трети марта, а по Дементьеву A950) в середине марта, но обычно пролет сильно
затягивается и главная масса выпей летит в апреле. Между 20 и 23 апреля
пролетные наблюдались и в вост. Иране (Зарудный, 1903). Вероятно пролет
затягивается еще дольше, так как в пустыне Кизыл-Кум у колодцев Дор-
Кудук и Аяк-Кудук отсталопролетные особи добывались Зарудным 21 мая
и 1 июня. На Сыр-Дарье в районе Кзыл-Орды малые выпи прилетают не ранее
5—12 апреля, и пролет идет весь апрель (Спангенберг). До средины мая пролет
идет в пределах Таджикистана (Иванов, 1940).
По нашим наблюдениям в вост. Закавказье (под Ленкоранью) в самом
начале апреля малые выпи уже не представляют редкость на лесных истылях.
Данные Сатунина A913) и Вильконского A894) о прилете на Черноморском
побережье Кавказа в конце первой трети мая, вероятно, ошибочны. В Крым
малые выпи прилетают уже в последней трети марта, а в различные даты
апреля появляются уже во многих частях Украины. Наиболее ранний прилет
в районе Мариуполя отмечен 2 апреля, но в другие годы значительно поздней,
например, 25 апреля (Боровиков, 1907). Под Харьковом первые особи приле-
тают в половине апреля, массовый прилет происходит приблизительно с 23
апреля и до 2 мая (Сомов, 1897). Не ранее половины апреля отмечалась эта
выпь под Уманью, а в Бессарабии в конце первой трети мая, в районе Куй-
бышева в конце апреля и 15 мая у Пскова (Зарудный, 1910).
Осенний отлет начинается в августе и затягивается до наступления настоя-
щей осени. В саду г. Барнаула молодая птица была поймана 1 сентября (Вели-
жанин, 1927). Пролет в районе Молотова происходит в течение всего августа
(Ушков, 1927). В конце августа интенсивный пролет идет на Рыбинском водо-
хранилище, в половине сентября по нашим наблюдениям попадаются еще под
Москвой. В Рязанской обл. несомненно пролетный экземпляр был добыт
18 сентября (Хомяков, 1901). В последней трети августа начинается пролет
у Тамбова. Шнитников A913) отмечает, что из-под Минска вероятно последние
птицы отлетают не ранее конца первой трети октября (экземпляр был добыт
8 октября). В августе летят через Крым, 15 августа экземпляр добыт
у Алушты. В районе Дзауджикау осеннее движение происходит между авгу-
стом и серединой октября (Беме, 1926). На Черноморском побережье Кавказа
малые выпи летят в последней трети сентября. Из района Куйбышева малые
выпи исчезают в начале октября, а на Сыр-Дарье редкие особи еще задер-
живаются в районе Кзыл-Орды до 17 октября (Спангенберг). Несомненно
пролетные особи у Ташкента отмечались 8 и 19 сентября (Сатаева, 1937),
В Туркмении пролет начинается в конце августа, сильный пролет идет в сен-
тябре, а в подгорной полосе Копет-Дага экземпляры добываются в октябре
(Дементьев, 1950). 20 сентября малые выпи наблюдались Северцовым в ущелье
Кзыл-Ата в пределах Таджикистана (Иванов, 1940), 20 ноября у Кульджи
(Шольцман, 1897) и до 30 октября в вост. Иране (Зарудный, 1903).
Во время пролетов малые выпи летят одиночками, никогда не образуя
стай. Летят в ночное время, а дневные часы птицы проводят в тростниковых
зарослях по берегам озер, в перелесках среди болот, а иногда и в мало подхо-
дящей обстановке. Так на южн. побережье Крыма малые выпи часто останавли-
ваются на дневку в мелких кустарниках дуба и бука, покрывающих сухие
холмы.
Биотоп. Гнездовый. Крупные и мелкие озера, пруды, водохранилища —
истыли в Закавказье, а на Украине и степные ставки с хорошо развитыми зарос-
лями тростника, камыша, ольхи, ивы, сосредоточенными то по берегам, то
внутри водоемов. Селятся также в речных старицах и разливах, если среди
последних есть участки, занятые тростниками, ольховым лесом, старыми
низкорослыми вербами, переплетенной хмелем, черемухой, или тугайными
зарослями—в Туркмении. На Рыбинском водохранилище заселяет также затоп-
ленные сосняки. Птицы избегают водоемов, расположенных среди сплошных
456 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
лесных массивов и предпочитают водоемы или среди открытого ландшафта
или где участки пойменного леса чередуются с открытыми местностями. Одним
словом, водоемы степи, лесостепи или площади, приобретшие лесостепной
характер вследствие вырубок, —коренные гнездовые места данного вида.
В Туркмении, однако, обитает не только по крупным рекам, но и по горным
ручьям (Шестоперов, 1937).
Кормовой биотоп малой выпи несколько шире гнездового. Если пищи
много, кормится в гнездовом участке, но часто вылетает за его пределы, посе-
щая канавы, отдельные наполненные водой ямки среди лугов, отмели озер
и рек. В конце размножения, когда многочисленное подросшее потомство
требует особенно много пищи, птицы предпринимают более далекие вылеты,
не превышающие однако 1 км. Местами (Сыр-Дарья, Аму-Дарья) малая выпь
поселяется и устраивает гнезда в глухих участках Камышевых крепей. Чаще
она встречается в сельскохозяйственных угодьях, где совсем не редко кор-
мится и строит гнездо в непосредственной близости от человеческого жилья.
Численность. Численность волчков кажется ограниченной. Но
такое впечатление во многих случаях ошибочно. Плотность населения местами
оказывается чрезвычайно густой. Волчок в большом количестве встречается
в Бессарабии (Остерман, 1915), во множестве населяет большую часть степной
и лесостепной Украины, где положительно каждый ставок, каждое подходя-
щее место реки оказывается заселенным хотя бы парочкой этих птиц, очень
обычна на гнездовье в Приазовье. На юге средней полосы Европейской части
Союза — она только обыкновенна, становясь более редкой ближе к сев. гра-
нице. На всем Кавказе в общем немногочисленна. Еще с меньшей плотностью,
исключая отдельных пятен, населяет водоемы Средней Азии и Зап. Сибири,
где местами становится положительно редкой. Несмотря на сравнительно
ограниченное распространение, которое значительно уже, чем у большой выпи,
общая численность этой цапли, вероятно.больше, чем названного вида.
Экология. Размножение. Половая зрелость в годовалом возрасте.
Отдельные пары всегда гнездятся обособленно, занимая большой гнездовый
участок, колоний не образует. Там, где птиц много, а площадь благоприятных
мест для устройства гнезд ограничена, как, например, на степных ставках
Украины, две —три пары поселяются на небольшом пруде. Гнездо от гнезда
в таких случаях бывают расположены метрах в 50, а иногда и ближе.
В зависимости от местных условий, гнезда сооружаются то в густых
тростниках и камышах среди водоемов, то на затопленных водой деревьях
ольхи, вербы или на кустарниковой иве, иногда на терновнике. Ряд авторов
указывает, что гнезда помещаются в тростниковых и камышовых зарослях,
другие находили их на вербе и иве, отмечают гнездование в одних и тех же
районах, и в тростниках, и на древесной или кустарниковой растительности.
Автор, много сталкивавшийся с гнездами малой выпи в различных частях
Союза, приходит к следующему выводу. Коренное место гнездовья малой выпи—
несомненно тростниковые и камышовые заросли, чем гуще они, тем более
привлекают птицу. Гнездо в этих случаях помещается низко над водной по-
верхностью. Оно то касается своим основанием воды, то устраивается на согну-
тых стеблях до полуметра над водою. Так устраиваются гнезда на степных
ставках Украины, на прудах средней полосы Союза. На указанных водое-
мах уровень изменяется относительно слабо и гнезда редко подвергаются за-
топлению.
В противоположность этому на обширных озерах Сыр-дарьинской ста-
рицы, в районе Кзыл-Орды, где уровень подвергается резким колебаниям,
часть птиц гнездится на кустарниках среди камышовых крепей и на прибреж-
ных ивовых порослях. Это уже вторичное явление. Оно особенно резко выра-
жено в плавнях Днепра и Волги. Подавляющее большинство гнезд малой
выпи помещается здесь на кустарниковой иве. на черной ольхе, обычно на
МАЛАЯ ВЫПЬ ИЛИ ВОЛЧОК 497
высоте в рост человека и значительно выше — до 2,5 м — на толстых, но низко-
рослых вербах. Так же гнездится большинство малых выпей в низовьях Куры,
на истылях под Ленкоранью, в Куйбышевской обл., где осмотренные гнезда
помещались на высоте от 65 см до 2 м* и в ряде других районов Союза.
При гнездовании в тростниках основной материал составляют стебли и
листья тростника. В начале гнездо имеет форму перевернутого конуса, но
позднее растаптывается и становится плоским. По данным Остермана A915)
в Бессарабии диаметр гнезда 23 см, высота 10 слс. В Харьковской обл. гнезда
имели поперечный диаметр в 25 см, высоту 11 см, диаметр лотка 10 см, глу-
бину 2,5 см (Сомов, 1897). Гнезда, построенные на кустарниках и деревьях
имеют некоторое сходство с гнездами горлинок. Их форма—хорошо выраженный
перевернутый конус. Материал —тонкие ольховые и ивовые прутики, расхо-
дящиеся в наклонном направлении из центра, в небольшом количестве ис-
пользуются также стебли камыша, и бывает ничтожная выстилка из зеленого
мха или из сухих толстых стеблей травы. Постройка чрезвычайно легка и
яйца видны сквозь боковые стенки. Диаметр вновь построенного гнезда от
17 до 21 см, диаметр лотка 5—б см, так что большая кладка не помещается
в нем, и боковые яйца, опираясь на гнездовые стенки, лежат несколько выше
центральных* Высота гнезда вначале около 12 см, позднее от 6,5 до 5 см. Гнездо
строится несомненно обеими птицами в течение 2—4 дней. Материал достав-
ляется и самкой и самцом. Птицы держат прутик в клюве не поперек, а по его
длине, что свойственно многим цаплям. Часть прутиков они находят близ
гнездового дерева, но часть приносят, видимо, издалека. Самка стоит внутри
начатой постройки и размещает принесенные веточки. При гнездовании в тро-
стниках гнездо, вероятно, подправляется птицами в течение насиживания и
подрастания молоди. К моменту вылета птенцов гнездо более массивно, чем
только что законченное.
Число яиц в кладках колеблется от 4 до 9. В Бессарабии их бывает б—9
(Остерман, 1915), в низовьях Днепра до 9 (Вальх, 1911), в районе Харькова
5—6 (Сомов, 1897), 5—6 в Куйбышевской обл. (Карамзин, 1901), в Турк-
мении 4—9 (Шестоперов, 1937). Яйца откладываются ежедневно. Форма яиц
в редких случаях правильная, яйцевидная, но обычно с одинаково острыми
концами. Скорлупа яиц шероховата и лишена блеска. Окраска скорлупы
вначале белая, несколько зеленоватая. К концу насиживания яйца сильно
загрязняются птицами. 6 июня 1923 г. в окрестностях ст. Водяной в 60 км
от Харькова автором была найдена кладка, свежие и едва насиженные
яйца которой оказались окрашены в темнокоричневый цвет, как у кобчика.
Такая окраска яиц —ненормальное явление. Размер яиц B7) 31,9--38,7 х
X 23,3—27,5 мм, среднее 34,17 х 25,32 мм.
Малая выпь появляясь рано весной приступает к размножению срав-
нительно поздно. В наиболее южных частях ареала птицы начинают гнездиться
приблизительно на месяц раньше, чем в наиболее северных. С другой стороны,
в одной и той же местности гнезда со свежими яйцами удается находить в тече-
ние всего лета. Это несомненно связано с возобновлением погибших кладок,
но не с нормальным повторением цикла размножения. Гебель A879) писал
о паре малых выпей, которые будто бы в течение одного лета с короткими
интервалами отложили три кладки. Первая кладка была найдена 2 июня,
вторая 11 июня и третья 23 июня. Несомненно в данном случае вкралась
ошибка. Непонятно и указание автора, что вторая кладка нормально содер-
жит большее количество яиц, нежели первая.
Брачный крик самца в районе Харькова удается слышать с десятых
чисел мая и до конца июля. В Бессарабии в последней трети мая птицы при-
ступают к постройке гнезд (Остерман, 1915). Следует отметить, что эта дата
несколько поздняя. В последней трети мая в гнездах в южн. частях ареала
удается находить законченные кладки; в районе Мариуполя кладка в 5 яиц
458 отряд голенастые птицы
была найдена 27 мая (Боровиков, 1907), у Запорожья в плавнях Днепра
несколько кладок с количеством яиц от 4 до 8 осмотрено мною между 17 мая
и 11 июнем, под Харьковом кладки в 5—6 яиц известны в последней трети
мая. В Куйбышевской обл. 10 мая и 5 июля в гнездах обнаружены кладки
в 6 и 5 яиц (Карамзин, 1901), а 12 июня гнездо с 6 сильно насиженными
яйцами (Волчанецкий и Яльцев, 1934). Когда начинают гнездиться малые
выпи в Закавказье —точно не установлено. На истылях под Ленкоранью
одно гнездо, осмотренное 24 мая, содержало б яиц в начальной стадии наси-
женности, другое, взятое 29 мая, 4 почти свежих яйца. Вероятно в это же время
заканчивается кладка в Минской обл., где 10 июня уже наблюдался вывод птен-
цов (Шнитников, 1913), и несколько позже (кладка найдена 2 июня) в районе
Умани (Гебель, 1879). Таким образом, во всяком случае в Европейской части
Союза кладка заканчивается на юге в последней трети мая, а в более северных
районах в первой трети июня. Значительно раньше, вероятно в конце апреля,
приступают к гнездованию малые выпи в Туркмении.
Гнезда, содержащие свежеснесенные яйца, удается находить в течение
июня, июля и до начала августа. Наиболее позднее гнездо, содержащее 3 све-
жих яйца, найдено 1 августа в Астраханском заповеднике (Воробьев, 1936).
Наиболее ранний вывод птенцов вероятно происходит в Туркмении, где
по сообщению Формозова на Мургабе 25 мая наблюдались подлетки, а вполне
летные птенцы в начале июня (Дементьев, 1950). В половине июня гнезда
содержат довольно крупных птенцов и на Сыр-Дарье, так как с этого времени
удается часто наблюдать взрослых птиц обоего пола, летающих за кормом
в дневные часы (Спангенберг, 1936). Под Херсоном 15 июня в гнезде были
обнаружены 2 птенца и 3 надклюнутых яйца (Браунер, 1898). В поло-
вине июня также появляются в гнездах птенцы в районе Харькова, а еще
не умеющая летать молодежь наблюдается здесь до половины августа. Под
Минском в гнезде, найденном 10 июня, оказалось 2 птенца и 3 яйца в различной
степени насиженности. Последнее, шестое яйцо кладки, лежало на земле под
гнездовым деревом (Шнитников, 1913). Под Весьегонском (Дарвиновский запо-
ведник) вывод птенцов в 1949 г. происходил 17 июля (Олигер и Немцов).
В средних числах июля в южн. частях ареала и в средней полосе уже часто
попадаются птенцы, почти достигшие роста взрослых. Летные в районе Ростова
отмечены в половине июля (Сарандинаки, 1908), в конце первой трети июля
в бывшем Донецком округе (Аверин, 1911), в Куйбышевской обл. в конце второй
трети июля. Несколько позднее происходит подъем птиц на крылья в более
северных и восточных частях ареала. В низовьях Урала близ устья Илека
еще нелетные птенцы из двух выводков были добыты Северцовым 24—25 июля.
В начале августа в Владимирской обл. (Плещеево озеро) добывалась молодая
птица (Птушенко и Гладков, 1933), плохо летающие молодые были найдены
1 августа в устье реки Великой в районе Пскова (Зарудный, 1910) и 1 ав-
густа еще нелетные под Барнаулом (Велижанин, 1910). Промежутки времени,
через которые откладываются отдельные яйца кладки, точно не выяснены. Пер-
вые три яйца птицей откладываются каждый день. Насиживание начинается
после отложения первого яйца, но самка сидит на первых яйцах небрежно,
много простаивая на гнезде. Однако в гнездах с птенцами хорошо заметна их
разница в величине, различны также и эмбрионы при вскрытии яиц одной
кладки (Спангенберг).
Насиживание продолжается по Остерману A915) 20 дней, но веро-
ятно несколько короче — 17—19 дней; начинается с первого яйца и произ-
водится обоими родителями. Только что вылупившиеся птенцы совершенно
беспомощны. Через неделю они уже стоят в гяезде и при приближении
человека принимают ту же позу, что и взрослые, т. е. вытягивают вверх
голову и шею и в таком положении остаются неподвижными. Кроме того,
чтобы отпугнуть врага, они широко открывают рты и раздвигают основание
МАЛАЯ ВЫПЬ ИЛИ ВОЛЧОК 459
нижней челюсти. Оперившиеся птенцы на 17—18-й день жизни при появлении
человека покидают гнездо и пытаются уйти, цепляясь за камышинки, и пере-
летают с ветки на ветку. В течение всего времени, пока птенцы остаются в гнезде,
самец и самка усиленно их кормят, летая за пищей и в дневные часы. На крыло
птенцы поднимаются в месячном возрасте.
После вылета из гнезд птенцы очень короткое время держатся выводками,
а затем разбиваются поодиночке.
Линька. Сезонного диморфизма нет, птицы имеют одну полную линьку
в году, начинающуюся в июле и не заканчивающуюся до отлета. Точные дан-
ные не установлены. Частичная линька гнездового наряда — смена мелкого
пера — протекает в первую же осень. Возрастные изменения незначительны,
и в годовалом возрасте, после первой полной линьки, молодые волчки надевают
окончательный наряд.
Питание. Волчки поедают животную пищу: мелкую рыбешку, лягу-
шек и головастиков, всевозможных водяных и наземных насекомых, червей и
улиток. Нередко поедают птенцов и разбивают яйца не только в гнездах
мелких воробьиных, гнездящихся в камышах, но нападают также на гнезда
родяных курочек, а при отсутствии самок и на гнезда уток и цапель. При
вскрытии птиц, убитых в районе Харькова, в их желудках найдены: остатки
ракушек, личинок, жуков-водолюбов и других насекомых (Сомов, 1897).
Под Ленкоранью малые выпи, гнездящиеся в лесных водохранилищах —
истылях, в большом числе поедают древесных лягушек — квакш, реже рыбу
и во множестве разнообразных насекомых. Незадолго до осеннего отлета и
весной главную пищу составляет мелкая рыбешка. У всех птиц, добытых на
осеннем пролете в Крыму, желудки оказались туго набиты остатками кобылок
и, видимо, богомолов (Спангенберг). В желудках птиц, убитых во время про-
летов на вост. берегу Каспия, также обнаружены остатки насекомых (Исаков
и Воробьев, 1940). Мелкую рыбу и водяных насекомых малая выпь искусно
ловит, подолгу просиживая на наклонившейся к воде ветке или пучке тростника.
Полевые признаки. Благодаря очень мелким размерам (птицы не круп-
нее кулика веретенника) и окраске (см. ниже) малую выпь в области ее распро-
странения невозможно смешать ни с одной нашей цаплей.
Птица ведет, главным образом, сумеречный и ночной образ жизни, но
в период вскармливания птенцов и во время спаривания деятельна днем.
Ее удается видеть, летящей над камышами, перелетающей с одного берега
реки на другой, но значительно чаще вспугивать из прибрежных зарослей и
тростниковых крепей водоемов. Несравненно реже, обычно у гнезда эту цаплю
удается наблюдать, сидящей на ветви дерева или в густых зарослях. Малая
выпь обладает исключительными способностями прятаться. Заметив прибли-
жение человека, она вытягивает вверх шею и голову и подолгу остается непо-
движной. В этом положении ее бывает трудно отличить от окружающих стеблей
тростника или сучьев дерева. По суше ходит крупными шагами, а при нужде
способна к быстрому бегу. Но особенно хорошо она передвигается в густых
тростниковых зарослях, охватывая растущие стебли тростника своими длин-
ными пальцами. Будучи вспугнутой, легко поднимается на воздух и, пролетев
сравнительно короткое расстояние, с разлета вновь бросается в заросли. При
более значительных перелетах летиг очень быстро. Она несколько вытягивает
шею, откидывает назад ноги и часто взмахивает крыльями. Ее полет имеет
нечто общее с полетом чирка. Спускаясь в камыши с большой высоты птица
некоторое время летит парящим полетом. Попав в воду, плавает, но делает это
очень неловко. В некоторых случаях подстреленная выпь пытается спастись
нырянием (Житков и Бутурлин, 1906).
В отличие от большинства наших цапель малая выпь держится обычно
в одиночку. Только в гнездовый период ее удается встречать парами и корот-
кое время после вылета из гнезд птенцов семьями.
460
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ.
Есть указания (Мензбир, 1918), что между самцами в период спарива-
ния бывают жестокие драки. Они происходят на земле, на деревьях и в воз-
духе. Драки прекращаются, когда птицы разбиваются на пары и присту-
пают к постройке гнезда.
Рис. 90. Волчок
Весенний крик самца малой выпи удается слышать ночью и в дневные
часы. Птица выкрикивает свои тумб» или «прумб», издавая их по два, по три раза
один за другим (Мензбир, 1918). Брачный крик малой выпи, по данным Со-
мова A897), звучит, «как отрывистые, несколько раз сряду повторяемые звуки
«бух» или «буэх». В другое время года птицы кричат совсем иначе, издавая
резкое и очень быстрое «ке-ке-ке-кек».
АМУРСКИЙ ВОЛЧОК
461
Описание. Размеры и строение. По складу — типичная цапля, с сжатым с бо-
ков телом, узкой головой и прямым острым клювом, коротким хвостом и широкими крыльями.
Перья на нижней части зоба удлинены. Формула крыла (не считая первого рудиментарного
махового): 1=2>3>4>5... Хвост прямой, из 12 рулевых. На затылке, как и у других выпей,
голое пятно, скрытое оперением. Голень обнажена в нижней трети. Вес около 136—145 г.
Длина крыла самцов A9) 150—160, самок B5) 140—150, в среднем соответственно 154,6
и 147,1 мм.
Окраска. Пуховой птенец охристо-желтоватый на спинной стороне, беловатый
на брюшной; вершины пушинок на спинной стороне шелковисто-блестящие. Клюв красный,
радужина зеленая.
Взрослые самцы: верхняя часть головы, спина, плечевые, надхвостье — черные с зе-
леноватым металлическим отливом; верхняя часть шеи сероватая, кроющие крыла желтые;
маховые и рулевые черные; брюшная сторона охристая с беловатыми основаниями перьев
и бурым продольным рисунком на груди; подкрылья белые. Клюв желто-зеленоватый,
голая уздечка желтая, ноги зеленые, рамфотека желтая.
Взрослые самки отличаются от самцов темнобурой окраской спины и плечевых; перья
на этих партиях окаймлены желтовато-охристым цветом; бока головы и шеи рыжевато-
бурые; на передней стороне шеи с брюшной стороны темный продольный рисунок. Клюв
желтый с бурым концом.
Птицы в гнездовом наряде (самцы и самки) сходны со взрослой самкой, но удлинен-
ных перьев на зобе нет и всюду по светлому фону темные пестрины, даже на больших крою-
щих крыла, темя буроватое с рыжеватыми отметинами; маховые и рулевые буроваты.
После Частичной линьки в первом зимнем наряде молодые волчки отличаются от взрос-
лых большим развитием продольных отметин на брюшной стороне, у самцов серый цвет
на шее менее чист; маховые и рулевые — от гнездового наряда, буроватые (иногда перелини-
вают только внутренние второстепенные маховые).
19. Амурский волчок Ixobrychus eurhythmus Swinhoe
Ardetta eurhythma. Swinhoe. The Ibis, 1873, стр. 74, Амой
Распространение. Ареал. Гнездовая область в СССР простирается от
южн. Забайкалья до Приморья включительно и, повидимому, до Сахалина.
За пределами Союза ССР — в Манчжурии, Корее и в Японии (Хоккайдо
и сев, половина Хондо), в Китае
(Сычуань, Чжензянь, Шаньдунь,
Чжили, к югу до Гуандуня и, быть
может, Фуцзяня). Детали распростра-
нения у нас таковы: на гнездовье
амурский волчок характерен для
большей части Приморья, где основ-
ной массой заселяет низменные терри-
тории, проникая местами по широким
речным долинам в область хребта
Сихотэ-алинь. Обнаружен на всем
протяжении долины Уссури и на мно-
гих правых притоках этой реки. Най-
ден в бассейне озера Ханка у Турьего
Рога, в истоках Сунгачи, в долине
Сиахе, на нижнем и среднем течении
Лефу, а также на реках Мо, Даубихе,
Улахе и Иману. Отмечался под Влади-
востоком у Сидеми и в ряде других
мест южн. Приморья. Обитает также
на берегах Японского моря, встречен
на п-ове Песчаном и о-ве Аскольде.
За пределами Приморья найден в ряде
мест, вероятно определяющих север-
ную и западную границу распростра-
нения вида. На восток от Борзи на
р. Аргуни (Цурухайтуй), на р. Дигун
Карта 84. Распространение амурского
волчка Ixobrychus eurhytmus
—граница гнездовий, б — не вполне выясненная
граница гнездовий, г — места зимовок.
462 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
и в Малом Хингане (Штегман, 1929; Шулышн, 1936). Гнездо с птенцами най-
дено на Зее под Благовещенском (устное сообщение Шапошникова), наблю-
дался также на Амуре (Давунда) под 49° с. ш. (Шренк, 1861) на озере Эворон.
Кроме того в Зоологическом музее Московского университета хран*ггся два
молодых экземпляра из коллекции Стаханова, с сев. Сахалина, добытые
6 августа и 13 сентября 1929 г. В зимнее время амурские волчки известны
из Южн. Китая, Индо-Китая (редки), с Малайского п-ова, с Филиппинских,
Больших Зондских о-вов и Целебеса.
Характер пребывания. В пределах Союза нормально гнездя-
щаяся перелетная птица. Весенний прилет совпадает с покрытием листвой
деревьев и подрастанием травы. В южн. Приморье (истоки Сунгачи, Турий
рог, Алтыновка) появляется 23—24 мая (Пржевальский, 1870; Шульпин,
1936). Несколько севернее —на нижнем течении р. Имана в 1938 г. первые
самки прилетели и были добыты мною 29 мая, первые самцы 31 мая. В наиболее
северные части своего ареала прилетают повидимому в самом начале июня.
О времени осеннего отлета мы пока можем судить только на основании добычи
в стране наиболее поздних экземпляров. В различных частях ареала, лежащего
в наших пределах (Амур 49° с. ш., Ханка, п-ов Песчаный) птицы добывались
со 2 по 8 сентября (Шульпин, 1936), на Сахалине 13 сентября (сборы Стаха-
нова). Запоздавшие особи задерживаются, вероятно, в южн. Приморье не-
сколько дольше.
Биотоп. Гнездовый—открытый ландшафт. Сухие и сырые луга и настоя-
щие болота, поросшие высокой травой, гривы среди пониженных луговых
участков, покрытые низкорослым густым кустарником монгольского дуба и
манчжурской лещины.
Пищу птицы добывают в лугах, в кочкарниках с открытой водой между
кочек, в вейниковых болотах, на берегах озер с редкими порослями камыша
и тростника на берегах канав и значительно реже на берегах крупных рек.
Птицы не избегают близости населенных пунктов и часто встречаются на забо-
лоченных участках огородов и по берегам водоемов, в самих селениях.
Численность. Довольно многочисленная птица в лугах широких
речных долин, расположенных на плоскости. В связи с сокращением соответ-
ствующих биотопов, в горной местности становится редкой и спорадичной.
Общая численность у нас, вследствие распространения на небольшой тер-
ритории, ограничена.
Экология. Размножение. Гнезда устраивает на земле среди очень
высокой и густой травы сухого или мокрого луга или среди густых мелких
кустарников дубняка и орешника. Строит гнезда амурский волчок и на болотах
(Мензбир, 1918). Небольшое углубление в почве выстилается тонким слоем
сухих стебельков, радиально расходящихся из центра гнезда. Имея форму
перевернутого конуса гнездо напоминает в этом отношении гнезда других
цапель, гнездящихся на деревьях и в камыше. Лоточек гнезда скрепляется
глиной. Ширина лотка внизу около 5,5 см, в верхней части гнезда 12—14 см.
Число яиц в законченных кладках от 3 до 5. На Имане полные кладки
с едва насиженными яйцами обнаружены в последних числах июня. Пови-
димому, они принадлежали поздно загнездившимся птицам, так как гнездо
с довольно сильно насиженными яйцами найдено там же 10 июня. Яйца
амурского волчка по окраске и характеру скорлупы очень похожи на
яйца малой выпи. Они белого цвета и белые на просвет. Скорлупа тонкая,
с хорошо видными порами и даже у насиженных яиц почти лишена блеска.
Форма несколько вытянутая, обычно эллипсоидальная. Размеры несколько
мельче, чем у малой выпи A5) 31,8—35,9 х 25,8—27,8 мм, в среднем 34,0 х
X 26,2 мм (р. Иман). Насиживание начинается с первого яйца, что доказы-
вается разновозрастностью эмбрионов и птенцов. Сроки появления птенцов
растянуты довольно сильно. На Имане я находил гнезда с маленькими птен-
АМУРСКИЙ ВОЛЧОК
463
дами в начале июля. На Ханке уже в сере-
дине июля появляются летные молодые
(Шульпин, 1936). Вскармливание птенцов
производится обоими родителями, но ос-
новная забота о доставке пищи лежит на
самце. В этот период самец часто попа-
дается на глаза не только с наступлением
сумерек, но и днем, он летает за пищей
в течение часа раза 3—4. В одних случаях
пища добывается поблизости от гнезда, но
чаще самец летит за ней сравнительно
далеко (от 0,5 до 1 км).
Линька. Схема линьки — как у
волчка. Самки приступают к линьке (смена
маховых) в июле. К началу августа махо-
вые полностью сменяются, тогда как руле-
вые и мелкое оперение продолжают интен-
сивно линять. В последних числах августа
самки имеют полное свежее оперение. Линь-
ка самцов не изучена. Во всяком случае
в июле они еще сохраняют брачное опе-
рение.
Ювенальная линька молодых—в июне.
Смена нарядов — пуховой, гнездовый —
первый годовой (переходный) с ювеналь-
ными маховыми рулевыми—второй годовой
(окончательный) и т. д.
Питание. В пищеводах и желудках
добытых на Имане волчков были обнару-
жены остатки насекомых и мелкая рыба
(вьюны до 76 мм длиной, бычки до 57 мм).
Можно предполагать, однако, что питание
птиц более разнообразно.
Полевые признаки и повадка. Амур-
ский волчок по своему общему сложению
очень похож на европейского волчка, но эти виды географически исключают
друг друга. От длинноногого волчка амурский волчок отличается несколько
иной окраской оперения.
Рис. 91. Амурский волчок
464 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Амурский волчок — одна из самых маленьких цапель, не крупнее водя-
ной курочки—камышницы. Образ жизни главным образом сумеречный;
в период спаривания активен ночью, а во время выкармливания птенцов
деятелен и днем. Птицу удается спугивать из высокой травы или наблюдать
летящей над лугами. При полете волчок быстро машет крыльями, но летит
относительно медленно, низко над землею и вскоре бросается в траву. Вторично
поднять пересевшую птицу очень трудно, так как она поспешно убегает в сто-
рону. Раненный в крыло волчок бежит с замечательной быстротой, делая
крупные шаги. Если нет выхода, птица смело защищается, нанося удары
клювом. Явно избегает древесной растительности, присаживаясь на нижние
ветви деревьев в исключительных случаях. Молчаливая птица. В момент
взлета издает грубый, глухой, отрывистый звук. В период спаривания часто
ночью и на зорях подает своеобразный голос, являющийся песней самца:
10—15 раз повторяется звук «гут «гуп-гуп-гуш, после чего следует корот-
кая пауза.
Описание. Окраска. Молодые птицы первого года жизни несколько похожи на
взрослую самку, но основной цвет верхней стороны бурее, а пятна не белые, а желтоватые.
Клюв зеленовато-желтый с бурым ребром верха, ноги желтовато-зеленые; радужина жел-
тая. У взрослого самца преобладают два цвета — каштановый сверху и охристый снизу.
Густо-каштановый цвет распространен на боках головы, верхней половине шеи и всей спине.
Верх головы и хвост более темной черновато-бурой окраски, причем на голове развит сла-
бый металлический блеск. Нижняя брюшная часть тела равномерно охристая, несколько
светлеющая на горле. От основания клюва вдоль горла, шеи и груди идет узкая бурая полоса,
несколько расширяющаяся на брюшке. Крыло в средней части серовато-охристое, по краю
обведено каштановой полосой, маховые свинцово-серые. Надклювье бурое, подклювье
значительно светлей (желтое). Голое кольцо вокруг глаза зеленовато-розовое, радужина
желтая. Ноги буро-зеленоватые, несколько желтеющие на задней стороне плюсны и нижней
стороне пальцев. Взрослая самка отличается более пестрой крапчатой окраской тела. Верх
головы темнобурый почти черный, бока головы каштановые. Верхняя часть шеи и спины
каштановой окраски с обильными пестринлми и пятнами белого цвета. Низ тела охристый
с продольными бурыми пятнами. Бурая полоса, идущая вдоль шеи и груди, хорошо заметна,
и на нижней стороне головы граничит со светлоохристой, почти белой, окраской. Крыло
пестрое из комбинации каштанового и охристого цвета. Маховые, как у самцов.
20. Длинноносый волчок Ixobrychus sinensis Gm.
Ardea sinensis. G m e 1 i n. §yst. Nat., 1789, стр. 642, Китай.
Распространение. Ареал. Южн. и ю.-в. Азия от Индии и Бирмы, Цей-
лона, Андаманских о-вов до Малайского п-ова, Индо-Китая, Китая и Японии,
Курильских о-вов и Сахалина; к югу до Филиппин, Б. Зондских о-вов,
Церама, Флореса, Тимора; кроме того на о-ве Палау, Каролинских и Мариан-
ских о-вах в Тихом и Сейшельских о-вах в Индийском океане.
Характер пребывания. В большей части ареала оседлый вид.
В Японии, Китае и в Корее перелетная птица.
Биотоп. Открытые пространства болот и берега водоемов.
Подвиды и варьирующие признаки. Незначительно варьирует в размерах и в окраске,
вероятно, и в деталях биологии. Известно пять подвидов: /. s. sinensis G me 1 i n, 1789 —
весь ареал за исключением областей следующих подвидов; /. s. astrologus W e t m о г е, 1918 —
Филиппинские о-ва; /, s. lepidus Н о г s f i e 1 d, 1821 — Суматра, Ява; /. s. bryani S e a 1 е,
1901 — Гуам; /» s. moorei Wetmore, 1919 — группа Каролинских о-вов. Самостоятель-
ность этих подвидов признана далеко не всеми авторами и вид быть может монотипичен
(Хартерт, 1915; Бутурлин, 1935).
Длинноносый волчок Ixobrychus sinensis sinensis G m.
Распространение. Ареал. Япония, Корея, весь вост. Китай, Индия,
Цейлон, Бирма, Малайя, Индо-Китай, Моллукские и Ново-Британские о-ва.
В наших пределах добыт на о-ве Кунасири на Курильской гряде (Синрио,
длинноносый волчок
465
1936), на озере Торайка на Сахалине несколько севернее залива Терпение
(Такахаси, 1937) и у Сакагаема на южн. Сахалине (Мунстерьельм, 1920) *.
15 мая 1879 г. взрослый самец добыт на о-ве Аскольд Янковским.
Характер пребывания. Вероятно гнездящаяся перелетная
птица о-ва Кунасири, а быть может и Сахалина, для Приморья случайно
залетная. Перелетен также в Китае, Корее и в Японии, где появляется в мае
и остается до октября.
Биотоп. Сходен с биотопом малой выпи в Европе.
Экология. Сведений по биологии в наших пределах нет. Гнезда устраивает
в зарослях камыша среди озер (Ян, 1942). Размеры яиц C0) 28,5 х 35,0 х
X 22,5—25,0, в среднем 32,0 х 24,5 мм (Китай, Латуш, 1934).
Карта 85. Распространение длинноносого волчка Ixobrychus sinensis
1. У. s. sinensis, 2. Y. s. astrologas, 3. Y. s. lepidus, 4. Y. s. bryani, 5. Y. s. moorei;
a — граница гнездовий, о — не вполне выясненная граница гнездовий.
Описание. Размеры и строение. Небольшая птица — несколько меньше евро-
пейского волчка, по своему общему сложению близок к другим видам волчков, но обладает
длинным клювом, который всегда значительно длиннее цевки. Длина крыла самцов из
Китая 140—146 мм, самок 130—136 мм; клюв 47—54 мм, цевка 45—48 мм (Латуш, 1934);
крыло птиц из Индии 129^143 мм, клюв 49—57 мм, цевка 44—51 мм (Бекер, 1929).
Окраска. У молодых птиц темя с рыжими пестринами, нижняя сторона пестрее
от более темных пестрин.
Общая окраска взрослых птиц землисто-бурая с рыжеватым оттенком. Край крыла
белый. Первостепенные маховые черные со слабым серым налетом; рулевые черные. Голова
сверху черная, иногда с серыми пестринами. Бока головы и шеи винного цвета. Горло белое
с продольной охристой полосой, переходящей в охристую окраску груди и брюха. С каждой
стороны груди пятно из черных с желтым краями перьев. Подхвостье белое. Голые лицевые
части и кольцо вокруг глаза зеленые; радужина желтая; верхняя челюстьтемнобурая, розо-
веющая к краям; нижняя челюсть розовато-бурая, зеленеющая к вершине; плюсна мясного
¦ Ошибочно определены были за Ixobrychus cinnamoneus, см. A Handlist of the
Japmese Birds. 1932, стр. 107.
30 Птицы СССР, т. II
466
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
цвета; пальцы и плюсневый сгиб желтый когти рогового цвета. Старые самки нередко носят
наряд старых самцов, но обычно отличаются от них более рыжим цветом верха, менее раз-
витым черным цветом головы, более развитым охристым пятном на середине зоба.
Рис. 92. Длинноносый волчок (самец и самка)
РОД ВЫПЬ BOTAURUS STEPHENS, 1819
Тип Ardea stellaris
21. Выпь Botauras stellaris L.
Ardea stellaris. Linnaeus. Syst. Nat, X, 1758, стр. 144, Швеция.
Русское название. Выпь — народное, этимология неясна. Иначе — бугай,
бухало, водяной бык (по характерному голосу птицы — «буханью»).
Распространение. Ареал. Европа от Англии (в настоящее время только
в Норфольке и Суффольке, раньше и в других местностях, в частности в южн.
Шотландии, Уэльсе, Ирландии), Испании и Португалии на юг до Средиземного
выпь
467
моря (но отсутствует на гнездовье в Греции и на островах, кроме Сардинии и
Майорки), на север до побережья Северного моря, Дании, южн. Швеции,
ю.-в. Финляндии; в Европейской части СССР. Африка—Марокко, Алжир,
Тунис и юг материка. Азия — умеренная полоса к северу, примерно до 58°
с. ш. у Тобольска, и в бассейне Енисея и около 62° в Якутии; на юг до Пале-
стины, Малой Азии, Ирана (быть может в Ираке и Афганистане), Джунгарии
и Кашгарии (озеро Лоб-нор), с.-з. Монголии, южн. Забайкалья и сев. излу-
Карта 86. Распространение выпи Botaurus stellar is
7. В. s. stellaris, 2. В. s. capensis;
a — граница гнездовий, б — не вполне выясненная граница гнездовий, г — область зимовок номи-
нальной формы.
чины Желтой реки; на восток до Сахалина, Японии и быть может провинции
Чжили в Сев. Китае. Зимовки северных выпей отмечены в сев. и тропи-
ческой Африке, Аравии, сев. Индии, Бирме, Южн. Китае (и северней,
см. ниже).
Характер пребывания. В сев. и средней Европе и Азии пере-
летная птица. Зимует, однако, местами, где не замерзают водоемы — уже
в Германии, южн. Швеции, Польше и т. д. Регулярно — у Средиземного моря,
в южн. Закавказье, в Туркмении у Черного моря. Южные популяции оседлы.
Биотоп. Водоемы и болота, заросшие тростником, камышом, ивня-
ком и т. д.
Подвиды и варьирующие признаки. Два подвида, различающихся деталями биологии,
а также размерами и окраской: В. s. stellaris Linnaeus, 1758 — в Европе, Азии и сев.
Африке; В, s. capensis S h 1 е g e 1, 1863 — южн. Африка.
Очень близкие виды распространены в Австралии и в Америке.
468 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
Северная выпь Botaurus stellaris stellaris L.
Синоним. Botaurus stellaris oriental is. Бутурлин. Наша Охота, 1938 (сент.),
стр. 9, Манчжурия.
Распространение. Ареал. Вне СССР указан выше, в очерке вида. Север-
ная граница в СССР пересекает южн. части Карело-Финской ССР, откуда идет
к Вологде и через район Кирова в Молотовскую обл., где выпь найдена на
широте 58г12° (Ушков, 1927). На Уральском хребте граница несколько пони-
жается, в Сибири с севера огибает Тюмень и подходит под 58° с. ш. к Тобольску,
В области Енисея, по данным Тугаринова A947) граница, примерно, проходит
по Подкаменной Тунгуске (под 64° с. ш. залеты) и резко поднимается к северу
в Якутии, проникая в бассейн Вилюя к Якутску. Гнездится ли в последнем
месте выпь или только залетает сюда, остается не выясненным, вернее послед-
нее. Далее на восток сев. граница плохо известна. На Охотском побережье
выпь гнездится К северу до Аяна, проникает на Сахалин, но отсутствует на
Камчатке. К югу от сев. границы выпь гнездится у нас в соответствующих
биотопах во всей Европейской и Азиатской частях Союза, а также далеко
проникает за южные границы Союза ССР.
Основная масса, вероятно, европейских птиц зимует в области Средизем-
ного моря и в Африке к югу до Абиссинии. Выпи, населяющие восточные части
ареала перезимовывают уже в Среднем и Южном Китае *, в Индии и изредка
в Бирме.
В наших пределах некоторое количество птиц зимует на берегах Черного
моря, во всем Закавказье, на Каспийском побережье к северу до устья Урала
и Волги и в Средней Азии к северу приблизительно до широты ст. Чийли на Сыр-
Дарье. В Мервском оазисе (Туркмения) выпи живут уже оседло. Залеты отме-
чены в Исландии, на Фарерские о-ва, Норвегию, Канарские о-ва, Мадейру.
Характер пребывания. У нас гнездящаяся, перелетная птица
северных и средних частей страны. На юге принадлежит местами к зимующим
и даже оседло живущим видам. Места наших зимовок таковы. Небольшое
число особей перезимовывает близ незамерзающих водоемов Крыма. Отдель-
ные особи всю зиму держатся на побережье Черного моря, зимует в Армении,
в ее более низких частях. Численность зимующих особей ограничена. В суро-
вые зимы, как например, зима 1924—25 г. выпь бывает под Ленкоранью крайне
многочисленной, вероятно это связано с перемещением сюда особей, нормально
зимующих несколько севернее. В январе 1925 г. южнее Ленкорани выпи
встречались положительно на каждом водоеме. Многие из них оказались
крайне истощенными и, будучи не в силах подняться на крылья, давались
в руки (Спангенберг). На Сев. Кавказе выпь отдельными особями перезимовы-
вает в районе Дзауджикау, по опросным данным зимует на текучей воде в устьях
Терека, где мною часто наблюдалась в декабре. Зимой в небольшом числе
встречается в приморской части дельты Волги. Одиночные экземпляры остаются
на зиму и в устьях Урала, хотя иногда эта зимовка кончается гибелью птиц
от голода. Нам часто приходилось наблюдать выпей в устье Урала в начале
декабря при наличии снегопадов и замерзании мелких водоемов. В районе
вост. побережья Аральского моря выпь не зимует, но южнее на Сыр-Дарье
(под Джулеком и у ст. Тюмень-арык) в теплые зимы перезимовывает отдель-
ными редкими особями (Спангенберг и Фейгин, 1936). Выше на Сыр-Дарье
вероятно зимует у Чиназа, где в 1923 г. наблюдалась автором в начале января.
Добывалась выпь также в зимнее время в долине Чирчика (Зарудный, 1910).
В другой работе там же авторA915) упоминает о редких зимовках птиц в пустыне
Кызыл-Кум в оазисе Тамды. В Туркмении выпь зимует регулярно, в частности—
в низовьях Атрека. По данным Дементьева A950) зимние находки известны
* Шань-Дунь, среднее и нижнее течение Яньцзе, Фуцзянь, Гуаньдун (Латуш, 1934).
выпь 469
в декабре 1888 г. у Ашхабада и в декабре 1890 г. у Мерва. В небольшом числе
зимует в пределах Таджикистана. Редкие особи вероятно перезимовывают
также в районе Джаркента, где по опросным сведениям известны случаи
добычи птиц в зимнее время.
Весенний прилет выпей очень сильно растянут. Первые особи появляются
рано, когда еще лежит снег, не редки метели и значительные ночные морозы.
Валовой пролет происходит недели на две, а то и на целый месяц, позднее
первоприлетных и часто совпадает со временем, когда южные особи уже при-
ступили к размножению. На юге Европейской части Союза первые выпи при-
летают в различные числа марта. В Бессарабии уже в феврале ст. ст. (Остер-
ман, 1912), в Подолии (вероятно не первые) в половине апреля. У Херсона
в последней трети марта, в районе Мариуполя в конце марта, иногда 23 марта
и в большем числе в самом начале мая (Боровиков, 1907). Первое «буханье»
у Ростова отмечено уже 13 марта (Сарандинаки, 1908). В районе Харькова
наиболее ранние особи отмечались 23 марта, валовой же пролет происходит
здесь в конце первой трети апреля, затягиваясь приблизительно до последних
чисел этого месяца (Сомов, 1897).
На Кавказе, где часть выпей перезимовывает, установить начало весен-
него прилета трудно. Во всяком случае в Закавказье в самом начале марта
численность птиц резко возрастает и сохраняется повышенной по меньшей мере,
в течение всего апреля. В последней трети марта выпи летят вдоль черномор-
ского побережья Кавказа, в конце марта достигают дельты Волги. Далее к се-
веру птицы появляются несколько позже: у Киева 10 апреля (Шарлемань, 1913),
на широте Минска, Тамбова, Куйбышева в различные даты второй половины
апреля, явно тяготея к концу этого месяца. Первый весенний крик в Моское-
ской обл. отмечен в последней трети апреля, под Горьким в середине апреля.
Под Казанью передовые птицы появляются в середине апреля, а массовый
пролет происходит в конце этого месяца. Весь весенний пролет у Пскова
укладывается между концом марта и концом апреля. В Ленинградскую обл.
выпи прилетают повидимому в конце апреля и приблизительно в те же сроки
в южные части Кировской обл. и на Белую в районе Уфы.
В Азиатской части Союза в марте наблюдается пролет птиц в районе Гассан-
кули и по внутренним водоемам Туркмении, в самом конце марта в окрест-
ностях Ташкента. На нижнем течении Сыр-Дарьи, к югу от станции Солс-
тюбе, передовые выпи регулярно появляются в начале марта, а к двадцатым
числам этого месяца весеннее буханье птиц удается слышать на всем протяже-
нии реки до ее устья и на озере Камышлы-баш. В эти же сроки несомненно
выпи появляются на вост. побережье Аральского моря. Однако, пролет здесь
всегда сильно затягивается и по нашим наблюдениям продолжается по меньшей
мере в течение всего апреля. В самом конце марта первоприлетные птицы
отмечались в устьях Урала, а также в районе Чкалова, где валовой пролет
начинается в последней трети апреля и длится до начала второй трети мая,
отсталые нормально летят в последних числах мая, а в 1880, как исключение,
даже до начала второй трети июня. Самое раннее буханье в 1890 г. отмечено
здесь 29 апреля.
В Сибири выпи также появляются или во второй половине апреля или
в первой половине мая. На озере Чаны в Зап. Сибири первый весенний голос
отмечен Даниловым 30 апреля, в бывшем Енисейском уезде 7 мая (Тугаринов
и Бутурлин, 1911). В районе Иркутска (в 50 км по Верхоленскому тракту)
самец был добыт 4 мая, а 9 мая в окрестных озерах выпи «бухали» напролет
все ночи (Спангенберг). На озерах Яблонового хребта весенний крик выпи
отмечен 2 мая (Дорогостайский, 1915). В Якутии в 1904 г. выпи наблюдались
2 мая и в 1905 14 мая (Иванов, 1929). Наблюдения по Якутии за 8 лет показы-
вают, что выпи прилетают между 3 мая и 2 июнем, чаще около середины этого
месяца (Скалой). Несколько раньше происходит прилет в Уссурийский край.
470 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
На Амурском заливе в урочище Славянска в 1912 г. впервые выпь была отме-
чена в зарослях Адими —23 марта (Медведев, 1913). Нормально же пролет
начинается в апреле и тяготеет ко второй половине этого месяца.
Таким образом, в конце апреля или в начале мая первые птицы появ-
ляются повсеместно, где они гнездятся. Массовый прилет происходит несколько
позднее, а полностью заканчивается значительно позже прилета передовых
особей. Особенно большой промежуток между прилетом ранних и запоздавших
птиц наблюдается в южн. полосе, где пролетные птицы состоят из различных
популяций. Начало осеннего отлета совпадает с наступлением осеннего не-
настья и похолодания и продолжается до наступления настоящих морозов.
В северных европейских частях ареала выпи начинают двигаться к югу вероят-
но в конце августа и в начале сентября. На Финском заливе весь осенний пролет
укладывается в промежуток между 23 сентября и 9 октября (Бихнер, 1884).
Начало движения к югу из под Пскова отмечено с половины сентября, вало-
вой отлет происходит ближе к концу этого месяца. Наиболее поздние птицы
задерживаются здесь до 20 ноября (Зарудный, 1910), т. е. до настоящей зимы.
Даты начала пролета в Бессарабии неизвестны, но известен случай добычи
экземпляра 8 декабря (Остерман, 1912). Под Минском осенний пролет выпей
протекает видимо, со второй половины сентября и в первой половине следую-
щего месяца. Приблизительно в те же сроки выпи отлетают из средней полосы
Союза — из-под Вышнего Волочка Калининской обл. в середине сентября,
в конце сентября из среднего Прикамья; в конце сентября летят по Еруслану,
не ранее середины сентября из района Уфы. Чем далее к югу, тем пролет
оказывается все более растянутым. На Черноморском побережье Кавказа, где
выпи не гнездятся, они появляются уже в середине сентября и бывают там до
начала января. В садах Алушты, где выпи также не гнездятся, мы наблюдали
их в 1949 г. уже 22 августа. В Харьковской обл. пролет тянется более двух ме-
сяцев с 8 сентября до 17 ноября (Сомов, 1897). Вероятно столь же продолжителен
пролет и в дельте Волги, где движение птиц отмечено в октябре и ноябре.
Далее на юге время пролета трудно отделить от частичных зимовок, так как
северные особи подлетают сюда рано, а одиночек можно наблюдать и в зимнее
время. У Запорожья на Днепре выпи еще не представляют большой редкости
в средине ноября (Спангенберг). По данным Вальха A899) в Днепропетров-
ской обл. пролет укладывается между серединой сентября и последней третью
ноября. Последние птицы летят при заморозках. На Сев. Кавказе у ст. Котля-
ровской один экземпляр был добыт 27 ноября (Беме, 1926). Под Кизляром во
второй половине ноября выпей бывает чрезвычайно много и несколько меньше
в первой половине декабря. Данные, касающиеся отлета выпей из азиатской
части Союза, отличаются большой скудностью. В Якутии основная масса птиц
отлетает в начале последней трети сентября, редко в конце его (Скалой).
В районе Тюмени осенний отлет несомненно начинается в самых последних
числах августа и хорошо выражен в первой трети сентября, в те же сроки
пролет птиц отмечался у Иркутска. На реке Алей на западной окраине Алтая
выпь наблюдалась 4 сентября (Аверин и Лавров, 1909). В Уссурийском крае
и на Охотском побережье отлет, видимо запоздавших особей, происходит в суро-
вых условиях, когда уже наступают морозы и нередко выпадает снег. Из-под
Аяна известен экземпляр, добытый 5 октября, а в Приморье из окрестностей
Троицкого даже 6 ноября (Шульпин, 1936). В низовьях Урала начало осеннего
отлета примерно совпадает с концом августа, в половине или в последней
трети сентября пролет достигает наибольшей силы. В теплые годы последние
выпи задерживаются до середины ноября. В верховьях Иргиза последние
птицы отмечены 16 октября (Сушкин, 1908). На Сыр-Дарье под Кзыл-Ордой
выпи попадаются часто во всяком случае до половины октября, в конце месяца
становятся крайне редкими и в начале ноября безусловно отлетают к югу и, если
встречаются, то лишь южнее ст. Соло-тюбе, где местами зимуют (Спангенберг).
выпь 47Т
В Семиречье, в районе Джаркента, пролетные особи отмечались 24 сентября
и несколько раз 7и8октября (Зарудный и Кореев 1900). У Кульджи пролет-
ная выпь была отмечена 20 ноября (Штольцман, 1897). У Гассан-кули пролет
происходит в течение всего ноября и декабря, а в вост. Иране 7 ноября (Заруд-
ный, 1903).
Пролет происходит в ночное время, изредка и днем в пасмурную погоду.
Как правило, птицы летят одиночными особями и редко парами, но иногда
десяток птиц тянется на небольшом расстоянии друг от друга. Сушкин
видел однажды двух выпей, присоединившихся к стае лебедей. В разгар весен-
него и осеннего пролета выпей часто удается поднимать десятками на совсем
маленьких участках, но каждая особь вылетает отдельно и, видимо, не связана
с соседней. Можно предполагать, что большие скопления птиц вызываются
не «общественными» инстинктами, а просто наличием благоприятных дневок.
Во время осеннего пролета выпи поднимаются с дневных стоянок в поздние
сумерки или даже после зари, когда птиц почти невозможно увидеть, но по
крику удается установить, что взлетевшая выпь делает большой круг над
озером и, лишь поднявшись на большую высоту, избирает известное направле-
ние. На дневку осенью выпи рассаживаются на утреннем рассвете, когда
бывает достаточно светло, чтобы видеть птицу. При этом также часто удается
слышать их голоса в воздухе. Места дневок пролетных птиц крайне разно-
образны. Выпи усаживаются на высокие вербы и другие деревья, чаще они
садятся осенью в камыши по берегам озер, а весной на груды камыша, черно-
быльника, дикой конопли, примятой снегом. На южном побережье Крыма
в октябре выпей удается поднимать во время перепелиной охоты из мелких
кустарников дуба и граба на сухой шиферной почве. Многие авторы отмечают
поразительную способность выпей прятаться на дневках в самой, казалось бы,
неподходящей обстановке.
Биотоп. Обширные водные или заболоченные пространства: пру-
ды, озера, речные заливы и плавни, сильно заболоченные луга и мелкие
ольховые поросли и заброшенные торфоразработки, если последние изобилуют
Камышевыми и тростниковыми крепями, зарослями высокой травы с рассеян-
ными здесь и там группами ивовых кустарников. Значительно реже, например
в Минской обл., выпи поселяются среди густых ивовых порослей. Обяза-
тельным условием обитания является наличие небольших участков чистой воды,
вкрапленных среди густой растительности. Повсеместно в период размноже-
ния выпь предпочитает стоячие водоемы, но изредка гнездится на мелких остров-
ках глухих речушек, обладающих слабым течением. В отличие от других
наших цапель кормовой биотоп выпей почти полностью совпадает с биотопом
гнездовым, и птицы не выходят за пределы гнездового участка. Несколько
разнообразные стации выпей после вылета из гнезд птенцов, в период пролетов
и на зимовках. Птиц удается вспугивать из тростников, осоковых и кустарни-
ковых зарослей водоемов самого разнообразного характера, а изредка и вдали
от водоема. Повсеместно выпи избегают сталкиваться с человеком, забиваясь
в наиболее глухие и труднодоступные заросли. В то же время они весьма часто
селятся близ рыбацких стоянок и около крупных населенных пунктов.
Численность. На обширной области распространения численность
выпей неодинакова. Весьма многочисленна выпь в Белоруссии в Полесье, на
большей части территории Украины по нижним и средним отделам рек, впа-
дающих в Черное море. В средней лесной полосе Европейской части Союза
численность значительно понижается, но к северу отсюда местами птицы вновь
встречаются в большом количестве, например в районе Пскова (Зарудный,
1910). В долине Волги многочисленна в среднем течении, только обыкновен-
на — в дельте. В нижнем отделе Урала выпь встречается в большом коли-
честве. В бывшей Оренбургской губ. в подходящих местах на пространстве
около полудесятины иногда можно найти до десяти гнезд птицы (Зарудный,
472 ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
1888). Напротив, во всем Закавказье и Средней Азии выпь немногочисленная
птица. Редка она и в Зап. Сибири, где численность несколько возрастает
лишь в редкие годы. В районе Байкала плотность населения выпей оказы-
вается весьма значительной. Далее на восток она вновь повсеместно должна
быть отнесена лишь к обычным гнездящимся птицам. Вследствие обширного
распространения и местами высокой плотности общая численность выпей
в нашей стране огромна. Этим объясняется большое скопление птиц в подхо-
дящих местах во время пролетов. В районе Кизляра в ноябре автору удавалось
поднимать до полусотни птиц за дневную экскурсию, в апреле на разливах
Илека 20—30 вспугнутых выпей—нередкое явление. Столь же большое скоп-
ление выпей (до 40 птиц за дневную экскурсию) наблюдалось осенью в устье
р. Великой близ Пскова (Зарудный, 1910).
Экология. Размножение. Половая зрелость в годовалом возрасте.
В противоположность многим другим нашим цаплям выпь не образует коло-
ниальных гнездовий. Каждая пара стремится обособить себя от близкого
соседства себе подобных и от других родственных видов. Однако, в наиболее
благоприятной для гнездования обстановке, отдельные пары поселяются
близко друг от друга, образуя пятно с большой плотностью населения. На
мелких водоемах гнезда строятся на выступающих из воды кочках среди зарос-
лей тростника, камыша или среди редких кустарников, иногда закладывается
на сыром грунте трясины. На глубоких озерах гнездо часто помещается на
застрявшей среди тростника куче отмершей растительности или, наконец,
на плесах, сплошь покрытых листьями кувшинки (Зарудный, 1888). В районе
Иркутска выпи избирают для гнездовья совсем маленькие островки — кочки,
сплошь покрытые высокой и густой осокой. Гнездо в этих случаях помещается
на твердом грунте, близ края чистой воды (Спангенберг). Гнездо представляет
собой небрежную постройку из стеблей и листьев надводных растений.
Законченное гнездо имеет округлую форму, но так как птица входит и
сходит с него в одном и том же месте, то один край гнезда бывает обмят и
утоптан (Шнитников, 1913). Поперечный диаметр свежевыстроенных гнезд
около 50 см, высота их около 35 см. По мере подрастания птенцов гнездо посте-
пенно погружается в воду и надстраивается стариками.
Вследствие этого гнезда, найденные летом, бывают несоразмерно велики,
достигая 90 см в поперечном диаметре, при высоте в 50 см (Зарудный, 1888).
Количество яиц в кладках колеблется от 3 до 5, как редкое исключение их
бывает б, изредка 7 (последнее число по западноевропейским авторам). Яйца
откладываются с промежутком в 2—3 дня.
Формой и особенно окраской яйца выпи отличаются от яиц всех осталь-
ных наших цапель. Форма их правильная, яйцевидная, окраска глинисто-серая.
Они имеют нечто общее с яйцами фазанов, но больших размеров и их скорлупа
лишена блеска. Относительно к величине птицы, яйца несколько малы. Размер
их: A2) 49,0—50,5 х 37,0 —38,1 мм, среднее 49,5 х 37,6 мм. У западно-
европейских A00) 47,5—58,2 х 35,5—41,5, в среднем 52,62 X 38,54 мм (Уай-
зерби, 1939).
Выпь приступает к размножению весьма скоро после прилета на гнездовья,
но и в одних и тех же районах гнездование происходит неодновременно. Одни
пары начинают гнездиться рано, другие, по сравнению с ранними, сильно запаз-
дывают. Причины этого не выяснены (разные возрасты?). Сроки начала размно-
жения в различных частях ареала изменяются почти так же, как и сроки прилета
первых особей. К сожалению конкретные данные по размножению крайне
скудны и не позволяют представить общей картины.
В Харьковской обл. кладка в конце первой трети мая или в послед-
них числах этого месяца, редко в более ранние сроки. Насиженные яйца
можно найти и позже. Самостоятельные птенцы попадаются со второй трети
июля, но чаще с середины июля и до первых чисел второй трети августа.
выпь 473
Приблизительно в те же сроки (около половины мая) гнезда содержат свежив
яйца в Минской обл. Вылупление происходит в первых числах июня, а в по-
следней трети этого месяца молодые оставляют гнезда. Даты нахождения
гнезд: 31 мая —насиженные яйца, 5 июня —2 яйца в гнезде надклюнуты,
13 июня —вполне оперившиеся птенцы (Шнитников, 1913). Там же по дан-
ным Федюшина A912) летные молодые бывают в первой трети июля. В Днепро-
петровской обл. летные молодые отмечались в конце июля. Под Псковом
9 апреля отмечено первое «буханье», 16—17 — целый концерт весенних голо-
сов выпей, а 17 мая у добытой самки обнаружено готовое яйцо — последнее
в кладке. Из двух гнезд, осмотренных 9 июня, в одном обнаружены три только
что вылупившихся птенца и 1 яйцо с «пискуном», другое содержало 5 птенцов
недельного возраста. 9 августа найдено еще одно гнездо с тремя птенцами,
величиной с взрослых. Во второй половине августа в устье р. Великой появ-
ляется множество летной молоди.
В районе Чкалова первые яйца удавалось находить в середине мая, а не-
давно вылупившихся птенцов — в середине июня. В Наурзумском заповед-
нике 24 августа пойман птенец —подлеток (Михеев, 1938). Видимо несколько
позднее происходит яйцекладка в Сибири. В свое время автору было достав-
лено свежеснесенное яйцо, взятое под Иркутском из кладки в 4 яйца 27 мая.
Южнее, на р. Тунке в 1933 г. несколько гнезд выпи были найдены Слудским.
Одно из них содержало 26 июня 6 птенцов, в других гнездах оказались
28 июня свежие и слабонасиженные яйца в количестве от 3 до 5 штук.
В Якутии 13 июня найдено гнездо с 5 яйцами, а нелетные птенцы
27 августа (Иванов, 1929). Крайне рано A2 июля) молодая птица в свежем
гнездовом пере была добыта при впадении в озеро Зайсан р. Колпянки
(Поляков, 1912). В Уссурийском крае первый весенний голос выпей отме-
чен 10 апреля. У самки, добытой 23 апреля, обнаружено много крупных фол-
ликул и среди них одна желточная в 10 мм (Шульпин, 1936). Даже за юж-
ными пределами Союза некоторые особи приступают к размножению поздно.
В вост. Иране сильно насиженные яйца Зарудный A903) находил 4 июня.
Насиживание, с первого яйца, длится 25—26 дней. Главная забота
по насиживанию лежит на самке, но вероятно и самец принимает участие.
В Псковской обл. известны случаи добычи самцов с наседными пятнами (Заруд-
ный, 1910). Самец иногда кормит сидящую на гнезде самку и приносит пищу
молодым. Самка начинает насиживать после отложения первого яйца, вслед-
ствие чего птенцы в одном гнезде сильно отличаются друг от друга ростом
(Зарудный, 1888). Сидящая на гнезде птица ведет себя крайне осторожно и при
опасности сходит с гнезда даже в момент вылупления птенцов (Шнитников,
1913). Вообще же насиживающая выпь подпускает к себе человека совсем
близко, а затем или уходит пешком или улетает, сравнительно редко описы-
вая круги над гнездом, как это делают другие цапли.
Птенцы при появлении у гнезда человека издают быстро друг за другом
следующие звуки или писк, широко разевают рты, раздвигая нижнюю челюсть,
и уже в возрасте 2—3 недель могут покидать гнездо. Попав в воду они вновь,
помогая крыльями, стремятся залезть в гнездо на кочки или камыш. Летными
становятся в 2-месячном возрасте. Подросшие птенцы относительно короткий
срок держатся вместе, семья вскоре разбивается и каждый ведет самостоятель-
ную жизнь. Размножающиеся самцы видимо продолжают свое весеннее
«буханье» в течение всего насиживания яиц самкой. В б. Оренбургской
губ. «буханье» удается слышать приблизительно до конца первой трети июля
(Спангенберг). В Уссурийском крае наиболее поздний весенний голос самца
отмечен 26 июля (Шульпин, 1936). На Вилюе в устье Мархи буханье удава-
лось слышать 21 июля (Воробьева, 1928).
Линька. Молодые сменяют мелкое оперение в первую осень с июля
до января. У взрослых — одна полная годовая линька между августом и
474
ОТРЯД ГОЛЕНАСТЫЕ ПТИЦЫ
январем, начинающаяся в конце периода размножения и кончающаяся на
зимовках. Подробности плохо изучены.
Питание. Главным образом животная, а по некоторым авторам (напри-
мер, Остерман, 1912; Исаков и Воробьев, 1940) и растительная пища. Основ-
ная —несомненно рыба: карась,
линь, окунь и, особенно часто,
небольшие щуки, достигающие
около 27 см длины (Шнитников,
1913), а в некоторых случаях и
около 35 см (Житков и Бутурлин,
1906). Помимо рыбы выпи в боль-
шом числе поедают лягушек,
головастиков, тритонов, червей,
а также связанных с водной сре-
дой насекомых и, видимо редко,
млекопитающих. Исследованные
желудки птиц, добытых под Лен-
коранью в зимнее время содер-
жали плавунцов (Macrodytes),
водолюбов (Hydrophilidae )игла-
дышей (Notonectes), а также
растительные остатки (Исаков и
Воробьев, 1940). В других слу-
чаях здесь же 27 февраля в же-
лудке была найдена медведка,
несколько плавунцов и тритон
(Тугаринов и Козлова-Пушка-
рева, 1935). В другое время в
желудке удавалось обнаружить
жуков-листогрызов и рыбу (Рад-
де, 1885). При вскрытии под-
летка, добытого 24 августа в
Наурзумском заповеднике, в пи-
щеводе найдена крупная водяная
крыса (Михеев, 1938). Пища
молодых, главным образом, со-
стоит из головастиков (Заруд-
ный, 1888). Поджидая добычу,
выпи подолгу стоят неподвижно
у уреза воды, а затем быстрым
движением схватывают ее клю-
вом. Реже в поисках пищи птицы
ходят по мелководью или по
листьям водяных растений подоб-
но пастушкам.
Полевые признаки. Размеры,
своеобразное строение и окраска
выпи позволяют безошибочно отличить ее в природе от других наших
цапель. Весной и в начале лета, не наблюдая самих птиц, можно безошибочно
установить их присутствие по их своеобразному буханью. Выпь ведет ночной
образ жизни. В дневные часы, будучи крайне осторожной, предпочитает дер-
жаться в густых тростниковых крепях и других зарослях, но во время пролета
часто проводит день в местах более открытых и доступных. В спокойном состоя-
нии птица втягивает шею, сгорбливается и стоит на одной ноге. Напротив,
потревоженная выпь вертикально вытягивает шею и голову, и, напоминает
Рис. 93. Выпь
выпь 475
в таком положении пучок сухих стеблей тростника, подолгу остается неподвиж-
ной. Заметить ее в этих случаях бывает очень трудно. Оживление наблюдается
с наступлением поздних сумерек и длится в течение всей ночи. Птица появ-
ляется на краях открытых плесов или лазает по камышу, охватывая стебли
своими длинными пальцами. Поздней осенью с наступлением темноты, выпи
часто поднимаются высоко в воздух и летают над камышами, издавая свое-
образные крики. Есть указания в литературе о том, что выпи садятся на деревья.
Увидеть выпь сидящей удается редко и в значительной степени случайно.
Трудно увидеть ее вследствие темноты и во время вечерних полетов. Чаще
птиц удается наблюдать, пробираясь по зарослям тростника, когда она, под-
пустив человека на самое близкое расстояние, наконец решается подняться
в воздух. Самый взлет почти вертикален. Он сопровождается частыми взма-
хами широких крыльев. С явным усилием выбившись из тростника, птица
летит уже спокойным полетом и, пролетев короткое расстояние, вновь опу-
скается в заросли. В летнее время выпь держится парами, затем короткое
время выводками, в другое же время одиночками. Есть указание, что иногда
во время пролета выпи образуют стайки до восьми-десяти особей в каждой
(Зарудный, 1910). В дневные часы даже при большом скоплении птиц в одном
месте, выпи всегда сидят одиночками, иногда близко друг от друга. Голос выпи
удается слышать после наступления темноты во время пролетов. Этот звук,
издаваемый на лету, может быть передан, как резкое «кау», отчасти напо-
минающее крик кваквы, но грубее и громче. «Иногда этот крик звучит как
«оу», реже слышится только одно грубое «о». Весенняя песня состоит из соче-
тания двояких звуков, которые можно передать следующим образом: «ып-ып-
ып-прумб, ып-прумб, ып-прумб, ып-прумб». По Мензбиру A918) самец, заме-
тив самку, начинает принимать в высшей степени оригинальные позы и, нако-
нец, как-то беспокойно, но вместе с тем и ловко, ударяет клювом плашмя по
воде. Затем, опустив клюв в воду, он жадно вбирает ее, отчего слышатся
повторяющиеся несколько раз звуки «у-у». Набрав достаточно воды, птица
резким движением забрасывает голову назад, так что затылок касается спины,
затем опять опрокидывает голову вперед и, опустив клюв в воду, производит
самые резкие, глубокие басовые звуки, вроде рева, которые можно передать
звуками, похожими на «прумб». За ревом следуют менее сильные звуки «бу-бу»,
происходящие от выбрасывания остающейся в зобе воды. Весной и в начале
лета выпь «бухает» на зорях, ночью и отнюдь не редко в дневные часы. По
данным Шульпина A936) и наблюдениям автора весенний крик удается слышать
и от пролетных особей.
Описание. Размеры и строение. Типичное «иаплиное» сложение. Темя пло-
ское, крылья длинные и широкие, хвост короткий; клюв сильный, прямой, слегка выпуклый
по надклювью, с мелкими зазубринами у вершины, длина его около 70—75 мм. Цевки отно-
сительно короткие (длина около 90—100 мм), пальцы длинные (средний палец с когтем
околоПО—120 мм). Форму л а крыла (без первого рудиментарного махового). 1=2=3>4>5...
Вырезок на первостепенных маховых нет. Рулевых 10, хвост закругленный, перья под-
хвостья достигают почти вершины хвоста. На затылке голое пятно. Дистальная часть го-
лени не оперена. Средний вес самцов 966 г, самок 867 г (Ленкоранские зимовки, Исаков и
Воробьев, 1940), но бывает и значительно выше — у зал.Посьет 7 октября добыт самец, весив-
ший 1702 г. (Горчаковская). Крыло самцов B1) 315—362, самок B0) 260—362, в среднем
309,3 и 343,3 мм.
Окраска. Пуховой наряд — на спинной стороне ржавчато-бурый, на брюшной
желтовато-бурый. Взрослые самцы и самки: темя черное, на затылке перья с охристыми
каемками; спина черная с желтовато-охристыми каймами перьев. Маховые ржавчато-
рыжие с неправильными темнобурыми, поперечными пятнами. Рулевые желтовато-бурые
с черноватым рисунком. Хорошо выраженные усы; шея желтоватая с темным поперечным
рисунком, горло беловатое с продольной ржавчатой полосой; брюшная сторона охристая
с бурым поперечным рисунком. Клюв зеленовато-желтый, бурый на вершине и у разреза
рта; голое пятно на уздечке и у шеи зеленоватое или бледно голубоватое; ноги бледно-
зелёные ; радужина желтая.
Птицы в гнездовом наряде сходны со взрослыми, но «усы» слабее выражены; темный
рисунок бледнее. Возрастные изменения таким образом весьма незначительны.
ОГЛАВЛЕНИЕ
Отряд голуби (Р. Н. Мекленбурцез)
Характеристика отряда 3
Таблица для определения видов голубей 6
Род голуби 6
1. Сизый голубь 6
Обыкновенный сизый голубь , 8
Туркестанский сизый голубь 11
2. Скалистый голубь 14
Северный скалистый голубь 15
Южный скалистый голубь 17
3. Белогрудый или белоспинный голубь fp
Западный белогрудый голубь 20
4. Бурый голубь 21
5. Клинтух 25
Европейский клинтух 27
Туркестанский клинтух 31
6. Вяхирь или витютень 33
Европейский вяхирь 34
Иранский вяхирь 38
Туркестанский вяхирь 39
Род горлицы 41
7. Горлица 41
Европейская горлица 42
Азиатская горлица 47
8. Большая или степная горлица 51
Таежная большая горлица 52
Туркестанская большая горлица 55
9. Кольчатая горлица 59
Западная кольчатая горлица 61
Центральноазиатская кольчатая горлица 62
10. Малая или египетская горлица 64
Туркестанская малая горлица 65
Род короткохвостые горлицы 69
11. Короткохвостая горлица 69
Китайская короткохвостая горлица 69
Отряд рябки (Г. П. Дементьев)
Характеристика стряда 71
Таблица для определения видов рябксв 73
Род рябки 73
1. Чернобрюхий рябок 73
Восточный чернобрюхий рябок 74
Западный чернобрюхий рябок 78
ОГЛАВЛЕНИЕ 477
2. Белобрюхий рябок 79
Восточный белобрюхий рябок 79
[Сенегальский рябок] 83
Род саджа 84
3. Обыкновенная саджа 84
4. Тибетская или горная саджа 88
Отряд трехперстки (Г. П. Дементьев)
Характеристика отряда 92
Род трехперстки 93
1. Пятнистая трехперстка 93
Восточная или большая пятнистая трехперстка 94
Отряд журавли (А. М. Судиловская)
Характеристика отряда 97
Таблица для определения видов журавлей 102
Семейство журавлей '. 102
Род журавли 102
1. Серый журавль 102
Западный серый журавль 1J4
Восточный серый журавль 110
2. Канадский журавль 114
Обыкновенный канадский журавль 114
3. Даурский журавль 117
4. Черный журавль или журавль-монах 121
5. Стерх или белый журавль 123
6. Манчжурский журавль 123
7. Австралийский журавль 131
[Индийский журавль] 133
Род журавли-красавки 133
8. Журавль-красавка или малый журавль 133
Отряд дрофы (Е. П. Спангенберг)
Характеристика отряда 139
Таблица для определения видов дроф 141
1. Стрепет 141
Восточный стрепет 142
2. Вихляй или джек 15J
Восточный вихляй 151
3. Дрофа 157
Европейская дрофа 158
Восточно-Сибирская дрофа 166
Отряд чистики (Г. П. Дементьев)
Характеристика отряда 169
Таблица для определения вида чистиковых 174
Род гагарки 175
1. Гагарка 175
Северная гагарка 176
Род люрики 179
2. Люрик, малая гагарка 179
Обыкновенный люрик 180
Большой люрик 182
Род кайры 184
3. Тонкоклювая или длинноклювая кайра 184
Североатлантическая тонкоклювая кайра 186
Тихоокеанская тонкоклювая кайра 189
4. Толстоклювая или короткоклювая кайра 190
Атлантическая толстоклювая кайра 191
Полярная толстоклювая кайра 196
Сибирская толстоклювая кайра 196
Чукотская толстоклювая кайра 197
Тихоокеанская толстоклювая кайра 197
478 ОГЛАВЛЕНИЕ
Род чистики 198
5. Чистик 198
Североатлантический чистик 201
Балтийский чистик 202
Полярный чистик 203
Таянов чистик 205
Берингов чистик, берингова каюрка 205
Курильский чистик, курильская каюрка 206
Командорский чистик, командорская каюрка 207
6. Очковый чистик 208
Род пыжики 210
7. Пестрый пыжик 210
Азиатский пестрый пыжик 211
Американский пестрый пыжик 212
8. Серый пыжик 213
(Алеутский пыжик] 214
Род старики 215
9. Старик 215
10. Хохлатый старик 217
Род конюга 218
11. Большая конюга 218
12. Малая конюга 221
13. Конюга-крошка 223
Род белобрюшки 225
14. Белобрюшка 225
Род тупики-носороги 227
15. Тупик-носорог 227
Род тупики 229
16. Тудик 229
Обыкновенный тупик 231
Полярный тупик 234
17. Ипатка 234
Род хохлатые топорики 237
18. Топорик, хохлатый топорик 237
Отряд гагары (Г. П. Дементьев)
Характеристика отряда 241
Таблица для определения видов гагар 244
Род гагары 244
1. Краснозобая гагара 244
Обыкновенная краснозобая гагара 245
Пестрая краснозобая гагара 249
2. Полярная гагара 250
Белонссая или белоклювая полярная гагара 250
Темноклювая полярная гагара 252
3. Чернозобая или полосатая гагара 253
Европейская чернозобая гагара 254
Восточносибирская чернозобая гагара , 258
Белошейная или тихоокеанская гагара 259
Отряд поганки (Г. П. Дементьев)
Характеристика отряда 261
Таблица для определения видов поганок 263
Род поганки 263
1. Большая поганка или чомга 263
Обыкновенная чомга 264
2. Серощекая поганка 268
Западная серощекая поганка 269
Восточная серощекая поганка 272
3. Рогатая или красношейная поганка 274
4. Черношейная или ушастая поганка 277
Обыкновенная черношейная или ушастая поганка 278
5. Малая поганка 282
Европейская малая поганка 283
Белокрылая малая поганка 284
Китайская малая поганка 286
ОГЛАВЛЕНИЕ 479
Отряд трубконосые или буревестники (А. М. Судиловская)
Характеристика отряда 287
Таблица для определения видов трубкокосых 293
Род альбатросы 294
1. Белоспинный альбатрос 2S4
2. Темноспинный альбатрсс 298
3. Черноногий альбатрос 301
Род тайфунник 3U4
4. Белолобый тайфунник 304
Бонинский белолобый тайфунник 305
Род глупыши 306
5. Глупыш 306
Атлантический глупыш 306
Тихоокеанский глупыш 313
Род буревестники 316
6. Серый буревестник 316
7. Тонкоклювый буревестник 319
8. Малый буревестник 323
Средиземноморский малый буревестник 324
9. Пестролицый буревестник 326
Род прямохвостые качурки 327
10. Малая прямохвостая качурка 327
Род вилохвостые качурки 330
11. Северная качурка 330
Северная большая качурка 331
12. Малая вилохвостая качурка 335
Азиатская малая вилохвостая качурка 336
13. Сизая качурка 336
Отряд фламинго (Е. П. Спангенберг)
Характеристика отряда 341
Род фламинго 343
1. Обыкновенный фламинго 343
Европейский фламинго или краснокрыл 343
Отряд голенастые птицы (Е. П. Спангенберг)
Характеристика отряда 350
Таблица для определения видов голенастых птиц 351
Семейство ибисы 357
Род колпицы 357
1. Колпица 357
Европейская колпица 358
Род каравайки 364
2. Каравайка 354
Европейская каравайка 364
Род белый ибис 372
3. Священный ибис 372
Материковый священьый и(л с 372
Род красноногий ибис 373
4. Красноногий ибис 373
Семейство аистов 376
Род аисты 376
5. Черный аист 376
6. Белый аист 383
Европейский белый аист ' 384
Туркестанский белый аист 390
Китайский белый аист 391
Семейство цаплевые 393
Род цапли 393
7. Серая цапля 393
Обыкновенная серая цапля 394
8. Рыжая цапля 404
Западная рыжая цапля 405
Восточная рыжая цапля 410
480 ОГЛАВЛЕНИЕ
Род белые цапли 412
9. Большая белая цапля 412
Северная большая белая цапля 413
Южная белая цапля 419
10. Средняя белая цапля 421
Азиатская средняя белая цапля 421
11. Чепура-нужда или малая белая цапля 422
Северная чепура-нужда 423
12. Желтоклювая чепура-нужда 427
Род малые чепуры 428
13. Египетская цапля 428
Западная египетская цапля 428
Восточная египетская цапля 433
Род косматые цапли 433
14. Желтая цапля 433
15. Белокрылая цапля 439
Род зеленые кваквы 441
16. Зеленая кваква 441
Амурская зеленая кваква 442
Род кваквы 445
17. Кваква 445
Обыкновенная кваква 446
Род волчки 453
18. Малая выпь или волчок 453
Северная малая выпь 453
19. Амурский волчок 461
20. Длинноносый волчок 464
Длинноносый волчок 464
Род выпь 466
21. Выпь 466
Северная выпь 468
Белогрудый голубь
Тибетская саджа
Северная качурка (вверху) и сизая, качурка (внизу)
Амурский волчок
Большеклювый волчок
Том II содержит описание одиннадцати отрядов птиц.
Отряды рябков, трехперсток, чистиков, гагар, поганок описаны Г. П. Д е-
ментьевым; голубей — Р. Н. Мекленбурцевым; журавлей и труб-
коносых — А. М. Судиловской; дроф, фламинго и голенастых —
Е. П. Спангенбергом.
Отв. редактор Н. И. Кузнецов
Рисунки художников: А. Н. Комарова, Н. Н. Кондакова и В. А. Ватагина
Ведущий редактор Л. П. Костина, художественная редакция К. И. Журинской
Технический редактор А. И. Прояева.
Т-10285. Подписано в печать 30/ХП 1950 г. Тираж 5.000 экз. Объем 30 п. л. + 5 цв.
вклеек. Уч.-авт. л. 47. Формат 70 X 1087ie- Цена в переплете 26 руб. Заказ 797.
2-я типография «Печатный Двор» им. А. М. Горького Главполиграфиздата при Совете
Министров СССР. Ленинград, Гатчинская, 26.