/






















































































































































































































Author: Флинт В.Е. Головкин А.Н.
Tags: aves птицы в целом орнитология птицы ссср
ISBN: 5-02-005232-9
Year: 1990




Text
ПТИЦЫ СССР
Чистиковые
академия наук ссср
Научный совет по проблеме
«Биологические основы освоения,
реконструкции
и охраны животного мира»
Институт
эволюционной морфологии
и экологии животных
им. А. Н. Северцева
Зоологический институт
ПТИЦЫ СССР
РЕДАКЦИОННАЯ КОЛЛЕГИЯ
В. Д. ИЛЬИЧЕВ
В. Е. ФЛИНТ
(ответственные редакторы)
Р. Л. БЁМЕ
Э. И. ГАВРИЛОВ
В. М. ГАЛУШИН
10. А. ИСАКОВ
Е. Н. КУРОЧКИН
Р. Л. ПОТАПОВ
С. Г. ПРИКЛОНСКИЙ
А. К. РУСТАМОВ
Л. С. СТЕПАНЯН
В. Е. ФОМИН
Ж
« и
Москва 1990
ПТИЦЫ СССР
Чистиковые
ic
Ответственные редакторы
доктор биологических наук
В. Е. ФЛИНТ
кандидат биологических наук
А. Н. ГОЛОВКИН
Москва 1990
УДК 598.2
Авторы
А. Н. ГОЛОВКИН, Н. Б. КОНЮХОВ, Н. Н. СКОКОВА,
Е. Э. СТОЦКАЯ, И. П. ТАТАРНИКОВА, С. П. ХАРИТОНОВ,
Ю. В. ШИБАЕВ, В. А. ЗУБАКИН (одуэнова чайка)
Птицы СССР. Чистиковые. М.: Наука, 1989. 207 с.
ISBN 5-02-005232-9
Очередной том серии «Птицы СССР» посвящен истинно морским птицам,
относящимся к подотряду чистиковых. Они населяют северные и дальневосточ-
ные моря Советского Союза, образуя многотысячные гнездовые колонии, или
«птичьи базары». В видовых очерках приведены современные сведения о
распространении, местах обитания, сроках и путях миграций, особенностях
гнездования, питания и размножения, поведения, о роли птиц в экосистемах.
Особое внимание уделено значению птиц для человека, их охране. Книга
снабжена цветными таблицами, на которых птицы изображены в разных воз-
растных и сезонных нарядах, а также их птенцы и яйца. Штриховые рисунки
иллюстрируют особенности строения и элементы поведения птиц. На картосхе-
мах изображены ареалы видов.
Для орнитологов, зоологов широкого профиля, работников охотничьего
хозяйства, любителей природы.
Цв. табл. 8. Ил. 49. Библиогр. 16 с.
Рецензенты
доктор биологических наук
А. В. МИХЕЕВ
член-корреспондент АН СССР
Е. Е. СЫРОЕЧКОВСКИЙ
Фото на суперобложке
Н. Б. КОНЮХОВА
Цветные иллюстрации
Ю. М. СМИРИНА
Черно-белые иллюстрации
В. М. ГУДКОВА и Ю. В. ШИБАЕВА
И
1907000000-025
055 (02)-89
582-89-Кн.
2.
© Коллектив авторов, 1989
ISBN 5-02-005232-9
Предисловие *
Очередной том сводки «Птицы СССР» посвящен обобщению материалов
по 20 видам подотряда Чистиковых, входящих в состав фауны птиц СССР,
т. е. достоверно обитающих на территории страны. В томе
сохранена схема построения видовых очерков, принятая в предыдущих
томах. Статус вида относится только к территории СССР. При состав-
лении описания, общей характеристики и полевых признаков специально
отмечается, какими особенностями отличается вид от других, похожих
на него представителей подотряда, и по каким полевым признакам его
можно узнать в природе.
Окраска нарядов приводится в соответствии с уже известными и опуб-
ликованными ранее описаниями видов, а также с некоторыми уточне-
ниями, которые авторы могли сделать в результате работы с коллек-
ционными материалами и в процессе собственных полевых наблюдений.
Названия частей тела и оперения птицы, а также способы измерения
соответствуют схеме, принятой для сводки в целом. Для ряда обычных
видов, по которым имеется массовый коллекционный материал, а также
опубликованные данные с большим числом промеров, авторы сочли воз-
можным провести совокупную среднюю с указанием границ изменчивости
и без ссылки на общее число измерений,,поскольку дальнейшее увеличе-
ние числа измерений практически не может изменить среднюю.
В связи с тем, что при составлении описания авторы широко использо-
вали материалы коллекций Зоологического института АН СССР и Зоо-
логического музея МГУ, авторы считают своей приятной обязанностью
поблагодарить сотрудников этих организаций за предоставленную воз-
можность работать с коллекциями.
Описание систематики подотряда базируется на фундаментальной
монографии Л. С. Степаняна (1975).
При подготовке видовых очерков авторы использовали не только опу-
бликованные данные, но и собственные наблюдения в природе, результаты
собственного анализа коллекционных материалов. Эти не опубликованные
ранее сведения специально оговариваются как данные автора, когда они
в той или иной степени имеют принципиальную новизну. Для того чтобы
настоящая сводка была более насыщена современными сведениями,
* Редколлегия рекомендует при цитировании сводки «Птицы СССР» ссылаться на авторов
конкретных очерков. Например: Скокова Н. Н. Тупик // Птицы СССР. Чистиковые.
М.: Наука, 1989.
5
Предисловие
в очерках использованы также личные сообщения ряда авторитетных
исследователей, касающиеся деталей распространения видов, особенностей
их поведения, численности в отдельных частях ареала, сроках основных
этапов жизненного цикла. В этих случаях обязательно указан автор сооб-
щения. Авторский коллектив тома выражает благодарность всем коллегам,
предоставившим возможность использовать их неопубликованные данные.
В конце книги помещен очерк об Одуэновой чайке, который не вошел
в предыдущий том, поскольку находка этого вида на территории СССР
была сделана уже после выхода тома о подотряде Чайковых.
Редакторы тома благодарят Н. Ю. Бурскую за большую помощь при
подготовке рукописи к печати.
Подотряд Чистиковые
Alcae
Чистиковые — истинно морские птицы, обитающие в шельфовой и пела-
гической зонах и связанные с сушей только в период гнездования. У совре-
менных видов размеры варьируют от мелких (около 100 г) до средних
(до 1,3 кг). Вымершая бескрылая гагарка, видимо, достигала веса 5 кг.
Они великолепно приспособлены к плаванию на поверхности воды
с помощью ног и в водной толще с использованием крыльев («подводный
полет»). Это наложило определенный отпечаток на морфологические
особенности чистиковых. Для большинства видов характерны несколько
удлиненное веретенообразное тело, плотное оперение, отнесенные назад
ноги, короткие заостренные крылья. Лапы снабжены плавательной пере-
понкой. Задний палец редуцирован. Череп схизогнатический, схизориналь-
ный со сквозными ноздрями. В развитых надорбитальных ямках имеются
большие носовые железы. Ноздри защищены кожистым клапаном. У взрос-
лых птиц базиптеригоидные сочленения не функционируют. Шейных
позвонков 15. Грудные позвонки не срастаются в грудную кость. Грудина
и таз узкие и удлиненные. Грудные мышцы хорошо развиты. Скелет
крыла уплощен, предплечье короче кисти. Весовая нагрузка на крыло зна-
чительно больше, чем у чаек. Задние конечности относительно короткие.
На суше чистиковые обычно опираются на всю плюсну и передвигаются
неуклюже. В целом скелет мало пневматизирован. Птерилии и аптерии
полностью покрыты пухом. Перья с побочным стволом.
Питаются только в море, добывая преимущественно в толще воды рыб
и мелких беспозвоночных (главным образом ракообразных). Язык мяси-
стый, удлиненный, иногда снабженный шипами в основной части, хорошо
приспособлен к удерживанию добычи. У взрослых люриков и конюг,
питающихся в основном зоопланктоном, образуется подъязычный мешок,
в котором они приносят корм птенцам. В связи с белковым питанием
железистый желудок развит хорошо, тогда как мускульный — мал
по объему.
Полет стремительный, у многих видов маломаневренный, с частыми
взмахами крыльев. Первостепенных маховых 11 (первое рудиментарно).
Рулевые короткие — 6—8 пар. Часто рулят с помощью растопыренных
лап. Воздушные мешки большие.
Окраска оперения двухцветная (белый низ и темный верх), темная,
реже пестрая. Полового диморфизма в окраске нет, но возрастные и
сезонные наряды различаются.
Чистиковые гнездятся, как правило, моно- и поливидовыми колониями,
часто крупными (более 10 тыс. особей), территориально приуроченными
к местам скопления корма. Моногамы. Гнездятся на скалах или в слое
торфа, открыто или в укрытиях (в щелях, полостях среди камней, в норах).
7
Подотряд Чистиковые
Настоящих гнезд, как правило, не строят. Только длинноклювый пыжик
гнездится на деревьях.
В кладке одно или два яйца. Насиживают оба родителя, у которых
образуются наседные пятна. Инкубация занимает 3—5 недель. Птенцы
вылупляются зрячими, одетыми густым пухом, который у открытогнездя-
щихся видов меняется на промежуточный наряд (мезоптиль).
В подотряде только одно семейство (Alcidae), объединяющее 13 родов,
в которых в общей сложности насчитывается 22 вида. Вероятный центр
формообразования чистиковых — Берингово море. Ископаемые остатки
известны из нижнего эоцена и более поздних слоев. По ним описано
11 вымерших видов. Современные виды найдены в плейстоценовых отло-
жениях. Один вид — бескрылая гагарка — в результате высокой степени
адаптации к подводному плаванию полностью утратил способность к по-
лету в воздухе. Он был истреблен в первой половине XIX в. Чистиковые
обособились от чаек в ходе приспособления к водному образу жизни и
освоению кормовых ресурсов водной толщи до глубины в несколько
десятков метров. Современные виды распространены в морях северных и
умеренных широт Северного полушария.
Для фауны СССР отмечено 20 видов, из которых 19 гнездятся и лишь 1
(алеутский пыжик) известен как залетный.
Головкин А. Н.
Род Alle Link, 1806
Люрик —
Alle alle Linnaeus, 1758
Little (англ.), Dovekie (амер.), Krabbentaucher (нем.). '
Син. (лат.): Plautus alle.
Статус. В пределах СССР гнездящийся, кочующий, зимующий вид.
Общая характеристика и полевые признаки. Один из самых мелких
чистиков. Окраска контрастная черно-белая: голова, шея и верхняя сторона
тела черные, низ белый..На крыльях зубчатый белый рисунок. Голова
крупная, при наполненном подъязычном мешке выглядит массивной. .Клюв
короткий, темный, слегка загнутый. Полет быстрый, с частыми взмахами
крыльев, довольно маневренный.. Летают люрики обычно стаями низко
над водой. Легко взлетают с камней и мелкой волны. На суше передви-
гаются неуклюже, опираясь на плюсну, но могут подниматься и на пальцы.
На камнях часто полулежат, приподнимаясь по тревоге. Прекрасно пла-
вают и ныряют. На поверхности воды гребут лапами, а для плавания под
водой используют крылья («подводный полет»), В период гнездования
образуют большие колонии в каменистых осыпях. Вблизи колоний крик-
ливы. При возвращении в колонию или при опасности издают повторяю-
щиеся свистящие крики «здии—зди-зди», затухающие к окончанию, а также
«уиррр-уир» и «крии-кри» — при взаимных контактах. Птенцы в гнезде
или в укрытии издают шуршащие звуки или слабое попискивание.
Подвидовая систематика. Известны два подвида, различающиеся раз-
мерами тела. Оба обитают на территории СССР.
8
Люрик
1. Alle alle alle
Alca alle Linnaeus, 1758, Syst. Nat., ed. 10, c. 131. Арктический океан
у Европы и Америки.
Мелкий подвид. Длина крыла самцов и самок НО—116 мм (сред-
нее 120,8). Распространен по всему ареалу вида, кроме Земли Франца-
Иосифа и, возможно, Северной Земли.
2. Alle alle polaris
Alle alle polaris Stenhouse, 1930, Scottish Naturalist, № 182, c. 47, Земля
Франца-Иосифа.
Крупный подвид. Длина крыла самцов и самок 123,5—138 мм (среднее
130,5). Населяет архипелаги Земли Франца-Иосифа и предположительно
Северной Земли (Степанян, 1975).
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла: . . Рулевых
перьев 12. Размеры самцов и самок: длина (в мм) крыла НО—138 (среднее
124,0) клюва 11,2—17,5 (среднее 14,8), плюсны 18,7—24,0 (среднее 21,1),
масса (в г) 127—224 (среднее 158,8) (Stonehouse, 1930, по Дементьеву,
1951; Johnson, 1944; Козлова, 1957; Vaurie, 1965; Norderhaug, 1980; Stemp-
niewicz, 1981; и др.). Клюв толстый с выпуклым надклювьем. Ноздри
овальные с кожистым утолщением. Контурное оперение плотное. В зимний
период размеры клюва уменьшаются.
Окраска. Самец и самка в брачном наряде. Полового диморфизма
в окраске нет. Голова, шея, верхняя часть груди, спина, крылья и хвост
черные, блестящие. На лицевой части и груди имеется коричневатый
оттенок. Спереди над глазом узкое белое пятнышко. Белые каемки на пле-
чевых перьях образуют зубчатый рисунок, а белые концы второстепенных
маховых — узкую белую полоску. Низ тела белый..На боках внутренние
дымчато-серые опахала кроющих образуют продольные размытые полосы.
Подкрылья буровато-серые. Нижние кроющие крыла с белыми вершинами.
Ноги серые с темно-бурыми пятнышками, темными плавательными пере-
понками и когтями. Клюв черный.. Радужина темно-коричневая.
Самец и самка в зимнем наряде. Горло и грудь белые, слегка испещрен-
ные серым за счет двухцветной серо-белой окраски кроющих. Белый цвет
распространяется также на щеку до затылка, окружая черное подглаз-
ное пятно полукольцом.
Пуховой наряд. Общий тон черный, дымчатый или коричневато-серый.
На брюшке пух более светлый, светлеет с возрастом.
Гнездовой наряд сходен по окраске со взрослым, но без блеска оперения
на спине. Среди темных перьев в верхней части груди и на зобе сохра-
няются белые пестринки. Клюв коричневатый, лапы серые.
Линька. Последовательность смены нарядов: пуховой — первый зим-
ний — брачный. Только что вылупившийся птенец покрыт густым длин-
ным мягким однотонным пухом. Через две недели пух становится более
редким и более светлым на брюшке. К концу третьей недели пух сменяется
перьевым покровом и к вылету из гнезда птенец почти полностью пере-
линивает в юношеский зимний наряд. Отдельные пушинки сохраняются
только на голове и крестце. Линька заканчивается уже в море. Взрослые
птицы меняют контурное оперение с мая по сентябрь. Линька маховых
и рулевых проходит в сентябре—октябре. К этому времени они приобре-
тают зимний наряд. С декабря по март зимний наряд меняется на брачный
(Горбунов, 1932; Cramp, 1985).
9
Семейство Чистиковые
Распространение. Гнездовой ареал. Высокие широты северных
морей, бассейнов Атлантического, Северного Ледовитого и Тихого океанов
(рис. 1, 2). Гнездится на западном и восточном побережьях Гренландии,
на островах Исландия, Ян-Майен, Медвежий, Шпицберген, Земля Франца-
Иосифа, Новая Земля, Северная Земля, Малый Диомид.
Зимовки. Большая часть люриков зимует вдали от мест гнездования.
Держатся неподалеку от кромки льдов, иногда в разводьях между ледовыми
полями или в открытом море. Птицы из Гренландии перемещаются к се-
веро-восточным берегам Америки и зимуют в проливе Дэвиса и южнее
вдоль Ньюфаундленда (Brown et al., 1975). Люрики из центральной и
восточной частей Северной Атлантики зимуют в Баренцевом, Норвежском,
Северном и Исландском морях (Белопольский, и др., 1982; Joiris, 1983;
Brown, 1984). Люрики со Шпицбергена найдены на зимовках у берегов
Гренландии (Salomonsen, 1971). Отмечена предпочитаемость районов
моря, покрытых льдом на 40—60 %, и избегание участков, покрытых более
чем на 90 % (Brown, 1984). Меньшая часть популяций зимует неподалеку
от мест гнездования или перемещается к северу (Рутилевский, Успенский,.
1957; Бутьев, 1959). К югу на зимовках доходят до Карибского моря и
Западного Средиземноморья. В восточных районах отмечены залеты
на о-в Св. Лаврентия (Bedard, 1966), о-в Врангеля (В. И. Придатко, личное
сообщ.), летом в 1987 г. — на о-в Талан в Тауйской губе Охотского моря
(С. П. Харитонов, личное сообщ.). С середины XIX в. известны много-
численные случаи встреч люриков у берегов и на суше в Великобритании,
куда, в частности в октябре-ноябре 1938 г., залетели тысячи люриков
(Fisher, 1954), а также на восточном побережье США. В небольшом числе
встречаются у берегов Нидерландов; среди 108 трупов люриков, найденных
на берегу, один был отнесен к северо-восточному подвиду A. a. polaris
(Camphuysen, 1986).
Миграции. Весенний прилет в районы гнездования начинается в конце
февраля-марте, когда берега еще покрыты льдами, а в местах расположения
колоний лежит снег (Горбунов, 1932; Рутилевский, 1957; Lovenskiold,
1964). В это время птицы не держатся здесь постоянно, а улетают и
возвращаются с интервалом в несколько дней. В середине апреля стайки
люриков появляются на суше, но массовый весенний пролет продолжается
до середины мая (Renaud et al., 1982). Направление миграций в основном
на восток и северо-восток. Осенние откочевки начинаются в августе—
начале сентября, становятся более интенсивными в сентябре и затухают
в октябре. В целом они продолжаются около трех месяцев (Рутилевский,
1957; Bateson, 1961; Norderhaug, 1967).
Численность. В связи со скрытностью гнездования общая численность
люриков в настоящее время может быть определена только очень приблизи-
тельно: вероятно, не менее 30 млн пар. Самая крупная популяция люриков
находится в Гренландии в проливе Ланкастер и близ западных побережий
Баффиновой Земли. На пролете в середине мая было учтено около 14 млн
особей, которые, скорее всего, гнездятся на северо-западном побережье
Гренландии (Renaud et al., 1982) . Предполагается, что всего в Гренландии
гнездится от 8 до 25 млн пар люриков (Evans, 1984). НаШпицбергене,
занимающем второе место по численности люриков, обитает от 10 до 15 млн
особей (Riippel, 1969) или гнездится 0,4—1,6 млн пар (Evans, 1984).
На о-ве Медвежий учтено около 10 тыс. пар, на о-ве Ян-Майен — 50 тыс.
пар. Реликтовая гнездовая колония в несколько пар сохранилась в Ислан-
10
60
60
Рис. 1. Область распространения люрика Рис. 2. Ареал люрика в СССР
а—граница гнездового ареала, б — область зимовок н миграций, а — гнездовой ареал, б — область миграций, в — предполагаемые места
в — залеты. Подвиды: / — А.а. alle, 2 — А.а. polaris гнездования и миграций. Подвиды: 1 — А.а. alle, 2 — А.а. polaris. ? — подвидовой
статус неизвестен
Семейство Чистиковые
дии (Evans, 1984). Небольшая колония обнаружена в Беринговом проливе
на о-ве Малый Диомид (Breckenridge, 1966). В СССР люрики гнездятся
на Земле Франца-Иосифа — около 250 тыс. пар (Успенский, 1959), в губе
Архангельской на Новой Земле — около 5,5 тыс. пар (Головкин, 1972).
Возможно, есть гнездовья в некоторых других пунктах Новой Земли,
а также на Северной Земле.
Местообитания. Типично морская птица, обитатель шельфовых районов
высокой Арктики. Вне шельфовой Зоны встречается редко. Гнездовое
местообитание — скалистые берега островов, каменистые осыпи у подно-
жия скал недалеко от уреза воды или расположенные в нескольких кило-
метрах от берега/Иногда населяют расселины скал до высоты 500 м над
уровнем моря. Возможно, существуют колонии, удаленные от моря на не-
сколько десятков километров (Bateson, 1961).
Размножение. Возраст первого размножения не известен, предположи-
тельно 2 года. Люрики гнездятся в колониях, насчитывающих от несколь-
ких до сотен тысяч пар (с плотностью до 1 гнезда/м2, иногда более).
Гнезда располагаются в полостях под камнями осыпей или — реже —
в трещинах скал на глубине 0,5— 1,0 м от поверхности. Они могут использо-
ваться одной и той же парой в течение нескольких лет. На дне гнездовой
камеры имеется тонкая (2—4 см толщиной) выстилка из ракушечника,
лишайников, тонких стеблей растений (Norderhaug, 1980). Сроки яйце-
кладки завйсят~от скорости стаивания снега в местах размещения гнезд.
На Земле Франца-Иосифа и Новой Земле кладка приходится на вторую
половину июня—начало июля (Горбунов, 1929; Рутилевский, 1957). При-
мерно в эти же сроки она происходит на Шпицбергене и в Гренландии,
тогда как в Исландии начинается в конце мая—начале июня (Cramp, 1985).
В кладке, как правило, одно яйцо, хотя у родителей имеется по два насед-
ных пятна (Norderhaug, 1980). Яйцо эллиптической формы, слегка утол-
щенное на одном конце. Окраска скорлупы бледная зеленовато-голубая
с редкими желтовато-бурыми, коричневыми, зелеными или розоватыми
точками или пестринами, рассеянными по всему полю или концентрирую-
щимися у тупого конца. Размеры яиц у подвидов сходны: 45—56X30—
39 мм (среднее 48,5X34,0); средняя масса 28 г. (Schonwetter, 1967; Norder-
haug, 1980).
Насиживают попеременно оба родителя, но сменяются они нерегулярно.
Срок инкубации 28—31 день (в среднем 29 дней) (Stempniewicz, 1981).
Только что вылупившегося птенца согревают и кормят оба родителя.
Частота кормления изменяется в зависимости от возраста птенца — от 4
до 14 раз в сутки и составляет в среднем 5,25—8,5 раза. От вылупления
до вылета птенцы проводят в гнезде 26— 29 дней (в среднем 27,1). Индекс
прироста птенцов наибольший среди чистиковых. Кривая прироста имеет
типичную S-образную форму. Максимальный прирост приходится
на 8-е сутки, а его уменьшение — на 17—25-й день, когда птенцы дости-
гают максимальной массы. После 21 дня частота кормлений снижается
(Brown, 1976; Norderhaug, 1980; Evans, 1981). Терморегуляция устанавли-
вается на 3—5-й день жизни. Перед выходом из гнезда птенцы имеют вес
в среднем 108—114 г (Norderhaug, 1980; Glutz, Bauer, 1982; по Cramp,
1985). Успешность инкубации на Шпицбергене составляет 65,3 %, а выжи-
ваемость птенцов — 80 % (Stempnievicz, 1981). В Гренландии успешность
размножения в целом составляет около 50 % (Evans, 1981).
Суточная активность, поведение. Ритм суточной активности в гнездовое
12
Люрик
время в Западной Гренландии близок к ритму вертикальных миграций
зоопланктона. Максимальная активность кормления приходится на вечер-
ние и утренние часы — на 22 и 8 ч. Наиболее активное кормление птенцов
происходит в период с 18 до 6 ч. При этом отмечена значительная синхрон-
ность полетов многих особей из колонии, по-видимому обусловленная
появлением массового корма (Evans, 1981). На Шпицбергене, например,
более активное кормление птенцов зарегистрировано в дневное время
с пиками активности рано утром и после полудня — с 15 до 18 ч (Norder-
haug, 1980). Ритм посещения мест гнездования и длительность пребывания
люриков в колонии меняются в течение сезона размножения (Stempniewicz,
1986). Длительность одновременного присутствия основной массы птиц
в колонии на Шпицбергене в период откладки яиц меняется от 80 ч в сере-
дине мая до 24 ч в конце мая и составляет лишь 12 ч в период вылупления
птенцов в конце июня. Туманы и сильные ветры, а также появление
хищников (песцов или бургомистров) заставляют тех люриков, которые
не находятся в норах, улетать в море.
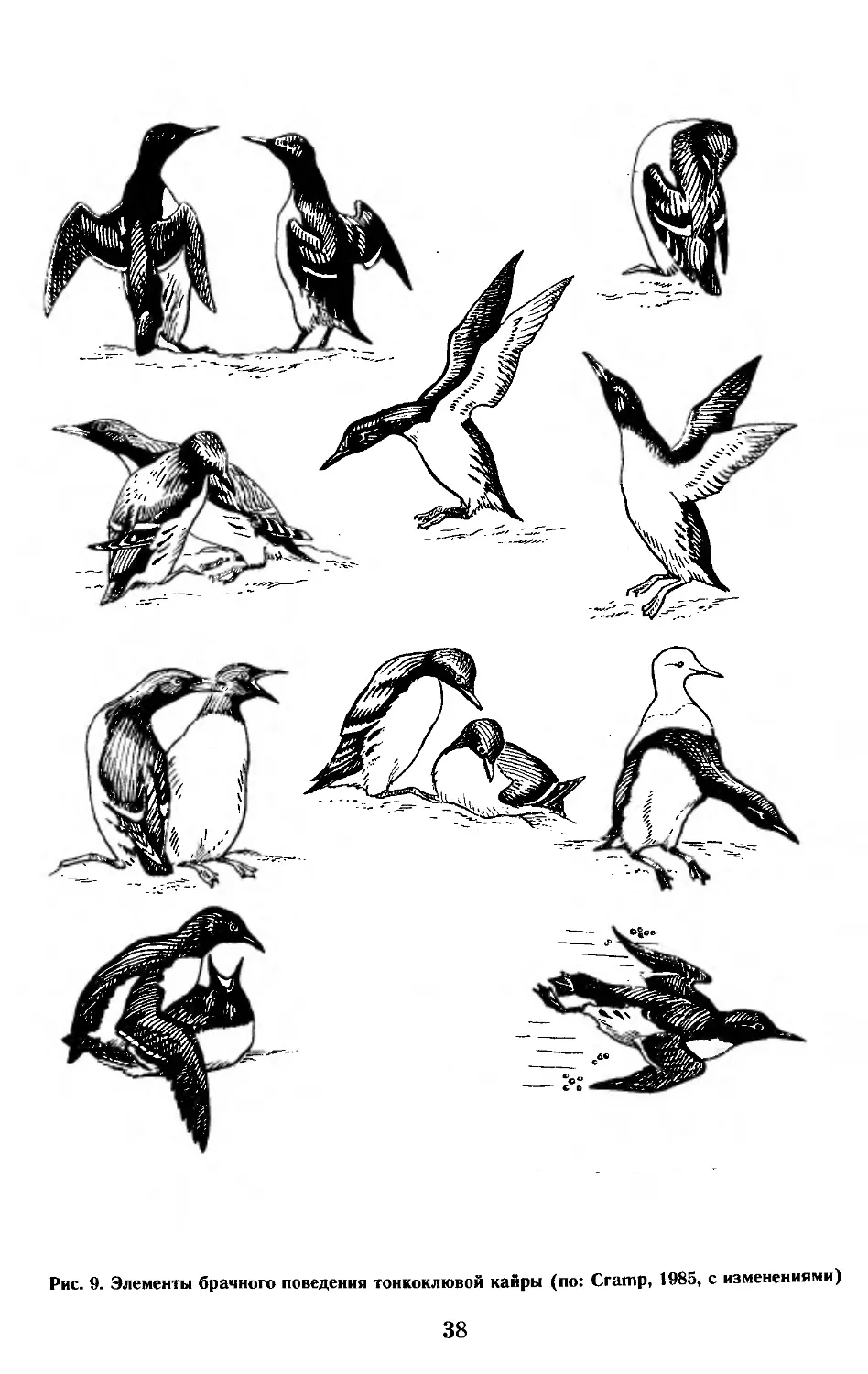
В период размножения люрикам свойственно сложное брачное пове-
дение, для которого характерны специфические позы, движения и крики.
Как элементы брачного поведения отмечены «импонирующий» полет с ред-
кими взмахами крыльев (3—5 взмахов в секунду против обычных 12—18),
«импонирующая» походка (2—3 шага в секунду против обычных 4—6),
«притоптывание», позы ухаживания, передача камешков от партнера
к партнеру. Кроме того, для различных периодов гнездового цикла из-
вестны позы тревоги и агрессии, полет «каруселью» при возвращении
в колонию, полеты с резкой сменой высоты, а также пять типов крика
(Горбунов, 1929; Рутилевский, 1957; Ruppel, 1969; Evans, 1981; и др.)
(рис. 3).
Питание. По характеру питания преимущественный планктофаг. В про-
бах из подъязычного мешка отмечены также мальки рыб, но они значи-
тельно уступают планктонным организмам как по встречаемости, так и
по количеству и общей массе. В период гнездования взрослые люрики
осуществляют сбор планктонных организмов в подъязычный мешок, ко-
торый представляет собой выпячивание дна ротовой полости. Его эла-
стичность обусловлена нарушением целостности подъязычной мышечной
выстилки. Список видов беспозвоночных, используемых люриками, вклю-
чает более 30 видов, в основном ракообразных (калянусы, эуфазииды,
гиперииды, мизиды, гаммариды, личинки декапод и др.). В летний период
в различных частях ареала основу питания люриков составляют рако-
образные, образующие плотные локальные скопления: Calanus finmarchi-
cus. С. glacialis, С. hyperboreus, Thysanoessa inermis, T. longicaudata,
Parathemisto spp. (Головкин и др., 1972; Zelickman, Golovkin, 1972; Evans,
1981). Калянусы, или амфиподы, являются основным кормом люриков и
во внегнездовое время в районе Баффиновой Земли (Bradstreet, 1982).
Количество ракообразных, обнаруженных в подъязычных мешках люри-
ков, варьирует от нескольких десятков до 3 тыс. особей. Зачастую люрики
последовательно облавливают моновидовые скопления различных рачков,
вследствие чего в подъязычном мешке накапливаются слои из особей раз-
ных видов. Птицы могут склевывать отдельных ракообразных, плавающих
у поверхности воды, но чаще кормятся, заныривая вглубь и захватывая
сразу большое количество животных в плотных скоплениях. Подсчитано,
что даже при очень высоких средних показателях плотности распреде-
13
Семейство Чистиковые
Рис. 3. Элементы брачного поведения люрика (по: Cramp, 1985, с изменениями)
ления ракообразных (биомасса до 1,9 г/м3) у берегов Новой Земли, но при
равномерном (не стайном) их размещении, люрики должны облавливать
не менее 50 м3 воды для получения суточной нормы корма. По-видимому,
практически нереально, чтобы люрики могли обеспечить себя кормом,
схватывая единичных мелких ракообразных. Это означает, что только очень
плотные скопления зоопланктона, где птицы захватывают сразу помногу
рачков, являются базой успешного питания люриков. Последовательные
порции корма в подъязычном мешке накапливаются и уплотняются в нем
за счет специфичных «поршневых» движений языка (Головкин и др.,
1972).
Кормятся люрики обычно в открытом море, улетая от берега на рас-
стояние от 2,5 до 9 км (Головкин и др., 1972; Zelickman, Golovkin, 1972;
Evans, 1981). По мнению Брауна (Brown, 1976), они могут совершать
кормовые перелеты и на расстояние 100—150 км от колонии, однако
энергетическая оправданность таких перелетов не известна. Они могут
кормиться также и в непосредственной близости от берега, о чем свидетель-
14
Люрик
ствует присутствие в их рационе литоральных беспозвоночных (Mathey-
Dupras, 1913; Демме, 1934; Hartley, Fisher, 1936; Lovenskiold, 1958).
Состав корма меняется по сезонам, годам и географическим районам.
Птицы имеют тенденцию избирательно добывать более крупных рако-
образных и более крупные их возрастные группы. При этом отмечена
значительная избирательность по сравнению с общим составом планктона
в водной толще (Bradstreet, 1982). Потребность люриков в пище не изучена,
но, по-видимому, вес потребляемого за сутки корма составляет не менее
50 % от веса птицы, т. е. в среднем около 80 г. планктона. Интенсивность
обмена у них велика. При голодании птицы гибнут на третий день, теряя
около половины веса тела (Головкин и др., 1972). .Многочисленность
люриков и специфичность их питания ставят их на одно из ведущих мест
среди высших позвоночных, потребляющих зоопланктон в экосистемах
морей высокой Арктики (Norderhaug, 1970; Головкин и др., 1972; Brad-
street, 1982). Подсчитано, что на Шпицбергене в период выкармливания
птенцов они переносят из моря на сушу от 280 до 1120 т зоопланктона.
Снабжая органическими веществами растительные сообщества в местах
гнездования, они способствуют увеличению продуктивности раститель-
ности участков высокоарктических тундр.
Враги, неблагоприятные факторы. Основными врагами люриков можно
считать бургомистров, которые нападают как на взрослых птиц, так и
на птенцов. Наибольший ущерб они причиняют колонии, уничтожая
слетков, совершающих первые полеты после выхода из укрытий. Этих
птиц, летающих еще неуверенно, чайки заставляют приземляться или
садиться на воду и убивают. Давление на колонию люриков оказывают
еще и песцы, разоряющие гнезда с яйцами или птенцами. В первую оче-
редь гибнет потомство в тех норах, которые расположены неглубоко
в осыпи. Хищничество песцов и бургомистров в колониях люриков известно
практически во всех местах их обитания (Демме, 1934; Evans, 1981;
Stempnievicz, 4981; и др.).
К числу неблагоприятных факторов следует отнести в первую очередь
нефтяное загрязнение моря. Гибель птиц от загрязнения в период зимовки
и предгнездовых кочевок отмечена у берегов Северной Америки и Грен-
ландии. Некоторое количество люриков гибнет, запутываясь в тонких став-
ных рыболовных сетях (Christensen, Lear, 1977; Brown, Nettleship, 1984).
В Гренландии аборигенное население охотится на люриков в местах их
гнездования, отлавливая сетями взлетающих птиц (Salomonsen, 1967).
В СССР антропогенное влияние на люриков в настоящее время мини-
мально, поскольку их колонии расположены в слабо освоенных районах.
Однако угроза нефтяного загрязнения растет с увеличением судоходства и
числа полярных станций в северных областях.
Хозяйственное значение, охрана. В настоящее время у нас в стране
в хозяйственных целях не используется. Наряду с другими морскими пти-
цами люрики расцениваются как важный биологический ресурс Крайнего
Севера и один из компонентов арктических экосистем. В связи с этим они
подлежат особой охране в соответствии с Указом Президиума Верховного
Совета СССР «Об усилении охраны природы в районах Крайнего Севера
и морских районах, прилегающих к северному побережью СССР», приня-
тым в ноябре 1984 г.
15
Семейство Чистиковые
Род Alca Linnaeus, 1758
Гагарка —
Alca torda Linnaeus, 1758
Rasorbill (англ.), Tordalk (нем.).
Статус. Гнездящийся перелетный вид.
Общая характеристика и полевые признаки. Наряду с кайрами самый
крупный представитель семейства чистиковых. Типично морская птица,
с сушей связана только в период гнездования. К передвижению по земле
приспособлена плохо. Положение тела при ходьбе вертикальное, обычно
ходит, опираясь на всю плюсну, реже — только на пальцах (в основном
птенцы). С ровной поверхности земли подняться не может, взлетает с обры-
вов, с высоких камней. С воды взлетает после длительного разбега, хлопая
крыльями по воде. Полет быстрый, с частыми взмахами крыльев, но мало-
маневренный; круто поворачивать на лету не может. Рулем при поворотах
служат перепончатые лапы, широко расправленные и несколько раздвину-
тые на лету. Отлично плавает и ныряет. Под водой передвигается с по-
мощью крыльев, которые держит в полусогнутом положении и производит
ими короткие редкие взмахи назад и вниз. Окраска двухцветная: спинная,
сторона черная, брюшная — белая. Клюв высокий, сжатый с боков, длин-
ный. Во время плавания характерно положение поднятого кверху хвоста.
Осторожна и молчалива. Только в период спаривания или при испуге
издает своеобразный хриплый скрипящий крик.
Подвидовая систематика. Описаны два подвида гагарок, различаю-
щиеся размерами.
1. Alca torda torda
Alca torda Linnaeus, 1758, Syst. Nat., ed. 10, c. 130, о-ва Стора Карлсё,
Балтийское море.
Более крупный подвид. Обитает на большей части ареала, в том числе
и в СССР.
2. Alca torda islandica
Alca torda islandica Briinnich, 1764 (Дементьев, 1951).
Мельче по размерам. Населяет Исландию, Великобританию, Ирлан-
дию, Фарерские острова, Бретань, Гельголанд.
Замечания по систематике. Ф. Соломонсон (Solomonsen, 1944) выска-
зал предположение, что гагарки из Норвегии, Гренландии и северных
частей СССР отличаются от балтийских большим числом борозд над-
клювья (обычно тремя). Однако у гагарок, обитающих на Мурмане,
количество борозд бывает разным. Г. П. Дементьев (1951) приводит дан-
ные по 26 птицам: у 19 было две борозды, у 1 — три, у 5 — одна, у 1 —
ноль. По-видимому, количество борозд на клюве гагарки, так же как
у тупиков, зависит от возраста птицы (Petersen, 1976а).
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла: .. Рулевых перьев
12—14. Размеры: длина (в мм) крыла самцов 180—220 (среднее 205,6),
самок 195—230 (среднее 210,4); масса (в г) самцов 524—890 (среднее
728,5), самок 600—910 (среднее 716) (Горчаковская, 1944; Дементьев,
1951; Кафтановский, 1961; Белопольский, 1957).
Половой диморфизм в размерах, по-видимому, отсутствует. Индиви-
16
Гагарка
дуальные колебания размеров значительны. Вершина клюва загнута
крючкообразно. Ноздри щелевидные.
Окраска. Полового диморфизма в окраске нет. Самец и самка
в брачном наряде. Спинная сторона, верх и бока головы и шеи, подбородок
и горло черные, с легким аспидным оттенком. Брюшная сторона, под-
крылья и вершины второстепенных махорых белые. От глаза к клюву про-
ходит белая полоса. Ноги черные. Клюв высокий, сжатый с боков, черный
с белой поперечной полосой и с одной-двумя темными поперечными бороз-
дами. В брачном наряде клюв вздутый у основания за счет развития
пластин. Ротовая полость ярко-желтого цвета.
Самец и самка в зимнем наряде. Спинная сторона черная. Подбородок,
горло и бока головы белые. Белой полосы от глаза к клюву нет.
Пуховой наряд. Верхняя сторона тела, бока и крылья коричневые:
окраска широко варьирует от светло- до темно-коричневого. На шее окраска
постепенно светлеет, переходя в белесовато-желтую, на голове — в почти
совершенно белую. Нижняя сторона шеи и горло сероватые, брюшко
белое. Ноги черные, клюв серовато-черный, по форме сходен с клювом
взрослой птицы, но поперечные борозды на нем едва заметны, белой по-
лоски нет. Яйцовый бугорок на клюве хорошо заметен и сохраняется
долго. Птенец в мезоптиле очень похож на взрослую птицу, отличаясь
лишь белесым цветом горла. Белой поперечной полосы на клюве нет.
Известно (Hudson, 1984), что некоторое количество птенцов (до 49 %)
могут иметь черную окраску горла. У белогорлых птенцов описано два
варианта окраски горла: 1) белые горло, подбородок и щеки, 2) белые
горло и щеки, но черный подбородок и черная полоса вокруг основания
горла.
Гнездовой наряд. Похожа на взрослую зимой, белой полосы поперек
клюва нет. Белая полоса на второстепенных меховых несколько уже,
чем у взрослых птиц. Поперечные борозды на надклювье развиты слабо.
Линька. Последовательность смены нарядов такова: эмбриональный
пух — мезоптиль — первый осенний (зимний) — летний предбрачный —
второй зимний — первый брачный — зимний послебрачный и т. д.
Пух птенца короткий, довольно жесткий. Густота пуха на брюшной и
спинной сторонах более или менее одинакова. Развитие оперения идет
очень быстро: на 3—4-й день показываются верхушки верхних кроющих
крыла, на 5—6-й появляются кроющие на всем теле. На 15—16-й день
эмбрионального пуха не остается, птенец весь покрыт пером (мезоптиль).
Но по структуре это перо отличается от пера взрослой птицы: оно короче
и мягче. Развитие мезоптиля позволяет птенцу в короткий срок одеться
густым перьевым покровом, предохраняющим его от переохлаждения
на воде. Рулевые и маховые развиваются позже, после спуска на воду.
Первый осенний наряд взамен мезоптиля развивается в августе-сентябре
(Witherby et al., 1941, цит. по Дементьеву, 1951).
Ко второму лету молодая птица отличается от взрослой в брачном
наряде серой с пестринками окраской горла. Линька начинается с конца
июня. Наряд второй зимы сходен с зимним нарядом взрослых птиц. Первый
брачный наряд развивается к третьему лету. У взрослых в году проходят
две линьки. Предбрачная неполная начинается в марте, а заканчивается
к концу мая. Маховые и рулевые не меняются. Послебрачная полная
линька начинается в конце августа с мелкого оперения на голове и шее,
затем меняются плечевые и кроющие ног, далее — перья брюшной стороны.
2 Птицы СССР
17
Семейство Чистиковые
Смена крупного пера проходит осенью в море. Маховые линяют одновре-
менно, поэтому птица временно теряет способность к полету (Дементьев,
1951).
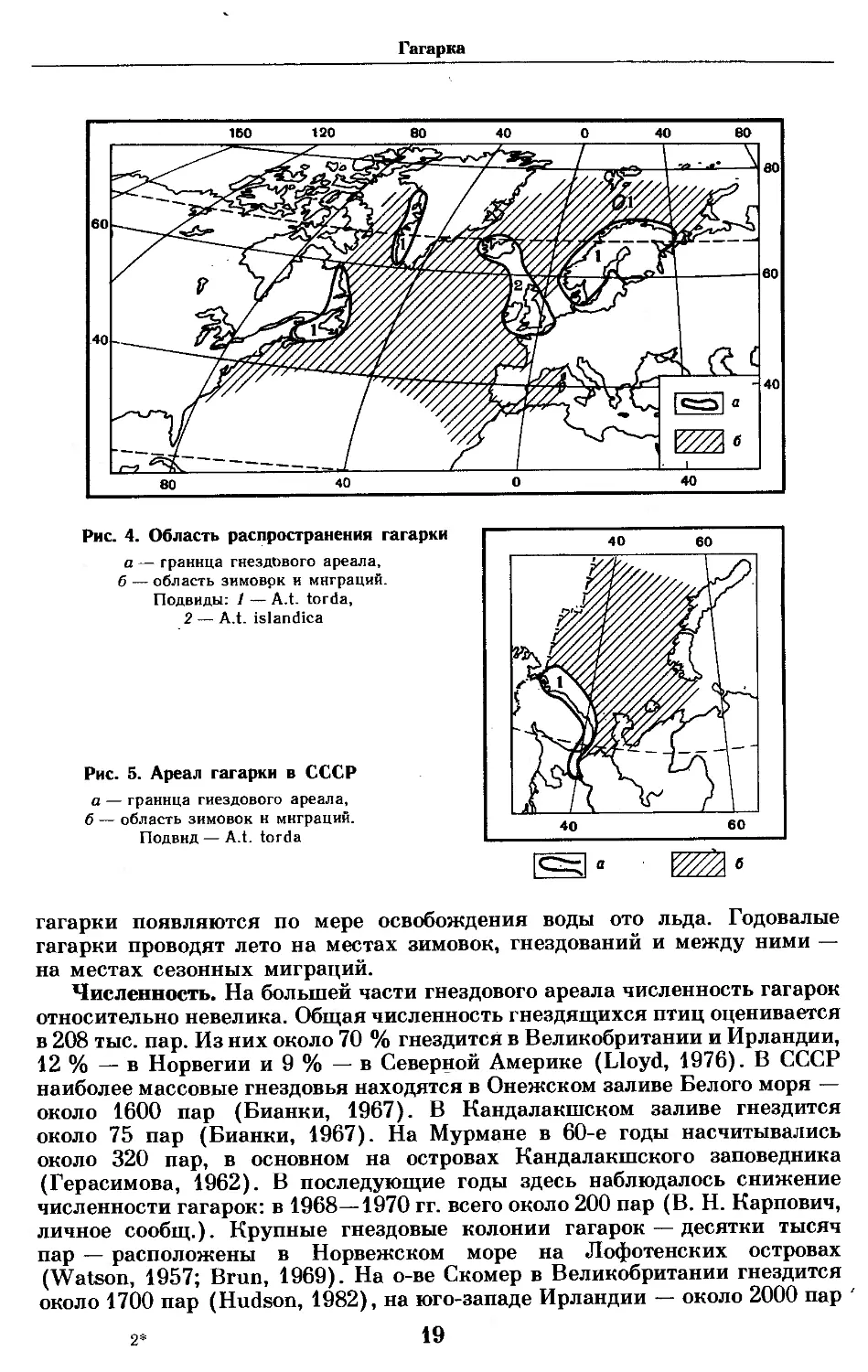
Распространение. Гнездовой ареал. По общему распростране-
нию гагарка относится к бореальным видам, встречаясь лишь в умеренных
широтах Северной Атлантики (рис. 4). Северный предел распространения
в Западной Атлантике совпадает с августовской изотермой 4° (Bedard,
1969) и проходит по юго-западному побережью Гренландии; на восточ-
ном побережье отсутствует. В Северной Америке населяет восточную часть
побережья от Нью-Брусвика, залива Св. Лаврентия, Ньюфаундленд и
Лабрадор. На юг доходила до берегов штата Мэн (Bent, 1919), в настоящее
время здесь не встречается. В Западной Европе населяет Исландию,
о-в Медвежий, острова у побережья Норвегии, Ирландии, Англии, Шот-
ландские острова. На юге распространена до побережья п-ова Бретань
во Франции и о-ва Гельголанд. На Балтийском море гнездится от северных
частей Ботнического залива до о-ва Борнхольм на юге. В Финляндии —
по берегам Финского залива.
В СССР гнездится на Ладоге, в Онежском и Кандалакшском заливах
Белого моря и на Мурмане (рис. 5).
Зимовки. В зимнее время встречается на море от Лабрадора
до о-ва Лонг-Айленд в Америке, в Европе — у западных берегов Голландии,
Франции, Португалии, в Северном и Балтийском морях. Наиболее южные
встречи зимой — на Средиземном море (Тунис, Италия). Случайные
залеты отмечены у Азорских и Канарских островов, на Шпицбергене и
Новой Земле (Дементьев, 1951).
Миграции. Миграции беломорских гагарок начинаются сразу же, как
только птенцы спустятся на воду — в первой половине августа (Бианки,
1967). Птенцы еще долго остаются с родителями; на Баренцевом море
в сентябре отмечали подросших птенцов, державшихся около взрослой
птицы. Места зимовок беломорских и баренцевоморских гагарок совпадают.
Большая часть птиц зимует у берегов Южной Норвегии, немногие отходят
к берегам Северного моря, встречаются в проливах Дании. Здесь же зимуют
и гагарки, гнездящиеся в Норвегии (Holgersen, 1953—1955). Гнездящиеся
на Балтике гагарки зимуют в основном в южной части Балтийского моря
(Nordstrom, 1957 — 1963). Видимо, здесь же зимуют гагарки, гнездящиеся
на Ладоге. В Финском заливе встречаются на пролете гагарки из финских
популяций (Йыги, 1975). Значительный пролет гагарок наблюдается
осенью и зимой у северо-восточных берегов Шотландии (Elkins, Williams,
1972). В Центральной Атлантике гагарки перемещаются шире, чем тонко-
клювые кайры, и регулярно встречаются зимой и весной в Средиземном
море (Mead, 1974), достигая в ноябре-декабре побережья Марокко (Robin,
1969). Среди необычных дальних залетов следует отметить находки не-
половозрелых гагарок во Флориде (Arnold, 1980). Зимой гагарки посещают
некоторые места гнездования колоний, причем в количествах, значительно
больших, чем их бывает в период гнездования. В частности, на о-ве Мэй
(Шотландия) зимой в колонию прилетает до 2000 гагарок, хотя летом
здесь гнездится лишь около 500 пар (Taylor; Reid, 1981).
Весенняя миграция половозрелых гагарок начинается, по-видимому,
в марте. Во второй половине апреля основная масса птиц появляется
на Западном Мурмане. К концу апреля массовый прилет гагарок отме-
чается на Восточном Мурмане (Белопольский, 1957). На Белом море
18
Гагарка
Рис. 4. Область распространения гагарки
а — граница гнездового ареала,
б — область зимовок и миграций.
Подвиды: 1 — A.t. torda,
2 — A.t. islandica
Рис. 5. Ареал гагарки в СССР
а — граница гнездового ареала,
б — область зимовок и миграций.
Подвид — A.t. torda
40 60
60
гагарки появляются по мере освобождения воды ото льда. Годовалые
гагарки проводят лето на местах зимовок, гнездований и между ними —
на местах сезонных миграций.
Численность. На большей части гнездового ареала численность гагарок
относительно невелика. Общая численность гнездящихся птиц оценивается
в 208 тыс. пар. Из них около 70 % гнездится в Великобритании и Ирландии,
12 % — в Норвегии и 9 % — в Северной Америке (Lloyd, 1976). В СССР
наиболее массовые гнездовья находятся в Онежском заливе Белого моря —
около 1600 пар (Бианки, 1967). В Кандалакшском заливе гнездится
около 75 пар (Бианки, 1967). На Мурмане в 60-е годы насчитывались
около 320 пар, в основном на островах Кандалакшского заповедника
(Герасимова, 1962). В последующие годы здесь наблюдалось снижение
численности гагарок: в 1968—1970 гг. всего около 200 пар (В. Н. Карпович,
личное сообщ.). Крупные гнездовые колонии гагарок — десятки тысяч
пар — расположены в Норвежском море на Лофотенских островах
(Watson, 1957; Brun, 1969). На о-ве Скомер в Великобритании гнездится
около 1700 пар (Hudson, 1982), на юго-западе Ирландии — около 2000 пар '
2*
19
Семейство Чистиковые
(Evans, 1970), в заливе Св. Лаврентий (Канада) — около 2000 пар (Bedard,
1969). На Балтийском море в Финляндии гнездится 1500—2000 пар (Meri-
kallio, 1958; Grenquist, 1961). В Швеции на о-ве Стора Карлсё в 1918 г.
насчитывалось более 7000 пар (Joppenthin, 1936; по: Kartaschew, 1960).
В Дании гнездится около 300 пар (Salomonsen, 1943; по: Fisher, Lockley,
1954).
Численность подвержена значительным колебаниям: в большинстве
районов происходит заметное ее снижение (Lloyd, 1976), но кое-где она
растет. В частности, в Шотландии на о-ве Мэй с 1921 по 1981 г. количество
гагарок увеличилось с 360 до 840—1010 пар, а в период с 1969 по 1981 г.
годовой прирост был наиболее существенным и составлял в среднем 16 %
(Harris, Galbraith, 1983). Тенденция увеличения численности гагарок
отмечена и на Оркнейских островах (Wanless et al., 1982) и в Ботническом
заливе Балтийского моря (Hilden, 1978).
Местообитания. В гнездовое время — прибрежные скалы морских побе-
режий и чаще — островов. Гнездится как на высоких, обрывистых остро-
вах, так и на скалистых берегах низких островов. На Белом море выводит
птенцов на скалистых маленьких островах, удаленных от материка.
Размножение. Половозрелой гагарка становится к третьему лету, в воз-
расте около двух лет (Дементьев, 1951), начинает размножаться, по послед-
ним данным, в 4—5 лет (Lloyd, Perrins, 1977).
К местам гнездования птицы подходят во второй половине апреля—
начале мая, когда на скалах еще лежит снег. Поэтому гагарки держатся
стайками на море или сидят на камнях на берегу. Брачные игры и копуля-
ция наблюдаются как на воде, так и на суше, у гнезд (рис. 6). Первое
спаривание на о-ве Скокхолм отмечено 27 марта (Plumb, 1965). В это
время часто слышатся хриплые скрипучие голоса гагарок.
Гнезда гагарки устраивают в укрытиях, в пустотах под обломками скал,
в нагромождениях валунов. Само гнездо обычно полуукрыто, яйцо ртклады-
вается у входа в пещеру и часто бывает хорошо видно сверху.. Выстилки
в гнезде гагарка не делает, хотя отдельные травинки в гнездо могут,
попадать. При возвращении к гнезду гагарка ориентируется по внешнему
окружению входа. При слабом изменении внешнего вида входа она беспо-
коится мало, тогда как сильное изменение приводит к длительному беспо-
койству. В пределах гнездовой полости гагарка имеет определенное гнездо-
вое место, куда и помещается яйцо. При перемещении яйца птица почти
всегда возвращает его на прежнее место (Ingold, 1976). На птичьих база-
рах, где гагарки селятся в непосредственном соседстве с кайрами и тупи-
ками, гагарка может загнездиться на открытых карнизах. На лудах Белого
моря гнезда иногда размещаются под нагромождением бревен плавника
(Бианки, 1967), и тогда яйцо бывает хорошо укрыто. Гнезда соседних
пар изолированы друг от друга. Гагарки, селятся, как правило, группами,
хотя одиночное расположение гнезд тоже встречается. Защиты гнездовой
территории у гагарок нет. Все птицы, гнездящиеся в колонии, вместе
отдыхают на определенных камнях — «клубах». Гнездовой консерватизм
у гагарок развит очень сильно: из года в год пары зайимают одни и те же
гнезда;
Откладка яиц на Мурмане проходит в конце мая—начале июня, на Бе-
лом море — чуть позже. В Англии гагарки начинают гнездиться почти
на месяц раньше — в конце апреля—начале мая, а в Норвегии — около
середины мая (Дементьев, 1951; Plumb, 1965). В кладке бывает одно
20
Рис. 6. Элементы брачного и гнездового поведения гагарки (по: Cramp, 1985, с изменениями)
Семейство Чистиковые
яйцо, очень редко — два. В последнем случае последнее яйцо бывает
неоплодотворенным. Экспериментально установлено, что гагарки не спо-
собны насиживать два яйца. При искусственном добавлении второго яйца
родители выкармливают обоих птенцов, но не достаточно эффективно.
В этих увеличенных выводках птенцы растут на 15—45 % медленнее, чем
в нормальных (Soikkeli, 1982). В случае потери яйца в начале насиживания
самка может отложить второе. Новое яйцо появляется через 12—18 дней
после исчезновения первого (Kartaschew, 1960).
Яйцо имеет удлиненную, приближающуюся к грушевидной, форму.
Окраска основного фона варьирует от чисто белой до зеленоватой или
желтоватой, редко — до коричневой. По основному фону разбросаны
черные и коричневые неправильной формы пятна, обычно сгущающиеся
у тупого конца. Расположение, размеры и цвет пятен сильно варьируют.
Некоторые пятна, находящиеся глубоко в толще скорлупы, просвечивают
слабо. В некоторых случаях пигмент оказывается на поверхности яйца и
может быть смыт. В гнездах часто бывает сыро и грязно, поэтому к концу
насиживания цвет яйца может измениться из-За загрязнения. Сцщшупа
яйца несколько шероховатая, пористая. Толщина ее в среднем 0,27 мм.
Размеры яиц гагарок на Мурмане и Белом море достоверно не разли-
чаются. Размеры 197 яиц с Семи островов: длина 60—86 мм (среднее 76,5),
ширина 44—59 мм (среднее 48,4). Вес свежих яиц 80—115 г (среднее 95,1).
В годы с неблагоприятными метеоусловиями размеры яиц бывают не-
сколько меньше. К концу сезона яйцекладки птицы Откладывают яйца
меньшего объема. Размер яиц увеличивается с возрастом родителей
до 15-летнего возраста. Потеря яиц достигает 30 % (Plumb, 1965; Lloyd,
1979) и происходит в основном в первые 10 дней после откладки яиц.
До 25 % потерь восполняется повторными кладками, обычно через 14 дней.
Однако повторные кладки могут быть лишь у птиц, отложивших яйца в на-
чале сезона яйцекладки (Lloyd, 1979).
В насиживании яиц принимают участие оба родителя. И у самки и
у самца имеются по два наседных пятна. Насиживание ведется со значи-
тельными перерывами, особенно в первые дни. По-видимому, развитие
эмбриона прерывистое, поскольку в инкубаторе эмбрион развивается всего
30 дней (Witherby et al., 1940—1941), а в гнезде период насиживания
длится дольше — 34—39 дней (Кафтановский, .1951; Bedard, 1969), в сред-
нем 34,3 дня (Paludan, 1960). Поведение насиживающей гагарки доволь-
но однообразно. Время от времени она сходит с гнезда, и если не летит
на кормежку, то прохаживается по соседним с гнездом скалам, как бы
разминаясь. Второй родитель отдыхает на скалах у моря или на «клубах»,
где собираются все не занятые в это время гагарки из близко расположен-
ных гнезд. Отдыхающая гагарка непрерывно поворачивает голову из сто-
роны в сторону и по временам делает несколько взмахов крыльями. На-
сиживающая гагарка осторожнее других чистиковых. При прибли-
жении человека покидает гнездо и начинает летать широкими кругами
над морем, берегом и гнездом. Только в конце насиживания и после вы-
лупления птенцов гагарка становится менее осторожной, при приближении
человека не вылетает из гнезда, а прячется в камнях или садится на скалу
вблизи гнезда и беспокойно кричит.
Процесс выплупления птенца длится двое-трое, иногда до шести суток,
по-видимому, из-за большой толщины скорлупы (Ingold, 1974). Гибель
вылупившихся птенцов составляет около 7 %. Чаще всего они гибнут
22
Гагарка
в первую неделю после вылупления, когда вес птенца находится в прямой
зависимости от веса яйца (Lloyd, 1979). Размеры птенца в первый день:
клюв 11 —12 мм, плюсна 24—25 мм, средний палец 32 —33 мм, масса 50—
70 г (Бианки, 1967), средняя масса 63—64 г (Кафтановский, 1951; Kart-
haschev, I960). В первые дни после вылупления птенец беспомощен, почти
не двигается, часто издает тонкий писк. Звуковое общение между роди-
телями и птенцом начинается еще во время вылупления (Ingold, 1973).
Родители обогревают птенца почти круглые сутки. Терморегуляция уста-
навливается на 4—6-й день, но взрослые птицы проводят в гнезде много
времени до самого выхода птенца. Даже при сильном беспокойстве —
отлове — гагарка не бросает птенца.
Выкармливают птенца рыбой, которую приносят в клюве. По сравнению
с другими чистиковыми птенец гагарки получает корм редко, в среднем
около двух раз в день. В некоторые дни наблюдений кормовых прилетов
родителей с рыбой не было отмечено вовсе, тогда как «без корма» гагарки
в гнездо прилетали 4—5 раз в сутки. Это заставляет предположить, что
гагарки выкармливают птенцов не только рыбой, но и еще какой-то мелкой
пищей, скорее всего ракообразными. В пользу этого предположения гово-
рит и наличие в гнезде красноватого помета с характерным запахом рако-
образных.
По характеру развития гагарка, как и кайры, занимает промежуточное
положение между выводковыми и полувыводковыми птицами (Sealy,
1973а). Рост птенца протекает интенсивно, кривая нарастания веса
по внешнему виду приближается к прямой линии, S-образный изгиб
почти отсутствует. Объясняется это, вероятно, тем, что развитие птенца
еще продолжается после выхода из гнезда. Возраст и вес выходящих
из гнезда птенцов в разных местах незначительно различается. На Белом
море птенцы выходят в возрасте 17—18 дней, при весе 170—190 г. (Бианки,
1967), на Мурмане — в возрасте 19—22 дня, при весе 250 г (Кафтановский,
1951), в Северной Америке — в возрасте 18 дней, при весе 140 г (Bedard,
1969), в Англии — в возрасте 18,5 дней, при весе 189 г (Plumb, 1965),
на Лофотенских островах — в возрасте 19,3 дня (Ingold, 1974). В послед-
ние дни перед выходом с птенцом остается одна взрослая птица, которая
проводит в гнезде много времени — до 13 ч в сутки. По-видимому, родители
прекращают кормить птенца, поэтому за день-два до того как покинуть
гнездо птенец часто пищит, отмечается некоторое снижение его веса.
Из гнезда он выходит обычно в утренние или дневные часы, на Лофотен-
ских островах — в вечерние часы (Ingold, 1974) в сопровождении взрослой
птицы. Они идут к воде рядом, оживленно «переговариваясь». Взрослая
птица издает хриплые звуки, птенец — негромкий мелодичный крик
«ви-ви-ви», произносимый с закрытым клювом. При опасности птенец
прячется под камни, взрослая птица пережидает опасность, сидя на камнях
сверху. Если потревоженная взрослая гагарка слетает на воду, птенец
начинает кричать резко и громко, широко открывая клюв. На воду птенец
спускается с высоких обрывистых скал, планируя при помощи своих
коротких крылышек. После спуска птенца гагарки сразу отходят от мест
гнездования. Дальнейшее развитие птенца проходит на море.
Эффективность размножения гагарок высокая — до 80 % от числа отло-
женных яиц. На Мурмане главной причиной гибели яиц и птенцов
является хищничество крупных чаек. В Англии птенцы гибнут больше
от недостаточного ухода родителей, чем от влияния хищников (Lloyd,
23
Семейство Чистиковые
1979). На Белом море во время массовой откочевки взрослых гагарок
с птенцами родители иногда бросают гнезда с поздними, как правило,
повторными кладками (Бианки, 1967).
Суточная активность. Поведение. Гнездовой период совпадает с пе-
риодом сплошного северного дня, поэтому определенного времени кор-
межки и отдыха у гагарок, по-видимому, нет. В ночное время — с 22 до 5 ч
птицы наименее активны. Большинства гагарок в это время у гнезд нет,
вероятно, они в море. В остальное время наблюдается два пика наибольшей
активности птиц — с 5 до 10 ч и с 13—14 до 18—19 ч.
Питание. Основой питания гагарки служат мелкие пелагические
рыбы — песчанка Ammodytes tobianus и мойва Mallotus villosus, мелкая
сельдь Clupea harengus, в единичных случаях — мальки крупных рыб —
трески и зубатки. Поедают гагарки и мелких ракообразных. В Баренцевом
море рыбы в питании гагарок встречаются значительно чаще (77,5 %), чем
ракообразные (1,5 %), полихеты (0,7 %) и моллюски (0,4 %) (Белополь-
ский, 1957). Питаясь теми же видами рыб, что и кайры и тупики, гагарки
частично избегают конкуренции за счет использования специфических
размерных групп своих жертв (Harris, 1970). Тем не менее временами
для гагарок больше, чем для других видов чистиковых, может сказываться
неблагоприятная кормовая ситуация, когда птицы очень сильно страдают
от бескормицы (Blake, 1984). Экспериментально установлено, что взрослые
гагарки в состоянии проглатывать рыб с высотой (т. е. максимальным
диаметром сечения) 23—26 мм и весом 16—18 г. Однако они предпочитают
поедать рыб, имеющих высоту около половины максимальной и длину
до 15—18 см. При выборе добычи ведущую роль играет зрение (Swennen,
Duiven, 1977).
В некоторых случаях (гагарки могут выступать как кормовой паразит,
отнимая добычу у летящих к гнезду тупиков (Ingold, Tschanz, 1970).
Враги, неблагоприятные факторы. Врагом взрослой гагарки может быть
кречет (на Семи островах), сапсан (на Белом море). Наиболее серьезными
врагами являются крупные чайки, особенно большая морская чайка.
С увеличением численности крупных чаек на островах Мурмана связано
резкое уменьшение численности гагарок (на Айновых островах с 75 пар
в 1960 г. до 5 пар в 1964 г.) (Коханов, Скокова, 1967). Чайки убивают
и поедают птенцов во время их выхода из гнезда и взрослых птиц в течение
всего гнездового периода. Гибель же яиц и птенцов в гнезде невелика.
Некоторое количество гагарок гибнет во время штормов как у мест
гнездования весной, так и на зимовках. На Балтике в суровые зимы гагарки
гибнут от тяжелых ледовых условий (Hilden, 1978).
По материалам массового кольцевания на о-ве Скокхолм (Уэльс) уста-
новлено, что до возраста 4—5 лет доживает лишь 18 % птенцов. Ежегодная
выживаемость взрослых птиц составляет 80—92 % (Lloyd, 1976; Lloyd,
Perrins, 1977).
В последние десятилетия иногда отмечается значительная гибель гага-
рок от нефтяного загрязнения моря (Olsson, 1974). В Норвегии отмечен
высокий уровень загрязнения яиц гагарок ртутью (до 10 мг/кг) (Fimreite
et al., 1974).
Известны случаи массового заболевания гагарок птичьей оспой, вызы-
ваемой вирусами и переносимой кровососущими паразитами (Bourne et aL,
1970).
Хозяйственное значение, охрана. Запасы гагарки в СССР невелики,
24
Тонкоклювая кайра
и поэтому хозяйственного значения она не имеет. Мясо гагарки без рыбного
запаха, яйца съедобны. Шкурка может идти на меховые изделия (Спанген-
берг, 1935). Питается сорной рыбой, рыболовству не вредит.
Специальных мер охраны гагарок не предусмотрено. На Белом и
Баренцевом морях запрещена охота на всю водоплавающую дичь, в том
числе и на гагарку.
Род Uria Brisson, 1760
Тонкоклювая кайра —
Uria aalge Pontoppidan, 1763
Guillemot (англ.), Common murre (амер.), Trottellumme (нем.).
Син. (лат): Colimbus aalge (Pontop).
Син. (рус.): длинноклювая кайра.
Статус. В пределах СССР гнездящийся перелетный, частично зимую-
щий вид.
Общая характеристика и полевые признаки. Птица средних размеров,
самая крупная из современных чистиковых, с плотным веретенообразным
телом, относительно длинной шеей, вытянутым почти прямым заострен-
ным клювом. Окраска контрастно двухцветная: голова, шея, верхняя сто-
рона тела темные, черновато-бурые, иногда с коричневатым оттенком;
нижняя сторона тела белая. Ноги сильно отставлены назад. На суше сидит,
опираясь на цевку и пальцы, тело держит обычно вертикально, реже —
наклонно или лежит на животе. Ходит медленно и неуклюже, слегка
раскачиваясь. Может подниматься на пальцы. Для взлета с суши необхо-
дим трамплин или обрыв. Полет быстрый, прямолинейный, с частыми
взмахами крыльев. Средняя скорость полета — около 60 км/ч, макси-
мальная— до 95 км/ч (Tuck, 1960). Не способна совершать крутой
вираж или резко менять высоту полета. Крылья узкие, заостренные,
короткие, в полете полусогнутые. В полете отличается от близких видов
по тонкому, стройному одноцветному клюву и относительно изящной
небольшой голове. Изредка летает с открытым клювом. Рулит в воздухе
и тормозит перед посадкой растопыренными лапами. Садится на воду
и приземляется часто с размаха, не до конца погасив скорость. Велико-
лепно плавает и ныряет; на поверхности воды гребет перепончатыми ла-
пами. При этом тело держит довольно высоко над водой или при опас-
ности почти все его погружает в воду, оставляя на поверхности голову, шею
и часть спины. Купаясь, может плавать на боку. Иногда привстает на
воде вертикально, часто хлопая крыльями. Нырнув, передвигается под
водой со скоростью до 20 км/ч с помощью крыльев («подводный полет»),
Заныривает на глубину в несколько десятков метров, по документирован-
ным данным — до 138 м (Burger, Simpson, 1986). С воды взлетает после
большого (10—12 м) разбега, чаще — оттолкнувшись крыльями на волне.
В период гнездования образует большие колонии на морских скалах
материка и островов, вблизи которых можно видеть их отдыхающими
и кормящимися на воде или на погруженных в воду обломках скал —
«клубах». Кайры на воде обычно молчаливы, но в колониях на гнездовых
карнизах, реже при подлете к колонии, а также на воде перед слетом
издают громкие хриплые крики. Какофония криков тысяч кайр послужила
основанием для того, чтобы назвать их гнездовья «птичьими базарами».
25
Семейство Чистиковые
Хорошо различимы крики «арра», разной продолжительности «уррр» и ко-
роткое «арр». Отмечено соответствие криков поведенческим реакциям.
Некоторые крики видоспецифичны и по сонограммам отличаются от кри-
ков толстоклювых кайр (Tschanz, 1972; Birkhead, 1976). У птенцов можно
различать тихий писк «уи-уи-уи» или звонкий и протяжный крик «фи-
лиип», который хорошо слышен даже на фоне общего шума птичьего
базара. Звонкий крик издают слегка подросшие птенцы в состоянии тре-
воги: потерявшие родителей, голодные, переместившиеся на непривычную
часть карниза, а также птенцы, собирающиеся прыгнуть с карниза.
Наибольшая активность кайр наблюдается в светлое время суток,
особенно в утренние и вечерние часы.
Подвидовая систематика. В настоящее время различают пять подвидов
(Степанян, 1975). Отмечены вариации размеров (длина крыла, длина
и высота клюва), оттенков окраски темной части оперения и развития
пестрин на боках тела и нижних кроющих крыла. В СССР обитают два
подвида.
1. Uria aalge hyperborea
Uria aalge hyperborea Salomonsen, 1932, Ibis, c. 130, о-в Медвежий.
Верхняя сторона тела черновато-бурая, нижние кроющие крыла с
пестринами, темными вершинными пятнами и наствольями. Самый круп-
ный атлантический подвид, но мельче тихоокеанского. Длина (в мм) клюва
46,0-48,0 (Tuck, 1960), 43,5- 50,3 (Красовский, 1937).
2. Uria aalge inornata
Uria aalge inornata Salomonsen, 1932, Ibis, c. 128, О-в Св. Матвея,
Берингово море.
Спинная сторона светлее, с более развитым сероватым оттенком.
Нижние кроющие крыла белые, без пестрин. Не имеют «очковой» морфы.
В размерах клюва размах вариации больше, чем у атлантического под-
вида: 42,0— 53,0 мм (Tuck, 1960).
Замечания по систематике. Дс| недавнего времени выделялось 7 —8 под-
видов тонкоклювой кайры (Дементьев, 1951; Иванов и др., 1953; Tuck,
1960). Обоснование самостоятельности многих подвидов сделано на сра-
внительно небольшом материале, не подвергалось статистическому ана-
лизу и поэтому спорно. В частности, нет четкого различия для таких
общепринятых подвидов, как U. a. aalge и U. a. hyperborea. Отмечено,
что в различных районах Норвегии, где обитает U. a. hyperborea, неко-
торые экземпляры подходят под описание номинального атлантического
подвида; различия в .размерах позволяют предположить клинальный ха-
рактер изменчивости (Mather, 1966, Gloe, 1977; Wus, 1981, и др.).
«Очковые» кайры сначала выделялись в самостоятельный вид Uria
ringwia Brunnich, 1764. Однако их видовая (или подвидовая) самостоя-
тельность по морфологическим показателям не подтвердилась (Кафтанов-
ский, 1951; Jefferies, Parslow, 1976). Известна тенденция клинального
увеличения доли «очковой» морфы в популяциях в Атлантике с юга на
север (Дементьев, 1951; Southern et al., 1965) и с запада на восток (Brun,
1970). В Северной Атлантике «очковые» кайры составляют около 1 %
популяции в южных районах и до 50 % — на севере (Southern et aL,
1965). Однако отмечено, что их процент изменяется по годам и особенно
непостоянен в мелких колониях. Возможно, это связано со спецификой
выбора биотопов «очковыми» кайрами, а также сильно развитым инстинк-
том хоминга, особенно у самок (Brun, 1970; Birkhead et al, 1980). Сохра-
26
Тонкоклювая кайра
нению этого признака, вероятно, способствует то, что кайры «очковой»
морфы в пределах общей колонии проявляют тенденцию гнездиться ближе
друг к другу. Тем не менее объединение их в группы последними иссле-
дованиями не подтвердилось (Harris, Wanless, 1986). «Очковость» —
рецессивный признак, определяемый геном аутосомы. У птенцов она отчет-
ливо проявляется на 54—66 день после вылупления (Jefferies, Parslow,
1976).
Близкородственные виды — толсто- и тонкоклювые кайры имеют много
общего во внешнем облике, экологии и поведении. При гнездовании в об-
щей колонии основными факторами, препятствующими скрещиванию, по-
видимому, являются видоспецифичные крики и сопровождающие их позы,
хотя известен случай межвидового скрещивания и благополучного выве-
дения гибридного потомства (Tschanz, 1972).
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла I<II>III>IV. .. Рулевых перьев
12—14. Размеры: длина (в мм) крыла самцов 200—228 (среднее 212,6),
самок 200—220 (среднее 210,4); клюва самцов и самок 42,5—51
(среднее 46,6); цевки самцов и самок 40—43 (среднее 42,4) ; масса (в г)
самцов 825—1285 (среднее 1060,1), самок 820—1260 (среднее 1047,8)
(Кафтановский, 1951; Гизенко, 1955; Успенский, 1956; Белопольский,
1957; Tuck, 1960; Cramp, 1985). Клюв вытянутый (прямой, к концу
слегка загнутый), чуть сжатый с боков, с узкими щелевидными ноздрями.
Контурное оперение плотное.
Окраска. Полового димррфизма в окраске нет. Самец и самка в брач-
ном наряде. Голова и шей буровато-черная с коричневым оттенком.
Верхняя их часть заметно темнее. От глаза кзади — отчетливая темная
бороздка. У «очковой» морфы вокруг глаза имеется белое кольцо, а за
глазом — белая полоска. Верхняя сторона тела, крылья и хвост темно-
бурые, иногда с сероватым оттенком. Вершины второстепенных маховых
имеют белую каемку, вследствие чего на крыле четко заметна белая
полоска. Нижняя сторона тела, начиная с груди, белая. На боках темные,
почти черные продольные полосы. Нижние кроющие крыла белые или
белые с темными вершинами. Подмышечные белые с бурыми пестринами.
Клюв черный или серовато-черный.
Ноги, плавательные перепонки темно-бурые, на передней стороне цевки
и внешней поверхности пальцев, светлее, с охристым просвечивающим
оттенком. Радужина темно-бурая.
Самец и самка в зимнем наряде. Вся передняя часть шеи становится
белой или испещренной мелкими пестринами. Иногда белый цвет полосой
распространяется и на заднюю часть шеи. За глазом остается широкая
бурая полоса, отделенная от темно-бурой «шапочки» белым оперением.
Темно-бурая окраска сохраняется на верхней части головы: лоб, уздечка
и под глазом. Несколько увеличивается количество пестрин. Клюв серова-
тый.
Известен случай находки полного альбиноса (самки) тонкоклювой
кайры с ярко-розовыми глазами и ногами, более светлым, чем обычно,
клювом (Arnold, 1950), а также случаи частичного меланизма, когда
кайры имели черные подхвостье, подмышечные и некоторые кроющие
груди (Кафтановский, 1951).
Пуховой наряд. Птенец покрыт коротким (5 —10 мм) жестким темно-
серым пухом с белой «проседью» на голове и белыми на груди и брюшке.
27
Семейство Чистиковые
В мезоптильном наряде голова, верх шеи, спина, кроющие хвоста, крылья
сверху и снизу черные. Нижняя часть шеи, грудь и брюхо белые. Рулевые
и маховые в этом наряде отсутствуют.
Гнездовой наряд. Сходен с зимним нарядом взрослых, но характери-
зуется несколько большим развитием белого цвета на шее и отсутствием
темных продольных пестрин на боках тела. На темном оперении заметны
черновато-бурые каемки перьев.
Линька. Последовательность смены нарядов: пуховой — мезоптиль —
первый зимний — неполный взрослый — зимний — первый взрослый
брачный — зимний (послебрачный). Смена пухового наряда птенцов начи-
нается вскоре после их обсыхания. Обычно в возрасте 4—5 дней стано-
вятся различимы верхушки кроющих крыла, на 6—7 день — кроющие
тела, на 11 — 12-й день — кроющие хвоста. В возрасте 15—20 дней, неза-
долго до спуска с гнездового карниза, птенец приобретает полный мезоп-
тильный наряд (Кафтановский, 1951). О ходе линьки из мезоптиль-
ного в первый зимний наряд известно очень мало. Она происходит в ав-
густе-сентябре, когда вместе со взрослыми птенцы откочевывают с мест
гнездования в районы зимовок. У взрослых птиц интенсивная послегнез-
довая линька в зимний наряд начинается со смены контурного оперения
на голове, шее и плечах еще в тот период, когда птицы находятся на
гнездовьях (Rees, 1969). Одновременно с контурным оперением интен-
сивно растут маховые перья. С потерей маховых перьев кайры теряют
способность к полету. Линька в зимний наряд сопровождается некоторым
увеличением размеров клюва и изменением его цвета до черного. Весной
молодые птицы вновь частично линяют, приобретая летний наряд, отли-
чающийся от брачного присутствием значительного числа пестринок на
горле. На следующую осень у молодых птиц происходит полная линька
во второй зимний наряд, а к середине зимы — в брачный наряд полово-
зрелых птиц. Завершив линьку в зимний наряд, кайры практически без
паузы линяют вновь и к началу февраля приобретают брачный дефини-
тивный наряд (Tuck, 1960).
По наблюдениям за кайрами, содержавшимися в неволе (Birkhead,
Taylor, 1977), установлено, что первостепенные маховые выпадают за
несколько дней, а их рост занимает от 42 до 90 дней (среднее 63). Рулевые
и второстепенные маховые выпадают почти одновременно, когда первосте-
пенные вырастают примерно наполовину. Рост рулевых происходит в те-
чение 35—86 дней (среднее 61), а второстепенных маховых — 14—25 дней.
Таким образом, кайры теряют способность к полету на 45—50 дней.
Линька контурного оперения на спинной стороне тела занимает 51 —
134 дня (среднее 96), а на брюшной она длится дольше. Послегнездовая
линька головы продолжается в среднем 62 дня. В северных широтах пред-
брачная линька головы начинается после паузы в 120 дней, а в южных ши-
ротах — практически без паузы. В зимний период, когда велики энерге-
тические затраты на линьку, а кормовые условия неблагоприятны,
особенно велика смертность птиц (Rees, 1969; Мегпе, 1969). Взрослые
птицы в брачном наряде, появляющиеся весной на гнездовых карнизах,
имеют уже сильно обтрепанное оперение. В Великобритании появление
кайр на гнездовых карнизах в летнем оперении наблюдали, начиная
со второй половины ноября, тогда как вдали от берегов птицы всю зиму
сохраняли зимний наряд. Это позволяет предполагать, что часть популя-
ции завершает годовую линьку еще осенью. Такое явление отмечено лишь
28
Рис. 7. Область распространения тонкоклювой кайры
а — граница гнездового ареала, б — область зимовок и миграций. Подвиды: 1 — U.a. aalge,2 — U.A. albionis,3 — U.a. ibericus,4 — U.a. hyperborea,
5 — U.a. inornata
Семейство Чистиковые
в последнее десятилетие, что, возможно, связано с общим потеплением
климата (Bourne, 1981).
Распространение. Гнездовой ареал. Арктические, субарктиче-
ские и умеренные моря (рис. 7, 8). В Атлантическом океане: западное
побережье Гренландии, побережье и острова Северной Америки от Север-
ного Лабрадора до Новой Англии, залива Св. Лаврентия, восточнее побе-
режья Ньюфаундленда. В Европе: побережье Исландии, острова Фарер-
ские, Гебриды, Оркнейские, Шетландские, Шотландия, Ирландия, Англия,
Гельголанд, острова и побережья Франции, Португалии и Испании. На юг
до островов Берленгас. Мелкие острова Балтийского моря и Ботниче-
ского залива. По северному побережью Скандинавии, острова и матери-
ковые берега Норвегии и Мурмана до мыса Святой Нос, острова Шпицбер-
ген, Медвежий, Новая Земля.
В Пацифике — западное побережье Америки от мыса Хоп на северо-
западе Аляски до Калифорнии, острова Алеутские, Прибылова, Корякское
побережье к югу от мыса Дежнева, побережье Камчатки, Охотского моря,
Северного Приморья до западного побережья о-ва Хоккайдо. Острова
залива Шелихова, Ямские, Шантарские, Сахалин, Тюлений, Командор-
ские, Курильские.
Зимовки. Изучены недостаточно полно. Значительная часть популя-
ций зимует в море относительно недалеко от гнездовых колоний (Cramp,
1985). Кайры, гнездящиеся в Баренцевом и Норвежском морях, пере-
мещаются на зимовку в районы, расположенные к западу и юго-западу,
избегая глубоководных районов и районов, полностью блокированных
льдами. Новоземельские кайры могут зимовать у берегов Мурмана и Скан-
динавии, а скандинавские птицы — в Северном море вплоть до Датских
проливов. Зимой кайры в значительных количествах отмечены в Норвеж-
ском море на банке Джорджес (Белопольский, и др., 1982). Птицы из
балтийских колоний зимуют неподалеку от мест гнездования. Северное
море, включая берега Южной Норвегии, и Атлантика к югу до Бискайского
залива используются для зимовки кайрами из колоний Исландии, Бри-
танских островов, Испании и Франции. Кайры с западного побережья Ат-
лантики зимуют в прилежащих водах, свободных ото льда, перемещаясь
к югу до залива Св. Лаврентия и Ньюфаундленда (Joiris, 1983). Районы
зимовки тихоокеанских кайр, видимо, также связаны с расположением
льдов и наличием корма. Поэтому они удаляются от мест гнездования
к югу настолько, насколько их оттесняют неблагоприятные условия
(Brown, et al., 1975). Зимовки кайр отмечены на юг до Орегона и Калифор-
нии, в массе — в юго-восточной части Берингова моря и вдоль побере-
жий Курильских островов и Охотского моря (Woodby, 1984).
Миграции. Весенний пролет к местам гнездований начинается в попу-
ляциях в различные сроки. В Британии и Ирландии первое появление
кайр на гнездовых карнизах приходится на начало марта. В феврале кайры
скапливаются неподалеку от берегов в открытых водах Гренландского,
Норвежского и Баренцева морей (Brown, 1984). На Балтике, в Норвегии
и на Мурмане они появляются на неделю позднее, а на Новой Земле —
к концу марта (Успенский, 1951; Белопольский, 1957; Cramp, 1985).
В зависимости от условий года и особенно от ледохода сроки прилета
могут запаздывать. В морях тихоокеанского бассейна прилет кайр проис-
ходит позднее, чем в Атлантике, в частности, для о-ва Св. Лаврентия ука-
30
Рис. 8. Ареал тонкоклювой кайры в СССР
Обозначения те же, что на рис. 7
Семейство Чистиковые
зывается конец апреля (Fay, Cade, 1959), а для бухты Провидения —
середина мая (Портенко, 1973).
Осенняя откочевка и миграция к местам зимовок начинаются практи-
чески сразу же после слета птенцов с гнездовых карнизов: на Британ-
ских островах — в конце июня—начале мая, на Мурмане и в Норвегии —
в последнюю неделю июля, в тихоокеанских колониях — в третью декаду
августа, на Новой Земле — в первую декаду сентября (Белопольский,
1957; Успенский, 1956; Гизенко, 1955; Tuck, 1960). В некоторых случаях
кайры могут возвращаться в районы гнездования в межгнездовой пе-
риод — октябре—январе. Иногда они держатся здесь в течение 1—2 мес.,
но потом улетают до начала нового гнездового сезона. В некоторых коло-
ниях зимой кайр собирается в несколько раз больше, чем гнездится летом
(Taylor, Reid, 1981). Молодые кайры в первые годы мигрируют дальше,
чем в последующие, и меньше времени проводят вблизи гнездовых колоний
(Perrins, 1978). Для птиц Норвежского и Баренцева морей наиболее веро-
ятное направление миграций — запад, юго-запад, для тихоокеанских мо-
рей — юг, юго-восток.
Численность. Современная численность гнездящихся тонкоклювых,
кайр составляет около 11,1 млн пар. По-видимому, примерно такой же
величины достигает число неполовозрелых и негнездящихся особей.
Наиболее многочисленны популяции Северо-Западной Европы — 3144 тыс.
пар (Evans, 1984), Аляски — 2750 тыс. пар (Lensink, 1984) и Охотского
моря — 1100 тыс. пар (Golovkin, 1984). В пределах СССР гнездится
около 1,6 млн пар тонкоклювых кайр, т. е. 14 % мировой популяции вида.
В Баренцевом море наибольшее количество тонкоклювых кайр гнездится
на Мурмане в колониях: мыс Май-Наволок — 1500 пар, мыс Городецкий —
2600, острова: Гавриловский — 850, Харлов — 4216, Кувшин — 1000, губа
Дворовая — 2800 (Герасимова, 1962; Головкин, Гаркавая, 1975; Шкляре-
вич, 1977). Небольшое количество кайр гнездится также в мелких коло-
ниях, а еще совместно с толстоклювыми кайрами на Новой Земле (Успен-
ский, 1956). Эти кайры относятся к северо-атлантическому подвиду
U. aalge hyperborea.
В колониях морей тихоокеанского бассейна, где обитает U. a. inornate,
на севере она уступает в численности толстоклювой кайре, тогда как на
юге доминирует. На Командорских островах гнездится несколько тысяч пар
тонкоклювых кайр. Только в одной из колоний на о-ве Арий Камень
(о-в Беринга) — 2,5 тыс. пар кайр; число тонкоклювых кайр больше, чем
толстоклювых (Карташев, 1961). На восточном побережье Камчатки в Ка-
рагинском заливе тонкоклювые кайры гнездятся на о-ве Карагинский —
около 30 тыс. пар (Вяткин и др., 1975) и на о-ве Верхотурова — около
10 тыс. пар (Вяткин, 1975).
Крупнейшие колонии вида в СССР расположены в Охотском море:
на Курильских островах — несколько десятков тысяч пар (Велижанин,
1969), в заливе Шелихова на о-ве Нансикан — около 50 тыс. пар (Яхон-
тов, 1975а), в Пенжинской губе в 20 колониях на мелких островах —
несколько сотен тысяч пар (Яхонтов, 1975). Самая крупная из известных
колоний тонкоклювой кайры в СССР расположена на Ямских островах,
где насчитывается 297 500 пар (Велижанин, 1975). Примерно по 50 тыс.
пар гнездится в колониях на о-ве Ионы и о-ве Тюлений (Головкин,
Георгиев, 1970; Бычков, 1975; Харитонов, 1975; Велижанин, 1975).
На побережье и островах Охотского моря также имеется несколько мелких
32
Тонкоклювая кайра
колоний. Численность вида в рассмотренных частях ареала довольно
стабильна. Существенное уменьшение колоний отмечено только для о-ва
Тюлений, где с 1940-х годов (Гизенко, 1955) численность тонкоклювых
кайр упала почти в 5 раз из-за того, что часть скал, где гнездились птицы,
была переоборудована человеком таким образом, чтобы расширились
площади лежбища котиков.
Местообитания. Типично морская птица, обитатель прибрежных
(шельфовых) морских районов Северного полушария. Вне шельфовой
зоны может встречаться в зимнее время и в период миграций. С сушей
связана только во время гнездования. Гнездовое местообитание — ска-
листые берега материка и островов, обрывающиеся в море или отде-
ленные от него узкой полоской суши, а также береговые каньоны в ска-
лах. Как правило, избегает участки скал, покрытые растительностью
и торфом. Высота расположения гнездовых участков — от нескольких
метров до нескольких сотен метров над уровнем моря. Гнездование
чаще происходит на узких и широких полках и карнизах, выступах
скал, в трещинах, а также на плоских, лишенных растительности вер-
шинах островков. Известны небольшие колонии, размещающиеся в пе-
щерах (Fogden, Greenwood, 1965; данные А. Н. Головкина). Гнездовые
колонии располагаются на участках прибрежья, неподалеку от мест отно-
сительно регулярного скопления пелагических рыб и беспозвоночных.
Размножение. Половая зрелость наступает в возрасте трех лет. В двух-
летнем возрасте птицы, одевшие окончательный брачный наряд, могут
занимать места на гнездовых карнизах и даже иногда откладывать
неоплодотворенные яйца, однако, их можно считать лишь «потенциально
зрелыми» (Слепцов, 1948). Моногамы. Пары, вероятно, сохраняются из
года в год. Приуроченность к территории прежнего гнездования очень
велика — до 95 % (Birkhead, 1976). Сроки прилета варьируют в зави-
симости от географического положения колонии, ледовой обстановки и
метеоусловий.. Близ мест размножения появляются за 5—8 дней до
начала откладки яиц (Белопольский, 1957), на Мурмане — в третьей де-
каде марта—начале апреля. Копуляция обычно происходит на скалах,
гнездовых карнизах, но случается также на воде или на льдине. Ей пред-
шествует период ухаживания со специфичным ритуальным поведением
партнеров: брачные полеты, «танцы» на воде, прекопуляционные позы
и крики. Копуляция осуществлятся в период за 5 дней до откладки яиц
и 14 дней после нее (Tuck, 1960). Кайры не строят гнезд, но иногда
подкладывают камешки рядом с яйцом. Гнездятся обычно колониями,
в которых насчитывается от нескольких пар до десятков тысяч пар.
Известны случаи гнездования единичными парами или объединениями
в 2 пары, которые располагались в колониях других видов птиц или вне
колоний (Antiainen, 1964; Марков, 1975; Стенченко, 1975; данные А. Н. Го-
ловкина). Яйца откладываются на плоскую или слегка наклонную по-
верхность карнизов скал, причем наклон может быть обращен к скале или
в сторону моря. (Наиболее крупная «плоскостная» колония (около 50 тыс.
пар), расположенная на относительно обширных плоских вершинах ост-
рова, в СССР известна на о-ве Тюлений). Соседствующие пары в период
откладки яиц и инкубации проявляют территориальную агрессивность,
однако столкновения из-за территории чаще случаются с особями, случайно
залетающими на чужой карниз, и непродолжительны. При совместном
гнездовании с толстоклювыми кайрами отчетливо выражена межвидовая
3 Птицы СССР
33
Семейство Чистиковые
конкуренция из-за удобных гнездовых карнизов. Тонкоклювые кайры
предпочитают более широкие карнизы и вытесняют толстоклювых кайр
на узкие и маленькие уступы, за исключением тех случаев, когда толсто-
клювые кайры намного преобладают в численности (Кафтановский,
1951; Белопольский, 1957; Williams, 1974). Зачастую расположение наси-
живающих птиц настолько плотное, что между ними не имеется проме-
жутка. В плотных колониях количество особей на 1 м2 достигает 45,0
(Головкин, Георгиев, 1970), а в относительно разреженных — 26,5—29,0.
Период откладки первых яиц на Мурмане приходится на 11—29 мая,
на о-ве Тюленьем — на 23 мая (Белопольский, 1957; Гизенко, 1955), а на
Новой Земле — примерно на три недели позднее (Успенский, 1956).
Между откладкой первых и последних яиц в колонии проходит от 22 (на
Мурмане), до 48 (на Новой Земле) дней, однако массовая кладка синхро-
низирована и занимает 9—10 дней (Белопольский, 1957). Растянутость
периода яйцекладки обусловлена' повторными кладками у птиц, потеряв-
ших яйца, а также неблагоприятными метеоусловиями (например, ранней
затяжной весной) (Белопольский, 1957). Сроки откладки яиц в Атлантике
примерно те же, что и на Мурмане, а на Тихоокеанском побережье Аме-
рики приходятся на начало июня (Tuck, 1960). В кладке одно яйцо, как
правило, грушевидной формы. Форма яиц варьирует. В колониях могут
быть найдены яйца от резко грушевидных до почти эллиптических. Груше-
видность яиц кайр имеет явно адаптивный характер, поскольку радиус
пути скатывающегося грушевидного яйца значительно меньше, чем оваль-
ного. Экспериментально установлено, что с карниза шириной в 20 см от
толчка падает вниз лишь 20 % яиц кайр. Вероятность падения растет
с увеличением силы толчка и уменьшается с увеличением степени наси-
женности (Tschanz et al., 1969). Скорлупа матовая, крупнопористая,
толстая, особенно в середине узкой части яйца и на остром конце. Средняя
толщина скорлупы варьирует от 0,31 мм (0,26—0,36) на Мурмане
(Кафтановский, 1941) до 0,43 мм на Ньюфаундленде (Mahoney, Tjrelfall,
1981). Окраска сильно варьирует от белой, кремовой, красноватой, ох-
ристой до зеленой и голубой. Скорлупа может быть совсем без пестрин
или иметь пестрины в виде точек, пятен, черточек или полосок, которые
обычно сгущаются к толстому концу яйца. Размеры яиц имеют до-
вольно большую изменчивость. При этом средние размеры яиц у U. а.
aalge и U. a. hyperborea практически одинаковы: соответственно 84,4Х
Х51,8 (п = 77) и 84,4X51,9 мм (п = 971), хотя размах колебаний у тихо-
океанского подвида больше: 74—96X45—56 против 79—93X49—55 (Каф-
тановский, 1951; Головкин, Георгиев, 1970). Другие подвиды в среднем
имеют яйца меньших размеров (Tuck, 1960). Вес ненасиженных яиц из
разных районов составляет 101 —144 г (среднее 112,9). Известны находки
необычно мелких яиц: 53X38 мм, вес 45 г (Кафтановский, 1951) и яиц-
гигантов: 105,6X61,0 мм, вес 191 г (Гизенко, 1955). В случае потери
яйца кайры могут откладывать второе и третье и даже четвертое яйцо
через 7 — 18 дней. Этот срок удлиняется в зависимости от степени наси-
женности потерянного яйца (Слепцов, 1948; Кафтановский, 1951; Бело-
польский, 1957). Первое яйцо формируется 14—15 сут, в том числе желток
в среднем — 11,5 сут. Второе яйцо формируется 12,7 сут, а его желток —
9,3 сут (Birkhead, Nevo, 1987). В норме первое яйцо сохраняется у 56 %
самок и более (Tuck, 1960). Яйца повторных кладок несколько мельче
первого.
34
Тонкоклювая кайра
Инкубация продолжается в течение 28—37 дней, чаще 30—34 дня,
что, возможно, зависит от метеоусловий и широты местности (Tuck, 1960).
Средняя потеря веса яйца при инкубации 0,55 г/сут (Johnson, West, 1975).
В течение первых 20 дней эмбрионы прибавляют в весе медленно, а затем
заметно быстрее. Средняя прибавка составляет 1,9 г/сут (Белопольский,
1957; Mahoney, Threlfall, 1981). Насиживают попеременно оба родителя,
которые имеют по одному большому вытянутому наседному пятну. Наси-
живающие птицы сменяются через 11—38 ч (среднее 17 ч) (Verspoor
et al., 1987). За время инкубации вследствие увеличения воздушной камеры
центр тяжести яйца смещается ближе к острому концу. Поэтому для
насиженного яйца уменьшается радиус окружности, по которой оно может
скатиться с карниза в отсутствие взрослой птицы. Тем самым снижается
опасность утери яйца на последних стадиях насиживания, когда возобнов-
ление кладки уже невозможно (Успенский, 1956).
Градиент температур верхней и нижней поверхности яйца, вероятно,
может превышать 30° при температуре в верхней части яйца 39° (Бело-
польский, 1957). Вылупление первых птенцов на Мурмане происходит
с 20 июня по 4 июля (среднее 27 июня), а массовый выклев — в первой
декаде июля. В эти же сроки вылупляются птенцы и в Охотском море
(Белопольский, 1957; Гизенко, 1955). Вес птенцов при вылуплении
составляет от 53 до 101 г (среднее 73,5 г) (Гизенко, 1955; Успенский,
1956; Cramp, 1985). Терморегуляция у птенцов начинает устанавливаться
между 4-й и 6-м днем жизни, к началу появления мезоптильного наряда,
когда отпадает яйцевой зуб, и заканчивается к 9—10-му дню (Рольник,
1948; Кафтановский, 1951; Johnson, West, 1975). Гомойотермия, по неко-
торым данным (Mahoney, Threlfall, 1981), устанавливается полностью
только после спуска птенца на воду. По характеру развития птенцов
тонкоклювые кайры отнесены к промежуточному типу между выводковыми
и полувыводковыми птицами (Sealy, 1973а). Получать пищу птенец начи-
нает со 2—3-го дня жизни, когда рассасывается желточный мешок. Увели-
чение его веса ускоряется с возрастом и в период с 1-го по 16-й день
жизни составляет в среднем 8,8 г в сутки (Mahoney, Threlfall, 1981). Для
увеличения веса на 1 г требуется в среднем 13,4 г рыбы (Tuck, Squires,
1955).
В первые дни жизни птенцов родители обогревают их, сменяясь
так же, как при инкубации яиц. Начиная с 10-го дня они оставляют их на
непродолжительное время. В холодные дни птенцы могут согреваться
под взрослыми соседями (по нескольку птенцов под одной птицей).
Родители и птенцы устанавливают звуковые контакты, обусловливаю-
щие взаимное опознавание еще на последних стадиях инкубирования,
за 90—100 ч до вылупления, когда птенцы начинают издавать писк в яйце
и родители им отвечают. Это звуковое общение развивается и усиливается
после вылупления птенца и играет решающую роль в отыскании его
родителями. Кормление усиливает прочность акустических связей
(Tschanz, 1968). Кормят птенца оба родителя, принося на карниз преиму-
щественно рыбу длиной от 52 до 129 мм (обычно около 100 мм) и массой
от 1 до 15 г. В питании птенцов отмечено более 20 видов рыб, но наиболее
часто встречаются песчанки (Ammodytes sp.) (Кафтановский, 1951;
Tuck, 1960; Головкин, Георгиев, 1970). Птицам свойственна врожденная
ориентация при схватывании рыбы из клюва родителей или с карниза.
Они заглатывают рыбу непременно с головы, ориентируясь на располо-
з*
35
Семейство Чистиковые
жение глаз (Oberholzer, 1975). Птенцы обладают также врожденным
«чувством края», т. е. стремлением к контакту со скалой в отсутствие
взрослых птиц, что предотвращает их непроизвольное падение с карниза.
В возрасте трех дней птенцы определяют расположение края уступа
визуально (Wehrlin, Tschanz, 1969). Жизнь птенцов на гнездовом карнизе
продолжается до 35 дней (чаще 20—25 дней). За это время они достигают
веса от 172 г (Новая Земля) до 296 г (Мурман), среднее 244 г. Птенцы
из повторных кладок к вылету набирают меньший вес, чем первые птенцы
(Успенский, 1956; Hedgren, Linnman, 1979; Карпович и др., 1980; Cramp,
1985). Начало спуска птенцов на воду происходит на Мурмане 20—
25 июля, а на Новой Земле и на Сахалине — в первую декаду августа
(Белопольский, 1957; Успенский, 1961; данные А. Н. Головкина). В по-
следние сутки перед сходом на воду родители (или один из них) перестают
кормить птенца. Птенцы, еще не умеющие летать, прыгая с карнизов,
попадают как на воду, так и на прибрежные камни. Для них наиболее
опасны прыжки с небольшой высоты, так как только с 15—20 м падение
приобретает характер планирования и сила удара о субстрат уменьшается.
Заполнение воздушных мешков также способствует смягчению приземле-
ния. Смертность птенцов при спуске составляет около 4,2 % (Карпович
и др., 1980). Спуск на воду обычно происходит в сумерки. Наибольшее
число птенцов спускается со скал с 20 до 23 ч. Во время массового слета
гибель птенцов от чаек наименьшая, поскольку количество хищничающих
птиц примерно постоянно и поэтому относительная доля добываемых
ими птенцов меньше. Отход птенцов зависит от удаленности гнездовых
скал от уреза воды. Если слетки прыгают непосредственно в воду, их
гибель от хищников существенно снижается (Williams, 1975). На воде
родители обнаруживают своего птенца по крикам и откочевывают с ним
от берега.
Успех размножения зависит от состояния кормовой базы, метеоусловий
года, удобства расположения и плотности колонии, степени беспокойства
колонии человеком и хищниками. При благоприятных условиях успех
размножения достигает 0,8 птенца на пару размножающихся птиц
(Hedgren, 1980). Наименьший успех имеют кайры, запоздавшие с кладкой,
или особи, впервые приступающие к размножению. При неблагоприятных
условиях гибель потомства (суммарно яиц и птенцов) может составлять
75 %. Однако расчеты показывают, что даже пополнения, составляющие
25 %, достаточны для сохранения стабильной численности в популяции
при условии незначительной смертности птенцов после их схода на воду.
При анализе изменений состояния популяций тонкоклювой кайры на
Аляске обнаружена функциональная зависимость между успехом размно-
жения и суммой отклонений температуры воздуха от 18 °C за период раз-
множения. На основании теоретической модели динамики численности
предполагается, что основным фактором падения численности является
смертность кайр в период зимовки (Murphy et al., 1986). В каждый
конкретный момент размножающаяся популяция представлена значитель-
ным числом (7—9) последовательных генераций (Leslie, 1966). Выжива-
емость взрослых птиц в норме составляет 0,87— 0,94 особи в год, средняя
продолжительность жизни одного поколения — 9 лет (Southern et al.,
1965), а средняя продолжительность жизни половозрелых кайр — 16 лет
(Birkhead, 1974).
Суточная активность, поведение. В гнездовое время в высоких широтах
36
Тонкоклювая кайра
в условиях полярного дня активны круглосуточно. Активность падает
в ночное время, особенно при темных ночах. Количество птиц на скалах
в апреле-мае зависит от времени суток и от уровня прилива. Наиболь-
шее количество присутствует вечером и при полном приливе. Утром на
карнизах находится около 60 % размножающихся кайр, а в середине дня —
лишь 30—42 % (Birkhead, 1978). Вероятно, это связано с условиями для
сбора корма: утром и днем при хорошей освещенности кайрам легче
ловить песчанку, выносимую течением во время отлива («малой воды»)
ближе к поверхностным слоям воды. Число кайр на карнизах снижаемся
также при дожде и сильном ветре, когда в колониях остаются только птицы,,
насиживающие яйца или согревающие птенцов, а все остальные откоче-
вывают в море (Slater, 1976). При плохой погоде, особенно при сильном'
волнении моря, родители кормят своих птенцов вдвое реже (Birkhead,
1976).
Температура тела кайр, находящихся в активном состоянии, достигает
42,5° (Iversen, Krog, 1972). Кайры исключительно выносливы к низким,
температурам при условии, что их оперение не намокает. Установлено,^
что при температуре —55° они в течение 18 ч поддерживают температуру
тела на уровне не ниже 38,9°. Это достигается за счет высокой потенциаль-
ной интенсивности метаболизма (Johnson, West, 1975).
Важнейшей особенностью социального поведения кайр является вы-
сокая степень колониальности их гнездования. Хотя изредка кайры могут
гнездиться отдельными парами, типичным для вида можно считать обра-
зование плотных колоний, в которых в крайнем варианте насиживающие
птицы соприкасаются друг с другом. Такой тип гнездования, вряд ли
связанный с недостатком пригодных для откладки яиц мест, несомненно
обусловленный условиями питания и особенно обилием корма и отчасти —
давлением хищников, привел к социальному стимулированию и синхрони-
зации размножения и дал определенные преимущества птицам, гнездя-
щимся в более плотных колониях. В плотных колониях гибель яиц и птен-
цов — наименьшая (Кафтановский, 1951; Birkhead, 1977; и др.).
Гнездовой консерватизм, характерный для кайр, в наибольшей степени
проявляется у пар, успешно размножавшихся в предыдущем году
(Hedgren, 1980).
Кайрам свойственно ритуальное ухаживание между партнерами, сопро-
вождаемое характерными позами и криками. Своеобразно также поведение
птиц в период захвата гнездовых участков, насиживания яиц и воспитания
птенцов. Различают 3 позы активной и 6 поз пассивной агрессии
(Birkhead, 1978) (рис. 9, 10).
Яйца кайр сильно варьируют по форме и окраске и могли бы служить
для распознавания своего гнезда. Тем не менее, возвращаясь после
отлучки на гнездовой карниз, кайры ориентируются в первую очередь
на место, где было оставлено яйцо, и лишь потом — на само яйцо, причем
только на ранней стадии насиживания, пока яйцо не изменило окраску
и форму от налипшего помета. Со второй недели насиживания кайры
насиживают любое яйцо или даже похожий на яйцо камень, положенный
на его место (Кафтановский, 1951; данные А. Н. Головкина). При
смене насиживающих партнеров кайры обычно не оставляют яйцо сво-
бодно лежать на уступе, а передают его от партнера к партнеру или
располагают яйцо таким образом, чтобы возможность скатиться была ми-
нимальной: прислоняют его к выступу, камешку и пр. (Ingold, 1980).
37
Рис. 9. Элементы брачного поведения тонкоклювой кайры (по: Cramp, 1985, с изменениями)
38
Рис. 10. Элементы гнездового поведения тонкоклювой кайры (по: Cramp, 1985, с изменениями)
39
Семейство Чистиковые
Звуковые контакты, устанавливаемые еще до вылупления, помогают роди-
телям распознавать своего птенца даже вне привычного места — при пе-
ремещении на соседний карниз или при его сходе на воду. Различают
9 типов крика птенца (Schommer, Tschanz, 1975), имеющих индивидуаль-
ную «окраску». Звуковое опознавание препятствует кормлению птенца
чужими родителями, хотя и известны случаи согревания и даже кормле-
ния птенцов кайрами, потерявшими собственных птенцов, а также объеди-
нения птенцов под одной взрослой кайрой наподобие «детских садов»
у пингвинов (Tschanz, 1979; данные А. Н. Головкина). Птенцы в возрасте
1 —2 дней согреваются родителями 96 % времени суток, а в возрасте
20 дней и старше — до 75 %. Рекогносцировочные перемещения птенцов
по карнизу отмечены лишь перед спуском на воду (Wehrlin, 1977).
Питание. По характеру питания тонкоклювая кайра — преимущест-
венно ихтиофаг. Состав кормов, встречающихся в желудках кайр, весьма
разнообразен. Он меняется по районам и сезонам года. В списках кормовых
объектов насчитывается более 40 видов, но доминирующее положение
как по частоте встречаемости, так и по доле в общем рационе занимают
рыбы. Беспозвоночные составляют менее 10 %. В питании кайр встре-
чаются как пелагические, так и донные рыбы, но последние добываются
в основном на мелководье. Большинство видов рыб, которыми кормятся
кайры, образуют скопления. Среди поедаемых кайрами ракообразных
также преобладают стайные виды. Наблюдения показывают, что кайры
предпочитают узких небольших рыб длиной 10—13 см и весом около 10 г.
К числу наиболее обычных кормовых объектов следует отнести такие виды,
как песчанки Ammodytes tobianus, A. hexapterus, A. lanceolatus, шпрот
Sprattus sprattus, мойва Mallotus villosus, минтай Theragra halcogramma,
треска Gad us morhua, сайка Boreogadus saida. Среди ракообразных
преобладают эуфаузииды Thysenoessa raschii, T. longipes, бокоплавы
Paratemisto libellula (Кафтановский, 1951; Белопольский, 1957; Головкин,
Георгиев, 1970; Ogi, Tsujita, 1973; Ogi, 1979; Blake, 1984; Woodby, 1984).
По-видимому, кайры в питании могут свободно переключаться с одного
вида на другой, достаточно массовый вид. В качестве случайных кормовых
объектов в желудках кайр встречаются полихеты, моллюски, растения
и даже насекомые (Белопольский, 1957).
Лишь небольшую часть рыб (около 1 %) кайры добывают на поверх-
ности, остальных — в толще воды, после непродолжительной погони.
Заглатывают обычно после выныривания и только с головы. Предельная
длина проглатываемых рыб — 22,5 см (Sandford, Harris, 1967). Экспери-
ментально установлено, что максимальный размер рыб, поедаемых кай-
рами, определяется не их длиной, а их высотой (максимальным диа-
метром). Кайры в состоянии проглатывать рыб с высотой 41—44 мм
и массой 62 — 96 г. Однако птицы предпочитают поедать рыб, имеющих
размеры вдвое меньшие максимальных (Swennen, Duiven, 1977). Пи-
таясь теми же видами рыб, что тупики и гагарки, кайры избегают конку-
ренции с родственными видами за счет использования преимущественно
других размерных группировок (Harris, 1970).
Суточная потребность в корме составляет около 28 % веса тела. Одно-
кратно кайра может съесть до 200 г рыбы, а после суточного голодания —
до 600 г за сутки. Скорость переваривания составляет 20—30 г в час. При
голодании в течение 6—8 дней кайры погибают, теряя до 39,4 % перво-
40
Тонкоклювая кайра
начального веса (Головкин, 1963; Sandford, Harris, 1967). Уровень стан-
дартного метаболизма— 140,4 ккал/сут (Johnson, West, 1975).
В районах кормежки в тех местах, где кормовые объекты — в пер-
вую очередь стайные рыбы (мойва, сайка и др.) — образуют аггрегации,
кайры объединяются в плотные скопления. По наблюдениям близ Ньюфа-
ундленда (Schneider, Piaff, 1986), такие скопления могут иметь протяжен-
ность от нескольких Десятков метров до 8,75 км. Они, как правило,
совпадают с расположением стай рыб, причем отмечена корреляция не
только по расположению скоплений, но и по количеству кайр и их жертв.
Аналогичные скопления со средней плотностью 26 птиц/км2 наблюдали
также в юго-восточной части Берингова моря, где кайры придерживались
наиболее продуктивной зоны между краем шельфа и серединой гидроло-
гического фронта по изобате 100 м (Woodby, 1984).
Обеспеченность кайр кормом меняется по годам, сезонам года и в тече-
ние одного сезона. Особенно значительным недостаток кормов бывает
зимой, когда часто можно встретить исхудавших и ослабевших птиц
(Blake, 1984). Малое количество корма весной приводит к растягиванию
кладки или уменьшению числа гнездящихся особей (Белопольский,
1957). Летом в поисках корма кайры могут перемещаться ежесуточно на
десятки километров.
Вследствие своей многочисленности кайры могут рассматриваться как
одна из существенных групп потребителей органического вещества на '
высших ступенях трофической пирамиды в экосистемах северных морей.
Особенно значимой их роль в круговороте веществ и энергии становится
в период гнездования, когда многие тысячи кайр, объединяясь в колонии,
выедают большое количество рыбы и удобряют воду своими экскремен-
тами на сравнительно небольшой по площади акватории в зоне разлета
на кормежку в непосредственной близости птичьих базаров (Успенский,
1959; Головкин, 1972, 1982). В связи с этим следует остановиться также
на экскреторной деятельности кайр (Головкин, Позднякова, 1965). При
оптимальном рационе питания, составляющем в среднем около 300 г рыбы
в сутки, кайры совершают 72—80 (среднее 74) дефекации. Масса экскре-
ментов, выделяемых при одной дефекации, варьирует в широких пределах
(от 0,35 до 5,87 г в сыром весе) при средней массе 2,5 г. Масса экскре-
ментов, продуцируемых в среднем за сутки, составляет 185 г, т. е. 67,1 %
от массы корма. Наблюдения в природе показали, что в период инкубации
и выкармливания птенцов лишь 14 % экскрементов кайр попадает на
скалы, тогда как остальная часть попадает в море, обогащая воду легко
растворимыми соединениями азота и фосфора. В летний период в усло-
виях дефицита растворенных биогенных веществ в поверхностных слоях
северных морей такое удобрение вод благоприятно воздействует на
развитие фитопланктона и способствует лучшему сохранению природного
комплекса гидробионтов. Размеры обогащаемых экскрементами зон в
прибрежье зависят от величины птичьих базаров и могут достигать де-
сятков и даже сотен квадратных километров (Головкин и др., 19726).
Враги, неблагоприятные факторы. Среди хищников, оказывающих
влияние на успешность размножения кайр, в первую очередь следует
отметить крупных чаек (серебристая, тихоокеанская, большая морская,
бургомистр), в популяциях которых некоторые особи или группы особей
специализируются на разорении колониально-гнездящихсЯ птиц (Моде-
стов, 1939; Tschanz, Barth, 1978). Хищничают в колониях кайр также ворон
41
Семейство Чистиковые
и ворона. Чаще всего хищники нападают на птенцов кайр в то время, когда
родители улетают с гнездовых карнизов за кормом. При этом в разре-
женных колониях ущерб, причиняемый хищниками, значительно больше,
чем в плотных колониях, где само присутствие на карнизах большой
массы птиц отпугивает хищника. В соответствие с этим в колониях,
где плотность гнездящихся кайр превышает 10 особей/м2, успех размно-
жения существенно выше (87,5 %), чем в разреженных колониях с плот-
ностью менее 3 особей/м2 (30,8 %) (Birkhead, 1977). Сильнее страдают
от влияния хищников периферийные участки колоний по сравнению с ее
центром. Воздействие хищников усиливается при наложении «фактора
беспокойства», особенно при посещении колоний людьми.
Существенное значение для успешного размножения кайр имеет обилие
доступного корма весной поблизости от птичьих базаров. При неблаго-
приятных кормовых условиях успех размножения и численность птиц
на карнизах снижается значительно (Blake, 1984). Недостаток кормов
осенью обусловливает резкое увеличение смертности кайр (Rees, 1969),
а ухудшение кормовой базы летом — увеличение гибели птенцов
(Tschanz, 1978). Губительное влияние на кайр могут оказывать резкие
изменения метеоусловий, в частности сильные шторма. В 1970 г. на
о-ве Унимак (Аляска) после четырехдневных штормов со скоростью ветра
до 135 км/ч число кайр, погибших от истощения, достигло 100 000 осо-
бей; на километр береговой линии — до 2000 особей (Bailey, Davenport,
1972). Значительное уменьшение популяций происходит при нерациональ-
ном промысле яиц и взрослых птиц в колониях. При нефтяном загрязне-
нии шельфовых вод кайры гибнут как из-за отравления нефтепродуктами,
так и в результате склеивания загрязненного оперения. До 16 % кайр
гибнет в условиях даже слабого нефтяного загрязнения, связанного
со сбрасываемыми с судов нефтепродуктами при очистке топливных ем-
костей (Hartwig, Ludtke, 1986). При нефтяных катастрофах в море кайры
составляют наибольшую долю из числа гибнущих морских птиц. Сотни
тысяч кайр гибнут ежегодно в ставных рыболовных сетях, особенно если
промысел ведется неподалеку от колоний. В Калифорнии в 1968—1970 гг.
было зарегистрировано уменьшение толщины скорлупы яиц (на 12,8 %)
и ее среднего веса (на 11,5 %), что, возможно, явилось следствием загряз-
нения пищевых объектов кайр пестицидами (Grees et al., 1971). На Балтике
отмечена отрицательная корреляция между толщиной скорлупы яиц и со-
держанием в яйцах метилртути. Однако имелась также и положительная
корреляция толщины скорлупы с изменением солености моря в период
яйцекладки. Поэтому утоныпение скорлупы лишь предположительно
связывается с загрязнением ртутью (Dyck, Kraul, 1984).
Хозяйственное значение, охрана. До начала 1960-х годов яйца кайр,
имеющие хорошие вкусовые и питательные качества, заготавливались
местным населением на Мурмане, в Охотском море и бригадой промысло-
виков на котиковом промысле на о-=ве Тюлений. В настоящее время
сбор яиц расценивается как браконьерство (Кафтановский, 1951; Гизенко,
1955; Карташев, 1963; Головкин, Георгиев, 1970). Охота на взрослых птиц
практиковалась ранее в разных районах в ограниченных масштабах.
В настоящее время кайра включена в список видов, на которые разрешена
охота только в Магаданской области. Несмотря на то что кайры потребляют
большое количество рыбы, в настоящее время они не являются конкурен-
тами рыболовному промыслу, поскольку питаются в основном непромысло-
42
Толстоклювая кайра
выми видами рыб (песчанка, сайка и др.,), а также многочисленной
молодью промысловых видов (мойва, молодь сельди и трески), не нанося
заметного ущерба их популяциям. В Гренландии птичьи базары рассмат-
риваются как индикаторы мест успешного рыболовства (Bibby, 1973).
За счет удобрения вод экскрементами кайр вблизи их колоний возни-
кают локальные зоны повышенной биологической продуктивности. Такие
зоны способствуют поддержанию высокого уровня продуктивности в при-
брежье северных морей.
Колонии птиц — место обитания иксодовых клещей Ixodes putus, пара-
зитирующих на кайрах. Эти клещи могут передавать человеку ряд инфек-
ционных заболеваний, однако контакты человека с птичьими базарами
очень невелики, поэтому эпидемиологическая опасность незначительна
(Флинт, Костырко, 1967; Карпович, 1970). Изредка встречаются кайры,
зараженные трематодами Cryptocotyle lingua, паразитирующими также на
рыбах. Роль кайр в распространении этих гельминтов ничтожно мала
(Подлипаев, Головкин, 1977).
В связи с большой уязвимостью колоний кайр к антропогенному
воздействию, их специфичной ролью в морских экосистемах птичьи базары
находятся под охраной. Они особо охраняются в Кандалакшском госу-
дарственном заповеднике на Мурмане и в государственном заповеднике
«Магаданский» в заливе Шелихова в Охотском море.
Увеличению численности кайр и особенно увеличению успешности их
размножения способствуют такие биотехнические мероприятия в гнездо-
вых колониях, как расчистка гнездовых карнизов и уничтожение хищ-
ников (Кафтановский, 1951; Corkhill, 1970). Кайр несложно содержать
в неволе. В вольерах при достаточном питании они могут образовывать
пары, откладывать яйца и выводить птенцов. В неволе добивались размно-
жения кайр во втором поколении (Marsault, 1975; Conway et al., 1977).
Толстоклювая кайра —
Uria lorn via Linnaeus, 1758
Briinnich’s Guillemot (англ.), Thick-billed Murre (амер.), Dickchnabellumme
(нем.).
Син. (рус.): короткоклювая кайра.
Статус. В пределах СССР гнездящийся перелетный или кочующий,
зимующий вид.
Общая характеристика и полевые признаки. Птица средних размеров,
по внешнему облику очень похожа на тонкоклювую кайру. Такое же
веретенообразное тело, но шея и клюв короче, отчего голова кажется более
крупной и округлой. Клюв острый, прямой, на конце слегка загнутый.
По нижнему краю надклювья от ноздрей к углу рта расположено вздутие
беловато-серой окраски, отличающееся от цвета основной рамфотеки и
хорошо различимое даже издали. По окраске этой полосы и более толстому
и короткому клюву данный вид можно, достаточно уверенно отличать
от тонкоклювой кайры, особенно если оба вида встречены одновременно.
Плотное жесткое оперение имеет контрастную двухцветную окраску.
Голова, шея, верхняя сторона тела черновато-бурые с темно-коричневым
оттенком. Нижняя сторона тела, грудь, живот, подкрылья белые без
пестрин на боках или с редкими пестринами. По сравнению с тонкоклювой
кайрой голова окрашена темнее, белая окраска на груди заходит на шею
43
Семейство Чистиковые
выше, белое зеркальце на крыльях несколько длиннее. Ноги сильно отстав-
лены кзади. На суше неуклюжа, стоит и передвигается так же, как тонко-
клювая кайра. Полет стремительный, маломаневренный с быстрыми взма-
хами крыльев. Прекрасно плавает и ныряет, но под водой менее маневренна,
чем тонкоклювая кайра (Spring, 1971). С воды или ровной поверхности
суши взлетает с трудом после толчка вверх с какой-либо неровности или
с гребня волны. В полете, на воде и на суше по движениям и позам очень
сходна с предыдущим видом. В период гнездования также образует
крупные колонии или «птичьи базары» — одновидовые или совместно
с тонкоклювыми кайрами, гагарками и моевками. На воде обычно молча-
лива. В колониях крики неумолчны. По первому впечатлению крики обоих
видов кайр неразличимы, хотя реально они различаются, что подтверждено
анализом сонограмм (Tschanz, 1972). Кроме общего крика «ар-ра», они
издают крик «ар-рр-р» с более грубо звучащей концовкой или «уррр-ррау-
ррау», а также «гррр». Особенно часто такой крик звучит в тех случаях,
когда на гнездовом карнизе возникают конфликты между соседями или
птицы обеспокоены появлением какой-либо опасности. В период брачной
активности и при копуляции издается двусложный крик «гау-гау», напо-
минающий лай собаки (Pennycuick, 1956). Взрослые птицы, собирающиеся
у подножия скал в период слета птенцов, издают раскатистые крики
«аррр» с повышением звука в конце. Крики соответствуют поведению
кайр. Птенцы, выпрашивая корм у родителей, или при поисках зоны
комфорта (например, при охлаждении) издают тихий писк «уи-уи».
Потерявшиеся подросшие птенцы при длительном отсутствии родителей
издают громкий крик «у-иип». Так же кричат птенцы, спустившиеся
с карнизов на воду. Этот крик служит сигналом для родителей к поискам
своего птенца. В условиях полярного дня кайры могут быть активны
в любое время суток.
Подвидовая систематика. В настоящее время различают четыре под-
вида, отличающиеся размерами и оттенками окраски оперения (Степанян,
1975). Все они обитают на территории СССР.
1. Uria lomvia lorn via
Uria lomvia Linnaeus, 1758, Syst. Nat, ed. 10, c. 130, Гренландия.
Верхняя сторона тела темная, черновато-бурая, со слабым сероватым
оттенком или без него. По окраске и размерам занимает промежуточное
положение между самой темной и сероватыми формами. Белая полоска
на крыле довольно широкая.
2. Uria lomvia eleonorae
Uria lomvia eleonorae Portenko, 1937, Mittel. Zool. Mus. Berlin, 22, c. 227,
о-в Преображения, Хатангский залив, Восточный Таймыр.
Верхняя сторона тела буровато-черная, с сильно развитым сероватым
или даже голубоватым оттенком. Светлый кофейный оттенок имеется на
передней стороне шеи. Пестрины на боках неясные или могут отсутство-
вать вообще. Белая полоса на крыле относительно широкая. Самый мелкий
из подвидов.
3. Uria lomvia heckery
Uria lomvia heckery Portenko, 1944, Докл. АН СССР, 43, c. 226, о-в Вран-
геля.
По окраске верха сходна с предыдущим видом. Размеры клюва больше.
4. Uria lomvia arra
Cepphys arra Pallas, 1811, Zoographia Rosso-Asiat., 2, c. 347, Камчатка.
44
Толстоклювая кайра
Окраска верхней стороны тела темная, буровато-черная, без сероватого
оттенка. На боках тела четкие пестрины. Клюв крупнее, чем у других под-
видов. Белая полоска на крыле узкая (Портенко, 1973).
Замечания по систематике. Самостоятельность подвида Uria lomvia
arroides, описанного Л. А. Портенко (Portenko, 1937) по материалам
с островов Франца-Иосифа, подвергалась сомнению (Дементьев, 1951;
Белопольский, 1957) и была признана недостаточно обоснованной (Коз-
лова, 1957).
Описание. Строение и размеры. Первостепенных маховых И,
первое рудиментарно. Формула крыла: .. Рулевых перьев
12—14. Размеры: длина (в мм) крыла самцов 205,8—234,0 (среднее
216,4), самок 192,5—240,0 (среднее 214,9); клюва самцов 35—48,5
(среднее 40,7), самок 31,1—48,0 (среднее 38,3); плюсны самцов (35—40
(среднее 37,8), самок 36 —40 (среднее 37,5). Масса (в г) самцов 760—
1160 (среднее 987,8), самок 730—1206 (среднее 997,9) (Кафтановский,
1941; 1951; Белопольский, 1957; Tuck, 1960; Cramp, 1985).
Клюв вытянутый, сходный с таковым у тонкоклювой кайры, но отно-
сительно более высокий, массивный, на конце менее загнутый. Контурное
оперение плотное, жесткое.
Окраска. Самец и самка в брачном наряде. Полового диморфизма
в окраске нет. Окраска спинной стороны черновато-бурая с большим
или меньшим развитием серого оттенка. При том же общем тоне, на боках
головы, подбородке и шее хорошо заметен коричневатый («шоколадный»)
оттенок, который меньше, чем у тонкоклювой кайры, распространяется
на спину и кроющие крыльев, но на боках головы более насыщен. Темная
полоска за глазом менее заметна. «Очковых» морф не бывает. Маховые
бурые. Внутренние опахала первостепенных маховых беловаты у основа-
ния. Вершины второстепенных имеют белую каемку. При сложенном и
раскрытом крыле они образуют хорошо заметную узкую белую полосу.
Нижняя сторона тела белая. Белая окраска с груди заходит на нижнюю
часть шеи в виде перевернутой буквы V, образуя более острый угол, чем
у тонкоклювой кайры. Бока белые, как правило, без пестрин. Нижние
кроющие крыла с редкими пестринами. По краю основания надклювья
беловато-серая (иногда с зеленоватым или желтоватым оттенком) роговая
полоска шириной до 3 мм. Ноги черные с просвечивающим желтовато-
оранжевым оттенком на передней стороне цевки и на наружной поверхно-
сти пальцев. Когти черные. Радужина темно-бурая.
Самец и самка в зимнем наряде. Белая окраска появляется на подбо-
родке и передней стороне шеи. Белые пестринки заметны и на щеках.
Клюв становится более светлым, сероватым и несколько менее вздутым,
чем в брачном наряде.
Пуховой наряд. Птенец покрыт коротким плотным пухом, коричневато-
бурым на голове и спинной стороне и белым на груди и животе. На голове
и шее видны светло-коричневые и беловатые пушинки или кончики
пушинок, делающие окраску слегка пестрой. Самый длинный пух (10—
11 мм) на голове, несколько короче (8—9 мм) на спине и боках, самый
короткий (5—6 мм) на животе (Успенский, 1956). Общий тон окраски
варьирует от светло-коричневой до почти черной. Клюв и ноги темно-
серые. Яйцевой бугорок большой (2—4 мм) на надклювье и несколько
меньше (0,4—2 мм) на подклювье (Кафтановский, 1951; Успенский,
1956). В мезоптильном наряде голова, верх шеи, спина, кроющие хвоста
45
Семейство Чистиковые
и крыльев насыщенно черные, без коричневого и серого оттенков, обычных
у взрослых птиц, нижняя часть шеи, грудь и брюхо белые. Рулевые и
маховые в этом наряде отсутствуют. На голове, бедрах и огузке сохра-
няются отдельные пушинки.
Первый зимний наряд. Напоминает зимний наряд взрослой птицы
и отличается большим развитием белого цвета на горле и щеках, наличием
пестринок на боках шеи. Известны альбиносы и меланисты (Кафтанов-
ский, 1951).
Линька. Последовательность смены нарядов такая же, как у тонкоклю-
вой кайры. Смена пухового наряда начинается в первые дни жизни птен-
цов. Верхние кроющие крыла появляются первыми на 2—3-й день.
На следующий день верхушки кроющих обнаруживаются и на других
частях тела, на 10—11-й день кроющие перья раскрываются наполовину,
а к 15-му дню — полностью. В это время птенец приобретает промежуточ-
ный, или мезоптильный наряд, в котором заметны лишь отдельные пу-
шинки. В таком наряде он спускается с гнездовой скалы в море, где
в течение первых двух месяцев происходит линька в первый зимний
наряд. Весной и в начале лета он меняется на неполный взрослый,
а осенью — на зимний наряд. Первый полный брачный наряд кайры
приобретают к середине второй зимы. У взрослых кайр линька из брачного
наряда в зимнее оперение начинается на гнездовых карнизах, а заканчи-
вается уже после откочевки в море. В первую очередь сменяется контурное
оперение на голове и шее. Во время откочевки вместе с птенцами из райо-
нов гнездования сменяется оперение на плечах, растут новые маховые
перья. К концу осени взрослые кайры теряют на время способность летать.
К середине—концу декабря некоторые птицы имеют еще не полностью
выросшие маховые перья. Закончив линьку в зимнее оперение, кайры
почти сразу же начинают линять вновь и в феврале приобретают брачный
наряд (Кафтановский, 1951; Tuck, 1960).
Распространение. Гнездовой ареал. Арктические и субаркти-
ческие моря. В Северной Америке распространена по западному побережью
вдоль берегов Аляски от о-ва Ванкувер на юге до мыса Лисбурн на севере,
на северном побережье в районе о-ва Бэнк, на восточном побережье к югу
до залива Св. Лаврентия (рис. 11, 12). На северном побережье Евразии —
Северная Норвегия, Кольский полуостров, восточное побережье Таймыра,
побережье Чукотского полуострова, Камчатки, Охотского моря до Север-
ного Приморья, к югу до устья р. Тумнин. Острова тихоокеанского бас-
сейна: Алеутские, Прибылова, Св. Матвея, Св. Лаврентия, Диомида,
Командорские, Карагинский, острова залива Шелихова, Ямские, Шантар-
ские, Ионы, Курильские, Сахалин, Тюлений. Острова атлантического
бассейна: Элсмир, Баффинова Земля, острова Гудзонова залива, Гренлан-
дия, Исландия, Ян-Майен, Медвежий, Лофотенские, Шпицберген, Земля
Франца-Иосифа, Новая Земля, острова Восточного Мурмана. Острова
бассейна Северного Ледовитого океана: Анжу, Преображения, Де-Лонга,
Врангеля, Геральд, Колючин.
Зимовки. После откочевки от гнездовий кайры постепенно переме-
щаются к местам зимовок, расположение которых диктуется границами
сплошных льдов. Районы зимовок находятся обычно недалеко от мест
гнездования. Кайры из северных районов Канады смещаются в южную
часть пролива Дэвиса, к берегам Гренландии и Ньюфаундленда. Значи-
тельные зимовки располагаются в заливе Св. Лаврентия, у южных берегов
46
Рис. 11. Область распространения толстоклювой кайры
а — граница гнездового ареала, б — область зимовок и миграций. Подвиды: / — U.l. lomvia, 2 — U.l. eleonorae, 3 — U.l. heckeri, 4 — U.l. arra
Семейство Чистиковые
Новой Шотландии и Ньюфаундленда, а также в открытом море в этих
районах в зоне шельфового склона (Tuck, 1960; Salomonsen, 1971).
В этих местах они встречаются с октября до начала мая, перемещаясь
вдоль границ плавающих ледяных полей и припая (Salomonsen, 1971;
Brown, 1968). Толстоклювые кайры с берегов Новой Земли и других
северных островов Баренцева моря зимуют, по-видимому, близ ледовых
полей, лежащих к северо-западу от мест гнездования (Успенский, 1956).
Места зимовки кайр Восточного . Мурмана, Северной Норвегии, видимо,
располагаются южнее. В зимний период отмечены случайные залеты
толстоклювых кайр к берегам Германии и Нидерландов (Goethe, Ringle-
ben, 1964; Tooren, 1967). Зимующих кайр наблюдали на банке Джорджес
(Белопольский и др., 1982). Кайры с островов и побережий морей тихо-
океанского бассейна также придерживаются границ ледовых полей и
границ шельфа. Они в массе встречаются и в юго-восточной части Берин-
гова моря, к середине зимы перемещаясь к югу и достигая широты
о-ва Хонсю (Kuroda, 1963а; Woodby, 1984). Предполагается, что в период
зимовки кайры совершают скорее кочевки, чем правильные миграции.
Миграции. Весенний прилет к местам гнездования в значительной
степени зависит от ледовой обстановки. Сроки прилета сильно различаются
в разных частях ареала и могут меняться от года к году. Зачастую кайры
появляются вблизи гнездовий значительно раньше, чем условия позволят
занять им гнездовые карнизы. В тех местах, где оба вида кайр гнездятся
в общих колониях, толстоклювые кайры появляются на гнездовых карни-
зах на 2—3 недели позднее, чем тонкоклювые (Кафтановский, 1951;
Успенский, 1956; Tuck, 1960). В феврале кайры концентрируются вблизи
берегов, но до начала мая могут быть также встречены и в открытом море,
хотя в меньших количествах (Brown, 1968; 1984).
Осенние миграции начинаются сразу же после слета птенцов с гнездо-
вых карнизов — в конце августа-сентябре. Взрослые птицы вместе с птен-
цами перемещаются со скоростью до 40 км в сутки (Gaston, 1982). Кайры
Баренцева и Норвежского морей мигрируют в западном или северо-запад-
ном направлении. В западных районах Атлантики основное направление
миграций — юго-восточное и южное. Прослежено, что толстоклювые кайры
с южной части Баффиновой Земли в конце августа мигрировали через
Гудзонов пролив, придерживаясь направления поверхностного течения.
Кайры из колоний в Гудзоновом проливе в начале сентября переместились
в открытое море и через несколько дней достигли Северного Лабрадора.
В конце сентября стаи кайр перемещались по морю Лабрадор к местам
зимовок вокруг Ньюфаундленда. Часть кайр с Баффиновой Земли мигри-
рует к западным берегам Гренландии (Orr, Ward, 1982). С азиатских
берегов Тихого океана осенний пролет направлен к востоку и юго-востоку,
а с американских — к западу.
Численность. Современная численность гнездящихся толстоклювых
кайр составляет около 5,3 млн пар (по материалам обзора «Status an Conser-
vation of the World’s Seabirds» 1984). Возможно, она несколько занижена,
поскольку еще далеко не все места гнездования кайр обследованы с доста-
точной полнотой. По мнению Л. Така (Tuck, 1960), толстоклювая кайра —
самая многочисленная морская птица Северного полушария в целом, хотя
и уступающая по численности в Северной Атлантике люрику, а в Северной
Пацифике — конюге-крошке. Общая численность вида оценена этим авто-
ром в 42 млн особей. Если допустить, что примерно половину популяции
48
Рис. 12. Ареал толстоклювой кайры в СССР
Обозначения те же, что на рис. 11
Семейство Чистиковые
составляют негнездящиеся особи, то количество гнездящихся пар,
по оценке Л. Така (Tuck, 1960) (около 10,5 млн пар), будет намного
отличаться от последних данных. Скорее всего, различия могут быть
объяснены более реалистичной оценкой, проводимой в последние годы
на базе учетов птиц в разных частях ареала.
Наиболее многочисленные популяции размещаются в Беринговом
море (около 2,5 млн пар), на северо-западе Атлантики (3,2 млн пар)
и в Баренцевом море (около 1 млн пар) (Lensink, 1984; Evans, 1984;
Golovkin, 1984).
В СССР гнездится около 1640 тыс. пар толстоклювых кайр, т. е. при-
мерно 1/5 часть мировой популяции. В том числе на побережьях Новой
Земли и Земли Франца-Иосифа гнездится около 911 тыс. пар (Успенский,
1956; 1959; Головкин, 1972), на о-ве Преображения и о-вах Анжу —
35 тыс. пар, на о-вах Де-Лонга и западном побережье о-ва Врангеля —
около 50 тыс. пар (Успенский, 1959; Успенский и др., 1963), на островах
и побережье Чукотского моря — около 53 тыс. пар (Успенский и др.,
1963; Головкин, Флинт, 1975; Кондратьев, 1975; Морозов, Томкович,
1981), на островах и восточном побережье Берингова моря, включая
восточное побережье Камчатки — около 280 тыс. пар (Успенский, 1959;
Мараков, 1963; Вяткин, 1975; Вяткин и др., 1975), на побережье и островах
Охотского моря, включая Курильские острова — около 310 тыс. пар (Ве-
лижанин, 1969, 1975; Бычков, 1975; Харитонов, 1975; Яхонтов, 1975а, б;
Велижанин, 1977а). Численность кайр более или менее стабильна.
Местообитания. Истинно морская птица, обитатель шельфовых зон
Северного полушария. В открытом море встречается зимой, а также
в период миграций. Гнездится на скалах на побережье материков и морских
островов. Использует скалы, обрывающиеся в море или отделенные от него
полоской суши шириной в несколько десятков метров. Высота гнездования
над уровнем моря варьирует от нескольких метров до нескольких десятков
и даже сотен метров. Гнездится как на широких, так и на узких карнизах
и выступах скал. Изредка образует «плоскостные» гнездовые колонии
на горизонтальных или слабо покатых скалистых вершинах небольших
островков. Обычно избегает участки суши, имеющие растительный покров
или покрытые слоем торфа. Гнездовые колонии приурочены к участкам
акватории с повышенной биологической продуктивностью вод.
Размножение. Биология размножения во многом сходна с таковой
тонкоклювой кайры. Половая зрелость наступает в возрасте двух лет
(Кафтановский, 1951). Моногамы, сохраняющие верность партнерам
и местам гнездования из года в год. При первом появлении вблизи гнездо-
вых карнизов в конце зимы кайры имеют еще недоразвитые гонады.
Гонады достигают максимального развития в третьей декаде_мая на Мур-
мане и в первой декаде июня на Новой Земле (Белопольский, 1957).
Сроки появления на гнездовых карнизах и начало размножения зависят
от широты местности и особенно от ледовых и метеорологических условий.
В районах, наиболее суровых в климатическом отношении, сроки между
появлением на карнизах и кладкой минимальны. Откладка яиц на Мурмане
происходит 11—29 мая, на юге Новой Земли 2—10 июня, на. севере —
20 июня—2 июля (Кафтановский, 1951; Успенский, 1956; Белопольский,
1957). Середина июня — типичный срок массовой откладки яиц в боль-
шинстве районов гнездования.
Копуляция происходит на гнездовых карнизах или вблизи них. Она
50
Толстоклювая кайра
продолжается и после начала откладки яиц. Ей предшествует период
ухаживания, сопровождающийся «танцами» на воде — плаванием кру-
гами, полетами, в которых выписываются замысловатые фигуры, предбрач-
ными позами на карнизах и вне их, а также специфичными криками.
Кайрам свойственна откладка яиц непосредственно на скалу без какого-
либо гнезда. Лишь изредка отмечаются атавистичные попытки гнездострои-
тельной деятельности: скобление субстрата, подкладывание под яйцо
примитивного гнездового материала (камней, травы, скорлупы яиц, кусоч-
ков засохшего помета, рыбы, перьев) (Williams, 1971). Сугубо типичным
следует считать колониальное гнездование. Гнездование отдельными
парами бывает крайне редко (Портенко, 1931). Колонии могут быть
одновидовыми или смешанными, где толстоклювые кайры соседствуют
с тонкоклювыми, другими видами чистиковых, моевками. При совместном
гнездовании с тонкоклювыми кайрами толстоклювые занимают менее
удобные краевые участки карнизов, узкие уступы (Кафтановский, 1951;
Белопольский, 1957; Tuck, 1960). На наиболее удобных широких карнизах
гнездящиеся толстоклювые кайры располагаются очень близко друг
от друга — по 32—37 насиживающих птиц на 1 м2 (Красовский, 1937;
Успенский, 1956). Наиболее крупная плоскостная колония на слабонаклон-
ной вершине скалистого островка известна в СССР в заливе Вилькицкого
на Новой Земле (Портенко, 1931; Головкин, 1972).
Массовая откладка яиц в колониях синхронизирована. Между ранними
и поздними кладками на Мурмане проходит 16 дней, а на Новой Земле —
4—6 дней (Белопольский, 1957). За счет повторных кладок взамен поте-
рянных первых общий период яйцекладки может растянуться до полутора
месяцев. В кладке одно яйцо грушевидной формы, с матовой, крупнопо-
ристой и бчен’ьтблсЯоЙ'скорлуной (0,22—0,26 мм на тупом конце, 0,32—
0,34 мм на остром конце) (Кафтановский, 1951). Окраска основного фона
чрезвычайно изменчива: от сочной темно-зеленой до светло-зеленоватой
с легким голубым оттенком. Коричневатые оттенки для яиц толстоклювой
кайры не характерны. Яйцо, как правило, испещрено коричневатыми
или черными пятнами и полосками, имеющими самую разнообразную
форму и расположение. Чаще всего они сгущаются и образуют «поясок»
у тупого конца. Размеры яиц в колониях сильно варьируют, но в среднем
для подвидов весьма сходны (Tuck, 1960). Можно считать типичной сред-
нюю величину 79,5Х50,8 мм. Известны гигантские яйца, у которых боль-
шой диаметр достигает 96 мм, и необычно мелкие яйца — 70 мм. Изменчи-
вость малого диаметра меньше — 14 мм (Успенский, 1956). Расхождения
в вариациях большого и малого диаметров определяют Изменчивость
формы от округло-грушевидной до удлиненно-грушевидной. Вес свеже-
отложенных яиц из разных районов варьирует в пределах 101 —118 г
и составляет около 11 % от веса тела самки. В случае потери первого
яйца самка может отложить второе, а при его утере — третье. Интервал
между кладками — 16—18 дней. Повторные яйца сходны с первыми
по форме, окраске и весу (Кафтановский, 1951). Средний объем яиц может
быть различным в разных колониях (Birkhead, Nettleship, 1981). Отме-»
чается, что в тех колониях, где кладка начинается раньше, и самки откла-
дывают более крупные яйца, успех размножения в расчете на одну пару
птиц выше. Птенцы, вылупившиеся из более крупных яиц, имеют больший
вес, но не отличаются быстрым развитием, а просто обладают большим
запасом питательных веществ. Поздно откладываемые яйца имеют мень-
4*
51
Семейство Чистиковые
ший вес. Предполагается, что это имеет адаптивное значение и связано
с необходимостью успеть отложить сформировавшееся яйцо к периоду,
когда еще достаточно корма для выращивания потомства (Nettleship,
1982).
Самка начинает насиживание сразу же после откладки яйца. В дальней-
шем насиживают оба родителя, сменяя друг друга с интервалом от несколь-
ких минут до нескольких часов. В первые дни инкубации яйцо иногда
остается без обогрева на 10—20 мин, но с 5—7-го дня насиживается
постоянно. Каждый из родителей имеет одно удлиненное наседное пятно.
Так же как у тонкоклювой кайры, по мере развития эмбриона центр
тяжести смещается к острому концу, что снижает вероятность падения
яйца с карниза при поспешном отлете родителей (Успенский, 1956).
Инкубационный период длится 32—35 дней. Различия в сроках инкубации
могут быть обусловлены способностью наклюнувшегося птенца полностью
освободиться от скорлупы. У некоторых птенцов на это уходит от 2 до
4 дней (Tuck, 1960). Средняя усушка яиц при инкубации составляет 0,55 г
в сутки (Johnson, West, 1975), а средняя скорость роста эмбриона — 3,4 г
в сутки (Nettleship, 1982). При вылуплении птенцы весят 65-^77 г (в сред-
нем 70,6 г) (Кафтановский, 1951). Птенцовый пух обсыхает в течение
суток. Птенцы начинают получать корм со 2—3-го дня после рассасывания
желточного мешка. Наиболее интенсивный прирост приходится на 4—10-й
день жизни. В это время у птенца устанавливается терморегуляция.
Первые дни жизни один из родителей согревает птенца. Кормят его преиму-
щественно рыбой, длиной 28—193 мм, изредка — ракообразными (Gaston,
Nettleship, 1981). Число кормлений изменяется от 3—4 в первые дни жизни
до 6—8 к концу выкармливания на карнизе. В среднем за сутки в зависи-
мости от возраста птенец получает от 40 до 100—120 г пищи. Для увели-
чения веса птенца на 1 г расходуется в среднем 13,4 г рыбы (Tuck,
Squires, 1955), а прибавка веса составляет 7—11 г в сутки (Nettleship,
1982). Длительность пребывания на карнизе варьирует от 18—20 сут
в наиболее суровых районах до 25 сут в местах с более мягкими климати-
ческими условиями. За это время вес птенца увеличивается соответственно
до 175—215 или 250—275 г (Кафтановский, 1951; Успенский, 1956;
Tuck, 1960). Установлено, что в более крупных колониях птенцы к моменту
спуска с гнездовых карнизов достигают меньшего веса (относительно
веса взрослой птицы), чем в колониях меньшего размера (Gaston et al.,
1983). Длина и ширина крыла птенцов, спускающихся с базаров, варьирует
значительно меньше, чем вес (Hatch, 1983).
Родители устанавливают звуковой контакт с птенцом еще в тот период,
когда он начинает только проклевываться (Pennicuick, 1956; Gaston,
Nettleship, 1981). Эти контакты многофункциональны. Они позволяют
взрослым птицам и птенцам распознавать друг друга, стимулируют роди-
телей к кормлению голодного птенца, предотвращают его нежелательные
перемещения по карнизу, служат стимулом птенцу для спуска на воду
в конце гнездового периода, помогают родителям отыскать своего птенца
на воде у подножия скал. Примерно за сутки до оставления гнездовых
карнизов родители прекращают кормить птенца. Птенцы в мезоптильном
наряде, еще не умея летать, прыгают со скал на воду или на прибрежные
камни. Наполнение воздушных мешков и быстрые взмахи маленьких
крыльев превращают падение в некоторое подобие планирования. Птенцы,
даже упав с большой высоты на твердый субстрат, как правило, не травми-
52
Толстоклювая кайра
руются и после непродолжительной паузы пешком добираются до кромки
воды. Преодолев полосу прибоя, они отплывают от берега на несколько
десятков метров, где происходит их встреча с родителями. К спустивше-
муся на воду птенцу одновременно подплывает множество взрослых птиц,
иногда образующих клиновидную стаю. Не признав птенца, первые отплы-
вают в сторону и уступают место следующим за ними родителям, которые,
обнаружив своего птенца, отплывают с ним в сторону моря. Если птенцы,
падая, попадают на сушу и им не удается сразу же добраться до моря, они,
не получая пищи, теряют в весе в среднем за сутки 17,5 г. В этом случае
собственный вес птенцов имеет большое значение для их выживаемости
(Hatch, 1983). Спуск птенцов в море имеет четко выраженную суточную
ритмику и наибольшее число птенцов спускается со скал в период между
20 и 24 ч. Планирующие и упавшие на сушу птенцы подвергаются напа-
дениям бургомистров. Синхронность схода птенцов с гнездовых карнизов
способствует тому, что большему числу птенцов удается избежать нападе-
ния хищников (Daan, Tinbergen, 1979). Гибель потомства составляет
около 60 %, в том числе около 50 % падает на период инкубации (Успен-
ский, 1956; Tuck, 1960).
Суточная активность, поведение. В течение суток четко выражена
активность в светлое время. На Шпицбергене и Новой Земле, где в начале
периода гнездования различия в условиях освещенности днем и ночью
незначительны, кайры активны круглые сутки (Pennycuick, 1956; Успен-
ский, 1956). Однако с наступлением периода темных ночей видно, что на
рассвете и перед закатом более 50 % кайр улетает из колонии на кормежку.
Характерно изменение активности в соответствии с фазами гнездового
сезона. Во время массового вылупления птенцов, а также незадолго до их
спуска частота отлетов и прилетов птиц резко возрастает. В полеты на кор-
межку дополнительно вовлекается до 35 % населения колонии. Особенно
это выражено в середине июля, когда отлеты принимают характер мас-
сового исхода (Tuck, 1960; Головкин, Позднякова, 1966).
Так же как у предыдущего вида, активность зависит от уровня прилива
и метеоусловий. Отлет на кормежку и для отдыха на воде происходит чаще
в период отлива, когда обычно уменьшается волнение. При сильном ветре,
дожде, волнении моря, а также при сплошном тумане на карнизах присут-
ствует большее количество кайр. В штормовых условиях кайры, согреваю-
щие яйца или птенцов, могут не покидать карнизы по нескольку дней.
Поведение на гнездовых карнизах тесно связано с колониальным типом
гнездования. В начале гнездового сезона, когда кайры занимают участки
на скалах, они агрессивны по отношению к соседям. Размеры защищаемой
территории невелики. Они ограничиваются зоной, куда может дотянуться
сидящая птица. На карнизах часто возникают драки, особенно при появле-
нии в сложившейся гнездовой группе птиц нового претендента на гнездо-
вую территорию. Позднее агрессивность снижается и плотность расположе-
ния птиц на наиболее удобных площадках достигает максимального
уровня. Птицы насиживают вплотную друг к другу.
Важным моментом' является социальная стимуляция активности
размножения, способствующая синхронности гнездования. Кайрам свойст-
венно ритуальное ухаживание, а также своеобразный комплекс поз и кри-
ков, соответствующий различным поведенческим ситуациям: тревоге,
смене насиживающих родителей, появлению птиц с кормом и др. (Wil-
liams, 1974; Gaston, Nettleship, 1981). При общем сходстве элементов
53
Семейство Чистиковые
поведения с таковыми у тонкоклювых кайр, отмечены и некоторые отличия,
позволяющие считать поведение видоспецифичным (Pennycuick, 1956;
Gaston, Nettleship, 1981' и др.). По-видимому, толстоклювые кайры менее
агрессивны, чем тонкоклювые, вследствие чего при совместном гнездовании
они оттесняются на менее удобные краевые участки карнизов (Кафтанов-
ский, 1951; Успенский, 1956; Tuck, 1960; Brun, 1965). Характерно усиле-
ние инстинкта сохранения потомства от начала к концу инкубации. Если
в первые дни после откладки яиц кайры покидают их при первой же опасно-
сти, то по прошествии нескольких дней они становятся менее пугливыми
и не вставляют яйцо, даже когда до птицы дотрагиваются рукой. Некото-
рые поведенческие реакции (возбуждение, отлет и прилет) проявляются
одновременно у групп особей, гнездящихся по соседству.
Питание. Преимущественный ихтиофаг. Состав корма может значи-
тельно варьировать как в течение года, так и в зависимости от географи-
ческого местоположения. В списке кормовых объектов насчитывается
более 30 видов рыб и 20 видов беспозвоночных. В гнездовой сезон явно
доминируют рыбы, составляя более 90 % как по встречаемости, так и по
удельному весу. Характерно, что в местах с тяжелыми ледовыми усло-
виями как в летний период, так и на зимовке доля беспозвоночных в пита-
нии увеличивается. Летом ряд видов встречается в питании особенно
часто. К ним относятся сайка Boreogadus saida, треска Cadus morrhua,
триглопс Triglops pingeli, люмпены Lumpenus spp. Песчанка Ammodytes
tobianus и мойва Mallotus villosus добываются в большом количестве
в южных частях ареала как во время гнездования, так и на зимовке (Каф-
тановский, 1951; Успенский, 1956; Белопольский, 1957; Tuck, 1960;
Birkhead, Nettleship, 1981; и др.). На зимовках у кромки льдов питаются
преимущественно криофиллами — сайкой и амфиподами Onisimus glacia-
lis (Bradstreet, 1979). Распределение кайр на зимовках в шельфовой зоне
также зависит от локальных скоплений их кормовых объектов — рыб и
крупного зоопланктона (Woodby, 1984). В зависимости от массовости
кормовых объектов в море ведущим кормом могут быть беспозвоночные —
амфиподы Parathemisto libellula (до 75 % весовой доли), а также кальмары
Gonatopsis borealis, Berrytenthis magister (до 72,6 % веса корма). В пита-
нии встречаются еще эуфаузииды Tyssanoessa raschii, Т. longipes и другие
виды (Ogi, Tsujita, 1973; Оги, 1979; Ogi, .1980; Ogi, Hamanaka, 1982).
В качестве дополнительных или случайных кормов могут быть бычки
(рр. Icelus, Myoxocephalus), сельдь Clupea harengus, декаподы, полихеты,
медузы. Беспозвоночные в целом составляют в питании толстоклювых
кайр большую долю, чем у тонкоклювых. Предполагается, что их ротовой
аппарат более приспособлен к добыванию планктонных организмов, чем
у предыдущего вида, а вследствие меньшей маневренности плавания
в толще воды им труднее ловить рыбу (Spring, 1971). Вероятно, большая
доля беспозвоночных в питании обусловлена также тем, что в высоких
широтах, где доминируют толстоклювые кайры, планктонные организмы
являются массовыми и более доступными, образующими к тому же круп-
ные скопления.
Обычно кайры кормятся в непосредственной близости от мест гнездова-
ния. Наибольшие скопления кормящихся кайр отмечали не далее 2 км
от колонии в Атлантике и 2—4 км — в Пацифике (Bedard, 1976), или
по другим данным — в радиусе 1,5—8 км (Tuck, Squires, 1955). Известны
54
Толстоклювая кайра
и дальние разлеты на кормежку на расстояние 100—200 км (Успенский,
1965; Cramp, 1985).
Обилие корма вблизи мест гнездования является одним из основных
факторов возникновения крупных колоний. При этом существенно
не только то, что колонии обычно располагаются в высокопродуктивных
районах моря (Белопольский, 1979), но и тот факт, что для крупных
колоний птицы обычно избирают мелкие острова или выступающие в море
скалистые мысы, где циркуляционные течения способствуют возникнове-
нию крупных скоплений кормовых организмов (Головкин, 1970; Голов-
кин и др., 19726; Brown, 1981). Предпочитаемые размеры рыб — 6—10 см,
при максимуме до 33 см (Успенский, 1956; Ogi, Tsujita, 1973), и беспозво-
ночных— 1,5—3,1 см (Ogi, Hamanaka, 1982). Суточная потребность
в корме взрослых птиц составляет в среднем 250—300 г. После суточного
голодания потребление может возрасти вдвое (Tuck, Squires, 1955; Голов-
кин, 1963). Уровень стандартного метаболизма 140,4 ккал/сут (Johnson,
West 1975).
Являясь одним из наиболее массовых видов истинно морских птиц
в Северном полушарии, толстоклювая кайра может быть отнесена к числу
важных потребителей биологической продукции на высшем трофическом
уровне в морских экосистемах. Колониальный образ жизни в период
гнездования способствует тому, что, добывая из моря большое количество
органического вещества, кайры возвращают вблизи колоний значительную
его часть в виде экскрементов, содержащих легко растворимые соедине-
ния, пригодные для использования их фитопланктоном. Тем самым
осуществляется трансформация и перераспределение веществ, имеющая
определенное значение для биогенного круговорота в прибрежной зоне
моря (Успенский, 1959; Tuck, 1960; Головкин, Позднякова, 1964; Голов-
кин, 1972, 1982; и др.).
Враги, неблагоприятные факторы. Обычно хищники не оказывают
существенного влияния на колонии кайр. Небольшое количество яиц
из разрозненных колоний крадут крупные чайки (бургомистр, серебристая
и большая морская). Упавших с карнизов птенцов поедают песец, ворон,
полярная сова. Взрослые кайры изредка становятся добычей кречета.
Чаще всего успеху хищников способствует фактор беспокойства, связан-
ный с появлением человека в колониях. Разорение колоний человеком —
наиболее существенный неблагоприятный фактор. Колонии страдают
также при разрушении скал вследствие эрозии. Недостаток корма, особенно
весной, сказывается на успешности размножения. Для толстоклювых
кайр, как и для тонкоклювых, с ростом интенсивности морского рыболов-
ства увеличивается опасность сокращения кормовой базы, что может
привести к катастрофическому уменьшению численности птиц (Головкин,
1982; Furnes, 1984). Колонии кайр считаются индикаторами мест, наиболее
успешных для промысла атлантического лосося (Salmo salar). В то же
время значительное число толстоклювых кайр (в частности, в Гренландии
от 100 до 500 тыс. особей) ежегодно гибнет в рыболовных сетях, выстав-
ляемых вблизи колоний. Помимо того, до 750 тыс. особей этого вида в год
добывают охотники Гренландии. Предполагается, что в западной части
Северной Атлантики смертность толстоклювых кайр превышает пополне-
ние популяции молодыми птицами (Tull et al., 1972; Bibby, 1973; Evans,
Waterston, 1978).
55
Семейство Чистиковые
Хозяйственное значение, охрана. В СССР до 1950-х годов промысел
яиц кайр производился на Новой Земле (Успенский, 1956; Белопольский,
1957). Позднее промысловый сбор яиц был прекращен. Случайные сборы
осуществляли только команды судов, проходивших мимо колоний.
В настоящее время сбор яиц расценивается как браконьерство. Использо-
вание яиц практиковалось и в других частях ареала толстоклювых кайр,
в частности в западной части Гренландии, где за год собирали до 750 тыс.
яиц (Salomonsen, 1944). Охота на взрослых птиц в море производится
для нужд местного населения на северо-востоке нашей страны. Масштабы
добычи невелики. Гнездовья кайр охраняются в Кандалакшском и Мага-
данском заповедниках и в ряде заказников.
Род Cepphus Pallas, 1769
Чистик —
Cepphus grylle Linnaeus, 1758
Black Guillemot (англ.), Gryllteiste (нем.).
Статус. Гнездящийся, оседлый, частично перелетный вид.
Общая характеристика и полевые признаки. Среднего размера, величи-
ной с голубя. Типично морская птица, с сушей связана только в период
гнездования. К движению по земле адаптирована плохо. При ходьбе
опирается на землю пальцами, но плюсну держит не вертикально,
а наклонно, под острым углом к земле. Походка кажется как бы приседаю-
щей, медленна и неуклюжа, тело находится в наклонном положении.
Иногда довольно быстро пробегает короткие расстояния, в таком случае
положение тела бывает вертикальным. При отдыхе присаживается на
землю всем корпусом и поджимает ноги. Полет быстрый, порхающий,
с частыми взмахами крыльев, как у всех чистиковых — маломаневренный,
хотя крылья относительно более широкие, чем у остальных чистиковых.
Прекрасно плавает и ныряет. Для взлета с воды нужен «разбег». С земли
взлетает с камней или обрыва. Окраска летом черная, зимой серебристая.
Клюв длинный, шиловидный. Лапы красные. Голос — тонкий резкий
свист.
Подвидовая систематика. Из пяти подвидов чистика три встречаются
на территории СССР. Подвиды различаются по размерам и интенсивности
развития белой окраски на первостепенных маховых и верхних кроющих
крыла (Степанян, 1975). *
1. Cepphus grylle grylle
Alca grylle Linnaeus, 1758, Syst. Nat., ed. 10, c. 130, Швеция.
Размеры мелкие. Длина крыла самцов 162—174 мм (среднее 168),
самок 157—170 мм (среднее 163). Первостепенные маховые от основания
до середины пера имеют белую окраску на внутренних опахалах. Большие
верхние кроющие крыла от основания до середины темно-бурые, вследствие
чего на белом зеркале различима темная полоса.
2. Cepphus grylle mandtii
Uriae mandtii Mandt., 1822, Observ. Hist. Nat. Itinere Groenlandico
Factae, c. 30, Шпицберген.
По размерам сходен с предыдущим подвидом. Внутренние опахала
первостепенных маховых белые от основания до вершины, которая имеет
56
Чистик
бурую окраску. Большие верхние кроющие крыла целиком белые или
буроватые у основания.
3. Cepphus grylle tajani
Cepphus grylle tajani Portenko, 1944, Докл. АН СССР, 43, c. 239, мыс
Пиллар, о-ов Врангеля.
Размеры крупные. Длина крыла самцов 168,5—180 мм (среднее 173,1),
самок 163—176 мм (среднее 171,2). По окраске сходен с предыдущим под-
видом.
Замечания по систематике. Подвидовое деление нуждается в дальней-
ших исследованиях. Необходимо более детальное изучение системати-
ческого положения как самых крупных чистиков, обитающих в восточной
части ареала (С. g. tajani), так и самых мелких, населяющих Белое море.
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла: . . Рулевых перьев
обычно 12, редко 13—14. Размеры самцов и самок: длина (в мм) крыла
130—180 (среднее 163,5); клюва 25—35 (среднее 31,2); плюсны 27—35
(среднее 32,9). Масса самцов и самок 198—500 г (среднее 407,3).
Клюв прямой, вытянутый, слегка сжатый с боков. Ноздри узкие,
щелевидные, прикрыты оперением. Цевка, сжатая с боков, сетчатая,
Контурное оперение плотное.
Окраска. Самец и самка в брачном наряде. Полового диморфизма
в окраске нет. Спинная сторона черная с более или менее заметным зеле-
новатым отливом. Брюшная сторона черновато-бурая. Маховые черные,
с белыми основаниями внутренних перьев. Рулевые черные. На крыле
белое зеркальце, образованное средними кроющими и вершинами больших
кроющих. Подмышечные и часть подкрыльев белые. Клюв черный, ноги
ярко-красные, когти черные. Радужина бурая. Ротовая полость ярко-
красная.
Самец и самка в зимнем наряде. Перья на спинной стороне тела черные,
с белыми каймами. Эти каемки особенно широки на голове, шее, плечевых
и пояснице. На уздечке и кроющих уха — черное пятно. Брюшная сторона
белая.
Пуховой наряд. Общий тон черный. Пух рыхлый, слабосцепленный,
на брюшной стороне заметно гуще. Клюв черный, розоватый у основания
надклювья. На клюве — острый белый яйцевой бугорок, внутренние части
рта и язык нежно-розовые.
Гнездовой наряд. Появляется к возрасту 30 дней. Спинная сторона
черная, с большей или меньшей примесью беловатых пестрин, особенно
на шее и плечевых. Щеки черноватые, с белыми пестринами. Маховые
и рулевые черные. Белое зеркальце на крыле испещрено черноватыми
пестринами. Брюшная сторона белая, с черноватыми каймами перьев.
Ноги лиловато-красные, клюв черный.
Линька. Последовательность смены нарядов: пуховой — гнездо-
вой — первый зимний — предбрачный — второй зимний — брачный. Пте-
нец вылупляется покрытым густым мягким пухом. На 6—7-й день появ-
ляются верхушки маховых и кроющих плеч, спины. На 8-й день показы-
ваются кроющие боков и брюха, на 10-й день — рулевые. Голова и шея
оперяются примерно на 15-й день. На 22—23-й день пух начинает исчезать
и полностью пропадает на 30—32-й день (Кафтановский, 1951). В пер-
вую же осень (между сентябрем и декабрем) птенцы линяют в зимнее
оперение, похожее на зимний наряд взрослых, но с меньшим развитием
белого цвета (Witherbi et al., 1941).
57
Семейство Чистиковые
У взрослых две линьки в году: полная послебрачная и неполная
предбрачная, охватывающая все оперение, кроме маховых и рулевых.
Весенняя пребрачная линька начинается в конце декабря—начале января
и заканчивается в мае. Со второй половины июля начинается послебрачная
линька — сначала на голове и шее, потом на спине. Эта линька продол-
жается до конца ноября. Смена маховых происходит одновременно (Де-
ментьев, 1951).
Распространение. Гнездовой ареал. В Атлантике распространен
в Северной Америке от побережья Мэна и Новой Шотландии до Южного
Лабрадора, в Исландии, на Фарерских островах, по берегам Англии и
Шотландии, на Гебридских, Оркнейских и Шетландских островах,
в Ирландии, Западной Швеции, Восточной Дании (по островам Каттегата),
по побережью Норвегии до Варангера (рис. 13). В высоких широтах
Северного Ледовитого океана на северо-востоке Америки ареал проходит
на юг до Северного Лабрадора и Гудзонова залива. Чистик встречается
на Среднем Лабрадоре, в Западной и Восточной Гренландии, о-ве Ян-Майен,
о-ве Медвежьем и Шпицбергене.
В СССР на Мурмане (Айновы острова, Гавриловские острова, Семь
островов, побережье материка), й Горле Белого моря (о-в Сосковец),
в Кандалакшском и Онежском заливах, на Балтике — в Ладожском озере,
на архипелаге Франца-Иосифа, о-вах Вайгач, Новая Земля, Северная
Земля, Новосибирских, Врангеля, Геральда, на северном побережье Чукот-
ского полуострова (рис. 14).
Зимовки. В негнездовое время чистики встречаются южнее, до ат-
лантического побережья Франции, Западной Германии, Бельгии и Голлан-
дии. Американские чистики на зимовке регулярно доходят до берегов
Массачусетса.
Миграции. Далекие миграции чистикам не свойственны. В южной
части ареала это более или менее оседлые птицы, в северной — кочующие
(Дементьев, 1951).
Беломорские чистики, как правило, кочуют вблизи мест гнездования
в Кандалакшском и Онежском заливах, вплоть до Терского берега. Зимуют
в полыньях Белого моря, большинство в дрейфующих льдах бассейна
Белого моря, в радиусе 300—400 км от мест гнездования (Бианки, 1967).
Так же ведут себя чистики на Балтике (Nordstrom, 1957—1963; Nilsson,
1982).
Чистики, гнездящиеся на островах Баренцева моря, частично кочуют
в прибрежье Мурмана, частично — к берегам Северной Норвегии. Самый
дальний возврат чистика, окольцованного на Семи островах, был получен
в феврале из Средней Норвегии с 65° с. ш. Кочующие чистики были
встречены в самых высоких широтах Северного Ледовитого океана, под
84- 86° с. ш. (Дементьев, 1951). На востоке СССР чистики встречаются
зимой на полыньях Чукотского моря (Портенко, 1973). В проливе Ланка-
стер и в западной части Баффинова залива во время весенних миграций
чистики предпочитают участки с умеренной и большой площадью ледового
покрытия и чаще размещаются вдоль кромки льда, чем вдоль свободного
ото льда побережья (McLaren, 1982). При этом средняя плотность распре-
деления составляет 1,07 особи/км2 по краю прибрежных льдов и
0,17 особи/км2 по границе паковых льдов (Renaud, Bradstreet, 1980).
Материалы кольцевания показывают, что дальность миграций чистиков
имеет большую индивидуальную и региональную изменчивость (Myrber-
get, 1973b).
58
Рис. 14. Гнездовой ареал чистика в СССР
а— граница гнездового ареала, б — область зимовок и миграций. Подвиды:
1 — C.g. grylle, 2 — C.g. mandtii, 3 — C.g. tajani
Рис. 13. Область распространения чистика
а — граница гнездового ареала, б — область зимовок и миграций
Семейство Чистиковые
Численность. На большей части гнездового ареала относительная
численность чистиков невелика, только в отдельных, видимо особо благо-
приятных,местах их колонии достигают крупных размеров. На Белом море
в Кандалакшском заливе гнездится около 300 пар чистиков, в Онежском
заливе — более 1200 пар (Бианки, 1967). На Мурмане Т. Д. Герасимовой
(1962) было учтено 2140 пар, в 1968—1970 гг. — около 2500 пар (В. Н. Кар-
пович, устное сообщение). На всем северном побережье СССР, по данным
С. М. Успенского (1969), обитает не менее 100 тыс. пар чистиков, а по рас-
четам А. Н. Головкина (1984) — около 60 тыс. пар. В Северо-Западной
Европе (без Норвегии) количество гнездящихся чистиков находится в пре-
делах от 60 до 160 тыс. пар (Evans, 1984а). На северо-востоке Северной
Америки (в проливе Ланкастер) — около 20 тыс. пар (Brown, Nettleship,
1984), на побережье США от штата Мэн до штата Виргиния — около 3 тыс.
пар (Buckley Р., Buckley F., 1984), в Гренландии — 23—60 тыс. пар
(Evans, 1984b).
Местообитания. В гнездовое время чистики неизменно связаны со ска-
лами на морских побережьях и морских островах, где гнездятся в расще-
линах, трещинах, среди валунов, иногда в торфе и плавнике. Зимой —
прибрежные и удаленные от берега свободные от сплошного льда участки
моря, разводья и полыньи.
Размножение. Некоторые исследователи считают, что размножаться
чистики начинают в возрасте двух лет (Кафтановский, 1951; Козлова,
1957). В. В. Бианки (1967) приводит случаи размножения годовалых птиц.
Основная масса приступает к гнездованию в 2—3-летнем возрасте.
По-видимому, начало гнездования растянуто. Там, где море зимой
покрывается льдом — на Белом море, на Земле Франца-Иосифа и северо-
востоке Новой Земли, — время подлета чистиков следует за появлением
полыней и разводий у мест гнездования (Антипин, 1953; Белопольский,
1957; Бианки, 1967). На Мурмане чистики подходят к островам в начале
или середине апреля, что определяется общим ходом весны. Поскольку
места гнездования еще долго бывают покрыты снегом и льдом, чистики
долгое время держатся на море вблизи берега или на камнях у воды. Этот
период времени назван предгнездовым и продолжительность его в разных
местах различная: на Белом море — около 30 дней (Бианки, 1967), на
Мурмане — 40—50 (Белопольский, 1957), на Новой Земле — 70 (Успен-
ский, 1956), на о-ве Врангеля — 80 дней (Портенко, 1973), на Земле
Франца-Иосифа — около 110 дней (Горбунов, 1932). Во время предгнездо-
вого периода на воде происходят брачные игры.
По степени укрытости гнезд чистик занимает промежуточное положе-
ние между тупиком, роющим норы в торфе, и гагаркой, устраивающей
гйездо в полуоткрытых нишах. Распределение гнезд по островам обуслов-
лено наличием удобных мест, убежищ — трещин в камнях, крупных валу-
нов и т. д. В Кандалакшском заливе таких мест чцстику не хватает, поэтому
он вынужден гнездиться под бревнами плавника, в торцовых дуплах
лежащих бревен. Охотно занимает искусственные укрытия, сделанные для
него из валунов и дерна. Гнездовая лунка чаще всего находится в отдален-
ной части укрытия. В Онежском заливе и на Мурмане чистики иногда
устраивают гнезда в торфе. На Белом море гнезда находятся обычно
близко от воды — в 5—10 м, на Мурмане — в 50—300 м, на Шпицбер-
гене — в 2—3 км (Бируля, 1910; Кафтановский, 1951). Известны случаи
гнездования чистика в постройках человека (Carnduff, 1981).
60
Чистик
Гнездятся чистики группами от 2 до 7 пар. Родители охраняют гнездо
в радиусе нескольких десятков сантиметров. В брачных играх принимают
участие все чистики группы. Гнездовой консерватизм развит очень сильно.
При уничтожении убежищ чистики ищут новые подходящие места вблизи
старых. После оттаивания гнездового участка птицы начинают заходить
в норы. Брачные игры еще больше оживляются, копуляция происходит
на воде. Первое яйцо откладывается через 6—10 дней после первого
посещения нор. Выстилки в гнезде нет, иногда лоток бывает покрыт
мелкими камешками или створками раковин мидий. Кладка чистиков
состоит из двух яиц, очень редко из одного. В кладке из трех яиц, по-види-
мому, смешаны яйца двух самок.
Начало размножения в разных частях ареала по срокам значительно
различается. На Белом море это последняя декада мая—первая пятидневка
июня (Бианки, 1967). В эти же сроки начинает размножаться чистик
на Балтике (Дементьев, 1951; Козлова, 1957). На Восточном Мурмане
начало кладки было отмечено с 13 мая по 10 июня, в среднем — 27 мая
(Белопольский, 1957), на Западном Мурмане — в среднем 31 мая
(13 мая — 15 июня), на Новой Земле — 18 июня—13 июля (Успенский,
1956), на о-ве Врангеля — начало июля (Портенко, 1973), на Земле
Франца-Иосифа — конец июня—конец июля (Горбунов, 1932). Растяну-
тость начала откладки яиц зависит от климатических условий; поздняя
весна отодвигает кладку на более поздние сроки.
Форма яиц яйцевидно-овальная. Окраска основного фона меняется
от чисто белой до слегка зеленоватой, желтоватой или. коричневатой.
По основному фону разбросаны мелкие коричневые пятна, сгущающиеся
к тупому концу, расположение их сильно варьирует, но меньше, чем у дру-
гих видов чистиковых. Толщина скорлупы в среднем 0,17 мм (Кафтанов-
ский, 1951). Размеры яиц (в мм) на Белом море: 52—64X35—41 (среднее
56,7X38,8), масса (в г) 43—54 (среднее 47,3) (Бианки, 1967); на Западном
Мурмане — 56—63,5X38—42 (среднее 59,7X40,1), масса 42—60 (сред-
нее 51,8); на Восточном Мурмане 57—65X38—44 (среднее 59,8X40,6),
масса 48—57 (среднее 52,2) (Кафтановский, 1951); на Новой Земле —
57—65X39—43 (среднее 61,2X41,0), масса 42,5—56 (среднее 53) (Успен-
ский, 1956).
В насиживании яиц принимают участие оба родителя. Насиживание
продолжается 26—31 день. При гибели кладки часть чистиков может
загнездиться повторно через 7—8 дней. Первое яйцо чистики обогревают
в ночные часы. Плотное насиживание начинается с появлением второго
яйца, которое откладывается через 2—3 дня после первого. В ночные часы
насиживание более плотное, чем в дневное. При беспокойстве насижи-
вающие чистики собираются на воде против своих гнезд, плавают, свистят.
Типичное движение, выражающее беспокойство, — полоскание в воде
клюва. В слабонасиженных брошенных кладках эмбрионы оставались
живыми до 10 дней, а незадолго до вылупления — до 2 дней (Bergman,
1971).
Птенцы в одном гнезде вылупляются с интервалом в 1—2 дня, сам
процесс вылупления продолжается 2—3 дня. Средние размеры птенцов
на Белом море: длина (в мм) клюва 10,5, плюсны 19,0 (Бианки, 1967);
на Западном Мурмане — длина клюва 12,3, плюсны 20. Средний вес
однодневных птенцов на Белом море, Семи островах и Новой Земле практи-
чески одинаков — от 34 до 34,5 г (Бианки, 1967; Кафтановский, 1951;
61
Семейство Чистиковые
Успенский, 1956), а на Западном Мурмане составляет 37 г. Первый раз
родители кормят птенца через 10—15 ч после вылупления. Вылупившиеся
птенцы очень спокойны и малоподвижны. Пух на теле покрыт беловатыми
чехликами, которые быстро разрушаются. На брюшной стороне пух гуще,
чем на спине. Пух очень мягкий, рыхлый. Он может предохранить птенца
от переохлаждения только в укрытом от ветра гнезде. Вначале родители
обогревают птенцов непрерывно. Терморегуляция устанавливается в воз-
расте 4 дней, потом постепенно стабилизируется (Кафтановский, 1951).
Птенцов старше недельного возраста родители не обогревают.
Кормят птенцов оба родителя. Частота кормления на Западном Мур-
мане в первые 10 дней — по 5—6 раз в сутки, в возрасте от 10 до 30 дней —
10—14, по истечении месяца частота кормления падает до 2—3 раз в сутки.
В среднем в период выкармливания птенцы получают корм 8 раз в сутки.
Такую же интенсивность кормления отмечали исследователи, работавшие
в районе Квебека (Канада) (Cairns, 1984) и в Финляндии (Bergman,
1971). Рост птенцов идет довольно интенсивно, кривая роста имеет обыч-
ный S-образный вид. Раньше других частей тела заканчивается рост
плюсны и среднего пальца. Клюв, крыло и хвост не успевают полностью
развиться до спуска птенца на воду. Вес птенца при выходе из гнезда
составляет от 83 до 106 % среднего веса взрослой птицы. Беломорские
молодые чистики весят 300—400 г (Бианки, 1967), на Семи островах —
360—460 г (Кафтановский, 1951; Белопольский, 1957), на Новой Земле —
около 400 г (Успенский, 1956).
В период выращивания птенцов чистики из близко расположенных
гнезд ежедневно в раннеутренние часы собираются на 3—4 ч в определен-
ных местах берега, на скалах у самой воды, стайками от 4 до 18 птиц.
Поведение птиц в таких группах спокойное, иногда они пересвистываются,
переходят с места на место. Так продолжается до начала выхода птенцов
из гнезд. Молодые покидают гнездо в возрасте 31—38 дней. При поздних
сроках гнездования время пребывания птенца в гнезде короче, чем при
ранних. На о-ве Гельголанд наблюдали спуск птенцов в воду в возрасте
24 дней (Steinbacher, 1982). Выходят птенцы из гнезд самостоятельно,
в сумерках, когда родителей близко нет. Младший птенец выходит на 2—
3 суток позже первого. Путь от гнезда до моря птенец совершает пешком,
в быстром темпе перебегая открытую скалистую полосу берега. Достигнув
литорали, иногда задерживается здесь, купаясь в лужах. Растянутость
выхода птенцов из разных гнезд велика (от 14 до 27 дней). В это время
в поведении взрослых чистиков наблюдаются интересные изменения (Тата-
рникова, 1977). Родители рано вышедших птенцов не покидают остров.
В утренние часы они продолжают проводить несколько часов с птицами
своей группы, подсаживаются к стайкам на берегу, потом подлетают
к гнездам, возбужденно свистят, ходят по камням и стараются заглянуть
в гнездо с оставшимися птенцами. Хозяева гнезда отгоняют их, демонстри-
руя угрожающие позы, но драк не бывает и посторонние чистики все-таки
прорываются в гнезда и осматривают их. Чем меньше остается гнезд
с птенцами, тем большее количество чистиков посещают их по утрам.
После осмотра гнезд часть чистиков улетает, потом поодиночке или груп-
пами подлетают другие. Меченье позволило установить, что это разные
птицы. Иногда посетители бывают из гнезд, расположенных на расстоянии
2 км (по береговой линии). Днем посторонних птиц у гнезд нет, не видно
их в море. Только родители прилетают с кормом для птенцов. «Инспекти-
. 62
Чистик
рование» продолжается до выхода последних птенцов, после чего чистики
отходят от острова и держатся одной стаей. В районе Айновых островов
в конце августа—начале сентября у берега материка возле Среднего
полуострова держатся крупные стаи чистиков — до нескольких сотен осо-
бей, молодые и взрослые птицы вместе. Ю. М. Кафтановский (1951)
отмечал, что видел молодых птиц отдельно, без взрослых.
Эффективность размножения чистиков, как правило, высокая —
65— 80 % от числа отложенных яиц (Бианки, 1967; Bergman, 1971; Ols-
son, 1974). Успех размножения в кладках из двух яиц выше, чем в кладках
с одним яйцом (Cairns, 1981). Гибель яиц увеличивается из-за фактора
беспокойства. В отдельные годы наблюдается большая гибель птиц при
сильных штормах, когда прибойная волна заливает гнезда. Г. Бергман
(Bergman, 1971) сообщает о случае гибели на Балтике в 1969 г. практи-
чески всех птенцов чистиков от голода, когда из-за резкого похолодания
моря во второй половине июля из района островов отошла рыба.
Суточная активность, поведение. В гнездовой период у чистиков
бывают хорошо заметны два пика активности — в утренние и вечерние
часы. В период инкубации в ночные часы птицы находятся на гнездах,
а с 6 до 10 ч активно кормятся или отдыхают на камнях вблизи гнезд.
При выкармливании птенцов в ночные часы чистики отдыхают на море
и на берегу, на удобных камнях. Наиболее активно кормят птенцов
с 5 до 10 ч утра. Затем кормовые прилеты становятся гораздо более ред-
кими, хотя дневное кормление не прекращается. В вечерние часы —
с 16 до 19—20 — активность кормовых прилетов вновь возрастает. После
вечернего кормления чистики группами отдыхают на скалах, а затем уле-
тают на море. Такая же суточная активность отмечается и у чистиков
в Канаде (Cairns, 1981), Финляндии (Bergman, 1971) и в Шотландии
(Slater Р., Slater Е., 1972).
Описаны характерные позы ухаживания в брачный период и при копу-
ляции, элементы оборонительного и агрессивного поведения, демонстра-
ций, интерференции между соседними парами (рис. 15). Некоторые формы
демонстраций могут быть вызваны различными причинами. В частности,
погружение клюва обычно предшествует заныриванию за добычей, а также
используется как демонстрация «смещенной активности» в некоторых
конфликтных ситуациях (Asbirk, 1979). При территориальных конфлик-
тах чистики, уже занявшие гнездовую территорию, как правило, изго-
няют чужаков без сопротивления. Настоящие драки происходят лишь
в тех случаях, когда обе конфликтующие птицы провели в потенциаль-
ном месте гнездования примерно одинаковое время. Если хозяин террито-
рии в сгорбленной позе издает свист низкого тона, это означает, что затем
последует атака на чужака (Nelson, 1984). К числу специфичных пове-
денческих реакций чистика можно отнести также достоверно установлен-
ное индивидуальное предпочтение к расположению рыбы в клюве родите-
лей. Птицы, приносящие корм птенцам, держат рыбу в клюве поперек,
располагая ее так, что голова оказывается с правой или с левой стороны
(Slater, 1974).
Питание. Корм добывают исключительно в море, ныряя в неглубоких
местах (5 — 20 м) в 1—4 км от гнездовой (Bergman, 1971). Под водой
находятся 30—35 с. (Бианки, 1967), не более 50 —60 с, проплывая за это
время до 75 м (Козлова, 1957).
Основным кормом служат придонные мелкие рыбы, молодь крупных
63
Рис. 15. Элементы брачного и агрессивного поведения чистика (по: Cramp, 1985, с изменениями)
64
Чистик
рыб, ракообразные и моллюски. По-видимому, предпочтение отдается
массовым кормам. Избирательность питания выражена слабо. На Белом
море около 50 % кормов чистика составляют бычки, 34 % — люмпенус,
в небольшом количестве встречаются треска и сельдь. Ракообразные
в пище молодых полностью отсутствуют (Бианки, 1967). На Баренцевом
море в питании преобладают другие виды рыб, причем в разные годы
на первое место по значению выходят различные виды. Так, на Айновых
островах в 1981 г. птенцы выкармливались почти одной только бельдюгой,
в 1983 г. в кормах 60 % составляла песчанка, бельдюга — только 30 %,
в 1984 г. в питании чистиков встречалась только песчанка. На Семи остро-
вах рыба составляет около 77 % в питании молодых чистиков, на долю
ракообразных приходится около 18 %, на Новой Земле рыба занимает
около 42 % и почти столько же — ракообразные. В апреле—мае в питании
взрослых птиц 2/3 кормов составляют морские ракообразные и другие
беспозвоночные (Белопольский, 1957). В питании дальневосточных чисти-
ков большое значение имеют не только мелкая рыба, но и креветки, крабы
и другие ракообразные, а также моллюски (Портенко, 1973). При наличии
прибрежного льда чистики добывают корм у нижней поверхности льда
и в придонном слое, а у ледовых полей в море — почти исключительно
у нижней кромки льда. При этом существенную роль в питании играют
гидробионты — криофиллы, в частности полярная тресочка (сайка)
Boreogadus saida и амфиподы Onisimus glacialis (Bradstreet, 1980).
В Шотландии при выкармливании птенцов заметное место (46,9 %)
в питании занимает маслюк Pholis gunnellus (Slater Р., Slater Е., 1972).
Враги, неблагоприятные факторы. Врагов у взрослых чистиков мало.
На Белом море на них могут охотиться сапсан, орлан-белохвост и ястреб-
тетеревятник. На насиживающих чистиков могут нападать лисицы и гор-
ностаи. Плохо скрытые гнезда могут разорять серые вороны и серебристые
чайки. Но в сумме гибель чистиков от этих хищников незначительна.
Больше всего их гибнет в рыбацких сетях. От общего числа находок
окольцованных чистиков птицы, попавшие в сети, составляют на Белом
море по 22 % сеголеток и годовалых особей (Бианки, 1967). Не меньше
птиц гибнет в сетях на Балтийском море и в датских водах. На Мурмане
в последние десятилетия врагами чистиков являются крупные чайки,
в основном большие морские, которые ловят как взрослых птиц, так и
выходящих из нор птенцов. На Баренцевом море врагами чистиков также
бывают кречет и белая сова, на Новой Земле и восточнее — песец (Успен-
ский, 1969).
Большую опасность для чистиков представляет загрязнение моря
нефтепродуктами (Magnar, Einar, 1977; Hallet, Miller, 1980).
Из наружных паразитов на птенцах чистиков нередко встречается
иксодовый клещ Ceratixodes putus, иногда в довольно большом коли-
честве.
Хозяйственное значение, охрана. Хозяйственное использование чистика
на большей части ареала незначительно. В небольшом количестве чистик
добывается в Исландии и в Гренландии. В СССР немного чистиков и их
яиц добывается на Чукотке. Шкурки этих птиц после обработки пригодны
для изготовления воротников, шляп и т. д. (Спангенберг, 1935).
Специальных мер охраны чистика не предусмотрено, но на Белом и
Баренцевом морях, в наиболее густонаселенных районах охота на них
запрещена.
5 Птицы СССР
65
Семейство Чистиковые
Тихоокеанский чистик —
Cepphus columba Pallas, 1811
Pigeon guillemot (англ.).
Син. (рус.): Cepphus columba columba — большой тихоокеанский чистик, камчатский чистик;
Cepphus columba kaiurka — малый тихоокеанский чнстик, командорский чистик, берингов
чистик; Cepphus columba snowi — курильский чистик.
Статус. Гнездящийся, кочующий, частично зимующий вид.
Общая характеристика и полевые признаки. Некрупный чистик. Опере-
ние черное с буроватым оттенком, зеленоватого отлива нет. Белое зеркало
на крыле рассечено клиновидным пятном, иногда его может не быть вовсе
или оно представлено 1—3 узкими белыми поперечными полосками. Ок-
раска зеркала индивидуально варьирует. Рисунок на правом и левом
крыльях может различаться (личное сообщ. В. А. Зубакина). Подмышеч-
ные и нижние кроющие крыла серо-дымчатые в отличие от чисто-белых
у атлантического чистика. Клюв черный, полость рта и лапы ярко-красные.
При плавании держится на воде довольно высоко, белые пятна на крыльях
хорошо заметны по бокам, хотя они выделяются меньше, чем в полете.
Рулевых перьев всегда 7 пар (у атлантического чистика их чаще 6). Летит
быстро над самой водой. Под водой плавает быстрее и легче, чем конюги
и кайры. Превосходно ныряет. При плавании часто обмакивает клюв
в воду, особенно если в клюве зажата добыча. Голос — набор пронзитель-
ных свистов и трелей (Бутурлин, 1934; Дементьев, 1951; Иванов и др., 1953;
Козлова, 1957; Gabrielson, Lincoln, 1959).
Подвидовая систематика. В настоящее время выделяют 3 подвида:
1. Cepphus columba columba
Cepphus columba Pallas, 1811, Zoographia Rosso Asiatica, 2, c. 348, Кам-
чатка.
Окраской сходен с С. с. kaiurka, отличается более крупными размерами
(длина крыла в среднем на 10 мм больше). От С. с. showi отличается белой
окраской вершинной половины или трети нижних кроющих крыла и нали-
чием одного белого опахала у нижних кроющих крыла.
2. Cepphus columba kaiurka
Cepphus columbus kaiurka Portenko 1937, Mitteil. Zool. mus Berlin, 22.
c. 228, Командорские острова.
Окраской сходен с С. С. columba, отличается более мелкими размерами.
3. Cepphus columba snowi
Cepphus columba snowi Steineger 1897. Auk. 14, c. 201, О-в Райкоке,
Центральные Курилы.
Отличается от других подвидов окраской кроющих перьев крыла.
Верхние кроющие крыла бурые с узкой вершинной белой полоской или
без нее, нижние кроющие целиком бурые. У молодых верхние кроющие
крыла, образующие зеркало, темные с белыми отметинами на краях в отли-
чие от С. с. columba и С. с. kaiurka, у которых предвершинная часть верх-
них кроющих крыльев белая с темными концами (Бутурлин, 1934; Степа-
нян, 1975). Для американской части ареала описано еще два подвида,
отличающихся общими размерами, длиной клюва и крыльев (С. с. eureka,
С. с. adianta) (Козлов, 1957).
Замечания по систематике. Самостоятельность подвида С. с. kaiurka,
населяющего Командорские острова, подвергалась сомнению Г. П. Де-
ментьевым (1951), мы присоединяемся к его мнению. Ограниченность
гнездового ареала, малая численность птиц, незначительность отмеченных
66
Тихоокеанский чистик
различий, наличие переходных экземпляров свидетельствуют в пользу
того, что мы имеем дело не с географической, а с популяционной измен-
чивостью. А. Г. Велижанин (1977а) обращает внимание на наличие пере-
ходных по окраске форм на Курильских островах, где совместно обитают
С. с. columba, С. с. snowi и Cepphus carbo. Он отмечает, что у очковых
чистиков размеры и форма белого пятна вокруг глаза варьируют от бесфор-
менного пятна длиной 2—2,5 см до очень узкого не более 2—3 мм кольца.
В то же время белые зеркальца на крыльях у курильских чистиков иногда
бывают очень широкими, приближающимися по размерам к таковым
у большого тихоокеанского чистика. Таким образом, подвидовая система-
тика тихоокеанского чистика заслуживает самого пристального внимания.
Описание. Размеры. Большой тихоокеанский чистик: длина (в мм)
крыла самцов 179—195 (среднее 185,6, п=32), самок 179—191 (среднее
184,3, n=23) (Drent, 1965; Портенко, 1972; Степанян, 1975; собств. измере-
ния*); клюва самцов 30,0—34,6 (среднее 32,4, п=10), самок 30,3—32,8
(среднее 31,2, п=30) (собств. измерения); длина плюсны 32,0—36,5
(Козлова, 1957). Малый тихоокеанский чистик: длина (в мм) крыла сам-
цов 169—182 (среднее 173,7, п=25), самок 168—184 (среднее 175,25,
n=16), (Drent, 1965; Портенко, 1972; Степанян, 1975; собств. измерения);
клюва самцов 29,2—32,0 (среднее 30,4, п=7), самок 24,9—32,0 (среднее
29,7, п=4) (собств. измерения); длина плюсны 29,0—34,5 (Козлова, 1957).
Курильский чистик: длина (в мм) крыла самцов 180—190 (среднее 183,8,
п=7), самок 184—192 (среднее 188,8, п=7); клюва самцов 31,7—36,8
(среднее 34,1, п=7), самок 30,0—37,2 (среднее 34,1, п=7) (собств.
измерения); длина плюсны 31,6—35,0 (Козлова, 1957). Масса (в г) самцов
курильского чистика 389— 446 (среднее 417, п=8), самок 415—481 (сред-
нее 453, п=5) (Карташев, 1979). Масса самцов без разделения на подвиды
360— 550 г (среднее 482, п=23), самок 422—580 г (среднее 509,6, п=5).
Окраска. Самец и самка в брачном наряде. Верхняя сторона тела
черно-стальной окраски. Бока головы и шеи, а также вся нижняя сторона
тела, крылья и хвост буровато-черные. У номинального и командорского
подвидов на белом зеркальце крыла резко очерченный буровато-черный
клин, у курильского подвида на темном крыле 1—3 узкие белые попереч-
ные полоски (или их может не быть), образованные вершинными белыми
пятнами или полосками средних и больших верхних кроющих крыльев.
Подмышечные бурые у курильского подвида и дымчато-буроватые у двух
северных подвидов. Нижние кроющие крыльев и маховые сплошь серовато-
бурые и курильского подвида; у двух северных подвидов нижние кроющие
имеют белые внутренние опахала, а внешние — дымчато-бурые; внутрен-
ние опахала и вершины первостепенных маховых у всех подвидов большей
частью белые, а внешние опахала и вершины маховых перьев буровато-
черные. Радужина бурая. Клюв черный, полость рта и ноги красные
(Бутурлин, 1934; Козлова, 1957).
Самец и самка в зимнем наряде. Верхняя сторона тела серовато-черная,
с белыми вершинными каймами перьев. Зашеек и надхвостье иногда
частично белые. Нижняя сторона тела белая. Бока белые с серовато-бурыми
продольными полосками. Белого на подкрыльях больше, но все же карпаль-
ный край дымчато-бурый, зеркальце — как в летнем наряде. Курильский
* Собственные измерения преведены по материалам Зоологического музея и Кафедры
биогеографии МГУ.
5* 67
Семейство Чистиковые
подвид в зимнем наряде сходен с северными подвидами, но резко отлича-
ется от последних окраской и рисунком верхних и нижних кроющих
крыльев и первостепенных маховых, которые окрашены так же, как и летом
(Бутурлин, 1934; Дементьев, 1951; Иванов и др., 1953; Козлова, 1957).
Пуховой наряд. Общая окраска угольно-черная. Пух однотонно-черный
со слабым дымчато-серым налетом, усиливающимся на брюшке, которое
выглядит темно-серым. Лапы черно-телесные: телесный цвет на перепон-
ках, черный с примесью телесного — на пальцах и цевке. Клюв черный
(с примесью телесного на подклювье), кончик белый. Глаз темный, зев
розовый (личное сообщ. В. А. Зубакина).
Гнездовой наряд. Перья гнездового наряда начинают пробиваться на
2—3-й день. Сначала появляются пеньки на плечевых птерилиях и перво-
и второстепенных маховых. На 5—6-й день пробиваются пеньки на брюш-
ной стороне тела, третьестепенные маховые, кроющие крыла, перья кры-
лышка, на 7—8-й — пеньки перьев спины, на 8—9-й — бедренные и руле-
вые, на 9—13-й — пеньки перьев головы. В возрасте 26—30 дней птенец
целиком оперен, пух остается на кончиках растущих перьев только по
бокам шеи, на бедрах и боках тела (Михтарьянц, 1974). Яйцевой зуб
теряется в 28—30 дней (Sealy, 1970). Общая окраска верха головы, спины
и плечевых перьев буровато-черная, на плечевых белые вершинные каймы,
зашеек и надхвостье белые с буровато-черными вершинными каймами.
Большие и средние верхние кроющие крыльев с бурыми основаниями и
вершинами и широкой предвершинной полосой, благодаря чему зеркальце
на крыле пестрое, сильно затемненное бурым. У молодых чистиков куриль-
ского подвида концы верхних кроющих не темные, а с белыми отметинами
на краях. Нижняя сторона тела белая, испещренная темными концами
перьев, образующими бурые поперечные полоски (Бутурлин, 1934; Ива-
нова и др., 1953; Козлова, 1957).
Первый зимний наряд. Сходен с нарядом взрослых птиц зимой. Верх
тела, включая плечевые, буровато-черный с белыми вершинными каймами
перьев. Нижняя сторона тела белая с буроватыми поперечными полосками,
но этих полос меньше, чем в юношеском наряде. Белое зеркальце затемнено
бурыми пестринами (Иванов и др., 1953; Козлова, 1957).
Первый брачный наряд. Сходен с таковым взрослых птиц, отличается
затемненными бурыми пестринами зеркальцем (Козлова, 1957).
Линька. Послебрачная линька взрослых чистиков наступает в конце
июля или в первой декаде августа, но проходит у разных особей не вполне
одновременно. Большинство птиц уже к ноябрю одеты в зимний наряд,
другие — к середине января еще не закончили линьку. Полная осенняя
линька взрослых птиц начинается со смены оперения нижней стороны
тела, затем выпадают маховые и одновременно появляются перья зимнего
наряда на спине, а позднее на надхвостье (Иванов и др., 1953; Козлова,
1957). По Л. А. Портенко (1972), смена перьев во время линьки происхо-
дит более или менее равномерно по всему телу. Молодые чистики закан-
чивают смену юношеского оперения на первый зимний наряд к середине
ноября (Иванов и др., 1953; Козлова, 1957). Предбрачная линька охваты-
вает все контурное оперение, но не маховые и рулевые, и тянется иногда
с февраля и даже с января до мая и даже июня (Дементьев, 1951). Первые
птицы в полном брачном наряде появляются в марте (Иванов и др., 1953).
Распространение. Гнездовой ареал (рис. 16, 17). Большой тихо-
океанский чистик: в пределах СССР — побережье Чукотского полуострова
68
Рис. 16. Область распространения тихоокеанского чистика
а — границы гнездового ареала, б — область зимовок и миграций.
Подвиды: 1 — С. с. columba, 2 — С. с. kaiurka, 3 — С. с. snowi
Семейство Чистиковые
Рис. 17. Гнездовой аре-
ал тихоокеанского чис-
тика
Обозначения те же, что
на рис. 16.
от Колючинской губы до залива Креста, о-в Малый Диомид, предполо-
жительно Анадырский лиман; на Корякском побережье — базары у мыса
Отвесного и мыса Барыкова, ограничивающих бухту Угольную, юго-за-
паднее Хатырки, бухты Северная и Глубокая; о-ва Верхотурова, Карагин-
ский, по восточному побережью Камчатки к югу от Кроноцкого полуост-
рова до южной оконечности, на западном побережье Камчатки к северу
от южной оконечности до р. Камбальной, предположительно — острова
Охотского моря у Аяна. Вне СССР — западное побережье Северной Аме-
рики от мыса Лисборн и залива Коцебу на севере Аляски к югу до северных
частей п-ова Калифорния, островов Санта-Барбара, Фараллон, Алеутские
острова, за исключением их западной половины, острова Берингова моря —
Большой Диомид, Кадьяк, Прибылова, Св. Лаврентия, Св. Матвея. Малый
тихоокеанский чистик: в пределах СССР — Командорские острова, вне
СССР — западная часть Алеутской гряды. В средних частях Алеутской
гряды предположительно интерградирует с предыдущим подвидом. Ку-
рильский чистик: только в пределах СССР — все острова Курильской
гряды. Наиболее обычен в центральной части от о-ва Оц^котан на севере
до о-ва Итуруп на юге. Менее обычен на о-ве Парамушир. На островах
Кунашир, Шикотан редок, на мелких островах Малой Курильской гряды
(в частности, Демина, Юрий) обычен. Наиболее типично окрашенные особи
встречаются в центральной части Большой Курильской гряды между
островами Шикотан и Черные Братья (Портенко, 1939; Козлова, 1957;
Карташов, 1961; 1979; Нечаев, 1969; Вяткин, 1975; Вяткин и др. 1975;
Степанян, 1975; Велижанин, 1977а, 1978; Кишинский, 1980).
Зимовки. Зимой кочует у берегов Камчатки, у Курильских островов
70
Тихоокеанский чистик
к югу до Японии (о-в Хоккайдо), вдоль берегов Северной Америки до
южных границ гнездового ареала (Козлова, 1957).
Миграции. Сезонные кочевки невелики, особенно в южных частях
ареала вида, где птицы круглый год остаются в районе гнездования. Весен-
нее передвижение на север происходит вслед за отступлением льдов
в начале—середине апреля. У мест будущего гнездования птицы появля-
ются в начале-середине мая. В Кроноцком заливе численность чистиков
возрастает с конца марта. На островах Диомида птицы появляются в конце
мая—начале июня. Осенние кочевки начинаются после спуска птенцов
на воду в августе и продолжаются до октября и даже до середины ноября.
Чистики с Командорских островов откочевывают в октябре (Дементьев,
1951; Гизенко, 1955; Портенко, 1972).
Численность. Немногочисленный вид, как правило не образующий
крупных скоплений. На Аляске известны 363 точки гнездования тихоокеан-
ского чистика. Суммарная учтенная численность здесь достигает 40 000
птиц. Девять мест имеют гнездовые популяции в 1—4 тыс. особей. В боль-
шинстве мест гнездится не более 100 птиц. Общая расчетная величина
Аляскинской популяции оценивается в 200 000 птиц (Sowls et. al., 1978).
На островах Фараллон в 1972 г. гнездилось около 2000 птиц (Ainley,
Lewis, 1974).
Среди колониальных морских птиц Дальнего Востока СССР тихоокеан-
ский чистик принадлежит к группе малочисленных видов. Его численность
достигает всего лишь несколько десятков тысяч. Отдельные колонии насчи-
тывают от 2—10 до нескольких десятков пар. Численность же популяций
отдельных островов, включающих несколько отдельных микроколоний
или колоний, может достигать 100—200 пар. На базаре мыса Халюсткин
в 1984 г. гнездилось около 70 пар тихоокеанских чистиков (личное сообщ.
В. А. Зубакина). На о-ве Верхотурова гнездилось 32 пары (Вяткин, 1975),
на о-ве Карагинский — отдельные пары и небольшие колонии (Вяткин
и др., 1975; Герасимов, 1979). На о-ве Сердце-Камень отмечены 2 колонии
в 15 и 30 пар (Головкин, Флинт, 1975), на о-ве Колючин учтено около
300 чистиков (Кондратьев, 1975), на о-ве Арий Камень — 16—20 пар, на
о-ве Топорков — 150—180 пар (Карташев, 1961, 1979). На всех Куриль-
ских островах, по подсчетам А. Г. Велижанина (1978), обитает около
10 000 тихоокеанских и очковых чистиков. При этом население лишь одного
о-ва Экарма составляет 500 тихоокеанских чистиков. Крупная колония
этого вида существует на о-ве Ловушка — около 300 птиц (Велижанин,
(1978). Что касается современных тенденций в изменении численности
тихоокеанского чистика, то, по мнению Фридмана (Fridman, 1934, цит. по
Бутурлину, 1934), нашедшего кости этой птицы в 16 археологических
раскопках, численность и распространение тихоокеанского чистика прак-
тически не йзменились за последние 2500 лет. На островах Фараллон, где
в 70-е годы нашего столетия гнездилось около 2 тыс. птиц, в начале века
численность была приблизительно такой же, но с началом нефтяного
загрязнения пошла на убыль и восстановилась лишь в 60-х годах (Ainley,
Lewis, 1974). Считается, что в настоящее время популяция достигла мак-
симальной величины, поскольку основная часть пригодных для устройства
гнезд мест оказалась занята.
Местообитания. Скалистые побережья и острова, валунные пляжи,
каменистые осыпи. Могут гнездиться не только на побережье, но и в глу-
бине островов. Часто собираются группами на небольших каменных усту-
71
Семейство Чистиковые
пах, в литоральной полосе моря, на воде недалеко от берега, кекурах и кам-
нях, выступающих из воды. Зимой иногда собираются на льдинах. Большая
часть птиц зимует в открытом море вдали от побережья (Козлова, 1957;
Vaurie, 1965; Sowls et al., 1978).
Размножение. Половозрелость достигается на третий год жизни. В ко-
лониях присутствует до 30 % молодых неполовозрелых птиц и холостую-
щих взрослых. Большинство пар постоянны (прослежены по 3—4 сезона),
но иногда могут распадаться даже при живых партнерах. Характерна
многолетняя привязанность каждой птицы к месту гнезда и отдыха
(Drent, 1965). Токовые игры и спаривание на Командорских островах
начинаются в начале июня (Карташев, 1979). В первой декаде апреля
чистиков можно видеть чаще на воде, чем на суше, но со второй и в третьей
декаде они все больше времени проводят в прибрежных камнях. Брачные
игры на воде, ухаживание и спаривание наблюдаются в течение всего июня,
а у некоторых пар — даже в июле. С середины июля птицы выбирают место
для откладки яиц и держатся на камнях, входящих в гнездовой участок
радиусом 8—10 м (Карташев, 1979).
Сведения по брачному поведению носят отрывочный характер. По лич-
ному сообщению В. А. Зубакина, наблюдавшего этих птиц на Чукотке
в 1984 г., чистики довольно часто и подолгу лежат на камнях перед входом
в свод щели. На воде плавают либо парами и одиночками, либо небольшими
непостоянными по составу стайками по 3—14 птиц. При этом наблюдаются
демонстрации такого рода: «свист клюв в клюв» — две птицы плавают,
почти соприкасаясь клювами друг с другом, и сначала одна, а затем другая
издают длинную свистовую трель, широко раскрыв клюв. В то время как
одна птица издает трель, другая слегка вибрирует клювом в горизонтальной
плоскости, изредка касаясь кончика клюва партнера. Потом другой парт-
нер свистит, а первый вибрирует клювом. Возможен дуэтный свист. Во
время выполнения этого ритуала обе птицы могут быть как неподвижными,
так и слегка вращаться вокруг общего центра. Подобная демонстрация
бывает не только на воде, но и на камнях, уступах скал. После «свистов
клюв в клюв» чистик нередко набрасывается на соседа-чужака, если тот
находится в пределах 1,5—2 м, гонится за ним по воде, а нередко и под
водой и прогоняет. Владельцы гнезд и территорий изгоняют всех приса-
живающихся неподалеку чистиков, издавая при этом свистовую трель,
взмахивая крыльями и двигаясь по направлению к вторженцу. Иногда при
стычках один из чистиков вцепляется в хвост другому, и оба летят вниз,
один на хвосте у другого.
Агрессивная демонстрация заключается в вытягивании шеи почти
вертикально вверх и легком вздергивании закрытого клюва. Иногда между
противниками разыгрывается своеобразная «дуэль свистов». Н. Н. Карта-
шев (1979) описывает позу угрозы иначе: вытянутая вперед шея держится
на одном уровне с туловищем, клюв направлен в сторону противника.
Дж. Габриельсон и Ф. Линкольн (Gabrielson, Linkoln, 1959) отмечают, что
на протяжении всего сезона размножения самцы постоянно вызывают друг
друга, при этом поднимают вверх хвост и раскрывают клюв, демонстрируя
широкий ярко-красный зев. Наибольшее число чистиков собирается в ко-
лонии на камнях или под скалами на воде в утренние часы, где устраивают
продолжительные хоровые свисты, после чего разлетаются за кормом.
Неразмножающиеся чистики ночуют в море (Drent, 1965).
Как правило, чистики не строят какого-либо подобия гнезда, яйца
72
Тихоокеанский чистик
откладывают прямо на камнях или нанесенной земле или торфе, иногда
затаскивая в гнездовую нишу или щель мелкую гальку, осколки камлей,
щепочки. Предпочитают большие ниши, выбитые морем (Gabrielson,
Lincoln, 1959) . Наличие скудной подстилки отмечали А. И. Гизенко (1955),
Э. А. Михтарьянц (1974), И. Н. Карташев (1979), В. А. Зубакин (личное
сообщ.).
Чистики гнездятся обычно невысоко над уровнем моря, начиная от
уровня заплеска волн при максимальном прибое (Fay, Cade, 1959). Гнез-
довая камера размещается в россыпях скальных обломков, под навесами
скал, в нишах, иногда в трещинах отвесных скал. Могут гнездиться в не-
занятых норах ипатки или даже выкапывать собственные норы (Sowls et
al., 1978). Описано 9 случаев использования построек человека для
гнездования. Яйца, как правило, откладывались между досками причалов,
и один раз кладка была обнаружена в штабеле досок (Campbell, 1977).
Нормальная полная кладка чистиков состоит из двух яиц, но кладка
одного яйца встречается также довольно часто. Из 20 гнезд, просмотрен-
ных Н. Н. Карташовым на о-ве Топорков 23.VI.1960 г., в 6 было по два
яйца, в 6 — по одному, 8 гнезд были пустыми (Карташев, 1979). По данным
Э. А. Михтарьянц (1974), в 23 из 33 обследованных на том же острове гнезд
находилось по два яйца, в 10 — по одному. По данным А. Г. Велижанина
(19776), в 5 из 13 гнезд было по два яйца и в 8 — по одному (Курильские
острова).
Период яйцекладки варьирует в зависимости от географической широты
и условий года и в пределах колонии может растягиваться на 18—42 дня
(Drent, 1965). Копуляция начинается за 23—28 дней и достигает макси-
мума за 12 дней до откладки яиц (Drent, 1965). В среднем массовая кладка
яиц происходит в пределах всего ареала в середине—конце июня (Де-
ментьев, 1951; Михтарьянц, 1974; Велижанин, 19776), самая ранняя
кладка найдена 7.VI.1963 г. на Арьем Камне (Карташев, 1979). По данным
В. А. Зубакина (личное сообщ.), 10—16 июля 1984 г. на мысе Халюсткина
было еще довольно много чистиков, не отложивших яйца, которые обра-
зовывали пары и защищали территории.
Форма яиц варьирует от заостренно-овальной до удлиненно-овальной.
Цвет фона изменяется от светло-зеленого до зеленовато-белого, голубо-
вато-белого (Gabrielson, Lincoln, 1959), беловатого или матово-голубоватого
и даже розоватого (Гизенко, 1955; Велижанин, 19776). Поверхность яйца
покрыта пятнышками, крапинами, пестринами различных оттенков — от
коричневого цвета до почти черного или сероватого. Некоторые яйца имеют
неотчетливые пятна. (Gabrielson, Lincoln, 1959; Велижанин, 19776). Раз-
меры яиц (в мм): 57—68,5X38,5—43,5 (среднее 60,5X41) (Дементьев,
1951), масса яиц 49,0—68,2 г (среднее 58,4±1,12) (Велижанин, 19776).
Второе яйцо откладывается через двое (Михтарьянц, 1972) или через трое
суток после первого (Drent, 1965). Интенсивное насиживание начинается
после откладки второго яйца (Drent, 1965) или спустя 2 суток (Михтарь-
янц, 1972).
Насиживают кладку оба партнера, согревая яйца большую часть (до
95 %) суток. Смена партнеров происходит довольно беспорядочно. Пере-
рывы в насиживании бывают от 10 мин до 10 ч. Температура обогреваемого
яйца 39,4°, насиживающей птицы — 40,6°. При отсутствии родителей
в течение 3—16 ч отмечено охлаждение яиц до 12—14° (Drent, 1965).
Насиживание длится 26—33 дня (Михтарьянц, 1972, 1974). Второй птенец
73
Семейство Чистиковые
вылупляется одновременно с первым, реже через 1—2 сут. Вылупление
происходит во второй половине июля (Дементьев, 1951), самое раннее
вылупление на Арьем Камне было зарегистрировано 13 июля, массовое —
21—24, самое позднее — 27 июля (Михтарьянц, 1972, 1974). Дневной обог-
рев птенцов осуществляется родителями только первые 1—2 дня после
вылупления. Едва оперившись, птенцы становятся очень подвижными, вы-
бегают из своих гнездовых укрытий, но спуск на воду происходит, лишь
когда у них отрастут крылья (Иванов и др., 1953), в возрасте 29—39 дней
(в среднем 35 дней). У однодневных птенцов температура тела равна
в среднем 37,9°, в возрасте 2—6 дней — 38,2—38,9°, после 20-го дня —
40,1-40,8° (Drent, 1965).
Суточная активность, поведение. Данных нет.
Питание. Родители выкармливают птенцов мелкой рыбой, которую ло-
вят на расстоянии 30—50 м от берега или в широких литоральных ван-
нах во время отлива (Михтарьянц, 1974). Спектр питания взрослых птиц
довольно широк: помимо мелкой рыбы, чистики добывают различных, в том
числе головоногих, моллюсков, мелких и крупных ракообразных, полихет,
которых в изобилии находят вблизи каменистых побережий, близ скал и
рифов, в глубоких морских тихих бухточках, где в больших количествах
произрастают морские водоросли (Гизенко, 1955; Gabrielson, Lincoln, 1959;
Drent, 1965; Портенко, 1972).
Враги, неблагоприятные факторы. Поскольку яйца чистиков бывают
хорошо укрыты в щелях, этот вид испытывает меньшее хищничество со
стороны чаек, чем, например, кайры. На Юго-Восточной Аляске их яйца
регулярно добывают вороны (Gabrielson, Lincoln, 1959). Хозяйственное
значение. Не имеет.
Очковый чистик —
Cepphus carbo Pallas, 1811
Spectacled Guillemot (англ.), Brillenteiste (нем.), Keima furi (яп.).
Статус. В СССР гнездящийся, мигрирующий и зимующий вид.
Общая характеристика и полевые признаки. Чистик средних размеров.
В брачном наряде серовато-черный с красными лапами и хорошо замет-
ным белым пятном вокруг глаза. Наличие пятна вокруг глаза и отсутствие
белого на крыле отличают его от других представителей рода Cepphus
(в частности, от С. columba). В зимнем наряде имеет темный верх и белый
низ. На воде держит голову высоко и приподнимает хвост. Во время охоты
опускает голову в воду «до глаз», высматривая добычу. Голос, который
чаще всего можно услышать в гнездовой период (в частности, во время
брачных демонстраций), — звонкая громкая трель, убыстряющееся к кон-
цу стаккато: «цсит-цси-цси-си-си-сисиси». Птицы издают ее в море и на
суше.
Подвидовая систематика. Монотипический вид.
Описание. Размеры. Длина (в мм) крыла самцов 186,0—193,0
(среднее 189,3), самок 184,0—192,0 (среднее 188,5) ; клюва самцов 34,4—
40,0 (среднее 38,5), самок 33,3—37,4) (среднее 35,7). Масса (в г) самцов
544—685 (среднее 598,5), самок 604—649 (среднее 633), самцов и самок
540— 655 (Козлова, 1957; Коллекционные материалы Биолого-почвенного
института ДВО АН СССР и ДГУ).
74
Очковый чистик
Окраска. Самец и самка в брачном наряде. Все оперение серовато-
черное, на границе с клювом небольшие участки белого. Снизу белое
постепенно переходит в серое горло. Вокруг глаза большое белое поле.
Передний край крыла (предплечье и кисть) светло-бурые. Нижние крою-
щие крыльев и подмышечные серовато-бурые. Радужина темно-бурая,
клюв и когти черные, ноги красные.
Самец и самка в зимнем наряде. Верхняя сторона тела серовато-черная,
нижняя — белая (проглядывает серая часть опахал, темные стержни).
Горло и шея спереди серые (интенсивность окраски варьирует). Участки
белого вокруг клюва заметно меньше, иногда практически отсутствуют.
Уменьшаются размеры белого поля вокруг глаза. Лапы не такие яркие,
как летом.
Пуховой наряд. Спинная сторона серовато-бурая (иногда серовато-чер-
ная) . На верхней стороне головы и у основания надклювья — темнее. Бока
головы, низ, бока тела и крылья светлее, более серые. Пух на брюшной
стороне густой и заметно более короткий, чем на спине. Язык и стенки
ротовой полости окрашены в оранжево-розовый цвет, постепенно перехо-
дящий к концу клюва в бледно-оранжево-фиолетовый. Клюв черно-бурый,
яйцевой зуб и маленькое пятнышко на подклювье перед вершиной белые.
Цевка и пальцы розовато-бурые, перепонки светлее. Когти черно-бурые,
светлеющие к основанию. Радужина темно-бурая (Назаров, Лабзюк, 1972).
Гнездовой наряд. Верхняя сторона тела, крылья, хвост буровато-чер-
ные; вокруг глаза небольшое белое кольцо, от него назад идет светлая
полоска. Нижняя сторона тела с многочисленными буровато-черными
поперечными полосами. К бокам и зобу интенсивность и густота темных
пестрин возрастает. «Подбородок» серый, горло и шея спереди черновато-
серые, бока головы темнее. Нижние кроющие крыльев серовато-бурые.
Лапы розовато-бурые. Клюв черный, на конце светлеющий, заметно более
низкий и короткий, чем у взрослой птицы.
Первый зимний наряд. Похож на зимний наряд взрослых, но на нижней
стороне тела на общем фоне выделяется некоторое число темно-бурых
поперечных пестрин. Крыло как в юношеском наряде.
Линька. Смена брачного наряда на зимнее оперение начинается у взрос-
лых птиц в августе, а возможно и раньше, и продолжается до декабря.
У птиц (3 экз.), добытых в конце августа и в сентябре, на нижней стороне
тела (зоб, бока, грудь, подхвостье) росли белые (зимние) перья. Большая
их часть, а иногда и все, еще скрыты под черным пером брачного наряда.
У части новых перьев опахала развернуты полностью, и они создают пят-
нистый рисунок нижней стороны тела (редкие белые пятна на черном
фоне). В первую очередь белые перья появляются на зобу и на наседных
пятнах. Кроме того, оперение зимнего наряда растет вокруг клюва, на ще-
ках, над глазами, на темени, зашейке, спине. В это время выпадают третье-
степенные и крайние первостепенные маховые. На месте последних отме-
чен рост нового пера. Махи и рули в сентябре истрепаны. Окраска лап
заметно бледнее, чем в брачном наряде.
В октябре линька идет, по-видимому, наиболее интенсивно, так как
птицы, добытые в середине ноября и середине декабря, имели уже зимний
облик. У них сменилось все контурное оперение, маховые и рулевые. Лапы
приобрели бурый оттенок. У одной особи линька в зимний наряд еще не
совсем закончилась: среди нового белого пера нижней стороны тела еще
остались редкие черные летние перья, у другой — уже началась линька
75
Семейство Чистиковые
в брачный наряд: под белым зимним пером зоба и груди растет новое
черное, свежие перья появились также на боках шеи и на темени. У тре-
тьей птицы зимнее оперение уже выросло, а брачное еще не начало расти.
Смена зимнего наряда на брачный происходит у взрослых птиц с декабря
до конца мая. В январе нижняя сторона тела становится пестрой. Рисунок
образован на 50 % перьями брачного наряда. Новое перо растет на зобу,
груди, боках, темени, зашейке, задней стороне шеи, в области лопаток и на
надхвостье. Горло и бока шеи еще не линяют и имеют зимнюю окраску.
В феврале сохраняются остатки белых зимних перьев на черной нижней
стороне тела. Активно растет перо брачного наряда, плечевые и все кон-
турные, в том числе на боках головы и горле, увеличивается белое поле
вокруг глаза и участки белого вокруг клюва. Однако лапы еще не приобре-
тают ярко-красного цвета, свойственного брачному наряду. В течение марта
линька в брачный наряд продолжается, а в апреле для некоторых популя-
ций*'по-видимому, заканчивается (Дементьев, 1951; Козлова, 1957). Од-
нако еще в мае попадаются особи с белыми перьями на нижней стороне
тела.
Линька молодых птиц из гнездового наряда в перый зимнии начи-
нается уже летом. У одной из просмотренных птиц на зобу были заметны
отдельные растущие перья зимнего наряда. У птиц, добытых в октябре,
всюду росли контурные перья (зоб, бока, грудь, брюхо, лоб, темя, затылок,
зашеек, спина, надхвостье, плечевые). Птицы уже имели «зимний» облик.
Оперение нижней части тела представляло собой смесь свежих белых
перьев и перьев с бурой каймой, оставшихся от гнездового наряда. Линька
молодых чистиков в зимний наряд заканчивается, по-видимому, к середине
зимы. В это время у них на груди появляются также отдельные перья
первого брачного наряда. По Е. В. Козловой (1957), у молодых птиц весен-
няя линька запаздывает и к концу мая бывает в полном разгаре.
Распространение. Ареал вида в целом. Северо-западная часть
Пацифики (от северо-восточной части Охотского моря до Корейского
полуострова и Японских островов). Гнездовой ареал. Основная
часть гнездового ареала находится в СССР (рис. 18). Охотское море:
северо-западное побережье Камчатки к северу от мыса Южный (Вяткин,
1986), Пенжинская губа (о-ва Добржанского, Второй, Крайний, Зубчатый)
(Дементьев, 1951; Яхонтов, 1975), п-ов Тайгонос (Дементьев, 1940),
Гижигинская губа (предположительно) (Велижанин, 1978), залив Шели-
хова (Кищинский, 1968), южное побережье полуострова Пьягина и Кони,
Ямские острова (Матыкиль, Баран, Хатемалью) (Велижанин, 1978),
Тауйская губа (Дементьев, 1951), район Аяна (Козлова, 1957), Шантар-
ский архипелаг (Дулькейт, Шульпин, 1937; Яхонтов, 1977; Росляков,
1986). В Сахалинском заливе и Амурском лимане отсутствует (данные
Ю. В. Шибаева). В Японском море — вдоль всего материкового побережья
к югу до пограничной реки Туманган (Шульпин, 1936; Белопольский,
1955, Лабзюк и др., 1971; Литвиненко, Шибаев, 1971; Лабзюк, 1975;
Елсуков, 1984; данные Шибаева Ю. В.), в частности на большинстве
островов залива Петра Великого (о-в Фуругельма, о-ва Римского-Корса-
кова, Стенина, Карамзина, Верховского, Циволько, Желтухина, Моисеева,
Сергеева, Клыкова, Наумова и др.). Берега Сахалина в основном непри-
годны для гнездования очкового чистика, поэтому там имеются лишь
отдельные поселения, приуроченные к скалистым мысам и редким остров-
кам: мыс Терпения, район мыса Анива, два мыса у южной оконечности
76
Очковый чистик
п-ова Крильон, о-в Монерон,
мыс Ламанон у г. Александров-
ска, две точки на п-ове Шмидта
(мыс Марии, мыс Горнера)
(Никольский, 1889; Гизенко,
1955; Нечаев, 1975а, 1986).
Курильские острова: по А. Г. Ве-
лижанину (1977а), имеет здесь
разорванный ареал: северная
часть — п-в Парамушир (юго-
восточное побережье) и о-в Дым,
южная часть — от о-ва Броуто-
на и о-вов Черные Братья до
конца Курильской гряды и да-
лее. Гнездование доказано для
о-ва Шикотан (Kobayashi, 1933;
Велижанин, 1978). В 1971 г. на
о-ве Шикотан очковый чистик
гнездился на ряде мелких остро-
вков и на скалах самого острова
в бухтах Крабовая, Безымян-
ная, Кабельная, на о-вах Девя-
тый вал и Средний. Кроме того,
птицы были отмечены в гнездо-
вое время у о-вов Юрий и Тан-
фильева (данные Ю. В. Шибае-
ва) . На Малой Курильской гря-
де очковый чистик, кроме того,
обитает на о-вах Лисьих, о-ве
Сторожевой (Велижанин, 1978)
и о-ве Анучин (Воронов, 1975;
Велижанин, 1978). Известны
залеты чистиков на Командоры
(Stejneger, 1885) и на северо-
востокКамчатки (Вяткин, 1975). Рис. 18.Областьраспространенияочковогочистнка
В целом гнездовое распростра-
нение очкового чистика в СССР
выяснено недостаточно хорошо,
особенно на Охотском побережье
и Курильских островах.
В Японии гнездится главным
образом на о-ве Хоккайдо: п-ов Сиретоко, побережье от п-ова Немуро
до Кусиро, о-ва Томосири, Юрури, Моюрури, Дайкоку (Фудзимаки, 1986),
мыс Эсан (Austin, Kuroda, 1953; Фудзимаки, 1986), о-в Кодзима, мыс Ся-
котан (Фудзимаки, 1986), о-в Теури (Austin, Kuroda, 1953; Фудзимаки,
1986). Предполагается гнездование на о-ве Ребун (Фудзимаки, 1986).
На о-ве Хонсю гнездится на п-ове Симокита (префектура Аомори) (Austin,
Kuroda, 1953). В 1978 г. гнездование на Хонсю было зарегистрировано
на мысе Сирия (префектура Аомори), о-ве Санкан (префектура Иватэ),
о-ве Футаго (префектура Мияги) и о-ве Тотиганэ в заливе Камаиси (пре-
фектура Иватэ) (Распространение гнездящихся птиц Японии, 1981).
1 — гнездовой ареал,
2 — область знмовок,
3 — залеты,
4 — отдельные колонии,
5 — сомнительное гнездование
77
Семейство Чистиковые
Корейский полуостров: о-в Кук (провинция Канвондо) (Austin, 1948;
Вон Хон-Гу, 1963), о-в Рэн (северо-восток провинции Хамгён Цукто)
(Austin, 1948).
Зимовки. В СССР — вдоль материкового побережья от района
Тернея и на юг до границы (Абрамов, и др., 1973; Елсуков, 1985; данные
Ю. В. Шибаева). Далее вдоль восточного и южного побережий Корейского
полуострова (Gore, Won, 1971). По А. Г. Велижанину (1972), обычный
зимующий вид Южных Курильских островов. Однако В. А. Нечаев (1969)
зимой его там не отметил. Не зарегистрирован он и на зимовке на охотском
побережье Хоккайдо (Tiede et al., 1973). Очковый чистик зимует в Японии
вдоль восточных берегов о-ва Хонсю. Как правило, птицы, не спускаются
южнее префектуры Фукусима. Залеты южнее — к о-вам Хатидзе, в залив
Суруга, к п-ову Сима. На западном побережье не добывался, но отмечался
на Цусиме (Austin, Kuroda, 1953).
Миграции.. Данных нет.
Численность. В СССР выяснена лишь для некоторых районов. Так,
в заливе Петра Великого в настоящее время гнездится вряд ли более
6000 пар очкового чистика (данные Ю. В. Шибаева), на Шантарских
островах — 19—20 тыс. пар (Росляков, 1986), на Ямских островах — более
4000 пар (Велижанин, 1978), на Сахалине В. А. Нечаев (1986) учел в обсле-
дованных колониях немногим более 200 пар (даже с учетом, вероятно,
пропущенного очковый чистик там, без сомнения, редок), немногочислен
на Курильских островах (Велижанин, 1978; данные Ю. В. Шибаева). На
материковом побережье были проведены два учета, которые дали следую-
щие результаты: Южное Приморье — на участке протяженность 50 км
летом 1973 г. было учтено 680 пар (Лабзюк, 1975); Северное Приморье —
на участке длиной 440 км летом 1977 г. учли около 2500 пар (Елсуков,
1984). В Японии в настоящее время численность вида, видимо, не превы-
шает 1000 пар (полного учета не было), но в прошлом она была выше
(Austin, Kuroda, 1953; Фудзимаки, 1986). В настоящее время можно лишь
весьма приблизительно оценить численность вида в целом, исходя, с одной
стороны, из реальных цифр учетов, имеющихся по ряду районов, с дру-
гой — из плотности гнездования на материковом побережье. По-видимому,
она находится между 50 000 и 400 000 пар.
Очковый чистик, как правило, нигде не создает значительных скопле-
ний. Он гнездится набольшими (от нескольких десятков до нескольких
сотен пар) колониями. Более крупные колонии редки и располагаются
только на островах. К настоящему времени известно лишь несколько
таких колоний: о-в Матыкиль (Ямские острова) — в 1974 г. 4000 пар
(Велижанин, 1978), о-ва Утичий и Птичий (Шантарские острова) — в
1982 г. 7000 и 3000 пар соответственно (Росляков, 1986), о-в Фуругельма
(залив Петра Великого), о-в Теури (Западный Хоккайдо) — в 1949 г.
по 3600 пар (по о-ву Фуругельма — данные Ю. Шибаева, по о-ву Теури —
Фудзимаки) (Austin, Kuroda, 1953), но в последние годы значительно
меньше (Фудзимаки, 1986).
Местообитания. Гнездится на скалистых побережьях островов и мате-
рика. Предпочитает каменистые осыпи, где при отсутствии хищников (что
бывает на островах) может достигать высокой плотности. Иа материковых
скалах обычно занимает щели в скалистых обрывах, в негнездовое время —
прибрежные участки моря.
Размножение. Колониальный вид. К размножеию приступают в воз-
78
Очковый чистик
расте 3—4 лет. Изредка гнезится отдельными парами. Характерная черта
биологии очкового чистика — гнездование только в естественных пустотах.
Поэтому избираемый ими гнездовой биотоп — скальные побережья с тре-
щинами и щелями и крупнообломочные осыпи с многочисленными пусто-
тами. Причем на островах, где нет наземных хищников, чистики предпо-
читают гнездиться в осыпях, а на побережье устраивают гнезда преиму-
щественно в трещинах труднодоступных скалистых обрывов. Высота
расположения гнезд зависит от наличия подходящих мест для их устрой-
ства. В заливе Петра Великого они гнездятся как у подножия скал неда-
леко от полосы прибоя, так и в верхней части скал или осыпей примерно
до высоты 100 м, однако большая часть гнезд располагается в 1,5—20 м
от полосы прибоя. Изредка устраивают гнезда в трещинах изолированных
скальных образований на вершинах островов среди зарослей кустарников
и мелколесья. В заливе Петра Великого гнездовые камеры располагались
на расстоянии 30—150 см от входа, в среднем — 60,7 см. Пол «норы» ка-
менный, реже земляной. Лоток обычно выстилается мелкими плоскими
камешками, иногда — сухой травой (на о-ве Фуругельма — рассученными
стеблями крестовника — Senecio — и злаками), в некоторых гнездах яйца
откладываются прямо на каменный пол (Назаров, Лабзюк, 1972; наблюде-
ния Ю. В. Шибаева).
В Южном Приморье откладка яиц начинается в конце апреля—начале
мая и продолжается до начала июня, массовая — во второй декаде мая
(Назаров, Лабзюк, 1972; данные Ю. В. Шибаева). По-видимому, примерно
в эти же сроки приступают к откладке яиц очковые чистики в Японии
(Fujimaki, 1961; Kuroda, 1963b). В некоторые годы начало гнездования
запаздывает: в районе бухты Ольги в 1973 г. первые кладки были обнару-
жены 2 июня (Лабзюк, 1975). На севере Татарского пролива сроки гнездо-
вания более поздние — по данным Л. И. Шренка (Schrenk, 1860), во всех
гнездах, осмотренных им 23 июня, было по одному слабо насиженному
яйцу.
В кладке обычно два яйца, а не одно, как это показано в некоторых
работах (Дементьев, 1951; Козлова, 1957; Гизенко, 1955). Из 60 гнезд,
просмотренных на островах в заливе Петра Великого в мае-июне, 60 %
содержали по два яйца (Назаров, Лабзюк, 1972). По два яйца откладывают
птицы в Японии (Austin, Kuroda, 1953).
Окраска яиц беловатая, иногда с кремовым или голубоватым оттенком.
На основном фоне разбросаны черные и темно-бурые поверхностные и
более глубокие серые пятна. У большинства яиц пятна сгущаются на тупом
конце, вокруг него. Форма яиц — правильно-яйцевидная или эллипсоидно-
удлиненная. Скорлупа матовая, мелко-зернистая. Размеры яиц (в мм):
из залива Петра Великого (п=24) 61,2—68,0X41,1—45,5, среднее 64,2Х
X43,2 (данные Ю. В. Шибаева). Масса (в г) трех свежих яиц 62,5; 66,4;
66,3 (Нечаев, Юдаков, 1968).
В насиживании участвуют оба родителя. Птицы имеют два наседных
пятна. Сроки насиживания не известны. В заливе Петра Великого вылуп-
ление начинается в конце мая—начале июня. В 1976 г. оно продолжалось
с 30 мая по 1 июля, в 1983 г. — с 8 июня до начала июля, массовое вылупле-
ние наблюдалось 22—23 июня. Примерно в те же сроки (конец первой—
начало второй декады июня) появляются первые птенцы на Сахалине
(Гизенко, 1955). В Японии (о-в Теури) в 1949 г. вылупление проходило
позднее — первые птенцы были зарегистрированы лишь 26 июня (Austin,
79
Семейство Чистиковые
Kuroda, 1953). На Шантарских острова^ начало вылупления отмечено
в конце июля (Яхонтов, 1977).
Масса однодневных птенцов из залива Петра Великого — 42 г; 47;
49,7 и 44,7 (данные Ю. В. Шибаева), 37,7 г (Назаров, Лабзюк, 1972).
Однодневные пуховики уже пытаются обороняться клювом. В возрасте
2—3 дней они уже довольно подвижны, забираются глубоко под камни
и становятся труднодоступными. Большая часть молодых в Южном При-
морье спускается на воду в середине июля. Птенцы покидают гнезда, имея
еще не доросшие крылья (Шульпин, 1936; Назаров, Лабзюк, 1972). Вылет
происходит, видимо, в основном в темное время суток. На о-ве Фуругельм
все птенцы, покинувшие гнезда, сразу же исчезают из пределов колонии.
У берегов острова остается лишь небольшая часть, вероятно ослабевших
слетков (ежегодно не более 2—3 десятков), которые самостоятельно кор-
мятся на мелководье, но на протяжении двух недель гибнут (все погибшие
молодые птицы были сильно истощены). К середине августа чистики
откочевывают из залива Петра Великого.
Суточная активность, поведение. Активность дневная. В период вы-
кармливания птенцов чистики наиболее заметны в колонии в утренние
часы, когда, вылетев с рассветом из гнезд, они до 9—10 ч сидят здесь же на
камнях или плавают у берега. После этого почти все птицы улетают на
кормежку и в течение дня на территории колонии или вблизи нее можно
видеть лишь прилетающих с кормом птиц или негнездящихся особей,
которые остаются здесь обычно до середины дня.
Демонстрации, наблюдаемые при конфликтных ситуациях, различны,
но для них чаще всего характерны такие признаки: птица пронзительно
кричит, широко открыв клюв, обращенный к сопернику, и поводя головой
из стороны в сторону. В зависимости от степени агрессивности, крылья
могут быть подняты вверх или слегка отставлены. Птица может двигаться
в направлении соперника или оставаться на месте (сидеть, стоять, лежать).
В некоторых ситуациях глаза прикрыты. Прогоняя соперника, чистик
иногда преследует его в воздухе или под водой. Драки наблюдаются редко.
Во время брачных церемоний самец и самка, сидя друг против друга,
покачивают из стороны в сторону головами и непрерывно покрикивают.
Птицы могут сидеть рядом, но тогда одна из них поворачивает голову
почти на 180°, чтобы клюв был обращен к партнеру. Временами обе птицы
принимаются перебирать перья на голове, шее и спине партнёра, а иногда
кладут шеи друг другу на спину и некоторое время остаются в такой позе.
Копуляция происходит на суше. Перед этим самец начинает ходить вокруг
самки, описывая небольшие круги, пока она не включается в демонстра-
цию. Похожее «кружение» отдельных пар наблюдается и на воде, но там,
возможно, оно имеет другое значение. Часто можно видеть групповые
демонстрации очковых чистиков на воде (реже — на суше), когда 20—30
птиц, собравшись в одну стайку и громко покрикивая, преследуют друг
друга в воздухе и под водой, причем к образовавшейся группе, как правило,
присоединяются другие птицы. Иногда такие стайки направляются к бе-
регу, выбираются на камни и некоторе время остаются на суше (наблюде-
ния Ю. В. Шибаева).
Питание. Кормятся чистики в 2—6 км от своих гнездовий, обычно
недалеко от берега на глубинах, не превышающих 15—20 м. В местах
кормежки держатся рассредоточенно, не ближе 50 м один от другого.
Насытившись, могут отдыхать здесь же, объединяясь в группы до 10 птиц.
80
Очковый чистик
Птенцов выкармливают в основном рыбой, а также десятиногими раками
и полихетами. В заливе Петра Великого в первые дни родители приносили
птенцам полихет (нередко длиной до 15 см), а в дальнейшем — мелких
бычков, полурылов Hemirhamphidae, маслюков Pholididae и других рыб,
реже креветок (Назаров, Лабзюк, 1972). На о-ве Манерой А. И. Гизенко
(1955) в качестве объектов питания птенцов обнаружил зубатую корюшку
Osmerus eperlanus, иваси Sardoniks sadax и тихоокеанскую сельдь Clupea
harengus pallasi. На о-ве Теури в 1949 г. чистики выкармливали птенцов
песчанкой Ammodytes hexapterus и анчоусом Engraulis japonicus (длина
рыб примерно 12 см). Желудок одной из птиц, добытых зимой у берегов
Северного Хонсю, содержал рыбу Platycephalus indicus, другой — остатки
осьминога, третьей — мелких крабов (Austin, Kuroda, 1953).
Враги, неблагоприятные факторы. В колониях Южного Приморья
основным врагом очкового чистика является большеклювая ворона, поедаю-
щая яйца и птенцов. Практически все птенцы рано или поздно становятся
ее жертвами (данные Ю. В. Шибаева). Взрослые птицы часто служат
добычей филина: в заливе Петра Великого (о-в Матвеева) очковый чистик
в добыче филина составлял 65 % (Назаров, Лабзюк, 1972), на побережье
Лазовского заповедника — 29 % (Коломийцев, Поддубная, 1985). Охо-
тится за ним сапсан (Гизенко, 1955; Назаров, Трухин, 1985), а на крупных
островах и на материковом побережье также четвероногие хищники —
лисица (Гизенко, 1955), колонок, харза (Лабзюк, 1975). В заливе Петра
Великого по отношению к чистику пиратствует чернохвостая чайка (Лит-
виненко, 1980). Обычно чайки поджидают чистиков, сидя на скалах, а за-
видя летящую птицу, начинают преследовать ее. Чистик, как правило, по-
ворачивает обратно к морю и, долетев до воды, тут же ныряет. Так может
повториться 2—3 раза, пока чистику не удастся проскочить в гнездо или
пока он не выронит добычу. Преследуют чистика 1—2 чайки, но когда он
роняет рыбку, тотчас присоединяются другие чайки.
Большой урон численности наносят неблагоприятные погодные усло-
вия. В 1983 г. после нескольких тайфунов, прошедших в июне с непродол-
жительными перерывами, частых штормов и затяжных дождей на о-ве
Фуругельма к 10 июля полностью погибла небольшая колония очкового
чистика (30 гнезд), в других колониях острова птенцы выжили лишь
в 10—15 % нор. Гибель яиц и птенцов наблюдалась в течение всего периода
непогоды с начала мая. Птенцы погибали как непосредственно от дождей
(норы были затоплены), так и от голода. Постоянное сильное волнение,
опреснение воды в прибрежной зоне, а также нарушение структуры дна
повлияло на распределение и численность кормовых объектов чистика
(вблизи острова наблюдалась массовая гибель моллюсков, креветок, мор-
ских ежей и звезд) (данные Ю. В. Шибаева).
Хозяйственное значение, охрана. В Приморском крае очковый чистик
под названием «краснолапка» служит объектом ружейной охоты местного
населения, хотя существенного значения в добыче не имеет. В последние
годы результаты антропогенного влияния на этот вид стали заметнее,
в частности, в заливе Петра Великого. Фактор беспокойства возрос много-
кратно ввиду широкого распространения маломерного флота и постоянного
присутствия отдыхающих на островах в колониях и вблизи них. Поэтому
важное значение в охране этого и других видов морских птиц в этом районе
имела организация в 1979 г. Дальневосточного морского заповедника.
Степень отрицательного влияния человека сейчас особенно хорошо за-
6 Птицы СССР
81
Семейство Чистиковые
метна — колонии очкового чистика процветают на охраняемых островах
и деградируют на всех остальных. Очковый чистик охраняется также
в нескольких заповедниках, выходящих к морю — Сихотэ-Алинском, Ла-
зовском (Приморский край), на Ямских островах в Магаданском заповед-
нике, а также в республиканском заказнике «Малые Курилы» (Малая
Курильская гряда). Существенным недостатком указанных природных
объектов является отсутствие охраны прилегающих акваторий, о чем писа-
лось в литературе (Жирмунский, 1981).
В Японии колония морских птиц на о-ве Теури, а среди них и очковый
чистик, охраняется как памятник природы (Austin, Kuroda, 1953).
Род Brachyramphus Brandt, 1837
Длинноклювый пыжик —
Brachyramphus marmoratus Gmelin, 1789
Marbled Murrelet (англ.). Marmelalk (нем.).
Син. (лат.): Colymbus marmoratus Gmelin.
Син. (рус.): пестрый пыжик.
Статус. В СССР гнездящийся и мигрирующий вид.
Общая характеристика и полевые признаки. Мелкий темно-окрашенный
пестрый (в брачном пере) чистик с относительно длинным и тонким
клювом. У плавающей птицы выделяется светлое горло. Зимой окраска
резко двухцветна — темный верх и белый низ. От старика в это время
отличается отсутствием заметной черной «шапочки» с белыми плечевыми
(две заметные полосы вдоль спины), от близкого короткоклювого пы-
жика — иными пропорциями головы (из-за разницы в длине клюва) и
одноцветными темными рулевыми (в природе их разглядеть трудно).
Голос громкий и оживленный, на воде и на лету они часто и отрывочно
повторяют односложное «миир, миир» (Козлова, 1957). А. П. Кузякин
(1963) и В. А. Нечаев (личное сообщ.) передавали его как резкий пронзи-
тельный свист.
Подвидовая систематика. Известны два подвида.
1. Brachyramphus marmoratus perdix
Cepphus perdix Pallas, 1811, Zoographia Rosso-Asiat, 2, c. c. 351, табл. 80.
Тауйский залив, Охотское море.
Более крупный (вес, длина крыла и клюва). Обитает на Азиатском
побережье Тихого океана. В СССР гнездится.
2. Brachyramphus marmoratus marmoratus
Colymbus marmorztus Gmelin, 1789, Syst. Nat., 1, c. 583. Бухта Принца
Вильяма, Аляска.
Размеры меньше. Обитает на американском побережье. В СССР изред-
ка встречается на кочевках у северных берегов Чукотского полуострова
и у островов Диомида в Беринговом проливе.
Описание. Размеры. На азиатском побережье длина (в мм) крыла
самцов 136,0—147,0 (среднее 141,2), самок 130,0—145,0 (среднее 138,3);
клюва самцов 18,9—22,2 (среднее 20,2), самок 18,0—21,0 (среднее 19,о,;
цевки самцов 17,0—18,7 (среднее 18,1), самок 16,8—19,0 (среднее 18,0).
Масса (в г) самцов и самок 258—357 (среднее 295,8) (коллекции Биолого-
почвенного института ДВНЦ АН СССР и Дальневосточного государствен-
ного университета; Кищинский, 1968).
82
Длинноклювый пыжик
В. m. marmoratus. Длина (в мм) крыла самцов 134,2±1,2 (128—140),
самок 132,6±1,8 (122—139); клюва самцов 15,5±0,3 (13,2—17,4), самок
15,3±0,4 (13,7—17,6); цевки самцов 16,2±0,2 (15,1—17,6), самок 15,9±
±0,3 (13,9—17,3). Масса (в г) самцов в период размножения 217,0
(196,2-252,5), самок 222,7 (188,1-269,1) (Sealy, 1975а).
Окраска. Самец и самка в брачном наряде. Вся верхняя сторона
тела буровато-черная с рыжеватыми или песочными каймами перьев.
В зависимости от интенсивности окраски и степени развития кайм верх
птицы может выглядеть более рыжеватым или более темно-бурым. На
темных участках пера чуть заметный металлический (фиолетовый) отлив.
Пестрые плечевые (сочетание темно-бурого с белым и рыжеватым) выде-
ляются светлой полосой (иногда выражена слабо) на общем темном фоне
спины. Эта полоса продолжается на боках надхвостья. Низ тела пестрый,
более светлый. Рисунок образован белыми контурными перьями с широ-
кими черно-бурыми и серо-бурыми каймами. Бока головы, шея спереди
и с боков, а также задняя часть брюха и подхвостье (не всегда) выглядят
как белые участки с темными пестринами. Остальной низ воспринимается
как темный с белыми пестринами. Наиболее интенсивно окрашен зоб.
Бока тела почти сплошь темно-бурые. Верхние кроющие крыльев темно-
бурые, иногда с узкими беловатыми каемками перьев. Подмышечные
буровато-серые. Нижние кроющие крыла — от сплошь буровато-серых до
почти белых. Рулевые буровато-черные. Иногда крайне рулевые с узкими
каймами или пестринами (белое с рыжеватым). Радужина у взрослых
птиц (у птенцов и молодых) темно-коричневая (Sealy, 1975а). Клюв
темно-роговой окраски, лапы красновато-розовые. Перепонка между паль-
цами и плюсна сзади черные.
Самец и самка в зимнем наряде. Верх темно-бурый с сизыми каймами
перьев, плечевые большей частью белые. Вокруг глаза бывает белое
кольцо. Бока головы, ожерелье на шее, доходящее почти до зашейка,
белые. Нижняя сторона тела белая, на боках примесь бурых перьев.
Пуховой наряд. Птенец (В. m. marmoratus) покрыт толстым слоем
желтоватого пуха с темными пестринами, сгущающимися на голове.
На животе густой светло-серый пух (Simons, 1980). Наряд В. не perdix
не описан.
Гнездовой наряд. Сверху однообразно темно-бурые, плечевые белые
с бурым (иногда присутствует рыжеватый цвет). Нижняя сторона тела
и бока головы белые с темно- или серо-бурыми поперечными пестринами.
Нижняя сторона и бока головы, задняя часть брюха и подхвостье заметно
светлее остальных участков низа. Нижние кроющие крыльев буровато-
серые с белым. Рулевые и маховые буровато-черные (внутренние опахала
светлее). На крайних рулевых могут быть белые черточки.
Первый зимний наряд. Верх в целом голубовато-серый (голова не-
сколько темнее). Беловатые узкие каймы не перьях образуют чешуйчатый
рисунок. В задней части спины каймы могут быть шире. Низ, в том
числе бока головы и шеи, белый, обычно с поперечными пестринами
на груди и в задней части брюха. Иногда пестрин очень мало. У части
экземпляров очень мелкие сероватые пестрины на горле. Остальное — как
у взрослых.
Линька. Сведения отрывочны. По Е. В. Козловой (1957), «полная
осенняя линька у некоторых взрослых пыжиков начинается уже в первой
декаде августа. У самки от 7 августа из северных частей Татарского
6*
83
Семейство Чистиковые
пролива (бухта Таба) интенсивно растут свежие перья на нижней стороне
тела. Маховые и рулевые еще не сменяются. У другой самки от 31
августа из Охотского моря выпали все маховые и рулевые, а мелкое
перо на нижней стороне тела большей частью еще в пеньках. Другие
взрослые особи несколько запаздывают с линькой и к последней декаде
августа (от 18 августа из пролива Литке и от 24 августа из Аяна)
находятся все еще в полном летнем наряде без признаков начала линьки».
У молодых птиц смена гнездового наряда на первый зимний проис-
ходит в течение сентября и октября. Птицы из южных районов Дальнего
Востока, добытые в августе (в том числе и в конце месяца), находились
в гнездовом наряде и еще не имели признаков линьки. Птицы из Южного
Приморья, добытые в последних числах октября и в ноябре, полностью
сменили гнездовой наряд на зимний, исключая оперение крыльев и
рулевые.
Распространение. Ареал вида в целом. Северная Пацифика.
Побережье материков, острова и прибрежные воды между 28 и 63° с. ш. на
азиатском континенте и между 32 и 60° с. ш. на американском (рис. 19).
Гнездовой ареал. На азиатском побережье описан преимущест-
венно на основании летних встреч птиц (рис. 20). Восточное побережье
Камчатки: пролив Литке, Авачинский залив (птицы в гнездовом наряде)
(Дементьев, 1951). Материковое побережье Охотского моря: устье р. Пен-
жины (Дементьев, 1951), залив Бабушкина и район Магадана (Кащин-
ский, 1968); у Охотска (гнездо) (Кузякин, 1963); в 200 км к юго-западу
от Охотска (добыта самка с яйцом) (Яхонтов, 1979). Сахалин: залив
Чайво (гнездо), визуально и по голосам отмечался в летнее время в ряде
мест на юге Сахалина; в районе залива Терпения; в бассейнах рек Тымь,
Поронай; на северо-восточном и северо-западном побережьях и на п-ове
Шмидта (В. А. Нечаев, личное сообщ.) Курильские острова:
Уруп, Итуруп (Велижанин, 19776), Кунашир (В. А. Нечаевым на берегу
Южно-Курильского пролива найдена мертвая птица в гнездовом наряде).
Материковое побережье Японского моря: Амурский лиман (отмечены
особи с кормом, улетавшие в глубь материкового побережья): бухта
Быки, севернее устья р. Тумнин (птица в гнездовом наряде добыта
в августе) (данные Ю. В. Шибаева), бухты Максимовка и Таежная на
севере Приморского края (Елсуков, 1984); бухта Ольги (гнездо)
(В. И. Лабзюк, личное сообщ.). Япония: район г. Мокото на охотоморском
побережье о-ва Хоккайдо (добыта насиживающая птица, гнездо не обнару-
жено) (Check-list of Japanese birds, 1974). Юго-Западное побережье
о-ва Хоккайдо — от мыса Зримо до Аккеси (Austin, Kuroda, 1953; Kuroda,
1955).
Зарегистрированы два залета внутрь материка молодой птицы на
озеро Ханка (добыта 3. Б. Сметаниной 26.Х 1967 г.) и взрослого пыжика
в район среднего течения р.Бикин (добыт Ю. Б. Шибневым 13. VII1980 г.).
, На побережье Северной Америки гнездится от восточных островов Алеут-
ской гряды и далее к югу вдоль материка до Северной Калифорнии
(Sealy, 1975а; Sowls et al., 1978). Известны залеты за пределы гнез-
дового ареала на о-ва Прибылова и о-в Св. Лаврентия (Sealy, 1975а).
Зимовки. Оба подвида зимуют за пределами СССР. В Японии —
обычный зимующий вид в водах о-ва Хоккайдо (исключая охотоморское
побережье), Северного Хонсю. Южнее редок (Кюсю, Центральные Рюкю,
о-в Амами) (Austin, Kuroda, 1953; Thiede et al., 1973; Check-list of
84
Длинноклювый пыжик
Рис. 19. Область рас-
пространения длинно-
клювого пыжика
а — районы гнездования,
б — области зимовок и
миграций,
в — места возможного
гнездования
Рис. 20. Места установ-
ленного (/) и возмож-
ного (2) гнездования
длинноклювого пыжи-
ка в СССР
Japanese birds, 1974). У берегов Корейского полуострова редкая (отдель-
ные встречи) зимующая птица (Gore, Won, 1971; Min et al., 1976).
В Китае В. m. marmoratus зимует вдоль материкового побережья Северной
Америки от п-ова Аляска до Южной Калифорнии (Birds of North Ameri-
ca, 1966).
Миграции. Данных нет.
Численность. Данные недостаточны. Объективные оценки численности
отсутствуют. По Бергману (1935), обычен в августе у восточных берегов
Камчатки. На Курильских островах редок (Велижанин, 19776). По-види-
мому, малочислен на северном побережье Охотского моря (Кищинский,
1968; Яхонтов, 1979). Относительно обычен летом в Амурском лимане
(данные Ю. В. Шибаева). Видимо, немногочислен в гнездовое время
у берегов Приморья (Елсуков, 1984; В. И. Лабзюк, личное сообщ.),
и на Сахалине (В. А. Нечаев, личное сообщ.).Районы с высокой числен-
85
Семейство Чистиковые
ностыо в СССР не известны. Зимой обычен в прибрежных водах Хоккайдо
и северного Хонсю, далее к югу — редок (Aistin, Kuroda, 1953; Check-list
of Japanese birds, 1974).
B. m. marmoratus многочислен у о-ва Кадьяк. У о-ва Принца Уэльского
летняя популяция насчитывает несколько сотен тысяч особей (Sowls et al.,
1978).
Местообитания. В период гнездования вид связан с распространением
на побережьях лесов таежного типа (Кищинский, 1968). Гнезда устраи-
вает в основном на деревьях, однако для В. m. marmoratus известны находки
гнезд на земле (Simons, 1980). Кормовые биотопы птиц — прибрежные
районы моря. Зимовка проходит в море в прилегающей к побережью
полосе.
Размножение. Колоний не образует. Гнездится отдельными изолиро-
ванными парами. К размножению приступает, по-видимому, в возрасте
3 лет. На о-ве Лангара (Британская Колумбия) в период гнездования
неполовозрелые птицы составляют примерно 15 % популяции (Sealy,
1975а). Гнездится главным образом в хвойных лесах. Не избегает горных
лесов (Кищинский, 1968; Sealy, 1974а). Продолжительное время считали,
что вид устраивает гнезда только на деревьях. В 1978 и 1979 гг. на
о-ве Баррен (Аляска) были обнаружены гнезда на земле (Simons, 1980).
Все найденные гнезда В. m. perdix помещались на лиственницах. Одно
гнездо в окрестностях г. Охотска находилось в лиственничном лесу в 6—
7 км от моря (Кузякин, 1963). Оно было устроено на высоте 7 м от земли,
на ветке с широкой плоской поверхностью из густого переплетения
мелких веточек, яйцо лежало на пучке лишайника Bryopodon. На скалис-
том берегу моря в бухте Ольга гнездо располагалось на лиственнице
в 2,5 мм от основания дерева и было устроено аналогично предыдущему.
Лоток помещался на плотном слое переплетенных ветвей, засыпанных
хвоей, сухими веточками, прядями лишайника (В. И. Лабзюк, личное
сообщ.). Еще одно гнездо было найдено па Сахалине (В. И. Нечаев,
личное сообщ.), в 2 км от залива. Чайво на обломанной вершине листвен-
ницы, в 5 м от земли.
В Северной Америке гнезда В. m. marmoratus были найдены на кедре
на высоте 15 м в 5 км от моря (Harris, 1971, цит. по: Sealy, 1974) и
на дугласии Pseudotsuga taxifolia на высоте 45 м (Simons, 1980). На
о-ве Баррен гнезда пыжика помещались на скалистом морском берегу
в 75 м от воды. Яйца лежали прямо на земле и на примятой траве
(Simons, 1980).
Сроки гнездования в различных частях ареала В. m. perdix близки. Яй-
ца и насиживающие птицы были обнаружены 17 и 23 июня в районе
г. Охотска и южнее (Кузякин, 1963; Яхонтов, 1979), 21 июня в Южном
Приморье (бухта Ольга) (В. И. Лабзюк, личное сообщ.), 19 июня
на Северном Сахалине (В. А. Нечаев, личное сообщ.), 15 июня на
Восточном Хоккайдо (Check list of Japanese birds, 1974). В Британской
Колумбии, судя по результатам вскрытия взрослых птиц, в 1970 и 1971 гг.
откладка яиц продолжалась с середины мая до конца июня (Sealy,
1974а), на о-ве Баррен (Аляска) в двух гнездах в 1978 и 1979 гг.
яйца были отложены в конце июня и в начале июля (Simons, 1980).
В кладке одно яйцо (Кищинский, 1968; Sealy, 1974). Окраска его
варьирует. У В. m. perdix по голубовато-зеленоватому фону разбросаны не-
многочисленные точки и мелкие пятна буроватого, орехового и темно-
86
Длинноклювый пыжик
песочного цвета. Форма удлиненно-овальная, почти цилиндрическая
(Кузякин, 1963); яйцо, извлеченное из яйцевода В. Д. Яхонтовым (1979),
было зеленовато-голубоватым с редкими мелкими буровато-серыми пятна-
ми. Яйцо у В, m. marmoratus светлое, оливково-зеленое с неравномерно
разбросанными коричневато-черными пятнами, едва заметно сгущающи-
мися вокруг тупого конца (Simons, 1980).
Размеры яиц (в мм): 63,6X39,3, масса 48—50 г (Кузякин, 1963);
63X38—39 (яйцо, готовое к откладке, было повреждено при вскрытии
птицы) (Яхонтов, 1979); 61,2X36,3 и 58,9X36,3, масса 41 и 38,5 г
(Simons, 1980).
Срок насиживания примерно 30 дней. У самца и самки развивается
по одному наседноМу пятну (Sealy, 1974). Насиживающие птицы меняются
через 24 ч. Смена происходит в сумерках и ночью (Simons, 1980).
Летящие к гнезду птицы издают громкий свист (Кузякин, 1963).
Размеры однодневного птенца В. m. marmoratus: длина (в мм) кры-
ла 25, цевки 19,5, клюва 10. Вес двух птенцов 34,5 г и примерно 32 т. Благо-
даря желтоватой с темными пятнами окраске птенец в гнезде почти не
заметен. Уже в первый день он реагировал на посторонние звуки вблизи
гнезда, а приближение человека замечал на расстоянии более 10 м. При
этом он поворачивался к опасности «лицом», широко раскрывая клюв и
делая им энергичные выпады. Изредка птенец издавал звуки, напомина-
ющие писк птенцов Fratercula, Lunda, Cerorhinca во время их кормления.
Густой пуховой покров сохраняется в течение почти всего периода
пребывания птенца в гнезде, скрывая развивающийся гнездовой наряд.
На 5-й день появляются пеньки первостепенных маховых, на 6-й —
всех маховых, плечевых и части кроющих спины. К 10-му дню кроющие
спины, брюха и второстепенные маховые выходят из пеньков. На 17-й день
появляются пеньки кроющих крыла. К этому времени оперяется голова
и пух на ней исчезает. На 21-й день рулевые наполовину выходят
из пеньков, сходит пух на брюхе. На 26—27-й день гнездовой наряд выгля-
дит полностью сформировавшимся (Simons, 1980).
Сроки вылупления птенцов В. m. perdix не известны. Период выкарм-
ливания у пыжиков, гнездящихся в районе устья Амура, приходится
на июнь. 14и 21 июля 1982 г. (перед заходом солнца и сразу после него)
здесь были зарегистрированы 3 птицы (одиночка и пара) с рыбой в клюве.
Поднявшись с воды и набрав высоту более 100 м, они направились
к берегу,, поросшему хвойным лесом (пихта, ель, лиственница), причем
пара птиц летела вместе, пока можно было следить за ними в бинокль
(данные Ю. В. Шибаева).
Судя по результатам взвешивания птенца В. m. marmoratus, родители
кормят его каждую ночь (Simons, 1980). Прилет взрослых с кормом
отмечался сразу после захода солнца, птицы прилетели поодиночке и
оставались в гнезде в среднем 7 мин. В одном случае удалось наблюдать
кормление птенца в возрасте 25 дней: прилетевшая птица с рыбкой
в клюве приземлилась рядом с гнездом, подошла к птенцу, бросила
добычу перед ним и улетела. Птенец тут же проглотил рыбу.
Срок пребывания птенцов в гнезде 27—28 дней (Simons, 1980).
Каким образом слетки преодолевают расстояние от гнезда до моря, до
сих пор не выяснено. Некоторые исследователи (Кузякин, 1963; Кищин-
ский, 1968) считают, что птенцы добираются пешком до ближайшего пото-
ка (ручья, реки) и по воде спускаются к морю. С. Сили (Sealy,
87
Семейство Чистиковые
1975а) полагает, что слетки преодолевают этот путь по воздуху, так как
крылья у них хорошо развиты (длина крыла составляет 86 % крыла
взрослой птицы) и недавно покинувшие гнездо птенцы хорошо летают:
при приближении лодки они легко взлетали и удалялись на расстояние
до 1 км. Слетевшие на воду птенцы, по-видимому, держатся самостоя-
тельно, так как, по наблюдениям С. Сили (Sealy, 1975а), большинство
из них встречается без взрослых птиц. Кроме того, за 50 ч наблюдений
в районе их кормежки вблизи о-ва Лангара этот автор ни разу не видел,
чтобы взрослые кормили молодых.
Суточная активность. Все прилеты птиц к гнезду, связанные с выращи-
ванием потомства (смена насиживающих птиц, кормление птенцов),
осуществляется в сумерках и ночью (Sealy, 1975а; Simons, 1980; данные
Ю. В. Шибаева). Свободные от «дежурства» птицы проводят день
в море.
Питание. Кормятся в прибрежных водах. В Британской Колумбии —
обычно в пределах 0,5 км от берега на глубине менее 30 м (Sealy,
1975а), в мелководном Амурском лимане кормившиеся пыжики встреча-
лись на расстоянии 5—10 км от суши. В период гнездования в местах
кормежки держатся главным образом парами и одиночками (Sealy,
1975а; данные Ю. В. Шибаева). В середине июля 1982 г. в Амурском
лимане из 104 встреченных там взрослых и, возможно, неполовозрелых
птиц 46,2 % составляли пары, 23 % — одиночные особи, остальные держа-
лись группами из 3—4 птиц, в одном случае из 8. Пыжики кормились
в мутной и сильно опресненной воде лимана на глубинах от 1 до 10 м.
Три птицы, улетевшие с кормом на берег, несли по одной рыбке.
Питается преимущественно рыбой. На о-ве Лангара основу питания
В. m. marmoratus в гнездовой период (май—август) составляли Ammody-
tes hexapterus и Cymatogaster aggregate. В апреле значительную часть в
питании занимали ракообразные Thysanoessa spinifera. Питание В. m. per-
dix не изучено.
Враги. В гнезде сапсана на о-ве Лангара найдены остатки крыльев
двух молодых пыжиков (Sealy, 1975а).
Хозяйственное значение, охрана. Хозяйственного значения не имеет.
Большая часть гнездового ареала В. m. perdix расположена в относи-
тельно малоосвоенной человеком зоне. Однако на северо-востоке Сахалина
в зоне нефтепромыслов отрицательное влияние человеческой деятельности,
видимо, может сказываться на состоянии местных популяций.
Короткоклювый пыжик —
Brachyramphus bravirostris Vigors, 1829
Kittlitz‘s Murrelet (англ.), Kurzsthnabelalk (нем.).
Син. (лат.): Uria brevirostris Vigors.
Статус. В СССР гнездится и зимует.
Общая характеристика и полевые признаки. Мелкий чистик в брачном
пере с темным верхом и белым низом. Зимой — с серым верхом и белым
низом. В природе может быть спутан с Brachyramphus marmoratus
и Aethia pusilia (в зимнем пере). От первого взрослые и молодые ко-
роткоклювые пыжики во всех нарядах отличаются коротким клювом
и белыми крайними рулевыми (особенно заметными при посадке на воду).
88
Короткоклювый пыжик
Кроме того, в брачном пере этот пыжик выглядит более пестрым, имеет
более светлую окраску верха и больше белого на нижней стороне тела;
в зимнем наряде отличается белыми боками головы (глаз — на белом фо-
не). От конюги-крошки его отличает большая величина и белые бока
головы (у последней, как и у длинноклювого пыжика, глаз — на темном
фоне) (Козлова, 1957; Birds of North America, 1966; Harrison, 1983).
Полет быстрый, легко поднимается с воды.
Подвидовая систематика. Монотипический вид.
Описание. Размеры самцов и самок: длина (в мм) крыла 126,0—
142,6; плюсны 15,5—18,0; клюва от переднего края ноздри до вершины
7,9—10,5 (Козлова, 1957).
Окраска. Самец и самка в брачном наряде. Верхняя сторона тела
темно-серая с многочисленными рыжеватыми вершинными пятнышками.
Бока головы, горло, область зоба и бока тела очень густо испещрены
черноватыми пестринами с палевыми предвершинными участками перьев
(рис. 21). Нижняя сторона белая. Переход от темного верха к белому
Рис. 21. Голова старика в брачном наряде (/), в зимнем наряде (2) и молодой птицы
в гнездовом наряде (3) (рисунок Ю. В. Шибаева)
низу постепенный. Маховые и верхние кроющие крыльев темно-бурые.
Нижние кроющие крыльев и подмышечные буровато-дымчатые. Несколько
крайних рулевых белые (количество их варьирует), остальные, включая
среднюю пару, серовато-бурые с белыми каемками. Радужина бурая,
клюв черный, ноги желтовато-серые.
Самец и самка в зимнем наряде. Окраска в целом контрастная.
Верхняя сторона тела темно-сизая. Голова в основном белая, исключая
узкую черноватую полосу сверху (передняя часть головы, темя, шея),
переходящую в окраску спины. Перед глазом — небольшое темное пятно
полулунной формы. На темном фоне верха — светлые пестрины. Белые
плечевые образуют продольную полосу. Низ белый, за исключением
узкого, прерванного посредине черноватого ошейника (ожерелья) в облас-
ти зоба. Верх крыльев черновато-серый.
Пуховой наряд. Не описан.
Гнездовой наряд. Молодые птицы в юношеском наряде весьма близки
по окраске оперения к молодым длинноклювого пыжика, но хорошо
отличаются от них маленьким коротким клювом и белыми крайними
рулевыми.
Линька. Время полной осенней линьки не установлено. Птица, добытая
в начале августа у северного побережья Чукотского полуострова, имела
89
Семейство Чистиковые
полный летний наряд. Весенняя частичная линька взрослых заканчи-
вается на южном берегу Чукотского полуострова в последних числах
мая. У пыжиков из залива Креста в летнем наряде от 28 мая много
перьев в пеньках было на голове, шее, спине и в области зоба. Неполовоз-
релые особи линяют позднее: у пыжика в зимнем наряде, добытого
20 мая на кочевке в бухте Карагинской (северо-восточный берег Камчат-
ки), сменилось только одно перо на зобе (Козлова, 1957).
Распространение. Ареал вида в целом. Побережья Аляски
к югу до 55° с. ш., Чукотский полуостров и северная часть Охотского
моря (рис. 22). Гнездовой ареал и область кочевок выяснены
недостаточно. Гнездование достоверно доказано (находки кладок) в 14
точках (одна — в СССР, остальные — в США). Другая информация, на
основании которой приходится судить о гнездовом ареале — встречи в лет-
нее время. В Азии гнездится по южному и арктическому побережьям
Чукотского полуострова, к северо-западу до о-ва Врангеля, а также на
о-вах Диомида в Беринговом проливе (Козлова, 1957). Обнаружение этого
вида на гнездовании (найдено гнездо) на охотском побережье в заливе
Шелихова (Кищинский, 1968) значительно расширяет пределы гнездового
распространения короткоклювого пыжика (до 61°, а возможно, и до 59°
с. ш.). Кроме того, А. А. Кищинский (1968) встретил этих птиц в гнездо-
вое время 24.VII 1964 г. в заливе Бабушкина. Птицы добыты также
28.V в заливе Креста, 6.VIII в бухте Провидения и 20.V в Карагинском
заливе (Козлова, 1957). Взрослая птица в летнем наряде добыта в заливе
Корфа (Кищинский, 1980). Отмечены еще встречи в летнее время в
Кроноцком заливе (Стенченко, 1975). Один пыжик добыт 12.VII на
кочевке у о-ва Парамушир (Yamashina, 1929). На Курильских островах
встречается очень редко (Козлова, 1957). Около 30 особей встречено
в проливе Лонга и 6 особей в Колючинской губе (Кондратьев, 1968).
Гнездовая область на американском побережье тянется от залива Ле-Конт
через северную часть залива Аляска к Алеутским островам. К северу —
вдоль побережья Берингова и Чукотского морей до участка между мысами
Лисбурн и Барроу (Boiley, 1973; Sowls et al., 1978; Day et al., 1983)
(см. рис. 22).
Зимовки. На кочевках зимой встречается в Азии лишь у побережий
Камчатки и Курильских островов (Козлова, 1957).
Численность. В СССР редок и спорадичен. На американском континен-
те наиболее многочислен в районе от юго-восточной части штата Аляска
(США) до о-ва Кадьяк. Обычен вдоль п-ова Аляска и на Алеутских
островах. В проливе Уильяма численность летом составляет, по-видимому,
десятки тысяч (Кищинский, 1968; Sowls et al., 1978). Крупные стаи
наблюдались во внегнездовое время в районе бухты Гласиер (Grinnel,
1909).
Местообитания. А. А. Кищинский (1968) отмечает совпадение общей
конфигурации гнездовой области короткоклювого пыжика с областью
распространения близ берегов гольцового ландшафта. Обнаруженные к
настоящему времени гнезда этого вида располагались в гольцах среди
россыпей и снежников (Thayer, 1914; Кищинский, 1968; Boiley, 1973;
Day et al., 1983). Высота гнездования над уровнем моря 230—1070 м
(средняя 570). В лесных районах гнездится выше. Расстояние между
гнездовым участком и морем — до 75 км (Day et al., 1983). В гнездовое
время держится в прибрежных районах моря.
90
Короткоклювый пыжик
Рис. 22. Область
распространения
короткоклювого пыжика
а — районы летних встреч,
б — места установленного
гнездования
Размножение. Гнездится отдельными изолированными парами. Гнездо,
обнаруженное А. А. Кищинским (1968) *, находилось в горном цирке
в 30 км от моря, на высоте 700 м над уровнем моря. Оно располагалось
на слегка пологой мелкокаменистой россыпи по днищу цирка, в гольцовой
обстановке, значительно выше стланника и в 8 км от ближайшего леса.
С россыпями чередовались пятна ягельников; в середине июля здесь
оставались еще не стаявшие снежники. Само гнездо представляло собой
площадку около 20 см в поперечнике, с которой был отброшен более
крупный щебень, а на оставшейся мелкой щебенке лежало без всякой
подстилки одно яйцо. Гнезда на Аляске (Thayer, 1914; Thompson et al.,
1966; Day et al., 1983) располагались в такой же обстановке. В гнезде,
найденном у залива Шелихова 16.VII 1963 г. (Кищинский, 1968), содер-
жалось одно яйцо. Птица насиживала. На Аляске гнезда с яйцами находи-
ли с 10 июня и до 26 июля (Day et al., 1983).
В кладке одно яйцо (Thayer, 1914; Кищинский, 1968; др.)» имеющие
эллипсоидную форму с почти одинаковыми концами. Основной цвет
его поверхности голубовато-зеленый. Фон густо покрыт пятнами двух
типов. Более глубокие и размытые пятна — от бледного серовато-фиолето-
вого до темного пепельно-серого оттенка. Поверхностные пятна более тём-
ные и резкие, как бы «наляпанные» кистью, окрашены в коричневые тона
от светло-бурого до темно-каштанового и темно-умбрового. Пятна сгуще-
ны к одному из концов, не образуют, однако, заметного венчика. Диаметр
глубоких пятен до 4—5 мм, поверхностных — до 7—8 мм (Кищинский,
1968). По Дею (Day et al., 1983), большинство яиц имеет оливково-
зеленый фон и лишь некоторые — голубовато-зеленый. Размеры: (п=90)
60,0±2,0 X 37,3±1,1 мм (Day et al., 1983).
Сведения об инкубации, развитии и выкармливании птенцов отсутству-
ют. По-видимому, в насиживании и выкармливании птенцов участвуют
обе птицы. Спуск на воду проходит в условиях близких к тем, в которых
проходит он у длинноклювого пыжика, и осуществляется, по мнению
А. А. Кищинского (1968), с использованием водотоков.
* Единственная находка в СССР.
91
Семейство Чистиковые
Суточная активность, поведение. Данных нет.
Питание. Кормятся в прибрежных водах. В пище найдены мелкие
ракообразные и моллюскоподобные организмы (Bent., 1919). А. А. Кищин-
ский (1968) обращает внимание на очень маленький клюв совершенно
не «рыболовного строения» и делает заключение о специализации этого
вида на питании беспозвоночными.
Враги, неблагоприятные факторы. Данных нет.
Хозяйственное значение, охрана. Относительная редкость вида, спора-
дичность его гнездования и труднодоступность во все сезоны года позво-
ляют пока быть спокойными за его судьбу. Потенциальной угрозой
его существованию является освоение шельфа, в частности загрязнение
моря нефтепродуктами. Вид занесен в Красную книгу РСФСР как очень
редкий, малоизученный периферийный (IV категория статуса). Изъятие
особей из природы на территории СССР возможно только по специальным
разрешениям.
Род Synthliboramphus Brandt, 1837
Старик —
Synthliboramphus antiquus (Gmelin, 1789)
Ancient Murrelet (анг.). Silberalk (нем.)
Син. (лат.): Alca antique Gmelin, 1789.
Статус. Гнездящийся мигрирующий и зимующий вид.
Общая характеристика и полевые признаки. Мелкий чистик с серым
верхом, белой брюшной стороной, черной широкой полосой (обычно
прикрыта крыльями), идущей вдоль боков тела, черной головой и коротким
светлым клювом. В брачном наряде выделяются широкие белые полосы
вдоль боков темени, смыкающиеся на затылке. Тело удлиненное, ноги
отнесены далеко назад.
В Южном Приморье и на Южных Курильских островах может быть
спутан с редким хохлатым стариком, от которого в природе трудно
отличим. Летает, как и другие мелкие чистики, быстро, прямолинейно,
низко над водой.
Взлетает с воды без разбега: сидящая птица, отталкиваясь ногами,
подпрыгивает на 50 см от поверхности воды и начинает полет. Сходным
образом взлетает с суши. В отличие от многих других чистиковых
старик на суше не сидит «столбиком», как его нередко изображают
в определителях, а лежит, опираясь грудью на субстрат. На суше птицы
большую часть времени неподвижно лежат на одном месте, хотя при необ-
ходимости перемещаются довольно резво (данные В. А. Зубакина,
С. П. Харитонова). Старики либо ползают, отталкиваясь лапами, либо
прыгают, перемещаясь за один прыжок на 20—30 см, либо передвигаются
короткими перебежками, во время которых держат туловище наклонно к
субстрату. Благодаря цепким когтям птицы хорошо лазают даже по почти
вертикальной поверхности.
В колониях старик очень криклив, особенно в начале размножения
и во время перехода птенцов из гнезда в море. Чаще всего слышна
громкая трель взрослых птиц «црррль». На воде старики обычно молча-
ливы, исключая время кормления птенцов. Птенцы в гнезде негромко
попискивают «пси-пси-пси. . .». Как только пуховик покидает гнездо, начи-
ная движение к морю, звучит новый сигнал — громкое настойчивое
92
Старик
«ции!. . .», повторяемый 4—6 раз подряд с постепенным усилением силы
звука к концу. Он слышен на расстоянии нескольких десятков метров.
С его помощью птенец поддерживает контакт с родителями на протяжении
перехода к морю. Ответный «призывный» крик взрослых — короткое «при-
рири» (или «црриври»).
Во время кормления птенцов родители, вынырнув, как правило, подают
сигнал «цк-цк-црррль» (или только первую часть). Птица кричит так
независимо от того, вынырнула она с добычей или без нее. Птенцы,
устремляясь навстречу, издают звонкое «цив-цив-цив». В остальное время
они негромко пищат, замолкая во время нахождения родителей под водой.
Подвидовая систематика. Описаны два подвида, различающиеся по
величине клюва и развитию рисунка на боках шеи (Степанян, 1975).
Оба подвида обитают на территории СССР.
1. Syhthliboramphus antiquus antiquus
Alca antique Gmelin, 1789, Syst. Nat.l, c. 554, запад Северной
Америки. Ареал вида, кроме Командорских островов.
На боках шеи широкие и многочисленные пестрины. Клюв массивный
и более высокий.
2. Synthliboramphus antiquus microrhynchos
Synthliboramphus antiquus microrhyhchos Stepanyan, 1972, Орнитоло-
гия, 10, c. 388. о-в Медный, Командорские острова.
Белый рисунок на боках шеи менее развит, пестрин меньше и они
заметно уже. Клюв менее массивный и высокий.
Описание. Размеры. Длина (в мм) крыла самцов (п=63) 136,5—
146,0 (среднее 140,3), самок (п=49) 131,0—149,0 (среднее 139,5); клюва
самцов (п=78) 12,5—15,1 (среднее 13,8), самок (п=69) 12,6—15,1
(среднее 13,8); цевки самцов (п=72) 25,1—29,0 (среднее 26,8); самок
(п=74) 24,6—29,0 (среднее 26,9). Масса (в г) самцов (п=87) 184,8—
267,2 (среднее 210,6), самок (п=86) 176,7—248,6 (среднее 206,4) (Sealy,
1976; Дементьев, 1951; данные Ю. В. Шибаева).
Окраска. Самец и самка в брачном наряде. Половой диморфизм
в окраске не выражен. Верх головы и зашеек черные. На боках темени
и затылке белая полоса в виде подковы, смыкающаяся сзади головы
(образована узкими удлиненными перьями). На межлопаточной области
(в большинстве случаев) и боках зоба — продольные пестрины из таких
же белых перьев. Иногда вокруг глаза не очень заметные светлые
полукольца. Бока головы, подбородок, горло, шея спереди буровато-чер-
ные. Бока шеи, зоб и вся нижняя сторона тела белые. Вдоль боков тела от
основания шеи до основания хвоста — широкая буровато-черная полоса,
контрастирующая с белым низом. Верх тела темно-серый. Маховые и руле-
вые буровато-черные. Внутренние опахала первостепенных в основной
части белые. Малые верхние кроющие крыльев черноватые, остальные
темно-серые. Нижние кроющие крыла белые, подмышечные пестрые
(сочетание белого с бурым). Клюв светлый, желтоватый. Вдоль гребня —
черная полоса. Ноги светло-серые, перепонки темные, когти
черные. Радужина у птиц всех возрастов темно-коричневая (Sealy, 1976).
Самец и самка в зимнем наряде. Верх головы, задняя часть шеи
и зашеек черные. Белых удлиненных перьев нет (или есть остатки).
Вокруг глаза узкие (около 1 мм) белые полукольца, иногда едва заметные.
Горло и шея спереди, как и весь низ, белые. Бока тела серые, часть
93
Семейство Чистиковые
перьев на боках двуцветные — сочетание белого с серым. Верх темно-
серый с голубизной. Остальное — как летом.
Пуховой наряд. Шапочка, зашеек, бока головы и подбородок черные.
Горло, шея спереди и низ тела белые. Пятно за ухом светло-серое. Верх
выглядит черновато-серым, он образован голубовато-серыми пушинками
с черными основаниями. На этом фоне выделяются три буровато-черных
пятна на крыльях и надхвостье. Бока буроватые. Подхвостье и область
бедер светло-серые. В целом распределение окраски повторяет таковое
взрослой птицы.
Гнездовой наряд. Черный или буровато-черный цвет верха головы
и зашейка постепенно переходит к черновато-серому цвету спины (у
взрослых птиц в брачном пере граница между зашейком и спиной резкая).
Бока головы светлее ее верхд, подбородок и горло буровато-серые, иногда
этот цвет заметен только на подбородке. У большинства птиц по бокам
темени штриховой рисунок различной степени интенсивности из очень
узких белых перышек (иногда он присутствует на затылке и на зашейке).
Передняя часть шеи, как и весь низ тела, белая. Бока буровато-серые
с чешуйчатым рисунком на границе с белым низом (рисунок образован
темными каймами на концах контурных перьев). Клюв меньше и темнее,
чем у взрослой птицы. Остальное — как у взрослой птицы в зимнем
оперении.
Первый зимний наряд. В целом птица очень похожа на взрослых
зимой, отличаясь меньшей высотой клюва и более темными боками.
Кроме того, в ноябре (а иногда и в декабре) на боках еще сохраняются
остатки «чешуйчатого рисунка».
Линька. Полная линька взрослых в наших водах бывает в сентябре
и в октябре, так как птицы материалов ЗИН АН СССР, добытые 22 августа
около Мальминских островов, находятся в летнем наряде без признаков
линьки, а старики от конца октября из Аяна, бухты Находки и от 21 октяб-
ря из Петропавловска-на-Камчатке — уже в зимнем оперении с дорастаю-
щими перьями (в пеньках) на горле и темени. Весенняя частичная
линька взрослых происходит в апреле. У самца с о-ва Хонсю в зимнем
наряде от 28 апреля интенсивно сменялось мелкое оперение на голове,
шее, спине, зобу и груди. Некоторые особи, по-видимому, линяют и раньше
апреля, так как у старика из залива Америка (район Владивостока)
от 30 мая летнее перо было уже сильно изношено, в особенности на
спине (Козлова, 1987).
От марта к августу (Приморье) идет процесс изнашивания оперения.
Это, в частности, очень заметно по деградации украшающих перьев головы
и зашейка. Рисунок становится менее заметным, белые перья исчезают на
затылке (из «подковы» получаются две заглазничпые белые полосы) и в
межлопаточной области. Процесс изнашивания оперения начинает особен-
но прогрессировать после спуска на воду молодых (в Приморье — с конца
мая до конца июня). Все июльские взрослые из Приморья по изношенности
оперения резко отличались от июньских. К этому времени значительно
деградируют украшающие перья, но особенно меняются маховые, которые
совершенно обтрепываются и становятся белесыми. Кульминацией яв
ляется полная потеря маховых, которые выпадают уже после распада семьи
(взрослая самка от 28 августа, добытая севернее устья р. Тумнин) (данные
Ю. В. Шибаева).
Распространение. Ареал в,® да в целом. Северная Пацифика,
94
Рис. 23. Область распространения старика
а — места гнездования, б — область зимовки
Рис. 24. Места установленного (/) и предполагаемого (2) гнездования старика в СССР
Семейство Чистиковые
побережья материков и островные гряды между 22 и 63° с. ш. (рис. 23, 24).
Гнездовой ареал. Северный предел гнездования на азиатском
побережье Тихого океана — Пенжинская губа. Гнездование известно со
времен Палласа. В. Д. Яхонтов (1974) пишет о гнездовании на мелких
островах в северной части губы и в южной ее половине (о-ва Добржан-
ского, Темчун, Второй, Крайний, Ровный). Нигде более на западном побе-
режье Камчатки не зарегистрирован. На восточном — достоверно гнездится
только на о-ве Старичков в районе Авачинской губы (Вяткин, 1989).
Об этой колонии писал ее Паллас (цит. по: Taczanowski, 1893). На
Командорских островах в настоящее время — один из самых редких
гнездящихся чистиков (Карташев, 1961; Мараков, 1966). В прошлом на
о-ве Медный был обычен (Иоганзен, 1934), гнездился на о-ве Арий
Камень (у о-ва Беринга) (Stejneger, 1885).
В незначительном числе населяет все Курилы (Yamashina, 1931; Вели-
жанин, 1972, 1978). Наиболее обычен на островах Птичьих (Базарный,
Бакланий, Две Гагары), у островов Парамушир, Чиринкотан, Ушишир,
Шикотан, Анучина (Велижанин, 1972, 1978). Молодые или яйца найдены
на о-вах Парамушир, Итуруп (Yamashina, 1931), Анучина (Воронов,
1975), на мелких островках у Шикотана (Кобаяси, 1933; Велижанин,
1978; данные Ю. В. Шибаева).
Для Сахалина доказано гнездование в двух точках — на о-ве Тюлений
(Нечаев, Тимофеева, 1980) и на о-ве Монерон. На мелких островах
у Монерона в конце июля 1976 г. была обнаружена скорлупа яиц и три
трупа старика (данные Ю. В. Шибаева). Помимо этого, в гнездовое
время старики встречались у берегов самого Сахалина (мыс Терпения —
28.VI 1947 г., одиночка; мыс Кузнецова — 29.V 1949 г., пара) (Гизенко,
1955).
На северо-западном побережье Охотского моря гнездование установ-
лено в районе Аяна (Мальминские острова, Козлова, 1957). Гнездится
на небольшом островке Талан Тауйской губы Охотского моря (данные
А. Я. Кондратьева и А. С. Китайского). Встречен на Шантарских островах.
Однако гнездование не доказано (Дулькейт, Шульпин, 1937). В. Д. Яхон-
тов (1977) на Шантарских островах этот вид вообще не отмечает.
На побережье Татарского пролива гнездование старика зарегистри-
ровано в заливе Де-Кастри на о-ве Обсерватории (Schrenk, 1861). С. В. Ел-
суков (1984), обследовавший большой (около 440 км) участок побережья
от Духовских озер (район Пластуна) и до р. Лудза (район мыса Золотой)
в течение лета 1977 г., учел в общей сложности 37 птиц. Он считает
старика гнездящимся в этом районе, однако не приводит никаких объек-
тивных доказательств.
В районе залива Ольги старик немногочисленный гнездящийся вид.
Наблюдались семьи с молодыми. Добыта птица с наседными пятнами
(Лабзюк, 1975). В заливе Петра Великого гнездится на островах Вер-
ховского, Карамзина (Нечаев, Юдаков, 1968; Лабзюк и др., 1971), Фуру-
гельма (Лабзюк и др., 1970), Стенина и Клыкова (Нечаев, Юдаков,
1968). На двух последних островах факт гнездования не подтверждается
конкретными находками. Обнаружение мертвого пуховика в мае 1979 г.
(данные Ю. В. Шибаева) на пляже о-ва Гильдебрандта позволяет гово-
рить о гнездовании на этом или на одном из близлежащих островков.
В целом старик в западной части гнездового ареала — спорадически
гнездящийся вид. Сложность обнаружения гнезд в связи с ночным
96
Старик
образом жизни позволяет полагать, что многие места гнездования его
еще не выявлены. В частности, это касается Курильских островов.
Известно также гнездование этого вида на пяти островках у берегов
Корейского полуострова, на двух — у берегов Японии и в одном месте
в Китае (Austin, 1948; Check-list of Japanese birds, 1974; Cheng, 1976).
В Северной Америке колонии этого вида размещаются на Алеутских
островах, на островах у п-ова Аляска, на побережье залива Аляска
и на юге архипелага Александра (США). Далее к югу вид обитает
на островах Королевы Шарлотты и по берегам залива Королевы Шарлотты
(Канада) (Sowls et al., 1978; Vermeer et al., 1983).
Зимовки. В СССР зимует у Командорских островов (Козлова,
1957), в прибрежных водах южной части Курильской гряды (Велижанин,
1972), у юго-восточного побережья Сахалина (Воронов, 1972) и в Южном
Приморье (нерегулярно), где доходит до бухты Ольги (Абрамов и др.,
1973; данные Ю. В. Шибаева).
В Японии зимой обычен вдоль всей гряды островов, распространяясь
на юг до Тайваня и побережья Китая (Check-list of Japanese birds,
1974). В Корее зимует у южных берегов полуострова, у островов Чеджудо
и Уллындо (Вон-Хон Гу, 1963; Gor, Won, 1971). В восточной части ареала
часть стариков зимой движется к югу вдоль побережья Калифорнии,
другие зимуют вблизи мест гнездования или подальше в море (Grinnel,
Miller, 1944; Gabrielson, Lincoln, 1959; Sealy, 1976).
Во время осенней миграции при сильных западных ветрах отмечены
многочисленные залеты стариков в глубь американского континента
(Munyer, 1965; Verbeek, 1966).
Миграции. Данных нет.
Численность. Оценка численности затруднена из-за скрытного образа
жизни. Данные по СССР отрывочны. На о-ве Старичков (восточное
побережье Камчатки) 12—14 тыс. птиц (Вяткин, 1983). На о-ве Талан
Тауйской губы Охотского моря гнездится примерно 10 тыс. пар стариков
(данные А. Я. Кондратьева и А. С. Китайского). Для Курильской гряды
численность оценивается в 3000 экз. (Велижанин, 1978). И. Ямасина
(1931) и А. Г. Велижанин (1972, 1978) считают, что старик — малочислен-
ный вид на Курильских островах. Однако А. Г. Воронов (1975) на
относительно коротком маршруте между островами Кунашир и Анучина
учел в июле более 2000 птиц. В заливе Петра Великого численность
в лучшем случае определяется несколькими тысячами. Стариков было
много на о-ве Обсерватории в заливе Де-Кастри в XIX в., однако сейчас
информация оттуда отсутствует.
Невелики и колонии, находящиеся за пределами нашей страны на
азиатском побережье Тихого океана. Японские острова: на о-ве Теури —
500 экз. (Murata, 1958). Н. Курода (Kuroda, 1963а), посетивший это
место позднее, колоний не обнаружил. Корейский полуостров: на о-ве
Рэн — «много»; на о-ве Ниси — несколько сот птиц; на о-ве Чибалдо, —
по-видимому, несколько сот (Austin, 1948). В целом в западной части
ареала, и в частности в нашей стране, старик — немногочисленный
вид. В восточной половине гнездового ареала численность вида определя-
ется примерно в 1 млн особей (Sowls et al., 1978; Vermeer et al.,
1983). В западной части ареала она, видимо, значительно ниже.
Местообитания. В гнездовое время — преимущественно небольшие
острова (с почвенным слоем и пустотами в разрушающихся скальных
7 Птицы СССР
97
Семейство Чистиковые
породах), расположенные в островных дугах или у материкового по-
бережья. В период кочевок и зимой — в основном районы моря, примыкаю-
щие к побережью.
Размножение. Колониальная птица. Обычно гнездится с другими
морскими птицами, образуя смешанные колонии. В Приморье селится
рядом с очковым чистиком (Cepphus carbo), тупиком-носорогом (Cerorhin-
са monocerata), вилохвостой качуркой (Oceanodroma monorhis), черно-
хвостой чайкой (Larus crassirostris). На Курилах обитает по соседству
с северной качуркой (Oceanodroma leucorrhoa), топорком (Lunda cirrata),
тупиком-носорогом и тихоокеанской чайкой (Larus schistisagus).
К размножению приступает, по-видимому, в возрасте трех лет. Не-
половозрелые птицы в возрасте 1—2 лет в период размножения посе-
щают гнездовые колонии. На о-ве Лангара (Британская Колумбия)
в середине мая они составляют около 20 % популяции (Sealy, 1976).
Рис. 25. Позы старика иа воде и на суше
Старику присущ гнездовой консерватизм. Один из помеченных стари-
ков использовал прошлогоднее гнездо, три пары птиц были пойманы на
своих гнездовых участках год спустя (Sealy, 1976). На островах Верхов-
ского в 1969 г. в нескольких гнездах было окольцовано по одной взрослой
птице, две из них были пойманы в следующем году рядом с прежними
норами (данные Ю. В. Шибаева).
Сведения о демонстративном поведении стариков отрывочны. Наиболее
часто на воде и на суше в конфликтных ситуациях и брачных кон-
тактах отмечаются две позы: первая — голова поднята под углом 45 °,
шея вытянута, вторая — шея как бы втянута в плечи. Брачные партнеры
исполняют некоторое подобие танца (billing), характерного для ряда
других чистиковых. Птицы располагаются клювами навстречу друг к
другу либо в первой, либо во второй позе, клювы почти соприкасаются
(хотя полного соприкосновения не наблюдали), и самец и самка при этом
исполняют песню. Отмечалась также распластанная поза: шея вытянута
параллельно земле или поверхности воды, клюв направлен вперед. Она
наблюдается как в территориальных конфликтах, так и в брачных взаимо-
действиях (рис. 25). При ссорах на воде, когда несколько птиц сближаются
в плотную группу. Уход стариков из зоны столкновения часто происходит
при помощи прыжков с раскрытыми крыльями. При этом может наблю-
даться короткий полет на 2—3 м и заныривание с воздуха. Но чаще
собственно полета не наблюдается, старики просто подпрыгивают на 20 см,
полностью раскрывают крылья (но не машут ими), пролетают по воздуху
0,5—0,7 м, падают на воду чаще всего крылом вперед и тут же заныривают.
98
Старик
Поэтому такие прыжки можно рассматривать как прерванный в самом
начале полет.
По наблюдениям в 1987 г., на о-ве Талан занятие территорий стариками
на колонии происходило в первую половину июня. Птицы, скорее всего
самцы, защищают территории диаметром 0,5—0,7 м (данные С. П. Харито-
нова). Территориальные конфликты стариков на островах залива Петра
Великого можно наблюдать в апреле и мае. Они дерутся на поверхности
земли и в норах, хватая соперника за основание клюва. Весной в колониях
часто встречаются птицы с выщипанными вокруг клюва перьями и со
следами крови на оперении. В одном случае драка была особенно ожесто-
ченной. В ней участвовали три птицы. Две из них 5 сидели, сцепившись
клювами, и, выворачивая шеи, пытались как бы опрокинуть соперника,
третья птица клевала обоих. Все они издавали стрекочущие трели. Клювы,
оперение птиц и камни вокруг были в крови. Драка продолжалась
без перерыва 30 мин., затем одна из птиц была вытолкнута соперником
из норы и улетела, но через некоторое время вернулась и драка возобно-
вилась. В общей сложности конфликт продолжался около двух часов.
Птицы дрались в полной темноте и на свет включаемого фонарика не
обращали внимания.
Спаривание стариков, по наблюдениям на о-ве Талан, происходит
главным образом на суше, в ночное время. Пик спаривания отмечен
здесь в третью декаду июня. Самец перед садкой приближается к самке
сзади, вытянув шею и направив клюв вперед и слегка вниз. Затем
он запрыгивает на самку, взмахивая развернутыми крыльями. Во время
спаривания самец лежит на спине самки, свесив наполовину развернутые
и мелко дрожащие крылья по бокам тела, иногда он опирается крыльями
на субстрат. После схода самца самка несколько раз вскидывает клюв.
Демонстрации, предшествующие спариванию, по-видимому, могут и отсут-
ствовать. Отмечались попытки спаривания с одной самкой одновременно
несколько самцов. А. С. Китайский однажды наблюдал садку на воде,
в полный штиль. Самец при этом располагается на самке «столбиком»,
вертикально (Данные В. А. Зубакина, С. П. Харитонова, А. С. Китайско-
го).
Гнездится преимущественно на небольших скалистых островках. Гнез-
да» располагаются в пустотах между камнями или в норах, вырытых
в_почв£. Иноща^анимад)т норы других.дтиц: в Приморье — вцдрддшстай
качурки, тупика-носорога. Норы встречаются как у подножия, так и на
вершине острововУНаостровах Верховского часть гнезд устроена между
валунами пляжа, в нескольких метрах от воды. Длина нор и основном
от 10 до 60 см (Вяткин, 1983; Murata, 1958). Гнездо помещаете ft в дальнем
конце норы^ лотокиногда слегка углублен. Гнезда без выстилки иди
с небольшим количеством сухой травы.
ИД ДЙНиЫМ 1987 Г. с о-ва Талан, среднее минимальное расстояние
между но рам ц ц рднЬне_пдо.т11огргнездования Нари?ор см (п==41),
минимальное — 10 см, распределение нор на поверхности случайное (дан-
ные С. 11. Харитонова).
В заливе Петра Великого старики занимают гнезда в первой декаде
апреля, к откладке яиц приступают в середине этого месяца. На Южных
Курилах сроки размножения более поздние — на о-ве Шикотан откладка
яиц в 1971 г. началась в первой декаде июня.
На о-ве Талан в 1987 г. первое яйцо старика найдено 23 июня,
7*
99
Семейство Чистиковые
массовое начало яйцекладки 28 июня (устное сообщ. А, С. Китайского).
В южных и центральных районах Корейского полуострова откладка
яиц начинается в середине марта, а на крайнем северо-востоке его — в
конце апреля (Austin, 1948), в Японии (о-в Теури) — в июне (Murata,
1958). В Британской Колумбии (о-в Лангара) первые яйца появляются
в последней декаде апреля (Sealy, 1976).
В кладке, как правило, два яйца (Козлова, 1957; Sealy, 1976). Во всех
осмотренных нами полных кладках в заливе Петра Великого (п=32)
было по два яйца. Однако 11 августа 1971 г. у о-ва Кунашир была
встречена семья стариков с тремя оперяющимися птенцами. Самка перед
откладкой каждого яйца несколько дней проводит в море, не возвращаясь
в колонию (Sealy, 1976). Между откладкой первого и второго яйца
проходит в среднем 5 (Ishizawa, 1983, цит. по: Sealy, 1976) или 7 дней
(Sealy, 1976).
Окраска яиц: по светлому сероватому, буровато-оливковому, охристому
или пятна.; ~и штрихи: — пцдерхност-
ные буроватые и коричневатые, и более глубокие серые. Пятна, щ много
сгущаются на тупом конце или распределены равномерно. Форма варьи-
рует от эллиптически- или пйлиндрически-овальной до удлиненно-яйце-
вйдной, Скорлуцкгладаад, .сдедк.а блестящая, мелкозернистая (Козлова,
1957)“
Размеры яиц. (в мм):Южное Приморье (н=72) 56,4—63,7X35,9—40,3,
в среднем 60,0X36,5; Британская^ Колумбия (n=200). в.„среднем 5X5 X
Х37,4 (Sealy, 1976); Северная Америка (п—51} в среднем 61,1X38,6
(Вепу, 1919), Масса(вдТ"снсжнх^1и|Гиз залива Петра Великого =_36,8;
40,6; 47,0 ^Нечаев,. 1968); ид.Бритдцркод/Крду.м6идДл==Д5) 40,4 —
46,2, в среднем 44 9 (Sealy. 1976).
Срок инкубации в среднем 35 дней (Sealy, 1976). Насиживание начи-
нается после откладки второго яйца. У насиживающих птиц два наседных
пятна. Партнеры меняются через трое суток (Sealy, 1976).
В Южном Приморье процесс вылупления в 1976 г. начался около
20 мая и продолжался до конца июня. Пик вылупления пришелся на
период с 28 по 31 мая. На о-ве Шикотан в 1971 г. вылупление и спуск
птенцов на воду наблюдались 15—16 июля, а к 1 августа все старики
покинули гнезда. На о-ве Старичкова (Камчатка) массовое появление
птенцов в 1972 г. зарегистрировано 1—2 августа: в 32 гнездах (из 42
осмотренных) шло вылупление (Вяткин, 1983). На о-ве Лангара (Британ-
ская Колумбия) и расположенном неподалеку от него о-ве Форрестер
(Аляска) птенцы появляются в период с конца мая до начала июля
(Sealy, 1976), т. е. примерно в те же сроки, что и в Южном Приморье.
Обращает на себя внимание сходство в сроках размножения стариков
в некоторых значительно удаленных по широте колониях: Южное Примо-
рье — 42° 30' с. ш., Британская Колумбия — 54°00' с. щ. Это, по-видимому,
связано с тем, что в обоих районах к началу апреля складываются благо-
приятные кормовые условия, что, вероятно, определяет сроки начала
гнездового сезона морских птиц.
Птенцы старика переселяются на воду вскоре после вылупления
(Heath, 1915; Sealy, 1975а, 1976; и др.). От времени вылупления до ухода
из гнезда проходит в среднем около 2,2 сут (Sealy, 1976). Все это время
птенцов не кормят, однако энергетических ресурсов, имеющихся у них,
достаточно для перехода из гнезда в море. Находясь в норе, птенец теряет
100
Старик
около 11 % своего веса (по Sealy, 1976—15 %). Непрерывность в обогре-
вании птенца является приспособлением к сохранению энергии. Этой же
цели служит манера птенца прятать лапы в пух, изолируя их таким обра-
зом от холодного пола норы, и его малоподвижность в гнезде.
Характер передвижения нескольких десятков семей старика, уходящих
из гнезд в море, был прослежен нами на островах Верховского в мае—ию-
не 1976 г. Птенец хорошо подготовлен к прохождению нелегкого маршрута.
Как у всех выводковых птиц, у него большие сильные лапы.
Цевка у вылупившегося пуховика составляет 93,5 % цевки взрослой птицы
(Sealy, 1976). На ровной поверхности он способен совершать прыжки до
40 см в длину. Плотный слой упругого пуха служит хорошим амортизато-
ром при падении. Двигаясь, птенец низко опускается на лапах и баланси-
рует крыльями. Его действия во время передвижения на зов родителей
энергичны и целеустремленны. Он прыгает с каменистых уступов, взби-
рается на большие глыбы камней, прыгая и карабкаясь по их крутой
поверхности, протискивается через узкие щели, неутомимо ищет обходные
пути, если препятствие непреодолимо, а в конце маршрута пересекает
полосу прибоя.
Процесс переселения птенцов в море (Шибаев, 1978) начинается после
наступления темноты. К вечеру, в день спуска, пуховики ведут себя актив-
нее, часто выбираются из-под обогревающей их птицы. После прилета
второго партнера родители издают громкую трель, которая сопровождается
писком птенцов. Через некоторое время взрослые начинают кричать по-
иному, после чего один из них выходит из норы и усаживается рядом,
непрерывно покрикивая и поджидая пока выйдут птенцы. Птенцы отвечают
также новым сигналом, отчетливо слышным за несколько десятков метров.
Оба сигнала непрерывно звучат во время перехода семьи к воде. Птенцы
выбираются из норы и в сопровождении взрослых птиц начинают движение
в сторону моря. Родители опекают птенцов на всем пути от гнезда
к воде. Если почему-либо птенец задерживается по дороге, он продолжает
«сигналить», взрослые птицы возвращаются и побуждают его к дальней-
шему движению (садятся рядом, покрикивают). Роль самца и самки
при передвижении птенцов не ясна — на суше их обычно сопровождала
одна птица, вторая чаще находилась в воде, причем обе птицы то и дело
покрикивали. В случае серьезного препятствия на пути (обрыв, появление
человека) вторая взрослая птица также улетает на воду, птенец же про-
должает движение самостоятельно, поддерживая контакт с родителями
при помощи звуковых сигналов. Птенцы из одного выводка, идущие на зов
взрослых птиц, не реагируют на крики друг друга, единственный ориентир
в это время — голос родителей. Скорость птенца меняется в зависимости
от сложности маршрута: в густой траве и среди крупных глыб каменистой
россыпи его продвижение вперед замедляется, на галечниковом пляже
он бежит очень быстро, а когда попадает в воду, то уносится подобно
моторной лодке.
С того момента, когда птенцы попадают в море, начинаются послегнез-
довые кочевки стариков. Родители сразу, буквально с момента встречи
семьи на воде, начинают уводить выводок от острова. Это хорошо просле-
живается по голосам, затихающим по мере удаления птиц. Наши попытки
обнаружить выводки в море вблизи острова наутро после их ухода из гнезд
не дали положительных результатов. Однако примерно через полтора
месяца после массового вылупления птенцов в районах гнездования начи-
101
Семейство Чистиковые
нают появляться молодые птицы, за редким исключением, уже без
сопровождения взрослых (Sealy, 1979). Исчезновение семейных групп ста-
риков из мест гнездования, по мнению С. Сили, связано с перемещением
их в кормовые районы. В заливе Петра Великого быстрая откочевка стари-
ков от гнездований, возможно, стимулируется также потенциальной
угрозой, исходящей от многочисленных чернохвостых чаек.
Образ жизни семьи старика после ухода ее из колонии до сих
пор оставался неизвестным. В 1980 г. в заливе Петра Великого (Приморье)
были встречены 4 семейные группы с птенцами в возрасте более 1 мес.
Одна из семей, состоящая из двух взрослых и двух оперяющихся птенцов
с пухом на спине и голове (размер птенцов — примерно 2/з взрослой
птицы), появилась здесь 29 июня и оставалась до 23 июля. Все это
время птенцы получали корм от родителей и ни разу не пытались охотиться
сами. В начале июля с птенцами осталась одна птица. Выводок держался
с западной и северной сторон острова, защищенных от летних ветров
и штормов, на акватории протяженностью не более 3 км. Птицы кормились,
как правило, в 50—200 м от берега в течение всего светлого времени
суток, а в сумерках удалялись в море, где проводили ночь (в середине июля
они прекращали охоту в 20 ч 20 мин—20 ч 45 мин). Дневная активность
стариков выражалась в последовательно чередующихся периодах охоты-
кормления (20—50 мин) и отдыха (15—30 мин). Во время отдыха семья
удалялась от места кормежки в сторону моря и, сделав кольцо, возвраща-
лась на прежнее место, где держалась плотной группой. В период кормеж-
ки птицы несколько рассредоточивались, причем взрослые птицы могли
удаляться от птенцов на расстояние 50—70 м. На дневной ритм влияли
обилие и доступность корма, погодные условия. Во время кормежки
птенцы постоянно следили за действиями родителей, поддерживая голосо-
вой и визуальный контакт (когда родители ныряли, опускали голову
в воду). В соответствии с поведением родителей они быстро меняли направ-
ление своего движения, чтобы к моменту появления птицы с добычей
оказаться поблизости. Корм доставался тому, кто успевал доплыть первым.
Отдав добычу птенцу, старик обычно тут же нырял снова. Если он задер-
живался, голодные птенцы побуждали его к дальнейшей охоте, с громким
писком подплывая к взрослой птице сзади и сбоку и касаясь клювом
подхвостья. Писк прекращался как только старик нырял.
До середины июля птенцы получали корм только на поверхности
воды и ныряли лишь в случае опасности. Позже они начали активно
нырять, гоняясь за родителями под водой, и там же часто получали корм.
К этому времени у них отросли маховые, что заметно повысило маневрен-
ность подводного плавания.
Суточная активность. Поведение. В колонии ведет ночной образ жизни.
Днем на суше остаются только насиживающие птицы. Перед прилетом
и после отлета из колонии птицы скапливаются в определенных зонах
вблизи гнездовий («stading area»); в вечернее время — за три часа до
наступления сумерек (Sealy, 1976).
У о-ва Верховского (залив Петра Великого) птицы скапливаются
в 300— 500 м от острова, в июне вечерний прилет их в колонию начинается
в сумерках и тянется 30—35 мин. Отлет из колонии происходит на
рассвете.
По наблюдениям, во второй половине июня—первой половине июля
на о-ве Талан скопления стариков первоначально располагаются
102
Старик
в 2,4—3 км от колонии. К 19—20 ч., т. е. за 3—4 ч до захода солнца, здесь
собираются тысячи стариков, птицы очень разреженной стаей (несколько
десятков метров между парами или группами птиц) начинают медленно
перемещаться в направлении колонии. К часу ночи, в глубоких сумерках
(примерно через два часа после захода солнца), стая подплывает к самому
берегу острова и старики перелетают на сушу. В отличие от других чисти-
ковых взлетевшие с воды старики летят вверх по склону низко над
землей, на высоте 1,5—5 м. Посадка напоминает падение вниз, по склону
они летят на той же высоте, переходя над пляжем на горизонтальный полет
и с ходу ныряя в воду. Примерно до трех часов ночи идет интенсивный об-
мен птицами между стаей на воде и колонией, затем поток стариков с
суши начинает преобладать и стая птиц постепенно отплывает от берега.
Слет последних птиц отмечен в 4 ч 48 мин, примерно за 15 мин до восхода
солнца (данные В. А. Зубакина, С. П. Харитонова).
Питание. В Британской Колумбии (о-в Лангара) старик в начале перио-
да размножения (апрель-май) питается главным образом двумя видами
эуфаузиид — Euphausia pacifica и Thysanoessa spinifera. Позже птицы пе-
реходят к кормлению рыбой — Ammodytes hexapterus (Sealy, 1975). В за-
ливе Петра Великого в июне 1980 г. пара стариков выкармливала птенцов
преимущественно молодью сельди — Clupea harengus. Таким образом,
основу питания составляют массовые планктонные ракообразные и стай-
ные виды рыб.
У о-ва Фуругельма (Приморье) в июне старик охотился за молодью
сельди (3—4 см), которая образовывала плотные косяки (0,5—1 м в диа-
метре) из нескольких сотен особей. Охота происходила в верхнем слое
воды, около 1,5 м от поверхности. Основной тактический прием состоял
в том, чтобы не допустить ухода косяка в глубину. В начале старик пла-
вал, часто опуская голову в воду в поисках объекта охоты. После обнару-
жения стаи рыб следовала серия очень коротких и неглубоких занырива-
ний, которые выглядели как рекогносцировка. Затем начинались более
глубокие заныривания, во время которых птица стремительно снизу или
сбоку врывалась в стаю, пересекала ее и выскакивала на поверхность,
иногда подпрыгивая над водой. В результате серии таких атак косяк под-
нимался к поверхности, а его структура и управление расстраивались на
0,5—2 мин. Часть рыб «брызгами» выпрыгивала из воды, другие плашмя
бились на поверхности. Старик, совершавший в это время свои охотничьи
атаки, ловил рыб только в толще воды, не трогая тех, которые были на ее
поверхности. Обычно к этому моменту рядом оказывалась группа черно-
хвостых чаек, которые следили за действиями старика. Они тут же под-
ключались к охоте. Наконец, рыбы восстанавливали строй и уходили в глу-
бину. После этого все могло повториться.
Враги, неблагоприятные факторы. В Южном Приморье старики часто
становятся жертвами сапсана (Falco peregrinus). Этот хищник охотится
за ними и в Британской Колумбии (Beebe, 1960). По сведениям П. С. Вят-
кина (1983), гнездовья стариков на о-ве Старичкова постоянно посещают-
ся тихоокеанскими чайками (Larus schistisagus), которые уничтожают
кладки в неглубоких норах. В колониях стариков залива Петра Великого
неоднократно приходилось наблюдать чернохвостых чаек (Larus crassiros-
tris), большеклювых и черных ворон (Corvus macrorhynchos, С. согопе),
которые, низко летая вдоль склонов или расхаживая по колонии, загля-
дывали в норы стариков. Добычей для этих «хищников» здесь могут быть
103
Семейство Чистиковые
только яйца стариков (в начальный период откладки, когда птицы еще
не приступили к насиживанию) и, возможно, пуховики, не успевшие
спуститься на воду ночью в период перехода их в море или плавающие
вблизи острова, что случается очень редко. Семья стариков (два взрослых
и два птенца с остатками пуха), за которой мы постоянно наблюдали в июле
1980 г., постоянно подвергалась нападениям чернохвостых чаек. Они
пикировали на птенцов и взрослых птиц, выныривающих с рыбкой, пыта-
ясь забрать добычу. Все нападения (около 30), кроме одного, были безус-
пешны.
На о-ве Талан небольшое число взрослых птиц гибнет, по-видимому,
по вине топорков, когда старики ночью забираются в их жилые норы.
Погибших стариков с ранами и кровоподтеками на шее неоднократно
находили на поверхности земли среди нор топорков и, реже, в самих норах
(устное сообщ. А. С. Китайского).
На о-ве Монерон кладки стариков может уничтожать серая крыса
(Rattus norvegicus сагасо). Известно о нескольких случаях уничтожения
кладок и птенцов старика на о-ве Лангара черной крысой (Rattans
rattus) (Campbell, 1968; Sealy, 1976).
Хозяйственное значение, охрана. На большей части гнездового ареала
не требует специальных мер охраны. Ряд гнездовых колоний в СССР,
в первую очередь расположенных вблизи крупных промышленных центров,
взяты под охрану. Колонии старика в заливе Петра Неликого вошли
в состав Дальневосточного морского заповедника, организованного в
1978 г., и двух памятников природы на островах Карамзина и Верховского,
действующих с 1984 г. Единственное место гнездования вида на Восточной
Камчатке — о-в Старичкова. В 1981 г. объявлен памятником природы.
Часть гнездовий на Малой Курильской гряде охраняется республиканским
заказником.
Хохлатый старик—
Synthliboramphus wumizusume Temminck, 1835
Japanese Murrelet, Crested Murrelet (анг.), Kanniuri-umisuzume (яп.). Japanalk (нем.).
Статус. Кочующий и, по-видимому, гнездящийся вид.
Общая характеристика и полевые признаки. Мелкий чистик. По общему
облику, повадкам и размерам очень похож на старика. В полевых условиях
взрослых птиц можно различить только на близком расстоянии. Отличается
от старика распределением черного и белого на голове, цветом и пропор-
циями клюва. Хохол в полевых условиях не всегда заметен, так как обычно
прилегает к голове.
Подвидовая систематика. Монотипический вид.
Описание. Размеры. Длина крыла, клюва и цевки самцов не известна.
Длина (в мм): крыла самок (п=3) 120,6—127,5, без определения пола
126 —136; клюва самок 15,3—18,0, без определения пола 16,5—18,0; цевки
самок 22,0—26,0. Масса самки 107 г (Hartert, 1921, 1922; Ю. И. Назаров,
личное сообщ.; данные Ю. В. Шибаева).
Окраска. Самец и самка в брачном наряде. Половой диморфизм
отсутствует. Лоб, горло, бока головы, шеи и зашеек чёрные. На лбу —
хохол из узких черных перьев длиной 2—3 см. По бокам темени тянутся две
широкие белые полосы, смыкающиеся на задней стороне шеи. Они образованы
узкими длинными перьями (рис. 26). Такие же перья, но едва заметные,
104
Хохлатый старик
иногда бывают на боках зоба (у старика эти места интенсивно испещрены).
Нижняя сторона тела белая, бока серые с тянущейся вдоль тела от зашей-
ка до хвоста черной каймой. Верх темно-серый с голубоватым оттенком.
Наружные опахала и концы первостепенных маховых буровато-черные,
внутренние опахала белые, буреющие к концу. Второстепенные маховые
имеют буровато-черные наружные и буровато-белые внутренние опахала.
Малые кроющие крыла буровато-черные, остальные темно-серые. Подмы-
шечные пестрые (сочетание белого с черноватым). Испод крыла белый.
Рулевые буровато-черные. Радужина темно-бурая. Лапы темно-серые.
Клюв голубовато-серый с черным гребнем.
Рис. 26. Голова хохлатого старика в брачном (/) и зимнем наряде (2) и молодой птицы
в гнездовом наряде (3) (рисунок Ю. В. Шибаева)
Самец и самка в зимнем наряде. Подбородок, горло, шея спереди и весь
низ белые. Верх головы, затылок черные. Хохол короче, чем летом. Укра-
шающих перьев нет (или есть остатки) *. Вокруг глаз черноватая полу-
маска, веки окаймлены белыми полукольцами шириной до 2 мм (см.
рис. 26). Черная полоса на боку тела отсутствует. Окраска остальной части
оперения — как в брачном наряде.
Пуховой наряд. Птенец сверху буровато-серый с беловатыми пестри-
нами. Низ белый.
Гнездовой наряд.Распределение окраски на голове в целом, как у взрос-
лой птицы в зимнем наряде (см. рис. 26), однако белый цвет в некоторых
местах замещен серым, а на боках темени и затылке есть полоса из редких
белых украшающих перьев (более узких, чем у взрослых). Хохол короткий
(до 1 см), состоит из перьев округлой формы. Низ тела белый, бока серые
с заметной поперечной исчерченностью. Спина, надхвостье темно-серые
с голубоватым оттенком. Наружные опахала маховых черные с едва замет-
ным металлическим блеском, внутренние буровато-черные на конце и
белые в основной части. Кроющие крыла (средние и большие) черноватые
с серой каймой. Плечевые буровато-серые с темными поперечными поло-
сами. Испод крыла белый. Рулевые черные. На концах буровато-серых
верхних кроющих хвоста поперечные полосы, как на плечевых. Лапы серые
с черной тыльной стороной и перепонками, когти черные. Клюв чернова-
тый, немного светлеющий в средней части.
* В некоторых определителях хохлатый старик в зимнем оперении изображается как
в брачном наряде, с широкой белой полосой вдоль темени. На самом деле украшающие
белые перья, образующие эту полосу, зимой исчезают (как и у старика) и верх головы
выглядит темным.
105
Семейство Чистиковые
Рис. 27. Область распространения хохлатого
старика
а — места гнездования.
Линька. Не изучена.
Распространение. Гнездо-
вой ареал. Прибрежные воды
Японского моря и тихоокеанского
побережья Японских островов
(Takahashi, 1937; Chesklist of Japa-
nese birds, 1974; Won Pyong-Oh,
1976; данные Ю. В. Шибаева).
Известные гнездовья сосредото-
чены в Японии в полосе между
33 и 35° с. ш. Северная граница
совпадает с южной границей гнез-
дового ареала старика (рис. 27).
В СССР встречается во время
кочевок в Южном Приморье.
Взрослые самки добыты 31.VIII
1959 г. в бухте Киевка и 6.IX
1971 г. у о-ва Карамзина (из
пары); одиночная взрослая птица
встречена В. И. Лабзюком (1975)
в районе залива Ольги; ослабевшая
половозрелая самка поймана 28.VI
1984 г. у о-ва Фуругельма; молодая
б — места встреч на зимовках в СССР,
в — область знмовок и миграций
лин (Takanashi, 1937) и на юге
самка в гнездовом наряде с остат-
ками пуха найдена мертвой 8.VII
1984 г. в бухте Бойсмана (Ю. Н. На-
заров, личное сообщ), у о-ва Саха-
Курильских островов (Судиловская,
1976; данные Ю. В. Шибаева). Находка молодой птицы с остатками пуха
в заливе Петра Великого позволяет предположить возможность гнездова-
ния вида в этом районе. Область гнездования в Японии образует два
хорошо очерченных гнездовых района — острова у восточного побережья
Хонсю от залива Сагами до п-ова Кии (наиболее известные гнездовья — на
о-вах Идзу) и крайний юго-запад Японских островов и Корейский пролив.
Зимовки. На кочевках в Японии встречается в основном вдоль ее
тихоокеанской стороны и в Корейском проливе (Austin, Kuroda, 1953).
На Корейском полуострове хохлатый старик зарегистрирован в период
кочевок в марте-апреле (Вон Хон Гу, 1963; Rare and Endangered species
of Animals and Plant of Republic of Korea, 1981) и зимой (Won Pyong-Oh,
1976). Зимой часть птиц, по-видимому, проникает в северную часть Восточ-
но-Китайского моря (Шунтов, 1972).
В районах гнездования птицы встречаются в течение всего года. В дру-
гих районах в период зимовок (у о-вов Хонсю и Кюсю) — с ноября по
апрель.
Миграции. Данных нет.
Численность. В целом, по-видимому, невысокая. Конкретные данные
имеются для двух островов в Японии (о-в Миминоама, пролив Миэ — 150—
200 гнездящихся особей; о-в Окиносима, пролив Фукуока — около 400 гнез-
дящихся особей) (Распространение гнездящихся птиц Японии, 1981).
В пределах СССР вид, вероятно, встречается чаще, чем это представля-
106
Алеутский пыжик
ется в настоящее время, но из-за сходства со стариком его не всегда удается
точно идентифицировать.
Местообитания. В гнездовое время — небольшие скалистые островки
(Austin, Kuroda, 1953). В период кочевок и зимовок — в основном при-
брежные районы моря.
Размножение. Гнездится небольшими колониями и отдельными парами.
Гнезда устраивает в нишах среди камней на плоских участках или склонах
островов в 15—60 м от воды. Лоток выстилается небольшим количеством
сухой травы. В кладке 1—2 яйца желтовато-коричневого или светло-серого
цвета со светло-коричневыми пятнами. Размеры яиц (в мм): 49,2—56Х
Х32-36,5 (Austin, Kuroda, 1953), 52-56,1X33,6-36, 53,37X35,05 (Har-
tert, 1921 — 22). Яйца и птенцы отмечались с февраля по май (Austin, Kuroda,
1953). Дж. Мойер (Moyer, 1957), посетивший гнездовья хохлатого старика
на скалах Санбондаке (о-в Мияке) в 1953 г., добыл вблизи колонии 10 самок
и 2 самца. В яйцеводах самок оказались яйца и фолликулы разной степени
зрелости (у одной — готовое яйцо; у восьми — фолликулы, развившиеся до
половины нормального яйца; у одной — менее развитые фолликулы). 26 мая
во время второго посещения острова автор обнаружил в нишах, где гнезди-
лись птицы, только скорлупу яиц, т. е. гнездовой сезон был закончен.
Суточная активность, поведение. Данных нет.
Питание. Данных нет.
Враги, неблагоприятные факторы. Данных нет.
Охрана. Внесен в Красные книги Японии (1975), Южной Кореи (1981)
и РСФСР (1983).
Род Ptychoramphus Brandt, 1837
Алеутский пыжик —
Ptychoramphus aleuticus Pallas, 1811
Cassin’s auk let (англ.).
Син. (лат.):Uria aleutica Pallas, Aethia aleutica Pallas.
Статус. Редкий залетный вид.
Общая характеристика и полевые признаки. Птицы мелких размеров,
хорошо плавают и ныряют. Полет быстрый, маневренный, при слете на воду
характерен полет зигзагом. При плавании под водой невозможно различить
отдельные взмахи крыльев. Плавая на поверхности воды, часто обмакивают
клюв в воду. Сложение плотное, компактное, шея короткая, клюв короткий
заостренный. Общая окраска за исключением брюха, темная, почти черная,
выделяются белые пятна над и под глазами. Голос: ночью в колонии алеут-
ских пыжиков слышен постоянный хор, напоминающий лягушачий или
визжание свиней; иногда приблизительно на 15 мин хор смолкает, но затем
возобновляется с новой силой. Описано 10 вариаций голоса пыжиков,
которые используются в различных ситуациях. При испускании звуков
голова выдвинута вперед, горло раздуто, все тело выбрирует (Thoresen,
1964).
Подвидовая систематика. Монотипический вид.
Описание. Размеры. Длина (в мм) крыла самцов и самок 110—130,
клюва 18—21 (Бутурлин, 1934); высота клюва самцов 10,89+0,44, самок
9,51+0,41; цевки самцов 30,9+1,01, самок 30,1+0,93 (n=71) (Nelson,
107
Семейство Чистиковые
1981). Масса самцов и самок в среднем 165 г (Roundybush et al., 1980;
Vermeer, Cullen, 1982).
Окраска. Самец и самка в брачном наряде. Полового диморфизма
нет. Верх черновато-аспидный, бока головы, подбородок, горло и зоб
несколько светлее, темно-дымчато-серые, подкрылья серо-коричневые,
грудь и брюхо белые (резкой границы нет). Над и под глазом по белому
пятну, в основании подклювья светлое пятнышко. Радужина белая, клюв
заостренный черный, ноги голубоватые или голубовато-бурые. Горловой
мешок у приступивших к размножению птиц начинает удлиняться, дости-
гая к моменту вылета птенцов 130 мм, осенью укорачивается до 65 мм (Бу-
турлин, 1934; Thoresen, 1964; Manuwal, 1978; Tuck, Heinzel, 1978).
Самец и самка в зимнем наряде. Окраска оперения сходна с брачной,
но горло светлое (Tuck, Heinzel, 1978).
Пуховой наряд. Верх варьирует от черного до буро-серого, низ — от
серого до палевого или бледно-серого. Участок кожи вокруг глаз не опушен.
Лапы розовые с черными коготками. Радужина темно-коричневая. До 10—
12 дней птенцы еще маленькие и пушистые. Сравнивая с другими видами,
их легко принять за 2—3-дневных. Цвет ног изменяется до темно-серых
с голубоватым оттенком на 10-й день. Первые перья появляются на 12—
16-й день, сначала — на брюхе, боках, средней линии спины и крыльях,
в 18 дней — на голове и шее. В 20—23 дня развиваются первостепенные
маховые. Юношеское оперение формируется к 30-дневному возрасту, когда
большая часть пуха заменяется пером, но участки пуха могут оставаться
на затылке и крестце до 42 дней. Перья хвоста начинают развиваться на
14-й день, в 18 дней достигают 10 мм и к вылету — 40 мм (Thoresen, 1964).
Птенцовый наряд. По окраске почти не отличается от взрослого, но
горло белое, клюв несколько короче (Gabrielson, Lincoln, 1959; Thoresen,
1964).
Первый брачный наряд. Очень сходен со взрослым, отличается более
коротким клювом и чуть беловатым горлом. Радужина темная или с тем-
ными пестринами. Предъязычного мешка обычно не имеется. До конца лета
оперение сохраняется свежим, черно-серым, как и у перелинявших взрос-
лых, без бурого оттенка (Gabrielson, Lincoln, 1959; Manuwal, 1978).
Линька. Линька гнездящихся на островах Фараллон алеутских пыжи-
ков частично совпадает с сезоном размножения и протекает на протяжении
его второй половины. Первостепенные маховые сменяются более быстро
и синхронно, чем оперение тела. Птица сохраняет способность летать на
протяжении всего периода линьки. Взрослые пыжики начинают послебрач-
ную линьку в то время, когда у них есть яйца и птенцы. Линька крыльев
происходит в июле—сентябре, линька тела — в июне—октябре. Скорость
линьки невелика. В июле птицы, уже выкармливающие птенцов, по
развитию линьки опережают насиживающих яйца, но те и другие сильно
отстают по сравнению с пыжиками, не отложившими яйца. Птицы, имею-
щие потомство, линяют на 40 % дольше по времени по сравнению с бездет-
ными (Payne, 1965). А. Торесен сообщает о птицах, утративших способ-
ность к полету в связи с обнашиванием оперения хвоста (вероятно, не-
размножающихся) (Thoresen, 1964).
Распространение. Гнездовой ареал. Острова западного побе-
режья Северной Америки от п-ова Аляска, Алеутских островов, о-вов
108
Алеутский пыжик
Рис. 28. Область распространения алеутского пыжика
а — граница гнездового ареала, б — места залетов в СССР
Кадьяк и Шумагин до 27° с. ш., п-ова Калифорния, о-ва Гуадалупе. Все
известные к настоящему времени колонии расположены к югу от 57° с. ш.,
но вид регулярно отмечается севернее пролива Аляска (Thoresen, 1964;
Степанян, 1975; Tuck, Heinzel, 1978) (рис. 28).
Зимовки. Зимует в пределах гнездового ареала, кочует там же (Бело-
польский, Шунтов, 1980). Популяция островов Фараллон оседла, вне
размножения пыжики посещают гнездовья весь год, но нерегулярно,
обычно ночью (Manuwal, 1974).
Алеутский пыжик был добыт на Курильских островах И. Г. Вознесен-
ским, который работал там в 1844—1845 гг. Лихтенштейн в 1854 г. упоми-
нает об экземпляре алеутского пыжика «из Камчатки» в Берлинском музее
(цит. по Бутурлину, 1934). Вероятно, те же случаи имеют в виду К. Вори
(Vaurie, 1965) и Л. С. Степанян (1975).
Численность. На островах Фараллон (Северная Калифорния) в 1969—
1970 гг. гнездилось 52 тыс. пар алеутских пыжиков с плотностью гнездова-
ния 0,02 — 1,09 пары/м2 (Manuwal, 1974). На о-ве Трайангл (Британская
Колумбия, Канада) в 1976—1977 гг. насчитывалось 360 тыс. пар, при
занятости нор на 36—58 % и плотности гнездования 0,1 —1,1 пары/м2, что,
вероятно, составляет 40 % от мировой популяции (Vermeer et al., 1979).
В Атласе морских птиц Аляски (Sowls et al., 1978) указывается, что распре-
деление и численность алеутских пыжиков на Аляске плохо изучены.
Известна 21 колония, но учет произведен лишь в 8. В этих 8 колониях
зарегистрировано 342 тыс. птиц. Некоторые из необследованных колоний
превышают по величине самые крупные из описанных. Предполагается, что
вся аляскинская популяция насчитывает не менее 600 тыс. птиц. Две
самые крупные колонии (не менее чем по 100 тыс. птиц каждая) располо-
жены на островах Унга и Нигруд. 47 тыс. особей гнездится на о-ве Кастль
Рок (Sowls et al., 1978).
109
Семейство Чистиковые
Местообитания. Гнездится колониями на скалистых побережьях, изоли-
рованных утесах, мысах, островах. Выделено 11 гнездовых биотопов.
Основное требование к месту для гнезда — крыша над головой и убежище
от чаек и других хищников. Предпочитают гнездиться в самостоятельно
вырытых норах среди невысокоисухой растительности или в редких зарос-
лях кустарника, роют норы на голой земле, под корнями деревьев й кустар-
нйкб'вГпод упавшими стволами и под камнями. Длительное существование
ш5бтнь1х~колоний приводит к исчезновению растительности и последова-
тельной эрозии почв (Thoresen, 1964; Vermeer et al., 1975; Sowls et al.,
1978). На островах Фараллон недостаток пригодных для рытья нор уча-
стков при высокой плотности вынуждает 51 % популяции гнездиться
в трещинах скал (Manuwal, 1974). Иногда роют норы в массивах гуано
бакланов Phalacrocorax penicillatus (Thoresen, 1964).
Размножение. Изучено практически на одной модельной популяции
на островах Фараллон (Северная Калифорния). Половозрелость насту-
пает в возрасте трех лет (Manuwal, 1974, 1978). В 1959 г. на острове
птицы появились в конце декабря, в 1960 г. — в начале января. Прилет
происходит после дождя, так как рытье нор возможно только во влажной
земле. Птицы прибывают ночью в большом количестве и сразу прини-
маются за расчистку старых нор, ухаживание, постройку новых нор.
В это же время образуются новые пары. Старые — сохраняются несколько
лет. Интенсивность заселения острова возрастает в феврале, достигая
пика в марте-апреле. При первом посещении колонии птицы задержи-
ваются на острове лишь на 3—4 ч. Первая кладка была найдена 2. II 1934 г.
(Van Kosen, 1939, цит. по: Thoresen, 1964). В основном яйца отклады-
ваются с апреля по июль, но свежие яйца находили также в середине
августа (Smith, 1934, цит. по: Gabrielson Lincoln, 1959). Молодые птицы,
готовые к вылету, отмечены в мае—июле. Птенцы из яиц, отложенных
в июле и августе, должны вылетать в сентябре-октябре (Thoresen, 1964;
Payne, 1965).
Устройство гнезд алеутского пыжика очень напоминает таковое у ста-
рика (Sowls et al1978). Гнездо помещается в конце норы, вырытой
самими пыжиками. Длина норы 45—120 см (в среднем около 1 м). Обычно
за ночь вырывается 5^7,ТГсм' 1Гщ)гда строительство норы затягиваемся:
так,- 2 н()рЬ|, Начатые 18 января, птицы рыли еще 17 марта. Глубилалти.х
нор была около 90 ем. Норы часто имеют слепые ответвления, бывают
изогнутыми или закрученными в. виде спирали (Thoresen, 1964). Ё норе
находится особый боковой тоннель, где_ птицы скапливают свои экскре-
менты (Gabrielson, Lincoln, 1959). Предполагается также, что насижи-
вающие птицы для испражнения выходят из .норы,., В результате чего
край жилой норы всегда забрызган розовым от криля пометом (Thoresen,
1964). ....... ~ .. .....‘ ..."....—........
Яйцо откладывается на голую почву или на редкую подстилку, для
которой птиць^.выщипывают траву вокруг норы. Кладка всегда ^состоит
йз'бднбго"яйца. У 84 % птиц, имеющих Первую кладку, образуется два
н'аседных пятна, которые редуцируются к концу насиживания. Наседные
пятна имеются или слабо развиты у 64 % птиц, потерявших первую кладку,
и у 68 % — отложивших вторую кладку (Manuwal, 1974). Яйцо имеет
форму промежуточную между овальной и эллиптической, обычно с совер-
шенно округлыми концами, но встречаются “яйца' заострен
или'с обоих концов. Цвет яйца матово-белый, молочно-белый, кремово
110
Алеутский пыжик
белый, щщща с.заметным голубоватым или зеленоватым оттенком (Gabri-
elsicmijLincoln, 1959). Вес яйца .в. среднем давен 29,74 г, .объем — 27,87 мл,
плотность — 1,08 г/мл, площадь поверхности — 45,7 см2, длина — 46,9 мм,
ширина — 33,9 мм (п=14) (Jloudybush et al., 1980). Д. Мануэль приводит
несколько иной вес - 27,4 г, указывая, что это составляет 16 % от веса
самки (Manuwal, 1974). Молодые и менее опытные птицы размножаются
позднее и кладут более мелкие яйца (Vermeer, Cullen, 1982). Насиживают
кладку оба партнера, сменяясь вечером после захода солнца. Насиживание
длится 37—42 дня, как правило, 38 дней (Thoresen, 1964; Manuwal, 1974;
Roudybush et al., 1980). Вылупление длится 2—3 дня, редко — сутки
и происходит утром (Thoresen, 1964). Средний вес вылупившегося птенца
равен 17,81 г (Thoresen, 1964), 21 г (Manuwal, 1974). Родители приносят
корм птенцам ночью. Через неделю после вылупления родители перестают
обогревать птенцов. С этого времени те получают пищу 2 раза в сутки.
В трехнедельном возрасте птенцы появляются у выхода из нор. В 30—
35 дней они начинают «тренировать крылья» около входа в нору (Tho-
resen, 1964; Manuwal, 1974). В возрасте 41—50, в среднем 44,7 (Thoresen,
1964), 35—46, в среднем 43,5 дней (Vermeer, Cullen, 1982) птенцы поки-
дают гнездо и улетают в море активным, но неуверенным полетом. Первые
тренировочные полеты происходят в 40 дней. Средний вес вылетевшего
птенца 117,5 г (Thoresen, 1964), 169 г (Vermeer, Cullen, 1982). Птенцы,
вылупившиеся в конце сезона размножения, при сходе на воду имеют
меньший вес, чем вылупившиеся в начале (Vermeer, Cullen, 1982). По дан-
ным А. Торсена (Thoresen, 1964), для 75 наблюдаемых нор успех размно-
жения составил 26,6 %, потеря яиц — 58,6, потеря птенцов от числа вылу-
пившихся 35,5 %. Однако автор отмечает, что это не естественное соот-
ношение, так как многие птицы бросили норы, не выдержав беспокойства
со стороны исследователей. По данным Д. Мануэля, из 52 тыс. пар, гнез-
дившихся на островах Фараллон в 1969—1970 гг., 36 тыс. пар успешно
вырастили птенцов, успех размножения составил 0,69 птенца на гнездо
(Manuwal, 1974; Speich, Manuwal, 1974). Для этой же популяции тем же
автором приводятся следующие данные: существует значительное коли-
чество неразмножающихся птиц — 40 тыс., которые на 56 % состоят из
потенциально половозрелых особей (начиная с трехлетнего возраста)
и на 44 % — из одно-двухлетних. Собственной территории эти птицы
не имеют. Из холостующих взрослых птиц 80 % ранее не размножалось.
Впервые ежегодно приступают к размножению 17 % популяции. Из 2600
пыжиков, пойманных у побережья Южной Калифорнии, 69 % были в воз-
расте не менее трех лет (Manuwal, 1978).
Суточная активность, поведение. Строго ночная птица. Появляются
в гнездовой колонии спустя 30 мин после захода солнца. Днем кормятся
и отдыхают в море. На море пыжики не демонстрируют никакой социаль-
ной активности. Все взаимодействия происходят на суше. Взаимные
демонстрации включают узнавание, приветствие, кружение друг с другом,
покачивание головой, поклоны, трясение головой из стороны в сторону,
поднимание крыльев. При территориальных конфликтах применяются
атакующие выпады, удары клювом, преследования. Спариванию предше-
ствуют поклоны, покачивание головой, имитация клевков, щебетание.
После спаривания обе птицы поправляют оперение, отряхиваются, взмахи-
вают крыльями. Активные бои в колонии развертываются, когда большин-
ство нор в колонии еще пустует. Описано несколько форм социальных
111
Семейство Чистиковые
взаимодействий, напоминающих игры: «пятнашки», «чехарда», «салочки»,
в которых могут принимать участие несколько птиц. Погружение клюва
в воду, характерное также для чистиков, объясняется несколькими причи-
нами: для питья, высматривания хищников, смачивания пищи, локализа
ции жертв перед нырком, ополаскивания солевых желез. Известно, что
активность погружений возрастает при беспокойстве (Thoresen, 1964).
Питание. Кормятся в море в нескольких километрах от побережья
(Payne, 1965), ныряя за кормом или склевывая его на поверхности воды
(Gabrielson, Lincoln, 1959). Взрослые и птенцы потребляют различных
планктонных ракообразных — эуфазиид, копепод, амфипод — 6—30 мм
длиной, реже — мальков рыб 15—45 мм (Thoresen, 1964; Manuwal, 1974).
Средний вес пищевого комка может быть 17,6 г при разбросе 8,6—45,6 г
(Vermeer, Cullen, 1982) или 27,8 г при разбросе 13—35 г (Manuwal, 1974).
Пробираясь с кормом по тоннелю к птенцу, родитель постоянно щелкает
клювом, а птенец отвечает ему частыми локационными щелчками (Thore-
sen, 1964).
Враги, неблагоприятные факторы. Большое количество птенцов уничто-
жают западные чайки Larus occidentalis, нападающие и на взрослых птиц
(Thoresen, 1964; Manuwal, 1974). На островах Фараллон на 52 тыс. пар
пыжиков приходится 10 тыс. пар западных чаек, которые успешно таскают
из неглубоких нор и трещин яйца и птенцов. Однако рост популяции
в первую очередь сдерживает недостаток мест, пригодных для гнездования,
а не влияние хищников (Manuwal, 1974). На Алеутских островах и остро-
вах у побережья Аляски численность алеутских пыжиков сильно подорвали
проникшие туда лисицы, но в настоящее время, когда лисы уничтожены,
популяция может восстановиться (Sowls et al., 1978). Со стороны человека
пыжикам грозят следующие опасности: как и всех ночных птиц, пыжиков
привлекает свет, поэтому они часто летят на искусственный источник света
и разбиваются о препятствия. Местные жители добывают пыжиков, прима-
нивая их светом костров. Кроме того, привлеченные теплом теплоэлектро
станции на Фараллоне, пыжика гибнут от угарного газа (СО) (Gabrielson,
Lincoln, 1959; Thoresen, 1964).
Хозяйственное значение, охрана. Хозяйственного значения не имеют.
Специальных мер охраны в СССР не предусмотрено.
Род Aethia Merrem, 1788
Большая конюга —
Aethia cristatella (Pallas, 1769)
Crested anklet (англ.), Schopfalk (нем.).
Син. (лат.): Alca cristatella Pallas, 1769.
Статус. Гнездящийся, кочующий, частично зимующий вид.
Общая характеристика и полевые признаки. Птица среднего размера,
темно-серого цвета, на воде кажется черной. На голове хохол из загибаю-
щихся вперед узких перышек того же цвета, что и оперение головы. Клюв
яркий, оранжево-красный с желтоватой вершиной. На гнездовой колонии
криклива. В весеннее время ее голос, похожий на лай, в ясные безветрен-
ные дни можно слышать на воде, где птицы обычно держатся стаями,
иногда очень большими. Одиночные птицы встречаются редко. Вообще для
большой конюги стайный образ жизни характерен в течение круглого года.
112
Большая конюга
Полет быстрый, над водой летает низко. Над колониями, в зависимости от
их положения на береговом склоне, может летать на высоте до полукило-
метра. От всех чистиковых птиц отличается однотонной окраской; от малой
конюги — большими размерами и, кроме того, имеет две (более короткие),
а не шесть, белые косицы на голове; в полете от белобрюшки — более
округлыми крыльями. Хохол в полете прижимается к голове и не заме-
тен.
Подвидовая систематика. Монотопический вид.
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла: . . Рулевых
перьев 14. Размеры самцов: длина (в мм) крыла 134—152 (среднее 140,4;
п=7), клюва 10,2—13,1 (среднее 12,2; п=7), цевки 26,2—29,7 (среднее
28,0; п=8); самок: длина крыла 125—144 (среднее 136,1; п=20); клюва
10,9—13,1 (среднее 12,0; п=11), цевки 25,5—28,6 (среднее 26,7; п=11).
Высота клюва у самцов равна 13,0+0,4 мм (п=30), у самок — 11,3±
+0,4 мм (п=30). Средний вес (в г) самцов и самок в начале сезона раз-
множения 300,3 (п=84) и в его конце — 265,1 (м=77) (Дементьев, 1951;
Bedard, 1969b; Cramp, 1985; материалы Зоологического музея МГУ).
Окраска. Половой диморфизм почти не выражен. Самец отличается
от самки более высоким клювом. В полевых условиях при соответствующем
навыке самца можно отличить по более массивной голове.
Самец и самка в брачном наряде. Вся спинная сторона тела почти чер-
ная, щеки, передняя часть шеи, грудь темного шиферного цвета, который на
брюхе и подхвостье несколько светлее. Сложенные крылья черные, внеш-
ние опахала маховых и концы первостепенных маховых сверху черные,
внутренние опахала маховых буровато-черные, снизу все маховые сереб-
ристо-серые. По мере изнашивания маховые светлеют и перед линькой
у некоторых птиц они могут приобретать светло-коричневый цвет. Верхние
кроющие крыла и рулевые черные, нижние кроющие крыла темно-шифер-
ного цвета. На лбу хохол из 14—30 загибающихся вперед перьев длиной
до 50 мм. От глаза назад — полоска из узких удлиненных перышек белого
цвета. В начале сезона размножения у больших конюг на голове разви-
ваются неразличимые в полевых условиях белые волосовидные перышки,
выпадающие после откладки яиц. Радужина белая или желтовато-белая.
Клюв яркий, оранжево-красного цвета с более светлой, желтоватой верши
ной. В этот период на клюве развиваются украшающие пластины: первая,
прикрывающая ноздри, вторая, лежащая под ноздрями вдоль режущего
края надклювья, третья находится в углу рта («розетка») и четвертая,
узкая пластинка лежит вдоль режущего края подклювья до угла рта.
Цевка и пальцы сверху серовато-голубые с черными прожилками между
щитками, цевка и пальцы снизу, а перепонки сверху и снизу черные.
Самец и самка в зимнем наряде. Аналогичен брачному, но хохол более
короткий и белая полоска за глазом менее выражена. Клюв желтовато-
коричневый, без украшающих пластин и по этой причине кажется более
длинным.
Пуховой наряд. Птенец покрыт густым пухом. На голове и брюхе его
длина 8 мм, спине 14 мм и на крыльях 11 мм. Цвет пуха на темени и
затылке может быть от черновато-серого до смолянисто-черного, на горле
черноватый, боках и крыльях черновато-серый и на брюхе от светло-серого
до черновато-серого. Радужина первоначально черная, но в возрасте около
8 Птицы СССР 113
Семейство Чистиковые
недели светлеет, приобретая черновато-серую окраску. Клюв серый с белым
яйцевым зубом. Цевка и пальцы светло-серые сверху и более темные снизу.
Яйцевой зуб отпадает в возрасте около 10 дней.
Гнездовой наряд. Надевают в месячном возрасте. У птиц в этом наряде
темя, затылок, уздечка, спина, надхвостье, рулевые и маховые перья,
а также кроющие крыла черновато-серые, кроющие уха и лопаточные
темно-серые, небольшой хохол, достигающий 5 мм длины, черно-серый,
однако он может и отсутствовать. Белая полоска за глазом неясно выражена
и прерывиста. Подбородок, горло, грудь и брюхо светло-серые, бока и
подхвостье серые. Клюв черновато-серый, его высота на уровне оперения
меньше длины на 3,2—2,6 мм, тогда как у взрослых птиц эта разность равна
1,0—1,2 мм, либо высота и длина клюва равны. Радужина жемчужно-серая.
Цевка и пальцы светло-серые сверху и более темные снизу.
Первый зимний наряд. Видимо, аналогичен гнездовому наряду, но
хохол у птиц в этом наряде несколько длиннее, чем в предыдущем.
Второй зимний наряд. У птиц в этом оперении уже хорошо выражена
белая полоска за глазом, хохол развит, но его длина меньше, чем у взрослых
птиц.
Второй летний наряд. Птицы в этом оперении похожи на взрослых
в брачном наряде, но украшающие пластинки на клюве у них меньшего
размера.
На о-ве Св. Лаврентия была встречена большая конюга с белыми грудью
и крыльями — частичный альбинос (Sealy, 1969).
Линька. Знания о ходе линьки у большой конюги неполны, несмотря
на то что ее изучали как в нашей стране, так и за рубежом (Козлова, 1957;
Bedard, 1969b; Bedard, Sealy, 1984). Как и у конюги-крошки, у большой
конюги, видимо, нет линьки из гнездового в первый зимний наряд, однако
после схода на воду у молодых птиц отрастает хохол. Птицы в первую зиму
отличаются от более старших птиц коротким хохлом, небольшим размером
клюва и отсутствием украшающих пластин на клюве.
В середине первого лета после вылупления птицы линяют из первого
зимнего сразу во второй зимний наряд, минуя первый летний. Линька
у этих птиц, видимо, начинается раньше, чем у взрослых. Следующей
весной они линяют из второго зимнего во второй летний наряд. От полово-
зрелых птиц они отличаются бледным цветом и меньшими размерами
украшающих пластин на клюве и более коротким хохлом, который, однако,
длиннее, чем у больших конюг в первом зимнем наряде.
Послебрачная линька протекает так же, как и у конюг-крошек, но более
интенсивно. У потерявших кладки или птенцов птиц смена как первосте-
пенных маховых, так и контурного пера начинается на 2—4 нед. раньше,
чем у успешно размножающихся птиц. В это же время линяет и хохол. Суще-
ствуют различия в сроках послебрачной линьки в разных частях ареала. На
севере ареала смена внутренних первостепенных маховых начинается
в начале июля, тогда как южнее, в районе Курильских островов, в июле—
августе. В 1987 г. на колонии мыса Уляхпэн (Чукотский полуостров)
смена первостепенных маховых у больших конюг началась в конце периода
насиживания (конец июля—начало августа), тогда как на о-ве Талан
птицы на этой же стадии линьки были ко времени слета птенцов (начало
сентября) (данные Н. Б. Конюхова). К сожалению, количество материала
в коллекциях зоологических музеев МГУ и ЗИН АН СССР недостаточно,
114
Большая конюга
Рис. 29. Область распространения большой конюги:
а — места гнездования, б — область зимовок и миграций, в — районы концентрации птиц иа зимовках
чтобы составить представление о различиях смены оперения у большой
конюги на всей территории ареала.
В конце сезона размножения отпадают украшающие пластины клюва:
сначала в углу рта, затем на подклювье и в последнюю очередь на над-
клювье. На о-ве Св. Лаврентия к концу репродуктивного периода в начале
сентября 10 % птиц уже сбросили украшающие пластинки. На о-ве Талан
в конце августе—начале сентября большая часть птиц сбросила украшаю-
щие пластинки и лишь у некоторых старая рамфотека на коньках над- и
подклювья. Клюв после линьки становится сероватым с оранжевым оттен-
ком, высота его уменьшается. Предбрачная линька у больших конюг
начинается в конце марта—начале апреля и длится около полутора меся-
цев. Вероятно, что в это время происходит смена оперения головы, шеи,
горла и верхней части спины.
Распространение. Гнездовой ареал. Побережье и острова Берин-
гова и Охотского морей (рис. 29); самые восточные колонии на Алеутской
гряде известны на островах Шумагина, хотя незначительное количество
птиц встречается и в акватории о-ва Кадьяк, но там не гнездится. В СССР
достоверно гнездится в следующих пунктах: о-в Ратманова, юго-восточное
побережье Чукотского полуострова (мысы Лысая Голова, Имтук, Уляхпэн,
Ягночымло и Кенилин), о-в Верхотурова, Командорские острова, Средние
Курильские острова: Чиринконтан, Ловушки, Райкоке, Матуа, Топорков,
Расшуа, Ушишир, Симушир и Черные Братья; восточное и западное побе-
8*
115
Семейство Чистиковые
режье Пенжинской губы, включая ее острова, о-в Ионы, о-в Сахалин,
о-в Талан (Тауйская губа, Охотское море), Ямские и, возможно, Шантар-
ские острова (Гизенко, 1955; Велижанин, 1978; Харитонов, 1980; Нечаев,
1986; данные А. С. Китайского, А. Н. Кондратьева, Н. Б. Конюхова).
В Чукотском море кочующие большие конюги могут встречаться от
мыса Шмидта на западе до мыса Барроу на востоке; на о-ве Врангеля отме-
чается как редкий залетный вид (Козлова, 1957; Стишов и др., 1985).
22 сентября 1937 г. на р. Юкон была найдена молодая птица. Место находки
расположено более чем в 600 км от устья реки и в 100 км от моря, отгоро-
женного горами (Geist, 1939). За пределы гнездового ареала известны
залеты в Исландию (Gramp, 1985) и в Калифорнию (округ Мэн) (Weyman,
1980).
Зимовки. Часть птиц остается зимовать в пределах гнездового
ареала там, где море не замерзает; круглогодично встречается на акватории
близ островов Прибылова и Алеутских островов. Часть птиц откочевывает на
юг до Приморья и Японии на западном побережье Тихого океана. На
восточном побережье важным районом зимовки является о-в Кадьяк
(Gabrielson, Lincoln, 1959; Sowls et al., 1978). В северо-западной части
Тихого океана большие конюги зимуют в районах Командорских и Куриль-
ских островов, островов Сахалин и Хоккайдо, образуя большие скопления
в акватории средней части Курильской гряды и у южной оконечности
о-ва Сахалин. Небольшое количество конюг встречено зимой по трещинам
и разводьям в ледовых полях Охотского моря, иногда далеко от открытой
воды. В суровые зимы откочевывает дальше на юг до 35—36° с. ш. Во время
метелей птицы, потеряв ориентацию, могут залетать в глубь суши, где, обес-
силев, гибнут (Yamashina, 1931; Austin, Kuroda, 1953; Гизенко, 1955; Ku-
roda, 1960; Kuroda, 1963; Мараков, 1965; Нечаев, 1969; Трухин, Косыгин,
1986).
Миграции. О миграциях больших конюг известно мало. В. П. Шунтов
(1965) отмечает, что конюги начинают лететь вдоль побережья Чукотки
со второй половины августа, массовый пролет идет в течение сентября. В это
время большие стаи конюг встречаются на юге Анадырского залива и в аква-
тории мыса Наварин. Пролет также идет и вдоль американского побережья,
но он менее интенсивен. По всей вероятности, в начале пролета летят
негнездящиеся птицы, поскольку вылет птенцов в северной части ареала
происходит в первой декаде сентября. Некоторая часть птиц задерживается
в водах Чукотки до третьей декады ноября (Козлова, 1957).
Численность. Общая численность вида оценивается в 4 млн особей,
в том числе в СССР не менее 2 млн, однако из-за недостаточной изученности
побережья и крайне приблизительных данных о численности вида во мно-
гих частях ареала общая численность, вероятно, занижена. По территории
гнездового ареала большая конюга встречается крайне неравномерно,
образуя огромные колонии при наличии обильной кормовой базы и мест для
гнездования. Наиболее крупные из них расположены на о-ве Св. Лаврен-
тия — 517 тыс. особей, мыс Ягночымло (Чукотский полуостров) —
500 тыс., о-в Булдырь (Алеутские острова) — 250 тыс., о-в Талан —
170 тыс., о-в Матыкиль (Ямские острова) — 220 тыс., о-в Горелый (Алеут-
ские острова) — 186 тыс., о-в Крузенштерна — 140 тыс., о-в Св. Матвея —
110 тыс., о-в Чиринконтан (Курильские острова) — 100 тыс., крупная
колония находится также на о-ве Ратманова (Портенко, 1973; Велижанин,
116
Большая коиюга
1978; Sowls et al., 1978; Golovkin, 1984; Lensink, 1984; данные А. С. Китай-
ского, А. Л. Кондратьева, H. Б. Конюхова).
Местообитания. Большая конюга — морская птица, встречается как
в открытом море, так и в прибрежных водах. С сушей связана только в пе-
риод размножения. В это время подавляющее большинство птиц придержи-
вается гнездовых участков и прилегающих к ним акваторий. Большие
конюги гнездятся в незадернованных осыпях, однако на о-ве Талан неболь-
шая часть птиц устраивает хдезда ,Д, .стар.зд.,НОЖх тор^^вГТТбдосодящие
для гнездования осыпи являются одним из лимитирующих факторов,
обусловливающих распределение колоний по побережью. Е. П. Кнудтсон и
Г. Б. Бёд (Knudtson, Byrd, 1982) отмечают, что при образовании подхо-
дящей для гнездования осыпи она сразу же занимается птицами. Места
гнездования могут располагаться как в непосредственной близости от моря,
так и в некотором удалении от него (Bedard, 1969а), а по вертикали — от
нескольких метров до полукилометра над уровнем моря. Большие конюги
могут гнездиться с плотностью до 99 особей на 200 м2, предпочитая участки
осыпи, на которых диаметр слагающих их камней равен 60—80 см (Bedard,
1969а) . Гнездовые камеры размещаются как на глубине В.несколько метров,
так и у самой"гГОверхпрсти. Отдельные пары могут гнездиться в щелях
скальных” обрывов изолированно от колонии, что, вероятно, случается
крайне редко.
Размножение. Половой зрелости достигают на третьем году жизни
(Sealy, Bedard, 1973). В район гнездовых колоний в южных частях ареала
прилетают в конце апреля, в северных частях — во второй декаде мая
(Searing, 1977; данные А. С. Китайского, Н. Б. Конюхова). Прилетевшие
птицы в течение 1—2 нед. держатся в море, а затем занимают гнездовые
колонии. Неполовозрелые особи, составляющие 30—35 % популяции, в те-
чение всего сезона размножения находятся в районе колоний (Bedard,
1969а).
Сведения по поведению больших конюг скудны. Согласно С. П. Харито-
нову (1980), большие конюги токуют на больших, возвышающихся над
поверхностью осыпи камнях. При спорах за место на камне птицы прини-
мают угрожающую позу (рис. 30), наклоняясь в сторону конкурента. При
большей степени возбуждения птицы взъерошивают перья на зашейке.
В ситуации, близкой к драке, большая конюга принимает S-образную позу.
Самец, начиная токовать, несколько раз встряхивает головой и взъеро-
шивает перья на шее. При каждом новом встряхивании он все выше и выше
поднимает голову, выпячивает вперед грудь и издает лающие звуки, делая
резкий выпад головой вверх при каждом новом крике. Кроме того, у боль-
шой конюги можно наблюдать токование парой — «биллинг». Самец и
самка, соприкасаясь клювами, сидят друг против друга, издавая низкую
трель. Также можно выделить позу ухаживания: птица распластана на суб-
страте, шея слегка выгнута вниз и перья на ней прижаты. Эта поза в основ-
ном адресуется самками самцам, токующим в одиночку, либо в паре. Как
правило, самцы агрессивно реагируют на ухаживание самок.
По наблюдениям В. А. Зубакина на о-ве Талан (Охотское море),
социальная структура поселений больших конюг довольно сложна. Пары,
поселяющиеся в отдельных норах под камнями, имеют небольшой индиви-
дуальньгй участок перед входом в нору? с кбтбрбго изгоняются Другие ОСоби.
п то ж^Тремя самцы-хозяева таких участков могут иметь большую терри-
117
Семейство Чистиковые
Рис. 30. Элементы брачного поведения большой конюги
торию для токования, в пределах которой их взаимоотношения построены
по типу линейной иерархии.;Пплчияенные~ самцы могут токовать здесь
только" при отсутствии самца-доминанта. В его присутствии они прижи-
мают перья на голове и шее (в результате чего принимают облик самки) и
начинают, как самки, перебирать клювом перья на шее доминанта. Самцов,
пытающихся токовать, доминирующий самец изгоняет.
Большой конюге свойствен гнездовой консерватизм. С. Сили (Sealy,
1975).пишет, что прилетевшие на колонию птицы размещаются по снежни-
кам, покрывающим гнездовую осыпь, не беспорядочно, а только там, где им
предстоит гнездиться. Они токуют над своими гнездовыми камерами и по
мере освобождения осыпи от снега занимают их. Сходную картину в 1987 г.
на о-ве Талан наблюдали А. С. Китайский и С. П. Харитонов.
Сроки откладки яиц на севере ареала зависят от погодных условий
весны и прошедшей зимы. При многоснежных зимах и холодных веснах
массовая яйцекладка происходит примерно на две недели позже, чем при
ранних веснах, хотя откладка первых яиц начинается примерно в одни и те
же сроки, поскольку даже в неблагоприятные сезоны существует неболь-
шое количество подходящих для гнездования мест, не покрытых снегом.
После особо снежных зим, когда снежные наносы лежат на колонии до сере-
дины лета, большие конюги могут откладывать яйца прямо на снег. Эти
яйца птицы бросают даже без попытки насиживания (Sealy, 1975b). В юж-
ных частях ареала, где осыпи освобождаются от снега к середине мая,
задержку яйцекладки могут вызвать только неожиданные снегопады
(Knudtson, Byrd, 1982). Откладка и насиживание яиц начинается через
118
Большая конюга
4—6 недлюсле прилета: на о-ве Св. Лаврентия — 13 июня—7 июля (в зави-
симости от погодных условий) (Sealy, 1975b) на побережье Чукотского
полуострова — во второй декаде июня (данные Н. Б. Конюхова), на Алеут-
ских островах (о-в Булдырь) — 24 мая—7 июня (Knudtson, Byrd, 1982),
на Курильских островах — в первой декаде июня (Гизенко, 1955), на
о-ве Талан в 1987 г. — в конце мая (данные А. С. Китайского).
Кладка состоит из одного яйца белого цвета. Яйцо овальное, поверх-
ностьТкорлупыТ^ёюкб^рництая, матбвдя. Размеры (в мм): 50—60X32,5—
42,5 (среднее 54.2X37,9; п=30) (Дементьев, 1951). Яйцо откладывается
без подстилки на грунт. Большая конюга, как самая крупная из конюг,
занимает все подходящие для гнездования пустоты, выгоняя оттуда мелкие
виды. Гнездясь в «пещерках», насиживающие птицы располагаются одна
от другой на расстоянии около метра и более. Если между ними находятся
камни, не позволяющие птицам видеть друг друга, это расстояние сокраща-
ется до 15 см (Knudtson, Byrd, 1982).
Насиживают кладку оба родителя, меняясь на яйце 1 раз в сутки, при-
чем в гнезде часто ночуют обе птицы (Byrd et al., 1983). Насиживание
на о-ве Булдырь длится 40—41 день, на о-ве Св. Лаврентия — 34—37 дней
(Sealy, 1968). Такой разброс в сроках насиживания С. Сили (Sealy,
1984) объясняет тем, что из-за неблагоприятных условий (заливания
гнезд дождевой водой) птицы могут оставлять гнезда, а затем возобнов-
лять насиживание. Время насиживания при этом увеличивается до 44 дней,
а вылупившиеся птенцы, как правило, не выживают. Первые птенцы на
Курилах отмечались в начале июля (Гизенко, 1955), в 1974 г. на о-ве
Ионы — 10 июля (Харитонов, 1980); массовое вылупление на о-ве Св.
Лаврентия — 29 июля—11 августа (Searing, 1977), на о-ве Булдырь —
5—12 июля (Lewbel, 1983).
Птенцов выкармливают оба родителя, пронося пищу в шейном мешке
(Портенко, 1973), а также и в зобе (Bedard, 1969b). За один раз взрослая
птица способна принести от 250 до 900 мелких ракообразных (Портенко,
1973), масса одной такой порции в среднем составляет 20—22 г; самцы
приносят 15—29 г пищи, самки 14—26,5 г. Так как в шейный мешок впа-
дают протоки слюнных желез, то его содержимое представляет собой
однородную массу (Bedard, 1969b). В период выкармливания птенцов
взрослых птиц на колонии можно видеть практически в любое время дня.
Согласно данным Г. В. Бед с соавторами (Byrd et al., 1983), двупиковая
активность на колонии во время выкармливания птенцов сохраняется;
в течение утреннего пика родители успевают принести птенцам корм
дважды, вечером корм приносится один раз. Кормиться больше конюги
могут как около берега, так и на расстоянии 30—50 км от него (Bedard,
1969b). Нами стаи кормящихся птиц были встречены за 100 км от бли-
жайшего места гнездования.
Вылет птенцов в разных частях ареала происходит в разные сроки:
на о-ве Булдырь первые слетки появляются к концу июля, а массовый
вылет зарегистрирован в первой декаде августа (Knudtson, Bwrd, 1982),
на о-ве Ионы — во второй декаде августа (Харитонов, 1980), на о-ве Св.
Лаврентия — с середины августа до второй декады сентября с пиком
3—9 сентября (Searing, 1977). У юго-восточного побережья Чукотского
полуострова слетки в море отмечались с конца августа до второй декады
сентября (данные Н. Б. Конюхова).
119
Семейство Чистиковые
Эмбриональная и постэмбриональная смертность соответственно равны
26 и 35 % при успешности размножения 43 % (Knudtson, Byrd, 1982).
Суточная активность, поведение. Большая конюга — дневная птица,
имеющая на гнездовых колониях в течение суток два пика активности —
в утреннее и вечернее время (Харитонов, 1980; Byrd et al., 1983; данные
Н. Б. Конюхова и др.). Для конюг во время пиков активности очень
характерно «роение». К гнездовым колониям южного побережья Чукот-
ского полуострова с кормежки птицы летят стайками от нескольких до
200—250 особей; на подлете к колонии стайки могут сливаться в непре-
рывные потоки. Прилетев в акваторию колонии, стайки птиц садятся на
воду, иногда птицы начинают кормиться. То одна, то другая стайка взле-
тает и направляется к колонии, но едва достигнув береговой линии, пово-
рачивает в море и опять садится на воду. З ак продолжается около часа.
За это время на воде уже собирается несколько тысяч птиц, сбившихся
в одну или две стаи. Вот, наконец, в воздух поднимается вся стая, вытяги-
ваясь в удлиненное облако. Она пересекает береговую линию и заходит
на посадку, но с первого раза садятся только несколько десятков птиц,
а вся стая, повернув, летит в море и, описав круг, опять заходит на посадку.
На этот раз садится около половины птиц. К этой «карусели» присоеди-
няются птицы, прилетающие с моря, и над колонией образуется непрерыв-
ное кольцо из летящих птиц. Время пиков активности зависит от этапов
репродуктивного периода и от состояния погоды. Интенсивность «роения»
уменьшается к концу гнездового сезона, а также во время сильных ветров
и туманов.
Как уже отмечалось выше, часть птиц проводит ночь в гнездах, но
в июле 1984 г. мы видели тысячные стаи больших конюг, ночующих в море
в 3—4 км мористее колонии. Возможно, это были неразмножающиеся
особи.
Питание. По данным Ж. Бедара (Bedard, 1969b), в питании больших
конюг существуют некоторые различия, связанные с циклом размножения.
С момента прилета и до вылупления птенцов в рационе птиц 48 % состав-
ляют представители семейства Gammaroidea, 25 % — Mysidacea, 17,1 % —
Parathemisto libellula и только 5,7 % — Thysanoessa spp., тогда как
в период выкармливания птенцов основной кормовой объект — Thysa-
noessa spp. (56 %), а также 28,7 % в рационе составляет Calanus finmar-
chicus. В акватории островов Прибылова основой в питании больших
конюг является также Thysanoessa spp. (68,7 %) и амфиподы (30 %),
среди которых большую часть составляет Parathemisto libellula (Hunt
et al., 1981). У берегов Чукотки пищей большим конюгам служат копеподы
(Copepoda) с небольшой примесью шизопод (Schisopoda). Вероятно, как
случайная пища могут потребляться моллюски (Портенко, 1973). Наи-
большая глубина, на которую ныряют большие конюги за планктоном, —
35-40 м (Bedard, 1969b).
Враги, неблагоприятные факторы. Естественными врагами больших
конюг являются сапсан, белоголовый и белоплечий орланы, бургомистр
и завезенный на многие острова Алеутской гряды песец (Murie, 1940;
Sowls et al., 1978; Knudtson, Byrd, 1982; данные А. С. Китайского).
В рационе белоголовых орланов большие конюги могут составлять 11,3 %
от всей потребляемой пищи. Бургомистры могут питаться большими
конюгами как случайно (6 % потребляемой пищи), так и специально
охотясь на них, и тогда они уже составляют 67 % в питании этих чаек
120
Малая конюга
(Knudtson, Byrd, 1982). Кладки и птенцы в ряде случаев подвергаются
нападению красной полевки Clethrionomys rutilus и полевки-экономки
Microtus oeconomus. В результате гибнет 1,9—3,4 % яиц и 5,3 % птенцов
(Sealy, 1982). Во время схода птенцов на воду при сильном встречном
ветре они сносятся на берег, где часть из них гибнет (Sealy, 1968). Пара-
зитирование клещей (до 11 штук на птенца) также увеличивает смертность
птенцов.
В незначительном количестве взрослые птицы добываются весной эски-
мосами. Небольшое количество птиц попадает в сети при коммерческом
лове рыбы (Ainley et al., 1981; Ogi, 1984). Ввиду перспективности Берин-
гова моря на разработку месторождений нефти предполагается, что воз-
можные разливы нефти будут наносить серьезный ущерб как кормовой
базе, так и самим птицам.
Хозяйственное значение, охрана. Прямого хозяйственного значения
не имеет. Вместе с другими морскими птицами охраняется в заповеднике
«Магаданский» (Ямские острова).
Малая конюга —
Aethia pygmaea Gmelin, 1789
Whiskered auklet (англ.), Bartalk (нем.).
Статус. Гнездящийся, кочующий, зимующий в некоторых местах,
возможно, оседлый вид.
Общая характеристика и полевые признаки. Мелкий чистик с плотным
веретенообразным телом, коротким и толстым ярко-красным клювом.
Верхняя сторона тела черная, нижняя — сероватая, более светлая на
брюшке. На суше сидит и передвигается, опираясь на цевку, реже подни-
мается на пальцы. Часто полулежит на камнях. Хотит медленно и неук-
люже. Полет быстрый, маневренный. Летает низко над водой или очень
высоко, поднимаясь на несколько сотен метров обычно стайками. Прек-
расно плавает и ныряет. На поверхности воды гребет лапами, для пере-
движения под водой пользуется крыльями (подводный полет). На воде
держит тело высоко над водой, голова обычно низко посажена. От других
видов конюг отличается наличием пучков длинных волосовидных перьев,
идущих от углов рта книзу вдоль щек («усов») и кверху к затылку. В пе-
риод гнездования совместно с другими конюгами образует колонии в ка-
менистых осыпях на скалистых побережьях островов. На воде обычно
молчаливы. На суше вблизи колоний .издают тихие щебечущие звуки,
а в полете — хрипловатый свист.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. При анализе географической изменчивости
конюг (Feinstein, 1959) были отмечены различия в размерах (особенно
длины крыла) и окраске птиц, обитающих на Алеутских и Курильских
островах. Изменчивость имеет клинальный характер, и конюги с Коман-
дорских островов занимают промежуточное положение, приближаясь
к особям курильской популяции. Высказано предположение, что алеут-
ская и курильская популяции могут иметь статус подвидов (Aethia pyg-
maea pygmaew Gmelin и A. camtschatica Lepechin). Однако это разделение
требует дальнейшего обоснования и пока не получило признания.
121
Семейство Чистиковые
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла: V. . . Рулевых перьев
14. Размеры: длина (в мм) крыла самцов 99,4—118 (среднее 106,9),
самок 97,3—119 (среднее 107,3); клюва самцов 8,4—11,2 (среднее 9,2),
самок 9,0—11,0 (среднее 9,5); плюсны самцов 19,2—23,1 (среднее 21,6),
самок 20,1 — 22,0 (среднее 21,1); масса (в г) самцов и самок 93—139 (сред-
нее 120,2) (Дементьев, 1951; Feinstein, 1959; Карташев, 1979; Knudtson,
Byrd, 1982; Материалы Зоологического музея МГУ).
Окраска. Самец и самка в брачном наряде. Полового диморфизма
в окраске нет. Голова, верхняя сторона тела, хвост черные или серовато-
черные. Шея спереди, грудь и бока тела черновато-серые. Низ груди,
брюшко, а также подхвостье беловатые или белые. Маховые, верхние
кроющие крыла заметно темнее подмышечных перьев. На лбу имеется
длинный хохолок из черных, загибающихся вперед полукольцом украшаю-
щих перьев. Над глазом светло-серая бровь. По бокам головы располо-
жены три пары пучков белых украшающих удлиненных нитевидных
перьев. Верхние и нижние пучки с каждой стороны головы начинаются
от общего белого пятна, отделенного от надклювья узкой черной полоской.
Нижний пучок, или «усы», начинается на уровне края рта и идет ниже
глаза вдоль щеки. Верхние пучки проходят над глазом к затылку и вы-
ступают над теменем в виде двух небольших хохолков. Средние пучки
белых перьев начинаются от заднего края глаза и спускаются книзу
на всю длину шеи. Клюв кораллово-красный с белой вершиной, выпуклый
по коньку, с узкими щелевидными ноздрями. Ноги светло-серые с черно-
ватыми плавательными перепонками и черными когтями. Радужина бело-
ватая.
Самец и самка в зимнем наряде. Окраска на горле становится белова-
той, а зоб и грудь серыми. Пропадает черный хохолок украшающих перьев,
а пучки белых перьев, хотя и сохраняются, но становятся значительно
короче. В результате смены рамфотеки окраска клюва меняется на жел-
товатую с черной вершиной.
Пуховой наряд. Сверху темно-бурый, середина брюшка светлее и серее
(Козлова, 1957).
Гнездовой наряд. Отсутствуют украшающие перья. Голова и верхняя
сторона тела черновато-серая и светлее, чем у взрослых. Окраска низа
сходна с таковой у взрослых птиц в зимнем наряде. Клюв темный.
Линька. Взрослые имеют две линьки: полная послебрачная и пред-
брачная. Полная линька начинается в июле, когда родители еще кормят
птенцов. В это время меняется контурное оперение, часть маховых и ру-
левые. Заканчивается она, по-видимому, в конце октября. Предбрачная
линька начинается в декабре и заканчивается в начале мая (Дементьев,
1951; Козлова, 1957). Птенцы спускаются в море практически полностью
оперившимися и способны к полету (Козлова, 1957). Редкие пушинки
сохраняются на шее и крестце (Карташев, 1979).
Распространение. Гнездовой ареал расположен в Беринговом
и Охотском морях; гнездится на Командорских островах (Мараков, 1957;
Карташев, 1979), в южной и западной частях Алеутской гряды (Lensink,
1984), в центральной части Курильских островов (Эскарма, Расшуа,
Среднева, Итуруп, Уруп, Симушир, Райкоке, Ушишир, Матуа) (Austin,
Kuroda, 1953; Гизенко, 1955; Велижанин, 1969; Клумов, личное ссобщ.),
на Ямских островах (Велижанин, 1975), о-ве Ионы (Харитонов, 1975),
122
Малая конюга
возможно, на островах Пенжинской губы (Яхонтов, 1975) и о-ве Монерон
(Нечаев, 1975) (рис. 31).
Зимовки. Изучены слабо. Предполагается, что в некоторых местах
(например, на Командорских островах) оседла (или совершает неболь-
шие кочевки) (Гизенко, 1955; Мараков, 1975). В ноябре малых конюг
отмечали у берегов Камчатки
(Лобков, 1980). В небольшом
количестве встречается зимой
у Южного Сахалина (Шунтов,
1965). Многочисленна зимой
в центральной части Куриль-
ского архипелага. Единичные
особи проникают к югу до япон-
ских островов Хондо и Сикоку
(Гизенко, 1955; Yamashina,
1931).
Миграции. Изучены недо-
статочно. Весной появляются
вблизи колоний в конце апреля.
Осенняя массовая откочевка
начинается в середине августа
(Knudtson, Byrd, 1982).
Численность. Общая числен-
ность на земном шаре не изве-
стна. В западных и центральных
частях Алеутских островов из-
вестно 10 гнездовых колоний
с общей численностью 6400 осо-
бей, предполагается, что их мо-
жет быть и больше. В восточной
части архипелага обитает от 10
до 15 тыс. особей. Величина
алеутской популяции в целом
оценивается в 50 000 птиц
(Byrd, Gibson, 1980; Lensink,
1984).
На Командорских островах
гнездится, по-видимому, не-
сколько тысяч особей. Учеты
гнездовий там не проводились.
Известны только учеты в море
в гнездовое время (июнь—ав-
густ) — 340 особей/50 км марш-
рута на о-ве Медном и 168 осо-
бей/50 км маршрута на о-ве
Беринга (Карташев, 1961). На
Курильских островах, по дан-
ным А. Г. Велижанина (1969),
в центральной части гряды гнез-
дится около 1 млн больших и
малых конюг. Доля малых КО-
Рис. 31. Область распространения малой конюги
а — места гнездования, б — область зимовок
и миграций
123
Семейство Чистиковые
нюг не указана. На Ямских островах (о-ва Матыкиль и Атыкан) отмечено
гнездование 2000 особей {Велижанин, 1975). В остальных местах гнездо-
вания численность не известна.
Местообитания. Типично морская птица, обитатель прибрежных мор-
ских районов. Вне шельфовой зоны может встречаться очень редко зимой..
Гнездовое местообитание — мелкие скалистые островки, щели в скалах
и каменистые осыпи на несколько метров выше уровня моря. Обычно
гнездится в колониях совместно с большими конюгами и конюгами-крош-
ками, качурками (Гизенко, 1955).
Размножение. Возраст наступления половой зрелости не известен.
Появление в местах гнездования происходит в различных районах при-
мерно в одинаковые сроки — в середине мая и варьирует по годам в зави-
симости от скорости таяния снега (Гизенко, 1955; Knudtson, Byrd, 1982).
Для гнездования занимают полости в скалах и между камнями объемом
всреднем 162,5 см3, т. е. заметно меньшим, чем у больших конюг, и сходным
с таковым у конюг-крошек. Предпочитают гнездиться в каменистых
осыпях, а не в почвенном субстрате. При недостатке удобных мест конку-
рируют из-за них с конюгами-крошками.
На Алеутских островах откладка яиц отмечается 24 мая—5 июня.
Инкубация занимает 35—36 дней. В. кладке .одноональноематошь-белое
яйцо. Размеры 45,2—48,5X31,5—33,5 мм (Козлова, 1957; Гизенко, 1955).
Насиживают яйцо попеременно оба родителя, сменяясь через сутки (Byrd
et al., 1983). Вылупление птенцов 24 июня—8 июля (Knudtson, Byrd,
1982). Спуск птенцов в море начинается в конце июля, массовый — в пер-
вые 10 дней августа. Выходят они из нор в темное время суток (Knudtson,
Bird, 1982). Вес птенцов при спуске может достигать 127 г (Карташев,
1979). Наибольшая гибель эмбрионов происходит на ранних стадиях наси-
живания. Успех размножения на Алеутских островах высок — 86 %
(Knudtson, Byrd, 1982).
Суточная активность, поведение. В течение всего сезона размножения
суточная активность характеризуется двумя пиками: утром с рассветом
и второй — перед сумерками. Оба родителя, видимо, ночуют на суше.
Период дневного покоя во время откладки яиц короче, чем во время инку-
бации. Насиживая яйцо, один из родителей вынужден голодать в течение
суток и потому должне кормиться дольше, чтобы компенсировать голо-
дание. Приуроченность конюг к колониям зависит от условий погоды,
распределения и обилия пищи в окрестных водах и от стадии гнездового
цикла (Knudtson, Byrd, 1982).
Питание. По характеру питания — типичный планктофаг. Основным
кормом служат мелкие ракообразные: амфиподы (гаммариды), изоподы,
декаподы, гастроподы (Козлова, 1957; Дементьев, 1951).
Враги, неблагоприятные факторы. Основные враги птенцов — серо-
крылые чайки Larus glaucescens и песцы. Взрослые птицы иногда стано-
вятся жертвами сапсана и белоголового орлана (3 % от общего числа
кормовых объектов) (Knudtson, Byrd, 1982).
Хозяйственное значение, охрана. Хозяйственного значения не имеет.
Вместе с другими природными объектами охраняется в заповеднике «Ма-
гаданский» (Ямские острова), в заказнике местного значения на Коман-
дорских островах.
124
Конюга-крошка
Конюга-крошка —
Aethia pusilia (Pallas, 1811)
Least auklet (англ.), Zwergalk (нем.).
Син. (лат.): Uria pusilia Pallas, 1811.
Син. (рус.): крошечная конюга.
Статус. Гнездящийся, кочующий, частично зимующий вид.
Общая характеристика и полевые признаки. Самая маленькая среди
чистиковых птиц. В море держится стаями, иногда можно встретить
и одиночных птиц. С воды взлетает легко, почти без разбега. Полет быст-
рый и более маневренный, чем у других чистиковых. В гнездовое время
«роится» над колонией и прилежащей акваторией в утренние и вечерние
часы. На камни в отличие от большой конюги садится легко. По суше
передвигается на цевке, однако может ходить и на пальцах. Голос — резкое
щебетание, похожее на крик волнистых попугайчиков.
Подвидовая систематика. Монотипический вид.
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла IV. . . Рулевых
перьев 14. Размеры самцов: длина (в мм) крыла 91 —101 (среднее 95,9;
п = 9), клюва 7,8—8,8 (среднее 8,5; и = 9), цевки 17,5—20,5 (среднее
19,0; и=9); самок: длина крыла 95—97 (среднее 95,6; п—1), клюва
8,0—8,8 (среднее 8,4; и=6), цевки 17,9—20,0 (среднее 18,8; и=8);
без определения пола: длина крыла 93—101 (среднее 96,8; п — 5), клюва
8,2—9,0 (среднее 8,5; и = 5), цевки 19,3—20,0 (среднее 19,6; и = 5).
Масса (в г): до сезона размножения 93,4 (и = 85), во время сезона раз-
множения 73,3—94,1 (среднее 84,5; п = 44) и после него — 83,4 (и = 52)
(Bedard, 1969b; Roby, Brink, 1986a; Материалы Зоологического музея
МГУ).
Окраска. Половой диморфизм не выражен.
Самец и самка в брачном наряде. Темя, затылок, задняя часть шеи,
спина, поясница, надхвостье черные. На лбу, уздечке, темени среди тем-
ного оперения разбросаны узкие белые перышки, которые на лбу и темени
длиннее, чем у основания клюва и на уздечке. За глазом эти белые перышки
сливаются в полоску. Очень узкая полоска вокруг основания клюва
коричневато-серая. Подбородок однотонны^ серый, горло чисто-белое.
Кроющие уха и бока шеи серые. В верхней части шеи спереди черно-
бурое «ожерелье», которое книзу становится пестрым. Грудь, брюхо и под-
хвостье пестрые: по белому фону разбросаны темные пятна, образованные
за счет темно-бурых каемок перьев. Пятнистость от «ожерелья» к под-
хвостью уменьшается и последнее почти чисто-белое, за исключением
нескольких пестрин. Крыло сверху черное, среди плечевых есть перья
с беловатыми каемками. Маховые и их кроющие буровато-черные. На
4—6 внутренних второстепенных маховых беловатые каемки. Снизу махо-
вые серые, большие кроющие серые с беловатыми концами, малые крою-
щие белые. Подмышечные коричневые. Рули черные. Клюв трехцветный.
Самая вершина надклювья серовато-белая, полупрозрачная. Вершинная
треть надклювья и половина подклювья красные. Весь режущий край
надклювья и вершинная половина режущего края подклювья темно-
красные. Остальная часть клюва, включая роговидный вырост на гребне
надклювья, развивающийся в период размножения, — черная. Цевка
и пальцы сверху серовато-голубые с черными прожилками между щитков;
125
Семейство Чистиковые
суставы более темные из-за более густой сети прожилок. Перепонка
сверху и снизу, цевка и пальцы снизу черные. Когти черные.
Самец и самка в зимнем наряде. Лоб, темя, затылок, задняя часть шеи,
спина, надхвостье, хвост, крылья черные. Щеки буровато-серые. Горло,
шея, грудь, бока, живот и подхвостье белые. Белые узкие перышки на го-
лове не выпадают. «Рог» на клюве становится меньше или исчезает
полностью. Плечевые перья белеют. Подбородок сероватый. Клюв чер-
новато-бурый со слабо-желтой вершиной.
Пуховой наряд. Птенец покрыт густым пухом, длина которого на голове
равна 6 мм, на брюхе — 13 мм, на крыльях — 7 мм. Пух на голове черный,
на спинной стороне его цвет варьирует от черновато-серого до черновато-
коричневого, на горле и брюхе серый, боках и крыльях черновато-серый.
Радужина черновато-серая, клюв серый с белым яйцевым зубом. Цевка
и пальцы светло-серые сверху и более темные снизу.
Гнездовой наряд. Это оперение птенцы приобретают в возрасте около
месяца. У птиц в этом наряде темя, затылок, уздечка, спина, поясница,
маховые и рулевые перья черные. Кроющие уха темно-серые. На передней
части головы находятся белые щетинковидные перья и неясная белая
полоса позади глаза. Подбородок, горло, грудь, бока, брюхо, плечевые,
нижние кроющие хвоста и вершины второстепенных маховых белые.
Подмышечные и нижние кроющие крыла белые с небольшим количеством
светло-серых перьев, образующих полосатый рисунок. Радужина жем-
чужно-серая, клюв черновато-серый без роговидного выроста на коньке
надклювья.
Первый зимний наряд. Птицы в этом пере отличаются от птиц в гнез-
довом наряде лишь более серыми боками шеи, от птиц во втором зимнем
и окончательном зимнем нарядах — меньшим размером клюва и отсут-
ствием роговидного выроста на надклювье.
Второй зимний наряд. По-видимому, практически неотличим от зим-
него наряда взрослых птиц.
Второй летний наряд. Приобретается в результате весенней линьки
на вторую весну после вылупления. У птиц в этом наряде подбородок
становится полосатым, грудь, бока и живот — пестрыми. Горло в отличие
от взрослых птиц темное. Радужина белая. Клюв бурый, на нем появля-
ется характерный роговидный вырост.
Дважды на о-ве Св. Лаврентия встречены конюги-крошки с аберрант-
ной окраской оперения: неполовозрелый самец с белыми рулевыми и
слеток — полный альбинос (Sealy, 1969).
Линька. Ход линьки у конюг-крошек изучен недостаточно полно.
Наиболее подробно этот процесс освещен в работе С. Сили и Ж. Бедара
(Bedard, Sealy, 1984). Некоторые данные имеются также в книге Е. В. Коз-
ловой (1957). По мнению первых двух авторов, молодые птицы проводят
свою первую зиму в гнездовом пере. Зимой или весной у них происходит
лишь частичная линька небольших участков контурного оперения, приво-
дящая к образованию сероватого налета на боковых сторонах шеи. В даль-
нейшем, в июле—сентябре, в результате полной линьки птицы меняют
первый зимний наряд фактически на зимний наряд взрослых птиц,
минуя первый летний наряд. На вторую весну после вылупления у птиц
происходит частичная линька, которая, вероятно, начинается позже, чем
у взрослых птиц, и завершается только тогда, когда они прилетают на
гнездовые колонии. Во время этой линьки заменяются рулевые и все
126
Конюга-крошка
контурное перо, за исключением области поясницы и бедер. Конюги-
крошки в этом наряде похожи на взрослых птиц, за исключением серого,
а не белого горла и большей пятнистости брюха. Узкие белые перышки на
голове короткие. «Рог» на клюве небольшой, клюв бурого цвета.
Зимний наряд взрослые птицы приобретают в результате почти полной
линьки. Ход линьки успешно размножающихся текущим летом конюг-
крошек отличается от такового неполовозрелых особей и птиц, потерпев-
ших неудачу при размножении. У гнездящихся конюг-крошек смена
первостепенных маховых начинается во второй половине июля, а через
месяц маховые меняют все размножающиеся особи. В начале сентября,
когда птицы покидают гнездовые колонии, они уже сменили или почти
сменили все первостепенные и начали смену второстепенных маховых,
которая начинается после выпадения VII первостепенного махового.
Линька контурного оперения начинается во второй половине июля или при-
мерно через две недели после выпадения XI первостепенного пера. У птиц,
неудачно размножавшихся в данном сезоне, смена контурного пера идет
более интенсивно, опережая этот процесс у размножающихся особей,
однако маховые у тех и других заменяются примерно в одни и те же сроки.
Послебрачная линька, по всей вероятности, заканчивается к концу октября.
Весенняя, предбрачная линька у взрослых птиц начинается в начале
апреля и завершается к концу мая. В это же время вершина клюва краснеет
и на надклювье появляется «рог». Смена рулевых перьев происходит
либо в течение этой линьки, либо сразу же после нее, так как в полную
осеннюю линьку птицы входят с новыми рулевыми.
Распространение. Гнездовой ареал охватывает в основном по-
бережье и острова Берингова моря, но есть колонии и в Охотском море
(рис. 32). На восток по Алеутской гряде конюга-крошка доходит до
островов Семиди. В СССР достоверно гнездится в следующих пунктах:
о-в Ратманова, мыс Уляхпэн, мыс Лысая Голова (юго-восточное побе-
режье Чукотского полуострова, о-в Верхотурова, Командорские острова,
район залива Камбальный (западное побережье Камчатки), о-в Талан
(единичные птицы), о-в Ионы и Ямские острова. Возможно гнездование
на мысах Имтук, Ягночымло и Кекилин (юго-восточное побережье Чукот-
ского полуострова), о-ве Ушишир (Средние Курильские острова), по по-
бережью и островам Пенжинской губы (Портенко, 1973; Велижанин,
1978; Лобков, 1986; данные А. С. Китайского и Н. Б. Конюхова) (рис. 33).
В летне-осеннее время периодически встречаются в водах о-ва Вран-
гель (Стишов и др., 1985), неоднократно добывались в акватории мыса
Барроу (Козлова, 1957).
Зимовки. В зимнее время конюги-крошки встречаются в акватории
мыса Лопатка (п-ов Камчатка), Алеутских и Командорских островов,
о-ва Сахалин, Приморья и северных островов Японии. Небольшие стайки
зимуют по разводьям и трещинам ледовых полей Охотского моря (Yama-
shina, 1931; Austin, Kuroda, 1953; Гизенко, 1955; Kuroda, 1960; Kenyon,
1961; Kuroda, 1963; Byrd et al., 1974; Трухин, Косыгин, 1986).
Миграции. Пути миграций сходны с таковыми большой конюги. На
Чукотском полуострове, по-видимому, могут оставаться до конца октября—
начала ноября (Портенко, 1973), у побережья Камчатки наиболее много-
численны в октябре-ноябре (Лобков, 1986), у берегов Сахалина отмеча-
ются с начала сентября, в акватории Командорских островов — с января
по март (Гизенко, 1955). К местам гнездования на юге ареала конюги-
127
Семейство Чистиковые
Рис. 32. Область
распространения
конюги-крошкн
а — места гнездования,
б — область зимовок и миграций
Рис. 33. Область
распространения
кон юг и-крошки в СССР
а — гнездовой ареал,
б — зимовки,
? — места предполагаемого
гнездования
крошки прилетают в конце апреля (Lewbel, 1983), на о-ве Св. Лаврентия
10—15 мая (Searing, 1977), на юго-восточном побережье Чукотского
полуострова в 1984 г. впервые отмечены 25 мая (данные Н. Б. Конюхова),
в районе о-ва Ратманова в 1958 г. — 23 мая (Портенко, 1973).
Численность. Самая многочисленная гнездящаяся птица Берингова
моря. Общая численность вида оценивается в 7 млн особей. Самые крупные
128
Конюга-крошка
колонии находятся в следующих пунктах: о-в Св. Лаврентия — 1003 тыс.,
о-в Крузенштерна — 980 тыс., о-в Св. Георгия (о-ва Прибылова) —
250 тыс., о-в Горелый — 402 тыс., о-в Киска — 250 тыс. и о-в Булдырь —
100 тыс. (Алеутские острова), о-в Св. Матвея — 175 тыс., Ямские ост-
рова — более 60 тыс., колония, насчитывающая более 100 тыс. особей,
есть на о-ве Ратманова (Портенко, 1973; Велижанин, 1978; Sowls et al.,
1978; Шунтов, 1986; данные Н. Б. Конюхова).
Местообитания. Морская птица, с сушей связана только в период
размножения. Единственным гнездовым биотопом являются не закреплен-
ные растительностью осыпи, где конюги-крошки гнездятся совместно
с большими и малыми конюгами. Гнездовые колонии, как правило,
расположены на берегу моря и обращены в его сторону, но могут нахо-
диться на расстоянии до полутора километров от уреза воды (Roby, Brihk,
1986b). Конюги-крошки обычно занимают верхние части осыпи, где обло-
мочный материал, слагающий осыпь, меньшего размера (в диаметре при-
мерно 30 см). Максимальная плотность гнездования на таких участках
составляет 129 особей на 200 м2 (Bedard, 1969а). Средний объем гнездовой
камеры равен 117,6 см3 (п = 40), причем из более обширных ниш конюги-
крошки в 70 % случаев изгоняются малыми конюгами и всегда — боль-
шими (Knudtson, Byrd, 1982).
Размножение. Почти всегда гнездится совместно с другими видами
конюг. Соотношение конюги-крошки и большой конюги в разных колониях
может колебаться от 1 : 10 до 20 : 1 (Fay, Cade, 1959; данные Н. Б. Коню-
хова).
Молодые конюги-крошки начинают посещать гнездовую колонию в пер-
вое же лето после вылупления. Незначительное количество особей в пер-
вом зимнем оперении встречено на о-ве Св. Лаврентия во второй половине
июня. Эти птицы были очень пугливы и не принимали участия в социаль-
ной активности колонии. Они встречались здесь не более месяца. Двухлет-
ние птицы во втором летнем наряде в июле составляли около 20 % от всех
птиц в колонии. Они уже были социально активны и некоторые из них,
вероятно, размножались, так как встречены особи, носившие корм; однако
успех размножения этих птиц не известен. Основная масса птиц присту-
пает к размножению в трехлетием возрасте, одев брачный наряд (Sealy,
Bedard, 1973; Bedard, Sealy, 1984).
Весной конюги-крошки, прилетевшие в район колоний, некоторое
время держатся в море, появляясь на гнездовых колониях в середине
мая на юге ареала и в конце месяца на севере.
Из года в год птицы занимают одни и те же гнездовые камеры. На о-ве
Св. Георгия 91 % птиц, находившихся под наблюдением, на следующий
год заняли свои старые гнезда (Roby, Brink, 1986а). В зависимости от
погодных условий откладка яиц на юге ареала происходит в конце мая—
начале июня, на севере — в июне—начале июля (Lewbel, 1983; Roby,
Brink, 1986а). Кладка состоит из одного яйца белого цвета. Скорлупа
мелкозернистая, матовая. Размеры яиц (в мм): 33,5—42,7 X26,7—30,9
(среднее 39,5 X 28,5; п = 122); масса яиц (в г) с о-ва Св. Георгия 14,95—
19,6 (среднее 17,4; и == 20) (Дементьев, 1952; Roby, Brink, 1986а). Яйцо
откладывается в гнездо без выстилки и насиживается обоими родителями
в течение 31,2 ± 2,7 дней (п = 15); в случае прерывания насиживания
птенцы могут вылупиться и через 39 дней, но, как правило, эмбрионы
гибнут (Seady, 1984). Родители на гнезде меняются примерно раз в сутки.
9 Птицы СССР
129
Семейство Чистиковые
Птенцы на о-ве Булдырь вылупляются 30 июня—9 июля (Knudtson, Byrd,
1982), на о-ве Св. Георгия — 3—13 июля (Roby, Brink, 1986а) и на о-ве
Св. Лаврентия — в конце июля—начале августа (Searing, 1977).
В зависимости от гидрологических условий и продуктивности приле-
жащей акватории за кормом конюги-крошки могут летать на расстояние
более 50 км. Корм птенцу приносится обоими родителями в шейном мешке,
а также и в зобе. Вес одной порции корма, приносимой самцом, равен
5,5—7,0 г, самкой — 3,5—8,5 г (Bedard, 1969b). Средний вес пищи, при-
несенной за один раз, — 5,28 г (и = 66); пища дается 4—7 раз в день
(в среднем 5,33; п = 12) (Roby, Brink, 1986а). Во время выкармливания
птенцов на о-ве Св. Георгия, по данным Д. Д. Роуби и К. Л. Бринка (Roby,
Brink, 1986а), на долю Calanus finmarchicus приходится 66,3 % от общего
количества пойманной добычи, а на долю копепод — 97 %; на о-ве Св. Лав-
рентия эти значения соответственно равны 88,9 и 90,7 %. Количество дру-
гих компонентов в пище в это время сокращается в несколько раз.
Птенцы покидают гнезда примерно через месяц после вылупления.
Сход птенцов на о-ве Св. Георгия происходит в утренние часы, причем
птенцы летят в море самостоятельно, без сопровождения родителей.
Вес слетков — 91,5 г (и = 11) (Roby, Brink, 1986а).
Эмбриональная смертность у конюг-крошек на о-ве Булдырь равна
32 %, постэмбриональная смертность достигает 25, успех размножения —
51 % от числа отложенных яиц; на островах Прибылова успех вылупле-
ния — 87 — 90, постэмбриональная смертность — около 24, общий успех
размножения — 66—74 % (Knudtson, Byrd, 1982; Roby, Brink, 1986a).
Суточная активность, поведение. Конюга-крошка — дневная птица.
Как и у большой конюги, у нее ярко выражена двупиковая активность
в колонии, которая в период выкармливания птенцов несколько сглажи-
вается. Взрослые птицы, потерявшие кладку, в гнездовой период продол-
жают посещать колонию; количество же неполовозрелых особей в это
время сокращается. На островах Прибылова основная масса конюг-крошек
прилетает в колонию вечером, в 20 ч 00 мин—22 ч 30 мин, 82 % из них
с кормом. Вечером же большая часть негнездящихся птиц улетает в море.
В гнезде ночует по крайней мере один из родителей, но, как правило, оба.
Утром, в 6—7 ч, они улетают за кормом, прилетая обратно в 8 ч 00 мин—
14 ч 30 мин (Roby, Brink, 1986а).
О поведении конюг-крошек известно немного. С. П. Харитонов (1980)
отмечает, что токующие партнеры сидят рядом друг с другом; при выра-
жении угрозы (рис. 34) птица вытягивает-шею и поднимает на ней перья.
В это время она может подавать голос.
Питание. Питается планктоном, за которым ныряет на глубину до
35— 40 м (Bedard, 1969b). Основа ее питания — копеподы. В разные годы
их количество и видовой состав могут изменяться. На о-ве Св. Лаврентия
копеподы составляют 74—95 % от объема всей пищи, на о-ве Св. Матвея —
85—89 и до 97 % на островах Прибылова. В первом случае самыми много-
численными видами были: в 1964—1966 гг. Calanus marshalae, в 1976 г. —
С. plumchrus, а в 1981 г. в пробах пищи С. plumchrus было в два раза
больше, чем С. marshalae, составлявшего 26 % от объема пищевых проб.
Во втором случае преобладал С. marshalae, а у птиц с островов Прибылова
в добыче по количеству особей доминировал С. marshalae, а по весу —
С. cristatus (Bedard, 1969b; Searing, 1977; Hunt et al., 1981; Springer,
Roseneau, 1985). Питание конюг-крошек на о-ве Св. Лаврентия в период
130
Белобрюшка
выкармливания птенцов отличается от питания птиц в другие периоды их
жизни. Во время прилета и насиживания яиц птицы едят Calanus finmar-
chicus и личинок десятиногих раков (триба Caridea), которые составляют
в их питании 39,2 и 33,3 % (Bedard, 1969b). Также в состав кормов,
потребляемых конюгами-крошками, входят эвфаузиды, амфиподы, глав-
ным образом Parathemisto libellula, представители сем. Gamaroidea разме-
ром менее 7 мм.
Враги, неблагоприятные факторы. Зарастание травянистой и кустар-
никовой растительностью осыпей приводит к сокращению численности
гнездящихся здесь конюг-крошек (Roby, Brink, 1986b). На о-ве Булдырь
конюги-крошки — главный компонент в питании живущих там сапсанов,
Рис. 34. Элементы
территориального
поведения конюги-
крошки
а в питании бургомистра она может составлять треть от всей потребляемой
пищи (Knudtson, Byrd, 1982). На островах Прибылова основной ее враг —
песец. Горностай, проникший на о-в Верхотурова, привел к сокращению
численности гнездящихся здесь конюг-крошек, а возможно, и других
видов мелких чистиковых птиц (Лобков, 1986). Кроме того, взрослые
птицы могут подвергаться нападению полевок Microtus oeconomus и
Clethrionomys rutilus, приводящему в некоторых случаях к их гибели.
Полевки же являются причиной гибели яиц, птенцов и слетков. Потери
среди этих возрастных групп, находившихся под наблюдением на о-ве
Св. Лаврентия в 1966—1967 гг., составляли 3,4—6,0; 1,0—9,3 и 2,3 %
(Sealy, 1982). Незначительное количество птиц гибнет, путаясь в сетях,
при коммерческом лове лосося. В будущем, при разработке месторожде-
ний нефти, птицы могут страдать от нефтяного загрязнения (Ainley et al.,
1981; Lensink, 1984; Ogi, 1984).
Хозяйственное значение, охрана. Прямого хозяйственного значения
не имеет. Вместе с другими морскими птицами охраняется в заповеднике
«Магаданский» (Ямские острова).
Род Cyclorrhynchus Каир, 1829
Белобрюшка —
Cyclorrhynchus psittacula (Pallas, 1769)
Parakeet auklet (англ.)» Papageialk (нем.).
Син. (лат.): Alca psittacula Pallas, 1769.
Статус. Гнездящийся, кочующий, частично зимующий вид.
Общая характеристика и полевые признаки. Среднего размера чисти-
ковая птица контрастной двухцветной окраски: нижняя часть груди,
брюхо и подхвостье белые, остальное оперение черное. Клюв красный
9*
131
Семейство Чистиковые
с загибающимся вверх подклювьем. Голос — вибрирующий свист, похож
на свист чистика. На суше, как и многие чистиковые, белобрюшка пере-
двигается, опираясь на цевку, но может непродолжительное время ходить
на пальцах. Хорошо плавает и ныряет, используя для передвижения под
водой крылья. С воды поднимается с разбега; при испуге улетает или
ныряет. Как в море, так и на суше в большие стаи не собирается.
Похожа на большую конюгу, на воде отличаясь от нее отсутствием
хохла и белым подхвостьем, в полете — более узкими, остроконечными
крыльями и меныпей частотой взмахов крыльев, при виде «сзади» —
белыми полосками по краям спины. От ипатки и кайры — меныпими
размерами, а также от первой — сплошь темным оперением головы, а от
второй — клювом красного цвета.
Подвидовая систематика. Монотипический вид.
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла: I < II > III > IV. . . Рулевых
перьев 14. Размеры самцов: длина (в мм) крыла 142—157 (среднее 151,8;
п = 4), клюва 15,1 — 16,1 (среднее 15,9; п — 3), цевки 28,0—30,4 (среднее
29,1; п = 4); самок: длина крыла 149—154 (среднее 150,6; и — 3); клюва
13,3—15,4 (среднее 14,3; п = 22), цевки 29,0—29,2 (среднее 29,1; п — 2);
без определения пола: длина крыла 142—157 (среднее 151,3; п = 14),
клюва 13,8—15,5 (среднее 14,8; п == 13), цевки 28,0—30,9 (среднее 29,2;
п = 14). Масса (в г) самцов и самок вскоре после прилета 297,4 (п = 11),
после сезона размножения 281,1 (п=29); самцов 245—347 (среднее
292,3; п=79), самок 255—337 (среднее 285,7; n=71) (Bedard, 1969;
Sealy, Bedard, 1973; Зоологический музей МГУ).
Окраска. Полового диморфизма нет.
Самец и самка в брачном наряде. Лоб, щеки, темя, затылок, задняя
часть шеи, спина, надхвостье серовато-черные либо графитно-черные, на
боках шеи некоторое количество белых перьев образует светлую струй-
чатость. Подбородок и горло темно-серые, причем на горле выражена
белая пятнистость, постепенно переходящая на передней части шеи в бе-
лый фон, на котором за счет серых концов перьев образуется крапчатость.
Грудь, брюхо, бока и подхвостье чисто-белые; под крыльями на боках
небольшое количество перьев с темными вершинами. Перья голени с внеш-
ней стороны коричнево серые. Наружные опахала маховых черновато-
серые, темнее внутренних. Последние темно-серые, слегка серебристые.
Большие кроющие черные, малые кроющие образуют чешуйчатый рисунок
за счет темных внешних и светлых внутренних опахал. Маховые и их
большие кроющие снизу серебристо-серые, малые кроющие и подмышеч-
ные дымчато-коричневые. Рулевые черные. Цвет оперения зависит и от
времени года: птицы в свежем пере более темные сверху, но к концу
периода размножения оперение изнашивается и становится более бурым;
маховые даже иногда приобретают коричневый цвет. За глазом полоска
из узких белых перьев. У основания надклювья развиваются украшающие
пластинки буроватого цвета. Мягкое вздутие вдоль нижнего края под-
клювья от ноздрей до разреза рта беловатое или розовато-белое. Радужина
белая, либо слегка желтоватая. Цевка и пальцы сверху светлые^ серовато-
голубые с темными прожилками между щитками. Суставы более темные
из-за большего количества щитков на них. Перепонки между пальцами
голубовато-серые, края перепонок черноватые. Цевка, пальцы и перепонки
снизу грязно-черные. Когти черные.
132
Белобрюшка
Самец и самка в зимнем наряде. Зимний наряд взрослых птиц сходен
с брачным, но весь низ тела, за исключением черновато-серого подбородка
и горла, белый. Украшающие пластинки у основания надклювья отпадают,
но форма клюва остается прежней.
Пуховой наряд. «У вылупившегося птенца голова и все туловище по-
крыты густым пухом. Окраска пуха на голове, передней стороне шеи,
спине и дорзальной стороне крыльев варьирует у разных птенцов от
черновато-бурых тонов до дымчато-серых. Низ груди и брюхо светлее:
буровато-серые или светло-серые. Длина отдельных пушин (в мм): на го-
лове — 8, на спине — 22, на бедрах — 23—24, на голени — 16, на брюхе —
15, кистевом отделе крыла — 7, локтевом отделе крыла — 17. Все над-
клювье и вершина подклювья темно-серые, остальная часть подклювья
и складки в углах рта светло-бурые или светло-серые. Вершина надклювья
изогнута под прямым углом. Подклювье ложкообразной формы, его
вершина изогнута кверху. В области ноздрей мягкое вздутие, ноздря
щелевидная длиной 3 мм. Яйцевой зуб белый, 1—2 мм. Ротовая полость
и язык розовые, у некоторых особей язык серый или розовый с небольшими
серыми пятнами или каймой. Радужина темно-коричневая. Окологлаз-
ничное кольцо голой кожи черное. Цевка спереди и пальцы сверху голу-
бовато-серые, иногда с розовым налетом, лапа снизу и когти темно-серые,
почти черные. Перепонка телесного цвета, иногда с серым налетом»
(Михтарьянц, 1981).
Гнездовой наряд. Птенцы приобретают его к концу первого месяца
жизни. Он, а также первый и второй зимние наряды очень похожи на
зимний наряд взрослых птиц. Различия между этими нарядами, возможно,
будут найдены в результате дальнейших исследований. Второй летний
наряд. Сходен с предыдущим, за исключением более развитой белой
полоски за глазом и окрашенных пластинок на клюве, отличаясь только
количеством темного оперения на подбородке и горле, а также цветом
клюва. Можно отметить, что у неполовозрелых птиц длина крыла, размер
клюва и вес тела меньше, чем у взрослых птиц.
Описан один случай отклонения от нормальной окраски: на о-ве Св.
Лаврентия встречена самка, у которой спинная сторона, крылья, хвост
и подбородок были желтовато-коричневыми. Птенец этой птицы имел
нормальную окраску (Sealy, 1969).
Линька. Ход линьки и смена нарядов у белобрюшки изучены недо-
статочно. Известно, что неполовозрелые птицы линяют из гнездового
наряда, по-видимому минуя первый зимний и первый летний наряды,
сразу во второй зимний наряд в июле—августе; в те же сроки происходит
линька птиц из второго летнего в окончательный зимний наряд. При
этом идет смена маховых и контурных перьев. Второй летний наряд
белобрюшки носят в течение нескольких недель во второе лето после
вылупления. У взрослых птиц послебрачная линька начинается со смены
контурного оперения в конце августа—начале сентября; смена первосте-
пенных маховых — с внутренних первостепенных и происходит несколько
позднее. Утрата украшающих пластин клюва по разным данным проис-
ходит в июле—сентябре со смены пластин подклювья. Линька в брачный
наряд происходит, вероятно, в марте-апреле (Козлова, 1957; Bedard,
1969b; Портенко, 1973; Bedard, Sealy, 1984).
Распространение. Гнездовой ареал. Побережье и острова север-
ной части Тихого океана (рис. 35) — от Берингова пролива на юг по
133
Семейство Чистиковые
Рис. 35. Область
распространения
белобрюшки
а — места гнездования,
б — область зимовок
и миграций
Рис. 36. Ареал
белобрюшки в СССР
Обозначения те же, что
на рис. 35
азиатскому побережью до о-ва Тюлений, по американскому побережью
до залива Принца Вильямса. В СССР гнездится: на о-ве Ратманова,
в окрестностях мыса Дежнева, по юго-восточному побережью Чукотского
полуострова — от мыса Чукотский до мыса Беринга, на островах Верхо-
турова, Карагинский, Командорских и Средних Курильских островах
134
Белобрюшка
(Райкоке, Ловушки, Чиринкотан), о-ве Матыкиль (Ямские острова),
о-ве Талан (Тауйская губа), островах Ионы, Сахалин и Тюленьем (Вась-
ковский, 1946; Белкин, Велижанин, 1965; Нечаев, 1969; Портенко, 1973;
Велижанин, 1978; Харитонов, 1980; Михтарьянц, 1981; Нечаев, 1986;
данные А. С. Китайского, А. Я. Кондратьева, Н. Б. Конюхова). В летне-
осеннее время встречается в Чукотском море — от мыса Сердце-Камень
на западе до мыса Барроу на востоке (Козлова, 1957) (рис. 36). За пре-
делы гнездового ареала известен залет в Швецию (Cramp, 1985).
Зимовки. Белобрюшки зимуют в свободной от льда южной части
Берингова моря, по западному побережью океана до северных островов
Японии, по восточному — почти до северных границ Мексики; в цент-
ральной части Тихого океана неоднократно отмечалась в северо-западной
части Гавайского архипелага в акваториях островов двух атоллов — Куре
и Мидуэй (Yamashina, 1931; Kenyon, 1937; Austin, Kuroda, 1953; Гизенко,
1955; Gabrielson, Lincoln, 1959; Kuroda, 1960; Kenyon, 1961; Kuroda,
1963; Clapp, 1986; Schreiber, 1987).
Миграции. Миграции белобрюшки изучены недостаточно полно. В ме-
стах гнездования на юге ареала они появляются в конце апреля, на
севере — во второй половине мая и примерно через неделю становятся
многочисленными. В особо благоприятные годы птицы могут прилетать
в начале мая (Sealy, Bedard, 1973). Отлет из северной части Берингова
моря в первой половине сентября, сразу же после вылета птенцов. Пролет
к местам зимовок идет вдоль азиатского и американского побережий
океана. На юге гнездового ареала белобрюшки встречаются круглый год.
Численность. Общая численность вида составляет не менее 900 тыс.
особей, причем основная часть популяции (около 90 %) обитает в амери-
канском секторе Северной Пацифики (Sowls et al., 1978). Незначительная
численность белобрюшки на азиатском побережье, возможно, объясняется
недостаточной изученностью данного региона. По мнению Ж. Бедара
(Bedard, 1969b), невысокая численность белобрюшек по сравнению с та-
ковой конюг обтясняется различиями в питании; питаясь хищным зоо-
планктоном, она занимает более высокое положение в пищевых цепях
по сравнению с конюгами.
Крупнейшие колонии белобрюшек находятся на островах Прибылова
(184 тыс.), о-ве Човиет (о-ва Семиди) (58 тыс.), о-ве Кинга (42 тыс.),
о-ве Крузенштерна (20 тыс.), о-ве Ратманова (20 тыс.), о-ве Горелый
(Алеутские острова) (20 тыс.), о-ве Св. Матвея (18 тыс.), о-ве Матыкиль
(Ямские острова) (15 тыс.), о-ве Талан (60 тыс.), о-ве Ионы (12 тыс.)
(Карташев, 1961; Портенко, 1973; Велижанин, 1978; Sowls et aL, 1978;
Харитонов, 1980; Михтарьянц, 1981; данные Н. Б. Конюхова).
Местообитания. По всему гнездовому ареалу белобрюшка связана со
скалистыми побережьями, обрывами, осыпями и прибрежными нагро-
мождениями валунов. Совместно с конюгами, как правило, не гнездится,
предпочитая осыпи, иногда очень пологие, покрытые слоем почвы и расти-
тельностью. Гнезда белобрюшки устраивают в естественных укрытиях:
расщелинах скал, пустотах и нишах под й между камнями, под кочками
на склонах и в старых порах топорков. Заселяя пологие склоны, бело-
брюшки конкурируют за места гнездования с топорком, но, гнездясь
в неглубоких либо с узким входом убежищах, ослабляют эту конкуренцию
(Bedard, 1969а; Портенко, 1973; Sealy, Bedard, 1973; Михтарьянц, 1981).
Размножение. Белобрюшки становятся половозрелыми на третий год
135
Семейство Чистиковые
жизни. На гнездовых колониях на север ареала появляются в зависи-
мости от погодных условий 4—13 мая. Большинство птиц, вернувшихся
в колонию, занимают свои старые гнездовые камеры.
Размножающаяся часть популяции на о-ве Топорков (Командорские
острова) составляет 30—40 %. На о-ве Св. Лаврентия самцов несколько
больше, чем самок, 53 и 47 % соответственно (n=159) (Sealy, Bedard,
1973; Михтарьянц, 1981). Токуют птицы рядом с убежищем, в котором
находится гнездо. После многоснежных зим, когда гнездовья еще покрыты
снегом, белобрюшки токуют на снегу над своими гнездовыми камерами
и по мере таяния снега занимают их (Fay, Cade, 1954; Sealy, Bedard, 1973).
При ограниченном количестве подходящих для гнездования мест размер
охраняемого участка может достигать 40—50 м, но, как правило, он бывает
меньше и в основном охраняется только вход в нору и прилежащая
к нему территория.
Спаривание происходит на воде. Откладка яиц начинается через
3—4 недели после массового прилета птиц в район гнездовых колоний.
В кладке только одно яйцо, но в случае его потери в начале периода раз-
множения возможна и повторная кладка; однажды новое яйцо было отло-
жено через 16 дней после гибели первого (Sealy, Bedard, 1973; Михтарьянц,
1981). Яйца откладываются без выстилки прямо на грунт, иногда в гнезде
присутствуют мелкие камешки. •
Яйца эллиптической формы, свежеотложенные — белые, иногда со
слабым'*5ёленоватым или голубоватым оттенком. Поверхность скорлупы
мелкозернистая, матовая. Размеры (в мм): 48,9—58,7 X 33,0—40,0 (сред-
псп 53,"8X37,4; п -= 88). Масса (в г): свежеотложенных яиц 36,6—39,1
(среднее_37,5;_ пмиц. навешенных незадолго до вылупления. 35.4
43,5~(среднее 37,6; п=6), масса проклюнутых яиц 28,4—34,5 (среднее
33,1; н=4^“^Дементье]в, 1951; Sealy, Bedard, 1973; Михтарьянц, 1981).
Насиживание начинается сразу же после откладки яйца и продол-
жается 34—37 дней. Насиживают оба родителя, причем у них развиваются
по два латеральных наседных пятна. Птенцы вылупляются в первой
половине июня на о-ве Булдырь (Алеутские острова), на месяц позже
на Командорских островах и с 25 июля до конца первой декады августа
на о-ве Св. Лаврентия и о-ве Ионы (Sealy, Bedard, 1973; Харитонов, 1980;
Михтарьянц, 1981; Knudtson, Byrd, 1982). Процесс вылупления птенца
длится 6—8 дней. Масса (в г) только что вылупившихся птенцов на
о-ве Св. Лаврентия — 25,5—34,4 (среднее 28,1; п = 4), на Командорских
островах — 24,7—31,0 (среднее 25,6; п = 5). Первые два дня после вылуп-
ления один из родителей обогревает птенца, а другой кормит его дважды
в день, утром и вечером. С 4—5-дневного возраста птенца начинают кор-
мить и днем. В недельном возрасте у птенца устанавливается терморегу-
ляция. С этого времени родители перестают его постоянно обогревать
и кормят вдвоем, причем один из родителей прилетает в гнездо с кормом
в 2,5—3 раза чаще другого. Интенсивность кормления в утренний пик
активности более чем в два раза выше, чем в вечерний. Время между при-
летами колеблется от 25 мин (1 — 3 мин в случае прилета обоих родителей)
до 3 ч; в дневные часы эти интервалы увеличиваются до 3—7 ч (Мих-
тарьянц, 1978, 1981). Основным кормом в этот период на о-ве Св. Лаврен-
тия являются Calanus cristatus (30,9 %), Thysanoessa spp. (23,9%),
Parathemisto libellula (17,5), а также небольшое количество рыбы (2,8 %)
(Bedard, 1969b). Пища птенцу приносится в шейном мешке и в зобе 6—
136
Белобрюшки
8 раз в сутки. Вес корма, приносимого самцами за один раз, составляет
13,5—28,0 г, самками — 15,5—23,5 г. При кормлении птенец пищит и те-
ребит оперение на шее родителя, слегка ударяя клювом в вершину шейного
мешка, провоцирует отрыгивание пищи. Пища выталкивается из шейного
мешка небольшими порциями. За время пребывания птенца в гнезде его
вес увеличивается в 7 —10 раз, достигая к вылету 194,1—259,1 г (среднее
222,6 г; п = 6) (Sealy, Bedard, 1973). Увеличение веса птенцов в течение
первых 25 дней происходит стабильно, а затем его значения начинают
колебаться. С возрастом у птенца изменяется цвет радужины от темно-
коричневого через буро-коричневый и светло-серый до беловатого с чуть
заметным серым оттенком перед вылетом. Яйцевой зуб отпадает в воз-
расте 14—16 дней.
Сход птенцов в море происходит в конце июля—первой половине
августа на Алеутских островах и в конце августа—начале сентября на
севере Берингова моря (Sealy, Bedard, 1973; Knudtson, Byrd, 1982). За
несколько дней до вылета птенцы выбираются из своих гнезд и тренируют
крылья. Слет происходит в утренние часы. Птенцы летят в море и садятся
на воду примерно в полукилометре от берега. В дальнейшем с родителями
они более не связаны.
Эмбриональная смертность у белобрюшки равна 32,3 %, постэмбрио-
нальная — 33,8, успех размножения колеблется в пределах 52—56 %
(Sealy, Bedard, 1973; Searing, 1977; Lewbel, 1983).
Суточная активность, поведение. Белобрюшка — дневная птица. По
материалам Э. А. Михтарьянц (1978), у белобрюшек на Командорских
островах на гнездовой колонии активность строго двупиковая, не завися-
щая от погодных условий: утром с5до12—13чс максимумом в 7 —10 ч
и вечером после захода солнца и до темноты (с 21—21 ч 30 мин до 22—
23 ч). Периодичность активности разных возрастных групп на колонии
неодинакова. У неразмножающихся и молодых птиц активность двух-
пиковая с дневной кормежкой в море. У размножающихся птиц ритм
суточной активности изменяется в связи с заботой о потомстве. С. Сили
и Ж. Бедар (Sealy, Bedard, 1973), проводившие исследования на о-ве Св.
Лаврентия, отмечали всего один, утренний пик активности, причем у не-
размножающихся и молодых птиц максимальная активность была смещена
к полудню, тогда как у размножающихся птиц максимум активности
приходился приблизительно на 10 ч утра. Возможно, что в период «белых
ночей» птицы могут кормиться и в ночные часы.
Ночуют белобрюшки, вероятно, в море недалеко от колонии, по крайней
мере ночью в норах остаются только насиживающие и обогревающие
птицы, а утром, в предрассветные часы, белобрюшки отмечаются на ко-
лонии в незначительном количестве, а затем начинается прилет птиц
с моря (Михтарьянц, 1978).
У белобрюшек отмечается своеобразная «карусель», характерная для
некоторых чистиковых (D. Manuwal, N. Manuwal, 1979). Прилетевшие
с кормом птицы сначала собираются на воде недалеко от колонии в стаю.
Затем эта стая поднимается в воздух и летит сначала в сторону моря,
а затем, повернув, к колонии. Прилетевшие птицы могут садиться прямо
к гнезду либо на склон, а затем пешком или перелетая с камня на камень
добираться до гнезда. Описанный выше порядок прилета в гнездовую
колонию у белобрюшек характерен в начале сезона размножения. К его
концу летящие с моря птицы уже не останавливаются в море перед
137
Семейство Чистиковые
колонией, а летят сразу к гнезду. Следует отметить, что в начале сезона
размножения, до вылупления птенцов, белобрюшки проводят в районе
гнездовых колоний как на скалах, так и на воде от 5 до 10 ч, затрачивая
на социальные контакты половину этого времени. Во время выкармливания
птенцов взрослые птицы у гнездовых колоний проводят не более трех
часов.
Поведение белобрюшек изучено недостаточно. Д. А. и Н. Дж. Мануэль
(D. Manuwal, N. Manuwal, 1979) отмечают у этого вида своеобразное
«пение». «Песня» исполняется как одиночными птицами, так и дуэтом,
причем в последнем случае поющие птицы не всегда являются членами
пары. Белобрюшки во время дуэта сидят друг против друга, иногда
соприкасаясь клювами. После интенсивного пения на воде отмечались
попытки спаривания. Угрожающие позы описаны Д. А. и Н. Дж. Мануэль
(1979) и С. П. Харитоновым (1980) (рис. 37). Наклонная угрожающая
Рис. 37. Элементы территориального поведения белобрюшки
поза принимается в том случае, когда расстояние между птицами доста-
точно велико; при сокращении расстояния между ними эта поза переходит
в другую, похожую на предыдущую, но клюв поднят вверх. Последняя
поза часто предшествует прямой атаке. На близких расстояниях может
также демонстрироваться вертикальная угрожающая поза. Все позы,
исключая последнюю, демонстрируются как на воде, так и на суше.
Питание. Белобрюшки планктоноядные птицы. Корм добывают в море
на расстоянии 1 — 6 км от берега, как склевывая его с поверхности, так
и ныряя на глубину до 50 м (Bedard, 1969b; Михтарьянц, 1981). О питании
взрослых птиц известно мало. По данным Ж. Бедара (Bedard, 1969b),
основным кормом белобрюшек вне периода выкармливания птенцов
в акватории о-ва Св. Лаврентия являются Parathemisto libellula и Lima-
cina, которые соответственно составляют 60,8 и 35,9 % от общего числа
пищевых объектов. Кроме того, в их рацион входят полихеты, мелкие
головоногие моллюски и мелкая рыба, составляющая 12 % от всей
потребляемой пищи, тогда как у большой конюги, имеющей сходные
размеры, на эту группу приходится только 1 %. В сравнении с большой
конюгой белоброшка предпочитает более крупную добычу. Доля круп-
ного хищного зоопланктона, имеющего размер более 7,0 мм, у первой
составляет около 10 % от веса потребляемой пищи, тогда как у второй
138
Тупик-носорог
в четыре раза больше. В акватории островов Прибылова на долю Thysa-
noessa inermis и Т. raschii от объема всей съеденной пищи приходится
10 и 19,4 %, на долю мелкой рыбы — 26,6 и полихет — 23,5 % (Hunt et al.,
1981). Л. А. Портенко (1973), исследуя содержимое шейных мешков
белобрюшки, обнаружил там остатки Calanus cristatus, Parathemisto
libellula и Policheta.
Планктон, которым питается белобрюшка, не так многочислен, как
тот, который служит пищей большой конюге и конюге-крошке, и более
равномерно распределен по акватории Берингова моря. Вероятно, что
по этой причине белобрюшки не образуют больших колоний, селятся
более равномерно, заселяя острова, на которых конюги не гнездятся.
По этой же причине на кормежке в море белобрюшки проводят на 60 %
времени больше, чем конюги.
Враги, неблагоприятные факторы. Как и у большой конюги, основные
естественные враги белобрюшки — сапсан, кречет, бургомистр, белоголо-
вый орлан, песец; также белобрюшек находили в желудках трески (Murie,
1940; Михтарьянц, 1981; Knudtson, Byrd, 1982). В репродуктивный период
потомству белобрюшек могут наносить некоторый ущерб полевки (Micro-
tus oeconomus, Clethrionomys rutilus), уничтожившие 3,2 % яиц и 20 %
птенцов, находившихся под наблюдением в 1967 г. (Sealy, 1982). Неболь-
шое количество птиц добывается эскимосами. Так же как и другие чисти-
ковые, белобрюшки гибнут в сетях при коммерческом лове лосося (Ainley
et al., 1981; Ogi, 1984). В местах разработки месторождений нефти
возможна массовая гибель птиц как от отравления нефтепродуктами,
так и от переохлаждения, связанного с ухудшением теплоизоляции за-
грязненного оперения (Lensink, 1984). Гибель птиц может происходить
при заглатывании большого числа (до 40) кусочков пластика (Мих-
тарьянц, 1981).
Хозяйственное значение, охрана. Прямого хозяйственного значения
не имеет. Вместе с другими морскими птицами охраняется в заповеднике
«Магаданский» (Ямские острова).
Род Cerorhinca Bonaparte, 1828
Тупик-носорог —
Cerorhinca monocerata Pallas, 1811
Rhinoceros Anklet Hornbilled Puffin (англ.), Utou (яп.).
Статус. В СССР гнездящийся и мигрирующий вид.
Общая характеристика и полевые признаки. Весьма крупный чистик
с темным (черноватым или бурым) верхом и белым (грудь, брюхо) низом.
Щеки, горло и зоб серовато-бурые. Клюв оранжевый, довольно высокий
с вертикальным выростом — «рогом» у основания; от углов рта и от глаз
назад идут белые косицы (птица в брачном наряде). Зимой похож на
молодого топорка, но с менее мощным клювом. Рог отсутствует.
В море молчалив. В колониях часто подает голос. Голос — мычащие,
гнусавые звуки двух типов: односложное «ууу, ууу, ууу. - »; двухсложное
«ук-ууу, ук-ууу. . .» (или «ак-ааа, ак-ааа»), У разных особей голос варьи-
рует по высоте.
Подвидовая систематика. Монотипический вид.
139
Семейство Чистиковые
Описание. Размеры. Длина (в мм) крыла самцов 181,0—195,0
(п=11) (среднее 188,5), самок 174,0—187,0 (п = 8) (среднее 178,9);
плюсны самцов 29,8—32,7 (п = 17) (среднее 31,5), самок 28,4—32,0
(п = 10) (среднее 30,1); клюва (от границы оперения) самцов 32,0—
40,0 (п = 17) (среднее 36,6); самок 30,7—36,3 (н=10) (среднее 33,5).
Масса (в г) самцов 550—670 (п = 10) (среднее 599), самок 491,5; 513,0;
533,0 (среднее 512,5) (Южное Приморье, о-в Монерон, Южные Куриль-
ские острова; коллекции БЦИ ДВНЦ АН СССР и Дальневосточного
государственного университета; данные Ю. В. Шибаева — промеры птиц
в природе).
Окраска. Самец и самка в брачном наряде. Верх головы и всего
тела, крылья, хвост и их верхние кроющие черновато-бурые. Мелкие
верхние кроющие крыльев по краю предплечья светло-бурые или бело-
ватые. Бока головы, шея, подбородок, горло, бока тела и нижние кроющие
крыльев серовато-бурые с различной интенсивностью окраски. Наиболее
темные бока головы. Переход от окрашенных боков тела и зоба к белым
груди и брюху постепенный. От углов рта под ухом отходят удлиненные
белые нитевидные перья. Такие же белые перья образуют узкую полосу
над глазом и по бокам затылка. В основании надклювья большой верти-
кальный вырост — «рог», между ним и оперением — узкая пластинка.
На киле подклювья — небольшой роговой валик. Радужина бледно-бурая,
слегка желтоватая, клюв оранжевый, конец надклювья черный, «рог»
желтоватый, вздутые края надклювья от ноздрей до угла рта желтые.
Ноги желтовато-бурые, плавательные перепонки черные.
Самец и самка в зимнем наряде похожи на взрослых в летнем наряде,
но окраска горла, шеи и зоба серая, почти без буроватого оттенка. Зимой
появляются белые удлиненные перья за глазами и по бокам клюва. Рого-
вого выроста в основании надклювья нет. Отсутствуют плоская узкая
поперечная пластинка в основании надклювья у грани оперения и малень-
кий роговой валик в основании киля подклювья.
Пуховой наряд. Сверху весь темно-бурый, снизу светлее, грудь и брюхо
серовато-бурые. На месте будущего «рога» в основании надклювья —
небольшой кожистый вырост (на тушках плохо заметен). Ноги и плава-
тельные перепонки желтоватые.
Гнездовой наряд. В целом похожи на взрослых в зимнем наряде.
Верх головы и всего тела, крылья, хвост и их верхние кроющие черные.
Бока головы темно-серые. Горло, шея, область зоба и бока буровато-
серые, более светлые. Грудь и брюхо белые, часть перьев с сероватыми,
а на подхвостье с черными вершинами. Клюв более короткий и низкий
с небольшим кожистым выступом в основании гребня надклювья.
Первый зимний наряд *. Похож на гнездовой наряд, но горло, шея
спереди и область зоба серые, без буроватого оттенка. В углах рта и за
глазами — по нескольку мелких, совсем коротких светло-бурых нитевид-
ных перышек. Клюв длиннее и выше, чем у молодых птиц осенью, но почти
вдвое ниже, чем у взрослых зимою. 1к<)сновании надклювья небольшой
вырост (описано по экземпляру от 12 декабря из Калифорнии; материалы
ЗИН АН СССР).
* По Е. В. Козловой (1957), гнездовой и первый зимний наряды не тождественны, как
считалось ранее.
140
Тупик-носорог
Линька. К декабрю, во время кочевки, заканчивается смена юноше-
ского наряда на первый зимний, или первый брачный, так как уже зимой
у молодой птицы появляются отдельные нитевидные светло-бурые перья
у углов рта и за глазами.
По Бенту (Bent, 1919), у молодой птицы в первую весну жизни линьки
не бывает, но появляются дополнительные роговые образования на клюве,
а темные каемки на перьях груди и брюха изнашиваются, так что эти
участки оперения становятся чисто белыми. Нитевидные перья на голове
меньших размеров, чем у взрослых. Клюв короче и ниже, чем у взрослых,
а киль вершинной части надклювья загибается книзу по более отлогой
дуге. Годовалые птенцы сменяют оперение очень рано — в середине июня.
Молодые птицы на втором году жизни начинают предбрачную линьку,
которая, видимо, охватывает большую часть мелкого оперения в начале
марта. К этому времени нитевидные белые перья только показываются
из пеньков (возможно, что сроки появления этих «украшающих» перьев
у разных особей различны), а на клюве роговых выростов еще нет.
Начало полной линьки взрослых в районе Южного Сахалина и на Юж-
ных Курильских островах отмечено в начале июля. К августу они теряют
значительную часть «украшающих» перьев и «рог», меняют часть кон-
турного оперения и приобретают новое оперение на наседных пятнах.
Предбрачная линька взрослых и рост белых нитевидных перьев, по
всей вероятности, происходят не раньше мая, судя по сообщению Даусона
(Dawson, 1940), который 14—17 апреля у о-ва Санта-Крус наблюдал
и добывал рогатых тупиков еще без дополнительных роговых образований
на клюве и без белых нитевидных перьев на боках головы.
К концу апреля в Южном Приморье (о-в Карамзина) тупики приоб-
ретают полный брачный наряд.
Распространение. Ареал вида в целом. Побережья и прибреж-
ные воды Северной Пацифики. Два района обитания — у берегов Азии
(между 40 и 46° с. ш.) и у берегов Северной Америки (между 48 и 59°
с. ш.) (рис. 38, 39).
Гнездовой ареал. Азия. В Южном Приморье: о-в Фуругельма
(Лабзюк и др., 1970; Назаров, Шибаев, 1984), острова Верховского, Карам-
зина, Клыкова (Лабзюк и др., 1971), о-в Чихаева (Лабзюк, 1975), вероятно,
о-в Стенина (Назаров, Шибаев, 1984). Для Шантарских островов тупик-
носорог приводится в качестве обычного вида (Дулькейт, Шульпин, 1937).
Однако в более поздних исследованиях он обнаружен не был (Яхонтов,
1977; Росляков, 1986).
На Сахалине: о-в Монерон (Takahashi, 1939; Гизенко, 1955; Нечаев,
1975), о-в Тюлений (Бычков, 1976; Нечаев, Тимофеева, 1980). Небольшое
гП^цовье на п-ове Крильон близ пос. Атласово, о котором писал А. И. Ги-
зенко (1955), в более позднее время найдено не было (Нечаев, 1936).
В. А. Нечаев (1986) предполагает возможность гнездования вида также
в некоторых других местах Сахалина.
Курильские острова: А. И. Гизенко (1955) в качестве мест гнездова-
ния приводит острова Анучина и Полонского (Малая Курильская гряда),
А. Г. Велижанин и А. Н. Белкин (1967) — о-в Пико (о-в Кунашир),
острова Анучина, Юрий, Демина, Лисьи, а также мелкие островки у Шико-
тана — Грига, Девятый Вал и Айвазовского (Малая Курильская гряда).
Ряд летних встреч у Итурупа позволяет предполагать здесь гнездование,
в частности, у о-ва Камень-Лев (Велижанин, Белкин, 1967). По данным
141
Рис. 38. Область распространения тупика-носорога
а — места установленного гнездования, б — места предполагаемого гнездования, в — места летних
встреч, г — область зимовок и миграций
Рис. 39. Область распространения тупика-носорога в СССР
Обозначения те же, что на рис. 38
142
Тупик-носорог
Ю. В. Шибаева, тупик-носорог гнездился в 1971 г. на мелких безымян-
ных островках у юго-восточного побережья Шикотана — в бухтах Маяч-
ной, Кабельной и Безымянной, кроме того, на о-ве Средний (бухта Димит-
рова) и на о-ве Девятый Вал. На о-ве Айвазовского в 1971 г. тупики не гнез-
дились.
На Командорских островах — залетный вид (Steijneger, 1985). Неясен
характер4 встреч на Средних Курилах — Черные Братья (Велижанин,
19776), Симушир, Райкоке (Yamashina, 1931).
В Японии вид гнездится на Хоккайдо (о-ва Рисири, вероятно, Ребун,
Теури, Кодзима, Дайкоку, Моюрури, Томосири) и на Северном Хонсю
(острова Цубаки и Аси) (Austin, Kuroda, 1953; Check-eist of Japanese birds,
1974; Фудзимаки, 1986). В заливе Муцу (Северный Хонсю) были добыты
птицы с яйцами, готовыми к откладке, однако колонии обнаружены
не были (Austin, Kuroda, 1953).
На Корейском полуострове, по Вон Хон Гу (1963—1965), вид гнездится
у западных и восточных берегов. Единственное конкретное место гнездо-
вания приводит Мори (цит. по: Austin, 1948) — о-в Джоб (провинция
Пхёпан-Пукто).
На американском побережье вид гнездится от Алеутских островов
(на о-ве Булдырь), вдоль п-ова Аляска и далее к югу вдоль побережья
материка до штата Калифорния (США). Крайняя южная точка гнездова-
ния — острова Фараллон (район Сан-Франциско) (Scott et al., 1974; Sowls
et al., 1978; Vermeer, 1979).
Любопытным фактом является смещенность к югу гнездового ареала
тупика-носорога в Азии по сравнению с Америкой. Отсутствие его в более
северных районах азиатского побережья (в частности, на большей части
Курильской гряды) может быть связано с воздействием четвероногих
хищников и возможной конкуренцией с топорком. По материалам, собран-
ным на Алеутской гряде, видимо, эти факторы для гнездования вида
существенны (Vermeer, 1979).
Зимовки. Выяснены недостаточно. В Азии зимует у берегов Япо-
нии — от Северного Хоккайдо (Thiede et al., 1973) к югу до Центрального
Хонсю, Кюсю и Цусимы (Check list of Japanese birds., 1974). Обычно
к югу от Фукусимы редок, однако в благоприятные в кормовом отноше-
нии годы доходит по тихоокеанскому побережью до Токио и залива Су-
руга (Austin, Kuroda, 1953). Зимует также у южных берегов Корейского
полуострова (Min, Won, 1976).
В Америке зимует в прибрежной полосе тихоокеанского побережья
до п-ова Калифорния (Birds of North America, 1966; Harrison, 1983).
Миграции. В гнездовые колонии Южного Приморья прилетает в сере-
дине апреля. Первые птицы на о-ве Карамзина в 1971 г. отмечены
12 апреля. На о-ве Монерон в 1938 г. тупики-носороги проявились 20 марта
(Takahashi, 1939), в 1949 г. — в первой декаде апреля (Гизенко, 1955).
В Японии (о-в Теури) весенний прилет начинается с конца февраля (Ku-
roda, 1963). По данным кольцевания птицы с о-ва Теури (Западный Хок-
кайдо) кочуют вдоль западных берегов Японии, птицы с о-ва Аси (Восточ-
ный Хонсю) — вдоль восточных берегов Хонсю (Japanese Birds Banding
in Becent Years, 1985).
Численность. В Южном Приморье репродуктивная часть популяции
вряд ли превышает 1000 пар. Эта оценка весьма приблизительна, так как
данные по численности недостаточны (Лабзюк и др., 1970, 1971; Лабзюк,
143
Семейство Чистиковые
1975; данные Ю. В. Шибаева). Численность на Сахалине находится в пре-
делах 2500 пар (Нечаев, 1975; Бычков, 1976; Нечаев, Тимофеева, 1980).
Основные гнездовья в СССР расположены на Южных Курильских остро-
вах (более 10 000 экз.) (Велижанин, Белкин, 1967; Велижанин, 1978).
В прошлом тупик-носорог, возможно, был более многочисленным. Во вся-
ком случае, на о-ве Монерон с 1938 по 1949 г. численность его сократилась
в несколько раз (с 14 069 до 1000 —1500 птиц) и осталась на низком уровне
(Takahashi, 1939; Гизенко, 1955; Нечаев, 1975).
Основная часть азиатской популяции обитает в Японии на о-ве Теури:
25 000 пар — в 1951 г. (Udagawa, 1953), не менее 50 000 — в 1963 г.
(Kuroda, 1963), 398 820 — в 1972 г., 302 560 пар в 1977 г. В других япон-
ских колониях гнездится, по-видимому, небольшое количество тупиков
(Фудзимаки, 1986). Состояние гнездовий на Корейском полуострове
не известно, но вряд ли там обитает много птиц. В целом репродуктивная
часть популяции в Азии, видимо, не превышает 500 000 пар.
Общая численность гнездящейся части популяции в Америке в 1976 г.
насчитывала 181 000 пар, из них примерно 56 000 пар — на Аляске,
104 000 — в Британской Колумбии, 21 000 — в штате Вашингтон, менее
100 пар в Орегоне и Калифорнии. 95 % популяции обитает на участке
побережья от о-ва Форрестер (Юго-Восточная Аляска) до о-ва Дистракшн
(т. е. на территории Канады, в Британской Колумбии) (Sowls et al., 1978;
Vermeer, 1979).
Местообитания. В гнездовое время — преимущественно небольшие
острова (с достаточно толстым почвенным слоем), расположенные в остро-
вных дугах или у материкового побережья. Непременным условием для
гнездования является отсутствие хищных млекопитающих. В период
кочевок и зимой — районы моря, примыкающие к побережью.
Размножение. Колониальная птица, гнездовья насчитывают от несколь-
ких десятков до нескольких сот тысяч пар. Гнездится рядом с другими
морскими птицами. В Приморье образует смешанные поселения со стари-
ком — Synthliboramphus antiquus, вилохвостой качуркой — Oceanodroma
monorhis, пестроголовым буревестником — Puffinus leucomelas, чернохво-
стой чайкой — Larus crassirostris, на Курилах — с топорком — Lunda cir-
rhata и тихоокеанской чайкой — Larus schistisagus.
Территориальное и брачное поведение почти не изучено. Серьезным
препятствием этому является ночной образ жизни птиц в гнездовых коло-
ниях. Фрагментарные наблюдения удалось сделать А. Торесену (Thoresen,
1983) на о-ве Теури, где этих птиц можно увидеть в дневное время.
Во время защиты территории у входа в нору хозяин гнезда обычно прини-
мает вертикальную позу, вытягивает вверх шею и голову с открытым клю-
вом, приподнимая при этом крылья и раздувая горло («upright huff»).
При появлении посторонней птицы тупик-носорог становится в наклонную
позу, вытягивает вперед шею и очень медленно (между отдельными ша-
гами иногда проходит 15 с) направляется к «врагу» («agressive hunched
walk»), последний обычно улетает или принимает оборонительную позу,
приподнимая крылья и открыв клюв («defensive winge-raise») (Thoresen,
1983).
Гнездится-м-цопах.-которые роет как на горизонтальной поверхности,
так и на склдяях рд?чимипй крутизны. Для приземления не нуждается
в“бсобых условиях — крутых склонах,. оголенных участках земли, как. это
требуёТсХ"” например," для топорка: в Восточной Пацифике обычно
144
Тупик-носорог
гнездование тупика-носорога у подножия деревьев или среди зарослей
кустарников; В таких биотопах прилетающие в колонию птицы часто
-на кусты и ветви деревьев, затем спрыгивают на землю и бегут
к норам (Vermeer, 1979). В Западной Пацифике занимает открытые
участию травяным покровом "Хколосняк — Elymus, вейник — Calamagros-
tis,''полынь — Artemisia), зарослями бамбука и оголенной почвой, в гус-
тых и высоких зарослях тупйкй-носброги протаптывают троиы (Гизенко,
1955; Fujimaki, 1961; данные Ю. В. Шибаева).
Распределение нор обусловливается в первую очередь наличием поч-
венного слоя, достаточно толстого и прочного для рытья нор (минималь-
ная толщина свода нор у птиц азиатской популяции. 10 см, максималь-
ная — 40 см). В заливе Петра Великого тупик-носорог гнездится как
в'Твёрхйёй'частй островов, так"и у подножия скал, куда не достигает прибой
и где сохраняются участки почвенного покрова. В колониях, где эти птицы
гнездятся вместо с тмюркбм, ОДи Занимают участки менее удобные для
взлета и посадки. В частности, на островках с плоской вершиной (некото-
рые колонии на Южных Курилах и Северо-Восточном Хоккайдо) топорок
поселяется у самой кромки обрыва, а тупик-носорог — в полосе за ним
(Fujimaki, 1961; данные Ю. В. Шибаева).
Ход норы обычно изогнут, а иногда и разветвлен, имеется 2—3 гнез-
довые камеры (реже одна), гнездо устраивается обычно в концевой камере.
Две” ЙТйногДа три норы могут соединяться. Длина нор птиц азиатской
популяции — от 30 до 230 см, чаще всего — 100— 500 см. Размеры
(в см) входа в нору: высота 6 —11, ширина 7—19, диаметр камеры
20— 30, высота 15—20. Лоток выстилается сухой травой (Гизенко, 1955;
Нечаев, Юдаков, 1968; Fujimaki, 1961; данные Ю. В. Шибаева).
В Южном Приморье приступает к откладке яиц в конце апреля—
начале мая. Примерно в те же сроки откладывают яйца птицы из других
колоний азиатской популяции — на Курилах, о-ве Монерон (Гизенко,
1955; данные Ю. В. Шибаева) и в Японии — на о-ве Теури (Kuroda, 1963а;
Thoresen, 1983). В Британской Колумбии (о-в Триангл) начинают откла-
дывать яйца на 3—4 нед позже (Vermeer, 1980).
В кладке одно яйцо гряящьбелого. цвета с неясными сероватыми или
буроватыми пятнами, иногда сгущающимися у тупого конца. Форма
правильно-яйцевидная. Скорлупа гладкая матовая (Нечаев, Юдаков, 1968;
ВёпТГТЭТ^'дай'йые’Ю.' В. Шибаева).
Рддмерьг лмц Дв. мм): Южное, Приморье — 61,5—75,0X42,1—50,0,
в среднем 68,7X47,2 (Austin, Kuroda, 1953; Гизенко, 1955; Нечаев,
Юдаков, 1"968г даййые Ю. В. Шибаева); Северная Америка — (п=39)
63,7 73,5X42,6 -50,0, в’среднем 68,5X46,2 (Beni, 1919).
В насиживаний принимают участие оба партнера, имеющие по два
наседных пятна. Период вылупления птенцов в Южном Приморье в 1976 г.
продолжался примерно с конца мая до конца июня (данные Ю. В. Ши-
баева). В Японии (на о-ве Теури) первые птенцы появляются в последнюю
неделю мая (Kuroda, 1963), в Британской Колумбии вылупление птенцов
в 1976 г. проходило с 1 по 22 июля, в 1978 г. — с 10 июня по 15 июля (Ver-
meer, 198(5).
Родители кормят птенца два раза в сутки. На о-ве Триангл с августа
1978 по 1979 г. птенцы получали в среднем соответственно по 64 и 66 г
пищи в сутки. Средняя продолжительность пребывания птенцов в гнезде на
о-ве Триангл — 51 день (1978 г.) и 55 дней (1976 г.), успех гнездования
10 Птицы СССР
145
Семейство Чистиковые
в этой колонии в 1975—1978 гг. был 31,9—61,7 % (Vermeer, 1979; Ver-
meer, Cullen, 1979).
В заливе Петра Великого птенцы покидают норы обычно во второй
половине июля. На Южных Курилах (о-в Шикотан) 1.VIII 1971 г. в норах
были обнаружены птенцы, готовые к вылету, а 9 и 11 августа в море вблизи
г. Малокурильск в стайках тупиков-носорогов встречены три молодые
птицы. Массовый вылет птенцов на о-ве Монерон в 1976 г. проходил
в последних числах июля.
Суточная активность, поведение. В гнездовое время на суше ведет
ночной образ жизни — прилетает в колонию после захода солнца и на рас-
свете улетает в море. В северных частях ареала это строго ночная птица —
появляется в колонии в полной темноте. В южных районах гнездования —
на американском континенте (в Орегоне и Калифорнии) и на юге Даль-
него Востока (Приморье, о-ва Монерон, Хоккайдо) птицы прилетают
на сушу в более светлое время — вскоре после захода солнца, а на о-ве
Теури в 1981 г. — за 1 ч до захода. В самых южных гнездовьях на американ-
ском побережье, на о-ве Теури в Японии и на о-ве Монерон в СССР птиц
можно увидеть на суше и в дневное время (Scott et al., 1974; Thoresen,
1983; Watanuki, 1983; данные Ю. В. Шибаева).
В заливе Петра Великого в июне—июле тупики-носороги начинают
прилетать в колонию через 30—40 мин после захода солнца, т. е. до наступ-
ления темноты, а остальные особи иногда появляются и перед заходом.
Обычно стаи птиц собираются у острова примерно за 1,5—2 ч до заката
в 100—150 м от побережья. Перелет на сушу происходит дружно —
птицы летят стайками, парами и одиночками. Часть из них, приблизившись
к острову, разворачивается над ним и снова улетает в море.
Утром птицы начинают покидать остров с первыми признаками рас-
света и примерно в течение 20 мин улетают. На Монероне вечерний прилет
наблюдался примерно в те же сроки, отлет же растягивался порой с рас-
света до 10 ч.
Ночная и сумеречная активность тупика-носорога, с одной стороны,
по-видимому, снижает влияние на его популяции пернатых хищников
(Sealy, 1973а), с другой — может быть связана с особенностями суточного
распределения его кормовых объектов (Vermeer, Cullen, 1979).
Питание. Питается в основном рыбой. Корм добывает обычно в откры-
том море, иногда удаляясь от мест гнездовий до 60 миль (Heath, 1915), реже
охотится у берега. На кормежке в любое время года держится в одиночку,
иногда разреженной стаей (Козлова, 1957).
На о-ве Тюлений с 1958 по'1965 г. у входов в норы тупика-носорога
в период выкармливания птенцов были собраны следующие виды: мойва
Mallotus villosus, песчанка Ammodytes sp., корюшка Osmerus eperlanus,
зубатки Anarhichas sp., морские лисички Agonidae, бельдюги Zoarces sp.,
ликоды Licodes sp., маслюки Pholis sp., а также молодь трески Gadus mor-
hua macrocephalus, горбуши Oncorhynthus gorbusha и желтоперой кам-
балы Pseudopleuronectes herzenstein (Бычков, 1976).
На Монероне птицы выкармливали птенцов тихоокеанской сельдью
Clupea harengus, иваси Sardinops sagax, молодью Oncorhynchus, песчанкой
и азиатской корюшкой (Гизенко, 1955).
В Японии, на о-ве Моюрури (Восточный Хоккайдо), в корме птенцов
обнаружены песчанка Ammodytes hexapterus, терпуги Hexagrammidae
и анчоус Engraulis japonica. За один раз птица приносит 10—16 рыб
146
Тупик-носорог
длиной около 7 см или 2—3 рыбы длиной около 14 см (Fujimaki,
1961).
В Восточной Пацифике в различных частях ареала — от Аляски до
штата Вашингтон одним из основных кормов птенцов служит песчанка —
Ammodytes hexapterus. Кроме того, в колониях на островах штата Вашинг-
тон в корме птенцов часто встречались: анчоус Engraulis mordax, корюшка
Hypomesus pretiosus и сельдь Clupea harengus; в Британской Колумбии:
сайра Cololabis saira, и нансения Nansenia candeda и сельдь, а на Аляске —
мойва. Длина рыб, приносимых птенцам в Британской Колумбии, —
от 4 до 23 см (Vermeer, 1979, 1980).
Враги, неблагоприятные факторы. Основными врагами тупика-носо-
рога в гнездовой период являются вороны и чайки. На о-ве Карамзина
(залив Петра Великого), где этот чистик гнездится в колонии чернохвостой
чайки — Larus crassirostris, птицы, приносящие корм птенцам, часто под-
вергаются нападению чаек (Литвиненко, 1980). Чаще всего чайки встре-
чают тупика-носорога у самой норы, когда тот приземляется и бежит
к гнезду, а не преследуют в воздухе, кай делают это в случаях нападения
на очковых чистиков. В местах, где тупик-носорог гнездится рядом с тихо-
океанской чайкой, ее добычей становятся птенцы во время слета на воду.
На о-ве Монерон у гнезд и на территории колонии этой чайки были найдены
остатки 16 тупиков-носорогов в гнездовом наряде (данные Ю. В. Шибаева).
На о-ве Теури птенцы тупика-носорога часто уничтожаются большеклювой
вороной Corvus macrorhynchos и тихоокеанской чайкой. Обе птицы тратят
много времени на отыскивание жертвы, так как птенцы тупика-носорога
в течение дня сидят в глубине норы и практически недоступны хищникам.
Обычно охота происходит рано утром, когда птенцы еще активны и под-
ходят близко к выходам нор. Зато успешность такой охоты высока —
у тихоокеанской чайки она составляет 93 % (14 из 15 попыток), у больше-
клювой вороны— 100 %', (17 из 17 попыток) (Watanuki, 1983).
На Аляске численность вида значительно снизилась после интродук-
ции песца. Скорее всего, это основная причина отсутствия тупика-носорога
на Алеутских островах. Лишь на о-ве Булдырь, где нет песца, сохранилась
небольшая колония этих птиц (Sowls et al., 1978).
На о-ве Теури большой вред тупику-носорогу наносят бродячие
собаки — ежегодно ими уничтожается около 6000 особей (Abe, 1967),
на о-велМонерон — серая крыса (данные Ю. В. Шибаева). В различные
сезоны года много тупиков-носорогов погибает в рыболовных сетях.
В, Японии в 1976, 1978 и 1984 гг. около 90 % возвратов колец (45 экз.)
было получено при извлечении птиц, запутавшихся в сетях (Japanese
Bird Banding in Recent Years, 1985; Report of the Bird Migration Research
Center, 1979, 1985).
Хозяйственное значение, охрана. Хозяйственного значения не имеет.
В СССР вид обитает на краю гнездового ареала и имеет невысокую числен-
ность, поэтому требуется специальная его охрана. В Приморье почти все
места гнездования тупика-носорога находятся под охраной: о-в Фуругельма
и о-в Стенина входят в состав Дальневосточного морского заповедника
с 1979 г.; острова Верховского и Карамзина объявлены памятниками при-
роды с 1984 г. (необходимо их включение в состав Дальневосточного мор-
ского заповедника). На Малой Курильской гряде часть гнездовий охраня
ется в заказнике республиканского подчинения «Малые Курилы». Нуж-
дается в охране колония на о-ве Монерон (острова Восточные), которая на
ю*
147
Семейство Чистиковые
протяжении многих лет разоряется местными жителями и командами
рыболовных судов (Гизенко, 1955; Б. Н. Вепринцев, личное сообщ.; дан-
ные Ю. В. Шибаева за 1976 г.).
В Японии тупик-носорог охраняется на островах Теури, Дайкоку (Хок-
кайдо) и на о-ве Аси (префектура Мияги) (Наканиси, 1953).
Род Fratercula Brisson, 1760
Тупик —
Fratercula arctica Linnaeus, 1758
Atlantic Puffin (англ.)» Papageitaucher (нем.)
Статус. В пределах ареала в СССР гнездящийся мигрирующий, изредка
зимующий вид.
Общая характеристика и полевые признаки. Птица средних размеров,
с компактным обтекаемым телом, крупной головой на короткой шее, конт-
растно окрашена: верх черный, низ белый. Вертикально сплюснутый мас-
сивный клюв почти треугольной формы яркого красно-оранжевого цвета.
Ноги тупика сильно отставлены назад, поэтому на суше в настороженной
позе он держит тело почти вертикально, опираясь на цевку и пальцы,
в спокойной позе — слегка под углом. Ходит довольно быстро, обычно
пригнувшись («сгорбившись»), по ровному месту легко бежит. Полет стре-
мительный — до 70 км/ч (Mills, 1981), с частыми взмахами крыльев, но
не маневренный; крылья узкие, относительно короткие и заостренные.
Короткая шея и массивная голова отличают тупика в полете от кайр, уток
и гагар. Рулит в воздухе растопыренными лапами, красный цвет которых
хорошо заметен. На воде сидит низко, плавает «по-утиному», прекрасно
ныряет и плавает под водой с помощью крыльев, а далеко отставленные
назад ноги выполняют функцию руля. В период гнездования тупиков чаще
всего можно видеть кормящимися на воде группами (от нескольких до
сотен птиц) или отдыхающими на краю обрывов или на скалистых пло-
щадках — «клубах». Встречается тупик у морских побережий материков,
но чаще — у морских островов. С сушей связан только в период размно-
жения. Большую часть года проводит в море, не очень далеко от берегов.
По суше ходит неохотно. При выходе из норы, а также при опасности
способен взлететь с места почти вертикально, но в местообитаниях с кру-
тыми склонами добирается до обрыва и бросается с него в воздух. Под-
летая к норе, тормозит в воздухе растопыренными лапами и, не присажи-
ваясь, вертикально опускается либо в трещину среди камней, либо вблизи
или точно у входа в нору и быстро скрывается в ней. С воды взлетает после
небольшого разбега.
Молчалив. Во время массового полета в стаях издает тихое низкое
«урчанье» («кхорр», «охрр»), в стаях на воде и в период спаривания
можно слышать громкий протяжный и хрипловатый звук. Активен в свет-
лые часы суток.
Подвидовая систематика. Различают три подвида.
1. Fratercula arctica arctica
Alca arctica Linnaeus, 1958, Syst. Nat. ed. 10, c. 130, Северная Норвегия.
Длина (в мм) крыла самцов 163—177 (среднее 167), самок 158—
176 (среднее 164). Длина клюва от границы оперения у самцов и самок
41,7—50,2 мм, высота клюва у основания 34,5—39,8 мм (Дементьев, 1951).
148
Тупик
2. Fratercula arctica naumanni
Fratercula arctica naumanni Norton, 1901, Proc. Portland Soc. Nat. Hist. 2,
c. 144, 145, табл. 2, фиг. 3, Шпицберген — предположительно.
Длина (в мм) крыла самцов 177 —195 (среднее 185), самок 175—
187 (среднее 183). Длина клюва от границы оперения самцов и самок
49,7—55,8 мм, высота клюва у основания 40,2—44,8 мм. Надклювье изо-
гнуто более круто, чем у другихподвидов.
3. Fratercula arctica grabae
Fratercula arctica grabae Brehm, 1831.
Наиболее мелкий южный подвид. Длина крыла самцов и самок Гебрид-
ских островов 144—170 мм (Harris, 1979).
Замечания по систематике. Географическая изменчивость тупика незна-
чительна, подвиды слабо различаются только весом и некоторыми разме-
рами (крыла, клюва). Однако величина клюва и крутизна гребня над-
клювья не могут служить надежным диагностическим признаком. Для
всех подвидов характерна большая индивидуальная изменчивость в ок-
раске оперения головы и щек. Во всех популяциях встречаются как очень
мелкие, так и очень крупные особи с перекрывающимися максимальными
размерами, что затрудняет отнесение их к какому-либо подвиду. В связи
с этим единого мнения о существовании трех подвидов тупиков и их разме-
щении внутри ареала вида нет. Е. Хартер (Hartert, 1903—1922) относит
к F. a. naumanni не только птиц, гнездящихся в северной половине Грен-
ландии и на Шпицбергене, но и на Ян-Майене, которых другие авторы
относят к F. a. arctica (Schaanning, 1933). Новоземельских тупиков
Е. Хартер (1903—1922) и М. А. Мензбир (1895) относят к F. a. naumanni,
а Б. М. Житков и С. А. Бутурлин (1901) — к номинальному подвиду
F. a. arctica. Л. О. Белопольский (1957) считает, что существует два под-
вида: более крупный полярный тупик F. a. naumanni, обитающий в Север-
ной Норвегии, на Западном и Восточном Мурмане, Новой Земле, Шпиц-
бергене, Ян-Майене и Северной Гренландии до 86° с. ш., и типичный более
мелкий тупик F. a. arctica, включая F. a. grabae, гнездящийся в Исландии,
на о-ве Медвежьем, в Средней и Южной Норвегии, Западной Швеции,
Фарерских островах, Британских островах, островах Ла Манша, в Бретани.
П. А. Мавродиади (1911) предполагает, что на Айновых островах, кроме
номинального подвида, гнездится также и F. a. naumanni. Массовые измере-
ния тупиков на Айновых островах показали, что птицы местной гнездя-
щейся популяции по средним размерам крыла и клюва превосходят соот
ветствующие показатели подвида F. a. arctica и ближе к F. a. naumanni
(данные Н. Н. Скоковой). Тупики из Южной Исландии по длине крыла,
цевки и клюва отличаются от тупиков Северной Исландии больше, чем
от птиц с Фарерских островов и, по принятым критериям деления тупиков
на подвиды птиц из Северной Исландии следует отнести к F. a. arctica,
а из Южной — к F. a. grabae. В связи с этим А. Петерсон (Petersen, 1976а)
предполагает, что тупики образуют размерную клину grabae—naumanni
и выделенные в настоящее время подвиды произвольны. Таксономия вида
нуждается в уточнении.
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла: I<II>III>IV. . ., у годовалых и
неполовозрелых П=Ш. Рулевых перьев 14. Самцы несколько крупнее
самок. Длина (в мм) крыла самцов 150—195 (среднее 176,6), самок 160—
195 (среднее 175,1); цевки самцов 26,0—39,0 (среднее 30,8), самок 27,0—
149
Семейство Чистиковые
36,0 (среднее 29,6); клюва самцов 40,5—60,0 (среднее 50,6), самок 40,7—
53,8 (среднее 48,9). Масса (в г) самцов 400 —610 (среднее 513,7), самок
400—572 (среднее 486,3) (Кафтановский, 1951; Н. Н. Горчаковская, лич-
ное сообщ.; Н. Н. Карташев, личное сообщ.; С. М. Успенский, цит. по Де-
ментьеву, 1951; данные Н. Н. Скоковой).
Окраска. Самец и самка в брачном наряде. Половой диморфизм
в окраске отсутствует. Лоб беловатый; темя, затылок темно-серые; оже-
релье на шее, спина, хвост черные; грудь, живот, подхвостье белые; бока
тела бледно-серые с постепенным переходом к белому животу; щеки от
беловатых до интенсивно серых и резко отграничены от окраски верха
головы и ожерелья на шее; подбородок темно-серый. Хвост и крылья
черные, внутренние опахала маховых бурые. Низ голени бурый. Контурное
оперение плотное, с легким блеском. Глаза небольшие, радужина серая,
вокруг глаза узкое кольцо карминного цвета, над глазом небольшая кожи-
стая лопасть, под глазом голая плоская бляшка аспидного цвета, переходя-
щая сзади в четкую уздечку в виде нитевидного углубления в оперении
щеки. Клюв высокий, уплощенный с боков, ярко окрашен; дистальная
часть рамфотеки карминно-красная с желтым концом, треугольные пла-
стинки в проксимальной части аспидно-серые, ограничивающие их в осно-
вании клюва рельефные валики — желтые, кожистая мягкая складчатая
розетка на надклювье и подклювье бурые. Лапы оранжево-красные, когти
черные.
Самец и самка в зимнем наряде. В окраске щек и на боках тела много
грязно-бурого и черноватого тона, окаймление кроющих спины сероватое,
ноги оранжево-желтые, клюв темно-бурый, более темный к основанию, без
красочной рамфотеки и цветной окраски голых участков около глаз.
Пуховой наряд. Покрыт длинным почти черным с буроватым оттенком
пухом с белым округлым пятном в нижней части груди и на животе, клюв
и ноги темно-серые, когти темно-бурые.
Гнездовой наряд (он же и первый зимний) схож с зимним нарядом
взрослых тупиков, но с еще большей примесью бурых тонов в оперении
щек, особенно вокруг глаз. Клюв темно-серый, с бурой вершиной, без попе-
речных бороздок, ноги голубовато-серые с буроватыми перепонками, когти
черные.
В полувзрослом наряде надклювье относительно ниже, чем у взрослых,
окраска рамфотеки менее интенсивная, поперечные борозды на клюве
почти не выражены или отсутствуют.
Число бороздок на клюве — возрастной признак. В годовалом возрасте
намечается след одной бороздки, в возрасте 2 лет — одна, 3 лет — одна-две
борозды, у половозрелых и старых — больше двух. В пределах каждого
возраста возможны значительные вариации. У неполовозрелых особей
меньше двух борозд (Harris, 1979, 1981). Размеры клюва, степень разви-
тия боковых пластинок и бороздок могут быть использованы для определе-
ния возраста тупиков от 2 до 6 лет (Petersen, 1967а).
Линька. Изучена слабо, однако предположительная последовательность
смены нарядов такова: пуховой — гнездовой (он же первый зимний) —
неполный взрослый (S/ad) — второй зимний (S/ad) — первый взрослый
брачный (S/ad) — второй зимний взрослый (S/ad)' — второй (дефинитив-
ный) взрослый брачный (ad?) — второй взрослый зимний (ad?) и т. д.
Пуховой наряд. Из мягкого густого эмбрионального пуха, длинного на
спине (30—40 мм) и более короткого на груди и животе (7—9 мм), к 44-му
150
Тупик
дню заменяется окончательным гнездовым нарядом в следующей последо-
вательности: на 9—10-й день появляются пеньки на спинной, брюшной
и боковых птерилиях; на 11 — 14-й — начинают развертываться пеньки кон-
турных перьев; на 15—17-й — появляются кисточки на пеньках маховых
и рулевых; на 18—20-й — интенсивно развертываются и растут перья по
всему телу; на 21—26-й — быстро выпадает пух на крыльях; на 27—31-й —
выпадает пух на спине и животе; на 35—37-й — остается пух только на
голове; на 44—45-й — единичные пушинки только на голове (Карташев,
1957, 1960; Корнеева, 1967). Оформляется гнездовой наряд,по структуре
очень незначительно отличающийся от оперения взрослых и сохраняю-
щийся в течение первой зимы. У взрослых две линьки. Частичная пред-
брачная линька, в основном мелкого пера, идет от головы к хвосту, у неко-
торых птиц — полная со сменой маховых и рулевых, начинается в апреле
и заканчивается к концу мая. Маховые выпадают в сжатые сроки, и тупики,
вероятно, утрачивают способность летать (Salomonsen, 1944). Линька
клюва начинается в середине апреля с покраснения надклювья, к середине
мая появляются выросты рамфотеки надклювья, к концу мая — попереч-
ные желтые полосы и борозды, роговые образования в основании клюва
и мягкие кожистые обрамления разреза рта (розетка), голые кожистые
образования у глаз. Ноги становятся оранжево-красными. Послебрачная
полная линька начинается по окончании гнездового периода у большин-
ства тупиков со смены кроющих головы и шеи, затем — рулевых и махо-
вых. Первостепенные и второстепенные маховые меняются почти син-
хронно, но первостепенные растут медленнее, и, видимо, в это время тупики
утрачивают способность к полету. К концу марта у взрослых и неполово-
зрелых особей маховые сменяются полностью. У некоторых птиц линька
начинается не раньше октября и сильно растянута. У годовалых птиц махо-
вые сменяются в конце лета (Harris, Yu be, 1977). С конца периода размно-
жения утрачиваются красочность рамфотеки и выросты клюва, розетки,
кожистые лопасти у глаз. Ноги становятся желтыми.
Распространение. Гнездовой ареал. Эндемик Северной Атлан-
тики. Широко распространен главным образом в ее бореальной части,
меньше — в арктической и умеренной (рис. 40). Восточное побережье
Северной Америки к северу от штата Мэн, заливы Фенди, Св. Лаврентия,
о-в Ньюфаундленд, п-ов Лабрадор, на север до залива Унгава, по западному
побережью Гренландии до пролива Смита, по восточному — до залива
Скорсби (71° с. ш.); острова Ян-Майен, Исландия, Шпицберген, Медве-
жий, западное побережье Новой Земли, северное побережье Кольского по-
луострова, северное и западное побережье Скандинавии, Фарерские остро-
ва, Британские, Норманские и другие острова Ла-Манша, побережье Бре-
тани.
F. a. naumanni — высокоширотный арктический подвид — гнездится
на Шпицбергене и в северо-западной части Гренландии от 76° с. ш. (север-
нее залива Мелвилла); F. a. arctica — южноарктический номинальный
подвид — обитает по западному побережью Северной Америки, западному
и восточному побережьям Гренландии до 71° с. ш., на островах Исландия,
Ян-Майен, Медвежий, в Норвегии, к северу от Бергена, на Мурманском
побережье Кольского полуострова и западном побережье Новой Земли;
F. a. grabae — самый южный подвид — гнездится на Фарерских островах,
спорадично — на западном побережье Шотландии и Англии, в Ирландии,
в Скандинавии, южнее Ставангера, на островах Ла-Манша, побережье
Бретани.
151
Семейство Чистиковые
В СССР тупик распространен только в Баренцевом море (рис, 41), где
гнездится номинальный подвид F. a. arctica; на островах Западного и Вос-
точного Мурмана — Айвовых, Арских, Большом Кие, Большом Гаврилов-
ском, Большом Гусинце, Харлове, Большом и Малом Зеленцах, Кувшине,
Малом Лицком, в некоторых губах западного побережья Новой Земли.
На севере — в районе Русская Гавань, мыс Желания, на юге — в районе
губ Грибовая, М. Кармакулы. На Айновых островах и Новой Земле воз-
можно обитание полярного подвида F. a. naumanni. Размещение тупика
внутри ареала спорадическое, преимущественно в районах стыка теплых
и холодных вод в зоне влияния теплых течений.
Зимовки. Районы обитания в негнездовое время охватывают как
гнездовой ареал, так и лежащие южнее районы Северной Атлантики.
Зимующие тупики многочисленны в районе Гебридских островов (Bourn,
Harris, 1979). Большинство молодых тупиков с о-ва Рёст и о-ва Ловунден
(Лофотенские острова) проводят первую зиму в водах Южной Норвегии
и Британии; тупики с о-ва Ловунден добыты в водах Шотландии и юго
восточных районах Северного моря (Myrberget, 1962а, 1973). В Север-
ной Атлантике немногие тупики зимуют в районе банки Джорджес (Бело-
польский и др., 1982). Два тупика, помеченные на о-ве Скомер (Юго-
Западный Уэльс), встречены в первую зиму у берегов Ньюфаундленда
(Saunders, 1966). В восточной части ареала тупики в зимнее время дости-
гают к югу 25° с. ш. Они встречены у берегов Марокко, в Западном Сре-
диземноморье — у Балеарских островов, Корсики, Сицилии, Мальты,
реже — в южных частях Адриатического моря. В Атлантике — у о-ва Ма-
дейры, Азорских и Канарских островов. В западной части ареала вдоль
берегов Северной Америки откочевывают к югу до 39° с. ш. до залива
Делавер (Hartert, 1921; Bent, 1919). Зимуют обычно вблизи берегов Север-
ной Америки, но иногда и в открытом море. Встречены в 450 милях от Ир-
ландии, в 400 милях от мыса Фарвел в Гренландии, в 300 милях от Фарер-
ских островов (Wynne-Edwards, 1935). Известны залеты в глубь материка
в Европе: в Эммерих и Гессен (Le-Boi, 1908), в ряд пунктов фин-
ской Лапландии (Плеске, 1887); 8.ХП 1961 г. один тупик встречен
на оз. Нейзидлер-Зе в Австрии (Leister, 1962), еще один — в устье Вислы,
два — в разные сроки — у балтийского побережья Польши (Lambert,
Nehls, 1965).
В СССР зимой в Баренцевом море у берегов Восточного Мурмана
тупики встречаются редко и, как правило, одиночные птицы (Кафтанов-
ский, 1951). Самый восточный залет — у северо-западных берегов
п-ова Канин: зимой добыт у пос. Шойна (Спангенберг, Леонович, 1969).
Одиночные птицы встречены в феврале—марте на Западном Мурмане
между Кольским заливом и Варангер-фьордом и у п-ова Рыбачий на рас-
стоянии до 20 км от берега (Курочкин, Герасимова, 1960), в марте —
в Варангер-фьорде уже, вероятно, на пути к местам гнездовий (Meinert-
zhagen, 1938). Единичные особи неежегодно зимуют в вершине Канда-
лакшского залива, куда попадают, видимо,пересекая Кольский полуостров
по водной системе: Кольский залив — р. Кола — оз. Имандра — р. Нива.
В октябре по одной молодой птице добыто у г. Кандалакша (Бианки,
1960а, б), в декабре — на Чунозере (Кафтановский, 1951).
Миграции. Мигрируют тупики вплавь, широко рассеиваясь в умеренной
и южной частях Северной Атлантики. Некоторые птицы остаются зимовать
в районе мест гнездования, другие уходят на разные расстояния. Напри-
152
Тупик
Рис. 40. Область распространения тупика
а — места гнездования, б — область зимовок
и миграций, в — места предполагаемого
гнездования. Подвиды: 1 — F.a. arctica,
2 — F.a. naumanni, 3 — F.a. grabae
Рис. 41. Распространение тупика в СССР
а — колонии, б — летние и зимние встречи
отдельных птиц
мер, тупики популяций восточных островов Великобритании практически
не мигрируют, а западных (в том числе гнездящиеся в Ирландии) мигри-
руют довольно широко (Elkins, Williams, 1972; Harris, 1982b). Дальность
миграций молодых птиц больше, чем взрослых (Mead, 1974). В пределах
СССР во время позднелетних кочевок и осенних миграций тупики через
горло Белого моря вплавь достигают вершины Кандалакшского залива.
Летом наиболее южные залеты неразмножающихся особей отмечены
в Белом море у 65° с. ш.: в августе 1957 г. — в Кандалакшском заливе (Би-
153
Семейство Чистиковые
анки, 1960а), в июне-июле 1961 г. — у Соловецкого архипелага (Карта-
шев, 1963). Сведения о размещении тупиков в негнездовое время очень
скудны и фрагментарны, миграции изучены слабо.
Численность. Общая численность в ареале в 50-е годы составляла 15 млн
особей (Lockley, 1953), в 60-е — 10,5—13,0 млн пар, из которых F. a. gra-
bae — 1 — 1,5 млн, F. a. arctica — 9,5—11,5 млн, a F. a. naumanni — около
15 тыс. (Harris, 1976а). Самые крупные очаги гнездования находятся
в Исландии — 8—10 млн пар (Harris, 1976а). На Фарерских островах
около 2,5 млн пар (Lockley, 1953), островах Северной Норвегии —
1,5—2 млн пар (Brun, 1966), на Британских островах — около 2,0 млн пар
(Lockley, 1953). Более многочисленны островные популяции: о-в Рёст —
700 тыс. пар и о-в Фуглёй — 500 тыс. пар в Северной Норвегии (Brun,
1966), о-в Сент-Килда (Внешние Гебриды) — около 250 тыс. особей (Flegg,
1972) и о-в Ловунден в Норвегии — около 250 тыс. особей (Myberget,
1959b), о-в Грейт-Айленд (Ньюфаундленд) — около 100 тыс. пар (Net-
tleship, 1972), о-в Шайант (Внутренние Гебриды) — около 90 тыс. жилых
нор (Brooke, 1972), о-в Фула (Шетлендские острова) — 70,2 тыс. пар
(Furness, 1981), о-в Веди (Лофотенские острова) — около 500 тыс. пар
(Watson, 1957), о-в Хорнёя (Варангер-фьорд) — около 20 тыс. пар («Vo-
gelfelsen», 1982), о-ва Фарн (Северо-Восточная Англия) — около 13,5 тыс.
пар (Hornug, 1978). Наименее многочисленны популяции тупиков в Грен-
ландии — 20— 200 пар (Salomonsen, 1944), на о-ве Мэй (Шотландия) —
50 пар (Harris, 1977), на побережье Гудзонова пролива (Канада) — 20 пар
(Gaston, Malon, 1980). В 60—70-е годы численность тупика почти повсе-
местно снижалась. Так, на о-ве Сент-Килда (Внешние Гебриды) в конце
XIX в. гнездилось 1 — 3 млн пар тупиков, в 50-е годы XX в. уровень чис-
ленности почти не изменился, в 60-е началось резкое снижение числен-
ности, и в 1971 г. на всех островах архипелага учтено около 163 тыс. пар
при-общей численности не более 250 тыс. особей (Flegg, 1972). На о-ве Ско-
мер (Юго-Западный Уэльс) за 40—50-е годы численность тупика снизи-
лась с 50 тыс. пар до 4 тыс. пар (Saunders, 1966). На о-ве Фуглёй в группе
Лофотенских островов число тупиков с 500 тыс. пар в 60-е годы сократи-
лось до 218000 в 70-е (Brun, 1966, 1971а, Ь). В 80-е годы на некоторых
островах шотландского побережья численность стабилизировалась и даже
возросла, в том числе на о-ве Мэй с 50 пар в 60-е годы до 3 тыс. пар в 1975 г.
(Harris, 1977). Резко уменьшилось число гнездящихся тупиков в Швеции
и Норвегии, в Бретани, и на Лабрадоре, в Англии (берега Кента и Сас-
секса), йа о-ве Гельголанде они перестали гнездиться (Карташев, 1951).
В пределах СССР в 20-е годы гнездилось не более 20 тыс. пар, т. е.
0,13 % мировой популяции вида. Самые многочисленные и стабильные по-
пуляции находятся на Айновых островах, насчитывавшие в конце 50-х гг.
12—15 тыс. пар (Скокова, 1967а). Остальные популяции Мурманского по-
бережья насчитывали от десятков до нескольких сотен пар: о-в Кий —
около 420 пар (данные Н. Н. Скоковой), о-в Гавриловский — 90 пар,
о-в Большой Гусинец — 10 пар, о-в Харлов — 100 пар, о-в Кувшин —
500 пар (Герасимова, 1962). Однако, как и всюду, численность тупика
на гнездовье в СССР неуклонно снижается: в 60-е гг. на Айновых островах
гнездилось 7—8 тыс. пар, к концу 70-х гг. — насчитано всего 3—4 тыс.
взрослых особей, т. е. за 50 лет численность уменьшилась более чем
в 10 раз (Карпович, 1984). На Новой Земле тупики гнездятся разреженно
и немногочисленны (Белопольский, 1957; Головкин, 1972).
154
Тупик
Показатели численности в гнездящихся парах, скорее всего, занижены
по сравнению с реальной численностью, поскольку в гнездовых колониях
обитает значительное число неразмножающихся особей (Lockley, 1953; Re-
gnell, 1956; Myrberget, 1959b; Скокова, 1967a).
Местообитания. Тупик — типичная морская птица, обитатель прибреж-
ных (шельфовых) морских пространств, не очень удаленных от матери-
ков, или вокруг островов. Во время миграций и зимовок держится на воде.
С сушей связан только в период размножения. Гнездовые местообита-
ния — морские побережья материков или крупных морских островов
в сочетании с местами скопления пелагических рыб.
Гнездятся только колониями — от нескольких пар до,десятков и сотен
тысяч. Стенотопный вид. Занимает два типа гнездовых биотопов: торфя-
ные пласты и крупнокаменистые россыпи, преимуществецнЬ на морских
островах. В торфе тупики активно роют норы с помощью клюва и сильных
лап с острыми,, и- загнутыми когтями. В южных частях ареала в очень
?едких случаях занимают норы кроликов (Naumann, 1897—1905; Brehm,
920—1921). Вход в нору обычно находится в небольшом углублении.
Старые норы имеют накатанные «подъезды» и «скаты». Одиночная простая
нЪра, как правило, изогнута почти под прямым углом или круто-дугооб-
разно. Длина ее не менее 1 м, ширина хода норы 22—25 см. В удаленном
конце находится гнездовая камера с подстилкой из сухих растений и еди-
ничных перьев (чаек). В зависимости от мощности торфяного пласта ход
и камера располагаются на глубине 40—90 см. Если нора вырыта на крутом
склоне, то камера оказывается на большей глубине, чем ход. В густонасе-
ленных процветающих колониях большинство нор сложные, с разветвле-
нием ходов, вырытых в разных горизонтах (этажах), соединяющихся друг
('Другом и образующих сложную сеть — «городок». В таких норах в гнездо
можно попасть через несколько входных отверстий несколькими путями,
причем один вход может вести в несколько гнезд, и, наоборот, одна гнездо-
вая камера — иметь несколько входов. Например, входы двух нор, раско-
панных в брошенной колонии на о-ве Б. Айнов, находились в одном общем
углублении. Мощность пласта торфа 1 м, наибольшая глубина располо-
жения ходов 90 см, средняя — 60 см; гнездовые камеры — на глубине 50—
90 см; общая длина ходов одной норы 4 м, второй — 7,4 м; средний диаметр
хода 20 22 см, толщина стенок между соседними ходами разных нор
йтбгда не превышала нескольких сантиметров. В одной норе — 6 гнездо-
вых камер и два выхода, в другой — одна гнездовая камера и три выхода.
Вариантов строения нор много, в том числе и кольцеобразные (Скокова,
1967а, б). Общая длина ходов в одной норе может достигать 15 м (Гераси-
мова, 1958). Обязательной принадлежностью жилой норы тупика является
«птенцовая уборная» — постоянное место испражнения, находящееся
обычно на крутом повороте хода норы, ближе к выходу; помет лежит ком-
пактными кучками с обеих сторон хода в небольших углублениях. В гнезде
всегда чисто.
'В крупнокаменистых россыпях гнездо находится в глубине, в трещинах
и пустотах под нависающими камнями. Вход в гнездо может быть сбоку
или сверху, даже через вертикальную щель, в которую тупик опускается
с лету или присев сначала на ближайший камень. «Промежуточный»
между этими типами биотоп представляет собой пласты торфа с погребен-
ными крупными камнями, где тупики также роют норы в обход этих кам-
ней с выходами в нишах под ними. Плотность гнездования в жилых коло-
155
Семейство Чистиковые
ниях Айновых островов 0,4—1,6 норы/м2 (Скокова, 1967а, б)., на Лофотен-
ских островах (о-в Рёст) — максимальная.— 2,72 пары/м2 (Brun, 1966),
на Британских островах (о-в Сент-Килда) — средняя 0,5 пары/м2 (Harris,
1980). Колонии тупиков либо сплошные, монолитные, либо состоят из отно-
сительно изолированных участков. В процессе жизнедеятельности птиц
гнездовой биотоп изменяется, разрушается и тупики постепенно покидают
непригодное место. В процветающей многолетней колонии торфяно|й пласт
настолько изрыт многочисленными и «многоэтажными» подземными хо-
дами, что в некоторых местах стенки обрушиваются. От тысячекратного
перемещения птиц по земле, особенно у края колонии, где они отдыхают
и откуда улетают в море, растительность полностью уничтожается. Эти
участки представляют собой голые торфяные площадки с отверстиями нор.
Роющая деятельность формирует зоогенную растительность: на Айновых
островах на свежих выбросах при расчистке нор поселяются пионерные
виды растений нарушенных почв — ромашка крупноцветковая, дрема лап-
ландская, которые пышно разрастаются на удобренной птичьим пометом
почве. На месте давно покинутых колоний растительность сменяется зо-
нальной морошково-вороничной ассоциацией с мелкокочковатым рельефом
(Скокова, 1967а; Коханов, Скокова, 1967; Парфентьева, 1969).
Колонии тупиков долговечны. На Айновых островах они существуют
не менее 120—150 лет (Мензбир, 1985; Пинегин, 19Q9; Мавродиади, 1911;
Emeis, 1929). На Гебридских островах в наиболее заселенных и уязвимых
участках колоний почвенный слой разрушается приблизительно за 100 лет
(Hornung, 1978). Своеобразие гнездовых биотопов, Их относительная ред-
кость определяет высокую степень колониальности и гнездовой консерва-
тизм тупиков при условии обязательного сочетания пригодных для гнездо-
вания мест с обилием пелагических кормов в прилегающих участках моря.
Размножение. Тупики — моногамы. Пары, видимо, постоянны, о чем
косвенно свидетельствует высокий процент возврата меченых тупиков
в те же колонии (99,6 %), конкретные участки колоний (83 %), пользова-
ние одними и теми же норами (Скокова, 1967а; Татарникова, Чемякин,
1970). На о-ве Скомер в Уэльсе только 6,8 % тупиков сменяли прошлогод-
ние норы и ежегодно распадалось около 8 % прошлогодних пар (Ashcroft,
1979). Потенциальная половозрел ость наступает, вероятно, в годовалом
возрасте, но реализуется в возрасте двух лет (Дементьев, 1951; Кафтанов-
ский, 1951), по более поздним исследованиям отдельные особи начинают
гнездиться на 3-й год, но основная масса образует пары на 4—5-й год
(Perrins, 1978; Ashcroft, 1979; Wehle, 1982); по другим данным, большин-
ство тупиков начинает гнездиться на 5—6-й год (Petersen, 1976b; Harris,
1981). Число неразмножающихся птиц в гнездовых колониях может быть
равно числу размножающихся или превышать его в 3—5 раз (Lockley,
1953; Regnell, 1956; Myrberget, 1959b) за счет обитания в колониях двух-
леток и годовиков (Myrberget, 1962а). На Британских островах 20—27 %
обитающих в колониях птиц в возрасте 2—3 лет не образуют пар (Ashcroft,
1979), а среди ежегодно отлавливаемых в Южной Исландии 100 тыс. тупи-
ков только 7,2 % — половозрелые (Petersen, 1976b). На Айновых остро-
вах не размножается около 20 % тупиков (Скокова, 1967а) . Соотношение
полов на Семи островах 1 : 1 (Кафтановский, 1951), на Айновых островах
на трех самцов приходилось две самки (в 1960 г. — 1,53:1, в 1961 —
1,44:1) (Скокова, 1967а, б). Это, вероятно, связано с тем, что самцы начи-
нают размножаться позднее самок (Petersen, 1976b).
156
Тупик
Продолжительность жизни тупиков, видимо, велика, хотя прямых дан-
ных об этом недостаточно. На Айновых островах трижды отлавливали
окольцованных тупиков, проносивших кольцо 11 лет. При условии гибели
не болей 5 % популяции в год (что установлено при массовом отлове и
мечении тупиков в гнездовых колониях) смена популяции этих островов
происходит, наверное, в течение 20 лет (Скокова, 1967а). Наибольшая
смертность приходится на первый год жизни (Wehle, 1982с). Хотя еже-
годная выживаемость взрослых гнездящихся тупиков достигает 95 %,
средняя продолжительность жизни, видимо, не превышает 6—7 лет, так
как до 4—5 лет доживает 10—15,6 % тупиков, помеченных птенцами
(Ashcroft, 1979). В неволе тупик прожил 20 лет (Wolley, 1905—1907).
К местам гнездования тупики прилетают, когда еще часть территории
находится под снегом, а норы забиты ледяными пробками. В зависимости
от экспозиции склона и времени оттаивания нор тупики занимают их
несинхронно, и период размножения в связи с этим растянут, разница
в сроках гнездования разных пар может составлять 20—25 дней. На Семи
островах этот период более растянут из-за более сложного рельефа, чем
на Айновых островах (Герасимова, 1951; Кафтановский, 1951). В Шотлан-
дии начало кладки на восточном и западном побережьях разнится
на 2—3 нед — с конца февраля до середины марта (Harris, 1982b) и в каж-
дой колонии зависит от просыхания почвы (Hornung, Harris, 1976). В Грен-
ландии тупики гнездятся на 1—2 мес позднее, чем на Британских остро-
вах, но при более синхронизированном цикле размножения (Evans, 1977).
На Мурмане прилетают во второй половине апреля, массовое появление
в гнездовой колонии 26.IV—18.V; первые кладки 15—30 мая; вылупление
первых птенцов 15.VI—4.VII; массовое вылупление 27.VII—1 .VIII; выход
первых птенцов из нор 28.VII —13.VIII, массовый — в середине августа.
Отдельные пары могут кормить птенцов до первой декады сентября (Ско-
кова, 1967а; Корнеева, 1967; Татарникова, Чемякин, 1970).
Плодовитость минимальная: самка откладывает одно яйцо, в редких
случаях (не более 1 %) — два (Bent, 1919; Dobson, 1937; Кафтановский,
1951). При утрате кладки потенциально может отложить три яйца
(М. Слепцов, по Белопольскому, 1957). Кроме того, установлено, что
из занявших норы тупиков 5—16 % не откладывают яиц (Ashcroft, 1979).
Размеры яиц (в мм) на Семи островах (п=55) 60—68X39—48, сред-
нее 63,7X44,4; вес 65,1 г. (Ю. М. Кафтановский, по Дементьеву, 1951).
На Айновых островах (п=14) 60,9—70,8X41,5—47,9; вес 70,5, максималь-
ный — 75 г! (Герасимова, 1958); (п=13) 60,3—70,5X42,0X53,7, среднее
64,8X45,8; вес свежего яйца 63 г, насиженного — 60—61 г. (Корнеева,
1967). Форма эллипсоидно-округлая, чуть заостренйая к одному концу.
Окраска простая, белая. В глубоком слое есть слабые лиловатые пятна и
разводы, хорошо видные на яйце, находящемся в яйцеводе. Последнее сви-
детельствует о депигментации скорлупы в связи с гнездованием в укры-
тиях (Кафтановский, 1951; Белопольский, 1957). Поверхность мелкозерни-
стая, матовая, при насиживании приобретает слабую глянцевитость и буро-
ватый налет, загрязняясь торфяной пылью. Толщина скорлупы одинакова
по всей поверхности яйца — 0,15 мм. Яйцо относительно крупное —
11,1 —14,2 % веса самки (Кафтановский, 1951). Аберрации формы очень
редки.
Обычно тупики прилетают к островам в сжатые сроки (2—5 дней),
первые дни живут в море, а гнездовые колонии начинают посещать по мере
157
Семейство Чистиковые
освобождения их от снега. В это, время происходят брачные игры и массо-
вое спаривание. С началом насиживания, которое длится 35—42 дня,
в колонии наступает затишье. Насиживают обе птицы, сменяя друг друга
через 6—8 ч (Корнеева, 1967), по другим данным — через 24—72 ч (Saun-
ders, 1966). Вылупление птенца продолжается 3—4 сут. Жизнь колонии
активизируется после массового вылупления птенцов, когда оба родителя
начинают летать за кормом.
Птенец тупика, защищенный от неблагоприятных условий погоды в за-
крытом гнезде, развивается по «птенцовому» типу и может быть отнесен
к группе «выводковых гнездарей», до конца постэмбрионального периода
зависящих от родителей (Portmann, 1935, 1950). Рождается зрячим. Гус-
той и длинный эмбриональный пух защищает птенца от переохлаждения
в норе, где температура не превышает 7—8 °C, так как норы иногда нахо-
дятся в промерзших и медленно оттаивающих пластах (Герасимова, 1951).
Первые три дня птенец малоподвижен, неуклюж, обычно сидит под обогре-
вающим его родителем, слабо пищит. Терморегуляция у птенца устанавли-
вается к 5—7-му дню (Рольник, 1948; Кафтановский, 1951; Карташев,
1960), по другим данным — на 3-й день (Tschanz, 1979). В возрасте 5—
10 дней птенец уже быстро перемещается внутри норы, в двухнедельном —
начинает испражняться в постоянной «уборной». Когти всех пальцев
однодневного птенца одинаковой величины, но с 10—14-дневного возраста,
когда птенец начинает, хотя бы немного, копать в гнездовой камере или
стенке хода, коготь второго пальца заметно увеличивается и становится
более загнутым. Клюв по мере роста птенца увеличивается в высоту (Кар-
ташев, 1957). К 25—26-му дню пытается приближаться к выходу из норы,
но при опасности быстро скрывается в ней. Хорошо выраженный отрица-
тельный фототаксис и рефлекс быстрого убегания в глубь норы или щели
имеет большое адаптивное значение, так как заставляет птенца в течение
всего гнездового периода оставаться в норе, не выходить преждевременно
из гнездового укрытия и уменьшает гибель от хищников и непогоды.
Интенсивность кормления птенцов нарастает к 25—26-му дню и посте-
пенно снижается к 40-му (Hudson, 1979). В 10—14 дней птенцы достигают
1/3 веса взрослых, в 20 дней — 2/3, в 44—45 дней — полного веса взрос-
лых (Карташев, 1960). В одних случаях родители кормят птенцов до самого
выхода из нор (Harris, 1976b), в других — прекращают кормление за 6—
9, в среднем за 3—5 дней до выхода из норы (Татарникова, Чемякин, 1970),
и птенцы теряют в весе от 5 до 40 г в сутки (Myrberget, 1962b), составляя
перед выходом 69—78 % веса взрослых (Корнеева, 1967; Ashcroft, 1979;
Harris, 1982а), максимально 81,6 % (Кафтановский, 1951). Однодневные
птенцы тупика на Семи островах имели размеры: длина (в мм) тела 120—
138, среднее 127,6, крыла 17,5—24,0, среднее 20,1; масса (в г) 40,0—
53,5, Среднее 48,0 (Кафтановский, 1951). Средняя максимальная масса
перед вылетом 404,5 г, средняя 332,3. Масса птенцов на Айновых островах
в возрасте 1—3 дня 33—54 г, 4—5 дней — 59—65 г, 7—8 дней — 90—
114 г, 9—10 дней — 116—167 г, 11—14 дней — 132—197 г, 15—17 дней —
195—218 г, 18—20 дней — 132—240 г, 21—26 дней — 225—265 г, 27—
31 день — 265—379 г, 32—35 дней — 315—397 г, 35—37 дней — 347—
410 г, 37—44 дня — 485—565 г. (Корнеева, 1967). Тупики продолжают
расти и набирают вес до пятого года жизни (Harris, 1981).
Побуждаемые голодом, первые птенцы могут покидать гнездо на 36—
37-й день, еще не всегда готовые к вылету. Массовый выход из нор проис-
158
Тупик
ходит на 42—46-й день (Saunders, 1966; Корнеева, 1967; Harris, 1978;
Ashcroft, 1979). Птенцы, вышедшие в начале массового выхода, крупнее
и активнее тех, которые покидают норы в последнюю очередь (Nettleshin,
1972). Одни птенцы оставляют гнездовую нору сразу, больше не возвра-
щаясь в нее (Козлова, 1957), другие — после нескольких выходов для
«разминки» (Татарникова, Чемякин, 1970). Большинство птенцов поки-
дают колонии в августе, в сумеречные часы и ночью, в одиночку (Saunders,
1966; Nettleship, 1969; Myrberget, 1981). При гнездовании на крутых скло-
нах, заканчивающихся обрывом, молодые тупики уходят в первый неуме-
лый планирующий полет, бросаясь с. края обрыва в воздух и падая на воду.
Из колоний, расположенных на плато и пологих склонах, добираются
до берега по суше пешком и уходят от островов вплавь, сразу же начиная
нырять. В послегнездовой период держатся отдельно от взрослых.
Успех размножения тупиков довольно высок. Эмбриональная смерт-
ность на островах побережья Уэльса составляет 20—25 %, постэмбрио-
нальная — 5 %, до вылета доживает 60—70 % птенцов (Perrins, 1978;
Ashcroft, 1979). Основной причиной эмбриональной гибели считается
хищничество малого буревестника, галки и чаек, разбивающих яйца и вы-
нуждающих тупиков бросать кладку. В гнездящейся популяции о-ва Рёст
(в группе Лофотенских островов) отмечено явление естественной флуктуа-
ции численности катастрофического характера с массовой гибелью птен-
цов: так, в 1964—1968 гг. размножение шло нормально, в 1969—1973 гг.
успех размножения был очень низким, в 1974 г. — относительно высоким,
в 1976 г. — ниже нормы, в 1975 г. и 1977—1980 гг. воспроизводства прак-
тически не было. Средний возраст погибших птенцов в 1975 г. составил
4,4 дня, в 1977 - 4,6, 1978 — 4,7, 1979 — 10,8, 1980 г. - 14—15 дней (Lid,
1981). Обычно на о-ве Рёст ежегодно выводится около 0,5 млн птенцов,
а в 1977—1979 гг. выжил один из тысячи, в 1980 — ни одного. Причиной
массовой гибели считается голодание (Mills, 1981). На Айновых островах
в неблагоприятные по погодным условиям годы (например, 1962 г.),
с дождливым и холодным летом, когда буйная растительность полегает
и забивает отверстие норы, большинство взрослых тупиков покидают коло-
нию до окончания гнездового периода, что влечет за собой массовую гибель
птенцов в норах от истощения (Скокова, 1967а).
Суточная активность, поведение. В суточной активности, несмотря на
полярный день, существует цикличность со снижением в «ночные часы».
Родители прилетают с кормом четыре (Perry, 1946; Lockley, 1953), шесть
(Myrberget, 1962а) раз в сутки, по другим данным в середине размноже-
ния — 1 — 9 раз, в конце — чаще всего 3—5 раз (Татаринкова, Чемякин,
1970). Ритм кормления разных пар и в разные дни может существенно
отличаться в зависимости от индивидуальных особенностей птиц. Но в це-
лом суточный ритм имеет два подъема: в 5—7 ч утром, хорошо выражен-
ный, и во второй половине дня — более растянутый; в конце периода раз-
множения — один пик в середине дня (Myrberget, 1962b; Скокова, 19676;
Татаринкова, Чемякин, 1970; Corkhill, 1973).
В синхронизации ритма жизни гнездовой колонии большое значение
имеют «карусели» — специфическая форма общественного поведения:
периодические (несколько раз в день) круговые полеты массы птиц над
колонией присущи каждой колонии, члены которой держатся обособленно,
даже при небольшом расстоянии между местами гнездовий. Во время
«карусели» тупики просушивают оперение после пребывания в морской
159
Семейство Чистиковые
воде, меняются местами с партнером, находившимся до этого в норе, воз-
можно, получают информацию о месте кормежки, более защищены
от нападений поморников (чем птицы, летящие с кормом в одиночку),
способствуют поддержанию продуктивности «своих», постоянных мест
кормежки и т. д. В пик активности в «карусели» обычно летает до 500—
1000 тупиков (Скокова, 1962а, 1967а). Тупики крупных колоний, отдель-
ных островов или групп островов представляют обычно отдельные попу-
ляции с выраженными особенностями экологии: постоянными местами
гнездования и кормежки, своими областями зимовок и миграций, сроками
размножения, стереотипом поведения, в том числе и местом «карусели»,
степенью стабильности численности, а также некоторыми морфометриче-
скими признаками (Скокова, 1967а; Perrins, 1978; Harris, 1978, 1979,
1982b) (рис. 42).
Питание. Тупик трофически тесно связан с морем. Добывает корм
исключительно в воде. При ловле добычи под водой ориентируется с помо-
щью зрения, поэтому даже в условиях полярного дня охотится только
в наиболее светлое время суток и, как правило, на некотором расстоянии
от берега, в прозрачной воде. Места кормежки находятся от нескольких
сотен метров (Скокова, 1962а, 1967а) до 2—3 км от гнездовой колонии
(Mills, 1981), максимально — 32 км (Corkhill, 1973).
В пище тупика доминируют пелагические формы (более 90 %) и го-
раздо реже встречаются донно-литоральные (менее 7 %). Тупики узко
специализированные ихтиофаги. Видовое разнообразие кормов невелико
и представлено преимущественно рыбами. Другие виды корма, обнаружен-
ные в содержании желудков тупиков, — пелагические моллюски (0,8 %),
ракообразные (4 %), полихеты (4 %), губки (0,8 %), насекомые (0,8 %),
растения (22,6 %) (Белопольский, 1957), скорее всего, добываются тупи-
ками попутно, случайно или попадают вместе с рыбой. В питании тупиков
чаще всего встречаются песчанки Ammodytes marinus, мойва Mallotus
villosus, шпрот Sprattus sprattus, атлантическая сельдь Clupea harengus,
морской налим Gaidropsarus sp., скумбрия Scombrus scombrus, мерланг
Merlangus merlangus, корюшка Osmerus eperlanus, тресочка Эсмарка
Trisopterus esmarkii, треска Gadus morhua, большая песчанка Hyperoplus
lanceolatus, угорь Anguilla anguilla (Кафтановский, 1951; Герасимова,
1951, 1958; Белопольский, 1957; Harris, 1970, 1982b; Swennen, Duiven 1977;
Harris, Hislop, 1978; Hudson, 1979; Lid, 1981). Во всех частях ареала основу
питания составляют песчанка, мойва, шпрот и сельдь. Так, по данным
Л. О. Белопольского (1957), в районе Семи островов в пище тупика рыбы
составляют 86,8 % (песчанка — 56,8 %, мойва — 21,0, сельдь — 19,7, тре-
ска — 2,5 %), в районе Айновых островов 82,3 % (песчанка — 6,3 %,
мойва — 43,7, сельдь—50 %). У берегов Уэльса песчанка составляет
56 %, шпрот — 39,7 % (Hudson, 1979), на Лофотенских островах домини-
руют те же виды, что и в питании тупиков на Айновых островах, — пес-
чанка, мойва и молодь сельди (Lid, 1981).
Размеры поедаемых животных более или менее одинаковы. В экспери-
ментальных условиях тупики были в состоянии проглотить рыб с макси-
мальной шириной 23—26 мм, но предпочитали рыб вполовину «тоньше»
и длиной до 15—18 см (Swennen, Duiven, 1977); Родители приносят в клюве
за один раз в среднем 5—14 мелких рыбок (Saunders, 1966) максимально —
более 60: в одном случае 62 малька песчанки и морского налима (Harris,
Hislop, 1978), в другом — 61 личинку угря (Mills, 1981). Взрослые либо
160
Рис. 42. Элементы брачного поведения тупика (по: Cramp, 1985, с изменениями)
11 Птицы СССР
161
Семейство Чистиковые
сразу скармливают рыб птенцам прямо в клюв, либо бросают на землю
в норе, а птенцы подбирают их сами (Corkhill, 1973; Hudson, 1979). Суточ-
ная потребность птенцов в пище возрастает от 10 г. в 1-й день до 120 г на
26-й и затем постепенно снижается, составляя перед выходом из норы около
20 г (Tschanz, 1979; Mills, 1981), по другим данным, она составляет 13,4—
64,0 г (Harris, Hislop, 1978). В неволе птенцы отвергают «излишний» корм
(Hudson, 1979). За весь период роста в норме птенец съедает около 3,7 кг
рыбы (Harris, 1978). Суточная потребность взрослой птицы в пище 100—
107 г (Harris, Hislop, 1978; Mills, 1981).
Враги, неблагоприятные факторы. Из пернатых хищников главным
врагом взрослых тупиков в период гнездования является большая морская
чайка, в меньшей степени — серебристая чайка. На Айновых островах
стереотип поведения этих чаек включает и специализацию питания тупи-
ками, нападения в постоянных местах, подстерегания на «трассах» «кару-
селей», между гнездовыми колониями и местами кормежки на море: в этих
местах обычны скопления останков погибших тупиков с расклеванными
грудными мышцами и вырванными внутренностями или скелетов с харак-
терно вывернутыми и не снятыми с головы шкурками (Скокова, 1962а).
По данным Т. Д. Герасимовой (1958), на о-ве Б. Айнов в 1950 г. тупики
встречались в питании крупных чаек в 5,3—15,6 % случаев. В 1960 г.
гибель окольцованных тупиков от чаек на этом острове составила 1,7 %
и не превышала 2,5 % популяции (Скокова, 1967а). На о-ве Сент-Килда
(Внешние Гебриды) в 1973—1979 гг. около 40 пар больших морских чаек
питались исключительно тупиками и уничтожали в год 1,5 % всей популя-
ции (Harris, 1980), а на некоторых островах резко снизили численность
тупиков (Flegg, 1972). Пресс хищничества крупных чаек в большей сте-
пени проявляется в колониях, расположенных на плато, чем на крутых
склонах, и обычно направлен на ограниченные участки, непосредственно
около которых гнездятся сами чайки (Скокова, 1962а; Nettleship, 1972;
Harris, 1980; Карпович, 1984). На взрослых тупиков иногда нападает лап
ландский кречет, в пищевых остатках которого на Семи островах они
встречались в некоторые годы в 62 % случаев (Карташев, 1949 а, б).
Отмечены нападения на тупика залетного ястреба-тетеревятника на Айно-
вых островах (Татаринкова, Чемякин, 1970). На о-ве Скомер в Уэльсе к их
врагам относят Сапсана и ворона (Saunders, 1966). Серебристая чайка,
малый буревестник и галка являются причиной эмбриональной смертности
тупиков (разбивают и поедают яйца) и постэмбриональной гибели птенцов
(Ashcroft, 1979; Nettleship, 1972). Гибель птенцов в период их выхода из
гнезда и спуска на воду на Айновых островах достигает 7—8 % (Татарин-
кова, Чемякин, 1970).
Среди птиц, обитающих вблизи гнездовых колоний тупиков (коротко-
хвостый поморник, большая морская чайка, серебристая чайка, клуша,
галка), развит клептопаразитизм. Теряя добытую рыбу при неожиданном
нападении этих птиц, тупик вынужден повторно, а иногда и несколько раз
возвращаться в море для охоты за рыбой, что нарушает режим питания
птенцов. Этот факультативный паразитизм особенно развит и широко
распространен у короткохвостого поморника и галки (Скокова, 1962а,
1967а,б; Saunders, 1966; Grant, 1971; Corkhill, 1973; Arnason, Grant, 1978;
Ashcroft, 1979; Mills, 1981).
Известен случай поедания тупика крупной рыбой (Legendre, 1926).
Решающим фактором резкого подавления численности тупиков в период
162
Тупик
гнездования являются наземные хищники — песец, лисица, выдра, гор-
ностай и особенно крыса. Так, на о-ве Лунди в 1952 г. численность тупика
была снижена крысами со 100 тыс. до 400 пар (Fisher, Lockley, 1954); резко
снижена численность на о-ве Сент-Килда (Flegg, 1972) и о-ве Скомер
(Saunders, 1966). Из-за песцов тупики не гнездятся в идеальных для них
условиях на побережье Сильефьорда в Северной Норвегии при соседствую-
щих колониях на о-ве Веди, насчитывающих более 100 тыс. пар (Watson,
1957). Лисица является причиной очень низкой численности тупиков на
побережье Беркшира в Шотландии по сравнению с их численностью на
ближайших островах (Prato S„ Prato Е., 1980).
В районах наибольшей численности (Исландия, Фарерские острова)
тупики находятся под постоянным прессом человека. Неумеренная заго-
товка птиц в гнездовых колониях с помощью силков, сетей, сачков, желез-
ных крючьев и даже специально дрессированных собак — постоянно сни-
жает их численность (Wolley, 1905—1907; Timmermann, 1938). Регуляр-
ный промысел приводит не только к уничтожению части популяции взрос-
лых птиц, но и к разрушению их местообитаний и норм (Saunders, 1966;
Petersen, 1976). В Великобритании и Ирландии смертность окольцованных
тупиков прямо или косвенно связана с человеком: 22 % добыты на охоте,
6 % погибли в орудиях рыбного промысла, 22 % — от загрязнения моря
нефтепродуктами (Mead, 1974), к которым тупики очень чувствительны
(Mills, 1981; Wehle, 1982с). В заливе Св. Лаврентия главной причиной
снижения численности тупиков является прямое преследование человеком
(Nettleship, Lock, 1973).
Численность тупиков может существенно снижаться описанными выше
катастрофическими циклами размножения вследствие неблагоприятных
метеорологических условий и ухудшения возможностей питания, необес-
печенности кормом, что приводит к голоданию птенцов в норах и их массо-
вой гибели (Скокова, 1962а, 1967а; Saunders, 1966; Harris, 1978; Lid, 1981;
Mills, 1981). Высокая смертность птенцов в 70-е годы в разных районах
Норвегии связывается с перепромыслом видов рыб, составляющих основу
питания тупиков (Lid, 1981; Mills, 1981).
Хозяйственное значение, охрана. За пределами СССР в местах высокой
численности (Исландия, Фарерские острова) тупик имеет промысловое
значение как дополнительный источник питания местного населения.
Используются грудные мышцы птиц, которые хранятся в копченом виде.
Высушенные остатки тушки иногда используются как топливо. Среди ры-
бацкого населения Исландии мясо тупика в 20—30-е годы подавалось на
обед не реже раза в неделю (Roberts, 1934). В 1913 г. в Исландии было
добыто около 232,6 тыс. особей, в 1923—212,4 а в 1933 г. — 91,3 тыс. особей
(Timmermann, 1938). В 70-е годы только на о-ве Вестманнаэйяр, у южного
побережья Исландии, местное население ежегодно отлавливало около
100 тыс. птиц (Petersen, 1976а). Продолжают промышлять тупиков на
Фарерских и Британских островах (Saunders, 1966). В связи с тенденцией
снижения численности промысловое значение тупика надает, а в южных
частях ареала он становится редок и входит в категорию охраняемых видов
(Бретань, острова Ла-Манша).
Роль тупиков в круговороте минеральных и органических веществ
в море может быть существенной в связи с тем, что они образуют много-
тысячные скопления в местах их постоянного массового гнездования,
кормежки и отдыха на воде. Потребляя огромное количество рыбы, они
н*
163
Семейство Чистиковые
обогащают поверхностные воды моря биогенными элементами через экс-
кременты, попадающие в воду непосредственно при дефекации или при
смыве из гнездовых колоний (Головкин, Позднякова, 1966; Головкин, 1982).
Однако это воздействие локально, учитывая точечный характер размеще-
ния гнездовий и мест скопления тупиков по сравнению с акваторией моря
(Портенко, 1931).
Роющая деятельность тупиков в гнездовых колониях в сочетании
с обогащением почвы экскрементами приводит к средообразующему эф-
фекту — формирует зоогенные растительные ассоциации, мелкокочкова-
тый микрорельеф и стимулирует пышное развитие растений (Скокова,
1967а; Парфентьева, 1969).
В СССР тупик не является объектом промысла или спортивной охоты,
самые значительные колониальные гнездовья этого вида заповеданы (Ай-
новы острова, Семь островов). В связи с малой плодовитостью, ограничен-
ностью пригодных для гнездования мест и низкой относительной числен-
ностью (0,13 % мировой популяции вида) тупик должен охраняться пов-
семестно и круглогодично.
Ипатка —
Fratercula corniculata Naumann, 1821
Horned Puffin (англ.).
Син. (лат.): Mormon corniculata Naumann, 1821.
Статус. В пределах СССР гнездящийся, кочующий, частично зимую-
щий вид.
Общая характеристика и полевые признаки. Размеры средние. Хорошо
плавает, ныряет, с воды взлетает с разбега. С горизонтальной поверхности
суши взлететь не может, взлетает только с крутых склонов. Полет прямой,
быстрый, слабо маневренный, с частыми взмахами крыльев. Верх тела
черный, низ — белый. Голова крупная, сжата с боков, бока головы белые.
Клюв массивный, сжатый с боков, имеет вид лопатки. Весной, летом и
в начале осени клюв ярко-желтый с красной вершиной, над глазами верти-
кальные кожистые неоперенные выросты, напоминающие рожки. От ту-
пика отличается белыми щеками, преимущественно желтым клювом и на-
личием надглазных кожных выростов. Шея короткая, когда птица летит
создается впечатление, что голова «втянута в плечи». Хорошо ходит по
скалам даже по наклонной плоскости, цепляясь когтями пальцев за неров-
ности. Сидит на скалах, опираясь на всю плюсну, но при ходьбе опирается
только на пальцы.
Молчаливая птица. В период размножения иногда издаёт негромкие
низкочастотные «ворчащие» звуки. В местах, где плотность гнездования
низка, ипатки почти не издают звуковых сигналов при взаимодействиях
между особями. Однако в районах с большой плотностью гнездования сиг-
налы ипаток слышны довольно часто и даже в значительной мере могут
составлять звуковой фон колоний морских птиц.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Морфологические различия ипатки и атлан-
тического тупика так значительны, что, несмотря на явную генеалогиче-
скую близость этих птиц и строгий географический викариат, их можно
считать только за особые виды (Демёнтьев, 1951). На основании исследова-
ния экземпляров, добытых на Охотском побережье, Камчатке, Курильских
164
Тупик
и Командорских островах, островах Прибылова, Анадырском лимане,
Чукотском побережье и Аляске, не отмечено дробление этого вида на
географические расы (Портенко, 1973).
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла: .. Рулевых перьев
14—16, изредка 18. Размеры: длина (в мм) крыла самцов 185—100 (ге=14,
среднее 191,7), самок 176—201 (п=12, среднее 185,7), годовалого самца
180 (п=1), неполовозрелой самки 176 (и=1); клюва летом: самцов 47,1 —
55,3 (п=14, среднее 50,6), самок 45,9—50,4 (п=12, среднее 47,9), годова-
лого самца 41,0 неполовозрелой самки — 42,2; цевки: самцов 29,0—33,2
(п=14, среднее 30,9), самок 26,9—31,8 (п=12, среднее 29,2), годовалого
самца 29,9, неполовозрелой самки 27,0 (промеры сделаны по материалам
Зоологического музея МГУ). Масса самцов 531—754 г, самок 499—691 г
(Портенко, 1973).
Цевка в месте соединения с голенью в задней части ноги уплощена, и
в этом месте, как у многих других видов чистиковых, имеется кожная
мозоль — утолщенный участок кожи, размером примерно 10X5 мм (про-
меры по материалам Зоологического музея МГУ). Мозоль переходит
в кожную оторочку высотой 1 мм, которая тянется по заднему краю цевки
до пальцев. Сидящая на цевке птица опирается на эту мозоль (данные
С. П. Харитонова).
Окраска. Половой диморфизм в окраске отсутствует.
Самец и самка в брачном наряде. Верх головы темно-бурый. Бока го-
ловы чисто-белые, от глаза кзади проходит узкая черноватая полоска.
Подбородок и горло буровато-черные и сливаются в один черный рисунок
с ожерельем на шее. Кожистый вырост над глазом длинный и узкий,
черного цвета, больших размеров, чем у тупика, достигает у некоторых
особей 12 мм в длину. Радужина буровато-серая. Голое кольцо вокруг глаз
красное. Вершина клюва красная, основная часть — желтая. На вершине
клюва имеется 3 вертикальные борозды. Розетка в углу рта ярко-оранже-
вая. Вся спинная сторона тела, крылья и хвост черные или буровато-чер-
ные. Подкрылья буровато-серые, чернеющие к кистевому сгибу. Брюшная
сторона тела белая. По задней части оперения голени идет буровато-черная
полоса, которая соединяется с кольцом такого же цвета, окаймляющим
голень в том месте, где кончается оперение. Это в природе не всегда за-
метно, так как голень бывает прикрыта белыми кроющими перьями. Ноги
яркие, оранжево-красные.
Самец и самка в зимнем наряде. Клюв менее высокий, основание клюва
бурое, вершина красноватая, имеет борозды. Базальный валик надклювья
и розетки отсутствуют. Бока лба, уздечка и передняя часть щек серовато-
бурые, остальные части боков головы беловато-серые. Узкая полоска
кзади от глаз черная. Выросты вокруг глаз отсутствуют, кольцо вокруг
глаз бурое. Остальное оперение как в брачном периоде. Ноги бледно-мясной
окраски.
Путовой наряд. Птенец весь темно-бурый, середина брюшка беловатая,
клюв невысокий, черный. Молодые птицы в первом зимнем наряде. Уздечка
и голое кольцо вокруг глаз черноватые. Бока головы над глазом и за глазом
дымчато-черные. Середина горла и шея спереди буровато-черные. Клюв
низкий, бурый, без борозд на надклювье. Остальная окраска,как у взрослых
зимой. Молодые птицы во вторую осень жизни как взрослые зимой, но
в вершинной части клюва никаких борозд нет. Имеется слабый кожный
165
Семейство Чистиковые
вырост над глазом. Первый брачный наряд. Розетка, базальный валик у
основания надклювья и вертикальные борозды на клюве только намечены,
щеки сероватые (Козлова, 1957, Дементьев, 1951).
Линька. Изучена слабо. Последовательность смены нарядов предполо-
жительно такова: пуховой — первый зимний — промежуточный — второй
зимний — первый брачный — зимний (окончательный) — второй брачный
(окончательный).
Послебрачная линька полная, охватывает рамфотеку, маховые, руле-
вые, кроющие. Смена рамфотеки начинается в конце сентября, оперение
сменяется, по-видимому, в октябре-ноябре. Предбрачная линька — частич-
ная, охватывающая мелкое оперение и рамфотеку, протекает, вероятно,
в конце зимы — начале весны (точные даты не установлены). Годовалые
птицы заканчивают полную осеннюю линьку рано: ипатка в возрасте одного
года, добытая 5 августа в бухте Провидения, находилась в совершенно
свежем наряде (Козлова, 1957; Дементьев, 1951).
Распространение. Гнездовой ареал. Северные части Тихого
океана (рис. 43). Гнездится на базарах восточной части о-ва Врангеля
(мыс Уэринг), на о-ве Колючип, далее по северному побережью Чукотского
полуострова к востоку от Колючинской губы, на восточном и южном
побережье Чукотского полуострова, островах Диомида, побережье Анадыр-
ского залива, о-ве Олюмка, восточном побережье Камчатки до пос. Жупа-
ново (Кроноцкий залив), на Командорских островах, о-ве Верхотурова,
о-ве Карагинский. На южном и западном побережьях Камчатки не най-
дена. Далее гнездится в Пенжинской губе, на п-ве Тайгонос, Гижигинской
губе, Ямских островах, Тауйской губе, на побережье у пос. Аян, о-ве Св.
Ионы, Шантарских островах, северном побережье Приморья, в ряде мест
на побережье Сахалина (мыс Терпения), о-ве Тюлений, северных и сред-
них Курильских островах, возможно, и на Южных Курилах (Козлова, 1957;
Портенко, 1973; Нечаев, 1969; Гизенко, 1955; Велижанин, 1978; Вяткин,
1975; Вяткин и др., 1975; Стенченко, 1975; Мараков, 1975; Марков, 1975;
Кондратьев, 1975; Харитонов, 1980; Яхонтов, 1975). Гнездование ипатки
на Олюторском и Охотском побережьях обследовано крайне слабо. На о-ве
Монерон ипатка отмечена в 1949 г. (Гизенко, 1955), но не найдена в 1973
(Нечаев, 1975) (рис. 44).
Вне СССР гнездится по всему побережью Аляски, южнее 68° 50' с. ш., .
возможно, есть на мысе Барроу (71° 20' с. ш.) — по крайней мере до о-ва
Форрестер на юге, на островах: Алеутских, Прибылова, Св. Лаврентия,
Нунивак, Св. Матвея (Sowls et aL, 1978). Маленькая колония ипаток обна-
ружена на островах Королевы Шарлотты (Британская Колумбия), взрос-
лые летом встречены возле островов Триангль и Ванкувер, что позволяет
предполагать, что южная граница гнездового ареала ипатки смещается
к югу (Harrison, 1983). Единичных негнездящихся ипаток иногда обнару-
живали летом в районе Киритаппу (о-в Хоккайдо) в гнездовой колонии
топорков (Massey et al., 1982).
Миграции. Мигрирует, по-видимому, вплавь. Зимует поблизости от
берегов, южнее границы льдов (Sealy, 1973b), но изредка остается на зиму
в северной части ареала при наличии открытой воды (Дементьев, 1951;
Портенко, 1973). Встречается зимой в водах у Курильских островов,
Южного Сахалина, Командорских островов, восточного берега Камчатки,
спускается на юг до Южных Курил, нерегулярно зимует в водах у Хок-
кайдо и Северо-Восточного Хонсю. Вдоль американского побережья про-
166
Ипатка
а — гнездовой ареал,
б — область зимовок
Рис. 44. Ареал ипатки
в СССР
двигается к югу до южных районов Калифорнии (Hoffman et al., 1975),
иногда до вод Мексики (Harrison, 1983).
Откочевка к местам зимовки начинается, по-видимому, в октябре, так
как до конца сентября даже в северных частях ареала птицы добываются
у колоний. Весной у мест гнездования на юге ареала появляются в апреле-
167
Семейство Чистиковые
мае, на севере — в мае—начале июня. На о-ве Крузенштерна в 1953 и
1958 гг. прилет отмечен 2 июня. В 1934 г. в районе Уэлена ипатки появи-
лись 3 июня (Портенко, 1973). В 1984 г. у пос. Сиреники (южное побережье
Чукотки) прилет ипаток наблюдали 30 мая (данные Н. Б. Конюхова).
В Анадырском лимане ипатки появляются в середине июня (Козлова,
1957). На о-ве Св. Лаврентия за 7 лет наблюдений прибытие первых ипаток
отмечено от 5 до 28 мая, на мысе Томпсон (Аляска) в 1960 г. — 6 июня,
в 1961 г. — 8 июня (Sealy, 1973b). На о-ве Монерон ипатки появлялись
в начале мая, на Курильских островах — в апреле (Гизенко, 1955).
Численность. Немногочисленна по всему ареалу, хотя в отдельных
местах колонии могут насчитывать до нескольких сот и даже более 1000
гнездящихся пар, в очень редких случаях численность достигает десятков
тысяч (о-в Талан). Данные по численности отрывочны, поскольку гнездо-
вой ареал изучен крайне неравномерно. Почти по всему ареалу, кроме са-
мой северной его части (к северу от мыса Чукотский), по
численности уступает топорку.
На о-ве Врангеля гнездится несколько пар ипаток (Портенко, 1973),
на о-ве Колючин — до 500 особей (Кондратьев, 1975). На побережье
Чукотского полуострова от мыса Сердце-Камень до мыса Энмелен, вклю-
чая прибрежные острова, кроме о-ва Ратманова и о-ва Балка, в 1985 г.
учтено более 14,5 тыс. ипаток, наиболее крупные гнездовья отмечены на
мысе Энмелен и мысе Кикелюн — по 3000 особей. Из прибрежных остров-
ков наиболее крупные поселения были на о-ве Кенкай — 1000 особей, на
о-ве Маркинкан — 700 особей (данные Л. С. Богословской, Б. М. Звонова,
Н. Б. Конюхова) и о-ве Балка — 500 особей (данные В. Д. Аниси-
мова) .
В пределах отдельных базаров Чукотского побережья численность
ипаток может значительно колебаться по годам в зависимости от характера
таяния снега. В годы, когда северные базары летом бывают покрыты сне-
гом, численность ипаток на более южных базарах может сильно увеличи-
ваться, хотя не ясно, все ли они там гнездятся. В такие годы численность
ипаток на о-ве Балка (залив Лаврентия) доходит до 2000 особей (данные
Л. С. Богословской).
На о-ве Олюмка имеется «многотысячная колония ипаток (Козлова,
1957). На о-ве Верхотурова примерно 500 ипаток (Вяткин, 1975), на
Командорских островах (Мараков, 1972), на о-ве Карагинском и далее на
юг по побережьям Камчатки и Охотского моря ипатка немногочисленна —
отдельные поселения не превышают нескольких пар или десятков особей
(Вяткин и др., 1975; Марков, 1975; Стенченко, 1975; Велижанин, 1978;
Яхонтов, 1975). На Ямских островах — 1000 пар ипаток (Велижанин,
1975), на о-ве Талан Тауйской губы Охотского моря — примерно 50—60
тыс. пар ипаток (данные А. Я. Кондратьева и А. С. Китайского), на о-ве
Св. Ионы — 20 пар (Харитонов, 1980). По оценке А. Г. Велижанина (1978),
на Курильских островах гнездится 3—4 тыс. ипаток. Общая численность
ипаток, по учетам в восточной части ареала вне пределов СССР — 768 тыс.
особей, гнездящихся в 435 местах; расчетная численность популяции для
Аляски — 1,5 млн особей (Sowls et al., 1978).
Местообитания. В гнездовое время населяет скалистые побережья и
острова. Гнезда устраивает в расщелинах и под камнями. Гнездится груп-
пами или отдельными парами в колониях других морских птиц, но изредка
на мелких островках образует и моновидовые колонии, например на о-ве
168
Ипатка
Рис. 45. Элементы территориального (а) и брачного (б) поведения ипатки
Меркинкан, в бухте Пэнкигнэй, и на восточном побережье Чукотки (дан-
ные Л. С. Богословской, Н. Б. Конюхова, Б. М. Звонова).
Размножение. Половой зрелости достигают на третий, изредка на
второй год жизни (Sealy, 1973b). Приступают к размножению, по-види-
мому, в возрасте 4—5 лет (Wehle, 1982а). В небольшом числе в районе
гнездовий могут присутствовать на скалах неполовозрелые особи (Sealy,
1973b). Моногамы. В паре самец, как правило, немного крупнее самки.
Весной в местах гнездований могут появляться, когда скалы еще находятся
под снегом. От прилета первых птиц до откладки первых яиц проходит
примерно 3 недели (Sealy, 1973b). Первую неделю после прилета ипатки
держатся стаями на воде возле мест будущей колонии, изредка посещая
скалы, на которых в дальнейшем проводят много времени (Wehle,
1982а).
Территориальное и брачное поведение изучено недостаточно. Ипатки
охраняют вход в занятые щели или норы. Самцы защищают самку, где бы
она ни находилась — и на скалах, и в воде. Партнеры в сформированных
парах часто держатся вместе, иногда совершают совместные полеты возле
скал. В период перед откладкой яиц в конце июня 1984 г. мы наблю-
дали множество пар ипаток на море вдоль восточного побережья Чу-
котки.
Демонстрационное поведение в основном сходно с таковым у тупика
и других чистиковых птиц. В качестве угрозы могут быть использованы,
по-видимому, две демонстрации: 1) распластанная поза — Forward threat
posture (Cramp, 1985): птица наклоняется вперед и выставляет клюв
в направлении другой особи (рис. 45), в отличие от тупика клюв при этом
обычно закрыт; 2) вскидывание клюва: птица поднимает клюв, откидывая
голову назад, клюв направлен вперед и вверх, иногда почти вверх и нахо-
дится в таком положении несколько десятков секунд (см. рис. 45), иногда
169
Семейство Чистиковые
при этом крылья полураскрыты и приподняты (данные С. П. Харитонова).
Наклонная угрожающая поза с раскрытым клювом — Frussel-gaping
(Svein, 1962), у ипатки, по-видимому, отсутствует (Wehle, 1982а). Настоя-
щие стычки и драки редки. Во время драк птицы хватают друг друга за
клюв или за зашеек (данные В. А. Зубакина). Демонстрируя свое присут-
ствие на территории, ипатки могут издавать довольно громкий сигнал:
«а-гаа-ха-ха-ха». Этот же сигнал нередко издается при территориальных
взаимодействиях двух самцов. Кроме того, для ипаток очень характерна
«демонстративная походка» в «клубах» на камнях. При этой демонстрации
во время ходьбы тело ипатки качается из стороны в сторону и создается
впечатление, что птица с силой ставит лапу при каждом шаге. Шаги при
этом резкие, мелкие, походка напоминает «ходьбу на месте».
В брачном поведении используются: а) статическое вскидывание клюва;
б) динамическое вскидывание клюва — Head-flicking (Cramp, 1985) —
голова ритмично вздергивается вверх, как при статическом вскидывании,
но к тому же при этом клюв периодически открывается и закрывается
(см. рис. 45). Иногда, при каждом вздергивании клюва вверх ипатка из-
дает короткий звук «оп». И при длительном динамическом вздергивании
клюва слышно ритмичное «оп-оп-оп-оп». . .». Демонстрация «биллинг» —
Billing (Cramp, 1985; Wehle, 1982а; Svein, 1962) — птицы стоят напротив
друг друга, почти соприкоснувшись клювами, и обе (или только самец)
делают быстрые движения клювом из стороны в сторону, при этом
раскрывая и закрывая клюв (см. рис. 45). Самец иногда хватает самку за
перья шеи спереди, звуков при этой демонстрации даже с близкого рас-
стояния не слышно (Харитонов, 1980). Между демонстрациями «бил-
линг» партнеры иногда демонстрируют поклоны друг другу — Bowing
(Svein, 1962) — как на суше, так и на воде (см. рис. 45) (данные В. А. Зу-
бакина). Иногда после демонстрации «биллинг» самец нагибается и хва-
тает самку за голень в том месте, где кончается оперение (см. рис. 45)
(Харитонов, 1980). Последняя демонстрация отмечалась нами у ипаток на
о-ве Св. Ионы и ни разу не наблюдалась на мысе Халюсткина (восточное
побережье Чукотки). «Биллинг» имеет место у птиц, которые сформиро-
вали пару в течение всего сезона размножения (Wehle, 1982а).
Брачное поведение ипаток имеет место как на воде, так и на суше,
обычно на плоских камнях, в так называемых «клубах», где в утренние
и вечерние часы собираются группы ипаток. Насиживающие птицы иногда
покидают гнездо и могут несколько минут провести в «клубе», где устраи-
вают «биллинг» со своими партнерами, или же парочка может сделать
1 — 2 круга возле колонии, после чего насиживающая птица возвращается
к яйцу. В «клубах» во время «биллинга» некоторые рядом находящиеся
ипатки пытаются участвовать в этой демонстрации вместе с токующей
парочкой. Они приближают свой клюв к клювам токующих птиц и
делают движения «биллинга». Токующая парочка агрессивно реагирует
на таких особей (данные В. А. Зубакина и С. П. Харитонова).
У ипаток отмечен очень интересный способ изгнания потенциального
соперника. Этот способ можно назвать демонстрацией отношений, кото-
рая, возможно, заключает в себе косвенную угрозу. Демонстрация отноше-
ний состоит в следующем: если к парочке приближается третья птица, то
партнеры (или только самец) начинают усиленно демонстрировать «бил-
линг», который прекращается, как только чужак удалится. На воде демон-
страция отношений исполняется, когда третья особь приближается на
170
Ипатка
расстоянии менее 3—4 м от парочки, на суше — при меньшем расстоянии
(данные В. А. Зубакина и С. П. Харитонова).
Перед спариванием на воде самец часто исполняет демонстрацию,
которую можно назвать «демонстративным плаванием»: передняя часть-
тела самца приподнята над водой по сравнению с нормальным положением
плавающей птицы, шея вытянута вертикально вверх, клюв направлен
вперед. Перед самым спариванием самец демонстрирует статическое
вскидывание клюва, затем делает садку. Спаривание обычно-на воде, из-
редка на суше. Избегающая спаривания самка заныривает при попытке
садки.
На севере ареала ипатки гнездятся в расщелинах скал, в каменистых
осыпях обычно в верхней части базаров, на юге — также в расщелинах,
но могут и в норах, иногда почти открыто среди травы (Гизенко, 1955).
Однако рытье нор ипатками никем не отмечалось. Скорее всего, они зани-
мают норы, вырытые топорками или длинноклювыми тупиками (Ги-
зенко, 1955). В качестве подстилки ипатки таскают сухую траву, перья,
иногда водоросли. Подстилка в гнездовых нишах может отсутствовать
вовсе, находиться возле яйца, но может быть и беспорядочно разбросанной
по нише или гнездовой камере. На мысе Халюсткина мы находили гнезда
ипаток, где вход в щель был густо устлан сухой травой, а яйцо лежало на
голом камне или торфе. Среднее минимальное расстояние между входами
в щели ипаток на о-ве Талан в 1987 г. было 1,5 м (п=24) (данные
В. А. Зубакина и С. П. Харитонова).
Откладка яиц у ипаток начинается в июне—начале июля, в зависимости
от географической широты и времени отхода льда. На о-ве Св. Лаврентия
семенники у самцов имеют наибольший размер в последнюю неделю
июня—начале июля (Sealy, 1973b). Начало яйцекладки на о-ве Св. Лав-
рентия в 1967 г. — 20 июня, пик — 29 июня, на о-ве Крузенштерна в
1966 г. — не ранее 25 июня, на островах Прибылова — 19 июня—1 июля
(Sealy, 1973b). На о-ве Балка в 1957 г. свежие яйца были 3 июля (Пор-
тенко, 1973). На мысе Халюсткина в 1984 г. яйцекладка началась не позд-
нее 7 июля, 4 августа вылупление птенцов еще не началось. На о-ве Талан
в 1987 г. откладка яиц началась 18 июня (данные В. И. Зубакина). На о-ве
Св. Ионы в 1974 г., судя по вылуплению, кладка яиц началась не позднее
15—16 июня (данные С. П. Харитонова).
Самец и самка ипаток имеют по 2 наседных пятна, размером 55X27 мм,
но в кладке одно яйцо (Sealy, 1973b; Wehle, 1982а). Приведенные
Л. А. Портенко (1973) со ссылкой на других исследователей случаи на-
ходки кладок из двух яиц сомнительны. Дело в том, что ипатки иногда
могут изгнать чистиков из занятых ими ниш и насиживать их кладку
(Divoky, 1982), что, возможно, и создает путаницу.
Яйцо грязно-белого цвета, матовое. На скорлупе, в основном на толстом
конце, просвечивают как бы замазанные буро-фиолетовые пятна пигмента.
Поверхность яйца часто испещрена мелкими бурыми пятнами, представля-
ющими собой высохшие сгустки крови. Причина появления этих пятен
не ясна, возможно они как-то связаны с эктопаразитами ипаток. Размер
яиц 57—76X41—50 мм (Козлова, 1957); вес 9 свежих яиц, собранных
на о-ве Балка — 62—81,7 г., в среднем 74 г. (Портенко, 1973; материалы
Зоологического музея МГУ). В случае исчезновения первого яйца повтор-
ная кладка возможна через 10—21 день (Wehle, 1983).
Насиживают оба партнера попеременно. Период насиживания, по дан-
ным с о-ва Св. Лаврентия, — 40—43 дня, в среднем 41, 1 дня (Sealy, 1973b).
171
Семейство Чистиковые
Вылупление — конец июля—начало августа. 12 августа 1985 г. на о-ве
Балка найдено 2 недавно вылупившихся птенца (данные В. Д. Анисимова,
Л. И. Барсовой, Т. Б. Голубевой), в 1974 г. на о-ве Св. Ионы вылупление
началось не позднее 26 июля (данные С. П. Харитонова).
Вес эмбрионов перед вылуплением 49—69 г, вес только что вылупив-
шихся 2 птенцов 48 и 49 г, вес суточного птенца 66 г (о-в Балка, данные
В. Д. Анисимова, Л. И. Барсовой, Т. Б. Голубевой). Птенцы полувыводко-
вые. Период выкармливания птенца 36 —42 дня (Sealy, 1973b). Птенцов
кормят 3—6 раз в день, перерывов в кормлении не отмечено. Во время
кормления птенцы принимают позу выпрашивания и пищат, взрослые
издают «ворчащие звуки» (Wehle, 1983). Терморегуляция устанавлива-
ется на 6-й день (Sealy, 1973b). Кормят птенцов практически только
рыбой, размер приносимых рыб 36—178 мм, в среднем 61—98 мм, вес
рыб 2,5—34 г. Рыбок ипатки держат поперек туловища; как и тупики,
могут приносить в клюве сразу по несколько штук (Wehle, 1983).
Характер роста птенца (по данным с о-ва Св. Лаврентия): вес одноднев-
ного птенца 58,6 г., 29-дневного 420,9, 37-дневного 400,2 г. Спускающийся
на воду птенец полностью оперен, имеет вес в две трети от веса взрослой
птицы. После схода на воду птенцы, по-видимому, совершенно самостоя-
тельны. Сход птенцов на воду происходит в сентябре (Портенко, 1973);
на о-ве Монерон проходил в первой декаде сентября (Гизенко, 1955), на
о-ве Талан в 1987 г. — в середине сентября (данные А. Я. Кондратьева
и А. Китайского).
Суточная активность, поведение. Ипатки ведут дневной образ жизни
(Велижанин, 1973). На колонии имеют пики активности в утреннее и ве-
чернее время. Часто сидят на камнях, образуя «клубы» от 2—3 до 10 и
более особей. Во время насиживания в «клубах» часто держатся свободные
от насиживания птицы. Утром, примерно к 10—12 ч по местному времени
«клубы» пустеют, птицы разлетаются на кормежку в море, вечером к 20—
22 ч вновь собираются в «клубы». На воде в гнездовое время держатся
парами, одиночками, либо небольшими группами из нескольких особей,
изредка у базаров в группы из 10 и более птиц (данные В. А. Зубакина,
С. П. Харитонова). В зимний период держатся, скорее всего, мелкими
группами (Портенко, 1973).
Питание. Пищу добывают под водой, при нырянии. В гнездовое время
кормятся в основном в 1 — 2 км от берега (Sealy, 1973b). Питаются мелкой
рыбой (песчанкой, мойвой, сельдью, треской, скумбрией), головоногими
моллюсками (кальмарами), мелкими рачками, многощетинковыми пелаги-
ческими червями. Основу питания (от 60 до 97 %) составляет рыба, рако-
образные и моллюски составляют до 20 и 15,4 % соответственно. Удельный
вес тех или иных объектов в питании меняется в зависимости от места,
сезона и состояния кормовых ресурсов (Wehle, 1982с).
Враги, неблагоприятные факторы. Поскольку ипатки гнездятся в норах
и на крутых склонах, их гнезда практически не разоряются хищниками
и успех размножения высок — до 91 % (Wehle, 1983). Ипатка относится к
числу видов, наиболее чувствительных к нефтяному загрязнению моря
(King, Sanger, 1979, цит. по: Wehle, 1982а), чувствительны к загрязнению
ядохимикатами (Wehle, 1982а). В желудках примерно четвертой части
птиц, кормящихся у побережья Аляски, обнаруживаются кусочки пластика
размером 1 —14 мм (Wehle, 1982с). Ипатки запутываются и гибнут в сетях,
выставленных на лососевых рыб (Ainley et al., 1981).
172
Топорок
Успех размножения сильно зависит от обилия кормовой базы. Скорость
роста птенцов в какой-то мере может служить показателем состояния рыб-
ных запасов в данном месте и в данном сезоне: если скорость роста птенца
составляет более 2,5 % веса взрослой птицы в сутки, то кормовая база
достаточна для нормального развития птенцов, если же менее 1,5 % —
имеется недостаток пищевых ресурсов. В случае обильной кормовой базы
ипатки могут вырастить двух птенцов в искусственно сдвоенных выводках
(Wehle, 1983).
Хозяйственное значение, охрана. Промыслового значения не имеет.
В ряде мест местное население проводит сбор яиц попутно с яйцами других
видов птиц, но яйца собираются в очень небольшом количестве. Специаль-
ных мер охраны не предусмотрено.
Род Lunda Pallas, 1811
Топорок —
Lunda cirrhata Pallas, 1811
Tufted Puffin (англ.).
Син. (лат.): Alca cirrhata Pallas, 1769. Stagmatorrhina lathami Bonaparte, 1851.
Син. (рус.): Топорик.
Статус, В пределах СССР гнездящийся, кочующий, частично зимующий
вид.
Общая характеристика и полевые признаки. Крупная птица темной
окраски. Хорошо плавает, ныряет, с воды взлетает с разбега, с поверхности
суши — только с крутых склонов или забравшись на камни высотой 1,5—
2,8 м, играющие роль «взлетных площадок» (Михтарьянц, 1977). Полет
прямой, быстрый, слабо маневренный, с частыми взмахами крыльев. Голова
крупная, сжата с боков, бока головы белые, позади глаз расположены пучки
желтых перьев — «косицы». Клюв массивный, сжатый с боков, имеет вид
лопатки. От ипатки отличается темным брюхом, красным клювом и нали-
чием косиц. Ходит на пальцах, может довольно быстро бегать. При ходьбе
и беге туловище наклоняется вперед. На цевку опирается редко — при
движении по наклонным камням и когда залезает в нору.
Молчаливая птица. В период размножения издает низкочастотные
«ворчащие» звуки. Характерно, что в местах, где плотность гнездования
топорков низка, они редко используют звуковые сигналы в контактах
между собой. В районах с большой плотностью гнездования звуковые
сигналы топорки употребляют значительно чаще.
Подвидовая систематика. Монотипический вид.
Описание. Строение и размеры. Первостепенных маховых 11,
первое рудиментарно. Формула крыла: 1<П>П1>1¥ . . . Рулевых перьев
14—16. Размеры: длина (в мм) крыла самцов 191 — 220 (п=19, среднее
202,4), самок 186—207 (п=16, среднее 195,7), молодых особей, добытых
в конце сентября—декабря (п=5), 134—172; клюва самцов 55,0—68,7
(п=20, среднее 59,9), самок 51,4—60,4 (п=15, среднее 56,2), молодых
особей (и=5) 37,0—46,5; цевки самцов 32,8—37,1 (и=20, среднее 34,5),
самок 31,7—36,0 (и=16, среднее 34,2), молодых особей (и=5) 29,0—35,2.
(Промеры сделаны по материалам Зоологического музея МГУ). Масса
взрослых особей 689—920 г (Sealy, 1973b).
173
Семейство Чистиковые
В задней части цевки в месте ее соединения с голенью, так же как и
у многих чистиковых, имеется кожистая мозоль размером примерно
11,2X6,0 мм; сзади цевки имеется кожистая оторочка от мозоли до паль-
цев (данные С. П. Харитонова).
Окраска. Половой диморфизм в окраске отсутствует. Самец и самка
в брачном наряде. Вся верхняя сторона тела, крылья, хвост и их верхние
кроющие буровато-черные. Уздечка, щеки, поле вокруг глаз и узкая по-
лоска вокруг клюва белые. Кзади от глаза — пучок беловато-желтых длин-
ных перьев. Горло, бока и вся нижняя сторона тела темно-бурые, немного
светлее, чем спина. Нижние кроющие крыла буровато-серые. Край крыла
вдоль предплечья имеет белые перышки. Радужина серовато-белая, голое
кольцо вокруг глаза красное. Вершина клюва красная, базальная часть
клюва бледно-оливково-зеленая или желтовато-оливковая. На вершине
клюва имеется до 4 вертикальных бороздок. Узкая полоска вокруг основа-
ния клюва на грани оперения красная. Лапы красные.
Самец и самка в зимнем наряде. Вся голова и нижняя сторона тела
темно-бурые, на брюшке бывают беловатые пестрины. За глазом более свет-
лое серовато-бурое пятно. Косиц нет. Верхняя сторона тела (кроме головы)
буровато-черная. Плоская пластинка в основании клюва над ноздрями,
роговой валик в основании гребня надклювья, поперечная пластинка
у грани оперения и розетка в углах рта отсутствуют. Основание клюва
бурое, вершина — красная.
Пуховой наряд. Птенец темно-бурый, снизу сероватый. Клюв невысо-
кий, буровато-серый, лапы буровато-черные. У части популяции (в от-
дельных районах до 6,8 %) у пуховых птенцов на брюшке бывают белые
пятна, диаметром от 20 мм и более, иногда все брюшко бывает беловатое
(Wehle, 1982b). Пух птенца густой и длинный — до 30 мм на спине.
Гнездовой наряд. Вслед за пуховым нарядом у птенцов развивается
мезоптильно-юношеский наряд, который меняется на первый зимний в пер-
вую осень жизни. У птиц в юношеском наряде верхняя сторона тела —
как у взрослых птиц зимой, за глазом намечается серая полоска. Горло, шея
спереди и частично область зоба буровато-серые; грудь и брюшко белые со
слабо выраженными серовато-бурыми вершинными каемками перьев. Ра
дужина буровато-серая, голое кольцо вокруг глаз черноватое. Клюв невы-
сокий, без борозд, бурого цвета, ноги светло-серые.
Первый зимний наряд. Молодые птицы в первом зимнем наряде сходны
с птенцами в гнездовом наряде, но серовато-бурые каемки белых перьев
на груди и брюшке всегда хорошо развиты. Молодые птицы в первую весну
жизни не линяют, но темные каемки белых перьев на груди и брюшке обна-
шиваются, и эти части оперения становятся белыми.
Второй зимний наряд. Молодые птицы на второй год жизни очень по-
хожи окраской оперения на взрослых, но нижняя сторона тела несколько
светлее и серее. От глаза кзади проходит сероватая полоска из волосовид-
ных, слегка удлиненных перьев. Основание клюва бурое, вершина оранже-
вая. Молодые птицы во вторую весну жизни после весенней частичной
линьки имеют белые перья по бокам головы, белое кольцо вокруг клюва
и короткие белые косицы за глазом. В базальной части надклювья разви-
ваются дополнительные роговые пластинки и валики. .На вершине над-
клювья намечаются борозды (Козлова, 1957; Дементьев, 1981).
Линька. Изучена слабо. Последовательность смены нарядов пред-
положительна такова: пуховой — мезоптиль — первый зимний — пер-
174
Топорок
вый летний — второй зимний — второй летний (предбрачный) — зимний
(окончательный) — летний (окончательный). В первый зимний наряд из
мезоптиля линяют в августе—октябре. В первую весну жизни молодые
не линяют, первый летний наряд формируется за счет снашивания опере-
ния первого зимнего наряда. Для всех других возрастов имеется полная
послебрачная линька в зимний наряд в конце сентября—октябре, возможно
и позднее: топорок, добытый 23 февраля у о-ва Беринга, находился в све-
жем зимнем наряде. Весенняя линька частичная, захватывает мелкое кон-
турное оперение, но не рулевые и не маховые. Тогда же развиваются
украшающие перья на голове и пластинки у основания клюва. Число бороз-
док на клюве увеличивается в первые годы жизни от 0 до 4. Весенняя
линька проходит не ранее марта и не позднее середины мая (Козлова, 1957;
Дементьев, 1951).
Распространение. Гнездовой ареал. Северные части Тихого
океана (рис. 46). Распространен по побережью Чукотского полуострова от
островов Врангеля и Колючин, на восток от мыса Сешан по северному, вос-
точному и южному побережью Чукотского полуострова (включая прибреж-
ные острова), о-в Ратманова, о-в Олюмка (Анадырский лиман), Корякское
побережье Камчатки (гнездование на побережье Олюторского залива и
северо-востоке Камчатки не доказано, но весьма вероятно), острова Верхо-
турова, Карагинский, Командорские острова, юго-западное побережье
Камчатки, островки вдоль западного побережья Камчатки (гнездование
не доказано), Пенжинская губа, весьма вероятно — губа Гижига, Ямские
острова, Тауйская губа, острова Талан, Нансикан, побережье у пос. Анн
(побережье Охотского моря изучено слабо), о-в Св. Ионы, Шантарские
острова, в ряде мест на побережье Сахалина, о-в Монерон, о-в Тюлений,
практически все Курильские острова, весьма вероятно — северное и вос-
точное побережье Приморского края (Козлова, 1957; Портенко, 1973;
Гизенко, 1955; Нечаев, 1969; Вяткин, 1975; Вяткин и др., 1975; Велижанин,
1978; Воробьев, 1954; Мараков, 1975; Марков, 1975; Кондратьев, 1975;
Стенченко, 1975; Нечаев, 1975; Стишов и др., 1985; Яхонтов, 1975).
В Приморье с начала столетия происходит сокращение ареала топорка.
В начале века этот вид гнездился на островах в заливе Петра Великого,
в 1949—1957 гг. отмечен на гнездовании в бухте Ольги, в настоящее время
в этих местах не гнездится; в заливе Петра Великого топорки не отмеча-
ются (Воробьев, 1954; Лабзюк, 1975; Лабзюк и др., 1971). Южная граница
гнездового ареала в Приморье в настоящее время не установлена (рис. 47).
Вне СССР гнездится на мелких островках на северо-востоке Хоккайдо:
побережье Киритаппу, Аккеси, острова Дайкоку, о-в Мойурури (Иванов,
1976; Massey et al., 1982). В Северной Америке гнездится от севера Аляски
(мыс Липсберн) на юг по побережью, на островах: Св. Лаврентия, Прибы-
лова, Алеутских, Крузенштерна, Нунивак, на побережье Британской
Колумбии, западном побережье США до острова Фараллон (37°40' с. ш.,
123°10' з. д.) и мысе Харрикайн-Пойнт в Калифорнии (Sowls et al., 1978;
Harrison, 1983). В прошлом и у американского побережья южная граница
ареала проходила южнее, чем в настоящее время, — топорки доходили до
Санта-Барбара, иногда гнездились на о-ве Сан-Николас (33° с. ш.) (Bent,
1919, по Дементьеву, 1951).
Зимовки. Зимует в открытом море, недалеко от мест гнездования
на свободных ото льда участках моря. Отмечен в районе Командор (добы-
вались в декабре—феврале), у берегов Камчатки, у Сахалина, на Куриль-
175
Семейство Чистиковые
Рис. 46. Область
распространения
топорка
а — гнездовой ареал,
б — отдельные колонии,
в — область зимовок и
миграций,
г — дальние залеты
Рис. 47. Ареал топорка
в СССР
а — отдельные колонии,
б — гнездовой ареал,
в — зимовки
ской гряде. На юг вдоль азиатского континента топорки спускаются до
севера Хонсю; посреди океана встречаются примерно до 35 параллели; у бе-
регов Калифорнии зимуют, видимо, лишь немного южнее границы гнездо-
вого ареала. Залеты отмечены у мыса Барроу, в море Бофорта, в штате Мэн
(Атлантика) (Козлова, 1957; Дементьев, 1951; Harrison, 1983).
176
Топорок
Откочевка с мест гнездования происходит в средней части Курильской
гряды в конце сентября —октябре, с Камчатки откочевывают в начале
сентября (Гизенко, 1955). С побережья Чукотки отлет происходит при-
мерно в это же время (Портенко, 1973).
Миграции. Появляются у гнездовых колоний на юге ареала в апреле—
мае, на севере — в конце мая—начале июня. На Командорских островах
прилет отмечен в конце апреля. В 1934 г. у Уэлена топорки впервые встре-
чены 3 июня (Портенко, 1973). В 1984 г. в районе пос. Сиреники (южное
побережье Чукотского полуострова) топорки появились 18 мая (данные
Н. Б. Конюхова). На о-ве Св. Лаврентия за 4 года наблюдений срок прилета
изменялся от 23 мая до 4 июня, на мысе Томсон (Аляска) за 2 года наблю-
дений — 8 и 16 июня, на о-ве Крузенштерна в 1953 и 1958 гг. — 2 июня
и 26 мая (Sealy, 1973b).
Численность. В самых северных частях ареала немногочислен, гнез-
дится отдельными парами или небольшими группами на птичьих базарах.
Подобное положение отмечается и на самом юге ареала. В средней части
ареала многочислен, если позволяют условия — гнездится большими ко-
лониями.
На о-ве Врангеля (мыс Уэринг) гнездится нерегулярно, отдельными
парами (Стишов и др., 1985). На о-ве Колючин насчитывается около
50 топорков. По Чукотскому побережью от Колючинской губы на восток,
далее по побережью до мыса Энмелен (включая прибрежные острова, кроме
островов Ратманова, Балка, Беннета) в 1985 г. учтено 20 700 топорков.
От мыса Чукотский по восточному и северному побережью Чукотки чис-
ленность топорков в пределах отдельных базаров ниже численности ипаток,
к западу от мыса Чукотский топорки преобладают над ипатками. Наиболее
крупные колонии на мысе Кикелюн — 6000 особей и мысе Укелюн —
3500 особей (данные Л. С. Богословской, Б. М. Звонова, Н. Б. Конюхова).
На о-ве Олюмка — от 50 до 100 особей (Козлова, 1957), о-ве Верхотурова —
500 особей (Вяткин, 1975), о-ве Карагинский — 5 тыс. особей, в том числе
до 80 % на маленьком островке Птичьем, возле о-ва Карагинский (Вяткин
и др., 1975). Вдоль восточного побережья Камчатки топорки встречаются
небольшими группами или отдельными парами (Марков, 1975; Стенченко,
1975). На Командорских островах основное население топорков сосредото-
чено на о-ве Топорков — 10—15 тыс. особей и на о-ве Арий Камень —
2500 особей (Карташев, 1979), на о-ве Медный, где на самом крупном ба-
заре отмечено до 2000 топорков (Велижанин, 1978). Курильские острова
практически все населены топорками, общая численность популяции здесь,
по оценкам Велижанина (1978), — 200 тыс. особей. На о-ве Топорков
(возле Матуа) имеется колония до 10 тыс. птиц (Гизенко, 1955). На о-ве
Талан Тауйской губы Охотского моря примерно 80 тыс. пар (данные
А. Я. Кондратьева и А. С. Китайского). На о-ве Св. Ионы примерно 300 пар
(Харитонов, 1980), на Ямских островах — 16 тыс. топорков (Велижанин,
1975). На северном и западном побережье Охотского моря, а также на
о-ве Сахалин топорки гнездятся преимущественно маленькими группами,
хотя на мелких островах их численность бывает значительна (Гизенко, 1955,
Велижанин, 1978). На о-ве Монерон отмечено несколько десятков пар
топорков (Нечаев, 1975). На Шантарских островах крупных колоний нет.
На побережье Приморья топорки немногочисленны (Велижанин, 1978),
немного их и в пределах Японии (Massey et al., 1982). На о-ве Мойурури
гнездятся примерно 230 особей (Fujimaki, 1961). В пределах штата Аляска
12 Птицы СССР
177
Семейство Чистиковые
учтено 2 108 535 топорков в 502 местах гнездования, расчетная численность
популяции здесь — 4 млн особей. Наиболее крупная колония на о-ве Кали-
гаган — от 250 тыс. до 500 тыс. топорков (Sowls et al., 1978). В Британской
Колумбии отмечено 31 тыс. топорков, 8000 — в пределах штата Вашингтон,
примерно 1000 — в пределах штатов Орегон и Калифорния (Vermeer, 1979).
Местообитания. В гнездовой период населяет побережья и острова.
На севере ареала гнездится преимущественно в расщелинах скал, южнее,
кроме того, и в норах, которые роет в слое торфа, иногда в песчаном и галеч-
ном грунтах (если почва закреплена корнями растений) (Гизенко, 1955;
Михтарьянц, 1977). Часто гнездится в колониях других морских птиц,
но может образовывать и самостоятельные многотысячные колонии,
последние — преимущественно на мелких островках.
Размножение. Половой зрелости топорки достигают, по-видимому,
на третий год жизни, приступают к размножению преимущественно
в возрасте 4—5 лет (Wehle; 1982а). Неполовозрелые особи могут присут-
ствовать в районе колонии в небольшом числе (Sealy, 1973b). Моногамы.
Период от прилета до откладки яиц примерно 3 недели (Sealy, 1973b).
Перед откладкой яиц топорки присутствуют на колонии периодически,
по 2—3 дня, после чего на 2—3 дня отлетают в море (Wehle, 1982а). Гнезда
располагаются в укрытиях (нишах или норах), но некоторые — почти
на поверхности почвы (Михтарьянц, 1977). Нору роют клювом, землю
выгребают обычно ногами. Входные отверстия открываются на поверхность
без особой маскировки, высота отверстия 13,5— 14,5 см, ширина — 16—17см
(Гизенко, 1955). Глубина норы 0,5—2 м, чаще — около 1 м, на 1 м2 почвы
может быть до 5 входных отверстий (Карташев, 1979). Среднее мини-
мальное расстояние между центрами нор, по данным с о-ва Талан Охот-
ского моря, 83 см (н=342), минимальное — 15 см. Плотность гнездоваТГШг
на о-ве Талан доходила до 182 жилых нор на 200 м2. Распределение втор
по площади колонии при плотности 66—182 норы на 200 м2 достоверно
равномерное (данные С. П. Харитонова).
Может гнездиться и в крупноблочных каменистых осыпях и завалах
камней вместе с другими укрыто гнездящимися чистиковыми. Занимают
ниши в камнях, размером примерно 30X50 см.
Территориальное и брачное поведение изучено крайне слабо (рис. 48).
На о-ве Топорков (Командорские острова) в конце мая 1969 г. почти все
норы уже были распределены между птицами, однако драки из-за них
наблюдались до середины июня. Драки происходили преимущественно
в утренние и вечерние часы. Топорки, сцепившись клювами, наносили друг
ДРУГУ Удары лапами и раскрытыми крыльями (Михтарьянц, 1977). Кон-
куренция между топорками бывает как за места,’ где в дальнейшем птицы
будут рыть норы, так и за прошлогодние норы. Нередко драки происходят
прямо в норах. На колонии топорки защищают участки диаметром при-
мерно около 1 —1,5 м, однако в плотных местах размеры участков меньше.
На крупных камнях среди колонии топорки могут образовывать
«клубы», где птицы сидят в 0,2—0,3 м друг от друга без каких-либо види-
мых взаимодействий. Однако на других камнях парочки и отдельные самцы
топорков могут образовывать подвижные охраняемые участки диаметром
0,7—1 м. Сигналом того, что участок уже занят парой топорков может слу-
жить так называемая «демонстрация отношений», когда пара при прибли-
жении возможного конкурента начинала демонстрацию «биллинг» между
собой.
178
Рис. 48. Элементы территориального и брачного поведения топорка
12*
179
Семейство Чистиковые
У топорков отмечены 3 позы угрозы. Слабая угроза: стоящий или си-
дящий топорок просто пригибает голову, наклоняя клюв вертикально вниз.
При этом хвост птица может поднять почти вертикально вверх. Эта поза
может иметь место и в движении; в последнем случае топорок может идти
в этой позе с раскрытыми крыльями. Более сильная угроза: клюв вытяги-
вается вперед и поднимается вверх под углом до 45°, клюв при этом раскрыт,
а косицы выпрямлены. Обычно топорки находятся в такой позе неподвижно
3—4 с. Кроме того, изредка отмечается и агрессивный «биллинг» одиноч-
ной особи (по-видимому, так демонстрируют угрозу некоторые самцы):
при приближении соперника птица наклоняет голову и начинает быстро
двигать клювом из стороны в сторону и вверх-вниз.
Колонии топорков между норами имеют множество тропинок, которые
ведут к «взлетным площадкам» — обрывистому берегу или высоким кам-
ням.
Демонстрация «биллинг» имеет место у топорков в течение всего сезона
размножения. Во время этой демонстрации самец и самка стоят друг перед
другом и двигают клювами из стороны в сторону, ударяя друг друга
по клювам. Во время «биллинга» самец может хватать клювом самку
за перья на любой части тела. Наблюдалось даже легкое хватание клювом
плавательной перепонки самки. Если «биллинг» демонстрируется парочкой
в «клубе», где есть еще топорки, то некоторые из них пытаются токовать
вместе с токующей парочкой. Особи, исполняющие «биллинг», агрессивно
реагируют на птиц, пытающихся пристроиться к току (данные С. П. Хари-
тонова) .
Спаривание у топорков происходит чаще на воде, но может быть
и на суше, возле нор. Перед копуляцией самцы топорков принимали позу,
аналогичную статическому вскидыванию клюва у ипаток. Спаривание
длится 30—60 с (Михтарьянц, 1977).
Хотя топорки, как и ипатки, имеют два наседных пятна, в кладке
у них обычно одно яйцо, два бывает очень редко (Дементьев, 1951).
Выстилка гнезда большей частью скудная, состоит из сухой травы и перьев,
только в редких случаях у топорков отмечается «хорошее» гнездо из сухой
травы (Михтарьянц, 1977). Вместе с тем у них ярко выражен инстинкт
собирания гнездового материала. На о-ве Топорков в период занятия нор
отмечался интенсивный сбор гнездового материала — птицы захватывали
клювами пучок сухой травы (при этом часто садились на цевку) и с силой
отрывали его от земли. Собранный материал птицы интенсивно носили
в норы (Михтарьянц, 1977).
Яйцо топорка типичной яйцевидной формы, грязно-белого, изредка
бело-кремового цвета с матовой поверхностью. Иногда на толстом конце
видны как бы замазанные белым пятна диаметром 1—4 мм. Но пигментиро-
ваны яйца топорков слабее, чем яйца ипаток. На скорлупе, так же как
у ипаток, имеются мелкие пятна засохшей крови. Размеры яйца (в мм):
длина 65,5—78, ширина 45,0—51,5 (Козлова, 1957). Масса четырех свежих
яиц (по материалам Зоологического музея МГУ) 70, 89, 89, 96 г.
Откладка яиц в зависимости от широты места начинается в мае—июне,
иногда в начале июля: на о-ве Триангль в среднем 16 мая (Burrel, 1980),
на Курильских островах — в конце мая—июне (Гизенко, 1955). В 1960 г.
на островах Топорков и Арий Камень (Командорские острова) первые
яйца появились в конце мая—начале июня (Карташев, 1979); на о-ве Та-
лан в 1987 г. 18 июня (данные А. М. Китайского). На о-ве Св. Ионы в 1974 г.,
180
Топорок
судя по вылуплению, яйца начали появляться не позднее 10—11 июня
(данные С. П. Харитонова). Яйцекладка растянута более чем на месяц.
В местах, где осуществляется массовый сбор яиц топорков, период откладки
яиц может быть растянут сильнее. В случае потери первого яйца, повторная
кладка происходит через 10—21 день (Wehle, 1982с).
Насиживание продолжается 38—46 дней (Михтарьянц, 1977; Burrel,
1980). Насиживают оба партнера, сменяя друг друга, по одним данным,
через 12 ч (Козлова, 1957), по другим — бессменное насиживание кладки
продолжается более суток (Михтарьянц, 1977).
Первые птенцы на о-ве Уруп в 1947 г. были обнаружены в начале
июля (Гизенко, 1955). В 1974 г. на о-ве Св. Ионы вылупление началось
не позднее 21 июля (данные С. П. Харитонова). В 1969 г. на о-ве Топорков
появление первых птенцов отмечено 25 июля (Михтарьянц, 1977). Период
выкармливания птенцов 43—59 дней, в среднем 50,4 дня (Burrel, 1980).
Птенцы полувыводковые. Кормят птенцов оба родителя 3—4 раза в день,
иногда на 1—2 дня делая перерывы. Кормят птенцов преимущественно
рыбой, но от 5 до 30 % приносимого корма составляют кальмары и рако-
образные (Vermeer, 1979; Wehle, 1983). Длина песчанок, приносимых
птенцам, варьирует в зависимости от года. На о-ве Триангль в 1977 г. она
была в среднем 84,9 мм, в 1978 г. (малокормный год) — 12,5 мм. В год
с оптимальными кормовыми условиями родители приносят птенцам в день
(в августе месяце) 75—80 г корма (Vermeer, 1979). На Командорских
островах в 1969 г. длина приносимых рыбок была в пределах 4—20 см,
крупных рыб топорки приносили в клюве по 1 — 2, мелких — до двух
десятков и более. Рыбок топорки держат поперек клюва, длинные рыбки
оказываются сложенными вдвое и прихваченными у хвоста и у головы
(Михтарьянц, 1977). Во время кормления птенцы принимают позу выпра-
шивания и издают писк, родители «урчат». При вылете из гнезда птенцы
имеют размер в 2/з взрослой птицы, маховые и рулевые перья слегка недо-
развиты (Гизенко, 1955, Козлова, 1957).
На о-ве Талан Охотского моря массовый спуск птенцов топорков
наблюдался в середине сентября (данные А. Я. Кондратьева и А. М. Китай-
ского). В Сахалинской области птенцы спускаются на воду в конце
августа, выведенные из повторных кладок — в середине или конце сентяб-
ря (Гизенко, 1955). Птенец с сильно недоросшими маховыми и рулевыми
и остатками пуха на брюшке добыт 15 октября на о-ве Беринга (Козлова,
1957). После схода на воду птенцы, по-видимому, совершенно самостоя-
тельны.
Суточная активность, поведение. Топорки наиболее активны в дневное
время суток (Велижанин, 1973). На колонии имеют утренний и вечерний
пики активности. На суше сидят возле нор или образуют «клубы» из не-
скольких птиц на камнях. Днем топорки улетают на кормежку в море,
ночуют в норах, и, по-видимому, на воде (Михтарьянц, 1977). У топорков
до начала откладки яиц, а также в периоды насиживания и выкармли-
вания птенцов имеют место колебания численности птиц в колонии. Спады
и нарастания численности отмечаются примерно раз в 5 сут. Период отсут-
ствия топорков (кроме насиживающих птиц) 1,5—2,5 суток, прилет после
спада численности происходит в вечернее время, после чего топорки бы-
вают многочисленны на колониях 2—3 дня, затем в утреннее время снова
наблюдается массовый отлет (Михтарьянц, 1978). На кормежку летают
181
Семейство Чистиковые
небольшими стаями, по 2—5 птиц; в море, видимо, кормятся тоже не-
большими группами (Михтарьянц, 1977, Гизенко, 1955).
Питание. Пищу добывают под водой при нырянии. В гнездовое время
обычно кормятся далеко от берегов — на расстоянии до 15 км (Sealy, 1973b)
или даже более 20 км (Михтарьянц, 1978). Мест кормежки достигают,
преимущественно, по воздуху. (Топорки чаще пользуются крыльями, чем
кайры и ипатки). Питаются мелкой рыбой: песчанкой, сельдью, мойвой,
треской, скумбрией, бычками, терпугом, в южных частях ареала — анчоу-
сом, поедают также головоногих моллюсков (кальмаров), пелагических
ракообразных и червей (Vermeer, 1979; Wehle, 1983). Есть указания, что
в рационе топорков встречаются морские ежи и губки (Гизенко, 1955).
Основу питания (от 60 до 97 %) составляет рыба, ракообразные и мол-
люски составляют до 20 и 15,4 % соответственно (Wehle 1983). Удельный
вес тех или иных объектов в питании сильно варьирует в зависимости
от места и сезона.
Враги, неблагоприятные факторы. На топорков охотится сапсан; яйца,
птенцов и взрослых птиц могут поедать попавшие на острова лисицы
(Гизенко, 1955). Иногда топорки страдают от клептопаразитизма со сто-
роны моевок, которые отбирают у топорков рыбу (Михтарьянц, 1977).
В желудках примерно четвертой части птиц, кормящихся у побережья
Аляски, содержатся мелкие кусочки пластика (Wehle, 1982с). Так же как
и другие чистиковые, топорки гибнут в сетях для лососевых рыб (Ainley
et aL, 1981). Топорки относятся к числу видов, весьма чувствительных
к загрязнению моря нефтепродуктами и ядохимикатами (King, Sanger,
1979, цит. по: Wehle, 1983).
Успех размножения — до 95 %, но часто бывает низким по причине
недостатка кормов (Vermeer, 1979). Скорость роста птенцов может служить
индикатором состояния кормовых ресурсов: оптимум — при скорости роста
птенцов 2,5 % веса взрослых в сутки, явный недостаток — при скорости
0,9—1,5 % веса взрослых в сутки. При обильной кормовой базе могут
выкормить двух птенцов (в искусственно сдвоенных выводках), причем
скорость роста птенцов в таких выводках не отличается от таковой в вывод-
ках с одним птенцом (Wehle, 1983).
Хозяйственное значение, охрана. Яйца и мясо топорков употребляются
в пищу. Яйца собирают в больших количествах, взрослых птиц ловят
сетями на колониях. Способ лова таков: перед летящими топорками охот-
ники неожиданно поднимают на палках сеть, в которую птицы запутыва-
ются. Из шкурок местное население изготовляет одежду — парки (Коз-
лова, 1957; Гизенко, 1955). Специальных мер охраны не предусмотрено.
Литература
Абрамов В. К., Косыгин Г. М., Тихоми-
ров Э. А. О зимовках некоторых видов
птиц в заливе Петра Великого // Вюл.
МОИП. Отд. биол. 1973. Т. 78, № 2.
С. 67 70.
Авилова К. В., Корнеева Т. М. Некоторые
эколого-морфологические особенности
глаза чайковых (Laridae) и чистиковых
(Alcidae) птиц // Зоол. журн. 1973.
Т. 52, № 9. С. 1521-1527.
Антипин В. М. Птичьи базары на северо-
востоке Новой Земли // Природа. 1953.
№ 3. С. 115-116.
Белкин А. Н., Велижанин А. Г. Распростра
нение белобрюшки на Курилах // Орни-
тология. 1965. Вып. 7. С. 257.
Белопольский Л. О. Птицы Судзухинского
заповедника // Тр. ЗИН АН СССР. 1955.
Ч. 2. Т. 17. С. 224-265.
Белопольский Л. О. Экология морских ко-
лониальных птиц Баренцева моря. М.;
Л.: Изд-во АН СССР, 1957а. 460 с.
Белопольский Л. О. Кормовые биотопы и
состав пищи морских колониальных птиц
Баренцева моря // Позвоночные Арк-
тики. Л., 19576. С. 19-31. (Тр.
АРКТНИИ; Т. 25).
Белопольский Л. О. Анализ факторов, опре-
деляющих массовые концентрации мор-
ских птиц в открытом море, на побе-
режьях и островах океана // Докл. АН
СССР. 1979. Т. 249, № 5. С. 43-45.
Белопольский Л. О., Бабарыка В. Н., Бу-
гаев Л. А., Смирнова Л. В. Экологиче-
ские особенности распределения морских
птиц в открытом море на протяжении
года // Там же. 1982. Т. 262, № 2.
С. 508- 512.
Бианки В. В. Материалы по миграциям
птиц в Кандалакшском заливе//Тр.
Третьей Прибалт, орнитол. конф. Виль-
нюс, 1957. С. 6.
Бианки В. В. Негнездящиеся виды чисти
ковых (Alcae) в Кандалакшском за-
ливе//Тр. Кандалакш. заповедника.
1960. Вып. 2. С. 113-121.
Бианки В. В. Кулики, чайки и чистиковые
Кандалакшского залива // Тр. Канда-
лакш. заповедника. 1967. Вып. 6. С. 259—
316.
Бируля А. А. Зоологические результаты
русских экспедиций на Шпицберген //
Ежегодник Зоол. музея Акад. наук.
1910. Т. 15. № 1/2. С. 167-206.
Бутурлин С. А. Кулики, чайки, чистики,
рябки, голуби // Бутурлин С. А., Де-
ментьев Г. П. Полный определитель
птиц СССР. М.; Л.: Когиз, ф-ка книги
«Красный пролетарий» в Москве, 1934.
Т. 1. С. 256.
Бутьев В. Т. Зимовка птиц на севере
Новой Земли // Орнитология. 1959.
Вып. 2. С. 99-101.
Бычков В. А. Состояние птичьих базаров
на о-ве Тюленьем (Сахалинская обл.)
в 1958—1965 г. // Колониальные гнез-
довья околоводных птиц и их охрана.
М.: Наука, 1975. С. 168-169.
Бычков В. А. О распространении шишко-
носой гагарки // Редкие, исчезающие и
малоизученные птицы СССР. Рязань,
1976. С. 184-186.
Васъковский А. П. Новые орнитологические
находки на северном побережье Охот-
ского моря // Зоол. журн. 1956. Т. 35,
№ 7. С. 1051 — 1058.
Велижанин А. Г. Морские колониальные
птицы Курильских островов // Орнито-
логия в СССР. Ашхабад, 1969. Т. 2.
С. 112-115.
Велижанин А. Г. Морские колониальные
птицы Курильских островов: Автореф.
дис. . . . канд. биол. наук. Новосибирск,
1972. 28 с.
Велижанин А. Г. Адаптивный характер
ночного образа жизни морских коло-
ниальных птиц // Проблемы эволюции.
Новосибирск: Наука, 1973. Т. 3. С. 249—
251.
Велижанин А. Г. Птичьи базары Ямских
островов // Охота и охотничье хоз-во.
1975. № 7. С. 18-19.
Велижанин А. Г. Новые сведения о морских
птицах Дальнего Востока // Зоол. журн.
1977а. Т. 16, № 7. С. 1077-1083.
Велижанин А. Г. О некоторых редких и
малоизвестных птицах Курильских ост-
ровов // Орнитология. 19776. Вып. 13.
С. 25—32.
Велижанин А. Г. Размещение и состояние
численности колоний морских птиц на
Дальнем Востоке // Актуальные вопросы
охраны природы на Дальнем Востоке.
Владивосток: ДВНЦ АН СССР, 1978.
С. 154 -172.
183
Литература
Велижанин А. Г., Белкин А. Н. Тупик-
носорог на Курильских островах // Ор-
нитология. 1967. Вып. 8. С. 336 — 337.
Вон Хон Гу. Птицы Кореи. Пхеньян,
1963. Т. 1. 129 с. На кор. яз.
Воробьев В. А. Птицы Уссурийского края.
М.: Изд-во АН СССР, 1957. 360 с.
Воронов В. Г. Орнитофауна о-ва Тюле-
ний // Итоги VI симпоз. по изуч. вирусов,
экологически связанных с птицами. М.,
1972. С. 52-58.
Воронов В. Г. Размещение морских колони-
альных птиц на о-ве Анучин и в аквато-
рии Южно-Курильского пролива в июле
1966 г. // Природные ресурсы Сахалина,
их охрана и использование. Южно-Са-
халинск, 1975. С. 111 — 114.
Вяткин II. С. Морские колониальные птицы
о-ва Верхотурова // Колониальные гнез-
довья околоводных птиц и их охрана.
М.: Наука, 1975. С. 169.
Вяткин II. С. Материалы по орнитофауне
о-ва Старичкова // Биология и промысел
охотничьих животных. Пермь, 1983.
С. 8—14.
Вяткин II. С. Кадастр гнездовий колониаль-
ных птиц Камчатской области // Мор-
ские птицы Дальнего Востока. Влади-
восток: ДВНЦ АН СССР, 1986. С. 20-36.
Вяткин II. С., Герасимов Н. Н., Мараков С. В.
Морские колониальные птицы о-ва Кара-
гинского / / Колониальные гнездовья око-
ловодных птиц и их охрана. М.: Наука,
1975. С. 170-171.
Герасимов Н. Н. Размещение и численность
колониальных птиц на северо-западном
побережье Карагинского острова//Ор-
нитология. 1979. Вып. 14. С. 189—190.
Герасимова Т. Д. Материалы по экологии
тупика Fratercula arctica L. // Учен. зап.
МГПИ им. В. П. Потемкина. 1951. Т. 18,
вып. 1. С. 115—124.
Герасимова Т. Д. К орнитофауне Айновых
островов // Тр. Кандалакш. заповедника.
1958. Вып. 1. С. 37-50.
Герасимова Т. Д. Состояние птичьих базаров
Мурманского побережья // Орнитология.
1962. Вып. 4. С. 11-14.
Гизенко А. И. Птицы Сахалинской области.
М.: Изд-во АН СССР, 1955. 328 с.
Головкин А. Н. О выедании рыбы кайрами и
моевками в гнездовой период в Баренце-
вом море // Зоол. журн. 1963. Т. 42, № 3.
С. 408-416.
Головкин А. Н. Гидрологические предпо-
сылки увеличения продуктивности сооб-
ществ близ птичьих базаров Баренцева
моря // Тез. докл. II съезда ВГБО. Ки-
шинев, 1970. С. 84.
Головкин А. Н. Птичьи базары севера
Новой Земли // Особенности биологиче-
ской продуктивности вод близ птичьих
базаров севера Новой Земли. Л.: Наука,
1972, С. 84-91.
Головкин А. Н. Роль птиц в морских
экосистемах // Роль птиц в экосистемах.
М.: ВИНИТИ, 1982. С. 92—157. (Итоги
науки и техники. Сер. Зоология позво-
ночных; Т. 11).
Головкин А. Н., Гаркавая Г. П. Удобрение
вод прибрежья Мурмана различными
типами колоний морских птиц // Биоло-
гия моря. 1975. № 5. С. 49—57.
Головкин А. Н., Георгиев А. А. Птичий
базар на о-ве Тюленьем (Дальний
Восток) // Бюл. МОИП. Отд. биол. 1970.
Т. 75, № 2. С. 53-59.
Головкин А. Н., Зеликман Э. А., Геор-
гиев А. А. Биология и пищевые связи
люриков Plotus alle с пелагическим сооб-
ществом на севере Новой Земли // Осо-
бенности биологической продуктивности
вод близ птичьих базаров севера Новой
Земли. Л.: Наука, 1972а. С. 74—83.
Головкин А. Н., Позднякова Л. Е. Влияние
морских колониальных птиц на режим
биогенных элементов в прибрежных
водах Восточного Мурмана // Новые ис-
следования планктона и бентоса Барен-
цева моря. М.; Л.: Наука, 1964. С. 88—
98.
Головкин А. В., Позднякова Л. Е. Влияние
морских колониальных птиц на режим
биогенных солей в прибрежных водах
Мурмана // Рыбоядные птицы и их зна-
чение в рыбном хозяйстве. М.: Наука,
1965. С. 210-230.
Головкин А. Н., Флинт М. В. Колониальные
гнездовья морских птиц в районе мыса
Сердце Камень (Чукотка) // Колониаль-
ные гнездовья околоводных птиц и их
охрана. М.: Наука, 1975. С. 171 — 173.
Головкин А. Н., Широколобое В. Н., Гарка-
вая Г. II. Особенности распределения
биогенных элементов в районе птичьих
базаров севера Новой Земли // Особен-
ности биологической продуктивности вод
близ птичьих базаров севера Новой
Земли. Л.: Наука, 19726. С. 46—62.
Горбунов Г. П. Материалы по фауне млеко-
питающих и птиц Новой Земли//Тр.
Ин-та по изуч. Севера. 1929. Вып. 40.
С. 169-239. '
Горбунов Г. П. Птицы Земли Франца-
Иосифа//Тр. Аркт. ин-та. 1932. Т. 4.
С. 1-244.
Горчаковская Н. Н. Орнитофауна Государ-
ственного заповедника «Семь островов»:
Дис. . . . канд. биол. наук. М., 1944.
204 с.
Дементьев Г. П. Отряд чистики // Птицы
Советского Союза. М.: Сов. наука, 1951.
Т. 2. С. 169- 240.
Демме Н. П. Птичий базар на скале Ру-
бини//Тр. Аркт. ин-та. 1934. Т. 11.
С. 55-86.
Дулькейт Г. Д., Шулъпин Л. М. Птицы
Шантарских островов // Труды Биол-
184
Литература
НИИ при Томском ун-те. Томск, 1937.
С. 113-136.
Елсуков С. В. Видовой состав и характер
пребывания птиц Среднего Сихотэ-
Алиня / / Сихотэ-Алинский биосферный
район: Экологические исследования. Вла-
дивосток, 1965. С. 104—113.
Елсуков С. В. К орнитофауне морского
побережья Северного Приморья // Фау-
нистика и биология птиц юга Дальнего
Востока. Владивосток: ДВНЦ АН СССР,
1984. С. 34-43.
Жирмунский А. В. Предисловие // Цветко-
вые растения островов Дальневосточного
морского заповедника. Владивосток:
ДВНЦ АН СССР, 1981. С. 3-5.
Житков Б. М., Бутурлин С. А. По северу
России // Землеведение. 1901. Кн. 3/4.
С. 29-206.
Иванов А. И. Каталог птиц СССР. Л.:
Наука, 1976. 276 с.
Иванов А. И., Козлова Е. В., Портенко Л. А.,
Тугаринов А. Я. Птицы СССР. М.; Л.:
Изд-во АН СССР, 1953. Ч. 2. 343 с.
Иоганзен Г. X. Птицы Командорских остро-
вов // Тр. Том. ун-та. 1934. Т. 86. С. 222—
266.
Йыги А. О миграциях чистиковых (Alcidae)
в Эстонской ССР // Сообщ. Прибалт,
комис. по изуч. миграций птиц АН ЭССР.
1975. № 8. С. 72-80.
Карпович В. Н. Птичьи базары как среда
обитания иксодового клеща Ceratixodus
putus Pick.-Camb.//Тр. Кандалакш. за-
поведника. 1970. Вып. 8. С. 356—368.
Карпович В. Н. Кандалакшский заповедник.
Мурманск, 1984. 155 с.
Карпович В. И., Пилипас И. И., Шкляре-
вич Ф. И. К экологии гнездового периода
тонкоклювой кайры // Экология птиц
морских побережий. М.: ЦНИЛ Главохо-
ты РСФСР, 1980. С. 5-16.
Карташев Н. Я. Хищные птицы на птичьих
базарах Восточного Мурмана по наблю-
дениям 1947 г. // Охрана природы. 1949а.
Вып. 6. С. 50—57.
Карташев В. Н. Птичьи базары Восточного
Мурмана//Там же. 19496. Вып. 7.
С. 115-122.
Карташев Н. И. Промысловое значение
чистиковых птиц и пути рациональной
эксплуатации птичьих базаров // Там же.
1951. Вып. 14. С. 67-82.
Карташев И. И. Материалы по постэмбрио-
нальному развитию некоторых видов
чистиковых птиц (отряд Alciformes) //
Зоол. журн. 1957. Т. 36. № 6. С. 909-921.
Карташев Н. В. О типах постэмбрионального
развития 1ггиц: Науч. докл. высш, школы.
Биол. науки. 1960. № 2. С. 33—38.
Карташев В. В. Птицы Командорских остро-
вов и некоторые предложения по их
рациональному использованию // Зоол.
журн. 1961. Т. 11. № 9. С. 1395—1410.
Карташев В. В. Количественная характери-
стика авиафауны Соловецких островов //
Орнитология. 1963. Вып. 6. С. 23—36.
Карташев В. В. Материалы к биологии
чистиковых птиц Командорских остро-
вов // Там же. 1979. Вып. 14. С. 144—149.
Кафтановский Ю. М. Опыт сравнительной
характеристики биологии размножения
некоторых чистиковых//Тр. Гос. запо-
ведника «Семь островов». 1941. Т. 1.
С. 53— 72.
Кафтановский Ю. М. Чистиковые птицы
Восточной Атлантики. М.: МОИП, 1951.
169 с.
Кищинский А. А. Птицы Колымского на-
горья. М_: Наука, 1968. 188 с.
Кищинский А. А. Птицы Корякского на-
горья. М.: Наука, 1980. 336 с.
Козлова Е. В. Ржанкообразные: подотряд
чистиковые // Фауна СССР. М.; Л., 1957.
Т. 2, вып. 3. С. 114—143.
Коломийцев В. И., Поддубная Н. Я. Мате-
риалы к биологии филина Bubo bubo (L.)
в Лазовском заповеднике // Редкие и ис-
чезающие птицы Дальнего Востока.
Владивосток: ДВНЦ АН СССР, 1985.
С. 81-84.
Кондратьев А. Я. О состоянии птичьих
базаров на острове Колючи н // Колони-
альные гнездовья околоводных птиц
и их охрана. М.: Наука, 1975. С. 174—176.
Кондратьев А. Я. Колонии морских птиц
на арктическом побережье крайнего
Северо-Востока СССР // Морские птицы
Дальнего Востока. Владивосток: ДВНЦ
АН СССР, 1986. С. 37-47.
Корнеева Т. М. О размножении тупика
на Айновых островах / / Тр. Кандалакш.
заповедника. 1967. Вып. 5. С. 178—184
Коханов В. Д., Скокова Н. В. Фауна птиц
Айновых островов // Там же. 1967.
Вып. 5. С. 185-267.
Красовский С. К. Этюды по биологии толсто-
клювой кайры//Тр. Аркт. ин-та. 1937.
Т. 77. С. 33-92.
Кузякин А. П. К биологии длинноклювого
пыжика//Орнитология. 1963. Вып. 6.
С. 315-320.
Курочкин Е. В., Герасимова Т. Д. Зимние
и ранневесенние наблюдения над мор-
скими птицами вблизи Мурманского по-
бережья // Тр. Кандалакш. заповедника.
1960. Вып. 3. С. 195—198.
Лабзюк В. И. Летняя авифауна морского
побережья в районе залива Ольги // Ор-
нитологические исследования на Дальнем
Востоке. Владивосток: ДВНЦ АН СССР,
1975. С. 279-284.
Лабзюк В. И., Назаров Ю. Н., Нечаев В. А.
Птицы островов северо-западной части
залива Петра Великого // Орнитологи-
ческие исследования на юге Даль-
185
Литература
него Востока. Владивосток: ДВНЦ АН
СССР, 1971. С. 52-78.
Лабзюк В. И., Назаров Ю. Н., Остапенко В. А.
Птичий базар на острове Фуругельма
. (Южное Приморье) // Материалы XV
Науч. конф, профес.-преподават. состава
Дальневост, ун-та. Владивосток, 1970.
С. 64-67.
Литвиненко Н. М. Чернохвостая чайка
Larus crassirostris Vieill. М.: Наука, 1980.
144 с.
Литвиненко Н. М., Шибаев Ю. В. К орнито-
фауне Судзухинского заповедника и до-
лины р. Судзухе // Экология и фауна
птиц юга Дальнего Востока. Владивосток,
1971. С. 127-186.
Лобков Е. Г. Материалы по миграциям
гагар, поганок, бакланов, чаек и чистиков
вдоль восточного побережья Камчатки //
Экология птиц морских побережий. М.,
1980. С. 74-86.
Лобков Е. Г. Гнездящиеся птицы Камчатки.
Владивосток: ДВНЦ АН СССР, 1986.
304 с.
Мавродиади II. А. Наблюдения над птицами,
усоногими раками и грегаринами Мур-
манского моря // Изв. Варшав. ун-та.
1911. № 3. С. 1-16; № 4. С. 17-37.
Мараков С. В. Звери и птицы острова
Медного // Бюл. МОИП. Отд. биол. 1957.
Т. 62, № 1. С. 117-118.
Мараков С. В. Птичьи базары острова Мед-
ного и возможности их практического
использования // НТИ Всесоюз. НИИ
животного сырья и пушнины. 1963.
Вып. 5(8). С. 51—64.
Мараков С. В. О зимних птицах Командор-
ских островов // Орнитология. 1965.
Вып. 7. С. 480.
Мараков С. В. Край непуганых птиц. М.:
Наука, 1966. 117 с.
Мараков С. В. Природа и животный мир
Командор. М.: Наука, 1972. 184 с.
Мараков С. В. К динамике колоний мор-
ских птиц на Командорских островах //
Колониальные гнездовья околоводных
птиц и их охрана. М.: Наука, 1975.
С. 176-178.
Марков В. И. Распределение и динамика
птичьих базаров в средней части тихо-
океанского побережья Камчатки // Там
же. 1975. С. 178-180.
Мензбир М. А. Птицы России. М., 1895.
Т. 1. 836 с.
Михтарьянц Э. А. Гнездование чистиковых
птиц на острове Топоркова (Коман-
доры) //Материалы IV совещ. зоологов
Сибири. Новосибирск: Наука, 1972.
С. 328-330.
Михтарьянц Э. А. К гнездовой биологии
малого тихоокеанского чистика Cepphus
columba kaiurka // Материалы VI Все-
союз. орнитол. конф. М.: Изд-во МГУ,
1974. Ч. 2. С. 87—88.
Михтарьянц Э. А. Заметки по экологии и
поведению топорка // Орнитология. 1977.
Вып. 13. С. 127-133.
Михтарьянц Э. А. Ритмы активности
белобрюшки (Cyclorrhynchus psittacula)
и топорка (Lunda cirrhata) и связанные
с ними регулярные флуктуации их чис-
ленности на гнездовых колониях // Эко-
логия и зоогеография некоторых позво-
ночных суши Дальнего Востока. Владиво-
сток: ДВНЦ АН СССР, 1978. С. 31-39.
Михтарьянц Э. А. Распространение и био-
логия белобрюшки Cyclorrhynchus psitta-
cula (Pall.) // Редкие птицы Дальнего
Востока. Владивосток: ДВНЦ АН СССР,
1981. С. 80-96.
Модестов В. М. Питание чаек Восточного
Мурмана и роль их в формировании
и жизни птичьих базаров // Научные
студенческие работы. М.: Изд-во МГУ,
1939. Вып. 9. С. 83-113.
Морозов В. В., Томкович П. С. Колонии мор-
ских птиц северо-восточной оконечности
Чукотского полуострова // Научные ос-
новы изучения колоний морских птиц.
М.: Наука, 1981. С. 93-95.
Назаров Ю. Н., Лабзюк В. И. Некоторые
данные о биологии очкового чистика
в Южном Приморье / / Биол. науки.
1972. № 3. С. 32-35.
Назаров Ю. Н., Трухин А. М. К биологии
сапсана и филина на островах залива
Петра Великого (Южное Приморье) //
Редкие и исчезающие виды Дальнего
Востока. Владивосток: ДВНЦ АН СССР,
1985. С. 70-77.
Назаров Ю. Н., Шибаев Ю. В. Список
птиц Дальневосточного государственного
морского заповедника // Животный мир
Дальневосточного заповедника. Влади-
восток, 1984. С. 175—195.
Наканиси Г. Острова естественных экспо-
натов//Кагаку-нодзиккен. 1953. № 10.
С. 60-63; № И. С. 40-46; № 12. С. 42-
46. На яп. яз.
Нечаев В. Н. Птицы Южных Курильских
островов. Л.: Наука, 1969. 246 с.
Нечаев В. А. Птицы острова Монерон //
Орнитологические исследования на Даль-
нем Востоке. Владивосток: ДВНЦ АН
СССР, 1975а. С. 5-25.
Нечаев В. А. Колониально гнездящиеся
птицы острова Монерон // Колониальные
гнездовья околоводных птиц и их охрана.
М„ 19756. С. 181-183.
Нечаев В. А. Новые сведения о морских
птицах острова Сахалин // Морские пти-
цы юга Дальнего Востока. Владивосток,
1986. С. 71-81.
Нечаев В. А., Тимофеева А. А. Птицы
186
Литература
острова Тюлений//Бюл. МОИП. Отд.
биол. 1980. Т. 85, № 1. С. 36-42.
Нечаев В. А., Юдаков А. Г. О гнездовании
морских птиц на островах залива Петра
Великого (Южное Приморье) // Изв. СО
АН СССР. 1968. Вып. 3, № 15. С. 93-96.
Никольский А. М. Остров Сахалин и его
фауна позвоночных животных // Зап.
Акад. наук. 1889. Т. 60, прил. № 5.
С. 334.
Оги X. Пища и питание кайр в Беринговом
море//XIV Тихоокеан. науч, конгр.,
Хабаровск, 1979, Ком. F, секция F II,
Общесекц. сомпоз. F II, I. Подсекцион-
ный F ПЬ: Тез. докл. М., 1979. С. 123—
124.
Парфентьева Н. С. Растительность Айновых
островов // Тр. Кандалакш. заповедника.
1969. Вып. 7. С. 413-424.
Пинегин Н. Айновы острова // Изв. Арханг.
о-ва изуч. Рус. Севера. 1909. № 13. С. 1 —
132.
Плеске Ф. Д. Критический обзор млекопи-
тающих и птиц Кольского полуострова //
Зап. Акад. наук. 1887. Т. 56, прил. № 1.
С. 1-536.
Подлипаев С. А., Головкин А. П. Роль
морских колониальных птиц Мурмана
в распространении гельминтов рыб
в связи с вопросами охраны птичьих
базаров // Научные основы охраны при-
роды. М., 1977. Вып. 5. С. 140-146.
Портенко Л. А. Производительные силы ор-
нитофауны Новой Земли. Л.: Изд-во
АН СССР, 1931. Прил. 2. 52 с. (Тр. био-
геохим. лаб.).
Портенко Л. А. Фауна Анадырского края //
Тр. НИИ поляр, земледелия, животно-
водства и промысл, хоз-ва. Сер. промысл,
хоз-во. 1939. Ч. 2, вып. 6. С. 4—198.
Портенко Л. А. Птицы Чукотского полу-
острова и острова Врангеля. Л.: Наука,
1972. Ч. 1. 423 с.
Портенко Л. А. Птицы Чукотского полу-
острова и острова Врангеля. Л.: Наука,
1973. Ч. 2. 324 с.
Распространение гнездящихся птиц Японии.
Токио: Упр. по делам окружающей среды,
1981. 556 с. На яп. яз.
Ролъник В. В. Развитие терморегуляции
у некоторых птиц Севера // Зоол. журн.
1948; Т. 27, № 6. С. 535-546.
Росляков Г. Е. Птицы морских побережий
Шантарских островов // Морские птицы
Дальнего Востока. Владивосток: ДВНЦ
АН СССР, 1986. С. 66-70.
Рутилевский Г. Л. О птицах острова Ру-
дольфа // Тр. Аркт. ин-та. 1957. Т. 205.
С. 87-96.
Рутилевский Г. Л., Успенский С. М. Фауна
млекопитающих и птиц Центральной
Арктики // Там же. 1957. Т. 205. С. 5—18.
Скокова Н. Н. Тупик на Айновых остро-
вах // Орнитология. 1962а. Вып. 5.
С. 7-12.
Скокова Н. Н. Некоторые факторы, опре-
деляющие состояние популяции тупи-
ков на Айновых островах // Материалы
III Всесоюз. орнитол. конф. Львов, 19626.
Кн. 2. С. 184-185.
Скокова Н. Н. О факторах, определяющих
состояние популяции тупиков в гнездовой
период / / Тр. Кандалакш. заповедника.
1967а. Вып. 5. С. 155-177.
Скокова Н. Н. К методике массового отлова
тупиков в гнездовых колониях / / Там же.
19676. Вып. 5. С. 367—373.
Слепцов М. М. Метод изучения интенсив-
ности размножения птиц по яичникам //
Охрана природы. 1948. Вып. 5. С. 119—
129.
Спангенберг Е. П. Меховая птица. М.;
Л.: КОИЗ, 1935. 68 с.
Спангенберг Е. П., Леонович В. В. Птицы
северо-восточного побережья Белого мо-
ря // Тр. Кандалакш. заповедника. 1960.
Вып. 2. С. 213-336.
Стенченко А. М. Колонии морских птиц
малых бухт Кроноцкого залива // Коло-
ниальные гнездовья околоводных птиц
и их охрана. М.: Наука, 1975. С. 183—
184.
Степанян Л. С. Состав и распределение
птиц фауны СССР: Неворобьиные Non-
Passeriformes. М.: Наука, 1975. 369 с.
Стишов М. С., Придатко В. И., Баранюк В. В.
Новые материалы о птицах острова
Врангеля // Бюл. МОИП. Отд. биол.
1985. Т. 90, № 2. С. 42-48.
Судиловская А. М. Некоторые интересные
поступления в коллекцию Зоологического
музея МГУ за 1962—1971 гг.//Орнито-
логия. 1976. Вып. 12. С. 255—259.
Татаринкова И. П. О поведении атланти-
ческого чистика в позднегнездовой пе-
риод//VII Всесоюз. орнитол. конф.:
Тез. докл. Киев: Наук, думка, 1977. Ч. 2.
С. 41.
Татаринкова И. П., Чемякин Р. Г. Харак-
теристика окончания гнездового периода
у тупика (Fratercula arctica) // Тр. Кан-
далакш. заповедника. 1970. Вып. 8.
С. 190-214.
Трухин А. М., Косыгин Г. М. Распределение
морских птиц во льдах Охотского моря
в зимний период // Морские птицы
Дальнего Востока. Владивосток: ДВНЦ
АН СССР, 1986. С. 48-56.
Успенский С. М. Современное состояние
запасов птиц на новоземельских птичьих
базарах // Охрана природы. 1951. Вып. 14.
С. 83-103.
Успенский С. М. Птичьи базары Новой
Земли. М.: Изд-во АН СССР, 1956. 180 с.
Успенский С. М. Морские колониально
гнездящиеся птицы северных и дальне-
187
Литература
восточных морей СССР, их размещение,
численность и роль как потребителей
планктона и бентоса // Бюл. МОИП.
Отд. биол. 1959. Т. 64, № 2. С. 39-52.
Успенский С. М. Жизнь в высоких широтах
на примере птиц. М_: Мысль, 1969.
463 с.
Успенский С. М., Беме Р. Л., Велижа-
нин А. Г. Авифауна острова Врангеля //
Орнитология. 1963. № 6. С. 57—67.
Флинт В. Е., Костырко И. Н. О биологии
клеща Ixodes putus Pick-Camb. // Зоол.
жури. 1967. Т. 44, № 8. С. 1253-1256.
Фудзимаки Ю. Колонии морских птиц
на острове Хоккайдо // Морские птицы
Дальнего Востока. Владивосток: ДВНЦ
АН СССР, 1986. С. 82-87.
Харитонов С. П. Морские колониальные
птицы острова Ионы // Колониальные
гнездовья околоводных птиц и их охрана.
М.: Наука, 1975. С. 184-185.
Харитонов С. П. Материалы по птицам
острова Ионы // Орнитология. 1980.
Вып. 15. С. 10—15.
Шибаев Ю. В. Спуск на воду и переход
к жизни в море у птенцов старика
Synthliboramphus antiquus (Gm) // Эко-
логия и зоогеография некоторых позво-
ночных суши Дальнего Востока. Влади-
восток: ДВНЦ АН СССР, 1978. С. 79-85.
Шкляревич Ф. Н. Об изменениях числен-
ности моевок и кайр на базарах острова
Харлова (Семь Островов, Восточный
Мурман) //VII Всесоюз. орнитол. конф.:
Тез. докл. Киев: Наук, думка, 1977. Ч. 2.
С. 344-345.
Шулъпин Л. М. Промысловые охотничьи
и хищные птицы Приморья. Владивосток:
Типография имени Волина, 1936. 436 с.
Шунтов В. П. О трубконосных и чистиковых
птицах Берингова моря//Орнитология.
1965а. Вып. 7. С. 276- 286.
Шунтов В. П. Сезонные аспекты в распре-
делении птиц в открытых водах Япон-
ского моря//Зоол. журн. 19656. Т. 44,
№ 3. С. 411-421.
Шунтов В. П. Морские птицы и биологи-
ческая структура океана. Владивосток,
1972. 376.
Яхонтов В. Д. Птичьи базары Пенжинской
губы и их состояние//Материалы VI Все-
союз. орнитол. конф. М.: Изд-во МГУ,
1974. С. 251-252.
Яхонтов В. Д. Остров Нансикан // Коло-
ниальные гнездовья околоводных птиц
и их охрана. М.: Наука, 1975а. С. 186—
187.
Яхонтов В. Д. Птичьи базары Пенжинской
губы и их состояние // Там же. 19756.
С. 185-186.
Яхонтов В. Д. Птицы Шантарских островов:
Некоторые вопросы экологии // Биогра-
фия Приамурья. Хабаровск, 1977.
Вып. 17. С. 155-171.
Яхонтов В. Д. Новое место гнездования
длинноклювого пыжика Brachyramphus
marmoratus perdix Pall. // Птицы Северо-
Востока Азии. Владивосток: ДВНЦ
АН СССР, 1979. С. 163.
Abe М. On the breeding habits of the
horn-billed puffin Cerorhinca monocerata
and its damage by ownerless in Teuri
Island // Tori. 1967. Vol. 18. P. 174-179.
Ainley D. G., De Gauge A. B., Jones L. L.,
Beach B. J. Mortality of seabirds in high-
seas salmon gill nets // Fish. Bull. 1981.
Vol. 79, N 4. P. 800—806.
Ainley D. G., Lewis J. The history of Faral-
lon Island marine bird populations,
1954-1972//Condor. 1974. Vol. 76.
P. 432-446.
Arnason E., Grant P. B. The signeficance
of kleptoparasitism during the breeding
season in colony of Arctic Skuas Stercora-
rius parasiticus in Iceland // Ibis. 1978.
Vol. 120, N 1. P. 38 -54.
Arnold J. B. An albino murre // Condor. 1950.
Vol. 52, N 3. P. 141.
Arnold K. A. Second Florida specimant of the
Razorbill // Fla Field Natur. 1980. Vol. 8,
N 2. P. 50.
Ashcroft В. E. Survival rates and breeding
biology of puffins on Skomer Islands,
Wales // Ornis scand. 1979. N 1. P. 100—
110.
Austin O. L. The birds of Korea // Bull. Mus.
Comp. Zool. 1948. Vol. 101, N 1.
P. 1-301.
Austin O. L., Kuroda N. The birds of Japan:
Their status and distribution // Ibid. 1953.
Vol. 109, N 4. P. 279-637.
Bailey E. P., Davenport G. H. Die-off of Com-
mon Murres on the Alaska Peninsula
and Unimak Island//Condor. 1972.
Vol. 74, N 2. P. 215.
Bateson P. P. Studies of less familiar birds.
112. Little Auk.//Brit. Birds. 1961.
Vol. 54. P. 272-277.
Bedard J. New records of alcids St. Lawrence
Island, Alaska//Condor. 1966. Vol. 68,
N 5. P. 503-506.
Bedard J. The nesting of the crested, least
and parakeet auklets on St. Lawrence
Island, Alaska // Ibid. 1969a. Vol. 71.
P. 386- 398.
Bedard J. Feeding of the least, crested,
and parakeet auklets around St. Laurence
Island, Alaska//Canad. J. Zool. 1969b.
Vol. 74. P. 1025-1050.
Bedard J. Coexistence, coevolution and conver-
gent evolution in seabird communities:
A comment // Ecology, 1976. Vol. 57, N 1.
P. 174-184.
Bedard J., Sealy S. G. Moults and feather
188
Литература
generations in the least, crested and
parakeet anklets //J. Zool. 1984. Vol. 202.
P. 461-488.
Beebe F. L. The marine peregrines of the
north-west Pacific coast // Condor. 1960.
Vol. 62. P. 145-180.
Bent A. C. Life histories of North-American
diving birds//Wash. (D. C.), 1919.
245 p. (Smiths Inst. Bull.; N 107).
Bergman G. Gryllteisten Cepphus grille in
einem Randgebiet: Nahrung, Brutresultat,
Tagesrhythmus und Ansiedlung // Com-
ment. Biol. Soc. Sci. Fenn. 1971. Vol. 42.
P. 26.
Bibby C. Net loss to auks // Birds. 1973. Vol. 4,
N 9. P. 248.
Birds of North America: A guide to field
identification. N. Y.: Golden press,
1966. 340 p.
Birkhead T. R. Movement and mortality rates
of British guillemots // Bird Study. 1974.
Vol. 21, N 4. P. 241-254.
Birkhead T. R. Effects of sea conditions
. on rates at which guillemots feed
chicks // Brit. Birds. 1976. Vol. 69, N 12.
P. 490-492.
Birkhead T. R. The effect of habitat and
density on breeding success in the common
guillemot (Uria aalge) //J. Anim. Ecol.
1977. Vol. 46, N 3. P. 751-764.
Birkhead T. R. Attendance patterns of guille-
mots Uria aalge breeding colonies on Sko-
mer Island // Ibis. 1978a. Vol. 120, N 2.
P. 219-229.
Birkhead T. R. Behavioural adaptations to
high density nesting in nestling in the
common guillemot Uria aalge // Anim. Be-
hav. 1978b. Vol. 26, N 2. P. 321-331.
Birkhead T. R., Biggins J. D., Nettleship
D. N. Non-random intra-colony distribu-
tion of bridled guillemots Uria aalge //
J. Zool. 1980. Vol. 192, N 1. P. 9-16.
Birkhead T. R., Del Neva A. J. Egg formation
and pre-laying period of the common
guillemot Uria aalge//Ibid. 1987. Vol. 211,
N 1. P. 83-88.
Birkhead T. R., Nettleship D. N. Reproductive
biology of thick-billed murres (Uria lom-
via) : an inter-colony comparison // Auk.
1981. Vol. 98, N 2. P. 258-269.
Birkhead T. R., Taylor A. M. Moult of the
guillemot Uria aalge // Ibis. 1977. Vol. 119,
N 1. P. 80-85.
Blake B. F. Diet and fish stock availability
as possible factors in the mass death of auks
in the North Sea // J. Exp. Mar. Biol,
and Ecol. 1984. Vol. 76. P. 89-103.
Boiley E. P. Discovery of a Kittlitz’s murrelet
nest // Condor. 1973. Vol. 75, N 4. P. 457.
Bourne W. R. P. Winter colony attendance
by auks and the danger of oil pollution //
Scot. Birds. 1981. Vol. 11, N 8. P. 254- 257.
Bourne W. R. P., Dixon T. Yule R. Note
of the occurrence of avian pox in auks,
with an appeal for the collection of sick
seabirds and their parasites // Seabird
Rep. 1970. P. 52-53.
Bourne W. R. P., Harris M. P. Birds of the
Hebrides: seabirds // Proc. Roy. Soc.
Edinburgh. 1979. Vol. 77. P. 445 —475.
Bradstreet M. S. W. Thick-billed murres and
black guillemots in the Barrow Strait area,
N. W. T., during spring: distribution and
habitat use // Canad. J. Zool. 1979. Vol. 57.
P. 1789-1802.
Bradstreet M. S. W. Thick-billed murres and
black guillemots in the Barrow Strait area
N. W. T., during spring: diets and food
availability along ice edges // Ibid. 1980.
Vol. 58, N 11. P. 2120-2140.
Bradstreet M. S. W. Pelagic feeding ecology
of dovekies, Alle alle in Lankaster sound
and Western Buffin Bay // Arctic. 1982.
Vol. 35, N 1. P. 126-140.
Breckenridge W. J. Dovekie on Little Diomede
Island, Alaska // Auk. 1966. Vol. 83, N 4.
P. 680.
Brook M. L. de. The Puffin population of the
Shiant Islands // Bird Study. 1972. Vol. 19,
N 1. P. 1-6.
Brown R. G. B. Sea birds in Newfoundland
and Greenland waters, April—May 1966 //
Canad. Field-Natur. 1968. Vol. 82, N 2.
P. 88-102.
Brown R. G. B. The foraging range of bree-
ding Dovekies, Alle alle. // Ibid. 1976.
Vol. 90, N 2. P. 166-168.
Brown R. G. B. Seabirds at sea // Ocean us.
1981. Vol. 24, N 2. P. 31-38.
Brown R. G. B. Seabird in the Greenland,
Barents and Norwegian Seas, February-
April 1982 // Polar Res. 1984. Vol. 2, N 1.
P. 1-18.
Brown R. G. B., Nettleship D. N. The
seabirds of Notheastern North America:
their present status and conservation
requirements // Status and conservation
of the world’s seabirds. Paston Press,
Norwich, England. 1984. P. 85—100.
Brown R. G. B., Nettleship D. N., German P.
et al. Atlas of eastern Canadian seabirds.
Ottawa, 1975. 220 p.
Brun E. Polarlomvien, Uria lomvia (L.) some
rugefugl i Norge//Sterna. 1965. Bd. 5.
S. 229-250.
Brun E. Hekkebestanden av lunde Fratercula
arctica (L.) i Norge//Ibid. 1966. Bd. 7,
N 1. S. 1-17.
Brun E. Utbredelse og hekkebestand av alke
(Alca torda) i Norge // Ibid. 1969. Bd. 8,
N 8. S. 345-359.
Brun E. Dimorph-ratio cline of bridled guille-
mots (Uria aalge) in Norway // Astarte.
1970. Vol. 3, N 1. P. 45-50.
Brun E. Populasjonsendringer hos noen
189
Литература
sjjlffuglarter i Sor-Norge//Sterna.
1971a. Bd. 10, N 1. S. 35-36.
Brun E. Census of puffin (Fratercula arctica)
on Nord-Fugloy, Troms //Astarte. 1971b.
Vol. 4, N 2. P. 41-45.
Buckley P. A., Buckley F. G. Seabirds
of the North and Middle Atlantic Coasts
of the United States: their status and
conservation // Status and conservation
of the world’s seabirds. Paston Press,
Norwich, England. 1984. P. 101 —134.
Burger A. E., Simpson M. Diving depths
of Atlantic puffins and common murres //
Auk. 1986. Vol. 103, N 4. P. 828-830.
Burrel G. C. Some observations on nesting
tufted puffins, Destruction Island, Wa-
shington // Murrelet. 1980. Vol. 61, N 3.
P. 92-94.
Byrd G. V., Day В. H., Knudtson E. P.
Patterns of colony attendance and censu-
sing of Anklets of Buldir Island // Con-
dor. 1983. Vol. 85, N 3. P. 274-280.
Byrd G. V., Gibson D. D. Distribution and
population status of Whiskered auklet in
the Aleutian Islands, Alaska//West.
Birds. 1980. Vol. 11, N 3. P. 135-140.
Cairns D. Breeding, feeding, and chick growth
of the black guillemot (Cepphus grylle)
in Southern Quebec / / Canad. Field-Natur.
1981. Vol. 95, N 3. P. 312-318.
Campbell B. W. Alexandrian rat predation
on ancient murrelet eggs // Murrelet.
1968. Vol. 49. P. 38.
Campbell B. W. Use of manmade structures
as nest sites by pigeon guillemots // Canad.
Field-Natur. 1977. Vol. 91, N 2. P. 193-
194.
Camphuysen C. J. Vondsten van Kleine Aiken
Alle alle en Papegaaiduikers Fratercula
arctica langs de Nederlandse kust // Li-
mosa. 1986. Bd 59, N 3. Biz. 138-141.
Carnduff D. Black guillemots breeding in the
inner Clyde estuary // Scot. Birds. 1981.
Vol. 11, N 6. P. 195-196.
Check-list of Japanese birds. Tokyo, 1974.
364 p.
Cheng Tso Hsing. Distributional list of Chinese
birds. 2nd rev. ed. Peking: Sci. Sinica,
1976. 1280 p.
Christensen O., Lear W. H. Bycatches in sal-
mon drift-nets at West Greenland in 1972 //
Medd. Grtfnland. 1977. Bd.' 205, N 5.
S. 1-38.
Clapp В. B. A summary of alcid records
from Hawaii // Colonial Waterbirds. 1986.
Vol. 9. P. 104-107.
Conway W. G., Bell J., Bruning D., Dolen-
se K. Alcidae at the New York Zoological
Park // Intern. Zoo Yb. 1977. Vol. 17.
P. 173-176.
Corkhill P. Improving guillemot ledges //
Seabird Rep. 1970. P. 46—47.
Corkhill P. Food and feeding ecology of
puffins // Bird Study. 1973. Vol. 20,
N 3. P. 207-220.
Cramp S. The birds of the Western Palearc-
tic. Oxford etc.: Oxford Univ, press,
1985. Vol. 4. 960 p.
Cramp S., Simmons К. E. L. The birds of the
Western Palearctic. Oxford etc.: Oxford
Univ, press, 1983. 913 p.
Daan S., Tinbergen J. Young guillemots
(Uria lomvia) leaving their arctic breeding
cliffs: A daily rhythm in numbers and
risk // Ardea. 1979. Vol. 67, N 3/4.
P. 96-100.
Day В. H., Oakley K. L., Barnard D. B.
Nest sites and eggs of Kittlitz’s and marbled
murrelets // Condor. 1983. Vol. 85, N 3.
P. 265-274.
Divoky G. J. The occurrence and behaviour
of non-breeding horned puffins at black
guillemot colonies in northern Alaska //
Wilson Bull. 1982. Vol. 94, N 3. P. 356-
358.
Dobson B. A. A 2-clutch of puffin // Oologist’s
Rec. 1937. По аннот.: Beitr. zur Fortpflan-
zungbiol. der Vogel, 1938.
Drent В. H. Breeding biology of the pigeon
guillemot Cepphus columba // Ardea. 1965.
Vol. 53. P. 99—160.
Dwight J. The gulls (Laridae) of the
World // Bull. Amer. Mus. Natur. Hist.
1925. Vol. 52, art. 3. P. 63-401.
Dyck J., Kraul 1. Environmental pollutants
and shell thinning in eggs of the guillemot
Uria aalge from the Baltic Sea and the
Faeroes, and a possible relation between
shell thickness and sea water salinity //
Dan. ornithol. foren. tidsskr. 1984. Bd. 78,
N 1/2. S. 1-14.
Elkins N., Williams M. H. Aspects of seabird
movement off north-east Scotland //
Scot. Birds. 1972. Vol. 7, N 2. P. 66-75.
Evans P. North to Greenland // Wildlife.
1977. Vol. 19, N 2. P. 82-85.
Evans P. G. H. The south-west Irish sea-bird
populations // Seabird Rep. 1970. P. 35—
40.
Evans P. G. H. Ecology and behaviour of the
Little auk Alle alle in West Greenland //
Ibis. 1981. Vol. 123, N 1. P. 1 — 18.
Evans P. G. H. Status and conservation
of seabirds in Northwest Europe (excluding
Norway and the USSR) // Status and
conservation of the world’s seabirds. Paston
Press, Norwich, England. 1984a. P. 293—
322.
Evans P. G. H. The seabirds of Greenland:
their status and conservation // Ibid.
1984b. P. 49—84.
Evans P. G. H., Waterston G. The decline
of the Brunnich’s guillemot in West
190
Литература
Greenland // Ibis. 1978. Vol. 120, N 1.
P. 131—132.
Fay F. H., Cade T. J. An ecological analysis
of the avifauna of St. Lawrence, Alaska //
Univ. Cal. Publ. Zool. 1959. Vol. 6, N 2.
P. 73-150.
Feinstein B. Geographic variation in the
Whiskered auklet//Auk. 1959. Vol. 76,
N 1. P. 60-67.
Fimreite N., Brun E., Frtfslie A. et al. Mercury
in eggs of Norwegian seabirds // Astarte.
1974. Vol. 7, N 2. P. 71-75.
Fisher J. Birds as animals. Pt 1. A history
of Birds. L„ 1954. 279 p.
Fisher J., Lockley R. M. Sea-birds: An intro-
duction to the natural history of the
sea-birds of the North Atlantic. L., 1954.
125 p.
Flegg J. J. M. The puffin on St. Kilda,
1969-1971 // Bird Study. 1972. Vol. 19,
N 1. P. 7-17.
Fogdea S., Greenwood J. Guillemots nesting
in deep cavity // Brit. Birds. 1965. Vol. 58,
N 11. P. 470.
Fujimaki Yu. Observations on sea birds of
Moyururi Island eastern Hokkaido // Tori.
1961. Vol. 16, N 78. P. 387 -398.
Furness B. W. Seabirds populations of
Foula//Scot. Birds. 1981. Vol. 11, N 8.
P. 237-253.
Furness R. W. Seabird biomass and food
consumption in the North Sea // Mar.
Pollut. Bull. 1984. Vol. 15, N 7. P. 244-
248.
Gabrielson J. N., Lincoln F. C. The birds
of Alaska. Wash. (D. C.), 1959. 922 p.
Gaston A. J. Migration of juvenile thick-billed
murres through Hudson Strait in 1980 //
Canad. Field-Natur. 1982. Vol. 6, N 1.
P. 30-34.
Gaston A. J., Chapdelaine G., Noble D. G.
The growth of thick-billed murre chicks
at colonies in Hudson Strait: Inter- and
intra-colony variation // Canad. J. Zool.
1983. Vol. 61, N 11. P. 2465—2475.
Gaston A. J., Malon M. Range extension of at-
lantic puffin and rasorbill in Hudson
Strait // Canad. Field-Natur. 1980. Vol. 94,
N 3. P. 328-329.
Gaston A. J., Nettleship D. N. The thick-
billed mu'rres of Prince Leopold Island —
a study of the breeding ecology of a colonial
high arctic seabird // Ottawa, 1981. 350 p.
(Canad. Wildlife Serv. Monogr. Ser.; N 6).
Geist O. W. Seabirds found far inland in
Alaska // Condor. 1939. Vol. 47. P. 68—70.
Gloe P. Bemerkungen zum Fund einer nicht
zu U. a. albionis (Witherby) gehorenden
Trottellumme (Uria aalge ssp) // Ornithol.
Mitt. 1977. Bd. 29, N 4. S. 86-88.
Goethe F., Ringleben H. Ein neuerer Nachweis
von Uria lomvia (L.) fur Deutschland//
J. OrnithoL 1964. Bd. 105, N 1. S. 54-56.
Golovkin A. N. Seabirds meeting in the USSR:
The status and protection of population //
Status and conservation of the world’s
seabirds. Paston Press, Norwich, England.
1984. P. 473-486.
Gor M. E. Y., Won P. O. The birds of Korea.
Seoul; Tokyo, 1971. 450 p.
Grant P. R. Interactive behaviour of puffins
(Fratercula arctica L.) and skuas (Sterco-
rarius parasiticus L.) // Behaviour. 1971.
Vol. 40, N 2. P. 263-281.
Grenquist P. Riistantutkimuslaitoksen meri-
tarkkailualuci den lintukannat vuosina
1949 ja 1959 // Suomen riista. 1961. N 14.
S. 15-34.
Gress F., Risebrough R. W., Sibley F. C.
Shell thinning in eggs of the common
murre, Uria aalge, from the Farallon
Islands, California // Condor. 1971. Vol. 73,
N 3. P. 368-369.
Grinnel J. Summer birds of Sitka, Alaska //
Auk. 1909. Vol. 15. P. 122-131.
Hallet D., Miller D. Effects of ingested crude
oil on black quillemots: a combined field
and laboratory study // AMBIO. 1980.
Vol. 9, N 1. P. 28-30.
Harris M. P. Differences in the diet of British
auks // Ibis. 1970. Vol. 112, N 4. P. 540-
541.
Harris M. P. The present status of the puffin
in Britain and Ireland // Brit. Birds. 1976.
Vol. 69, N 7. P. 239-261.
Harris M. P. Puffins on the Isle of May //
Scot. Birds. 1977. Vol. 9, N 6. P. 285-290.
Harris M. P. Supplementary feeding of young
puffins, Fratercula arctica //J. Anim. Ecol.
1978. Vol. 47, N 1. P.' 15—23.
Harris M. P. Measurements and weights
of British puffins//Bird Study. 1979.
Vol. 26, N 3. P. 179-186.
Harris M. P. Breeding performance of puffins
Fratercula arctica in relation to nest den-
sity, laying date and year // Ibis. 1980.
Vol. 122, N 2. P. 193-209.
Harris M. P. Age determination and first
breeding of British puffins // Brit. Birds.
1981. Vol. 74, N 6. P. 246-256.
Harris M. P. Seasonal variation in fledging
weight of the puffin Fratercula arctica //
Ibis. 1982a. Vol. 124, N 1. P. 100-103.
Harris M. P. The breeding seasons of British
puffins // Scot. Birds. 1982b. Vol. 12, N 1.
P. 12-17.
Harris M. P., Gabbraith H. Seabird populations
of the Isle of May // Ibid. 1983. Vol. 12,
N 6. P. 174-180.’
Harris M. P., Hislop J. R. G. The food of young
puffins Fratercula arctica //J. Zool. 1978.
Vol. 185, N 2. P. 213-236.
Harris M. P., Yube R. F. The moult of the puffin
Fratercula arctica // Ibis. 1977. Vol. 119,
N 4. P. 535-541.
Harris M. P., Murray S. Monitoring of puffin
191
Литература
numbers at Scottish colonies / / Bird Study.
1981. Vol. 28, N 1. P. 15—20.
Harris M. P., Wanless S. A comparison of the
biology of the normal and bridled forms
of the guillemot Uria aalge at a single
colony//J. Zool. 1986. Vol. 210, N 1.
P. 121-130.
Harrison P. Seabirds: An identification guide.
Boston, 1983. 448 p.
Hartert E. Die Vogel der Palaarktischen
Fauna. B. 1921-1922. Bd. 3. S. 1765-
2328.
Hartley C., Fisher J. The marine food of birds
in inland-fiord region in West Spitzbergen.
Pt 2 // Anim. Ecol. 1936. Vol. 5. P. 115 —
121.
Hartwig E., Liidtke S. Auswirkungen «schlei-
chender» verolung auf die Vogelwelt
im Bereich der Nordenseeinsel Sylf im Win-
ter 1984/85 // Angew. Ornithol. 1986.
Bd. 6, N 1/2. S. 1-16.
Hatch S. A. The fledging of common and
thick-billed murres on Middleton Island,
Alaska //J. Field Ornithol. 1983. Vol. 54,
N 3. P. 266-274.
Heath H. Birds observed on Forrester Island,
Alaska, during the summer of 1913 // Con-
dor. 1915. Vol. 17. P. 20-41.
Hedgren S. Reproductive success of guillemots
Uria aalge on the Island of Stora Karlso //
Ornis fenn. 1980. Vol. 57, N 2. P. 49-57.
Hedgren S., Linnman A. Growths of guillemot
Uria aalge chicks in relation to time
of hatching // Ornis scand. 1979. Bd. 10,
N 1. S. 29-36.
Hilden O. Merenkurkun ruokkikannan Kehi-
tyksesta viime aikoina // Ornis fenn. 1978.
Vol. 55, N 1. P. 42-43.
Hoffmann W., Elliot W. P., Scott J. M.
The occurrence and status of the horned
puffin in the Western United States //
West. Birds. 1975. Vol. 6, N 3. P. 87—94.
Holgersen H. Bird-banding in Norway 1950—
1951 // SmSskr. Zool. Ser. 1951/1952. N 3.
S. 5.
Holgersen H. Bird-banding in Norway 1952—
1954//Sterna. 1953/1955. Vol. 1, N 11.
S. 16-21.
Hornung M. Puffin-soil interactions on the
Fame Islands Nortumberland (U. K.) //
Ibis. 1978. Vol. 120, N 1. P. 111.
Hornung M., Harris M. P. Soil water-levels
and delayed egg-laying of puffins // Brit.
Birds. 1976. Vol. 69, N 10. P. 402-408.
Hottola P. Bird records in Georgian SSR //
Dutch Birding. 1987. Vol. 9, N 3. P. 123.
Hudson P. J. The parent-chick feeding rela-
tionship of the puffin, Fratercula arctica //
J. Anim. Ecol. 1979. Vol. 48, N 3. P. 889-
898.
Hudson P. J. Nest site characteristics and
breeding success in the Razorbill Alca
torda // Ibis. 1982. Vol. 124, N 3.
P. 355-359.
Hudson P. J. Plumage variation of Razorbill
chicks // Brit. Birds. 1984. Vol. 77, N 5.
P. 208-209.
Huges M. H. Some observations on ion and
water balance in the puffin, Fratercula
arctica // Canad. J. Zool. 1970. Vol. 48,
N 3. P. 479—482.
Hunt C. L. (Jr.), Burgeson B., Songer G. A.
Feeding ecology of sea-birds of the eastern
Bering Sea // The eastern Bering Sea shelf
oceanography and resources. Seattle
(Wash.): Univ. Wash, press, 1981. Vol. 2.
P. 629-647.
Ingold P. Zur lautlichen Beziehung des Elters
zu seinem Kucken bei Tordalken (Alca
torda) // Behaviour. 1973. Vol. 45, N 1/2.
P. 154-190.
Ingold P. Brutverhaeltnisse bei Tordalken
(Alca torda) auf der Vogelinsel Ves/y
(Lofoten) //Sterna. 1974. Bd. 13, N 4.
S. 205-210.
Ingold P. Zur Beziehung des Tordalken Alca
torda zur Nestumgebung // Ornithol. Beob.
1976. Bd. 73, N 1. S. 1-10.
Ingold P. Anpassungen der Eier und des Brut-
verhaltens von Trottellummen (Uria aalge
aalge Pont.) an das Briiten auf Felssim-
sen // Ztschr. Tierpsychol. 1980. Bd. 53,
N 4. S. 341-388.
Ingold P., Tschanz B. Tordalken, Alca Torda,
als Nahrungsschmarotzer//Sterna. 1970.
Bd. 9, N 4. S. 201-206.
Iversen J. A., Krog J. Body temperatures in
active and resting Charadriiforme birds
(murres, puffins and auklets) at different
ambient temperatures // Norw. J. Zool.
1972. Vol. 20, N 2. P. 145-146.
Japanese bird banding in recent years (1961 —
1983). Tokyo, 1985. 202 p.
Jefferies D. J., Parslow J. L. F. The genetics
of bridling in guillemots from a study
of hand-reared birds //J. Zool. 1976.
Vol. 179, N 3. P. 411-420.
Johnson H. A. Weight records for some
Atlantic Alcidae // Wilson Bull. 1944.
Vol. 56. P. 161-168.
Johnson S. B., West G. C. Growth and develop-
ment of heat regulation in nestlings and
metabolism of adult common and thick-
billed murres //Ornis scand. 1975. Bd. 6,
N 1. S. 109-115.
Joiris C. Seabirds recorded in the Fladenground
area nothern North Sea, in April, May
and June 1976 // North Sea dynamics. B.;
Heidelberg: Springer, 1983. P. 676—686.
Kartaschev N. N. Die Aiken-Vogel des Nordat-
lantiks: Die neue Brehm-Bucherei. Witten-
berg; Lutherstadt, 1960. 154 S.
Kenyon K. W. Two sea-bird record for sou-
192
Литература
them California//Condor, 1937. Vol. 39.
P. 257—580.
Kenyon K. W. Birds of Amchitka Island,
Alaska //Auk. 1961. Vol. 78. P. 305-326.
Knudtson E. P., Byrd V. G., Breeding biology
of crested, least, and whiskered auklet
on Buldir Island, Alaska // Condor. 1982.
Vol. 84. P. 197-202.
Kobayashi K. Summer birds in Shikotan
Island, South Kuriles // Tori. 1933. Vol. 8,
N 36. P. 10-22.
Kuroda N. Analysis of seabird distribution
in the northwest Pacific Ocean//Pacif.
Sci. 1960. Vol. 14, N 1. P. 55-57.
Kuroda N. A winter sea-bird census between
Tokyo and Kushiro, Hokkaido // Misc.
Rep. Yamashina’s Inst. Ornithol. 1963a.
Vol. 3, N 4. P. 227-238.
Kuroda N. A survey of sea birds of Teuri
Island, Hokkaido, with notes on land
birds // Ibid. 1963b. Vol. 3, N 5. P. 58—67.
Lambert K., Nehls H. W. Masconur, Fratercula
arctica (L.) u ujscia Wisly//Acta orni-
thol. 1965. Vol. 9, N 3. P. 165.
Legendre R. Un poisson qui peche des
oiseaux // Nature. 1926. N 2725. P. 16—21.
Leister B. Papageiteucher (Fratercula arctica)
am Neusiedlersee//Egretta. 1962. Bd. 5,
N 1. S. 1-3.
Lensink С. T. The status and conservation
of seabirds in Alaska // Status and conser-
vation of the world’s seabirds. 1984. Paston
Press, Norwich, England. 1984. P. 13—27.
Le-Roi P. Ornithologosghes aus Rheinpro-
vinz // Ornithol. Monatsber. 1908. N 7/8.
27 p.
Leslie P. H. The intrinsic rate of increase
and the overlap of successive generations in
a population of guillemots (Uria aalge
Pontop.) //J. Anim. Ecol. 1966. Vol. 35,
N 2. P. 291-301.
Lewbel G. 8. Bering Sea biology: An evolution
of the environmental data base related
to Bering Sea oil and gas exploration
and development. Anchorage, 1984. 180 p.
Lid G. Reproduction of the puffin on R/st
in the Lofoten Islands in 1964—1980 //
Fauna norw. C. 1981. Bd. C4, N 1, S. 30—
39.
Lloyd C. S. An estimate of the world breeding
population of the razorbill // Brit. Birds.
1976. Vol. 69, N 8. P. 298—304.
Lloyd C. S. Factors affecting breeding of razor
bills Alca torda on Stockholm // Ibis. 1979.
Vol. 121, N 2. P. 156-176.
Lloyd C. S., Perrins С. M. Survival and age
at first breeding in the razorbill (Alca
torda) // Bird-Band. 1977. Vol. 48, N 3.
p 239______262
Lockley R. M. Puffins. L., 1953. 186 p.
Lovenskiold H. L. Diaries: Vestspitsbergen,
1954 and 1956. Bjoroya, 1958. 215 p.
Lovenskiold H. L. Avifauva Svalbardensis
with a discussion on the geographical distri-
bution of the birds of Spitsbergen and
adjacent Islands // Norsk Polarinst. skr.
1964. N 129. S. 1-460.
Magnar N., Einar B. Barentshavets sj/ fugler
essurser // Medd. Norsk Polarinst. 1977.
N 104. S. 119.
Mahoney 8. P., Threlfall W. Notes on the eggs,
embryos and chick growth of common
guillemots Uria aalge in Newfound-
land // Ibis. 1981. Vol. 123, N 2. P. 211 —
218.
Makatsch W. Studies of less familiar birds. 154.
Audouin’s gull // Brit. Birds. 1969. Vol. 62,
N 6. P. 230-232.
Manuwal D. A. The incubation patches of Cas-
sin’s Auklet // Condor. 1974. Vol. 76, N 4.
P. 481-484.
Manuwal D. A. Criteria for aging Cassin’s
Auklet//Bird-Band. 1978. Vol. 49, N 2.
P. 157-161.
Manuwal D. A., Manuwal N. J. Habitat spe-
cific behaviour of the Parakeet Auklet in
the Barron Islands, Alaska //West. Birds.
1979. Vol. 10. P. 189-200.
Marsault В. M. Breeding guillemots Uria aalge
and rasorbills Alca torda in captivity //
Intern. Zoo Yb. 1975. Vol. 15. P. 105-108.
Massey I. A., Matsui S., Suzuki T. et al.
A field guide to the birds of Japan. Tokyo,
1982. 336 p.
Mather J. R. The occurrence of the nothern
guillemot (Uria aalge aalge) in Yorkshire
waters// Naturalist. 1966. N 898. P. 81 —
84.
Mathey-Dupras A. Ornithologiques receueillies
au coers de la croisieres du «Grosser
Kuzfiirst» an Spitzbergentre le 18 juillet
et 16 aout 1911//Neuchatel. Soc. sci.
natur. 1913. Vol. 39. 161 p.
Mayol J. Observaciones sobre la gaviota de
Audouin, Larus audouinii Rayr., en el
Miditerraneo occidental (primavera de
1978) // Natur. hisp. 1978. N 20. P. 3-34.
McLaren P. L. Spring migration and habitat
use by seabirds in the eastern Lancaster
Sound an Western Baffin Bay // Arctic.
1982. Vol. 35, N 1. P. 88-111.
Mead C. J. The results of ringing auks
in Britain and Ireland // Bird Study.
1974. Vol. 21, N 1. P. 45-86.
Meinertzhagen R. Winter in Arctic Lapland //
Ibis. 1938. Vol. 4. P. 18-56.
Merikallio E. Finnish birds, their distribution
and numbers // Soc. fauna et flora fenn.
1958. Vol. 3. P. 132.
Merne O. The auk kill in country Wexford,
September 1969 // Seabird Rep. 1969. N 12.
22 p.
Mills 8. Graveyard of the puffin //New Sci.
1981. Vol. 91, N 1260. P. 10-13.
Min Byung Yoon, Won Pyong Oh. A winter
seabird survey on the shore south coast
13 Птицы СССР
193
Литература
of Korean Peninsula (Mogro-Wando-
Yeosu area) // Misc. Rep. Yamashina
Inst. Ornithol. 1976. Vol. 18, N 1 (45).
P. 53-67.
MunyerE. A. Inland wanderings of the ancient
murrelet//Wilson Bull. 1965. Vol. 77.
P. 235-242.
Murata E. The breeding of the ancient auk
Synthliboramphus antiquus (Gm.) of the
Island of Teuri, Hokkaido//Tori. 1958.
Vol. 14, N 70. P. 22-26.
Murie 0. J. Food habits of the northern
Bald eagle in the Aleuntian Island,
Alaska // Condor. 1940. Vol. 42. P. .198—
202.
Murphy E. C., Springer A. M., Roseneaw D. G.
Population status of common guillemots
Uria aalge at a colony in western Alaska:
results and simulations // Ibis. 1986.
Vol. 128, N 3. P. 348-363.
Myrberget S. Vekslinger i antall lundefugl
inne Ved Kolonien // Sterna. 1959. Bd. 3,
N 6. S. 12-24.
Myrberget S. Lundefugllens vandringer//
Fauna (Norge). 1962a. Bd. 15, N 3.
S. 1957-1964.
Myrberget S. Inders ^kelser over forplatnings-
biologien til lunde (Fratercula arctica
L.) // Ibid. 1962b. S. 1970-1988.
Myrberget S. Merking av toppskarv og lunde
рй R/st//Sterna. 1973a. Bd. 13, N 5.
S. 307- 315.
Myrberget S. Ringmerking av teiste langs
den skandinaviske vestkyst // Ibid. 1973b.
Bd. 12, N 1. S. 33-40.
Myrberget S. The fledging period of puffins
Fratercula arctica on Rost, northern
Norway // Fauna norv. 1981. Bd. C4, N 1.
S. 27-29.
Naumann J. F. Naturgeschichte der Vogel
Mitteleuropas, Mera—Untermhaus. Kohler.
1905. Bd. 10.
Nelson D. A. Sexual differences in measure-
ments of Casein’s Auklet //J. Field Or-
nithol. 1981. Vol. 52, N 3. P. 233-234.
Nelson D. A. Communication of intentions in
agonistic context by pigeon guillemot,
Cepphus columba // Behaviour. 1984.
Vol. 88, N 1/2. P. 145-189.
Nettleship D. N. Trapping common puffin
fledgings // Bird-Band. 1969. Vol. 40, N 2.
P. 139-144.
Nettleship D. N. Breeding success of the com-
mon puffin (Fratercula arctica L.) on
different habitats at Great Island, New-
foundland // Ecol. Monogr. 1972. Vol. 42,
N 2. P. 239-268.
Nettleship D. N. The adaptive significance
of egg size and laying date in thick-billed
murres Uria lomvia // Ecology. 1982.
Vol. 63, N 2. P. 300-306.
Nettleship D. N., Lock A. R. Tenth census
of seabirds in the sanctuaries of the north
shore of the Gulf of St. Lawrence // Canad.
Field-Natur. 1973. Vol. 87, N 4. P. 395-
402.
Nilsson L. Tobisgrisslans Cepphus grylle
vinterutbredning utmed den svenska
ostkusten // Anser. 1982. Bd. 21, N 2.
S. 89-92.
Norderhaug M. Triekkforhold, stedstrohet og
pardanelse hos alkekonge рй Svalbard //
Fauna (Norge). 1967. Bd. 20, N 4.
S. 236-244.
Ndrderhaug M. Breeding biology of the little
auk (Plautus alle) in Svalbard//Norsk
Polarinst. skr. 1980. N 173. S. 1—45.
Oberholzer A. Angeborene Orientierungsvor-
gange im FreBablauf bei Trottellummen
(Uria aalge aalge Pont.) // Ztschr. Tier-
psychol. 1975. Bd. 39. S. 150—172.
Ogi H. Пища и питание кайр в Беринговом
море//XIV Тихоокеан. науч, конгр.
Хабаровск, 1979. Ком. F II в.: Тез. докл.
М., 1979. С. 123-124. На рус. яз.
Ogi Н. The pelagic feeding ecology of thick-
billed murres in the North Pacific,
March-June // Bull. Fac. Fish. Hokkaido
Univ. 1980. Vol. 31, N 1. P. 50—72.
Ogi H. Seabird mortality incidental to the
Japanese salmon gillnet fishery // Status
and conservation of the world’s seabirds.
Paston Press, Norwich, England, 1984.
P. 717-720.
Ogi H., Hamanaka T. The feeding ecology of
Uria lomvia in the northwestern Bering
Sea region // Misc. Rep. Yamashina Inst.
Ornithol. 1982. Vol. 14, N 2/3. P. 270- 280.
Ogi H., Tsujita T. Preliminary examination
of stomach contents of murres (Uria spp.)
from the eastern Bering Sea and Bristol
Bay, June—August, 1970—1971 //Jap. J.
Ecol. 1973. Vol. 23, N 5. P. 201—209.
Olsson V. Forandringar inom en population
av tordmule Alca torda och tabisgrissla
Cepphus grylle i Oster gotlands skargard
1954-1973 // Var fagelvarld. 1974. Bd. 33,
N 1. S. 3-14.
Orr C. D., Ward R. M. P. The autumn migra-
tion of thick-billed murres near Southern
Baffin Island and Northern Labrador //
Arctic. 1982. Vol. 35, N 4. P. 531-536.
Pennycuick C. J. Observations on a colony
of Briinnich’s guillemots, Uria lomvia,
in Spitsbergen//Ibis. 1956. Vol. 98.
P. 80-89.
Perrins С. M. Life tables of auks // Ibid.
1978. Vol. 120, N 1. P. 128-129.
Perry R. Lundy: Isle of puffins. L., 1946.
P. 340.
Petersen A. Size variables puffins Fratercula
arctica from Island, and bill features as
criteria of age // Ornis scand. 1976a. Bd. 7,
N 2. S. 185-192.
Petersen A. Age of first breeding of puffin
194
Литература
Fratercula arctica (L.) // Astarte. 1976b.
Vol. 9, N 2. P. 43-50.
Plumb W. J. Observations on the breeding
biology of the razorbill // Brit. Birds.
1965. Vol. 58, N 11. P. 449-456.
Portenko L. A. Einige neue Unterarten palae-
arktischer Vogel. T. 2 // Mitt. Zool. Mus.
Berlin. 1937. Bd. 22, N 2. S. 219-229.
Portmann A. Die Ontogenese der Vogel als
Evolutionsproblem // Acta biotheor. 1935.
Vol. 1. 312 S.
Portmann A. Development postembryonnaire.
Traite de zoologie. Anatomie, systematique,
biologie // Oiseaux France. 1950. Vol. 15.
1164 p.
Prato S. R. D., Prato E. S. The seabirds
of Berwickshire // Scot. Birds. 1980.
Vol. 11, N 1. P. 13-20.
Rees E. J. S. Guillemot kill in North Wales —
autumn 1969 // Seabird Rep. 1969. P. 7—
11.
Regnell S. Lunnetagelns (Fratercula arctica)
nackning pa Soteskar // Vfir fAgelvSrld.
1956. Bd. 15, N 4. P. 280-281.
Renaud И7. E., Bradsteet M. S. W. Late
winter distribution of black guillemots
in Nothern Baffin Bay and the Canadian
High Arctic // Canad. Field-Natur. 1980.
Vol. 94, N 4. P. 421-425.
Renaud W. E., McLaren P. L., Johnson S. R.
The dovekie, Alle alle, as a spring migrant
in Eastern Lancaster Sound and Western
Baffin Bav // Arctic. 1982. Vol. 35, N 1.
P. 118-125.
Report of the Bird Migration Research
Center (February 1, 1978—January 31,
1979). Tokyo, 1979. 246 p.
Report of the Bird Migration Research Center
(February 1, 1984-January 31, 1985).
Tokyo, 1985. 212 p.
Roberts B. Notes on the birds of central and
southeastern Iceland // Ibis. 1934. P. 18—
24.
Robin P. Alca torda 31°30/ N, 9°48' ouest,
durant 1’hiver 1968—1969 a Essaouira//
Alauda. 1969. Vol. 37, N 2. P. 162.
Roby D. D., Brink K. L. Breeding biology
of least anklets on the Pribilof Islands,
Alaska // Condor. 1968a. Vol. 88. P. 336—
346.
Roby D. D., Brink K. L. Decline of breeding
least auklets on St. George Island,
Alaska //J. Field Ornithol. 1986b. Vol. 57,
N 1. P. 57—59.
Roudybush T., Hoffman L., Rahn H. Conduc-
tance, pore geometry and water loss of eggs
of cassin’s auklet // Condor. 1980. Vol. 82,
N 1. P. 105-106.
RUppell G. Beitrage zum Vorhalten den Krab-
bentauchCrs (Plautus alle alle) // J. Orni-
thol. 1969. Bd. 110, N 2. S. 161-169.
Salomonsen F. The Atlantic Alcidae. The
seasonal and geographical variation of the
auks inhabiting the Atlantic Ocean and
the adjacent waters // Sjatte foljden.
B. 1944. Bd. 3, N 5. S. 138.
Salomonsen F. Grenlands fugle // The birds
of Greenland: In 3 vol. K/benhavn, 1950—
1951. 124 p.
Salomonsen F. Fuglene p§ Gr^nland. Copen-
hagen: Rhodos, 1967. 341 s.
Salomonsen F. Recoveries in Greenland
of birds ringes abroad // Medd. Gr/rdand.
1971. Bd. 191, N 2. S. 1—52.
Sanford R. C., Harris S. W. Feeding behaviour
and food-consumption rates of a captive
California murre // Condor. 1967. Vol. 69,
N 3. P. 298—302.
Saunders D. R. Clowns on Skomer Island //
Animals. 1966. Vol. 8, N 7. P. 170-173.
Schaanning H., Tho L. A contribution to the
bird fauna of Jan-Mayen. Oslo, 1933. 49 p.
Schenk H., Meschini E. Gabbiano corso Larus
audouinii Payraudeu, 1826 // Distribu-
zione e popolazione dei Laridi e Sternidi
nidificanti in Italia. 1986. P. 41—51.
। (Bologna Suppl. Ric. Biol. Selvaggina;
Vol. 11).
Schneider D. C., Piatt J. F. Scale-dependent
correlation of seabirds with schooling fish
in a coastal ecosystem // Mar. Ecol. Progr.
Ser. 1986. Vol. 32, N 2/3. P. 237-246.
Schommer M., Tschanz B. LautauBerungen
junger Trottellummen (Uria a. aalge)
als individuelle Merkmale//Vogelwarte.
1975. Bd. 28, N 1. S. 17-44.
Schreiber R. W. Parakeet auklet at Midway
Aioli//Elepaio. 1987. Vol. 47. P. 16.
Schrenk L. Reisen und Forschungen im Amur-
Lande in den Yahren 1854—1856 // Vogel
Amur-Landes. 1860. Bd. 1. S. 217—567.
Scott J. M., Hoffman W. E., Ainley D.,
Zeillemaker C. F. Range expansion and
activity patterns in Rhinoceros Auklets //
West. Birds. 1974. Vol. 5. P. 13-20.
Sealy S. G. Color aberration in some alcids
on St. Lawrence Island, Alaska //Wilson
Bull. 1969. Vol. 81, N 2. P. 213-214.
Sealy S. G. Egg teeth and hatching methods
in some alcids // Ibid. 1970. Vol. 82, N 3.
P. 289-293.
Sealy S. G. Adaptive significance of post-
hatching developmental patterns and
growth rates in the Alcidae // Ornis scand.
1973a. Bd. 4, N 2. S. 113-121.
Sealy S. G. Breeding biology of the horned
puffin on St. Lawrence Island, Bering Sea
with zoogeographical notes on the North
Pacific puffins // Pacif. Sci. 1973b. Vol. 27,
N 2. P. 99-119.
Sealy S. G. Breeding phenology and clutch
size in the marbled murrelet // Auk.
1974. Vol. 91, N 1. P. 10—23.
Sealy S. G. Feeding ecology of the ancient
. and marbled murrelets near Langara Is-
13*
195
Литература
land, British Columbia// Canad. J. Zool.
1975a. Vol. 53. P. 418- 433.
Sealy S. G. Influence of snow on egg-laying
in auklets // Auk. 1975b. Vol. 92. P. 528—
538.
Sealy S. G. Biology of nesting ancient murre-
lets // Condor. 1976. Vol. 78. P. 294-306.
Sealy S. G. Voles as a source of egg and
nestling loss among nestling auklets //
Murrelet. 1982. Vol. 63. P. 9—14.
Sealy S. G., Bedard J. Breeding biology of the
parakeet auklet (Cyclorrhynchus psitta-
cula) on St. Lawrence Island, Alaska //
Astarte. 1973. Vol. 6. P. 59- 68.
Searing G. F. Some aspects of the ecology
of cliff-nesting seabirds at Konckok Bay,
St. Lawrence Island, Alaska, during 1976:
Final rep. // Environ, assess. Alaskan con-
tin. shelf, Annu. rep. Nat. Oceanic Atoms
Admin, and Bur. Land Mgkht., Outer con-
tin. shelf environ, assess. Program.
Boulder (CoL), 1977. Vol. 5. P. 263-412.
Simons T. B. Discovery of ground-nesting
marbled murrelet // Condor. 1980. Vol. 82.
P. 1-9.
Slater P. J. B. Orientation of fish in the
Tystie’s beak // Bird Study. 1974. Vol. 21,
N 4. P. 238- 240.
Slater P. J. B. Tidal rhythm in a seabird //
Nature. 1976. Vol. 264, N 5587. P. 636-
638.
Slater P. J. B., Slater E. P. Behaviour of the
tystie during feeding of the young // Bird
Study. 1972. Vol. 19, N 2. P. 105-113.
Soikkeli M. The razorbill Alca torda does not
incubate two eggs // Ornis fenn. 1982.
Vol. 59, N 2/3. P. 155-156.
Southern H. N., Carrick B., Potter W. G.
The natural history of a population
of guillemots (Uria aalge Pont.) // J.
Anim. Ecol. 1965. Vol. 34, N 3. P. 649-665.
Sowls A. L., Hatch S. A., Lensink C. J.
Catalog of Alaskan seabirds colonies.
Anchorage, Alaska. 1978. 356 p.
Spring L. A comparison of functional and
morphological adaptations in the common
murre (Uria aalge) and thick-billed murre
(Uria lomvia) // Condor. 1971. Vol. 73.
P. 1-27.
Springer A. M., Boseneau D. G. Copepod-based
food webs: auklets and oceanography
in the Bering Sea // Mar. Ecol. Progr.
Ser. 1985. Vol. 21. P. 229-237.
Stejneger L. Results of ornithological explora-
tion in the Commander Islands and
Kamtschatka // Bull. US Nat. Mus. 1885.
N 29. P. 7-362.
Stempniewicz L. Breeding biology of the little
auk Plautus alle in the Hornsund region,
Spitsbergen // Acta ornithol. 1981. Vol. 18,
N 2. P. 141-165.
Stempniewicz L. Factors causing changes in
the rhythm of attendance of the little auks,
Plautus alle (L.) at a colony during the
breeding season in Svalbard // Ecol. pol.
1986. Vol. 34, N 2. S. 247-263.
Svein M. Lundens seksuelle og so siale ad-
ferd // Fauna (Norge). 1962. Bd. 15, N 1.
S. 1-10.
Swennen C., Duiven P. Size of food objects
of three fish-eating seabird species: Uria
aalge, Alca torda, and Fratercula arctica
(Aves, Alcidae) // Netherl. J. Sea Res.
1977. Vol. 11, N 1. P. 92-98.
Taczanowski L. Fauna ornithologique de la Si-
berie Orientale. St. Petersburg, 1891 —1893.
1278 p. (Mem. Acad. Sci. St. Petersburg.
Ser. VII; Vol. 39).
Takahashi T. On the unrecorded birds from
South Saghalien // Tori. 1937. Vol. 9, N 45.
P. 36- 52.
Takahashi T. О птицах острова Кайбато
и гнездовании на нем Cerorhinca mono-
cerata (Pallas) // Rep. Saghalien Gov.
Mus. 1939. Vol. 3, N 3. P. 159-214.
На яп. яз.
Taylor K., Beid J. B. Earlier colony attendance
by guillemots and razorbills // Scot. Birds.
1981. Vol. 11, N 6. P. 179-180.
Thayer J. E. Nesting of the kittlitz’s murre-
let // Condor. 1914. Vol. 16, N 3. P. 31—36.
Thiede W., Takedatsu M., Thiede U. Bird
life in winter at the Ochotsk Sea coast
of Hokkaido // Tori. 1973. Vol. 22,
N 93/94. P. 1-13.
Thompson M. C., Hines J. Q., William-
son F. S. L. Discovery of the downy young
of Kittlitz’s murrelet // Auk. 1966. Vol. 83,
N 3. P. 349-351.
Thoresen A. C. The breeding behaviour of the
Cassin’s auklet // Condor. 1964. Vol. 66,
N 6. P. 456-476.
Thoresen A. C. Diurnal activity and social
displays of Rhinoceros auklets on Teuri
Island, Japan // Ibid. 1983. Vol. 85, N 3.
P. 373—375.
Timmermann G. Die Vogel Islands. Visinda-
felag // Islendinga. 1938. Bd. 21. 109 p.
Tooren D. Ein nieuwe Vondst van de dikbek-
zeekoet in Nederland // Limosa. 1967.
Bd. 40, N 4. Biz. 175-181.
Tschanz B. Trottellummen (Uria aalge aalge
Pont.) // Ztschr. Tierpsychol. 1968. Bd. 4.
S. 287—301.
Tschanz B. Beobachtungen an Dickschnabel-
und Trottellummen (Uria lomvia und
aalge) auf Vedoy (Lofoten, Norwegen) //
Ornithol. Beob. 1972. Bd. 69, N 3/4.
S. 169-177.
Tschanz B. Untersuchungeu zur Entwicklung
des Trottellummenbestandes auf Vedoy
(Rost, Lofoten) // J. Ornithol. 1978.
Bd. 119, N 2. S. 133-145.
Tschanz B. Zur Entwicklung von Papagaiteu-
cherkiiken Fratercula arctica in Freiland
und Labor bei unzulanglichem und ausrei-
196
Литература
chenden I' ulterangebot // Fauna norv.
1979. Bd. C2, N 2. S. 70-94.
Tschanz B., Barth E. K. Svingninger i lomvi-
bestanden рй Ved/y рй R/st//Fauna
(Norge). 1978. Bd. 31, N 4. S. 205-219.
Tschanz B., Eymann H., Ingold P. et al.
Familieliv hos lomvi, Uria aalge // Ibid.
1969. Bd. 22, N 1. S. 1-14.
Tschanz B., Ingold P., Lengacher H. Eiform
und Bruterfold bei Trottellummen Uria
aalge aalge Pont // Ornithol. Beob. 1969.
Bd. 66, N 2. S. 25-42.
Tuck G., Heinzel H. A field guide to the sea-
birds of Britain and the world. L., 1978.
292 p.
Tuck L. M. The murres. Ottawa, 1960. 260 p.
(Canad. Wildlife Serv. Monogr.; N 1).
Tuck L. M., Squires H. J. Food and feeding
habits of Brunnich’s murre (Uria lomvia
lomvia) in Akpatok Islands // J. Fish.
Res. Board Canada. 1955. Vol. 12, N 5.
P. 781-792.
Tull С. E., Germain P., May A. W. Mortality
of thick-billed murres in the West Green-
land salmon fishery // Nature. 1972.
Vol. 237, N 5349. P. 42-44.
Udagawa T. The avifauna of Teuri and
Jagishiri Island, Hokkaido//Tori. 1953.
Vol. 13, N 63. P. 68-84.
Vaurie C. The birds of Palearktic fauna.
1. A systematic reference non-passerifor-
mes. L., 1965. 763 p.
Verbeek N. A. Wanderings of the ancient
murrelet: some additional comments //
Condor. 1966. Vol. 68. P. 75-87.
Vermeer K. Nesting requirements food and
breeding distribution of Rhinoceros
auklets, Cerorhinca monocerata, and Tufted
puffins Lunda cirrhata//Ardea. 1979.
Vol. 67. P. 101 — 110.
Vermeer K. The importance of timing and
type of prey to reproductive success
of Rhinoceros auklets Cerorhinca mono-
cerata // Ibis. 1980. Vol. 122, N 3.
P. 343-350.
Vermeer K., Cullen L. Growth comparison
of a plankton- and a fish-feeding alcids //
Murrelet. 1982. Vol. 63, N 2. P. 34-39.
Vermeer K., Robertson J., Campbell R. W.
et al. Distribution and densities of marine
birds on the Canadian west coast. Ottawa,
1983. 73 p.
Verspoor E., Birkhead T. R., Nettleship D. N.
Incubation and brooding shift in the com-
mon murre Uria aalge // Canad. J. Zool.
1987. Vol. 65, N 2. P. 247-252.
Vogelfelsen // Kosmos. 1982. N 7. S. 42—49.
Wunless S., Erench D. D., Harris M. P.,
Langlow D. R. Detection of annual changes
in the number of cliff-nesting seabirds
in the Orkney 1976—1980 // J. Anim. Ecol.
1982. Vol. 51, N 3. P. 785-795.
Watanuki Y. Predation and anti-predation
behaviour in seabirds on Teuri Island,
Hokkaido // J. Yamashina Inst. Ornithol.
1983. Vol. 15. P. 167-174.
Watson A. Notes on birds in Arctic Norway //
Sterna. 1957. Bd. 2, pt 3, N 28. P. 82-88.
Wehle D. H. S. Color phases in the downy
and juvenal plumages of Tufted puffins //
Condor. 1982a. Vol. 84. P. 444-445.
Wehle D. H. S. Food of adult and subadult
Tufted and Horned puffins // Murrelet.
1982b. Vol. 63, N 2. P. 51-58.
Wehle D. H. S. Puffins: East and west //
Sea Front. 1982c. Vol. 28, N 6. P. 324-329.
Wehle D. H. S. The food, feeding and develop-
ment of young Tufted and Horned puffins
in Alaska // Condor. 1983. Vol. 85, N 4.
P. 427-442.
Wehrlin J. Verhaltensan-passungen junger
Trottellummen (Uria aalge aalge Pont.)
aus Felsklippen- und Kolonierbriiten //
Ztschr. Tierpsychol. 1977. Bd. 44, N 1.
S. 45-49.
Wehrlin Tschanz B. Cliff-Response bei
Trottellummen (Uria aalge) // Rev. suisse
zool. 1969. Vol. 76, N 4. P. 1132-1144.
Williams A. J. Laying and nest-building
behaviour in the larger auks (Aves,
Alcidae)//Astarte. 1971. Vol. 4, N 2.
P. 61-67.
Williams A. J. Site preferences and interspe-
cific competition among guillemots Uria
aalge (L.) and Uria lomvia (L.) on Bear
Island // Ornis scand. 1974. Bd. 5.
S. 113-121.
Williams A. J. Guillemot fledging and preda-
tion on Bear Island // Ibid. 1975. Bd. 6,
N 2. S. 117-124.
Witherby H. F. A practical handbook of British
birds. Pt 2. High Holborn. L., 1924.
498 p.
Witherby H. F., Gourdain F. C. R., Tice-
hurst N. F. et al. Handbook of British birds.
L„ 1941. Vol. 5. 434 p.
Witt H. Zur Biologic der Korallenmawe Larus
audouinii — Brut und Ernahrung // J. Or-
nithol. 1977a. Bd. 118, N 2. S. 134—155.
Witt H. Zur Verhaltenbiologie der Korallen-
mowe Larus audouinii // Ztschr. Tierpsy-
chol. 1977b. Bd. 43, N 1. S. 46-67.
Wolley J. Some observations on the birds
of Faroe Islands // Ootheca Wolleyana.
London, 1905—1907. Appl. 41.
Won Pyong Oh. Checklist of the birds
of the Republic of Korea. Seoul, 1976.
30 p.
Woodby D. A. The april distribution of murres
and prey patches in the southeastern
197
Литература
Bering Sea // Limnol. and Oceanogr. 1984.
Vol. 29, N 1. P. 181 — 188.
Wus W. J. R. de. Uria aalge hyperborea
in Nederland? // Limosa. 1981. Bd. 54,
N 2. Biz. 63-64.
Wynee-Edwards V. C. On the habits and
distribution of birds on the North Atlan-
tic // Proc. Boston Soc. Natur. Hist. 1935.
Vol. 40. P. 233-246.
Yamashina Y. Notes on a collection of birds
from Southern Sakhalin. 2 // Tori. 1928.
Vol. 5, N 25. P. 419-453.
Yamashina Y. On a collection of birds from
Paramushir Islands, N. Kuriles // Ibid.
1929. Vol. 6, N 27. P. 22-56.
Yamashina Y. Die Vogel der Kurilen //J. Or-
nithol. 1931. Bd. 79, N 4. S. 491-541.
Zelickman E. A., Golovkin A. N. Composi-
tion, structure and productivity of neretic
plankton communities near the bird colo-
nies of the nothern shores of Novaya
Zemlya // Mar. Biol. 1972. Vol. 17, N 3.
P. 265-274.
Дополнение к тому
«Птицы СССР.
Чайковые»
Одуэнова чайка
Larus audouinii Payraudeau, 1826
Audouin’s Gull (англ.), Korallenmowe (нем.), Goeland d’Audouin (фр.).
Статус. Залетный вид.
Общая характеристика и полевые признаки. Белоголовая чайка, круп-
нее сизой и несколько мельче серебристой. Окраска взрослых особей
сходна с окраской средиземноморского подвида серебристой чайки, от кото-
рой одуэнова чайка отличается кораллово-красным с черной перевязью
и желтым кончиком клювом (издали клюв выглядит черным), оливково-
зеленой окраской лап и меньшим числом белых «зеркалец» на черных
концах крыльев. Кроме того, одуэнова чайка выглядит несколько более
длиннокрылой и короткохвостой, чем серебристая. Птицы в гнездовом
наряде более «светлолицые», чем серебристые чайки, и с более четкими
пестринами на спине. В отличие от серебристой чайки хвост в этом наряде
почти целиком черный, [резко контрастирующий с белым надхвостьем.
Промежуточные наряды светлее, чем у серебристой чайки, с большим раз-
витием черного цвета в дистальной части крыла (Cramp, Simmons, 1983).
Голос неприятный, хорошо отличается от голосов серебристой и сизой
чаек. Для него характерны хриплые тона (Makatsch, 1969).
Подвидовая систематика. Монотипический вид.
Описание. Размеры.
Длина (в мм): крыла самцов 401—423 (среднее 408, п=14), самок
384—406 (среднее 393, и=12); клюва самцов 46,5—51,9 (среднее 49,6,
и=13), самок 40,3—48,2 (среднее 44,5, п=12); цевки самцов 57,3—
62,1 (среднее 59,1, п=13), самок 52,5—57,8 (среднее 55,8, n=12) (Cramp,
Simmons, 1983).
Окраска. (Dwight, 1925; Cramp, Simmons, 1983).
Самец и самка в брачном наряде. Голова, шея, центр груди, брюшко,
подхвостье, надхвостье, поясница и хвост белые. Задняя часть шеи, верх-
няя часть мантии, бока груди и бока очень светло-серые. Мантия, плечевые
перья и спина светло-серые, несколько темнее, чем у озерной чайки; плече-
вые перья с белыми кончиками. Второе (первое видимое) первостепенное
маховое черное с серым основанием внутреннего опахала, небольшим
овальным белым предвершинным пятном на внутреннем опахале и белым
кончиком. Следующие четыре первостепенных черные с белыми кончи-
ками, причем серый цвет основания пера распространяется по обоим опа-
халам все ближе к вершине. Седьмое первостепенное серое с черной пред-
вершинной полосой и белым кончиком, остальные четыре — серые с белым
кончиком. Второстепенные маховые серые с белыми вершинами. Наруж-
ные кроющие первостепенных маховых беловатые, остальные верхние
199
Дополнение
кроющие крыла светло-серые. Клюв красный с предвершинной черной
перевязью (иногда двойной) и желтым кончиком. Лапы оливково-зеленые.
Радужина темно-коричневая, края век красные.
Самец и самка в зимнем наряде. По бокам головы, спереди и за глазами,
на затылке и задней стороне шеи редкие буроватые пестрины. Остальное
оперение, клюв и лапы как в предыдущем наряде.
Пуховой наряд. Основной тон окраски верхней части тела желтовато-
серый, на голове более светлый. Мелкие черные пестрины на голове фор-
мируются в полоски. Спина в темно-коричневых пестринах, обычно со-
ставляющих три прерывистые полоски. Нижняя часть тела светлее верх-
ней, брюшко беловатое. Клюв темно-серый с красноватым кончиком, лапы
темно-серые, радужина темно-коричневая.
Гнездовой наряд. Голова, шея, подбородок, горло и грудь светлые
серо-коричневые, с более светлым лбом и оперением у основания под-
клювья. На голове и горле небольшие продольные, а на подбородке и груди
поперечные пестрины. Перья мантии темно-коричневые с белыми оконеч-
ностями, плечевые перья темно-коричневые с широкими светлыми каем-
ками. Поясница и верхние кроющие крыла белые, каждое перо с предвер-
шинной бурой полосой. Нижняя часть тела белая с серо-коричневым оттен-
ком на брюшке, бока с четкими темно-коричневыми пестринами. Хвост
коричневато-черный с белой вершинной полосой, основания рулевых
перьев белые. Первостепенные маховые черные, второстепенные — корич
невато-черные с белыми вершинами и белым полем на основании внут-
реннего опахала. Большие верхние кроющие крыла светло-коричневые,
остальные верхние кроющие крыла как плечевые. Нижние кроющие крыла
темно-коричневые. Клюв черный с серовато-розовым основанием, лапы
серые.
Первый зимний наряд. Как гнездовой, но перья мантии и плечевые
серые с большими бурыми отметинами.
Первый летний наряд. Оперение крыльев и хвоста остается от гнездо-
вого наряда. Перья мантии и плечевые серые с коричневыми пятнами.
Голова и грудь беловатые с коричневыми пестринами. Нижняя часть туло-
вища в основном белая. Подкрылья темно-коричневые.
Второй зимний наряд. Голова, мантия, поясница и нижняя часть тела
как у взрослых птиц в зимнем наряде, но пестрины на голове и шее более
густые. Грудь с коричневатым налетом, на спине и плечевых небольшое
количество бурых пятен. Хвост белый с черной предвершинной полосой.
Второе—восьмое первостепенные маховые тускло черные, большинство
из них с белыми кончиками (белого кончика нет на втором первостепен-
ном, его может не быть также на третьем и четвертом первостепенных).
Начиная с шестого, на основании внутреннего опахала появляется серое
поле, которое постепенно растет, так что внутренние опахала девятого—
одиннадцатого первостепенных становятся сплошь серыми с широкой
белой вершинной полосой. Второстепенные маховые черные с широкой
белой вершинной полосой и каймой по краю внешнего опахала, внутрен-
ние опахала их серые. Верхние кроющие первостепенных маховых темно-
бурые со светлыми центрами и белыми кончиками. Малые верхние крою-
щие крыла с темно-бурыми центрами и белыми или светло-серыми кай-
мами, средние кроющие серые с тонкими темными пестринами, большие
кроющие крыла, кроющие третьестепенных маховых и сами третьестепен-
ные серые с темно-бурыми центрами. Черный цвет на клюве сокращается
2(Ю
Дополнение
до черной предвершинной полосы, светлое основание клюва начинает кра-
снеть. Лапы становятся оливковыми (возможно, это происходит раньше —
к первой зиме).
Второй летний наряд. Как предыдущий, но голова и нижняя часть тела
белые, мантия и плечевые чисто-серые.
Третий зимний и третий летний наряды. Как соответствующие окон-
чательные наряды, но с большим развитием черного цвета на первостепен-
ных маховых, более густыми пестринами на голове и шее зимой, с остат-
ками бурых пятен на второстепенных маховых и верхних кроющих крыла,
остатками темной полосы на хвосте. У некоторых птиц, возможно, третий
наряд неотличим от окончательного. У большинства окончательными ста-
новятся четвертый зимний и четвертый летний наряды.
Линька. Линька из гнездового в первый зимний наряд частичная
(сменяется оперение головы, нижней части тела, мантии, а также плече-
вые перья), происходит, очевидно, с конца сентября по ноябрь. В после-
дующем линьки бывают дважды в году, полная осенняя и частичная весен-
няя. Послебрачная (осенняя) линька взрослых птиц протекает с середины
мая по октябрь, первостепенные маховые сменяются в июне—октябре.
Сроки частичной предбрачной линьки, при которой сменяется оперение
головы и туловища, не известны. Линьки неполовозрелых птиц протекают
в несколько более ранние сроки, чем взрослых (Cramp, Simmons, 1983).
Распространение. Гнездовой ареал. Ограничен исключительно
Средиземноморьем (рис. 49). Гнездовые колонии известны на Балеарских
островах и островах Колумбретес у берегов Испании, на северо-восточном
побережье Испании, на Корсике, на мелких островках у побережья
о-ва Сардиния, на островах Тосканского архипелага, по северному побе-
режью Марокко, Алжира и Туниса, на ряде островов Эгейского моря, Кипре
и у южного побережья Турции (Cramp, Simmons, 1983; Schenk, Meschini,
1986).
Зимовки. Большинство птиц, по-видимому, зимует в Средиземном
море, дисперсно распределяясь главным образом в его южной части и
наиболее часто встречаясь в этот период у северного побережья Марокко,
Алжира, Туниса и Западной Ливии. Значительная часть птиц, однако,
покидает Средиземное море через Гибралтарский пролив и зимует на атлан-
тическом побережье Марокко, причем тенденция покидать на зиму Сре-
диземное море, по-видимому, сильнее у молодых птиц, чем у взрослых.
Миграции. В Средиземном море, очевидно, носят характер кочевок.
Стаи одуэновых чаек отмечены осенью у юго-восточного побережья Ис-
пании, зимой эти птицы встречаются здесь редко. Часть чаек мигрирует
через Гибралтарский пролив и далее в южном направлении вдоль атланти-
ческого побережья Марокко, причем отдельные особи долетают до Сене-
гала. Не ясно, имеет ли здесь место направленная миграция, или же это
просто крайняя форма дисперсии особей во внегнездовой сезон. Взрослые
птицы отмечаются на атлантическом побережье Марокко только в зимний
период, годовалые особи, кроме того, и в летнее время.
Отмечены залеты птиц в Португалию, Швейцарию, Иорданию, на
Канарские острова (Cramp, Simmons, 1983). Изредка одуэновы чайки зале-
тают в Черное море. Достоверный залет этого вида на территорию СССР
зарегистрирован 3 октября 1984 г., когда взрослая одуэнова чайка была
встречена финскими орнитологами на берегу Черного моря у Батуми
(Hottola, 1987). Зимой 1979—1980 гг. чайка, напоминающая одуэнову,
201
Дополнение
Рис. 49. Область распространения
чайки Одуэна
а — гнездовые поселения,
б — область зимовок,
в — залет на территорию СССР
отмечена на Южном берегу Крыма
(Ю. В. Костин, личное сообщ.). Однако
уверенности в достоверности определе-
ния вида не было, и Юлий Витальевич
не включил эту чайку в свою сводку по
птицам Крыма. По-видимому, одуэнова
чайка периодически, хотя и очень редко,
залетает на территорию нашей страны.
В связи с ростом численности этого вида
частота подобных залетов будет, вероят-
но, возрастать.
Численность. Общая численность
мировой популяции вида составляет
ныне, вероятно, несколько более 5 тыс.
пар. В Западном Средиземноморье
она, по-видимому, возрастает (Cramp,
Simmons, 1983; Schenk, Meschini,
1986).
Местообитания. В сезон гнездова
ния — небольшие каменистые необитае
мне островки, прибрежные скалы сла-
бой или средней крутизны высотой
обычно не более 50 м над уровнем моря, прилегающие морские акватории.
Во внегнездовой сезон — морские побережья.
Размножение (Makatsch, 1969; Witt, 1977а, b; Mayol, 1978; Cramp,
Simmons, 1983; Schenk, Meschini, 1986). Время наступления половой зре-
лости не известно. Моногам, факультативно-колониальный вид. Некото-
рые колонии из года в год располагаются на одних и тех же местах, другие
нестабильны.
Пары формируются в «клубах», где холостые самцы удерживают
охраняемую территорию. Демонстрационное поведение — как у других
крупных белоголовых чаек, однако имеются и видовые особенности.
Позы при долгом крике сходны с подобными позами сизой чайки, однако
у одуэновой чайки последовательность поз завершается серией вскидыва-
ний клюва, происходящих в более быстром темпе, чем во время ритуаль-
ного кормления. В агрессивной прямой позе клюв нередко опущен почти
вертикально. «Квохтание» и отворачивание клюва ритуализованы слабо,
распластанная поза не отмечена. Как и у других крупных белоголовых
чаек, выпрашивая корм у самца, самка принимает сгорбленную позу
и вскидывает клюв. Вскидывание клюва обоих партнеров обычно предшест-
вует спариванию. В целом демонстрационное поведение одуэновой чайки
больше всего напоминает такое же поведение сизой чайки (подроб-
ности демонстрационного поведения см.: Witt, 1977b; Cramp, Simmons,
1983).
Гнездится небольшими колониями (средний размер — около 50 гнезд),
однако изредка встречаются колонии величиной более 100 гнезд. Отмеча-
ется и одиночное гнездование. Под колонии занимаются обычно неболь-
шие каменистые островки (в том числе расположенные неподалеку
от побережья материка или крупного острова), покрытые травянистой рас-
тительностью, нередко с участками низкорослых кустарников, а также при-
брежные скальные обнажения. Гнезда помещаются на земле среди камней
202
Дополнение
и разреженной растительности. Среднее расстояние между центрами бли-
жайших соседних гнезд в разных колониях варьирует от 1,66 до 7,75 м,
в плотных участках колонии между гнездами обычно 1—4 м. Индекс
плотности гнездования, вычисленный по имеющимся данным (п=291),
равен в среднем 0,03 (от 0,11 до 0,02 в разных колониях). Диаметр гнезда
(в мм) по измерениям на Балеарских островах составляет (п—7) 230—
360, в среднем 310; диаметр лотка — 160—190, в среднем 180. В качестве
материала для гнезда используется сухая трава, веточки, листья и отдель-
ные перья.
Откладка яиц начинается с середины или с последней декады апреля,
что на 6—7 недель позже, чем у L. argentatus michahellis. В полной кладке
2—3, реже 1 яйцо. Средняя величина кладки в разных колониях и по годам
варьирует от 2,32 до 2,54. Размеры яиц (в мм): 58,9—68,1X40,9—46,6,
в среднем (п=63) 63,6X44,5; средний вес 67,5 г (Makatsch, 1969). Окраска
яиц напоминает окраску яиц сизой чайки. Насиживают кладку оба парт-
нера, начиная С первого яйца. Длительность инкубации от 26 до 33 дней,
в среднем 28,5 дней (первое яйцо кладки насиживается в среднем 29,8 дней,
второе — 27,8, третье — 27,0). Промежуток между откладкой первого и
третьего яйца в среднем составляет 4,1 дня, интервал между вылуплением
первого и третьего птенца — 1,7 дня.
Первые птенцы появляются с середины—второй половины мая. Вылу-
пившийся птенец весит в среднем 46 г (п=13). После вылупления роди-
тели выносят скорлупу яйца из гнезда (Makatsch, 1969). В течение пер-
вого дня жизни птенец остается в гнезде, на второй день он начинает поки-
дать его на некоторое время, а с третьего дня не возвращается в гнездо.
Кормят птенцов оба родителя. На крыло молодые птицы поднимаются,
по-видимому, в возрасте 35—40 дней; родители некоторое время продол-
жают кормить уже летающих птенцов. Точных данных по успеху размно-
жения нет. В Эгейском море в 1974 г. вылупилось 33 птенца из 66 отложен-
ных яиц. На островах у побережья Марокко в 1062 гнездах погибло 420 яиц
и было найдено мертвыми 560 птенцов. В пяти колониях на Балеарских ост-
ровах в 382 гнездах погибло 30 яиц и найдено 16 мертвых птенцов.
Суточная активность, поведение. Активность, по-видимому, в основном
дневная, хотя птицы могут кормиться и ночью. В сезон гнездования брач-
ные партнеры ночуют в колонии на своей территории. Вне сезона размно-
жения отмечаются коллективные ночевки на каменистых островках. Во все
сезоны обычно держится стайками от 3—4 до 20 птиц, но может образовы-
вать стаи до 200 птиц (Cramp, Simmons, 1983).
Питание. Кормовые объекты — главным образом рыба, гораздо реже
водные и наземные беспозвоночные, мелкие птицы и растения. Кормятся
одуэновы чайки в основном в море, гораздо реже над сушей неподалеку
от моря, предпочитая в последнем случае пашни и сжатые поля. Могут
ловить в воздухе летающих насекомых. Отмечено поедание отбросов рыбо-
ловного промысла. В целом одуэнова чайка далеко не столь явно выражен-
ный пелагический вид, как предполагалось ранее.
Способ кормежки одуэновой чайки над спокойной поверхностью моря
достаточно своеобразен. Птица круто снижается к поверхности, а затем,
опустив лапы, очень медленно летит над самой водой. Быстрыми движе-
ниями головы, часто не замедляя полета, она выхватывает из воды добычу,
причем воды касается только клюв. Через некоторое время чайка взмывает
выше, затем вновь снижается и летит над самой водой в обратном направ-
203
Дополнение
лении. Время от времени птица может при этом присаживаться на воду
(Cramp, Simmons, 1983).
Враги, неблагоприятные факторы. Негативное воздействие на гнездова-
ние одуэновых чаек оказывает фактор беспокойства со стороны человека,
сбор яиц, хищничество серебристых чаек-хохотуний, ворон и крыс (Mayol,
1978; Schenk, Meschini, 1987).
Хозяйственное значение, охрана. Одуэнова чайка занесена в Красную
книгу Международного союза охраны природы и природных ресурсов.
Поправки к предыдущим томам сводки
«Птицы СССР»
ИСТОРИЯ ИЗУЧЕНИЯ. ГАГАРЫ, ПОГАНКИ, ТРУБКОНОСЫЕ.
М.: Наука, 1982.
Автор очерка «Приенисейская Сибирь, Саяны, Тува» в разделе «Региональные очерки
истории изучения фауны птиц СССР» — Р. Л. Наумов.
На первой странице суперобложки тома — белошейная гагара (фото А. Г. Сорокина),
на последней странице суперобложки — чомга (фото Г. М. Русанова).
КУРООБРАЗНЫЕ. ЖУРАВЛЕОБРАЗНЫЕ.
Л.: Наука, 1987.
Черно-белые иллюстрации журавлей, пастушковых и дроф выполнены художником
Ю. М. Смириным.
На первой странице суперобложки тома — стерх (фото 3. В. Назарова), на последней
странице суперобложки — дикуша (фото 3. В. Назарова).
ЧАЙКОВЫЕ.
М.: Наука, 1988.
Автор очерка «Чернохвостая чайка» — Н. М. Литвиненко.
На цветной таблице VII под номером 10 изображена вилохвостая чайка в гнездовом
(а не в первом зимнем) наряде.
Из списка литературы выпали две процитированные в тексте работы:
Андрусенко Н. Н. О весеннем пролете птиц в юго-западной части оз. Кургальджин //
Миграции птиц в Азии. Душанбе: Дониш, 1980. С. 191 — 201.
Слепцов М. М. Южно-полярный поморник в водах СССР // Орнитология. 1963. Вып. 6.
С. 482.
Таблица I
1—3, 12 — гагарка (2 — взрослая в летнем наряде, 2 — взрослая в зимнем наряде (голова), 3 —
в летнем наряде в полете, 12 — в летнем наряде на воде)
4—7, 11 — тупик (4 — взрослый в летнем наряде, 5 — взрослый в зимнем наряде, 6 — молодой летом,
7 — взрослый в летнем наряде в полете, 11 — то же на воде)
8, 9, 10 — люрик (8 — взрослый в летнем наряде, 9 — в зимнем наряде,//) — в летнем наряде на воде)
Таблица II
1—4 кайра толстоклювая (7 — взрослая в летнем наряде, 2 — то же в полете, 3 - взрослая в зим-
нем наряде, 4 — взрослая в летнем наряде, подвид Uria lomvia eleonorae)
5—9 — кайра тонкоклювая (5 — взрослая в летнем наряде, 6 — взрослая в зимнем наряде, 7 — голова
очковой морфы, 8 — птенец после вылупления, 9 — птенец перед слетом с карниза)
10 — 12 — чистик (10 — взрослый в летнем наряде, 11 — взрослый в зимнем наряде, 12 - молодой)
Таблица III
7, 2, 12 — топорок (7 — взрослый в летнем наряде, 2 — взрослый в полете, 12 — в летнем
наряде на воде)
7, 77 — ииатка (3 — взрослая в летнем наряде, 4 — то же в полете, 77 — то же на воде)
5, 14 — тупик-носорог
6—8, 13 — тихоокеанский чистик (6 — взрослый в летнем наряде, 7 — взрослый в зимнем
наряде, 8 — молодой, 13 — взрослый в летнем наряде на воде)
9, 10 — очковый чистик (9 — взрослый в летнем наряде, 10 — молодой)
Таблица IV
1—3 — большая конюга (/, 2 — взрослая в летнем наряде, 3 — взрослая в зимнем наряде
на воде)
4, 5 — малая конюга (4 — взрослая в летнем наряде, 5 — то .же на воде)
6—9 — конюга-крошка (6 — взрослая в летнем наряде, 7 — то же на воде, 8 — взрослая
в зимнем наряде, 9 — то же на воде)
10, 11 — белобрюшка (10 — взрослая в летнем наряде, 11 — то же на воде)
Таблица V
1,2 — старик (1 — взрослый в летнем наряде, 2 — взрослый в зимнем наряде)
3 — хохлатый старик
4, 5 — пыжик короткоклювый (4 — взрослый в летнем наряде, 5 — взрослый в зимпем
наряде на воде)
6,7 — длинноклювый пыжик (6 — взрослый в летнем наряде, 7 — взрослый в зимпем наряде
на воде)
8 — алеутский пыжик взрослый в летнем наряде
Таблица VI
1 — птенцы гагарки, 2 — гагарка у гнезда, 3 — белобрюшки у гнезда, 4 — пуховой птенец
люрика, 5 — пуховой птенец короткоклювого пыжика, 6 — группа ипаток на гнездовье,
7 — голова тупика с кормом для птенца, 8 — пуховой птенец тупика, 9 — пуховой птенец
тихоокеанского чистика, 10 — голова птенца тихоокеанского чистика
Таблица VII
7—5 — вариации окраски яиц тонкоклювой кайры, 6—8 — толстоклювой кайры. Слева и
внизу — кайры на гнездовье
Таблица VIII
1—3 — яйца гагарки (вариации окраски), 4, 5 — тихоокеанского чистика (варианты
окраски), 6 — старика, 7 — длинноклювого пыжика, 8 — топорка, 9 — ипатки, 10, 11 —
туника (варианты окраски), 12 — большой конюги, 13 — тупика-носорога, 14 — люрика.
Внизу — топорки на гнездовье
Оглавление
ПРЕДИСЛОВИЕ
5
ПОДОТРЯД ЧИСТИКОВЫЕ
А. Н. ГОЛОВКИН
7
СЕМЕЙСТВО ЧИСТИКОВЫЕ
Род Alle Link, 1806 . ...... 8
Люрик ........................ 8
А. Н. ГОЛОВКИН
Род Alca Linnaeus, 1758 ..... 16
Гагарка...................... 16
И. П. ТАТАРИНКОВА,
А. Н. ГОЛОВКИН
Род Uria Brisson, 1760 ...... 25
Тонкоклювая кайра.............25
А. Н. ГОЛОВКИН
Толстоклювая кайра............43
А. Н. ГОЛОВКИН
Род Cepphus Pallas, 1769 .... 56
Чистик........................56
И. П. ТАТАРИНКОВА,
А. Н. ГОЛОВКИН
Тихоокеанский чистик ....... 66
Е. Э. СТОЦКАЯ
Очковый чистик............... 74
Ю. В. ШИБАЕВ
Род Brachyramphus Brandt, 1837 . 82
Длинноклювый пыжик............82
Ю. В. ШИБАЕВ
Короткоклювый пыжик...........88
Ю. В. ШИБАЕВ
Род Synthliboramphus Brandt, 1837 92
Старик .......................92
Ю. В. ШИБАЕВ
Хохлатый старик..............104
Ю. В. ШИБАЕВ
Род Ptychoramphus Brandt, 1837 . 107
Алеутский пыжик...............107
Е. Э. СТОЦКАЯ
Род Aethia Merrem, 1788 ..... 112
Большая конюга................112
Н. Б. КОНЮХОВ
Малая конюга..................121
А. Н. ГОЛОВКИН
Конюга-крошка ................125
Н. Б. КОНЮХОВ
Род Cyclorrhynchus Каир, 1829 . . 131
Белобрюшка....................131
Н. Б. КОНЮХОВ
Род Cerorhinca Bonaparte, 1828 . . 139
Тупик-носорог.................139
Ю. В. ШИБАЕВ
Род Fratercula Brisson, 1760 . . . 148
Тупик ........................148
н. Н. СКОКОВА
Ипатка........................164
С. И. ХАРИТОНОВ
Род Lunda Pallas, 1811........173
Топорок ......................173
С. П. ХАРИТОНОВ
Литература ...................183
Дополнение к тому «Птицы СССР.
Чайковые». Одуэнова чайка.....199
В. А. ЗУБАКИН
Поправки к предыдущим томам
сводки «Птицы СССР»...........205
Научное издание
ПТИЦЫ СССР
*
Чистиковые
Утверждено к печати
Институтом
эволюционной морфологии
и экологии животных
им. А. Н. Северцова
Академии наук СССР
Редакторы издательства
Т. А. Хот у лева, 3. А. Вишнякова
Художественный редактор
Н. Н. Михайлова
Технические редакторы
М. Ю. Соловьева, Н. Н. Плохова
Корректоры
Л. А. Иванова, К. И. Келаскнна
ИБ № 39573
Сдано в набор 10.04.89.
Подписано к печати 7.08.89.
Т-00003. Формат 70X100’/i6.
Бумага офсетная № 1
Гарнитура обыкновенная.
Печать’ офсетная.
Усл. печ. л. 16,9. Усл. кр. отт. 22.26. Уч.-изд.л. 18,
Тираж 10 400 экз. Тип. зак. 1479.
Цена 1 р. 60 к.
Ордена Трудового Красного Знамени
Издательство «Наука»
117864 ГСП-7, Москва В-485
Профсоюзная ул., 90.
1-я типография издательства «Наука»
198034, Ленинград, В-34, 9-я линия, 12
1 р. 60 к.
•НАУКА-