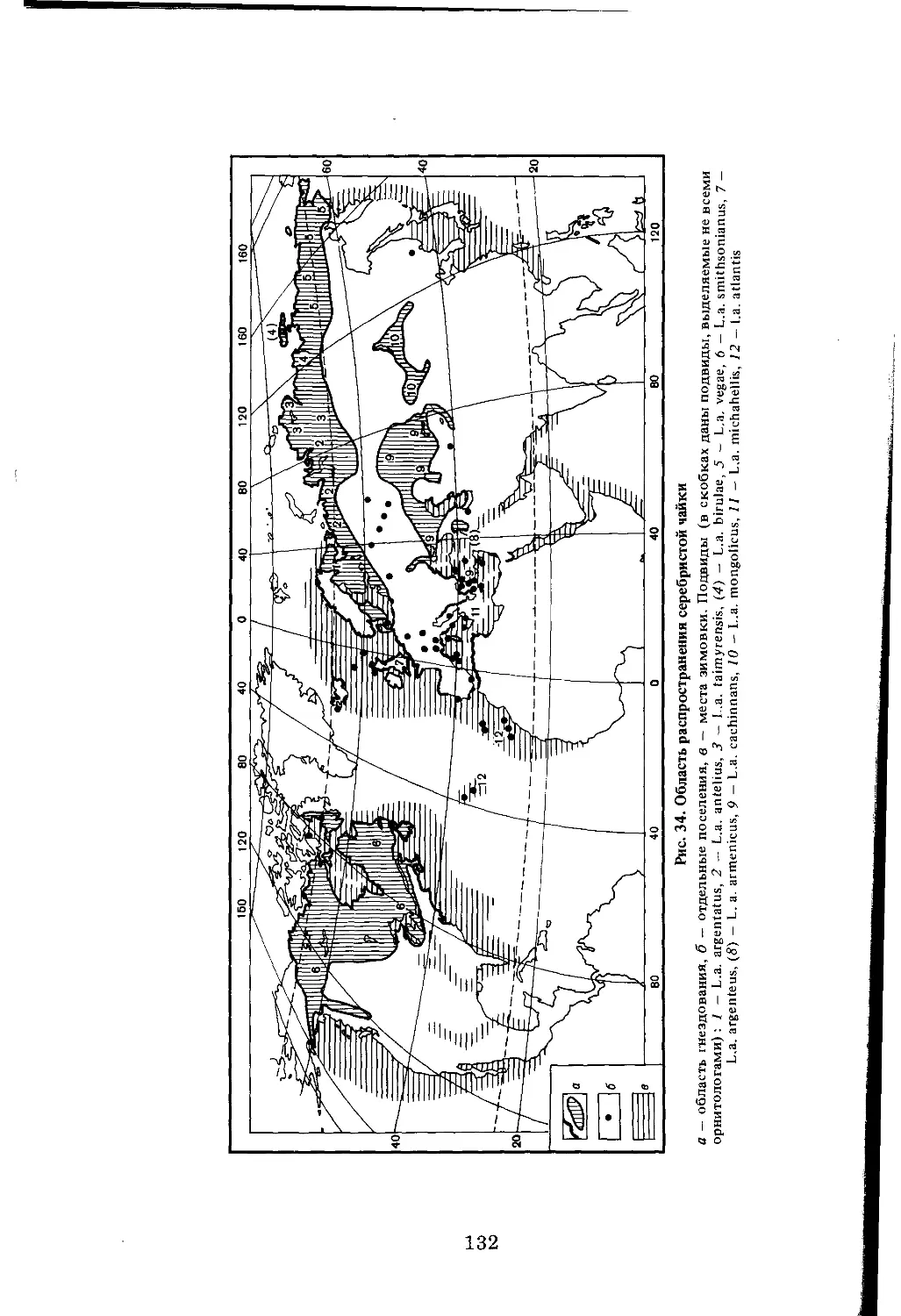
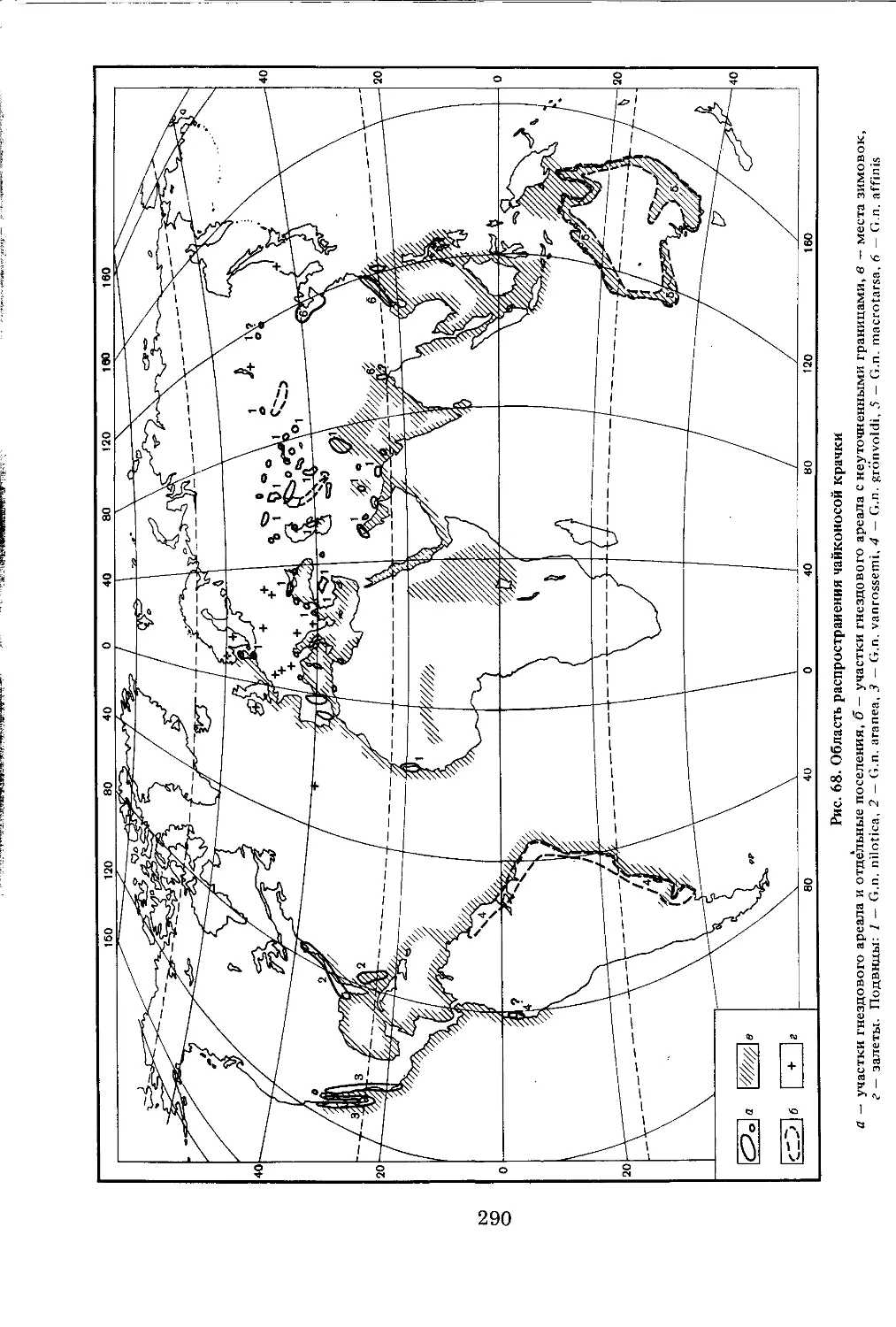
/










































































































































































































































































































































































































































Author: Флинт В.Е. Зубакин В.А.
Tags: aves птицы в целом орнитология птицы ссср
ISBN: 5-02-005255
Year: 1988




Text
ПТИЦЫ СССР
Чайковые
ПТИЦЫ СССР
РЕДАКЦИОННАЯ КОЛЛЕГИЯ
В.Д. ИЛЬИЧЕВ
В.Е. ФЛИНТ
(ответственные редакторы)
Р.Л. БЁМЕ
Э.И. ГАВРИЛОВ
В.М. ГАЛУШИН
Ю.А. ИСАКОВ
Е.Н. КУРОЧКИН
Р.Л. ПОТАПОВ
С.Г. ПРИКЛОНСКИЙ
А.К. РУСТАМОВ
Л.С. СТЕПАНЯН
В.Е. ФОМИН
Москва 1988
ПТИЦЫ СССР
Чайковые
*
Ответственные редакторы
доктор биологических наук
В.Д. ИЛЬИЧЕВ
кандидат биологических наук
В.А. ЗУБАКИН
Ж
го №ЙИ| со
'*АУ^
Москва 1988
УДК 598.2 (47 + 57)
Авторы
В.О. АВ ДАНИН, Я.А. ВИКСНЕ, В.А. ЗУБАКИН,
А.А. КИЩИНСКИЙ, Н.М. ЛИТВИНЕНКО, Е.Г. ЛОБКОВ,
В.А. НЕЧАЕВ, Л.В. ФИРСОВА, В.Е. ФЛИНТ, К.А. ЮДИН
Птицы СССР. Лайковые. М.: Наука, 1988. 416 с. ISBN 5-02-005255 - 8.
Серия книг ’’Птицы СССР” рассказывает о распространении и образе жизни
всех видов птиц, населяющих территорию Советского Союза. В данном томе при-
ведены очерки о всех видах птиц подотряда чайковых, отмеченных в СССР. Опи-
саны все возрастные и сезонные наряды каждого вида, географическая изменчи-
вость окраски и размеры птиц. Рассмотрены современные данные о гнездовом
ареале, зимовках, местообитаниях в разные сезоны года, экологии гнездования,
различных аспектах поведения, о питании и других деталях биологии, а также о
хозяйственном значении и проблемах охраны этих важных для человека групп
птиц. Монография снабжена цветными таблицами, картами и рисунками.
Для орнитологов, зоологов широкого профиля, работников охотничьего хо-
зяйства, любителей природы.
Цв. табл. 12. Ил. 86. Табл. 20. Библиогр. 43 с.
Рецензенты
доктор биологических наук
А.В. Михеев
доктор биологических наук
Л.П. Познании
2005000000-339
п------------------ 246-87 - IV
042 (02)-88
© Издательство ’’Наука”, 1988 г.
ISBN 5-02-005255-8.
Предисловие
Очередной том сводки ’’Птицы СССР” посвящен описанию 37 видов подотряда Чайко-
вых, отмеченных на территории СССР. Схема изложения видовых очерков принята той
же, что и в предыдущих томах; в ряде случаев в нее внесены незначительные изменения.
Для некоторых разделов видового очерка необходимо привести уточнения и разъяс-
нения.
Синонимы. Указаны наиболее употребительные русские и латинские синонимы назва-
ния вида, которые можно встретить в крупнейших отечественных и зарубежных свод-
ках и монографиях.
Статус. В очерке приводится статус вида на территории СССР.
Описание. Окраска нарядов приводится по материалам коллекций ЗИН АН СССР,
Зоологического музея МГУ, а также по монографии Дж. Дуайта (Dwight, 1925), свод-
кам Г.П. Дементьева (1951) и С. Крэмпа (Cramp, Simmons, 1983; Cramp, 1985), реже по
специально указанным источникам. Названия частей тела и оперения птицы, а также спо-
собы измерения приняты по ’’Определителю птиц СССР” (Гладков и др., 1964). Нумера-
ция первостепенных маховых осуществляется от дистального (наружного) конца крыла
к проксимальному, второстепенных маховых — от первостепенных к проксимальному
концу крыла, рулевых — от центральной пары к краям хвоста. Первостепенные махо-
вые обозначаются римскими цифрами (I—XI), второстепенные и рулевые — арабскими.
Промеры птиц, кроме особо оговоренных случаев, сделаны по музейным экземплярам.
Длина клюва измерялась до границы лобного оперения, длина крыла — при прижатом
и выпрямленном крыле. Цвет неоперенных частей тела дан по живым или по только
что добытым экземплярам. Описания нарядов, линьки, а также промеры особей белой,
серебристой, тихоокеанской, серокрылой, полярной, морской, сизой и вилохвостой
чаек, клуши, бургомистра, моевки и красноногой моевки сделано Л.В. Фирсовой по
материалам Зоологического института АН СССР, обработанным в свое время К.А. Юди-
ным. Поэтому редколлегия сводки включила покойного Константина Алексеевича в
число авторов видовых очерков перечисленных видов. В число авторов очерка о розо-
вой чайке включен покойный А.А. Кищинский, материалы которого использованы при
написании очерка.
Распространение. Гнездование вида считалось достоверно установленным, если найде-
ны его нелетные птенцы или гнездо. В очерках К.А. Юдина и Л.В. Фирсовой факт гнездо-
вания считался установленным и в случае добычи птицы с наседным пятном.
Размножение. Позы, крики и ритуалы демонстрационного поведения выделяются в
соответствии с терминологией Н. Тинбергена (Tinbergen, 1959) и С. Крэмпа (Cramp,
Simmons, 1983; Cramp, 1985), реже других, специально оговоренных в тексте авторов.
Английские названия демонстраций даны ниже. Индекс плотности гнездования вида,
приведенный в большинстве очерков, вычисляется по В.А. Зубакину (19756): средний
диаметр лотка гнезда делится на среднее расстояние между центрами ближайших сосед-
них гнезд. Индекс позволяет сравнивать плотность гнездования разных видов вне зави-
симости от величины особи вида. Описание окраски яиц, если оно проведено авторами
видовых очерков, сделано по Ю.В. Костину (1977). Все даты, кроме особо оговорен-
ных, приведены по новому стилю.
5
Предисловие
Цветные иллюстрации выполнены Ю.М. Смириным, штриховые рисунки в тексте —
В.М. Гудковым.
При написании видовых очерков, помимо литературных источников, были использо-
ваны данные ряда исследователей, особо оговоренные в тексте. Авторы тома пользу-
ются случаем поблагодарить всех орнитологов, любезно предоставивших в их распоря-
жение материалы своих наблюдений, особенно В.В. Леоновича, а также В.М. Лоскота,
П.С. Томковича и И.Н. Добрынину за оказанную возможность ознакомиться с орнито-
логическими коллекциями Зоологического института АН СССР и Зоологического му-
зея МГУ и с архивами Центра кольцевания АН СССР. Редакторы тома благодарят
И.А. Харитонову, О.Н. Волошину и Е.В. Степаницкую за помощь при подготовке руко-
писи к печати.
Редколлегия рекомендует при цитировании сводки ’’Птицы СССР” ссылаться на ав-
торов конкретных видовых очерков. Например: Виксне Я.А. Озерная чайка // Пти-
цы СССР. Чайковые. М.: Наука, 1987.
В тексте использованы сокращения: ЗИН — Зоологический институт АН СССР;
ЗМ МГУ — Зоологический музей МГУ; ЦКАН — Центр кольцевания АН СССР.
Английские названия наиболее обычных поз, криков и других демонстраций чайко-
вых птиц (Tinbergen, 1959; Cramp, Simmons, 1983; Cramp, 1985).
Чайки
Тревожный крик — Alarm call
Долгий крик — Long call
Наклонная поза долгого крика - Oblique-cum-Long-calL
Агрессивная прямая поза — Aggressive upright
Испуганная прямая поза — Intimidated upright
’’Квохтание” (’’кашляние”) - Choking
Распластанная поза — Forward
’’Огрызание” (ложное клевание) — Jabbing
Дерганье травы — Grass pulling
Клевание земли — Ground pecking (Pecking-into-the-ground)
Мяукающий крик — Mew call
Отворачивание клюва (головы) — Facing away (Head flagging)
Вскидывание клюва (головы) — Head tossing
Взаимное вскидывание клюва — Mutual head tossing
Сгорбленная поза — Hunched posture
Церемония встречи партнеров — Meeting ceremony
Выпрашивание корма — Food begging
Ритуальное кормление — Courtship feeding
Крачки
’’Рекламный крик” — Advertising call
’’Высотный полет” — High flight
’’Низкий полет” — Low flight
Согнутая поза — Bent posture
Агрессивная прямая поза — Aggressive upright
Выпрямленная поза — Erect posture
Наклонная поза — Stoop posture
ПОДОТРЯД ЧАЙКОВЫЕ
LARI
Подотряд Lari включает околоводных птиц мелкого и среднего размера (масса тела от
40—50 г до 2,5 кг) с характерным обликом чайки или крачки. Он разделяется на три
семейства.
1. Семейство Поморниковые (Stercorariidae), которое объединяет один-два рода и от
четырех до семи видов. Представители этого семейства в сезон гнездования распростра-
нены биполярно. В фауне СССР пять видов поморников, из которых три гнездятся.
2. Семейство Чайковые (Laridae) объединяет 14—17 родов и 86—87 видов чаек и кра-
чек (разными систематиками выделяется большее или меньшее число родов и видов).
Виды семейства распространены по всем материкам, кроме центральных частей Антарк-
тиды, и по многим океаническим островам. В фауне СССР 32 вида, из которых 30 гнез-
дятся.
3. Семейство Водорезовые (Rynchopidae), которое включает один род Rynchops с
тремя видами, обитающими в тропических зонах Азии, Африки и Америки.
Ископаемые представители подотряда, относимые к семейству Laridae, известны из
эоцен-олигоценовых отложений Франции. Поморники (р. Stercorarius) появляются по
крайней мере в среднем миоцене (США), ископаемые виды современного р. Larus —
в нижнем миоцене (Франция). Прежнее предположение о принадлежности ископаемых
остатков из миоцена Грузии крачке р. Sterna ныне подвергается сомнению (Olson,
1985). По-видимому, формирование подотряда шло на рубеже мелового и третичного
периодов, а разделение на семейства произошло уже к эоцену.
Чайковые имеют удлиненное туловище, длинные крылья и среднего размера хвост —
либо прямой (большинство чаек), либо выемчатый и вильчатый (крачки), либо с уд-
линенными средними рулевыми (поморники). Клюв удлиненный, сильно сжат с боков,
обычно с хорошо заметным выступом на нижней челюсти. Конец надклювья или заги-
бается вниз крючком (чайки и поморники), или заострен (крачки, водорезы). Ноздри
сквозные, схизоринальные. Череп схизогнатический, с хорошо развитым сошником.
Шейных позвонков 15. Ноги умеренной длины, передние пальцы соединены плаватель-
ной перепонкой. Задний палец мал, расположен выше уровня передних; у некоторых
видов (моевка) он недоразвит. Гортань трахеобронхиальная, с хорошо развитыми голо-
совыми мышцами. Пищевод сильно растяжим, однако зоба обычно нет. Мускульный же-
лудок с толстыми стенками, слепые кишки рудиментарны (лучше развиты у поморни-
ков).
Первостепенных маховых 11, первое обычно недоразвито. Формула крыла: II > III >
> IV... Рулевых перьев 12. Для большинства видов во взрослом состоянии отмечается
преобладание белой окраски. Половой морфизм окраски не выражен, однако у отдель-
ных видов (например, у черной крачки) самку можно отличить от самца по интенсив-
ности окраски. Характерна резкая возрастная и менее резкая у большинства видов, но
отчетливая сезонная смена окраски. Для поморников во взрослом состоянии характер-
но наличие нескольких цветовых морф.
У птенцов один пуховой наряд: у большинства видов пестрый, криптический, у неко-
торых — однотонный: либо бурый (поморники), либо светлый (черноголовый хохотун,
моевка). Взрослые птицы линяют дважды в году: после сезона размножения бывает
полная линька (в том числе маховых и рулевых перьев), перед брачным сезоном —
7
Подотряд Чайковые
частичная, при которой сменяется мелкое контурное оперение. Для крачек характерна
относительно большая независимость смены контурного оперения и первостепенных
маховых, причем у большинства видов еще до завершения полной линьки первостепен-
ных начинается их повторная смена (а у некоторых еще и третья смена), которая преры-
вается перед сезоном размножения. В результате не всегда возможно разграничить ко-
нец послебрачной и начало предбрачной линьки. Линька первостепенных маховых идет
от проксимальных перьев к дистальным (т.е. от XI к II), смена рулевых начинается с
центральной пары. В период линьки птицы не теряют способности к полету.
Чайковые - моногамы, хотя в последние годы у чаек все чаще отмечают полигинные
трио (Cramp, Simmons, 1983; Fitch, Shugart, 1983; Conover, 1984). После сезона размно-
жения пары, по-видимому, распадаются, однако в следующий сезон птицы обычно обра-
зуют пару с прошлогодними партнерами. Половая зрелость даже у мелких видов насту-
пает не ранее, чем в конце второго года жизни, у крупных — часто в возрасте 3—5 лет, у
некоторых Особей — и в более старшем возрасте. Характерна моноциклия размножения,
однако отдельные виды (например, Larus novaehollandiae) могут выращивать два вывод-
ка за сезон (Nicholls, 1974; Woller, Dunlop, 1979). Цикл размножения тропических кра-
чек может несколько не совпадать с годовым (Diamond, 1976; Dunlop, 1985).
В начале сезона размножения отмечается ритуальное кормление самцом самки. Де-
монстрационное поведение хорошо развито у большинства видов, особенно у чаек.
Спаривание происходит на суше — как на гнездовой территории, так и вне ее. Гнезда
чаще всего устраиваются на земле или сплавинах, реже на уступах скал, еще реже на
деревьях или кустарниках. Отдельные виды (Gygis alba) вообще не строят гнезд, другие
ограничиваются выкапыванием ямки в грунте, большинство видов устраивают более
или менее оформленное гнездо из растительной ветоши. Расположение гнезд открытое,
только Larosterna inca гнездится в укрытиях. Полная кладка у большинства видов чаек
и крачек состоит из 2—3 яиц, у некоторых — из 1 яйца или 1—2 яиц; у поморников —
из 2 яиц, у водорезов — из 4. Яйца пестро окрашенные, обычно слабогрушевидной фор-
мы. Насиживают оба партнера. Птенцы полувыводкового типа: вылупляются зрячими,
опушенными. Родители выкармливают птенцов до подъема молодых на крыло и до-
кармливают их некоторое время после этого. Птенцов кормят либо отрыгнутой пи-
щей (чайки, крачки рода Anous), либо принесенной в клюве (большинство крачек).
Становление взаимного индивидуального опознавания родителей и птенцов у колони-
альных видов происходит после оставления выводками гнезда. У видов, птенцы кото-
рых не покидают гнезда вплоть до подъема на крыло (например, моевки), родители
весь этот период не отличают своих птенцов от чужих.
Большинство видов гнездится колониально, лишь для большинства поморников бо-
лее характерно одиночное гнездование. Различается два типа колониальности: облигат-
ный и факультативный. Облигатная колониальность характеризуется исключительно
высокой плотностью гнездования, отсутствием охраняемой гнездовой территории, на-
личием морфологических и поведенческих компонентов, демаскирующих гнездо
(уменьшение маскирующих свойств окраски яиц и птенцов, дефекация насиживающей
птицы на край гнезда, отсутствие выноса скорлупы после вылупления птенца), наличи-
ем у большинства видов птенцовых ’’ясель”, формированием птенцовых табунков во
время тревоги. Охрана потомства у облигатно-колониальных видов, к которым отно-
сятся черноголовый хохотун, реликтовая чайка, морской голубок, крачки рода Tha-
lasseus и, возможно, еще некоторые виды, осуществляется главным образом с помощью
’’эффекта плотности” — отпугивающего воздействия на хищника плотной массы птиц.
Факультативный тип колониальности сохраняет элементы одиночно-территориального
гнездования: охраняемую гнездовую территорию (которая, однако, может быть очень
мала), способность гнездиться отдельными парами, маскировку гнезда (дефекации на
8
Подотряд Чайковые
гнездо нет, скорлупа удаляется) и др. Внутри этого типа колониальное™ выделяется
диффузное и плотное гнездование. Диффузно гнездящиеся факультативно-колониаль-
ные виды (большинство видов подотряда) сочетают рассредоточение криптически окра-
шенных кладок и птенцов с активной зашитой потомства. Плотно гнездящиеся факуль-
тативно-колониальные виды (черноголовая и буроголовая чайки, чеграва и некоторые
другие виды) наряду с активной защитой используют и ’’эффект плотности” (Зубакин,
1976в,1983).
Чайковые птицы хорошо летают, пользуясь гребным полетом. Многие виды (как
правило, чайки) способны парить, однако парящий полет не используется в качестве
поискового. Чайки хорошо плавают, на воде держатся высоко. Крачки, как правило,
на воду садятся крайне редко. Пищу собирают обычно в полете, пикируя на добычу
(многие крачки и часть чаек способны нырять с высоты нескольких метров, полностью
погружаясь в воду). Чайки могут добывать пищу и бродя по суше или мелководью, а
также при плавании. Пища либо исключительно, либо главным образом животная.
Большинство видов трофически связаны как с сушей, так и с водоемами, меньшее
число — только с водоемами, отдельные виды (черноголовая чайка, чайконосая крач-
ка) — главным образом с сушей. Значительное место среди кормов занимает мелкая
и средней величины рыба. Крупные виды поедают также мелких наземных позвоноч-
ных — грызунов, птиц и их птенцов, ящериц. Большое значение имеют водные и назем-
ные насекомые, особенно в питании мелких видов. Поедаются также земляные черви,
креветки, моллюски и другие водные и наземные беспозвоночные. Многие виды чаек
охотно поедают падаль и отбросы. Среди растительных кормов большое значение имеют
ягоды. Узкая специализация в кормовом отношении не свойственна. Характерно пере-
ключение на появляющиеся массовые корма.
Подотряд в целом и особенно его центральное семейство Laridae — процветающая
группа. Многие виды обладают большой экологической пластичностью, что позволяет
им хорошо адаптироваться к происходящим ныне в природе антропогенным изменени-
ям. Это особенно характерно для чаек. Переход на антропогенные корма (в частности,
на питание отбросами) и повсеместная охрана гнездовых колоний способствовали рез-
кому увеличению численности ряда видов чаек в Европе (в том числе и в нашей стране),
особенно озерных, сизых и серебристых чаек, многие популяции которых стали почти
синантропными. Наблюдается постепенное освоение лайковыми птицами прежде не-
свойственных им гнездовых биотопов, в частности крыш домов. Число гнездящихся на
зданиях особей возрастает, растет и число видов, устраивающих здесь свои гнезда
(Cramp, 1971; Fisk, 1978). В Болгарии больше половины всей размножающейся популя-
ции серебристых чаек ныне гнездится на крышах приморских городов и сел, причем в
последние годы серебристые чайки стали поселяться в материковой части страны, за-
селяя города (Нанкинов, 1981, 1982). В конце 1970-х годов отмечены первые гнезда
чаек на зданиях и в нашей стране — в Риге и Таллине. Ныне на крышах здесь гнездятся
серебристые и сизые чайки, клуши и речные крачки (Рандла, 1985; Strazdins et al.,
1987). Все больше чаек остается в городах на зимовку, причем в последние годы от-
мечены многочисленные зимовочные скопления озерных и сизых чаек и единичные зи-
мующие особи серебристых чаек и клуш в городах, удаленных от моря, таких, как
Киев и Брест (Разгонов и др., 1984; Шокало и др., 1984). Создается впечатление, что
мы становимся свидетелями начальных этапов мощного глобального процесса синантро-
пизации многих видов чайковых птиц, последствия которого сейчас вряд ли предска-
зуемы.
В силу своей массовости чайковые птицы имеют большое хозяйственное значение,
которое, видимо, с течением времени будет возрастать. Практическое значение разных
видов и разных популяций одних и тех же видов неоднозначно. Виды, питающиеся глав-
9
Семейство Поморииковые
ным образом насекомыми (черноголовая чайка, чайконосая крачка), могут приносить
большую пользу сельскому хозяйству. Отдельные популяции рыбоядных видов могут
в ряде случаев наносить локальный ущерб (в рыбхозах). В то же время гнездование
чайковых птиц (озерная чайка) на прудах рыбхозов способствует концентрации здесь
же на гнездовье многих ценных охотничье-промысловых водоплавающих птиц, кладки
и выводки которых чайки эффективно охраняют от серых ворон. Крупные виды чаек,
и в первую очередь серебристая, нередко сами ведут себя как хищники, уничтожая по-
томство других водных и околоводных видов. При возрастании их численности подоб-
ное воздействие на других птиц становится настолько ощутимым, что в ряде стран прак-
тикуется искусственная регуляция численности крупных чаек. У нас такие мероприятия
приходится проводить в Кандалакшском и Черноморском заповедниках. Следует, од-
нако, иметь в виду, что разного рода ущерб могут причинять лишь отдельные виды
чайковых и далеко не во всех пунктах своего ареала, поэтому огульное зачисление всех
чайковых птиц в число ’’вредных”, как это имело место несколько десятилетий назад,
совершенно недопустимо. Многие виды чайковых хозяйственного значения не имеют.
Несмотря на процветание группы в целом, отдельные виды имеют невысокую числен-
ность, а некоторые находятся под угрозой вымирания. В Красную книгу МСОП занесе-
ны два вида чаек (Larus audouinii, L. relictus), а также два вида и один подвид крачек
(Sterna balaenarum, S. zimmermanni, S. albifrons browni). Во второе издание Красной кни-
ги СССР (1984) занесено шесть видов чайковых птиц: белая чайка, черноголовый хо-
хотун, реликтовая, буроголовая и розовая чайки, алеутская (камчатская) крачка.
СЕМЕЙСТВО ПОМОРИИКОВЫЕ STERCORARIIDAE
Семейство Поморниковых представляет собой небольшую, но хорошо обособленную и
монолитную в морфологическом и экологическом планах группу видов подотряда
чаек. Это среднего размера птицы (масса тела от 0,4 до 2,0 кг) с длинными, сравнитель-
но узкими, обычно заостренными крыльями и коротким хвостом, в котором средняя
пара рулевых более или менее удлинена и выдается за его задний обрез. Лапы относи-
тельно- короткие, три передних пальца заключены в плавательную перепонку, задний
палец слабо развит и приподнят. Пальцы снабжены сильными, загнутыми и острыми
когтями. Клюв недлинный, но крепкий, несколько сжатый с боков в дистальной части,
надклювье заканчивается острым крючком. Рамфотека надклювья состоит из четырех
обособленных пластин. Окраска поморников преимущественно темная, у некоторых
видов нижняя сторона тела светлая. Для ряда видов характерен морфизм окраски.
Лапы и клюв темные, от буровато- или синевато-черного до черного цвета. Половой ди-
морфизм окраски отсутствует, однако самки обычно несколько крупнее самцов. В
году две линьки: неполная предбрачная и полная послегнездовая.
В экологическом плане поморники обитатели открытых пространств тундры, мор-
ских побережий и островов высоких и низких широт. Они прекрасно летают и хорошо
плавают, крупные виды быстро бегают, а более мелкие из-за коротких лап передвига-
ются по земле с некоторым трудом. Питание смешанное, с отчетливым преобладанием
в рационе животных кормов. Моногамы. Гнезда примитивного строения, располагаются
на земле. В кладке, как правило, два яйца, окраска птенцов однородная, темно-бурая.
В семействе 7 видов, относящихся к единственному роду Stercorarius, из них 4 вида
(S. skua, S. pomarinus, S. parasiticus, S. longicaudus) характерны только для северного по-
лушария и 3 вида (S. maccormicki, S. antarcticus, S. chilensis) — для южного. Крупные ви-
ды, гнездящиеся в южном полушарии, нередко объединяют с S. skua и относят к само-
стоятельному роду Catharacta, но правильнее рассматривать их в качестве подрода.
В фауне СССР зарегистрировано три гнездящихся и два залетных вида.
10
Большой поморник
Род Stercorarius Brisson, 1760
Большой поморник — Stercorarius skua
(Brunnich, 1764)
Great Skua (англ.), Grosse Raubmowe (нем.), Grand Labbe (фр.).
Статус. Залетный вид.
Общая характеристика и полевые признаки. Самый крупный из палеарктических
поморников. Общая длина до 66 см, размах крыльев 145—155 см. Сложение плотное.
По сравнению с другими палеарктическими поморниками клюв и ноги более длинные,
крылья шире и менее острые. Хвост несколько клиновидный, центральная пара руле-
вых почти не выдается за его обрез, особенно у молодых птиц. Окраска в целом темно-
коричневая, на крыльях в основании первостепенных маховых хорошо заметные белые
пятна, которые, однако, видны только у летящей птицы. Размерами и внешним обли-
ком в полете несколько напоминает неполовозрелых серебристых чаек (рис. 1), но
Рис. 1. Поморники в полете (по: Lofgren, 1984, с изменениями)
1 — длиннохвостый, 2 — короткохвостый, 3 — средний, 4 — большой
отличается от них более темной окраской, белыми пятнами на крыльях, тяжелым сло-
жением при относительно более коротких крыльях и хвосте. От среднего поморника,
к которому иногда бывает близок по размерам, отличается более длинным клювом,
широкими, слегка закругленными крыльями и коротким хвостом (а также белыми
пятнами на крыльях), однако на значительном расстоянии и в плохую погоду эти приз-
наки улавливаются с трудом, что делает определение ненадежным.
Спокойный полет большого поморника напоминает полет крупных чаек с глубо-
кими взмахами крыльев, но в ветреную погоду и при преследовании добычи он стано-
11
Семейство Поморниковые
вится, как и у более мелких поморников, очень маневренным, с резкими подъемами
и спусками и крутыми виражами. По земле в отличие от других поморников нашей
фауны большой поморник бегает довольно быстро. Во внегнездовое время держится
поодиночке нли небольшими группами по 3—4 птицы, в основном в море или вблизи
островов. Во время кочевок и миграций молчалив, хотя на местах гнездования от-
носится к крикливым птицам. Голос, как и у других поморников, очень разнообра-
зен. У взрослых птиц различают более 13 различных звуковых сигналов, выполняющих
самостоятельные функции, и более 3 у птенцов (Дементьев, 1951; Heinzel et al., 1974;
Glutz v. Blotzheim, Bauer, 1982; Cramp, Simmons, 1983; Nat. Geogr.Soc., 1983).
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Систематические взаимоотношения в подроде Catharacta
весьма сложны и недостаточно ясны, поэтому взгляды систематиков на таксономичес-
кую структуру подрода до настоящего времени остаются разноречивыми. Часть иссле-
дователей рассматривает его как единый вид с 5 (Дементьев, 1951) или 6 подвидами
(Степанян, 1975), другие выделяют 2 самостоятельных вида (S. skua с 6 подвидами и
S. maccormicki) (Gruson, 1976). Третьи насчитывают в составе подрода 3 самостоятель-
ных вида, выделяя, помимо уже названных, еще S. chilensis (Devillers, 1977, 1978; Cramp,
Simmons, 1983; Howard, Moor, 1984). Наконец, есть последователи более дробного де-
ления, которые рассматривают подрод в составе 4 самостоятельных видов (S. skua,
S. maccormicki, S. chilensis и S. antarcticus с 3 подвидами) (Brooke, 1978; Harrison, 1983).
В коллекциях Советского Союза нет материалов, достаточных для критического рас-
смотрения упомянутых выше концепций, однако сравнительный анализ литературных
данных по географическому распространению, окраске, гибридизации и этологии пред-
ставителей подрода Catharacta позволяет провизорно принять точку зрения Р.К. Бруке
(Brooke, 1978), поддерживаемую также П. Хэррисоном (Harrison, 1983), и рассматри-
вать S. skua как монотипический палеарктический вид.
Описание. Размеры (мм) и масса тела (г) (Cramp, Simmons, 1983):
Длина крыла:
<Jd - 382-414 (среднее 399);
99 — 398—428 (среднее 413).
Длина клюва:
dd — 47—52 (среднее 50,1);
99 —49—53 (среднее 51,2).
Длина цевки:
dd —64—70 (среднее 67,1);
99 — 66—72 (среднее 68,7).
Масса тела:
dd - 1210-1410 (среднее 1338);
99 — 1390—1630 (среднее 1525).
Окраска. Самец и самка в брачном наряде. Морфизма окраски нет. Лоб и темя
темно-коричневые (’’шапочка”), верхняя сторона тела несколько светлее; перья на
верху головы с узкими красновато-коричневыми наствольями, а начиная с передней
части спины с широкими красновато-коричневыми пятнами на концах. Бока шеи и
затылок более светлые из-за ланцетовидных перьев с контрастными соломенно-жел-
тыми наствольями, образующих нечто вроде ’’гривы”. Горло и передняя сторона шеи
12
Большой поморник
темно-бурые с размытыми красно- или желто-коричневыми наствольями. Остальная
нижняя сторона тела бледно-бурая с более или менее выраженной красновато-коричне-
вой волнистостью. Перья на боках и подхвостье с коричневатыми или красноватыми
наствольями. Рулевые черно-бурые, маховые черно-бурые с белыми полями у основа-
ния, которые у второстепенных маховых закрыты кроющими крыла, а у пяти наруж-
ных первостепенных маховых выступают из-под кроющих на 30—60 мм? Кроющие кры-
ла и кисти буровато-черные со светлыми красноватыми или коричневатыми оконча-
ниями. Подмышечные и нижние кроющие крыла темно-коричневые с красновато-ко-
ричневыми окончаниями. Лапы черные, иногда со свинцовым оттенком. Радужина тем-
но-коричневая. Клюв черный, у основания слегка оливковый или синеватый.
Самец и самка в зимнем наряде. Зимний наряд похож на брачный, но в целом более
однороден, особенно на голове и на шее, где ланцетовидные перья с соломенно-желты-
ми наствольями сменяются на закругленные и одноцветные.
Пуховой птенец. Верхняя сторона тела одноцветная, красновато-коричневая с шел-
ковистым блеском, нижняя сторона несколько светлее, середина брюшка буровато-
серая. Клюв свинцово-серый, лапы синеватые, радужина черно-коричневая.
Гнездовой наряд. Голова черновато-серая, более темная сверху и светлеющая к гор-
лу. Шея и мантия от черновато-серого до серо-коричневого цвета, на каждом пере блед-
но-желтоватое U-образное предвершинное пятно, размеры которого варьируют. Крою-
щие крыла и верхние кроющие хвоста тускло-черные, обычно с такими же пятнами,
как на перьях мантии. Вся нижняя сторона тела светло-коричневая, часто с легким
розоватым или сиренево-серым оттенком, более однородная и менее испещренная, чем
во взрослом наряде. Горло темно-серое. Рулевые и маховые, как у взрослых, но белое
поле на первостепенных маховых меньше и менее контрастно. Первый предбрачный
наряд (весна и лето второго года жизни). Похож на зимний взрослый наряд, но белое
поле на первостепенных маховых меньше (как в гнездовом наряде). Голова и шея
сравнительно одноцветные, черновато-коричневые, часть перьев со светло-коричневы-
ми окончаниями, все перья мантии с рыжеватыми вершинными пятнами. Второй пред-
брачный наряд (старше двух лет). Как взрослый наряд, но оперение сменено лишь
частично, и в нем наряду со свежим пером присутствует много обношенных перьев.
У неполовозрелых птиц на лапах сохраняются синеватые пятна.
Линька. Формирование окончательного наряда происходит на 4—5-м году жизни.
Пуховой наряд образован длинными, мягкими, шерстеподобными пушинками с утон-
ченными шелковистыми окончаниями. Гнездовый наряд сменяется первым промежу-
точным, линька полная, начинается в декабре с оперения головы и нижней части тела,
завершается к середине марта — апрелю. Оперение крыла (включая маховые), спины
и хвоста начинает сменяться в марте-апреле, и смена длится до конца июля — середины
августа. Последующие линьки промежуточных нарядов в основном повторяют схему,
характерную для линьки взрослых птиц, однако смена внутренних маховых и оперения
головы и тела начинается иногда уже в мае.
У взрослых птиц в году две линьки — полная послегнездовая и неполная предбрач-
ная. Во время послегнездовой линьки первостепенные маховые сменяются центробеж-
но, последовательно. Начало линьки маховых с августа по первую декаду сентября,
окончание в феврале-марте. Остальное оперение крыльев, голова, тело и хвост обычно
линяют в эти же сроки, но иногда на месяц раньше. Линька птиц, потерявших кладки
или незагнездившихся, нередко растягивается с июля до января. Предбрачная линька
начинается сразу после послегнездовой и заканчивается по возвращении на места гнез-
дования. Детали линьки неизвестны. Во всяком случае, она захватывает голову и шею,
иногда нижнюю часть тела и мантию, но не крылья и хвост (Fjeldsa, 1977; Cramp, Sim-
mons, 1983).
13
Семейство Поморииковые
Распространение. Гнездовой ареал. Исландия, Фарерские, Шетландские,
Оркнейские и другие острова и северное побережье Шотландии, о-в Медвежий, Шпиц-
берген, некоторые мелкие острова у северного побережья Норвегии (рис. 2). В СССР
не гнездится, но зарегистрированы залеты на Мурман, южнее о-ва Колгуева и к запад-
ным берегам Новой Земли, в 1949 и 1950 гг. — на Айновы острова, в 1967, 1977 и
1983 гг. — в Латвию, в 1979 г. — в Литву (Pleske, 1928; Дементьев, 1951; Белопольс-
кий, 1957; Cramp et al., 1974; Sharrock, 1976; Harrison, 1982; Синкявичюс, Палтанави-
чюс, 1982; Виксне, 1983; Skuja, 1984).
Зимовки. Северная половина Атлантики, ограниченная южной оконечностью
Гренландии, западными побережьями Европы и Африки (до Гвинейского залива) и
восточными’берегами Северной и Южной Америки (до Бразилии) (Harrison, 1983).
Миграции. Большой поморник связан с сушей только в период гнездования, и по
сути дела вся его жизнь вне этого периода, а также во время, предшествующее наступле-
нию половой зрелости, проходит в миграциях в пределах указанной выше области
зимовок. Установлено, однако, что у разных возрастных групп характер размещения
различен. После подъема на крыло в августе молодые птицы из Шотландии в течение
двух первых лет жизни частично разлетаются исключительно широко к югу, тогда как
часть нх остается в районах гнездования до декабря, а иногда залетает во внутренние
континентальные районы (известны залеты в Польшу, Чехословакию, Венгрию,
Австрию и другие страны Европы). Видимо, в этой возрастной группе существуют се-
зонные перемещения с юга на север в летние месяцы и в обратном направлении зимой.
На третьем году жизни дисперсность размещения достигает максимума, причем прояв-
ляется тенденция к летнему проникновению в районы гнездования или лежащие к севе-
ру от них (Гренландия). Летом четвертого года жизни наблюдается еще большая кон-
центрация в северных районах и близ мест гнездования, хотя часть птиц в этом процессе
не участвует. Зимой все большие поморники этой возрастной группы скапливаются в
водах вдоль побережий Пиренейского полуострова и северо-западной части Африки.
С пятого года жизни тенденция к проникновению в северные воды исчезает и птицы,
достигшие половой зрелости, еще более отчетливо собираются к весне в конкретных
районах гнездования. Внегнездовой период, т.е. с сентября по апрель, они проводят ли-
бо в водах, окружающих места гнездования, где в это время нет молодых птиц, либо
спускаются несколько к югу в восточную Атлантику и к водам Пиренейского полу-
острова, возвращаясь весной на места гнездования. Птицы с Исландии в значительно
большей степени тяготеют к западной Атлантике и прибрежным водам Канады и США,
присутствуя, в частности,на Ньюфаундлендской банке в течение круглого года (Cramps,
Simmons, 1983).
Численность. В Исландии численность в настоящее время относительно стабилизи-
ровалась, здесь насчитывают около 6 тыс. гнездящихся пар, и на Фарерских островах
учтено около 500 пар, но численность в последние годы сокращается. На севере Велико-
британии (Шетландские и Оркнейские острова) численность быстро растет и к 1977 г.
достигла примерно 6 тыс. гнездящихся пар (по сравнению с 3172 парами, учтенными
в 1969-1970 гг.).
Местообитания. В гнездовое время населяет открытые равнинные или всхолмленные,
лишенные древесной и кустарниковой растительности участки, покрытые заболочен-
ной мохово-осоковой тундрой, в основном в прибрежной зоне, часто прилегающие к
скальным обнажениям или окруженные ими. В Исландии заселяет обширные галечные
пляжи. В негнездовой период держится в пелагических или прибрежных водах и избе-
гает суши.
Размножение. Половой зрелости большие поморники достигают в возрасте 5, чаще
6—7 и даже 9 лет. Моногамы, пары сохраняются на протяжении всей жизни, хотя в пе-
14
Рис. 2. Область распространения большого поморника (7—5) н южно-полярного поморника (4—6)
1, 4 — область гнездования; 2, 5 — область кочевок; 3,6 - залеты на территорию СССР
f
Семейство Поморниковые
риод кочевок и миграций птицы живут порознь. Известны брачные союзы из трех птиц
(самец и 2 самки). В этих случаях обе самки откладывают яйца (обычно 3) в одно
гнездо и насиживают их совместно. В зависимости от условий гнездятся как одиноч-
ными парами (реже), так и в колониях, которые достигают значительных размеров
(до многих сотен пар). Расстояние между гнездами составляет 25—130 м, иногда до
10 м. В крупных колониях плотность поселения выше. Индивидуальные гнездовые
территории очень постоянны и занимаются из года в год. Площадь территорий от
0,25 до 1,36 га. Территория поддерживается самцом, который возвращается на нее
первым и ожидает самку. В тех случаях, когда самка не появляется, самец образует
другую пару на той же гнездовой территории.
На места гнездования взрослые большие поморники прибывают в начале-середине
апреля, и к 20 апреля все гнездовые территории обычно заняты. Откладка яиц в Ис-
ландии начинается в середине мая, реже в конце месяца или даже в начале июня, на
Шетландских островах — в конце апреля, разгар откладки — во второй декаде мая.
Образование новых пар происходит вне территории гнездовых колоний, на так на-
зываемых клубах — местах, где собираются неполовозрелые птицы. Оно сопровожда-
ется сложными демонстрациями (рис. 3), куда входят элементы агрессии, а на более
поздних стадиях образования пары — ’’выпрашивание” и ’’имитация птенца”. Предко-
пуляционное поведение включает ’’вертикальную позу”, долгий крик, ’’выпрашивание”,
ритуализированное кормление, ’’копуляционный крик”, особый ’’танец” и другие эле-
менты. Копуляция происходит на занятой гнездовой территории, которую птицы охра-
няют от соседей.
Гнезда располагаются на приподнятых местах, более сухих и с более развитым расти-
тельным покровом. Гнездо примитивной конструкции и изначально состоит из ямки
диаметром 23—30 см и глубиной 4,5—8,0 см, которую делают обе птицы, выскребая
грунт лапами и выравнивая вращательными движениями груди. В процессе насижива-
ния в ямку попадают кусочки лишайников, мха, сухие листочки, так что образуется
более или менее выраженная выстилка. Кладка состоит из двух (реже одного) яиц.
Кладки в три яйца, по-видимому, всегда отложены двумя самками. В году нормально
одна кладка, однако при утере ее птицы откладывают повторную. Интервал между
откладкой обычно 2-3 дня, в среднем 2,6 сут. Яйца имеют эллипсоидную, нормально-
яйцевидную форму, скорлупа свежих яиц матовая, в процессе насиживания приобре-
тает блеск; окраска основного фона коричневатая, оливковая или серо-зеленоватая,
изредка голубая, наружные пятна коричневые, обычно концентрируются на широком
конце, иногда встречаются яйца без пятен. Размеры (мм) (и = 1547) : 61-82 X 46-55,
среднее 71 X 50. Масса 88—93 г. Насиживание начинается с первого яйца, длительность
инкубации 26-32, в среднем 29 дней. Насиживают кладку оба родителя, однако глав-
ным образом самка. Вылупление птенцов несинхронное, интервал между вылуплением
в одной кладке составляет около 36 ч. Вскоре после вылупления птенцы покидают
гнездо и держатся поодиночке в пределах гнездовой территории. Самка в основном ох-
раняет их, а самец приносит корм, хотя кормят птенцов оба родителя, отрыгивая пищу.
В возрасте 40-50 дней птенцы поднимаются на крыло, после чего вскоре переходят
к самостоятельной жизни. У гнезда и птенцов взрослые птицы чрезвычайно агрессив-
ны и нападают на всех появляющихся животных и человека.
Суточная активность, поведение. Большие поморники активны в светлое время
суток. На Шетландских островах первые вылеты в поисках корма отмечены между
3 ч 30 мин и 5 ч 42 мин и заканчивались в интервале между 22 ч 19 мин и 23 ч 30 мин.
На фарерских островах, лежащих на 2° севернее, период ночного покоя длится 2,5—
4,0 ч. Ночные часы птицы проводят в пределах гнездовых территорий, где спят в от-
личие от чаек лежа. Больше, чем другие виды поморников северного полушария, прово-
дят времени на воде. Часто купаются в пресноводных озерках, особенно в середине дня.
16
Большой поморник
Во внегнездовый сезон держатся обычно поодиночке, иногда в группах по 2—3 пти-
цы, реже образуют небольшие стайки, айв отдельных случаях даже заметные скоп-
ления у рыболовных баз или флотилий.
Питание. Основу пищи больших поморников составляет рыба, которую они само-
стоятельно добывают в море или отнимают у птиц других видов. Рыбу хватают с по-
верхности или слегка погружаясь в воду, но не ныряя. Помимо рыбы, поедают яйца
и охотятся на птенцов и даже взрослых морских птиц, кормятся отбросами зверобой-
Рис. 3. Демонстрационное поведение большого пцмориика (по: Cramp, Simmons, 1983,
с изменениями)
1 — долгий крик, 2 — агрессивная поза
ного промысла, трупами морских животных. При отнимании добычи у других видов
птиц обычно атакуют жертву вдвоем и преследуют ее, пока она не отрыгнет пищу. Таким
образом большие поморники отбирают добычу у чистиковых птиц, крачек, моевок
и олуш, нередко даже атакуют птиц своего вида. Большие поморники — сильные и лов-
кие хищники. Поодиночке или вдвоем они легко ловят и убивают молодых и взрослых
птиц как в полете, так и на воде, где часто и поедают добычу. Нередки случаи канни-
бализма, в том числе и поедание своих же птенцов. Среди жертв больших поморников
известны взрослые тупики, моевки, серебристые чайки, чистики, обыкновенные гаги,
многие виды куликов, включая кулика-сороку и большого кроншнепа, хохлатые
бакланы, короткохвостые поморники и многие другие виды птиц. Кроме того, обыч-
ной добычей на островах являются дикие кролики и даже зайцы-беляки. Интересно,
что разные особи применяют различные способы охоты. Например, тупиков большие
поморники ловят в воздухе, на воде, подкарауливают у входов в норы или даже вытас-
кивают из нор, проникая туда достаточно глубоко. Птенцов большие поморники кор-
мят в основном рыбой, но нередко приносят мелких грызунов и птиц, реже беспозво-
ночных (типулид). Во внегнездовое время основу пищи также составляет рыба, морс-
кие пелагические беспозвоночные, отбросы зверобойного промысла и рыболов-
ства.
Враги, неблагоприятные факторы. Взрослые большие поморники практически не
имеют естественных врагов. Однако успешность размножения вида в целом исключи-
тельно низка: около 15,6% яиц гибнет от хищников, а из оставшихся неоплодотворен-
ные яйца составляют 12%, погибшие эмбрионы 6%, погибшие при вылуплении птенцы
3,2% и птенцы с тератологическими признаками 0,4%. Предполагается, что это является
следствием химического загрязнения среды, поскольку в яйцах и тканях взрослых
птиц обнаружены в высоких концентрациях тяжелые металлы (кадмий, ртуть). Смерт-
ность в первые 6 мес. жизни составляет у больших поморников 16%, в последующие
17
Семейство Поморииковые
12 мес. падает до 8% и дня взрослых птиц колеблется около 7%. Известный возраст-
ной предел в природе 16 лет 2 мес.
Хозяйственное значение, охрана. В местах гнездования как очевидный ’’хищник”
в прошлом подвергался официальному преследованию, однако сейчас оно прекращено.
В СССР как редкий залетный вид никакого хозяйственного значения не имеет и мер
по его охране не предусмотрено.
Южно-полярный поморник —
Stercorarius maccormicki Saunders, 1893
South Polar Skua (англ.), MacCormick Raubmowe (нем.), Skua Antarctique (фр.).
Статус. Залетный вид.
Общая характеристика и полевые признаки. Несколько мельче большого поморни-
ка, общая длина до 53 см, размах крыльев 127—132 см. Сложением, формой крыльев
и хвоста очень похож на большого поморника, но клюв кажется более тонким и корот-
ким. Окраска трех типов — темного, светлого и промежуточного. Взрослые птицы тем-
ного типа однотонные, черновато-бурые, с заметной темной ’’шапочкой”, но без крас-
новатых и рыжеватых тонов и без светлых желтоватых пестрин на верхней стороне
тела. У птиц светлого типа окраски верх однотонно бурый, нижняя сторона тела и го-
лова грязно-серые, беловатые, ’’шапочки” нет. Птицы промежуточного типа окраски
напоминают птиц светлого типа, но низ тела желто-коричневатый, темя несколько
темнее. Неполовозрелые птицы окрашены, как взрослые, но голова и низ серее, верх-
няя сторона тела с заметными желтоватыми пестринами. В основании первостепенных
маховых у птиц всех типов окраски контрастные белые пятна, как у большого помор-
ника. Поведением, характером полета и голосом практически не отличается от большо-
го поморника.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. В большинстве отечественных работ S. maccorcmicki
рассматривается как подвид большого поморника (Дементьев, 1951; Нечаев, 1969;
Степанян, 1975; и др.). Однако в подавляющем большинстве современных зарубежных
и некоторых новейших отечественных сводках (Белопольский, Шунтов, 1980) южно-
полярный поморник имеет статус самостоятельного вида. Мы придерживаемся этой же
точки зрения (см. предыдущий вид).
Описание. Строение и размеры. Размеры (мм): длина крыла (оба пола)
390—410, среднее 397,5; хвоста 140—152, среднее 147,4; цевки 62,4— 65,6, среднее
64,1. Масса тела (г) : 1224 (среднее из 4, пол неизвестен (Murphy, 1936). Добытый в
территориальных водах СССР южно-полярный поморник (самец в возрасте 1 года)
имел следующие размеры: длина крыла 410, хвоста 145, клюва 50, цевки 60 мм
(Слепцов,1963).
Окраска. Самец и самка в брачном наряде. Окраска полиморфная. Темная
морфа: окраска сравнительно однотонная; голова в 'основном темно-бурая с чернова-
той ’’шапочкой” и более светлым зашейком. У основания клюва, особенно у нижней
челюсти, беловатое поле. Верхняя сторона тела однотонная черновато-бурая, низ тела
несколько светлее, темно-бурый. Крылья сверху черно-бурые, основания маховых бе-
ловатые, на первостепенных белое поле больше, чем на второстепенных. Хвост чернова-
тый. Светлая морфа: окраска контрастная; голова обычно от бледного розовато-буро-
го до серовато-белого цвета с более светлым зашейком, смотрящимся как незамкну-
тый светлый ошейник. На темени и боках шеи золотистые пестрины, менее выражен-
ные, чем у большого поморника. Верхняя сторона тела однотонная буровато-черная
с небольшим количеством светлых пестрин. Нижняя сторона тела от светлого розовато-
18
Южно-полярный поморник
бурого до серовато- или грязно-белого цвета. Крылья и хвост, как у темной морфы.
Промежуточная морфа: похожа на светлую морфу, но голова, задняя сторона шеи и
нижняя сторона тела соломенно- или темно-желтые. Встречаются экземпляры, переход-
ные от светлой к темной морфам. Лапы и клюв черновато-серые или черные, радужина
темно-коричневая.
Самец и самка в зимнем наряде. Зимний наряд похож на брачный, но голова и ниж-
няя сторона обычно желтее и серее.
Пуховой птенец. По структуре пухового наряда похож на пухового птенца большого
поморника, но значительно светлее, верхняя сторона тела бледно-желтая или голубовато-
серая. Гнездовой наряд. Нет данных. Промежуточные наряды. Темная морфа: в целом,
как зимний наряд взрослых птиц, но верх тела более пестрый из-за соломенно-желтых
окончаний перьев спины и кроющих крыла, на нижней стороне тела волнистый рисунок.
Светлая морфа: как взрослый, но верх пестрее (как у темной фазы), нижняя сторона
тела более грязного тона, на нижней стороне шеи и груди узкие продольные пестрины.
У птиц светлой морфы клюв светло-синий с черным кончиком, лапы синие. Относитель-
но возрастных изменений промежуточных нарядов данных нет (Devillers, 1977; Harri-
son, 1983).
Линька. Полная линька происходит в негнездовой период, во время северного лета.
Детали неизвестны, но, вероятно, общая последовательность смены нарядов не отли-
чается от таковой у большого поморника (Devillers, 1977).
Распространение. Гнездовой ареал. Побережья Антарктиды и прилежащие
острова (см. рис. 2). Темная морфа преобладает в северных частях Антарктического
полуострова, светлая во всех районах гнездования на континенте (Harrison, 1983). Вы-
деляются три главных места гнездования: 1. Побережье Земли Грейама с прилежащими
островами (Южные Шетландские, Южные Оркнейские и др.). 2. Побережье и острова
моря Росса. 3. Побережье Земли Королевы Мод (Spellerberg, 1967; Salomonsen, 1976).
Зимовки. Основные места зимовки охватывают Тихий океан к северу до Куриль-
ских островов, Атлантический океан до широты Ньюфаундленда в Северной Америке
и тропика Козерога в Африке, южные части Индийского океана. В СССР добыт 25 ав-
густа 1954 г. в территориальных водах недалеко от мыса Южно-Курильский (море
Немуро). В 1954 г. наблюдался в Южно-Курильском проливе в непосредственной бли-
зости от островов Шикотан и Кунашир. В южной части Охотского моря отмечен у север-
ной оконечности Кунашира и в проливе Лаперуза, а в Японском море - у мыса Моне-
рон (Слепцов, 1963).
Миграции. Разлет с колоний и миграции начинаются в феврале—апреле, взрослые
птицы перемещаются в ограниченных пределах, тогда как неполовозрелые мигрируют
широко по Тихому, Атлантическому и Индийскому океанам. Часть их находится в
состоянии миграции на протяжении круглого года. У берегов Японии большие поморни-
ки проводят май—июль, на севере Тихого океана появляются в июле-августе, у запад-
ных берегов Северной Америки на широте Калифорнии - в сентябре-октябре, на се-
вере Атлантики - в июне—сентябре. Отдельные особи залетают к берегам Западной
Европы и Гренландии (Harrison, 1983).
Численность. В секторе моря Росса на гнездовье отмечено около 2000 пар, на Земле
Адели не более 50 пар, на континентальной части полуострова Земля Грейама, Южных
Шетландских и Южных Оркнейских островах около 700 пар (Croxall et al., 1984). Таким
образом, по неполным данным общая гнездовая численность вида составляет примерно
2750 пар. Вероятно, в три раза большее число составляют неполовозрелые особи, про-
водящие весь год вдали от районов гнездования, и общую численность вида с известны-
ми оговорками можно оценить в 10—15 тыс. особей. Местами численность относительно
стабильна, местами (например, на Земле Адели) в последнее десятилетие увеличива-
ется (Croxall et al., 1984).
19
Семейство Поморниковые
Местообитания. В гнездовое время заселяет ровные участки и пологие склоны пред-
горных шлейфов в прибрежных районах континента и островов, освобождающиеся от
льда и снежного покрова; вне гнездового сезона держится в открытом море.
Размножение. На местах гнездования южнополярные поморники прилетают в конце
октября, и в начале ноября число их быстро растет, по мере того как начинается отклад-
ка яиц в гнездах пингвина Адели (Pygoscelis adeliae). Гнездятся как одиночными пара-
ми, так и в колониях на участках, свободных от льда, обычно на склонах солнечной
(северной) экспозиции, нередко по соседству с колониями пингвинов Адели, но иногда
и вдалеке от них. В более центральных участках континента гнездовья южно-полярных
поморников, как правило, связаны с колониями трубконосых. Гнездовые территории
обычно постоянны и занимаются одной и той же парой на протяжение ряда лет, хотя
место расположения гнезда может меняться. Поддерживается территория преимущест-
венно самцами, которые появляются на местах гнездования раньше самок. В колониях
обычно бывает 10—20 гнезд, но иногда насчитывается до 100—200 гнезд на площади
около 10 га, минимальное расстояние между гнездами 10—20 м, но иногда и меньше,
в среднем 25 м. Гнезда располагаются на относительно ровных местах с каменистым
грунтом и скудной растительностью и представляют собой ямку практически без вся-
кой выстилки.
В демонстрационном поведении важную роль играют позы агрессии. Агрессивное
поведение складывается из тех же элементов, которые известны дня чайковых птиц,
в том числе и для большого поморника. Предкопуляционное поведение включает
агрессивную прямую позу со стороны самца, на которую самка отвечает угрожающей
прямой и сгорбленной позами. Копуляция не происходит до тех пор, пока эти позы не
сменятся отрыгиванием пищи и ритуализированным кормлением со стороны самца
(Young,, 1963а; Spellerberg, 1971).
Откладка яиц начинается в последних числах ноября, массовая откладка падает на
декабрь. В случае гибели первой кладки птицы откладывают повторную. В кладке
2 яйца. Окраска, как у большого поморника. Размеры (мм) (и = 27): 66,7—76,4Х
X 47,1—52,1, среднее 71,3 X 49,6. Интервал между откладкой яиц около 48 часов. Наси-
живают оба родителя, но самка значительно дольше. Длительность инкубации состав-
ляет 27—30 дней. Птенцы вылупляются с интервалом 2,1—2,3 сут. Вылупление в по-
пуляции происходит с первой декады января до середины февраля, в среднем массовое
вылупление падает на 17—19 января. Вылупившиеся птенцы нередко проявляют агрес-
сию по отношению друг друга. Масса тела новорожденного пуховика 60-63 г. На крыло
птенцы поднимаются примерно в возрасте 50 дней. Гнездовая и послегнездовая смерт-
ность южнополярных поморников относительно высока, и успешность гнездования
составляет в среднем 0,4—0,6 птенцов на гнездящуюся пару. В отдельные годы, однако,
эти показатели могут быть как ниже (до 0,13), так и значительно выше (Murphy,
1936; Spellerberg, 1971; Wood, 1971; Young, 1972; Hemmings, 1984).
Суточная активность, поведение. Суточная активность и сезонность социальной
структуры популяции, вероятно, мало отличается от того, что характерно для большо-
го поморника.
Питание. По-видимому, южнополярные поморники в значительно меньшей степени
хищники, нежели другие представители подрода Catharacta (например, S. lonnbergi).
По данным Хеммингса (Hemmings, 1984), практически единственным кормом в гнез-
довое время служит рыба Pleuragramma antarcticum из семейства нототениевых. Этим
же кормом выкармливаются птенцы. Несмотря на то что поблизости от колоний южно-
полярных поморников всегда имеются колонии пингвинов или бакланов (Phalacroco-
rax atriceps), клептопаразитизма, как и прямого хищничества, на этих колониях не от-
мечено (Young, 1963).
20
Средний поморник
Средний поморник -Stercorarius pomarinus
(Temminck, 1815)
Pomarine Skua (англ.), Spateliaubmowe (нем.), Labbe pomarin (фр.).
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Самый крупный из гнездящихся на тер-
ритории СССР поморников, хотя по величине лишь незначительно превосходит коротко-
хвостового поморника (молодые особи почти равны взрослым короткохвостым помор-
никам) . Сложение типично для поморников - сравнительно длинные острые крылья,
удлиненный заостренный хвост, в целом темная окраска (особенно верха). Общая дли-
на около 50 см, размах крыльев 122-127 см. У взрослых птиц средние рулевые далеко
выдаются за обрез хвоста, закруглены и в дистальной части повернуты под прямым
углом, отчего при наблюдении птицы в полете создается впечатление, что они имеют
концевое расширение. У молодых птиц центральные рулевые слабо выступают за обрез
хвоста и не повернуты. В полете на исподе крыла у основания первостепенных махо-
вых хорошо заметно белое поле, хотя оно и не так отчетливо, как у большого помор-
ника. По сравнению с короткохвостым и особенно длиннохвостым поморниками сло-
жение кажется более тяжелым и грубым, крылья более широкие, клюв массивнее.
В целом средние поморники более молчаливы, чем другие наши поморники. Среди
голосовых сигналов различают: 1) ’’короткий крик”, при появлении или нападении
других поморников, чаек или какого-либо четвероногого хищника, звучит как низ-
кое ”гакк” или ”гекк”, напоминая голос ястреба; 2) долгий крик — серия громких
гнусавых звуков, издаваемая при решении территориальных конфликтов или при об-
щении брачных партнеров на гнездовой территории; 3) ’’дрожащий крик” — высокий
тремолирующий звук ”уа-уа-уа-уа...”, издаваемый при нападении на хищника; 4) Крик
тревоги — двухсложное визгливое ”вии-вииф”, обращаемое обычно на человека. Извес-
тен и ряд других голосовых сигналов (Cramp, Simmons, 1983).
Полет, как и у других поморников, сильный, свободный, с глубокими взмахами
крыльев. В полете птицы часто и умело используют восходящие токи воздуха и ветер
и способны искусно маневрировать при почти неподвижных крыльях, легко и быстро
пикируя почти до земли и резко взмывая в воздух на значительную высоту. Такая
система полета чаще всего используется при поисках добычи. Из-за сравнительно корот-
ких лап по земле передвигается мелкими шагами, медленно и неохотно. Во внегнез-
довое время держится парами, тройками или небольшими группами, но нередко в мес-
тах, обильных кормом, собирается в скопления (например, на мусорных свалках у
поселков, в местах забоя северных оленей и т.д.). Голос исключительно разнообразен,
преобладают резкие мяукающие звуки, слабо отличающиеся от криков других помор-
ников нашей фауны.
Подвидовая систематика. Монотипический вид.
Описание. Размеры (мм) (Cramp, Simmons, 1983) и масса тела (г) (Дорогой,
1981):
Длина крыла:
33 — 354-374 (среднее 363);
99 - 363-382 (среднее 373).
Длина клюва:
33 - 38-42 (среднее 39,8);
99 - 39-44 (среднее 40,9).
21
Семейство Поморннковые
Длина цевки:
<3<3 - 50-56 (среднее 53,7);
99 - 53-58 (среднее 55,1).
Масса тела:
<3<3 (и = 18) - 520—700 (среднее 631);
99 (п = 16) — 610-750 (среднее 687).
Окраска. Самец и самка в брачном наряде. Окраска диморфна; соотношение
светлой и темной морфы примерно 20:1. Светлая морфа: на голове буровато-черная
’’шапочка”, остальные части головы и задняя сторона шеи желто-белые. Верхняя сторона
тела черновато-коричневая, нижняя — в основном желтовато-белая, часто с поперечной
темной полосой на груди и темным бурым подхвостьем. Полоса на груди в разгар брач-
ного сезона отсутствует. Крылья сверху черновато-бурые, основания первых маховых
беловатые, снизу крыло такое же, но белые основания маховых более заметны. Хвост
черноватый, боковые рули сероватые. Темная морфа: основная окраска черновато-
бурая, за исключением коричневато-желтых щек и воротника. Белые основания перво-
степенных маховых, как у светлой морфы. Клюв темный коричнево-роговой или свин-
цово-серый, с темным концом, ноги черные.
Самец и самка в зимнем наряде. В целом зимний наряд почти не отличается от брач-
ного, но шапочка на голове несколько серее, особенно это заметно у светлой морфы.
Пуховой птенец. Общая окраска темно-коричневая, переходящая в тускло-серый
бледный цвет на подбородке, вокруг глаз, на концах крыльев и на животе. Основания
пушинок серые, что придает пуху легкий сероватый оттенок. Клюв свинцово-серый
с темным концом, ноги светло-голубые.
Гнездовой наряд (первый зимний). Окраска в гнездовом наряде сильно варьирует
и может быть условно разделена на три типа. Светлая морфа: голова и шея беловатые,
сильно испещренные темными серо-коричневыми крапинками на темени, затылке
и уздечке, переходящими в пестрины на шее и горле. Верхняя сторона тела и крыльев
темная шиферно-бурая, перья с желтоватыми каймами. Нижняя сторона белая с бурым
на боках и подхвостье, на груди и животе коричневые крапинки. Полосатая морфа:
голова и шея от темного серо-бурого до темного шиферно-серого цвета, перья на верху
головы с узкими желтоватыми концами, на задней стороне шеи, боках головы и гор-
ле — с желтоватым окаймлением. Верх тела и верхние кроющие крыла от тускло-
черного до грязно-бурого, перья с желтоватыми окончаниями или окаймлением. Ниж-
няя сторона тела с поперечными серо-бурыми и желтоватыми полосами, особенно
контрастными на животе и подхвостье. Голова часто одноцветно-темная. Темная морфа:
окраска целиком грязно-черная, и только основания первостепенных маховых, как и
у других морф, белые или беловатые. Клюв коричневый, оливково-серый или желто-
ватый с черным кончиком, лапы светло-голубоватые, пальцы и перепонки черные,
радужина темно-коричневая.
Первый полувзрослый весенне-летний (промежуточный) наряд (весна и лето второ-
го календарного года жизни). Светлая и полосатая морфы: похож на гнездовый наряд,
но нижние кроющие крыла однотонные, темные, без желтоватых окончаний. У темной
морфы первый полувзрослый наряд идентичен по окраске гнездовому. Первый полу-
взрослый зимний наряд (вторая зима). Светлая морфа (полосатая морфа переходит
в этот же тип окраски): похожи на взрослых негнездящихся птиц, но нижняя сторона
тела и крыльев с отчетливой полосатостью. Темная морфа: окраска одноцветная свин-
цово-черная, ’’шапочка” на голове слегка темнее оперения спины. Второй полувзрос-
22
ьо
со
160 120 80 40 0 40 80 120 160 160
Рис. 4. Область распространения среднего поморника
1 - гнездовой ареал, 2 — южная граница зимне-весенних кочевок, 3 — южная граница весенне-летних кочевок, 4 — северная граница весенне-
летних кочевок
Семейство Поморннковые
лый летний наряд (лето третьего календарного года жизни). Светлая морфа: ’’шапочка”
грязно-черная, резко контрастирующая со светлыми боками головы, горлом и задней
стороной шеи, которые обычно испещрены грязно-черными крапинками. На боках
головы появляется желтоватый оттенок. Верх тела и крыльев свинцово-черный, перья
мантии с узкими белыми окончаниями. Надхвостье черное с белыми полосами. Нижняя
сторона тела и крыльев с буро-черными и белыми полосами, маховые, как у взрослых.
Темная морфа: как первый полувзрослый зимний наряд. Второй полувзрослый зимний
наряд (третья зима). Светлая морфа: как второй полувзрослый летний наряд, но живот
сравнительно светлее, на боках головы меньше желтого цвета. Темная морфа: как пер-
вый полувзрослый зимний. Третий полувзрослый летний наряд (лето четвертого кален-
дарного года). Светлая морфа: как второй полувзрослый летний наряд, но желтый
цвет на боках головы отчетливее и опятнение слабее, нижняя сторона тела, за исклю-
чением подхвостья, приобретает однородную окраску. Темная морфа: как взрослые
птицы, но еще отсутствуют заостренные перья по бокам головы. Формирование полного
брачного наряда заканчивается на шестом календарном году (Fjeldsa, 1977; Cramp,
Simmons, 1983; Harrison, 1983).
Линька. Чередование летних и зимних полувзрослых (промежуточных) нарядов
продолжается до возраста шести лет, когда птицы одевают окончательный брачный
наряд, сменяющийся, в свою очередь, взрослым зимним. Пуховой наряд птенцов обра-
зован волосовидными мягкими пушинками с утонченными шелковистыми окончани-
ями. Гнездовой наряд сменяется на первый полувзрослый весенне-летний (первый
промежуточный) в результате полной линьки, начинающейся с головы, тела и хвоста
(ноябрь—апрель) и заканчивающейся сменой оперения крыльев (февраль—июль).
Далее (с июля) следует неполная линька, в результате которой формируется первый
полувзрослый зимний наряд (второй промежуточный), который соответственно сме-
няется в процессе полной линьки на второй полувзрослый летний. Эта схема сохраня-
ется до образования окончательного брачного наряда, однако у полувзрослых птиц
четкое разграничение полных и частичных линек несколько нарушено и они нередко
смещаются во времени, налегая отчасти друг на друга.
Линька взрослых птиц более строго связана с сезонами года. Полная послегнездо-
вая линька начинается со смены оперения тела еще в гнездовом ареале в конце августа—
сентябре, у негнездившихся или потерявших кладки птиц с июля, заканчивается на зи-
мовках. Первым сменяется оперение на верху головы, шее, мантии, затем на груди,
боках, надхвостье, после чего последовательно линяют остальные части головы, брюш-
ная сторона, спина, поясница. Нередко линька захватывает большую часть тела одно-
временно, заканчивается в ноябре—декабре. Маховые линяют последовательно с ок-
тября — начала ноября и полностью сменяются к середине мая — началу апреля. Опе-
рение хвоста линяет в октябре—феврале, линька начинается с центральных рулевых.
Верхние и нижние кроющие крыла, оперение спины и поясницы сменяются только
раз в год — в то же время, когда и маховые. Предгнездовая линька взрослых птиц
неполная, захватывает голову, часть оперения тела (кроме спины и поясницы), а также
центральную пару рулевых; протекает обычно с февраля по апрель. Голова, шея, ман-
тия, подхвостье и грудь линяют в последнюю очередь. Завершается линька на мигра-
циях, и в местах гнездования птицы появляются в полностью сформировавшемся
наряде.
Распространение. Гнездовой ареал. Ареал циркумполярный, охватывает
северные побережья и острова северо-восточной части Европы, Азии, Северной Амери-
ки, Гренландию (рис. 4). Гнездовой ареал не стабилен во времени, и наличие гнездя-
щихся популяций определяется запасом кормов (прежде всего высокой численностью
леммингов).
24
Средний поморник
В СССР зона гнездования (рис. 5) охватывает п-ов Канин, Малоземельскую и Боль-
шеземельскую тундры, Югорский полуостров, Ямал к югу до р. Щучьей, Гыданский
полуостров, Таймыр к северу до 74° 30' с.ш., северные части междуречий Хатанги,
Лены, Яны, Индигирки, Колымы, побережья Чаунской и Колючинской губ и Чукот-
ский полуостров, побережье Анадырского залива (Дементьев, 1951). Гнездится на ост-
ровах Новая Земля и Вайгач (Успенский, 1960), на Новосибирских островах (Pleske,
1928), на о-ве Врангеля (Портенко, 1973; и др.), на о-ве Беринга (Иогансен, 1934).
Отсутствует на гнездовье на Кольском полуострове, не гнездится в Корякском на-
горье (Кищинский, 1980), на Курильских островах (Нечаев, 1969) и на Камчатке
(Лобков, 1978), хотя нередок в этих районах на пролете и кочевках.
Вне СССР гнездится на Аляске, в ряде районов Канадской Арктики, на западном
побережье Гренландии. На Шпицбергене, в Исландии и на других островах Северной
Атлантики не гнездится, хотя и отмечен в гнездовое время (Cramp, Simmons, 1983).
Зимовки. Средний поморник зимует как в открытом море, так и вдоль побе-
режий материков. Основные места зимовки — северная Атлантика, северная Пацифи-
ка, отчасти Северный Ледовитый океан, южнее зимует у восточных побережий Север-
ной Америки и в Карибском море, вдоль западного побережья Африки, в Индийском
океане у побережий Аравийского полуострова, у берегов Австралии, о-ва Новая Гви-
нея и Индонезии, у берегов Японии. Вместе с тем часть птиц зимует дисперсно, вероят-
но, по всем океанам (Harrison, 1983).
Миграции. Основных миграционных путей в СССР, по-видимому, два. Часть птиц
осенью движется вдоль морских побережий на запад, в воды Атлантики, другая часть
с восточных участков гнездового ареала — в восточном направлении, а затем к югу,
в воды Тихого океана, к побережьям Австралии и в восточные части Индийского оке-
ана (Дементьев, 1951). Птицы с Аляски и из канадских тундр огибают с обеих сторон
североамериканский континент и летят вдоль западного побережья в южноамерикан-
ские воды, а вдоль восточного побережья - в Карибское море (Harrison, 1983). На Ба-
ренцевом море миграция средних поморников проходит вдали от побережий и лишь
25
Семейство Поморннковые
весной часть птиц подходит на 3—5 км к материку, образуя большие скопления (стаи
до 600—750 особей) в местах концентрации рыбы. Общее количество мигрантов очень
значительно: имеются указания, что при определенных обстоятельствах (подход льдов)
16 мая 1966 г. у Лицких островов на маршруте в 10 км зарегистрировано 5 тыс. птиц,
отдыхающих на воде, и кроме того за 2 ч 20 мин отмечено 7490 особей, мигрирующих
с запада (Коханов, 1970).
Незначительная часть птиц с северо-европейских и азиатских тундровых побережий
летят через материк на юг, этим объясняется значительное число залетов во внутрен-
ние моря Советского Союза и встречи средних поморников вообще вдали от крупных
водоемов. Обзор таких залетов за первую половину XX в. сделан Г.П. Дементь-
евым (1951). В последние десятилетия накопился дополнительный материал, под-
тверждающий регулярность появления средних поморников в центральных и южных
областях европейской и азиатской частей СССР. Такие залеты зарегистрированы
в 1957-1976 гг. в Крыму (Костин, 1983), в 1966 г. в Армении (Айрумян и др., 1968),
на северо-западе Прикаспия в Калмыкии в 1974 г. (Савицкий, Пославский, 1977),
в 1973 г. на Каспийском побережье в Туркмении (Щербина, 1977), в окрестностях
Иркутска в 1952 г. (Гагина, 1962), в Тувинской котловине в 1961 г. (Берман, Забелин,
1963) и в ряде других мест.
Мигрируют средние поморники в одиночку или небольшими группами по 4—5 птиц,
реже стаями до нескольких десятков. Однако, как показано выше, бывают и исклю-
чения. Нередко поморники на пролетных путях образуют довольно крупные скопле-
ния в местах, богатых кормом (в местах забоя морского зверя, у рыболовных флоти-
лий и пр.).
Численность. Средний поморник относится к обычным птицам, хотя, по-видимому,
заметно уступает по численности короткохвостому и длиннохвостому поморникам.
Вместе с тем размещение популяции среднего поморника исключительно лабильно
и целиком определяется наличием основного корма — мелких млекопитающих. Поэто-
му в одних и тех же местах вид может быть зарегистрирован и как обычный, и как
крайне редкий, что создает картину резких колебаний численности (чего в действи-
тельности нет). Так, в 1975 г. в период нарастания численности леммингов на о-ве Вран-
геля на двухсоткилометровом маршруте в начале августа встречена одна неразмно-
жающаяся пара средних поморников, тогда как в 1976 г., в пик численности леммин-
гов, плотность поселения этих птиц составляла 1, 2 гнездящихся пар на 1 км2 (Доро-
гой, 1981), а на Аляске даже 6-8 пар на 1 км2 (Pitelka et al., 1955). Определить общую
численность средних поморников на территории СССР не представляется возможным.
Местообитания. В гнездовый период средний поморник населяет типичные и аркти-
ческие тундры с равнинным или слабохолмистым рельефом, в большей или меньшей
степени заболоченные. В годы депрессии леммингов прослеживается связь с долинами
крупных рек и морскими побережьями, тогда как в периоды подъема численности
грызунов большинство птиц держится на более возвышенных и сухих участках тундры.
В негнездовой период средние поморники ведут преимущественно пелагический образ
жизни и лишь в небольшой мере связаны с побережьями.
Размножение. Возраст достижения половой зрелости точно не известен - по-видимо-
му, не менее 5 лет, причем такие птицы размножаются только при пике численности
леммингов и не каждый год. Моногамы, однако длительность брачного союза не из-
вестна. На местах гнездования часть средних поморников появляется уже в парах.
На всей территории гнездового ареала весенний прилет протекает примерно в одни
и те же сроки и лишь в годы с особенно поздней весной запаздывает на несколько дней.
Как правило, основная масса птиц появляется между 25 и 30 мая, концентрируясь в
местах, богатых кормом (обычно у поселков), а к 7—10 июня уже рассредоточивается
26
Средний поморник
в тундре по гнездовым участкам. Негнездящиеся особи некоторое время находятся
там же, а к концу июня постепенно исчезают, хотя часть их нередко держится в потен-
циально пригодных для гнездования местах до отлета в середине августа - - сентябре.
Гнездовые стации среднего поморника довольно разнообразны. На о-ве Врангеля
большинство птиц гнездится в равнинных среднеувлажненных дриадово-мохово-разно-
травных тундрах и лишь 15% птиц предпочитают щебнистые, слегка увлажненные мохо-
во-лишайниково-дриадовые тундры по склонам холмов, причем гольцовый пояс и силь-
ноувлажненные понижения явно избегаются (Дорогой, 1981). В районе Колючинской
губы гнездование средних поморников отмечено на сухих участках кочкарниковой
тундры (Кречмар и др., 1978). Вместе с тем в районе Уэллена на Восточной Чукотке и на
побережье Хромской губы в Северной Якутии местом массового гнездования были
обширные заболоченные низины, занятые мохово-осоковой тундрой (Успенский
и др., 1962; Томкович, Сорокин, 1983). Примерно такая же разнообразная картина
наблюдается и на Аляске (Pitelka et al., 1955; Maher, 1974).
Средние поморники относятся к числу строго территориальных птиц, причем в от-
личие от короткохвостого и длиннохвостого поморников активно защищают гнездо-
вую территорию от вторжения как особей своего вида, так и других поморников. Раз-
меры гнездовой территории в целом определяются величиной кормовых запасов и мо-
гут варьировать в очень широких пределах. Так, на Аляске в годы массового размноже-
ния леммингов на 1 км2 гнездится 6-8 пар и размеры гнездовой территории составля-
ют 390—425 м в диаметре, тогда как при более низкой численности грызунов на 1 км2
гнездится 0,07—1,5 пар (при еще более низкой численности леммингов средние помор-
ники к гнездованию вообще не приступают). На о-ве Врангеля в годы высокой числен-
ности леммингов площадь гнездовой территории составляет около 1 км2, а расстояние
между гнездами не превышает 800—900 м. На побережье Хромской губы расстояние
между гнездами еще меньше — 400-500 м (Успенский и др., 1962; Дорогой, 1983;
Pitelka et al., 1955; Maher, 1974).
Территориальное поведение начинает демонстрироваться за 1—2 недели до откладки
яиц. Охрана гнездовой территории осуществляется двумя принципиально различными
путями. В одних случаях птицы патрулируют границы территории в особом полете,
включающем редкие, как бы замедленные, глубокие взмахи крыльев и короткие
отрезки парения с V-образно поставленными крыльями. При нарушении границы сле-
дует прямое нападение на нарушителя. Второй путь охраны — демонстрация занятости
территории, когда владелец территории постоянно находится на каком-нибудь воз-
вышенном месте и при появлении нарушителя принимает наклонную позу с подняты-
ми крыльями, сопровождая ее долгим криком. В охране территории принимают участие
оба члена пары. В общей схеме демонстрационных поз и акустических сигналов просле-
живается значительное сходство с большим поморником (Andersson, 1973).
Гнезда средних поморников располагаются на относительно более сухих мохово-
лишайниковых бугорках, плоских кочках и гривках и представляют собой лишь слабо
выраженное углубление со скудной выстилкой из стеблей сухих трав, листочков ивы
или лишайников (Thamnolia vermicularis). Диаметр гнезда 200-270, глубина до 50-
70 мм. В постройке гнезда участвуют оба члена пары. Период откладки яиц растянут
(до двух недель и более), начало его зависит от местных погодных условий. В годы
с ранней и дружной весной откладка яиц начинается в первую пятидневку июня, а раз-
гар падает на середину месяца, тогда как в холодные поздние весны начало откладки
сдвигается на середину июня, а массовая откладка наблюдается в третьей декаде этого
месяца. Яйца откладываются с интервалом около 48 ч. В кладке, как правило, 2 яйца,
значительно реже 1 (для Аляски кладки с 1 яйцом составляли 5%, для о-ва Вранге-
ля 2%) или как исключение 3 яйца (за последние десятилетия таких кладок не найде-
27
Семейство Поморииковые
но). В годы с холодной и затяжной весной кладки с 1 яйцом встречались чаще. За сезон
в норме одна кладка, но при утрате ее в начале насиживания возможна вторая, как
правило, в 1 яйцо. Яйца обычно нормально-яйцевидной, реже укороченно-яйцевидной
формы с гладкой, слегка блестящей скорлупой. Основной фон окраски скорлупы
оливково-коричневый, иногда с зеленоватым оттенком, поверхностные пятна темно-
коричневые, ржавчато-бурые или даже черноватые, разбросаны беспорядочно, чаще
группируются на тупом конце. Размеры (мм): с о-ва Врангеля (п = 11) 60,8—
66,2 X 40,9—45,5, среднее 63,2 X 43,5 (Дорогой, 1981), с п-ова Таймыр (и = 7) 58,5—
65,0 X 42,7—46,3, среднее 61,8 X 44,3 (ЗМ МГУ), из зарубежных частей ареала (п -
= 37) 57,2—72,6 X 41,5—47,5, среднее 63,9 X 45,0 (Witherby et al., 1941). Масса яиц
(и = 7) 58,3—68,3, среднее 62,8 г. Насиживание начинается с первого яйца, однако в
первые сутки оно не постоянно. В насиживании принимают участие оба партнера, но на
долю самки приходится до 75% времени пребывания птицы на гнезде.
Поведение средних поморников при появлении человека в окрестностях гнезда
разнотипно. Иногда оба партнера ведут себя довольно индифферентно и летают по-
одаль, издавая ’’короткий” или ’’дрожащий” крики (см. выше) и не атакуя нарушите-
ля. В других случаях птицы активно пикируют на человека, иногда даже ударяя ее
лапами. В непосредственной близости от гнезда они прекращают атаки и приступа-
ют к отвлекающей демонстрации, имитируя раненую птицу, а при более напряжен-
ной ситуации имитируя и птенца, что сопровождается тонким писком. Если в гнезде
идет вылупление или уже появились птенцы, птицы никогда не бывают агрессивны
и летают поодаль или садятся на землю далее 50—70 м (Томкович, Сорокин, 1983).
Длительность инкубации 25—27 сут, массовое вылупление птенцов в зависимости
от погодных условий весны приходится на первую декаду или середину июля. После
вылупления птенцы в течение 2—4 сут остаются в гнезде, а затем расходятся и прячут-
ся поодиночке вплоть до подъема на крыло в окрестностях гнезда, не удаляясь от не-
го более 100 м. Птенцы агрессивны по отношению друг к другу, и старший может
напасть на младшего. Выкармливают оба родителя, принося пищу и отрыгивая ее птен-
цам. Летать птенцы начинают в возрасте 31—32 дней, иногда несколько позже, но по
крайней мере еще две недели спустя после подъема на крыло подкармливаются роди-
телями.
Успешность гнездования среднего поморника сильно варьирует и зависит от общей
плотности поселения и наличия пищи. На северной Аляске в поселениях средней плот-
ности она составляет 30—35%. В поселениях с большой плотностью успешность гнездо-
вания может быть как высокой (до 55%), так и весьма низкой (до 4%) (Maher, 1974).
Высокая степень гибели кладок и смертность птенцов в плотных поселениях, по-види-
мому, может быть связана с возникающим дефицитом кормов в июле, что влечет за
собой необходимость более далеких и длительных отлучек взрослых птиц в поисках
корма и способствует увеличению каннибализма. В разреженных же поселениях вы-
сокая смертность, по всей вероятности, связана со слабой защищенностью гнезд и птен-
цов против четвероногих хищников. Следует, однако, заметить, что на о-ве Врангеля
в сравнительно разреженных поселениях среднего поморника, несмотря на сильный
пресс песцов, гибель кладок и птенцов в целом была незначительной: из 74 гнездящих-
ся пар, находившихся под наблюдением, у 47 (64%) птенцы благополучно поднялись
накрыло (Дорогой, 1981).
Суточная активность, поведение. В период гнездования в условиях круглосуточной
освещенности средние поморники активны в любое время суток, однако большинство
птиц в ночные часы отдыхает, сидя на кочках или сухих бугорках. Вне гнездового пери-
ода, на море или побережьях активны в светлое время суток, спят обычно на воде или на
прибрежных скалах. Летом большинство негнездящихся птиц, как уже говорилось,
28
Средний поморник
держится группами по три-четыре птицы, реже образуют небольшие скопления в местах
с обильным кормом.
Питание. В гнездовое время в тундрах основу питания среднего поморника состав-
ляют мелкие грызуны, преимущественно лемминги разных видов: на азиатской терри-
тории сибирский (Lemmus sibiricus) и копытный (Dicrostonux torquatus), а на американ-
ской — бурый (L. trimucronatus). Из других грызунов чаще всего используются полевки
рода Microtus. Добычу птица высматривает с возвышения, замечая лемминга на рассто-
янии до 100 м, после чего взлетает, настигает зверька и с лету наносит ему сильный
удар клювом. Нередко, однако, поморник разыскивает добычу в полете, кружа с ко-
роткими остановками в воздухе над гнездовой территорией, а затем, при обнаружении
лемминга, пикирует и хватает зверька клювом. Когда после выпадения снега лемминги
реже выходят на поверхность, поморники караулят их'у входов в норы, а иногда пы-
таются разрывать норы. Есть наблюдения, что на Аляске поморники разрывают верх
надземного гнезда леммингов. Пойманного лемминга птица как правило съедает на
месте или относит партнеру, сидящему на гнезде.
Обычно поморник расклевывает зверька и съедает его по частям, оставляя только
желудок и кишечник. Однако он может и проглотить целиком лемминга весом до
100 г, что чаще бывает при некотором дефиците корма или при нападении на поморника
других птиц. При обилии корма один поморник удовлетворяется в сутки тремя лем-
мингами массой по 60 г. В азиатских тундрах копытные лемминги, по-видимому, ча-
ще становятся добычей, нежели сибирские. Во всяком случае, на о-ве Врангеля встреча-
емость копытных леммингов в желудках и погадках средних поморников составляла
50—80%, тогда как сибирских 17,6—19,2%. Из других видов корма в питании среднего
поморника зарегистрированы мелкие птицы (в частности, плосконосые плавунчики),
насекомые (жужелицы, гусеницы бабочек белянок, личинки двукрылых). Отмечены
случаи нападения поморников на гнезда и птенцов белого гуся (Дорогой, 1981).
Негнездящиеся птицы концентрируются и подолгу задерживаются в местах забоя
северных оленей, у трупов морских животных, на свалках. По данным, полученным
на Аляске, рацион негнездящихся средних поморников в тундре составляли мыше-
видные грызуны (41%), птицы (25%), яйца птиц (20%), падаль (14%), рыба (12%),
насекомые (9%) и морские беспозвоночные (3%) (Maher, 1974). Эти соотношения,
вероятно, могут изменяться в широких пределах.
Во время миграций и на зимовках средние поморники в основном питаются рыбой, ко-
торую ловят в поверхностных слоях воды. Несколько меньшую роль в питании играет по-
едание падали, нападение на чаек и других птиц, клептопаразитизм (Cramp, Simmons,
1983).
Враги, неблагоприятные факторы. Главный неблагоприятный фактор для средних
поморников — депрессии численности леммингов, что влечет за собой негнездование
значительной части популяции, хотя и не приводит непосредственно к снижению ее чис-
ленности. Крайне неблагоприятны также погодные условия некоторых лет, когда
весна бывает затяжной и холодной, снежный покров сохраняется до июня-июля, а озе-
ра долго покрыты льдом. Из хищников только песец может причинить серьезный ущерб
кладкам и нелетным птенцам. Местное население при случае использует яйца среднего
поморника наряду с другими в пищу, однако это не имеет регулярного характера.
Хозяйственное значение, охрана. Хозяйственное значение средних поморников нич-
тожно. Поедая отбросы и падаль у поселков, они играют определенную положительную
роль. Вместе с тем, повреждая наравне с чайками попавшую в сети рыбу, нападая на
гнезда и птенцов промысловых птиц, средние поморники причиняют незначительный
ущерб человеку. Гораздо важнее их роль в балансе энергетических процессов в тундро-
вых биоценозах. В годы массового размножения леммингов средние поморники стано-
29
Семейство Поморииковые
вятся важным регулятором их численности. По наблюдениям на Аляске, например,
пара поморников для выкармливания птенцов изымает 31 лемминга с площади 0,5 га
(Maher, 1974). На о-ве Врангеля средние поморники уничтожают 3—5% среднелетней по-
пуляции копытных и сибирских леммингов (Чернявский, Дорогой, 1981). В годы, бед-
ные леммингами, резко возрастает нагрузка на популяции мелких птиц тундры.
Никаких специальных мер по охране средних поморников не предусмотрено.
Короткохвостый поморник — Stercorarius parasiticus
(Linnaeus, 1758)
Arctic Skua (англ.), Parasitic Jaeger (амер.), SchmarotzerraubmSwe (нем.), Labbe parasite (фр.).
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Среднего размера поморник, лишь
немногим уступающий по величине среднему, но обычно заметно крупнее длинно-
хвостого. Окраска либо темная, либо двухцветная: верх темный, низ светлый. Общая
длина около 45 —48 см (с хвостом), размах крыльев 88—125 см. Сложение, в принципе,
как у других наших поморников, однако птицы менее массивны, чем средний помор-
ник, и относительно более ширококрылы нежели длиннохвостый. В полете на нижней
стороне крыла заметно белое поле у оснований первостепенных маховых, но оно еще
менее контрастно, чем у среднего поморника. У взрослых птиц средняя пара рулей
шиловидно заострена и на 8—14 см выдается за обрез хвоста, однако эти перья у корот-
кохвостого поморника всегда относительно короче, чем у длиннохвостого, и в отличие
от него в полете не изгибаются от ветра. У молодых птиц выступающие концы цент-
ральных рулевых едва заметны; в отличие от длиннохвостого поморника все рулевые
заострены (Walter, 1962) (рис. 6).
Полет, как и у других поморников, быстрый, сильный и маневренный, с довольно
глубокими взмахами крыльев, причем машущий полет часто и легко сменяется сколь-
жением на неподвижных крыльях, что позволяет птице в случае обнаружения добычи
стремительно пикировать с высоты к самой земле, а затем так же быстро и легко взмы-
вать вверх. По земле птицы передвигаются медленно и неохотно, предпочитая переле-
тать даже на самые короткие расстояния. Гнездящиеся птицы держатся парами, а не
участвующие в размножении — обычно небольшими группами по 3—4 птицы. Сразу по
прилете, когда снег в тундре еще не стаял или в годы с затяжной холодной весной, ко-
роткохвостые поморники концентрируются в более крупные (иногда сотенные) скоп-
ления в местах обилия корма, часто на свалках у поселков, где объединяются со сред-
ними и длиннохвостыми поморниками и серебристыми чайками. Голос — визгливые,
гнусавые или мяукающие очень разнообразные звуки, слабо отличающиеся от кри-
ков других поморников нашей фауны.
Насчитывают более 12 специфических звуковых сигналов, главными из которых
являются: 1. ’’Короткий крик”. Ряд ритмически повторяющихся коротких звуков
вроде ”кекк-кекк-кекк...” Демонстрируется в конфликтных ситуациях при агрессии
и нападении; 2. Долгий крик. Громкое носовое ”и-и-и-е ...и-и-и-е...”, повторяющееся
обычно 3—4 раза (до 12 раз) почти без пауз. Функционально эквивалентен долгому
крику других чайковых птиц; 3. Тревожный крик. Различные жалобные мяукающие
звуки, издаваемые при появлении человека у гнезда или при попытках поймать птенца;
4. ’’Лающий крик”. Высокие пронзительные звуки, отдаленно напоминающие лай
собаки. Издается птицей при атаке на нее с воздуха или при приближении человека,
функционально заменяя тревожный крик.
Подвидовая систематика. Монотипический вид.
30
Короткохвостый поморник
Замечания по систематике. Количественное соотношение различных цветовых морф
а популяциях короткохвостого поморника представляет несомненный интерес. Доми-
нирование или преобладание темных птиц отмечено в Исландии, на Фарерских островах,
титтяпе Гренландии, Лабрадоре и Командорах. Примерно равное соотношение темной
I светлой морф характерно для Соловецкихостровов, Мурмана и Камчатки. На северо-
востоке Гренландии, Шпицбергене, Ян-Майене, о-ве Медвежьем, на Аляске, п-ове Канин,
с-ве Колгуев, Новой Земле, в Тиманской тундре и по всей полосе тундр Сибири птицы
fee, 6. Вершины крыла и обрез хвоста молодых поморников: короткохвостого (I), длиннохвос-
того (2) (по: Glutz v. Blotzheim, Bauer, 1982, с изменениями)
светлой морфы составляют подавляющее большинство, а местами темная морфа не
встречается вовсе. Таким образом, как будто подтверждается предположение С.А. Бу-
турлина (1934) о том, что по мере продвижения на север удельный вес в популяции
пип темной морфы падает, а выше 68° с.ш. она перестает встречаться вовсе. Следует
также заметить, что намечаются как бы два очага распространения темных птиц: один
и севере Европы (Исландия, Фареры, Мурман), а другой на востоке Азии (Камчатка,
Командоры). Вопрос нуждается в дополнительных исследованиях, но априорно можно
сказать, что систематического значения, как это предполагалось в прошлом, поли-
морфизм короткохвостых (как и других) поморников не имеет.
Описание. Строение и размеры.
Размеры (мм) и масса тела (г) птиц с Восточного Мурмана (Дементьев, 1951) и из
Западной Палеарктики (Cramp, Simmons, 1983) :
31
Семейство Поморннковые
Длина крыла:
(и= 123) - 290-345 (среднее 322);
9 2 (п = 119) — 290—340 (среднее 326).
Длина клюва:
<3<3 (п = 56) — 25-37 (среднее 30,6);
9 9 (я = 27) — 27—34 (среднее 31,4),
Длина цевки:
<3<3— 41—47 (среднее 44,3);
9 9 — 42—47 (среднее 44,4).
Масса тела:
<3<3 (я = 108) - 306-585 (среднее 429);
9 9 (п = 125) — 315—636 (среднее 462).
Окраска. Самец и самка в брачном наряде. Окраска полиморфна, различают
три морфы: светлую, промежуточную и темную. Светлая морфа: верх головы (’’шапоч-
ка”) темная черновато- или серовато-бурая, спина, поясница, кроющие крыла, брюхо
и подхвостье серовато-бурые, но светлее ’’шапочки”. Бока головы, задняя сторона
шеи, горло и вся грудь белые, но боках и задней стороне шеи золотисто-желтый отте-
нок. Промежуточная морфа: в типе окраска, как у светлой морфы, но на груди попе-
речная (иногда неполная) буроватая полоса, оперение верха тела несколько темнее,
чем у светлой морфы, бурый цвет на брюхе распространен дальше к груди и заходит
на бока. Темная морфа: окраска однородная, серовато-бурая, более темная и насыщен-
ная на верху головы (на ’’шапочке”). У всех морф маховые и их кроющие, рулевые
и нижние кроющие крыла черновато-бурые, почти черные. Клюв черный с оливковым,
зеленоватым или серым оттенком у основания, лапы черные.
Самец и самка в зимнем наряде. В целом зимний наряд похож на брачный, но у
светлой и промежуточной морф темные участки оперения (за исключением кроющих
крыла) несколько светлее из-за беловатых окончаний свежего пера, на боках и задней
стороне шеи, горле и боках появляются темно-коричневые пятнышки, а на пояснице
и животе — темные и светлые поперечные полосы.
Пуховой птенец. Общая окраска серовато-бурая, желтовато-коричневая или желто-
вато-серая, наиболее темная на затылке и спинной стороне тела, более бледная вокруг
глаз, на подбородке, концах крыльев и животе. Клюв розовато-серо-синий, с черным
кончиком, лапы бледные серо-голубые с розоватыми пятнышками.
Гнездовой наряд. Окраска в гнездовом наряде сильно варьирует и может быть, как и
у среднего поморника, условно подразделена на три типа (морфы), которые, однако,
не коррелируют с морфизмом взрослых птиц. Светлая морфа: голова и шея грязно-
темно-желтые с легкой узкой исчерченностью. Мантия, спина, надхвостье и поясница
черные, причем каждое перо имеет широкое темно-желтое окаймление. Рулевые и ма-
ховые черные. Грудь и живот или черно-бурые с примесью красно-бурого (особенно
на брюхе), или покрыты густыми пятнами и поперечными полосами темно-бурого и
ржавчато-желтого (иногда с примесью белого) цвета. Темная морфа: общая окраска
грязноватая черно-бурая, похожа на окраску взрослых птиц темной морфы, но перья
на животе и надхвостье, а также нижние кроющие часто имеют ржавчато-бурое окай-
мление. Иногда такое же окаймление есть и на перьях мантии, головы и всей нижней
стороны тела. От взрослых птиц молодых в гнездовом наряде отличают короткие,
32
Зак.826
СО
со
Рис. 7. Область распространения короткохвостого поморника
1 — гнездовой ареал, 2 — места зимовок, 3 — южная граница осенних кочевок
Семейство Поморииковые
слабо заостренные центральные рулевые. Полосатая морфа: голова и шея испещрены
тускло-черными, сероватыми с легким желтоватым оттенком продольными пятнами,
более темными на верху головы и светлыми на горле. Черные перья верхней стороны
тела имеют широкие и контрастные беловатые окаймления, нижняя сторона тела (влю-
чая нижние кроющие крыла) покрыта поперечными черно-бурыми и белыми полосами.
Между этими морфами встречается ряд переходов с большими или меньшими уклоне-
ниями в сторону светлой или темной морфы. Клюв у всех птиц в гнездовом оперении,
как у взрослых, на ногах заметно постепенное потемнение либо сверху к пальцам, либо
наоборот. Первый полувзрослый (промежуточный) наряд (весна и лето второго кален-
дарного года жизни) сильно варьирует и в целом похож на гнездовой. Центральные
рулевые более удлинены и заострены. Птицы в этом наряде в основном остаются в
местах зимовки. Светлая и полосатая морфы: голова в целом темнее, чем в гнездовом
наряде, мантия и спина менее полосаты, свежее перо либо имеет желтоватое окаймле-
ние, либо однотонное свинцово-черное. Верхние кроющие крыла однотонные грязно-
бурые. Нижняя часть тела, кроющие хвоста и нижние кроющие крыла, как в гнездовом
наряде, но основной тон несколько светлее. Темная морфа: птицы по окраске практи-
чески неотличимы от поморников в гнездовом наряде.
Второй полувзрослый (промежуточный) наряд (зима второго и лето третьего кален-
дарного года жизни) : светлая и полосатая морфы по окраске выравниваются. Некото-
рые полосатые перья, похожие на перья гнездового наряда, сохраняются до конца , зимы.
В целом наряд близок к взрослому и отличается от предыдущих менее развитым жел-
тым оттенком. Наряд сохраняется в течение всего лета, за исключением некоторых
участков на голове, шее и мантии. Темная морфа: по окраске, как взрослые птицы,
но на верхней стороне тела имеются как новые, так и старые перья, чего у взрослых
не наблюдается. Третий полу взрослый (промежуточный) наряд (зима третьего и лето
четвертого календарного года) : похож на взрослый зимний наряд, но зимой часть ниж-
них кроющих крыла имеет бледные окончания. У большинства на цевке сохраняются
голубовато-серые пятна. В летнее время у птиц светлой морфы, по общей окраске
неотличимых от птиц в брачном наряде, на желтых участках оперения шеи часто сохра-
няются мелкие темные крапины и светлые окаймления на перьях мантии, а также по-
перечная полосатость на груди и брюхе. В четвертом полувзрослом наряде птицы прак-
тически неотличимы от взрослых в весеннем наряде. Радужина во всех возрастах темно-
коричневая.
Линька. Полный брачный наряд окончательно формируется на пятом-шестом году
жизни.
Общий ход линьки практически не отличается от такового среднего поморника,
т.е. в году две линьки: полная послегнездовая и неполная предгнездовая, захватываю-
щая голову, шею, мантию, нижнюю сторону тела. Центральная пара рулевых в отличие
от среднего поморника линяет только раз в году. Линька протекает быстрее, чем у
среднего поморника, и обе линьки (послегнездовая и предгнездовая) заканчиваются
в пределах осенне-зимнего сезона.
Распространение. Гнездовой ареал (рис. 7). Зона тундр Европы, Азии, Север-
ной Америки (циркумполярный ареал). В Западной Европе: Исландия, прибрежные
районы Норвегии, Швеции, Финляндии, север Шотландии, острова Гебридские, Оркней-
ские, Шетландские, Фарерские, Шпицберген, Медвежий, Ян-Майен. В Северной Америке:
прибрежные районы п-ова Аляска, северное побережье и острова Канады, Западная и
Восточная Гренландия. В СССР (рис. 8): прибрежные районы Кольского полуострова
и прилежащие острова, п-ов Канин, Малоземельская тундра, Большеземельская тундра,
Югорский полуостров, Ямал, Гыданский полуостров, Таймыр к северу до 75-й и к югу
до 72-й параллели, тундры низовий Анабара, Лены, Яны, Индигирки, Колымы, Чаунская
34
Короткохвостый поморник
низменность, прибрежные районы Чукотского полуострова, Анадырская низменность,
приморские районы Корякского нагорья, Камчатка, северное побережье Охотского
моря, острова Колгуев, Новая Земля, Земля Франца Иосифа, Командорские. Не гнез-
дится на о-ве Вайгач, Северной Земле, Новосибирских островах, о-ве Врангеля и Север-
ных Курильских островах, хотя встречи в гнездовое время регистрировались в этих
местах неоднократно (Pleske, 1928; Карпович, Коханов, 1967; Нечаев, 1969; Дорогой,
1981; и др.).
Зимовки. Зимует в океанических водах Северной Атлантики, в прибрежных
водах Южной Америки, у восточных побережий Северной Америки, Южной Австралии,
у западного побережья Африки, восточного побережья Азии, в Красном море и Персид-
ском заливе. Незначительное количество птиц зимует на Черном и Средиземном морях.
Миграции. Как и у среднего, у короткохвостого поморника на территории СССР
имеются, по-видимому, два основных пролетных пути. Птицы западных популяций
осенью движутся побережьями или прибрежными водами северных морей на запад в
обход Мурмана или через Белое и Балтийское море, пападая в воды, омывающие север-
ное побережье Европы, откуда следуют далее вдоль побережий Франции и Испании в
Атлантику. Птицы восточных популяций мигрируют осенью на восток, спускаясь затем
в воды Тихого океана. Однако граница между западными и восточными популяциями
пока недостаточно ясна. Часть короткохвостых поморников, уклоняясь к югу, проле-
тает над сушей, и таких птиц осенью, зимой и весной нередко отмечают вдали от круп-
ных водоемов в различных частях страны от Белоруссии, Украины и центральных об-
ластей европейской части СССР до Средней Азии (обзор залетов см.: Деменьев, 1951).
В Сибири залеты известны до оз. Малый Чан (Чернышов, Юрлов, 1986). Как правило,
это молодые птицы. Однако несмотря на регулярность встреч короткохвостых помор-
ников в центральных районах, основной областью пролета в СССР следует считать
прибрежные воды, острова и побережья северных морей (Балтийского, Баренцева,
Карского, Лаптевых, Восточно-Сибирского, Чукотского, Берингова, а также Охотского
и Японского).
35
Семейство Поморииковые
Вне СССР залеты зарегистрированы в Чехословакии, Югославии, Румынии, Иране,
Ираке, Сирии, Алжире, Ливии, Ливане и на ряде островов Средиземного моря (Scott
et al., 1975; Cramp, Simmons, 1983).
Период весеннего пролета короткохвостых поморников охватывает вторую поло-
вину моя, осеннего — август — сентябрь. На пролете птицы держатся обычно небольши-
ми группами, хотя нередко концентрируются в значительные скопления в местах,
богатых кормом. На зимовках в отличие от других поморников большие скопления
образует редко.
Численность. В тундрах в период гнездования обычная, а местами многочисленная
птица. В европейской части СССР, несомненно, самый многочисленный вид из помор-
ников, в азиатской части заметно уступает в численности длиннохвостому поморнику,
причем к востоку от Чаунской низменности снижается не только удельный вес, но и
общая численность. Локальные колебания численности по годам хотя и выражены,
но значительно менее отчетливо, чем у среднего поморника. По самым приблизитель-
ным экспертным оценкам общая численность на территории СССР может быть опреде-
лена в 100—200 тыс. особей.
Местообитания. В гнездовый период основным местообитанием являются типичные
и кустарниковые тундры. Наиболее характерны сильно заболоченные мохово-осоко-
вые равнинные тундры с густой сетью разной величины озер и проток, с кустарниковы-
ми зарослями по берегам рек. В более сухих и лучше дренированных осоково-пушице-
вых кочкарниковых тундрах, в гористых ландшафтах и на водораздельных плато
распространение связано с более или менее обширными понижениями вокруг озер,
в связи с чем оно приобретает отчетливый пятнистый характер. Во время послегнез-
довых кочевок короткохвостые поморники часто концентрируются по широким,
заросшим кустарниками долинам рек, на пролете и зимовках ведут пелагический
образ жизни и с сушей связаны мало.
Размножение. Половой зрелости достигают в возрасте 3—5 лет (есть указание, что
иногда начинают гнездиться и в годовалом возрасте, еще в первом промежуточном
наряде, однако оно нуждается в подтверждении). Моногамы, пары обычно постоянны,
и на места гнездования птицы прибывают чаще всего в парах. Прилет в Западной Европе
падает на конец апреля - начало мая. В местах гнездования в. СССР короткохвостые
поморники появляются почти одновременно на всей площади ареала в третьей декаде
мая и только в годы с затяжной и холодной весной сроки прилета несколько сдвигают-
ся. По прилете птицы держатся в местах, богатых кормом, обычно вместе с крупными
чайками, а по мере схода снежного покрова в тундре распределяются по гнездовым
территориям, что происходит в среднем к 5—10 июня.
В зависимости от характера местообитаний короткохвостые поморники гнездятся
полуколониально или одиночными парами, причем первый тип гнездования свойствен
гористым ландшафтам, где удобные территории ограничены по площади (в частности,
на Мурмане и островах), а второй — низинным мохово-осоковым тундрам (тундры
Сибири, Канадского Севера и Аляски). При полуколониальном гнездовании расстояние
между гнездами в среднем составляет около 100 м, а величина охраняемой территории
несколько превышает 1 га. В равнинных тундрах расстояние между гнездовыми парами
колеблется от 0,5 до нескольких километров и гнездовые территории обычно даже
не соприкасаются. Гнездовые территории довольно постоянны и занимаются из года
в год, вселение новых пар сопровождается территориальными конфликтами, хотя в це-
лом короткохвостые поморники при защите гнездовых территорий менее агрессивны,
нежели средние, Плотность гнездования в тундрах Европейского Севера СССР и Сиби-
ри сравнительно мало колеблется по годам.
Гнездо короткохвостые поморники всегда устраивают на сухом приподнятом ме-
36
Короткохвостый поморник
сте - на торфяных плоских кочках, на выступающих из воды валиках полигональных
структур, на сухих шлейфах пологих холмов. Гнездо представляет собой небольшое
углубление в грунте диаметром 120—150 мм и глубиной до 50 мм, выстилка состоит
из сухих листочков карликовой березки или ив и кусочков лишайников, иногда она
практически отсутствует.
Начало откладки яиц в Западной Европе (о-в Фэр-Айл, Оркнейские острова) прак-
тически стабильно и падает на вторую половину мая, хотя отдельные пары задерживают
откладку до середины июня. В тундрах Сибири и на Шпицбергене, где весна начинается
позже, а погодные колебания выражены более резко, начало откладки яиц смещается
на первую декаду — середину июня и иногда задерживается до конца июня. Интервал
между откладкой первого и второго яйца обычно составляет 1—2 дня, иногда больше.
В кладке в норме 2 яйца (87%), реже 1 (13%) (в неблагоприятные годы, при утрате
первой кладки, у молодых птиц) и как исключение 3 яйца (0,4%) (поскольку у по-
морников два симметричных наседных пятна, такие аномальные кладки обречены на
гибель). Яйца нормально-яйцевидной формы. Скорлупа гладкая, слегка блестящая,
что особенно заметно у насиженных яиц. Окраска ее сложная, основной фон оливко-
во-коричневый или зеленовато-оливковый, изредка голубоватый, поверхностные пятна
и точки темно-бурого или грязно-коричневого, черноватого цвета, распределяются
неравномерно по всей поверхности яйца, число их сильно варьирует. Размеры (мм):
с территории СССР (кроме Семи Островов) (п = 13) 55,2—62,2 х 39,7—42,8, среднее
58,3 х 41,4; с Семи Островов (п = 28) 55,0—60,7 х 40,0—45,0, среднее 57,1 х 41,2;
из зарубежных частей ареала (л = 100) 50—62 х 37—44, среднее 57 х 40. Масса яиц
49,0-50,2 г.
Насиживают кладку оба партнера начиная с первого яйца; период инкубации 25-28,
в среднем 26 дней. Вылупление асинхронно. Птенцы после вылупления первые дни
остаются в гнезде, а затем расходятся в пределах гнездового участка, но обычно не
далее 100—200 м. Родители кормят их отрыжкой. В возрасте 25—30 дней птенцы подни-
маются на крыло, но еще в течение 2—5 недель подкармливаются родителями, после
чего переходят к самостоятельной жизни.
В период инкубации короткохвостые поморники исключительно агрессивны по
отношению к любым врагам, приближающимся к гнезду. В непосредственной бли-
зости от гнезда оба члена пары переходят к отвлекающей демонстрации, имитируя
раненую птицу в сочетании с имитацией птенца. После вылупления птенцов активность
отвлекающей демонстрации резко снижается.
Успешность размножения зависит от возраста родителей: у пар, гнездящихся два и
более года, она составляет в среднем 1,51, у гнездящихся впервые — 0,65 вставших
на крыло молодых на пару птиц. Кроме того, в парах, где самец относится к темной
морфе (такие птицы начинают гнездиться несколько раньше), успешность размноже-
ния выше, чем у пар с самцом светлой морфы. По-видимому, на успешности размноже-
ния сказывается также характер весны и общая обеспеченность кормами (Дементьев,
1951; Perdeck, 1963; Maher, 1974; Glutz v. Blotzheim, Bauer, 1982; Cramp, Simmons,
1983;.O’Donald, 1983).
Суточная активность, поведение. В пределах гнездового ареала в условиях кругло-
суточной освещенности короткохвостые поморники активны в любое время суток,
однако между 23.00 и 2.00 ч ночи наблюдается отчетливый спад активности, когда боль-
шинство не занятых насиживанием птиц спокойно сидит невдалеке от гнезд на сравни-
тельно постоянных местах, обычно несколько возвышенных (кочках, бугорках и т.п.).
В это же время прекращают поиски пищи и негнездящиеся птицы.
Негнездящиеся птицы летом в тундрах держатся в большинстве случаев небольшими
группами по 3—4 особи, которые нередко скапливаются в период подъема на крыло
37
Семейство Поморннковые
птенцов мелких воробьиных птиц в заросших кустарниками речных долинах. Образуют
скопления и в других местах с обильным кормом, например у трупов тюленей, север-
ных оленей. Вне гнездового периода чаще образуются концентрации в районах птичьих
базаров, рыбных и зверобойных промыслов. Активны в этот период в светлое время.
Питание. По типу питания короткохвостые поморники — типичные эврифаги: они
легко переходят с одного вида корма на другой. Вместе с тем отчетливо намечаются
два основных направления пищевой специализации, проявляющиеся в гнездовой период
и связанные с характером гнездовых местообитаний. При гнездовании на морских побе-
режьях и островах, особенно вблизи птичьих базаров, основу кормового поведения
составляет клептопаразитизм и добывание пищи на воде и в воде. Чаще всего поморники
отбирают корм у моевок, полярной и других крачек, тупиков, гагарок, чистиков,
причем наиболее успешным бывает нападение вдвоем, втроем и даже впятером. В море
поморники добывают рыбу и беспозвоночных животных в основном с поверхности,
причем видовой состав жертв чрезвычайно разнообразен. На птичьих базарах поморники
часто поедают яйца и птенцов чайковых и чистиковых птиц. Весной и осенью в рацио-
не заметную роль играют ягоды, в основном вороники.
При гнездовании в материковых тундрах и во внутренних частях крупных островов
короткохвостые поморники проявляют себя как типичные хищники. В годы высокой
численности леммингов и других мелких млекопитающих они становятся основным
кормом поморников, однако в отличие от среднего поморника короткохвостый в слу-
чае отсутствия грызунов легко переключается на питание птицами. Основной его добы-
чей служат пуховые птенцы и кладки куликов, слетки воробьиных птиц, яйца гагар,
уток, чаек, белых куропаток. Помимо позвоночных животных, короткохвостые помор-
ники поедают значительное число членистоногих, моллюсков и других беспозвоночных.
Весной и осенью в питании важную роль игра.эт ягоды вороники, брусники,голубики,
а также падаль и отбросы. Однако в целом короткохвостого поморника следует рас-
сматривать как неспециализированного орнитофага.
Враги, неблагоприятные факторы. К числу неблагоприятных факторов можно от-
нести погодные аномалии, задерживающие начало откладки яиц, и территориальную
конкуренцию со средним поморником, однако серьезного влияния на численность
короткохвостого поморника это не оказывает. Из хищников только песец в отдельные
годы может причинить известный ущерб кладкам и нелетным птенцам этого вида.
Местное население при случае использует яйца поморников в пищу, но это не имеет
характера регулярного промысла.
Хозяйственное значение, охрана. Хозяйственное значение короткохвостого поморни-
ка такое же, как и среднего. В гагачьих хозяйствах может причинить серьезный ущерб.
Никаких специальных мер по' его охране не принимается, и в них нет нужды.
Длиннохвостый поморник —
Stercorarius longicaudus Vieillot, 1819
Long-tailed Skua (англ.), Lond-tailed Jaeger (амер.), Kleine Raubmowe (нем.), Labbe a longue
queue (фр.).
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Самый мелкий из гнездящихся на терри-
тории СССР поморников. Окраска сверху темная, снизу — светлая, белая. Длина взрос-
лых птиц (с хвостом) около 50—58 см, размах крыльев 105—117 см. Общий облик как
у других поморников — сравнительно длинные и узкие, заостренные крылья, длин-
ный, хорошо заметный издали хвост, у взрослых птиц значительно более длинный,
38
Длиннохвостый поморник
нежели у короткохвостого поморника; причем обычно хорошо заметно, как при боко-
вом ветре центральные рулевые, выступающие за обрез хвоста на 15 — 18 см и образую-
щие вершину хвоста, сгибаются и как бы относятся в сторону, У молодых птиц в воз-
расте 1—2 лет концы центральных рулевых едва выступают за обрез хвоста; все руле-
вые, включая центральные, в отличие от короткохвостого поморника слегка закругле-
ны. Телосложение в целом заметно более легкое и элегантное, нежели у короткохвосто-
го и среднего поморников, клюв значительно короче, тоньше и слабее.
Полет, как и у других поморников, сильный, легкий, с глубокими взмахами крыль-
ев, еще более стремительный, чем у короткохвостого поморника. Птица может мгновен-
но сбросить высоту, спикировав к самой земле, а затем легко взмыть снова вверх.
Нередко длиннохвостые поморники в полете ’’зависают” подобно пустельге и высмат-
ривают добычу, а затем ныряют в пустоту подобно крачкам, но, не долетев до земли,
снова поднимаются почти по вертикали, Поймав же добычу, птица легко садится на
землю. Прекрасно плавают, но не ныряют, по земле передвигаются медленно и неохотно,
предпочитая перелететь с места на место. Голос, как и других поморников, исключи-
тельно разнообразен, выразителен и в целом неприятен. Обычно это визгливые, мяу-
кающие, плачущие жалобные резкие звуки. Как правило, если в тундре слышишь
незнакомый голос, он оказывается принадлежащим длиннохвостому поморнику.
Насчитывают около 10 звуковых сигналов взрослых птиц и 3 птенцов. Наиболее
важными и частыми являются: 1. ’’Короткий крик”, звучащий как быстро повторяемое
”крикк.„крикк,„” и издаваемый при появлении наземной опасности, в том числе
человека. 2. Долгий крик составляет комбинацию элементов ’’короткого крика”, пов-
торяющегося несколько (2—4) раз, с серией протяжных криков, звучащих как
”фиюю...”, и издается во время территориальных конфликтов, при появлении коротко-
хвостого поморника, реже при приближении человека (наиболее обычный крик в
гнездовий период). 3, ’’Замирающий крик” — короткий двухсложный мелодичный
крик, звучащий как ”клии-о” или ”клии-у”, издается при атаке на крупных чаек или
пикировании на приближающегося человека, 4. Крик тревоги, напоминающий лай
мелкой собаки. 5, Агрессивный крик, звучащий как ”куу-лии-о” или ”кииро-а”, изда-
ваемый при атаке на крупных чаек и хищных птиц. 6. Тонкий жалобный писк, издавае-
мый во время отвлекающей демонстрации на земле или при ритуализированном кор-
млении во время предкопуляционной церемонии (Drury, 1960; Andersson, 1971).
Подвидовая систематика. Два слабо различимых подвида.
1. Stercorarius longicaudus longicaudus
Stercorarius longicaudus Vieillot, 1819. Nouv. Diet. d’Hist., nouv. ed., 32, c. 157, север Евро-
пы и частично Азии (на восток до Таймыра).
Отличается более темным низом тела.
2. Stercorarius longicaudus pallescens
Stercorarius longicaudus pallescens Loppenthin, 1932, Meddelelser om Gronland, (91), c. 85.
Гренландия, север Азии к востоку от Таймыра, Северная Америка.
Отличается более светлым низом тела.
Замечания по систематике. Самостоятельность названных подвидов неоднократно
подвергалась сомнению, и в частности в последней сводке Л.С. Степаняна (1975).
В целом вопрос нуждается в дополнительных исследованиях, однако сочтено целесооб-
разным провизорно принять точку зрения, утверждающую поли типичность вида.
Описание. Строение и размеры. S. 1. longicaudus (мм) : длина крыла самцов
(п = 21) 283—326 (среднее 309,6), самок (п = 14) 296-325 (среднее 312,1); клюва
самцов и самок 27—30; цевки самцов и самок 40—46 (Дементьев, 1951); из Западной
Европы (Скандинавия) (самцы и самки, и = 17): длина крыла 292—320 (среднее 307);
39
Семейство Поморннковые
длина клюва 27—30 (среднее 28,5); длина цевки 40—45 (среднее 42,5) (Cramp, Sim-
mons, 1983). Размеры S.l. pallescens (мм): длина крыла самцов (п = 10) 298—321
(среднее 308,9), крыла самок (и = 9) 297-326 (среднее 311,7) (Дементьев, 1951);
из Гренландии (самцы и самки): длина крыла (и = 95) 291—327 (среднее 308); длина
клюва (и = 91) 26—31 (среднее 28,5); длина цевки (и = 86) 39—46 (среднее 42,3)
(Cramp, Simmons, 1983). Масса тела самцов S.l. longicaudus 269—317, среднее 274,5 г;
самок 260—319, среднее 297,3 г (Дементьев, 1951). Масса тела самцов SJ. pallescens
с Аляски (и = 26) 236—343, среднее 280; самок (и = 18) 258—358, среднее 313r (Maher,
1974). Существенный разницы в размерах по полу и подвидам нет.
Окраска. Самец и самка в брачном наряде Верх головы до глаз черный (’’ша-
почка”), остальные части головы и шея белые, окончания перьев на задней стороне и
боках шеи желтые, иногда желтый цвет заходит на горло. Мантия, спина, поясница и
надхвостье шиферно-серые, рулевые несколько темнее к концам. Грудь белая, брюхо
и бока тела серые, у птиц, обитающих в Азии к востоку от Таймыра, в Северной Аме-
рике и Гренландии белый цвет заходит и на брюхо. Первостепенные маховые черные,
второстепенные и нижние кроющие крыла однородно-серые. Клюв черный, оливково-
серый или коричневатый у основания, лапы черные.
Самец и самка в зимнем наряде. В целом, как гнездовой наряд, но верх головы
темно-бурый, остальная часть головы и шея белые с мелкими буроватыми пестринами
различной формы, желтый цвет отсутствует. Верх тела как в гнездовом наряде, но
окончания перьев мантии несут желтоватую или светло-серую кайму. Нижняя сторона
тела белая с темно-серой поперечной полосой через грудь.
Пуховой наряд. Общая окраска от желтовато-коричневой до желтовато-серой, бока
головы и нижняя сторона тела светло-серые. В целом пуховой наряд длиннохвостого
поморника светлее и серее, чем у короткохвостого, менее контрастен. Клюв светло-
голубой с красноватым основанием и черноватым концом, лапы светло-синие с крас-
новатым оттенком на перепонках.
Гнездовой наряд. Имеются две морфы - светлая и полосатая, связанные множест-
вом переходов. Светлая морфа: верх головы пестрый, темно-серый с белым, задняя
сторона шеи, бока головы и горло белые со слабыми серыми пестринами. Верхняя
сторона тела черно-коричневая, каждое перо с беловатым окаймлением. Вся нижняя
сторона белая, на боках и подхвостье темные полосы, еще более контрастная полоса-
тость есть на надхвостье. Рулевые грязно-черные с узкой белой окантовкой на дисталь-
ном конце. Полосатая морфа: верх головы темно-бурый, края перьев светлые, бока
головы, шея, горло темно-серые с темными пестринами. Верхняя сторона тела и нижние
кроющие крыла как у светлой морфы, но окаймления перьев уже и светлее. Нижняя
сторона тела резко полосатая, серо-белая, полосы более отчетливы на боках и под-
хвостье. Полосатая морфа короткохвостого поморника довольно сходна с длинно-
хвостым, но окаймление контурных перьев на голове и шее желтовато-коричневое.
Клюв серо-голубой с черным концом, лапы серо-голубые, проксимальные участки пере-
понки розовые.
Первый полувзрослый (промежуточный) наряд (зима первого и лето второго кален-
дарного года жизни). В отличие от гнездового наряда окраска более однородна и разли-
чия в морфах сглаживаются. Голова, как в гнездовом наряде, темя темнее, чем осталь-
ные части головы. Горло светлее, иногда равномерно белое, обычно появляются желтые
окончания перьев на боках головы и задней стороне шеи. Верхняя сторона тела черная,
перья окаймлены узкой белой полоской, имеется обычно примесь одноцветных шифер-
но-серых перьев. На надхвостье сохраняется полосатый рисунок, рулевые черные с ши-
ферно-серым основанием. На груди темная серовато-бурая поперечная полоса, на боках
и подхвостье контрастная белая и буроватая полосатость, брюхо белое, иногда с серым
40
Рис. 9. Область распространения длиннохвостого поморника
1 — гнездовой ареал, 2 — южная граница знмне-весенннх кочевок, 3 — то же весенне-летних кочевок
Семейство Поморниковые
оттенком. Второстепенные маховые и нижние кроющие крыла как гнездовом наряде.
Лапы серо-голубые, как в гнездовом наряде, но клюв черный, как у взрослых птиц.
В отличие от среднего и короткохвостого поморников, длиннохвостые в этом наряде
уже появляются иногда вблизи мест гнездования.
Второй полувзрослый (промежуточный) наряд (вторая зима). Похож на взрослый
зимний наряд, но обычно сохраняется полосатость на нижних кроющих крыла и второ сте-
пенных маховых. Третий полувзрослый (промежуточный) наряд (лето третьего календар-
ного года жизни). Похож на брачный наряд, но в оперении черной ’’шапочки” присутст-
вуют серые или бурые перья. В оперении мантии еще сохраняются отдельные черные перья
с белыми окантовкой и точками. Сохраняются также темные точки и пестрины на боках
головы и горле и остатки поперечной темной полосы на груди. Нижние кроющие крыла
и второстепенные маховые темно-серые, кое-где сохраняется полосатость. Есть указа-
ния, что в этом наряде отдельные птицы приступают к размножению. Четвертый полу-
взрослый (промежуточный) наряд (третья зима). Практически неотличим от взрослого
зимнего и только иногда сохраняется остаточная полосатость на нижних кроющих
крыла. Пятый полувзрослый наряд (лето четвертого года жизни). Неотличим от взрос-
лого гнездового, для многих птиц его, по-видимому, можно считать окончательным.
Цвет лап постепенно формируется по мере смены промежуточных нарядов, проходя
ряд стадий от серо-голубого в первом полувзрослом до черного в пятом полувзрослом.
Радужина во всех возрастах темно-коричневая (Дементьев, 1951; Fjeldsa, 1977;
Glutz v. Blotzheim, Bauer, 1982; Cramp, Simmons, 1983).
Линька. В году две регулярные линьки: полная послегнездовая и неполная пред-
гнездовая. У взрослых размножавшихся птиц послегнездовая линька контурного опере-
ния головы и большей части тела падает на октябрь—ноябрь, у негнездившихся или
потерявших кладку начинается иногда уже в июле, Спина, поясница, крылья и хвост
линяют в ноябре, смена иногда затягивается до февраля-марта и почти смыкается
тогда с предгнездовой линькой. Предгнездовая линька захватывает голову, центральные
рулевые и контурное оперение тела, кроме спины, лопаток и поясницы. В основном
она падает на март—апрель; смена центральных рулевых, которая, как и у коротко-
хвостого поморника, происходит дважды в год, осуществляется в течение 30—
40 дней. Линька из гнездового наряда в первый промежуточный полная и проходит на
местах зимовки. Первостепенные маховые и рулевые меняются с января по июнь,
оперение тела — в октябре—феврале. Линька полувзрослых птиц протекает по той же
схеме, что и взрослых, но отличается деталями и несколько сдвинутыми сроками.
Распространение. Гнездовой ареал. Зона тундр Европы, Азии, Северной
Америки (циркумполярный ареал), местами далеко выходит к югу за пределы собствен-
но зоны тундр. В Западной Европе: Скандинавия (Северная Швеция, Финляндия, Нор-
вегия), Шпицберген, Ян-Майен (гнездился в прошлом). На о-ве Медвежьем наблюдался
в летнее время, но гнездование не установлено. В Северной Америке: п-ов Аляска,
материковые тундры и острова (Баффинова Земля и др.) севера Канады, северная
оконечность Лабрадора, западное и восточное побережья Гренландии (рис. 9). В СССР:
вся полоса тундр на севере европейской части и в Азии, побережье Кольского полу-
острова, п-ов Канин, Тиманская, Малоземельская и Большеземельская тундры, Ямал,
Гыданский п-ов, Таймыр, полоса тундр от Хатанги до Чукотского полуострова, где
длиннохвостый поморник заходит и во внутренние части полуострова, Анадырская
низменность, побережье Берингова моря, Камчатка, северное побережье Охотского
моря. Гнездится также на Новой Земле, Новосибирских островах и о-ве Врангеля
(рис. 10). По-видимому, не гнездится на Вайгаче, Земле Франца-Иосифа, Северной
Земле, Командорских островах, Сахалине и Курилах, хотя встречи птиц в летнее время
регистрировались неоднократно.
42
Длиннохвостый поморник
Зимовки. Зимует в океанических водах Северной Атлантики, спускаясь далеко
к югу вдоль побережий Африки и Америки, в северной половине Тихого океана, где
также спускается далеко на юг вдоль западного побережья Америки; зимует в значи-
тельном числе в водах и у побережий Северного Ледовитого океана (особенно по се-
верной окраине Американского континента), а также у северных побережий Европы.
Миграции. Изучены недостаточно. По-видимому, как и у среднего и короткохвостого
поморников, на территории СССР имеются два основных пролетных пути. Птицы запад-
ных популяций осенью двигаются побережьями или прибрежными водами северных
морей на запад, через Мурман, Белое и Балтийское моря и попадают к северным бере-
гам Западной Европы, вдоль которых следуют далее на юго-запад, в Атлантику. Птицы
восточных популяций, вероятно, мигрируют осенью на восток, спускаясь затем в воды
Тихого океана. Граница между популяциями неясна, и видимо, она не носит абсолютно-
го характера, так как и на зимовках и даже во время миграций птицы двух разных
подвидов, гнездящиеся географически разобщенно, в значительной мере перемешивают-
ся. Как и у короткохвостых поморников, часть птиц залетает во внутренние районы СССР,
однако такие встречи значительно реже, чем у предыдущего вида. Вне СССР залеты
длиннохвостого поморника отмечены в Бельгии, ГДР, Польше, Чехословакии, Австрии,
Венгрии, Швейцарии, Италии, Румынии, Югославии, Турции, Португалии (Cramp, Sim-
mons, 1983).
Весенняя миграция длиннохвостого поморника у берегов Европы и Азии проходит
в мае, несколько смещаясь иногда по точным срокам в зависимости от климатических
условий. Осенью основной пролет падает на август—октябрь. Во время миграций птицы
держатся как поодиночке, так и группами, обычно небольшими, но иногда в местах
с обильным кормом образуются довольно значительные скопления, в основном на
ранних этапах пролета.
Численность. В гнездовой период в тундрах Европы и Азии обычная, местами даже
многочисленная птица. Отчетливо прослеживается нарастание численности на террито-
рии нашей страны в направлении на восток, т.е. обратное тому, что наблюдается для
43
Семейство Поморннковые
короткохвостого поморника. Если в западных частях ареала короткохвостый поморник
доминирует по численности, то в восточных тундрах Сибири картина бывает об-
ратной.
Численность и плотность поселения длиннохвостого поморника в разных частях
ареала весьма различна. Так, на косе Беляка (Колючинская губа) в благоприятный год
зарегистрирована плотность гнездования в 0,1 пары на 1 км2, в Яно-Индигирской
тундре в среднем она составляет от 0,8 до 1,2 (по: Успенский и др., 1962, даже до
10,0) пары на 1 км2, на о-ве Врангеля в разные годы от 0,08 до 0,4 пары на 1 км2,
в Северной Швеции варьировала от 0,02 до 0,63 пары на 1 км2; для Аляски и Севера
Канады известны колебания от 0,068 до 0,94 пары на 1 км2 (Mahler, 1974; Andersson,
1976; Кречмар и др., 1978; Дорогой, 1981). В целом в сухих тундрах относительно
более редок, в сильно увлажненных — заметно более многочислен. Численность по срав-
нению с другими видами поморников несколько более стабильна, особенно в матери-
ковых равнинных заболоченных тундрах. Численность длиннохвостого поморника в
Скандинавии равна около 12-15 тыс. особей. На территории СССР по самым прибли-
женным экспертным оценкам она может составлять не менее 200—250 тыс. птиц.
Местообитания. В период гнездования характерными местообитаниями длиннохвос-
того поморника являются типичные и кустарниковые тундры. Наиболее предпочитаемы
сильно увлажненные равнинные или слегка холмистые мохово-осоковые тундры с гус-
той сетью мелких и крупных озер, соединенных протоками, с обширными болотистыми
депрессиями с мелкими торфяными бугорками, островками карликовой березки и
полигональными структурами, с кустарниковыми зарослями по берегам рек и проток.
В более сухих и хорошо дренированных тундрах, в сильно бугристых или увалистых
ландшафтах и на водораздельных плато население длиннохвостых поморников реже, но
в отличие от короткохвостых они распределены сравнительно равномерно. В целом
в тундрах они предпочитают участки с наиболее пестрыми растительным покровом и
микрорельефом. В гористых местностях гнездятся в субальпийском поясе. Во время
послегнездовых кочевок, как и короткохвостый поморник, длиннохвостый концен-
трируется по широким закустаренным долинам рек, где в это время в массе соби-
раются молодежь воробиных птиц и мелкие грызуны. На пролете и зимовках боль-
шинство птиц ведет пелагический образ жизни и с сушей почти не связано.
Размножение. Половой зрелости длиннохвостые поморники в норме достигают в
возрасте 4—5 лет, однако, как указывалось выше, иногда начинают гнездиться на вто-
ром году жизни, еще в промежуточном наряде. Моногамы, брачные пары поддержи-
ваются по несколькулет.В места гнездования прибывают в парах или образуют их сразу
по прилете. Прилет на гнездовья в середине — третьей декаде мая, еще по снегу, в наиболее
северных районах — в начале июня; сроки прилета иногда смещаются в связи с холод-
ной затяжной весной, но в целом довольно стабильны. Сразу после прилета птицы дер-
жатся в местах, богатых кормом, в основном у поселков, а по мере исчезновения сне-
гового покрова распределяются по гнездовым территориям.
Характер гнездовых стаций во многом определяется ходом весны. В годы с ранней и
теплой весной гнездовые территории распределяются сравнительно равномерно, хотя
определенное преимущество отдается окраинам крупных приозерных депрессий. Птицы
охотно гнездятся и по сухим пологим склонам холмов. В гористом рельефе предпочи-
тают увлажненные понижения с моховым покровом. В годы с холодной затяжной вес-
ной и поздним снеготаянием птицы вынуждены начать откладку яиц еще до того, как
наиболее пригодные места выйдут из-под снега. В такие годы места гнездования как бы
сдвигаются вверх по склонам холмов (в основном солнечной экспозиции) и ближайшее
окружение гнезда чаще всего составляет осоково-пушицевый кочкарник. Но о-ве Вран-
геля гнезда размещаются в сухой щебнистой мохово-дриадовой тундре, как на равнине,
44
Длиннохвостый поморник
Рис. 11. Демонстрационное поведение длин-
нохвостого поморника — долгий крик
(по: Glutz v. Blotzheim, Bauer, 1982, с
изменениями)
так и на склонах. В равнинных приморских тундрах, заливаемых иногда нагонной во-
» дрй, длиннохвостые поморники практически не гнездятся.
। На территории СССР длиннохвостый поморник всегда гнездится отдельными пара-
j ми, хотя иногда плотные поселения создают видимость колониального гнездования. Рас-
I стояние между гнездами в таких случаях составляет 200-300 м, но никакой коллектив-
ной защиты территории при приближении опасности не бывает. Вместе с тем, хотя гнез-
довые территории часто соприкасаются, а иногда и перекрываются, пограничные кон-
фликты исключительно редки. Размеры гнездовых территорий и расстояния между
ближайшими гнездами сильно варьируют.
На Аляске среднее расстояние между гнез-
дами в двух различных местообитаниях со-
ставляло 0,9 и 1,3 км; на Земле Элсмира
(Канада) при высокой численности по-
морников - в среднем 2,52 км, на Восточ-
ной Чукотке в окрестностях Уэллена —
от 0,8 до 2,5 км (Maher, 1974; Томкович,
Сорокин, 1983). Площадь гнездовой терри-
тории составляет в среднем около 1 км2,
хотя может заметно варьировать (An-
dersson, 1971; Maher, 1974; Дорогой,
. Территории весьма стабильны и зани-
мают из года в год, хотя птицы могут и не
гнездиться. Вместе с тем известны случаи
незначительного смещения территории в
разные годы. Поведение длиннохвостых поморников при охране гнездовых территорий
и образовании новых пар похоже на соответствующее поведение других видов поморни-
ков (рис. 11).
Гнездо длиннохвостые поморники всегда делают на сухом месте. В одних случаях
это плоская торфяная кочка или бугорок, в других — выступающие из воды валики
полигональных структур, ровная площадка среди осоково-пушицевых или дриадовых
кочек или просто небольшой участок щебенки. Гнездо делает самка; оно представляет
собой ямку с диаметром 10—15 см и глубиной от 2 до 5 см. Гнездовая выстилка со-
стоит из небольшого количества сухих листочков ивы или карликовой березки, кусоч-
ков лишайника или iyixa, иногда выстилки нет совсем.
Откладка яиц у длиннохвостых1 поморников в Западной Европе начинается с середи-
ны мая; на Мурмане, в тундрах европейской части СССР, Сибири и на о-ве Врангеля на-
чало откладки обычно падает на первую декаду — середину июня, хотя иногда затяги-
вается до начала июля (возможно, однако, что это повторные кладки). Интервал между
откладкой яиц 1-2, изредка 3 сут. Полная кладка в норме состоит из 2 яиц. Кладки
с 1 яйцом встречаются редко, обычно в годы с худшими кормовыми условиями (от-
сутствие или низкая численность леммингов) или у молодых птиц. На о-ве Врангеля
кладки с 1 яйцом в ’’безлемминговые” годы составляют до 60% (Дорогой, 1961).
В Северной Швеции в годы с низкой численностью грызунов средняя величина кладки
составляла от 1,37 до 1,73 (Andersson, 1976). Крайне редко встречаются кладки в
3 яйца, но успешная инкубация их невозможна.
Яйца у длиннохвостого поморника нормально-яйцевидной формы, скорлупа на-
сиженных яиц гладкая, маслянисто-блестящая. Окраска скорлупы сложная, основной
фон — оливково-коричневый или зеленовато-оливковый, очень редко встречаются
яйца с ярким чисто-голубым основным фоном. Поверхностные пятна и крапинки
темно-бурого, грязно-коричневого или черноватого цвета распределяются равномерно
45
Семейство Поморииковые
или расположены беспорядочно, нередко гуще сгруппированы на тупом конце яйца,
число их и густота сильно варьируют, иногда они почти отсутствуют. Размеры (мм):
S.l. pallescens (и = 31) 48,7—59,6X35,6—40,8, среднее 53,9X38,1; по другим данным
(и = 79) 47,1—59,8X36,7—41,2, среднее 54,8X39,1 (Дорогой, 1981); оба подвида (и =
= 73) 49,0—65,7X33,0—42,2, среднее 55,4X38,5. Средняя масса ненасиженных яиц
(и = 6) 40,0 (33,6-47,6) г (ЗМ МГУ).'
Кладку насиживают оба родителя начиная с первого яйца, однако самке принадле-
жит главная роль. Во время инкубации яйца лежат сверху на перепонках лап. Длитель-
ность инкубации каждого яйца 23-25, всей кладки 25-27 сут. Вылупление асинхронно,
птенцы после вылупления остаются в гнезде 1—2 сут затем расходятся, но обычно не
далее 20—100 м. В это время роль родителей различна: самка в основном обогревает и
кормит молодых, самец охраняет ее и снабжает кормом, хотя также принимает участие
в кормлении. В возрасте 24—26 сут птенцы поднимаются на крыло, а еще 10—20 дней
спустя переходят к самостоятельной жизни. В случае гибели ненасиженной кладки
птицы могут отложить повторную.
Длиннохвостые поморники активно защищают гнездо от пернатых и четвероногих
хищников. При приближении к гнезду человека обе птицы с криком пикируют на него,
однако практически никогда не ударяют, хотя проносятся над самой головой. В лите-
ратуре есть указания на то, что длиннохвостые поморники в экстремальной ситуации
переходят к имитации птенца и раненой птицы; для территории нашей страны таких
наблюдений нет. На тревожные крики птиц, беспокоящихся около гнезда, нередко
прилетают поморники из соседней пары, однако они как правило летают кругами
выше хозяев и непосредственного участия в защите не принимают.
Успешность размножения во многом зависит от возраста родителей, кормовых и
погодных условий. В благоприятные годы она может достигать 96%, в неблагоприятные
падать до 31,2% (Дорогой, 1981). В годы, особенно бедные кормом, откладка яиц
иногда вовсе не происходит (Дементьев, 1951; Perdeck, 1963; Mahler, 1974; Andersson,
1976; Glutz v. Blotzheim, Bauer, 1982; Cramp, Simmons, 1983).
Суточная активность, поведение. Суточная активность длиннохвостых поморников
в условиях круглосуточной освещенности складывается так же, как и у короткохвос-
тых: птицы практически активны все время, но в полуночные часы (примерно с 23 до
2 ч ночи) активность заметно спадает. Большинство поморников в это время неподвиж-
но сидят или лежат на брюхе на различных повышениях рельефа. Свободные от насижи-
вания птицы отдыхают в это время неподалеку от гнезд. Ночью птицы менее осторожны,
подпускают человека очень близко.
Негнездящиеся и потерявшие кладку длиннохвостые поморники летом держатся,
как правило, группами по 3—5 птиц. Эти группы широко кочуют в поисках мест, бога-
тых кормом. Около падали обычно очень скоро образуются смешанные скопления раз-
личных чаек и поморников разных видов, в том числе и длиннохвостых. Вне гнездового
периода нередко собираются в районах птичьих базаров, рыбных и зверобойных про-
мыслов. Спят часто вместе с чайками на воде, на скалах, на песчаных ко^ах или на кам-
нях в отливной зоне.
Питание. Питание длиннохвостого поморника исключительно разнообразно и вклю-
чает практически все виды доступного корма — млекопитающих, птиц, рыб, беспозво-
ночных (особенно насекомых), ягоды, падаль, отбросы. Вместе с тем основу питания
(до 99%) в гнездовое время все же составляют при высокой их численности мелкие
млекопитающие, и прежде всего грызуны (лемминги разных видов, серые и лесные
полевки), хотя значительный удельный вес составляют и землеройки. Лемминги
вообще занимают особое место в рационе длиннохвостого поморника, он поедает
даже их трупы, вытаивающие из-под снега. В годы высокой численности леммингов
46
Длиннохвостый поморник
гнездящихся птиц больше, размер кладок и выживаемость птенцов выше, птицы упи-
таннее, к гнездованию приступают молодые особи в. промежуточном наряде. Однако
низкая численность леммингов не оказывает такого катастрофического воздействия
на гнездование, как у среднего поморника. Птицы легко переходят на другие корма,
в основном на насекомых и в несколько меньшей степени на яйца и птенцов, ягоды.
В большинстве случаев даже при полном отсутствии мышевидных грызунов длинно-
хвостые поморники в тундрах Сибири продолжают гнездиться, хотя известны случаи
на о-ве Врангеля и в Северной Швеции, когда гнездования при этих условиях не было.
В целом же длиннохвостый поморник может быть охарактеризован как типичный по-
лифаг, и именно полифагией объясняется его относительно стабильная локальная чис-
ленность, особенно на обширных территориях равнинных, сильно заболоченных мохо-
во-осоковых тундр, где население птиц и беспозвоночных исключительно богато. Здесь
в отличие от сухих и более бедных кормом тундр низкая численность леммингов вооб-
ще не отражается на гнездовании длиннохвостых поморников.
Список видов животных, поедаемых длиннохвостым поморником, очень широк.
Из млекопитающих это норвежский, сибирский и копытный лемминги (а в Америке -
Lemmus trimucronatus), серые полевки рода Microtus (М. gregalis, М. oeconomus),
лесные полевки (Clethrionomys rufocanus, С. rutilus), землеройки (Sorex arcticus
и др.), из птиц, прежде всего, яйца и птенцы воробьиных, куликов, уток, даже белых
гусей и полярных сов. Насекомые представлены в питании личинками и имаго дву-
крылых (типулиды), шмелями, жуками, гусеницами бабочек; из ягод основную роль
в питании играет вороника, но используются также брусника, морошка. Из рыб часто
поедается девятииглая колюшка, многочисленная в мелких водоемах тундры, мальки
щуки, хотя в целом рыба в гнездовый период серьезного значения не имеет.
Питание длиннохвостого поморника в период миграций и на зимовках изучено
слабо. У птичьих базаров он паразитирует на чистиках, чайках, глупышах, отбирая у
них добычу в полете. Нередко птицы охотятся самостоятельно, подхватывая с поверх-
ности воды всплывающих морских беспозвоночных, прежде всего различных рако-
образных. Как уже упоминалось, определенную роль в питании играет падаль и
отбросы.
Способы охоты у длиннохвостого поморника различны. Лемминга он в основном
подкарауливает, сидя на каком-либо возвышении, а затем стремительно летит к нему
и хватает или бьет клювом. Более универсальный способ охоты — обнаружение добычи
с полета. Птица патрулирует территорию на сравнительно быстром полете на высоте
30—50 м и при виде добычи резко меняет направление полета и устремляется к земле
примерно под углом 30—45°, хватая или ударяя добычу клювом, после чего обычно са-
дится на землю и добивает жертву. Так поморники добывают леммингов, нелетных
птенцов, яйца, крупных насекомых. Поднявшихся на крыло птенцов они ловят в поле-
те или преследуют до тех пор, пока птенцы не сядут на землю. Леммингов и птенцов
целиком заглатывают редко, обычно разрывают на куски или сначала приносят к
гнезду, а затем разделывают, заглатывают и отрыжкой кормят своих птенцов.
Враги, неблагоприятные факторы. Гибель гнезд и птенцов длиннохвостого поморни-
ка от песцов и других хищников — явление достаточно обычное, однако она не носит
массового характера. Не влияют катастрофически на численность длиннохвостого
поморника и погодные отклонения.
Хозяйственное значение, охрана. В хозяйственном плане длиннохвостый поморник
для человека безразличен. Экологическая роль его в принцйпе не отличается от таковой
для других видов поморников. Специальных мер охраны длиннохвостых поморников
не предусмотрено.
47
Семейство Чайковые
СЕМЕЙСТВО ЧАЙКОВЫЕ - LARIDAE
Центральное семейство подотряда Lari. Сформировалось путем адаптивной радиации
в околоводных местообитаниях; предковые формы обособились, по-видимому, от при-
митивных генерализованных ржанок (Юдин, 1965). Ископаемые остатки известны с
эоцен-олигоценовых отложений Франции, в том числе род Larus - из нижнего миоцена
Франции (Olson, 1985). Очевидно, чайковые птицы возникли в северном полушарии.
Точно определить центр происхождения семейства не представляется возможным (Дар-
лингтон, 1966); в качестве предполагаемого центра назывался океан Тетис (Williams,
1952, цит. по: Белопольский, Шунтов, 1980). Ныне представители семейства населяют
все материки (кроме большей части Антарктиды) и большинство океанических остро-
вов.
Современные виды — птицы мелких и средних размеров (масса тела от 40—50 г до
2,5 кг) характерного облика чайки или крачки. В отличие от поморников в окраске
взрослых особей большинства видов преобладает белый и светло-серый цвета; ха-
рактерна только возрастная и сезонная изменчивость окраски. Рамфотека надклювья
сплошная. Экологически группа связана с побережьями морей и внутренних водое-
мов и прибрежными водами, вне сезона размножения некоторые виды держатся в
открытом море.
Систематика чайковых птиц, несмотря на хорошую изученность этой группы, до
конца не разработана. Разногласия возникают в основном по вопросу количества родов
в семействе и в отношении статуса некоторых форм, главным образом из числа круп-
ных белоголовых чаек. В основе разногласий чаще всего лежат не столько разные
фактические данные, сколько приверженность авторов к широкой или узкой трактовке
рода или вида. Наиболее последовательные приверженцы первой концепции приходят к
выводу о существовании в семействе только 4 родов (Larus, Anous, Larostema,
Sterna), не считая родов Stercorarius и Rynchops, которые они также включают в се-
мейство чайковых (Moynihan, 1969). Приверженцы второй концепции (Wolters, 1975)
включают в семейство 24 рода (по 12 родов чаек и крачек). Большинство других
орнитологов принимают 12—17 родов. Самым сложным остается вопрос о взаимо-
отношении форм внутри комплекса L. argentatus-L. fuscus (Stegmann, 19346; Ти-
мофеев-Ресовский, Штреземан, 1959; Goethe, 1963; Майр, 1968; и др.). Накапливается
все больше сведений,которые противоречат представлению о подвидовом статусе, по
крайней мере, некоторых из этих форм, относимых обычно к L.argentatus. В частности,
обнаружено перекрывание гнездовых ареалов и совместное гнездование (вплоть до
смешанных колоний) при отсутствии смешанных пар и гибридов ’’светлоспинных”
(L.a. argentatus) и ’’темноспинных” (L.a. antelius, другое название — L.a.heuglini)
чаек на Кольском полуострове (Коханов, 1981; Михайлов, Фильчагов, 1984; Фильча-
гов,устное сообщ.) и п-ове Канин (Спангенберг, Леонович, 1960; Леонович, устное
сообщ.). Есть некоторые данные о совместном обитании двух форм серебристых чаек
в западной части Таймыра (Колюшев, 1933). Во Франции не отмечается смешанных
пар и гибридов при совместном гнездовании L.a. argentatus и L.a.cachinnans (Marion
et al., 1985).
Все это говорит о том, что в данном случае мы, скорее всего, имеем дело с самостоя-
тельными, хотя и близкими видами. Даже если предположить, что формы antelius
и taimyrensis - это подвиды клуши (Cramp, Simmons, 1983), что достаточно пробле-
матично, видовая самостоятельность cachinnans сомнений почти не вызывает. Не исклю-
чено, что в конце концов придется вернуться к слегка откорректированной точке
зрения С.А. Бутурлина (1934), выделявшего в качестве отдельных видов L.argenta-
tus, L.vegae, L.cachinnans, L.fuscus и L.taimyrensis (включая L.t.antelius). Однако
48
Семейство Чайковые
в настоящее время в связи с практически полным отсутствием полевых наблюдений за
взаимоотношениями большинства форм в зоне их предполагаемых контактов нет воз-
можности ни уверенно говорить о количестве отдельных видов в комплексе argenta-
tus-fuscus, ни четко определить границы форм, видовой статус которых представляет-
ся несомненным. Поэтому, чтобы не усугублять путаницы, царящей в этом вопросе,
мы решили в данной сводке описывать вид L.argentatus в его прежнем широком объе-
ме, сознавая всю условность и явную устарелость подобной трактовки.
Семейство чайковых обычно разделяют на два подсемейства, которых многие систе-
матики возводят в ранг семейств.
1. Подсемейство Чайки (Larinae). Объединяет роды Pagophila, Larus, Rissa, Rho-
dostethia, Gabianus, Creagrus, Хета, Hydrocoloeus (последние 4 рода нередко включают
в род Larus) - в общей сложности 45 видов1 , из которых в СССР встречается 22 и
гнездится 20 видов.
2. Подсемейство Крачки (Sterninae). Объединяет роды Chlidonias, Phaetusa, Gelo-
chelidon, Sterna, Larostema, Procelsterna, Anous, Gygis, Hydroprogne, Thalasseus (по-
следние два рода изредка включают в род Sterna) — всего 43 вида1, из которых в
СССР встречается и гнездится 10.
Виды семейства Чайковых мировой фауны
(по: Белопольский, Шунтов, 1980; Howard, Moor, 1984; с изменениями)
Подсемейство Чайки - Larinae
1. Большеклювая чайка - Gabianus pacificus
2. Магелланова чайка - G. scoresbii
3. Белая чайка — Pagophila eburnea
4. Темная чайка - Larus fuliginosus
5. Серая чайка - L. modestus
6. Чайка Хеермана - L. heermanni
7. Белоглазая, или красноморская, чайка - L. leucophthalmus
8. Аденская чайка - L. hemprichii
9. Ацтекская, или смеющаяся, чайка - L. atricilla
10. Чайка Франклина - L. pipixcan
11. Черноголовый хохотун - L. ichthyaetus
12. Реликтовая чайка - L. relictus
13. Черноголовая чайка - L. melanocephalus
14. Буроголовая чайка - L. brunnicephalus
15. Сероголовая чайка - L. cirrocephalus
16. Андская чайка - L. serranus
17. Австралийская чайка - L. novaehollandiae
18. Буллеровачайка - L. bulled
19. Патагонская чайка - L. maculipennis
20. Озерная чайка - L. ridibundus
21. Морской голубок - L. genei
22 ; Бонапартова чайка - L. Philadelphia
23. Китайская чайка - L. saundersi
24. Чайка Симеона, или Симеонова чайка, - L. belcheri
25. Чернохвостая чайка - L. crassirostris
26. Одуэнова чайка, или чайка Одуэна, - L. audouinii
27. Делавэрская чайка - L. delawarensis
28. Сизая чайка - L. canus
29. Клуша - L. fuscus
30. Серебристая чайка - L. argentatus
1 Отдельные систематики принимают как несколько большее, так и несколько меньшее число видов
чаек и крачек.
49
Семейство Чайковые
31. Чайка Тэйера, или тэйерова чайка, - L. thayeri
32. Калифорнийская чайка - L. californicus
33. Западная чайка - L. occidentals
34. Доминиканская чайка - L. dominicanus
35. Тихоокеанская чайка - L. schistisagus
36. Серокрылая чайка - L. glaucescens
37. Полярная чайка - L. glaucoides
38. Бургомистр - L. hyperboreus
39М орская чайка - L. marinus
40. Галапагосская чайка - Creagrus furcatus
41. Вилохвостая чайка - Хета sabini
42. Моевка - Rissa tridactyla
43. Красноногая моевка - R. brevirostris
44. Малая чайка - Hydrocoloeus minutus
45. Розовая чайка - Rhodostethia rosea
Подсемейство Крачки - Sterninae
1. Черная крачка - Chlidonias niger
2. Белокрылая крачка - Ch. leucopterus
3. Белощекая крачка - Ch. hydbridus
4. Большеклювая крачка - Phaetusa simplex
5. Чайконосая крачка - Gelochelidon nilotica
6. Чеграва - Hydroprogne caspia
7. Крачка Берга - Thalasseus bergii
8. Королевская крачка - Th. maximus
9. Бенгальская крачка - Th. bengalensis
10. Китайская крачка - Th. bernsteini (=zimmermanni)
11. Гвианская крачка - Th. eurygnatha
12. Элегантная крачка - Th. elegans
13. Пестроносая крачка - Th. sandvicensis
14. Индийская крачка - Sterna aurantia
15. Южноамериканская крачка - S. hirundinacea
16. Речная крачка - S. hirundo
17. Полярная крачка - S. paradisaea
18. Антарктическая крачка - S. vittata
19. Кергеленская крачка - S. virgata
20. Крачка Форстера, или форстерова крачка, - S. forsteri
21, -КрачкаТрудео - S. trudeaui
22. Розовая крачка - S. dougallii
23. Белолобая крачка - S. striata
24. Аравийская крачка - S. repressa
25. Светлая, или лысая, крачка - S. sumatrana
26. Чернобрюхая крачка - S. melanogaster
27. Камчатская крачка - S. camtshatica
28. Очковая крачка - S. lunata
29. Бурокрылая крачка - S. anaethetus
30. Темная крачка - S. fuscata
31. Австралийская крачка - S. nereis
32. Чернолобая крачка - S. albistriata
33. Амазонская крачка - S. superciliaris
34. Дамарская, или китовая, крачка - S. balaenarum
35. Чилийская крачка - S. lorata
36. Малая крачка - S. albifrons
37. Мекранская крачка - S. saundersii
38. Инкова крачка — Laro sterna inca
39. Серая глупая крачка - Procelsterna cerulea
40. Обыкновенная глупая крачка - Anoiis stolidus
41. Малая глупая крачка - A. tenuirostris
42. Белошапочная глупая крачка - A. minutus
43. Белая крачка - Gygis alba
50
Белая чайка
Род Pagophila Каир, 1829
Белая чайка — Pagophila ebumea
(Phipps, 1774)
Ivory Gull (англ.), ElfenbeinmSwe (нем.), Goeland senateur (фр.).
Син. (лат.): Larus eburneus Phipps, 1774; Pagophila alba (Gunnerus, 1767)
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Чайка средних размеров, от других ви-
дов отличается чисто белой окраской взрослых, сильно оперенной голенью и грубыми
покровами ног. Голос напоминает звуки, издаваемые речной крачкой и вилохвостой чай-
кой. Самая северна” из всех чаек, гнездится на арктических островах, в течение всей
жизни связана со льдами. Кроме обычной для всех чаек пищи, состоящей из рыбы и
беспозвоночных, может кормиться экскрементами белых медведей.
Подвидовая систематика. Монотипический вид.
Описание. Размеры (мм).
Длина крыла:
66 (и = 21) - 320—355 (среднее 338);
9? (и =20) —310—342 (среднее 327).
Длина клюва:
66 (и = 21) - 33,0—49,0 (среднее 36,2);
9? (п = 20) - 31,0-36,0 (среднее 32,8).
Длина цевки;
dd (и = 21) - 43,0-47,0 (среднее 39,4);
99 (и =20) - 35,0-40,0 (среднее 37,7).
Окраска. Самец и самка в летнем и зимнем наряде. Все оперение чисто белое со
слабым желтоватым оттенком. Две основные трети клюва голубовато-серого цвета с зе-
леноватым оттенком; вершинная треть его желтая с примесью красного цвета на концах
челюстей. Радужина черновато-бурая, края век карминно-красные, ноги черные.
Пуховой наряд (Cramps, Simmons, 1983). Белый с серой спиной, серыми неясными
полосами на крыльях и серой серединой брюшка.
Гнездовой наряд. Лоб, бока головы, подбородок и горло, а иногда и прилежащие час-
ти шеи темные дымчато-серые со светлыми пестринами. Остальное оперение чисто-белое
с темно-бурыми пятнами, развитыми на темени, затылке, межлопаточной области и
верхних кроющих крыла. На крупных плечевых имеется стреловидный рисунок, вытя-
нутый в поперечном направлении; на всех верхних кроющих хвоста пятна имеют капле-
видную форму. Прочее оперение верхней стороны туловища белое без пестрин. На ниж-
ней его стороне редкие темные пятна развиты по бокам груди, брюшка и на кроющих
хвоста. Первостепенные маховые и их верхние кроющие, а также рулевые имеют буро-
вато-черные вершины, окаймленные (за исключениемдрех проксимальных первостепен-
ных) узкой белой полосой. На второстепенных маховых развиты буровато-черные на-
ствольные предвершинные пятна. Клюв черный с беловатой вершиной и килем нижней
челюсти, радужина черновато-бурая, края век и ноги черные.
Первый зимний наряд похож на гнездовой, но отличается от него меньшим развитием
темной лицевой маски и темных пестрин на туловище и крыльях. Степень развития это-
го темного рисунка, однако, сильно индивидуально варьирует. У всех линяющих в дан-
51
ел
ьэ
Рис. 12. Область распространения
белой чайки
1 — область гнездования, 2 —
отдельные гнездовые поселения;
3 — места зимовок
Рис. 13. Ареал белой чайки в СССР
Обозначения те же, что иа рис. 12
Белая чайка
ный наряд экземпляров обнаружены только чисто-белые растущие перья. Клюв с жел-
тыми тонами в области вершины, ноги и края век черные.
Линька. Подробности линьки изучены К.А. Юдиным (1960), который установил, что
линька в первый зимний наряд у некоторых особей начинается до того, как гнездовое
перо полностью сформируется. Свежие перья всегда чисто-белые; сроки этой линьки
сильно варьируют индивидуально. Первая весенняя (предбрачная) линька в отличие от
большинства других представителей Laridae полная и начинается в конце апреля или на-
чале июня. Об этом свидетельствуют особи, добытые на Шпицбергене, Новой Земле и
Анадыре. Вторая и последующие предбрачные линьки у птиц старше одного года, в том
числе и у половозрелых гнездящихся, также полные. Они Начинаются в апреле (иногда в
марте) и заканчиваются в июне, затягиваясь у некоторых особей до июля и первой по-
ловины августа (Юдин, 1960). По материалам П.С. Томковича (1986), собранным на
Земле Франца-Иосифа в 1981 г., подавляющее большинство готовых к размножению
птиц не успевают закончить эту линьку к началу гнездования и потому прерывают ее на
стадии смены IV—VI первостепенных. Часть особей, видимо, продолжает линять и во вре-
мя размножения. Вопрос о сроках и полноте послегнездовой линьки взрослых птиц и
птиц второго года жизни пока остается открытым.
Распространение. Гнездовой ареал располагается циркумполярно на аркти-
ческих островах между 72° и 82° с.ш. (рис. 12). Белая чайка размножается на Шпицбер-
гене, у северных берегов Гренландии и на некоторых островах Канадского Арктическо-
го архипелага (Dalgety et al„ 1931; Dalgety, 1932; Salomonsen, Gitz-Johansen, 1944;
Snyder, 1957; Voous, I960; Salomonsen, 1961; Lovenskiold, 1964; Birkenmajer, 1969).
В СССР гнездование известно на о-ве Виктория, Земле Франца-Иосифа, Новой Земле и
Северной Земле (Горбунов,1932; Антипин, 1938; Лактионов,1946; Говоруха, 1970;
Беликов,Рандла, 1984; Булавинцев, 1984; Томкович, 1984) (рис. 13).
Зимовки. Размещаются в Северной Атлантике к югу до моря Баффина (у запад-
ных берегов Гренландии), Исландии, Фарерских островов и Скандинавского полуостро-
ва; часть особей зимует, по-видимому, поблизости от районов размножения — в Грен-
ландии, на Шпицбергене, Земле Франца-Иосифа и Новой Земле (Salomonsen, Gitz-Jo-
hansen, 1944; Withserby et al., 1941; Бутьев, 1959; Успенский, 1972; Lovenskiold,
1964). Зимовки белых чаек отмечены также в Беринговом море (Шунтов, 1972; Divo-
ky, 1976). Интересен факт добычи в январе 1984 г. на юго-западном побережье Анадыр-
ского залива самца белой чайки, окольцованного взрослой птицей 17.VI 1981 г. на Земле
Франца-Иосифа (Томкович, устное сообщ.).
Во время кочевок нередко залетают далеко за пределы ареала как в высокие широты
Арктики (до 87—88° сш.), так и в глубь материков на юг до таежных районов Енисея,
Западной Сибири и центральных областей Камчатки (Bergman, 1935; Портенко, 1946;
Дементьев, 1951; Успенский, 1969; Рогачева и др., 1978; Раевский, 1982). Вне СССР
известны залеты во Францию, Голландию, ГДР и ФРГ, Швейцарию; в Северной Амери-
ке — в Британскую Колумбию и на оз. Онтарио; в Японию (Bent, 1921; Withserby et
al., 1941; Hand-list of the Japanese Birds 1958; Faanes, 1981; Stribemy, 1981).
Миграции. Изучены слабо. Весной к местам размножения на Земле Франца-Иосифа
первые птицы прилетают в первой-второй декадах марта, в массе — в первой половине
апреля (Горбунов, 1932). Молодые особи в возрасте одного года появляются здесь
позднее - после 15 апреля (Успенский, 1972). В северные районы Новой Земли прибы-
вают в начале апреля (Антипин, 1938). У восточных берегов Баренцева моря, а также в
горле Белого моря и у северных его побережий пролетные и кочующие, очевидно, не
размножающиеся птицы встречаются во второй половине апреля и первой половине мая
(Смирнов, 1926; Скалинов, 1960). В восточную часть ареала - на Северную Землю -
прилетают в середине мая — первой декаде июня (Лактионов, 1946). Л.А. Портенко
53
Семейство Чайковые
(1973) предполагает, что белые чайки движутся сюда от Чукотки и о-ва Врангеля в те-
чение апреля и мая. На восточном побережье Камчатки в районе пос. Жупаново единич-
ных птиц наблюдали 2.IV 1976 г. и 11 .V 1975 г. (Лобков, 1980). В открытых водах Чу-
котского моря брачные игры отмечены в первой декаде июня (Косыгин, 1975). Послед-
ние пролетные или кочующие особи встречены 2.VI 1905 г. и 13.VI1931 г. в Анадырском
крае (Белопольский, 1934; Портенко, 1939), 11.VI 1973 г. в Колючинской губе (Креч-
мар и др., 1978) и в начале июня на Новосибирских острорах (Рутилевский, 1958). По
недавним сведениям из западной части ареала (Orr, Parsons, 1982) большое число птиц
концентрируется в марте—июне у кромки льдов между Гренландией и п-овом Лабрадор.
Отмечена значительная концентрация белых чаек в апреле—июне в Анадырском заливе
и прилегающих участках Берингова моря (Косыгин, 1985).
Осенью у берегов Новой Земли птиц наблюдали до конца ноября (Антипин, 1938;
Бутьев, 1959). В восточном секторе Арктики (от Таймыра до Берингова пролива) на
осеннем пролете встречаются с начала сентября до начала ноября (Бируля, 1907; Scha-
aning, 1928; Воробьев, 1963; Успенский, 1969; Портенко, 1946, 1973; Засыпкин,
1981). В Беринговом море известны октябрьские встречи на Анадыре и восточном по-
бережье Камчатки в районе пос. Жупаново (Портенко, 1939; Лобков, 1980). У северных
берегов Аляски (мыс Барроу, Уэйнрайт) в некоторые годы эта чайка нередка в сентяб-
ре-октябре (Bailey, 1925, 1948; Divoky, 1976). В канадском секторе Арктики сотни
птиц концентрируются в это же время у островов Элсмир и Девон, откуда в середине
октября перемещаются в западную часть моря Баффина (Renaud, McLaren, 1982).
Численность. Повсеместно немногочисленна. Общая численность в западной части
Шпицбергена составляет 334 пары и повсеместно уменьшается (Birkenmajer, 1969), что
связывают с интенсивным таянием ледников (Bateson, Plowright, 1959а) или с умень-
шением численности белых медведей (Birkenmajer/; 1969). На о-ве Греэм Белл (Зем-
ля Франца-Иосифа) общая численность птиц в 1981 г. не превышала 170—200 пар (Том-
кович, 1984), на о-ве Большевик (Северная Земля) обитает около 100 пар (Булавин-
цев, 1984). Крупные негнездовые скопления птиц (до 35 тыс. особей) недавно найде-
ны весной в проливе Дэвиса у западных берегов Гренландии (Orr, Parsons, 1982).
Местообитания. Летом это арктические тундры на побережьях архипелагов. Белые
чайки предпочитают селиться по соседству с участками моря, покрытыми льдами, а
также на побережьях с материковыми льдами или хотя бы частично покрытых снегом.
При отступлении ледников птицы могут покидать обжитые места (Bateson, Plowright,
1959а; Lovenskiold, 1964). Зимой так же тесно связаны со льдами как на суше, так и
на море, нередко тяготеют к человеческому жилью и местам охоты на морского зверя
(Бутьев, 1959; Lovenskiold, 1964; Успенский, 1972; Divoky, 1976; Orr, Parsons,
1982).
Размножение. К местам гнездования прибывают за один-два месяца до начала размно-
жения; в различных регионах это происходит с марта по июнь (Горбунов, 1932; Лак-
тионов, 1946; Bateson, Plowright, 1959а, b; Lovenskiold, 1964; McDonald, 1976; Том-
кович, 1986).
Демонстративное поведение подробно изучалось лишь на Шпицбергене (Bateson,
Plowright, 1959b). Установлено, что набор демонстраций у белой чайки в целом тот же,
что и у других чаек. Некоторые из них — ’’квохтание”, вскидывание клюва, ’’огрыза-
ние” — сходны с соответствующими демонстрациями моевок, хотя в деталях позы мо-
гут и отличаться (рис. 14) . Однако у белой чайки есть особая угрожающая демонстра-
ция (”Roor”), которая не встречается у других чаек. Это наиболее часто применяемая
демонстрация угрозы. Исполняя ее, птица держит голову и шею вытянутыми вперед и
вверх, клюв обычно направлен в сторону оппонента, но может быть и отвернут от него.
Принимая эту позу, чайка издает низкое ворчание. Опознавательный сигнал (Note call)
54
Белая чайка
напоминает крик тревоги речной крачки, долгий крик — соответствующий крик озер-
ной чайки.
Гнездится преимущественно отдельными парами или небольшими рассредоточенны-
ми поселениями, величина которых колеблется от нескольких до 50 пар (Bateson, Plow-
right, 1959а; Hakala, 1975; Frisch, Morgan, 1979; Булавинцев, 1984; Томкович, 1984).
Самая крупная колония (170 пар) была обнаружена в западной части Канадского Аркти-
ческого архипелага на островке Силур к северу от о-ва Баттерст (McDonald, 1976). Ко-
Рис. 14. Демонстрационное поведение белой чайки (по: Bateson, Plowright, 1959b, с изменениями)
1 — ’’квохтание”, 2 — ”Roor” 3—4 — последовательные позы долгого крика
лонии обычно располагаются как на утесах во внутренних частях или побережьях остро-
вов, так и на плоских каменистых участках побережий, небольших плоских островках
или на покатых вершинах и склонах холмов; расстояние от поселения до побережья мо-
жет достигать 60 км (Salomonsen, Gitz-Johansen, 1944; Bateson, Plowright, 1959a, b;
Lovenskiold, 1964; Hakala, 1975; McDonald, 1976; Frisch, Morgan, 1979; Wright, Mat-
thews, 1980; Булавинцев, 1984; Томкович, 1984). Гнезда помещаются на широких кар-
низах и уступах скал, в выемках нависающих ледников, на плоских участках — в выем-
ках микрорельефа. На самых плотных участках поселения минимальное расстояние
между ними 1,0 м, обычно же гнезда располагаются на расстоянии 2—5 м друг от друга
(Bateson, Plowright, 1959а, b; McDonald, 1976; Булавинцев, 1984; Томкович, 1986).
Среднее расстояние между центрами ближайших соседних гнезд на Земле Франца-Ио-
сифа в 1981 г. составляло 3,29 и (и = 105) (Томкович, 1986). Гнездостроение протека-
ет обычно в июне—июле, в сжатые сроки (Горбунов, 1932; Белопольский, 1957; Mc-
Donald, 1976), так что первые яйца появляются иногда уже через 30 ч после начала по-
стройки гнезда.
Строительным материалом для гнезд служат мох, сухие стебли и листья камнеломок,
иногда используются также водоросли, лишайники, перья; при этом материал часто при-
носится издалека (Dalgety et al., 1931; Dalgety, 1932; Hakala, 1975; Булавинцев, 1984).
Известны случаи поселения белых чаек в гнездах гаг и среди выбросов на берегу (Анти-
пин, 1938; Makatsch, 1974).
Откладки яиц на Земле Франца-Иосифа начинается в середине июля (Горбунов,
1932; Белопольский, 1957), по данным П.С, Томковича (1984), в начале этого месяца и
продолжается до 21-22 числа; средний размер кладки (и = 51) здесь был 1,31. Нор-
55
Семейство Чайковые
мальная кладка на Шпицбергене состоит из 2 яиц, реже встречаются кладки из 1 или 3
яиц (Dalgety, 1932; Bateson, Plowright, 1959а, b; Lovenskiold, 1964). Размеры яиц
(мм) в разных частях ареала следующие. В Северной Америке (Beht, 1921) 57,0-69,3 X
X 40,0—44,6, в среднем 60,5 X 43,6 (и = 32); в Гренландии (McNeile, цит по: Makatsch,
1974) в среднем (и = 3) 60,0 X 42,7; на Шпицбергене (Jourdan, Collett, 1888, цит. по:
Makatsch, 1974 ; Schalow, 1905; Dalgety, 1932; Sladen, 1932) 53,0-69,3 X 40,0-46,2,
в среднем (и = ПО) 60,71 X 42,96; на Земле Франца-Иосифа (Томкович, 1986) 55,5—
63,8 X 40,2—43,9, в среднем (и = 24) 59,1 X 42,1.
Окраска яиц (Dalgety, 1932) варьирует от темно-коричневой до светлой серовато-
бурой с желтоватым или зеленоватым оттенками; встречаются серовато-зеленые яйца
со следами коричневых тонов. Пятна на скорлупе свинцовые или лавандово-серые, ок-
раска их в поверхностных слоях скорлупы варьирует от почти черной или коричневой
до светлых оттенков цвета хаки. В общем, как считает Макач (Makatsch, 1974), яйца
белой чайки практически неотличимы от яиц сизой чайки. Насиживание начинается, ве-
роятно, с первого яйца и длится 24-25 дней, участвуют в нем оба родителя (Dalgety,
1932; Bateson, Plowright, 1959а, b; Томкович, 1984), На Северной Земле и островах
Канадского Арктического архипелага найдены повторые кладки (Лактионов, 1946;
McDonald, 1976).
Птенцы на Земле Франца-Иосифа вылупляются в конце июля — начале августа (Бело-
польский, 1957; Томкович, 1984). Птенцы выкармливаются обоими партнерами, при
этом частота кормлений обычно небольшая. При искусственном выкармливании в
экспериментальных условиях они проявляют интерес к пище только через 8—10 ч пос-
ле кормления (Булавинцев, 1984). В естественных условиях птенцы берут пищу из
клюва родителей или с края гнезда, куда она отрыгивается взрослыми (Bateson, Plow-
right, 1959b.). По наблюдениям последних исследователей птенцы достаточно подвиж-
ны и в очень раннем возрасте способны покидать гнездо и затаиваться в его окрестнос-
тях. В.И. Булавинцев (1984), напротив, указывает, что в возрасте 1—3 дней они боль-
шую часть времени проводят в полудремотном неактивном состоянии. Способность к
полету молодые приобретают в возрасте 4—5 недель ( Bateson, Plowright, 1959а,b; Була-
винцев, 1984). Гнездовья на Земле Франца-Иосифа чайки покидают в начале сентября,
отлет с мест размножения начинается в середине этого месяца и заканчивается к его
концу или к началу октября (Горбунов, 1932; Белопольский, 1957). На Северной Зем-
ле зто происходит в разные годы с середины сентября до конца октября в зависимости
от ледовых условий (Лактионов, 1946).
Суточная активность, поведение. У гнезд птицы, как правило, очень агрессивны
(Lovenskiold, 1964) и подобно крачкам активно защищают их, яростно пикируя на чело-
века, часто задевая его при этом ногами или ударяя клювом. Агрессивность характерна
для белых чаек и по отношению к особям своего вида — соседям по колонии (Bateson,
Pbwright, 1959b; McDonald, 1976; Томкович, 1986).
Питание. Летом основу рациона составляют рыба (преимущественно сайка Boreoga-
dus saida) и морские беспозвоночные ( ракообразные Amphipodae, моллюски Clione),
которых птицы добывают из поверхностных слоев воды (Бируля, 1907; Bateson,Plow-
right, 1959b; Voous, 1960; Lovenskiold, 1964) слёта или посредством поверхностного пог-
ружения (Divoky, 1976); собирают пищу и плавая у кромки льдов ( Lovenskiold, 1964;
Laybourne, 1978), предпочитая места, где материковые льды достигают моря. Именно
летом особенно проявляются хищнические наклонности этих чаек. В колониях они
отбирают пищу у люриков и кайр, а также ловят раненых люриков (Горбунов, 1929;
Salomonsen, Gitz-Johansen, 1944; Lovenskiold, 1964). На Земле Франца-Иосифа отмечено
расклевывание яиц в соседних гнездах своего вида (Томкович, 1984). Зимой белая чай-
ка — преимущественно мусорщик: собирает корм на промыслах морского зверя,
56
Черноголовый хохотун
поедает трупы животных, выброшенные морем, питается отходами человеческого жилья
ва зимовках полярников и у дрейфующих судов (Бируля, 1907; Смирнов, 1926; Пор-
тенко, 1946; Бутьев, 1959; Успенский, 1972; Беликов, Рандла, 1984; Булавинцев,
1984). Многие авторы упоминают о тесной связи белых чаек с белыми медведями, кото-
рых они сопровождают в скитаниях среди льдов и поедают остатки их трапез и экскре-
менты. Однако такая связь существует, видимо, только в самое трудное для птиц вре-
мя и не везде. Так,по наблюдениям у берегов Аляски (Divoky, 1976), большинство чаек
кормятся на воде у кромки паковых льдов вне связи с белыми медведями, возможно,
используя в пищу организмы, поедающие фитопланктон, цветение которого происходит
на поверхности льдов и в поверхностных слоях воды вдоль южной их кромки.
Враги, неблагоприятные факторы. Естественных врагов у белой чайки мало. На Зем-
ле Франца-Иосифа, по-видимому, только бургомистр может в незначительной степени
разорять их гнезда. В Канадской Арктике успех размножения белой чайки в большой
степени зависит от хищничества песцов (McDonald, 1976). Основными причинами гибе-
ли и уменьшения численности вида считают ухудшение кормовых условий и уменьше-
ние площади ледников в местах гнездования (Lovenskiold, 1964; Birkenmajer, 1969;
Томкович, 1984).
Хозяйственное значение, охрана. Хозяйственного значения не имеет. Малочисленный
узкоареальный и малоизученный у нас в стране вид. Занесен в Красную книгу СССР.
Род Larus Linnaeus, 1758
Черноголовый хохотун —
Larus ichthyaetus Pallas, 1773
Great Black-headed Gull (англ.), Fischmowe (нем.), Goeland ichthyaete (фр.).
Статус. Гнездящийся мигрирующий, на Каспии частично оседлый вид.
Общая характеристика и полевые признаки. Одна из самых крупных и красивых
чаек нашей страны и мировой фауны, по размерам близка к бургомистру и большой
морской чайке. От других крупных чаек хорошо отличается бархатисто-черной голо-
вой, от темноголовых чаек - очень большими размерами. По рисунку первостепенных
маховых хохотун сходен с реликтовой чайкой, от которой легко отличается размерами
и трехцветным клювом (оранжево-желтое основание, красный кончик и черная пере-
вязь у вершины). Летает, медленно взмахивая крыльями, может парить. Хорошо пла-
вает, на воде сидит высоко, не ныряет. Очень осторожен, вне колонии молчалив. Голос
очень низкий: в колонии и ее окрестностях можно слышать низкое ”гау” или ”ау”,
долгий крик напоминает глухой рев. Вопреки названию и некоторым литературным
данным (Дементьев, 1951) и в отличие от серебристой чайки-хохотуньи черноголовый
хохотун не хохочет — если не считать хохотом короткий тревожный крик ”ха-га-га”.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Большинство систематиков подчеркивают определенную
обособленность черноголового хохотуна в ряду остальных чаек, выделяя его в моно-
типический подрод (Dwight, 1925; Бутурлин, 1934) и даже род Ichthyaetus (Wolters,
1975). Однако имеются основания для сближения хохотуна с реликтовой чайкой. На
родство (хотя, по-видимому, не очень близкое) этих видов указывает большое сходст-
во в окраске первостепенных маховых, а также близость пуховых и промежуточных
нарядов. Вместе с тем демонстрационное поведение этих двух чаек различается в боль-
шей степени, чем у видов внутри группы серебристой чайки (крупные белоголовые
чайки) и даже внутри группы озерной чайки.
57
Семейство Чайковые
Описание. Строение и размеры.
Размеры (мм) (Cramp, Simmons, 1983; ЗМ МГУ) и масса тела (г) (Дементьев,-1951;
Долгушин, 1962; Gramp, Simmons, 1983):
Длина крыла:
< 3 <3 (п = 17) — 485—517 (среднее 503);
9 9 (п = 16) — 462—493 (среднее 472).
Длина клюва:
< 3 <3 (п = 18) — 59,1-70,3 (среднее 64,2);
9 9 (п = 15) — 47,5—65,3 (среднее 56,7).
Длина цевки:
< 3 <3 (п = 18) — 77,6—88,6 (среднее 81,4);
9 9 (п = 16) - 68,4-84,9 (среднее 73,6).
Масса тела:
< 3 <3 ( п = 8) - 1130-2000 (среднее 1600);
9 9 (и = 8) - 960-1500 (среднее 1220).
Окраска (Dwight, 1925; Дементьев, 1951; Долгушин, 1962; ЗИН, ЗММГУ).
Самец и самка в брачном наряде. Голова бархатисто-черная, причем черный цвет спере-
ди слегка захватывает верх шеи, а сзади весь затылок. Над и под глазами узкие белые
полоски. Шея, грудь, брюхо, бока, надхвостье, подхвостье и рулевые белые. Спина,
плечевые и верхние кроющие крыла бедные сизовато-серые, задние плечевые с белыми
вершинами. Первостепенные маховые белые, кроме серых трех последних. II (первое
видимое) первостепенное с черным на большом протяжении наружным опахалом и
черным предвершинным пятном, иногда не доходящим до внутреннего края опахала.
На III расположено почти такое же пятно, не доходящее до внутреннего края опахала,
и, кроме того, маленькое черное пятнышко почти у самой вершины. Черные предвер-
шинные пятна (полосы) есть на IV—VII. Второстепенные маховые серые с белыми вер-
шинами. Клюв оранжево-желтый с предвершинной черной перевязью, конец клюва
красный. Лапы зеленовато-желтые. Радужина бурая, края век ярко-красные.
Самец и самка в зимнем наряде. Лоб, темя, затылок, уздечка темно-бурые, с густы-
ми белыми пестринами (могут быть и белыми с бурыми пестринами), кроющие уха
однотонные темно-бурые; горло и подбородок белые. Остальное оперение, как в брач-
ном наряде.
Пуховой наряд. Птенец целиком серебристо-белый или кремово-белый, нередко
сверху с охристым оттенком. Изредка на спине и крыльях неясные светло-серые пятна.
Лапы черновато-серые, клюв черный с желтоватой вершиной.
Гнездовой наряд. Лоб, темя, затылок покрыты серо-бурыми перьями с серыми каем-
ками, в результате чего голова кажется сероватой с неясными более темными пестри-
нами. Такие же, но более крупные перья есть на шее и спине. Плечевые и кроющие кры-
ла, кроме больших, серовато-бурые с охристыми каймами; большие кроющие крыла
серые. Серовато-бурый цвет перьев может быть разной интенсивности у разных особей,
нередко с коричневатым оттенком. Подбородок, горло, бока, брюхо, надхвостье белые,
грудь белая с бурыми пестринами. Первостепенные маховые темно-бурые, начиная с
III на внутреннем опахале в средней части пера появляется светлое поле. Второстепен-
ные маховые бурые с белыми вершинами. Рулевые белые с темно-бурой верхней третью.
58
Черноголовый хохотун
Клюв буроватый у основания с почти черной вершинной частью. Лапы серовато-бурые,
радужина темно-бурая, края век черные.
Первый зимний наряд. Лоб, подбородок, горло, грудь, бока, низ шеи, брюхо, над-
хвостье белые. Темя белое с бурыми пестринами, которые становятся густыми на
затылке и задней части шеи. Перед глазом и на кроющих уха бурое пятно. Перья спи-
ны и плечевые становятся сероватыми с более темными бурыми центрами. Крылья и
рулевые сохраняются от гнездового наряда.
Рис. 15. Область распространения черноголового хохотуна
1 — область гнездования, 2 — отдельные поселения, 3 — предполагаемые места гнездования, 4 — мес-
та зимовок
Первый летний наряд. Как первый зимний, но на голове появляются темные бурова-
тые перья.
Второй зимний наряд. Лоб, темя, горло, подбородок, надхвостье и нижняя часть ту-
ловища белые. Затылок белый с темно-бурыми пестринами, шея белая с отдельными
крупными темно-серыми пестринами. Перед глазом и на кроющих уха нечеткое темное
пятно. Спина серая с буроватым оттенком и отдельными бурыми пестринами. Перво-
степенные маховые черные с белыми основаниями и белыми полями на внутренних
опахалах; на II и III первостепенном обычно появляются небольшие белые предвершин-
ные пятна. Рулевые белые с широкой черной предвершинной полосой, иногда отсутст-
вующей на паре боковых перьев.
Второй летний наряд. Голова черно-бурная с белыми пестринами, густота которых
индивидуально варьирует. Остальное оперение, как в предыдущем наряде.
Третий зимний и третий летний наряды, как окончательные, но на первостепенных
маховых (особенно на II и III) черный рисунок развит больше, а на рулевых у части
особей сохраняются остатки черных предвершинных пятен.
Линька. Во время частичной линьки в первый зимний наряд в сентябре—декабре
сменяется оперение головы и туловища, оперение крыла и рулевые остаются от гнездо-
вого наряда. Последующие линьки бывают дважды в году: частичная предбрачная, при
которой не меняются рулевые и оперение крыла, и полная послебрачная. Предбрачная
линька взрослых бывает, очевидно, между январем и мартом, послебрачная — между
59
<7>
О
Рис. 16. Ареал черноголового хохотуна в СССР
1 — область гнездования, 2 — постоянные поселения, 3 — временные поселения, 4 — предполагаемые места гнездования,
5 — северная граница области регулярных кочевок, б — направление осенних миграций, 7 — направление весенних миграций,
8 — позднелетние скопления, 9 — залеты, 10 — места зимовок
Черноголовый хохотун
июнем и декабрем (возможно, январем). Полные и частичные линьки в промежуточные
наряды проходят, по-видимому, в несколько более ранние сроки (Дементьев, 1951;
Cramp, Simmons, 1983).
Распространение. Гнездовой ареал. Область гнездования простирается от
северных районов Крыма до котловины Больших озер в МНР и оз. Кукунор (по неко-
торым данным до среднего течения р. Хуанхэ) и от оз. Чаны до южной части Каспия.
Сплошного ареала вид не образует, для него характерны отдельные гнездовые поселе-
ния, нередко удаленные на несколько сот кило метров < друг от друга (рис. 15, 16).
Одни поселения существуют в течение десятков лет, другие возникают на короткое
время при улучшении гидрологического режима водоемов. На севере Крыма хохоту-
ны гнездятся на Сиваше и Лебяжьих островах (Каркинитский залив Черного моря).
На Сиваше гнездование известно с начала 30-х годов: в 1932—1934 гг. на островах
у Чонгарского моста (Воронцов, 1937) и, по крайней мере, с конца сороковых годов
по настоящее время на о-ве Китай (Киселев, 1949; Солдатенко, 1956; Зубакин, 1975а;
Сиохин, 1981а). На Лебяжьих островах гнездование регистрируется с 1947 г. (Костин,
1983). По материалам ЦКАН, в 1977 г. существовала колония в Кирпильском лимане
Приморско-Ахтырского района Краснодарского края. В 1983 г. отмечено гнездование
на Кривой косе в Донецкой области (Букреев и др., 1986).
С начала 1960-х годов хохотун стал регулярно отмечаться на гнездовье на озерах
и водохранилищах долины Маныча: на оз. Маныч-Гудило, Пролетарском и Чограйском
водохранилищах, на оз. Казинка и Меклетинских озерах Калмыкии (Петров, Мино-
ранский, 1962; Кривенко, Любаев, 1975, 1981; Языкова, 1975; Казаков и др., 1981;
Кривенко, 1981; Любаев, Кривенко, 1981), а также на расположенных неподалеку рыб-
хозах (Бичеров, 1981). В то же время на Сарпинских озерах, где хохотун гнездился
ранее (Мензбир, 1895), он ныне не обнаружен (Кривенко, 1983). На Каспийском море
в настоящее время гнездится лишь на севере Каспия: на о-ве Жемчужном, в дельте
Урала и на островах северо-восточного побережья Каспия от Гогольской косы до
п-ова Дурнаев (Кривоносов, 1976). Крайне нерегулярно гнездится в дельте Волги
(Модестов, 1943; Кривенко, 1975). В 1952 г. отмечено гнездование на Тюленьих (Ман-
гистауских) островах (Гладков, Залетаев, 1956), современное состояние этого поселе-
ния неизвестно. На юге Каспия, где прежде гнездование отмечалось в заливе Кирова
(Азербайджанское побережье), и, по-видимому, в Красноводском заливе, на Челекене
и о-ве Огурчинском (Дементьев, 1951, 1952) ныне не гнездится (Виноградов, Черняв-
ская, 1965; Васильев и др., 1975; Щербина и др., 1975). Гнездовое поселение в зали-
ве Кара-Богаз-Гол, существование в 1979—1980 гг. (Панов и др., 1980, 1982), исчезло
после перекрытия пролива, соединявшего залив с морем. В Казахстане постоянно
гнездится на озерах Тенгиз (Целиноградская обл.), Алаколь, Балхаш (Долгушин,
1962; Азаров, 1975а; Ауэзов, Грачев, 1975; Волков, 1975; Андрусенко, 1981; Ауэзов,
Сема, устное сообщ.). В 1948 г. отмечена колония в 3 тыс. особей на о-ве Комсомоль-
ский в Аральском море (Гладков, 1949). Последующая судьба этой колонии, как и
колоний в устье р. Сырдарьи и по восточному побережью Аральского моря (Спанген-
берг, Фейгин, 1936), неизвестна. Судя по исчезновению колонии на о-ве Барсакельмес
(Гисцов, 1974), поселения хохотунов на Аральском море сейчас испытывают период
депрессии. Подобное наблюдается на оз. Зайсан, где эти птицы ныне перестали гнез-
диться (Самусев, 1975), хотя прежде, по-видимому, были многочисленны (Долгушин,
1962). В 1962 г. найдена колония на оз. Жалаулы в Павлодарской области (Гаврилов
и др., 1983). Севернее известно лишь одно постоянное поселение хохотунов — на оз. Ча-
ны в Новосибирской области (Ходков, 1981).
Весьма вероятно (хотя и не доказано) гнездование черноголовых хохотунов на
озерах в низовьях Тургая, а также на оз. Карасор Карагандинской обл. (Долгушин,
61
Семейство Чайковые
1962). В Киргизии гнездование этих птиц возможно на оз. Иссык-Куль у мыса Кара-Бу-
лун (Янушевич и др., 1959; Шукуров, 1981).
Среди возникающих на короткое время поселений отмечены гнездовья на оз. Сор-
куль восточнее пос. Джаныбек (Уральская обл.) в 1968—1971 гг. (Линдеман, Зале-
таев, 1977) ив 1981 г. (Леонович, устное сообщ.), на оз. Жарколь (Кустанайская обл.)
в 1966—1968 гг. (Самородов, 1975), на оз. Таволжанном (юг Тюменской обл.) в 1959—
1961 гг. (Азаров, 19756). На основании опросных данных высказано предположение
о гнездовании хохотунов на оз. Джулукуль на юго-востоке Алтая в 1962—1964 гг. (Ири-
сов, 1972); в 1977 г. хохотуны здесь не гнездились (Стахеев, 1981). 24.V 1985 г. оди-
ночное гнездо черноголового хохотуна найдено в колонии серебристых чаек и речных
крачек на о-ве Таволжанском Запорожского водохранилища (Клестов, Лепешков,
устное сообщ.).
Вопреки первоначальным предположениям (Дементьев, 1951; Долгушин, 1962),
хохотун не гнездится на оз. Маркаколь (Березовиков, 1981), на Памире (Потапов,
19666) и, по-видимому, на Камыш-Самарских озерах (Шевченко, 1975), хотя в послед-
нем случае возможны нерегулярные поселения.
Вне СССР достоверно гнездится на озерах Хара-Ус-Нур, Толбо-Нур и Убсу-Нур (Сте-
панян, Болд, 1983; Болд, устное сообщ.), очень вероятно гнездование на оз. Хяргас-Ну-
ре (В.Е. Фомин, устное сообщ.). Гнездование указывалось также для озер Урэг-Нура,
Ачит-Нура и Бон-Цаган-Нура (Stubbe, Bolod, 1971), однако неясно, основываются
ли эти сведения на находках гнезд или же, как в случае Ачит-Нура, на встречах птиц
в гнездовом наряде (что для хохотуна не может служить доказательством гнездова-
ния). Вопреки прежним представлениям (Дементьев, 1951), на оз. Хубсугул (Косо-
гол) хохотун не гнездится (Долгушин, 1962; Скрябин, 1982). Для Китая отмечается
гнездование на оз. Кукунор, на озерах истока р. Хуанхэ и в среднем течении этой реки
(Внутренняя Монголия) (Cheng Tso-hsin, 1976; Schauensee, 1984) также без указания
на находки гнезд. Что касается оз. Кукунор, то, судя по недавним работам (Melville,
1984; Heinz, 1985), гнездование хохотунов здесь можно считать доказанным (в замет-
ке Хейнца приведена цветная фотография колонии хохотунов, правда, с подписью,
что это колония буроголовых чаек). Мнение же о гнездовании этих птиц на озерах
верховий р. Хуанхэ, высказанное впервые Н.М. Пржевальским (1888), до сих пор
находками гнезд не подтверждено (Cheng Tso-hsin et al., 1983).
В отношении динамики ареала хохотуна можно, очевидно, пока говорить лишь о
перераспределении колоний внутри области гнездования, а не о каком-либо измене-
нии ее границ. Исключение составляет лишь юг Каспия, где эти птицы перестали гнез-
диться, скорее всего, вследствие изменения уровня Каспия. Однако то, что хохотун
оказался способным гнездиться на водохранилищах, показывает потенциальную воз-
можность расширения ареала в будущем, — о чем, в частности, говорит недавняя попыт-
ка гнездования на Запорожском водохранилище. В последние годы участились случаи
встреч чаек этого вида на Куйбышевском водохранилище (Аюпов и др., 1983; Водолаж-
ская, 1986).
Зимовки. Основной район зимовки - Каспийское море (главным образом его
южная часть), откуда получено 12 (40%) из 30 зимних (декабрь—февраль) возвратов
хохотунов, окольцованных в Казахстане; 23,3% зимних возвратов поступило с внутрен-
них водоемов Ирана (в основном из озерно-болотной местности Заболь), 13,3% - с
водоемов юга Средней Азии (Хаузханское водохранилище, Каракумский канал, Айдар-
ские разливы, Амударья в Сурхандарьинской обл.), 6,7% — из Индии и по 3,3% — с внут-
ренних водоемов Калмыкии и Азербайджана, Эфиопии (оз. Тана), Кувейта и южного
побережья Аральского моря (Гаврилов и др., 1983). Анализ данных, поступивших в
ЦКАН, показывает, что с юга Каспия (включая разливы р. Артек) получено 34,4% из
62
Черноголовый хохотун
32 зимних возвратов хохотунов, окольцованных на севере Каспия, из северных и севе-
ро-западных районов Каспия — 21,9%, с внутренних водоемов Волгоградской и Сара-
товской областей — 15,6%, из внутренних районов Ирана и Эфиопии (оз. Тана) — по
63%, и по 3,1% с внутренних водоемов Азребайджана, северного Ирака, побережья
Красного моря в ЙАР, Омана (Салала) и из г. Батуми. От хохотунов, окольцованных
на водоемах Маныча, получено по два зимних возврата из Астраханской обл. и южных
районов Каспия, по одному — с внутренних водоемов Ростовской обл. и Сирии, побе-
режья Израиля, Персидского залива, из Эфиопии (оз. Тана) и Индии. Основное место
зимовок хохотунов северного Причерноморья, видимо, тоже Каспийское море (Кос-
тин, 1983).
В целом с учетом данных кольцевания и визуальных наблюдений в область зимовки
этого вида следует включить южную часть Средиземного моря (а не только египетское
побережье, как указано в сводке Cramp, Simmons, 1983), Красное море, внутренние
водоемы Эфиопии, морские побережья от Адена и Персидского залива до Бирмы,
внутренние водоемы Сирии, Ирака, Ирана и Пакистана, незамерзающие участки Кас-
пийского и изредка Аральского морей, внутренние водоемы Волгоградской, Саратов-
ской и Ростовской областей, юга Средней Азии и Казахстана (Ali, Ripley, 1969; Гаври-
лов и др., 1983; Ходков, 1983; Cramp, Simmons, 1983). Зимующие особи отмечены
на оз. Иссык-Куль (Янушевич и др., 1959). В Черном море у берегов Крыма зимовки
носят, по-видимому, нерегулярный характер (Костин, 1983).
Миграции. Покидает места зимовок в конце февраля — начале марта, хотя отдель-
ные особи появляются в окрестностях гнездовий, расположенных по соседству с райо-
нами зимовок, уже в феврале: 11—16.11 на Лебяжьих островах (Костин, 1983), 11.11 на
о-ве Барсакельмес (Исмагилов, Бурамбаев, 1973). Пролет взрослых особей завершает-
ся в сжатые сроки: к концу марта — началу апреля на оз. Чаны, однако, первые пти-
цы появляются лишь в середине — конце апреля (Ходков, Тотунов, 1979). Неполово-
зрелые птицы начинают миграцию одновременно со взрослыми, но их пролет сильно
растянут и завершается в конце апреля — первой половине мая (Гаврилов и др., 1983).
Пролет части казахстанской популяции хохотунов, по-видимому, может идти через
Центральную Азию — оз. Лобпор и озера Тибета. На зто как будто указывает выражен-
ный пролет в северном направлении через Джунгарские ворота в конце марта — апреле
(Гаврилов и др., 1983). Первые хохотуны на оз. Лобнор отмечены 19.11, на оз. Куку-
нор— в начале — середине марта (Пржевальский, 1875, 1883, 1888).
После подъема молодых на крыло (июнь—июль) хохотуны начинают широко коче-
вать, нередко залетая на сотни километров за пределы области гнездования вида. Пре-
обладающее направление. кочевок казахстанских хохотунов в июле - августе запад-
ное и северо-восточное; направленный пролет отмечается уже в августе, в последующие
месяцы он приобретает все более выраженное западное и юго-западное направ-
ление (некоторые особи долетают до Азовского моря). В большинстве райо-
нов Казахстана пролет завершается в октябре (Гаврилов и др., 1983). Сходная
картина наблюдается у хохотунов с оз. Чаны (Ходков, 1983). У птиц с Лебяжьих остро-
вов направление осеннего пролета восточное, в сторону Каспия (Костин, 1983). Боль-
шинство молодых хохотунов, окольцованных на севере Каспия, держатся в районе
рождения, часть кочуют довольно далеко в западном, юго-западном и северном (вплоть
до Татарской АССР) направлениях (Кривоносов и др., 1978). По данным ЦКАН, хохо-
туны с водохранилищ системы Маныча до сентября держатся главным образом в Рос-
товской обл. (66% возвратов), часть откочевывает на север до Волгоградской и Сара-
товской областей, другие появляются западнее — на побережье Азовского моря, юго-за-
паднее и южнее — в Краснодарском и Ставропольском краях; на побережье Каспия от-
мечаются лишь единичные особи. В октябре — ноябре птицы распределены по тому же
63
Семейство Чайковые
обширному району, но более равномерно; встречи на севере Каспия становятся боле:
частыми.
Широкие послегнездовые кочевки особей из разных поселений, захватывающие почт?:
всю область гнездования, наряду с общими местами зимовки позволяют предположить
что все черноголовые хохотуны Казахстана представляют единую популяцию, внутр?:
которой не исключена возможность перемещения птиц из одних гнездовых колоний
в другие (Гаврилов и др., 1983). По тем же причинам к этой популяции должны отно-
ситься птицы с Каспия и оз. Чаны. Не исключено, что все хохотуны, гнездящиеся е
СССР, представляют собой единую популяцию.
В период гнездовых кочевок взрослые и молодые птицы нередко в массе появляются
на богатых рыбой озерах, где отсутствовали на гнездовье: оз. Маркаколь на Южном
Алтае, оз. Зоркуль и Яшилькуль на Памире, оз. Малый Чан в Новосибирской области
и др. (Потапов, 19666; Ходков, Тотунов, 1979; Березовиков, 1981; наши данные)
Как в сезон гнездования, так и вне его отмечены многочисленные залеты черного-
ловых хохотунов за пределы гнездового ареала и области зимовок. К дальним зале-
там можно отнести встречи птиц в Англии, Бельгии, Нидерландах, Дании, Швеции, Поль-
ше, Италии, Греции, Румынии, на о-ве Мадейра (Cramp, Simmons, 1983), на Западной
Украине (Бокотей, устное сообщ.), в восточной части Латвии (Виксне, 1983), в Том-
ской обл. (Миловидов, Стрелков, 1978), Борзинском районе Читинской обл. (ЦКАН).
Гонконге (Melville, 1977), на оз. Виктория в Уганде (Britton, Brown, 1974).
Численность. На севере Причерноморья гнездится до 290 пар (Сиохин, 1981а; Кос-
тин, 1983), на водохранилищах системы Маныча — около 1800 пар, возможно несколь-
ко больше (Казаков и др., 1981; Кривенко, 1981; Кривенко, Любаев, 1981; Любаев.
Кривенко, 1981). На о-ве Жемчужном в разные годы гнездится от 3500 до 10 тыс. пар.
а всего на севере Каспия — около 10 тыс. пар или несколько больше (Кривоносов.
1976; Гаврилов, Кривоносов, 1981). На Кара-Богаз-Голе в 1979 г. гнездилось 628 пар
(Панов и др., 1980). На оз. Тенгиз в последние годы гнездилось 343-875 пар (Андру-
сенко, 1981), на оз. Алаколь - 2250—3700 пар (Ауэзов, 1980), на оз. Балхаш - 2500-
3000 пар (Ауэзов, Сема, устное сообщ.), на оз. Чаны — 72—190 пар (Ходков, 1981)
Всего в СССР гнездится, видимо, 20—25 тыс. пар. Общая тенденция изменения числен-
ности не ясна в связи со значительными несинхронными колебаниями ее в разных
частях ареала. Постоянное увеличение численности отмечено для северных районов
Причерноморья; на оз. Алаколь после роста в 1970-х годах она, видимо, стабилизи-
ровалась; на оз. Тенгиз в целом снижается. Численность вне СССР неизвестна.
Местообитания. В сезон гнездования — крупные соленые и солоновато-водные озе-
ра, морские мелководья и реже водохранилища в степной полупустынной и пустынной
зонах. Для гнездования необходимы изолированные от наземных хищников острова,
для кормежки, — по-видимому, мелководья с непостоянным уровнем воды либо иные
участки, где хохотуну легко добывать рыбу не ныряя или собирать ее в большом коли-
честве полуснулой. Во внегнездовое время — морские побережья, долины рек, озера,
водохранилища, пруды рыбхозов.
Размножение. К размножению хохотуны приступают, по-видимому, на четвертое лето,
отдельные особи, возможно, на год раньше. Черноголовый хохотун — облигатно-коло-
ниальный вид; моногам. Характерны резкие колебания численности птиц в колониях
по годам, связанные с перераспределением гнездящихся пар в пределах одного озера
или ближайших озер и, возможно, с негнездованием части особей. Не исключено к
перераспределение пар между поселениями в разных частях ареала в зависимости от из-
менения гидрологического режима водоемов.
Черноголовые хохотуны гнездятся только на островах (как на останцах, так и плос-
ких косах), причем нередко удаленных на несколько километров от берега. Под гнез-
64
Черноголовый хохотун
довья выбираются лишенные растительности возвышенные участки. Хохотуны практи-
чески всегда гнездятся вместе с серебристыми чайками, которые поселяются по краю
их компактных колоний или в непосредственной близости от них. По соседству с коло-
ниями хохотунов могут гнездиться и другие чайковые птицы (Модестов, 1943; Глад-
ков, 1949; Долгушин, 1962; Зубакин, 1975а; Языкова, 1975; Линдеман, Залетаев,
1977; Ауэзов, 1980).
Рис. 17. Демонстрационное поведение черноголового хохотуна
а—г — последовательность поз долгого крика, д — ’’огрызание”, е — мяукающий крик
На территории колонии можно наблюдать различные формы демонстрационного
поведения, которое у хохотунов имеет ряд своеобразных черт (Зубакин, 1975г) .Харак-
терная особенность хохотунов во время выполнения различных демонстраций — взъеро-
шивание перьев спины, плечевых и третьестепенных маховых, в ряде случаев — развора-
чивание веером хвоста. Долгий крик хохутуна достаточно своеобразен. Его можно раз-
делить на четыре фазы (рис. 17,а-г); в первых двух издается протяжный ревущий
крик. Общая продолжительность ритуала 3—5 с. Как и у других чаек, долгий крик хохо-
туна полифункционален.
Основная агрессивная демонстрация — ’’огрызание”, или ложное клевание, — выпад
раскрытым клювом в сторону противника (см. рис. 17,д). К сфере брачного поведения
относятся ’’кашляние” или ’’квохтанье”,и мяукающий крик. В позе мяукающего крика
(см. рис. 17, е) хохотун издает протяжное ”охх-ггооо” (или ”ухх-ггууу” при закрытом
клюве) . Данная демонстрация отмечается во время брачных церемоний, при смене парт-
неров на гнезде и кормлении птенцов. ’’Квохтание” похоже на аналогичную демонстра-
цию серебристой чайки.
Как и у других чаек, у хохотунов в период формирования пар и реже во время наси-
живания можно наблюдать кормление самки отрыгнутой самцом пищей; выпрашива-
ние корма сопровождается вскидыванием клюва. Прямая поза и отворачивание головы
ритуализованы слабо, ’’распластанная” поза и ’’клевание травы” отсутствуют.
Величина колоний хохотунов варьирует от нескольких десятков до нескольких сот
пар. Реже отмечаются колонии в несколько пар (Кривенко, Любаев, 1981; Костин,
1983), гнездование отдельными парами бывает исключительно редко и всегда в коло-
3. Зак. 826
65
Семейство Чайковые
нии других видов чайковых (Киселев, 1949; Клестов, Лепешков,устное сообщ.). Гнез-
да в колониях располагаются пространственно разделенными группами — субколония-
ми, в которые входит от нескольких до нескольких сот гнезд (Гладков, 1949; Долгушин,
1962), по пространственной конфигурации различаются плотные многорядные,
плотные линейные и разреженные линейные колонии (Панов и др., 1980). Плотность
гнездования очень велика. Среднее расстояние между центрами ближайших соседних
гнезд в колониях севера Причерноморья составляет 1,0 м( «=200), индекс плотности
гнездования 0,32 (Зубакин, 19756). Со сходной плотностью хохотуны гнездятся и в
других частях ареала: так, в заливе Кара-Богаз-Гол среднее минимальное расстояние
между краями гнезд составляло в разных субколониях от 0,39 до 2,13 м, в большинстве
случаев 0,4—0,9 м (Панов, Зыкова, 1982), на оз. Чаны среднее расстояние между центра-
ми ближайших соседних гнезд («=72) было 0,92 м (Ходков,Тотунов, 1979).
В качестве материала для гнезда используются сухие водоросли, сухие стебли травя-
нистой растительности, отдельные перья. В начале насиживания в гнезде может не быть
выстилки; достройка гнезда продолжается весь период насиживания. Наружные стенки
гнезд в период насиживания всегда запачканы пометом. Размеры гнезд (мм) : на оз. Тен-
гиз диаметр гнезда 500—700, диаметр лотка около 260, глубина лотка 70—80 (Долгу-
шин, 1962); на оз. Жарколь диаметр гнезда («=18) 290—582, средний 445, диаметр лотка
250—354, средний 299, глубина лотка 56—83, средняя 72 (Самородов,Рябов, 1972); на
Лебяжьих островах (п=16) диаметр гнезда 350—560, средний 449, диаметр лотка
220—280, средний 253, глубина лотка 65—110, средняя 88 (Костин, 1983). •
Откладка яиц в Крыму начинается в конце марта — начале апреля, в Казахстане на I
оз. Жарколь 13—19. IV, на оз. Тенгиз 16. IV, на о-ве Барсакельмес 20.IV. В случае гибели •
гнезд могут быть повторные кладки. Полная кладка содержит 1—3, чаще всего 3 яйца;
на Лебяжьих островах очень редко отмечается 4 яйца (Долгушин, 1962; Самородов, Ря-
бов, 1972; Костин, 1983). Окраска яиц, по И.А. Долгушину (1962), в целом гораздо свет-
лее, чем у серебристой чайки. По беловатому с кремовым или желтоватым налетом,
реже серовато-оливковому фону разбросаны серовато-фиолетовые глубокие и бурова-
тые или черноватые поверхностные пятна. Размеры яиц (мм): с Лебяжьих островов
(« = 200)70,5—88,3 х 48,3—56,7, в среднем 77,3 х 53,8 (Костин, 1983); сСиваша («=226)
72,0—90,7 х 49,2—58,4, в среднем 78,6 х 54,1 (наши данные); с оз. Тенгиз («=50) 72-
86 х 51—57, в среднем 78 х 54,5 (Долгушин, 1962); с оз. Жарколь («=36) 70—83 х 40-
59, в среднем 77,9 х 53,8, масса свежих яиц («=3) 120—127 г,в среднем 123 г (Саморо-
дов, Рябов, 1972); с Аральского моря («=10) 66,7—82 х 52—55,2,в среднем 77,9 х 54,1
(Зарудный, 1916).
Насиживают оба партнера. На оз. Жарколь смена происходила несколько раз в день,
самки сидели без смены по 3—7 ч и более, самцы — по 1—4 ч (Самородов, Рябов, 1972).
На Сиваше, по наблюдениям за 4 гнездами в конце периода насиживания, смены проис-
ходили 2-3 раза в сутки, причем промежутки между ними достигали 17 ч42 мин (воз-
можно, из-за удаленности мест кормежки) ; ночью насиживают и самки и самцы. Наси-
живание с первого яйца, первый птенец вылупляется через 25—29 сут, в среднем через
27 сут («=13, Лебяжьи острова). Между появлением трещины на яйце и вылуплением
проходит 1—5 сут, чаще всего 3 или 2 сут (50% и 29% соответственно; «=78),
между появлением отверстия в скорлупе и вылуплением — главным образом (75%)
1 сут. Между вылуплением первого и последнего птенца в гнезде с кладкой из трех
яиц проходит 1—6 сут, чаще всего 2 сут (48,5% случаев), реже 1 сут(21,2%) ' и 3 сут
(15%) (данные В.А. Зубакина) . После вылупления птенцов родители не уносят скорлу-
пу, ее либо затаптывают, либо выталкивают на край гнезда (Чекменев, 1964; Зубакин.
19766).
На Лебяжьих островах первые птенцы появляются 11—19.IV (птенцы повторных кла-
66
Черноголовый хохотун
зок 17— 23.VI), на Сиваше 17IV, на оз. Тенгиз с 6.V по 1—5. VI, на оз. Жарколь
'.5.V, в заливе Кара-Богаз-Гол 30.IV. Вылупление птенцов в каждой отдельной коло-
нии протекает обычно в достаточно сжатые сроки — от 11 до 21 дня (Чекменев, 1964;
Самородов, Рябов, 1972; Зубакин, 1975а; Панов, Зыкова, 1982; Костин, 1983). Масса
тела однодневных птенцов на оз. Тенгиз составляет 40—80 г (Чекменев, 1964), на оз.
Жарколь 82—90 г (Самородов, Рябов, 1972) . На 4—6-е сут после вылупления старшего
гтенца выводки оставляют колонию и кочуют по берегу в стайке с другими выводками.
В эти же сроки (на 4—5-е сут) формируется взаимное опознавание родителей и птенцов.
Во время тревоги птенцы в первые дни жизни затаиваются в гнезде или непосредствен-
ной близости от него, а после оставления колонии сбиваются в плотный табунок (Зуба-
кин, 1976а). Поднимаются на крыло в возрасте около 45 дней (Долгушин, 1962) .
Первая попытка кормления птенца отмечена через 3 ч после вылупления. По наблю-
дениям на Сиваше, в первый день жизни птенцов кормят 11 раз в сутки (данные по 1
гнезду), в возрасте 1—2 сут — 27—30 раз (2 гнезда), 5—^суточных — 15—27 раз
(2 гнезда), 13—17-суточных — 6—16 раз (2 гнезда). Родители кормят птенцов
круглосуточно, чаше всего с 9 до 21 ч (76% всех кормежек). Самцы кормят
в среднем в 1,4 раза чаще самок (и = 132). Родители обогревают (в жару за-
теняют) птенцов до 11—13-суточного возраста, при этом смена партнеров проис-
ходит 4—7 раз в сутки, в среднем (п = 7) 4,7 раза (данные В.А. Зубакина). После
становления индивидуального опознавания родители агрессивно относятся к чужим
птенцам своего вида, нередко забивая их насмерть. В разных колониях количество
погибших по этой причине птенцов варьирует от 1% (Зубакин, 1976в) до21,3%иболее
от числа вылупившихся (Панов и др., 1980). Отмечены случаи забивания родителями
насмерть своих птенцов, чаще всего младших в выводке (Костина, Панов, 1982).
Стратегия защиты потомства у хохотуна носит в основном пассивный характер на
основе ’’эффекта плотности” (отпугивающее воздействие плотной массы сидящих птиц)
и направлена против неспециализированных пернатых хищников типа серебристой чай-
ки (Зубакин, 1983). Черноголовые хохотуны значительно менее агрессивны, чем более
мелкие серебристые чайки, однако в редких случаях способны пикировать на челове-
ка, находящегося в колонии (Долгушин, 1962; Самородов, Рябов, 1972).
Эмбриональная и постэмбриональная смертность варьирует в разных поселениях. На
оз. Жарколь в 1967 г. через месяц после начала вылупления осталось в живых 47,5%
птенцов от числа отложенных яиц, на Сиваше в 1973 г. — 23,2%, на оз. Тенгиз в 1976—
1978 гг. — 7,4—21,8%, в заливе Кара-Богаз-Гол в 1979 г. — 45 —50%;• процент неоплодот-
воренных яиц в различных колониях варьирует от 1,4% до 33,3% (Самородов, Рябов,
1972; Зубакин, 1975а; Панов и др., 1980). На Сиваше из 102 погибших птенцов 45,1%
съедены серебристыми чайками, 3,0% раздавлены родителями в гнезде, 17,6% оставле-
ны родителями и погибли от голода, 3,0% убиты взрослыми хохотунами, 31,3% исчез-
ли (очевидно, также съедены серебристыми чайками). Здесь же разорено серебристыми
чайками от 21,9% до 32,8% кладок (Зубакин, 1975а). На оз. Тенгиз в 197&-1978 гг. в
условиях дефицита корма в качестве основной причины гибели птенцов указывалось
уничтожение их взрослыми хохотунами, в том числе каннибализм (Панов и др., 1980) ;
не исключено, однако, что в последнем случае оказалась неучтенной хищническая
деятельность серебристых чаек.
По данным кольцевания, смертность молодых в первый год жизни составляет 80%,
в последующие примерно по 50% ежегодно. Максимально известный возраст черноголо-
вого хохотуна 16 лет 3,5 месяца (Гаврилов и др., 1983).
Суточная активность, поведение. В сезон гнездования активность, по-видимому,
круглосуточная,хотя в светлое время суток хохотуны несколько более активны, чем в
темноте. На оз. Жарколь и на Сиваше неоднократно наблюдали ночные смены партнеров
67
Семейство Чайковые
на гнезде, кормления птенцов, прилет птиц в колонию, их отлет; на оз. Жарколь отме-
чена ночная кормежка (Самородов,Рябов, 1972; данные В.А.Зубакина).
В сезон гнездования черноголовые хохотуны очень общественные птицы, однако в
период кочевок и зимовки они обычно встречаются одиночно или небольшими груп-
пами (Долгушин, 1962). Большие стаи ( до нескольких сот особей) изредка могут
образовывать отдыхающие птицы на пролете и зимовках (Березовиков, 1981; Cramp,
Simmons, 1983).
Питание. Трофически в большей степени связан с водоемами, чем с сушей. В боль-
шинстве случаев в пищевом рационе хохотуна преобладает рыба; мелкие грызуны,
насекомые, птенцы водоплавающих птиц имеют подчиненное значение (Мензбир,
1895, Гладков, 1949; Бородулина, 1960,1965; Долгушин, 1962; Заблоцкий, Заблоц-
кая, 1963; Чекменев, 1964; Залетаев, 1960, 1965; Сиохин, 1980, 19816). На поедание
хохотунами наряду с рыбой большого количества саранчи и жуков во время массового
появления этих насекомых указывали Е.П. Спангенберг и ГА. Фейгин (1936). В 94
погадках, собранных в 1973 г. у гнезд хохотунов на о-ве Китай (Сиваш), рыба встречена
в 84,0%, грызуны — 8,5%, птицы— 1,1%, наземные насекомые — 2,1%, речной рак -
1,1%, падаль —9,6%, остатки растений — 3,2%. В 62 погадках, найденных в 1974 г. у
гнезд этого вида на Лебяжьих островах, рыба составляла 93,6% встреч, череп степно-
го хорька — 1,6%, пух птенца — 1,6%, наземные насекомые — 3,2%, растения — 24,2%
(Зубакин,1976в).
Однако есть данные, что животные суши могут играть большую роль в питании
хохотуна в некоторых районах в случае ухудшения кормовых качеств водоемов. Ана-
лиз 521 погадки на оз. Жарколь за 1966—1968 гг. и содержимого 9 желудков показал,
что млекопитающие (из которых преобладает водяная полевка) встречены в 81,8%
погадок, рыбы (в основном карась) — в 43,9%, птицы и их яйца — в 15,8%,насекомые
(главным образом, жуки-водолюбы) — в 28,6% (Самородов, Рябов, 1971). Не исклю-
чено, однако, что погадки черноголовых хохотунов оказались немного” разбавлен-
ными” погадками серебристых чаек-хохотуний, гнездившихся поблизости. Во всяком
случае, в 9 желудках хохотунов была встречена только рыба (14 карасей и 2 окуня) и
3 жука. Что касается нередко цитируемой работы И.С. Солдатенко(1956), из которой
следует, что остатки наземных животных встречаются в 91,44% погадок хохотунов с о-ва
Китай, а рыбы — только примерно в 20%, то псчти наверняка за погадки хохотунов бы-
ли в большинстве случаев приняты погадки серебристых чаек. За это говорит большое
сходство в спектре питания этих двух видов, обнаруженное И.С. Солдатенко, большое
количество (634) погадок, собранное всего от 51 пары птиц за малый период времени,
и сбор погадок в период, когда выводки хохотунов уже покинули колонию и перемеща-
лись по острову.
Наблюдений за кормовым поведением хохотуна мало. По-видимому, он способен
улетать кормиться за десятки километров. По Д.И. Чекменеву, здоровую рыбу в глубо-
кой воде хохотун Поймать не может — разве что в местах ее больших скоплений или на
высыхающих водоемах, где подвижность рыбы ограничена. Нередко хохотун добывает
рыбу из сетей или поедает отбросы промысла (Долгушин, 1962). На озерах кормится
как в одиночку (Потапов, 19666), так и стаями (Березовиков, 1981) . Отмечена ловля
насекомых (жуков) в воздухе в сумерках (Спангенберг, Фейгин, 1936) .
В норме хохотун в отличие от серебристой чайки крайне редко поедает птенцов свое-
го вида (Чекменев, 1964; Зубакин, 1976в), но в отдельных случаях в условиях недостат-
ка корма каннибализм, по-видимому, может принимать значительные размеры (Панов
и др., 1980).
Враги, неблагоприятные факторы. Серьезное воздействие на колонии черноголового
хохотуна оказывает серебристая чайка-хохотунья, особенно при усилении фактора
68
Реликтовая чайка
беспокойства. Яйца и птенцов чаек этого вида в отдельных случаях могут уничтожать
лисицы, барсуки и кабаны (Самородов, Рябов, 1972). Отмечалось затопление колоний
во время нагонных ветров и штормов (Модестов, 1943; Костин, 1983) .
Из-за своих крупных размеров и рыбоядности хохотун до сих пор еще нередко попа-
дает под выстрел, на что указывает достаточно высокий процент возврата колец (Зуба-
кин, 1977; Ходков,1983).
Хозяйственное значение, охрана. Несмотря на рыбоядность, отрицательного хозяй-
ственного значения в силу малочисленности не имеет. Вид занесен в Красную
книгу СССР.
Реликтовая чайка —
Larus relictus Lonnberg, 1931
Relict Gull (англ.), Lonnbergmowe (нем.), Goeland relique (фр.).
Син. (англ.) : Mongolian Gull.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Чайка с темной головой, среднего раз-
мера, величиной с сизую чайку. Рисунком крыла напоминает черноголового хохотуна,
но отличается от него значительно более мелкими размерами. От всех темноголовых
чаек нашей страны в полевых условиях хорошо отличается широкими белыми поло-
сками над и под глазами, которые нередко смыкаются за глазом, образуя полукольцо
(’’очки”). По этому признаку реликтовая чайка сходна с американской L. pipixcan.
Полет, плавание, как и у других чаек сходного размера; не ныряет. Вне колонии обыч-
но молчалива. Тревожный крик — трехсложное короткое ”кав-кав-кав” — сходен
с таким же криком других чаек. Кроме него, в колонии обычно слышны хриплые кри-
ки типа ”аррр”, ’’арррииу”, короткие визгливые ”рвиу”, разнообразные урчащие
и визгливые звуки, напоминающие скуление щенка или повизгивание поросенка.
Подвидовая систематика. Монотипический вид.
Замечания по сйстематике. Реликтовая чайка была описана Е. Лённбергом в качест-
ве подвида черноголовой чайки Larus melanocephalus relictus по одному экземпляру,
добытому 24.IV 1929 г. во Внутренней Монголии в нижнем течении р. Эдзин-Гол (Lon-
nberg, 1931). Позже были высказаны предположения, что это уклоняющийся от обыч-
ной окраски экземпляр буроголовой чайки (Дементьев, 1951) или гибрид меж-
ду черноголовым хохотуном и буроголовой чайкой (Vaurie, 1962). 14.V 1963 г. и
12.V 1965 г. на Торейских озерах в Читинской области А.Н. Леонтьевым были до-
быты две реликтовые чайки, определенные как буроголовые (Ларионов, Чельцов-
Бебутов, 1972). 4.VI 1967 г. им же здесь была найдена колония этих чаек, которых
вследствие неправильного определения он продолжал считать буроголовыми (Леонть-
ев, 1968). Первое сообщение о видовой самостоятельности реликтовой чайки бы-
ло сделано Э.М. Ауэзовым (1970, 1971) после находки в 1968—1969 гг. колонии на
оз. Алаколь и сравнения добытых экземпляров с чайками других видов в коллекци-
ях Зоологического музея МГУ н ЗИН АН СССР, проведенного А.Ф. Ковшарем. Почти
одновременно с этим предположение о видовой самостоятельности реликтовой чай-
ки высказали М. Штуббе и А. Болд (Stubbe, Bolod, 1971).
Описание. Строение и размеры (табл. 1).
Окраска. Взрослые птицы в брачном оперении (ЗМ МГУ; ЗИН АН СССР). Под-
бородок и лоб светло-кофейного или насыщенного серого цвета, который быстро
темнеет и превращается в темно-бурый и черный на темени, горле, затылке и шее.
Граница ’’капюшона” сзади проходит по затылку, спереди черный цвет захватывает
и верх передней части шеи. Над и под глазом белые полоски шириной 4—7 мм, ко-
торые в ряде случаев смыкаются за глазом. Шея, грудь, бока, брюхо, хвост, надхвостье,
69
Семейство Чайковые
Таблица 1. Реликтовая чайка. Размеры особей (мм) и масса тела (г)
Признак Пол Птицы из Казахстана (ЗМ МГУ; ЗИН; Ауэзов, 1971; Ковшарь, 1974) Птицы из Забайкалья (ЗМ МГУ; ЗИН; Ларионов, Чельцов-Бебу- тов, 1972; Васильченко, Голо- вушкин, Осипова, устное сообщ.)
п lim Среднее п lim Среднее
Длина крыла dd 7 338—356 348 18 337—362 347 99 8 322-347 328 4 334-354 343 Длина клюва 66 7 35,0-38,0 36,9 18 32,3-42,5 36,9 99 8 32,5-36,1 34,4 4 32,2-36,6 34,1 Длинацевки 66 7 53,0-64,6 60,2 18 53,1-65,4 57,9 99 8 52,5-59,0 56,2 4 49,0-58,5 54,4 Массатепа dd 7 470-575 505,8 _ _ _ 99 5 420-488 462,8 - - dd„ 99 - - - 7 499-665 573,7
крылышко, подмышечные и нижние кроющие крыла белые; спина и верхние крою-
щие крыла светло-серые. II (первое видимое) первостепенное маховое белое со свет-
ло-серым основанием, черным наружным опахалом почти до вершины пера и черным
предвершинным пятном. Третье первостепенное с черным предвершинным пятном,
черным наружным опахалом в дистальной части пера и вторым черным пятном у самой
вершины; серый цвет основания постепенно переходит в белый примерно в средней
части пера. На IV пере черное предвершинное пятно и черный участок наружного опа-
хала сдвинуты к вершине, светло-серый цвет основания распространяется примерно
на две трети пера. V и VI первостепенные маховые с черными предвершинными пятна-
ми у самой вершины; серый цвет доходит почти до самого пятна. Остальные перво-
степенные маховые серые, на VII может быть черное предвершинное пятно. Второсте-
пенные маховые светло-серые с белой дистальной частью. Клюв и лапы темно-красные.
По А.Ф. Ковшарю (1974), радужина темно-коричневая, края век ярко-красные.
Зимний окончательный наряд не описан.
Пуховой наряд (ЗИН; живые птенцы с Торейских озер). Голова, шея, грудь, бока
и брюшко серебристо-белого цвета. Лоб, подбородок, спина и крылья светло-серые
с отдельными темно-серыми пушинками; на спине, верхней части шеи и крыльях тем-
ные пушинки нередко объединяются в неясные темно-серые пятна. Клюв черно-серый,
с кончиком цвета слоновой кости, лапы серовато-бурые.
Гнездовой наряд (Ковшарь, 1974). Лоб, щеки и горло белые, темя и затылок с не-
ясным темным рисунком. Перья зашейка белые с широкими предвершинными буры-
ми полосами; перья спины и верхних кроющих крыла сизые, с широкими бурыми
предвершинными полями и широкими беловатыми окончаниями. Надхвостье, бока
и весь низ белые. Хвост белый с широкой предвершинной черно-бурой полосой. Пер-
вое и второе маховые сплошь черные, остальные с постепенно увеличивающимися
в проксимальном направлении белыми полями на внутренних опахалах; все маховые
с белыми каплевидными пятнами на вершинах. Клюв темно-бурый, более светлый
у основания подклювья, лапы темно-серые. Радужина темно-коричневая, края век
черные.
Первый зимний наряд (экземпляры колекции ЗИН № 157 118 и 157 119 из Китая,
провинция Хэбей, г. Дату, 8.IV 1935 г. и 29.X 1934 г.). Крылья и рулевые, как в гнез-
довом наряде. Лоб, горло, уздечка, нижняя часть шеи, брюхо, бока, надхвостье и под-
70
Реликтовая чайка
хвостье - белые, на темени мелкие темно-бурые пестрины, которые постепенно укруп-
няются на затылке и задней части шеи и превращаются в редкие каплевидные пестри-
ны. Грудь белая, в мелких редких бурых пестринах или без них. Спина серая.
Первый летний и второй зимний наряды не описаны.
Второй летний наряд (экземпляр № 157 117 коллекции ЗИН от 9.IV 1935 г. из Ки-
тая, провинция Хэбей, г. Дату). Как окончательный, но II (первое видимое) и III ма-
ховые черные с белыми предвершинными пятнами. На внутренних опахалах II—V ма-
ховых широкое светло-серое поле, длина которого увеличивается от II к V перу; у
последнего оно не доходит до края крыла 2—3 см. VI—VII первостепенные маховые
серые с бурым предвершинным пятном, остальные серые.
Третий зимний и третий летний наряды, по-видимому, окончательные, хотя отдель-
ные черты промежуточных нарядов у некоторых особей могут сохраняться.
Линька. Почти не изучена. По данным М.А. Осиповой (1987а), линька взрослых
птиц в зимний наряд носит растянутый характер и протекает во второй половине ию-
ля — августе, завершаясь, по-видимому, уже на зимовках. Первым начинает меняться
контурное оперение, последними — рулевые и кроющие рулевых. Линька маховых
и их кроющих начинается с внутренних первостепенных. Сроки линьки промежуточ-
ных нарядов не установлены. В коллекции ЗИН АН СССР имеются 2 экземпляра в
первом зимнем наряде, добытые 29.X и 8.IV, и особь во втором летнем наряде, добы-
тая 9.IV.
Распространение. Гнездовой ареал. Гнездование достоверно отмечено лишь
в трех пунктах (рис. 18, 19) : на оз. Алаколь (Талды-Курганская обл. Казахской ССР),
на оз. Барун-Торей (Читинская область РСФСР) и в 1984 г. на оз. Балхаш (Ауэзов,
1986). Резкие колебания численности гнездящихся чаек по годам и крайне незначи-
тельное число встреч окольцованных взрослых птиц в колонии при почти сплошном
кольцевании птенцов говорят о наличии еще каких-то неизвестных колоний (Ауэзов,
1980). В сезон размножения взрослые птицы добыты в июне 1957 г. на оз. Ихзс-Нур
в Гобийском Алтае (Piechocki et al., 1981) и 15.V 1966 г. на оз. Байан-Нур южнее
оз. Буйр-Нур в восточной части Монголии (Stubbe, Bolod, 1971). Одиночная птица
встречена 1—2.V 1975 г. на берегу р. Булган-Гол в западной части Монголии (Piechocki
et al., 1981); 20 пар отмечены 24.IV—5.V 1977 г. на оз. Орок-Нур и 3 пары около
оз. Таатзин-Цаган-Нур в монгольской Долине озер, 3 птицы добыты 5.VII 1977 г. на
оз. Хух-Нур в северо-восточной части Монголии (Kitson, 1980). Две залетных особи
были отмечены на северном склоне горного массива Мунх-Хайрхан в западной части
Монголии (Кищинский и др., 1982). В других местах Монголии, в том числе в Котло-
вине больших озер, пока не встречена (Kitson, 1980). Из перечисленных мест гнез-
дование (по-видимому, не ежегодное) наиболее вероятно в окрестностях озер Бай-
ан-Нур, Хух-Нур и в Долине озер (Stubbe, Bolod, 1971; Kitson, 1980). Возможно, ко-
лонии реликтовых чаек есть на территории Китая по соседству с озерами Алаколь
и Барун-Торей.
Зимовки реликтовых чаек расположены, по-видимому, в Юго-Восточной Азии.
29.Х 1934 г., 8 и 9.IV 1935 г. реликтовых чаек добывали в заливе Бохайвань Желто-
го моря (ЗИН), a 30.IX 1971 г. молодая чайка, окольцованная птенцом 3.VI того же
года на оз. Алаколь, была добыта на оз. Байты-Лонг в провинции Куангнин Вьетнама
(Ауэзов, 1974, 1977).
Миграции. Пути и сроки миграции реликтовых чаек практически не изучены. Мож-
но предположить, что осенний и весенний пролет торейских чаек идет через залив Бо-
хайвань. Возможно, от этого залива идет на запад и весенний миграционный путь ала-
кольских чаек, который, вероятно, проходит затем через пустыню Гоби, через Долину
озер и далее через Джунгарские ворота, в окрестностях которых первая реликтовая
71
Семейство Чайковые
чайка в 1973 г. отмечена 31 марта (Ауэзов, 1980). Птицы летят одиночно, парами
и группами до 9 особей (Ауэзов, 1980). По данным трех возвратов колец (Ауэзов,
1977), одни молодые птицы после подъема на крыло до конца сентября держатся
неподалеку от места гнездования (до 250- 300 км к северо-западу от оз. Алаколь),
другие в этот же период совершают перелет за тысячи километров до мест зимовок.
Поздним летом и осенью реликтовые чайки отмечались на оз. Хур-Нур в Восточной
Гоби (9 и 12.VIII 1970 г.) и по южным предгорьям Хангая, где птицы 15—17.IX 1982 г.
Рис. 18. Область распространения
реликтовой чайки
1 — гнездовые поселения, 2 — встре-
чи птиц в сезон размножения, 3 —
предполагаемая граница участков
гнездового ареала, 4 — предполагае-
мые места зимовок, 5 — предпола-
гаемые направления весенних миг-
раций
80 100 120
80
Рис. 19. Ареалы реликтовой {1—2} и буроголовой (^-5) чаек в СССР
1> 3 — известные гнездовые поселения, 2, 4 — районы легких кочевок, 5 — залет буро-
головых чаек
кормились полевками Брандта и исчезли 20.IX с установлением морозов (Е.Н. Куроч-
кин, устное сообщ.).
Численность. На оз. Алаколь в 1968 г. гнездилось 15—20 пар, в 1969 г. — 25—30,
в 1970 г. — 118, в 1971 г. — 35, в 1972 г. — более 120, в 1973 г. — не загнездились, в
1974 г. — 40 пар, в 1975 г. — около 500 пар, в 1976 г. — около 800, в 1977 г. — око-
ло 1200, в 1978 г. — около 350, в 1979 г. — около 300, в 1980 г. — 414, в 1981 г. —
252, в 1982 г. — 350, в 1983 г. — 700 и в 1984 г. — 700 пар (Ауэзов, 1975; Ауэзов и др.,
1981; Ауэзов, Сема, устное сообщ.). На оз. Барун-Торей в 1967 г„ отмечено не менее
100 гнездящихся пар, в 1970 г. - 81, в 1975 г. - 322, в 1976 г. - 493, в 1977 г. - 86,
в 1979 г. — 612, в 1980 г. — 312, в 1981 г. — 280, в 1982 г. — 653, в 1983 г. — не загнез-
72
Реликтовая чайка
лились, в 1984 г. — 320, в 1985 г. — 1025 пар (Леонтьев, 1968; Потапов, 1971; Го-
ловушкин, 1977; Зубакин, 1978; Васильченко, 1986). На оз. Балхаш в 1984 г. встре-
чена 1 пара птиц с птенцом в возрасте 15-17 дней (Ауэзов, 1986). В целом в СССР,
по-видимому, может гнездиться до 2,2 тыс. пар; мировая популяция вряд ли превыша-
ет 10 тыс. взрослых особей.
Местообитания. В сезон гнездования - соленые степные озера с непостоянным уров-
нем воды. В период пролета реликтовые чайки держатся по речным долинам и внутрен-
ним водоемам, во время зимовки, — по-видимому, по морским побережьям.
Размножение. К размножению приступает, видимо, в возрасте 2-3 лет. Моногам,
облигатно-колониальный вид. Формирование пар происходит вне территории колонии.
В этот период и позже в колонии можно наблюдать разнообразные агрессивные и брач-
ные демонстрации..
У реликтовых чаек отмечена большая часть демонстраций, характерных для дру-
гих видов чаек (Зубакин, 1979; Zubakin, Flint, 1980). Наклонная поза с долгим кри-
ком состоит из четырех фаз (рис. 20, а—г); во второй фазе птица издает крик из 3—5
слогов — своеобразное похрюкивание со взвизгиванием, напоминающее визг поросен-
ка. Длительность всей демонстрации 3-5,5 с, в среднем 3,9 с (п = 6).
К агрессивным демонстрациям относятся прямая поза, ’’квохтание”, ’’огрызание”
и демонстрация угрозы на близком расстоянии. Прямая поза (рис. 20,д) производит
впечатление менее ритуализованной демонстрации по сравнению с прямой позой озер-
ной и серебристой чаек и употребляется в агрессивных столкновениях редко — глав-
ным образом вне колонии. ’’Квохтание” (рис. 20,е) довольно часто можно видеть
в колонии, оно сходно с аналогичной демонстрацией других чаек. В большинстве слу-
чаев ’’квохтание” имеет агрессивный характер по отношению к соседним птицам. Вне
колонии оно отмечается во время брачной церемонии. Демонстрация угрозы на близ-
ком расстоянии (рис. 20,ж), по-видимому, видоспецифична для реликтовой чайки.
Это наиболее часто встречающаяся агрессивная демонстрация в колонии. ’’Огрызание”
(выпады открытым клювом в сторону противника) у реликтовой чайки в отличие
от других облигатно-колониальных чаек отмечается лишь изредка.
К сфере брачных демонстраций принадлежат: мяукающий крик (рис. 20,з), отво-
рачивание клюва и вскидывание клюва, которые употребляются в тех же ситуациях,
что и у других видов чаек.
На оз. Алаколь первые реликтовые чайки появляются 4—13.IV (Ауэзов, Сема, уст-
ное сообщ.), на Торейских озерах — в третьей декаде апреля (Васильченко, устное
сообщ.) или в первой половине мая (Леонтьев, 1972). Колонии располагаются либо
на невысоких галечниковых и глинисто-щебнистых островах-косах, появившихся
в результате падения уровня воды (о-в Хухан на оз. Барун-Торей, острова урочища
Чубар-Тюбек на оз. Алаколь), либо на высоких (до 60-65 м) небольших островах-
останцах (о-в Арал на оз. Барун-Торей и о-в Средний на оз. Алаколь). Выбранные
под колонию острова удалены от берега не менее чем на 100 м, чаще на несколько
километров. Колонии каждый год образуются на новом месте, хотя нередко в тече-
ние ряда лет в пределах одного и того же острова. Выбираются ровные участки либо
лишенные растительности, либо с низкой редкой растительностью (Ауэзов, 1980;
Зубакин, 1981а; Осипова, 19876). Величина колонии варьирует от 15-20 до 1200 пар,
нередко колонии состоят из 2—3 субколоний размером от нескольких гнезд до не-
скольких сот, удаленных друг от друга на расстояние до 100 м (Ауэзов, 1975; Головуш-
кин, 1977; Zubakin, Flint, 1980). Внутри субколоний, в свою очередь, можно неред-
ко различить отдельные микроколонии, гнезда в которых расположены близко друг
к другу. Вылупление птенцов в большинстве гнезд такой микроколонии происходит
почти одновременно (Головушкин, 1977; Zubakin, Flint, 1980). Колонии реликто-
73
Семейство Чайковые
вых чаек практически всегда примыкают к колониям чеграв или чайконосых крачек
(Ауэзов, 1980; Zubakin, Flint, 1980).
На месте будущей колонии реликтовые чайки до откладки яиц появляются лишь
периодически. Строительство гнезд начинается за 1—2 дня до откладки яиц одновре-
менно группой прилетевших на остров птиц, причем с каждым днем число загнездив-
шихся чаек возрастает. Основная масса птиц приступает к размножению в сжатые
сроки, но весь период формирования колонии может занимать 20-25 дней (Ауэзов,
Рис. 20. Демонстрационное поведение реликтовой чайки (по: Зубакин, 1979, с изменениями)
а—г — последовательность поз при долгом крике, д — прямая поза, е — ’’квохтание”, ж — демонстра-
ция угрозы на близком расстоянии, 3 — мяукающий крик
1977, 1980). Откладка яиц на о-ве Среднем начинается в первой декаде мая; в 1970 г.
полная кладка из 3 яиц найдена 11.V, начало массовой откладки яиц отмечено 14.V
(Ауэзов, 1977). На Торейских озерах яйцекладка начинается примерно на неделю
позже. Отдельные колонии могут формироваться и позже — в начале июня.
Плотность гнездования очень высокая. На оз. Барун-Торей среднее расстояние меж-
ду центрами ближайших соседних гнезд составляет 0,45 м (и = 134), индекс плотности
гнездования равен 0,40 (Zubakin, Flint, 1980). На оз. Алаколь гнезда располагаются
в 0,3—0,5 м, редко дальше 1 — 1,5 м друг от друга, общая площадь колонии обычно
не превышает 40-50 м2 (Ауэзов, 1977, 1980). Размеры гнезд на оз. Барун-Торей (мм):
диаметр гнезда 230—250, в среднем 300; диаметр лотка 160—195, в среднем 180;
глубина лотка 35—50, в среднем 40 (п = 25) (Zubakin, Flint, 1980). Гнезда построены
из сухих стеблей растений и перьев, в ряде случаев они могут представлять собой ям-
ки в грунте без всякой выстилки; края гнезд запачканы пометом.
Величина кладки 1—4 яйца (крайне редко 5), чаще всего 3 яйца. На о-ве Алаколь
средняя величина кладки колебалась от 2,4 до 2,8 яиц (Ауэзов, 1980), на оз. Барун-
74
Реликтовая чайка
Торей — от 2,0 (поздно загнездившаяся группа птиц) до 3,1 яиц (Головушкин, 1977;
Zubakin, Flint, 1980). Размеры яиц (мм) на оз. Алаколь (п = 100): 55,8—65,0 X 38,5—
46,0, в среднем 59,7 Х42,8. Масса (г) ненасиженных яиц (п = 14): 52,7-70,7, в сред-
нем 60,1; яиц.перед вылуплением (и = 11): 43,5-54,0, в среднем 49,8 (Ауэзов, 1977).
Размеры яиц (мм) на оз. Барун-Торей (и = 56): 55,5—65,6 X40,6—46,0, в среднем
60,1 X43,4 (Головушкин, 1977; Zubakin, Flint, 1980). Форма яйцевидная, окраска
сложная. Фон яйца светлый: светло-бежевый, светло-оливковый, светло-зеленоватый,
кремовый. Рисунок в виде достаточно редких четких пятен, крапин и черточек, глубо-
ких — темно-серого цвета, поверхностных — от коричневато-черного до темно-оливко-
вого цвета (Ковшарь, 1974; Zubakin, Flint, 1980). Насиживание начинается с перво-
го яйца и продолжается 24—26 дней, насиживают попеременно самец и самка (Ауззов,
1977). Первые птенцы вылупляются на оз. Алаколь 27—28 мая — 3 июня, на оз. Ба-
рун-Торей в первую декаду июня (Потапов, 1971; Головушкин, 1977; Zubakin, Flint,
1980). После вылупления птенцов чайки-родители не уносят скорлупу яиц из гнезда.
Она либо затаптывается, либо выталкивается на край гнезда. Основная масса птен-
цов вылупляется в течение нескольких дней.
Масса тела только что вылупившихся птенцов (и = 12) 36—45, в среднем 42,1 г;
обсохших — 27—30 г. В первые 5—7 дней после вылупления один из родителей постоян-
но находится с птенцами, согревая их или защищая от солнца. Птенцов кормят пищей,
отрыгнутой из зоба. На оз. Алаколь в 50 из 88 кормлений пуховичков пищей служили
звонцы (сем. Chironomidae), в 21 — жуки, в 6 — прямокрылые и клопы, в 7 — неопре-
деленные насекомые, в 4 - рыба. В корме для птенцов более старших возрастов также
преобладали звонцы (Ауэзов, 1977). В возрасте 3-4 дней птенцы покидают гнезда
и собираются в группы из 15—30 особей, которые находятся под охраной нескольких
взрослых птиц (Головушкин, 1977). Во время тревоги в первые дни жизни птенцы
затаиваются в гнезде’, затем, как птенцы черноголовых хохотунор и морских голуб-
ков, сбиваются в плотный табунок (Ауэзов, 1977; Zubakin, Flint, 1980). В первые
дни после вылупления родители не отличают своих птенцов от чужих (Потапов, 1971).
Более старшие птенцы подвергаются жестоким нападениям (вплоть до смертельно-
го исхода), если пытаются присоединиться к чужому выводку. Молодые поднимают-
ся на крыло в возрасте 40—45 дней (Ауззов, 1977). После подъема на крыло моло-
дые и взрослые птицы начинают откочевывать из района колонии, где в начале ав-
густа остаются лишь единичные особи.
Доля неоплодотворенных яиц в колонии на оз. Алаколь составляет 2—4,4%, число
поднявшихся на крыло молодых в пересчете на пару птиц колеблется по годам от
0,3 до 2,0; в 1974, 1980, 1982, 1983, 1984 гг. колонии погибали целиком, причем в 4
последних года они были смыты штормами (Ауэзов, 1980; Ауэзов, Сема, устное со-
общ.). На оз. Барун-Торей доля неоплодотворенных яиц в колонии в 1982 г. состави-
ла около 4%, погибло около 20% вылупившихся птенцов. В 1970 и 1976 гг. птенцы
в колонии на о-ве Хухан погибли полностью (Потапов, 1971; Зубакин, 1979). При-
чины гибели яиц и птенцов, помимо штормов, — хищничество серебристых чаек, на-
падения взрослых птиц на птенцов, пытающихся присоединиться к чужим выводкам,
каннибализм. Гибель птенцов от этих причин резко увеличивается при возрастании
фактора беспокойства (Ауэзов, 1980; Zubakin, Flint, 1980).
Суточная активность, поведение. Активность дневная, однако есть указания на не-
которую активность ночью, в частности на прилет чаек в колонию до рассвета (Ауэ-
зов, 1977). Характерная черта поведения реликтовых чаек в колонии — их малая пуг-
ливость. В период вылупления птенцов чайки подпускают человека к колонии на 10—
20 м, после чего меньшая часть птиц взлетает, а большинство начинают отходить от
своих гнезд (Потапов, 1971; Ауззов, 1977; Zubakin, Flint, 1980).
75
Семейство Чайковые
Питание. Кормовые стации реликтовых чаек — мелководья у берега и зона запле-
ска, где чайки поедают сдутых в воду и прибитых к берегу насекомых, снулых рыб,
а также ракообразных; кроме того, целинные степи и поля (Журавлев, 1975; Головуш-
кин, 1977; Ауэзов, 1980; Осипова, 19876). На оз. Алаколь в период гнездования основ-
ным кормом служат насекомые, которые отмечены в 100% погадок взрослых птиц
(где составляют 98,5% общего числа кормовых объектов) и в 97,1 — 100% отрыжек
птенцов. Изредка среди пищевых объектов отмечена рыба, мелкие млекопитающие,
воробьиные птицы, мелкие ракообразные и пауки; в отдельные годы часто встречают-
ся зерна ячменя. Среди насекомых главный корм взрослых птиц и птенцов — комары —
звонцы (хирономиды). В отличие от озерных и серебристых чаек реликтовых чаек
не отмечали на свалках населенных пунктов, у звероферм и рыбоприемных пунктов
(Ауэзов, 1980).
По данным анализа погадок, собранных на Торейских озерах 11—16.VI 1976 г.
(п = 163) и в июле 1982 г. (и = 120), зерна культурных злаков содержались в 97,5%
погадок в 1976 г. и в половине погадок в 1982 г. Насекомые (главным образом жу-
ки) в 1976 г. встречены в 55,8% погадок, водные ракообразные — в 24,6%, рыбы —
в 18,4%, воробьиные птицы — в 0,6%, полевки Брандта - в 1,2%, тушканчик-прыгун -
в 0,6%. В 63,2% погадок отмечены гастролиты. В 1982 г. встречены остатки 2 полевок
Брандта; среди насекомых более 98% составляли жуки, как правило, массовые ви-
ды (чернотелки, долгоносики, жужелицы). В 1976 г. отмечено поедание яиц чеграв,
а также яиц и, возможно, птенцов своего вида (Zubakin, Flint, 1980; Осипова, 19876).
Враги, неблагоприятные факторы. Колонии реликтовых чаек недоступны для на-
земных хищников. Из пернатых хищников наибольшее воздействие на их поселения
оказывают серебристые чайки, в отдельных случаях нацело уничтожая яйца и птен-
цов (Зубакин, 1979). Важное значение имеют погодные факторы. Установлено, что
численность загнездившихся реликтовых чаек увеличивается в засушливые и теплые
годы, а в холодные и дождливые снижается (Ауэзов, 1980). Неоднократно отмечалась
гибель колоний во время штормов.
Особую роль в гибели потомства у реликтовых чаек играет фактор беспокойства.
Исключительная чувствительность к нему объясняется, во-первых, сочетанием канни-
бализма по отношению к кладкам с высокой плотностью гнездования и, во-вторых,
совместным поселением с серебристыми чайками. В отсутствие беспокойства реликто-
вые чайки в отличие, например, от более мелких черноголовых могут противосто-
ять хищничеству серебристых чаек. Однако общие взлеты птиц колонии во время
беспокойства ее человеком не только чрезвычайно облегчают серебристым чайкам
добычу яиц и птенцов, но и дают возможность особям-каннибалам расклевать клад-
ки чаек, опускающихся в колонию позже других. Достаточно частые тревожные взле-
ты поэтому приводят к изреживанию колонии, которая становится неспособной про-
тивостоять натиску серебристых чаек и гибнет. В качестве фактора беспокойства,
запускающего подобный механизм уничтожения колонии, могут выступать не толь-
ко посещения колонии человеком или четвероногими хищниками, но и некоторые
погодные условия: дождь с сильным ветром, шторма. По-видимому, именно особая
чувствительность к фактору беспокойства ответственна за крайнюю малочисленность
этого вида, и описанная выше малая пугливость реликтовых чаек в колонии — эво-
люционная попытка как-то снизить его катастрофическое воздействие (Зубакин, 1979;
Zubakin, Flint, 1980).
Хозяйственное значение, охрана. Из-за крайней редкости хозяйственного значе-
ния вид не имеет. Реликтовая чайка занесена в Красную книгу МСОП и Красную книгу
СССР. Сохранение этой чайки от вымирания зависит от обеспечения ей возможности
гнездования в условиях, исключающих всякое беспокойство со стороны человека.
76
Черноголовая чайка
Черноголовая чайка —
Larus melanocephalus Temmmck, 1820
Mediterranean Gull (англ.), Schwarzkopfmowe (нем.), Mouette melanocephale (фр.).
Син. (лат.) : Hydrocoloeus melanocephalus (Temminck, 1820).
Син. (рус.): средиземноморская чайка.
Син. (англ.): Adriatic Gull.
Статус. Гнездящийся перелетный, частично зимующий вид.
Общая характеристика и полевые признаки. Темноголовая чайка средней величины,
с озерную чайку или чуть крупнее. От последней взрослые особи в сезон гнездования
отличаются белыми концами крыльев, угольно-черной головой и более мощным клю-
вом. Молодые птицы в гнездовом наряде отличаются от озерных чаек более темным
цветом головы и верхней части тела, а также преобладанием темного цвета на первых
(дистальных) первостепенных маховых. Полет, как и у других чаек сходного размера.
Хорошо плавает, на воде сидит высоко, не ныряет. Голос не похож на голос озерной
чайки: совершенно отсутствуют хриплые каркающие крики. Основной видовой крик —
гнусавое ”эва”, его часто можно услышать в сезон размножения как в колонии, так и
вдали от нее. Тревожный крик в колонии — трехсложное ”ка-ка-ка”; реже он состоит
из 2—4 слогов. Вне сезона размножения черноголовая чайка, по С. Крэмпу (Cramp,
Simmons, 1983), молчаливая птица.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Статус черноголовой чайки в подсемействе Larinae не
совсем ясный. По окраске и размерам она близка к ’’капюшонным” чайкам, рисунок
первостепенных маховых двухлетних птиц сходен с расцветкой маховых взрослых
черноголовых хохотунов и реликтовых чаек, массивным клювом (Dwight, 1925) и не-
которыми деталями демонстрационного поведения (Mauersberger, 1970) черноголо-
вая чайка напоминает крупных белоголовых чаек. Отмечены смешанные пары и гиб-
риды черноголовых чаек с озерными чайками (Kohler et al., 1983). Дж. Дуайт помеща-
ет эту чайку в один род с озерной, хотя и с оговорками (Dwight, 1925). Сходных взгля-
дов придерживается Г. Волтере (Wolters, 1975), предлагая, однако, выделить черного-
ловую чайку в особый подрод. Т. Мойнихан (Moynihan, 1959) относит ее в сборную
группу ’’примитивных капюшонных чаек”.
Описание. Размеры (мм) из Тендровского залива Черного моря (ЗМ МГУ):
Длина крыла:
33 (и = 9) — 295—311 (среднее 305);
99 (и = 4) — 282—307 (среднее 296).
Длина клюва:
33 (и = 9) - 32,5-37,0 (среднее 35,1) ;
99 (и = 4) — 31,5—37,0 (среднее 33,7).
Длина цевки:
33 (п = 8) — 48,9—54,7 (среднее 51,7):
99 (и = 4) — 46,9—52,0 (среднее 49,7).
Окраска (Dwight, 1925; Дементьев, 1951; Cramp, Simmons, 1983; ЗИН АН
СССР; ЗМ МГУ). Самец и самка в брачном наряде. Голова черная, причем черный цвет
захватывает не только затылок и горло, но и верхнюю часть передней и задней стороны
шеи. Над и под глазами небольшие белые пятнышки (полоски). Шея, грудь, бока, брю-
хо, поясница, надхвостье, подхвостье, рулевые и подкрылья белые. Спина, кроющие
77
Семейство Чайковые
крыла и плечевые светло-серые. Первостепенные и второстепенные маховые светло-се-
рые, но несколько светлее спины, к вершине постепенно белеющие. По краю наружного
опахала II (первого видимого) первостепенного тонкая черная полоска, не доходящая
до вершины пера. Клюв ярко-красный, нередко с бурой предвершинной полосой;
кончик клюва может быть желтым. Лапы красные, радужина коричнево-черная, края
век ярко-красные.
Самец и самка в зимнем наряде. Голова белая с серовато-бурыми пятнами перед
глазом и на кроющих уха, на затылке отдельные серо-бурые пестрины. Остальное опе-
рение, как в брачном наряде. Клюв и лапы темно-красные, иногда оранжевые.
Пуховой наряд. Общий тон окраски серый или рыжевато-серый, брюшко белое.
Лоб и уздечка обычно черные, на темени, затылке, задней части шеи и крыльях — много-
численные густые черные крапины, нередко группирующихся в полоски. На спине пят-
на сливаются в две-три неясные широкие продольные темные полосы, чередующиеся
со светлыми полосами. Горло охристое или белое, подбородок и грудь темно-серые
(пушинки с темно-серым основанием и светлым концом). В отличие от птенцов других
видов чаек фауны СССР у пуховичков черноголовых чаек вершины нескольких пуши-
нок соединены вместе и заканчиваются одной длинной остью; в результате создается
впечатление, что птенец еще не обсох (подобное строение пуха и у птенцов пестроносых
крачек). Клюв темный, серо-коричневый с кончиком телесного цвета, лапы серо-телес-
ные (ЗИН АН СССР; живые пуховички с Чонгарских островов).
Гнездовой наряд. Лоб, темя, затылок и задняя часть шеи серо-бурые с мелкими
поперечными охристыми пестринами (каемки перьев); уздечка и кроющие уха нес-
колько темнее, без пестрин. Подбородок, горло, передняя часть шеи, брюхо и под-
хвостье белые. Грудь серая с неясными светлыми поперечными пестринами, бока белые
с нечеткими серо-бурыми пестринами. Поясница серая, надхвостье светло-серое с неяс-
ными темными пестринами. Большие кроющие крыла серые, остальные кроющие
крыла, третьестепенные маховые и перья спины бурые с охристыми каемками. Перво-
степенные маховые темно-бурые, начиная с III (второго видимого) на внутренних опа-
халах появляется белое поле, не доходящее 3—6 см до вершины пера. Иногда небольшое
белое поле заметно уже на II первостепенном. Начиная с VII на наружном опахале отме-
чается серое поле. Второстепенные маховые серые с широкой темно-бурой предвершин-
ной полосой и белыми или охристыми вершинами. Рулевые белые с черным предвер-
шинным пятном, менее развитым на двух крайних парах. Клюв черный с более свет-
лым основанием, ноги мясного или темно-телесного цвета, радужина бурая.
Первый зимний наряд. Голова белая, на затылке, задней стороне шеи и кроющих
уха буроватые пестрины и пятна, перед глазом черное полулунное пятно. Спина светло-
серая, иногда с коричневатым оттенком. Крылья и хвост, как в предыдущем наряде.
Шея и нижняя часть туловища белые.
Первый летний наряд. Верхняя часть головы до затылка черная с белыми пестрина-
ми, подбородок и горло белые. Спина и поясница светло-серые. Остальное оперение,
как в предыдущем наряде.
Второй зимний и второй летний наряды отличаются от окончательного зимнего и
летнего нарядов расцветкой первостепенных маховых: они с белыми вершинами, чер-
ными или буроватыми предвершинными полосами и большим или меньшим развитием
темного цвета на наружных опахалах и пристволовой части внутренних опахал. Темный
рисунок на первостепенных маховых весьма изменчив. Клюв буровато-красный, ноги
красноватые.
Линька. Линька из гнездового в первый зимний наряд (август—ноябрь) частичная,
сменяется оперение головы и туловища. В последующем линька бывает дважды в году:
полная осенняя и частичная весенняя. Предбрачная линька взрослых птиц протекает
78
т
Черноголовая чайка
о декабря-января по март-апрель (сменяются перья головы, туловища, часть кроющих
крыла и 1—2 второстепенных маховых), полная послебрачная — с июня по октябрь
(декабрь). Линька в промежуточные наряды происходит, видимо, в более ранние сро-
ки (Cramp, Simmons, 1983).
Распространение. В прошлом веке гнездовой ареал вида включал, очевидно,
только северное и северо-восточное побережья Черного моря и, возможно, Эгейское
море (рис. 21). Здесь и сейчас сосредоточена почти вся мировая популяция вида. В на-
Рис. 21. Область распространения черноголовой
чайки
1 — постоянные поселения, 2 — временные посе-
ления, 3 — область регулярных кочевок, 4 — мес-
та зимовок, 5 — залеты
стоящее время в этом регионе гнездовые
поселения отмечены в Греции, Румынии,
на внутренних водоемах Турции (Cramp,
Simmons, 1983). В советском Причер-
номорье основные колонии располагают-
ся на островах Орлов, Бабин, Смаленый
и некоторых других Тендровского зали-
ва, в меньшем количестве черноголовая
чайка гнездится на о-ве Каланчакском,
недалеко от г. Скадовска, в Картказац-
ком заливе западнее Красноперекопска,
на Чонгарских островах и о-ве Коянлы
Сиваша (Ардамацкая, 1975а, в, 1981;
Костин, 1975; Зубакин, Костин, 1977;
Сиохин, 1981а). Одна пара в 1975 г. гнез-
дилась в Тилигульском лимане (Черни-
ченко, Стойловский, 1981).
С 60-х годов нашего столетия (в Ни-
дерландах и Венгрии несколько раньше —
с 1933 г. и 1940 г. соответственно) не-
большие колонии в несколько пар и от-
дельные пары птиц стали гнездиться на
побережьях Северного и Балтийского мо-
рей, а также внутренних водоемах почти
всех стран Западной и Центральной Евро-
пы. Небольшие поселения этих птиц по-
явились и в западной части Средиземного
моря. В Англии черноголовая чайка нере-
гулярно гнездится с 1968 г. (Cramp, Sim-
mons, 1983), отдельные пары пытались
гнездиться в 1970, 1973 и 1975 гг. в Да-
нии (Moller, 1978), 1 пара в 1981 г.-и 3—4 пары в 1982 г. гнездились в Польше (Ni-
tecki, 1984; Wesolowski et al., 1984). Нерегулярно гнездится в Бельгии с 1964 г., поч-
ти ежегодно с 1953 г. в Венгрии, с 1968 г. в Нидерландах, с 1963 года в ГДР, с 1965 г.
в ФРГ и во Франции. Отмечено гнездование в 1966 г. в Чехословакии, в 1969, 1973,
1976 гг. в Швейцарии (1 пара), в 1970 г. в Швеции, в 1977 г. в Австрии (1 пара), в
1978 и 1981 гг. в Италии (Cramp, Simmons, 1983); в 1962 и 1967 гг. черноголовая
чайка загнездилась в Эстонии (Манк, 1970). Одновременно выявлено расшире-
ние ареала в восточном направлении (Кривенко и др., 1973): в 1972 г. отме-
чены одиночное гнездование на Молочном лимане (Запорожская область) и
две колонии в 67 и 30 пар на Чограйском водохранилище (Восточный Маныч)
(рис. 22).
По-видимому, еще рано квалифицировать массовое выселение отдельных особей
79
Семейство Чайковые
Рис. 22. Ареал черноголовой чайки в СССР
Обозначения те же, что на рис. 21
и групп птиц в 1960—1970-е годы за пределы основного ареала в западном и северо-за-
падном направлениях как расширение ареала вида. Поселения в Западной и Централь-
ной Европе неустойчивы, увеличения их размеров в большинстве случаев не проис-
ходит. Это, вероятно, говорит о неподходящих условиях для обитания здесь черного-
ловых чаек. В то же время в случае некоторых изменений в экологии вида (в частнос-
ти, при переходе на корма антропогенного характера) и при должной охране колоний
можно ожидать в будущем расселения этой чайки по всей Европе. На водохранилищах
системы Маныча черноголовая чайка
нашла себе подходящие условия обита-
ния, ныне она в большом количестве
гнездится на оз. Маныч-Гудило, Чог-
райском водохранилище и Меклетин-
ских озерах (Кривенко, Любаев, 1975,
1981; Кривенко, 1981; Любаев, Кри-
венко, 1981). Закрепилась черноголо-
вая чайка и на Молочном лимане (Сио-
хин, 1981а). Таким образом, правомер-
но говорить о расширении области гнез-
дования вида в восточном направлении
почти на 900 км.
Зимовки. По данным кольцева-
ния, главные места зимовок чаек черно-
морских колоний — средиземномор-
ские побережья Греции, юга Италии,
Сицилии и Туниса; большое число чаек
концентрируется также на средиземно-
морском побережье северной Италии,
Франции, Испании и Марокко. В мень-
ших количествах черноголовые чайки
зимуют вдоль Черноморского побережья Кавказа и Южного берега Крыма, а также в
устье Нила и на озерах Нижнего Египта (Шеварева, 1955; Ардамацкая, 19756, 1977в).
Зимующие птицы отмечены также на атлантических побережьях Марокко (залетают
до Сенегала и Гамбии), Португалии, Испании; с 1950-х годов в небольшом, но посте-
пенно возрастающем числе встречаются на побережьях западной и северо-западной Ев-
ропы от Франции до ГДР (Cramp, Simmons, 1983).
Миграции. Начинают покидать места зимовок, видимо, в конце февраля — марте.
На севере Крыма появляются в середине марта, к 20-м числам становятся обычными
(Костин, 1983); в Тендровский залив первые птицы прилетают в первой половине
марта, иногда в конце февраля (Ардамацкая, 1977в). В апреле основная масса поло-
возрелых чаек уже не встречается в области зимовок, тогда как неполовозрелые птицы
продолжают кочевать здесь всю весну и лето (Шеварева, 1955).
Послегнездовые кочевки, начинающиеся с середины июля, постепенно переходят
в осенний пролет, который заканчивается в ноябре. Основное направление пролета
черноморских чаек — западное, причем часть птиц затем поворачивает на юг и летит
вдоль западного побережья Черного .моря в Грецию. Другие, по-видимому, напрямик
пересекают Балканский полуостров и с августа появляются в северной части Адриати-
ческого моря. Часть птиц летит на восток от места гнездования — вдоль побережий
Крыма, Азовского моря, затем вдоль Черноморского побережья Кавказа. Небольшое
количество молодых чаек пересекают материк в северо-западном направлении, появ-
ляясь на побережьях Балтийского и Северного морей уже в августе (Шеварева, 1955;
80
Черноголовая чайка
Ардамацкая, 19756). Некоторые чайки летят с Черного моря в Средиземное, пересе-
кая азиатскую часть Турции (Cramp, Simmons, 1983).
В Тендровском заливе птицы покидают окрестности колонии во второй половине
августа (Шеварева, 1955). Из северных районов Крыма исчезают в разные годы в кон-
це августа — октябре (Костин, 1983).
Черноголовые чайки, особенно молодые, нередко залетают в глубь материка и за пре-
делы областей пролета и зимовки; чаще всего это бывает в период кочевок. Известны
залеты в Норвегию, Ирландию, Швецию, Ирак, Кувейт, восточную часть Саудовской Ара-
вии, Азорские и Канарские острова, о-в Мадейру (Cramp, Simmons, 1983), в Латвию
во второй половине мая 1970 г. (Виксне, 1983), Московскую область 26.V. 1982 г.
(Зубакин, Харитонов, 1983), в дельту Амударьи 11.III 1974 г. (Ажимуратов, 1984).
В 1950-е годы отмечены регулярные залеты черноголовых чаек на Восточный и Северо-
Восточный Каспий (Залетаев, 1960).
Численность. Непрерывно возрастает, хотя в каждом конкретном поселении может
резко колебаться по годам. На островах Тендровского залива в 1935 г. гнездилось
около 19 тыс. пар, в 1965—1970 гг. — от 100 тыс. до 170 тыс. пар, в 1982 г. отмечено
335 тыс. пар (Ардамацкая, 1975а, 1984). На о-ве Каланчакском в 1972 г. гнездилось
около 5 тыс, пар, в 1973 г. — примерно 15 тыс, пар, в Картказацком заливе в 1973 г. —
450 пар, на Сиваше в 1977—1979 гг. — 987—1457 пар, на Молочном лимане в эти же го-
ды — 1—302 пары, на озерах и водохранилищах Маныча в 1974—1979 гг. — примерно от
2 тыс. до 13,4 тыс. пар (Ардамацкая, 1975в; Костин, 1975; Сиохин, 1981а; Кривенко,
1981; Кривенко, Любаев, 1981; Любаев, Кривенко, 1981). За пределами нашей стра-
ны гнездится вряд ли больше 2 тыс. пар (Cramp, Simmons, 1983), Мировая популяция
черноголовых чаек насчитывает, таким образом, 340—370 тыс. размножающихся пар,
из которых более 99% гнездится в СССР и более 90% - в Тендровском заливе Чер-
ного моря.
Местообитания. В сезон гнездования — морские побережья, заливы, лиманы; в по-
следние годы также соленые, солоноватые и реже пресноводные озера и водохранили-
ща степной и полупустынной зон. Необходимо сочетание недоступных наземным хищни-
кам и редко посещаемых человеком небольших островов (гнездовой биотоп) и при-
брежных степей и полей, богатых массовыми видами насекомых (кормовой биотоп).
Вне сезона размножения — морские побережья, нередко гавани и окрестности городов.
В последние годы отмечается склонность к соседству с человеком, особенно во вне-
гнездовое время, и тенденция к гнездованию в местах, достаточно резко отличающихся
по климату, растительности и кормовым характеристикам от стереотипных для вида
(Cramp, Simmons, 1983).
Размножение. К размножению в большинстве случаев приступает в трехлетием воз-
расте, отдельные особи — годом раньше. Моногам. Плотно гнездящийся факульта-
тивно-колониальный вид. Одни и те же острова могут заниматься чайками в течение
десятков лет (о-в Орлов), в то же время известны поселения, существующие только
один сезон или недолгие годы. Характерны резкие колебания численности в различных
поселениях по годам, что связано, вероятно, с перераспределением пар между коло-
ниями и негнездованием части особей. По крайней мере часть пар образуется не на тер-
ритории колонии (Cramp, Simmons, 1983), однако, по-видимому, формирование пар
происходит и в колонии, поскольку здесь отмечаются холостые самцы, охраняющие
гнездовую территорию (Taverner, 1976).
Репертуар демонстрационного поведения, по наблюдениям Г. Мауэршбергера и
Дж. Тавернерав Западной и Центральной Европе (Mauersberger, 1970; Taverner, 1976)
и В.А. Зубакина на Сиваше, включает многие демонстрации, характерные для других
чаек. Наиболее своеобразна последовательность поз при долгом крике. В первой фазе
81
Семейство Чайковые
(рис. 23 fl) чайка принимает позу, как при ’’квохтании”: задняя часть туловища припод-
нята, грудь опущена, шея вытянута наклонно вперед и вверх и изогнута, клюв направ-
лен наклонно вниз. Затем, почти не меняя положения головы и шеи по отношению
к туловищу, чайка опускает задний конец тела, так что туловище становится горизон-
тальным, а грудь выпячивается вперед. В конце этой (второй) фазы чайка может слег-
ка вскинуть клюв вверх, но в ряде случаев этого не происходит и сразу начинается
третья фаза: птица вытягивает шею наклонно вперед и вверх и, широко раскрыв клюв,
Рис. 23. Демонстрационное поведение черноголовой чайки
а—в — последовательность долгого крика, г — ’’изогнутая” поза, д — прямая поза
издает свой характерный слегка гнусавый крик ”эва”. В первой-второй фазе слышны
негромкие урчащие звуки, из которых первые 1—5 слогов, как и последний слог, ко-
роткие, а предпоследний — длинный и протяжный. В отдельных случаях третьей фазы
может и не быть. Запястные суставы крыльев во время этой и большинства других
демонстраций слегка опущены и отодвинуты от туловища. Чаще всего долгий крик
используется в агрессивных ситуациях. Возможно, он характерен только для самцов
(Cramp, Simmons, 1983).
К другим агрессивным демонстрациям относятся хорошо ритуализованная агрес-
сивная прямая поза, ’’огрызание” — выпады открытым или закрытым клювом в сто-
рону противника и ’’клевание травы”, которое отмечается при столкновениях на гра-
нице участка, но реже, чем у крупных белоголовых чаек.
К демонстрациям сферы брачного поведения относятся мяукающий крик, сгорблен-
ная поза, вскидывание клюва, а также ’’изогнутая” поза. При мяукающем крике шея
вытянута вперед и слегка вверх и изогнута, клюв полуоткрыт; сам крик — нечто вроде
протяжного гнусавого ’’эээууу” или ’’эээуууа”. Данную демонстрацию нередко можно
видеть при смене партнеров на гнезде, но чаще всего в этом случае можно наблюдать
’’изогнутую” позу (рис. 23,г), в которой шея вытянута вверх и слегка изогнута, клюв
направлен наклонно вниз, перья головы и шеи прижаты, так что голова кажется мень-
ше, а шея тоньше, чем обычно; звуки в этой позе не издаются. ’’Изогнутую” позу часто
принимают оба партнера, идя бок о бок и отворачивая клювы друг от друга. Отворачи-
82
Черноголовая чайка
вание клюва можно нередко наблюдать и после агрессивных столкновений. Сгорблен-
ную позу, часто со вскидыванием клюва, принимает самка, выпрашивая корм у сам-
ца перед ритуальным кормлением. Вскидывание клюва можно наблюдать и из ’’изог-
нутой” позы. Г. Мауэршбергер (Mauersberger, 1970) в числе брачных демонстраций
отметил и ’’квохтание”.
Черноголовая чайка гнездится колониями, насчитывающими обычно сотни и тыся-
чи пар, реже десятки и десятки тысяч пар. Еще реже отмечено гнездование отдельными
парами, причем только в колониях с другими видами чайковых (озерная и сизая чайки,
речная крачка и др.); при малой численности возможны смешанные пары с озерной чай-
кой. Заселяет как материковые островки-останцы, так и острова наносного происхож-
дения, как правило, вдали от берега. Под колонии выбирает участки, заросшие не-
густой растительностью — полынью, сведой, лебедой и др., совершенно голых участков
избегает. По соседству с колониями черноголовых чаек могут располагаться колонии
чайконосых крачек, морских голубков, пестроносых и речных крачек, шилоклювок.
Соседства серебристых чаек избегает и на островах, где они гнездятся, не поселяется
(Клименко, 1950а; Кривенко и др., 1973; Ардамацкая, 1975в, 1977а,б, 1981; Ta-
verner, 1976; Зубакин, Костин, 1977; Milbled, Apchain, 1978; Сиохин, 1980; Cramp,
Simmons, 1983; Kohler et al., 1983).
Плотность гнездования высокая. На островах Тендровского залива расстояние между
гнездами, по М.И. Клименко (1950а), варьирует от 0,3 до 1,5 м. На Сиваше расстояние
между центрами ближайших гнезд изменяется от 0,30 до 4,0 м (и = 268), составляя
в среднем 0,63 м; индекс плотности гнездования в среднем равен 0,29, варьируя по ко-
лониям от 0,20 до 0,34. Несмотря на большую плотность, вокруг каждого гнезда есть
небольшая охраняемая территория (Зубакин, 1976а). В Греции (Isenmann, 1975) рас-
стояние между центрами ближайших гнезд 0,32—0,60 м (в среднем 0,44 м;. п = 16).
В качестве материала для гнезд используются сухие стебли травы, сухие водоросли,
отдельные перья. На Сиваше диаметр (мм) гнезда составляет (и = 35) 235—350, сред-
ний 298; лотка 155—200, средний 182; глубина лотка 25—65, средняя 38 (данные
В.А. Зубакина).
Откладка яиц в Тендровском заливе начинается 2—14.V, кладка проходит дружно,
в 8—10 дней (Клименко, 1950а; Бородулина, 1960; Ардамацкая, 19776,в). В отдель-
ных колониях яйцекладка может происходить на месяц позже. В полной кладке 1—4 яй-
ца, чаще всего 2—3, в поздних и повторных кладках 1—2. Размеры яиц (мм): 48,0—
59,8 X 32,6—41,5, в среднем (и = 97) 54,7 X 38,0 (Костин, 1983). Форма яйцевидная,
окраска сложная: по светло-зеленоватому, светло-палевому, сероватому или светло-
оливковому фону разбросаны негустые пятна, крапинки или штрихи, глубокие — се-
рого цвета, поверхностные — бурого. Окраска яиц в целом темнее, чем у морских
голубков, но светлее, чем у озерных чаек. Насиживают оба партнера. Длительность
инкубации 23—26 дней (Ардамацкая, 1977в; Cramp, Simmons, 1983).
Вылупление в Тендровском заливе начинается в первой декаде июня (Ардамацкая,
1977в) и происходит очень дружно (Кузякин, 1959). В 1974 г. на Сиваше вылупление
в разных колониях началось 7 и 8.VI и продолжалось 13—15 дней; в 1973 г. оно завер-
шилось за 12 дней. Длительность вылупления в одном гнезде с 3 яйцами здесь состав-
ляла 1—6 сут, чаще всего 3,2 и 4 сут (в 44,8%, 24,1% и 20,7% случаев соответственно;
и = 29), между появлением первой трещины на яйце и вылуплением проходило 1—3 сут
(чаще всего 3 сут — в 39,3%; и = 33), между появлением отверстия и вылуплением
в 96,8% случаев — 1 сут (и = 31). После вылупления птенцов родители выносят скор-
лупу из гнезда (данные В.А. Зубакина).
Птенцов выкармливают оба родителя, отрыгивая пищу из зоба. По наблюдениям
в Тендровском заливе, кормление бывает 10—18 раз в сутки, а смена партнеров, обо-
83
Семейство Чайковые
гревающих птенцов, 2—3 раза в день (Бородулина, 1960). По наблюдениям на Сиваше,
во время тревоги птенцы в возрасте до 2 сут лежат в гнезде, начиная с 3-суточного воз-
раста выбегают из гнезда и затаиваются по соседству. Примерно через 5 сут после вы-
лупления первого птенца выводок оставляет гнездо и держится среди травы в районе
колонии. В возрасте 15 сут и старше птенцы во время тревоги начинают выбегать из рас-
тительности на открытый берег, еще через неделю в подобной ситуации формируется
плотный табунок, аналогичный табунку морских голубков и черноголовых хохотунов.
После образования табунка к нему могут присоединяться и более молодые птенцы.
Индивидуально опознавать птенцов родители начинают примерно через 2 сут после
вылупления первого птенца, после чего птицы крайне агрессивно относятся к чужим
птенцам, нередко убивая их ударами клюва. По этой причине может погибать до 11%
вылупившихся птенцов (данные В.А. Зубакина). Птенцы поднимаются на крыло че-
рез 35—40 дней после вылупления, в Тендровском заливе это происходит 5—10.VII
(Ардамацкая, 1977в).
В Тендровском заливе в разные годы гибнет от 9 до 23% кладок, главным образом
от наводнений, в меньшей степени от пернатых хищников; гибель среди птенцов 5—
7%, на сухих участках она может достигать 22—30%, по-видимому от перегрева (Кли-
менко, 1950а). Одна из важных причин гибели кладок — расклевывание чайками яиц
своего вида при беспокойстве колонии, особенно на ранних стадиях насиживания (Кли-
менко, 1950а; Зубакин, Костин, 1977). Это происходит в результате сочетания высо-
кой плотности гнездования с хищническими наклонностями вида. Непосредственные
причины гибели гнезд, по наблюдениям на Сиваше, — неодновременное возвращение
чаек на гнезда после тревоги (некоторые птицы возвращаются к гнезду на 10 мин и
более позже остальных) и наличие в колонии неразмножающихся особей, кормящихся
на ее территории и быстро расклевывающих оставленную кладку. После нескольких
тревожных взлетов плотность колонии может настолько уменьшиться, что оставшимся
парам станет не хватать социальной стимуляции для продолжения насиживания. Птицы
оставляют гнезда без видимой причины, и колония окончательно гибнет. Расклевыва-
ния чайками в подобной ситуации своих собственных кладок (Бородулина, 1960) не
отмечалось (данные В.А. Зубакина).
В результате повышенной чувствительности к фактору беспокойства в 1973 г. на Чон-
гарских островах в разных колониях погибло от 52,5 до 98% гнезд (Зубакин, Костин,
1977). В 1974 г. по этой причине погибло 22,7% яиц (и = 225), еще 6,2% составили не-
оплодотворенные яйца; в целом эмбриональная смертность оказалась равной 31,1%
(данные В.А. Зубакина).
Максимальный возраст черноголовой чайки, по данным кольцевания, 15 лет (Арда-
мацкая, 1977в).
Суточная активность, поведение. Активность в основном дневная (Авилова, 1972),
хотя ночью в районе колонии неоднократно приходилось слышать голоса летящих ча-
ек. Черноголовая чайка в высокой степени общественная птица, она держится группа-
ми и стаями во все сезоны. Нередко отмечаются стаи в несколько сот особей;
в Крыму скопления чаек могут достигать 6—8 тыс. особей (Костин, 1983). У черного-
ловых чаек, очевидно, существуют прочные группировки, состоящие из одновозраст-
ных птиц, которые держатся вместе во время пролета и на зимовках, а также совместно
возвращаются в район гнездовий. Стайки определенного состава могут сохраняться
в течение ряда лет, возможно и на протяжении всей жизни данных особей (Шеварева,
1955). В колониях черноголовая чайка более агрессивна и более осторожна, чем чай-
ковые сходного размера (например, морской голубок). На человека у гнезда пики-
рует крайне редко.
84
Озерная чайка
Питание. В сезон гнездования черноголовая чайка трофически связана главным об-
разом с сушей. Основные кормовые стации в это время - целинные приморские степи
и поля. Главные кормовые объекты — насекомые, обычно самые массовые виды. По-
едает также мелких грызунов, яйца и птенцов морских голубков, пестроносых крачек
и других птиц, ящериц, изредка рыбу и зерна культурных злаков (Шевченко, 1937;
Клименко, 1950а; Шеварева, 1955; Бородулина, 1960; Сиохин, 1980, 19816). Анализ
159 погадок, собранных в 1973—1974 гг. на Чонграских островах, показал, что назем-
ные членистоногие (главным образом насекомые) встречаются в 89,3% случаев, мелкие
грызуны — в 37,1% случаев (в том числе полевки Microtus sp. 32,1%), птицы (мелкие
воробьиные, птенцы чаек и крачек) — в 5,0%, яйца птиц — в 1,3%, растения — в 14,5%
случаев. Кроме того, отмечено поедание яиц и птенцов своего вида (Зубакин, 19766).
В 458 пищевых пробах, собранных у птенцов на островах Тендровского залива,
содержалось 36 100 пищевых объектов; 96,6% объектов составляли остатки живот-
ных, из них 96,0% — насекомые (чешуекрылые 25,1%, двукрылые 24,1%, перепончато-
крылые 19,2%, жесткокрылые 16,3%, полужесткокрылые 6,9%, прямокрылые 4,3%).
Остатки позвоночных животных (морские иглы, бычки, ящурки разноцветные, птен-
цы пестроносых и речных крачек и черноголовых чаек, общественные полевки) встре-
чены в 3,7% пробах (0,15% всех экземпляров). Водные животные составляли0,6%кор-
мовых объектов; виды, образующие скопления, — 67,0%. Корм черноголовые чайки
собирают с растений и, видимо, с поверхности почвы. Радиус разлета за кормом чаще
всего 5—45 км, но может достигать 50—60 км (Ардамацкая, Вакаренко, Петрусенко,
устное сообщ.). В Греции он может достигать 28 км (Isenmann, 1975).
Вне сезона размножения основной корм морская рыба и моллюски, а также отбро-
сы рыболовного промысла и пищевые отходы (Cramp, Simmons, 1983).
Враги, неблагоприятные факторы. Из пернатых хищников ущерб колониям черного-
ловых чаек, по-видимому, в наибольшей степени наносит серебристая чайка-хохотунья.
Среди врагов черноголовых чаек указывались также болотный лунь, ворон, серая
ворона. Из неблагоприятных абиотических факторов большое значение имеют нагон-
ные ветры и штормы, смывающие колонии на низких островах, а также ливни с пони-
жением температуры и град (Клименко, 1950а). Сочетание хищнических тенденций
с высокой плотностью гнездования делает колонии этого вида крайне чувствитель-
ными к беспокойству в период насиживания.
Хозяйственное значение, охрана. Уничтожением большого количества насекомых-
вредителей и мышевидных грызунов приносит пользу сельскому хозяйству. Занесе-
на в Красную книгу Эстонской ССР. Колонии охраняются в Черноморском заповедни-
ке (около 90% мировой популяции вида).
Озерная чайка —
Larus ridibundus Linnaeus, 1766
Black-headed Gull (англ.), Lachmowe (нем.), Mouette rieuse (фр.).
Син. (лат.): Hydrocoloeus ridibundus (Linnaeus, 1766), Chroicocephalus ridibundus (Linnaeus, 1766).
Син. (рус.): речная чайка, обыкновенная чайка.
Статус. Гнездящийся, мигрирующий вид, в южной части ареала — зимующий.
Общая характеристика и полевый признаки. Чайка средних размеров (немного
меньше черноголовой чайки и морского голубка, примерно на 25% меньше сизой
чайки). В брачном наряде имеет темно-коричневую голову (издали выглядит черной),
граница коричневой окраски на шее у стоящей птицы косая — идет от затылка к горлу
(у малой и черноголовой чаек головы в брачном наряде черные, включая затылок,
85
Семейство Чайковые
в связи с чем граница черного цвета почти горизонтальная). Концы крыльев черные
(у черноголовой и малой чайки белые), нижняя сторона крыльев относительно свет-
лая (у малой чайки темно-серая). Хорошим отличительным признаком во всех на-
рядах служит клинообразно расширяющийся в дистальной части ярко-белый передний
край крыла, образованный первостепенными маховыми и их кроющими. В зимнем
наряде по окраске очень похожа на морского голубка, но отличается от него относитель-
но более короткими шеей и клювом, более высоким лбом. Молодые птицы по харак-
терному рисунку верхней стороны крыльев хорошо отличимы от молодых птиц дру-
гих видов чайковых (у похожего морского голубка верхняя сторона более светлая).
Во всех возрастах на озерную очень похожа американская чайка Larus Philadelphia,
которая среди американских чайковых выделяется наиболее' частыми залетами в Ев-
ропу (темно-графитовый рисунок головы в брачном наряде по форме похож на та-
ковой озерной чайки, концы крыльев снизу светлые, в то время как у озерной чайки
они темнее остальной части низа крыла). В Центральной Азии следует при определении
учесть сходство с несколько более крупными буроголовой чайкой и реликтовой чайкой.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Выделение дальневосточных птиц в подвид Larus ridi-
bundus sibiricus Buturlin, 1911 на основе несколько более крупных размеров приз-
нано необоснованным (Kozlova, 1932, цит. по: Дементьев, 1951).
L. ridibundus,. L. cirrocephalus, L. maculipennis очень близкородственны и вместе с
L. hartlaubii, L. novaehollandie, L. buileri, L. serranus, а также с несколько более от-
даленными L. brunnicephalus, L. saundersi, L. Philadelphia и L. genei образуют хорошо
обособленную группу (Cramp, Simmons, 1983).
Описание. Строение и размеры. Данные по размерам взрослых птиц в пе-
риод гнездования для трех пунктов ареала в СССР обобщены в табл. 2.
Окраска. Самец и самка в брачном наряде. Голова до затылка, подбородок
и горло темно-коричневые. Глаза сверху и снизу окаймлены узкой белой полоской.
Передняя и средняя часть спины и верхняя часть крыла серые. В дистальной части кры-
ла клинообразное белое пятно, расширяющееся к концу крыла. Концы II—VII перво-
степенных маховых черные, на конце VIII субтерминальное черно-серое пятно. Осталь-
ная часть оперения — шея, нижняя часть тела, хвост, надхвостье — белые. Клюв, края
век, ноги — темно-красные, радужина бурая. Взрослые самец и самка в зимнем наря-
де. Как в брачном наряде, однако голова белая. Глаз спереди окаймлен черным. Под
глазом (иногда и за ним) и в области ушных перьев черно-серые пятна. Пятна у глаз
(менее выражены пятна у ушных отверстий) могут быть связаны темной полосой,
проходящей поверх головы. Клюв светло-красный с темным концом, ноги светло-
красные.
Пуховой наряд. Охристо-бурый (варьирует в довольно широких пределах) с круп-
ными черно-бурыми пятнами различной формы. Клюв грязно-мясного цвета с темным
концом, ноги тоже грязно-мясные.
Гнездовой наряд. Голова светло-бурая, более светлая в передней и нижней части.
На ней можно различить более темное пятно перед глазом, полосу под глазом и пятно
в области уха, более светлое пятно за глазом. Задняя сторона шеи и передняя часть
спины серовато-бурая с желтоватыми концами перьев. Задняя часть спины серая с
переходом в белое в области надхвостья. Хвост белый с темно-бурой предвершинной
полосой шириной 15—25 мм, за который следует узкая желтоватая каемка на самом
конце перьев (на крайней паре рулевых предвершинное пятно может отсутствовать
или быть небольшим). Плечевые перья бурые с более светлым желтоватым окаймле-
нием и серые. Второстепенные маховые с темными концами. Первостепенные махо-
вые черновато-бурые с белым (II—V) и серым (VI—VII) клинообразным рисунком,
86
Озерная чайка
Таблица 2. Озерная чайка. Размеры (мм) и масса тела (г)
взрослых особей озерной чайки в период гнездования
Показатель Самцы Самки
М lim п м lim п
Длина крыла 311,0 Патвия, оз. Эн1 299-320 уре (данные i 6 . Виксне) 296,3 284-312 17
ПЛЮСНЫ 46,7 42-49 И 44,2 42-52 25
клюва 35,0 35 2 — — —
черепа 84,0 80-86 13 77,2 72-80 45
Масса 293,8 265-300 11 281,1 215-310 21
Московская обл., оз. Киёво (Исаков и др., 1947)
Длина
крыла 319,1 309-340 66 303,1 . 288-332 91
ПЛЮСНЫ' 47,3 40-63 65 43,2 40-46 90
клюва 36,2 33-39 65 33,0 30-37 90
Масса 293 23 257 37
Оз. Байкал (Скрябин, 1977а)
Длина
крыла 310,2 32 . 295,7 24
плюсны 46,3 32 44,0 24
клюва 35,9 32 34,3 24
Масса 282 32 246 24
который на VIII—XI маховых становится расплывчатым. Верхние кроющие крыла бе-
лые, серые и разных оттенков бурого и черновато-бурого цвета. Нижние кроющие
серые, на переднем краю крыла белые. Нижняя часть тела белая со светло-буроватой
расплывчатой полосой поперек груди. Клюв грязно-желтый с темным концом, ноги
грязно-желтые.
Первый зимний наряд. Как зимний наряд взрослых, но хвост и крылья, а иногда
и плечевые сохранены от гнездового наряда.
Первый летний наряд. Как первый зимний наряд, но голова у большинства птиц
бурая с белыми пестринками, особенно на лбу, горло часто почти белое. Ювенильный
рисунок на крыльях и хвосте стал более светлым в результате износа. Во втором зим-
нем наряде птицы иногда отличаются от взрослых, по темным отметинам на крыль-
ях (в основном на кроющих наружных первостепенных маховых). Птицы в брачном
наряде с белыми пестринками на голове, по-видимому, двухлетние. Более подроб-
ные описания приводятся в литературе (Glutz v. Blotzheim, Bauer, 1982; Cramp, Sim-
mons, 1983).
Линька. Послебрачная линька у взрослых полная (конец мая—ноябрь), предбрачная —
частичная, сменяется оперение головы и туловища (январь—март, иногда затягивается
до мая). Линька в первый зимний наряд частичная: сменяется оперение головы, туло-
вища, часть кроющих крыла (июль—декабрь). Предбрачная после первой зимовки
частичная: сменяется оперение головы, туловища, часть кроющих крыла, иногда цент-
ральная пара рулевых (январь—май). Послебрачная годовалых: как послебрачная у
взрослых, но на 3—4 недели раньше (Cramp, Simmons, 1983).
Распространение. Гнездовой ареал. Сильное расширение ареала началось
87
00
Рис. 24. Область распространения озерной чайки
1 — гнездовой ареал, 2 — неуточненная граница ареала, 3 — места зимовок
Озерная чайка
с XIX в., но особенно проявилось в XX в., что связано предположительно с потепле-
нием климата, улучшением охраны во многих странах и освоением чайками новых
антропогенных источников кормов (Glutz v. Blotzheim, Bauer, 1982; Cramp, Simmons,
1983).
Граница современного ареала на севере вне СССР влючает о-в Ньюфаундленд (пер-
вое гнездование в 1977г.), юго-запад Гренландии (с 1969 г.), Исландию, Фарерские
острова, Британские острова, проходит по северу Скандинавского полуострова и Фин-
ляндии (Glutz V. Bloezheim, Bauer, 1982; Cramp, Simmons, 1983). В СССР северная
граница ареала включает Онежское озеро и проходит около Архангельска (Степанян,
1975), включает верховья Вычегды (Естафьев, 1981а), проходит на Урале около 60°
с.ш., около 67° с.ш. на Оби, 65° с.ш. на Енисее (Степанян, 1975), 65° с.ш. на притоке
Вилюя — Мархе, 68° с.ш. на Колыме (Дегтярев и др., 19816; Перфильев, 1981). Гнез-
дится на Камчатке и к северу от полуострова в Корякском нагорье, а также на о-ве Ка-
рагинском (Лобков, 1975, 1981а). Отсутствуют данные о гнездовании на обширных
территориях на западном берегу Охотского моря, но гнездится на его южном побе-
режье, например в нижнем течении Амура (Росляков, Рослая, 1981). По Г.П. Дементье-
ву (1951), гнездится на Сахалине, но более поздние авторы этого не подтверждают
(Гизенко, 1955).
Южная граница гнездового ареала на западе включает отдельные гнездовья в Испа-
нии, на юге Франции, на островах Сардиния и Сицилия, долину р. По в Италии, север
Югославии и Болгарии, отдельные гнездовья в центральной и восточной частях Тур-
ции (Giutz V. Blotzheim, Bauer, 1982; Cramp, Simmons, 1983). В СССР граница ареа-
ла проходит по северному побережью Черного моря (отсутствует в Крыму — Кос-
тин, 1983), включает Закавказские республики, огибает с севера Каспий, проходит
по дельте Волги (Луговой, 1958), через Камыш-Самарские озера, Актюбинск, спуска-
ется к Аральскому морю, включает его северную часть, Сырдарью, равнинные районы
Южного и Восточного Казахстана (Долгушин, 1962), озера Иссык-Куль и Сон-Кёль
в Киргизии (Кыдыралиев, 1981). Гнездовые поселения отмечены также на оз. Айдар-
куле (Мухина, 1983). Вне СССР южная граница далее проходит по Монголии, где
озерная чайка гнездится на озерах и реках западной, северо-западной и центральной
частей страны (Glutz v. Blotzheim, Bauer, 1982; Cramp, Simmons, 1983), на оз. Буйр-
Hyp -и Восточной Монголии, включает северо-восток Китая и советское Приморье
(Дементьев, 1951; Поливанова, 1971) (рис. 24).
Зимовки. Популяции, гнездящиеся к северу и востоку от январской изотермы
— 2,5 ° С, являются мигрантами, гнездящиеся на Британских островах и в бассейне
Средиземного моря оседлы, у населяющих промежуточные области молодые птицы
в большинстве случаев мигрируют, а у взрослых в направлении с востока на запад
прослеживается возрастающая тенденция к оседлому образу жизни. В массовом коли-
честве зимуют к югу и западу от изотермы 0 °C (Glutz v. Blotzheim, Bauer, 1982).
Таким образом, в область зимовки озерной чайки входят почти все страны Европы,
где это позволяет ледовая обстановка зимой, Средиземноморье, побережья Черного,
Каспийского морей, а также морей Индийского и Тихого океанов, омывающих юж-
ные и восточные (к югу от 45° с.ш.) берега Азиатского материка. За последние деся-
тилетия чайка стала зимовать на побережье Северной Америки от о-ва Ньюфаундленд
до Нью-Йорка, на западном побережье Африки к югу до Нигерии, в Мали и Нигере
(в последние, вероятно, попадает, пересекая Сахару), а также в странах восточного
побережья Африки к югу даже до Кении и Танзании, где рост числа зимующих птиц
связывают с ростом численности озерной чайки в СССР (Glutz v. Blotzheim, Bauer,
1982; Cramp, Simmons, 1983).
89
Семейство Чайковые
Для чаек, гнездящихся в СССР, зимовки и миграции наиболее подробно изучены
у восточнобалтийской популяции, объединяющей птиц Эстонии, Латвии, Литвы и Ка-
лининградской обл. (Schiiz, Weigold, 1931; Тауринып и др., 1953; Йыги, 1957; Викс-
не, 1961, 1962, 1968а; Шеварева, 1965; Вайткявичюс, Скуодис, 1965; и др.). Эти пти-
цы зимой встречены на обширной территории от Канарских островов и северо-запад-
ных берегов Африки до восточного побережья Черного моря (отдельные встречи —
на Багамских островах и на Каспии), однако наиболее характерными местами зимов-
ки этой популяции следует считать западное побережье Балтийского моря — юг Шве-
ции, Дания, север ГДР, побережье Северного моря, включающее северо-запад ФРГ,
Нидерланды, Бельгию, северный берег Франции и южную половину Британских остро-
вов, атлантическое побережье Франции и Пиренейский полуостров, озера и реки внут-
ри континента (в основном в ФРГ, Швейцарии, Венгрии), а также западное Средиземно-
морье, особенно северное побережье Адриатического моря, долину р. По, юг Фран-
ции. Указанные зимовки достигаются в основном двумя путями: 1) чайки пересе-
кают Балтийское море в западном направлении и попадают в южную Швецию и Данию,
откуда достигают североморских и атлантических побережий Европы и Африки, а
пересекая материк, попадают в Швейцарию и в западную часть Средиземного моря;
2) птицы двигаются вдоль восточного берега Балтийского моря, проходят через Ка-
лининградскую обл. (летевшие по первому пути в это время уже в Дании), попа-
дают в Польшу и ГДР, где пересекают материк и достигают Адриатического моря.
Чайки, гнездящиеся в северной части Прибалтийского региона, более склонны к пер-
вому из описанных выше путей, гнездящиеся в южной части — ко второму.
Чайки центральных областей европейской части СССР — Московской, Ивановской,
Рязанской и Ярославской — зимой встречаются от юга Франции на западе до Каспия
на востоке, однако их основными зимовками являются побережье Черного и юго-вос-
точное побережье Азовского морей, побережья Италии, Югославии, Греции, восточ-
ное Средиземноморье (устье Нила, Ливан, Кипр и др.), среднее и верхнее течение Ду-
ная, Швейцария (Птушенко, 1948; Сапетина, 1959; 1962; Шеварева, 1965). В ходе
миграции к местам зимовок эти чайки примерно на 3 месяца задерживаются в среднем
и нижнем течении Днепра, в низовьях Дона и на Азовском море, что дало основание
выделить у них так называемый промежуточный перелет (Шеварева, 1965).
Озерные чайки Казахстана и Западной Сибири (Долгушин, 1962; Ходков, 1977а)
зимуют на Каспии и предположительно также на берегах Персидского залива и Аравий-
ского моря. Кольцеванием установлена зимовка камчатских озерных чаек в Японии
(Sugawa et al., 1982).
Миграции. Оставление гнездовых колоний начинается сразу после подъема моло-
дых птиц на крыло, по срокам оно широко варьирует в зависимости от широты и
местных условий в данном году и происходит в основном с третьей декады июня до
начала августа. Озерной чайке свойственны послегнездовые кочевки, которые у птиц,
гнездящихся вдали от границы ареала, имеют ненаправленный характер (Птушенко,
1948 и др.), а вблизи границы ареала могут иметь выраженную направленность и свя-
заны с расширением ареала (Виксне, 1968а). Наиболее отдаленных зимовок озерные
чайки достигают в середине декабря, в феврале их число там сокращается. На ближай-
ших зимовках (например, Адриатика, юго-запад Балтики) чайки пребывают примерно
с конца октября — начала ноября до середины марта. Сроки прилета сильно варьи-
руют в зависимости от хода весны, но прилетает озерная чайка, как правило, несколь-
ко раньше освобождения водоемов ото льда. Например, в Эстонии в 1948—1966 гг.
первые озерные чайки прилетали в среднем 7 апреля, наиболее ранняя встреча 23.III
(Rootsmae, Rootsmae, 1976); в Калининской обл. за 36 лет наблюдений 26.III-23.IV,
в среднем 7-8.IV, массовый пролет 1-17.IV (Зиновьев и др., 1981); на оз. Киёво
90
Озерная чайка
первые особи 24.Ill—8.IV, массовое появление ЗО.Ш—27.IV (Исаков и др., 1947);
в Барабинской степи первые особи в первой декаде апреля, пролет до двадцатых чи-
сел мая, наиболее интенсивный — в конце апреля — начале мая (Ходков, 1977а). На
Байкале (южная часть) озерная чайка появляется в середине апреля (Скрябин, Раз-
махнина, 1978). В нижнем течении Сырдарьи первые птицы появляются в конце фев-
раля, пролет в основном в марте, в Алма-Ате 10—19.III, под Семипалатинском 7—17.IV
(Долгушин, 1962).
Численность. Начавшиеся уже в XIX в. рост численности и расширение ареала в За-
падной Европе продолжаются во многих странах и в настоящее время. В 1970-х го-
дах численность в странах Европы, кроме СССР, оценивалась минимум в 1400 тыс.
гнездящихся пар (Glutz v. Blotzheim, Bauer, 1982). Более поздние публикации по
отдельным странам (Saurola, in: Hyytia, Kellomaki, Koistinen, 1983) дают основание
предположить, что в этой части ареала гнездится от 1515 тыс. до 1820 тыс. пар озер-
ных чаек. В том числе в Финляндии гнездится 150 тыс., в Швеции — 270 тыс., Дании —
210 тыс., ФРГ — 67 тыс., ГДР — 90 тыс., Польше — 84,5 тыс., Чехословакии — 200—
350 тыс., Венгрии — 12 тыс., Нидерландах — 200 тыс., на Британских островах —
150-300 тыс. пар.
Определение общей численности озерной чайки в СССР пока проведено для ряда
западных территорий. В 1970-х годах она составляла: для Эстонии 80 тыс., Латвии
97 тыс., Литвы 30 тыс. пар (Виксне и др., 1981), Белоруссии 104 тыс. (Наумчик, 1981),
Московской обл. 30—32 тыс. пар (Зиновьев и др., 1981). Прослеживается и рост чис-
ленности на упомянутых территориях: в Эстонии к 1960 г. гнездилось 20 тыс. пар,
в 1967-1969 гг. — 30 тыс. пар (Оппо, 1966, 1971), в Латвии в конце 1930-х — начале
1940-х годов — 10 тыс. пар (Berzins, 1946), в 1964—1966 гг. — 30 тыс. пар (Виксне,
Балтвилкс, 1966), в Литве в начале 1970-х годов гнездилось 15—18 тыс. пар (Valius,
1974). Рост численности озерной чайки в последние десятилетия имел место также в
других областях на северо-западе и в средней полосе европейской части СССР (Зи-
новьев и др., 1981; Мальчевский, Пукинский, 1983), а также вблизи населенных пунк-
тов на Камчатке (Лобков, 1981а).
Местообитания. В гнездовое время преимущественно мелководные внутренние
водоемы — озера, пруды, водохранилища, дельты рек и т.д. — со сплавинами из корне-
вищ надводной растительности или луговыми островами, торфяные карьеры, болота
(преимущественно низинные), заросшие надводной растительностью морские бухты
и морские острова вблизи берега. Реже гнездится на прибрежных лугах, дюнах, топ-
ких прибрежных кочкарниках и т.п. Местами добычи корма служат преимущественно
открытые ландшафты — пахотные земли, луга и пастбища или водоемы. С усилением
синантропизации местами кормежки все чаще становятся свалки, зверофермы, рыбо-
консервные заводы и другие предприятия пищевой промышленности, рыболовецкие
гавани, города. В период миграции больше связана с морским побережьем и поймами
крупных рек, в период зимовки — с морским побережьем, сельскохозяйственными
угодьями и населенными пунктами вблизи водоемов.
Размножение. Начинает размножаться в возрасте 1—4 лет (т.е. после первой—чет-
вертой зимовки), при этом самки начинают гнездиться в возрасте 1, 2 (в основном)
и 3 лет, самцы — в возрасте 2, 3 (в основном) и 4 лет (Виксне, 19686). Участие годо-
валых самцов в размножении, по-видимому, очень редкое. Моногам, диффузно-гнез-
дящийся факультативно-колониальный вид (Зубакин, 1976в). Колонии весьма посто-
янны и могут существовать в течение многих десятилетий, их перемещения вызваны
обычно сукцессионными изменениями, продолжительным воздействием какого-либо
неблагоприятного фактора (например, присутствие хищного млекопитающего), экст-
ремальными условиями (например, затопление прежнего места колоний). По край-
91
Семейство Чайковые
ней мере часть чаек возвращается с зимовок парами (Cramp, Simmons, 1983), у боль-
шинства пары образуются после прилета вне колонии или в ней, но, как правило, не
на месте будущего гнезда (Glutz v. Blotzheim, Bauer, 1982; Харитонов, 1983a). Мно-
гие чайки меняют несколько партнеров, прежде чем приступят к постройке гнезда.
До постройки гнезда, а также после вылупления птенцов чайки нередко меняют поло-
жение своего участка в колонии и переселяются в другое место. Первые прилетевшие
чайки занимают будущий центр колонии, затем в течение нескольких дней занимается
вся площадь, но с малой плотностью. С прибытием новых птиц и увеличением плот-
ности наблюдаются центростремительные тенденции — чайки стараются переселиться
в более предпочитаемые области колонии (Patterson, 1965; Харитонов, 19836; и др.).
Поведение озерной чайки изучено весьма подробно (Moynihan, 1955; и др.), деталь-
ные его описания приводятся также в обобщающих работах (Glutz v. Blotzheim, Bauer,
1982; Cramp, Simmons, 1983). Антагонистической демонстрацией в воздухе можно
считать полет преследования и ’’налет и планирование” (Swoop and Soar Display):
резкое увеличение скорости полета преследователя приводит к сходной реакции прес-
ледуемого, преследующая птица издает долгий крик и вибрирующий крик, после че-
го внезапно взмывает вверх, планируя на поднятых и отведенных назад крыльях,
ноги при этом опущены, шея и голова вытянута вперед и вниз. Среди антагонистичес-
ких демонстраций на земле следует выделить долгий крик в наклонной позе, являю-
щийся предупреждением другой птице, приближающейся по воздуху, земле и воде
(рис. 25,а). При этом голову птица держит поднятой высоко вверх и вперед, клюв
более или менее горизонтально, красный зев хорошо заметен во время крика, запяст-
ные суставы отодвинуты от туловища; во время демонстрации птица обычно смотрит
в сторону противника. Распластанная поза демонстрируется на воде и на суше (см.
рис. 25, б). Агрессивная прямая поза — голова поднята вверх, оперение на шее и голо-
ве приподнято, клюв опущен, крылья немного приподняты. В этой позе птица либо
стоит, либо двигается в сторону противника. Свидетельством готовности к бегству
может служить испуганная прямая поза — шея также вытянута вверх, но оперение
в ее верхней части прижато, клюв направлен более или менее горизонтально, запястные
суставы немного приподняты, птица смотрит в сторону от оппонента (см. рис. 25, в).
В территориальных спорах часто можно видеть демонстрацию мяукающего крика,
которая обычно наблюдается после драки или атаки (см. рис. 25, г). ’’Квохтанье”
(см. рис. 25, д) особенно характерно в затянувшихся территориальных спорах (оно
достоверно чаще наблюдается между соседями, чем между владельцем территории
и чужаком — Харитонов, 1983а), его часто можно видеть в плотных колониях. К анта-
гонистическим демонстрациям относится также клевание земли. Отворачивание головы
(см. рис. 25, ж), по-видимому, служит позой умиротворения.
Брачное поведение включает многие антагонистические демонстрации. При встре-
че самки самец издает долгий крик в наклонной позе, затем следует распластанная
поза, которая отличается от агрессивного варианта приподнятым вверх клювом при
опущенной горизонтально, на уровне туловища, шее (кроме того, в неагрессивных
распластанных позах птицы обычно стоят бок о бок). В церемонию приветствия вхо-
дит также отворачивание головы. Формирование пары завершается ритуальным корм-
лением. Самка выпрашивает корм в сгорбленной позе, издавая своеобразный звук.
Гнездовые колонии варьируют по величине от нескольких и нескольких десятков
пар (в основном вблизи границ ареала — Дегтярев и др., 19816; Кыдыралиев, 1981;
Жордания, 1981; Естафьев, 1981а и др.) до многих тысяч (в средней полосе в условиях
культурного ландшафта, где чайки освоили антропогенные источники кормов). Такие
крупные колонии имеются на оз. Киёво (16500 пар; Харитонов, 1983а), на водохра-
нилищах на Днепре (до 10 тыс.; Булахов, Мясоедова, 1975), на озерах Бабите и Энгу-
92
Озерная чайка
ре в Латвии. Самая крупная колония в Западной Европе находится в Нидерландах,
в районе устья р. Шельда — 21 тыс. пар в 1976 г. (Glutz v. Blotzheim, Bauer, 1982).
В условиях Прибалтики из крупных поселений (тысячи пар), занимающих несколь-
ко десятков квадратных километров, около 36% молодых птиц расселяются на рас-
стояние до 500 км; самки с самцы одинаково часто. В пределах таких поселений (оз. Эн-
гуре, 35 км2) 45,6% оставшихся гнездятся на расстоянии 1,5 км от места рождения
(Виксне, 1970). Самцы более часто гнездятся в родной колонии (40,8%), чем самки
а — долгий крик, б — распластанная поза, в — испуганная прямая поза, г — мяукающий крик, д —
’’квохтание”, е — ритуальное кормление, ж — отворачивание головы
(24,3%). В условиях оз. Энгуре около 20% выживших взрослых меняют гнездовую
колонию; повторные кладки (бывают у птиц с возраста 3 лет) после отлова отмеча-
лись всегда в другой колонии, до 5 км от прежнего места. После неуспешного гнездо-
вания в следующем году чайки предпочитают гнездиться в другой колонии на том
же или другом водоеме (Виксне, 1968в, г).
В колониях озерной чайки охотно селятся поганки (особенно черношейная, серо-
щекая и красношейная), ряд уток (особенно хохлатая чернеть, также красноголовый
нырок, серая утка, кряква и др.), речная крачка, малая чайка, пестроносая крачка
и другие виды.
Озерная чайка охраняет территорию 32—47 см в диаметре (Харитонов, 1978), оп-
тимальное расстояние между гнездами примерно вдвое больше (Glutz v. Blotzheim,
93
Семейство Чайковые
Bauer, 1982). Меньшее расстояние между гнездами бывает в случае, если что-нибудь
препятствует визуальным контактам между насиживающими птицами. В Латвии на
оз. Энгуре на 79 пробных площадках в 1974—1984 гг. плотность гнезд составляла
1,4—13,1 гнезд на 10 м2, в среднем 5,3 гнезд на 10 км2; в радиусе 1 м'25,5% гнезд
соседей не имело, 25,5% имело > 1; 21,3% — 2; 14,7% — 3,..., 0,3% — 7 соседей (Викс-
не, 1981а). На оз. Киёво к 1980 г. плотность колебалась в разных участках от 3—5
до 67 гнезд на 100 м2 (Зубакин, Харитонов, 1981). На Украине (Булахов, Мясоедо-
ва, 1975) в крупных колониях (1500—10000 пар) плотность достигает 2—5 гнезд на
1 м2, в мелких (100—250 пар) 8—10 гнезд на 100 м2. На Байкале в дельте Селенги в
колониях величиной 16—1034 пар плотность гнезд составляла 0,5—7,2 на 100 м2 (Ги-
левич, 1977), на Северном Байкале в колониях величиной 5—107 пар — 0,3—27,5 пар
на 100 м2 (Садков, 1977). В Северном Казахстане плотность достигает 32 гнезд на
100 м2 (Самородов, 1979).
Гнездо отличается от гнезд других чаек, населяющих сходные места, использовани-
ем более грубых материалов и некоторой небрежностью постройки (Glutz v. Blotzheim.
Bauer, 1982). Его размеры сильно варьируют в зависимости от расположения: напри-
мер, на оз. Киёво (Исаков и др., 1947) гнезда на сухих и мокрых участках имели соот-
ветственно диаметр у основания 22 и 50 см, а высоту 4,5 и 30 см; в мокрых местах
гнезда могут иметь еще большие размеры — диаметр 90 см, высоту 40 см (Glutz v.
Bloezheim, Bauer, 1982).
Сроки размножения варьируют на одних и тех же водоемах по годам в зависимости
от хода весны, особенно уровня воды, поэтому выборочно приведем лишь самые
общие сведения. В Латвии на оз. Энгуре первые яйца появляются в последней декаде
апреля, средняя .дата вылупления в 1974—1978 гг. на пробных площадках колебалась
от 30.V до 9.VI (Viksne, Janaus, 1980), в целом по озеру на несколько дней раньше.
В Литве в 1966—1973 гг. на оз. Жувинтас начало строительства гнезд отмечено 9—28.IV.
первые птенцы появились 15—30.V (Недзинскас, 1975). На водохранилище — охлади-
теле, отличающемся повышенной температурой, первое яйцо в половине отложенных
кладок в среднем появляется до 29.IV, т.е. на две недели раньше, чем в целом по рес-
публике (Виткаускас, 1981а). На оз. Киёво в 1933—1936 гг. первые яйца отмечены
17—24.IV, первые птенцы появились 18—23.V, начало массового вылупления в сухих
рогозовых участках 22.V—2.VI (Исаков и др., 1947); в 1975, 1977 и 1978 гг. на озе-
ре первые птенцы вылупились соответственно 3.V, 9—10.V, 13.V (Зубакин, Харито-
нов, 1981). Под Казанью первые яйца в 1974 г. отмечены 30.IV, массовая кладка
5—14.V (Водолажская, 1979). В Северном Казахстане первые яйца найдены 8—24.IV,
первые птенцы — 16—18.V (Самородов, 1979). На Байкале в дельте Селенги в 1973—
1975 гг. в разных колониях начало яйцекладки зарегистрировано 7—13.V, массовая
кладка проходила в период с 8 по 20.V в течение 3—8 дней, начало вылупления птен-
цов 1 —12.VI (Гилевич, 1977); на Северном Байкале в 1972—1976 гг. начало яйцеклад-
ки отмечено 19.V—22.V, первые птенцы появляются 7.VI, обычно 15—18.VI (Садков,
1977). Более старые особи гнездятся раньше, чем трехлетние и двухлетние; при задерж-
ке начала размножения паводком различия в сроках сокращаются (Виксне, 19686).
В кладке 1—3 (чаще всего 3) яйца, возможность снесения одной самкой кладки
из 4 яиц не доказана (Glutz v. Blotzheim, Bauer, 1982). Встречаются совместные клад-
ки двух и нескольких самок, содержащие до 14 яиц (Кумари, 1953); подобные клад-
ки чаще всего наблюдаются при дефиците мест гнездования при высоком паводке.
Средняя величина кладки за 5 лет в Латвии составляла 2,47—2,84 (обследовано 261—
415 кладок в год; Viksne, Janaus, 1980), в Литве — 2,68 ± 0,03 яиц (Виткаускас,
1981а); в ходе сезона наблюдается уменьшение среднего размера кладки. В Северном
Казахстане средний размер кладки 2,7 (обследовано 2140 яиц; Самородов, 1979).
94
I
j
I Озерная чайка
i На Байкале в дельте Селенги в 1973-1975 гг. в разных колониях из 14-822 гнезд
| средний размер кладки составлял 2,03—2,70 яиц (Гилевич, 1977), на Северном Бай-
’ кале в 1973—1976 гг. в отдельных колониях из 13—91 гнезд — 2,43—2,90 яиц (Сад-
ков, 1977). Если первое и последующие яйца вынимаются из гнезда в течение 12 ч
после откладки, одна самка может снести в гнездо до 7 яиц (Weidmann, 1956; Викс-
не, 1968в). В размерах яиц не прослеживается географических вариаций (Glutz v.
Biotzheim, Bauer, 1982; Cramp, Simmons, 1983). На оз. Киёво длина яиц составляла
50,54 ± 0,09 мм (и = 1348), ширина — 35,32 ± 0,07 мм (п = 1211) (Исаков и др., 1947),
под Ижевском - 54,2 ± 0,17 X 35,9 ± 0,12 (л = 175) (Рысьева и др., 1984), на Северном
Байкале в двухколониях 52,2 ± 0,36 X 36,9 ± 0,4 (п = 52) и 51,9± 0,18 X 36,6± 0,15
(п = 60) (Садков, 1977). Яйца сильно варьируют по окраске; чаще всего их основной
фон буровато- или оливково-зеленоватый со сравнительно равномерно распределенны-
ми! пятнами неправильной формы сероватого цвета (глубокие) и разных оттенков
бурого (поверхностные). Наряду с нормально окрашенными встречаются также очень
* светлые голубоватые яйца, почти полностью лишенные рисунка.
В насиживании участвуют оба пола, однако данные о их относительной роли проти-
> воречивы. По одним данным, по крайней мере в светлое время суток, в среднем боль-
‘ ше насиживает самец (53,5%, наблюдения за 9 гнездами, всего 669 ч, см.: Ytreberg,
I 1956). По другим данным гораздо больше насиживает самка, при этом во время яйце-
; кладки, ночью и в конце периода инкубации ее участие в насиживании особенно ве-
' лико (Водолажская, 1979; Самородов, 1979; Виткаускас, 19816). Время от снесения
j до вылупления для 1-го яйца, по данным трех исследований (Ytreberg, 1956; Виткаус-
t кас, 1981а; Климова, 1984), составляло соответственно 24,88 ±0,81, 24,81 ±0,07,
| 24,2 ±0,15 сут, для 2-го яйца — 23,59 ± 0,80, 23,87 ± 0,06 и 23,1 ±0,14 сут, для 3-го
‘ яйца — 22,82 ± 0,86, 22,73 ± 0,05 и 22,4+ 0,16 сут.
J От появления первых трещин на яйце до вылупления птенца проходит 2—6, в сред-
, нем 3,3 дня. Последовательность вылупления в большинстве случаев соответствует
(последовательности снесения яиц. Обычно между вылуплением первого и второго
птенца проходит 12 ч, между вылуплением второго и третьего — 24 ч (Ytreberg, 1956).
После обсыхания птенца скорлупа яйца удаляется; взрослая птица выбрасывает ее,
отлетев на несколько метров (Tinbergen, 1967). Масса только что вылупившихся
и однодневных птенцов на оз. Энгуре в 1978 и 1979 гг. соответственно составляла
26,1 г (п = 510) и 25,4 г (п = 582) (Янаус, 1981). Масса птенцов в день вылупления
на оз. Киёво (и = 85) 26,3 г (Исаков и др., 1947), на севере Казахстана (п = 43) 24,8 г
(Самородов, 1979). В выкармливании птенцов участвуют оба родителя, по данным
Т.И. Водолажской (1979), по крайней мере первые 5 дней жизни более активно птен-
цов кормит самка. В среднем птенцов кормят 5,7 раза в день (Isenmann, 1977), по
другим данным (Исаков и др., 1947), — 4—5 раз, что составляет 100—150 г корма.
Родители и птенцы узнают друг друга как визуально, так и по голосу через 3—4 дня.
В первый день жизни птенцы обычно не проявляют тенденции к бегству, с 2—4 дня
при опасности все чаще покидают гнездо, после 6-го дня бегство в определенные убе-
жища — участки густой растительности — становится нормой; с возрастом длительность
пробежек увеличивается. В убежищах птенцы разных выводков не проявлют взаим-
ной враждебности, но их путь обратно к своему гнезду приводит к нападениям взрос-
лых птиц — владельцев территорий и подросших (пятидневных и старше) птенцов
(Glutz V. Biotzheim, Bauer, 1982). Примерно с 25—30-дневного возраста молодые
ничинают летать (Исаков и др., 1947), однако у части птенцов подъем на крыло затя-
гивается на 2—3 недели. Как правило, взрослые оставляют колонию раньше молодых,
самец часто раньше самки (Исаков и др., 1947; Makatsch, 1952); по другим наблю-
дениям, родители нередко продолжают кормить и летных птенцов, часто взрослых
95
Семейство Чайковые
отлетает из гнездовых колоний вместе со своими птенцами, и их контакт некоторое
время сохраняется (Самородов, 1979; наблюдения Я. Виксне).
Размеры гибели яиц и птенцов варьируют в широких пределах. Например, в Латвии
на оз. Энгуре в 1974-1981 гг. процент вылупления сосатавлял 70,7-86,0% (1,75-2,34
птенца на пару), выжило до 25 дня жизни 37,2—66,4% из числа вылупившихся или
41,1—57,1% из числа снесенных яиц (0,94—1,55 птенца на пару). Смертность птенцов
до 25 дня жизни в среднем составляла 42,9% (в отдельные годы 33,6—54,3%). В сред-
нем 60,5% птенцов погибало до 4-дневного возраста, около 80% — до 10-го дня жиз-
ни (Виксне, 1981а, б, 1982; Янаус, 1981; Viksne, Janaus, 1980). Новый пик смертнос-
ти имеет место в момент перехода птенцов к самостоятельной жизни. На водохра-
нилище Литовской ГРЭС за 3 года успешность вылупления составляла 81,8%, в сред-
нем 2,19 птенца на кладку (Виткаускас, 1981а). В Белоруссии на Минском море за
два года отход яиц составил 12,1%, постэмбриональная смертность — 57,5%, выросли
в среднем 1,12 молодых на пару взрослых (Никифоров, Шкляров, 1983). На Байкале
в дельте Селенги (Гилевич, 1977) и на севере этого озера (Садков, 1977) в нормаль-
ных условиях отход яиц составлял соответственно 20,0—32,5% и до 16% кладок,
при наводнениях могут погибнуть свыше 90% кладок.
Отмечены различия в успешности размножения в зависимости от сроков гнездова-
ния — наивысшие показатели соответствуют либо самым ранним, либо средним сро-
кам (Viksne, Janaus,, 1980; Виткаускас, 1981а; Никифоров, Шкляров, 1983; и др.).
Погодные условия оказывают относительно меньшее влияние на смертность птенцов
до 10-го дня жизни, чем на более старших птенцов, требующих больше корма (Яна-
ус, 1983).
На первом году жизни после подъема на крыло смертность составляет ориентиро-
вочно 38%, взрослых особей 15% в год (Виксне, 19816, 1982). Считается что на рост
численности озерной чайки большее влияние оказало сокращение смертности взрос-
лых птиц, чем изменение успешности размножения (Lebreton, Isenmann, 1976).
Максимальный возраст, по данным кольцевания, 32 года 1 месяц (Rydzewski, 1978;
цит. по: Cramp, Simmons, 1983).
Суточная активность, поведение. Активна как днем, так и ночью (Самородов, 1979),
в том числе и вне периода размножения; мигрирует также не только днем, но и ночью.
Имеется два дневных пика активности — утро и ранний вечер (Glutz v. Blotheim,
Bauer, 1982). Общественна круглый год. В Западной Европе в местах ночевок зимой
скапливается до 100 тыс. особей (Glutz v. Blotzheim, Bauer, 1982).
Питание. Употребляет весьма разнообразные, преимущественно животные корма,
легко переключается на наиболее доступные виды. В странах Западной и Средней Ев-
ропы (Cramp, Simmons, 1983) в период гнездования доминируют дождевые черви
(до 50% общей массы) и насекомые (около 15%); летом и осенью значение дождевых
червей уменьшается, чайки переходят на другие виды беспозвоночных, рыбу, фрукты
и. семена растений, пищевые отбросы; на зимовках насекомые поедаются редко, доми-
нирует рыба и пищевые отбросы. Вследствие большого разнообразия условий в пре-
делах ареала озерной чайки от этой общей схемы имеется много отклонений, которые
в значительной степени определяются все расширяющейся и углубляющейся синантропи-
зацией вида в значительной части ареала.
Анализы содержимого желудков, собранных в мае—сентябре 1947—1959 гг. в Латвии
(Тима, 1961), показали доминирующую роль беспозвоночных в рационе. В частности,
дождевые черви доминировали в мае, стрекозы, жуки и двукрылые — в июне, июле,
августе; в сентябре увеличилось количество потребляемой рыбы, однако в целом
она составляла лишь 3% по встречаемости. В рационе птенцов озерной чайки оз. Энгуре
(побережье Рижского залива) встречаемость явно антропогенных кормов (отбросы
96
Озерная чайка
пищевых продуктов, зерна злаков) и рыбы (добытой преимущественно в местах по-
грузки) менялась следующим образом: 1959 г. — 0% и 89$%, 1963 г. — 23,3% и 52,0%,
1971 г. — 52,1% и 35,2% (Виксне, 1975), что иллюстрирует радикальное изменение
питания чаек этого водоема.
В центре европейской части СССР основательные исследования питания озерных
чаек проведены в 1930-1936 гг. на оз. Киёво (Исаков и др., 1947). В ранневесендий
период в рационе чаек преобладали мышевидные грызуны (в основном серая полев-
ка) — 62,8%, насекомые и другие беспозвоночные, связанные с водой, — 26,1% встре-
чемости. В период насиживания доминировали личинки жуков (41,9%), наземные
насекомые (26,2%); дождевые черви (28,3%), личинки жуков, живущие в почве
(34,9%), и наземные насекомые (18,5%) преобладали также в период выкармлива-
ния птенцов. Животные, вредные для сельского хозяйства (майские жуки, их личинки,
личинки щелкунов, серая полевка и др.), в период гнездования составляли в рационе
62,2% по встречаемости, полезные животные - 18,2%. Во время послегнездовых ко-
чевок в рационе доминировали рыба (45,2%), насекомые (14,5%) и щитни (30,7%),
а на зимовках на Каспии и Черном море рыба, добытая преимущественно у заводов
и причалов. В 1970-е годы состав кормов чаек киевской колонии сильно изменился,
так как основным местом кормежки стала свалка бытовых отходов (Зубакин, Хари-
тонов, 1978). Для ряда других колоний средней полосы европейской части СССР боль-
шую роль по сравнению с киевской колонией в период гнездования в рационе играет
рыба (Зиновьев и др., 1981).
Преобладание насекомых в рационе в период гнездования констатировано также
в Новосибирской обл. (Бородулина, 1960); в Казахстане (Долгушин, 1962) весной
в значительном количестве поедает мышевидных грызунов, летом — разных степных
насекомых, в том числе пруса, осенью увеличивается потребление рыбы. На Байкале
озерная чайка питается исключительно животными кормами, основу которых состав-
ляют беспозвоночные (Скрябин, Размахнина, 1978). Весной 50,3% содержимого же-
лудков по объему составляли гаммариды, 30% — насекомые, моллюски, 11,8% —
рыба. Летом насекомые (поденки, стрекозы, ручейники, двукрылые и др.) составляли
94% объема, рыба — 3,9%.
Корм озерные чайки добывают разнообразными способами: плавая и передвигаясь
пешком, в полете — с поверхности воды, земли или растений; ловят в воздухе.
Дальность полетов за кормом зависит от численности птиц в колонии (группе ко-
лоний) и количества корма в окрестности. Она колеблется от нескольких до 70 км;
для большинства птиц в крупных колониях все же не превышает 40 км (Исаков и др.,
1947; Виксне, Янаус, 1986).
Враги, неблагоприятные факторы. Основную опасность представляет довольно
широко практиковавшийся в прошлом неконтролируемый сбор яиц, что иногда при-
водило к исчезновению даже крупных колоний (Berzins, 1946; и др.), и всякая хо-
зяйственная деятельность (выпас скота и пр.) в местах колоний в период гнездования.
Естественные враги те же, что и у других водоплавающих. Особую опасность пред-
ставляют млекопитающие — лисица, енотовидная собака, американская норка, домаш-
няя собака, кабан и др., однако часто гнездовья для большинства из них недоступны.
Из пернатых значительный вред озерной чайке причиняют ворона, серебристая чайка,
ворон, камышевый лунь, локально - филин, ястреб-тетеревятник и др. При этом успех
хищничества вороны и серебристой чайки значительно увеличивается при беспокойстве
колонии человеком.
Резкий подъем уровня воды (например, во время штормов, на водохранилищах
ГЭС) может полностью уничтожить все кладки в колонии. Основная причина гибели
птенцов — недостаток кормов, наступающий, как правило, при продолжительной дождли-
4. Зак. 826 97
Семейство Чайковые
вой прохладной погоде. При беспокойстве колонии гибель птенцов может достигнуть
серьезных масштабов в результате агрессивного поведения взрослых по отношению к
чужим птенцам. В литературе (Glutz v. Blotzheim, Bauer, 1982) приводятся сведения о
повышенной гибели птенцов, вызванной пастереллезом, ботулизмом, сальмонеллеза-
ми, а также о разных гельминтозах.
Хозяйственное значение, охрана. Как массовый вид имеет весьма ощутимое значение
в разных отраслях. Играет положительную роль в охотничьем хозяйстве, так как в гнез-
довых колониях озерной чайки плотность утиных кладок и их сохранность выше, чем в
прилегающих сходных угодьях вне колоний (Fabricius, 1937; Haartmann, 1937; Mihel-
sons et al., 1976; Bergman, 1982; и др.). Легкое переключение на наиболее массовые
виды кормов и большой радиус разлета за кормом определяют активное участие озер-
ной чайки в уничтожении и ограничении численности вредителей сельскохозяйственных
культур — насекомых и полевок — при массовом их появлении (Исаков и др., 1947;
Долгушин, 1962 и др.). Доказана возможность целенаправленного привлечения чаек
на кормежку в определенных местах путем выставления макетов (Харитонов, 19806),
что открывает возможность использования чаек для подавления локальных вспышек
численности вредителей. Имеет определенное значение как санитар, собирая кормовые
отбросы в звероводческих фермах и т.д. В Дании разрешен в установленные сроки сбор
яиц озерной чайки (Bloch-Nielsen, 1975), в ряде стран она считается охотничьей пти-
цей (в Дании, ФРГ, Бельгии, частично в Австрии — Lampio, 1983).
Наряду с положительной ролью необходимо отметить создаваемую большими скоп-
лениями чаек опасность для авиации (Якоби, 1974), а также локально и кратковремен-
но проявляющееся отрицательное воздействие чаек на прудовое рыболовство (уничто
жение молоди) (Koubek, 1982). Однако вред, причиняемый рыбному хозяйству, силь-
но преувеличен.
При необходимости регуляции численности рекомендуется ограничение доступа к
антропогенным кормам, изменение гидрологического режима для устранения возмож-
ностей гнездования (Glotz v. Blotzheim, Bauer, 1982).
Основные мероприятия по охране — обеспечение покоя в гнездовых колониях в пе-
риод размножения.
Буроголовая чайка —
Larus brunnicephalus Jerdon, 1840
Indian Black-headed Gull (англ.), Tibetlachmowe (нем.) , Mouette de Tibet (фр.)
Син. (лат.): Hydrocoloeus brunneicephalus (Jerdon, 1840).
Син. (рус.): тибетская чайка, тибетская буроголовая чайка.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Чайка среднего размера, несколько
крупнее озерной, на которую очень похожа. От последней, однако, хорошо отличается
сильным развитием черного цвета на дистальных первостепенных маховых, в результа-
те чего в полете на конце крьиа видна широкая черная перевязь. Крыло кажется более
широким и тупым, чем у озерной чайки, клюв — более массивным. По полету сходна с
озерной чайкой и другими чайками среднего размера. Хорошо плавает; как и другие
чайки, на воде сидит высоко. Не ныряет, но во время кормежки может, на мгновение
взлетев с воды, с ходу наполовину погрузиться в воду. Голос похож на голос озерной
чайки, но несколько грубее. Основной видовой крик — дребезжащее карканье. В коло-
нии и вне ее буроголовая чайка гораздо молчаливее и спокойнее озерной.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. В начале нашего века видовая самостоятельность бурого-
ловой чайки сомнений не вызывала (Dwight, 1925), но затем Б.К. Штегманом было
98
Буроголовая чайка
предложено считать эту чайку подвидом озерной на основании разобщенности ареалов и
находки на Кашгарской равнине предположительно гибридных особей с рисунком махо-
вых промежуточного характера (Stegmann, 1935, цит. по Дементьеву, 1951). Эта точка
зрения приобрела сторонников (Moynihan, 1959), тем не менее большинство орнитоло-
гов ее не поддержали. В частности, Е.В. Козлова (1952) объясняла нетипичность окрас-
ки кашгарских особей возрастной изменчивостью. Подтвердили видовую самостоятель-
ность буроголовой чайки и позднейшие работы (Потапов, 1966а; Зубакин, 1982). Одна-
ко повторный внимательный осмотр экземпляров из Кашгарии, хранящихся
в ЗИН АН СССР, показал, что их нельзя отнести к озерным чайкам с остатками ювениль-
ного наряда. Упомянутые экземпляры занимают четкое промежуточное положение меж-
ду озерной и буроголовой чайками как по размерам, так и по окраске головы и перво-
степенных маховых (рисунок последних сильно варьирует, причем по степени развития
черного цвета можно построить почти непрерывный ряд от озерной до буроголовой чай-
ки) . Все это как будто действительно указывает на гибридный характер кашгарских
особей, подтверждая точку зрения Б.К. Штегмана. Однако известные ныне различия в
поведении, окраске пуховых птенцов и характере гнездования этих видов таковы, что
не позволяют усомниться в видовой самостоятельности буроголовой чайки. Таким об-
разом, вопрос о статусе кашгарских чаек пока остается открытым. Наиболее вероятны
два варианта решения проблемы. Либо это подвид буроголовой чайки, гнездящийся в
совершенно нехарактерных для вида как по биотопу, так и по абсолютной высоте мес-
тах (на гнездование с большой долей вероятности указывает добыча в Марал-Ба-
ши 16.V.1909 г. самца с наседным пятном), либо, что более вероятно, мы имеем дело
с гибридной популяцией двух хороших видов, входящих в комплекс типа superspe-
cies (Степанян, 1983). Следует отметить, что из Кашгарии нет коллекционных экземп-
ляров типичных для буроголовых чаек размеров и расцветки.
Описание. Строение и размеры (табл. 3).
Окраска (особи с Памира и Тибета — ЗИН, ЗМ МГУ).
Самец и самка в брачном наряде. Голова бледно-бурая, светло-бежевая у клюва, на лбу
и подбородке и постепенно темнеющая к затылку и шее, от которой резко отделяется
черно-бурым ожерельем. Последнее сзади не спускается ниже верхней части затылка,
а спереди захватывает верх передней части шеи. Над и под глазом узкие белые полоски
шириной 1—1,5 мм. Шея, грудь, бока, брюхо, хвост, надхвостье, подхвостье, крылышко,
кроющие кисти и подмышечные белые; подкрылья светло-серые, спина и кроющие кры-
ла серые. II (первое видимое) и III первостепенные маховые черные (темно-бурые) с
белым основанием, белым предвершинным пятном и черной вершиной; IV — с черной
верхней половиной, белым основанием и бурым краем внутреннего опахала; V и VI —
с черной вершиной, бурым краем внутреннего опахала и серым полем на внутреннем
опахале, более широким на VI пере. VII первостепенное маховое с черной вершиной,
белым наружным и серым внутренним опахалами с бурым краем, VIII — серое с бурым
краем внутреннего опахала и бурым предвершинным пятном. Остальные первостепен-
ные маховые серые, IX нередко с неясным бурым краем внутреннего опахала. Второсте-
пенные маховые серые. Клюв и лапы темно-красные, радужина светлая, желтоватая.
Самец и самка в зимнем наряде. Голова белая с серовато-бурыми пятнами у глаза
и на кроющих уха; неясные серовато-бурые пятна могут быть на затылке и шее на
месте ожерелья. Остальное оперение, как в брачном наряде.
Пуховой наряд (описание живых птенцов с оз. Каракуль). Основной тон окраски
гораздо светлее, чем у озерной чайки, и лишь немного темнее, чем у морского голубка:
светло-серый, светло-бежевый, светлый рыжевато-серый. По этому фону на темени,
верхней стороне шеи, спине и крыльях разбросаны контрастные черные или темно-
бурые пятна, величина и густота которых сильно варьирует. Черный пух разной густо-
99
Семейство Чайковые
Таблица 3. Буроголовая чайка. Размеры (мм)
Географический район Пол Длина крыла
п Lim сред нее
Памир (ЗИН, dd 7 341-352 346
ЗМ МГУ) 99 1 333 —
dd и 12 328-352 344
99*
Тибет (ЗИН, dd 20 325-354 342
ЗМ МГУ, Vaurie, 1972) 99 13 315-337 327
dd и 315-354 336
99* 34
Кашгарская равнина и dd 5 320-336 326
окрестности оз. Лобнор 99 1 319 —
(ЗИН) dd и 7 319-336 324
99*
*Просуммированы данные о всех промеренных самцах, самках и птицах неизвестного пола.
ты присутствует также на подбородке и уздечке, в ряде случаев — на лбу. Иногда
отдельные черные пушинки равномерно покрывают всю голову, в результате чего она
кажется темнее туловища. Клюв и лапы розовато- или телесно-серые, кончик ключа
темный. Радужина темная.
Гнездовой наряд. Сходный с соответствующим нарядом озерной чайки, однако
II—IV первостепенные маховые темно-бурые, белый цвет есть только у основания перьев.
В первом зимнем наряде (Dmight, 1925) спина серая, иногда с небольшими бурыми
пестринами. Голова белая с темно-серым затылком и темными пятнами на кроющих
уха и у глаз. Крылья и хвост, как в гнездовом наряде. В первом летнем наряде сохра-
няется оперение крыльев и хвоста от гнездового наряда. Голова становится бледно-
бурой с примесью белых перьев, особенно на лбу. Второй зимний и второй летний наря-
ды — окончательные, однако у части особей могут сохраняться темные пятна на рулевых
и большее развитие черного цвета на первостепенных маховых.
Линька. Частичная линька в первый зимний наряд (оперенье крыльев и рулевые не
сменяются) заканчивается, по-видимому, к октябрю (хотя 19 сентября 1956 г. добыта
самка в гнездовом наряде — Потапов, 19666). Частичная линька в первый летний наряд
(ювенильные маховые и рулевые по-прежнему сохраняются) происходит в апреле—мае.
Полная линька во второй зимний наряд происходит с июля до октября—ноября, частич-
ная линька во второй летний наряд — в марте—мае. В последующем линьки бывают
дважды в году: частичная предбрачная и полная послебрачная. Как и у других видов
чаек, наблюдается широкая индивидуальная изменчивость сроков линьки (Dwight,
1925; Дементьев, 1951).
Распространение. Гнездовой ареал. Высокогорья Центральной Азии — Ти-
бетское нагорье, Ладакх, Памир. На Тибетском нагорье заселяет в основном западный,
южный и северо-восточный участки (включая оз. Кукунор) (Vaurie, 1972), а также
Цайдамскую котловину (Cheng Tsohsin, 1976) (рис. 26). По данным последнего авто-
ра, гнездовой ареал захватывает и западную часть Кашгарской равнины, что, видимо,
соответствует ареалу кашгарской популяции. В СССР отмечена на Центральном (Во-
сточном) Памире, где в настоящее время достоверно гнездится лишь на оз. Каракуль
100
Буроголовая чайка
Длина клюва Длина цевки
п Lim сред- нее п Lim сред- нее
8 36,6-43,6 39,8 8 49,9-57,9 53,6
2 36,2-37,3 36,8 2 52,6-53,1 52,9
14 34,0-48,0 39,9 14 49,9-59,3 54,2
2 39,2-40,2 39,7 2 50,8-54,4 52,6
3 36,8-37,7 37,3 3 48,0-51,2 49,7
6 36,8-40,2 38,4 6 48,0-54,4 50,7
5 35,8-38,5 36,4 5 44,7-52,3 48,1
1 34,8 — 1 44,4 —
7 34,8-38,5 36,2 7 44,4-52,3 47,4
(Потапов, 19666; Зубакин, 1982). Встречи буроголовых чаек в сезон гнездования
и сразу после него на памирских озерах Зоркуль и Яшилькуль, а также в долине р. Али-
чур (Северцов, 1879; Абдусалямов, 1971) явно относятся к летним бродячим или
кочующим особям (см. рис. 19).
Зимовки. Морские побережья Южной и Юго-Восточной Азии от Адена на восток,
по крайней мере до Гонконга (Baker, 1929; Peters, 1934; Melville, 1977). Зимовки
отмечены также на материковой части Китая в провинции Юньнань (Cheng Tsohsin,
1976; Schauensee, 1984), отдельные птицы встречаются по р. Брахмапутре (Baker,
1929) и у г. Лхасы (Vaurie, 1972).
Миграции. На Памире буроголовые чайки появляются в конце апреля, когда свобод-
ные ото льда участки озер есть только в устьях рек (Потапов, 19666). В южном Тибете
(Гианцзе) первые птицы отмечаются раньше - в середине марта (Baker, 1929). В это
же время первые чайки отмечены на оз. Кукунор в 1873 г. (Пржевальский, 1875).
Неизвестно, летят ли чайки в Тибет через Гималаи широким фронтом или же по доли-
нам рек, прорезающим Гималайский хребет (Инд, Брахмапутра), либо стекающим с
восточной части нагорья (Меконг, Салуин и др.). Во всяком случае, в марте (как и
в ноябре) на р. Брахмапутре отмечены стаи буро голо вых чаек (Baker, 1929).
В августе—сентябре, после окончания гнездового сезона на Памире, птицы кочуют
по высокогорным озерам и долинам памирских рек. Отлетают в октябре (Потапов,
19666). Тибетские птицы покидают места гнездования, очевидно, в сходные сроки -
в октябре они уже появляются на равнинах восточной Индии и Бирмы (Baker, 1929).
В то же время Н.М. Пржевальский в октябре встречал буроголовых чаек еще в преде-
лах нагорья (экземпляр № 55 288 коллекции ЗИН с оз. Кукунор).
В сезон гнездования бродячие птицы встречены во Внутренней Монголии на оз. Сого-
Нур 13.VII 1928 г. (Lonnberg, 1932), на оз. Ханка в Приморском крае 7.VII 1949 г.
(Лебедев, 1950)3 и 30.VIII 1973 г. (Поливанова, Глущенко, 1974). В период пролета
3Предположение А.И. Иванова (1976), что в последнем случае это была реликтовая чайка, не
подтвердилось при повторном осмотре тушки в Зоомузее МГУ.
101
Семейство Чайковые
40 80 120
Рис. 26. Область распространения буроголовой
чайки
1 — область гнездования, 2 — район встреч гиб-
ридных (?) особей, 3 — места зимовок, 4 —
залеты
чайки отмечены в китайских провинциях Шаньси, Хэбэй и Внутренняя Монголия (Cheng
Tsohsin, 1976; Schauensee, 1984). Зарегистрирован залет в северную Танзанию (Britton,
Вготп, 1975).
Численность. Общая численность мировой популяции неизвестна. В 1977 г. на остро-
вах оз. Каракуль найдено 331 гнездо (Зубакин, 1982), в 1983 г., по данным
П. Мераускаса, здесь гнездилось только 25—30 пар; общая численность памирской по-
пуляции с учетом холостых бродячих особей, по-видимому, не превышает 1—2 тыс.
взрослых птиц.
Местообитания. В сезон гнездования -
крупные высокогорные озера на высотах
от ~ 3200 м (оз. Кукунор) до ~ 4860 м
(Cheng Tsohsin et al., 1983). На Памире
гнездится только на островах (Потапов,
19666; Зубакин, 1982), в других местах
отмечено гнездование и по берегам озер
(Baker, 1929; Ali, Ripley, 1969). На про-
лете и зимовках держится по морским
побережьям, нередко в гаванях вместе с
другими видами чаек (Ali, Ripley, 1969),
по долинам рек и внутренним водоемам.
Размножение. Приведенные ниже све-
дения взяты в основном из работ Р.Л. По-
тапова (1966а, б) и В.А. Зубакина
(1982). Половая зрелость достигается,
по-видимому, на второе, а у части осо-
бей, возможно, на третье лето. Бурого-
ловая чайка — моногам; это плотно гнез-
дящийся факультативно-колониальный
вид. Колонии в одних случаях могут
существовать на одном месте в течение ряда лет (гнездование на одном и том же остро-
ве оз. Каракуль отмечено в 1962 и 1977 гг.), так и менять свое расположение (Зубакин,
1982).
Демонстрационное поведение сходно с таковым озерной чайки, однако различается
в деталях (Зубакин, 1982). Наиболее часто наблюдаемая демонстрация в колонии -
поза долгого крика, которая используется в агрессивных ситуациях и при встрече
партнеров (рис. 27, а, б). Она состоит из двух фаз; в первой из них птица издает
несколько громких звуков, похожих на хриплое карканье. Общая длительность долгого
крика 2,2-3,8 с. Остальные демонстрации еще более сходны с такими же демонстра-
циями других чаек, прежде всего озерной. Прямая поза (рис. 27, в) и ’’квохтание”
(рис. 27, г), так же как и у озерной чайки, используются при территориальных
конфликтах. ’’Квохтание” обычно отмечается на границе двух гнездовых территорий,
при этом чайки-соперники стоят параллельно друг другу по обе стороны от границы.
Агрессивная прямая поза в отличие от такой же демонстрации озерной чайки чаще
используется не на охраняемой территории у гнезда, а на нейтральной территории у
края колонии. Для буроголовых чаек, как и для озерных, очень характерна распластан-
ная поза (рис. 27, д). Как и прямая поза, она может использоваться и в агрессивных
ситуациях, и при встрече брачных партнеров. В качестве агрессивной демонстрации на
близком расстоянии используется ’’огрызание”, или ложное клевание, — выпады закры-
тым или полуоткрытым клювом в сторону противника (рис. 27, е). При этом издается
102
Буроголовая чайка
негромкий урчащий или скулящий звук. Эта демонстрация у озерной чайки наблю-
дается несравненно реже.
К обычным для буроголовой чайки неагрессивным демонстрациям относятся вски-
дывание клюва, отворачивание клюва и мяукающий крик. По характеру исполнения
они близки к аналогичным демонстрациям озерной чайки. Очень похожа у этих двух
видов и церемония встречи брачных партнеров: чайки идут навстречу друг другу в
распластанных позах, затем одновременно или одна за другой резко выпрямляются,
Рис. 27. Демонстрационное поведение буроголовой чайки (по: Зубакин, 1982, с изменениями)
а—б — последовательность поз долгого крика, в — прямая поза, г — ’’квохтание”, д — распластан-
ная поза, е — ’’огрызание”
принимая прямую позу, после чего отворачивают клювы друг от друга. Отворачивание
клюва отмечается и при смене партнеров на гнезде. Вскидывание клюва обычно
предшествует ритуальному кормлению. По-видимому, как и у других видов чаек,
вскидывание клюва может предшествовать и спариванию. Мяукающий крик издает
птица, сменяющая партнера на гнезде, а также чайка, кормящая пуховичка.
На Памире отмечено гнездование в двух биотопах: на плоских, высотой не более
1—1,5 м, покрытых коркой соли голых островках и на крутых каменистых островах
площадью 0,7—1,5 га. Совместно с буроголовыми чайками здесь гнездятся горные гуси
и речные крачки. На низких солончаковых островках чайки занимают наиболее высокие
участки, на каменистых островах — как вершины, так и склоны. Гнезда могут распола-
гаться на открытых местах и среди камней, реже по краю зарослей травянистой расти-
тельности. Среди растительности гнезд не обнаружено. В Ладакхе гнезда находили также
по болотистым берегам на наполовину затопленных кочках (Ali, Ripley, 1969), в Тибете
буроголовые чайки гнездятся как на островах озер, так и на высокогорных болотах
(Cheng Tsohsin et al., 1983). Величина памирских колоний варьирует от 4 до 173 гнезд.
В литературе есть упоминание о крупных колониях в Ладакхе (Baker, 1929); в Тибете
известны колонии от 10 до нескольких сот особей, в одной колонии отмечено около
30 тыс. особей (Cheng Tsohsin et al., 1983) Минимальное расстояние между центрами
103
Семейство Чайковые
ближайших соседних гнезд чаек составляет в среднем 1,08 м (п = 177; 0,27—17 м),
средний по четырем колониям индекс плотности гнездования равен 0,18 (в разных
колониях варьирует от 0,04 до 0,29). Размеры гнезд (мм) памирских птиц (п = 41):
диаметр гнезда 250—410, в среднем 310; диаметр лотка 160—215, в среднем 190;
глубина лотка 20—70, в среднем 45. В качестве материала для гнезда используются
сухие стебли злаков, выбросы водных растений, перья.
Откладка яиц на Памире происходит в последней декаде мая — начале июня, однако
возможны и более поздние (не исключено, что повторные) кладки: свежие и слабо
насиженные яйца найдены 26.VI 1961 г. (Потапов, 19666). В Ладакхе гнездится в июне-
июле (Ali, Ripley, 1969). В кладке 1—4 яйца, чаще 3 яйца (55,8%), средний размер
кладки 2,4 яйца (п = 52). Размеры яиц памирской популяции (п = 28); 56,1-65,1 X
X 38,1—43,5, в среднем (п = 41) 61,4 X 40,8. Масса (г) : 37,6-57,6, в среднем (п= 11)
46,6 (Потапов, 19666; Зубакин, 1982). Размеры яиц из Ладакха (п= 100): 57,1—66,9 X
X 39,1—45,6, в среднем 61,3 X 42,6 (Baker, 1929). Форма яйцевидная. Окраска сложная.
Фон бледный: зеленовато-оливковый, зеленоватый, бежевый или серовато-бежевый.
Рисунок в виде редких, четких, разбросанных пятен и крапин, глубоких — серого цве-
та, поверхностных — бурого. Скорлупа мелкозернистая, матовая.
Насиживают оба родителя, по наблюдениям за 2 гнездами в светлое время суток
происходит 3—6 смен партнеров. Длительность периода насиживания неизвестна.
В 1977 г. вылупление началось 12—16 июня, последний птенец вылупился 8 июля. Про-
межуток от появления наклевки до вылупления птенца равен в среднем 2 сут (п = 9;
1,5—3 сут), между вылуплением первого и третьего птенцов в гнезде проходит в сред-
нем 2,8 сут (п = 3; 1,5—4 сут). В отличие от большинства факультативно-колониальных
видов чайковых буроголовая чайка не уносит скорлупу яиц из гнезда после вылупле-
ния, а либо затаптывает ее, либо выталкивает на край гнезда.
Масса тела (г) обсохших пуховичков в день вылупления (и = 9) 28,7—36,7, в среднем
33,3. Вес птенцов в возрасте 2 сут (п = 5) 34,5—51,1, в среднем 44,7; в возрасте 4 сут
(п = 3) 46,2—77,0, в среднем 63,5; в возрасте 6 сут (л = 1) 97,2. Пеньки контурного
оперения начинают пробиваться на 3—4 сут, в первую очередь на спине и груди. Пеньки
первостепенных маховых появляются примерно на 6—7 сут.
Первое кормление происходит через 2,5—3 ч после вылупления птенца. По наблюде-
ниям за 3 гнездами, в которых старшим птенцам было 2—2,5 сут, в светлое время суток
родители кормили птенцов 15—21 раз (в среднем 17,3 раза), причем самец 10—12 раз
(в среднем 11), самка 4—9 раз (в среднем 6,3). Наиболее интенсивно кормление проис-
ходит в утренние часы. Родители постоянно обогревают птенцов примерно до
10-суточного возраста, причем самки более длительное время, чем самцы, 10—14-суточ-
ных птенцов родители согревают только ночью, а оперяющихся птенцов в возрасте бо-
лее 2 недель перестают обогревать, несмотря на ночные морозы.
Выводок остатется в гнезде 3—4 сут после вылупления старшего птенца, затем роди-
тели уводят птенцов. К этому времени, судя по всему, происходит становление взаимно-
го опознавания птенцов и родителей. После ухода из гнезда выводок не имеет постоян-
ной территории. Во время тревоги птенцы первое время затаиваются, а с возраста
3—4 сут бегут к воде и уплывают. Плотные табунки, характерные для облигатно-ко-
лониальных видов, не образуются. Молодые поднимаются на крыло в возрасте больше
месяца, после этого они еще некоторое время Держатся вместе с родителями.
Точных данных по величине эмбриональной и постэмбриональной смертности нет.
Доля неоплодотворенных яиц и яиц с погибшими зародышами может быть достаточно
высокой — около 16%. Птенцы в первые дни жизни нередко гибнут, отстав от выводка.
Большинство из них просто замерзает, некоторые, видимо, гибнут от побоев взрослых
птиц, отгоняющих их от своих выводков. Изредка отмечаются случаи поедания яиц и
104
Буроголовая чайка
птенцов своего вида. Птенцы более старших возрастов могут гибнуть во время сильных
ночных морозов. Средняя величина вставшего на крыло выводка 1,4 птенца (п = 12).
Суточная активность, поведение. Активность сдвинута в сторону светлого времени
суток, хотя, по-видимому, может быть круглосуточной. По крайней мере, в гнездовой
колонии ночью жизнь не прекращается, хотя и несколько затихает: слышны крики
пролетающих чаек, голоса птенцов н их родителей. Спать чайки могут как днем, так и
ночью, причем в светлое время чаще в ранние утренние и вечерние часы, обычно не бо-
лее 30-45 мин без перерыва. Ночью партнер насиживающей птицы, как правило, не оста-
ется на своем гнездовом участке.
Питание. В сезон размножения основу питания составляет мелкая рыба, водные
беспозвоночные и насекомые, в том числе имаго ручейников (Молчанов, Зарудный,
1914; Потапов, 1966а, б). Анализ погадок (п = 81), собранных 18.VI-13.VII 1977 г.
на островах оз. Каракуль, показал, что мелкая рыба—голец (Nemachilus lacusnigri) —
встречается в 81,5% случаев, беспозвоночные — в 59,3% (наземные примерно в 1,5 раза
чаще, чем водные), птицы — в 4,9% (мелкие воробьиные, вертишейка, пуховички чаек),
яйца воробьиных птиц — в 1,2%. В 50,6% случаев отмечены растительные остатки (в
основном водные растения), в 34,6% — мелкие камешки — гастролиты (Зубакин,
1982). Осенью на памирских озерах питаются рыбой (Потапов, 19666), во время про-
лета изредка поедают червей, насекомых и других наземных беспозвоночных, на зимов-
ках на морских побережьях кормятся рыбой, креветками, отходами рыболовного
промысла, нередко кухонными отбросами (Ali, Ripley, 1969).
Характерные места кормежки на памирских озерах — берега озер у зоны заплеска,
мелководья у берега, устья впадающих в озера речек, соленые лужи и озерки в
непосредственной близости от озера. Кормежек на суше, в том числе на помойках на-
селенных пунктов, в сезон гнездования не отмечено. На воде птицы собирают корм
с глубины в несколько сантиметров (иногда 20—30 см), погружая в воду голову и
шею, реже — с поверхности воды (поедая хирономид) . Наблюдаются кормежки на бе-
регу в прибойной полосе. Изредка отмечается поедание яиц и маленьких птенцов свое-
го вида.
Враги, неблагоприятные факторы. В настоящее время буроголовая чайка на Пами-
ре почти не имеет естественных врагов. Колонии их недоступны наземным хищникам,
а из пернатых, по-видимому, только ворон может наносить некоторый ущерб, поедая
кладки или птенцов (Абдусалямов, 1971). Гораздо чаще гибель птенцов, как указы-
валось выше, зависит от погодных факторов или происходит по вине взрослых чаек.
Прямого преследования со стороны человека не отмечается, однако часть кладок
может гибнуть в результате браконьерского сбора яиц горных гусей. Данные о величи-
не смертности взрослых птиц отсутствуют.
Хозяйственное значение, охрана. Хозяйственного значения не имеет. Как малочислен-
ный периферийный вид фауны СССР подлежит охране. Занесена в Красную книгу СССР.
Морской голубок -
Larus genei Breme, 1839
Slender-billed Gull (англ.), Diinnschnabelmowe (нем.), Goeland railleur (фр.).
Син. (лат.): Hydrocoloeus genei (Breme, 1839), Larus gelastes Thienemann, Brehm, 1838.
Син. (рус.): тонкоклювая чайка.
Статус. Гнездящийся перелетный, частично зимующий вид.
Общая характеристика н полевые признаки. Чайка среднего размера, величиной с
озерную или слегка крупнее. Окраской крыльев, в том числе и первостепенных махо-
вых, очень похожа на озерную чайку; в брачный сезон отличается от нее белой головой
105
Семейство Чайковые
и хорошо заметным розовым оттенком нижней части туловища. В гнездовом и зимнем
нарядах эти виды трудноразличимы в полевых условиях, но клюв и шея у морского
голубка заметно длиннее, а темные пятна перед глазом и на кроющих уха гораздо
светлее; нередко они совсем отсутствуют. Сидящий голубок несколько более высо-
коногий и заметно более длинношеий, чем озерная чайка. Радужина у взрослых птиц
в отличие от озерной чайки светлая. Полет и плаванье, как у чаек сходного размера.
Голубки чаще, чем другие чайки, бродят по брюхо в воде по мелководью, как кулики.
Голос похож на голос озерной чайки. Основной видовой крик — дребезжащее карканье,
которое звучит немного мягче, чем у озерной чайки. Тревожный крик — короткое
”ка-ка-ка”.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Большинство систематиков сближают морского голубка
с озерной чайкой и близкими к ней видами. Для выделения его в монотипический род
Gelastes, как это сделано Ш. Бонапартом, нет достаточных оснований (Dwight, 1925).
Описание. Размеры особей (мм) (ЗМ МГУ) и масса тела (г) (Бородулина,
1960; Долгушин, 1962):
Длина крыла:
66 (п = 22) — 287 — 319 (среднее 308);
9 9 (п = 12) - 275 - 304 (среднее 291).
Длина клюва:
66 (п = 22) — 35,6 — 45,2 (среднее 41,9);
9 9 (и = 13) — 35,6 — 39,9 (среднее 37,5).
Длина цевки:
66 (п = 23) — 50,7 — 55,7 (среднее 53,1);
9 9 (и = 13) — 45,8 — 53,1 (среднее 48,8).
Масса тела:
66 (л = 15) — 276 — 375 (среднее 330);
9 9 (п = 15) — 230 - 360 (среднее 288).
Окраска (ЗМ МГУ). Самец и самка в брачном наряде. Голова, грудь, брюхо,
бока, надхвостье, подхвостье, рулевые и подмышечные перья белые, причем нижняя
часть туловища, особенно брюхо, с ярким розовым оттенком. Спина, поясница, плече-
вые и верхние кроющие крыла сизо-серые, довольно светлые. Испод крыла серый,
темнее, чем у озерной чайки (почти такого же цвета, как и верхние кроющие крыла),
передний край крыла белый. II (первое видимое) первостепенное маховое белое с
черной вершиной и узкой черной полосой по внешнему краю наружного опахала;
у части особей по внешнему краю внутреннего опахала может быть узкая серо-бурая
каемка. Ill—V первостепенные белые, с черной вершиной, бурой каймой по краю внут-
реннего опахала и серым полем на внутреннем опахале; площадь серого поля и шири-
на каймы увеличиваются к V перу. VI—VIII первостепенные серые с черной вершиной
и бурой каймой по краю внутреннего опахала, остальные серые, у ряда особей с неясной
бурой каймой на внутреннем опахале. Второстепенные маховые серые. Клюв и лапы
у живых птиц почти черные, реже темно-красные, радужина белесая или желтоватая.
Края век ярко-красные.
106
Морской голубок
Самец и самка в, зимнем наряде. На темени и затылке серый налет, перед глазом
и на кроющих уха серо-бурые пятна. Розовый цвет развит слабее. Остальное оперение,
как в брачном наряде. Клюв и лапы более светлые (Cramp, Simmons, 1983), красные.
Пуховой наряд (ЗИН, живые пуховички с Чонгарских островов). Общий фон свет-
лый, гораздо светлее, чем у озерной чайки: светло-серый, кремовый или светло-охрис-
тый, иногда почти рыжий. По верхней части головы, шее, спине, крыльям и бокам —
контрастные темно-бурые или черные пятна различной густоты. Изредка лоб, темя и
затылок могут быть в густых черных пушинках, в результате чего голова кажется тем-
нее туловища. Подбородок белый, на горле неясные темные пестрины. Грудь и брюхо
белые, причем брюхо может быть с отдельными темными пестринами. Клюв и ноги
серо-телесные.
Гнездовой наряд. Лоб, темя, затылок, задняя часть шеи — буровато-серые, интенсив-
ность цвета -может варьировать у разных особей; кроющие уха бурые. Подбородок,
горло, передняя часть шеи, грудь, брюхо, надхвостье и подхвостье белые, иногда с ро-
зоватым оттенком, по бокам груди буроватый налет. Перья спины, плеч, верхние крою-
щие крыла (кроме больших) бурые (разной интенсивности цвета у разных особей)
с серыми основаниями и светлыми вершинами. Большие кроющие крыла серые. II (пер-
вое видимое) — V первостепенные маховые белые с черно-бурой вершиной и широки-
ми каймами такого же цвета по внешним краям наружного и внутреннего опахал;
на внутреннем опахале V пера развит серый цвет. Внутренние первостепенные серые
с черно-бурыми концами и бурыми внутренними опахалами; на самых кончиках перьев
белые пятнышки. Второстепенные маховые серые с большими бурыми предвершинны-
ми пятнами. Рулевые белые с черным вершинным пятном. По С. Крэмру (Cramp, Sim-
mons, 1983), клюв и лапы оранжево-телесные, радужина темная.
Первый зимний наряд. Голова, шея и грудь белые, на кроющих уха серое пятно.
Спина и плечевые сизо-серые. Остальное оперение, как в предыдущем наряде. Первый
брачный наряд отличается от первого зимнего белой головой и появлением розового
налета на белом оперении (Дементьев, 1951).
Линька. Линька из гнездового в первый зимний наряд (август—октябрь) частичная,
сменяется оперение головы и туловища. Затем линьки бывают дважды в году: полная
осенняя и частичная весенняя. Предбрачная линька взрослых птиц (сменяется оперение
головы и туловища) протекает в апреле—мае, послебрачная (сменяется все опере-
ние) — в июле—октябре—ноябре (Дементьев, 1951). Линька промежуточных нарядов
происходит, возможно, в более ранние сроки (Cramp, Simmons, 1983).
Распространение. Гнездовой ареал. Первоначально область гнездования вида,
по-видимому, охватывала северное и северо-западное побережья Черного моря, Малую
Азию, Каспийское море, западную, центральную и южную часть Казахстана, внутренние
водоемы Ирака, Ирана (возможно, и иранское побережье), побережье Пакистана.
В республиках Средней Азии эти чайки на гнездовье встречались лишь на побережье
Каспия (Дементьев, 1951, 1952). Примерно с 60-х годов нашего столетия небольшие
поселения голубков стали отмечаться в западной части Средиземного моря, а также
на атлантическом побережье Марокко, Мавритании и Сенегала (Latour, 1973; цит. по:
Isenmann, 1976; Cramp, Simmons, 1983). Впрочем, не исключено, что эти чайки и преж-
де нерегулярно гнездились в западном Средиземноморье, в частности в южной Испании
(Dwight, 1925; Wallace, 1964). В 1960—1970-е годы голубки стали поселяться на возни-
кающих в это время крупных водохранилищах Средней Азии, чуть раньше на водоемах
Предкавказья, заселив территорию, ранее входившую лишь в область пролета вида.
Как и черноголовый хохотун, сплошного гнездового ареала морской голубок не
107
Семейство Чайковые
образует; отдельные постоянные или временные гнездовые поселения удалены нередко
на сотни километров друг от друга (рис. 28. 29). Не менее 50 лет существуют посе-
ления в Тендровском заливе Черного моря (Ардамацкая, 1975а), не менее 90 лет —
на островах Сиваша (Браунер, 1898; Сеницкий, 1898; Молчанов, 1906; Воронцов,
1937; Зубакин, Костин, 1977; Сиохин, 1981а), с 1968 г. — на Молочном лимане (Фило-
нов, Лысенко, 1971; Филонов и др., 1974; Сиохин, 1981а), с 1953 г. — на водоемах
системы Маныча: оз. Маныч-Гудило, Пролетарском водохранилище, Меклетинских
озерах (Бакеев и др., 1957; .Кривенко, Любаев, 1975,1981; Языкова, 1975; Кривенко,
1981; Любаев, Кривенко, 1981). Колонии морских голубков отмечались также в
1959 г. на лиманах Таманского полуострова (Кищинский, 19606) и в 1973 г. в Картка-
зацком заливе Черного моря в районе Перекопского перешейка (Костин, 1975).
На Каспии морской голубок периодически гнездится в авандельте Волги, в приустье-
вой части дельты р. Урал (по-видимому, не ежегодно), на о-ве Осушном в Красноводс-
ком заливе, на разливах низовья р. Атрек; в прежние годы отмечались колонии в устье
Эмбы, на Тюленьих (Мангистауских) островах (1951—1952 гг.) и в устье пролива Ка-
ра-Богаз-Гол (1973 г.), однако современных данных по этим территориям мы не имеем
(Гладков, Залетаев, 1956; Долгушин, 1962; Бондарев, 1975; Васильев и др., 1975;
Кривоносов, 1976; Караваев, 1981; Гаузер, 19836).
В Казахстане неизвестны постоянные поселения морских голубков. Очевидно, это
связано с непостоянным гидрологическим режимом казахстанских озер. Колонии го-
лубков известны с 1959 г. на оз. Тенгиз в Целиноградской области, однако птицы гнез-
дились здесь не ежегодно (Чекменев, 1964; Азаров, 1975а; Волков, 1975; Андрусенко,
1981). В период обводнения озер появляются колонии на оз. Челкар-Тенгиз и на других
озерах низовий Тургая, на озерах и сорах Приаральских Каракумов и л отдельных мес-
тах северного побережья Аральского моря (Сушкин, 1908; Спангенберг, Фейгин, 1936;
Долгушин, 1962; Ауэзов, Хроков, 1975; Варшавский и др., 1977). В 1961 г. морской
голубок гнездился на пресноводном оз; Казоты недалеко от низовий р. Таласе (Долгу-
шин, 1962); дальнейшая судьба этой колонии неизвестна. В 1968-1969 гг. и 1981 г.
отмечено гнездование на оз. Соркуль восточнее пос. Джаныбек Уральской обл. (Линде-
ман, Залетаев, 1977; Леонович, устное сообщ.). Предположение о гнездовании голубков
на Балхаше (Долгушин, 1962) не подтвердилось (Ауэзов, 1986). На Камыш-Самарс-
ких озерах эта чайка в настоящее время, очевидно, не гнездится (Шевченко, 1975), хотя
голубков периодически отмечают здесь в сезон гнездования (Леонович, устное сообщ.).
Самая северная колония морских голубков зарегистрирована на оз. Термакуль
близ курорта Карачи в Новосибирской области в 1925 г. (Рузский, 1926). В последую-
щие годы эта чайка здесь не гнездилась (Долгушин, 1962).
В Средней Азии морской голубок с 1962 г. гнездится на Хаузханском водохранили-
ще Туркмении (Ташлиев и др„ 1967), с 1966 г. — на Кайраккумском водохранилище
Таджикистана (Абдусалямов, 1981). В Узбекистане отмечено гнездование на оз. Айдар-
куль (Мухина, 1983; Остапенко и др., 1984).
Вне СССР крупные гнездовые поселения (несколько сот пар) расположены в настоя-
щее время на побережье Мавритании, о-ве Сардиния, на внутренних водоемах Турции
и Ирака, в дельте Нила. Небольшие колонии в несколько пар отмечаются, обычно не
ежегодно, вдоль атлантического побережья Марокко, в Испании (устья рек Гвадалкви-
вир и Эбро), с 1948 г. (регулярно с 1971 г.) в Камарге во Франции, в северо-восточной
части Италии, в Греции в дельтах рек Аксиос и Нестос (Cramp, Simmons, 1983). Гнездо-
вые поселения найдены в Сенегале, в Персидском заливе (Бахрейн) и в заливе Сонмия-
ни в Пакистане; возможно гнездование и в других пунктах побережья Пакистана
(Wallace, 1964; Ali, Ripley, 1969; Isenmann, 1976). Современных данных о распростра-
нении голубка в Иране нет.
108
Морской голубок
Рис. 28. Область распространения морского голубка
1 — постоянные поселения, 2 — временные или периодически возникаю-
щие поселения, 3 — места зимовок, 4 — область регулярных кочевок,
5 — залеты
Рис. 29. Ареал морского голубка в СССР
Обозначения те же, что на рис. 28
109
Семейство Чайковые
Появление новых поселений морских голубков в западном Средиземноморье, воз-
можно, говорит о том, что начинается расширение области гнездования этого вида в
западном направлении. Однако, как и в случае с черноголовой чайкой, с уверенностью
говорить об этом пока преждевременно, поскольку устойчивые поселения в большинст-
ве новых мест не сформировались (за исключением колоний у побережья Мавритании,
время возникновения которых точно не известно). Обращает на себя внимание, что ны-
нешнее расселение морских голубков и черноголовых чаек происходит в целом в те же
регионы (западное Средиземноморье, южные побережья Северного и Балтийского мо-
рей) , куда в более отдаленные времена расселилась пестроносая крачка — еще один вид
черноморско-каспийского бассейна. Не исключено, что вслед за пестроносой крачкой
морской голубок и черноголовая чайка закрепятся в этих регионах и даже попытаются
образовать поселения в Америке. Видимо, в этом же направлении следует ожидать рас-
ширения, области гнездования черноголового хохотуна в случае возрастания его чис-
ленности.
Зимовки. Черноморские ^орские голубки зимуют, по данным кольцевания,
главным образом в двух областях Средиземного моря: в Нижнем Египте и на северном
и северо-восточном побережьях Туниса н прибрежных участках о-ва Сицилия. Меньшая
часть птиц зимует в Черном море у берегов южного Крыма и Кавказа и в Азовском
море; в некоторые годы стайки голубков отмечаются в водах Черноморского заповед-
ника. Отдельные птицы зимуют в Персидском заливе (Семенов, Сабиневский, 1957).
Голубков, окольцованных в дельте Волги, добывали зимой на южном побережье Каспия
и в Омане (Кривоносов и др., 1978); на юго-западном Каспии этот вид зимует не еже-
годно (Заблоцкий, Заблоцкая, 1963). Зимовки отмечены также на побережье Маврита-
нии, средиземноморском побережье Испании, в Красном море и по морскому побе-
режью от Персидского залива до Бомбея (Ali, Ripley, 1969; Cramp, Simmons,
1983).
Миграции. Весенние миграции начинаются, по-видимому, в конце февраля — первых
числах марта. Увеличение численности голубков в водах Черноморского заповедника
наблюдается в конце февраля — середине марта, массовый прилет — 22—28.III (Семе-
нов, Сабиневский, 1957). Перемещения птиц со средиземноморских зимовок идет,
видимо, в основном по западному побережью Черного моря, поскольку на Черноморс-
ком побережье Кавказа голубок в весеннее время редок (Вронский, Томкович, 1975).
В начале апреля он пролетает в северо-восточном направлении вдоль северного побе-
режья Азовского моря (Лысенко, 1975), на внутренних водоемах Приазовья появля-
ется в значительных количествах во второй декаде марта, пролет идет до конца мая
(Казаков, Языкова, 1982). Что касается Средней Азии и Казахстана, то в низовьях
р. Сурхандарьи в 1975 г. пролет голубков отмечался 27.11—15.IV (Остапенко и др.,
1978), в среднем течении р. Мургаб в 1974 г. — в марте (Эминов, 1978), первые особи
на Айдаркуле появляются 18-27.III, основной пролет проходит в конце апреля — на-
чале мая (Калабин, 1986), в дельте Амударьи в 1974 г. интенсивный пролет голубков
шел в 2—4-ю пятидневках апреля (Ажимуратов, 1978), на нижней Сырдарье слабый
пролет отмечался в середине апреля (Спангенберг, Фейгин, 1936). В окрестностях мест
гнездования в Красноводском заливе голубки появляются 18.III—4.IV (Гаузер, 19836),
на оз. Тенгиз первых особей наблюдали 9.V, к 15.V голубки были уже обычны (Чек-
менев, 1964).
Послегнездовые кочевки начинаются в июле, в августе—сентябре они приобретают
характер пролета. Черноморские голубки летят в основном на запад, затем повора-
чивают на юг вдоль западного побережья Черного моря. В августе и сентябре часть птиц
уже появляются на средиземноморских зимовках, в ноябре зимовок достигают практи-
чески все особи. Меньшее число голубков летят из Тендровского залива в восточном
110
Морской голубок
направлении — на зимовки в Азовское море и на побережья южного Крыма и Кавказа.
Отмечены позднеосенние встречи голубков на северном и юго-западном Каспии (Семе-
нов, Сабиневский, 1957). В Северном Приазовье пролет идет в основном в западном
и юго-западном направлениях двумя волнами: в конце июля и в конце октября — пер-
вых числах ноября (Лысенко, 1975). Осенние миграции в азиатской части СССР просле-
жены слабо. Известно, что на Айдаркуле массовый пролет проходит в сентябре;
в 1981 г. его пик приходился на 15-25.IX (Остапенко и др., 1984). На юго-западном
Каспии пролет идет с августа до начала декабря (Заблоцкий, Заблоцкая, 1963).
Залеты морских голубков отмечены в Великобританию, ФРГ, Австрию, Португалию,
Югославию (Cramp, Simmons, 1983), на Канарские о-ва (Семенов, Сабиневский, 1957),
в Березинский заповедник (Долбик, 1970), в г. Оренбург, г. Вольск Саратовской обл.
(Водолажская, Залетаев, 1977), на оз. Севан в Армении (Маргарин, 1981) и на о-ве
Шри-Ланка (Berg et al., 1982).
Численность. Возрастает, хотя и не столь стремительно, как у черноголовой чайки.
В Тендровском заливе в 1935 г. гнездилось 11 тыс. пар, в 60-е годы — 7,4—22,6 тыс. пар,
в 70-е годы — 10,3—24,8 тыс. пар, в начале 80-х— 26,2—37,3 тыс. пар (Ардамацкая,
1975а, 1984), на Сиваше в 1973—1979 гг. - 270-3913 пар, на Молочном лимане -
221-1128 пар (Зубакин, Костин, 1977; Сиохин, 1981а), на озерах и водохранилищах
системы Маныча в 1974—1979 гг. —0,7—11 тыс. пар (Кривенко, 1981; Любаев, Кривен-
ко, 1981), на Северном Каспии — около 12 тыс. пар (Кривоносов, 1976), на о-ве Осуш-
ной в Красноводском заливе - от 7 пар в 1972 г. до 1340 пар в 1985 г. (Гаузер, 1986),
на разливах Атрека в 1978—1979 гг. — 212—314 пар (Караваев, 1981), на оз. Соркуль
в Уральской обл. в 1968-1969 гг. - 300-800 пар (Линдеман, Залетаев, 1977), на оз.
Тенгиз в 1977 г. - 450—470 пар (Андрусенко, 1981), на оз. Казоты в 1961 г. - 150-180
пар (Долгушин, 1962). Общая численность морских голубков в СССР составляет в на-
стоящее время не менее 52 тыс. гнездящихся пар (возможно, до 60 тыс. пар). Вне СССР
гнездится, по приблизительной оценке, 5—10 тыс. пар (Cramp, Simmons, 1983)
Местообитания. В сезон размножения — соленые, солоноватоводные и реже пресно-
водные озера и водохранилища, морские заливы, лиманы и побережья (особенно участ-
ки в устьях рек) степной, полупустынной и пустынной зон. В Казахстане в многовод-
ные годы может поселяться на залитых водой сорах и других пересыхающих водоемах.
Для гнездования необходимы изолированные от наземных хищников островки, для
кормежки — мелководья. Вне сезона размножения — морские побережья, чаще всего
мелководные заливы и лиманы; в период пролета также — соленые и реже пресные озе-
ра, еще реже долины рек.
Размножение. Приступает к размножению, по-видимому, в возрасте двух лет.
Моногам. Облигатно-колониальный вид. Одни и те же острова в благоприятных услови-
ях голубки могут заселять в течение десятков лет (однако места расположения коло-
ний на острове из года в год меняются), в то же время отдельные поселения могут
существовать один или несколько сезонов. Характерны значительные колебания числен-
ности по годам в каждом конкретном поселении.
В демонстрационном поведении морского голубка, по наблюдениям на Сиваше и в
Камарге (Франция), много своеобразия, хотя сходство с озерной чайкой прослежи-
вается (Зубакин, 19766; Isenmann, 1976). Голубка отличает преобладание ’’вертикаль-
ных” демонстраций, когда птица вытягивается ’’столбиком”; позы, в которых туло-
вище горизонтально или задний конец тела приподнят (распластанная, ’’квохтанье”,
мяукающий крик) отсутствуют. Не отмечено также ’’клевание травы”. Долгий крик
полифункционален, исполнение этой демонстрации включает три фазы (рис. 30,а—в).
Он звучит как ”ува-ка-ка-ка-кау” (или ”ува-кра-кра-кра-крау”), причем его первый
и последний слога произносятся в первой и третьей фазе, а остальные (их может быть
111
Семейство Чайковые
3—10) — во второй. Длительность всего крика 2—5 с. Как и при большинстве других
демонстраций, сложенные крылья слегка отодвинуты от боков туловища.
Одна из наиболее частых демонстраций у голубков — вертикальная поза, которая,
видимо, гомологична неагрессивной прямой позе озерных чаек. В фигуре птицы в этой
позе есть что-то змеиное (см. рис. 30,г). Вертикальную позу голубки принимают при
смене партнеров на гнезде, встрече партнеров и перед столкновениями вне колонии.
С данной демонстрацией тесно связано отворачивание клюва, которое, как и у озерной
Рис. 30. Демонстрационное поведение морского голубка
а—в — последовательность поз долгого крика» — г — вертикальная поза
чайки, входит в ритуал встречи партнеров. Основная агрессивная демонстрация в коло-
нии — ’’огрызание”, или выпад раскрытым клювом в сторону противника; при этом изда-
ется резкий крик ’’уввааа”. Угольно-черная окраска ротовой полости, видимо, усилива-
ет отпугивающий эффект. ’’Огрызание” используется также и против пернатых хищни-
ков типа серебристой чайки; в этом случае одновременно демонстрируют черную
’’пасть” все птицы колонии и их голоса сливаются в мощный хор.
У морских голубков отмечены также сгорбленная поза и вскидывание клюва —
демонстрации, с помощью которых самки выпрашивают корм у самца во время ри-
туального кормления.
Морские голубки гнездятся почти исключительно на небольших островках, лишь
в Казахстане в редких случаях отмечалось гнездование на топких побережьях. Остро-
ва могут быть либо материкового, либо, что гораздо чаще, наносного происхождения.
Под колонию голубки выбирают участки, лишенные или почти лишенные растительнос-
ти; нередко колонии тяготеют к повышениям микрорельефа. По соседству с морски-
ми голубками обычно гнездятся чайконосые и пестроносые крачки, чегравы, реже чер-
ноголовые чайки, однако в таких смешанных колониях голубки в большинстве слу-
чаев образуют компактные одновидовые группировки. Соседства с серебристой чайкой
они явно избегают. Величина колоний варьирует от нескольких до нескольких сот и да-
же тысяч гнезд (в Черноморском заповеднике известны колонии в 13 тыс. гнезд),
случаи гнездования одиночными парами крайне редки, отмечаются лишь во вновь
образующихся поселениях и только в колониях других видов чайковых. Колонии не-
редко состоят из 2—18 субколоний, которые включают от нескольких до нескольких
десяткой гнезд, реже сто и более гнезд (Долгушин, 1962; Чекменев, 1964; Wallace,
112
Морской голубок
1964; Ali, Ripley, 1969; Ардамацкая, 19776, 1981; Зубакин, Костин, 1977; Гаузер,
19836; Cramp, Simmons, 1983).
Формирование поселений на о-ве Орлов Тендровского залива начинается с конца пер-
вой декады апреля, в Красноводском заливе — в конце апреля. Образуется несколько
стай (клубов), которые держатся, нередко перемещаясь, в определенных районах ост-
рова. Особи клуба агрессивно относятся к появлению поблизости другой стаи. Переме-
щения клубов прекращаются с появлением гнезд (Семенов, Сабиневский, 1957; Гау-
зер, 19836).
Строительство гнезда и охрана его начинаются только за 2—3 дня до откладки яиц
(Isenmann, 1976), по другим данным — накануне откладки (Семенов, Сабиневский,
1957). Гнездо — обычно массивная постройка из сухих водорослей, веточек солянок
и другой растительной ветоши. Края гнезд в период насиживания всегда запачканы по-
метом. В Тендровском заливе средний диаметр гнезда (мм) 310, лотка 185, средняя
глубина лотка 40; на Сиваше (п = 22) 293 (255—340), 185 (155—240), 34 (25—52);
в Красноводском заливе 257, 163, 56; на оз. Тенгиз 250,140, 50 соответственно (Боро-
дулина, 1960; Чекменев, 1964; Гаузер, 19836; данные В.А. Зубакина). Плотность раз-
мещения гнезд очень высокая, нередко их края соприкасаются. Среднее расстояние
между центром ближайших соседних гнезд (м) на Сиваше равно (п =544) 0,42 (0,2—3,0),
в Камарге (п = 13) - 0,44 (0,26-0,78) . На Сиваше индекс плотности гнездования по
8 колониям составляет 0,45 (0,35-0,51), в Красноводском заливе - 0,67. В других
местах голубок гнездится со сходной плотностью (Зубакин, 1976в; Isenmann, 1976;
Гаузер, 19836).
Первые кладки в Тендровском заливе отмечаются 28.IV—5.V (Семенов, Сабиневс-
кий, 1957), по более поздним данным - 13-15.IV (Ардамацкая, 19776,в). Судя по сро-
кам вылупления, на Сиваше первые яйца появляются около 20.IV (Зубакин, Костин,
1977). В то же время в отдельных, обычно вновь образовавшихся поселениях, гнездо-
вание может начинаться почти на месяц позже (Филонов и др., 1974; Isenmann, 1976).
На оз. Маныч-Гудило кладки появляются с первой декады мая (Казаков, Языкова,
1982). В Казахстане откладка яиц начинается позже, видимо в середине мая, в некото-
рых поселениях и в июне (Долгушин, 1962). Однако в 1981 г. на оз. Соркуль Уральс-
кой обл. отдельные гнезда имели полные кладки из 3 яиц уже 6.V (Леонович, устное
сообщ.). Откладка яиц в каждой конкретной колонии происходит в сжатые сроки —
обычно не более 3—5 дней (Клименко, 1950а; Семенов, Сабиневский, 1957). В случае
гибели гнезд бывают повторные кладки (Ардамацкая, 19776). В полной кладке 1—5,
чаще всего 2—3 яйца. Окраска яйца сложная: основной фон белый, с легким кремовым
или розоватым оттенком, реже с охристым налетом. Глубокие фиолетово-серые и по-
верхностные контрастные черно-бурые пятна разной величины и формы относительно
равномерно распределены по поверхности скорлупы (Долгущин, 1962). На Сиваше
изредка попадаются яйца с бледно-зеленоватым фоном. В целом яйца голубков резко
отличаются окраской от яиц большинства видов чайковых и напоминают яйца пестро-
носых крачек.Размеры яиц (мм): с Тендровского залива (п = 37) 50,2—59,5 X 33,6—42,5,
в среднем 54 X 39 (Бородулина, 1960); с Молочного лимана (п = 86) 48,8—57,7 X
X 34,0—43,0, в среднем 53,9 X 38,3 (Филонов и др., 1974); с оз. Тенгиз (п = 39)
48-59 X 36-42, в среднем 55 X 39 (Чекменев, 1964); с оз. Маныч-Гудило (п = 24)
в среднем 54,0 X 39,3 (Казаков, Языкова, 1982). Масса (г) свежих яиц: с оз. Тенгиз
(п = 30) 41-52, среднее 44,6 (Долгушин, 1962); с оз. Соркуль (п =.27) 35,1-49,7, сред-
нее 43,5 (данные В.В. Леоновича); с оз. Маныч-Гудило (и = 24) 42-49, среднее 44 (Ка-
заков, Языкова, 1982).
Насиживают кладку оба партнера, но в большей степени самка. Длительность перио-
да насиживания 21-22 дня (Семенов, Сабиневский, 1957; Бородулина, 1960), по дру-
113
Семейство Чайковые
гим данным 25-28 дней (Isenmaim, 1976). По наблюдениям на Сиваше, смена на гнезде
происходит 3-4 раза в сутки (и = 2). Охраняемая территория у гнезда отсутствует,
охраняется только само гнездо. Между вылуплением первого и последнего птенца в
гнезде с 3 яйцами проходит 1—5 сут (и = 110), чаще всего 3 сут (в 50% случаев), реже
2 сут (27,3%), еще реже 4 сут (14,5%). Как и другие облигатно-колониальные виды,
голубки не уносят скорлупу из гнезда после вылупления (наши данные). Средняя мас-
са (г) птенцов: в день вылупления 28,8 (и = 55), в суточном возрасте 31,6 (и = 47),
в возрасте 3 сут 41,1 (и = 13), в 5 сут (и = 2) 68,0 (Гаузер, 19836).
Голубки кормят птенцов отрыжкой, в первый раз через 3-4 ч после вылупления.
В выкармливании принимают участие оба партнера (на Сиваше примерно в равной
степени), кормление пуховичков в Тендровском заливе происходит 18—22 раза в сут-
ки, а смена птиц на гнезде - всего 2-3 раза. На Сиваше 1-5 суточных птенцов родители
кормят 6-13 раз в сутки; смена птиц, обогревающих птенцов, происходит 2-9 раз в
сутки (и = 4) (Семенов, Сабиневский, 1957; Бородулина, 1960; наши данные). Вывод-
ки обычно оставляют гнездо через 3 сут после вылупления старшего птенца. В это же
время происходит становление взаимного опознавания родителей и птенцов, после чего
взрослые птицы начинают агрессивно относиться к чужим птенцам, приближающимся
к их выводку. После оставления колонии выводки собираются в так называемые
ясли — большие стаи птенцов, в которых постоянно держится и некоторое количество
взрослых птиц. Эти стаи кочуют по берегу, объединяясь, вновь разделяясь и постепенно
вбирая в себя выводки разных колоний. Такие ясли нередко состоят из нескольких сот
и даже тысяч птенцов, однако родители находят в них своих отпрысков и кормят толь-
ко их. Во время тревоги птенцы яслей сбиваются в плотный табунок. Подобная реак-
ция на хищника характерна с 4—5-суточного возраста, более младшие птенцы затаивают-
ся в гнезде или там, где их застала тревога (Зубакин, 1976а,в; Гаузер, 19836). На кры-
ло молодые поднимаются в возрасте 33—37 дней (наблюдения В.А. Зубакина), по дру-
гим данным - 30-35 дней (Isenmann, 1976). Родители докармливают птенцов и после
оставления колонии (Костин, 1983) .
Как и у черноголового хохотуна, стратегия защиты потомства у морского голубка
основана на ’’эффекте плотности” (Зубакин, 1976в, 1983) , Элементы активной защиты,
в частности пикирование на хищника, голубками используются реже, чем, например,
озерными чайками. На человека голубок пикирует очень редко.
По наблюдениям в Красноводском заповеднике, в отсутствие хищников средний
отход яиц за 4 года составил 5,9%, достигая в отдельных поселениях 20,9%; причем
46,9% всех погибших яиц приходилось на долю неоплодотворенных. В качестве важней-
шей причины эмбриональной смертности указывается также оставление родителями
последних яиц в кладке в результате раннего увода птенцов в ясли. На Сиваше в 1973 г.
9% гнезд погибло от наводнений, еще 18,5% гнезд было брошено птицами после оставле-
ния колонии основной массой выводков. Процент найденных в колонии погибших птен-
цов оказался сходным на Сиваше и в Красноводском заливе — 9% и 10,6% от числа вы-
лупившихся птенцов, соответственно, причем 29,3% и не менее 34% этих птенцов были
убиты взрослыми птицами. Кроме того, определенное количество птенцов на Сиваше
погибло в результате хищничества серебристых и черноголовых чаек (Зубакин, 1976в;
Зубакин, Костин, 1977; Гаузер, 19836).
Суточная активность, поведение. Активность преимущественно дневная (Авилова.
1972, 1973), однако, судя по наблюдениям на Сиваше, ночью жизнь в колонии оконча-
тельно не замирает: постоянно слышны крики пролетающих птиц, писк птенцов, отдель-
ные голубки покидают колонию, другие возвращаются; в одном случае в темноте от-
мечена смена партнеров на гнезде (данные В.А. Зубакина). Есть данные о миграциях
голубка в ночное время (Андрусенко, 1980).
114
Морской голубок
Голубок — общественная птица, держится группами и стаями как в сезон гнездова-
ния, так и вне его. Состав стай, судя по всему, может быть постоянным в течение ряда
лет, причем такие стаи могут основывать новые колонии на большом расстоянии от
прежних (Бакеев и др., 1957; Семенов, Сабиневский, 1957).
Питание. Морской голубок трофически в большей степени связан с водоемами,
чем с сушей: основу питания его во все сезоны составляют водные животные, главным
образом ракообразные и мелкая рыба. Наряду с доминированием водных кормов от-
мечалось поедание и наземных насекомых (Сушкин, 1908; Клименко, 1950а; Семенов,
Сабиневский, 1957; Долгушин, 1962; Заблоцкий, Заблоцкая, 1963; Чекменев, 1964;
Wallace, 1964; Ali, Ripley, 1969; Зубакин, 1976в; Isenmann, 1976; Сиохин, 19816;
Казаков, Языкова, 1982).
Основные места кормежек: мелководные лагуны, где голубки добывают мелкую
•рыбу (хамсу, атеринок, бычков, морских игл) и беспозвоночных (креветок, морских
тараканов), пресноводные болотца, где они собирают мотыля и других личинок насе-
комых, а также очень соленые водоемы, где голубки поедают листоногих рачков Аг-
temia salina. Отмечены кормежки и в прибрежной степи саранчевыми и другими насе-
комыми. Радиус разлета за кормом может достигать 15—20 км. Рыбу и водных бес-
позвоночных голубки ловят, либо бродя по мелководью, как кулики, либо плавая
по воде, вытянув вперед шею и делая броски головой за добычей. Нередко они взле-
тают или слегка выпрыгивают из воды и ныряют, погружаясь в воду по крылья.
Видимо, подобными способами питания определяется большая, чем у других чаек,
относительная длина шеи и лап у морских голубков (Бородулина, 1960; Долгушин,
1962; Кистяковский, 1967; Isenmann, 1976).
Несмотря на агрессивное отношение взрослых птиц к чужим птенцам, которых
нередко забивают до смерти, поедание птенцов (как и яиц) своего вида для голуб-
ка не характерно. За 2 года наблюдений на Сиваше лишь единственный раз была отме-
чена попытка расклевать яйца брошенной кладки (данные В.А. Зубакина).
Врага, неблагоприятные факторы. Определенный ущерб колониям морских го-
лубков могут наносить серебристые чайки-хохотуньи и черноголовые чайки, поедаю-
щие яйца и птенцов, особенно в условиях повышенного беспокойства колоний со сто-
роны человека. Гнезда нередко гибнут от затопления в результате нагонных ветров
и штормов, птенцы - от холодных дождей с градом (Клименко, 1950а; Зубакин,
Костин, 1977). В качестве причины гибели птенцов определенное значение имеют со-
циально обусловленные факторы, в частности уничтожение птенцов взрослыми птица-
ми и оставление младших птенцов в момент увода выводков из колонии (Гаузер,
19836). Отмечена массовая гибель взрослых и птенцов в колонии на оз. Маныч-Гу-
дило либо от отравления, либо в результате эпизоотии (Бакеев и др., 1957).
Хозяйственное значение, охрана. Хозяйственного значения вид не имеет. Колонии
охраняются в Черноморском, Красноводском и Кургальджинском заповедниках и
ряде заказников.
Китайская чайка -
Laras saundersi (Swinhoe, 1871)
Saunders’s Gull (англ.), Kappenmove (нем.), Mouette de Saunders (фр.).
Син. (лат.): Hydrocoloeus saundersi (Swinhoe, 1871).
Син. (рус.) : китайская малая чайка.
Статус. Залетный вид.
Общая характеристика и полевые признаки. Мелкая чайка с черной головой, круп-
нее малой чайки. Рисунок первостепенных маховых и окраска крыла в целом при-
115
Семейство Чвйковые
мерно такая же, как у озерной чайки. Отличается от последней, помимо размеров,
коротким высоким, сжатым с боков, черным клювом.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. По рисунку первостепенных маховых близка к видам
группы озерной чайки, особенно к L. bulleri и L. maculipennis, что, по-видимому, ука-
зывает на некоторое филогенетическое родство с этой группой. Однако различия в
строении клюва и лап таковы, что Дж. Дуайт выделил китайскую чайку в отдельный
монотипический подрод Saundersia (Dwight, 1925).
Описание. Размеры (мм) (Dwight, 1925) :
Длина крыла:
<5<5 (и = 9) - 277-293 (среднее 284);
99 (и = 6) — 268—282 (среднее 275).
Длина клюва:
<3<3 (п = 9) — 28—29 (среднее 28,3);
99 (п = 6) — 23-27 (среднее 25,2).
Длина цевки:
с5с5 (п = 9) - 42-44 (среднее 43,2);
99 (п = 6) - 39-41 (среднее 40,5).
Клюв очень короткий, сжатый с боков и высокий, цевка и пальцы тонкие, а плава-
тельные перепонки с более глубокой выемкой, чем у других видов чаек.
Окраска. Самец и самка в брачном наряде (Dwight, 1925). Голова черная, над
и под глазами белые отметины. Шея, нижняя часть тела и рулевые — белые. Спина и
кроющие крыла светлые голубовато-серые. Дистальные первостепенные маховые бе-
лые с широкой черной каймой на внутреннем опахале; по краю наружного опахала II
(первого видимого) первостепенного махового, как правило, проходит узкая черная
полоса; на III—VI пере — предвершинное черное пятно, иногда такое пятно есть и на
II. VI первостепенное обычно серое с узкой темной каймой на внутреннем опахале,
иногда белое; остальные первостепенные маховые серые с беловатыми вершинами.
Второстепенные маховые в значительной части белые. Клюв черный, лапы красные,
радужина темно-коричневая, края век черные. Окончательный зимний наряд отли-
чается от брачного белой головой с серым налетом на затылке; перед глазом и на
кроющих уха черное пятно.
Пуховой наряд не описан. Гнездовой наряд (экземпляр № 157 114 коллекции ЗИН
из Китая, Циндао, 20.VIII 1929 г.). Лоб, подбородок и горло беловатые, у глаза и на
кроющих уха темные пятна. Темя и затылок коричневато-бурые с охристыми пестри-
нами, спинная сторона тела такая же, но несколько светлее. Грудь, брюхо, бока, над-
хвостье, подхвостье и подмышечные - белые. Большие кроющие крыла и нижние
кроющие серые, кроющие первостепенных маховых серые с отдельными буроваты-
ми пятнами, остальные верхние кроющие крыла коричневато-бурые с охристыми каем-
ками. Темно-бурые каймы внутренних опахал II (первого видимого) — IV первосте-
пенных на большом протяжении доходят до стержня, отступая от него лишь в дисталь-
ной части пера; черные каемки имеются и на наружных опахалах, вершины перьев
также черные, хотя на конце может быть светлый кончик. Остальные первостепенные
маховые серые с темной каймой по внутреннему опахалу; вершины V—VIII темно-
бурые или с предвершинным пятном того же цвета. Второстепенные маховые серые
с небольшими темными пятнами на наружных опахалах, третьестепенные - серые с
116
Китайская чайка
бурыми вершинами. Рулевые белые с неширокими бурыми каемками на вершине.
Клюв (Dwight, 1925) черный или бурый, лапы темные, красно-коричневые.
Первый зимний наряд (экземпляр № 55 345 коллекции ЗИН из юго-восточного
Китая, Фу-Чао, 4X1 1883 г.). Голова белая с неясной темной поперечной полосой на
затылке, темными кроющими уха и темным пятном перед глазом. Шея, грудь, брю-
хо, бока, надхвостье и подхвостье белые. Спина серая (нередко с буроватым
налетом - Dwight, 1925), крылья и хвост, как в гнездовом наряде.
Первый летний наряд (экземпляры № 157 115 и
157 116 коллекции ЗИН из Китая, 30 и 25.IV 1931 г).
Как предыдущий, но затылок черно-бурый с белыми
пестринами.
Линька. Линька из гнездового в первый зимний на-
ряд частичная (сменяются перья головы и туловища),
протекает, видимо, уже в начале августа (экземпляр
№ 157 114, 8.VIII 1929 г., коллекция ЗИН). В последую-
щем она бывает дважды в году: полная осенняя и час-
тичная весенняя. Детали линьки не известны. Есть дан-
ные, что в феврале—марте большинство взрослых птиц
уже одевают брачный наряд (Melville, 1977).
Распространение. Гнездовой ареал. До сих
пор остается практически не известным (рис. 31).
Прежние предположения о гнездовании китайской чай-
ки на внутренних водоемах северного Китая (Dwight,
1925; Peters, 1934) пока не подтверждаются (Cheng
Tsohsin, 1976; Melville, 1984). Не гнездится эта чайка
также в Моноголии (Stubbe, Bolod, 1971), Японии
(Hand-list of Japanese Birds, 1958) и на юге о-ва Ко-
рея (Gore, Won Pyong-Oh, 1971; Won Pyong-Oh, 1976).
Причем в последних двух странах отмечается как залет-
ная, а в Монголии не встречена ни разу. Единственное
определенное указание на гнездование имеется для
КНДР, где эта чайка якобы гнездится на морских остро-
вах в северо-восточной и юго-западной частях Восточно-
го
Рис. 31. Область распростра-
нения китайской чайки
1 — предполагаемые места
гнездования, 2 — места зимо-
вок, 3 — залеты на террито-
рию СССР
Корейского залива (Won Hong Коо, 1964).
Зимовки. Юго-восточный Китай от Шанхая до о-ва Хайнань (провинции Чжэцзян,
Фуцзянь, Гуандун), включая о-в Тайвань (Cheng Tsohsin, 1976). Регулярно встреча-
ется в Гонконге с ноября по март, главным образом в феврале—марте. Здесь отме-
чаются в основном взрослые особи, встречи птиц в возрасте меньше года единичны
(Melville, 1977). Крайне редко китайская чайка отмечается зимой в южной части Ко-
рейского полуострова. Отсюда известны только три находки: 23.11 1927 г. в провин-
ции Чолла-Намдо, 18.1 в провинции Кенгидо и 20.1 1967 г. в дельте р. Нактон (Gore,
Won Pyong-Oh, 1971).
На территории СССР достоверно зарегистрирована дважды: 18.IV 1882 г. в Сидеми
близ Владивостока (по-Видимому, годовалый самец) (Taczanowski, 1891-1893)
и на оз. Благодатном в Сихотэ-Алинском заповеднике 16.V 1970 г. (Елсуков,
1974а).
Миграции. Практически не изучены. Пролетные особи отмечались в северо-восточном
и восточном Китае в провинциях Гирин, Ляонин, Хэбэй, Шаньдун (Cheng Tsohsin, 1976)
и по западному побережью п-ова Корея (Won Hong Коо, 1964).
Численность. Неизвестна, однако, судя по редкости встреч, явно невелика.
117
Семейство Чайковые
Местообитания. В сезон гнездования — неизвестно. Во внегнездовое время — морские
побережья и внутренние водоемы.
Размножение. Не изучено.
Суточная активность, поведение. Не изучены.
Питание. Не изучено.
Врага, неблагоприятные факторы. Данных нет.
Хозяйственное значение, охрана. В настоящее время это, по-видимому, наименее
изученный вид чаек мировой фауны. Если предположения о крайней узости его ареала
и невысокой численности подтвердятся, китайская чайка безусловно заслуживает за-
несения в Красную книгу МСОП.
Клуша —
Laras fascus Linnaeus, 1758
Lesser Black-backed Gull (англ.), Heringsmowe (нем.), Goeland brun (фр.).
Статус. Гнездящийся мигрирующий и частично зимующий вид.
Общая характеристика и полевые признаки. Крупная чайка. По окраске оперения
похожа на морскую чайку, но мельче, более легкого телосложения, с относительно
более длинными крыльями и желтыми ногами. Поселяется на поросших травой скалис-
тых побережьях и внутренних озерах, везде предпочитает острова. Голос грубее, чем
у серебристой чайки, несмотря на более мелкие размеры.
Подвидовая систематика. Разные исследователи принимают от 2 до 4 подвидов,
распространение которых показано на рис. 32 (Hartert, 1912—1921; Dwight, 1925;
Vaurie, 1965; Barth, 19756; Степанян, 1975). В СССР обитает один подвид:
Larus fuscus fuscus
Larus fuscus Linnaeus, 1758, Syst. Nat., ed. 10, c. 136, Швеция. Самый крупный и
темный подвид.
Замечания по систематике. Систематика крупных белоголовых чаек комплекса
L. argentatus—L. fuscus разработана недостаточно. Некоторые авторы относят к виду
L. fuscus западно-сибирскую форму antelius и таймырскую taymirensis (Dwight, 1925;
Cramp, Simmons, 1983). Однако Б.К. Штегман относил эти формы к серебристой чайке
(Stegmann, 1934b). Этой же точки зрения придерживаются и авторы настоящего
очерка.
Описание. Строение и размеры (табл. 4).
Окраска. Самец и самка в брачном наряде. Голова, шея, зашеек, весь низ и бока
туловища, а также рулевые и кроющие хвоста белые. Спина, поясница, плечевые, верх-
ние кроющие крыла аспидно- или буровато-черные. Межлопаточная область несколь-
ко светлей остальных частей мантии с более явственным аспидным налетом. Длинные
Таблица 4. Клуша. Размеры (мм)
Подвид Пол Длина крыла
п Lim среднее
L. f. fuscus dd 8 398-438 419
99 6 405-430 412
L. f. graellssii dd 3 408-423 416
99 2 390-420 405
118
Клуша
плечевые с белыми вершинами. Первостепенные маховые черные, с более или менее
заметным буроватым оттенком, слегка и постепенно светлеющие к краям внутренних
опахал. II (первое видимое) маховое обычно с белой каемкой на конце и того же цве-
та предвершинным пятном, иногда затемненным на внутреннем опахале серовато-
аспидным налетом. III маховое с белым кончиком и изредка с зачатком белого пред-
вершинного пятна. Проксимальные первостепенные с аспидным оттенком, белыми
концами и неясно очерченными черноватыми предвершинными пятнами. Второсте-
пенные окрашены как мантия, концы их белые, внутренние опахала у некоторых осо-
бей сероватые при основании. Перья крылышка черновато-аспидные, с белыми осно-
ваниями и наружными опахалами. Ноги и клюв ярко-желтые. Радужина светло-жел-
тая. Края век и пятно на подклювье красные или карминовые.
Взрослые птицы в зимнем наряде. На затылке и зашейнике развиваются мелкие,
но резко очерченные бурые продольные пятна. Остальное оперение окрашено как
в предыдущем наряде. Неоперенные части менее яркие, чем летом.
Пуховой наряд у птиц из западной части ареала (Cramp, Simmons, 1983) очень похож
на соответствующий наряд номинального подвида серебристой чайки, но пестрины на
голове и спине несколько более отчетливые и окрашены в чуть более холодные тона.
На спине они к тому же более редкие.
Гнездовой наряд. В общем, очень похож на соответствующий наряд западно-сибир-
ского подвида серебристой чайки, но основной тон окраски верха обычно более черно-
ватый, светлые охристо-сероватые каемки на перьях развиты сильней, поперечный
рисунок на плечевых и верхних кроющих крыла едва намечен или совсем отсутствует.
Мраморный рисунок в основании центральных рулевых крупней, обычно образует
грубые, более или менее правильные поперечные полосы. От светлых форм серебрис-
той чайки клуша в гнездовом наряде отличается, кроме того, равномерно бурой окрас-
кой IV первостепенного.
Первый зимний наряд (Cramp, Simmons, 1983). Голова и нижняя сторона туловища
часто более светлые, чем в гнездовом пере. Свежие перья мантии и плечевые обычно
более темно-серые или серовато-бурые и с более размытыми предвершинными пятна-
ми, чем у номинального подвида серебристой чайки.
Первый летний наряд. У птиц из западной части ареала он также похож на соответ-
ствующий наряд номинального подвида серебристой чайки (Cramp, Simmons, 1983),
но голова, поясница и нижняя сторона туловища светлее, оперение мантии и межло-
паточной области (включающее гнездовые перья, перья от первого зимнего наряда
и темно-серые перья как у взрослых) — темнее.
Второй зимний наряд. У особей из западной части ареала (Cramp, Simmons, 1983)
темя и шея испещрены больше, чем у взрослых в соответствующем наряде, но осно-
вание шеи более темно-коричневое. Мантия, межлопаточные и спина грифельно-серые,
Длина клюва Длина цевки
п Lim среднее п Lim среднее
8 46,0-56,0 51,0 8 60,0-65,0 62,1
6 45,0-55,0 48,2 6 58,0-63,0 59,8
3 45,0-51,0 48,7 3 58,0-69,0 64,0
2 45,0-57,0 51,0 2 62,0-67,0 64,5
119
Семейство Чайковые
но более тусклые, чем у взрослых, и имеют более или менее развитый коричневый
рисунок. Плечевые также с темным рисунком и обычно более коричневые, со свет-
лыми вершинами. Поясница и верхние кроющие хвоста белые с коричневыми поло-
сами и пятнами. Нижняя часть туловища сильно испещрена, бока и нижние кроющие
хвоста с коричневыми полосами. Первостепенные похожи на гнездовые, но прокси-
мальные перья имеют гораздо более светлые вершины и светло-коричневые полосы.
Кроющие кисти коричневато-черные с более светлыми вершинами. Самые крупные
Рис. 32. Область распространения клуши
а — область гнездования, б — отдельные по-
селения и гнездование узкой полосой вдоль
побережий, в — неуточненная граница области
гнездования, г — места зимовок. Подвиды:
1 — L.f. graellssii, 2 — L.f. intermedins, 3 — L. f.
fuscus
из них светло-коричневые, с более тем-
ным наружными опахалами и светло-ко-
ричневыми каймами. Средние и малые
кроющие кисти имеют более однотонные
темно-коричневые центры, чем в первом
зимнем наряде. Второстепенные — с чер-
но-коричневыми наружными опахалами
и коричневато-белыми вершинами и кай-
мами, самые проксимальные из них
с предвершинной коричневато-белой мра-
морной полосой; иногда некоторые из
них грифельно-серые. Отдельные перья
того же цвета встречаются среди кроющих
крыла. Рулевые очень похожи на соответ-
ствующие перья в гнездовом наряде, но
обычно с более узкой черной предвершин-
ной полосой и более белыми основаниями;
они гораздо темнее, чем в соответствую-
щем наряде у номинального подвида
серебристой чайки.
Второй летний наряд (Cramp, Sim-
mons, 1983). Голова и шея в основном
белые с большим или меньшим числом
светлокоричневых пестрин. Мантия более
однотонная грифельно-серая, чем зимой.
Крестец белый, некоторые верхние крою-
щие хвоста с коричневыми полосами.
Нижняя часть туловища в основном белая
с немногочисленными коричневыми пят-
нами или полосами. Кроющие крыльев
сильно изношены, вершины у них более
светлые, чем зимой.
Третий зимний наряд (Cramp, Simmons, 1983) похож на соответствующий наряд
взрослых, но на задней стороне шеи более развиты коричневые тона. На перьях ман-
тии, плечевых и кроющих крыла, а также на груди и боках остатки более или менее
развитого коричневого рисунка. Хвост иногда белый, иногда в значительной степени
испещрен коричневато-черными пятнами или полосами. II первостепенное часто с бе-
лой предвершинной полосой, III иногда с белым зеркалом. На кроющих кисти в той
или иной степени сохраняются коричневые тона. Второстепенные иногда как у взрос-
лых, но чаще с черными штрихами; иногда они очень похожи на соответствующие
перья второго зимнего наряда, но с более развитыми серыми тонами.
Третий летний наряд (Cramp, Simmons, 1983). Голова, шея и нижняя часть тулови-
ща белые. На верхней стороне тела серые перья имеют менее развитый коричневый
120
Клуша
рисунок, чем в предыдущем наряде. Четвертый зимний и летний наряды (Cramp, Sim-
mons, 1983) в большинстве случаев как у взрослых, но с черными штрихами на крою-
щих первостепенных.
Линька. У птиц с территории СССР изучена слабо. Линька в первый зимний наряд
частичная, охватывает оперение зашейка, начинается уже в конце июля и продолжа-
ется до октября. По С. Крэмпу (Cramp, Simmons, 1983), у номинального подвида
линька протекает, вероятно, позднее, чем у западного.
Последующие линьки неполовозрелых
птиц, по данным тех же авторов, изучены
слабо: у некоторых особей на втором и треть-
ем году жизни в феврале меняются прокси-
мальные первостепенные (X, IX) и в апреле —
VIII—VII первостепенные.
Линька взрослых в зимний наряд полная.
У птиц с Белого моря (Бианки, 1967) или
с Северной Норвегии (Barth, 1975а) смена
первостепенных маховых начинается в июне-
июле, линька мелкого оперения туловища,
по данным В.В. Бианки (1967), - в июне.
Оба исследователя указывают на значитель-
ные индивидуальные колебания сроков этой
линьки. По С. Крэмпу (Cramp, Simmons,
1983), эта линька у номинального подвида
протекает гораздо раньше, чем у западного,
обычно в октябре—апреле. Первостепенные
начинают меняться в октябре—ноябре, т.е.
уже на зимовках; но два внутренних пера
обьино сменяются в районах гнездования.
Голова становится пестрой, как правило, в
октябре. Линька первостепенных. заканчива-
ется иногда в феврале, но чаще — в марте-
апреле, на зимовках или уже на весеннем
пролете (Stresemann, Stresemann, 1966;
Stresemann, 1971, цит. по: Cramp, Simmons,
1983; Bart, 1975a;. экземпляры из европей-
ских музеев).
Линька взрослых в летний наряд (Cramp, Simmons, 1983) частичная, охватывает
голову и шею. Она протекает, вероятно, в феврале—апреле. Иногда некоторые внут-
ренние первостепенные сменяются в январе—феврале одновременно с концом преды-
дущей послегнездовой линьки дистальных первостепенных (Stresemann, Stresemann,
1966).
Распространение. Гнездовой ареал простирается от южных побережий Ис-
ландии через Британские острова и Северную Норвегию до южных берегов Белого
моря (рис. 32, 33). Южная граница размножения проходит по южным берегам Север-
ного и Балтийского морей от северо-западных побережий Франции, через Голландию,
Данию, ГДР и ФРГ до берегов Финского залива и Онежского озера (Gudmundsson,
1955; Бианки, 1967; Hentzel, Fitter, Parslow, 1972; Зимин, Ивантер, 1974; Makatsch,
1974; Monaghan, Coulson, 1977; Коханов, 1981; Хохлова, 1981; Мальчевский, Пукин-
ский, 1983). Единичные случаи гнездования отмечены недавно на северо-западном
побережье Испании, в бухте Виго (Вагсепа, 1977).
121
Семейство Чайковые
Зимовки занимают побережья самой юго-западной части Баренцева моря, а так-
же берега Норвежского, Северного и Балтийского морей. Вдоль атлантического по-
бережья Европы они простираются до Гибралтара и охватывают бассейн Средиземно-
го и Черного морей и берега Аравийского полуострова (Исаков, Воробьев, 1940; Де-
ментьев, 1951; Heinzel, Fitter, Parslow, 1972; Строков, 1974; Gorski, 1981). В Афри-
ке зимует вдоль северных, западных и восточных побережий на юг до Нигерии и Ке-
нии, обычна и на больших озерах в центре континента (Currey-Lindahl, 1981). По круп-
ным рекам и озерам восточного побережья на юг доходит до Ботсваны и Южной Аф-
рики. Известны также зимовки на побережьях Пакистана и Западной Индии (АН,
Ripley, 1969).
Отмечены залеты (в основном во внегнездовое время или неполовозрелых особей)
в Канаду, Панаму и южные штаты США (Buckley, Капе, 1975; Stevenson, 1975; Webb,
Conry, 1978; Powers, 1979; Anderson, 1980; Smith, 1982). В Европе единичных птиц
встречали в Австрии (Biirkli, 1981) и на Шпицбергене (Bruns, 1981).
Миграции. Весной африканские, пакистанские и индийские зимовки последние осо-
би покидают в мае (Meinertzhagen, 1930, 1954; Ali, Ripley, 1969). На территории СССР
на северном побережье Черного моря в районе Тилигульского лимана небольшое число
пролетных особей встречается в апреле-мае (Черничко, устное сообщ.). На Днепре
у Херсона и Голой Пристани первые птицы появляются обычно с 1 по 7 марта, пролет
'длится здесь почти до конца мая (Клименко, 19506). В районе Харькова мигрантов
отмечали во второй половине марта (Сомов, 1897), а в окрестностях Киева (Шар-
леман, 1913) — в апреле и второй половине мая. Единичные особи зарегистрированы
весной в устье реки Белой (Теплов и др., 1959). В район Волжско-Камского водо-
хранилища первые клуши прилетают в разные годы в период с 8 по 30 апреля, в сред-
нем 15 апреля (Водолажская, Залетаев, 1977). В Белоруссии и Литве пролет наблю-
дается обычно во второй половине этого месяца (Уилсон, 1977).
Осенний пролет на Белом море начинается уже в начале сентября (Бианки, 1960),
в середине следующего месяца отмечаются лишь отдельные стаи (Скокова, 1960).
В Карелии, по данным Т.Ю. Хохловой (1977), последняя взрослая особь встречена
в 1974 г. 11.IX. В Ленинградской обл. осенние миграции начинаются в первой декаде
августа, пик численности мигрантов приходится на конец сентября, многие остаются
на весь октябрь, а отдельные особи - до начала ноября (Мальчевский, Пукинский,
1983). В Эстонии интенсивные миграции в районе Пухту (Йыги и др., 1961) и на се-
вере республики (Манк, 1959; Renno, 1978) протекают с середины сентября до сере-
дины октября, отдельные особи наблюдаются в это же время и на северо-западном по-
бережье Чудского озера (Роотсмяе, 1959). В те же сроки пролет регистрировался и
на Куршской косе (Вайткявичюс, 1961). В Горьковской обл. мигрантов встречали
в разные годы с 11 по 26 сентября (Воронцов, 1967). В районах Киева и Харькова
первые пролетные клуши отмечались с 9 по 20 августа, в середине следующего ме-
сяца миграции становятся здесь хорошо заметными (Сомов, 1897; Шарлеман, 1913);
иногда массовый пролет в этих районах регистрируется в октябре, последние
особи встречаются здесь до начала ноября. В низовьях Днепра первые птицы на-
блюдаются обычно 17-20.IX и держатся здесь до замерзания Днепра (Клименко,
19506).
Основные направления осенних миграций у птиц с Белого моря юго-западное и
юго-восточное (Бианки, 1967). Большинство сеголетков летит при этом к бассей-
нам Западной Двины, Днепра и Волги. В этом же направлении перемещаются сканди-
навские клуши (Valicongas, Nordstrom, 1957; Nordstrom, 1957—1963, цит. по.: Биан-
ки, 1967). На востоке основные миграционные пути ограничиваются бассейнами рек
Северной Двины и Волги. Неполовозрелые особи в возрасте 1—3 лет и сеголетки ветре-
122
Клуша
чаются преимущественно по Восточно-Европейской равнине и к юго-востоку от нее.
На юг залетают до Кустаная и Омска (Долгушин, 1962), известен даже один возврат
из Бирмы (Бианки, 1967). На восток птицы проникают до бассейна Печоры (Тепло-
ва, 1957). Гораздо меньше возвратов дает, как указывает В.В. Бианки, (1967), юго-
западное направление (Польша, Дания, Франция, Италия). В Ленинградской области
основной поток мигрантов направляется вдоль побережий Финского залива. Как по-
казывают данные кольцевания (Мальчевский, Пукйнский, 1983), здесь пролетают
в основном птицы из Финляндии (19 возвратов) и Швеции (2 возврата).
На пролете птицы держатся обычно небольшими стаями, часто вместе с другими
крупными чайками. Охотно посещают в зто время места лова рыбы в море, рыбо-
разводные пруды, городские свалки.
Численность. На гнездовьях крупных скоплений не образует, селится отдельными
парами или небольшими (до нескольких десятков пар) колониями. Вероятно, самая
крупная в СССР колония на о-ве Малом в Финском заливе насчитывает (Renno, 1978)
более 700 пар. В Карелии (Хохлова, 1977) самая крупная колония в 1976 г. состоя-
ла из 73 гнезд. На о-ве Мэй у восточных берегов Шотландии известна колония чис-
ленностью 2500 пар (Duncan, 1981а). В южной части карельского берега Белого моря
обитает всего 50 пар, в Онежском заливе — около 300 (Бианки, 1967). На о-вах Фин-
ского залива в 1971 г. в пределах Эстонии было учтено всего 295 пар (Ренно, 1972),
у западных берегов этой республики в период с 1959 по 1971 г. насчитывалось от 32
до 167 пар (Аумезс, 1967, 1972). По последним данным О. Ренно (Renno, 1978), на
островах Ухтью у северо-восточных берегов Эстонии гнездится до 390 пар, у полу-
острова Кугальский уже в Ленинградской обл. — около 150 пар.
Общая тенденция изменения численности почти повсеместно одинакова. В запад-
ной части ареала на Британских островах и островах в южной части Балтийского моря
численность неуклонно увеличивается и в некоторых районах ежегодный прирост
ее достигает 20—29% (Paludan, 1951; Brown, 19676; Parslow, 1967; Harris, 1970b;
MacRoberts M., MacRoberts B., 1972; Greenhalgh, 1974; Davis, Dunn, 1976; Monaghan,
Coulson, 1977; Sandeman, 1982). Здесь предпринимаются специальные меры, ограни-
чивающие дальнейший рост популяции (Duncan, 1981а, б; Jones, White, 1982). В от-
дельных районах на Британских о-вах численность в некоторые годы локально умень-
шается из-за эпизоотий (Mudge, Frust, 1981; Fems, 1982). Несколько иная картина
динамики численности вырисовывается в последнее время в Ленинградской обл. и
Финляндии. До начала 60-х годов в Финляндии численность птиц увеличивалась или
оставалась постоянной (Lemmetyinen, 1980), позднее она стала уменьшаться из-за
конкуренции с серебристой чайкой, численность которой к этому моменту резко воз-
росла (Kilpi et al., 1980; Bergman, 1982; Kilpi, 1983). В Ленинградской обл. резкое
падение численности отмечено с середины 50-х годов (Мальчевский, Пукинский, 1983).
Причиной его, кроме усилившейся конкуренции с серебристой чайкой на местах гнездо-
вания, эти авторы считают возросшее беспокойство птиц людьми во время размно-
жения.
Местообитания. Летом — скалистые побережья, прибрежные острова и острова на
крупных внутренних озерах. В бассейне Белого и Балтийского морей, а также на круп-
ных озерах предпочитает мелкие островки, расположенные по возможности далеко
от берега, не избегает при этом (в отличие от серебристой чайки) понижений рельефа
и сплошного ковра растительности (Аумезс, 1967; Бианки, 1967; Пеэдосаар, Онно,
1970; Хохлова, 1977; Renno, 1978; Мальчевский, Пукинский, 1983). В Англии в по-
следние десятилетия успешно осваивает антропогенные биотопы (Salmon, 1958; Owen,
1975; Monaghan, Coulson, 1977). В пределах Ленинграда несколько лет существовала
небольшая колония на намывном участке берега в районе Угольной гавани (Пантелеев,
123
Семейство Чайковые
1984). Зимой птицы держатся по побережьям и в прибрежных участках моря, сопро-
вождая суда, посещая рыбообрабатывающие предприятия и свалки.
Размножение. Половая зрелость у большинства особей, судя по нарядам, наступает
в возрасте 4—5 лет. На местах гнездования в Эстонии появляются, по многолетним
данным, 6 апреля, самая ранняя встреча известна 29 марта (Rootsmae, Rootsmae, 1976).
По данным других авторов (Renno, 1978), на западном побережье республики в ран-
ние весны они появляются в начале марта, а в поздние — на месяц позднее. В Ленингра-
де первых клуш отмечают в середине или в 20-х числах апреля, у Выборга в разные
годы с 27.III по 6.V, в среднем 19.IV (Бианки, 1967; Мальчевский, Пукинский, 1983).
В Карелию на крупные озера прибывают в 20-х числах мая (Владимирская, 1948);
по более поздним данным Т.Ю. Хохловой (1981), на западном побережье Онежского
озера первые клуши в 1976 г. наблюдались 10—12.V. На Белое море птицы прилетают
в середине мая (Бианки, 1967); на Западном Мурмане, по старым сведениям (Goebel,
1901, цит. по Pleske, 1928), появляются в разные годы от 30.Ш до l.V. В.Н. Коханов
и Н.Н. Скокова отметили одиночную особь на Айновых островах 4.V 1959 г.
На места гнездования прибывают парами или образуют их в клубах поблизости
от колоний. В это время наиболее отчетливо выражено демонстративное поведение,
которое подробно изучено на Британских островах. Сравнительные исследования
в смешанных колониях клуш и серебристых чаек показали, что, в общем, демонстра-
тивное поведение у этих видов сходно, но смешанные пары редки (Brown, 1967а, б;
Harris, 1969, 1970а). Число их резко увеличилось после эксперимента по взаимной
замене кладок и воспитанию чайками птенцов другого вида. Оказалось, что в смешан-
ных парах большинство птиц было выкормлено воспитателями другого вида. При
этом самцы образовывали пары как с партнерами своего, так и чуждого вида,
самки же выбирали в основном партнеров, сходных по окраске с птицами-воспи-
тателями.
На Белом море клуша в отличие от серебристой чайки гнезда устраивает обычно
на вороничнике, среди приморско-луговой растительности (Бианки, 1967). Сходные
данные получены в смешанных колониях этих двух видов в Англии (Hosey, Goodrige,
1980), хотя здесь отдельные пары клуш могут помещать их и на утесах среди гнезд
серебристых чаек. На западном побережье Ладожского озера (Мальчевский, Пукин-
ский, 1983) гнезда строятся преимущественно на островах, лишенных растительности,
хотя отдельные пары могут устраивать их на скалах в шхерах или на островках, по-
росших ивовыми кустами и соснами. В Англии в условиях растущей численности и
недостатка привычных для размножения мест в густонаселенных районах клуши мо-
гут строить гнезда на крышах строений. В последние десятилетия тенденция эта зна-
чительно усиливается (Cramp, 1971; Owen, 1975; Monaghan, Coulson, 1977), в 1981 г.
первый случай такого рода отмечен в нашей стране, в г. Таллине (Рандла, 1985).
Гнезда клуш менее массивные, чем у серебристых чаек, и напоминают, скорее, гнез-
да сизых чаек (Bergman, 1951; Paludan, 1951; Бианки, 1967). На Белом море и в Ле-
нинградской обл. строительным материалом служат мох, сухая прошлогодняя трава,
кусочки тростника, мелкие щепочки (Бианки, 1967; Пантелеев, Маклашова, устное
сообщ.). Размеры (мм) 12 гнезд, промеренных В.В. Бианки были: диаметр лотка
200-260, в среднем 230, глубина лотка 45-60, в среднем 50; 11 гнезд из Ленингра-
да, промеренных А.В. Пантелеевым, имели размеры (мм): диаметр гнезда 200-800.
в среднем 416, диаметр лотка 170—250, в среднем 215, высота стенок гнезда от зем-
ли 15—120, в среднем 41, глубина лотка 40—75, в среднем 51. Расстояние между гнез-
дами в одной карельской колонии колебалось от 0,5 до 5 м (Хохлова, 1977).
Откладка яиц в Ленинградской обл. начинается в конце мая - начале июня (Маль-
чевский, Пукинский, 1983). На Белом море массовая откладка яиц отмечалась с 20-х
124
Клуша
чисел мая до начала июня (Бианки, 1967), на побережье южной Финляндии — в сере-
дине мая (Bergman, 1951).
17 кладок (27,8%) из 61, находившихся под наблюдением в Ленинградской обл.
(Мальчевский, Пукинский, 1983; Пантелеев, Маклашова, устное сообщ.), содержали
по три яйца, 28 (45,9%) - по два и 16 (26,2%) - по одному яйцу; средний размер
кладки составил здесь, таким образом, 2,01 яйца на грездо. В Карелии этот показа-
тель был равен 1,88 в колониальных поселениях и 2,33 в одиночных гнездах (Хох-
лова, 1981). В Онежском заливе Белого моря в 19 (90%) осмотренных кладках было
по три яйца и в 2 (10%) — по два; средний размер кладки составил здесь 2,90 яйца
на пару (Бианки, 1967). В Эстонии обнаружены повторные кладки, содержавшие,
как правило, лишь одно или два яйца (Renno, 1978). Насиживают кладку оба партне-
ра в течение 22—28 дней (Paludan, 1951), первое яйцо в среднем (и = 65) 24,3 дня,
второе — 24,7 и третье (и = 55) - 26,1 дня.
Яйца имеют овальную или короткоовальную форму и крупнозернистую матовую
скорлупу (Makatsch, 1974). Окраска основного фона яиц варьирует от темно-корич-
невой до зеленовато-серой или оливковой и от песчаной до блеклой зеленовато-серой.
Темно- или светло-коричневые неравномерно расположенные поверхностные пятна
или разводы сконцентрированы обычно на тупом конце яйца. Во внутренних слоях
скорлупы более или менее развиты светло- или темно-серые, а также коричневато-
фиолетовые пятна. Средние размеры яиц из Скандинавии 66,77 X46,21 (и = 22); из
Финляндии (и = 12) 66,63X46,63; из Швеции (и =96) 66,58X46,58 (Makatsch, 1974).
Размеры яиц (мм) эстонской клуши 61,0—77,0X46,0—55,0 (Renno, 1978); из Ле-
нинградской обл. (Пантелеев, Маклашова, устное сообщ.) 54,0—70,0X41,3—46,1,
в среднем (п = 17) 63,6 X 44,6; 140 яиц из Карелии имели размеры 59,0—73,9 X 42,2—
51,2, в среднем 66,1 Х46,7 (Хохлова, 1981).
Вылупление птенцов в Ленинградской обл. протекает во второй половине июня —
начале июля (Мальчевский, Пукинский, 1983). В Карелии в 1976 г. первые птенцы
в колонии появились 12.VI (Хохлова, 1977). В Онежском заливе Белого моря в 1960
и 1961 гг. вылупление птенцов происходило с 10.VI по 20.VII (Бианки, 1967).
В первые сутки после вылупления чайчата сидят в гнезде или держатся в радиусе
1 м от него. В возрасте 4—5 дней и более подвижность их резко возрастает; в возрас-
те 10 дней они могут отходить от гнезда на 30 м, в возрасте 15 дней - на 50 м (Paludan,
1951).
Первые лётные молодые в Ленинградской обл. (Мальчевский, Пукинский, 1983)
встречаются с конца июля, к 1 .VIII большинство их уже хорошо летает. На Белом море
в районе Кемьских шхер первых летных молодых наблюдали 19.VII 1960 г. (Бианки,
1967), но большинство молодых начинает летать на Белом море, по данным того же
автора, в 20-х числах июля — первой половине августа.
Питание. У птиц с территории СССР изучено слабо. В погадках, собранных в июле
на Белом море (Бианки, 1967) 83% (по встречаемости) составили семена вороники,
насекомые — 43%, рыба — 35%, моллюски (в основном мидии) - 23%, полихеты (Ne-
reis virens) — 5%, ракообразные (Hyas araneus, Crangon crangon) — 3,3%. В желудках
добытых взрослых птиц обнаружены остатки мелкой рыбы и насекомых. Пища птен-
цов (материал из 4 отрыжек) состоит из мелкой рыбы (песчаники Ammodytes hexap-
terus marinus, мойвы Mallotus villosus). Кроме рыбы в отрыжках встречаются остат-
ки миног Lampetra japonica, полихеты Nereis pelagica, насекомые: усачи, щелкуны,
жужелицы, летающие формы муравьев Formica, мухи журчалки и оводы.
В окрестностях Киева в желудках, добытых на осеннем пролете клуш, находили
уклеек, пескарей; однажды была обнаружена крупная полевка Microtus arvalis (Шар-
леман, 1913). Этот же автор наблюдал, как клуши поедали подранков речной чайки
125
Семейство Чайковые
и чибиса. В Швеции они в больших количествах поедают миног, когда те заходят на
нерест в реки (Sjbberg, 1973). В Англии отмечен случай поедания гусениц зимней
пяденицы Operohptera brumata на вересковых пустошах во время массового размно-
жения этих насекомых (Picozzi, 1981). В Рижском заливе во время весеннего проле-
та клуши вместе с другими крупными чайками кормятся в море у рыболовных се-
тей, а на берегу - у рыбозаводов (Леинь, Каспарсон, 1961).
Как и другие чайки, клуши в течение всего года охотно кормятся на свалках в круп-
Hbix городах и портах, но в отличие от серебристых чаек они все же меньше тяготе-
ют к питанию отходами (Brown, 1967в; Harris, 1969). На свалках, как показали спе-
циальные исследования (Verbeek, 1977), большинство клуш специализируется на кра-
же уже найденного серебристыми чайками корма.
Врага, неблагоприятные факторы. Как и у других крупных чаек, отрицательное
влияние на успех размножения оказывают наземные и воздушные хищники, беспо-
койство людьми во время размножения, а также неблагоприятные кормовые или
погодные условия. В местах больших скоплений на кормежках и местах отдыха стра-
дают от эпизоотий (Mudge, Frust, 1981; Ferns, 1982).
Хозяйственное значение, охрана. Хозяйственного значения вид не имеет, охрана
не предусмотрена. В некоторых регионах, где крупные скопления птиц становятся
эпидемиологически опасными для людей, с ними ведется борьба (Jones, White, 1982).
Серебристая чайка —
Larus argentatus Pontoppidan, 1763
Herring Gull (англ.), Silbermowe (нем.), Goeland argente (фр.).
Син. (лат.) : Larus cachinnans Pallas, 1811, Larus vegae Palmen, 1887 , Larus taimyrensis Buturlin, 1911
Син. (рус): хохотунья, сибирская клуша.
Статус. Гнездящийся мигрирующий, частично зимующий вид.
Общая характеристика и полевые признаки. Крупная белоголовая чайка с типичными
признаками рода Larus. Мантия от светлосерого до аспидно-сизого цвета, на маховых хо-
рошо заметен черный рисунок. Клюв желтый с красным пятном на изгибе подклювья.
От бургомистра и серокрылой чайки отличается наличием черного цвета на первостепен-
ных маховых, от клуши и морской чайки — более светлой окраской мантии. Набор зву-
ков и демонстраций, как у других крупных чаек, но голос выше, чем у клуши, и ниже,
чем у бургомистра.
Подвидовая систематика. Разработана не достаточно. В мировой фауне различные ис-
следователи признают от 4 до 18 подвидов* 1 (Hartert, 1912—1921; Dwight, 1925; Peters,
1934; Stegmann, 1934; Vaurie, 1965; Степанян, 1975; Cramp, Simmons, 1983), различаю-
щихся в основном по окраске верхней стороны тела и ног. В СССР обитает, по представ-
лениям разных авторов, от 6 до 11 подвидов (Тимофеев-Ресовский, Штреземан, 1959;
Долгушин, 1962; Vaurie, 1965; Степанян, 1975). Относительно хорошо дифференциро-
ванными из них можно считать лишь 6 (первоописания и диагнозы большинства подви-
дов приведены по: Степанян, 1975):
1. Larus argentatus argentatus
Larus argentatus Pontoppidan, 1763, Danske Atlas, 1, c. 622, Дания.
Спина бледная, серовато-сизая, светлее, чем у других рас, и сходная с таковой у
cachinnans. Ноги красновато-розовые.
*В последние годы накапливается все больше данных, свидетельствующих, что комплекс подвидов
L. argentatus на самом деле может быть комплексом близких видов. Подробнее об этом смотри
в характеристике семейства Чайковых.
126
Серебристая чайка
2. Larus argentatus antelius2
Larus fuscus antelius Iredale, 1913, B.B.O.C., 31, c. 69, низовья Оби.
Спина темная, шиферно-серая. Ноги желтые.
3. Larus argentatus taimyrensis
Larus affinis taimyrensis Buturlin, 1911, Орнитол. вести., 2, с. 149; p. Глубокая, восточ-
ное побережье Енисейского залива.
Спина темно-серая, светлее чем у antelius и темнее, чем у vegae. Окраска ног варьиру-
ет от желтой до светло-розовой.
4. Latus argentatus vegae
Larus argentatus Brunn, var. vegae Palmen, 1887, Vega-Exped. Vetensk. lakttag, 5, c. 370.
Питлекай, Чукотский полуостров.
Спина серовато-сизая, светлее, чем у предыдущего подвида, но темнее, чем у номи-
нального. Окраска ног варьирует от серовато-розовой до серовато-желтой.
5. Larus argentatus cachinnans
Larus cachinnans Pallas, 1811, Zoographia Rosso-Asiat., 2, c. 318, Каспийское море.
Спина бледная, как у номинального подвида, но менее серая. Ноги желтые.
6. Larus argentatus mongolicus
Larus argentatus mongloicus Suschkin, 1925. Список и распределение птиц Русского
Алтая, с. 63, оз. Урюгнор, северо-западная Монголия.
Спина серовато-сизая, как у vegae, темнее, чем у cachinnans. В зимнем наряде отлича-
ется от vegae менее развитыми темными пестринами на голове. Окраска ног варьирует
от серовато-розовой до желтой.
Пять других ранее описанных подвидов (L. a. omissus, L.a. birulae, L.a. ponticus, L.a.
armenicus, L.a. barabiensis) либо уже сведены в синонимы, либо настолько слабо диффе-
ренцированы или описаны на столь малом материале, что реальность их вызывает сом-
нение3 .
Замечания по систематике. В настоящее время не вполне установился не только объ-
ем группы серебристых чаек, но и взгляды на историю возникновения и родственные от-
ношения внутри ее. В литературе эта группа неоднократно приводилась как пример
кольцевого ареала, иллюстрирующего географическое видообразование. В одной из по-
следних работ Майр (1968), анализируя и обобщая исследования последних десятиле-
тий, касающиеся этой группы птиц (Voous, 1960; Тимофеев-Ресовский, Штреземан,
1959; Goethe, 1960; Smith, 1960; Macpherson, 1961), вынужден признать, что реальная
ситуация в этом кольце оказалась сложнее, чем это представлялось раньше. Рассматри-
вая историю возникновения группы, он вслед за некоторыми из указанных авторов
предполагает, что в плейстоцене ареал серебристой чайки был разбит на несколько ре-
фугиумов, существовавших как в Палеарктике, так и в Неарктике. Группа cachinnans,
характеризующаяся желтыми ногами, эволюционировала в Арало-Каспийской области
и позднее дала начало атлантической группе fuscus. Группа vegae и родственные ей фор-
мы с розовыми ногами развивались на тихоокеанском побережье Азии и дали начало
близкородственной форме smithsonianus в Северной Америке, которая относительно
недавно проникла в Западную Европу, где образовала номинальную форму argentatus.
Там, где argentatus или vegae встречаются с желтоногими формами, между ними в ряде
случаев происходит обмен генами. С другой стороны, на побережье Европы, где обитают
вместе argentatus и fuscus, они ведут себя как хорошие виды, практически не образую-
щие подвидов. Изоляты в Северной Америке дали формы thayeri и glaucoides.
’Применяющееся в последних сводках (Дементьев, 1951; Vaurie, 1965; Степанян, 1975) название
для этой формы ’’heuglini” не валидно (Goethe, 1973), так как типовой экземпляр, как выясни-
лось после повторного исследования, принадлежит форме taimyrensis и описан из района зимовок.
’Реальность подвида L.a. armenicus у большинства систематиков сомнений не вызывает. (Прим, ред.)
127
Семейство Чайковые
Таблица 5. Серебристая чайка. Размеры (мм)
Подвид Пол Длина крыла
n lim среднее
L.a. argentatus dd 26 430-472 451
99 24 395-440 420
L.a. antelius dd 23 420-466 440
99 15 406-442 420
L.a. taimyrensis dd 12 435-467 454
99 12 405-433 425
L.a. birulae dd 27 433-466 449
99 12 414-436 425
L.a. vegae dd 17 441-466 449
99 23 402-443 422
L.a. cachinnans dd 18 445-462 454
99 14 395-445 424
L.a. mongolicus dd 16 430-476 451
99 6 419-448 434
Описание. Строения и размеры (табл. 5).
Окраска. Самец и самка в брачном наряде. Голова, шея, зашеек, бока, нижняя
сторона тела и хвост белые. Спина, плечевые, верхние кроющие крыла серые различных
оттенков у разных форм (от очень светлого голубовато-серого до темного аспидно-
сизого или стального). Вершины длинных плечевых и кончики всех маховых белые.
Первостепенные маховые до VI—IX с черным рисунком, по крайней мере в вершинных
частях. В предвершинной части II (первого видимого) обычно развито белое пятно, от-
деленное от того же цвета вершины черной поперечной полосой. На III первостепенном
белое предвершинное пятно развито значительно слабей, иногда его нет совсем. На IV
и последующих первостепенных серый цвет все больше оттесняет черный к вершине
пера. Окраска ног варьирует от серовато-мясного цвета до розоватого или ярко-желтого
с оранжевым оттенком. Клюв зеленовато- или лимонно-желтый с красным пятном на
изгибе подклювья. Радужина песочная, серебристо-серая или светло-желтая. Края век
киноварно- или оранжево-красные, у некоторых форм желтые. Углы рта оранжевые или
к расно-о ранжев ые.
Самец и самка в зимнем наряде. Отличаются от птиц в летнем наряде более или менее
густыми темными пестринами на голове, а иногда и шее. Исключение составляет L.a.
cachinnans, многие особи которой зимой имеют чисто-белую голову и шею.
Пуховой наряд (L.a. argenteus по: Cramp, Simmons, 1983). Голова и верхняя сторона
туловища буровато-серые с крупными черновато-коричневыми пятнами. На темени они
более округлые и резко очерченные, на спине более продолговатые и размытые. На го-
лове и шее пятна располагаются обычно в виде более или менее определенного рисун-
ка, на спине они беспорядочны. Нижняя сторона тела буровато-белая. Клюв черный с ро-
зоватым кончиком, ноги телесно-розовые, радужина черно-коричневая.
Гнездовой наряд. Лоб, бока головы, подбородок и горло беловатые с продольным
рисунком в виде наствольных полос или пятен, иногда равномерно бурые. Перья теме-
ни, затылка и боков шеи с белыми основаниями, бурыми предвершинными пятнами и
беловатыми или охристыми каемками на конце. Спина, плечевые и верхние кроющие
крыла бурые со светлым рисунком в виде грязно-охристых или беловатых каемок.
128
Серебристая чайка
Длина клюва Длина цевки
п lim среднее п lim среднее
26 53,0-61,0 56,0 26 65,0-73,0 69,7
24 48,0-55,0 50,9 24 61,0-67,0 63,8
23 50,0-63,0 57,3 23 67,0-75,0 70,0
15 49,0-61,0 52,6 15 62,0-73,0 66,2
12 54,0-58,0 56,1 12 67,0-75,0 71,5
12 51,0-57,0 53,2 12 64,0-72,0 67,0
27 52,0-62,0 56,6 27 64,0-76,0 70,0
12 50,0-58,0 52,8 12 62,0-68,0 65,0
17 54,0-61,0 57,9 17 66,0-76,0 70,7
23 50,0-58,0 52,9 23 63,0-72,0 66,2
18 55,0-66,0 60,8 18 67,0-76,0 72,9
14 50,0-61,0 55,9 14 62,0-73,0 66,2
16 50,0-59,0 55,7 16 62,0-74,0 68,4
6 50,0-55,0 53,0 6 64,0-70,0 66,8
Крупные плечевые и верхние кроющие крыла более или менее поперечно-пятнистые.
Основной фон поясницы обычно беловатый, рисунок образован стреловидными буро-
ватыми предвершинными пятнами и поперечными полосами. Верхние кроющие хвоста
окрашены сходно с поясницей, но белый цвет здесь чище, а бурый поперечный рисунок
резче и правильней. Нижняя сторона тела от белой с немногочисленными размытыми
бурыми пятнами до почти равномерно светло-бурой; длинные перья на боках туловища
с темным поперечным рисунком. На зобе у светлоокрашенных форм развита попереч-
ная бурая перевязь. Первостепенные маховые черновато-бурые, второстепенные — бу-
рые, вершины их окаймлены беловатой или грязно-охристой полосой. Рулевые белые
при основании, буровато-черные в вершинной половине с беловатой каемкой на конце.
Между белым основанием и темной вершиной хвоста — переходная зона с мраморным
рисунком. Ширина темной предвершинной полосы резко сокращается от центральной к
крайним парам рулевых. Клюв черно-коричневый, у основания подклювья обычно те-
лесно-розовый; ноги коричневато-розовые. Радужина черно-коричневая.
Первый зимний наряд. Отличается от предыдущего значительным сокращением раз-
меров темных пестрин на голове и по всему низу туловища, а также большим развитием
светлых поперечных пятен и полос на спине, плечевых и верхних кроющих крыла.
Первый летний наряд. Голова, шея и низ туловища еще белее, чем в предыдущем на-
ряде. Мантия выглядит значительно менее пестрой, благодаря тому что границы темного
рисунка нерезки, а сам рисунок значительно сокращен. Межлопаточные равномерно бу-
ровато-серые или сизые с большей или меньшей примесью буроватого оттенка. Сизые
тона здесь достигают максимального развития, с одной стороны, у L.a. vegae и L.a. biru-
lae, с другой — у L.a. cachinnans. Поясница остается пестрой. Рулевые, маховые и часть
верхних кроющих крыла сохраняются от гнездового наряда.
Второй зимний наряд. Спина от межлопаточной области до поясницы включительно
более или менее равномерно сизая разных оттенков, обычно с буроватым налетом.
Мелкие верхние кроющие хвоста хотя бы частично белые. На II первостепенном иногда
уже развито белое предвершинное пятно или его зачатки. Рисунок на проксимальных
первостепенных, начиная с V—VI, в общем похож на таковой у взрослых птиц, но свет-
s. Зак. 826 129
Семейство Чайковые
лые поля на рх внутренних опахалах обычно буровато-серые или охристо-беловатые и
более или менее испещрены мелким бурым крапом. Второстепенные маховые светлые
серовато- или палево-бурые с широкими охристыми или беловатыми каймами на кон-
цах и по краям наружных опахал, темными предвершинными пятнами и иногда мелким
бурым крапом на внутренних опахалах. Предвершинная белая перевязь на хвосте уже,
а мраморный рисунок в основании центральных рулевых мельче и гуще, чем в преды-
дущих нарядах.
Второй летний наряд. Маховые, большая часть верхних кроющих крыла и рулевые от
второго зимнего наряда. Мантия более или менее равномерно сизая. Верхние кроющие
хвоста обычно белые. Голова, шея, весь низ тела, а также концы крупных плечевых —
белые. На темени, шее, верхней части груди и брюшка иногда сохраняются светло-бурые
пестрины. Среди плечевых и верхних кроющих крыла остается значительное количество
перьев второго зимнего наряда.
Третий зимний наряд. Спина, плечевые, надхвостье, а иногда и верхние кроющие
крыла сизые. Крупные плечевые с белыми каймами и остатками буровато-черного ри-
сунка в виде продольных пятен. Буровато-черная предвершинная полоса на хвосте зна-
чительно уже, чем в предыдущих нарядах; нередко она разбивается на отдельные пятна
или становится сплошь мраморной, иногда рулевые могут быть чисто-белыми. Рисунок
первостепенных маховых, как у взрослых, но белое предвершинное пятно на II махо-
вом значительно меньше и не столь резко очерчено. Второстепенные маховые с белы-
ми концами и с более или менее развитым буровато-черным пятном.
Третий летний наряд. Отличается от всех предыдущих нарядов и окончательного
летнего тем, что окраска маховых, рулевых и больших наружных кроющих кисти, как
в предыдущем наряде, а голова, шея, весь низ и бока туловища, а также кроющие
хвоста белые. Серый цвет на маховых благодаря выгоранию становится более бледным.
Четвертый зимний и летний наряды. Отличаются от соответствующих нарядов взрос-
лых птиц несколько большим развитием черного рисунка на первостепенных и нали-
чием черного продольного рисунка на больших верхних кроющих кисти и I (редуциро-
ванном) первостепенном.
Линька. Линька в первый зимний наряд частичная, у большинства экземпляров
охватывает незначительное число перьев в межлопаточной области, среди мелких плече-
вых и зашейка. У некоторых особей, кроме того, линяет часть оперения по всей спине
и нижней стороне туловища. Начинается эта линька в конце июля в межлопаточной
области, затем распространяется на зашеек, плечевые и спину и заканчивается в ок-
тябре. Линька в первый летний наряд также частичная, она затрагивает оперение меж-
лопаточной области, груди и мелкие плечевые перья. При этом у некоторых особей
сменяются лишь единичные перья, у других — свежие перья на участках, затронутых
линькой, преобладают. Протекает эта линька в апреле—мае. Линька во второй зимний
наряд полная, начинается во второй половине июня обычно со смены проксимальных
первостепенных. Второстепенные маховые начинают линять с дистального пера в момент
смены VII—VI первостепенных. К моменту смены IV—III первостепенных второстепен-
ные, вероятно, полностью обновляются. Рулевые начинают линьку с центральной пары
в момент смены VIII—VI первостепенных и заканчивают ее заменой крайней пары
перьев во время линьки VII—VI первостепенных. Линька контурного оперения тулови-
ща у большинства особей начинается и заканчивается во время смены первостепенных.
Но у некоторых птиц начало ее опережает или запаздывает по отношению к началу сме-
ны первостепенных маховых. Заканчивается эта линька в августе—начале сентября.
Линька во второй летний наряд частичная, охватывает небольшое число перьев на спине
и среди мелких плечевых. Протекает она в феврале—мае.
Линька в третий зимний наряд полная. Порядок смены оперения в общих чертах тот
130
Серебристая чайка
же, что и в ходе линьки во второй зимний наряд. Дистальное второстепенное растет во
время смены VI- V первостепенных, а иногда во время роста VII первостепенного все
второстепенные уже свежие. Рулевые также начинают меняться с центральной пары,
у одних особей во время линьки VII—V первостепенных, у других - позднее, во время
роста IV—III первостепенных. Начало линьки кроющего оперения туловища у разных
особей также может совпадать с началом смены первостепенных, опережать ее или силь-
но запаздывать. Начинается эта линька в июне и заканчивается во второй половине ав-
густа. Линька в третий летний наряд частичная, ход и сроки ее не ясны из-за отсутствия
материала. Линька в четвертый зимний наряд полная, начинается в июле (меняются XI
и X первостепенные; у некоторых особей они в это время уже свежие). Заканчивается
эта линька в октябре (дорастает II первостепенное).
Линька в четвертый летний наряд частичная, сроки не ясны из-за отсутствия материа-
ла. Линька из четвертого летнего в пятый (окончательный) зимний наряд полная, начи-
нается в июле—августе (растут XI и IX первостепенные). Сроки окончания этой линьки
на нашем материале не улавливаются, ясно лишь, что они также сильно варьируют инди-
видуально. Об этом можно судить по состоянию линьки у июльских и августовских осо-
бей. Линька из пятого (окончательного) зимнего в пятый (окончательный) летний на-
ряд частичная, протекает в марте—апреле. Линька из пятого (окончательного) летнего
в окончательный зимний наряд полная, представлена серией из 136 экз. Начальные ее
этапы (смена X—XI) на севере ареала от Мурманского побережья до Анадырского края
протекают с 18.VI по 31.VII. На юге ареала это происходит с 1.VI по 27.VII. Окончание
этой линьки протекает с 1.XI до 13.XII. Таким образом, осенняя линька у взрослых
особей длится 6 месяцев с июня по декабрь.
Распространение. Гнездовой ареал. Охватывает Голарктику циркумполярно
и располагается во всех растительно-климатических зонах от тундровой до средиземно-
морской включительно (рис. 34); северная граница его лежит между 70° и 80° с.ш.,
южная — между 30° и 40° с.ш. (Voous, 1960). В пределах СССР, как впрочем и в За-
падной Европе, гнездовой ареал довольно четко распадается на северную и южную зоны
(рис. 35). Северная зона у нас в стране располагается между 59° и 76° с.ш., южная —
между 56° и 39° с.ш., конфигурация границ этих зон сложная, каждая граница может
отклоняться на расстояние до 10°. Так, южная граница северной зоны проходит между
59° и 69° с.ш., северная — между 70° и 76 с.ш. Северная граница южной зоны прохо-
дит между 45° и 56° с.ш., южная — между 39° и 49° с.ш.
В последние годы в европейской части СССР отмечено появление отдельных поселе-
ний серебристых чаек за пределами границ основного гнездового ареала. В небольшом
количестве они ныне гнездятся на Запорожском, Днепродзержинском и, возможно,
Киевском водохранилищах (Клестов, 1983; Мельничук, Головач, 1984; Клестов, Ле-
пешков, устное сообщ.). В 1982 г. В.В. Ивановским и А.В. Наумчиком зарегистрирова-
но гнездование 11—13 пар на верховом болоте в Миорском районе Витебской обл.
БССР (Бирюков и др., 1982); в этом же году 3 пары чаек гнездились на Ситников-
ских торфяных карьерах в Борском районе Горьковской обл. (С.В. Бакка, устное со-
общ.). Численность серебристых чаек на Рыбинском водохранилище, где эти птицы
гнездятся с 1949 г., за 1970-е годы возросла в шесть раз (Немцев, 1980). Изолирован-
ное гнездование серебристой чайки отмечено для Приханкайской низменности (Глу-
щенко, 1981).
Зимовки. Зимний ареал, как и у других чаек, занимает обширные акватории
(приблизительно между 60° и 20° с.ш.) как в Атлантике, так и в Тихом океане
(Austin, Kuroda, 1953; Meinertzhagen, 1954; Шунтов, 1972; Sanger, 1973; Tuck, Heinzel,
1980). В пределах СССР одиночные особи в теплые зимы остаются на Баренцевом и
Белом морях (Кафтановский, 1941; Бианки, 1959), в большем или меньшем числе
131
132
Рис. 34. Область распространения серебристой чайки
а - область гнездования, б - отдельные поселения, в - места зимовки. Подвиды (в скобках даны подвиды, выделяемые не всеми
орнитологами): 1 - L.a. argentatus, 2 - L.a. antelius, 3 - L.a. taimyrensis, (4) - L.a. birulae, 5 - L.a. vegae, 6 - L.a. smithsonianus, 7-
L.a. argenteus, (5) - L. a. armenicus, 9 - L.a. cachinnans, 10 - L.a. mongolicus, 11 - L.a. michahellis, 12 - La. atlantis
133
Рис. 35. Ареал серебристой чайки в СССР
1 —область гнездования, 2 — изолированные поселения, 3 — места зимовок
Семейство Чайковые
у побережий Прибалтики (Манк, 1957; Йыги, 1965, 1969; Леинь, Каспарсон, 1960;
Пятрайтис, 1974). Регулярно зимует по всему северному побережью Черного и Азов-
ского морей, а также на Черноморском побережье Кавказа (Винокуров и др., 1960;
Андоне и др., 1965; Винокуров, 1965; Строков, 1974; Бескаравайный, 1978). На
Каспии регулярно, хотя обычно и в небольшом числе, проводит зиму у северных побе-
режий от Калмыкии до залива Кара-Богаз-Гол, на юге — в районе Кызыл-Агача (за-
лив Кирова) и устья Атрека (Исаков, Воробьев, 1940; Доброхотов, 1962; Залетаев,
1963; Кривоносов, 1963; Виноградов, Чернявская, 1965; Самородов, Самородов,
1969а,б). В восточной части ареала единичные зимующие особи отмечены на юге При-
морья и южных Курильских островах (Нечаев, 1969; Панов, 1973).
Миграции. На юге ареала в Черноморском заповеднике, на Сиваше, а также на юго-
западном побережье Каспия в районе залива Кирова первые чайки появляются в февра-
ле (Дюнин, 1948; Киселев, 1951; Бородулина, 1949; Ардамацкая, 1977в); на Ле-
бяжьих островах в последние 10 лет птицы появляются в колонии уже в середине
января (Костин, 1983). На северное побережье Азовского моря и в Восточное Пред-
кавказье они прибывают в первой половине марта (Филонов и др., 1974; Казаков,
Языкова, 1982). На северном побережье Черного моря в районе Тилигульского ли-
мана наиболее интенсивные перемещения наблюдаются в апреле—мае (Черничко, устное
сообщ.). На юго-западном побережье Каспия в заливе Кирова пролет усиливается в
конце февраля — начале марта, к концу этого месяца он заметно ослабевает и закан-
чивается в первой половине апреля (Заблоцкий, Заблоцкая, 1963). На огромной терри-
тории Казахстана (Долгушин, 1962) сроки начала весенних миграций колеблются от
начала марта на полуострове Мангышлак на Каспии до начала июня на Иртыше; в бас-
сейне Иргиза пролет заканчивается в конце апреля — середине мая. На озерах Бара-
бинской низменности самое раннее появление зарегистрировано 4.IV 1973 г., хорошо
выраженные миграции наблюдались в последней декаде апреля — начале мая; закан-
чивались они здесь в конце второй декады мая, иногда даже в начале июня (Гынгазов,
Миловидов, 1977; Ходков, 1977). На юго-восточный Алтай чайки прилетают 14—20.IV,
на южный Байкал — 28.III—12.IV, на северный Байкал — 12—22.IV (Кучин, 1976; Скря-
бин, 19776). На Байкале массовые перемещения мигрантов зарегистрированы в доли-
не Селенги с 15 по 22.IV и в районе Верхней Ангары с 22.IV по 7.V; заканчивается про-
лет в этих регионах в конце апреля — первой декаде мая (Скрябин, Шароглазов, 1974).
В Приханкайскую низменность первые чайки прилетают во второй половине марта
(Глущенко, 1981), а пролетные птицы в южном Приморье отмечались во второй поло-
вине апреля — начале мая (Черский, 1915; Панов, 1973). На Сахалине движение на
север начинается в первой декаде апреля (Гизенко, 1955).
На севере ареала раньше всего (в разные годы с 22 по 26.III) серебристые чайки
прилетают на незамерзающие побережья Баренцева моря (Модестов, 1967), позднее
(с 26.V по 13.VI) — в прибрежные районы Сибири и Таймыра до Индигирки (Бируля,
1907; Pleske, 1928; Успенский и др., 1962; Матюшенков, 1979).
В Эстонию по многолетним данным первые особи прибывают в среднем 3.IV (Root-
smae, Rootsmae, 1976). Интенсивная миграция в Прибалтике отмечалась с 16 по 30.V
(Леинь, Каспарсон, 1961), на Белом море - с середины апреля до 9.V (Бианки, 1959.
1967; Коханов, Скокова, 1960). На Баренцевом море пролет наблюдается до первой
половины мая (Pleske, 1928; Курочкин, Скокова, 1960; Скалинов, 1960; Коханов.
1965), наиболее интенсивно он протекает здесь в марте—апреле. В среднем течении Оби
у поселка Нарым и на среднем Тыме первые птицы отмечены 2—14.V (Гынгазов, Мило-
видов, 1977). На Среднем Енисее в районе пос. Мирный и на р. Оленек массовые мигра-
ции зафиксированы в третью декаду мая (Ларионов, Седалищев, 1978; Рогачева и др..
1978). На Вилюе Б.Н. Андреев (1974) наблюдал хорошо выраженный пролет с 5 по
134
Серебристая чайка
7.V. На юго-восточном побережье Камчатки, по данным Е.Г. Лобкова (1980), мигра-
ции наблюдаются с середины апреля до конца мая. На севере этого полуострова в устье
р. Апуки регулярный пролет зарегистрирован в 1960 и 1961 гг. с 4 по 26.V (Кищин-
ский, 1980), а на Анадыре в районе пос. Марково — с 11 по 22.V (Портенко, 1939).
В миграционных скоплениях молодые могут составлять от 20 до 80% от числа взрос-
лых, при этом к концу пролета их число увеличивается, что свидетельствует о том, что
взрослые особи начинают и заканчивают пролет раньше молодых (Сушкин, 1908; Ку-
рочкин, Герасимова, 1960; Ходков, 19776, 1981а; Кречмар и др., 1978; Кищинский,
1980). В районах морских побережий и крупных рек чайки часто летят вдоль них,
но могут пересекать и большие пространства суши вдали от побережий. На Белом море
во время миграций часто держатся у становищ или в открытом море, в районах промыс-
ла тюленей (Курочкин, Герасимова, 1960; Скалинов, 1960).
Осенним миграциям обычно предшествует период кочевок, который длится в разных
регионах от 7—10 дней до 2,5 месяцев и характеризуется большим разбросом направле-
ний (Модестов, 1967; Бианки, Бойко, 1972, 1975; Курочкин, Скокова, 1960; Вино-
куров, 1965; Ходков, 1967). Позднее кочевки постепенно переходят в настоящий про-
лет. На Баренцевом море он начинается в середине августа — первой четверти сентября
и заканчивается в середине сентября (Коханов, Скокова, 1960; Модестов, 1967). На
Белом море пролет проходит с конца июля до конца октября (Благосклонов, 1960;
Скокова, 1960; Флеров, Скалинов, 1960), массовые миграции в Кандалакшском заливе
в конце 1960-х годов отмечались во второй декаде сентября (Бианки, Бойко, 1972,
1975), что на 10—15 дней раньше, чем в 1950-х годах. Далее на восток на полуострове
Канин в районе устья Кулоя Б. Житков (1904) заметил первую отлетную стаю 18.VII.
На Новой Земле кочевки и пролет в разные годы наблюдались с 6.VIII по 19.IX (Горбу-
нов, 1929). На остальной территории северной части ареала пролет отмечается пример-
но с середины августа по октябрь. На юго-восточном побережье Камчатки слабый про-
лет проходит с середины сентября до первых чисел ноября (Лобков, 1980).
В Прибалтике миграции отмечаются с августа по ноябрь, особенно интенсивно они
протекают с середины сентября до середины октября. На территории Волжско-Камско-
го междуречья пролет идет с конца сентября до начала ноября (Водолажская, Залетаев,
1977); на черноморском побережье в устье Дуная — с октября по декабрь (Андоне и
др., 1965); на Тилигульском лимане (окрестности Одессы) заметное увеличение числа
пролетных птиц наблюдается уже в июле (Черничко, устное сообщ.). На восточном по-
бережье Азовского моря интенсивные миграции протекают в октябре (Винокуров,
1965) ; на Каспии в районе Гасан-Кули — с октября до второй половины ноября (Иса-
ков, Воробьев, 1940). На Барабинских озерах активные миграции отмечены во второй
половине сентября — октябре (Ходков, 19776, 1983). В юго-западном Забайкалье про-
лет проходил со второй декады сентября до конца этого месяца (Измайлов, 1967).
Птицы, гнездящиеся в европейской части СССР, Казахстане и на юге Западной Сиби-
ри, осенью мигрируют в западном и юго-западном направлениях, в бассейн Атлантичес-
кого океана и Средиземного моря. Чайки, размножающиеся к востоку от Западного
Таймыра, движутся к юго-востоку в бассейн Тихого океана. Как и весной, птицы при-
держиваются морских побережий или долин крупных рек, но могут и пересекать боль-
шие пространства суши и крупные внутренние водоемы напрямик (Луговой, 1958;
Йыги и др., 1961; Вайткявичюс, 1968). На Белом море, Гыданском полуострове, в За-
падной Сибири молодые покидают места размножения раньше взрослых (Наумов,
1931; Курочкин, Скокова, 1960), в других (Вилюй, Байкал, Магаданская обл.) на-
против, взрослые улетают раньше молодых (Андреев, 1974; Кречмар и др., 1978;
Шкатулова, 1981). Так или иначе эти исследования показывают, что основная масса
молодых и взрослых особей в процессе миграций разобщена во времени.
135
Семейство Чайковые
Численность. Общая численность гнездящихся птиц выяснена лишь для некоторых
регионов страны. Так, на Мурманском побережье, по оценкам Т.Д. Герасимовой (1962)
и И.П. Татарниковой (1970, 1975), размножается 6—7 тыс. пар, в заповедной части Кан-
далакшского залива — не менее 1,3 тыс. пар (Бианки, 1967); на западном побережье
Эстонии (Пеэдосаар, Окно, 1970) и южном берегу Финского залива (Ренно, 1972) —
640 и 658 пар соответственно. На Лебяжьих островах в Черном море в 1979 г. гнезди-
лось 9417 пар (Костин, Тарина, 1981); на Молочном лимане Азовского моря в
1975-1979 гг. учитывали от 481 до 630 пар (Сиохин, 1981); в устье Дуная в 1976—
1979 гг. регистрировали около 500 пар (Петрович, 1981); в Восточном Предкавказье
в 1968—1980 гг. учитывали от 240 до 3270 размножающихся пар (Кривенко, Любаев,
1975, 1977, 1981; Языкова, 1975; Казаков и др., 1981; Казаков, Языкова, 1982).
На Каспии в районе Бакинского архипелага в 1961 — 1967 гг. гнездилось от 2750 до
3500 пар (Туаев и др., 1972). На Байкале в районе Верхней Ангары и Кичеры заре-
гистрировано 270 пар (Попов, 1979; Попов, Садков, 1981); в районе Малого моря,
устья Ангары и на островах Чивыркуйского залива размножалось 560, 90 и 320 пар со-
ответственно. Позднее в Малом море учитывали до 870, а в дельте Ангары — до 1200 пар
(Литвинов и др., 1977; Скрябин и др., 1977). На Торейских озерах в 1976 г. гнездилось
3,7 тыс. пар (Зубакин, 1981а).
Во многих районах СССР численность неуклонно растет, например на Баренцевом мо-
ре, в Восточной Прибалтике, Черном море и Сиваше, Рыбинском водохранилище, в
Восточном Предкавказье и на Байкале (Аумеэс, 1972; Ренно, 1972; Костин, 1975;
Кривенко, Любаев, 1975, 1977, 1981; Немцев, 1980; Костин, Тарина, 1981; Кривенко,
1981; Скрябин и др., 1977; Татаринкова, 1975, 1981; Кумари, 1978; Попов, 1979;
Попов, Садков, 1981; Сиохин, 1981а), в других (верховые болота Прибалтики, отдель-
ные острова Вайкайского заповедника у побережья Эстонии, о-в Жемчужный у север-
ного побережья Каспия) — падает (Аумеэс, 1967; Кумари, 1978; Бауманис, 1980;
Гаврилов, Кривоносов, 1981; Петринып, 1982). Вне СССР, особенно в Западной Ев-
ропе и на атлантическом побережье США, судя по литературным данным, наблюдается
резкое увеличение численности. Причины столь резкого и быстрого увеличения числен-
ности за последние 40—50 лет многие исследователи видят в приспособлении к кор-
мам антропогенного происхождения. Следствием резкого увеличения численности мо-
жет быть переход от одиночного к колониальному гнездованию (Bergmann, 1982).
Местообитания. Очень разнообразны, особенно летом. Во время размножения во всех
ландшафтно-географических зонах от тундровой до полупустынной поселяются как на
морских побережьях (скалистых или сглаженных), так и во внутренних районах мате-
рика, везде предпочитая острова: морские, на крупных реках и озерах, различного типа
болотах и больших водохранилищах. С конца прошлого столетия наметилась тенденция
к освоению антропогенных биотопов, которая выразилась в приспособлении к гнез-
дованию на крышах различного рода строений в Болгарии, на Британских островах, на
территории ГДР и ФРГ, Финляндии и США (Reiser, 1894; Paynter, 1963; Cramp, 1971;
Kosonen, Makinen, 1978). За последние десятилетия эта тенденция значительно усили-
лась (Kumerloeve, 1957, Goethe, 1960; Mountfort, Ferguson, 1961; O’Meara, 1975:
Monaghan, Coulson, 1977; Fisk, 1978; Hoyer, Hoyer, 1978; Monaghan, 1982; Нанки-
нов, 1981, 1982). В СССР гнездование на зданиях зарегистрировано в Риге с конца
1970-х годов (Strazdins et al., 1987). Зимой серебристые чайки держатся в прибрежных
районах моря и на побережьях вблизи от источников пищи.
Размножение. Большинство особей приступает к размножению в возврасте 5—6 лет
(Татаринкова, 1970). Серебристые чайки — моногамы, но иногда в колониях наблюдает-
ся полигиния (Shugart, 1980; Fitch, Shugart, 1983). Образование пар часто происходит
зимой или уже на гнездовых участках. Прибывают на места размножения птицы за
136
Серебристая чайка
20--40 дней до начала гнездования, когда на крупных водоемах появляются промоины и
забереги. На морских побережьях сразу после прилета чайки держатся стаями в лито-
ральной зоне, на прибрежных скалах или во внутренних районах тундры (например, на
Западном Таймыре); они в это время тяготеют к жилью человека, где легче добыть
корм (Белопольский, 1957; Модестов, 1967; Кречмар, 1966). После появления неболь-
ших проталин в тундре и оттаивания мелких водоемов большинство птиц рассредоточи-
вается по местам гнездования и приступает к размножению.
Рис. 36. Демонстрационное поведение серебристой чайки (в-д, ж - по: Cramp, Simmons, 1983, с
изменениями)
а—б — последовательность поз долгого крика, в — мяукающий крик, г — ’’квохтание”, д — дерганье
травы, е — прямая поза, ж — сгорбленная поза
Именно в это время наблюдается ярко выраженное демонстративное поведение,
которое довольно подробно изучено на западно-европейских колониях (Tinbergen,
1953, 1959, 1960,1962, 1964; Goethe, 1937, 1963; Brown, 1967b; Drury, Smith, 1968;
McLannahan, 1973; и др.). Хороший обзор зарубежных работ по этому вопросу дан в
сводке С. Крэмпа (Cramp, Simmons, 1983). У нас в стране поведение серебристой
чайки изучалось в основном в плане взаимоотношений взрослых птиц и птенцов в се-
мейных группах и в колониях (Зубакин, 1976в; Панов, Зыкова, 1981). У серебристой
чайки описаны многие демонстрации, имеющиеся у других крупных чаек (рис. 36):
долгий крик, мяукающий крик, ’’квохтание”, или ’’кашляние”, вскидывание клюва,
дерганье травы, прямая поза, отворачивание клюва, сгорбленная поза, ритуальное кор-
мление (Courtship feeding) и др., а также тревожный крик, опознавательный крик
(Note call) и другие типы криков; более или менее известны их функции и происхож-
дение (см.: Cramp, Simmons, 1983). Как и у других чаек, демонстративное поведение
137
Семейство Чайковые
распадается на агрессивный и брачный комплексы. При этом подавляющее большинство
указанных выше демонстраций и поз применяется в обоих.
Серебристая чайка — диффузно гнездящийся факультативно-колониальный вид
(Зубакин, 1976в, 1983). Гнездится как колониями, так и одиночными парами. Наибо-
лее крупные поселения в пределах страны известны на Айновых островах в Баренцевом
море, где размножаются 4,0—4,5 тыс. пар (Татарникова, 1975, 1981), на сивашских
островах Китай и Мартынячий — 1,6 и 2,3 тыс. пар соответственно (Сиохин, 1981а),
на каспийском о-ве Огурчинском и одном из островов Торейских озер — по 2,5 тыс. пар
(Васильев и др., 1975; Гаузер и др., 1977; Зубакин, 1981). В других частях ареала
от Эстонии до Казахстана, Западной Сибири и Байкала самые крупные поселения насчи-
тывают только до нескольких сот пар, обычно всего по нескольку десятков пар. На се-
вере и востоке ареала от п-ова Канин до Анадыря и Корякского нагорья встречаются
лишь мелкие поселения от нескольких десятков до единичных пар (Портенко, 1939;
Рутилевский, 1958; Успенский и др., 1962; Кищинский, 1980).
При выборе места для гнезда серебристые чайки проявляют большую пластичность,
но во всех биотопах гнезда помещаются на открытых местах, так что птицы имеют хо-
роший обзор: на скалах, косах, отмелях, по краям сплавин, на мысах рядом с обшир-
ными плесами или на отдельных кочках на мелководьях. Лишь иногда в очень жарких
и засушливых районах в дельте Волги, на Каспии, Арале и в Восточном Предкавказье
(Исаков, 1948; Модестов, 1943; Исмагилов, 1955; Луговой, 1958; Языкова, 1975;
Казаков, Языкова, 1982), редко в лесной и тундровой зонах (Коханов, Скокова, 1960;
Болотников и др., 1979) гнезда располагаются в густых зарослях травянистой или
кустарниковой растительности, защищающей птиц от прямых лучей солнца или силь-
ных ветров.
Гнездо строят, как и у всех чаек, самец и самка. В качестве строительного материала
птицы обычно используют растения, которые они находят в непосредственной близости
от гнезда. На севере ареала в тундре и лесотундре, как показывают многочисленные
литературные данные, это сухие листья и стебли злаков, осок, мхи, лишайники, ве-
точки вороники, карликовых форм ивы, березы; иногда в гнездах встречаются перья
птиц, оленья шерсть, у побережий — водоросли, особенно фукусы, зостера (Бируля,
1907; Thayer, Bangs, 1914; Михель, 1935; Портенко, 1939; Белопольский, 1957;
Коханов, Скокова, 1960; Успенский и др., 1962; Бианки, 1967; Остапенко, 1973;
Кищинский, 1980; Данилов и др., 1984). В лесной зоне на Байкале и в Витебской обл.,
кроме осок, злаков и мхов, используются прутики тальника, веточки багульника (Гу-
сев, 1960; Малышев, 1960; Наумчик, устное сообщ.). На юге ареала, помимо злаков
(пырея, ковыля, типчака), осок и других травянистых растений, применяются рогоз,
тростник, тамариск; попадаются также в гнездах шерсть, перья птиц и водоросли (Модес-
тов, 1943; Дюнин, 1948; Исаков, 1948; Исмагилов, 1955; Луговой, 1958; Бородули-
на, 1960; Долгушин, 1962; Измайлов, 1967; Самородов, Рябов, 1968; Ардамацкая,
1977в; Ходков, 1981а; Казаков, Языкова, 1982). Выстилка лотка обычно выполняет-
ся из более мягкого материала, чем стенки гнезда: мелких сухих листьев, стеблей и ко-
решков трав. Однако иногда встречаются гнезда совсем без выстилки, в виде ямки
среди песка и ракушечника (Долгушин, 1962). В этом случае доля яиц с трещинами
достигает 56,5%.
Обычно гнезда помещаются на земле, но изредка они могут оказаться приподняты-
ми над ней или над водой на высоту от 0,5 до 2,0 м (Рашкевич, 1959; Коханов, Скоко-
ва, 1960). Нередко гнезда используются многократно, что особенно характерно для
южных колоний, но иногда встречается и в лесной зоне. Плотность расположения гнезд
в колонии сильно варьирует и зависит от многих конкретных условий: площади поселе-
ния, микрорельефа местности, численности птиц и ее динамики в данной колонии,
138
Серебристая чайка
взаимоотношений с сопутствующими видами и т.д. Расстояние между гнездами в самых
плотных колониях равно 1-3 м, в разреженных 25—30 м (Дюнин, 1948; Кречмар, 1966;
Бианки, 1967; Самородов, Рябов, 1968; Татаринкова, 1975; Ардамацкая, 1977в; Хо-
дков, 1981а; Бианки, Плюснин, 1983; Данилов и др., 1984). Индекс плотности гнез-
дования для колоний Сиваша равен 0,09, для колоний Торейских озер — 0,1 (данные
В.А. Зубакина). Одиночное гнездование так же обычно, как и колониальное; при оди-
ночном поселении на Ямале чайки гнездятся не ближе, чем в 3—5 км друг от друга (Да-
нилов и др., 1984). Внутренняя структура мелких и более крупных колоний, как по-
казали специальные исследования на Белом море, существенно различны (Бианки,
Плюснин, 1983). Мелкие колонии (10—15 пар) состоят из одиночных гнезд и групп
в 2-3 гнезда, которые распределяются в пространстве равномерно друг относительно
друга. Более крупные колонии (20—25 пар) содержат обычно группы по 7—9 гнезд и
одиночные гнезда. При этом распределение гнезд в пространстве становится случайным
и наблюдается деформация гнездовых территорий у большинства пар. Заселение коло-
ний идет от периферии к центру острова.
Размеры гнезд в разных частях ареала также сильно варьируют (табл. 6).
По многолетним наблюдениям откладка яиц на Баренцевом и Белом морях начи-
нается в среднем 9—10. V (Белопольский, 1957; Бианки, 1967). Далее на востоке в
окрестностях Воркуты, на Новосибирских островах, побережье Анабарской губы,
в низовьях Колымы и на р. Канчалан свежие кладки находили с 6 по 24.VI (Плеске,
1928; Успенский, 1958, 1965; Воробьев, 1963; Кищинский и др., 1983). В устье р. Чаун
на Чукотке массовая откладка яиц протекала в 1975 и 1976 гг. с 2 по 22.VI (Кондрать-
Таблица 6. Размеры гнезд (мм) серебристой чайки
в разных частях ареала
Место наблюдений, автор п Диаметр гнезда Диаметр лотка Высота стенки гнезда от зем- ли Глубина лотка
Белое море (Бианки, 1967) — 350-500 220-250 60-170 50-70
Белое море (Коханов, личное 20 240-395 188-230 — 46-78
сообщ.) Витебская обл. (Наумчик, личное (302) * (308) (209) 200-240 (58) (50)
сообщ.) Дельта Волги (Луговой, 1958) — 300-400 170-250 (70) 50-130
Дельта Волги (Модестов, 1943) — (350) 20-800 (220) 240-460 320-390 (80)
Южные моря СССР (Бородулина, — 400-610 210-250 — 80-100
1960) Восточное Приазовье (Казаков, 150 (519) 340-960 (218) 190-380 10-400 (91) 20-140
Языкова, 1982) (520) (280) (140) (70)
Казахстан (Долгушин, 1962) — 200-400 — 90-300 —
Юго-западный Каспий (Дюнин, 1948) — 300-400 — 30-90 55-120
Юг западной Сибири, оз. Чаны 19 (361) 300-1200 180-250 (61) 70-300 (81) 23-100
(Ходков, 1981а) (419+46) (209+3) (162,5 + 13,7) (65+5)
Байкал (Малышев, 1960) — 400-500 — 100-150 —
Байкал (Гусев, 1960) 11 390-400 240-260 — (100)
Устье р. Чаун (Остапенко, 1973) - (300) — — (70)
*В скобках приведены средние значения.
139
Семейство Чайковые
Таблица 7. Серебристая чайка. Размер кладок
в различных частях ареала
Место наблюдений, автор Число кладок Доля кладок (%) из числа яиц Средний раз мер кладки
1 2 3 4
Баренцево море (Белопольский, 1957; Модестов, 1967) 597 0 35,51 63,99 0,5 2,65
Белое море (Бианки, 1967) 183 3,8 15,9 79,8 0,5 —
Юго-западный Ямал (Данилов и др., 1984) 10 20,0 10,0 70,0 0 2,5
Западный Таймыр (Кречмар, 1966) 48 6,2 29,1 66,8 0 2,58
Новосибирские острова (Бируля, 1907) 5 20,0 40,0 40,0 0 2,2
Низовья Лены (Лабутин и др., 1982) — — — — — 2,74
Дельта Селенги (Богородский, 1981) 14 — — — — 2,70
Северный Байкал (Попов, 1979) 262 9,37 18,76 71,83 0,4 2,50
Байкал (Скрябин и др., 1974) Сиваш, о-в Китай (Зубакин, 1975а) 1250 12,7 26,4 60,0 0,9 2,24-2,59
Центральная часть острова 93 0 9,7 89,2 1,1 2,91
Периферийная ” 53 13,2 30,2 56,6 0 2,43
Восточное Приазовье (Казаков, Языкова, 1982) 128 8,6 25,8 65,6 0 -
Дельта Волги (Модестов, 1943) 54 11,1 40,7 48,1 0 2,37
Дельта Волги (Бородулина, 1960) — 0 38,1 68,2 0 —
Юго-западный Каспий (Дюнин, 1948) 23 8,6 39,1 52,1 0 2,46
Арал, о-в Барсакельмес (Исмагилов, 1955) 343 11,7 20,1 68,3 - 2,56
Западная Сибирь, оз. Чаны (Ходков, 18 - — — - 2,50+0,16
1981а)
ев, 1979). На юге ареала на Черном море (Лебяжьи острова), в Присивашье, восточной
части Предкавказья, на Каспии (дельта Волги, залив Кирова) первые кладки в разные
годы отмечали с конца марта по 25.IV (Модестов, 1943; Дюнин, 1948; Бородулина,
1949, 1960; Киселев, 1951; Филонов и др., 1974; Ардамацкая, 1977в; Казаков и др.,
1980; Казаков, Языкова, 1982; Костин, 1983). На территории Казахстана от восточ-
ных побережий Каспия, аральского о-ва Барсакельмес, низовий Сырдарьи до озер
Алакольской котловины и Зайсана свежие яйца в разные годы находили с 20-х чисел
марта до 20.V (Зарудный, 1916; Спангенберг, Фейгин, 1936; Исмагилов, 1955; Долгу-
шин, 1962). На озерах Барабинской низменности в Западной Сибири откладка яиц
начинается в середине мая (Гынгазов, Миловидов, 1977). В средней части Байкала
гнездование начинается 5 —10.V, на севере озера — 10—15.V (Скрябин, 19776).
Средний размер кладки, а также численное соотношение разновеликих кладок
сильно колеблются в разных частях ареала (табл. 7). Этот показатель зависит также
от погодных и пищевых условий в разные годы в одном и том же регионе (Кречмар,
1966; Модестов, 1967; Скрябин, Шароглазов, 1974). Средний размер кладки колеб-
лется от 2,2 до 2,91 яйца. Крупные кладки из 4-7 яиц, обнаруженные на Каспии и на
Ямале (Дюнин, 1948; Данилов и др., 1984), откладываются, вероятно, двумя самками.
Это доказано для серебристых чаек, гнездящихся на Великих озерах в США (Shugart,
1980). Повторные кладки отмечены на севере ареала от Баренцева моря до Чукотки
(Белопольский, 1957; Спангенберг, Леонович, 1960; Кречмар, 1966; Бианки, 1967;
140
Серебристая чайка
Таблица 8. Серебристая чайка. Размеры яиц (мм) в разных частях ареала
Место наблюдений, автор п Длина Ширина Средние раз- меры
Баренцево море (Модестов, 1967) % 65,0-81,0 41,0-54,0 72,7 X 49,6
Белое море (Бианки, 1967) 180 67,0-81,0 46,0-59,0 73,2 X 50,7
Таймыр и Новосибирские острова (Бируля, 1907) 11 66,0-75,8 46,1-52,3 72,3 X 51,0
Анадырский край (Портенко, 1939) 15 67,0-80,0 45,0-52,0 72,9 X 48,7
Байкал, Чевыркуйский залив (Гусев, 1960) 6 75,4-81,2 50,2-53,3 78,1 X 51,6
Черное море, Лебяжьи острова (Костин, 1983) 185 63,3-80,5 44,6-58,5 72,8 X 50,4
Восточное Приазовье (Казаков, Языко- ва, 1982) 311 79,6-63,0 46,0-53,5 70,9 X 50,4
Дельта Волги (Луговой, 1958) 66 65,7-81,5 44,4-52,8 71,2 X 49,6
Юго-западный Каспий (Дюнин, 1948) 56 61,0-79,0 40,0-52,0 72,0 X 49,0
Юго-восточный Каспий, залив Кара-Богаз-Гол (Щербина, 1979) 18 66,0-77,0 43,5-52,2
Арал, о-в Барсакельмес (Исмагилов, 1955) 705 64,0-84,0 44,0-55,0 71,0 X 50,0
Кустанайская обл., оз. Жарколь (Само- родов, 1970) 47 67,0-82,0 45,0-57,0 72,8 X 50,3
Юг западной Сибири, оз. Чаны (Ходков, 1981а) 42 66,6-73,7 44,5-51,3 70,0 X 48,1
Портенко, 1973). По наблюдениям Л.О. Белопольского (1957), только 28% самок,
потерявших кладки, могут их восстанавливать.
Размеры яиц серебристой чайки в разных частях ареала приведены в табл. 8. Форма
яиц в большинстве случаев варьирует от овальной до короткоовальной, иногда от
удлиненно-овальной до остроовальной; скорлупа грубозернистая, глянцевитая. Окраска
фона яиц и их рисунка очень изменчива. Обычно окраска фона — от тускло-коричне-
вого до светлого зеленовато-голубого. Поверхностные пятна коричневого цвета различ-
ной интенсивности; обычно они большие, округлой формы, но могут быть пятна
неправильной формы и извилистые линии. Глубокие пятна от сероватого до коричне-
вато-фиолетового цвета. Очень редко яйца могут быть красноватыми или чисто-голу-
быми без пятен, Яйца подвидов по окраске неразличимы (Makatsch, 1974).
Интервал между откладкой яиц одной самкой 1—2, реже 3—4 дня. Насиживают
кладку оба родителя начиная с первого или второго яйца. Плотное насиживание начи-
нается обычно со второго или третьего яйца, при этом самки сменяются через 2—5 ч
(Бианки, 1967). По наблюдениям Ю.А. Самородова (1970) в Казахстане, самцы могут
насиживать кладку без перерыва от 40 мин до 3 ч, самки — от 3 до 6 ч. Длительность
насиживания кладки в разных частях ареала колеблется от 26 до 32 дней (Модестов,
1943, 1967; Дюнин, 1948; Исмагилов, 1955; Бианки, 1967; Ардамацкая, 1977в).
По данным Ю.А. Самородова (1982), масса яиц (и = 9) в процессе насиживания умень-
шается в среднем на 0,4—0,9 г в сутки; средняя масса яиц на стадии вылупления птен-
цов равна 81,0 г (и = 14). За весь период насиживания яйцо теряет в массе в среднем
16,6 г (и = 89), что составляет 18,8% его первоначальной массы (Казаков, Языкова,
1982),
Первые птенцы на Баренцевом море (Айновы острова, Семь островов) отмечались
в разные годы с 20.V по 15.VI (Успенский, 1941; Белопольский, 1957; Татарникова,
141
Семейство Чайковые
1958; Коханов, Скокова, 1960; Модестов, 1967); массовое вылупление на Айновых
островах происходило с 25 по 30.VI, а на Семи островах — 9.VI. На Белом море (Канда-
лакшский и Онежский заливы, восточное побережье) первые птенцы и массовое вылуп-
ление наблюдались 5—25.VI и с 15.VI по первые числа июля соответственно (Кузякин,
Леонович, 1958; Спангенберг, Леонович, 1960; Бианки, 1967). В северотаежной части
долины р. Лены появление первых птенцов в разные годы отмечали с третьей декады
июня до первой декады июля (Лабутин и др., 1982), а в среднем течении Омолона —
в первой половине июля (Кречмар и др., 1978), в низовьях р. Чаун — 5—25.VII (Конд-
ратьев, 1979). На восточном побережье Чукотки (мыс Халюсткина) в 1984 г. вылупле-
ние началось 28—29.VI (Зубакин, устное сообш.).
На юге ареала на Сиваше первые птенцы появляются примерно 10—15.IV (Зубакин,
1975а); на Маныче в Восточном Предкавказье — в конце апреля — начале мая, массо-
вое вылупление происходит здесь в середине мая (Казаков, Языкова, 1982). В заливе
Кара-Богаз-Гол первые пуховички найдены 10.V 1938 г. (Исаков, 1948). На территории
Казахстана (п-ов Мангышлак, о-в Барсакельмес, оз. Тенгиз, оз. Жарколь из группы
Наурзумских озер) первые птенцы зарегистрированы во второй половине мая (Дол-
гушин, 1962; Самородов, Рябов, 1968; Самородов, 1970), на оз. Соркуль в Уральской
обл. в 1981 г. — 6.V (Леонович, устное сообщ.). На озерах Барабинской низменности
вылупление пуховичков отмечалось примерно с середины июня до 20.VII (Гынгазов,
Миловидов, 1977; Ходков, 1981а).
На Байкале первых пуховичков находили с 10 по 21.VI (Гусев, 1960; Скрябин,
Шароглазов, 1974), на Торейских озерах в Забайкалье первые птенцы появились в
последних числах мая (Зубакин, 1981а).
Порядок вылупления птенцов соответствует порядку откладки яиц, при этом на
Белом море вылупление всех пуховичков в одном гнезде длится 1—2 дня, а в популя-
ции этот процесс растягивается в среднем на 40 дней (Бианки, 1967). Отход яиц в раз-
ных частях ареала составляет от 10 до 38%. Основные его причины — действие хищни-
ков, гибель кладок при неблагоприятных погодных условиях (перегрев на юге, высо-
кие приливы и шторма на севере), гибель зародышей на разных стадиях эмбриогенеза
и наличие неоплодотворенных яиц.
В первые сутки после вылупления птенцы беспомощны, находятся в гнезде и не
получают пищи. При опасности, однако, они могут покидать гнездо и затаиваться
около него даже в день вылупления (Бианки, 1967). На Баренцевом море терморегу-
ляция у чайчат устанавливается в возрасте 1,5—2 сут (Белопольский, 1957). Со второго
дня жизни птенцов уже кормят (самым маленьким пищувзрослыеподносятв клюве,
позднее пуховички могут подбирать отрыжку с земли), они способны вставать на
ноги и начинают двигаться (Дюнин, 1948; Модестов, 1967). Кормят птенцов оба роди-
теля, в Казахстане это происходит не только днем, но и ночью (Самородов, 1970, 1971,
1973). В возрасте 3—4 дней пуховички свободно бегают и плавают; они могут уходить
от гнезда на расстояние до нескольких десятков метров, хотя обычно держатся на
гнездовом участке, куда родители приносят корм (Бианки, 1967). Окончательно поки-
дают гнездо птенцы в возрасте не более двух дней и при опасности уходят в воду.
При отсутствии беспокойства они могут находиться в непосредственной близости от
гнезда до 8-10 дней (Бородулина, 1960; Долгушин, 1962; Самородов, 1970), а с этого
времени предпочитают держаться на воде. На юге ареала подросшие птенцы при опас-
ности с воздуха иногда выходят на открытые участки и сбиваются в разреженные та-
бунки-ясли, которые находятся под охраной летающих взрослых птиц (Дюнин, 1948;
Щербина, 1979; Панов, Зыкова, 1981). Родители активно нападают на хищников и
нарушителей спокойствия. Птиц атакуют в воздухе и длительно преследуют. При появ-
лении в колонии человека вылетают ему навстречу с беспокойными криками, вблизи
142
Серебристая чайка
от гнезд или птенцов активно пикируют на него, задевают ногами или ударяют клю-
вом, поливают пометом; другие наземные животные (овцы, коровы, собаки, лисы)
также атакуются с разной интенсивностью (Patten, Southern, 1977, цит. по: Cramp,
Simmons, 1983).
Рост и развитие птенцов изучены довольно хорошо. На Баренцевом море только что
вылупившиеся пуховички имеют массу от 49 до 66 г, в среднем 57 г, что составляет
около 5% массы взрослых особей (Белопольский, 1957; Модестов, 1967). Заметное
нарастание веса птенцов здесь начинается с третьего дня их жизни и продолжается до
25-го дня, затем приостанавливается, а к 35-му дню иногда наблюдается снижение веса.
На Черном море и в дельте Волги средняя масса только что вылупившихся птенцов
составляет 60,7 г (Бородулина, 1960). На Каспии и в Казахстане в первый день жизни
птенцы теряют в массе от 2 до 6 г, в последующие три дня их масса увеличивается на
3 г в сутки, с 4-го дня ежесуточный привес составляет от 10 до 30 г, а максимума (око-
ло 50 г) он достигает к 12—13-дневному возрасту. Веса взрослых птенцы достигают к
35-45-дневному возрасту (Дюнин, 1948; Долгушин, 1962). На Баренцевом и Белом
морях пеньки перьев над поверхностью кожи на плечевой птерилии появляются на
4—5-й день жизни, пеньки первостепенных маховых и кроющих спины — на 5—7-й день,
рулевых — на 12-й день; опахала плечевых и первостепенных маховых перьев начинают
разворачиваться соответственно на 5—7-й и 8—10-й день, рост оперения на плечевой
птерилии заканчивается к 28—30-му дню жизни птенца (Бианки, 1967; Модестов,
1967).
Лётными птенцы становятся, по данным разных авторов, в возрасте от 38 до 45 дней
(Дюнин, 1948; Белопольский, 1957; Бородулина, 1960; Долгушин, 1962; Бианки,
1967; Модестов, 1967; Ардамацкая, 1977в). На Баренцевом и Белом морях это про-
исходит в разные годы с конца июня по 15.VIII, средняя многолетняя дата на Барен-
цевом море 23.VII (Успенский, 1941; Белопольский, 1957; Благосклонов, 1960;
Малышевский, 1962). На островах Колгуев, Вайгач и в Тиманской тундре лётных
молодых отмечали с конца июля по 15.VIII (Толмачев, 1927; Успенский, 1958, 1965;
Гладков, 1951); в низовьях Енисея и на Западном Таймыре — с 10 по 28.VIII (Бируля,
1907; Тугаринов, Бутурлин, 1911; Кречмар, 1966). В среднем течении Омолона и в
низовьях Чауна лётные птенцы обнаружены 5.VIII и с 20 по 30.VIII соответственно
(Кречмар и др., 1978; Кондратьев, 1979). На Восточной Чукотке и в Корякском на-
горье их видели с 31.VII по 21.VIII (Портенко, 1983; Кищинский, 1980). На юге ареа-
ла подъем молодых на крыло происходит с начала—середины июня по конец июля.
На Байкале первых лётных птенцов находили в конце июля — начале августа (Скрябин,
Шароглазов, 1974).
После приобретения способности к полету молодые в течение по крайней мере 30
дней еще полностью зависят от родителей, которые их активно кормят. Самосто-
ятельно кормиться птенцы начинают в возрасте не менее 70 дней, после чего частота
кормлений их взрослыми резко падает. В это время молодые постепенно переходят к
самостоятельному образу жизни, выводки группируются в стаи (Burger, 1981). На
Белом море в стаи объединяются чайки всех возрастов от сеголетков до взрослых
(Бианки, 1967), на Каспии молодые собираются в стаи, обособленные от взрослых
(Дюнин, 1948). Стаи ведут кочевой образ жизни, концентрируясь в местах скопления
корма.
На Баренцевом море чайки откочевывают с колоний на литораль в конце августа
(Модестов, 1967), на Белом море — в конце июля — первой декаде августа (Благо-
склонов, 1960; Бианки, Бойко, 1975). В Корякском нагорье и на Чукотке взрослые
покидают места размножения с середины или конца августа, в октябре здесь встре-
чаются в основном сеголетки (Портенко, 1973; Кищинский, 1980). На Черном море
143
Семейство Чайковые
молодые оставляют колонии во второй половине июля или начале августа (Бородули-
на, 1949; Киселев, 1951), на водоемах Восточного Предкавказья — во второй половине
августа (Казаков, Языкова, 1982), на Каспии и Аральском море — в июле (Дюнин,
1948; Степанян, Галушин, 1962), на озерах Барабинской низменности — в начале августа
(Ходков, 1977), анаБайкалев дельте Селенги —с середины августа (Богородский, 1981).
Успех размножения серебристой чайки в разных частях ареала колеблется от 0,2
до 1,9 птенца на пару размножавшихся птиц (Бианки, 1967; Богородский, 1981; Панов,
Зыкова, 1981), В неблагоприятные по кормовым условиям годы или в условиях высо-
кого уровня численности птицы не гнездятся вовсе или частично, либо успех их размно-
жения практически сводится к нулю (Сдобников, 1971; Кречмар и др., 1979; Татарни-
кова, 1970, 1981). В благоприятные годы отход птенцов составляет от 10 до 57%
(Андреев, 1940; Клименко, 1950а; Бианки, 1967; Попов, 1979). Много молодых
гибнет от каннибализма и хищничества чаек (Клименко, 1950а; Коханов, Скокова,
1960).
Максимальная продолжительность жизни, по данным кольцевания, 31 год И меся-
цев (Rydzewski, 1978, цит. по: Cramp, Simmons, 1983).
Суточная активность, поведение. В некоторых регионах активны не только днем,
но и ночью (Самородов, 1971, 1973; Goethe, 1937). Описано несколько типов осенних
и зимних территорий вне гнездовых колоний и в их пределах, которые сохраняются в
течение нескольких лет (Cramp, Simmons, 1983). Члены одной семейной группы могут
контактировать и зимой, особенно если дисперсия молодых незначительна, что харак-
терно для урбанизированных местообитаний (Drury, Smith, 1968; Bourne, 1979;
Holley, 1982, цит. no: Cramp, Simmons, 1983). Вне колоний поддерживается индивиду-
альная дистанция, равная приблизительно 0,3 м (Greebb, 1974; цит. по: Cramp, Sim-
mons, 1983), В период размножения и во внегнездовое время описаны синхронные
полеты потревоженных особей на высоте до 1000 м (Goethe, 1937; Brown,
1967b; Cook, Ross, 1972, цит. no: Cramp, Simmons, 1983; McRoberts, McRoberts,
1972).
Питание. Серебристая чайка — типичный эврифаг, использующий водные и наземные
корма, способный к сезонной их смене, быстрому переходу от одного массового корма
на другой в течение одного сезона и применяюший разнообразные способы добычи пищи.
В состав водных кормов входят как донно-литоральные, так и пелагические морские и
пресноводные животные: рыбы, моллюски, ракообразные, иглокожие, черви. Наземные
объекты питания не менее разнообразны: грызуны, яйца птиц и птенцы, ящерицы,
черепахи, различные насекомые и их личинки, черви, а также ягоды. Особый тип кормов
составляют отходы рыбного и охотничьего промыслов, пищевые отбросы, скапливаю-
щиеся на свалках у населенных пунктов.
Соотношение различных пищевых, объектов в данный момент зависит от место-
обитания, сезона, наличия массовых кормов. Так, на Баренцевом и Белом морях в
гнездовое время преобладают морские корма (Белопольский, 1957; Герасимова,
1958, 1965; Благосклонов, 1960; Коханов, Скокова, 1960; Бианки, 1967; Модестов,
1967), которые составляют, по подсчетам Л.О. Белопольского (1957), 82% по встре-
чаемости (из них 56% приходится на донно-литоральных и 44% — на пелагических жи-
вотных). Наземные корма составляют в этом регионе лишь 12%. Вдали от моря в
тундре (Таймыр, низовья Енисея, Лены и Индигирки) основой питания в гнездовое вре-
мя служат наземные корма: лемминги, серые полевки и водяные крысы, птенцы и
яйца птиц, отходы охотничьего промысла; рыба занимает здесь подчиненное положе-
ние (Тугаринов, Бутурлин, 1911; Михель, 1935; Кречмар, 1966; Лабутин и др., 1982).
На юге ареала наземные корма составляют основу рациона даже в колониях, располо-
женных в непосредственной близости от моря. Так, на Лебяжьих островах Черного
144
Серебристая чайка
моря, как сообщают Т.Л. Бородулина (1949) и Ф.А. Киселев (1951), чайки питаются
преимущественно вредными для сельского хозяйства грызунами (сусликами, полевка-
ми), которые составляют до 72% встреч в погадках и желудках птиц. На долю ящериц
здесь приходится 10-17%, насекомых - 11-29% встреч. Сходный состав кормов об-
наружен в Присивашье (Ишунин, 1963), на Каспии (Дюнин, 1948; Исаков, 1948;
Луговой, 1961) и в Казахстане (Долгушин, 1962). Во всех этих регионах рыба исполь-
зуется только как дополнительный корм, в больших количествах птицы едят ее только
во время путины, потребляя в основном отходы промысла или рыбу, погибшую при
заморах.
Изменчивость соотношения кормов в разные сезоны зависит от пола и возраста чаек,
а также их способности быстро переключаться с одного массового корма на другой.
Эти особенности отмечались многими исследователями, работавшими как на севере,
так и на юге ареала (Бируля, 1907; Исмагилов, 1955; Белопольский, 1957; Герасимо-
ва, 1958, 1965; Коханов, Скокова, 1960; Луговой, 1961; Бородулина, 1965; Бианки,
1967; Модестов, 1967; Скрябин и др., 1981; Ходков, 1981а; Казаков, Языкова, 1982).
Разнообразию объектов питания соответствует разнообразие кормовых биотопов
и способов добычи корма. Рыбу серебристые чайки ловят сидя на воде, погружая в нее
голову и шею и хватая добычу в поверхностных слоях воды. Могут пикировать за ней
с воздуха, иногда зависая перед броском, как крачки (Бойко и др., 1970). Ловко
достают рыбу из сетей с глубины до 50 см, выбирая их клювом (Михель, 1935; Дунае-
ва, Кучерук, 1941; Долгушин, 1962; Воробьев, 1963; Заблоцкий, Заблоцкая, 1963;
Кречмар, 1966; Ходков, 1981а,б). Иногда охотятся совместно с другими морскими
животными (белухами, бакланами), подбирая распуганную или раненную ими рыбу
(Банников, 1934; Заблоцкий, Заблоцкая, 1963). Раковины двустворчатых моллюсков,
панцири крабов и морских ежей разбивают, бросая их с высоты на землю (Тинберген,
1974).
На суше чайки также обнаруживают большое разнообразие способов охоты и прояв-
ляют при этом удивительную сообразительность. Насекомых могут ловить в воздухе,
собирать их с растений и земли и даже находить в земле, разрывая их норы (Дюнин,
1948). Мелких млекопитающих размером до суслика, ящериц, молодых черепах актив-
но ловят, подкарауливая у нор или высматривая с воздуха (Бородулина, 1949). По
наблюдениям А.Г. Дюнина (1948), во время осенних палов идут непосредственно за
линией огня и ловят выходящих на поверхность грызунов. Во время паводков в Запад-
ной Сибири вылавливают грызунов на незатопляемых участках и в прошлогодних
стогах сена (Ходков, 1981а). В антропогенных ландшафтах добывают пищу, следуя
за сельскохозяйственными машинами (Бородулина, 1949; Ходков, 1981а).
На протяжении всего ареала большое место среди способов добычи корма занимают
клептопаразитизм и хищничество в колониях. Объектами клептопаразитизма служат
другие чайки (озерная, сизая), крачки, тупики, поморники, утки, бакланы, причем
не только летом, но и зимой (Модестов, 1967; Скокова, 19676; Бойко и др., 1970;
Краснов, 1981; Краснов, Николаева и др., 1982; Дурнев, 1983). В колониях обычно
грабят оставленные без присмотра гнезда с яйцами или маленькими птенцами как
своего, так и других видов. Такие нападения бывают особенно успешными во время
тревоги, спровоцированной хищником или человеком (Краснов, Головацкая и др.,
1982); нередко источником беспокойства являются сами чайки-грабители, иногда перед
нападением они активно сгоняют с гнезд птиц-хозяев. В северных колониях или вблизи
от них чайки-хищники ловят не только птенцов, но и ослабленных, больных и даже
здоровых взрослых особей, размером от мелких воробьиных до уток; птенцов гаги
чайки хватают за голову и держат их под водой, пока они не перестают сопротивляться
(Бойко и др., 1970). На южном Байкале в последние десятилетия отмечается значитель-
145
Семейство Чайковые
ное усиление хищничества как способа кормодобывания, что вызвано, возможно,
резким сокращением естественных кормов, ростом численности самих чаек и увели-
чением доступных пищевых отходов, добываемых на свинофермах, зверофермах и
птицефермах (Мельников, Лысиков, 1983; Мельников, Шинкаренко и др., 1983).
Каннибализм отмечается по всему ареалу как в северных, так и в южных колониях
(Клименко, 1950а; Долгушин, 1962; Чекменев, 1964; Бойко и др., 1970; Мельников,
Лысиков, 1983; Мельников, Шинкаренко и др., 1983). Однако активно хищничают
лишь отдельные особи, максимальное их число в популяции может достигать 10—12%
(Мельников, Лысиков, 1983).
Большую часть кормов чайки добывают все же не путем активной охоты, а соби-
ранием на литорали во время отлива или среди отходов (Белопольский, 1957; Бианки,
1967). У крупных экземпляров мертвых рыб, которых они не могут заглотить цели-
ком, птицы проделывают клювом отверстие в коже и через него выедают внутренности
и мышцы; так же они поступают при разделке трупов китообразных и тюленей (Бойко
и др., 1970). На юге ареала чайки также подбирают падаль и насекомых, которые скап-
ливаются на ней (Сушкин, 1908; Долгушин, 1962). На Аральском море подросшие
птенцы чаек держатся в колониях цапель и бакланов, подбирая оброненную ими рыбу
(Рашкевич, 1969).
Враги и неблагоприятные факторы. Естественные враги серебристых чаек — лисицы,
ястреба, совы, вороны. Некоторые колонии сильно страдают от сбора яиц людьми.
Отдельные птенцы могут разбиваться, падая со скал. Однако большая часть молодняка
погибает в отдельные годы от неблагоприятных кормовых или погодных условий.
На успех размножения значительное воздействие может оказывать каннибализм
(Brown, 1967b; Parsons, 1971; Зубакин, 1975а).
Хозяйственное значение, охрана. Уничтожением грызунов серебристые чайки при-
носят пользу сельскому хозяйству. Однако хищничество этого вида в ряде мест может
наносить значительный ущерб другим птицам. Так, успех размножения уток на южном
Байкале снижается чайками по крайней мере на 35—40%; в Коми АССР в 1979 г. из-за
хищничества чаек погибло 70% кладок у полярных крачек (Шкарин и др., 1982; Мель-
ников, Лысиков, 1983; Мельников, Шинкаренко и др., 1983). Поэтому в некоторых
заповедниках и на других охраняемых природных территориях проводится регуляция
численности серебристых чаек (Аумеэс, 1967; Бианки, 1967; Татаринкова, 1975, 1981).
В ряде мест за рубежом численность их также искусственно уменьшается путем отстре-
ла или другими способами (Coulson et al., 1982; Шедцен, 1982). Специальные меры
охраны не предусмотрены.
Тихоокеанская чайка —
Larus schistisagus Stejneger, 1884
Slaty-backed Gull (англ.) JCamtchatkamove (HeM.),Goeland du Kamtchatka (фр.)
Син. (рус.): тихоокеанская морская чайка, тихоокеанская клуша.
Статус. Гнездящийся мигрирующий и частично зимующий вид.
Общая характеристика и полевые признаки. Крупная чайка. От серебристой и серо-
крылой чаек отличается более темной окраской верха, от морской чайки и клуши,
совместно с которыми не встречается, — более светлой окраской верхней стороны тела.
Весь гнездовой ареал располагается в пределах СССР. Размножается в непосредственной
близости от моря, в глубь материка не идет. Экология и демонстративное поведение,
как у других крупных белоголовых чаек. Голос значительно выше, чем у морской чай-
ки и клуши, и немного выше, чем у серокрылой чайки.
Подвидовая систематика. Монотипический вид.
146
Тихоокеанская чайка
Замечания по систематике. Взгляды различных исследователей на систематическое
положение и таксономический ранг этой чайки до сих пор окончательно не установи-
лись. А. Бент (Bent, 1921) и Дж.Дуайт (Dwight, 1925) считали ее самостоятельным
видом, близким к серебристой чайке. Б.К.Штегман (Stegmann, 19346) придавал ей
статус подвида морской чайки, Л.А.Портенко (1963, 1973) — восточного подвида
серебристой чайки. Более поздние исследования возрастной и сезонной изменчивости
оперения (Фирсова, 1975а, 1986), выполненные на новом серийном материале, подтвер-
дили высказанное ранее К.А.Юдиным предположение о принадлежности тихоокеанской
чайки к особому комплексу крупных белоголовых чаек, обитающих в тихоокеанском
бассейне. Ни серебристая, ни морская чайки в этот комплекс не входят.
Описание. Размеры (мм):
Длина крыла:
33(и = 46) - 408-467 (среднее 438);
9$ (и = 31) - 391-455 (среднее 417).
Длина клюва:
33 (п = 46) - 53,7—64,1 (среднее 62,0);
99 (и = 31) — 48,0—59,4 (среднее 53,8).
Длина цевки:
33 (и = 46) - 64,6-86,0 (среднее 73,9);
99 (и = 31) - 59,8-78,9 (среднее 67,7).
Окраска (Фирсова, 1975а). Самец и самка в брачном наряде. Голова, шея,
зашеек, верхние кроющие хвоста и рулевые, а также весь низ и бока туловища белые.
Спина, плечевые и верхние кроющие крыла темно-аспидные. Крупные плечевые с широ-
кой белой каймой на концах, все первостепенные маховые с белыми вершинами.
II—III первостепенные черные с аспидно-серым клиновидным полем на внутреннем
опахале; на II белая предвершинная перевязь до 4 см шириной, на III обычно разви-
вается белое округлое предвершинное пятно. Далее по направлению к проксимальному
концу кисти область аспидно-серой окраски расширяется, оттесняя черный цвет к вер-
шине пера. На VI—VII от него остается лишь узкая предвершинная перевязь, которая
на V—VII отграничена от аспидно-серой области белым пятном. VIII—XI аспидные
с белыми вершинами. Второстепенные маховые аспидно-серые с широкими белыми кай-
мами на вершинах. Перья крылышка того же цвета с белыми основаниями и наружны-
ми опахалами. Клюв желтый с красным пятном в вершинной части подклювья, ноги
телесно-розовые.
Самец и самка в зимнем наряде. Голова, шея, зашеек, а иногда и верхняя часть груди
с коричневато-бурым продольным рисунком. Радужина золотисто-желтая, края век ро-
зовые, Прочее оперение, как в предыдущем наряде.
Пуховой наряд. Окраска верха пятнистая; на серовато-буром фоне черные пятна,
более мелкие и резко очерченные на голове. Передняя часть шеи и' грудь также пят-
нистые, брюшко чисто белое. На предплечье, пояснице и брюшке развит охристый налет.
Клюв светлый на вершине и темный на две трети от основания, ноги серовато-телесные,
пальцы и коготки чуть более темные, чем перепонки.
Гнездовой наряд. Верхняя сторона тела темно-бурая, на голове мелкие светлые
пестрины, на шее они более крупные и редкие. Зашеек темно-бурый однотонный с про-
глядывающими кое-где беловатыми основаниями перьев. На перьях межлопаточной
области, плечевых и большинстве верхних кроющих предплечья хорошо развиты
147
Семейство Чайковые
грязновато-белые краевые каемки, которые создают чешуйчатый рисунок этих участков
оперения; на спине эти каемки развиты несколько хуже. Задняя часть спины и верхние
кроющие хвоста более пестрые, чем передняя часть спины, так как здесь на перьях
развиты крупные бурые предвершинные и белые дистально-латеральные пятна. Нижняя
сторона тела темно-бурая, но заметно светлее, чем спина. Подбородок и верхняя часть
шеи беловатые, с мелкими темными наствольными пестринами. Шея и верхняя часть
груди с мелким беловатым поперечным рисунком, который образован очень узкими
неясными краевыми каемками на перьях. На брюшке эта поперечная испещренность
выражена слабее. Подхвостье более пестрое. II—VI первостепенные маховые однотон-
ные, темно-бурые, чуть более темные, чем основной тон окраски верхней стороны тела,
и светлеющие по направлению к проксимальному концу кисти. На VII— XI развиты
узкие краевые каемки, расширяющиеся в этом же направлении. Второстепенные махо-
вые бурые с более светлыми внутренними опахалами и ясной буровато-охристой каем-
кой на концах и по краю наружных опахал. Рулевые окрашены как дистальные перво-
степенные, с узкими беловатыми краевыми каемками; на наружных опахалах крайней
пары развиты узкие беловатые пятна, почти сливающиеся в каемки. В основании всех
рулевых также заметны беловатые пестрины, особенно развитые на крайних парах.
Клюв однотонно-черный с более светлой вершинной частью.
Первый зимний наряд. От предыдущего отличается более светлым оперением лба,
затылка, шеи и подбородка. Все оперение более выгоревшее и обношенное, отчего
краевые каемки на перьях верхней стороны тела выделяются не так отчетливо.
Первый летний наряд. От предыдущего и последующего нарядов отличается более
светлой окраской оперения. От первого зимнего наряда отличается кроме того тем, что
на некоторых свежих перьях межлопаточной области появляется шиферный налет.
Второй зимний наряд. От первого зимнего отличается более светлым коричневатым
тоном окраски свежих перьев и примесью на них шиферных тонов, а также меньшей
отчетливостью рисунка. Окраска первостепенных в описываемом наряде шиферно-бу-
рая, заметно более темная, чем в свежем гнездовом пере.
Второй летний наряд. От предыдущего отличается наличием шиферно-серого
треугольника в верхней части спины и выгоревшими и обношенными маховыми и
рулевыми.
Третий зимний наряд. От второго зимнего отличается наличием белого предвершин-
ного пятна на II и более серой окраской остальных первостепенных маховых, а также
преимущественно шиферно-серой окраской верхних кроющих крыла.
Третий летний наряд. От предыдущего отличается чисто-белой головой и сильно обно-
шенными маховыми и рулевыми.
Четвертый зимний наряд. От третьего зимнего отличается отсутствием бурых тонов
на проксимальных первостепенных и на верхних кроющих предплечья, от окончатель-
ного зимнего — остатками бурых пятен на I (редуцированном) первостепенном, неко-
торых верхних кроющих кисти и перьях крылышка.
Четвертый летний наряд. От предыдущего отличается чисто-белой головой и сильно
выгоревшими и обношенными маховыми и рулевыми.
Линька. В первый зимний наряд частичная, захватывает немногочисленные перья на
голове, шее, зашейке и мантии, а у некоторых экземпляров, кроме того, ею затраги-
ваются единичные перья на надхвостье, груди и боках. Линька в первый летний наряд
также частичная, охватывает оперение головы, шеи, зашейка и межлопаточной области.
Сроки протекания не ясны, в июне наряд уже полностью сформирован. Линька во вто-
рой зимний наряд полная, начинается уже в июне, Сроки окончания этой линьки на на-
шем материале не ясны. Линька во второй летний наряд частичная, охватывает опере-
ние головы, шеи, зашейка, мантии и груди. Сроки ее протекания не ясны. Линька
148
Тихоокеанская чайка
в третий зимний наряд полная, начинается в конце мая: в это время выпадает XI перво-
степенное. Сроки окончания этой линьки не ясны. Линька в третий летний наряд частич-
ная, охватывает оперение головы, шеи, зашейка, кроющих крыльев, а также груди и
брюшка. Сроки протекания ее не ясны, по Дуайту (Dwight, 1925) она происходит пред-
положительно в апреле —мае. Линька в четвертый зимний наряд полная, протекает
в июне — ноябре. Линька в четвертый летний наряд частичная, охватывает в основном
оперение головы, шеи, зашейка, мелких плечевых и груди. Протекает она, судя по на-
Рис. 37. Область распространения тихоокеанской чайки (1-4) и серокрылой чайки в СССР (5-7)
1, 5 — область гнездования, 2 — отдельные поселения тихоокеанской чайки, 3 — места зимовок
тихоокеанской чайки, 4, 7 — залеты на территории СССР, 6 — случайные гнездования серокрылой
чайки
шим материалам, в сжатые сроки в апреле. Линька в пятый (окончательный) зимний
наряд полная, протекает в июне — октябре.
Распространение. Гнездовой ареал. Почти целиком расположен в пределах
СССР (рис. 37), он охватывает побережья Охотского моря, Камчатки (на север до бух-
ты Угольной), Курильских островов и о-ва Сахалин, простираясь на юг до Приморья
включительно (Allen, 1905; Гизенко, 1955; Нечаев, 1975; Яхонтов, 1975а,б; Кищин-
ский, 1980; Литвиненко, 1980; Харитонов, 1980а; Фирсова и др., 1981; ЗИН).
Вне СССР известны две колонии у северо-западных и юго-восточных берегов о-ва Хок-
кайдо (Fennell, 1953; Fujimaki, 1961; Watanuki, 1982), На этом же острове зарегистри-
рован единичный случай гнездования на мысе Сякотан (Mazda, Higuchi, 1976).
Зимовки. Зимний ареал охватывает южные районы размножения от незамерзаю-
щих побережий Берингова и Охотского морей, юго-западных и восточных побережий
149
Семейство Чайковые
Камчатки до островов Хоккайдо, Рюкд и Тайвань (Черский, 1915; Austin, Kuroda,
1953; Gore, Won, 1971; Шунтов, 1972), Во время кочевок эту чайку наблюдали и
добывали гораздо севернее и восточнее районов размножения: на Чукотке (Томкович,
Морозов, 1982), о-ва Врангеля (Стишов и др,, 1985), Алеутских островах и у побере-
жий Аляски (Nelson, 1887; Clark, 1910; Hersey, 1917; Swarth, 1934; Gabrielson, Lincoln
1959; Murie, 1959).
Миграции. Весной в северной части Тихого океана (к северу от 30° с.ш.) начало миг-
раций приходится на март. Так, в Восточно-Китайском море, у южных берегов Кореи и
Японии весенние перемещения птиц на север становятся заметными в первой половине
марта (Austin, Kuroda, 1953; Gore, Won, 1971). В Японском и Охотском морях начало
пролета также приходится на март, но основная масса мигрантов здесь проходит в апре-
ле и мае (Шунтов, 1972). На восточном побережье Камчатки в районе Кроноцкого запо-
ведника миграции обычно начинаются 9—И.IV и продолжаются до начала июня, при
этом неполовозрелые особи в возрасте от 1 до 3 лет летят позднее (с начала мая),
чем взрослые (Фирсова и др,, 1982). На Командорских островах пролетные чайки
в большом числе появляются в начале марта и обычны здесь до конца мая, одиночные
встречаются до середины июня (Иогансен, 1934). В восточной части Берингова моря
пролет начинается только в первых числах апреля (Шунтов, 1972). По наблюдениям
Шунтова (1972), в Беринговом море пролет идет преимущественно вдоль побережий,
но часть птиц вместе с другими чайками остается в открытых водах в течение всего
периода размножения. Через Кроноцкий заповедник мигранты летят поодиночке, пара-
ми или небольшими стайками по 4—8 особей на высоте до 250 м, иногда движутся
строго вдоль береговой кромки, используя восходящие потоки воздуха (Фирсова и
др., 1982).
Интенсивность пролета невелика; в отдельные дни в 1975 г. здесь регистрировали
от 266 до 499 птиц, за весь период весенней миграции в том же году было учтено около
22 тыс. особей.
Массовые осенние кочевки молодых в прибрежной зоне моря в районе Кроноцкого
заповедника и на севере Охотского моря начинаются в начале сентября (Allen, 1905;
Фирсова и др., 1982). С середины этого месяца кочевки приобретают характер массово-
го и направленного пролета, который в окрестностях заповедника продолжается до
середины или конца октября. Последние мигранты отмечались здесь в середине ноября.
Западное побережье Камчатки большинство птиц покидает во второй половине декабря,
когда появляется береговой припай льда, позже здесь встречаются лишь одиночные
особи. На северном охотском побережье у Гижиги пролет хорошо выражен в начале ок-
тября (Allen, 1905), к концу этого месяца здесь остаются лишь единичные птицы.
В Кроноцком заповеднике осенние миграции протекают более интенсивно, чем весен-
ние. Так, в 1975 г. за 1 ч у пос. Жупаново в отдельные дни массового пролета регистри-
ровали от 300 до 1200 особей (Фирсова и др., 1982). Картина осенних миграций,
наблюдаемая в отдельных пунктах побережья, хорошо согласуется с данными, получен-
ными во время морских исследований. Перемещения чаек в Беринговом, Охотском
и Японском морях становятся заметными в сентябре, в октябре они приобретают
хорошо выраженное южное направление. В Японском море во второй половине октября
и в ноябре идет интенсивный пролет через Татарский пролив и пролив Лаперуза.
В это же время происходит явное увеличение численности чаек в северной части Тихого
океана. В Восточно-Китайское море чайки прибывают в конце октября, здесь мигранты
придерживаются в основном более холодной западной части моря. В Беринговом море
и у восточных побережий Камчатки тихоокеанская чайка в отличие от серебристой чай-
ки и бургомистра придерживается западной половины моря и почти не выходит за пре-
делы шельфа, в Охотском и Японском морях птицы широко рассеиваются по всей аква-
150
Тихоокеанская чайка
тории, хотя основные потоки мигрантов проходят также вдоль побережий (Шун-
тов, 1972).
Численность. Лучше всего изучена на территории Камчатской обл, (Фирсова и др.,
1982). В этом регионе известно более 200 поселений общей численностью не менее
47 тыс. пар. Численность отдельных колоний колеблется здесь от 10 до 1500 пар.
Преобладают мелкие (до 100 пар) поселения, средних (100—500 пар) — почти вдвое
меньше, а крупных (500—1000 пар) всего три. Все они обнаружены у восточных побе-
режий Камчатки (мыс Шипунский, мыс Столбовой, устье р. Жупанова), Самая круп-
ная колония (1500 пар) находится на острове Птичьем у северо-восточных берегов
острова Карагинского. На охотском побережье, Курильских островах и Сахалине,
а также на территории Приморья крупных поселений, судя по литературе, нет (Гизен-
ко, 1955; Лабзюк и др., 1971; Нечаев, 1975; Яхонтов, 1977). На о-ве Хоккайдо най-
дены небольшие колонии численностью в 250 и 144 пары (Watanuki, 1982).
Об общих тенденциях изменения численности судить трудно из-за отсутствия мате-
риалов прошлых лет. На острове Монерон (Нечаев, 1975) за последние 25 лет числен-
ность несколько уменьшилась.
Местообитания. Летом предпочитает скалистые островки в устьях рек и на лиманах
или кекуры на морском побережье. В глубь материка далеко не заходит. Зимой при-
держивается в основном прибрежных шельфовых акваторий.
Размножение. Как и другие крупные чайки, половозрелости достигают, судя по опе-
рению, в возрасте 4—5 лет. Гнездовые колонии на северных Курильских островах
начинают посещать в середине мая (Подковыркин, 1956). На севере Охотского моря
концентрируются в колониях к l.VI (Allen, 1905). Демонстрации взрослых птиц,
по наблюдениям в колониях на севере Камчатской обл, неотличимы от демонстраций
серокрылой чайки, лишь голос у тихоокеанской чайки несколько выше.
Гнезда помещаются на береговых утесах (на высоте от 15 до 200 м),на закрепленных
травянистой растительностью склонах или на вершинах кекуров, а также на плоских
островках в устьях рек. На кекурах гнезда располагаются в нишах, на карнизах и в
углублениях среди камней. Нередко чайки устраивают их и среди нагромождения валу-
нов прямо в литоральной зоне, однако избегают горизонтальных участков на примор-
ской террасе. Вне зоны скалистых побережий птицы занимают также берега и острова
тундровых озер.
В Кроноцком заповеднике тихоокеанские чайки приступают к гнездостроению
в начале мая — через несколько дней после появления в колонии; большинство птиц
заканчивает строительство гнезд к концу этого месяца или в начале июня (Фирсова и
др., 1982). На плоских вершинах скал или кекуров гнезда располагаются довольно
плотно, в среднем одно гнездо на 0,5 м2, на крутых склонах — более разреженно, в сред-
нем одно гнездо на 70 м2. В плоскостных и тундровых поселениях плотность
расположения гнезд колеблется от 0,3 до 100 гнезд на 1 га. Гнезда на сырых участках
более массивные (диаметр у основания до 1200 мм, высота до 310 мм), чем на сухих
островках, где диаметр гнезд колеблется от 380 до 1100, составляя в среднем (и = 92)
526,4 + 9,5; диаметр лотка равен 200—320, в среднем 262,0 ±3,0; глубина лотка
50-100, в среднем 65,0 + 1,0; высота гнезда от земли 50-200, в среднем 82,0 + 4 мм.
В качестве строительного материала на сухих островках используются листья и стебли
злаков, веточки шикши, мох. На сырых участках тундры основу гнезда часто состав-
ляет мох, в небольших количествах добавляются сухие злаки и веточки кустарничков.
Спаривание в истоках реки Кроноцкой и в бухте Гека на Камчатке наблюдалось
с 24. V по начало июля (Фирсова и др., 1982). Откладка яиц на восточном побережье
Камчатки между Семячикским лиманом и р. Шумной начинается обычно в последних
числах мая — начале июня, в массе это происходит в конце первой — начале второй де-
151
Семейство Чайковые
кады июня. В некоторые годы откладка яиц сильно задерживается. На севере охотского
побережья в районе Гижиги пик откладки яиц отмечен 10.VI (Allen, 1905).
Из 128 кладок, находившихся под наблюдением в бухте Гека, 1,6% содержали по од-
ному, 18,8%-по два и79,7%-по три яйца; в среднем в каждой кладке было 2,8 яйца. .
Размеры (мм) яиц из этой колонии (п = 270) 69,0—82,0 X 47,8—56,7, в среднем
74,6 X 51,5. На Курильских островах средняя величина кладки была 2,1 яйца, а разме-
ры (мм) составили (и = 13) 68,5—80,1 X 49,1—55,2 в среднем 71,7 X 52,2 (Подковыр-
кин, 1956). Изучение 27 яйц, собранных на Камчатке, показано, что наиболее типичная
окраска основного фона яиц изменяется от бежевой до желтовато-серой (по шкале
цветов Бондарцева, 1964), реже встречаются яйца с коричневатым или зеленоватым
фоном. Окраска поверхностных пигментных пятен варьирует от темно-умбровой до
темно-песочной, внутренние пятна обьино более тусклые. Пигментные пятна образуют
пятнистый рисунок, иногда с элементами точечного и линейного рисунков. Распределе-
ние элементов рисунка по поверхности яйца изменяется от очень редкого до среднегус-
того, при этом на тупом конце яйца густота рисунка обьино на 30—40% больше, чем
на остром (Фирсова и др., 1982).
Насиживают кладку, как и у других чаек, оба родителя. Длительность инкубации
(от появления первого яйца до вылупления последнего птенца) по наблюдениям
на Камчатке была 28—30 дней, в среднем (и = 56) 29 дней (Фирсова и др., 1982),
на Курильских островах — 31—33 дня (Гизенко, 1955). Массовое вылупление птенцов
в бухте Гека в 1977 г. протекало с 25 июня по 7 июля. На о-ве Ионы в западной части
Охотского моря оно начинается, по С.П.Харитонову (1980а), 20 июня.
Птенцов выкармливают оба партнера в течение 40—45 дней. Первые летные молодые
в бухте Гека в 1977 г. отмечены 5 августа, через 5 дней большинство птенцов уже лета-
ло, но пределы колонии в это время покидали лишь немногие из них и на короткое
время. В Кроноцком заповеднике лётные молодые появляются в середине августа,
но часть особей не летает еще в первой декаде сентября. В конце августа — начале сен-
тября молодежь в массе покидает колонии (Фирсова и др., 1982).
Суточная активность, поведение. Тихоокеанские чайки ведут преимущественно днев-
ной образ жизни. Защитное поведение у птенцов и взрослых, как у большинства других
чаек. Птенцы при опасности прячутся в укрытиях или спасаются бегством на воду.
Взрослые особи вылетают навстречу нарушителю границ колонии, окрикивают его или
активно атакуют, пикируя и задевая при этом ногами.
Питание. Во все сезоны основу рациона составляет рыба и морские беспозвоночные
(моллюски, ракообразные, морские ежи и звезды). Во время массового нереста лосо-
севых поднимаются вверх по рекам (изредка до 30 км, обычно не более 10), где
поедают отнерестившуюся рыбу, или сопровождают вылавливающих рыбу медведей и
подбирают остатки их трапез (Кищинский, 1968; Велижанин, 1972; данные Л.В.Фир-
совой) . В течение всего года, и особенно зимой, хорошим подспорьем в питании служат
отходы рыболовного и зверобойного промыслов. В конце лета птицы охотно кормятся
на приморских ягодниках (Фирсова и др., 1982). В колониях не упускают случая пожи-
виться чужими яйцами или птенцами. У побережья о-ва Хоккайдо птенцы чернохвостой
чайки составляют 4,1% (по весу) рациона (Watanuki, 1982).
Враги, неблагоприятные факторы. Как и другие гнездящиеся на земле чайки, тихо-
океанские чайки страдают от наземных хищников. На севере Камчатки островные коло-
нии в устьях рек подвергаются разорению медведями, домашними оленями. Однакс
основные причины гибели молодняка — неблагоприятные кормовые или погодные
условия.
Хозяйственное значение, охрана. Хозяйственного значения вид не имеет, специальные
меры охраны не предусмотрены.
152
Серокрылая чайка
Серокрылая чайка —
Larus glaucescens Naumann, 1840
Glaucous-winged Gull (англ.), Beringsmowe (нем.), Goeland £ ailes glauques (фр.).
Статус. Гнездящийся мигрирующий, частично зимующий вид.
Общая характеристика и полевые признаки. Крупная чайка. От бургомистра отли-
чается более мелкими размерами и темными стержнями первостепенных маховых.
От других чаек, обитающих в тех же районах, — отсутствием черного рисунка на этих
перьях. Гнездится обычно в непосредственной близости от моря. Экология, демонстра-
тивное поведение, как у других крупных чаек. Голос немного ниже, чем у тихо-
океанской чайки.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Дж.Дуайт (Dwight, 1925) сближал серокрылую чайку
с серебристой и придавал ей статус вида. Б.К. Штегман (Stegmann, 1934b) считал ее
подвидом бургомистра. Позднее К.А.Юдин на основании изучения возрастной изменчи-
вости оперения пришел к выводу о принадлежности серокрылой чайки к комплексу
морских тихоокеанских чаек (L.schistisagus, L. occidentalis, L.dominicanus), в кото-
рый не входят ни серебристая чайка, ни бургомистр. В дальнейшем эта идея получила
подтверждение (Фирсова, 1975а) на новом более полном материале с Командорских
островов. Связи серокрылой чайки с западной чайкой L.occidentalis, а через нее
с доминиканской чайкой L. dominicanus подтверждаются работами американских иссле-
дователей (Hoffman et al., 1978; Hand, 1981), изучавших вокализацию этих видов,
а также гибридизацией серокрылой и западной чаек на побережье США.
Описание. Размеры (мм):
Длина крыла:
dd (п = 7) - 415-435 (среднее 429);
29 (л =4) - 405-428 (среднее 415);
dd и 99 (п = 7) — 410—443 (среднее 430) .
Длина клюва:
dd (и = 7 ) — 55,0—64,0 (среднее 61,0) ;
99 (п = 4) — 48,0—57,0 (среднее 52,7);
dd и 99 (л = 7) — 52,0—66,0 (среднее 59,1).
Длина цевки:
dd ( п = 7 ) — 67,0—73,0 (среднее 69,8) ;
99 (л =4) — 61,0—74,0 (среднее 66,7);
dd и 99 (л = 7) — 69,0—76,0 (среднее 73,1).
Окраска. Самец и самка в брачном наряде. Голова, шея, зашеек, весь низ и бока
туловища, а также все рулевые и кроющие хвоста белые. Все маховые и крупные плече-
вые имеют более или менее развитые белые вершины. Спина, плечевые, верхние крою-
щие крыла и крылышко грифельно-серые. II (первое видимое) первостепенное
аспидно-серое, заметно темней спины, с белым предвершинным пятном и грифельно-се-
рым полем на внутреннем опахале. На следующих пяти перьях площадь, занимаемая
аспидно-серой окраской, постепенно уменьшается, а грифельно-серой увеличивается.
На VI и VII первая имеет вид узких предвершинных полосок, примыкающих к белой
вершине. Предвершинное белое пятно так же прогрессивно сокращается по направлению
153
Семейство Чайковые
к проксимальному концу кисти. На VI и VII эти пятна превращаются в узкие полоски,
подчеркивающие снизу аспидно-серые предвершинные пятна. VIII, IX и все второсте-
пенные маховые грифельно-серые с белыми вершинами. Рисунок на крыле очень похож
на таковой у тихоокеанской чайки, но черный цвет заменен аспидно-серым, а светлые
части рисунка у тихоокеанской чайки становятся бледно-грифельно-серыми у серокры-
лой. Крылышко грифельно-серое, бледнеющее по краям наружных опахал. Клюв жел-
тый с красным пятном на подклювье, ноги розовато-телесные.
Самец и самка в зимнем наряде. Вся голова, шея, зашеек, а иногда и верхняя часть
груди в густом буром крапе. Прочее оперение, как в предыдущем наряде. Клюв бурова-
то-желтый, с темной перевязью в области красного пятна на подклювье.
Пуховой наряд. Верхняя сторона тела серовато-бурая с более темными черноватыми
пятнами, которые на голове мельче и отчетливее, чем на спине. Бока головы и шеи
с едва заметным охристым налетом. Грудь и верхняя часть брюшка почти однотонные
грязно-белые. Подбородок, шея, большая часть брюшка и бока туловища серовато-бу-
рые, но более светлые, чем окраска на верхней стороне тела. На подбородке крупные
удлиненные черноватые пятна, на брюшке и по бокам туловища они мельче и более
отчетливые. По бокам шеи, на груди и в нижней части брюшка охристый налет.
Ноги темно-серые, блестящие, с чуть более светлыми перепонками. Клюв черный на
две трети от основания, вершинная его часть бежевая с чуть более темным кончиком.
Гнездовой наряд. Верхняя сторона тела буровато-серая. На голове, шее и зашейке
развиты беловатые пестрины, более мелкие, частые и резко очерченные на голове.
Межлопаточные, плечевые и верхние кроющие предплечья также буровато-серые, но с
чешуйчатым беловатым рисунком, который образован краевыми каемками на перьях.
Нижняя сторона тела более однотонная буровато-серая. Светлые пестрины на ней осо-
бенно отчетливо выражены на подбородке; на шее и верхней части груди число их
уменьшается. Нижняя часть груди и брюшко почти однотонные серовато-бурые;
неясные пестрины появляются на самой нижней части брюшка, по направлению
к хвосту отчетливость и число их увеличиваются. Нижние кроющие хвоста с резко
очерченными поперечными беловатыми пестринами в виде пятен и узких полос.
Рисунок здесь дополняется широкими бурыми полосками в предвершинной части
перьев. Первостепенные маховые однотонные буровато-серые, постепенно светлеющие
к основаниям перьев, особенно на внутренних опахалах. Кроме того, на них развиты
очень узкие светлые каемки, хорошо заметные с IV первостепенного и далее к прокси-
мальному концу кисти. Верхние кроющие кисти без пестрин, более серые, чем кроющие
предплечья. Рулевые также буровато-серые с мелким беловатым крапом, особенно
развитым на наружных опахалах и увеличивающимся от центральных к наружным па-
рам перьев. Клюв и ноги однотонно черные.
Первый зимний наряд. Очень похож на гнездовой, но голова и шея становятся более
светлыми за счет сокращения бурых пятен на свежих перьях. На остальных участках,
где гнездовое перо не меняется, оно сильно выгорает и обнашивается.
Первый летний наряд. По Дж.Дуайту (Dwight, 1925), мало отличается от предыду-
щего. Свежие перья на голове, шее, груди и спине, хотя и светлее, чем в предыдущем
наряде, но кажутся более темными на фоне выгоревшего оперения предыдущих ге-
нераций. Голова и шея почти белые, на спине иногда некоторое число серых перьев.
Второй зимний наряд. От всех предыдущих отличается менее заостренной формой
дистальных концов рулевых и маховых, а также более однотонной серовато-бежевой
окраской всего оперения. Кое-где отчетливо заметна примесь серых тонов. Клюв чер-
ный с небольшими желтыми участками у основания и вершины.
Второй летний наряд. От предыдущего хорошо отличается почти чистой (без примеси
бурых тонов) грифельно-серой межлопаточной областью, спиной и плечевыми, образую-
154
Серокрылая чайка
щими отчетливый треугольник на верхней стороне туловиша. Клюв буровато-желтый
в основании и буровато-черный в вершинной части.
Третий зимний наряд. От предыдущего отличается большим развитием бурых тонов
на голове, шее и зашейке. В окраске маховых и рулевых серые тона преобладают над
бурыми, на маховых развиваются беловатые вершинные пятна.
Третий летний наряд. От второго летнего отличается примесью серых тонов в окраске
маховых и рулевых и развитием белых вершинных пятен по крайней мере на VI—VII
первостепенных. Кроме того, на кроюших предплечья в описываемом наряде в большей
или меньшей степени развито оперение грифельно-серого цвета, чего нет во втором лет-
нем наряде.
Четвертый зимний наряд. От третьего зимнего отличается полным отсутствием
бурых тонов на кроющих предплечья и почти полным исчезновением их на кроющих
кисти и крылышке. Брюшко у птиц в описываемом наряде чисто-белое в отличие от
третьего зимнего. От окончательного зимнего отличается бурым крапом, развитым на
некоторых рулевых.
Четвертый летний наряд. От предыдущего отличается чисто белыми головой, шеей,
зашейком и нижней стороной туловища, от окончательного летнего — остатками бурого
рисунка на рулевых, а иногда и на I первостепенном.
Линька. Линька в первый зимний наряд неполная, охватывает сравнительно ограни-
ченные участки тела: голову, шею; перья груди и боковых ветвей брюшной птерилии
сменяются только отчасти и, видимо, не у всех особей. Число сменяющихся перьев
бывает обычно незначительным. Начинается эта линька сразу после окончания фор-
мирования гнездового наряда (или чуть раньше), в августе—сентябре, и заканчивается
к декабрю. Линька в первый летний наряд, по Дж. Дуайту (Dwight, 1925), неполная,
протекает в марте—апреле. Линька во второй зимний наряд полная, протекает с конца
мая — начала июня до ноября. Линька во второй летний наряд неполная, хотя и охва-
тывает многие участки оперения. Кроме оперения головы, шеи, зашейка и верхней сто-
роны туловища, сменяется практически все оперение межлопаточной области, спины,
а также плечевые. Сроки не известны.
Линька в третий зимний наряд полная, начинается в мае со смены XI и X первосте-
пенных, в августе идет смена рулевых, дистальных первостепенных, мелкого оперения
туловиша. Конец линьки на нашем материале не прослеживается. Линька в третий
летний наряд неполная, охватывает оперение головы, шеи, зашейка и некоторых других
участков. Сроки не известны. Линька в четвертый зимний наряд полная, начинается во
второй половине июня со смены XI и X первостепенных, оперения межлопаточной
области, брюшка и боков туловиша и заканчивается в конце декабря. Линька в четвер-
тый летний наряд неполная, протекает в марте, охватывает оперение передней половины
туловиша от головы до мелких плечевых и груди, а также оперение надхвостья.
Линька в пятый зимний наряд полная, в середине июля она идет полным ходом,
сменяются V, 20—21-е второстепенные, средние кроющие предплечья, средние и малые
кроющие хвоста, а также мелкое оперение плечевых, груди и нижней стороны шеи.
К началу декабря она уже закончена.
Линька в пятый летний наряд неполная, протекает в феврале—мае, охватывает опере-
ние головы, шеи, зашейка, межлопаточной области и груди.
Как показывают изложенные материалы, сроки и продолжительность линьки перво-
степенных маховых в значительной степени колеблются в зависимости от возраста
птиц, при этом длительность предвзрослых полных линек может быть как больше, так
и меньше, чем у взрослых особей (Фирсова, 19786).
Распространение. Гнездовойареал. В СССР в массе гнездится только на Коман-
дорских островах (см. рис. 37). На юге Корякского нагорья Л.В. Фирсовой недавно
155
Семейство Чайковые
Рис. 38. Область распространения серокрылой чайки
/ — область гнездования, 2 — область массовых зимо-
вок, 3 — область зимовок небольшого числа птиц
установлено размножение одной особи серокрылой чайки в паре с обычной здесь тихо-
океанской чайкой (Кищинский, 1980). У самки, добытой 28. VI 1978 г. на восточном
побережье Чукотки, обнаружены интенсивно зарастающие наседные пятна (Томкович.
Морозов, 1982). Авторы предполагают гнездование этой особи где-то поблизости.
За пределами Советского Союза обитает на Алеутских островах и островах Прибылова
(Murie, 1959), а также на тихоокеанском побережье и прилежащих островах Север-
ной Америки - от мыса Пирс в
Бристольском заливе на Аляске
(Strang, 1977) до островов Кол-
вилл и Дестракшн в штате Вашинг-
тон (Dawson, 1908; Gillet et al.,
1975) (рис. 38). Молодые и холос-
тые взрослые птицы в гнездовое
время залетают гораздо севернее
районов размножения в Беринго-
вом море (ЗИН АН СССР; Кищин-
ский, 1980).
Зимовки. Часть птиц зи-
мует в гнездовой области. Осо-
бенно многочисленны чайки в
незамерзающей юго-восточной час-
ти Берингова моря, вдоль всех
Алеутских и Командорских остро-
вов и у побережий Аляски (Шун-
тов, 1972). Кроме того, зимой
чайки широко кочуют по всей акватории северной части Тихого океана на юг до о-ва
Хоккайдо (Austin, Kuroda, 1953) и штата Калифорния в США (Cooke, 1915), а также
у северных берегов Мексики, в Нижней Калифорнии (Blake, 1953). Одиночки достигают
Гавайских островов. При этом взрослые птицы предпочитают более северные районы
зимней акватории, молодые — более южные. Последние, кроме того, рассеиваются го-
раздо шире (Шунтов, 1972; Harrington, 1975; Butler et al., 1980). Во внегнездовое
время чаек отмечали гораздо севернее упомянутых районов размножения и зимовок:
на Чукотке у поселков Уэлен и Лаврентия (Томкович, Морозов, 1982), в Анадырском
заливе и заливе Креста (Белопольский, 1934), в бухте Провидения (Портенко, 1973)
и на о-ве Св. Лаврентия (Fay, Cade, 1959). Единственной достоверной находкой на се-
верной Аляске, по мнению Бейли (Bailey, 1948), остается обнаружение костей этого
вида при археологических раскопках на мысу Принца Уэльского (Fridmann, 1934).
Миграции. Весеннее движение птиц к местам размножения начинается в марте, мас-
совые миграции наблюдаются в апреле. Во второй половине этого месяца, по наблюде-
ниям В.П. Шунтова (1972), в глубоководных районах океана эти чайки почти не встре-
чаются, основной пролет идет в мелководной юго-восточной части Берингова моря.
У восточного побережья Камчатки в районе пос. Жупаново массовые миграции отмеча-
лись до последней декады апреля, самых поздних птиц видели здесь в разные годы
с 15 по 24.V (Лобков, 1980). У восточного побережья Чукотки единичные особи обыч-
ны до середины июля (Томкович, Морозов, 1982) .
Осенью на Командорских островах, как показало кольцевание (Фирсова, 19786).
некоторые сеголетки держатся, по крайней мере, до декабря. На восточном побережье
Камчатки (Лобков, 1980) осенний пролет наблюдается с середине сентября до середины
ноября. По данным кольцевания на юге Канады (Vermeer, 1963), сеголетки покидают
156
Серокрылая чайка
окрестности колоний во второй половине сентября. Эти факты хорошо согласуются с
наблюдениями В.П. Шунтова (1972) в открытых водах Берингова моря, где первые
единичные мигранты появляются в августе, в сентябре здесь преобладают (до 75%)
именно молодые серые особи, а в следующие два месяца доля взрослых увеличивается
до 40%.
Численность. На Командорских островах общая численность гнездящихся птиц оцени-
вается приблизительно в 1000 пар, из них около 500 пар размножается на двух неболь-
ших островках у северной части о-ва Беринга (Фирсова, 19786, 1983). Пять самых
крупных колоний (от 2 до 6 тыс. пар) находятся у побережий Аляски и восточной
части Алеутской гряды (Gabrielson, Lincoln, 1959; Murie, 1959; Hunt et al., 19816).
Общая численность популяции на восточном побережье Берингова моря оценивается
в 42 тыс. пар (Sowls et al., 1978). В СССР численность в последние десятилетия ос-
тается приблизительно на одном уровне, в заливе Аляска у побережий одноименного
полуострова она, по мнению американских исследователей, увеличивается (Baird
et al., 1979).
Местообитания. Летом излюбленные биотопы — небольшие' скалистые островки и
кекуры в прибрежной зоне или береговые утесы крупных островов и материка (Daw-
son, 1908; Jones, 1908; Gabrielson, Lincoln, 1959; Vermeer, 1963; Фирсова, 19786,
1983). Чайки предпочитают селиться в непосредственной близости от моря, лишь иногда
гнездятся на островках внутренних озер на Аляске и в штате Вашингтон (США) на
расстоянии до 25—30 км от морского побережья (Osgood, 1904; Walker, 1920; Wil-
liams, 1947; Wahl, 1972). В последнее время успешно используют антропогенные биото-
пы, поселяясь в США на крышах домов на расстоянии до 1 км от берега (Cramp, 1971).
Зимой вместе с другими чайками держатся в основном в прибрежных шельфовых участ-
ках моря в пределах 100—150 миль от берега,’ в открытом океане встречаются реже
(Шунтов, 1972; Sanger, 1973). Часто концентрируются в районах промысла, сопро-
вождают торговые и пассажирские суда, подбирая отходы, выброшенные за борт.
На берегу также скапливаются на участках, богатых кормом, в окрестностях круп-
ных портовых городов, на свалках, у рыбообрабатывающих предприятий.
Размножение. К местам гнездования на Командорских островах птицы прилетают
в начале марта, в апреле занимают гнездовые участки (Иогансен, 1934). В Бри-
танской Колумбии (западное побережье Канады) первые чайки прилетают в райо-
ны колоний в первой половине февраля, и только во второй половине марта они
занимают гнездовые участки (Vermeer, 1963). Наблюдения этого исследователя за
индивидуально помеченными особями показали, что большинство птиц поселяется
ежегодно на одних и тех же гнездовых участках, многие пары сохраняются с прошлого
сезона, а образование новых пар происходит в колонии.
Именно в это время наблюдается ярко выраженное демонстративное поведение.
Относительно подробно оно изучено у нас в стране (Фирсова, 19786, 1983) и у птиц
из американской части ареала (Vermeer, 1963; Moyle, 1966; Stout, Brass, 1969; Stout,
Wilcox, Creitz, 1969). Установлено, что набор поз и демонстраций у серокрылой чайки
тот же, что и у других крупных белоголовых чаек (рис. 39). В систему демонстраций
агрессивного комплекса входят долгий крик, ’’квохтание”, мяукающий крик, дергание
травы, прямая поза. В конфликтных ситуациях возможны самые разнообразные после-
довательности выполнения этих демонстраций, они определяются степенью возбуждения
взаимодействующих птиц. Демонстрации агрессивного комплекса можно наблюдать
не только на гнездовых территориях, но и на местах кормления и отдыха. Комплекс
брачного поведения включает церемонию встречи партнеров, состоящую из последова-
тельно выполняемых демонстраций долгого и мяукающего криков, отворачивания
157
Семейство Чайковые
головы и собственно предкопуляционного поведения, которое, в свою очередь, вклю-
чает дружественное, или парное ’’квохтание”, вскидывание головы, выпрашивание
корма, взаимное вскидывание головы. Последовательность демонстраций в брачном
комплексе поведения строго фиксирована, и этим брачное поведение отличается от
агрессивного. Кроме того, у серокрылой чайки известны звуки и позы, непосредст-
венно не связанные с агрессивным или брачным демонстративным поведением, а отра-
жающие скорее эмоциональное состояние птицы. К ним относятся крики тревоги и лег-
Рис. 39. Демонстрационное поведение серокрылон чайки (по: Фирсова, 1983, с изменениями)
а — долгий крик, б — прямая поза, в — дерганье травы, г — ’’квохтание”, д — мяукающий крик,
е — вскидывание клюва
кого беспокойства, призывный крик (Note call), атакующий крик (Charge call), поза
настороженности, поза разглядывания ног.
Гнезда помешаются на крутых каменистых обрывах и сглаженных участках, лишен-
ных травянистой растительности, задерненных склонах и плоских вершинах островов
(Gabrielson, Lincoln, 1959; Murie, 1959; Фирсова, 19786, 1983). На юге ареала, где
острова зарастают древесно-кустарниковой растительностью, гнезда могут располагать-
ся и под кустами (Jones, 1908; Williams, 1947). На о-ве Богослов (Алеутская гряда)
гнезда устраивались на плоском песчаном участке в кратере потухшего вулкана
(Bent, 1921; Gabrielson, Lincoln, 1959). На юго-западном побережье США в г. Сиэтл
отмечается гнездование на крышах домов (Cramp, 1971).
Гнездостроение на Командорских островах заканчивается в первой — второй де-
кадах июня (Фирсова, 19786, 1983). В строительстве гнезда принимают участие оба
158
Серокрылая чайка
члена пары, обычно самец приносит строительный материал (который собирается,
как правило, поблизости от гнезда), а самка его укладывает. Поэтому в гнездах, рас-
полагающихся среди травы, основу строительного материала составляют сухие листья
и стебли колосняка вместе с небольшими добавками зеленых частей растений и мха,
а в гнездах, устроенных среди голых камней недалеко от воды, — водоросли. Размеры
гнезд зависят от их расположения. Наиболее массивные постройки находили в прибреж-
ных камнях недалеко от воды, средняя толщина стенок здесь была 147±6 мм, тогда
как у гнезд, расположенных вдали от берега среди травы, — лишь 110 + 11 мм (Фир-
сова, 19786, 1983). Размеры (мм) 42 гнезд на Командорских островах следующие:
диаметр гнезда 300—600, в среднем 488; диаметр лотка 150—230: в среднем 202;
глубина лотка 65—159, в среднем 88.
Сроки откладки яиц на Командорских островах в 1969—1971 гг. были растянуты с
конца мая почти до конца июня, что объясняется, очевидно, сбором яиц жителями
близлежащего поселка. Интенсивная откладка яиц в последние два года происходила
с 3 по 22.V. По всему ареалу кладки состоят обычно из 2—3 яиц, реже 1—4
(Grinnell, 1898: Vermeer, 1963; Фирсова, 19786, 1983). Средняя величина кладки
на Командорских островах, по данным последнего автора, была 2,5, в Канаде
(Vermeer, 1963) — 2,82, в США на юге ареала (штат Вашингтон) (Gillet, Hayward,
1975) — 2,33, на севере ареала в заливе Аляска (Nysewander, Barbour, 1979) — от
2,49 до 2,64 яйца на гнездо. В повторных кладках на Командорских островах (Фир-
сова, 19786, 1983) преобладали кладки из двух яиц, возрастало их число с одним
яйцом и резко падало — с тремя; около трети птиц не возобновляли кладки вообще.
Средние размеры (мм) яиц с Командорских островов (Фирсова, 19786, 1983) (и = 54)
и из американских музеев (Bent, 1921) (и = 47) равны 76,5 X 53,0 и 72,8 X 50,8 соот-
ветственно. Изучение размеров яиц из канадских колоний (Verbeek, Richardson, 1982)
показало, что они положительно коррелируют с размерами самок и имеют тенденцию
снижаться от первого к последнему яйцу в кладке. Наиболее типичная окраска (по
шкале Бондарцева, 1954) фона 29 собранных на Командорских островах яиц (Фирсо-
ва, 19786, 1983) — средняя между темно-песочной и бледно-бурой. Часть яиц уклоняет-
ся в сторону зеленоватых тонов, другая — в сторону коричневатых. Поверхностные
пигментные пятна чаще всего бледно-умбровые, образующие пятнистый рисунок, иног-
да дополняющийся элементами точечного рисунка. Густота рисунка варьирует от очень
редкого до средне густого, при этом на тупом конце она обычно на 30—40% больше,
чем на остром. На Алеутских островах изредка встречаются однотонные бледно-голу-
бые яйца (Murie, 1959).
Насиживают кладку, по наблюдениям на Командорских островах, поочередно самец
и самка в течение 26—31 дня, считая от появления первого яйца до вылупления послед-
него птенца. У большинства пар (70%, и = 70) длительность насиживания составила
28—29 дней. (Фирсова, 19786, 1983). В Канаде насиживание длится в среднем 27 дней,
считая с момента откладки второго яйца (Vermeer, 1963). Отход яиц на Командорских
островах в первых кладках составил 12,5% (и = 382), в Канаде (и =210) - 17%
(Vermeer, 1963). Основные причины гибели яиц — беспокойство птиц людьми, особенно
на ранних стадиях насиживания, и наличие неоплодотворенных яиц.
Вылупление птенцов на Командорских о-вах в 1970—1971 гг. протекало C25.VI
по 1 .VIII, но в большинстве гнезд пуховички появлялись в гораздо более сжатые сроки.
Так, в 1970 г. в 90% гнезд они вылупились с 5 по 9.VII, в 1971 г. в 77% гнезд — с
27.VI по 10.VII.B повторных кладках в 1971 г. вылупление птенцов протекало более
дружно: с 22 по 27.VII, а в большинстве гнезд с 22 по 25.VII (Фирсова, 19786, 1983).
В первые сутки после вылупления пуховичков родители их усиленно обогревают.
Замерзшие или голодные птенцы слабо попискивают, затаивание и оборонительная
159
Семейство Чайковые
реакция у них еше не выражены. К концу третьих суток жизни у всех птенцов эти
реакции хорошо развиты: при опасности они затаиваются прямо на гнезде, а при попыт-
ке взять их в руки клюются и издают особые дребезжащие звуки. Кормят их в этом
возрасте приблизительно один раз в час, пишу они берут только из клюва взрослых.
К концу первой недели жизни птенцы осваивают ближайшие окрестности гнезда, при
опасности прячутся среди травы или камней. Пищу уже могут брать и с земли, окон-
чательно вырабатывается реакция выпрашивания корма: пуховички не только клюют
красное пятно на подклювье родителей, но и преследуют их, кланяются, принимают
сгорбленные выпрашивающие позы и издают высокий свист. Кормят их теперь прибли-
зительно один раз в два часа, а обогревают в основном по утрам и вечерам или в холод-
ную погоду. К концу второй недели жизни птенцы не только хорошо знают свой гнез-
довой участок, о чем свидетельствует их умение быстро прятаться в одних и тех же
укрытиях, но и защищают его от чужих птенцов. К концу третьей недели жизни оборо-
нительная реакция у некоторых чайчат слабеет, а на 29—32 день — полностью исчезает.
При опасности они теперь прячутся, а затем бегут к линии прибоя, но в воду пока не
идут; в возрасте 40 дней в таких ситуациях обычно уходят в море.
В 1970 и 1973 гг. первые лётные молодые были замечены в конце июля — начале ав-
густа, в первой декаде сентября большинство птенцов уже летали (Фирсова, 19786,
1983). Молодые поднимаются на крыло обычно в возрасте 45 дней. До 60—70-дневного
возраста птенцы не могут добывать корм самостоятельно, они выпрашивают его у роди-
телей, непрестанно преследуя их на воде и в воздухе. В возрасте 70—80 дней молодые
большую часть времени вместе со взрослыми проводят вне колонии, возвращаясьтуда
только на ночь, а затем постепенно покидают ее совсем. Обычно это происходит в се-
редине сентября.
Успех размножения в 1971 и 1973 гг. на Командорских островах составил 0,5—0,6
птенца на пару размножавшихся птиц (Фирсова, 19786, 1983), что значительно ниже,
чем соответствующие показатели (1,0—1,7) на юге Канады (Vermeer, 1963), и несколь-
ко ниже, чем в заливе Аляска (0,8-1,39; Nysewander, Barbour, 1979). Основная причи-
на гибели птенцов, по наблюдениям на Командорских островах (Фирсова, 19786,
1983), — беспокойство птиц людьми, особенно в первой половине июля, когда малень-
кие птенцы в панике забегают на чужие участки, где их часто убивают взрослые особи.
В плохую погоду пуховички, не успевшие быстро вернуться на свой участок, часто за-
мерзают.
Суточная активность, поведение. Серокрылая чайка ведет преимушественно дневной
образ жизни. Формы защитного поведения у птенцов и взрослых те же, что и у боль-
шинства других чаек: птенцы затаиваются или убегают от опасности, взрослые вылетают
навстречу вторженцу, окрикивают его или активно атакуют, задевая во время пикиро-
вания ногами или ударяя клювом.
Питание. На протяжении всего года и по всему ареалу серокрылая чайка всеядный
санитар, питающийся отходами рыбного, китобойного и котикового промысла, посе-
щающий портовые свалки крупных городов и подбирающий пишевые отходы с кораб-
лей (Stejneger, 1885; Bent, 1921; Gabrielson, Lincoln, 1959; Murie, 1959; Фирсова,
19786; Hunt et al-, 19816). Вовремя хода лососевых рыб на нерестилища Командорс-
ких, Алеутских островов и Аляски птицы в массе питаются мертвой рыбой и ее икрой,
скатывающимися в низовья речек и ручьев после нереста; не брезгуют и остатками
трапезы медведей, добывающих идущую на нерест рыбу (Murie, 1959; Gabrielson,
Lincoln, 1959; Isleib, Kessel, 1973). В период размножения на Командорских островах
почти половину (по числу встреч) пищевых объектов у взрослых птиц составляют
литоральные животные, которых они собирают во время отлива: морские ежи р. Stron-
gylocentrotus; моллюски из родов Modiolus, Mytilus, Collisella, Protothaca, Amonia,
160
Полярная чайка
Nucella, Littorina, Buccinum (Фирсова, 19786). Другую половину составляют непромыс-
ловые виды рыб из семейств Gadidae, Liporidae, Agonidae, Cottidae, остатки млекопи-
тающих, птиц (кайры, белобрюшки), а также их яиц. Птенцов на Командорских ост-
ровах (Фирсова, 19786) серокрылая чайка кормит главным образом непромысловым
северным одноперым терпугом, а промысловые виды Gadus morhua macrocephalus,
Thegagra chaicogramma, судя по крупным остаткам костей, берет, очевидно, в основном
из отходов промысла. Большого разнообразия достигают и способы кормодобывания.
Содержимое панцирей морских ежей и моллюсков с прочными раковинами чайки
добывают, бросая их с высоты и разбивая о твердый субстрат (Bent, 1921; Barash et al.,
1975; Rockwell, 1982). При добыче рыбы в море пользуются двумя способами: хватают
ее, плавая по поверхности воды, и ныряют, подобно моевкам, с воздуха (Seargy, 1978).
Питаясь лососевыми на Аляске, также используют два способа добычи корма. На отме-
лях и по берегам расклевывают и заглатывают скатывающихся после нереста мертвых
и ослабленных рыб или подбирают остатки от кормежки медведей. Плавая на глубоких
местах и стремнинах, кормятся икрой, приносимой течением, выхватывая ее из по-
верхностных слоев воды (Moyle, 1966).
Враги, неблагоприятные факторы. К врагам серокрылых чаек относятся песцы,
лисицы, другие чайки. Основной отход молодняка происходит вследствие беспокойст-
ва птиц во время размножения, реже из-за неблагоприятных погодных или кормовых
условий.
Хозяйственное значение, охрана. Хозяйственного значения не имеет. О необходимос-
ти особой охраны неоднократно упоминалось в связи с тем, что в пределах СССР это
узкоареальный и малочисленный вид (Фирсова, 1977, 19786, 1979). Занесен в Красную
книгу РСФСР.
Полярная чайка
Larus glaucoides Meyer, 1822
Iceland Gull (англ.), Polarmowe (нем.), Goeland a ailes blanches (фр.)
С н н. (лат.) : Larus leucopterus Vieillot, 1820.
Син. (рус.) : малая полярная чайка.
Статус. Залетный вид. Дважды добывался на Новой Земле (один из экземпляров
находится в ЗИН АН СССР).
Общая характеристика и полевые признаки. Чайка средних размеров, очень похожа
на бургомистра, но гораздо мельче, с более округлой головой и относительно слабым
клювом. Гнездится на скалистых побережьях, часто вместе с моевками.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Некоторые систематики различают (Vaurie, 1959; Сте-
панян, 1975) два подвида (L.g. glaucoides из Гренландии и более темнокрылый
L.g. kumlieni из Канадской Арктики). Большинство исследователей (Hartert, 1912—
1921; Dwight, 1925; Peters, 1934; Voous, 1960; Cramp, Simmons, 1983) полагают,
что различия между этими формами не достаточны для выделения подвидов. Такой
знаток чаек, как Дж. Дуайт, считал канадскую форму гибридом между L. glaucoides
и L.a. thayeri, обитающей здесь же, на севере Канады (Dwight, 1925). Окончательно
вопрос о ее статусе до сих пор не решен.
Описание. Размеры (мм):
Длина крыла:
dd(n = 1) — 415;
99 (п = 4) — 390—450 (среднее 395),
6.Зак.826
161
Семейство Чайковые
Длина клюва:
<3<$(п= 1) — 43,0;
2? (п = 4) - 38,0—42,0 (среднее 39,7).
Длина цевки:
66 (и = 1)-62,0;
2? (п = 4) — 55,0—59,0 (среднее 56,7).
Окраска. Самец и самка в брачном наряде. Голова, шея, зашеек, весь низ и бока
туловища, а также рулевые и кроющие хвоста белые. Спина, плечевые, поясница, махо-
вые и верхние кроющие крыла очень светлые голубовато-серые. Вершины длинных пле-
чевых и всех маховых белые. Перья крылышка белые с нежным голубовато-серым на-
летом.Границы белого и серого цветов на спине и маховых нерезкие. В общем,
по окраске и рисунку оперения эта чайка выглядит, как уменьшенная копия бурго-
мистра.
Самец и самка в зимнем наряде. Голова, шея и зашеек испещрены очень бледными и
размытыми буроватыми продольными пятнами, прочее оперение, как в брачном наряде.
Пуховой наряд. Не описан.
Гнездовой наряд. Голова, горло и прилежащие части шеи беловатые с сильно размы-
тым бледно-бурым рисунком в виде продольных или поперечных вершинных пятен.
У переднего угла глаза темно-бурое полукольцо. Зашеек почти однотонный, бледно-
бурый. Спина, плечевые, верхние кроющие предплечья, а также поясница и верхние
кроющие хвоста того же цвета, но с беловатым рисунком в виде неправильных попереч-
ных пятен и полос и местами мраморных пестрин. Грудь и брюшко также бледно-бурые.
Длинные перья боков и нижние кроющие хвоста с поперечными светлыми полосами.
Первостепенные маховые беловатые с отчетливым буровато-палевым оттенком, причем
проксимальные со стреловидным бурым пятном у вершины и того же цвета крапом.
Верхние кроющие кисти одноцветные бледно-бурые, самые крупные из них с беловаты-
ми каемками на вершинах. Второстепенные бледно-бурые с беловатой вершинной кай-
мой и неясным, но более темным, чем фон, мраморным рисунком. Перья крылышка
бледно-бурые с остатками темного рисунка. Рулевые окрашены, как мантия, беловатый
мраморный рисунок на них развит на концах и по краям перьев.
Первый зимний наряд. Как у бургомистра, но свежие перья более бурые (Cramp,
Simmons, 1983).
Первый летний наряд (Cramp, Simmons, 1983). Как у бургомистра, все старые перья
выцветают почти до белой окраски. Свежие перья на мантии и межлопаточной области
коричневые, некоторые из них с серым налетом.
Второй зимний наряд. Голова, шея, зашеек и верхняя часть зоба белые. На темени и
затылке бледные продольные пятна буровато-серого цвета. У переднего угла глаза
черноватое полукольцо. Спина и плечевые очень бледные сероватые с буровато-палевым
оттенком, плечевые с бледно-бурым крапом в виде поперечных полос или каемок. Верх-
ние кроющие хвоста и граничащая с ними часть оперения поясницы бледно-бурые с попе-
речными полосами или пятнами. Низ и бока туловища бледные, буровато-палевые.
Длинные перья боков и нижние кроющие хвоста светло-бурые с беловатым поперечным
рисунком. Дистальные первостепенные однотонные серовато-палевые, проксимальные -
того же цвета, но с бледно-бурым крапом у вершин. Сходно с ними окрашены второ-
степенные и крылышко. На верхних кроющих крыла преобладает бледный буровато-
палевый тон, равномерный или с мраморным рисунком. Рулевые бледно-бурые, заметно
темней спины и со сплошным мелким мраморным рисунком от вершин до оснований
перьев. По С.Крэмпу (Cramp, Simmons, 1983), мантия в этом наряде по сравнению с
162
Полярная чайка
соответствующим нарядом бургомистра в большей степени или тхлшущественно серая.
В отличие от первого зимнего наряда нет предвершинного темного рисунка на перво-
степенных.
Второй летний наряд (Cramp, Simmons, 1983). Как у бургомистра, но более серый
на спине.
Третий зимний и летний наряды (Cramp, Simmons, 1983) похожи на соответствую-
щие наряды у бургомистра. Четвертый зимний и летний наряды, по С. Крэмпу (Cramp,
Simmons, 1983), похожи на окончательные, но с отдельными признаками третьего
наряда.
Линька. Изучена недостаточно. Линька в первый зимний наряд (Cramp, Simmons,
1983) очень ограничена, хотя и охватывает оперение головы, мантии, плечевых, спины,
поясницы и нижней стороны туловища. На этих участках встречаются часто лишь единич-
ные растущие перья. Протекает осенью и зимой, хотя у некоторых особей ее еще нет в
марте. Линька в первый летний наряд (Cramp, Simmons, 1983) частичная, охватывает
оперение головы, шеи и туловища, но, как и предыдущая, очень неполная, протекает в
апреле — мае. Линька во второй зимний наряд полная, в ноябре дорастают III и II перво-
степенные и палевые перья межлопаточной области. По С. Крэмпу (Cramp, Simmons,
1983), эта линька протекает, как у взрослых, в более ранние сроки; все последующие
линьки неполовозрелых особей протекают так же, как у взрослых, но раньше. После-
гнездовая линька взрослых птиц полная, у самца от 10.XI1908 г. из Исландии она закан-
чивается: дорастает II первостепенное, прочее оперение уже свежее. По С. Крэмпу, эта
линька протекает с мая по ноябрь, в течение 6—7 месяцев (Hedgren, Larsson, 1973; ма-
териалы европейских музеев). Предгнездовая линька взрослых (охватывает оперение
головы и шеи) протекает, вероятно, в апреле—мае (Dwight, 1925).
Распространение. В гнездовой ареал входят южное побережье Гренландии
(приблизительно до 70° с.ш. по восточному и до 75°, с.ш. по западному побережьям),
южное (от 75° с.ш.) и восточнее (до 68° с.ш.) побережья Баффиновой Земли. Отдель-
ные очаги размножения известны на п-ове Лабрадор и островах в Гудзоновом проливе
(Cramp, Simmons, 1983) (рис. 40). В конце прошлого века гнездование было зарегист-
рировано на о-ве Ян-Майен (Clark, 1890, цит. по: Cramp, Simmons, 1983) и в 1939 г.
у берегов Исландии (Einersson, цит. по: Makatch, 1974).
Зимовки. Область регулярных зимовок охватывает Северную Атлантику (меж-
ду 50° и 70° с.ш.) и простирается от западных берегов Гренландии до северных побере-
жий Норвегии. На юге эта область ограничена линией, проходящей от Ирландии к севе-
ро-восточным берегам Великобритании и далее к северным берегам Дании (Salomonsen,
1935; Bannerman, 1962; Williamson, 1970; Haftorn, 1971; Hedgren, Larsson, 1973; цит.
no: Cramp, Simmons, 1983). Во время кочевок залеты зарегистрированы в Западной
Европе по побережьям Польши, ГДР и ФРГ. Нидерландов, Бельгии, Португалии, Фран-
ции, Италии, Румынии, а также во внутренних районах Австрии, Венгрии и на Азорских
островах (Witherby et al., 1941; Cramp, Simmons, 1983).
Миграции. Подробнее всего изучены у западногренландской, популяции, для которой
методом кольцевания установлено, что подавляющее большинство птиц с августа по
ноябрь кочует в пределах того побережья, где они были помечены (Tuck, 1971; цит. по:
Cramp, Simmons, 1983). Молодые особи, по данным упомянутого исследователя, после
подъема на крыло перемещаются из внутренних районов фьордов к побережью и широ-
ко рассеиваются по нему. Позднее район кочевок несколько локализуется, при этом
большинство птиц держится южнее мест кольцевания. Однако некоторые совершают и
дальние миграции, достигая побережий США и северо-западной Норвегии. Чайки из вос-
точных районов Гренландии, по предположению Саломонсена (Salomonsen, 1950), дви-
жутся осенью в основном к берегам Исландии и далее к побережьям Норвежского и Се-
163
Семейство Чайковые
верного морей. На местах зимовок птицы держатся с декабря по февраль, в марте—ап-
реле начинается миграция к местам размножения. Но у берегов Исландии они еще обыч-
ны в мае; задерживаются здесь, вероятно, в основном неполовозрелые особи (Banner-
man, 1962).
Численность. Размеры колоний на побережье Гренландии варьируют от нескольких
пар до нескольких сот гнезд. Самая крупная колония насчитывала 1000 пар (Salomon-
sen, 1950). Общая тенденция изменения численности не известна.
Рис. 40. Область распространении полярной чайки
1 — область гнездования, 2 — места зимовок, 3 — залеты в СССР
Местообитания. В период размножения — внутренние части фьордов и далеко вдаю-
щихся в сушу заливов; незащищенных участков побережий избегают. Зимой — побе-
режья и прибрежные участки моря.
Размножение. Лучше всего изучено на западном побережье Гренландии (Salomonsen,
Gitz-Johansen, 1944; Salomonsen, 1950; Cramp, Simmons, 1983). Сезон размножения
длится здесь с мая по август. Гнезда помещаются на крутых утесах на высоте от 100 до
800 м, часто вместе с моевками или выше их, но при этом полярные чайки занимают бо-
лее широкие и плоские уступы. Там, где птиц не беспокоят, они могут селиться и на
плоских низких островках. Гнезда строят в основном из мха и травы, в кладке 2—3 яй-
ца. По форме и окраске яйца похожи на яйца бургомистра, но заметно мельче. Средние
размеры яиц (мм) следующие: в Северной Америке (Bent, 1921) (и = 54) 68,0 X 54,0;
в Гренландии (и = 14) 67,8 X 47,7;в разных частях ареала (и = 61) 68,6 X 48,3 (Makatsch,
1974). Длительность насиживания не установлена. Воспитывают птенцов, как и у других
чаек, вероятно, оба родителя. Детали гнездовой и послегнездовой жизни мало известны.
Питание. Основу рациона составляют мелкая рыба (Salmo, Sprattus, Clupea), ракооб-
разные, моллюски (Witherby et al., 1941; Salomonsen, 1950). Охотно поедают мертвую
рыбу, погибших птенцов моевок, люриков. В желудках встречаются также семена (Ног-
deum, Avena), ягоды (Empetrum). В период гнездования места кормежки могут быть
удалены на десятки километров от колоний. Способы добычи корма, как и у других
чаек, достаточно разнообразны. Могут брать пищу с поверхности воды, сидя на ней, и
слету. Подобно крачкам, способны бросаться в воду с высоты до 5 м с частичным или
164
Бургомистр
полным, но кратковременным погружением. Там, где их не беспокоят, кормятся и на
берегу. Зимой часто посещают свалки, места рыбного промысла.
Враги, неблагоприятные факторы. Не изучены. Вероятно, как и у моевок, отрицатель-
ное влияние на успех размножения оказывают в основном неблагоприятные кормовые
или погодные условия.
Хозяйственное значение, охрана. Хозяйственного значения не имеет, охрана не преду-
смотрена.
Бургомистр
Larus hyperboreus Gunnerus, 1767
Glaucous Gull (англ.), Eismowe (нем.), Goeland bourgmestre (фр.).
Син. (лат.) : Larus glaucus Brunhich, 1764.
Син. (рус.): большая полярная чайка, полярная чайка.
Статус. Гнездящийсямигрирующий, частично зимующий вид.
Общая характеристика и полевые признаки. Одна из самых крупных чаек. От боль-
шинства других видов отличается размерами, светлой окраской мантии и отсутствием
черного рисунка на концах первостепенных маховых; от серокрылой и малой поляр-
ной чаек — лишь первыми двумя признаками. Гнездится циркумполярно как на ска-
листых, так и на низких тундровых побережьях, обычно отдельными парами или неболь-
шими колониями. Голос, как у других крупных чаек, но выше, чем можно было бы
ожидать у птицы таких крупных размеров.
Подвидовая систематика. В настоящее время большинство авторов признает три под-
вида (Peters, 1934; Портенко, 1939; Vaurie, 1965; Степанян, 1975; Cramp, Simmons,
1983), которые различаются по средним размерам и окраске верхней стороны тела.
В СССР обитают два из них.
1. Larus hyperboreus hyperboreus.
Larus hyperboreus Gunnerus, 1767, Leem, Beskrivelse over Finmarkens Lapper, pt. 1,
c. 226, Лапландия в северной Норвегии.
Общая окраска темнее, размеры мельче.
2. Larus hyperboreus pallidissimus
Larus hyperboreus pallidissimus Portenko, 1939, Ibis, с. 266, Наукан и Уэлен, Чукот-
ский полуостров.
По сравнению с номинальным подвидом он крупнее и светлее.
Описание. Строение и размеры (табл. 9).
Окраска. Самец и самка в брачном наряде. Голова, шея, зашеек, весь низ и бока
туловища, а также рулевые и кроющие хвоста белые. Спина, плечевые, большая часть
верхних кроющих крыла, а также нижняя часть спины светло-сизые. Плечевые с белы-
ми. вершинами. Первостепенные маховые также светло-сизые, бледнеющие в вершинной
части. Второстепенные того же цвета с широкой белой каймой на вершине. Перья кры-
лышка белые с более или менее развитым голубовато-серым налетом. Клюв желтый
с красным пятном у вершины нижней челюсти. Окологлазное кольцо желтое или оран-
жевое. Ноги розовато-телесного цвета.
Самец и самка в зимнем наряде. Голова, шея, зашеек, а иногда и зоб белые с размы-
тыми буро-коричневыми пятнами, имеющими округлую форму на перьях зашейка и
продольно-вытянутую на голове и шее. Прочее оперение, как в предыдущем наряде.
Пуховбй наряд. (Cramp, Simmons, 1983 ). Как у морской чайки, но чуть более серый
и бледный, с менее резкими и крупными пятнами на голове, на спине пятна также бо-
лее бледные и менее резкие.
Гнездовой наряд. Верх и бока головы, а также зашеек и шея продольно-полосатые.
Каждое перо здесь имеет бурый центр и беловато-охристые края. Подбородок и гор-
165
Семейство Чайковые
Таблица 9. Бургомистр. Размеры (мм)
Подвид Пол Длина крыла
п lim среднее
LJi. hyperboreus Uh. pallidissimus „ бб рр бб рр 22 440-496 468 17 422-480 450 15 461-498 472 13 441-468 452
по светлей прилежащих частей головы, обычно беловатые с очень размытым, более
темным рисунком, поперечным или продольным. У переднего угла глаза неясно ограни-
ченное буровато-черное пятно. Спина, плечевые, а также верхние малые и средние крою-
щие предплечья светло-бурые с неправильным беловато- или серовато-охристым попе-
речным рисунком в предвершинной части каждого пера. Верхние кроющие хвоста по
окраске сходны с оперением спины, но поперечный рисунок здесь более резкий и пра-
вильный. Низ шеи и туловища буроватый с неясным поперечным рисунком из светлых
супротивных пятен. Рулевые светло-бурые с более или менее развитым сероватым от-
тенком и мраморным беловатым рисунком, образованным многочисленными пятнами
у вершин и по краям опахал. Первостепенные маховые светло-бурые с сероватым или
палевым оттенком, беловатыми кончиками и бурыми предвершинными пятнами стре-
ловидной формы. Второстепенные — несколько светлей, с белой вершинной каймой
и бурыми предвершинными пятнами, а иногда с бурым крапом по краям обоих опахал.
Крылышко окрашено, как маховые, но почти без темного рисунка. Большие верхние
кроющие предплечья и все верхние кроющие кисти светлые охристо-буроватые с неяс-
ным в виде теней бурым крапом по краю наружных опахал. Самые крупные из верхних
кроющих кисти с беловатой вершиной и темным предвершинным пятном. Окраска и
рисунок кроющего оперения крыльев и мантии в значительных пределах варьируют ин-
дивидуально, отчего птицы одного возраста выглядят то очень светлыми, то довольно
темными. Кроме того, по мере изнашивания все оперение значительно светлеет, а махо-
вые могут выцветать почти до белой окраски. Клюв сероватый при основании, бурова-
то-черный в вершинной трети. Ноги серовато-розовые, радужина коричневая.
Первый зимний наряд. Отличается от предыдущего более светлой головой, белова-
тыми горлом и передней частью низа шеи. Темная окраска на свежих перьях головы зна-
чительно сокращается, а охристые каемки светлеют.
Первый летний наряд. От двух предыдущих отличается более однотонными светло-
бурыми зашейком и верхней частью спины, которые почти лишены продольно-полоса-
того рисунка. Среди малых и средних верхних кроющих крыла появляются одноцвет-
ные охристо-бурые перья. В остальном этот наряд неотличим от изношенного гнездово-
го или первого зимнего нарядов.
Второй зимний наряд. Белый с нежным палевым оттенком, реже - чисто белый.
Характеризуется сильным сокращением поперечного рисунка. Продольный светло-бу-
рый рисунок заметен на белых перьях лба, темени и затылка. На зашейке, прилежа-
щих частях спины, плечевых и верхних кроющих предплечья развит того же цвета ри-
сунок в виде более или менее округлых пятен. Большие верхние кроющие предплечья
беловато-палевые с более темным мраморным рисунком в вершинной части и по краям
опахал. Маховые, крылышко и верхние кроющие кисти белые с охристым или кремо-
вым оттенком. На дистальных маховых (у некоторых экземпляров и на второстепен-
ных) иногда развит неясный светло-бурый рисунок в виде стреловидных предвершинных
166
Бургомистр
Длина клюва Длина цевки
’ п lim среднее п 1 lim среднее
22 56,7-69,0 63,3 22 70,0-81,0 74,5
17 50,0-61,0 56,6 17 63,0-74,0 69,2
15 58,0-70,0 64,1 15 71,0-81,0 75,4
13 57,0-63,0 59,6 13 67,0-75,0 71,0
пятен и крапа на обоих опахалах. Нижняя часть спины и верхние кроющие хвоста с яс-
ным поперечным темным рисунком, менее резким, чем в предыдущих нарядах. Руле-
вые, как в гнездовом наряде, но темная окраска на них менее насыщена и занимает
меньшую площадь, чем белая. Окраска низа туловища пестрая, так как перья груди,
брюха и боков тела белые в основании, бледно-бурые в вершинной трети и с белыми
каемками.
Второй летний наряд. От предыдущих нарядов отличается тем, что в нем впервые
развиваются сизые перья в межлопаточной области, среди плечевых и больших верхних
кроющих предплечья. У наиболее полно перелинявших в этот наряд особей голова, шея
и грудь белые, зашеек того же цвета с немногочисленными буровато-палевыми ок-
руглыми пятнами. Верх спины, плечевые, часть локтевых и большие верхние кроющие
крыла светлосизые с охристо-беловатыми нерезкими каемками. Брюшная сторона
пестрая, часть перьев здесь белая, другая с беловатой вершиной. Прочее оперение от пре-
дыдущего наряда. Основание надклювья серовато-желтое, спереди от ноздрей желтею-
щее, у вершины темное пятно. Подклювье у основания серовато-розовое, с красным
пятном на выступе киля и бурой вершиной. Ноги светло-розовые, радужина светло-
желтая.
Третий зимний наряд. От второго зимнего отличается более темными коричневато-
бурыми маховыми, на которых уже развиты беловатые вершины, от второго летнего
наряда — отсутствием сизых тонов на верхней стороне тела.
Третий летний наряд. Легко узнается по комбинации буровато-палевых маховых и
верхних кроющих крыла со светло-сизой мантией и белыми головой и низом туловища.
Четвертый зимний наряд. В этом наряде впервые к зиме появляются сизые перья
на мантии и наружных кроющих крыла. Первостепенные маховые также светло-сизые,
хотя и имеют буроватый оттенок. Этими двумя признаками четвертый зимний наряд
отличается от третьего зимнего; от третьего летнего наряда он хорошо отличается про-
дольно-полосатой головой и пестрым низом туловища.
Четвертый летний наряд отличается от предыдущего белым цветом головы и низа ту-
ловища, а также более чистой светло-сизой окраской мантии. От второго и третьего лет-
них нарядов отличается сизыми второстепенными маховыми и сокращением темного
рисунка на хвосте.
Линька. В первый зимний наряд неполная, охватывает оперение головы (лоб, щеки)
и зашейка. Протекает в сентябре—октябре. По С. Крэмпу (Cramp, Simmons, 1983), очень
неполно, но захватывает, кроме того, оперение мантии, плечевых, спиныи надхвостья;
протекает осенью и зимой. Линька в первый летний наряд также неполная, охватывает
оперение зашейка, межлопаточной области, груди и брюшка. По С. Крэмпу (Cramp,
Simmons, 1983), захватывает, кроме того, оперение мантии, плечевых и надхвостья;
протекает предположительно в феврале—апреле. Линька во второй зимний наряд пол-
ная, в первой декаде декабря уже закончена. Линька во второй летний наряд частичная,
167
168
Рис. 41. Область распространения бургомистра
а — область гнездования, б — отдельные поселения и гнездование узкой полосой на побережье, в — область массовых зимовок, г — область нерегу-
лярных зимовок. Подвиды: 1 — L.h. hyperboreus, 2 — L.h. pallidissimus, 3 - L.h. barrovianus
Бургомистр
у особей из западной части ареала она протекает с конца апреля до середины июня;
сменяется оперение на голове, зашейке, в области плечевых, межлопаточных, на спине,
надхвостье и среди верхних кроющих крыла. По С. Крэмпу (Cramp, Simmons, 1983),
эта и все последующие линьки протекают как у взрослых, но, вероятно, раньше.
Линька в третий зимний наряд полная, в июле—сентябре. Линька в третий летний на-
ряд частичная, в апреле—мае.
Линька в четвертый зимний наряд полная. У особи от 10.IX 1900 г. с Таймыра (за-
лив Миддендорфа) она почти закончена: дорастает II, мелкое кроющее оперение на
спине, зашейке и по всему низу туловища. Линька из четвертого зимнего в окончатель-
ный брачный наряд частичная, у самца, добытого 13.VI 1935 г. на о-ве Белый (у побе-
режья Ямала), она уже закончена. Линька из четвертого летнего в окончательный зим-
ний наряд полная, в августе она заканчивается: дорастают II и Ill, старые перья есть
среди верхних кроющих хвоста и крыльев, прочее оперение свежее; в середине ноября
эта линька уже закончена.
Линька взрослых птиц в окончательный зимний наряд полная. Начинается в июне-
июле (меняются IX—X первостепенные), в сентябре находится лишь на средних ста-
диях развития (растут IV первостепенное, все рулевые сразу, интенсивно линяет мел-
кое оперение туловища) или (реже) приближается к завершению. По данным П.С. Том-
ковича (1984), на Земле Франца-Иосифа линька маховых начинается уже с третьей
декады июня — первой декады июля, хотя встречаются особи, в середине июля еще не
приступившие к линьке. Линька взрослых птиц в окончательный летний наряд частич-
ная, по мнению Скалинова (1960), начинается в марте—апреле. Три самца и две самки,
добытые им в Белом и Баренцевом морях 23.III—21.IV, уже ее закончили, а две
самки, добытые здесь же 23.III и 8.IV еще не приступили к ней.
Распространение. Гнездовой ареал. Располагается циркумполярно в аркти-
ческой климатической зоне (Voous, 1960), лишь в некоторых районах, например на
полуострове Лабрадор, выходит за ее пределы (рис. 41). Северная граница проходит
почти везде по северным арктическим островам (Гренландии, Шпицбергену, Земле
Франца-Иосифа, Северной Земле, Новосибирским островам, о-ву Врангеля и островам
Канадского Арктического архипелага), южная — по приморским арктическим тундрам
от п-ова Лабрадор, через южную часть Гренландии, западное побережье Исландии, остро-
вам Ян-Майену, Медвежьему. В СССР эта граница проходит вдоль тундровых побере-
жий от Кольского полуострова в районе устья Поноя (Коханов, 1981) до устья Анады-
ря и далее на юг до устья р. Хатырки (Кищинский, 1980) (рис. 42). На Аляске она
идет через о-в Св. Матвея и устье Юкона (Strang, 1977; Sowls et al., 1978).
Зимовки. Как у большинства других чаек, занимают обширные акватории в Ат-
лантическом и Тихом океанах. При этом часть птиц зимует в южных районах размно-
жения севернее кромки льдов до 70° стп., кочуя среди разводий у южных и западных
побережий Новой Земли (Горбунов, 1929), у Мурманских берегор (Спасский, 1925;
Кафтановский, 1941; Коханов, Скокова, 1967), в горле Белого моря (Смирнов, 1926)
и у южных берегов Чукотки (Портенко, 1973). Но большая часть особей отлетает к
югу от ледовой кромки, рассеиваясь широко и равномерно по всей акватории прибли-
зительно до 30° с.ш. По наблюдениям В.П. Шунтова (1972), в Тихом океане зимую-
щие чайки преобладают в прибрежной шельфовой зоне, где концентрируются в районах
рыболовного промысла. Молодые и неполовозрелые особи, по данным этого же автора,
склонны к большей дисперсии и встречаются не только в бореальной, но и в северной
части тропической акватории, что подтверждается и другими исследователями (Bent,
1921; Voous, 1960). На зимовках держатся, судя по срокам осенних и весенних мигра-
ций, с ноября—декабря по февраль-март.
169
Семейство Чайковые
Миграции. Весной протекают в разных регионах с марта по июнь. На Белом и Ба-
ренцевом морях весенний пролет прослеживается до первой декады мая включительно
(Смирнов, 1926; Кафтановский, 1941), а во внутренних районах южного Ямала миг-
рирующие особи встречаются в конце мая (Данилов, и др., 1984), иногда до середины
июня. В устье р. Апуки на юге Корякского нагорья пролет отмечали с 30.IV по 6.VI
1960 г., наиболее интенсивным он был здесь с 6 по 20 мая и шел в направлении на
восток и северо-восток (Кищинский, 1980). На восточном побережье Камчатки в
Рис. 42. Ареал бургомистра в СССР
1 — область гнездования и отдельные поселения, 2 — места зимовок
районе пос. Жупаново интенсивные миграции наблюдали во второй половине апреля,
последних пролетных птиц зарегистрировали здесь в период с 16 до 21 мая (Лобков,
1980). На западном побережье этого полуострову в окрестностях Усть-Большерецка
вторая волна мигрантов проходит в конце мая — начале июня, в это время здесь наблю-
даются и неполовозрелые особи (Остапенко и др., 1975). У побережья о-ва Уруп (Ку-
рильская гряда) интенсивные перемещения птиц отмечали 25—26.IV 1974 г. (Велижа-
нин, 1975). Направление весеннего пролета в районе Хромской губы (Яно-Индигирская
низменность) западное (Успенский и др., 1962). Того же направления придерживают-
ся, очевидно, и птицы, размножающиеся на территории от Чукотки до Новосибирских
островов. Об этом свидетельствуют даты появления первых особей на местах гнездо-
вания.
Осенние миграции взрослых особей, размножающихся на Земле Франца-Иосифа,
начинаются с середины сентября, у молодых — с конца сентября до второй половины
170
Бургомистр
октября (Горбунов, 1932); на о-ве Вайгач — со второй половины августа у взрослых,
с конца этого месяца и начала следующего — у молодых (Карпович, Коханов, 1967).
На Новой Земле (Антипин, 1938) сеголетки кочуют в районах гнездования в сентябре,
в октябре они начинают мигрировать на зимовки, в конце ноября улетают последние
особи. На Северной Земле и Новосибирских островах миграция протекает в сентябре-
октябре (Бируля, 1907; Лактионов, 1946; Рутилевский, 1958). В устье Яны послед-
них бургомистров (преимущественно молодых) видели 26 сентября (Bunge, 1887).
Вне районов размножения на северо-востоке Охотского моря в окрестностях Гижиги
осенний пролет наблюдали в первую неделю октября (Allen, 1905), на юге Корякского
нагорья в устье р. Апуки (Кищинский, 1980) — 13 октября и 9 ноября; а на восточ-
ном побережье Камчатки в районе пос. Жупаново он отмечался с середины октября до
первой декады ноября (Лобков, 1980).
В период кочевок птицы доходят на север до 83° с.ш. (Портенко, 1946), на юг до
20° слп. (Bent, 1921; Snyder, 1956; Fay, Cade, 1959: Curry-Lindahl, 1981). Известны
также залеты во внутренние районы материка на Украину (К1стяк1вский, 1957, Гаври-
ленко, 1960), в бассейн Дона (Барабаш-Никифоров, Семаго, 1963) и Волги (Житков,
Бутурлин, 1960; Водолажская, Залетаев, 1977). Вне СССР зафиксированы встречи на
Боденском озере (Suter, Wicht, 1981) и в Швейцарии (Winkler, 1981;Sermet, Muriset,
1982).
Численность. В СССР самые крупные поселения найдены на Новой Земле, где в гу-
бах Грибовой и Безымянной обитает 200 пар (Успенский, 1957); на о-ве Вайгач найде-
на колония в 70 пар (Карпович, Коханов, 1967), а по данным С.М. Успенского (1965)
на мысу Стакан на северо-востоке этого острова есть колония в 150 пар. На о-ве Вран-
геля (мыс Пиллар) обнаружено несколько десятков гнездящихся пар (Успенский,
1963). На о-ве Большевик (Северная Земля) обитает всего 25—50 пар (Булавинцев,
1984). Общие тенденции изменения численности для большинства регионов неизвест-
ны. В Исландии (Ingolfsson, 1970) численность прогрессивно уменьшается из-за сильной
конкуренции с проникшей сюда серебристой чайкой.
Местообитания. Летом — скалистые и низкие тундровые побережья и прибрежные
острова, предпочтительно поблизости от птичьих базаров или колоний гаг (Lovenskiold,
1964; Карпович, Коханов, 1967). На низменных участках тундры охотнее всего посе-
ляются на косах и островках в лагунах, устьях рек и на внутренних озерах (Pleske,
1928; Тугаринов, Толмачев, 1934; Гладков, 1951; Рутилевский, 1958; Успенский и
др., 1962; Карпович, Коханов, 1967; Винокуров, 1981). Зимой предпочитает прибреж-
ные шельфовые акватории.
Размножение. Весной к местам гнездования прибывает вскоре после появления в
окрестностях колоний открытой воды. На Земле Франца-Иосифа, Новой Земле, о-ве
Вайгач и в Тиманской тундре это происходит в разные годы с середины марта до се-
редины апреля, в зависимости от ледовой обстановки и общего хода весны (Горбу-
нов, 1932; Антипин, 1938; Гладков, 1951; Карпович, Коханов, 1967). На северном
побережье Таймыра, в устье Лены и на Новосибирских островах первые птицы появля-
ются в последней пятидневке мая (Бируля, 1907; Pleske, 1928, Рутилевский, 1958);
восточнее в устье Хромской губы (Яно-Индигирская низменность) первые птицы
наблюдались 23 мая (Успенский и др., 1962). В район от Медвежьих островов до мыса
Шелагского чайки начинают прибывать во второй половине мая (Schaanning, 1928;
Остапенко, 1973; Портенко, 1973; Кречмар и др., 1978), на мыс Шмидта, в Колючинс-
кую губу и на о-в Врангеля, по данным тех же авторов, — в первой половине мая
и, наконец, на восточное побережье Чукотки (Портенко, 1973) с начала апреля до на-
чала мая.
Демонстративное поведение сходно с аналогичным поведением серебристой чайки.
171
Семейство Чайковые
Судя по описаниям в сводке С. Крэмпа (Cramp, Simmons, 1983), в агрессивных, при-
ветственных и брачных ситуациях, а также в процессе смены партнеров на гнезде и в
ходе выкармливания птенцов используются те же демонстрации, что и у других круп-
ных чаек: долгий крик, прямая поза, ’’квохтание”, вскидывание клюва, клевание тра-
вы, кормление при ухаживании и др.
Гнездостроение на Земле Франца-Иосифа начинается в начале июня (Демме, 1934),
на Новой Земле — в конце апреля (Антипин, 1938), на о-ве Вайгач — в конце мая — на-
чале июня (Карпович, Коханов, 1967). Гнездятся бургомистры обычно отдельными па-
рами или небольшими колониями до нескольких десятков пар. Гнезда устраивают на
широких скальных карнизах, избегая ниш или навесов (Горбунов, 1929а,б; Портен-
ко, 1973; Головкин, Флинт, 1975); в тундре помещают их в защищенных от четверо-
ногих хищников местах: на островах, косах; в равнинных тундрах — на сильно заболо-
ченных берегах озер ( Pleske, 1928; Тугаринов, Толмачев, 1934; Гладков, 1951; Рути-
левский, 1958; Успенский и др., 1962; Карпович, Коханов, 1967); иногда - на каме-
нистых россыпях под береговыми скалами (Кречмар и др., 1978). На Шпицбергене
(Lovenskiold, 1964) отмечены попытки гнездиться на крышах заброшенных строений.
В постройке гнезда, как и у других чаек, принимают участие оба члена пары, В качест-
ве строительного материала используются сухие стебли и корни травянистых растений,
мох, водоросли (Успенский и др,, 1962, Карпович, Коханов, 1967; Портенко, 1973).
Спаривание, по данным последнего автора, на восточном и южном побережьях Чукот-
ки происходит в последней декаде мая — первой декаде июня.
Откладка яиц на Мурманском побережье в разные годы наблюдается от 14.V до
3.VI (Гёбель, 1902). На Земле Франца-Иосифа это происходит с 10 по 12 VI (Гор-
бунов, 1932), По наблюдениям В.Д, Коханова на Белом море (устное сообщ.), в север-
ной части Терского берега 28.V 1979 г. 12 пар из 30 уже плотно насиживали кладки,
а у остальных яиц еще не было. На Новой Земле в 1947 г. откладка яиц происходила
с 9.V по 31.VII (Белопольский, 1957), а на о-ве Вайгач — в конце мая — начале июня
(Успенский, 1965; Карпович, Коханов, 1967). Далее на восток на территории от Ново-
сибирских островов и устья Лены до Медвежьих островов и устья Колымы кладки раз-
ной степени насиженности находили в разные годы с 11 по 22 июня (Bunge, 1887; Thay-
er, Bangs, 1914; Pleske, 1928; Schaanning, 1928; Успенский и др., 1962). На о-ве Вран-
геля и северном побережье Чукотского полуострова свежие кладки были обнаружены
20 и 29.VI соответственно (Портенко, 1973).
Кладка содержит от 1 до 3 яиц (Портенко, 1973). Окраска основного фона яиц
варьирует от светлой желтовато-коричневой до темной оливково-желтой или от блед-
ной оливково-зеленоватой до тусклой голубовато-зеленой. Рисунок в поверхностных
слоях скорлупы образуется пятнами неправильной формы, штрихами и завитушками
от светло- до черновато-коричневого цвета. Рисунок этот обычно сгущается на тупом
конце яйца и нередко образует здесь венчик. Более редкие пятна в глубоких слоях
скорлупы окрашены в серовато- или коричневато-фиолетовые, а также в черновато-
коричневатые тона. Форма яиц овальная или продолговатоовальная, скорлупа круп-
нозернистая, матовая. Яйца бургомистра очень похожи на яйца других крупных чаек,
особенно большой морской, от которых они с уверенностью не отличимы (Makatsch,
1974). Размеры яиц (мм): с Новосибирских островов (п = 5) 74,0—81,0 X 51,0—55,0,
в среднем 80,2 X 53,3 (Pleske, 1928); с о-ва Врангеля (и = 2) 77,6-78,3 X 52,6-54,3,
в среднем 77 Д X 53,4 (Портенко, 1973).
Насиживают кладку оба партнера, на Шпицбергене (Lovenskiold, 1964) 29—30 дней,
на Аляске (Strang, 1973) от 22 до 28, в среднем 27,6 дня. Первые птенцы на Земле
Франца-Иосифа появляются между 8 и 18 июля (Горбунов, 1932; Томкович, 1984),
172
Бургомистр
на северном побережье Белого моря (Коханов, устное сообщ.) - 12 июля: в южной
части Новой Земли они зарегистрированы 11 июля, на севере этого острова вылупле-
ние протекает несколько позднее (Горбунов, 1929). В устье Лены и на Новосибирских
островах оно отмечалось с 11 по 18 июля (Bunge, 1887; Pleske, 1928); в дельте Колы-
мы пуховичков находили с 5 июля по 1.2 августа (Thayer, Bangs, 1914, Портенко, 1973).
В устье Амгуемы и на о-ве Колючин недавно вылупившиеся птенцы обнаружены
Л.А. Портенко 3—6 июля, а на северном побережье Чукотки в районе мыса Сердце-
Камень он же видел пуховичков в гнезде 29 июня. Родители активно защищают свое
потомство. Способы активной и пассивной защиты гнезда и птенцов от пернатых и чет-
вероногих хищников у бургомистров те же, что и у других крупных чаек (окрикива-
ние, пикирование, задевание ногами и удары клювом).
Птенцы приобретают способность к полету через 49 дней после вылупления (Loven-
skiold, 1964). На Земле Франца-Иосифа это происходит между 20 августа и 1 сентября
(Горбунов, 1932; Демме, 1934). По данным С.Е. Беликова и Т.Э. Рандла (1984), на
о-ве Гукера (скала Рубини) первые молодые покинули гнезда между, 15 и 20 августа.
П.С. Томкович (1984) на о-ве Греэм-Белл видел летного птенца в сопровождении
взрослых 8—10.IX 1981 г. В Тиманской тундре Н.А. Гладков (1951) наблюдал много
хорошо летавших молодых в конце июля — начале августа 1938 г. На Новой Земле и
о-ве Вайгач вылет молодых начинается в первые числа августа (Антипин, 1938; Успенс-
кий, 1965; Карпович, Коханов, 1967) . На Чукотке и о-ве Врангеля летающие молодые
встречаются обычно в последней декаде августа (Портенко, 1973), хотя уже 8.VIII
1938 г. на мысу Шмидта отмечен лётный сеголеток. В Корякском нагорье в устье р. Ха-
тырки А.А. Кищинский (1980) наблюдал молодых на гнездах накануне вылета 8 ав-
густа, а 9—10 августа в бухте Угольной он видел птенцов, уже сошедших на воду.
После оставления гнезд молодые и взрослые держатся некоторое время вместе не-
большими стайками (Горбунов, 1929; Демме, 1934; Lovenskiold, 1964),.в которых
родители еще подкармливают молодых. Позднее взрослые отделяются от сеголетков
и улетают на зимовки раньше их, молодые же, сбившись в стаи, кочуют в районах раз-
множения до ледостава (Salomonsen,Gitz-Johansen, 1944; Портенко, 1973).
Выводки на о-ве Вайгач содержали от 1 до 3 птенцов, в среднем 2,5 птенца (Карпо-
вич, Коханов, 1967). Успех размножения на побережье Аляски к югу от устья Юкона
составил в 1972—1973 гг. 1,21 птенца на пару размножавшихся птиц (Strang, 1973),
а в заливе Нортон, расположенном значительно севернее, он колебался от 0,5 до 0,67
птенца (Drury, Steele, 1977; Biederman, Drury, 1978).
Питание. Как и большинство других крупных чаек, бургомистры всеядны. Кормо-
вые объекты и способы кормодобывания у них весьма разнообразны, но в основном
это хищники и санитары-мусорщики. Охотятся, как правило, на берегу или в непосред-
ственной близости от него. Основу питания составляет рыба (Boreogadus saida, Salmo
alpinus), морские беспозвоночные, обитающие в поверхностных слоях воды в прибреж-
ной зоне (амфиподы и другие низшие ракообразные), и литоральные животные: морс-
кие ежи и звезды, моллюски, крабы (Salomonsen, Gitz-Johansen, 1944; Белопольский,
1957; Lovenskiold, 1964; Карпович, Коханов, 1967; Ingolfsson, 1970; Портенко, 1973;
Strang, 1982).
В гнездовое время живут в основном за счет обитателей птичьих базаров или гусе-
образных, гнездящихся в тундре; в больших количествах поедают яйца, птенцов, ловят
также и взрослых птиц размерами от люрика до моевки и кайры (Бутурлин, 1906;
Житков, Зензинов, 1915; Горбунов, 1925; Демме, 1934; Антипин, 1938; Salomonsen,
Gitz-Johansen, 1944; Портенко, 1973; Кречмар и др„ 1978; Strang, 1982). В некоторых
регионах сильно вредят гнездовьям гаг и сводят успех их размножения почти на нет
173
Семейство Чайковые
(Lovenskiold, 1964; Карпович, Коханов, 1967). В Гренландии охотятся на только что
родившихся тюленей (Lovenskiold, 1964). В низменных районах тундры чаще питаются
грызунами (Успенский и др., 1962; Карпович, Коханов, 1967; Винокуров, 1981; Strang,
1982). Осенью охотно едят ягоды (Salomonsen,Gitz-Johansen, 1944; Велижанин, 1972).
Во все времена года, а особенно зимой, часто кормятся на свалках у поселков, под-
бирают отходы рыбообрабатывающих и зверопромысловых предприятий (Демме,
1934; Антипин, 1938; Скалинов, 1960; Карпович, Коханов, 1967; Портенко, 1973). На
Ямале отмечено поедание трупов оленей (Данилов и др., 1984), а на Курильских остро-
вах, по наблюдениям А.Г. Велижанина (1972), всю осень в массе кормятся отнерестив-
шимися и погибшими лососевыми.
По данным Л.О. Белопольского (1957), в рационе бургомистра корма, добываемые
на суше, занимают больше места, чем у других крупных чаек. Соотношение различ-
ных кормов подвержено, кроме того, как указывает этот автор,географической измен-
чивости Так, на Западном Мурмане рыба составляет 39% (по встречаемости), на Новой
Земле — лишь 14%, ракообразные, наоборот, чаще встречаются на Новой Земле (43%),
чем на Западном Мурмане (5%).
Из редко используемых способов кормодобывания можно отметить поверхностное
’’ныряние” с воздуха или воды, которое наблюдали у молодой особи, кормившейся
водными животными (Brown, 1949). На Новой Земле бургомистры вытаскивают рыбу
из сетей и приманку из песцовых капканов (Горбунов, 1929).
Враги, неблагоприятные факторы. Естественных врагов мало, птенцы гибнут чаше
всего из-за неблагоприятных кормовых или погодных условий.
Хозяйственное значение, охрана. Большого хозяйственного значения не имеет. Спе-
циальные меры охраны не предусмотрены. На некоторых охраняемых природных тер-
риториях, где бургомистры оказывают сильное отрицательное воздействие на других
птиц, проводится регуляция их численности.
Морская чайка —
Larus marinus Linnaeus, 1758
Great Black-backed Gull (англ.), Mantelmbwe (нем.), Goeland marin (фр.).
Син. (лат.): Larusmaximus Brehm, 1822.
Син. (рус.): большая морская чайка, большая чайка.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Одна из самых крупных чаек в фауне
СССР, имеет очень темную черновато-аспидную окраску мантии и ноги розовато-мясно-
го цвета. От сходной по этим признакам тихоокеанской чайки отличается крупными
размерами, от клуши — окраской ног и размерами. Голос немного ниже, чем у сереб-
ристой чайки. Вследствие крупных размеров более склонна к хищничеству, чем дру-
гие чайки.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Б.К. Штегман (Stegmann, 1934b) сближает морскую
чайку с тихоокеанской, что, на наш взгляд (Фирсова, 1975а), неверно из-за существен-
ных отличий в окраске промежуточных нарядов.
Описание. Размеры (мм) и масса тела (г):
Длина крыла:
dd (и = 17) — 460—500 (среднее 478);
99 (п = 14) - 400-485 (среднее 457).
174
Морская чайка
Длина клюва:
< 5<3 (п = 17) - 56,0-63,7 (среднее 61,2);
99 (п = 14) - 50,0-58,0 (среднее 56,3).
Длина цевки:
< 5(5 (и = 17) - 75,0-83,0 (среднее 77,5);
99 (п = 14) — 68,0-77,6 (среднее 74,6).
Масса тела:
< 5<5(и = 6) - 1662-1980 (среднее 1811);
99 (п = 3) - 1326-1490 (среднее 1434).
Окраска. Самец и самка в брачном наряде. Мантия, спина, плечевые и верхние
кроющие крыла очень темные, черновато-аспидные. Крупные плечевые с широкими бе-
лыми краевыми каймами. Основной фон окраски первостепенных маховых от буро-
вато-черного на дистальных (И (первое видимое) — IV) перьях до серовато- или бело-
вато-аспидного на средних (V—VII) и чуть более темного черновато-аспидного тона на
внутренних (VIII—XI) перьях. Все первостепенные имеют белые вершины. На II эта вер-
шина образована обширным белым полем (6—7 см по высоте вдоль стержня), кото-
рое иногда разделяется на две части узкой поперечной чернодатой полосой. На III раз-
виты, кроме того, темная предвершинная полоса и белая поперечная перевязь. На V—
VII также имеются черноватые предвершинные поля или перевязи. На II—IV, а также
на V—XI белая окраска от вершины перьев спускается вдоль стержня вниз, заходя в
пределы темноокрашенных участков. Перья крылышка черновато-аспидные с белой
каймой по наружному опахалу. Все остальное оперение белое. Клюв лимонно-желтый
с красным пятном на дистальном конце нижней челюсти. Ноги розовато-мясного цве-
та. Радужина серебристо-белая или очень светло-желтая. Края век киноварно-красные.
Самец и самка в зимнем наряде. Бока головы и зашеек с бурыми узкими и немно-
гочисленными пестринами. Остальное оперение и неоперенные участки тела окрашены,
как в предыдущем наряде.
Пуховой наряд. Похож (Witherby et al., 1941; Fjeldsa, 1977; Cramp, Simmons, 1983)
на соответствующий наряд у британских серебристых чаек, но имеет более светлый
и чистый серый общий тон окраски. Голова также светлее, с более редкими мелкими
и округлыми пятнами; спина тоже испещрена меньше. Однако все эти различия не
являются абсолютно надежными (Fjeldsa, 1977). Уверенно различать двух-трех дневных
пуховичков морской и серебристой чаек позволяет, как показал этот автор, лишь
индекс отношения длины цевки (измеренной по вентральной стороне) к длине нижней
челюсти (от вершины клюва до заднего угла нижней челюсти), который у морской
чайки равен 0,60—0,75, у серебристой - 0,47-0,58.
Гнездовой наряд. Спина и брюшная сторона туловища бурые с буровато- или серо-
вато-охристым светлым рисунком. На спинной стороне он резко выраженный дольча-
тый, на брюшной — неясный, образован тусклыми поперечными полосами. Наибольше-
го развития рисунок достигает на крупных плечевых перьях и по бокам тела. Голова
гораздо светлее спины, беловатая с продольным бурым рисунком, степень развития
которого сильно варьирует индивидуально. Подхвостье немного светлее брюшной
стороны, с бурыми поперечными пятнами и полосами. Дистальные первостепенные
маховые и все верхние кроющие кисти темно-бурые, первые из них светлеют по краям
внутренних опахал, вторые имеют светлые кончики. Проксимальные первостепенные
более светлые, буровато-охристые с темными вершинами и беловатыми каемками на
175
Семейство Чайковые
самых кончиках. Второстепенные маховые и верхние кроющие предплечья бурые.
Рисунок на дистальных второстепенных образован резкими беловатыми каймами на
их концах и (частично) по краю наружных опахал. На проксимальных второстепен-
ных появляются охристо-беловатые предвершинные пятна и мраморный рисунок на
внутренних опахалах. Рулевые с беловатыми каемками на концах перьев и черновато-
бурыми предвершинными полосами, более широкими на центральной паре и резко
суживающимися на краевых перьях. В основании рулевых, кроме того, развит мра-
морный рисунок. Клюв темно-рогового цвета, ноги буровато-телесного, радужина ко-
ричневая.
Первый зимний наряд. Отличается от предыдущего большим развитием поперечно-
го рисунка на спине и верхних кроющих крыла, а также сокращением бурого рисунка
на голове, шее и нижней стороне туловища. У переднего угла глаза темно-бурое полу-
кольцо, клюв буровато-черный. К началу первой весенней линьки все оперение сильно
выцветает, бурый тон светлеет, охристый становится грязно-белым.
Первый летний наряд. Отличается от двух предыдущих светлым серовато-бурым
цветом мантии, почти белой головой и сокращением поперечного рисунка на межло-
паточной области. Посветление верха туловища обусловлено, с одной стороны, силь-
ным выцветанием перьев предыдущих нарядов, с другой — бледной окраской свежего
пера. Этот наряд надевается разными особ^ями и с различной степенью полноты и носит-
ся сравнительно недолго.
Второй зимний наряд .(Cramp, Simmons, 1983). Похож на первый зимний, но темя
и задняя часть шеи светлее и менее пестрые: перья верхней части спины и плечевые
более серые и полосатый рисунок на них менее выражен, чем в первом зимнем наря-
де. Нижняя часть спины и верхние кроющие хвоста испещрены сильнее, а на нижней
стороне тела рисунок из пятен и полос выражен гораздо слабее, чем в первом зим-
нем наряде. Первостепенные также более серые, чем в первом зимнем наряде, светле-
ющие по направлению к проксимальному концу кисти. Они имеют белые вершины бо-
лее округлой формы. На пяти дистальных перьях обычно различимы светлые предвер-
шинные пятна или поля на внутренних опахалах. На остальных перьях, как правило,
развиты темные коричневые пристержневые и предвершинные полосы. Второстепен-
ные имеют более выраженный, чем в первом зимнем наряде, червеобразный темно-
коричневый рисунок, а их большие кроющие — отчетливый мелкий серовато-бурый
или темно-коричневый крап; средние и малые кроющие предплечья полосатые, подоб-
но верхней части спины и плечевым. Окраска рулевых гораздо более контрастна, чем
в первом зимнем наряде, так как серый оттенок на перьях, а также белые и темные
участки на них развиты сильнее.
Второй летний наряд (Cramp, Simmons, 1983). Голова и нижняя часть тела менее
пестрые, а перья верхней части спины и плечевые более темные, чем в предыдущем
наряде.
Третий зимний наряд. Голова, шея, низ и бока туловища в основном белые, только
на темени, затылке и зашейке есть небольшие бурые продольные пятна, а на брюшке
и боках иногда развиты немногочисленные бледно-бурые пестрины. Верхняя часть спи-
ны и кроющие крыльев черновато-аспидные, как у взрослых, но с бурым оттенком.
При этом большая часть кроющих крыла имеет остатки бурого рисунка в виде полос,
пятен ,или мелкого крапа. На верхней части спины он развит преимущественно на круп-
ных перьях. Исключение составляют самые длинные плечевые, на которых бурые тона
преобладают. Первостепенные и второстепенные маховые имеют белые вершины, нс
предвершинные белые пятна на первостепенных развиты еще слабо и обычно имеют
серый налет или несколько затемнены бурым крапом. Основной фон окраски на этих
перьях также имеет бурый оттенок. На второстепенных маховых вдоль наружных опа-
176
Морская чайка
хал и в предвершинной части перьев развиты светлые крапчатые зоны. Рулевые с бе-
лыми вершинами и основаниями, а также мраморным рисунком в предвершинной
части перьев. Все кроющие хвоста белые.
Третий летний наряд. Голова, шея, низ и бока туловища чисто белые. Свежее опере-
ние межлопаточной области, большинства плечевых, а также нижней части спины — тем-
ные серовато-аспидные, но не такого чистого тона, как у взрослых. Среди локтевых
и наружных кроющих крыла также есть группы свежих перьев, окрашенных, как
спина. Прочее оперение верхней стороны тела, так же как и большая часть оперения
крыльев и хвоста, остается от предыдущего наряда и сильно выцветает до серовато-
бурого или грязно-белого цвета. Клюв желтый с красным пятном на подклювье и
черноватым — на сгибе крючка надклювья. Ноги бледно-розовые, радужина желто-
ватая.
Линька. В первый зимний наряд неполная, охватывает лишь оперение туловища
от подбородка до груди и от зашейка до межлопаточной области и мелких плечевых,
протекает в сентябре. По С. Крэмпу (Cramp, Simmons, 1983), она длится с августа по
ноябрь. Частичная линька в первый летний наряд охватывает оперение зашейка, верх-
ней части спины, у некоторых особей частично и оперение нижней стороны туловища.
Протекает она со второй половины марта до второй половины мая, но у некоторых
экземпляров в конце апреля еще не начинается. По С. Крэмпу (Cramp, Simmons, 1983),
она длится с января по апрель. Линька во второй зимний наряд полная, в начале июля
идет полным ходом: у самца с о-ва Харлов 5 июля сменились четыре внутренних перво-
степенных, растущие перья были среди плечевых, наружных кроющих крыла и на шее.
По С. Крэмпу (Cramp, Simmons, 1983), эта линька протекает с апреля по октябрь-
ноябрь. Линька во второй летний наряд частичная, охватывает оперение головы и ту-
ловища, длится с декабря по март (Witherby et al., 1941) или апрель (Dwight, 1925).
Следующие две линьки в третий зимний и летний наряды протекают, как у взрослых
(Cramp, Simmons, 1983), но, вероятно, раньше: первая из них идет с мая или апреля
по ноябрь. Линька в четвертый (окончательный) зимний наряд полная, в июне—июле
находится на начальных стадиях: меняются три проксимальных первостепенных пера.
У взрослых птиц (в возрасте 4 лет и старше) осенняя линька также полная, охваты-
вает все оперение тела, начинается в июне. В этом месяце меняются X—XI первосте-
пенные, а также оперение туловища в области зашейка, шеи, груди, брюха, межлопа-
точных и мелких плечевых. Вместе с тем в это же время встречаются особи, у кото-
рых линька еще не началась.
По С. Крэмпу (Cramp, Simmons, 1983), осенняя линька взрослых птиц идет с сере-
дины мая до декабря или начала января. Весенняя линька взрослых птиц частичная,
охватывает оперение головы, шеи, отчасти спины и кроющих крыльев; протекает
во второй половине марта. Так, у самца, добытого (Скалинов, 1960) в Белом море,
линяло оперение на спине и верхние кроющие крыльев. По литературным данным
(Dwight, 1925; Witherby et al., 1941), эта линька протекает с января по апрель.
Распространение. Гнездовой ареал. Занимает побережья Северной Атлан-
тики от п-ова Лабрадор и берегов Северной Каролины на западе до побережий Барен-
цева моря на востоке (рис. 43). Северная граница размножения проходит через южные
побережья Гренландии, Исландию, Шпицберген, южная — вдоль южных берегов Север-
ного, Балтийского, Белого и Баренцева морей (Voous, 1960; Lovenskiold, 1964; Ingolfs-
son, 1970). В СССР гнездится на Мурманском побережье Баренцева моря от Варангер-
фьорда до устья р. Поной и мыса Данилова, а также в Кандалакшском, Онежском
заливах и вдоль Карельского берега на Белом море (Татаринкова, 1970; Бианки,
1967; Коханов, устное сообщ.). Единичные случаи размножения отмечены у о-ва Вай-
гач (Карпович, Коханов, 1967) и на Новой Земле в губе Грибовой (Белопольский,
177
Семейство Чайковые
1957). Гнездится также на островах у берегов Эстонии (Салури, 1959; Аумеэс, 1967;
Ренно, 1972) и на Березовых островах Выборгского залива (Храбрый, 1984).
В нашем столетии наблюдается (Voous, 1960) интенсивное расширение ареала как на
север, что связывает с общим потеплением климата северной Атлантики, так и на юг.
В 20—30-х годах нашего столетия началось продвижение вида на север до о-ва Мед-
вежьего и Шпицбергена, а на побережье Северной Америки за последние 50 лет граница
гнездового ареала продвинулась на юг более чем на 1000 км.
Рис. 43. Область распространения морской чайки
Обозначения те же, что на рис. 42
Зимовки. Расположены в Северной Атлантике между 70° и 30° с.ш., массовые —
между 60° и 40° с.ш. (Peters, Burleigh, 1951; Heinzel, Fitter, Parslow, 1972; Curry-Lindahl,
1981). В СССР в небольших количествах зимует у берегов Западного Мурмана, Эстонии
и Литвы (Успенский, 1941; Йыги, 1969; Пятрайтис, 1974). Как показало кольцевание
(Татаринкова, 1970), птицы, размножающиеся в берегов Мурмана, проводят зиму в
основном на побережьях Северного и Балтийского морей: в Англии, южной Сканди-
навии и вдоль побережий от Франции до Польши.
Миграции. Весенние миграции в Эстонии (Rootsmae, Rootsmae, 1976) по многолетним
данным начинаются с 23 марта, средняя дата появления птиц в этой республике 1 апре-
ля. Изредка в апреле и мае встречаются во внутренних районах вдали от моря
Так с 10 апреля по 25 мая 1956 г. наблюдали одиночных птиц в устье р. Белой (Теплов
и др., 1959), а 27 мая 1973 г. (Уилсон, 1977) - в Белоруссии.
Осенние миграции на Баренцевом море происходят с августа по ноябрь. При этом,
как доказано с помощью кольцевания (Татаринкова, 1970), сеголетки с Мурманского
побережья разлетаются по трем основным направлениям: на юго-восток (40% возвра-
тов), в Архангельскую, Кировскую области, Коми АССР и Чувашию; на запад к бере-
гам Норвегии и Англии; на юго-запад через Кольский полуостров, Белое море и Фин-
ский залив в Балтийское и Северное моря. При этом большая часть особей совершает
эти миграции постепенно, задерживаясь в кормных местах, некоторые совершают пря-
мые перелеты и быстро достигают зимовок.
В Эстонии осенние перемещения отдельных птиц начинаются уже в первой половине
августа и кончаются в середине октября (Кумари, 1961). В районе Пухту и на севере
178
Морская чайка
республики интенсивный пролет идет с середины сентября до середины октября (Манк,
1959; Йыги и др., 1961). Последние особи встречаются в Эстонии (Rootsmae, Rootsmae,
1978) до 7 декабря. Изредка отмечаются на осеннем пролете вдали от моря: в сентяб-
ре 1928 г. самка была добыта в районе г. Кирова (Водолажская, Залетаев, 1977), один
экземпляр добыт 23.Х 1883 г. под Львовом (Страутман, 1963).
Неполовозрелые особи Мурманской популяции (Татаринкова, 1970) достигают зи-
мовок на Северном море в ноябре. При этом они, как показало кольцевание, придер-
живаются кормных мест в районе зимовок в течение нескольких лет, со временем
постепенно приближаясь к местам размножения.
Численность. В СССР самые крупные колонии известны на Айновых и Семи островах
Баренцева моря, где в 1969—1970 гг. было зарегистрировано около 5000 пар, причем на
одном только о-ве Большой Айнов в 1970 г. гнездилось 3000 пар (Татринкова, 1970,
1975). Подсчитано, что за последние десятилетия численность крупных чаек, в том чис-
ле и морской, на этих островах увеличилась в 5 раз, на островах Северного архипелага
в Белом море — в 3 раза (Бойко и др., 1970).
У западных берегов Эстонии в 1967—1969 гг. учтено 210 пар (Пеэдосаар,Окно, 1970).
По данным одних авторов (Аумеэс, 1972), численность птиц здесь уменьшается в ре-
зультате борьбы с этим видом, другие исследователи считают, что она растет (Самури,
1959).
Вне СССР по всему ареалу от побережий Канады и США до Финляндии численность
неуклонно увеличивается (Voous, 1960; Parslow, 1967; Ingolfsson, 1970; Miller, 1978a;
Kilpi et al., 1980; Lemmetyinen, 1980), что связывают обычно (Voous, 1960) с потеплени-
ем климата и улучшением питания птиц в зимнее время. В некоторых районах на Бри-
танских островах численность их достигает такого уровня, что здесь, по сообщению
английских исследователей (Jones, White, 1982), вынуждены уничтожать до 37% чаек
ежегодно.
Местообитания. В сезон размножения — скалистые побережья и прибрежные острова.
На Баренцевом и Белом морях, а также у побережий Эстонии предпочитает небольшие
островки подальше от материка (Коханов, Скокова, 1967; Бианки, 1967; Аумеэс,
1967). В Канаде, Гренландии, Исландии и Швеции известно гнездование на островах
крупных внутренних озер (Salomonsen, Gitz-Johansen, 1944; Peters, Burleight, 1951;
Voous, 1960). Вне сезона размножения — морские побережья; охотно посещают свалки
портовых городов, бухты, заливы и устья рек, где есть рыбообрабатывающие пред-
приятия.
Размножение. Движение к местам гнездования в Белом и южной части Баренцева
моря начинается уже в конце февраля — начале марта (Скалинов, 1960). В это время
первые птицы начинают появляться в районе Семи островов, в середине марта они уже
посещают побережье, и в апреле их можно видеть на местах будущих гнездовий (Бе-
лопольский, 1957; Модестов, 1967). В Кандалакшский залив Белого моря они приле-
тают 1—13апреля,в среднем за 10 лет — 7 апреля (Коханов, устное сообщ.). Гнездовые
участки занимают обычно в апреле — первых числах мая (Бианки, 1967; Мо-
дестов, 1967).
В этот период во время территориальных конфликтов и формирования пар можно
наблюдать различные формы демонстрационного поведения. Набор демонстраций,
криков и поз, которые их сопровождают, а также их функциональное значение у мор-
ской чайки сходны с соответствующими демонстрациями и криками, описанными у
других крупных белоголовых чаек (серебристой, клуши, серокрылой). Но в деталях
различия все же имеются. Так, в третьей фазе долгого крика морская чайка держит го-
лову и шею перпендикулярно спине, тогда как другие виды — под углом 45° к ней.
Указанное различие трактуется некоторыми авторами (Brown, 1967а,b; Goethe,
179
Семейство Чайковые
1963а,б; Harris, 1969, 1970а) как видовое. Наши наблюдения в смешанных колониях
морских и серебристых чаек показали, однако, что различия такого рода определяют-
ся скорее физиологическим состоянием птиц в момент демонстрации.
На Айновых островах в Баренцевом море гнезда устраиваются чаще всего на купо-
лообразных вершинах холмов, заросших вороникой и папоротником, а также на сгла-
женных участках и отдельных камнях на берегу (Коханов, Скокова, 1967; Татарнико-
ва, 1975). На Семи островах их помещают обычно (Модестов, 1967; наши наблюдения)
на вершинах сопок и в верхней трети их склонов, поросших травянистой раститель-
ностью, или на прибрежных скалах поблизости от воды. Средние участки склонов за-
нимают преимущественно серебристые чайки.
В Гренландии занимают общие с бургомистрами биотопы, но в отличие от них не
гнездятся на уступах птичьих базаров, хотя и могут строить гнезда на их плоских верши-
нах (Salomonsen, Gitz-Johansen, 1944).
С 1970 г. на Британских островах началось освоение нового биотопа - крыш забро-
шенных строений (Cramp, 1971; Monaghan, Coulson, 1977); эта тенденция усиливается.
Гнездятся морские чайки в основном отдельными парами, если и образуют колонии,
то гораздо более разреженные и малочисленные, чем серебристые чайки. Так, на Семи
островах в Баренцевом море (Модестов, 1967) в наиболее плотных участках колоний
минимальное расстояние между гнездами составляло 5 м, а на Айновых островах, где
общая численность вида гораздо выше, на 1 га приходилось (Татаринкова, 1975) 30—
40 гнезд, что значительно меньше, чем у серебристых чаек, размножающихся здесь же.
Образование пар происходит, вероятно, как на пролете (Курочкин, Герасимова.
1960; Модестов, 1967), так иша гнездовых участках (Коханов, Скокова, 1967). К гнез-
достроению приступают обычно сразу после распределения по гнездовым участкам.
Первые постройки на Мурманском побережье в 1939—1941 гг. были отмечены 10-
12 мая, массовое гнездостроение наблюдалось здесь 15 мая (Белопольский, 1957; Мо-
дестов, 1967). Отдельные пары приступают к строительству за 5—10 дней до откладки
яиц. В колониях, по наблюдениям Модестова (1967), оно начинается несколько рань-
ше, чем у одиночно-гнездящихся особей. В качестве строительного материала использу-
ются сухие стебли злаков, дремы, вороники, мхов, папоротников, лишайников, водо-
рослей (Белопольский, 1957; Коханов, Скокова, 1967; Модестов, 1967). Размеры
14 гнезд, промеренных нами на Семи островах, были следующими (мм): диаметр
гнезда 300—530, в среднем 398; диаметр лотка 230—280, в среднем 260; глубина лот-
ка 60—90, в среднем 67. Размеры (мм) 5 гнезд из Кандалакшского залива (Коханов,
устное сообщ.): 360—500, в среднем 416; 260—285, в среднем 279; 89—110, в сред-
нем 96 соответственно. На Айновых островах после многоснежных зим птицы строят
обычно более массивные гнезда, чем в годы со средним уровнем зимних осадков (Ко-
ханов, Скокова, 1967).
Первые кладки на Айновых островах в разные годы находили 16—30 апреля, массо-
вая откладка яиц здесь происходила 2—15 мая; при этом в годы с плохими кормовыми
условиями она несколько задерживалась (Коханов, Скокова, 1967). На Семи остро-
вах в нормальные по погодным условиям годы откладка яиц происходит в первой
и второй декадах мая (Белопольский, 1957; Модестов, 1967). Погибшие кладки пти-
цы восстанавливают. Специальные гистологические исследования, проведенные на Се-
ми островах, показали, что повторные кладки имеют 20% размножавшихся самок (Бе-
лопольский, 1957). Кладки содержат от 1 до 3 яиц.
Окраска яиц (Дементьев, 1951) от серовато-охристой до оливково-бурой с глубо-
кими серыми и поверхностными темно-бурыми пятнами. По мнению этого автора, они
отличаются от яиц серебристой чайки, кроме размеров, более округлой формой, мато-
вым тоном окраски и крупными пестринами. Размеры (мм) 32 яиц с о-ва Харлов
180
Морская чайка
следующие: 67,5-83,5 X 50,0-55,6 в среднем 76,4 X 52,6 (Модестов, 1967); 27 яиц
с о-ва Вешняк (наши данные) имели размеры: 69,5—80,0 X 48,6—54,7, в среднем 76,5 X
X 52,1; 13 яиц из Кандалакшского залива имели размеры: 73,6-82,3 X 52,1—56,5, в сред-
нем 78,5 X 54,3 (Коханов, устное сообщ.).
Насиживание кладки длится от 28 до 33 дней, чаще 29—30 дней (Модестов, 1967).
Первые птенцы на Семи островах по многолетним наблюдениям (Белопольский, 1957;
Модестов, 1967; наши данные) появляются в разные годы в период с 19 мая по 28 ию-
ня, в среднем 8 июня. На Айновых островах первые птенцы отмечены 18—28 мая (Гера-
симова, 1958; Коханов, Скокова, 1967), массовое вылупление их здесь протекает
с 31 мая по 12 июня. На Белом море вылупление птенцов длится (Бианки, 1967) с се-
редины июня до конца июля.
Только что вылупившиеся, еще не обсохшие пуховички весят 80—90 г (Модестов,
1967). Со следующего дня родители начинают кормить их отрыгиваемой рыбой. У трех-
дневных птенцов уже выражена оборонительная реакция: при опасности они отрыги-
вают пищу и пытаются клевать. На Айновых островах (Коханов, Скокова, 1967) в воз-
расте около 7 дней птенцы сидят в основном в гнездах. Позднее, вплоть до подъема
на крыло, держатся в радиусе нескольких метров от них. По В.М. Модестову (1967),
этот период длится приблизительно до 40-дневного возраста птенцов. На Айновых
островах в средние по погодным условиям годы молодые в массе поднимаются на кры-
ло в конце июля — начале августа (Коханов, Скокова, 1967). На Семи островах это
происходит, как сообщает Л.О. Белопольский (1957), с 6 июля по 6 августа, в среднем
20 июля. На Айновых островах (Коханов, Скокова, 1967) в первые дни после оставле-
ния колоний птенцы еще плохо летают, часто отдыхают на воде и держатся вместе со
взрослыми на вороничниках в районе гнездовья. Позднее птенцы свободно летают
и кормятся вместе с родителями на литорали и в открытом море. Часто образуют от-
дельные стаи. В этот период полусамостоятельной жизни, как отмечают указанные
авторы, много молодых гибнет вследствие каннибализма в тундре у мест гнездовий,
где взрослые убивают ослабевших от голода птенцов. Острова птицы покидают к сере-
дине сентября. Кандалакшский залив Белого моря они оставляют (Бианки, Бойко,
1975) в сентябре—октябре. На Айновых островах, как сообщают Коханов и Скокова
(1967), перед отлетом молодые и взрослые держатся отдельными стаями. На Белом
море (Бианки, Бойко, 1975) места размножения покидают сначала молодые, затем
старые особи.
В плотных популяциях с растущей численностью, особенно на небольших островках,
резко возрастает количество негнездящихся особей. На Айновых островах их число
в разные годы колебалось от 40 до 80% (Коханов, Скокова, 1967; Татаринкова, 1975,
1981). Сходный эффект обнаруживается в годы с плохими кормовыми условиями.
Так, в 1979 г., по нашим наблюдениям, размножалось лишь 8% имевших гнезда птиц,
причем ни один птенец не дожил до лётного состояния, большинство их погибло в пер-
вую неделю жизни.
Питание. Довольно подробно изучено на Мурманском побережье. Основной корм
здесь рыба: треска, сельдь, мойва. Обычно птицы кормятся на плотных косяках у по-
верхности моря. Способы добычи рыбы весьма разнообразны. Чайки ловят добычу,
сидя на поверхности воды или слету. В последнем случае во время броска тело птицы
частично погружается в воду. Часто сопровождают рыболовные суда, подбирая отхо-
ды промысла, или посещают ставные сети. Охотно кормятся на свалках рыбообраба-
тывающих предприятий, а также подбирают рыбу, выброшенную морем или потерян-
ную другими морскими птицами. Нередко участвуют в охоте тюленей Halichoerus grypus
и подбирают распугиваемую ими рыбу (Спангенберг, 1941; Герасимова, 1958; Мо-
дестов, 1967; Бойко и др., 1970). В условиях эксперимента чайки съедали 150—800 г
181
Семейство Чайковые
рыбы в сутки, в среднем 450 г, что составляет 36% от среднего веса взрослой особи
(Бианки и др., 1975).
Вторая большая группа пищевых объектов — птицы и их яйца. Значение этих кор-
мов резко возрастает в годы плохого хода рыбы. Нападают на птенцов кайр, моевок,
обыкновенных гаг, турпанов, длинноносых крохалей, тупиков и полярных крачек;
поедают также ослабленных или больных взрослых особей этих и других птиц (крякв,
гоголей), а также птенцов своего вида. Успешно охотятся в воздухе и на вполне здо-
ровых взрослых тупиков и моевок. Яйца и птенцов морских птиц чайки добывают
нередко прямо в колониях, прогоняя при этом хозяев с гнезд (Герасимова, 1958;
Модестов, 1967; Бойко и др., 1970).
Из литоральных животных используют в пищу моллюсков, в основном мидий Ми-
tylis, ракообразных Gammarus sp., морских ежей Strongylocentropus; однако все это
дополнительные корма. Здесь же собирают личинок мух с водорослей, в тундре -
насекомых с растений и ягоды (Бойко и др., 1970; Бианки и др., 1975).
Из других способов кормодобывания следует отметить пищевой паразитизм (клеп-
топаразитизм). Чайки активно отбирают корм у гаг (особенно зимой) и других уток,
а также у крупных чаек, моевок, поморников и даже у серых тюленей (Бойко и др..
1970; Краснов, 1981; Каверкина, Давыдова, 1982; Краснов и др., 1982). Но полностью
обеспечить себя пищей за счет этого способа добычи корма, как считают указанные
выше авторы, чайки все же не могут.
Состав кормов подвержен географической, сезонной, половой и возрастной измен-
чивости (Белопольский, 1957). Так, на Западном Мурмане рыба составляет 72% встреч,
на Восточном — лишь 45%, при этом во втором регионе в рацион, кроме рыбы, вклю-
чаются насекомые, иглокожие, мшанки, полихеты, ракообразные, млекопитающие и
отбросы, отсутствующие в меню у птиц на Западном Мурмане. В июне и августе по-
требление рыбных кормов увеличивается, в июле - остается на одном уровне; потреб-
ление ягод также максимально в июне и августе. В период откладки яиц на Восточном
Мурмане рыба в рационе самцов составляет от 40 до 72%, у самок только 23—33%.
Зато у последних в это время лучше представлены литоральные корма: моллюски 26%.
ракообразные 21%, иглокожие 4% (против 9, 12 и 2% у самцов). У птенцов состав
кормов менее разнообразный, чем у взрослых. Основу их рациона составляет рыба
(треска 61%, сельдь 28%) и отчасти, насекомые. Литоральные корма, птицы и их яй-
ца, ягоды встречаются в очень небольших количествах; насекомых птенцы потребля-
ют, напротив, чаще, чем взрослые. Зимой в питании значительно повышается роль пи-
щевых отходов (Jones, White, 1982).
Враги, неблагоприятные факторы. Гибель молодняка обусловлена в основном небла-
гоприятными климатическими и кормовыми условиями, беспокойством людьми гнез-
дящихся птиц.
Хозяйственное значение, охрана. В заповедниках, где хищничество морских чаек
наносит существенный ущерб успеху размножения других видов, осуществляется
регламентация их численности (Аумеэс, 1957; Бойко и др., 1970).
Сизая чайка —
Larus canus Linnaeus, 1758
Common Gull (англ.), Mew Gull (амер.), Sturmmowe (нем), Goeland cendre (фр.).
Статус. Гнездящийся мигрирующий, частично зимующий вид.
Общая характеристика и полевые признаки. Чайка средних размеров с относитель-
но длинными ногами и узкими крыльями. От крупных белоголовых чаек, кроме раз-
меров, отличается лимонно-желтым клювом и отсутствием на нем красного пятна.
182
Сизая чайка
от моевок — желтыми ногами. Населяет разнообразные ландшафты от тундровых
до полупустынных. Голос очень высокий, с преобладанием визгливых звуков.
Подвидовая систематика. В настоящее время большинство исследователей признают
четыре подвида сизой чайки (Vaurie, 1965; Степанян, 1975; Cramp, Simmons, 1983).
1. Larus canus canus
Larus canus Linnaeus, 1758, Syst. Nat., ed. 10, c. 136, Швеция.
Самый светлый подвид.
2. Larus canus major (= L. c. heinei)
Larus canus L. var. major Middendorff, 1853, Sib. R. 11, 2, c. 243, от Станового хребта
до Охотского моря.
Более крупный и темный подвид, обладающий более сильным клювом.
3. Larus canus kamtschatschensis
Larus kamtschatschensis Bonaparte, 1857, Consp. Avium, 2, c. 224, Камчатка.
Еще более темный, чем предыдущий подвид, с еще более сильным клювом.
4. Larus canus brachyrhynchus
Larus brachyrhynchus Richardson, 1931, Wilson and Bonaparte, Amer. Ornith., Jameson
ed., 4, c. 352. Большое Медвежье озеро, Форт Франклин, Маккензи, Канада.
По окраске занимает промежуточное положение между L.c. canus и L.c. major; наибо-
лее короткоклювый подвид.
Замечания по систематике. Материалы ЗИН АН СССР подтверждают изложенное вы-
ше разделение на подвиды, предложенное Б.К. Штегманом еще в 1935 г. (Stegmann,
1935). Однако границы распространения типичных форм и границы переходных зон
между подвидами выяснены недостаточно. Л.А. Портенко (1939), например, считает,
что на Анадыре, Колыме и даже Индигирке встречаются особи, переходные между кам-
чатским и сибирским подвидами. Г.П. Дементьев (1940), основываясь на материалах
Зоомузея МГУ, утверждает, что на Колыме и Алазее обитает камчатский подвид. Не
везде ясна граница между номинальным и сибирским подвидами.
Описание. Строение и размеры (табл. 10).
Окраска. Самец и самка в брачном наряде. Голова, шея, зашеек, весь низ и бока
туловища, а также рулевые и кроющие хвоста белые. Спина, плечевые, верхние крою-
щие крыльев, все второстепенные маховые, проксимальные первостепенные маховые,
а также крылышко сизые разных оттенков. Все маховые с более или менее развитыми
белыми вершинными пятнами, длинные плечевые с белыми вершинными каймами.
II—VII (иногда и VIII) первостепенные с черным рисунком, на II он преобладает на
обоих опахалах. На III на наружном опахале появляется сизый цвет, на внутреннем он
доходит до середины его или более. Далее от IV к VIII сизый цвет все более оттесняет
черный к вершине пера: на VII остается лишь узкая черная первязь, на VIII — неболь-
шие парные пятна. Кроме того, на II и III всегда есть довольно большие резко очер-
ченные белые предвершинные пятна, иногда развивающиеся и на внутреннем опахале
IV. Расплывчатые белые пятна имеются обычно и на V—VII у нижней границы черного
предвершинного рисунка. Ноги желтые, клюв лимонно-желтый.
Самец и самка в зимнем наряде. Верх, бока головы и шеи белые с бурыми пятнами,
продольными на темени, щеках и затылке и округлыми или угловатыми на шее. За-
шеек обычно белый без пестрин. У переднего угла глаза черноватое полукольцо. Прочее
оперение, как в брачном наряде. Окраска неоперенных частей менее яркая, чем
летом.
Пуховой наряд (L.c. canus; Cramp, Simmons, 1983). Голова и верхняя часть тела
бледно-бурые или серовато-бурые, сильно испещренные резко очерченными точками
или пятнами. На затылке, в нижней части шеи, а также на груди и спине пятна часто
сливаются в полосы. Нижняя сторона тела очень светло-бурая, иногда почти белая, без
183
Семейство Чайковые
Таблица 10. Сизая чайка. Размеры (мм)
Подвид Пол Длина крыла
n lim среднее
L.c. canus d d 10 333-367 356
9 9 12 325-355 338
L.c. major d d 46 355-402 381
9 9 36 350-400 371
L.c. kamtschatschensis d d 10 350-390 377 '
9 9 16 342-382 360
L.c. brachyrhynchus d d 3 356-367 361
9 9 2 332-341 336
резкого рисунка. Клюв черный с розоватым концом, ноги серо-телесные, радужина
черно-коричневая.
Гнездовой наряд. Лоб, бока головы под глазами, а также подбородок й горло бело-
ватые с более или менее развитым буроватым налетом. На перьях щек наствольные блед-
но-бурые пятна. На темени и затылке довольно густые продольные пестрины охристо-
или серовато-бурового цвета. Перья зашейка и верхней части спины, а также малые
и средние верхние кроющие предплечья бурые с охристо-сероватыми светлыми каемка-
ми. Большие верхние кроющие крыльев буровато-серые одноцветные или с неясными
охристо-буроватыми каемками. Задняя часть спины и поясница буровато-охристые
или сероватые однотонные, но несколько светлей, чем оперение межлопаточной облас-
ти или плечевых. Верхние кроющие хвоста белые с охристыми кончиками, иногда с
бурыми пятнами. Нижняя часть шеи, грудь, брюшко и бока белые. Поперек зоба идет
пестрая перевязь, образованная расплывчатными бурыми пятнами, такие же пятна есть
и на боках туловища. Нижние кроющие хвоста белые, иногда с редкими бурыми пят-
нами. Рулевые белые с серовато-охристыми каемками на концах и черноватой пред-
вершинной перевязью, более широкой на центральной паре. II—VI первостепенные
маховые черновато-бурые с охристо-сероватым клином на внутренних опахалах, VII—
XI охристо-сероватые с темно-бурыми предвершинными пятнами и светлыми конце-
выми каемками. Второстепенные маховые черновато-бурые со светлыми сероватыми
внутренними опахалами или сероватые с черновато-бурыми пятнами; на вершинах
и по краю наружных опахал развита белая кайма. Перья крылышка окрашены, как
первостепенные. Клюв черный с телесным основанием, ноги телесно-розовые (Cramp.
Simmons, 1983).
Первый зимний наряд. От предыдущего отличается сизыми межлопаточными и
плечевыми перьями, последние уже с белыми вершинами. Перья задней части спины и
поясницы тоже сизые, но с буроватым оттенком, иногда с неясным поперечным рисун-
ком. Прочее оперение от гнездового наряда, но более обношенное и выгоревшее, без
охристых тонов.
Первый летний наряд. Отличается от предыдущего сокращением, а иногда и полным
отстутствием бурого рисунка на голове, шее, груди. У некоторых особей среди верхних
кроющих крыла один-два ряда сизых перьев.
Второй зимний наряд. Голова, шея и низ туловища окрашены, как у взрослых зимой,
но пестрины на голове и шее обычно более развиты. Мантия сизая, но со значительно
развитым буровато-охристым оттенком, особенно заметным на верхних кроющих пред-
плечья; самые мелкие из этих перьев могут быть сплошь буровато-охристыми. Большие
184
Сизая чайка
Длина клюва Длина цевки
п lim среднее п lim среднее
10 12 32,0-38,0 30,0-35,0 35,1 32,7 10 12 49,0-54,0 43,0-51,0 51,3 48,6
46 32,0-43,0 38,4 46 49,0-63,0 55,9
36 31,0-40,0 34,8 36 49,0-61,0 53,5
10 36,0-44,0 40,8 10 52,0-61,0 56,6
16 33,0-45,0 38,6 16 48,0-60,8 64,0
3 33,0-35,0 34,3 3 48,0-52,0 50,3
2 32,0-35,0 33,5 2 48,0-50,0 49,0
верхние кроющие кисти темно-бурые с серыми пристержневыми пятнами или с того же
цвета широкими каймами. Маховые окрашены, как у взрослых, но темный рисунок
развит больше, а серый цвет имеет охристый оттенок; белое предвершинное пятно на
III обычно менее выражено. Крылышко серовато-бурое. Мелкое оперение края кисти
с мелким чешуйчатым бурым рисунком. Рулевые обычно белые, иногда с остатками
черноватого рисунка в предвершинной части перьев.
Второй летний наряд. От предыдущего отличается отсутствием бурого рисунка
на голове и шее.
Третий зимний и летний наряды отличаются от соответствующих окончательных
нарядов у взрослых только продольными черновато-бурыми пятнами на больших
верхних кроющих кисти и I первостепенном.
Линька. Линька в первый зимний наряд частичная, охватывает оперение головы,
шеи, зашейка, межлопаточных и плечевых. Протекает с конца июля до второй поло-
вины октября. У некоторых особей, однако, во второй половине августа и в сентябре-
октябре эта линька уже закончена. У других экземпляров в это же время она еще не
начинается. Линька в первый летний наряд частичная, охватывает оперение головы,
шеи, зашейка, межлопаточной области, груди и боков туловища. Протекает в апре-
ле—мае. У некоторых особей она заканчивается гораздо раньше — в марте—апреле.
Линька во второй зимний наряд полная, начинается в мае—июне (растет IX перво-
степенное, большие верхние кроющие предплечья и кисти частично сменились) и закан-
чивается в сентябре (дорастает II первостепенное, а также оперение на верхней части
спины, горле и нижней стороне шеи). Линька во второй летний наряд частичная. У
самца от 8.IV 1894 г. из бассейна Эмбы оперение туловища частично перелиняло, голо-
ва еще в зимнем пере; экземпляр, добытый 11.V, эту линьку уже закончил.
Линька в третий зимний наряд полная, начинается во второй половине июня (растет
XI, прочее оперение старое) и заканчивается в сентябре. Самка от 29.IX 1854 г. из Ни-
колаевска-на-Амуре эту линьку уже закончила. Линька в третий летний наряд частич-
ная, у номинального подвида протекает, как у взрослых, но, вероятно, раньше (Cramp,
Simmons, 1983); сроки изучены недостаточно. Линька взрослых в окончательный зим-
ний наряд полная, начинается в мае (растет IX, прочее оперение еще старое), заканчи-
вается в сентябре —октябре (дорастают I и II, прочее оперение уже свежее; у двух са-
мок, добытых 9—18.Х в низовьях р. Урал, эта линька уже полностью закончена). Линь-
ка взрослых в окончательный летний наряд частичная, охватывает оперение головы,
шеи, груди и боков, протекает в апреле—мае. По литературным данным, предбрачная
линька у большинства взрослых на Восточном Мурмане и Белом море (Горчаковская,
185
186
Рис. 44. Область распространения сизой чайки
а — область гнездования, б — примерная граница области гнездования, в — отдельные поселения, г — места зимовок. Подвиды: 1 - L.c. canus,
2 — L.c. major, 3 — L.c. kamtschatschensis, 4 — L.c. brachyrhynchus
Сизая чайка
1948; Бианки, 1967) к середине мая заканчивается, но у некоторых следы ее обнару-
живаются в конце этого месяца или начале следующего. В Ленкорани эта линька отме-
чается уже в феврале (Заблоцкий, Заблоцкая, 1963).
Распространение. Гнездовой ареал. Охватывает в основном северную часть
лесной зоны Голарктики (Voous, 1960) (рис. 44). Лишь в некоторых регионах (север
Скандинавии, Мурманское побережье СССР, побережья Аляски) он выходит за преде-
лы лесной зоны в лесотундру и тундру или в степную зону и даже зону полупустынь
(северная часть Казахстана, юг Западной и Восточной Сибири). В СССР гнездится от
Мурманского побережья и Прибалтики до низовий Анадыря и берегов Камчатки
(рис. 45). Северная граница гнездового ареала в нашей стране проходит приблизитель-
но между полярным кругом и 70° с.ш., опускаясь к югу от полярного круга лишь на
крайнем северо-востоке (от устья Колымы до мыса Наварин на Камчатке). Южная гра-
ница проходит через южную Белоруссию (около 47°с.ш.), на Днепре опускается при-
близительно до 50° с.ш. в районе Каневского водохранилища (Клестов, Осипова, 1981)
и далее проходит между 50° и 53° с.ш. до станции Жарма (к северо-западу от Усть-Ка-
меногорска) в Казахстане (Долгушин, 1962), откуда опять поднимается до 50° с.ш.
в районе оз. Убсу-Нур в Монголии. Далее на восток до Байкала граница не прослеже-
на. От восточного его побережья она идет до Зейского водохранилища, а дальше на
восток опять не прослеживается. Упоминавшееся (Гизенко, 1955) предположитель-
ное гнездование у южных берегов Сахалина и на средних Курильских островах до сих
пор документально не подтверждено. В Северной Америке обитает в северо-западной
части материка: на восток до Больших озер и 110°в.д. (Voous, 1960; Bent, 1921),
на юг до побережий Британской Колумбии. С конца прошлого — начала текущего столетия
наблюдается расширение гнездового ареала в Западной Европе на север и запад (Voous,
1960). В Нидерланды, на Фарерские острова и в Исландию эта чайка проникла в пер-
вые десятилетия нашего столетия.
В западной части ареала от Британских островов и Исландии до Финляндии и Мур-
манского побережья в СССР обитает L.c. canus; L.c. major живет на северо-востоке Ев-
ропы, в Казахстане и Сибири (до Забайкалья и Яблоневого хребта). На Камчатке, в
Корякском нагорье и на Анадыре обитает L.c. kamtschatschensis, американскую часть
ареала занимает подвид L.c. brachyrhynchus, залеты которого отмечены на Командор-
ские острова (Степанян, 1975).
Зимовки. Зимует у восточных побережий Атлантики, вокруг Европы — от запад-
ных берегов Баренцева моря, северной Скандинавии и побережий Балтики до север-
ных берегов Средиземного моря (Горчаковская, 1948; Манк, 1957; Йыги, 1958;
Бианки, 1967; Vande Weghe, 1971; Heinzel et al., 1972; Пятрайтис, 1974; Kastepold,
Kabal, 1982; Харитонов, 1982). Во внутренних районах материка сизая чайка доходит
на север до Швейцарии (Kanel, 1977; Sermet, Muriset, 1982), Австрии и Чехословакии
(Cerny, 1965). Небольшое число птиц проводит зиму на Черном, Азовском и Каспий-
ском морях (Тугаринов, Козлова-Пушкарева, 1935; Исаков, Воробьев, 1940; Федорен-
ко, 1959; Винокуров и др., 1960; Залетаев, 1963; Заблоцкий, Заблоцкая, 1963; Андо-
не и др., 1965; Самородов, Самородов, 1969а,б; Бескаравайный, 1978; Банникова и
др., 1984). Область зимовки на африканском побережье на восток простирается от Гиб-
ралтара до Египта, на юг — до 20° с.ш., изредка встречается на Канарских островах
(Curry-Lindahl, 1981; Lensch, 1981). Зимует также на побережье Персидского залива
и у берегов Пакистана (Meinertzhagen, 1930; Tuck, Heinzel, 1980). В восточной части
ареала в это время года встречается у берегов Сахалина, на юге Приморья, на юг до
Японии и Кореи (Austin, Kuroda, 1953; Воробьев, 1954; Гизенко, 1955; Gore, Won,
1971; Лабзюк и др., 1971; Панов, 1973). На восточном побережье Тихого океана зи-
мует от южной оконечности о-ва Ванкувер (Канада) до южной Калифорнии (Bent,
187
188
Рис. 45. Ареал сизой чайки в СССР
1 — область гнездования, 2 — предполагаемая граница области гнездования, 3 — изолированные гнездовые поселения, 4 — места
зимовок
Сизая чайка
1921). Во внегнездовое время встречается иногда у восточных побережий Северной
Америки в районе о-ва Род-Айленд (Finch, 1975).
Миграции. По данным кольцевания (Kastepold, Kabal, 1982), птицы, встреченные
в Эстонии, покидают западноевропейские зимовки в марте, с зимовок на Черном и
Каспийском морях улетают в феврале—марте; молодые на Каспии задерживаются до
апреля (Заблоцкий, Заблоцкая, 1963; Кривоносов, 1963; Строков, 1974). На места
гнездования первые птицы прилетают обычно до ледохода, когда появляются промои-
ны на реках, озерах и в морских заливах. В Эстонии это происходит в среднем между
18.III и 2.IV (Rootsmae, Rootsmae, 1976; Аумеэс и др., 1977), на Белом море — с 20.IV
по 7.V, в среднем 28.IV (Бианки, 1967), а на Баренцевом море — в последних числах
апреля или в первой декаде мая (Белопольский, 1957; Герасимова, 1958; Коханов,
Скокова, 1967; Модестов, 1967). В Московской обл. сизые чайки появляются в конце
марта — первых числах апреля (Леонович, устное сообщ.); в верховьях Камы в районе
Чердыни и на Камском водохранилище первые птицы в разные годы отмечались с 14
по 22.IV, массовый пролет протекал здесь с 20 по 25.IV, заканчивалась миграция в се-
редине мая (Болотников и др., 1979). В верховьях Печоры первые чайки появляются
в разные годы с 9 по 26.V, а немного севернее — в среднем течении р. Большая Сыня —
лишь 26—29.V (Естафьев, 1982). В Северный Казахстан сизые чайки прилетают обычно
в первой половине апреля,, реже в конце марта (Долгушин, 1962). На юг Западной
Сибири в Омск, Новосибирскую обл., Томск прибывают с 9 по 16.IV (Гынгазов, Мило-
видов, 1977; Ходков, 1975в, 19776). На Южном Ямале в районе Яр-Сале ина р. Хады-
таяхе чайки отмечаются с 13 по 16 и с 22 по 31.V соответственно (Данилов и др., 1984),
а в Кондо-Сосьвинском заповеднике в северном Зауралье (Раевский, 1982) — с 24.IV
по 21.V, в среднем 7.V. В предгорьях Алтая на р. Катунь в районе Образцовки птицы
появляются 21.IV (Кучин, 1976). В среднем течении Енисея в районе пос. Мирный пер-
вые птицы отмечаются 4—20.V, массовые миграции начинаются здесь на 10 дней позд-
нее, заканчивается пролет в первой половине июня (Рогачева и др., 1978). На Западном
Таймыре в истоках р. Рыбной первых особей наблюдали в 1955 г. только 1. VI (Креч-
мар, 1966). На Байкале в районе островов Чивыркуйского залива и на оз. Рангутай
сизые чайки появляются 20— 21.IV, а севернее, в бухте Давше, лишь 10—12.V (Гусев,
1960). В среднем течении Лены первых мигрантов видели в 1973 и 1974 гг. 7—12.V,
массовые перемещения птиц зафиксированы здесь 11—19.V (Дегтярев и др., 1978; Ла-
бутин, Поздняков,1978).
Зимовки в Японии птицы покидают с начала апреля по начало мая (Austin, Kuroda,
1953), при этом взрослые начинают движение к местам размножения раньше, чем моло-
дые. На южном побережье Приморья первые особи отмечены 22 марта 1960 и 1961 гг.,
пролет здесь в эти годы продолжался до второй декады апреля (Панов, 1973). В Кро-
ноцком заповеднике на восточном побережье Камчатки в 1943 и 1946 гг. первые чайки
появились 18.V (Аверин, 1948), а в 1971 — 1978 гг. первая пролетная волна проходила
здесь с 6 по 10.V, вторая, образованная в основном взрослыми особями, — 26—30.V
(Лобков, 1980). На Шантарских островах в Охотском море первые птицы появляются
около середины мая, севернее — в Магадане, в районе Гижиги, а также на территории
Корякского нагорья и на Анадыре — во второй его половине (Allen, 1905; Дулькейт,
Шульпин, 1937; Портенко, 1939; Кишинский, 1980).
Начало осеннего отлета с Айновых и Семи островов на Баренцевом море начинается
в конце августа — начале сентября (Горчаковская, 1948; Коханов, Скокова, 1967; Мо-
дестов, 1967). В Кандалакшском заливе Белого моря сизые чайки оставляют колонии
во второй и третьей декадах августа; при этом в последние годы это происходит на 10—
15 дней раньше, чем 15—20 лет назад (Бианки, 1959,1967; Бианки, Бойко, 1972, 1975).
Как показало кольцевание, миграции сизых чаек на Белом море протекают с августа по
189
Семейство Чайковые
ноябрь. При этом последние сеголетки начинают ее в октябре, когда некоторые особи
уже достигают зимовок в Дании (Бианки, 1967) .Некоторые молодые чайки из Эстонии
появляются на этих зимовках уже во второй декаде июля, большинство достигает их,
по данным кольцевания, в ноябре (Rootsmae, Rootsmae, 1978; Kastepold, Kabal, 1982).
Из бассейна Печоры птицы отлетают около 20.IX (Естафьев, 1982). В северном Зауралье
последние чайки отмечены 11.Х.1940 г. (Раевский, 1982). В низовьях Оби пролет наб-
людается в первых числах сентября (Дерюгин, 1898), а в среднем течении его регистри-
руют до конца октября (Гынгазов, Миловидов, 1977). На юге Западной Сибири в райо-
не оз. Малый Чан интенсивные миграции отмечаются со второй декады октября, а в
конце его и начале ноября часть особей уже достигает зимовок или находится на пути к
ним на восточном побережье Каспия, в низовьях Дона или на Азовском море (Ходков,
1975в, 19776, 1983). На территории Кроноцкого заповедника на Камчатке интенсивный
пролет наблюдается в сентябре, заканчивается в начале октября (Лобков, 1980). У побе-
режий Южного Приморья хорошо выраженные миграции регистрировались со второй
половины сентября до 20-х чисел ноября (Панов, 1973). На Шантарских островах пос-
ледние особи держатся до ноября (Дулькейт, Шульпин, 1937), из района Гижиги отле-
тают в середине сентября (Allen, 1905). В Корякском нагорье А.А. Кищинский (1980)
отмечал последних птиц 5.IX на р. Ачайваям, в низовьях Апуки 27.IX, до З.Х здесь оста-
вались только сеголетки. На зимовку на о-в Хоккайдо первые особи прилетают в конце
сентября (Austin, Kuroda, 1953).
Направления осеннего пролета относительно хорошо изучены с помощью кольцева-
ния лишь в некоторых регионах. Установлено, что часть мурманской популяции движет-
ся на юго-запад вдоль Скандинавии, достигая берегов Балтийского и Северного морей
(Горчаковская, 1948). Из Кандалакшского залива Белого моря птицы разлетаются по
трем направлениям (Бианки, Бойко, 1975). Самый мощный миграционный поток идет
на юго-запад к берегам Балтийского и Северного морей в район Дании. Второе направ-
ление — восточное, к вершинам Онежского и Двинского заливов и далее к Черному
морю. И третий, северный поток мигрантов направляется к оз. Имандра и затем пово-
рачивает на запад и юго-запад, попадая к Ботническому заливу Балтийского моря.
Двигаясь вдоль его побережий на юг, птицы соединяются с юго-западным потоком миг-
рантов. Небольшая часть чаек продолжает лететь на север, попадает на Западный Мур-
ман и далее огибает северное побережье Скандинавии. Беломорские птицы залетают
иногда во Францию, Италию и до побережий Средиземного моря (Бианки, 1967; Биан-
ки, Бойко, 1975). Эстонские и финские чайки летят теми же маршрутами, что и бело-
морские птицы, но беломорские сеголетки прибывают на зимовки на 2,5—3 мес позд-
нее, чем эстонские (Йыги, 1958; Nordstrom, 1963). В Карелии на северо-восточном по-
бережье Ладожского озера пролет идет вдоль береговой линии на северо-запад
(Зимин, 1965).
Птицы, размножающиеся в Казахстане, а также особи, попадающие сюда из более
северных районов, летят почти строго в западном направлении (Долгушин, 1962).
Чайки, гнездящиеся на юге Западной Сибири, во время миграции придерживаются,
кроме западного, еще и южного направлений (Ходоков, 1977в, 1983).
Во время пролета чайки движутся обычно медленно, с длительными остановками
на отдых и кормежку. В это время они связаны в основном с литоральной зоной
побережий или с крупными внутренними водоемами (Онно, 1957; Йыги, 1958; Бианки.
1960; Йыги и др., 1961; Кумари, 1961; Зимин, 1965). Часто концентрируются в местах
скопления корма: на свалках у портовых городов, у зверосовхозов и зверокомбинатов
(Носков и др., 1985). Пролетая над континентальными районами, чайки придерживают-
ся, как правило, речных долин, но могут двигаться и напрямик через водоразделы ши-
роким фронтом (Сомов, 1897; Бианки, Бойко, 1975).
190
Сизая чайка
Численность. В СССР самые многочисленные поселения известны в Западной Си-
бири (оз. Малый Чан) — 1800 пар (Ходков, 1981), на о-ве Туренец Камского водохра-
нилища — 500—700 пар (Скрылева, 1977; Болотников и др., 1979); в Уральской обл.
в окрестностях пос. Джаныбек в 1974 г. была известна колония численностью 250 пар
(Линдеман, Залетаев, 1975). В Московской обл. самые крупные колонии, как сообща-
ет В.А. Зубакин (19816), достигают численности 100—150пар. Абсолютная численность
чаек на Айновых островах не более 100 пар (Коханов, Скокова, 1967), на Белом море
их не более 2000 пар (Бианки, 1967). В Эстонии, по последним данным, численность
достигла 40 тыс. пар (Kumari, 1976), в Латвии — 300 пар (Виксне, 1978), в Москов-
ской обл. учтено 500—550 пар и предполагается гнездование не менее 1 тыс. пар (Зуба-
кин, 19816).
Общие тенденции изменения численности в разных регионах различны. В Эстонии,
Латвии, Московской и Ярославской областях и на Камском водохранилище она растет
(Онно, 1958, 1965, 1967, 1970; Аумеэс, 1972; Ренно, 1972, 1976; Виксне, 1978; Зуба-
кин, 19816; Белоусов, 1985), а на Байкале (Гусев, 1960; Попов, Садков, 1981) падает.
Вне СССР резкое увеличение численности наблюдается в ФРГ (Mende, Sparr, 1971),
Финляндии (Kilpi et al., 1980) и на северном побережье Швеции (Grenmyr, Sundin,
1981). В Дании и некоторых других странах Западной Европы, а также на некоторых
островах Балтийского моря в последние 40—50 лет отмечается резкое ее падение
(Franzmann, 1974; M011er, 1978а; Breife, 1981). В ГДРуменьшение численности вызва-
но осуществлением’с 1971 г. специальных мероприятий по сокращению числа птиц
(Nehls, 1979).
Местообитания. Летом предпочитают лесные реки, озера и болота во внутренних
районах материков. Не избегает, однако, и морских побережий и прибрежных островов.
В Скандинавии заселяет даже горные озера, расположенные на высоте от 1300 м над
уровнем моря (Voous, 1960), вдоль южной границы ареала — степные и полупустынные
озера (Сушкин, 1908; Долгушин, 1962). В последние годы наблюдается тенденция к
освоению антропогенных биотопов: карьеров, рыборазводных прудов, построек чело-
века. Зимой держится у побережий, на косах и мелководьях, а также во внутренних
районах на полях и в степях (Радде, 1884; Никольский, 1891; Тугаринов, Козлова-
Пушкарева, 1935; Meinertzhagen, 1954; Федоренко, 1959; Заблоцкий, Заблоцкая,
1963; VandeWeghe, 1965; Kanel, 1977).
Размножение. Сизая чайка — моногам; возраст первого размножения, степень гнездо-
вого консерватизма, а также процесс образования пар подробно изучены у эстонских
птиц (Онно, 1965, 1967—1969). Самцы достигают половой зрелости в возрасте 2—3,
а самка — 3—4 лет. Самец и самка из одной пары часто зимуют отдельно и весной встре-
чаются на месте гнездования. При этом самец во время образования пары более акти-
вен, он же выбирает и охраняет гнездовой участок. Молодые птицы чаще образуют пары
между собой. Поскольку самки достигают половой зрелости обычно на год позднее
самцов, нередки пары, в которых они на год старше самцов. В 72% случаев пары обра-
зуют те же партнеры, что и в предшествующем году, в 28% — новые, при этом у 40%
особей прежний партнер жив. Новые пары образуются часто между птицами, гнездив-
шимися в прошлом сезоне недалеко друг от друга.
Гнездовой консерватизм развит очень сильно. При отсутствии беспокойства до 90%
чаек селится на прошлогодних гнездовых территориях и только 2% покидают их и
строят гнезда в радиусе 500 м от прошлогоднего места. У самцов гнездовой консер-
ватизм выражен больше, чем у самок. Зависит он также от возраста птиц: сильнее все-
го проявляется у особей, гнездившихся на одном месте в течение трех или более лет.
Диффузно-гнездящийся факультативно-колониальный вид (Зубакин, 1976в; 1983).
Гнездятся чайки на большей части ареала обычно небольшими колониями (до 50 пар)
191
Семейство Чайковые
или одиночно (Горчаковская, 1948; Герасимова, 1958; Онно, 1958; Кищинский, 1960а;
Спангенберг, Леонович, 1960; Карташев, 1975; Хохлова, 1977; Бианки, Плюснин,
1983). Крупные колонии редки. Гнезда располагаются на плоских участках, но часто и
на небольших возвышенностях (буграх, бревнах, камнях). Чайки помещают их на
островах, берегах или косах у водоемов. Иногда гнезда могут быть построены среди
густой травы, в сырых местах — на торцах пней, сломанных вершинах деревьев или на
их стволах на высоте от 0,2 до 6 м, изредка под прикрытием пней или валежин (Туга-
ринов, Бутурлин, 1911; Успенский и др., 1962; Бианки' 1967; Андреев, 1974; Стрел-
ков, 1974а; Татарникова, 1975; Виксне, 1978; Болотников и др., 1981; Дегтярев,
Ларионов, 1981а,б; Лабутин и др., 1981). Некоторые исследователи (Кречмар и др.,
1978) считают, что гнездование на деревьях на северо-востоке страны — обычное явле-
ние. Нередко гнезда устраиваются также на плоских участках скал (Bergman, 1939;
Haarnmann, 1980; Хохлова, 1981), иногда на скалистых обрывах (Гусев, 1960; Ки-
щинский, 1980) или плавнике (Спангенберг, Олигер, 1949). В последние годы как в
СССР, так и за его пределами наблюдается тенденция к освоению новых мест гнездова-
ния. Так, в Московской обл. и Белоруссии птицы заселяют залитые водой карьеры,
оставшиеся после торфоразработок (Авданин и др., 1980; Зубакин, 19816; Самородов.
1981а; Наумчик, устное сообщ.). В Прибалтике они все больше осваивают верховые
болота и рыборазводные пруды (Kumari, 1976; Пукас и др., 1981; Петриньш, 1982).
В Англии, Швеции, Финляндии, Латвии и Эстонии известны единичные случаи гнездова-
ния на крышах (Cramp, 1971; Monaghan, Coulson, 1977; Рандла, 1985; Strazdins
et al., 1987).
В начале сезона размножения наблюдаются различные формы демонстративного по-
ведения. Оно довольно подробно изучено в Шотландии (Weidmann, 1955) и в общем
сходно как с поведением крупных белоголовых чаек (Tinbergen, 1953, 1959; Goethe,
1937), так и мелких темноголовых чаек. По сравнению с серебристой и озерной чай-
ками имеются лишь незначительные отличия в форме выполнения, мотивации проис-
хождения некоторых демонстраций.
В гнездостроении, как и у других чаек, принимают участие оба партнера. На постройку
гнезда уходит от 2 до 5 дней, иногда старые постройки подновляются, ремонтируются и
используются в течении нескольких лет (Горчаковская, 1948; Гусев, 1960; Бианки,
1967; Литвинов и др., 1978; Самородов, 1978а; Болотников и др., 1979). Строитель-
ным материалом служит обычно растительность, окружающая гнездо. В тундре это зла-
ки, веточки вороники, ивы, мхи, лишайники (Горчаковская, 1948; Кищинский, 1960а;
Спангенберг, Леонович, 1960), на луговых лудах Белого моря — сухие стебли травяни-
стых растений, реже мох и водоросли (Бианки, 1967). В лесной зоне, кроме стеблей и
корневищ злаков, используются сухие древесные веточки, кукушкин лен и другие мхи
(Гусев, 1960; Болотников и др., 1979), в Казахстане — мелкие обломки тростника
(Долгушин, 1962).
Размеры гнезд также во многом зависят от окружающей их обстановки. На сухих
местах это, как правило, невысокие с тонкими стенками и дном постройки, на
сырых — толстостенные и высокие сооружения. В различных точках ареала были зафик-
сированы следующие размеры гнезд (мм). В Эстонии диаметр гнезда 203—310, в сред-
нем (п = 40) 262; диаметр лотка 140—185, в среднем 158; глубина лотка 25—65, в сред-
нем 39 (Аумеэс и др., 1977). В Белоруссии (Наумчик, устное сообщ.) средние размеры
гнезд (п = 18): диаметр гнезда 261, диаметр лотка 176, глубина лотка 74. На юго-вос-
токе Корякского нагорья (бухта Гека) диаметр гнезда 220—300, в среднем 247; диа-
метр лотка 125—250, в среднем 185 (и = 19; данные Л.В. Фирсовой).
Плотность гнезд в эстонских колониях,по С. Онно (19676), выше на мелких, чем на
крупных островах: 32,1 против 15,5 гнезд на 1 га соответственно; среднее расстояние
192
Сизая чайка
между гнездами при самой большой их плотности в среднем равно 5 м, что больше,
чем у других колониальных чайковых в этом регионе. В центре крупной колонии на
Камском водохранилище плотность составила 20—25 гнезд на 1 га, и на периферии это-
го же поселения — лишь 5—10 гнезд (Болотников и др., 1979). Индекс плотности гнез-
дования для Северного Казахстана равен 0,02 (Зубакин, 1976в).
Откладка яиц на Баренцевом море (Айновы острова) в разные годы начинается
13—17.V, в очень поздние годы с 6 по 10.VI; массовая откладка яиц проходит здесь
в разные годы с 18.V по 2.VI (Герасимова, 1958; Коханов, Скокова, 1967). На Семи ос-
тровах, по многолетним наблюдениям, первые кладки отмечаются с 20.V по 7.VI, в
среднем 28.V (Белопольский, 1957). При этом в годы с поздними веснами откладка
яиц начинается позднее и проходит в более сжатые сроки. На Белом море (Канда-
лакшский залив) первые кладки появляются с И по 28 .V, в среднем (за 12 лет) 18.V;
массовая откладка яиц протекает обычно с 20 по 25 .V (Бианки, 1967). На восточном
побережье Белого моря свежие кладки находили с 29.V по 3.VI (Спангенберг, Леоно-
вич, 1960). На Камском водохранилище откладка яиц в разные годы проходит с 7 по
20.V (Болотников и др., 1979), в Московской обл. — первых числах мая (Леонович,
устное сообщ.). На Наурзумских озерах на севере Казахстана откладка яиц начинает-
ся обычно в конце первой декады апреля (Самородов, 1978а), в окрестностях Омска и
на Байкале — во второй декаде мая (Гусев, 1960; Гынгазов, Миловидов, 1977). На се-
веро-востоке Камчатки (бухта Гека в заливе Корфа) первые кладки обнаружены нами
14.VI 1977 г., но еще 21.VI мы находили здесь свежие неполные кладки.
На Мурманском побережье и Белом море откладка яиц у одной пары длится от 3 до
7 дней, очень редко до 14 дней (Горчаковская, 1948; Бианки, 1967; Модестов, 1967).
Интервалы между откладкой яиц в гнезде здесь колеблются от 1,5 до 2 дней, реже яй-
ца откладываются ежедневно. В Эстонии средний промежуток между откладкой яиц
немного короче 48 ч; на откладку яиц, особенно в начале гнездования, сильно влия-
ют метеорологические факторы: температура воздуха, воды (Онно, 19676). В плотных
популяциях откладка яиц начинается раньше и часто (но не всегда) протекает быстрее.
Сроки начала откладки яиц у каждой самки, по С. Онно, наследственно закреплены и
варьируют в разные годы не более чем на 4 дня по отношению к среднему для всей по-
пуляции сроку начала этого процесса. Этот показатель не зависит от положения гнезда
в центре или на периферии колонии. Молодые особи в возрасте 2—3 лет откладывают
яйца в среднем на 4 дня позднее среднего для всей популяции срока (Онно, 19676)
В популяции откладка яиц занимает в разные годы от 40 до 48 дней, в среднем 43 дня
(Аумеэс и др., 1977).
О размерах кладок в разных частях ареала дает представление табл. 11. Повторные
кладки на Баренцевом море, по Л.О. Белопольскому (1957), имеют 11% самок,
Н.Н. Горчаковская (1948) их здесь не обнаружила. Процент восстанавливаемых кла-
док, как показали исследования в Эстонии (Онно, 19676), зависит от стадии насижи-
вания первых кладок в момент их гибели, возраста самок и степени привязанности
партнеров друг к другу. Легче всего восстанавливаются кладки, погибшие в первые
четыре дня насиживания, их возобновляет 80,5% самок. Этот показатель резко увели-
чивается при совместном размножении партнеров от 1 до 4 лет и у самок, достигших
возраста от 6 до 11 лет. Перерыв между гибелью первой и появлением повторной клад-
ки колеблется у 90% самок от 3 до 15 дней, в среднем 9,51± 0,18 дня; повторные клад-
ки в среднем на 5 г легче первых, и средняя величина их также заметно меньше за счет
увеличения числа кладок из двух яиц.
Окраска яиц на Семи островах Баренцева моря (Горчаковская, 1948) светлая серо-
ватая или серовато-оливковая с неравномерно разбросанными пятнами, штрихами и
черточками коричневого цвета; темный рисунок нередко образует венчик у тупого кон-
7. Зак. 826
193
Семейство Чайковые
Таблица 11. Сизая чайка. Величина кладок
в разных частях ареала
Место, автор п Доля кладок (%) при числе яиц 1 —4 Средний размер кладки
1 2 3 4
Мурманское побережье (Горчаковская, 1948) 65 10,77 32,31 56,92 0 2,46
Белое море (Бианки, 1967) — 8,8 20,7 70,2 0 2,3
Карелия (Хохлова, 1981) — — — — — 2,59
Эстония (Онио, 19676) — 0 8,0 91,0 1,0 —
Белоруссия (Наумчик, не- опубликованные данные) - 0 70,0 30,0 0 -
Казахстан (Самородов, 1978а) 58 8,4 11,3 80,3 0 -
Корякское нагорье (наши данные) 36 5,5 8,3 86,2 0 2,8
Таблица 12. Сизая чайка. Размеры (мм) яиц
в разных частях ареала (в скобках - средние величины)
Место, автор п Размеры яиц
Баренцево море, Семь островов (Горчаковская, 1948) 139 51,5—61,8 X 38,0-43,0 (56,7 X 40,5)
Кандалакшский залив Белого моря (Бианки, 1967) 425 (58,5 X 41,7)
Карелия, острова Заонежья (Хохлова, 1977) 200 53,3-65,6 X 38,6-45,6 (58,1 X 41,2)
Белоруссия (Наумчик, неопубликованные дан- ные) 54 (59,7 X 41,3)
Казахстан, Наурзумские озера (Самородов,1978а' 35 56,7-65,0 X 39,5-46,4 (59,5 X 43,6)
Байкал, Чивыркуйский залив (Гусев, 1960) 16 57,5-69,0X 41,3-45,3 (63,0 X 43,2)
Камчатка, Олюторский залив, бухта Гека (наши данные) 68 54,4-69,0 X 38,4-45,2 (62,3 X 43,0)
ца яйца. Изредка встречаются темно-коричневые яйца с черным рисунком. На Белом
море (Бианки, 1967) окраска фона яиц варьирует от голубовато-зеленой до коричне-
вой. Большинство яиц имеет серовато-оливковый фон с равномерно разбросанными по-
верхностными пятнами и штрихами того же цвета, но более темными. Коричневые яйца
составляют отдельные кладки.
Данные о размерах яиц из различных точек ареала приводятся в табл. 12.
Масса яиц (г) с Баренцева моря была 44,5—58,3, в среднем (и = 70) 51,0; вес скор-
лупы 2,91—3,3 (Горчаковская, 1948). На Белом море вес 149 свежих яиц был равен в
среднем 54,8, длина яиц превышала их ширину в 1,35—1,5 раза, обычно не более чем в
1,45 раза; более округлые и вытянутые яйца составляют всего 9% (Бианки, 1967). По
данным эстонских орнитологов (Онно, Бугаев, 1976; Онно и др., 1977), объем яиц, от-
кладываемых одной и той же самкой, в разные годы может изменяться. Этот показа-
тель, а также форма и вес яиц зависят от возраста самок; зависимость эта имеет слож-
ный нелинейный характер. Длина и масса яиц, как показано этими же исследователями,
194
Сизая чайка
увеличиваются с севера на юг. Масса яиц изменяется, кроме того, в цикле кладки,
достигая максимума у второго яйца; третье яйцо меньше по всем показателям для
всех географических популяций. В процессе насиживания масса яиц уменьшается в сред-
нем (п = 20) на 0,3—0,9 г в сутки (Самородов, 1982), средняя масса яиц (и = 5), в кото-
рых началось вылупление, 55 г.
Обогревают кладку оба партнера. Плотное насиживание на Белом море, по наблюде-
ниям Н. Чернышовой (Бианки, 1967), начинается, как правило, с момента появления
второго или третьего яйца. Но в особых условиях, — например, когда кладки сильно
страдают от хищничества других чаек, — родители могут плотно насиживать и с первого
яйца (Kantak, 1954). Самки проводят на гнездах больше времени, чем самцы, особенно
на первых и последних этапах инкубации (Калинин, 1975). Средняя длительность насижи-
вания после окончания откладки яиц, по данным разных авторов, варьирует от 25 до
27 дней (Белопольский, 1957; Бианки, 1967; Аумеэс и др., 1977). При этом длитель-
ность насиживания первого яйца может быть 27—29 дней, второго — 25—28 и третьего —
24-26 дней. В Камском Предуралье эти цифры составляют в среднем 25,8,24,6, и 23,6
дней (Болотников и др., 1979).
Первые птенцы на Айновых островах Баренцева моря отмечаются 15—22.VI, массо-
вое вылупление их здесь происходит 27.VI—12.VII (Герасимова, 1958; Коханов, Скоко-
ва, 1967). На Семи островах, по многолетним наблюдениям, первые пуховички появля-
ются с 14.VI по 2.VII, в среднем 22.VI (Белопольский, 1957). На Белом море вылупле-
ние пуховичков начинается в ранние по погодным условиям годы 5—10.VI, в поздние —
10-20.VI; массовое вылупление их здесь отмечается с 20-х чисел июня (Бианки, 1967).
На п-ове Канин (р. Чижа) маленьких пуховичков наблюдали 20.VI (Спангенберг, Лео-
нович, 1960), а на юге Ямала в среднем течении р. Хадытаяхи - 8.VII (Данилов и др.,
1984). На Наурзумских озерах первых птенцов находили в разные годы с 6 по 24.V,
массовое вылупление их здесь наблюдалось в 1967 г. в конце мая — начале июня, по-
следние пуховички в этом же году были обнаружены 16.VI (Самородов, 1978а). В юж-
ной части Западной Сибири вылупление начинается в первых числах июня (Стрелков,
1974а; Гынгазов, Миловидов, 1977). На Байкале О.К. Гусев (1960) отмечал первых
птенцов 15—18 июня. На юго-востоке Корякского нагорья (бухта Гека в заливе Кор-
фа) только что вылупившихся и еще не обсохших чайчат находили 7.VII 1977 г. (дан-
ные Л.В. Фирсовой); А.А. Кищинский (1980) видел пуховичков на р. Ачайваям 12.VII
1960 г.
Длительность вылупления одного птенца в< Эстонии (Аумеэс и др., 1977) колеблется
от 62 до 99 ч, в среднем.составляет 79 ч 41 мин. При этом очередность появления накле-
вов на скорлупе соответствует очередности откладки яиц, но из последних яиц птенцы
вылупляются обычно быстрее (в среднем за 65 ч 7 мин) , чем из первых (в среднем за
82 ч 15 мин). На Белом море скорость вылупления отдельных птенцов варьирует от 2
до 7 дней, обычно равна 3 дням; в одном гнезде пуховички появляются в течение 2—3
дней (Бианки, 1967). При этом два первых птенца чаще освобождаются от скорлупы
в один день с интервалом в несколько часов, третий — обычно на сутки позднее; реже
все чайчата появляются с суточным интервалом или в один день. На Камском водохра-
нилище все пуховички в одном гнезде вылупляются в течение 1,9—2,0 дней (Болотни-
ков и др„ 1979). После появления птенцов взрослые обычно уносят скорлупу из гнезда.
Вылупление птенцов в популяциях на Белом море и в Казахстане растянуто, как пра-
вило, на 30—35 дней (Бианки, 1967; Самородов Д 978а). В Эстонии оно длится от 37 до
45 дней, в среднем 40 дней (Аумеэс и др., 1977), Указанные авторы объясняют растя-
нутость этих сроков наличием повторных кладок.
Отход яиц на о-ве Харлов в 1939 г. составил 30,72% (Горчаковская, 1948). На Белом
море этот показатель увеличивается с увеличением среднего размера кладки (Бианки,
195
Семейство Чайковые
1967): при средней кладке в 2,3 яйца он равен 13,1%, при кладке в 2,7—18,1%, а при
кладке в 2,83 яйца — 30,4%. На северном побережье Байкала отход яиц в 1978 г. соста-
вил 20% (Попов, 1979). В Эстонии ежегодно гибнет 27—36% яиц, большей элиминации
подвергаются яйца молодых птиц (Мянд, 1981).
В первые три дня после вылупления птенцы обычно большую часть времени находят-
ся в гнезде и обогреваются родителями; терморегуляция у пуховичков устанавливается
на второй-третий дни жизни. Но в случае опасности они способны покидать гнездо и
удаляться от него на 10—20 см уже через 3,5 ч после вылупления (Самородов, 1978а).
В возрасте 3—5 дней птицы, как правило, оставляют гнезда и держатся на гнездовом
участке, который охраняется ими и родителями (Ivask, Lilleleht, 1978). По наблюде-
ниям на Байкале, уже через неделю после окончания вылупления птенцы в колонии не
встречаются (Попов, 1979) . На Белом море родители иногда уводят 1—2-дневных пухо-
вичков на литораль за десятки метров от гнезда (Бианки, 1967). На Семи островах Ба-
ренцева моря некоторые семьи уже через 5—7 дней после появления птенцов имеют ох-
раняемые взрослыми особями подвижные гнездовые участки (вне гнездовой террито-
рии) радиусом от 8 до 20 м (Модестов, 1967). Время пребывания родителей на гнездо-
вом участке в период выкармливания птенцов уменьшается, но в начале периода самцы
проводят здесь больше времени, чем самки, а в середине этого периода — наоборот; к
концу выкармливания различия сглаживаются. Пребывание партнеров на гнездовой
территории хорошо согласовано и не зависит от времени суток; члены колонии, так
же как и птенцы одного выводка, знают друг друга ”в лицо” (Ivask, 1978).
Маленьких пуховичков родители кормят из клюва полупереваренной массой, позд-
нее отрыгивают ее на землю, откуда птенцы берут ее сами. Как правило, родители узна-
ют своих чайчат с 4—6-дневного возраста и кормят только их, хотя изредка наблюдаются
случаи смешивания выводков (Ivask, Lilleleht, 1978). На Семи островах Баренцева мо-
ря чужие взрослые особи часто подбирают остатки пищи после кормления птенцов или
даже преследуют птенцов и заставляют отрыгивать уже проглоченный корм, что, вероят-
но, связано с недостатком пищи (Модестов, 1967; наши наблюдения). На Баренцевом
море, по Л.О. Белопольскому (1957), относительная масса только что вылупившихся
пуховичков составляет около 9% массы взрослых птиц (около 27 г). Наиболее интен-
сивно растут птенцы с 11-го по 15-й день жизни, относительный прирост массы достига-
ет в это время 4,9% массы взрослых. В Кандалакшском заливе Белого моря птенцы рас-
тут быстрее, чем на Баренцевом море, и достигают массы взрослых в среднем к 20-му
дню жизни. Перед самым подъемом на крыло вес молодых несколько падает, хотя и
не у всех особей (Горчаковская, 1958; Бианки 1967). Средняя масса пуховичков эс-
тонских чаек в момент вылупления равна 36,6 ± 1,2 г, максимальной массы они дости-
гают только к 26-му дню жизни (Laidna, 1978). При этом быстрее всего растут птенцы
из ранних и средних по срокам кладок. Для Эстонии установлена также высокая поло-
жительная корреляция между массой свежих яиц и массой и размерами вылупившихся
из них птенцов. В первые 8 дней птенцы из крупных яиц растут быстрее, затем разница
сглаживается (Аумеэс и др., 1977).
На крыло большинство птенцов поднимается, по данным разных авторов, в возрасте
от 29 до 40 дней. На Мурманском побережье Баренцева моря первые лётные птенцы от-
мечаются в конце июля — начале августа, на Белом море — в середине июля, на севере
Казахстана — в конце июня, в Нарымском крае Западной Сибири — в середине июля,
в Корякском нагорье, на Шантарских островах и на Анадыре — во второй декаде ав-
густа. Общение с родителями продолжается после подъема на крыло еще в течение 2—3
недель, когда взрослые еще кормят лётных молодых (Дулькейт, Шульпин, 1937; Пор-
тенко, 1939; Горчаковская, 1948; Белопольский, 1957; Кищинский, 1980; Долгушин,
1962; Бианки, 1967; Стрелков, 1974а; Ivask, 1978; Самородов, 1978а, 1981а).
196
Сизая чайка
Успех размножения на Белом море равен в среднем 1,5 птенца на пару, отход яиц и
птенцов составляет здесь 45—50% от числа отложенных яиц (Бианки, 1967). На Айно-
вых островах в благоприятные годы поднимается на крыло лишь 0,25 птенца на пару
размножавшихся птиц, чаще все птенцы гибнут (Татарникова, 1975). В Эстонии этот
показатель достигает 1,0 птенца на пару взрослых, наибольший отход птенцов наблюда-
ется в течение первых двух недель их жизни. Смертность первого птенца в выводке ни-
же, чем у двух других; наибольшая смертность зарегистрирована у чайчат из поздних
выводков (Laidna, 1978). Низкий успех размножения у молодых особей, как показали
многолетние наблюдения за индивидуально помеченными птицами (Раттисте, 1981),
компенсируется высоким значением этого показателя у особей, гнездящихся шестой
раз и более старых.
Максимальная продолжительность жизни, по данным кольцевания, 24 года 2 месяца
(Rydzewski, 1978,цит. по: Cramp, Simmons, 1983).
Суточная активность, поведение. Сизые чайки ведут преимущественно дневной образ
жизни. Защитные формы поведения, как у большинства других чаекюблеты с тревож-
ными криками, пикирование и обливание пометом наземных хищников и человека;
воздушных хищников атакуют в воздухе. Иногда применяют отвлекающие демонстра-
ции (Cramp, Simmons, 1983).
Питание. Кормовые стации, способы кормодобывания и набор пищевых объектов
весьма разнообразны. Охотно собирают корм на морских побережьях и литорали, а
также во внутренних районах островов и материка на озерах и реках. Охотятся и на от-
крытом зеркале воды, добывая рыбу у ее поверхности, разыскивают корм, плавая
или расхаживая по мелководью, опуская голову в воду; высматривают добычу с возду-
ха и могут нырять за ней до глубины 1 м как с воды, так и с воздуха. В последнем слу-
чае зависают перед броском в воздухе, как крачки (Горчаковская, 1948; Белополь-
ский, 1957; Кищинский, 1960а; Герасимова, 1965; Бианки, 1967; Коханов, Скокова,
1967; Модестов, 1967; Самородов, 1978а). Большое место среди объектов питания за-
нимают различные отходы и остатки сельскохозяйственной продукции, которые птицы
собирают на свалках у поселков и рыбозаводов, на рыборазводных прудах, у зверо-
ферм и птицефабрик, а также на полях (Горчаковская, 1948; Белопольский, 1957;
Зимин, 1965; Модестов, 1967; Самородов, 1978а; Хохлова, 1977; Vauk-Hentzelt,
Schumann, 1979, 1980; Мельников, Лысиков, 1983; Мельников и др,, 1983; Liittrin-
ghaus, Vauk-Hentzelt, 1983).
Хищничают в колониях по сравнению с крупными представителями рода мало, но все
же иногда поедают яйца куликов, уток, крачек, а также птенцов мелких птиц (Бело-
польский, 1957; Wadewitz,1978; Мельников, Лысиков, 1983; Мельников и др., 1983).
В дельте Селенги на южном Байкале нередко отмечается каннибализм (Мельников,
Лысиков, 1983) . На юге ареала в степях подобно хищникам подолгу сидят на возвыше-
ниях, высматривая добычу, или подстерегают мелких грызунов у нор (Заблоцкий, Заб-
лоцкая, 1963; Самородов, 1978а). Насекомых хватают в воздухе или собирают на
полях и лугах с растений, а также следуя за сельскохозяйственными машинами (Со-
мов, 1897; Тугаринов, Бутурлин, 1911; Тугаринов, Козлова-Пушкарева, 1935; Ewing,
Ewing, 1975; Самородов, 1978а; Picozzi, 1981).
Заметное место среди способов добывания пищи на всем ареале занимает клептопара-
зитизм. На Баренцевом море сизые чайки отбирают корм у птенцов других видов чаек
и у тупиков (Белопольский, 1957; Краснов и др., 1982). В Норвегии похищают песко-
жилов у малых веретенников (Vader, 1979а, Ь), В Швейцарии зимующие на крупных
озерах чайки специализируются на питании моллюсками, отбирая их у лысух и нырко-
вых уток (Kanel, 1977). В ГДР описаны удачные и неудачные попытки клептопарази-
тизма на чернозобых гагарах, больших бакланах, серых воронах и озерных чайках.
197
Семейство Чайковые
Количественные соотношения различных пищевых объектов, их сезонная и возраст-
ная динамика, а также изменение состава кормов по годам довольно подробно изучены
на Баренцевом и Белом морях (Горчаковская, 1948; Белопольский, 1957; Кищинский,
1960а; Герасимова, 1965; Бианки, 1967: Коханов, Скокова, 1967; Модестов, 1967;
Бианки и др., 1975). Так, на Баренцевом море, по Л.О. Белопольскому (1957), расте-
ния по частоте встречаемости составляли 34,9%, насекомые 24,2%, рыба 22%, моллюски
9,2%, ракообразные 5,5%, отбросы 2,2%, прочие корма (полихеты, иглокожие, птицы,
млекопитающие) от 0,2 до 0,5%. При этом около 50% кормов добывалось на суше;
среди морских кормов преобладали пелагические животные (70%), а донно-литораль-
ные встречались в минимальных количествах. На Белом море, напротив, чайки кормят-
ся в основном на литорали и животные объекты в их рационе преобладают над расти-
тельными (Бианки, 1967). Основу рациона здесь составляют трехиглая колюшка, ми-
дии, нереисы (Nereis pelagica, N. virens Sarv.). Из растительных кормов это вороника
и зостера. Морские корма в отличие от Баренцева моря составляют здесь 69% от общего
числа встреч; из них литоральные — 60% и пелагические — только 9%. Наземные корма
в этом регионе также богаче и разнообразнее, чем на Баренцевом море.
Экспериментальные исследования питания сизой чайки на Белом и Баренцевом мо-
рях показали, что средняя суточная норма потребления пищи у нее составляет 170—210 г
(45—57% от массы взрослой особи) при кормлении смешанной пищей из рыбы, мидий,
пескожилов и ягод (Бианки и др., 1975). При избыточном кормлении рыбой птицы по-
едают ее от 66 до 300 г, в среднем 181 г за один прием; ягод они съедают от 12,5 до
64 г в сутки. В августе последний вид корма может составлять до 35—40% суточного
рациона и является одним из основных источников витаминов. Расчетное количество
энергии, необходимое подопытным чайкам, оказалось равным 111 ккал в сутки.
Сезонная динамика питания на Баренцевом море (Белопольский, 1957) выражается
в увеличении объема потребляемых насекомых от весны к лету; в июле этот показа-
тель становится максимальным и затем снижается. Сходным образом меняется и потреб-
ление рыбы. Потребление ягод, наоборот, здесь максимально ранней весной и с конца
лета. На Белом море, по В,В, Бианки (1967), потребление грызунов резко возрастает
в июне, что связано с повышенной активностью в это время молодняка данного года
рождения; в конце мая—июне повышается процент встреч в погадках скорлупы яиц.
В июне—июле увеличивается также процент встреч в погадках рыбы, что связано с мас-
совым ее подходом; количество моллюсков и ракообразных при этом уменьшается,
так как рыба более предпочитаемый корм. Изменение состава кормов по годам упомя-
нутые выше авторы объясняют изменением их доступности. Так, на Баренцевом море
в 1946 и 1947 гг. преобладали (60—70%) наземные корма, а в 1935 и 1948 гг. — морские
(Белопольский, 1957). Повышение потребления грызунов на Белом море в 1958 г. бы-
ло обусловлено массовым их размножением в предыдущие годы (Бианки, 1967). У
птенцов по сравнению со взрослыми наблюдается обедненный состав корма.
Гораздо меньше материала по питанию сизой чайки из других частей ареала. В Латвии
основу рациона, по анализу 15 желудков (Тима, 1961), составляла рыба (63,3% по
встречаемости) и насекомые (33,3%). В условиях сильно урбанизированного ландшафта
Московской обл. сизая чайка питается в основном рыбой, погибшей от заморов и ин-
вазий, изредка посещает садовые участки, добывает дождевых червей (Самородов.
1981а). На юге ареала, в частности в Казахстане, рацион довольно разнообразен (Дол-
гушин, 1962). В него входит рыба (которая, однако, не всегда составляет его основу).
ракообразные, водные насекомые. Чайки охотно кормятся грызунами, особенно ранней
весной, а в годы массового размножения — и летом. При вспышках численности саран-
човых длительное время питаются ими. На полях собирают зерна злаков (Самородов.
1978а). По данным этого же автора, исследовавшего погадки, в 1967—1971 гг. млеко-
198
Сизая чайка
питающие в них составляли 68,3% встреч, насекомые — 41,8%, рыбы — 14,9%, птицы и
их яйца - 12,2% и растительные корма — 17,2%. В низовьях Индигирки сизые чайки
питались в основном полевками (Михель, 1935), на северном Байкале весной и летом
рыба составляла 79—84% рациона по объему (Размахнина, 1979; Скрябин, Размахнина,
1979). В Корякском нагорье и на Курильских островах осенью в больших количествах
едят погибших после нереста лососевых (Нечаев, 1969; Кищинский, 1980).
Враги, неблагоприятные факторы. На Баренцевом море основными причинами ги-
бели птёвдов сизой чайки считают истощение от недоедания, переохлаждение под густым
пологом растительно'сти и затрудненный выход из-под него на открытые места (Коханов,
Скокова, 1967). На Белом море гнезда сизых чаек и их птенцы страдают от хищдичества
горностая, лисицы, серой вороны. Взрослые и молодые особи иногда становятся жертва-
ми сапсана, орлана-белохвоста (Бианки, 1967).
Хозяйственное значение, охрана. Хозяйственного значения вид не имеет, специальные
меры охраны не предусмотрены.
Чернохвостая чайка —
Larus crassirostris Vieillot, 1818
Japanese Gull (англ.), Japanmowe (нем.), Gotland й queue noire (фр.).
Син. (англ.) : Black-tailed Gull, Temminck’s Gull.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Чайка средней величины. Взрослые пти-
цы хорошо отличаются от других чаек черной предвершинной полосой на хвосте. Серый
цвет мантии и кроющих крыла значительно темнее, чем у сизой и серебристой чаек, и по
тону близок к окраске тихоокеанской чайки. Конец крыла издали выглядит черным,
так как белые пятна на маховых очень малы. Общая окраска первогодков бурая, тем-
нее, чем у серебристой и тихоокеанской чаек. С большого расстояния такие птицы ка-
жутся черными. Молодых чернохвостых чаек в переходных нарядах можно спутать с
близкими по размеру молодыми сизыми чайками (оба вида встречаются вместе в райо-
нах миграций и зимовки). Однако чернохвостая чайка отличается тяжелыми пропорция-
ми, особенно головы и клюва, и более темной окраской всех нарядов.
Вокализация в общих чертах, как у чаек из группы серебристой, но голос звучит ни-
же, чем, например, у тихоокеанской и серебристой чаек. Весной в Приморье черно-
хвостая чайка очень криклива — из смешанных стай пролетных чаек доносится главным
образом ее голос. Крик тревоги — многократно и быстро повторяющиеся звуки
”ка-ка-ка-ка. . . ка-ка” или короткие носовые звуки ”ау. . . ау. . Нападающая птица
в момент пикирования издает громкий крик ”ку-у-а-ау!”.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Дж. Дуайт (Dwight, 1925) сближал чернохвостую чайку
с L. belcheri, L. hemprichii, L. heermanni, L. fuliginosus и некоторыми другими чайками
с темной окраской туловища, выделяя их в подрод Adelarus. Однако особенности де-
монстрационного поведения, характер возрастных нарядов и линьки указывают на
родство чернохвостой чайки с чайками группа серебристой (Литвиненко, 1980).
Описание (коллекции ЗИН, ЗМ МГУ, Дальневосточного университета, а также эк-
земпляры, добытые автором очерка). Размеры (мм) и масса тела (г):
Длина крыла:
88 (п = 15) - 366-385 (среднее 375);
99 (n = 11) — 345—393 (среднее 364).
199
Семейство Чайковые
Длина клюва:
6<5 (и = 19) — 46—59 (среднее 49,5);
9 9 (и = 16) — 43—49 (среднее 46,5).
Длина цевки:
<5 <5 (л = 19) - 52-59 (среднее 55,1);
9 9 (и = 16) — 48—57 (среднее 52,2).
Масса тела:
<5<5 (и = 6) — 523—636 (среднее 591);
9 9 (и = 12)- 437-615 (среднее 522).
Окраска. Самец и самка в брачном наряде. Голова, шея, весь низ и бока туло-
вища, а также кроющие хвоста белые, иногда с теплым розоватым оттенком. Спина,
плечевые и верхние кроющие крыла сизо-серые, на концах двух-трех самых крупных
нижних плечевых белая кайма. Первостепенные маховые со II (первое видимое) по
VI черные или темно-бурые, слегка сереющие на внутреннем опахале, начиная с III или
IV — с белым концом. Остальные 5 первостепенных аспидно-серые, светлеющие в прок-
симальном направлении и тоже с белым концом, на VII—IX — предвершинное черное
пятно или перевязь. Второстепенные маховые того же цвета, что и спина, с белой каймой
на конце и по краю внутреннего опахала. Рулевые белые с черной предвершинной пе-
ревязью, сужающейся по направлению к крайним рулевым. На 6-й паре рулевых от по-
лосы остается лишь черное предвершинное пятно на внутреннем опахале и черный
штрих вдоль стержня пера. Ширина черной полосы обычно 4—5 см, белой, идущей по
краю, 1 см. Клюв желтый с черной предвершинной перевязью. Пятна на клюве, кожа в
углах рта и края век ярко-оранжевые или киноварно-красные, радужина кремовая.
Ноги желтые, перепонка снизу с телесным оттенком, когти черные.
Самец и самка в зимнем наряде. Окраска оперения, как у взрослых птиц в брачном
наряде, но на темени, затылке, боках головы и зашейке темно-серые размытые пестри-
ны, которые сгущаются на зашейке, образуя темную перевязь, переходящую иногда
в узкое пестрое ожерелье под горлом. У некоторых птиц пестрины сливаются, образуя
равномерно окрашенный серый капюшон. Белыми остаются лоб, горло и кольцо вокруг
глаз. Клюв голубовато-серый, с желтым оттенком или без него, цвет пятен менее яркий,
чем летом. Радужина, как в брачном наряде, кремовая, края век черные, ноги желтова-
то-серые, реже голубовато-серые; иногда передние пластины на цевке буроватые.
Пуховой наряд. Основной тон светло-охристый, у разных птенцов он варьирует от
охристо-серого до охристо-рыжего. На верхней части тела и на горле буровато-черные
пятна. Низ однотонный, лишь на верхней части груди выделяется охристо-белое пятно.
Клюв голубовато-белый с беловато-розовым концом, яйцевой зуб белый. Радужина
бурая. Ноги серые с фиолетовым оттенком, когти черные с розовато-белыми концами.
Гнездовой наряд. Голова буровато-серая, более темная сверху, светлеющая с боков
и к основанию надклювья. Перед глазом темно-серое или черное полукольцо. Перья
затылка и шеи серовато-бурые с чуть заметной рыжеватой каймой, на спине и плечах
они бурые с резкой серовато-охристой или беловатой каймой. Грудь и верхняя часть
брюха дымчатая, нижняя часть белая. Маховые буровато-черные, слегка светлеющие к
краю внутреннего опахала. Рулевые тоже буровато-черные с белыми основаниями и
белой или бледно-охристой каймой шириной 1—3 мм. Клюв темно-розовый с черным
концом и бурым налетом по коньку, радужина темно-бурая. Ноги светло-коричневые
с бурым налетом на передних щитках цевки.
200
Чернохвостая нанка
Первый зимний наряд. Лоб, темя, уздечка и бока светло-серые, перед глазом серое
полукольцо, затылок и зашеек буровато-серые. Перья спины и плечевые буровато-аспид-
ные с широкой светлой охристо-серой каймой, поясница пепельная. Подбородок, сере-
дина горла, зоба и груди беловатые или совсем белые, брюхо белое.Маховые, рулевые
и большая часть кроющих крыла остаются от гнездового наряда, меняются лишь неко-
торые средние кроющие крыла, что почти не изменяет буроватую окраску обтрепанных
крыльев. Клюв, ноги, как в гнездовом наряде. На конце клюва появляется светлое
пятно.
Первый летний наряд. Выглядит почти так же, как первый зимний, отличаясь свет-
леющей головой и белым низом. Маховые и рулевые остаются от гнездового наряда
и имеют очень выгоревший, изношенный вид, спина и плечи одеты в основном в перья
первого зимнего наряда. Клюв становится кремовым с голубизной у основания. Края
век и углы рта желтоватые или бледно-оранжевые. Ноги кремовые или серые с голубо-
ватым оттенком, когти черные. Радужина бурая.
Второй зимний наряд. Лоб, темя, уздечка светло-серые, реже белые. Щеки и область
уха у многих птиц с буровато-серыми пестринами. Затылок, зашеек и передняя часть
спины буровато-аспидные и у некоторых птиц особенно хорошо выделяются на фоне
довольно светлой мантии. Спина, плечевые и поясница сизовато-серые, почти как у
взрослых птиц, резко отличаются от буроватых крыльев. Верхние кроющие хвоста бе-
лйе. Весь низ белый или с буровато-серым налетом на шее и на боках груди. Первосте-
пенные маховые со II (первого видимого) по VII темно-бурые, а с VIII по XI серовато-
бурые с белым концом. Второстепенные — серовато- или черновато-бурые с белой кай-
мой на вершине и по краю наружного опахала. Рулевые в основании на треть белые,
дальше темно-бурые с узкой (2—5 мм) белой каймой на конце. Среди рулевых появля-
ются контрастные черно-белые перья, характерные для последующих нарядов. Ноги
и светлая часть клюва принимают зеленовато-голубой оттенок, на конце надклювья
появляется оранжевое пятно; радужина бурая.
Второй летний наряд. У большинства чаек исчезают темные пестрины на голове и
шее; остальное оперение, как в предыдущем наряде. Клюв принимает различные оттен-
ки от зеленовато-голубого до зеленовато-желтого. Увеличивается размер оранжевого
пятна на конце надклювья, появляются еще два таких же пятна на подклювье. Радужина
кремовая, края век и углы рта приобретают оранжевый цвет. Цвет ног варьирует от
бледно-зеленого или голубоватого до бледно-желтого с голубым или зеленым от-
тенками.
Третий зимний наряд. Как окончательный зимний, однако первостепенные маховые
с бурым оттенком, концевые белые пятна есть лишь на нескольких проксимальных
перьях (обычно VII—XI) и на закрытом крыле не видны. Белая кайма второстепенных
маховых значительно уже, чем у взрослых птиц. Черная полоса на рулевых шире, чем у
взрослых (в среднем 7 см). Эти же ’’юношеские” черты сохраняются и в третьем лет-
нем наряде.
Линька. Частичная линька в первый зимний наряд (конец июля—январь) охватывает
перья головы, шеи, туловища, малые верхние кроющие крыла. Птенцы приступают к
ней до того, как сформируется гнездовой наряд. В июле—августе меняются перья го-
ловы, шеи, передней части туловища, в сентябре — задней части туловища. Частичная
линька в первый летний наряд (январь—май) перекрывается с началом полной линьки
во второй зимний наряд. Меняются перья головы (лоб, темя, подбородок), горла и
отдельные перья туловища.
Полная линька во второй зимний наряд (конец апреля—январь) — самая продолжи-
тельная линька чернохвостой чайки. В мае начинают меняться первостепенные маховые,
плечевые, средние кроющие второстепенных маховых; в начале июня — второстепенные
201
Семейство Чайковые
маховые, перья головы, шеи; в июле — рулевые. К концу августа все маховые, рулевые,
а также их кроющие меняются, а в сентябре—октябре большинство из них достигает
нормального размера. Линька перьев головы, туловища и проксимальных второстепен-
ных продолжается до декабря. Последними меняются кроющие хвоста. Средние крою-
щие второстепенных маховых в процессе этой линьки меняются дважды — в начале ее
(май—июнь) и в конце (сентябрь—октябрь). Последующие линьки бывают дважды в
году: частичная предбрачная и полная послебрачная; по срокам и последовательности
смены оперения они почти не отличаются от линьки взрослых.
Предбрачная линька взрослых птиц частичная, меняются перья головы и часть перьев
туловища. Начинается она, по-видимому, в январе (у добытой в феврале 1911 г. птицы
голова полностью вылиняла, на спине, плечах, груди и животе продолжалась смена
перьев) и завершается к началу марта. Послебрачная линька полная, протекает со вто-
рой половины июня по конец октября. В июне начинают меняться первостепенные махо-
вые и перья туловища, в конце июля — налаче августа — второстепенные маховые,
перья головы, шеи, в августе — рулевые. В конце сентября — начале октября маховые,
рулевые и их кроющие полностью сменяются, однако среди них еще много недорос-
ших; в основном заканчивается линька контурных перьев.
Распространение. Гнездовой ареал. Побережья Японского и Желтого морей,
а также тихоокеанское побережье Японии (рис. 46). В СССР колонии сосредоточены
главным образом в заливе Петра Великого (Южное Приморье) — на островах Фуру-
гельма, Карамзина, Стенина и Антипенко, небольшие группы птиц гнездятся также на
о-ве Де-Ливрона и на скале, расположенной у о-ва Матвеева. Кроме того, колонии есть
на о-ве Монерон (Гизенко, 1955), о-ве Попова в Татарском проливе (сообщ. Ю.В. Ши-
баева) и на южных островах Курильской гряды (острова Демина) (Велижанин, 1977).
В Японии гнездится на побережьях островов Хоккайдо и Хонсю, наиболее крупные
колонии расположены на островах Теури, Кодзима, Кабудзима и Эносима (Наканиси,
1953; Распространение гнездящихся птиц Японии, 1981; Austin, Kuroda, 1953; Fuji-
maki, 1961; Check-list of Japanese birds, 1974). На Корейском полуострове обитает на
восточном и западном побережьях, в южной его части колоний, по-видимому, нет
(Won Hong Коо, 1964; Austin, 1948). О гнездовании вида в КНР сведений мало. Единст-
венное известное место, где есть одна или две колонии, находится у побережья Шандунс-
кого полуострова (Swinhoe, 1875; Cheng, 1976). Сведения Ж. Латуша (La Touche,
1931—1934) о гнездовании чернохвостой чайки в провинции Фуцзянь вызывают сомне-
ние, так как местность лежит значительно южнее известного ареала этого вида. Кроме
того, небольшой размер яиц (56 X 41,5 и 59,5 X 41,5), найденных там, позволяет пред-
положить, что произошла ошибка в определении.
Зимовки. Зимует на юге Японского моря, в Корейском проливе, в северной час-
ти Восточно-Китайского моря вдоль гряды Японских островов (см. рис. 46). В не-
большом числе встречается и южнее (самая южная точка зимовки — о-в Сянган
(Cheng, 1976; King et al., 1976). Основной район зимовки - Корейский пролив, где в
зимний период концентрируются сардина, ставрида, анчоус, сайра и кальмар и работает
многочисленный рыболовный флот (Шунтов, 1972). В Восточно-Китайском море, по
данным этого автора, распространение чернохвостой чайки ограничивается областью
холодных вод, выходящих из Желтого моря. У юго-западной оконечности Корейского
полуострова эта чайка становится уже немногочисленной (Min, Won, 1976). На тихо-
океанском побережье Японии численность зимующих чернохвостых чаек невелика, и,
по-видимому, колеблется, возрастая в годы массового хода иваси (Komatsu, 1935).
Из 6 зимних находок птиц первого года жизни, окольцованных в Приморье, 5. относят-
ся к берегам Японии, из них 3 чайки встречены на о-ве Кюсю. Еще одна зимняя встреча
в заливе Терпения на Сахалине, по-видимому, случайна, так как залив покрывается
202
Чернохвостая чайка
льдом. Неполовозрелые чернохвостые чайки зимуют в более южных районах, чем
взрослые (Шунтов, 1972); в Восточно-Китайском море зимой среди чаек преобладают
молодые птицы, в Японском — взрослые.
По результатам кольцевания, основной район зимовки молодых чаек Южного При-
морья — воды, омывающие западные берега островов Кюсю, Сикоку и южного Хонсю
(Литвиненко, Шибаев, 1972); чаек, гнездящихся на Северном Хонсю (о-в Кабудзи-
ма), — восточные берега тех же островов (Narita,
1979).
Миграции. В августе—сентябре начинаются после-
гнездовые кочевки чаек,гнездящихся в заливе Петра
Великого. Судя по результатам кольцевания (220
возвратов от 23 550 окольцованных птиц), большая
часть птиц летит в северо-восточном направлении,
придерживаясь линии побережья, и скапливается в
Татарском проливе. Многие чайки остаются в Юж-
ном Приморье, и лишь небольшое число птиц на-
правляется на юго-запад, вдоль побережья Корей-
ского полуострова, к местам зимовки. Основной
причиной откочевки к северу является, скорее
всего, недостаток пищи в районе гнездования. Из-
вестно, что Татарский пролив — наиболее продук-
тивный район Японского моря в летний период
(Шунтов, 1972). В конце сентября, когда субтро-
пические виды рыб (сайра, анчоус, сардина, скум-
брия) отходят в южную часть моря (Шунтов,
1972), начинается обратное движение чернохвостых
чаек. Отлет к югу проходит в сжатые сроки, и к
концу октября этих птиц можно встретить в не-
большом числе лишь вблизи населенных пунктов
Рис. 46. Область распространения
чернохвостой чайки
1 — колонии, 2 — область зимов-
ки, 3 — залеты, 4 — сведений о
зимовке нет
Приморья. Последние чайки из Южного Приморья
улетают в конце ноября. Наиболее ранняя встреча на зимовке (юг Корейского полу-
острова) чайки первого года жизни, окольцованной в заливе Петра Великого, зареги-
стрирована 16.Х. Послегнездовые кочевки японских чернохвостых чаек имеют аналогич-
ный характер (Narita, 1979).
Массовый весенний отлет чернохвостых чаек из районов зимовок проходит в мар-
те-апреле, наиболее мощный поток мигрантов идет в западной половине Восточно-
Китайского моря и вдоль берегов Кореи (Шунтов, 1972). В Южном Приморье первые
чернохвостые чайки появляются в начале марта, но основная часть птиц прилетает
в конце этого месяца.
Залет отмечен на о-в Атту (Алеутские острова) в мае 1980 г. (Harrison, 1983).
Численность. Общая численность репродуктивной части вида, по-видимому, не превы-
шает 500 тыс. особей. В СССР гнездится примерно 100 тыс., из них около 96 тыс. в зали-
ве Петра Великого: на о-ве Фуругельма около 80 тыс. (учет 1983 г., устное сообщ.
Ю.В. Шибаева), на о-ве Карамзина около 11 тыс. (учет 1972 г.), на о-ве Стенина не более
5 тыс., на о-вах Антипенко несколько сотен (сообщ. Ю.В. Шибаева), несколько десят-
ков пар — на о-ве Де-Ливрона и на небольшой скале у о-ва Матвеева. В последние 10 лет
численность чаек на о-ве Фуругельма выросла примерно в 1,5 раза, что связано, по-ви-
димому, в основном с организацией в заливе Петра Великого Дальневосточного госу-
дарственного морского заповедника, в состав которого вошел этот остров. Примерно
2 тыс. птиц гнездятся на о-ве Монерон (Гизенко, 1955), 600—700 особей на о-ве Попова
203
Семейство Чайковые
в Татарском проливе (устное сооб. Ю.В. Шибаева) и около 800 птиц на южных островах
Курильской гряды (Велижанин, 1977).
Число птиц, населяющих 4 наиболее крупные колонии Японии, не превышает 150 тыс.
(о-в Теури — 40—50 тыс. птиц, о-в Кодзима — 5—7 тыс., о-в Кабудзима — около 10 тыс.,
о-в Эносима — несколько десятков тысяч) (Наканиси, 1953; Austin, Kuroda, 1953;
Udagawa, 1953; Kuroda, 1963; Watanuki, 1982). Если предположить, что в других,
менее многочисленных колониях (их около 20) гнездится такое же количество птиц,
то и тогда общее число чернохвостых чаек, населяющих Японию, составит не более
300 тыс. О численности этого вида в Корее и КНР сведений нет.
Местообитания. Гнездится на морских островах. Колонии располагаются на скалис-
тых обрывах или склонах, покрытых травой. Кормится в море, на пляжах, прибрежных
лугах, пресных озерах, лагунах. Иногда проникает в глубь материка, но обычно не далее
10 км. Во время миграции и зимовок предпочитает морские побережья и прибрежные
морские воды. Нередко отмечается на городских свалках и зверофермах.
Размножение. Приступает к размножению в возрасте около 3 лет (2 года 11 меся-
цев) , когда наряд еще сохраняет ’’юношеские” черты. Гнездится колониями от несколь-
ких десятков до 40 тыс. пар. Развит гнездовой консерватизм — 70% чаек, окольцован-
ных на о-ве Кабудзима (Austin, Kuroda, 1953), были пойманы там на следующий год,
48% из них устроили гнезда на прежнем месте. В Приморье в непосредственной близос-
ти от гнезд чернохвостой чайки гнездятся уссурийский и берингов бакланы, тонкоклю-
вая кайра, очковый чистик, тупик-носорог, старик, пестролицый буревестник, малая
качурка, серая цапля.
В агрессивном и брачном поведении очень много общего с чайками из группы сереб-
ристой (Литвиненко, 1980). Наиболее распространенная демонстрация при территори-
альных конфликтах — прямая поза. Варианты позы различной степени агрессивности
и сопутствующие элементы этой демонстрации очень похожи на таковые у серебристой
чайки. Долгий крик состоит из двух, реже — трех фаз (рис. 47). После одного-двух
глухих звуков, которые птица издает чуть вытянув шею вперед (первая фаза), она
резко опускает голову вниз и назад, часто при этом прижимая подклювье к брюху и
сопровождая это движение двумя-тремя хрипловатыми криками (II фаза). Затем пти-
ца так же резко вскидывает голову и издает серию громких и чистых, постепенно зати-
хающих звуков (III фаза), при этом крылья в локтевых сгибах всегда слегка отстав-
лены. I фазу птицы часто пропускают. Продолжительность долгого крика 6—8 с, изда-
ют его птицы, сидящие, стоящие, плавающие и летящие. Из других демонстраций, харак-
терных для крупных белоголовых чаек, отмечен мяукающий крик, ’’кашляние”, или
’’квохтанье”, вскидывание клюва. Спаривание, включая предшествующее ему ритуаль-
ное кормление, своеобразное вскидывание клюва и копуляцию, выглядит, в общем,
как у других чаек. Инициатор спаривания — всегда самка. Копуляционный крик сам-
ца — быстрое аритмичное ”ко-ко-ко-кох-кооо-о-ко-ко-кго-кого...”.
На территории колоний чайки Южного Приморья появляются обычно в начале ап-
реля. К середине месяца большинство птиц занимает гнездовые участки. Сроки размно-
жения чернохвостых чаек залива Петра Великого и чаек из японских колоний (остро-
ва Теури, Кабудзима), расположенных приблизительно в одних широтах, совпадают
(Komatsu, 1935; Kuroda, 1963), на о-ве Киодзима (южный Хонсю) гнездование начи-
нается на 2 месяца раньше (Wada, 1932).
После 20.IV птицы приступают к постройке гнезд. При выборе участка важным ус-
ловием является наличие укрытия для птенцов (от солнца, хищников) и хороший об-
зор. Излюбленные места — травянистые склоны с выступающими грядами камней;
отвесных скал и участков с высокой густой травой чайки избегают. Состав раститель-
204
Чернохвостая чайка
ности на территориях колоний довольно однообразен: лебеда, крестовник, полынь,
колосняк.
Основной строительный материал гнезд — сухая прошлогодняя растительность по со-
седству с гнездом. Гнезда, расположенные ближе к воде, могут быть выстроены из
зостеры, а гнезда у границы леса — из прошлогодних листьев дуба, липы. Средние раз-
меры гнезд (мм): диаметр гнезда 301, диаметр лотка 177, глубина лотка 44 (п = 18).
Средняя плотность гнездования (учет на 17 пробных участках, площадью 100 м2)
Рис. 47. Долгий крик чернохвостой чайки
д— П фаза, б — III фаза
0,29 на 1 м2 (устное сообщ. Ю.В. Шибаева). Среднее расстояние между двумя соседними
гнездами 1,75 м (о-в Карамзина), 1,52 м (о-в Фуругельма), наименьшее — 30 см.
Откладка яиц у чернохвостых чаек залива Петра Великого начинается в первых чис-
лах мая. реже в середине мая. В кладке 2—3 яйца, изредка (0,1—0,2%) — 4 (и = 800).
Птицы часто подкладывают в гнезда небольшие (размером с яйцо или крупнее) камни.
Они попадаются в 20% гнезд. Окраска яиц: по коричневато- или серовато-оливковому
фону разбросаны темно-бурые и зеленоватые пятна (поверхностные и глубокие), ре-
же — линии. Пятна сгущаются на тупом конце, иногда образуя ’’тонзуру”. Реже фон
бывает голубовато-оливковым, а еще реже — палевым. Средние размеры яиц (мм)
63,07 X 43,58 (и = 108) (Литвиненко, 1980).
Насиживание длится 24—25 дней, редко 27 (Komatsu, 1935). Насиживающие птицы
меняются обычно через 6—7 ч. В начале ритуала смены партнеров на гнезде обе птицы
обмениваются долгим криком. Иногда этого бывает достаточно, чтобы насиживающая
птица сошла с гнезда. Но чаще вслед за этим обе птицы издают мяукающий крик. Если
после неоднократного его повторения насиживающая птица не уходит с гнезда, партнер
прибегает к демонстрации ’’кашляния”, приносит к гнезду строительный материал или
(1 случай) пытается столкнуть насиживающую птицу с гнезда.
Птенцы появляются в конце мая — начале июня. Процесс вылупления занимает
2—3 дня. Скорлупа выбрасывается обычно в день вылупления. Масса только что вы-
лупившихся пуховичков (и = 5) 36—44,5, в среднем 40,2 г. У большинства 3—4-дневных
пуховиков уже ясно намечаются птерилии. На 5—6-й день появляются первые пеньки на
плечевой и грудной птерилиях. В это же время обычно отпадает яйцевой зуб. С 7-го на
8-й день пробиваются второстепенные, а затем первостепенные маховые и кроющие
крыла.
Через несколько часов после вылупления птенец получает первую порцию корма.
Кормление птенцов обычно сопровождается мяукающим криком. Заслышав сигнал
205
Семейство Чайковые
опасности, птенец уже в первый день покидает гнездо и прячется за его краем, а через
день-два убегает в ближайшие кусты травы. У каждого выводка или отдельного птен-
ца на гнездовом участке есть постоянные укрытия, часто в таких местах прячутся
вместе птенцы из соседних выводков.
В возрасте 35—40 дней птенцы поднимаются на крыло. Массовый вылет молодых
чаек в заливе Петра Великого происходит в середине июля. В августе чайки Южного
Приморья покидают гнездовую колонию.
Выживаемость птенцов в 1970 г. на о-ве Карамзина составила примерно 70%; в
1976 г. на о-ве Фуругельма — 63%, здесь на 1 гнездящуюся пару приходилось 1,2 слет-
ка (55 гнезд). Причины смертности: раны, нанесенные взрослыми чайками, — 39,4%,
переохлаждение во время сильного додждя — 13,1%, неизвестные причины — 1,6%,
птенцы не были найдены учетчиком - 31,6% (некоторые из этих птенцов, возможно,
выжили). Основная причина гибели в Приморье — преследование взрослыми чайками
птенцов, убежавших от родителей из-за паники в колонии или непогоды. В местах,
где чернохвостая чайка гнездится рядом с тихоокеанской, часть птенцов (на о-ве Теури
4% от вылупившихся) гибнет из-за хищничества последней (Watanuki, 1982).
Питание. Основу питания составляет рыба, насекомые, ракообразные и отбросы, реже
в корме встречаются моллюски, морские ежи, многощетинковые черви, случайно —
мелкие воробьиные и грызуны. Ловит рыбу, как все чайки, с поверхности воды или
ныряя на небольшую глубину (лишь иногда полностью скрываясь под водой). Охотно
кормится в местах рыбных промыслов, следуя за рыболовными судами. Во время ноч-
ного лова сайры охотится ночью при свете судовых прожекторов. Собирает водных
беспозвоночных на мелководьях и во время отлива. Добычу с жестким панцирем
(крабов, морских ежей) разбивает о камни, бросая ее с высоты нескольких метров;
у колюшки, перед тем как ее проглотить, ломает колючки, ударяя ею о твердый грунт.
Насекомых ловит, преследуя их в воздухе, выпугивая (при этом она летает ’’челно-
ком” над самой землей) или расхаживая пешком и разыскивая их в траве. Собирает
отбросы на свалках, в море — у выходов городских коллекторов, на зверофермах и
рыбокомбинатах.
Склонна к пиратству (клептопаразитизму) — в гнездовых колониях Южного При-
морья отбирает добычу у очкового чистика, тупика-носорога. На зимовках отбирает
рыбу у моевки, озерной чайки, речной крачки, уссурийского баклана; отмечен случай
нападения ее на летящего с добычей черного коршуна (Nakamura, 1972).
По материалам, собранным в 1968—1976 гг., в Южном Приморье предпочитаемым
кормом чернохвостой чайки является рыба, при недостатке ее в питании возрастает
значение насекомых. В плохую погоду (шторм, дождь), когда добыча этих объектов
затруднена, многие чайки переключаются на сбор отбросов.
В марте—апреле, когда массовые виды рыб еще не проникли в северную часть Япон-
ского моря, а большинство прибрежных водоемов и морских мелководий покрыты
льдом, чайки кормятся главным образом отбросами. В конце апреля — начале мая
в лагуны Южного Приморья заходит на нерест трехиглая колюшка (Gasserosteus acu-
leatus), с этого времени здесь кормятся тысячи чернохвостых чаек. Встречаемость
колюшки в корме в апреле составляет 84%. В мае, кроме колюшки (32,8%), чайки
кормятся также анчоусом Engraulis japonicus (35%) и насекомыми Pentatomidae
и Carabidae (около 20%).
В конце июня количество рыбы в рационе чаек значительно снижается, одновремен-
но возрастает значение насекомых, встречаемость которых превышает 50%. В основ-
ном это различные Carabidae (31,2%), Scarabeidae (14,5%), Diptera (15,6%). В июле,
когда к берегам Приморья подходит сайра (Cololabis saira) и тихоокеанская сельдь
(Clupea pallasi), чайки переключаются на питание этими видами (встречаемость их со-
206
Вилохвостая чайка
ответственно 56,6% и 23,2%). В начале августа сайра все еще остается основным кор-
мом чернохвостой чайки (встречаемость 54%), но к середине месяца снова начинает
расти доля насекомых. Теперь это в основном крылатые особи муравьев — Lasius,
Camponotus (16,3%), Curculionidae (18,4%) и Lepidoptera (18,4%).
В октябре — начале ноября набор кормов разнообразен, но главным образом это
рыба: мелкая навага Eleginus gracilis, красноперка Leuciscus brandti, колюшка Нуро-
mesus olidus, Н. pteriosus, а также ракообразные: Crangon septemspinosa, Pandalus
latirostris, Anisogammarus kugi и отбросы.
Сравнение характера питания чернохвостых чаек Южного Приморья за десятилет-
ний период с 1968 по 1977 г. показало отсутствие различий как в составе питания,
так и в сезонной смене кормовых спектров. Однако в последние годы (1982—1984 гг.)
состав корма изменился — в частности, главенствующую роль в летнем питании чаек
теперь занимает не сайра, а иваси Sardinops sagax. Связано это с изменениями в гидро-
логическом режиме моря, вызвавшими изменения состава ихтиофауны.
Хозяйственное значение, охрана. Чернохвостая чайка нуждается в охране как ло-
кально распространенный вид, ареал которого совпадает с наиболее освоенными райо-
нами стран Дальнего Востока. В настоящее время состояние вида благополучное. В на-
шей стране колонии островов Фуругельма и Стенина включены в состав Дальневосточ-
ного государственного морского заповедника, а о-в Карамзина объявлен памятником
природы. Таким образом, примерно 95% чернохвостых чаек, обитающих в СССР, взяты
под охрану. В Японии охраняются 5 колоний, расположенных на островах Эносима,
Фумидзима, Кабудзима, Тобисима и Цубакидзима (Наканиси, 1953).
Род Хета Leach, 1819
Вилохвостая чайка -
Хета sabini (Sabine, 1819)
Sabine’s Gull (англ.), Schwalbenmbwe (нем.), Mouette de Sabine (фр.).
Син. (лат.): Larus sabini Sabini, 1819.
Син. (рус.): малая чернокрылая чайка. ,
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Сравнительно мелкая, довольно длин-
нокрылая чайка с темной головой и черным полем по переднему краю крыла, клино-
образно расширяющемся к его концу. Кроме того, от всех прочих отличается сильно
вырезанным, вильчатым хвостом (глубина вырезки достигает 1/4 его длины) и двух-
цветным черным с желтой вершиной клювом. По типу окраски первостепенных махо-
вых, пропорциям ног, полету и способам охоты приближается к крачкам. Голос также
напоминает голос крачек, в нем преобладают трескучие звуки.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. В.К. Штегман (Stegmann, 1934) выделил подвиды X. s.
sabini и X. s. palaearctica, а Л.А. Портенко (1939) внутри палеарктических птиц разли-
чал чукотскую X. s, tschuktscherum. Кроме того, он же описал X. s. woznesenskii
из Западной Аляски, самую темную и мелкую. Однако различия, характеризующие
эти формы, слишком малы и постепенны, чтобы служить основанием для выделения
подвидов.
Описание. Строение и размеры (табл. 13).
Окраска. Самец и самка в летнем наряде. Голова, горло и верхняя часть шеи
сизовато-аспидные. Нижняя часть шеи, весь низ и бока туловища, все кроющие хвоста
и рулевые белые. Темное оперение головы и белое шеи разделены узкой черной поло-
207
Семейство Чайковые
Таблица 13. Вилохвостая чайка. Размеры (мм)
Географический район Пол Длина крыла
п lim среднее
Гренландия <5 <5 2 273-280 277
9 9 2 262-263 262,5
Территория от Таймыра до Чукотки <5 <5 12 270-287 280
9 9 12 257-285 268
Чукотка <3 <5 17 262-273 271
Западная Аляска <3 <3 1 264 -
9 9 1 263
сой, образующей ошейник. Спина, плечевые и верхние кроющие предплечья светлые
аспидно-сизые. Шесть дистальных первостепенных маховых буровато-черные с белыми
кончиками и того же цвета широкими полосами по краю внутренних опахал. Остальные
первостепенные и дистальные второстепенные белые. Крылышко и верхние кроющие
кисти черные. Клюв черный с желтой вершиной, ноги темно-серые с черными когтями.
Радужина темно-бурая, края век ярко-красные.
Самец и самка в зимнем наряде (Cramp, Simmons, 1983) похожи на птиц в летнем
пере, но передняя часть и бока головы, а также подбородок и верхняя часть груди
белые. Перед глазом и в области уха черные пятна, затылок и верхняя часть шеи черно-
ватые. От глаза или уха к затылку идет темно-серая полоса. Верхняя часть мантии, бока
шеи и груди светло-серые.
Пуховой наряд (Cramp, Simmons, 1983). Верхняя сторона тела теплого золотисто-
бурого тона с черными пятнами и полосами. Одиночные пятна располагаются над клю-
вом и по бокам подбородка, много их в нижней части спины. Узкая полоса имеется
над глазом (иногда и под ним), на темени — три продольные, на затылке — одна попе-
речная. В межлопаточной области две широкие продольные полосы, переходящие в
испещренность в нижней части спины. Нижняя сторона тела бурая с белой или беловатой
серединой груди и брюшка.
Гнездовой наряд. Верх головы, шеи, плечевые и большая часть верхних кроющих
крыла, спина, надхвостье и бока зоба буровато-серые с охристыми вершинными каем-
ками на перьях. На плечевых и верхних кроющих крыла развиты, кроме того, чернова-
то-бурые предвершинные поперечные полоски. Лоб и надбровная полоса грязно-белые
с охристым налетом, у переднего угла глаза черноватое пятно. Подбородок, горло,
весь низ и бока туловища белые. Пять дистальных первостепенных буровато-черные
с широкой белой полосой по краям внутренних опахал. На двух следующих перьях
появляются светлые вершинные пятна. Остальные первостепенные, так же как и ди-
стальные второстепенные, белые. На проксимальных второстепенных появляется буро-
вато-серый налет, занимающий на самых внутренних из них большую часть пера. Боль-
шие верхние кроющие предплечья буровато-серые в основании и белые на вершинах,
средние и малые — буровато-серые. Кроющие хвоста и рулевые белые, на рулевых раз-
виты буровато-черные вершины и узкие охристые концевые каемки. Клюв черноватый
с розовато-серым основанием подклювья, ноги серовато-телесные.
Первый зимний наряд (Dwight, 1925). Голова, шея, спина и вся нижняя часть туло-
вища, как у взрослых в зимнем пере. Крылья и хвост от гнездового наряда.
Первый летний наряд (Cramp, Simmons, 1983). Похож на соответствующий взрос-
208
Вилохвостая чайка
Длина клюва Длина цевки
п lim | среднее п lim среднее
2 26,0-26,0 2 33,0-33,0 —
2 23,0-24,0 23,5 2 30,0-32,0 31,0
12 24,0-27,0 25,9 12 33,0-36,0 34,2
12 24,0-27,0 25,9 12 30,0-36,0 33,0
17 24,0-29,0 25,4 17 30,0-35,0 33,2
1 25,0 — 1 35,0 —
1 25,0 — 1 34,0 —
лый, но на темной шапочке заметна примесь белых перьев или же лоб белый. Задняя
часть и бока шеи, а также бока.груди серые. На рулевых отдельные черные пятна.
Линька. Изучена слабо. Линька в первое зимнее перо частичная, охватывает оперение
туловища и головы, полнота ее точно не установлена, о сроках известно мало (Cramp,
Simmons, 1983). У экземпляров, добытых в середине сентября в Калифорнии (Dwight,
1925), а также в сентябре и в декабре во Франции (Bureau, 1901), она только начинает-
ся. Линька в первый летний наряд полная, рулевые и маховые от гнездового наряда
меняются в феврале—мае (Stresemann, Stresemann, 1966). Линька во второй зимний
наряд также полная, сроки ее протекания неясны (Cramp, Simmons, 1983). Линька
взрослых в зимний наряд. На местах гнездовий экземпляры, добытые от Таймыра
до о-ва Врангеля, меняют лишь мелкое перо на груди и брюшке. В конце августа про-
должают встречаться как птицы, у которых линяет мелкое оперение туловища (в част-
ности, на верхней части спины), так и совсем не линяющие. Линька маховых происхо-
дит уже на зимовках (Lambert, 1975) с конца ноября, у большинства — в первой поло-
вине декабря. Линька оперения головы и туловища начинается в августе или сентябре и
заканчивается к октябрю. Линька взрослых в летний наряд частичная, охватывает
оперение головы и туловища, протекает, вероятно, в марте—апреле (Cramp, Simmons,
1983).
Распространение. Гнездовой ареал. Располагается циркумполярно, но
отдельными пятнами, приблизительно между 60° и 81° с.ш. (рис. 48), охватывает
прибрежные материковые тундры (на Аляске к югу до залива Квичак) и острова Шпиц-
берген, Гренландию, Канадский Арктический архипелаг (Salomonsen, Gitz-Johanson,
1944; Snyder, 1957; Gabrielson, Lincoln, 1959; Lovenskiold, 1964). В СССР гнездова-
ние доказано на Таймыре (ЗИН АН СССР; Middendorff, 1851; Тугаринов, Толмачев,
1934; Scalon, 1938), Новосибирских островах (ЗИН), в низовьях Индигирки (к западу
до р. Хромы) (Успенский и др., 1962), Колымы, Чауна и в прилежащих районах
(Schaanning, 1928; Воробьев, 1963; Остапенко, 1973; Засыпкин, 1981), а также на
крайнем северо-востоке Чукотского полуострова и о-ве Врангеля (ЗИН; Портенко,
1973; Дорогой, 1984) (рис. 49) .
Зимовки. Размещаются у западных и южных побережий Африки, наиболее мас-
совые скопления обнаружены здесь между 25° и 10° с.ш. и 17—34° с.ш. (Lambert,
1975; Curry-Lindal, 1981). В тихоокеанском бассейне отмечалась в это время года от
южных берегов Британской Колумбии в Канаде до берегов Перу (Chapman, 1969).
Наиболее массовые зимовки зарегистрированы, однако, в водах холодного Перуанско-
го течения между 4° и 21° ю.ш. На зимовках у берегов Африки появляются в октябре,
209
210
Рис. 48. Область распространении вилохвостой чайки
/ область гнездования и отдельные поселения, 2-- места зимовок
Вилохвостая чайка
достигая максимальной численности в декабре—феврале. Отдельные стаи насчитывают
здесь в это время до 1000 особей (Loutendyk, 1965, 1968; Lambert, 1975). Покидают
зимовки в марте—апреле. На тихоокеанских зимовках наблюдаются с ноября по апрель,
наибольшей численности достигают здесь в декабре (Chapman, 1969) и, так же как в
Атлантике, держатся крупными стаями.
Кочующие особи встречаются далеко за пределами гнездового и зимнего ареа-
лов. Так, в СССР отдельные встречи известны в Горьковской обл., на Енисее
Рис. 49. Ареал вилохвостой чайки в СССР
1 — области гнездования, 2 — отдельные поселения
(около 62° 20' с.ш.) и даже к северу от Новосибирских островов (Портенко, 1946;
Воронцов, 1967; Рогачева, и др., 1978). Вне СССР одиночные кочующие особи реги-
стрировались в Швейцарии, ГДР, ФРГ, Скандинавских странах, Польше, Венгрии, Румы-
нии, Италии, Египте, Израиле, а также у юго-восточных берегов Кореи (Gore, Won,
1971; Cramp, Simmons, 1983).
Миграции. Весной единичные птицы встречаются на пролете в июне (с 5 по 19) в райо-
не Кроноцкого заповедника у восточного побережья Камчатки (Лобков, 1980), на Ко-
мандорских островах (Иогансен, 1934), в Корякском нагорье (Кищинский, 1980) ина
Анадыре (Портенко, 1939). На о-ве Врангеля пролет наблюдается в первой декаде июня
(Портенко, 1973; Дорогой, 1984). В Чаунской низменности в 1975 г. массовый пролет
зарегистрирован с 4 по 6 июня (Кондратьев, 1979). Вне СССР небольшое число мигри-
рующих особей замечено в апреле в юго-восточной части Берингова моря (Шунтов,
1961).
Осенью на Новосибирских островах единственный взрослый экземпляр добыт
1.IX 1939 г. (Рутилевский, 1958). На северном побережье о-ва Врангеля и в Колючин-
ской губе взрослые и молодые мигрирующие особи были встречены в третьей декаде
августа, а в первых числах сентября зарегистрирован массовый пролет в районе Уэлле-
на, одиночные птицы отмечались здесь до 1 октября (Портенко, 1973; Кречмар и др.,
1978). В заливе Анива на Сахалине одна молодая особь была замечена с борта судна
26.Х 1974 г. (Остапенко и др., 1977).
В Атлантике осенний пролет идет вдоль восточных побережий океана, где массовые
скопления птиц наблюдали у Британских островов (Ricard, 1966) и Испании (Biermann,
1966). Иногда в это время года встречаются также в бассейне Балтийского и Северного
морей (Witherby et al., 1941; Дементьев, 1951; Ренно, 1968; Lambert, 1975; Sermet,
Muriset, 1982). В Тихом океане пролет идет в основном вдоль американских берегов
(Bent, 1921; Murphy, 1936; Портенко, 1939, 1973; Gabrielson, Lincoln, 1959; Wolford,
1972; Taylor, 1972; Cottam et al., 1975).
211
Семейство Чайковые
Численность. В период размножения в СССР нигде не бывает большой. На зимовках,
как упоминалось, птицы концентрируются в крупных стаях.
Местообитания. Низменные прибрежные болотистые тундры и плоские островки,
покрытые мохово-травянистой растительностью. Во внегнездовой период — морские
побережья.
Размножение. На места гнездования прибывает в разные годы с конца мая — начала
июня (Чукотка, о-в Врангеля) до середины июля (восточная часть Таймыра), когда на
реках и озерах появляются лишь первые промоины (Middendorff, 1851; Бутурлин,
1905; Scalon, 1938; Остапенко, 1973; Портенко, 1973; Кондратьев, 1979). Иногда
поселяется на мелких островках на озерах и реках, чаще среди болотистой тундры или
по берегам озер, как исключение — на галечных морских косах, нередко совместно с
полярными крачками (Middendorff, 1851; Лебедев, Филин, 1959; Воробьев, 1963;
Портенко, 1973; Кищинский и др., 1983; Дорогой, 1984; Кондратьев, Кондратьева,
1984), обычно не далее 20—25 км от берега моря, но известны случаи гнездования
и в 50—100 км и более от морских побережий.
Демонстративное поведение имеет некоторые особенности, редко встречающиеся или
вообще отсутствующие у других чаек (рис. 50). К ним относятся: выразительность и
частое применение ’’сутулой” (Crouch) и ’’распластанной” поз, в том числе при долгом
крике, своеобразная форма прямой позы (Quarter upright). Характерны демонстра-
ция ’’изгиб с поклоном” (Arch-and-Bow) и редкое применение вскидывания клюва в
процессе ритуального кормления и перед спариванием; прямое кормление (а не отрыж-
кой) самки самцом в начале сезона размножения; привлечение самки на свободную
территорию с помощью демонстрации ’’изгиб с поклоном”, а не с помощью долгого
крика (Brown et al., 1967).
Гнездится обычно отдельными парами, небольшими группами или разреженными
колониями численностью до 12 пар (Лебедев, Филин, 1959; Воробьев; 1963; Brown
et al., 1967). Гнезда представляют собой углубления во мху, выстланные сухими листья-
ми и стеблями злаков, осок и другой травянистой растительностью. В качестве строи-
тельного материала используются также водоросли и перья птиц. Размеры гнезд (мм)
в разных частях ареала варьируют: диаметр лотка 80—135, глубина его от 10 до 30;
наружные стенки гнезда обычно плохо выражены (Middendorff, 1851; Schaanning,
1928; Воробьев, 1963; Портенко, 1973; Дорогой, 1984).
Откладка яиц на о-ве Врангеля начинается в конце второй декады июня (Дорогой,
1984), в Чаунской губе и на о-ве Айон, по-видимому, в первую декаду июня (Schaan-
ning, 1928; Остапенко, 1973). Свежие кладки в низовьях реки Коньковой найдены
в третьей декаде июня (Воробьев, 1963); на Таймыре, в устье р. Гусихи, слабо наси-
женные яйца обнаружены 12 июля (Scalon, 1938).
Кладка содержит обычно 2—3 яйца. На побережье Чаунской губы полные кладки
состояли из 1—3 яиц, в среднем (п = 49) 2,6 ± 0,42 яйца (Кондратьев, Кондратьева.
1984). На о-ве Врангеля средняя величина кладки была (п = 32) 2,7 яйца (Дорогой.
1984). При этом 11 кладок содержали по 2 яйца, 20 — по 3 и 1 кладка — 4 яйца. Пов-
торные кладки зарегистрированы К.А. Воробьевым (1963) на северо-востоке Якутии
в низовьях р. Коньковой и М.Ю. Засыпкиным (1981) на западе Чукотки в низовье
р. Чаун. Возможны они, по мнению А. Миддендорфа (1851), и на Таймыре.
Яйца (Makatsch, 1974) коричневато- или зеленовато-оливковые с расплывчатым
рисунком из более темных оливково-коричневых поверхностных и серовато- или зе-
леновато-коричневых внутренних пятен. На тупом конце яйца рисунок, как правило,
более густой. По окраске яйца вилохвостых чаек неотличимы от яиц розовых чаек.
Размеры яиц (мм) в разных частях ареала следующие: на о-ве Врангеля (п = 59) 40,2-
212
Вилохвостая чайка
47,7 X 30,5-33,5, среднее 44,1 X 31,8 (Дорогой, 1984); в Чаунской губе (и = 9) 40,5-
47,0 X 31,3-34,0, среднее 45,1 X 32,9 (Schaanning, 1928); в Якутии (и = 9) 40,8
47,3 X 30,1—32,5, среднее 44,7 X 31,7 (Воробьев, 1963). В сводке В. Макача (Makatsch,
1974) приводятся следующие размеры яиц. Из Восточной Сибири (сборы В.Е. Флинта):
средние (п = 14) 44,7 X 32,3, максимальные 47,6 X 32,0 и 42,5 X 33,2, минимальные
42,5 X 33,2 и 44,4 X 31,3; с Аляски и северо-восточной Сибири: средние (п = 9) 43,7 X
X 31,6, максимальные 45,5 X 30,2 и 42,2 X 33,1, минимальные 41,4 X 31,1 и 45,5 X 30,2;
Рис. 50. Демонстрационное поведение вилохвостой чайки (по: Brown et al., 1967, с изменениями)
а-д — последовательность поз долгого крика, е — прямая поза (Quarter upright), ж— з— ’’изгиб с
поклоном”
из Северной Америки (Bent, 1921): средние (и = 56) 45,5 X 32,0, максимальные 49,5 X
X 31,5 и 47,0 X 34,0, минимальные 39,5 X 30,0. Масса (г) 26 слабо насиженных яиц
на о-ве Врангеля (Дорогой, 1984) 23,0—25,6; яйца средней степени насиженности имели
массу 19,5—22,5,в среднем (п = 33) 21,2.
Насиживают кладку оба партнера в течении 23—26 дней (Makatsch, 1974). На о-ве
Врангеля на откладку трех яиц самки затрачивают от 3 до 5 сут, насиживание начинается
с первого яйца (Дорогой, 1984). У гнезд птицы обычно очень агрессивны, активно за-
щищают их, пикируя на нарушителя, ударяя его клювом и лапами и обливая пометом
(Воробьев, 1963; Лебедев, Филин, 1959; Портенко, 1973; Кондратьев, Кондратьева,
1984).
Вылупление птенцов в разных частях ареала протекает с конца июня - первой поло-
вины июля. На Чукотке (р. Канчалан) 13 июля видели маленького пуховичка, которого
еще грели родители (Кищинский и др., 1983). На о-ве Врангеля пуховичек был найден
19 июля (Успенский и др., 1963), а в устье р. Чаун В.А. Остапенко (1973) обнаружил
только что вылупившегося птенца 27 июня. 29 июля на Таймыре А. Миддендорф (1851)
отметил уже подросших пуховичков. Многие родители при опасности отводят от гнезд
213
Семейство Чайковые
с кладками на стадии вылупления или с маленькими птенцами, хотя и не так активно,
как это делают некоторые кулики или поморники (Brown et al., 1967).
Успех вылупления птенцов на западе Чукотки (низовья р. Чаун) в разные годы
составлял 80—82% (Кондратьев, Кондратьева, 1984), при этом основной причиной от-
хода яиц была гибель эмбрионов на ранних (до 2 недель) стадиях развития. Период
вылупления всех птенцов выводка длится, по данным этих же исследователей, от 1,5
до 4 сут. По наблюдениям на Аляске (Brown et al., 1967), сразу после вылупления
птенцов родители удаляют скорлупу из гнезда.
По наблюдениям в низовьях р. Чаун на западе Чукотки, средняя масса только что
вылупившихся птенцов (п = 13) составляет 16,2 г. Первую порцию корма пуховички
получают, вероятно, уже через 2—4 ч после вылупления и к концу третьих суток разви-
тия удваивают массу. В возрасте от 4 до 14 сут прирост массы у них остается постоян-
ным, но более высоким, чем в первые трое суток; у некоторых особей он достигает
20—25 г в сутки. Позднее этот показатель несколько снижается, но к 16—18-дневному
возрасту птенцы весят (п = 16) от 180 до 225 г, что составляет 110% от массы взрослых
птиц. К моменту поднятия на крыло масса молодых несколько снижается, затем снова
немного увеличивается. Развитие оперения у птенцов идет достаточно интенсивно, хотя
и не так быстро, как у розовых чаек. Уже к 20-дневному возрасту оперение крыльев
и хвоста настолько развито, что птенцы могут подлетывать, хотя полностью рост перво-
степенных и рулевых у них завершается лишь к 30—35-дневному возрасту. Плюсна
достигает окончательных размеров уже к двухнедельному возрасту (Кондратьев, Кон-
дратьева, 1984).
Маленькие пуховички очень подвижны и уже через 6 ч после освобождения от скор-
лупы уверенно двигаются и даже пытаются склевывать комаров. При беспокойстве
родителей даже только что обсохшие чайчата отбегают от гнезда и затаиваются, а еще
через 2—3 сут спасаются от опасности вплавь. При этом они способны долго оставаться
активными в холодной воде, терморегуляция окончательно устанавливается у них в
возрасте около 20 дней. Семьи остаются у гнезд не более суток после вылупления по-
следнего птенца, позднее они кочуют по кормным водоемам в окрестностях коло-
ний. Во время этих кочевок выводки одиночно-гнездившихся и колониальных пар
держатся на одних и тех же участках. Добывать пищу птенцы начинают очень рано:
уже в первые сутки жизни они пытаются склевывать насекомых, а через 2—3 дня актив-
но ищут и добывают корм. Однако родители все же подкармливают их, вероятно, до
20-дневного возраста (Кондратьев, Кондратьева, 1984).
На о-ве Врангеля и на западном побережье Чаунской губы лётными птенцы стано-
вятся в конце июля (Лебедев, Филин, 1959; Успенский и др., 1963). Семьи в отличие
от розовой чайки при подъеме молодых на крыло не распадаются и места гнездовий
покидают семейные группы (Кондратьев, Кондратьева, 1984) .
В низовьях р. Чаун средняя величина выводка вскоре после вылупления птенцов
в разные годы составляла 1,8—2,1 птенца, в момент поднятия молодых на крыло —
1,5—2,0 птенца. Пуховички гибнут больше всего в период вылупления, так как в это
время резко возрастает агрессивность взрослых особей и они затаптывают малышей
в ходе столкновений (Кондратьев, Кондратьева, 1984) .
Послегнездовая жизнь изучена слабо. На о-ве Врангеля места размножения в 1938 г.
птицы покинули между 20 и 27 августа. Во время послегнездовых кочевок вилохвос-
тые чайки в отличие от розовых придерживаются низких берегов, лагун и озер в при-
брежной тундре и избегают открытого моря (Портенко, 1973).
Питание. Летом основу его составляют наземные и водные насекомые и их личинки
(Chironomidae, Tipulidae), моллюски, листоногие раки (Branchinecta), щитни, пауко-
образные, перепончатокрылые, бабочки, морские ракообразные (Gammarus, Amphipo-
214
Моевка
dae, Chmocetosopilio и др.); из позвоночных — мелкая рыба: сайка, колюшка (Scalon,
1938; Успенский и др., 1962; Воробьев, 1963; Кречмар и др., 1978; Рогачева и др.,
1978; Остапенко, 1973; Портенко, 1973; Кондратьев, Кондратьева, 1984). В Гренлан-
дии и на Аляске отмечены случаи поедания яиц полярных крачек и плавающих трупов
животных, а также клептопаразитизма вилохвостой чайки на полярной крачке (Voous,
1960; Brown et al,, 1967). На африканских зимовках основу рациона составляют отхо-
ды рыболовного промысла (Lambert, 1975), на тихоокеанских — морские беспозво-
ночные (Chapman, 1969).
Способы кормодобывания достаточно разнообразны. Птицы могут склевывать пищу
с поверхности воды и ныряя с высоты подобно крачкам; собирают беспозвоночных,
бегая по мелководью, как кулики; сопровождают суда и подбирают промысловые и
бытовые отходы; грабят кладки полярных крачек и отбирают у них добычу
(Brown et al., 1967; Chapman, 1969; Портенко, 1973; Lambert, 1975; Кищинский,
1980).
Враги, неблагоприятные факторы. В природе основными врагами являются песцы,
поморники, крупные чайки, но главная причина гибели пуховичков, как упомина-
лось, — агрессивность взрослых, которые давят их во время столкновений. Характер
изменений численности не известен.
Хозяйственное значение, охрана. В связи с ’’разорванным” ареалом и малочислен-
ностью вида у нас в стране предложено занести его в Красную книгу СССР (Зубакин,
1977).
Род Rissa Stephens, 1826
Моевка —
Rissa tridactyla (Linnaeus, 1758)
Kittiwake (англ.), Black-legged Kittiwake (амер.), Dreizehenmowe (нем.), Mouette tridactyle (фр.).
Син. (лат.): Larus tridactylus Linnaeus, 1758.
Син. (рус.): обыкновенная моевка, черноногая моевка, трехпалая чайка, говорушка.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Чайка средних размеров . От всех других
чаек отличается недоразвитым задним пальцем, от красноногой моевки — черными но-
гами и белой окраской нижней стороны крыльев, от сизой чайки в полете — сплошными
черными, без белых пятен, концами крыльев. Гнездится циркумполярно, обычно на ска-
листых утесах, где образует вместе с другими морскими птицами птичьи базары. Во вре-
мя размножения очень крикливы, основной звук — часто повторяющееся ”кити-уэйк”.
Питается мелкой рыбой, морскими беспозвоночными. Часто ныряет за кормом с воз-
духа.
Подвидовая систематика. Различают два подвида (Bent, 1921; Hartert, 1919-1921;
Dwight, 1925).
1. Rissa tridactyla tridactyla
Larus tridactylus Linnaeus, 1758, Syst. Nat., ed. 10, c, 136, Северная Европа.
Более мелкий подвид, характеризующийся меньшим развитием черного рисунка на
первостепенных маховых.
2. Rissa tridactyla pollicaris
Rissa tridactyla pollicaris Stejneger, 1884, Baird, Brewer and Ridgw., Water B.N.
Amer. II, c. 202.
Крупнее предыдущего, черный рисунок на первостепенных маховых более развит.
Описание. С т р о е н и е и размеры (табл. 14).
215
Семейство Чайковые
Таблица 14 Моевка. Размеры (мм)
Пол Длина крыла Длина клюва Длина цевки
п lim среднее п lim среднее п lim среднее
<5 <5 28 298-330 314 R.t. 28 tridactyla 34,0-38,0 35,5 28 33,0-37,0 35,0
9 9 8 297-310 306 8 32,0-34,0 33,3 8 33,0-35,0 33,8
<5 <5 23 303-337 323 R.t. pollicaris 23 35,0-42,0 38,7 23 34,0-39,0 35,9
9 9 25 302-333 317 25 32,0-42,0 37,0 25 31,0-37,0 34,5
Окраска. Самец и самка в брачном наряде. Голова, шея, зашеек, верхние крою-
щие хвоста, рулевые и весь низ тела белые. Мантия и крупные плечевые сизые, послед-
ние — с белыми вершинами. Первостепенные маховые в основном сизые. II (первое
видимое) из них с черным концом и наружным опахалом, III—V с черными вершинами
и небольшими белыми или светло-сизыми пятнами на самых концах. VI с черной пред-
вершинной поперечной полосой и светлой вершиной, иногда сходный рисунок развит
и на следующем VII пере. Остальные первостепенные, так же как и все второстепенные,
сизые с беловато-сизыми вершинами. Крылышко и все верхние кроющие крыла сизые.
Клюв желтый, углы и полость рта ярко-оранжевые. Края век красные, ноги черные.
Самец и самка в зимнем наряде. На голове от задней половины темени на верхней
стороне шеи и по ее бокам интенсивный сизый налет. На зашейке развивается иногда
аспидно-сизая поперечная полоса, соединяющаяся с того же цвета ушными пятнами.
Перед глазом и под ним черное полукольцо. Все прочее оперение, как в брачном наряде.
Пуховой наряд (Cramp, Simmons, 1983). Голова, шея и весь низ тела блестяще-
белые. Спина серая с неясно выраженными светлыми серовато-коричневыми полосами
или пятнами. Крылья серые с белыми основаниями и вершинами.
Гнездовой наряд. Лоб, темя, бока головы, верхние кроющие хвоста и весь низ тела
белые. Зашеек и верхняя часть спины белые с равномерным сизым налетом. По бокам
и на задней части шеи черно-бурая поперечная перевязь, охватывающая шею сверху
полукольцом. В области ушей черно-бурые округлые пятна, иногда сливающиеся с тем-
ным оперением перевязи. Вокруг глаза темное кольцо. Мантия сизая, мелкие и средние
плечевые того же цвета и с неясными охристыми каемками, крупные плечевые сизые
с белыми вершинами. П—IV первостепенные буровато-черные с широкой белой полосой
по краю внутреннего опахала. V в вершинной части того же цвета, ниже область черной
окраски сужается, идет только вдоль стержня и постепенно сходит на нет, основание
пера белое. Наружное опахало этого пера сизое. VI и VII сизые с белыми концами и чер-
новатыми поперечными предвершинными полосами. Остальные первостепенные и все
второстепенные сизые с белыми вершинами. Крылышко, верхние кроющие II—V,
а также ряд мелких кроющих кисти и сгиба крыла буровато-черные. На предплечье
широка;: продольная темная полоса, образованная буровато-черными предвершинными
полосами или пятнами на малых кроющих. На локтевых также более или менее развиты
бурые тона. Крайняя пара рулевых белая, остальные — белые с буровато-черными вер-
шинами. Клюв черный, ноги буровато-черные, но более светлые, чем у взрослых.
Первый зимний наряд. Очень похож на предыдущий, но сизый цвет на перьях темени
и затылка концентрируется главным образом вдоль стержней, поэтому эти части опере-
ния выглядят неясно продольно-полосатыми. Кроме тощ, сизый налет на шее и у перед-
ней границы темной перевязки становится гуще, а сам перевязь суживается, и среди
216
Моевка
перьев ее образующих появляются перья цвета спины. Клюв при основании и в области
ноздрей светлеет. Ноги как у птиц в гнездовом пере.
Первый летний наряд. Голова и шея, как в предыдущем наряде, но темная перевязь
исчезает полностью или (реже) частично. Оперение спины и плечевые сизые, как у
взрослых. Маховые, все кроющие крыла и рулевые сохраняются от гнездового наряда.
Клюв бледно-желтый с темными пятнами в вершинной части, ноги бурые.
Второй зимний наряд. От окончательного зимнего отличается продольной полоса-
тостью на темени и затылке, а также наличием бурых пятен или полос на крылышке,
больших и средних верхних кроющих кисти, а также верхних кроющих I (рудиментар-
ного) первостепенного. Клюв грязно-желтый с темным пятном на сгибе надклювья,
ноги бурые.
Второй летний наряд. Сходен с предыдущим, но сизый налет на голове и шее слабее.
От взрослых птиц в летнем наряде отличается неясными сизыми пятнами в области
ушей и темным рисунком на крылышке и некоторых верхних кроющих кисти. Клюв
грязно-желтый без темных пятен, ноги бурые.
Третий зимний и летний наряды отличимы от соответствующих окончательных
нарядов только по окраске верхнего кроющего рудиментарного махового: у взрослых
птиц оно сизое, у особей в описываемом наряде — сизое с черным пятном.
Линька. Линька в первый зимний наряд неполная, охватывает оперение головы и
шеи. У некоторых особей с Чукотки и из Северного Ледовитого океана в сентябре она
уже закончена, тогда как у других она еще идет в октябре. По С. Крэмпу (Cramp,
Simmons, 1983) эта линька частичная и очень ограниченная: охватывает некоторые
перья на голове и шее. Протекает в августе—ноябре.
Линька в первый летний наряд также неполная, но охватывает гораздо больше опе-
рения, чем предыдущая; протекает в мае. Идет смена оперения на голове, шее, груди
и по бокам тела, сменяются некоторые партии перьев среди мелких плечевых и средних
кроющих предплечья, межлопаточной области, верхней и нижней части спины. В коллек-
ции ЗИН АН СССР есть один весенний экземпляр, сменивший центральную пару руле-
вых. Вероятно, в этом случае мы имеем дело с аномальной линькой рулевых.
Линька во второй зимний наряд полная, охватывает все оперение. У птиц из восточ-
ной части ареала в конце июня она находится на начальных стадиях (меняются IX пер-
востепенное, 19-е второстепенное, рулевые, оперение туловища от шеи до хвоста).
У птиц из западной части ареала в первой половине октября она заканчивается: единич-
ные растущие перья есть по всему телу, прочее оперение, включая маховые и рулевые,
свежее. В сводке С. Крэмпа (Cramp, Simmons, 1983) указывается, что эта линька пол-
ная, протекает с мая по ноябрь, раньше, чем у взрослых. Линька во второй летний наряд
частичная, но оперение туловища охватывает довольно полно. Кроме головы и шеи,
ею затрагиваются мелкие и средние плечевые, межлопаточные, оперение спины до
хвоста; сменяется (по крайней мере частично) оперение от подбородка до груди.
У птиц из восточной части ареала линька заканчивается в начале мая. У особей западно-
го подвида во второй половине мая эта линька уже закончена.
Линька в третий зимний наряд полная. В восточной части ареала начинается в первой
половине июля — второй половине августа со смены XI первостепенного и мелкого
оперения туловища. У птиц западного подвида эта линька начинается во второй полови-
не июня, в это время растут IX—XI, 21-е, единичные перья на боках и брюшке. Линька
в третий летний наряд неполная, охватывает в основном оперение головы, шеи, частично
туловища; на спинной стороне до нижней части спины включительно, на брюшной -
от подбородка до груди. У птиц восточного подвида протекает в апреле, у особей запад-
ного подвида в конце марта она уже закончена.
Линька в окончательный (четвертый) зимний наряд полная, у птиц восточного
217
218
Рис. 51. Область распространения моевки
а — гнездовые поселения и гнездование узкой полосой вдоль побережья, б — область массовых знмовок, в — область нерегулярных зимовок.
Подвиды: 1 : R.t. tridactyla, 2 — R.t. pollicaris
Моевка
подвида начинается в первой половине августа (линяют средние плечевые, межлопаточ-
ные, оперение на груди, боках и брюшке). У птиц западного подвида эта линька также
начинается в августе и заканчивается зимой. По С. Крэмпу (Cramp, Simmons, 1983),
смена маховых происходит в мае—декабре. II первостепенное растет обычно в сентяб-
ре-ноябре; все первостепенные часто обновлены уже в ноябре. Линька в окончатель-
ный летний (брачный) наряд частичная. У особей из восточной части ареала к 20 апреля
уже закончена. В западной части ареала она заканчивается во второй половине этого
же месяца, в начале мая уже закончена. По С. Крэмпу (Cramp, Simmons, 1983), линька
эта частичная, охватывает голову и шею, протекает в марте—апреле.
Распространение. Гнездовой ареал располагается циркумполярно в аркто-
бореальной климатической зоне (рис. 51) (Voous, 1960). Северная его граница почти
везде проходит по северным островам Арктического бассейна (Гренландия, Шпицбер-
ген, Земля Франца-Иосифа, Новая и Северная Земля и о-в Врангеля), лишь в районе
Канадского Арктического архипелага опускается до 74° с.ш. Южная граница размноже-
ния в Атлантике проходит через залив Св. Лаврентия у побережий Северной Америки
и северо-западные берега Франции и даже Испании (Silvar, Rabago, 1975); в Тихом
океане — через Сахалин и Южные Курильские острова к устью р. Медной в Канаде
(Gabrielson, Lincoln, 1959). В СССР, помимо указанных пунктов, гнездится на Мур-
манском побережье, о-ве Вайгач, на Новосибирских островах, у побережья Таймыра,
а также на Медвежьих островах, у побережий Чукотского полуострова, на севере и юго-
западе охотского побережья, на восточном побережье Камчатки, Командорских и
Курильских островах (Бируля, 1907; Гизенко, 1955; Герасимова, 1958; Карпович,
Коханов, 1967; Модестов, 1967; Портенко, 1973; Яхонтов, 1979; Шунтов, 1985 и др.).
В пределах СССР от Баренцева моря до Таймыра и Северной Земли живет номинальный
подвид, на восток от Чаунской губы в Чукотском море — R. t.pollicaris. Птицы с терри-
торий от Новосибирских островов до Чаунской губы демонстрируют промежуточные
признаки двух подвидов.
Зимовки. Размещаются в основном между 60° и 40° с.ш. как в Атлантическом,
так и в Тихом океане, единичные особи встречаются до 20°—15° с.ш. (Gore, Won, 1971;
Curry, Lindahl, 1981; Ralph, Pyle, 1981; Pyle, 1983). В СССР немногочисленные нере-
гулярные зимние встречи известны на Черном море (Бернацкий, 1954; Кгстяк1вский,
1957; Строков, 1974; Костин, 1983). В восточной части ареала известны три основных
района зимних скоплений (Шунтов, 1972). Первый находится в юго-восточной части
Японского моря у п-ова Корея и о-ва Хонсю, второй — в северной части Тихого океана,
к югу от Алеутской гряды (приблизительно между 40° и 50° с.ш., 150° в.д. и 150° з.д.);
третий — у побережий штатов Вашингтон, Орегон и Калифорния в США. Есть указания,
что зимовки неполовозрелых особей расположены в общем дальше от мест размноже-
ния, чем у взрослых (Дементьев, 1955; Willett, 1915; Isleib, Kessel, 1973). Методом
кольцевания доказано, что особи, гнездящиеся в южных частях ареала, и зимуют
южнее, хотя наблюдается значительное перекрывание (Bannermann, 1962; Coulson,
1966).
Миграции. Судя по датам прибытия на места размножения, зимовки покидают в
феврале—марте. Об этом же свидетельствуют прямые наблюдения на центрально-атлан-
тических зимовках (между 50° и 40° с.ш.), о которых упоминается в сводке С. Крэмпа
(Cramp, Simmons, 1983). На тихоокеанских зимовках весеннее движение птиц на север
начинается по мере прогрева поверхностных вод со второй половины марта в южных
районах и первых чисел апреля в северных (Шунтов, 1972; Harrington, 1975). Дальней-
шее продвижение мигрантов зависит от ледовой обстановки. Основное направление
пролета в юго-восточной части Берингова моря, по данным В.П. Шунтова (1972), севе-
ро-восточное, при этом неполовозрелые особи начинают мигрировать позднее, чем
219
Семейство Чайковые
взрослые. На Курильских островах в районе о-ва Итуруп первые мигранты замечены
20 апреля, а у расположенного к северу от него о-ва Уруп они появились на шесть
дней позднее (Велижанин, 1975). У западных берегов более холодного Охотского моря
в районе Шантарских островов первые чайки отмечены лишь 15 17 мая (Дулькейт,
Шульпин, 1937); у западных побережий Камчатки в районе Болыперецка в конце
этого месяца и в начале следующего наблюдали вторую волну пролета, образованную
в основном неполовозрелыми особями (Остапенко и др., 1975). На Командорских
островах, где не бывает сплошного ледового покрова, заметное увеличение числен-
ности мигрантов начинается в начале апреля (Stejneger, 1885; Иогансен, 1934). На
восточном побережье Камчатки весенний пролет, по мнению Е.Г. Лобкова (1980), выра-
жен плохо.
Здесь в первой половине апреля 1975 г, зарегистрировано всего несколько
дар птиц, пролетевших на север. В более северных регионах, на о-ве Св. Лаврентия и
в прилежащих районах Чукотки, первые моевки появляются 6—11 мая (Fay, Cade,
1959; Портенко, 1973) , а на о-ве Малый Диомид и мысу Принца Уэльского, располо-
женных еще севернее, - 15-17 мая (Bailey, 1925; Kenyon, Brooks, 1960).
Осенние кочевки моевок, обитающих на севере ареала от Земли Франца-Иосифа
и Баренцева моря до о-ва Врангеля, начинаются в конце августа — сентябре и продолжа-
ются до октября, когда они постепенно переходят в хорошо выраженный пролет (Горбу-
нов, 1925, 1929, 1932; Демме, 1934; Михель, 1937; Антипин, 1938; Успенский, 1941;
Модестов, 1967; Томкович, 1984) . По данным кольцевания, как взрослые, так и моло-
дые моевки с Мурманского побережья летят на запад вдоль берегов Скандинавии в
Северную Атлантику до Ньюфаундленда, западной части Гренландии, а также побережий
Британских островов, Дании и Норвегии (Дементьев, 1955). В восточной части ареала
начало осенних кочевок приходится на сентябрь (Шунтов, 1972). В этом и следующем
месяце моевки становятся многочисленными по всей акватории Берингова моря и в
глубоководной части Охотского моря, но основные потоки мигрантов в последнем
случае направлены вдоль шельфов по западным и восточным его берегам, к проливу
Лаперуза, Курильским островам и о-ву Хоккайдо. С середины октября они становятся
многочисленными у его берегов и у побережья Приморья. В конце октября мигранты
достигают Корейского пролива, позднее — Желтого и Восточно-Китайского морей.
У восточного побережья Камчатки активные миграции наблюдаются с середины до
конца сентября, хотя отдельные особи встречаются здесь и в ноябре (Лобков, 1980).
Во время пролета моевка обычна или многочисленна в открытом океане на огромной
акватории между Японией, Камчаткой и восточной частью Алеутских островов. В севе-
ро-восточной части Тихого океана, к востоку от о-ва Унимак, в открытом океане она,
напротив, немногочисленна. Основные пути пролета здесь проходят вдоль шельфа
залива Аляска, Британской Колумбии и далее к Калифорнии (Kenyon, 1949; Шунтов,
1972).
Во время кочевок моевки могут встречаться далеко за пределами гнездовой области:
в высоких широтах Арктики до 86° с.ш. (Успенский, 1969), на Днестре, Днепре, Дону,
Волге и Каспии, в Средней Азии (Дементьев, 1934, 1955; К1стяк1вский, 1957; Бара-
баш-Никифоров, Семаго, 1963; Страутман, 1963; Баранов, 1978), в Западной Сибири
(Раевский, 1982) и на оз. Байкал (Гагина, 1962). Вне СССР осенние и зимние встречи
известны в Тироле (Landmann, 1981) и во внутренних районах США и Канады (Smith,
Kuyt, 1983).
Численность. В СССР самые крупные колонии (18—19 тыс. пар) известны на о-ве
Харлов (Восточный Мурман) и мысу Городецком (Западный Мурман) (Герасимова,
1962; Шкляревич, 1977). На скале Рубини (о-в Гукера, Земля Франца-Иосифа) обитает
5 тыс. пар (Беликов, Рандла, 1984), на Новой Земле в губах Грибовой и Безымянной —
220
Моевка
4,5 и 2,2 тыс. пар соответственно (Успенский, 1941; Головкин, 1972). На Чукотке в
районе пос. Уэлен недавно найдено поселение численностью около 1,9 тыс. пар (Моро-
зов, Томкович, 1981); на юго-восточном побережье о-ва Сахалин В.А. Нечаев (устное
сообщ.) обнаружил недавно колонию в 2,5 тыс. пар. Известна общая численность птиц
на отдельных островах. Так, на о-ве Карагинском у восточных берегов Камчатки гнездит-
ся более 120 тыс. пар (Вяткин и др., 1975), на островах Нансикан, Ямских и Ионы в
Охотском море — 50, 13,8 и 5 тыс. пар соответственно (Харитонов, 1975,1980а; Яхон-
тов, 1975; Шунтов, 1985); на Командорских островах (Фирсова, 19756) — 8,0—
8,5 тыс. пар, на о-ве Тюлений у берегов Сахалина — 0,75 тыс. пар (Гизенко, 1955),
на о-ве Колючин в Чукотском море — 7,0 тыс. пар (Кондратьев, 1975), на Курильских
островах — 45 тыс. пар (Шунтов, 1985).
Вне СССР самые крупные колонии (около 100 тыс. пар) известны в заливе Диско
на западном побережье Гренландии (Salomonsen, Gitz-Johansen, 1944) и на северо-
восточном побережье Бристольского залива и юго-восточной части Берингова моря
(Sowls et al., 1978, цит. по: Hunt et al., 1981b).
В последние десятилетия во многих регионах отмечается неуклонный рост числен-
ности этого вида: на Британских островах (Coulson, White, 1956,1958; Coulson, 1963;
Doran, 1967; Harris, Galbraith, 1983), в Дании (Paludan, 1955; Zontrillo, 1982), на
севере Норвегии (Brun, 1971), на о-ве Ньюфаундленд (Maunder, Threlfall, 1972) ина
многих островах Берингова моря (Gabrielson, Lincoln, 1959; Huntetal., 1981b). Та же
тенденция наблюдается и в СССР на Мурманском побережье (Карпович, Шкляревич,
1975) и чукотском о-ве Колючин (Кондратьев, 1975).
Местообитания. В сезон гнездования — скалистые береговые утесы, обращенные
в сторону моря, или берега фьордов, часто поблизости от источников пресной воды
(Salomonsen, Gitz-Johansen, 1944; Lovenskiold, 1964). Необходимыми условиями
для образования колонии являются: наличие постоянно возникающих открытых вод-
ных пространств, обеспечивающих весной доступ к местам размножения; высокая
продуктивность ближайших акваторий; защищенность места колонии от хищников и
его пригодность для инкубации и выращивания молодняка (Nettleship, 1982). В усло-
виях растущей численности при недостатке мест для размножения птицы могут занимать
несвойственные им местообитания: плоские островки, заросшие травой или покрытые
валунами (Howell, 1948; Paludan, 1955), постройки человека (Thorburn, 1935; Coulson,
White, 1958; Coulson, Macdonald, 1962).
Во внегнездовое время — морские акватории, по сравнению с другими видами пред-
почитает более пелагические районы (Bent, 1921; Шунтов, 1972). В Тихом океане,
по В.П. Шунтову, район зимовок совпадает с областью, имеющей зимние поверхностные
изотермы в пределах от +1 до +15°С.
Размножение. По данным кольцевания и гистологических исследований достигают
половой зрелости и гнездятся в первый раз в возрасте 3—4 лет, некоторые особи пыта-
ются размножаться в возрасте 2 лет (Белопольский, 1957; Слепцов, 1948; Coulson,
White, 1959).
К местам гнездования прибывают, как только в ближайших окрестностях колоний
появляется открытая вода. На Мурманском побережье по многолетним данным это
происходит с 5 по 22,111, в среднем 13.III (Белопольский, 1957; Модестов, 1967).
На Земле Франца-Иосифа первые птенцы появляются с И по 24.IV (Jackson, 1899;
Горбунов, 1932; Демме, 1934), на Новой Земле с 20 IV по 11.V (Горбунов, 1932;
Антипин, 1938), на Северной Земле в середине мая, а иногда и позднее (Лактионов,
1946). В восточной части ареала у мест размножения первые особи появляются с нача-
ла апреля (Командорские острова) до середины мая (Охотское море и Чукотка),
о чем свидетельствуют данные многих авторов (Stejneger, 1885; Bailey, 1925; Иоган-
221
Семейство Чайковые
сен, 1934; Дулькейт, Шульпин, 1937; Fay, Cade, 1959; Kenyon, Brooks, 1960; Портенко,
1973).
К гнездованию приступают обычно через 1—2 месяца после прилета, когда скалы
полностью освобождаются от снега. В это время птицы еще ведут зимний образ жизни:
держатся на воде большими стаями, очень шумливы, места будущих колоний посещают
лишь изредка; постепенно эти посещения учащаются (Hagerup, 1891; Salomonsen,
Gitz-Johansen, 1944; Модестов, 1967).
Рис. 52. Демонстрационное поведение моевки (по: Cramp, Simmons, 1983, с изменениями)
а—б — ’’воркование”, в—д— приветственная церемония партнеров
В начале периода размножения, до появления кладок, интенсивно проявляются
различные формы демонстрационного поведения. Оно подробно изучалось в Дании,
наБританскихи Командорских островах (Paludan, 1955; Cullen, 1957; McLannahamm,
1973; Фирсова, 19786) и, в общем, сходно с демонстративным поведением, описанным
для красноногой моевки (тот же набор демонстраций, поз и звуков). При наблюдении
обоих видов на Командорских островах (Фирсова, 1978а,б) подмечены следующие
различия, Во время долгого крика клюв у обыкновенной моевки открывается мень-
ше, ’’воркование” (Cooing) у нее гораздо более интенсивное: поклоны более низкие,
клюв почти касается земли, крылья приподняты (рис. 52). Звуки, которые при этом
издаются, более длительные и громкие.
На Земле Франца-Иосифа интенсивное гнездостроение в 1931 г. отмечено 6. VI и
длилось до 16.VI (Демме, 1934). На Семи островах Баренцева моря в 1939 г. оно нача-
лось в первых числах мая (Белопольский, 1957; Модестов, 1967). На севере Чукотки
в районе поселка Наукан 21,VI гнездостроение у большинства птиц еще продолжалось
(Портенко, 1973), а на Командорских островах в 1970—1973 гг. к середине июня у
большинства пар оно уже закончилось (Фирсова, 1978а,б).
Гнезда помещаются на узких карнизах и выступах скал, предпочтительно на стенках
с отрицательным уклоном, в нишах, защищающих постройки от падающих сверху кам-
ней и дождя (Salomonsen, Gitz-Johansen, 1944; Lovenskiold, 1964). В Западной Европе
при дефиците мест для размножения птицы могут строить гнезда в несвойственной для
них обстановке: среди валунов и на участках, заросших травой, а также наподоконни-
222
Моевка
ках и крышах домов (Thorburn, 1935; Howell, 1948; Paludan, 1955; Coulson, White,
1958; Coulson, Macdonald, 1962).
В строительстве гнезда принимают участие оба члена пары. В качестве строительного
материала используется мох, сухие злаки, водоросли; все это цементируется влажной
почвой и за счет фекалий, попадающих на стенки постройки. На Мурманском побережье
в старых колониях гнезда из года в год только достраиваются, так что некоторые из
них достигают веса до 10 кг; длительность строительных работ здесь у отдельных пар
колеблется от 3 до 10 дней, а в популяции от 25 до 30 дней (Белопольский, 1957;
Модестов, 1967).
Размеры гнезд (мм) с Командорских островов (п - 23): диаметр гнезда 285—385,
в среднем 336 ± 10,2; диаметр лотка 94-145, в среднем 131 ± 3,8; глубина лотка
20—55, в среднем 38 ± 1,9 (Фирсова, 19786).
Откладка яиц в разных колониях на Земле Франца-Иосифа наблюдалась с 20.VI
по 1.VII (Горбунов, 1932); на о-ве Греэм-Белл, по данным П.С. Томковича (1984),
первые кладки были отмечены 24—25 .VI, последние — в начале второй декады июля,
но большинство из них было завершено к началу этого месяца. На Семи островах Барен-
цева моря, по многолетним наблюдениям (Б ело польский, 1957), откладка яиц происхо-
дит с 5 по 27.V, в среднем 17.V; в 1968 г. она началась здесь на две недели раньше
средней многолетней даты — 3.V (Карпович, Шкляревич, 1975). На Новой Земле отклад-
ка яиц отмечена с 5 по 11.VI (Антипин, 1938; Белопольский, 1957), в среднем 9. VI.
В более суровых условиях Северной Земли яйца откладываются во второй половине
июля (Лактионов, 1946). В Чукотском море на о-ве Колючин свежие кладки Й. Курин
добывал 10.VII, а Л.А. Портенко там же видел гнезда с полными кладками 19—26.VII
(Портенко, 1973). На о-ве Врангеля, по данным того же автора, в 1960 г. моевки начали
откладывать яйца 30.VI. На Командорских островах в 1970—1973 гг. первые кладки бы-
ли обнаружены с 27.V по 7 июня (Фирсова, 19786); на о-ве Тюлений в Охотском мо-
ре — 18.VI, массовая откладка яиц отмечалась здесь с 22 по 30.VI (Гизенко, 1955).
Наблюдения за индивидуально меченными птицами на Британских островах показа-
ли (Thomas, 1982), что в разные сезоны каждая самка стремится откладывать яйца
в одно и то же время. Сдвиг этих сроков может быть вызван сменой самца. Если он
старше предыдущего, откладка яиц начинается раньше.
Полная кладка может содержать от 1 до 3 яиц, очень редко — 4 (Данилов, 1948).
О соотношении больших и маленьких кладок и их средних размерах в разных частях
ареала дает представление табл. 15. Даже в одной колонии в разные годы средняя величи-
на кладки может в значительной степени колебаться. Так, на Семи островах в 1937—
1939 гг. были зафиксированы (Успенский, 1941; Модестов, 1967) средние размеры
кладок в 2,9; 1,33 и 1,3 яйца соответственно.
Размеры яиц (мм): с Мурманского побережья (Модестов, 1967) 52,0—61,0 х 40,0—
44,0, в среднем 57,2 х 41,7; с Командорских островов (л = 64) 50,0—63,0 х 38,0-48,0,
в среднем 55,6 х 41,8 (Фирсова, 19786). Подробные сведения о размерах и весе яиц
из разных частей ареала приведены в сводке В. Макача (Makatsch, 1974). Наблюдения
на Британских островах за индивидуально помеченными самками показали, что объем
яиц у каждой самки увеличивается с возрастом, достигая максимума у птиц в воз-
расте 15—23 лет (Thomas, 1982). Основной фон окраски яиц с Командорских островов
средний между беловато-серым по шкале цветов Бондарцева (1954) и чисто-белым.
Поверхностные пятна имеют окраску среднюю между темно-песочной и ореховой,
разбросаны они по поверхности яйца более или менее равномерно.
Насиживают кладку поочередно самец и самка в течение 24—25 дней (Модестов,
1967), по другим данным (Фирсова, 19786; Hunt et al., 1981а), - 26—28 дней. Мини-
мальный отход яиц на Командорских островах в 1973 г. составил 23% (Фирсова,
223
Семейство Чайковые
Таблица 15. Моевка. Величина кладки
в разных частях ареала
Число кладок Доля кладок (%) из числа ЯИЦ Среднее значение Доля повтор- ных кладок, %, Место наблюдения, автор
I 2 3
117 25,64 71,79 2,56 1,76 0 Командорские о-ва (Фирсова, 19756)
295 13,43 74,21 12,34 1,92 0 Британские о-ва (Coulson, White, 1958, 1961)
225 16,00 82,00 2,00 1,85 32 О-в Ньюфаундленд (Maunder, Threlfall, 1972)
577 23,00 59,00 18,00 2,00 17 Баренцево море (Белополь-
ский, 1957)
19786). Вылупление птенцов на Семи островах Баренцева моря, по многолетним дан-
ным (Белопольский, 1957), происходит с 4 по 28.VI, в среднем 19.VI, по В.М. Модесто-
ву (1967), — только 30.VL На Земле Франца-Иосифа оно протекает с 14 по 25 .VII
(Горбунов, 1932), но в самых поздних кладках на о-ве Греэм-Белл 6.VIII1981 г. птенцы
еще не вылупились (Томкович, 1984). На Северной Земле пуховички появляются
только в первой половине августа (Лактионов, 1946). На Командорских островах
первые птенцы были отмечены в 1970—1971 и 1973 гг. 26, 14 и 22.VII соответственно,
при этом 92% птенцов в 1973 г. вылупились в период с 1 по 16.VII (Фирсова, 19786).
На юге Охотского моря (о-в Тюлений у берегов Сахалина) в 1949 г. вылупление было
отмечено 27—28 июля (Гизенко, 1955), на о-ве Ионы в 1974 г. оно продолжалось с
12 по 20.VII (Харитонов, 1980а).
Пуховички в первые сутки жизни весят от 28 до 40 г (Фирсова, 1978). Средняя нор-
ма ежедневного прироста массы птенцов (по отношению к весу взрослых) на Мурман-
ском побережье составляет 2,6%; максимума в 3,7% этот показатель достигает между
16-м и 20-м днями жизни птенцов (Белопольский, 1957). В возрасте от 1 до 3 дней
птенцы большую часть времени сидят в гнезде и обогреваются родителями, даже во
время кормлений пуховички еще не встают на ноги. В возрасте 5 дней они уже уверен-
но стоят на ногах, позже птенцы большую часть времени проводят, стоя в гнезде. Разви-
тие голосовых и двигательных реакций у них протекает так же и в те же сроки, что и
у красноногой моевки. Частота кормления птенцов в возрасте от 1 до И дней состав-
ляет в среднем 0,4—0,8 в 1 ч на одного птенца и, по всей вероятности, не зависит от их
возраста. По данным, полученным в шотландских колониях, этот показатель увеличи-
вается по мере роста птенцов (Galbraith, 1983), особенно резко в возрасте от 12 до
20 дней, и не зависит от размеров выводка.
На Семи островах Баренцева моря первые молодые покидают гнезда в возрасте
33—34 дней в середине июля, через месяц большинство их уже летает (Модестов, 1967).
По наблюдениям на Командорских островах (Фирсова, 19786) это происходит позднее.
Так, в 1973 г. первые лётные птенцы здесь отмечены 9 .VIII, поздние особи научились
летать только к 5.IX, но большинство (80%) приобрело способность к полету в период
с 15 по 29.VIII в возрасте от 37 до 47 дней, из них 62% — в возрасте 39—42 дней. На о-вах
Прибылова молодые начинают летать (Hunt et al., 1981) в возрасте 42—44 дней. В Охот-
ском море на островах Тюленьем (Гизенко, 1955) и Ионы (Нечаев, Тимофеева, 1973)
птенцы становятся лётными к концу августа.
Наблюдения на Командорских островах показали (Фирсова, 19786), что в течение
224
Моевка
по крайней мере 12—15 дней после приобретения способности к полету молодые, прого-
лодавшись, возвращаются на гнезда, где их находят и подкармливают родители. Веро-
ятно, птенцы окончательно покидают гнезда, когда полностью переходят на самостоя-
тельное питание.
На Семи островах Баренцева моря суммарный отход яиц и птенцов в нормальные по
погодным и кормовым условиям годы составляет 20—25%, тогда как в годы с плохими
условиями - более 80% (Карпович, Шкляревич, 1975). Кроме неблагоприятных кормо-
вых и погодных условий причиной гибели птенцов моокет быть хищничество крупных
чаеки поморников (Anderson, 1976; Barrett, Runde, 1980). На Командорских островах
отход яиц и птенцов в 1973 г. составил 58%, а успех размножения здесь в этом же году
был 0,76 птенца на пару размножавшихся птиц (Фирсова, 1978а,б). В колониях юго-
восточной части Берингова моря этот показатель колеблется от 0,4 до 1,23 (Hunt et
al., 1981а,б), а наиболее стабильным, хотя и не очень высоким (0,29—0,51), он оказы-
вается на о-вах Прибылова. На успех размножения здесь также влияют погодные и
кормовые условия.
Суточная активность, поведение. Защитные реакции на воздушных и наземных хищ-
ников у моевок по сравнению с чайками, гнездящимися на земле, практически отсутст-
вуют (Cullen, 1957). Но все же, по наблюдениям на Баренцевом море (Тихонов, Фокин,
1982), при появлении у колонии кречета чайки все вместе снимаются с утесов и выле-
тают ему навстречу.
Питание. Основу его везде составляют планктонные организмы: ракообразные
Isopodae, Amphipodae; крылоногие моллюски Clione, Limacinae; реже головоногие
моллюски Cephalopodae; крупные черви Polychaetae, Annelidae и мелкая рыба (мойва
Mallotus vilosus, М. arcticus, песчанка Ammodytes, сайда Bodeogadus saida, мальки
тресковых и сельдевых рыб) (Salomonsen,Gitz-Johansen, 1944; Белопольский, 1957;
Lovenskiold, 1964; Модестов, 1967; и др.) . Не брезгуют моевки также отходами рыбно-
го промысла, а во вне гнездовое время используют их в больших количествах (Hersey,
1917; Prebl, McAtee, 1923; Шунтов, 1972; Карпович, Шкляревич, 1975). В условиях
недостатка привычной пищи могут кормиться на трупах морских зверей и использо-
вать отходы зверобойного промысла (Lovenskiold, 1964; Портенко, 1973). Соотноше-
ние различных кормов в рационе подвержено как географической, так и сезонной
изменчивости (Белопольский, 1957; Hunt et al., 1981а). Как и другие чайки, моевки
легко переходят с одного массового корма на другой. Экспериментальными исследо-
ваниями установлено (Головкин, 1963, 1972), что норма суточного потребления пищи
у взрослых составляет 120 г, у птенцов — 60 г.
Основной кормовой биотоп — поверхность открытого моря, но могут добывать
корм и на мелководьях около берега и даже на литорали (Белопольский, 1957; Drury,
Biederman, 1978; цит, по: Hunt et al., 1981а). На севере ареала излюбленные места кор-
межки-верховья ледовых фьордов и кромка плавучих льдов (Salomonsen, Gitz-
Johansen, 1944; Lovenskiold, 1964; Беликов, Рандла, 1984), где за счет разности темпе-
ратур и солености воды создаются сильные вертикальные течения, выносящие пищу на
поверхность.
Способы кормодобывания достаточно разнообразны. Планктон и рыбу добывают
на поверхности или с небольших глубин (до 0,5 м), погружая в воду голову и шею
или ныряя в нее с воздуха (Nelson, 1887; цит. по: Bent, 1921; Prebl, McAtee, 1923;
Nicholson, 1930; Демме, 1934; Белопольский, 1957; Gabrielson, Lincoln, 1959; Hunt
et al., 1981a). Могут отбирать рыбу друг у друга или у других птиц: кайр, тупиков,
топорков, полярных крачек (Гизенко, 1955; Lovenskiold, 1964; Михтарьянц, 1972;
Краснов, Николаева, Чернышова, 1982). В Гренландии (Salomonsen, Gitz-Johansen,
1944) и на Аляске (Nelson, 1887, цит. по: Bent, 1921) моевки охотно сопровождают
8. Зак. 826 225
Семейство Чайковые
китов и тюленей, вылавливая рыбу, которую звери подгоняют к поверхности. Сопровож-
дают рыболовные и пассажирские суда, подбирая отходы. На Британских островах в
условиях роста численности все чаще наблюдаются кормежки в антропогенных ланд-
шафтах (Coulson, Macdonald, 1962). Здесь моевки ловят рыбу в пресной воде, подни-
маясь вверх по рекам на расстояние до 10 миль, берут пищу из рук людей, использу-
ют отходы из сточных вод.
Дальность кормовых полетов может достигать 100 км (Демме, 1934; Salomonsen,
Gitz-Johansen, 1944; Модестов, 1967; Hunt etal., 1981а,б).
Враги, неблагоприятные факторы. Пуховички и молодые особи часто гибнут из-за
неблагоприятных кормовых или погодных условий, а также вследствие хищничества
крупных чаек и поморников. Причиной почти повсеместного увеличения численности,
по-видимому, является, как и у других чаек, улучшение питания за счет освоения от-
ходов и других антропогенных кормов, особенно в зимнее время.
Хозяйственное значение, охрана. Большого хозяйственного значения не имеет. Спе-
циальные меры охраны не предусмотрены.
Красноногая моевка — Rissa brevirostris Bruch, 1853
Red-legged Kittiwake (англ.), Klippenmowe (нем.), Mouette des brumes (фр.).
Син. (рус.) : красноногая говорушка.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Чайка средних размеров. От всех прочих
чаек отличается слабо развитым задним пальцем, от моевки — красными ногами, более
темной окраской мантии и нижней стороны крыльев. Гнездовой ареал ограничен Берин-
говым морем и прилежащими районами Тихого океана. Экология, как у моевки, но
данный вид предпочитает более открытые участки океана, особенно во время миграций
и на зимовках. Голос заметно выше, чем у моевки.
Подвидовая систематика. Монотипический вид.
Описание. Размеры (мм):
Длина крыла:
dd (и =1) 99 (и = 3) 306; 295—316 (среднее 306);
dd и 99 (и = 3) 283-314 (среднее 307).
Длина клюва:
dd (и =1) 99 (и = 3) Длина цевки: 26,0; 27,0—29,0 (среднее 28,0).
dd (п = 1) 99 (и = 3) 32,0; 30,0—32,0 (среднее 30,6);
ddn99 (и = 3) 31,0-31,0.
Окраска. Самец и самка в брачном наряде. Голова, шея, рулевые и их верхние
кроющие, а также весь низ тела белые. Межлопаточная область, плечевые, спина, все
верхние кроющие крыльев аспидно-сизые. На крупных плечевых хорошо развиты белые
концевые каемки. Первостепенные на большем протяжении аспидно-сизые, причем на
226
Красноногая моевка
дистальных (II—VI) более или менее развита черная окраска. На II она занимает боль-
шую часть наружного опахала и вершинную — внутреннего. На последующих перьях
область черного пигмента прогрессивно уменьшается (до небольшого пятна на VI),
а аспидно-сизой окраски увеличивается. Кроме того, на V—VI перьях развиты вершин-
ные серые пятна, увеличивающиеся по направлению к проксимальному концу кисти.
Проксимальные первостепенные (VII—XI) аспидно-сизые с белыми вершинами и бе-
лыми полосами вдоль латерального края внутренних опахал. Ширина этих полос увели-
чивается по направлению к проксимальной части кисти. Второстепенные аспидно-сизые
с большими белыми вершинами, переходящие в узкие краевые каемки в дистальной
части предплечья. Клюв лимонно-желтый; ноги, края век и внутренняя полость рта
ярко-красные.
Самец и самка в зимнем наряде. Щеки, затылок и граничащие с ними участки темени
белые с темными пестринами. Вокруг глаза черноватое кольцо, более широкое снизу
и спереди. В области ушей аспидно-сизые пятна. По верху и бокам шеи развита аспидно-
сизая перевязь, почти сливающаяся с ушными пятнами. Верхняя часть шеи, расположен-
ная каудально от перевязи, белая с сизым налетом. Все прочее оперение, как у птиц
в брачном пере. Клюв желтый, ноги красные.
Пуховой наряд. Голова, шея и весь низ тела блестяще-белые. Спинная сторона темно-
серая с седоватым налетом, на предплечьях он выражен сильнее. Кисти белые в основа-
нии, с небольшими серыми пятнами на верхней стороне. На границе серой и белой
окраски по всему телу, а особенно на предплечьях, развиты едва заметные охристые
тона. Клюв однотонный темно-серый, яйцевой зуб белый. Цевка и пальцы серые, пере-
понки между пальцами более светлые, телесные, коготки черные. Радужина темно-ко-
ричневая, зрачок голубоватый.
Гнездовой наряд. Голова, зашеек, рулевые и их кроющие, а также весь низ туловища
чисто-белые. Перед глазом черное полукольцо, в области уха того же цвета пятно, а по
бокам головы неясная поперечная темная испещренность. На верхней стороне шеи и
особенно по ее бокам развит широкий черный полуошейник. Плечевые, межлопаточные,
перья спины и кроющие крыльев чистого аспидно-сизого цвета, чуть более светлого,
чем у взрослых. Кроме того, в отличие от них на дистальных концах кроющих пред-
плечья, мелких и средних плечевых есть узкие беловатые краевые каемки, которые
образуют неясный чешуйчатый рисунок на этих участках тела. II (первое видимое)
первостепенное однотонное, черновато-бурое, постепенно светлеющее в проксимальной
части, особенно на внутреннем опахале. На внутреннем опахале в проксимальной
части III появляется серое поле, размеры которого увеличиваются на IV, на V серая
окраска появляется и на наружном опахале, а самый кончик пера становится белова-
тым. На VI и VII серый цвет преобладает, черновато-бурые тона остаются лишь в дис-
тальной их части в виде отчетливых неправильной формы полос или пятен; на вершинах
этих перьев развиваются крупные белые пятна. Остальные первостепенные серые с белы-
ми вершинами, без черных тонов. Второстепенные также серые с белыми концами,
на некоторых из них развиты узкие пристержневые бурые полоски, на перьях крылыш-
ка и мелких кроющих кисти — отчетливые бурые пятна. Клюв однотонный буровато-
черный, ноги буровато-коричневые с более темными когтями.
Первый зимний наряд. По Дж.Дуайту (Dwight, 1925), по сравнению с предыдущим
нарядом черный полуошейник становится уже.
Первый летний наряд. От гнездового и первого зимнего нарядов отличается, вероят-
но, полным отсутствием черного полуошейника и темных пестрин по бокам головы.
Ноги и клюв желтовато-бурые.
Второй зимний наряд. От гнездового и первого зимнего нарядов отличается полным
отсутствием черного полуошейника, от первого летнего — свежими маховыми и
227
Семейство Чайковые
аспидно-сизым налетом на верхней стороне шеи. Клюв желтовато-зеленый, ноги в отли-
чие от взрослых желтовато-оранжевые. В природе птицы этого возраста легко опознают-
ся именно по цвету ног и остаткам бурого оперения на мелких кроющих предплечья и
кисти. В сводке Дж.Дуайта (Dwight, 1925) этот наряд описан как окончательный.
Второй летний наряд. В материалах ЗИН АН СССР ни одного экземпляра в этом на-
ряде нет, но, судя по особям в предыдущем и последующем нарядах, он, вероятно,
отличается от предыдущего меньшим развитием аспидно-сизого полуошейника или
полным его отсутствием.
Третий зимний наряд. Похож на второй зимний, отличается от него отсутствием бу-
рых тонов на перьях крылышка и кроющих кисти. Третий летний наряд отличается от
предыдущего отсутствием аспидно-сизого полуошейника, темных пестрин по бокам го-
ловы и того же цвета полуколец вокруг глаз. Эти наряды отличаются от соответствую-
щих окончательных лишь наличием бурых тонов на рудиментарном первом первосте-
пенном маховом, о периодичности линьки которого ничего не известно. Поэтому дан-
ные наряды, возможно, различимы не у всех особей.
Линька. Частичная линька в первое зимнее перо охватывает, вероятно, только опере-
ние шеи и зашейка. Линька в первый летний наряд также частичная, захватывает опере-
ние по бокам головы, на верхней стороне шеи и зашейке. О сроках ее протекания
известно очень мало. У самки от 30.VI 1913 г. с Берингова моря уже началась линька
в следующий наряд. Линька во второй зимний наряд полная. У упомянутой самки от
30.VI сменились четыре внутренних первостепенных и часть проксимальных второ-
степенных. Растущие перья есть среди верхних кроющих рулевых, среди плечевых,
на груди, по бокам тела и на брюшке. Во второй половине августа эта линька уже близ-
ка к завершению: сменилось по крайней мере восемь первостепенных, половина второ-
степенных, идет смена 4-й — 5-й пар рулевых; мелкое оперение туловища в основном
уже свежее. Линька во второй летний наряд неполная, охватывает в основном оперение
на верхней стороне шеи и зашейке, сроки ее протекания неизвестны. Линька в третий
зимний наряд полная. У особи, добытой 8.Х 1906 г. в Анадырском лимане она только
начинается; растет 17-е второстепенное, мелкие плечевые, а также оперение на зашейке
и груди. Линька в третий летний наряд частичная, охватывает оперение головы, шеи,
зашейка. Сроки протекания неясны. Линька в окончательный зимний наряд полная,
сроки ее протекания неизвестны. У экземпляра, добытого на Командорских островах
23.VIII, она еще не началась. Линька в окончательный летний (брачный) наряд частичная,
сроки ее протекания также неизвестны.
Распространение. Гнездовой ареал ограничен островами Алеутской гряды
на севере Тихого океана (рис. 53). Колонии известны на Командорских островах и о-вах
Прибылова, а также на островах Богослов и Булдырь из Алеутской группы (Elliot, 1882;
Stejneger, 1885; Byrd, 1878; Hunt et al., 1981 a,b). На других Алеутских островах отмеча-
лись лишь на пролете (Gabrielson, Lincoln, 1959;Murie, 1959). Вне Алеутской гряды, по
данным последних авторов, единичные особи встречены в Бристольском заливе (Hurley,
1931, 1932, цит. по: Gabrielson, Lincoln, 1959), на о-ве Св.Лаврентия (Fay, Cade, 1959),
на Чукотке в районе Уэллена (Bailey, 1925). В материалах ЗИН АН СССР есть осенний
экземпляр с Анадырского лимана. Известны также встречи в Охотском море у о-ва Са-
халин и Курильских островов, у восточного побережья Камчатки, которые В. П. Шунтов
(1972) не считает случайными.
Зимовки. Изучены слабо. По мнению американских исследователей (Gabrielson,
Lincoln, 1959), они предположительно находятся в открытых водах Берингова моря
между Алеутскими островами и о-вами Прибылова. В.П. Шунтов (1972) считает, что
большинство птиц покидает на зиму Берингово море и обитает в это время несколько
южнее, в глубоководных северо-восточных частях Тихого океана.
228
Красноногая моевка
Рис. 53. Область распространения красно-
ногой моевки
7 — гнездовые поселения, 2 — предполагае-
мые места основных зимовок, 3 — залеты
Миграции. Массовый весенний пролет наблюдается во второй половине апреля — нача-
ле мая и заканчивается в конце мая — начале июня (Шунтов, 1961, 1963). Осенью после
оставления колоний на Командорских островах часть птиц держится поблизости в море
до ноября (Иогансен, 1934), а на о-вах Прибылова — даже всю зиму (Elliott, 1882).
Одиночных особей наблюдали 30.IX и 1.Х 1973 г. на восточном побережье Камчатки
в районе пос. Жупаново (Лобков, 1980). В открытых водах Берингова моря осенняя
миграция прослеживается с конца сентября до ноября, общее ее направление юго-вос-
точное (Шунтов, 1963, 1972). При этом, как
и весной, птицы предпочитают глубоковод-
ные районы моря в отличие от черноногой
моевки, придерживающейся преимуществен-
но его прибрежных участков.
Численность. Как показано американски-
ми исследователями (Byrd, 1978, Kessel,
Gibson, 1978; Hunt et al., 1981b), большая
часть (88%) мировой популяции гнездится на
о-вах Прибылова. Самая крупная колония
(ПО тыс. пар) расположена здесь на о-ве
Св. Георгия, более мелкие (300 и 500 пар)
на островах Св. Павла и Бобровом. На о-ве
Богослов (Byrd, 1978) учтено 200 пар, о-ве
Булдырь — 2 тыс.
В пределах СССР на Командорских остро-
вах размножается не более 2,0—2,5 тыс. пар (Фирсова, 19756),по С.В. Маракову (1975,
1976), — только тысяча пар. Общая численность мировой популяции этого вида состав-
ляет, таким образом, около 115 тыс. гнездящихся пар. По другим оценкам, только
в юго-восточной части Берингова моря обитает 125 тыс. пар (Sowls et al., 1978).
Местообитания. В гнездовое время — скалистые участки побережий, на пролете и зи-
мой — глубоководные районы недалеко от области размножения.
Размножение. В районы гнездования на Командорские острова и о-ва Прибылова
птицы начинают прилетать в конце марта — начале апреля, в начале мая они становятся
здесь многочисленными (Elliott, 1882; Stejneger, 1885; Иогансен, 1934; Hanna, цит. по:
Gabrielson, Lincoln, 1959; Мараков, 1976). На местах размножения они занимают
отвесные береговые утесы, отдельные камни-кекуры, или небольшие островки в при-
брежной зоне (Elliott, 1882; Stejneger, 1885; Фирсова, 1978а). В любом случае гнезда
помещаются на карнизах, небольших выступах и в нишах скал, которые лишены травя-
нистой растительности и настолько узкие, что непригодны для других обитателей
птичьих базаров: кайр, бакланов, крупных чаек и даже обыкновенных моевок (Фирсо-
ва, 1978а; Hunt et al., 1981 a,b). Часто все же красноногие и обыкновенные моевки гнез-
дятся на одних и тех же участках скал, но внутри такой смешанной колонии обычно
выделяются скопления того или другого вида, величина которых, очевидно, опреде-
ляется шириной карнизов. В очень крупных смешанных поселениях красноногие моев-
ки часто оттесняются другими видами на самые нижние карнизы, почти на уровень при-
боя (Gabrielson, Lincoln, 1959;Фирсова, 1978а).
В начале сезона размножения в колониях можно наблюдать различные формы демон-
страционного поведения. Оно подробно изучено (Фирсова, 1978а) на Командорских
островах (рис. 54). Агрессивный комплекс демонстраций включает долгий крик,
’’квохтание”, воркование (Cooing), прямую позу угрозы, ложное клевание. Все эти де-
монстрации обеспечивают защиту места для гнезда и самого гнезда от других особей.
Комплекс брачных демонстраций состоит из церемоний встречи партнеров, вскидыва-
229
Семейство Чайковые
ния клюва и выпрашивания корма самкой, кормления самки самцом и взаимного
вскидывания клюва. Интенсивность выполнения всех демонстраций снижается к концу
сезона, некоторые из них могут выпадать вовсе. В целом демонстрационное поведение
красноногой моевки сходно с описанным у обыкновенной моевки (Paludan, 1955;
Cullen, 1957; McLannahan, 1973), но голос у первой из них значительно выше (Фир-
сова, 19786).
Рис. 54. Демонстрационное поведение красноногой моевки (по: Фирсова, 1978а, с изменениями)
а, г — долгий крик, б — ’’воркование”,в — ’’квохтание”,д — парное ’’квохтание”, е — выпрашивание
корма, ж— взаимное вскидывание клюва
Недавние наблюдения на о-вах Прибылова (Souibb, Pitts, 1982) показали, что неде-
монстративные формы копулятивного поведения самцов у этих двух видов различны.
Самцы красноногих моевок во время спаривания раньше начинают взмахивать
крыльями и удерживаются на спине самки с помощью клюва, чего не наблюдается
у самцов обыкновенных моевок. Эти особенности, по мнению авторов, связаны с тем.
что красноногие моевки занимают более мелкие крутые уступы и потому вынуждены
затрачивать больше усилий на сохранение равновесия.
В строительстве гнезда принимают участие оба партнера (Elliott, 1882; Фирсо-
ва, 1978а), при этом самец чаще всего приносит строительный материал, а самка егс
утрамбовывает, а затем формует стенки гнезда точно так же, как это описано у обыкно-
венной моевки (Paludan, 1955). В качестве основы для гнезда используется влажная
почва, которую птицы берут тут же на острове; стенки гнезда строятся из водорослей.
230
Красноногая моевка
сухих травинок, перемешанных с влажным грунтом (Elliott, 1882; Фирсова, 19786).
На Командорских островах в 1970—1971 и 1973 гг. гнездостроение у основной массы
птиц закончилось к середине июня, вспышки его (вероятно, у неполовозрелых или
утративших кладки птиц) наблюдались также в конце августа. Размеры (мм) 13 гнезд
с Командорских островов (Фирсова, 19786) следующие: диаметр гнезда 140—220,
в среднем 186; диаметр лотка 80—110, в среднем 94; глубина лотка 20—40, в сред-
нем 27.
Первые кладки на Командорских островах обнаружены 7 и 17 июня в 1970—1971 и
1973 гг. соответственно. Размеры кладки здесь минимальные. Все 75 кладок, находив-
шихся под наблюдением, содержали только по одному яйцу, тогда как на о-вах Прибы-
лова раньше (Elliott, 1882) нормальная кладка состояла из двух яиц, встречались клад-
ки и из трех яиц. Позднее (Kenyon, Phillips, 1965) там встречались кладки из одного,
очень редко из двух яиц. В самых недавних публикациях (Hunt et al., 19816) указывает-
ся, что кладка на этих островах состоит только из одного яйца. По наблюдениям на
о-вах Прибылова, восстановление утраченной кладки возможно через 7—10 дней
(Elliott, 1882). По нашим данным, на Командорских островах потерянные кладки не
восстанавливались. Более того, в 1973 г. 18% пар здесь не отложили яиц вообще, хотя и
имели нормальные гнезда (Фирсова, 19786).
Размеры яиц (мм) с Командорских островов (Фирсова, 1978а,б) (и = 21)
52,0—65,0 X 38,0—44,0, в среднем 56,8 X 40,5. Основной фон их окраски бледно-серый
с небольшими вариациями. Пигментные пятна на поверхности темно-коричневые,
глубинные — более светлые, приглушенного коричневого цвета; размеры пятен от 1
до 10 мм, на тупом конце яйца они расположены, как правило, гуще.
Насиживают кладку поочередно самец и самка, продолжительность насиживания на
Командорских островах, по нашим данным (15 гнезд), 28—30 дней; на о-вах Прибыло-
ва 24—26 (Elliott, 1882) или 29 дней (Hunt et al., 19816). На Командорских островах
в 1973 г. из 44 яиц вылупилось 27 птенцов (3 яйца оказались болтунами, 10 кладок бы-
ли брошены, судьба 4 осталась невыясненной); минимальный отход яиц составил,
таким образом, 29% (Фирсова, 19786). Первые птенцы в этом году вылупились 6 июля,
последние 31 июля, большинство (80%) — в период с 12 по 22 июля.
Наши наблюдения у гнезд показали, что в первые трое суток после вылупления птен-
цы малоподвижны, большую часть времени сидят в гнезде. Передвигаться могут только
на цевках, иногда помогая себе при этом клювом. Взрослые птицы все время обогре-
вают их, прерывая это занятие только на время кормления или смены партнеров
на гнезде. Оборонительная реакция или реакция отворачивания клюва у птенцов в этом
возрасте еще не выражены. Замерзшие или голодные птенцы слабо пищат или ритмично
поскрипывают. Выпрашивая корм они клюют подклювье родителей и склевывают пищу
прямо из их клюва, так что подстилка гнезда пищей не пачкается. Частота кормления
птенцов, по нашим данным, не зависит от их возраста и составляет в среднем 0,4 корм-
ления в час.
В возрасте 3—5 дней у некоторых пуховичков появляется слабая реакция отворачива-
ния клюва. К концу первой недели жизни или чуть позже они хорошо стоят на ногах,
но большую часть времени проводят все же сидя. Реакция отворачивания клюва выраже-
на уже у всех птенцов, иногда очень сильно: у некоторых появляется слабая оборони-
тельная реакция. К концу второй недели жизни птенцы становятся заметно более актив-
ными и подвижными, хотя подвйжность их и ограничена пределами гнезда. Большую
часть времени они стоят на нем, в обогреве почти не нуждаются. Некоторые птенцы
-издают звук, промежуточный между поскрипыванием самых маленьких птенцов и кри-
ком ”ой-ой-ой”, издаваемым взрослыми в момент непосредственной опасности. На чет-
вертой неделе жизни, перед тем как при опасности отрыгнуть корм, птенцы начинают
231
Семейство Чайковые
издавать очень тихий и высокий односложный писк. Еще через неделю сигнал опасности
”ой-ой-ой” становится у них очень похожим на соответствующий крик взрослых птиц.
Наблюдения в колонии за 21 меченым птенцом показали (Фирсова, 19786), что боль-
шинство из них начинают нормально летать в возрасте 45—47 дней, когда заканчивают
развитие основные категории оперения. Некоторые особи могут оставлять гнезда в воз-
расте 38—39 дней, но такие вылеты, как правило, бывают спровоцированными, обычно
птенцы оказываются неспособными вернуться в гнезда и гибнут. Проголодавшись,
лётные птенцы по крайней мере в течение 15 дней возвращаются в гнезда, где их под-
кармливают родители. На о-вах Прибылова взрослые кормят лё'тных молодых по край-
ней мере неделю (Hunt et al., 19816). Между кормлениями птенцы проводят время
на гнездах, в непосредственной близости от них в клубах или вне колонии. Первые лёт-
ные молодые в 1973 г. на Командорских островах были отмечены 22.VIII.4.IX 19
(из 21) птенцов уже летали, большинство (61%) поднялось на крыло в период
25—31.VIII, только 13% их стало лётными 22—24.VIII и 26% — 1—6.IX.
Успех размножения в 1973 г. составил 0,47 птенца на пару размножавшихся птиц,
отход яиц и птенцов достиг 52% (Фирсова, 1978а) ; на о-вах Прибылова первый показа-
тель оказался равным 0,38 (Hunt et al., 19816).
Питание. Анализ 29 проб корма птенцов с Командорских островов показал (Фирсо-
ва, 19786), что основу их рациона составляет полупереваренная рыба песчанка, которая
встречалась в 96% проб, причем 47% проб состояло только из этой рыбы. Ракообразные
(Amphipodae, сем. Lusianassidae) встречены в 36% проб, мелкие головоногие мол-
люски (Cephalopodae) диаметром 15—20 мм — в 24% проб, икра (вероятно, рыбья) —
в 6% проб.
Недавние исследования на о-вах Прибылова (Hunt et al., 1981а,b) показали, что
по сравнению с 20-ми годами нашего века объем потребляемых головоногих моллюс-
ков в этом регионе снизился, а потребление рыбы (преимущественно сем. Myctophidae)
увеличилось, особенно в конце сезона. Наличие в рационе терпуга (Theragra chalco-
gramma) в июне отражает, по мнению авторов, тот факт, что красноногие моевки пи-
таются у края шельфа в большей степени, чем обыкновенные. Об этом же косвенно
свидетельствует и меньшее использование красноногими моевками в начале сезона
обитающих на небольших глубинах амфипод Parathemisto libellula. По данным этих же
авторов (Hunt et al., 1981 а,b), красноногие моевки питаются мелкой рыбой и круп-
ным зоопланктоном, корм они добывают на глубине до 0,25 м от поверхности.
При этом птицы могут погружаться на небольшую глубину как с поверхности воды,
так и бросаясь в нее с воздуха. Большая часть их рациона состоит из рыб сем.
Myctophidae, обитающих на больших глубинах и редко встречающихся в пределах
шельфовых вод. Именно этим указанные авторы объясняют тот факт, что колонии
этого вида моевок обнаружены только на островах, расположенных близко к склону
шельфа, а также то, что большинство моевок этого вида кормится ночью.
Враги, неблагоприятные факторы. В природных условиях естественных врагов
мало, основная причина гибели молодняка — это неблагоприятные погодные или кор-
мовые условия. Общие тенденции изменений численности неизвестны.
Хозяйственное значение, охрана. Хозяйственного значения вид не имеет. Вследствие
очень узкого распространения и малочисленности красноногой моевки у нас в стране
были неоднократные предложения о внесении вида в Красную книгу СССР (Фирсо-
ва, 19756, 1977, 1979; Мараков, 1976; Зубакин, 1977); в настоящее время занесена
в Красную книгу РСФСР и подлежит строгой охране. На Командорских островах
организован областной заказник, включающий гнездовья красноногой моевки и других
редких видов морских птиц.
232
Малая чайка
Род Hydrocoloeus Каир, 1829
Малая чайка -
Hydrocoloeus minutus (Pallas, 1776)
Little Gull (англ.), Zwergmowe (нем.), Mouette pygmce (фр.).
Син. (лат.): Larus minutus Pallas, 1776.
Статус. Гнездящийся мигрирующий, частично зимующий вид.
Общая характеристика и полевые признаки. Самая мелкая чайка мировой фауны.
От других темноголовых чаек, помимо размеров, отличается легким порхающим поле-
том, сравнительно короткими и округлыми крыльями, темным исподом крыла.
От озерной чайки, кроме того, — светлыми концами крыльев, в сезон размножения —
угольно-черной, а не коричневой головой и иной формой границы между темной голо-
вой и белой шеей: черный цвет у малой чайки захватывает не только темя и затылок,
но и верхнюю часть шеи. Птицы в гнездовом наряде характеризуются темным, черно-
вато-бурым цветом спины и такого же цвета зигзагообразной полосой вдоль крыла.
Малые чайки хорошо плавают, не ныряют. По земле передвигаются мало и неохотно,
как крачки. Хорошо летают, характерны разнообразные воздушные демонстрации.
Голос малых чаек мелодичен и напоминает голос крачек, особенно чайконосых.
Чаще всего можно слышать крики типа ”кей, кей, кей”, трели ”ке-кей, ке-кей,
ке-кей ... ”, ”ке-кой, ке-кой, ке-кой ... ”. Крик тревоги в колонии — быстрая сухая
трелька ”ке-ке-кек”, которая в отличие от подобных криков большинства других чаек
содержит различное (2—5) количество слогов. В тревожных ситуациях в колониях
можно слышать резкие визгливые крики типа ”уик” или ”уить”.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Дж.Дуайт (Dwight, 1925) с некоторыми оговорками
объединил малую чайку в один род Hydrocoloeus с группой видов, родственных озерной
чайке. После сведения ряда родов семейства чайковых до уровня подродов рода Larus
малую чайку продолжали в большинстве случаев помещать либо рядом с черноголовой,
либо по соседству с озерной чайкой (Бутурлин, 1934; Дементьев, 1951; Степанян, 1975).
Однако целый ряд морфологических особенностей (почти равная длина цевки и средне-
го пальца, характер расцветки первостепенных маховых у взрослых птиц, темный цвет
верхней части тела и характер окраски крыльев в гнездовом наряде и др.) вместе с осо-
бенностями демонстрационного поведения (см, ниже) говорит о необходимости выде-
ления малой чайки в отдельный род и возвращения ей прежнего родового названия
Hydrocoloeus Каир, 1829, как это было сделано в одном из недавних списков птиц
мира (Wolters, 1975).
За подобное выделение говорит и рост доказательств некоторой филогенетической
близости малой и розовой чаек. Предположение о родстве этих видов, высказанное
более четверти века назад (Moynihan, 1959), получает все большее подтверждение.
У этих видов много общего в окраске яиц, птенцов и гнездового наряда. Для обоих ви-
дов характерны выселения за пределы гнездового ареала с образованием неустойчивых
поселений. Голоса этих чаек тоже достаточно сходны, как и ряд черт демонстрацион-
ного поведения, — в частности, у обоих видов отсутствует четко оформленный.долгий
крик, столь характерный для других чаек. Последний признак, по-видимому, следует
считать примитивным в подсемействе Larinae. Интересно, что по нему, как и по харак-
теру расцветки крыла в гнездовом наряде, малая чайка сближается и с моевками
(р. Rissa ) . Не исключено, что все эти ныне далеко разошедшиеся виды,различия между
которыми достигли родового ранга, связаны общим происхождением и стоят несколько
особняком среди других чаек.
233
Семейство Чайковые
Описание. Строение и размеры. Размеры особей (мм) (ЗМ МГУ) и масса
тела (г) (Бородулина, 1960):
Длина крыла:
33 (и = 28) — 218—238 (среднее 225) ;
99 (и = 12) - 215-227 (среднее 222).
Длина клюва:
33 (и = 24) - 20,6-23,7 (среднее 22,2);
99 (п = 11) — 20,7—23,9 (среднее 21,7).
Длина цевки:
33 (и = 27) - 24,8-27,7 (среднее 26,2);
99 (л = 12) — 24,8—26,8 (среднее 25,8).
Масса тела:
33 (л = 22) - 103-127 (среднее 115);
99 (и = 5) - 93-122 (среднее 107).
Окраска (Дементьев, 1951; Cramp, Simmons, 1983; ЗМ МГУ; ЗИН). Самец и
самка в брачном наряде. Голова бархатисто-черная, причем черный цвет захватывает
также и верхнюю часть шеи. Белые полоски над и под глазами в большинстве случаев
отсутствуют. Шея, грудь, брюхо, бока, надхвостье, подхвостье и рулевые белые, причем
низ тела может быть с легким розоватым оттенком. Нижние кроющие крыла дымчато-
серые. Первостепенные и второстепенные маховые сизо-серые с белыми вершинами;
нижняя поверхность первостепенных черновато-серая. Клюв темно-красный, ноги
ярко-красные. Радужина темно-бурая, края век красные.
Самец и самка в зимнем наряде. Лоб, уздечка, подбородок и горло белые; темя и
затылок темно-серые, кроющие уха и пятно перед глазом несколько темнее. Остальное
оперение, как в брачном наряде. Клюв черноватый или черный, ноги розовато-ко-
ричневые.
Пуховой наряд (живые птенцы из Московской обл.). Верх головы и туловища серо-
вато-бежевый, серовато-рыжий разной густоты. Горло такого же цвета или более
рыжеватое, брюшко несколько светлее, нередко с охристым налетом. На зашейке,
реже на крыльях и между лопатками рыжий оттенок различной интенсивности. На голо-
ве, задней стороне шеи, спине и крыльях черные пятна и крапины; нередко они форми-
руют неясные полоски. Отмечены индивидуальные вариации как в цвете и интенсив-
ности тона, так и в размере, рисунке и густоте черных пятен. В целом птенцы малых
чаек темнее птенцов большинства других видов чаек. Клюв телесного цвета с темным
кончиком, лапы телесные, радужина черная.
Гнездовой наряд. Лоб, уздечка, подбородок, горло, брюхо, бока, грудь, надхвостье,
подхвостье, испод крыла белые. На темени и затылке черно-бурая шапочка с неясными
мелкими охристыми пестринами, такого же цвета кроющие уха, пятно перед глазом
и пятна по бокам зоба. Спина темно-бурая с узкими охристыми пестринами, Большие
верхние кроющие крыла серые, малые и средние — черно-бурые с охристыми каемками,
внутренний ряд малых кроющих беловатый с темно-бурыми основаниями перьев.
II (первое видимое) — VI первостепенные маховые черные с широкими белыми поля-
ми на внутренних опахалах. Внутренние первостепенные серые с темными стволами,
белыми полями на внутренних опахалах и широкой белой вершинной каймой. Второ-
степенные маховые сероватые с белыми каемками и вершинами и черными пристволь-
234
Малая чайка
аыми пятнами. Рулевые белые с черной вершинной частью; крайняя пара может быть
целиком белой. Клюв черный или черновато-бурый, ноги темно-телесные, радужина
бурая.
Первый зимний наряд. Темя и затылок темно-серые, кроющие уха и пятно перед
глазами темно-бурые. Исчезают темные пятна по бокам зоба. Спина, плечевые и верхняя
щсть шеи серые. Остальное оперение, как в предыдущем наряде.
Рис. 55. Область распространения малой чайки
1 — область гнездования, 2 — неуточненная граница области гнездования, 3 — нерегулярные поселе-
ния вне области гнездования, 4 — предполагаемые места гнездования, 5 — места зимовок
Первый летний наряд. Как первый зимний, но голова серовато-бурая с белыми пест-
ринами, наиболее густыми в области лба.
Второй зимний наряд. Как окончательный зимний, но на концах дистальных перво-
степенных маховых сохраняются остатки темно-бурого цвета (признак этот сильно
варьирует) ; нижние кроющие крыла серовато-белые, светлее, чем у взрослых птиц.
Второй летний наряд. Как предыдущий, но голова черная.
Линька. (Дементьев, 1951; Cramp, Simmons, 1983). Неполная линька в первый зим-
ний наряд (сменяется оперение головы, нижней части туловища, передней части спины
и большинство плечевых перьев) происходит в августе—октябре; по-видимому, у неко-
торых могут сменяться рулевые. Частичная линька в первый летний наряд (сменяется
оперение туловища и головы) происходит, видимо, в марте—апреле, а полная линька
во второй зимний наряд — с июня по сентябрь (возможно, октябрь); линька перво-
степенных маховых может заканчиваться в августе. Последующие линьки бывают
дважды в году: частичная предбрачная (оперение головы и туловища) и полная после-
брачная. Линька в брачный наряд у взрослых особей происходит в марте—мае, полная
линька в зимний наряд — в июле—ноябре. Темные перья головы сменяются в основном
к августу, первостепенные маховые заканчивают линьку в сентябре—ноябре. Птицы
в возрасте двух лет линяют, видимо, в несколько более ранние сроки, чем взрослые.
Сроки линьки во все наряды индивидуально варьируют.
Распространение. Гнездовой ареал. Для малой чайки наряду с наличием
более или менее сплошного гнездового ареала характерны периодические выселения
235
Семейство Чайковые
за несколько сот и даже тысяч километров за его пределы, однако без четкой направ-
ленности на запад и северо-запад, отмеченной у черноголовой чайки и морского голуб-
ка (рис. 55). В большинстве случаев подобные поселения существуют только год,
иногда гораздо дольше. В 1962 г. малая чайка неожиданно загнездилась на американ-
ском континенте, в Канаде (Scott, 1963). С тех пор маленькие колонии и отдельные
пары периодически отмечаются в различных штатах Канады и США (McRae, 1984;
Godfrey, 1986) . Крайне нерегулярное гнездование или попытки его отмечены в Англии
(1975 и 1978 гг.), ФРГ (1965 и 1979 гг.), ГДР (1951 г.), Австрии (1963 г.), Румынии
(1866, 1966 гг. и, возможно, 1964 г.). Более или менее постоянное гнездование отме-
чается в Нидерландах (с 1942 г.), Дании (с 1902 г.), Польше (по крайней мере
с 1821 г.), Финляндии (с 1879 г.) и в Швеции на островах Эланд и Готланд (Cramp,
Simmons, 1983).
Основной гнездовой ареал малой чайки лежит в пределах СССР (рис. 56).Центр его,
очевидно, находится в богатой озерами степной зоне юга Западной Сибири и Северного
Казахстана. Здесь малые чайки находят оптимальные условия для гнездования, тут они
наиболее многочисленны, и их колонии распределены относительно равномерно.
Северная граница ареала в этом регионе проходит, видимо, по северу Курганской обл.,
в Тюменской обл. несколько севернее Тобольска, по северной части Омской и Ново-
сибирской областей; в долине р. Оби поднимается до Обь-Чаинского междуречья
(Калинин, Болотников, 1975; Кошелев, 1975; Ходков, 1975; Адам, Сурнаев, 1979;
Сулимов, 1981). Что касается распространенной точки зрения о гнездовании малой чай-
ки по всей Оби (Дементьев, 1951; Гынгазов, Миловидов, 1977), то, кроме Том-
ской обл., достоверные гнездования отмечены лишь в ее низовьях (см. далее) и у
устья Иртыша (Ясков, 1981). Встречи же особей и стаек малых чаек (Гордеев, 1963;
Равкин, 1978) далеко не всегда свидетельствуют о близости гнездовой колонии.
По-видимому, между гнездовьями по верхней и нижней Оби существует большой раз-
рыв, о чем косвенно свидетельствует редкая встречаемость малых чаек на средней
Оби по сравнению с нижней Обью и особенно с более южными районами (Равкин,
1978).
Южная граница основного ареала проходит в Казахстане через север Уральской обл.,
низовья Тургая, далее примерно по широте 49° до долготы Усть-Каменогорска; в кот-
ловине оз. Зайсан отсутствует (Долгушин, 1962; Дебело, Чибисов, 1975; Саму-
сев, 1975).
Западнее, в европейской части СССР, малая чайка становится гораздо более редкой,
ее колонии распределяются внутри ареала неравномерно и нередко меняют свое распо-
ложение. В сущности, о сплошном ареале здесь можно говорить лишь с некоторой на-
тяжкой. Северная граница его огибает с юга Урал, проходит через Башкирию, подни-
мается до Камского водохранилища Пермской обл. и через Удмуртию идет до Рыбин-
ского водохранилища и Центральной части Вологодской обл.; в Кировской обл. не гнез-
дится. Далее в гнездовой ареал входит Ленинградская обл., Ладожское озеро и Прибал-
тика (Карташев, 1975; Попов, 1975; Плесский, 1976; Бутьев, 1978; Ильичев, Фо-
мин, 1979; Болотников и др., 1981; Гражулявичюс, 1981; Пукас и др., 1981; Виксне,
1983; Мальчевский, Пукинский, 1983). Южная граница ареала проходит по южной Бело-
руссии, спускается к Киевскому и Каневскому водохранилищам; далее на восток
достоверно известно гнездование в Калининской, Московской, Рязанской, Горьковской
(С.В.Бакка, устное сообщ.) областях, Татарской АССР, Куйбышевской и Волгоград-
ской областях. Временные поселения отмечены также на севере Воронежской и юге Пен-
зенской обл. (Птушенко, Иноземцев, 1968; Кистяковский, Мельничук, 1975; Семаго,
1975; Водолажская, Залетаев, 1977; Наумчик, 1979; Зиновьев, 1980; Аюпов, 1981;
Денисов, Фролов, 1981; Зубакин, 19816; Клестов, Осипова, 1981; Кубанцев, Чернобай.
236
237
Рис. 56. Ареал малой чайки в СССР
1 — область гнездования, 2 — область наиболее высокой численности, 3 — неуточненная граница области гнездования, 4 — нерегуляр-
ные поселения вне области гнездования, 5 — предполагаемые места гнездования, 6 — примерная северная граница области регуляр-
ных кочевок, 7— залеты, 8 — места зимовок
Семейство Чайковые
1982; Долб1к, Навумчык, 1983; Мельничук, Головач, 1984). Гнездования малых чаек
в Брянской и Калужской областях и в Мордовии не отмечено (Луговой, 1975; Кудле-
нок, 1981; Баранов, Марголин, 1981).
В Восточной Сибири сплошного ареала вид не образует. Отмечены гнездовые поселе-
ния в северной Хакасии (Прокофьев, 1981) и южной Туве (Баранов, 1981; Леонович,
устное сообщ.), Прибайкалье и Забайкалье (Садков, 1977; Шкатулова, 1979; Егоров,
1981; Мельников, 1984; Васильченко, 1986), долине р. Лены от Якутска примерно до
впадения Вилюя и в долине среднего Вилюя (Андреев, 1953, 1974; Лабутин, Поздня-
ков, 1978; Дегтярев, 1985). Вероятно, хотя и не доказано, гнездование малых чаек
где-то на среднем, а возможно, и нижнем Енисее (Рогачева и др., 1978; Сыроечковский,
Рогачева, 1980; Рогачева, Вахрушев, 1983). В 1961 — 1964 гг. около 10 пар малых чаек
гнездились на оз. Джулукуль на юго-востоке Алтая; в последующие годы они здесь
встречены не были (Ирисов, 1972; Стахеев, 1981).
Отмечалось еще несколько временных поселений (выселений) вдали от основного
ареала. Возможно, малая чайка изредка гнездится в Волынской обл. (Сребродольская
и др., 1981). Орнитологи конца прошлого — начала нашего столетия единодушно сооб-
щали о гнездовании малых чаек на Сиваше (Шатилов, 1874; Никольский, 1891;
Браунер, 1898; Сеницкий, 1898; Молчанов, 1906). В настоящее время малая чайка
здесь не гнездится, а поскольку условия Сиваша достаточно несхожи с характерным
для вида гнездовым биотопом, возникает предположение, что за гнездящихся птиц
могли быть приняты пролетные особи, которые нередко в больших количествах скап-
ливаются весной на Сиваше (Костин, 1983).
Бесспорно гнездование малых чаек на севере европейской части СССР, где поселения
отмечались в устье Печоры (Дмоховский, 1933), на п-ове Канин в окрестностях Семжи
в 1965 г. (Леонович, устное сообщ.); указывалось гнездование и для района Архан-
гельска (Мензбир, 1895). Достоверное гнездование отмечено на южном Ямале и нижней
Оби у с. Мужи (Данилов, 1965; Данилов и др., 1984). Не исключено, что в данном слу-
чае речь может идти не о периодическом выселении отдельных групп на север, а еще об
одном, северном участке ареала от Архангельска до низовий Оби, на протяжении кото-
рого чайки образуют непостоянные поселения. Возможно также, что таких участков
два: североевропейский и нижнеобский.
Вне СССР з Азии возможно гнездование на оз. Буйр-Нур в Монголии (Stubbe, Во-
lod, 1971);
Зимовки. Малая чайка на территории Палеарктики зимует в Балтийском море,
на атлантическом побережье Западной Европы и Марокко, Азорских островах, в Среди-
земном море, на крымском и Кавказском побережье Черного моря, южном побережье
Каспия (ЗИН; ЦКАН; Строков, 1974; Костин, 1983; Cramp, Simmons, 1983). На вос-
токе зимовки отмечены в северо-западном, северо-восточном и восточном районах Ки-
тая (Schauensee, 1984) . По крайне скудным результатам кольцевания можно предпо-
ложить зимовку чаек из Прибалтики и Ленинградской обл. на атлантическом побе-
режье Западной Европы и в западном Средиземноморье; по крайней мере часть сибир-
ских чаек летит зимовать на юг Каспия и далее на запад: чайка, окольцованная птен-
цом 14.VII 1974 г. на севере Байкала, встречена 15.X того же года в г.Махачкале;
неполовозрелая чайка, окольцованная в августе 1970 г. в Бельгии, добыта в июне
1973 г. в Барабинской степи (Виксне, 1962; Мальчевский, Пукинский, 1983; Cramp.
Simmons, 1983; ЦКАН).
Миграции. В связи с крайне незначительным количеством возвратов колец изучены
недостаточно. Весенний и осенний пролет чаек, гнездящихся в Прибалтике и Ленин-
градской обл., изучен лучше других, он проходит вдоль южного побережья Балтийского
238
Малая чайка
и Северного морей; часть птиц (чаще весной, чем осенью) пересекают материк между
Балтийским и Средиземным морями (Виксне, 1962; Cramp, Simmons, 1983).
Первые чайки появляются весной в конце апреля —начале мая, в Ленинградской обл.—
чаще всего во вторую пятидневку мая. Откочевка части птиц начинается уже в июне,
осенний пролет со второй половины июля до середины — конца августа (Носков и др.,
1981; Мальчевский, Пукинский, 1983). Последние пролетные птицы в Латвии отмечены
21.Х (Виксне, 1962).
Меньше ясности с птицами из Центра европейской части СССР. Прилетают малые
чайки сюда в конце апреля (в Московскую обл. в среднем 23.IV, в Калининскую
20.IV —2.V), начинают отлетать со второй половины июля и исчезают в августе
(Птушенко, Иноземцев, 1968; Зиновьев, 1980) . Отсутствие данных об осеннем пролете
малых чаек непосредственно южнее Московской и Рязанской областей породило
предположение о перемещениях этих птиц либо вниз по Оке и далее по Волге (Поля-
ков, 1910), либо через водоемы верховий Волги в западном направлении (Птушенко,
Иноземцев, 1968). В то же время выраженный осенний пролет во второй половине
августа — начале сентября отмечается на Днепре под Каневом (Юстякивскш, 1957) и
в конце июля — начале сентября в Белоруссии (Федюшин, Долбик, 1967). Что касается
весенней миграции, то она, скорее всего, идет с Черного моря через материк. Об этом
свидетельствует пролет малых чаек по Днепровским водохранилищам со второй поло-
вины марта по первую половину апреля (Булахов, Мясоедова, 1978), весенний пролет
на р.Мокше в Мордовии (Луговой, 1975) и более ранние сроки появления чаек в Мос-
ковской обл. по сравнению с Ленинградской обл. и Татарской АССР (Водолажская,
Залетаев, 1977).
По-видимому, узловым пунктом как весенней, так и осенней миграции служит север-
ный Крым, где первое весеннее появление малых чаек отмечается 16.Ш— 9. IV, а пик
пролета — в апреле. После сезона размножения птицы начинают пролетать здесь
в начале — середине июля; в начале августа в Присивашье образуются скопления до
30 тыс. птиц (Костин, 1983). Большая величина скоплений говорит о том, что здесь
пролетают не только европейские, но и, видимо, часть азиатских чаек. В отношении
миграции последних данных еще меньше. На севере Казахстана и в Западной Сибири
основное направление весенних миграций восточное. В низовьях Иргиза и Тургая пер-
вые птицы отмечались 2—10.V, в Наурзумском заповеднике 30.IV—4.V, в окрест-
ностях Караганды 15 —19.V, северо-восточнее Семипалатинска 13—20.V, у оз.Чаны
2—3.V, в окрестностях Нарыма Томской обл. 5—15.V, у Ханты-Мансийска 4—24.V
(в среднем 17.V) (Долгушин, 1962; Елкин, 1975; Дубовик и др., 1977; Ходков,
19776; Гордеев, 1979). Незначительный весенний пролет идет и через Аральское море
(Березовский, 1983). Осенний пролет в Казахстане и Западной Сибири начинается в
конце июля и идет, по-видимому, в западном направлении. Последние малые чайки из
Б арабы и западной части Казахстана исчезают в конце августа — первых числах сентяб-
ря; в среднем течении р. Урал пролет длится весь сентябрь и первую половину октября.
Судя по появлениям первых чаек в окрестностях Улан-Удэ (22.IV—2.V), восточно-
сибирские чайки летят из иных мест зимовок, по-видимому из Китая. В Якутии в доли-
не р.Лены первых малых чаек отмечают 11—12.V, пролет идет с запада на восток.
На среднем Вилюе чаек встречают с 20.V. Первые малые чайки на северном Байкале
отмечаются 12—18.V, осенние миграции начинаются 26.VII—4.VIII, последние птицы
исчезают в начале сентября (Андреев, 1974; Садков, 1977; Дегтярев и др., 1978;
Лабутин, Поздняков, 1978; Шкатулова, 1979). По данным кольцевания, байкальские
малые чайки отлетают осенью в западном направлении (Скрябин и др., 1981) .
Залеты малых чаек отмечены в Исландию, на Фарерские острова, Канарские острова,
239
Семейство Чайковые
в Норвегию, Мавританию, Гамбию, Нигерию, Сьерра-Леоне, Анголу, Кению, Ирак,
Иорданию, Кувейт (Cramp, Simmons, 1983), в окрестности Бомбея и Ладакх (АН,
Ripley, 1969), на Айновы острова (Коханов, Скокова, 1967), западный Таймыр
(Кречмар, 1966), в Яно-Индигирскую низменность в июне 1976 г. (пос. Берелях) и
окрестности Магадана в июне 1972 г. (Леонович, 1981, устное сообщ.), на Памир в ав-
густе 1977 г. (данные В.А.Зубакина), в бассейн р.Сарыджаз (высота 3500 м) на восто-
ке Киргизии (Янушевич и др., 1959), окрестности Верхоянска (ЗИН).
Численность. За исключением степной зоны Казахстана, везде малочисленна или
редка. В связи с резкими колебаниями численности в разных поселениях по годам
общая оценка численности затруднена. В Латвии гнездится 1000—2000 пар (Виксне,
1983), в Ленинградской обл. — около 2000 пар (Мальчевский, Пукинский, 1983),
в Белоруссии — 1600—2300 пар (Наумчик, 1979; Бирюков и др., 1982), в Московской
обл. в начале 1980-х годов —300—400 пар (данные В.А.Зубакина), в Омской обл. —
около 1 тыс. пар (Сулимов, 1981), на оз. Чаны — около 200 пар (Ходков, 1981),
в дельте р. Селенги — не менее 600 пар (Мельников, 1984). Общая численность вида
в СССР, по-видимому, несколько десятков тысяч гнездящихся пар; вне СССР — вряд
ли более 1,5 тыс. пар (Cramp, Simmons, 1983). Тенденции изменения численности
мировой популяции не ясны. В Финляндии, Вологодской, Ленинградской и Московской
областях отмечен рост численности; в Дании, ряде колоний Латвии и в дельте Селенги—
ее снижение.
Местообитания. В сезон гнездования — пресноводные равнинные озера, старицы, пой-
менные озера и болота в долинах рек степной зоны, зоны смешанных лесов и тайги.
В период миграции — морские побережья и разнообразные внутренние водоемы;
на материке, видимо, предпочитает мигрировать по долинам крупных рек. В период
зимовки — морские побережья, где чайки держатся как в прибрежных мелководьях,
у городов и устьев рек, так и в 1—7 км от берега (Долгушин, 1962; Костин, 1983;
Cramp, Simmons, 1983).
Размножение. В норме малая чайка приступает к размножению, видимо, на третий
год жизни (Cramp, Simmons, 1983). Моногам. Диффузно-гнездящийся факультативно-
колониальный вид. Пары формируются как в период пролета, так и вскоре после при-
лета, вне территории колонии. Колонии, как правило, каждый год возникают на но-
вом месте. Характерны резкие колебания величины поселения по годам вплоть до
периодического прекращения гнездования. По-видимому, происходит интенсивное
перераспределение гнездящихся пар на обширном пространстве, измеряемом десятками
тысяч, а возможно и сотнями тысяч квадратных километров. Окольцованная в Латвии
в июне 1974 г. птенцом чайка была найдена мертвой в июне 1981 г. примерно в 700 км
северо-восточнее — в Олонецком районе Карельской АССР (ЦКАН).
Демонстрационное поведение малых чаек очень своеобразно. Воздушные демонстра-
ции преобладают над наземными, что обычно характеризует крачек. Начиная с середины
периода насиживания демонстрации чаек на своей гнездовой территории отмечаются
редко — этим малые чайки напоминают розовых чаек. Среди наземных демонстраций
можно выделить лишь прямую позу, напоминающую такую же позу черноголовых чаек,
’’квохтанье”, вскидывание клюва перед ритуальным кормлением и спариванием и
наклоны головы в сторону вместо обычного для ряда чаек отворачивания клюва.
Все эти демонстрации относятся к сфере брачного поведения. Агрессивные намерения
чаще всего передаются криками — быстрой трелькой цокающих звуков, которую
издает сидящая на гнезде птица по отношению к противнику, и резкими визгливыми
криками типа ”уик” или ”уить”, с которыми взлетевшая птица гонится за другой чай-
кой. Воздушные демонстрации представляют собой чередования активного полета,
более глубоких и более редких взмахов крыльев и планирования, сопровождающиеся
240
Малая чайка
серией быстро повторяющихся криков ”ке-кей, ке-кей, ке-кей ... ”, ”ке-кой, ке-кой,
ке-кой ... ” и звучащих реже предыдущих и только после них криков ”ке-кееу, ке-кееу,
ке-кееу ... ”. В последнем случае шея и закрытый или полуоткрытый клюв вытянуты
наклонно вверх, крылья распростерты горизонтально, хвост слегка поднят вверх;
вся поза напоминает позу при долгом крике в полете у серебристой или озерной чай-
ки (рис. 57,а). Изредка такой крик можно слышать и у сидящей птицы (рис. 57,в).
По-видимому, данный крик, а возможно и весь описанный демонстративный полет,
Рис. 57. Демонстрационное поведение малой чайки
а — поза при долгом крике в полете (крик ”ке-кееу, ке-кееу”), б — крик ”ке-кой, ке-кой” на земле,
в — поза при долгом крике на воде, г — наклон головы в сторону
можно считать гомологом долгого крика, однако своей неоформленностью, длитель-
ностью и рядом других деталей он сильно отличается от долгого крика большинства
других чаек (Moynihan, 1955; Veen, 1980, цит. по: Cramp, Simmons, 1983; данные
В. А. Зубакина).
Характерный гнездовой биотоп малой чайки — пойменные мелководные болотца,
заросшие манником, осокой, телорезом и другой водной и околоводной растительно-
стью; заболоченные берега пойменных и иных равнинных озер и водохранилищ с топ-
кими сплавинами; прибрежные заболоченные луга; реже низкие сырые острова,
заросшие водяной сосенкой (дельта р. Селенги). Крайне редко колонии располагаются
на сухих островках и солончаковых берегах. В ряде случаев чайки могут оказаться
на сухом островке в результате резкого падения уровня воды после паводка. В послед-
ние годы отмечено постепенное заселение рыборазводных прудов. Колонии обычно
невелики — 5—30 пар, нередко до 50 пар, более крупных мало (в Московской обл.
до 170 пар, в Казахстане до нескольких сот, в дельте р.Селенги свыше 100 пар).
Гнездование отдельными парами отмечалось (Сушкин, 1908), но, видимо, оно бывает
лишь в колониях других видов чайковых. Моновидовые колонии образуются редко
(в 7,3% случаев в дельте Селенги и в 22,2% в Московской обл.). В зависимости от места
241
Семейство Чайковые
гнездования предпочитают гнездиться либо с белокрылыми крачками (Московская обл.,
Казахстан), либо с озерной чайкой и речной крачкой (Латвия', Белоруссия, Ленинград-
ская обл., дельта Селенги, Северный Байкал). Гнезда располагаются на плавающих
растениях, кучках растительного мусора, тонких сплавинах, заломах тростника, реже
на осоковых кочках, еще реже — прямо на земле. В последнем случае выстилка гнезда
бывает крайне скудной, обычно же чайки строят хорошо оформленные гнезда из кусоч-
ков растительного мусора. Диаметр гнезда (мм) по измерениям в Московской обл.
составляет (и = 47) 160—240, средний 190; диаметр лотка (и = 87) 95—135, сред-
ний 115; глубина лотка (и = 48) 20—40, средняя 25 (Бородулина, 1960; Долгушин,
1962; Садков, 197-7; Зубакин, 19816; Бирюков и др., 1982; Виксне, 1983; Мальчев-
ский, Пу кинский, 1983; Мищенко, 1983; наши данные).
Плотность гнездования невысокая. На Северном Байкале она варьирует от 1,2 до
9,0 гнезд на 100 м2. Среднее расстояние между центрами ближайших соседних гнезд на
оз. Энгуре в Латвии составляет 4,3 м (1,4—10 м; п = 36), в Московской обл. (и = 225)
4,7 м, в дельте Селенги, по данным Д.М.Очагова, — 6,6 м (2—39 м; и = 128), в Якутии—
4,1 м (1,3—10 м; п = 19). Индекс плотности гнездования вида, таким образом, ра-
вен 0,02. Первые яйца появляются в Ленинградской обл. обычно в середине третьей де-
кады мая, в Вологодской — в первых числах июня, в Московской — в конце второй или
начале третьей декады мая, в Казахстане в конце мая — первых числах июня, в Томской
обл. в первой половине июня, на Северном Байкале с 29—30.V, в Якутии 30—31.V.
В отдельных колониях откладка яиц может начаться на месяц позже обычного срока;
по-видимому, такие колонии образуют птицы, потерявшие первые кладки. Величина
кладки 1—5 яиц, в большинстве случаев 3 яйца, реже 2. Размеры яиц (мм) : из Москов-
ской обл. (и = 100) 37,2—44,1 X 27,7—31,7, в среднем 40,8 X 29,8; из Вологодской обл.
(и = 7) 39,1—42,0 X 28,9—31,2, в среднем 40,5 X 30,1; из Томской обл, (и = 48) в сред-
нем 41,2 X 30,0; Из Казахстана (и = 9) 39,8—43,3 X 29,4—31,3, в среднем 41,2 X 30,2.
По данным В.В.Леоновича, масса свежих яиц (г) с п-ова Канин (и = 12) 15,9—20,0,
в среднем 18,4; из Московской обл. (и = 6) 18,3—203, в среднем 19,3. Окраска яйца
сложная. Фон темный: оливковый, оливково-зеленый, коричневато-оливковый.
Темно-бурые поверхностные пятна обычно мелкие; рисунок редкий, сгущающийся у
тупого конца или образующий поясок на тупом конце. Насиживание начинается с пер-
вого яйца, птенец вылупляется через 19—24 дня, обычно через 20—22 дня (Долгушин,
1962; Садков, 1977; Адам, Сурнаев, 1979; Ковшарь, Левин, 1982; Мальчевский,
Пукинский, 1983; Дегтярев, 1986; наши данные).
В период откладки яиц насиживают только самки, затем оба партнера, но самка си-
дит на гнезде больше — 51 %—85% времени суток. Ночью чаще насиживают самки.
Температура в гнезде в начале периода насиживания составляет 34,0—39,6°С (Болотни-
ков, Калинин, 1975; Калинин, 1975). Смена, как правило, происходит быстро:
услышав крик прилетевшей птицы, насиживающая чайка взлетает, и партнер опускается
прямо в гнездо. Смененная птица затем нередко несколько раз приносит материал для
гнезда. Между появлением трещины на яйце и вылуплением проходит 1—3 сут
(и - 21), чаще 2 сут (в 47,6% случаев), реже 3 сут (в 33,3% случаев); между появле-
нием отверстия и вылуплением — чаще всего 1 сут (83,3% случаев). Вылупление птен-
цов в гнезде из 3 яиц обычно длится 2—3 сут. После появления птенца родители уносят
скорлупу из гнезда. В Московской обл. первые птенцы появляются 7—10.VI, в Ново-
сибирской обл. — в начале второй декады июня. Птенец в день вылупления весит (л = 7)
12,5—16,9 г, в среднем 15,1 г (Московская обл.). В первые дни жизни родители обогре-
вают птенцов — сначала на гнезде, а после ухода выводка на третий день вне его.
10—11-суточных птенцов родители не обогревают даже в дождь и при сильном похолода-
242
Малая чайка
нии. Выводок может как остаться вплоть до подъема на крыло в окрестностях гнезда,
так и переселиться в другую часть колонии. Выкармливают птенцов оба родителя,
10—11-суточных птенцов кормят 11 — 13 раз за светлое время суток. Корм — отрыгнутые
разнообразные насекомые. Во время тревоги в колонии птенцы разбегаются и за-
таиваются. Родители активно преследуют серых ворон, сорок, серых цапель и других
явных и потенциальных врагов. На человека, однако, пикируют редко. Летать птенцы
начинают в возрасте 23—25 дней (Бородулина, 1960; Мальчевский, Пукинский, 1983;
наши данные).
Обращает на себя внимание сходство сроков размножения малых чаек в разных
частях ареала. Кладки появляются в конце мая — середине июня, а лётные птенцы —
в середине июля — первых числах августа как в центре европейской части СССР, так и
на севере Байкала, в Якутии и даже на юге Ямала и в низовьях р.Полуй (Бойков, 1965;
Данилов и др., 1984).
Точных данных по величине эмбриональной и постэмбриональной смертности мало.
В годы с благоприятным гидрорежимом на севере Байкала отход яиц составляет 17,8%,
пуховиков — 3,2%. В многоводные годы летние паводки губят до 80% кладок (Сад-
ков, 1977). В Якутии до подъема на крыло доживает 1,32—1,38 птенца на выводок
(Дегтярев, 1986). Максимально известная длительность жизни 7 лет (ЦКАН).
Суточная активность, поведение. Ярко выраженная дневная активность, ночью жизнь
в колонии замирает. Общественная птица, обычно встречается стайками в несколько
особей. На кормежке может образовывать скопления в несколько сот и даже тысяч
особей.
Питание. В сезон гнездования - главным образом водные и наземные беспозвоноч-
ные, насекомые. В Новосибирской области основной корм — хирономиды, личинки пла-
вунцов, гладыши, гребляки, мелкие стрекозы и ручейники; остатки позвоночных жи-
вотных не встречены (Бородулина, 1960). В Кустанайской обл., помимо беспозвоноч-
ных, малая чайка в небольшом количестве поедает рыбу — остатки небольших карасей
и сазанов встречены в желудках у 23,5% от 17 птиц. Из беспозвоночных преобладают
жуки-плавунцы и стрекозы (Самородов, 19786). В других районах Казахстана пита-
ется личинками двукрылых, водяными клопами и их личинками, личинками водяных
жуков, ракообразными (Artemia salina). На суше поедает мелких саранчовых, жуков
(Долгушин, 1962). Главнейшие компоненты питания на севере Байкала - хирономиды
и ручейники (Скрябин, Размахнина, 1978), то же отмечено для Дарвинского заповед-
ника (Зиновьев, 1980). В Латвии основным кормом служат насекомые, изредка рыба
(Тима, 1961). В осенний период, по В.С. Залетаеву (1955, 1960), главный корм — мел-
кая рыба, моллюски, реже насекомые. Весной иногда кормится на свалках (Воробьев,
1981).
По способам добывания корма малые чайки напоминают болотных крачек. Кормят-
ся обычно небольшими стайками, чаще всего над заросшими растительностью мелко-
водьями, лугами, травяными болотами. Летают низко над водой или травой, склевы-
вая насекомых с воды или растений. Не ныряют. Нередко ловят летающих насекомых
в воздухе; рачков Artemia salina добывают, бродя по мелководью (Бородулина, 1960;
Тима, 1961; Долгушин, 1962; Самородов, 19786).
Враги, неблагоприятные факторы. Гнезда малых чаек нередко разоряют болотные
луни и серые вороны. Кладки могут гибнуть при изменениях уровня воды в водоеме,
птенцы погибают от переохлаждения среди густой травы в холодные и дождливые дни.
Отрицательное воздействие могут оказывать и антропогенные факторы (Бирюков и
др., 1982); выпас скота, сельхозработы, фактор беспокойства.
Хозяйственное значение, охрана. Хозяйственного значения вид не имеет, занесен в
Красную книгу БССР.
243
Семейство Чайковые
Род Rhodostethia MacGillivray, 1842
Розовая чайка
Rhodostethia rosea (MacGillivray, 1824)
Ross’s Gull (англ.). Rosenmowc (нем.). Mouette de Ross (фр.).
Син. (лат.): Larus roseus MacGillivray, 1824; Larus rossii Richardson, 1825.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Мелкая, исключительно красивая чайка
легкого изящного склада, с отчетливо клиновидным хвостом и коротким тонким клю-
вом. В гнездовое время безошибочно отличается от других видов чаек розовой окрас-
кой нижней стороны тела и черным узким ’’ожерельем” на шее. В негнездовое время
розовая окраска незаметна: по размерам и отсутствию черного цвета на концах крыльев
розовая чайка в негнездовом наряде может быть спутана с малой чайкой, но отличается
от нее более светлым подбоем крыла, клиновидным хвостом и более коротким клю-
вом. Полет очень легкий, маневренный и грациозный, несколько напоминающий полет
крачек; при поисках пищи над водоемом птицы нередко зависают на месте, после чего
с плеском бросаются в воду, погружаясь почти целиком. При полете на значительные
расстояния взмахи крыльев ровны и глубоки. Розовые чайки очень любят сидеть на
льду, предпочитая опуститься на льдину, даже когда участки льда единичны. Легко и
грациозно плавают, но на воде этих птиц приходится видеть не очень часто. Голос ро-
зовой чайки негромок, нежен и в каком-то аспекте мелодичен. Чаще всего от пролетаю-
щих птиц можно слышать серию криков ”э-вей, э-вей” или ”ке-век, ке-век, ке-век”.
Этот крик также издает птица, подлетающая к своему гнезду. Используется он и в аг-
рессивных демонстрациях. В возбуждении птицы издают мягкие, слегка гнусавые зву-
ки ”вав-вав-вав-вав...” и визгливые крики ”кья, кья...”, которые иногда звучат как
”кийя”. С этими криками чайка может преследовать поморника или птицу своего вида.
Характерный для чаек трехсложный крик тревоги у розовой чайки звучит как ”кэ-вэ-
вэ” или ”квэ-вэ-вэ”. Однако слышать его можно только в гнездовой колонии и исполь-
зуется он редко. В период брачных демонстраций в скоплениях чаек слышны сухие
трескучие трельки. В целом голос розовой чайки несколько напоминает голос неко-
торых крачек и малой чайки.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Розовая чайка по морфологическим характеристикам
стоит несколько особняком среди других видов чаек (Dwight, 1925). Обычно ее выде-
ляют в отдельный род Rhodostethia (Peters, 1934; Бутурлин, 1934; Vaurie, 1965; Степа-
нян, 1975; Howard, Moore, 1984), реже объединяют с другими чайками в род Larus
(Moynihan, 1959). Филогенетически розовая чайка ближе всего, по-видимому, к ма-
лой чайке, а по ряду признаков близка также к родам Rissa и Хета (Dwight, 1925;
Юдин,1965; Фирсова, 19786).
Описание. Размеры (мм) (ЗМ МГУ) и масса тела (г) (ЗМ МГУ, Воробьев,
1963):
Длина крыла:
dd (п = 21) - 242-275 (среднее 263);
99 (п = 16) — 240—282 (среднее 258).
Длина клюва:
dd (п = 20) - 16,3-20,5 (среднее 18,2) ;
99 (п = 16) - 16,2-19,4 (среднее 17,9).
244
Розовая чайка
Длина цевки:
33 (и = 21) - 29,1-33,7 (среднее 31,7);
99 (и = 16) - 29,7-32,8 (среднее 31,0).
Масса тела:
33 (и = 4) - 157-190 (среднее 171);
99 (и = 2) - 142-145 (среднее 144).
Окраска. (ЗМ МГУ, ЗИН, сборы А.А. Кищинского).
Самец и самка в брачном наряде. Голова, шея, вся нижняя сторона тела и рулевые
розового цвета различной интенсивности. На верхней части головы, надхвостье и руле-
вых розовый цвет наименее ярок; последние иногда почти белые. Розовая окраска у
самцов обычно более интенсивна. Розовый цвет уже вскоре после добычи и препаровки
птицы начинает терять интенсивность, а при хранении в коллекциях со временем пол-
ностью пропадает. Вокруг шеи узкая черная полоска, замыкающаяся в своеобразное
’’ожерелье”. Спина, плечевые и верхние кроющие крыла мягкого сизо-пепельного
цвета. Второстепенные маховые того же цвета, но с белыми вершинами. Первостепен-
ные маховые сизые, слегка темнеющие по направлению к дистальному концу крыла.
II (первое видимое) первостепенное имеет черное наружное опахало; черный цвет не
доходит до вершины пера. Вершины трех проксимальных первостепенных маховых
белые. Клюв черный, полость рта, края век и ноги ярко-красные, когти черные. Радужи-
на коричневая.
Самец и самка в зимнем наряде. Спина, плечевые и кроющие крыла сизо-серые, не-
сколько светлее, чем в предыдущем наряде. Прочее контурное оперение, как и руле-
вые, белое. Черного ожерелья нет. Слабо заметные черноватые пятнышки, образованные
темными окончаниями перьев, расположены книзу от глаза и в ушной области. На
темени едва заметный сероватый налет. Остальное оперенье, как в предыдущем наряде.
Ноги, по Г.П. Дементьеву (1951), оранжево-красные.
Пуховой наряд. Основной цвет серовато-желтый разного оттенка — от серно-желтого
до охристо-буроватого или ржавчатого. По основному фону разбросаны многочислен-
ные темные пестрины различной формы, образующие продольный рисунок на туло-
вище, поперечный на затылке и суживающийся клином на темени. Пестрины на тулови-
ще буровато-серые и нерезкие, на голове — почти черные, резкие. По нескольку округ-
лых пестрин есть по бокам горла и шее и одна длинная в виде бровной полосы — над
глазом. Зоб охристый, со слабыми черными пестринками; охристый налет есть на бо-
ках и гузке. Середина груди и брюха беловатые, без пестрин. Ноги и клюв багрово-
телесного цвета, когти и кончик клюва буроватые.
Гнездовой наряд. Верхняя сторона тела черновато-бурая с поперечными охристыми
пестринами. Лоб и уздечка сизовато-бурые. Перед глазом нерезкое серо-бурое или чер-
новато-бурое пятно, распространяющееся и на ушную область. На задней стороне шеи
неясный беловатый ошейничек. Верхние кроющие хвоста белые с неясными охристыми
вершинами и небольшими аспидно-бурыми предвершинными пестринами. Малые и боль-
шие кроющие крыла, кроме кроющих кисти, белые со светло-охристыми вершинами,
средние и кроющие кисти черно-бурые; второстепенные маховые белые. Три дисталь-
ных первостепенных маховых пера (не считая редуцированного) черно-бурые с белой
внутренней частью внутреннего опахала, не достигающей вершины. Последующие пер-
востепенные маховые все более светлые, черно-бурый цвет остается лишь на вершинах
перьев, и 2—3 проксимальных первостепенных уже белые. В целом развернутое крыло
белое, с темной зигзагообразной полосой. Рулевые белые, с узкими охристыми вершин-
245
Семейство Чайковые
ними каемками; 3—4 средних пары рулевых с черно-бурыми вершинами. Хвост клино-
видный, но разница между средними и крайними рулевыми меньше, чем у взрослой пти-
цы. Нижняя сторона тела белая, с сильным сизовато-глинистым налетом и серо-бурой
пестристостью на зобе и груди- Нижние кроющие крыла вдоль края кисти буроватые.
Глаза и края век черно-бурые. Ноги багрово-телесного цвета, когти черноватые. Клюв
серовато-бурый, бледнее к основанию, с телесным основанием нижней челюсти.
Первый зимний наряд. Спина бледно-серая, светлее, чем в брачных нарядах. Голова
и шея белые с серовато-сизым налетом на темени и зашейке. В ушной области и у глаз
буровато-черные пятна, вокруг них черноватые пестрины. Нижняя сторона тела белая с
розоватым оттенком. Оперение крыльев и рулевые перья сохраняются от гнездового
наряда. Окраска клюва и ног та же, что в гнездовом наряде.
Первый летний наряд. Спина серовато-сизая, как в окончательном наряде. Голова и
верхняя сторона шеи белые с розовым оттенком, светлее, чем у взрослых птиц. На
темени и затылке сероватый оттенок. У большинства птиц заметны черноватые пятна
вокруг глаз, изредка и в ушной области. Нижняя сторона тела розового цвета, чуть свет-
лее, чем в последующих брачных нарядах. Черный ошейник имеется, но иногда преры-
вается снизу. Большие кроющие второстепенных маховых,серовато-сизые с широкими
белыми краями внешних опахал, остальные кроющие крыла черновато-бурые со свет-
лыми каймами. Концы и внутренние опахала больших кроющих первостепенных махо-
вых, начиная с 6-го, белые. Первостепенные маховые черно-бурые; на II—IV большая
часть внутренних опахал белая, но белый цвет не доходит до вершин перьев. V первосте-
пенное буровато-серое с белыми внутренними опахалом и предвершинной полосой и
черной вершиной; на следующих первостепенных маховых белые части еще более уве-
личиваются, а IX—XI совсем белые. Второстепенные маховые белые с сизым оттенком
на внутренней части; на внешних опахалах последних двух черноватые участки. Плече-
вые — чернобурые. Рулевые — белые; хвост клиновидной формы. Клюв и кольцо во-
круг глаз черные, ноги буровато-красные, не столь яркие, как у взрослых птиц.
Второй зимний наряд. Отличается от окончательного зимнего наличием большого
количества черно-бурого цвета на стержне и наружном опахале I (редуцированного)
первостепенного махового пера и на одном или нескольких кроющих перьях. В ушной
области заметны отчетливые черноватые пятна. Чернота вокруг глаз сохраняется от
1-го брачного наряда. Темя и затылок с пепельно-серым налетом. Наряд приобретается
на вторую осень жизни; он ошибочно описан Дж. Дуайтом (Dwight, 1925) в качестве
окончательного зимнего наряда.
Второй летний наряд. Отличается от окончательного брачного наличием бурого или
черно-бурого цвета на стержне и опахале I первостепенного махового и на 1—3-м боль-
ших кроющих крыла.
Третий зимний наряд. Отличается от окончательного наличием буроватого цвета на
I первостепенном маховом. Есть серый налет вокруг глаз и едва заметный сероватый от-
тенок в ушной области.
Третий летний наряд. Отличается от окончательного наличием серовато-бурого цвета
на I (редуцированном) первостепенном маховом или на 1-м большом кроющем.
Линька. Первый зимний наряд одевается в августе—сентябре после частичной линьки,
в результате которой сменяется контурное оперение головы, шеи и туловища; оперение
крыльев и рулевые перья сохраняются от гнездового периода (Dwight, 1925). Частичная
линька в первый летний наряд идет медленно и постепенно, так что в начале лета могут
быть добыты птицы в разной ее стадии. В отличие от большинства других чаек первая
смена рулевых у розовых чаек происходит весной (а не осенью) второго года. Большая
часть оперения крыльев сохраняется от гнездового наряда. Линька во второй зимний
наряд начинается в июне, со средних и малых кроющих крыла. В сентябре она заверша-
246
247
Рис. 58. Область распространения розовой чайки
1 — гнездовой ареал, 2 — изолированные поселения вне основного гнездового ареала, 3 — область зимних кочевок, 4 — область летне-осенних коче-
вок , 5 — направления миграций, 6 — залеты, 7 — предполагаемые направления миграций и места гнездования
Семейство Чайковые
ется; смены рулевых не происходит. В последующие годы бывает две линьки в году:
полная и послебрачная, которая по С. Крэмпу (Cramp, Simmons, 1983) протекает с
июля августа1 по сентябрь — начало октября, и, видимо частичная, предбрачная, о де-
талях которой почти ничего не известно.
Распространение. Гнездовой ареал. Охватывает Яно-Индигирскую и Колым-
скую низменности от Омолоя до Колымы (Бутурлин, 1905, 1934; Дементьев, 1951;
Воробьев, 1963, 1967; Переломов, 1976; Рощевский, 1976; Томкович, 1980). К северу
он доходит до южной границы арктических тундр и только в дельтах Индигирки и Ко-
лымы проникает в южную часть этой подзоны. Большая часть области гнездования ле-
жит в подзоне субарктических тундр, на юге она захватывает открытые озерно-болот-
ные равнины предтундровых редколесий (рис. 58, 59).
Спорадическое гнездование розовых чаек отмечается и вне основного гнездового
ареала. Факты достоверного гнездования известны для Чаунской низменности (Андре-
ев, Кондратьев, 1981; Засыпкин, 1981), дельты Лены (Лабутин и др., 1985), низовий
р. Б. Балахня на восточном Таймыре (Павлов, Дорогов, 1976) , западной (залив Диско)
и северо-восточной Гренландии (Hjort, 1980; Kampp, Kristensen, 1980; Evans, 1984b).
для островов Королевы Елизаветы Канадского Арктического архипелага и окрест-
ностей Черчилла в канадской провинции Манитоба (Balch et al., 1979; Blomqvist, Elan-
der, 1980; Chartier, Cooke, 1980; Bledsoe, Sibley, 1985; Godfrey, 1986). По-видимому,
возможно периодическое гнездование розовых чаек на р. Баранихе (Schaanning, 1928):
попытки гнездования отмечены также на Шпицбергене (Lovenskiold, 1964). Хотя боль-
шинство попыток гнездования вне основного гнездового ареала отмечено в 1970-х го-
дах, говорить о расширении области гнездования вида в последние годы, видимо, преж-
девременно. Скорее всего, это периодическое выселение части особей за пределы ос-
новного гнездового ареала. Так, гнездование розовых чаек в заливе Диско отмечалось
и в 1885 г., а на канадском о-ве Миен — в начале XX столетия (Glutz v. Blotzheim, Bauer.
1982). В Гренландии и Северной Америке, по-видимому, поселяются птицы, ’’запер-
тые” на зимовках в Северной Атлантике (подробнее - при описании миграций).
Зимовки. О местах зимовок розовых чаек до сих пор приходится говорить в
достаточной степени предположительно, хотя работы последних лет внесли некоторую
ясность в этот вопрос (Богословская, Вотрогов, 1981; Meltofte et al., 1981; Андреев.
1985; Герасимов, 1985; Косыгин, 1985; Bledsoe, Sibley, 1985; Трухин, Косыгин, 1986).
Можно определенно утверждать, что гипотеза о зимовках розовых чаек в Северном Ле-
довитом океане севернее мест гнездования (Бутурлин, 1934; Дементьев, 1951), в част-
ности в Великой Сибирской полынье в районе Новосибирских островов (Успенский.
1973), не подтвердилась. Большинство розовых чаек, размножающихся в пределах
основного гнездового ареала, проводит зиму, по всей видимости, в северной части
Берингова моря, где птицы держатся на обширных незамерзающих полыньях (в част-
ности, вдоль южного побережья Чукотского полуострова), а также в Охотском море,
особенно у восточного Сахалина. Часть берингийских птиц откочевывает (или откосится
штормами) южнее и восточнее, где их периодически отмечают в декабре—марте на
о-ве Св. Лаврентия, Командорских и Курильских островах (Sealy et al., 1971; Нечаев.
1969; Портенко, 1973) и в качестве крайне редких залетов на японском о-ве Хоккайдо
(Mori, 1976).
Зимние залеты в Западную Европу (Densley, 1977а) заставляют предполагать нали-
чие зимовок где-то в северной Атлантике, возможно у кромки льдов в Гренландском
море. Некоторое число птиц зимует в море Баффина и Девисовом проливе (Glutz v.
‘Однако П.С. Томкович (устное сообщ.) в 1981 г. встретил на Земле Франца-Иосифа активно линя-
ющую взрослую птицу с зарастающим наседным пятном уже 1/VII.
248
Розовая чайка
Blotzheim, Bauer, 1982). Зимовки в атлантическом секторе Арктики и соседних субарк-
тических водах имеют второстепенное значение.
Миграции. Изучены далеко недостаточно, поскольку проходят преимущественно
вдали от берегов. У арктических побережий и островов розовые чайки появляются не-
регулярно, в зависимости от погодных условий и ледовой обстановки, оказывающих
большое влияние на характер и направление миграционных потоков (Придатко, 1981).
На основании анализа разрозненных наблюдений общая картина миграций розовых
Рис. 59. Ареал розовой чайки в СССР
1 — гнездовой ареал, 2 — изолированные гнездования вне основного ареала, 3 — область зимних
кочевок, 4 — область летне-осенних кочевок, 5—7 — направления миграций: 5 — весенних, 6 — лет-
них, 7 — позднелетних и осенних, 8 — направления зимних кочевок, 9 — предполагаемые места
гнездования и направления миграций
чаек представляется следующим образом. Покидая в конце июля - первых числах ав-
густа места гнездования (Приклонский и др., 1962; Андреев, 1985), взрослые и моло-
дые птицы широко разлетаются к северу, северо-западу и северо-востоку, достигая к
середине августа островов Врангеля и Геральда и окрестностей Новосибирских остро-
вов (Рутилевский, 1958; Портенко, 1973). О возможности разлета некоторых птиц
даже до Земли Франца-Иосифа говорит встреча там Ф. Нансеном в конце прошлого
века молодой птицы 27.VII (Collett, Nansen, 1900; Бутурлин, 1934) (если это не по-
томство птиц, загнездившихся по соседству). Западнее молодых птиц в период мигра-
ций не отмечали (Meltofte et al., 1981).
Неразмножающиеся, а также потерявшие кладку птицы начинают покидать места
гнездовий уже в середине июня и почти исчезают к началу июля. Они разлетаются на бо-
лее дальние расстояния. Во второй декаде июня чайки могут достигать Колючинской
249
Семейство Чайковые
губы, где в конце июня 1974 г. в районе косы Беляка отмечено не менее 350 взрослых
и годовалых особей (Кречмар и др., 1978),и окрестностей пос. Уэлен (Томкович, Соро-
кин, 1983). С первых чисел июля такие особи отмечаются на Земле Франца-Иосифа
(Томкович, 1984). В дальнейшем розовые чайки летят в сторону Гренландии. 7—11. VII
1980 г. севернее Шпицбергена встречено несколько сот особей, большинство из которых
мигрировали в западном направлении (Meltofte et al., 1981).В августе — начале сентября
большое количество розовых чаек держится среди пакового льда между Шпицбергеном
и северной Гренландией около 82° с.ш. Эта область, как и район Шпицбергена и Земли
Франца-Иосифа, где большое количество розовых чаек наблюдал еще Ф. Нансен (Col-
lett, Nansen, 1900), по-видимому, служит важным местом летней кормежки и линьки
для неразмножающихся птиц. Возможно, часть птиц в это время проникает в море Баф-
фина к западу от Гренландии (Kampp, Kristensen, 1980). В середине сентября севернее
Шпицбергена уже идет обратный пролет в восточном или юго-восточном направлении
(Meltofte et al., 1981), однако часть птиц остается в северной Атлантике на зимовку.
Продвигаясь на восток, мигрирующие чайки достигают в сентябре Северной Земли
(Лактионов, 1946), окрестностей Новосибирских островов (Бируля, 1907), а затем
и о-ва Врангеля, где заметный пролет отмечен 15.IX—24.X (Придатко, 1981). По-види-
мому, осенью чайки разных возрастов придерживаются полосы вновь образующихся
льдов. В конце сентября (иногда уже с 6.VIII) розовые чайки появляются у мыса Бар-
роу на Аляске, пролет продолжается в октябре и заканчивается к началу ноября. В
отдельные годы здесь отмечают тысячи птиц, причем их появление связано с северо-за-
падным ветром (Murdoch, 1899, цит. по: Портенко, 1973; Densley, 1977b). От мыса
Барроу розовые чайки отлетают в северо-восточном направлении и далее следы их те-
ряются, обратного пролета здесь никто не наблюдал. Возможно, часть чаек через Ка-
надский Арктический архипелаг долетает до моря Баффина и Девисова пролива, и имен-
но эта пролетная ветвь ответственна за загадочные залеты розовых чаек в ноябре—де-
кабре в глубь американского материка (Miliotis, Buckley, 1975; Balch et al., 1979; Bled-
soe, Sibley, 1985). Однако большинство розовых чаек в начале зимы оказывается уже
в Беринговом море. По словам эскимосов, у о-ва Св. Лаврентия они появляются в кон-
це ноября — декабре, когда подходят большие льды из Чукотского моря (Fay, Cade,
1959, цит. по Портенко, 1973). Видимо, тысячные стаи чаек, пролетающих на северо-вос-
ток мимо мыса Барроу, затем, перемещаясь вдали от берегов, возвращаются в Чукот-
ское море (28.XII 1979 г. одна особь встречена на о-ве Врангеля — Стишов и др., 1986),
откуда после окончательного замерзания моря двигаются на юг (Андреев, 1985).
Возможно, какая-то часть птиц летит из моря Бофорта в Берингово море напрямик,
пересекая Аляску.
В течение зимы часть популяции держится вдали от берегов в северной части Берин-
гова моря, другие розовые чайки вместе со льдами постепенно опускаются к югу мимо
Камчатки вплоть до южных Курильских островов. В апреле отмечены значительные
скопления птиц в юго-западной части Охотского моря. Так, по сообщению А.А. Кибаль-
чича, 13.IV 1977 г. стайки розовых чаек по 4—5 птиц встречены среди льдин в северной
части залива Терпения. 11.IV 1982 г. большое количество розовых чаек отмечено непо-
далеку, восточнее южной части Сахалина; в конце апреля — мае эти птицы, видимо,
перемещаются в северную и северо-восточную части Охотского моря (Косыгин, 1985).
Примерно с середины мая начинается направленный пролет розовых чаек через мате-
рик к местам гнездования. Чайки из Анадырского залива, залива Шелихова и Пенжин-
ской губы, окрестностей Карагинского и Олюторского заливов летят на северо-запад
и север в сторону Колымской и Яно-Индигирской низменностей, в том числе долинами
рек Омолона и Колымы (Портенко, 1939, 1973; Дементьев, 1940; Велижанин, 1976;
Кречмар и др., 1978; Кищинский, 1980; Фирсова, Левада, 1982; Андреев, 1985). При
250
Розовая чайка
изменении погодных условий розовые чайки в отдельные годы в большом числе отме-
чены на побережье и даже во внутренних районах Камчатки (Герасимов, 1985). Часть
птиц задерживается в Охотском море, Анадырском заливе и в Беринговом море южнее
мыса Наварин до июня (Косыгин, 1985). На северном побережье Чукотского полу-
острова весенний пролет не наблюдается и здесь лишь изредка появляются отдельные
особи и стайки.
На всем пространстве основного гнездового ареала розовые чайки появляются в
последнюю декаду мая - первые числа июня, в период ледохода на реках, причем преж-
де всего — в долине Колымы. Массовый пролет начинается через 1—7 дней после появ-
ления первых птиц и продолжается несколько дней. В низовьях Колымы выражены се-
верное, северо-восточное и восточное направления пролета, в среднем течении Алазеи —
северное, в дельте Индигирки, междуречье Хромы и Индигирки — западное, северо-
западное и юго-западное, на южном побережье губы Буор-Хая и в бухте Тикси — се-
веро-западное (Schaanning, 1928; Воробьев, 1967; Григорьев, 1976; Томкович, 1980;
Андреев, 1985; наши данные). Следует заметить, что прилетевшие розовые чайки при
резких похолоданиях нередко бывают вынуждены отлетать южнее. Они возвращаются
сразу же при потеплении, что создает ложную картину массового пролета, но направле-
ние этих перемещений может отличаться от направления сезонной миграции.
Часть розовых чаек, летующих и линяющих у края паковых льдов севернее Земли
Франца-Иосифа, Шпицбергена и у побережья Гренландии, по неясным причинам (воз-
можно, просто ’’отсеченная” замерзанием моря) остается зимовать в северной части Ат-
лантики у края льдов. Весной эти чайки оказываются запертыми здесь сплошными
ледяными полями замерзших сибирских и канадских арктических морей. Не находя
подходящих мест для гнездования, они могут загнездиться в несвойственных биотопах.
Так, по-видимому, возникают упомянутые выше поселения в Гренландии, на Шпицбер-
гене, в Канадском секторе Арктики и на Северо-Американском материке южнее поляр-
ного круга. Разлетом с атлантических зимовок можно объяснить и появление розовой
чайки в стае моевок 8.VI на Новой Земле (Антипин, 1938).
Отражением зимних кочевок птиц в северной Атлантике являются частые встречи ро-
зовых чаек в декабре—феврале в Великобритании, а в период весеннего миграционного
беспокойства они учащаются в апреле—мае (Densley, 1977а). Немногочисленность зим-
них встреч в Европе говорит о невысокой численности розовых чаек на зимовке в се-
верной Атлантике. Однако за последние 10 лет число зимних встреч этих птиц в Европе
и Северной Америке резко возросло (Densley, 1977а; Bledsoe, Sibley, 1985), что отра-
жает не только рост числа наблюдателей, но и свидетельствует о реальном увеличении
числа розовых чаек на североатлантических зимовках.
Годовалые птицы встречаются в течение лета в тех же местах, что и неразмножающие-
ся взрослые особи, пролетая через области гнездования и разлетаясь затем по аквато-
рии Северного Ледовитого океана. В дельте Индигирки в июне они составляют до 20%
всей весенней популяции, в низовьях Колымы в 1979 г. — менее 5%. В прииндигирских
тундрах единичные розовые чайки в первом летнем наряде и их группы задерживаются
до конца июня, а иногда и до третьей декады июля. В июле они отмечаются на Земле
Франца-Иосифа (Томкович, устное сообщ.) и обычны на кочевках севернее Шпицберге-
на, где составляют около 12% всех розовых чаек (Meltofte et al., 1981).
В Европе залеты розовых чаек отмечены на юг Норвегии и Швеции, в Великобрита-
нию, Данию, ФРГ, Нидерланды, Францию, Италию (на о-в Сардинию), в Азии — в Япо-
нию, Китай, а также в глубь Северо-Американского материка (Bledsoe, Sibley, 1985).
Численность. В междуречье Хромы и Индигирки, где розовая чайка очень обычна на
гнездовье, в среднем плотность гнездования составляет от 0,15 до 0,83 пары на 1 км2.
Размножающаяся часть популяции в СССР насчитывает несколько десятков тысяч,
251
Семейство Чайковые
возможно не менее 50 тыс. гнездящихся пар. В Гренландии и Северной Америке чис-
ленность крайне незначительна.
Местообитания. В сезон размножения — низинные болотистые тундры с многочис-
ленными озерами, в меньшей степени — болотистые низины в дельтах значительных
сибирских рек и открытые озерно-болотные равнины в области предтундровых редко-
лесий. Приморских луговых тундр избегает. Во внегнездовое время — арктические и
субарктические моря, где розовые чайки держатся у края многолетних льдов, среди
битого льда и в полыньях. Открытой воды без льдин и сплошных ледяных полей без
разводий избегают. В период весеннего пролета — обычно долины вскрывшихся рек.
Размножение. Половой зрелости достигает в возрасте двух лет, однако регулярно
размножаться начинает в трех-четырехлетнем возрасте. Годовалые птицы изредка
образуют пары (иногда со взрослыми), но в размножении не участвуют. Моногам.
Диффузно-гнездящийся факультативно-колониальный вид. Численность гнездящихся
птиц резко варьирует в зависимости от погодных условий. Большинство розовых чаек
прилетают на места гнездования уже сформированными парами. Птицы сначала дер-
жатся в поймах вскрывшихся рек, кормясь у края льда и на льду пойменных озер,
затопляемых талыми водами. Здесь могут образовываться скопления от нескольких
десятков до 200 особей, в которых присутствуют как сформировавшие пару птицы,
так и формирующие пару холостые особи.
К особенностям демонстрационного поведения розовых чаек относится его четкая
приуроченность к ранневесеннему периоду. Большинство брачных и агрессивных де-
монстраций можно видеть в короткий период после прилета до начала гнездования,
главным образом в скоплениях на льду. В период насиживания территориальные кон-
фликты соседних пар, а также церемонии встречи и смены партнеров на гнезде, харак-
терные для большинства видов чаек, у розовых практически отсутствуют (Зубакин,
Авданин, 1983).
Демонстрационное поведение розовых чаек изучено недостаточно,'в целом оно на-
поминает такое же поведение малых чаек. Четко оформленный долгий крик, который
у большинства чаек служит самой распространенной и универсальной демонстрацией
в сезон размножения, у розовой чайки отсутствует. По-видимому, долгим криком
у этого вида следует считать своеобразную демонстрацию, когда птица, стоя со втяну-
той шеей и слегка опущенным передним концом тела, быстро приподнимает задний ко-
нец туловища и хвост наклонно, а затем почти вертикально вверх, изгибая при этом
шею назад и направляя клюв наклонно вверх (рис. 60, а-г). В это время издается
визгливый вскрик и трескучая трелька, при исполнении которой птица поводит голо-
вой из стороны в сторону, после чего принимает ту же позу, в какой была в начале
демонстрации. Чаще всего эту демонстрацию можно видеть у членов пары, которые
находятся в скоплении среди других розовых чаек или при постройке гнезд, а также
у особей, охраняющих кормовую территорию в скоплении птиц. Долгий крик можно
наблюдать в полете, при этом шея вытянута вперед и наклонно вниз, чайка скользит
на неподвижных крыльях, которые сначала подняты V-образно, а потом распластыва-
ются горизонтально. На земле или на льду взаимно адресованную демонстрацию произ-
водят одновременно до трех птиц.
Наиболее часто встречающаяся агрессивная демонстрация в скоплениях чаек напо-
минает начальную позу при долгом крике и немного похожа на распластанную позу
других чаек, но голова направлена не вперед, а вперед и слегка вниз (см. рис. 60, д).
В этой позе чайка бежит в сторону противника с агрессивным стрекотом — быстро пов-
торяющимися криками ”э-вей, э-вей, э-вей...”, либо с серией быстро повторяющихся
слогов ”вав-вав-вав-вав-вав...”. Такие же крики часто можно слышать в полете, когда
одна чайка гонится за другой. При интенсивной погоне они переходят в звонкое визг-
252
Розовая чайка
ливое ”кья, кья, кья...”. Нередко агрессивный стрекот заканчивается долгим криком.
Прямой позы у розовых чаек не отмечено. Весной нередко наблюдается ритуальное
кормление самки самцом, особенно перед копуляцией. Самка выпрашивает корм в
сгорбленной позе, но характерного для чаек вскидывания клюва в этой ситуации ни
разу не отмечалось. Наблюдалась также поза, сходная с выпрямленной позой крачек:
партнеры вытягивали шею и закрытый клюв наклонно вверх, почти соприкасаясь клю-
Рис. 60. Демонстрационное поведение розовой чайки
а—г — последовательность поз долгого крика, д — поза при агрессивном крике
вами. Эту позу птицы принимали после того, как один из членов пары прогонял сопер-
ника.
Гнездовой биотоп однотипен по всему основному гнездовому ареалу: это открытые,
лишенные кустарника и заросшие осокой низинные болота по берегам озер, заливаемые
весенними водами, с отдельными грядами и моховыми кочками-островками, или же
залитые весенними водами заболоченные термокарстовые низины, также заросшие
осокой и имеющие бугорки-острова. В низинной мокрой тундре с мелкими лужами и
несколькими крупными озерами розовые чайки гнездятся и в Канаде, в окрестностях
Черчилла (Chartier, Cooke, 1980). В то же время в других местах вне основного гнездо-
вого ареала эти птицы могут изредка поселяться в совершенно несвойственных им мес-
тах: в губе Буор-Хая на приморском мелководном озерке (Томкович, 1980); на пес-
чаной косе, отделяющей приморскую лагуну от моря на одном из островов в заливе
Диско (Kampp, Kristensen, 1980); на ровном, продуваемом ветрами каменистом берегу
северо-восточной Гренландии (Hiort, 1980).
Чайки появляются на территории будущих колоний за несколько дней до откладки
первых яиц, иногда за день или даже в день откладки, хотя до этого обычно держатся на
льду озера по соседству. Появляющиеся на месте колонии пары перелетают с одного
островка на другой, ходят по ним, делают во мху ямки. Копуляция происходит здесь же
или на льду ближайшего озера. В этот период чайки вступают в столкновения (иногда
жестокие драки) с другими парами, уже занявшими территории и изгоняющими сопер-
ников с расстояния 10—20 м от своих гнезд.
253
Семейство Чайковые
В большинстве колоний розовых чаек гнездятся полярные крачки в количестве
1—4 пар, нередко по соседству поселяются морянки и другие утки, гагары, многие ку-
лики, вилохвостые и серебристые чайки, бургомистры. Гнезда розовых чаек располага-
ются на моховых (обычно сфагновых) или осоковых островках-кочках, на примыкаю-
щих к берегу валиках полигональных структур либо просто на берегу. Как исключение
гнезда могут располагаться на валике растительной ветоши и заломах стеблей арктофи-
лы, на плоском илистом островке и на куче растительных остатков на льдине (Томко-
вич, 1980).
Места расположения некоторых колоний (обычно наиболее крупных) постоянны,
и птицы гнездятся там даже в самые неблагоприятные годы, хотя и в ничтожном коли-
честве (6—8% от нормального числа). В то же время некрупные колонии по 5—8 пар воз-
никают в благоприятные годы в самых разных местах, где чайки до этого не гнездились,
и, просуществовав 1—2 года, снова исчезают. В особо благоприятные годы с ранней и
дружной весной без возврата холодов небольшая часть розовых чаек гнездится вне ко-
лоний одиночными парами даже среди сухой тундры, где есть небольшая лужа. В годы
с высокой весенней водой гнезда смещены к периферии озерной депрессии и по мере
спада воды оказываются на совершенно сухом месте довольно далеко от воды. В нор-
мальные годы колония распределяется относительно равномерно по залитым водой
участкам.
Число гнезд в колониях различно. В низовьях Колымы* 2 оно составляет от 2 до 27.
в среднем 13 гнезд. В междуречье Хромы и Индигирки3 в 12 из 22 колоний было по
2—5 пар, в 6 — по 6—10 гнезд, в 4 — по 14—18 гнезд. В тех же местах, но в другие годы
величина некоторых колоний доходила до 40—50 гнезд.
Плотность колонии невелика. В хромо-индигирских тундрах среднее расстояние
между двумя ближайшими гнездами (по 85 измерениям в 12 колониях) составляет
43 м, минимальное отмеченное здесь расстояние равно 4,8 м, хотя в другие годы бывает
и до 0,8 м. В низовьях Колымы среднее расстояние между соседними гнездами в плот-
ных колониях составляет 14—20 м, в рассеянных и агрегированных — 33—48 м, от 15
до 55% птиц гнездится на удалении 200 м и более от соседней. Индекс плотности гнездо-
вания розовых чаек в разных колониях варьирует от 0,01 до 0,001, составляя в сред-
нем 0,003.
Откладка яиц происходит в первой декаде — середине июня, в особо холодные весны
даже позже (в Яно-Индигирской низменности в разные годы 6—19 июня, в низовьях Ко-
лымы — 1 — 16 июня), откладка в популяции длится от 4 до 20 дней. В сходные сроки
отмечено начало откладки яиц в западной Гренландии и в Канаде у Черчилла (Chartier.
Cooke, 1980; Kampp, Kristensen, 1980). Гораздо позднее (около 1 .VII) началась в 1973 г.
кладка на Таймыре (Павлов, Дорогов, 1976).
Гнездо устроено примитивно — это ямка во мху или дернине, выстланная сухой осо-
кой или пушицей, изредка слоевищами лишайников. Подавляющее большинство гнезд
(83 из 87) не имеет выраженного гнездового валика. Материал для гнезда птица соби-
рает либо в непосредственной близости от гнезда, либо приносит его в клюве с некото-
рого расстояния. Диаметр лотка (мм) (гнездовой ямки) 100—170, в среднем (и = 87)
131; глубина лотка — 20—60, в среднем 45. В кладке 2-3, изредка 1 яйцо. В низовьях
Колымы средняя величина кладки изменяется по годам от 2,2 до 2,7 яиц, процент кла-
док из 3 яиц варьирует от 31% до 73%, из 1 яйща — от 7% до 33%. В междуречье Хромы
и Индигирки в 1978 г. кладки из 3 яиц отмечены в 19% из 68 гнезд, из 2 яиц — в 63%.
’Данные по низовьям Колымы здесь и далее взяты из работ А.В. Андреева и А.Я. Кондратьева
(1981) , А.В. Андреева (1985).
3По междуречью Хромы и Индигирки здесь и далее использованы материалы авторов очерка.
254
Розовая чайка
из 1 — в 18%; в 1979 г. — в 36%, 36% и 27% соответственно (и = 22). По наблюдениям
в Колымской низменности, средняя величина кладки больше в годы с ранней и друж-
ной весной.
Окраска яиц (Бутурлин, 1905; Дементьев, 1951; Воробьев, 1963) сравнительно
темная, основной фон оливково-зеленый или серовато-оливковый различного оттенка.
По этому фону разбросаны негустые и нечеткие буроватые или шоколадно-коричневые
поверхностные и сероватые глубинные пятна, гуще сконцентрированные на тупом кон-
це яйца. Размеры яиц (мм): с низовий Колымы (п = 36) 38,6—45,8 X 30,0—32,9, в сред-
нем 43,3 X 31,6 (Бутурлин, 1905); из междуречья Хромы и Индигирки (и = 172) 34,5—
48,3 X 29,5—34,1, в среднем 42,8 X 31,3; с Таймыра (и = 10) 38,0—44,3 X 29,5—32,5,
в среднем 41,2 X 30,9 (Павлов, Дорогов, 1976). Яйца розовых чаек в целом более
округлы, чем яйца других чаек. Масса ненасиженных яиц (г) с низовий Колымы равна
в среднем (п = 29) 22,2 ± 0,4, из междуречья Хромы и Индигирки - 19,4-22,7, в сред-
нем (п = 19) 21,1.
Откладка яиц у каждой пары продолжается до 4 сут, насиживание начинается с пер-
вого яйца. Повторных кладок взамен погибших, вопреки мнению С.А. Бутурлина
(1905), у розовых чаек, по-видимому, не бывает. Инкубация первого яйца продолжает-
ся в среднем 20 дней (и = 18), второго — 19,5 (и = 17), третьего — 19 дней (и = 8). В
случае длительных холодов, по данным А.В. Андреева и А.Я. Кондратьева (1981), срок
инкубации затягивается до 22—28 сут. В насиживании принимают участие оба родителя.
По наблюдениям в междуречье Хромы и Индигирки смена партнеров происходит в сред-
нем через 2,5 часа, очень быстро и просто: услышав крик подлетающего партнера, наси-
живающая птица взлетает и в большинстве случаев немедленно улетает из колонии, а
сменяющая ее практически с лету садится на гнездо. Нередко смена происходит без во-
кализации. Свободные от насиживания партнеры крайне редко находятся на гнездовом
участке и вообще в колонии (иногда не более 2 мин в сутки). Лишь в первые дни наси-
живания партнеры ненадолго задерживаются, чтобы принести материал для гнезда. В
результате в колонии розовых чаек в отсутствие беспокойства совершенно нет шума,
характерного для поселений других видов чайковых. Самец насиживает в сутки на
1—3 ч больше, чем самка. Максимальная длительность непрерывного насиживания сам-
ца 5 ч, самки 3,5 ч. Ночью насиживает как самка, так и самец. Переворачивание яиц в
кладке, по наблюдениям на Колыме, в среднем происходит каждые 19 мин, среднесу-
точная температура яйца составляет в период с 6-го по 15-й день насиживания 29,6±1,5°,
ее колебания (605 измерений) равны 23,0—34,6°.
Самое раннее вылупление птенцов в низовьях Колымы зарегистрировано 21.VI
1981 г., самое позднее начало вылупления отмечено 6.VII 1982 г. В междуречье Хромы
и Индигирки первые птенцы появились в 1978 и 1979 гг. 4.VII. После появления трещи-
ны на яйце до вылупления проходит 1—3 сут, чаще всего 2 и 3 сут (по 40% случаев;
п - 25), от появления проклева до вылупления чаще всего проходят сутки (73% случа-
ев, п = 22), реже 2 сут (18%) и меньше суток (9%). Родители выносят и выбрасывают
в отдалении скорлупу яйца. Первая неудачная попытка кормления птенца наблюдалась
через 4 ч 40 мин после вылупления, первое удачное кормление — через 9 ч. В колонии
из 5 гнезд вылупление в 1978 г. продолжалось 5 сут, в колонии из 17 гнезд — 6 сут.
Масса только что вылупившегося птенца 12,0—16,6 г, в среднем (и = 18) 15,1 г. К
двухнедельному возрасту птенцы обычно достигают максимальной массы, которая в
ряде случаев немного превосходит массу взрослых чаек. В возрасте около суток птенцы
способны при опасности покидать гнездо и затаиваться неподалеку. Вскоре после вы-
лупления последнего птенца выводок оставляет район гнезда и птенцы перемещаются
по колонии обычно в пределах 40—80 м от гнезда, но могут удаляться от него и на не-
сколько сот метров. В первые день-два после вылупления с птенцами находятся оба ро-
255
Семейство Чайковые
дителя, постоянно их кормя и согревая. Уже на 5— 7-й день жизни птенцов родителей
можно видеть лишь во время кормления птенцов, которое в этот период бывает в сред-
нем 21 раз в сутки. В возрасте 17—19 дней птенцов кормят только 4 раза в сутки. Уже
с 3-го дня жизни они начинают самостоятельно отыскивать пищу. Основной корм птен-
цов — комары и их личинки, брюхоногие моллюски, в меньшей степени жуки и рако-
образные. Установление постоянной температуры тела у молодых птиц происходит не-
посредственно перед подъемом на крыло. До этого у птенцов наблюдается хорошо вы-
раженная гипотермия, они способны активно передвигаться и разыскивать корм при
температуре тела 29—30°. Лётными птенцы становятся на 18—20-й день жизни. Вскоре
после подъема на крыло, в последних числах июля — первых числах августа, молодые
птицы покидают тундру. Взрослые успешно отгнездившиеся чайки покидают места
гнездовий на 4—7 дней раньше, в среднем 24.VII (Андреев, Кондратьев, 1981; Зубакин,
Авданин,1983; Андреев, 1985).
Стратегия охраны потомства основана на рассредоточении криптически окрашенных
кладок и птенцов, уменьшении заметности колонии для четвероногих хищников в со-
четании с активным нападением на хищника. Характерная особенность защитного по-
ведения розовых чаек-родителей — наличие отвлекающих демонстраций (Зубакин.
Авданин, 1983).
Успех размножения розовых чаек в целом весьма невелик — от 0 до 76%, в среднем
25%. В низовьях Колымы прослежена судьба 165 кладок, из которых 28% были разоре-
ны хищниками, а 7,2% брошены родителями. Успешно инкубировано 280 яиц в 107
кладках, из которых вылупилось 272 птенца. Поднялось на крыло только 99 птенцов.
Таким образом, основные потери приходятся на период роста птенцов: погибает почти
64% (Андреев, 1985). В междуречье Хромы и Индигирки в 1978 и 1979 гг. поднялось
на крыло соответственно чуть больше 15% и 29,3% птенцов от числа отложенных яиц.
причем в первый год основная гибель потомства была за счет птенцов (погибли 81,4%
из 43 вылупившихся птенцов), а во второй — за счет брошенных кладок при похолода-
нии (погибла половина из 36 отложенных яиц). Гибель гнезд в отдельные годы состав-
ляет до 65,4% (и = 81).
Особенно губительно сказываются на воспроизводстве популяции аномально небла-
гоприятные метеорологические условия весны. В затяжные и холодные весны чайки не
только гнездятся исключительно в сравнительно немногих постоянных колониях и в
ничтожно малом количестве, но и сокращают величину кладки до 1, реже 2 яиц, причем
около половины их оказываются неоплодотворенными. Кроме того, одиночные гнезда
в подавляющем большинстве случаев гибнут по вине хищников. Таким образом, вос-
производство в основном гнездовом ареале фактически становится равным нулю. Часть
негнездящихся птиц в такие годы до начала июля держится в стаях по 80—100 птиц
на нерастаявшем льду крупных озер, а затем покидает гнездовую область, часть же еше
с ранней весны смещается южнее, где гнездится в подходящих местообитаниях в преде-
лах северных частей таежной зоны (Лабутин, Ларионов, 1976). Успешность гнездования
во время таких сдвигов ареала неизвестна.
Суточная активность, поведение. В период полярного дня активна круглосуточно, с
некоторым спадом в ночные часы. Общительные птицы, хотя больших стай в период
перелетов обычно не образуют, мигрируя парами и стайками до 20, редко до 50 особей.
На осеннем пролете изредка отмечаются более крупные стаи и скопления. Так, одна
стая на мысе Барроу содержала более 4500 особей (Kessel, Gibson, 1978). В предгнездо-
вой период в пределах области гнездования также могут образовываться скопления до
нескольких сот птиц.
Питание. В сезоны гнездования — наземные и водные насекомые и их личинки, вод-
ные беспозвоночные (ракообразные, моллюски), реже мелкая рыба и изредка зеленые
256
Розовая чайка
части и семена растений, которые могут быть случайными примесями (Бутурлин, 1905;
Буякович, 1960; Успенский и др., 1962; Воробьев, 1963; Рощевский, 1976; Андреев,
1985). В целом розовые чайки способны поедать любые мелкие пищевые объекты, по-
являющиеся у поверхности воды, на мелководье, на льду, а также крупных летающих
насекомых. В течение гнездового сезона они многократно меняют места и способы до-
бычи корма, постоянно переключаясь на наиболее доступные и массовые группы пище-
вых объектов, главную роль среди которых играют различные Diptera. Вместе с тем
прослеживается определенная сезонность как в использовании главных кормов, так и
в способах добычи. Сразу после прилета чайки концентрируются на помойках и свалках
около поселков или в местах замора девятииглой колюшки в освободившихся ото
льда лужах. Во второй половине весны (первая декада июня) основными кормами
служат наземные беспозвоночные (брюхоногие моллюски, жуки, пауки), которых пти-
цы собирают, подолгу расхаживая по тундре. Наконец, с вылетом комаров и выходом
на поверхность их личинок птицы переключаются в основном на питание этими кормо-
выми объектами. Вне сезона размножения розовая чайка тесно связана со льдами,
поскольку питается животными, обитающими под ними. В этот период большое значе-
ние в питании имеет сайка (в 79% желудков встречены отолиты) и амфиподы (54%),
особенно Apherusa glacialis. Кроме них, отмечено поедание различных креветок, гамма -
русов, полихет, мелких головоногих моллюсков (Collett, Nansen, 1900; Портенко,
1973; Divoky, 1976).
В море розовые чайки кормятся обычно в 2—3 м от края льда, присаживаясь на воду
и склевывая корм с поверхности или ныряя с воздуха и частично погружаясь в воду.
Нередко собирают корм, расхаживая пешком по краю льдины или плавая и склевывая
корм с поверхности воды, как плавунчики. Отбросы с кораблей, как и экскременты
моржей на льду, их привлекают мало (Divoky, 1976; Densley, 1979). В пределах гнез-
дового ареала используются те же способы кормодобывания, но чайки на воде прово-
дят больше времени, кормясь, как плавунчики, или глубоко опуская в воду голову.
На большей глубине добычу схватывают, опрокидываясь, как речные утки, или ны-
ряют, взлетев с воды, изредка погружаясь почти целиком. Кроме того, могут ловить
насекомых в воздухе. В местах концентрации корма нередко собираются стаями до
нескольких десятков особей. В менее кормных местах могут охранять временные кор-
мовые участки радиусом до 30 м, обычно 5—15 м, что особенно характерно для первой
половины июня.
Враги, неблагоприятные факторы. Наибольший ущерб потомству розовых чаек из
пернатых наносят поморники, серебристые чайки и бургомистры, причем крупные чай-
ки чаще похищают птенцов, чем яйца. Из млекопитающих угрозу для колоний может
представлять песец и, возможно, лисица, а также стада домашних северных оленей и
сопровождающие оленеводов собаки. Крайне неблагоприятны для размножения розовых
чаек поздняя холодная весна и возврат холодов в начальный период насиживания.
Хозяйственное значение, охрана. Хозяйственного значения розовая чайка не имеет.
Занесена в Красную книгу СССР.
9. Зак. 826
257
Семейство Чайковые
Род Chlidonias Rafinesque, 1822
Черная крачка —
Chlidonias niger (Linnaeus, 1758)
Black Tern (англ.), Trauerseeschwalbe (нем.), Guifette noire (фр.).
Син. (лат.) : Hydrochelidon nigra (Linnaeus, 1758).
Син. (рус.): черная болотная крачка.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Мелкая крачка, от белокрылой крач-
ки в брачном наряде отличается малоконтрастной однотонной темно-серой окраской
(белое только подхвостье), от белощекой — отсутствием контрастных белых ”щек”
и черным клювом. От других крачек — общей темной окраской, небольшими размера-
ми, относительно коротким хвостом с неглубокой вырезкой. В зимнем оперении и
гнездовом наряде от белокрылых и белощеких крачек отличается наличием темных
пятен по бокам зоба. Полет легкий, порхающий; может зависать на одном месте, тре-
пеща крыльями. Подобно другим болотным крачкам, как правило, не ныряет, на во-
ду не садится. По земле ходит неохотно и мало, туловище держит горизонтально. Го-
лос — негромкое ”кир”, ”крир”, иногда звучит как ”крер”. В окрестностях колонии
слышны крики ”кирр-ли”, ”кирр-лит” или ”стирр-ли”, ”стирр-лит”, с которыми крач-
ки возвращаются к своему гнезду.
Подвидовая систематика. Различают два подвида: палеарктический Ch.n. niger и се-
веро-американский Chai, surinamensis (Peters, 1934; Степанян, 1975). В СССР обитает
только один подвид:
Chlidonias niger niger
Sterna nigra Linnaeus, 1758, Syst. Nat., ed. 10, c. 137, Швеция.
Общая окраска светлее, а размеры несколько больше, чем у американского под-
вида.
Описание. Размеры (мм) (ЗМ МГУ) и масса тела (г) (Бородулина, 1960):
Длина крыла:
3 3 (п = 46) — 208—228 (среднее 217);
9 9 (п = 29) - 207-226 (среднее 215).
Длина клюва:
6 6 (и = 47) - 24,7-29,6 (среднее 27,2);
9 9 (п = 29) — 22,8—29,2 (среднее 25,9).
Длина цевки:
38 (п = 47) — 15,0—18,6 (среднее 16,4);
9 9 (п = 29) — 14,4—18,2 (среднее 15,9).
Масса тела:
6 6 (п = 76) — 53-76 (среднее 65);
9 9 (п = 41) - 49-76 (среднее 63).
Окраска (Дементьев, 1951; Cramp, 1985). Самец и самка в брачном наряде.
Голова и шея черные, подбородок и горло могут быть несколько светлее, особенно у
самок. Плечевые перья и верхние кроющие крыла аспидно-серые. Передняя часть спины
у самцов тускло-черная, постепенно переходящая в аспидно-серую, у самок обычно
аспидно-серая. Грудь, брюхо и бока у самцов от тускло-черных до черновато-аспидных.
258
Черная крачка
у самок аспидно-серые. Первостепенные маховые аспидно-серые, дистальные с белыми
стволами (кроме их вершинной части), внутренний край внутренних опахал беловатый.
Второстепенные маховые серые с бледным краем внутренних опахал. Рулевые серые,
надхвостье серое, подхвостье белое. Клюв черный, ноги красновато-бурые, радужина
темно-коричневая.
Самец и самка в зимнем наряде. Лоб, передняя часть темени, подбородок и горло
белые; перед глазом черное пятно; кроющие уха, задняя часть темени и затылок чер-
новатые, причем на перьях темени и затылка могут быть узкие светлые каемки. Спина,
маховые, плечевые, верхние кроющие крыла, надхвостье и рулевые серые, светлее,
чем летом. Грудь, бока, брюхо, подхвостье белые, по бокам груди серые пятна. Ниж-
ние кроющие крыла бледно-сизо-серые. Клюв и ноги, как в брачном наряде.
Пуховой наряд. Спинная сторона желтовато-коричневая с небольшими черными пят-
нами на темени, шее, спине, крыльях и пояснице; пятна нередко группируются в неяс-
ные полоски. Передняя часть лба, уздечка, подбородок, кольцо вокруг глаз и нередко
концы крыльев белые. Нижняя часть щек и горло от тускло-черного до светло-рыже-ко-
ричневого цвета. Брюшная сторона охристо-серовато-бурая, центральная часть брюшка
беловатая. Клюв черноватый, у основания серо-телесный, ноги серо-телесные. Радужина
темно-коричневая.
Гнездовой наряд. Лоб и уздечка светло-охристые, скоро выцветающие до белого,
темя и затылок буровато-черные с узкими светлыми пестринами в свежем пере. Перед
глазом черное пятно, кроющие уха буровато-черные. Подбородок, шея, грудь, бока,
брюхо белые. На задней стороне шеи буроватое ожерелье, заходящее на бока зоба и
образующее здесь такого же цвета пятна. Спина темно-бурая с прикрытыми сероваты-
ми основаниями перьев и узкими охристо-бурыми каемками, плечевые серые с широ-
кими темно-бурыми предвершинными пятнами и буровато-охристыми каемками на вер-
шине; в целом цвет верхней части тела весьма изменчивый, особенно при износе пера.
Поясница и надхвостье беловатые с серовато-охристым налетом и неявными охристы-
ми каемками перьев. Малые верхние кроющие крыла черноватые с белыми каймами
перьев, верхний ряд малых кроющих белый; средние и большие кроющие серые с тем-
но-бурыми предвершинными пятнами и охристо-буроватыми вершинами. Первостепен-
ные маховые, как у взрослых, но стволы обычно буровато-серые, а не белые, а свет-
лое поле на крае внутренних опахал белое. Дистальные второстепенные, как у взрос-
лых, проксимальные — с серой основной частью, темно-бурым пятном перед вершиной
и охристо-бурой вершинной каймой. Хвост серый, почти без вырезки, с закругленными
вершинами на рулевых. Клюв черный с розоватым или желтоватым основанием под-
клювья, ноги желтовато- или розовато-коричневые.
Первый зимний наряд. Как окончательный зимний, но первостепенные маховые
остаются от гнездового наряда и сменяются между началом января — началом июля.
Предполагается, что первый зимний наряд замещается сразу вторым зимним (Cramp,
1985), по-видимому, не отличимым от окончательного зимнего; первый полный брач-
ный наряд черные крачки надевают ранней весной третьего календарного года. Неко-
торые птицы возвращаются с зимовок в смешанном наряде: голова, часть шеи и грудь
в зимнем наряде, оставшиеся участки нижней части тела темные, как в брачном на-
ряде. Предполагается, что это либо годовалые птицы с ускоренной линькой в брачный
наряд, либо, что более вероятно, двухлетние птицы с запаздывающей предбрачной
линькой; они не размножаются (Cramp, 1985).
Линька из гнездового в первый зимний наряд полная, проходит поздно, уже на
зимовках. Обычно начинается в октябре—ноябре сменой оперения головы, передней
части спины и плечевых; в декабре—январе выпадают XI первостепенные и средние
(1-е) рулевые. К началу марта голова, тело, хвост и большинство верхних кроющих
259
Семейство Чайковые
крыла в оперении первого зимнего наряда. К июню—августу второго календарного
года все маховые уже новые, у некоторых птиц в мае—июне начинается повторная
линька первостепенных маховых. Первый зимний наряд замещается сразу вторым
зимним нарядом примерно в то же время, когда происходит линька взрослых в зим-
ний наряд; линька первостепенных маховых начинается примерно в июле.
У взрослых птиц бывает две линьки в году: полная послебрачная и частичная пред-
брачная. Послебрачная линька начинается в конце мая — начале июня сменой опере-
ния у основания клюва и вокруг глаз. К середине июля обьино голова, шея и зоб в све-
жем пере, начинается смена плечевых и оперения спины, брюшка. К середине августа
голова, туловище, 1-я, 2-я, реже и 3-я пары рулевых и многие средние и малые верхние
кроющие крыла в свежем наряде. Линька первостепенных маховых начинается со сме-
ны XI в начале июля — середине августа, прерывается в период осенней миграции на ста-
дии новых IX (VIII)—X. Завершается линька на зимовках сменой II (первого видимо-
го) первостепенного в январе—феврале. Последовательность смены пар рулевых: 1-, 2-,
3-, 6-, 4-, 5-я. В предбрачной линьке сменяется оперение головы, туловища, внутренних
первостепенных и кроющих крыла, кроме части средних и малых кроющих крыла
и изредка 4—5-й пары рулевых. Повторная смена внутренних первостепенных маховых
начинается с XI между концом ноября и началом февраля, часто до завершения преды-
дущей (послебрачной) линьки первостепенных. Линька прерывается в марте обычно
на стадии новых VII—VIII. Третьей линьки внутренних первостепенных у палеарктичес-
кого подвида Ch.n. niger в отличие от Char, surinamensis не отмечено. Завершается
линька к середине марта — началу апреля (Cramp, 1985).
Распространение. Гнездовой ареал. Номинальный подвид образует обшир-
ный ареал от Франции и южной части Испании на западе до оз. Байкал на востоке. Ареал
американского подвида занимает центральную часть Северной Америки (рис. 61). В
Евразии черная крачка гнездится почти во всех странах Западной, Центральной и Юж-
ной Европы; в 60-е годы нашего столетия загнездилась в Великобритании, Ирландии и
Финляндии (Cramp, 1985). Большая часть ареала лежит в пределах СССР (рис. 62).
Северная граница его, не во всех случаях точно установленная, проходит через При-
ладожье и юг Карелии, далее, по-видимому, через центральную часть Вологодской обл.,
Костромскую и северную часть Горьковской обл., центральную часть Кировской обл.,
центральную часть Удмуртии, Камское водохранилище в Пермской обл. (Хохлова,
1972; Попов, 1975; Плесский, 1976; Водолажская, Залетаев, 1977; Зимин, 1977;
Бутьев, 1978; Болотников и др., 1981; Мальчевский, Пукинский, 1983; Литун, Мака-
ров, 1984). Далее северная граница проходит, видимо, в окрестностях Свердловска,
по югу Тюменской и Омской областей, северу Барабинской степи; по Оби поднимает-
ся до окрестностей с. Молчаново, Обь-Чаинского междуречья и Нарыма (Дементьев.
1951; Стрелков, 19746; Москвитин и др., 1977; Ходков, 1977; Адам, Сурнаев, 1979;
Сулимов, 1981). Возможно, поселения черной крачки есть и несколько севернее ука-
занных районов — так, этих птиц ежегодно отмечали на р. Конде (Гордеев, 1977). По
долинам рек Оби и Томи проходит, по-видимому, восточная граница сплошного ареала,
которая затем огибает с запада и юга Алтай (Кучин, 1975). Далее на восток указыва-
лось гнездование под Минусинском (Дементьев, 1951), однако в последние годы здесь
отмечены лишь залетные особи (Прокофьев, 1975). Отдельные гнездовые поселения
недавно найдены на юге Тувы (Баранов, 1971; В.В. Леонович, устное сообщ.) и в При-
байкалье в устье р. Иркут (Мельников, 1985).
Южная граница области гнездования огибает с юга котловину оз. Зайсан, поднимает-
ся к Семипалатинску, далее проходит южнее Караганды, поднимается к оз. Тенгиз,
опускается к низовьям рек Иргиз и Тургай. Далее черная крачка гнездится по долинам
и устьям рек Эмбы, Урала, Уила, Волги (Луговой, 1958; Долгушин, 1962; Пославский.
260
Черная крачка
1974; Самусев, 1975; Кривоносов, 1976; Неручев, Макаров, 1982). В гнездовой ареал
входит Предкавказье, Восточное Приазовье (Бородулина, 1960; Казаков, Языкова,
1982). Затем эта граница области гнездования идет по югу Украины (исключая Крым)
до устья Дуная (Булахов, Мясоедова, 1975; Ардамацкая, 1981; Костин, 1983; и др.).
Южнее границы сплошного ареала черные крачки образуют гнездовые поселения в
ряде мест Казахстана и Средней Азии: В Талды-Курганской обл., в устье р. Или, в доли-
нах рек Чу и Сырдарья, в многоводные годы могут населять пересыхающие степные и
Рис. 61. Область распространения черной крачки
а — область гнездования (пунктиром показана неуточненная граница) , б— отдельные гнездовые по-
селения, в — места зимовок, г — залеты. Подвиды: 1 — Ch. n. niger, 2 — Ch. n. surinamensis
пустынные водоемы (Спангенберг, Фейгин, 1936; Долгушин, 1962; Кисленко, 1972;
Ауэзов, Грачев, 1975; Умрихина, 1975; Шукуров, 1981). Без ссылок на находки гнезд
указано гнездование для низовьев Сурхандарьи (Остапенко и др., 1978) и трассы Кара-
кумского канала, низовьев Теджена и Мургаба (Ташлиев и др., 1967). Отмечено гнез-
дование в низовьях Амударьи (Кенжегулов, 1972) и в Армении на оз. Гилли (Ляйстер,
Соснин, 1942). Что касается предположения о гнездовании черных крачек в низовьях
Атрека в юго-западной Туркмении, то новейшие сведения его не подтвердили (Щерби-
на, 1975; Караваев, 1981). В архиве Центра кольцевания есть данные о добыче в Индии
7.ХП 1970 г. черной крачки, окольцованной 18.VII того же года птенцом в низовьях
Атрека. Однако зимовки в Индии характерны для белощекой крачки, и, по-видимому,
261
262
Рис. 62. Ареал черной крачки в СССР
1 — область гнездования (пунктиром показана неуточненная граница), 2 — отдельные поселения, 3 — места предполагаемого гнез-
дования, 4 — залеты
Черная крачка
з данном случае произошла ошибка в определении. Об этом же говорят и размеры до-
бытой особи: длина крыла 221 мм, масса 72 г (Abdulali, Ambedkar, 1984), что больше
подходит для молодой белощекой крачки.
Предположения о гнездовании черных крачек в Монголии и Тибете (Stubbe, Bolod,
1971; Vaurie, 1972; Schauensee, 1984) не подтверждены находками гнезд. Однако по-
пытка гнездования черных крачек в северной части оз. Убсу-Нур, отмеченная В.В. Лео-
новичем в 1975 г., не исключает возможности гнездования этого вида на других озерах
Монголии. Сообщалось о гнездовании черных крачек в восточной части Тянь-Шаня
(Козлов, 1899), что сомнительно.
Зимовки (Cramp, 1985). Для палеарктического подвида - побережья западной
Африки от Мавритании до Намибии и отдельные участки побережья в ЮАР, а также
долина Нила в южном Египте и Судане; иногда залетает до оз. Виктория (Britton,
Brown, 1974). Одиночных птиц отмечали в зимнее время у Черноморского побережья
Кавказа и в Красноводском заливе Каспия (Самородов, Самородов, 1972; Строков,
1974); по-видимому, это случайные встречи. Американский подвид зимует на атлан-
тическом и тихоокеанском побережье севера Южной Америки.
Миграции. По данным ЦКАН и А.Й. Йыги (1968), пути миграции прибалтийских
и, видимо, белорусских черных крачек отличаются от путей пролета крачек остальной
части ареала. Они летят по южному побережью Балтики и далее по атлантическому
побережью Западной Европы: возможно, часть птиц пересекает материк и оказывает-
ся у средиземноморского побережья Франции. Весенние миграции, видимо, идут тем
же путем вдоль атлантического побережья Европы.
Черные крачки, гнездящиеся в центральных и южных частях европейской части
СССР, после сезона размножения летят на юг, к Черному и Азовскому морям, в част-
ности через Сиваш и северный Крым, где они появляются в середине июля, достигают
большой численности в конце июля и первой половине августа и затем дружно исче-
зают (Костин, 1983). В середине августа—сентябре крачки, окольцованные в Окском
заповеднике, уже отмечались в Алжире и Испании, хотя часть особей в это же время
встречены еще в Донецкой обл. Из Средиземноморья черные крачки летят к Гибралта-
ру и далее на атлантическое побережье Африки. Пролет через Сахару, если и имеет мес-
то, то весьма слабый. Мест зимовок достигают в основном в ноябре (Cramp, 1985).
Судя по немногочисленным возвратам колец, к Черному и Азовскому морям в авгус-
те стягиваются крачки из Зауралья и Западной Сибири.
Весенний пролет, который начинается в конце марта (Cramp, 1985) идет, по-види-
мому, по тому же маршруту, что и осенний. Через Средиземное море пролетают пти-
цы, которые затем распределяются по всему ареалу в пределах СССР, кроме При-
балтики. Это показывают возвраты колец от крачек, окольцованных на пролете на
северо-западном побережье Италии, близ г. Пизы (Таманцева, 1955; ЦКАН). Интерес-
но, что все крачки, от которых получены возвраты, окольцованы здесь не ранее первых
чисел мая, обычно в середине мая—начале июня. Поскольку в большинстве мест гнездо-
вания первые птицы появляются в конце апреля — самом начале мая, остается предпо-
ложить, что через северо-западную Италию летят более молодые птицы, сроки размно-
жения которых более поздние, а старые птицы пролетают где-то южнее. Не исключено,
что существует и какой-то более короткий путь, которым следуют особи, первыми по-
являющиеся в местах гнездований. Возможно, это крачки, зимовавшие в долине Нила,
или транссахарские мигранты.
Поскольку пролет черных крачек отмечается вдоль Черноморского побережья
Кавказа (Строков, 1974), на водохранилищах Азербайджана (Туаев,Васильев, 1972),
на восточном побережье Каспия (Залетаев, 1960) и восточном побережье Аральского
моря (Березовский, 1983), можно предположить еще один миграционный поток, пере-
263
Семейство Чайковые
секающий п-ов Малая Азия и разделяющийся на черноморскую и каспийскую ветви.
Неясными остаются места зимовок черных крачек, пролетающих весной на север по
долине Мургаба (Эминов, 1978), встреченных 29.IV 1975 г. в низовьях Сурхандарьи
(Остапенко и др., 1978), а также карчек, отмеченных в период пролета в высокогорьях
Центрального Тянь-Шаня, в Монголии и Цайдаме (Козлов, 1899; Дементьев, 1951; Кы-
дыралиев, 1973).
Первые особи черных крачек появляются весной в Ленинградской обл. 10—15.V
(Мальчевский, Пукинский, 1983), в Белоруссии в конце апреля — первой половине
мая (Федюшин, Долбик, 1967), а Крыму в 20-х числах апреля (Костин, 1983), на Запад-
ной Украине во второй половине апреля — начале мая (Страутман, 1963), в Ворошилов-
градской обл. в конце апреля (Панченко, 1975; Сулик, Енакиев, 1978), в Сумской обл.
12.V (Кныш, Пулях, 1975). В Московскую обл. прилетают 19.IV—12.V, в среднем 3.V
(Птушенко, Иноземцев, 1968), в Горьковскую обл. - в первых числах мая (Водолаж-
ская, Залетаев, 1977), в Предкавказье — во второй декаде апреля (Казаков, Языкова.
1982). В низьвьях Сырдарьи первых крачек отмечали с начала второй декады апреля
(Спангенберг, Фейгин, 1936), в Наурзумском заповеднике — 25—30.IV (Елкин, 1975).
у Семипалатинска и Павлодара —«в середине мая (Долгушин, 1962), под Карагандой -
15—25.V, в среднем 20.V (Ленхольд, Гаврилов, 1978), на оз. Малый Чан — 12—17.V
(Ходков, 1978), на верхней Оби - 15—20.V (Блинова, 1986). Пролет весной идет обыч-
но стайками 6—20 штук, реже парами, одиночками и стаями до 40 особей и еще реже -
до 70—100 особей (Долгушин, 1962; Ходков, 1978).
Осенний отлет на протяжении всего ареала начинается в середине июля — начале ав-
густа; к концу августа птицы обычно исчезают. Отдельные особи изредка задержива-
ются до сентября и даже октября.
Крачки в возрасте 1-го года не появляются в местах гнездования и держатся в об-
ласти между местами гнездования и зимовки (Таманцева, 1955). Залеты черных крачек
отмечены в Исландию, Норвегию, на Фарерские острова и о-в Мадейра, в Ирак, Кувейт.
Иорданию (Cramp, 1985), Индию (All, Ripley, 1969), на Айновы острова (Коханов, Ско-
кова, 1967), на Енисей возле Нижнешадринска (Бурский, Вахрушев, 1983), в юго-за-
падное Забайкалье (Измайлов, Боровицкая, 1973).
Численность. В Западной Европе, по-видимому, 22—25 тыс. гнездящихся пар (Cramp.
1985). В Латвии насчитывается 1000—1500 пар (Виксне, 1983), в Белоруссии -
43 тыс. пар (Долбж, Навумчык, 1983), в Московской обл. — не менее 300 пар (Зуба-
кин, 19816). Очень многочисленна крачка в Казахстане (Долгушин, 1962) и на юге
Западной Сибири (Дементьев, 1951). Численность вида в СССР можно оценить в нес-
колько сот тысяч пар; тенденция ее изменения не ясна. Отмечен рост численности в Ле-
нинградской и Калиниской областях (Зиновьев, 1980; Мальчевский, Пукинский.
1983), снижение (в основном в результате мелиорации) — в Западной Украине, Волго-
градской обл., Мордовии (Луговой, 1975; Сребродольская, 1975; Талпош, 1975;
Сребродольская и др., 1981; Кубанцев, Чернобай, 1982).
Местообитания. В сезон гнездования - заросшие околоводной и водной раститель-
ностью пресные, часто неглубокие, равнинные водоемы со стоячей или медленно теку-
щей водой; в Казахстане черная крачка изредка гнездится в тростниковых зарослях по
берегам рек со спокойным течением и еще реже на осоковый болотах (Долгушин.
1962). В лесной зоне предпочитает поймы крупных рек. В последние десятилетия успеш-
но заселяет пруды рыбхозов (Пукас и др., 1981; Мищенко, 1983).
Во время миграций — озера, водохранилища, морские побережья, заливы, эстуа-
рии; пролет идет как над сушей, так и над морем. На зимовках крачки главным об-
разом держатся по морским побережьям, эстуариям и лагунам, изредка на внутренних
водоемах (Cramp, 1985).
264
Черная крачка
Размножение. Размножение начинается не ранее, чем в двухлетнем возрасте. Моно-
гам., Диффузно гнездящийся факультативно-колониальный вид. Нередко меняет места
гнездования, однако в случае неизменности условий может поселяться в одних и тех
же местах в течение нескольких лет. Из болотных крачек обладает, по-видимому, наи-
большим гнездовым консерватизмом. Крачка, окольцованная взрослой 10.VI 1964 г.
в Шацком районе Рязанской обл., отмечена в этом же районе пять лет спустя (ЦКАН).
Численность поселений значительно колеблется по годам, но в несколько меньшей сте-
пени, чем у белокрылой крачки. Очень редко образует смешанные пары с белокрылой
крачкой (Зарудный, 1910; Мельников, 1985).
Для черных крачек, как и большинства других видов крачек, характерны воздуш-
ные формы демонстрационного поведения; по сравнению с чайками наземные демон-
страции отличаются гораздо меньшим разнообразием - (Baggerman et al., 1956; Cramp,
1985). Сразу после прилета в места гнездования можно наблюдать ’’высотный полет”,
при котором 2 20 птиц кругами поднимаются в воздух на высоту до 200 м, глубоко
и резко взмахивая крыльями. Затем начинается планирующий спуск, нередко по ши-
рокой спирали. ’’Низкий полет”, или ’’полет с рыбкой” (Fish-flight), можно наблюдать
над будущей территорией колонии. Самец, поймав какую-нибудь добычу, летает круга-
ми на высоте 10—15 м над колонией. К нему присоединяется самка, которая приземля-
ется после нескольких кругов. Самец опускается вслед за ней и кормит ее. Кормле-
ние самцом самки продолжается и после формирования пары, обычно при смене на
гнезде. При посадке самца с кормом партнеры обычно принимают ’’выпрямленную”
позу, при которой передняя часть тела приподнята, шея вытянута наклонно вверх,
клюв образует 40-70“ с горизонталью, крылья отодвинуты от боков туловища
(рис. 63,а). Если самец опустился без корма, чаще всего принимается ’’наклонная”
поза, при которой задняя часть тела приподнята так, что спина образует угол пример-
но в 30° с горизонталью (см. рис. 63,6).
Величина колоний черных крачек обычно составляет 5—20 гнезд, реже меньше 5 или
20—50 пар, еще реже отмечаются колонии в 100 и более пар — до 800 пар в Казахстане;
отдельными парами гнездится лишь в виде исключения (Долгушин, 1962; Зиновьев,
1980; Зубакин, 19816; Виксне, 1983; Мальчевский, Пукинский, 1983; Мищенко,
1983). Колония в несколько тысяч гнезд наблюдалась в Новосибирской обл. (Боро-
дулина, 19536), но судя по описанию, это был комплекс из нескольких колоний.
В крупных колониях крачек гнезда располагаются группами в несколько, реже в нес-
колько десятков гнезд, удаленными на некоторое расстояние друг от друга.
Гнезда обычно помещаются по краю зарослей тростника или рогоза, на тонких ро-
гозовых сплавинах, скоплениях плавающих растительных остатков, листьях плавающих
водных растений, плавающих кучах растительного мусора, реже на кочках среди топ-
ких мест, еще реже на осоковых кочках. В Московской обл. отмечена четкая приуро-
ченность колоний к озерам, где есть сплавины. Часто поселяется вместе с малыми
чайками, черношейными и красношейными поганками, нередко по соседству с коло-
ниями озерных чаек, реже белокрылых крачек. Для гнезда черных крачек характерно
небольшое количество гнездового материала — оно построено из отмерших кусочков
растений, собранных рядом с гнездом; в отличие от белокрылой крачки листья и стеб-
ли зеленых растений практически не используются. Размеры гнезд (мм) из Москов-
ской обл. (п = 14): диаметр гнезда 140—190, в среднем 170; диаметр лотка 90—100,
в среднем 95; глубина лотка 10—45, в среднем 20. Среднее минимальное расстояние
между центрами ближайших соседних гнезд на оз. Энгуре Латвийской ССР составляет
(и = 29) 0,50—7,0 м, в среднем 2,2 м; в Московской обл. — в среднем 4,3 м (п = 21),
причем в отдельных колониях уменьшается до 1,3 м (п = И); в Новосибирской обл. —
в среднем 1,5 м (наименьшее — 0,43 м). В других частях ареала крачки гнездятся со
265
Семейство Чайковые
сходной плотностью (Бородулина, 19536; Долгушин, 1962; наши данные). Индекс
плотности гнездования, рассчитанный по имеющимся данным, равен 0,03.
Сроки начала откладки яиц сходны в разных частях ареала: середина или конец
второй декады мая; лишь в Новосибирской обл. первые яйца отмечаются в самом
конце мая, а в дельте р. Или — уже 11.V. Для черной крачки характерен сильно растя-
нутый период откладки яиц — свежие кладки можно встретить даже в первых числах
июля. Возможно, позже начинают гнездиться более молодые птицы и пары, потерявшие
Рис. 63. Демонстрационное поведение черной крачки (а, б - по: Cramp, 1985, с изменениями)
а — выпрямленная поза, б — наклонная поза, в — кормление самкн самцом во время смены парт-
неров на гнезде
первые кладки. В кладке 1—4, чаще всего 3 яйца, несколько реже 2. Окаска яиц слож-
ная, обычно темная. Фон от охристого до темно-коричневого, нередко оливковый или
оливково-коричневый, реже зеленоватый. Рисунок в виде редких четких пятен и кра-
пин. Глубоких сероватых пятен немного, они малозаметны. Поверхностные пятна свет-
ло-бурые, бурые или почти черные. Обычно они сгруппированы у тупого конца, нередко
образуя венчик, реже равномерно разбросаны по всей поверхности скорлупы (Спан-
генберг, Фейгин, 1936; Бородулина, 19536; Долгушин, 1962; Мальчевский, Пукинский,
1983; наши данные). Размеры яиц (мм): из европейской части СССР (и = 20) 33,0-
36,5 X 24,0-25,8, в среднем 34,5 X 25,2; из низовий Сырдарьи (и = 18)32,9—36,5 X 24,1-
26,1, в среднем 34,8 X 25,2; из Западной Сибири (и = 13) 31,2—37,4 X 23,8—25,6, в сред-
нем (и = 6) 34,0 X 24,9; из Казахстана (п = 35) 33,5—39,8 X 23,2—26,5, в среднем 35,7 X
X 25,1. Масса (г) слабонасиженных яиц из низовий Сырдарьи (и = 12) 10,4—12,3, в
среднем 11,3 (Дементьев, 1951; Долгушин, 1962; Кисленко, 1972; Гынгазов, Милови-
дов, 1977; Ковшарь, Левин, 1982).
Насиживание начинается с первого яйца, длительность его 18—22 дня (Карпович
и др., 1958; Бородулина, 1960; Cramp, 1985; наши данные); указание на 14—17 дней
266
Черная крачка
(Дементьев, 1951; Долгушин, 1962), по-видимому, ошибочно. Насиживают оба парт-
нера, самка несколько больше, чем самец (Калинин, 1975). По наблюдениям 31.V
1.VI 1983 г. в Московской обл., смены партнеров происходят 16—21 раз в сутки (и =
= 3), только в светлое время (первая смена наблюдалась в 5 ч 46 мин, последняя —
в 21 ч 25 мин, время летнее). В Кустанайской обл. 31.V—1.VI 1971 г. отмечено 10 смен
партнеров за сутки (п = 1), первая и последняя смена были примерно в те же сроки (на-
ши данные).
Вылупление птенцов на большей части ареала начинается с первой декады июня,
в Новосибирской обл. — в конце второй декады июня. Вылупление в гнезде с 3 яйца-
ми длится 1—4 сут (и = 29), чаще всего 2 сут (48,3% случаев) и 1 сут (34,5%). После
вылупления крачки выносят и выбрасывают скорлупу из гнезда. В первые дни жизни
родители постоянно обогревают пуховичков. Кормят птенцов оба родителя 65—85 раз
в сутки. Корм (обычно водных и околоводных насекомых и их личинок) приносят
только в клюве. По измерениям в Кустанайской обл. птенцы в день вылупления весят
6,8—9,5 г, в среднем (и = 8) 8,3 г. В возрасте 2 сут начинают прорезываться пеньки
перьев на груди и плечевых, 3 сут — пеньки первостепенных маховых. У пятисуточных
птенцов появляются пеньки рулевых и начинают разворачиваться опахала плечевых,
у шестисуточных — опахала первостепенных маховых. Уже в суточном возрасте птенцы
могут оставлять гнезда при тревоге, в последующем в тревожной ситуации они всегда
разбегаются и затаиваются в растительности. В возрасте 2—3 сут после вылупления
первого птенца выводки оставляют гнездо и, как правило, уходят в другой участок
колонии, но могут держаться и на своей гнездовой территории. В этом же возрасте
родители начинают отличать своих птенцов от чужих и нападать на чужих птенцов,
оказавшихся по соседству (Бородулина, 1960; Мальчевский, Пукинский, 1983; наши
данные). На пернатых хищников, появляющихся в колонии, крачки активно нападают
и обычно быстро прогоняют их. На человека пикируют, нередко ударяя клювом (осо-
бенно после вылупления птенцов).
На крыло молодые поднимаются в возрасте 18—21 день и, как правило, около неде-
ли держатся на территории колонии. К самостоятельному питанию, по одним данным,
переходят в возрасте 25 дней, по другим не менее чем через 2 недели после подъема
на крыло. С летными молодыми могут держаться как оба родителя, так и один (Кар-
пович и др., 1958; Бородулина, 1960). Родители продолжают докармливать молодых
и в период послегнездовых кочевок (Кныш, Пулях, 1975).
По наблюдениям в Окском заповеднике, до подъема молодых на крыло гибнет,
главным образом от хищников, 36% птенцов от числа вылупившихся или 46% от числа
отложенных яиц. Число неоплодотворенных яиц здесь было крайне незначительным
(Карпович и др., 1958). В 1971 г. в Кустанайской обл. в одной колонии из 19 гнезд
погибло 66,7% всех яиц, главным образом из-за затяжного ненастья в начальный пе-
риод насиживания. В другой колонии отход яиц составил лишь 6,7% (и = 150), причем
50% погибших яиц пришлось на долю неоплодотворенных, еще 20% было, видимо,
уничтожено хищниками и 20% — брошено родителями. Отход птенцов в этой коло-
нии составил 25% от числа вылупившихся или 30% от числа отложенных яиц (наши дан-
ные) .
Максимально известная продолжительность жизни черной крачки, по данным кольце-
вания, 17 лет и 2 месяца (Cramp, 1985).
Суточная активность, поведение. Активность дневная, хотя дольше, чем другие виды,
может охотиться в сумерках (Авилова, 1973). Ночью жизнь в колонии замирает. Наси-
живающие и обогревающие птенцов крачки спят на гнездах, свободные от насиживания
партнеры в колониях, как правило, не ночуют (наши данные). Общественная птица
как в сезон гнездования, так и вне его. В период миграций на морских побережьях
267
Семейство Чайковые
крачки могут образовывать тысячные стаи, ночевочные скопления включают до 40 тыс.
особей (Cramp, 1985).
Питание. Основной корм в сезон гнездования — водные и околоводные насекомые:
стрекозы и их личинки, личинки плавунцов и водолюбов, водяные клопы, личинки
хирономид. Кроме того, черная крачка поедает других водных беспозвоночных (пия-
вок, пауков и др.), а также сухопутных насекомых: бабочек, кобылок, кузнечиков,
жуков, за которыми летает в степь или на луга; в отдельных случаях доля сухопут-
ных кормов может даже превышать долю водных. В ряде мест, например в дельте Вол-
ги и особенно в низовьях Кубани, большое значение в питании черных крачек могут
иметь мальки рыб и головастики. В других местах рыба в питании не отмечается или
составляет незначительный компонент корма (Бородулина, 1960; Тима, 1961; Дол-
гушин, 1962; Ширяева, 1965; Птушенко, Иноземцев, 1968; Самородов, 1976; Костин.
1983; Мальчевский, Пукинский, 1983). Черная крачка почти не кормится над открытой
водой — главным образом над водными и околоводными растениями и над травой
на лугах и в степи. Высота поискового полета обычно не превышает 2—3 м. Заметив
добычу, крачка скользит вниз и склевывает ее с поверхности воды или с растений.
Гораздо реже можно наблюдать броски в воду с высоты 1,5-2 м, при которых крач-
ка погружается почти целиком, кроме концов крыльев. Высматривая корм, птицы
часто трепещут на одном месте. Черные крачки могут ловить насекомых и в воздухе.
Радиус разлета из колонии за кормом обычно не первышает нескольких километров
(Бородулина, 1960; Долгушин, 1962; Авилова, 1973. 1975; наши данные). На зимов-
ках, по-видимому, большое значение в питании имеет рыба (Cramp, 1985).
Враги, неблагоприятные факторы. По-видимому, наибольший ущерб колониям кра-
чек наносит болотный лунь, изредка возможно хищничество озерных чаек, серых цапель
и серых ворон. Нередко гнезда гибнут в результате сильных ветров и летних паводков.
Хозяйственное значение, охрана. Хозяйственного значения вид не имеет, специальные
меры охраны не предусмотрены.
Белокрылая крачка —
Chlidonias leucopterus (Temminck, 1815)
White-winged Black Tern (англ.), Weissflilgelseeschwalbe (нем.), Guifette leucoptere (фр.).
С и n. (лат.) : Hydrochelidon leucoptera (Temminck, 1815).
Син. (рус.): светлокрылая крачка, белокрылая болотная крачка.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Мелкая крачка, отличается от других
болотных крачек в сезон размножения контрастной черно-белой окраской (черное
туловище и голова, белое надхвостье и хвост, белый верх сгиба крыла). От других кра-
чек отличается темной окраской, небольшими размерами и неглубокой вырезкой
хвоста; от черной в зимнем оперении — отсутствием темных пятен по бокам зоба.
На воду не садится, не ныряет. Полет легкий, порхающий; нередко птицы зависают
на одном месте, трепеща крыльями (в этом случае издали видны их ярко-белые хвост
и надхвостье). По земле передвигается мало и плохо, туловище держит горизонтально.
Голос несколько грубее, чем у черной крачки. Основной видовой позыв — короткое
чирканье ”тчир” или ”чвир”, который можно слышать как вне колонии, в том числе
от пролетающих стаек, так и в колонии. В частности, этот крик издают крачки, под-
летая к гнезду с кормом для птенцов. Крик ”чвир-лит”, напоминающий крик ”кирр-
лит” черной крачки и, видимо, гомологичный ему, в этой ситуации употребляется
гораздо реже. Трескучее щебетание типа ”ки-ти-кирр”, ”ки-ти-ки-кирр” можно слы-
шать во время внутривидовых столкновений, особенно в воздухе. Звонкая цокающая
268
Белокрылая крачка
трелька ”цти-ти-ти-ти” используется во время погонь за пернатыми хищниками. Крик
тревоги — звонкое громкое ”чвик”.
Подвидовая систематика. Монотипический вид.
Описание. Строение и размеры (табл. 16).
Окраска (Дементьев, 1951; Cramp, 1985). Самец и самка в брачном наряде.
Голова, шея, грудь, брюхо, бока интенсивно-черные. Спина и плечевые тускло-черные
или грифильно-серые. Нижние кроющие крыла черные, кроме бледно-серых нижних
больших кроющих. Поясница, надхвостье, подхвостье и рулевые белые. Малые верхние
кроющие крыла белые, средние и большие кроющие -- серые. Первые 2- 3 дистальных
первостепенных черноватые, слегка окрашенные в серебристо-серый цвет весной и бу-
ровато-черный осенью. Остальные первостепенные со светло-серыми внутренними
опахалами и светло-серебристо-серыми или почти белыми внешними; самые внутрен-
ние первостепенные могут быть сплошь светлого серебристо-серого цвета. Передние
второстепенные маховые светло-серые с беловатой вершинной каймой, задние — серова-
то-черные. Клюв темно-красный или чаще черный с красноватым оттенком; ноги ярко-
красные. Радужина темно-коричневая.
Самец и самка в зимнем наряде. Лоб, горло, уздечка, передняя часть шеи, грудь,
бока, брюхо и подхвостье белые. Темя и затылок дымчато-серые с белыми пестрина-
ми, задняя часть шеи белая с сероватыми пятнами. Пятно перед глазом черное, кроющие
уха черноватые. Спина, плечевые и верхние кроющие крыла светло-серые, поясница и
надхвостье несколько светлее. Рулевые светло-серые, крайние две пары белые или почти
белые. В сентябре—декабре дистальные первостепенные и второстепенные маховые
черные (иногда со светло-серым налетом), проксимальные первостепенные светло-
серые. В декабре—феврале самые крайние первостепенные серые с серебристо-серым
налетом, проксимальные и второстепенные светло-серые, иногда с более темной цен-
тральной частью. Клюв черный, ноги темно-красные.
Пуховой наряд (живые птенцы из Московской обл.). Как у черных крачек, но в це-
лом темнее. Верх тела от серовато-коричневого до черновато-коричневого цвета, лоб
от темно-бурого до угольно-черного. Вокруг основания клюва и глаз белые кольца,
уздечка также ‘белая. Подбородок и горло темно-серые или почти черные. Брюшко
и грудь темно-серые. На спинной стороне крупные и мелкие темные пятна. Клюв чер-
ный, у основания серо-телесный, лапы серо-телесные. Радужина почти черная.
Гнездовой наряд. Похож на соответствующий наряд черной крачки, но по бокам
груди нет темных пятен, поясница почти белая, а не сероватая, серый цвет плечевых,
хвоста, маховых и верхних кроющих крыла светлее. Спина от бурой до темно-бурой;
темя с большим количеством белых пестрин, чем у черных крачек. Клюв черновато-
бурый, ноги буровато-красные.
Первый зимний наряд. К декабрю—марту становится похожим на окончательный
зимний наряд, но дистальные первостепенные сохраняются от гнездового наряда, они
темнее, чем у взрослых птиц. Как у черных крачек, первый зимний наряд замещается
летом вторым зимним, минуя брачный наряд (Cramp, 1985). Второй зимний не отличим
от окончательного зимнего, но дистальные маховые осенью и ранней зимой имеют более
свежий вид. Брачный наряд одевается лишь весной третьего календарного года.
Линька (Cramp, 1985). Сроки полной линьки в первый зимний наряд варьируют:
у одних она начинается в сентябре, у других — только в ноябре. Завершается обычно к
марту, но у отдельных особей остаются некоторые ювенильные перья до начала мая.
Первостепенные маховые начинают линять с XI в конце ноября — начале февраля, их
линька завершается в июне—августе. С мая—июля второго календарного года начинается
повторная линька первостепенных, в это же время оперение головы, туловища, хвоста и
кроющих крыла замещается перьями второго зимнего наряда.
269
Семейство Чайковые
Таблица 16. Белокрылая крачка. Размеры особей (мм) (ЗМ МГУ)
и масса тела (г) (Бородулина, 1960; Долгушин, 1962)
Географический район Пол Длина крыла среднее
п Lim
Западная часть гнездового ареала dd 18 202-224 212
99 10 200 -212 206
Забайкалье, Хабаровский край dd 4 210-220 214
99 3 204-211 208
У взрослых птиц две линьки в году: полная послебрачная и частичная предбрачная.
Послебрачная начинается с июня и завершается сменой первостепенных маховых в де-
кабре-марте. Ее протекание напоминает ход линьки черной крачки, но в отличие от по-
следнего вида повторная смена первостепенных маховых, начавшись в конце ноября -
начале января, прерывается лишь в конце апреля или начале мая обычно на стадии
новых IV—V. В это же время начинается повторная линька рулевых, немного позднее —
повторная смена второстепенных маховых. Предбрачная линька стартует в начале
февраля — начале марта и завершается в конце апреля — начале мая; сменяется опере-
ние головы, туловища, хвоста, кроющие крыла и внутренние первостепенные. Третья
линька первостепенных (бывает примерно у 80% особей) начинается между первыми
числами февраля “и серединой апреля, прерывается в мае, обычно на стадии но-
вых IX—XI.
Распространение. Гнездовой ареал состоит из двух крупных участков,
один из которых протянулся от Венгрии и восточной Польши (Chmielewski, 1984;
Keller, 1984) до верхней Оби, а другой охватывает северо-восточный Китай и приле-
гающие территории СССР. В декабре-феврале 1974—1975 гг. одна пара загнездилась
в Новой Зеландии (Robertson, 1985). В недавнее время изолированные поселения
появились в Прибайкалье и Лено-Амгинском междуречьи в Якутии (рис. 64).
Западная часть ареала почти целиком лежит в пределах СССР (рис. 65). Вне нашей
страны гнездование в настоящее время зарегистрировано лишь в восточной Польше.
Венгрии, Румынии; небольшие поселения отмечены в Италии, Югославии и, возможно,
в Болгарии. Обычна на гнездовье белокрылая крачка также в Ираке (Cramp, 1985).
В пределах СССР западная граница сплошного ареала проходит по Белоруссии, где крач-
ка заселяет Гродненскую, Брестскую, Могилевскую и Витебскую области (Долбж,
Навумчык, 1983). В Западной Украине достоверно гнездилась в Волынском Полесье
(Страутман, 1963) , вновь появилась там на гнездовье в 1984—1985 гг. (Горбань, устное
сообщ.). Отмечается на гнездовье во многих местах Молдавии (Аверин и др., 1981),
на Украине по Днепровским водохранилищам (Булахов, Мясоедова, 1975; Мельни-
чук, Головач, 1984).
Северная граница гнездового ареала ныне проходит через южную и восточную Литву
(Валюс и др., 1976; Пукас и др., 1981; Мачикунас, 1982), северную Белоруссию, Смо-
ленскую обл., где она постепенно сдвигается к северу (Андриевский, 1974), южную
часть Калининской обл. (Зиновьев, Шапошников, 1978), Ярославскую обл. близ Яро-
славля, где гнездование постоянно отмечается с 1971г. (Белоусов, Макковеева, 1981).
юг Костромской обл. и Горьковскую обл. севернее Горького (Хохлова, 1972; Водо-
лажская, 1981), Удмуртию (Попов, 1975), видимо, значительно севернее Тюмени (Ша-
270
Белокрылая крачка
Длина клюва Длина цевки Масса тела среднее
п Lim среднее п Lim среднее п Lim
18 20,3-26,0 23.4 16 17,0 21,8 19,2 43 59 -79 67
11 21,8 - 24,8 23,1 и 17.0-21,6 19,2 28 53-76 63
4 23.0 26.4 24.5 4 19.3 20.3 19,8 - - -
3 23,8-25.3 24,5 3 19,0-21.1 19,7 -
ронов, 1954; Гынгазов, Миловидов, 1977), несколько севернее Омска (Сулимов, 1981)
и по северной границе Барабинской степи (Ходков, 1978) . По долине Оби белокрылая
крачка гнездится до Обь-Чаинского междуречья (Адам, Сурнаев, 1979). В пределах
Томской и Новосибирской областей проходит восточная граница данного участка
ареала, которая огибает с запада Алтай и входит в пределы Казахстана в окрестностях
Семипалатинска. Южная граница ареала поднимается по долине Иртыша, спускается к
Караганде, далее идет к низовьям Тургая и Иргиза, затем по северной части Уральской
обл. до Волги (Долгушин, 1962). Гнездится в низовьях р. Уил (Пославский, 1974).
Отмечена на гнездовье в Волгоградской обл. (Коленов, 1981; Кубанцев, Чернобай,
1982), в дельте Волги (Воробьев, 1936; Кривоносов, 1976), дельте Терека, на разли-
вах р. Кумы и Восточном Маныче (Волчанецкий, 1959) , в дельтах Дона, Кубани, на во-
дохранилищах Западного Маныча (Казаков, Языкова, 1982). Далее граница проходит
по югу Украины севернее Крыма (Букреев и др., 1986: Костин, 1983).
С 1957 г. известно поселение белокрылых крачек в восточной Латвии на оз. Лубана
(Виксне, 1983). В 1982 г. 1 пара загнездилась в Эстонии (Cramp, 1985), с этого же
года отмечено гнездование в центре Кировской обл. (Литун, Макаров, 1984; Савельев,
Чарушина, 1984), с 1968 г. — на Оби ниже устья Иртыша (Ясков, 1981) . Изолированные
поселения отмечены в низинных частях Грузии (Жордания, 1977), в Кызылагачском за-
ливе (Заблоцкий, Заблоцкая, 1963), в низовьях р. Атрек в юго-западной Туркмении
(Караваев, 1981), в Зайсанской и Алакольской котловинах (Грачев, 1968; Сурвилло,
1968; Ауэзов, Грачев, 1975; Самусев, 1975), в северной Хакасии (Прокофьев, 1981),
южной Туве (Баранов, 1981), на Торейских озерах (Васильченко, 1986) и, возможно,
в дельте Амударьи (Кенжегулов, 1972). Восточный участок гнездового ареала бело-
крылых крачек ранее заходил на территорию СССР лишь в окрестностях оз. Ханка
(Воробьев, 1954; Поливанова, 1971) и, возможно, в юго-восточном и восточном Забай-
калье (Дементьев, 1951). Однако во второй половине нашего столетия началось резкое
расширение в северном направлении области гнездования белокрылой крачки на восто-
ке. Крачка заселила почти весь нижний Амур, достигнув в 1972 г. оз. Чукчагирского
(Росляков, 1977, 1979)1; по-видимому, гнездится по всей Зейско-Бурейской равнине
(Дымин, 1975). В начале 1960-х годов эта птица стала гнездиться на Байкале в устье
Селенги, на перешейке п-ова Святой Нос и в устье Верхней Ангары и Кичеры (Скрябин,
1967), в 1967 г. — в Иркутской обл. в нижнем течении р. Иркут (Безбородов, 1979), в
1974—1975 гг. найдены колонии в Забайкалье на озерах Б. Еравное, Сосновское, Харта,
Исинга и Михайловское, а также в окрестностях г. Улан-Удз (Шкатулова, 1979). В
1 Впрочем, по В.А. Нечаеву (1963), она была не редка на гнездовье на оз. Эворон еще в 1959 г.
271
272
Рис. 64. Область распространения белокрылой крачки
1 — область гнездования (пунктиром показана неуточненная граница), 2 — изолированные гнездовые поселения, 3 — предполагаемые
места гнездования, 4 — места зимовок, 5 — залеты
273
Рис. 65. Ареал белокрылой крачки в СССР
у _ область гнездования (пунктиром показана неуточнеииая граница), 2— изолированные гнездовые поселения, 3 — предполагаемые
места гнездования, 4 — залеты
Семейство Чайковые
1965 г. отмечен залет белокрылой крачки в Лено-Амгинское междуречье. В 1968 г. она
найдена на гнездовье в среднем течении р. Вилюй, в 1971—1973 гг. — в Лено-Амлинском
междуречье, где с тех пор постоянно гнездится, и в районе устья Вилюя. Залетные особи
и стайки белокрылых крачек, отмеченные в июле 1975 г. в истоках р. Тюкян (66 ° с.ш.)
и 8— 10.VI 1977 г. в среднем течении р. Оленек (68° с.ш.) говорят о возможности даль-
нейшего продвижения на север этого вида (Андреев, 1974; Дегтярев и др., 1981а).
Есть указания на гнездование белокрылой крачки в Монголии на озерах Ачит-Нур,
Орог-Нур, Буир-Нур, реках Селенга и Тола (Vaurie, 1965; Stubbe, Bolod, 1971).
Рассматривая ареал белокрылой крачки в целом, необходимо отметить проявившую-
ся в конце 1950-х—начале 1970-х годов тенденцию к расширению области гнездования
к северу, особенно заметную на востоке ареала, где крачка продвинулась примерно на
1,5 тыс. км. В европейской части СССР отмечены сдвиги границы ареала к северу на
150—250 км в Прибалтике, Ярославской и Кировской областях.
Зимовки. Область зимовки, как и гнездовой ареал, разделена на две части. Евро-
пейские, западносибирские и казахстанские крачки зимуют, видимо, во внутренних
районах Африки от 18° с.ш. до юга Африки. Другой район зимовок включает острова
Юго-Восточной Азии, северное побережье Австралии, Новую Зеландию, юго-восток
Китая, страны Индокитая, Бирму, Бангладеш, Шри-Ланка. Небольшое количество
птиц отмечается в Персидском заливе и на побережье Пакистана, крайне редкие встре-
чи зарегистрированы на западном побережье Индии (АН, Ripley, 1969; Cramp, 1985;
Robertson, 1985). Отдельные особи встречены зимой у Черноморского побережья
Кавказа (Строков, 1974).
Миграции. Изучены хуже, чем у черной крачки. В отличие от последнего вида пролет
через северную и западную часть Европы слабый, в Центральной и Юго-Восточной Евро-
пе белокрылые крачки отмечаются чаще на весеннем, чем на осеннем пролете. Места
зимовок, по крайней мере в Африке, птицы начинают покидать в марте. Пролет идет
по Нилу, в меньшей степени через Сахару. Далее часть птиц летит через Балканский п-ов
и Турцию в Восточную Европу, другие — в Закавказье (Cramp, 1985). Затем часть
птиц направляется вдоль Кавказского побережья Черного моря, где пролет достаточно
интенсивен (Строков, 1974; Вронский, Томкович, 1975). Другие крачки летят к Кас-
пийскому морю, причем, видимо, долиной Аракса или через северный Иран: отмечен
пролет у Еревана (в апреле 1978 г. добыта белокрылая крачка, окольцованная в ЮАР
19.XII 1972 г.; ЦКАН), а на Мингечаурском водохранилище крачка на пролете отсут-
ствует (Туаев, Васильев, 1972). В дальнейшем пролет идет через Каспийское море
(Залетаев, 1960; Заблоцкий, Заблоцкая, 1963), далее, очевидно, часть птиц летит вверх
по Волге, другие поворачивают на восток и летят через Северный Казахстан в Западную
Сибирь. Незначительный пролет отмечен по восточному побережью Аральского моря
(Березовский, 1983). Возможно, часть птиц с зимовок на побережье Индийского океана
летит через Иран, Пакистан и Афганистан. Об этом может свидетельствовать встреча
стайки из 6 особей 3.V 1975г. в низовьях р. Сурхандарьи (Остапенко и др., 1978) и
появление белокрылых крачек в последние годы в апреле—мае около Алма-Аты (Еро-
хов, 1986).
В Крыму первые особи появляются не ранее 29.IV, наиболее интенсивный пролет
идет в мае, однако белокрылая крачка здесь явно малочисленнее черной (Костин,
1983). В Предкавказье крачки появляются во второй половине апреля, массовый про-
лет проходит в начале мая (Казаков, Языкова, 1982). В конце апреля первые белокры-
лые крачки отмечаются в Ворошиловградской обл. (Панченко, 1975; Сулик, Енакиев.
1978), с 7.V — в Сумской обл. (Кныш, Пулях, 1975), с 2—15.V — в Московской обл.
(Птушенко, Иноземцев, 1968; наши данные), 1-3.V в г. Вольске Саратовской обл.,
19.V в Татарской АССР (Водолажская, 1981), в Мордовии — не ранее середины мая
274
Белокрылая крачка
(Луговой, 1975). На юго-западном Каспии первые птицы появляются 2.V (Заблоцкий,
Заблоцкая, 1963); близ устья Урала прилет отмечен 8.V, восточнее Караганды — 13—
16.V, северо-восточнее Семипалатинска — 19.V(Долгушин, 1962) .В Нарзуме первые
крачки отмечаются 23—27.IV (Елкин, 1975), на оз. Малый Чан — 10—17.V (Ходков,
1978), в долине верхней Оби — 15—25.V (Блинова, 1986).
На оз. Ханка белокрылые крачки прилетают 13—15.V (Поливанова, 1971),под Ха-
баровском первых птиц отмечают 15—18.V, на оз. Эворон — 25—30.V (Росляков, 1977) .
В дельте Селенги первые крачки появляются 16—21.V, на Верхней Ангаре — 22—25.V
(Мельников, 1977); в окрестностях Улан-Удэ в 1974—1979 гг. первые птицы отмечены
21—25.V, пролет шел до 29.V—7.VI (Шкатулова, 1984). В Лено-Амгинском междуречье
в 1978—1983 гг. прилет зарегистрирован 23—27 .V,в среднем 26.V, в устье р. Вилюй —
26.V 1974 г. и 31.V 1975 г., в районе устья р. Наманы — 29.V 1973 г.; миграции про-
должались до 10.VI (Дегтярев и др., 1981а; Дегтярев, 1986).
Осенние миграции, как и у черной крачки, начинаются в конце июля—начале августа.
К концу августа большинство птиц исчезает, отдельные особи изредка отмечаются и в
сентябре. Осенний пролет идет, видимо, по тому же маршруту, что и весенний, но в об-
ратном направлении. Величина пролетных стай белокрылых крачек примерно такая же,
как у черных. В дельте Волги крачки концентрируются перед отлетом в тысячные
скопления (Бородулина, 1960).
Залеты отмечены в Исландию, Ирландию, Великобританию, Бельгию, Норвегию,
Швецию, Финляндию, Португалию, на Фарерские острова, о-в Мадейру (Cramp, 1985),
о-в Барбадос, США (Дементьев, 1951), на юг Кореи (Gore, Won, 1971), Шантарские
острова (Дулькейт, Шульпин, 1937), о-в Сахалин (Гизенко, 1955), заполярную Якутию
(Дегтярев и др., 1981а).
Численность. В европейской части СССР в целом малочисленнее черной крачки. В Бе-
лоруссии гнездится 5400 пар (Долбж, Навумчык, 1983), в Латвии около 20—40 пар
(Виксне, 1983), по-видимому, столько же в Литве; в Московской обл. гнездится до
400—500 пар (наши данные). Много белокрылых крачек в степном Казахстане и Бара-
бинской степи (Долгушин, 1962), в Курганской обл. (Калинин, Болотников, 1975).
В Омской обл. гнездится 260 пар (Сулимов, 1981). На оз. Эворон в 1969 г. гнездилось
более 1000 пар, затем численность уменьшилась (Росляков, 1979); в дельте Селенги
гнездится 10—12,5 тыс. пар (Мельников, 1978). На оз. Ханка отмечены десятки тысяч
птиц в 1961, 1962 и 1964 гг., несколько сот пар — 2,5 тыс. пар в 1980—1981 гг. (Поли-
ванова, 1971; Глущенко, 19846). В Западной Европе зарегистрировано несколько сот
гнездящихся пар (Cramp, 1985). Всего в СССР гнездится, по-видимому, несколько
сот тысяч пар; общая тенденция изменения численности не ясна. В ряде мест числен-
ность снижается в связи с мелиорацией, в других местах (в частности, в Мордовии,
Московской обл.) — возрастает.
Местообитания. В сезон гнездования — мелководья и заболоченные берега поймен-
ных и водораздельных озер и водохранилищ, низинные осоковые и манниковые боло-
та и болотистые луга, дельты рек с заболоченными участками, плесы степных речек;
в последние годы — пруды рыбхозов, но белокрылая крачка заселяет их менее охотно,
чем черная.
Вне сезона размножения (Cramp, 1985) — морские побережья, лагуны, мангровые
заросли, но чаще — долины рек и берега озер; может встречаться и вдали от водоемов.
Размножение. Птицы впервые приступают к размножению не ранее двухлетнего
возраста. Белокрылая крачка моногам, факультативно-колониальный вид. Характерна
частая смена мест гнездования и резкие колебания численности по годам, вплоть до
негнездования в отдельные годы (Глущенко, 19846; наши данные). Демонстрационное
поведение, по наблюдениям в Московской обл., сходно с поведением черной крачки.
275
Семейство Чайковые
Формирование пар происходит после прилета и не на территории будущей колонии.
Отмечен ’’высотный” и ’’низкий” полеты, кормление самки. Детали не изучены. В ред-
ких случаях может образовывать смешанные пары с черной и белощекой крачками
(Зарудный, 1910; Мельников, 1985) .
В большинстве регионов величина колоний белокрылой крачки составляет 2—20 пар,
реже до 50 пар и еще реже 100 и более пар; в Барабинской степи отмечались колонии
в 300—400 гнезд (Шнитников, 1913; Долгушин, 1962; Мельников, 1977; Дегтярев и
др., 1981а). На оз. Ханка рыхлые поселения крачек состоят из групп в 10—40 гнезд или
могут тянуться на многие километры (Поливанова, 1971; Глущенко, 19846). Отмечено
гнездование отдельными парами (Безбородов, 1979; Букреев и др., 1986). В отличие от
черных крачек белокрылые предпочитают гнездиться на мелководье, устраивая гнезда
чаще всего на осоковых кочках, полегших или подмятых стеблях околоводных расте-
ний, на кучах растительного мусора, осоковых или вейниковых сплавинах, плавающих
островках мха или камыша; в дельте Селенги — на обсыхающих грязевых отмелях и
иногда на Твердых, Слегка увлажненных берегах. Может образовывать моновидовые
колонии, но нередко селится вместе с другими видами птиц: чаще всего с малыми чай-
ками, реже — с речными и черными крачками, озерными чайками и черношейными по-
ганками (Долгушин, 1962; Мельников, 1977; Дегтярев и др., 1981а; Виксне, 1983;
наши данные).
В качестве материала для гнезда используются отмершие и зеленые стебли окру-
жающей растительности. Размеры гнезд (мм): диаметр гнезда (и = 32) 130—180, в
среднем 155; диаметр лотка (и = 69) 90-110, в среднем 100; глубина лотка (п =
= 43) 10—30, в среднем 18 (Московская обл.). Гнездится более разреженно, чем чер-
ная крачка. Среднее минимальное расстояние между центрами ближайших соседних
гнезд в Московской обл. (« = 97) составило 7,3 м (0,75—36 м). В Новосибирской
обл. среднее расстояние между гнездами было равным 5,8 м, минимальное — 3,3 м,
в Татарии между гнездами было 0,9—16,4 м, в Якутии — 8—50 м, в Бурятии —1,5—25 м,
в среднем 3,3 м (Бородулина, 1960; Водолажская, Мингазова, 1979; Шкатулова, 1979;
Дегтярев и др., 1981а; наши данные). Индекс плотности гнездования, рассчитанный по
имеющимся данным, равен 0,01.
Откладка яиц происходит на несколько дней позже, чем у черной крачки. В Москов-
ской обл. первые яйца отмечаются в третьей декаде мая, в отдельные годы 18—19.V
(наши данные), в Татарской АССР, видимо, в эти же сроки (Водолажская, Мингазова.
1979). В дельте Селенги первые яйца отмечаются 3—6.VI, на Верхней Ангаре — 8-
9.VI (Мельников, 1977), в Якутии — 10.VI (Дегтярев и др., 1981а), на оз. Ханка —
5.VI (Поливанова, 1971) . Массовая откладка яиц происходит на несколько дней позже.
Как и для черной крачки, для белокрылой характерен растянутый период яйцекладки,
для дельты Селенги он составляет 40-44 дня (Мельников, 1977). В кладке от 1
до 6 яиц, чаще всего 3 яйца. В повторных кладках, которые бывают после гибели пер-
вой, количество яиц меньше. По наблюдениям на р. Селенге, отдельные птицы могут
загнездиться в третий раз (Мельников, 1977).
Окраска яиц сложная, в целом довольно темная. Фон оливково-коричневый разной
интенсивности, оливковый, оливково-зеленый. Рисунок в виде редких четких пятен и
крапинок различной величины и густоты, которые часто группируются у тупого конца,
нередко образуя венчик. Поверхностные пятна коричнево-бурые до почти черных. В це-
лом цвет яиц сильно варьирует в разных гнездах и нередко в одном гнезде. Размеры
яиц (мм): из Московской обл. (п = 86) 32,7—39,3X24,1—27,2, в среднем 35,7X25,8:
из Западной Сибири (п = 17) 31,0—37,0X24,0—26,5, в среднем 34,4X25,0; с оз. Ханка
(п = 36) 26,5—36,8X23,6—32,1, в среднем 34,5X24,9 (Поливанова, 1971; Гынгазов.
276
Белокрылая крачка
Миловидов, 1977; наши данные). Масса (г) яиц с оз. Ханка (и = 16) 10,0-13,0, в сред-
нем 11,9 (данные В.В. Леоновича).
Насиживание начинается с первого яйца и продолжается 18—22 дня (Карпович и др.,
1958; Бородулина, 1960; Мельникова, 1977; Водолажская, Мингазова, 1979). Наси-
живают оба партнера, причем самка в среднем больше, чем самец (Калинин, 1975).
По наблюдениям за одним гнездом в Московской обл., смена партнеров на гнезде
бывает 16 раз в сутки, причем только в светлое время (наши данные).
Вылупление птенцов в Московской обл. и Татарии начинается в конце первой-
начале второй декады июня, в Новосибирской обл. и на Байкале — в последнюю пяти-
дневку июня—начале июля. По наблюдениям на Селенге, со времени появления первых
трещин на яйце до полного вылупления проходит обычно 2—3 дня, вылуцление в одном
гнезде продолжается 1—4 сут, чаще всего 2—3 сут. Скорлупу яйца родители выносят и
выбрасывают. Кормят птенцов оба партнера, в первый раз спустя 1,5—2 ч после вы-
лупления. По наблюдениям в Московской обл. за гнездом с двухсуточными птенца-
ми, самец кормил 115 раз, самка 174 раза за сутки. Корм родители приносят в клю-
ве. По измерениям в Московской обл., в день вылупления птенцы весят (и = 8) 7,7—
9,5 г, в среднем, 8,5 г. В возрасте 2—3 сут появляются пеньки плечевых перьев, в воз-
расте 3—4 сут — пеньки первостепенных маховых. Выводки покидают гнездо на 2—3-е
сут после вылупления первого птенца. Родители активно нападают на хищников, появ-
ляющихся в колонии. Птенцы при этом разбегаются и затаиваются (Бородулина, 1960;
Мельников, 1977; Водолажская, Мингазова, 1979; наши данные) .
На крыло молодые поднимаются в возрасте 18—22 сут (Карпович и др., 1958; Бо-
родулина, 1960; Водолажская, Мингазова, 1979) или 24—25 сут (Мельников, 1977) . Ко-
лонию покидают через 3—7 дней после подъема на крыло. Родители продолжают кор-
мить выводок; обычно с молодыми держится только одна птица, как правило самец
(Карпович и др., 1958; Мельников, 1977). Докармливание молодых продолжается и в
период по еле гнездовых кочевок (Кныш, Пулях, 1975) .
До подъема на крыло в Рязанской обл. доживает около 70% птенцов от числа отло-
женных яиц; число неоплодотворенных яиц здесь было крайне незначительным (Карпо-
вич и др., 1958). В дельте Селенги в многоводные годы птенцы вылупляются только
из 5,25% яиц, в обычные годы — из 34%, в годы с низким уровнем воды — из 44% яиц.
Гибнет от 65% до 75% птенцов. Основные причины гибели яиц — колебание уровня
воды (гибнет от 4,3% до 94,4% яиц), хищничество сизых чаек (2,3—100%), в маловод-
ные годы вытаптывание скотом (6,4—100%). Отход птенцов увеличивается в затяжное
ненастье (Мельников, 1977).
Суточная активность, поведение. Активность сугубо дневная, ночью жизнь в колонии
замирает. Однако отмечались кормежки в сумерках (Бородулина, 1960) . Общественная
птица как в сезон гнездования, так и вне его. Отмечены массовые ночевки этих птиц на
оз. Ханка, где скапливается до 15 тыс. особей. Птицы концентрируются в плотную стаю
на участке острова размером 40X50 м (Поливанова, 1971; Глущенко, 19846).
Питание. Основной корм в сезон размножения — водные и околсводные насекомые:
взрослые стрекозы и их личинки, водяные клопы, взрослые жуки-плавунцы и их личин-
ки и др. В разных регионах состав кормов несколько меняется. На Байкале белокрылая
крачка питается только беспозвоночными, причем почти исключительно насекомыми —
главным образом ручейниками, стрекозами, двукрылыми и прямокрылыми (Скрябин,
Размахнина, 1978). Сходная картина отмечена в Новосибирской обл., где главным кор-
мом были взрослые стрекозы (52,2% встреч) и личинки плавунцов (39% встреч).
В дельте Волги, при доминировании в питании насекомых, отмечено поедание рыбы
до 45,1% встреч и головастиков до 21,4% (Бородулина, 1960). В Тургайской долине
277
Семейство Чайковые
(Самородов, 1976) в состав кормов крачек также входили водные и околоводные
насекомые (стрекозы — до 55,5% встреч) и рыба (до 25% встреч). В Якутии основной
корм — жуки (78,6% встреч) и их личинки (35,7%), главным образом плавунцы и жу-
желицы. Роль рыбы незначительна — 7,1% встреч (Дегтярев и др., 1981а).
Кормятся белокрылые крачки, используя ту же технику поимки добычи, что и чер-
ные, однако более активно ловят летающих насекомых (Бородулина, 1960). Места
кормежки — болота и заросшие мелководья, берега водоемов, луга, степь. Чаще, чем
черные крачки, кормятся группами и стаями — в Московской обл. до 18 особей (наши
данные), на оз. Ханка до 500 особей (Поливанова, 1971) .
Спектр питания в зимний период зависит от места зимовки, главным образом это
мелкая рыба и насекомые (Cramp, 1985).
Враги, неблагоприятные факторы. Ущерб кладкам и птенцам могут наносить болот-
ные луни, серые и большеклювые вороны, сизые чайки, американские норки. Много
гнезд гибнет в результате колебания уровня воды. Урон колониям наносит и фактор
беспокойства (Поливанова, 1971; Мельников, 1977; Водолажская, Мингазова, 1979;
Росляков,1979).
Хозяйственное значение, охрана. Хозяйственного значения не имеет, специальные
меры охраны не предусмотрены.
Белощекая крачка —
Chlidonias hybridus (Pallas, 1811)
Whiskered Tern (англ.), Weissbartseeschwalbe (нем.), Guifette moustac (фр.).
Син. (лат.): Hydro chelid on leucopareia Temminck, 1820, Hydrochelidon hybrida (Pallas, 1811).
Син. (рус.): белощекая болотная крачка.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Крупнее черной и белокрылой кра-
чек, более длиннокрылая. В брачном наряде отличается от них белыми щеками и под-
бородком, резко контрастирующими с черной шапочкой и темно-серой грудью. Ха-
рактерен также очень светлый подбой крыла и красный клюв. В целом окраской на-
поминает крачек рода Sterna, но гораздо темнее. Хвост, однако, как у типичной болот-
ной крачки, с неглубокой вырезкой. В зимнем наряде белощекая крачка очень похо-
жа на белокрылую, нечетко отличаясь более длинным клювом и более серой поясни-
цей. Полет легкий, как у других болотных крачек, но временами напоминает полет
речной крачки. На воду не садится. Как и речная крачка, может нырять с разлета. Го-
лос громче и грубее, чем у других болотных крачек; характерны каркающие звуки.
Подвидовая систематика. Разными авторами различается 3—7 подвидов, измен-
чивость проявляется в интенсивности окраски оперения и размерах (Peters, 1934;
Дементьев, 1951; Vaurie. 1965; Степанян, 1975; Cramp, 1985). В СССР гнездятся
два подвида (Степанян, 1975).
1. Chiidonias hybridus hybridus
Sterna hybrida Pallas, 181 l.Zoographia Rosso-Asiat., 2, c. 338,низовья Волги и Сарпинские
озера.
Нижняя сторона тела светлее, чем у следующего подвида, размеры крупнее.
2. Chlidonias hybridus javanicus
Sterna javanica Horsfield, 1822, Trans.Linn. Soc. London, 13, pt. 1, c. 198, Ява.
Нижняя сторона тела немного темнее, чем у предыдущего подвида, размеры меньше.
Замечания по систематике. Обладая большим сходством с черной и белокрылой
крачкой и явно находясь с этими видами в филогенетическом родстве (отмечены сме-
шанные пары с белокрылой крачкой — Мельников, 1985), белощекая крачка некото-
278
Белощекая крачка
рыми деталями морфологии напоминает и крачек рода Sterna. По этой причине от-
дельные авторы выделяют белощекую крачку в особый подрод Peiodes рода Chlido-
nias (Wolters, 1975).
Описание. Строение и размеры (табл. 17).
Окраска (Дементьев, 1951; Cramp, 1985). Самец и самка в брачном наряде.
Лоб, верхняя часть уздечки, темя, затылок и задняя часть шеи черные. Подбородок
и щеки белые, граница черного и белого цвета проходит через глаз. Спина серая; пле-
чевые, поясница и верхние кроющие крыла и надхвостье светло-серые; рулевые та-
кого же цвета, боковая пара их с белым наружным опахалом. Светло-серый цвет горла
постепенно переходит в темно-серую окраску груди и черно-серый цвет брюха и бо-
ков. Надхвостье белое. Внешнее опахало II (первого видимого) первостепенного махо-
вого серое с серебристым налетом, у остальных первостепенных более светлое, сереб-
ристо-серое. Внутреннее опахало II темно-серое в пристволовой части и белое по краю
опахала, причем белый цвет не доходит до вершины. Внутренние опахала других перво-
степенных такие же, но размер белого поля постепенно уменьшается. Второстепенные
маховые светло-серые с узкой белой каемкой на вершине и более светлыми внутренни-
ми опахалами. Окраска оперения самки может быть слегка светлее, чем у самца. Клюв
темно-красный, ноги красные, радужина от красновато-бурой до темно-коричневой.
Самец и самка в зимнем наряде. Лоб, уздечка, щеки, подбородок и горло белые.
Темя белое с черными пестринами, затылок черный с белыми пестринами; перед
глазом черное пятно, такого же цвета кроющие уха. Спинная сторона несколько блед-
нее, чем в брачном наряде, брюшная сторона белая. Клюв черный, ноги красновато-
бурые.
Пуховой наряд. Очень похож на аналогичный наряд черных и белокрылых крачек,
но отличается меньшим количеством белого цвета на концах крыльев, едва намечен-
ным белым кольцом вокруг глаза, черным пятном на лбу, доходящим до основания
клюва, и белой нижней стороной тела (кроме темно-коричневого пятна в области груд-
ной клетки и охристых боков). Клюв темно-серый с серовато-телесным основанием,
ноги серовато-розовые, радужина темно-коричневая.
Гнездовой наряд. Напоминает окончательный зимний наряд. Лоб и уздечка с охрис-
тым налетом в свежем пере. Перед глазом черное пятно, черный цвет на кроющих уха
занимает большее пространство, чем у взрослых крачек в зимнем наряде. Перья спи-
ны, плечевые, третьестепенные маховые, внутренние верхние кроющие крыла и руле-
вые тускло-серые с отчетливой черноватой предвершинной полосой и узкой охристо-
беловатой каемкой. Оперение поясницы, надхвостья и остальных верхних кроющих
крыла тускло-серое со светлыми беловатоохристыми каемками, иногда с более серы-
ми предвершинными пятнами. Щеки и брюшная сторона белые. Первостепенные махо-
вые несколько темнее, чем у взрослых, серебристый оттенок развит слабо, вершины
буроватые. Клюв красновато-коричневый с черным кончиком, ноги красновато-корич-
невые, радужина темно-коричневая.
Первый зимний наряд. Как окончательный зимний, от которого с трудом отличает-
ся обношенными дистальными первостепенными маховыми, оставшимися от гнездо-
вого наряда.
В отношении последующих нарядов нет полной ясности (Cramp, 1985). По-види-
мому, некоторые птицы надевают после первого зимнего первый брачный наряд, не-
отличимый от взрослого. У других, возможно, первый зимний наряд переходит пря-
мо во второй зимний, как у черных и белокрылых крачек.
Линька (Cramp, 1985). Линька у белощекой крачки значительно более изменчивая
по срокам и скорости протекания, чем у других крачек, в большей степени увязана с
условиями обитания данной особи в данный момент. Большинство видов крачек,
279
Семейство Чайковые
Таблица 17. Белощекая крачка. Размеры особей (мм) (ЗИН, ЗМ МГУ)
и масса тела (г) (Бородулина, 1960)
Пол Длина крыла Длина клюва
п | lim среднее п lim | среднее
dd 25 218-243 Ch. h. hybridus 232 26 27,2-31,4 29,1
99 19 211-238 225 21 25,1-30,2 26,9
dd 1 228 Ch. h. javanicus 1 29,8
99 1 226 - 1 26,1 -
как правило, не завершают прерванную сезоном гнездования линьку первостепенных
маховых, а вновь начинают ее с XI пера. Белощекая крачка, однако, может как про-
должить прерванную серию смены маховых, так и начать ее заново. Линька в первый
зимний наряд полная, начинается сменой перьев передней части спины, плечевых, XI
первостепенного и 1-й пары рулевых иногда вскоре после подъема на крыло, однако
чаще в октябре — начале ноября. К ноябрю — январю, а иногда и к августу линька
завершается, однако остаются несмененными дистальные первостепенные и второ-
степенные. Повторная смена первостепенных начинается в январе—апреле, после за-
вершения первой линьки маховых. По-видимому, с февраля начинается линька в пер-
вый брачный наряд. Часть особей (вероятно, позже начавших линьку) линяют прямо
во второй зимний наряд.
У взрослых особей две линьки в году: частичная предбрачная и полная послебрач-
ная. Послебрачная начинается в июне—июле сменой оперения темени, уздечки, плече-
вых и перьев спины. Последовательность смены рулевых, как у черной крачки. Перво-
степенные начинают линять между концом июля — концом сентября; линька преры-
вается на период миграции на стадии новых V—XI и продолжается на зимовках. За-
вершается линька в большинстве случаев к концу ноября, иногда в середине октяб-
ря или в декабре—январе. Смена первостепенных заканчивается между январем —
началом марта, а в конце октября — начале февраля начинается их повторная смена.
Предбрачная линька (конец января — начало апреля) захватывает голову, тело, крою-
щие крыла, рулевые и внутренние первостепенные. Повторная смена первостепенных
прерывается в сезон размножения обычно на стадии новых V—VI. Третья смена перво-
степенных бывает лишь у 34% птиц, она стартует между концом декабря и началом
марта.
Распространение. Гнездовой ареал включает 5 изолированных участков
(Vaurie, 1965; Cramp, 1985): европейско-среднеазиатский (от Испании на западе до
примерно 67°-68° в.д. на востоке), восточноазиатский (от Байкала и оз. Ханка на се-
вере до Вьетнама на юге), африканский (южная и восточная Африка), индийский (до-
лина р. Ганг и нижнее течение р. Брахмапутры) и австралийский. Эти участки в основ-
ном соответствуют областям гнездования подвидов (рис. 66).
Номинальный подвид Ch. h. hybridus не образует сплошного ареала. Постоянные
поселения этого вида располагаются обычно в нескольких сотнях километров друг от
друга. В то же время для подвида (по-видимому, и для вида в целом) характерно об-
разование временных поселений, нередко вдали от основной области гнездования.
Этим белощекая крачка весьма напоминает малую чайку. В Западной Европе и Перед-
ней Азии постоянные поселения отмечены в Испании, на юге и в центральной части
280
Белощекая крачка
Длина цевки Масса тела
п lim среднее п Lim среднее
25 21,0-24,7 22,6 22 76-93 86
21 19,8-24,3 21,8 26 60-101 83
1 21,9 — — —
1 20,5 — — - —
Франции, на севере Италии, в Венгрии, Югославии, Румынии, Греции, Турции и Ираке.
В отдельные годы начиная с конца прошлого столетия отмечены гнездования в Пор-
тугалии, Бельгии, Нидерландах, ФРГ, Польше, Чехословакии, Австрии, в Марокко,
на севере Туниса, в Алжире (Cramp, 1985).
На территории СССР постоянные поселения крачек этого подвида существуют на ли-
манах восточного Приазовья, в том числе в Ахтарско-Гривенских плавнях (Бороду-
лина, 1960; Казаков, Языкова, 1982), в пойме Нижней Волги от Волгограда до дельты
(Воробьев, 1936; Луговой, 1958; Кривоносов, 1976; Николаев и др., 1977; Кубанцев,
Чернобай, 1982), в долине нижней Сырдарьи, примерно от широты г. Туркестана до
устья (Зарудный, 1916; Спангенберг, Фейгин, 1936; Долгушин, 1962). Без указаний
на находки гнезд отмечено гнездование в низовьях Амударьи (Кенжегулов, 1972),
в .низовьях Мургаба и по трассе Каракумского канала (Ташлиев и др., 1967). Вероят-
но, к постоянным можно отнести поселения в низовьях р. Атрек на юго-западе Турк-
мении и в окрестностях залива Кирова на юго-востоке Азербайджана (Дементьев и
др., 1955; Заблоцкий, Заблоцкая, 1963; Щербина, 1975; Караваев, 1981), а также по-
селение в низовьях Дуная, часть которого находится на территории СССР (Петрович,
1981; Жмуд, Балацкий, 1986) (рис. 67).
Помимо постоянных, белощекая крачка образует нерегулярные временные посе-
ления. В одних случаях их появление можно связать с обводнением прежде пересох-
ших водоемов, в других — причины возникновения новых колоний не ясны. Подоб-
ного типа поселения известны для низовий Днестра (Черничко, устное сообщ.), ниж-
него течения Днепра (Орлов, 1957; Булахов, 1968; Клестов, Осипова, 1981), север-
ной части Харьковской обл. (Лисецкий и др., 1978), оз. Маныч-Гудило, Веселовско-
го и Усть-Манычского водохранилищ (Кривенко, 1981; Казаков, Языкова, 1982),
Воронежского водохранилища (Семаго и др., 1974), низовий рек Урала и Эмбы, Ка-
мыш-Самарских озер, окрестностей г. Уральска, р. Хобды, низовий рек Иргиза и Тур-
гая, по-видимому для Зайсанской котловины и окрестностей Семипалатинска (Дол-
гушин, 1962; Дебело, Шевченко, 1986). Относительно двух последних случаев, по-
жалуй, можно говорить о дальних выселениях за пределы основной области гнездо-
вания. Как дальние выселения можно квалифицировать и появление колонии вели-
чиной чуть больше 10 пар на оз. Жувинтас в Литве в 1959 г. (Валюс и др., 1976), коло-
ний в Зборовском районе Тернопольской обл. в 1974 г. (Талпош, 1977), на прудах
большинства областей Западной Украины в 1983 г. и в Тернопольской обл. в 1985 г.
(данные И.М. Горбаня), на Каунасском водохранилице в 1978 г. (Мичикунас, 1981)
и Куйбышевском водохранилище в 1980 г. (Аюпов и др., 1983).
Гнездование в СССР подвида Ch. h. javanicus отмечается с 1963 г. на оз. Ханка (По-
281
282
Рис. 66. Область распространения белощекой крачки
а — постоянные гнездовые поселения (пунктиром показана неуточненная граница), б — временные поселения, в — предполагаемые места гнез-
дования. г — места зимовок, д— залеты. Подвиды: 1 Ch.h. hybridus. 2 — Ch. h. javantcus, 3 — Ch. h. sclateri, 4 — Ch. h. fluviatilis
283
Рис. 67. Ареал белощекой крачки в СССР
1 — постоянные гнездовые поселения (пунктиром обозначена неуточненная граница), 2 — временные поселения, 3 — предполагаемые
места гнездования, 4— граница области наиболее частого гнездования, 5— залеты
Семейство Чайковые
Ливанова, 1971; Глущенко, 19846). По-видимому, этот же подвид ныне гнездится на
Байкале в дельте р. Селенги. Первая колония здесь найдена в 1977 г. (Мельников,
1979). Нерегулярное гнездование отмечено на Торейских озерах (Шкатулова, 1972;
Васильченко, 1986). Южнее пос. Архара Амурской обл. С.М. Смиренский встречал
беспокоящихся белощеких крачек и птиц, летающих с кормом в колонию белокры-
лых крачек.
Зимовки. Крачки подвида Ch. h. hybridus из европейской части СССР, очевидно,
зимуют в Восточной Африке от Судана до Кении, в Ираке, Иране и Пакистане, а также
в дельте Нила; птицы из Средней Азии и Прикаспия, — по-видимому, в Иране и Па-
кистане, возможно достигают Индии, Шри-Ланки и Восточной Африки. Небольшое
число птиц отмечено на зимовке у берегов Румынии, изредка в Турции (Cramp, 1985).
В мягкие зимы отдельные особи и стайки задерживаются до середины декабря на
юго-восточном Каспии (Дементьев и др., 1955) и на восточном побережье Азовского
моря (ЦКАН). Подвид Ch. h. javanicus, очевидно, зимует в Индокитае и на островах
Юго-Восточной Азии.
Миграции. Белощекая крачка покидает места зимовок, по-видимому, в марте
(Cramp, 1985). Весенний пролет в СССР, вероятно, нигде не бывает значительным.
Он отмечен на Черноморском побережье Кавказа (Вронский, Томкович, 1975), на
юго-западном Каспии с 15.IV до конца этого месяца (Заболоцкий, Заболоцкая, 1963),
на юго-восточном Каспии во второй половине апреля (Дементьев и др., 1955). По-ви-
димому, эти птицы летят с иранских и иракских зимовок, достигая восточного При-
азовья в конце апреля (пролет идет здесь весь май), дельты Волги и Урала в начале
мая (Долгушин, 1962; Казаков, Языкова, 1982). Дунайские, днестровские и днепро-
вские крачки мигрируют, видимо, западным берегом Черного моря. Об этом говорит
полное отсутствие белощеких крачек на весеннем пролете в Крыму (Костин, 1983).
На нижнем Днепре первые птицы появляются 3—11.V (Орлов, 1957).
В Среднюю Азию белощекие крачки летят, видимо, с пакистанских и индийских
зимовок долинами Мургаба, где пролет проходит в апреле (Эминов, 1978), долинами
Амударьи, Сурхандарьи, где мигрирующие особи отмечались 20.IV—14.V (Остапенко
и др., 1978). Птицы могут пересекать и обширные безводные пространства; есть све-
дения о транссахарских перелетах (Cramp, 1985). Крачки с оз. Байкал и оз. Ханка про-
летают, видимо, через восточную часть Китая и появляются в окрестностях гнездовий
в конце второй — середине третьей декады мая (Поливанова, 1971; Мельников, 1979;
Леонович, устное сообщ.). Размер весенних стай — 30—80 особей или несколько круп-
нее (Заблоцкий, Заблоцкая, 1963; Остапенко и др., 1978).
Послегнездовые кочевки начинаются в августе, затем они постепенно переходят
в не всегда выраженный осенний пролет. Крачки, окольцованные молодыми в начале
августа в дельте Волги, в начале сентября того же года отмечались как в Волгоград-
ской обл., так и в Марийской обл. Туркмении. В то же время крачка, окольцованная
птенцом 2.VII 1959 г. в дельте р. Атрек, встречена 13.VIII 1961 г. в окрестностях г. Геор-
гиевска Ставропольского края. Все это указывает на широкие послегнездовые ко-
чевки вида. В августе—сентябре проходит пролет вдоль побережья Болгарии (Нанки-
нов, 1980). На востоке Приазовья осенние миграции начинаются в конце августа и про-
должаются весь сентябрь, отдельные птицы задерживаются до конца октября — первых
чисел ноября (Казаков, Языкова, 1982). В конце августа — сентябре проходит пролет
на юго-западном и юго-восточном Каспии; Е.Л. Шестоперовым отмечен осенний пролет
по Узбою (Дементьев и др., 1955; Заблоцкий, Заблоцкая, 1963). Во второй полови-
не августа слабо выраженный пролет проходит на Аральском море (Долгушин, 1962).
По-видимому, некоторые среднеазиатские крачки пролетают во второй половине сен-
284
Белощекая крачка
тября через Ладакх и Западный Тибет (Vaurie, 1972). С оз. Ханка птицы исчезают
в августе (Поливанова, 1971).
Годовалые птицы, по-видимому, остаются летом в районе зимовок (Cramp, 1985).
Залеты белощеких крачек отмечены в Великобританию, Ирландию, Люксембург, Да-
нию, Норвегию, ГДР, на острова Мадейра и Барбадос (Cramp, 1985), в Псковскую
(Зарудный, 1910), Новгородскую (Бианки, 1915), Московскую (Харитонов, 1986)
и Новосибирскую (Рузский, 1932) области.
Численность. В советской части дельты Дуная найдена колония до 100 пар (Петро-
вич, 1981), в дельте Волги располагаются колонии до тысячи и более пар (Маркузе,
1975; Кривоносов, 1976), на юго-западном Каспии в 1950-х годах гнездилось более
500 пар (Заблоцкий, Заблоцкая, 1963). В Казахстане численность достаточно высо-
ка и стабильна только на Сырдарье (Долгушин, 1962). В низовьях Атрека гнездится
до 5 тыс. пар (Караваев, 1981), на оз. Ханка, — вероятно не менее 500 пар (Полива-
нова, 1971), в дельте Селенги численность возрастала до 1979 г. (800 особей) затем
резко упала (Мельников, 1984). Всего в СССР гнездится, по-видимому, несколько
десятков тысяч пар; общая численность мировой популяции неизвестна.
Местообитания. В сезон гнездования — стоячие и слаботекучие равнинные водоемы
степной и пустынной зон, густо заросшие водной и околоводной растительностью,
(клубнекамышом, рогозом, ситником, водяной лилией) и имеющие небольшие плесы:
относительно крупные озера, водохранилища, заводи в дельтах крупных рек и т.д.
Предпочитают прозрачную воду (Спангенберг, Фейгин, 1936, Бородулина, 1960; Заб-
лоцкий, Заблоцкая, 1963; Маркузе, 1975; Мачикунас, 1982). На пролете и зимовках —
внутренние водоемы, морские побережья.
Размножение. Сроки достижения половой зрелости неизвестны, по-видимому не ра-
нее двухлетнего возраста. Моногам. Диффузно гнездящийся факультативно-колониаль-
ный вид. Характерны резкие изменения количества гнездящихся пар в поселениях по
годам, вплоть до негнездования в отдельные годы (Караваев, 1981; Мельников, 1984).
Есть данные о перераспределении гнездящихся пар между колониями, удаленными на
расстояние около 2 тыс. км: окольцованная в Испании крачка встречена через 2 года в
мае в Венгрии (Schmidt, 1978, цит. по: Cramp, 1985).
Демонстрационное поведение сходно с таким же поведением черной крачки, однако
в ’’высотном полете” участвует обычно меньше птиц — до 5 особей, а ’’низкий полет”
менее развит (Cramp, 1985). Видоспецифические особенности демонстраций разобра-
ны в специальных работах (Swift, 1960; Szekely, 1984). Белощекие крачки гнездятся
как моновидовыми колониями (видимо, чаще всего), так и совместно с речными
крачками (Орлов, 1957), черными крачками (Долгушин, 1962; Аюпов и др., 1983),
черными крачками и черношейными поганками (Талпош, 1977), озерными чайками
(Поливанова, 1971), белокрылыми крачками (Мельников, 1979), белокрылыми крач-
ками и черношейными поганками (Долгушин, 1962). Величина колонии варьирует от
нескольких пар до нескольких сот пар, в отдельных случаях — до тысячи гнезд (такие
колонии образуются в результате постоянного подселения все новых пар); обычно
10—30 пар. В колониях гнезда часто располагаются отдельными группами от 2—4 до
6—10 гнезд (Орлов, 1957; Бородулина, 1960; Заблоцкий, Заблоцкая, 1963; Поливано-
ва, 1971; Маркузе, 1975; Глущенко, 19846). Плотность гнездования белощеких кра-
чек, по-видимому, несколько выше, чем у черных крачек. Так, в дельте Волги между
гнездами было обычно 1,5—2 м (Бородулина, 1960), в низовьях Атрека — 1—3 м (Ка-
раваев, 1979), в центре колонии на оз. Ханка гнезда располагались на расстоянии около
1 м друг от друга, на периферии — 2—5 м (Поливанова, 1971). В пойме Днепра между
гнездами отмечалось от 3 до 10-12 м (Орлов, 1957). В Западной Европе средние рас-
285
Семейство Чайковые
стояния между ближайшими соседними гнездами в разных колониях варьируют от
2,2 до 8 м и более (Cramp, 1985) .
Колонии располагаются на заросших водной растительностью водоемах. Гнезда час-
то помещаются на плавающих листьях кувшинок, нимфейника, рдестов и т.д., среди
редкого рогоза, ежеголовника, камыша, на сплавинах среди жесткой надводной рас-
тительности. На юго-востоке Азербайджана нередки плавучие гнезда, в низовьях Атре-
ка отдельные гнезда располагаются на старых гнездах чомг, на оз. Ханка отмечены гнез-
да крачек на пустых гнездах озерных чаек и на поросшей редким травостоем илистой
отмели. Гнезда белощеких крачек нередко представляют собой более массивную по-
стройку, чем у черных и белокрылых крачек. Диаметр гнезда (и = 40) равен 300—
640 мм, в среднем 430, диаметр лотка 80—200, в среднем 120, глубина лотка 15—30,
в среднем 20; общая высота гнезда 100—260, в среднем 150; высота над водой 25—50,
в среднем 30 мм (дельта Волги). В качестве материала для гнезда используются от-
мершие и зеленые стебли и листья водных и околоводных растений. Лоток гнезда су-
хой (Луговой, 1958; Бородулина, 1960, Заблоцкий, Заблоцкая, 1963; Поливано-
ва, 1971, Караваев, 1979, Глущенко, 19846).
Первые яйца в дельте Волги и Предкавказья появляются в конце мая (Бородулина,
1960), в низовьях Днепра — 9—10.V1 (Орлов, 1957), на юго-востоке Азербайджана -
23—25.V (Заблоцкий, Заблоцкая, 1963), в низовьях Сырдарьи — примерно 10.V1
(Спангенберг, Фейгин, 1936). Слабо насиженные яйца в Тернопольской обл. встречены
19.VI (Талпош, 1977). На оз. Ханка в 1964 г. кладка с 3 яйцами отмечена 31 .V (Леоно-
вич, устное сообщ,). В дельте Селенги в 1977 г. первые яйца были отложены 10—12.V1
(Мельников, 1979). Полная кладка содержит от 1 до 5 яиц, чаще всего 3 яйца, несколь-
ко реже 2. Из-за частой гибели гнезд характерен очень растянутый период откладки
яиц; свежие яйца отмечаются в ряде случаев до конца июля (Воробьев, 1936).
Окраска яиц сложная. Фон обычно светлее, чем у других болотных крачек: зелено-
ватый, сероватый, реже охристо-буроватый: пестрин также несколько меньше (Дол-
гушин, 1962). На оз. Ханка яйца, как правило, светло-зеленого цвета с глубинными
серыми и поверхностными темно-коричневыми крапинами, сгущающимися к тупому
концу. Основной фон яйца здесь может варьировать от светло-зеленого до густо-темно-
зеленого цвета, иногда он оливково-охристый, бурый или серый. Крапинки чаще всего
крупные. Цвет и форма яиц нередко варьируют даже в одном гнезде (Поливанова,
1971). Размеры яиц (мм) : из Предкавказья (и = 123) 34,8—44,3 X 26,2—32,3, в сред-
нем 38,7 X 28,3; из дельты Волги (и = 93) 34,7—41,0 X 26,7—29,8, в среднем 38,3 X
X 28,1; из юго-восточной части Азербайджана (п = 73) 35—42 X 25—30, в среднем 36,9 X
X 28,7: из юго-западной части Туркмении (и = 78) 35,1-43 X 25,7-29,6, в среднем
38,3 X 27,8; с оз. Байкал (и = 20) 35,9—43,5 X 26,7—30,0, в среднем 39,3 X 28,4: с оз.
Ханка (п = 16) 28,9—41,7 X 26,0—35,0, в среднем 38,6 X 27,2. Масса (г) свежих яиц
из юго-западной части Туркмении (и = 9) 14,2-17, в среднем 15,6 (Луговой, 1958:
Заблоцкий, Заблоцкая, 1963; Поливанова, 1971, Караваев, 1979; Мельников, 1979^
Казаков, Языкова, 1982). Длительность насиживания 18—20 дней. В насиживании
участвуют оба партнера, птицы сменяются на гнезде через 10—40 мин. Вес только что вы-
лупившегося птенца равен 10—10,7 г. Пеньки маховых появляются у трехдневных птен-
цов, рулевых — у пятидневных. Подлетывать птенцы начинают в возрасте 20 дней, пол-
ностью лётными становятся на 23—25-й день (Орлов, 1957: Заблоцкий, Заблоцкая.
1963).
Гибель гнезд белощеких крачек часто бывает очень значительной, главным образом
от подъема воды вследствие сильных ветров, дождя или паводков, а также в резуль-
тате падения уровня воды и из-за штормов. Гибнет от 35% до 100% кладок (Заблоц-
кий, Заблоцкая, 1963; Поливанова, 1971; Мельников, 1979; Караваев, 1981).
286
Чайконосая крачка
Максимально известный возраст, по данным кольцевания, 7 лет 5 месяцев (ЦКАН).
Суточная активность, поведение. Активность, по-видимому, дневная, хотя точные
данные отсутствуют. В меньшей степени общественная птица, чем белокрылая крачка;
крупных стай и ночевочных скоплений не образует (Cramp, 1985).
Питание. Среди болотных крачек белощекие наиболее всеядны, соотношение кормов
зависит от условий обитания. В дельте Волги вдали от рыбхозов в питании преоблада-
ли наземные насекомые, особенно жужелицы, кобылки и сверчки, а из водных форм —
личинки плавунцов. Лягушки и рыба были встречены тойько у 13,5% добытых птиц.
На рыбхозах дельты Волги лягушата и головастики встречены у 92,7% добытых птиц,
рыба — у 48,0%, беспозвоночные — у 70,4%; по весу эти корма составляли соответст-
венно 68,3%, 20,4%, 11,3% (Бородулина, 1960). На юго-востоке Азербайджана со време-
ни прилета до последней трети мая пища представлена исключительно насекомыми
(чаще всего личинками стрекоз), с конца мая крачки преимущественно питаются
мальками рыб (чаще всего сазана и атеринки), в июле—августе значительную долю кор-
ма составляют лягушки (Заблоцкий, Заблоцкая, 1963). В Предкавказье в питании бе-
лощеких крачек отмечена рыба, лягушки, личинки плавунцов, стрекоз и наземные
беспозвоночные (Казаков, Языкова, 1982). На оз. Ханка кормовыми объектами слу-
жат мелкие бычки, бокоплавы, стрекозы и личинки плавунцов (Поливанова, 1971).
Способы добывания корма более разнообразны, чем у других болотных крачек;
белощекие крачки чаще кормятся над сушей. Птицы собирают корм, летая над степью,
сырым лугом или водными растениями на высоте до 0,5 м, время от времени снижа-
ясь и схватывая насекомых. Нередко ныряют в воду с высоты около 2,5 м, как реч-
ные крачки. Способны лосить летающих насекомых (Бородулина, 1960).
Враги, неблагоприятные факторы. Наибольшее отрицательное воздействие на коло-
нию оказывают абиотические факторы: изменение уровня воды, сильные ветры, за-
тяжное ненастье. Нередко крачки бросают гнезда из-за беспокойства людьми (Маркузе,
1975). Из хищников наибольшее воздействие оказывает, по-видимому, болотный лунь
(Swift, 1960). Наблюдалось разорение гнезд лисицами и шакалами после высыхания
водоема. Отмечено поедание птенцов болотной черепахой (Караваев, 1979).
Хозяйственное значение, охрана. Хозяйственного значения не имеет. Специальные
меры охраны не предусмотрены,
Род Gelochelidon C.L. Brehm, 1830
Чайконосая крачка —
Gelochelidon nilotica (Gmelin, 1789)
Gill-billed Tern (англ.), Lachseeschwalbe (нем.), Sterne hansel (фр).
Син. (лат,) : Sterna nilotica Gmelin, 1789; Gelochelidon anglica (Montagu, 1813).
Син. (рус) : черноносая крачка.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Относительно крупная (крупнее реч-
ной) крачка. От других крачек отличается размерами, от пестроносой — более корот-
ким и массивным черным клювом, более коротким хвостом с неглубокой вырезкой,
несколько более длинными и широкими крыльями, а также отсутствием хохолка.
Цевка относительно длинная, поэтому на земле птица выглядит выше в ногах, чем
другие крачки. По земле ходит мало, хотя чувствует здесь себя более уверенно, чем
болотные или речные крачки. На воду в норме не садится, ныряет за добычей крайне
редко. Голос разнообразный, более мелодичный, чем у других крачек; в нем совер-
шенно отсутствуют скрипучие и каркающие звуки. В сезон размножения чаще всего
287
Семейство Чайковые
Таблица 18. Чайконосая крачка. Размеры особей (мм)
Географический район Пол Длина крыла
п lim Среднее
Центральная и Южная Европа, d3 35 309-341 326
Персидский залив(Сгатр, 1985) 99 24 307-333 319
Северное Причерноморье (ЗМ МГУ) 33 5 318-335 325
99 5 305-338 315
Каспий, Средняя Азия, Казахстан 33 19 302-325 315
(ЗМ МГУ) 99 11 299-320 312
можно слышать двухсложные крики ”че-век, че-век”, ”че-кво,че-кво”. С этими кри-
ками чайконосые крачки возвращаются в колонию с кормом, их можно слышать во
время формирования пар и просто от пролетающих стаек. Крик тревоги — немного
гнусавая трелька ”кэ-ве-ве”. В агрессивных столкновениях с особями своего вида
употребляется сухая стрекочущая трелька, с подобным стрекотом птицы преследуют
серебристых чаек и других пернатых врагов.
Подвидовая систематика. Различают обычно шесть подвидов (Vaurie, 1965; Cramp,
1985). Изменчивость проявляется в окраске оперения верхней стороны тела и в раз-
мерах клюва. В СССР один подвид (Степанян, 1975) ;
Gelochelidon nilotica nilotica
Sterna nilotica Gmelin, 1789, Syst. Nat., 1, c. 606, Египет.
Верхняя сторона тела, надхвостье и рулевые темнее, а клюв длиннее и тоньше, чем у
восточнокитайского подвида G.n. affinis.
Описание. Строение и размеры (табл. 18).
Масса тела (г) (Бородулина, 1960; Долгушин, 1962; Заблоцкий, Заблоцкая, 1963):
33 (п = 39) -- 130—261 (среднее 211);
99 (и = 15)— 172—260 (среднее 210).
Окраска (Дементьев, 1951; Cramp, 1985). Самец и самка в брачном наряде.
Лоб, верхняя часть уздечки, темя, затылок и верхняя часть задней стороны шеи черные:
граница черного цвета проходит по нижнему краю глаза. Спина, плечевые, третьесте-
пенные маховые и верхние кроющие крыла светлые, сизо-серые. Поясница, верхние
кроющие хвоста и рулевые такого же цвета или слегка светлее (особенно крайние
рулевые) , но никогда не бывают белого цвета, контрастирующего со спиной и крылья-
ми, как у пестроносой крачки. Подбородок, щеки, горло и нижняя часть туловища
белые. Наружные опахала первостепенных маховых, кроющих первостепенных и все
второстепенные маховые серые, слегка светлее, чем оперение спины, с серебристым
налетом по свежему перу; ношенные первостепенные постепенно темнеют, становясь
к осени почти тускло-черными. Внутренние первостепенные и все второстепенные с
белыми вершинами. Внутренние опахала первостепенных маховых светло-серые с ши-
роким белым полем по краю. Клюв и ноги черные, радужина темно-коричневая.
Самец и самка в зимнем наряде. Голова белая, на темени и затылке с сероватым
оттенком и нерезкими тонкими черноватыми штрихами. Перед глазом темно-серые
или черноватое пятно, такого же цвета пятно за глазом и кроющие уха. Остальное опе-
рение, как в брачном наряде, но спинная сторона обычно несколько светлее.
Пуховой наряд (живые птенцы с Сиваша). Пух довольно длинный и мягкий. Окрас-
ка чрезвычайно изменчива. Спинная сторона варьирует от желтовато-серого до тем-
288
Чайконосая крачка
i Длина клюва Длина цевки
* п lim среднее п Кт среднее
36 38-42 39,8 34 33-38 34,8
28 35-40 37,7 27 31-35 33,4
5 37,8-40,7 39,2 5 35,2-36,6 36,0
5 34,4-38,8 36,1 5 32,3-37,2 34,5
21 34,5-40,8 37,3 20 30,2-37,8 34,3
10 33,4-36,3 35,0 11 31,3-36,3 33,9
ного, коричневато- и буровато-серого цвета и от светлого, почти белого, до серовато-
бежевого разных оттенков. На темени, затылке, шее, спине, пояснице и крыльях чер-
ные пятна, величина и густота которых сильно варьирует: нередко они формируют
неясные полоски. У некоторых птенцов пятен практически нет, причем это могут быть
как очень светлые, так и темные пуховички. Подбородок и горло обычно серого цве-
та разной интенсивности с белым пятном под клювом, реже почти черные, еще реже
белые; как правило, у более светлых особей они светлее. Брюшко белое. Клюв и ноги
серые, телесно-серые, серовато-розовые или серовато-оранжевые.
Гнездовой наряд. Лоб беловатый, темя и затылок от беловатого до светло-серого,
нередко с охристым налетом, у некоторых особей с мелкими дымчатыми продольными
пестринами, всегда имеющимися на кроющих уха. Перед глазом и за ним небольшие
серые пятна. Задняя часть шеи серая, перья спины, плечевые, кроющие крыла серые
с охристыми вершинами: интенсивность окраски их индивидуально варьирует. На зад-
них плечевых, а иногда и на других перьях верхней стороны тела буроватые предвер-
шинные пятна. Нижняя часть тела белая. Первостепенные маховые окрашены, как
у взрослых, но в общем темнее, с менее выраженным серебристым налетом и со сла-
бо развитыми светлыми полями по заднему краю внутренних опахал. Внутренние
первостепенные с беловатыми вершинами. Клюв светло-оранжевый с темной верши-
ной, осенью постепенно чернеющий. Ноги розовато-бурые или красновато-серые. Раду-
жина серовато-коричневая или темно-коричневая.
Первый зимний наряд. Как зимний наряд взрослых, но дистальные первостепен-
ные и их кроющие, некоторые второстепенные и часто рулевые и третьестепенные
сохраняются до весны второго календарного года. Окончательный брачный наряд
к лету второго календарного года не приобретается: темя и затылок в это время года
могут быть с крупными черными пестринами.
Линька (Cramp, 1985). Линька в первый зимний наряд полная, начинается в конце
августа — конце сентября сменой оперения головы. К декабрю—февралю все оперение
головы, туловища, большинство кроющих крыла и третьестепенных маховых свежие.
Рулевые линяют в октябре—феврале, последними сменяются 4-я и 5-я пары. Смена
первостепенных начинается с XI между началом декабря и концом марта, их линька
часто задерживается в,начале лета и завершается во время линьки во второй зимний
наряд в июле — начале сентября, когда оперение головы и туловища первого зимнего
наряда замещается оперением второго зимнего наряда, а внутренние первостепенные
и рулевые начинают новую серию линьки. В дальнейшем бывает две линьки в году:
полная послебрачная и частичная предбрачная. Послебрачная начинается со смены са-
мых внутренних первостепенных в конце июня — середине августа; линька преры-
вается в период миграции на стадии новых VIII—XI. Смена оперения головы и туло-
10. Зак.826
289
290
Рис. 68. Область распространения чайконосой крачки
а — участки гнездового ареала и отдельные поселения, б — участки гнездового ареала с неуточненными границами, в — места зимовок,
г — залеты. Подвиды: 1 — G.n. nilotica, 2 — G.n. aranea, 3 — G.n. vanrossemi, 4 — G.n. gronvoldi, 5 — G.n. macrotarsa, 6 — G.n. affinis
Чайконосая крачка
вища начинается в конце июля — середине августа (обычно с оперения темени, а не
лба, как у многих других крачек) и завершается в октябре—декабре, линька руле-
вых — к ноябрю. Смена первостепенных маховых заканчивается в середине декабря -
конце января, у некоторых крачек — в феврале—марте. Предбрачная линька начина-
ется с внутренних первостепенных в период между концом декабря и концом февра-
ля, обычно после завершения послебрачной линьки дистальных первостепенных. Сме-
на первостепенных прерывается с января по март обычно на стадии новых IX—XI:
прерванная линька не продолжается в конце сезона размножения. Около трети евро-
пейских крачек вообще не сменяют первостепенных маховых весной. Рулевые ли-
няют в конце января — апреле, обычно полностью. Перья головы, нижней части туло-
вища, верхней части спины и плечевые сменяются с середины февраля по март.
Распространение. Гнездовой ареал. Заселяет все континенты, кроме Ан-
тарктиды, но, по-видимому, везде гнездится спорадически (Vaurie, 1965; Cramp, 1985).
В Палеарктике от Испании до Забайкалья обитает номинальный подвид G.n. nilotica;
он же гнездится в Иране, Пакистане, западной и северной Индии и, по-видимому, в
Монголии (Stubbe, Bolod, 1971). На юге и востоке Китая и в низовьях Ганга (АН,
Ripley, 1969) гнездится G.n. affinis. Отдельный подвид заселяет Австралию, три дру-
гих подвида — Северную и Южную Америку (рис. 68) .
Номинальный подвид в Западной и Южной Европе ныне гнездится в Испании, на
юге Франции, на северо-востоке и в южной части Италии, в Греции, Болгарии, Дании
и ФРГ. Отмечены также постоянные поселения в Мавритании, Алжире, Тунисе и Тур-
ции. В отдельные годы чайконосая крачка гнездилась в Великобритании, Нидерлан-
дах, Австрии, ГДР, Чехословакии, Венгрии, Португалии, Марокко, Сирии и Ираке
(Cramp, 1985). На территории СССР вид не образует сплошного ареала, а гнездится
сп 'радически отдельными участками и поселениями, удаленными друг от друга на
сотни километров (рис. 69). Область постоянного гнездования лежит между север-
ным Причерноморьем на западе и оз. Алаколь на востоке. В Причерноморье и Приазовье
крачки гнездятся только в приморской полосе, в Казахстане заселяют внутренние
водоемы на север примерно до 51°—52° с.ш. Изолированный участок ареала находит-
ся в Забайкалье, где ныне чайконосные крачки гнездятся на Торейских озерах (Зуба-
кин, 1981а). Недавно гнездовое поселение этого вида обнаружено в Южной Туве (Ба-
ранов, 1981).
Поселения чайконосых крачек нередко располагаются относительно близко друг
к другу — в нескольких десятках километров, формируя пространственно изоли-
рованные участки ареала. Не исключено, что по крайней мере часть таких группиро-
вок представляют собой отдельные популяции. Несмотря на дальние выселения от-
дельных пар и репродуктивных групп за пределы области гнездования вида (см. ниже),
доказательства обмена гнездящимися особями между такими участками ареала от-
сутствуют.
Существует несколько пространственно-изолированных участков ареала чайконосых
крачек. Два из них — в бассейне р. Илек и в окрестностях Челкар-Тенгиза (включая
низовья рек Иргиза и Тургая) — выделил еще П.П. Сушкин (1908). Можно выделить
также азово-черноморский участок, включающий поселения в Тилигульском лимане
(Черничко, Стойловский, 1981); в Тендровском и Ягорлыцком заливах Черномор-
ского заповедника, на о-ве Каланчакском (Ардамацкая, 1975а, в, 1984), в Картка-
зацком заливе (Костин, 1975), на Сиваше (Зубакин, Костин, 1977; Сиохин, 1981а),
Молочном лимане (Филонов и др., 1974) и, возможно, поселения на Таманском полу-
острове (Кищинский, 19606), и в устье Дуная (Жмуд, Балацкий, 1986). В отдельные
участки ареала, очевидно, группируются поселения долины Маныча, включая озера
Калмыкии и, возможно, низовья Терека (Волчанецкий, 1959; Языкова, 1975; Кривен-
291
292
Рис. 69. Ареал чайкоиосой крачки в СССР
1 — участки гнездового ареала (пунктиром показаны неуточнениые граница) , 2 — отдельные поселения, 3 — отдельные нерегулярные
поселения, 4 — область нерегулярных зимовок, 5 — область регулярных кочевок, 6 — залеты
Чайконосая крачка
ко, 1981; Кривенко, Любаев, 1981; Любаев, Кривенко, 1981; Кукиш, 1986), поселе-
ния низовий Сырдарьи и северной части Аральского моря (Спангенберг, Фейгин, 1936;
Долгушин, 1962), среднего течения Сырдарьи, включая Кайраккумское водохранили-
ще (Долгушин, 1962; Абдусалямов, 1981), Амударьи от устья, возможно, до границ
Таджикистана (Гладков, 1932; Дементьев, 1952; Эминов, 19746, 1975). В Таджики-
стане гнездование чайконосых крачек по Амударье и Пянджу предполагается, но не
доказано (Иванов, 1969; Абдусалямов, 1971). По-видимому, еще одним участком
ареала ныне можно считать трассу Каракумского канала, где колонии найдены в ни-
зовьях Теджена, на Хаузханском водохранилище и в ряде других мест (Ташлиев и др.,
1967; Эминов,1975).
Отдельные поселения чайконосых крачек отмечены на Каспии; на Тюленьих (Ман-
гистауских) островах (Залетаев, 1960), островах в заливе Кара-Богаз-Гол (Щербина,
1979), островах Красноводского залива (Гаузер, 19836), о-ве Огурчинском (Щерби-
на и др., 1975), не ежегодно — в низовьях Атрека (Караваев, 1981). Гнездится эта
птица в Зеравшанской долине (Сагитов, Салимов, 1975), на Айдаркуле (Остапенко
и др., 1984), есть указание на гнездование по Сурхандарье (Остапенко и др., 1978).
На основании летних встреч предполагалось гнездование в Киргизии (Янушевич и др.,
1959), что требует проверки. Поселения чайконосых крачек есть в дельте Волги (Кри-
воносов, 1976) , на Камыш-Самарских озерах (Леонович, устное сообщ.) и по р.М. Узень
(Дебело, Шевченко, 1986), по рекам Иргизу, Тургаю, Чу, Или, Талассу, в низовьях
р. Сарысу, на Балхаше, на озерах между Карагандой и Баян-Аулом (Долгушин, 1962),
на Алаколе (Ауэзов, Грачев, 1975). Севернее чайконосая крачка гнездится уже не-
регулярно: на Соркуле у пос. Джаныбек (Линдеман, Залетаев, 1977), озерах Наурзум-
ского заповедника (Чельцов-Бебутов, 1958); на оз. Тенгиз (Чекменев, 1964; Азаров,
1975а; Волков, 1975) колонии и гнездящиеся пары отмечаются не каждый год. В 1975 г.
гнездо чайконосой крачки найдено на оз. Чаны (Юрлов и др., 1977; Ходков, 1980).
Это, по-видимому, один из случаев нерегулярного выселения крачек за пределы об-
ласти гнездования. Видимо, подобное выселение произошло и в 1952 г. в район юж-
ной Кулунды (Юрлов, 1959).
Зимовки. По данным ЦКАН, крачки из северного Причерноморья зимуют на
побережье Западной Африки (Сенегал), в долине р. Нигер (Мали) и на Черноморском
побережье Кавказа. Предполагаются также зимовки птиц этой популяции в верхнем
течении Нила, Эфиопии и на озерах Восточной Африки (Cramp, 1985). Каспийские
крачки, возможно, зимуют в Персидском заливе, на побережье Ирана, Пакистана и в
Индии. Здесь же, очевидно, зимуют казахстанские и среднеазиатские птицы. Места зи-
мовок забайкальских крачек не известны, скорее всего это Юго-Восточная Азия.
Миграции. По данным ЦКАН, у птиц, окольцованных на севере Причерноморья,
в первую осень жизни существует четкая направленность перемещений в западном,
а затем и юго-западном и южном направлениях. В середине августа часть птиц дости-
гает Румынии, в начале сентября — Испании, другие в конце сентября еще встречают-
ся в Одесской обл. В декабре крачки отмечаются на местах зимОвок. Однако у птиц
в возрасте двух и более лет столь же четко в августе—сентябре преобладает восточное
направление пролета: они отмечаются в Крыму и Восточном Приазовье, лишь неко-
торые особи — в Одесской обл. Перемещения к местам зимовок у более старых птиц
начинаются несколько позже, чем у молодых.
Места зимовок птицы покидают, видимо, в марте. Первые крачки на севере Кры-
ма отмечены 27.III, с начала апреля они уже обычны в восточной части Каркинитского
залива и Присивашье (Костин, 1983). В Черноморском заповеднике и на островах
Маныча птицы регистрируются в первых числах апреля (Ардамацкая, 19776, Каза-
ков, Языкова, 1982). С 7.IV отмечается сильно растянутый пролет вдоль Черноморско-
293
Семейство Чайковые
го побережья Кавказа (Вронский, Томкович, 1975), в апреле чайконосая крачка про-
летает вдоль юго-западного побережья Каспия, с 26.III — начала апреля — вдоль юго-
восточного берега (Дементьев и др., 1955; Заблоцкий, Заблоцкая, 1963). В Средней
Азии пролет также проходит в апреле. На Сурхандарье в 1975 г. птицы отмечались
с 3.IV по 15.V, пролет был достаточно выраженным (Остапенко и др., 1978). На Ай-
даркуле первые птицы появляются в последних числах марта -- начале апреля, мас-
совый пролет идет в середине месяца (Остапенко и др., 1984; Калабин, 1986) . В Таджи-
кистане птицы летят по долинам рек Вахша и Яхсу с 31.Ill по 1 .V (Абдусалямов, Ле-
бедев, 1977, 1978; Абдусалямов и др., 1980). Незначительный весенний пролет отме-
чен на Памире (Абдусалямов, 1971). В Киргизии весной и осенью чайконосые крач-
ки встречаются в долинах рек Атбаши, Нарын, Кочкорка, а также на оз. Сонкёль, где
иногда летуют (Кыдыралиев, 1973). В Туркмении пролет идет по рекам Мургаб и
Амударья в апреле (Дементьев, 1952; Карташев, 1958; Эминов, 1978). В Казахста-
не на о-ве Барсакельмес первых птиц отмечали в конце марта, на нижней Сырдарье —
в середине апреля. На Эмбе, близ Темира, первые особи отмечены 18.IV, в низовьях
Илека пролет идет с конца апреля до середины мая (Долгушин, 1962). В низовьях
Тургая первые птицы были встречены 14.IV, в Наурзуме — 25.IV—3.V (Елкин, 1975:
Ауэзов, Хроков, 1977).
Осенний пролет относительно поздний, не всегда выраженный, проходит в боль-
шинстве случаев через те же пункты, что и весенний. Заметный пролет отмечается
у устья Сырдарьи 10—12.VIII, последние особи в Казахстане встречаются до конца
октября (Долгушин, 1962). На Айдаркуле наиболее интенсивный пролет идет во вто-
рой половине сентября, последняя птица встречена 5.Х (Остапенко и др., 1984). Сла-
бый осенний пролет, видимо, проходит через Памир: 11.VIII 1977 г. на оз. Булункуль
встречена стайка из 7 взрослых и 4 молодых птиц (наши данные).
На юго-западе Каспия осенний пролет отмечен в августе—сентябре (Заблоцкий,
Заблоцкая, 1963). В Присивашье и на севере Крыма крачки обычны весь сентябрь,
реже встречаются в октябре; самая поздняя осенняя встреча — 22.XI (Костин, 1983).
Чайконосые крачки летят одиночками и стайками по 15—36 особей. Годовалые
птицы проводят лето, видимо, в районах зимовки (Остапенко и др., 1978, 1984; Андру-
сенко, 1980; ЦКАН).
Залеты чайконосых крачек отмечались в Ирландию, Бельгию, Люксембург, Нор-
вегию, Швецию, Швейцарию, Польшу, Югославию, на Азорские острова (Cramp, 1985),
на юг Приморья, в окрестности Киева и Чернигова (Дементьев, 1951) и дельту Селен-
ги (Богородский, 1976).
Численность. На севере Причерноморья и в Приазовье ныне гнездится, очевидно,
2,2—3 тыс. пар; численность несколько снизилась с конца 1940-х годов, когда только
в Черноморском заповеднике гнездилось до 3,5 тыс. пар (Ардамацкая, 1975а, в; Сио-
хин, 1981а; Черничко, Стойловский, 1981; Костин, 1983). На водохранилищах Ма-
ныча и озерах Калмыкии гнездится не менее 3,5—5 тыс. пар (Языкова, 1975; Кри-
венко, 1981; Кривенко, Любаев, 1981; Любаев, Кривенко, 1981), на юго-восточном
Каспии — видимо, не более 100 пар (Щербина и др., 1975; Щербина, 1979). На оз. Сор-
куль Уральской обл. отмечалось до 120 гнездящихся пар (Линдеман, Залетаев, 1977),
на оз. Алаколь — около 2 тыс. пар (Ауэзов, Грачев, 1975), на оз. Тенгиз — до 15 пар
(Волков, 1975), на Хаузханском водохранилище — около 400 пар (Эминов, 1975),
на Кайраккумском водохранилище — 70—75 гнезд в 1966 г. и 110—115 в 1979 г. (Аб-
дусалямов, 1981). Многочисленна эта птица в низовьях Сырдарьи, на Аральском море,
в низовьях Тургая и Иргиза (Долгушин, 1962). На Торейских озерах найдена колония
в 248 гнезд (Зубакин, 1981а). Общая численность чайконосой крачки в СССР, по-ви-
димому, не превышает 30—40 тыс. гнездящихся пар; тенденции изменения численнос-
294
Чайконосая крачка
ти не ясны. В зарубежной Европе, Африке и Западной Азии предполагается гнездо-
вание 10 тыс. пар; численность резко уменьшилась с начала столетия (Miller,
1975).
Местообитания. В сезон гнездования на территории СССР — морские побережье,
заливы, лиманы; голые и слабо заросшие растительностью побережья равнинных со-
леных, солоноватоводных и изредка пресных внутренних водоемов, а также долины
крупных рек пустынной, полупустынной и реже степной зон. Кормовые биотопы —
Рис. 70. Демонстрационное поведение чайконосой крачки (по: Cramp, 1985, с изменениями)
а — агрессивная прямая поза, б — выпрямленная поза крачек-партнеров
прилегающие к водоемам участки степи и полупустыни, реже прибрежные мелко-
водья. На горных озерах, по-видимому, не гнездятся, хотя может отмечаться здесь
во время пролета и летних кочевок; постоянно кочующее из сводки в сводку утверж-
дение о гнездовании чайконосой крачки в Армении (Ляйстер, Соснин, 1942) лишено
фактических доказательств и по меньшей мере сомнительно (как и указания на гнездо-
вание здесь целого ряда других чайковых птиц). Вне сезона размножения — морские
побережья, внутренние водоемы в тропической зоне (Cramp, 1985).
Размножение. Чайконосые крачки приступают к размножению, по-видимому, не ра-
нее двухлетнего возраста; по наблюдениям в Дании, — не ранее пятилетнего возраста
(М011ег, 1975а, цит. по.: Cramp, 1985). Моногамы. Формирование пар по крайней мере
у части птиц происходит до прилета на места гнездования, пары сохраняются наизмен-
ными в течение ряда лет (Cramp, 1985). Диффузно гнездящийся факультативно-ко-
лониальный вид. Поселения нередко меняют свое месторасположение, величина их
сильно варьирует по годам.
Демонстрационное поведение чайконосой крачки характеризуется относительно
большим удельным весом наземных демонстраций, чем у многих других крачек. Пти-
цы интенсивно защищают территорию вокруг гнезда, при территориальных конфлик-
тах нередко принимают агрессивную прямую позу, в которой издается агрессивный
стрекочущий крик (рис. 70, а). Формирование пар включает как воздушные (’’вы-
сотный полет”, ’’низкий полет”), так и наземные демонстрации; среди последних
различают ’’выпрямленную” позу, при которой клюв может быть направлен горизон-
тально, вертикально вверх или наклонно вниз (рис. 70, б). В таких позах крачки-
партнеры ходят бок о бок и друг вокруг друга, расходятся и вновь сближаются. При
этом очень часто птицы поворачивают головы вправо-влево вдоль продольной оси
295
Семейство Чайковые
(Head-turning). Перед откладкой яиц самец нередко.кормит самку, в период наси-
живания это бывает гораздо реже (Lind, 1963; Cramp, 1985) .
Величина колоний варьирует от нескольких гнезд до нескольких сот гнезд. В зару-
бежной Европе колонии чаще всего состоят из нескольких пар и нескольких десят-
ков гнезд (Lind, 1963; М011ег, 1975, 1978b; Нанкинов, Даракчиев, 19806; Cramp,
1985). В СССР также достаточно обычны колонии до 20—50 пар (Бородулина, 1960;
Кищинский, 19606; Чекменев, 1964; Филонов и др., 1974), однако часто встречают-
ся и более крупные колонии — 50—200 гнезд; отмечались колонии до 500 и даже
1000 пар (Долгушин, 1962; Абдусалямов, 1971; Ауэзов, Грачев, 1975; Зубакин, Кос-
тин, 1977; Зубакин, 1981а). По-видимому, не составляет большой редкости и гнездо-
вание отдельными парами (Залетаев, 1960; Кищинский, 19606; Ходков, 1980; Лео-
нович, устное сообщ.). Гнездятся крачки чаще всего на островах, реже — на солонча-
ковых берегах водоемов. Предпочитают намывные песчаные и ракушковые острова-
косы, заросшие невысокой и негустой растительностью (обычно различными видами
солянок); здесь гнезда нередко располагаются на невысоких грядах — выше, чем у
речных и малых крачек. На материковых островах, наоборот, отмечено гнездование
в солончаковых понижениях. На реках гнездится исключительно на песчаных бере-
гах островов. Гнезда обычно располагаются среди невысокой растительности, реже
открыто. Чайконосая крачка часто поселяется вместе с пестроносой, речной и малой
крачками, чегравой, морским голубком, озерной чайкой, шилоклювкой, тиркушка-
ми, однако может образовывать и одновидовые колонии (Бородулина, 1960; Кищин-
ский, 19606; Долгушин, 1962; Зубакин, Костин, 1977; Казаков, Языкова, 1982) .
Гнездо — ямка в грунте, либо без выстилки, либо со скудной выстилкой из сухих
стеблей травянистых растений и выброшенного водой растительного мусора; в отно-
сительно сырых местах гнездовая постройка может быть более массивной. Диаметр
гнезда (мм) в Черноморском заповеднике равен 190—290 (в среднем 227), диаметр
лотка 120—160 (в среднем 134), глубина лотка 25—40 (в среднем 36); на Чонгарских
островах соответственно 225—320 (270; и = 21), 135—210 (179; л = 37), 15—40 (24;
п = 37). Гнезда в колониях Черноморского заповедника обычно располагаются на рас-
стоянии 1,5—2 м, минимальное отмеченное расстояние — 0,83 м; однако в конце 20-х го-
дов расстояние между гнездами здесь было 10—20 м (Кистяковский, 1967). На ли-
манах Таманского полуострова расстояние между гнездами составляло 1—20 м, обыч-
но 3—5 м. На Чонгарских островах между центрами ближайших гнезд было 0,7—5,3 м
(и = 161), в среднем 1,9 м, на Торейских озерах в среднем 1,22 м (и = 50); на Кайрак-
кумском водохранилище гнезда располагались в 0,2—0,8—1,5 м друг от друга (Боро-
дулина, 1960; Кищинский, 19606; Абдусалямов, 1971; наши данные). Индекс плот-
ности гнездования около 0,1.
Крачки появляются на территории колонии, по одним данным, в среднем за 19 дней
до откладки яиц; самцы устанавливают гнездовую территорию, которую затем жест-
ко охраняют оба партнера (М011ег, 1975а, цит. по: Camp, 1985). По другим данным,
охрана гнездовой территории начинается за 1 —2 дня до откладки первого яйца (Sears,
1978). Сроки начала откладки яиц в разных частях ареала на территории нашей стра-
ны довольно близкие: конец апреля — начало второй декады мая (Бородулина, 1960:
Долгушин, 1962; Филонов и др., 1974; Ардамацкая, 19776; Зубакин, Костин, 1977:
Щербина, 1979; Казаков, Языкова, 1982). Сообщение о находке 30.IV колонии, в ко-
торой, помимо кладок, были подросшие птенцы (Спангенберг, Фейгин, 1936), — ско-
рее всего, опечатка. Почти все исследователи отмечают крайне растянутый период
яйцекладки — до конца июня, а иногда и середины июля.
В кладке 1—6 яиц, чаще всего 2—3 яйца. Крайне редко в одном гнезде отмечают-
ся кладки нескольких птиц: гнездо с 12 яйцами найдено на Молочном лимане (Фило-
296
Чайкоиосая крачка
нов и др., 1974), с 17 яйцами — на нижней Сырдарье (Спангенберг, Фейгин, 1936); в
последнем случае кладку насиживали несколько птиц. Окраска яйца сложная. Основ-
ной тон ее желтовато-охристый, в некоторых случаях с зеленоватым или краснова-
тым оттенком. Немногочисленные глубокие пятна фиолетово-серые, поверхностные
пятна буроватые, блеклые, нерезко очерченные; их также немного, обычно несколь-
ко больше на тупом конце (Долгушин, 1962). Яйца чайконосых крачек отличаются
от яиц других видов слабой грушевидностью и слабой удлиненностью (Костин, 1983).
Размеры яиц (мм): с Молочного лимана (и = 44) 45,5—53,0 X 31,5—36,8, в среднем
48,8 X 34,8; из Предкавказья (и = 110) 43,2—52,0 X 33,0—39,9, в среднем 47,6 X 34,9;
из Казахстана (и = 29) 42,1—51,1 X 30,1—36,3, в среднем 48,0X 33,8; с Хаузханского
водохранилища (и = 104) 44,0—53,0 X 34,0—37,5, в среднем 47,6 X 34,5 (Ташлиев и др.,
1967; Филонов и др., 1974; Казаков, Языкова, 1982; Ковшарь, Левин, 1982). Мас-
са (г) свежих и слабо насиженных яиц (и = 33) 25,0—33,2, в среднем 29,2 (Дементьев,
1951; данные В.В. Леоновича) .
Насиживание начинается с первого яйца и продолжается 20—23 дня (Дементьев, 1951;
Эминов, 19746; Cramp, 1985). Насиживают оба партнера. Смена партнеров в Черно-
морском заповеднике отмечается 3—7 раз в сутки, причем самка насиживала больше,
чем самец (Бородулина, 1960), в Туркмении — 4—5 раз за 12 ч (Эминов, 19746),
на Сиваше (и = 3) — 5—11 раз в сутки. Изредка при смене партнеров самец кормит
самку, после чего принимает ’’выпрямленную” позу с клювом, направленным вверх.
Свободный от насиживания партнер много времени проводит на охраняемой гнездо-
вой территории. От появления первой трещины на яйце до вылупления проходит 1-
4 сут (и = 61), чаще всего 2 и 3 сут (39,3% и 36,1% случаев соответственно); от появ-
ления отверстия в яйце до вылупления — от нескольких часов до 2 сут (« = 49) , чаще
всего сутки и меньше суток (53,1% и 38,8%). Вылупление птенцов в гнезде с 3 яйца-
ми длится 2—3 сут (наши данные) .
После вылупления птенцов родители уносят из гнезда скорлупу, но тенденция эта
выражена менее четко, чем у многих других крачек (Зубакин, 1976в; Sears, 1978).
Масса птенцов в день вылупления 24—29 г (Бородулина, 1960; Ташлиев и др., 1967).
По наблюдениям на Сиваше, птенцов начинают кормить через несколько часов после
вылупления и в течение первых суток жизни кормят 13—37 раз (и = 2). Смена птиц,
согревающих птенца, в это время происходит 6-15 раз в сутки, обогревает птенцов
в основном самка, кормит — самец. Маленьких птенцов родители кормят мелкими
насекомыми и мелкими морскими иглами (наши данные). По наблюдениям в Черно-
морском заповеднике, на одного 2—3-дневного птенца в сутки приходится 9—19 корм-
лений, в зависимости от размера кормовых объектов. Среди приносимого птенцам
корма в 1957 г. (п = 240) здесь на первом месте стояли креветки (30%), затем шли
ящерицы (14,3%), насекомые (7,4%), пауки (6,5%), рыбы (1,3%); остальные пище-
вые объекты остались неопределенными (Бородулина, I960). На Сиваше (Зубакин,
1976в) подросших птенцов в 1973—1974 гг. в 45,2% случаев кормили наземными
насекомыми, в 1,9% — многоножками и паукообразными, в 17,8% — рыбами, в 24,8% —
земноводными, в 3,8% — ящерицами, в 2,5% — птенцами воробьиных и чайковых птиц
(в том числе в 0,6% случаях — пуховичками чайконосых крачек), в 3,8% — мыше-
видными грызунами (и = 157).
По наблюдениям на Сиваше, первые дни выводок обычно находится в гнезде, по-
кидая его лишь во время тревоги. Когда старшему птенцу исполняется 3 (реже 2)
суток, взрослые птицы обычно уводят птенцов за границы охраняемой территории.
К этому времени родители начинают отличать своих птенцов от чужих и агрессивно
относиться к чужим птенцам (Зубакин, 1976в). Летными птенцы становятся в воз-
расте 28—35 дней (Дементьев, 1951; Эминов, 19746; Cramp, 1985).
297
Семейство Чайковые
Данных по успеху размножения мало. По наблюдениям на Сиваше, в 1973 г. птен-
цы вылупились из 78,6% яиц (п = 98), в 1974 г. — из 71,8% (и = 71), причем неопло-
дотворенные яйца и яйца с погибшими зародышами составили соответственно 6,1%
и 7,0%, яйца, брошенные после вылупления первых птенцов — 3,1% и 9,9%, яйца пол-
ностью брошенных кладок — 2,0% и 11,3%, яйца, расклеванные чайками, — 10,2% и
0% (наши данные). Из 37 вылупившихся в начале июня птенцов здесь осталось в жи-
вых к подъему на крыло лишь 12 (32,4%). Один птенец погиб от побоев взрослых
птиц, другой найден мертвым без видимых повреждений, остальные 23 исчезли, —
по-видимому, были съедены особями-каннибалами или погибли от хищников. Из
72 птенцов поздних выводков удалось выжить только 2, подавляющее большинство
остальных были съедены самими чайконосыми крачками (Зубакин, 1975в, 1976в).
В Дании поднимается на крыло 62,9% птенцов от числа отложенных яиц (М011ег, 1975а,
цит. по: Cramp, 1985).
Максимальная известная продолжительность жизни, по данным кольцевания, 15 лет
9 месяцев (Cramp, 1985).
Суточная активность, поведение. Активность в основном дневная, хотя в лунные
ночи жизнь в колонии полностью не замирает — неоднократно отмечались смены парт-
неров на гнездах. Свободные от насиживания птицы ночуют либо на гнездовом участ-
ке, либо вблизи колонии на голых днищах высохших луж и в солончаковых пониже-
ниях. Первые вылеты за кормом отмечены за 50—60 мин до восхода солнца (наши
данные). В меньшей степени общественная птица, чем большинство крачек. Крупных
стай не образует даже в период миграций и на зимовках — обычно до 30—40 особей
(Britton, Brown, 1974). Кормится в основном в одиночку (Erwin, 1978).
Питание. В отличие от других крачек трофически связана главным образом с су-
шей, причем основным кормом служат наземные насекомые. Кроме них, крачки ло-
вят ящериц, амфибий, мелких мышевидных грызунов, птенцов других чайковых,
куликов и воробьиных птиц (Мензбир, 1895; Спагенберг, Фейгин, 1936; Шевченко,
1937; Клименко, 1950а; Дементьев и др., 1955; Залетаев, 1960; Долгушин, 1962;
Эминов, 19746; Зубакин, 1975в; Самородов, 1976; М011ег, 1976; Сиохин, 1981а).
Рыба и водные беспозвоночные в питании занимают подчиненное положение, хотя
в ряде случаев, видимо, могут быть главным кормом (Деревягин, 1948; Dragesco,
1961; Абдусалямов, 1971).
В Черноморском заповеднике основным кормом, по результатам анализа 76 же-
лудков и погадок, служат насекомые (жуки, крупные стрекозы и медведки) и яще-
рицы (63,2% и 34,1% встреч соответственно), реже креветки (14,5%) и мышевидные
грызуны (13,3%), еще реже рыбы, птенцы и яйца птиц (5,2%, 1,3%, 1,3%). На севере
Крыма основу питания также составляли насекомые (певчие цикады — 28% встреч,
хлебные жуки — 27%, кукурузные навозники — 27%), ящерицы встречались реже (12%),
рыбы и морские беспозвоночные не встречены вовсе. На кубанских рыбхозах основ-
ной корм — лягушки (Бородулина, 1960). На Сиваше наземные насекомые встрече-
ны в 95,8% из 97 погадок, паукообразные — в 3,2%, рыбы — в 2,2%, земноводные —
в 3,2%, ящерицы — в 2,2%, полевки рода Microtus — в 3,2%, птенец крачки рода
Sterna — в 1,1%, остатки растений в 2,2% погадок. Здесь же отмечены частые случаи
поедания птенцов своего вида и отнимания добычи у пестроносых крачек (Зубакин,
1975в, 1976в). На зимовках чайконосые крачки питаются ракообразными, лягуш-
ками, мелкой рыбой, насекомыми. Отмечено поедание отбросов из сточных вод (АН,
Ripley, 1969).
За кормом птицы могут улетать далеко от мест гнездования, иногда до 30 км. В от-
личие от ряда других видов крачек крайне редко ныряет за кормом, обычно подхва-
298
Чеграва
тывает его с поверхности воды, земли или с растений; может ловить насекомых в
воздухе (Бородулина, 1960; Долгушин, 1962; Cramp, 1985).
Враги, неблагоприятные факторы. Из врагов чайконосых крачек следует отметить
серебристую чайку-хохотунью, черноголовую чайку и болотного луня, которых крач-
ки активно, но не всегда успешно прогоняют с территории своей колонии (Долгу-
шин, 1962; Эминов, 19746; наши данные). Колонии могут страдать от беспокойства
со стороны человека (Зубакин, Костин, 1977) и лисиц (Эминов, 19746), от затопле-
ния при нагонных ветрах; птенцы нередко гибнут во время продолжительного не-
настья (Клименко, 1950а). Большое воздействие на успех размножения может ока-
зывать каннибализм — поедание птенцов своего вида (Зубакин, 1975в).
Хозяйственное значение, охрана. Уничтожением насекомых чайконосые крачки
приносят пользу сельскому хозяйству. Колонии охраняются в Черноморском, Кур-
гальджинском и в ряде других заповедников и заказников; специальные меры охра-
ны вида не предусмотрены. В силу малочисленности чайконосая крачка заслуживает
занесения в некоторые республиканские Красные книги, особенно Украинской ССР.
Род Hydroprogne Каир, 1829
Чеграва -
Hydroprogne caspia (Pallas, 1770)
Caspian Tern (англ.), Raubseeschwalbe (нем.), Sterne caspienne (фр.).
Син. (лат.) : Hydroprogne tschegrava (Lepechin, 1770), Sterna caspia Pallas, 1770.
Син. (рус.) : крачка-чеграва
Статус. Гнездяшийся мигрирующий вид.
Общая характеристика и полевые признаки. Очень крупная крачка, размером лишь
немного уступает серебристой чайке. От всех крачек отличается размерами, от чаек —
длинным красным клювом, вырезанным хвостом и характерной для крачек черной
’’шапочкой”. Полет сильный, с медленными, как у крупных чаек, взмахами крыльев.
В то же время чегравы способны останавливаться в воздухе, трепеща крыльями, как
другие крачки. На воду в норме, по-видимому, не садится. Добычу ловит, ныряя в воду
с разлета, причем обычно погружается целиком. Неплохо ходит по земле, чувствует
себя здесь гораздо увереннее многих крачек.
На территории колонии очень шумная птица. Голос громкий, резкий; в нем преоб-
ладают каркаюшие и скрежещущие звуки. Крик тревоги - повторяющиеся звуки типа
”рак, рак, рак”. Агрессивные столкновения сопровождаются стрекочущим криком,
который представляет собой серию быстро повторяющихся слогов ”ра-ра-ра-ра-ра”.
В период инкубации именно этот крик создает шумовой фон колонии. Характерен
громкий скрежещущий крик, который можно приблизительно передать буквами как
’’чкарркакаррриу”. Его издает чеграва, возвращающаяся (обычно с добычей) в ко-
лонию.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Иногда чеграв Австралии и Новой Зеландии выделяют в
особый подвид H.c.strenua (Peters, 1934; Howard, Moore, 1984). C.A. Бутурлин (1910)
выделил в отдельный подвид Н.с. leptorhyncha крачек из Уссурийского края. Однако
большинство систематиков считают чеграв монотипическим видом (Vaurie, 1965;
Степанян, 1975; Cramp, 1985). Не решен окончательно и вопрос о выделении чегравы
в отдельный род. Часть авторов (Moynihan, 1959; Cramp, 1985) относят ее к роду
Sterna, однако большинство систематиков выносят чеграву в монотипический род
299
Семейство Чайковые
Hydroprogne (Peters, 1934; Дементьев, 1951; Vaurie, 1965; Степанян, 1975; Wolters,
1975) , что, очевидно, более правильно.
Описание. Размеры (мм) (ЗМ МГУ) и масса тела (г) (Бородулина, 1960; Долгу-
шин, 1962; Заболоцкий,Заболоцкая, 1963).
Длина крыла:
3 3 (п = 11) — 389—432 (среднее 417) ;
$ $ (п = 13) — 394—423 (среднее 409) .
•Длина клюва:
3 3 (и = 14) — 62,8—72,3 (среднее 67,3) ;
9$ (п = 14) — 63,1—69,5 (среднее 65,9) .
Длина цевки:
6 6 (п = 14) — 43,9—50,0 (среднее 46,7);
2 $ (и = 13) — 43,9—48,5 (среднее 45,4) .
Масса тела:
3 <3 (п = 10) — 640—700 (среднее 673);
$ $ (и = 14) — 482—730 (среднее 623).
Окраска (Дементьев, 1951; Cramp, 1985). Самец и самка в брачном наряде.
Лоб, темя, затылок, верхняя часть уздечки, окологлазная область и кроющие уха чер-
ные; перья затылка слегка удлинены и образуют хохол. Спина, плечевые, кроющие
крыла светлые серебристо-сизые; задняя часть шеи, поясница, верхние кроющие хвоста
и рулевые белые, последние^ нередко с сероватым налетом. Нижняя часть тела белая.
Наружные опахала дистальных и оба опахала проксимальных первостепенных маховых
светлые, серебристо-серые, концы и край внутренних опахал 5—6 дистальных перво-
степенных черновато-серые. Нижняя поверхность дистальных первостепенных темно-
серая. Клюв ярко-красный, ноги черные, радужина темно-бурая.
Самец и самка в зимнем наряде. Лоб и уздечка белые, темя и затылок в белых и чер-
ных пестринах; перед глазом черное пятно, такого же цвета кроющие уха. Остальное
оперение, как в предыдущем наряде. Окраска клюва несколько светлеет, кончик
становится темным.
Пуховой наряд (Cramp, 1985; живые птенцы с Лебяжьих островов). Окраска сильно
варьирует от серебристо-серой, почти белой, до желтовато -серой, причем различную
окраску нередко имеют и птенцы одного выводка. По верхней стороне тела разбро-
саны отдельные мелкие темные пестрины, иногда они практически отсутствуют. Нижняя
часть тела светлая, подбородок и брюшко белые, уздечка и нижняя часть горла нередко
серые. Клюв оранжево-красный с темным пятном на вершине, ноги оранжево-желтые,
радужина темно-коричневая.
Гнездовой наряд. Верхняя часть головы белая с густыми черными продольными
пестринами, кроющие уха и пятно перед глазом черные или темно-серые. Перья верх-
ней части спины, плечевые и третьестепенные маховые тускло-серые с беловатыми или
охристыми каймами. Многие из них имеют предвершинные V-образные тускло-черные
пятна; количество перьев с такими пятнами индивидуально варьирует. Задняя часть спи-
ны, поясница и большинство верхних кроющих крыла, как у взрослых птиц, но верши-
ны перьев нередко с охристым налетом. Часть малых кроюших более темного цвета
в предвершинной части, они образуют неявную темную полосу вдоль крыла. Задняя
часть шеи, нижняя часть тела и нижние кроющие крыла белые. Рулевые серые с белыми
300
Чеграва
внутренними опахалами, светлыми концами и темными предвершинными пятнами.
Первостепенные маховые, как у взрослых, но темнее; проксимальные первостепен-
ные с белой вершиной. Клюв оранжево-желтый или оранжево-красный с темным кон-
чиком, ноги желтые с черноватым налетом, радужина темно-коричневая.
Первый зимний наряд. Как окончательный зимний, но маховые, рулевые и часть
кроющих крыла остаются от гнездового наряда. Весной и летом второго календарного
года первый зимний наряд прямо переходит во второй зимний (Cramp, 1985). Летом
третьего и, возможно, четвертого календарного года часть птиц имеют белое пятно или
пестрины в передней части лба.
Линька (Cramp, 1985). Линька в первый зимний наряд полная, начинается в ноябре
(иногда октябре) — декабре, завершается к марту—апрелю, однако первостепенные
маховые часто не успевают смениться до июня. Повторная линька первостепенных на-
чинается с самых внутренних в марте—апреле. Завершается она сменой II (первого ви-
димого) первостепенного в сентябре—октябре, когда оперение головы, туловища, хвос-
та и кроющих крыла из первого зимнего наряда линяет непосредственно во второй
зимний наряд. Последующие линьки проходят, как у взрослых птиц, но послегнездо-
вые могут начинаться раньше или продолжаться дольше, а предгнездовые вовлекать
большее число внутренних первостепенных, чем линьки взрослых птиц. Кроме того,
у неполовозрелых особей могут несколько смещаться сроки линьки первостепенных
маховых.
У взрослых особей бывает две линьки в году: полная послебрачная и частичная
предбрачная. Послебрачная начинается линькой оперения лба и темени между концом
июля и концом сентября. Проксимальные первостепенные либо начинают сменяться в
это же время, либо после появления птиц на зимовках. Линька оперения тела, хвоста и
крыльев завершается в январе—феврале. Предбрачная линька включает смену оперения
головы и части оперения туловиша (февраль—март), хвоста и обычно нескольких прок-
симальных первостепенных маховых (их смена начинается после завершения смены
первостепенных маховых послебрачной линьки). Линька первостепенных прерывается
на стадии новых 2—4 проксимальных, а не продолжается после сезона размножения.
Птицы, поздно завершившие послебрачную линьку, не приступают к повторной смене
первостепенных до сезона размножения.
Распространение. Гнездовой ареал. Обитает на всех материках, кроме Южной
Америки и Антарктиды, однако везде поселяется спорадично (рис. 71). В зарубежной
Европе в настояшее время гнездится в Швеции и Финляндии. В отдельные годы гнезда
этих птиц отмечаются в Норвегии, ГДР, Польше и Италии; в прошлом столетии и начале
нынешнего чеграва гнездилась во Франции, Дании и на территории ФРГ (Cramp,
1985).
В СССР чегравы гнездятся отдельными изолированными поселениями, удаленными
друг от друга на десятки и сотни километров (рис. 72). Этим они очень напоминают
черноголовых хохотунов, с которыми часто поселяются на одних и тех же водоемах
и даже островах. Современное размещение гнездовых поселений следующее. В Эстонии
чегравы гнездятся в Матсалуском заповеднике и окрестностях о-ва Вильсанди (Аумеэс,
1972; Кастепыльд, 1972); впервые гнездование в Эстонии отмечено в 1910 г. (Никлус,
1957). В 1976 г. одна пара вывела птенцов на оз. Энгуре в Латвии (Виксне, 1983). На
севере Причерноморья чеграва гнездится в Тендровском заливе (Черноморский за-
поведник) , на Лебяжьих островах Каркинитского залива и Чонгарских островах Сива-
ша (Зубакин, Костин, 1977; Костин, 1983; Ардамацкая, 1982, 1984). В Тендровском
заливе поселения чеграв известны по крайней мере с 1935 г., на Лебяжьих островах —
с начала 1950-х годов. На Чонгарских островах гнездование этой крачки обнаружено в
1901 г. (Молчанов, 1906), однако затем чегравы, видимо, переселились на о-в Китай,
301
302
Рис. 71. Область распространения чегравы
1 — гнездовые поселения, 2 — места зимовок, 3— залеты
303
Рнс. 72. Ареал чегравы в СССР
1 — постоянные гнездовые поселения, 2 — временные гнездовые поселения, 3 — предполагаемые места гнездования, 4 — область
осеннего пролета балтийских чеграв, 5 — область регулярных кочевок черноморских чеграв, 5 — область регулярных кочевок казах-
станско-среднеазиатских чеграв, 7 — область нерегулярных зимовок, 8 — залеты, 9 — направление осеннего пролета, 10 — направ-
ление весеннего пролета
Семейство Чайковые
где гнездились в 1934-1937 гг. и 1953 г. (Иваненко, 1936; Андреев, 1940; Солдатенко,
1956). По крайней мере с 1973 г. поселения чеграв опять отмечаются на Чонгарских
островах (Сиохин, 1981а; Костин, 1983).
Далее на восток известны нерегулярные гнездования отдельных пар и небольших
групп на водохранилищах Маныча (Петров, Миноранский, 1962; Языкова, 1975). Круп-
ные поселения чеграв расположены на северном Каспии, где птицы гнездятся в авандель-
те Волги на островах Жемчужный и Морской Очиркин, в дельте Урала (по-видимому,
не регулярно) и на островах прибрежных мелководий восточной части северного Кас-
пия (Бондарев, 1975; Кривоносов, 1976). В начале 1950-х годов чегравы гнездились
на Тюленьих (Мангистауских) островах (Гладков, Залетаев, 1956), современные дан-
ные об этом поселении отсутствуют. На юго-востоке Каспия гнездование отмечено лишь
в заливе Кара-Богаз-Гол (Васильев и др., 1975; Щербина, 1979) .
В Казахстане чеграв'а гнездится на оз. Челкар-Тенгиз и в низовьях Тургая и Иргиза,
на Балхаше, озерах Бийлюкульской и, возможно, Теликульской группы, в низовьях
р. Чу (Долгушин, 1962; Ауэзов, 1986) , на Алаколе (Ауэзов, Грачев, 1975) и оз. Тенгиз
(Чекменев, 1964; Волков, 1975; Андрусенко, 1981); с 1983 г. — на Капчагайском
водохранилище (Сема и др., 1986). Гнездование отмечалось в ряде мест на Аральском
море (Зарудный, 1916; Исмагилов, 1955; Долгушин, 1962; Степанян, Галушин, 1962),
однако в связи с прогрессирующим усыханием Арала по крайней мере часть поселений
в настояшее время испытывают депрессию (Гисцов, 1964). Неизвестно современное
состояние колоний низовий Амударьи (Гладков, 1932). В Средней Азии чеграва, по-
мимо Кара-Богаз-Гола, достоверно гнездится ныне лишь на Хаузханском водохранили-
ще (Эминов, 1975) иАйдаркуле (Мухина, 1983; Остапенко и др., 1984).
Колонии чеграв отмечены также на оз. Чаны (Ходков, 1981), с 1977 г. в дельте
р. Селенги на Байкале (Мельников, 1979) и на Торейских озерах (Зубакин, 1981а).
На оз. Ханка чеграва, вопреки прежним указаниям (Бутурлин, 1934; Дементьев, 1951;
Иванов, 1976), не гнездится, по крайней мере, в настоящее время (Воробьев, 1954;
Поливанова, 1971; Глущенко, 19846). В Туве статус чегравы пока неясен (Баранов,
1981; Савченко, 1983), по соседству достоверное гнездование отмечено для оз. Хара-
Ус-Нур на северо-востоке Монголии (Степанян, Болд, 1983). Нерегулярно гнездится на
озерах северной Кулунды (Гынгазов, Миловидов, 1977).
Зимовки. Эстонские, финские, шведские и черноморские чегравы имеют, видимо,
сходные места зимовок: побережье Западной Африки (особенно Гвинейский залив),
долина р. Нигер, долина р. Нил в Египте и Судане, Средиземное море (побережье Туниса).
Чегравы с Каспия зимуют в Иране (провинции Систан и Гюрген), Ираке, Саудовской
Аравии и Индии. Казахстанские и среднеазиатские чегравы отмечены на зимовках
главным образом во внутренних районах Ирана (Систан, Заболь), а также в Индии,
Пакистане, Бангладеш, Персидском заливе, Омане, во Вьетнаме, Кампучии и Судане.
Имеются два зимних возврата из Дании от чеграв, окольцованных птенцами в Эстонии
и на оз. Алаколь в Казахстане (Таманцева, 1955; Шеварева, 1962; Кривоносов и др.,
1978; Костин, 1978, 1983; Cramp, 1985; Сема и др., 1986; ЦКАН). Изредка чеграв от-
мечают зимой на юго-западном Каспии (Виноградов, Чернявская, 1965).
Таким образом, птицы из нашей страны зимуют на обширной территории от побе-
режья Западной Африки до Вьетнама. Области зимовок разных популяций перекры-
ваются, но птицы западных регионов зимуют, как правило, в западной части зимовоч-
ного ареала, восточных — в восточной. Отдельные птицы разных популяций могут про-
водить зиму на побережье Западной Европы. Места зимовок чеграв с Байкала и из За-
байкалья не установлены.
Миграции. В период миграций чегравы большей частью летят напрямик через конти-
нент, пересекая обширные безводные пространства. В августе—сентябре у европейских
304
Чеграва
и азиатских чеграв происходят послегнездовые кочевки, когда птицы отмечаются как
в окрестностях места гнездования, так и в нескольких десятках и сотнях километров
от него, причем нередко в направлении, противоположном миграционному. Однако у
чеграв, гнездяшихся в Финском заливе, в это время уже наблюдается осенний пролет,
который проходит через Ленинградскую обл., республики Прибалтики, Калининградс-
кую обл., Белоруссию, западную половину Украины, далее через дельту Дуная (Кор-
зюков, 1981) и Румынию в Средиземное море; часть птиц затем пересекают Сахару,
достигая Гвинейского залива. Птицы, гнездящиеся севернее -- в Ботническом заливе,
пролетают через территорию нашей страны тем же маршрутом в сентябре и частично
октябре. В октябре—ноябре большинство чеграв достигают мест зимовок, причем
северные птицы прилетают сюда в целом несколько раньше. Весенний пролет финно-
скандинавских чеграв проходит в апреле—мае напрямик через Чехословакию, Венгрию,
Польшу, лишь слегка захватывая Львовскую и Калининградскую области. На Черное
и Азовское моря чегравы возвращаются, видимо, вдоль побережий, достигая Крыма
в первых числах марта (Шеварева, 1962; Смогоржевский, 1970; Костин, 1978, 1983).
На Балтике в районе г. Паланги первых чеграв отмечают в начале второй декады апре-
ля (Петрайтис, 1975), в Швеции первые чегравы появляются 10—15.IV, в Финляндии —
несколькими днями позже (Cramp, 1985). На Ладоге в районе Свирской губы пролет
идет в первой половине мая (Носков и др., 1981).
В азиатской части ареала весенний пролет отмечен на южном Каспии с начала марта
до первой половины апреля (Дементьев и др., 1955; Заблоцкий, Заблоцкая, 1963),
в долине р. Мургаб — в апреле (Эминов, 1978), на Айдаркуле — с 14.Ш (Остапенко
и др, 1984), В дельте Волги чегравы появляются в апреле (Луговой, 1958), в районе
Кургальджина — 27.IV—13.V, у Семипалатинска — в третьей декаде апреля (Долгушин,
1962). на Торейскихозерах в первой половине мая (Леонтьев, 1972), в дельте Селен-
ги — с 30.IV, чаше с 17—19.V (Мельников, 1979). Осенний пролет на юго-западе Каспия
отмечен в сентябре—октябре; отдельные птицы задерживаются здесь до первой трети
ноября (Заблоцкий, Заблоцкая, 1963). В других местах осенний пролет не выражен,
птицы исчезают незаметно. Северную половину Казахстана птицы покидают в сентябре,
южную — в октябре (Сема и др., 1986). На Айдаркуле последние чегравы отмечаются
22.IX (Остапенко и др., 1984; Калабин, 1986), на Торейских озерах — во вторую поло-
вину сентября (Леонтьев, 1972).
Судя по данным кольцевания, чегравы с оз. Алаколь осенью пролетают через южную
часть Туркмении и Таджикистана. По-видимому, сушествует также пролет через Джун-
гарские ворота и Китай. Весенние пути пролета, вероятно, повторяют осенние. Аральс-
кие чегравы попадают на зимовки в Персидский залив через долину Мургаба. Чегравы
с оз. Тенгиз осенью, по-видимому, летят в Персидский залив через Каспий (Сема и др.,
1986).
Часть неполовозрелых чеграв в возрасте 1—2 года отмечаются летом в местах зимо-
вок (Шеварева, 1962; Костин, 1978,1983). Для этого вида характерны широкие летние
кочевки бродячих птиц, которые захватывают всю область гнездования вида и нередко
выходят за ее пределы. В частности, кочующие особи постоянно наблюдаются по всему
Казахстану (Долгушин, 1962), встречаются в Омской обл. (Сулимов, 1981). Пролет
чеграв идет одиночными особями или небольшими группами в 2—7 особей (Долгушин,
1962; Cramp, 1985).
Залетных чеграв встречали в Великобритании, Ирландии, Норвегии, на Фарерских
островах (Cramp, 1985), на Семи островах (Герасимова и др., 1967), в Московской
обл. (Птушенко, Иноземцев, 1968), в окрестностях Барнаула (Велижанин, 1928),
южном Приморье (Taczanowski, 1893; Глущенко, Шибнев, 1984), на о-ве Сахалин
(Гизенко,1955).
305
Семейство Чайковые
Численность. В целом невысокая. В Эстонии число гнездящихся пар возрастает:
в 1971 г. гнездилось 350 пар (Кастепыльд, 1972), в конце 1970-х годов — около
500 пар (Cramp, 1985). Численность черноморско-сивашской популяции сильно ко-
леблется по годам: в отдельные годы она превышала 1 тыс. гнездящихся пар (1970 г.),
в середине 1970-х годов держалась на уровне 500 пар, затем снизилась до примерно
300 пар (Сиохин, 1981а; Костин, 1983; Ардамацкая, 1984). Возможно, это связано
как с негнездованием части особей, так и существованием пока неизвестных колоний.
На севере Каспия гнездится, по-видимому, не менее 5 тыс. пар (Кривоносов, 1976),
в заливе Кара-Богаз-Гол — 50—90 пар (Щербина, 1979), на оз. Тенгиз в 1970-х годах-
75—840 пар (Волков, 1975; Андрусенко, 1981), на оз. Алаколь — до 3 тыс. пар (Ауэ-
зов, Грачев, 1975; Ауэзов, 1980), на Аральском море не менее нескольких сот пар
(Долгушин, 1962). На оз. Чаны в 1978 г. отмечено 124 гнездяшиеся пары (Ходков,
1981), на Хаузханском водохранилище в 1963 г. — 25 пар (Эминов, 1975), на Торейс-
ких озерах в 1976 г. — 635 пар (Зубакин, 1981а), в дельте Селенги в 1977—1981 гг. —
300—400 особей (Мельников, 1984) .
Всего в СССР в благоприятные годы гнездится, по-видимому, 12,5—15 тыс. пар.
Обшая тенденция изменения численности не ясна: она либо стабильна, либо медленно
возрастает. В Швеции и Финляндии численность растет; в конце 1970-х годов здесь
гнездилось по 1 тыс. пар (Cramp, 1985) .
Местообитания. В сезон гнездования - морские побережья и крупные внутренние
водоемы, как правило соленые, реже пресные. Чегравы явно предпочитают естествен-
ные водоемы, водохранилища их привлекают в меньшей степени. Поселяются как на
рыбных, так и безрыбных водоемах; необходимые условия гнездования: достаточно
удаленные от берега небольшие островки с ровными, лишенными растительности участ-
ками и наличие в относительной близости от гнездовья — по И.А. Долгушину (1962).
не далее 40—50 км — водоемов с рыбой. Вне сезона размножения — морские побережья,
крупные озера, долины крупных рек.
Размножение. К размножению птицы приступают в возрасте не менее 3 лет (Ludwig,
1965, цит. по: Cramp, 1985). Чеграва — моногам, плотно гнездящийся факультативно-
колониальный вид. Степень постоянства пар, возможно, связана не с успехом размноже-
ния в предыдущей год, а со стабильностью гнездовых биотопов. Так, на оз. Мичиган
пары сохранялись достоверно чаще, если место гнездования прошлого года не было за-
топлено (Cuthbert, 1985). Для пространственной структуры вида характерно, видимо,
сушествование более или менее изолированных популяций, объединяющих поселения,
удаленные друг от друга на несколько десятков, а иногда и сотен километров. Между
поселениями внутри популяции существует заметный обмен особями. В настоящее
время можно выделить балтийскую популяцию, включающую чеграв Швеции, Фин-
ляндии и Эстонии (Кастепыльд, 1972), и черноморскую, куда входят поселения север-
ного Причерноморья и Сиваша (Шеварева, 1962; Ардамацкая, 1982; Костин, 1983).
Колонии одного поселения могут как менять свое местоположение по годам (однако
обычно в пределах группы близлежащих островов), так и, реже, находиться на одном
и том же месте.
Демонстрационное поведение чегравы имеет черты сходства с поведением других
видов крачек и специфические особенности (Cramp, 1985; наши данные). Несмотря
на плотное гнездование, вокруг гнезд существуют небольшие охраняемые территории
(по-видимому, с нечеткими границами). Высокая внутривидовая агрессивность при-
водит к постоянным территориальным конфликтам. Выделяют агрессивную прямую
позу, сходную с такой же позой у чайконосой крачки: шея вытянута наклонно вперед
и вверх, хохол приподнят, клюв открыт и направлен вперед (рис. 73, л). В этой позе
чеграва издает стрекочущий крик и, то вытягивая шею по направлению к другой птице.
306
Чеграва
то вновь приподнимая ее, медленно движется к противнику. Продолжая стрекотать,
птицы сближаются на границе участков (см. рис. 73,6), после чего либо вспыхивает
драка, либо одна из птиц начинает медленно оттеснять другую вглубь ее территории.
После изгнания противника, чеграва нередко принимает выпрямленную позу, при кото-
рой шея и закрытый клюв вытянуты вертикально или наклонно вверх, крылья полу-
раскрыты, иногда целиком развернуты, и держатся параллельно земле (см. рис. 73,в).
По-видимому, к агрессивной сфере относится демонстрация, напоминающая долгий
Рис. 73. Демонстрационное поведение чегравы (в, д - по: Cramp, 1985, с изменениями)
а — агрессивная прямая поза, б — агрессивный стрекот чеграв-соседей на границе гнездовых участ-
ков, в — выпрямленная поза, г — ’’долгий крик”, д — ’’поклон со вскидыванием головы”, е — вып-
рямленная поза партнеров
крик чаек (см. рис. 73,г): шея вытянута вертикально, широко раскрытый клюв на-
правлен горизонтально, крылья развернуты и приподняты, при этом чеграва несколько
раз взмахивает ими с малой амплитудой. Крик, который птица издает в этой позе —
однократное или двукратное ”ррау”. Функции данной демонстрации не ясны.
Процесс формирования пар включает как воздушные, так и наземные демонстрации.
Отмечен ’’высотный полет”, который, однако, бывает реже, чем у речной крачки; участ-
вуют в нем только две птицы. Нередко отмечается "низкий полет”, при котором самец
с рыбкой в клюве летает, сопровождаемый самкой; периодически он издает очень не-
приятный скрежещущий крик типа "чкаррчкакаррриу”, как бы ныряя в это время в
воздухе. С подобным криком самец предлагает самке рыбку и на земле, при этом он
сначала приподнимает задний конец тела и вытягивает шею параллельно земле, затем
резко вскидывает шею и клюв почти вертикально (см. рис. 73,6). Отдав рыбку сам-
ке, самец принимает выпрямленную позу, сходную с описанной (см. рис. 73,е), однако
307
Семейство Чайковые
крылья при этом не развернуты, а голова поворачивается вдоль продольной оси впра-
во-влево. Самка, выпрашивая корм, издает негромкие стонущие звуки.
В большинстве случаев на территории колонии чегравы появляются уже сформиро-
ванными парами (Cramp, 1985). Для гнездования птицы выбирают, как правило, низ-
кие песчаные, галечниковые, ракушечниковые и, реже, каменистые островки, нередко
удаленные от берега на несколько километров. Колонии располагаются на лишенных
растительности участках. Величина варьирует от нескольких пар до более тысячи гнезд,
причем, по-видимому, наиболее обычны колонии от нескольких десятков до 100 —
200 гнезд. Гнездование одиночными парами также достаточно характерно для вида,
особенно на севере. По соседству с чегравами гнездятся другие виды чайковых (Заруд-
ный, 1916; Долгушин, 1962; Эминов, 1969; Кастепыльд, 1972; Зубакин, Костин,
1977; Bergman, 1980; Зубакин, 1981а; Виксне, 1983; Костин, 1983; Ардамацкая,
1984).
Гнездо — обычно ямка в грунте, чаще всего без всякой выстилки. Гнездовой валик
нередко плохо выражен, состоит из ракушек, камешков, реже кусочков растений.
Размеры гнезд (мм) на Сиваше и Лебяжьих островах; диаметр гнезда (и = 27) 350-
460, в среднем 396; диаметр лотка (гнездовой ямки) 210—330, в среднем 267 (п =
= 27); глубина лотка (п = 7) 40—50, в среднем 45. Гнездование очень плотное. Среднее
расстояние между центрами ближайших соседних гнезд на Сиваше и на Лебяжьих остро-
вах составляло (п = 203) 0,7 м (0,4—3,1), на оз. Чаны (и = 124) — 0,8 м (0,4—1,2),
на Торейских озерах (п = 94) — 0,79. Со сходной плотностью вид гнездится и в других
частях ареала. В Казахстане между гнездами отмечается 0,5—4 м, на Кара-Богаз-Голе -
0,6-4 м. В дельте Селенги на площади 30 X 18 м отмечено 100 гнезд (Долгушин, 1962;
Зубакин, 1976в, 1981а; Мельников, 1979; Ходков, Тотунов, 1979; Щербина, 1979;
наши данные). Индекс плотности гнездования равен 0,36.
На Балтике откладка яиц отмечается с начала мая (Cramp, 1985). Первые яйца на се-
вере Причерноморья обычно появляются в третью декаду апреля — начале мая (Костин,
1983; Ардамацкая, 1984), в Казахстане, по-видимому, в эти же сроки; на севере рес-
публики чуть позже — в первую декаду мая (Кузякин, 1959; Долгушин, 1962). В пер-
вой декаде мая отмечается начало гнездования на юге Туркмении (Эминов, 1969).
На оз. Чаны в 1978 г. и в дельте Селенги в 1977 г. первые кладки появились, видимо,
в третьей декаде мая (Мельников, 1979; Ходков, Тотунов, 1979); на Торейских озе-
рах в 1976 г. - в первых числах июня (Зубакин, 1981а). Характерна сильная растяну-
тость гнездового периода чегравы. Свежие кладки встречаются до середины июля,
причем поздно загнездившиеся птицы часто присоединяются к существующей колонии.
Размер кладки 1-5 яиц, чаще всего 2-3 яйца. В одних колониях преобладают кладки
из 3 яиц, в других, особенно сформировавшихся в более поздние сроки, — из 2.
Окраска яйца сложная. Цвет основного фона яйца варьирует от светлого охристо-
беловатого до более темного, светло-коричневого. По этому фону разбросаны глубокие
серые и поверхностные темные (светло-бурые, бурые и черноватые) пятна (Долгушин,
1962). Несмотря на вариации окраски яиц, они в целом светлее яиц гнездящихся по
соседству серебристых чаек. Размеры яиц (мм): с Лебяжьих островов (п = 215) 57,4—
72,0 X 40,5—49,1, в среднем 65,4 X 44,6 (Костин, 1983); из Казахстана (п = 41) 48,5-
71,3 X 30,4—47,3, в среднем 64,6—44,1 (Ковшарь, Левин, 1982); с Байкала (п = 20)
60,9-70,2 X 42,0-46,2, в среднем 64,5 X 44,8 (Мельников, 1979). Форма яиц, как и
окраска, сильно изменчива. Масса (г) свежих яиц (п = 90) 58,3-70,2, в среднем 65,7
(Эминов, 1969; Костин, 1983).
Насиживание начинается с первого яйца и продолжается 20—22 дня (Cramp, 1985)
или 26—29 дней (Эминов, 1969). Насиживают оба партнера, по наблюдениям на Сиваше
смена на гнезде происходит 3-5 раз в сутки (п = 5), при этом самец изредка кормит
308
Чеграва
самку. Вылупление птенцов в одном гнезде длится 1- 2 сут (по другим данным —
2-3 сут, см.: Эминов, 1969), между появлением первой трещины на яйце и вылупле-
нием проходит 3—4 сут, появлением отверстия и вылуплением — 1 сут. Суточного
птенца, по наблюдениям на одном гнезде, кормили 3 раза в сутки мелкой рыбой (на-
ши данные). После вылупления птенцов родители, как правило, уносят скорлупу из
гнезда, но иногда она остается лежать в нем или по соседству. При беспокойстве коло-
нии взрослые птицы уводят птенцов из гнезда через 2—5 сут после вылупления старшего
птенца и держатся с ними вне колонии. В отсутствие, беспокойства подобный увод
выводков происходит, видимо, позже. Во время тревоги птенцы в возрасте до 5 сут
замирают там, где их застала тревога. Нередко так же ведут себя и более старшие птен-
цы — в возрасте 10—12 сут. В более старшем возрасте птенцы при появлении опасности
стремятся убежать, объединяясь в стайки. В крупных колониях птенцы во время тре-
воги сбиваются в табунки, которые, однако, не такие плотные, как у черноголовых
хохотунов. Родители активно нападают на хищников и изгоняют их за пределы колонии.
Часто пикируют и на человека (Чекменев, 1964; наши данные). На крыло молодые
поднимаются в возрасте 30—35 сут (Cramp. 1985) или 35—38 сут (Эминов, 1969).
Сведения об успехе размножения скудные. На Хаузханском водохранилище общий
отход яиц и птенцов в 1963 г. составил около 35% (Эминов, 1969) .
Максимальная продолжительность жизни, по данным кольцевания, 26 лет 2 месяца
(Clapp et al., 1982, цит. по; Cramp, 1985). Смертность молодых чеграв в первый год
жизни, по-видимому, значительно меньшая, чем у чаек (Шеварева, 1962).
Суточная активность, поведение. Активность преимущественно дневная, но отдель-
ные чегравы могут возвращаться в колонию или улетать из нее в полной темноте. Сво-
бодные от насиживания партнеры обычно ночуют у гнезд или на краю колонии. Мас-
совый вылет птиц из колонии начинается примерно за 1 ч — 1 ч 10 мин до восхода солн-
ца (наши данные). Несмотря на гнездование плотными крупными колониями, вне се-
зона размножения чегравы держатся одиночно, парами или небольшими группами до
7 особей, лишь во время отдыха и ночевок могут собираться вместе более 100 птиц.
Кормятся чегравы обычно также в одиночку или парами (Cramp, 1985). Данные коль-
цевания (ЦКАН) как будто указывают на постоянство состава мигрирующих групп
в первый год жизни (птиц, окольцованных в одной колонии, добывали в один и тот
же день и в одном месте на зимовках).
Питание. Чеграва питается почти исключительно рыбой 15—20, иногда 30 см длиной,
гораздо реже — водными беспозвоночными, в частности раками (Мензбир, 1895; Bent,
1921; Шевченко, 1937; Деревягин, 1948; Гладков, Залетаев, 1956; Бородулина, 1960,
1965; Чекменев, 1964; Koli, Soikkeli, 1974; Cramp, 1985). По-видимому, иногда способ-
на поедать и наземных насекомых, например саранчу в период ее массового появления
(Спангенберг, Фейгин, 1936). Кочующее из сводки в сводку утверждение о том, что
чеграва иногда поедает яйца и птенцов других птиц (Дементьев, 1951; Долгушин,
1962 и др.) , по-видимому, впервые было высказано еще М.А. Мензбиром (1895) и фак-
тическими данными не подтверждено. Подобные факты (если они были) можно тракто-
вать только в качестве редчайшей случайности. Видовой состав добычи зависит от мес-
та обитания конкретной особи. В северном Крыму это преимущественно бычки раз-
ных видов (Костин, 1983), на юго-западе Каспия — кутумы, вобла и атеринка (Заблоц-
кий, Заблоцкая, 1963), на Кургальджине — плотва и карась, в Балхаше и Алаколе —
маринка (Долгушин, 1962), в южной Туркмении — аральская плотва, закаспийская
храмуля и сазан (Эминов, 1969).
Основной способ охоты чегравы напоминает поисковый полет речной крачки без тре-
петания. Птица, задержавшись в воздухе на высоте 3—20 м, вертикально бросается в
воду, часто скрываясь в ней целиком. За кормом чегравы способны улетать на рассто-
309
Семейство Чайковые
яние до 30 км от колонии, в отдельных случаях, видимо, до 100 км (Спангенберг,
Фейгин, 1936; Бородулина, 1960; Cramp, 1985).
Враги, неблагоприятные факторы. Гнезда чеграв нередко разоряют поселяющиеся
по соседству серебристые чайки, особенно при усилении фактора беспокойства (наши
данные). На Балтике кладки страдают от хищничества серых ворон (Cramp, 1985).
Отмечена гибель гнезд от затопления (Эминов, 1969). Крупные размеры и рыбоядность
нередко приводит к тому, что птица попадает под браконьерский выстрел. Косвенное
свидетельство тому — большее по сравнению с другими крачками число возвратов
от окольцованных особей.
Хозяйственное значение, охрана. В силу малочисленности хозяйственного значения
не имеет. Занесена в Красную книгу Эстонской ССР. Без сомнения, чеграва заслуживает
занесения в Красные книги Украины и РСФСР, а возможно, и в Красную книгу СССР.
Род Thalasseus Boie, 1822
Пестроносая крачка —
Thalasseus sandvicensis (Latham, 1787)
Sandwich Tern (англ.) , Brandseeschwalbe (нем.) , Sterne cauger (фр.) .
Син. (лат.): Sterna sandvicensis Latham, 1787, Sterna cantiaca Gmeiin, 1789.
Син. (рус.): пестроклювая крачка.
Статус. Гнездящийся мигрирующий, возможно, частично оседлый вид.
Общая характеристика н полевые признаки. Крупная крачка, сходная по размерам
с чайконосой, от которой отличается длинным и более тонким двухцветным клювом
(черным с желтой вершиной), светлыми, при взгляде снизу, концами крыльев (у чай-
коносой они темно-серые) и голосом. Как у большинства крачек, в брачный период
на голове черная шапочка, причем перья затылка образуют заметный хохолок, более вы-
раженный, чем у чегравы. Полет легкий и быстрый, добычу ловит, ныряя в воду с разле-
та. Несмотря на относительную коротконогость, по земле передвигается довольно актив-
но, однако лишь в колонии и ее окрестностях и, реже, в местах отдыха. На воду в норме
не садится. Голос напоминает голос черной крачки, но он более резкий и в нем больше
скрипучих тонов. В окрестностях гнездовья можно слышать крик ’’киррит” (или ”кир-
лит”), с которым крачки возвращаются в колонию; этот же крик используется в
брачных демонстрациях. Во время тревоги издаются отрывистые крики ”кир, кир.
кир”, при агрессивных конфликтах в колонии слышны сухие стрекочущие трельки
типа ”кр-кр-кр-кр-кр”, а также скрипучее ”кррр”. Более подробно звуковое общение
пестроносых крачек разобрано у И.Д. Никольского (1985).
Подвидовая систематика. Обычно различают два подвида: палеарктический
Th. s. sandvicensis и американский Th. s. acuflavidus (Peters, 1934; Vaurie, 1965). Некото-
рые авторы сводят до уровня подвида пестроносой крачки обитающую на восточном
побережье Южной Америки Th. eurygnathus (Howard, Moor, 1984; Cramp, 1985). В СССР
только один подвид (Степанян, 1975) :
Thalasseus sandvicensis sandvicensis.
Sterna sandvicensis Latham, 1787, Gen. Synopsis Birds,suppl. 1, c. 296, Кент, Англия.
Белая окраска на внутренних опахалах первых трех первостепенных маховых рас-
пространена шире, чем у Th. s. acuflavidus, и достигает вершины пера. Размеры больше.
Замечания по систематике. Ряд систематиков относят пестроносую крачку к роду
Sterna, выделяя ее лишь в особый подрод Thalasseus (Бутурлин, 1934; Vaurie, 1965;
Степанян, 1975). Однако различия в морфологии и поведении дают право выделить
пестроносую крачку (вместе с Th. bergii, Th. maximus, Th. bengalensis, Th. zimmermanii,
310
Пестроносая крачка
Th. eurygnathus и Th. elegans, с которыми она образует компактную филогенетическую
группу) в отдельный род (Peters, 1934; Wolters, 1975; Howard, Moore, 1984).
Описание. Размеры (мм) (ЗМ МГУ) и масса тела (г) (Бородулина, 1960; Дол-
гушин, 1962; Заблоцкий,Заблоцкая, 1963),
Длина крыла:
66 (п = 16) - 295-324 (среднее 309);
99 (и = 11) — 292-318 (среднее 305) .
Длина клюва:
<5<5 (и = 18) — 49,7-60,2 (среднее 54,8) ;
99 (п = 11) - 50,3-56,5 (среднее 53,7).
Длина цевки:
с5с5 (и = 19) - 25,4-29,9 (среднее 28,2);
99 (п = 11) - 25,3-29,7 (среднее 27,4).
Масса тела:
<5<5 (и = 26) - 214-311 (среднее 254);
99 (и = 15) — 156-294 (среднее 234).
Окраска (Дементьев, 1951; Cramp, 1985). Самец и самка в брачном наряде.
Лоб, верхняя часть уздечки, темя, затылок и кроющие уха шелковисто-черные; перья
затылка удлинены и образуют хохол. Спина, плечевые перья, третьестепенные маховые
и кроющие крыла бледные сизо-серые — светлее, чем у речной и полярной крачек.
Третьестепенные маховые и часть длинных плечевых с белыми вершинами. Нижняя
часть уздечки, горло, шея, вся брюшная сторона, надхвостье и рулевые белые; наруж-
ные опахала наружных рулевых могут быть сероватыми. II (первое видимое) — VI пер-
востепенные маховые серебристо-серые на наружных опахалах и с белыми полями на
краях внутренних опахал, доходящими до вершин перьев. Проксимальные первосте-
пенные, начиная с VII, светлые, почти белые. Второстепенные маховые, как кроющие
крыла, с белыми вершинами и внутренними опахалами. Клюв черный с желтой верши-
ной, ноги черные, радужина темно-коричневая.
Самец и самка в зимнем наряде. Лоб и уздечка белые, темя белое с черными пестри-
нами. Перед глазом черное пятно, такого же цвета кроющие уха и затылок, последний
может быть с белыми пестринами. Остальное оперение, как в предыдущем наряде.
Пуховой наряд. Строение пуха своеобразное: на голове, шее, спине и верхней части
зоба он имеет волосовидный характер, поскольку отдельные бородки объединяются
у вершин общим чехликом. В результате птенец кажется необсохшим. Окраска пуха
варьирует от кремовой и светло-охристой до коричневато-серой, по верхней стороне
тела разбросаны многочисленные черноватые пятна и крапинки, иногда группирующие-
ся в неявные полосы. В целом окраска верхней части тела имеет отчетливый криптичес-
кий характер, пожалуй, в большей степени, чем у других наших чаек и крачек. Нижняя
часть тела беловатая или кремовая, горло может быть несколько темнее. Клюв голубо-
вато-серый, часто с черным предвершинным пятном. Ноги серые, иногда с розоватым
или желтоватым оттенком.
Гнездовой наряд. Лоб, верхняя часть уздечки, темя, затылок и кроющие уха черные
с охристыми каемками перьев и такими же пестринами, особенно густыми на лбу и
уздечке. Задняя сторона шеи белая с черноватыми отметками. Перья спины, плечевые,
третьестепенные маховые серые с охристой вершиной и темно-бурым предвершинным
311
Семейство Чайковые
V-образным пятном. Поясница и надхвостье белые, иногда с такими же пестринами.
Рулевые с белыми основаниями, средняя пара их с черноватыми предвершинными пят-
нами и охристой каймой, остальные с темно-серыми наружными опахалами, буровато-
серой широкой предвершинной полосой и беловато-охристыми вершинами. Нижняя
часть тела белая. Большие верхние кроющие крыла и кроющие маховых серые с белы-
ми вершинами, средние и часть малых кроющих могут быть либо светло-серыми, либо
такими, как плечевые. Остальные малые кроющие крыла темно-серые. II—IV первосте-
пенные маховые темнее, чем у взрослых, с меньшим развитием серебристого налета;
белое поле на внутреннем опахале не доходит до вершины пера. Проксимальные перво-
степенные с широкой беловатой каймой на вершине. Второстепенные маховые серые
с бурыми пятнами. Клюв обычно целиком черный, он короче, чем у взрослых птиц.
Первый зимний наряд. Голова и верхняя часть тела окрашены как у взрослых птиц
в зимнем наряде, однако маховые, рулевые и темно-серые малые кроющие крыла
остаются от предыдущего наряда. Летом второго календарного года птицы надевают
второй зимний наряд, минуя первый брачный. Весной третьего календарного года на-
девается частичный брачный наряд, при котором среди черных перьев лба, уздечки и
темени остаются белые перья предыдущего наряда.
Линька (Cramp, 1985). Линька в первый зимний наряд полная, начинается в большин-
стве случаев в конце августа — середине сентября (иногда в конце июля). К середине
зимы птица почти целиком в свежем пере, за исключением первостепенных маховых
и их кроющих и, редко, 4-й или 5-й пары рулевых. Смена первостепенных маховых
начинается в декабре—январе, завершается в мае—июле второго календарного года;
в июне—июле начинается повторная смена первостепенных (с XI) . Летом второго кален-
дарного года первый зимний наряд замещается вторым зимним. Последующие линьки,
как у взрослых птиц, но сроки их иногда смещены.
У взрослых птиц две линьки в году: полная послебрачная и частичная предбрачная.
Послебрачная начинается между серединой июня и серединой июля сменой оперения
на лбу, уздечке и темени. Линька оперения головы завершается в конце августа — кон-
це сентября. Рулевые начинают сменяться обычно во второй половине июля и заверша-
ют линьку к началу октября; приблизительная последовательность смены пар рулевых:
2, 1,3, 6, 4, 5-я. Оперение туловища и кроющие крыла завершают линьку в конце сен-
тября — конце октября. Начало смены первостепенных маховых отмечается в середине
июля — конце августа, завершается она к концу октября — декабрю. Предбрачная линь-
ка включает смену оперения головы, внутренних первостепенных и, возможно, всего
туловища и хвоста. Линька головы происходит обычно с середины февраля по конец
марта; смена внутренних первостепенных начинается в ноябре—декабре и прерывается
в конце января — феврале на стадии новых VI—VIII первостепенных. Смена оперения
туловища и хвоста, возможно, происходит в этот же период.
Распространение. Гнездовой ареал. Сплошного ареала пестроносая крачка
не образует, везде гнездится спорадично, отдельными поселениями (рис. 74). В Запад-
ной Европе пестроносые крачки гнездятся в Великобритании, Ирландии, на атлантичес-
ком и средиземноморском побережьях Франции, в Нидерландах, Дании, ФРГ (по край-
ней мере с начала XIX столетия), ГДР (с 1957 г.), южной Швеции (с 1911 г.) , на среди-
земноморском побережье Испании (с 1961 г.), в северо-восточной Италии и Румынии
(дельта Дуная). Нерегулярное гнездование отмечалось в Норвегии, Польше, Греции
(Cramp, 1985). В 1981 г. после длительного перерыва загнездилась в Болгарии (Нан-
кинев, Даракчиев, 1984). В СССР можно выделить три, по-видимому, достаточно изо-
лированных популяции: балтийскую (точнее, западноевропейскую, область гнездования
которой заходит на территорию СССР лишь своим крайним восточным участком) , азо-
во-черноморскую и каспийскую (рис. 75). Гнездование балтийских пестроносых кра-
312
Пестроносая крачка
чек отмечено лишь в Эстонии, где эти птицы гнездятся начиная с 1962 г. в небольшом
количестве и, видимо, не ежегодно (Манк, 1970; Пеэдосаар, Онно, 1970; Thomas, 1982,
цит. по: Cramp, 1985). Крачки азово-черноморской популяции заселяют северные побе-
режья Черного и Азовского морей от дельты Дуная до Кривой косы. Гнездовые посе-
ления этой популяции в настоящее время отмечены в дельте Дуная (Петрович, 1981),
не ежегодно в лиманах Бурнас и Телигульском Одесской обл. (Корзюков, 1981; Чер-
ничко, Стойловский, 1981), в Тендровском заливе (Черноморский заповедник),
Рис. 74. Область распространения пестроносой крачки
а — гнездовые поселения, б — места зимовки, в — залеты. Подвиды: 1 — Th.s, sandvicensis, 2 — Th.s.
acuflavidus
на о-ве Каланчакском в окрестностях г. Скадовска (Ардамацкая, 1975а,в, 1984),
в Картказацком заливе (Костин, 1975), на Чонгарских островах и о-ве Коянлы Сиваша,
Обиточном заливе, Молочном лимане (Филонов и др., 1974; Сиохин, 1981а), Кривой
и Белосарайской косах (Молодая, Сиренко, 1981; Букреев и др., 1986). По-видимому,
к этой популяции относится и поселение на Таманском полуострове (Кищинский,
19606).
Поселения каспийских пестроносых крачек отмечены в авандельте Волги, на
о-ве Морской Очиркин, в дельте Урала, на островах прибрежных мелководий восточ-
ной части северного Каспия (Бондарев, 1975; Кривоносов, 1976; Пославский, Криво-
носов, 1976). В 1951 г. отмечено гнездование на Тюленьих (Мангистауских) островах
(Гладков, Залетаев, 1956). До 1955 г. пестроносая крачка в небольшом количестве
гнездилась в заливе Кирова (Заблоцкий, Заблоцкая, 1963). В 1970-е годы на южном
313
Семейство Чайковые
Каспии колонии найдены в заливе Кара-Богаз-Гол, на о-ве Осушном Красноводского
залива и на о-ве Огурчинском (Васильев, Гаузер, 1975; Васильев и др., 1975; Щерби-
на и др., 1975; Щербина, 1979) .
В настоящее время можно констатировать расширение области гнездования западно-
европейской популяции на восток, в результате чего пестроносая крачка загнездилась
в Эстонии (Aumees et al., 1983) и все чаще встречается в период послегнездовых коче-
вок в Латвии (Виксне, 1983). Отмечено также расширение области гнездования азово-
Рис. 75. Ареал пестроносой крачки в СССР
1 — отдельные поселения, 2 — области гнездования разных популяций, 3 — места нерегулярных зи-
мовок, 4 — главные направления весенних миграций, 5 — главные направления осенних миграций,
6 — залеты
черноморской популяции на северо-восток вдоль северного побережья Азовского мо-
ря. Так, в конце прошлого — начале нашего века пестроносая крачка в восточной части
Азовского моря не отмечалась (Боровиков, 1907; Алфераки, 1910), хотя была доста-
точно обычной на Сиваше (Молчанов, 1906). В 1933 г. она гнездилась на о-ве Бирючем
(Шевченко, 1937), с 1952 г. загнездилась на Молочном лимане (Филонов, Лысенко,
1971), ныне отмечается на гнездовье вплоть до Кривой косы (Донецкая обл.). В отно-
шении каспийской популяции можно, по-видимому, говорить не об изменении границ
области гнездования, а о перераспределении поселений внутри нее (подробнее см.:
Пославский, Кривоносов, 1976).
Зимовки. Западноевропейская популяция зимует в основном на западном и юж-
ном побережьях Африки от Мавритании до мыса Доброй Надежды, часть птиц остает-
314
Пестроносая крачка
ся в Западной Европе (М011ег, 1981, цит. по: Cramp, 1985). Зимовки азово-черномор-
ской популяции расположены в Средиземном море (главным образом в его централь-
ной части: Греция — юг Италии - о-в Сицилия — Тунис; кроме того, в устье Нила, Се-
верной Италии, на средиземноморских побережьях Франции, Испании, Марокко и Ал-
жира) , реже на Черном и Азовском морях (юг Крыма, Черноморское побережье Кав-
каза, окрестности г. Темрюка), у западных побережий Португалии и Африки на юг до
Берега Слоновой Кости (Ардамацкая, 19756, 1977в; Бескаравайный, 1978). Каспий-
ские пестроносые крачки, по данным ЦКАН, встречаются зимой на Каспии по восточно-
му побережью от г. Шевченко до Красноводского залива, дагестанскому и азербай-
джанскому побережью, а также на крайнем юге Индии (г. Нагаркоил) и западном по-
бережье о-ве Шри-Ланка. Одна птица в возрасте 3 лет 7 месяцев отмечена в Намибии.
Каспийские птицы зимуют также в Персидском заливе, на юге Аравии и по побережью
Ирана и Пакистана (Кривоносов и др., 1978; Cramp, 1985).
Миграции. Пути миграций трех популяций различны. Западноевропейские крачки
после кочевок в июле—августе мигрируют вдоль атлантического побережья Европы
и Африки. Некоторые особи достигают мест зимовок в Африке уже в начале сентября
(отдельные молодые птицы — даже в начале августа), в ноябре большинство молодых
крачек отмечаются в тропической Западной Африке и лишь некоторые в Европе. Взрос-
лые птицы начинают покидать места зимовок уже в феврале (Cramp, 1985) .
Миграции азово-черноморских крачек изучены в основном на птицах Черноморского
заповедника (Ардамацкая, 1977в). В июле—августе часть крачек держится в окрест-
ностях мест размножения, другие откочевывают в западном и восточном направлениях.
Крачки, мигрирующие из Черноморского заповедника на восток, отмечаются в южном
Крыму, на северном и восточном побережьях Азовского моря. В сентябре продолжает-
ся пролет птиц вдоль западного побережья Черного моря на юг и по побережью Среди-
земного моря на запад; крачки достигают побережий Испании, Франции и Португалии.
В конце сентября — начале октября проходит основная волна пролета пестроносых
крачек по северному побережью Азовского моря (Лысенко, 1975), — по-видимому,
идет обратная миграция части птиц, откочевавших на восток. В октябре происходит
дальнейшее смещение основной массы птиц к западу и юго-западу. Последние птицы
покидают окрестности Черноморского заповедника и северный Крым (Костин, 1983)
в середине — конце октября. С ноября—декабря птицы держатся в местах зимовок,
которые начинают покидать в марте. В окрестностях Черноморского заповедника пер-
вые крачки появляются обычно в третьей декаде марта, иногда в конце февраля — на-
чале марта (Ардамацкая, 1977в), в северном Крыму — 27—31.III, с 10-х чисел апреля
здесь отмечаются регулярно (Костин, 1983). Вдоль северного побережья Азовского
моря пролет идет в начале апреля (Лысенко, 1975) .
О миграциях каспийских пестроносых крачек можно судить на основании возвратов
колец от птиц, помеченных в Красноводском заповеднике (ЦКАН) . В июле крачки дер-
жатся в основном в окрестностях мест размножения, в августе отмечаются по западно-
му и восточному побережьям Каспия, главным образом в Дагестане и Азербайджан-
ской ССР; единичные долетают до Астрахани. В сентябре распределение птиц сохра-
няется примерно прежним, но часть особей достигают иранского побережья Каспия
и южной оконечности Индии. В октябре—ноябре количество возвратов резко умень-
шается, но распределение птиц остается прежним. В октябре — начале ноября отмечен
пролет в окрестностях пос. Гассан-Кули на юго-востоке Каспия (Дементьев и др., 1955),
осенний пролет на юго-западном Каспии идет с конца июля до первых чисел ноября
(Заблоцкий, Заблоцкая, 1963). Мест зимовок птицы достигают, видимо, в ноябре—де-
кабре (часть особей остается зимовать на Каспии), покидают их, очевидно, в марте,
так как в конце марта — апреле проходит пролет на юго-западном, и, более выражен-
315
Семейство Чайковые
ный, на юго-восточном Каспии (Дементьев и др., 1955; Заблоцкий, Заблоцкая, 1963).
В Красноводском заливе пестроносые крачки появляются в разные годы 22—31.III
(Гаузер, Васильев, 1979).
Птицы в возрасте 1—2, реже до 3 лет отмечаются летом в местах зимовок, иногда
в окрестностях ближайших к местам зимовок поселений (Ардамацкая, 1977в; Криво-
носов и др., 1978; Cramp, 1985; ЦКАН). По данным ЦКАН, окольцованная птенцом
3.VII 1974 г. в Запорожской обл. крачка 11.VI 1977 г. встречена в Дании. Летят птицы
обычно группами от 2 до 20 особей. Залеты пестроносых крачек отмечались в Финлян-
дию, Чехословакию, Венгрию, на Азорские острова, острова Мадейра, Ян-Майен (Cramp,
1985), в Танзанию (Britton, Brown, 1974), в Харьковскую обл. (Ардамацкая, 1977в),
к Оренбургу (Зарудный, 1888) , на Аральское море на о-в Кыз-Арал и в устье Сырдарьи
(Зарудный, 1916; Спангенберг, Фейгин, 1936), в Рязанскую и Омскую области (Кри-
воносов и др., 1978).
Численность. В Эстонии в конце 1960-х годов нерегулярно гнездилось несколько
пар (Манк, 1970; Пеэдосаар, Онно, 1970), в 1973—1977 гг. — около 100 пар (Thomas,
1982, цит. по: Cramp, 1985). Численность азово-черноморской популяции растёт, что
видно по динамике численности птиц Черноморского заповедника (1935 г. — 5,6 тыс.
пар, 1946 г. — 10,2 тыс. пар, 1966 г. — 6,5 тыс. пар, 1976 г. — 23,6 тыс. пар) . Ныне азово-
черноморская популяция насчитывает, видимо, около 40 тыс. гнездящихся пар, причем
до 30,8 тыс. (1981 г.) гнездится в Черноморском заповеднике (Ардамацкая, 1977в,
1984; Корзюков, 1981; Молодая, Сиренко, 1981; Петрович, 1981; Сиохин, 1981а;
Черничко, Стойловский, 1981). На севере Каспия в начале — середине 1970-х годов
гнездилось 9—10 тыс. пар, на юго-востоке Каспия в 1973 г. — 9,5 тыс. пар (Васильев
и др., 1975; Кривоносов, 1976). Затем численность поселения на о-ве Осушной в Крас-
новодском заливе стала интенсивно расти, достигнув в 1985 г. 31,1 тыс. пар (Гаузер,
1986). Данных о численности колоний на северном Каспии в этот год мы не имеем;
если она осталась прежней, каспийскую популяцию можно оценить примерно в 40 тыс.
гнездящихся пар. Однако даже еслй большая часть каспийских крачек переселилась
в Красноводский залив, численность каспийской популяции не будет ниже 33 тыс. пар.
Общая численность вида в СССР составляет, таким образом, 75—80 тыс. гнездящихся
пар. В Западной Европе в конце 1970-х годов гнездилось 38—40 тыс. пар (Cramp, 1985).
Повсеместно отмечается довольно быстрый рост численности.
Местообитания. Во все сезоны — морские побережья, заливы, лиманы; для гнездо-
вания необходимы небольшие плоские островки. Внутренних водоемов в сезон раз-
множения, как правило, избегает, хотя в Ирландии отмечалось гнездование на островах
пресных озер (Cramp, 1985).
Размножение. Птицы приступают к размножению в возрасте 3—4 лет, изредка 2 лет
(Cramp, 1985). Пестроносая крачка — моногам, пары могут сохраняться до 4 лет.
птицы предпочитают образовывать пары с особями сходного возраста (Cramp, 1985).
Облигатно-колониальный вид. Гнездовой консерватизм почти отсутствует, характерна
постоянная смена мест гнездования (Пославский, Кривоносов, 1976). Даже если пти-
цы из года в год гнездятся на одном и том же острове, колонии ежегодно возникают
в новых его участках (Гаузер, 19836,в). Характерно резкое колебание численности
в поселениях по годам, вплоть до исчезновения их в отдельные годы (Филонов и др..
1974; Ардамацкая, 1975а, 1977в; Сиохин, 1981а; Гаузер, 19836,в; и др.) . Это связано,
по-видимому, как с перераспределением особей между поселениями, так и с негнездо-
ванием части птиц.
В демонстрационном поведении пестроносых крачек много общего с другими вида-
ми крачек (Cramp, 1985; наши данные). Однако в наземных демонстрациях они в боль-
шей степени, чем другие виды, разворачивают крылья, распластывая их параллельно
316
Пестроносая крачка
земле (рис. 76). Основная агрессивная наземная демонстрация - агрессивный стрекот
(быстрая сухая трелька ”кр-кр-кр-кр-кр”), который птица издает часто не сходя с гнез-
да, то вытягивая шею по направлению к противнику, то поднимая ее почти вертикально
(см. рис. 76,а). В отличие от чеграв, данный крик не составляет основного звукового
фона колонии. У пестроносой крачки, как и у чегравы, отмечена демонстрация, напоми-
нающая позу при долгом крике у чаек (см. рис. 76Д) . Крачка вытягивает шею и широ-
ко раскрытый клюв наклонно вверх и издает сухой скрипучий крик ”кррр”. Крылья
Рис. 76. Демонстрационное поведение пестроносой крачки
а — агрессивный стрекот, б — поза при крике ”кррр” (’’долгий крик”), в—г — брачные демонстра-
ции партнеров
при этом могут быть развернуты и подняты (но птица не взмахивает ими, как чеграва) .
Чаще, однако, крылья прижаты к телу. В последнем случае этот крик можно слышать
в ситуациях, когда птица чем-то обеспокоена или настроена агрессивно; нередко он пе-
реходит в агрессивный стрекот или стрекот завершается этим криком. Возможно,
данный крик гомологичен крику ’’киррряя” речной крачки.
Брачное поведение включает как воздушные, так и наземные демонстрации. Среди
первых характерен ’’высотный полет”, ’’низкий полет” не развит. Как и у других кра-
чек, в наземных демонстрациях часто используется ’’выпрямленная поза”, при которой
клюв может быть направлен как горизонтально, так и почти вертикально. В последнем
случае крачки слегка поворачивают головы то в одну, то в другую сторону вдоль про-
дольной оси. Как и у других крачек, в период брачных демонстраций и, реже, во вре-
мя насиживания самец кормит самку. Подношение рыбки обычно сопровождается
криком ’’кирлит”. Выпрашивая корм, самка бегает перед самцом, вытянув вверх
шею, взъерошив хохол и держа клюв горизонтально; время от времени она коротко
вскрикивает, слегка вскидывая клюв. После передачи рыбки самец принимает ’’вы-
прямленную позу” с клювом, направленным вверх (см. рис. 76,в). Перед спариванием
самка принимает ’’сгорбленную” позу: втягивает голову в плечи, слегка приподни-
мает раскрытый клюв и тонко пищит (см. рис. 76,г).
317
Семейство Чайковые
Формирование пары происходит вне территории колонии. Птицы, гнездившиеся
в предыдущем сезоне, появляются весной раньше и быстрее приступают к размноже-
нию, чем впервые размножающиеся особи (Cramp, 1985). По наблюдениям на
о-ве Осушной в Красноводском заливе, колония формируется следующим образом.
В последней трети апреля птицы образуют компактные предгнездовые скопления от
нескольких сот до 1,5—2 тыс. особей, приуроченные к отмелям, расположенным вблизи
места будущей колонии. Отдельные пары из состава предгнездовой группировки на-
чинают перемещаться вглубь острова и рыть ямки для гнезд; затем они вновь воз-
вращаются на берег. Это повторяется до тех пор, пока некоторое ’’критическое” число
пар не обоснуется окончательно на месте гнездового поселения. Гнезда первых птиц
не всегда располагаются максимально компактно, другие пары, вероятно, могут посе-
ляться между первыми гнездами и вокруг них. Гнездовая группа привлекает других
птиц, и колония быстро растет. Если предгнездовое скопление птиц по каким-либо при-
чинам покидает район начавшей формироваться колонии, птицы обычно оставляют толь-
ко что отложенные кладки (Гаузер, 1983а). Образование гнездовой группировки
может быть инициировано колониями других видов: на Чонгарских островах Сива-
ша — чайконосой крачкой, на Молочном лимане — шилоклювкой и отчасти морским го-
лубком, в Красноводском заливе — морским голубком (Сиохин, 1978; Гаузер, 1983в).
Размер колонии варьирует от нескольких пар до нескольких тысяч гнезд (Гладков,
Залетаев, 1956; Филонов и др., 1974; Зубакин, 1976в; Пославский, Кривоносов, 1976;
Гаузер, 19836; Ардамацкая, 1984). В некоторые годы колония в Нидерландах дости-
гала 35 тыс. пар (Cramp, 1985). Гнездование отдельными парами виду не свойственно,
хотя крайне редко и отмечается (Манк, 1970) . Колонии обычно состоят из нескольких
пространственно разобщенных участков — субколоний. Их величина также может до-
стигать нескольких тысяч гнезд, однако чаще всего составляет от 11 до 50 гнезд. В по-
добных субколониях все репродуктивные процессы идут, как правило, гораздо син-
хроннее, чем в колонии в целом: так, разница в откладке яиц птиц одной субколонии
обычно составляет 3—4 дня, тогда как во всей колонии она может достигать 40 дней
(Langham, 1974; Пославский, Кривоносов, 1976; Campredon, 1978; Гаузер, 19836).
По наблюдениям за меченными особями, разделение птиц на группы, которые затем
станут субколониями, может происходить еще до занятия территории колонии, в пред-
гнездовом скоплении (Smith, 1975).
Пестроносые крачки гнездятся только на островах, предпочитая невысокие песчаные
или ракушечниковые островки наносного происхождения, реже небольшие материковые
острова. Под колонии птицы занимают плоские, лишенные растительности наиболее
возвышенные районы острова, в отдельных случаях — участки с полегшей раститель-
ностью (Клименко, 1950а; Бородулина, 1960; Зубакин, 1976в; Пославский, Криво-
носов, 1976; Ардамацкая, 1981, 1984; Гаузер, 19836). Вместе с пестроносыми крач-
ками на островах могут гнездиться чайконосые, речные и малые крачки, морские го-
лубки, шилоклювки; в Западной Европе нередко и озерные чайки (Клименко, 1950а;
Fuchs, 1977; Сиохин, 1978). Плотность гнездования очень высока. Среднее расстояние
между центрами ближайших соседних гнезд на Сиваше и в Черноморском заповеднике
составляет (и = 326) 0,35 м (0,21—2,65 м), на о-ве Осушном в Красноводском зали-
ве — 0,32 м, в ФРГ (и = 39) 0,35 м, варьируя в пределах 0,24—0,45 м (Dircksen, 1932.
цит. по: Cramp, 1985; Гаузер, 1976; Зубакин, 1976в). В других местах пестроносые
крачки гнездятся ,со сходной плотностью (Гладков, Залетаев, 1956; Бородулина.
1960; Кищинский, 19606; Пославский, Кривоносов, 1976; Campredon, 1978; Щер-
бина, 1979). Индекс плотности гнездования в субколониях может быть очень высо-
ким — до 0,49—0,51 (Гаузер, 19836); рассчитанный по имеющимся данным в целом
для вида он равен 0,48.
318
Пестроносая крачка
Гнездо — простая' ямка в грунте без всякой выстилки, ее края сцементированы
пометом насиживающей птицы; в ряде случаев может быть выражен гнездовой валик.
В Черноморском заповеднике диаметр гнезда (мм) равен 200-285, в среднем 260,
диаметр лотка (ямки) 106-200, в среднем 140, глубина лотка 18-45, в среднем 31
(Бородулина, 1960); на Сиваше и Молочном лимане соответственно 260—350, в сред-
нем (и = 27) 300, 145—200, в среднем (п = 60) 169, 20—40, в среднем (и = 30) 30 (на-
ши данные).
Откладка первых яиц в разных популяциях происходит почти одновременно - в пер-
вую декаду мая. В Черноморском заповеднике это 1 —10.V, причем массовая откладка
яиц здесь отмечается во второй декаде мая (Ардамацкая, 1984), на Молочном лимане —
5—7.V (Сиохин, 1978), в Красноводском заливе — 30.IV—8.V (Гаузер, Васильев, 1979),
во Франции - первые числа мая (Campredon, 1978), в Шотландии - 6-8.V (Fuchs, 1977) .
В то же время, отдельные колонии могут начать формироваться значительно позже —
в середине и даже в конце июня, а свежие кладки можно встретить вплоть до июля (По-
славский, Кривоносов, 1976; Гаузер, Васильев, 1979). В полной кладке 1—2, реже 3,
крайне редко 4 яйца (Conover, 1984). В ранних кладках чаще всего бывает 2 яйца, в
поздних или повторных — 1 яйцо (Ардамацкая, 1977в) . Окраска яиц сложная, в целом
как у морского голубка. Фон очень светлый: светло-оливковый, светло-бежевый, свет-
ло-охристый, светлый голубовато-серый, почти белый. Рисунок в виде четких редких
разбросанных пятен и крапин. Глубокие пятна сероватые, поверхностные - контраст-
ные, темно-бурые. Размеры яиц (мм) : с Молочного лимана (п = 50) 48—57 X 32—38,
в среднем 54,3 X 37,4 (Филонов, Лысенко, 1971); из Черноморского заповедника 47—
60 X 34—39, в среднем 52 X 36 (Бородулина, 1960) ; с залива Кара-Богаз-Гол; 48,5—
63,6 X 35,7—42,5 (Щербина, 1979); из Западной Европы (и = 250) 44—59 X 33—40,
в среднем 51 X 36 (Cramp, 1985). Масса (г) насиженных яиц (и = 20) 32,0—43,0, в сред-
нем 36,8 (Филонов и др., 1974).
Насиживание начинается с первого яйца, длительность его 22—23 дня, иногда до
25 дней (Ардамацкая, 1977в) ; по другим данным 21—29, в среднем 25 дней (Langham,
1974; Smith, 1975). Насиживают оба партнера, по наблюдениям на Сиваше, смена партне-
ров бывает 4—12 раз в сутки (и = 4). Вылупление птенцов в гнезде из 2 яиц длится 2—
4 сут (и = 24), чаще всего (в 45,8% случаев) — 3 сут, несколько реже (37,5%) — 2 сут.
Скорлупу яиц после вылупления птенцов родители из гнезда не уносят (наши данные) .
В Красноводском заливе интервал вылупления в одном гнезде равен 1—4 сут (и = 38),
в среднем 2,3 сут (Гаузер, 19816). Вес вылупившегося птенца 22—24 г. Родители кор-
мят птенцов 8—12 раз в сутки (Бородулина, 1960). В возрасте 2—5 (в среднем 3,1 сут;
п = 59) крачки уводят птенцов из колонии в так называемые ясли — скопления птенцов,
с которыми держится некоторое количество взрослых птиц. Во время перехода через
колонию второй птенец выводка, которому в это время бывает 1—2 сут от роду, неред-
ко отстает, подвергается нападениям со стороны чужих взрослых птиц и, как прави-
ло, гибнет. В колонии бывают брошены 86,5% младших птенцов (Гаузер,
19816).
Во время тревоги в колонии птенцы в первые дни жизни затаиваются в гнезде, а
после оставления гнезда — в том месте, где их застигнет опасность. На 15—17-е сут подоб-
ная реакция на тревогу у птенцов исчезает и они, как и птенцы морских голубков,
в тревожной ситуации начинают сбиваться в плотный табунок (Зубакин, 1976в), кото-
рый, однако, может формироваться не во всех колониях (Smith, 1975). Стратегия
защиты потомства у пестроносых крачек двоякая. С одной стороны, используется
эффект плотности (отпугивающее воздействие плотной массы сидящих птиц), с дру-
гой стороны — активная защита потомства: пикирование на врага, удары клювом.
Активная защита может быть весьма действенной: пестроносые крачки совместно с
319
Семейство Чайковые
озерными чайками способны насмерть забить ворону (Fuchs, 1977). Родители опозна-
ют своих птенцов в яслях (Smith, 1975) .
Полностью лётными птенцы становятся в возрасте 32—35 дней (Ардамацкая, 1984),
по другим данным — 28—30 дней (Smith, 1975). Родители продолжают докармливать
их и в августе — в период осенних миграций (Резанов, 1978) .
По наблюдениям в Красноводском заливе, в отсутствие наземных и пернатых хищ-
ников выживает 77,7% птенцов от числа отложенных яиц, или 1,3 птенца на пару взрос-
лых птиц. Отход яиц незначительный, колеблется по годам от 1,9% до 13,8%, причем
основная причина эмбриональной смертности (62,5% случаев) — оставление вторых
яиц после вылупления первого птенца. Гибель птенцов за все сезоны здесь составляет
в среднем около 15,0%, причем в 96,6% случаев птенцов забивают взрослые пестроно-
сые крачки (Гаузер, 1983а). В Черноморском заповеднике смертность птенцов незна-
чительная. Основные причины отхода яиц и птенцов здесь — затопление колоний и
разорение гнезд хищниками (Ардамацкая, 1984). Детальные сведения о связи успеха
размножения с плотностью гнездования, синхронностью откладки яиц, размером и фор-
мой колонии и т.д. изложены в работах Н. Лангхама (Langham, 1974), Дж. Вина (Veen,
1977) и М.Е. Гаузер (1981а,б, 1983а-в).
Максимальная продолжительность жизни, по данным кольцевания, 23 года 7 меся-
цев (Rydzewski, 1978, цит. по: Cramp, 1985).
Суточная активность, поведение. Активность дневная, но птицы могут добывать
корм при сравнительно низкой освещенности (Авилова, 1972). По наблюдениям на Си-
ваше, в лунные ночи крачки улетают из колонии и возвращаются в нее на протяжении
всей ночи. Свободные от насиживания птицы могут ночевать как на краю колонии, так
и вне ее (наши данные).
Пестроносая крачка — общественная птица как в сезон гнездования, так и вне его.
Кормится обычно в одиночку, хотя в местах обилия корма иногда собирается несколь-
ко сот особей. На ночевке и отдыхе могут образовываться скопления в несколько ты-
сяч особей (Ардамацкая, 1977в; Cramp, 1985) . По данным кольцевания, у пестроносых
крачек существуют постоянные по составу стайки из птиц одного возраста, которые
держатся вместе во время пролета, на зимовках и, возможно, одновременно возвра-
щаются к местам гнездования. В то же время по сравнению с черноголовой чайкой
у пестроносой крачки чаще встречаются смешанные разновозрастные группы (Ар-
дамацкая, 1977в).
Питание. Пестроносая крачка — почти абсолютный интиофаг, изредка она может
поедать мелких ракообразных (креветок) и очень редко — снесенных в воду насе-
комых (Шевченко, 1937; Клименко, 1950а; Гладков, Залетаев, 1956; Смогоржевсь-
кий, 1959; Бородулина, 1960, 1965; Зубакин, 1976в; Сиохин, 19816). Спектр пита-
ния зависит от наличия в данном месте тех или иных видов рыб. На Лебяжьих остро-
вах, в Присивашье и Черноморском заповеднике это главным образом бычки, на Азов-
ском море — атеринка, реже тюлька (Шевченко, 1937; Бородулина, 1960; Костин,
1983), на юго-западе Каспия — годовики сазана, сельди, кильки и кефали (Заблоц-
кий, Заблоцкая, 1963), в районе Мангышлака — килька (Залетаев, I960) . Крачка обыч-
но кормится в неглубоком (1,5—2 м) морском заливе, летая на высоте 10 м и более
над водой. Заметив рыбу, ныряет с разлета, совершенно скрываясь под водой (Бороду-
лина, 1960). Есть сведения о кормовых разлетах крачек на 60—70 км от колонии (За-
летаев, 1960).
Враги, неблагоприятные факторы. Потомство пестроногих крачек нередко уничто-
жают серебристые, черноголовые и озерные чайки, врановые птицы; из наземных
хищников - изредка горностай (Fuchs, 1977; наши данные). Определенный урон мо-
жет наносить степная гадюка (Ардамацкая, 1984). Значительное количество гнезд и
320
Речная крачка
птенцов гибнет в результате штормов и затопления от нагонных ветров (Костин, 1983;
Ардамацкая, 1984). Большое влияние на успех размножения могут оказывать соци-
ально обусловленные факторы — агрессивность взрослых птиц по отношению к птен-
цам, оставление второго яйца в кладке и младшего птенца после вылупления старшего
ит.д. (Гаузер, 1981а,б, 1983а—в).
Хозяйственное значение, охрана. Хозяйственного значения пестроносая крачка не име-
ет. Занесена в Красную книгу Эстонской ССР. Колонии охраняются в Черноморском
и Красноводском заповедниках (около 50% всей палеарктической популяции).
Род Sterna Linnaeus, 1758
Речная крачка —
Sterna hirundo Linnaeus, 1758
Common Tern (англ.), Flussseeschwalbe (нем.), Sterne pierregarin (фр.).
Син. (лат.): Sterna fluviatilis Naumann, 1819.
Син. (рус.) : обыкновенная крачка.
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Среднего размера крачка, мельче пестро-
носой и чайконосой, но крупнее болотных крачек. Общая окраска спинной стороны си-
зо-серая, брюшной — сероватая. На голове черная шапочка. Крылья очень длинные и
узкие, хвост вильчатый, глубоко вырезанный. От полярной крачки отличается более
коротким хвостом (у сидящей речной крачки хвост не выдается за сложенные концы
крыльев). Речные крачки, обитающие на Северо-Востоке СССР, в сезон гнездования
отличаются от полярных черным клювом (у последних он красный). Различение евро-
пейских речных и полярных крачек по цвету клюва ненадежно. Молодые птицы в поле-
вых условиях практически неотличимы.
Голос разнообразный, преобладают скрипучие тона. Характерен громкий крик
’’киррр” или ’’киаррр” (по И.Д. Никольскому (1975) — .’’кирря”), который можно
слышать как в колонии, так и вне ее. По-видимому, это крик тревоги или беспокойст-
ва. Во время тревоги в колонии часто слышны короткие крики ”кик”. В период брач-
ных демонстраций, а также во время агрессивных столкновений можно слышать ’’рек-
ламный” крик, который в разных вариациях звучит как ”киир”, ”кии-ири”, ”кри-ир”,
а нередко и более продолжительно: ”ки-ки-ир, ки-киир”. С этим же криком крачка
возвращается в колонию; его можно слышать также от самца во время ритуальной
кормежки (самка при этом тонко пищит ”ки-ки-ки-ки”). Более подробно о криках
речных крачек смотри у С. Крэмпа (Cramp, 1985). Спектральные характеристики
криков приведены у И.Д. Никольского (1975, 1978).
Подвидовая систематика. Обычно различают 4 подвида (Peters, 1934; Vaurie, 1965).
Изменчивость проявляется в окраске оперения верхней и нижней стороны тела, в окрас-
ке клюва и ног, в размерах крыла и клюва. Все четыре подвида обитают на территории
СССР (Степанян, 1975).
1. Sterna hirundo hirundo
Sterna hirundo Linnaeus, 1758, Syst. Nat., ed. 10, c. 137, Швеция.
Наиболее светлая раса, без буроватого налета в оперении. Клюв красный с черной
вершиной, ноги красные.
2. Sterna hirundo minussensis
Sterna hirundo minussensis Sushkin, 1925. Список и распространение птиц Русского Ал-
тая, с. 64, Новоселове на Енисее, Минусинский район.
И.Зак.826 321
Семейство Чайковые
Таблица 19. Речная крачка. Размеры особей (мм) (ЗИН, ЗМ МГУ) и масса тела (г)
(Дементьев, 1951; Бородулина, 1960; Долгушин, 1962; Скрябин, 1977)
... Подвид Пол Длина крыла Длина клюва
п lim среднее п lim среднее
S.h. hirundo <5 <5 31 260-288 271 29 31,6-38,5 35,4
(европейская часть 9 9 25 252-286 270 27 31,0-36,9 34,1
СССР, Западная Сибирь) S.h. minussensis dd 16 262-287 272 17 30,8-36,2 33,6
(Алтай, окрестности Минусинска, Байкал) 9 9 7 250-272 264 6 30,2-35,3 32,7
S.h. tibetana dd 9 265-279 273 9 29,9-37,8 33,2
(Памир, Тибет) 9 9 7 265-288 278 9 28,1-33,9 31,3
S.h. longipennis dd 22 258-291 273 25 31,3-37,1 34,5
(Якутия, северо-восток Сибири, Дальний Восток) 99 7 266-277 271 7 27,8-35,2 32,6
Темнее сверху и снизу, чем номинальная раса. Черная окраска на клюве распростра-
нена шире. Окраска ног варьирует от красной до буровато-красной.
3. Sterna hirundo longipennis
Sterna longipennis Nordmann, 1835, Erman, Verz. Thieren und Pflanzen, c. 17, дельта реки
Кухтуй у Охотска.
Верхняя сторона тела, как у предыдущего подвида, нижняя сторона темнее. Черная
окраска клюва распространена еще шире, и у многих особей (особенно у восточных
популяций) клюв целиком черный. Окраска ног варьирует от красновато-бурой до
черно-бурой.
4. Sterna hirundo tibetana
Sterna tibetana Saunders, 1876, Proc. Zool. Soc. London, c. 649, Тибет.
Наиболее темная раса, на верхней стороне тела развит буроватый налет, на ниж-
ней — сероватый. Клюв красный с черной вершиной, ноги красные.
Замечания по систематике. С.А. Бутурлин (1934) считал SJi. longipennis отдельным
видом, указывая на совместное обитание этой расы с речной крачкой на территории от
Абаканской степи до восточной Монголии. Впоследствии крачки этого региона были
отнесены к расе S.h. minussensis, которую в силу неоднотипности принадлежащих к
ней птиц обычно рассматривают в качестве гибридной между S.h. hirundo и S.h. longi-
pennis (Дементьев, 1951; Vaurie, 1965). В этой связи некоторые авторы даже не выде-
ляют S.h. minussensis в качестве подвида (Cramp, 1985).
Описание. Строение и размеры (табл. 19).
Окраска (Дементьев, 1951; Cramp, 1985). Самец и самка в брачном наряде. Лоб,
верхняя часть уздечки, темя и затылок черные; граница черного цвета проходит по ниж-
нему краю глаза. Нижняя часть уздечки, подбородок, щеки, горло, нижние кроющие
крыла и хвоста белые. Белый цвет передней части шеи и надхвостья постепенно перехо-
дит в сероватую окраску груди и брюшка, которая слабее всего развита у европейских
и западносибирских птиц и более интенсивная у крачек на востоке нашей страны и в
горах Центральной Азии. Иногда на груди и брюшке заметен слабый розовый оттенок.
Задняя часть шеи сероватая; спина, плечевые, третьестепенные маховые и верхние крою-
щие крыла светло-серые, темнее брюшка; они более темные у крачек из Восточной Си-
бири и с буроватым налетом у крачек с Памира и Тибета. Поясница и верхние кроющие
322
Речная крачка
Длина цевки Масса тела
п lim | среднее п lim среднее
32 29 18,8-21,7 20,3 18,5-22,2 20,1 57 39 W1-146 97—175 122 128
17 18,7-21,8 20,4 41 120,9 ± 1,31
7 19,3-20,8 20,1 20 — 121,5 ± 0,60
9 19,4-21,8 20,3 — — —
9 18,8-21,8 19,9 — — —
22 17,6-21,1 19,7 4 95-118 110
7 17,4-21,2 19,6 — — —
хвоста белые. Средние рулевые перья белые или светло-серые, прочие рулевые с белы-
ми стволами и внутренними опахалами и серыми наружными опахалами, причем серый
цвет темнее на двух крайних парах. II—V первостепенные маховые с темно-серым на-
ружным опахалом и вершиной; внутренние опахала с обширным белым полем, серый
цвет остается здесь на вершине пера и полосой в 3—6 мм вдоль стержня пера. Стволы
этих перьев белые. VII и последующие первостепенные серые с беловатыми узкими
каймами по краю внутренних опахал. В свежем пере все маховые с серебристым нале-
том, слабеющим по мере обнашивания пера. Второстепенные маховые серые с белыми
вершинами и белым краем внутренних опахал. Клюв ярко-красный с черной вершиной,
в некоторых случаях целиком красный; по мере продвижения к восточным границам
ареала черный цвет вершины клюва распространяется на весь клюв и восточнее Забай-
калья клюв становится целиком черным. Ноги красные, радужина темно-коричневая.
Самец и самка в зимнем наряде. Лоб и уздечка белые, темя белое с темно-серыми
или черноватыми пестринами, перед глазом черное пятно, под глазом черные пестрины.
Задняя часть шеи белая, иногда с серыми вершинами перьев. Серая окраска спинной
стороны бледнее, чем в брачном наряде. Нижняя часть тела белая. Поясница и верхние
кроющие хвоста серые, рулевые с серым налетом. Клюв черноватый с более или менее
развитым красным оттенком у основания или же черный. Ноги красновато-бурые.
Пуховой наряд. Основной тон спинной стороны довольно светлый, бледно-охристый,
охристо-буроватый или серый; на темени, затылке, спине и крыльях немногочисленные
черноватые крапинки, на голове и спине нередко группирующиеся в неявные полоски.
Горло, уздечка, щеки и реже лоб серовато-бурые или черно-бурые. Грудь, брюшко,
подбородок и пятно у основания клюва белые. Клюв розовый или оранжевый с черно-
ватой вершиной, ноги оранжевые или розоватые, радужина темно-коричневая.
Гнездовой наряд. Окраска головы и верхней части туловища сильно варьирует. Лоб
и уздечка белые, обычно с охристым налетом. Темя и затылок матово-черные, на теме-
ни с беловатыми или охристыми пестринами. Перед глазом черное пятно, кроющие
уха сероватые или черноватые. Перья задней стороны шеи с белыми основаниями,
серыми предвершинными пятнами и охристыми каемками. Перья спины, плечевые,
третьестепенные маховые и большинство верхних кроющих крыла светло-серые с бе-
ловатыми, охристыми или рыжеватыми каемками и обычно с буроватой поперечной
323
Семейство Чайковые
предвершинной полосой. Верхний ряд малых кроющих крыла белый, остальные —
темно-бурые, образующие на крае крыла темную карпальную полосу. Перья поясницы
и верхние кроющие хвоста светло-серые с белыми каемками. Горло, щеки, передняя
часть шеи, грудь и брюхо белые, последние нередко с охристым оттенком. Рулевые
светло-серые, с охристыми вершинными каемками и нередко темными предвершинны-
ми пятнами. Наружные опахала 2—3 крайних пар рулевых черноватые. Маховые серые,
темнее, чем у взрослых (особенно внутренние второстепенные), с белыми вершинами.
Клюв черновато-бурый с желтоватым основанием, ноги розовато-оранжевые или желто-,
вато-оранжевые, радужина темно-коричневая.
Первый зимний наряд. Напоминает зимний наряд взрослых птиц, но маховые, руле-
вые, задние плечевые и многие кроющие крыла остаются от гнездового наряда. После
первого зймнего крачки надевают второй зимний наряд, минуя стадию первого брач-
ного наряда. Во втором зимнем наряде лоб и часть темени белые, на крыле сохраняется
темная карпальная полоса, нижняя часть тела белая, хвост короткий, клюв черный.
Первостепенные маховые более тусклые и темнее, чем у взрослых. Клюв обычно цели-
ком черный, ноги красновато-бурые. Второй брачный наряд очень изменчив. В одних
случаях он напоминает второй зимний с элементами брачного наряда, в других похож
на брачный наряд взрослых птиц, но с белым лбом, белой нижней частью тела и остат-
ками темной карпальной полосы; некоторые птицы во втором брачном наряде очень
похожи на взрослых, но сероватый цвет нижней части тела светлее. Клюв либо чер-
ный, либо как у взрослых. В третьем брачном наряде у некоторых птиц сохраняются
отдельные белые перья на лбу и уздечке и темные — на месте карпальной полосы.
Линька (Cramp, 1985). Линька в первый зимний наряд полная. Иногда начинается
перед осенней миграцией, обычно же — на зимовках, в ноябре—декабре (иногда в фев-
рале). Смена оперения головы, туловища, хвоста и большинства кроющих крыла завер-
шается к февралю—марту; линька первостепенных маховых начинается в январе—фев-
рале (реже в декабре) и завершается в июне—августе; у некоторых особей приостанав-
ливается на стадии новых III—V перьев. Повторная смена первостепенных начинается
в мае—июле второго календарного года, протекает она медленно, часто приостанавли-
вается летом и завершается в марте—июне третьего календарного года. В июне—августе
второго календарного года первый зимний наряд головы, туловища и хвоста замещает-
ся непосредственно вторым зимним нарядом. Третья серия линьки первостепенных на-
чинается в декабре—феврале третьего календарного года. Она прерывается в мае—июне
у тех птиц, которые посещают места гнездования, и лишь изредка приостанавливается
у особей, которые остаются на лето в местах зимовки; завершается в августе—октяб-
ре. Частичный брачный наряд (40—90% оперения головы и нижней части туловища,
обычно меньшее количество перьев верхней части тела и крайне изменчивое число
кроющих крыла) надевается в феврале—июне третьего календарного года. Последую-
щие линьки, как у взрослых птиц.
У взрослых бывает две линьки в году: полная послебрачная и частичная предбрач-
ная. Линька оперения крыльев начинается с XI первостепенного в начале июля — конце
августа, реже в конце мая — июне или в начале сентября. Она приостанавливается во
время миграции в конце августа — сентябре на стадии 1 — 4 новых внутренних перво-
степенных. Остальные первостепенные сменяются на зимовке к концу января — началу
марта. Линька рулевых начинается примерно в то же время, что и маховых, завершается
она на зимовках. Линька головы, туловища и кроющих крыла проходит главным обра-
зом на зимовках в октябре—январе; небольшая часть оперения может смениться до
осенней миграции. У некоторых особей послебрачная линька может начинаться лишь на
зимовках с октября — начала ноября. В предбрачную линьку сменяется оперение голо-
вы, туловища, хвоста, верхних кроющих крыла, проксимальных первостепенных и дис-
324
Речная крачка
тальных второстепенных. Она начинается с XI первостепенного и рулевых в декаб-
ре-феврале, заканчивается в марте—апреле, когда завершается линька головы, тулови-
ща, хвоста; линька первостепенных маховых прерывается главным образом на стадии
VII—VIII новых первостепенных. У некоторый особей, раньше приступивших к линьке,
может начаться повторная смена внутренних первостепенных, прерывающаяся на стадии
новых X—XI перьев.
Общее течение линьки у крачек из восточных частей ареала, видимо, такое же, как и
из западных. Однако, возможно, послебрачная линька и линька в первый зимний наряд
проходят здесь в более ранние сроки, поскольку во второй половине сентября отмеча-
лись взрослые особи, закончившие послебрачную смену мелкого оперения, а 22.IX в
Приморье добыта молодая птица в первом зимнем наряде (Дементьев, 1951). О линьке
тибетских и памирских крачек сведений нет.
Распространение. Гнездовой ареал. Широко распространена по всей Пале-
арктике, кроме Крайнего Севера, где ее заменяет полярная крачка. Населяет также
Северную Америку (рис. 77). Обитает во всех странах зарубежной Европы, за исключе-
нием Португалии, где гнездится лишь изредка. Заселяет все норвежское побережье,
заходя по нему севернее 70° с.ш. На юге отдельные поселения известны в Нигерии,
Сенегале, Мавритании, на Азорских островах и о-ве Мадейра, в Тунисе, Израиле; нере-
гулярные - в Марокко, Ливии, Сирии, на Кипре. Кроме того, изолированные участки
гнездового ареала расположены в Турции, Ираке, Иране, Афганистане и Пакистане
(Cramp, 1985).
Северная граница сплошного ареала вида на территории СССР (рис. 78) проходит
по северу Карелии (Зимин, Ивантер, 1974), по-видимому по низовьям р. Онеги (Кор-
неева и др., 1984), севернее р. Вычегды (Гурьев, 1982), вероятно по среднему течению
р. Печоры (Естафьев, 19816), пересекает Урал в окрестностях Денежкина Камня (Де-
ментьев, 1951), по долине Оби и ее притокам поднимается к северу до широты Салехар-
да (Бойков, 1965; Данилов и др., 1984), в бассейне р. Таз проходит севернее 64° с.ш.
(Скалой, Слудский, 1941). На Енисее граница гнездового ареала проходит где-то между
66° и 68° с.ш. (Рогачева, Вахрушев, 1983; Рогачева и др., 1983), далее ареал включает
бассейн Нижней Тунгуски (Ткаченко, 1937, цит. по: Дементьеву, 1951). По долине
р. Лена граница ареала достигает 69° с.ш. (Дегтярев и др., 1981а), в восточной части
Якутии она проходит, по-видимому, в районе полярного круга (Воробьев, 1963),
причем по Колыме поднимается к северу до 67,5° с.ш. (Бутурлин, 1934). Далее к
востоку гнездовой ареал включает долину среднего течения р. Анадырь (Портенко,
1939), его граница проходит по северному краю Корякского нагорья (Кищинский,
1980); в бассейне Канчалана речных крачек не отмечали (Кищинский и др., 1983).
Речные крачки относительно равномерно заселяют Камчатку (Лобков, Головина,
1978), в том числе о-в Карагинский (Каверкина, 1982), а также о-в Сахалин (Гизенко,
1955) и острова заливов Петра Великого и Ольги на юге Приморья (Лабзюк и др.,
1971; Лабзюк, 1975). Достоверных доказательств гнездования на Курильских островах
нет (Нечаев, 1969). Почти неизвестен статус речной крачки на северном и северо-запад-
ном побережье Охотского моря. Гнездование здесь отмечалось в окрестностях Магадана
(Васьковский, 1956) и в Пенжинском районе (Яхонтов, 1979); в Колымском нагорье
речной крачки нет (Кищинский, 1968), на Шантарских островах и о-ве Ионы она отме-
чалась лишь изредка в период весеннего и осеннего пролета (Дулькейт, Шульпин, 1937;
Харитонов,1980а).
Южная граница гнездового ареала проходит между реками Хуанхэ и Янцзы, далее по
южной границе Тибета, включая Ладакх (Ali, Ripley, 1969; Vaurie, 1972; Cheng Tsoh-
sin, 1976). В Корее и Японии не гнездится (Hand list of the Japanese Birds, 1958; Won
Pyong-Oh, 1976). Далее на запад южная часть гнездового ареала речной крачки ста-
325
326
160 120 80 40 0 40 80 120 160 160
Рис. 77. Область распространения речной крачки
а — область гнездования и отдельные поселения, —неуточненная граница области гнездования,в — места зимовок, г — залеты.
Подвиды: L - S.h. hirundo, 2 — S.h. minussensis, 3 — S.h. longipennis, 4 — S.h. tibetana
327
Рис. 78. Ареал речной крачки в СССР
Пунктиром обозначена неуточненная граница
Семейство Чайковые
ловится разорванной: крачки селятся по долинам рек и отдельным водоемам и отсут-
ствуют на обширных безводных пространствах. В Средней Азии речные крачки гнез-
дятся в Киргизии, кроме ее юго-западной части (Янушевич и др., 1959; Кыдыралиев,
1975; Шукуров, 1981), заселяют северный, центральный, равнинную часть южного и
юго-западного Таджикистана, Памирское нагорье, долины рек Зеравшан, Сырдарья,
Амударья, Сурхандарья, оз. Айдаркуль и трассу Каракумского канала (Дементьев,
1952; Потапов, 19666; Ташлиевидр,, 1967; Иванов, 1969; Абдусалямов, 1971; Кенже-
гулов, 1972; Абдреимов, 1975; Сагитов, Салимов, 1975; Эминов, 1975; Остапенко и
др., 1978, 1984). На юго-восточном Каспии поселения отмечены у пролива Кара-Богаз-
Гол (Щербина, 1979), в Красноводском заливе, на о-ве Огурчинском (Васильев и др.,
1975). Нерегулярное гнездование отмечается в низовьях Атрека (Дементьев, 1952;
Караваев, 1979).
Речная крачка достоверно гнездится в республиках Закавказья, хотя сведения о
конкретном распространении здесь этого вида немногочисленны (Ляйстер, Соснин,
1942; Виноградов, 1967; Туаев, Васильев, 1969; Жордания, 1975; Туаев, 1975). В
Крыму гнездится в его северной и западной частях (Костин, 1983).
Номинальный подвид заселяет Северную Америку, Европу и Азию на восток до
средней части Западной Сибири. Подвид Sh. minussensis распространен от средних
частей Западной Сибири к востоку до долины Аргуни и оз. Буир-Нур. Долину Нижней
Тунгуски и всю восточную часть ареала от юго-восточного Забайкалья заселяет S.h. lon-
gipennis. Подвид S.h. tibetana распространен на Памире, в Ладакхе, Тибете к востоку
до Цайдама, Куконора и Ганьсу (Дементьев, 1951; Степанян, 1975).
Тенденции изменения границ ареала не ясны; в ряде районов, возможно, имеет
место некоторое расширение ареала к северу.
Зимовки. Область зимовки палеарктических крачек включает все побережье
Африки, Красное море, Персидский залив, морские побережья от Адена до Бангладеш,
а также побережья п-ова Малакка, островов Суматра, Ява, Новая Гвинея и ряда других,
западное побережье Австралии. Изредка крачек отмечают в зимнее время также на
атлантическом побережье Европы и в Средиземном море (Дементьев, 1951; Cramp,
1985).
Крачки из Прибалтики, по данным кольцевания, отмечаются зимой в южной части
Африки (Ангола, ЮАР), здесь же находятся зимовки птиц из северного Причерноморья.
Возможно, часть черноморских крачек зимует в Средиземном море (отдельные особи,
видимо, и в Черном море: у южного берега Крыма и в Керченском проливе), а неко-
торые птицы из Прибалтики — в Западной Европе (Таманцева, 1955; Виксне, 1962;
Лебедева, 1962; Вайткявичюс, Скуодис, 1965; Ардамацкая, 1977в; Мальчевский,
Пукинский, 1983; Стойловский, 1986). Зимовки крачек северного Каспия отмечены
в Кении и Саудовской Аравии (Кривоносов и др., 1978). Видимо, изредка крайне
незначительная часть каспийских крачек остается зимовать на юге Каспия (Доброхотов,
1962, 1963), отдельные случайные птицы отмечаются даже у п-ова Мангышлак (Послав-
ский, и др., 1964). Крачки западной Сибири, Казахстана и Средней Азии, по-видимому,
зимуют на побережьях Ирана, Пакистана и Индии. Возможны зимовки и на внутренних
водоемах юга Средней Азии. Так, речных крачек в декабре — январе 1975 — 1976 гг.
отмечали в долине р. Пяндж на юге Таджикистана (Абдусалямов и др., 1980). Область
зимовок S.h. minussensis, по-видимому, находится где-то в Южной Азии. SJi. longipen-
nis зимует, видимо, в Индонезии, на побережьях о-ва Новая Гвинея и п-ова Малакка
(Дементьев, 1951). S.h. tibetana зимует в Индии, Бангладеш, достигая на юге о-ва
Шри-Ланка (Ali, Ripley, 1969).
Миграции. Прибалтийские крачки мигрируют по южному побережью Балтийского
моря и далее вдоль атлантического побережья Европы и Африки. Отлет начинается в
328
Речная крачка
первых числах августа или в конце июля, большинство птиц покидает регион в конце
августа — начале сентября. Последних птиц в Латвии отмечали 5. IX, в Ленинградской
обл. — 20.IX. Весной речные крачки отмечаются на побережье в Прибалтике с конца
апреля (Михельсон, 1953; Петрайтис, 1975), на оз. Жувинтас — 18—23.IV (Станявичюс,
1983); в Ленинградском обл. - с 4-5.V, пролет здесь идет до конца мая (Носков и
др., 1981; Мальчевский, Пукинский, 1983). Крачки, окольцованные в Черноморском
заповеднике, начинают кочевать во второй половине июля — августе. Кочевки имеют
как западное (основное), так и восточное направления. В августе птиц отмечают от
восточного побережья Азовского моря до Болгарии. В сентябре—октябре кочевки как
к западу, так и к востоку от Черноморского заповедника продолжаются. К ноябрю
речные крачки достигают Сицилии, откуда затем направляются в Африку (Лебедева,
1962; Ардамацкая, 1977в). Крачки Центра европейской части СССР, по-видимому,
мигрируют в южном направлении (Таманцева, 1955), зимуя, скорее всего, там же,
где 11 черноморские.
Осенний пролет крачек северных, центральных и южных областей европейской
части СССР начинается почти одновременно — в конце июля — начале августа; в север-
ных и центральных областях он завершается в конце августа — начале сентября (Страут-
ман, 1963; Птушенко, Иноземцев, 1968; Плесский, 1976; Матвиенко, 1978; Зиновьев,
1980; Корнеева и др., 1984), а в южных растягивается на весь сентябрь и завершается
в начале (реже в середине) октября (Лысенко, 1975; Булахов, Мясоедова, 1978;
Аверин и др., 1981; Казаков, Языкова, 1982; Костин, 1983; Ардамацкая, 1984). Вес-
ной первые крачки появляются в Северном Причерноморье в конце марта — начале
апреля, в большом количестве в 20-х числах апреля (Ардамацкая, 1977в; Стойловский,
1981; Костин, 1983), в Приазовье пролет идет в первую - вторую декаду апреля (Лы-
сенко, 1975; Казаков, Языкова, 1982). На Западной Украине первых птиц отмечают
в начале апреля — начале мая (Страутман, 1963), в Молдавии — в конце апреля —
начале мая (Аверин и др., 1981), в Белоруссии — с первых числе мая (Федюшин, Дол-
бик, 1967), под Киевом — 28.IV—16.V (в среднем за 11 лет 5.V), в Харьковской обл. —
22.IV— 11.V (в среднем за 7 лет 4.V) (Кютякивскш, 1957), в Московской обл. —
17.IV—10.V (в среднем 2.V); массовый пролет здесь идет 9—20.V, в это же время от-
мечен пролет крачек в Калужской обл. (Птушенко, Иноземцев, 1968; Баранов, 1978).
В Кировскую обл. первые крачки прилетают в последних числах апреля — начале мая,
массовое появление приходится на вторую половину мая (Плесский, 1976); в низовьях
р. Онеги пролет идет в начале мая (Корнеева и др,, 1984).
Крачки северной части Каспия в период послегнездовых кочевок отмечаются вблизи
места рождения, а также в Волгоградской обл. и Ставропольском крае; осенью они
кочуют на юге Каспия (Кривоносов и др., 1978). В дельте Волги осенний пролет и отлет
заканчивается в конце сентября, причем первыми исчезают взрослые птицы. Осенний
пролет на южном Каспии проходит с августа до начала октября. Весенний пролет на юго-
восточном Каспии прослеживается с середины марта и идет весь апрель, на юго-западном
Каспии — с 19—20.IV по начало мая; в дельте Волги прилет отмечается со второй поло-
вины апреля (Дементьев и др., 1955; Луговой, 1958; Заблоцкий, Заблоцкая, 1963;
Караваев, Белоусов, 1983).
По крайней мере часть западносибирских крачек летят к местам зимовок через
Каспийское море (Ходков, 1983), пути миграций казахстанских и среднеазиатских
птиц не прослежены, но часть из них (возможно, большинство) летят по долинам рек
и далее напрямик на юг через материк. Нередко птицы в это время отмечаются в высо-
когорье (Кыдыралиев, 1973), Осенний пролет на всей территории Западной и Средней
Сибири, Казахстана и Средней Азии идет уже в августе и продолжается обычно до
середины сентября; нередко он слабо выражен. Последних птиц отмечают до октября
329
Семейство Чайковые
(З.Х — в Самарканде, 6.Х — на Аральском море, в конце октября — в низовьях Сыр-
дарьи и на Айдаркуле). Весенний пролет в Средней Азии и южной части Казахстана
проходит в апреле — первых числах мая (Спангенберг, Фейгин, 1936; Иванов, 1969;
Абдусалямов, 1971; Эминов, 1978). На Айдаркуле первых птиц отмечают в начале
апреля (Остапенко и др., 1984), на Иссык-Куле — 14.IV (Янушевич и др., 1959), в до-
лине р. Или — 10.IV (Долгушин, 1962). В Наурзуме первые крачки появляются 18—
28.IV, валовый пролет проходит в конце апреля — начале мая (Елкин, 1975). На боль-
шей части Казахстана первых крачек отмечают в середине — конце апреля (Долгушин,
1962). В южной Барабе первые птицы появляются 8—14.V, пролет идет до конца мая
(Ходков, 19776), в окрестностях Томска первых крачек регистрируют в среднем
10.V (Гынгазов, Миловидов, 1977), у Нарыма — 21—27.V (Дубовик и др., 1977), у Хан-
ты-Мансийска - 10—28.V, в среднем 23.V (Гордеев, 1979). На Енисее под Туруханском
речные крачки появляются в конце мая — начале июня (Рогачева, Вахрушев, 1983).
На Памире первых речных крачек отмечали 7 и 14.V (Мекленбурцев, 1946; Абдуса-
лямов, 1971). Осенний отлет здесь не прослежен, в конце сентября большие стаи птиц
еще отмечались на оз. Зоркуль (Потапов, 19666). На юге Тувы на оз. Убсу-Нур в 1980 г.
первые птицы встречены 2.V (Савченко, 1983). Первые речные крачки на Байкале
в дельте Селенги появляются 16—19.V, реже 10—12.V, на северном Байкале и Братском
водохранилище — 17—21.V. Через 2—7 дней наблюдается массовый пролет, который
длится 3—8 дней. Отлет начинается в последних числах августа, массовый пролет прохо-
дит в первой половине сентября, отдельные птицы встречаются до 5 —10.Х (Мельников,
Садков, 1977). В Забайкалье пролет отмечен с третьей декады августа по 12.IX (Измай-
лов, Старков, 1960).
Миграции черноклювых речных крачек (Sh. longipennis) проходят как по морским
побережьям, так и через материк. На оз. Ханка птицы появляются 7—8.V (Поливанова,
1971; Леонович, устное сообщ.), массовый пролет на оз. Болонь проходит 7—15.V
(Росляков, 1977), на Сахалине - 18—30.V (Гизенко, 1955). На юго-восточном побе-
режье Камчатки первые крачки отмечаются 16—27.V, в среднем 18.V (Лобков, Голо-
вина, 1978), на западном побережье - 16.V (Остапенко и др., 1975), на северном побе-
режье Охотского моря — 24.V (Васьковский, 1956), на юге Корякского нагорья —
27—29.V, а в окрестностях Хатырской котловины — только 16.VI (Кищинский, 1980).
В пос. Марково на р. Анадырь в 1970 г. крачки наблюдались с 1.VI (Леонович, устное
сообщ.). В Центральную Якутию речные крачки прилетают раньше — 14—24.V, в сред-
нем 19.V. Осенние миграции здесь начинаются в десятых числах августа, наиболее вы-
ражен пролет 15—25.VIII, а последних птиц отмечали 23.IX (Дегтярев и др., 1981а).
Из района Корякского нагорья крачки исчезают в конце августа, вдоль юго-восточного
побережья Камчатки пролетают в массе во второй половине августа, пролет на Саха-
лине начинается с первой декады сентября (Гизенко, 1955; Лобков, Головина, 1978;
Кищинский, 1980).
На весеннем пролете крачки нередко летят парами и одиночками (Ходков, 19776;
Мальчевский, Пукинский, 1983; Остапенко и др., 1984), а также небольшими группами
и стайками до 20 птиц (Гизенко, 1955; Долгушин, 1962; Кищинский, 1980). На осен-
нем пролете стаи обычно крупнее (Кищинский, 1980; Мальчевский, Пукинский, 1983).
Годовалые птицы крайне редко отмечаются летом в местах гнездования, тогда как
встречи двухгодовалых птиц здесь достаточно часты (Cramp, 1985).
Залеты отмечены в Исландию и на острова Кабо-Верде (Cramp, 1985).
Численность. В большинстве мест обитания — многочисленный вид. В Латвии гнез-
дится около 2 тыс. пар (Виксне, 1983), на островах Западной Эстонии - 2,9 тыс. пар
(Пеэдосаар, Онно, 1970), всего в Эстонии в 1970-х годах — около 5 тыс. пар (Thomas,
1982, пит. по: Cramp, 1985), в Белоруссии — 18,6 тыс. пар (Наумчик, 1979), в Москов-
330
Речная крачка
ской обл. - несколько более 200 пар (Зубакин, 19816; Мищенко, 1983), на севере
Причерноморья и Приазовье — не менее 25 - 30 тыс. пар (Костин, 1975, 1983; Молодая,
Сиренко, 1981; Петрович, 1981; Сиохин, 1981а; Черничко, Стойловский, 1981; Арда-
мацкая, 1984), на севере Каспия - не менее 20 тыс. пар (Кривоносов, 1976), на юго-
восточном Каспии — около 20 тыс. пар (Васильев и др., 1975; Щербина, 1979; Гаузер,
19836), на Торейских озерах — не менее 250 пар (Зубакин, 1981а), на оз. Ханка -
около 770 пар (Глущенко, 19846). Для Камчатки предполагается гнездование около
80 тыс. пар (Лобков, Головина, 1978). Численность тибетского подвида на Памире
невелика, не превышает немногих сот пар. В 1977 г. на оз. Каракуль было найдено 133
гнезда (наши данные).
Общую численность вида на территории СССР можно оценить в несколько сот тысяч
гнездящихся пар. Глобальные тенденции изменения численности не ясны, но в ряде
регионов — в частности, в Центре европейской части СССР (Луговой, 1975; Зиновьев,
1980) - численность заметно сократилась по сравнению с прошлым веком. В Западной
Европе и на Ближнем Востоке гнездится около 120 тыс. пар, в большинстве стран
численность крачек снижается (Cramp, 1985).
Местообитания. Наиболее эвритопная из наших крачек. В сезон гнездования заселяет
долины и дельты рек, разнообразные пресные, солоноватоводные и соленые озера, торфя-
ные карьеры, рыборазводные пруды, лиманы, морские заливы и побережья. Обитает как
на равнинных, так и на горных водоемах, причем на Памире гнездится по озерам до высо-
ты 4150 м, а в Тибете -- вплоть до 4800 м (Потапов, 19666). На Корякском нагорье мо-
жет поселяться и по горным рекам (Кищинский, 1980), хотя в целом явно предпочитает
стоячие водоемы или реки с небыстрым течением. Населяет природные зоны от север-
ной тайги до пустыни. В ряде мест проникает в лесотундровую зону и даже в зону тунд-
ры, но гнездится здесь лишь по долинам рек; типичных тундровых ландшафтов в
отличие от полярной крачки явно избегает.
Размножение. Речные крачки приступают к размножению обычно в возрасте 3 -4 лет,
реже годом раньше. Моногамы, 79% пар сохраняются по крайней мере два сезона под-
ряд (Cramp, 1985). Диффузно гнездящийся факультативно-колониальный вид (Зуба-
кин, 1976в; 1983). В нестабильных местообитаниях (зарастающие островки, размывае-
мые косы и т.п.) часто меняют места гнездования, однако в более стабильных условиях
поселения могут существовать десятилетиями. В последнем случае в колонию возвра-
щаются для гнездования около 14,3% от поднявшихся на крыло птенцов, только 1—2%
выбирают для гнездования другие места. Обмен взрослыми особями между колониями
здесь незначителен (DiCostanzo, 1980). Однако, по наблюдениям в Эстонии, примерно
15% переживших зиму особей переселяются на новое место, иногда на расстояние
10 км и более; по-видимому, это более характерно для молодых птиц (Онно, 1965;
Мянд, 1979). Численность поселений колеблется по годам, особенно в нестабильных
местообитаниях. Вопросы постоянства пар, гнездового консерватизма и демографии
речной крачки разобраны в работах О. Остина (Austin, 1940, 1945, 1947, 1949; Austin,
Austin, 1956). Изредка речная крачка образует пары с розовой крачкой (S. dougallii),
причем такие пары могут быть постоянными и, видимо, давать жизнеспособное потом-
ство (Robbins, 1974; Hays, 1975; Cramp, 1985). По-видимому, крайне редко возможны
смешанные пары речной и малой крачек (Очагов, 1982; Мельников, 1985).
Демонстрационное поведение речной крачки сходно с поведением других видов
крачек, особенно полярной. К наземным агрессивным демонстрациям относится ’’сог-
нутая” поза, которая используется в территориальных столкновениях (рис. 79,а).
При наиболее интенсивной демонстрации клюв опущен почти вертикально. Нередко в
этой ситуации издается ’’рекламный” крик ”кии-ри” или ’’криири”. Побежденный
противник принимает ’’выпрямленную” позу с клювом, поднятым почти вертикально
331
Семейство Чайковые
(см. рис. 79,6). Для наземных демонстраций речной крачки, как и большинства других
видов рода Sterna, характерен вздернутый вверх хвост (Cramp, 1985). В агрессивных
столкновениях нередко используется сухая трескучая трелька, с которой крачка может
преследовать особь своего вида в воздухе; часто такая же трелька слышна и во время
преследования чаек и других пернатых хищников.
Брачное поведение включает как воздушные, так и наземные демонстрации. К пер-
вым относятся ’’высотный” и ’’низкий” полеты (Cullen, 1960), некоторые авторы
Рис. 79. Демонстрационное поведение речной крачки (по: Cramp, 1985, с изменениями)
а — согнутая поза, б — выпрямленная поза
выделяют еще ’’змеевидный” полет (Каверкина, 1983). К наземным брачным демонст-
рациям относятся ритуальное кормление самки, ’’выпрямленная” поза, при которой
клюв направлен вертикально или наклонно вверх и голова слегка поворачивается то в
одну, то в другую сторону вдоль вертикальной оси (в этой позе обычно происходит
встреча партнеров), а также разнообразные ’’танцы”, или кружения и повороты парт-
неров друг перед другом (Каверкина, 1983). Формирование пар обычно происходит
вне территории колонии, нередко довольно далеко от нее (Palmer, 1941, цит. по: Cramp,
1985; Austin, 1947; Каверкина, 1983).
Величина колоний варьирует от нескольких пар до нескольких тысяч гнезд. Крупные
колонии (сотня и более гнезд) расположены обычно на морских островах или остро-
вах крупных озер, в других местах характерны колонии из нескольких пар или из
немногих десятков гнезд. Гнездование отдельными парами также не составляет ред-
кости (Долгушин, 1962; Зубакин, Костин, 1977; Лобков, Головина, 1978; Erwin,
1978; Блинов, Ходков, 1979; Bergman, 1980; Молодан, Сиренко, 1981; Виксне, 1983;
Савченко, 1983; Корнеева и др., 1984; и др,). В выборе места для гнездования речные
крачки показывают большую эвритопность, чем остальные наши крачки. Наиболее
характерные места гнездования — расположенные в морских заливах и лиманах, на
крупных соленых озерах, крупных реках и в их дельтах наносные островки из песка,
ракушечника, ила и изредка гальки, заросшие невысокой растительностью или (что
реже) совершенно голые. Иногда речная крачка поселяется на такого же характера
берегах водоема. На относительно высоких островках речная крачка гнездится по
их краю, недалеко от воды; низкие острова занимает целиком (Луговой, 1958; Боро-
дулина, 1960; Филонов и др., 1974; Мельников, Садков, 1977; Блинов, Ходков, 1979;
Ардамацкая, 1981; Зубакин, 1981а; Титавнин, 1981).
Реже речная крачка устраивает гнезда на сплавинах, плавающей растительной ветоши,
на наносах прошлогоднего тростника (Долгушин, 1962; Мельников, Садков, 1977;
332
Речная крачка
Зиновьев, 1980; Виксне, 1983). В Корякском нагорье крачки гнездятся по галечным
и галечно-песчаным берегам и островам горных рек (Кищинский, 1980), в Центральной
Якутии - как по песчаным, так и по заболоченным берегам озер, помещая в последнем
случае гнезда на кочках, кучах плавника, моховых гривах (Дегтярев и др., 1981а),
на Камчатке - на песчаных и галечных островах и косах, в травянистых тундрах, иногда
даже под пологом ивняка на островах (Лобков, Головина, 1978) . На оз. Ханка колонии
располагались по песчаным островкам; кроме того, к.рачки устраивали полуплавучие
гнезда среди залитых водой зарослей вейника (Поливанова, 1971).
Речная крачка может образовывать самостоятельные колонии, но нередко гнездится
вместе с другими видами птиц: малой и чайконосой крачками, малой и озерной чайка-
ми, морским голубком, шилоклювками и др. Плотность гнездования сильно варьирует,
причем наиболее разреженные колонии отмечаются на доступных наземным хищникам
берегах водоемов (Блинов, Ходков, 1979). В Московской обл. среднее расстояние
между ближайшими соседними гнездами составляет 2,0 м (0,76 11 м; п = 34), в коло-
ниях на Сиваше - 2,0 м (0,40-11 м; п = 130), на Памире - 6,5 м (п = 43), варьируя от
1,75 м до 33 м (наши данные). В Красноводском заливе расстояние между ближайши-
ми соседними гнездами составляет 0,7—13,0 м, в среднем (и = 612) 3,65 м (Гаузер,
19836), на островах Молочного лимана - 0,3—23,7 м, в среднем (и = 59) 1,6 м, на о-ве
Карагинском — 8,5—60,0 м, в среднем (п = 4) 28,3 м (Каверкина, 1983), в дельте Селен-
ги 1,0—102 м, в среднем (и = 66) 10,0 м (данные Д.М. Очагова). Индекс плотности
гнездования вида, вычисленный по имеющимся данным, равен 0,03.
Вокруг гнезда всегда есть охраняемая территория с нечеткими границами, которую
защищают обе птицы. Привязанность к ней, по-видимому, зависит от характера гнездо-
вого биотопа. Нередки случаи, когда пары занимали один и тот же гнездовой участок в
течение нескольких лет (Cramp, 1985). В других местах отмечалось, что речные крачки
в отличие от полярных на следующий год часто занимали новую территорию (Онно,
1965).
В сухих местах гнездо обычно представляет собой ямку в грунте со скудной выстил-
кой или без нее. В более сырых местах гнездо хорошо оформлено и иногда бывает
достаточно массивным. Материал гнезда — сухие стебли и листья травянистых растений,
сухие водоросли; он в значительной степени определяется окружающей раститель-
ностью (Сиохин, Скрипко, 1978). Средние размеры гнезд (мм): в дельте Волги и
Черноморском заповеднике диаметр гнезда 240, диаметр лотка 115, глубина лотка
28 (Бородулина, 1960); на Хаузханском водохранилище в Туркмении (и = 10) 225,
105 и.29 (Эминов, 1974а); на Байкале 162 (и = 43), 96 (п = 43) и 25 (и = 38) соот-
ветственно (Мельников, Садков, 1977). В Московской обл. (и = 4) диаметр гнезда
равен 240-290, в среднем 270, лотка 110—145, в среднем 130, глубина лотка 20—25,
в среднем 23; на Сиваше соответственно 155-230, в среднем (и = 7) 195, 110—170 в
среднем (и = 37) 131, 20—40, в среднем (и = 10) 29; на Памире 190—240, в среднем
(я = 7) 211, 105 — 130, в среднем (и = 12) 125, 20—40, в среднем (и = 12) 33 (наши
данные).
Сроки откладки первых яиц зависят от географической широты местности: разница
между южной и северной частями ареала в пределах СССР может быть больше месяца.
На севере Причерноморья, в Приазовье и Предкавказье первые кладки появляются в
первой — начале второй декады мая (Филонов и др., 1974; Ардамацкая, 19776; Зуба-
кин, Костин, 1977; Казаков, Языкова, 1982), на Западной Украине — в начале второй
декады мая (Страутман, 1963), в Московской обл. — в середине мая (наши данные),
в Ленинградской обл. - в конце второй — начале третьей декады мая (Мальчевский,
Пукинский, 1983), в Кировской обл. — в середине первой декады июня (Плесский,
1976). Птицы, гнездящиеся на водохранилище-охладителе Литовской ГРЭС, приступают
333
Семейство Чайковые
к откладке яиц раньше (2—7.V), чем на водоемах по соседству (Виткаускас, Гражуля-
вичюс, 1976; Виткаускас, 1977).
В заливе Кара-Богаз-Гол первые яйца появляются, судя по датам подъема птенцов
на крыло, в конце апреля — первых числах мая (Щербина, 1979), в дельте Волги, на
Хаузханском водохранилище Туркмении, в северном Таджикистане - в середине мая
(Луговой, 1958; Абдусалямов, 1971; Эминов, 1974а), на оз. Иссык-Куль и в Казахста-
не - во второй половине мая (Янушевич и др., 1959; Долгушин, 1962), на юге Запад-
ной Сибири - в конце мая - начале июня (Гынгазов, Миловидов, 1977), в низовьях
Оби - видимо, в середине июня (Данилов и др., 1984).
В Восточной части ареала гнездование начинается примерно на месяц позже, чем в
западной, и почти одновременно как на севере, так и на юге. На юге Тувы первые клад-
ки отмечены в конце мая (Леонович, устное сообщ.), на Байкале — 4—10.VI (Мельни-
ков, Садков, 1977), в Центральной Якутии - в первых числах июня (Дегтярев и др.,
1981а), на оз. Ханка — в конце мая - начале июня (Поливанова, 1971), на о-ве Саха-
лин - в конце первой — середине второй декады июня (Нечаев, 1977; Леонович, устное
сообщ.), на Камчатке — в самом конце мая — начале июня (Лобков, Головина, 1978),
в Корякском нагорье - видимо, в середине июня (Кищинский, 1980). Крачки на Пами-
ре, судя по срокам вылупления, приступают к откладке яиц в последних числах мая -
первых числах июня (Потапов, 19666; наши данные) .
Массовая откладка начинается через несколько дней после появления первых яиц.
Для речной крачки характерен сильно растянутый период откладки яиц — вплоть до
20-х чисел августа (Ардамацкая, 1984), однако на северо-востоке ареала кладка прохо-
дит в достаточно сжатые сроки — 13—20 дней (Дегтярев и др., 1981а). В полной кладке
1-3 яйца, чаще всего 2-3, редко 4-5 и крайне редко 6 яиц (Щербина, 1979; Conover,
1980). В более ранних кладках преобладает 3 яйца, в поздних и повторных — 1—2
(Бородулина, 1960; Лобков, Головина, 1978; Блинов, Ходков, 1979; Дегтярев и др.,
1981а; Костин, 1983 и др.). Зависимость размера кладки от величины колонии разоб-
рана в работе Ю.К. Рощевского и Л.Н. Лаухиной (1980).
Окраска яиц сложная. Основной-фон блеклый палевый, серовато-бежевый или свет-
ло-коричневый, реже зеленовато-серый или оливково-серый. По этому фону в беспо-
рядке разбросаны глубокие фиолетово-серые и поверхностные бурые и черноватые
пятна и крапинки. Пятна обычно равномерно покрывают яйца, реже группируются
ближе к тупому концу или образуют поясок (Долгушин, 1962; Лобков, Головина,
1978). Размеры яиц (мм): из Крыма и Присивашья (п = 178) 38,2—46,9 х 27,2-32,9,
в среднем 41,7 х 30,4 (Костин, 1983); из дельты Волги (л = 217) 37,1-46,2 х 28,9—
34,0, в среднем 41,6 х 31,1 (Луговой, 1958); из Московской обл. (и = 9) 39,3—43,5 х
х 29,7—31,7, в среднем 41,5 х 30,7 (наши данные); из Казахстана (л = 42) 36,7—45,1 х
х 27,1—32,5, в среднем 40,9 х 30,7 (Ковшарь, Левин, 1982); из Западной Сибири (и =
= 137) 36,6—46,4 х 27,6—32,5, в среднем 41,1 х 30,2 (Стрелков, цит. по: Гынгазов,
Миловидов, 1977); с Байкала (и =112) в среднем 40,7 х 29,5 (Мельников, Садков,
1977); из Центральной Якутии (л = 135) в среднем 41,7 х 30,1 (Дегтярев и др., 1981а) ;
с Камчатки (и = 101) 37,0—45,3 х 23,4-31,6, в среднем 42,1 х 28,5 (Лобков, Головина,
1978). Яйца крачек с Памира (S.h. tibetana) несколько крупнее, чем из других участ-
ков ареала (п = 44): 41,3—46,3 х 29,0—31,6 в среднем 43,4 х 30,6 (Потапов, 19666;
данные В.А. Зубакина). Масса (г) свежих яиц: из Предкавказья (п = 106) 17—21, в
среднем 19 (Казаков, Языкова, 1982); с Камчатки (л = 35) 16,2—21,0, в среднем
18,5 (Лобков, Головина, 1978) .
Насиживание начинается с первого яйца и продолжается 18—20 дней (Бородулина,
1953а; Янушевич и др., 1959; Эминов, 1974а) или 20—24 дня (Лобков, Головина,
1978; Дегтярев и др., 1981а; Ардамацкая, 1984; Cramp, 1985). Насиживают оба роди-
334
Речная крачка
теля, но самка несколько больше, чем самец, - особенно в начале инкубации и в период
вылупления. Во время откладки яиц насиживает только самка (Болотников, Калинин,
1975; Калинин, 1975), по другим данным — и самец, но в меньшей степени. По наблю-
дениям на Сиваше, смена партнеров в конце насиживания происходит 4—10 раз в сутки
(п = 3); на Памире в начальный период насиживания отмечено 15 смен партнеров в
сутки (п = 1), причем изредка наблюдалось кормление Самки самцом (наши данные).
После вылупления родители уносят из гнезда скорлупу яйца. Масса птенцов в день
вылупления — 12—14 г (Бородулина, 1960; Эминов, 1974а; Лобков, Головина, 1978).
Первый раз родители пытаются кормить птенца через 2—3 ч после вылупления; нередко
сначала птенцу не удается схватить и проглотить корм. В зависимости от размера кор-
мовых объектов (главным образом рыба, мелкие ракообразные, реже насекомые),
выводок кормят 10—41 раз в сутки (Бородулина, 1960; наши данные). В первые дни
родители постоянно согревают птенцов, затем делают это все реже. На Памире, в суро-
вых условиях высокогорья, родители продолжают согревать даже 10—12-суточных
птенцов (по наблюдениям за одним гнездом их обогревают 12 ч. 54 мин. в сутки, глав-
ным образом в ночное время). Кормят выводок родители в этот период 13'раз в сутки
(« = 1), только в светлое время (наши данные).
Выводки могут находиться на гнездовой территории вплоть до подъема на крыло
и даже дольше (Cramp, 1985), однако в большинстве колоний они, по-видимому, по-
кидают ее в первые дни жизни (Бородулина, 1960; Мельников, Садков, 1977; Дегтя-
рев и др., 1981а). Становление взаимного индивидуального опознавания родителей и
птенцов отмечено на 7—10-й день жизни птенцов, до этого главную роль в опознавании
своего потомства у крачек играет место гнезда и индивидуальная территория (Наза-
ренко и др., 1977).
Во время тревоги птенцы разбегаются и затаиваются, взрослые птицы активно
нападают на хищника, пикируя и ударяя его клювом. Наиболее активны в этом отно-
шении пары, гнездящиеся по периферии колонии (Рощевский, 1978). Лётными птенцы
становятся в возрасте 22—28 дней (Nisbet, Drury, 1972; Мельников, Садков, 1977;
Ардамацкая, 1984). Родители продолжают кормить лётных молодых и в период осен-
них миграций (Резанов, 1978).
Величина эмбриональной и постэмбриональной смертности колеблется в значитель-
ных пределах по годам и между колониями. В дельте Селенги до подъема на крыло
доживает 55—60% вылупившихся птенцов, в устье Верхней Ангары — 30—35%; эмбрио-
нальная смертность составляет в среднем, соответственно, 29,3% и 11,5% (Мельников,
Садков, 1977). В Красноводском заливе эмбриональная смертность в разные годы
равна 16,1—36,9%, гибель птенцов — 28,9—45,9%; до подъема на крыло доживает 1,5
птенца на пару птиц (Гаузер, 1983а). Основные причины гибели кладок — затопление
в результате подъема уровня воды и фактор беспокойства. Птенцы гибнут главным
образом от недостатка корма, причем чаще самые младшие и реже — самые старшие
птенцы выводка (подробнее см.: Langham, 1972; Гаузер, 1983а,б). На успех размноже-
ния оказывают воздействие степень зарастания территории колонии, величина и плот-
ность колонии (Рощевский, 1974, 1977; Ковалев, Рощевский, 1977).
Максимальная продолжительность жизни речной крачки, по данным кольцевания,
25 лет (Cramp, 1985). До начала размножения доживает не менее 16% от числа вставших
на крыло молодых; ежегодная смертность взрослых птиц 8,0% (DiCostanzo, 1980).
Суточная активность, поведение. Активность дневная, хотя птицы могут добывать
корм и при сравнительно низкой освещенности (Поливанова, 1971; Авилова, 1972).
По наблюдениям на Сиваше, партнеры насиживающих птиц ночуют, как правило, вне
колонии — на берегах островов или здесь же в солончаковых понижениях. По наблю-
дениям на Памире, свободные от насиживания птицы проводят ночь на своих гнездо-
335
Семейство Чайковые
вых участках. Общественная птица во все сезоны года. Кормится как одиночно, так
и стаями до 200 особей (Erwin, 1978). Есть некоторые данные о постоянстве состава
пролетных группировок речных крачек (Стойловский, 1981).
Питание. Основным кормом во все сезоны служит мелкая рыба, в меньших коли-
чествах крачки добывают водных беспозвоночных (креветок, гаммарид, водных насе-
комых) , наземных насекомых, изредка — земноводных и ящериц. Соотношение кормов
зависит от конкретного места обитания крачек (Шевченко, 1937; Клименко, 1950а;
Бородулина, 1953а, 1960; Гладков, Залетаев, 1956; Янушевич и др., 1959; Тима,
1961; Долгушин, 1962; Ширяева, 1965; Абдусалямов, 1971; Самородов, 1976; Сиохин,
19816). В дельте Волги, по данным анализа погодок и содержимого желудков, рыба
составляет 82,8% встреч (чаще всего бычок кругляк и сазан — 20,4% и 19,3% встреч
соответственно). В Крыму рыба встречена в 50,5% случаев, насекомые — в 70%; в
Черноморском заповеднике крачки питаются главным образом креветками (Бороду-
лина, 1960). На юго-западном Каспии основу питания речных крачек также составляет
рыба - в основном сеголетки сазана (Заблоцкий, Заблоцкая, 1963), на северо-восточ-
ном Каспии — килька, атеринка, бычки (Залетаев, 1960), на Хаузханском водохрани-
лище — плотва, храмуля, быстрянка, пескарь и др.; насекомые (стрекозы, жуки, кло-
пы, личинки плавунцов) по встречаемости составляют 59,5% (Эминов, 1974а). На
водохранилищах системы Маныча встречаемость в корме насекомых (личинок стрекоз,
гладышей, жуков-кузек) равна 79,0%, причем крачки кормятся только на водоемах,
наземных склевывают с воды (Казаков, Языкова, 1982). На Байкале летом основу
питания составляют насекомые (главным образом летающие), весной и осенью —
мелкая рыба (Скрябин, Размахнина, 1978). Большое значение имеют насекомые (жест-
кокрылые, стрекозы, ручейники, прямокрылые) в питании крачек из Центральной
Якутии (74,5% встреч). Крачки добывают их в воздухе, реже собирают с поверхности
воды у берега. Рыба (обычно гольян и елец) составляет 45,5% встреч (Дегтярев и др.,
1981а). На Памире крачки поедают мелких рыб и водных беспозвоночных (Потапов,
19666; Абдусалямов, 1971).
Места кормежки — открытые плесы водоемов, мелководья; кроме того, крачки
могут ловить насекомых на лету, как ласточки. Изредка можно наблюдать поисковый
полет крачек в степи неподалеку от водоема. За рыбой птицы ныряют с разлета, завис-
нув перед этим в воздухе; могут подхватывать добычу с поверхности воды (Бородули-
на, 1960; Поливанова, 1971). В гнездовой период дальность разлета за кормом обычно
не превышает 10 км, однако в ряде мест характерны дальние полеты на 20—26 км и
более (Бородулина, 1960; Носков и др., 1981; Cramp, 1985). Нередко крачки отнима-
ют добычу у особей своего вида, причем клептопаразитизм может иногда принимать
большие размеры и способствовать гибели птенцов от голода (Гаузер, 1983а; Панов и
др., 1985),
Враги, неблагоприятные факторы. Ущерб колониям речных крачек наносят крупные
чайки (чаще всего серебристые и сизые), врановые (как правило, серая ворона), из
млекопитающих — ласки, горностаи, колонки, лисицы, енотовидные собаки, кабаны;
в некоторых местах (Черноморский заповедник) птенцы страдают от степных гадюк
(Бородулина, 1960; Lemmetyinen, 1971; Мельников, Садков, 1977; Fuchs, 1977).
Один из главнейших неблагоприятных факторов — повышение уровня воды в результа-
те нагонных ветров, паводков, затяжных дождей и т.д., которое может приводить
к гибели всей колонии или большей части кладок. Затяжные дожди могут способст-
вовать также гибели птенцов (Мельников, Садков, 1977) и уменьшать плодовитость
крачек, если дожди предшествуют откладке яиц (Becker et al., 1985). Отмечалась по-
вышенная смертность птенцов в результате высокой зараженности клешами и, видимо,
от арбовирусной инфекции (Щербина, 1979). Большую роль в гибели кладок может
336
Полярная крачка
играть беспокойство колонии человеком, сбор яиц, выпас скота в местах гнездования
птиц (Бородулина, 1960; Виткаускас, 1977; Наумчик, 1982; Гаузер, 1983а).
Хозяйственное значение, охрана. В южных регионах страны в силу своей многочис-
ленности речная крачка может приносить некоторый локальный ущерб, поедая маль-
ков промысловых рыб на территории рыбхозов. На большинстве территории страны
хозяйственного значения не имеет. Специальные меры охраны не предусмотрены.
Полярная крачка —
Sterna paradisaea Pontoppidan, 1763
Arctic Tern (англ.), Kustenseeschwalbe (нем.), Sterne arctique (фр.).
Син. (лат.): Sterna macrura Naumann, 1819.
Син. (рус.): длиннохвостая крачка.
Статус. Гнездящийся мигрирующий вид.
Обшдя характеристика и полевые признаки. Крачка среднего размера, с речную, на
которую очень похожа. Отличается более длинным хвостом (у сидящей птицы он вы-
дается за концы сложенных крыльев); от S.h. hirundo, кроме того, более темной окрас-
кой нижней части туловища, а от S.h. logipennis — красным клювом. Молодые птицы
в полевых условиях практически неразличимы. Характер полета, как у речной крач-
ки. За добычей птица ныряет с разлета. По земле передвигается мало и неохотно; у си-
дящей птицы обращают на себя внимание короткие hoih (короче, чем у речной крачки).
Голос очень сходен с голосом речной крачки, но несколько выше. Крик тревоги зву-
чит глуше, чем у речной крачки, — как скрипучее’’керрр” или ”кррр”.Во время тревоги
в колонии часто слышны крики ”кия”, которые издают птицы, летающие над нарушите-
лем спокойствия. Крик крачки, возвращающейся в колонию (Advertising-call по:
Cramp, 1985), звучит как ’’криир” или ”пиир”; почти всегда он переходит в звонкое
стрекотание типа ”кити-ки-кийер, кити-ки-кийер ...” или ”кити-ки-кири . ..”. Подоб-
ный крик издает самец, кормящий самку (последняя, выпрашивая корм, тонко пищит
’’пи-пи-пи-пи. . .” или ”ти-ти-ти-ти. . .”), а также крачки во время агрессивных столкно-
вений. В последнем случае можно нередко слышать сухую трескучую трельку (она ис-
пользуется также во время погонь за пернатыми хищниками) и звонкие щелкающие
или цокающие звуки (подробнее см.: Анзигитова и др., 1980; Cramp, 1985).
Подвидовая систематика. Монотипический вид.
Описание. Размеры особей (мм) (ЗМ МГУ) и масса тела (г) (Бианки, 1967):
Длина крыла:
dd (и ?44) —257—286 (среднее 268);
99 (и =20) - 246-276 (среднее 265).
Длина клюва:
dd (и = 41) — 26,2—33,8 (среднее 30,3);
9 9 (и =20) — 26,7—31,1 (среднее, 28,8),
Длина цевки:
dd(n =43) — 13,7—16,7 (среднее 15,3);
9 9 (и =21) - 13,8-16,7 (среднее 15,1).
Масса тела:
dd (п = 56) — 82—135 (среднее 104);
9 9 (п = 37) — 89—153 (среднее 107).
337
Семейство Чайковые
Окраска (Дементьев, 1951; Cramp, 1985). Самец и самка в брачном наряде.
Окраска оперения почти такая же, как у речной крачки, но черная шапочка спускается
с боков головы немного ниже, окраска верхней части туловища более голубовато-се-
рая и менее пепельная, а сероватый цвет нижней части туловища более интенсивный, чем
у речной крачки, и поднимается вверх до подбородка и нижней части щек. Длинные пле-
чевые перья с более явственными белыми каемками, рулевые обычно все белые, только
наружные опахала двух крайних пар серые, причем у внешней пары серый цвет темнее.
Первостепенные маховые, как у речной крачки, но белое поле внутренних опахал более
широкое, между ним и стержнем пера остается серая полоска шириной лишь в 1,5—
2,5 мм. Белый цвет на вершинах и внутренних опахалах второстепенных маховых раз-
вит больше. Клюв ярко-красный, иногда с черным кончиком, лапы красные, радужина
темно-коричневая.
Самец и самка в зимнем наряде. Очень похожи на речных крачек в соответствующем
наряде; отличаются окраской первостепенных и второстепенных маховых (см. выше), а
также меньшим развитием серого цвета на пояснице, верхних кроющих хвоста и руле-
вых.
Пуховой наряд. Очень сходен с пуховым нарядом речной крачки; пуховички этих
двух видов отличаются с трудом и не надежно. Общий тон окраски верха варьирует от
светло-серого до рыжевато-коричневого; по этому фону разбросаны темные пятна и
крапины. Лоб, уздечка и Горло от бурого до темно-бурого цвета, подбородок бывает бе-
лым довольно редко. Нижняя часть тела белая, на боках и брюшке с серым или бурым
налетом. Клюв, радужина и ноги, как у речной крачки.
Гнездовой наряд. Окраска головы и туловища, как у речной крачки, но поясница и
верхние кроющие хвоста белые. Наружные опахала рулевых серые, концы и внутренние
опахала их белые. Окраска крыльев несколько иная, чем у речной крачки: карпальная
полоса светлее и уже, второстепенные маховые светлее больших кроющих крыла (а не
темнее, как у речной крачки), белый цвет на их концах более развит, внутренние опа-
хала первостепенных маховых с более широким белым полем. Клюв черный с розова-
тым или оранжевым основанием, к сентябрю обычно чернеет целиком; ноги оранжево-
красные, розовато-серые или серовато-красные; радужина темно-коричневая.
Первый зимний наряд. После полной линьки похож на окончательный зимний наряд,
однако на крыле остается карпальная полоса. Весной и летом второго календарного го-
да крачки не надевают брачный наряд, сохраняя зимний. Отдельные особи в это время
могут появляться в северном полушарии; они отличаются от речных крачек в сходном
наряде по тем же признакам, что и взрослые зимние птицы, а также характером линьки
первостепенных маховых. В третьем календарном году крачки надевают брачный на-
ряд, однако у части птиц (около 11%) остаются отдельные перья предыдущего зимне-
го наряда на крыльях, лбу, уздечке и брюхе.
Линька (Cramp, 1985). Линька в первый зимний наряд полная, начинается на зимов-
ках. Однако оперение головы, низа тела, спины и плечевые перья могут иногда начать
сменяться в октябре, во время миграции. К февралю линька мелкого оперения и руле-
вых заканчивается, смена маховых начинается в декабре—январе и завершается, види-
мо, к маю. У части птиц, возможно, линька первостепенных маховых проходит в более
ранние сроки, как у взрослых. Линька во второй зимний наряд происходит в те же сро-
ки, что и у взрослых. Линька во второй брачный наряд начинается позже, чем у взрос-
лых, и захватывает меньшую часть оперения: не сменяются все верхние кроющие крыла,
часть перьев спины и отдельные перья лба и брюха. Крайне редко в это же время могут
сменяться 1—2 внутренних первостепенных маховых.
Последующие линьки бывают дважды в году: полная послебрачная и частичная пред-
брачная. Послебрачная линька начинается обычно на зимовках. Точные сроки ее начала
338
Полярная крачка
неизвестны — видимо, конец сентября — начало ноября. В январе птицы уже в свежем
зимнем мелком оперении; первостепенные маховые сменяются к началу февраля —
началу марта. Предбрачная линька протекает в конце февраля — марте, завершается к
началу весенней миграции. Сменяются перья головы, туловища, хвоста и кроющие кры-
ла; в отличие от речной крачки смены внутренних первостепенных и наружных второ-
степенных маховых не происходит.
Распространение. Гнездовой ареал. Гнездится циркумполярно, заселяя при-
мыкающие к Северному Ледовитому океану территории Евразии и Северной Америки,
острова и побережья северной Атлантики и северной Пацифики (рис. 80). В Западной
Европе гнездование отмечено в Исландии, на о-ве Ян-Майен, о-ве Медвежьем, Шпиц-
бергене, по побережьям Великобритании, Ирландии, Нидерландов, Дании, ФРГ, ГДР,
Норвегии; заселяет все балтийское побережье Швеции и Финляндии, а на севере этих
стран — и внутренние водоемы. Нерегулярные поселения отмечались во Франции, Бель-
гии и Польше (Cramp, 1985).
На территории СССР (рис. 81) гнездовые поселения известны в Прибалтике, главным
образом на островах на западе и севере Эстонии (Пеэдосаар, Онно, 1970; Аумеэс, 1972;
Ренно, 1972; Aumees et al., 1983). В 1978 г. доказано гнездование полярной крачки в
окрестностях г. Риги (Страздс, 1981; Страздс, Страздс, 1982), предполагается ее появ-
ление в Латвии после 1950-х годов (Виксне, 1983), В небольшом количестве полярная
крачка гнездится на Березовых островах у устья Выборгского залива (Храбрый, 1984);
в других местах Ленинградской обл. ныне не гнездится, хотя в 1940-х годах колония
была обнаружена на восточном побережье Ладожского озера (Мальчевский, Пукинский,
1983). Севернее полярная крачка заселяет баренцевоморское и беломорское побережья
Кольского полуострова, включая Айновы острова, Семь островов и другие острова (Ус-
пенский,1941; Благосклонов, 1960; Кищинский, 1960а; Малышевский, 1962; Бианки,
1967; Коханов, Скокова, 1967), все остальное побережье Белого моря, включая Соло-
вецкие острова (Спангенберг, Леонович, 1960; Карташев, 1963; Корнеева и др., 1984).
Гнездование отмечено на крупных озерах Кольского полуострова (Владимирская,
1948), на озерах южной Карелии не гнездится (Нейфельдт, 1970).
Далее на восток южная граница ареала отступает от побережья и более или менее
точно совпадает с южной границей зоны тундры, иногда спускаясь в лесотундру и даже
северную тайгу (Дементьев, 1951; Успенский, 1960). Северная граница материкового
ареала проходит по побережью Северного Ледовитого океана и близлежащим островам.
Крачки населяют Малоземельскую и Большеземельскую тундры (Гладков, 1951; 1962;
Лобанов, 1975; Минеев, 1982), гнездятся по всему Ямалу (Данилов и др,, 1984) ; далее
южная граница ареала проходит, видимо, в окрестностях полярного круга, на Енисее -
около Игарки (Скалой, Слудский, 1941; Рогачева и др., 1983). Есть сведения о гнездо-
вании этого вида гораздо южнее — на средней Оби в окрестностях Сургута и по средне-
му течению р. Вах (Вдовкин, 1941; Шаронов, 1951; ЗИН); по-видимому, это изолиро-
ванное гнездование, так как на Нижней Оби полярная крачка не отмечена южнее г. Ла-
бытнанги (Данилов, 1965). Далее к востоку полярная крачка заселяет Таймыр, хотя и
не везде равномерно: в ряде мест полуострова она на гнездовье не отмечена (Кречмар,
1966; Зырянов, Ларин, 1983; Кокорев, 1983; Матюшенков, 1983; Павлов и др., 1983;
Якушкин, 1983; Морозов, 1984). В бассейне Хатанги граница, видимо, проходит в
окрестностях 68° с.ш. (Иванов, 1976).
На р. Лене южная граница ареала лежит севернее 68°30z с.ш. (Лабутин и др., 1981),
на Индигирке — южнее 69° 30 с.ш. (Успенский и др., 1962), на Колыме — между 67° и
67°30' с.ш. (Бутурлин, 1934; Лабутин и др., 1981). Отмечено гнездование полярных
крачек на Алазее (Воробьев, 1967), в Чаунской губе и на о-ве Айон (Лебедев, Филин,
1959; Засыпкин, 1981) , востоке Чукотки (Томкович, Сорокин, 1983), во всему бассей-
339
340
160 120 80 40 0 40 80 120 160 160
Рис. 80. Область распространения полирной крачки
1 — область гнездования (пунктиром показана неуточненная граница), 2 — гнездование на узкой прибрежной полосе и отдельные
поселения, 3 ~ предполагаемые места гнездования, 4 — область пролета, 5 — места зимовок, 6 — залеты
341
Рис. 81. Ареал полярной крачки в СССР
1 — область гнездования (пунктиром показана неутомленная граница), 2 — гнездование в узкой прибрежной полосе, 3 — отдельные
поселения, 4 — места предполагаемого гнездования, 5 — залеты, 6 — направления весенних миграций, 7 — то же осенних миграций
Семейство Чайковые
ну р. Канчалан (Кищинский и др., 1983). Южная граница сплошного ареала проходит
по среднему течению р. Анадырь и северному краю Корякского нагорья, образуя край-
не узкую зону симпатрии с речной крачкой (Кищинский, 1980) . По-видимому, населяет
всю Чукотку, но гнездится здесь спорадично (Портенко, 1973). Южнее границы сплош-
ного ареала известно несколько изолированных гнездовых поселений: в Парапольском
долу (Дементьев, 1940: Лобков, 1983), в низовьях р. Караги (Лобков, 19816), в бухте
Гека в низовьях р. Гатымынваям (Фирсова, Левада, 1982), на о-ве Карагинском (Гера-
симов, 1979а), на западном побережье Камчатки в устье р. Тигиль (Остапенко и др.,
1977) и у пос. Кировский (Лобков, 1985). Предполагается гнездование в низовьях
р. Пенжины и на побережье Пенжинской губы (Яхонтов, 1979), а также на юго-запад-
ном побережье Камчатки в Усть-Большерецком районе (Глущенко, 1984а).
Полярные крачки1 населяют также острова арктического бассейна. Гнездование отме-
чено на Земле Франца-Иосифа (Горбунов, 1932; Паровщиков, 1963; Успенский, 1972;
Томкович, 1984), на Новой Земле (по крайней мере на ее западном и северо-западном
побережьях), о-ве Вайгач (Белопольский, 1957; Успенский, 1960; Карпович, Коханов,
1967); нет точных сведений о гнездовании этого вида на о-ве Колгуев (Дементьев,
1951). Далее к востоку гнездование отмечено на о-ве Большевик (Булавинцев, 1984),
достоверных сведений о гнездовании на других островах Северной Земли нет (Лак-
тионов, 1946). Гнездится полярная крачка также на Новосибирских островах и о-ве
Врангеля (Дементьев, 1951; Рутилевский, 1958; Портенко, 1973).
Зимовки. Полярные крачки зимуют в зоне пакового льда морей, омывающих
Антарктиду, проникая на юг в море Уэдделла до 74°ю.ш., а в море Росса, по-видимому,
до 76° 52' ю.ш. Основной район зимовок европейских птиц лежит между 30° и 150°в.д.,
с наибольшей концентрацией птиц между 50° и 110°в.д. (Дементьев, 1951; Cramp,
1985).
Миграции, Полярные крачки Белого и Баренцева морей, а также, видимо, птицы с
побережий Карского моря, Таймыра (возможно, и из более восточных регионов) летят
осенью на запад, затем перемещаются вдоль северных и западных берегов Европы и за-
падного побережья Африки, достигая мест зимовок в ноябре—декабре. Сходным путем
летят птицы из западной половины Северной Америки, соединившись с западно-пале-
арктическими крачками у побережий Западной Европы. Полярные крачки Берингова
моря и Аляски летят на юг вдоль западного побережья Северной и Южной Америк.
По-видимому, этим же путем летят крачки восточных регионов СССР (Cramp, 1985).
Наиболее изучены миграции беломорских птиц (Бианки, 1967). Массовый отлет по-
лярных крачек из Кандалакшского залива начинается в середине двадцатых чисел ию-
ля и заканчивается в начале—середине августа; в конце 1960-х годов у птиц этой попу-
ляции проявилась тенденция отлетать в более поздние сроки — примерно на 20 дней
позднее, чем прежде (Бианки, Бойко, 1972). Начиная с августа крачки перемещаются в
юго-западном направлении, пролетая через Балтийское море и побережье Западной Ев-
ропы. В сентябре большинство птиц еще отмечается в Европе, однако передовые уже
достигают западного побережья тропической Африки. В октябре—ноябре крачки про-
должают движение на юг вдоль западного побережья Африканского континента и в де-
кабре достигают мест зимовок в приантарктических водах. Обратное движение начина-
ется, видимо, в марте, и в конце второй декады мая первые птицы появляются в Канда-
лакшском заливе (за 17 лет наблюдений сроки появления первых крачек варьировали
с 6 по 23.V, средняя дата 16.V), Так же как и осенью, птицы весной не огибают Кольско-
го полуострова, а летят через Балтийское море, Финляндию и Ленинградскую обл. Не-
значительный весенний пролет идет через юго-восточную часть Ладожского озера в кон-
це мая—начале июня (Носков и др., 1981).
Некоторые птицы, особенно молодые, могут сбиваться с магистрального пути проле-
342
Полярная крачка
та, их обнаруживают в глубине материка. Так, молодые птицы 27.VIII 1958 г. и 30.VIII
1960 г. были встречены в Челябинской обл, и на Западной Украине (Хмельницая обл.);
отмечали их и на Черном море (Бианки, 1967).
На Айновых островах (Западный Мурман) первые птицы появляются 8—25.V, в сред-
нем за 21 год 18.V (Анзигитова и др., 1980), на Семи островах (Восточный Мурман) —
24—31.V, в среднем 28.V (Белопольский, 1957), на озерах Лапландского заповедника —
21.V—6.VI, в среднем за 11 лет 29.V (Владимирская, 1948), на Земле Франца-Иосифа —
7—24.VI, в среднем 18.VI или немного раньше (Горбунов, 1932; Паровщиков, 1963;
Томкович, 1984). В Малоземельской тундре первые полярные крачки отмечаются 25—
31.V, в Большеземельской тундре — 31.V—3.VI (Минеев, 1982), на юге Ямала — 28-V—
8.VI, обычно в начале июня (Данилов и др., 1984), на Западном Таймыре в разные годы
и в разных пунктах — с 3 по 21.VI (Кречмар, 1963, 1966), в низовье Енисея севернее
Игарки — в первой декаде июня (Рогачева и др., 1983). Перечисленные сроки, несмотря
на то, чю они сильно варьируют по годам в зависимости от хода весны, четко указыва-
ют на продвижение полярных крачек весной с запада на восток вплоть до Таймыра. На
Восточный Таймыр, видимо, прилетают крачки, которые двигаются с востока, с Чукот-
ского и Берингова морей; они появляются здесь 11 —15.VI и в августе отлетают также
на восток (Матюшенков, 1979, 1983). Восточнее Таймыра полярные крачки появляют-
ся в местах гнездовий раньше: в Приколымской тундре 27.V, на Алазее 31.V, в Яно-
Индигирской тундре 30.V— 1.VI (Воробьев, 1963, 1967), в Чаунской низменности 1.VI
(Кондратьев, 1979), в Уэлене 31.V, в заливе Креста 1 .VI, на о-ве Врангеля — 12.VI
(Портенко, 1973). Заслуживает внимания, что сроки появления крачек в тундрах севе-
ро-востока Якутии несколько более ранние, чем на побережье Чукотки. Если это не
случайное следствие более теплых и ранних весен в период наблюдений, можно предпо-
ложить миграцию крачек через материк где-нибудь в районе залива Шелихова и Пен-
жинской губы. Во всяком случае, на восточном побережье Камчатки в Тигильском рай-
оне крачек отмечали уже во второй половине мая (Остапенко и др., 1975), а в 1972—
1973 гг. пролетных птиц встречали 22—26.V на р. Омолон (Кречмар и др., 1978).
Осенью из большинства районов гнездования полярные крачки исчезают в течение ав-
густа. Задержки до начала—середины сентября отмечались лишь на юге Ямала (Данилов
и др., 1984),в Большеземельской тундре (Минеев, 1982) и на Земле Франца-Иосифа
(Паровщиков, 1963 ; Томкович, 1984) . В отношении направления осеннего пролета раз-
ных популяций пока еще нет ясности, можно лишь предположить, что осенью птицы
мигрируют, в общем, по тем же маршрутам, что и весной, но в обратном направлении.
Пролетные стаи до 100—350 особей в окрестностях Уэлена появляются в третьей декаде
августа (Томкович, Сорокин, 1983) .
В летние для северного полушария месяцы годовалые крачки кочуют по всей огром-
ной территории от Антарктики до мест гнездования в Арктике. То же свойственно, ви-
димо, и части двухгодовалых птиц (Бианки, 1967). В период весенних миграций поляр-
ные крачки обычно летят группами в несколько особей, реже стаями в 100—150 птиц
(Минеев, 1982; Данилов и др., 1984). Осенние стаи и стаи птиц на зимовках обычно
крупнее (Cramp,1985).
Залеты полярных крачек, помимо упомянутых, отмечены в Псковскую область
(Зарудный, 1910), Чехословакию, Австрию, Швейцарию, Италию, Турцию, Алжир, на
Кипр (Cramp, 1985). Экспедиция ’’Фрама” добыла полярную крачку 27.VII 1895 г: под
84°32'с.ш. (Дементьев, 1951).
Численность. Для большинства регионов СССР не определена. В Латвии гнездится
10—25 пар (Страздс, 1981; Страздс, Страздс, 1982), примерно столько же — на Березо-
вых островах Финского залива (Храбрый, 1984), в Эстонии — около 10 тыс. пар (Пеэ-
досаар, Онно, 1970; Ренно, 1972), по другим данным 12,5 тыс. пар (Thomas, 1982,
343
Семейство Чайковые
цит. по: Cramp, 1985) . На Белом море в 1960-х годах гнездилось, как минимум, 25 тыс.
пар, на Мурманском побережье — около 10 тыс пар (Бианки, 1967). Численность бело-
морской популяции с тех пор уменьшилась (Бианки, Хляп, 1970: Бианки, Бойко,
1972); видимо, то же произошло с популяцией крачек западного Мурмана (Анзигито-
ва и др., 1980). Немногочисленна полярная крачка на Земле Франца-Иосифа — в 1981 г.
на о-ве Греэм-Белл гнездилось не более 30 пар (Томкович, 1984), малочисленна на вос-
токе Таймыра (Матюшенков, 1983), редка в восточной части Чукотки (Томкович, Со-
рокин, 1983) и, в общем, малочисленна на Чукотском полуострове и о-ве Врангеля
(Портенко, 1973). Достаточно обычна эта крачка в тундрах Якутии (Воробьев, 1963)
и в ряде других мест: в Чаунской низменности и на о-ве Айон (Лебедев, Филин, 1959),
в Колючинской губе (Кречмар и др., 1978), в низовьях р. Канчалан (Кищинский и др.,
1983). На о-ве Карагинском гнездится, видимо, несколько сот пар полярных крачек
(Герасимов, 1979а). В целом наиболее многочисленны полярные крачки в западной, ат-
лантической части палеарктического ареала: так, только в Исландии гнездится свыше
100 тыс. пар, в Норвегии — 21 тыс. пар (Cramp, 1985). Общая численность вида в СССР
составляет, по-видимому, несколько сот тысяч гнездящихся пар.
Местообитания. В сезон гнездования — морские побережья, озера и долины рек тунд-
ровой зоны; арктические острова как тундровой, так и, реже, зоны арктических пус-
тынь. По внутренним водоемам крачки заходят в лесотундру и даже северную часть
тайги, по морским побережьям в Западной Европе — в зону смешанных лесов. Вне се-
зона размножения — открытое море, на зимовках - зона антарктического пакового
льда.
Размножение. Полярные крачки впервые гнездятся обычно в возрасте 3—4 лет, из-
редка в возрасте 2 лет и как исключение в годовалом возрасте (Бианки, 1967). В Кан-
далакшском заливе крачки в возрасте 1 — 2 лет составляют 4,9% размножающихся птиц,
3—4 лет — 30,7%, 5—6 лет — 27,0%, 7—8 лет —20,9%, 9 лет и старше — 16,5% (Бианки,
1967); в Великобритании соответственно 0%, 30,2%, 29,6%, 18,5%, 21,2% (Coulson,
Horobin, 1976). Моногамы, пары имеют тенденцию сохраняться из года в год, распада-
ются они чаще у молодых, чем у старых птиц. Крачки предпочитают образовывать пары
с птицами своего возраста: только у 4 из 29 пар возраст партнеров различался больше
чем на 2 года (Coulson, Horobin 1976). Нет доказательств, что пары сохраняются на
зиму; по-видимому, их восстановление происходит в стаях, которые формируются в
окрестностях колонии перед началом гнездования (Cramp, 1985). В это время можно
наблюдать различные формы демонстрационного поведения, которое у полярных
крачек очень сходно с поведением речных крачек (подробнее см.: Cullen, 1960; Cramp,
1985).
Полярная крачка — диффузно гнездящийся факультативно-колониальный вид (Зу-
бакин, 1976в, 1983). Места расположения колоний могут быть постоянны в течение
многих лет (Бианки, 1967; Coulson, Horobin, 1976); в то же время неоднократно от-
мечалась смена мест гнездовий (Карташев, 1949; Белопольский, 1957). По данным
кольцевания, после благоприятного по питанию года 47—81% взрослых беломорских
полярных крачек возвращаются на следующий год в ту же колонию, после неблаго-
приятного - только 15-25% (Бианки, Хляп, 1970). Лишь около трети молодых крачек
гнездятся в той же колонии, где они вывелись, примерно столько же поселяются в
1—5 км от родной колонии, еще 27,2% — в 5—100 км от нее; 2,2% молодых птиц пе-
реселяются более чем на 100 км. Так, в Кандалакшском заливе Белого моря поселяют-
ся птицы, окольцованные птенцами в Ботническом заливе Балтийского моря и на
Айновых островах Баренцева моря (Бианки, 1967). По наблюдениям в Эстонии, око-
ло 15% переживших зиму крачек ежегодно переселяются на новое место. Большинство
вернувшихся особей гнездится на прошлогодней гнездовой территории (Онно, 1965).
344
Полярная крачка
Первыми в колонии появляются птицы старше 8 лет, опережая трехлетних особей при-
мерно на 17 дней, а двухлетних на месяц (Coulson, Horobin, 1976).
Величина колонии колеблется от нескольких гнезд до нескольких тысяч пар. В Кан-
далакшском заливе число гнезд в колониях обычно не превышает 100 и только как
исключение бывает больше 150. Самые крупные колонии на Белом море (Соловецкие
острова) содержат 1,5—3 тыс. пар (Бианки, 1967). Колония о-ва Большой Айнов в
1970-х годах состояла из 200—650 пар (Анзигитова и др., 1980), колонии Семи остро-
вов в разные годы — из 15—159 пар (Успенский, 1941, Ковалев, 1981). В Великобри-
тании на Оркнейских островах отмечена колония в 17,5 тыс. пар (Cramp, 1985). Восточ-
нее Баренцева моря размер колоний резко уменьшается — обычно он не превышает де-
сяти пар, лишь изредка достигая 15—20 гнезд (Успенский и др., 1962; Кречмар, 1966;
Засыпкин, 1981, Данилов и др., 1984). Столь же невелики поселения полярных кра-
чек на о-ве Греэм-Белл Земли Франца-Иосифа (Томкович, 1984). Помимо колониаль-
ного гнездования, для этого вида очень обычно поселение отдельными парами (Малы-
шевский, 1962; Воробьев, 1963; Кречмар, 1966; Bergman, 1980; Кищинский и др.,
1983; Томкович, Сорокин, 1983; Данилов и др., 1984).
Плотность гнездования также весьма изменчива. На островах крачки гнездятся, как
правило, гораздо плотнее, чем по берегам тундровых рек и озер и в заболоченных озер-
ных котловинах. На о-ве Большой Лицкий расстояние между ближайшими соседними
гнездами составляет в среднем 8,3 м (1,8—23 м), на 100 м2 здесь приходится 0,50—
1,35 гнезда (Ковалев, 1981). Сходные данные для этого же острова получены Н.П. Ка-
веркиной (1983): рассчитанное по ее данным (п = 118) среднее расстояние между бли-
жайшими соседними гнездами составляет 8,1 м (0,75—53 м). На Айновых островах,
по В.А. Зубакину (1976в), это расстояние равно 3,1 м (1,6—5,4 м; п = 41). На остро-
вах Кандалакшского залива 80,4% всех гнезд (п = 102) располагаются не далее 10 м
друг от друга, причем 29,4% - не далее 3 м (Бианки, 1967) . В Я но-И нди гирек ой тунд-
ре среднее расстояние между ближайшими соседними гнездами (л = 9) составляет 172 м
(наши данные), в низовьях р.Канчалан отмечалось расстояние между гнездами в 100—
200 м (Кищинский и др., 1983). Индекс плотности гнездования, вычисленный по
имеющимся данным, для островных колоний равен 0,01, для поселений в тундре —
0,0004.
Крачки образуют как самостоятельные колонии, так и совместные поселения с
куликами (зуйки, камнешарки, кулики-сороки и др.), сизыми и розовыми чайками,
малыми, камчатскими, а в местах симпатрии — изредка и с речными крачками (Во-
робьев, 1963, Бианки, 1967; Остапенко и др., 1977, Герасимов, 1979а; Страздс, 1981).
Характерный гнездовой биотоп на островах Баренцева и Белого морей — приморс-
кие луга, участки вороничной тундры, реже обнажения скальных пород с небольшими
дерновинами (Кищинский, 1960а; Бианки, 1967; Анзигитова и др., 1980). В тундре
крачки поселяются по берегам и островам озер и рек, устраивая гнезда как на сухих
песчаных и галечных косах, так и в заболоченных местах на моховых островах-кочках,
торфяных буграх (Кречмар, 1966, Портенко, 1973; Лобанов, 1975; Кречмар и др.,
1978; Минеев, 1982; Кищинский и др„ 1983; Данилов и др., 1984). Гнездо - ямка
в грунте или во мху, со скудной выстилкой из сухих кусочков растений, собранных в
непосредственном окружении гнезда. Гнездовой валик не выражен. Средний размер
гнездовой лунки (мм) : на Белом море 120, глубина ее 20; на Айновых островах соот-
ветственно 73 (60—80, п = 5), и 15—25; в Большеземельской тундре 103 (89—120),
26(20—40). Полная кладка содержит 1—3 яйца, крайне редко 4 и 5 яиц (Conover,
1984); чаще всего в кладке 2 яйца (Кречмар, 1966, Бианки, 1967; Зубакин, 1976в;
Белопольский и др., 1977; Анзигитова и др., 1980; Минеев, 1982; Каверкина, 1983;
Данилов и др., 1984).
345
Семейство Чайковые
Раньше всего на территории СССР откладка яиц начинается в Латвии (видимо,
10—15.V) и в Эстонии (начало третьей декады мая), примерно на неделю позже в Кан-
далакшском заливе Белого моря - в среднем l.VI (17.V-7.VI). На Баренцевом море
первые яйца появляются позже:на Айновых островах в среднем 7.VI, на Семи островах
10—22.VI, в среднем 17.VI. В 1970 г., однако, откладка яиц на Семи островах началась
29.V (Б ел о польский, 1957; Бианки, 1967; Белопольский и др., 1977; Анзигитова
и др., 1980; Страздс, 1981). На озерах Лапландского заповедника кладка начинается
10—20.VI (Владимирская, 1948). На Земле Франца-Иосифа первые яйца появляются,
видимо, в конце первой — начале второй декады июля (Паровщиков, 1963; Томкович,
1984), на полуострове Канин — в первых числах июня (Спангенберг, Леонович, 1960).
В Малоземельской и Большеземельской тундрах первые кладки отмечены 14—16.VI,
в отдельные годы - только 16.VII (Минеев, 1982), на южном Ямале - 9-25.VI (Да-
нилов и др., 1984), в низовьях Енисея — в третьей декаде июня — начале июля (Рогаче-
ва и др., 1983), на Таймыре, видимо, во второй декаде июня (Кречмар, 1966; Матюшен-
ков, 1983). В Приколымской тундре первые кладки с 1 яйцом регистрировались уже
6.VI, в Яно-Индигирской тундре свежая кладка из 2 яиц встречена 12.VI (Воробьев,
1963). В Чаунской губе на о-ве Айон откладка яиц начинается, видимо, в конце пер-
вой — начале второй декадыиюня (Лебедев, Филин, 1959), в Колючинской губе — 14—
20.VI (Кречмар и др., 1978), на Восточной Чукотке и в низовьях р. Канчалан, судя по
датам вылупления птенцов, — в начале второй декады июня (Кищинский и др., 1983;
Томкович, Сорокин, 1983). В западной части Камчатки 7.VI уже отмечались гнезда
(Остапенко и др., 1977), на о-ве Карагинский откладка первых яиц зарегистрирована
12—16.VI (Каверкина, 1982). Массовая яйцекладка начинается обычно через несколь-
ко дней после появления первых яиц. Растянутость периода откладки яиц относительно
невелика — обычно не более месяца; в более суровых климатических условиях она про-
ходит в более сжатые сроки (Бианки, 1967; Белопольский и др., 1977; Кищинский
и др., 1983). У ряда особей отмечаются повторные кладки после гибели первой (Бе-
лопольский, 1957; Бианки, 1967).
Обращает на себя внимание, что в тундрах по всему побережью Ледовитого океана
откладка яиц начинается почти одновременно — примерно во вторую декаду июня,
с небольшим сдвигом сроков в ту или иную сторону в зависимости от характера весны.
Лишь в тундрах северо-востока Якутии полярные крачки начинают откладку яиц,
видимо, на несколько дней раньше.
Окраска яиц сложная. Фон яйца сильно варьирует от чисто-голубого до коричневого;
наиболее часто встречаются яйца серовато-зеленого, оливкового и буроватого цветов.
По этому фону разбросаны бурые и черновато-бурые поверхностные пятна, обилие и
характер распределения которых также чрезвычайно разнообразны. В одном гнезде
могут быть яйца как одного цвета, так и, реже, различной окраски (Кищинский, 1960а;
Бианки, 1967; Анзигитова и др„ 1980). Размеры яиц (мм) : с Айновых островов (и =
= 349) в среднем 39,4 X 28,9 (Анзигитова и др., 1980); с Семи островов (и = 167) в
среднем 40,1 X 29,4 (Каверкина, 1983); с Кандалакшского залива Белого моря (п =
= 395) в среднем 40,1 X 29,4 (Бианки, 1967); из Латвии (п = 35) 38,3—42,6 X 28,2—
30,7, в среднем 40,4 X 29,3 (Страздс, 1981); из Большеземельской тундры 35,5-42 X
X 27,0-30,0, в среднем 39,1 X 28,4 (Минеев, 1982); из южной части Ямала (п = 14)
35,8—41,9 X 27,2- 30,1, в среднем 39,2 X 28,9 (Данилов и др., 1984). Масса свежесне-
сенного яйца (п = 198) 17,5 г (Анзигитова и др., 1980).
Насиживание начинается с первого яйца и продолжается обычно 18—24 дня, его
длительность изменяется в зависимости от погоды и условий питания (Бианки, 1967;
Анзигитова и др., 1980; Данилов и др., 1984; Cramp, 1985). Насиживают оба роди-
теля. По данным 7-суточных наблюдений, самец находится на гнезде в среднем
346
Полярная крачка
8 ч 30 мин, а самка — 13 ч. 40 мин, 1,5—2 ч в сутки кладка не обогревается. Смена
партнеров на гнезде может происходить с ритуальным подношением рыбки (Анзиги-
това и др., 1980).
Первые трещинки на яйцах в районе Белого моря чаще всего появляются за 2 сут
до вылупления, на Айновых островах — за 1 сут. Промежуток между вылуплением
первого и последнего птенца в гнезде с 2—3 яйцами обычно равен суткам. После вы-
лупления родители уносят из гнезда скорлупу. Вес однодневного птенца равен 10—
13,5 г, в среднем 13,0 г. Птенцы сидят в гнезде около 2 сут, затем выводок покидает
гнездо и либо держится на гнездовом участке вплоть до подъема на крыло, либо остав-
ляет его и перемещается в другой участок колонии. Начиная с 1— 1,5-суточного воз-
раста во время тревоги птенцы разбегаются и затаиваются (Бианки, 1967, Анзигитова
и др., 1980). Крачки-родители активно атакуют хищника и нарушителя спокойствия,
пикируя на него, ударяя клювом и обливая пометом. Защитная активность достигает
максимума во время вылупления птенцов. В крупных колониях наблюдается большая
групповая, но меньшая индивидуальная защитная активность (Каверкина, Рощевский,
1984).
Заботятся о птенцах оба родителя, но самец чаще кормит, а самка больше времени
проводит с птенцами. В первый день жизни родители постоянно обогревают птенцов,
в 2—4-суточном возрасте на обогрев тратится около 10 ч в сутки, к двухнедельному
возрасту обогрев прекращается. Количество прилетов родителей с кормом сильно
варьирует даже в гнездах с близкими по возрасту птенцами; в среднем птенцов кормят
около 20 раз в сутки (п = 10), к концу периода выкармливания несколько реже, чем
в начале. Помимо родителей, птенцов изредка кормят крачки-”помощники” — возмож-
но, это птицы, потерявшие свой выводок (Анзигитова и др., 1980). Родители начинают
отличать своих птенцов от чужих в норме, видимо, уже на вторые сутки после вылупле-
ния. Однако в условиях высокой густой растительности, полностью скрывающей птен-
цов (Айновы острова), отмечено значительное запаздывание сроков становления ин-
дивидуального опознавания (Анзигитова, Зубакин, 1975). Птенцы поднимаются на
крыло в возрасте 21—24 дней (Бианки, 1967; Cramp, 1985) или 25—28 дней (Анзиги-
това и др., 1980).
В Кандалакшском заливе в разные годы поднимается на крыло 1,0—1,2 молодых
на пару птиц (Бианки, 1967), на Айновых островах в благоприятные годы — только
0,7, причем в 10 из 21 сезона гнездование всех крачек колонии здесь было неудачным
(Анзигитова и др., 1980). На юго-западе Финляндии поднимается на крыло 0,53—1,05
молодых на пару птиц (Lemmetyinen, 1973), на Шпицбергене - 0,52 (Bengtson, 1971).
В Кандалакшском заливе гибнет от 18 до 37% яиц и 10—25% птенцов от числа вылупив-
шихся, на Айновых островах в относительно благоприятные годы — 20—26% яиц и 33%
птенцов. Основные причины гибели кладок и птенцов — затяжные шторма и хищ-
ничество.
Ежегодная смертность взрослых птиц на Кандалакшском заливе составляет в сред-
нем 19,7% (Бианки, Хляп, 1970), в Великобритании — 12—13% (Coulson, Horobin,
1976). Максимальная продолжительность жизни, по данным кольцевания, 34 года
(Clappetal., 1982,цит.по: Cramp, 1985).
Суточная активность, поведение. Общественная птица во все сезоны года. Часто кор-
мится большими скоплениями, однако нередки и одиночные кормежки. В период зи-
мовки полярные крачки держатся крупными стаями (Erwin, 1978; Cramp, 1985).
Питание. Главной пищей полярных крачек служат мелкие виды рыб и мальки более
крупных рыб, а также ракообразные. Кроме того, крачки поедают полихет и насеко-
мых. Видовой состав кормов зависит от конкретных условий обитаний крачек. Так,
в Кандалакшском заливе основной корм, от подхода которого зависит успех гнездо-
347
Семейство Чайковые
вания, — трехиглая колюшка. На Восточном Мурмане поедается преимущественно
песчанка и сельдь (Белопольский, 1957; Бианки, 1967). На юге Ямала в желудках
крачек находили гольянов, колюшек, мальков сиговых рыб, а также водных беспозво-
ночных — жуков-плавунцов и бокоплавов (Данилов и др., 1984), в Яно-Индигирекой
низменности — водных жуков, стрекоз, щитней и рыбу (Успенский и др., 1962). Для
полярных крачек на Таймыре характерно питание насекомыми, плавающими на по-
верхности воды или неглубоко в воде, рыба в питании здесь также играет значитель-
ную роль (Кречмар, 1966). Помимо водных насекомых (в частности, куколок хиро-
номид, которые имеют большое значение в питании крачек в тундре), птицы поедают
и наземных насекомых, снесенных ветром в водоемы (Чернов, 1967).
Враги, неблагоприятные факторы. Основные враги полярных крачек из наземных
хищников — песцы, лисицы, горностаи; из пернатых — крупные чайки (включая сизую,
которая выступает по отношению к крачке как главный хищник на Айновых островах),
в ряде мест — серые вороны. В отдельных случаях ущерб колониям могут наносить
орланы-белохвосты и сапсаны. Отмечено выпивание яиц крачек камнешаркой. Из абио-
тических факторов наибольшее отрицательное воздействие оказывают затяжные штор-
ма, во время которых крачки нередко бросают насиженные кладки и птенцов. В дожд-
ливую погоду птенцы часто гибнут от переохлаждения (Благосклонов, 1960; Бианки,
1967; Анзигитова и др., 1980; Данилов и др., 1984).
Хозяйственное значение, охрана. Хозяйственного значения вид не имеет. Специаль-
ные меры охраны не предусмотрены. Колонии охраняются в Кандалакшском, Вильсан-
дийском и в некоторых других заповедниках.
Камчатская крачка —
Sterna camtschatica Pallas, 1811
Aleutian Tern (англ.), Aleutenseeschwalbe (нем.), Sterne aleoute (фр.).
Син. (лат.) : Sterna aleutica Baird, 1869.
Син. (рус.) : алеутская крачка, белолобая крачка.
Статус. Гнездящийся мигрирующий, возможно частично зимующий вид.
Общая характеристика и полевые признаки. Крачка среднего размера. Похожа на
речную и полярную крачек, но отличается от них более массивным телосложением,
более темной окраской верха тела, треугольным белым пятном на лбу, относительно
коротким белым хвостом и черными ногами. Кроме того, отличается по голосу, напо-
минающему чириканье (Лобков, Головина, 1978; Нечаев, 1981): ”черь, черь...” (’’чирк,
чирк...”), или ’’свирь, свирь, свирьи...” (”цирь, цирь”). Часто в полете птицы посви-
стывают ”цвит, цвит...” (’’цирк, цирк...”), иногда издают торопливые звуки: ”цвит.
цвит-пи-и” (”псии, пени...”). В момент агрессивных бросков у гнезд с птенцами слыш-
но негромкое кряканье ’’крек, крек...”, отсутствующее у птиц в другое время.
На земле держится горизонтально, ходит мало и неохотно. Полет легкий, порхаю-
щий, маневренный, с глубокими взмахами крыльев. На воду присаживается редко
и сразу же взлетает. Не ныряет, пикирующие броски с погружением не отмечены. До-
бычу схватывает с поверхности воды или из ее верхних слоев. Постоянно держится
в море; на суше отмечается только в период гнездования. Гнездится колониями, в
период миграций образует многочисленные стаи.
Подвидовая систематика. Монотипический вид.
Замечания по систематике. Камчатская крачка описана дважды. Первое описание,
несомненно, относящееся к крачке этого вида, дается под названием Sterna camtscha-
tica Pallas, 1811, но это описание скудное, неточное и в некоторых деталях противоре-
чивое. Так, в первых строках текста совершенно верно указываются такие характер-
348
Камчатская крачка
ные черты вида, как черный клюв, белый лоб и белая надглазничная полоса. Далее
сообщается, что эта крачка многочисленна в морях у Камчатки и на прилежащих остро-
вах, что также соответствует распространению камчатской крачки. В то же время
П.С. Даллас ошибочно пишет о красной окраске ног (на эту неточность справедливо
указывал еще С.А. Бутурлин, 1934) — признак нетипичный для данного вида. Однако
ниже в том же диагнозе П.С. Даллас, более подробно описывая окраску, упоминает
еще один отличительный признак этой крачки — белый без каких-либо темных кае-
мок хвост. Здесь же приводится и окраска ног; они буровато-красные с черным по-
крытием (’’обмазаны” черным). Следовательно упомянутый автор дважды приводит
описание окраски ног, причем во второй раз совершенно правильно. Действительно,
у живых и недавно добытых камчатских крачек ноги черные или буровато-черные,
а у экземпляров, длительно хранившихся в музеях, приобретают красновато-бурый
цвет. В данном случае П.С. Паллас описал камчатскую крачку по сборам Г.В. Степле-
ра, пролежавшим в коллекциях более 50 лет. К сожалению, некоторые орнитологи
не обратили внимания, что Паллас в двух местах своего текста приводит описание
камчатской крачки. Он дает противоречивое описание окраски ног, но в то же вре-
мя правильно указывает на ее такие характерные особенности, как белые лоб, бро-
ви и хвост.
Позднее (через 58 лет) полное описание этой крачки под названием Sterna aleutica
дает С. Бэрд (Baird, 1869), причем приводит рисунок. Подробное и ясное описание
крачки С. Бэрдом послужило причиной считать его первооткрывателем этого вида.
С тех пор название ’’алеутская” крачка получило широкое распространение (Vaurie,
1965; Степанян, 1975; и др.). Однако по правилу приоритета, как это справедливо
полагал С.А. Бутурлин (1934) и отметил Л.А. Портенко (1973), ’’алеутскую” крач-
ку все же следует называть камчатской — Sterna camtschatica.
Описание. Размеры (мм) и масса тела (г) (ЗИН; сборы В.А. Нечаева и Е.Г. Лоб-
кова) :
Длина крыла:
dd (п = 20) — 250—284 (среднее 268);
99 (п = 10) — 255—277 (среднее 266).
Длина клюва:
dd (п = 10) — 30,0-36,5 (среднее 32,2);
99 (п = 5) - 30,0-35,0 (среднее 32,1).
Длина цевки:
dd (п = 10) — 17—22 (среднее 19,5);
99 (п = 5) — 16—20 (среднее 17,6).
Масса тела:
dd (и = 11) — 117—130 (среднее 124);
99 (п = 8) — 117—138 (среднее 128).
Окраска. Самцы и самки в брачном наряде. Лоб, короткая бровь, щеки, под-
бородок, горло, нижняя сторона крыльев, подхвостье, надхвостье и хвост белые. Верх
головы (темя, затылок), уздечка и кольцо вокруг глаз черные. Спина, поясница, пле-
чевые и верхние кроющие маховых свинцово-серые. Грудь, брюшко и бока тела серые
с легким буроватым, в свежем пере розоватым оттенком. Первостепенные маховые
со II (первого видимого) по VII темно-серые с серебристым налетом и белыми стерж-
349
Семейство Чайковые
нями, наружное опахало II махового черное с серой вершиной. На внутренних опаха-
лах II—VII маховых протягивается широкая продольная белая полоса. Остальные пер-
востепенные серые. Второстепенные маховые также серые, но с белыми каемками на
вершинах наружных опахал и большими белыми полями на внутренних опахалах.
Клюв, ноги и когти черные. Радужина темно-коричневая.
Пуховой наряд. Птенец покрыт густым и длинным (до 17 мм) пухом. Условно
выделяется светлый (более редкий) и темный типы окраски, но встречаются особи
и с переходной окраской (Лобков, Головина, 1978). У птенцов темной окраски го-
лова и верхняя сторона тела коричнево-оливковые (лоб и темя покрыты крупными
черными пятнами); уздечка, пух вокруг глаз, бока подбородка черные, но середина
подбородка белая, горло темно-серое. У птенцов светлой окраски голова и спина свет-
ло-коричневые, реже желтые с некрупными пестринами грязно-бурого цвета; уздеч-
ка, бока подбородка и пух вокруг глаз грязно-бурые, середина подбородка белая,
горло серое. Грудь, брюшко, нижняя поверхность крыльев белые, на груди нередко
выступает желтоватый налет. Основание клюва от бледно-розового до оранжевого,
надклювье темное, вершина клюва от темно-розового до черного. Яйцевой зуб серый,
иногда с желтоватым оттенком. Складки в углах рта и ротовая полость розовые или
розовато-оранжевые. Ноги бледно-розовые или розовато-желтые с серым оттенком
на передней стороне, когти черные. Радужина буроватая. Первоописание пухового
наряда камчатской крачки (Nelson, 1887) относится к птенцам с темной окраской
пуха.
Гнездовой наряд (Нечаев, 1981). Лоб, уздечка, углы рта и шея серовато-бурые.
Темя такого же цвета, но с черными продольными пестринами. На затылке черная
изогнутая полоса (шириною до 15 мм) в виде полукруглой короны, передние концы
которой подходят к заднему краю глаза. Вблизи переднего края глаза черное пятно^
Щеки и подбородок белые с серовато-бурым налетом. Горло и брюшко белые, по бело-
му фону груди и боков тела серовато-бурый оттенок. Спина и верхние кроющие крыль-
ев черные с охристыми каемками. Поясница серовато-черная с охристыми полоска-
ми. Верхние кроющие хвоста сизые с черной предвершинной полосой и охристой верши-
ной, нижние — белые с охристым оттенком. Первостепенные маховые пепельно-серые.
Наружное опахало II (первого видимого) первостепенного черное, остальных — се-
рое. На внутренних опахалах белые полоски. На вершинах первостепенных маховых
(кроме II—IV) охристые (или беловатые) каемки. Второстепенные серые с белыми
или буроватыми вершинами, образующими сплошную полосу. Наружные рулевые
серовато-белые, у остальных — серые наружные и белые внутренние опахала. На всех
рулевых черные предвершинные полосы и охристые вершины. Подкрылья белые.
Ноги оранжево-серые, когти черные. Надклювье темно-серое, подклювье розовато-
оранжевое, на вершине клюва потемнение.
Характер возрастной и сезонной изменчивости окраски не изучен.
Линька. Сведения о характере линьки камчатской крачки отсутствуют. По всей
вероятности, последовательность смены нарядов, как у речной крачки. Взрослые пти-
цы, участвовавшие в размножении, начинают полную линьку во второй половине ав-
густа — начале сентября и заканчивают ее на зимовках. Самка от 23 августа (северная
часть Сахалина) начала смену мелких перьев на спине, где отмечались пеньки и раз-
вернувшиеся кисточки.
Распространение. Гнездовой ареал. Охватывает отдельные участки на по-
бережье Тихого океана, Охотского и Берингова морей (рис. 82). В СССР гнездование
отмечено на побережье Корякского нагорья, п-ова Камчатка и о-ва Сахалин, а так-
же на северо-западном побережье Охотского моря (Лобков, Нечаев, 1982) (рис. 83).
На п-ове Камчатка — от устья р. Апука (Портенко, 1964; Кищинский, 1980) до дель-
350
Камчатская крачка
ты р. Авача на восточном побережье (Лобков, 1976) и от устья р. Тигиль (Остапенко
и др., 1977) до низовьев р. Опала на западном побережье (данные Е.Г. Лобкова), а так-
же на о-ве Карагинский (Герасимов, 1979а, б); взрослые птицы встречаются на море
до мыса Лопатка (п-ов Камчатка). На о-ве Сахалин — на побережье залива Анива и
оз. Лебяжье, на островах оз. Невское, на северо-восточном побережье и островах за-
ливов Набиль, Ныйво, Даги, Чайво и Пильтун, на побережье п-ова Терпения и на севе-
ро-западном побережье острова от залива Помрь на севере до окрестностей пос. Трам-
Рис. 82. Гнездовой ареал камчатской крачки
Рис. 83. Гнездовой ареал камчатской крач-
ки в СССР
баус на юге (Hesse, 1915; Munsterhjelm, 1922; Гизенко, 1955; Нечаев, 1981; данные
В.А. Нечаева). Гнездование установлено на северо-западном побережье Охотского
моря (Тауйская губа) вблизи рек Ойра и Широкая (Леонович, 1976). Вне СССР гнез-
дится на западном побережье Аляски от залива Коцебу (лагуна Тасейчик) на севере
до залива Якутат и немного южнее до бухты Литуя (залив Аляска) на юге, включая
п-ов Аляска, а также на о-ве Кадьяк и Алеутских островах: Унимак, Умнак, Адак,
Амчитка, Алаид, Атту (Gabrielson, Lincoln, 1959; Kessel, Gibson, 1978; Sowls et al.,
1978).
Зимовки. Места зимовок неизвестны. Предполагается, что большинство кам-
чатских крачек зимует в северо-западной части Тихого океана (Gabrielson, Lincoln,
1959; Sowls et al., 1978). Есть указания (Бутурлин, 1934; Гизенко, 1955) на зимов-
ки этого вида у берегов о-ва Сахалин, но коллекционными экземплярами это не под-
тверждается. По данным О. Остина и Н. Курода (Austin, Kuroda, 1953), камчатская
крачка не мигрирует к югу, а держится в северных морях круглый год. В Японии
это нерегулярно залетный вид (Check-list of Japanese birds, 1974).
Миграции. Весной на о-ве Сахалин камчатские крачки появляются в мае, пролет
продолжается до первой декады июня. На озерах Лебяжье и Невское в 1978—1980 гг.
первые птицы появились 9—15 мая (Воронов, Еремин, 1982). На побережье залива
Терпения (устье р. Владимировка) 23 мая — 3 июня 1977 г. регулярно наблюдали
стаи пролетных птиц, состоящие из 30—40, редко из 8—20 и 50—60 особей. Наиболее
интенсивно они летели в конце третьей декады мая. На юго-восточное побережье п-ова
Камчатка возле Семячинского лимана крачки прилетают 19—29, в среднем за 6 лет
22 мая (Лобков, Головина, 1978). Больших скоплений на образует: максимум до
200 особей. В заливе Корфа первые птицы появились 28 мая, а в устье р. Апука —
29 мая 1960 г. (Кищинский, 1980). Весной на Аляске крачки раньше всего (с 30 ап-
реля по 13 мая) появляются в заливе Аляска (на о-ве Кадьяк, в дельте р. Коппер и др.),
затем (с 16 мая по 7 июня) в западных и северных районах (Kessel, Gibson, 1978).
351
Семейство Чайковые
Сроки осенних миграций не прослежены. На о-ве Сахалин с озер Лебяжье и Невское
птицы улетают в сентябре (Воронов, Еремин, 1982). В Кроноцком заповеднике (Во-
сточная Камчатка) они исчезают к середине сентября: самая поздняя встреча — 13 сен-
тября 1973 г. (Лобков, Головина, 1978). На Аляске отлет начинается вскоре после
того, как молодые поднимутся на крыло; пролет наблюдали с 12 августа по 19 сен-
тября (Gibson, Kessel, 1978). На о-ве Св. Михаила (в заливе Нортон-Саунд) их отме-
чали до 10 сентября (Nelson, 1887).
В период пролета крачки найдены у берегов п-ова Камчатка (Лобков, 1976) и
о-ва Сахалин (Нечаев, 1981). Миграции у берегов Аляски не отмечены; предполагает-
ся, что птицы прилетают на места гнездования с моря и улетают осенью в открытые
воды (Kessel, Gibson, 1978). Залетных крачек регистрировали в августе 1879 г. в
заливе Св. Лаврентия на юго-восточном берегу Чукотского полуострова (Newcomb,
1888), 7 сентября 1911 г. на о-ве Медном (Командорские острова) и 24 августа
1930 г. вблизи г. Охотска (Дементьев, 1951).
Численность. На о-ве Сахалин наиболее многочисленные колонии этого вида рас-
полагаются на северо-восточном побережье: на о-ве Лярво в заливе Даги (в 1976 г.
около 500 пар, в 1984 г. до 200 пар птиц) и на о-ве Чайка в заливе Набиль (600—
700 пар, по данным учета в 1981 и 1984 гг.). Колонии численностью до 200 пар най-
дены на юге п-ова Терпения. На оз. Невское гнездится 250—300 птиц (Воронов, Ере-
мин, 1982). Колонии из 20—40 пар отмечены на побережьях заливов Набиль, Чайво
и Пильтун, на северо-западном побережье о-ва Сахалин вблизи мыса Погиби, заливов
Тык и Виахту, окрестностей пос. Трамбаус, на оз. Лебяжье и побережье залива Ани-
ва (данные В.А. Нечаева). Общая численность камчатской крачки в обследованных
колониях на о-ве Сахалин, по данным 1979—1984 гг., составляет 2,1 тыс.парптиц.
На северо-западном побережье Охотского моря в Тауйской губе в 1972 г. обна-
ружена колония численностью до 30 пар (Леонович, 1976). На п-ове Камчатка чис-
ленность этого вида составляет около 4 тыс. пар. Общая численность камчатской крач-
ки в азиатской части ареала (по данным на 1984 г.) немногим более 6 тыс. пар.. По
материалам, собранным на Аляске до 1977 г., описано более 60 поселений общей числен-
ностью примерно 3,4 тыс. пар (Kessel, Gibson, 1978). Общая (предполагаемая) числен-
ность этого вида в американской части ареала оценивалась в 10 тыс. особей (Sowls
et al., 1978). Однако в последние годы отмечен рост численности: в дельте р. Коппер
до 1973 г. было известно несколько колоний общей численностью 300—500 особей
(Isleib, Kessel, 1973), в 1978 г. там было учтено 34 колонии численностью 2428 осо-
бей (Holtan, 1980). Некоторое увеличение численности отмечено ина Камчатке. Общая
численность мировой популяции камчатской крачки достигает 11—12 тыс. пар, при-
чем на долю СССР приходится не менее половины этого количества.
Местообитания. В гнездовой период камчатские крачки населяют полосу морского
побережья шириною 3—6, редко до 20 км, изрезанную лагунными заливами с низмен-
ными травянистыми островами, приустьевыми участками рек, протоками и озерами,
а также прибрежные заболоченные, сильно разреженные леса (подобно лесотундре)
из низких и искривленных ветрами лиственниц; реже склоны и вершины возвышен-
ностей, покрытые зарослями из вересковых кустарничков, шикши сибирской и разно-
травными лугами. В период миграций держится в основном на-морском побережье.
Размножение. Половая зрелость наступает, по-видимому, в возрасте 1 года; но боль-
шинство особей нормально размножается в возрасте 2 лет. Камчатская крачка - мо-
нетам. Пары образуются на местах гнездования. Брачное поведение отчетливо выра-
жено в- июне, в период формирования пар (через неделю после прилета) и колоний,
а также во время откладки яиц. У птиц, потерявших первые кладки и способных к
повторным, брачное поведение можно наблюдать и в июле.
352
Камчатская крачка
Весенний ток начинается с групповых полетов и переклички. Для воздушных ритуа-
лов характерен полет с рыбкой. Движения партнеров синхронны. Полет с рыбкой сме-
няется одновременным купанием. Из наземных ритуалов у этого вида известны не-
сколько поз основных демонстраций, ритуальная чистка оперения, своеобразный ри-
туальный танец и ритуал подношения рыбки, которые мало чем отличаются от тако-
вых у других видов крачек (данные Е.Г. Лобкова и Н.П. Каверкиной). В период фор-
мирования пар можно наблюдать выпрямленную позу, когда один из партнеров (или
оба) резко вытягивают шею вверх, вздергивают голову, опускают крылья и поднима-
ют хвост под углом около 45°.
Выражено и территориальное поведение. Камчатские крачки охраняют приколо-
ниальную территорию в радиусе до 500 м; для них характерны ’’птицы-сто рожа” (Ка-
веркина, 1983). Камчатские крачки раньше других видов крачек реагируют на появ-
ление опасности. Голосовая активность доминирует над двигательной, однако звуко-
вая активность камчатской крачки в 5-6 раз ниже активности речной крачки (Кавер-
кина, Давыдова, 1982). Колонии формируются примерно через 10—15 дней после при-
лета (Лобков,1976).
На северо-восточном побережье о-ва Сахалин и оз. Невское гнездовые биотопы вида—
низинные острова, расположенные в лагунных заливах (Набиль, Даги, Пильтун) и по-
крытые мохово-осоковыми болотами, разнотравными лугами и зарослями кустар-
ничков (шикши сибирской, брусники, ивы буреющей и т.д.), а также некоторые участ-
ки побережья этих заливов (Нечаев, 1981). На северо-западном побережье о-ва Саха-
лин (окрестности мыса Погиби, заливов Тык и Виахту) и на побережье залива Анива
и оз. Лебяжье птицы гнездятся на кочковатых осоково-пушицевых болотах с клюк-
вой и морошкой среди разреженных и заболоченных лиственничных лесов с кедро-
вым стланником на расстоянии от 1 до 20 км от берега моря. На п-ове Терпения места
обитания вида — мохово-травянистые, слабо кочковатые болота, а также сухие скло-
ны и вершины возвышенностей (высотой до 300 м), покрытые шикшей, брусникой
и разнотравными лугами с одиночными кустами кедрового стланника, и участки на
песчаных дюнах среди редкой травянисто-кустарниковой растительности.
На побережье Корякского нагорья (в пойме р. Апука) населяют низинные острова
в устьевой части реки, густо заросшие вейником и осокой (Кищинский, 1980). На
побережье Тауйской губы колонии обнаружены на островке в устье р. Ойра и на при-
легающих участках приморской косы (Леонович, 1976). На п-ове Камчатка выделя-
ются три основных типа гнездовых местообитаний: 1) заросшие растительностью пес-
чаные и песчано-галечные косы и острова в устьях рек, в лагунных заливах и лиманах;
2) заболоченные участки тундр (марши) с мозаикой небольших озер в низовьях рек,
главным образом в дельтах и у лиманов; 3) сырые кочкарники на невысоких равни-
нных приморских террасах с озерами. На Аляске населяют мохово-травянистые бо-
лота, колосниковые луга, заболоченные участки тундры среди озер (Gill, Dick, 1977;
Holtan, 1980).
Камчатские крачки гнездятся колониями, небольшими поселениями, иногда от-
дельными парами. На п-ове Камчатка из 27 колоний в 21 было до 50 пар, в двух —
по 50—100 пар, еще в двух — от 100 до 200 пар и еще в двух колониях было учтено
более чем по 200 пар. Самые крупные колонии на п-ове Камчатка описаны в 1976—
1977 гг. в устье р. Новый Семячик (150 пар), в 1981 г. в устье р. Воровская (250—
300 пар) и в 1983 г. в лимане устья р. Ича (200—250 пар). На о-ве Сахалин самая круп-
ная колония в заливе Набиль (около 700 пар). Из 34 колоний в дельте р. Коппер 16 бы-
ли численностью до 50 особей, 10 — от 50 до 100 особей, 6 — от 100 до 200 особей,
2 — от 200 до 500 особей (Holtan, 1980). Самые большие колонии известны на Аляс-
ке: они достигают 800—1200 особей в заливе Моллер и 600 особей в заливе Гудньюс
12. Зак.826
353
Семейство Чайковые
(Sowls et al., 1978; Kessel, Gibson, 1978). В последние годы в связи с увеличением
численности происходит укрупнение мелких колоний и возникновение новых по-
селений.
Постройка гнезд начинается в первой половине июня и у разных пар продолжается
до середины июля. Гнезда представляют собою углубления без выраженных валиков
по краям. Они располагаются среди мха, кустарничков и трав, реже на заломах сухих
стеблей колосняка и листьев осоки. На островах найдены гнезда, устроенные на ку-
чах древесного мусора, веток и травы, намытых волнами. На приморских косах и бе-
регах — на песчано-галечном грунте среди редких стеблей колосняка. Обычно же гнез-
да помещаются на возвышенных участках — буграх и кочках (Леонович, 1976; Лоб-
ков, Головина, 1978; Кищинский, 1980; Нечаев, 1981). Лоток выстилается неболь-
шим количеством сухих листьев осок, злаков, карликовой ивы, морошки, сабель-
ника и других растений, стебельками шикши, кусочками корешков и веточек мха.
Толщина подстилки достигает 8—10 мм. Иногда гнезда вообще лишены выстилки и
представляют собою углубления в песке или гальке.
На п-ове Камчатка гнезда, построенные на болотах, имеют диаметр лотка 105 — 150,
в среднем (и = 26) 125 мм, глубину 32—65, в среднем (п = 17) 47 мм; а на колосни-
ковых лугах соответственно 100—170, в среднем (и = 12) 126, 20—58, в среднем
(и = 11) 41 (данные Е.Г. Лобкова). На о-ве Сахалин в густонаселенной колонии на
о-ве Лярво поперечник лотков составлял 60—120X80—130, в среднем (и = 17)
92 X 95 мм, глубина 15—50, в среднем (и = 15) 30 мм; в разреженной колонии вбли-
зи мыса Погиби соответственно 100—120 X 105 —125, в среднем (н = 12) 111X118,
25—57, в среднем (п = 12) 42 (Нечаев, 1981). В дельте р. Коппер средний диаметр лот-
ка (п = 126) 111 мм и глубина 38 мм (Holtan, 1980). Лоток имеет овальную, реже
округлую форму, а в период насиживания яиц он становится немного шире и глубже.
Необычно массивные гнезда были на валах выброшенной водой травы в устье р. Any-
ка: 250—300 мм в поперечнике и около 100 мм высотой, размеры лотков 100—120,
а глубина 30—40 мм (Кищинский, 1980).
В поселениях крачек отмечено рассеянное размещение гнезд. На о-ве Сахалин в
’’плотных” колониях гнезда располагаются на расстоянии 1—8 м одно от другого
(п = 17; 4,88 ± 0,4), в разреженных — от 10 до 100 м (п = 12; 57,83 ± 8,3). Неред-
ко селятся изолированными парами и группами из 2—3 пар. На п-ове Камчатка самая
высокая плотность в колониях 2 пары на 10 м2, при наименьшем расстоянии между
гнездами 1,6—1,9 м (данные Е.Г. Лобкова). В дельте р. Коппер средняя плотность
от 36,9 до 151,5 гнезд на гектар, а среднее (по колонии) ближайшее расстояние меж-
ду ними от 3,22 до 9,09 м (Holtan, 1980).
В азиатской части ареала вида первые яйца отмечены в конце первой — во второй
декадах июня. Массовая откладка в конце второй — начале третьей декад июня на
о-ве Сахалин (Нечаев, 1981) и с 8—10 по 19—21 июня на п-ове Камчатка (Лобков,
Головина, 1978). Поздние (повторные) свежие кладки найдены в последних числах
июля. На Аляске (дельта р. Коппер) в раннюю весну 1978 г. пик откладки яиц был с
12 по 17 мая (Holtan, 1980).
В кладке обычно 2, редко 1 и еще реже 3 и 4 яйца (Лобков, Головина, 1978; Кова-
лев и др., 1980; Нечаев, 1981; Gill, Dick, 1977; и др.). Интервалы между откладка-
ми яиц составляют от 1 до 4 сут (Лобков, Головина, 1978). Средняя величина клад-
ки для о-ва Сахалин и п-ова Камчатка 1,6 яиц (Каверкина, 1983). В дельте р. Коппер
из 164 гнезд в 10 колониях 139 содержали по 2 яйца и 25 — по одному; за весь сезон
1978 г. была найдена лишь одна кладка из 3 яиц (Holtan, 1980).
Размеры яиц (мм): на п-ове Камчатка: 39,1—48,9X27,4—30,5, в среднем (и =88)
43,9X29,1; масса свежих яиц (г) 17—21,9, в среднем (п = 30) 19,6 (Лобков, Голо-
354
Камчатская крачка
вина, 1978). На о-ве Сахалин: размеры 41,0—48,0X26,0—32,0, в среднем (и = 121)
43,8 Х29,6 (Нечаев, 1981); масса свежих яиц (п = 15) в среднем 20,1 (Воронов, Ере-
мин, 1982). На побережье Тауйской губы: 40,2—45,7 X 28,0—30,6, в среднем (и = 20)
42,9 X29,0; масса 16,3—21,9, в среднем 19,4 (Леонович, 1976). На Аляске: размеры
яиц (п = 83) в среднем 43,5 X 29,6; масса 19,6 (Holtan, 1980) .
Форма яиц в основном продолговато-овальная. Окраска сложная и сильно варьиру-
ет как в пределах одной, так и в разных кладках. На о-ве Сахалин выделено шесть ос-
новных типов окраски основного фона яиц (и =125): песочно-зеленоватый (51,2%),
зеленоватый (13,0%), песочный (10,4%), буровато-зеленый, голубовато-зеленый и
буроватый (Нечаев, 1981). Рисунок четкий, густой (реже редкий) из крупных пятен,
чаще удлиненной формы: глубокий рисунок — серый, серовато-бурый; поверхност-
ный — темно-бурый, бурый, каштановый. Пятна разбросаны по всей поверхности скор-
лупы (80%), реже образуют сгущения на тупой половине, еще реже — в виде венчика
возле тупого полюса или поперек середины яйца. Единично встречаются яйца с мел-
кой пятнистостью. Скорлупа тонкая, на просвет зеленоватая. Ее поверхность мелко-
зернистая, слабошероховатая, матовая. При выцветании зеленый цвет буреет. На о-ве Са-
халин из 40 кладок с 2 яйцами в 27 окраска была более или менее одинаковая.
Яйца насиживают оба партнера, но самка значительно больше (Лобков, Головина,
1978; Каверкина, 1983; Gill, Dick, 1977; Holtan, 1980). Насиживание начинается с пер-
вого яйца. На о-ве Сахалин и п-ове Камчатка птенцы вылупляются в конце первой
декады июля, в ранние весны — в конце июня; массовое вылупление — в середине
июля, самое позднее — в конце июля (Лобков, Головина, 1978; Нечаев, 1981). В до-
лине р. Коппер в 1978 г. птенцы вылуплялись в течение всего июня, пик отмечен 4—9.VI
(Holtan, 1980).
В первые сутки птенцы обычно находятся в гнезде, но на вторые у них появляется
стремление покинуть его в случае опасности. В возрасте 2—3 сут они убегают из гнезда
и затаиваются, но потом возвращаются обратно. В первые два дня взрослые птицы
почти не кормят птенцов, но интенсивно обогревают (Лобков, Головина, 1978). Мас-
са птенцов (и = 6) в это время 12,2—14,8 г, в среднем 13,3 г. В возрасте 10 сут они
прячутся под кочками, среди травянистой растительности, а при опасности быстро
убегают, реже уплывают (Нечаев, 1981). Молодые поднимаются на крыло в возрасте
около 25 сут (Holtan, 1980). На о-ве Сахалин первые слетки отмечены в начале треть-
ей декады июля, летные молодые — в конце июля — первой декаде августа, а на
п-ове Камчатка — в третьей декаде июля — первой декаде августа. Докармливание
молодых в разных выводках продолжается в течение августа.
Успех размножения невысок. Отмечена значительная эмбриональная и постэмбрио-
нальная смертность. В долине р. Коппер в 1978 г. доля гнезд, в которых благополуч-
но вывелись птенцы, варьировала от 56,3 до 86,7% в разных колониях; здесь же было
зарегистрировано от 12,5 до 28,1 мертвых птенцов на 100 гнезд (Holtan, 1980). Одна
из причин, влияющих на успех размножения, — частое посещение колоний человеком
и отрицательное влияние хищников, а также неблагоприятной погоды (затяжные дож-
ди, похолодание и т.д.), когда птенцы погибают от переохлаждения. Из врагов на
п-ове Камчатка указываются ворон, черная ворона, поморники, лисицы и бурый мед-
ведь (Лобков, 1976), на о-ве Сахалин — черная и большеклювая вороны, филин, ли-
сица, бурый медведь , росомаха (Нечаев, 1981) .
Суточная активность, поведение. Камчатские крачки активны в светлое время су-
ток. Характерна склонность к ’’спонтанным взлетам” и образованию так называемых
клубов — постоянных скоплений в стороне от колонии (Каверкина, 1983). В гнездо-
вое время птицы отдыхают и спят на гнездовых участках, а негнездящиеся — группа-
ми на отмелях и косах.
355
Семейство Чайковые
Питание. Крачки отыскивают пищу в прибрежных водах моря, в солоноватых водое-
мах (озерах, лиманах), в устьях рек, реже на пресных водоемах и еще реже на мохо-
во-травянистых и травянистых болотах. За добычей обычно улетают на ближайшие
водоемы в 3-10 км от колоний, но нередко и значительно дальше: до 30 км на
о-ве Сахалин, до 50—70 км на Аляске, до 70—80 км на п-ове Камчатка (Нечаев, 1981;
Gill, Dick, 1977; данные Е.Г. Лобкова). В первые дни после прилета основу питания
составляют мелкие рыбы: трехиглая и девятииглая колюшка, молодь некоторых ло-
сосевых, корюшка, мойва, песчанка, молодь сельди, наваги, красноперки и др. В гнез-
довой период, помимо рыбы, поедают насекомых (жуков, клопов, веснянок, ручей-
ников, двукрылых, крупных чешуекрылых — совок, перепончатокрылых) и дру-
гих беспозвоночных. Птенцов кормят главным образом рыбой — трехиглой колюш-
кой, песчанкой и др., а также мелкими ракообразными — амфиподами (Holtan, 1980;
Нечаев, 1981; данные Е.Г. Лобкова). В питании молодых птиц преобладает мелкая
рыба, но нередки и насекомые, в частности стрекозы (Holtan, 1980). Добычу птицы
выхватывают из воды, нередко ’’зависают” в трепещущем полете. При этом опуска-
ют в воду клюв или часть тела, но не погружаются полностью (Лобков, 1976; Нечаев,
1981). Насекомых ловят в воде, на поверхности земли и листьях растений.
Враги, неблагоприятные факторы. Помимо хищников, существенным фактором
прямого уничтожения колоний камчатских крачек являются люди — сборщики яиц.
На п-ове Камчатка известны случаи выпаса коров в пределах рассеянных колоний.
На о-ве Сахалин колонии, расположенные вблизи населенных пунктов, уже исчезли
или находятся на грани исчезновения. Причины смены мест гнездования изучены не-
достаточно. Известно, что крачки покидают участки, зарастающие кустарниками
(Holtan, 1980) или заливаемые высокими паводками (данные Е.Г. Лобкова).
Хозяйственное значение, охрана. Камчатская крачка прямого хозяйственного зна-
чения не имеет, добыча ее запрещена. Как редкий и спорадично распространенный
вид она занесена в Красную книгу СССР. На о-ве Сахалин в заливе Даги (о-в Лярво)
организован заказник постоянного срока действия; это место также объявлено памят-
ником природы Сахалинской обл. На п-ове Камчатка 20—25% популяции этого вида
охраняется в Кроноцком заповеднике и в 7 комплексных заказниках областного под-
чинения.
Малая крачка —
Sterna albifrons Pallas, 1764
Little Tern (англ.), Zwergseeschwalbe (нем.), Sterne naine (фр.).
Син. (лат.): Sterna minuta Linnaeus, 1766; Sternula albifrons (Pallas, 1764).
Статус. Гнездящийся мигрирующий вид.
Общая характеристика и полевые признаки. Самая мелкая из крачек, размером ту-
ловища примерно со скворца. От речной крачки отличается меньшим размером (при-
мерно на одну треть), пропорциями тела (птица кажется более длинноклювой и боль-
шеголовой) , белым лбом во всех перьевых нарядах, в сезон размножения — желтым
с черным кончиком клювом. В отличие от крачек среднего размера полет, как пра-
вило, более прямолинейный и менее грациозный, с быстрыми и неглубокими взма-
хами крыльев; порой он больше напоминает полет кулика, чем крачки. За добычей
ныряет с разлета, полностью или частично погружаясь в воду. На земле держится сво-
бодно, в колонии может довольно быстро бегать. Обычно при этом туловище распо-
ложено горизонтально, голова втянута в плечи.
В голосе малой крачки отсутствуют громкие скрипучие тона и нередки булькаю-
щие и журчащие звуки. Наиболее часто как в колонии, так и вне ее можно слышать
отрывистые крики ’’квит, квит”, иногда они звучат как ”квут” или ’’кивит”. По
356
Малая крачка
С. Крэмпу (Cramp, 1985), ’’рекламный” крик, который издает птица, возвращаясь
в колонию, а также во время брачных демонстраций, звучит как ”че-ре-ти-тет” или
”пе-ри-ти-ти-тит” с ударением на последнем слоге. В число агрессивных криков вхо-
дит, быстрая трелька ”кит-кит-кит-кит. ..” (более подробно о голосе малой крачки
см.: Cramp, 1985).
Подвидовая систематика. Разными авторами выделяется 7—9 подвидов; изменчи-
вость проявляется в размерах, а также в окраске клюва, оперения спины, поясницы,
маховых и рулевых перьев (Peters, 1934; Vaurie, 1965; Howard, Moore, 1984; Cramp,
1985). В СССР обитает два подвида (Степанян, 1975).
1. Sterna albifrons albifrons
Sterna albifrons Pailas, 1764, Vroeg, Cat. Adumbratiunculae, c. 6, Голландия.
Ствол II (первого видимого) первостепенного махового пера темно-бурый. Вер-
шина клюва черная. Длина наружных рулевых перьев 71—95 мм.
2. Sterna albifrons sinensis
Sterna sinensis Gmelin, 1789, Syst. Nat., 1, c. 608, Китай.
Ствол II первостепенного махового пера белый. Окраска вершины клюва варьи-
рует от черной до желтой. Длина наружных рулевых перьев 95—140.
Замечания по систематике. S. albifrons, S. saundersi1, S. superciliaris, S. lorata
и S. nereis образуют хорошо очерченную группировку надвидового ранга (Cramp,
1985), которой разные исследователи придают различный таксономический статус.
С.А. Бутурлин (1934) выделял ее в отдельный род Sternula, однако большинство
авторов считают существующие морфологические и поведенческие отличия малой
крачки и близких к ней видов от настоящих крачек не достигающими родового ран-
га. В поддержку последней точки зрения говорит и предположительная возможность
гибридизации малой и речной крачек (Очагов, 1982; Мельников, 1985). По-видимо-
му, группу малой крачки следует выделить в отдельный подрод Sternula рода Sterna,
как это сделано Волътерсом (Wolters, 1975).
Описание. Строение и размеры (табл. 20).
Окраска (Дементьев, 1951; Cramp, 1985). Самец и самка в брачном наряде.
Лоб и узкая полоса над глазом, доходящая до его заднего края, белые. Темя, затылок,
кроющие уха и полоса, проходящая от надклювья по уздечке через глаз, черные. Щеки,
горло и вся нижняя сторона тела белые. Спина, плечевые, третьестепенные маховые
и все верхние кроющие хвоста светло-серые, в свежем состоянии с голубоватым от-
тенком. Поясница, верхние кроющие хвоста и рулевые варьируют у разных особей
от белого до тускло-серого цвета, более светлого, чем спина. Поясница белая с серо-
ватым налетом; такой же налет присутствует на центральных перьях верхних крою-
щих хвоста, на центральной паре рулевых и внешних опахалах 2-й и 3-й пары руле-
вых. II (первое видимое) — III первостепенные маховые с черным наружным опаха-
лом, черным стволом (у S. a. sinensis ствол II, реже II—III и даже IV первостепенно-
го белые, иногда сероватые), черной прилежащей к стволу частью внутренного опа-
хала и белым полем вдоль внутреннего края этого опахала. IV первостепенное серое
или почти черное с черным стволом и белым пятном вдоль края внутреннего опаха-
ла. Остальные первостепенные серые с узкой белой каймой по краю внутреннего опа-
хала. Второстепенные маховые серые с белой вершинной каймой и широким белым
полем на внутренних опахалах. Клюв ярко-желтый, обычно с темным кончиком; ноги
от желтых до оранжево-красных; радужина темно-коричневая.
Самец и самка в зимнем наряде. Лоб, уздечка и нижняя часть тела белые, темя белое
с темными пестринами. Пятно перед глазом, кроющие уха и затылок черные, послед-
Не исключено, что S. saundersi только подвид малой крачки (Cramp, 1985).
357
Семейство Чайковые
Таблица 20. Малая крачка. Размеры особей (мм) (ЗИН, ЗМ МГУ)
Географический район Пол Длина крыла
п lim среднее
Европейская часть СССР dd 21 166-182 175
99 12 165-179 173
Каспийское море dd 13 163-180 174
99 4 163-177 171
Казахстан dd 26 170-198 178
и Средняя Азия 99 16 ’ 165-185 174
Дальний Восток dd 6 180-187 184
(S. a. sinensis) 99 3 170-179 175
Масса тела (г) (Дементьев, 1951; Бородулина, 1960; Долгушин, 1962):
dd (п = 13) — 43 — 54 (среднее 50) ;
99 (п = 23) — 43 — 60 (среднее 50) .
ний может быть с белыми пестринами. Перья спины, плечевые, верхние кроющие крыла
такие же, как в предыдущем наряде, но с большим голубоватым оттенком. Поясница,
верхние кроющие хвоста и рулевые несколько серее, чем в предыдущем наряде. Цвет
первостепенных маховых зависит от времени, прошедшей с линьки: свежие перья
серебристо-серые, затем они постепенно темнеют и чернеют. Клюв черноватый; ноги
желтовато-коричневые.
Пуховой наряд. Пух короче, чем у других крачек. Спинная сторона желтовато-се-
рая или охристо-песочная с мелкими черными или буроватыми крапинами, которые
либо диффузно покрывают голову, туловище и крылья, либо группируются в три пре-
рывистые линии на темени и две на спине. Нижняя часть тела, в том числе подбородок
и горло, однотонная, белая или кремовая. Вариации в окраске пуховичков незначи-
тельные. Клюв розоватый или серовато-телесный с темным кончиком; ноги телесно-
розовые; радужина темно-коричневая.
Гнездовой наряд. Лоб и уздечка охристые или серовато-желтые. Темя такого же
цвета, но с черными пестринами. Вокруг глаза черное пятно, распространяющееся
на кроющие уха. Затылок черный с охристыми каемками перьев. Спина, плечевые,
третьестепенные маховые охристые с черными предвершинными v-образными пятнами
и беловатыми каймами, придающими рисунку чешуйчатый характер. Верхние кроющие
крыла светло-серые и охристыми или беловатыми каймами, из них средние и часть
малых с черными предвершинными пятнами. Остальные малые кроющие крыла и
кроющие первостепенных маховых серовато-черные с узкой серой каймой; они обра-
зуют четкую карпальную полосу. Поясница, верхние кроющие хвоста светло-серые
с широкими беловатыми каймами: крайние рулевые белые, остальные бледно-серые
с охристым оттенком и темно-бурыми предвершинными пятнами. Нижняя часть тела
и нижняя часть задней стороны шеи белая. Внутренние (проксимальные) первостепен-
ные светло-серые, постепенно темнеющие к дистальному концу крыла; II (первое
видимое) почти черное. Стержни проксимальных первостепенных сверху белые, ди-
стальных — черные или темносерые. Внутренние опахала первостепенных маховых
белые с широкой серой полосой вдоль стержня. Вершины второстепенных маховых
и проксимальных первостепенных с большим развитием белого цвета, чем у взрослых
птиц. Радужина темно-коричневая, клюв от серо-коричневого до почти черного, основа-
358
Малая крачка
Длина клюва Длина цевки
п lim среднее п lim среднее
21 25,1-31,9 29,3 20 15,7-18,4 16,9
12 25,1-29,6 27,6 12 15,9 - 1 7,4 16,6
13 25,4-31,7 29,4 13 16,5-18,2 17,3
4 27,9-29,4 28,8 4 16,5-17,4 17,1
26 26,9-34,1 30,0 26 16,0-19,0 17,6
16 26,0 29,9 28,0 16 15,7-18,2 17,3
6 27,9-31,1 28,9 6 16,3-17,9 17,2
3 26,7-30,3 28,6 3 17,3-18,3 17,7
ние клюва розовато-желтое или желтоватое. Ноги от серо-телесного до желтовато-
коричневого цвета.
Первый зимний наряд. Напоминает окончательный зимний наряд; по С. Крэмпу
(Cramp, 1985), отличается характером линьки первостепенных маховых и степенью
их обношенности: в сентябре—октябре они светлее и свежее, чем у взрослых, к нояб-
рю-январю становятся темнее по сравнению со свежими светлыми перьями у взрослых.
Брачный наряд в первую весну жизни не надевается, но иногда на темени появляются
отдельные черные перья, а на темной карпальной полосе — серые перья.
Второй зимний наряд отличается от окончательного зимнего более свежими дисталь-
ными первостепенными маховыми. Иногда задержка линьки первостепенных маховых
отмечается на иной стадии, чем у взрослых птиц; кроме того, новая серия линьки
проксимальных первостепенных может начинаться раньше, чем у взрослых.
Второй летний наряд. Как окончательный брачный наряд, но на темени могут оста-
ваться отдельные белые перья, а в районе карпальной полосы — отдельные черные
перья.
Линька (Cramp, 1985). В отличие от большинства других видов крачек характерно
наличие трех последовательных серий линьки первостепенных маховых, каждая из
которых начинается с XI пера и приостанавливается на различной стадии завершенности
в начале сезона размножения.
Линька в первый зимний наряд полная, начинается в начале августа — конце сентября
сменой перьев верхней части спины и плечевых. В конце августа — начале октября боль-
шая часть контурного оперения уже свежая; линька маховых и рулевых между концом
сентября — началом декабря. Первая серия смены первостепенных маховых завершает-
ся в апреле—мае, вторая серия начинается в феврале—марте. В мае у ряда особей начи-
нается третья серия смены первостепенных маховых, в это же время иногда появляются
отдельные перья брачного наряда на темени, спине, плечах и кроющих крыла. По-види-
мому, данную смену оперения можно считать частичной предбрачной линькой. В конце
мая—июне вторая и третья серии смены первостепенных маховых приостанавливаются.
Дальнейшая ситуация не ясна. В июле птицы могут начать новую серию линьки перво-
степенных, не завершая предыдущие. В то же время птицы с более поздними сроками
начала линьки могут с июля продолжить линьку из первого зимнего прямо во второй
359
360
Рис. 84. Область распространения малой крачки
а — гнездовые поселения (пунктиром обозначена неуточненная граница) , б — места зимовок. Подвиды: 1 — S.a. albifrons, 2 — S.a. sinensis, 3 -
S.a. pusilia, 4 - S.a. antillarum, 5 - S.a. athalassos, 6 — S.a. browni, 7S.a. mexicana, 8 — S.a. staebleri, 9 — S.a. guineae
Малая крачка
зимний наряд, не приостанавливая второй серии смены первостепенных маховых и
в то же время начиная новую серию. На некоторых особях можно наблюдать одновре-
менно четыре серии линьки превостепенных.
У взрослых особей бывает две линьки в году — полная послебрачная и частичная
предбрачная. Послебрачная линька начинается со смены проксимальных первостепенных
маховых в конце июня — конце августа. Вскоре после этого начинается смена оперения
головы и верхней части тела. Линька приостанавливается во время миграции и завер-
шается на зимовках, причем первостепенные маховые сменяются к середине декабря.
После смены оперения головы, туловища и хвоста (середина октября — середина де-
кабря, реже ранее) начинается повторная линька первостепенных маховых, которая
прерывается на стадии новых IV—V в начале сезона размножения. У некоторых особей
(возможно, у всех) отмечается начало повторной смены рулевых.
Предбрачная линька протекает с конца февраля до начала апреля: сменяется опере-
ние головы, туловища, кроющих крыла и хвоста (в последнем случае, видимо, про-
должается начавшаяся ранее повторная смена рулевых). В феврале — середине марта
начинается третья серия смены первостепенных маховых, которая прерывается перед
началом инкубации, обычно на стадии новых VIII—X первостепенных.
Распространение. Гнездовой ареал. Европа (кроме северной ее части),
Передняя, Средняя, Южная и Юго-Восточная Азия, Северная Африка, центральная часть
Западной Африки, атлантическое и тихоокеанское побережье южной части Север-
ной Америки, бассейн р. Миссисипи, острова Карибского бассейна, восточное побе-
режье Австралии. В большинстве регионов сплошного ареала не образует и гнездится
спорадично (рис. 84). В зарубежной Европе заселяет побережья Великобритании (на
север до Оркнейских островов), Ирландии, Нидерландов, Дании, ФРГ, ГДР, южное
побережье Швеции. На внутренних водоемах этих стран крачки поселяются лишь из-
редка. Во Франции, Испании, Португалии, Италии, Румынии отмечается гнездование
как на побережье, так и на материковых водоемах. С начала 1960-х годов малая крачка
гнездится на западном побережье Финляндии. Отдельные поселения отмечаются на
внутренних водоемах Венгрии, возможно гнездование в Югославии. До 1955 г. малая
крачка гнездилась в Чехословакии и до 1962 г. — в Австрии. Спорадичные поселения
существуют на побережьях почти всех стран Средиземноморья (Cramp, 1985). Гнездо-
вание отмечено также в Болгарии (Нанкинов, Даракчиев, 1980а).
В Европе, Северной Африке, Сибири, Казахстане и Средней Азии обитает форма
S.a. albifrons (Vaurie, 1965; Howard, Moore, 1984). Этот же подвид населяет Турцию,
Иран, Ирак, Афганистан, Пакистан, западную часть Индии (Paludan, 1959: АН, Ripley,
1969; Cramp, 1985) и, по-видимому, Монголию, где гнездовья малой крачки обнаруже-
ны на озерах Орог-Нур и Бон-Цаган-Нур и весьма вероятны в котловине Больших озер
(Stubbe, Bolod, 1971). Форма S.a. pusilia, признаваемая не всеми систематиками, оби-
тает на Яве, Суматре, по рекам северной Индии и Бирмы и, возможно, на п-ове Малакка
(Vaurie, 1965). Советское Приморье и Приамурье, восточный и юго-восточный Китай,
п-ов Корею, Японию, о-в Шри-Ланка, большую часть Юго-Восточной Азии и Австралию
заселяет S.a. sinensis (Дементьев, 1951; Gore, Won, 1971; Cheng, 1976; Hasegawa,
1984; Schauensee, 1984; Cramp, 1985). S.a. guineae гнездится в Экваториальной Афри-
ке, еще 5 подвидов заселяют Северную и Центральную Америку (Vaurie, 1965) .
В СССР малая крачка распространена довольно широко, но гнездится спорадично,
населяя морские побережья и долины крупных рек (рис. 85). В Прибалтике гнездова-
ние отмечено в долинах Немана, Западной Двины, на Балтийском побережье, на неко-
торых озерах и рыбхозах (Красная книга Литовской ССР, 1981; Пукас и др., 1981;
Страздс, 1981; Виксне, 1983; Lipsbergs et al., 1985), на островах западной и северной
части Эстонии (Пеэдоссар, Онно, 1970; Аумеэс, 1972: Ренно, 1972; Aumees et al.,
361
362
Рис. 85. Ареал малой крачки и СССР
1 — гнездовые поселения (пунктиром обозначена неуточненная граница), 2 — граница области регулярных кочевок европвйско
сибирско-казахстанских малых крачек, 3 — граница области регулярных кочевок прибалтийских малых крачек, 4 — залеты
Малая крачка
1983); в Ленинградской обл. — на Березовых островах и островах южного побережья
Финского залива, в западном Приладожье, устье Свири (Носков и др., 1981: Маль-
чевский, Пукинский, 1983; Храбрый, 1984).
В Причерноморье и Приазовье малая крачка гнездится по северному побережью
Черного моря от устья Дуная до Северного Крыма, на Сиваше, по северному и восточ-
ному побережьям Азовского моря и на лиманах Таманского полуострова (Кищинский,
19606; Молодан, Сиренко, 1981; Петрович, 1981; Сиохин, 1981а; Черничке, Стой-
ловский, 1981; Казаков, Языкова, 1982; Костин, 1983; Ардамацкая, 1984). Днепр
крачка заселяет от южной Белоруссии до устья; гнездится также по Припяти, по при-
токам Днепра поднимается выше Чернигова, до Сумской и Брянской областей (К1стя-
кивскш, 1957; Орлов, 1959; Булахов, Мясоедова, 1975; Кистяковский, Мельничук,
1975; Клакоцкий, 1975; Марисова, Самофалов, 1977; Клестов, 1981; Клестов, Оси-
пова, 1981; Кудденок, 1981; Долб1к, Навумчык, 1983; Мельничук, Головач, 1984:
Грищенко, устное сообщ.). Зарегистрировано гнездование в Закарпатской обл. по
р. Тисе (Юстякивскш, 1957) и в Самборском районе Львовской обл. (Горбань, устное
сообщ.): в Молдавии малые крачки не отмечены (Аверин и др., 1981).
По Волге эти птицы гнездятся от устья вверх до Калининской обл. (в том числе
на Рыбинском водохранилище); поселяются также на притоках Волги — Оке (вверх
до границы Калужской обл.), Мокше, Суре (вплоть до Пензенской обл.), Каме (по-
видимому, только до границ Пермской обл., см.: Воронцов, 1949; Болотников и др.,
1981), Самаре. В бассейне Волги малые крачки гнездятся также по р. Цне в Моршан-
ском районе Тамбовской обл., по р. Вятке вверх до устья р. Моломы, в Башкирии
(Третьяков, 1940; Воронцов, 1967; Луговой, 1975; Щеголев, 1975; Плесский, 1976;
Николаев и др., 1977; Зиновьев, Шапошников, 1978; Ильичев, Фомин, 1979; Аюпов,
1981; Водолажская, 1981; Денисов, Фролов, 1981; Рощевский, Маслов, 1981; Титав-
нин, 1981; Кубанцев, Чернобай, 1982). По-видимому, крачки заселяют и долину Дона,
однако достоверные сведения об их гнездовании мы имеем лишь из Воронежской обл.
(Барабаш-Никифоров, Семаго, 1963; Семаго и др., 1974) . В бассейне Северского Донца
гнездование отмечено по Осколу и на прудах Печенежского рыбхоза (Юстякивскш,
1957; Лисецкий и др., 1978).
В Предкавказье малая крачка гнездится по всем водохранилищам системы Маныча,
разливам Кумы, низовьям Терека и дагестанскому побережью Каспия (Волчанецкий,
1959; Пишванов, 1975; Казаков и др., 1981; Кривенко, 1981; Кривенко, Любаев,
1981; Казаков, Языкова, 1982). В Закавказье гнездование ныне отмечено в Азербайд-
жане — на островах Бакинского архипелага (Туаев, Васильев, 1969) и оз. Аггель в
долине р. Куры (Виноградов, 1967; Туаев, 1975); до 1956 г. гнездилась в Кызыл-
Агачском заповеднике (Виноградов, Чернявская, 1965). В Армении эти птицы отме-
чались в летнее время, однако гнездование не доказано (Ляйстер, Соснин, 1942). На
Каспийском море, помимо упомянутых выше, известны поселения в авандельте Волги,
на островах Малый Жемчужный и Морской Очиркин, в устьях рек Урала и Эмбы, на
островках восточной части северного Каспия (Кривоносов, 1976), Тюленьих о-вах
(Мангистауских) и прилегающих побережьях Мангышлака (Залетаев, 1960), в Красно-
водском заливе (Васильев и др., 1975), проливе Кара-Богаз-Гол (Щербина, 1979) и в
низовьях р. Атрек (Караваев, 1979, 1981).
В Казахстане малая крачка гнездится по рекам Уралу, Илеку, Эмбе, Иргизу, Тургаю,
Сырдарье, Сарысу, Чу, Таласу, Или, Караталу, на всем протяжении Иртыша, в окрест-
ностях Кургальджина, на Аральском море, Балхаше и Алаколе (Долгушин, 1962;
Неручев, 1968; Ауэзов, Грачев, 1975; Саму сев, 1975; Неручев, Макаров, 1982) . В Сред-
ней Азии она заселяет долину Амударьи, по-видимому на всем ее протяжении, долину
Сурхандарьи, Айдаркуль, низменные участки северного и юго-западного Таджикистана,
363
Семейство Чайковые
долины Мургаба и Теджена, трассу Каракумского канала (Дементьев, 1952; Ташлиев
и др., 1967; Абдусалямов, 1971, 1981; Кенжегулов, 1972; Эминов, 1975; Остапенко
и др., 1978; Мухина, 1983; Бидос, 1985). Гнездование отмечено также на оз. Иссык-
Куль (Кыдыралиев, 1973; Кыдыралиев, Султанбаева, 1977). В Западной Сибири гнез-
дится по Иртышу в пределах Омской обл. (Сулимов, 1981) и, по-видимому, по Тоболу
и Ишиму (Гынгазов, Миловидов, 1977). Малые крачки заселяют также долину Оби
от окрестностей Барнаула до устья р. Тым (Велижанин, 1928; Велижанин, Велижанин,
1929; Стрелков, 1974а,б; Блинов, Блинова, 1979).
На Дальнем Востоке гнездование отмечается на оз. Ханка (Поливанова, 1971; Глу-
щенко, 19846), по долине Амура от Хабаровска до оз. Эворон (Росляков, 1981), на
о-ве Путятин в заливе Петра Великого и оз. Дорицени в Хасанском районе (Лабзюк
и др., 1971). Возможно, гнездится по р. Уссури.
За последние 20—30 лет отмечено расширение ареала вида в северном направлении
в Прибалтике, где малая крачка заселила Ленинградскую обл. (Мальчевский, Пукин-
ский, 1983), по долине Оби (Миловидов, Стрелков, 1978) и, видимо, на Дальнем
Востоке (Росляков, 1981; Глущенко, 19846). В пределах области гнездования вида
крачки нередко поселяются на новых водохранилищах в местах, где их ранее не отме-
чали (Семаго и др., 1974; Щадилов, 1980).
Зимовки. Западноевропейские малые крачки зимуют в Западной Африке (глав-
ным образом в Гвинейском заливе), а также, по-видимому, в Южной и Восточной
Африке (на север до Кении). Зимовки птиц из Восточной Европы расположены в Крас-
ном море и на южном побережье Аравии, из Арало-Каспийского региона — в Персид-
ском заливе и на западном побережье Индии. Подвид S.a. sinensis зимует на островах
Малайского архипелага (Cramp, 1985).
Миграции. Западноевропейские популяции малых крачек покидают места зимовок
в первых числах апреля. Птицы летят вдоль атлантических побережий Северной Афри-
ки и Европы, а затем вдоль побережий Северного и Балтийского морей. Миграции че-
рез континент почти не прослеживаются. Осенние миграции начинаются с конца (реже
с начала) июля и завершаются в октябре. Крачки в это время в основном летят вдоль
побережий, однако часть птиц пересекает континентальную Европу в юго-западном на-
правлении (Cramp, 1985). Крачки, гнездящиеся в Эстонии, Латвии, Литве и Ленинград-
ской обл., мигрируют, по-видимому, как западноевропейские птицы (Йыги, 1968).
В Эстонии, по данным Э.В. Кумари, они появляются весной во второй трети мая, отлет
происходит с середине июля по середину августа (Дементьев, 1951).
Малые крачки номинального подвида из других регионов нашей страны мигрируют
через материк, придерживаясь, где это возможно, речных долин и побережий внутрен-
них морей. Восточноевропейские крачки пересекают весной Египет, Иорданию и Сирию;
затем часть птиц, видимо, мигрирует через восточную Турцию, другие летят к Черному
морю через Босфор (Cramp, 1985). Первый из этих миграционных потоков идет затем
на северо-запад вдоль Черноморского побережья (Вронский, Томкович, 1975), другой,
видимо, по западному побережью Черного моря. В окрестностях Черноморского запо-
ведника (Тендровский залив) первые крачки отмечаются уже 13.IV; массовое их по-
явление регистрируется в начале третьей декады апреля (Ардамацкая, 19776, 1984).
В Северном Крыму, на Молочном лимане Азовского моря и Предкавказье первые ма-
лые крачки отмечаются во второй половине апреля—начале мая (Филонов и др., 1974;
Казаков, Языкова, 1982; Костин, 1983). На Украине под Уманью первых птиц встреча-
ли 22 и 26.IV под Киевом - 28.IV-23.V (в среднем за 10 лет 10.V), на Западной Украи-
не — в конце апреля—начале мая (К;стякивскш, 1957; Страутман, 1963). В сходные
сроки (29.IV—14.V, в-среднем 8.V) малая крачка появляется в Рязанской обл. (Пту-
шенко, Иноземцев, 1968). В Белоруссии первые, птицы появляются весной в конце
364
Малая крачка
апреля (Шнитников, 1913), по другим данным - в третьей декаде мая (Федюшин,
Долбик,1967).
Осенние миграции крачек центра европейской части СССР отмечаются в конце июля—
августе (Воронцов, 1967; Птушенко, Иноземцев, 1968; Луговой, 1975; Плесский,
1976), в более южных районах — в августе—сентябре (Юстякивскш, 1957; Страут-
ман, 1963). В Крыму основная масса птиц исчезает в середине сентября, отдельные
особи отмечаются до середины октября (Костин, 1983), на Молочном лимане послед-
няя волна пролета прослеживается в конце сентября — -начале октября (Лысенко,
1975). В период осенних миграций малые крачки встречаются по черноморскому по-
бережью Болгарии (Нанкинов, 1980).
Пролет малых крачек, гнездящихся в Средней Азии и Казахстане, идет по
побережьям Каспийского моря и долинам рек, в частности по Амударье, Сырдарье,
Сурхандарье, Вахшу, Мургабу и, видимо, Теджену. Весенний пролет начинается здесь
в конце апреля (в отдельные годы во вторую декаду апреля) и заканчивается в начале
мая (Спангенберг, Фейгин, 1936; Дементьев и др., 1955; Заблоцкий, Заблоцкая, 1963;
Эминов, 1964, 1978; Ташлиев и др., 1967; Абдусалямов, 1971; Абдусалямов, Лебе-
дев 1977; Остапенко и др., 1978, 1984; Сухинин, 1979; Абдусалямов и др., 1980).
Весной и осенью отмечен пролет по восточному побережью Аральского моря (Заруд-
ный, 1916; Березовский, 1983). Осенний пролет в Казахстане проходит в конце июля—
начале сентября (Долгушин, 1962), в Таджикистане - в начале сентября (Абдусалямов,
1971). На Иссык-Куле крачек отмечали до середины сентября (Кыдыралиев, Султан-
баева, 1977), на Айдаркуле — до 24.IX; наиболее заметен здесь пролет в начале сен-
тября (Остапенко и др., 1984; Калабин, 1986). В сходные сроки заканчивается пролет
в дельте Волги (Воробьев, 1936). В августе—сентябре проходят миграции на южном
Каспии; последних особей здесь отмечают до середины октября (Исаков, Воробьев,
1940; Дементьев и др., 1955; Заблоцкий, Заблоцкая, 1963; Караваев, Белоусов, 1983).
На нижнем Амуре первые крачки появляются весной в середине мая, последние
птицы здесь встречены 20.IX (Росляков, 1981). 21.X 1968 г. одиночная малая крачка
отмечена на льду оз. Благодатное в Сихоте-Алинском заповеднике (Елсуков, 19746).
Крачки мигрируют поодиночке, парами или небольшими группами в 3—5 особей
(Остапенко и др., 1978, 1984; Костин, 1983). Годовалые птицы, по-видимому, остают-
ся летом в местах зимовки. Двухлетние особи отмечаются в пределах гнездового ареала
вида, но нередко вдали от места появления на свет (Cramp, 1985).
Залеты малых крачек отмечены на Соловецкие острова (Карташев, 1963), в низовья
Енисея (Рогачева, Вахрушев, 1983) и дельту Селенги (Мельников, 1979).
Численность. Почти везде невелика. В относительно большом количестве малая
крачка гнездится лишь на островах северного Каспия (несколько тысяч пар — Криво-
носов, 1976) и в Северном Причерноморье и Приазовье (видимо, не менее 4 тыс. пар, —
см.: Зубакин, Костин, 1977; Молодан, Сиренко, 1981; Петрович, 1981; Сиохин, 1981а;
Черничко, Стойловский, 1981; Ардамацкая, 1984; Букреев и др., 1986). В Литве гнез-
дится около 20 пар (Красная книга Литовской ССР, 1981), в Латвии — не менее 240 пар
(Страздс, 1981), в Эстонии - около 200 пар (Пездосаар, Онно, 1970; Ренно, 1972),
в Ленинградской обл. — не более нескольких десятков пар (Носков и др., 1981; Маль-
чевский, Пушкинский, 1983; Храбрый, 1984), в Белоруссии — не более 600—800 пар
(Долб1к, Навумчык, 1983), в Московской обл. — не более 100—150 пар (Зубакин и
др., 1986). На водохранилищах системы Маныча гнездится, видимо, несколько более
1 тыс. пар (Казаков и др., 1981; Кривенко, 1981; Кривенко, Любаев, 1981), на Даль-
нем Востоке — около 1500 пар (Росляков, 1981; Глущенко, 19846). В целом в СССР
гнездится вряд ли больше 50 тыс. пар. В Прибалтике и на Дальнем Востоке отмечается
рост численности (Росляков, 1981; Виксне, 1983; Мальчевский, Пукинский, 1983;
365
Семейство Чайковые
Страздс, Страздс, 1983), во многих местах европейской части СССР — ее снижение
(Щеголев, 1975; Клестов, Осипова, 1981; Титавнин, 1981).
Численность малой крачки в Западной Европе и Средиземноморье ныне оценивается
примерно в 10—11 тыс. пар; отмечено значительное ее снижение в текущем столетии
(Cramp, 1985). Численность американских подвидов также невелика: S.a. athalassos
около 625 гнездящихся пар (Downing, 1980) , S.a. browni 600-700 пар (King, 1981) .
Местообитания. В сезон гнездования — морские побережья, главным образом мелко-
водные заливы и лиманы. В глубине материка — долины крупных рек, водохранилища
и значительные по величине озера. В последние годы отмечается гнездование на рыб-
хозах и прудах — отстойниках сточных вод (Пукас и др., 1981; Бухаринов, Олигер,
1984). В период зимовки — морские побережья (Cramp, 1985). В целом малая крач-
ка — птица низменностей, однако в Средней Азии гнездится на оз. Иссык-Куль на
абсолютной высоте 1608 м, а на пролете отмечается на озерах Сонкель и Чатыркель на
высоте 3016 м и 3530 м (Кыдыралиев, 1973).
Размножение. Малые крачки впервые приступают к размножению в среднем в воз-
расте 3 лет, иногда 2 лет. Моногамы. Пары могут сохраняться более 1 сезона размно-
жения, однако они распадаются, по-видимому, чаще, чем у ряда других чайковых
(Cramp, 1985). Малая крачка — диффузно гнездящийся факультативно-колониальный
вид (Зубакин,1976в,1983).
Процесс формирования пары включает воздушные и наземные демонстрации (Cramp,
1985). Как и у других крачек, характерен ’’высотный полет”, в котором обычно участ-
вуют 2—3 птицы. После исполнения ’’высотного полета” крачки спускаются, планируя
по спирали, но более быстро и зигзагообразно, чем другие виды крачек. В это время
слышен повторяющийся ’’рекламный” крик ”че-ре-ти-тет”. Этот же крик сопровождает
кормление самцом самки во время Наземных демонстраций. После кормления крачка
принимает выпрямленную позу (рис. 86, а). При территориальных столкновениях на
земле у малых крачек, как и у многих других крачек, можно видеть агрессивную
прямую позу, при которой шея вытянута наклонно вверх, клюв направлен на против-
ника или слегка наклонен, перья черной шапочки взъерошены. В этой позе издается
быстрая агрессивная трелька из щелкающих звуков (см. рис. 86, б).
Колонии малых крачек, как правило, невелики — обычно меньше 50 пар, чаще
всего до 10—15 пар. Реже отмечаются колонии до 100 гнезд, еще реже — в несколько
сот пар. Часто крачки поселяются отдельными парами (Зарудный, 1916; Бородулина,
1960; Кищинский, 19606; Долгушин, 1962: Эминов, 1964; Абдусалямов, 1971; Зу-
бакин, Костин, 1977; Костин, 1983; Ардамацкая, 1984; Wesolowski et al., 1985).
Характерные гнездовые биотопы — намывные песчаные, илистые, ракушечные, солон-
чаковые, реже галечниковые острова, отмели, косы и материковые пляжи, голые
либо поросшие редкой невысокой травянистой растительностью. Гнезда нередко рас-
полагаются вблизи воды. В отличие от речной малая крачка на сплавинах и в заболо-
ченных местах либо вовсе не гнездится, либо селится в исключительных случаях.1
Подобная стенотопность объясняет жесткую приуроченность малых крачек в сезон
размножения либо к морским заливам и лиманам, либо к руслам крупных рек и водо-
хранилищам. Эфемерность островков, кос и отмелей, то появляющихся, то исчезающих
в зависимости от полноводности рек и преобладания сгонных или нагонных ветров,
привела к выработке у малых крачек способности часто менять места гнездования.
Этот вид характеризуется слабым гнездовым консерватизмом (McNicholl, 1975), его
колонии часто меняют свое расположение и величину. Изредка поселения крачек отме-
1 Указание на такого рода гнездование в Белоруссии (Шнитников, 1913), по-видимому, основано
на ошибке.
366
Малая крачка
чаются в необычных местах,, хотя биотопически часто сходных с предпочитаемыми
этим видом отмелями. Так, колонии крачек встречены на сильно обмелевшем пруду —
отстойнике сточных вод (Бухаринов, Олигер, 1984) и на пашне после позднего схода
весенних паводковых вод (Гусев, 1978). В США зарегистрировано гнездование на кры-
шах зданий (McNicholl, 1975; Fisk, 1978; Cramp, 1985). Малые крачки могут образо-
вывать как одновидовые колонии, так и (чаще) совместные поселения с речной крач-
кой. Нередко вместе с крачками гнездятся малые или морские зуйки, реже ходулочни-
Рис. 86. Демонстрационное поведение малой крачки
а — выпрямленная поза,б — агрессивная прямая поза
ки, тиркушки, и некоторые другие кулики (Луговой, 1958; Долгушин, 1962; Кислен-
ко, 1972; Зубакин, Костин, 1977; Глущенко, 19846). Близкого соседства с чайконо-
сой крачкой малая обычно избегает (Зубакин, Анзигитова, 1975).
Гнезда представляют собой ямку в грунте, часто без всякой выстилки или с не-
большим количеством сухих веточек растений, мелких камешков или ракушек. Гнез-
довой валик часто не выражен. Нередко под гнездовую ямку используется естественная
выемка в грунте, след человека, коровы и т.д. (Луговой, 1958; Бородулина, 1960;
Долгушин, 1962; Поливанова, 1971). Диаметр гнезда (мм) на Сиваше (п = 13) в сред-
нем 161 (140—180), диаметр лотка, т.е. гнездовой ямки (п = 25), 104 (90—130), глу-
бина лотка (п = 25) 22 (15—30); в Московской обл. диаметр лотка в среднем (п =9)
109 (95—125), его глубина 27 (20—30) (наши данные).
В Черноморском заповеднике расстояние между гнездами составляет 0,7—2 м, в
среднем (п = 150) 1,8 м (Ардамацкая, 1984); на лиманах Таманского полуострова —
2—15 м, обычно 5—10 м (Кищинский, 19606); в заливе Кара-Богаз-Гол — 0,8—4 м
(Щербина, 1979); в Таджикистане от 0,15—0,20 до 0,60-0,65 м (Абдусалямов, 1971);
на оз. Ханка — обычно 0,2—0,5 м (Поливанова, 1971). Среднее расстояние между цен-
трами ближайших соседних гнезд на Сиваше (и = 123) равно 1,2 м (0,35—5,0 м) (Зуба-
кин, 1976в), в Московской обл. (и = 19) —10,7 м (4—50 м) (наши данные). Индекс
плотности гнездования, рассчитанный по имеющимся данным, равен 0,05.
Наиболее ранние сроки начала откладки яиц — первые числа мая — отмечены для ни-
зовий Атрека (Караваев, 1979). На внутренних водоемах Туркмении первые яйца
появляются в первой трети мая (Эминов, 1964), в Тендровском заливе Черного моря,
на Молочном лимане и Сиваше — с конца первой декады мая до середины мая (Филонов
и др., 1974; Ардамацкая, 19776; 1984; Зубакин, Костин, 1977), на оз. Иссык-Куль —
10—12.V (Кыдыралиев, Султанбаева, 1977), на водохранилищах системы Маныча — во
второй декаде мая (Казаков, Языкова, 1982). С конца мая — начала июня начинается
откладка яиц в Мордовии (Луговой, 1975) и Рязанской обл. (Птушенко, Иноземцев,
367
Семейство Чайковые
1968), а также в Казахстане, где кладка в основном происходит в июне (Долгушин,
1962). В Кировской обл. откладка яиц начинается 8—14.VI (Плесский, 1976), на сред-
ней Оби - видимо, в эти же сроки (Миловидов и др., 1979). S.a. sinensis на территории
СССР (оз. Ханка) начинает гнездиться в начале июня (Поливанова, 1971). Массовая
кладка обычно происходит через несколько дней после откладки первых яиц. Сроки
откладки яиц сильно растянуты (до середины июля) в связи с поздним и неодновре-
менным освобождением от паводковых вод гнездовых биотопов крачек и частой
гибелью первых кладок (Долгушин, 1962; Филонов и др., 1974).
В полной кладке бывает 1—3, чаще всего 3 яйца, редко 4. Отмечены гнезда с 5 и 9
яйцами — по-видимому, в данных случаях в одной гнездовой ямке были яйца несколь-
ких кладок (Кищинский, 19606; Самородов, 19816). Отмечена тенденция возрастания
средней величины кладки с увеличением числа гнездящихся пар в колонии (Рощев-
ский, Лаухина, 1980). Окраска яиц сложная. Фон ее обычно песочный, песочно-охри-
стый, серовато-песочный. На оз. Ханка отмечаются также бледно-оливковые и изредка
бледно-зеленоватые яйца (Поливанова, 1971). По этому фону разбросаны мелкие
округлые, реже неправильной формы или изогнутые в виде запятой пятна; цвет глу-
бинных пятен серо-фиолетовый, поверхностных - коричневый или коричнево-бурый.
Пятна могут как равномерно распределяться по яйцу, так и скапливаться у его ту-
пого конца, образуя неявный поясок. Размеры яиц (мм): из Предкавказья (и = 151)
28,9—34,9X21,5—28,1, в среднем 31,5X23,6 (Казаков, Языкова, 1982): со среднего
Днепра (и = 23) 27,6—34,5X22,1—24,5, в среднем 31,8X23,5 (Самородов, 19816); из
Московской обл. (и = 25) 30,4—35,1X22,8—24,7, в среднем 31,9X23,6 (наши данные);
из Азербайджана (и = 6) 29,5—31,1X23,0—23,8, в среднем 30,5X23,4 (данные В.В. Лео-
новича) ; из Казахстана (и = 17) 30,0—38,5X22,2—24,2, в среднем 33,2X23,3 (Ковшарь,
Левин, 1982); со средней Оби (и = 18) 28,8—32,6X22,3—25,7, в среднем 30,5X23,4
(Миловидов и др., 1979). Масса свежих яиц (г): из Крыма (п = 11) 8,4—10,6, в сред-
нем 9,5 (Костин, 1983); с низовий Сырдарьи (и = 9) 8,1—8,8, в среднем 8,5 (Кислен-
ко, 1972).
Насиживание начинается с первого яйца и продолжается 18—22 дня (Бородулина,
1960; Эминов, 1964; Птушенко, Иноземцев, 1968). Насиживают кладку обе птицы;
смена партнеров бывает около 5 раз в день (Бородулина, 1960), по другим данным —
25—39 раз в сутки (Гусев, 1978); при этом самец изредка кормит самку. Продолжи-
тельность стадии наклева от 7 ч 35 мин до 23 ч 40 мин, вылупления от 1 ч 14 мин до
2 ч 36 мин (Гусев, 1978). Вес вылупившегося птенца равен в среднем (и = 30) 5,2 г
(Ардамацкая, 1984). По наблюдениям на Сиваше, между вылуплением первого и по-
следнего птенца в гнезде проходит 1—2 сут. Первые два дня один из родителей постоян-
но обогревает птенцов; смена партнеров в этот период происходит 12—14 раз в день.
В трехдневном возрасте птенцы оставляют гнездо и держатся по соседству. С этого
времени родители, по-видимому, начинают индивидуально опознавать своих пухович-
ков. Птенцы, оказывающиеся поблизости от чужого выводка, подвергаются нападению
взрослых птиц (Бородулина, 1960; Cramp, 1985).
В день в гнезде бывает 51—63 кормления птенцов (Бородулина, 1960). Во время
тревоги в колонии птенцы разбегаются и затаиваются; подобное поведение проявляет-
ся уже в суточном возрасте. Молодые малые крачки поднимаются на крыло в возрасте
15—21 дня, после чего молодые отмечаются в окрестностях колонии еще 5—10 дней.
Птицы держатся рассредоточено, каждая лётная молодая крачка занимает определенный
(обычно постоянный) участок берега острова, однако участки эти, по-видимому, не
охраняются (Эминов, 1964; Птушенко, Иноземцев, 1968; наши данные). Родители
продолжают кормить молодых и после оставления колонии, в том числе и в августе, в
период миграции (Резанов, 1978).
368
Малая крачка
Успех размножения сильно варьирует в разных поселениях. В целом гибель потом-
ства довольно значительна. Так, в пойме р. Припять гибнет 80—100% гнезд (Наумчик,
1982). На Молочном лимане в 1969 г. выход пуховиков составил около 60%, а их
смертность в гнездах достигала 30% (Филонов и др., 1974). В 1974 г. на Сиваше в ко-
лонии из 46 гнезд поднялись на крыло только 3 птенца (0,07 молодых на пару птиц или
2,8% от числа отложенных яиц), гибель птенцов происходила главным образом в ре-
зультате хищничества чайконосых крачек (наши данные). В Западной Европе в разных
колониях поднимается на крыло 0-2,5 птенцов на пару птиц (Cramp, 1985). Одна из
главных причин гибели потомства — затопление колонии во время паводков, штормов
и нагонных ветров. Например, от колебания уровня Волги в Куйбышевской обл. гибнет i
до 60% кладок (Титавнин, 1981); постоянно затапливаются колонии во время штор- !
мов в Черноморском заповеднике (Ардамацкая, 1984).
Максимальная продолжительность жизни малых крачек, по данным кольцевания,
21 год 1 месяц (Clapp et al., 1982, цит. по: Cramp, 1985).
Суточная активность, поведение. Активность дневная, однако птицы способны кор-
миться и в густых сумерках (Бородулина, 1960; Авилова, 1972). Малая крачка — об-
щественная птица во все сезоны года, однако больших стай, как правило, не образует;
мигрирует и охотится нередко в одиночку или небольшими группами. Тем не менее
в местах скопления корма ее стаи могут состоять из нескольких тысяч птиц (Исаков,
Воробьев, 1940). .
Питание. Трофически связана главным образом с водоемами. В Крыму основу |
питания составляет мелкая рыба (91% встреч), креветки и мелкие рачки отмечены |
в 27% желудков. У малых крачек, добытых в дельте Волги, в желудках была только j
рыба (Бородулина, 1960). Мальками рыб (атеринкой, воблой, сазаном, килькой,
сельдью, бычками) питаются эти птицы и на Каспии (Залетаев, 1960; Заблоцкий, За- I
блоцкая, 1963). В Присивашье и на севере Приазовья основу питания (62,3%) состав-
ляют водные ракообразные (Сиохин, 19816), в Таджикистане — мальки рыб и водные
беспозвоночные (Абдусалямов, 1971), в Туркмении — мелкая рыба и насекомые I
(Ташлиев и др., 1967). На Пролетарском водохранилище Ростовской обл., малые j
крачки питаются в основном насекомыми (79,0% по встречаемости); наибольшее
значение имеют личинки стрекоз (44,4%) гладыша (11,5%) и муравьи (14,5%), рыба
обнаружена лишь в 25,2% проб (Казаков, Языкова, 1982). Питание наземными на- '
секомыми (наряду с рыбой и водными беспозвоночными) отмечено в Казахстане
(Долгушин, 1962), в Рязанской обл. (Птушенко, Иноземцев, 1968) и на оз. Ханка
(Поливанова, 1971).
Малые крачки, как и речные, ныряют за кормом с высоты нескольких метров,
часто скрываясь целиком под водой. Высота поискового полета составляет 5—7 м (Ави-
лова, 1975). Для этого вида характерно частое и продолжительное трепетание на месте.
Нередко крачка охотится на небольшом кормовом участке у берега, куда не допускает
других особей (Бородулина, 1960). Отмечено склевывание насекомых со стеблей и
листьев околоводных растений (Поливанова, 1971).
Враги, неблагоприятные факторы. Из абиотических факторов наибольший урон, как
уже упоминалось, наносит подъем воды в результате паводков, штормов и нагонных
ветров. Колонии, расположенные на отмелях и низких островках, в этом случае неред-
ко гибнут целиком. В жаркие солнечные дни на лишенных растительности островах
возможна гибель птенцов от перегрева (Кищинский, 19606). Серьезное воздействие
на колонии малых крачек может оказывать хищничество некоторых видов, среди ко-
торых разные авторы называют серебристых и других чаек, пустельг, куликов-сорок,
лисиц, шакалов, кабанов (Спангенберг, Фейгин, 1936; Бородулина, 1960; Караваев,
1979; Cramp, 1985). На Чонгарских островах Сиваша, как уже отмечалось, птенцов
369
Семейство Чайковые
малых крачек почти полностью выедали чайконосые крачки. Значительное отрица-
тельное воздействие оказывает и деятельность человека, особенно на поселения по ре-
кам и водохранилищам. Гнезда здесь смываются волнами от проходящих судов (Клес-
тов, Осипова, 1981; Росляков, 1981), гибнут в результате постоянного посещения ко-
лоний купальщиками, рыболовами, туристами (Наумчик, 1982; Зубакин и др., 1986).
Фактор беспокойства наряду с антропогенной перестройкой местообитаний был важней-
шей причиной резкого уменьшения численности малых крачек в нашем столетии в За-
падной Европе (Cramp, 1985). Он же, по-видимому, оказывает ныне одно из главней-
ших отрицательных воздействий на популяции крачек в местах с большой плотностью
населения и в нашей стране, в том числе в курортных районах Северного Причерно-
морья и Приазовья. Однако слабая гнездовая консервативность малых крачек, их спо-
собность быстро менять места гнездования в случае ухудшения условий обитания,
попытки гнездования в новых, ранее не свойственных им биотопах (в частности, на
крышах домов) вселяют надежду на возможность сохранения этого вида в антропоген-
ном ландшафте при условии создания зон покоя хотя бы на некоторых территориях,
пригодных для гнездования.
Хозяйственное значение, охрана. В силу малой величины и невысокой численности
хозяйственного значения малая крачка не имеет. Занесена в Красные книги Белоруссии,
Латвии, Литвы, Эстонии. Колонии охраняются в Вильсандийском, Красноводском,
Черноморском и в ряде других заповедников.
* * *
В начале нашего столетия в литературе были указания на нахождение в пределах
СССР мекранской крачки Sterna saundersi, Hume, 1877 (Сушкин, 1908; Зарудный,
1910; 1916). Однако, по Г.П. Дементьеву (1951), они относятся к нетипичным особям
S. albifrons. Подробные сведения об отличительных признаках этих двух видов приведе-
ны у С. Крэмпа (Cramp, 1985).
7.V 1975 г. над р. Зеравшан у Пенджикента была встречена птица, определенная
визуально как индийская крачка Sterna aurantia Gray (Wiist, 1976). Залеты этого вида,
гнездящегося по побережьям Индии и Пакистана (Ali, Ripley, 1969), на территорию
Советской Средней Азии никем ранее не отмечались, несмотря на хорошую изученность
региона. Мы считаем преждевременным включение индийской крачки в фауну СССР,
однако призываем орнитологов, работающих в Средней Азии, учитывать возможность
подобных залетов и обращать внимание на нетипичных особей речных крачек. Индий-
ская крачка отличается от речной более крупными размерами, более длинным хвостом
и более, мощным желтым клювом.
Литература
Абдреимов Т. К биологии некоторых колониаль-
ных птиц низовьев Амударьи // Колониаль-
ные гнездовья околоводных птиц и их охра-
на. М.: Наука, 1975. С. 117.
Абдусалямов И.Л. Птицы. Душанбе: Дониш,
1971. Ч. 1. 398 с. (Фауна Таджикской ССР;
Т. 19).
Абдусалямов ИА. Современное состояние гнез-
довых колоний околоводных птиц Таджи-
кистана // Размещение и состояние гнездовий
околоводных птиц на территории СССР.
М.: Наука, 1981. С. 124-125.
Абдусалямов ИА., Лебедев ЮА. К изучению
перелета птиц в Вахшской долине Таджики-
стана // Миграции птиц в Азии. Новосибирск:
Наука, 1977. С. 170-180.
Абдусалямов ИА., Лебедев ЮА. Миграция птиц
в низовьях р. Яхсу в Таджикистане // Мигра-
ции птиц в Азии. Ташкент: Фан, 1978. С. 34-
59.
Абдусалямов И.А., Лебедев ЮА., Муратов Р.Ш.
Миграции птиц в системе Дарвазского и Ка-
ратегинского хребтов (Таджикистан) // Ми-
грации птиц в Азии. Душанбе: Дониш, 1980.
С. 9-62.
Абдусалямов И.А., Муратов Р.Ш., Лукьянов Г.Н.
К изучению зимней орнитофауны на юге
Таджикистана // Миграции птиц в Азии. Ду-
шанбе: Дониш, 1980. С. 307-310.
Авданин В.О., Гарушянц К.Ю., Зубакин В А.,
Мищенко А.Л. О некоторых интересных на-
ходках в Московской области // Орнитоло-
гия. М.: Изд-во МГУ, 1980. Вып. 15. С. 191-
192.
Аверин Ю.В. Наземные позвоночные восточной
Камчатки // Тр. Кроноцкого заповедника.
М., 1948. Вып. 1. 223 с.
Аверин Ю.В., ГаняИ.М., Зубков И.И., Мунтя-
ну А.И., Успенский ГА. Животный мир
Молдавии: Птицы. Кишинев: Штиинца, 1981.
336 с.
Авилова Ю.В. Суточная активность чайковых
птиц по наблюдениям в Черноморском и
Дарвинском заповедниках // Вести. МГУ.
Сер. 16. Биология. 1972. № 2. С. 25-29.
Авилова К.В. Эколого-морфологические особен-
ности глаза чайковых птиц: Автореф. дис. ...
канд. биол. наук. М.: МГУ, 1973. 24 с.
Авилова К.В. Некоторые особенности пищедо-
бывательного поведения крачек // Коло-
ниальные гнездовья и их охрана. М.: Наука,
1975. С. 7-8.
Адам А.М., Сурнаев В.Н. К распространению и
экологии малой чайки (Larus minutus Pall.)
в Томском Приобье // Вопр. зоологии Сиби-
ри. Томск: Изд-во Том. ун-та, 1979. С. 121-
122.
Ажимуратов X. О миграции чайковых птиц в
дельте Амударьи // 2-я Всесоюз. конф, по
миграциям птиц: Тез. сообщ. Алма-Ата:
Наука, 1978. Ч. 2. С. 4-5.
Ажимуратов X. Влияние синоптических процес-
сов на динамику миграций птиц в дельте
Амударьи // Миграции птиц в Азии. Таш-
кент: Фан, 1984. С. 89-98.
Азаров В.И. Гнездовые колонии околоводных
птиц на озерах Кургальджинского заповед-
ника (по наблюдениям в 1966 и 1967 гг.)
// Колониальные гнездовья околоводных
птиц и их охрана. М.: Наука, 1975а. С. 119 —
121.
Азаров В.И. Колониальные гнездовья некото-
рых околоводных птиц на юге Тюменской
области // Там же, 19756. С. 117-118.
Айрумян К.А., Адамян М.С., Маргарян НА. О на-
хождении среднего поморника в Армении //
Орнитология. М.: Изд-во МГУ, 1968. Вып. 9.
С. 335-336.
Алфераки С.Н. Птицы восточного Приазовья //
Орнитологический вестник. 1910. № 1. С. 11-
35; № 2. С. 75-93; № 3. С. 162-170; № 4.
С. 245-252.
Андоне Г., Пашковский С., Алмэшан X., Андо-
не Л. Материалы по зимовке водоплавающих
и болотных птиц в дельте Дуная в 1959—
1961 гг. // Миграции птиц и млекопитающих.
М.: Наука, 1965. Вып. 4. С. 133-145.
Андреев А.В. Успех размножения розовой чай-
ки Rhodostethia rosea (McGill.) в Нижнеко-
лымской тундре и определяющие его факто-
ры // Редкие и исчезающие птицы Дальнего
Востока. Владивосток: ДВНЦ АН СССР,
1985. С. 110-131.
Андреев А.В., Кондратьев А.Я. Новые данные по
биологии розовой чайки (Rhodostethia rosea)
// Зоол. журн. 1981. Т. 60, вып. 3. С. 418—
425.
Андреев Б.Н. Птицы Среднего Вилюя. Якутск:
Кн. изд-во, 1953. 127 с.
Андреев Б.Н. Птицы Вилюйского бассейна.
Якутск: Кн. изд-во, 1974. 312 с.
Андреев И.Ф. О биоценозе о-ва Китай (Сиваш)
// Пращ науково-дослщного зоолого-бюло-
пчного шституту Харювск. держ. ун-ту.
371
Литература
Харюв: Изд-во Харк. ун-та. 1940. № 8/9.
С. 343-362.
Андриевский И.В. Некоторые особенности ави-
фауны Смоленской области // География и
экология наземных позвоночных. Владимир:
Владимир, пед. ин-т, 1974. Вып. 2. С. 3-4.
Андрусенко Н.Н. Динамика численности коло-
ниальных околоводных птиц в гнездовой
период на оз. Тенгиз // Размещение и со-
стояние гнездовий околоводных птиц на
территории СССР. М.: Наука, 1981. С. 118-
120.
Анзигитова Н.В., Зубакин ВЛ. Об индивидуаль-
ном опознавании у полярных крачек (Sterna
paradisaea Pontopp.), гнездящихся на острове
Большой Айнов // Вопр. зоопсихологии,
этологии и сравнительной психологии. М.:
Изд-во МГУ, 1975. С. 77-79.
Анзигитова НВ., Зубакин В.А., Татарин-
кова И.П. Экологический и этологический
аспекты гнездования полярной крачки // Эко-
логия птиц морских побережий. М.: ЦНИЛ
Главохоты РСФСР, 1980. С. 27-45.
Антипин В.М. Фауна позвоночных северо-восто-
ка Новой Земли // Пробл. Арктики. 1938.
№2. С. 153-171.
Ардамацкая Т.Б. Изменение численности чай-
ковых птиц на островах Тендровского за-
лива // Колониальные гнездовья околовод-
ных птиц и их охрана. М.: Наука, 1975а.
С. 54-56.
Ардамацкая Т.Б. Миграции некоторых чайко-
вых в Северном Причерноморье // Материа-
лы Всесоюз. конф, по миграциям птиц. М.:
Изд-во МГУ, 1975 б. С. 107-108.
Ардамацкая Т.Б. Расширение гнездового ареала
черноголовой чайки в Северном Причерно-
морье // Колониальные гнездовья около-
водных птиц и их охрана. М.: Наука, 1975в.
С. 57-58.
Ардамацкая Т.Б. Межвидовые отношения чайко-
вых птиц на островах Тендровского залива //
Управление поведением животных: Докл.
Всесоюз. конф, по поведению животных.
М.: Наука, 1977а. С. 8-10.
Ардамацкая Т.Б. Особенности гнездования раз-
личных видов чайковых птиц на островах
Тендровского залива в 1976 г. // 7-я Всесоюз.
орнитол. конф.: Тез. докл. Киев: Наук,
думка, 19776. Ч. 1. С. 192-194.
Ардамацкая Т.Б. Сезонное размещение и мигра-
ции чайковых, гнездящихся в Черноморском
заповеднике // Сообщ. Прибалт, комис. по
изуч. миграций птиц. Тарту: АН ЭССР. 1977в.
№ 10. С. 87-113.
Ардамацкая Т.Б. Методика обследования коло-
ниальных гнездовий околоводных птиц
Северного Причерноморья // Научные основы
обследования колониальных гнездовий
околоводных птиц. М.: Наука, 1981. С. 26-30.
Ардамацкая Т.Б. Размещение и численность че-
гравы в Северном Причерноморье // Вести,
зоологии. 1982. № 2. С. 65-66.
Ардамацкая Т.Б. Гнездование утиных и ржанко-
образных на островах Тендровского залива
Черного моря // Орнитология. М.: Изд-во
МГУ, 1984. Вып. 19. С. 41-49.
Аумеэс Л. Об орнитофауне морских островов
западного побережья Сааремаа // Ornitol.
kogumik. Tartu, 1967. К. 4. Lk. 40-57.
Аумеэс Л. Итоги деятельности Вайкаского гос-
заповедника в 1959-1971 гг. // Тез. докл.
8-й Прибалт, орнитол. конф. Таллин: АН
ЭССР. 1972. С. 5-7.
Аумеэс Л., Горяйнова Г.П., Бугаев Л.А. Характе-
ристика гнездового периода сизой чайки в
Вильсандиском государственном заповедни-
ке ЭССР // Тез. докл. 7-й Всесоюз. орнитол.
конф. Киев: Наук, думка. 1977. Ч. 1. С. 197 -
198.
Ауэзов Э.М. О находке колонии реликтовой
чайки Larus relictus Lonnb. // Вести. АН
КазССР. 1970. № 1. С. 59-60.
Ауэзов Э.М.Таксономическая оценка и система-
тическое положение реликтовой чайки Larus
relictus Lonnb. // Зоол. журн. 1971. Т. 50,
№ 2. С. 235-242.
Ауэзов Э.М. Северный Вьетнам - новое место
встречи реликтовой чайки (Larus relictus)
// Зоол. журн. 1974. Т. 53, № 1. С. 139.
Ауэзов ЭМ. Реликтовая чайка на озере Ала-
коль // Колониальные гнездовья около-
водных птиц и их охрана. М.: Наука, 1975.
С. 58-59.
Ауэзов Э.М. О биологии реликтовой чайки //
Редкие и исчезающие звери и птицы Казах-
стана. Алма-Ата: Наука, 1977. С. ПЭ-
ВО.
Ауэзов Э.М. Биология реликтовой чайки и меро-
приятия по ее охране: Дис. ... канд. биол.
наук. Алма-Ата, 1980. 108 с.
Ауэзов Э.М. Колониально-гнездящиеся птицы
островов озера Балхаш // Всесоюз. совещ.
по пробл. кадастра и учета животного мира:
Тез. докл. М„ 1986. Ч. 2. С. 229-231.
Ауэзов ЭМ., Гаврилов Э.И., СемаА.М. Дина-
мика численности реликтовой чайки на озе-
ре Алаколь // Размещение и состояние гнез-
довий околоводных птиц на территории
СССР. М.: Наука, 1981. С. 23-25.
Ауэзов ЭМ., Грачев В.А. Колониальные гнез-
довья околоводных птиц Алакольских озер
// Колониальные гнездовья околоводных
птиц и их охрана. М.: Наука, 1975. С. 121
123.
Ауэзов Э.М., Хроков В.В. Значение островов для
околоводно-гнездящихся птиц // Колониаль-
ные гнездовья околоводных птиц и их охра-
на. М.: Наука, 1975. С. 8-9.
Ауэзов Э.М., Хроков В.В. Весенний пролет птиц
372
Литература
в низовьях Тургая. // Миграции птиц в
Азии. Новосибирск; Наука, 1977. С. 115
121.
Аюпов А.С. Современное состояние колониаль-
ных околоводных птиц Куйбышевского во-
дохранилища // Размещение и состояние
гнездовий околоводных птиц на территории
СССР. М.: Наука, 1981. С. 90-91.
Аюпов А.С., Прохоров Е.В., Горшков Ю.А., Ив-
лев В.Г. О встречах редких птиц в Татарской
АССР // Орнитология. М.: Изд-во МГУ, 1983.
Вып. 18. С. 163-164.
Бакеев Н.Н., Скалой О.И., Чугунов Ю.Д. Новое
в распространении и экологии морского
голубка // Тр. Бюро кольцевания. М.: Изд-во
Минсельхоз, 1957. Вып. 9. С. 133-136.
Банников А.Г. Заметки по орнитофауне полу-
острова Канина // Тр. Аркт. ин-та. 1934.
Т. И. С. 87 - 94.
Банникова А.А. Калякин М.В., Соколова К.И.,
Цеханская М.Ю. О зимующих птицах Ша-
ховской косы // Орнитология. М.: Изд-во
МГУ. 1984. Вып. 19. С. 173-174.
Барабаш-Никифоров И.И., Семаго Л.Л. Птицы
юго-востока Черноземного Центра. Воронеж:
Изд-во Воронеж, ун-та, 1963. 211 с.
Баранов А.А. Эколого-фаунистический анализ
птиц южной Тувы: Автореф. дис. ... канд.
биол. наук. М.: МГПИ, 1981. 16 с.
Баранов Л.С. Весенний пролет чаек на р. Оке //
2-я Всесоюз. конф, по миграциям птиц: Тез.
сообщ. Алма-Ата: Наука, 1978. Ч. 2. С. 12-
13.
Баранов Л.С., Марголин В.А. К фауне и разме-
щению колониальных околоводных птиц
Калужской обл. // Размещение и состояние
гнездовий околоводных птиц на территории
СССР. М.: Наука, 1981. С. 86-87.
Бауманис Я. Изменения орнитофауны верхо-
вого болота Ценас в течение 10 лет (1968—
1978) // Редкие растения и животные. Рига:
ЛатНИИНТИ. 1980. С. 33.
Безбородов В.И. О редких и новых птицах Ир-
кутской области // Орнитология. М.: Изд-во
МГУ, 1979. Вып. 14. С. 186-187.
Беликов С.Е., Рандла Т.Э. К орнитофауне остро-
ва Гукера (Земля Франца-Иосифа) // Там
же, 1984. Вып. 19. С. 174-175.
Белопольский Л.О. К орнитофауне Анадырско-
го края // Тр. Аркт. ин-та. 1934. Т. 11. С. 23-
44.
Белопольский Л.О. Экология морских .коло-
ниальных птиц Баренцева моря. М.; Л.:
Изд-во АН СССР, 1957. 460 с.
Белопольский Л.О., Горяйнова Г.П., Миловано-
ва Н.И. и др. Особенности размножения по-
• лярной крачки на Баренцевом. Белом и
Балтийском морях // Орнитология. М.:
Изд-во МГУ. 1977. Вып. 13. С. 95-99.
Белопольский Л.О., Шунтов В.П. Птицы морей
и океанов. М.: Наука, 1980. 185 с.
Белоусов ЮЛ. Влияние антропогенных факто-
ров на формирование и структуру орнито-
ценоза Ляпинских водоемов // Влияние ант-
ропогенных факторов на структуру и функ-
ционирование биоценозов. Калинин: Кали-
нин, ун-т, 1985. С. 130-132.
Белоусов Ю.А., Макковеева И.И. Новые данные
по географическому распространению коло-
ний белокрылой крачки в Ярославской об-
ласти // Размещение и состояние гнездовий
околоводных птиц на территории СССР. М.:
Наука, 1981. С. 11.
Березовиков Н.Н. О размещении и численности
черноголового хохотуна на оз. Маркаколь //
Размещение и состояние гнездовий около-
водных птиц на территории СССР. М.: Наука,
1981. С. 20-22.
Березовский В.Г. Весенний пролет водоплаваю-
щих и околоводных птиц на восточном по-
бережье Аральского моря в 1978-1979 гг. //
Миграции птиц в Азии. Алма-Ата: Наука,
1983. С. 26-34.
Берман Д.И., Забелин В.И. Новые материалы по
орнитофауне Тувы // Орнитология. М.:
Изд-во МГУ, 1963. Вып. 6. С. 153-160.
Бернацкий Г.И. Зимовки моевок в Черном мо-
ре // Природа. 1954. № 2. С. 119.
Бескоровайный М.М. О зимней фауне водо-
плавающих морской акватории Южного бе-
рега Крыма // 2-я Всесоюз. конф, по мигра-
циям птиц: Тез. сообщ. Алма-Ата: Наука,
1978. Ч. 1.С. 12-13.
Бианки В.В. Материалы по миграции птиц в Кан-
далакшском заливе // Тр. 3-й Прибалт, ор-
нитол. конф. Вильнюс: АН ЛитССР, 1959.
С. 21-30.
Бианки В.В. Пролет птиц в окрестностях с. Вирь-
мы осенью 1958 г. // Тр. Кандалакш. запо-
ведника. Мурманск: Кн. изд-во, 1960. Вып. 3.
С. 175-190.
Бианки В.В. Кулики, чайки и чистиковые Канда-
лакшского залива // Там же, 1967. Вып. 6.
366 с.
Бианки В.В., Бойко Н.С. Динамика осеннего
пролета морских птиц в вершине Кандалак-
шского залива // Тез. докл. 8-й Прибалт, ор-
нитол. конф. Таллин: Ин-т зоологии и бота-
ники. 1972. С. 14-16.
Бианки В.В., Бойко Н.С. Динамика осеннего
пролета морских птиц в вершине Кандалак-
шского залива // Сообщ. Прибалт, комис. по
изуч. миграций птиц. Тарту: АН ЭССР, 1975.
№9. С. 14-24.
Бианки В.В., Карпович В.Н., Пилипас Н.И., Та-
таринкова И.П. О суточной потребности в пи-
ще некоторых морских птиц Севера // Тр.
Кандалакш. заповедника. Мурманск: Кн.
изд-во, 1975. Вып. 9. С. 100 -128.
Бианки В.В., Плюснин ЮМ. Организация коло-
ний и проблема родства в сообществах се-
верных морских птиц // Материалы Всесоюз.
373
Литература
конф, по поведению животных. М.: Наука,
1983. Т. 2. С. 115 117.
Бианки В.В., Хляп Л.А. Опыт изучения гнездово-
го консерватизма и смертности взрослых
полярных крачек // Материалы 7-й Прибалт,
орнитол. конф. Рига, 1970. Кн. 1. С. 25-30.
Бианки В.Л. Первое дополнение к статье ’’Наши
сведения о птицах Новгородской губернии”//
Ежегодник Зоол. музея Академии наук.
1915. Т. 20, №3. С. 1-4.
Бидос В.С. Материалы о летней орнитофауне ант-
ропогфзного ландшафта Яванской долины //
Природные ресурсы и заповедный фонд. Ду-
шанбе: Дониш, 1985. С. 157-174.
Бируля А. Очерки из жизни птиц полярного по-
бережья Сибири // Научные результаты рус-
ской полярной экспедиции в 1900-1903 гг.
под начальством барона Э.В. Толля. СПб..
1907. Т. 1, вып. 2. С. 1-157.
Бирюков В.П., Блажевич Р.Ю., Вынаев Г.В. и др.
Охраняемые растения и животные БССР:
Обзор, информ. Минск: БелНИИНТИ, 1982.
51 с.
Бичеров А.П. Орнитофауна рыбоводных прудов
и рисовых чеков на Ставрополье // Экология
и охрана птиц. Кишинев: Штиинца, 1981. С.25.
Благосклонов К.Н. Птицы Кандалакшского за-
поведника и окрестностей Беломорской био-
логической станции Московского универси-
тета. // Тр. Кандалакш. заповедника. Мур-
манск: Кн. изд-во, 1960. Вып. 2. С. 5-104.
Блинов В.Н., Блинова Т.К. О гнездовании малой
крачки на реке Обь у северной границы ареа-
ла // Тез. Всесоюз. конф, молодых ученых
’’Экология гнездования птиц и методы ее
изучения”. Самарканд: Самарканд, ун-т,
1979. С. 36-38.
Блинов В.Н., Ходков Г.И. К характеристике ко-
лоний речной крачки в разных местообита-
ниях Приобья // Миграции и экология птиц
Сибири. Якутск: ЯФ АН СССР, 1979. С. 123-
125.
Блинова Т.К. Сезонные миграции чайковых в
долине верхней Оби // Миграции птиц в Азии.
Новосибирск: Наука, 1986. С. 212-218.
Богородский Ю.В. Орнитологические находки в
Прибайкалье // Орнитология. М.: Изд-во
МГУ, 1976. Вып. 1ч2. С. 223-224.
Богородский Ю.В. Серебристая чайка на южном
Байкале // Размещение и состояние колоний
околоводных птиц на территории СССР. М.:
Наука, 1981. С. 31-32.
Богословская Л.С., Вотрогов Л.М. Массовые
зимовки птиц и китов в полыньях Беринго-
ва моря // Природа. 1981. № 1. С. 42-43.
Бойко Н.С., Коханов ВЛ., Татаринкова Н.П.
О способах добывания корма большой мор-
ской и серебристой чайками на Мурмане и в
Кандалакшском заливе // Тр. Кандалакш.
заповедника. Мурманск: Кн. изд-во, 1970.
Вып. 8. С. 120-148.
Бойков ВЛ. Материалы по фенологии птиц се-
верной лесотундры (низовья о. Полуя) //
Тр. Ин-та биологии УФ АН СССР. Сверд-
ловск: УФ АН СССР, 1965. Вып. 38. С. 111 -
140.
Болотников А.М., Дьяконов Ю.В., Еремчен-
ко М.И. и др. Количественное соотношение
и биотопическое размещение колониальных
околоводных птиц Камского Предуралья //
Размещение и состояние колоний околовод-
ных птиц на территории СССР. М.: Наука,
1981. С. 91-93.
Болотников А.М., Калинин С.С. Время отклад-
ки яиц и характер насиживания в этот период
у чайковых (Laridae) // Гнездовая жизнь
птиц. Пермь: Перм. пед. ин-т, 1975. С. 64-69.
Болотников А.М., Литвинов Н.А., Скрыле-
ва Л.Ф., Тарасов В.А. К вопросу о гнездовой
жизни сизой чайки в Камском Предуралье //
Там же, 1979. С. 3-11.
Бондарев Д.В. Околоводные колониально-гнез-
дящиеся птицы и изменения условий среды
(на примере Северного Каспия) // Колони-
альные гнездовья околоводных птиц и их
охрана. М.: Наука, 1975. С. 11-13.
Бондарцев А.С. Шкала цветов; Пособие для био-
логов при научных и научно-исследователь-
ских учреждениях. М.; Л.: Изд-во АН СССР,
1954. 27 с.
Боровиков Г.А. Материалы для орнитологии
Екатеринославской губернии // Сб. студ.
биол. кружка при Новорос. ун-те. Одесса,
1907. №2. С. 272.
Бородулина Т.Л. Сельскохозяйственное значение
чайки-хохотуньи в прибрежных степях Север-
ного Крыма // Охрана природы. М.: ВООП,
1949. №7. С. 34-40.
Бородулина Т.Л. Биология и рыбохозяйственное
значение речной крачки // Тр. Ин-та морфо-
логии животных им. А.Н. Северцова. М.:
Изд-во АН СССР, 1953а. Вып. 9. С. 118-139.
Бородулина Т.Л. К биологии болотных крачек //
Там же, 19536. Вып. 9. С. 100-117.
Бородулина Т.Л. Биология и хозяйственное зна-
чение чайковых птиц южных водоемов
СССР // Там же, 1960. Вып. 32. С. 3-130.
Бородулина Т.Л. Рыбохозяйственное значение
чаек и крачек Азово-Черноморского и Кас-
пийского бассейнов // Рыбоядные птицы и
их значение в рыбном хозяйстве. М.: Наука,
1965. С. 34-54.
Браунер А.А. Заметки о птицах Крыма. // Зап.
Новорос. о-ва естествоиспытателей. Одесса,
1898. Т. 23, вып. 1. С. 1-44.
Букреев С.А., Молодая Г.Н., Залевский ВЛ- К
современному состоянию фауны чайковых
птиц в Северо-Восточном Приазовье // Всесо-
юз. совещ. по пробл. кадастра и учета живот-
ного мира: Тез. докл. М., 1986. Ч. 2. С. 240-
241.
Булавинцев В.И. Птицы острова Большевик, ар-
374
Литература
хипелаг Северная Земля // Орнитология. М.:
Изд-во МГУ, 1984. Вып. 19. С, 175 176.
Булахов В.Л. Формирование орнитофауны Днеп-
родзержинского водохранилища // Там же,
1968. Вып. 9. С. 178 187.
Булахов В.Л., Мясоедова О.М. К характеристике
колониальных поселений чаек на Днепро-
дзержинском и Запорожском водохранили-
щах // Колониальные гнездовья околоводных
птиц и их охрана. М.: Наука, 1975. С, 60-61.
Булахов В.Л., Мясоедова О.М, Сезонные мигра-
ции птиц в районе Днепровских водохрани-
лищ степной зоны Украины // 2-я Всесоюз.
конф, по миграциям птиц: Тез. сообщ Алма-
Ата: Наука, 1978. Ч. 1. С. 87—88.
Бурский О.В., Вахрушев А.А. фауна и население
птиц енисейской южной тайги // Животный
мир енисейской тайги и лесотундры и при-
родная зональность. М.: Наука, 1983. С. 106-
167.
Бутурлин С.А. Гнездовья розовой чайки // Псо-
вая и ружейная охота. М., 1905. № 11/12.
С. 41-65.
Бутурлин С.А. Охотничьи птицы низовья Колы-
мы // Псовая и ружейная охота. Тула, 1906.
№ 1. С. 12-13; № 2. С. 28-29.
Бутурлин СА. Птицы Приморской области //
Наша охота. 1910. Кн. 5. С. 43-56.
Бутурлин С.А. Кулики, чайки, чистики, рябки
и голуби // Птицы СССР. М.; Л.: КОИЗ, 1934.
Т. 1.225 с.
Бутьев В.Т. Зимовки птиц на севере Новой Зем-
ли // Орнитология. М.: Изд-во МГУ, 1959.
Вып. 2. С. 99-101.
Бутьев В.Т. К авифауне Вологодской области //
Фауна и экология позвоночных животных.
М.: МГПИ им. В.И. Ленина, 1978. С. 3-19.
Бухаринов С.В., Олигер И.М. Формирование ко-
лоний чайковых птиц на отстойниках сточных
вод // Эколого-морфологические особеннос-
ти животных Среднего Поволжья. Казань,
1984. С. 132-135.
Буякович Н.Г. К охране розовой чайки // Охрана
природы. М.: ВООП, 1950. № 15. С. 119—
121.
Вайткявичюс А.П. Миграция птиц вдоль Курш-
ской косы осенью 1958 г. // Ornitol. Kogumik.
Tartu, 1961. К. 2. Lk. 130-145.
Вайткявичус А.П. Видимые миграции водопла-
вающих и других водяных птиц в Литве //
Сообщ. Прибалт, комис. по изуч. миграций
птиц. Тарту: АН ЭССР, 1968. № 5. С. 44-66.
Вайткявичюс А.П., Скуодис В.К. Перелет птиц.
Вильнюс: Минтис, 1965. 194 с.
Валюс М., Калиндрене А., Пятрайтис А., Скуо-
дис В. Исторические и современные материа-
лы по орнитологии Литовской ССР // Эколо-
гия птиц Литовской ССР. Вильнюс: Ин-т зо-
ологии и паразитологии АН ЛитССР, 1976.
Ч. 1.С. 5-32.
Варшавский С.Н., Варшавский Б.С., Гарбу-
зов В.К. Некоторые редкие и исчезающие
птицы Северного Приаралья // Редкие и ис-
чезающие звери и птицы Казахстана. Алма-
Ата: Наука, 1977. С. 146-153.
Васильев В.И., Гаузер М.Е. Пестроносая крачка
на южном Каспии // Колониальные гнездовья
околоводных птиц и их охрана. М.: Наука,
1975. С. 64.
Васильев В.И., Гаузер М.Е., Щербина А.А. Совре-
менное состояние колониальных гнездовий
чайковых птиц на юго-восточном Каспии //
Там же. С. 62-63.
Васильченко А.А. Учет колониальных птиц на
озере Барун-Торей (Юго-Восточное Забай-
калье) // Всесоюз. совеш. по пробл. кадастра
и учета животного мира: Тез. докл. М., 1986.
Ч. 2. С. 244-246.
Васьковский А.П. Новые орнитологические на-
ходки на северном побережье Охотского мо-
ря // Зоол. журн. 1956. Т. 35, вып. 7. С. 1051-
1058.
Вдовкин М.С. Новые данные к распространению
птиц в низовьях реки Оби // Заметки по фау-
не и флоре Сибири. Томск, 1941. Вып. 4.
С. 1-6.
Велижанин А.Г. Курильские острова как путь
пролета птиц // Трансконтинентальные связи
перелетных птиц и их роль в распространении
арбовирусов. Новосибирск: Наука, 1972.
С. 87-90.
Велижанин А.Г. О весеннем пролете водных и
околоводных птиц на Курильских островах //
Материалы Всесоюз. конф, по миграциям
птиц. М.: Изд-во МГУ, 1975. Ч. 2. С. 9-11.
Велижанин А.Г. Краткое сообщение о розовой
чайке в Беринговом море // Редкие, исчезаю-
щие и малоизученные птицы СССР. Рязань,
1976. С. 177. (Тр. Окского заповедника;
Вып. 13).
Велижанин А.Г. О некоторых редких и малоиз-
вестных птицах Курильских островов // Ор-
нитология. М.: Изд-во МГУ, 1977. Вып. 13
С. 25-32.
Велижанин А.П., Велижанин Г.А. Список птиц
Барнаульского округа // Uragus. 1929. Кн. 9,
№ 1. С. 5-14.
Велижанин Г.А. Добавление к орнитофауне Бар-
наульского округа // Там же. 1928. Кн. 6,
№ 1. С. 12-20.
Виксне Я.А. Некоторые вопросы миграции лат-
вийской обыкновенной чайки // Экология
и миграции птиц Прибалтики. Рига: Изд-во
АН ЛатвССР, 1961. С. 227-233.
Виксне Я.А. Миграции и сезонное размещение
чайковых птиц Латвии по данным кольцева-
ния // Миграции водоплавающих и прибреж-
ных птиц Латвийской ССР. Рига: Изд-во
АН ЛатвССР, 1962. С. 77-145.
ВикснеЯ.А. О значении послегнездовых кочевок
в территориальном размещении гнездующих
озерных чаек (Larus ridibundus) (на примере
375
Литература
восточно-прибалтийской' популяции) // Эко-
логия водоплавающих птиц Латвии. Рига:
Зинатне, 1968а. С. 167-205.
Виксне Я.А. О структуре гнездовой популяции
озерной чайки (Laius ridibundus L.) на при-
морских озерах Латвии // Там же, 19686.
С. 207-228.
Виксне Я.А. Связь с гнездовой территорией, рас-
селение молодых птиц и формирование но-
вых колоний у озерной чайки (Larus ridibun-
dus L.) // Миграции животных. Л.: Наука,
1968в. Вып. 5. С. 116-133.
Виксне Я.А. Территориальные связи и демогра-
фия популяции озерной чайки (Larus ridibun-
dus L.) Восточной Прибалтики: Автореф.
дис.... канд. биол. наук. Таллин, 1968 г.
22 с.
Виксне Я.А. Связь с местом рождения у озерной
чайки (Larus ridibundus L.) // Материалы 7-й
Прибалт, орнитол. конф. Рига: Зинатне,
1970. Ч. 1. С. 41-44.
Виксне Я.А. Численность и распространение
озерной чайки в Латвии // Колониальные
гнездовья околоводных птиц и их охрана. М.:
Наука, 1975. С. 64-66.
ВикснеЯ.А. Численность и территориальное раз-
мещение гнездовых колоний чайковых в Лат-
вии // Сообщ. Прибалт, комис. по изуч. миг-
раций птиц. Тарту: АН ЭССР, 1978. № И.
С. 76-89.
Виксне ЯА. Успешность размножения озерной
чайки (Larus ridibundus L.) в зависимости от
плотности гнезд // 10-я Прибалт, орнитол.
конф.: Тез. докл. Рига: Ин-т биологии АН
ЛатвССР, 1981а. Т. 2. С. 27-30.
Виксне Я.А. Численность, распространение и де-
мография озерной чайки (Larus ridibundus L.)
в Латвии // Сообщ. Прибалт, комис. по изуч.
миграций птиц. Тарту: АН ЭССР, 19816.
№ 12. С. 95-115.
Виксне Я.А. Долгосрочное изучение демогра-
фии популяции озерной чайки (Larus ridi-
bundus L.) в Латвии // 18-й Междунар. орни-
тол. конгр., Москва, 1982: Тез. докл. и стенд,
сообщ. М.: Наука, 1982. С. 246-247.
Виксне ЯА. Семейство Laridae // Птицы Лат-
вии: Территориальное размещение и чис-
ленность. Рига: Зинатне, 1983. С. 98-112.
Виксне ЯА., Балтвилкс Я.П. Распространение
и численность чайковых птиц в водоемах
Латвии // Материалы 6-й Прибалт, орни-
тол. конф. Вильнюс, 1966. С. 27-28.
Виксне ЯА., Недзинскас В.С., Ренно О.Я. Чис-
ленность и распространение озерной чайки
в Прибалтике и ее динамика // Распростра-
нение и численность озерной чайки. М.:
Наука, 1981. С. 5-14.
Виксне Я.А., Янаус М. Кормовые полеты озер-
ной чайки оз. Энгурес // Орнитология, М.:
Изд-во МГУ, 1986. Вып. 21. С. 31-37.
Виноградов В.В. Биологические ресурсы водно-
болотных охотничьих угодий Мильской сте-
пи, их производительность и перспективы
хозяйственного использования // Тр. заповед-
ников Азербайджана. М.: Лесн. пром-сть,
1967. Вып. 2. С. 143.
Виноградов В.В., Чернявская С.И. Материалы по
орнитофауне Кызыл-Агачского заповедник^/
Там же, 1965. Вып. 1. С. 22-79.
Винокуров АА. Позднелетние и осенние мигра-
ции некоторых болотных и водоплавающих
птиц в северо-западном Предкавказье // Миг-
рации птиц и млекопитающих. М.: Наука,
1965. №4. С. 39-55.
Винокуров А.А. Серебристая чайка и бурго-
мистр на центральном Таймыре // Размеще-
ние и состояние колоний околоводных птиц
на территории СССР. М.: Наука, 1981. С. 37.
Винокуров А А., Залетаев В.С., Кищинский А А.
и др. Зимовки водоплавающих птиц на по-
бережье Черного и Азовского морей зимой
1957-1958 гг. // Миграции животных. М.:
Изд-во АН СССР, 1960. Вып. 2. С. 45-54.
Виткаускас Н.В. К биологии размножения реч-
ной крачки на островах водохранилища //
7-я Всесоюз. орнитол. конф.; Тез. докл.
Киев: Наук, думка, 1977. Ч. 2. С. 115-116.
Виткаускас Н.В. Биология размножения озер-
ной чайки в центральной Литве // 10-я При-
балт. орнитол. конф. Рига: Ин-т биологии
АН ЛатвССР, 1981а. Т. 2. С. 31-38.
Виткаускас Н.В. Характер насиживания во вре-
мя яйцекладки у озерной чайки // Там же,
19816. Т. 2. С. 33-36.
Виткаускас Н.В. Устойчивость насиживания у
речной крачки // Тез. докл. 11-й Прибалт,
орнитол. конф. Таллин: Ин-т зоологии и бо-
таники АН ЭССР, 1983. С. 71-72.
Виткаускас Н., Гражулявичюс Г. К сравнитель-
ной экологии размножения речной крачки //
Материалы 9-й Прибалт, орнитол. конф.
Вильнюс: Ин-т зоологии и паразитологии АН
ЛитССР, 1976. С. 63-65.
Владимирская М.И. Птицы Лапландского запо-
ведника // Тр. Лапланд. заповедника. М.:
Гл. упр. по заповедникам, 1948. Вып. 3.
С. 171-245.
Водолажская Т.Н. О режиме насиживания озер-
ной чайки // Орнитология. М.: Изд-во МГУ,
1979. Вып. 14. С. 206-207.
Водолажская Т.Н. К размещению крачек в Волж-
ско-Камском крае // Размещение и состояние
гнездовий околоводных птиц на территории
СССР. М.г Наука, 1981. С. 12-13.
Водолажская Т.Н. Материалы по изменению фау-
ны птиц Куйбышевского водохранилища //
Всесоюз. совещ. по пробл. кадастра и учета
животного мира: Тез. докл. М., 1986. Ч. 2.
С. 249-250.
Водолажская Т.И., Залетаев В.С. Отряд чайко-
образные // Птицы Волжско-Камского края:
Реворобьиные. М.: Наука, 1977. С. 199-220.
376
Литература
Водолажская Т.И., Мингазова Н.М. К экологии
гнездования белокрылой крачки (Chlidonias
leucoptera Temm.) // Экология гнездования
птиц и методы ее изучения: Тез. Всесоюз.
конф, молодых ученых. Самарканд: Самар-
канд. ун-т, 1979. С. 45-46.
Волков Е.Н. Динамика численности и размеще-
ния колониальных птиц на оз. Тенгиз (Цент-
ральный Казахстан) // Колониальные гнез-
довья околоводных птиц и их охрана. М.:
Наука, 1975. С. 125-129.
Волчанецкий И.Б. Очерк орнитофауны восточ-
ного Предкавказья // Тр. НИИ биологии и
Биол. факультета ХГУ. Харьков: Изд-во
Харьк. ун-та, 1959. Т. 28. С. 7-38.
Воробьев Г.П. Массовые налеты чаек на свалку
в холодные весны // Экология и охрана
птиц. Кишинев: Штиинца, 1981. С. 45-46.
Воробьев К.А. Материалы к орнитологической
фауне дельты Волги и прилежащих степей //
Науч. тр. госзаповедников. Сер. 1. Астра-
ханский заповедник. 1936. Вып. 1. С. 3-52.
Воробьев К.А. Птицы Уссурийского края. Вла-
дивосток: ДВ ФАН СССР, 1954. 351 с.
Воробьев К.А. Птицы Якутии. М.: Изд-во АН
СССР, 1963. 336 с.
Воробьев К.А. Орнитологические исследования
на Алазее (Северо-Восточная Якутия) // Ор-
нитология. М.: Изд-во МГУ, 1967. Вып. 8.
С. 150-159.
Воронов Г.А., Еремин Ю.П. О гнездовании алеут-
ской крачки Sterna camtschatica на Сахалине//
Эколого-фаунистические исследования неко-
торых позвоночных Сахалина и Курильских
островов. Владивосток, 1982. С. 26-35.
Воронцов ЭМ. До шзнання орнитофауни При-
сивашия и Сиваш1в // Праш наук.-досл. зо-
ол.-бюлог. ин-ту. XapKiB: Изд-во Харк. ун-ту,
1937. Т. 4. С. 83-125.
Воронцов ЕМ. Птицы Камского Приуралья.
Горьк. ун-т, 1949. 114 с.
Воронцов ЕМ. Птицы Горьковской области.
Горький: Волго-Вят. кн. изд-во, 1967. 168 с.
Вронский Н.В., Томкович П.С. К фауне пролет-
ных птиц приморской Колхиды // Материалы
Всесоюз. конф, по миграциям птиц. М.: Изд-
во МГУ, 1975. Ч. 2. С. 13-15.
Вяткин П.С., Герасимов Н.Н., Мараков С.В.
Морские колониальные птицы острова Кара-
гинского // Колониальные гнездовья около-
водных птиц и их охрана. М.: Наука, 1975.
С. 170.
Гавриленко Н.И. Находки арктических и боре-
альных птиц на Полтавщине // Орнитология.
М.: Изд-во МГУ, 1960. Вып. 3. С. 436-439.
Гаврилов Н.Н., Кривоносов Г.А. Колониальные
гнездовья чайковых птиц на о-ве Жемчужном
(Северный Каспий) // Размещение и состоя-
ние гнездовий околоводных птиц на террито-
рии СССР. М.: Наука, 1981. С. 15-16.
Гаврилов Э.И., Ауэзов ЭМ., Сема AM., Вол-
ков Е.Н. Сезонные миграции черноголового
хохотуна в Казахстане // Миграции птиц в
Азии. Алма-Ата: Наука, 1983. С. 102-125.
Гагина Т.Н. Залетные птицы Восточной Сиби-
ри // Орнитология. М.: Изд-во МГУ. 1962.
Вып. 4. С. 367-372.
Гаузер М.Е. Массовые гнездовые колонии пест-
роносых крачек на островах Красноводско-
го залива как возможный очаг арбовирус-
ных инфекций на Восточном Каспии // Эко-
логия вирусов. М.: Изд-во АМН СССР. 1976.
Вып. 4. С. 99-101.
Гаузер М.Е. Социально обусловленная смерт-
ность птенцов в колониях пестроносой крач-
ки Thalasseus sandvicensis (Laridae) на остро-
вах Красноводского залива. 1. Общая харак-
теристика ювенильной Смертности и ее воз-
можные причины // Зоол. журн. 1981а. Т. 60,
вып. 4. С. 530-539.
Гаузер М.Е. Социально обусловленная смерт-
ность птенцов в колониях пестроносой крач-
ки Thalasseus sandvicensis (Laridae) на островах
Красноводского залива. 2. Роль структуры и
организации колонии // Там же. 19816. Т.60.
Вып. 6. С. 879-886.
Гаузер М.Е. Адаптивная ценность различных ти-
пов колониального гнездования у чайковых
при отсутствии пресса хищничества // Коло-
ниальность у птиц: структура, функции, эво-
люция. Куйбышев: Куйбышев, ун-т, 1983а.
С. 121-143.
Гаузер М.Е. Этологическая структура популяций
и успех размножения у чайковых птиц с раз-
личными типами колониального гнездова-
ния (на примере Sterna hirundo L., Sterna
sandvicensis Lath., Larus genei Br.): Дис. ...
канд. биол. наук. Красноводск, 19836. 220 с.
Гаузер М.Е. Этологическая структура популяции
пестроносой крачки Thalasseus sandvicensis
Lath. (Laridae) на островах Красноводского
залива // Изв. АН ТССР. Сер. биол. наук.
1983в. № 1. С. 43-49.
Гаузер М.Е. Численность пестроносой крачки
и морского голубка в Красноводском запо-
веднике и обусловливающие ее факторы //
Всесоюз. совещ. по пробл. кадастра и учета
животного мира: Тез. докл. М., 1986. Ч. 2.
С. 254-256.
Гаузер М.Е., Васильев В.И. Фенологические ас-
пекты биологии пестроносой крачки на Вос-
точном Каспии // Экология гнездования
птиц и методы ее изучения: Тез. Всесоюз.
конф, молодых ученых. Самарканд: Самар-
канд. ун-т, 1979. С. 58-60.
Гаузер М.Е., Васильев В. И., Караваев А.А. Дина-
мика численности чайковых на Восточном
Каспии // Тез. докл. 7-й Всесоюз. орнитол.
конф. Киев: Наук, думка, 1977. Ч. 1. С. 226-
227.
Гёбель Г.Ф. Материалы по орнитофауне Лаплан-
дии и Соловецких островов // Тр. Санкт-Пе-
377
Литература
тербург, о-ва естествоиспытателей. Отд. зоо-
логии и физиологии. 1902. Т. 33, вып. 2.
С. 97-137.
Герасимов Н.Н. Размещение и численность ко-
лониальных птиц на северо-западном побе-
режье Карагинского острова // Орнитоло-
гия. М.: Изд-во МГУ. 1979а. Вып. 14. С. 189—
190.
Герасимов Н.Н. Эколого-географический анализ
авифауны острова Карагинского: Автореф.
дис. ... канд. биол. наук. М.: ЦЛОП. 19796.
С. 1-24.
Герасимов Н.Н. Встречи розовой чайки на Кам-
чатке // Редкие и исчезающие птицы Дальне-
го Востока. Владивосток: ДВНЦ АН СССР,
1985. С. 132-134.
Герасимова Т.Д. К орнитофауне Айновых остро-
вов // Тр. Кандалакш. заповедника. Вологда:
Кн. изд-во, 1958. Вып. 1. С. 37-50.
Герасимова Т.Д. Состояние птичьих базаров
Мурманского побережья // Орнитология. М.:
Изд-во МГУ, 1962. Вып. 4. С. 11-14.
Герасимова ТД. Питание чаек мурманского по-
бережья // Рыбоядные птицы и их значение в
рыбном хозяйстве. М.: Наука, 1965. С. 194—
209.
Герасимова Т.Д., Горчаковская Н.Н., Карпо-
вич В.Н., Рахилин В.К. Дополнение к фауне
птиц Семи Островов // Тр. Кандалакш. запо-
ведника. М.: Лесн. пром-сть, 1967. Вып. 5.
С. 339-344.
Гизенко А.И. Птицы Сахалинской области.
М.: Изд-во АН СССР, 1955. 328 с.
Гилевич А.Л. Размножение озерной чайки в
дельте р. Селенги на Байкале // Экология
птиц Восточной Сибири. Иркутск: Иркут,
ун-т, 1977. С. 37-58.
Гисцов А.П. Некоторые изменения фауны птиц
острова Барсакельмес за последние десяти-
летия // Материалы 6-й Всесоюз. орнитол.
конф. М.: Изд-во МГУ, 1974. Ч. 1. С. 189-190.
Гладков Н.А. Орнитологические результаты
поездки на Амударью летом 1931 г. // Бюл.
МОИП. Отд. биол. Н.С. 1932. Т. 41, вып. 3/4.
С. 351-398.
Гладков Н.А. Наблюдение над птицами остро-
ва Комсомольского (Аральское море ) //
Охоана природы. М.: ВООП, 1949. № 8.
С. 29-43.
Гладков Н.А. Птицы Тиманской тундры //
Сб. трудов Зоомузея МГУ. М.: Изд-во МГУ,
1951. Т. 7. С. 15-89.
Гладков Н.А. Материалы по птицам окрестнос-
тей Воркуты (восток Большеземельской
тунды) // Орнитология. М.: Изд-во МГУ,
1962. Вып. 4. С. 15-28.
Гладков Н.А., Дементьев Г.П., Птушенко Е.С.,
Судиловская А.М. Определитель птиц СССР.
М.: Высш, шк., 1964. 536 с.
Гладков Н.А., Залетаев В.С. О фауне птиц Ман-
гышлака и Мангистауских островов // Тр.
Ин-та биологии АН ТССР. 1956. Т. 4.
С. 120-164.
Глущенко Ю.Н. К фауне гнездящихся птиц При-
ханкайской низменности // Редкие птицы
Дальнего Востока. Владивосток: ДВНЦ АН
СССР, 1981. С. 25-33.
Глущенко Ю.Н. Материалы к орнитофауне
юго-западной Камчатки // Орнитология. М.:
Изд-во МГУ, 1984а. Вып. 19. С. 177.
Глущенко Ю.Н. Состояние гнездовий чайковых
птиц на побережье озера Ханка в период его
низкого уровня // Фаунистика и биология
птиц юга Дальнего Востока. Владивосток:
ДВНЦ АН СССР, 19846. С. 79-86.
Глущенко Ю.Н., Шибнев Ю.Б. К орнитофауне
заповедника "Кедровая падь” и сопредель-
ных территорий // Фаунистика и биология
птиц юга Дальнего Востока. Владивосток:
ДВНЦ АН СССР, 1984. С. 44-48.
Говоруха Л.С. Остров Виктория // Советская
Арктика: Природные условия и естествен-
ные ресурсы СССР. М., 1970. С. 359-362.
Головкин А.Н. О выедании рыбы кайрами и
моевками в гнездовой период в Баренце-
вом море // Зоол. журн. 1963. Т. 42, № 3.
С. 408-416.
Головкин А.Н. Птичьи базары севера Новой
Земли // Особенности биологической про-
дуктивности вод близ птичьих базаров севера
Новой Земли. Л.: Наука, 1972. С. 84-91.
Головкин А.Н, Флинт В.Е. Колониальные гнез-
довья морских птиц в районе мыса Сердце-
Камень (Чукотка) // Колониальные гнездо-
вья околоводных птиц и их охрана. М.: Нау-
ка, 1975. С. 171-173.
Головушкин М.И. О колонии реликтовых чаек
в Читинской области // 7-я Всесоюз. ориитол.
конф.: Тез. докл. Киев: Наук, думка, 1977.
Ч. 2. С. 207-209.
Горбунов Г.П. Птичьи базары Новой Земли //
Тр. НИИ по изуч. Севера. 1925. Вып. 26,
ч. la. С. 48.
Горбунов Г.П. Материалы по фауне млекопи-
тающих и птиц Новой Земли // Там же.
1929. Вып. 40. С. 169-239.
Горбунов Г.П. Птицы Земли Франца-Иосифа //
Тр. Аркт. ин-та. 1932. Т. 4. 244 с.
Гордеев Ю.Н. Некоторые данные по птицам
Казыма (Тюменская обл.) // Орнитология.
М.: Изд-во МГУ, 1963. Вып. 6. С. 469-
470.
Гордеев Ю.И. Материалы по распространению
птиц в Ханты-Мансийском округе // Орнито-
логия. М.: Изд-во МГУ, 1977. Вып. 13. С 33-
39.
Гордеев Ю.И. Прилет птиц в окрестности
г. Хаиты-Мансийска // Ритмы природы Си-
бири и Дальнего Востока. Иркутск: Ин-т
Сибири и Дальнего Востока, 1979. С. 93-105.
Горчаковская Н.Н. Экология сизой чайки по
наблюдениям на Восточном Мурмане // Ох-
378
Литература
рана природы. М.: ВООП, 1948. Вып. 5.
С. 77-83.
Гражулявичюс Г.Б. Околоводные колониальные
птицы заказника Биржулис // Размещение
и состояние 'гнездовий околоводных птиц
на территории СССР. М.: Наука, 1981.
С. 83-84.
Грачев В.А. Белокрылая крачка на Алакульских
озерах // Новости орнитологии Казахстана.
Алма-Ата: Наука, 1968. С. 220.
Григорьев В.Р. Краткое сообщение о розовой
чайке на севере Сибири // Редкие, исчезаю-
щие и малоизученные птицы СССР. Рязань,
1976. С. 178. (Тр. Окского заповедника;
Вып. 13).
Гурьев В.Н. Видовой состав, численность и
биомасса птиц среднего течения р. Вычег-
ды // фауна и экология птиц и млекопитаю-
щих европейского северо-востока СССР.
Сыктывкар: Коми ФАН СССР. 1982.
С. 10 20.
Гусев А.А. Случай гнездования малых крачек
в нетипичных условиях // Научные основы
охраны и рационального использования птиц.
Рязань, 1978. С. 353- 355. (Тр. Окского
заповедника; Вып. 14).
Гусев О.К О гнездовании птиц на островах
Чивыркуйского залива Байкала и оз. Ран-
готуй // Тр. Вост.-Сиб. фил. СО АН СССР.
Зоология. 1960. Вып. 23. С. 69-88.
Гынгазов А.М., Миловидов С.П. Орнитофауна
Западно-Сибирской равнины. Томск: Изд-во
Том. ун-та, 1977. 351 с.
Данилов Н.Н. О гнездовом паразитизме у чаек //
Природа. 1948. № 12. С. 62.
Данилов Н.Н. Птицы нижней Оби и изменения в
их распространении за последние десятиле-
тия // Тр. Ин-та биологии УФ АН СССР.
1965. Вып. 38. С. 103-109.
Данилов Н.Н., Рыжановский В.Н., Рябицев В.К.
Птицы Ямала. М.: Наука, 1984. 334 с.
Дарлингтон Ф. Зоогеография. М.: Прогресс,
1966. 519 с.
Дебело П.В., Чибисов Н.Г. Видовой состав и
численность некоторых водных птиц Чижин-
ских и Балыктинских разливов // Колониаль-
ные гнездовья околоводных птиц и их охра-
на. М.: Наука, 1975. С. 131-132.
Дебело П.В., Шевченко В.Л. Распространение
и численность некоторых крачек в Северном
Прикаспии // Всесоюз. совещ. по пробл.
кадастра и учета животного мира: Тез. докл.
М., 1986. Ч. 2. С. 271-272.
Дегтярев А.Г, Дегтярев В.Г, Ларионов А.Г.
К экологии крачек центральной Якутии //
Миграции и экология птиц Сибири. Новоси-
бирск: Наука, 1981а. С. 127-134.
Дегтярев А.Г., Дегтярев В.Г., Ларионов А.Г.
Озерная чайка в Центральной Якутии //
Распространение и численность озерной чай-
ки. М.: Наука, 19816. С. 94-98.
Дегтярев А.Г., Ларионов Г.П. Опыт учета числен-
ности чайковых бассейна среднего течения
р. Лены // Научные основы обследования
гнездовий околоводных птиц. М.: Наука.
19816. С. 44-46.
Дегтярев А.Г., Ларионов Г.П., Гермогенов Н.И.
Миграции водно-болотных птиц в районе
устья реки Наманы // Водно-болотные виды
птиц долины Средней Лены. Якутск: Яф АН
СССР, 1978. С. 29 48.
Дегтярев В.Г. Внутривидовые и межвидовые от-
ношения в гнездовых поселениях чайковых на
севере Лено-Алданского междуречья // Тео-
ретические аспекты колониальности у птиц.
М.: Наука, 1985. С. 32 -35.
Дегтярев В.Г. Чайковые птицы северной части
Лено-Алданского междуречья: Дис. ... канд.
биол. наук. Якутск, 1986. 213 с.
< Дегтярев В.Г., Ларионов А.Г. К биологии сизой
чайки Лено-Алданского междуречья // Разме-
щение и состояние колоний околоводных
птиц на территории СССР. М.: Наука, 1981а.
С. 34-35.
Дементьев Г.П. Залеты моевок в глубь азиат-
ского материка в пределах СССР // Сб. тр.
Зоол. музея МГУ. М.: Изд-во МГУ, 1934.
Вып. 1. С. 161-162.
Дементьев Г.П. Птицы полуострова Канина. //
Там же. 1935. Вып. 2. С. 23-35.
Дементьев Г.П. Материалы к авифауне Коряк-
ской земли. М.: МОИП, 1940. 83 с.
Дементьев Г.П. Отряд Чайки (Lariformes) // Пти-
цы Советского Союза. М.: Сов. наука, 1951.
Т. 3. С. 373-603.
Дементьев Г.П. Птицы Туркменистана. Ашхабад:
Изд-во АН ТССР, 1952. 547 с.
Дементьев Г.П. Миграции Rissa tridactyla // Тр.
Бюро кольцевания. М.: Изд-во МСХ СССР,
1955. Вып. 8. С. 22-32.
Дементьев Г.П., Караев М.К., Карташев Н.Н.
Птицы юго-западной Туркмении // Уч. зап.
МГУ. 1955. Вып. 171. С. 53-172.
Демме Н.П. Птичий базар на скале Рубини (ост-
ров Гукера, Земля Франца-Иосифа) // Тр.
Аркт. ин-та. 1934. Т. 11. С. 55-86.
Денисов В.П., Фролов В.В. Распространение око-
ловодных колониальных птиц в Пензенской
области // Размещение и состояние гнездо-
вий околоводных птиц на территории СССР.
М.: Наука, 1981. С. 88-90.
Деревягин П.Я. Материалы по экологии некото-
рых видов крачек // Тр. Алма-Атинского
зоопарка. Алма-Ата, 1948. Вып. 1. С. 55-64.
Дерюгин К.М. Путешествие в долину нижнего
и среднего течения р. Оби и фауна этой облас-
ти // Тр. О-ва естествоиспытателей. СПб.,
1898. Т. 24, вып. 2. С. 140.
Дмоховский А.В. Птицы Средней и Нижней
Печоры // Бюл. МОИП. Отд. биол. Н.С. 1933.
Т. 42, вып. 2. С. 214-242.
Доброхотов Б.П. Современное состояние зимо-
379
Литература
вок птиц на юго-восточном берегу Каспия //
Орнитология. М.: Изд-во МГУ. 1962. Вып. 5.
С. 362-367.
Доброхотов Б.П. Современное состояние зимов-
ки птиц в Кызыл-Агачском заповеднике //
Там же, 1963. Вып. 6. С. 355-359.
Долбик М.С. Птицы Березинского заповедни-
ка// Березинский заповедник: Исследования.
Минск: Ураджай. 1970. Вып. 1. С. 149 155.
Долбгк М.С., Навумчык А.В. Чайкавыя птушкг
Беларуси асаблгвасц i ix распаусюджання i
размяшчэння // Весц! АН БССР. Сер. бгял.
н. 1983. №3. С. 86 -90.
Долгушин Н.А. Отряд Чайки - Laiiformes // Пти-
цы Казахстана. Алма-Ата: Изд-во АН КазССР,
1962. Т. 2. С. 246-327.
Дорогой И.В. Экология поморников // Экология
млекопитающих и птиц острова Врангеля.
Владивосток: ДВНЦ АН СССР, 1981. С. 38-
55.
Дорогой И.В. Материалы по биологии вилохвос-
той чайки // Орнитология. М.: Изд-во МГУ,
1984. Вып. 19. С. 198.
Дубовик А.Д., Миловидов С.П., Стрелков В.Е.
Фенология весеннего прилета птиц в Томской
области // Миграции птиц в Азии. Новоси-
бирск: Наука, 1977. С. 108-115.
Дулькейт ГД., Щульпин Л.М. Птицы Шантар-
ских островов // Тр. Биол. НИИ Том. ун-та.
1937. Т. 4. С. 114 136.
Дунаева Т.Н., Кучерук В.В. Материалы по эколо-
гии наземных позвоночных тундры Юж-
ного Ямала // Материалы к познанию фауны
и флоры СССР. Отд. зоол. Н.С. М.. 1941.
Вып. 4 (19). С. 5-80.
Дурнев Ю.А. К изучению клептопаразитических
отношений у птиц в условиях Сибири //
Материалы 3-й Всесоюз. конф, по поведе-
нию животных. М.: Наука, 1983. Т. 2. С. 197
198.
Дымин В.А. Новые данные о распространении
некоторых птиц в Верхнем Приамурье //
Орнитологические исследования на Дальнем
Востоке. Владивосток: ДВНЦ АН СССР,
1975. С. 277 -278.
Дюнин А.Г. Чайка-хохотунья на юго-западном
Каспии // Чайка-хохотунья и фламинго на
Каспийском море. М.: Гл. упр. по заповед-
никам, 1948. С. 3 31.
Егоров В.Г. Колониальные птицы Чивыркуй-
ского залива оз. Байкал // Размещение и
состояние гнездовий околоводных птиц на
территории СССР. М.: Наука, 1981. С. 33-34.
Елкин К.Ф. Весенняя миграция водоплавающих
и болотных птиц в средней части Тургайской
депрессии (Северный Казахстан) // Сообщ.
Прибалт, комис. по изуч. миграций птиц.
Тарту: Изд-во АН ЭССР, 1975. № 9. С. 61 --82.
Елсуков С.В. К авифауне Северо-Восточного
Приморья // Материалы 6-й Всесоюз. орни-
тол. конф. М.: Изд-во МГУ, 1974а. Ч. 1.
С. 199-200.
Елсуков С.В. Некоторые дополнительные све-
дения о птицах Среднего Сихотэ-Алиня //
Орнитология. М.: Изд-во МГУ, 19746. Вып. 11.
С. 371.
Ерохов С.Н. формирование русел пролета птиц
в пустынной зоне юго-востока Казахстана
в связи с изменением экологических усло-
вий // Миграции птиц в Азии. Новосибирск:
Наука, 1986. С. 24-32.
Естафьев А.А. Распространение и численность
озерной чайки на европейском северо-восто-
ке СССР // Распространение и численность
озерной чайки. М.: Наука, 1981а. С. 3-5.
Естафьев А.А. Современное состояние, распре-
деление и охрана авифауны таежной зоны
бассейна р. Печоры. Сыктывкар: Коми
ФАН СССР. 19816. 54 с.
Естафьев А.А. Сроки прилета, размножения
и отлета гнездящихся птиц таежной зоны
бассейна р. Печоры // Фауна Урала и приле-
жащих территорий. Свердловск: Урал, ун-т,
1982. С. 25 -34.
Житков Б.М. В тундре п-ова Канин // Зап. Рус.
геогр. о-ва. 1904. Т. 41, № 1. С. 170.
Житков Б.М., Бутурлин С.А. Материалы для ор-
нитофауны Симбирской губернии // Зап.
И.Р.Г. о-ва по общей географии. СПб., 1906.
Т. 41, № 2. 275 с.
Житков Б. Зензинов В. Орнитологическая фауна
крайних северных частей Сибири // Журн.
Отд. зоол. о-ва любителей естеств. истории,
антропологии и этнографии Н.С. 1915. Т. 3,
№2. С. 1-19.
Жмуд М.Е., Балацкий К.Л. Современное состоя-
ние колоний чайковьгх и куликов в примор-
ской части Дунай-Днестровского между-
речья // Всесоюз. совещ по пробл. кадастра
и учета животного мира: Тез. докл. М.,
1986. Ч. 2. С. 290-292.
Жордания Р.Г. Колониальные гнездовья около-
водных птиц в Грузии // Колониальные
гнездовья околоводных птиц и их охрана.
М.: Наука, 1975. С 136.
Жордания Р.Г. Орнитогеография Грузинской ССР:
(Зоогеографическое районирование на ави-
фаунисгичеекой основе и анализ типов фау-
ны) // 7-я Всесоюз. орнитол. конф.: Тез.
докл. Киев: Наук, думка, 1977.4.1. С. 60-61.
Жордания Р.Г. Озерная чайка в Грузии // Распро-
странение и численность озерной чайки.
М.: Наука, 1981. С. 72-73.
Журавлев М.Н. К наблюдениям за реликтовой
чайкой // Колониальные гнездовья около-
водных птиц и их охрана. М.: Наука, 1975.
С. 165-167.
Заблоцкий В.И., Заблоцкая Л.И. Эколого-
фаунистический обзор чайковых птиц юго-
западного Каспия и их рыбохозяйственное
380
Литература
значение // Тр. Астрах, заповедника. 1963.
Вып. 8. С. 309-348.
Залетаев В.С. Малая чайка на Восточном Кас-
пии // Природа, 1955. № 8. С. 116-117.
Залетаев В.С. Материалы по биологии рыбояд-
ных птиц Восточного Каспия // Охрана приро-
ды и озеленение. М.: ВООП, 1960. Вып. 4.
С. 11-44.
Залетаев В.С. Зимовки водоплавающих птиц
на северном, восточном и юго-восточном
Каспии // Тр. Астрах, заповедника, 1963.
Вып. 8. С. 349-372.
Залетаев В.С. Распространение и численность
рыбоядных птиц на восточном побережье
Каспийского моря // Рыбоядные птицы и
их значение в рыбном хозяйстве. М.: Наука,
1965. С. 141-150.
Зарудный НА. Орнитологическая фауна Орен-
бургского края // Зап. Акад. наук. СПб.,
1888. Т. 57, прил. № 1. С. 1-338.
Зарудный НА. Птицы Псковской губернии //
Зап. Акад, наук по физ.-мат. отд. Сер. 8.
СПб., 1910. Т. 25. № 2. С. 182.
Зарудный НА. Птицы Аральского моря // Изв.
Туркм. отд. И.Р.Г. о-ва. 1916. Т. 12, вып. 1.
С. 1-229.
Засыпкин М.Ю. Распространение птиц на запад-
ной Чукотке и зоогеографический анализ
ее авифауны // Орнитология. М.: Изд-во
МГУ, 1981. Вып. 16. С. 100-114.
Зимин В.Б. Пролет птиц на восточном берегу
Ладожского озера осенью 1962 г. // Сообщ.
Прибалт, комис. по изуч. миграций птиц.
Тарту: АН ЭССР, 1965. № 3. С. 28-49.
Зимин В.Б. О структуре периферийной зоны
ареала у птиц // 7-я Всесоюз. орнит. конф.:
Тез. докл. Киев: Наук, думка, 1977. Ч. 1.
С. 62-63.
Зимин В.Б., Ивантер Э.В. Птицы (Животный
мир Карелии). Петрозаводск: Карелия, 1974.
256 с.
Зиновьев В.И. Птицы лесной зоны европейской
части СССР (Ржанкообразные). Калинин:
Калинин, ун-т, 1980. 83 с.
Зиновьев В.И., Молодовский А.В., Зубакин В.А.,
Белоусов Ю.А., Измаилов А.И. Размещение
и численность озерной чайки в центральных
районах Нечерноземья // Распространение и
численность озерной чайки. М.: Наука, 1981.
С. 36-42.
Зиновьев В.И., Шапошников Л.В. Материалы
по орнитофауне Калининской обл. // Гео-
графия и экология наземных позвоночных.
Владимир, 1978. № 3. С. 40-53.
Зубакин ВА. Гнездование чайковых на острове
Китай в 1973 году // Колониальные гнездовья
околоводных птиц и их охрана. М.: Наука,
1975а. С. 72-74.
Зубакин ВА. Индекс плотности гнездования
некоторых видов чайковых птиц и способ
его вычисления // Зоол. журн. 19756. Т. 54,
№9. С. 1386-1389.
Зубакин В А. Каннибализм у чайконосой крачки
(Gelochelidon nilotica Gm.) // Вести. МГУ.
Сер. 16, Биология. 1975в. № 3. С. 32-36.
Зубакин ВА. О демонстрационном поведении
черноголового хохотуна (Larus ‘ichthyaetus
Pall.) // Вопр. зоопсихологии, этологии и
сравнительной психологии. М.: Изд-во МГУ,
1975 г. С. 55-57.
Зубакин ВА. Некоторые вопросы индивидуаль-
ного опознавания у чайковых птиц (Laridae)
// Бюл. МОИП. Отд. биол. 1976а. Т. 81,
№ 3, С. 31-37.
Зубакин ВА. О демонстрационном поведении
морского голубка // Групповое поведение
животных: Докл. 2-й Всесоюз. конф, по по-
ведению животных. М.: Наука, 19766.
С. 140-142.
Зубакин В А. Сравнительная экология колони-
ального гнездования чайковых птиц. Дис. ...
канд. биол. наук. М.: МГУ, 1976в. 173 с.
Зубакин ВА. О необходимости включения в
Красную книгу СССР некоторых редких ви-
дов чаек // Редкие животные и их охрана в
СССР. М.: Изд-во ЦЛОП МСХ СССР, 1977.
С. 77-80.
Зубакин ВА. Реликтовая чайка на Торейских
озерах // Докл. МОИП. Зоология, ботаника.
2-е полугодие 1976 г. М.: Изд-во МГУ, 1978.
С. 6-8.
Зубакин ВА. Некоторые аспекты демонстра-
ционного и гнездового поведения реликто-
вой чайки, гнездящейся на Торейских озе-
рах // Бюл. МОИП. Отд. биол. 1979. Т. 84,
вып. 2. С. 15-20.
Зубакин ВА. Колониальные птицы Торейских
озер // Размещение и состояние гнездовий
околоводных птиц на территории СССР.
М.: Наука, 1981а. С. 132-134.
Зубакин В А. Чайковые птицы Московской
области и их адаптация к антропогенному
ландшафту // Научные основы обследова-
ния колониальных гнездовий околоводных
птиц. М.: Наука, 19816. С. 51-56.
Зубакин ВА. Экология и поведение буроголо-
вой чайки (Larus brunnicephalus) на Пами-
ре // Бюл. МОИП. Отд. биол. 1982. Т. 87,
вып. 4. С. 45-56.
Зубакин ВА. Роль различных факторов в воз-
никновении и развитии колониальности у
чайковых птиц // Колониальность у птиц:
структура, функции, эволюция. Куйбышев,
1983. С. 37-64.
Зубакин ВА., Авданин В.О. Особенности коло-
ниального гнездования розовой чайки (Rho-
dostethia rosea) // Зоол. журн. 1983. Т. 62,
вып. 11. С. 1754-1756.
Зубакин В.А., Анзигитова И.В. Межвидовые от-
ношения некоторых видов чайковых и ку-
381
Литература
ликов юга европейской части СССР // 2-я
Всесоюз. конф, молодых ученых по вопро-
сам сравнит, морфологии и экологии живот-
ных. М.: Наука, 1975. С. 162 163.
Зубакин ВЛ., Костин Ю.В. Гнездящиеся птицы
Чонгарских островов // Орнитология. М.:
Изд-во МГУ, 1977. Вып. 13. С. 49-55.
Зубакин В А., Мищенко А.Л., Абоносимова Е.В.,
Волошина О.Н. и др. Современное состояние
некоторых редких видов птиц Московской
области: Неворобьиные // Орнитология. М.:
Изд-во МГУ, 1986. Вып. 21. С. 77-93.
Зубакин В.А., Харитонов С.П. Современное со-
стояние колонии озерных чаек на озере
Киёво // Растительность и животное населе-
ние Москвы и Подмосковья. М.: Изд-во МГУ,
1978. С. 102-104.
Зубакин ВА., Харитонов С.П. Гнездование озер-
ных чаек на оз. Киёво (Московская обл.) //
Распространение и численность озерной чай-
ки. М.: Наука, 1981. С. 45-49.
Зубакин В А., Харитонов С.П. Залет черноголо-
вой чайки в Московскую область // Орнито-
логия. М.: Изд-во МГУ, 1983. Вып. 18. С. 167.
Зырянов В.А., Ларин В.В. Видовой состав птиц
гор Путорана // Птицы Таймыра: (Экология,
охрана и хозяйственное использование). Но-
восибирск: СО ВАСХНИЛ, 1983. С. 3-9.
Иваненко И.Д. Бюлогия i сшьскогосподарське
значения чайки-реготухи // Зб1рник наук,
праць. Азово-С)ваш. заповедник. Киш: Хар-
KiB, 1936. С. 109-131.
Иванов А.И. Птицы Памиро-Алтая. Л.: Наука,
1969. 448 с.
Иванов А.И. Каталог птиц СССР. Л.: Наука,
1976.276 с.
Измайлов И.В. Птицы Витимского плоскогорья.
Улан-Удэ: Бурят, кн. изд-во, 1967. 304 с.
Измайлов И.В., Боровицкая Г.К Птицы юго-
западного Забайкалья. Владимир: Владимир,
пед. ин-т, 1973. 315 с.
Измайлов И.В., Старков И.А. Наблюдение за
пролетом птиц в озерной долине Еравны
(Бурятская АССР) // Орнитология. М.:
Изд-во МГУ, 1960. Вып. 3. С. 405-409.
Ильичев В.Д., Фомин В.Е. Орнитофауна Башки-
рии и ее изменения в XX веке // Там же,
1979. Вып. 14. С. 83-96.
Иогансен Г.Х. Птицы Командорских остро-
вов // Тр. Том. ун-та. 1934. Т. 86. С. 222-266.
Ирисов В.А. Особенности распространения неко-
торых птиц в юго-восточном Алтае // Орни-
тология. М.: Изд-во МГУ, 1972. Вып. 10.
С. 248-251.
Исаков ЮА. О колониальном гнездовании чаек-
хохотуний на острове Тараба // Чайка-хохо-
тунья и фламинго на Каспийском море. М.:
Гл. упр. по заповедникам, 1948. С. 33-43.
Исаков Ю.А., Воробьев КА. Обзор зимовок и
пролета птиц на южном Каспии // Тр.
Всесоюз. орнитол. заповедника Гассан-Кули.
М.: Гл. упр. по заповедникам, 1940. Вып. 1.
С. 5-159.
Исаков ЮА., Крумина М.К., Распопов М.П.
Материалы по экологии обыкновенной чайки
(Larus ridibundus L.) //Очеркиприроды Под-
московья и Московской области. М.: МОИП,
1947. С. 104-187.
Исмагилов М.И. Чайки острова Барсакелъмес
(Аральское море) // Тр. Ин-та зоол. АН
КазССР. Алма-Ата: Изд-во АН КазССР,
1955. Т. 4. С. 166-172.
Исмагилов М.И., Бурамбаев К.Б. Условия су-
ществования и характер пребывания птиц
в заповеднике Барсакельмес // Биологиче-
ские науки. Алма-Ата, 1973. Вып. 5. С. 119—
126.
Ишунин Г.И. Питание чайки-хохотуньи в При-
сивашье // Орнитология. М.: Изд-во МГУ,
1963. Вып. 6. С. 471-472.
Йыги А.И. Миграция эстонской обыкновенной
чайки по данным кольцевания // Тр. 2-й
Прибалт, орнитол. конф. М.: Изд-во АН
СССР, 1957. С. 193-201.
Йыги А. Перелеты эстонской сизой чайки по
данным кольцевания. (Eesti kalakajakate ran-
dest fongasmatise andmeil) // Ornitol. kogumik.
Tartu, 1958. К. 1. Lk. 192-212.
Йыги А. Зимовка водоплавающих и других
водяных птиц в Эстонии за 1960- 1963 гг. //
Сообщ. Прибалт, комис. по изуч. миграций
птиц. Тарту: АН ЭССР, 1965. №3. С. 115-132.
Йыги А.И. О результатах мечения птиц в Мат-
салуском гос. заповеднике // Тр. гос. заповед-
ников ЭССР. Таллин: Валгус, 1968. Вып. 1.
С. 107-131.
Йыги А. Результаты учета зимующих водопла-
вающих птиц в Эстонии в январе 1967 года //
Сообщ. Прибалт, комис. по изуч. миграций
птиц. Тарту: АН ЭССР, 1969. № 6. С. 94-96.
Йыги А.И., Линг Р.Г., Онно С. Видимые перелеты
птиц в окрестностях Пухтуской орнитологи-
ческой станции осенью 1957 г. // Ornitol.
kogumik. Tartu, 1961. К. 2. Lk. 42-78.
Каверкина Н.П. Гнездование настоящих крачек
на о-ве Карагинском (Камчатка) // Эколо-
гические исследования и охрана птиц При-
балтийских республик: Тез. докл. Прибалт,
конф, молодых орнитологов. Каунас, 1982.
С. 86-88.
Каверкина Н.П. Сравнительная экология и по-
ведение крачек в репродуктивный период:
Дис. ... канд. биол. наук: Ленинград: Изд-во
ЛГУ, 1983. 294 с.
Каверкина Н.П., Давыдова Г.Ю. Динамика за-
щитной активности в колонии алеутской
крачки // Экология и охрана животных:
Межвуз. сб. Куйбышев: Куйбышев, ун-т,
1982. С. 98-100.
Каверкина Н.П., Рощевский Ю.К Защитное
поведение полярной крачки (Sterna paradi-
382
Литература
saea Brun.) // Весты. ЛГУ. Биология. 1984.
№ 15. С. 19-25.
Казаков БД., Ломадзе Н.Х., Гончаров В.Т.
Состояние колоний околоводных птиц дель-
ты Дона и Западного Маныча // Фауна, эколо-
гия и охрана животных Северного Кавказа.
Нальчик, 1980. С. 112-129.
Казаков Б.А., Ломадзе Н.Х., Гончаров В.Т.
Голенастые и крачки Веселовского водохра-
нилища // Размещение и состояние гнездовий
околоводных птиц на территории СССР.
М.: Наука, 1981. С. 69-71.
Казаков Б.А., Ломадзе Н.Х., Языкова ИМ.
Чайки и голенастые оз. Казинка Ц Разме-
щение и состояние колоний околоводных
птиц на территории СССР. М.. Наука, 1981.
С. 71-73.
Казаков Б.А., Языкова ИМ. Отряд Ржанко-
образные // Ресурсы живой фауны. Ч. 2.
Позвоночные животные суши. Ростов н/Д:
Изд-во Рост, ун-та, 1982. С. 204-230.
Калабин С.Ю. Пролет чайковых птиц в районе
озера Айдаркуль в 1980-1982 гг. // Мигра-
ции птиц в Азии. Новосибирск: Наука, 1986.
С. 163-167.
Калинин С.С. Степень участия семейных партне-
ров в насиживании у чайковых // Гнездовая
жизнь птиц. Пермь: Перм. пед. ин-т, 1975.
С. 128-134.
Калинин С.С., Болотников А.М. Гнездования
чайковых в Зауралье // Колониальные гнез-
довья околоводных птиц и их охрана. М.:
Наука, 1975. С. 76-77.
Караваев А.А. Материалы по фауне гнездящихся
водно-болотных птиц низовьев р. Атрек //
Природная среда и птицы побережий Кас-
пийского моря и прилежащих низменностей.
Баку: Азерб. гос. изд-во, 1979. С. 62-82.
Караваев А.А. Численность и распространение
чайковых и голенастых птиц в низовье р. Ат-
рек (Юго-Западная Туркмения) // Размеще-
ние и состояние гнездовий околоводных
птиц на территории СССР. М.: Наука, 1981.
С. 125-128.
Караваев А.А., Белоусов ЕМ. Миграции птиц
на юго-восточном Каспии осенью 1977 года //
Миграции птиц в Азии. Алма-Ата: Наука,
1983. Вып. 8. С. 52-65.
Карпович В.Н., Коханов В.Д. Фауна птиц остро-
ва Вайгач и северо-востока Югорского полу-
острова // Тр. Кандалакш. заповедника. М.:
Лесн. пром-сть, 1967. Вып. 5. С. 268-338.
Карпович В.Н., Соловьева-Волынская Т.Н.,
Шехт Н.Н. К экологии болотных крачек
поймы среднего течения р. Оки // Тр. Окско-
го заповедника. М.: Гл. упр. охотничьего
хоз-ва и заповедников. 1958. Вып. 2. С. 78-
101.
Карпович В.Н., Шкляревич Ф.Н. Причины и
размеры гибели яиц и птенцов моевок в ко-
лониях острова Харлова (Восточный Мур-
ман) // Колониальные гнездовья околовод-
ных птиц и их охрана. М.: Наука, 1975.
С. 173-174.
Карташев Н.Н. Об изменении границ гнездовых
ареалов и проявлении периодических циклов у
северных птиц // Охрана природы. М.: ВООП,
1949. Вып. 8. С. 50-58.
Карташев Н.Н. Наблюдение над весенним проле-
том птиц в Кара-Кумах // Уч. зап. МГУ. М.:
Изд-во МГУ, 1958. Вып. 197. С. 113-124.
Карташев Н.Н. Количественная характеристика
авифауны Соловецких островов // Орнито-
логия. 1963. Вып. 6. С. 23-36.
Карташев Н.Н. Годовые изменения численности
чайковых птиц Моложского отрога Рыбин-
ского водохранилища // Колониальные гнез-
довья околоводных птиц и их охрана. М.:
Наука, 1975. С. 76-77.
Кастепыльд Т. О гнездовом консерватизме
чегравы // Тез. докл. 8-й Прибалт, орнитол.
конф. Таллин: Изд-во АН ЭССР, 1972. С. 46.
Кафтановский Ю.М. Зимовки птиц в заповед-
нике ’’Семь островов ”//Тр. гос. заповедника
’’Семь островов”. М.: Гл. упр. по заповедни-
кам. 1941. Вып. 1. С. 47-52.
Кенжегулов К. К экологии голенастых и чай-
ковых птиц дельты Амударьи//Экология
важнейших млекопитающих и птиц Кара-
калпакии. Ташкент: Фан. 1972. С. 158-192.
Киселев Ф.А. В государственном сивашском
заповеднике на острове Китай//Охрана при-
роды. М.: ВООП, 1949. № 8. С. 115-117.
Киселев ФА. Сельскохозяйственное значение
чайки-хохотуньи в северо-западном Крыму//
Тр. Крым. ФАН СССР. Симферополь: Крым-
издат, 1951. Т. 2. С. 21-30.
Кисленко Г.С. Материалы по биологии птиц
низовьев Сырдарьи//Орнитология. М.: Изд-
во МГУ, 1972. № 10. С. 337- 339.
К1стякивськш О.Б. Птахи: Загальна характе-
ристика птах1в. Курин!. Голуби. Рябки.
Пастушки. Журавль Дрофи. Кулики. Мар-
тини. Киш: Изд-во АН УССР, 1957. 432 с.
Кистяковский А.Б. О пластичности экологии
птиц//Орнитология. М.: Изд-во МГУ, 1967.
Вып. 8. С. 357-358.
Кистяковский А.Б., Мельничук В.А. Измене-
ния колониальных гнездовий птиц после
образования Киевского водохранилища//Ко-
лониальные гнездовья околоводных птиц
и их охрана. М.: Наука, 1975. С. 138-139.
Кищинский АД. К фауне и экологии птиц
Териберского района Мурманской облас-
ти//Тр. Кандалакш. гос. заповедника. Мур-
манск: Кн. изд-во, 1960а. Вып. 2. С. 122—
212.
Кищинский АД. Современная авифауна ли-
манов северо-восточного Причерноморья
и биология гнездящихся здесь чайковых
птиц//Птицы водоемов/Охрана природы и
383
Литература
озеленение. М.: ВООП, 19606. Вып. 4.
С. 69-75.
Кищинский А.А. Птицы Колымского нагорья.
М.: Наука, 1968. 188 с.
Кищинский А.А. Птицы Корякского нагорья.
М.: Наука, 1980. 336 с.
Кищинский А.А., Томкович П.С., Флит В.Е.
Птицы бассейна Канчалана//Распространение
и систематика птиц. М.: Изд-во МГУ, 1983.
С. 3-76.
Кищинский А.А., Фомин В.Е., Болд А., Цэвэн-
мядаг В. Птицы горного массива Мунх-
Хайрхан (МНР) //Зоологические исследова-
ния в МНР. М.: Наука, 1982. С. 62-81.
Клакоцкий В.П. Колониальное гнездование не-
которых околоводных птиц по среднему
течению реки Припять//Колониальные гнез-
довья околоводных птиц и их охрана. М.:
Наука, 1975. С. 139-140.
Клестов Н.Л. Колониальные околоводные птицы
Кременчугского водохранилища//Размеще-
ние и состояние гнездовий околоводных
птиц на территории СССР. М.: Наука, 1981.
С. 104-106.
Клестов Н.П. Орнитофауна среднего Днепра
и ее изменение под влиянием гидрострои-
тельства: Автореф. дис. . . . канд. биол.
наук. Киев: Ин-т зоологии АН УССР, 1983.
26 с.
Клестов Н.Л., Осипова М.А. О формировании
колониальных поселений околоводных птиц
на Каневском водо.хранилище//Научные осно-
вы обследования колониальных гнездовий
околоводных птиц. М.: Наука, 1981. С. 57—58.
Клестов Н.Л., Севастьянов В.И., Чуприн С.Л.
О гнездовании Larus argentatus cachinnans
Pall, на Киевском водохранилище//Вестн.
зоологии. 1986. № 1. С. 86.
Клименко М.И. К экологии чайковых северного
Черноморского побережья//Тр. Черномор,
гос. заповедника. Киев: Изд-во Киев, ун-та,
1950а. Вып. 1. С. 53-69.
Клименко М.И. Материалы по фауне птиц района
Черноморского заповедника//Там же, 19506.
Вып. 1.С. 3 -52.
Климова И.Г. Темп развития эмбрионов озерной
чайки в Камском Предуралье//Огражение
достижений орнитологической науки в учеб-
ном процессе средних школ и вузов и народ-
ном хозяйстве. Пермь: Перм. пед. ин-т, 1984.
С. 51-53.
Кныш Н.П., Пулях В.П. Пролет водно-болотных
птиц на искусственных водоемах возле
г. Сумы//Материалы Всесоюз. конф, по миг-
рациям птиц. М.: Изд-во МГУ, 1975. Ч. 2.
С. 23 -25.
Ковалев А.Н. О гнездовании полярной крачки
на острове Б. Лицкий (Баренцево море)//
Экология и охрана птиц. Кишинев: Штиинца,
1981. С. 107.
Ковалев А.Н., Каверкина Н.П., Давыдова Г.Ю.,
Коротицкая Л.Н., Рощевский Ю.К. О гнездо-
вании алеутской крачки на озере Невском
(Сахалин)//Орнитология. М.: Изд-во МГУ,
1980. Вып. 15. С. 196-197.
Ковалев А.Н., Рощевский Ю.К. Зарастание
колонии и выживаемость птенцов речной
крачки//7-я Всесоюз. орнитол. конф.: Тез.
докл. Киев: Наук, думка, 1977. Ч. 1. С. 257-
258.
Ковшарь А.Ф. Реликтовая чайка//Птицы Казах-
стана. Алма-Ата: Наука, 1974. Т. 5. С. 407-411.
Ковшарь А.Ф., Левин А. С. Каталог оологической
коллекции Института зоологии АН КазССР.
Алма-Ата: Наука, 1982. 103 с.
Козлов П.К. Труды экспедиции Императорского
Русского географического общества по Цент-
ральной Азии. СПб., 1899. Ч. 2. 296 с.
Козлова Е.В. Авифауна Тибетского нагорья,
ее родственные связи и история//Тр. Зоол.
ин-та АН СССР. 1952. Т. 9. № 4. С. 964-1028.
Кокорев Я.И. Орнитофауна бассейна Пуры
(западный Таймыр)//Птицы Таймыра: (Эко-
логия, охрана и хозяйственное использова-
ние) . Новосибирск: СО ВАСХНИЛ, 1983.
С. 15-19.
Коленов В.В. Материалы по размещению коло-
ний околоводных птиц в Волгоградской
области//Размещение и состояние гнездовий
околоводных птиц на территории СССР.
М.: Наука, 1981. С. 98-99.
Колющее И.П. Материалы по фауне и промыслу
западного угла Таймырского полуострова//
Материалы по изучению Сибири. Томск:
Изд-во Том. ун-та, 1933. Т. 4. С. 68-106.
Кондратьев А.Я. О состоянии птичьих базаров
на острове Колючин//Колониальные гнез-
довья околоводных птиц и их охрана. М.:
Наука, 1975. С. 174-175.
Кондратьев А.Я. Сезонные явления в жизни
птиц Чаунской низменности//Птицы северо-
востока Азии. Владивосток: ДВНЦ АН СССР,
1979. С. 95-105.
Кондратьев А.Я., Кондратьева Л.Ф. Рост и разви-
тие птенцов вилохвостой чайки//Орнитоло-
гия. М.: Изд-во МГУ, 1984. Вып. 19. С. 81-88.
Корзюков А.И. Миграции некоторых околовод-
ных и хищных птиц прибалтийских популя-
ций в северо-западном Причерноморье//10-я
Прибалт, орнитол. конф.: Тез. докл. Рига:
Инт-т биологии АН ЛатвССР, 1981. Т. 1.
С. 136-139.
Корнеева Т.М., Быков А.В., Речан С.П. Наземные
позвоночные низовьев реки Онеги. М.: Наука,
1984. 89 с.
Костин Ю.В. Колониальные гнездовья около-
водных птиц Крыма//Колониальные гнез-
довья околоводных птиц и их охрана. М.:
Наука, 1975. С. 140-141.
Костин Ю.В. О методике ооморфологических
исследований и унификации описаний оологи-
ческих материалов//Методика исследования
384
Литература
продуктивности и структуры видов птиц в
пределах их ареалов. Вильнюс: Мокслас,
1977. С. 14-22.
Костин Ю.В. Сезонное размещение черноморс-
ких чеграв по данным кольцевания//2-я
Всесоюз. конф, по миграциям птиц. Алма-
Ата: Ин-т зоологии АН КазССР, 1978. Ч. 2.
С. 69-71.
Костин Ю.В, Птицы Крыма. М.: Наука, 1983.
240 с.
Костин Ю.В., Тарина Н.А. Новые сведения о сос-
тоянии гнездовий околоводных колониаль-
ных птиц Крыма//Размещение и состояние
колоний околоводных птиц на территории
СССР. М.: Наука, 1981. С. 113-115.
Костина Г-H., Панов Е.Н. Смертность пуховиков
и характер персональных связей между птен-
цами и родителями у черноголового хохо-
туна Larus ichthyetus//Зоол. журн. 1982. Т.61,
вып. 10. С. 1531-1542.
Косыгин Г.М. Белая чайка на Чукотском полу-
осгрове//Колониальные гнездовья околовод-
ных птиц и их охрана. М.: Наука, 1975.
С. 176.
Косыгин Г.М. Регистрация белой, вилохвос-
товой и розовой чаек в Чукотском, Беринго-
вом и Охотском морях//Редкие и исчезаю-
щие птицы Дальнего Востока. Владивосток:
ДВНЦ АНСССР, 1985. С. 135-137.
Коханов В.Д. Весенние миграции птиц в районе
Айновых островов.//Тр. 5-й Прибалт, орни-
тол. конф_ Таллин: Валгус, 1967. С. 175 —
183.
Коханов В.Д. Материалы по миграции среднего
поморника на Мурмане//Тр. Кандалакш. гос.
заповедника. Мурманск: Кн. изд-во, 1970.
Вып. 8. С. 182-189.
Коханов В.Д. Современные ареалы чаек на Бе-
лом море и Мурмане//Научные основы обсле-
дования колониальных гнездовий околовод-
ных птиц. М.: Наука, 1981. С. 63-64.
Коханов В.Д., Скокова Н.Н. Наблюдения за ве-
сенней миграцией птиц в Кандалакшских
шхерах в 1958 году//Тр. Кандалакш. гос.
заповедника. Мурманск: Кн. изд-во, 1960.
Вып. 3. С. 121-132.
Коханов В.Д., Скокова Н.Н. Фауна птиц Айно-
вых островов//Тр. Кандалакш. гос. заповед-
ника. М.: Лесн. пром-сть, 1967. Вып. 5.
С. 185-267.
Кошелев А.И, Смешанная колония на оз. Петра-
ковское (Западная Сибирь) //Колониальные
гнездовья околоводных птиц и их охрана.
М.: Наука, 1975. С. 141-142.
Красная книга Литовской ССР. Вильнюс: Мокс-
лас, 1981. 84 с.
Краснов Ю.В. Клептопаразитизм чаек рода
Larus в районе Семи островов (Восточный
Мурман)//Экология и охрана птиц. Кишинев.
Штиинца, 1981. С. 117-118.
Краснов Ю.В,, Головацкая И.Ф., Соловьева Т.Ю.
13. Зак. 826
Сравнительная характеристика хищнического
поведения крупных чаек в колониях кайр и
моевок Семи островов//Экология и морфо-
логия птиц на крайнем северо-западе СССР.
М.: ЦНИЛ Главохоты РСФСР, 1982. С. 80-91.
Краснов Ю.В,, Николаева Н.Г, Чернышева НМ-
Особенности кормового поведения коротко-
хвостовых поморников в колониях морских
птиц Восточного Мурмана//Тез. докл. Прибалт,
конф, молодых орнитологов. Каунас, 1982.
С. 119-121.
Кречмар А.В. Сезонные явления в жизни птиц
района Норильских озер//Орнитология. М.:
Изд-во МГУ, 1963. Вып. 6. С. 37-48.
Кречмар А.В, Птицы западного Таймыра//Био-
логия птиц. М.: Л.: Наука, 1966. С. 185-312.
Кречмар А.В., Андреев А.В., Кондратьев А.Я.
Экология и распространение птиц на Северо-
Востоке СССР. М.: Наука, 1978. 196 с.
Кривенко В,Г. Экологические сукцессии и вод-
ные птицы дельты Волги//Охотоведение.
Биологические и экономические основы ве-
дения охотничьего хозяйства. М.: Лесн. пром-
сть, 1975. С. 211-234.
Кривенко В.Г. Закономерность динамики чис-
ленности гнездящихся птиц в водоемах до-
лины р. Маныча//Научные основы обследо-
вания колониальных гнездовий околовод-
ных птиц. М.: Наука, 1981. С. 68-75.
Кривенко В.Г. Черноголовый хохотун//Красная
книга РСФСР. М.: Россельхозиздат, 1983.
С. 273-275.
Кривенко В.Г, Лысенко В.И., Филонов К.П.
Расширение гнездового ареала черноголовой
чайки (Larus melanocephalus) // Зоол. журн.
1973.№4.С.618-619.
Кривенко В.Г, Любаев В.Л. Колониально гнез-
дящиеся птицы Восточного Маныча//Коло-
ниальные гнездовья околоводных птиц и их
охрана. М.: Наука, 1975. С. 142-143.
Кривенко В.Г., Любаев В.Л. Изменения чис-
ленности гнездящихся птиц Восточного Ма-
ныча//”Тез. докл. 7-й Всесоюз.орнитол. конф.
Киев: Наук, думка, 1977. Ч. 1. С. 72-74.
Кривенко В.Г,, Любаев В.Л. Изменение числен-
ности гнездящихся птиц на Чограйском водо-
хранилище//Размешение и состояние гнездо-
вий околоводных птиц на территории СССР.
М^Наука, 1981. С. 93-96.
Кривоносов Г.А. Зимовки птиц на Северном
Каспии в 1960-1961 гг. (по наблюдениям в
районе о-ва Искусственного)//Тр. Астрах,
заповедника. Астрахань: Волга, 1963. Вып. 8.
С. 377-391.
Кривоносов Г.А. Очаги массового гнездования
чайковых птиц на Северном Каспии и факто-
ры, определяющие длительность их сушесгво-
вания//Экология вирусов :(Сб. трудов Ин-та
вирусологии АМН СССР). М.: Изд-во АМН
СССР. 1976. С. 46-51.
Кривоносов ГА., Бондарев Д.В., Гаврилов Н.Н.
385
Литература
Некоторые результаты кольцевания чайко-
вых птиц в дельте Волги и на Северном Кас-
пии//2-я Всесоюз. конф, по миграциям птиц.
Алма-Ата: Наука, 1978. Ч. 1. С. 131-132.
Кубанцев Б.С., Чернобай В.Ф. Птицы северных
районов Нижнего Поволжья. Волгоград: Вол-
гоград. пед. ин-т, 1982. 72 с.
Кудленок И.А. Водные угодья и колониальные
околоводные птицы Брянской области//Раз-
мещение и состояние гнездовий околоводных
птиц на территории СССР. М.: Наука, 1'981.
С. 84-86.
Кузякин А.П, Материалы по биологии колониаль-
но гнездящихся птиц//Уч. зап. Моск. обл.
пед. ин-та. 1959. Т. 71, вып. 4. С. 4-21.
Кузякин А.П,, Леонович В.В. Птицы беломорс-
ко-мезенской лесотундры//Там же. 1958.
Т. 65, вып. 3. С. 119-140.
Кукиш А.И. Состояние колоний птиц на внутрен-
них водоемах Калмыкии//Всесоюз. совещ. по
проблемам кадастра и учета животного
мира: Тез. докл. М., 1986. Ч. 2. С. 325-326.
. Кумари Э.В. Пролет и зимовка птиц на террито-
рии Эстонской ССР и задачи их исследова-
ния//Перелеты птиц в европейской части
СССР. Рига: Изд-во АН ЛатвССР, 1953. С.
44-55.
Кумари Э.В. О позднелетних скоплениях чаек
на северном берегу Чудского озера// Ornitol.
kogumik. Tartu, 1961. К. 2. Lk. 164-171.
Кумари Э.В, Верховые болота Эстонии как мес-
тообитания птиц//Орнитология. М.: Изд-во
МГУ, 1965. Вып. 7. С. 36-43.
Кумари Э.В. Динамика распространения сереб-
ристой чайки в Восточной Прибалтике в тече-
ние последних 125 лет// Ornitol. kogumik. Tal-
lin: Valgus, 1978. К. 9. Lk. 7-43.
Курочкин E.H., Герасимова Т.Д. Зимние и ранне-
весенние наблюдения над морскими птицами
вблизи Мурманского побережья//Тр. Канда-
лакш. гос. заповедника. Мурманск: Кн.
изд-во, 1960. Вып. 3. С. 195-198.
Курочкин Е.Н,, Скокова Н.Н. Миграция птиц
в Кандалакшских шхерах осенью 1958 года//
Тр. Кандалакш. гос. заповедника, Мурманск:
Кн. изд-во, 1960. Вып, 3. С. 133-144.
Кучин А.П. Распространение и экология чаек и
крачек на Алтае // Колониальные гнездовья
околоводных птиц и их охрана. М.: Наука,
1975. С. 81.
Кучин А.П. Птицы Алтая. Барнаул: Алт. кн.
изд-во, 1976. 231 с.
Кыдыралиев А.К. Птицы водоемов Центрально-
го Тянь-Шаня. Фрунзе: Илим, 1973. 117 с.
Кыдыралиев А.К. О гнездовых колониях озер-
ных чаек и речных крачек на озерах
Иссыккуль и Сонкуль (Тянь-Шань) // Коло-
ниальные гнездовья околоводных птиц и их
охрана. М.: Наука, 1975. С. 82.
Кыдыралиев А.К. О гнездовых колониях озер-
ной чайки на озерах Иссыккуль и Сонкуль
(Тянь-Шань) // Распространение и числен-
ность озерной чайки. М.: Наука, 1981.
С. 91-94.
Кыдыралиев А.К., Султанбаева З.Б. Наблюдения
за малоизвестными птицами Киргизии // Миг-
рации птиц в Азии. Новосибирск: Наука,
1977. С. 218-222.
Лабзюк В.И. Летняя авифауна морского побе-
режья в районе залива Ольга // Орнитологиче-
ские исследования на Дальнем Востоке. Вла-
дивосток: ДВНЦ АН СССР, 1975. С. 279 —
284.
Лабзюк В.И., Назаров Ю.Н, Нечаев В.А. Птицы
островов северо-западной части залива Пет-
ра Великого // Орнитологические исследова-
ния на юге Дальнего Востока. Владивосток:
ДВНЦ АН СССР, 1971. С. 52-78.
Лабутин Ю.В., Гермогенов Н.И., Поздняков В.И.
Серебристая чайка северотаежной части доли-
ны р. Лены // Миграции и экология птиц Си-
бири. Новосибирск: Наука, 1982. С. 112-127.
Лабутин Ю.В., Дегтярев А.Г., Блохин Ю.Ю. Пти-
цы // Растительный и животный мир дельты
реки Лены. Якутск: ЯФ АН СССР, 1985.
С. 88-110.
Лабутин Ю.В., Ларионов Г.П. О расширении
ареалов й новых видах птиц фауны Якутии //
Природные ресурсы Якутии и их охрана.
Якутск: ЯФ АН СССР, 1976. С. 46-52.
Лабутин Ю.В., Поздняков В.И, Миграции водно-
болотных птиц в долине среднего течения
р. Лены // Водно-болотные виды птиц долины
средней Лены. Якутск: ЯФАН СССР, 1978.
С. 5-28.
Лабутин Ю.В., Поздняков В.И, Гермогенов НИ,
Дегтярев А.Г. Чайки приполярных районов
долин рек Лены и Колымы // Размещение и
состояние колоний околоводных птиц на тер-
ритории СССР. М.: Наука, 1981. С. 35-36.
Лактионов А.Ф. Северная Земля. М.; Л.: Глав-
севморпуть, 1946. 150 с.
Ларионов В.Ф., Чельцов-Бебутов А.М. Находка
реликтовой чайки на Торейских озерах (За-
байкалье) // Орнитология. М.: МГУ, 1972.
Вып. 10. С. 277-279.
Ларионов Г.П, Седалищев В.Т. О весеннем про-
лете пластинчатоклювых и чаек в районе
среднего течения р. Оленек // Тез. докл.
2-й Всесоюз. конф, по миграциям птиц.
Алма-Ата: Наука, 1978. Ч. 2, С. 86-88.
Лебедев В.Д. Залет буроголовой чайки в При-
морье // Природа. 1950. № 11. С. 62.
Лебедев В.Д., Филин В.Р. Орнитологические на-
блюдения в западной Чукотке // Орнитоло-
гия. М.: Изд-во МГУ, 1959. Вып. 2. С. 122-
129.
Лебедева М.И. Миграции речной крачки // Мигра-
ции животных. М.: Изд-во АН СССР, 1962.
Вып. З.С. 87-91.
Леинь Г., Каспарсон Г. Перелет птиц на мысе
Колкасрагс в Латвийской ССР весной 1958г.
386
Литература
//Оrnitol.kogumik.Tartu, 1961.Sej. 2. Lpp.. 94 -
102.
Ленхольд B.A., Гаврилов Э.И. Сроки пролета
птиц в Караганде и ее окрестностях // Мигра-
ции птиц в Азии. Ташкент: Фан, 1978.
С. 90-97.
Леонович В.В. Новое место гнездования алеут-
ской крачки // Редкие, исчезающие и мало-
изученные птицы СССР. Рязань, 1976. С. 181 —
182. (Тр. Окского заповедника; Вып. 13).
Леонович В.В. Новые данные по распростране-
нию птиц в Магаданской области // Орнитоло-
гия. М.: Изд-во МГУ, 1981. Вып. 16.
С. 154-155.
Леонтьев А.Н. Гнездовая колония чаек и бакла-
нов на Торейских озерах // Изв. Забайк. фил,
Геогр. о-ва СССР. 1968. №6. С. 79-80.
Леонтьев А.Н, Фенология пролета птиц на Торей-
ских озерах // Охрана природы и воспроиз-
водство естественных ресурсов. Чита, 197 2.
С. 60-62. (Зап. Забайк. фил. Геогр. о-ва
СССР; Вып. 73).
Линдеман Г.В., Залетаев В.С. О периодических
изменениях видового состава чаек в гнездо-
вой колонии // Колониальные гнездовья око-
ловодных птиц и их охрана. М.: Наука,
1975. С. 84.
Линдеман Г.В., Залетаев В.С. К распределению
редких видов чаек в Западном Казахстане //
Редкие и исчезающие звери и птицы Казахста-
на. Алма-Ата: Наука, 1977. С. 199-201.
Лисецкий А.С., Кривицкий И.А.,КуниченкоА.А.,
Шурубура П.В. Заметки о некоторых редких
и исчезающих птицах Харьковской области //
Вести. Харьк. ун-та. 1978. № 164. С. 97-101.
Литвиненко НМ. Чернохвостая чайка. М.: Наука,
1980, 144 с.
Литвиненко Н.М., Шибаев Ю.В. Результаты пер-
вых лет кольцевания птенцов чернохвостой
чайки (Larus crassirostris Vieill.) на юге При-
морья // Трансконтинентальные связи пере-
летных птиц и их роль в распространении
арбовирусов. Новосибирск: Наука, 197 2.
С. 92-93.
Литвинов НА., Мирсаитова Г.И., Карабулин В.В.
Насиживание и инкубация яиц сизой чайки
в Камском Предуралье // Гнездовая жизнь
птиц. Пермь; Перм. пед. ин-т, 1978. С. 11-
16.
Литвинов НИ., Скрябин Н.Г., Сонин В.Д. Птицы
островов Малого моря // Тез. докл. 7-й Все-
союз. орнитол. конф. Киев: Наук, думка,
1977. Ч. 1. С. 81.
Литун В.И., Макаров В.А. Орнитологические
находки на территории Кировской области //
Проблемы охоты, воспроизводства и охраны
промысловых зверей и птиц. Пермь, 1984.
С. 23-26.
Лобанов В.А. Колониальные гнездовья полярной
крачки и куликов в восточной части кустар-
никовой подзоны Большеземельской тунд-
ры // Колониальные гнездовья околоводных
птиц и их охрана. М.: Наука, 1975. С. 143.
'Лобков Е.Г, Экология колоний озерных чаек на
Восточной Камчатке // Колониальные гнез-
довья околоводных птиц и их охрана. М.:
Наука, 1975. С. 85.
Лобков Е.Г. Распространение и экология кам-
чатской крачки (Sterna camtschatica) на
Камчатке // Зоол. журн. 1976. Т. 55, вып. 9.
С. 1368-1374.
Лобков Е.Г. Краткий обзор птиц центральных
районов Восточной Камчатки // Птицы и
пресмыкающиеся: Исследования по фауне
Советского Союза. М., 1978. С. 52-72. (Сб.
гр. Зоол. музея МГУ; Т. 17).
Лобков Е.Г. Материалы по миграции гагар, пога-
нок, чаек, бакланов и чистиков вдоль восточ-
ного побережья Камчатки // Экология птиц
морских побережий. М.: ЦНИИ Главохоты
РСФСР, 1980. С. 74-86.
Лобков Е.Г. Озерная чайка на Камчатке // Рас-
пространение й численность озерной чайки.
М.: Наука, 1981а. С. 105-113.
Лобков Е.Г. Фаунистические находки на севере
Камчатки // Орнитология. М.: Изд-во МГУ,
19816. Вып. 16. С. 155-156.
Лобков Е.Г. Материалы по фауне птиц Параполь-
ского дола // Там же, 1983. Вып. 18. С. 13-22.
Лобков Е.Г. Совместное гнездование камчатской
крачки (Sterna camtschatica) с речной (St.
hirundo longipennis) и полярной (St. para-
disaea) на Камчатке // Редкие и исчезающие
птицы Дальнего Востока. Владивосток:
ДВНЦ АН СССР, 1985. С. 146-147.
Лобков Е.Г., Головина Н.М. Сравнительный
очерк биологии камчатской и речной крачек
на Камчатке // Бюл. МОИП. Отд. биол. 1978.
Т. 83, вып. 6, С. 27-37.
Лобков Е.Г., Нечаев В.А. Современное состояние
популяций камчатской крачки (Sterna cam-
tschatica Pallas) на территории СССР // Тез.
докл. и стенд, сообщ. 18-го Междунар. орни-
тол. конгр. М.: Наука, 1982. С. 189-190.
Луговой А.Е. Чайковые птицы низовий дельты
Волги и примыкающих участков Северного
Каспия // Тр. Астрах, заповедника. Астрахань:
Волга, 1958. Вып. 4. С. 269-294.
Луговой А.Е. Питание чаек-ихтиофагов в ни-
зовьях дельты Волги // Там же, 1961. Вып. 5.
С. 220-231.
Луговой А.Е. Птицы дельты реки Волги // Там
же, 1963. Вып. 8. С. 9-186.
Луговой А.Е. Птицы Мордовии. Горький: Горьк.
пед. ин-т, 1975. 299 с.
Лысенко В.И. Миграции гидрофильных птиц в Се-
верном Приазовье // Материалы Всесоюз.конф
по миграциям птиц. М.: Изд-во МГУ, 1975.
Ч..2. С. 26-29.
Любаев В.Л., Кривенко В.Г. Колониальные гнез-
довья птиц на Меклетинских озерах Калмы-
кии // Размещение и состояние гнездовий око-
387
Литература
ловодных птиц на территории СССР. М.: На-
ука, 1981. С. 96-98.
ЛяйстерА.Ф., Соснин Г.В. Материалы по орнито-'
фауне Армянской ССР. Ереван: Изд-во
АФАН СССР, 1942. 402 с.
Майр Э. Зоологический вид и эволюция. М.: Мир,
1968. 597 с.
Малышев Л.И. Птицы северо-восточного побе-
режья Байкала // Первая Всесоюз. орнитол.
конф. Л.: Изд-во АН СССР, 1960. С. 81-91.
(Тр. проблемат. и темат. совещаний ЗИН
АН СССР; Т. 9).
Малышевский Р.И. Летние наблюдения над пти-
цами Терского берега Белого моря // Орнито-
логия. М.:Изд-во МГУ, 1962. Вып. 5. С. 13-
27.
Мальчевский А.С., Пукинский Ю.Б. Птицы Ле-
нинградской области и сопредельных терри-
торий. Л.: Изд-во ЛГУ, 1983, Т. 1. 480 с.
Манк А.Я. Пролет птиц в окрестностях Талли-
на//Тр. 2-й Прибалт. орнитол. конф. М.:
Изд-во АН СССР, 1957, С. 100-107.
Манк А.Я. Миграция птиц на морском побережье
Северо-Западной Эстонии осенью 1956 года//
Тр. 3-й Прибалт, орнитол. конф. Вильнюс,
1959, С. 187-195.
Манк А.Я. Новые залетные и новые гнездовые
птицы Эстонии // Материалы 7-й Прибалт,
орнитол. конф. Рига: Зинатне, 1970. Кн. 3.
С. 25-30.
Мараков С.В. Красноногая говорушка//Редкие,
исчезающие и малоизученные птицы СССР.
Рязань, 1976. С. 171-174. (Тр. Окского гос.
заповедника; Вып. 13).
Маргарян Н.А. О нахождении в Армении мор-
ского голубка // Орнитология. М.: Изд-во
МГУ, 1981. Вып. 16. С. 156.
Марисова И.В., Самофалов М.Ф. Изменения ор-
нитофауны Нежинщины за последние 50 лет
// 7-я Всесоюз. орнитол. конф.: Тез. докл.
Киев: Наук, думка, 1977. Ч. 1. С. 83.
Маркузе В.К. Особенности гнездования, числен-
ность и биомасса чайковых птиц в рыбхозах
дельты Волги // Колониальные гнездовья
околоводных птиц и их охрана. М.: Наука,
1975. С. 86-88.
Матвиенко М.Е. Осенние миграции птиц на севе-
ро-востоке Украины // 2-я Всесоюз. конф,
по миграциям птиц: Тез. сообщ. Алма-Ата:
Наука, 1978. Ч. 1. С. 140-141.
Матюшенков Н.В. Орнитофауна бассейна реки
Бикады // Науч.-техн. бюл. НИИ сельского
хозяйства Крайнего Севера. Новосибирск:
СО ВАСХНИЛ, 1979. Вып. 21. С. 33-35.
Матюшенков Н.В. Малочисленные и залетные
птицы бассейна Бикады (восточный Таймыр)
// Птицы Таймыра; (Экология, охрана и хо-
зяйственное использование). Новосибирск:
СО ВАСХНИЛ, 1983. С. 19-23.
Мачикунас А. Новое гнездовье белощекой крач-
ки в Литве // 10-я Прибалт, орнитол. конф.:
Тез. докл. Рига; Ин-т биологии АН ЛатвССР,
1981. Т. 1. С. 49-50.
Мачикунас А. Некоторые особенности гнездова-
ния болотных и водоплавающих птиц на
Каунасском водохранилище // Орнитология.
М.: Изд-во МГУ, 1982. Вып.. 17. С. 101-105.
Мекленбурцев Р.Н. Зимующие и пролетные пти-
цы Восточного Памира // Бюл. МОИП. Отд.
биол. 1946. Т. 51, № 1. С. 87-110.
Мельников Ю.И. Экология белокрылой крачки
Восточной Сибири // Экология птиц Восточ-
ной Сибири. Иркутск: Иркут, ун-т, 1977.
С. 59-92.'
Мельников Ю.И. Формирование стай и осенний
отпет белокрылой крачки на южном Байкале
// 2-я Всесоюз. конф, по миграциям птиц:
Тез. сообш. Алма-Ата: Наука, 1978. Ч. 2.
С. 102-104.
Мельников Ю.И. Новые сведения о птицах юж-
ного Байкала // Экология птиц бассейна
оз. Байкал. Иркутск: Иркут, ун-т, 1979.
С. 148-152.
Мельников Ю.И. Численность и распределение
редких и малоизученных птиц дельты р. Се-
ленги // Орнитология. М.: Изд-во МГУ, 1984.
Вып. 19. С. 58-63.
Мельников Ю.И. О гибридизации крачек // Бюл.
МОИП. Отд. биол. 1985. Т. 90, № 4. С. 32-36.
Мельников Ю.И, Лысиков С.И. О хищничестве
чайковых птиц на южном Байкале // Бюл.
МОИП. Отд. биол. 1983. Т. 88, № 5. С. 21-28.
Мельников Ю.И., Садков В.С. Материалы по био-
логии размножения речной крачки оз. Бай-
кал // Экология птиц Восточной Сибири.
Иркутск: Иркут, ун-т, 1977. С. 92-109.
Мельников Ю.И, Шинкаренко А.В., Подковы-
ров В.А., Мельников И.И. Чайки и антропо-
генный ландшафт // Материалы 3-й Всесоюз.
конф, по поведению животных. М.: Наука,
1983. Т. 3. С. 111-114. .
Мельничук В.А., Головач О.Ф. Распределение
колониальных птиц в северной части Киев-
ского водохранилища в период его стабилиза-
ции // Вести, зоологии, 1984. № 5. С. 85-86.
Мензбир М.А. Птицы России. М., 1895. Т. 1.
836 с.
Миловидов С.П., Стрелков В.Е. Предваритель-
ный список птиц Томского Приобья //
Проблемы экологии позвоночных Сибири.
Кемерово, 1978. С. 65-78.
Миловидов С.П., Стрелков В.Е., Гуреев С.П,
Рудковский В.П„ Петров С.Ю. К биологии и
распространению некоторых птиц Томского
Приобья // Вопр. зоологии Сибири. Томск:
Изд-во Том. ун-та, 1979. С. 126-130.
Минеев Ю.Н. Околоводные птицы тундр европей-
ского Северо-Востока СССР // Фауна и эколо-
гия птиц и млекопитающих европейского Се-
веро-Востока СССР. Сыктывкар: Коми
ФАН СССР, 1982. С. 36-38.
Михайлов К.Е., Фильчагов А.В. Особенности рас-
388
Литература
пространения и расселения некоторых видов
птиц в тундре Кольского полуострова // Ор-
нитология. М,: Изд-во МГУ, 1984. Вып. 19.
С. 22 -29.
Мищенко А.Л. Значение рыборазводных Прудов
для сохранения орнитофауны в условиях
антропогенного ландшафта Московской об-
ласти // Бюл. МОИП. Отд. биол. 1983. Т. 88,
вып. 4. С- 15-25.
Михель Н.М. Материалы по птицам Индигир-
ского края // Тр. Аркт. ин-та. 1935. Т. 31.
С. 1-101.
Михель Н.М. Наблюдения над птицами и млеко-
питающими района Карской губы // Изв. Гос.
геогр. о-ва. 1937. Т. 69, вып. 3. С. 443-447.
Михельсон Г.А. Наблюдение над пролетом птиц
на южном побережье Рижского залива //
Перелеты птиц в европейской части СССР.
Рига: Изд-во АН ЛатвССР, 1953. С. 141-
154.
Михтарьянц Э.А. Гнездование чистиковых птиц
на о-ве Топорков (Командоры) // Зоологи-
ческие проблемы Сибири. Новосибирск:
Наука, 1972. С. 328-330.
Модестов В.М. К экологии гнездового периода
чаек в дельте Волги // Зоол. журн. 1943.
Т. 22, № 2. С. 92-101.
Модестов В.М. Экология колониально-гнездя-
щихся птиц (по наблюдениям на Восточном
Мурмане и в дельте Волги) // Тр. Кандалакш.
заповедника. М.; Лесн. пром-сть. 1967.
Вып. 5. С. 49-154.
Молодая Г.Н., Сиренко В.А. Современное состоя-
ние гнездовий околоводных птиц северо-вос-
точного побережья Азовского моря // Разме-
щение и состояние гнездовий околоводных
птиц на территории СССР. М.: Наука, 1981.
С. 115-116.
Молчанов Л.А. Список птиц Естественно-истори-
ческого музея Таврического губернского
земства (в г. Симферополе) // Материалы
к познанию фауны и флоры Рос. империи.
Отд. зоол. 1906. Вып. 7. С. 248-301.
Молчанов Л.А., Зарудный Н.А. К авифауне Па-
мира // Ежегодник Зоол. музея Акад. наук.
СПб., 1914. Т. 19, №4. С. 439-477.
Морозов В.В. Орнитофауна окрестностей озера
Капчук, плато Путорана // Орнитология. М.:
Изд-во МГУ, 1984. Вып. 19. С. 30-40.
Москвитин С.С., Дубовик А.Д., Горд Б.Я. Птицы
долины р. Кеть // Фауна и систематика позво-
ночных Сибири. Новосибирск: Наука, 1977-
С. 245-279.
Мухина Е.Н, Гнездящиеся околоводные и водо-
плавающие птицы Арнасайского заповедни-
ка // Экология растений и животных запо-
ведников Узбекистана. Ташкент: Фан, 1983.
С. 66-82.
Мянд Р. Связь гнездового консерватизма с пока-
зателями гнездовой экологии у крачек //
Экология гнездования птиц и методы ее изу-
чения: Тез. Всесоюз. конф, молодых ученых.
Самарканд: Самарканд, ун-т, 1979. С. 151.
Мянд Р.А. Влияние естественной элиминации на
размеры и форму яиц сизой чайки // Эколо-
гия и охрана птиц. Кишинев: Штйинца, 1981.
С. 160.
НазаренкоЛ.Ф., Черничка И.И., Стойловский В.П.
Территориальные механизмы распознавания
кладок у птенцов взрослыми речными крач-
ками // 7-я Всесоюз. орнитол. конф.: Тез.
докл. Киев: Наук, думка, 1977. Ч. 2. С. 30-
32.
Наканиси Г. Острова естественных экспонатов
// Кагаку-но-дзиккен. 1953, № 10. С. 60-63;
№ 11. С. 40-46; № 12. С. 42-46. На яп. яз.
Нанкинов Д.Н. Места концентрации мигрирую-
щих водоплавающих и околоводных птиц
у болгарского побережья Черного моря //
Орнитология. М.: Изд-во МГУ, 1980. Вып. 15.
С. 36-41.
Нанкинов Д.Н. Стадии в урбанизацията и синан-
тропизацията на Българските птици Ц Орни-
тол. информ, бюл. София, 1981. № 10.
С. 9-29.
Нанкинов Д.Н. Птиците на град София // Там же,
1982. № 12. 386 с.
Нанкинов Д., Даракчиев А. Върху биологията на
белочелата рибарка (Sterna albifrons Pall.) в
България / / Науч. тр. Пловдив, ун-та
им. Паисий Хилендарски. 1980а. Т. 18, кн. 4.
С. 131-151.
Нанкинов Д., Даракчиев А. Разпространение и
екология на дебел ок люната рибарка (Geloche-
lidon nilotica Gm.) в България // Там же.
19806, Т. 18, кн. 4. С. 103-120.
Нанкинов Д., Даракчиев А. Гривестата рибарка
(Thalasseus sandvicensis Latham) отново
гнезда по българското черноморско крайбре-
жие // Там же. 1984. Т. 22, кн. 1. С. 143-147,
Наумов С.П. Млекопитающие и птицы Гыданско-
го полуострова (Северо-Западная Сибирь) //
Тр. Полярной комис. АН СССР. Л., 1931.
Вып. 4. С. 1-106.
Наумчик А.В. Распределение чайковых птиц в
Белоруссии // 7-я Всесоюз. зоогеогр. конф.:
Тез. докл. М.: Наука, 1979. С. 152-153.
Наумчик А.В, Численность и особенности геогра-
фического распределения озерной чайки в Бе-
лоруссии // Распространение и численность
озерной чайки. М.: Наука, 1981. С. 25-31.
Наумчик А.В. Чайковые птицы Белоруссии и не-
которые факторы, влияющие на их числен-
ность // Экологические исследования и охра-
на птиц Прибалтийских республик. Каунас:
Комитет ЛитССР по охране природы, 1982.
С. 62-63.
Недзинскас В.С. Колонии обыкновенной чайки
в Литве // Колониальные гнездовья около-
водных птиц и их охрана. М.: Наука, 1975.
С. 91-93.
Нейфельдт И.А. Обзор орнитологических иссле-
389
Литература
дований в Карелии // Тр. ЗИН АН СССР.
1970. Т. 47. С. 67-105.
Немцев В.В. Состояние численности некоторых
видов птиц Рыбинского водохранилища за
последние тридцать пет // Комплексное изуче-
ние и рациональное использование природных
ресурсов. Калинин. 1980. С. 192-193.
Неручев В.В. Новые данные о птицах нижней
Эмбы и приэмбинских пустынь // Орнитоло-
гия. М.: Изд-во МГУ, 1968. Вып. 9. С. 137-141.
Неручев В.В., Макаров В.И. Материалы по гнез-
довой фауне и населению птиц нижней Эмбы
// Там же, 1982. Вып. 17. С. 125-129.
Нечаев В.А. Новые данные о птицах Нижнего
Амура Ц Там же, 1963. Вып. 6. С. 177-181.
Нечаев В А. Птицы южных Курильских остро-
вов. Л.: Наука, 1969. 246 с.
Нечаев В.А. Колониально-гнездящиеся птицы
острова Монерон // Колониальные гнездовья
околоводных птиц и их охрана. М.: Наука,
1975. С. 181-183.
Нечаев В.А. Сравнительная характеристика гнез-
дования камчатской и обыкновенной крачек
на о-ве Сахалин // 7-я Всесоюз. орнитол.
конф.: Тез. докл. Киев: Наук, думка, 1977.
Ч. 1. С. 291-293.
Нечаев В.А. Распространение и биология кам-
чатской крачки Sterna camtschatica Pall, на
острове Сахалин // Редкие птицы Дальнего
Востока. Владивосток: ДВНЦ, 1981. С.47-55.
Нечаев В.А., Тимофеева А.А. О птицах острова
Ионы // Бюл. МОИП. Отд. биол. 1973. Т. 78,
вып. 1. С. 35-39.
Никифоров М.Е., Шкляров Л.П. Факторы смерт-
ности и успех размножения озерной чайки
в Центральной Белоруссии // Тез. докл. 11-й
Прибалт, орнитол. конф. Таллин: Ин-т зооло-
гии и ботаники АН ЭССР, 1983. С. 154-156.
Никлус М. Гнездование и перелеты крачки-чег-
равы в Эстонской ССР. // 3-я Прибалт, орни-
тол. конф.: Тез. докл. Вильнюс: Ин-т биоло-
гии АН ЛитССР, 1957. С. 60-61.
Николаев В.В., Кошелев А.И., Чернышев В.М.,
Тотунов В.М., Акулинин В.Н. Оологическая
и индологическая коллекция Зоологи-
ческого музея Биологического института
СО АН СССР (Новосибирск) // Фауна и
систематика позвоночных Сибири. Новоси-
бирск: Наука, 1977. С. 214-244.
Никольский А.М. Позвоночные животные Кры-
ма. СПб, 1891. 484 с.
Никольский ИД. Некоторые акустические сиг-
налы в поведении речной крачки (Sterna
hirundo) II Зоол. журн. 1975. Т. 54, № 11.
С. 1734-1737.
Никольский И.Д. Роль звуковых сигналов в ре-
гуляции группового защитного поведения
птиц: Автореф. дис. ... канд. биол. наук. М.:
Изд-во МГУ, 1978. 21 с.
Никольский ИД. Значение голосовых сигналов
в поведении пестроносой крачки // Вести,
зоологии. 1985. № 4, С. 84-87.
Носков Г.А., Гагинская Е.Р., Каменев В.М.,
Хааре А.О., Большаков КВ. Миграции птиц
в восточной части Финского залива // Сообщ.
Прибалт, комис. по изуч. миграций птиц.
Тарту: АН ЭССР, 1965. № 3. С. 3-27.
Носков Г.А., Зимин В.Б., Резвый С.П., Рымке-
вич Т.А., Лапшин Н.В., Головань В.И. Птицы
Ладожского орнитологического стационара и
его окрестностей // Экология птиц Прила-
дожья. Л.: Изд-во ЛГУ, 1981. С. 3-86.
Онно С.Х. Летние и раннеосенние передвижения
. некоторых видов водяных и прибрежных
птиц на водоемах Эстонской ССР Ц Тр. 2-й
Прибалт, орнитол. конф. М.: Изд-во АН СССР,
1957. С. 166-170.
Онно С.Х. ( Оппо S. ) Vekogudega seotud haude-
lindude levikust Eestis. О распространении
гнездовых птиц, связанных с водоемами Эс-
тонии// Ornitol. kogumik. Tartu, 1958. К. 1.
Lie. 52-79.
Онно С.Х. О гнездовом консерватизме сизой
чайки, речной и полярной крачек // Сообщ.
Прибалт, комис. по изуч. миграций птиц.
. Тарту: АН ЭССР, 1965. № 3. С. 140-162.
Онно С.Х. Смертность сизой чайки по данным
кольцевания // Там же, 1967а. № 4. С. 109—
127.
Онно С.Х. Экология гнездования сизой чайки //
Ornitol. kogumik. Tallinn, 19676, К. 4.
L.k. 144-146.
Онно С.Х. Возрастной состав и продолжитель-
ность жизни сизой чайки в Эстонии // Сообщ.
Прибалт, комис. по изуч. миграций птиц.
Тарту: АН ЭССР, 1968, № 5. С. 81-109.
Онно С.Х. Гнездовые пары у сизой чайки: (К изу-
чению экологии популяции) // Орнитология
в СССР. Ашхабад: Ин-т зоологии АН ТССР,
1969. Кн. 2. С. 468-470.
Онно С.Х. Изучение популяционной экологии
прибрежных птиц Эстонии // Птицы по двум
сторонам Финского залива. Тарту: АН ЭССР,
1970. С. 111-127.
Онно С.Х., Бугаев Л.А. Корреляция между фи-
зическими характеристиками яйца (длина,
ширина, индекс формы и вес) и возрастом
самок сизой чайки // Материалы 9-й Прибалт,
конф. Вильнюс: Ин-т зоологии и паразитоло-
гии АН ЛитССР, 1976. С. 194-196.
Онно С., Бугаев Л.А., Горяйнова Г.П. Изменчи-
вость физических характеристик яиц сизой
чайки // Тез. докл. 7-й Всесоюз. орнитол.
конф. Киев: Наук, думка, 1977. Ч. 1.
С. 294-295.
Орлов П.П. Про пйздування б!лощокого кряч-
ка (Chlidonias hybrida Pall.) в плавнях нижне-
го Днепра // Наук. зап. Мелыопол. пед. ин-ту.
Мелитополь: Пед.ин-т, 1957. Т.4. С.111-120.
Орлов П.П. Изменения в орнитофауне Нижнего
390
Литература
Днепра в районе строительства Каховского
гидроузла // Тр. НИИ биологии и биол. ф-та
Харьк. ун-та. Харьков: Изд-во Харьк. ун-та,
1959. Т. 28. С. 101-114.
Осипова М.А. К изучению линьки реликтовой
чайки (Laius relictus Lonnb.)// Веста, зооло-
гии. 1987а. №2. С. 15-17.
Осипова М.А. Состояние забайкальской популя-
ции реликтовой чайки (Larus relictus LOnnb.)
в 1982 г.//Тамже. 19876, № 1. С. 84-86.
Остапенко В.А. Авифауна дельты реки Чаун //
Биол. проблемы Севера. Магадан, 1973.
Вып. 2. С. 59-73.
Остапенко В.А., Гаврилов В.М., Ефремов В.Д.
Характер и особенности весенних миграций
птиц западной Камчатки // Материалы Все-
союз. конф, по миграциям птиц. М.: Изд-во
МГУ, 1975. Ч. 2. С. 32-35.
Остапенко В.А., Гаврилов В.М., Ефремов В.Д.,
Голубева Т.Б. О некоторых интересных на-
ходках птиц на Камчатке и о-ве Кунашир //
Орнитология, М.: Изд-во МГУ, 1977. Вып. 13.
С. 192-193.
Остапенко М.М., Кашкаров Д.Ю., Лановен-
ко Е.Н., Третьяков Г.П., Филатов А.К. Весен-
ний пролет птиц в низовьях р. Сурхандарьи //
Миграции птиц в Азии. Ташкент: Фан, 1978.
С. 5-33.
Остапенко М.М., Шерназаров 3., Гончаров Г.Ф.,
Зинченко Т.Н., Калабин С.Ю., Парамо-
нов С.Е. О пролете некоторых птиц на Айдар-
куле // Миграции птиц в Азии. Ташкент:
Фан, 1984. С. 103-113.
Очагов Д.М. О возможном гибриде Sterna albi-_
frons sinensis X Sterna hirundo из дельты Се-
ленги // Бюл, МОИП. Отд, биологии. 1982.
Т. 87, вып. 5. С. 39-42.
Павлов Б.М., Бейльман А.А., Крашевский О.Р.
К орнитофауне бассейна Верхней Таймыры //
Птицы Таймыра-. (Экология, охрана и хозяй-
ственное использование). Новосибирск:
СО ВАСХНИЛ, 1983. С. 9-14.
Павлов Б.М., Дорогое В.Ф. Розовая чайка на Тай-
мыре // Орнитология. М.: Изд-во МГУ, 1976.
Вып. 12. С. 240-241.
Панов Е.Н. Птицы Южного Приморья. Новоси-
бирск: Наука, 1973. 376 с.
Панов Е.Н., Гаузер М.Е., Зыкова Л.Ю. Катастро-
фическое воздействие переуплотнения на по-
пуляцию речной крачки Sterna hirundo// Тео-
ретические аспекты колониальности у птиц.
М.: Наука, 1985.
Панов Е.Н., Зыкова ЛЮ. Поведение хохотуньи
(Laius argentatus cachinnans) на поздних ста-
диях репродуктивного цикла // Зоол. журн.
1981. Т. 60, № 11. С. 1658-1669.
Панов Е.Н., Зыкова Л.Ю. Социально обусловлен-
ная смертность в колониях черноголового
хохотуна (Larus ichthyaetus). 2. Динамика и
размах ювенильной смертности в колониях
с различной пространственной структурой //
Зоол. журн. 1982. Т. 61, вып. 9. С. 1396-
1411.
Панов Е.Н., Зыкова Л.Ю.. Костина Г.Н., Андру-
сенко Н.Н. Социально обусловленная смерт-
ность птенцов и каннибализм в колониях
черноголового хохотуна (Larus ichthyaetus ).
1. Масштаб и причины ювенильной смертно-
сти // Там же. 1980. Т. 59, вып. И. С. 1694-
1705.
Пантелеев А.В. Гнездование клуши в городе Ле-
нинграде // Птицы и урбанизированный
ландшафт. Каунас: Ин-т зоологии и парази-
тологии АН ЛитССР, 1984. С. 109-110.
Панченко С.Г. Миграции птиц на востоке Ук-
раины // Материалы Всесоюз. конф, по миг-
рации птиц. М.: Изд-во МГУ. 1975. Ч. 2.
С. 35-36.
Паровщиков В.Л. О новых птицах Земли Алек-
сандры // Орнитология. М.: Изд-во МГУ,
1963. Вып. 6. С. 481.
Переломов Г.Б. Краткое сообщение о розовой
чайке в дельте Яны // Редкие и малоизучен-
ные птицы СССР / Тр. Окского заповедника.
Рязань, 1976. Вып. 13. С. 178-179.
Перфильев В.И. Озерная чайка в Северной Яку-
тии // Распространение и численность озерной
чайки. М.: Наука, 1981. С. 98-99.
Петрайтис А.К. Весенняя миграция птиц на побе-
режье Балтийского моря // Материалы Все-
союз. конф, по миграции птиц. М.: Изд-во
МГУ, 1975. Ч. 2. С. 36-38.
Петриныи А. Орнитофауна болота Лиелайс Ке-
меру Тирелис // Редкие растения и животные.
Рига; ЛатНИИНТИ, 1982. С. 33. На латыш, яз.
Петров В.С.. Миноранский В.А. Летняя орнито-
фауна озера Маныч-Гудило и прилежащих
степей // Орнитология. М.: Изд-во МГУ,
1962. Вып. 5. С. 266-275.
Петрович З.О. Колонии околоводных птиц в со-
ветской части дельты р. Дунай // Размещение и
состояние гнездовий околоводных птиц на
территории СССР. М.: Наука, 1981. С. 106-
109.
Пеэдосаар Ю., Онно С. Гнездовая фауна птиц на
островах Западной Эстонии // Материалы
7-й Прибалт, орнитол. конф. Рига: Зинатне,
1970. Ч. 3. С. 59-63.
Пишванов Ю.В. Некоторые данные к вопросу о
колониальных гнездовьях околоводных птиц
в Дагестане // Колониальные гнездовья око-
ловодных птиц и их охрана. М.: Наука, 1975.
С. 151-153.
Плесский ИВ. Птицы // Животный мир Киров-
ской обл. Киров: Гос. пед. ин-т, 1976. Вып.З.
С. 49-138.
Подковыркин Б.А. Материалы по биологии
размножения некоторых птиц северных Ку-
рильских островов // Зоол. журн. 1956.
Т. 35, вып. 12. С. 1892-1901.
391
Литература
Поливанова Н.Н. Птицы озера Ханка: (Охот-
ничье-промысловые водоплавающие и коло-
ниальные) // Тр. заповедника "Кедровая
Падь”. 1971. Ч. 1. С. 239.
Поливанова Н.Н., Глущенко Ю.Н. Новые данные
о некоторых редких и малочисленных птицах
Приморья // 7-я Всесоюз. орнитол, конф.:
Тез. докл. Киев: Наук, думка, 1977. Ч. 1.
С. 95-96.
Поляков Г,И. К орнитологической фауне Мос-
ковской губ. Птицы отрядов Pygopodes, Lon-
gipennes, Lamellirostres, Steganopodes // Мате-
риалы к познанию фауны и флоры Рос. импе-
рии. Отд. зоол. М., 1910. Вып. 10. С. 1-211.
Попов Ю.К. Материалы о колониальном гнездо-
вании чайковых у г. Ижевска // Колониаль-
ные гнездовья околоводных птиц и их охра-
на. М.: Наука, 1975, С. 93-94.
Попов В.Д. К экологии размножения чаек се-
верного Байкала // Тез. Всесоюз. конф, мо-
лодых. ученых. Самарканд: Самарканд, ун-т,
1979. С. 171-172.
Попов В.Д., Садков В.С. Чайки северного Байка-
ла // Экология и охрана птиц, Кишинев:
Штиинца, 1981. С. 183.
Портенко Л.А. Фауна Анадырского края. Птицы
// Тр. Ин-та поляр, земледелия, животновод-
ства и промысл, хоз-ва. Сер. Промысл, хоз-во.
1939, Вып. 5. С. 5-209; Вып. 6. С. 5-197.
Портенко Л.А. Птицы высоких широт Северного
Ледовитого океана. М.; Л.: Г лав сев мор путь,
1946. С. 21-29.
Портенко Л.А. Таксономическая оценка и систе-
матическое положение тихоокеанской мор-
ской чайки (Larus argentatus schistisagus
Stejn) // Фауна Камчатской области. М.; Л.:
Изд-во АН СССР, 1963. С. 61-64.
Портенко Л.А. Очерк фауны птиц Корякского
нагорья // Проблемы орнитологии: Тр. 3-го
Всесоюз. орнитол. конф. Львов: Изд-во
Львов, ун-та, 1964. С. 57-66.
Портенко Л.А. Птицы Чукотского полуострова и
острова Врангеля. Л.: Наука, 1973. Ч. 2.
323 с.
Пославский А.Н. К орнитологической характе-
ристике Северного Прикаспия // Орнитоло-
гия. М.: Изд-во МГУ, 1974. Вып. 11. С. 238-
252.
Пославский А.Н., Кривоносов Г.А. Экология
пестроносой крачки (Thalasseus sandvicensis
Lath.) на границе ареала // Экология. 1976.
№3. С. 51-56,
Пославский А.Н., Постников Г.Б., Самарин Е.Г.
О зимовках птиц в Северном Прикаспии и на
Мангышлаке // Охотничьи птицы Казахстана:
(Фауна, экология н практическое значение).
Алма-Ата: Изд-во АН КазССР, 1964. С. 157—
180.
Потапов Р.Л. Гнездование буроголовых чаек
(Larus brunnicephalus Jerd.) в СССР //
ДАН СССР, 1966а. Т. 167. №6. С. 1409-1410.
Потапов Р.Л. Птицы Памира // Тр. Зоол. ин-та
АН СССР. М.; Л.: Наука, 19666.Т.39.С.3-119.
Потапов Р.Л. Находка на Торейских озерах //
Природа. 1971. № 5. С. 77-81.
Пржевальский Н.М. Монголия и страна тангутов.
СПб.: Изд-во Рус. геогр. о-ва, 1875. Т. 1.
381 с.
Пржевальский Н.М. Из Зайсана через Хами в
Тибет и на верховья Желтой реки. Там же,
1883. 473 с.
Пржевальский Н.М. От Кяхты на истоки Желтой
реки, исследование северной окраины Тибета
и путь через Лоб-нор по бассейну Тарима.
Там же, 1888. 536 с.
Придатко В.И. Ледовый режим моря и миграции
розовых чаек в 1978-1980 гг. // Экология и
охрана птиц. Кишинев: Штиинца, 1981. С. 186.
Приклонский С.Г, Беме Р.Л., Успенский С.М.
Материалы по пролету птиц в дельте Ин-
дигирки // Миграции животных. М.: Изд-во
АН СССР, 1962. Вып. 3. С. 145-159.
Прокофьев С.М, О гнездовании чайковых в степ-
ной зоне Хакасии // Колониальные гнез-
довья околоводных птиц и их охрана. М.:
Наука, 1975. С. 94-95.
Прокофьев С.М. К характеристике колониаль-
ных поселений чаек в Ширинской озерно-кот-
ловинной степи (Хакасия) // Размещение и
состояние гнездовий околоводных птиц на
территории СССР. М.: Наука, 1981. С. 29-31.
Птушенко Е.С. Миграции озерной чайки Larus
ridibundus ridibundus//Tp.Центр. бюро кольце-
вания М., 1948. Вып. 7. С. 195-267.
Птушенко Е.С., Иноземцев А.А. Биология и хо-
зяйственное значение птиц Московской об-
ласти и сопредельных территорий. М.: Изд-во
МГУ, 1968. 461 с.
Пукас А.А., Мераускас П.П., Гячис В.П. Чайко-
вые птицы рыбхоза Арненис и особенности их
гнездования // Размещение и состояние гнез-
довий околоводных птиц на территории
СССР. М.: Наука, 1981. С. 6-7.
Пятрайтис А. К экологии зимующих водопла-
вающих птиц в Литве // Материалы конф, по
изуч. миграций птиц и охраны птицБалт. бас-
сейна. 1974. С. 47-49.
Равкин Ю.С. Птицы лесной зоны Приобья. Ново-
сибирск: Наука, 1978. 288 с.
Радде Г. Орнитологическая фауна Кавказа. Тиф-
лис, 1884. 450 с.
Раевский В.В. Позвоночные животные северного
Зауралья. М.: Наука, 1982. 145 с.
Разгонов С.А., Лопарев С.А., Клестов Н.Л. Зи-
мующие водоплавающие и околоводные пти-
цы урбанизированных ландшафтов Украины
// Птицы и урбанизированный ландшафт.
Каунас, 1984. С. 113-114.
Размахнина О.В. Характеристика питания чайко-
вых птиц Северного Байкала // Тез. Всесоюз.
конф, молодых ученых. Самарканд: Самар-
канд. ун-т, 1979. С, 175-177.
392
Литература
Рандла Т. О зимней орнитофауне г. Таллина //
Сообщ. Прибалт, комис. по изуч. миграции
птиц. Тарту: АН ЭССР, 1985. Вып. 17.
С. 137-141.
Распространение гнездящихся птиц Японии. Уп-
равление по делам окружающей среды.
Токио, 1981. 556 с. На яп. яз.
Раттисте К. О стратегии воспроизводства популя-
ций у сизой чайки // Экология и охрана
птиц. Кишинев: Штиинца, 1981. С. 190,
Рашкевич Н.А. Колония рыбоядных птиц на юж-
ном берегу Аральского моря // Орнитология
в СССР. Ашхабад: Ин-т зоологии АН ТССР,
1969. Кн. 2. С. 530-533.
Резанов А.Г. Докармливание молодых крачек
в период осенней миграции // 2-я Всесоюз.
конф, по миграции птиц: Тез. докл. Алма-Ата:
Наука, 1978. Ч. 1. С. 56-57.
Ренно О.Я. Список птиц Матсалуского госзапо-
ведника // Тр. гос. заповедников ЭССР. Тал-
лин: Валгус, 1968. Т. 1. С. 139-150.
Ренно О.Я. К гнездовой орнитофауне островов
южной полосы Финского залива // Тез. докл.
8-й Прибалт, орнитол. конф. Таллин: Ин-т
зоологии и ботаники АН ЭССР, 1972.
С. 96-97.
Ренно О. Авифаунистические исследования в
Эстонской ССР Ц Материалы 9-й Прибалт,
орнитол. конф. Вильнюс: Ин-т зоологии и
паразитологии АН ЛитССР, 1976. С. 256-227.
Рогачева Э.В., Вахрушев А.А. Фауна и население
птиц енисейской северной тайги // Животный
мир енисейской тайги и лесотундры и природ-
ная зональность. М.: Наука, 1983. С. 47-106.
Рогачева Э.В., Равкин Е.С., Сыроечковский Е.Е.,
Кузнецов Е.А. фауна и население птиц енисей-
ской лесотундры // Животный мир енисей-
ской тайги и лесотундры и природная зональ-
ность. М.: Наука, 1983. С. 14-47.
Рогачева Э.В., Сыроечковский Е.Е., Бур-
ский О.В., Ангизитова Н.В., Готфрид А.Б.
Птицы среднетаежного Енисея: фауна, ее
охрана и рациональное использование //Охра-
на фауны Крайнего Севера и ее рациональное
использование. М.: ЦНИЛОП МСХ СССР,
1978. С. 30-165.
Роотсмяе Л.Т. Осенняя миграция водяных
и прибрежных птиц на северо-западном
побережье Чудского озера в 1954-1956 гг.//
Тр. 3-й Прибалт, орнитол. конф. Вильнюс,
1959. С. 253-259.
Росляков Г.Е. Водоплавающие и околоводные
птицы Нижнего Приамурья и их участие
в циркуляции арбо- имиксовирусов: Дис. ...
канд. биол, наук. Хабаровск, 1977. 178 с.
Росляков Г.Е. Колониальные поселения бело-
крылой крачки на озерах Эворон и Чукча-
гирское // Орнитология. М.: Изд-во МГУ,
1979. Вып. 14. С. 196.
Росляков Г.Е, Краткие сведения о некоторых
редких и малоизученных птицах нижнего
Приамурья //Редкие птицы Дальнего Востока.
Владивосток: ДВНЦ АН СССР, 1981. С. 112-
115.
Росляков Г.Е., Рослая И.Г. Озерная чайка на
Нижнем Амуре // Распространение и числен-
ность озерной чайки, М.: Наука, 1981.
С. 102-105.
Рошевский Ю.К. Динамика группового эффекта
в гнездовом поведении речной крачки //
Материалы 6-й Всесоюз. орнитол. конф.
М.: Изд-во МГУ, 1974. Ч. 1. С. 166,
Рошевский Ю.К. Краткое сообщение о розовой
чайке в Яно-Индигирской низменности //
Редкие, исчезающие и малоизученные птицы
СССР // Тр. Окского заповедника. Рязань.
1976. Вып. 13. С. 177-178.
Рошевский Ю.К. Формула соседских взаимоот-
ношений у настоящих крачек //7-я Всесоюз.
орнитол. конф.: Тез. докл. Киев: Наук,
думка, 1977. Ч. 2. С. 34-35.
Рошевский Ю.К. Особенности группового пове-
дения животных. Куйбышев: Куйбышев,
ун-т, 1978. 99 с.
Рошевский Ю.К., Лаухина Л.Н. Влияние плот-
ности населения на размер кладки у настоя-
щих крачек // Орнитология, М.: Изд-во
МГУ, 1980. Вып. 15. С, 138-141.
Рощевский ЮГС., Маслов С.К. Население крачек
р. Самары в Куйбышевской обл. // Научные
основы обследования колониальных гнез-
довий околоводных птиц. М.: Наука, 1981,
С. 110-111.
Рузский М.Д, О морском голубке и о других
чайках на курорте ’’Карачинское озеро” //
Изв. Том. ун-та. Биология. Отд. оттиск.
1926. Т. 77, вып. 2. С. 113-114.
Рузский М.Д. Материалы по фауне курорта
Карачи. О залетных и редко или периоди-
чески гнездящихся птицах Карачинского
курорта и его окрестностей // Тр. Том.
ун-та. Биология. 1932. Т. 85 С 150—
159.
Рутилевский Г.Л. Птицы острова Большого
Ляховского// Пробл. Арктики. 1958. Вып. 4.
С. 79-90.
Рысьева Т.Г., Стремоусова Н.А., Третьякова Е.Ю.
К биологии размножения озерной чайки
в окрестностях города Ижевска // Отраже-
ние достижений орнитологической науки в
учебном процессе средней школы и вузов
и народном хозяйстве, Пермь: Перм. пед.
ин-т, 1984. С. 100-101.
Савельев А.П,, Чарушина АН. Авифаунисти-
ческие изменения на искусственных водое-
мах Кировской области // Там же. С. 101 —
103.
Савицкий Б.П., Пославский А.Н. О залете сред-
него поморника в Северо-Западный Прикас-
пий // Орнитология. М.: Изд-во МГУ, 1977.
Вып. 13. С. 194.
Савченко А.П. Заметки о птицах оз. Убсу-Нур //
393 •
Литература
Птицы Сибири. Горно-Алтайск: Горно-Алт.
пед. ин-т, 1983, С. 100.
Сагитов А.К., Салимов Х.В. К экологии крачек
в пойме реки Зеравшан // Колониальные
гнездовья околоводных птиц и их охрана.
М.: Наука, 1975. С. 95.
Садков В.С. Материалы по экологии озерной
и малой чаек Северного Байкала и сведения
о залетах чайковых птиц // Экология птиц
Восточной Сибири. Иркутск: Иркут, ун-т,
1977, С. 109-128.
Салури Р. О гнездовой фауне птиц морских
островков западной Эстонии // 2-я Всесоюз.
орнитол. конф.: Тез. докл. М.: Изд-во МГУ,
1959. Ч. 2. С. 76-77.
Самородов А.В., Самородов ЮА. О встрече
зимой на восточном побережье Каспийского
моря черной крачки // Орнитология. М.:
Изд-во МГУ, 1972. Вып. 10. С. 387.
Самородов ЮА. Биология и экология серебри-
стой чайки-хохотуньи, гнездящейся в Се-
верном Казахстане // Вести, МГУ. Сер. 16,
Биология. 1970. Вып. 6. С. 22-28.
Самородов ЮА. Ночное поведение некоторых
чайковых Кустанайской области // Там же.
1971. Вып. 6. С. 22-26,
Самородов ЮА. Ночное поведение чаек // При-
рода. 1973. № 1. С. 88-90.
Самородов ЮА. Динамика численности чайко-
вых птиц на озере Жарколь (Наурзумский
заповедник) // Колониальные гнездовья око-
ловодных птиц и их охрана. М.: Наука, 1975.
С. 97-99.
Самородов ЮА. О питании черной, белокрылой,
чайконосой и речной крачек Тургайской до-
лины (Северный Казахстан). М., 1976. 15 с.
Деп. в ВИНИТИ, № 940-76.
Самородов ЮА. К экологии западносибирской
сизой чайки Larus canus heinei Homeyer,
гнездящейся на озерах Наурзумского за-
поведника Кустанайской области // Проб-
лемы экологии позвоночных Сибири. Кеме-
рово, 1978а. С. 3-36.
Самородов ЮА. О питании малой чайки на во-
доемах Тургайской ложбины. М., 19786.
5 с. Деп. в ВИНИТИ, № 526-78.
Самородов ЮА. Экология озерной чайки Наур-
зумского заповедника (Северный Казах-
стан) // Экология птиц бассейна оз. Байкал.
Иркутск: Иркут, ун-т, 1979. С. 101-
125.
Самородов ЮА. О новом гнездовании сизой
чайки на Храпуновских болотах Московской
области // Размещение и состояние колоний
околоводных птиц на территории СССР.
М,: Наука, 1981а. С. 9-11.
Самородов ЮА. О совместном гнездовании
малых крачек и зуйков на Днепре // Орнито-
логия. М.: Изд-во МГУ, 19816. Вып. 16.
С. 182-183.
Самородов ЮА. Изменение веса яиц чайковых
птиц Наурзумского заповедника // Там же,
1982. Вып. 17. С. 188-189.
Самородов ЮА. Изменение веса яиц во время
насиживания и рост птенцов чайковых птиц
в Северном Казахстане // Наземные и водные
экосистемы. Горький: Горьк. ун-т, 1983.
Вып. 6. С. 35-36.
Самородов Ю.А., Рябов В.Ф. К экологии сереб-
ристой чайки (Larus argentatus Pont) Куста-
найской области // Вести. МГУ. Сер. 16,
Биология. 1968. Вып. 4. С. 99-102.
Самородов ЮА., Рябов В.Ф. Питание черного-
лового хохотуна Larus ichthyaetus Pall. Куста-
найской области // Там же. 1971, Вып. 2.
С. 29-33.
Самородов Ю.А., Рябов В.Ф. О черноголовом
хохотуне в Кустанайской области // Орнито-
логия. М: Изд-во МГУ, 1972. Вып. 10. С. 266-
276.
Самородов ЮА., Самородов А.В. Зимующие
птицы Калмыцкого побережья Каспия //
Вести, МГУ. Сер. 16, Биология. 1969а, Вып. 4,
С. 31-33.
Самородов Ю.А., Самородов А.В. Зимующие
птицы Калмыкии //Тамже. 19696. Вып. 6.
С. 39-43.
Самусев И.Ф. К гнездованию чайковых в Восточ-
ном Казахстане // Колониальные гнездовья
околоводных птиц и их охрана. М.: Наука,
1975. С. 99-100.
Сапетина ИМ. Результаты двухлетнего кольце-
вания озерной чайки в Ивановской области //
Тез. докл. 2-й Всесоюз. орнитол. конф.
М„ 1959. Вып. 2. С. 89-90.
Сапетина ИМ. Результаты трехлетнего кольце-
вания озерной чайки (Larus ridibundus) в
Ивановской и Рязанской обл. // Тр. Окско-
го заповедника, Вологда, 1962. Вып. 4.
С. 193-224.
Сдобников ВМ. Материалы по фауне и экологии
птиц Ленско-Хатангского края (по сборам
и наблюдениям А.А. Романова) // Тр. Ин-та
биологии ЯФ АН СССР. 1959. Т. 6. С. 119-
143.
Сдобников ВМ. О причинах перерывов в гнездо-
вании у арктических птиц // Зоол. журн.
1971. Т. 50, вып. 5. С. 734-740.
Северцов Н.А. Заметка о фауне позвоночных
Памира // Зап. Туркест. отд-ния О-ва люби-
телей естествознания, антропологии и этно-
графии. 1879. Т. 1, вып. 1. С. 58-89.
Сема AM., Гаврилов Э.И., Ауэзов Э.М. Мигра-
ции чегравы по материалам кольцевания в
Казахстане // Миграции птиц в Азии. Новоси-
бирск: Наука, 1986. С. 174-182.
Семаго Л.Л. Колониальные чайки н цапли в бас-
сейне Среднего Дона // Колониальные гнез-
довья околоводных птиц и их охрана. М.:
Наука, 1975. С. 156-157.
Семаго Л.Л., Уварова Т.Б., Шевцов А. Орнито-
логическая обстановка на Воронежском во-
394
Литература
дохранилище летом 1972 года // Материалы
6-й Всесоюз. орнитол. конф. М.: Изд-во
МГУ, 1974. Ч. 2. С. 132 -134.
Семенов С.М., Сабиневский Б.В. Сезонное
размещение и миграции морского голубка
(Laius genei Вт.) по данным кольцевания и
визуальных наблюдений // Тр. Бюро кольце-
вания. М.: МСХ СССР, 1957. Вып. 9. с. 86-
132,
Сеницкий А.И. Птицы Тархан-Сунака. Симфе-
рополь, 1898. 99 с.
Синкявичюс С., Палтинавичюс С. Новый вид в
орнитофауне Литвы // Орнитология. М.:
Изд-во МГУ, 1982. Вып. 17. С. 173.
Сиохин ВД. О межвидовых отношениях пест-
роносой крачки ( Sterna . sandvicensis Lath.) в
смешанных колониях // Вести, зоологии.
1978. Вып. 3. С. 73-74.
Сиохин ВД. Экология чайковых птиц северо-
западного побережья Азовского моря и
Сиваша: Автореф. дис. ... канд. биол. наук.
Днепропетровск, 1980. 24 с.
Сиохин ВД. Распределение и численность чай-
ковых птиц на северном побережье Азовско-
го моря и Сиваше // Размещение и состоя-
ние гнездовий околоводных птиц на терри-
тории СССР. М.: Наука, 1981а. С. 17-20.
Сиохин ВД. Трофические связи чайковых птиц
в наземных и водных экосистемах Приси-
вашья // Эколого-морфологические особен-
ности животных и среда их обитания. Киев:
Наук, думка, 1981. С. 61-63.
Сиохин ВД, Скрипка Г.С. О некоторых связях
колоний речных крачек ( Sterna hirundo L.)
с растительными ассоциациями островов
// Веста, зоологии. 1978. Вып. 1. С. 77-79.
Скалинов С.В. Ранневесенние, наблюдения над
птицами в северной части и Горле Белого
моря // Тр. Кандалакш. заповедника. Мур-
манск: Кн. изд-во, 1960. Вып. 3. С. 191-194.
Скалой В.Н., Слудский А.А. Птицы Елогуйско-
Тазовского бассейна // Природа и соц. хоз-во.
1941. Т. 8, ч. 2. С. 421-434.
Скокова Н.Н. Пролет птиц осенью 1958 г. в
окрестностях с. Поньгомы // Тр. Кандалакш.
заповедника. Мурманск: Кн. изд-во, 1960.
Вып. 3. С. 152-174.
Скрылева Л.Ф. Морфологическая разнокачест-
венность яиц сизой чайки // Гнездовая жизнь
птиц. Пермь: Перм. пед. ин-т, 1977. С. 41-45.
Скрябин НГ. К орнитофауне Прибайкалья //
Орнитология. М.: Изд-во МГУ, 1967. Вып. 8.
С. 386-387.
Скрябин Н.Г. Морфофизиологическая характе-
ристика чайковых птиц // Экология птиц
Восточной Сибири. Иркутск: Иркут, ун-т,
1977а. С. 128-176.
Скрябин Н.Г. Экология серебристой и сизой
чаек на Байкале // Там же, 19776. С. 4-36.
Скрябин Н.Г. Околоводные птицы экосистем
крупнейших озер Азии (на примере оз. Бай-
кал, СССР и оз. Хубсугул, МНР) // 18-й
Междунар. орнитол. конгр.: Тез. докл. и
стенд, сообщ. М.: Наука, 1982. С. 233.
Скрябин Н.Г., Мельников Ю.И., Садков В.С.
Численность и распределение чаек и крачек,
гнездящихся на Байкале // Тез. докл. 7-й
Всесоюз. орнитол. конф. Киев: Наук, думка,
1977. Ч. 1. С. 104-105.
Скрябин Н.Г., Размахнина О.В. Питание чаек
н крачек Байкала // Роль птиц в биоценозах
Восточной Сибири. Иркутск: Иркут, ун-т,
1978. С. 4-52.
Скрябин Н.Г., Размахнина О.В. Роль основных
кормов в питании чаек и крачек Байкала //
Экология птиц бассейна озера Байкал. Ир-
кутск: Иркут, ун-т, 1979. С. 77—90.
Скрябин Н.Г, Садов В.С., Пыжьянов С.В.,
Гилевич А.Л., Сафронов Н.Н, Попов В.Д.,
Мельников Ю.И. Результаты кольцевания
чаек и крачек на Байкале // Экология и
охрана птиц. Кишинев: Штиинца, 1981. С. 209.
Скрябин Н.Г., Шароглазов В.И. К эколого-
морфологической характеристике серебри-
стой чайки на озере Байкал // Материалы
6-й Всесоюз. орнитол. конф. М.: Изд-во
МГУ, 1974. Ч. 2. С. 135-136.
Слепцов ММ. Метод изучения интенсивности
размножения птиц по яичникам // Охрана
природы. М.: ВООП, 1948. Вып. 5. С. 119-
129.
Смирнов НА. Весенние орнитологические наб-
людения в Горле Белого моря // ДАН СССР.
1926, Сер. А. С. 32-34.
Смогоржевський Л.О. Рибощш птахи Украши.
Кигв: Вид-во Киш. ун-ту, 1959. 122 с.
Смогоржевский Л.А. Пролет чеграв на терри-
тории Украины // Вести, зоологии. 1970.
Вып. 4. С. 86-87.
Солдатенко И.С. К экологии чайковых о-ва Ки-
тай // Тр. Одес. ун-та. 1956. Т. 146, вып. 4.
С. 119-123.
Сомов Н.Н. Орнитологическая фауна Харьков-
ской губернии. Харьков: Тип. А. Дарре,
1897. 680 с.
Спангенберг Е.П. Состав авифауны острова
Харлова и прилегающего побережья в гнездо-
вой период 1932 г. // Тр. заповедника ’’Семь
островов”. М.: Гл. упр. по заповедникам,
1941. Вып. 1. С. 74-84.
Спангенберг Е.П. О птицах низовьев Колымы //
Орнитология. М.: Изд-во МГУ, 1960. Вып. 3.
С. 106-111.
Спангенберг Е.П., Леонович В.В. Птицы северо-
восточного побережья Белого моря // Тр.
Кандалакш. заповедника. Мурманск: Кн.
изд-во, 1960. Вып. 2. С. 213-336,
Спангенберг Е.П., Олигер И.М. Орнитологи-
ческие исследования в Дарвиновском запо-
веднике в 1946 и 1947 годах // Тр. Дарвинов,
заповедника. 1949. Вып. 1. С. 245-
302.
395
Литература
Спангенберг Е.П., Фейгин Г.А. Птицы нижней
Сырдарьи и прилежащих районов // Сб.
тр. Зоол. музея при МГУ. М.: Биомедгиз,
1936. Вып. 3. С. 41—184.
Спасский Н.Н. Список птиц Кольского залива
по работам 1922-1923 гг. // Работы Мурман,
биол. станции. 1925. Т. 1. С. 55-90.
Сребродольская Н.И. Колониальные гнездования
черных крачек в Волынском Полесье //
Колониальные гнездовья околоводных птиц
и их охрана. М.: Наука, 1975. С. 101.
Сребродольская Н.И., Мисюк Т.В., Татух С.И.,
Штыркало Я.В. Околоводные колониальные
птицы Западноукраинского Полесья // Раз-
мещение и состояние гнездовий околоводных
птиц на территории СССР. М.: Наука, 1981.
С. 103-104.
Станявичюс В. Весенний прилет птиц в заповед-
ник Жувинтас // Тез. докл. 11-й Прибалт,
орнитол. конф. Таллин: Ин-т зоологии и
ботаники АН ЭССР, 1983. С. 204-206.
Стахеев В.А. Состояние колониальных гнездо-
вий околоводных птиц в Алтайском запо-
веднике // Размещение и состояние гнездо-
вий околоводных птиц на территории СССР.
М.: Наука, 1981. С. 131-132.
Степанян Л.С. Состав и распределение птиц
фауны СССР: Неворобьиные Non-Passetifor-
mes. М.: Наука, 1975. 370 с.
Степанян Л.С. Надвиды и виды-двойники в
авифауне СССР. М.: Наука, 1983. 294 с.
Степанян Л.С., Болд А. Материалы по гнездо-
вой экологии птиц Тувинской АССР и Мон-
гольской народной республики // Орнито-
логия. М.: Изд-во МГУ, 1983. Вып. 18.
С. 33-39.
Степанян Л.С., Галушин ВМ. Материалы по
авифауне заповедника Барса-Кельмес // Там
же, 1962. Вып. 4. С. 200-207.
Стишов М.С., Придатке В.И., Баранюк В.В.
Новые материалы о птицах острова Вранге-
ля // Бюл. МОИП. Отд. биол. 1985. Т. 90,
вып. 2. С. 42-48.
Стойловский В.П. Фенология пролета речной
крачки в низовьях Тилигульского лимана //
Экология и охрана птиц. Кишинев : Штиинца,
1981, С. 214.
Стойловский В.П. Трансконтинентальные мигра-
ции речных крачек северо-западного Причер-
номорья // Кольцевание и мечение птиц
в СССР, 1979-1980 годы. М.: Наука, 1986.
С. 21-23.
Страздс АД, Гнездовые колонии чайковых птиц
в Рижском и Лимбажском районах Латвий-
ской ССР // 10-я Прибалт, орнитол. конф.
Рига: Ин-т биологии АН ЛатвССР, 1981.
Т. 1. С. 65-68.
Страздс А., Страздс М. Орнитофауна водно-
болотного угодья Даугавгрива (г. Рига) //
Экологическое исследование и охрана птиц
Прибалтийских республик: Тез. докл. При-
балт. конф, молодых орнитологов. Каунас,
1982. С. 170-171.
Страздс М„ Страздс А. Изменения орнитофауны
г. Риги за период 1940-1982 гг. // Тез. докл.
9-й Прибалт, орнитол. конф. Таллин: Ин-т
зоологии и ботаники АН ЭССР, 1983.
С. 43-44.
Страутман Ф.И. Птицы западных областей УССР.
Львов: Изд-во Львов, ун-та, 1963. Т. 1. 200 с.
Стрелков В.Е. К экологии размножения кули-
ков и чаек в бассейне Средней Оби // Мате-
риалы 6-й Всесоюз. орнитол. конф. М.: Изд-во
МГУ, 1974а. Ч. 2. С. 141-143.
Стрелков В.К. Распространение куликов и
чаек в бассейне Средней Оби // Тр. НИИББ
при ТГУ. Томск: Изд-во Том. ун-та, 19746,
Т. 4. С. 31-37.
Строков В.В. Зимовки водоплавающих птиц у
Черноморских берегов Кавказа // Орнито-
логия. М.: Изд-во МГУ, 1974. Вып. 11.
С. 274-278.
Сулик В.Г., Енакиев С.В. Пролет птиц в Луган-
ском госзаповеднике // 2-я Всесоюз. конф,
по миграциям птиц: Тез. сообщ. Алма-Ата:
Наука, 1978. Ч. 1. С. 165.
Сулимов АД. О колониальных околоводных
птицах Омского Прииртышья // Размещение
и состояние гнездовий околоводных птиц на
территории СССР. М.: Наука, 1981. С. 129-
130.
Сурвилло А.В. О гнездовании малой чайки и
белокрылой крачки в Зайсанской котлови-
не // Новости орнитологии Казахстана, Алма-
Ата: Наука, 1968. С. 220-221.
Сухинин А.Н. Видовой состав птиц Бадхыза//Ох-
рана природы Туркменистана. Ашхабад:
Ылым, 1979. Вып. 5. С. 36-45.
Сушкин П.П. Птицы Средней Киргизской сте-
пи//Материалы к познанию фауны и фло-
ры Рос. империи. Отд. зоологии. М., 1908.
Вып. 8. 803 с.
Сушкин П.П. Птицы Советского Алтая и при-
лежащих частей северо-западной Монголии.
М.; Л.: Изд-во АН СССР, 1938. 316 с.
Сыроечковский Е.Е., Рогачева Э.В. Животный
мир Красноярского края. Красноярск: Кн.
изд-во, 1980. 359 с.
Талпош В.С. Черная крачка в Закарпатской об-
ласти//Колониальные гнездовья околовод-
ных птиц и их охрана. М.: Наука, 1975.
С. 101-102.
Талпош В.С. О гнездовании крачки белощекой
(Chlidonias hybrida Pall.) на западе УССР //
Вести.’ зоологии. 1977. Вып. 4. С. 83-86.
Таманцева Л.С. Сезонные миграции крачек//Тр.
Бюро кольцевания. М.: МСХ СССР, 1955.
Вып. 8. С. 91-100.
Татаринкова И.П. Результаты кольцевания боль-
ших морских и серебристых чаек на Мурма-
не//Тр. Кандалакш. заповедника. Мурманск:
Кн. изд-во, 1970. Вып. 8. С. 149-181.
396
Литература
Татаринкова И.П. Влияние крупных чаек на раз-
мещение колоний некоторых колониальных
птиц//Колониальные гнездовья околоводных
птиц и их охрана. М.: Наука, 1975. С. 24-25.
Татаринкова И.П. Особенности экологии круп-
ных чаек на Айновых островах как возмож-
ный механизм регуляции численности попу-
ляции//Научные основы обследования коло-
ниальных гнездовий околоводных птиц. М.:
Наука, 1981. С. 113-114.
Тауриныи Э.Я., Вилкс К.А., Михельсон Г.А.
Сезонное размещение и миграция некото-
рых видов птиц по данным кольцевания
орнитологической станции Академии наук
Латвийской ССР//Перелеты птиц в европейс-
кой части СССР. Рига: Изд-во АН ЛатвССР,
1953. С. 57-102.
Ташлиев А.О., Сухинин А.Н., Бельская Г.С.,
Кекилова А.Ф., Эминов А. Птицы культурно-
го ландшафта. Ашхабад: Ылым, 1967. 140 с.
Теплое В.П., Карпович В.Н., Приклонский С.Г.,
Сапетин Я.В. Сравнительная характеристика
пролета птиц весной и осенью 1956 г. в райо-
не среднего течения р. Оки и в устье р. Бе-
лой//Тр. 3-й Прибалт, орнитол. конф. Виль-
нюс, 1959. С. 295-304.
Теплова Е.Н. Птицы района Печоро-Илычского
заповедника//Тр. Печоро-Илыч. заповедника.
Сыктывкар, 1957. Вып. 6. С. 5-115.
Тима Ч.Б. Состав корма чайковых Латвии//Эко-
логия и миграции птиц Прибалтики. Рига:
Изд-во АН ЛатвССР, 1961. С. 141-146.
Тимофеев-Ресовский Н.В., Штреземан Е. Видо-
образование в цепи подвидов настоящих
чаек группы серебристая-хохотунья-клу-
ша//Тр. Урал. отд. МОИП. Свердловск, 1959.
Вып. 2. С. 99-115.
Тинберген Н. Мир серебристой чайки. М.: Мир,
1974. 272 с.
Титавнин А.П. Распространение чайковых и ант-
ропогенное влияние на них в Куйбышевской
области//Размещение и состояние гнездовий
околоводных птиц на территории СССР. М.:
Наука, 1981. С. 13-15.
Тихонов А.В., Фокин С.Ю. Защитные реакции
ржанкообразных птиц в гнездовой период//
Орнитология. М.: Изд-во МГУ, 1982. Вып. 17.
С. 54-65.
Толмачев А.И. К авифауне острова Колгуе-
ва//Ежегодник Зоол. музея АН СССР. Л.,
1927. Т. 28, вып. 3. С. 355-365.
Томкович П.С. Необычное гнездование розовых
чаек//Орнитология. М.: Изд-во МГУ, 1980.
Вып. 15. С. 215.
Томкович П.С. Птицы острова Греэм-Белл,
Земля Франца-Иосифа//Там же, 1984. Вып. 19.
С. 13-21.
Томкович П.С. Материалы по биологии белой
чайки на о-ве Греэм-Белл (Земля Франца-
Иосифа) //Актуальные проблемы орнитоло-
гии. М.: Наука, 1986. С. 34-39.
Томкович П.С., Морозов В.В. Фаунистические
находки на востоке Чукотского полуостро-
ва//Орнитология. М.: Изд-во МГУ, 1982.
Вып. 17. С. 173-175.
Томкович П.С., Сорокин А.Г. Фауна птиц Восточ-
ной Чукотки//Распространение и систематика
птиц. М.; Изд-во МГУ, 1983. С. 77-
159.
Третьяков А.В. Орнитофауна Калининской об-
ласти//Учен. зап. Калинин, пед. ин-та. Кали-
нин: Калинин, пед. ин-т, 1940. Т. 9, вып. 11.
С. 3-58.
Трухин А.М., Косыгин Г.М. Распределение морс-
ких птиц во льдах Охотского моря в зимний
период//Морские птицы Дальнего Востока.
Владивосток: ДВНЦ АН СССР, 1986.
С. 48-56.
Туаев Д.Г. Материалы по фауне и экологии
водоплавающих и болотных птиц Кура-Ара-
ксинской низменности в современных усло-
виях//Материалы по фауне и экологии назем-
ных позвоночных Азербайджана. Баку: Элм,
1975. С. 151-188.
Туаев Д.Г., Васильев В.И. Колониально-гнездя-
щиеся птицы водоемов Азербайджана//Орни-
тология в СССР. Ашхабад: Ин-т зоологии
АН ТССР, 1969. Кн. 2. С. 649-652.
Туаев Д.Г., Васильев В.И. К видовому составу
и биологии птиц Мингечаурского и Варва-
ринского водохранилищ//Орнитология. М.:
Изд-во МГУ, 1972. Вып. 10. С. 260-265.
Туаев Д.Г., Васильев В.И, Абушев Ф.А. О слу-
чае массовой гибели серебристой чайки в
гнездовой колонии на острове Глиняный
Азербайджанской ССР//Там же. С. 394-395.
Тугаринов А.Я., Бутурлин С.А. Материалы по
птицам Енисейской губернии. Красноярск:
Тип. М.И. Абалакова, 1911. 440 с.
Тугаринов А.Я., Козлова-Пушкарева Е.В. Зимов-
ка птиц в Талыше//Тр. Азерб. фил. АН СССР.
Баку, 1935. Т. 23. С. 1-119.
Тугаринов А.Я., Толмачев А.И. Материалы для
авифауны Восточного Таймыра//Тр. Поляр,
комис. АН СССР. 1934. Вып. 16. С. 1-47.
Уилсон М. Орнитологические наблюдения на
Минском море//Орнитология. М.: Изд-во
МГУ, 1977. Вып. 13. С. 195-196.
Умрихина Г.С. Колонии черных крачек в Чуйс-
кой долине//Колониальные гнездовья около-
водных птиц и их охрана. М.: Наука, 1975.
С. 103-104.
Успенский В.С. Птицы заповедника ’’Семь ост-
ровов”//Тр. заповедника ’’Семь островов”.
М.: Гл. упр. по заповедникам, 1941. Вып. 1.
С. 5-46.
Успенский С.М. Применение массового кольце-
вания, маркирования и учета численности
морских колониальных птиц при изучении
их экологии//Тр. 2-й Прибалт, орнитол.
конф. М.: Изд-во АН СССР, 1957. С. 349-354.
Успенский С.М. Некоторые виды птиц на севе-
397
Литература
ро-востоке европейской части СССР//Учен,
зап. МГУ. 1958. Вып. 197. С. 35 48.
Успенский С.М. Широтная зональность авифау-
ны Арктики//Орнитология. М.: Изд-во МГУ,
1960. Вып. 3. С. 55-70.
Успенский С.М. Птицы востока Болыпеземельс-
кой тундры, Югорского полуострова и остро-
ва Вайгач//Тр. УФ АН СССР. Свердловск,
1965. Т. 38. С. 65-102.
Успенский С.М. Жизнь в высоких широтах:
На примере птиц. М.: Мысль, 1969. 463 с.
Успенский С.М. К авифауне Земли Франца-Иоси,-
фа//Орнитология. М.: Изд-во МГУ, 1972.
Вып. 10. С. 123-129.
Успенский С.М. Загадка розовой чайки//При-
рода. 1973. №8. С. 102-106.
Успенский С.М., Беме Р.Л., Велижанин А.Г.
Авифауна острова Врангеля//Орнитология.
М.: Изд-во МГУ, 1963. Вып. 6. С. 58-67.
Успенский С.М., Беме Р.Л., Приклонский С.Г.,
Bexoe Н.В. Птицы Северо-Востока Якутии//
Там же, 1962. Вып. 4. С. 64-86.
Федоренко А.П. К вопросу о зимовках птиц в
районе дельты Дуная//Тр. 3-й Прибалт,
орнитол. конф. Вильнюс, 1959. С. 317-328.
Федюшин А.В., Долби к М.С. Птицы Белоруссии.
Минск: Наука и техника, 1967. 520 с.
Филонов К.П., Лысенко В.И. О гнездовании
морского голубка и пестроносой крачки на
Молочном лимане//Науч. докл. высш. шк.
Биол. науки. 1971. Вып. 7. С. 31-33.
Филонов К.П., Лысенко В.И., Сиохин В.Д. Осо-
бенности гнездования куликов и чаек на ост-
ровах Молочного лимана//Вестн. зоологии.
1974. Вып. 5. С. 52-58.
Фирсова Л.В. Возрастные наряды серокрылой
. чайки Larus glaucenscens Naum, и тихоокеанс-
кой морской чайки Larus schistisagus Stejneg.//
Тр. Биол.-почв. ин-та ДВНЦ АН СССР. 1975а.
Т. 29(132). С. 40-62.
Фирсова Л.В. Гнездование моевок Rissa tridac-
tyla (L.) и Rissa brevirostris (Bruch) на Коман-
дорских островах//Тез. докл. отчет, науч,
сес. Зоол. ин-та АН СССР. Л.: ЗИН АН СССР.
19756. С. 21.
Фирсова Л.В. Красноногая моевка и серокрьшая
чайка - виды фауны СССР, нуждающиеся в
особой охране//Тез. докл. 8-й Всесоюз.
орнитол. конф. Киев: Наук, думка, 1977.
Ч. 2. С. 248-249.
Фирсова Л.В. Гнездование красноногой и обык-
новенной моевок Rissa brevirostris (Bruch),
Rissa tridactyla (Linnaeus) на Командорских
островах//Систематика и биопогия редких и
малоизученных птиц. Л.: ЗИН АН СССР,
1978а. С. 36-45.
Фирсова Л.В. Чайки Командорских островов:
Дис. . . . канд. биол. наук. Л., 19786.
214 с.
Фирсова Л.В. Экология чаек Командорских
островов и необходимость их охраны //
Тез. докл. 1-й Тихоокеан. науч, конгр. 1979.
Комитет А, сектор А. Вып. 11. С. 91-
92.
Фирсова Л.В. Размножение и поведение серо-
крылой чайки (Larus glaucescens) на Ко-
мандорских островах I/ Бюл. МОИП. Отд.
биол. 1983. Т. 88, вып. 4. С. 39-52.
Фирсова Л.В. К вопросу об изменчивости опе-
рения у тихоокеанской чайки // Морские
птицы Дальнего Востока. Владивосток:
ДВНЦ АН СССР, 1986. С. 135-143.
Фирсова Л.В., Левада А.В. Орнитологические
находки на юге Корякского нагорья // Ор-
нитология. М.: Изд-во МГУ, 1982. Вып. 17.
С. 112-118.
Фирсова Л.В., Лобков Е.Г., Вяткин П.В. Разме-
щение, численность и размножение тихо-
океанской чайки на Камчатке // Размеще-
ние и состояние колоний околоводных
птиц на территории СССР. М.: Наука, 1981.
С. 43-44.
Фирсова Л.В., Лобков Е.Г., Вяткин П.В. Ти-
хоокеанская чайка (Larus schistisagus) в Кам-
чатской области // Бюл. МОИП. Отд. биол.
1982. Т. 87, вып. 1.С. 30-35.
Флеров А.И., Скалинов С.В. Осенняя мигра-
ция птиц в районе Кемьлудских островов
в 1958 г. // Тр. Кандалакш. заповедника.
Мурманск: Кн. изд-во, 1960. Вып. 3. С. 145-
151.
Харитонов С.П. Морские колониальные пти-
цы о-ва Ионы Ц Колониальные гнездовья
окоповодных птиц и их охрана. М.: Нау-
ка, 1975. С. 184-185.
Харитонов С.П. К вопросу о территории и ре-
гуляции плотности колонии обыкновенной
чайки (Larus ridibundus) // Lindude Kaitu-
mine. Ornitol. kogumik. Tallinn: Valgus. 1978.
K. 8. Lk. 82-98.
Харитонов С.П. Материалы по птицам острова
Ионы // Орнитология. М.: Изд-во МГУ,
1980а. Вып. 15. С. 10-15.
Харитонов С.П. Некоторые пути адаптации озер-
ных чаек к антропогенному ландшафту //
Фауна Нечерноземья, ее охрана, воспроизве-
дение и использование. Калинин, 19806.
С. 134-141.
Харитонов С.П. О влиянии погоды на поведе-
ние зимующих птиц // Тез. докл. Прибалт,
конф, молодых орнитологов. Каунас, 1982.
С. 134-135.
Харитонов С.П. Пространственные связи и
групповые взаимоотношения озерных чаек
в гнездовой период: Автореф. дис. ... канд.
биол. наук. М.: Изд-во МГУ, 1983а. 16 с.
Харитонов С.П. Структура колонии и динами-
ка переселений озерных чаек (Larus ridi-
bundus) в сезон размножения // Зоол. журн.
19836. Т. 62, вып. 7. С. 1068-1076.
Харитонов С.П. Залеты бепощеких крачек
и черноголовой чайки в Московскую об-
398
Литература
ласть // Орнитология. М.: Изд-во МГУ, 1986.
Вып. 21. С. 142-143.
Ходков Г.И. Пролет серебристой и озерной
чаек в районе оз. Малые Чаны (Западная
Сибирь) в 1972 г. // Материалы 6-й Все-
союз. орнитол. конф. М.: Изд-во МГУ, 1974.
Ч. 4. С. 209-210.
Ходков Г.И. К экологии гнездования озер-
ной и малой чаек в районе озера Чаны (За-
падная Сибирь) // Колониальные гнездовья
околоводных птиц и их охрана. М.: Наука,
1975а. С. 106-107.
Ходков Г.И. Материалы по гнездованию сереб-
ристой чайки на озере Петраковский мыс
(Барабинская низменность) // Колониаль-
ные гнездовья околоводных птиц и их охра-
на. М.: Наука, 19756. С. 107-108.
Ходков Г.И. Особенности и фенология мигра-
ций некоторых видов чайковых на юге
Барабинской низменности в 1972-1973 гт.//
Материалы Всесоюз. конф, по миграциям
птиц. М.: Изд-во МГУ, 1975в. Ч. 1. С. 152—
155.
Ходков Г.И. Некоторые предварительные ито-
ги кольцевания озерной чайки (Larus ri-
dibundusL.) на юге Барабы // Миграции
птиц в Азии. Новосибирск: Наука, 1977а.
С. 100-101.
Ходков Г.И. Характер видимых миграций
чайковых на юге Барабинской низменно-
сти // Там же, 19776. С. 56-70.
Ходков Г.И. Генеральные направления сезон-
ных миграций серебристой и озерной чаек
в 1974 году в районе озера Чаны (Западная
Сибирь) // Трансконтинентальные связи пе-
релетных птиц и их роль в распростране-
нии арбовирусов: Докл. симпоз. Новоси-
бирск, Академгородок. Новосибирск: Нау-
ка, 1978а. С. 84-86.
Ходков Г.И. Сезонные миграции болотных кра-
чек в районе оз. Малый Чан (Барабинская
низменность) // 2-я Всесоюз. конф, по ми-
грациям птиц. Алма-Ата: Наука, 19786.
Ч. 2. С. 159-161.
Ходков Г.И. Расширение ареала чайконосой
крачки (Gelochelidon nilotica Gm.) в Запад-
ной Сибири // Фауна и экология позвоноч-
ных Сибири. Новосибирск: Наука, 1980.
С. 240-243.
Ходков Г.И. Материалы по размещению и био-
логии серебристой и озерной чаек на юге
Барабинской низменности // Экологические
и биоценотические связи перелетных птиц
Западной Сибири. Новосибирск: Наука,
1981а. С. 87-101.
Ходков ГИ. Современное размещение, чис-
ленность и состояние охраны колоний чай-
ковых птиц на оз. Чаны // Размещение и со-
стояние гнездовий околоводных птиц на тер-
ритории СССР. М.: Наука, 19816. С. 25-29.
Ходков Г.И. Миграции и сезонные размеще-
ния чайковых птиц Барабинской низмен-
ности по данным кольцевания // Миграции
птиц в Азии. Алма-Ата: Наука, 1983. Вып. 8.
С. 143-156.
Ходков ГИ., Тотунов В.М. Черноголовый
хохотун и чеграва на озере Чаны (Запад-
ная Сибирь) // Миграция и экология птиц
Сибири: Тез. докл. орнитол. конф. Якутск:
Изд-во ЯФ АН СССР, 1979. С. 189-191.
Хохлова Н.А. Экологический анализ разме-
щения и численности птиц побережий Горь-
ковского водохранилища // Экология и
проблема внутривидовой дифференциа-
ции животных среднего Поволжья. Горь-
кий: Горьк. ун-т, 1972. С. 11-21.
Хохлова Т.Ю. Орнитофауна островной части
Заонежья: Дис. ... канд. биол. наук. Л., 1977.
262 с.
Хохлова Т.Ю. Материалы по численности н
биологии чайковых птиц Онежского озе-
ра // Размещение и состояние колоний око-
ловодных птнц на территории СССР. М.:
Наука, 1981. С. 7-9.
Храбрый В.М. Птицы Березовых островов //
Материалы по фауне Выборгского заказ-
ника. Л.: ЗИН АН СССР, 1984. С. 116-146.
Чекменев Д.И. Размещение и численность коло-
ниальных птиц на озере Тениз (Централь-
ный Казахстан) // Материалы 3-й Всесоюз.
орнитол. конф. Львов: Изд-во Львов, ун-та,
1962. С. 229-230.
Чекменев Д.И. Очерки по биологии колони-
ально-гнездящихся птиц озера Тениз // Охот-
ничьи птицы Казахстана: (Фауна, экология
и практическое значение). Алма-Ата: Нау-
ка, 1964. С. 65-82.
Чельцов-Бебутов А.М. О пульсации ареалов
некоторых видов птиц в районе Тургай-
ской меридиональной депрессии // Пробле-
мы зоогеографии суши. Львов: Изд-во Львов,
ун-та, 1958. С. 325-334.
Черничка И.И., Стойловский В.П. Колониаль-
ное гнездование ржанкообразных птиц в
низовье Тилигульского лимана // Размеще-
ние и состояние гнездовий околоводных
птиц на территории СССР. М.: Наука, 1981.
С. 110-113.
Чернов Ю.И. Трофические связи птиц с насе-
комыми в тундровой зоне // Орнитоло-
гия. М.: Изд-во МГУ, 1967. Вып. 8. С. 133-
149.
Чернышов В.М., Юрлов А.К. Новые и редкие
виды птиц Причановского участка Барабы //
Миграции птиц в Азии. Новосибирск: Нау-
ка, 1986. С. 260-262.
Чернявский Ф.Б., Дорогой И.В. О роли хищ-
ников в динамике численности леммингов
(на примере острова Врангеля) // Эколо-
гия млекопитающих Северо-Восточной Сиби-
ри. М.: Наука, 1981. С. 32-50.
Черский А.И. Орнитологическая коллекция
399
Литература
музея Общества изучения Амурского края
во Владивостоке// Зап. О-ва изучения Амур,
края. 1915. Т. 14. С. 143-276.
Шарлеман Э.В. Заметки о некоторых чайко-
вых (Lariformes) окрестностей Киева //
Отдел орнитологии Русск. о-ва акклима-
тизации животных и растений. 1913. Вып. 3,
№47. С. 1-10.
Шаронов А.Д. Некоторые результаты изучения
птиц в таежной зоне Западной Сибири //
ДАН СССР. 1951. Т. 78, № 5. С. 1057-
1059.
Шаронов А.Д, Новые данные о распростране-
нии некоторых видов птиц в Западной Си-
бири // Там же. 1954. Т. 96. № 3. С. 669-
671.
Шатилов И.Н. Каталог орнитологического собра-
ния птиц Таврической губернии, пожерт-
вованного Зоологическому музею Москов-
ского университета И.Н. Шатиловым //
Изв. О-ва любителей естествознания, ан-
тропологии и этнографии. 1874. Т. 10,
вып. 2. С. 82-96.
Шеварева Т.П. Об изучении некоторых сторон
биологин средиземноморской чайки (La-
rus melanocephalus Ternm.) методом коль-
цевания Ц Тр. Бюро кольцевания. М.: МСХ
СССР, 1955. Вып. 8. С. 46-90.
Шеварева Т.П. Новые данные о встречах окольцо-
ванных чеграв (Hydroprogne tschegrava Le-
pechin) // Миграции животных. М.: Изд-во
АН СССР, 1962. Вып. 3. С. 92-105.
Шеварева Т.П. Изучение сезонного размещения
и миграций птиц СССР с помощью коль-
цевания // Миграции птиц и млекопитающих.
М.: Наука, 1965. С. 56-104.
Шевченко В.В. До екологп чайок i крячктв
Присивашшя И Пр. н.-д. зоолого-бюлопч-
ного ш-ту. Харкшський ун-т. Харкгв: Вид-во
Харкш. ун-ту, 1937. Т. 4. С. 127-152.
Шевченко В.Л. Современное состояние колони-
альных гнездовий околоводных птиц Камыш-
Самарской озерной депрессии // Колониаль-
ные гнездовья околоводных птиц и их охра-
на. М.: Наука, 1975. С. 161-162.
Шедден К. Загрязнение водных запасов чайками
и одно решение// 18-й Междунар. орнитол.
конгр.: Тез. докл. и стенд, сообщ. М.: На^ка,
1982. С. 228.
Ширяева С.С. Питание некоторых видов чайко-
вых // Новости орнитологии. АлмагАта,
1965. С. 422-423.
Шкарин В.С., Родимцев А.С., Иванов С.А., Овсян-
ников И.Н. Взаимоотношения полярных кра-
чек и серебристых чаек в гнездовой период //
Природа и экономика Кузбасса: Тез. докл.
к предстоящей науч. конф. Новокузнецк,
1982. С. 77-78.
Шкатулова А.П. Орнитофауна Торейских озер
и ее охрана // Охрана природы и воспроиз-
водство естеств. ресурсов. Чита: Забайк.
фил. Геогр. о-ва СССР, 1972. Вып. 2. С. 58 -
60.
Шкатулова А.П. Материалы по орнитофауне
Бурятской АССР Ц Орнитология. М.:
Изд-во МГУ, 1979. Вып. 14. С. 97-107.
Шкатулова А.П. Численность и поведение чай-
ковых птиц в период весенних и осенних
миграций на территории Забайкалья // Эколо-
гия и охрана птиц. Кишинев: Штиинца, 1981.
С. 243-244.
Шкатулова А.П. Миграции чайковых на террито-
рии Забайкалья // Фауна и экология птиц
Восточной Сибири. Иркутск: Иркут, ун-т,
1984. С. 150-158.
Шкляревич Ф.Н. Об изменениях численности
моевок и кайр на базарах о-ва Харлова
(Семь островов, Восточный Мурман) //
Тез. докл. 7-й Всесоюз. орнитол. конф.
Киев: Наук, думка, 1977. Ч. 1. С. 344-
345.
Шнитников В.Н. Птицы Минской губернии// Маг
териалы к познанию фауны и флоры Рос.
империи Отд. зоол. М., 1913. Вып. 12. С. 475.
Шокало С.И., Гайдук В.Е., Шокало Б.И. Харак-
теристика скоплений птиц города Бреста //
Птицы и урбанизированный ландшафт. Кау-
нас, 1984. С. 142-143.
Шукуров Э.Д. Птицы Киргизии. Фрунзе: Мек-
теп, 1981. Ч. 1. 110 с.
Шунтов В.П. Миграции и распределение морских
птиц в юго-восточной части Берингова моря
в весенне-летний период // Зоол. журн. 1961.
Т. 40, вып. 7. С. 1058-1069.
Шунтов В.П. Летнее распределение моевок в Бе-
ринговом море // Орнитология. М.: Изд-во
МГУ, 1963. Вып. 6. С. 325-330.
Шунтов В.П. Морские птицы и биологическая
структура океана. Владивосток: Дальневост,
кн. изд-во, 1972. 376 с.
Шунтов В.П. Биологические ресурсы Охотско-
го моря. М.: Агропромиздат, 1985. 224 с.
Шухов И.Н. Птицы Обдорского края // Ежегод-
ник Зоол. музея Акад. наук. СПб., 1915.
Т. 20, №2. С. 167-238.
Щадилов Ю.М. Формирование населения назем-
ных позвоночных животных на малых водо-
хранилищах Подмосковья // Комплексное
изучение и рациональное использование при-
родных ресурсов. Калинин: 1980. С. 196.
Щеголев В.И. Гнездование крачек в Чернозем-
ной полосе // Колониальные гнездовья око-
ловодных птиц и их охрана. М.: Наука, 1975.
С. 110-111.
Щербина А.А. Колониальные гнездовья крачек
на разливах Атрека // Колониальные гнез-
довья околоводных птиц и их охрана. М.:
Наука, 1975. С. 163-164.
Щербина А.А. О нахождении среднего поморни-
ка в Туркмении ,'/ Орнитология. М.: Изд-во
МГУ, 1977. Вып. 13. С 200.
Щербина А.А. Новые данные по гнездовой фауне
400
Литература
колониальных и околоводных птиц островов
Кара-Богаз-Гола // Природная среда и птицы
побережий Каспийского моря и прилежащих
низменностей. Баку: Азерб. изд-во, 1979.
С. 89-100.
Щербина А.А., Караваев А.А., Андреев В.П.
Колониальные гнездовья чайковых птиц ост-
рова Огурчцнский // Колониальные гнездовья
околоводных птиц и их охрана. М.: Наука,
1975. С. 112-113.
Эминов А. К биологии размножения малой крач-
ки // Изв. АН ТССР. Сер. биол. 1964. № 6.
С. 84-86.
Эминов А. К экологии обыкновенной чергавы
в южной Туркмении // Орнитология в СССР.
Ашхабад: Ин-т зоологии АН ТССР, 1969.
Кн. 2. С. 747-749.
Эминов А. К экологии речной крачки в южной
Туркмении // Материалы 6-й Всесоюз. орни-
тол. конф. М.: Изд-во МГУ, 1974а. Ч. 2.
С. 156-157.
Эминов А. Экология чайконосой крачки в юж-
ной Туркмении // Фауна и экология птиц
Туркмении. Ашхабад: Ылым, 19746. Вып. 1.
С. 149-161.
Эминов А. О гнездовых колониях крачек юж-
ной Туркмении // Колониальные гнездовья
околоводных птиц и их охрана. М.: Наука,
1975. С. 113-114.
Эминов А. О сроках весенней миграции неко-
торых птиц долины р. Мургаба // 2-я Всесоюз.
конф, по миграциям птиц: Тез. сообщ. Алма-
Ата: Наука, 1978. Ч. 1. С. 171-172.
Юдин КА. Линька белой чайки // Тр. проблем-
ных и тематических совещаний ЗИН АН
СССР. Л.: ЗИН АН СССР, 1960. Вып. 9.
С. 298-303.
Юдин КА. Филогения и классификация ржанко-
образных М.; Л.: Наука, 1965. 262 с. (Фауна
СССР. Птицы; Т. 2, вып. 1, ч. 1).
Юрлов КТ. О распространении некоторых птиц
в Юго-Западной Сибири. // Орнитология.
М.: Изд-во МГУ, 1959. Вып. 2. С. 176-179.
Юрлов К.Т., Чернышов В.М., Кошелев А.И.,
Сагитов В.А. и др. Новые и редкие птицы
в районе озера Чаны // Миграции птиц в Азии.
Новосибирск: Наука, 1977. С. 205—209.
Языкова И.М. Чайки и крачки Пролетарского
водохранилища // Колониальные гнездовья
околоводных птиц и их охрана. М.: Наука,
1975. С. 115-116.
Якоби В.Э. Биологические основы предотвра-
щения столкновений самолетов с птицами.
М.: Наука, 1974. 164 с.
Якушин Г.Д. Гнездование птиц на островах
в устье Бикады // Птицы Таймыра: (Эколо-
гия, охрана и хозяйственное использование).
Новосибирск: СО ВАСХНИЛ, 1983. С. 23-29.
Ямасина Е. Белая чайка Pagophila ebumea
(Phipps), добытая на Хоккайдо // Tori. 1934.
Т. 8, № 39. С. 326-327. На яп. яз.
Янаус М. Динамика веса птенцов озерной чайки
на оз. Энгуре в 1978 и 1979 гг. // 10-я При-
балт. орнитол. конф. Рига: Ин-т биологии
АН ЛатвССР, 1981. Т. 2. С. 205 - 209.
Янаус М. Влияние метеорологических условий
на смертность птенцов озерной чайки // Тез,
докл. 11-й Прибалт, орнитол. конф. Таллин:
Ин-т зоологии и ботаники АН ЭССР, 1983.
С. 241-243.
Янушевич А.И., Тюрин П.С., Яковлева И.Д.,
Кыдыралиев А., Семенова Н.И, Птицы Кир-
гизии. Фрунзе: Изд-во АН КиргССР, 1959.
Т. 1. 229 с.
Ясков В.А. Новые и залетные виды птиц в ниж-
нем Приобье (Западная Сибирь) // Экология
и биоценотические связи перелетных птиц
Западной Сибири. Новосибирск: Наука, 1981.
С. 180-181.
Яхонтов В.Д. Остров Нансикан // Колониальные
гнездовья околоводных птиц и их охрана.
М.: Наука, 1975а. С. 186-187.
Яхонтов В.Д. Птичьи базары Пенжинской
губы и их состояние // Там же, 19756.
С. 185-186.
Яхонтов В.Д, Птицы Шантарских островов:
некоторые вопросы экологии // Вопросы
географии Дальнего Востока. Хабаровск,
1977. №17. С. 150-171.
Яхонтов В.Д. Птицы Пенжинского района //
Птицы Северо-Востока Азии. Владивосток:
ДВНЦ АН СССР, 1979. С. 135-164.
Abdulali Н., AmbedkarV.C Occurrence of the black
tern Chlidonias niger (Linn.), in India // J. Bom-
bey Natur. Hist. Soc. 1984. Vol. 80, N 3. P. 640.
AU S., Ripley S.D. Handbook of the birds of India
and Pakistan. L. etc.: Oxford Univ, press, 1969.
Vol. 3. 325 p.
Allen J.A. Report on the birds collected in North-
Eastern Siberia by the Jesup North Pacific expe-
dition, with field notes by the collectors //
Bull. Amer. Mus. Natur. Hist. 1905. Vol. 21,
art. 13. P. 219-257.
Anderson B.R. Summary of highest counts of indi-
viduals for Canada // Amer. Birds. 1980. Vol. 34,
N4. P. 708-710.
Andersson M. Breeding behaviour of the long-
tailed skua // Ornis scand. 1971. Vol. 2.
P. 35-54.
Andersson M. Behaviour of the pomarine skua
Stercorarius pomarinus Temm. with compara-
tive remarks on Stercorariinae // Ibid. 1973.
Vol. 4, N l.P. 1-16.
Andersson M. Predation and kleptoparasitism by
skuas in a Shetland seabird colony // Ibis.
1976. Vol. 118, N 2. P. 208-217.
Aumees V., Soots H., Kullapere A. Vilsandi loodu-
skaitseala linnustikust // Vilsandi - looduskait-
seala Eesti NSV laanerannikul. Tallin: Valgus,
1983. S. 70-80.
Austin O.L. Some aspects of individual distribution
401
Литература
in the Cape Cod tern colonies // Biid-Band.
1940. Vol. 11. P. 155-169.
Austin O.L. The role of longevity in the successful
breeding by the common tern (Sterna hirundo) //
Ibid. 1945. Vol. 16. P. 21-28.
Austin O.L. A study of the mating of the common
tern (Sterna h. hirundo) // Ibid. 1947. Vol. 18.
P. 1-16.
Austin O.L. The birds of Korea // Bull. Mus. Com-
par. Zool. 1948. Vol. 101, N 1. P. 1-301.
Austin O.L. Site tenacity, a behaviour trait of
the common tern (Sterna hirundo L.) // Bird-
Band. 1949. Vol. 20. P. 1-39.
Austin O.L., Austin O.L. (JR.) Some demographic
aspects of the Cape Cod populations of common
tern (Sterna hirundo) // Ibid. 1956. Vol. 27.
P. 55-66.
Austin O.L., Kuroda N. The birds of Japan, their
status and distribution // Bull. Mus. Compar.
Zool. 1953. Vol. 109, N 4. P. 279-637.
Baggerman B., Baerends G.P., Heikens H.S.,
Mook J.H. Observations on the behaviour of
the black tern, Chlidonias n. niger, in the bre-
eding area // Ardea. 1956. Vol. 44, N 1/3.
P. 1-71.
Bailey A.M. A report on the birds of Northeastern
Alaska and regions adjacent to Bering Strait //
Condor. 1925. Vol. 27. P. 20-32.
Bailey A.M. Birds of Arctic Alaska // Colo. Mus.
Natur. Hist. Pop. Ser. 8. 1948. 317 p.
Baird S.F. On additions to the bird-fauna of North
America, made by the scientific corps of the
Russo-American telegraph expedition // Trans.
Chicago Acad. Sci. 1869. Vol. 1. 321 p.
Baird P.A., Barbour D.B., DeGange A.R. et al.
Population dynamic and trophic relationships
of marine birds in the Gulf of Alaska //
Environ. Assess. Alaskan Continent. Shelf.
NOAA/OCEAP. Annu. Rep. 1979. Vol. 2.
P. 7-20.
Baker S.E.C. The fauna of British India, including
Ceylon and Burma // Birds. L., 1929. Vol. 6
500 p.
Balch L.G., Bohlen H.D., Rosenband G.B. The
Illinois Ross’s gull // Amer. Birds. 1979. Vol. 33
N 2. P. 140-142.
Bannerman D.A. The birds of the British Isles.
Edinburg; L.: Oliver and Boyd, 1962. 366 p
Barash D.P., Donovan P., Myrick S. Clam dropping
behaviour of the glaucous-winged gull (Larus
glaucescens) // Wilson Bull. 1975. Vol. 87 N 1
P. 60-64.
Barcena de Limia F. Colonias de aves marinas en
las islas Gies // Natur. hisp. 1977. N 9. P. 1-24.
Barrett R.T., Runde O. Growth and survival of nest-
ling Kittiwakes Rissa tridactyla in Norway //
Ornis scand. 1980. Vol. 11, N 3. P. 228-235.
Barth E.K. Moult and taxonomy of the Herring
gull Larus argentatus and Lesser black-backed
gull. L. fuscus in Northwestern Europe // Ibis
1975a. Vol. 117. P. 384-387.
Barth E.K. Taxonomy of Larus argentatus and La-
rus fuscus in North-Western Europe // Ornis
scand. 1975b. Vol. 51. P. 49-63.
Bateson P.P., Plowright R.G. The breeding biolo-
gy of the Ivory gull in Spitsbergen // Brit. Birds.
1959a. Vol. 52, N 4. P. 105-114.
Bateson P.P., Plowright R.G. Some aspects of the
reproductive behaviour of the Ivory gull //
Ardea. 1959b. Vol. 47, N 3/4. P. 157-176.
Becker PH., Firtkk P., Anlauf A. Rainfall preceding
egg-laying - a factor of breeding success in com-
mon terns (Sterna hirundo) // Oecologia. 1985.
Bd. 65. S. 431-436.
Bengtson S.-A. Breeding success of the Arctic
tern, Sterna paradisaea (Pontoppidan), in the
Kongsfjord Area, Spitsbergen in 1967 // Norw.
J. Zool. 1971. Vol. 19, N 1. P. 77-82.
Bent A.C. Life histories of North American gulls
and terns. Wash.: Gov. Print, off., 1921. 340 p.
Berg A.B., Bosman C.A.W., Rozendaal F.G. First
record of Slender-billed gull, Larus genei, in Sri
Lanka // Ardea. 1982. Vol. 70. P. 82-83.
Bergman G. Unterschiede von SilbermSve Larus
a. argentatus, H eringmove L. fuscus in Leben-
weise und Stimme // Vogelwarte. 1951. Bd. 16.
S. 17-18.
Bergman G. Single-breeding versus colonial breeding
in the Caspian tern Hydroprogne caspia, the com-
mon tern Sterna hirundo and the Arctic tern
Sterna paradisaea // Ornis fenn. 1980. Vol. 57.
P. 141-152.
Bergman G. Interrelationships between ducks
and gulls // Managing wetlands and their birds.
SUmbridge: IWRB, 1982a. P. 241-247.
Bergman G. Population dynamics, colony forma-
tion and competition in Larus argentatus, L. fus-
cus and L. marinus in the archipelago on Fin-
land // Ann. zool. fenn. 1982b. Vol. 19, N 3.
P. 143-164.
Bergman St. Zur Kenntnis Nordasiatischer Vogel.
Ein Beitrag zur Systematik, Biology und Ver-
britung der Vogel Kamtschatkas und Kurilen.
Stockcholm: Bonniers, 1935. 268 S.
Berzins B. Nagot om Lettlands masfaglar // Var
fagelvarld. 1946. N 3.
Biederman J.O., Drury W.H. Ecological studies
in the Northern Bering Sea: Studies of sea-
birds in Bering Strait // Environ. Assess. Alas-
kan Continent. Shelf. NOAA/OCEAP. Annu.
Rep. 1978. Vol. 2. P. 751-838.
Bierman W.H. Sabine’s gull (Larus sabini) in the
North Atlantic // Ardea. 1966. Vol. 54. P. 271.
Birkenmajer K. Obserwacie na mewa modrodzioba
Pagophilla eburnea (Phipps), w poludniowej
cz^sci Zachodniego Spitsbergenu// Acta ornithol.
1969. Vol. 11, N 13. P. 461-476.
Blake E.R. Birds of Mexico. Chicago: Univ. Chicago
press, 1953. 644 p.
Bledsoe A.H., Sibley D. Patterns of vagrancy of
Ross’s guU // Amer. Birds. 1985. Vol. 39, N 2.
P. 219-227.
402
Литература
Bloch-Nielsen Н. Jagtloven og beslaegtede love //
Jeg er jaeger. K^benhavn: Politikens fori., 1975.
459 s.
Blomqvist S., Elander M. The masfSglar i Arktis //
Fauna och flore. 1980. Bd. 75, N 5. P. 217—
230.
Bourne W.R.P. Prolonged parental care in Herring
gulls nesting in towns in Eastern Scotland //
Bird Study. 1979. Vol. 26. P. 196 197.
Braun B., Hunt G.L. Brood reduction in black-
legged kittiwakes // Auk. 1983. Vol. 100, N 2.
P. 469-476.
Breife B. Bird fauna from Knissa mosse then and
now // Calidris. 1981. Vol. 10, N l.P. 23-24.
Britton P.L., Brown L.H. The status and breeding
behaviour of East African lari // Ostrich. 1974.
Vol. 45,N2.P. 63-82.
Brooke R.K. The Catharacta skuas occuring in
South African waters // Durban Mus. Novit.
1978. Vol. 11. P. 295-308.
Brown D.H. Glaucous gulls diving for food // Brit.
Birds. 1949. Vol. 42. P. 95.
Brown R.G.B. Courtship behaviour in the Lesser
black-backed gull Larus fuscus // Behaviour.
1967a. Vol. 29. P. 122-153.
Brown R.G.B. Breeding success and population
growth in a colony of the Herring and Lesser
black-backed gulls Larus argentatus and L. fus-
cus // Ibis. 1967b. Vol. 109, N 4. P. 502-515.
Brown R.G.B. Species isolation between the Her-
ring gull Larus argentatus and Lesser black-
backed gull L. fuscus // Ibis. 1967c. Vol. 109,
N3.P.310-317.
Brown R.G.B., Blurton J.N.J., Hussel D.J. The
breeding behaviour of Sabine’s gull Хеша
sabini // Behaviour. 1967. Vol. 28. P. 110—
140.
Brun E. Breeding distribution and population of
kliff-breeding seabirds in Sor-Varanger, North
Norway // Astarte. 1971a. Vol. 4, N 2. P. 53-60.
Bruns H. Ornithologische Beobachtungen auf einer
Kreutzfahrt in die Arctis (Nordpolarmeer,
Bareninsel, Spitzbergen, Jan Mayen) // Ornithol.
Mitt. 1981. Bd. 33, N 7. S. 171-190.
Buckley P.A., Kane R. The winter season De-
cember 1, 1974 - March 31, 1975 // Amer.
Birds. 1975. Vol. 29, N 3. P. 667-672.
Bunge A. Brief au den lestandigen Sekretar der
Akademie uber den Gang der von ihr veranstal-
teten. Expedition nach den Neusibirischen
Inseln und dem Jana-Lande // Beitr. Kennt.
Russ. Reiches. Ser. 3. 1887. Bd. 3. S. 119-139.
Bureau L. Sur les plumages de la mouette de Sabi-
ne // Ornis. 1901. Vol. 2. P. 285-306.
Burger J. On becoming independent in Herring
gulls: parent-young conflict // Amer. Natur.
1981. Vol. 117, N 4. P. 444-456.
Bilrkli W. Heringsmove Larus fuscus im Oberen-
gadin // Ornithol. Beob. 1981. Bd. 78, N 1.
S. 51.
Butler R.W., Verbeek N.A.M., Foottit R.G. Mor-
tality and dispersal of the glaucous-winged
gulls of Southern British Columbia // Canad.
Field.-Natur. 1980. Vol. 94, N 3. P. 315-320.
Byrd G.V. Red-legged kittiwake colonies in the
Aleutian Islands, Alaska // Condor. 1978. Vol. 80.
P. 250.
Campredon P. Reproduction de la Sterne caugek
Thalasseus sandvicensis Lath, sur le Banc d’Ar-
guin (Gironde): Aperqu de sa distribution hiver-
nale // Oiseau et Rev. fran<$. ornithol. 1978.
Vol. 48,N3.P. 263-279.
Z / </
Cerny W. Tan a prezimovani raska Bournina (Larus
canus) v Ceskoslovensku // Sylvia. 1965. Vol. 17.
P. 95-105.
Chapman St.E. The Pacific winter quaters of Sabi-
ne’s gull (Xema sabini) // Ibis. 1969. Vol. Ill,
N4.P. 615-617.
Chartier B., Cooke F. Ross’s gulls (Rhodostethia
rosea) nesting at Churchill, Manitoba, Canada //
Amer. Birds. 1980. Vol. 34, N 6. P. 839-841.
Check-list of Japanese birds. Tokyo: Gakken, 1974.
364 p.
Cheng Tso-hsin. Distributional list of Chinese birds.
Peking: Sci. Sin., 1976. 1218 p.
Cheng Tso-hsin, Li Dehao, Wang Zuxiang et al.
The aviafauna of Xizang. Peking, 1983. 360 p.
Chmielewski S. Gniezdzenie siq rybitwy biafoskr-
zydfej (Chlidonias leucopterus) pod Siedlcami //
Not. ornitol. 1984. Wol. 25, N 1/4. S. 63-64.
Clarck A.H. The birds collected and observed du-
ring the cruise of United States fisheries steamer
’’Albatross” in the North Pacific Ocean and
Bering, Okhotsk, Japan and Eastern Seas from
April to December 1906 // Proc. US Nat. Mus.
1910. Vol. 38. P. 25-74.
Clarke W.E. The birds of Jan Mayen Islands //
Zoologist. 1890. Vol. 14,N3.P. 1-16,41-51.
Collett R., Nansen F. An account of the birds //
The Norwegian North polar expedition 1893—
1896: Scientific results / Ed. F. Nansen. L.,
1900. Vol. 1. P. 1-53.
Conover M.R. Occurrence of supernormal clutches
in the Laridae // Wilson Bull. 1984. Vol. 96,
N2. P. 249-267.
Cooke IP. IP. Distribution and migration of North
American gulls and their allies //US Dep. Agr.
Bull. 1915. N 292. P. 1-70.
Cottam Cl., Bolen E.G., Zink R. Sabine’s gull on
South Texas Coast // South-West. Natur. 1975.
Vol. 20, N l.P. 134-135.
Coulson J.C. The status of the kittiwake in British
Isles // Bird Study. 1963. Vol. 10. P. 147-179.
Coulson J.C. The movements of the kittiwake //
Ibid. 1966. Vol. 13. P. 107-115.
Coulson J.C. The changing status of the kittiwake
Rissa tridactyla in the British Isles, 1969-
1979 // Ibid. 1983. Vol. 30, N 1. P. 9-16.
Coulson J.C., Duncan N., Thomas C. Changes in
the breeding biology of the Herring gull (Larus
argentatus) induced by reduction in the size
403
Литература
and density of the colony // J. Anim. Ecol.
1982. Vol. 51, N 3. P. 739-756.
Coulson J.C., Horobin J. The influence of age on
the breeding biology and survival of the Arctic
tern Sterna paradisaea // J. Zool. 1976. Vol. 178.
P. 247-260.
Coulson J.C., Macdonald A. Recent changes in habits
of the kittiwake // Brit. Birds. 1962. Vol. 55.
P. 171-177.
Coulson J.C., White E. A study of colonies of the
kittiwake Rissa tridactyla (L.) // Ibis. 1956.
Vol. 98, N l.P. 69-79.
Coulson J.C., White E. Observation of the breeding
of the kittiwake // Bird Study. 1958. Vol. 5,
N 2. P. 74-83.
Coulson J.C., White E. The post-fledging mortality
of kittiwake // Ibid. 1959. Vol. 6. P. 97-102.
Cramp S. Gulls nesting on buildings in Britain and
Ireland // Brit. Birds. 1971. Vol. 64. P. 476-
487.
Cramp S. The Birds of the Western Palearctic.
Oxford, London, New York: Oxford Univ.
Press. 1985. Vol. 4. 960 p.
Cramp S., Bourne W.R.P., Saunders D. The seabirds
of Britain and Ireland. L.: Collins, 1974. 287 p.
Cramp S., Simmons K. The Birds of the Western
Palearctic. Oxford, London, New York: Oxford
Univ. Press. 1983. Vol. 3. 913 p.
Cullen E. Adaptations in the kittiwake of cliff
nesting // Ibis. 1957. Vol. 99. P. 275-302.
Cullen J.M. The aerial display of the Arctic tern
and other species // Ardea. 1960. Vol. 48,
N 1/2. P. 1-37.
Curry-Lindahl K. Bird migration in Africa. L.;
N.Y.: Acad, press, 1981. Vol. 1. 444 p.
Cuthbert F. Mate retention in Caspian terns// Con-
dor. 1985. Vol. 87, N 1. P. 74-78.
Dalgety C.T. The Ivory gull in Spitsbergen // Brit.
Birds. 1932. Vol. 26. P. 2-7.
Dalgety C.T., McNeile J.H., Ingram M.J. Notes
on birds observed in Spitsbergen during the
spring 1930 // Ibis. 1931. Vol. 73. P. 243-254.
Davis G.W.F., Dunn E.K. Intraspecific predation
and colonial breeding in Lesser black-backed
gulls Larus fuscus // Ibid. 1976. Vol. 118, N 1.
P. 65-77.
Dawson W.L. The new reserves of the Washington //
Condor. 1908. Vol. 10. P. 45.
. Densley M. Ross’s gulls in Britain // Scot. Birds.
1977a. Vol. 9, N 7. P. 334-342.
Densley M. The Ross’s gull (Rhodostethia rosea)
in Arctic Alaska // Polar Rec. 1977b. Vol. 18.
P. 603-605.
Densley M. Ross’s gulls in Alaska // Brit. Birds.
1979. Vol. 72. P. 23-28.
Devillers P. The skuas of the North American Paci-
fic coast II Auk. 1977. Vol. 94. P. 417-429.
Devillers P. Distribution and relationships of South
American skuas // Gerfaut. 1978. Vol. 68.
P. 374-417.
Diamond A.W. Subannual breeding and moult
cycles in the Bridled tern Sterna anaethetus
in the Seychelles // Ibis. 1976. Vol. 118. P. 414-
419.
DiCostanzo J. Population dynamics of a common
tern colony // J. Field Ornithol. 1980. Vol. 51,
N3.P. 229-243.
Divoky G.J. The pelagic feeding habits of Ivory
and Ross’s gulls // Condor. 1976. Vol. 78,
N l.P. 85-90.
Doran G. Possible effect of kittiwake and guillemot
colonies on Fulmar colony // Irish Natur. J.
1967. Vol. 15. P. 337-339.
Downing R.L. Survey of Interior Least tern nesting
populations // Amer. Birds. Vol. 34, N 2.
P. 209-211.
Dragesco J. Observations ethologiques sur les
oiseaux du Banc d’Arguin // Alauda. 1961.
Vol. 29, N 2. P. 6-98.
Drury W.H. Breeding activities of Longtailed jaeger,
Herring gull and Arctic tern on Bylot Island,
Northwest Territories, Canada // Bird-Band.
1960. Vol. 31. P. 63-79.
Drury W.H., Smith W.J. Defence of feeding areas
by adult Herring gulls and intrusion by young //
Evolution. 1968. Vol. 22. P. 193-201.
Drury W.H., Steele B.B. Studies of populations,
community structure and ecology of marine
birds at King Isl., Bering Strait region, Alaska //
Environ. Assess. Alaskan Continent. Shelf.
NOAA/OCEAP. Annu. Rep. 1977. Vol. 5.
P. 75-149.
Duncan N. The Lesser black-backed gull on the
Isle of May // Scot. Birds. 1981a. Vol. 11,
N6.P. 180-188.
Duncan N. The Abbeystead and Mallowdale gull
colony before control // Bird Study. 1981b.
Vol. 28, N 2. P. 133-138.
Dunlop J.N. Reproductive periodicity in a popu-
lation of Crested terns, Sterna bergii Lichtens-
tein, in South-Western Australia // Austral.
Wildlife Res. 1985. Vol. 12. P. 95-102.
Dwight J. The gull (Laridae) of the world; Their
plumages, moults, variations, relationships and
distribution // Bull. Amer. Mus. Natur. Hist.
1925. Vol. 52,art. 3.P. 63-408.
Elliott H.W. A monograph of the Islands of Alaska //
Spec. Bull. US Commiss. Fish and Fisheries.
1882. N 176. P. 125-136.
Erwin R.M. Coloniality in terns: the role of social
feeding // Condor. 1978. Vol. 80, N 2. P. 211-
215.
Evans P.G.H. The seabirds of Greenland: their
status and conservation // Status and conser-
vation of the world’s seabirds. 1984. P. 49-84.
(ICBP Techn. Publ.; N 2.)
Ewing L.S., Eving A.W. Common and black-headed
gulls flight-feeding over ragwort // Brit. Birds.
1975. Vol. 68, N l.P. 44.
Faanes Cr. Birds of the St. Croix River Valley:
Minnesota and Wisconsin // North Amer. Fauna.
1981. Vol. 73. P. 1-196.
404
Литература
Fabricius Е. Nagra iak-tagelser rorande viggens,
Nyroca fuligula (L.) beroende av masfiglarna
sasom hackfagel i skargarden // Ornis fenn.
1937. Vol. 14. P. 115 124.
Fay F.H., Cade T.J. An ecological analysis of the
aviafauna of St. Lawrence Island, Alaska //
Univ. Cal. Publ. Zool. 1959. Vol. 6, N 2. P. 73-
150.
Fennell ChM. Notes of the birds of Daikokujima,
Hokkaido, Japan // Condor. 1953. Vol. 55.
P. 38-42.
Ferns P.N. Changes fortunes in the Bristol Channel //
Bull. Gull. Sci. Group. 1982. N 4. P. 20-21.
Finch D. W. The spring migration April 1 - May 31,
1974, North-Eastern maritime region // Amer.
Birds. 1975. Vol. 29, N 1. P. 125-129.
Fisk E.J. The growing use of roofs by nesting birds //
Bird-Band. 1978.Vol. 49.N2.P. 134-141.
Fitch M.A., Shugart G.W. Comparative biology
and behaviour of monogamous pairs and one
male - two female trios of Herring gulls //
Behav. Ecol. and Socio biol. 1983. Vol. 14,
N l.P. 1-7.
Fjeldsa J. Guide to the young of European Pre-
cocial birds. Tisvildeleje: Skarv Nat. Publ.,
1977. 285 p.
Franzmann N.E. Greesholm - en status pSskoert //
Feltornithologen. 1974. Bd. 16, N 1. S. 58-
59.
Friedmann H. Birds bones from Eskimo Ruins
on St. Lawrence Island // J. Wash. Acad. Sci.
1934. Vol. 24. P. 230-237.
Frisch T., Morgan W.C. Ivory gull colonies in Sout-
heastern Ellesmere Island, Arctic Canada //
Canad. Field.-Natur. 1979. Vol. 93, N 2.
P. 173-174.
Fuchs E. Predation and anti-predator behaviour
in a mixed colony of terns Sterna sp. and black-
headed gulls Larus ridibundus with special
reference to the Sandwich tern // Ornis scand.
1977. Vol. 8,N l.P. 17-32.
Fujimaki Yu. Observations on seabirds of Moyururi
Island, Eastern Hokkaido // Tori. Bull. Ornithol.
Soc. Jap. 1961. Vol. 16, N 78. P. 387-398.
Gabrielson I.N., Lincoln F.C. The birds of Alaska.
Wash., 1959. 922 p.
Galbraith H. The diet and feeding of breeding kit-
tiwakes Rissa tridactyla // Bird Study. 1983. Vol.
30, N 2. P. 109-120.
Gill K., Dick M. A book at current breeding dis-
tribution and abundance of the Aleutian tern
in North America (abstract) // Pacif. Seabird
Group Bull. 1977. Vol. 41, N 1. P. 22-
23.
Gillett W.H., Hayward J.L., Stout J.F. Effects of
human activity on eggs chick mortality in a
glaucous-winged gull colony // Condor. 1975.
Vol. 77, N 4. P. 492-495.
Glutz von Blotzheim UJV., Bauer KM. Handbuch
der Vogel der Mitteleuropas. Wiesbaden: Akad.
VerL, 1982. Bd. 8/1.699 S.
Godfrey W.E. The birds of Canada. Ottawa: Queen’s
Printe, 1966. 428 p.
Godfrey W.E. The birds of Canada: Rev. ed. Ot-
tawa: Nat. Mus. Canada, 1986. 595 p.
Goethe F. Beobachtungen und Untersuchungen zur
Biologie der Silbermowe (Larus argentatus Pont.)
auf der Vogelinsel Memmertsand // J. Ornithol.
1937. Bd. 85. S. 1-119.
Goethe F. Felstbrutertum und weitere beachtens-
werte Tendenzen bei der Silbermowe // XII
Intern. Ornithol. Congr. Proc. Helsinki, 1960.
S. 252-258.
Goethe F. Zur Taxonomie der Silbermowe (Larus
argentatus) im siidlichen deutschen Nordseege-
biet // Vogelwarte. 1961. Bd. 21, H. 1.
S. 1-24.
Goethe F. Verhantelsunterschiede zwischen europa-
ischen Formen der Silbermowengruppe (Larus
argentatus-cachinnans-fuscus) // J. Ornithol.
1963. Bd. 104. S. 129-141.
Goethe F. Zim ’’Typus” von Larus argentatus heug-
lini Bree im Staatlichen Museum fur Naturkunde
zu Stuttgart //Stuttgart Beitr. Naturk, A. 1973.
N 261. S. 1-8.
Gore M.E.J., Won Pvong-Oh. The Birds of Korea.
Seoul,Tokyo- 1971.450 t>.
Gorski W. Awifazna w legowa dolinach i pradoli-
nach rzek Pobrzeza Pomorskiego // Acta zool.
Crac. 1981. Vol. 26, N 3. P. 95-147.
Greenhalgh M.E. The pennine gullery // Bird Stu-
dy. 1974. Vol. 21, N 2. P. 146-148.
Grenmyr U., Sundin J.A. The bird fauna of the
Vasterbooten coast-changes since 30’s. //
V&r fagelvarld. 1981. Bd. 40, N 2. S. 91-104.
Grinnell J. Summer birds of Sitka, Alaska // Auk.
1898. Vol. 15. P. 122-131.
Gruson E.S. Checklist of the Birds of the World. L.:
Collins. 1976. 212 p.
Gudmundsson F. Inslen fuglar XII, Silamafur (La-
rus fuscus) // Natturufroedingurinn. 1955. Bd.
25. S. 215-226.
Haartman L. von. Bidrag till kannedomen om vig-
gens (Nyroca fuligula L.) hacknigspsykologi i
skargarden // Ornis fenn. 1937. Vol. 14.
P. 125-134.
Haartman L. von. Nest sites of the common gull
Larus canus in relation to ice age geology and
other factors // Ibid. 1980. Vol. 57, N 1.
P. 11-16.
Hakala A. An Ivory gull colony of Lardyfjellet,
Spitsbergen // Sterna. 1975. N 2. P. 91-94.
Hand J.L. A comparaison of vocalization of
Western gulls (Larus occidentalis occidenta and
L. o. livens) // Condor. 1981. Vol. 83, N 4.
P. 289-301.
Hand-list of the Japanese birds: 4th ed. Tokyo:
Ornithol. Soc. Jap., 1958. 264 p.
Hanna D.G. The summer birds of the St. Matthew
Island bird reservation // Auk. 1917. Vol. 34.
P. 403-410.
Harrington B.A. Pelagic gulls in winter off Southern
405
Литература
California // Condor. 1975. Vol. 77, N 3.
P. 346-350.
Harris M.P. Species separation in gulls // Animals.
1969. Nol. 12, N l.P. 9-12.
Harris M.P. Abnormal migration and hybridization
of Larus argentatus and Larus fuscus after unter-
species fostering experiments // Ibis. 1970a.
Vol. 112, N 4. P. 488-498.
Harris M.P. Rates and causes of increases of some
British gull populations // Bird Study. 1970c.
Vol. 17. P. 325-335.
Harris M.P., Galbraith H. Seabirds populations of
the Isle of May // Scot. Birds. Vol. 12, N 6.
P. 174-180.
Harrison C. An atlas of the birds of the Western Pa-
learctic. London: Collins, 1982. 322 p.
HarrisonP. Seabirds: An identification guide. Boston:
Houghton Mifflin, 1983. 448 p.
Hartert E. Die Vogel der palaarctischen Fauna. B.:
Friedlander, 1912-1921. Bd. 2. 1764 S.
Hasegawa H. Status and conservation of seabirds
in Japan, with special attention to the short-
tailed albatross // Status and conservation of the
world's seabirds. 1984. P. 487-500. (ICBPTechn.
Publ.; N 2).
Hays H. Probable common X roseate tern hybrids //
Auk. 1975. Vol. 92. N 2. P. 219-234.
Hedgren S., Larsson L. Vittrut Larus hyperboreus,
vitvingad trut L. glaucoides eller missfargad
annan trut-sharigheter vid faltbestamning //
Var fagelvard. 1973. Bd. 32. S. 173-198.
Heim T. Der Kukunor oder Qinghoi-See vom Austroc-
kenen bedroht // Gefiederte Welt. 1985. N 2.
S. 49-50.
Heinzel H., Fitter R., Parslow J. The birds of Bri-
tain and Europe with North Africa and Middle
East. L.: CoUins, 1974. 320 p.
Hemmines A.D. Aspects of the breeding biology
of McCormick’s Skua Catharacta maccormicki
at Signy Island, South Orkney Islands // Brit.
Antarct. Surv. BuU. 1984, N 65. P. 65-79.
Hersey F.S. The present abundance of birds in the
vicinity of Fort St. Michael, Alaska // Auk. 1917.
Vol. 34. P. 147-158.
Hesse E. Neuer Beitrag zur Ornis von Sachalin //
J. fur Ornithol. 1915. Bd. 63. S. 341-402.
Hjort C. Ross's gull Rhodostethia rosea breeding in
Peary Land, North Greenland, 1979 // Dan.
ornithol. foren. tidsskr. 1980. Bd. 74, N 1/2.
S. 75-76.
Hoffman W., Wiens J.A., Scott J.M. Hybridization
between gulls (Larus glaucescens and Larus occi-
dentalis) in the Pacific North-West // Auk. 1978.
Vol. 95, N3. P. 441-458.
Holley A.J.F. Post-fledging interactions on the terri-
tory between parents and young Herring Gulls
Larus argentatus // Ibis. 1982. Vol. 124, N 2.
P. 198-203.
Holtan L.H. Nesting habitat and ecology of Aleu-
tian terns on the Copper River Delta, Alaska //
Unpublished Final Report US Forest Service,
Pacif. North-West Forest and Range Exp. Stat.
Portland (Oregon). 1980. N 56. P. 1-85.
Hosey G.R., Goodrige F. Estableshment of territo-
ries in two species of gull on Walney Island,
Cumbria // Bird Study. 1980. Vol. 27, N 2.
P. 73-80.
Howard R., Moore A. A complete check-list of
the birds of the world. L., 1984. 732 p.
Howell J.C. Observation on certain birds of the
region of Kodiak, Alaska // Auk. 1948. Vol. 65.
P. 352-358.
Hoyer H., Hoyer G. Beobachtungen in den Bulga-
rischen Salinengebieten // Falke. 1978. Bd. 25.
S. 224-228.
Hunt G.L., Burgeson B., Sanger G. Feeding ecolo-
gy of seabirds of the Eastern Bering Sea // The
Eastern Bering Sea shelf. Seattle: Univ. Wash,
press, 1981a. P. 629-647.
Hunt G.L., Eppley Z., Drury W.H. Breeding dist-
ribution and reproductive biology of marine
birds in the Eastern Bering Sea // Ibid. P. 649-
687.
Hyytia K„ Kellomaki E., Koistinen J. Suomen
lintuatlas. Helsinki: Lintutieto Oy, 1983.
520 p.
Ingolfsson A. Hybridization of Glaucous gulls Larus
hyperboreus and Herring gulls L. argentatus
in Iceland // Ibis. 1970. Vol.l 12, N 3. P. 340-362.
Isenmann P. Contribution a 1’etude de la biologie
de la reproduction et de 1’ecologie de la mouette
melanocephale Larus melanocephalus // Nos
oiseaux. 1975. Vol. 33. P. 66-73.
Isenmann P. Contribution a 1’e'tude de la biologie
de la reproduction et de I’etho-^cologie du
Gueland Railleur, Larus genei // Ardea. 1976.
Vol. 64, N 1/2. P.48—61.
Isenmann P. Donnees sur la biologie de la reproduc-
tion de la mouette rieuse en Camargue // Nos
oiseaux. 1977. Vol. 34. P. 143-154.
Isleib M.E., Kessel B. Birds of the North Gulf Coast,
Prince Wiliam Sound region, Alaska // Biol.
Pap. Univ. Alaska. 1973. Vol. 14. P. 1-149.
Ivask M. Kalakajakate kaitaumisest poegade hool-
damise perioodil // Ornitoloogiline kogumik.
Tallinn: Valgus, 1978. K. 9. Lk. 124-145.
Ivask M., Lilleleht V. Kalakajaka (Laius canus)
vanemlindude ja poegade vahelistest suhetest //
Ornitol. kogumik. Tallinn: Valgus. 1978. Bd. 8.
S. 149-169.
Jones F., White W.R. Gull control on a water cat-
chment // BuU. GuU Sci. Group. 1982. N 4.
P. 16.
Jones L. June with the birds of Washington Coast //
Wilson BuU. 1908. Vol. 20. P. 19-57.
Kampp K., Kristensen R.M. Ross’s guU Rhodostethia
rosea breeding in Disko Bay, West Greenland,
1979 // Dan. ornithol. foren. tidsskr. 1980.
Bd. 74, N 1/2. S. 65-74.
Kanel A. Die Mittwinterverbreitung der Sturmmowe
Larus canus in der Schweiz // Ornithol. Beob.
1977. Bd. 74, N2.S. 55-61.
406
Литература
Kantak F. Sturmmowen auf Langenwerder. Wit-
tenberg; Lutherstadt: Ziemsen, 1954. 135 S.
Kastepold T. Loodusevaatlusi 1980. Tallinn: Valgus,
1983. 136 1k.
Kastepold T., Kabal R. Vajaspool eesti NSV-d
Rongastatud lindude Taasleiud eesti NSV-s
1956-1980. Tallinn: Valgus. 1982. К. 1.
1401k.
Keller M. Gniezdzenie sie rybitwy bialoskrzydlej
(Chlidonias leucopterus) na Polesin Lubelskim //
Not. ornithol. 1984. Vol. 25, N 1/4. S. 62-63.
Kenyon K. W. Distribution of the Pacific kittiwake
in November and December of 1948 // Condor.
1949. Vol. 51, N 4. P. 188.
Kenyon K.W., Brooks J.W. Birds of Little Diomide
Island Alaska // Ibid. 1960. Vol. 62, N 6.
P. 457-463.
Kenyon K.W., Phillips R. Birds from the Pribilof
Islands and vicinity // Auk. 1965. Vol. 82,
N 4.P. 624-635.
Kessel B., Gibson D. Status and distribution of
Alaskan birds. Lawrence (Kansas): Cooper
Ornithol. Soc. 1978. N 1. 100 p.
Kilpi M. Populations trends and selection of nest-
sites in Larus argentatus and Larus fuscus on
the Finish coast // Ornis fenn. 1983. Vol. 60.,
N 2. P. 45-50.
Kilpi M., Puntti H., Toivonen T. Numbers of gulls
nesting on the Northern coast of Finland //
Ibid. 1980. Vol. 57, N 4. P. 153-160.
King B., Woodcock M., Dickinson E.C. A field
guide to the birds of South-East Asia. Glas-
gow: Collins, 1976. 480 p.
King W.B. Endangered birds of the world // The
ICBP Bird Red Data Book. Wash., 1981.
Kitson A.R. Larus relictus - a review // Bull. Brit.
Ornithol. Club. 1980. Vol. 100, N 3. P. 178—
184.
Kohler D., Kneis P„ Mauersberger G. Gestalt und
Verhalten zweier Bastarde Larus melanocephalus
X L. ridibundus // Mitt. Zool. Mus. Berlin.
Suppl. Ann. Ornithol. 7. 1983. Bd. 59. S. 27-46.
Komatsu M. On the habits of Larus crassirostris
on Kabushima in Pref. Aomori // Tori. Bull.
Ornithol. Soc. Jap. 1935. Vol. 8, N 40. P. 446-
461.
Koli L., Soikkeli M. Fish prey of breeding Caspian
terns in Finland // Ann. zool. fenn. 1974.
Vol. 11, N 4. P. 304-308.
Kosonen L„ Makinen R. Harmaalokin posinta
rakennukessa // Ornis fenn. 1978. Vol. 55.
P. 87.
Koubek P. Hospodarsky vyznam racka chechtaveho
(Larus ridibundus L.) // Vertebratologicke zpravy.
1982. S. 47-50.
Kumari E. The increase in the numbers of the
common gull Larus canus and its colonization
of Estonian peat-bogs in recent decades // Ornis
fenn. 1976. Vol. 53, N 2. P. 33-39.
Kumerloeve H. Silbermowenbruten auf Hausda-
chern // Ornithol. Mitt. 1957. Bd. 9. S. 112.
Kuroda N. A survey of the seabird of Teuri Island
Hokkaido, with notes on land birds // Miscell.
Rep. Yamashina’s Inst. Ornithol. and Zool.
1963. Vol. 3, N4.P. 227-238.
Laidna A. Kalakajaka poegade kasv ja suremus //
Ornitoloogiline kogumik. Tallinn: Valgus, 1978.
K. 9. Lk. 108-123.
Lambert K. Schwalbermowen-Beobachtungen, Xe-
masabini (Sabine 1819) im Sudsommer 1972-
1973 im siidafrikanischen Winterquartier //
Beitr. Vogelk. 1975. Bd. 21, N 6. S. 410-
415.
Lampio T. Waterfowl hunting in Europe, North
America and some African and Asian countries
in 1980-81 // IWRB Spec. Publ. N 3. 1983.
35 p.
Landmann A. Zur Bedeutung. der Gewasser Nord-
tirols als Rast - und Uberwinterungsstatten
fur Wasservogel (Gaviidae, Podicipedidae, Ana-
tidae, Rallidae, Laridae) // Egretta. 1981. Bd. 24,
N 1. S. 1-40.
Langham N.P.E. Chick survival in terns (Sterna spp.)
with particular reference to the common tern //
J. Anim. Ecol. 1972. Vol. 41, N 2. P. 385-
395.
Langham N.P.E. Comparative breeding biology of the
Sandwich tern // Auk. 1974. Vol. 91, N 2.
P. 255-277.
La Touche J.D.D. A handbook of the birds of Eas-
tern China. L.: Taylor and Fransis, 1931-1934.
Vol. 2.566 p.
Laybourne S. Ivory gull bathing setting on water //
Brit. Birds. 1978. Vol. 71, N 1. P. 39.
Lebreton J.D., Isenmann P. Dynamique de la popu-
lation camarguaise de Mouettes rieuses: in mo-
dele mathe'matique // Terre et vie. 1976. Vol. 30
P. 529-549.
Lemmetyinen R. Breeding success in Sterna para-
disaea Pontopp. and S. hirundo L. in Southern
Finland // Ann. zool. fenn. 1973. Vol. 10
N 4. P. 526-535.
Lemmetyinen R. Vesti - ja lokkilintjen kannan-
muutoksista gullkronan selalla, Turun saaristos-
sa vuosina 1948-1977 // Suomen riista. 1980.
N 28. S. 42-48.
Lensch A. Zum Vorkommen einigen Vogelarten
auf der Insel La Palma (Kanaren) // Ornithol.
Mitt. 1981. Bd. 33, N 9. S. 229-232.
Lind H. The reproductive behaviour of the gull-
billed Tern, Sterna nilotica Gmelin // Vid. medd.
Dan. naturhist. foren. 1963. Bd. 125. S. 407-
448.
Lipsbergs J., Lipsbergs U., Strazds A., Strazds M.
Retie un aizsargajamie putni Rigas jukas lica
ziemelaustrumu piekraste // Retie augiun dzivnie-
ki. Riga: LatZTIZPI, 1985. Lpp. 63-70.
Lofgren L. Ocean birds. L.; Canberra, 1984. 240 p.
Lonnberg E. A remarkable gull from the Gobi
Desert // Ark. zool. B. 1931. Bd. 23, H. IB.
N 2. S. 1-5.
Lonnberg E. A contribution to the bird fauna
407
Литература
of Southern Gobi // Ibid. 1932. Bd. 23, H. 2B.
N 12. S. 118.
Loutendyk P. The occurence of Хеша sabini of
the coast of Southern Africa // Ostrich. 1965.
Vol. 36, N 1. P. 15-16.
Loutendyk P. The occurence of Sabine’s gull Хеша
sabini of the Cape Peninsula // Ibid. 1968.
Vol. 39, N l.P. 9-11.
Lovenskiold H.L. Avifauna Svalbardensis with
a discussion on the geographical distribution
of the birds in Spitsbergen and adjacent islands //
Norsk Polarinst. skrift. 1964. Bd. 129. 460 S.
Luttringhaus Ch., Vauk-Hentzelt E. Ein Beitrag zur
Ernahrung auf Mullplatzen gesammelter Silber-,
Sturm- und Lachmowen (Larus argentatus,
L. canus, L. ridibundus) von Emden und Leer //
Vogelwelt. 1983. Bd. 104, N 3. S. 95-107.
MacDonaid S.D. Phantoms of the pack ice // Audu-
bon. 1976. Vol. 78, N 3. P. 3-19,
Macpherson A.H. Observations on Canadian Arctic
Larus gulls, and on the taxonomy of L. thayeri
Brooks // Techn. Pap. Arct. Inst. North. Amer.
1961. N 7. P. 1-40.
MacRoberts B.R., MacRoberts M.H. Social stimu-
lation of reproduction in Herring and Lesser
black-backed gulls // Ibis. 1972. Vol. 114, N 4.
P. 495-506.
MacRoberts M.H., MacRoberts B.R. The relation-
ship between laying date and incubation period
in Herring and black-backed gulls // Ibid. N 1.
P. 93-97.
Magi E. Jarelkurn ja munemisoime kalakajakal //
Ornitoloogiline kogumik. Tallinn: Valgus, 1978.
K. 9. Lk. 81-107.
Maher W.J. Ecology of Pomarine, parasitic and long-
tailled jaegers in Northern Alaska // Cooper Orni-
thol. Soc. 1974. N 37. P. 1-148.
Makatsch W. Die Lachmowe. Leipzig: Geest und
Portig, 1952. 98 S.
Makatsch W. Die Eier der Vogel Europas. Leipzig:
Neumann, 1974. 476 S.
Marion L., Yesou P., Dubois Ph.J., Nicolau-Guillau-
met P. Coexistence progressive de la reproduction
de Larus argentatus et de Larus cachinnans sur les
cotes atlantiques frantjaises // Alauda. 1985.
Vol. 53, N 2. P. 81-89.
Mauersberger G. Verhalten und taxonomische
Stellung der Schwarzkopfmbwe (Larus melano-
cephalus Temminck) // Beitr. Vogelk. 1970.
Bd. 15, H. 4/5. S. 210-319.
Maunder J.E., Threlfall W. The breeding biology of
the black-legged kittiwake in Newfoundland //
Auk. 1972. Vol. 89, N 4. P. 789-816.
Mazda M., Higuchi Y. Редкий случай гнездования
тихоокеанской чайки на Хоккайдо// Tori Bull.
Ornithol. Soc. Jap. 1976. Vol. 25, N 100.
P. 81-82.
McLannahan H.M.C. Some aspects of the ontogeny
of cliff nesting behaviour in the kittiwake Rissa
tridactyla and Herring Gull Larus argentatus //
Behaviour. 1973. Vol. 44, N 1/2. P. 36-88.
McNicholl M. Larid site tenacity and group adhe-
rence in relation to habitat // Auk. 1975. Vol. 92,
N l.P. 98-104.
McRae R.D. First nesting of the little gull in Manito-
ba // Amer. Birds. 1984. N 3. P. 368-396.
Meinertzhagen C.R. Nicoll's birds of Egypt. L.:
Hugh press LTD, 1930. Vol. 11. P. 350-700.
Meinertzhagen C.R. Birds of Arabia. Edinburg; L.:
Oliver and Boyd, 1954. 624 p.
Meltofte H, Edelstam C, Granstrom G., Ham-
mar J., Hjort Ch. Ross's gulls in the Arctic
pack-ice // Brit. Birds. 1981. Vol. 74, N 8.
P. 316-320.
Melville D, New Laridae records from Hong Kong//
Bull. Brit. Ornithol. Club. 1977. Vol. 97, N 2.
P. 34-37.
Melville D.S. Seabirds of China and surrounding
seas // Status and conservation of the world’s
seabirds. 1984. P. 501-511. (ICBP Techn. Publ.;
N 2).
Mende A., Sparr G. Ungewohnlicher Neststandort
der Sturmmbwe (Larus canus) // Corax. 1971.
Bd. 3,N4. S. 184-185.
Middendorff A.Th. Sibirische Reise. St.-Petersburgh,
1851. Bd. 2, T. 2: Sangethiere, Vogel und Amphi-
bien. 256 S.
Mihelsons H, Viksne J., Mednis A. The improvement
of waterfowl nesting sites // Proc. Intern. Conf.
Conservation of Wetland and Waterfowl. Heili-
genhafen, FRG, 2-6 Dec. 1974. Slimbridge:
IWRB, 1974. S. 345-348.
MilbledT., Apchain C. Nidification et migrations de
la mouette melanocephale Larus melanocephalus
sur le littoral du Nord de la France // Alauda.
1978. Vol. 46, N3.P. 235-256.
Miliotis P„ Buckley P.A. The Massachusetts Ross’s
gull // Amer. Birds. 1975. Vol. 29, N 3.
P. 643-646.
Min Byung-Yoon, Won Pyong-Oh. A winter seabird
survey on the offshore South Coast of Korean
Peninsula // Misc. Rep.Yamashina's Inst. Ornithol,
1976. Vol. 8, N l.P. 53-67.
Mpllcr A.P. Ynglebestanden af Sandterne Gelocheli-
don n. nilotica Gm. i 1972 i Europa, Africa og
Vestasien, met et tilbageblik over bestandsaend-
ringer i dette arhundrede // Dan ornithol. foren
tidsskr. 1975. Bd. 69, N 1/2. S. 1-8.
Mfiller A.P. Magernes Larinae yngleudbredelse, bes-
tandsstorrelse og aendringer i Danmark, med
supplerende oplysninger om forholdene i det
tfvrige Europa // Ibid. 1978a. Bd. 72, N 1/2.
S. 15-40.
Mfiller A.P. Skiftende Koloniplaceringer hos danske
Sandterner Gelochelidon n. nilotica Gm. // Ibid.
1978b. Bd. 72, N3.S. 119-126.
Monaghan P. The breeding ecology of urban nesting
gulls // Urban ecology: The Second European
ecological symposium. Berlin, 8-12 Sept., 1980.
Lee; Seaward: Bornkamm, 1980. P. Ill—
121.
Monaghan P., Coulson J.C. Status of the large gulls
408
Литература
nesting on buildings // Bird Study. 1977. Vol. 24,
N 2. P. 89-104.
Mori Sh.Встреча розовых чаек на Хоккайдо / /
Tori Bull. Ornithol. Soc. Jap. 1976. Vol. 25,
N 100. P. 83-84.
Mountford G., Ferguson L.I.J. Observations on the
birds of Bulgaria // Ibis. 1961. Vol. 103a.
P. 443-471.
Moyle P. Feeding behaviour of the Glaucous-winged
gull on the Alaskan salmon streams // Wilson
BuU. 1966. Vol. 78, N 2.P. 175-190.
Moynihan M. Some aspects of reproductive beha-
viour in the black-headed Gull and related spe-
cies // Behaviour. Suppl. 1955. Vol. 4. 201 p.
Moynihan M. A revision of the family Laridae //
Amer. Mus. Novit. 1959. N 1928. P. 1-42.
Mudge Cr„ Frust W. Breeding guU populations of
the inner Bristol Channel // GuU Sci. Group
BuU. 1981.N 3.P. 10-11.
Munsterhjelm L. Some ornithological notes from
a journey to SaghaUen in 1914 // Medd. Gbte-
borgs mus. zool. avd. 1922. Bd. 13. S. 1 — 112.
Murie O.J. Fauna of the Aleutian Islands and Alas-
ka Peninsula // North Amer. Fauna. 1959.
Vol. 61.406 p.
Murphy R.C. Oceanic birds on South America. N.Y.:
Amer. Mus. Nat. Hist., 1936. Vol. 2. P. 642-
1245.
Nakamura K. A black-taUled guU: Obtaining food in
flight by piracy // Tori. BuU. Ornithol. Soc. Jap.
1972. Vol. 21, N 91-92. P. 15-19.
Narita K. Some results from banding of the black-
taHled guU, Larus crassirostris in Kabu-shima //
J. Aomori Biol. Soc. 1979. Vol. 17. P. 21-26.
National Geografic Society. Field guide to the birds
of North America. Wash., 1983. 464 p.
Nehls H.W. Notwendigkeit und Ergebnisse der
Bestandsregflierung bei Mo wen (Larus) // Beitr.
Vogelk. 1979. Bd. 25, N 1/2. S. 41-49.
Nelson E. W. Report upon natural history coUections
made in Alaska between the у ears 18 7 7 and 1881//
Arct. Ser. Publ. Sign. Serv. US Army. Wash.,
1887. Vol. 3, pt 1: Birds of Alaska. P. 19-226.
Nettleship D.N. The causes and consequences of co-
loniahty in Arctic seabirds // XVIII Congr. Intern.
Ornithol.: Abstr. Symp. and Poster Present. Mos-
cow: Nauka, 1982. P. 149-150.
Newcomb R.L. Our lost explorers, the narrative of
the Jeannette Arctic expedition. Hartford, 1888.
479 p.
Nicholls C.A. Double brooding in a Western Austra-
lian population of the sUver guU, Larus novaehol-
landiae Stephens // Austral. J. Zool. 1974.
Vol. 22. P. 63-70.
Nicholson E.M. Field-notes on Greenland birds //
Ibis. 1930. Vol. 12, N 6. P. 395-428.
Nisbet I.C.T., Drury W.H.Post-fleging survival in her-
ring guUs in relation to brood-size and date of
hatching // Bird-Band. 1972. Vol. 43, N 3.
P. 161-172.
Nitecki C. Pierwsze stwierdzenie gniazdowania mewy
czarnogfowej (Larus melanocephalus) w Polsce //
Not. ornitol. 1984. Wol. 25, N 1/4. S. 57-
58.
Nordstrom G. Einige Ergebnisse der Vogelberingung
in Finland in den Jaren 1913-1962. TeU 1 //
Ornis fenn. 1963. Vol. 40, N 3. P. 81-123.
Nysewander D.R., BarbourD.B. The breeding biology
of marine birds associated with Chiniak Bay,
Kodiak Island, 1975-1978 // Environ. Assess.
Alaskan Continent. Shelf. NOAA / OCEAP. An-
nu. Rep. 1979. Vol. 2. P. 21-106.
O’Donald P. The Arctic skua: a study of the ecology
and evolution of a seabird. Cambridge: Cambrid-
ge Univ, press, 1983. 324 p.
Olson S.L. The fossil record of birds // Avian biolo-
gy. N.Y.: Acad, press, 1985. Vol. 8. P. 79-
238.
O’Meara M. Building-nesting Herring gull (Larus ar-
gentatus Pontoppidan) in Co Waterford // Irish
Natur. J. 1975. Vol. 18. P. 152-153.
Onno S. Faunistical and ecological researches // Or-
nithological researches in Estonia. Tallinn: Val-
gus. 1966. 40 p.
Onno S. Veranderungen im Bestane der in Estland
briitenden Wasservogel und ihre Griinde // Beitr.
Vogelk. 1971. Bd. 17, N 4/5. S. 339-348.
Orr C.D., Parsons J.L. Ivory gulls Pagophilla eburnea,
and ice edges in David Strait and the Labrador
Sea // Canad. Field.-Natur. 1982. Vol. 96, N 3.
P. 323-328.
Osgood W.A. A biological recognaissance of the base
of the Alaska Peninsula // North. Amer. Fauna.
1904. N 24. P. 9-86.
Owen D.F. Inland breeding by gulls in Gloucester //
J. Glouc. Natur. Soc. 1975. Vol. 24, N 9.
P. 249-250; Vol. 26, N 3. P. 35-36. N 8/9.
P. 107-108.
Pallas P.S. Zoographia Rosso-Asiatica. Petropoli,
1811. Vol. 2.374 p.
Paludan K. Contribution to the breeding biology of
Larus argentatus and Larus fuscus. Copenhagen:
Kommiss. Ejenar Munksgaard, 1951. 142 p.
Paludan K. Some behaviour patterns of Rissa tfida-
cryla // Vid. medd. Dan. naturhist. foren. 1955.
Bd. 117. S. 1-21.
Paludan K. On the birds of Afghanistan. K4>benhavn,
1959. 332 p.
Parmelee D.F, Rimmer C. Status of known-age birds
banded as chicks near Palmer Station in the
1970’s // Antarct. J.U. 1984. N 5. P. 164-165.
Parslow J.L.F. Changes in status among breeding
birds in Britain and Ireland // Brit. Birds. 1967.
Vol. 60, N 5. P. 177-202.
Parsons J. Cannibalism in Herring gulls // Ibid. 1971.
Vol. 64, N 12. P. 528-537.
Patterson I.J. Timing and spacing of broods in the
black-headed gull Larus ridibundus // Ibis. 1965.
Vol. 107, N 4. P. 433-459.
Paynter R.A. North American Herring gulls nesting
on a building // Wilson Bull. 1963. Vol. 75.
P. 88.
409
Литература
Perdeck А.С. The early reproductive behaviour of
the Arctic Skua // Ardea. 1963. Vol. 51. P. 1-
15.
Peters H.S., Burleigh T.D. Birds of Newfoundland.
St. John’s, 1951. 431 p.
Peters J.L. Check-list of birds of the world. Camb-
ridge: Harvard Univ, press. 1934. Vol. 2. 401 p.
Picozzi N. Common gull predation of winter moth
larvae // Bird Study. 1981. Vol. 28, N 1. P. 68-
69.
Piechocki R., Stubbe M., Uhlenhaut K., Sumjaa D.
Beitrage zur Aviafauna der Mongolei. Teil 3. Non-
Passeriformes // Mitt. Zool. Mus. Berlin. Suppl.
Ann. Ornithol. 5. 1981. Bd. 57. S. 71-128.
Pitelka F.A., Tomich P.Q., Treichel G.W. Ecologi-
cal relations of jaeggers and owls near Barrow,
Alaska // Ecol. Monogr. 1955. Vol. 25. P. 85-
117.
Pleske Th. Birds of the Eurasian Tundra. Boston,
1928.485 р.
Powers K.D. Lesser black-backed gull Larus fuscus
in Labrador Waters // Canad. Field-Natur. 1979.
Vol. 93, N 4. P. 445-446.
Prebl E.A., McAtee W.L. A biological survey of the
Pribilof Islands, Alaska // Birds and mammals
of the Pribilof Islands, Alaska. 1923. 128 P.
(US Bur. Biol. Surv. North Amer. Fauna.
Vol. 46.)
Pyle R.L. Hawaiian Islands region // Amer. Birds.
1983. Vol. 37,N3.P. 340-342.
Ralph C.J., Pyle R.L. Hawaiian Islans region // Ibid.
1981. Vol. 35, N 3. P. 337-338.
Renaud W.E., McLaren P.L. Ivory gull (Pagophila
eburnea) distribution in later summar and au-
tumn in Eastern Lancaster Sound and Western
Baffin Bay // Arctic. 1982. Vol. 35, N l.P. MI-
MS.
Reiser O. Materialen zu einer Ornia Balcanica. II.
Bulgarien. Wien, 1894. 204 S.
Renno O. Tommukajkas eestis // Ornitol. kogumik.
Tallinn: Valgus, 1978. K. 9. Lk. 44-50.
Ricard M. Observation des mouettes de Sabine //
Oiseaux et Rev. framj. ornithol. 1966. Vol. 36.
P. 64-65.
Robbins Ch.S. Probable interbreeding of common
and roseate terns // Brit. Birds. 1974. Vol. 67.
P. 168-170.
Robertson C.J.R. Reader’s digest complete book of
New Zealand birds Sydney, 1985. 320 p.
Rockwell E.D. Intraspecific food robbing in Glau-
cous-winged gulls // Wilson Bull, 1982. Vol. 94,
N 3. P. 282-288.
Rootsmae J., Rootsmae L. Randlindude saabumine
Eestisse 1957-1966. II // Abiks Loodusevaat-
lejale, Tartu, 1976. N 69. 96 lk.
Rootsmiie K.L, Rootsmae L. Randlindude lahku-
mine Eestist 1967-1976 // Ibid. 1978. N 73.
Lk. 3-104.
Salmon H.M. Lesser black-backed and Herring gulls
nesting on a factory roof inland in Glamorgan //
Brit. Birds. 1958. Vol. 51. P. 339-401.
Salomonsen F. Aves // Zool. Faroes. 1935. Vol. 3,
N 64, pt 2. 269 p.
Salomonsen F. Grinlands Fugle. K</>benhavn: Mun-
ksgaard, 1950. 608 S.
Salomonsen F. Ismagen (Pagophila eburnea) som
yngfuge, Gronland // Dan. ornithol. foren.
tidsskr. 1961. Bd. 55, N 4. S. 177-180.
Salomonsen F. South Polar skua Stercorarius mac-
cormicki Saunders in Greenland // Ibid. 1976.
Bd. 70. S. 81-89.
Salomonsen F., Gitz-Johansen. The birds of Green-
land. Copenhagen: Munksgaard, 1944. Pt. 2.
P. 159-347.
Sandeman P. W. Inland colonies of Lesser black-
backed gull // Scot. Birds. 1982. Vol. 12, N 4.
P. 119-120.
Sanger G.A. Pelagic records of Glaucous-winged and
Herring gulls in the North Pacific Ocean // Auk.
1973. Vol. 90. P. 384-393.
Savile B.O. Spring observations of Sabine’s gull //
Canad. Field-Natur. 1972. Vol. 86, N 4.
P. 389.
Scalon W.N. Les oiseaux du Sud de Taimyr // Ger-
faut. 1938. Vol. 28. P. 165-175.
SchaanningH.T.Z. Birds from the North-Eastern Si-
berian Arctic Ocean // Medd. Zool. Mus. Oslo.
1928. N 15. P. 1-16.
Schalow H. Die Vogel der Arktis // Fauna arct.
1905. Bd. 4. S. 79-288.
Schauensee R.M. de. The birds of China. Oxford:
Oxford Univ, press, 1984. 602 p.
Schiiz E., WeigoldH. Atlas des Vbgelzuges. B., 1931.
160 S.
Scott D.A., Hamadani H.M., Mirhosseyni A.A.
The birds of Iran. Teheran: Dep. Environ., 1975.
406 p.
Scott G. First nesting of the little gull Larus minutus
in Ontario and the New World // Auk. 1963.
Vol. 80. P. 548-549.
Sealy S.G., Bedard J., Udvardy M.D.F., Fay EH.
New record and zoogeographical notes on the
birds of St. Lawrence Island, Bering Sea // Con-
dor. 1971. Vol. 73, N 3. P. 322-336.
Seargy W.A. Foraging success in three age classes of
Glaucous-winged gulls// Auk. 1978. Vol. 95, N 3.
P. 586-588.
Sears H.F. Nesting behaviour of the gull-billed tern //
Bird-Band. 1978. Vol. 49, N 1. P. 1-16.
Sermet E., Muriset J.C. Le passage et 1’hivernage des
Laridaes sur le Haut - lac de Neuchatel // Nos
oiseaux. 1982. Vol. 36, N 386. P. 197-232.
Sharrok J.T.R. The atlas of breedign birds in Britain
and Ireland. Hertfordshire: Poyser, 1976. 477 p.
Shugart G. W. Frequency and distribution of polygy-
ny in Great Lakes Herring gulls in 1978 11 Con-
dor. 1980. Vol. 82, N 4. P. 426-429.
Sibley F.C., McFarlane R.W. Gulls in the Central
Pacific // Pacif. Sci. 1968. Vol. 22. P. 314-
321.
Silvar R., Rabago D. Primera nidificacion de la ga-
viota tridactyla (Rissa tridactyla) en el SW de
410
Литература
Europa // Ardeola. 1975. Vol. 21, N 1. P. 409-
414.
Sfobarg K. Fiskatande faglar: ett stromekosystem //
Zool. revy. 1973. Bd. 35, N 3. S. 131-134.
Skuja V. Jauns lielas klijkaijas - Stercorarius skua
Brunn. - sastap^anas gadijums Latvija // Retie
augi un dzivnieki. Riga: LatZTIZPI, 1984.
Lpp. 43-44.
Sladen A.G.L. Notes upon the eggs of the Ivory
Gull (Pagophila eburnea) // Bull. Brit. Ornithol.
Club. 1932. Vol. 52. P. 95-97.
Smith A.J.M. Studies of breeding Sandwich terns //
Brit. Birds. 1975. Vol. 68, N 4. P. 142-156.
Smith A.R., Kuyt E. Occurences of the black-legged
kittiwake, Rissa tridactyla, in the Prairie Pro-
vinces and North-Central United States // Canad.
Field.-Natur. 1983. Vol. 97, N 1. P. 111-113.
Smith N.G. Visual isolation in gulls // Sci. Amer.
1966. Vol. 217, N 4. P. 95-102.
Smith N.G. Lesser black-backed gull in Panama //
Amer. Birds. 1982. Vol. 36, N 3. P. 336-337.
Snyder L.L. Arctic birds of Canada. Toronto:
Univ. Toronto press, 1957. 310 p.
Souibb R.C., Pitts M.E. Copulatory behaviour of the
cliff-nesting red-legged kittiwake // Anim. Behav.
1982. Vol. 30, N 3. P. 931.
Sowls A.L., Hatch S.A., Lensink C.J. Catalog of
Alaskan seabird colonies. Fish and Wildlife Serv.
US. Dep. Inter. 1978. 32 p. 153 maps, tables.
Spellerberg I.F. Distribution of the McCormick
skua (Catharacta maccormicki) // Notornis.
1967. Vol. 14. P. 201-207.
Spellerberg I.F. Breeding behaviour of the McCor-
mick skua Catharacta maccormicki in Antarc-
tica // Ardea. 1971. Vol. 59, N 3/4. P. 189-
230.
Stegmann B.K. Eine neue Form von Xema sabini
Sabine // Ornithol. Monatsber. 1934a. Bd. 42,
Nl.S. 25-26.
Stegmann B.K. Uber die Formen der grossen Mo-
wen ’’subgenus Larus” und ihre gegenseitigen
Beziehungen // J. Ornithol. 1934b. Bd. 72, N 3.
S. 340-380.
Stegmann B.K. Die palaarktishen Formen der Sturm-
rnowe // Ornithol. Monatsber. 1935. Bd. 43, N 1.
S. 20-21.
Stevenson H.M. Lesser black-backed gull summering
in Florida// Florida Field.-Natur. 1975. Vol. 3,
N l.P. 22.
Stout J.E., Brass M.E. Aggressive communication by
Larus glaucescens. Part. 2. Visual communication
// Behaviour. 1969. Vol. 34, N 1/2. P. 42-54.
Stout J.F., Wilcox K.R., Creitz L.E. Aggressive com-
munication by Larus glaucescens. Part 1. Sound
communication // Ibid. P. 29-41.
Strang C.A. The Alaskan Glaucous Gull (Larus hy-
perboreus barrovianus Ridgway): autecology,
taxonomy, behaviour // Progress report 1973.
West Lafayette: Dep. Forest. Conserv. Purdue
Univ., 1973. 60 p.
Strang C.A. The Alaskan Glaucous Gull (Larus hy-
perboreus barrovianus Ridgway); autecology,
taxonomy, behaviour // Progress report 1974.
West Lafayette: Dep. Fortest. Nat. Resour.
Purdue Univ., 1974. 45 p.
Strang C.A. Variation and distribution of Glaucous
Gulls in Western Alaska // Condor. 1977.Vol. 79,
N 2.P. 170-175.
Strang C.A. Diet of Glaucous Gulls in Western Alas-
ka // Wilson Bull. 1982. Vol. 94, N 3. P. 369-
372.v
Strazdins G., Strazds A., Strazds M. Par kaijveidigo
ligdosanu uz eku jumtiem un citiem augsti ur-
banizetas vides elementiem Riga // Putnu no-
verojumi - 1987. Riga: Zinatne, 1987. S. 20-
24.
Stresemann E., Stresemann V. Die Mauser der Vo-
gel // J. Ornithol. 1966. Bd. 107. 447 S.
Stribemy W. Elfenbeinmbwe (Pagophila eburnea)
auf Sylt // Ornithol. Mitt. 1981. Bd. 33, N 2.
S. 36-40.
Stubbe M., Bolod A. Mowen und Seeschwalben (La-
ridae, Aves) der Mongolei // Mitt. Zool. Mus.
Berlin. 1971. Bd. 47, N 1. S. 51-62.
Sugawa H., Otsuki S., Gerasimow N. An attempt of
the research on the biology of the black-headed
gulls by ornithologists of Japan and USSR //
XVIII Congr. Intern. Ornithol.: Abstr. Symp.
and Poster Present. Moscow: Nauka, 1982.
P. 296.
Suter W., Wicht U. Erstes Auftreten einer Eismowe
Larus hyperboreus am Bodensee // Ornithol.
Beob. 1981. Bd. 78, N l.S. 50.
Swarth H.S. Birds of Nunivak Island // Pacif. Coast
Avifauna. 1934. Vol. 22. P. 1-64.
Swartz L.G. Sea-cliff birds // Environment of the
Cape Thomson region, Alaska. Oak-Ridge: US
Atom. Energy Comm., 1966. P. 611—678.
Swift J.J. Notes on the behaviour of whiskered
terns // Brit. Birds. 1960. Vol. 53, N 12. P. 559-
572.
Swinhoe R. Ornithological notes made at Chefoo
(North China) // Ibis. 1875. Vol. 5. P. 114—
139.
Szekely T. A fattyuszerk'd (Chlidonias hybrida)
kotlasvaltasanak leirasa es elemzese // Aquila.
1984. Vol. 91. P. 157-161.
Taczanowski L. Faune ornithologjque de la Siberie
oriental // Memoirs 1’Academie des Sci. St. Pe-
tersb. 1891-1893. T. 39. Serie VII. P. 1-1278.
Taverner J.H. Voice, behaviour and display of Me-
diterranean gulls // Brit. Birds. 1976. Vol. 69,
N l.P. 4-8.
Taylor Ph.S. Sabin’s gull sighting in Alberta // Ca-
nad. Field-Natur. 1972. Vol. 86, N 4. P. 388.
Thayer J.E., Bangs O. Notes on the birds and mam-
mals of the Arctic Coast of East Siberia // Proc.
New England Zool. Club. 1914. Vol. 5. P. 1-48.
Thomas C.S. Individual differences in aspects of the
breeding biology of female kittiwake gulls Rissa
tridactyla // Bull. Gull Sci. Group. 1982. N 4.
P. 25.
411
Литература
Thomas C.S. The relationships between breeding
experience, egg volume and reproductive suc-
cess of the kittiwake Rissa tridactyla // Ibis.
1983. Vol. 125, N 4. P. 567-574.
Thorburn A. Kittiwakes nesting on empty warehouse
// Brit. Birds. 1935. Vol. 29. P. 89.
Tinbergen N. The Herring gull’s world: A study of
the social behaviour of birds. L.: Collins, 1953.
255 p.
Tinbergen N. Comparative studies of the behaviour
of gulls (Laiidae) // Behaviour. 1959. Vol. 15,
N 1/2. P. 1-70.
Tinbergen N. The evolution of behaviour in gulls //
Sci. Amer. 1960. Vol. 203, N 6. P. 118-130.
Tinbergen N. An itroduction to the behaviour of
British gulls // The British birds of Isles by
D.A. Bannerman. Edinburg; L.: Oliver and Boyd,
1962. P. 191-200.
Tinbergen N. On adaptive radiation in gulls (Tribe
Larini) // Zool. meded. 1964. Bd. 39. S. 209-
223.
Tinbergen N. Adaptive features of the black-headed
Gull Larus ridibundus L. // Proc. XIV Intern.
Ornithol. Congr. Oxford, 24-30 July 1966.
Oxford; Edinburgh, 1967. P. 43-59.
Tuck G„ Heinzel H. Die Meeres Vogel der Welt.
Hambug; B.: Parey, 1980. S. 11-336.
Tuck L.M. The occurence of Greenland and Euro-
pean birds in Newfoundland // Bird-Band. 1971.
Vol. 42. P. 184-209.
Udagawa T. The avifauna of Teuri and Yagishiri
Island, Hokkaido // Tori Bull. Ornithol. Soc.
Jap. 1953. Vol. 13, N 63. P. 68-84.
Vader W. Makespor i blotbunnesfjaera: Gull tracks
on sandflats // Var fuglefauna. 1979a. Bd. 2,
N4. S. 206-212.
Vader W. Riskemaker raner beitende lappspover //
Fauna (Norge). 1979b. Bd. 32, N 2. S. 62-65.
Valius M. Uber die Wasservogelforschung in der
Litauischen SSR // Falke. 1974. N 5. S. 150-
. 157.
Vande Weghe J.P. Adult-juvenile ratio in the black-
headed (Larus ridibundus) and common gull (La-
rus canus) during winter // Gerfaut. 1971.
Vol. 61. P. 111-124.
Vauk-Hentzelt E., Schumann K. Zur Winterriahrung
durchziehender und rastender Sturmmowen
(Larus canus) aus dem Bereich der Insel Helgo-
land // Angew. Ornithol. 1979-1980. Bd. 5,
N4. S. 178-184.
Vaurie Ch. The status of Larus relictus and of other
hooded gulls from Central Asia // Auk. 1962.
Vol. 79, N 3. P. 303-309.
Vaurie Ch. The birds of the Palearctic fauna: A sys-
tematic reference Non-Passeriformes. L., 1965.
763 p.
Vaurie Ch. Tibet and its birds. L., 1972. 407 p.
Veen J. Functional and causal aspects of nest distri-
bution in colonies of the Sandwich tern. (Sterna
s. sandvicensis L. ) // Behaviour. Suppl. 1977.
Vol. 20. 196 p.
Verbeek N.A.M. Interactions between Herring
and Lesser black-backed gulls feeding on refu-
se // Auk. 1977. Vol. 94. N 4. P. 726-
735.
Verbeek N.A.M. Timing of primary molt and egg-
laying in glaucous-winged gulls // Wilson Bull.
1979. Vol. 91, N 3. P. 420-425.
Verbeek N.A.M., Richardson H Limits to egg size
in gulls: another point of view // J. Field Orni-
thol. 1982. Vol. 53, N 2. P. 168-170.
Vermeer K. The breeding ecology of the Glaucous-
winged gull (Larus glaucescens) on Mandrate Is-
land, B.C. // Univ. Brit. Columbia Press. 1963.
Vol. 6. 110 p.
Viksne J., Janaus M. Breeding success of the black-
headed gull Larus ridibundus in relation to the
nesting time // Ornis fenn. 1980. Vol. 57, N 1.
P. 1-10.
Voous K.N. Atlas of European Birds. London:
1960.284 р.
Wada K. On migratory seasons of the black-tailed
gull at Kabushima // Tori Bull. Ornithol. Soc.
Jap. 1932. Vol. 7, N 33/34. P. 300-301.
Wadewitz O. Die Sturmmbwe auf der Fahinsel //
Falke. 1978. Bd. 25, N 7. S. 230-236.
Wahl T.R. Glaucous-winged gull nesting at a Lake
Whatcom, Bellingham // Murrelet. 1972. Vol. 53,
N 3.P. 51.
Walker E.P. Southerly nesting records of the Arctic
Tern in South-Eastern Alaska // Condor. 1920.
Vol. 22, N 2. P. 72-73.
Wallace D.I.M. Studies of less familiar birds. 128.
Slender-billed gull // Brit. Birds. 1964. Vol. 57,
N6.P. 242-247.
Walter H. Vergleichende Untersuchungen an den
Raubmowen Stercorarius parasiticus und longi-
caudus // J. Ornithol. 1962. Bd. 103. S. 166-
179.
Walters M. The complete birds of the world. L.:
David and Charles, 1980. 340 p.
Watanuki Y. Size selective hunting by Slaty-baked
gulls (Larus schistisagus) and influence on fled-
ging success of black-tailled gulls (L. crassiros-
tris) // J. Yamashina Inst. Ornithol. 1982.
Vol. 14. P. 25-34.
Webb B.E., Conry J.A. First record of the Lesser
black-backed gull in Colorado // West. Birds.
1978. Vol. 9, N 4. P. 171-173.
Weidmann U. Some reproductive activities of the
common gull Larus canus L. // Ardea. 1955.
Vol. 43. P. 85-132.
Weidmann U. Observations and experiments on egg-
laying in the black-headed gull // Brit. J. Anim.
Behav. 1956. Vol. 4. P. 150-161.
Wesolowski T, Gfazewcka E., Gfazewski L. et al.
Rozmieszczenie i liczebnosc ptakow siewkowa-
tych mew i rybitw gniazdujacych na wyspach
Wisly srodkowej // Acta ornithologica. 1984.
Vol. 20.N2. P. 159-185.
Wesolowski T, Glazewska E., Giazewski L. et al.
Size, habitat distribution and site turnover of
412
Литература
gull and tern colonies on the middle Vistula //
Ibid. 1985, Vol. 21, N 2. P. 45-67.
Willett G. Summer birds of Forrester Island, Alas-
ka // Auk. 1915. Vol. 32. P. 295-305.
Williams R.B. Notes on the Arctic tern in Alexander
Archipelago, South-Eastern Alaska // Ibid. 1947.
Vol. 64. P. 143-144.
Williamson K. The Atlantic Islands. L., 1970.
Winkler K. - Der Schweizer ’’Polarmowen” - Beleg
erweist sich als Eismowe Larus hyperboreus //
Ornithol. Beob. 1981. Bd. 78, N 1. S. 50-51.
Witherby H.F., Jordain F.C.R., Ticehurst N.F.,
Tucker B.W. The handbook of British birds. L.,
1941. Vol. 5. 382 p.
Wolford J.W. Sabin’s gulls in Alberta // Canad.
Field-Natur. 1972. Vol. 86, N 4. P. 372.
Woller R.D., Dunlop J.N. Multiple laying by the
silver gull Larus novaehollandiae Stephens, on
Carnac Island, Western Australia // Austral.
Wildlife Res. 1979. Vol. 6. P. 325-335.
Wolters H.E. Die Vogelarten der Erde. Bd. 1. Lie-
ferung - Boden 1-5. Hamburg: B., 1975. S. 1-
80.
Won Hong Koo. The birds of Korea. Pyongyang,
1964. Vol. 2. 344 p.
Won Pyong-Oh. Check-list of the birds of the Repub-
lic of Korea. Seoul, 1976. 30 p.
Wood R. C. Population dynamics of breeding South
Polar skua of unknown age // Auk. 1971.
Vol. 88, N 4. P. 805-814.
Wooler R.D. Repead laying by kittiwakes Rissa tri-
dactyla // Ibis. 1980. Vol. 122. N 2. P. 226-229.
Wright N.J.R., Matthews D. W. New nesting colonies
of the Ivory gull Pagophila eburnea in Southern
East Greenland // Dan. ornithol. foren. tidsskr.
1980. Bd. 74, N 1/2. S. 59-64.
Wust W. Verlauf und Ergebnisse zweier Fruhjahrs-
fahrten in den Jahren 1974 und 1975 durch
Zentralasien: Usbekistan, Tadschikistan und (nur
1974) Kasachstan // Anz. Ornithol. Ges. Bayern.
1976. Bd. 15, N2/3. S. 121-160.
Young E.C. The breeding behaviour of the South
Polar skua Catharacta maccormicki // Ibis.
1963a. Vol. 105, N 2. P. 203-233.
Young E.C. Feeding habits of the South Polar skua
// Ibid. 1963b. Vol. 105, N 3. P. 301-318.
Young E.C. Territory establishment and stability
in McCormick’s skua // Ibid. 1972. Vol. 114,
N 2. P. 234-244.
Ytreberg N.J, Contribution to the breeding biology
of the black-headed gull (Larus ridibundus L.)
in Norway // Nytt mag. zool. 1956. Bd. 4. S. 5-
106.
Zelickman E.A., Golovkin A.N. Composition, struc-
ture and productivity of neretic plankton com-
munities near the bird colonies of the Notern
shores of Novaya Zemlya // Mar. Biol. 1972.
Vol. 17, N 3. P. 265-274.
Zonfrillo B. Isle of May bird observatory and field
station report for 1981 // Scot. Birds. 1982.
Vol. 12. P. 43-49.
Zubakin V.A., Flint W.E. Okologie und Verhalten
der Relictmiiwe (Larus relictus L’dnnb.) // Beitr.
Vogelk. 1980. Bd. 26, N 5. S. 253-275.
Оглавление
ПРЕДИСЛОВИЕ
5
ПОДОТРЯД ЧАЙКОВЫЕ
В.А. ЗУБАКИН
7
СЕМЕЙСТВО ПОМОРННКОВЫЕ
В.Е. ФЛИНТ
10
Род Stercorarius Brisson, 1760 .... 11
Большой поморник ...................11
Южно-полярный поморник .............18
Средний поморник................21
Короткохвостый поморник.........30
Длиннохвостый поморник..........38
СЕМЕЙСТВО ЧАЙКОВЫЕ
В.А. ЗУБАКИН
48
Род Pagophila Каир, 1829 ........ 51
Белая чайка.......................51
К.А. ЮДИН, Л.В. ФИРСОВА
Род Larus Linnaeus, 1758 .........57
Черноголовый хохотун..............57
В.А. ЗУБАКИН
Реликтовая чайка..................69
В.А. ЗУБАКИН
Черноголовая чайка . . . .........77
В.А. ЗУБАКИН
Озерная чайка ....................85
Я.А. ВИКСНЕ
Буроголовая чайка.................98
В.А. ЗУБАКИН
Морской голубок..................105
В.А. ЗУБАКИН
Китайская чайка..................115
В.А. ЗУБАКИН
Клуша............................118
К.А. ЮДИН, Л.В. ФИРСОВА
Серебристая чайка .............126
К.А. ЮДИН, Л.В. ФИРСОВА
Тихоокеанская чайка ...........146
К.А. ЮДИН, Л.В. ФИРСОВА
Серокрылая чайка ..............153
К.А. ЮДИН, Л.В. ФИРСОВА
Полярная чайка ................161
К.А. ЮДИН, Л.В. ФИРСОВА
Бургомистр.....................165
К.А. ЮДИН, Л.В. ФИРСОВА
Морская чайка..................174
К.А. ЮДИН, Л.В. ФИРСОВА
Сизая чайка....................182
К.А. ЮДИН, Л.В. ФИРСОВА
Чернохвостая чайка ............199
Т.Б. ЛИТВИНЕНКО
Род Хеша Leach, 1819...........207
Вилохвостая чайка..............207
К.А. ЮДИН, Л.В. ФИРСОВА
414
Оглавление
Род Rissa Stephens, 1826..........215
К.А. ЮДИН, Л.В. ФИРСОВА
Моевка .......................... 215
Красноногая моевка................226
Род Hydrocoloeus Каир, 1829 ..... 233
Малая чайка.......................233
В.А. ЗУБАКИН
Род Rhodostethia MacGillivray, 1842 . . . 244
Розовая чайка.....................244
В.А. ЗУБАКИН, А.А. КИЩИНСКИЙ,
В.Е. ФЛИНТ, В.О. АВДАНИН
Род Chlidonias Rafinesque, 1822 . 258
В.А. ЗУБАКИН
Черная крачка ....................258
Белокрылая крачка ................268
Белощекая крачка..................278
Род Gelochelidon C.L. Brehm, 1830 . . . 287
Чайконосая крачка................287
В.А. ЗУБАКИН
Род Hydroprogne Каир, 1829 ..... 299
Чеграва ........................ 299
В.А. ЗУБАКИН
Род Thalasseus Boie, 1822 ...... 310
Пестроносая крачка ..............310
В.А. ЗУБАКИН
Род Sterna Linnaeus, 1758 ...... 321
Речная крачка....................321
В.А. ЗУБАКИН
Полярная крачка .................337
В.А. ЗУБАКИН
Камчатская крачка................348
В.А. НЕЧАЕВ, Е.Г. ЛОБКОВ
Малая крачка.....................356
В.А. ЗУБАКИН
ЛИТЕРАТУРА
371
Научное издание
ПТИЦЫ СССР
ЧАЙКОВЫЕ
Утверждено к печати
Институтом
эволюционной морфологии
н экологии животных
им. АЛ. Северцова
Академии наук СССР
Редактор
В.М. Гаврилов
Редактор издательства
Г.М. Орлова
Художник
Н.И. Шевцов
Художественный редактор
НЛ. Власик
Технические редакторы
Г.П. Каренина, BJB. Лебедева
Корректоры
Е А. Мишина, Н.И. Харламова
Набор выполнен в издательстве
на наборно-печатающих автоматах
ИБ № 35141
Подписано к печати 27.08.87. Т — 15765
Формат 70 X 100 1/16. Бумага офсетная № 1
Гарнитура Пресс-Роман. Печать офсетная
Усл. печл. 33,8 +1,0 вкл. Усл. кр.-отт. 41,6
Уч.-изд-л. 41,2. Тираж 12900 экз.
Тип.зак.826
Цена 3 р. 30 к.
Ордена Трудового Красного Знамени
издательство ’’Наука”
117864 ГСП-7, Москва В-485
Профсоюзная ул., д. 90
2-я типография издательства ’’Наука”
121099, Москва, Г—99
Шубинский пер .,10
Таблица I. Поморники
/—3—большой поморник (/— взрослая птица, 2 — взрослая птица в полете. 3 — молодая птица);
4 — южно-полярный поморник (взрослая птица); 5—8— средний поморник (5 — взрослая птица свет-
лой морфы, 6 — взрослая птица темной морфы, 7 — взрослая птица светлой морфы в полете, 8 —
молодая птица); 9—12 — короткохвостый поморник (9— взрослая птица светлой морфы; 10 — взрос-
лая птица темной морфы, // — взрослая птица светлой морфы в полете, 12—молодая птица в
полете); 13—/5 — длиннохвостый поморник (13— взрослая птица светлой морфы, 14 — взрослая
птица светлой морфы в полете, 15 — молодая птица)
Таблица II. Чайки
1—2 —полярная чайка (1— в брачном наряде, 2— в полете летом), 3—8 — бургомистр (3— в брач-
ном наряде, 4— в полете летом, 5 — голова взрослой птицы зимой, 6 — молодая птица, 7 — птица
в первом зимнем наряде, 8 — птица во втором зимием наряде) 9—14 — серокрылая чайка (9 —
в брачном наряде, 10 — в брачном наряде в полете, 11—голова взрослой птицы зимой, 12 —'Взрос-
лая птица зимой в полете, 13— молодая птица, 14 — молодая птица в полете); 15—19 — черно-
хвостая чайка (15 — в брачном наряде, 16 — в полете летом, 17 — голова взрослой птицы зимой,
18 — молодая птица, 19 — молодая птица в полете)
Таблица Ш. Чайки
/—//—серебристая чайка (/—2 — L. a. argentatus в брачном наряде, 3 — голова птицы зимой,
4 L. a. antelius, 5 — L. a. taimyrensis, 6— L. a. armenicus, 7 — L. a. mongolicus, 8 — L. a. vegae,
9 — L. a. cachinnans, 10 — молодая птица, II— молодая птица в полете); 12—16 — сизая чайка
(12 — в брачном наряде, 13 — в полете летом, 14 —голова взрослой птицы зимой, 15 — в первом
зимнем наряде, 16 — во втором зимнем наряде); /7—19— белая чайка (17— в брачном наряде,
18 — летом в полете, 19 — в первом зимнем наряде)
Таблица IV. Чайки
/—5—клуша (1— в брачном наряде, 2— в полете летом, 3 —голова взрослой птицы зимой, 4— мо-
лодая птица, 5 —молодая птица в полете); 6—5 — морская чайка (6—в брачном наряде, 7 —летом
в полете, 8 — голова взрослой птицы зимой, 9 — молодая птица); 10—13 — тихоокеанская чайка
(10— в брачном наряде, // — летом в полете, 12 — голова взрослой птицы зимой, 13— молодая
птица); 14—19— черноголовый хохотун (14— в брачном наряде, /5 — летом в полете, 16 — голова
взрослой птицы зимой, 17 — молодая птица, 18 — молодая птица в полете, 19—птица во втором
зимнем наряде)
Таблица V. Чайки
1—5 — реликтовая чайка (/ — в брачном наряде, 2 — летом в полете, 3 — голова птицы зимой,
4 — молодая птица, 5 — молодая птица в полете); 6—11— черноголовая чайка (6 — в брачном на-
ряде, 7 — летом в полете, 8 — голова взрослой птицы зимой, 9 — молодая птица, 10 — молодая
птица в полете, И— птица во втором зимнем наряде); 12—18 — малая чайка (12 — в брачном на-
ряде, 13—14 — летом в полете, 15 — голова взрослой птицы зимой, 16 — молодая птица, 17 — моло-
дая птица в полете, 18 — птнца в первом летнем наряде в полете)
'/2 И Зак. 826
Таблица VI. Чайки
1—5 — озерная чайка (/ — в брачном наряде, 2 — летом в полете, 3 — голова взрослой птицы зимой.
4 — молодая птица. 5—птица в первом зимнем наряде в полете); 6—9—-морской голубок
(6 — в брачном наряде, 7 — летом в полете, 8 — молодая птица, 9— птица в первом зимнем наряде
в полете); 10—14— буроголовая чайка (10— в брачном наряде, 11— летом в полете, 12 — голова
взрослой птицы зимой, 13 — молодая птица, 14— молодая птица в полете); 15—/7 — китайская
чайка (15— в брачном наряде, 16— летом в полете, 17—голова взрослой птицы зимой)
Таблица VII. Чайки
1—5— розовая чайка (/ — в брачном наряде, 2 — летом в полете, 3—голова взрослой птицы зимой.
4 — молодая птица, 5 — птица в первом летнем наряде); 6—10— вилохвостая чайка (6 — в брачном
наряде, 7 — летом в полете, 8 — голова взрослой птицы зимой, 9— молодая птица, 10 — птица в
первом зимнем наряде); 11—15 — моевка (11 — в брачном наряде, 12 — летом в полете, 13—голова
взрослой птицы зимой, 14 — молодая птица, 15 — молодая птица в полете); 16—19—красноногая
моевка (16 — в брачном наряде, /7 — летом в полете, 18 — молодая птица, 19 — молщ&я птица в
полете)
Таблица VIII- Крачки
/—4 — черная крачка (/ — в брачном наряде, 2 — летом в полете, 3 — зимой в полете, 4 —молодая
птица в полете); 5—8— белокрылая крачка (5 — в брачном наряде, 6— летом в полете, 7 — зимой
в полете, 8 — молодая птица в полете); 9—12 — белошекая крачка (9 — в брачном наряде, to —
летом в полете, // — зимой в полете. 12 — молодая птица в полете); /3—/6—малая крачка (13 —
в брачном наряде, /4 — летом в полете. 15—голова взрослой птицы зимой, 16— молодая птица в
полете); 17—20 — камчатская крачка (/7 — в брачном наряде, 18 — летом в полете, 19 — голова
взрослой птицы зимой, 20 — молодая птица в полете)
Таблица IX. Крачки
1—4 — чеграва (/ — в брачном наряде, 2 — летом в полете, 3 — голова взрослой птицы зимой. 4 —
молодая птица в полете); 5—8 — чайкой оса я крачка (5 — в брачном наряде. 6 — летом в полете.
7— голова взрослой птицы зимой, 8— молодая пгица в полете); 9—12 — пестроносая крачка
(9— в брачном наряде, 10— летом в полете, 11— голова взрослой птицы зимой, 12—молодая
птица в полете); 13—17 — речная крачка (13—S. h. hirundo в брачном наряде, 14— летом в по-
лете, 15 — S. h. longipennis летом в полете, 16 — голова речной крачки зимой, 17 — молодая птица
в полете); 18—20 — полярная крачка (18 — в брачном наряде, 19 — летом в полете, 20— молодая
птица)
Таблица X. Птенцы
/ — короткохвостый поморник, 2 — моевка, 3— белая чайка, 4 — черноголовый хохотун, 5 — релик-
товая чайка, 6— черноголовая чайка, 7 — бурогсловая чайка, 8 — морской голубок. 9 — озерная
чайка, 10 — серебристая чайка, 11 — клуша, 12 — бургомистр, 13 — сизая чайка, 14 — морская чайка
Таблица XI. Птенцы
1 — южно-полярный поморник, 2 — тихоокеанская чайка, 3 — малая чайка, 4 — вилохвостая чайка,
5 —розовая чайка. 6—7 — черная крачка, 8—9 — белокрылая крачка, 10 — белощекая крачка, 11 —
чеграва, 12 — чайконосая крачка, 13 — пестроносая крачка, 14—15 — малая крачка, 16—17 — речная
крачка, 18—полярная кпачка
?O. C L
Таблица XII. Яйца
/—серебристая чайка, 2 — черноголовый хохотун, 3 — черноголовая чайка,
вилохвостая чайка, 6 — короткохвостый поморник, 7 — моевка, 8 — чеграва,
10—// — черная крачка, 12—13 — речная крачка
4 — розовая чайка, 5 —
9 — чайконосая крачка,
•НАУКА*