/





















































































































































































































































































Author: Семенов-Тян-Шанский О. И. Гилязов А.С.
Tags: aves птицы в целом орнитология биология зоология птицы
ISBN: 5-02-005396-1
Year: 1991




Text
А.С.ГИЛЯ30В
АКАДЕМИЯ НАУК СССР
МОСКОВСКОЕ ОБЩЕСТВО ИСПЫТАТЕЛЕЙ ПРИРОДЫ
О.И.СЕМЕНОВ-ТЯН-ШАНСКИЙ
А.С.ГИЛЯЗОВ
ПТИЦЫ
ЛАПЛАНДИИ
МОСКВА"НАУКА”
1991
УДК 598.2
Птицы Лапландии / О.И. Семенов-Тян-Шанский, А.С. Гилязов -
М.; Наука, 1991. 288 с. ISBN 5-02-005396-1
Эколого-фаунистическая сводка по птицам внутренней части Мурманской
области. Описано 192 вида, обнаруженных за время существования Лапланд-
ского заповедника с 1930 по 1987 г. Данные по экологии, динамике популя-
ций приведены за 40—50 лет.
Для зоологов и специалистов, интересующихся вопросами охраны при-
роды.
Табл. 18, ил. 59, библ. 97 назв.
Birds of Lapland / O.l. Semyonov-Tyan-Shansky,- A.S. Guylyazov. M.: Nauka,
1991,288 p. ISBN 5-02-005396-1
The book presents an ecological and faunistic survey of birds of the interior part of the
Murmansk Region.
It includes characteristics of 192 species, observed since the foundation of the Lapland
Reserve in 1930 and up to the 1987. Data on ecology and population dynamics covers the
period of 40—50 years.
Might be of interest for zoologists and specialists involved in nature conservation.
Tabl. 18, fig. 59, bibl. 97.
Ответственный редактор
доктор биологических наук Т.В. Кошкина
Рецензенты:
доктор биологических наук Р.П. Потапов,
кандидат биологических наук Ю.Б. Пукинский
Издание научное
[СЕМЕНОВ-ТЯН-ШАНСКИЙ Олег Иэмайлович|
ГИЛЯЗОВ Александр Сабирович
ПТИЦЫ ЛАПЛАНДИИ
Утверждено к печати
Московским обществом испытателей природы
Редактор издательства Г.М. Орлова. Художественный редактор Н.Н. Михайлова
Технический редактор Г.И. Астахова. Корректор З.Д. Алексеева
Набор выполнен в издательстве на наборно-печатающих автоматах
ИБ № 48112
Подписано к печати 09.01.91. Формат 60 X 90 1/16. Бумага офсетная № 1
Гарнитура Пресс-Роман. Печать офсетная. Усл.печ.л. 18,0. Усл.кр.-отт. 18,0
Уч.-издл. 22,3. Тираж 1500 экз. Тип. зак. 1023. Цена 4р. 80к.
Ордена Трудового Красного Знамени издательство ’’Наука”
117864, Москва В-485, Профсоюзная ул., д. 90
Ордена Трудового Красного Знамени 1-я типография издательства ’’Наука”
199034, Ленинград В-34, 9-я линия, 12
С 1907000000-«оз 551 J полугодие © издательство ’’Наука”, 1991
U*rZ^ vZ/ 1
ISBN 5-02-005396-1
ПРЕДИСЛОВИЕ
Составленный В.В. Бианки обзор истории орнитологических исследо-
ваний на Кольском полуострове (1982, с. 20-25) показал, что старые
работы имели преимущественно фаунистическое направление и в боль-
шей мере охватывали морские побережья, чем внутреннюю горно-лесную
территорию полуострова. Организация двух государственных заповедни-
ков (в 1930 г. - Лапландского, комплексного, в бассейне оз. Имандра и в
1932 г. - Кандалакшского, преимущественно орнитологического, состоя-
щего из ряда участков, разбросанных по островам и побережьям Белого и
Баренцева морей) создала необходимую базу для стационарных экологи-
ческих исследований. При этом Кандалакшский заповедник в силу
своего географического положения изучает в основном морских и при-
брежных птиц, включая и равнинную тундру, а Лапландский - авифауну
лесной зоны и горных тундр.
Настоящая работа характеризует только внутренние, материковые
части Кольского полуострова, в основном по материалам Лапландского
заповедника, как ранее опубликованным, так и рукописным (важнейшие
из них - ’’Летопись природы” за 30 лет (1958-1987), картотечный фонд и
фенологические журналы, ведущиеся с 1930 г.) Хотя в тематике Лапланд-
ского заповедника преобладало териологическое направление, достаточ-
но внимания уделялось и птицам. По орнитологической тематике здесь
работали О.И. Семенов-Тян-Шанский (1930-1988), В.Ф. Седова (1934),
П.С. Мальчевский (1937), Г.А. Новиков (1937-1939), М.И. Владимирская
(1937-1941), Ю.Г. Гаев (1958-1965), А.Б. Брагин (1966-1983), С.Н. Баккал
(1971-1978), А.С. Гилязов (1976-1988), Д.В. Карелин (1980) и Л.В. Кирен-
ков (1986-1988). Практически в сборе материалов участвовал весь кол-
лектив заповедника - от лесников и проходивших производственную
практику студентов до директоров.
В деятельности заповедника было два перерыва: первый в годы
Великой Отечественной войны (1941-1945), второй - с 1951 г., когда
заповедник был закрыт, до 1958 г., когда он был восстановлен на той же
территории. Несмотря на эти разрывы, заповеднику удалось сохранить
преемственность работ и почти все свои научные фонды. Благодаря этому
фенологические ряды по некоторым явлениям уже превышают 50 лет и
стали уникальными для всей Субарктики.
На протяжении этого времени методика исследований сильно измени-
лась: до 1941 г. значительная часть животных добывалась путем отстрела,
а с 1948 г. приоритет получило изучение жизни птиц в природной обста-
новке с применением дистанционных самописцев, ружье уступило место
фотоаппарату. Раньше основным источником сведений о питании птиц
был анализ содержимого желудков и зобов, а теперь для этой цели
служат погадки и остатки добычи хищных птиц, временная перевязка
пищевода гнездовых птенцов певчих и визуальные наблюдения над
всеми остальными.
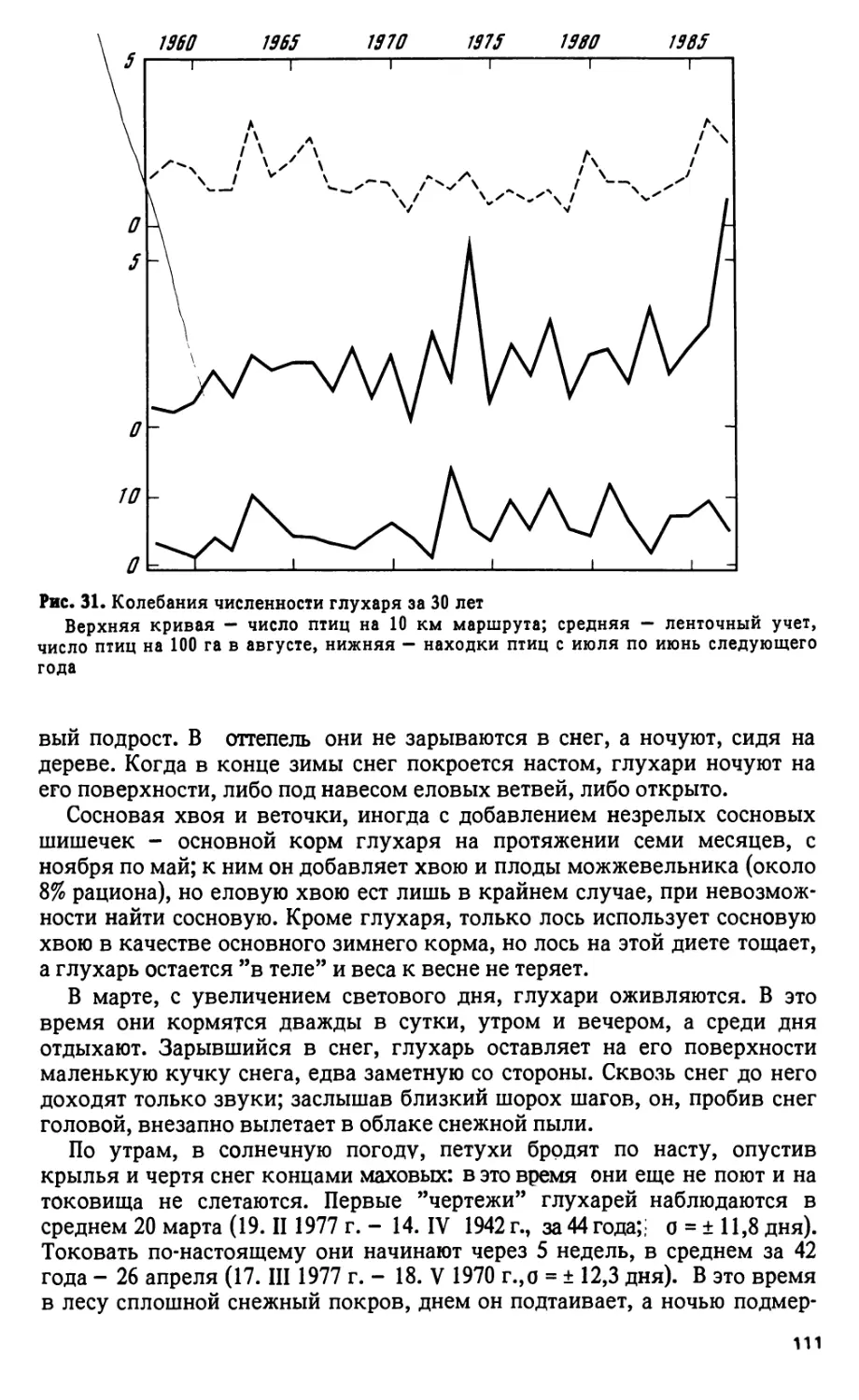
В заповеднике ежегодно проводятся учеты птиц, с 1936 г. - тетереви-
ных на ленточных пробах общей площадью 560 га, с 1966 г. - водопла-
вающих на береговых маршрутах протяженностью ПО км (А.Б. Брагин,
з
А.С. Гилязов), с 1977 г. - певчих птиц на стационарных площадках
(А.С. Гилязов).
Коллекционный материал для первой эколого-фаунистической сводки
’’Птицы Лапландского заповедника” (Владимирская, 1948) в объеме
320 экз. был определен проф. Г.П. Дементьевым до подвида. Ревизовать
эту работу ввиду отсутствия новых сборов не имеет смысла, в связи с
чем в настоящей работе, преимущественно экологической, мы ограничи-
лись бинарной номенклатурой. Приведенные в тексте промеры птиц
заимствованы из картотеки биосъемки заповедника.
Система птиц и транскрипция названий даются по ’’Каталогу птиц
СССР” А.И. Иванова (1976). Местные названия приводятся, только если
они отличаются от общепринятых, с использованием словаря ’’Живая
речь Кольских поморов” И.С. Меркурьева (1979). Саамские названия
записаны нами со слов работавших в заповеднике саамов Ф.К. и А.К. Ар-
хиповых (Екостровский погост), А.С. Сергина (Бабинский погост), и
И.П. Герасимова и И.П. Мошникова (Нотозерский погост) и др.
Наша задача состояла в возможно полном использовании оригиналь-
ных материалов, а не в составлении литературной сводки, поэтому
список цитированной литературы далек от полноты. То же относится и к
результатам кольцевания птиц. Особое внимание мы уделили характе-
ристике численности птиц в период исследования и их изменениям, как
периодическим, так и вековым. Сроки прилета и отлета птиц даются
сокращенно - средняя дата, предельные и число лет наблюдений; для
большинства видов приводятся также среднее квадратическое отклоне-
ние (сигма) (этот показатель лучше характеризует изменчивость дат, чем
амплитуда) и температура дня наступления явления, снятая с графика
средних декадных температур метеостанции Хибины за 25 лет (1960-
1984). Местами даны коэффициенты корреляции между датой явления по
годам и температурой воздуха соответствующего месяца или соседними
периодическими явлениями. Даты прилета и отлета некоторых птиц за
каждый год опубликованы в другом издании (Аблаева, Семенов-Тян-
Шанский, 1983).
Книга представляет собой своего рода отчет об орнитологических
исследованиях Лапландского заповедника за 58 лет его существования,
это надо иметь в виду, поскольку мы не могли назвать автора каждого
наблюдения поименно. При составлении книги А.С. Гилязов обработал
отряд воробьиных, а О.И. Семенов-Тян-Шанский все остальные отряды,
им же написаны предисловие и заключение. Авторы фотографий указаны
в подписях к ним.
При подготовке рукописи к печати большую техническую помощь
оказали нам сотрудники Лапландского заповедника Е.В. Гилязова,
Р.И. Катаева и Н.В. Костюкович. Пользуемся возможностью поблагода-
рить их за участие и содействие.
ОТРЯД ГАГАРЫ - GAVIIFORMES
Краснозобая гагара - Gavia stellata (Pont.)
Местные названия: саамское - лашш-тбут (болотная гагара).
Встречается и гнездится по всей территории Кольского полуострова,
хотя и реже чернозобой гагары: в Лапландском заповеднике зарегистри-
рованы 84 визуальные встречи краснозобой и 3494 встречи чернозобой
гагары, в соотношении 1:42. Такая разница объясняется не только относи-
тельной редкостью, но и более скрытным образом жизни первого вида:
чернозобая держится на открытом пространстве озер и легко доступна
для наблюдения, тогда как краснозобая гнездится на маленьких уеди-
ненных озерках в глубине лесов и болот, ввиду чего редко попадается на
глаза. В 30-х годах она довольно регулярно встречалась и, вероятно,
гнездилась на Кокоринском озерке в юго-восточной части заповедника
(годы 1931, 1934, 1937, 1938, 1941) и на Лебяжьем озерке в долине р. Чуна
(1932, 1934, 1938), но эти места часто посещаются людьми и в послевоен-
ные годы гагар на них уже не встречали. Эпизодически их наблюдали на
Садозере, Райкор-яурынче, безымянных озерках под Ягельной тундрой и
Ельявруайвенчем, на Нижнем Гарьюсном озере; водная площадь этих
озер в пределах 1-25 га. Из более крупных озер (50-150 га) их встречали
на Румель-, Улынч-, Тулп-, Чингльс-явре, Островском и Купись, но на
таких больших, как Чунозеро или Пиренгское, попадались только стайки
холостых особей. С 1931 по 1941 г., и в 1948-1950 гг. кразнозобых гагар
отмечали почти ежегодно, в среднем по 4,0 птицы за сезон, но с 1951 г. по 1968 г.
не было ни одной встречи; позднее, с 1968 по 1984 г., их видели почти
каждое лето (по 4,4 особи), а в 1985-1987 гг. не наблюдали ни разу. В
1972-1984 гг. пара гагар регулярно гнездилась на заболоченном, с водной
площадью не более 3 га, Гагарьем озерке у подножия Чуньявр-вараки.
Потаенных лесных озерков, где может гнездиться эта птица, в заповедни-
ке немало: численность ее можно оценить в 2-5 пар.
Прилетает краснозобая гагара позднее чернозобой, в среднем за 18 лет
28 мая (17. V 1977 г., 12. VI 1985 г.); среднее квадратичное отклонение
даты прилета + 7, 9 дней, средняя температура 6,5е. Отлет не прослежен,
самая поздняя встреча - 17. IX 1937 г. Зимуют краснозобые гагары на
морях у берегов Западной и Южной Европы.
Гнездовые участки они занимают сразу по прилете, заявляя о своем
прибытии далеко слышным брачным криком ”ку-куии, ку-куии, ку-
куии...” По этим крикам 23. V 1931 г. мы вышли на Кокоринское озерко
и впервые увидели брачные игры двух пар гагар; в сентябре 1934 г.
встретили выводок (1 птенец), 25. V 1938 г. токующую пару наблюдал
Г.А. Новиков, а в июле 1968 г. А.Б. Брагин нашел разбитое насиженное яйцо.
31. V 1972 г. он же описал брачную игру гагар на заберегах оз. Улын-
чявр: ’’Длительные, звонкие, согласованные и разнообразные крики,
напоминающие голоса лебедей; крича, гагара загибала клюв к воде”.
31. V 1978 г. мы наблюдали на Гагарьем озерке молчаливое ухаживание,
одна гагара ложилась на воде перед партнером на бок, демонстрируя
белое брюхо и черные лапы; там же 27. V 1981 г. слышали брачные крики,
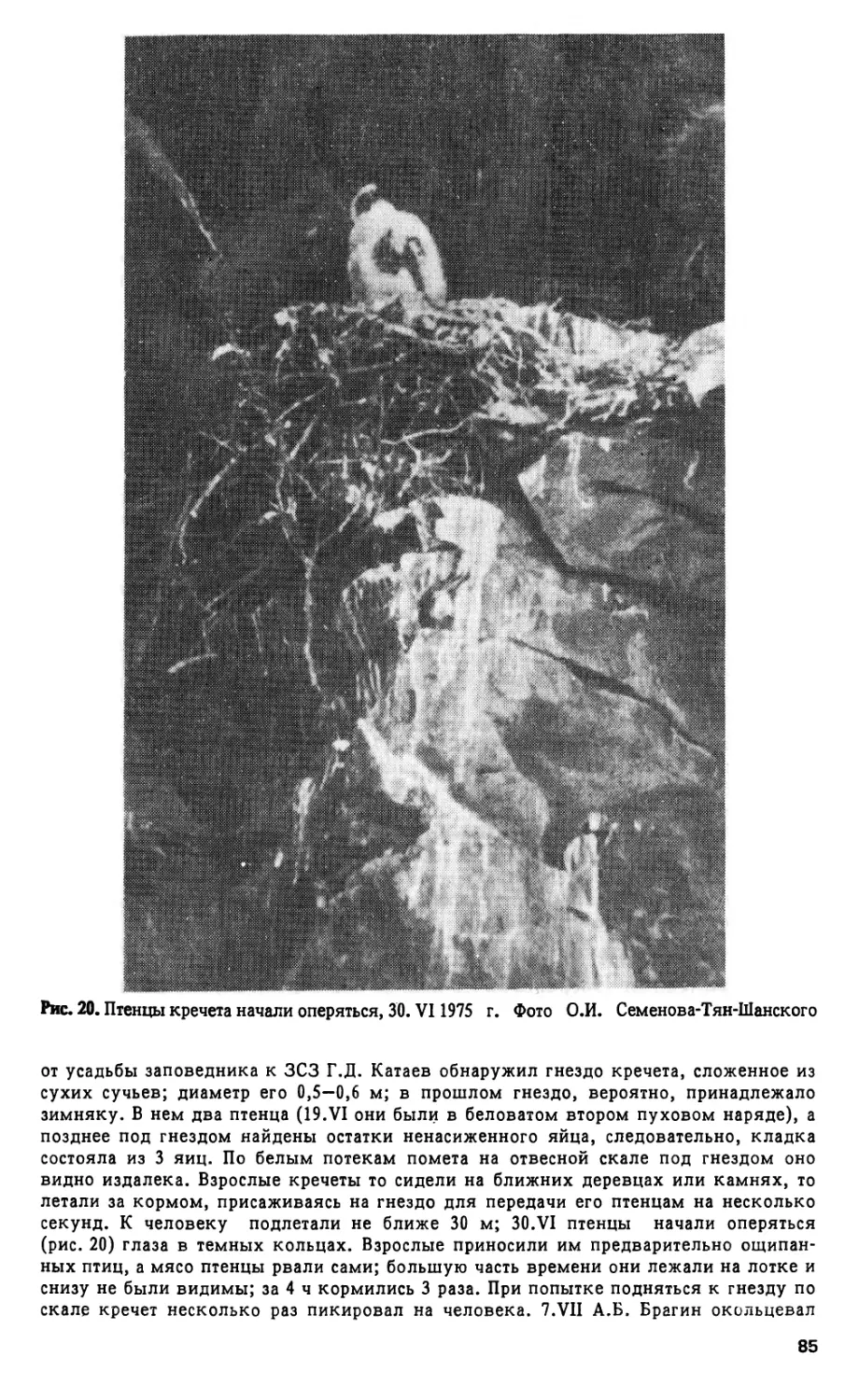
5
a 4. VII 1974 г. А.Б. Брагин нашел гнездо, устроенное на кочке, заросшей
водной осокой (Carex aquatilis), посреди плеса, в 30 м от ближайшего
берега: в нем было 2 яйца. Потом гагары гнездились на той же самой
кочке в 1979, 1981, 1982 и 1985 гг. - в последнем случае 7 июля в гнезде
находились пуховой птенец и яйцо с погибшим эмбрионом. В промежу-
точные годы ни гнезда, ни птенцов гагары здесь не находили.
Даты встреч гнезда или насаживающей птицы - 14, 16, 19, 20 июня,
1 и 4 июля (срединное число 20. VI). Принимая продолжительность инку-
бации 28 дней, а интервал между откладкой яиц 2 дня (Флинт, 1982,
с. 254), находим ориентировочные сроки: откладка первого яйца 5. VI,
вывод последнего птенца 5. VII.
Яйца краснозобой гагары отличаются от яиц чернозобой только мень-
шей величиной: размеры одного 74 х 47 мм, масса 73,5 г. При опасности
наседка незаметно соскальзывает в воду и бесшумно ныряет. В отличие
от чернозобой гагары отвлекающей демонстрации у краснозобой мы не
наблюдали. Около гнезда она плавает, более или менее погрузившись в
воду, выставив над ней только голову и шею.
С прилета до откладки яиц гагары держатся парами, но на озерке с
гнездом обычно находится только одна: по-видимому, свободный парт-
нер летает кормиться на другой, более кормный водоем. Птенцов, за
одним исключением (сентябрь 1937 г.), нам не приходилось наблюдать.
Своих материалов по питанию краснозобой гагары у нас нет, но по лите-
ратурным сведениям она кормится мелкой рыбой, которую ловит как на
озерках, так и на реках; пищу для птенцов она приносит в клюве, иногда
издалека, чем и объясняется нахождение гнезд на безрыбных водоемах.
Кроме территориальных пар, летом мы иногда встречали стаи до
6 (9. VI 1975 г.), 8 (18. VII 1969 г.) и даже 13 гагар (18. VIII 1950 г.). Очевид-
но, это были холостые птицы, так как гагары по второму году еще не
гнездятся, но к ним могли примкнуть и птицы, потерявшие кладку.
Гагары иногда гибнут в рыболовных сетях: весной 1935 г. в ставную
сетку на Садозере попались 4 краснозобые гагары.
Размеры двух взрослых птиц: длина 650-675, крыло 302-306, хвост
58-67 мм; масса тела самца (июнь 1941 г.) - .1987 г, птицы неизвестного
пола (август 1936 г.) - 1873 г.
Чернозобая гагара - Gavia arctica (L.)
Местные названия: саамское - й£фьм-тоут (йырьм-голомень, т. е.
открытое пространство моря или озера, тоут-гагара). Русское население
видов гагар не различает.
Ареал этой гагары сходен с ареалом краснозобой, но не заходит так далеко
на север. На Кольском полуострове встречается повсеместно. В Лапландском
заповеднике чернозобые гагары довольно регулярно гнездятся на следующих
озерах: Чунозеро (площадь 2096 га; обычно 2 пары), Охтозеро (1350 га), Нявка
(764 га), Вайкись (475 га), Купись (224 га), Чингльс-явр (139 га), Ельявр (114 га) и
по одной паре на смежных, Румель- и Улынчьявр (110 и 95 га), Тулп- и Кен-
зисъявр (58 и 116 га), Витте и Островское (140 и 147 га), Красная и Матренины
ламбины (в сумме 100-150 га). Эпизодически гнездятся также на Куркозере
(223 га) и озерах площадью менее 100 га - Сейд озере, Верхнем Ташким,
Мясном, Верхнем Гарьюсном, Пельозере, Куропачьем, трех Райкоряурынчах, а
6
также на плесах рек Купись и Нявка; всего на заповедной площади 1600 км2
гнездится до 16 пар в год.
Кроме территориальных, встречаются летом и холостые особи, иногда
стайками до 5-7 голов, так что общая численность этой птицы в заповед-
нике оценивается в 50 особей, примерно по 130 га водной площади на
каждую. Большие стаи наблюдались только в отлетное время, например
5. X 1966 г. над Охтозером пролетели 25-30, а 11. X 36 гагар.
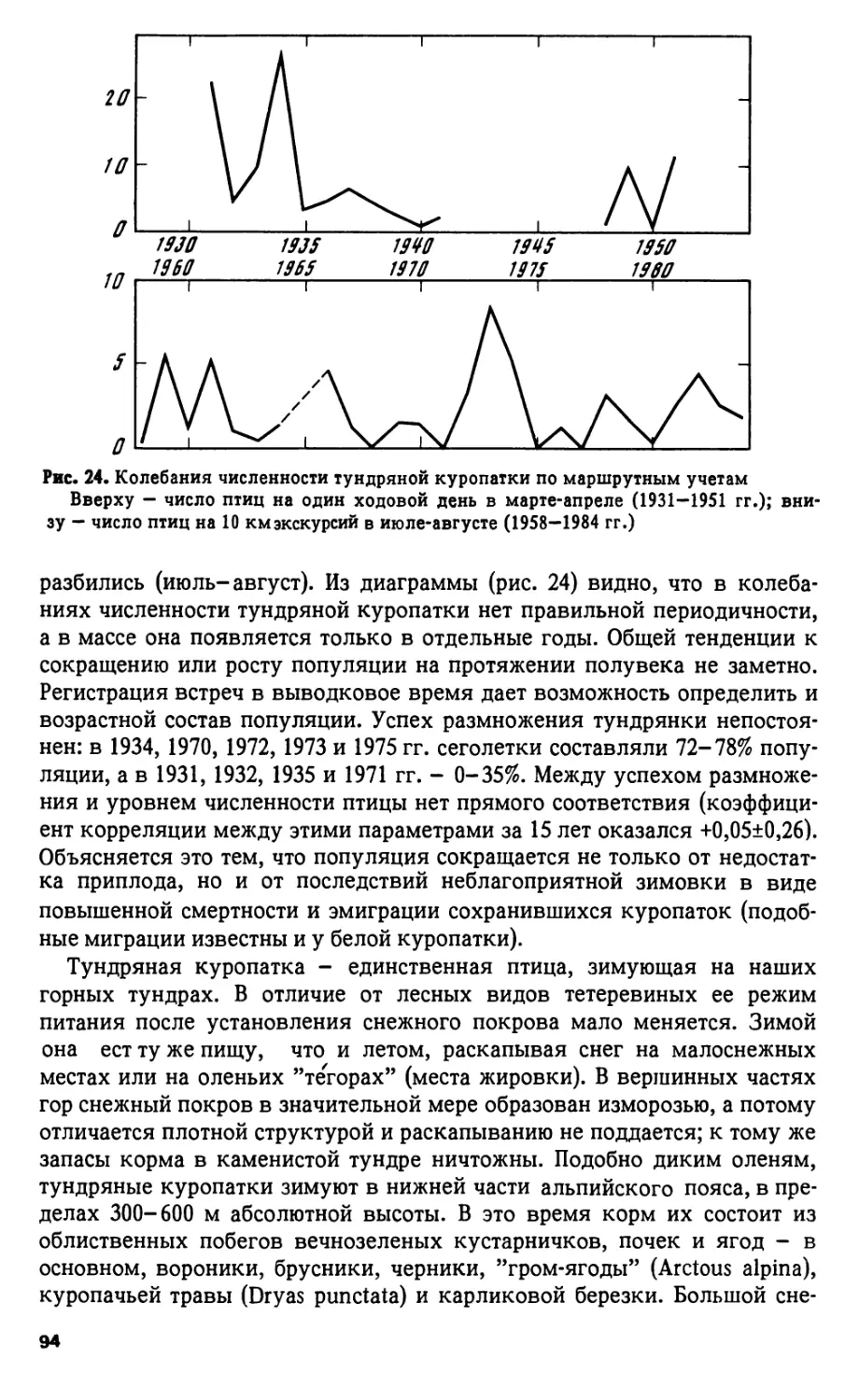
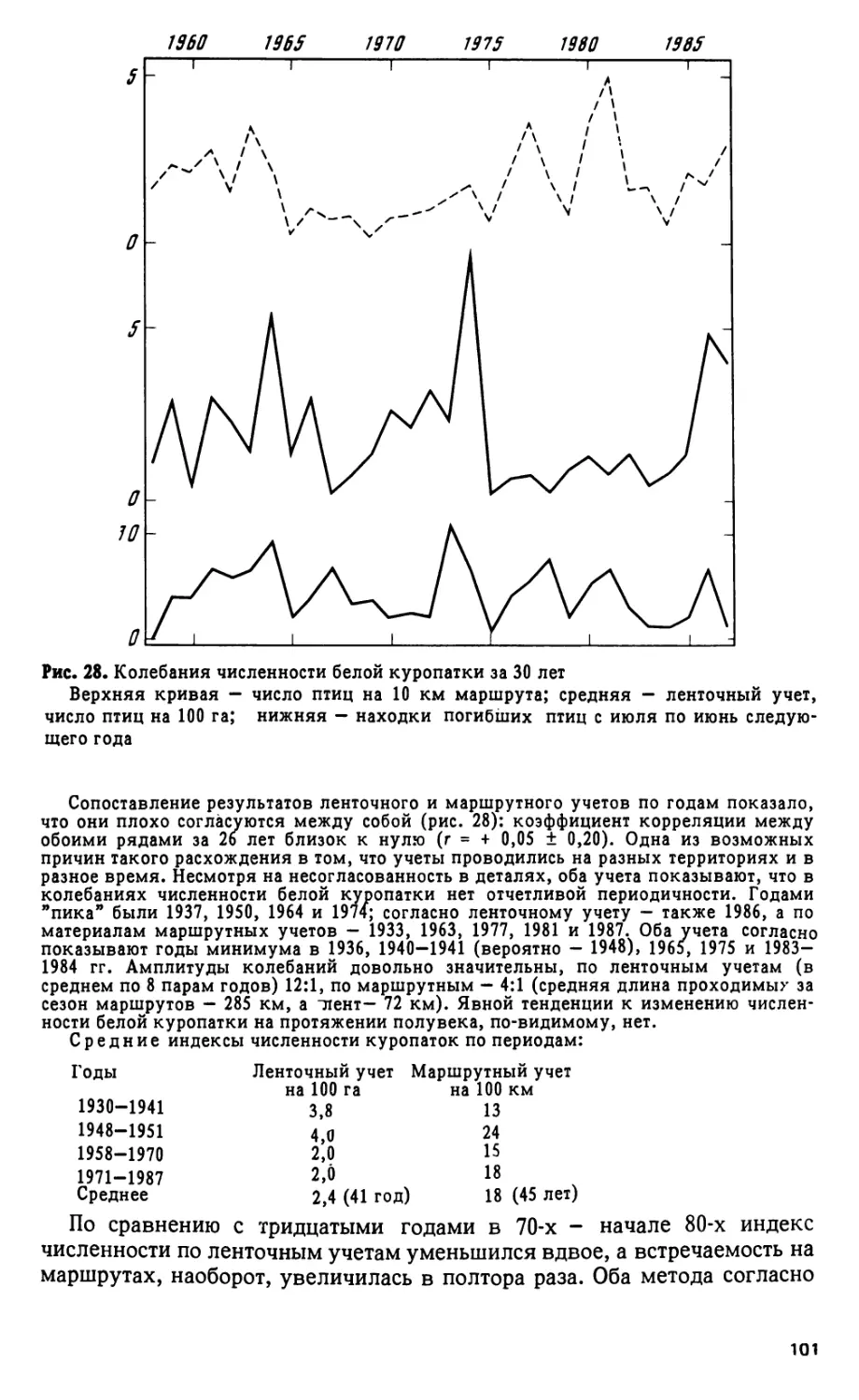
Регистрация гагар по годам обнаружила тенденции к снижению чис-
ленности: в 40-х годах по 15,5 встречи на 1 наблюдателя, в 50-х - 9,9, в
60-х - 12,0, в 70-х - 7,7, в 80-х - 8,3. За последние 23 года заметны слабо
выраженные колебания численности с периодом в 3 года, с максимумами
в 1966, 1969, 1971, 1974, 1978, 1981 и 1984 гг.
Чернозобая гагара прилетает (в среднем за 50 лет) 20 мая, крайние
сроки: 12. V 1934 и 1. VI 1985 г., 6 ± 5,2 дня, средняя температура дня
прилета 4,7е. Жизнь этой птицы тесно связана с открытой водой, появ-
ляется она обычно через 4 дня после образования заберегов и за 12 дней
до вскрытия больших озер. Коэффициенты корреляции между датами
наступления этих явлений равны + 0,53 ± 0,11 (забереги) и + 0,46 ± 0,13
(вскрытие озера); корреляция прилета с температурой мая невелика:
- 0,38 ± 0,13. Видимо, прилет гагары мало зависит от конкретных гидро-
метеорологических условий, у других явлений соответствующие коэф-
фициенты корреляции значительно выше (например, между температу-
рой мая и вскрытием Чунозера г = - 0,80 ± 0,05).
Гагара очень заметная, а весной к тому же и крикливая птица, поэтому
прилет ее замечается своевременно и точно. Значительно труднее опреде-
лить время отлета. В среднем за 38 лет последняя встреча приходится на
12 октября (20. IX 1977 г. - 2. XI 1933 г.), 6 ± 15,3 дня, температура + 1,Г.
Дата отлета заметно коррелирует с замерзанием глубоких озер (г =
= 0,48 ± 0,13 за 36 лет), а отчасти и с температурой осенних месяцев (для
сентября г = * 0,34 ±0,14, для октября ♦ 0,37 ± 0,14).
Гагары прилетают готовыми парами, и брачные песни их раздаются со
дня прилета. Наиболее заметные позы ухаживания: самец ложится на
воду боком, выставляя белое брюхо и помахивая в воздухе лапой; или
закидывает шею назад и раскрывает обращенный кверху клюв. 31. V и
1. VI 1976 г. мы наблюдали, как одна гагара (вероятно, самка) вылезала
на отлогий берег, делая вид, что садится на гнездо, а ее партнер припод-
нимался на воде взмахивая крыльями.
Гнезда с яйцами находили самое раннее 1 июня, в 1976 г. - через 10, а в
1983 г. - через 17 дней после прилета. Из 75 зарегистрированных гнезд 8
найдены в первой декаде июня, 25 - во второй, 28 - в третьей, 10 - в
первой декаде июля, 3 - во второй и 1 - в третьей (22. VII 1936 г.); самые
поздние кладки, очевидно, вторичные, после гибели первых.
Гагара гнездится на защищенном от прибоя участке берега, обычно
гнездо ее открыто, но иногда помещается под краем еловой кроны или
можжевелового куста, на расстоянии 20-200 см от края воды (в среднем
60 см) и на 10-50 см выше уровня; оно представляет собой утоптанную
площадку диаметром около 30 см (рис. 1). Иногда гагара поднимает
уровень лотка, подкладывая кусочки торфа. Она предпочитает гнездить-
ся на острове (75% гнезд), на стороне, обращенной к материку, где нет
волнения; если же на материковом берегу - то в бухточке или за грядой
7
Рис. 1. Гнездо чернозобой гагары, 19. VI 1963 г. Фото О.И. Семенова-Тян-Шанского
Рис. 2. Смена партнеров на гнезде чернозобой гагары. VI 1965 г. Фото А.В. Кречмара
камней (18%); иногда гнездится на берегу речного плеса (6%), а одно
гнездо находилось на плавучем острове у устья р. Нявки (июль 1970 г.).
Гагара не может ходить, с гнезда в воду она соскальзывает, а из воды
на гнездо всползает, поэтому дорожка к воде всегда гладко укатана. Из
найденных нами гнезд 45 содержали по два яйца, 24 - по одному, в 6 -
число яиц неизвестно. Яйца крупные, удлиненно-эллипсоидальной
8
формы, скорлупа очень темная, оливково-бурая с черными пятнами.
Средние размеры 8 яиц 87,2 х 52,8 (83 - 95 х 51-57) мм, масса 15 яиц в
разных стадиях инкубации от 122 до 139,5, а в среднем 130,1 г.
Успех размножения низок: из 78 яиц, судьба которых прослежена,
вывелись только 29 птенцов (37%), 7 яиц погибли, затопленные подъемом
воды, 10 расклеваны вброном, вороной или чайкой, 5 съедены норкой,
8 исчезли неизвестным путем; в 2 яйцах эмбрион погиб в процессе
инкубации, 2 повреждены самой гагарой, 15 брошены ею по причине
беспокойства. Если кладка погибает в начале инкубации, гагара строит
новое гнездо по соседству.
В спокойной обстановке гагара сидит на гнезде, положив голову на
крыло, встревоженная - вытягивает голову в сторону воды и несколько
вверх, а потом соскальзывает в воду и бесшумно ныряет. Если человек
подойдет к гнезду берегом, гагара шумно несется по воде, хлопая крыль-
ями, а в 20-30 м от гнезда вздымается над водой ’’свечкой”, кричит и
ныряет. Потревоженная, возвращается на гнездо не скоро, так что яйца
не менее часа остаются открытыми. В нормальных условиях яйца обогре-
ваются непрерывно, так как самец подменяет самку на гнезде. Свобод-
ный партнер подплывает к гнезду, всползает на берег, а наседка однов-
ременно соскальзывает в воду (рис. 2). Вне времени гнездования гагара
может совсем не выходить на сушу, она даже спит на воде, положив
голову на крыло (наблюдение 11. VI1981 г.).
Обеспокоенная гагара так легко бросает гнездо, что нам не удавалось
исследовать режим инкубации ее яиц даже с помощью самописцев. По
литературным указаниям, промежуток между откладкой яиц составляет
1-2 дня, и насиживать начинает с откладки первого. В гнезде, содержав-
шем в день находки 6. VI 1977 г. одно яйцо, 4. VII оказались 2 пуховичка,
следовательно, инкубация продолжалась 27-28 сут; в трех других
гнездах - не менее 24-26 сут; в согласии с литературными указаниями.
Птенцы остаются в гнезде около суток. Первое время они не могут
нырять и кормят их родители. В каком возрасте они начинают летать, не
прослежено. Из 61 учтенного выводка в 35 был один птенец, в 25 - два,
в 1 - три (соответственно 57:41:2).
Чернозобая гагара питается исключительно рыбой и ловит ее в откры-
той воде; она проныривает до 60-70 м, оставаясь под водой до 65 с, а
возможно, и долее. 6. VI 1981 г. лесник А.А. Царев видел, как гагара в
течение 5 мин. заглатывала сига, очевидно слишком большого для нее. В
желудке гагары, добытой в июне 1938 г., были остатки налима и камни, в
желудке другой - только 19 камешков.
У гагары мало природных врагов, и слабый темп размножения компен-
сируется большой продолжительностью жизни: кольцеванием установ-
лен максимальный возраст не менее 27 лет (Миграции птиц Восточной
Европы и Северной Азии: Гагарообразные, 1978, с. 27-37). Среди остатков
добычи орлана нами найдены две гагары; среди жертв других хищников
ее не встречали. Охотники редко добывают гагару, тем более что она
очень крепка на рану, но один раз, в 1960 г., на берегу Нявка-озера мы
нашли кости гагары, по-видимому, убитой и брошенной охотником.
Иногда гагара гибнет, попав в рыболовную сетку. 13. IX 1966 г. на Куро-
пачьем озере едва гагару освободили из сетки, в которой она запуталась,
как она тут же снова в нее попала. Другая гагара, попавшая в сетку на
Пиренгском озере 22. VI1971 г., была окольцована и выпущена.
9
Плывущая гагара держит клюв не горизонтально или наклонно,
подобно уткам, а несколько вверх, хвост опускает в воду. Обычно она
сидит на воде не глубже, чем нырковые утки, но может выпустить
воздух из воздушных мешков и тогда погружается, как подводная
лодка, оставляя над водой только голову и шею. Ныряя, иногда вскрики-
вает ”ой”. Взлетает с воды с разбега, медленно набирая высоту; частота
взмахов крыльев меньше, чем у гусей, но больше, чем у крохалей. На
лету вытягивает ноги дальше хвоста, а шею держит низко, на уровне
брюха, так что спина несколько горбится.
Самка немного меньше самца, не отличается от него по окраске.
Размеры двух самцов, добытых в июне 1938 г.: длина 720 и 744, крыло 330
и 340, хвост 99 и 87 мм, масса 3340 и 3482 г.
ОТРЯД ПОГАНКИ - PODICIPEDIFORMES
Серощекая поганка - Podiceps griseigena (Bodd.)
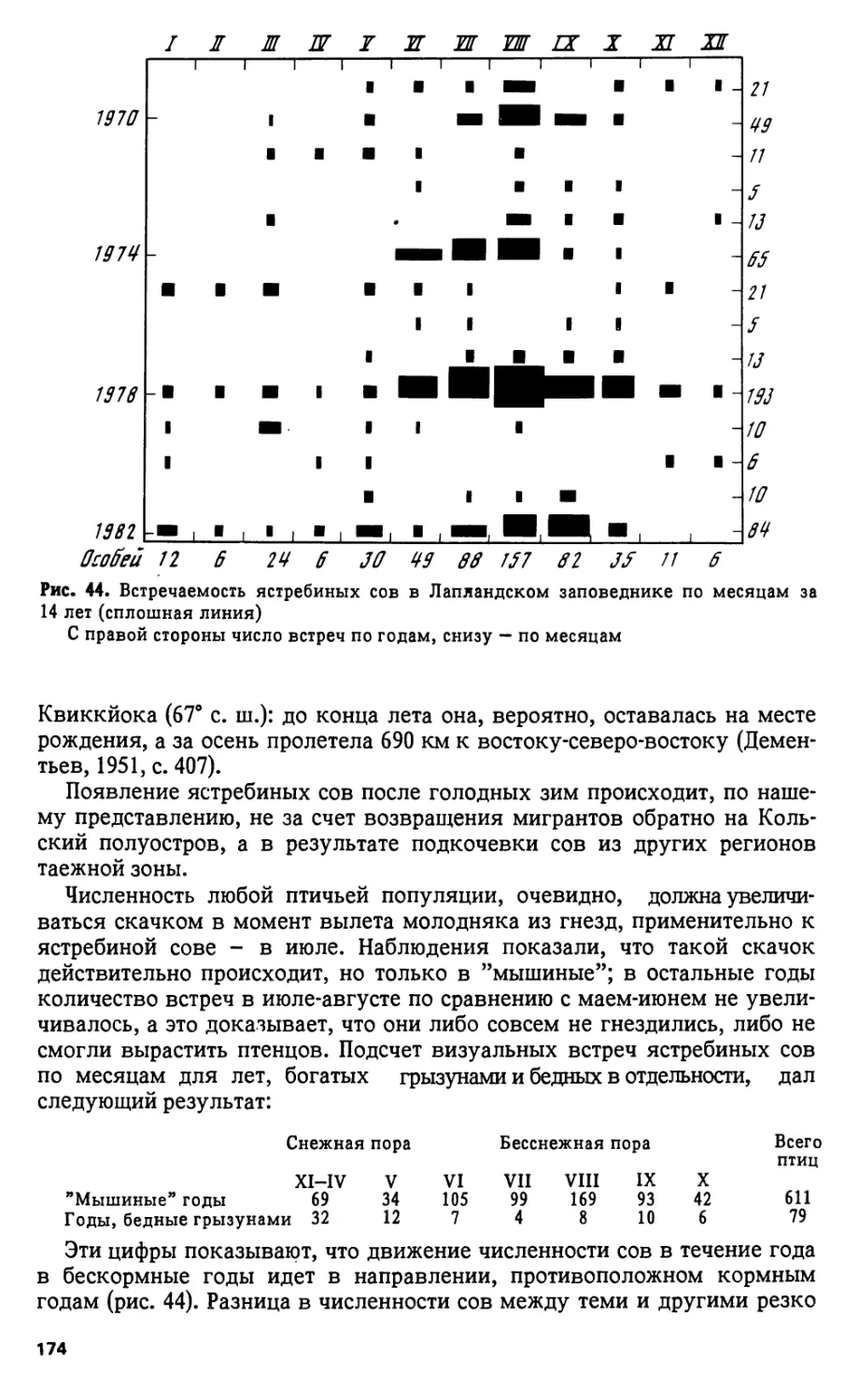
В середине XIX в. в Финляндии эта птица доходила к северу до Китти-
ля и даже озера Инари (Плеске, 1887, с. 506). В Лапландском заповеднике,
на Чунозере, летом 1937 г. М.И. Владимирская видела выводок, а А.А. На-
симович - одиночную поганку, но оба наблюдения сделаны на большом
расстоянии и без уверенности в правильности определения (Владимир-
ская, 1948, с. 175). Новых сведений о паганке в принятой нами области не
поступало, но она по-прежнему гнездится на островах Кандалакшского
залива, где впервые ее наблюдал Плеске.
ОТРЯД ВЕСЛОНОГИЕ - PELECANIFORMES
Большой баклан - Phalacrocorax carbo (L.)
На Мурманском берегу баклан гнездится, хотя и в небольшом коли-
честве, но в глубь Кольского полуострова только залетает (Плеске, 1887,
с. 331-333). , В Лапландском заповеднике бакланов неоднокртно наблю-
дали на Пиренгском озере: 18. VI 1937 г. А.Б. Васильев видел одного,
сидевшего на камне; 28. V 1949 г. лесник А.С. Сергин видел баклана,
летевшего над водой; 26. VIII и 10. IX 1962 г. - одного баклана видел и
подробно описал лесник Я.Р. Януше; в 1968 г. он же видел на Пиренге
3. VII четырех бакланов, а 5. VIII - трех.
ОТРЯД ГОЛЕНАСТЫЕ - CICONIIFORMES
Серая цапля - Ardea cinerea L.
Весной 1983 г., по словам охотника Н. Копина (пос. Куцколь), с 29 апре-
ля по 1 мая залетную цаплю несколько раз видели между озерами Ольче
и Куцколь (30 км к северу от границы заповедника). Ближайшие гнез-
довья этого вида находятся в сотнях километров к югу, в Псковской
области, или к западу - на побережье Норвегии. Между 1963 и 1979 г.
одиночных цапель наблюдали на Айновый островах^ близ Кандалакши и
на Терском берегу у мыса Орловского (Коханов, 1987, с. 22).
ю
ОТРЯД ПЛАСТИНЧАТОКЛЮВЫЕ - ANSERIFORMES
Лебедь-кликун - Cygnus cygnus (L.)
Местные названия: саамское - нюхчь (взрослая птица) и
йетмь (молодая, с серым оперением).
Ареал кликуна в прошлом занимал всю бореальную зону Европы
и Азии. В настоящее время в большинстве европейских стран он
вымер, сохранившись только на Севере материка. В Мурманской
области распространен повсеместно, не гнездится только в глухих
безлюдных местах. На территории Лапландского заповедника
численность лебедей подсчитывается ежегодно. По материалам
’’Летописи природы” на площади 1600 км2 с 1960 по 1983 г. она
колебалась от 8 до 21, в среднем 12,5 особей (7,8 птиц на 1000 км2);
в 1984-1987 гг. на площади 2784 км2 - от 26 до 43, в среднем 33
птицы (11,9 на 1000 км2). До 1960 г. абсолютного учета не велось, но
регистрация встреч показывает, что численность их довольно
стабильна, хотя в 30-х годах они встречались несколько чаще, а в
50-х (годы отмены заповедного режима) - реже, чем в другие
десятилетия:
Годы: 30-е 40-е 50-е 60-е 70-е 80-е Среднее
Индекс
встреч: 13,3 8,8 5,5 7,6 8,4 9,8 9,2
Лебеди нашего края зимуют сравнительно недалеко, у южных берегов
Балтийского и Северного морей, прилетают очень рано, задолго до
вскрытия озер, а потому не меньше месяца живут и кормятся на неболь-
ших участках открытой воды, в зашейках озер и на речных быстринах.
Благодаря этому обстоятельству* а так же крупным размерам и импозант-
ной внешности прилет лебедей замечается легко и своевременно. В
старинном календаре саамов первый месяц года назывался ’’нюхчь” -
месяц прилета лебедей.
В район заповедника лебедь прилетает, в среднем за 50 лет - 13 апре-
ля, крайние сроки 25. III 1935 г. и 30. IV 1940 г.; среднее квадратичное
отклонение даты прилета (сигма) ± 8,0 суток, средняя температура - 2,9е.
Корреляция между датой прилета и средней температурой марта равна
- 0,06 ± 0,15, а с температурой апреля - 0,14 ± 0,15, следовательно, время
прилета не зависит от температуры, но оно довольно точно привязано к
календарю: две трети дат укладываются в двухнедельный интервал 6-20
апреля. В южной и западной частях Мурманской области лебедь появ-
ляется раньше, чем в Лапландском заповеднике; так, в 1963 г. на его
территории первый лебедь замечен 25. IV, а у пос. Аллакурти (130 км к
юго-западу) - 11. III; в 1971 г. в заповеднике - 22. IV, а на р. Ковда
(100 км к югу) - 11. IV; в 1972 г. в заповеднике - 17. IV, а на р. Гирвас
(60 км к западу) - 2. IV.
С момента прилета лебеди держатся парами, но на пролете летят
стаями; так, 13. IV 1950 г. над Чунозером пролетели на восток 10 лебедей;
22. IV 1950 г. на льду того же озера сидело 10 птиц; 17. IV 1961 г. через
Купись-озеро на восток летели 7; 18. IV 1965 г. над Пиренгским озером
пролетели на восток 6; 16. IV 1967 г. через Чунозеро на север - 6;
17. IV 1983 г. через Чунозеро на северо-восток - 6. Путь лебяжьих стай
11
указывает, что летят они к нам с зимовок кратчайшим путем через
Ботнический залив и Финляндию.
Молодые лебеди сохраняют серый ювенильный наряд всю зиму и
весну, до летней линьки. Лебеди образуют пары задолго до наступления
половой зрелости, и летом холостых пар всегда бывает больше, чем
гнездящихся. Интересно, что весной изредка наблюдали разновозрастные
пары: один лебедь белый, другой серый.
Эффектные брачные церемонии лебедей редко случается наблюдать.
22. V 1979 г. в 19 ч. 30 мин1: привлеченные громкими кликами обоих
партнеров в унисон, мы увидели самца на спине самки, крылья его были
полуразвернуты, а ось тела приподнята под углом почти 45е; шеи обеих
птиц были вытянуты вверх параллельно друг другу, но голова самки
была в горизонтальном положении, а голова самца продолжала линию
шеи. В этом положении птицы находились не более 2 с, потом разъедини-
лись и некоторое время оставались на воде лицом друг к другу, почти
вплотную. На следующий день в 20 ч. 15 мин. на том же месте мы снова
наблюдали брачную игру: со звонким криком ”оуйи, оуйи..лебеди
плыли навстречу друг другу и, приподнявшись над водой, одновременно
взмахивали крыльями (рис. 3). Заметив нас, лебеди прервали церемонию
и улетели. В другой раз, около полудня, церемония началась с вокально-
го дуэта. Обратясь лицом друг к другу, лебеди вскрикивали поочередно,
продолжая сближаться, крики становились все громче и эмоциональнее,
наконец, обе птицы приподнялись на воде и несколько секунд взмахива-
ли друг на друга крыльями. Приветственный ритуал можно наблюдать и
позднее, когда самка уже насиживает. Вечером 17. VI 1967 г. один
лебедь кормился на оз. Румельявр, а в 22 ч. 45 мин. в устье реки появил-
ся второй, очевидно самка с гнезда; первый поднялся, подлетел ко
второму, оба кричали, вытягивая шеи вдоль поверхности воды; некото-
рое время они плыли рядом, потом вместе улетели на болото (А. Брагин).
Гнездо лебедя представляет большую кучу сфагнового торфа в форме
усеченного конуса, диаметром в основании 1,3-2,0 м, высотой 0,5-0,7 м,
с лотком глубиной до 20 см и диаметром 55-70 см. Как правило, гнездо
строится на твердом основании, на маленьком островке диаметром
3-5м, иногда поросшем карликовой березкой, посреди глухого, зарас-
тающего с берегов сплавиной, озерка глубиной 0,9-1,5 м, или среди
трясины. Только одно гнездо находилось на берегу р. Куудас, в густой
заросли лапландской ивы и карликовой березки. Одно гнездо может
служить лебедю повторно. Гнездо, впервые обнаруженное нами 31. V
1976 г. на заболоченном лесном озерке (получившем название Лебяжье-
го), в 400 м к северу от оз. Улынчъявр, позднее было занято в 1979, 1980 и
1984 гг. В 2,8 км к ЗЮЗ от этого гнезда есть другое, построенное также на
болоте, метрах в 200 от южного берега оз. Румельявр - в нем лебеди
вывели птенцов в 1939 и 1975 гг. В 1987 г. лебеди гнездились на крошеч-
ном островке под северным берегом того же озера, но неудачно.
В том же районе, но в долине р. Чуна, известно еще два гнезда: в
одном, на Гагарьем озерке, в 1,9 км к ВЮВ от первого гнезда, лебеди
гнездились до 1974, в 1985, 1986 и 1987 гг.; другое (4,5 км от первого к
ВЮВ) было построено и брошено в 1932 г. и с тех пор больше не использо-
хВо всех наблюдениях указано местное среднее солнечное время: на долготе
заповедника оно равно гражданскому зимнему —50'мин,.
12
валось. Расстояние между крайними гнездами этой группы не превышает
7 км. Лебеди нередко гнездятся и в районе окруженного болотами
оз. Нявка. Гнездо с 4 яйцами было найдено 25. VI 1939 г. на топком болоте
к северу от оз. Нявки и в 2 км к западу от Купись-озера; два старых
гнезда обнаружены в июле 1960 г., одно на островке среди зыбуна, не
далее 1 км к западу от Нявки, другое на безлесном берегу речки Куудас,
впадающей в то же озеро с юга. Расстояние между этими тремя гнездами
не более 9 км.
Несколько раз гнездились лебеди и в долине р. Курка. В июне 1939 г.
старое гнездо со скорлупами яиц обнаружено к югу от Тульпъявра на
плавучем островке среди болотного озерка, глубиной 1,2 м; в июле
1941 г. старое гнездо среди зарослей ивняков и осоки на острове посре-
ди Лебяжьего озерка, к югу от Чингльс-явра; в июне 1960 г. кладка
лебедя обнаружена на болоте к северу от Куркозера. Здесь также дистан-
ция между гнездами не превышала 9 км.
Гнезда с яйцами или насиживающей самкой находили 22, 24, 28,
31 мая, 1, 9, 10, 15 июня, а 1 июля - пуховичка, только что оставившего
гнездо. Первые выводки с пуховичками встречали между 30.VI 1940 г. и
8.VII 1978 г., а в среднем за 7 лет - 3 июля. Принимая продолжительность
инкубации в 34 дня (Palmer, 1976, р. 53), находим, что лебеди строят гнез-
до с середины мая, в последней декаде начинают насиживать и выводят
птенцов около 1 июля.
Насиживающая самка укладывает шею на спину, издали она похожа на
белую подушку, лежащую на плоской кочке. Оставляя гнездо, самка не-
которое время топчется на нем, укрывая яйца рыхлым слоем торфа,
благодаря чему они сохраняют тепло на протяжении нескольких часов.
15.VI 1932 г. на озерке в долине Чуны Г.М. Крепе и О.И. Семенов-Тян-
Шанский заметили лебедя в тот момент, когда он сходил с гнезда. Потом,
когда лебеди улетели, они положили на трясину жерди, добрались до
гнезда, нашли в нем 5 укрытых торфом яиц, сфотографировали их и не-
медленно ушли, убрав за собой жерди. Лебеди видели эту операцию с
воздуха и к гнезду не вернулись, а яйца впоследствии расклевал ворон.
После этого случая сотрудники заповедника не подходили к гнезду, если
заставали на нем лебедя. В гнезде, найденном 25.VI 1939 г. близ оз. Няв-
ка, лесник И. Зайков в бинокль сосчитал 4 яйца, но и эта кладка была
брошена: впоследствии здесь видели только пару взрослых лебедей без
птенцов. Гнездо, найденное геологами в июне 1960 г. близ Куркозера,
также было брошено и разорено. Гнездо, найденное в 1985 г., было осмот-
рено после того, как оба лебедя с одним птенцом его оставили, в нем
оказалось укрытое торфом теплое живое яйцо и скорлупа другого. Через
15 дней лебедей на озерке не было, гнездо оказалось пустым, а через
2 мес, когда выводок нашли, в нем был только один птенец. Еще в одном
гнезде по скорлупам определено, что кладка состояла из 3 яиц.
Лебеди не агрессивны, как правило, они не пытаются защищать ни
гнездо, ни птенцов, ни самих себя (при отлове линяющих для кольцева-
ния); только один раз, 27.VII 1980 г., когда лесник Г.Н. Заводчиков застал
на Жемчужном ручье выводок лебедей с тремя птенцами, старики нале-
тели на него, били крыльями и заставили отступить. К потомству они
относятся заботливее, чем утки. Встречи выводков до 20 июля редки,
видимо, потому, что родители прячут маленьких птенцов на уединенных
болотах, в глубине леса. На воде обычно одна из взрослых птиц плывет
впереди, за ней гуськом птенцы, вторая взрослая - в арьергарде. Если
выводок застигнут лодкой на реке, взрослые взлетают (если они не линя-
ют), а птенцы рассеиваются, прячутся под нависшими кустами, иногда
ныряют (по скорости плавания как над водой, так и под нею они уступа-
ют уткам), иногда вылезают на берег и пытаются убежать или спрятаться.
Изредка встречаются выводки без взрослых птиц: одиночного, еще пухо-
вого, лебеденка видели в Пора-губе на Имандре 22.VIII 1963 г.; с 3.VIII по
24.IX 1970 г. на Красной ламбине жил выводок из трех птенцов, позднее
оставались только два (родители, возможно, были убиты); 26.VIII 1977 г.
на оз. Нявка плавали 4 лебеденка отдельно от взрослых, находившихся
на том же озере.
Молодые поднимаются на крыло очень недружно, первые взлетавшие
лебедята отмечены 13.VIII 1974 г., 28.VIII 1946 г. и 5.IX 1973 г.; последние
неспособные летать - 24.IX 1970 г., 26.IX 1978 г. и 12.IX - в 1938 и 1983 гг.
Большинство, вероятно, начинают летать во второй декаде сентября в
возрасте около 75 дней. Лебеденок, оставивший гнездо 1.VII 1985 г.,
13 сентября, в возрасте 74 дней, по-видимому, еще не летал.
Если выводок обнаружен человеком на небольшом озерке, старики
14
немедленно переводят в другое место, иногда даже в другой речной бас-
сейн, что требует перехода по лесу пешком. Семья из пары взрослых с
одним птенцом, обнаруженная 30.VI 1940 г. на Чунозере, с 21.VII по З.Х.
проживала в том же составе на озере Румельявр: расстояние до него по
прямой линии 7 км, включая 3 км сухопутьем через перевал; или следо-
вало преодолеть перешеек между Чунозером и Охтозером шириной
0,4 км и проплыть не менее 20 км кружным путем. Другая семья такого
же состава, встреченная 25.VII 1972 г. в устье р. Куудас, на оз. Нявка, к
11. VIII переселилась на Купись-озеро, проплыв 7 км и пройдя 2 км по
суше.
Выводки лебедей наблюдали не только в окрестностях описанных
выше гнезд, но и в других местах заповедника, в частности в районе Сад-
озера и речки Пельйок, по всей долине р. Купись и ее притока Северный
Ташким, в долине Ливы и других местах, где, очевидно, они также
гнездятся.
Темп размножения лебедей невысок. За 47 лет наблюдений выводки
встречались на протяжении 33 и не попадались за 14 сезонов. Молодняк
составляет только 21% всех зарегистрированных с 1.VII до отлета особей,
однако учет лебедей, о котором речь идет ниже, дает более надежный
результат. Учтенные нами 49 выводков по количеству птенцов распреде-
ляются так:
Число птенцов 1 2 3 4 5 6 В среднем 2,8
Число выводков 10 11 13 И 3 1
Взрослые лебеди линяют очень недружно, в один и тот же день можно
встретить лебедей, неспособных взлететь и прекрасно летающих. Нелет-
ных встречали в разные сроки, с 17.VII до 25.VIII. Во время линьки они
живут скрытно и редко попадаются на глаза. Встречи лебедей распреде-
ляются по месяцам так (в %): апрель - 10, май - 22, июнь - 22, июль - 10,
август - 6, сентябрь - 21, октябрь - 8, ноябрь и март - по 0,5.
Излюбленное место линьки лебедей - десяток озерков с берегами,
заросшими осочником и ивняком в лесотундре, под восточным склоном
Сальной тундры (эта местность в 1983 г. вошла в состав заповедника). На
них 20.VII 1964 г. мы видели 11, а 23.VII 1985 г. 9 взрослых лебедей. Не-
способных летать птиц и выпавшие при линьке белые перья встречали и
в других местах, на оз. Нявка и окружащих его болотах, на Румельявре,
в пойменных березняках по среднему течению р. Чуна, на Лебяжьем
озерке в долине Курки. Линяющий лебедь не чувствует себя в безопас-
ности на воде, обычно от людей он прячется на суше, затаившись в кус-
тах или траве, при этом прижимает голову к земле, а середина шеи оста-
ется приподнятой горбом. Неспособность летать продолжается, вероятно,
не более двух недель. Из 84 лебедей, встреченных за 15 дней в разгаре
линьки (26.VII - 9.VIII), 26% не могли летать, 25% взлетели, состояние
остальных 49% не установлено.
Молодые лебеди начинают летать примерно за месяц до отлета. В это
время выводки и холостые птицы нередко собираются на кормных озе-
рах большими стаями. 23.IX 1962 г., пролетая на самолете Як-12 над бога-
15
Рис. 4. Кормежка лебедей на глубокой воде,'22. V 1978 г.Фото О.И. Семенова-Тян-
Шанского
тым водной растительностью Пече-озером, мы подняли с воды 50 лебедей,
взлетавших большей частью попарно, 28.IX 1982 г. лесники А. Царев и
В. Беднягин насчитали на оз. Нявке 30, a 15.IX 1986 г. над Островским
озером пролетели 27 лебедей. Стаи по 10-18 птиц встречали неоднократ-
но: на Чунозере - в октябре 1947 и 1948 гг., на оз. Нявка - в октябре 1965
и 1978 и сентябре 1983 и 1984 гг. По сообщению ботаника Е.Г. Чернова
(1950), на озерах восточной части Кольского полуострова осенью собира-
ются стаи до полусотни лебедей. Как только начнут замерзать озерные
мелководья, лебеди отлетают. Последние встречи приходятся, в среднем
за 44 года, на 18 октября, крайние сроки 20.IX 1966 г. и 30.XI 1950 г., сигма
±13,0 дней. Средняя температура дня отлета -0, Г, корреляция даты
отлета с температурой октября г = +0,72±0,08, а с датой замерзания мел-
ких озер +0,54±0,11 (за 41 и 39 лет).
Лебедь - птица преимещественно травоядная, но в отличие от гусей
кормится он на воде, опуская клюв до дна, на всю длину шеи. Чтобы дотя-
нуться до дна при высоком уровне воды во время половодья, ему иног-
да приходится опрокидываться в воде, хвостом вверх, балансируя лапа-
ми для сохранения равновесия (рис. 4). Во время кормежки голова оста-
ется под водой 10-18 с, над водой только 3-4 с. Лебедь с погруженной голо-
вой ничего вокруг себя не видит, поэтому в паре они обычно кормятся
поочередно, пока один достает корм, другой его страхует. Весной и
осенью лебеди кормятся исключительно водными растениями, судя по
остаткам на местах жировки - главным образом полушником (Isoetes
lacustris), затем хвощом (Equisetum fluviatilis), водяной сосенкой (Hippuris
vulgaris), урутью (Myriophyllum alterniflorum), водяным лютиком (Batra-
chim peltatum) и шильником (Subularia aquatica). Летом в помете лебедей
16
обнаружены нитчатые водоросли, листья осоковых и ягоды морошки,
следовательно, в это время они кормятся не только на воде, но и на
суше. Экскременты их имеют вид тёмных ’’колбасок” диаметром 17-
18 мм; судя по количеству помета, птицы они прожорливые.
В августе 1934 и 1935 гг. мы держали по одному лебеденку в неволе.
Кормили их смесью из ягод черники, вороники, голубики или брусники,
накрошенного хлеба, овса или гречневой крупы и нарубленной рыбы или
мяса, залив все водой; лебеденок ел эту смесь, щелокча клювом, как
утка. Давали ему также пучки лесного хвоща или луговика.
Лебедь - самая крупная птица нашей фауны, и врагов у него немного.
В материалах по питанию хищников остатков его не находили, но в
августе 1964 г. на берегу оз. Нявка нашли скелет крыла и перья линного
лебедя, пойманного хищником год назад, а в августе 1982 г. на берегу
р. Нявка - перья с кровью птенца, пойманного хищной птицей. Нелетно-
го лебедя может поймать любой хищный зверь: медведь, росомаха, волк,
возможно и лиса; а из птиц для него опасны только орел и орлан. 12.IX
1963 г. на Куропачьем озере И.А. Паракецов видел, как орел (или орлан?)
несколько раз бросался на пару взрослых лебедей, но кончилось тем, что
они поднялись с воды и благополучно улетели.
Кладки лебедей нередко гибнут от гнездовых хищников - ворона или
вороны. Недаром говорится в саамской пословице ”и в гнезде ворона
можно найти лебединое яйцо” (Саамско-русский словарь, 1985, с. 102).
9.VI 1976 г. мы снимали в кино с большого расстояния лебедей у гнезда.
Самка сошла с гнезда, вероятно, от блеска кинокамеры, но продолжала
плавать поблизости вместе со своим партнером. К гнезду с карканьем
подлетела ворона, самец быстро подплыл, поднялся на островок, но не сел
на яйца, а дождался самки, которая сменила его и возобновила насижи-
вание. Расклеванные яйца находили в гнездах неоднократно: 27.VI
1932 г. на гнезде в долине Чуны остатки 4 яиц, 24.V 1979 г.на лотке гнезда
близ Улынчъявра - 1, а остатки яиц, похищенных из гнезда, были найде-
ны 4.IX 1938 г. в долине р. Курка и 26.VII 1977 г. в долине ручья Лылчи.
Больше всего потерь лебедь несет от человека. Формально охота на
лебедей в нашей стране давно запрещена, но в ’’Красную книгу РСФСР”
пока внесен только один вид - малый лебедь. Ареал кликуна гораздо
шире, и статус его считается благополучным. По учету, проведенному в
январе 1967 г., общая численность этого вида в Старом Свете оценивалась
в 36 900 голов (Palmer, 1976, с. 54), а в Мурманской области, по учетам
Кандалакшского заповедника, в 1200-1400 голов, включая <400-450
гнездящихся пар (Редкие и нуждающиеся в охране животные и растения
Мурманской области, 1979, с.41-42). В действительности перспективы
лебедя внушают тревогу.
В лесной зоне Северной Америки нашего кликуна заменяет близкий
вид, лебедь-трубач. В начале XX в. он был близок к полному истребле-
нию по той причине, что в XIX в. компания Гудзонова залива вела заго-
товку лебяжьих шкур и за 57 лет продала в Лондоне 108 тыс. Теперь в
Канаде и США трубач гнездится только в отдельных точках его прежнего
ареала; сохранился он главным образом на Аляске, где численность его в
2. Зак. 1023
17
1968 г. оценивалась в 3500-4000 голов, в остальных штатах - в 900, в Ка-
наде - в 100 (Palmer, 1976, с.69). Та же опасность угрожает и нашим лебе-
дям с тех пор, как они стали объектом браконьерского промысла. При
авиаучете диких оленей на Таймыре в июне 1982 г., налетав на самолете
Ан-2 более 42 ч, мы едва насчитали в тундре 5 лебедей (малых). Факты
подтверждают, что пассивная охрана лебедей недостаточна и истребле-
ние их продолжается. В 1956 г. в заповеднике Ред Рок Лэйк, в бассейне
Маккензи, Канада, поймали и проверили на флуороскопе 103 линяющих
американских лебедя и в 15 птицах обнаружили дробь (Palmer,1976, с.69).
При аналогичном обследовании 303 малых лебедей, отловленных на
зимовке в Слимбридже, Англия, в 1970-1973 гг. в теле 103 особей обнару-
жена дробь, причем встречаемость ее пропорциональна возрасту, у годо-
валых птиц - 12%, в пятилетних - 67% (Evans et al., 1973; цит. по: РЖ
’’Биология”, 1974, 7И437)2. Практически вся популяция малого лебедя
гнездится в наших тундрах, в Англии он только зимует.
Сохранению лебедя на Кольском полуострове и в Карелии в прошлом
помогали пережитки древнего культа этой птицы. Один из героев Кале-
валы жизнью поплатился за убийство Туонельского лебедя. Среди саа-
мов еще в 30-х годах бытовало поверье, что убой лебедя влечет за собой
гибель охотника или одного из членов его семьи.
Лебеди страдают не только от охоты, но и от других форм деятельности
человека в природе. Строительство гидроэлектростанций, рубки леса,
мелиорация земель сокращают пригодную для обитания этой птицы пло-
щадь. Ахиллесова пята лебедя - его гнездо: видное на открытом болоте
издалека большое гнездо с сидящей на нем белой птицей привлекает
внимание не только браконьеров, но и лояльных людей, а лебедь даже на
любопытство реагирует болезненно и бросает кладку. В то время как
лебедь-шипун выводит детей даже в городских парках, кликун успешно
размножается только в ’’пустыне” (принимая это слово в его старинном
значении первобытной природы), а найти пустынный утолок с каждым
годом становится все труднее, в самых глухих местах работают экспеди-
ции, проникают туристы и рыболовы.
Собранный в ’’Летописи природы” материал позволяет составить при-
близительную модель возрастного состава популяции лебедей (использо-
ваны только учеты на старой территории заповедника за 1960-1984 гг.)
(табл. 1).
Среднее число лебедей в заповеднике 12,52, в том числе семейных пар
1,16, 3,28 птенца и 6,92 холостых особей; следовательно, на одну семей-
ную пару приходится три холостых. Сеголетки составляют 26% популя-
ции - это первая возрастная когорта (0 + лет); каждая следующая когор-
та, очевидно, должна быть меньше на величину естественной смертности.
Экстраполируя данные учетов Лапландского заповедника на всю
Мурманскую область, мы оцениваем запас этого вида в ИЗО голов, в том
числе 105 пар с выводками - в 4 раза ниже оценки мурманской ”Крас-
2Водоплавающие птицы часто заглатывают дробь с кормом, однако в цитированных
работах речь идет о ранении, а не заглатывании дроби.
18
Таблица 1. Учет лебедя в Лапландском заповеднике
Плс щадь 1613 км2
Год Год
лебедей выводков птенцов
1960 8 1 1 4
1961 18 2 2 + 4
1962 19 2 3 + 4
1963 18 2 3 + 4
1964 11 1 2
1965 8 — —
1966 8 — —
1967 13 1 2
1968 11 1 2
1969 14 1 4
1970 12 2 3 + 3
1971 9 1 3
1972 10 2 2 + 1
1973 8 —
1974 10 1 3
1975 14 2 2 + 2
1976 12 1 2
1977 12 1 4
1978 17 2 4 + 3
1979 9 —
1980 14 2 3 + 3 1984
1981 8 — — 1985
1982 13 2 1 + 3 1986
1983 16 2 3 + 3 1987
1984 21 — — 1988
Сумма 313 29 82
Среднее 12, 52 1, 16 3,28
Площадь 2784 км2
лебедей выводков птенцов
27 — —
35 3 1 + 3 + 2
26 — —
43 3 1 + 2 + 1
131 6 10
32, 75 1, 5 2,5
ной книги”. Возможно, и эта оценка завышена, так как условия гнездо-
вания лебедей в угодьях общего пользования, вероятно, хуже, чем в за-
поведнике.
По словам охотника, добывавшего старого лебедя в конце 30-х годов,
масса тела его равнялась 30 фунтам (12 кг). Остальные лебеди измерены
живыми: длина двух взрослых холостых 134 и 140 см, хвосты 23 и 21 см,
масса тела двух птенцов - 1,52 кг (4.VIII 1935 г.) и 4,60 кг (25.VIII 1934 г.,
самец). Промеры одного яйца 113 х 75 мм, его масса тела, по расчету,
около 318 г. След лебедя, без заднего пальца, который обычно не печа-
тается, 16-17 см в длину и 15-16 в ширину. Длина двойного шага на сне-
гу 70 см, а на взлете - от 160 до 330 см к концу разбега. Лебедь машет
крыльями реже, чем другие водоплавающие: на кинопленке один цикл
занимает 6 кадров, т.е. 0,38 с при скорости съемки 16 кадров в секунду.
С 1960 по 1985 г. заповедником окольцовано 5 взрослых линяющих ле-
бедей и 15 птенцов (возвратов не поступало).
19
Гусь-гуменник — Anser fabalis (L.)
Местное название: саамское - чуань
Ареал гуменника обширнее, чем других гусей; в нашей стране он за-
нимает две зоны, тундровую и таежную, включая и всю Мурманскую
область. В Лапландском заповеднике на пролете обычен, на гнездовье
сравнительно редок, как это видно по сезонному распределению встреч.
Встречаемость гуся гуменника по месяцам (%):
IV V VI VII VIII IX х XI
4 26 21 1 6 32 9 1
В среднем за сезон в картотеке регистрируется по 95 гусей, или по
12 особей на одного сотрудника, однако количество это сильно колеблет-
ся по годам, без заметной периодичности. Много гусей (по 25-35) отме-
чены в 1932, 1961, 1964, 1965, 1969 и 1987 гг.; очень мало (0-10) в .1931,
1937,1940-1941,1948-1951, 1958-1959,1966-1968,1977, 1982-1986 гг. Под-
счет встречаемости по десятилетиям не обнаруживает явной тенденции
популяции к росту или сокращению с годами:
Годы 30-е 40-е 50-е 60-е 70-е 80-е
Число гусей на 1 наблюде- 17 6 2 18 11 18
ние в год
Прилетают гуси небольшими стайками, в среднем по 4,5 особей, на
отлете стаи втрое крупнее, в среднем по 14 птиц. Группируя встречи по
размерам стай, мы нашли, что малые (по 1-10 особей) в апреле-июне дают
91% всех встреч, а в сентябре-ноябре - 55%; средние (по 11-20) соответ-
ственно 6 и 27%; крупные (21-40 птиц) - 3 и 12%; стай более 40 голов вес-
ной не встречали, осенью они составляют 6% встреч.
Гуси прилетают позже лебедей, а отлетают раньше. Средняя дата при-
лета 30 апреля (крайние сроки 12.IV 1969 г. и 1984 г. - 16.V 1945 г.; о ±7,8
дней за 48 лет наблюдений); дата отлета 7 октября (17.IX 1976 г. - 4.XI
1948 г.; 6 ± 13,9 за 29 лет). Время прилета приурочено к моменту перехода
суточных температур через 0,4° и появлению воды на льду озер, хотя
корреляция с температурой апреля и невысока, г = -0,20 ± 0,14. От-
лет происходит при температуре +2, Г, примерно в одно время с вы-
падением первого снега и коррелирует как с температурой сентября
(г= +0,43±0,15), так и с датой первого снежного покрова в лесу (г= +0,45±0,15);
осенью связь с погодой слабее выражена, чем у лебедя. Места зимовки и
пути перелета наших гусей еще не документированы кольцеванием, но,
по всей вероятности, они летят через Финляндию, Швецию, приморские
районы ГДР и зимуют в Нидерландах (Миграции птиц Восточной Европы и
Северной Азии: Аистообразные - Пластинчатоклювые. М., 1979, с. 1.60-
161).
В районе заповедника гуси гнездятся редко и, вероятно, не ежегодно.
Три гнезда с полной ненасиженной кладкой были найдены на болотах к
югу от Чунозера и Охтозера 15.V 1936 г., 18 и 22.V 1938 г.: в двух было по
5 яиц, в третьем гусыня 4 июня уже сидела на яйцах. Еще одно найдено в
том же районе уже после вывода птенцов, в сентябре 1939 г. на сухой
20
гривке среди сфагнового болота. На территории заповедника гусиных
гнезд не находили, но нелетные выводки встречали неоднократно: 24.VI
1975 г. - на осоковом болоте у берега Тулпъявра видели пару с одним
птенцом, 4.VII там же слышали их голоса, а 20.VIII три гуся пролетели над
Куркозером (3. Кондратьева, А. Якуничев и Я. Януше); 3.VII 1976 г. - на
Улынчьявре в осочнике выводок из самки и двух пуховых гусят, раз-
мером не менее чирка (А. Гилязов); 7.VIII 1980 г. - в южном течении
р. Чуна встречен выводок из трех гусят, без взрослых; потом их видели
13.VIII вместе с маткой (А. Гилязов, Д. Карелин); 18.VII 1984 г. на Палие-
вом озерке, севернее Красной ламбины, гусыня с тремя гусятами (Н. Бе-
лоусов).
Вне заповедника, на болотах к югу от Чунозера, 28.VI 1949 г. охотник
Н. Михайлов поймал трех пуховых гусят. Объединенные летные выводки
наблюдались в том же районе 18.VIII 1932 г. - 22 гуся, 23.VIII 1933 г. - 17,
3.IX 1960 г. - 23, 27.VIII и 10.IX 1987 г. - 39. В заповеднике выводок из
шести гусей поднят на р. Чуна 11.VIII 1974 г.; семья из двух взрослых и
трех молодых, по-видимому еще нелетных, встречена на р. Нявка 17.VIII
1974 г. Семья из шести птиц - там же 20.VIII и 18.IX 1987 г. Гнездование
гусей в заповеднике или его окрестностях зарегестрировано 12 раз за
48 лет наблюдений. На протяжении лета гусей встречали еще в 19 сезо-
нах, но это могли быть и холостые птицы; еще 17 лет гусей наблюдали
только в пролетное время.
В гнездовое время гуси ведут очень скрытный образ жизни, их выводки
гораздо труднее обнаружить, чем лебединые, хотя в некоторые годы в
заповеднике обитало, вероятно, больше гусей, чем лебедей.
Гнездятся гуси рано, в 1936 и 1938 гг. полные кладки были найдены
через 24-26 дней после прилета первых особей. Принимая, что инкубация
занимает 27-29 дней и молодые поднимаются на крыло в двухмесячном
возрасте (Harrison, 1975, с. 76), нормальными сроками размножения
можно считать: кладка - с 14.V (через 2 недели после прилета), инкуба-
ция с 20. V, вылупление птенцов 17.VI, подъем на крыло 14.VIII (за 7-8 не-
дель до отлета). На гнездовьях гуси проводят около 160 дней.
Сведения о линьке фрагментарны. Летающих гусей встречали на про-
тяжении всего лета, но во второй половине июля реже всего. 18.VII 1962 г.
найдены выпавшие при линьке маховые и контурные перья гусей. 18.VII
1984 г. самка при выводке не могла взлететь, но взрослые при выводках,
встреченных 24.VI 1975 г., 3.VII 1976 г. и 13.VIII 1980 г., летали.
Кормятся гуси преимущественно на суше или на болотах. В желудке
молодого гуся, добытого 23.VIII 1933 г. на оз. Мерьт-явр, оказались толь-
ко семена морошки, вороники и песок; помет гусей, собранный 20.VIII
1974 г., состоял из вегетативных частей растений, черники и других ягод
(А. Брагин).
Крупные хищные птицы иногда добывают гусей: 1 .VIII 1935 г. на юж-
ном побережье Чунозера нашли обклеванный остов взрослого гуся, a 2.XI
1939 г. обнаружены остатки другого в гнезде беркута в долине р. Витте.
Масса тела молодого гуся, добытого в августе 1933 г., 2820 г, длина -
750, крыло - 423, хвост - 128 мм. След гуся около 11 см в длину и ши-
рину.
21
Пискулька - Anser erythropus (L.)
Молодой экземпляр этого гуся добыт в начале сентября 1939 г. близ
Тик-губы (Екостровская Имандра) А.Б. Васильвым и определен М.И. Вла-
димрской и О.И. Семеновым-Тян-Шанским.
Пискулек не раз наблюдали визуально: 25.V 1937 г. пару на р. Чуна,
27.VI 1969 г. трех на болоте у оз. Румельявр, 25.V 1982 г. пара пролетела на
высоте 150-200 м над Чунозером на север.
В пролетное время еще несколько раз наблюдали мелких гусей: писку-
лек или казарок, отличавшихся от гуменников и по голосу (в 1948 г; в
сентябре, в 1960,1967,1969, 1971,1982 и 1985 гг. в мае или июне).
В лесной части Кольского полуострова пискулька появляется нерегу-
лярно, на пролете, но на тундрах Мурманского побережья изредка гнез-
дится (Фильчагов, Черенков, 1984; Коханов, Скокова, 1967).
Белощекая казарка - Branta leucopsis (Bechst.)
Редкая птица с ограниченным ареалом (гнездится в Гренландии, на
Шпицбергене, Новой Земле и Вайгаче, зимует на южном побережье Север-
ного моря). На Кольском полуострове - редкая пролетная птица.
Белощеких казарок несколько раз наблюдали с 9.V по 17.VI 1967 г. в
верхнем конце Чунозера А.Б. Брагин и О.И. Семенов-Тян-Шанский, по
1-2 птицы, a 21.V - стайку в 5 экз. В дальнейшем в том же районе 28.V
1971 г. видели одну, 18-19 V1973 г. 2-4 птицы (А. Брагин, М. Владимир-
ская); 7.VI 1974 г. на Турухтанном болоте (побережье Охтозера) одну
видел О.И. Семенов-Тян-Шанский, 5.V 1975 т. одна пролетела над Чун-
озерской усадьбой, 9.V 1977 г. семь птиц наблюдал в устье р. Чуна И. Па-
ракецоЪ, a 2.V 1984 г. трех летевших над Чунозерской усадьбой видели
О. Макарова и А. Хохлов.
Белощекая казарка легко отличается от монотонно-серых настоящих
гусей контрастной окраской, напоминающей белую трясогузку.
Черная казарка - Branta bemicla (L.)
Черная казарка гнездится по азиатским и американским побережьям
Ледовитого океана, а зимует южнее, у берегов Атлантического и Тихого
океанов. В Лапландском заповеднике - редкий пролетный вид. Пару ка-
зарок, по-видимому черных, наблюдал 7.VI 1938 г. на оз. Кензис-явр
лесничий заповедника Н. Благовещенский; 24.V 1960 г. Ю. Гаев видел
шесть черных казарок, пролетевших над Чунозерской усадьбой; в 1970 г.
А. Брагин заметил 12.V четырех над Мончегорском, a 18.V - трех над
р. Чуна; в 1971 г. он же наблюдал 10.V шесть над Мончегорском и 15.V
трех над Нижней Чуной; 28.V 1977г. над устьем Верхней Чуны пролетели
8 мелких темных казарок (О. Семенов-Тян-Шанский), а 23.IV 1983 г.
9 черных гусей летели над Чунозером (И. Паракецов). К сожалению, ни
одно из этих наблюдений не документировано ни добытой птицей, ни
фотографией.
22
Кряква - Anas platyrhynchos L.
Местное название: саамское - вадж-лоньт (приблизительный
перевод ’’матерая птица”).
В Лапландии, по сообщению Плеске (1887, с.325), кряква гнездилась к
северу до оз. Инари, р. Тулома и Териберка (примерно до 69° с.ш.). В Лап-
ландском заповеднике встречается и гнездится регулярно, но немного-
численна. По материалам ежегодных учетов водоплавающих за 22 года на
10 км течения реки в среднем приходится 0,9 крякв, а на 10 км береговой
линии озер - только 0,1. При таком низком уровне численности по уче-
там трудно судить о колебаниях ее по годам; для этой цели лучше воспользо-
ваться регистрацией встреч кряквы в картотеке биологических наблю-
дений. Вероятность встреч зависит, очевидно, не только от численности
птиц, но и от количества лиц, принимавших участие в наблюдениях;
чтобы привести сведения за разные годы к одному знаменателю, мы
делим число встреченных особей на число лиц, заполнявших карточки.
Так, например, в сборе материалов по крякве в 1931-1938 гг. в среднем
участвовало по 1,4 наблюдателя, а в 1968-1979 - по 13,7.
На протяжении последних 30 лет намечаются колебания численности
кряквы с периодом около 4 лет и амплитудой порядка 1:2; с другой
стороны, на протяжении полувека заметна тенденция к росту популяции:
один наблюдатель встречал в сезон в среднем в 30-х годах - по 5,2, в 40-х -
6,9, в 60-х - 7,0, в 70-х - 7,5, в 80-х - по 9,6 особей.
Подобно другим речным уткам, кряква нуждается не столько в воде,
сколько в траве. На больших озерах с чистой водой, Чунозере, Охтозере
или Пиренгском, летом эту утку можно встретить только в защищенных
от волны мелких заливах с подводной растительностью. Кряква предпо-
читает спокойные участки рек и мелкие лесные озера. На территории
заповедника распространена довольно равномерно, но общая числен-
ность ее даже в конце лета вряд ли превышает сотню. Малая плотность
северных популяций кряквы объясняется, очевидно, бедностью кормо-
вой базы.
Кряква прилетает раньше других речных уток, сразу вслед за гого-
лем, в среднем за 40 лет, 5 мая (предельные сроки 20.IV в 1964 и 1968 гг. и
21.V в 1935 г.); соответственно и отлетает поздно, около 16 октября
(28.IX 1958 г. - 16.XI 1930 г. за 24 года наблюдений). Отдельные особи или
даже стайки крякв иногда остаются у нас на зиму. Одиночных селезней
наблюдали 7-14 февраля 1951 г. на речке Сейд и 17-18 февраля 1981 г. на
Нижней Чуне. В начале 1966 г. на протяжении двух месяцев (20.11 - 20.1V)
пару крякв шесть раз встречали в районе оз. Ельявр. Зимой 1970/71 г.
группы крякв видели в разных местах: 29.ХП на Ельявруае - пару, 30.1
на р. Нявка - 4, 31 .III на Чалмозере - 8 особей. 1 ОЛП 1973 г. на Ёрмпороге
(р. Коложно) насчитали 21 крякву; с 21.11 до 28.Ш 1975 г. в истоке р. Няв-
ка, на речке Колнса и в Мончегорске видели по 2-7 крякв одновремен-
но; 22.ХП 1977 г. при температуре -30° в истоке Ельявруая подняли оди-
ночную самку; в 1982 г. 21.III близ Мончегорска видели 8 крякв, а 13.IV
на р. Вайкись - пару. По сообщению главного госохотинспектора области
А.М. Хохлова зимой 1982/83 г. в Мурманске, на полынье, подогреваемой
23
сточными водами, зимовало не менее 200 крякв - явление, аналогичное
регулярной зимовке их в Хельсинки, Ленинграде, Берлине и других го-
родах, где образовались оседлые городские популяции крякв.
В предвоенные годы, когда численность крякв была ниже современ-
ной, они прилетали относительно позже, а отлет их не удавалось просле-
дить. Приводим средние сроки по десятилетиям:
Годы 30-е 40-е 50-е 60-е 70-е 80-е
Прилет 17.V 8.V — 2.V 2.V 1.V
Последняя встреча — — — 15.Х 16.Х 17.Х
С прилета до начала июня, когда самки садятся на яйца, кряквы дер-
жатся парами. Сроки размножения известны только приблизительно, так
как гнезда находили редко.
14.VI 1967 г. - под елкой на краю Турухтанного болота (Охтозеро) по взлетевшей
самке найдено ее гнездо (подробностей нет).
3.VI 1971 г. - на берегу Парусной ламбины (р. Нявка) в бинокль замечена сидя-
щая на гнезде кряква; осмотреть его не удалось.
9.VI 1974 г. — на Гагарьем озерке (долине Чуны) под кустом карликовой березки,
на островке, найдено гнездо с 8 яйцами и скорлупой девятого; 2.VII утка еще наси-
живала, а 4.VII выводок ушел, следовательно, инкубация продолжалась не менее
24 суток.
6.VI 1982 г. — на берегу Парусной ламбины найдено гнездо с 5 яйцами.
Все 4 гнезда были устроены на земле, непосредственно около воды. В Лаплад-
ском заповеднике кряквы никогда не пытались занять одно из искусственных гнезд,
предназначенных для гоголей и крохалей.
Первые выводки кряквы появляются около 1 июля, самое раннее 20.VI
(1937, 1947, 1976), самое позднее - 10.VII 1965 г. (за 16 лет). По литератур-
ным указаниям, кряква насиживает 27-28 дней (Palmer, 1976. Vol. 2,
р. 301). Учитывая, что выводок обычно замечают, когда утятам уже не-
сколько дней, можно считать, что нормально птенцы выводятся 27-30.VI,
насиживание начинается в первых числах июня, а кладка яиц - в послед-
ней декаде мая. Утята ранних выводков поднимаются на крыло около
13 августа (4.VIII 1982 г. - 27. VIII 1966, за И лет), а поздние - в начале
сентября: так, 1.IX 1977 г. на Ельявре встретили выводок из 9 нелетных
утят.
Число утят в 108 учтенных в разные годы выводках колебалось от
1 до 10 , в среднем 5,0; маленькие семьи, по 1-2 утенка, составляют 14%
всех встреченных, большие, по 8-10 утят, - 11%. Уменьшение числа
птенцов за время их роста на нашем материале не прослеживается: в
конце июня в среднем 4,8, в июле - 5,3, в августе - 4,6, в начале сентяб-
ря - 5,8 утят в семье.
Известно, что пока самки насиживают, самцы собираются в стаи и отле-
тают к местам линьки, но прямых наблюдений за этим явлением мало:
9.VI 1949 г. видели стаю в 9 птиц, 12.VI 1976 г. - 8, 4.VII 1987 г. - 9, 4-
27.VI 1982 г. замечено несколько стаек по 4-11 селезней. Самки, очевид-
но, линяют, пока водят нелетных утят; изредка встречали и холостых
самок, неспособных взлететь (9.VII 1976 г.).
Лапландский заповедник находится так близко к границе гнездового
ареала кряквы, что весной мы видим крякв, прибывших на место гнездо-
24
вания, а не пролетных. По той же причине и осенний пролет едва заметен:
большие отлетные стаи наблюдались только дважды: З.Х 1979 г. в Монче-
горске (около сотни птиц) и 21.Х 1983 г. на Воче-ламбине (около двухсот).
Сотрудники заповедника А.Б. Брагин, В.М. Бусуек и О.А. Макарова
проследили зарождение городской популяции крякв в Мончегорске, на
Нюдозере и речке Нюде, подпертой дамбой при устье. Образовавшийся в
результате мелководный и богатый кормами пруд стал убежищем уток,
куликов и чаек. В августе 1971 г. здесь учли 6 выводков кряквы, всего
37 птиц,в июле 1975 - не менее 50 особей в 6 выводках, в июле 1981 г. -
7 выводков и несколько холостых уток, всего около 75 голов. В середине
июля 1979 г. дважды наблюдали, как матка переводила пуховичков-
утят через улицу: машины останавливались, чтобы их пропустить. Ребя-
та подкармливали утят хлебом, но в основном они питаются здесь при-
родными кормами - мотылем и взрослыми комарами. В июле 1981 г. на
утенка, отставшего от выводка при переходе его через грязевую отмель,
напали три вороны; утка безуспешно пыталась их отогнать, но они уле-
тели, как только наблюдавший эту сцену А.Б. Брагин взял в руку ка-
мень.
В городе кряквы гнездятся на 2-3 недели раньше, чем в заповеднике:
в 1977 г. первые выводки были отмечены соответственно 12 и 28 июня, в
1979 г. - 14. VI и 2.VII, в 1981 г. - 22. VI и 14. VII. Городские выводки обычно
многочисленнее природных, насчитывая в среднем по 7,0 утят.
В заповеднике окольцовано только несколько случайно пойманных
крякв - и все же известны два возврата: взрослая самка, окольцованная
в ноябре 1956 г. на севере Голандии, в июле была убита рабочим геологи-
ческой партии в южном конце Нотозера; другая кряква, окольцованная
птенцом в июле 1978 г. на Румельявре, убита 18. X в Хартиле, Финляндия
(6Г 30' с.ш., 26° в.д.), раньше, чем окончился отлет ее сородичей из запо-
ведника (21. X 1978 г.). Видимо, кряквы отлетают в юго-юго-западном
направлении и зимуют у южных берегов Балтийского и Северного морей.
Остатки крякв, ставших жертвой хищников, находили неоднократно:
2. VI 1939 г. на берегу Чунозера; 13. V 1964 г. при устье р. Мавра (самец,
добыча норки); 19. VII 1978 г. наУлынчъявре (птенец); 20. VIII 1980 г. на
Ондатровом озерке при устье р. Чуна (птенец, съеден зверем). В гнездах
хищных птиц остатков кряквы не находили.
Размеры двух взрослых (самца и самки), добытых в мае (в мм): длина
540 и 530, крыло 287 и 266, хвост 100 и 95, масса тела 1000 и 974 г. соот-
ветственно. Молодые ко времени подъема на крыло почти сравниваются
со взрослыми, масса тела молодой самки, добытой 13. VIII 1983 г., равна
955 г.
Чирок-свистунок - Anas сгесса L.
Местные названия: саамское - чйивж; русское - чирок - не веер
да означает эту утку, так как поморы на Белом море называют чирком по-
лярную крачку (Карпович, 1984, с. 124).
На Кольском полуострове встречается повсеместно, и в заповеднике
он многочисленнее других речных уток. На больших открытых водоемах
25
чирок не встречается и селится преимущественно по рекам и ручьям, из-
бегая только порожистых участков, а также по лесным и болотным озер-
кам, вплоть до верхнего предела леса. По материалам ежегодного учета
водоплавающих (50 км) береговой линии озер и 60 км течения рек) он
почти в 4 раза многочисленнее кряквы: на 10 км речного русла приходит-
ся 4,0 чирка. Результаты учета в начале августа плохо согласуются с
регистрацией встреч, проводимой на протяжении всего сезона, поэтому
вопрос о колебаниях численности неясен. По регистрации встреч больше
всего чирков было в 1948-1950 гг. - по 27-69 на одного наблюдателя в
сезон; меньше всего, по 4-5 встреч, в 1962-1963, 1965, 1968 и 1984 гг. По
материалам учетов в 1972-1973 и в 1978 гг. на 10 км рек приходилось по
8-9 чирков, а в 1966, 1969, 1974, 1977, 1979-1984 и 1986 гг. только по 1-3.
В вековом масштабе популяция обнаруживает тенденцию к сокраще-
нию: на одного наблюдателя в сезон приходилось чирков, в 30-х годах, -
14,9 в 40-х - 22,7, в 50-х - 13,2, в 60-х - 7,7, в 80-х - 9,1. Утиные попу-
ляции зависят не только от успеха размножения и других местных при-
чин, но и от тех условий, которые они встречают на путях перелетов и
местах зимовок, поэтому их колебания трудно интерпретировать.
Чирок прилетает дней на десять позже кряквы, в среднем 15 мая, при
средней суточной температуре около 3, 6е (25. IV 1951 г. - 29. V 1968 г. за
43 года); улетает раньше других уток, около 12 сентября при температу-
ре 7,0° (25 VII 1936 г., 1987 - 3. X 1940 г. за 39 лет). Сроки прилета доволь-
но стабильны (л ± 6,1) и зависят от температуры: корреляция с темпера-
турой апреля, г = - 0,49 + 0,12, и мая, г = - 0, 22 + 0,15. Время отлета силь-
но варьирует (о ± 11,4) и от температуры сентября практически не зави-
сит (г = + 0, 03 + 0,16).
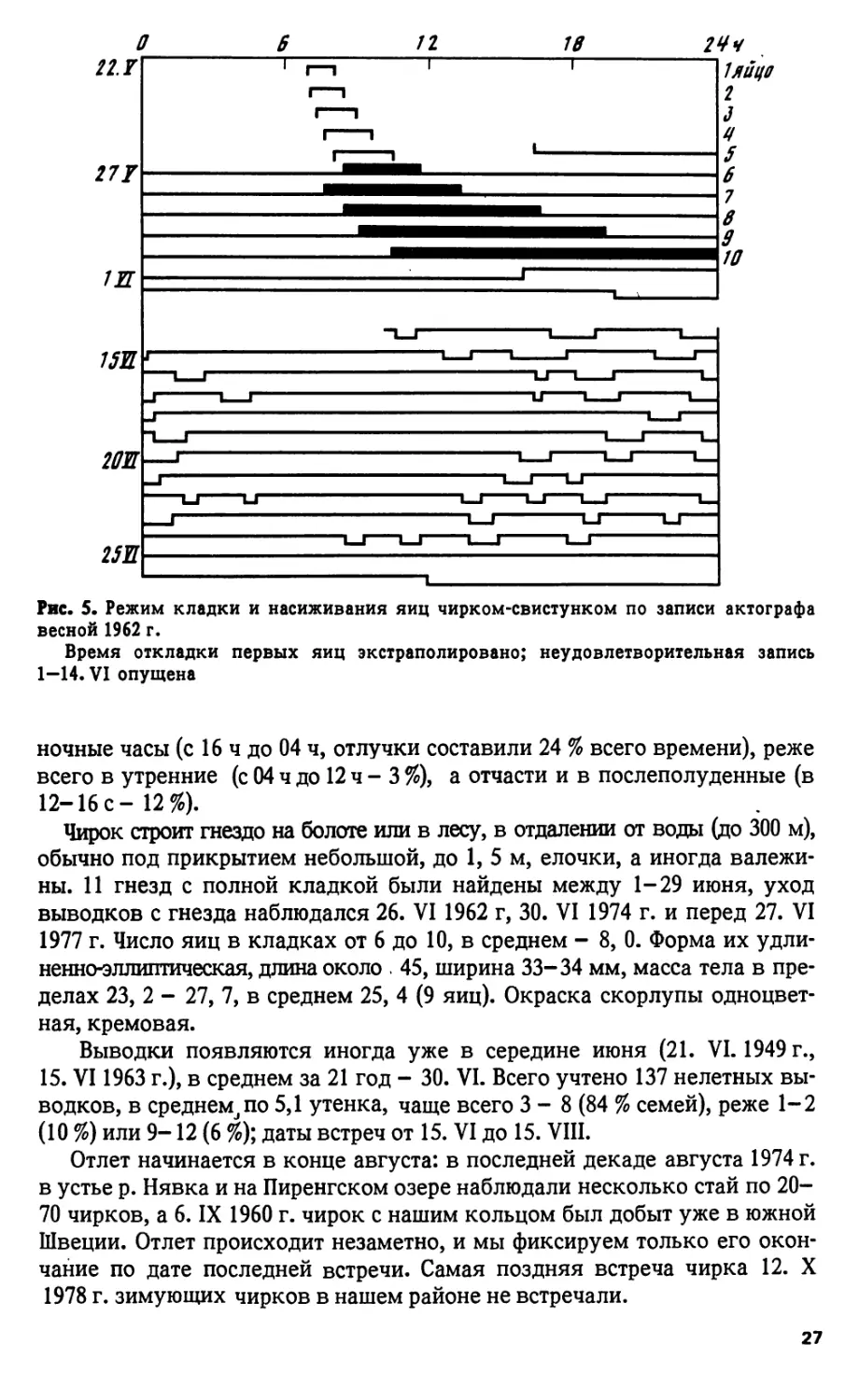
Гнездование чирка подробно прослежено только один раз, в 1962 г.
По срокам наступления фенологических явлений этот год был нормаль-
ным, так что приведенные здесь даты близки к средним многолетним.
24 мая около 9ч утра на выдвинутой в болото лесной гривке, в сотне
метров от ручья, вылетевший из-под елочки чирок обнаружил этим свое
гнездо, содержавшее 3 яйца. При установке актографа 26. V в 14 ч 30 мин
утка отсутствовала, а в гнезде, под слоем моха с примесью пуха, было
уже 5 яиц. Утка неслась ежедневно, появляясь около 8-10 ч утра и
оставаясь на гнезде с каждым днем все дольше: при шестом яйце 3, 5 ч,
а при десятом - 14 ч. К 24 ч 31 мая кладка была закончена (рис. 5), а
1 июня в 16 ч утка села на яйца. Согласно предложенной на диаграмме
экстраполяции, первое яйцо снесено утром 22 мая. Птенцы начали выводиться
25 VI перед полуднем, матка целые сутки не оставляла гнезда, авпол-
день 26. VI увела выводок. От начала инкубации яиц до оставления
гнезда птенцами прошло 24, 8 сут - на 2-4 дня больше, чем указывается
в литературе. В другом гнезде (1974 г.) интервал между первым прокле-
вом скорлупы и уходом выводка равнялся 37 ч.
Во время инкубации наседка оставляла гнездо 2-5 раз в сутки, про-
должительность прогулок варьировала от 0,5 ч до 3, 7 ч. Фактически утка
грела кладку 85 % всего времени, а 15 % яйца оставались одни, прикры-
тые пуховым одеялом. Кормилась она преимущественно в вечерние и
26
Рис. 5. Режим кладки и насиживания яиц чирком-свистунком по записи актографа
весной 1962 г.
Время откладки первых яиц экстраполировано; неудовлетворительная запись
1—14. VI опущена
ночные часы (с 16 ч до 04 ч, отлучки составили 24 % всего времени), реже
всего в утренние (с 04 ч до 12 ч - 3 %), а отчасти и в послеполуденные (в
12-16 с - 12%).
Чирок строит гнездо на болоте или в лесу, в отдалении от воды (до 300 м),
обычно под прикрытием небольшой, до 1, 5 м, елочки, а иногда валежи-
ны. И гнезд с полной кладкой были найдены между 1-29 июня, уход
выводков с гнезда наблюдался 26. VI 1962 г, 30. VI 1974 г. и перед 27. VI
1977 г. Число яиц в кладках от 6 до 10, в среднем - 8, 0. Форма их удли-
ненно-эллиптическая, длина около . 45, ширина 33-34 мм, масса тела в пре-
делах 23, 2 - 27, 7, в среднем 25, 4 (9 яиц). Окраска скорлупы одноцвет-
ная, кремовая.
Выводки появляются иногда уже в середине июня (21. VI. 1949 г.,
15. VI 1963 г.), в среднем за 21 год - 30. VI. Всего учтено 137 нелетных вы-
водков, в среднем^о 5,1 утенка, чаще всего 3 - 8 (84 % семей), реже 1-2
(10 %) или 9-12 (6 %); даты встреч от 15. VI до 15. VIII.
Отлет начинается в конце августа: в последней декаде августа 1974 г.
в устье р. Нявка и на Пиренгском озере наблюдали несколько стай по 20-
70 чирков, а 6. IX 1960 г. чирок с нашим кольцом был добыт уже в южной
Швеции. Отлет происходит незаметно, и мы фиксируем только его окон-
чание по дате последней встречи. Самая поздняя встреча чирка 12. X
1978 г. зимующих чирков в нашем районе не встречали.
27
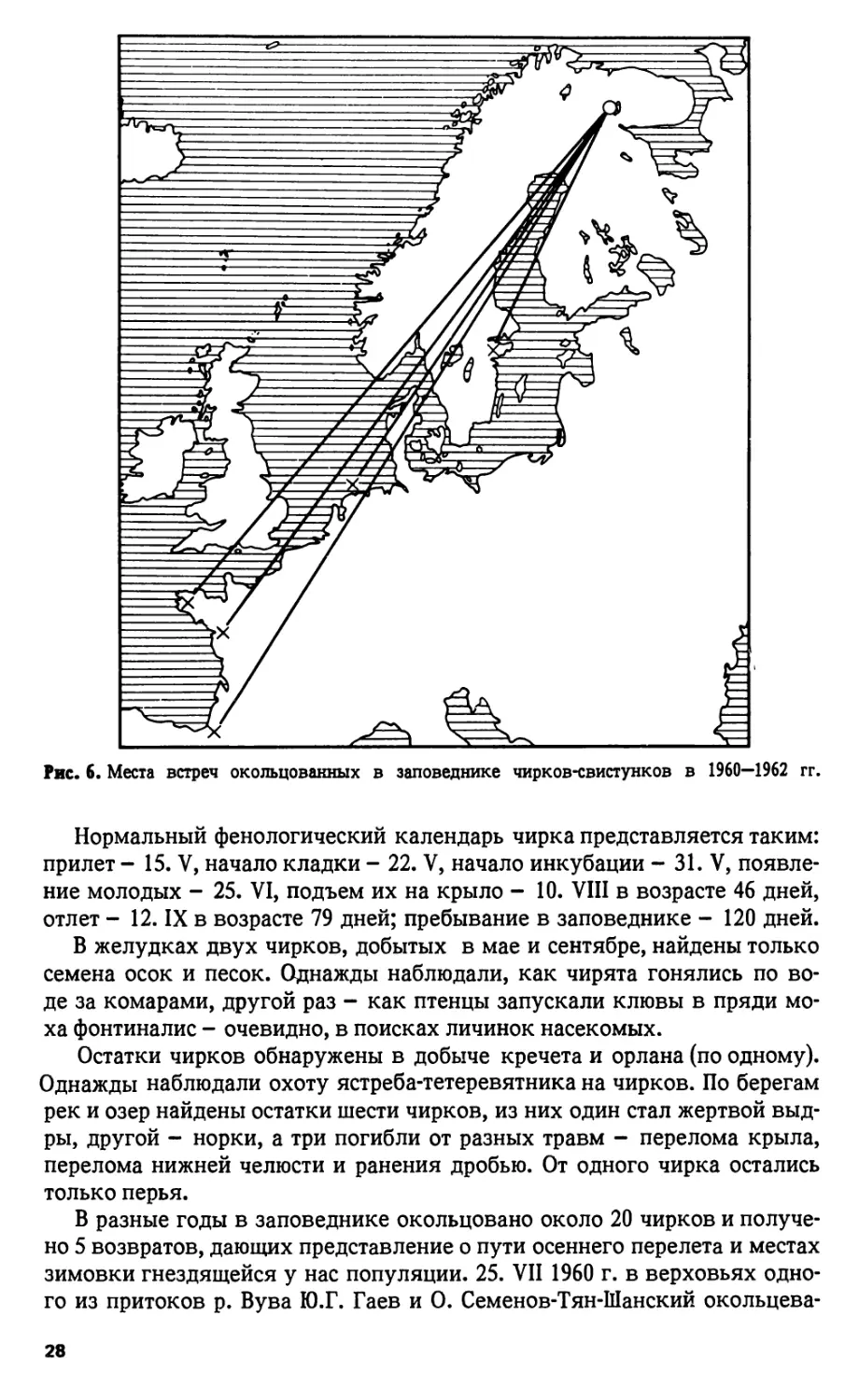
Рис. 6. Места встреч окольцованных в заповеднике чирков-свистунков в 1960—1962 гг.
Нормальный фенологический календарь чирка представляется таким:
прилет - 15. V, начало кладки - 22. V, начало инкубации - 31. V, появле-
ние молодых - 25. VI, подъем их на крыло - 10. VIII в возрасте 46 дней,
отлет - 12. IX в возрасте 79 дней; пребывание в заповеднике - 120 дней.
В желудках двух чирков, добытых в мае и сентябре, найдены только
семена осок и песок. Однажды наблюдали, как чирята гонялись по во-
де за комарами, другой раз - как птенцы запускали клювы в пряди мо-
ха фонтиналис - очевидно, в поисках личинок насекомых.
Остатки чирков обнаружены в добыче кречета и орлана (по одному).
Однажды наблюдали охоту ястреба-тетеревятника на чирков. По берегам
рек и озер найдены остатки шести чирков, из них один стал жертвой выд-
ры, другой - норки, а три погибли от разных травм - перелома крыла,
перелома нижней челюсти и ранения дробью. От одного чирка остались
только перья.
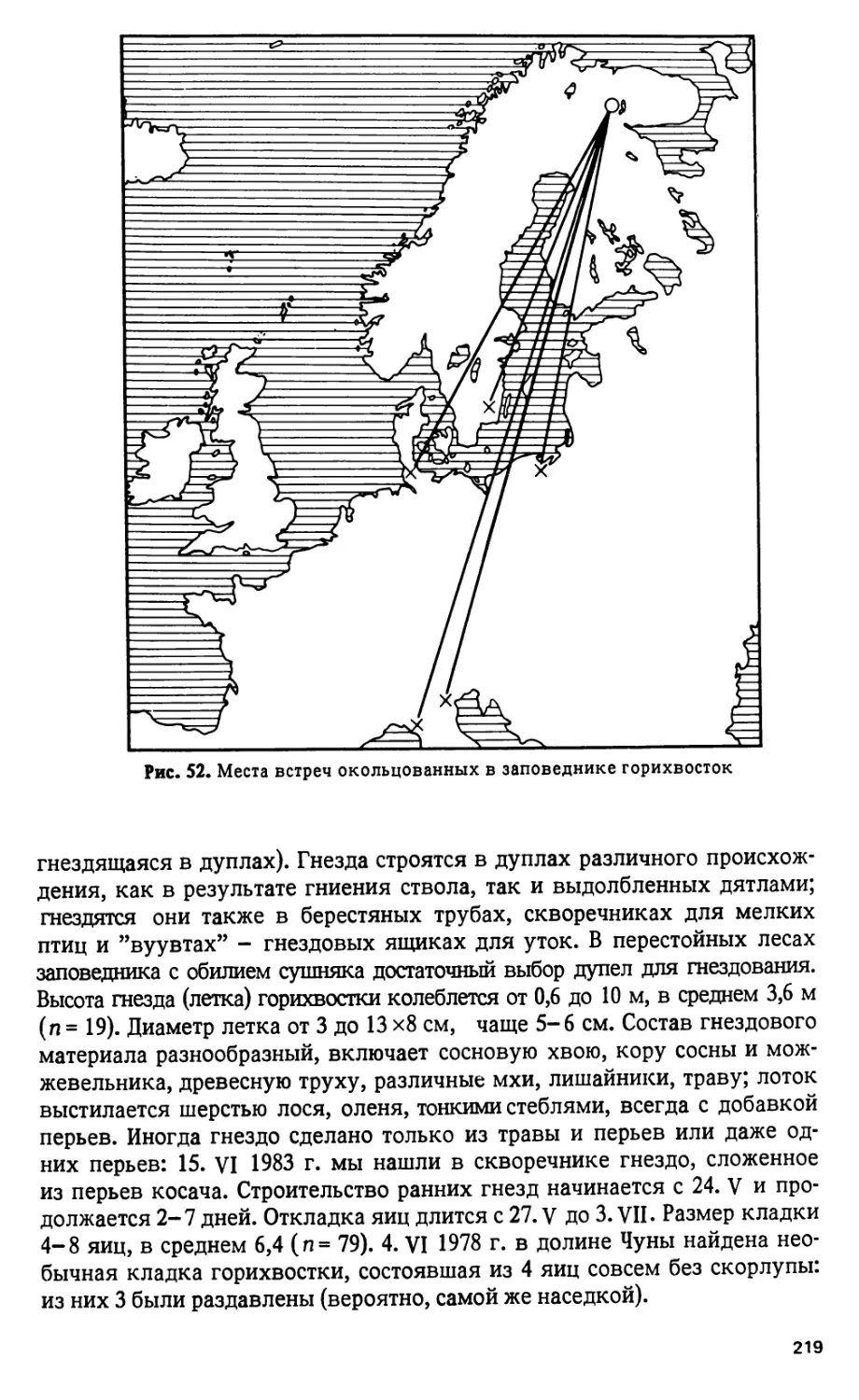
В разные годы в заповеднике окольцовано около 20 чирков и получе-
но 5 возвратов, дающих представление о пути осеннего перелета и местах
зимовки гнездящейся у нас популяции. 25. VII 1960 г. в верховьях одно-
го из притоков р. Вува Ю.Г. Гаев и О. Семенов-Тян-Шанский окольцева-
28
ли 8 чирят - весь выводок. Уже 6. IX один был добыт на балтийском по-
бережье южной Швеции (58° 33 с.ш., в 1270 км к юго-западу от места рож-
дения), другой 5. X на о-ве Амеланд у северного побережья Голландии
(53° 27 с.ш., 2150 км на юго-запад). Третий и четвертый убиты в феврале-
марте 1962 г. на зимовках, на атлантическом побережье Франции - при
устье Луары и в Гасконских Ландах (3000-3300 км на юго-запад от роди-
ны). Еще один чирок, окольцованный 1. VIII 1961 г. наПельозере, добыт
4. II 1962 г. в Бретани (3000 км; рис. 6).
Размеры 5 взрослых самцов (в мм): длина 340-390, крыло 173-185,
хвост 70-80; масса тела 285-328 г, в среднем 305 г; размеры самок, соот-
ветственно - 330-345, 162-178 и 62-70 мм (масса тела не измерена).
Масса тела 5 молодых чирков неизвестного пола в августе-сентябре в
среднем 288 г.
Свиязь - Anas peneloge L.
Местные названия: саамское - наартель.
В мурманской области распространена повсеместно, по мелким лес-
ным озеркам и участкам рек со спокойным течением, но немногочислен-
на. В Лапландском заповеднике, по материалам учетов 1966-1987 гг.,
на 10 км береговой линии озер приходится в среднем 0,4 свиязи, на 10 км
рек - по 0,5.
В среднем за сезон наблюдатели отмечали около 12 свиязей: в 30-х го-
дах - по 13, в 40-х - по 23, в 50-х - по 4, в 60-х - по 10, в 70-х - по 12, в
80-х - по 10. Снижение запасов в 50-х годах не вполне убедительно, так
как в наблюдениях пробел с 1952 по 1957 г., но повышение численности
в 1940-1949 гг. (когда был такой же пробел), вероятно, реально. На про-
тяжении последних 24 лет заметны регулярные колебания численности с
периодом 4 года и амплитудой примерно 1:4; максимумы падают на годы
1966, 1971,1974, 1978,1982 и 1985.
Свиязей легко наблюдать только весной, пока они держатся парами и
кормятся на виду, по разливам и в устьях рек. Летом они встречаются
гораздо реже, а выводки отмечались в заповеднике не каждый год. Брач-
ные игры начинаются сразу после прилета. Своеобразный, громкий и
чистый свист самцов облегчает своевременную регистрацию даты прилета
(рис. 7).
Свиязь прилетает на две недели позднее кряквы, в среднем за 47 лет
19 мая, при средней температуре 4,5е (крайние сроки - 5. V. 1948 г. и
31. V 1932 г., 1971 и 1972 гг.). Время прилета коррелирует с температурой
мая (г = — 0,55 + 0,15 ), но сравнительно мало варьирует по годам (о +
7,2 дня). На протяжении полувека заметны колебания средних сроков
прилета: в 30-х годах - 25. V, в 40-х - 11. V, 50-х - 15. V, 60-х и 70-х -
21.V, 80-х- 19. V.
За все годы в заповеднике обнаружено только 5 гнезд:
16. VI 1933 г. — на северном берегу Чунозера, в 10 м от воды, под еловым кусти-
ком; на подстилке из опавшей хвои, стеблей вероники, зеленых мхов и трухи лежа-
ли 6 яиц, окруженных барьером из пуха и трухи. Выводок ушел между 6—9 июля.
1. VI 1936 г. — в сосновом редколесье близ Чунозерской ламбины (мелкий залив
29
или старица), под трехметровой елочкой, на ягельнике, на подстилке из опавшей
хвои и пуха, 7 яиц. Утка поднята с гнезда. К 27. VI оно опустело, остался только
"болтун”.
15. VI 1961 г. — у берега р. Курка, на кочке мокрого болота (без подробностей).
26. VI 1974 г. — на высоком берегу р. Купись ниже впадения в нее Урды, под
кустом можжевельника в 5 м от воды гнездо с 8 яйцами (утка слетела заранее). Ло-
ток диаметром 17, глубиной 8 см, выстлан мхом и сухой травой, смешанной с тем-
ным пухом.
9. VI 1977 г. — на северном берегу Улынчъявра, в 1 м от воды, под сосновой ва-
лежиной, по взлетевшей утке найдено ее гнездо с яйцами; спугнутая вторично,
свиязь бросила гнездо.
Самая ранняя кладка с яйцами найдена 1. VI, самая поздняя - 6. VII,
средняя дата 19. VI (п = 8). При допущении, что свиязь насиживает столь-
ко же времени, как и чирок (25 сут), кладка яиц начинается, вероятно,
в последней пентаде мая. инкубация - с 7. VI до 2. VII.
Число яиц в кладке от 6 до 9, их размеры: длина 58- 65, ширина 38-
42 мм, масса 9 яиц в начальной стадии инкубации 39, 9 - 47,0 г, в сред-
нем 47,7; скорлупа одноцветная, сливочно-белая.
Первые выводки встречаются обычно в первой декаде июля, самое
раннее 25. VI 1987 г., самое позднее 14. VII 1971 г.; дата встречи последне-
го нелетного выводка - 11. VIII 1986 г., а первого способного летать -
5. VIII 1949.
Нелетные выводки насчитывают от 1 до 8 утят, в среднем 4,8, при сред-
30
нем размере кладки - 7, 5 яиц. Из 60 учтенных выводков по 1-2 птенца
имели 10 %, 3-6 птенцов - 65 %, 7-8 утят - 25 %.
Самцы свиязи отлетают на места линьки уже в первой половине июня,
как только самки сядут на гнезда. Селезней группами по 2-3 встречали
между 25. V и 24. VI, последнее наблюдение взрослого самца - 30. VII
1934 г. Начиная с середины июня не слышно и свиста селезней. Молодняк и
самки отлетают в сентябре довольно незаметно, самых последних встре-
чали в первой половине октября. Стаи свиязей втречаются у нас редко
и только в пролетное время, например 3. VI 1941 г. в устье р. Нявка-
28 птиц; 8. IX 1948 г. там же - 15; 23. IX 1948 г. на оз. Нюхчи - 50; 4. IX
1949 г. над Чунозером пролетела на запад большая стая. Средняя дата
последней встречи 26. IX (13. IX 1987 г. - 15. X 1970 г. за 16 лет; стандарт-
ное отклонение = ± 9, 8). Как правило, свиязи не пытаются зимовать на
местах гнездования, но зимой 1946/1947 г. она продержалась на р. Витте
до февраля (М.С. Тютин).
Судя по отрывочным наблюдениям, свиязь питается главным образом
растительной пищей. В желудке самца, добытого 2. X 1932 г. близ Чунозе-
ра, были семена вороники и песок, в желудках двух птенцов 27. VII
1950 г. - перетертая зелень и песок. Весной свиязи ищут корм среди
плывущего по реке, крутящегося в водовороте или прибитого течением
к кромке льда мусора (обломанные ростки водных растений, семена
осок и др.). В июне 1978 г. А. Брагин наблюдал, как свиязь рвала на бе-
регу молодую траву, а потом поплыла и собирала корм с поверхности во-
ды; в июле А. Гилязов видел, как утята, плывшие вдоль пояса осоки,
подпрыгивали, хватая насекомых с листьев.
Сведений о врагах мало: в июле 1940 г. один птенец задавлен на
Островском озере кошкой, 1 VII 1982 г. утенок обнаружен в желудке щу-
ки, пойманной в долине р. Чуна.
Размеры самца и самки (мм): длина 450 и 440, крыло 260 и 239, хвост
95 и 93 см, масса тела 700 и 617 г (осенью). Масса двух молодых 27. VII
1950 г.: самца - 442 г, самки - 420 г.
Шилохвость — Anas acuta L.
Местное название: саамское - чуарш или чуэрш.
В Лапландском заповеднике гнездится заметно реже, чем предыдущие
три вида, и не ежегодно. При учете водоплавающих в августе на 10 км
реки приходится в среднем 0,10, а на таком же протяжении озерного бе-
рега - только 0,01 шилохвость. Однако учет визуальных встреч дает
на одного наблюдателя, в среднем за 46 лет, по 5, 0 шилохвостей за сезон,
при этом повышение численности отмечалось в 1938-1939, 1949-1951,
1969-1971 и в 1977 гг. Обнаружена тенденция к сокращению популяций:
в первые годы существования заповедника (1931-1951) встречали в год
пр 8,4 особи, во второй период (1958-1987) только по 3,2.
Шилохвость появляется небольшими стайками, состоящими из 1-4
пар, примерно в одно время с чирком, в среднем 15 мая (крайние даты
21.IV 1938 и 1939 гт. - 29. V 1968 и 1976 гт. за 28 лет ; среднее квадратичное
отклонение ± 10,8 дней).
31
Рис. 8. Самка шилохвости у выводка, 22. VII 1975 г. Фото О.И. Семенова-Тян-Шан-
ского
Единственное гнездо найдено 27. V 1940 г. на Турухтанном болоте у бере-
га Охтозера, в корнях сосны, в 1 м от воды: утка слетела с гнезда, в
котором было 3 яйца. Пиренгское водохранилище, в состав которого
входит Охтозера, в это время накапливало воду: через несколько дней
вода затопила гнездо, в котором было уже 7 яиц (рис. 8).
Нелетные выводки шилохвостки встречали самое раннее 16. VI
1936 г., самое позднее - 9. VIII 1970 г.; первая встреча летных утят - 10.
VIII 1949 г. В учтенных 11 выводках было от 3 до 10 птенцов, в среднем
6,3 (рис. 8). Встречали их преимущественно на тихих участках рек и
ручьев с высокими, поросшими ивняком или торфяными берегами, реже
на травянистых озерках с водной площадью в несколько гектаров (прото-
ки на Пасма-ламбине, р. Нявка, Купись, Лылчи, Жемчужный ручей и
речка Нюдовка в Мончегорске; озера Ондатровое, Окуневое (близ Ель-
явра) и Румельявр; болота Лосиное и Турухтанное). Наряду с выводками
в тех же местах все лето встречаются и холостые особи. Необычайно позд-
ний нелетный выводок наблюдали 7. X 1981 г. в Мончегорске.
Отлет начинается в сентябре и заканчивается в октябре. В это время
шилохвости иногда образуют большие стаи, например 4. IX 1949 г. над
Чунозером пролетела на запад стая в 65 особей; 22. IX 1950 г. на Румельяв-
ре плавали 47, а 30. X 1961 г. в Травяной губе Верхнего Пиренгского
озера поднялись из осоки 20 шилохвостей. Средняя дата последних
встреч - 29 сентября (14. IX 1949 г. ~ 31. X 1961 г. за 10 лет, о = ± 15,3).
Зарегистрированы остатки четырех погибших :шилохвостей: две раз-
бились о провода электропередачи (25. VIII1961 г. и 20. VIII 1983 г.), одна
стала жертвой хищной птицы (8. IX 1973 г.), одна погибла от ранения
(6. VIII 1949 г.).
Масса тела одной взрослой самки - 760, молодого самца - 730 и сам-
ки - 695 г (август).
32
Чирок-трескунок - Anas querquedula L.
Эта утка характерна для зон смешанных лесов и степей, а на Кольский
полуостров залетает только изредка. В Лапландском заповеднике чирок-
трескунок появлялся дважды: 26. V 1978 г. пару видел на р. Чуна А.С. Ги-
лязов, а 21-22. V 1981 г. другую пару наблюдал на полынье в устье
р. Мавра А.М. Хохлов.
В окрестностях г. Кандалакши трескунок изредка гнездится, выводки
были встречены в 1968 и 1978 гг. в августе (Коханов, 1987, ’с. 27).
Широконоска - Anas clypeata L.
Граница ареала этой утки проходит в Карелии, откуда она залетает на
Кольский полуостров, а изредка даже гнездится.
В Лапландском заповеднике широконоску наблюдали пять раз: 20-
21. V 1967 г. А.Б. Брагин и О.И. Семенов-Тян-Й1анский наблюдали пару на
р. Чуна; 23. VI 1970 г. лесник В.И. Владимирский видел пару в устье
р. Нявка; 28. VII 1981 г. А.Б. Брагин встретил одиночную самку в Монче-
горске, на речке Нюдовка; 22. V 1982 г. А.С. Гилязов и О.И. Семенов-Тян-
Шанский видели в устье Верхней Чуны селезня широконоски в обществе
свиязей; 28. VII 1986 г. на оз. Красная Ламбина, по сообщению лесника
А. Сапунова, плавал выводок, самка с 9 утятами. В 1960, 1962 и 1967 гг.
залетные широконоски гнездились на северо-западной оконечности Коль-
ского полуострова, Айновых островах (Коханов, 1987, с. 28).
Гага обыкновенная - Somateria mollissima (L.)
Ареал гаги ограничен морским побережьем Северной Атлантики. В
Мурманской области гнездится по Мурманскому, Терскому и Карельско-
му берегам. Согласно исследованиям Кандалакшского заповедника, бе-
ломорские гаги зимуют на разводьях среди льдов, преимущественно в
Онежском заливе; мурманские частично остаются близ мест гнездования,
а частью сдвигаются вдоль побережья к западу. В глубь материка гаги
редко залетают, по крайней мере в Лапландском заповеднике летом их
не встречали ни разу.
26.1 1980 г. лесник П.П. Кохов заметил над кордоном Нявка летевшую
строем клина стаю черных уток в 50-70 голов. В тот же день в 40 км к
ССВ от этого места лесник В.И. Станюлис наблюдал над оз. Вайкись про-
летевший на север, на высоте около 200 м, клин ’’гусей”, около 160 осо-
бей. Загадочные птицы почти наверное были гагами, возможно даже, что
оба наблюдателя видели одну и ту же стаю. Перелет морских птиц на
север в середине зимы при очень коротком световом дне, очевидно, был
вызван сильными морозами: на Чунозере средние суточные температуры
23, 24 и 25 января были ниже -30е, а 26 - 28°. По всей вероятности, раз-
водья на Белом море покрылись льдом и гаги направились кратчайшим
путем к незамерзающему Баренцеву морю. При скорости полета уток
около 75 км/ч перелет от ближайшей открытой воды на Белом море, Горо-
децкого и Купчинского порогов и о-ва Великого, до Мурмана потребовал
бы около 4 ч.
3. Зак. 1023
33
Хохлатая чернеть - Aythia fuligula (L.)
Местные названия: саамское - видт или видый.
В Лапландском заповеднике хохлатая чернеть впервые достоверно
наблюдалась в 1962 г., и с тех пор она довольно регулярно появляется на
пролете и в небольшом количестве гнездится.
При ежегодном учете водоплавающих хохлатая чернеть отмечена
только за 4 сезона из 19. В среднем на 10 км береговой линии озер при-
ходится О, 4 чернети. В картотеке биологических наблюдений за 45 лет в
заповеднике зарегистрировано 307 хохлатых чернетей, в это число
входит несколько больших стай, встреченных в пролетное время: 14. X
1962 г. в устье р. Мавра плавала стая до сотни особей, а 3. VI 1983 г. на
Румельявре - 25 чернетей.
Чернеть появляется сравнително поздно, с незначительными колеба-
ниями по годам: в среднем 23 мая, от 15. V 1980 г. до 2. VI 1971 г. за 21
год, среднее квадратичное отклонение ± 4, 6 дней. Средняя температура
дня прилета около 5,4е.
Единственное гнездо хохлатой чернети найдено на Гагарьем озерке в
долине р. Чуна 28. VI 1982 г. А.Б. Брагиным; оно находилось на небольшой
кочке, среди воды, и содержало девять сильно насиженных серо-зеленых
яиц.
Выводки чернетей встречали на протоке, соединяющей озера Улынч-
и Румельявр, на озерках в долинах Чуны и Ливы и 8 раз в г. Мончегорске.
Число утят в И выводках варьировало от 1 до 7, в среднем 3,5. Нелетные
выводки встречали между 7. VII - 18. VII, в среднем 28. VII: очевидно,
птенцы хохлатой чернети выводятся и поднимаются на крыло позже, чем
у других уток.
Хохлатая чернеть предпочитает мелкие озерки с относительно бога-
той растительностью и речные протоки с медленным течением. С 1977 г.
чернети регулярно гнездятся в г. Мончегорске, в обществе крякв и дру-
гих речных уток. 7. VII 1981 г. на крошечном, не более 0,2 га, пруду
плавали две кряквы с выводками и две самки чернети; одна из крякв жесто-
ко била и топила обеих чернетей, отгоняя их от своих птенцов. При одной
из чернетей был пуховый утенок, остальные, видимо, погибли или
разбежались; позднее и этот утенок исчез.
Хохлатая чернеть кормится как речные утки, опрокидываясь в воде
(не ныряя).
4. VI 1980 г. среди группы деревьев у берега р. Нюдовки нашли остат-
ки самца, пойманного за несколько дней до того хищной птицей.
Самые поздние встречи уток этого вида 14. X 1962 г. и 6. IX 1981 г.; в
другие годы отлета не прослежено.
Морская чернеть - Aythia mania (L.)
Местные названия: саамское - видт или видный (одинаково с хох-
латой чернетью).
Морская чернеть, как и хохлатая, распространена, очевидно, по всему
Кольскому полуострову и примерно так же немногочисленна. В Лапланд-
34
ском заповеднике с 1933 по 1979 г. зарегистрированы 270 птиц этого вида,
причем попадались они не ежегодно, а с 1980 г. не было ни одной встре-
чи. За 22 года учетов на стационаре приходится в среднем по 0, 15 морс-
ких чернетей на 10 км береговой линии озер и 0,16 на 10 км рек.
Прилетает она немного позже хохлатой чернети, в среднем за 18 лет -
26 мая (20. V 1940 г. - 6. VI 1933 г.; среднее квадратичное отклонение
± 4,8 дня), при переходе средней температуры воздуха через 6, 0е.
В отличие от хохлатой чернети морская предпочитает более открытые
водоемы и попадается не только на маленьких лесных озерах, но и на
больших и глубоких, каковы Чунозеро, Охтозеро и Пиренгское.
В разные годы учтено 10 нелетных выводков, в каждом от 2 до 10 птенцов,
в среднем 4,0. Эта утка гнездится очень поздно, средняя дата встреч не-
летных выводков - 14. VIII самая поздняя - 3. IX 1941. Семьи встреча-
ли на Румельявре (4 раза), старицах . р. Чуна (2), озерах в долине Витте
(2) и по одной на Охтозере, Куркозере и в г. Мончегорске. Поскольку мо-
лодняк поднимается на крыло только в сентябре, морские чернети и от-
летают поздно, средняя дата последних встреч за 5 лет - 12 октября
(29. IX 1966 г. — 17. X 1974 г.), при температуре воздуха +1,Г.
2 и 3. VII 1972 г. лесник В. Михалев наблюдал пару уток этого вида на
р. Нижняя Чуна: возможно, они принадлежали к позднему выводку, по-
чему и не улетели вовремя.
Размеры взрослой самки, добытой на осеннем пролете: длина 438,
крыло 215, хвост 64 мм, масса тела 899 г.
Турпан - Melanitta fusca (L.)
Местное название: саамское - куалнас.
В Мурманской области встречается по озерам повсеместно, но в 6 раз
реже синьги (в картотеке за 1931-1987 гг. зарегистрированы 3444 синьги
и 587 турпанов). На учетном стационаре турпанов видели только 3 раза:
в 1980 г. один выводок на Улынчьявре, в 1983 и 1985 гг. по одному вывод-
ку на Румельявре. В среднем за 22 года на 10 км береговой линии озер
приходится только 0,15 турпана.
Приблизительное время прилета турпана - 3 июня (15. V 1983 г. - 20.
V 1969 г, за 29 лет), а отлета - 5 октября (12. IX 1967 г. - 30. X 1931 г.;
10 лет). Турпан встречается не часто, поэтому прилет мог бы отмечаться
с запозданием, а последняя встреча не всегда, соответствует моменту от-
лета. •
Турпан встречался нам преимущественно на озерах системы р. Нявка -
Румельявре, Улынчьявре, Окуневом, Нявка-озере и на ламбинах в
нижнем течении Нявки (65 % всех встреч); реже - на больших озерах -
Пиренгском, Улынчьявре и Екостровской Имандре (32 %). Большинство
встреч приурочено к весеннему пролету (в мае - 18 %, июне - 49 % наб-
людений), но часть особей живет в заповеднике все лето и гнездится.
4. VII 1948 г. под елочкой в 50 м от восточного берега Румельявра по
взлетевшей утке обнаружено гнездо турпана с 6 яйцами; к 16. VII из 5
яиц вывелись птенцы, шестое оказалось разбитым; 9. VIII - 4. IX там же
наблюдали выводок турпана.
35
В разные годы зарегистрировано 12 нелетных выводков, содержавших
от 2 до 8 утят, в среднем 5,1; встречали их с 13 июля по 15 августа. Ново-
рожденные плавают тесной кучкой и не ныряют, но уже через несколько дней
ныряют вместе с маткой. 20. VII1978 г. А. Гилязов видел, как птенец ненадол-
го сел на спину самки. Время подъема молодых турпанов на крыло не прос-
лежено.
Синьга - Melanitta nigra (L.)
Местные названия: саамское - нёрг, русское - чернуха.
В Мурманской области встречается по озерам повсеместно, но в общем
немногочисленна. На стационаре Лапландского заповедника по учету
водоплавающих встречается не каждый год; средняя плотность этой утки
1,4 птицы на 10 км береговой линии озер и 0,2 на 10 км рек. Осенью на
всей территории заповедника (1600 км2) проживает примерно сотня
синьги.
Летом синьга приурочена к открытым озерам, но выводки нередко
встречаются на речных плесах со спокойным течением. Численность ко-
леблется от года к году, без ясно выраженной периодичности, так как
интервал между ее подъемами варьирует от 2 до 8 лет. Тенденция к росту
или сокращению популяции в вековом масштабе также не обнаружена.
Количество встреч на 1 наблюдателя в год, в среднем по десятилетиям,
следующее:
Годы 30-е 40-е 50-е 60-е 70-е 80-е Среднее за
все годы
Число встреч 20 16 15 22 13 15 17
Прилетает синьга сравнительно поздно, примерно, за неделю до вскры-
тия озер, в среднем за 49 лет - 25 мая (7. V 1938 г. - 16. VI 1958 г.; о ±8,1),
при средней температуре мая около 5,8°. Даты прилета коррелируют со средней
температурой мая (г = -0,37 ± 0,13) и с моментом вскрытия озер (г = +0,33±0,13),
хотя соответствующие коэффициенты невелики.
Синьга появляется парами или группами до 5-7 пар, а позднее - по-
больше: так, 22. V 1948 г. на плесе р. Нявка видели стаю синьги в 33 осо-
би, а 5. VI 1978 г. на Чунозере - около 45. В состав таких стай входят,
вероятно, не только производители, но и годовалые птицы, не участвую-
щие в размножении. До середины июня синьги держатся парами, потом
начинают встречаться одиночные особи или группы птиц одного пола,
очевидно, в связи с началом отлета самцов к местам линьки; в. начале
июля 1974 г. на Чунозере неоднократно видели стаю из 15 взрослых
самцов.
Брачные игры синьги наблюдали 21. VI 1959 г. и 1966 г., а драку сам-
цов - 17. VI 1977 г. У самки, попавшей 19. VI1961 г. в рыболовную сетку,
яичники были заметно увеличены (фолликулы размером с горошину, т.е.
около 5 мм), но желтых тел в них не обнаружено. Хотя гнезд синьги в заповед-
нике не находили, не подлежит сомнению, что гнездится она поздно.
Первые выводки пуховичков встречали в разные годы между 7 и 29
июля, а в среднем за 20 лет - 19. VII. Учитывая, что первых утят замечают
не в первый день их жизни, а с некоторым запозданием, и и считая продол-
36
жительность инкубации в 29 суток (27-31-й день по Гаррисону, 1975,
с. 89), синьга, вероятно, насиживает со второй декады июня, а нестись
начинает не ранее первой декады. Пуховичков синьги встречали 15. VII
1977 г. и 18. VII 1978 г. на маленькой ламбине между озерами Нявка и
Купись (2. VII 1977 г. здесь же подняли на болоте самку, вероятно с
гнезда); 19. VII 1973 г. на болотном озерке у южного берега Чунозера;
22. VII 1968 г. на Травяных ламбйнах близ Куропачьего озера; 13. VII 1980 г.
7. VII 1983 г на речке Туреск у кордона ’’Купись”. Отсюда следует,
что синьга гнездится на болотах и переводит утят на ближайшее озеро,
когда они несколько подрастут.
За 45 лет наблюдений зарегистрированы 80 выводков синьги, больше
половины их - на Чунозере и по нескольку на озерах Купись, Нявка и Ру-
мельявр. Одиночные семьи отмечались также на озерах Куропачьем, Кен-
зис, Пагель и Островском. В 1981 и 1986 гг. на Чунозере жило по 4 вывод-
ка, в 1966 и 1984 гг. - по 3, в четыре лета - по 2, 12 лет - по одному, И -
выводков не было.
Количество утят в семье варьирует от 1 до И, в среднем 4,9 (выводок
из 18 птенцов, встреченный на Нявка-озере в августе 1937 г., явно был
сборным). Подсчет состава семей по месяцам: в июле - 4,8, в августе -
5,3, сентябре - 5,1 утят. Видимо, детская смертность у синьги незначи-
тельна. Интересно, что в 2-3 случаях неподалеку от выводка пухович-
ков наблюдали и самца. Птенцы остаются нелетными весь август, некото-
рые не поднимаются на крыло и в начале сентября, но большинство в пер-
вой декаде месяца уже летает.
Наряду с выводками в заповеднике все лето держится много холостых
особей, вероятно годовалых, еще неполовозрелых. Они образуют стайки
до десятка птиц, а иногда и больше. Подсчет встреч показал, что в июле и
августе сеголетки составляют только 42% всех особей, а в сентябре боль-
шинство уже не отличимы от взрослых. В это время синьга сбивается в
стаи; так, 11. IX 1971 г. в низовьях Нявки видели стаю в два десятка, а в
конце сентября и октября неоднократно встречали по 2-4 десятка. Такие
стаи держатся в ’’голомени” больших озер, вне заповедника.
Отлетает синьга позже других уток, недели за три до замерзания
больших озер,: в среднем за 35 лет - 22 октября, , при температуре воздуха
- 0,9° (крайние сроки 4.Х в 1970,1971 гг. и 1978 гг. - 14.XI 1938 г.; о ± 10,9)
Дата отлета слабо коррелирует как с температурой октября (г = + 0,15±
±0,17), так и с временем замерзания озер (г = + 0,26 ± 0,16; средняя дата
ледостава - 9.XI).
Питание синьги в заповеднике не изучено: известно, что она кормится
преимещественно бентосом.
Синьга превосходно ныряет, оставаясь под водой около 30 с, а в одном
случае - даже до 55 с; хорошо ныряют и пуховые птенцы. Летает обычно
низко над водой.
В добыче хищников остатков синьги мы не находили. Часто она гиб-
нет в рыболовных сетках: из 7 таких случаев, описанных в картотеке,
4 синьги погибли, а 3 были своевременно освобождены и выпущены.
Размеры 3 взрослых птиц, добытых весной (в мм): длина 477-516,
37
Рис. 9. Самка морянки на взлете, 1. VII 1978 г. Фото А.С. Гилязова
крыло 220-238; масса тела самцов 1020 и 1140 г, самок - 970 и 1040, мас-
са тела одной молодой самки в сентябре - 750 г.
Саамское название синьги - звукоподражательное, ее обычный позыв
действительно напоминает слог ”нёрг”. При брачных играх самец издает
мелодичный негромкий свист. При взлете птицы иногда слышен звук,
напоминающий трель среднего кроншнепа.
Морянка - Clangula hyemalis (L.)
Местные названия: саамское - англоньт,^русское - аллейка,
аулейка.
Морянка гнездится по пресным озерам тундровой зоны Евразии и
Северной Америки, в основном к северу от Полярного круга. На Коль-
ском полуострове гнездится только в тундре, вдоль Мурманского по-
бережья (Плеске, 1987, с.370; Фильчагов, Черенков, 1984, с. 215).
В Лапландском заповеднике морянка нередка на весеннем (33% всех встреч)
и осеннем (60%) пролетах; холостые особи изредка попадаются и летом
(7% особей зарегистрированы в июле и августе). За 48 лет наблюдений
отмечено 535 особей и добыто 7 (рис. 9).
У самки, добытой 6.VI 1935 г., диаметр самого крупного яйца в яични-
ке достигал 30-40 мм: очевидно, до места гнездования ей осталось лететь
недалеко.
Морянка прилетает последней среди уток, в среднем за 18 лет - 4 июня
(23.V 1974 г. - 17.VI 1960 г.; о ±6,9 сут; температура дня прилета +7,9°).
Птицы появляются обычно парами, реже стайками или поодиночке. В
стайках число самцов не всегда соответствует числу самок; так,18. V 1983 г.
38
на Чунозере видели стаю из 12 самцов и 9 самок. В районе заповедника
морянок встречали только на больших озерах - Имандре, Пиренге, Охт-
озере и Чунозере, хотя гнездятся они (например, на Таймыре) по мелко-
водным тундровым озеркам, площадью порядка нескольких гектаров.
Осенью морянки встречаются поодиночке или стайками до 16 птиц; толь-
ко один раз, осенью 1941 г., на Имандре видели большие, до сотни голов,
стаи.
В желудках двух морянок, добытых в октябре, найдены остатки боко-
плавов^ третьем желудке - домики ручейников и несколько мальков; в
двух весенних желудках - только песок.
Осенний пролет начинается в сентябре ( в среднем за 5 лет - 17.1Х)и
продолжается около месяца; морянка отлетает окончательно вместе с
синьгой, как только начнут замерзать озерные мелководья, - обе эти
утки питаются бентосом и не могут кормиться на глубокой воде. Средняя
дата последней встречи - 20 октября, при средней суточной температуре
около - Г (предельные сроки за 23 года - 4.Х 1960 и 1975 гг. - 4.XI 1961 г.;
о ±8,3 дня).
Это довольно крикливая птица, голос ее похож на ”аулёу... аулёу...”
- отсюда и местное название.
Размеры 3 самцов (в мм): длина 490-570, крыло 225-237, хвост 220—
229 мм, масса тела 927-1007 г; размеры 4 самок соответственно - 395-
415, 208-222 и 67-84 мм, масса тела 600-753 г.
Гоголь - Bucephala clangula (L.)*
Местные названия: саамское - чуадьк, русское (поморское)-
звонок (самец), звонуха, звончйха (самка).
Повадка гнездиться в дуплах деревьев ограничивает распространение
гоголя лесной зоной Европы, Азии и Северной Америки. Распространение
его в нашей области сто лет назад Ф.Д. Плеске, характеризовал так: ”В
лесной части Лапландии гоголь, без сомнения, самая обыкновенная ут-
ка” (Плеске, 1887, с.366). Это определение верно и теперь для Лапланд-
ского заповедника. За 39 лет регистрации встреч животных в заповедни-
ке учтено более 27 тыс. гоголей, в среднем по 677 особей в год, или по 46
на одного наблюдателя в сезон. Этот индекс мало колеблется по годам и
десятилетиям: в 40-х - 53, 50-х - 38, 60-х - 51, 70-х - 50, 80-х - 38. При
ежегодном учете водоплавающих птиц за 22 года (1966-1987) гоголь не-
изменно занимает первое место по численности, на его долю приходится
55% всех уток, встреченных на озерах, и 40% - на реках. Средняя плот-
ность популяции - 12,0 гоголей на 10 км береговой линии озер и 8,8 - на
таком же протяжении рек. Амплитуда колебаний численности по годам
едва превышает 1:2. Максимумы наблюдались в 1968, 1974, 1981 и 1985 гг.,
минимумы - в 1966, 1971, 1980 и 1984 гг.: правильной периодичности
здесь не видно. Оба источника информации, регистрация встреч и учет на
стационаре, констатируют заметное снижение численности гоголя в
последние годы. Суммируя данные по озерам и по рекам, находим, что
средняя плотность на 10 км составляла: в 60-х годах - 10,2, 70-х - 11,8,
80-х - 8,8 гоглей.
39
Прилетает гоголь раньше всех других уток, только на 2-3 недели
позднее лебедя, нормально - 29 апреля (крайние сроки - 15.IV 1985 г. -
16.V 1942 г., за 52 года; о±8,0сут).. Он появляется в момент переходов
средних суточных температур через +0,2° и за месяц до вскрытия озер.
Корреляция между сроками прилета гоголя и другими фенологическими
явлениями едва выражена, но он явно зависит от температуры апреля:
г = - 0,47 ± 0,11 (п = 49 лет).
В момент прилета гоглей в природе еще мало открытой воды, им при-
ходится кормиться на порожистых реках, на промоинах в устьях рек, на
протоках между озерами, (рис. 10). Первые гоголи появляются обычно
парами, стайки (до десятка пар) появляются только через неделю. Брач-
ные игры можно наблюдать с середины мая до начала июня. Токуя, селе-
зень распушает оперение головы и дергает ею взад и вперед, потом вытя-
гивает шею вверх и быстро откидывает голову назад, касаясь затылком
крестца: в этот момент он издает негромкий звук, напоминающий шипе-
ние, - ”фффф. . .”. Иногда самец приподнимает переднюю часть тела над
водой и хлопает крыльями. Самка, принимая приглашающую позу, втя-
гивает и опускает шею. Позднее, когда часть уток уже сидит на гнездах,
случается, что к паре гоголей подсаживаются до четырех свободных селез-
ней и все наперебой ухаживают за одной самкой; ее ’’законный” кавалер
этому обычно не препятствует. Иногда один самец погонится за другим,
но настоящих драк между ними мы не замечали.
Старая самка обычно занимает дупло, где гнездилась раньше. Гого-
люшке необходимо дупло с достаточно широким летком, не менее
8-10 см в диаметре. Из всех дятлов только желна делает дупла такого
размера, но бывают летки и другого происхождения, например в стволе с
40
сердцевинной гнилью - от выпавшего сука; если такое дерево сломано
ветром, гоголь иногда занимает полость, открывшуюся на месте слома
(торцовое дупло). Внутренний диаметр жилого дупла около 20 см, глу-
бина от нижнего края летка до дна 25-45 см. В заповеднике найдено
всего 12 занятых гоголями естественных дупел в стволах сухих или жи-
вых сосен и 4 торцовых дупла (Брагин, 1974). Саамы и карелы с давних
пор вели своеобразный яичный промысел, выставляя по берегам озер и
рек искусственные дуплянки (саамское название их ’’вуувт”): утки,
преимущественно гоголюшки, в них несутся, а предприимчивый охотник
собирает яйца (Энгельгардт, 1897, с.258). Лапландский заповедник ис-
пользует ’’вуувты” для экологических исследований. С этой целью
А.Б. Брагин расставил в заповеднике около 200 гнездовых ящиков из
досок. Гоголюшки охотно гнездятся как в долбленых ’’вуувтах”, так и в
дощатых искусственных гнездах (занимают их на 35%).
Кладка яиц начинается в среднем 17 мая (4.V 1937 г. - 30.V 1961 г. за
35 лет, о±6,1 дней). Утка несется ежедневно или через день (средний
интервал кладки 32-37 ч). Для этого она остается в гнезде 10-30 мин в
начальной стадии кладки, но постепенно время ее пребывания в гнезде
увеличивается, достигая к концу кладки 11ч. Если гнездо находится у
открытой воды, утка держится около него и в свободное время, в какой-
то мере охраняя кладку; если же вода перед гнездом покрыта льдом или
дупло находится в глубине леса, гнездо подолгу остается без присмотра,
что создает благоприятные условия для коллективных кладок. Нередко
в одно гнездо несутся две или даже три утки, иногда разных видов
(гоголь и луток или гоголь и большой крохаль). Так, 28.V 1951 г. в вуувт
на берегу озера Ельявр за 12 ч было снесено три гоголиных яйца, 29.V -
ни одного, 30.V - снова 3 за 18 ч: очевидно, в одно гнездо неслись три
гоголюшки. 1.VI кладка пополнилась еще 2 яйцами, а вечером 2.VI одна
из уток начала насиживать, но, просидев 6 сут, бросила гнездо, вероятно,
в связи с холодной и ненастной погодой (Семенов-Тян-Шанский, 1960,
с. 281). В других случаях утка насиживает смешанную кладку до благополуч-
ного конца и становится для чужих детей мачехой - первый шаг к воз-
никновению гнездового паразитизма. По оценке Брагина, 39% всех гого-
линых кладок принадлежат 2-3 уткам; число яиц в смешанных кладках
доходит до 20, а общая масса кладки - до 1200 г, вдвое больше массы
тела самой утки. Прогреть такую массу яиц она не может, поэтому боль-
шие кладки отличаются высокой эмбриональной смертностью, а чаще
утка бросает такую кладку либо до насиживания, либо в процессе ин-
кубации.
Беспорядочная откладка яиц разными самками в одно гнездо или
одной самкой в разные гнезда приводит к тому, что размер кладки варь-
ирует от 3 до 20 яиц. 62% кладок состоят из 6-12 яиц, из них 9% были бро-
шены; из кладок по 3-5 яиц (20% общего числа) брошены 56%; из больших
по 13-20 яиц (18%), 31% брошены до насиживания (Брагин, 1974, табл. 17).
Успех размножения гоголя варьирует по годам, как показал Брагин,
главным образом в результате ’’дезертирства” наседок, либо вследствие
конкуренции между ними, либо под влиянием ненастной или слишком
холодной погоды. Количество брошенных кладок составляет 19% в бла-
41
гоприятные весны и 30% в сезоны с неблагоприятной погодой. Для кла-
док, снесенных одной уткой, Брагин определил, что двухлетние гнездя-
щиеся впервые самки несут в среднем по 7,8 яйца (22 кладки), а самки
3 лет и старше - 9,6 (18 кладок); при этом молодые самки гнездятся и
выводят утят позже старых.
Насиживание начинается после окончания кладки, и в первые сутки
наседка не оставляет гнезда (непрерывно сидит в среднем 25 ч). В даль-
нейшем гоголюшка затрачивает на прогулки и кормежку от 10 до 22%
всего времени инкубации, сходя с гнезда от 2 до 6 раз в сутки, на срок от
40 м до 3 ч. Перед уходом утка тщательно укрывает яйца пухом, что за-
медляет их остывание. Она греет яйца главным образом лапами, темпера-
тура которых во время насиживания достигает, по измерениям Брагина,
4Г. Температура внутри яиц, измеренная термистором, равнялась в сред-
нем от 32,7 до 34,2° (в разных гнездах), варьируя в пределах 28,1-37,4°,
пока наседка на гнезде, а в ее отсутствие иногда падала до 12,3°, что не
повлияло на развитие эмбрионов.
Продолжительность инкубации до наклева яиц составляет 28,0-28,5
суток, а считая до ухода выводка с гнезда - 30,9-31,2 суток. Утята оста-
ются в гнезде не менее суток без пищи, используя энергетический ресурс
желточного мешка. Выскакивают они из дупла сами, на зов матки,
ожидающей птенцов под гнездом. Утята поочередно подпрыгивают вверх
(до 60 см), цепляясь когтями за край летка, потом спрыгивают на землю.
Собрав выводок, матка ведет его ближайшим путем к воде. Гоголята
родятся хорошо развитыми, с первого же дня они самостоятельно кор-
мятся, плавая и ныряя, оставаясь до 10 с под водой.
Эмбриональная смертность в благополучных кладках незначительна,
в среднем утята выходят из 95% яиц, (п = 638). ’’Неурожай” молодняка в
отдельные годы объясняется главным образом гибелью целых кладок,
либо от ’’дезертирства” наседок, либо от хищников, так как гнезда,
особенно в искусственных дуплянках, им легко находить. По расчету
Брагина, из 1771 яйца за 8 лет 22% яиц были брошены наседками, 21% -
стал добычей хищников, 2% - взяты браконьерами, в 3% - эмбрионы по-
гибли в процессе инкубации и только из 52% яиц вывелись утята. Разо-
ряют гнезда главным образом норки и куницы, при этом нередко погибает
и наседка. Вороны достают только отдельные яйца, если могут дотянуть-
ся до них через леток. Несколько раз дуплянку ломал или сбрасывал
медведь, а иногда росомаха.
Выводки гоголей появляются на водоемах раньше, чем других водо-
плавающих птиц, в среднем за 46 лет - 24 июня (15.VI 1937,1948 гг. - 2.VII
1966 г.; о±4,4); вместе с тем сроки размножения сильно варьируют,
вследствие чего в середине июля наряду с полуоперенными гоголятами
встречаются и маленькие пуховички. Гоголята живут дружно и уже в
месячном возрасте нередко кормятся самостоятельно. В природе при хо-
рошем питании они начинают летать через 51-52 дня после выхода из
гнезда; при воспитании в вольере - на 64-е сутки. Первые летные вывод-
ки мы встречали в среднем за 35 лет - 14 августа, самое раннее - 30.VII
1939 г.; поздние поднимаются на крыло в сентябре, а один нелетный вы-
водок наблюдался 12.IX 1938 г.
42
После того как утки сядут на яйца, селезни собираются стаями и отле-
тают к местам линьки. Последних самцов встречали обычно в последней
декаде июня, в некоторые годы - 10-15.VI (1948, 1951, 1964л 1966), в дру-
гие они задерживаются до начала или середины июля. Самые поздние
встречи отмечены 27.VII 1974 г. (6 самцов на оз. Витте) и 21.VII 1978 г.
(17 самцов). Задерживаются с отлетом, по-видимому, годовалые, еще
неполовозрелые самцы. Ранний отлет селезней, возможно, объясняется
ограниченностью кормовых фондов рек и озер: улетая, самцы не конку-
рируют за пищу со своим потомством.
Годовалые самки прилетают позже взрослых гоголей, пар не образуют,
но явно интересуются гнездовьями. С середины июня до 10 июля моло-
дые утки летают над лесом, разыскивая дупла, заглядывают в ’’вуувты”,
залетают в поселки, садятся на дымовые трубы и даже залезают в них.
8.VI 1978 г. в дымоходе одного из домов на Чунозерской усадьбе нашли
обгорелый труп годовалой гоголюшки, которая забралась в трубу, но не
смогла из нее выбраться. Летние стаи гоголей, по всей вероятности,
состоят из холостых годовалых уток.
В сентябре выводки объединяются в стаи и кочуют в районе гнездо-
вания. Несколько гоголей, окольцованных в заповеднике, были добыты
между 7-20 сентября на расстоянии 10-120 км от места мечения, из них
четыре - к востоку (оз. Имандра и Малый Вудьявр), по одной - к югу
(близ Кандалакши), к юго-западу (р. Толва) и к северу, за станцией Ло-
парской. В сентябре же начинается и отлет. Одна из окольцованных в
заповеднике гоголюшек была убита 4.IX 1982 г. в Финляндии, близ Оулу
(450 км к юго-западу); другая - 4.Х 1975 г. в Швеции близ Упсалы, за
1100 км; третья добыта 10.Х 1976 г. в Финляндии, за 485 км; четвертая - в
октябре 1966 г. в Финляндии же, за 590 км; пятая, сеголеток, - 15.X
1977 г. в Финляндии, за 970 км; шестая - 18.Х 1982 г. на Вуоксе Ленин-
градской области, в 800 км; седьмая - 26.Х 1982 г. в Норвегии, район
Тронхейма, в 1160 км от заповедника - все на пролете, в направлении от
Ю до ЗЮЗ от места мечения.
В октябре в районе заповедника не раз встречали большие стаи про-
летных гоголей: 29.IX 1973 г. через Монче-губу на запад пролетело до
200 голов, а 11.Х 1972 г. на оз. Купись видели три стаи, в сумме около
120 птиц; стаи по нескольку десятков уток наблюдали также в 1940, 1947,
1973,1980 и 1982 гг.
Средняя за 47 лет дата последних встреч - 13 октября (21.IX в 1945 и
1959 гг. - ЗО.Х 1938 г.), о ±10,0 дней. Средняя температура воздуха в день
отлета 0,9°, время отлета коррелирует со средней температурой октября
(г = +0,54±0,11), а в меньшей степени и сентября (г = +0,32±0,14). Одиноч-
ные, отсталые гоголя попадаются иногда даже в ноябре: в 1972 г. одного
или двух видели 10 и 27.XI на р. Вайкись, потом 2 и 7.XII - на Нижней
Чуне; в 1982/83 г. один гоголь продержался на р. Витта всю зиму: 13.1 ви-
дели двух, 4.II и 27.Ш поднимали только одного.
Результаты кольцевания показали, что наши гоголи зимуют в Запад-
ной и Центральной Европе (рис. 11). Зимние возвраты поступили из Дании
(5 колец, с 15.Х по 18.11, расстояние 1700-1800 км к ЮЗ), Южной Швеции
(2 кольца в XI—XII, расстояние 1350-1600 км к ЮЗ), по одному кольцу
43
Рис. 11. Места встреч окольцованных в заповеднике самок гоголей
поступило из Норвегии (район Тронхейма, 1160 км на ЗЮЗ; конец октяб-
ря), Ирландии (ЮВ побережье, 1700 км на ЗЮЗ), Швейцарии (район Бо-
денского озера, 2600 км на ЮЗ, февраль), Венгрии (оз. Балатон, 2450 км на
ЮЮЗ, декабрь) и Украины (Киевское водохранилище, 1900 км к югу,
октябрь).
В общей сложности в заповеднике окольцовано 180 взрослых самок,
отловленных в ”вуувтах”,и 92 пуховичка; получен 141 возврат от 94 осо-
бей - в основном повторные отловы уток в той же или другой дуплянке.
Поскольку гоголюшки впервые гнездятся в возрасте 2 лет, большинство
взрослых уток были окольцованы именно в этом возрасте, после чего их
отлавливали повторно, до пяти раз. По нашим материалам, максималь-
ный возраст самки гоголя 17+ лет: утка, окольцованная в дуплянке на
оз. Тулпъявр 1.VII 1967 г., была добыта 4.Х 1982 г. близ Оулу. Еще две
прослежены до десятилетнего возраста, одна, окольцованная на р. Чуна в
июне 1973 г., ловилась нами неоднократно, последний раз в июне 1981 г. в
’’вуувте” на Кокоринском озерке, в 28 км к ЮВ от первого места; другая,
помеченная на р. Чуна в июне 1976 г., трижды ловилась в других ”вуув-
44
тах” по той же речке, а последний раз - 6.VI 1984 г., в 1 км от места
мечения. Наблюдения подтверждают сильную привязанность гоголей к
своему гнезду - улетая зимовать за 1400-1900 км, они обычно возвраща-
ются в ту же самую дуплянку. Несомненно также, что естественная
смертность гоголей старше 2 лет незначительна.
Гоголи кормятся в основном бентосом - беспозвоночными, живущими
на дне рек и озер. В желудках двух особей обнаружены остатки личинок
поденок (или других насекомых), в других - зеленые части растений, а в
одном - 5 отолитов рыб. При кольцевании гоголей в июне А. Брагин
собрал 44 пробы экскрементов; исследовавший их В.Н. Михеев (кафедра
беспозвоночных МГУ) определил в них 117 кормовых объектов, среди
которых преобладали личинки ручейников (34%), далее - растительные
остатки (27%), поденки (16%), жуки (8%) и бокоплавы (8%); единично
встречались веснянки, раковины моллюсков, личинки комаров и мошек.
Весь свой корм гоголь достает со дна водоема, ныряя. Гоголь остается
под водой по 7-50, а в среднем около 30 с; передышки короче, по 6-14, в
среднем - 9 с. Даже при таком жестком режиме времени жировка гоголя,
судя по хронометражу насиживающих самок, занимает 2-5 ч в сутки.
Материалы кольцевания показали, что естественная смертность у го-
голя невысока, хотя врагов у них много. Американская норка и куница,
залезая в дупла за яйцами, нередко ловят при этом утку. В заповеднике
зарегистрированы остатки 12 гоголей, пойманных хищными зверями,
половина их - птенцы, остальные - взрослые самки (в том числе три
окольцованные). Одну гоголюшку добыла в августе 1938 г. выдра, дру-
гую весной 1979 г. поймала в дуплянке куница, остальные 10 гоголей, по
всей вероятности, стали жертвами норок: остатки трех птиц найдены
либо в норе этого хищника, либо рядом с ней, в остальных случаях ее
участие наиболее вероятно. А.А. Насимович и О.И. Семенов-Тян-Шан-
ский, изучая питание хищных зверей по экскрементам и остаткам добы-
чи, часто находили уток, в том числе и гоголей, в йище лисы, куницы,
норки, а иногда и выдры. Находили остатки уток также под гнездами
или в погадках орлана, ястреба-тетеревятника и кречета. 11.IX 1938 г.
Семенов-Тян-Шанский, a 18.VIII 1974 г. М. Владимирская наблюдали, как
ястреб охотился на гоголей на р. Чуна - оба раза безуспешно; 21.V 1983 г.
на той же реке ястреб поймал гоголя и унес к гнезду.
Утят ловят также щуки. В июле 1970 г. из вольера, где А. Брагин
содержал вышедших из инкубатора гоголят, пропали два утенка. Заме-
тив в воде около вольера щуку, Брагин поймал ее и нашел в желудке
остатки гоголенка, птенца улита и перья еще какой-то птицы. Интересно,
что эта щука массой 1,7 кг, сумела продернуть утенка сквозь ячею сетки.
Гибнут гоголи и от несчастных случаев: одна самка задохнулась в ды-
мовой трубе, три утонули, запутавшись в рыболовной сетке (май 1940 г.,
август 1941 г., июнь 1978 г.), четвертую успели освободить из сетки и вы-
пустить (июль 1971 г.). Летом 1977 г. на Чунозере поймали гоголюшку,
повредившую шею при ударе о провод; рану ее вылечили, и через 9 дней
утку выпустили на волю.
В кишечниках гоголя часто встречаются гельминты, а в их гнездах
45
довольно обычны блохи, преимущественно Ceratophyllus gallinae (опреде-
лены Н.Ф. Дарской). Зимуют эти блохи, по-видимому, в земле, так как в
образцах подстилки, взятых из гнезд зимой, блох не оказалось.
В полете крылья селезня издают своеобразный свист или звон, от кото-
рого и происходит его поморское название.
Средние размеры 5 селезней, добытых в мае: длина 455 (445-476), кры-
ло 226 (215-234), хвост 94 (90-102) мм, масса тела 896 (817-955) г. Разме-
ры 4 самок: длина 402 (400-405), крыло 197 (188-204), хвост 81 (76-86) мм,
масса тела 594 г. (532-681). Средняя масса гоголюшек, отловленных на
гнезде в начале инкубации, равна 621 ±7,0 г (550—690)(п = 36), к концу
инкубации она уменьшается на 8-9% (Брагин, 1975).
Яйца гоголя относительно крупные, средняя масса их перед насижи-
ванием 59,6 ±0,2 г (49,8-72,2; п = 323); средние размеры 58,2 x41,6 мм,
окраска скорлупы однородная, светло-голубовато-зеленая. Масса тела
новорожденных утят в гнезде 35,5 г (п = 33).
Луток - Mergus albellus L.
Местное название: саамское - нюннель.
Луток, хотя и широко распространен в лесной зоне Старого Света, на
большей части ареала довольно редок. На Кольском полуострове встре-
чается сравнительно часто, по регистрации встреч за 35 лет - в 60 раз
реже гоголя, по материалам учетного стационара, за 22 года, - в 29 раз.
Регулярно наблюдается только весной, прилет зарегистрирован 34 раза за
48 лет наблюдений, выводки отмечены за 22 сезона, время отлета про-
слежено 6 раз (рис. 12).
Луток прилетает парами или небольшими стайками сравнительно
поздно, в среднем 21 мая (5.V 1984 г. - 8.VI 1935 г.; о ±9,1 дней), по гра-
фику суточных температур, при 4,9°.
Гнездится, как и гоголь, на деревьях, в естественном или искусствен-
ном дупле и начинает нестись вскоре после прилета. Наблюдения по гнез-
дованию немногочисленны, и мы приводим их полностью:
17.VI 1949 г. — гнездо в естественном дупле на о-ве Пиренгского озера;
23.V 1950 г. — там же — гнездо в искусственной дуплянке;
26.VI1951 г. — в том же месте луток вывел утят;
5.VI 1969 г. — одно яйцо лутка найдено в кладке гоголя в дуплянке на берегу
оз. Румельявр;
25.V 1972 г. - в дуплянке на лесном озерке в долине р. Чуна, луток снес первое
яйцо; 6.VI в гнезде 8 яиц, под ним на земле два разбитых, вытащенных, вероятно,
вороной. Тут же на летке укрепили козырек для защиты от ворбн. Это отпугнуло
лутка, и он бросил кладку;
19.VI 1978 г. — в долине р. Нявка луток насиживал в дуплянке смешанную
кладку из 6 своих яиц и 5 гоголиных; 6.VII в гнезде 5 птенцов гоголя и 3 - лутка, 1 мерт-
вый птенец и 1 испорченное яйцо.
Согласно этим наблюдениям, луток начинает нестись в последней
декаде мая, насиживать - около 1 июня, выводит птенцов около 1 июля.
Средняя дата 21 встречи нелетных выводков лутка - 18 июля, самые ран-
ние отмечены 26.VI 1936 и 1951 гг. и 29.VI 1975 г. В семье от 1 до 7 птенцов,
в среднем 4,4 ± 0,4. Выводки попадались как на речных плесах (низовья
Нявки и Чуны, проток Меркенч), так и на озерах (Румельявр, Улынчьявр,
46
Рис.12. Пара лутков в дни прилета, 18. V 1982 г.Фото А.С. Гилязова
Куркозеро, Чунозеро и болотные озерки - Щучье, Гагарье, Полмех). В
трех выводках были одни птенцы, без матки, а в одном случае (7.VII
1974 г.) поблизости от самки с 7 утятами, по мнению наблюдателя, плавал
и селезень. Летом 1948 г. в низовьях р. Нявка жил выводок, потерявший
матку: 6.VII в нем было 6 птенцов, 1.VIII - 5, 21 и 26.VIII — только 3. 1.IX
1948 г. впервые отмечено, что два молодых лутка взлетели.
В сентябре лутки иногда собираются стаями: 4.IX 1949 г. над Чунозером
пролетели 35 лутков, a 6.IX на Румельявре видели 12 особей. Средняя за
6 лет дата последней встречи - 6 октября (20.IX 1949 г. - 17.Х 1970 г.), при
средней температуре воздуха 2,4°.
По литературным источникам, луток питается в основном раститель-
ным кормом и беспозвоночными, ныряя за ними до дна. По наблюде-
ниям Брагина, он остается под водой 5-15 с.
Размеры одного самца (V. 1938 г.): длина 415, крыло 199, хвост 91 мм.
Яйца, как и у других крохалей, удлененно-эллиптические, кремово-
белого цвета, средняя масса тела 41,1 г (39,3 - 42,5 = 11).
Средний, или длинноносый, крохаль - Mergus senator L.
Местные названия: саамское - шбпперьг, русское - малый кро-
халь, а чаще - просто крохаль (местные русские обычно виды крохалей
не различают).
В Мурманской области средний крохаль - обычная утка, уступает по
численности только гоголю и большому крохалю. В записях сотрудников
заповедника вид крохаля нередко назван неверно или совсем не указан,
47
поэтому по материалам картотеки трудно судить о количественном
соотношении и колебаниях численности обоих видов. Несомненно
только, что в довоенные годы средний крохаль встречался гораздо чаще
большого, а позднее соотношение переменилось в пользу последнего
вида. По надежным материалам ежегодного учета водоплавающих,
соотношение среднего и большого крохаля на последние годы составля-
ло: в 1966-1969 гг. - 51:49, в 1970-1974 и 1975-1979 - 37:63, в 1980-1984 -
43:57, в 1985-1987 - 50:50. Учеты показывают, что средний крохаль живет
преимущественно на озерах (в том числе глубоких, тектонического про-
исхожденияе, а большой крохаль - на реках. На 10 км береговой линии
озер, в среднем за 22 года, приходится 4,5, а на 10 км рек - 0,9 средних
крохалей. Колебания численности за эти годы нерегулярны: максимумы
отмечены в 1969, 1980 и 1985 гг., минимумы - в 1966, 1970, 1973 и
1981 гг.; амплитуда колебаний от 1:3 до 1:10.
Средний крохаль прилетает дней на 10 позже большого, в среднем за
44 года, 19 мая, при средней температуре воздуха около 4,5° (крайние
сроки 4.V 1983 г. - 4.VI 1939 г.; о ±7,0 дней).; Даты прилета слабо коррели-
руют со средней температурой мая, г-0,25±0,15.
Крохали прилетают парами и вскоре принимаются за брачные игры. В
1973 г. средние крохали прилетели 18 мая, а 20 А. Брагин уже наблюдал
спаривание.
Большинство гнезд с неполной кладкой найдены нами во второй
декаде июня (4-22. VI), а с насиженной кладкой - в третьей декаде; самая
ранняя находка - 18.VI1931 г, самая поздняя, с наклюнутыми яйцами, -
3.VIII 1941 г. По календарному графику находок гнезд мы нашли, что
кладка яиц нормально приходится на 10-20 июня, а инкубация их - на
20.VI - 20.VII, позже, чем у других уток.
Величина полных кладок варьирует от 4 до 14 яиц, чаще 7-10, а в сред-
нем 3,3±0,3 (л = 39). Средний крохаль устраивает гнездо на берегу озера,
на расстоянии от 0,7 до 15 м от уреза воды, обычно на береговом валу,
под густыми нижними ветками береговых елок, иногда под прикрытием
куста можжевельника. Реже утка гнездится в щели между камнями,
подобно большому крохалю, но в отличие от него никогда не несется в
дуплянке. Охотнее всего утка гнездится на острове, выбирая берег, об-
ращенный к материку и потому защищенный от прибоя (по Брагину, 1975,
82% гнезд находится на островах), иногда на материковом берегу (14%
гнезд), а одно гнездо было устроено на берегу маленького, пересыхающе-
го летом лесного озерка. Гнездовые ямки под елками, с остатками скор-
лупы, сохраняются годами, и крохали занимают их повторно. Постоян-
ство крохалей при выборе места для гнезда облегчает их нахождение, а
потому хищники разоряют их чаще, чем гнезда других уток. Из 22 гнезд, учтен-
ных Брагиным до 1975 г., и 36, найденных в 1976-1987 гг., утята вывелись
только в 25 (43%), уничтожены хищниками 17 кладок (30%), брошены
наседкой 3 (5%), судьба остальных 13 неизвестна. Разоряют гнезда
ворон, серая ворона, а из зверей - куница, норка и горностай.
Первое яйцо крохаль откладывает в пустую ямку, пух появляется
позднее, в процессе кладки; у среднего крохаля он темно-серый, у боль-
48
шого - почти белый - по этому признаку можно определить видовую
принадлежность гнезда. Во время насиживания яйца окружены валиком
из пуха; уходя на прогулку, утка сдвигает его к центру, закрывая им
яйца от холода и посторонних глаз.
Режим инкубации среднего крохаля был исследован с помощью само-
писцев Семеновым-Тян-Шанским - в 2 гнездах на протяжении 21 суток
(Семенов-Тян-Шанский, 1960), Брагиным - в 2 гнездах в течение 10 суток
(Брагин, 1974). Одна кладка в 1951 г. была уничтожена горностаем, дру-
гая - в 1966 г. - норкой, в двух других (1951 и 1968) благополучно выве-
лись птенцы. В 1951 г. удалось установить продолжительность инкуба-
ции: утка села на яйца ночью 21-22.VI, 21.VII в 12 ч из яиц слышался
писк и стук, в тот же час 22.VII появились проклевы, 23.VII в гнезде
находились 6 утят и 1 испорченное яйцо, 24.VII в 7 ч актограф отметил
уход выводка. Таким образом, инкубация продолжалась около 31 сут и
еще не менее 1,3 сут утята оставались в гнезде. Все 4 утки за время
инкубации кормились только днем, между 5 и 20 часами.
Крохали питаются исключительно рыбой, а ловля ее требует хорошего
освещения и много времени. Нормально наседка оставляет гнездо 2 ра-
за в день, утром и вечером, реже - 1 или 3 раза. В гнездах 1951 г. продол-
жительность прогулки варьировала от 13 мин до 4 ч 10 мин (в среднем
1 ч 45 мин). Общая продолжительность отлучек составляла 4,1 ч, или 17%
суток (по наблюдениям автора в 1951 г.) (1,35 ч. т.е. 6% суток, по данным
Брагина, 1968 г.). В одном гнезде регистрировалась также температура
подкладного яйца, снабженного термистором: в часы насиживания она
колебалась в пределах 30,9°-38,2°, составляя в среднем 35,8°, снижаясь
во время прогулок наседки до 29,5е (при температуре окружающего воз-
духа +5, Г... +13,5°).
Два гнезда, найденные нами 20.VI 1951 г. на о-ве Баран-суол, находи-
лись на расстоянии 1 м одно от другого и содержали по 8 яиц. Наседки
оставляли гнездо для прогулки поочередно. Регистрация велась только в
одном гнезде, а после его разорения (9.VII) прибор был подключен к дру-
гому. Видимо, недостаток подходящих мест заставляет крохалей тесни-
ться, образуя подобные гнездовые колонии: на том же Баран-суоле, остров-
ке площадью около 1 га, в 1982 и 1981 гг. было учтено по 6, а в 1984 г. -
даже 8 гнезд средних крохалей.
Первые выводки среднего крохаля появляются, в среднем за 25 лет, 19
июля (6.VII 1948 г. - 4.VII 1967, 1976 гг.). Крохали ловят рыбу довольно
своеобразным способом. Выводок во главе с маткой плывет вдоль бе-
рега озера, при этом часть птиц держит клюв и глаза погруженными в
воду. Едва один крохаль заметит добычу и бросится за ней - все осталь-
ные опускают головы в воду и с шумом, в брызгах, гонят стайку голья-
нов или мальков к берегу. Поймав рыбу, крохаль поднимает голову, пе-
ребирает добычу клювом, поворачивая ее головой к себе, и тут же про-
глатывает. Один и тот же выводок в разные дни можно встретить в раз-
ных местах озера, из чего следует, что у крохалей нет индивидуальных
участков и озеро они осваивают сообща. Для отдыха семья выбирается на
выступающий из воды плоский камень, птенцы ложатся на него грудью и
4. Зак. 1023
49
засыпают. Камни, на которых отдыхают крохали, можно узнать по при-
сохшему к нем белому помету. Экскременты удобряют субстрат, в ре-
зультате чего на таких камнях пышно развивается водоросльТгеп!ероЬНа
Jolitha (пользуемся случаем поблагодарить сотрудника БИН АН СССР
Н.В. Сдобникову за определение этого растения); смоченное волнами и
освещенное солнцем ее слоевище становится ярко-красным, похожим на
пролитую кровь (этот мотив отражен в некоторых саамских сказках).
Размеры выводков сильно варьируют, в них может быть от одного
птенца до четырех десятков; большие образуются путем объединения не-
скольких семей вокруг одной самки. 4.VIII 1974 г. лесник А. Безруков
наблюдал на оз. Островском драку двух крохалюшек, при которых
находилось около 18 птенцов. Возможно, что и в других случаях сильная
утка создает сборную семью насильственным путем. Интересно, что такая
искусственная семья оказывается неожиданно прочной; одну, включав-
шую 35 крохолят, мы наблюдали на Чунозере не менее месяца, с 11 ав-
густа по 11 сентября. Большие сборные выводки (от 13 до 38 утят) состав-
ляют 6% общего числа семей, большие ординарные (9-12 птенцов) - 14%,
средние (3-8 молодых) - 75%, маленькие, по 1-2 птенца, - 5% общего
числа. Детская смертность велика только при рождении утят и в первые
дни их жизни: при средней величине кладки 8,3 яйца средний размер
выводка в июле - 6,0, в августе - 6,1, в сентябре - 5,8 (учтены только
нелетные выводки).
8. VIII 1966 г. на Чунозере мы поймали и взяли на воспитание отстав-
шего от выводка крохаленка, примерно трехдневного возраста (без
яйцевого зуба). Первое время его кормили только живыми гольяна-
ми пущенными в таз с водой. Крохаленок самостоятельно нырял за ними,
ловил, заглатывал головой вперед, а насытившись 2-3 рыбками, выпры-
гивал из таза на пол и устраивался на отдых. Рос он быстро, масса тела
10.VIII была 53 г, а 24.VIII 260 г, следовательно, суточный прирост состав-
лял 12%. До трехнедельного возраста он был одет пухом, а достигнув
250 г начал оперяться.
В природе ранние выводки поднимаются на крыло около 15 сентября,
другие задерживаются, так что нелетных крохалят встречали не только в
последней декаде сентября, но даже в начале октября (8.Х 1971 г., 2.Х
1972 г.). При опасности нелетный выводок дружно убегает по воде,
действуя главным образом лапами, поднимая тучу брызг; моторная лод-
ка при скорости 10-12 км/ч не может их догнать. К 1 октября молодые
крохали по массе тела сравниваются со взрослыми.
Взрослые селезни крохалей, как и других уток, отлетают от нас уже
летом: последних, в среднем за 8 лет, наблюдали 6 июля (20.VI - 20.VII).
Матки с молодняком отлетают поздно: осенью они ловят довольно круп-
ную рыбу и не зависят от образования льда на мелководьях. Время отле-
та обоих видов крохалей, равно как и сведения о смертности, рассматри-
вается в следующем очерке.
Единичные стайки крохалей, вероятно запоздалые выводки, иногда
остаются у нас и на зиму По словам Ф.К. Архипова, семь средних крохалей про-
жили на р. Монча всю зиму 1935/36 г. Крохалей неизвестного вида неод-
50
некратно наблюдали и в последующие годы: 30.1 1970 два крохаля про-
летели над оз. Ельявр, (студент В. Матвеев); одиночного крохаля лесни-
ки видели на р. Вайкись 27.XII 1972 г.; также одиночного видел научный
сотрудник Г. Катаев в устье речки Колнаса 21.Ш 1974 г.; двух крохалей
поднимали в истоке р.Нявка 14.П 1977 г. и 24.11 1982 г.
Сотрудниками заповедника окольцовано несколько средних кроха-
лей, большею частью освобожденных из рыболовных сеток, но от них
возвратов не поступало. Самка, пойманная 12.VII 1960 г. на гнезде на
островке оз. Ельявр, ровно через год вторично поймана на гнезде на
островке Баран-суол, Чунозеро, на расстоянии 2 км к ЮЗ. Другая самка,
окольцованная на Баран-суоле 30.VI 1960 г., добыта 15.Х того же года на
Северском Донце, на расстоянии 2120 км к ЮЮВ.
По питанию среднего крохаля есть только случайные наблюдения.
Весной 1962 г. лесник Е. Черных видел, как эта утка поймала и съела
крупную ряпушку; в 1979 г. А. Брагин наблюдал самку, поймавшую боль-
шого (около 100 г) окуня, а два других крохаля пытались отнять у нее
добычу; весной 1985 г. в истоке Ельявруая самец вынырнул с пойманной
им форелью или гольяном, самка бросилась к нему, чтобы отнять добычу,
а потом она сама поймала такую же рыбку.
Размеры взрослых птиц (в мм): самец - длина 555-630, крыло 232-257,
хвост 78-95, масса тела 960-1325, в среднем ИЗО г (п= 11); самка соот-
ветственно - 540, 225 и 85 мм, масса тела 2 экз. 827 и 980 г. Яйца эллипти-
ческой формы, кремово-беловатые, масса 56, 4-70 ,5, в среднем 65,8±0,7 г
(п = 21).
Большой крохаль - Mergus merganser L.
Местные названия: саамское - куальс, русское - крохаль, ре-
же большой крохаль.
На Кольском полуострове большой крохаль обычен, а в Лапландском
заповеднике занимает по численности второе место среди уток, вслед за
гоголем. По материалам учета водоплавающих, в августе, на 10 км бе-
реговой линии приходится на озерах 1,4, на реках - 6,8 больших кроха-
лей: в отличие от среднего крохаля он явно предпочитает реки озерам.
В 30-х годах большой крохаль встречался значительно реже среднего,
в послевоенные годы они примерно сравнялись по численности, а в 70-х
годах соотношение изменилось в пользу большого крохаля (5 больших
на 3 средних крохалей). По данным учета, заметны колебания численнос-
ти с периодом 5-7 лет: максимумы отмечены в 1966, 1973, 1980 и 1985—
1987 гг., минимумы - в 1968, 1974 и 1984 гг.
Большой крохаль прилетает вскоре после гоголя, в среднем за 44 го-
да-8 мая, при температуре воздуха - 2, Г (17. IV 1967 г. - 27. V 1942 и
1958 гг.; о ± 9,3 дня). Дата прилета коррелируется с температурой апреля
(г = - 0, 51 ± 0,12), но слабо связана с наступающими примерно в то же время
фенологическими явлениями, например с образованием заберегов на
озерах (г =+0,28± 0,15).
Кладка яиц начинается в конце мая: в яйцеводе самки, добытой 1. VI
1938 г., было почти готовое яйцо, а в яичнике три пустых фолликула -
51
следовательно, первое яйцо было снесено не позднее 30 мая. В одной
дуплянке на берегу р. Мавра 24. V 1964 г. обнаружены 3, а 24. V 1965 г. -
4 крохалиных яйца: в обоих случаях самка начала нестись не позднее
21 мая. Полные кладки больших крохалей находили с 7. VI до 25.VII
(средняя дата 30 июня). Из зарегистрированных 24 гнезд 11 находились
на островах озер, 10 - на берегах рек и ручьев, одно гнездо - на мате-
риковом берегу озера и два брошенных постройках. Большой крохаль,
как и средний, предпочитает строить гнездо под низкой еловой кроной на
береговом валу (46 % гнезд), но часто занимает искусственную дуплянку
(33 %) или скважину между камней (8 %); по одному гнезду - под еловой
валежиной, под полом сарая и на чердаке барака.
Характер размещения гнезд и ничтожная площадь пригодных для них
мест делают крохалиные яйца легкой добычей хищников, что снижает
успех размножения крохалей. Из состоявших на учете 24 гнезд большо-
го крохаля птенцы вывелись только в 12 (50 %), разорены хищниками -
3 (12 %), брошены наседкой - 5 (21 %), судьба четырех (17 %) не выяснена.
Для крохалей характерна изменчивость в сроках размножения, индиви-
дуальная и по годам.
Режим инкубаций О.И. Семенов-Тян-Шанский и А. Брагин изучали при по-
мощи самописцев в трех гнездах. В гнезде, найденном 8. VII 1951 г. под
покрытием елочки на крошечном островке посреди быстрого ручья Ель-
явруай, получено 13 сут записи поведения наседки. Утка оставляла гнез-
до 2-3 раза в день: на первую прогулку выходила между 5 ч и 12 ч, с
последней возвращалась не позднее 20 ч, однажды - уже в 13 ч. Она кор-
милась только в самые светлые часы суток, а на протяжении 11-17 ’’ноч-
ных” часов не оставляла гнезда. Средняя продолжительность прогулки -
62 мин, а всего кладка оставалась без обогрева 2 ч 32 мин, или 0,106 сут.
Оставляя гнездо, она укрывала яйца пухом, затрачивая на эту операцию
не более 1 мин. Птенцы начали выводиться вечером 24. VII; матка более
18 ч не оставляла гнезда, на следующий день в 13 ч ушла кормиться, ос-
тавив под слоем пуха 9 спящих утят, а 26. VII в 6 ч увела их с гнезда
(Семенов-Тян-Шанский, 1960, с. 284-285).
Другое гнездо, с 8 яйцами, было найдено под ’’елью в юбке” на берегу
Чунозера 26. VI 1968 г. в первый день инкубации. С этого гнезда удалось
получить только автоматическую запись температуры яиц через каждые
6 мин за двое суток, 6-8 июля. Пока утка сидела на гнезде, она колеба-
лась в пределах 33,8-35,9°, в среднем 34,5°; без наседки, покрытые пу-
хом яйца остывают медленно: разница между температурой яйца и тем-
пературой окружающего воздуха после 6 мин экспозиции составляла
0, 987 исходной разницы температур. 6 июля в 21 ч после отсутствия ут-
ки в течение 3 ч 24 мин при температуре воздуха 7,7° температура яиц сни-
зились до 23, 2°: это было в конце инкубации, когда эмбрионы сами про-
дуцируют тепло. Под наседкой яйца нагреваются быстрее, чем остывают
в ее отсутствие. В ночь на 8 июля, в 1 ч 22 мин на крохалюшку напала ку-
ница; утка спаслась, но яйца исчезли, даже скорлуп не осталось.
3. VI 1978 г. на о-ве Баран-суол, под большим камнем, найдено гнездо
большого крохаля с 4 яйцами; 6. VI в нем полная кладка из 6 яиц, утки
52
не было; 11. VI она насиживала; 11. VII в 15 ч яйца были наклюнуты, а
утром 14. VII матка увела выводок. Продолжительность инкубации,
включая и время пребывания птенцов в гнезде, составляла от 33 до
37 сут.
23 гнезда с полной кладкой содержали в среднем по 6, 8 яйца, макси-
мум - 11. Из 7 кладок большого крохаля, находившихся в ’’вуувтах”,
3 были смешанными, крохалю принадлежали соответственно 6,3 и 5
яиц, гоголю - 2, 1 и 3. Насиживали все три кладки крохалюшки и кончи-
ли тем, что увели на воду вместе с собственными птенцами и гоголят.
Среднее число птенцов в учтенных нелетных выводках больших кро-
халей оказалось в июне 5,7, июле - 6,4, августе - 6,3, сентябре - 5,6
(учтено 219 ординарных семей; 14 больших сборных выводков исключе-
ны из подсчета). По-видимому, отход птенцов в выводковый период нез-
начителен, различия по месяцам так малы, что их можно признать слу-
чайными.
Как только самки начнут насиживать, пары распадаются, самцы со-
бираются стайками и отлетают к местам линьки. Отлет происходит в пос-
ледней декаде июня, но иногда селезни задерживаются до середины ию-
ля: средняя дата последней встречи самцоа за 17 лет - 1. VII, самые позд-
ние - 27. VII 1965 г. и 26. VII 1986 г.
Выводки появляются на воде в среднем за 37 лет 9 июля (20. VI 1975 г. -
31. VII1931 и 1967 гг.). В довоенные годы большой и средний крохали
гнездились практически одновременно (средние даты встреч первых вы-
водков 20 и 19 июля); за последние 40 лет выводки больших крохалей
появляются на 15 дней раньше средних, соответственно 5 и 20. VII. На
озерах молодняк обоих видов цедет себя одинаково; на реках выводки
больших крохалей кочуют вверх и вниз по течению, проходя до 9 км за
сутки.
Птенцы начинают летать в возрасте 60 дней, так как в среднем за 30
лет первые летные выводки мы встречали 8 сентября (20. VIII 1974 г. -
29. IX 1968 г.). Сроки гнездования крохалей сильно варьируют, поэтому
даже в начале октября иногда встречаются нелетные выводки.
Отлет плохо прослеживается, так как осенью крохали держатся на
открытой воде, а с большого расстояния утки не поддаются уверенному
определению. Средняя дата встреч крохалей (оба вида) 20 октября (29.
IX 1968 г. - 8. XI 1949 г. за 45 лет; о ± 8,7 дней), нормальная температу-
ра этого дня +0,5°'. Время отлета зависит от температуры воздуха, корре-
ляция его со средней месячной октября г = + 0,37 ±0,13, а сентября г =
= + 0,52 ± 0,11 (за 45 лет). Случаи зимовки крохалей в районе Лапландско-
го заповедника описаны в предыдущем разделе.
Оба вида крохалей питаются преимущественно рыбой, но большой,
будучи крупнее, ловят и добычу покрупнее: 14. X 1967 г. на порожистом
участке р. Чуна А. Брагин наблюдал 4 крохалей, один из них поймал и
старался заглотать взрослого сига (в это время они идут на нерест). Хвост
сига торчал из клюва утки, а тело заполнило пищевод, ввиду чего она
плыла в неестественной позе. Через полчаса три крохаля взлетели, а
четвертый оторвался от воды, только когда отрыгнул сига. Передняя
53
половина рыбы была уже переварена, но по размерам остатка масса тела
оценивается в 1/3 кг. 9. VI 1968 г. в устье р. Чуна самка большого крохаля
на наших глазах с трудом заглатывала крупную, около 1/4 кг, рыбу
13. VI. 1975 г., поднимаясь вверх по Чуне, мы встретили двух селезней, один
сразу взлетел от лодки, другой бежал по воде, хлопая крыльями, он не
мог от нее оторваться. -На месте взлета первого крохаля мы нашли
крупного (около 0, 3 кг) сига без головы. Когда мы нагнали второго кро-
халя, он подпустив лодку, отрыгнул сига (поменьше первого) и только
тогда взлетел.
На глубоких местах крохали ловят рыбу ныряя, а на мелководье до-
гоняют ее, глиссируя по поверхности с опущенной в воду головой, ра-
ботая крыльями в воздухе. 13. IX 1970 г. В. Фролов наблюдал, как два
еще нелетных молодых крохаля ловили рыбу в подречье Ельявруая. Тут
же плавали две серебристые чайки. Если крохаль выныривал с рыбкой, -
чайка тут же налетала на него, принуждая бросить добычу. Одна из уток
пыталась отогнать чайку, но та только поднимала крылья вверх и громко
кричала. За 10 мин чайки отобрали у крохалей 2-3 рыбки.
Желудки убитых крохалей обычно содержат только немного песка,
но в желудке птенца, добытого 26.VIII 1934 г., оказался щуренок весом
100-150 г, а у взрослого самца 5. IX 1937 г. - зубастая челюсть и другие
кости (вероятно, форели). У молодого самца, застреленного 25. IX 1949 г.
на ручье, в желудке обнаружены икра кумжи, остатки переваренной
рыбы и шерсть полевки, а желудок и пищевод молодой самки 30. IX
1967. были наполнены мелкими окуньками и щурятами, общей массой до
300 г.
Материалы по причинам смертности обоих видов крохалей мы рас-
смотрим здесь совместно. В картотеке заповедника учтены 103 случая
гибели или находок остатков, в том числе 9 больших и 13 средних кроха-
лей были добыты на охоте и еще 2 погибли в результате ранения пулей
или дробью. От рыболовного промысла крохали страдают больше всех
других птиц: 29 крохалей попались в озерные мережи, 20 - погибли в
ставных сетях, а еще 5, попавших в сети, были своевременно освобожде-
ны и выпущены.
Из хищных птиц уток ловят орлан, орел, ястреб-тетеревятник и кре-
чет, но крохали достоверно обнаружены только в добыче орлана (не ме-
нее 3 экз.). Из крохалей, остатки которых найдены на месте поимки, 4
стали жертвой хищных птиц, а 6 - хищных зверей, преимущественно
норок. По наблюдению А.С. Сергина, в августе 1948 г. одного крохаленка
схватила зубами щука, но почему-то не смогла проглотить.
В июне 1984 г. один большой крохаль, по-видимому, разбился о про-
вод высоковольтной линии электропередачи, труп потом расклевали во-
рбны. Причину смерти 6 пуховых птенцов, 2 молодых и 2 взрослых птиц
не удалось установить.
Яйца крохалей гораздо чаще становятся добычей хищников, чем сами
птицы. Часть гнезд находят и разоряют вброн и серая ворона. На бере-
гах и даже в глубине леса не раз мы находили скорлупы яиц, унесенных
и расклеванных этими хищниками. В июне 1950 г. дважды видели вбро-
54
нов, летевших с яйцом в клюве. Пять гнезд были разорены мелкими хищ-
ными зверьками, судя по записи актографов, происходило это ночью:
2. VII 1968 г. в 1 ч 22 мин кладка большого крохаля была уничтожена ку-
ницей, другая крохалюшка 29. VI 1966 г. в 23 ч 22-43 мин подверглась
нападению норки, сама спаслась, а яйца погибли; еще 3 кладки разорили
норка и горностай.
Из нескольких больших крохалей, окольцованных в заповеднике, од-
на самка, помеченная 20. VI 1964 г. в дуплянке на берегу р. Мавра, пов-
торно ловилась в другой дуплянке, в 1 км от первой, в июле 1965, 1966.
1967, 1968,1969 гг. и в 1970 г., когда ей было не менее 8 лет.
Большой крохаль в полтора раза крупнее среднего по массе тела. Раз-
меры трех самцов (в мм): длина 655 (620-700), крыло 277 (265-288),
хвост 111 (100-125) мм; масса тела 1440-1740 г; размеры 7 самок соот-
ветственно - 606 (560-640), 238 (200-266), ПО (80—137) мм; масса тела
1430(1160-1650) г.
Средняя масса 8 яиц из 3 кладок 72,0 ± 0,4 г (70, 2 - 73,3).
В заключение обзора отряда приведем количественное соотношение
птиц отряда гусеобразных по материалам учетов в Лапландском заповед-
нике за 22 года (1966-1987) (в %):
Гоголь 464 Луток 16
Большой крохаль 193 Хохлатая чернеть 8
Средний крохаль 129 Морская чернеть 7
Чирок-свистунок 93 Турпан 4
Синьга 36 Шилохвость 2
Кряква 23 Лебедь-кликун 4
Свиязь 20 Гусь-гуменник 1
ОТРЯД ХИЩНЫЕ - FALCONIFORMES
Скопа- Pandionhaliaetus(L.)
Местные названия: саамское ’’чиихч” и ”нурр-коацкемь (= моло-
дой орлан).
На Кольском полуострове скопа - широко распространенная и регу-
лярно гнездящаяся, но редкая хищная птица; на зиму улетает в Африку,
к югу от экватора. Прилетает она одновременно со вскрытием рек и неде-
ли за две до вскрытия озер, в среднем за 33 года 16 мая (25. IV 1949 г. -
5. VI 1966 г.; о ± 10, 0 дня), при средней суточной температуре + 3, 8°. От-
летает относительно рано, в среднем 8 сентября (22. VII 1940 г. - 25. IX
1980 г.) за 29 лет; о ± 12, 2 дня, за полтора-два месяца до замерзания озер,
при температуре около 7, 7°. Скопа не так часто попадается на глаза, по-
этому прилет ее иногда наблюдался с запозданием, а последние особи
могли отмечаться до действительного отлета. Несмотря на это, время
прилета заметно коррелирует со сроками вскрытия рек (г = ± 0, 49 ± 0, 14
за 30 лет) и температурой мая (г = - 0, 36 ± 0, 15 за 33 года). Отлет с други-
ми фенологическими явлениями не коррелирует, а с температурой ав-
густа связан таким образом, что, чем теплее, тем раньше она улетает
55
(r= -0,29 ± 0,17 за 29 лет). На своих лапландских гнездовьях скопа про-
водит в среднем 115 дней.
За 45 лет наблюдений в картотеке зарегистрированы 346 встреч скопы
в среднем по 8 раз в год, но с колебаниями от 0-1 в 1986 г. и 1969 г. до
29 - в 1986 г. С годами уровень численности, по-видимому, не снижается:
на 1 наблюдателя в год приходилось в среднем в 30-х годах по 4,1 встре-
чи, в 40-х - 70-х - по 2,2 -2,6, в 80-х - по 2,5.
В поисках добычи скопа подолгу летает или кружит над озером на
высоте 20-40 м, иногда зависает на месте, трепеща крыльями, подобно
пустельге, а заметив жертву, падает, подняв крылья над спиной, с плес-
ком уходит в воду и тут же взмывает кверху. Бросок не всегда бывает
удачным, только в 5 случаях из 19 скопа поднималась с добычей в ког-
тях. Примерно такая результативность охоты наблюдалась и в Дарвинс-
ком заповеднике, где из 42 бросков было удачных 8 (Белко, 1985, с. 120).
Пойманную рыбу она держит обеими лапами вдоль линии полета, голо-
вой вперед. Добычу уносит на гнездо или съедает сама, сидя на йершине
дерева и отрывая клювом кусок за куском; закончив еду, она нетороп-
ливо чистит клюв о кору и улетает. Мы не находили под гнездами скопы
ни остатков добычи, ни погадок, так что о составе ее пищи приходится
судить по случайным наблюдениям. Она ловит преимущественно некруп-
ную рыбу, весом 100-300 г - сигов, хариусов и кумжу. Мелкую рыбу длиной
10-15 см иногда держит одной лапой и съедает на лету (единичное наблю-
дение). По ошибке иногда хватает и такую крупную рыбу, что не может
поднять ее на воздух. 27 июля 1932 г. в полный штиль мы заметили ка-
кое-то движение на середине Чунозера и в бинокль различили, что на
воде лежала скопа. По временам взмахивала крыльями, пытаясь взле-
теть, наконец поднялась и полетела к берегу, держа рыбу в когтях и во-
лоча ее по воде. Птица еще два раза ложилась на воду отдыхать, но нако-
нец выпустила добычу и улетела.
Нам не раз попадались кумжи и щуки со свежими или заживши-
ми ранами от когтей на боках. У одной кумжи в заросший шрам вросли
два пилорических придатка кишечника: только скопа или орлан могли
нанести такие раны.
15 сентября 1961 г. на оз. Нюхчи (в системе р. Витте) скопа схватила
попавшего в сеть сига, запуталась сама и погибла. Скопа нападает иногда
и на птиц. 8. IX 1974 г. скопа, сидевшая на сосне на берегу р. Нявка, бро-
силась на плывшего по воде чирка, но он успел нырнуть и спасся (Н. Сер-
гачев); 17. VI 1975 г. в среднем течении р. Чуна самка гоголя демонстра-
тивно кричала и ныряла, стараясь отвлечь человека от гнезда с утятами:
скопа на нее бросилась, но схватить не успела (А. Брагин). 9. VII 1975 г.
под гнездом скопы близ Куртваренча мы нашли бедренную кость утки.
Ондатру скопа, по-видимому, не трогает: в июне 1933 г. мы наблюдали
ондатру, занятую своим туалетом, сидя на бревне: на пролетавшую мимо
скопу она не обратила внимания. Некоторые птицы сами преследуют ско-
пу, принимая за врага или конкурента: дважды наблюдали, как скопу
атаковал дербник, раз - пара крачек, четырежды - сизые и серебристые
чайки. С другой стороны, в 1961 и 1962 г. видели скопу, атакующую орла-
на.
56
Скопа строит гнездо на вершине сосны или ели непосредственно
над стволом, почему добраться до него очень трудно. Гнездо использует-
ся многократно и сохраняется десятки лет. В одном гнезде, построен-
ном на сосне близ вершины Куртваренч, у восточной границы заповед-
ника, впервые обнаруженном 28. V 1973 г., скопы потом гнездились в
1967, 1974, 1975 и 1976 гг., а к весне 1979 г. оно развалилось. По площади
оно сходно с гнездами орлана или орла, уступая им по высоте; лоток поч-
ти плоский. Откладку яиц мы не наблюдали, но известно, что она начи-
нается вскоре после прилета. В высоких широтах (67° с.ш. и выше) Фин-
ской Лапландии в середине прошлого века Воллей и другие исследовате-
ли нашли 7 гнезд с яйцами между 19 мая и 24 июня (в среднем 31 мая);
размер этих кладок 2-3 яйца (Плеске, 1987, с. 329-330).
В Лапландском заповеднике известны 3 гнезда, в одном из ко-
торых скопа выводила птенцов 5 раз.
24. VIII 1931 г. — на северном берегу Чунозера гнездо с 3 птенцами, вылетевшими
через несколько дней. Это гнездо летом 1932 г. пустовало, а потом исчезло, вероят-
но упало вместе с деревом.
28. V 1937 г. - под северным склоном Куртваренча (яйца ?); 14. VI и 4. VII матка
на гнезде (птенцы ?); 11. VIII гнездо v-же пусто.
30. VII 1967 г. — скопа несла рыбу к этому гнезду; 3. VIII видели птенцов; 19.VIII —
2 слётка и 2 взрослые птицы.
6. VIII 1974 г. — в том же гнезде 1 птенец и 2 взрослые птицы.
21. VI 1975 г. — то же гнездо, одна скопа насиживает, другая принесла добычу;
9. VII птенцы вывелись, под гнездом свежая скорлупа; взрослая кормилица птенца,
вероятно отрыгивая пищу; 14. VIII в гнезде один вполне оперенный птенец.
25< VI 1976 г. — гнездо вновь занято (результаты гнездования неизвестны).
4. IX 1986 г. — на северном склоне Рентваренча (к югу от Чунозера) около гнез-
да, построенного на вершине сосны, 2 слетка скопы.
Скопиные выводки наблюдались и в других местах заповедника: 1.
VII 1939 г. на Куписьозере летали одновременно 3 скопы; 12. VIII, 1964 г.
к западу от озера Нявка в бору по склону вараки встретили 2 слетков;
1. IX 1971 г. на Парусной ламбине по р. Нявка ловили рыбу 3 скопы (вы-
водок). В июле 1950 г. на Туресквараке и в июне 1977 г. в бору близ
Улынчъявра, судя по беспокойному поведению пары взрослых скоп,
также было гнездо, но разыскать его не удалось.
По литературным сведениям скопа насиживает яйца 35-38 дней и
птенцы остаются в гнезде еще 51-59 дней (Harrison, 1975).Увязав эти дан-
ные со своими наблюдениями, определяем нормальные сроки размноже-
ния скопы в Русской Лапландии: кладка яиц в третьей декаде мая,
появление птенцов около 1 июля, вылет их из гнезда во второй и третьей
декадах августа, а иногда и раньше.
Число слетков уступает количеству яиц в кладке: только в одном вы-
водке было 3 птенца, в трех - по 2, в двух - по 1. Остатки пищи в гнез-
де скопы привлекают в него насекомых: в гнезде 1967 г. ползали му-
равьи, а в гнезде 1975 г. было много мух, и серая мухоловка их ловила.
Судя по нашим наблюдениям, скопа мало плодовита;.; возможно, и
гнездится она не ежегодно. Популяция скопы зависит от состояния рыб-
ных запасов не только в водоемах области ее гнездования, но и на путях
пролета и на зимовках. Страдает скопа и от сплошных рубок леса, так
57
как при этом гибнут перестойные и сухие сосны и ели, на которых она
гнездится. В Западной Европе (Швеция, Великобритания) замечено сни-
жение плодовитости скопы в результате накопления в яйцах сельскохо-
зяйственных ядов, ртути и хлор-органических соединений.
Скопу легко отличить от других хищных птиц. Она крупнее большин-
ства их, уступая в размерах только орлану и орлу. Взрослый самец, до-
бытый 22. V 1935 г., имел в размахе крыльев 188 см, длину тела 56 см,
крыла - 50, 3, хвоста - 21, 5 см, масса тела 1356 г. Крылья длинные, но
сравнительно узкие, угловатые. Полет легкий, взмахи крыльев переме-
жаются со скольжением. . На охоте молчалива, крик у гнезда - высокий
по тону клекот ”тив-тив-тив” или ”кли-кли-кли”, напоминающий го-
лос ястреба. Голос птенцов - тонкий прерывистый писк с интервалами в
1/4 секунды.
Осоед- Pemisapivorus(L.)
Северная граница гнездового ареала осоеда достигает Северной
части Финляндии, где в прошлом веке его гнезда находили до
66° с.ш. (Кеми Лаппмарк, Кусамо, Торнио; Плеске, 1887, с. 316). В
Мурманскую область он только залетает, и в Лапландском заповед-
нике зарегистрирован 4 раза.
24. X 1938 г. - осоед парил над восточным склоном Чуна-тундры;
18. VII 1939 г. на болото к северу от Курт-варенча налетел сначала
один осоед, потом пара пролетела на север; 18. IV 1972 г. - один
летал над Чунозерской усадьбой кругами, набирая высоту; 17. V 1983 г. -
пара осоедов кружила над долиной Верхней Чуны; 25. V пара пролетала
над лесом, с Охтозера к Чунозеру; 31. V один летел над устьем Чуны, а
1. VI там же охотилась пара.
Появление осоедов в Лапландском заповеднике парами (1939 и
1983 гг.) указывает на возможность их гнездования. В Средней Карелии
он настолько обычен, что в годы Великой Отечественной войны в Мед-
вежьегорском районе (около 63° с.ш.) мы регулярно наблюдали его прилет и
отлет: прилет - 25. IV 1942 г., 29. IV 1943 г., 27. IV 1944 г., в среднем 27 ап-
реля; последние встречи - 19. IX 1942 г., 31. VIII 1943 г., 1. IX 1944 г., в
среднем 7 сентября.
Сходный по сложению и характеру полета с зимняком, осоед отличает-
ся от него темной окраской нижней стороны крыльев (кроме оснований
маховых), более длинным хвостом с темной каймой на конце и двумя
полосами поперечными полосами при основании, способностью трепе-
тать на месте, как пустельга, и молчаливостью. Осоед меньше зимняка:
размеры одного самца из района Повенца (в мм): длина 585, крыло 405,
хвост 268, масса тела 725 г.
Черный коршун - Milvus korshun (Gm.)
Регулярно перелетная хищная птица, гнездовой ареал которой зани-
мает большую часть Европы и Азии, кроме северных частей. На Кольский
полуостров только залетает, самая северная известная нам встреча - на
о-ве Хар лове - 22. VIII 1963 г. (Коханов, 1987, с. 25).
58
В районе Лапландского заповедника одиночных коршунов наблюдали
восемь раз: 29. V 1950 г., 26. V 1958 г., 30. V 1964 г. и 11. V 1986 г. - на Чун-
озере, 21. V и 7. VI. 1974 г. - в долине р. Чуна (Н. Караваев), 15. IV 1960 г.
- на Имандре, у Екостровского пролива и (29. IX 1967 г. - на оз. Купись
(А. Брагин). Все встречи происходили в пролетное время, что позволяет
определить среднюю дату прилета - 17 мая.
Внешность коршуна настолько характерна (парящий полет, темная ок-
раска, выемка на конце хвоста), что определение птицы не вызывает ни-
каких сомнений. Голос коршуна - протяжный свист, к концу переходя-
щий в трель.
Орлан-белохвост - Halliaeetus albicilla (L.)
Местные названия: как саамы, так и русские не отличают орла-
на от орла и называют этих птиц одним именем; русские - ’’орел”, са-
амы - ’’коацкемь”, ’’куацкемь” или (в отличие от скопы) ’’вуамькоац-
кемь” (большой орел).
Орлан распространен по всей Мурманской области, как по морским
побережьям, так и внутри страны, по берегам озер и рек. В отличие от
беркута регулярно отлетает на зиму, возвращаясь на гнездовья рано, ког-
да земля еще покрыта глубоким снегом, но солнце уже пригревает и вос-
ходящие потоки теплого воздуха создают необходимые условия для па-
рящего полета.
Средняя дата прилета орлана - 15. IV. (4. III 1977 г. и 7. V 1968 г. за 41 год
о ± 12,7 дней), отлета - 3. X (10. IX 1939 г. - 26. X 1985 г. за 41 год; о ± 11,3
дня); в области гнездования он проводит в среднем 171 день. Время при-
лета совпадает с переходом средних суточных температур через -2,6°,
отлет происходит при 3,0°. Сроки перелетов практически не зависят от
других фенологических явлений и средних месячных температур и соот-
ветствующие коэффициенты корреляции не превышают +0,24 +15 (для
пары дата отлета - температура сентября).
В Лапландском заповеднике зарегистрировано 579 встреч орланов за
40 лет. Численность на протяжении полувека остается на одном уровне:
на одного наблюдателя в год в 30-е годы приходилось по 3,3 встречи,
40-е - 4,2, в 50-е - 2,0, 60-е - 2,5, 70-е - 2,6, 80-е - 2,6.
Большинство встреч приурочено к озерам и рекам, гораздо реже орла-
на видят парящим над лесом или горной тундрой. Первое время после
прилета орланы питаются падалью, остатками крупных зверей, добытых
медведем или волком или погибших зимою от других причин. В марте,
апреле и мае орланов или их следы наблюдали на трупах лосей 8 раз, на
остатках оленей - 5 раз. Орланы патрулируют оленьи стада и во время
отела важенок поедают трупы погибших пыжиков (4 наблюдения), отго-
няя воронов и других конкурентов. Вероятно, в это раннее время орлан
охотится также на глухарей и зайцев; самый процесс охоты нам не случа-
лось наблюдать, но 16 мая 1982 г. орлан подлетел к остаткам глухаря, ве-
роятно им же и пойманного, а под гнездами орланов остатки глухарей
находили часто.
После вскрытия рек щуки выходят на залитые полой водой осочники
59
и мечут икру - тут их и ловят орланы. 7 июня 1951 г. лесник А.С. Сергин
видел, как орлан за полчаса поймал на озере Пасма-ламбина 2 щук и
обеих унес в лес, очевидно к гнезду. В разное время найдены остатки
еще 7 щук, расклеванных орланом на месте добычи, и 40 - под гнездом.
Из костей скелета лучше всех сохраняется крупная нижнечелюстная
кость (os dentale). В 1950 г. мы составили номограмму для определения
длины и веса щуки по размерам этой кости. Среди пойманных орлана-
ми щук масса четырех была в пределах 1-2 кг, четырех - от 2 до 4 кг,
а четырех самых крупных определена в 5,03, 5,5, 5,9 и 6,1 кг (длина их
от 91 до 112 см). Очевидно, орлан способен выхватить из воды добычу,
несколько превышающую его собственный вес. При случае он ловит ры-
бу и летом: 10. VIII 1986 г. на оз. Румельявр орлан расклевал огромную
щуку3 длиной 126 см; 5. VIII 1980 г. другого спугнули с остатков пой-
манной им кумжи массой около 3 кг; 5. VII 1949 г. на оз. Кензисъ-явр най-
ден поклеванный орланом сиг массой 1, 5 кг, а 9. VIII 1969 г. на р. Нявка
был подобран сиг в 0, 6 кг, погибший от нанесенных когтями ран.
На воде орлан ловит также ондатру и уток. Броски орлана на плава-
ющих уток наблюдали неоднократно: 14. X 1967 г. - на одиночного гого-
ля, 5. VIII 1976 г. - на выводок из 4 гоголей, 9. VIII 1984 г. - на 4 молодых
больших крохалей, 8. VIII 1981 г. - на 6 утят того же вида, с маткой. Во
всех четырех случаях утки ловко ныряли и орлану ни одну не удалось
схватить. 10. IX 1972 г. наблюдали нападение орлана на выводок белых
куропаток, а 9. IX 1978 г. лесник О. Петровская на берегу оз. Купись спуг-
нула орлана с куропатки, которую он расклевывал.
Под семью гнездами мы собрали все остатки добычи и погадки орланов
и несколько нашли на местах кормежек; с добавлением визуальных наб-
людений мы учили 321 животное, пойманное или съеденное орланом
(табл. 2).
’’Спектры питания”, составленные на основе анализа погадок - с од-
ной стороны, и по остаткам добычи (включая и визуальные наблюдения) -
с другой, существенно различаются. В погадках полевки и лемминги
составляют 34 % всех учтенных объектов и занимают в добыче хищника
второе место; среди остатков жертв и мелкие грызуны редки (всего 3%).
С другой стороны питаясь падалью или рыбой, орлан рвет и глотает поч-
ти чистое мясо, которое переваривается полностью, поэтому среди ос-
татков рыба составляет 37 %, падаль - более 14 %, а встречаемость их в
погадках составляет только 15 и 8 % соответственно. Объединение мате-
риалов, полученных разными методами, надо полагать, даст наиболее
близкий к истине результат (см. последнюю колонку табл. 2).
Примерно половину своего рациона орлан добывает на воде (рыба,
ондатра, норка, утки и другие водоплавающие); другую половину на су-
ше (падаль, тетеревиные птицы и большая часть зверей).
Добычу свою орлан либо высматривает с воздуха, паря высоко над
землей, либо подкарауливает, сидя на дереве. Во время перелетов слу-
чается, что орланы задерживаются и концентрируются на местах, бога-
3Этой щукой орлан кормился три дня; от нее остался только позвоночник с частью
головы.
60
Таблица 2. Питание орлана-белохвоста
Вид корма, добычи Остатки пищи Погадки (122 образца) Состав пищи, % То же, обоб- щен., %
Млекопитающие
Заяц 1 3 13 1
Белка — 1 3 / I1/2
Ондатра 5 7 37 4
Полевки* — 38 118
Лесные полевики — 9 28 101/2
Водяная крыса — 1 3
Норвежский лемминг 4 10 44
Лесной лемминг — 1 3
Лиса 1 — 3 1
Норка 1 1 6 ) 1
Лось (падаль) 6 2 25) 101/2
Олень (падаль) 14 12 81/
Птицы* 6 19 78 8
Гагара 2 — 6
Утки 21 34 171 -
Чирок 1 — 3 19
Крохаль 4 — 13
Тетеревиные* 1 3 13
Белая куропатка 7 5 37 11
Глухарь 15 3 56 1 X
Тетерев 2 — 6
Рептилии (ящерица жи- — 1 3 1/2
вород.)
Рыбы* — б 19
Форель и кумжа 1 — 3
Сиг 2 1 9 25
Хариус 1 3 13
Щука 50 16 206
Всего, особей 145 176
*Ближе не определено
тых кормом (так, во время нереста семги в верховьях р. Печора 5. X
1953 г. видели 5 орланов у остатков расклеванного ими лоха). Парящий
орлан использует для полета внешний источник энергии, конвективные
потоки, образующиеся при нагреве земной поверхности солнцем. Зимой,
при отсутствии восходящих потоков, орлан поневоле переходит на актив-
ный полет, это ограничивает длину его маршрута и шансы найти добычу:
поэтому он и отлетает на зиму к югу. Впрочем, при наличии доступного
корма орлан иногда остается в Мурманской области и на зиму. В январе
1967 г. на Терском берегу А.И. Флеров видел двух орланов на туше дико-
го оленя, а в начале марта 1974 г. охотники, занятые промыслом диких
оленей на озерах в бассейне Р. Пиренга, неоднократно встречали орланов
на местах разделки оленьих туш.
Гнездо орлана представляет массивную постройку из сучьев в диамет-
ре до 1,8 м и до 2 м в высоту, с плоским лотком. Восемь известных нам
61
гнезд находились не далее 1 км от озера или реки, на соснах, близ вер-
шины, но сбоку от ствола. Высота лотка над землей от 9 до 15 м. Пара
орланов занимает гнездо и начинает его ремонтировать вскоре после
прилета, во второй половине апреля. В гнезде, обнаруженном в 1937 г.
в долине Чуны, уже 10 мая было одно яйцо; после осмотра гнезда птицы
его бросили. 20 августа яйцо оставалось на том же месте в целости. В
1938 г. гнездо было снова занято, и 10 июля в нем находился 1 оперен-
ный птенец; 4 августа он уже вылетел, но держался поблизости. На про-
тяжении следующих 10 лет гнездо пустовало и в 1949 г. стало разрушать-
ся. В том же лесном квартале в 1959 г. обнаружено другое такое же ста-
рое гнездо. В этих гнездах орланы выводили птенцов и раньше: 25. VIII
1935 г. здесь наблюдали летного молодого, а в 1936 г. пару орланов встре-
чали неоднократно на протяжении всего лета, очевидно, она гнездилась
здесь четыре сезона подряд.
Другое гнездо было найдено на сухой сосне близ южного берега оз.
Мерть-явр 4. VIII 1934 г. - в день, когда слетка поймала и растерзала
семья медведей. Вторично это гнездо было занято в 1941 г., к 13 августа
оно опустело, но поблизости видели взрослую птицу и одного птенца.
Орланы жили и охотились в этом же районе до 17 сентября.
В 1935 г. в бору на берегу Мавра-губы было обнаружено старое гнездо.
24 апреля 1941 г. оно было занято и подремонтировано; на лотке лежали
зеленые ветки (результат гнездования не установлен, но 1-2 орлана наб-
людались здесь до начала октября). В 1950 г. гнездо снова занято, в нем
вывелся один птенец; 6 июля он был окольцован, ^20 августа гнездо уже
опустело. В 1951 г. орланы снова его заняли, вывели одного птенца, и
10 августа он вылетел. После того заповедник был закрыт, наблюдения
прекратились, лес вокруг Мавра-губы был вырублен, и гнездо исчезло.
Пятое гнездо у южной оконечности оз. Витте было занято в 1939 и
1946 гг.
В 1961 г. в смешанном лесу у берега Железной губы Екостровской
Имандры обнаружены два старых гнезда в 1 км друг от друга. 22 апреля
одно из них было занято, но позднее брошено. Орланов и следы их пре-
бывания находили в этом районе также в 1959,1960 и 1962 гг.
В июне 1968 г. в бору на юго-восточном берегу Куропачьего озера наш-
ли занятое гнездо орлана. Весной 1975 г. птицы заняли его снова и выве-
ли одного птенца; 16 июля он был окольцован (рис. 13), а 9 августа уже
летал. По соседству с этим гнездом оказались еще три старых.
Одно гнездо известно в бору по левому берегу р. Купись; оно было
занято 25 июня 1973 г., а потом - 13 мая 1982 г.; наблюдений за ним не
велось, но 23. IX здесь летали 3 орлана - вся семья.
10. VIII 1986 г. на вершине сосны в долине Нявки найдено гнездо, с
которого слетели 2 молодых орлана; семья держалась там же до 22. VIII.
Под гнездом собраны остатки 8 птиц, 5 щук, 1 норки и 24 погадки, состо-
явшие в основном из сухих листьев осок и злаков, с примесью эпифит-
ных лишайников, лосиной и оленьей шерсти и костей жертв.
В 1967 г. на горном склоне к р. Вайкись обнаружено гнездо, постро-
енное не на дереве, а в скалах: 11. VII в нем был один птенец.
52
Рис. 13. Молодой орлан-белохвост, 16. VII 1975 г. Фото О.И. Семенова-Тян-Шанского
На мелководных и рыбных озерах Нявка и Купись орланы встречаются
чаще, чем в других местах: в 1938 г, 1960 и 1977 гг. в этом районе находи-
ли старые гнезда, но жилых еще не встречали.
Судя по нашим наблюдениям, орлан гнездится на одном месте до
четырех лет подряд, потом гнездо на протяжении нескольких лет пус-
тует. На гнездовом участке может быть до 2-4 гнезд на расстоянии ме-
нее 1 км между соседними: очевидно, все они принадлежат в данный мо-
мент одной паре. Расстояние между гнездовыми участками гораздо
больше, не менее 4,5 км, обычно же 14-34 км. Гнездовые участки приу-
рочены либо к южной, либо к северной границе заповедника, где нахо-
дятся большие озера.
Молодые орланы оставляют гнездо в первой половине августа, вылет
наблюдали 4, 9, 10 и 11 августа, а 13, 20 и 25 у трех других гнезд видели
летных молодых. Из одного гнезда вылетели дваЪтенца, из семи - по
одному, и еще один орленок погиб в день вылета; два гнезда с яйцами
брошены, судьба остальных трех не прослежена.
63
Рис. 14. Орлан у гнезда, 16. VII 1975 г. Фото О.И. Семенова-Тян-Шанского
В каком возрасте орланы начинают гнездиться - неизвестно; во всяком
случае, размножаются они медленно. Судя по тому, что за последние 30
лет в заповеднике найдены трупы или останки 5 орланов, смертность
довольно высока. Одна самка была застрелена в июне 1963 г. при устье
Чуны браконьером (следы дроби на теле), в остальных случаях найдены
только кости и перья (долина Сылпуя 14. XI 1967 г.; берег Чунозера
17. VIII1971 г.; Сейдапахта 12. VII 1975 г.; берег р. Нявка 19. VII 1978 г.).
Природных врагов у орлана мало, только раз слетка поймал медведь;
возможно, яйцо может похитить вброн. Чайки, крачки и скопа нередко
преследуют летящего орлана, а дербник и зимняк нападают, если он
приблизится к их гнездам.
Орлан - самая крупная хищная птица в нашей области. Размеры сам-
ки, найденной в 1963 г. (в см): размах крыльев 220, длина 85,5, хвост
31,4, крыло 62; масса тела несколько подсохшего трупа 3, 03 кг. Длина
хвоста другого, вероятно самца, 27,5 см.
Орлан обычно парйт, сильно растягивая крылья, вследствие чего пе-
редние и задние края их представлены прямыми линиями, без выступа
на кистевом сгибе (рис. 14). Хвост короткий, округлый, у старых птиц
весь белый, у молодых серовато-бурый, как и остальное оперение; при
этом крылья заметно темнее туловища, а голова светлее. Голос довольно
высокий по тембру, напоминает ”кре-ке-ке-ке-ке-ке...”.
64
Ястреб-тетеревятник - Accipiter gentilis (L.)
Местное название: саамское ”кынц-лоньт” (саам Екостров-
ского погоста Ф.К. Архипов).
По сведениям Ф.Д. Плеске, ’’тетеревятник, хотя и в ограниченном
числе, живет оседло в весьма северных широтах и гнездится.. .”
(Плеске, 1887, с. 314). По наблюдениям Лапландского заповедника,
у нас зимуют только единичные особи в отдельные годы, а боль-
шинство ястребов откочевывает на зиму к югу. В теплую половину
года, с мая по октябрь включительно, на один месяц приходится в
среднем по 0,5— 0,6 встречи, в зимнее - с ноября по апрель, -
только по 0,1.
Вероятность встреч ястреба тетеревятника по месяцам:
I п ш IV V VI VII VIII IX х XI хп
0,1 0,1 0,1 0,1 0,5 0,6 0,6 0,6 0,5 0,5 0,1 0,1
Только в одну зиму 1938/39 г. ястреб попадался довольно часто, в
зимы 1940/41 и 1982/83 гг. их видели по два раза, по одной встрече прихо-
дится на 10 других, а на протяжении остальных 31 зимы ястребов в
природе совсем не встречали, но в 1984/85, 1985/86 и 1986/87 гг. они
зимовали в г. Мончегорске, питаясь голубями. В 80-х годах тетеревятни-
ки регулярно зимовали в Кандалакше, где охотились на голубей и ворон
(Коханов, 1985, с. 103-104).
Из 6 ястребов, окольцованных птенцами, один добыт в декабре 1966 г.
в возрасте полутора лет у деревни Корос-озеро (район Балтийско-Бело-
морского канала), в 480 км к ЮЮВ от места рождения и мечения.
За 44 г. в заповеднике зарегистрировано 176 визуальных встреч ястре-
ба-тетеревятника. На одного наблюдателя в год приходится: в 30-х годах
по 2,1 птицы, в 40-х - 50-х, 60-х, 70-х и 80-х годах по 1,5.
Из приведенных сведений можно заключить, что на Кольском полуост-
рове тетеревятник сравнительно редок и основная масса птиц около
1 ноября откочевывает на зиму к югу, возвращаясь обратно около 1 мая;
только при наличии благоприятных условий часть ястребов зимует на
месте. Популяция не обнаруживает тенденции ни к росту, ни к сокраще-
нию, но численность ее заметно колеблется с периодом в 4 года, причем
минимум приходится на високосные годы:
После високоса 1-й 2-й 3-й 4-й
Число встреч в год 1,5 7,3 4,3 2,8
Естественно предположить, что колебания численности ястреба отра-
жают соответствующие колебания численности его основной добычи,
тетеревиных птиц, однако корреляция между индексом численности
ястреба и суммарной плотностью четырех видов тетеревиных птиц на
1 км2 леса (по материалам учетов на ленточных пробах) оказалась невы-
сокой,! = + 0,35 ±0,14 (за 39 лет). Четырехлетний цикл колебаний числен-
ности характерен для полевок, однако связь между индексом встречае-
мости ястреба и численностью мелких млекопитающих на учетном
стационаре оказалась еще ниже, = + 0,24 ±0,15 (39 лет). Правда, пери-
одичность циклов полевок непостоянна, увеличиваясь иногда до 5-6
лет, а популяции тетеревиных птиц не обнаруживают правильной перио-
дичности.
5. Зак. 1023 65
Таблица 3. Питание ястреба тетеревятника
(по визуальным наблюдениям, остаткам добычи и погадкам с 5 гнезд)
Вид Число особей Процент Обоб- щен. %
Добыча Погадки (97 шт.) В добыче в погадках
Млекопитающие — — — — 1 33
Насекомоядные (зем- — 1 — 1
леройки)
Грызуны — — — —
Белка 1 9 2 7
Полевка лесная 4 24 7 19
Полевка серая — 1 — 1
Полевки* — 6 — 4
Хищные (горностай) — 1 — 1
Птицы* 1 7 2 5 67
Водоплавающие (гоголь) 1 1 2 1
Хищные (ястреб) — 1 — 1
Тетеревиные* — 4 — 3 12%
Глухарь И 1 19 1
Тетерев 6 — 10 —
Белая куропатка 25 9 43 7
Рябчик 1 1 2 1
Кулики и ржанки* — 3 — 2
Голуби (домашние) 1 — 2 —
Совы (ястребиная) — 1 — 1
Стриж — 1 — 1
Дятлы (большой и — 4 — 3
трехпалый)
Певчие птицы* — 37 — 29 41%
Кукша 2 1 3 1
Дрозды 3 12 5 9
Овсянка, вьюрок, 2 3 3 2
чечетка
Насекомые — 44 — —
Растительные остатки
Ягоды брусники, во- — 22 х — —
роники и др.
Сосновая хвоя — 18 — —
Всего позвоночных, 58 128
особей
*Ближе не определены.
Ястреб-тетеревятник-специализированный хищник орнитофаг. На охоте
он летити по лесу низко над землей, стараясь застать птицу врасплох и
схватить ее на взлете. Осенью 1934 г. О. Семенов-Тян-Шанский наблюдал,
как ястреб летел перед носом охотничьей лайки с явным намерением
ловить спугнутую ею дичь, то же самое наблюдал в 1938 г. и Г. Новиков.
Сотрудниками заповедника отмечены нападения ястреба на чирков,
гоголей, глухарей, тетеревов, белых куропаток, рябчиков, ворон и на
выводок кукш. 14 августа 1970 г. на наших глазах глухарка летела через
66
болото в долине ручья Сылп-уай, следом за ней ястреб. Через несколько
секунд хищник догнал жертву, сбил на землю и тут же стал расклевы-
вать. Видевшая это собака помчалась к ястребу, он бросил добычу и
улетел, держась низко над землей. У глухарки в районе правой лопатки
были расклеваны мускулы и сломано клювом ребро.
Крупную добычу ястреб оставляет на месте, возвращаясь к ней повтор-
но: так, 16. III 1941 г. Владимирская и А. Васильев подняли ястреба с
промерзших остатков добытого им раньше глухаря самца, а 19. IX 1941 г.
другого спугнули с живого глухаря. Таким же образом спаслась куропат-
ка, которую ястреб сбил на лед Чунозера 12 мая 1981 г. (М. Заваруев).
Обычно ястреб в первую очередь отрывает и съедает голову жертвы, а
тушку, перед тем как есть, ощипывает. Куропатку, масса тела которой
около 650 г. ястреб уносит легко, тетерку поднимает с трудом; вместе с
тем на гнездах не раз находили остатки даже взрослых глухарей, масса
тела которых превышает таковую ястреба втрое; вероятно, хищник
съедал часть мяса на месте, а птенцам приносил только остатки добычи.
Если выводок при виде ястреба затаится, он садится на дерево и караулит,
выжидая удобный момент для броска.
Под пятью гнездами мы собрали остатки жертв и погадки птенцов. Как
и в случае с орланом, анализ погадок дает несколько иной спектр пита-
ния, чем прямые наблюдения: в погадках обнаружены полевки, которых
ястреб, очевидно, заглатывает целиком, так как среди остатков добычи
их не удавалось обнаружить. В материале, собранном под гнездами,
заметно возрастает роль мелких птиц (певчих, дятлов, куликов); в одной
погадке оказался стриж. Большинство мелких птиц на зиму улетает, и в
это время года ястреб охотится преимущественно на куропаток, глуха-
рей, тетеревов и рябчиков (табл. 3).
Кроме птиц и млекопитающих в погадках нередко попадаются остатки
жуков, крупных перепончатокрылых, мух и других насекомых, семена
вороники, брусники, ”гром-ягоды”, свежая сосновая хвоя и другие
примеси. Часть этих объектов происходит из желудков его жертв, но сосновую
хвою птенцы, вероятно, срывают с зеленых веток, лежащих на лотке.
Остатки ястреба в одной погадке 1965 г. - свидетельство каннибализма:
5. VI в гнезде было 3 птенца, а через 10 дней остались только два (старшие
съели младшего).
Зарегистрировано 9 гнезд ястреба-тетеревятника:
20. V 1951 г. — гнездо с яйцами на северном побережье Чунозера. Построено
в смешанном лесу, на ветвях сосны с южной стороны ствола, в 6 м от земли;
диаметр 80 X 90 см, толщина 40 см. В лотке, выстланном зелеными
хвойными веточками, 3 яйца. 3. VI самка убита; в яйцах вполне сформиро-
ванные птенцы, одетые пухом, должны были вывестись 5-8, VI.
9. V 1959 г. - в том же районе другое гнездо, также на ветвях сосны на
высоте 7 м, содержит 1 яйцо. После осмотра гнездо брошено.
10. VI 1963 г. - там же третье гнездо, на сосне по южному склону холма, в
9 м над землей; в нем 2 яйца. После второй проверки 12. VI гнездо разорено
или брошено (27. VI оказалось пустым).
5. VI 1964 г. — в ельнике в долине речки Ташким, в боковой лощине
гнездо ястреба в кроне густой ели на высоте 5—6 м, с 4 пуховыми птенцами;
15 VII оно уже пусто, но один слеток найден неподалеку.
4. VI 1965 г. — на склоне вараки к берегу Охтозера, на юго-восточной
стороне большой сосны в 8 м над землей гнездо ястреба с 3 птенцами; 11. VI
матка греет птенцов на гнезде; 5 VII они наполовину оперены, средняя масса
тела птенцов 750 г. 15. VII остались два птенца (младший съеден старшими);
22. VII гнездо пусто, оба слетка в 100—200 м от него.
27. V 1973 г. — то же гнездо занято вновь, самка насиживает; 8.VI на
гнезде матка и птенцы; 14.VI три пуховых птенца без взрослых; 22.VI
67
полуоперенные птенцы окольцованы; 8. VII старший уже вне гнезда, осталь-
ные два взлетели от человека. В гнездовой подстилке найдено цельное яйцо
без признаков эмбрионального развития. К весне 1978 г. гнездо наполовину
обвалилось.
25. V 1983 г. — гнездо занято в третий раз, самка насиживает; 9.VI осмотр
гнезда, в нем только 1 яйцо, после чего ястреба бросили гнездо; 22.VI яйцо
взято, в нем почти готовый к вылуплению птенец.
23. VI 1986 г. — в верховьях р. Конья найдено гнездо с птенцами, которые
через несколько дней погибли.
24. VI 1987 г. - в том же гнезде ястреба вывели трех птенцов. Гнездо
построено на боковых ветвях сосны на высоте 6 м, размер 142 X 135, высота
45 см, на лотке свежие сосновые и березовые ветки. Птенцы окольцованы.
Летом 1988 г. ястреба шездились в нем в третий раз, вывели 4 птенцов,
вылетевших к концу июля.
Приблизительные сроки размножения ястреба-тетеревятника по этим
наблюдениям: кладка яиц и начало насиживания - не позднее первой
декады мая; вывод птенцов - первая декада июня; их вылет - 8-
22 июля.
Среднее число яиц по 7 гнездам - 2,6 (от 1 до 4); они крупные, шаро-
видно-эллиптической формы, белые со слабым зеленоватым оттенком.
Размеры и масса трех яиц: 45,2 х 59,3 мм, 64,0 г (1959); 68,0 г (1973);
45x57 м, 56,7 г (1983).
Ястреб может пользоваться тем же гнездом несколько лет сряду, в
других случаях он меняет место гнездования, возвращаясь к прежнему
через 4-10 лет, но можно предполагать, что это уже другая пара.
На лотке занятого гнезда всегда лежат свежие ветки, обычно сосны:
возможно, они камуфлируют гнездо сверху, кроме того, хвоя может
служить источником витамина С для птенцов.
Потревоженные в начале инкубации ястреба бросают гнездо после
первого же или второго посещения его: птенцов, а также яйца накануне
вывода птенцов не оставляют, даже если птицу караулят для фотографи-
рования. По нашим наблюдениям, успех гнездования ястребов невысок,
но надо учитывать, что его снижало вмешательство человека. Из девяти
гнезд с 25 яйцами четыре (с 7 яйцами) были брошены или разорены, в
остальных пяти из 18 яиц вывелось 17 птенцов, причем один погиб в
гнезде и был съеден, так что вылетели только 16 молодых ястребов; из их
числа один погиб в то же лето вблизи гнезда. Еще в одном гнезде погиб-
ли все птенцы.
Стремительный бросок ястреба на добычу иногда становится причиной
его гибели. В Печоро-Илычском заповеднике дважды (в марте 1939 г. и
апреле 1954 г.) находили трупы ястребов, насквозь проткнутых острым
сучком: очевидно, хищник напарывался на сук в тот момент, когда
выбрасывал лапы вперед, пытаясь схватить спрятавшегося в кроне
дерева рябчика или тетерева. Во втором из описанных случаев сук
вонзился в брюхо, а конец его вышел наружу в области лопаток (видимо,
в момент ранения ястреб летел брюхом вперед), сук обломился, смер-
тельно раненный хищник продолжал лететь, пока не упал на речной лед,
где его и нашли (Семенов-Тян-Шанский, 1959, с. 194). Зимой 1985/86 г. на
берегу Монче-губы Имандры нашли труп молодого ястреба, напоровшего-
ся на сук. В Лапландском заповеднике в августе 1962 г. молодой ястреб
был убит пропеллером ветряка электрогенератора на кордоне ’’Мавра”. В
марте 1987 г. в Мончегорске ястреб при поимке голубя разбил окно.
Остатки ястребов, погибших от разных причин, находили в заповедни-
ке еще 6 раз. В то же время три ястреба были застрелены в окрестностях
68
заповедника, а один окольцованный добыт в декабре 1966 г. в Карелии.
Природных врагов у ястреба почти нет, но в жизни его два критических
момента - при вылете птенца из гнезда, пока он не научится самостоя-
тельно ловить добычу, и зима - голодное время, когда хищник принуж-
ден откочевывать в незнакомые места. Смертность несомненно высока,
так как число убитых или найденных в районе заповедника мертвыми
составляет 7% от числа встреченных живых особей; при этом 5 ястребов
из 12 погибли слетками, летом, остальные взрослыми, в разные сезоны.
Крик взрослых птиц - звонкие ”кья-кья-кья. . .” с периодом около
0,3 с, голос птенца ”ти-ти-тириририри...”. В отличие от канюков, соколов
и скопы, ястреб у гнезда очень осторожен, близко не налетает, а прячется
за деревьями.
Размеры взрослой самки, добытой 3. VI 1951 г.: размах 121 см, длина
тела 62, хвоста 25,4, крыла 37 см, масса тела 1514 г. Масса тела двух
молодых неизвестного пола (октябрь 1961 г. и август 1962 г.) 1005 и 1162 г.
Ястреб-перепелятник - Accipiter nisus (L.)
Этот ястреб, как и тетеревятник, встречается в лесной части Мурман-
ской области, но заметно реже его. В Лапландском заповеднике за все
годы наблюдений зарегистрировано только 59 перепелятников на 176
тетеревятников. Подсчет числа визуальных встреч на одного наблюдате-
ля в год показал, что популяция перепелятника за половину века сильно
сократилась, очевидно в связи с устойчивым снижением численности всех
видов перелетных певчих птиц. Индекс встречаемости этого ястреба
изменялся по десятилетиям следующим образом: 30-е годы - 2,9; 40-е -
1,8; 50-е - 0,3; 60-е - 1,1; 70-е - 0,5; 80-е - 0,4.
Перепелятник никогда не зимует на Крайнем Севере; в среднем за 6
лет он прилетает 10 мая (самое раннее 16. IV в 1939 г.), последняя встре-
ча - 27 сентября (самая поздняя - 10 X в 1938 г.).
В отличие от тетеревятника, этот ястреб чаще попадается в ельниках
по склонам горных тундр, чем в борах. По способу охоты он не отличает-
ся от тетеревятника, но добывает преимущественно певчих птиц: в число
его жертв входят кукша, дрозд (белобровик и певчий), юрок, чечетка,
гаичка, 3 певчие птички, не определенные точнее, и 1 кулик-перевозчик.
В сентябре 1932 г. наблюдали нападение этого ястребка на выводок
рябчиков, по-видимому без успеха. Весной 1961 г., когда научный сот-
рудник заповедника Ю.Г. Гаев ловил лучком мелких птиц для кольцева-
ния, на точок с манными птицами несколько раз налетали перепелятни-
ки; двух удалось поймать и закольцевать. Взрослый самец, окольцо-
ванный 20 V 1961 г. через 3 года, 15 III 1964 г. добыт на месте зимовки у
Фарентуна в Швеции (близ Стокгольма), на расстоянии 1160 км к юго-за-
паду от Лапландского заповедника.
24. VII 1937 г. П.С. Мальчевский нашел на юго-восточном склоне Чуна-
тундры, в кроне густой елки, только что оставленное выводком гнездо
перепелятника и около него видел одного из ястребов. Под гнездом были
остатки добычи - кукша и юрок. 1. IX 1972 г. в том же районе и также на
елке, на высоте 4,5 м от земли найдено еще гнездо, старое, и в ней погад-
ка, содержавшая желудок и перья мелкой птички. 20 и 21 августа 1939 г.
в бору черничнике под западным склоном Чуна-тундры наблюдали
выводок, не менее 3 птиц.
69
Единственный экземпляр, взрослая самка, добыт 1. VIII 1938 г. в
долине р. Нявка, между гор, в тенистом ельнике с покровом из чернични-
ка, лесной герани, плаунов, мелких папоротников и зеленых мхов.
Длина птицы 387, хвоста 192, крыла 241 мм, вес 257 г; в желудке ястреба
остатки чечетки и еще одной птички.
Мохноногий канюк (зимняк) - Buteo lagopus (Pont.)
Местные названия: саамские ”рехпь-6угти” (от рехпь - белая
куропатка) и ”кёммесь-чиихч”.
Мохноногий канюк распространен циркумполярно, гнездится в зонах
тундры, лесотундры и северной тайги, на зиму отлетая к югу, в мало-
снежные регионы. По всей Лапландии это самая многочисленная хищная
птица.
,От 42 зимняков, окольцованных в Лапландском заповеднике в 1959—
1983 г., поступило 4 возврата из зарубежных стран: один добыт в первую
же зиму в горах центральной Греции, другой найден мертвым весной
1964 г. в восточной Австрии, третий в мае 1969 г. добыт в возрасте 6 лет в
восточной Венгрии, четвертый найден мертвым в начале 1978 г. близ
Рованиеми, в Финляндии. Расстояния от места рождения до места гибели
этих птиц - 3200, 2350, 2250 и 350 км в направлении от южного до юго-за-
падного. Видимо, у зимняка нет определенной области зимовки, а в
зависимости от запасов живого корма и состояния снежного покрова он
отлетает на разные расстояния, иногда достигая берегов Средиземного
моря.
Как это свойственно всем кочующим птицам, время возвращения его
на родину сильно варьирует, так что амплитуда дат прилета достигает
двух месяцев - от 23. III 1974 г. до 23. V 1965 г., в среднем 27 апреля
(41 год, б ± 12,5 дней, средняя температура - 0,2°). Корреляция между
временем прилета и средней температурой апреля низкая, г = - 0,13 ± 0,12;
это показывает, что прилет зависит не только от времени таяния снега, но
и от других причин, в первую очередь от обеспеченности кормом. Время
отлета варьирует между 2. IX 1976 г. и 17. XI 1939 г., в среднем 25 сентяб-
ря (27 лет, б ± 15,4, средняя температура + 4,5°). От температуры сентября
отлет не зависит (г= - 0,02 ± 0,19).
За 43 года наблюдений в заповеднике зарегистрировано 1374 встречи
зимняка. В зависимости от обеспеченности кормом численность его
колеблется по годам: коэффициент корреляции между учетным индек-
сом полевок и числом зимняков, встреченных одним наблюдателем за
год, г = + 0,78 ± 0,08. Вместе с тем тенденции к сокращению или увеличе-
нию численности за все время наших наблюдений не замечено: за послед-
ние 7 полных или неполных десятков лет число встреч на одного наблю-
дателя в год остается стабильным, от 5,1 - в 60-х, до 5,6 - в 30-х годах.
По роду питания мохноногий канюк, как и другие виды канюков,
миофаг. Питание его в заповеднике изучалось по составу погадок, соб-
ранных под гнездами, а отчасти по остаткам добычи и анализу желудков
добытых птиц; эту работу вели в 1938-1941 гг. научные сотрудники
А.А. Насимович и М.И. Владимирская, в 1959-1987 гг. - О.И. Семенов-
Тян-Шанский и др.
В табл. 4 не включены объекты, попавшие в погадки случайно или
находившиеся в желудках жертв, как то: семена из ягод, насекомые,
70
Таблица 4. Состав пищи мохноногого канюка
Вид добычи 1938-1941 гг. Число особей 1959-1987 гг. Число особей Состав пищи, %о Суммар- H 0 "%о
Млекопитающие — — ~ — ' 932
Насекомоядные (земле- ройки) — 2 2 2
Грызуны — — — 929
Полевки* 83 19 107 —
Лесные полевки 155 437 621 —
Серые полевки 34 112 153 —
Водяная крыса 7 — 7 —
Норвежский лемминг 14. 15 31 —
Лесной лемминг 1 8 10 —
Хищные (ласка) — 1 1 1
Птицы* (не определенные) 12 12 25 64
Тетеревиные** 8 6 15 —
Кулики — 1 1 —
Певчие 5 17 23 —
Пресмыкающиеся — — — 3
Живородящая ящерица 2 — 2 —
Гадюка — 1 1 1
Земноводные (лягушка травяная) — 1 1 1
Всего особей 321 632 953 1000%о
Всего погадок 215 367 582
*Ближе не определено.
**Большей частью птенцы.
собственные перья хищника, скорлупа собственных яиц. Соотношение
отдельных видов полевок в пище зимняка, вычисленное по всему мате-
риалу, следующее (в %):
Красно-серая полевка 59
Рыжая полевка 14
Красная полевка 2
Темная полевка 9
Экономка. 10
Водяная крыса 1
Норвежский лемминг 4
Лесной лемминг 1
В исследованном нами материале часть полевок определена только до
рода, а в некоторых случаях не указан даже род. Поэтому при составле-
нии таблицы особи, не определенные до вида, разверстаны по видам
пропорционально их численности в точно определенной части пробы. По
всей вероятности, видовой состав полевок в добыче зимняка отражает
количественное соотношение мелких млекопитающих в природе. Срав-
нивая видовой состав проб, полученных из разных гнезд, мы обнаружили
существенные различия между ними во времени и пространстве.
Норвежский и лесной лемминги доступны зимняку только в годы
массового размножения, в нашем материале это 1938, 1974 и 1977 гг.
71
Рис. 15. Зимняк, 29. VII 1963 г. Фото О.И. Семенова-Тян-Шанского
’’Удельный вес” их в лемминговые годы составлял для первого вида
13%, для второго - 2% добычи; в остальные годы он равен нулю. В части
Лапландского заповедника, находящейся к востоку от Чуна-тундры, в
последнее время от загрязнения воздуха промышленными отходами
(главным образом двуокисью серы) деградировали и вымерли эпифитные
лишайники, вследствие чего резко сократилась численность питающихся
ими рыжей и красной полевок и повысился ’’удельный вес” красно-серой
полевки. Анализ погадок зимняка, собранных в разных районах заповед-
ника, показал, что их можно использовать как индикатор видового
состава полевок. Соотношение трех видов лесных полевок, красно-серой,
рыжей и красной, в сборах под гнездами к востоку от Чуна-тундры
оказалось 95:5:0, а в районе к западу от горного хребта, где эпифитные
лишайники, при сравнительной чистоте воздуха, продолжает расти, оно
равно 71:26:3 (сборы относятся к 1959-1982 гг.) (рис. 15).
Зимняк высматривает добычу, медленно летя против ветра или кружа
на неподвижных крыльях; иногда он задерживается на месте, трепеща
крыльями и опустив хвост, подобно пустельге. Кроме полевок, он ловит
и птиц, например птенцов дроздов, куропаток и глухарей, изредка и
72
других мелких животных, землероек, ящериц и лягушек. Роль птиц в его
рационе увеличивается в ’’неурожайные” на полевок годы (в такие годы
он редко гнездится). После ’’пика” полевок в 1950 г. наступила резкая
депрессия по численности. 23 июня 1951 г. нашли под Курт-варенчем
гнездо зимняка с 3 пуховыми птенцами; в нем лежал свежий труп крас-
но-серой полевки; 29. VI найден труп глухаренка массой 39 г; 2. VII в
гнезде оставался один птенец4, 11. VII в гнезде и под ним найдены крас-
но-серая полевка, лягушка, птенец певчего дрозда и перья взрослой
куропатки.
В наших материалах по питанию мохноногого канюка остатки белых
куропаток встречены 8 раз, тундряной - 1 раз, глухарят - 4 раза, 1. X
1938 г. плотники, строившие избу при устье Сылпуая, подняли с раскле-
ванного, еще теплого трупа куропатки двух хищников: вероятно, это
были молодые зимняки, так как летели они с ’’писком”. В августе 1981 г.
на оз. Ташким лесники заповедника видели, как канюк поднял на
воздух, а потом уронил куропатку. В обоих случаях правильность
определения хищников остается под сомнением. В июле 1969 г. двое
юннатов наблюдали, как зимняк, летевший на высоте около 3 м, сделал
бросок и поймал зверька, по-видимому лемминга. В августе 1986 г. под
гнездом зимняка вместе с погадками найден чисто обклеванный скелет
крупной гадюки.
Канюки нередко конфликтуют с другими хищными птицами: как
правило, они преследуют более крупных птиц (орлан, филин), а сами
подвергаются атакам птиц меньшего роста - дербника, пустельги, крачек
и др.
Зимняк сравнительно редко строит новое гнездо (11% всех занятых
гнезд), а чаще занимает старое гнездо другого хищника или вброна.
Обычно оно строится на толстых ветках, примыкая к стволу, на высоте от
6 до 15 м, предпочтительно на сосне (46% гнезд), реже - на елке (32%);
дерево может быть сухим (39%), засыхающим (11%) или здоровым, зеле-
ным (28%). Иногда гнездо располагается на толстом суку в отдалении от
ствола (21% всех гнезд). Диаметр гнезд колеблется от 50 до 120 см в
зависимости от того, кем оно построено и сколь давно. Новое, только что
построенное зимняком, гнездо всегда меньше старого. Строится оно из
сухих веток. Лоток несколько углублен, устлан стеблями злаков и
эпифитными лишайниками с добавлением нескольких зеленых веток
сосны, ели, иногда плауна. Иногда зимняк гнездится в камнях, на карни-
зе скалы или в нише (22% гнезд), но не выше верхнего предела леса; в
гольцовом поясе мы не встречали гнезд. Помимо зимняка, на уступах
скал охотно гнездятся и другие птицы, в частности кречет, пустельга и
вброн; на удобных ’’полочках” всегда можно найти остатки старых гнезд
этих птиц и остатки их добычи.
Как правило, мохноногий канюк гнездится только в ’’мышиные” годы,
а в промежутках между ними старые гнезда по 2-3 сезона пустуют. Из 45
сезонов наблюдений зимняки гнездились только в 19. Даты встреч гнезд с
яйцами варьируют от 10 мая до 27 июня, в среднем 3 июня; пуховых
птенцов встречали с 25 мая по 15 июля, в среднем 27 июня, а оперенных
или полуоперенных - с 7 по 29 июля, в среднем 17 июля; вылет птенцов
4Возможно, что старший птенец убил и съел младших, но доказать это не удалось,
так как собранные под гнездом погадки не были исследованы.
73
Рис. 16. Гнездо зимняка ”на отлете”. Фото О.И. Семенова-Тян-Шанского
наблюдался, в среднем за 9 лет, 29 июля (20. VII 1962 - 9. VIII 1970 г.). По
литературным указаниям, мохноногий канюк насиживает 28-31 день, и
птенцы вылетают в возрасте 41 дня ( Harrison, 1975). Можно считать, что
зимняки у нас несут яйца около 15-20 мая, птенцы выводятся 15-20
июня и вылетают 29 июля, с отклонениями ± 10 дней.
Птенцов в гнезде от 1 до 5, с среднем по 27 гнездам 3,0. Систематичес-
ких наблюдений за гнездами мы не проводили и можем только прибли-
зительно определить успех размножения.Две кладки из 31 были брошены
или погибли. В момент находки в 20 гнездах учтено 8 яиц и 57 птенцов
(всего 65); из 2 яиц птенцов не вывелось, 7 молодых погибли в гнездах, 54
74
прослежены до или почти до вылета. Отсюда эмбриональная смертность
не менее 4%, детская - не менее 12%.
Мы уже упоминали о предположительной гибели двух птенцов от
каннибализма в голодном 1951 г. В бедном полевками 1969 г. в том же
районе 27 июня обнаружено гнездо с яйцами, 9 VII в нем были три пухо-
вичка, 18. VII в гнезде оставался один, а два лежали на земле мертвыми.
Оперенные птенцы иногда оставляют гнездо преждевременно, когда они
еще не могут взлететь с земли и в это время легко становятся жертвой
хищника, что подтверждено и прямыми наблюдениями: 31. VII 1969 г. и
15. VII 1982 г. в ельниках под Чуна-тундрой и 1. X 1987 г. при устье р. Чуна
обнаружены свежие остатки молодых зимняков, пойманных другой
хищной птицей, а 23. IX 1982 г. на склоне Нявка-тундры нашли перья
слетка и 11 его погадок - доказательство, что родители кормили его
несколько дней на земле, а потом его нашел и сожрал, вероятно, мед-
ведь; 3. X 1982 г. на Охтозере и 11. VIII 1983 г. близ Чунозера найдены
остатки слетков, пойманных куницей или лисой.
Взрослые зимняки наряду с ястребиными совами чаще других хищных
птиц становятся жертвами беспутных охотников: двух подстреленных
зимняков мы нашли на берегах Чунозера в сентябре 1968 и в октябре
1981 г., к ним можно добавить и окольцованных, добытых в Греции,
Австрии и Венгрии. Цельные трупы молодых зимняков находили также
в сентябре 1971 г. близ Чингльс-явра и в июне 1980 г. на Чуна-тундре;
последний погиб, вероятно, еще осенью от голода.
Зимняк - хищник среднего размера, покрупнее обыкновенного
канюка и осоеда. Размеры 4 добытых нами экземпляров: размах крыльев
124-143 см, длина тела 565-587 мм, крыла 373-450, хвоста 220-250 мм,
масса тела самцов 707 -896 г, самок 990, ИЗО и 1506 г.
Зимняк довольно крикливая птица, особенно у гнезда; его голос
жалобный, протяжный (1-2 с), как у всех канюков, - их и назвали так по
характерному просящему крику.
Орел беркут - Aguila chrysaetos (L.)
Местные названия: саамские ’’коацкемь” или ’’вуамь коац-
кемь” (большой орел), применяется и. к орлу, и к орлану.
Беркут, крупнейший из орлов, распространен по всей таежной терри-
тории Европы, Азии и Америки; южнее встречается только в горах. По
сведениям Плеске, в прошлом веке он гнездился в Финляндии и Норве-
гии к северу до оз. Инари и р. Тана (70° с. ш.). На Кольском полуострове,
по-видимому, не выходит за пределы лесной зоны. В Лапландском
заповеднике редок, но встречается и гнездится довольно регулярно: за
44 года учтены 94 встречи, обнаружены 4 гнезда, добыты или найдены
мертвыми 4 экз. Численность довольно стабильна: на одного наблюдателя
в год приходилось в 30-х годах по 1,5 особи, в 40-х по 2,8, в 50-х по 1,1, в
60-х и 70-х по 1,2, в 80-х по 1,5 птицы в год. Повышение встречаемости в
сороковых годах, вероятно, зависело от благоприятных кормовых
условий: в те годы на территории заповедника было обилие тетеревиных
птиц и зимовало много диких оленей. Когда добыча легко доступна, орел
живет у нас оседло, но при недостатке корма откочевывает на зиму,
возвращаясь в апреле. Вероятность встреч в светлую половину года по
0,3 особи в месяц, в темную - только 0,05.
75
Таблица 5. Питание орла беркута
Вид добычи Количество особей ; Соотно- шение ВИДОВ, %о
под гнездом в погад- ках полевые наблюдения
Млекопитающие:
Заяц 5 3 1 7
Белка 1 5 — 5
Ондатра — 1 — 1
Полевки и лемминги — 9 — 7
Лиса 4 3 — 6
Куница 2 2 — 3
Норка американская 2 — — 2
Горностай 1 1 — 2
Олень-пыжик* 3 2 1 5
Олень взрослый* 1 4 9 11
Лось взрослый* — — 1 1
Птицы, не определенные 1 4 — 4
точнее
Гусь 1 — 1 2
Утки (разные виды) 2 3 — 4
Глухарь 25 1 3 24
Куропатка 8 6 1 12
Ястребиная сова — 2 — 2
Ворон — — 1 1
Скорлупа яиц — (2) — —
Рыбы—щука — 1 — 1
Количество объектов 56 47 18 100%
пищи
*Падаль.
Сильные лапы орла, вооруженные мощными когтями, дают ему воз-
’можность ловить самых крупных птиц и зверей, масса тела которых
превышает его собственную. Известно, что орлы в отличие от соколов и
ястребов ловят птиц не на лету, а хватают на земле, так же как и зверей.
Состав пищи орла в районе заповедника впервые описан по остаткам
его жертв (44 образца) и погадкам (17), собранным в 1939 г. под гнездом в
долине р. Витте (Владимирская, 1948, с. 195). Новые материалы найдены
под тем же гнездом (1946) и под двумя другими (1959, 1962), а также при
наблюдениях за охотой орла: всего исследовано 74 остатка добычи и 25
погадок (табл. 5).
Прямые наблюдения за хищными птицами обычно ведутся при пита-
нии падалью и при охоте на крупных птиц, а ловля млекопитающих
редко замечается; добыча, принесенная в гнездо целиком, состоит из
сравнительно крупных зверей и птиц, но мелких млекопитающих (и
птиц?) орел заглатывает целиком, и остатки их обнаруживаются только в
погадках. Крупную добычу хищник предварительно ощипывает от
перьев, потом рвет масо по кусочкам, оставляя почти весь костяк,
поэтому в погадках можно обнаружить только единичные перья, волосы
и осколки костей. Таким образом, крупный объект (например, лисица)
регистрируется по меньшей мере дважды: в виде костяка и непереварен-
ных остатков в погадках.
Орлы, как и орланы, питаются главным образом зимой и по этой при-
чине нередко попадаются в капканы, поставленные на волка, лису или
росомаху.
76
Взаимоотношения между орлом и такими хищниками, как лиса и ку-
ница, двойственные: с одной стороны, охотясь на те же самые виды дичи,
на зайцев, куропаток и глухарей, они являются прямыми конкурентами,
с другой стороны, орел включает этих хищников в число своих жертв.
Наши наблюдения показывают, что в пище орла представители отряда
хищных мало уступают по значению другим отрядам млекопитающих -
грызунам и копытным.
Под гнездами орлов мы находили не только черепа, но и почти полные
костяки зайцев, лис и глухарей: видимо, птицы приносят их на гнездо
целиком; однако от оленьих телят мы находили только кости ног. Мас-
са тела взрослого зайца в Мурманской области в пределах 3,0-3,8 кг,
лисы - 3,6-5,8, глухаря самца - 2,5-4,6 (в зависимости от возраста),
новорожденных оленьих телят - 5-7 кг (Семенов-Тян-Шанский, 1959,
1982). Несомненно, что орел убивает животных даже крупнее самого
себя, но, способен ли он лететь с таким грузом, неизвестно. Возможно,
что в гнездо он приносит только остатки жертвы, а часть съедает на месте
добычи.
Приведем несколько наблюдений, иллюстрирующих приемы охоты
орла. В сентябре 1963 г. И. Паракецов был свидетелем нападения орла на
пару лебедей, севших на оз. Куропачье. После нескольких атак лебеди
поднялись и улетели. В октябре 1981 г. А. Гилязов видел, как орел,
летевший над вершинами березняка на старой гари, спикировал под
елку. Оттуда с криком и сильным треском вылетел самец куропатки,
поднялся высоко над лесом и скрылся из виду. Орел его не преследовал
и полетел дальше. В апреле 1936 г. на вараке близ устья р. Чуна О. Семе-
нов-Тян-Шанский нашел (около 17 ч) еще теплые остатки взрослого самца
глухаря. По следам на снегу было видно, что преследуемый хищником
глухарь сел на снег и забежал под елку, ветви которой спускались до
земли; хищник также сел, пробежал по снегу, сквозь ветки бросился на
глухаря, вырвав хвост и другие перья. Потом обе птицы выскочили на
поверхность наста, хищник убил глухаря, съел мясо с шеи, спины и
одной ноги (от таза до плюсны), сердце, обклевал копчик и плечевые
кости. Судя по форме и размерам следа (15-18 см), а также количеству
мяса, съеденного за один прием, это был не ястреб и не филин, а именно
орел. В августе 1935 г. лесник П. Корнилов на берегу оз. Мерьт-яурынч
нашел остатки взрослого гуся гуменника, ощипанного и объеденного
сходным образом. В апреле 1951 г. на льду р. Чуна О. Семенов-Тян-Шан-
ский обнаружил занесенные снегом остатки глухарки - клюв, перья и
сосновую хвою из зоба. Она была поймана хищной птицей, которая съела
на месте только голову и шею, а большую часть туши унесла в когтях.
Двумя неделями раньше в том же районе орел попался пальцем в волчий
капкан, но сумел освободиться: вероятно, он и поймал глухарку.
25 апреля 1959 г. в 15 ч близ Гольцового озера, на еловой гари О. Семенов-Тян-
Шанский и П. Тютин подняли сидевшего на снегу орла: он клевал оленью голову. Эту
голову вырыла из-под занесенной снегом валежины лиса, шронесла сотню метров, в
этот момент на нее напал орел. Снег сохранил следы драки — отпечатки крыльев орла
по обе стороны от следов лисицы; она отказалась от дальнейшей борьбы, уступила
добычу орлу и спаслась бегством, сначала скачками, потом рысью (рис. 17).
77
Рис. 17. Следы нападения орла на лису, 25. IV 1959 г. Фото О.И. Семенова-Тян-Шан-
ского
Орлов, клюющих падаль, наблюдали в апреле 1941 г. на трупах оленей, задранных
волками (дважды), и в августе 1947 г. на лосе, добытом медведем. При этом лесник
А. Сергин видел в бинокль, как орел сел на снег недалеко от туши и подошел к ней
"пешком”.
В июле 1972 г. на Сальной тудре О. Семенов-Тян-Шанский нашел труп двухмесяч-
ного олененка, поклеванного орлом (определен по выпавшему перу). В отличие от
ворона, в первую очередь съедающего глаза и внутренности, орел ел только мясо.
20.IV 1941 г. мы наблюдали брачную игру орлов над южным склоном Чуна-
тундры. Сперва они летали вместе, потом один спустился и исчез в лесу; второй
сложил крылья и спикировал к нему под углом 45° и только в лесу расправил
крылья; тут же из-под ели вылетел и сел на берегу озера первый орел.
На том же месте 30.III 1986 г. А. Гилязов и В. Курганский встретили пару орлов и
нашли построенное ими гнездо. Оно помещалось на "ведьминой метле”, на толстой
ветке сосны, в 2 м от ствола и в 10 м над землей. Потом орлов видели или слышали у
гнезда 11.V и 5.VII, но никаких признаков птенцов не было, а под ним, кроме кос-
тей глухаря, никаких остатков не обнаружено. Попытка гнездования оказалась
неудачной (рис. 18).
14.VI 1939 г. лесники М. Тютин и М. Малодушен нашли на большой сосне, в сме-
шанном лесу близ Мясного озера (долина р. Витте) занятое гнездо орла — огромная
старая постройка, опиравшаяся на толстые ветви и примыкавшая к стволу. Диаметр
гнезда 1,2—1,4 м, толщина 0,9 м, лоток плоский, на высоте 14 м над землей. В гнез-
де находились два начавших оперяться птенца, но уже 17.VI оставался только один,
остатки другого лежали под гнездом, впоследствии погиб и он. До 1949 г. орлы не
селились в этом гнезде, но посещали его неоднократно: весной 1940 и 1941 гг. на сне-
гу под ним оказались выпавшие орлиные перья, а в 1946 г. собраны остатки добычи.
26.VII 1959 г. научный сотрудник заповедника Ю. Гаев у берега Гольцового озера
нашел на сосне гнездо с одним молодым орлом, слетком, который был окольцован.
На следующее лето это гнездо стало разваливаться, но ближе к озеру было постро-
ено, а потом брошено второе.
16.VII 1962 г. лесник Я. Януше нашел в 4 км от северо-западного берега Верхнего
Пиренгского озера, в густом смешанном лесу на берегу ручья орлиное гнездо, по-
строенное, как и предыдущее, на большой сосне, сбоку от ствола, на высоте 16 м. На
78
Рис. 18. Гнездо орла в долине р. Витте. Фото О.И. Семенова-Тян-Шанского
краю гнезда сидел птенец, почти полностью оперившийся. Погадок под гнездом не
оказалось, собраны только кости 6 глухарей (3 самца и 3 самки) и половина тушки
норки. Взрослые птицы не появлялись. До 1975 г. гнездо не занималось орлами и
начало разваливаться.
Судя по встречам сеголеток, орлы гнездились в районе заповедника
также в 1931,1971 и 1979 гг.
По-видимому, беркуты выводят только одного птенца, даже если
самка отложит яйца (второй птенец в гнезде 1939 г., вероятно, был убит
или сброшен на землю старшим). Молодые оставляют гнездо в конце
июля, но еще месяца два не могут самостоятельно жить и в это время
79
нередко погибают. 23.IX 1931 г. директор заповедника Г.М. Крепе застре-
лил на берегу р. Нижняя Чуна молодого самца, шкурку которого передал
в ЗИН АН СССР. 23.IX 1979 г. на Чунозере в воде найден несвежий труп
молодой самки; она была истощена и, вероятно, погибла от голода. При
попытках истребления волков и росомах капканами, поставленными у
падали, зимой 1940/41 гг. были пойманы два орла, оба самца (16.11 и
22. IV). Еще один беркут попал в капкан в начале апреля 1951 г., но выр-
вался, оставив коготь. Охотник-промысловик Н.А. Михайлов, ловивший
лис капканами близ оз. Куропта в конце 1960 г., поймал двух орлов.
Несомненно, капканный промысел наносит большой урон популяции
орлов. Чтобы обеспечить эффективную охрану этого редкого хищника,
занесенного в Красную книгу СССР, необходимо разработать и внедрить
жесткие правила установки капканов, предотвращающие возможность
попадания в них хищных птиц - орла и орлана.
Размеры орлов в Лапландии: размах крыльев 2 самцов - 199 и 204 см,
молодой самки - 190; длина тела соответственно - 87, 83 и 89 см; крыла
3 самцов - 580, 615, 603 и 1 самки - 641 мм, длина хвоста самцов - 355,
340, 325, самки - 350 мм. Масса тела упитанного самца, пойманного в
феврале 1941 г., 3692 г, истощенной самки в сентябре 1979 г. - 3250 г.
След орла крупнее следов других хищных птиц, на песчаном грунте он
достигает 15-18 см в длину и И см в ширину; внешний палец характерно
оттопырен. Шаг широкий, расстояние между отпечатками той же лапы
75- 85 см. Орел чаще и охотнее ходит, чем другие хищники, он и добычу
иногда ловит ’’пешком”.
Крик напоминает высокий, отрывистый лай собаки ”кья. . . кья...
кья. . . кья. . .” с интервалом около 1 с. Орлы очень осторожны и
гнездо свое не обороняют.
Полевой лунь - Circus cyaneus (L.)
В бассейне оз. Имандра полевой лунь - редкая залетная птица: 17.VII
1947 г. близ Полярно-альпийского ботанического сада (г. Кировск)
А.А. Насимович видел белого луня, охотившегося на лугу и по кустам
около ручья, а 9.VI 1978 г. Г.Д. Катаев наблюдал другого луня в Лаплан-
дском заповеднике под юго-восточным склоном Чуна-тундры. Залетных
луней встречали и в других местах Кольского полуострова, вплоть до
Айновых островов на север (19.V 1962 г.; Коханов, Скокова, 1967), с. Пя-
лица на востоке (июнь-июль 1961 г.; Малышевский, 1963, с.475).
Ближайшие находки гнезд полевого луня в окрестностях Муонио и
Торнио, у границы Финской и Шведской Лапландии (Плеске, 1887, с. 304)
находятся в 350-450 км к западу и юго-востоку от Лапландского заповед-
ника. Приведенные факты не дают права включать в ареал луня всю
южную часть Кольского полуострова, как это сделали Г.П. Дементьев
(1951, с. 200) и В.Е. Флинт (1968, с. 156).
При пользовании опросными сведениями необходимо иметь в виду,
что местное население (например, на Поное) лунем называет белую сову.
80
Пустельга - Cerchneis tinnunculus (L.)
Пустельга широко распространена по всему Старому Свету, кроме
Крайнего Севера; только на Скандинавском и Кольском полуостровах ее
ареал заходит к северу за Полярный круг. На Кольском полуострове ее
впервые наблюдал в июне 1883 г. в Хибинах Энвальд (Плеске, 1887,
с. 327).
В Лапландском заповеднике она появляется не ежегодно, из 44 сезо-
нов наблюдений на протяжении 16 ее совсем не встречали, в течение 19
попадались одиночные залетные птицы, и только в 18 сезонах наблюда-
лись’’территориальные” пустельги, обычно парами: всего зарегистриро-
вано 118 особей.
Появление и частота встреч в нашем районе тесно связаны с циклич-
ностью мелких грызунов (рис. 19). В годы депрессии полевок пустельга
либо совсем не появляется, либо только залетает; пары занимают терри-
тории и гнездятся либо в год ’’пика” (все годы до 1975), либо в следую-
щий за ’’пиком” год (1976-1987 гг.). Коэффициент корреляции между
индексом численности полевок и количеством встреч пустельги в том
же году+0,62±0,18 (26 лет; 1936-1975 гг.); между индексом полевок и
встречами пустельги в следующем году составил +0,76±0,12 (12 лет,
1976-1987 гг.). Такая же зависимость гнездования пустельги от числен-
ности полевок и в Кандалакшском заповеднике (Бианки, Бойко, 1985).
Достоверно пустельга гнездилась в Лапландском заповеднике 5 раз:
1938 г. — 10.VIII в ельнике под скалистым обрывом Второго Ельнюна встречены
две молодые птицы, близко подпустившие наблюдателя; 9.IX здесь же летали вместе
3 пустельги;
1962 г. — 12.VIII под Сейда-пахтой найден труп слетка-самца;
1963 г. — 28.V в смешанном лесу в 1,5 км к ЮВ от усадьбы заповедника Ю.Г. Га-
ев нашел гнездо пустельги с 5 яйцами. Гнездо старое, построенное на сосне на вы-г
соте 8 м. При проверке гнезда 26.VI птица слетела с него, в лотке оказалось только
одно яйцо (судьба его не прослежена).
1978 г. - 10.VIII на южном склоне Сейднотчорра летал выводок, три птицы вместе.
1979 г. - 11.VIII в долине р. Чуна выше бывшего лесоучастка А.Б. Брагин встре-
тил плохо летавшего слетка, сидевшего на елке вместе со взрослой пустельгой.
Территориальные пары наблюдались на определенных местах, где
пустельга, вероятно, гнездилась: в 1938, 1941, 1950, 1959 и 1976 гг. - на
Ельнюне, юго-восточной оконечности Чуна-тундры; в 1962, 1970, 1978, 1982
и 1983 гг. - на южном склоне Чуна-тундры; в 1937, 1938 и 1970 гг. - в рай-
оне Нявка-тундры, а также в сосновых лесах, в долинах Сылпуая (1969),
р. Купись (1974). Чуны (1973, 1979) и на берегу Чунозера (гнездо 1963 г.).
Охотится пустельга чаще всего на горных тундрах, а если в лесу, то
предпочтительно в сосновом.
Пустельга прилетает к нам позже остальных хищных птиц, в среднем
за 16 лет 26 мая (8.V 1967 г. - 15.VI 1960 г.; средняя температура +0,6°),
соответственно рано и отлетает - около 29 августа (13.VIII 1986 г. - 1.Х
1938 г., при температуре +9,4°). Время прилета и отлета отмечалось, ве-
роятно, недостаточно точно, так как пустельга сравнительно редко
попадается на глаза. Молодые вылетают из гнезд около 10-12 августа
(3 наблюдения).
6. Зак. 1023
81
1935 1990 1960 1965 1970 1975 1980 1985
Рис. 19. Число встреч пустельги за год (сплошная линия) и индекс учета полевок
осенью (пунктир)
Зависимость пустельги от циклов мелких грызунов показывает, что в
Мурманской области она является по преимуществу миофагом; дейст-
вительно, в желудке слетка, найденного мертвым в августе 1962 г., ока-
зались остатки лесной полевки и трех жуков.
Дербник - Aesalon columbarius (L.)
Местное название: саамское ’’еЪткынч”.
Дербник распространен циркумполярно, гнездится в таежной зоне, а
на зиму отлетает в южные области Европы и Азии. На Кольском полуострове
это обычная хищная птица, уступающая по численности только зимняку.
Дербник прилетает к нам в одно время с коньками, но после дроздов и
овсянок, когда снег наполовину стаял, в среднем 17 мая (25.1 V 1978 г. -
6.VI 1950 г., при температуре +0,4°); отлетает одновременно с пеночками,
варакушками и другими певчими птичками, до начала листопада, около
7 сентября (15.VIII 1964 г. - 22.Х 1934 г., температура +7,9°). Дербник про-
водит у нас около 114 дней (от 85 дней в 1964 г. до 134 в 1974 г.).
Не считая повторных наблюдений у гнезд, за 44 года зарегистрировано
269 визуальных встреч. Численность дербника колеблется по годам,
повышаясь через каждые 3-6 лет, в зависимости от колебаний числен-
ности полевок: годы частых встреч и находок гнезд - 1933, 1938, 1950,
1958, 1963, 1969, 1977, 1986 и 1987. Независимо от периодических флукту-
аций численность дербника за полвека уменьшилась вдвое. Число ви-
зуальных встреч дербника на 1 наблюдателя в год составляло в 30-х го-
дах - 5,1, в 40-х - 2,0, 50-х - 3,3, 60-х - 2,5, в 70-х - 2,6, в 80-х - 2,5.
О питании дербника в Лапландии приходится судить по случайным
наблюдениям: в двух желудках обнаружены остатки полевок, в третьем -
стрекозы. В августе 1941 г. мы наблюдали, как дербник, сделав бросок,
схватил летящую стрекозу. В гнезде в июле 1963 г. найдены остатки
дрозда, а в мае 1971 г. на месте охоты дербника обнаружены перья пой-
82
манного им конька. В августе 1977 г. пара дербников, а в августе 1960 г. -
один охотились на белых трясогузок над р. Чуна. В августе 1932 г. дерб-
ник безуспешно гонялся за желной, лавировавшей между деревьями.
14 июня 1974 г. в 6 ч утра дербник дважды нападал на сидевшую на гнез-
де глухарку: наседка вскочила с взъерошенным оперением, отбила атаки
и через 14 мин уселась снова на гнездо. Глухарка в 8-10 раз больше
дербника и никак не могла стать его добычей; возможно, он напал на нее,
как на воображаемого конкурента (его гнездо находилось не далее 200 м
от гнезда глухарки), а скорее всего, хищник намеревался сбить глухарку
с выводка и схватить одного из птенцов; обнаружив, что под глухаркой
были не цыплята, а яйца, он оставил ее в покое.
По-видимому, дербник довольно универсальный хищник, но птиц
предпочитает другой добыче; ловит он их на открытых местах - в гор-
ной тундре, на старых гарях, болотах, по берегам озер и рек.
За все годы найдено 15 гнезд дербника. Обычно он занимает старое
гнездо другого хищника, построенное на большой сосне в верхней части
кроны, на высоте 10-18 м, в негустом бору или смешанном лесу (80%);
реже гнездится на земле (13%) или в скалах (7%).
26.VI 1933 г. на бровке юго-восточного склона хр. Райкорчорр, у границы леса
(380 м над ур. м.) из зарослей карликовой березки вылетел дербник: на земле под
кустами оказалось его гнездо в виде ямки без подстилки с 5 яйцами; 6.VII в гнезде
4 птенца, одетые белым пухом, и одно наклюнутое яйцо; 17.VII все 5 птенцов были в
сером втором пуховом наряде, у них начался рост маховых и рулевых.
22.VI 1938 г. пара дербников вылетела из щели в скале на вершине Пель-тундры
(590 м над ур. м.), но добраться до гнезда нам не удалось. В тот же день в елово-
березовом редколесье на южном склоне Пель-тундры, на земле, под еловой вале-
жиной, найдено гнездо другой пары: на подстилке из сухих еловых веточек и бере-
зовых листьев кладка из 4 яиц.
2.VIII 1938 г. в бору, почвенный покров которого выгорел 25 лет назад, в долине
Сылпуая нашли 3 оперенных птенцов дербника, сидевших кучкой под сосной. От
человека они уходили прыжками или перепархивая, а один пытался влезть на сосну,
цепляясь за кору когтями и крыльями. Один из птенцов, взятых на воспитание,
10.VIII уже летал.
25.V 1962 г. на сосне у края болота в 0,5 км к западу от усадьбы заповедника на
Чунозере Ю.Г. Гаев нашел гнездо дербника. 3.VI самка насиживала, 6.VII в гнезде
2 пуховых птенца в возрасте около 10 дней; 17.VII оба окольцованы. 31.V 1963 г. то
же гнездо снова занято, в нем 4 яйца; 19.VII — 3 слетка (первый, возможно, вылетел
раньше ?), все спланировали на землю; двое разысканы и окольцованы.
20.VI 1967 г. — на вырубке между озером Улынчъявр и Беличьей избушкой гнездо
на вершине сосны; в нем А.И. Флеровым 16. VII обнаружены 5 испорченных яиц.
22. V 1974 г. пара дербников заняла старое гнездо на сосне в 13—15 м над землей, на
сосновой вараке при устье р. Чуна; лоток еще пуст. 9.VI в гнезде полная кладка из
5 яиц, самка слетела, подпустив человека на 1,5 м; 24-25.VI гнездо, начавшее раз-
валиваться, брошено, в нем 4 насиженных яйца (С. Баккал).
22.VII 1975 г. у гнезда в бору при устье р. Мавра летали два молодых дербника.
Пара дербников вновь прилетела к этому гнезду 25.IV 1978 г.
14.VII 1977 г. в бору у края лесосеки, между реками Чуна и Нявка, почти у верши-
ны высокой сосны на боковой ветке, в 18 м над землей и 1,5 м от ствола С. Баккал
нашел гнездо; пара взрослых дербников летала вокруг.
29.V 1986 г. в бору оз. Улынчъявр дербниками занято старое гнездо ворона на
сосне на высоте 12 м над землей; 16.VII в гнезде птенцы.
22.V 1987 г. пара дербников заняла старое воронье гнездо в смешанном лесу близ
Чунозерской усадьбы заповедника; 27.V в нем 3 яйца, самка насиживала; 9.VIII виде-
ли 2 летных птенцов.
83
Из этих материалов заключаем, что кладка дербника состоит из 3-
5 яиц, в среднем 4,3; среднее число птенцов в гнезде 3, 6. Средняя дата
находки 6 полных кладок - 9 июня (27.V - 26.VI), а гнезд с пуховыми
или оперенными птенцами - 14 июля (2.VII - 2.VIII); птенцы вылетали
между 19.VII и 10.VIII. Принимая срок инкубации равным 28 сут, нахо-
дим, что кладка яиц происходит в последней декаде мая или первой
декаде июня, птенцы выводятся в конце июня - начале июля и вылетают
из гнезд в конце июля - начале августа. Прослежена судьба 6 кладок,
содержавших 27 яиц: вылетели 15 молодых (56%), брошена 1 кладка с
4 яйцами (15%), исчезли 3 яйца и в 5 погибли эмбрионы (11 и 18%).
У гнезда дербник довольно осторожен, перелетает с беспокойным
криком, присаживаясь ненадолго на деревья, но на человека не налетает.
Дербники часто вступают в конфликт с другими птицами. Нами отме-
чены нападения дербника на ворона (2 раза), ворону (4 раза), сороку,
зимняка, бородатую неясыть и одну из чаек. С другой стороны, дербника
преследовали золотистая ржанка, полярные крачки (3 раза), свиристели,
чечетки и белые трясогузки (3 раза); ястребиная сова и дербник нападают
друг на друга взаимно. В добыче других хищных птиц дербник нам не
попадался, но в июне 1978 г. в ельнике под Волчьей тундрой найдены
перья дербника, вероятно съеденного хищником.
С 1933 по 1977 г. окольцовано 16 птенцов и получен 1 возврат: дербник,
окольцованный в Лапландском заповеднике 19.VII 1963 г., убит 26.Х того
же года на зимовке во Франции, близ Авиньона, в 3100 км к юго-западу
от места рождения.
Размеры взрослого самца, добытого в июне 1938 г.: длина 285, крыло
223, хвост 122 мм, масса тела 168 г; размеры 2 молодых самок, добытых в
июле-августе, соответственно - 315-320, 200-213 и 125-132 мм, масса
тела 218 и 228 г.
Кречет - Falco gyrfalco L.
Местные названия: саамское (по Плеске) ’’валль” или ”фалли”.
Кречет распространен в тундре, лесотундре и северной тайге обоих
полушарий, но по всему обширному ареалу - одна из самых редких
хищных птиц. В русской Лапландии может встретиться всюду, особенно
по Мурманскому побережью. В Лапландском заповеднике за 44 года
зарегистрирован визуально 81 кречет, но неравномерно, на протяжении
19 лет его совсем не видели. Встречается он как летом, так и зимой (при-
мерно в два раза реже, чем летом), поэтому его можно считать условно
оседлым. На год приходится в среднем по 1,8 встречи. Заметной тенден-
ции к росту или сокращению численности не замечено. Гнездование
наблюдалось нами четыре раза:
5.V 1938 г. в смешанном лесу на СВ побережье Чунозера, в 1 км к юго-востоку от-
усадьбы заповедника найдено гнездо кречета с насиживающей самкой. Гнездо старое,
диаметр его 1 м, высота до 1,5 м, построено на сосне на высоте 6-7 м, с южной сто-
роны ствола на толстых сучьях. 21 июня в нем два больших, но еще одетых пухом
птенца, к 25 июля они покинули гнездо (Владимирская, 1948, с. 188—189).
4.VI 1975 г. на уступе скалы на северо-восточном склоне Ельнюна Второго, в 1 км
84
от усадьбы заповедника к ЗСЗ Г.Д. Катаев обнаружил гнездо кречета, сложенное из
сухих сучьев; диаметр его 0,5—0,6 м; в прошлом гнездо, вероятно, принадлежало
зимняку. В нем два птенца (19.VI они были в беловатом втором пуховом наряде), а
позднее под гнездом найдены остатки ненасиженного яйца, следовательно, кладка
состояла из 3 яиц. По белым потекам помета на отвесной скале под гнездом оно
видно издалека. Взрослые кречеты то сидели на ближних деревцах или камнях, то
летали за кормом, присаживаясь на гнездо для передачи его птенцам на несколько
секунд. К человеку подлетали не ближе 30 м; 30.VI птенцы начали оперяться
(рис. 20) глаза в темных кольцах. Взрослые приносили им предварительно ощипан-
ных птиц, а мясо птенцы рвали сами; большую часть времени они лежали на лотке и
снизу не были видимы; за 4 ч кормились 3 раза. При попытке подняться к гнезду по
скале кречет несколько раз пикировал на человека. 7.VII А.Б. Брагин окольцевал
85
Рис. 21. Кречет у гнезда, 7. VII 1985 г. Фото О.И. Семенова-Тян-Шанского
одного птенца, а другой убежал по карнизу и спрятался в щель. 18.VII птенцы сидели
врозь, вне гнезда, а 19.VII их уже не удалось обнаружить, только матка летала
вокруг с беспокойным криком.
15.VII 1986 г. на уступе скалы на южном склоне Сейда-пахты, Чуна-тундра А.С. Ги-
лязов нашел гнездо кречета диаметром 0,6—0,7 м, толщиной 0,3 м; птица только
добавила слой сосновых веточек к старому гнезду. Единственный птенец уже выле-
тел, но оставался поблизости до 30.VII, а к 3.VIII вся семья исчезла. Раньше пару
кречетов наблюдали 23.VI в 7 км, a 1.V - одного, в 8 км от гнезда. К востоку от него,
не далее 10 км, в мае—июне были обнаружены остатки 3 куропаток, пойманных этой
парой. Семью кречетов видели в том же районе еще 4.XI и 7.XII 1986 г.
29.VII 1986 г. на скалистом юго-западном склоне Падун-уайвенча, Нявка-тундра
А. Гилязов и Л. Киренков заметили слетка кречета в сопровождении обеих взрослых
птиц, но не смогли разыскать их гнездо.
Кречет насиживает 35 дней, а птенцы оставляют гнездо в возрасте
46-49 дней (Palmer 1988, с. 396). По нашим наблюдениям, птенцы вылета-
ли около 20 июля, следовательно, яйца были отложены около 1 мая, а
птенцы выводились в первой декаде июня. В 1975 г. кречетов встречали в
районе гнезда еще 20.IV (один), 18.V (пара) и 24.IX (один молодой).
По всей вероятности, кречеты гнездились и в другие годы в южной
части Чуна-тундры и на Нявка-тундре, где их неоднократно встречали
парами (1934,1938,1939,1977 и 1980 гг.).
Под гнездами в 1938,1975 и 1986 гг. были собраны все остатки добычи и
погадки. Часть птиц представлена более или менее полными скелетами,
без головы, с обклеванным мясом, другие были расчленены. Во избежа-
ние повторного учета той же особи мы считали птиц по килям, а остатки
млекопитающих по черепам. Взрослые приносят птенцам ощипанных
птиц, и в их желудки попадает почти чистое мясо, без перьев и костей;
86
Таблица б. Питание кречета (по материалам 3 гнезд и визуальным наблюдениям)
Вид добычи 1938 г 1975, 1986 гг. Состав пищи
остатки погадки (28 шт). остатки погадки, (42 шт.) по остат- кам, % по погад- кам, %
Млекопитающие* — — — —
Грызуны — — — — — —
Полевки лесные и 3 20 — 42 2 51
серые
Лемминг норвеж- 6 6 — — 4 5
ский
Птицы* — 14 — 1 — 12
Водоплавающие — — 1 — 1 —
Чирок 1 — — — 1 —
Гоголь — — 7 — 4 —
Хищные (зимняк?) — — 1 — 1 —
Тетеревиные 1 3 1 — 1 3
Тундряная куропат- 1 — 5 — 4 —
ка
Белая куропатка 34 9 28 1 40 8
Глухарь — 2 11 — 7 2
Тетерев 3 — 10 — 8 —
Кулики — — 2 — 1
Чайки (3 вида) — — 9 — 6 —
Крачка полярная — — 9 13 6 11
Голуби (сизарь) — — 1 — 1 —
Совы (3 вида) 2 — 1 — 2 —
Кукушки и дятлы 2 — — — 1 —
Воробьиные* 5 1 — — 4 1
Дрозды 1 — 2 — 2 —
Кукша 4 — — — 3 —
Ворона серая — — 2 — 1 —
Яичная скорлупа — 1 — — — 1
Рыба** — 3 — — — 2
Насекомые — 5 — — — 4
Всего 63 64 90 57 153 121
*Ближе не определено.
*Возможно, из старых погадок прежнего владельца гнезда.
грызунов, наоборот, они заглатывают целиком. Поэтому подсчет состава
пищи по остаткам добычи, с одной стороны, и по погадкам - с другой,
дает разные результаты (см. табл. 3-6).
Кречеты, гнездящиеся на Мурманском берегу, кормят птенцов глав-
ным образом моевками и тупиками (Шкляревич, Краснов, 1980, с.17-26),
а живущие в тундре и лесотундре охотятся преимущественно на белых
куропаток. В долине р. Колвилл (Аляска) они составляют 91% рациона
кречета (White, Cade, 1971); в долине Утсйоки, Финская Лапландия, - 71%
(Mikkola, Sulkava, 1972); в Мурманских тундрах куропатка также занима-
ет
ет в его добыче первое место (Кищинский, 1958). В Лапландском заповед-
нике в 1938 г. куропатки составляли 56% всей добычи кречета, в 1975 г. -
25%, в 1986 г. - 55%. Кроме куропаток, кречет часто ловит тетерок (на 8
тетерок - 2 взрослых косача) и даже глухарок, масса тела которых
превышает массу тела самого хищника.
В апреле 1941 г. лесник А.С. Сергин в долине Сылпуая спугнул кречета с добычи
еще теплых остатков глухаря самца. Малочисленность тетеревиных в 1975 г. застав-
ляла кречета охотиться на озере. В числе его жертв определены 3 сизые чайки, 1 се-
ребристая, 1 речная, ‘8 полярных крачек и 6 гоголей, из них 1 самец. На болотах он
поймал турухтана и среднего кроншнепа, а близ усадьбы — 2 ворон. В его добыче
преобладали взрослые птицы.
Под гнездом 1938 г. найдены череп мохноногого сыча и лапа воробьиного сыча,
под гнездом 1975 г. - остатки ястребиной совы. В добыче 1986 г. обнаружен домаш-
ний голубь, принесенный, очевидно, из ближайшего города, Мончегорска или Зашей-
ка, расстояние от которых до гнезда не менее 30 км по прямой.
19.XII 1982 г. в редком елово-березовом лесу к югу от Чунозера А.С< Гилязов
поднял кречета из-под ели, где он рвал добычу: он взлетел, держа в правой лапе
остатки тетерки. Судя по следам, он ударил тетерку, когда она кормилась на березе,
упал вместе с нею в снег и скатился по склону под елку.
24.1 1939 г. на Нявка-тундре А.Б. Васильев поднял с поверхности снега кречета,
расклевывавшего тундряную куропатку, остатки которой унес в когтях.
6.Х 1961 г., опускаясь по р. Чуна на лодке, О.И. Семенов-Тян-Шанский услышал
тревожное кряканье и увидел посреди реки гоголя, а под берегом двух крякв:
переполох был вызван кречетом, сидевшим на прибрежной березе, увидев лодку,
кречет улетел.
13.V 1975 г. на Чунозерской усадьбе под сосной нашли костяк левого крыла
годовалой глухарки с частью маховых и грудиной; очевидно, это была жертва
кречета, гнездо которого находилось не далее 1 км от усадьбы. Интересно, что из
остатков крачек, найденных в том же году под гнездом, две сохранили оба крыла, а
у пяти оставались только левые крылья: это свидетельство, что кречет разделывает
добычу по определенному шаблону.
Мы привели здесь только часть наблюдений за охотничьими приемами кречета,
именно те, когда хищника и жертву удалось рассмотреть и определить.
23.XII 1950 г. в 8 ч работники заповедника, на лошади ехавшие по льду Чунозера,
подняли с берега белую сову. На снегу осталась ее добыча — еще теплый труп
взрослой самки кречета; сова успела оторвать и съесть голову, обклевать мясо с
шеи и передней части груди, у вилочки. Желудок кречета пуст, но сама птица упи-
танная, с подкожным жиром. Длина крыла 412, хвоста 236, размах крыльев 1290 мм,
вес оставшейся части 1345 г. Очевидно, что живой вес ее был не менее 1500 г. Скелет
и шкурка поступили в коллекцию ЗИН АН СССР. Обеспечили победу сове два
обстоятельства: инициатива была на ее стороне и нападение произошло в темноте,
едва начало рассветать.
Кречет довольно молчаливая птица; крик хриплый, отрывистый,
напоминает ”кре..или ”кве”, а голос птенцов - ”пэе... пэе..
Сокол-сапсан - Falco peregrinus Tunst.
Сапсан распространен по всему земному шару, но распределен по тер-
ритории очень неравномерно, на Кольском полуострове он встречается
реже кречета. В Лапландском заповеднике сокола впервые достоверно
наблюдали 2 августа 1963 г. над Каменным ущельем на Нявка-тундре: он
3-4 раза с криком пикировал на О.И. Семенова-Тян-Шанского, в этот
момент был сфотографирован, а потом исчез. Судя по поведению, у него
88
Рис. 22. Сапсан пикирует, 2. VIII 1963 г. Фото О.И. Семенова-Тян-Шанского
были птенцы, но разыскать их или гнездо не удалось. На снимке (рис. 22)
отчетливо видны черные ”усы” - признак, отличающий сапсана от кре-
чета.
В начале августа 1967 г. студент А. Печенюк нашел в бору между р. Чу-
ной и оз. Улынчъявр остатки хищной птицы, съеденной лисой; судя по
острой форме и длине крыла (около 30 см), а также окраске кроющих,
белой с серыми поперечными полосами, это был сапсан. Без уверенности
в определении сапсанов встречали еще раз десять - в районе озер Нявка
и Купись, на Нявка-тундре, в долине Чуны и над Чунозером, в разные
годы, с мая по октябрь.
Гнезда сапсана находили только в долине Поноя в 1976 и 1977 гг.
(Редкие и нуждающиеся в охране животные..., 1979, с. 26).
Гнездование сокола в заповеднике вполне вероятно. В отличие от
кречета сапсан из северных областей отлетает на зиму к югу.
Регистрация встреч хищных птиц в заповеднике дает некоторое пред-
ставление об относительной численности видов. Птицы парящего полета,
высматривающие добычу с воздуха, легче замечаются, чем летающие
низко над землей или в лесу, поэтому в подборке мы поместили их в
разные колонки (в %<>):
Птицы парящего полета Птицы активного полета
Зимняк 439 Дербник 86
Орлан 185 Ястреб-тетеревятник 56
Скопа 110 Ястреб-перепелятник 19
Пустельга 38 Кречет 26
Орел беркут 30 Сапсан 4
Осоед 3 Лунь полевой 1
Коршун 3
Итого: 808°/оо Итого: 1927<
ОТРЯД КУРИНЫЕ - GALUFORMES
Пять видов тетеревиных птиц в составе нашей авифауны занимают
особое место как по относительно высокой численности, так и по нали-
чию специальных адаптаций, дающих им возможность пережить без
потерь длинную, холодную и снежную зиму на родине. Летом тетереви-
ные кормятся листьями, цветами и ягодами растений, образующих на-
почвенный покров, и в это время являются наземными птицами. Снеж-
ный покров закрывает этот источник питания на полгода. Глухарь, тете-
рев, рябчик и белая куропатка разрешили проблему зимнего питания
полным изменением кормового режима, довольствуясь легкодоступны-
ми и в это время древесными кормами. Этот природный ресурс широко
используют многие млекопитающие - лось, бобр, заяц-беляк и некото-
рые полевки, но никакая другая птица, кроме тетеревиных, не имеет
необходимых для этого адаптаций. К числу их относится сильный клюв с
режущими роговыми кромками рамфотеки и сильной мускулатурой
нижней челюсти, позволяющими стричь хвою и ветки; объемистый зоб,
вмещающий достаточный запас корма для постепенной переработки его
на протяжении долгой зимней ночи; необычайное развитие слепых
кишок, служащих для длительного усвоения питательных веществ из
концентрированного химуса, после освобождения его от крупных дере-
вянистых частиц, составляющих основную массу зимнего кала. Большую
часть суток тетеревиные проводят, зарывшись в толщу снежного покро-
ва, избегая таким путем потери тепла через излучение и от контакта тела
с холодным воздухом, что обеспечивает экономичное использование
энергетических ресурсов при низких температурах окружающей среды.
Диагностические признаки семейства тетеревиных птиц - ноздри,
прикрытые перьями, оперенная плюсна (а у белых куропаток и пальцы),
роговые выросты на краях пальцев зимой, сильное развитие побочного
опахала на контурных перьях - явно адаптивного происхождения; они
повышают термоизоляционные свойства оперения, облегчают ходьбу по
снегу и его раскапывание, а оперение ноздрей предохраняет дыхатель-
ные пути от попадания снега, когда птица находится в лунке.
Поскольку тетеревиные птицы и летом, и зимой питаются массовыми
растительными кормами, продуктивность которых во много раз превы-
шает потребности консументов, рост популяции лимитируется, очевид-
но, не кормовыми ресурсами, а какими-то другими причинами, влияющи-
90
ми на успех размножения, либо на смертность. Уже в первые годы работы
в Лапландском заповеднике мы заметили, что успех размножения глуха-
ря резко меняется по годам в зависимости от температуры воздуха в
период размножения. Исследование этого вопроса на протяжении не-
скольких десятилетий раскрыло механизм влияния температуры на раз-
множение как глухаря, так и других видов тетеревиных (Семенов-Тян-
Шанский, 1938,1959,1983 и др.).
С 1936 г. в Лапландском заповеднике ежегодно в августе проводится
количественный учет четырех лесных видов тетеревиных птиц на ленточ-
ных пробах, общая площадь которых первоначально равнялась 227 га, с
1961 г. - 354, а в последнее время доведена до 561 га. У этого метода ряд
преимуществ: он дает непосредственно число особей всех четырех
видов на единицу площади в удобный момент, когда численность птиц
стабилизировалась после окончания ранней детской смертности, и перед
началом охотничьего сезона. Недостатки метода - заметная трудоем-
кость (для отработки 354 га требуется не менее 24 человеко-дней, без
учета простоев из-за непогоды и затраты времени на переезды) и некото-
рая зависимость результатов от погоды и личных свойств исполнителей.
По этим причинам его результаты на малых пробах недостаточно отража-
ют колебания численности из года в год и количественное соотношение
видов; при обобщении цифр за ряд лет эти недостатки сглаживаются. Как
видно из диаграммы (рис. 23), за истекшие 50 лет средняя плотность
популяций глухаря, рябчика и белой куропатки существенно (примерно
в пять раз) уменьшилась. Колебания численности тетерева более сложны,
и мы рассмотрим их отдельно.
В одной из прежних работ мы сделали попытку приблизительно опре-
делить биомассу разных экологических групп лесных птиц и пришли к
выводу, что ’’тетеревиные, составляющие по числу особей 5-25% всего
птичьего населения тайги, по весу составляют около 75% его” (Семенов-
Тян-Шанский, 1959, с. 187). Сходную по масштабу биомассу дают и две
основные группы лесных млекопитающих: с одной стороны, мелкие гры-
зуны, с другой - олени. Эти три группы в сумме являются главными
потребителями первичной продукции биогеоценоза тайги и основным
Таблица 7. Количественное соотношение тетеревиных птиц (в %)
Вид Ленточный учет 1936- 1988 Маршрут- ный учет 1931-1983 Учет добы- тых птиц 1930-1951 Остатки по- гибших птиц, 1931-1987 Регистрация в картотеке, 1979-1987
Глухарь 40 1 42 ’ 46 ' 29 1 ‘ 20
Тетерев 5 19 3 12 8
Рябчик 27 5 14 2 1
Белая куропатка 28 34 32 55 70
Тундряная куро- — — 5 2 1
патка
91
ГоВьГ. 30-35 36-01 08-51 58-60 65-69 70-70 75-78 80-80 85-88
на 100
км
50
О
на 100
ta
15
Ю
5
О
Нелая куропатка
юухарь
тетереВ
рябчик
Рис. 23. Движение численности тетеревиных птиц по пятилетиям по учетам: марш-
рутному (вверху) и ленточному (внизу)
источником ее вторичной продукции, животного белка, за счет которого
существует большая часть хищных зверей и птиц, а через охоту, прямо
или косвенно, получает свою долю и человек. В этом и состоит биоцено-
тическая роль тетеревиных птиц, выдвигающая их в экономическом
отношении на первое место в авифауне нашего региона.
По причинам, не вполне понятным, учет на ленточных пробах преуве-
личивает относительную численность рябчика. Во всяком случае, про-
стая регистрация встречаемости отдельных видов птиц на охоте и осен-
них экскурсиях (с августа по октябрь, 1931-1983 гг.) дает иное количест-
венное соотношение видов. Мы подсчитали это соотношение также по
карточкам биосъемки добытых птиц за 1930-1951 гг., по числу найден-
ных в природе остатков погибших птиц (1931-1983) и, наконец, по реги-
страции встреченных птиц в картотеке биологических наблюдений запо-
ведника за последние девять лет (1979-1987). Эти подсчеты дают некото-
рое представление и об относительной численности тундряной куро-
патки.
92
В экологическом отношении тетеревиные изучены лучше всех других
птиц Лапландского заповедника, к тому же результаты этих исследова-
ний опубликованы; поэтому обзоры по этим видам даются конспектив-
ные, и только новые материалы изложены несколько подробнее.
Тундряная куропатка Lagopus \iutus (Mont.)
Местные названия: саамское ’’кйсрымь” (поговорка ’’кйерымь-
чаппись нюн сие'рымь” - ’’тундрянка - черные нос и ноздри”); русское -
тундрянка или горная куропатка.
Тундряная куропатка - преимущественно горная птица с циркумпо-
лярным ареалом; в Гренландии и на Земле Франца Иосифа она заходит и
севернее 80° широты, а в горных регионах местами опускается к югу до
50°, в Японии даже до 40°. На Кольском полуострове она приурочена к
горным тундрам, на Мурманском берегу встречается на равнинной тунд-
ре, но в лес даже не залетает. Летом самцы и холостые самки живут
высоко в горах, среди каменной пустыни и помет их мы встречали даже
на самых высоких вершинах (Эбрчорр - 1100 м над ур. м.).
Учет тундряной куропатки на пробных площадях не проводится, но
некоторое представление о колебаниях численности дает регистрация
встреч на экскурсиях, а соотношение количества зарегистрированных
белых и тундряных куропаток позволяет оценить и ее запасы. За 1977-
1987 гг. в картотеке учтены 15 562 белые куропатки и 335 тундряных,
количественное соотношение 46:1. Этот индекс несомненно занижен, так
как биотопы тундрянки в меньшей мере охвачены маршрутами охраны
и научных сотрудников, чем биотопы белой куропатки. Соотношение
двух видов куропаток среди найденных остатков погибших птиц составляет
23:1, а среди птиц, добытых в 1930-1951 гг. для исследования, 6,3:1. С
1958 г. на летне-осенних экскурсиях (июль-сентябрь) мы регистрирова-
ли встреченных птиц с учетом длины маршрута. В среднем за сезон мы
проходили тундрой по 46 км и встречали по 2 тундрянки на 10 км мар-
шрута. Принимая ширину полосы обзора 100 м (обоснование приведено
ниже в разделе о белой куропатке), находим среднюю плотность тундря-
ной куропатки равной 2 птицам на 1 км2, а так как горные тундры в за-
поведнике занимают около 450 км2, общий запас этого вида можно
оценить в 900 голов. Цифра эта, вероятно, завышена, так как тундры
заселены неравномерно и площадь пригодных биотопов, очевидно,
меньше общей площади тундр.
Без большой ошибки можно принять, что общая численность тундря-
ных куропаток в заповеднике в 10-20 раз меньше, чем белой.
Учет визуальных встреч на экскурсиях дает некоторое представление
и о колебаниях численности этой птицы. До 1951 г. для учета диких оле-
ней мы ежегодно в конце зимы (март- апрель) объезжали горные тундры
на оленьих упряжках и параллельно вели учет встреченных тундряных
куропаток: полученные цифры характеризуют статус этого вида перед
сезоном размножения (Семенов-Тян-Шанский, 1959, с. 283-284). С 1959 г.
заповедник перешел на учет оленей с воздуха и тундряных куропаток,
как и других птиц, стали регистрировать в конце лета, пока выводки не
93
Рис. 24. Колебания численности тундряной куропатки по маршрутным учетам
Вверху - число птиц на один ходовой день в марте-апреле (1931-1951 гг.); вни-
зу — число птиц на 10 км экскурсий в июле-августе (1958—1984 гг.)
разбились (июль-август). Из диаграммы (рис. 24) видно, что в колеба-
ниях численности тундряной куропатки нет правильной периодичности,
а в массе она появляется только в отдельные годы. Общей тенденции к
сокращению или росту популяции на протяжении полувека не заметно.
Регистрация встреч в выводковое время дает возможность определить и
возрастной состав популяции. Успех размножения тундрянки непостоя-
нен: в 1934, 1970, 1972, 1973 и 1975 гг. сеголетки составляли 72-78% попу-
ляции, а в 1931, 1932, 1935 и 1971 гг. - 0-35%. Между успехом размноже-
ния и уровнем численности птицы нет прямого соответствия (коэффици-
ент корреляции между этими параметрами за 15 лет оказался +0,05±0,26).
Объясняется это тем, что популяция сокращается не только от недостат-
ка приплода, но и от последствий неблагоприятной зимовки в виде
повышенной смертности и эмиграции сохранившихся куропаток (подоб-
ные миграции известны и у белой куропатки).
Тундряная куропатка - единственная птица, зимующая на наших
горных тундрах. В отличие от лесных видов тетеревиных ее режим
питания после установления снежного покрова мало меняется. Зимой
она ест ту же пищу, что и летом, раскапывая снег на малоснежных
местах или на оленьих ’’тегорах” (места жировки). В вершинных частях
гор снежный покров в значительной мере образован изморозью, а потому
отличается плотной структурой и раскапыванию не поддается; к тому же
запасы корма в каменистой тундре ничтожны. Подобно диким оленям,
тундряные куропатки зимуют в нижней части альпийского пояса, в пре-
делах 300-600 м абсолютной высоты. В это время корм их состоит из
облиственных побегов вечнозеленых кустарничков, почек и ягод - в
основном, вороники, брусники, черники, ”гром-ягоды” (Arctons alpina),
куропачьей травы (Dryas punctata) и карликовой березки. Большой сне-
94
гопад, если он не сопровождается ветром, лишает куропаток доступа к
наземным кормам. В таких случаях тундряные куропатки клюют почки и
побеги берез на верхней границе леса, для чего иногда взлетают на дере-
вья. В условиях континентального климата, например на Аляске и в
Северо-Восточной Сибири, в областях стационарного зимнего антицикло-
на с характерным для него безветрием, наземные корма зимой вообще
малодоступны, и тундряные куропатки по необходимости питаются,
подобно белым, ивовыми и березовыми побегами, а отчасти и цветочны-
ми сережками ольхи, подобно рябчику (Андреев, 1980, с. 28-30).
Клюв тундряной куропатки меньше и слабее, чем у белой, для пита-
ния веточным кормом он менее пригоден. В многоснежные зимы тун-
дрянки обычно переходят вслед за дикими оленями на наветренные
северные оконечности горных массивов. По причине относительной
сухости северных ветров здесь выпадает меньше снега, чем в других
местах, и выдувы занимают большее пространство. Пользуясь концентра-
цией тундряных куропаток на северном склоне Монче-тундры в апреле
1938 г., один промысловый охотник добыл там около 50 куропаток.
В снежное время тундрянки держатся стайками до 6 птиц или пооди-
ночке, но в самом начале зимы иногда сбиваются в большие стаи, до трех
десятков, становятся осторожными и взлетают далеко вне выстрела.
Сильно уплотненный метелями снег на открытой горной тундре куро-
патке трудно разрыть; северный олень легче справляется с этой задачей,
острая кромка его передних копыт хорошо приспособлена для разламы-
вания наста и разгребания снега. На копанках куропатки легко находят
ягоды и другой подснежный корм, почему они охотно пасутся вместе с
оленями, иногда бегая буквально у них под ногами. В плотном снегу
куропатке трудно выкопать и лунку для ночлега. В метель она садится с
подветренной стороны камня, и ветер быстро заносит ее снегом, так что
она оказывается почти в такой же лунке, какую при других обстоятель-
ствах выкопала бы сама. Днем в ветреную погоду тундрянки смирны и
легко подпускают к себе метра на четыре, а потом убегают, не пытаясь
взлететь.
Самцы начинают токовать еще до начала брачной линьки - в середи-
не или конце апреля. При тихой и сравнительно теплой погоде они
токуют всю светлую ночь. Раскрыв ярко-красные брови, петушок бегает
по снегу, останавливаясь на мгновение для исполнения своей брачной
песни: в этот момент он опускает шею и хвост, так что спинная сторона
образует дугу, и кричит ”кррраэ-кре-ке-крэээ-эээ-э-э...”, поднимая шею
вверх. Брачная песня тундряной куропатки несколько напоминает крик
коростеля. Токует он перед курочкой, но держится она тихо, и в белом наряде
ее трудно заметить на фоне снега. Призывный крик самки - тихое
”мя... мя... мя. ..”. Днем они не токуют, самец обычно сидит на камне,
прижавшись к нему грудью, головой на ветер, а самка прячется у подно-
жия камня. При подходе человека самец взлетает, но, не решаясь оста-
вить подругу, садится неподалеку, а когда самка взлетит, следует за нею,
и оба скрываются вдали. Если куропатки убегают, самца легко отличить
по приподнятому полуразвернутому хвосту, самка же бежит, сгорбив-
шись. При появлении соперника, куропач налетает на него и с криком
гонит прочь.
95
Весенняя линька начинается около 10 мая. У самца первые летние
перья появляются на затылке, после чего наступает пауза, и он долго
остается в белом оперении с черноватым теменем и затылком - это и есть
его брачный наряд (рис. 25). Линька возобновляется только через месяц,
когда самка уже начинает насиживание. Некоторое время самец держит-
ся поблизости от гнезда, а в середине июня бросает наседку. Петухи
заканчивают линьку и проводят лето на каменистых вершинах в одино-
честве или вместе с другими самцами и холостыми самками, по 2-5
куропаток в стае. Выпадение белых перьев зимнего наряда на спинной
стороне заканчивается у самок около 5 июня, у самцов - 8 июля, а на
нижней стороне тела еще позже. Практически тундряная куропатка
линяет все теплое полугодие без перерыва. Рисунок и окраска летних
перьев, вырастающих на месте выпавших зимних, зависит от времени их
роста: растущие в мае и июне темные, почти черные, с редкими и сравни-
тельно крупными охристыми или беловато-охристыми пятнышками,
июльские светлее, крапинки на них мельче и группируются в частые
поперечные полоски (рис. 26).
По этой причине летний наряд тундрянок представляет сочетание
перьев двух типов, причем в оперении самок преобладают перья первого,
весеннего типа, а у самцов - второго (они замещают белые перья, выпав-
шие в июне и июле, и преобладают по всей верхней стороне, исключая
голову и шею). У обоих полов перья, выросшие в начале сезона, сохра-
няются до сентября и сменяются только белыми. Белые перья нижней
стороны, рулевые, маховые и большая часть кроющих крыла сменяются
только один раз в году и во всяком наряде бывают белыми - кроме се-
ми пар рулевых, черных с белой каемкой на конце. Ф.Саломонсен( ^^обна-
ружил, что часть выпавших белых перьев не замещается перьями летней
генерации и перьевой сосочек остается свободным до осени когда из него
вырастает белое же перо. Вследствие этого летнее оперение тундряной
(и белой) куропатки реже и рыхлее зимнего.
fC концу августа нижняя сторона куропатки уже одета новыми перьями, а к
10-15 сентября и на спине появляются первые белые перья. Зимняя линька
продолжается около месяца. Последние летние перья выпадают в середине
октября, и с этого момента на протяжении почти 7 мес. тундряная куропатка
носит полный зимний наряд. Сроки зимней и весенней линьки в среднем почти
совпадают со сроками образования и разрушения снежного покрова на горных
тундрах.
Тундряная куропатка начинает насиживать сразу по окончании весен-
ней линьки. Ее летнее оперение - прекрасный пример камуфляжной
окраски: пока птица неподвижна, ее невозможно заметить на фоне
выветрившихся и покрытых накипными лишайниками камней.
Из 11 найденных в Лапландском заповеднике гнезд - шесть располага-
лись в каменных россыпях, либо открыто, либо под прикрытием большо-
го камня; одно - на злаково-ягельной лужайке между россыпей (рис. 27);
четыре - в кустарничковой тундре, среди вороничника, два - под
ветками елового стланника, одно - под веткой можжевельника и одно -
совсем открыто. Гнезда находились на разной высоте: от 380 м, немного
выше верхней границы леса, почти до 800 м, в каменной пустыне близ
вершины хребта. Лоток представляет собой неглубокую ямку в грунте,
почти без подстилки. В 11 кладках было от 3 до 8 яиц, в среднем 5,5. По
96
Рис. 26. Самка тундряной куропатки, перелинявшая наполовину, 19. V 1983 г. Фото
А.С. Гилязова
окраске они сходны с яйцами белой куропатки, но размерами поменьше;
средняя масса 19,0 г (п = 8). Процесс кладки не наблюдался. Даты находок
гнезд с яйцами от 10. VI 1980 г. до 4. VII 1982 г. (в среднем по 7 гнездам -
26 июня), а средняя дата встреч пуховичков - 2 июля (9 лет); срединная
дата начала насиживания 7-8. VI, а вывода птенцов - 30. VI.
Курочка сидит на гнезде исключительно крепко. 4. VII 1982 г., спус-
каясь с вершины Сейднотчорра (300 м над ур. м.), О. Семенов-Тян-Шан-
ский едва не наступил на сидевшую на гнезде куропатку; она не шевели-
лась, пока ее фотографировали, и вскочила только при попытке взять ее
рукой. Заметить гнездо, закрытое такой ’’шапкой-невидимкой”, как
самка куропатки, практически невозможно, поэтому и находят их редко.
Другое гнездо было найдено 10. VI 1980 г., когда наседка взлетела из-за
собаки.. Видевшим это студенткам едва удалось найти гнездо вторично
21. VI, когда одна из них заметила куропатку под ногами. Гнездо тут же
было оборудовано самописцами для регистрации поведения наседки и
температуры яиц. Наседка оставляла гнездо для кормежки и моциона по
4-7 раз в сутки, каждый сеанс 8-17 мин; яйца оставались открытыми
5% всего времени инкубации. Погода в те дни была солнечной и ветре-
ной, средняя суточная температура воздуха + 11,2°, а внутри яиц 32,Г,
с колебаниями от 38,0 до 13,5° в момент возвращения наседки с прогул-
ки. В отсутствие птицы яйца остывали почти вдвое быстрее, чем яйца
белой куропатки при тех же обстоятельствах, очевидно по той причине,
что в открытой тундре они обвевались ветром.
По расчетам, птенцы должны были вылупиться около 1-3 июля, но они
погибли в яйцах на последней стадии развития, уже оперенные; куропат-
ка продолжала насиживание еще 40 дней и только 11 августа бросила
кладку. За это время ее поведение изменилось, в последние 10 дней она
98
оставляла гнездо по 8-10 раз, затрачивая на прогулки 2-3 ч в сутки. В
последний день она дважды возвращалась только через 3 ч, а на третий
раз ушла окончательно. Причиной гибели птенцов, вероятно, была
чрезмерная потеря влаги яйцами в солнечные и ветреные дни 28. VI -
2. VII. В связи с открытым расположением гнезд эмбриональная смерт-
ность у тундряной куропатки выше, чем у других тетеревиных птиц.
Всего нами учтено 60 яиц, птенцы вывелись в 20% случаев, погибли в
процессе инкубации 38%, судьба остальных 42% не прослежена. Летом
1934 г. нашли два гнезда, в них все 11 яиц были с мертвыми эмбрионами.
Выводки встречаются обычно в нижней части альпийского пояса, на
травянистых луговинах, в кустарничковых зарослях по долинам ручьев,
на тундровых гарях, поросших политриховыми мхами, иногда в скалис-
тых ущельях, где сохраняются снежники. Летом тундряные куропатки
питаются побегами, листьями, цветами и ягодами черники, голубики,
брусники, ”гром-ягоды”, вороники, цветами и листьями разных видов
ив, куропачьей травы, камнеломок, андромеды, альпийской ястребинки,
семенами осок, ситников, ожик, карликовой березки, луковичками
живородящего горца, спорангиями политрихумов; птенцы, сверх того,
едят слизней, пауков, жуков, гусениц и других насекомых. В отличие от
белой куропатки самец тундряной не принимает участия в воспитании
птенцов.
Оперение птенцов сходно с оперением матки, смешанное, с преоблада-
нием перьев первого летнего наряда, Маховые первой генерации (с
серыми опахалами) отрастают рано, так что уже в середине июля птенцы
перепархивают. Смена этих маховых на белые, дефинитивного наряда,
начинается в июле и заканчивается в августе. Два дистальных первосте-
пенных маховых ювенильного наряда отличаются от остальных белыми
опахалами; они вырастают последними, остаются на зиму и сменяются
только на следующее лето. В зимнем наряде они отличаются от остальных
заостренными вершинами с темными крапинами и служат удобным
признаком для определения возраста птицы.
Существование популяции с таким слабым темпом воспроизводства,
как у тундряной куропатки (небольшая кладка и высокая эмбриональ-
ная смертность), возможно только при малой естественной смертности взрос-
лых птиц. За 45 лет зарегистрированы остатки только 23 тундряных куропаток,
в большинстве ставших жертвой хищных птиц и зверей. Из них первое место
принадлежит кречету: среди остатков добычи, собранных под тремя гнездами,
определены 4 тундрянки, а в январе 1939 г. на Нявка-тундре видели кречета,
терзавшего еще одну. В мае 1933 г. на Чуна-тундре наблюдали белую сову, а
позднее нашли ее погадку, содержавшую череп тундряной куропатки. От од-
ной тундрянки остался почти целый скелет, признак, что она погибла от
несчастного случая (например, замурована в снежной лунке), но не от
хищника.
Зимнее оперение тундряной куропатки чисто-белое, только семь пар
рулевых и клюв - черные. Она заметно меньше белой, клюв ее короче и
слабее, лапки оперены беднее, и площадь их вдвое меньше, чем у послед-
ней. Самец отличается черной уздечкой (полоска перьев от клюва до
глаза и за ним). Между обеими куропатками есть существенные анатоми-
ческие различия: у тундряной слепые кишки значительно короче, сум-
марная длина их равна 0,9 длины тонкого отдела кишечника, а у белой
99
куропатки превышает ее в 1,5-1,6 раза. Большая разница и в размерах
сердца - у первой оно равно 2,05%, у второй 1,45% массы тела.
Общий тон летнего наряда тундряной куропатки ближе к нейтрально-
серому, белой куропатки - к ржавчато-охристому. У обоих видов круг-
лый год остаются белыми маховые, часть кроющих крыла, нижняя часть
груди и брюхо; у тундрянки иногда и центральная пара рулевых (у белой
они на лето заменяются рябыми).
Все отмеченные различия адаптивного свойства. Тундряная куропатка
мало использует веточный корм, а потому и не нуждается в таком мощном
клювые,как белая. Жизнь среди открытой тундры, почти постоянно на
ветру, предъявляет повышенные требования к полету: тундряная куро-
патка летает не только быстрее белой, но и легче набирает высоту, подни-
маясь вдоль крутого склона, поэтому сердце ее несет большую нагрузку,
чем у лесных тетеревиных птиц. Бегает она по уплотненному ветром
снегу, а потому не нуждается в большой опорной площади лап.
Средние размеры тундрянки: длина самца 383, самки - 368, крыло -
204 и 198, хвост - 115 и 105 мм; зимняя масса тела 530 и 496 г. Эта куро-
патка отличается от белой большой географической изменчивостью (описа-
но около 20 подвидов по всему ареалу), в частности варьирует и по
размерам. Континентальные тундрянки северо-восточной Сибири по
массе, примерно на 10%, меньше Кольских (средняя масса тела самца
486, самки 444; Андреев, 1980, с. 30), а островные, шпицбергенские, на
35% крупнее (самец 738 г, самка 701; Salomonsen, 1941, с. 1-30).
Белая куропатка - Lagopus lagopus (L.)
Местные названия: саамское ”рехпь”; самец в брачном наряде -
’’лякстак”, самка в брачную пору - ’’коамгынч”; русское - к^ропать,
куропатка или лесная куропатка.
Общее распространение циркумполярное с очень широким ареалом,
включающим зоны тундры (до 75° с. ш. на Новосибирских островах),
тайги и степи Казахстана, к югу почти до 50°. В Лапландии встречаются
повсеместно, в равнинной тундре, лесотундре, тайге, а отчасти и на
горных тундрах, однако не поднимаясь так высоко, как тундряная
куропатка. По общей численности занимает первое место среди тетереви-
ных птиц.
В заповеднике, по результатам ленточного учета за 31 год (1958-1988),
средняя плотность популяции в лесах различных типов составляет 20 птиц,
на 10 км2. При общей площади заповедника, равной 1613 км2, стации,
заселенные белой куропаткой, - леса, криволесья и болота - занимают
66,7 %, горные тундры, заселенные ею частично, - 27,9%, наконец, водо-
емы - 5,3%. Если принять, что куропатка занимает 70% территории запо-
ведника (1129 км2), средний запас белых куропаток на ней оценивается в
2258 особей. Территория всей Мурманской области составляет 144900 км2;
допуская, что куропаткой заселено 75% всей площади и средняя плот-
ность ее такая же, как и в лесах Лапландского заповедника, численность
ее в области приближается к 217 тыс. голов. Эта оценка скорее занижена,
чем завышена, так как в лесотундре, где другие виды тетеревиных птиц
не встречаются, плотность ее может быть выше, чем в хвойных и смешан-
ных лесах.
100
1960 1965 1970 1975 1980 1985
Рис. 28. Колебания численности белой куропатки за 30 лет
Верхняя кривая — число птиц на 10 км маршрута; средняя — ленточный учет,
число птиц на 100 га; нижняя — находки погибших птиц с июля по июнь следую-
щего года
Сопоставление результатов ленточного и маршрутного учетов по годам показало,
что они плохо согласуются между собой (рис. 28): коэффициент корреляции между
обоими рядами за 26 лет близок к нулю (г = + 0,05 ± 0,20). Одна из возможных
причин такого расхождения в том, что учеты проводились на разных территориях и в
разное время. Несмотря на несогласованность в деталях, оба учета показывают, что в
колебаниях численности белой куропатки нет отчетливой периодичности. Годами
"пика” были 1937, 1950, 1964 и 1974; согласно ленточному учету — также 1986, а по
материалам маршрутных учетов - 1933, 1963, 1977, 1981 и 1987. Оба учета согласно
показывают годы минимума в 1936, 1940—1941 (вероятно — 1948), 1965, 1975 и 1983-
1984 гг. Амплитуды колебаний довольно значительны, по ленточным учетам (в
среднем по 8 парам годов) 12:1, по маршрутным — 4:1 (средняя длина проходимых за
сезон маршрутов — 285 км, а “лент— 72 км). Явной тенденции к изменению числен-
ности белой куропатки на протяжении полувека, по-видимому, нет.
Средние индексы численности куропаток по периодам:
Годы Ленточный учет на 100 га Маршрутный учет на 100 км
1930-1941 3,8 13
1948-1951 4,0 24
1958-1970 2,0 15
1971-1987 2,0 18
Среднее 2,4 (41 год) 18 (45 лет)
По сравнению с тридцатыми годами в 70-х - начале 80-х индекс
численности по ленточным учетам уменьшился вдвое, а встречаемость на
маршрутах, наоборот, увеличилась в полтора раза. Оба метода согласно
101
констатируют повышение численности белой куропатки около 1950 г. К
вопросу о причинах колебаний численности куропаток мы еще вернемся.
В отличие от глухаря, тетерева и рябчика обе куропатки (белая и
тундряная) всегда кормятся на поверхности земли или снега. Из полез-
ных адаптаций куропаток самая наглядная - сезонный диморфизм
оперения. В ржавчатом с черными крапинами летнем наряде, сходном с
окраской опавших листьев, она теряется на фоне напочвенного покрова
точно так же, как крапчато-серая тундряная куропатка, сливается с
окраской выветрившихся камней; ко времени образования снежного
покрова оба вида приобретают чисто-белое оперение и на фоне снега
становятся невидимками.
Зимой куропатки держатся небольшими стайками смешанного состава
(вероятно, это выводки). Для добывания корма им достаточно самого
скудного освещения: едва забрезжит рассвет, что соответствует освеще-
нию около 0,1 люкса и положению центра Солнца на 8-10° ниже гори-
зонта, куропатки уже выходят из снежных лунок, перелетают и начи-
нают кормиться. Даже в темную пору, когда солнце не появляется над
горизонтом (с 9. XII по 3. I), куропатки деятельны до 7 ч в сутки, с 8 ч
30 мин до 15 ч 30 мин солнечного времени. Перелеты куропаток в утрен-
них сумерках сопровождаются брачными криками самцов.
На жировке куропатки перебегают по снегу от куста к кусту, с легким
хрустом отламывая клювом от веток кусочки длиной около 8 мм, весом по
20-30 мг. Для наполнения зоба нужно много времени, так как количест-
во палочек в одном зобу достигает 2500 (Новиков, 1981). Всего охотнее они
кормятся тальником по берегам рек и ручьев, предпочитая молодую
поросль Salix phylicifolia; на болотах они едят веточки карликовой
березки (Betula nanaj, а в лесу, особенно на гарях, кормятся прикорне-
вой порослью берез (Betula subarctica, В. verrucosa)* За одну жировку
куропатка проходит около 1 км. В середине дня они отдыхают, обычно на
поверхности снега, переваривая корм, вечером кормятся вторично, а на
ночь зарываются в снег, перелетев с жировки в другое место. Прямо с
посадки птица роет наклонный туннель, откидывая снег назад лапами,
пока не окажется в снежной камере с потолком толщиной около 10 см, и
наглухо заделанным входом. Все члены стайки ночуют по соседству,
каждая птица в отдельной лунке.
В толще снежного покрова большой температурный градиент: поверх-
ность снега в ясную ночь холоднее окружающего воздуха вследствие
радиационной потери тепла, а температура под снегом остается лишь
немного ниже нуля благодаря непрерывному притоку тепла из глубины
земли. Температурный градиент обеспечивает циркуляцию воздуха в
толще снега, благодаря чему птице не грозит опасность задохнуться.
Изолированная от внешней атмосферы слоем рыхлого снега, куропатка
нагревает свою камеру дыханием, запас корма в ее зобу оттаивает, и за
ночь она постепенно переваривает его. Мускульный желудок птицы
перетирает почки и кору, а палочки проходят через кишечник почти
целыми. Перетертая пульпа из тонкой кишки поступает для дальнейшей
переработки в парные слепые (у белой куропатки они превышают по
емкости остальные отделы кишечника и желудок), а кусочки древесины
переходят непосредственно в прямую кишку, формируются там в экскре-
менты и выходят через клоаку. За ночь на дне лунки скапливается
102
большая куча таких экскрементов, но слепые кишки опорожняются
только раз в сутки, утром, при выходе птицы из лунки.
Питание веточным кормом, доступным птице всю зиму, ночлег в
тепле, под снегом, хорошие изоляционные свойства и защитная окраска
оперения, большая опорная поверхность лап, способность кормиться при
минимальном освещении обеспечивают существование куропатки в
суровых условиях полярной зимы.
С наступлением ’’весны света”, уже в марте, стаи куропаток распа-
даются, все чаще они встречаются парами или поодиночке; ночами самцы
кричат свою брачную песнь и ухаживают за самками. Поведение куропа-
ток изменяется постепенно, и стайки их, наряду с парами, встречаются
вплоть до начала, а иногда и конца апреля. К этому времени большая
часть ивовой и березовой поросли скрыта в толще снега, а оставшиеся на
виду ветки уже подстрижены зайцами и куропатками; в это время корм
приходится разыскивать и куропатки залетают вверх по склонам до
березового криволесья и даже в горную тундру. В марте и апреле они
нередко садятся на березы, обстригая побеги и цветочные сережки,
подобно тетеревам. Еще охотнее они кормятся на первых проталинах, по
солнопекам, бугристым болотам и берегам озер - как раз в это время в
их рационе появляются перезимовавшие ягоды вороники, брусники и
зеленые побеги черники.
В середине апреля самцы начинают линять. Первые красно-коричневые
перья брачного наряда появляются на горле, р среднем за 45 лет, 19
апреля. Начало линьки коррелирует с появлением первых проталин
(г = + 0,41 ± 0,14 за 36 лет) и средней температурой апреля" (г = - 0,32 ±0,15),
но связь эта сравнительно слабая. Когда голова и шея перелиняют пол-
ностью, в линьке наступает пауза до последней декады мая; в это время
петух носит брачный наряд, отличающийся от зимнего ржавчато-красным
оперением головы и шеи, с сурико-красной, отороченной гребешком
’’бровью” над глазом (рис. 29).
Куропатки умело пользуются своей покровительственной окраской,
как зимой, так и летом они затаиваются, а заметить неподвижную птицу,
неотличимую по окраске от окружающей среды, почти невозможно. В
брачном наряде петушок теряет это преимущество, но в брачную пору он
и не пытается затаиваться, а, наоборот, ведет себя демонстративно,
взлетает с громким криком, садится на открытых местах, на камни,
валежины, даже на вершины елок.
У самца линька толовища начинается в конце мая, и к 23 июня он
получает летний наряд: перья спинной стороны ржавчато-охристые с
черновато-бурыми пестринами, включая кроющие хвоста и большей
части крыльев. Маховые, малые и большие кроющие крыла, а также
брюхо все лето остаются белыми; рулевые черные, с белой каемкой.
Самка линяет очень быстро, в среднем с 11 по 29 мая, приобретая летний
наряд прежде, чем сядет на яйца, в то время как самец только начинает
смену оперения туловища. Летнее оперение курочки несколько светлее,
чем у самца, ржавчатый цвет заменен охристым, а перья шеи у нее не
одноцветные, а такие же рябые, как и на спине.
Куропатка предпочитает строить гнездо на открытом месте - на
болоте, в редколесье, реже - в ельнике или в бору; если на болоте, то на
кочке, под прикрытием стеблей пушицы, осоки, андромеды, голубики,
багульника и др.; на лесных полянах - или открыто, среди мхов и ягеля,
103
Рис. 29. Самец белой куропатки в брачном наряде, 30. V 1982 г. Фото О.И. Семенова*
Тян-Шанского
либо под прикрытием елочки, сосенки или березки не более 1,5 м ростом.
Лоток глубокий, со скудной подстилкой из сухих былинок и перьев
матки. Яйца по кремовому фону густо испещрены черновато-бурыми
пятнами; масса свежих яиц в пределах 19,7-21,2 г. Полная кладка содер-
жит обычно 6-12 яиц, в среднем - 9. Самки начинают нестись в послед-
ней декаде мая, садятся на гнезда около 8 июня, первые выводки появ-
ляются, в среднем за 38 лет, 30. VI (20. VI - 12. VII). В двух, состоявших
под наблюдением, гнездах инкубация продолжалась 22,3 сут.
Насиживает только самка, но самец держится поблизости, в пределах
104
200 м от гнезда, и активно охраняет его от врагов. Наседка сидит очень
крепко, позволяя сфотографировать себя с 2 м, иногда взлетает букваль-
но из-под ног. По этой причине гнезда куропаток редко удается находить.
Будучи спугнутой с гнезда, она возвращается к нему через 0,5-2,0 ч, но в
первые дни насиживания возвращение ее может задержаться на несколь-
ко часов - до 10.
Порядок кладки яиц нам не удалось проследить, но вряд ли куропатка
отличается в этом отношении от глухарки или рябчика. Э. Пуллиайнен
(Финляндия) определил средний интервал между откладкой яиц в 32
часа (Pulliainen, 1978). Режим насиживания изучался нами с помощью
актографов в шести гнездах (2152 ч записи). Наседка оставляет гнездо для
кормежки и моциона 3-5 раз в сутки, каждый раз на срок от 8 до 22 мин,
в результате яйца остаются открытыми на протяжении 0,035-0,052
времени инкубации (50- 75 мин в сутки). Время прогулок не приурочено
к определенным часам и день за днем смещается в ту или другую сторо-
ну. Объединив материал по всем гнездам, мы нашли два максимума
суточной активности: утренний (01-11 ч) и дневной (15-17 ч); в вечерние
и ночные часы (17-01) она снижается, но не прерывается.
Температура окружающего гнездо приземного слоя воздуха за время
инкубации варьирует в широких пределах, от -6° до + 39°, в среднем
+ 9,4°. Средняя температура яиц значительно ниже температуры тела
наседки, так как она греет их только сверху, а снизу и с боков они
охлаждаются от соприкосновения с грунтом и окружающим воздухом.
Температура яиц в среднем по трем гнездам + 33,0°, снижаясь до + 28,2° к
моменту возвращения наседки с прогулки: минимальная температура
яиц за все время инкубации оказалась + 19,7°.
Эмбриональная смертность у белой куропатки довольно велика: из 28
состоявших под наблюдением кладок с 244 яйцами десять, содержавшие
78 яиц, погибли целиком, три брошены наседками в начале инкубации
или до нее, шесть уничтожены хищниками (на одной погибла наседка) -
при этом потеряно 31% всех яиц; еще 7% погибли за время насиживания в
благополучных кладках, и только из 62% яиц вывелись цыплята.
Выводок водят обычно обе взрослые птицы. Кормятся они преиму-
щественно на болотах разных типов, особенно морошковых, затем на
старых гарях в стадии преобладания в их напочвенном покрове ягодных
кустарничков и верхоплодных мхов (Polytrichum sp.) и в ельниках.
Куропачьи выводки встречаются по склонам гор до верхней границы
леса и даже выше ее, в субальпийском поясе, в зарослях ивняков с
разнотравьем. Иногда два выводка объединяются, тогда с одного места
взлетает до двух десятков куропаток, в числе которых 3-4 взрослых.
Летний рацион белых куропаток включает листья голубики, стебли
лесного хвоща, побеги и цветы черники, ягоды вороники, черники,
реже - голубики и брусники, семена марьянника. Взрослые птицы
только изредка едят насекомых и других беспозвоночных; птенцы,
напротив, в июле питаются преимущественно животным кормом -
личинками пилильщиков, гусеницами и другими насекомыми, а также
пауками. Из растительных кормов птенцы отдают предпочтение самым
калорийным - семенам (осок, марьянника, андромеды), спорангиям
мхов, а также грибам. В августе ягоды, зелень и цветы преобладают над
семенами и насекомыми, а в сентябре состав пищи птенцов приближается
к рациону взрослых птиц. Во второй половине сентября куропатки
105
переходят с болот на лесные гривы и вараки и с этого времени, вплоть до
декабря, питаются в основном зелеными побегами черники, ягодами
черники и вороники.
У птенцов маховые первой генерации серые, линька их начинается с кисте-
вого сгиба крыла одновременно в двух направлениях, и на месте выпавших
серых вырастают уже белые. Два дистальных первостепенных маховых
первой генерации имеют белые опахала и остаются на зиму, сменяясь
только на следующее лето. Смена маховых и рулевых у молодых птиц
происходит с середины июля до конца сентября. Около 21 сентября на
спинах куропаток показываются первые белые перья, а через месяц
(20. X в среднем) они получают полный зимний наряд. Нормально оконча-
ние линьки совпадает с установлением в лесу снежного покрова, но
прямой связи между этими явлениями нет: соответствующий коэффи-
циент корреляции за 41 год оказался + 0,02 ± 0,16. Первый снегопад
обычно застает куропаток в разгаре линьки, но иногда они получают
белый наряд еще до выпадения снега и на время лишаются преимуществ
охранительной окраски. Время окончания . линьки зависит от температу-
ры октября, коэффициент корреляции за 45 лет оказался = + 0,43 ±0,12.
Пока снежный покров еще рыхлый и неглубокий, куропатки кормятся
на черничниках, разгребая снег, и постепенно переходят на веточный
корм. В декабре, при средней высоте снежного покрова около 50 см,
ветки ив и берез составляют около 55% рациона, черничные побеги - 44%,
и ягоды - 1%.
Осенью куропатки отдыхают и спят в естественных убежищах под
шатром еловых ветвей, а после выпадения снега - в открытой ямке,
полулунке. Ночлег в снежной камере возможен при толщине снежного
покрова не менее 30-40 см. В лесах Лапландского заповедника снег
достигает 30 см 20. XI и 40 см - 30. XI (в среднем за 23 года), а куропатки
впервые зарываются в снег 24. XI (в среднем за 17 лет), но в этих датах
бывают большие отклонения по годе 1. В 1966 г. снег выпал необычайно
рано, 10. X глубина его достигла 43 см, и в этот день куропатки ночевали
уже в лунках. Следующая зима 1967/68 г. оказалась самой малоснежной
за последние годы, высота снега на 31. XII едва достигла 32 см, ввиду
чего куропатки смогли зарываться в снег только со 2 января.
Численность куропаток осенью зависит в первую очередь от успеха
размножения, а последний - от метеорологических условий весны.
Успех размножения характеризуется возрастным составом осенней
популяции. По карточкам биосъемки добытых птиц и регистрации ви-
зуальных встреч в июле, августе и начале сентября на протяжении 29 лет,
доля молодняка в популяции варьировала от 37% в 1971 г. до 89% в
1974 г., в среднем 65%, что соответствует приблизительно 4 птенцам на
пару взрослых (размер проб от 20 до 480 куропаток за сезон, в среднем по
135). Индекс молодняка у куропатки коррелирует с температурой июня
(г= + 0,38 ± 0,17), а также с численностью куропаток на ленточных пробах
в августе (г = + 0,38 ± 0,17). Невысокие значения приведенных коэффи-
циентов - признак сложности связей, о которых идет речь: количество
приплода зависит не только от эмбриональной смертности, контролируе-
мой температурой среды, но также от выживаемости птенцов, пресса
хищников и других причин; численность популяции - как от ее очеред-
ного пополнения, так и от количества перезимовавших производителей,
их смертности и миграций. Имеют значение и ошибки самих учетов. В
106
результате вариации различных популяционных индексов сходны лишь
в общих чертах, но расходятся в деталях.
Резкое сокращение численности куропаток в 1967 и 1975 гг. при нор-
мальном успехе размножения объясняется эмиграцией их из-за неблаго-
приятных условий зимовки. Ранние и обильные снегопады в 1966 г.
уменьшили запас доступных куропаткам веточных кормов по всему
Кольскому полуострову, принудив их в массе откочевать к Терскому берегу,
где их успешно промышляли охотники: по сведениям Госохотинспекции зи-
мой 1966/67 г. там было заготовлено около 25 тыс. куропаток. Зимой 1974/75 г.
высота снега была нормальной, ни частые оттепели (не менее 12 за 5
месяцев) привели к образованию на нем ледяных корок. Куропатки не
могли раскапывать снег ни для кормежки, ни для ночлега и двинулись к
Терскому берегу, где в ту зиму было заготовлено 12 тыс., а в следую-
щую- еще 6-7 тыс. штук. В обычные зимы куропатки живут оседло,
охотятся на них любители и размер добычи не поддается учету.
О миграциях куропаток мы узнаем из случайных источников. По
словам научного сотрудника Полярно-альпийского Ботанического сада
Е.Г. Чернова, массовая концентрация куропаток на Терском побережье
наблюдалась также в 1938/39 г.; местные жители ловили их сетями и
силками. По-видимому, куропатки летели и дальше, на Зимний берег,
так как в горле Белого моря находили на льду сотни замерзших куропа-
ток (Семенов-Тян-Шанский, 1959, с. 43). В сборнике материалов по исто-
рии Кольского полуострова в XVI-XVII вв. есть упоминание о пошлине,
взысканной за 54 пуда куропачьего пера, вывезенного с Поноя в
”1767” году (1659 г.; Андреев, 1930, с. 16): для этого надо было ощипать
27 тыс. куропаток (Семенов-Тян-Шанский, 1959, с. 6).
В старину куропатка была одним из главных промысловых животных
саамов, занимая, вероятно, второе место после дикого оленя: ’’Досельные
лопари были знаменитые охотники и промышляли больше всего дикарей,
мясом которых питались, еще брали куропаток и рыбу”, - пишет
В.В. Чарнолуский (Чарнолуский, 1930, с. 26).
Множество куропаток гибнет от различных хищников. Среди 982
зарегистрированных нами остатков погибших тетеревиных птиц куропат-
кам принадлежит первое место - 55%; из них 40% стали добычей хищных
птиц, 20% - зверей, 29% - от неизвестных хищников, 3% погибли от
ранения или случайных причин, в 8% случаев причина смерти неясна. В
добыче кречета найдены 62 куропатки, ястреба-тетеревятника - 25,
зимняка - 8, орла беркута - 9, орлана - 7. Одна куропатка стала жертвой
совы - вероятно филина, белой совы или неясыти. Здесь указаны только
остатки добычи, не учитывая погадок.
При исследовании питания хищных зверей по экскрементам остатки
птиц редко удается определить до вида, а число особей в таблицах
питания не учитывается. Тетеревиные птицы имеют существенное
значение в питании лисы и куницы; зимой они составляют 21% добычи
лисы и 16% - куницы; летом соответственно - 8% и 3%. Оба хищника
активно ловят куропаток (и других птиц), преимущественно ночью,
когда они спят в снежных лунках. Охотятся на тетеревиных птиц и
другие звери: норка (13% зимнего рациона), горностай (5% зимой), росома-
ха (19% зимой), но по крайней мере часть птиц достается им в виде остат-
ков чужой добычи, хотя ловят они и сами. В феврале 1982 г. одну куро-
патку поймала даже выдра.
Рис. 30. Труп куропатки, погибшей зимой, в снежной лунке, 26. V 1970 г. Фото
О.И. Семенова-Тян-Шанского
Учет остатков погибших птиц показал, что огромное большинство их
кончает насильственной смертью, очень немногие, вероятно не более 5%,
умирают от травм, несчастных случаев и болезней. Приведем примеры.
В 1962 г. найдены остатки 3 куропаток, разбившихся о провода теле-
фонной линии; позднее, на протяжении 15 лет, зарегистрированы 3
куропатки, сбитые машинами на автодороге Мурманск-Ленинград. В
1979 г. птенец куропатки погиб, попав в ловчую канавку для учета
мелких Млекопитающих.
В июне 1977 г. на берегу оз. Ельявр поймана отощавшая от голода
годовалая куропатка, самка, весом всего 300 г; при осмотре обнаружена
сквозная рана в ротовой полости, в которую попал язык, вследствие чего
птица не могла кормиться.
24 мая 1970 г. на берегу ламбины в верхнем конце Чунозера найден
недавно вытаявший из снега труп куропатки, самки, в зимнем наряде,
лежавший в кусте карликовой березки на груди с поднятой вверх шеей.
Судя по этой позе и пустому пищеварительному тракту, птица погибла в
снежной лунке, не сумев пробиться через наметенный ветром плотный
сугроб либо образовавшуюся на поверхности снега ледяную корку
(рис. 30).
К смерти от неизвестных причин мы относим все находки более или
менее цельных трупов или скелетов птиц, не съеденных (а следователь-
но, и не пойманных) хищниками: в их число попадают и погибшие под
настом.
Количество находимых останков куропаток варьирует по годам,
повышаясь в год максимума или в следующий за ним год. Величина
смертности не только зависит от численности популяции, но и сама
108
влияёт на ее колебания, хотя и в меньшей мере, чем успех размножения
или миграции.
Сезонные наряды белой куропатки описаны выше, в связи с линькой.
Характерный признак белой и тундряной куропаток, от которого и проис-
ходит латинское название рода (Lagopus - заячья лапка), - богатое
оперениб лапок, включая и пальцы. В зимнем наряде перышки боковых
сторон пальцев загибаются вниз, полностью закрывая подошву, а когти
отрастают в длину и ширину. Площадь следа белой куропатки около
25 см2, а статическая весовая нагрузка на обе лапы - около 12 г/см2
(Новиков, 1981, с. 66) - меньше, чем у других бегающих птиц. Весной
оперение лап и когти линяют; летние перышки лап едва прикрывают
кожу.
Размеры наших куропаток (в среднем): длина самца - 417, крыло -
210, хвост - 127 мм, масса тела - 659 г; самка соответственно - 392, 197,
115 мм и 583 г. Масса тела взрослых птиц сильно меняется по сезонам,
достигая максимума в декабре-январе (самец 727, самка 628г); в феврале
масса тела уже снижается, у самцов достигает минимума в июне-июле
(597 г, или 82% от максимального); у самок в мае: во время кладки масса
тела повышается, а в период насиживания и вождения птенцов снижает-
ся, примерно, до 500 г (80% от максимального). Еще резче меняется масса
тела пищеварительных органов, особенно мускульного желудка, вес
которого в мае-июне составляет всего 60% от зимнего веса, очевидно в
связи с сокращением объема работы этих органов в летний сезон.
Глухарь - Tetiao urogallus L.
Местные названия: саамское - ’’ч^ухч” (самец) и ”ку£ппель” (самка),
русское - глухарь (самец) и кбппала (самка).
Глухарь, характерный элемент фауны европейской и сибирской тайги,
на Кольском полуострове приурочен к сосновым лесам и распространен
главным образом в юго-западной части его; граница ареала проходит
примерно от г. Никеля через оз. Хутоявр на Колу, дальше в юго-восточ-
ном направлении на Ловозеро и Поной, доходя по его долийе до Каневки,
а по Терскому берегу к востоку до р. Пялица.
В Лапландском заповеднике глухарь многочисленнее других тетере-
виных птиц, но в целом по области занимает второе место после белой
куропатки. С момента организации в 1936 г. ленточного учета до 60-х
годов численность глухаря в заповеднике сокращалась, а потом стабили-
зировалась и последние два десятка лет держится на уровне 25 особей на
10 км2 (табл. 8).
Оценим запас глухаря в заповеднике в последние годы. Леса занима-
ют 53,2% его территории; из них сосновые и смешанные (биотоп глухаря)
составляют не менее половины: глухарь встречается и в ельниках, но
гораздо реже, чем в борах. Принимая заселенную глухарем территорию в
500 км2 (31% площади заповедника), а плотность его в 20 голов на 10 км2
(1958-1987), получаем среднюю численность 1000 голов. Площадь Мур-
манской области равна 144 900 км2, леса занимают в ней 23%; считая, что
из них половина сосновых, оцениваем площадь последних в 16 664 км2, а
запас глухаря в них около 33 тыс. голов. Однако плотность глухаря в
лесах госфонда должна быть ниже, чем в заповеднике, где нет охоты;
кроме того, значительная часть боров в настоящее время представлена
109
Таблица 8. Движение численности глухаря и тетерева по десятилетиям/
Ленточный учет Маршрутный учет
Десятилетия На 1000 га на 100 км / 1
глухарь тетерев глухарь у тетерев
1930-1939 117 1 41 / 1
1940-1949 58 8 20 / 24
1950-1959 24 11 16 16
1960-1969 15 7 18 14
1970-1979 21 1 9 6
1980-1987 29 1 17 10
Среднее, все годы: 44 5 20 12
лесосеками и гарями. По этим причинам ближе к действительности будет
оценить запас глухаря ниже 20-30 тыс. голов.
Численность сильно варьирует по годам, судя по ленточным учетам,
амплитуда достигает 1:25, но в ее колебаниях нет правильной периодич-
ности (рис. 31). Хотя цифры ленточного и маршрутного учетов плохо
согласуются (коэффициент корреляции между ними + 0,12 ±0,19),, все же
годами максимумов можно признать 1963, 1966, 1972, 1974, 1978, 1980 и
1987 гг., а минимумов - 1962,1967,1971,1975,1979 и 1982 гг.
Подобно куропатке, в середине зимы глухарь большую часть суток
проводит под снегом. В сумерках он активен, пока солнце не опустится
на 5° ниже горизонта, что соответствует освещенности в 10 люкс; поэтому
во второй половине декабря, при полном отсутствии солнца, он деятелен
не более 4-5 ч в сутки. Утром, выйдя из лунки, глухарь извергает из
слепых кишок особый кал, консистенцией и цветом напоминающий
столовую горчицу, взлетает на сосну и начинает кормиться. На жировке он
отстригает клювом кончики побегов с хвоей, роняя на поверхность снега
хвоинки и обычные экскременты, состоящие из перемятой хвои. Зимой
глухари держатся поодиночке или группами по 2-3, а иногда до 6 птиц одного
пола. Самки предпочитают спелые сосновые леса со сравнительно густым дре-
востоем, самцы - более открытые места, сфагновые болота с низкими редкими
сосенками или разреженные боры на выходах скал. В годы высокой численнос-
ти случалось видеть большие стаи глухарей; так, в декабре 1934 г. на большом
болоте мы насчитали с одного места 26 петухов открыто рассевшихся по
низким соснам, до 2-3 на одном дереве. Зимой они живут оседло. Самки
прячутся в глубине кроны, используя преимущества покровительственной
окраски, сходной по тону со старой, потрескавшейся сосновой корой, тогда как
охристо-рыжий зоб не отличается по цвету от молодой коры вершин и ветвей.
Наполнив зоб, вмещающий у самцов до 250, у самок до 150 г. корма
(более 8% от массы тела птицы), птица снова зарывается в снег, обычно
рядом со старой лункой. Случается на одной поляне находить до 30-50
лунок разной свежести с большой кучей хвойного помета на дне каждой.
Глухарь остается в лунке до 19-20 ч. сряду.
В сильный мороз глухари избегают взлетать и кормятся на поверхнос-
ти снега, как куропатки, подстригая кусты можжевельника или сосно-
110
Рис. 31. Колебания численности глухаря за 30 лет
Верхняя кривая - число птиц на 10 км маршрута; средняя - ленточный учет,
число птиц на 100 га в августе, нижняя — находки птиц с июля по июнь следующего
года
вый подрост. В оттепель они не зарываются в снег, а ночуют, сидя на
дереве. Когда в конце зимы снег покроется настом, глухари ночуют на
его поверхности, либо под навесом еловых ветвей, либо открыто.
Сосновая хвоя и веточки, иногда с добавлением незрелых сосновых
шишечек - основной корм глухаря на протяжении семи месяцев, с
ноября по май; к ним он добавляет хвою и плоды можжевельника (около
8% рациона), но еловую хвою ест лишь в крайнем случае, при невозмож-
ности найти сосновую. Кроме глухаря, только лось использует сосновую
хвою в качестве основного зимнего корма, но лось на этой диете тощает,
а глухарь остается ”в теле” и веса к весне не теряет.
В марте, с увеличением светового дня, глухари оживляются. В это
время они кормятся дважды в сутки, утром и вечером, а среди дня
отдыхают. Зарывшийся в снег, глухарь оставляет на его поверхности
маленькую кучку снега, едва заметную со стороны. Сквозь снег до него
доходят только звуки; заслышав близкий шорох шагов, он, пробив снег
головой, внезапно вылетает в облаке снежной пыли.
По утрам, в солнечную погоду, петухи бродят по насту, опустив
крылья и чертя снег концами маховых: в это время они еще не поют и на
токовища не слетаются. Первые ’’чертежи” глухарей наблюдаются в
среднем 20 марта (19. II 1977 г. - 14. IV 1942 г., за 44 года;; о = ± 11,8 дня).
Токовать по-настоящему они начинают через 5 недель, в среднем за 42
года - 26 апреля (17. III 1977 г. - 18. V 1970 г.,о = ± 12,3 дня). В это время
в лесу сплошной снежный покров, днем он подтаивает, а ночью подмер-
111
Рис. 32. Глухарка бродит ночью по поселку, 10. VI 1974 г. Фото О.И. Семенова-Тян-
Шанского
зает; ночи уже светлые, вечерняя заря переходит в утреннюю. Первые
проталины образуются на солнопеках около 7 апреля, а с 3 мая (19. IV -
13. V за 23 года) глухарки уже кормятся на обтаявших буграх болот
перезимовавшими ягодами вороники и брусники, цветочными колоска-
ми пушицы, листьями андромеды и зелеными побегами черники. Видимо,
летние корма, особенно генеративные органы растений (цветы), создают
энергетический ресурс, необходимый для откладки яиц. В то же время
глухарки продолжают кормиться и на соснах (особенно петухи, в ночные
часы).
Токовище обычно располагается в перестойном бору, на грядах или
пригорках, перемежающихся со сфагновыми болотами, а если на них есть
сосны - то токуют и на болоте. Глухари предпочитают пересеченную
местность, где рано появляются проталины; на наш человеческий
взгляд - это самые живописные места.
Первый кордон заповедника на берегу Чунозера был построен в 1930 г.
рядом с токовищем, и, поскольку глухарей никто не трогал, они продол-
жали токовать не прежнем месте, так что песню можно было слышать с
порога кордона. На новой усадьбе заповедника и на кордоне ’’Нявка”
глухари также токовали поблизости от жилых построек. В последние
годы прямо из окна можно видеть, как глухарка кормится на проталинах
по южному склону, а ночью бродит между жилыми домами (рис. 32).
Токование полигамных птиц можно определить как ритуализирован-
ный парад самцов, на котором устанавливаются иерархические отноше-
112
ния Между ними, а самки выбирают партнеров для продолжения рода.
Глухари слетаются на токовище едва начинает смеркаться, шумно садят-
ся на сосны и первое время сидят молча, а в 21-23 ч начинают петь.
Вначал^ слышатся только отдельные отрывистые звуки, похожие на
падение^тяжелых капель, потом они сдваиваются в ”те-ке. . . те-ке.. .те-
ке” и, постепенно учащаясь, переходят в короткую трель, которая закан-
чивается 1 глухим звуком, напоминающим хлопок от вытащенной из
бутылки Пробки (сходный звук издает и домашний индюк): слышен он
только на коротком расстоянии, не далее 10 м. У западноевропейских
глухарей этот хлопок звучит громче остальных элементов песни, почему
немцы и называют его ’’главным ударом” (Hauptschlag). За хлопком
следует заключительное колено песни, напоминающее громкий лязг
железа о точильный камень. Хорошо распевшийся глухарь исполняет до
9 песен в минуту (троекратное ”те-ке” - 3 с и ’’точенье” - 3 с). Услышав
или увидев глухарку, петух токует энергичнее, слетает с дерева и про-
должает петь уже на земле - тут он вступает и в контакт с самкой.
Вместо последнего колена песни глухарь иногда взлетает, шумно хлопая
крыльями. Драки между соперниками если и бывают, то очень редко.
После восхода солнца, между 2 ч - 3 ч 30 мин солнечного времени,
глухари перестают петь, но некоторое время еще сидят по деревьям,
время от времени ’’скиркая” - так охотники называют резкий вскрик
”3к-ке-ке”, напоминающий икоту: этот звук - признак окончания тока.
Поют на току преимущественно вполне взрослые петухи, трех лет и
старше; годовалые и двухлетние, хотя и слетаются на ток, в присутствии
доминантных самцов не отваживаются токовать (половозрелость их не
подлежит сомнению, при содержании в неволе годовалый самец успешно
оплодотворяет самку). Положение меняется, если на току нет ни одного
взрослого петуха; 20 мая 1934 г. на позднем току, в отсутствие самок и
старых глухарей, Семенов-Тян-Шанский убил под песню, 3 самцов, и все
они оказались годовалыми. В районе Лапландского заповедника конку-
ренция между самцами носит скорее символический характер, птиц с
травмами от драк мы никогда не встречали. При низкой плотности
популяции глухари токуют редко и вяло, на токах, где бывало пело до
10-12 самцов (Семенов-Тян-Шанский, 1938, с. 229) в последние годы мы
находили только 1-2; тем не менее глухари и теперь оплодотворяются и
несут яйца, как прежде. Сократилась и продолжительность периода
токования: так, в 1931-1941 тг. последнюю песню глухаря отмечали в
среднем 5. VI, а в 1972-1987 гг. - 20. V; весь период токования сократил-
ся с 39 дней до 29, но и в это время токование зависит от погоды; глухари
энергично поют в тихую теплую, пасмурную ночь, в морозную же или
ветреную, а тем более в метель сидят по деревьям молча.
Около 20 мая глухарки уже оплодотворены и начинают нестись. В это
время в борах еще много снега и место для гнезда приходится подыски-
вать на проталине, в пределах небольшого участка, где в это время
глухарка кормится и живет (наблюдения 24-29 мая 1974 г. и 18-22 мая
1982 г.).
По наблюдениям, проведенным на пяти гнездах, интервал между
ноской яиц составляет 27-29 ч, благодаря чему момент откладки посте-
пенно сдвигается с раннего утра (2-3 ч) на послеполуденные часы (14-
17 ч), потом глухарка один день пропускает, а на второй несется рано
утром. Полная кладка состоит из 4-9 яиц, в среднем по 85 гнездам - 6,1,
8. Зак. 1023 ИЗ
Рис. 33. Глухарка на гнезде, 10. VI 1974 г. Фото О.И. Семенова-Тян-Шанского
для откладки которых требуется 7-8 дней. Наседка начинает наси-
живать вечером (около 21-22 ч), садясь на еще неполную кладку: послед-
нее яйцо она откладывает на следующее утро (рис. 33). Глухариное яйцо
размером с куриное, средняя масса его 48,6 г (л = 73), окраска светло-ко-
ричневато-желтоватая с мелкими коричневыми крапинами.
Гнездо чаще всего располагается у подножия сосны или другого
дерева или под прикрытием валежины или большого камня, в общем
довольно открыто. Оно представляет неглубокую ямку среди живого
растительного покрова, со скудной подстилкой из опавших листьев и
хвоинок - практически яйца лежат на земле. Наседка греет их сверху, а
нижняя сторона остывает от контакта с землей и окружающим воздухом.
Инкубация, включая несколько часов пребывания птенцов в гнезде,
до ухода выводка, продолжается в среднем 25,8 суток. Глухарка сидит на
яйцах около 23 ч в сутки, отлучаясь для кормежки обычно 2 или 3 раза в
день: в это время яйца остаются открытыми. С гнезда на место жировки
и обратно она летит, не оставляя следа, по которому зверь мог бы обнару-
жить гнездо. Продолжительность прогулки чаще всего 18-32 мин. Темпе-
ратура яиц все время колеблется, в среднем за время инкубации она
составляет от + 32,7° до + 36,4° (в разных гнездах), максимальная в преде-
лах + 35,9°. . . + 41,4°, минимальная, в присутствии наседки + 25,2°. .. +
+ 32,6°, а за время ее прогулок падает до + 12,8°.. . + 23,7°, в зависимости
от температуры воздуха и продолжительности отлучки. Спугнутая с
гнезда глухарка возвращается к нему только через несколько часов ( от
0,5 до 10 ч), и за это время температура яиц может снизиться до + 5°... +
+ 10°, без вреда для дальнейшего развития эмбрионов. Во время вылуп-
ления птенцов наседка сидит на гнезде непрерывно около суток, ста-
раясь увести выводок среди дня и в хорошую погоду. У одной глухарки
птенцы вывелись из яиц во время позднего снегопада 20-22 июня 1977 г.:
матка оставалась с ними в гнезде 48 ч, выжидая пока растает снег, и
только в полдень 22. VI, когда выглянуло солнце и земля обнажилась,
увела их в лес.
114
Глухарка выводит птенцов раньше, чем другие виды тетеревиных, в
среднем за 39 лет 26 июня (17. VI 1940 г. - 8. VII 1938 г., о = ± 5,7).
Описанный режим инкубации обеспечивает развитие яиц в большом
диапазоне погодных условий - и тем не менее успех размножения
зависит рт температуры воздуха во время насиживания. Корреляция
между средней температурой июня и относительным количеством молод-
няка в популяции осенью характеризуется коэффициентами г = + 0,78 ±
± 0,12 (1931—1941 гг.) или + 0,45 ± 0,15 (1958-1987 гг.). Различие в величи-
не этого коэффициента объясняется, вероятно, тем, что в 1931-1941 гг.
колебания возрастного состава популяции были ярче выражены, чем в
позднейшие годы; кроме того, до 1941 г. возрастной состав определялся
по добытым на охоте птицам, а с 1958 г. - преимущественно визуально.
Первоначальное предположение, что в ’’неурожайный” год эмбрионы
погибают от холода до или в процессе насиживания, не подтвердилось.
Из прослеженных нами 62 гнезд, содержавших 353 яйца, только в одном
случае глухарка насиживала 4 яйца с мертвыми эмбрионами, причем
погибли они не от холода, а. в середине инкубации, в жаркие дни, вероят-
но от потери влаги. Неожиданно выяснилось, что яйца менее чувстви-
тельны к холоду, чем наседка, которая легко бросает кладку при небла-
гоприятной для насиживания погоде, особенно если ее в это время
побеспокоят. Три гнезда были брошены во время кладки (29. V 1966 г. - 4
яйца, 6.VI1981 г. - 3 яйца, 18.V 1982 г. - 1 яйцо), три глухарки бросили гнезда в
начале инкубации (11.VI 1951 - 5 яиц, 13.VI 1951 г. - 6 яиц, 3.VI 1976 г. - 7 яиц).
Последний случай хорошо документирован, так как гнездо было оборудовано
актографом и электротермометром, датчик которого находился внутри одного
из яиц. Гнездо было найдено 27 мая, в первый день инкубации; глухарка
насиживала только неделю, вечером 2.VI резко похолодало, и через сутки, 3
июня в 21 ч, она покинула гнездо за несколько часов до сильного заморозка,
во время которого температура брошенных яиц опустилась ниже нуля.
Из 63 глухариных кладок с 360 яйцами, судьба которых известна, 18 с
91 яйцом погибли целиком (8 разорены хищниками, 8 брошены, 2 сгоре-
ли). В 45 благополучных кладках, содержавших 269 яиц, в 17 яйцах погиб
эмбрион, 8 разбиты или похищены, 13 птенцов погибли при вылуплении,
уведены с гнезд 231 цыпленок. Таким образом, успех гнездования -
64%, потери от хищников - 12%, брошены глухаркой - 10%, погибли в
пожаре - 3%, отход в процессе инкубации - 11%.
В выводковую пору (июль-сентябрь) глухари кормятся на поверхнос-
ти земли в основном листьями и цветами травянистых растений и вечно-
зеленых кустарничков (у взрослых птиц 49%, у птенцов - 37% рациона) и
ягодами , (у взрослых - 31%, у птенцов - 38%), отчасти различными
семенами (соответственно 7 и 12%), насекомыми и другими мелкими
животными (6 и 10%), с небольшой добавкой грибов и древесных кормов
(галлы на еловых побегах). Птенцы явно предпочитают животный корм и
семена, а взрослые - зелень^ особенно листья голубики. Основу летнего
питания глухаря составляют два растения, черника и голубика,- в
сумме они дают 75% рациона взрослых птиц или 43% - молодых. Интерес-
но, что птенцы без разбора едят ягоды обоих видов, а взрослые предпочи-
тают чернику: на 10 ягод голубики в пище молодых приходится 6, а в
пище взрослых - 6.0 ягод черники. Очевидно, умение выбирать лучший
корм приобретается опытом.
115
Летом глухари выбирают места с хорошими ремизными условиями;
лишенных укрытий, хорошо просматриваемых, чистых боров они в это
время избегают, а предпочитают пересеченную местность, гряды впере-
межку с болотами, речные долины, разновозрастные смешанные леса с
наличием валежин, подроста и елей, кроны которых спускаются до
земли. Помимо укрытий, им нужны обнажения песчаного грунта, чтобы
’’порхаться”, пересыпая оперение пылью (средство защиты от пухоедов и
кровососущих насекомых?). Они находят такие места под корнями
вывороченных бурей деревьев, перед челом медвежьей берлоги, на
оленьих тропах, кострищах или в сухой трухе сгнившего древесного
ствола. Глухари начинают ’’порхаться”, как только просохнет земля,
около 30 мая (13. V - 18. VI за 27 лет), а кончают в самое различное
время,в зависимости от характера осени (сухая или дождливая), от 2. VIII
1980 г. до 10. X 1960 г. (в среднем 5. IX).
Взрослые птицы линяют на протяжении всего лета и осени: в мае
начинается линька шеи и маховых (первостепенные сменяются начиная
от внутренних к внешним, второстепенные в обратном направлении).
Смена рулевых происходит, начиная с внешних к внутренним, у сам-
цов - со второй половины июня, у самок - на месяц позже, заканчиваясь
в сентябре. Линька контурного пера кончается в октябре. Последними
вырастают роговые бахромы подотеки и оперение лап.
У птенцов маховые первой генерации начинают рост с рождения, а с 20
дней они постепенно сменяются перьями второй генерации, кроме двух
наружных первостепенных маховых, которые сменяются только на
второе лето. Они отличаются от маховых второй генерации крапчатыми и
заостренными вершинами: по этому признаку молодых птиц легко
отличить от более старых на протяжении целого года. В трехнедельном
возрасте птенцы полностью оперены и самцов можно отличить от самок
по рисунку перьев шеи, серых с тонкими поперечными полосками у
первых, охристых с черными пестринами у вторых. В первой декаде
августа на шее петушков появляются черные перья второй генерации, а к
середине октября они отличаются от взрослых петухов главным образом
коротким хвостом и меньшим весом. К этому времени выводки распа-
даются и самцы держатся отдельно от сЬмок.
Осеннее токование глухарей на Кольском полуострове наблюдается
редко, в разные сроки между 19. VIII и 14. X.
Переход глухарей с подножных летних кормов на древесные, зимние,
происходит постепенно, с конца сентября до начала ноября. Хвоя и
древесные веточки перемалываются в мускульном желудке с помощью
камешков (гастролитов). Глухари заглатывают камешки (преимущест-
венно кварц) с сентября и продолжают собирать их на озерных пляжах,
лесных дорогах и других обнажениях грунта, вплоть до выпадения снега.
Галечный сезон продолжается около месяца (27. IX - 30. X в среднем). В
начале зимы желудок птицы содержит несколько сот гастролитов, и вес
их достигает 1% веса тела; к весне запас сокращается, и уже в марте
глухари заходят под вывороты, выбирая камешки, застрявшие в переп-
лете корней.
В начале зимы глухари ночуют на деревьях, а в сильный мороз - на
земле, в снежной ямке. Минимальная высота снежного покрова, при
которой глухарь зарывается полностью - 40 см - для самок и 50 см - для
самцов. Снежный покров достигает 50 см, в ельниках - около 14. XII, в
116
борах - 28. XII; средняя дата первых ночевок глухарей под снегом
23. XII.
В Лапландии глухарь живет оседло, таких концентраций, как белая
куропатка, не образует и объектом промысловой охоты не служит.
Сведений о количестве глухарей, добываемых охотниками, у нас нет.
Глухари несут большие потери от хищных зверей и птиц. Из пернатых
хищников первое место принадлежит беркуту. Этот сильный и энергич-
ный хищник без труда ловит и уносит в свое гнездо даже крупных
петухов. Орлан белохвост, хотя и крупнее орла, уступает ему в вооруже-
нии и ловкости, к тому же он предпочитает охотиться на воде, а не в
лесу. В добыче кречета мы находили только глухарок и молодняк. В
гнездах ястреба тетеревятника неоднократно находили кости взрослых
глухарей; вероятно, хищник съедал часть добычи на месте поимки и
приносил в гнездо только остаток. Филин и белая сова, хотя и ловят
глухарей, не приносят большого ущерба, так как редко появляются в
лесной части Кольского полуострова. Летом они, как и другие совы,
охотятся преимущественно на грызунов, а птиц добывают только зимой
(несколько случаев нападений хищных птиц на глухарей описано выше).
Филин, по всей вероятности, добывает тетеревиных птиц спящими,
хватая их в лунке броском, сквозь снег таким же образом, как бородатая
неясыть ловит мелких грызунов (Mikkola, 1981, с. 41).
Из зверей активно преследуют глухарей куница и лиса. Летом они
кормятся главным образом полевками, но с наступлением зимы ловить
их становится тем труднее, чем больше снега, и оба хищника переклю-
чают свою энергию на поиски крупной добычи, прежде всего тетеревиных
птиц. По сообщению Насимовича (1948 б, с. 88), летом их остатки обнару-
жены в 6% всех данных по питанию куницы, зимой - в 33%, а из остатков
тетеревиных в добыче куницы за 1930-1941 гг. на долю глухаря прихо-
дится 81% (Там же, с. 93). Сходные результаты А.А. Насимович получил и
при исследовании питания лисы: ”В экскрементах лисицы, собранных
зимой 1938/39 г. в борах по р. Чуна, остатки глухарей обнаружены в 50%
просмотренных данных. . . Большинство тетеревиных лисица добывает
не самостоятельно, а в виде остатков на следах лесной куницы” (Насимо-
вич, 1948а, с. 55).
Куница, охотясь преимущественно ночью, замечает спящую под
снегом дичь еще издали, вероятно чутьем или, возможно, на слух, обхо-
дит стороной для точной локализации и в несколько прыжков накрывает
сверху, хватая птицу в момент вылета из снега (Семенов-Тян-Шанский,
1959, с. 210). Тем же приемом охоты пользуется лисица (наблюдение
Н. Заводчиковой, 31.1 1982 г.) и другие хищники. Иногда зверь скрады-
вает птицу на жировке5, а 11. V 1933 г., по наблюдению Л. Кокорина,
куница поймала токующего глухаря на дереве и вместе с ним упала на
снег.
Справиться с такой крупной птицей, как глухарь, нелегко, жертве
удается иногда вырваться, отделавшись потерей хвоста.
Иногда глухари гибнут от несчастных случаев. 5. IV 1977 г. на берегу
р. Нижняя Чуна взрослый самец погиб от удара о высоковольтный провод;
11. X 1973 г. залетевшая в г. Мончегорск глухарка, пробив стекло, разби-
5Дважды на наших глазах охотничь и лайки ловили взрослого глухаря, жировавше-
го на черничнике (7. X 1949г. и 11. IX 1955 г.).
117
Таблица 9. "Удельный вес” глухаря в питании хищных птиц и зверей
Птица Число глуха- рей Доля в рацио- не, % Объем матери- ала по питанию Зверь* Число глуха- рей Доля в рацио- не, % Объем материа- ла по пи- танию
Орел беркут 27 37 73 1 Куница, зима 1 48 1 1 16 ' 1 592
Орлан бело- 15 10 145 Куница, лето 32 3 2034
хвост Ястреб-тетере- 11 19 58 Лиса, зима 65 15 433
вятник Кречет 11 7 153 Лиса, лето 28 7 406
Зимняк 4 0,4 953 Росомаха, 15 13 113
Филин 5 71 7 зима Норка, зи- 1 1 77
Белая сова 1 25 4 ма Горностай 3 2 133
Всего: 74 — 1393 Медведь 1 193 0.5 150 3918
*Хищные птицы и куница, как правило, едят собственную добычу; часть, глухарей
в пище других зверей представляет остатки чужой добычи.
лась насмерть в помещении детского сада. В утренних сумерках 19. XII
1970 г. на Чунозерской усадьбе заповедника глухарка налетела на элект-
ропровода, вызвав короткое замыкание. 3. X 1981 г. на берегу Охтозера
собака поймала молодого глухаря, который не мог взлететь из-за трав-
мы: между лучевой и локтевой костями левого крыла застрял еловый
сучок длиной 12 см. В последние годы были добыты также два глухаря со
сросшимися переломами бедреной кости и плюсны.
Табл. 9 дает представление о значении глухаря в питании различных
хищников: для ее составления использованы материалы настоящей
работы (хищные птицы) и опубликованные работы А.А. Насимовича(1948)
и О.И. Семенова-Тян-Шанского (1959,1982).
Из 291 глухаря, остатки которых найдены в природе с 1931 по 1987 г.,
хищными птицами добыты 39 %, зверями - 37%, неопределенными
хищниками - 12%, от несчастных случаев, болезней и огнестрельных ран
погибли 4% и от неизвестных причин - 8%. Приведенные материалы
показывают, что от хищных зверей глухари терпят примерно такой же
урон, как и от птиц, и что около 90% их становятся жертвой хищников,
так что у глухаря почти нет шансов умереть от старости. Последний
вывод подтверждается и анализом возрастного состава популяции.
Медленное окостенение черепа у глухаря дает возможность опреде-
лять возраст самцов до 4 лет, самок - до 3 лет (Кириков, 1944, с. 16-25;
Семенов-Тян-Шанский, 1959, с. 169-176). За 1946-1983 гг. мы отпрепари-
ровали с этой целью 85 черепов взрослых особей из общего числа 332, ис-
пользованных для биосъемки: на основе этого материала и составлена
табл. 10.
Уменьшение числа глухарей с возрастом дает представление о смерт-
ности и вероятности выживания птиц на протяжении года. В стабильной
популяции, очевидно, годовая смертность должна быть равна средне-
118
Таблица 10. Возрастной состав и выживаемость глухаря
Показатель Возраст, годы
0 + > 1 1 + 2 + 3 + 4 + 5 + 6 + 7 + 8 + 9 +
Число, особей, экз. Возрастной сос- тав, % 174 158 34 28 17 6 — — — — —
факт. 524 (476) 190 157 95 34 — — — — —
расч. 524 191 114 69 41 25 1 9 5 7
Вероятность доживания 1000 476 285 171 102 61 36 21 12 7
му ежегодному приросту. Нами исследовано 174 сеголетка и 158 глуха-
рей старше года, это дает возрастное соотношение 524 молодых: 476 взрос-
лых и сооответствует среднему приплоду 110:100 производителей.
В первой строке таблицы дан исходный материал (число особей), во
второй - возрастной состав в промилле, по фактическим данным. На ос-
нове этих цифр находим количественное соотношение соседних возраст-
ных групп: 0, 36 - 0, 83 - 0, 60 - 0, 36. Цифры здесь, как говорится, ’’ска-
чут” по недостаточности статистической выборки (85 черепов), а возмож-
но, и отдельных ошибок в определении возраста черепов. Полагая, что
смертность на первом году жизни должна быть выше, чем в последую-
щие, мы оставляем коэффициент выживания за первый год без измене-
ний (0,36), а для последующих лет принимаем арифметическое среднее
из трех коэффициентов (0,60). Рассчитанный на основе этих коэффициен-
тов выровненный возрастной состав популяции приведен в третьей стро-
ке. В четвертой показана вероятность доживания глухаря до соответ-
ствующего возраста, исходя из первоначального контингента в 1000 птиц
2-3-месячного возраста, т.е. переживших пору детской смертности. По
идее глухарь - долговечная птица, в неволе он достигает 18 лет, а ре-
кордный возраст, по материалам кольцевания, в природе (Южная Финлян-
дия) - 10 + лет (1950 - IX 1960 г.; Grzimek, 1969, с. 370). На деле у него
очень мало шансов дожить до такого возраста: по нашему расчету, до 4
лет доживает только 10 % глухарей, до 8-9 лет - 1 %, а до 13 лет - только
один из тысячи.
Размеры взрослых самцов в среднем: длина 903, крыло 388, хвост 321
мм, масса тела 3930 г; соответственные размеры самок - 600, 295 и 186
мм, масса тела 1888 г. Молодые не успевают вырасти в первое лето, зимой
рост приостанавливается, а на следующее лето они подрастают, сравни-
ваясь со взрослыми. Размеры молодых самцов в первую зиму - 804, 374 и
247 мм, масса тела -3000 г, масса тела самок - 1700 г (по линейным раз-
мерам они мало отличаются от взрослых). Сезонные изменения массы те-
ла такие же, как и белой куропатки, но слабее выражены.
119
Тетерев - Lyrurus tetrix (L.)
Местные названия: саамское - ’’косач” (самец) и ’’тетёрка” (сам-
ка).
Отсутствие собственного названия этой птицы у саамов показывает,
что тетерев заселил Кольский полуостров сравнительно недавно. В конце
прошлого века граница его ареала проходила по южному побережью
Имандры (Плеске, 1887, с. 406). В начале 30-х годов нашего века он встре-
чался в южной части Лапландского заповедника, но жившие к северу от
его границы саамы Нотозерского погоста его еще не знали. В окрестнос-
тях Мончегорска выводок тетеревов впервые был замечен в 1937 г., а
в 1957 г. тетерев доходил уже до Кольского залива на широте Мурманска
(Семенов-Тян-Шанский, 1959, с. 10). Расширению его ареала к северу спо-
собствовали рубки лесов и пожары, с замещением хвойных насаждений
березовыми. В районе Лапландского заповедника в 30-х годах тетерев
был самым малочисленным видом тетеревиных птиц (около 1% добытой
боровой дичи), и только в обжитой полосе вдоль Мурманской железной
дороги он встречался часто. Рост численности начался с 1940 г.: в декабре
уже видели стаю в 39 птиц. Одновременно с сокращением популяции
глухаря в 40-х и 50-х годах, популяция тетерева сильно увеличилась,
по материалам ленточных учетов примерно в 5 раз, по учету визуальных
встреч - до 24 раз. В то время в мае на льду озер токовало до 20-30 ко-
сачей, а 11. V 1951 г. лесник А.С. Сергин, проезжая на оленях по Пиренг-
скому озеру насчитал 98 токующих птиц. Согласно ленточным учетам,
тетерева в то время составляли около 10 % всей боровой дичи, а по марш-
рутным учетам - даже 30 %. Потом наступила катастрофа: летом 1963 г.
было массовое размножение осенней пяденицы (Oporinia autumnata).
Весной ее гусеницы объели едва распустившиеся листья березы и других
лиственных древесных пород. В июле гусеницы окукливались, а в ав-
густе и сентябре бабочки в массе летали по лесу, напоминая снежную ме-
тель. Летом гусеницы, а осенью бабочки стали едва ли не главным кормом
глухариных и тетеревиных выводков. Нашествие пядениц продолжалось
еще два года (до 1965). От ежегодного уничтожения листвы березы стали
усыхать, около половины деревьев погибли, а оставшиеся на протяжении
нескольких лет не цвели и не приносили семян. Цветочные сережки бе-
рез - основная зимняя пища тетерева, поэтому гибель берез стала для него
бедствием. Основная масса особей, по-видимому, мигрировала на юг: с ок-
тября до начала декабря 1965 г. сотрудники Кандалакшского заповед-
ника наблюдали, как стаи тетеревов летели через г. Кандалакшу и за-
лив на Карельский берег. Численность тетерева в Лапландском заповед-
нике резко сократилась, и вплоть до 1972 г. токования почти не было
слышно (Семенов-Тян-Шанский, 1975, с. 106, 161). Средняя встречаемость
тетерева в 1959-1965 гг. составляла 24 на 100 км, в 1966-1972 гг. только
3. Позднее популяция начала восстанавливаться, и в 1973-1987 гг. сред-
няя встречаемость составляла 10 на 100 км.
Зимой тетерева, как и глухари, кормятся на деревьях, а ночуют в
лунках под снегом. В отличие от глухарей самцы вместе с самками об-
разуют большие смешанные стаи до 30- 40 птиц и в поисках корма совер-
120
шают большие перелеты. Питаясь хвоей, глухарь обладает практически
неограниченным запасам зимних кормов, правда малопитательных. Зим-
ний рацион тетерева включает полноценные генеративные органы деревь-
ев, преимущественно цветочные сережки берез, запас которых в лесу
довольно устойчив от года к году. Кроме сережек, тетерев, подобно
белой куропатке, стрижет и березовые веточки с лиственными почками.
На соснах он кормится не столько хвоей, как ’’озимью” неспелыми
шишечками около 1 см в диаметре. Семена сосны созревают на второй
год после цветения, поэтому зимой на ней бывают шишки двух возрас-
тов - большие, содержащие спелые семена, и маленькие, с опыленными,
но еще неоплодотворенными яйцеклетками. В зобу одного тетерева мы
нашли 176 таких шишечек. Сосна цветет и плодоносит не ежегодно, поэ-
тому ’’озимь” доступна птицам только в некоторые годы. У можжевель-
ника тетерев ест главным образом плоды (’’ягоды”) и только отчасти
хвою.
Запасы цветочных сережек, сосновой ’’озими” и можжевеловых
’’ягод” невелики, быстро иссякают, и вторично в. ту же зиму на одном
дереве тетерев не может прокормиться: этим объясняются как большие пе-
релеты их на кормежку, так и зависимость их численности от состояния
кормовых фондов.
Веточным кормом тетерева кормятся с конца октября до середины
мая, около 194 дней в году. Летом он по сравнению с глухарем ест боль-
ше ягод и семян, но меньше зелени. Уже в апреле в поисках подснежных
ягод брусники, вороники и ”гром-ягоды” тетерева залетают в горную
тундру кормиться на первых проталинах. В мае их пища состоит из бе-
резовых сережек, перезимовавших ягод, зеленых побегов черники, а в
конце лета и осенью - из ягод черники и голубики, плодов осок, семян
марьянника и других растений, с добавкой листьев голубики, андромеды
и других. Животный корм потребляют преимущественно птенцы.
Тетерева токуют энергичнее, демонстративнее и дольше, чем глухари.
Песня их состоит из двух элементов: продолжительного ’’бормотания”,
напоминающего воркование голубя вяхиря, и короткого ’’чуфыканья”.
Оба звука разносятся на большое расстояние, на открытом месте (через
озеро) до 4 км, так что начало токования замечается легко и без опозда-
ния. В среднем тетерева начинают петь 14 апреля (15. III 1967 г. - 10. V
1935 г., за 50 лет; о ± И, 3 дня). Первое время тетерева токуют на деревь-
ях и только днем, позднее - на поверхности снега; петь начинают до вос-
хода солнца, а позднее, с вечера, токуя всю ночь. В Лапландии токовища
располагаются на льду озер или на обширных болотах. Интенсивность
токования зависит как от погоды, так и от численности птиц: в 1972,
1979, 1980 и 1983 гг. тетеревов было мало, на ток вылетали 1-2 петуха,
и слышали песню всего 4-6 раз за весну; напротив, в 1974, 1978, 1981 и
1986 гг. за весну отмечено по 20-26 дней токования, и на лед вылетало
до 15-30 птиц. Особенно хорошо токовали тетерева в 1949-1951 гг. Пос-
леднее пение наблюдается в среднем 9 июня (13. V 1979 г. - 24. VI 1971 г.
за 36 лет). В позднюю пору косачи снова поют на деревьях.
В мае на лед вылетают и тетерки, в их присутствии самцы оживляются
121
и нередко ожесточенно дерутся: черные перья, вырванные в драке,
часто попадаются на токовищих, а однажды (20. V 1951 г.) мы добыли
косача с заживающими ранами на затылке и в области зоба, с разрежен-
ным в этих местах оперением.
Поза токующего самца очень характерна, хвост поднят вертикально
и развернут до предела, так что концы боковых рулевых касаются грун-
та, а нижние кроющие образуют яркое белое зеркало, видимое не только
сзади, но и спереди, поскольку эти кроющие длиннее средних рулевых;
крылья опущены, концы их чертят борозды на снегу, шея вытянута впе-
ред, и оперение на ней оттопырено.
Тетерки гнездятся либо в смешанном сосново-елово-березовом или
елово-березовом лесу, либо на открытом сфагновом болоте с сосною
(в чистом бору гнезд мы не находили). Два гнезда находились в 1-4 м
от торной пешеходной тропинки (обе кладки разорили хищники), а два
других - в 6-8 м от строившейся автодороги, на которой работали трак-
торы и другая техника, - и в обоих вывелись птенцы. Гнездо устраивает-
ся под прикрытием елки или березы, а на болоте - открыто. Лоток глу-
бокий, так что насиживающую тетерку сбоку не видно за растительнос-
тью. Сидит она крепко, подпуская на 2-3 м прежде, чем взлететь. Кладка
содержит от 5 до 9 яиц, в среднем 7, 3; средняя масса ненасиженного яйца
31,6 г. Процесс кладки мы не наблюдали; время инкубации определено
только в одном гнезде - 24, 3 сут. Режим насиживания регистрировался
самописцами в 4 гнездах, в общей сложности на протяжении 63 сут.
Наседка оставляет гнездо для прогулки 2-5 раз в день, в среднем 3, 0
раза, на срок от 5 до 40 мин, в среднем 20, 4. Вне гнезда она проводит око-
ло 60 мин в сутки, или 0, 04 всего времени инкубации. Будучи спугнутой
с гнезда, она возвращается через 0, 5-0, 3 ч. Температура яиц, по автома-
тической записи на протяжении 107 ч в одном гнезде, варьировала в пре-
делах 20, 5 - 39, Г, в среднем, пока наседка сидит на яйцах, 34, 4, пони-
жаясь в ее отсутствие в среднем до 25°. Сроки кладки и насиживания,
как у глухарки или чуть позже, в среднем за 11 лет первые выводки
встречались 29 июня (24. VI 1937 г. - 7. VII 1963 г.) Тетеревиные выводки
предпочитают сравнительно открытые стации - болота, гари, вырубки.
Относительное количество молодняка в популяции варьирует по го-
дам, как у глухаря, от 20 % в 1977 г. до 81 % в 1970 г., и также зависит от
температуры воздуха во время насиживания: коэффициент корреляции
между температурой июня и процентом сеголетков осенью, г = + 0, 48 ±
±0,17 (за 21 год).
Осеннее токование наблюдается только в годы достаточно высокой
численности, но регулярнее, чем у глухаря, в широких пределах, от се-
редины августа (15. VIII 1950 г, 19. VIII 1949 г.) до 29. XI в 1966 г. (в сред-
нем за 12—14 лет с 16. IX до 24. X). Поют не только взрослые петухи, но
и сеголетки, обычно они только ’’бормочут”, сидя на дереве. Начинается
ток рано утром, в августе с 4 ч, в сентябре - с 6-8 ч (молодые поют и сре-
ди дня). После замерзания озер тетерева иногда токуют на льду, как вес-
ною: 15. XI 1948 г. на льду Охтозера токовали 24 косача.
122
Во второй половине сентября, с понижением температуры, насекомые
и другие беспозвоночные прячутся на зимовку, и находить животный
корм все труднее. В это время тетерева раскапывают муравейники, едят
муравьев, а возможно, и'”порхаются”, оставляя на раскопе перышки, по
которым можно определить вид птицы (белые куропатки и глухари
также копаются в муравейниках, но реже): это явление мы наблюдали
между 8. IX - 4. X
В октябре и ягоды и черничные побеги уходят под снег; с этого момен-
та тетерева переходят на древесное питание, а на поверхности или в тол-
ще снега только ночуют.
Естественная смертность относительно выше, чем у глухаря: с 1931 г.
зарегистрированы остатки 113 тетеревов, из них 50 % добыты хищными
птицами, 29% - зверями, 16% - хищниками, точнее не определенными, 5%
погибли от случайных или неизвестных причин. В добыче кречета найде-
но 13 особей, ястреба-тетеревятника - 6, орлана - 2. Остатки тетеревов
обнаружены в питании лисы и куницы, хотя и реже, чем у глухарей и
белых куропаток; среди жертв росомахи, орла, зимняка и крупных сов не
встречены.
16.1 1962 г. лесник Я.Р. Януше на лесосеке у Мавра-губы руками пой-
мал 4 косачей которые не могли взлететь из-за обмерзания крыльев и
задней части тела. Освобожденные от льда, они улетели. Каким образом
птицы намокли в морозную погоду установить не удалось.
Половой диморфизм в размерах слабее выражен, чем у глухаря: дли-
на тела взрослого самца 616, крыла 272, хвоста 216 мм, масса тела - 1426 г,
самца сеголетка соответственно - 569, 268, и 174 мм, масса тела 1242;
самки - 472,240 и 126 мм, масса тела 968 г.
Рябчик - Tetrastes bonasia (L.)
Местное название: саамское”пынг”.
Рябчик - типичная таежная птица, распространения по всей’ Пале-
арктике, от Скандинавии до Тихого океана. На Кольском полуострове
он приурочен к хвойным лесам, и в 30-х годах его ареал доходил к севе-
ру до г. Колы и с. Краснощелье (на Поное), но теперь, вероятно, сократил-
ся в связи с вырубкой лесов. По материалам ленточного учета за 41 год,
средняя плотность рябчика в лесах Лапландского заповедника - 24 пти-
цы на 1000 га, равная плотности белой куропатки: это обстоятельство
ставит под сомнение репрезентативность ленточного учета, так как по
данным маршрутного учета, регистрации визуальных встреч и другим
косвенным показателям, численность рябчика много ниже (табл. 7); нап-
ример, по маршрутным учетам он стоит на последнем месте, составляя
только 5 % всей боровой дичи. Линии ленточного учета пересекают мно-
го ручьев, сбегающих с Чуна-тундры, с ольшанником по берегам, а мар-
шрутный учет обычно сочетался с охотой и проводился в лесах и на боло:
тах к югу от Чунозера и Охтозера, где мало пригодных для рябчика
стаций. Принимая, что рябчик составляет 5 % всей учтенной на лентах
дичи, получаем среднюю плотность его 4 на 1000 га; при площади лесов
123
в заповеднике, равной 852 км2, численность рябчика оценивается по пер-
вому расчету в 2060, по второму - в 380 особей.
Численность сильно колеблется по годам, учеты указывают на повы-
шение ее в 1933-1938, 1959-1962, 1970-1971, 1976 1979,1986 гг. и сниже-
ние в 1939-1940, 1958, 1966-1969, 1972, 1978 и 1982-1983 гг. Эти кратко-
временные колебания происходят на фоне постепенного сокращения
популяции: на протяжении полувека она уменьшилась, по крайней мере,
в пять раз. По этой причине в Мурманской области с 1983 г. охота на него
запрещена.
Зимой рябчики живут парами или поодиночке, преимущественно по
долинам ручьев или лощинам, где растет серая ольха, встречаются они
и в густых ельниках с примесью лиственных пород. Рябчик довольствует-
ся малой территорией, при условии, что она удовлетворяет его жизнен-
ным потребностям: ели дают укрытие, а ольха, ивы и березы - пищу.
Открытых мест - болот, гарей, а особенно горной тундры - он избегает.
Под снегом он не только ночует, но долго продолжает доставать из-под
него зеленые побеги черники в качестве добавки к древесному корму -
цветочным сережкам, почкам и побегам ольхи и березы, составляющим
основу его диеты на протяжении семи зимних месяцев (с октября по
апрель). Хотя по сравнению с глухарем и белой куропаткой рябчик ис-
пользует более питательный корм, зимой его энергетический баланс отри-
цательный. Максимальной упитанности он достигает в ноябре-декабре
(средняя масса тела самца 432, самки 414 г): к марту масса тела самок
уменьшается на 9%, но к началу сезона размножения они восстанавли-
вают потери, и средняя масса тела 4 самок, добытых в мае, оказалась
406 г (по самцам материала нет).
В апреле рябчики находят на проталинах ягоды брусники и вороники,
а в мае питаются в основном наземными кормами (побеги черники и
перезимовавшие ягоды), к этому добавляются начинающие в это время
цвести сережки ив и берез (около 40 % рациона). В июне и июле он
охотно ест цветы, а потом завязи черники и других ягодных кустарничков с
добавлением листьев голубики и древесных пород. В августе-сентябре основу
рациона составляют ягоды черники, вороники и другие, затем сухие пло-
ды и семена осок, герани, марьянника и берез. В сентябре он начинает
есть ольховые и березовые веточки и сережки; с октября до нового года
’’удельный вес” цветочных кормом постепенно возрастает, а ягод и по-
бегов кустарничков снижается. Птенцы летом кормятся главным обра-
зом ягодами, семенами и беспозвоночными (последние в июле состав-
ляют треть их рациона). В летнем рационе взрослых птиц насекомые, пау-
ки и слизни составляют 3-7 %.
Сведения о размножении рябчика в Лапландском заповеднике скуд-
ны. Кладка яиц происходит в последней декаде мая: 22. V 1934 г. добыта
самка с формирующимся яйцом в яйцеводе, а 31. V 1947 г. найдено гнез-
до с 7 яйцами; другое найдено в ельнике у берега оз. Ельявр летом 1964 г.
третье в разреженном ельнике на восточном склоне Ельнюна в сентябре
1987 г.; в обоих скорлупы яиц, из которых вывелись птенцы, в одном 8,
124
Рис. 34. Рябчик самец, 3. 1973 г. Фото С. Баккала
в другом 5. Первые выводки рябчиков встречали, в среднем за 12 лет,
30 июня (22. VI 1937 г. - 6. VII1971 г. и 1981 г.).
Процесс кладки и насиживания описаны нами по одному гнезду,
найденному в Печоро-Илычском заповеднике в 1955 г. Инкубационный
период - 25,4 сут. Самец не подменяет самку на гнезде, но держится
поблизости, а иногда вместе с ней находится при выводке. Птенцы уже
в раннем возрасте взлетают на деревья и там затаиваются.
Рябчики обнаружены в добыче ястреба-тетеревятника, лисы и куницы
реже, чем другие куриные птицы. Остатки погибших (22 находки) состав-
ляют 2% от всех зарегистрированных остатков тетеревиных птиц; кроме
одной особи, все были жертвами хищников. В октябре-декабре 1973 г.
рябчики (до 7-13 птиц одновременно) нередко кормились на усадьбе
заповедника (рис. 34), обрывая ивовые почки, ольховые и березовые се-
режки: по-видимому, в ту зиму в лесу им не хватало корма.
Размеры самцов: длина 378, крыло 167, хвост 123 мм; самки соответ-
ственно - 370, 165 и 116 мм. Средняя масса тела 36 самцов и 42 самок
одинаковая - 398 г.
Рябчик вполне оседлая птица. Вылет у него шумный, но летает он ма-
ло и только на короткое расстояние, в связи с чем сердце у него малень-
кое - всего 0,5 % живой массы тела птицы: это в 4 раза меньше, чем у тун-
дряной куропатки, которая летает больше, чем остальные тетеревиные.
125
ОТРЯД ЖУРАВЛЕОБРАЗНЫЕ - GRUIFORMES
Серый журавль - Grus grus (L.)
Журавль, по сообщению Плеске, ’’лопарям на Имандре хорошо извес-
тен, хотя, по их словам, он появляется в указанной местности не ежегод-
но” (Плеске, 1887, с. 413). В наше время ареал журавля доходит до линии
пос. Верхнетуломский-Ревда- Краснощелье-Сосновка, но регулярно
гнездится только на юге Мурманской области (Коханов, 1982, с. 45-48).
По сообщению В.С. Дребенцова, журавль встречался на Поное, у Красно-
щелья и в конце 40-х годов.
В районе Лапландского заповедника их впервые наблюдали 10. VIII
1950 г.: стая около 10 журавлей летела на юг. С 1966 г. их видели почти
ежегодно, обычно на лету, одиночных, парами, иногда по 3- 4, чаще вес-
ной (со второй декады мая по вторую декаду июня) или осенью (трижды
в августе и дважды в октябре). Сроки осенних встреч совпадают либо
с началом осенних кочевок журавлей (август), либо с окончанием их от-
лета в более южных широтах (Мальчевский, Пукинский, 1983, Т.Т, с. 244-
245). Средняя дата первой весенней встречи журавлей в Лапландском за-
поведнике - 22 мая (13. V - 3. VI, за 13 лет).
Регулярные весенние залеты журавлей напоминают такие же залеты
чибиса и некоторых других птиц, но возможно, что изредка они гнездят-
ся в нашем регионе. 17. VI 1977 г. на болоте в долине Пельйок лесник
Н.А. Сергачев видел одного журавля, который подпустил его метров на
сто и не взлетел, а только тревожно кричал: создалось впечатление, что
его партнер сидел на гнезде где-то поблизости. В 1982 г. журавлей виде-
ли или слышали в долинах рек Нявка, Вува и на Чунозере пять раз, с
22. V по 15. VII.
Несколько раз журавли появлялись стаями: 17. VI 1971 г. над кордо-
ном Нявка пролетел на северо-восток косяк в 18 птиц; 21. VII 1976 г. на
поле близ станции Тайбола сидели, а потом улетели 13 журавлей; в
конце августа 1980 г. на болоте близ поселка Уполокша Н.А. Сергачев
видел стаю около 30 птиц; 26 июня 1983 г. в 3 ч среднего солнечного вре-
мени 23 журавля пролетели над Чунозером на юго-запад.
Всего с 1966 по 1987 г. в заповеднике зарегистрировано визуально 109
журавлей. Подсчет встреч на одного наблюдателя в год показал заметный
рост численности журавля за последние годы:
Годы 30-е 40-е 50-е 60-е 70-е 80-е
Журавлей на 1 0,0 0,0 1,0 0,5 2,9 3,7
наблюдение в'
год
27. VI1965 г. Ю.Г. Гаев видел на болоте близ Кокоринского озерка (под
Курт-варенчем) пару крупных птиц, белых с черными концами крыльев,
по его мнению - залетных стерхов. Ближайшие гнездовья этого журав-
ля находятся в низовьях Оби, в 1500 км к востоку от Кольского полу-
острова, поэтому залет его маловероятен; легко допустить, что наблю-
датель видел белых аистов, птицу более обычную и гнездящуюся не да-
лее 1000 км к ЮЮЗ от нас.
126
Лысуха - Fulica atra L.
На северо- западе нашей страны, в Приладожье, лысуха гнездится до
62° с.ш., но изредка залетает и в Лапландию. В 1857 и 1875 гг. одиночные
лысухи были добыты в районе Варангер - фиорда (Плеске, 1887, с. 412),
а в наше время их встречали на Айновых островах (Курочкин, Кошелев,
1987, с. 445), и близ с. Ловозера (Михайлов, Фильчагов, 1984, с. 26).
3. VII 1969 г. пару лысух наблюдали в Лапландском заповеднике на
оз. Кензисьявд московские юннаты Стопалов, Луценко и Добрушин; к
сожалению, в карточке не указаны признаки, по которым они идентифи-
цировали вид, поэтому не исключено, что за лысуху была принята синьга,
птица, сходная с ней по размерам и окраске.
ОТРЯД РЖАНКООБРАЗНЫЕ - CHARADRIIFORMES
ПОДОТРЯД КУЛИКИ - CHARADRII
Тулес- Squatarola squatarola (L.)
Тулес гнездится на равнинных тундрах Азии и Северной Америки; в
нашей стране от п-ова Канина до Чукотки. Зимует на морских побережь-
ях Британских островов, Западной Европы и Африки. На Кольском полу-
острове встречается нерегулярно и только в пролетное время.
18. IX 1934 г. на Железном наволоке (Екостровская Имандра) Семенов-
Тян-Шанский наблюдал смешанную стайку пролетных куликов, красно-
зобиков вместе с двумя десятками тулесов и одну молодую самку до-
был. Одиночного тулеса, кормившегося ягодами на вершине Ельявруай-
венча, добыл 18. IX 1939 г. А.А. Насимович. В начале августа 1959 г. двух
тулесов видел на Пельтундре Ю.Г. Гаев; 15. VIII 1961 г. на дороге между
Екостровским проливом и поселком Тикгуба также двух видел Семенов-
Тян-Шанский.
Наблюдали тулесов и весной: 17 и 18. V 1966 г. А.И. Флеров видел пару
на р. Нявка (они сидели на льду у края воды), а 2. VI 1975 г. пару летев-
ших над южной оконечностью Нявка-тундры видел А.Б. Брагин. Всего
за 45 лет 6 встреч, около 30 особей.
Размеры молодой самки, добытой в 1939 г., длина - 282, крыло - 204,
хвост - 78 мм, масса тела 165 г.
Золотистая ржанка - Pluvialis apricarius (L.)
Местные названия: поморское - семенуха, семенушка(поПлес-
ке); саамское нам не известно.
Золотистая ржанка гнездится по горным и равнинным тундрам, от
Скандинавского до Таймырского полуострова. В Лапландском заповед-
нике в 30-х годах эту ржанку встречали всего два раза - 28. VI 1936 г.
одиночный самец добыт А.Н. Жуковым на Ельнюне (Чуна-тундра), а 27.
VIII1937 г. одну птицу видел там же Семенов-Тян-Шанский: в то время
считалось, что она в заповеднике не гнездится (Владимирская, 1948,
127
с. 202). С 1960 г. она стала обычной гнездящейся птицей горных тундр.
Регистрация ее встреч, в пересчете
следующий результат:
Годы:
Ржанок на 1
наблюдателя
в год
Число птиц
Прилетают к
16 лет - 24 мая (13. V 1977 г. - 8. VI 1967 г.). Последних ржанок отмечали
в разные сроки, от 10. VIII. 1982 г. до 20. IX 1964 г., а в среднем за 10 лет -
27 августа (даты эти приблизительны, так как стации ржанок посещались
нерегулярно).
Типичная стация золотистой ржанки - горная тундра. Скалистые мес-
та ее не привлекают, она предпочитает ровные, плоские участки с преоб-
ладанием ягельно-кустарничкового или злаково-лугового покрова, но
селится и ниже, в сухом березовом криволесье ' со сходным напочвен-
ным покровом (на горные тундры приходится 64 % встреч). Часть ржанок
гнездится в лесном поясе, преимущественно на обширных сухих кочкар-
ных болотах, богатых вороникой и морошкой (20 % встреч), реже - на
старых гарях с редким сосновым древостоем или вторичным березняком
(6 % птиц). Остальные 10 % ржанок учтены вне характерных стаций, на
лету.
Золотистые ржанки занимают гнездовые участки сразу после прилета.
В заповеднике найдены три гнезда ржанок и несколько раз находили
на одного наблюдателя
в год, дала
30-е
0,2
40-е
0,0
50-е
0,0
60-е
9,8
70-е
7,5
80-е
6,2
Всего
4,0
2
нам
о
ржанки
о
обычно
276
164
третьей декаде
206
мая, в
648
среднем за
в
пуховых птенцов.
Вечером 12. VI 1967 г. на вырубке ягельного бора в долине р. Чуна, в
1,5 м от края заброшенной дороги, по взлетевшей птице обнаружено
гнездо ржанки с 4 яйцами. Инкубация, вероятно, началась только что,
так как 10. VI проходившие по дороге лица никого не спугнули (8. VI
неподалеку видели летевшую ржанку). С помощью актографа, поставлен-
ного в гнездо 14/VI в 18 ч., установлено, что насиживают кладку оба
партнера, сменяясь два раза в сутки, утром, между 4 ч 50 мин и 8 ч
29 мин (в среднем за 8 дней - в 7 ч 09 мин), и вечером, между 19 ч 15 мин
и 21 ч 12 мин (в среднем за 5 дней - в 20 ч 35 мин). Ночная вахта продол-
жалась около 10 ч 38 мин, дневная - 13 ч 26 мин. Смена происходит поч-
ти мгновенно, только один раз яйца оставались открытыми до 3 мин.
По наблюдениям, в горах Норвегии днем на гнезде золотистой ржанки
сидит самец, ночью - самка (Byrkyedal, 1985, р. 486-501).
Люди и звери, проходя по дороге, не раз спугивали наседку, но она
возвращалась и продолжала насиживание в среднем через 20 мин (от 7
до 37 мин: 7 случаев). Если птицу спугивали незадолго до урочного сро-
ка смены партнеров - на гнездо садилась уже другая птица. Вечером 21.
VI, после 7 сут отчетливой записи, регулировка прибора нарушилась, и в
дальнейшем удалось получить только две суточные ленты; 24-25 июня
партнеры сменялись в прежнем порядке, а 28. VI режим резко изменился:
насиживание продолжила только одна птица, причем за сутки она пять
раз оставляла гнездо для кормежки (в 7-ом, 9-ом, дважды в 21-м и в 24-м
128
часу), на срок от 5 до 16 мин: на протяжении 24 ч она отсутствовала 48
мин (3, 3 % всего времени). 29 июня в 2 ч 46 мин запись прекратилась; по-
видимому, хищник согнал ржанку и уничтожил кладку. Гнездо состоя-
ло под контролем немного более 16 сут, тогда как по литературным ука-
заниям золотистая ржанка насиживает 27-28 дней. Предполагая, что ин-
кубация началась 10-11. VI, находим, что самец и самка насиживали по-
очередно от 14 до 17 сут, после чего одна из птиц отступилась, предоста-
вив другой заканчивать инкубацию (Семенов-Тян-Шанский, Брагин, 1969,
с. 61-63).
21. VII 1969 г. на Волчьей тундре А.И. Флеров встретил самку с пуховыми птен-
цами.
9. VI 1979 г. в ягельном березовом редколесье (лесотундре) к северу от оз. Нявка
А.С. Гилязов нашел гнездо с 3 яйцами и сидевшей на них ржанкой.
20. VII 1980 г. на Ельнюнлаге, Чуна-тундра, у озера, встречен выводок в составе
2 взрослых птиц и 3 летных птенцов (Семенов-Тян-Шанский).
8. VI 1982 г. на склоне Рейтлагуая, Нявка-тундра, лесник В. Данченко нашел сре-
ди мха и брусничника гнездо ржанки с 4 яйцами.
Из этих наблюдений следует, что золотистая ржанка начинает насижи-
вание в первой или второй декаде июня, а птенцы выводятся с конца
июня до 20 июля.
Желудок самца, добытого в июле 1960 г. на болоте у Нявка-озера, со-
держал ягоды вороники, морошки и одну гусеницу.
Главными врагами ржанки надо считать различных соколов. 4.Х. 1961
на Большой Нялл-тундре найден общипанный костяк ржанки с сохранив-
шейся головой, а отчасти и грудными мускулами: очевидно, она стала
жертвой мелкого хищника, например дербника. 7. VI 1979 г. в долине
р. Лива А.С. Гилязов нашел перья погибшей ржанки, а 3. VIII 1983 г.
О.А. Макарова нашла у конторы Северного лесничества, на берегу Монче-
озера, крылья молодой ржанки, пойманной кошкой. Под гнездом кре-
чета летом 1975 г. обнаружены остатки двух крупных куликов. У гнезда
сапсана в июле 1982 г. на берегу р. Пясина (Таймырская тундра) мы наш-
ли остатки одной из ржанок (золотистой или тулеса).
Золотистая ржанка - крупный кулик с большой головой; верхняя сто-
рона крапчатая, золотисто-желтая с черным, нижняя - черная. Белые над-
бровные полоски продолжаются по бокам шеи и туловища, резко отделяя
золитистую окраску верхней стороны от черной груди: на птице, обращенной к
наблюдателю в профиль, она образует белую букву S. Голос - протяж-
ный, минорный свист, слегка напоминающий крик зимняка, но в 2-3 ра-
за короче. Кричащая птица поворачивает голову в разные стороны, от-
чего кажется, что птиц несколько и они кричат со всех сторон; вдобавок
она все время перебегает, замирая на месте в момент остановки, так что
ее трудно разглядеть. Брачную песню можно передать словами ’’тюрлй-
тирлй”.
Размеры двух добытых нами экземпляров: длина 277-290, крыло 185-
191, хвост 74-76 мм, масса тела 175 г (самец) и 211 г (пол неизвестен).
9. Зак. 1023
129
Галстучник - Charadrius hiaticula L.
Местные названия: саамское ’’пулдыг” , русское - зуек.
Галстучник гнездится по берегам морей, рек и озер в зонах тундры,
лесотундры и северной тайги; южнее его замещает близкий вид - малый
зуек. В нашем регионе галстучник распространен по песчаным галечни-
ковым пляжам повсеместно, но немногочислен. При учете уток на Чун-
озере 6. VIII 1979 г. на 36 км береговой линии мы встретили только 13 гал-
стучников (6 взрослых и 7 молодых); не больше особей попадалось и в
другие годы.
Заметна тенденция к росту численности с годами: на одного наблюда-
теля в сезон приходилось в 30-х и 40-х по 2,8 встречи, в 50-х - 1,6, в 60-х -
3,8, в 70-х - 4,0, в 80-х - 8,4.
Галстучники появляются обычно в третьей декаде мая, в случае
поздней весны - в первой декаде июня, а в среднем за 32 года - 28 мая
(крайние сроки 18. V 1986 г. - 13. VI 1935 г.). Они прилетают парами, с
первого же дня начинают токовать и приступают к гнездованию, как
только пляжи освободятся от полой воды. Брачная игра галстучника сос-
тоит из песни, которую он издает на лету, подобно песочнику или поле-
вому жаворонку, и ’’пляски”, которая начинается с погони партнеров
друг за другом; потом они топчутся на месте, выкидывая попеременно
ноги в ускоряющемся темпе. Игра кончается тем, что самка приседает, под-
гибая ноги, а самец вскакивает на нее и копулирует (запись А.С. Гиля-
зова 9. VI 1977 г., 23 ч.).
Гнездо представляет мелкую ямку в песке, около 8 см в диаметре и
3 см в глубину, со скудной подстилкой из травинок, кусочков коры и
другого мусора. Чаще всего оно располагается в наносной полосе мусора
на пляже, под плавником или другим укрытием, нарушающим однооб-
разие песчаного фона. Поблизости от занятого гнезда, иногда находится не-
сколько пустых ямок: так, 16. VI1982 г. Гилязов нашел около действующего
гнезда 6 пустых. Из 12 полных кладок, найденных в сжатые сроки, с 7
по 18 июня, одно содержало 3 яйца, остальные - по 4. Особняком стоит
тринадцатое гнездо, найденное на берегу Чунозера 20. VII 1979 г. с 3 яйца-
ми: очевидно, это была вторая либо дополнительная кладка взамен по-
гибшей. Яйца в сравнении с величиной птицы большие, с крупными бу-
рыми пятнами по кремовому или бледно-оливковому фону. Масса 12
яиц из 3 свежих кладок от 10, 1 до 11, 8 г, в среднем 10,8 ± 0,2 г, так что
масса полной кладки составляет 79 % массы тела наседки. Размеры 4
яиц: длина 33—35, ширина 25-26 мм.
Галстучник, спугнутый со свежей кладки (вероятно, самка), выпол-
няет ’’отвлекающую демонстрацию”: бегает, волоча крыло и хвост по
песку, иногда поднимает одно крыло вверх, стрекочет, приближаясь к
человеку ближе чем на метр (рис. 35). Его партнер держится осторожнее,
беспокойно кричит, бегает по пляжу, но ближе 2-4 м не подходит. Начи-
ная со следующего дня на протяжении всей инкубации, птицы ведут се-
бя совсем иначе. Наседка уходит с гнезда заранее и незаметно, потом обе
птицы с криком бегают по пляжу в нескольких десятках метров от гнез-
да. Во время вывода птенцов галстучник возвращается к прежней так-
130
Рис. 35. С .влекающая демонстация галстучника, 29. VI 1978 г. Фото О.И. Семенова-
Тян-Шанского
тике и ’’отводит” от гнезда, прибегая к такой же ’’агонистической демон-
страции”, как при только что снесенных яйцах.
Самая ранняя находка гнезда 3. VI 1987 г.: между 14 и 17 часам сне-
сено второе яйцо. В гнезде, найденном 8.VI. 1978 г. со свежей полной кладкой,
27. VI одно яйцо было уже наклюнуто, в другом - птенец пищал; 28 и 29
все яйца наклюнуты и пищат, но вывод птенцов задерживается, вероятно
по причине слабости их от потери влаги. 30. VI в гнезде обнаружен один
слабенький птенец, остальные исчезли, так же как и скорлупы; объеден-
ный муравьями труп одного птенца найден неподалеку.
В гнезде, найденном 16. VI 1938 г., вечером 4 июля оказались 1 птенец,
1 наклюнутое и 1 цельное яйцо (старший птенец, вероятно, покинул гнез-
до ?). 18. VII птенцы уже начали оперяться.
20. VII 1979 г. на пляже у мели Кошк-чёльм найдено гнездо с полной
кладкой из 3 яиц; через 17 дней, 6 августа, на том же месте встретили од-
ного галстучника с 3 пуховичками.
Около гнезда с 4 яйцами, обнаруженного на пляже в западном конце Чун-
озера 13. VI 1980 г., 20 дней спустя, 3 июля, встречены 2 взрослых галс-
тучника с 2 пуховичками.
Согласно нашим наблюдениям, вероятный срок инкубации у галстуч-
ника 20 дней - меньше, чем обычно указывается в литературе (23-26
дней, по: Harrison, 1975, с. 140).
В большинстве гнезд птенцы выводятся в первой декаде июля. Пухо-
вых или частью оперенных, но еще не летающих птенцов мы встречали на
протяжении всего июля (с 28. VI до 8. VIII), а в среднем из 20 наблюдений
- 22 июля. Птенцы оставляют гнездо почти сразу после вылупления, но
131
живут и кормятся на том же пляже, пока не научатся летать. Обычно
встречают 1-2 птенцов при одной взрослой птице: вероятно, родители
делят птенцов между собой, как это делает малый зуек (Мальчевский,
Пукинский, 1983, т. 1).
Успех размножения, видимо, невысок: из 52 яиц, находившихся в 14
гнездах, вывелось не более 22 птенцов (число их в выводке не всегда
удавалось определить): 2 погибли при вылуплении, 4 - уничтожены хищ-
ником, одно гнездо с 1 яйцом залито прибоем, 8 мертвых яиц остались в
гнездах, судьба 15 яиц не выяснена. Принимая средний размер кладки
в 4 яйца, при такой смертности в осенней популяции может быть не более
44 % молодых птиц. Фактически, в числе 30 галстучников, встреченных
в августе в разные годы, было 14 взрослых и 16 молодых (53 %).
Откочевка с мест гнездования начинается как только молодые подни-
мутся на крыло. Последних галстучников мы встречали в разных числах
августа, самое позднее 11 и 13. IX 1966 г., самое раннее - 2. VIII 1983 г.,
4. VIII в 1939 и 1977 гг.; в среднем за 16 лет - 12 августа. Таким образом,
галстучник живет на гнездовьях в среднем 75 дней.
В заповеднике окольцовано 10 птенцов, возвратов не поступало, но
на фотографии одного галстучника, снятого у гнезда 12. VI 1975 г., вид-
но кольцо на правой лапке.
Размеры двух самцов: длина 185, крыло 132-134, хвост 63-65 мм, мас-
са тела 50 и 60 г.
Крикливый зуек - Charadrius vociferus
4 августа 1970 г. на Лебяжьем озерке в верховьях р. Курка студент
МГУ В. Матвеев видел вместе с парой фифи пару куликов с красным
надхвостьем. В нашей фауне нет куликов с таким признаком, но он свой-
ствен американскому крикливому зуйку, изредка залетающему в Запад-
ную Европу. Его легко отличить от галстучника по двум черным пояскам
поперек белой шеи и громкому крику ”килл-ди, килл-ди”, откуда и про-
изошло его американское название ”killdeer”.
Хрустан - Eudromias mormellus (L.)
Эта ржанка гнездится только на горных и сухих арктических тундрах
Европы и Азии, образуя несколько изолированных колоний; одна из них
занимает Скандинавский и Кольский полуостров. Зимует хрустан в Се-
верной Африке и Юго-Западной Азии.
На Кольском полуострове хрустан гнездится преимущественно по гор-
ным тундрам - Хибинам, Чуне, Монче, Нявке, Сальной, Туадаш и др.,
а в восточных частях Мурманского и Терского берегов - от Иоканьги до
Пялицы - и по равнинным.
По встречам хрустана трудно судить о его численности, так как мес-
та его обитания посещаются людьми нерегулярно; с другой стороны, хо-
лостые особи много летают и, по-видимому, не привязаны к определен-
ной территории. На Нявка-тундре (6 200 га) вряд ли проживает больше
10-15 пар хрустанов, т.е. не более 3-4 птиц на 10 кма. Численность замет-
132
но колеблется по годам; больше всего встреч зарегистрировано в I960,
1967 и 1976 гг.: по 10-15 на одного наблюдателя в год; в другие годы
встречали только по 1-2 птицы за сезон. На одного наблюдателя в год
приходится в 30-х годах по 4, 5 птицы, в 60-х - по 8, 2, в 70-х - по 6, 0, в
80-х - по 5, 6 (всего зарегистрировано 320 особей).
Прилетает хрустан поздно, в среднем 3 июня (крайние сроки за 15 лет
24. V 1937 г. - 10. VI 1979 г.), а отлетает рано, в среднем 20 августа (13.
VIII1931 и 1939 гг. - 26. VIII 1930 и 1973 гг., за 13 лет). На местах гнездова-
ния он проводит около 78 дней, или 0,21 года.
В 1967 г. на Ельнюнлаге и Втором Ельнюне найдено несколько гнезд
хрустана, 29 июня два, с 3 яйцами в каждом, 1 июля еще одно, с наклю-
нутыми яйцами, и выводок с 3 птенцами.
25 июня 1983 г. в том же районе А.Д. Карпенко нашел гнездо с 3 яйца-
ми: наседка отбежала от него так незаметно, что ее и не видели. 27. VI
мы поставили в гнездо актограф. Гнездо расположено на северо-восточ-
ном склоне близ вершины Второго Ельнюна, на абсолютной высоте око-
ло 450 м, в каменистой тундре с редким растительным покровом из кар-
ликовой березки, вороники, брусники, снежной цетрарии и др. Лоток
около 10 см в поперечнике, едва выражен, подстилка скудная, из сухих
листьев. Яйца оливково-бурые с неправильными черными пятнами,
масса их 14, 0, 14, 4 и 14, 7 г. Спугнутая с гнезда наседка молча бегала
вокруг, приближаясь до 7 м, потом села на яйца. Запись актографа пока-
зала, что насиживал только один из членов пары (по литературным ука-
заниям самец). Он оставлял гнездо для кормежки 8-9 раз в сутки, на
короткий срок, от 3 до 18 мин, затрачивая на прогулки 5-6 % инкуба-
ционного времени. В дневные часы отлучки продолжительнее (в 9-15 ч,
10 % всего времени), чем в ночные (в 21-03 ч, 2 % времени). ’’Агонисти-
ческую демонстрацию” хрустан показал только один раз и на короткое
время; обычно он молча сходил с гнезда и тут же кормился, на ходу
склевывая корм, вероятно насекомых. Быстро привыкнув к человеку,
он подбегал на 3, 5 м. Птенцы вывелись 4 июля, в 12 ч хрустан двух увел
с гнезда, а третий погиб в яйце.
Пуховых птенцов мы встречали также с 1. VII 1967 г. по 23. VII 1972 г.,
1. VII и 22. VII (средняя дата 13 июля); полуоперенных и хорошо бегающих
-16. VII 1964 г. - 30. VII 1976 г. (средняя дата 24 июля),слетков - в среднем
28 июля. Таким образом, птенцы появляются на свет около 2 июля, ос-
тавляя гнездо почти сразу после рождения, развиваются быстро и начи-
нают летать в возрасте около 24 дней. В третьей декаде июля мы встреча-
ли как пуховичков (1967), так и летающих птенцов (1972, 1979, 1980 гг.):
очевидно, сроки размножения в разные годы и у разных пар варьируют
в пределах 3 недель.
Летом хрустан питается, по-видимому, насекомыми: в клюве одного
птенца мы нашли комара-долгоножку Tipula sp. При опасности птенцы
затаиваются поодиночке, а самец активно демонстрирует, стараясь
увести врага подальше (рис. 36).
Хрустаны избегают каменистых россыпей, зарослей ивняков и высо-
котравья; их любимый биотоп - ровная злаково или лишайниково-кус-
133
Рис. 36. Бегущий хрустан, 24. VII 1985 г. Фото О.И. Семенова-Тян-Шанского
тарничковая тундра с дриадой, овсяницей, вороникой и брусникой. Вра-
ги у хустана те же, что и у других ржанок - сокола и, возможно, горнос-
тай. В агусте 1970 г. на Нявка-тундре мы нашли расклеванные остатки
молодого хрустана.
В разные годы в заповеднике окольцовано 14 птенцов, но ни одного
возврата не поступило.
Повадками хрустан напоминает золотистую ржанку, но голос тише,
несколько дрожащий, подобный свисту снегиря; издает он и другой звук,
вроде ”кхек”. Размеры 2 самок и 1 самца: длина 225-240, крыло 145-
150, хвост 65-80 мм, масса тела самца 107 г.
Чибис - Vanellus vanellus (L.)
Чибис - птица степной и лесной зон Европы и Западной Сибири,
гнездится к северу до Петрозаводска; зимует в Западной Европе и
Передней Азии. Во второй половине прошлого века дважды залетал на
Варангер-фиорд (Плеске, 1887, с. 424).
В Лапландском заповеднике до 1959 г. не встречался. Впервые 4
чибисов видели на Охтозере у протока Меркенч 1. VIII 1959 г. Ю.Г. Гаев
134
и Б. Травкин; после этого залеты участились и стали настолько регуляр-
ны, что по ним можно определить время прилета, в среднем 11 мая (13.
IV1977 г. - 31.V1972 г.; 13 лет).
Чибисы поодиночке или небольшими стайками до 6 птиц попадаются
все лето; самые поздние встречи - 11. VIII 1962 г., 5. VIII 1973 г. и 22. VIII
1980 г. В бассейне Имандры они регулярно наблюдались с 1965 по 1983 г. -
за эти годы нами учтено 107 особей; позднее, в 1984-1988 гг., они совсем
не попадались. В Лапландском заповеднике их видели в зоне затопления
Пиренгского водохранилища (25% всех встреч), затем на Нижней Чуне
(14%) и при устье Верхней Чуны (6%). В те же годы чибисы часто появля-
лись в окрестностях Мончегорска (36%), на совхозных полях у Апатит
(12%), а также и в других местах Кольского полуострова - на Айновых
островах (Татарникова, Чемякин, 1965), на р. Кица (в 1978 г., по сооб-
щению А.Б. Брагина), в 1975-1976 гг. в среднем течении Поноя, а в 1980 г.
у Ревды (Михайлов, Фильчагов, 1984, с. 25).
Чибисы изредка гнездились в нашем районе - в 1976 г. у с. Чальмны-
Варрэ, а в 1982 г. у поселка Тик-губа, где Е.М. Васильева (научный сот-
рудник Полярного отделения Всесоюзного института растениеводства)
наблюдала две семьи и видела, как на одного птенца напали вороны, а
взрослый чибис их отгонял.
Кулик-сорока - Haematopus ostralegus L.
На побережьях Баренцева и Белого морей кулик-сорока обычная
гнездящаяся птица, но на внутренние водоемы Мурманской области
залетает только изредка. В Лапландском заповеднике его наблюдали
пять раз: 31. V 1969 г. в Нявка-губе видели трех, 28. V 1970 г. - пять,
7. VII1971 г. на Охтозере - шесть, 12. VI 1982 г. на Чунозере - 18 и 6. VI
1987 г. там же - 12.
Кулик-сорока питается преимущественно моллюсками, в частности
мидиями, раковины которых он легко раскрывает своим сильным клю-
вом. На морских побережьях он хорошо обеспечен кормом, но на наших
реках и озерах не находит себе пищи.
Фифи - Tring a glareola L.
Летом фифи широко распространен по всей лесной полосе СССР и
Северной Европы, зиму проводит в Африке, к югу от Сахары. В материко-
вой части Кольского полуострова фифи наряду с перевозчиком самый
многочисленный кулик; по этой причине регистрации его не уделялось
должного внимания, и дать такие же показатели численности фифи, как
для ржанок, мы не можем.
Фифи прилетает в среднем 20 мая (10. V 1983 г. - 1. VI 1935 г., за 39 лет;
температура дня прилета, 4,7е); отлетает рано, в среднем 12 августа (1. VIII
1939 г. - 22. VIII 1958 г., за 33 года; температура +12,0°). На местах гнездо-
вания он проводит около 84 дней (0,23 года). Сроки прилета и отлета мало
варьируют по годам, среднее квадратическое отклонение даты прилета
±4,9, отлета ±5,4 дня. Время прилета связано корреляцией с температу-
135
Рис. 37. Фифи, июнь 1972 г. Фото С. Баккала
рой двух первых декад мая ( г =-0,54 + 0,15). Явно зависит от температу-
ры и время отлета, но здесь связь обратная: чем первые две декады
августа теплее, тем раньше он отлетает ( г = -0,53±0,17). Естественно
предположить, что время отлета в какой-то мере зависит от времени
прилета, однако и тут корреляция оказалась обратной, в годы позднего
прилета фифи отлетает раньше (г = - 0,46±0,15 за 27 лет).
Первое время после прилета фифи кормятся преимущественно по
берегам, патрулируя вдоль кромки воды и собирая насекомых, выполза-
ющих на лед или на отмель. Через 1-7 дней после прилета они уже току-
ют и спариваются.
Брачную песню фифи буквами можно изобразить как ”тю-ри, тю-ри,
тю-ри...”. Токующий самец трепещет крыльями, подняв их вертикально
над спиной. На лету он иногда щебечет, подобно ласточкам. Обычный
крик звучит как ”фи-фи” - отсюда и название птицы.
К соседям фифи относится миролюбиво, и только один раз, 29 мая 1968 г.,
мы наблюдали отчаянную драку двух самцов: они били друг друга
крыльями, стараясь ухватить соперника клювом за перья затылка, потом
оба упали в воду, и тут один старался утопить другого.
Гнезда с полной кладкой мы находили на протяжении всего июня, са-
мое раннее-4. VI 1978 г. и 7. VI 1981 г., самое позднее - 4. VII 1969 г. и
11. VII 1967 г. Гнездо помещается обычно на осоковом или морошко-
вом комплексном болоте на кочке и представляет собой ямку во мху,
выстланную листьями осоки или пушицы, полуприкрытую окружаю-
щей растительностью - морошкой, вороникой, андромедой, багульником
или осоковыми. В десяти гнездах было по 4 яйца, в трех - по 3. Окраска
136
скорлупы кремовая или зеленоватая, с коричневыми пятнами. Длина
40-41, диаметр 27-28 мм, вес яйца от 11,5 до 15,0, в среднем 13,2 г.
30 мая 1978 г. в 10 ч 30 мин мы нашли гнездо с одним холодным яйцом
на осоковой кочке среди болота по левому берегу р. Чуна; у гнезда были
обе птицы, самец отлетел первым, самка ’’отводила”. На следующий день,
в тот же час, гнездо оборудовано актографом. Второе яйцо снесено 31. V
между 17 ч 21 мин и 17 ч 52 мин, третье - 2. VI между 01 ч 23 мин и 01 ч 50
мин; через 3 ч, в 4 ч 51 мин, самка начала насиживание, после чего снесла
четвертое яйцо. 23 июня в 12 ч три яйца были наклюнуты и слышно было
дыхание птенцов, а 24. VI в гнезде находились три птенца (четвертое
яйцо повреждено, и эмбрион в нем погиб). Запись актографа показала,
что время, потребное для откладки очередного яйца, - около 30 мин;
интервал между откладкой последовательных яиц - 32 ч; насиживание
начинается до откладки последнего яйца; продолжительность инкубации
22 сут. Первое яйцо было снесено, по всей вероятности, 30 мая около 9 ч,
четвертое - 3 июня около 10 ч.
Эта кладка была самой ранней из всех учтенных нами. Пуховых
птенцов мы находили также 27. VI 1979 г., 2. VII 1940 г., 4. VII 1977 г. и
5. VII 1978 г. (средняя дата 2. VII), а птенцов из запоздалых кладок - 18.
VII 1964 г. и 22. VII 1977 г. Принимая во внимание среднюю дату находок
13 гнезд с полной кладкой, 20 июня, находим, что нормальное время
начала инкубации 9-10 VI, а ее окончание - 1-2 VII. Летных птенцов
встречали между 13-25 июля, в среднем 20. VII.
Любимая стация фифи во время гнездования - комплексные сфагно-
вые болота с угнетенными соснами; он также охотно садится на деревья,
как улит или щеголь. Из наших куликов фифи наряду с плавунчиком
самый доверчивый, весной к нему нетрудно подойти на 4-5 м, а сам он
может приблизиться к неподвижному наблюдателю даже на 3 м (рис. 37).
Врагами фифи можно считать всех хищных птиц, в добыче которых
попадались кулики: ястребов, тетеревятника и перепелятника, кречета и,
вероятно, дербника, но большого урона от них он не терпит: только один
раз, в июне 1951 г., мы нашли остатки фифи, пойманного хищником.
Размеры 9 добытых особей: длина 195-223, крыло 118-133, хвост
50- 64 мм, масса тела двух самцов в 49,6 и 56 г, двух самок 53 и 58,5 г (в
среднем 54,0 г).
В 1964-1978 гг. в заповеднике окольцовано 6 птенцов, но возвратов не
поступало.
Черныш - Tringa ochropus L.
Ареал черныша в сравнении с фифи смещен к югу и лишь немного
заходит к северу от Ботнического залива. В последние годы он регуляр-
но залетает в район Лапландского заповедника, а возможно и гнездится.
Впервые черныша видел А.С. Гилязов 6. V 1981 г. на Северном ручье,
впадающем в оз. Ельявр, затем А.Б. Брагин, 7. VIII1981 г. на р. Чуна. С тех
пор одиночных чернышей встречали в тех же местах ежегодно, а в июне
1986 г. видели токующую пару. Средняя дата первой встречи за 7 лет 22
мая. Добывать их не было необходимости, так как приведенные на
137
карточках наблюдений признаки не оставляют сомнений в правильности
определения.
Черныш немного крупнее фифи, держится осторожнее его и отличается
более контрастной окраской оперения.
Большой улит - Tringa nebularia (Gunn.)
Местное название: саамское ’’янкьлоньт” (дословно - болот-
ная птица).
Обычный болотный кулик, хотя численно и уступает фифи. В
среднем за 25 лет на одного наблюдателя приходится 7 встреч
улитов в год; индекс численности колеблется, заметно повыша-
ясь через каждые 3-4 года (в одном случае 5 лет). Намечаются и
волны второго порядка: с 1959 до 1967 г численность постепенно
возрастала, при среднем уровне 5 встреч, в 1968-1975 гг. остава-
лась на высоком уровне, в среднем по 10,5 встреч (до 18,7 в 1969 г.),
с 1976 г. снизилась до 5 встреч в год в среднем.
Улит прилетает к нам раньше всех других куликов, в среднем за
47 лет 12 мая (4. V 1960 г. - 22. V 1968 г.; о ±4,2 дня, температура 2,9°);
спустя 4-8 дней он уже токует.
Весной улиты кормятся преимущественно по берегам рек и озер, у
края воды, где легко доступны наблюдению (на май приходится 48% всех
встреч). Летом они держатся преимущественно по большим комплексным
болотам с соснами по буграм. Интересно, что из пяти известных нам
гнезд, безусловно принадлежащих улитам, ни одного не было на болоте,
все находились на сухих возвышенных местах, в бору, чаще на старой
гари:
5. VI 1939 г. — в долине Чуны, в редком бору, горевшем 40 лет назад, с напочвен-
ным покровом из вереска, трубчатых кладоний и начинающего отрастать ягеля.
Гнездо на земле, под обугленной валежиной и древесной трухе; в кладке 4 яйца без
признаков насиживания (размер одного 50x34,5 мм).
30. V1950 г. — на той же старой гари гнездо с полной кладкой из 4 яиц; как и
предыдущее, найдено по отводившей от него наседке.
29. V 1977 г. - на лесосеке в ягельном бору, в долине Чуны, в сотне метров от
болота, у ствола поваленного дерева, гнездо с яйцами на подстилке из хвои и
березовых листьев.
31. V 1978 г. — в долине Змеиного ручья на бугре, на 38-летней сосновой rapHj
гнездо с 4 яйцами. При установке актографа 12. VI птица побежала, подпустив на 2 м;
вернулась первый раз через 9 мин, второй раз через 2 мин. 13. VI гнездо разорено
зверем, яйца съедены, гнездовой контакт выкинут.
7. VI 1986 г. в редком бору-скальнике с черничным покровом, на склоне Ельяв-
руайвенча лаборант Л.В. Киренков нашел около полудня (по взлетевшей птице]
гнездо улита; оно находилось в углублении под березовым пеньком в 30 см от
тропинки и содержало 4 яйца. Актограф, установленный в отсутствие наседки, в 16
9. VI, работал до 27. VI - в то утро в 3 ч 38 мин улита, по-видимому, согнал мел
кий хищник, менее чем за час утащивший все яйца; в 17 ч 45 мин в гнезде и вокруг
него не осталось даже скорлуп, не обнаружено и улитов на ближних болотах.
Режим инкубации оказался своеобразным. Насиживала только одш
птица (вероятно, самка); для кормежки она оставляла гнездо от 3 до S
раз в сутки, на срок от 12 мин до 8,5 ч (в среднем на 54 мин), так что яйце
оставались открытыми и без обогрева четверть всего времени регистра
138
ции. Улит придерживался строгого суточного ритма: в ночные часы
(22-02) он ни разу не оставлял гнезда, на протяжении 7 утренних часов
насиживал 85% всего времени, за 8 дневных часов (09-17) сидел только
45%, так что яйца более половины времени лежали открыто, за 5 вечер-
них часов (17-22) сидел на протяжении 84%. По этой причине средняя
температура воздуха за время прогулок (16,5°) оказалась значительно
выше температуры за все время инкубации (13,0°). Продолжительность
прогулок не зависит ни от солнца и облачности, ни от температуры суток:
корреляция между этими величинами близка к нулю.
Все 5 гнезд найдены, видимо, в первые дни насиживания, очевидно
оно начинается в последних числах мая. Одиночных нелетных птенцов
находили 1. VII 1964 г. и 13. VII 1980 г., а способных летать - 27. VII 1938 г.
и 19. VII 1979 г. (3 птенца). Летные выводки остаются на гнездовьях не
больше месяца. Отлет заканчивается, в среднем за 32 года, 17 августа (7.
VIII 1967 г. - 3. IX 1962 г.; о ± 6,3.дня, температура +11,3°). Улиты живут
у нас около 97 дней; дата прилета отчетливо коррелирует со средней
температурой апреля ( г = -0,56±0,11 ) и мая ( г = -0,53±0,11 за 41 год);
время отлета не контролируется температурой (г = -0,09±0,19 ).
Для улита опасны те же хищники, что и для других куликов. Остатки
одного мы обнаружили в добыче ястреба-тетеревятника, остатки птенца
А.Б. Брагин нашел 22. VII 1970 г. в желудке щуки (массой 1,7 кг).
Размеры двух взрослых птиц: длина 338-340, крыло 184-193, хвост
89-90 мм, масса тела самца 160, самки - 156 г.
Травник - Tringa totanus L.
В Лапландии травник приурочен к морскому побережью (Плеске, 1887,
с. 454; Михайлов, Фильчагов, 1984, с. 25), в глубь страны он только зале-
тает. В Лапландском заповеднике достоверно его наблюдали летом 1979 г.
- 6 июня О.И. Семенов-Тян-Шанский видел одного на Турухтанном
болоте по берегу Охтозера, а 26 июля студент МГУ Д.В. Карелин наблю-
дал травника в 2-3 км к северу, на ерниковом болоте у оз. Улынчъявр.
Щеголь - Triga erythropus (Pall.)
Щеголь гнездится в тундре, лесотундре и северной тайге от Скандинав-
ского полуострова до Анадырской губы; зимует главным образом на
побережьях Средиземного моря. В Лапландском заповеднике гнездится
регулярно, хотя и реже улита (за 30 лет в картотеке зарегистрированы
1010 улитов и только 226 щеголей). Число встреч колеблется по годам
параллельно с колебаниями численности улита (максимумы в 1964 , 1969,
1972, 1974,1978 и 1982 гг.).
Прилетает щеголь позже улита, в среднем за 25 лет 21 мая (14. V 1940 и
1978 гг. - 31. V 1969 г.; а ± 5,1 дня, температура дня прилета + 4,9°).
Щеголь в большей мере привязан к болотам, чем улит, и редко встре-
чается на берегах рек и озер (рис. 38). Найдено всего три гнезда: 3. VI 1972 г.
в бору близ оз. Румельявр, в 150 м от лесного болотца, под маленькой
сосенкой, по взлетевшей птице А.Б. Брагин нашел гнездо с 4 яйцами на
139
Рис. 38. Щеголь за кормежкой, июнь 1972 г. Фото С. Баккала
подстилке из листьев ивы и березы. Масса одного, почти ненасиженного,
21,3 г.
12. VI 1972 г. в бору-скальнике к югу от оз. Улынчъявр поднят с гнезда щеголь,
подобно предыдущему, оно устроено под маленькой (1 м) сосенкой; в нем 4 яйца,
оливково-зеленых с размытыми темно-коричневыми пятнами. Окружающая расти-
тельность — альпийская кладония, вереск, багульник, черника и брусника.
17. VI 1977 г. на болоте между озерами Островским и Витте, на кочке возле
сосны, лесник В.М. Бусуек нашел гнездо кулика с 4 наклюнутыми яйцами.
20. VI 1981 г. на берегу Гагарьего озерка между реками Чуна и Нявка найдена
скорлупа яйца со следами кровеносных сосудов; щеголь с криком летал вокруг.
Судя по этим наблюдениям, щеголь насиживает примерно с 1 по 20
июня. Летний выводок щеголей (4 птенца при 2 взрослых) наблюдали на
болоте севернее Румельявра 19. VII, 1978 г., а взрослых щеголей, активно
отводивших от птенцов, - 22. VII 1964 г., 1. VII 1972 г. и 24. VI 1981 г.
Щеголи гораздо агрессивнее улита и других куликов, на человека они
налетают в упор, подобно крачкам или поморникам (наблюдения
С. Баккала и А. Печенюка).
Последних щеголей отмечали между 8-21 августа, в 1973 г. даже
23. IX, а в 1960 г. явно отсталую птицу О.И. Семенов-Тян-Шанский видел
на Воче-ламбине 3 октября.
Крик щеголя напоминает большого улита, но тоньше и звонче. Брач-
ную песню можно передать буквами ’’тюййри, тюййри”; издает он также
трель, напоминающую среднего кроншнепа. Размеры 9 взрослых самцов:
длина 315-325, крыло 162-165, хвост 65-75 мм, масса тела 134-152 г;
длина самки 320, крыло 170, хвост 68 мм (масса тела не определена).
В желудках двух щеголей, добытых в июле, обнаружены только насе-
комые - мелкие жуки-водолюбы, личинки ручейников и стрекоз.
140
Перевозчик - Actitis hypoleucos (L.)
Местные названия: саамские ’’пулдыш” или ’’пубвдынч”.
На материковой части ' Кольского полуострова перевозчик самый
многочисленный или, по крайней мере, самый доступный наблюдению
кулик. Живет он по берегам рек и озер и постоянно попадается на глаза:
за 30 лет учтено около 2500 особей, или по 15 встреч в год на наблюдателя
(в 2,4 раза больше, чем улитов.) Определенной тенденции к росту или
сокращению популяции, по-видимому, нет (повышение встречаемости в
80-х годах, скорее всего, следствие более внимательной регистрации птиц
в последние годы). Перевозчик обнаруживает нерегулярные колебания
численности с повышениями ее через каждые 2-6 лет.
Прилетает перевозчик в среднем 19 мая (10. V 1983 и 1987 гг. - 1. VI
1935 г. за 45 лет; о ± 5,4 дня), обычно сразу после вскрытия рек, при сред-
ней суточной температуре воздуха +4,5°. Момент прилета зависит от
температуры апреля ( г = - 0,24±0,15 ) и мая ( г = — 0,48±0,12; 44 года); еще
лучшую корреляцию дают первая и вторая декады мая ( г = - 0,58±0,14; 23
года).
С первого дня перевозчики держатся парами и через 3-7 дней уже
токуют и спариваются. Цитируем карточку наблюдений О.И. Семенова-
Тян-Шанского: ”2 июня 1969 г. в 23 ч 45 мин в устье р. Чуна наблюдал
токование. Два перевозчика сели на кромку снега и с криком взлетали
друг перед другом, а иногда друг через друга; у одного крылья на секун-
ду становились вертикально вверх. Потом оба легли на снег, навстречу
один другому, но под углом. Один весь распушился и в таком виде
побежал прочь, раскрыв хвост веером и немного его опустив. Другой
оставался в обычной позе, а потом улетел прочь. Все представление
заняло не более 1 мин”.
Гнездо устраивается обычно на сухом месте, в бору, среди ягеля и
ягодных кустарничков, метрах в 20-60 от воды. Лоток углублен в расти-
тельный покров, так что наседку или яйца невозможно заметить даже на
близком расстоянии. Полная кладка, как правило, содержит 4 яйца (41
гнездо), иногда 3 (4 гнезда). Первое время самка сидит крепко, взлетает
чуть не из-под ног, обнаруживая этим гнездо; позднее она оставляет его
заранее, незаметно, и в этом случае его трудно обнаружить.
Масса свежих яиц варьирует от 10,7 до 12,5 г, в среднем 11,6±0,2 (п=11),
так что масса полной кладки примерно равна весу самой наседки. Мы
получили надежную запись только за одни сутки 24-25 июня 1963 г.; она
показала, что кладка ни минуты не оставалась открытой; очевидно,
насиживали самка и самец попеременно, а смена партнеров происходила
без задержки.
В общей сложности нами зарегистрировано 47 гнезд перевозчиков.
Приведем здесь лишь наиболее результативные наблюдения:
16. VI 1940 г. в ельнике-черничнике у берега Второго ручья найдено гнездо с 2
яйцами; 18. VI птица сидела на гнезде, в нем 3 яйца, а 20. VII — 4. 8 июля в 22 ч 30
мин из одного яйца птенец вывелся, другое наклюнуто; 9. VII гнездо уже пусто,
скорлупы удалены, птенцы прятались поблизости.
Около 7. VI 1940 г. в лесу у берега Ельявруая найдено другое гнездо с полной
141
кладкой из 4 яиц, впоследствии разоренное. Около того места 12 июля обнаружена
вторая кладка; вечером 19. VII в ней было 4 яйца, а 20. VII в 15 ч — 4 птенца; днем
21. VII выводок уже оставил гнездо.
21. VI 1949 г. у берега Парусной ламбины гнездо перевозчика с 4 наклюнутыми
яйцами; 22. VI в нем птенцы, 23. VI гнездо пусто.
16. VI 1974 г. в 12 ч в бору при устье Чуны по взлетевшей птице найдено гнездо
перевозчика с 3 яйцами; 17. VI в 12 ч в нем 4 яйца; 7. VII — 3 обсохших пуховичка и
крышечка от одного из яиц.
28. VI 1974 г. в 6 ч 30 мин там же, на открытом южном склоне обнаружено другое
гнездо с 4 яйцами; 5 июля три яйца наклюнуты; 7. VII в гнезде осталось одно яйцо с
мертвым эмбрионом, а птенцы и скорлупы исчезли.
15. VI 1977 г. в бору на берегу р. Чуна у первого порога — гнездо перевозчика с 3
яйцами, под прикрытием сосенки в 0,7 м. 22. VI гнездо окружено свежевыпавшим
снегом, но насиживание продолжается; 25. VI слышно дыхание птенцов в яйцах;
26. VI птенцы вывелись и окольцованы; 27. VI гнездо пусто; 28. VI в 23 ч 45 мин при
температуре воздуха + 5°... +6° два птенца вернулись и сидели в гнезде, очевидно
спасаясь от холода (третий не найден).
13. VII 1979 г. в янгельном бору близ берега Чуны найдено гнездо с 4 пуховыми
птенцами и обогревавшим их перевозчиком: насиживание яиц в этом случае
началось только 23. VI.
10. VI 1983 г. в ягельно-ягодниковом бору на южном берегу Чунозера найдено по
взлетевшей самке гнездо перевозчика: в 12 ч в нем было 3 яйца, в 15 ч — уже 4
(глубина лотка 45, диаметр 70—75 мм); 29 июня в 15 ч в гнезде по-прежнему 4 яйца,
без наклевов, а 3 июля в 12 ч остались только 2 яйцевые крышечки.
Сопоставляя эти наблюдения, находим, что инкубация яиц продол-
жается около 20 сут (до ухода выводка 20-21 сут); птенцы выводятся
дружно и хорошо развитыми, в возрасте нескольких часов они уже
бегают и прячутся (от первого проклева до оставления гнезда выводком
проходит немногим более суток); скорлупы взрослая птица удаляет из
гнезда; в случае холодной погоды пуховички иногда возвращаются в
гнездо на ночь.
Прослежена судьба 24 кладок из 47: в 22 вывелись птенцы, 2 погибли
целиком. Эмбриональная смертность в ’’благополучных гнездах”, видимо,
незначительна: из одного гнезда два яйца бесследно исчезли во время
инкубации, в третьем погиб эмбрион, так что вывелся только один
птенец; в другой кладке из 4 яиц вывелись 3 птенца. Достоверно из 84
яиц погибло 5, но на самом деле потерь было больше, так как не все
птенцы были найдены и подсчитаны.
Средняя дата оставления гнезда выводком 3 июля (от 23. VI до 13. VII;
22 случая, включая два, когда этот момент рассчитан по началу насижи-
вания); вычитая 21 день, получаем дату начала инкубации - 12 июня.
Отдельные поздние кладки, вероятно дополнительные, в этот расчет не
включены: одна насиживалась с 1 по 21 июля 1941 г., две другие полные
кладки найдены были 21. VII 1940 г. и 10. VII 1978 г. Как видно, сроки
размножения перевозчика сильно растянуты.
Нелетных птенцов встречали в разные годы между 24. VI и 27. VII
(средняя дата 13 июля, п = 15), а летних - с 15. VII по 17. VIII (в среднем 29
июля, п = 6). Птенцы растут быстро, в возрасте примерно 20 дней они уже
летают. В первой декаде августа перевозчиков можно встретить стайка-
ми по 7-13 птиц; вероятно, это объединенные выводки (наблюдения 4.
VIII 1976 г. и 4. VIII 1980 г.). Отлет проходит незаметно, последних особей
мы отмечали между 11. VIII и 13. IX, а в среднем за 45 лет - 20 августа,
142
при средней температуре воздуха +10,8°. Время отлета, по-видимому, не
зависит от температуры: корреляция между средней температурой
августа и датой отлета равна -0,03±0,16 (38 лет). Поздно встреченные
кулички могут быть и отсталыми, обреченными на гибель особями.
Перевозчики кормятся, по всей вероятности, насекомыми, по берегам
у края воды. Весенние возвраты холодов лишают их естественного
корма: в такие дни они, подобно воробьям, подбегают к человеческому
жилью в поисках всяких остатков, подбирают остатки рыбы, хлеб, вермишель
и т.д. Летняя смертность перевозчиков, видимо, незначительна: остатки
одного, пойманного хищной птицей, найдены в июне 1952 г., летом 1940 г.
взрослого поймал ястреб-тетеревятник, а птенца задавила собака. Цель-
ные трупы перевозчиков найдены в июле 1971 г. в верховьях р. Нявка и
в августе 1986 г. на берегу Чунозера.
На зиму перевозчики отлетают в Африку. Пути перелета тех, которые
гнездятся у нас, неясны, возврата колец не поступало.
Перевозчик - самый мелкий из улитов. Размеры одного самца: длина
185, крыло 109, хвост 52 мм, масса тела 33,8 г, новорожденного
птенца - 7,2 г.
Мородунка - Xenus cinereus (Giild.)
Мородунка гнездится в лесной и лесотундровой зонах Сибири и
Европейской России, к западу до берегов Белого и Балтийского морей; в
Скандинавию и на Кольский полуостров она залетает только изредка. В
окрестностях Лапландского заповедника наблюдалась дважды. Встречу
на р. Нота 22. VII, 1962 г. Семенов-Тян-Шанский описал так: ’’Около
надувной лодки летал куличок, сверху темно-коричнево-серый, почти
без белого в хвосте, с желтыми или оранжевыми ногами и слегка загну-
тым вверх клювом”. Много лет спустя, 25. V 1986 г., Л.В. Киренков видел
одиночную мородунку, с криком летевшую над Чунозерской усадьбой в
восточном направлении.
Круглоносый плавунчик - Phalaropus lobatus (L.)
Гнездится в зонах тундры и лесотундры Европы, Азии и Америки, а в
виде исключения и в лесной зоне (Эстония, Вологодская обл.), и даже
степной (Северный Казахстан, в XIX в.). Зимуют круглоносые плавунчи-
ки на Аравийском море и на других морях у берегов. Во время миграции
они летят через Казахстан, пересекая Евразию. Вероятно, значительную
часть года они проводят в миграциях.
В Лапландском заповеднике плавунчик впервые встречен и добыт 5
июня 1939 г. в нижнем течении р. Чуна (одиночная самка). Начиная с 1963 г.
плавунчики почти ежегодно наблюдались и неоднократно гнездились
на болотах между реками Нявка, Чуна и Охтозером, а также на водое-
мах среди лесотундры и редкостойных лесов, к северу от озер Туреск и
Купись (1964), в долине речки Лылча (1977) и в долине р. Лива (1979): весь
этот район в 1983 г. включен в территорию Лапландского заповедника.
143
Рис. 39. Круглоносый плавунчик, 4. VI 1969 г. Фото О.И. Семенова-Тян-Шанского
Кроме того, их наблюдали в городской черте Мончегорска (1976, 1979,
1980, 1981 гг.) и на Монче-озере (1980 г.).
За 25 лет нами зарегистрированы 220 плавунчиков, в том числе 8
пуховых птенцов; больше всего в 1974 г. - 32 особи. Даты первых встреч
от 19. V 1987 г. до 23. VI 1963 г., в среднем за 21 год 3 июня. Здесь самые
поздние даты отбракованы, но и остальные нельзя считать точными: по
причине скрытого образа жизни, специфике занимаемых стаций и мало-
численности, первых плавунчиков не раз отмечали с запозданием.
Последних наблюдали между 14 и 26 июля, вероятно задолго до настоя-
щего отлета.
Гнездится плавунчик на больших осоковых болотах, образующихся в
результате зарастания озер сплавиной: этот биотоп характерен для
краснозобой гагары, турухтана, полярной крачки, озерной чернети и
’’благородных” уток. На больших озерах и реках с медленным течением
он появляется только на весеннем пролете, иногда стайками до 10-15
птиц (27. V 1967 г., 9. VI 1974 г.). Позднее плавунчики встречаются парами
или поодиночке. Плавунчик - малозаметная птица, держится скрытно,
громко не кричит, а только ’’чмокает”; вместе с тем он очень доверчив, к
человеку подлетает или подплывает нередко на 3- 4 м.
На воде он напоминает миниатюрную уточку (рис. 39), посадка у него
легкая, как у чайки, хвост держит высоко над водой, но при быстром
движении опускает его в воду, как нырковая утка. Питаются плавунчи-
ки насекомыми, ловят их иногда опуская голову глубоко в воду, а чаще
хватают с поверхности, при этом и сами плавают бросками, как клопы-
водомерки. Кормятся как на открытой воде, так и в зарослях осоки.
Во время брачной игры плавунчик по несколько секунд крутится на
144
воде, как волчок, иногда он машет поднятыми вверх крыльями, точно
обвевает ими партнера. 5. VI 1977 г. видели, как самец топтал самочку на
воде.
Одно гнездо с 4 яйцами было найдено 16. VI 1987 г. в Мончегорске
В.М. Бусуйком. Пуховых птенцов наблюдали неоднократно: 2. VII 1967 г.
А. Печенюк видел одного; 28. VI 1982 г. А. Брагин нашел два выводка, в
одном два, в другом четыре птенца при одной взрослой птице (очевидно,
это были самцы); 29. VI 1975 г. он же наблюдал самца, отводившего от
птенцов, и нашел скорлупу яйца. Птенец, встреченный А. Гилязовым
21. VII 1978 г., хорошо плавал, и на подрастающих маховых уже лопнули
чехлики.
Принимая, что плавунчики подобно перевозчику и фифи, насиживают
20 дней, время инкубации яиц можно определить с 5-10 по 25-30 июня.
Размеры одной самки: длина 180, крыло 114, хвост 48 мм, масса
тела 31,3 г.
Камнешарка - Arenaria interpres (L.)
Камнешарка встречается по морским берегам почти всего света, в
одних местах летом, в других зимой. На Кольском полуострове обычна
по берегам Белого и Баренцева морей, откуда изредка залетает и в глубь
страны. В районе Лапландского заповедника пару камнешарок впервые
наблюдал 3. VII 1977 г. лесник В. Бусуек, на островке в зашейке Монче-
озера. У птиц, судя по их поведению, могли быть птенцы, но обнаружить
их не удалось. Позднее камнешарок наблюдали в г. Мончегорске, 26. V
1982 г. пару видел В. Бусуек, а 30. VIII 1986 г. - Г. Катаев (также пару).
Турухтан - Philomachus pugnax (L.)
Местные названия: саамское ”рыймь-рауд”, ’’раудынч”, ”рим-
ровг”.
В отличие от улитов турухтан распространен по Кольскому полуостро-
ву не равномерно, а небольшими колониями - на одних болотах гнез-
дится, на других полностью отсутствует. Хотя такой характер распростра-
нения по территории и затрудняет учет численности турухтанов по
регистрации встреч, попытаемся все же оценить численность этого вида
тем же путем, как сделали это в отношении улитов. За 30 лет нами заре-
гистрировано 1188 турухтанов, в среднем по 40 за сезон, или по 14,0 на
одного наблюдателя в год - в 2 раза больше, чем больших улитов. Встре-
чаемость сильно варьирует по годам, больше всего их встречалось в 1968
-1972 гг. (по 32 птицы на 1 наблюдателя в год) и в 1979-1981 гг. (по 20
птиц); мало турухтанов в 1959-1967, 1973-1978, 1982-1987 гг. (по 2-10 в
год на человека). В отдельные годы (1959, 1960, 1961) не видели ни одного
турухтана. За 11 довоенных лет нами зарегистрированы только 54 птицы.
Прилетает турухтан поздно, в среднем за 26 лет 23 мая (19. IV 1984 г. -
6. VI 1941 и 1964 гг.; 6±6,7 дней, средняя температура прилета +5,4°).
Корреляция между датой прилета и средней температурой мая
-0,54±0,15.
10. Зак. 1023
145
Гнездится турухтан в тундровой, лесотундровой и таежной зонах
Европы и Сибири; зимует преимущественно в Африке.
Самая значительная колония турухтанов в Лапландском заповеднике
расположена на большом (не менее 100 га) осоковом болоте с обширными
’’окнами” и островами на них, на северном побережье Охтозера, в зоне
затопления Пиренгского резервного водохранилища. Воду из него
зимой спускают для поддержания уровня оз. Имандра, а перед поло-
водьем шлюз закрывают, уровень воды быстро повышается и достигает
максимума к 1 июля, затопляя упомянутое болото, которое, таким
образом, становится ловушкой для гнездящихся на поверхности земли
птиц, в первую очередь куликов.
На этом болоте, которое так и назвали ’’Турухтанным”, подсчитывали
турухтанов в бинокль. Приводим максимальные цифры по годам:
1949 1950 1968 1969 1970 1972 1974 1977 1978 1979
67 56 50 20 20 38 15 22 4 8
Сразу после прилета самцы занимают свои индивидуальные террито-
рии, ’’точки”, обычно это островок или кочка на болоте. В то время как
самки оживленно бегают по грязевым отмелям, разыскивая корм, самцы
большую часть дня стоят неподвижно и молча, каждый на своем посту,
распушив пестрое оперение, с втянутой шеей и наклоненным книзу
клювом. В этой позе турухтана можно скорее принять за выросший на
болоте фантастический цветок, чем за живую птицу. Время от времени он
оживляется и токует, поднимая крылья вверх и трепеща ими несколько
секунд. Если на точок сядет соперник, оба самца взлетают и трепещут в
воздухе друг против друга. Иногда они вступают в драку, один схваты-
вает другого за длинные перья воротника, подпрыгивает и треплет
соперника, но чаще несколько петушков мирно токуют по соседству,
каждый сам по себе, обратясь лицом все в одну сторону - на ветер. При
подходе самки, а иногда соперника петушок обычно ложится на землю
как был, с распушенным оперением. Изредка на точок ’’стационарного”
самца налетает группа из 3- 5 посторонних петушков, все они мгновенно
сбегаются, и несколько секунд на болоте кружится пестрый клубок, в
котором невозможно ничего разобрать. Все представление происходит в
полной тишине; турухтаны очень молчаливы, и только на лету самец
иногда издает приглушенный звук ”лок-логгй, лок-логгй, лок-логгй...”,
слыша который, трудно понять, с какой стороны он доносится. Видимо,
от этого трехсложного крика происходит и название птицы - его можно
представить и другими буквами ”ту-рух-тан, ту-рух-тан...”.
7 июня 1966 г. на Турухтанном болоте найдены два гнезда с полными
кладками, одно под прикрытием сухой прошлогодней осоки, другое за
корягой; самки ’’отводили” молча, не далее 15-20 м, а потом скрыва-
лись. Окраска яиц светло-оливково-зеленая с размытыми буроватыми
пятнами.
В середине июня самцы продолжают сидеть на своих точках, но уже
почти не токуют. Последних самцов в брачном наряде наблюдали в
последней декаде июня, самое позднее 4. VII 1981 г.; исчезают они
незаметно.
146
Одного нелетного птенца мы поймали 21. VII 1971 г. на Кымм-озере, а
летных наблюдали в разные годы между 18 и 29 июля. Самки и молодые
отлетают около 17 августа, самая поздняя встреча 12. IX 1967 г. и 14. X
1980 г. А. Гилязов видел стаю в две сотни куликов, по-видимому турух-
танов, летевших над долиной Чуны к югу, - это были явно не местные
птицы, а летевшие издалека.
Зимуют турухтаны в Африке и Южной Азии.
Турухтаны в брачном наряде окрашены крайне пестро и разнообразно:
большие пучки перьев на голове (”уши”) и длинные перья ’’воротнич-
ков” представляют всевозможные комбинации белого, охристо-
желтого, золотисто-красного, ржаво-коричневого, серого и черного
цветов; последний может быть с зеленым или другим металлическим
блеском, а перья других цветов одноцветными, крапчатыми или попе-
речно-полосатыми. Некоторых самцов мы наблюдали на том же болоте из
года в год; например, петушка с охристыми ’’ушами” и ’’воротником” и
пестрой, рыжей с черным спиной мы видели в 1969, 1970 и 1971 гг.; друго-
го, с черными ’’ушами” и золотисто-красным ’’воротником”, - в 1971,
1972, 1974, 1976 и 1977 гг.; третьего, с золотистыми ’’ушами” и черным
’’воротником”, - в 1969,1970 и 1971 гг.; четвертого, с черными ’’ушами” и
крапчатым серым ’’воротником”, - в 1969, 1970, 1971, 1972, 1974, 1977 гг.,
а в 1978 г. - на другом болоте, в 4 км к северу от первого. Пятого, с
серыми ’’ушами”, черным ’’воротником” и крапчатой, серой с черным
спиной, - в 1970,1971 и 1972 гг. Иногда самца с определенной расцветкой
вторично замечали после перерыва в несколько лет (белые ”уши” и чер-
ный ’’воротник” замечен в 1972 и 1977 гг.) - в таких случаях легче допустить
случайное повторение той же комбинации цветов у другой особи. Самки
турухтанов и молодые оперены скромнее, верхняя сторона вместе с
надхвостьем буро-серые со светло-охристыми каемками перьев, низ
беловатый. Самка в полтора раза меньше самца. От улитов и других
куликов сходного размера турухтана можно узнать по темному над-
хвостью (основания рулевых, однако, белые), молчаливому поведению и
тому, что он не садится на деревья.
Остатки турухтана найдены летом 1975 г. у гнезда кречета и весной
1941 г. - в желудке щуки. Из двух найденных в 1966 г. гнезд одно через
несколько дней было затоплено подъемом воды в водохранилище.
Размеры добытых нами турухтанов: самки: длина 315, крыло 186-188,
хвост 69 мм, масса тела взрослого 182, молодого (конец июля) - 153 г;
молодой самки 98 г. В желудке молодого самца обнаружены только
остатки насекомых, пауков и моллюсков.
Белохвостый песочник - Calidris temminckii (Leisl.)
Этот мелкий куличек гнездится в тундрах от Норвегии до Чукотки. В
Лапландском заповеднике одиночная самка добыта М.И. Владимирской
4. VII 1938 г. на островке в устье р. Чуна; яичник и яйцевод были увели-
чены, что дало основание признать белохвостого песочника редкой
гнездящейся птицей. Стайку песочников в то лето наблюдали еще дваж-
ды, 30. VII на р. Чуна и 4. VIII в ее устье. Позднее песочников видели: трех
147
- 23. VIII1961 г. на каменистой корге в Воче-ламбине, одного 24. VI 1969
г. на р. Нижняя Чуна, одного - 29. VII 1971 г. на болоте между реками
Чуна и Нявка; и в Мончегорске - с десяток 20. VIII 1979 г. и трех - 7. VI
1987 г. (птицы определены по фото В. Бусуйка).
Пару песочников и их токование наблюдал Семенов-Тян-Шанский на
Турухтанном болоте 31. V и 12. VI в 1972 г. и 5. VI 1977 г. Оба партнера
поднимались в воздух, как коньки или жаворонки, песня песочника
напоминала щебетание ласточки. Нет уверенности во всех случаях, что
наблюдались именно белохвостые песочники, так как характерные
признаки этого вида (белые крайние рулевые и светлая окраска ног) не
всегда удавалось рассмотреть. Учитывая, что белохвостого песочника
неоднократно находили в Шведской, Финской и Русской Лапландии в
прошлом веке (Плеске, 1887, с. 439-442), и вероятнее всего, и у нас встре-
чается этот вид.
Размеры единственной добытой самки: длина 141, крыло 93, хвост 47
мм, масса тела 27,5 г.
Куличок-воробей - Calidris minuta (Leisl.)
Этот куличок гнездится исключительно в тундрах и на север
заходит дальше, чем предыдущий вид.
Пролетных куличков-воробьев в районе Лапландского заповедника,
по всей вероятности, наблюдали в августе 1960 г.: 7. VIII М.И. Владимир-
ская видела двух очень доверчивых куличков с золотистой окраской
спины на сплавине в Пасма-ламбине, а 22-го Семенов-Тян-Шанский
наблюдал на корге в Воче-ламбине стайку из семи куличков, отличав-
шихся от белохвостого песочника темными рулевыми и охристым оттен-
ком зоба.
Морской песочник - Calidris maritima (Brunn.)
Морской песочник населяет побережье Ледовитого океана и принад-
лежащих ему морей. На Мурманском берегу он довольно обычен, как
летом, так и зимой, но на Терском встречается только на пролете (Биан-
ки, 1958, с. 57). В нашем районе его видел только Ю.Г. Гаев 27. IX 1958 г. в
Тик-губе: стая в два десятка морских песочников пролетела над полем
по направлению к оз. Имандра.
Краснозобик - Calidris ferruginea (Pontopp.)
Краснозобик во множестве гнездится в Сибирских тундрах к востоку
от Енисея, зимует по морским побережьям Африки; в Европе появляет-
ся только на пролетах.
В районе Лапландского заповедника наблюдался трижды. 18 сентября
1934 г. О.И. Семенов-Тян-Шанский видел на Железном Наволоке Екост-
ровской Имандры смешанную стаю куликов двух видов, тулесов и крас-
нозобиков, определив последних по загнутому клюву; летом 1938 г.
148
Рис. 40. Краснозобик, 12. IX 1973 г. Фото С. Баккала
А.А. Насимович наблюдал этих куличков на островке в устье Чуны;
12 сентября 1973 г. С.Н. Баккал видел и сфотографировал на расстоянии
около 1 м одиночного краснозобика на берегу р. Чуна в среднем ее тече-
нии (рис. 40).
Чернозобик - Calidris alpina (L.)
Чернозобик гнездится в тундрах Европы, Азии и Америки, зимует по
морским побережьям Западной Европы, Африки, Азии и Америки. Иног-
да залетает внутрь Кольского полуострова, а изредка и гнездится.
Одного чернозобика Семенов-Тян-Шанский наблюдал в устье р. Чуна
на протяжении трех дней, 27-29 мая 1965 г. (рис. 41). Позднее он же 27. V
1967 г. видел кормившуюся на Турухтанном болоте стайку из 12 черно-
зобиков, а 6. VI 1976 г. - снова стайку на том же месте. На этот раз одна
пара кормилась вблизи и самец токовал, сначала он кричал ’’кливливл, клив-
ливл, кливливл...”, затем издавал трель, напоминавшую песню полево-
го жаворонка, или кричал ”вит-вит-вит-вит-вит”.
1. VII 1979 г. юннаты Луценко и Стопалов видели, по-видимому, черно-
зобика на болоте у берега Кензисъявра. 17. VI 1976 г. Гилязов заметил на
берегу р. Чуна летавшего с тревожным криком чернозобика, стал искать
гнездо, а нашел выбежавшего с писком из-под кочки пухового птенца;
на его крик отзывались еще 2-3 птенца и матка. Убегая от человека, пте-
нец, которому было не более 3 дней, переплыл протоку шириной около
2 м.
149
Рис. 41. Чернозобик, 28. V 1965 г. Фото О.И. Семенова-Тян-Шанского
Грязовик - Limicola falcinellus (Pontopp.)
Редкий кулик, гнездится на болотах тундровой и таежной зон, найден
на полуостровах Скандинавском и Кольском и в нескольких пунктах
Северной Сибири (границы ареала до сих пор неясны).
18. VII 1964 г. на зыбучем осоковом болоте6, в лесотундре на водораз-
деле рек Вува и Купись, О. Семенов-Тян-Шанский и А. Печенюк встре-
тили вместе с щеголем и парой фифи пару грязовиков с птенцами, одно-
го из которых поймали и закольцевали, а потом еще троих взрослых,
один из которых был добыт, а другой сфотографирован. 22 июля на дру-
гом болоте, дальше к северу, Печенюк встретил еще шесть грязовиков,
судя по поведению - с птенцами.
18. VII 1970 г. на топком болоте в долине р. Нявка Семенов-Тян-Шан-
ский снова встретил пару грязовиков, а 25. VII 1972 г. в третий раз пов-
стречал этого куличка на берегу озерка под восточным склоном Саль-
ной тундры, вместе со щеголем и фифи. В начале июня 1988 г. Гилязов
наблюдал семь грязовиков на Турухтанном болоте.
бБолото необычное, со склоном к северу; вода с поверхности стекает в провал глу-
биной 2,5 м, с отвесными торфяными стенками и минеральным грунтом на дне.
150
Гаршнеп - Lymnocryptes minimus (Brunn.)
Гаршнеп населяет лесотундру и болота северной части лесной зоны
Евразии; зимует преимущественно в Средиземнорморье. На Кольском
полуострове - редкая гнездящаяся птица.
13. VIII 1934 г. А. Ильин добыл одного гаршнепа, сеголетка, на Южном
берегу Чунозера, а раньше он же поймал руками птенца из-под собаки
(Владимирская, 1948, с. 206).
6. IX 1937 г. Г.А. Новиков поднял гаршнепа в заболоченном елово-бере-
зовом лесу с кочками осоки на берегу оз. Тулпъявр.
Токование гаршнепа слышали 23. VI 1963 г. на Турухтанном болоте;
14. VI 1976 г. на болоте между Чуной и Улынчъявром; 21-23. VI 1979 г.
и 18. VI 1984 г. - при впадении р. Нявка в оз. Румельявр (А. Гилязов).
Бекас - Gallinago gallinago (L.)
Местное название: саамское ’’ёммиль-л^мпсынч” (дословно’’бо-
жий барашек”).
Ф.Д. Плеске в свое время пришел к выводу, что в ’’Русской Лаплан-
дии” бекас весьма редок и гнездящимся еще с достоверностью не най-
ден” (Плеске, 1887, с. 432). Возможно, сто лет назад бекас действительно
был редок в нашей области, зато теперь мы считаем его одним из обыч-
ных болотных куликов.
Представление о численности бекаса дает регистрация его встреч, как
визуальных, так и акустических. За 30 лет (1958-1987) в картотеке учте-
но 833 бекаса, в среднем по 5, 4 на одного наблюдателя в год - немного
меньше, чем улитов. Встречаемость варьирует по годам, достигая мак-
симума через каждые 4-5 лет: в годы максимумов средний индекс чис-
ленности составлял 7, 6. В 1959-1963 гг. средний индекс численности был
заметно ниже (3, 5), чем в последующие 24 года (5, 9), вероятно по причи-
не неполной регистрации встреч в те годы.
Бекас прилетает и начинает токовать в одно время с большинством
наших куликов, а отлетает позже всех. Первое характерное ’’блеяние”
токующего в воздухе бекаса отмечали, в среднем за 44 года, 21 мая
(5. V 1983 г. - 5. VI 1950 г.; о± 1, 4, температура + 4, 9е); последнее ’’блея-
ние”, в среднем за 27 лет, - 29. VI (16. VI 1972 г. - 12. VII 1966 г.). Сезон
токования продолжается около 39 дней, максимум - 54 (19. V - 12. VII
1966 г.).
Бекасы азартно токуют в тихую ясную погоду или ночью, после теп-
лого солнечного дня; под дождем, при снегопаде или в ветреную погоду
не токуют в воздухе по нескольку дней. В зависимости от погоды они на-
чинают ’’блеять” или сразу после прилета, или через несколько дней, а
иногда и недель: так, например, в 1960 г. первого бекаса видели 2 мая,
а ’’блеяние” услышали только 30; в 1962 г. прилет 8 мая, начало токования
- 26-го; в 1982 г. на кордоне Мавра губа первое ’’блеяние” слышали уже
4. V, а в других пунктах наблюдений - 17, 20 или 22. V.
Даты последних встреч бекаса от 20. VIII 1941 г. до 11. X 1970 г., в сред-
нем за 26 лет - 10 сентября. Бекас ведет скрытный образ жизни и на гла-
151
за попадается не так часто, поэтому дата последней встречи не всегда
совпадает с днем отлета (по этой причине все ’’последние” встречи, да-
тированные до 20 августа, мы отбраковывали). Приняв во внимание, что
прилетает он обычно дней за 9 до того, как начнет токовать, продолжи-
тельность его пребывания на гнездовьях около 120 дней.
Брачная игра бекаса в воздухе происходит на высоте 25- 75 м, по приб-
лизительно круговому маршруту; на протяжении его птица попеременно
то набирает высоту частыми взмахами крыльев, то пикирует, развернув
хвост веером, - в это время и слышится ’’блеяние”. Нередко несколько
бекасов (до пяти) токуют над одним болотом одновременно: встретив-
шись в воздухе, два соседних бекаса некоторое время летят параллель-
ным курсом, потом расходятся, каждый в свою сторону. У бекаса есть и
другая форма токования, сидя неподвижно на вершине дерева (обычно
сухостойного) или на кочке, он подолгу громко кричит ”т£-ки, т£-ки,
тй-ки, тй-ки, т£-ки. . . ” ; эту песню можно слышать во всякую погоду.
Обе формы токования заканчиваются обычно тем, что птица слетает на
кочку и принимается добывать корм, глубоко запуская клюв в моховую
подушку или иловатую грязь среди осочника.
Гнездится бекас, как правило, на болотной кочке, под прикрытием
пучка осоки, кустика ивы или карликовой березки; лоток глубокий. Из
десяти найденных гнезд восемь содержали по 4 яйца, два - по 3. Средняя
дата находок насиженных кладок 21 июня, самое раннее 29. V 1987 г.,
самое позднее - 23. VII 1966 г. Птенцы выводятся (в среднем из 7 наблю-
дений) 7. VII, крайние сроки 22. VI и 25. VII, но в действительности бы-
вают и более ранние выводки: 26. VI 1948 г. встретили 4 едва летавших
птенцов, которые могли родиться не позднее 8 июня.
Бекасы, вероятно, мало страдают от хищников: перья бекаса, пойман-
ного хищной птицей, мы нашли только один раз, 9. VIII 1978 г., на южном
берегу Чунозера.
Размеры одной самки: крыло 123, хвост 56 мм, масса тела 103 г.
Вальдшнеп - Scolopax rustocola L.
Вальдшнеп распространен к северу до южного побережья Белого моря
и на Кольский полуостров редко залетает (Плеске в своей сводке о нем
даже не упоминает).
В Лапландском заповеднике впервые встречен 4. VII 1938 г. в ивняке
при устье р. Чуна М.И. Владимирской; другого 5. VII 1941 г. поднял в
ельнике-черничнике близ Гарьюсного озера А.А. Насимович. Ночью 23.
VI 1966 г. во вторичном березовом лесу в долине речки Колнса-йок
А.И. Флеров видел протянувшего с характерным хорканьем и циканьем
самца.
Средний кроншнеп - Numenius phaeopus (L.)
Местное название: саамское ’’пбтчельг”.
Средний кроншнеп широко распространен в лесной полосе Евразии, от
Норвегии до Тихого океана, но всюду малочислен; зимует в Африке,
Южной Азии и Австралии. В Лапландском заповеднике нередкая гнездя-
152
щаяся птица. Наш кроншнеп относится к номинальному подвиду Nume-
nius phaeopus phaeopus (Владимирская, 1948, с. 203).
В 1931-1941 гг. средний кроншнеп был обычной птицей, за 11 лет в
заповеднике учтено 217 встреч - по 6,3 на одного наблюдателя в год; в
1958-1987 гг. он попадался заметно реже, за 30 лет отмечено 395 птиц, по
3, 4 на человека в год. На фоне общего снижения численность этого вида
нерегулярно, но сильно колеблется по годам: на протяжении 13 лет
отмечали по 21-40 кроншнепов в год, за 3 года - по 15-20, а в остальные
25 лет только по 1-14 за сезон (по 1-3 на 1 наблюдателя). Следует заме-
тить, что кроншнеп - крупный, заметный кулик, обладающий характер-
ным, слышным на большом расстоянии криком, вследствие чего регист-
рируется он легче и точнее, чем другие кулики.
Время прилета среднего кроншнепа мало варьирует погодам, и эти
колебания, видимо, не зависят от температуры: средняя дата прилета -
18 мая (7. V 1986 г. - 2. VI 1959 г. за 41 год; о ± 5, 8 дней, температура
4, 3°, корреляция между датой прилета и средней температурой мая
+ 0, 10 ± 0,15). Отлетает он около 17 августа (6. VIII 1948 г. - 5. IX 1970 г.
за 23 года; о ±8, 6, температура + 11, 3°). На гнездовьях он проводит око-
ло 92 дней. Холодная погода несколько ускоряет, а теплая - задерживает
отлет: коэффициент корреляции между температурой августа и датой от-
лета г = + 0,44 ± 0,17 (23 года).
В прилетное и отлетное время кроншнеп часто встречается на берегах
озер, а гнездятся они либо на обширных открытых комплексных болотах
с морошкой, черникой, голубикой по кочкам или в субальпийском поясе
горных тундр, на сыроватых участках с озерками, зарослями ивняков и
карликовой березки, а иногда и на открытых старых лесных гарях. За
время существования Лапландского заповедника распределение средне-
го кроншнепа по стациям заметно изменилось, как это видно по резуль-
татам регистрации встреч этих куликов (%):
1931-1941 гг. 1959—1983 гг.
Горные тундры 60 4
(260—450 м над ур.м.) Болота в лесу или 19 71
лесотундре Старые лесные гари 2 2
Берега озер 12 15
Культурный ландшафт (поля,города) 7 8
Гнездятся кроншнепы поодиночке или небольшими колониями, до
5 пар. 14. VI 1937 Т.П. Некрасова нашла гнездо кроншнепа с 3 яйцами на
Ельнюнлаге, Чуна-тундра, на высоте около 450 м, определив его по вив-
шимся над ним птицам. На том же Ельнюнлаге в июне 1931 г. видели 7
кроншнепов, а 25. VII встретили летные выводки, всего 19 птиц. С 1932
по 1940 г. ежегодно здесь встречали по 1-2 пары кроншнепов, а 21. VII
1934 г. наблюдали летный выводок. Последний раз семью в составе 6
кроншнепов встретил здесь А.А. Насимович в августе 1960 г.
Выводок кроншнепов наблюдали так же на Ягельной тундре 28. VII
153
Рис. 42. Средний кроншнеп, 18. VII 1985 г. Фото О.И. Семенова-Тян-Шанского
1941 г., а одиночных птиц или пары - на Ельявруайвенче, Туйпл-пахк,
Поадз-уайвенч (в 30-х годах) и на Сальной тундре (в 1969-1972 гг.) В
лесном поясе выводки попадались нам на небольших болотах к югу от
оз. Воче-ламбины и Чунозера, 18. VIII 1932 г. из 5 встреченных кроншне-
пов добыт один молодой; 5. VIII 1973 п также видели 5 птиц. На таком
же болоте по левому берегу Нижней Чуны с 1965 г. почти ежегодно встре-
чали пару кроншнепов, а 2. VIII 1967 г. и 26. VII 1978 г. видели летные вы-
водки.
29. VI 1968 г. над Лосиным болотом в долине р. Мавра летали 5 пар
кроншнепов: выводков здесь не отмечали, вероятно, только потому, что
место это редко посещается. 11. VIII 1966 г. над болотом в долине р. Лива
кружили с криком 4 кроншнепа, очевидно выводок. На обширных боло-
тах у Нявка-озера 23. VII I960 г. мы встретили два летных выводка: мо-
лодые отличались от взрослых коротким и почти прямым клювом. Лес-
ники заповедника нашли гнезда кроншнепов, одно в районе Матрениных
ламбин 27. V 1978 г., другое, разоренное, между озерами В. Ташким и
В. Гарьюсное 20. VI 1979 г. (обе записи без подробностей).
Средний кроншнеп легко осваивает и культурный ландшафт, его неод-
нократно наблюдали на полях Хибинского опытного поля (ПОВИР) и сов-
хоза ’’Индустрия” в г. Апатиты, а с 1968 г. - в г. Мончегорске, на берегах
озер и разливах речки Нюдовка (выводков здесь не замечали).
Кроншнепы горячо защищают птенцов и даже яйца, атакуя собак,
(18. VIII 1932 г.) ворон и чаек.
Подобно золотистой ржанке, кроншнеп охотно ест ягоды, и во всех
исследованных нами желудках обнаружены семена: 4. VI 1933 г. - брус-
154
ники и ”гром-ягоды” (Arctous alpina),, 27. VII 1934 г. - морошки и ворони-
ки, 18. VIII 1933 г. - вороники.
Летом 1975 г. остатки кроншнепа найдены в гнезде кречета.
Кроншнепа легко узнать по длинному, загнутому вниз клюву, разме-
рам и голосу - своеобразной продолжительной трели, напоминающей
трель кукушки, козодоя или степной жабы. Размеры двух взрослых сам-
цов: длина 430-445, крыло 246-255, хвост 100 мм, масса тела 300 и 330 г.
18. V 1975 г. Семенов-Тян-Шанский из окна автомашины заметил
крупную птицу, взлетевшую с пахотного поля совхоза ’’Индустрия”
(Апатиты), признав ее за большого кроншнепа (Numenius arquata L.).
Большой кроншнеп гнездится вплоть до северного конца Ботнического
залива и южного побережья Белого моря, но залетает и на Кольский
полуостров, вплоть до Айновых островов (Коханов, Скокова, 1967).
Малый веретенник - Limosa lapponica (L.)
Малый веретенник распространен по тундре, лесотундре и северной
тайге от Норвегии до Чукотки. В 1880 г. Ф.Д. Плеске наблюдал веретен-
ников у Зашейка, на Колозере и в низовьях р. Тулома; упоминает он так-
же о находках этого кулика в трех пунктах Русской Лапландии и 6 -
Финской (Плеске, 1887, с. 461-462); он признал малого веретенника
характерной птицей субальпийского, а отчасти и лесного поясов Кольс-
кого полуострова.
В Лапландском заповеднике веретенник встречается редко. 3. VI 1932 г.
на старице р. Чуна Г.М. Крепе и Г.Г. Шубин увидели плывущего вере-
тенника, признав его по загнутому вверх клюву.
Летом 1934 г. студент ЛГУ А. Ильин 1. VH добыл на заболоченных про-
токах в Пасма-ламбине самку, а через два дня видел там же еще одного
веретенника; 12. VII на Гарьюсном озере он же и В.Ф. Седова встретили
самку с хорошо летавшим птенцом; 9. VII Семенов-Тян-Шанский видел
одного на той самой старице, где веретенника наблюдали в 1932 г. 27.
V 1938 г. Семенов-Тян-Шанский наблюдал пару веретенников на старой га-
ри в бору по среднему течению р. Чуна, отметив загнутый вверх клюв и
ржавчато-рыжую окраску птицы.
На основе этих сведений М.И. Владимирская пришла к выводу, что
веретенник ’’гнездится в небольшом количестве на осоковых болотах”
(Владимирская, 1948, с. 203-204).
После восстановления заповедника веретенников встречали только
в 1969 г. юннаты Добрушин и Бурский - 23. VII одного подняли с берега
Чунозера, 31. VII пара кормилась в зарослях хвоща при устье р. Чуна.
Сопоставление всех этих данных показывает, что численность мало-
го веретенника за последние сто лет сильно сократилась: был он обычной
гнездящейся птицей, потом редкой, а в последнее время, вероятно, толь-
ко залетной.
155
ПОДОТРЯД ЧАЙКОВЫЕ - LARI
Средний поморник - Stercorarius pomarinus (Ternm.)
Этот поморник гнездится в тундрах от п-ова Канина до Чукотского;
на Кольский полуостров только залетает.
В Лапландском заповеднике залетная молодая самка добыта 1. X
1949 г. на Чунозере, ее размеры: длина 485, крыло 343, хвост 147 мм (длин-
ные рулевые еще не развиты), масса тела 535 г. В желудке найдено нем-
ного рыбьих костей (шкурка не сохранилась).
Взрослого среднего поморника легко определить по хвосту, длинные
средние рулевые скручены по оси так, что концы опахал находятся в
вертикальной плоскости.
Длиннохвостый поморник - Stercorarius longicaudus Vieill.
Распространен кругополярно, в тундрах Евразии и Америки, включая и
Мурманское побережье, откуда залетает и в глубь Кольского полуостро-
ва.
25. V 1937 г. три длиннохвостых поморника кружились над вершиной
Оарекпакх, Нявка-тундра (Семенов-Тян-Шанский; 4. IX 1938 г. мертвый
поморник этого вида найден на пляже Чунозера около Управления за-
поведника; 5. VI 1962 г. одна птица пролетела низко над Чунозером
(М. Львовский); 29. V 1975 г. пара летала над Чунозером (Семенов-Тян-
Шанский).
Длиннохвостый поморник меньше других представителей этого рода:
добытая птица имела в длину 400, крыло 305, хвост 150 мм, масса тела
222 г: у этого молодого экземпляра центральные рулевые только на 20 мм
превышали другие,, тогда как у взрослых они более чем вдвое длиннее
остальных.
Короткохвостый поморник - Stercorarius parasiticus (L.)
Ареал сходен с ареалом двух предыдущих видов, при этом на Мурман-
ском побережье этот поморник самый многочисленный.
Залетного короткохвостого поморника наблюдал А.И. Флеров на
Сальной тундре 4. VI 1969 г. К тому же виду, вероятно, принадлежали
почти все залетавшие в заповедник поморники без длинных перьев в
хвосте, замеченные 26. V 1964 г. (оз. Верхняя Пиренга, один), 15. VI
1964 г. (Екостровская Имандра, стая), 14. X 1978 г. и 25. V 1948 г. (Чунозе-
ро), 14. VI 1985г.(В. Пиренга) и 26.X 1585 г. (Воче-ламбина).
Центральные рулевые короткохвостого поморника заостренные, не
длиннее, чем у среднего, прямые. У молодых птиц всех трех поморников
центральные рулевые едва превышают остальные, а оперение одинако-
вое, темное - поэтому при визуальном наблюдении не всегда удается
определить вид птицы.
156
СЕМЕЙСТВО ЧАЙКОВЫЕ - LARIDAE
В Лапландском заповеднике и его окрестностях нормально гнездятся
четыре вида чаек и один - крачек; еще два вида чаек и один - крачек
только залетают. За 1936-1983 гг. в картотеке было зарегистрировано
около 5 800 чаек, из них почти половина без указания вида. Допуская, что
в остальных случаях наблюдатели называли вид чайки правильно, мы
нашли соотношение видов таким (в %): сизая - 35, серебристая - 40, клу-
ша - 22, речная - 2, прочие виды - 1. Часть чаек, вероятно, была опреде-
лена ошибочно: окраска спинной стороны серебристой чайки варьирует,
почему темная особь легко может быть принята за клушу, а светлая -
за сизую чайку (если нет масштаба для сравнения размеров). По этой при-
чине мы не видим смысла в описании движения численности каждого ви-
да в отдельности.
Суммарная численность чаек всех видов на протяжении полувека из-
менялась таким образом: до 1941 г. чаек встречали мало, хотя в то вре-
мя заповедник сообщался с внешним миром только через оз. Имандра и
летом сотрудники проводили на воде больше времени, чем теперь; на
одного наблюдателя в год приходилось от 2 до 9 встреч чаек. В 1948-
1951 гг. чаек стало заметно больше и мы отмечали по 5-27 птиц в год. К
1958 г. в результате перелова ценных пород рыб состав ихтиофауны оз.
Имандра изменился и основным видом в уловах стала ряпушка; эта
мелкая рыбка держится у поверхности воды и потому служит самым дос-
тупным для чаек кормом. В 1958-1965 гг. чайки в массе гнездились на
каменистых коргах по всей Имандре, в августе они вместе с поднявшим-
ся на крыло молодняком объединялись в стаи по 100-300 птиц, сохраняв-
шиеся до отлета. В зависимости от размещения больших стай по аквато-
рии и дислокации наблюдателей число учтенных птиц варьировало от
20 до 210 особей на человека. Как только вошла в эксплуатацию автома-
гистраль Мурманск-Ленинград (1966), заповедник перешел с водного
транспорта на автомобильный и перенес резиденцию управления с Чун-
озера в г. Мончегорск. В результате акватория Имандры выпала из поля
зрения сотрудников заповедника. На территории Мончегорска находит-
ся несколько озер и болот; в прошлом олиготрофные, они в результате
загрязнения бытовыми отходами стали эутотрофными, привлекательны-
ми для многих видов уток, куликов и чаек. Примерно в то же время за-
пасы ряпушки на Имандре настолько сократились, что промысел ее с 1967 г.
был л прекращен (Дребенцов, Сурков, 1974, с. 49). В результате этих пе-
ремен экология чаек резко изменилась, и теперь они кормятся преиму-
щественно отбросами человека, стали конкурентами ворон и голубей.
Уже в 60-х годах сотенные стаи чаек кормились на городских свалках
Мурманска, Кировска, Мончегорска, а с 70-х годов главным образом на
полях около зверосовхозов ’’Кольский” и ’’Мурманск”.
Не только в портовом г. Мурманске, но и в Мончегорске можно видеть
чаек, сидящих вместе с голубями и воронами на крышах и трубах домов
и даже парящих над трубами комбината ’’Североникель” в атмосфере,
насыщенной ядовитым сернистым газом.
Городских чаек мы не включали в подсчеты численности. На природ-
157
ных водоемах начиная с 1967 г. в год на 1 наблюдателя мы регистрирова-
ли по 10-30 чаек разных видов; не обнаруживая правильной периодич-
ности, число их колеблется с интервалом между максимумами от 2 до
5 лет. Общий уровень численности всех видов чаек был ниже всего в
1936-1941 гг. (около 5 встреч на наблюдателя в год), выше в 1948-1951
(10 встреч), достиг максимума в 1958-1965 гг. (28 встреч) и стабилизиро-
вался на уровне около 18 встреч в 1966-1983 гг.
Сизая чайка - Larus canus L.
Местные названия: саамское ”к£йи”, молодые чайки ’’чабар”,
русское ’’чайка” (названия эти общие для сизой и серебристой чаек).
Сизая чайка широко распространена по рекам и озерам лесной полосы
Европы и Сибири; зимует у морских берегов Европы и в других местах.
В районе Лапландского заповедника, по-видимому, уступает по числен-
ности серебристой чайке.
Прилетает немного позже серебристой чайки, в среднем за 29 лет 15
мая (3. V 1984 г. - 31. V 1940 г.); отлетает, вероятно, вместе с другими чай-
ками.
Брачные игры наблюдали мы в последней декаде мая. В начале самка
и самец сидят на льду рядом и пронзительно кричат; иногда самец взле-
тает и пикирует над сидящей самкой с полусложенными крыльями, раз-
вивая большую скорость (23. V 1980). Игра заканчивается совокуплением:
самец вскакивает на спину самки, которая при этом стоит на ногах, а
иногда шагает с самцом на спине. Вечером 5. VI 1975 г. две чайки подра-
лись; сцепившись клювами, они кричали, хлопали крыльями, пытаясь
утопить противника; драка продолжалась не менее 1-2 мин, а потом со-
перники разлетелись.
За все годы найдено только четыре гнезда:
9. VII 1969 г. на болотистом Кокоринском озерке, на торчащем из воды камне
гнездо, сложенное из тонких веток и соломинок; в нем два новорожденных птен-
ца и одно наклюнутое яйцо. Обе взрослые птицы атаковали человека.
11. VI 1974 г. — лесное озерко в долине р. Чуна, на травяной кочке среди воды,
в 30 м от берега, гнездо с 2 яйцами; наседка слетела с него заранее.
22. VI 1974 г. — в том же районе, на другом болотистом озерке, на кочке среди
воды гнездо с двумя сильно насиженными яйцами (в воде всплывают); самка сле-
тела заранее. 4 июля здесь найден один пуховой птенец. В 10 м от этого гнезда на-
ходилось гнездо кряквы.
19. VI 1980 г. — болотце у автодороги Мурманск—Ленинград, в 3 км к северу от
р. Витте, в 30 м от дороги, на кочке среди воды, замечена насиживающая чайка. Она
находилась на том же месте 20. VI и 4. VII, сидела открыто, на проезжающие автома-
шины не реагировала. 30 июля здесь видели слетка.
Пуховых птенцов встречали в разные годы между 23-31 июля; опе-
ренных, но еще неспособных летать 2-4 августа, а первых летных моло-
дых чаек между 30. VII - 7. VIII (в среднем за 9 лет - 4 августа).
По литературным данным, насиживание продолжается 22-27 дней и
молодые поднимаются на крыло в возрасте 4 недель (Harrison, 1975,
с. 175); отсюда находим, что сизые чайки насиживают яйца с 5-15. VI по
30. VI - 9. VII.
Отдельные наблюдения дают представление о питании сизой чайки: в
158
желудке самки, добытой 28. VII 1969 г., найдены два сижка примерно
по 50 г; 12. VI 1972 г. чайка выуживала из воды выброшенные рыбьи киш-
ки; в погадках обнаружены 1. VI 1973 г. кости лягушки и остатки бого-
плавов, 1. VII 1974 г. - кости лягушек и собственные перья, 14. VIII 1981 г.
- кости и чешуя окуня; 9. IX 1978 г. чайка отрыгнула на песок до 20 снет-
ков; 8. VII 1981 г. - подбирала остатки собачьего корма и достала из по-
мойной ямы кусок хлеба.
Сизая чайка сходна с серебристой по окраске оперения, но ростом мень-
ше ее. Размеры одной самки: длина 420, крыло 355, хвост 135 мм, масса
тела 370 г.
Серебристая чайка - Larus argentatus Pontopp.
Местные названия те же, что и сизой чайки.
Самая обычная по морским побережьям, озерам и большим рекам
СССР чайка. Преимущественно перед другими чайками живет и кормится
в городах и вообще в культурном ландшафте, в связи с чем численность
ее сильно увеличилась в последние десятилетия.
Прилетает к нам раньше других чаек, в среднем за 43 года 7 мая (4.
IV 1988 г. - 27. V 1934 г.; 6 ± 13, 8 дней, температура дня прилета 1,9°).
В связи с происходящим на наших глазах изменением экологии (пере-
ход с рыбного питания на отбросы человека) серебристая чайка все мень-
ше нуждается в открытой воде и стала прилетать гораздо раньше, чем
прежде. Рассмотрим средние сроки прилета ее по десятилетиям:
Годы 30-е 40-е 50-е 60-е 70-е 80-е За весь период наблюдений
Прилет 22. V 17. V 11. V 9. V 4. V 21. V 7. V
Отлет 20.Х 22.Х 18.Х 15.Х 18.Х 29.Х 20.Х
Современные поколения серебристых чаек появляются в материковой
Лапландии на 31 день раньше, чем прилетали их предки полвека назад.
Отлетают все наши чайки около 20 октября (29. IX 1966 г. - 9. XI1961 г.
за 42 года; 6 ± 8, 8 дней, температура - 0,5е). Средние сроки отлета на
протяжении 30 лет смещались на более ранние, а потом на более поздние.
В результате эволюции сроков миграций серебристые чайки проводили
в бассейне оз. Имандры в 30-х годах 151 день, а 80-х 191.
Независимо от общей тенденции изменения сроков миграций колеба-
ния их от года к году явно зависят от сроков вскрытия и замерзания вод,
а следовательно, и от температуры весенних и осенних месяцев. При вы-
числении соответствующих коэффициентов корреляции оказалось, что
полувековое смещение дат прилета подавляет зависимость годичных ко-
лебаний от явлений окружающей среды; поэтому для анализа прилета
мы ограничились более коротким рядом в 20 лет (1968-1987):
Коррелирующие пары
Коэффициент Число лет
Температура апреля — прилет чайки — 0,56± 0,15
Появление воды на льду озер — прилет чайки + 0,67 ± 0,12
Температура октября — отлет чаек + 0,49 ± 0,12
Замерзание мелких озер — отлет чаек + 0,47 ± 0,12
20
20
42
40
159
Сдвиг сроков прилета на более ранние хорошо выражен и у сизой чай-
ки, только разница между 30-ми и 80-ми годами у нее меньше, всего 18
дней.
Гнездовых колоний серебристой чайки в бассейне Имандры мы не на-
ходили, на территории Лапландского заповедника гнездятся только оди-
ночные пары. Те молодые чайки, что появляются в Мончегорске в сен-
тябре, могут быть не только местными, но и подкочевавшими с морского
побережья, на кормное место. На Чунозере площадью 21 км2 обычно
выводят птенцов одна-две пары. Брачные игры мы наблюдали только
один раз, 21. V 1972 г. в устье р. Чуна. 17. VI 1978 г. в полусотне метров от
Ленинградской автодороги А.Б. Брагин заметил чайку, сидевшую на гнез-
де, устроенном на кочке среди болота. Другое гнездо с 3 яйцами нашел
30.V 1987 г. на окраине Мончегорска В.М. Бусуек; 13. VI в нем вывелись
птенцы. На озерах заповедника пуховых птенцов встречали несколько
раз между 27.VI и 24.VH, а слетков - между 28. VII и 6. VIII (в среднем
за 8 лет 3 августа). В общем по срокам размножения серебристая чай-
ка почти не отличается от сизой.
Лесник Г.Н. Заводчиков подсчитал возрастной состав серебристых
чаек в нескольких стаях на Пиренгском озере, в октябре: в 1977 г. из 111
чаек молодыми были 23, а 1986 г. из 28-9. Низкий процент молодняка
в популяции объясняется тем, что эти чайки начинают гнездиться в воз-
расте 3-4 лет, так что среди взрослых по цвету оперения птиц особи 2-
3 поколений являются неполовозрелыми.
В природных условиях серебристая чайка питается преимущественно
рыбой или отходами рыболовного промысла, что подтверждают и наши
наблюдения: раз чайка достала со дна сига-заглотыша из желудка выпот-
рошенной щуки; две чайки поочередно клевали мертвую рыбу; на воде
чайка долбила мертвого крупного сига, придерживая его лапами; летев-
шая чайка выронила из клюва окуня; видели, как чайка заглатывала
лягушку; в желудке погибшей чайки обнаружены остатки гольяна и жу-
ка. Чайки постоянно подбирают рыбьи потроха, а иногда расклевывают
попавшую в сеть рыбу. Одна чайка, наблюдавшая, как ловят налимов на
ставные крючки, подошла к проруби и стала вытягивать из нее леску,
пока крючок не зацепился за лед; другая заглотала наживку и попалась
на крючок перемета.
13. V 1982 г. в устье р. Чуна пара серебристых чаек летела за скопой,
вероятно, с целью поживиться ее добычей (А. Гилязов). Крачки часто
атакуют крупных чаек, защищая свои гнезда, иногда в этом им помогают
и сизые чайки. При совместной кормежке разных птиц на промоинах ран-
ней весной М.И. Владимирская заметила определенную иерархию в их
отношениях: крохали боятся крупных чаек, а гагары, напротив, их про-
гоняют.
Из чаек, гнездящихся на внутренних водоемах региона, серебристая
- самая крупная; окраска ее белая, но спина и крылья сверху серые раз-
ной насыщенности, от светло-сизого до очень темного. Таких темных чаек
часто принимают за клуш, но внимательный наблюдатель может отличить
первых по более узким и длинным крыльям и легкому полету. На Мур-
160
мане гнездится номинальный подвид, серебристой чайки Latus argenta-
tus argentatus Pont, с светло-серой мантией, а восточнее, начиная с Тер-
ского берега и по-ва Канина, распространена западносибирская Larus а.
antelius Iredale с очень темной мантией (Юдин, Фирсова, 1988, с. 126). Эк-
земпляр последней формы был добыт на Имандре в 1926 г. Г.М. Крепсом
(Владимирская, 1948, с. 207).
Размеры взрослого самца: длина 610, крыло 454, хвост 167 мм, масса
тела 1098 г.
Клуша - Larus fuscus L.
Ареал этой чайки ограничен Северо-Западной Европой, от британских
островов и Исландии до Мурманской области и Карелии, зимует она в
основном на Средиземном море.
Действительный статус клуши в бассейне Имандры трудно определить
из-за того, что ее часто путают с серебристой чайкой западносибирского
подвида: возможно, она составляет не 22 % общего числа чаек (см. выше),
а несколько меньше. Индекс встреч клуш был выше всего в 1965-1976 гг.
- 3,6; значительно ниже как до того (в 1936-1964 гг. - 0,9), так и после
(в 1977-1982 гг.) - 1,0 на 1 наблюдателя в год).
Прилетает клуша почти одновременно с серебристой чайкой, в сред-
нем за 18 лет 10 мая, а иногда гораздо раньше: 31. III 1976 г. лесники виде-
ли 11 клуш на промоине в зашейке Монче-озера.
Факт гнездования клуши на внутренних водоемах Русской Лапландии
впервые установил Ф.Д. Плеске, ’’нашел несколько гнездящихся пар на Вели-
ком острове в оз. Имандра, наблюдал ее около Разноволока и добыл 15 и
18 августа 1880 г. из двух пар, гнездившихся на Колозере, около Масель-
ги, трех старых чаек” (Плеске, 1887, с. 177-178). Продолжает она гнездит-
ся и теперь, сто лет спустя.
Брачные игры клуш мы дважды наблюдали в верхнем конце Чунозе-
ра: 22. V 1972 г. в 11-12 ч. две пары чаек исполняли ритуал, сидя на льду
близ промоины на мелководье, 1. VI 1976 г. в 22 ч, пара совершала его,
сидя на затопленной коряге.
Гнезд не находили, но гнездование клуш доказывают находки птен-
цов: 4. VII 1972 г. трех пуховичков видел на Сейд-озере у зашейка лесник
Безруков, а на Чунозере неоднократно встречали слетков, 4. VIII 1976 г. -
двух при паре взрослых у Кооньт-нярка, 16. VIII 1976 г. - одного с родите-
лями в устье Чуны, 6. VIII 1978 г. - двух у Кооньт-нярка, 16. VIII 1980 г. -
трех и пару взрослых в Ельлухт.
Как видно, по срокам размножения клуша не отличается от сизой и
серебристой чайки; с последней она сходна и по составу пищи и по взаи-
моотношениям с другими птицами.
Вечером 20. V 1974 г. прилетели к устью р. Чуна и сели на лед 2 клуши,
в бинокль было видно, как одна заглатывала принесенную с собой рыбу;
29. VI 1974 г. там же на пляже нашли остатки расклеванного клушей оку-
ня; 13. VI 1978 г. налетевшая клуша выхватила из воды выброшенные
рыбьи кишки; 6 и 7. VI 1966 г. клуши расклевали попавших в сетку двух
сигов, хариуса и окуня.
11. Зак. 1023 161
19. IX 1974 г. - клуша атаковала парившего навстречу ветру орлана;
23. VII 1975 г. над Купись-озером и 24. VI 1977 г. над Кокоринским озерком
видели, как клуша гнала зимняка, но он оторвался от преследования,
быстро набрав высоту; 5. VIII 1978 г. там же пара клуш пикировала на
О.А. Макарову, пока на сцене не появилась ястребиная сова и обе
чайки не бросились на нее. В свою очередь, клуш атаковали: 26. V 1978 г.
- крачки и кроншнепы, 27. VI 1978 г. - сизая чайка и полярная крачка;
24. VI 1980 г. - десяток крачек.
Клуша немного меньше серебристой чайки, но по форме крыла и ха-
рактеру полета больше напоминает сизую. Оперение белое, спина и
крылья почти черные, неотличимые от черных концов маховых.
Бургомистр (полярная чайка) - Larus hyperboreus Gunn.
Кругополярная птица, гнездится по берегам Ледовитого океана и зи-
мует в Северной Атлантике; в глубь Кольского полуострова только за-
летает. 24. V 1938 г. молодую полярную чайку видел на льду Чунозера
В.К. Эверт; в 1981 г. А.Б. Брагин трижды наблюдал полярных чаек в Мон-
чегорске, 4. VII - четырех на разливе речки Нюдовка, 17. VII - более де-
сятка на берегу Нюдозера, 23. VIII - двух в стае других чаек, кормивших-
ся отбросами под ближайшим к Нюдовке домом; 30. V 1985 г. одиночного
бургомистра видел А. Гилязов на Охтозере.
От всех гнездящихся у нас чаек взрослая полярная отличается круп-
ными размерами и очень светлой окраской, у взрослых крылья бледно-
серебристо-серые до концов маховых, молодые на вторую зиму чисто-бе-
лые.
Озерная чайка - Larus ridibundus L.
Пресноводная чайка распространена по всей территории СССР: север-
нее Ботнического залива, Ладожского и Онежского озер до недавнего
времени не гнездилась, а на Кольский полуостров только залетала.
В районе Лапландского заповедника впервые появилась в 1970 г. -
6 июня трех озерных чаек наблюдали в устье р. Чуна, а 29 июля с десяток
в г. Мончегорске (Семенов-Тян-Шанский, А. Брагин). На следующий год
(1971) 11 мая их видели уже на Кольском заливе, 12.V - в Княжной салме,
на Имандре, 25. V пару - на нижней Чуне, 5. VI одну на Пиренгском озере,
12 и 19. VII - одну на Чунозере, all. VIII в Мончегорске А. Брагин наблю-
дал летный молодняк. С тех пор озерные чайки регулярно гнездятся в
Мончегорске, откуда залетают и на территорию Лапландского заповедни-
ка, где ни гнезд, ни нелетных птенцов пока еще не встречали.
Озерная чайка прилетает немного раньше сизой, в среднем 9.V (24.IV1982 г-
6. VI 1970 г. за 16 лет; 6 + 11,1 дня, температура + 2,3°); отлетает немного
раньше серебристой чайки.
Спонтанное продвижение озерной чайки на север в связи с освоением
его нового биотопа, городской среды - явление того же порядка, что и
экспансия кольчатой горлицы, заселившей после 1950 г. всю Западную
Европу, или ’’завоевание” Америки египетской цаплей в первой поло-
162
вине XX в. Подобно другим птицам, расширяющим свой ареал (например,
чибису), первое время озерная чайка прилетала поздно (в 1970-1978 гг.
в среднем 20. V), а укоренившись на новом месте - все раньше и раньше
(в 1979-1988 гг. 3.V).
Гнездование озерных чаек в Мончегорске наблюдал А.Б. Брагин, оце-
нивший численность этой колонии в 1980 г. в 100-200 особей. 23. VI 1979 г.
на разливе - запруженной речки Нюдовка он нашел несколько гнезд, в том
числе одно с наклюнутыми яйцами и два разоренных, яйца из которых
были выкинуты, но не расклеваны. Гнезда располагались на кочках,
были выстланы сухой осокой и напоминали гнезда крачек. В 80-х годах
гнезда и птенцов ежегодно наблюдает бывший лесник заповедника
В.М. Бусуек. Самая ранняя находка неполной кладки 13. V 1987 г., вы-
лупление птенцов наблюдалось 10. VI 1985 г., пуховичков встречали 29.
VI 1980 г и 11. VI 1987 г., оперенных, но еще нелетных птенцов - 7. VIII
1978 г., 28. VII 1981 г. и 7. VIII 1987 г., а летающих - 10. VIII 1981 г., 22 и 25.
VII 1982 г.
Общая численность Мончегорской колонии озерных чаек оценивает-
ся теперь примерно в 200 особей; кроме нее, на Кольском полуострове
найдены еще две - большая близ поселка Мурмаши (в 1974 г. 160 пар) и
маленькая, у Зашейка (в 1975 г. 20 гнезд; Коханов, 1987, с. 29).
Озерные чайки отлетают уже в зимнем наряде, который отличается
от брачного отсутствием коричневого капюшона на голове.
Зимуют они на Средиземном море и в городах Западной и Центральной
Европы, например в Праге и Берлине.
Малая чайка - Larus minutus PalL
В фауне СССР это сравнительно редкая птица, гнездится рассеянными
колониями в Прибалтике, в бассейнах Волги, Оби и других местах; бли-
жайшие к нам пункты - Ладожское озеро и север Ботнического залива.
Одну малую чайку, летевшую вдоль берега Воче-ламбины на юг, ви-
дел 17. X 1960 г. Ю.Г. Гаев.
Полярная крачка - Sterna paradisaea Pontopp.
Местные названия: саамское ’’цёррик”, русское (у поморов) -
’’чирок” (Карпович, 1984, с. 124).
Вид циркумполярный, гнездится от лесной зоны до ближайших к
полюсу пределов суши - Земли Франца-Иосифа, Шпицбергена, Гренлан-
дии; зимует в Южном полушарии, на морях, омывающих Антарктический
материк. По длине сезонных перелетов полярная крачка не имеет конку-
рентов в мире птиц.
Одна из обыкновенных птиц по всем водоемам Мурманской области,
от самых больших (Имандра) до мелких болотных луж.
Полярная крачка прилетает сравнительно поздно, в среднем за 51 год
26 мая (19. V 1963,1975 гг. - 6. VI 1932 г; о ± 4,0 дня температура + 6,0°). Время
прилета коррелирует с температурой мая (г = - 0,60 ± 0,09) и с близкими
по времени фенологическими явлениями, например со вскрытием озе-
163
pa (г = +0,59 ± 0,09). Сравнительно небольшие амплитуда (18 дней) и квад-
ратическое отклонение дат прилета показывают, что момент прилета за-
висит не только от времени наступления благоприятных для жизни усло-
вий, но и от наличия собственного годичного ритма организма, завися-
щего от астрономического ритма (изменения долготы дня или полуден-
ной высоты солнца).
Крачка - очень заметная птица, и прилет ее регистрируют наблюдате-
ли на большинстве кордонов заповедника. В 1969-1983 гг. раньше всего
они появлялись на южной границе его (Мавра-губа - 24. V, устье Нявки -
25. V), затем в устье Верхней Чуны (27. V), на Нижней Чуне (27. V), в райо-
не Монче-губы и долинах рек Ярва и Витте (27-28. V). Это свидетельст-
вует, что они прилетают к нам с юго-запада, с Ботнического залива.
Правда, в отдельные годы на северной или северо-восточной окраине
заповедника они появлялись раньше, чем на южной: в 1975 г. на Купись-
озере 11. V на 15 дней раньше, чем на Чунозере; в 1976 г. в Мончегорске
20. V (за 3 дня до прилета на Чунозеро); в 1977 г. в Мончегорске 13. V (на
14 дней раньше, чем на Чунозере). Расценивая эти случаи как прежде-
временные залеты крачек с Мурманского берега, мы исключили их из
фенологического ряда. За полвека наблюдений замечено, что крачки
стали прилетать раньше, чем прежде, - средняя дата прилета за 30-е го-
ды 29 мая, а за 80-е - 25 мая.
Полярные крачки гнездятся либо на песчаных пляжах, подобно гал-
стучнику, либо на осоковых кочках болот (при наличии на них открытых
’’окон”). Одна колония гнездилась на песчаном пляже островка в дельте
р. Чуна, в удобном для наблюдений месте, с 1931 по 1951 г. Численность
колонии колебалась от 12-15 пар в 1931, 1932, 1938 и 1950 гг. до 2-3 пар
в 1934, 1936, 1940 и 1948 гг.; в 1935, 1947 и 1951 гг. на островке не было
ни одного гнезда.
После закрытия Лапландского заповедника в 1956 г. в долине Чуны
начались рубки леса; для обеспечения его сплава уровень Чунозера был
поднят плотиной почти на 2 м, и вода затопила островок с колонией
крачек. В 1961 г. на озере восстановлен прежний гидрологический режим,
но колония крачек на островке не восстановилась. С другой стороны, для
превращения Пиренгского озера в запасное водохранилище гидроэлек-
тростанций Нивского каскада его уровень в 1937-1939 гг. был поднят
на 5 м, и вода затопила обширные болота по берегам Пиренги и соединен-
ного с ним Охтозера. Пласты торфа местами всплыли, образовав зыбкие
плавучие острова; на одном из них в устье р. Нявка в 1949 г. начали гнез-
диться крачки. 28. VI1951 г. лесник А.С. Сергин учел на нем 46 гнезд, но
острова оказались недолговечными, и к 1958 г. колония пропала.
1977-1980 гг. В.М. Бусуек и А.Б. Брагин наблюдали колонию крачек
(около 40 пар) на двух островках в зашейке Монче-озера, на границе ох-
ранной зоны заповедника. Гнезда размещались частью на песчаном
пляже, частью на каменистой отмели, частью среди травы.
Одиночные пары или небольшие группы крачек гнездятся на болотных
озерках в разных местах, в частности около озер Нявка, Островское,
Кокоринское, на Турухтанном болоте, на болотах между реками Нявка и
164
Чуна, а также на разливах речки Нюдовка в Мончегорске. В таких местах
они помещают гнездо на кочке среди осоки и устилают лоток сухими сте-
бельками. Гнезда на песке представляют неглубокую ямку почти без
подстилки.
Из учтенных 216 гнезд 19 % содержали по одному яйцу, 71 % - по два,
10 % - по три. Окраска скорлупы оливково-буроватая с темно-коричне-
выми пятнами; по размерам и темной окраске они напоминают яйца хрус-
тана и некоторых других куликов, но узкий конец не заострен. Масса
яйца 17, 8 г, новорожденного птенца 12, 8 г.
Судя по разнице в сроках вылупления птенцов, насиживание начи-
нается с откладки первого яйца. В Чунской колонии крачек 18. VI 1931 г.
было 12 гнезд с полной кладкой, а 7. VII вывелся первый птенец - значит,
инкубация продолжалась не менее 19 суток; в 1950 г. в устье Нявки
первые полные кладки замечены 9. VI, а 1. VII в большинстве гнезд были
птенцы, что дает срок инкубации около 22 сут. Первые кладки с яйцами
мы находили, в среднем за 10 лет, 13 июня (9. VI 1950 и 1963 гг. - 18. VI
1931 г.); вылупление птенцов - в среднем 5 июля (1. VII 1950 г. - 11. VII
1969 г.); летать они начинают (в среднем за 9 лет) 25 июля (23. VII 1936 г. -
2. VIII 1938 г.). Отсюда находим вероятный срок инкубации 22 сут и
возраст слетков 20 дней.
Птенцы остаются в гнезде не более суток, позднее они прячутся в
траве или другом укрытии. При появлении врага они бегут к воде и
отплывают на десятки метров от берега. В августе молодые птицы отли-
чаются от взрослых коротким хвостом, белым лбом и более темной
окраской оперения (рис. 43).
Крачки питаются преимущественно мелкой рыбой, на охоту вылетают
парами или небольшой группой, чаще всего в ночные часы, когда ветер
стихает. Заметив рыбу, крачка складывает крылья над спиной, падает в
воду, исчезая на мгновение, и взмывает кверху с добычей в клюве. Пе-
ред броском она иногда трепещет на одном месте, подобно пустельге.
Свою добычу - ряпушку, колюшку или малька крупных пород - она не-
сет, держа в клюве только голову, выставив сверкающую рыбку напоказ,
и летает с нею над болотом, как будто хвастаясь; иногда за ней с криком
летят еще 2-4 крачки. Крачка заглатывает рыбку целиком, сидя на зем-
ле или на дереве. Часть добычи передается насиживающей самке, а позд-
нее птенцам. В картотеке зарегистрировано не менее 20 крачек с рыбкой
в клюве. Кроме рыбы, они ловят и насекомых, летающих над водой, -
стрекоз, веснянок, поденок, ручейников, мух и других. При исследова-
нии желудков взрослых крачек и погибших птенцов, а также экскремен-
тов и при визуальных наблюдениях учтено 33 кормовых объекта, из них
на долю рыб приходится 79 %, насекомых 21 %.
Способ охоты полярной крачки ставит ее в зависимость от погоды: при
сильном ветре и волнении крачка не в состоянии добыть корм и неизбеж-
но голодает. Во время сильной северной бури 6-8 июля 1950 г. в коло-
ниях крачек при устьях Чуны и Нявки погибла от холода и голода боль-
шая часть птенцов: после бури Т.В. Кошкиной и нами найдено 15 мерт-
вых птенцов, у одних желудки были пусты, у других содержали остатки
165
Рис. 43. Полярная крачка, 31. V 1977 г. Фото О.И. Семенова-Тян-Шанского
рыбы. Подобный случай был и в 1932 г. При обследовании колонии в 11
пар крачек 28 июля мы обнаружили только одного здорового птенца и
шесть гнезд, содержавших 11 яиц с мертвыми эмбрионами. Одиночных
погибших птенцов находили также в июле 1933, 1938 и 1978 гг. Гибнут от
летних бурь и взрослые крачки. После ’’морянки” со снегопадом 20-22
июня 1977 г. близ оз. Вайкись лесники нашли мертвую крачку, другую
подобрал 24. VI возле гнезда на Кокоринском озерке А.С. Гилязов.
Немало крачек гибнет от хищников. Под гнездами кречетов в 1975 и 1986 гг.
обнаружено 9 костяков крачек, остатки еще двух пойманных хищными птицами
найдены на берегах рек и озер в 1980 и 1984 гг. Причиной смерти бывает и несчастный
случай. В июле 1961 г. близ Екостровского пролива нашли взрослую крачку со сло-
манным крылом: либо она ударилась о провод, либо попала под выстрел браконье-
ра. В июле 1982 г. на набережной в Мончегорске летевшая через дорогу крачка была
сбита автомашиной УАЗ и застряла в радиаторе: водитель освободил и выпустил
птицу, но пальцы на одной лапке были травмированы. 26. VI 1985 г. на оз. Кензисъ-
явр нашли крачку, умершую от внутреннего кровоизлияния; вероятно, при
неудачном броске за добычей она ударилась о камень.
Весной, до начала кладки, и в конце лета, когда молодняк уже под-
нялся на крыло, крачки не агрессивны, но около своих гнезд и птенцов
они яростно атакуют всякого нарушителя, чаще всего это бывает ворон
(10 случаев) или ворона (6 раз). Сидящий на земле ворон не реагирует на
атаки, даже если на него пикируют одна за другой 8 крачек. Более успеш-
но они отгоняют чаек, серебристую, клушу и сизую, (10 наблюдений).
Трижды. видели атаки крачек на пролетавшую скопу и по одному разу -
на зимняка, тетеревятника, дербника и сидевшую на сухой елке ястре-
биную сову. В июле 1966 г. А.Б. Брагин видел, как крачка пикировала на
норку, плывшую с берега оз. Кензисъявр к островку, на котором крачки
гнездились: она добралась до цели, хотя там на нее с криком бросались
уже пять крачек.
166
В июне 1951 г. М.И. Владимирская наблюдала, как крачка налетала на
самца куропатки, сидевшего на верхушке елки, стараясь ударить его в
голову: дело происходило на островке, где крачка гнездилась. В июне
1974 г. несколько крачек напали на пару лебедей, пролетавших над боло-
том, где они гнездились, а в июле 1985 г. крачки атаковали плавающих
лебедей (А. Гилязов). Все эти атаки крачек наблюдались не раньше 3
июня и не позже 28 июля.
Крачки готовятся к отлету, как только молодняк научится летать.
Пролет проходит незаметно. Средняя дата последней встречи 17 августа
(3. VIII1961 г. - 1. IX. 1963 и 1986 гг. за 45 лет; о ±7,6 дней температура
+ 11,3е). На гнездовьях полярные крачки проводят около 83 дней, так что
77 % года они кочуют по морям и океанам. Отлет стимулируется не похо-
лоданием, температура в это время на 5° выше, чем в момент прилета,
а корреляция даты отлета с температурой августа мала и к тому же име-
ет отрицательный знак (г= - 0,25 ± 0,15); нет корреляции и с отлетом дру-
гих птиц, например перевозчика (г= + 0,15 ± 0,16).
После отлета местных крачек в некоторые годы снова появляются
большие стаи (сотни птиц). Их наблюдали на озерах, преимущественно
Пиренгском, между 2. IX 1971 г. и 18. X 1967 г., в среднем за 7 лет 25. IX.
Возможно, так поздно пролетают полярные крачки, гнездившиеся на
Крайнем Севере Сибири.
Крачки отличаются от чаек своеобразным, изящным полетом с редки-
ми взмахами крыльев; в отличие от других птиц удар крыла вниз зани-
мает у них меньше времени, чем обратное движение его вверх. Крик
”кирри, кирри...” звучит приятнее резких голосов чаек.
Размеры взрослых птиц - 3 самцов и 3 самок: длина 361-370, крыло
264-276, хвост 162-178, плюсна 15-16 мм, масса тела самцов 101 и 106 г,
самок 96 и 106 г.
Речная крачка - Sterna hirundo L.
Речная крачка широко распространена по территории СССР и в запад-
ной части его территории гнездится до Белого моря, а залетает и к севе-
ру от Полярного круга.
В бассейне Имандры впервые обнаружена Б.К. Штегманом, опреде-
лившем среди нескольких полярных крачек, -доставленных нами в ЗИН
АН СССР, одну речную (самец), добытую 6. VIII 1931 г. на Воче-ламбине.
В коллекции Лапландского заповедника также нашлась тушка речной
крачки (без этикетки). 12. VI 1982 г. А.С. Гилязов подобрал на берегу оз.
Улынчъявр труп самца речной крачки, погибшего, очевидно, в дни сне-
гопада 6-10 июня. Его размеры: крыло 271, хвост 176, плюсна 18 мм, мас-
са тела 85, 4 г, размер семенников 4,5 х 5,5 мм. В желудке - личинки вод-
ных насекомых.
Все три птицы были определены, когда находились в руках. Вероятно,
речная крачка залетает на Кольский полуостров гораздо чаще, но в поле-
вых условиях ее не отличают от очень похожей на нее полярной крачки.
20. VI 1980 г. В.Д. Коханов нашел гнездо речной крачки у южного берега
Имандры; там же они гнездились в 1983 и 1984 гг. (Коханов, 1987, с. 28).
167
СЕМЕЙСТВО ЧИСТИКОВЫЕ -ALCIDAE
Тонкоклювая кайра -Uria aalge (Pontopp.)
Кайры гнездятся большими колониями (птичьи базары) на уступав
скал по берегам северных морей, в том числе и на Мурманском побе-
режье; вне сезона гнездования живут в открытом море, не выходя на бе-
рег.
Залетных кайр несколько раз находили в глубине Кольского полу-
острова, в бассейне Имандры. 28.1 1941 г. студенты МГУ нашли погибшую
от истощения кайру в лесу у берега р. Нижняя Чуна. М.И. Владимирская
определила ее как тонкоклювую кайру.
20.1 1936 г. Я.Р. Януше заметил трех кайр на льду Пиренгского озера:
при подходе они поднялись и полетели на юг; в тот же день он нашел на
льду Чунозера мертвую кайру (определил ее Ю.Г. Гаев); 4. II 1964 г. Янусш
поднял с переката на р. Мавра, по-видимому, кайру.
18. I 1976 г. Мончегорский охотник застрелил кайру, летевшую над
Монче-губой оз. Имандра (А.Б. Брагин); 2.1 1986 г. в зашейке Монче-озе-
ра В.В. Кузьмин отбил у воронов кайру, вероятно налетевшую на провод;
ее взяли, кормили рыбой, но вскоре она погибла (А.С. Гилязов, Г.Д. Ка-
таев).
Интересно, что все пять залетов произошли в узкий отрезок времени:
2.1 - 4. II, в самый холодный период зимы, сразу по окончании ’’поляр-
ной ночи” (анализ погодных условий в дни залетов не проводился).
Размеры двух экземпляров (пол неизвестен): длина 400-432, крыло
203-227, хвост 52-55 мм, вес 1100 (1986 г.) и 692 г (1941).
Люрик - АНе alle (L.)
Люрик гнездится на островах атлантического сектора Арктики и у
Мурманского побережья встречается лишь во внегнездовое время.
10. X 1976 г. З.Х. Аблаева заметила на Чунозере у берега непрерывно
нырявшую птичку, ростом со скворца, приняв ее за нелетного гоголенка;
рассмотрев птицу в бинокль, М.И. Владимирская и О.И. Семенов-Тян-
Шанский определили ее как люрика. Возможно, того же самого люрика
видели вторично 22. I 1977 г. А.А. Хохлов и И.А. Паракецов на льду
Чунозера у большой промоины.
18. XI 1984 г. на южном берегу Чунозера А.С. Гилязов нашел остатки
люрика, расклеванного хищной птицей. Судя по длине крыла (128 мм),
этот экземпляр принадлежал к северному подвиду Alle alle polaris Stenh.,
гнездящемуся на Земле Франца-Иосифа.
Тупик - Fratercula arctica L.
Морская птица Северной Атлантики и Ледовитого океана; гнездится в
норах, вырытых в торфяном грунте, а иногда в щелях между камнями.
На Кольском полуострове гнездовые колонии есть на Айновых островах
и Семи островах (Кандалакшский заповедник).
Известны два залета в район Лапландского заповедника, оба в середи-
168
не зимы: 26. XII1936 г. живого тупика в зимнем пере поймал на льду Чун-
озера Л.Г. Кокорин; 18.1 1975 г. на разливе речки Нюдовка, в Мончегор-
ске, школьники поймали еще живого тупика; на следующий день птица
погибла, и труп ее был передан в Лапландский заповедник (В.Г. Федото-
ва).
ОТРЯД ГОЛУБИ - COLUMBIFORMES
Сизый голубь - Columba livia L.
Полудомашний сизый голубь проник на Кольский полуостров вслед
за человеком, но значительно позже, чем воробей. Плеске (1887), а потом
Владимирская (1948) о голубе вообще не упоминают. Мы впервые замети-
ли пару голубей в поселке Апатиты 3. XII 1939 г., а в Мурманске - 15. XII
1957 г.: три голубя кормились при электрическом освещении у колонн
здания Облисполкома (ул. Воровского). В апреле 1958 г. на том же месте
жили 4 голубя, пара - в Мончегорске и еще пара в пос. Монча. К 1970 г. в
городах Мурманской области голуби по численности уступали только
воробьям.
Единичные пары голубей селятся и в отдаленных от железной дороги
поселках, например в Краснощелье (зимы 1966/67 и 1975/76 гг.) или на
лесоучастке Куцколь (1984/85 г.). Начиная с 1972 г. одиночные голуби или
пары неоднократно залетали на Чунозерскую усадьбу Лапландского
заповедника, всегда осенью (5. VIII - 20. X), но через несколько дней
исчезали; только один, появившийся в июне 1973 г., прожил почти год.
Обилие голубей привлекает в города и хищников: о случаях поимки
их ястребом-тетеревятником и кречетом мы упоминали в своем месте.
Существование голубей к северу от Полярного круга стало возмож-
ным только благодаря благоустройству современных городов. В мороз-
ную погоду голуби сидят на улице только там, где грунт подогрет труба-
ми теплоцентралей, а дневное освещение им заменяет электричество.
24. XII 1981 г. в Кандалакше в 8 ч солнечного времени, когда заря на
востоке едва занималась, голуби уже кормились при свете уличных
фонарей.
Вяхирь - Columba palumbus L.
На Кольском полуострове редкая залетная птица. 27. V 1969 г. В.И. Вла-
димирский и Р.Е. Сухов видели 6 лесных голубей над устьем р. Нявка, а
7. VIII на глазах А.Б. Брагина один перелетел Чунозеро. В этих случаях
вид голубей не был точно определен, но 12. VI 1975 г. на Чунозерской
усадьбе Брагин снова видел и рассмотрел вяхиря. В июне 1977 г. лесни-
чий С.Е. Сорокин заметил пролетевшего над усадьбой вяхиря.
Есть сведения о гнездовании вяхиря на юге Кольского полуострова,
между Зашейком, Кандалакшей и Турьим мысом (Коханов, 1987, с. 29).
Горлица - Streptopelia turtur (L.)
В наше время горлица активно расширяет свой ареал в северо-запад-
ной части СССР. В начале столетия она гнездилась только до Пскова, а в
60-х годах заселила Ленинградскую область (Мальчевский, Пукинский,
1983, т. 1, с. 390) и стала залетать на Кольский полуостров. Летом 1963 г.
169
научный сотрудник Мурманского краеведческого музея В.С. Дребенцов
добыл пару горлиц в долине Поноя (личное сообщение). Известны также
залеты этой птицы в Кандалакшу, устье Чапомы и на Семь островов
(Коханов, 1987, с. 25).
1. VII 1986 г. на Чунозерской усадьбе Лапландского заповедника
одиночную горлицу наблюдал на близком расстоянии лесничий С.В. Шес-
таков.
ОТРЯД КУКУШКООБРАЗНЫЕ - CUCULIFORMES
Кукушка - Cuculus canorus L.
Одна из обыкновенных перелетных птиц по всей лесной части Коль-
ского полуострова. Замечают кукушку главным образом на слух -
самцы кукуют, самки издают трель, и оба пола при возбуждении громко
’’хохочут”. Сравнительно редко удается увидеть кукушку; так, в карто-
теке Лапландского заповедника в 1968-1983 гг. зарегистрировано в
среднем по 19 кукушек в сезон, или по 2,1 особи на одного наблюдателя в
год. Численность их от года к году колеблется, при этом в годы максиму-
мов (1968, 1971, 1976, 1979) отмечали в среднем по 24 кукушки, в годы
минимумов - по 11 за сезон. В последние годы численность кукушек
сократилась так же, как и других певчих перелетных птиц: в 1984-
1987 гг. регистрировали только по 6 встреч, или по 1,1 на наблюдателя.
Кукушки возвращаются со своих африканских зимовок сравнительно
поздно - это по календарю; по сравнению с фенологической обстановкой
во время прилета кукушек в средней полосе в Лапландию они прилетают
рано, обычно за неделю до начала зеленения берез. В среднем за 53 года
кукушки начинают куковать 27 мая (17. V 1949 г. - 11. VI 1941 г.; 6 ± 4,9
дня, температура + 5,2°). Кукушка начинает куковать не сразу после
прилета, но, пока она молчит, ее трудно заметить: в 1967 г. первую виде-
ли за 1 день, в 1980 г. - за 20, а в среднем за 8 лет за 8 дней до первого
кукования.
Дата начала кукования с течением времени постепенно меняется
(годы):
30-е 40-е 50-е 60-е 70-е 80-е
Дата первого 27 29 30 25 24 26 мая
кукования
Корреляция между датой первого кукования и средней температурой
мая характеризуется коэффициентом г = - 0,48 ± 0,11. Связь с другими
близкими по времени фенологическими явлениями выражена слабее,
например, с началом зеленения березы г = + 0,38 ± 0,12.
Активность кукушки зависит от погоды, и куковать она начинает
обычно в теплый солнечный день. Приведенная выше средняя температу-
ра дня первого кукования (+ 6,2°) снята с графика декадных температур
за 26 лет; в это время весны она повышается на 0,2° в сутки. Фактическая
температура этого дня, выбранная из таблиц средних суточных темпера-
тур за каждый день с 1965 по 1982 г., значительно выше (рис. 48). Интерес-
но, что, начав, кукушка продолжает куковать и в ненастную погоду,
даже при северном ветре и температуре около нуля. Однако при длитель-
ном похолодании с образованием снежного покрова кукушки остаются
170
без корма и замолкают на несколько дней. Так, при позднем снегопаде
21-22. VI 1977 г. кукушки еще куковали при температуре + 2°, но по
окончании снегопада и температуре+4°. . .+9° несколько дней молчали и
вновь закуковали только 2. VII при + 15°.
Севернее Полярного круга кукушки активны круглые сутки; в первой
половине июля их слышно все реже, а во второй они совсем умолкают.
Последнее кукование отмечалось, в среднем за 46 лет, 20. VII (3. VII
1984 г. - 7. VIII 1982 г.; 6 ± 6, 8 дней, температура + 14,5°). С течением
времени окончание кукования сдвигается на более ранние сроки: в 30-х
годах оно датировалось 21. VII, в 80-х - 17. VII. Взрослые кукушки,
возможно, отлетают, как только перестанут петь, так как в августе мы
встречали главным образом молодых, иногда вместе с приемными
родителями.
Кукушки кукуют тем дольше, чем теплее лето: корреляция между
продолжительностью сезона кукования и температурой шести декад
(21. V - 20. VII) оказалась + 0,35 ± 0,14 (за 38 лет).
Отлет кукушки трудно проследить, потому что молодые не подают
голоса и держатся скрытно. Средняя дата последней встречи 13. VIII
(24. VII 1937 г. - 19. IX 1965 г. за 34 года; 6 ± 12,1, температура + 11,9°).
Большая амплитуда и среднее квадратическое отклонение объясняют-
ся неточностью наблюдений; в действительности кукушки отлетают
несколько позже указанной нами даты.
Гнезд певчих птиц с яйцом или птенцом кукушки мы до сих пор не
находили.
Брачные игры кукушек и конфликты между ними с участием 2-3 птиц
наблюдались преимущественно в начале сезона размножения, с 28. V до
10. VI. В годы с поздней весной кукушек иногда встречали парой даже в
июле (5. VII 1949 г., 14. VII 1939 г.).
Певчие птицы относятся к кукушке враждебно, особенно яростно ее
преследуют белые и желтые трясогузки, иногда группой до 2-3 пар;
реже - юрки, белобровики и веснички. Эпизодически нападают чечетки,
большие синицы, а также птицы, в гнезда которых кукушка не подкла-
дывает яиц, - щеголь, сорока и ворона.
Летных кукушат встречали с 21 июля по 15 августа, нередко в сопро-
вождении приемных родителей - в этой роли чаще всего фигурируют
юрки (4 случая), затем веснички, мухоловки и горихвостки.
Кукушки не приурочены к определенному биотопу и встречаются
повсеместно: в борах, ельниках, березняках, на старых гарях, в речных
долинах, на болотах с редкими деревцами, залетают даже высоко в
горную тундру (в южной части Чуна-тундры, на высоте около 500 м над
ур. м. 13. VI 1950 г. видели одну, а 9. VI 1939 г. - пару).
Кормятся они обычно на земле, в лесу или на открытом болоте с
редкими соснами. 15. VIII 1971 г. А.Б. Брагин наблюдал, как кукушка
ловила насекомых на лету. Когда уровень водоемов поднимается от
паводка или таяния снега в горах, сухой мусор на берегах всплывает, в
нем скопляется много насекомых. В такие дни, обычно в июне, кукушки
вылетают на берега и кормятся в полосе наносов. Считается, что кукуш-
ки питаются преимущественно волосатыми насекомыми. В Лапландии
бабочек гораздо меньше, чем, например, в средней полосе России, поэто-
му гусеницы становятся массовым кормом только в отдельные годы,
как было, например, при массовом размножении осенней пяденицы в
171
начале 60-х годов. Желудки кукушек обычно содержат перетертые
хитиновые остатки разных насекомых и пауков, по материалам Г.А. Но-
викова - личинки пилильщиков (лжегусеницы), гусеницы бабочек,
различных жуков (долгоносики, усачи, щелкуны, короеды, листоеды,
божьи коровки и др.), муравьев и пауков; единично попадаются семена
вороники и брусники (Новиков, 1952). В ненастную погоду кукушкам
приходится и голодать: лесник О.М. Петровская наблюдала кукушку,
клевавшую кашу из собачьей миски.
Размеры 6 взрослых кукушек из окрестностей заповедника (3 самца, 2
самки, 1 неизвестного пола): длина 341-375, крыло 223- 250, хвост 175-
204 мм; масса тела 5 взрослых особей 96-124, в среднем 112 г, двух моло-
дых - 98,5 и 113,4 г.
ОТРЯД СОВЫ - STRIGIFORMES
Филин - Bubo bubo (L.)
Местное название: саамское - лыеблыкль.
Редкая птица, но, вероятно, все же гнездящаяся на Кольском полу-
острове. Встречается здесь во все времена года, то оседло, то кочуя,
залетая при этом даже на тундры Мурманского побережья, до г. Полярно-
го и о-ва Хар лова (Карпович, 1985, с. 164).
В Лапландском заповеднике за 46 лет наблюдений филина видели 16
раз, слышали брачный крик (уханье) 11 раз, нашли останки одного погиб-
шего и 5 раз находили выпавшие перья.
Судя по наблюдениям, зимой филин охотится преимущественно на
глухарей (найдены остатки 3 самок и 2 самцов) и мелких грызунов: в 4
погадках, собранных у остатков добычи, обнаружены 4 глухарки и 5
полевок - 1 красно-серая, 2 рыжие и 2 экономки. Местным саамам филин
хорошо знаком.
Белая сова - Nyctea scandiaca (L.)
Местные названия: саамское - тундр-лыёблыкь (тундровый
филин), русское (на Поное) - лунь.
Ф.Д. Плеске ошибочно признал белую сову оседлой птицей горных
тундр Кольского полуострова: ’’Свою станцию (имеется в виду тундра на
границе снеговой линии. - О. СТШ.) она покидает только зимой и появля-
ется тогда на открытых равнинах и низменных местах” (Плеске, 1887,
с. 296). В позднейшей литературе нет конкретных указаний на гнездова-
ние белой совы на Кольском полуострове, и А.И. Иванов не включил его
в гнездовой ареал вида (Иванов, 1976, с. 122). Размножение этой совы на
равнинных тундрах Мурманского побережья впервые доказал сотрудник
Кандалакшского заповедника Ю.В. Краснов: 27. V 1982 г. в районе Даль-
них Зеленцов он нашел 3 гнезда с яйцами (Краснов, 1985, с. 110).
На горных тундрах Лапландского заповедника белая сова определен-
но не гнездится и только изредка залетает на его территорию во время
зимних кочевок, с декабря по май. За 46 лет наблюдений зарегистрирова-
но 8 встреч.
172
1933 — 6. V в долине Чоквуоменч на Чуна-тундре О. Семенов-Тян-Шанский заметил
сидевшую на камне белую сову, а летом на вершине Реутчокки нашел ее погадку;
1950 — 28. XII на рассвете, ехавшие по Чунозеру работники заповедника спугнули с
берега белую сову и отобрали ее добычу — самку кречета; сова успела съесть
голову, обклевать шею и переднюю часть грудных мускулов. Перед тем, 18. XII, в
бору на побережье Воче-ламбины найдены остатки глухарки со следами крыльев
и лап крупной совы на снегу; вероятно, ее поймала та же сова, поскольку расстоя-
ние между обеими находками не превышало 7 км;
1962 — 10. VIII на Нявка-тундре О. Семенов-Тян-Шанский нашел остатки белой совы,
пойманной кречетом, ястребом или орлом; сохранился почти весь костяк, кроме
головы и шеи, и перья;
1969 — 18. I на лесника М. Брускова, ловившего на Чунозере рыбу на крючки, сзади в
упор налетела белая сова;
1980 — 27. III около 6 ч сотрудник заповедника В.Э. Берлин заметил поднявшуюся со
снега в лесу близ Чунозера белую сову; 20. IV Д.М. Шкляров при учете диких
оленей с самолета заметил летевшую ниже его белую сову в районе юго-западной
границы заповедника;
1985 — 5. V на берегу Нявка-озера лесники А. Царев и А. Беднягин подняли белую
сову с остатков пойманной ею белой куропатки;
1986 — 8. I на Воче-ламбине егерь А. Андреев видел белую сову, севшую на лед; в
том же районе ее видел 25. V лесник П. Пакулин.
Белая сова выкармливает птенцов преимущественно леммингами и
полевками, а зимой, когда этот ресурс становится недоступным под
снегом, кочует и ловит птиц. В числе ее жертв нами отмечены тундрянка,
белая куропатка, глухарка и кречет; с другой стороны, она сама может
стать добычей кречета. Залеты белой совы в леса Кольского полуострова,
по-видимому, не зависят от численности мелких грызунов: залеты 1933,
1969, 1962 и 1986 гг. происходили в фазе подъема их популяций, в 1950 г. -
в фазе пика, в 1980 г. - при депрессии.
Ястребиная сова - Surnia ulula (L.)
Местные названия: саамское - куньлоньт (приблизительный
перевод ’’срамная птица”), также чузень-вуоппе (чузень - искра, вуоп-
пе - сова; от поверья, что сова глотает искры от костра).
Ястребиная сова населяет таежную зону обоих полушарий. На Коль-
ском полуострове распространена по лесам всех типов: сосновым, ело-
вым и березовым, но предпочитает старые гари. Охотясь на грызунов,
залетает и на горные тундры, где нередко находили ее погадки и помет.
Встречается значительно чаще других сов, чему способствует и дневной
образ жизни, и демонстративное поведение. Подсчет визуальных встреч
сов по картотеке за все годы дал такое соотношение видов (в %0):
Ястребиная сова 819
Болотная сова 81
Мохноногий сыч 27
Бородатая неясыть 28
Длиннохвостая неясыть 16
Воробьиный сыч 12
Филин И
Белая сова 6
Ястребиная сова попадается во все месяцы ] года, но зимой приблизи-
тельно в 8 раз реже, чем летом, а в некоторые зимы отсутствует. Видимо,
в снежное время они кочуют, и только при благоприятных условиях
часть популяции зимует на местах гнездования (годы 1975, 1978, 1979,
1982,1983,1987). В конце 1969,1973, 1980 и 1983 гг. одиночные ястребиные
совы встречались до декабря, а затем откочевывали. В осени, бедные
грызунами (1968,1972,1975,1976 и 1984), совы исчезали уже в октябре, а в
годы депрессии полевок (1971, 1979 и 1985) и в августе. Вызванные бес-
кормицей откочевки уводят ястребиных сов далеко от места их рожде-
ния. В ’’лемминговый” год 3. XI 1946 г. близ Мурманска была добыта
ястребиная сова, окольцованная 24. VI 1946 г. в Шведской Лапландии, у
173
Рис. 44. Встречаемость ястребиных сов в Лапландском заповеднике по месяцам за
14 лет (сплошная линия)
С правой стороны число встреч по годам, снизу — по месяцам
Квиккйока (67° с. ш.): до конца лета она, вероятно, оставалась на месте
рождения, а за осень пролетела 690 км к востоку-северо-востоку (Демен-
тьев, 1951, с. 407).
Появление ястребиных сов после голодных зим происходит, по наше-
му представлению, не за счет возвращения мигрантов обратно на Коль-
ский полуостров, а в результате подкочевки сов из других регионов
таежной зоны.
Численность любой птичьей популяции, очевидно, должна увеличи-
ваться скачком в момент вылета молодняка из гнезд, применительно к
ястребиной сове - в июле. Наблюдения показали, что такой скачок
действительно происходит, но только в ’’мышиные”; в остальные годы
количество встреч в июле-августе по сравнению с маем-июнем не увели-
чивалось, а это доказывает, что они либо совсем не гнездились, либо не
смогли вырастить птенцов. Подсчет визуальных встреч ястребиных сов
по месяцам для лет, богатых грызунами и бедных в отдельности, дал
следующий результат:
Снежная пора Бесснежная пора Всего
птиц
XI-IV V VI VII VIII IX X
"Мышиные” годы 69 34 105 99 169 93 42 611
Годы, бедные грызунами 32 12 7 4 8 10 6 79
Эти цифры показывают, что движение численности сов в течение года
в бескормные годы идет в направлении, противоположном кормным
годам (рис. 44). Разница в численности сов между теми и другими резко
174
выражена только летом (соотношение 25:1 - 10:1), а зимой незначительна
(соотношение 2:1). Это обстоятельство наводит на мысль, что ястребиная
сова не способна добывать полевок из-под снега и во время зимних
кочевок питается случайной добычей, зверьками, выбегающими на
поверхность снега, и птицами, а потому уровень численности полевок
зимой не имеет такого большого значения, как летом.
Зависимость успеха размножения от обеспеченности кормом и зим-
ние миграции, перераспределяющие совиную популяцию по территории,
подчиняют колебания численности ястребиных сов циклам мелких
грызунов. Годы минимумов грызунов и сов, как правило, совпадают,
но ’’пик” сов наступает иногда в год ’’пика” полевок (1938, 1950, 1958,
1970, 1987), а чаще на следующий за ним год (1964, 1974, 1978, 1983;
рис. 49). Коэффициент корреляции между индексом учета полевок в
начале сентября и количеством встреченных сов, в пересчете на 1 наблю-
дателя в год, составляет за 1958-1987 гг. + 0,30 ± 0,17, если брать вариан-
ты за тот же самый год, или + 0,60 ± 0,12, если индекс сов брать за следую-
щий после учета грызунов год.
Судя по встречаемости выводков, ястребиная сова гнездится как в
сосновых, так и в еловых лесах, но в борах мы нашли 6 гнезд, а в ельни-
ках только 2; из этих гнезд 4 найдены в 1983 г.
Три гнезда располагались в дуплистой сосновой сушине с природным или выдолб-
ленным желной боковым входом.
6. VI 1947 г. — дупло в сухой сосне, на высоте 4 м, в елово-сосновом лесу в долине
р. Витте; в нем 5 близких к вылету птенцов (М.С. Тютин).
15. VI 1978 г. — в старой сушине в 8 отверстиями, из которых сова выбрала
верхнее, обращенное на юг, на высоте 10 м. В момент находки в гнезде были птенцы,
но при осмотре его А. Гилязовым 21. VI они уже вылетели. Найдено в бору на южном
склоне Чунъявр-вараки.
19. V 1982 r. — в бору ша северном склоне той же вараки, в дупле сухой сосны на высоте
5,5 м, отверстием на ЮЗ; совы расширили леток, выломав клювами кусочки трухля-
вого дерева с краев; внутренний диаметр дупла до 26 см, глубина 10 см; в кладке 7
яиц на подстилке из трухи. В момент находки самка насиживала, самец находился
вблизи. В 1987 г. совы гнездились в этом дупле вторично, 18. VI в нем был полуопе-
ренный птенец.
Другой тип дупла — на торце сломанного ветром гнилого дерева:
7. VII 1978 г. — в ельнике на берегу оз. Ельявр А.Б. Брагин обнаружил гнездо совы
на торце сломанной ветром ели, в 7—8 м над землей; к 16. VII птенцы вылетели.
16. V 1983 г. — в елово-березовом лесу на берегу Чуны А.Гилязов нашел гнездо на
торце гнилой березы, сломанной на высоте 4 м. Влезть по шаткому стволу было
невозможно, но с соседнего дерева в гнезде видны 2 яйца.
7. VI 1983 г. — в бору в долине Чуны, на торце сосны, расшатанной в корне и
сломанной ветром на высоте 5 м А. Гилязов обнаружил гнездо с 3 птенцами.
Два гнезда были устроены необычным образом:
2. VI 1983 г. — в гнездовом ящике для уток, на берегу озерка в разреженном
сосняке в долине Чуны А. Гилязов нашел гнездо, в нем 2 оперенных совенка и 2
яйца-болтуна.
5. VI 1983 г. — в бору на берегу оз. Красная Ламбина о. Макарова нашла гнездо
ястребиной совы на дереве, в углублении между сучьями, построенное из сухих
веточек (осмотреть его не удалось).
Ястребиные совы образуют пары и начинают брачные игры в апреле
(11-17. IV 1983 г.), иногда даже в марте (15.111 1987 г., 22. III 1973 г.). Нам
не удалось наблюдать откладку яиц, а гнезда с насиженными яйцами
были найдены 19. V 1982 г. и 16. V 1983 г.; 24. V 1934 г. добыта самка с
наседным пятном (Владимирская, 1948, с. 212); 30. V 1982 г. наблюдали
вылупление птенцов (2 птенца и 5 яиц в гнезде). Гнезда с птенцами
встречались между 2. VI - 7. VII (в среднем 12. VI; п = 6).
По литературным указаниям ястребиная сова насиживает 25-30 дней и
. птенцы вылетают в возрасте 23- 27 сут (Harrison, 1975, с. 202). Эти сведения
175
дают, возможность рассчитать сроки размножения, зная время вылета
птенцов из гнезд. Первых слетков мы встречали между 5. VI 1983 г. -
30. VI 1941 г., в среднем за 9 лет 20 июня. Отсюда следует, что нормально
птенцы выводятся в последней декаде мая, а кладка яиц начинается
около 30 апреля. В гнезде 1982 г. первый птенец появился на свет, види-
мо, 28-29 мая, последний - 7 июня, вылетели они соответственно 19 и 30
июня в возрасте 22-23 дней; следовательно, яйца были снесены в первой
декаде мая. Сроки размножения заметно варьируют по годам и индиви-
дуально. Возможно, ястребиная сова иногда гнездится и выводит вторич-
но, по крайней мере А.А. Насимович однажды встретил слетка необычай-
но поздно, 2. IX 1946 г. (Владимирская, 1948, с. 213).
Размер кладки сильно варьирует, в найденных нами гнездах было от 2
до 7 яиц, в среднем 4,2.
Масса тела двухнедельного птенца 21. V 1982 г. была 20,3 г, другого
птенца в 21-дневном возрасте, перед самым вылетом, 226 г. Средняя
суточная прибавка в массе вгнездовый период 13% в сутки. В 1982 г. при
хорошем питании совята росли очень быстро, 13. VI они своей массой
заполнили дупло, так что обогревавшей их самке пришлось выставить
наружу хвост, 17-го выставила и крыло, но к этому времени птенцы
полностью оперились, и она перестала их обогревать. Первые совята
вылетали дружно, вероятно с интервалом в сутки, потом через двое,
последний через трое суток, так что вылет семи птенцов занял 11 дней:
причиной этого может быть такая же растянутость кладки яиц или же
отставание в росте младших птенцов по сравнению со старшими. В ре-
зультате слетки рассеиваются и выводок не удается сосчитать полностью:
только в одном, встреченном 23. VI 1938 г. Г.А. Новиковым, было 5 совят,
во всех остальных от 1 до 3.
Ястребиная сова активна ночью и днем. Осенью она бесшумно подле-
тает к бивачному костру как в сумерки, так и в полночь, неожиданно
пугая путника своим неописуемым диким криком: от этой повадки
происходят оба саамских названия ястребиной совы. Ночуя в лесу
23- 24. IX 1982 г., мы слышали ее крик задолго до рассвета.
Днем ее обычно видят на наблюдательном посту сидящей на макушке
дерева, иногда на лету, в момент охоты. Ястребиная сова довольно
доверчива, легко подпускает на 20-30 м, а потому служит соблазнитель-
ной мишенью для ’’воскресных охотников” и часто становится жертвой
их ружей.
В 1982 г. мы изучали суточный цикл этой совы, закрепив на пороге
дупла датчик актографа. Взрослые навещали гнездо довольно равномер-
но на протяжении суток: активность распределяется по часам следующим
образом (сравните с мохноногим сычом, см. ниже):
Часы суток 09 13 17 21 01 05 09 За 24 ч.
При птенцах 13 18 18 17 17 17 100%
Суточная активность достигала минимума в полуденные часы (0,36
посещений в час), а максимума в вечерние (в 18-20 ч по 0,74 в час). В
разные дни совы прилетали к гнезду от 6 до 22 раз в сутки. Частота
визитов, по-видимому, не зависит ни от осадков, ни от температуры (она
за три недели колебалась в пределах - Г. . . + 27°, при средней + 5,9°), но
явно зависит от возраста птенцов: в первую неделю наблюдений по 10,9,
во вторую - 11,3, в четвертую - по 18,0 посещений гнезда в сутки. Если
176
сова каждый раз приносила по полевке, то в первую неделю каждый из 7
птенцов получал по 1,5, в последнюю же по 2,5 передачи в сутки.
Совята вылетают с остатками пуха в оперении и неокрепшими маховы-
ми; в первые 1-2 дня птенец сидит неподвижно на сучке или камне, под
которым скопляется его помет и 1-2 погадки; в это время его можно
взять руками. Совята постарше подпускают к себе не ближе 15 м, летят
низко над землей и садятся на нижние ветки деревьев, иногда баланси-
руя крыльями и хрипло посвистывая, как собака, когда просится в дверь.
Движения крыльев служат не для сохранения равновесия, как нам
показалось вначале, а сигналом требования корма. Слетка легко отли-
чить от взрослой совы по темному оперению и полному комплекту
маховых и рулевых: старики в середине лета линяют и потому в их
крыльях и хвосте заметна недостача больших перьев. К концу июля
совята становятся самостоятельными, с августа в подкормке уже не
нуждаются, по крайней мере встречаются по одиночке.
Из всех наших сов ястребиная - самая агрессивная. У гнезда она
яростно нападает на собаку или человека, стараясь застать врага врас-
плох. Она не носится над головой с криком, подобно крачке, поморнику
или улиту, а садится в засаду на одну из нижних веток дерева и, уловив
момент, когда на нее не смотрят, срывается и, быстро набирая скорость,
несется по прямой, целясь в голову нарушителя; в последний момент она
взмывает, проносясь со свистом над самой головой, или наносит удар
грудью и когтями в голову, руку или спину. В июне 1982 г. самка ударила
одного из нас в лоб, оставив три кровоточащие царапины над бровью. В
следующий раз мы подходили к гнезду, держа над головой бамбуковую
палку - сова ударила ее с такой силой, что могла бы сломать себе крыло
или шею. Самец был осторожнее, атаковал раз в десять реже, никогда не
ударял; свое беспокойство он выражал главным образом криком. Агрес-
сивность сов варьирует индивидуально, а не в зависимости от пола.
Самец другой пары, защищая гнездо в мае 1983 г., ударил одного из нас
сзади, сорвав с головы вязаную шапку. Одна из сов, занимавших в 1983 г.
утиную дуплянку, ударила человека в голову, тогда как другая пара,
гнездившаяся на сломанной сосне, вела себя мирно. Некоторые совы
атакуют и после того , как выводок оставит гнездо.
В июне 1937 г. Л. Кокорин видел, как пара ястребиных сов напала на
глухаря, одна ударила его в спину так, что он свалился в дерева, но
падая все же выровнялся и улетел.
Ястребиные совы проявляют агрессивность и во внегнездовое время: в
марте 1939 г. одна бросилась на глухаря в лесу близ Воче-ламбины,
другая в марте 1979 г. атаковала норку, бежавшую по снегу к р. Курка.
Однажды мы наблюдали ’’воздушный бой” совы с дербником и неодно-
кратно - нападения ястребиной совы на ворона. С другой стороны, ястре-
биную сову нередко преследуют мелкие птицы, чаще всего дрозды, затем
чечетки, клесты и кукши; иногда большой сорокопут, серая ворона,
полярная крачка, гаички и пеночки-веснички.
Собранный нами материал по питанию ястребиной совы характеризует
преимущественно бесснежный период: он состоит из 35 желудков, добы-
тых главным образом в довоенные годы (из них 5 пустых), 76 погадок,
собранных под 5 гнездами в 1978, 1982 и 1983 гг., и 13 подобранных
случайно (табл. 11).
12. Зак. 1023
177
Таблица 11. Питание ястребиной совы
Вид добычи 1931-1951 гг., желудки 1978—1983гг., погадки 5 гнезд Случайные сборы По всему материалу. %
Млекопитающие Насекомоядные 2 1
(землеройки) Грызуны* 7 3 1 6
Лесные полевки* 1 20 4 14
Красно-серая по- 6 51 7 36
левка Рыжая полевка 4 3 2 5
Красная полевка 2 2 — 2
Серые полевки* — 8 5 7
Пашенная полевка — 4 3 4
Экономка — 3 — 2
Норвежский лем- 8 21 3 18
минг Лесной лемминг 3 — 2
Птицы (конек, овсян- 1 1 2 2
ка и др.) Насекомые — 1 1
Всего объектов: 34 117 27 178
Число желудков, 30 76 16 122
погадок.
*Ближе не определены.
Мелкие грызуны составляют 97% летней пищи ястребиной совы.
Погадки и помет птенцов, а иногда и недоеденные остатки добычи в
гнезде образуют плотную, сравнительно сухую подстилку, толщина
которой достигает 5 см.
Ястребиная сова одна из самых обычных хищных (по образу жизни)
птиц нашей фауны, поэтому остатки погибших особей этого вида находят
довольно часто. В погадках орла под гнездом 1939 г. найдены остатки
двух, под гнездами кречетов - трех, в добыче ястреба-тетеревятника -
одного. Из 15 сов, найденных на месте гибели в разные годы, 5 съедены
лисами (судя по огрызенным очинам маховых), 2 пойманы хищными
птицами, 1 разбилась о высоковольтный провод, 1, по-видимому, утону-
ла, и труп ее выброшен на берег волной, 2 - погибли зимой от голода;
причины смерти остальных 4 не определены. Больше всего сов гибнет в
годы падения численности грызунов, когда хищники-миофаги голодают и
конкуренция между ними обостряется.
Размеры ястребиных сов: самцы - длина 384 (мм) (370-400), крыло 238
(233-242), хвост 188 (180-196; всего 6 экз.); самки соответственно - 366
(344-380), 246 (232-277) и 175 (165-190); 3 экз.). Масса тела заметно
меняется по сезонам, летом (VI-IX), средняя масса тела самца 310 ± 8,5 г
(и = ГО), самки - 340 ± 8,5 (и = 11); в снежное полугодие (X-V) самец
277 ± 13,7 (п = 3); самка - 321 ± 8,1 (п = 7). Предельные веса самцов 250-
178
348 самок 277-385 г. Средняя масса тела самки на 12% выше массы тела
самца, а зимняя масса тела обоих полов на 8% ниже летней; сеголетки,
видимо, по массе не отличаются-от взрослых. Вес двух яиц с погибшими
эмбрионами 19,0 и 19,6 г.
Воробьиный сыч (сычик) - Glaucidium passerinum (L.)
В прошлом веке сычик еще не был обнаружен на Кольском полуостро-
ве, и Ф.Д. Плеске сообщает только о нескольких находках его в Финской
Лапландии, к северу до Каресуандо (68° 27' с. ш.; Плеске, 1887, с. 303). В
Лапландском заповеднике его встречали неоднократно, во все месяцы
года, кроме января. Гнездование сычика в заповеднике пока не доказа-
но, но оно очень вероятно, так как он является относительно оседлой и
не очень редкой птицей: за 46 лет отмечено 17 встреч и добыто 6 экз.
1931 г. — на Старой базе на Чунозере наблюдался О. Семеновым-Тян-Шанским на слух
с XII. 1930 г. до IV. 1931 г.; 4. II один пролетел ночью около дома.
1935 г. — 1. X на берегу Чунозера на вершине сосны видели сычика.
1937—1938 гг. — в районе Чунозера П.С. Мальчевским и Г.А. Новиковым добыто 5
воробьиных сычей - два в ельнике, до верхней границы леса на склоне Чуна-Тун-
дры, два - в бору, один - в смешанном лесу. Даты добычи 30. VII - 15. VIII. Крик
сычика дважды слышали на усадьбе заповедника в октябре, а в сентябре 1938 г. в
ельниках между озерами Ельявр и Райкоряурынч.
1948 г. — в бору близ Чунозерской усадьбы заповедника в апреле найден труп
сычика, погибшего, видимо, от истощения.
1950 г. — в сентябре, в сумерках, сычик пролетел над усадьбой.
1971 г. — 20. VI в редком елово-березовом лесу между вараками Ельявруайвенч и
Курт-варенч О. Семенов-Тян-Шанский видел пару сычиков, но не смог найти их
гнездо.
1974 г. — 10 и 21 августа, в вечерних сумерках, И.А. Паракецов видел сычика в бору
между Чунозером и Охтозером; в утренних сумерках 3. XII сычика наблюдали на
усадьбе заподведника.
1977-1978 гг. - лесники А.В. Белозеров и Ю.А. Куприянов неоднократно наблюдали сычиков
на берегу оз. Вайкись: в мае 1977 г. одного видели на вершине сосны, дважды в
ноябре на гари между озерами Пагель и Матрениной лабминой, в августе 1978 г.
еще 2 раза в том же районе.
Встречи воробиных сычей не коррелируют с численностью полевок: в
1930-1931, 1937-1938, 1950, 1974, 1977-1978 гг. она была наибольшей, а в
1935,1948 и 1971 гг. находилась в фазе депрессии.
Сведения по питанию сычика фрагментарны: в желудках 3 особей
обнаружены остатки полевок, по одной в каждом, 3 желудка были пусты.
В августе 1981 г. под Ельявруайвенчем найдена погадка из перьев певчей
птицы, еще погадка с остатками красно-серой полевки обнаружена летом
1975 г. в скворечнике для синиц (его леток узок для мохноногого сыча).
Весной 1984 г. в 4 скворечниках найдены мумии большой синицы, лап-
ландской гаички, лемминга и красно-серой полевки: по всей вероятнос-
ти, это были запасы сычика.
На сычике, погибшем в 1948 г., пойманы 2 блохи, по определению
В.В. Дубинина и И.Г. Иоффа - Ceratophyllus gallinae.
Сычиков встречали в разное время суток, ночью на лету, днем -
обычно сидящих на дереве. Они довольно доверчивы, садятся на виду и
подпускают метров на десять, иногда и ближе. Сычик значительно
меньше других наших сов, голова относительно маленькая, с плохо
выраженным лицевым диском, хвост очень короток. Размеры 6 добытых
экземпляров: длина 148-181, крыло 98-108, хвост 55-72 мм, масса тела
49-79, в среднем 70,7 г (экземпляр, павший от истощения, исключен из
подсчета).
179
Бородатая неясыть - Strix nebulosa Forst
Местные названия: саамское - хиппик или ый-хгеппи.
Бородатая неясыть распространена в бореальной зоне обоих полуша-
рий. На Кольском полуострове она предпочитает сосновые и сосново-ело-
вые леса; открытых мест, например горной тундры, избегает, но иногда
пролетает над озерами. В районе Лапландского заповедника встречается
чаще и регулярнее других крупных сов. К сожалению, наши наблюдатели
не всегда отличали бородатую неясыть от длиннохвостой, что затрудняет
обработку наблюдений: за 46 лет зарегистрированы визуально 63 неясы-
ти, в том числе 25 бородатых, 14 длиннохвостых и 24 - неизвестного
вида. В середине зимы, на протяжении 100 дней, неясытей не встречали
(самая поздняя встреча бородатой неясыти 8. XI, самая ранняя - 15. II);
вероятно, в это время они кочуют и при этом смещаются немного к югу. В
Печоро-Илычском заповеднике, расположенном на 6° южнее Лапландско-
го, бородатая неясыть считается оседлой, по крайней мере встречается
там на протяжении всего года.
Рассмотрим наблюдения за этим видом по годам.
10. X 1941 г. О. Семенов-Тян-Шанский впервые встретил и рассмотрел с близкого
расстояния бородатую неясыть в бору на южном берегу Воче-ламбины. 25. VIII 1947 г.
на Чунозерской усадьбе видели крупную сову ”с кругами у глаз”. Летом 1951 г. в том
же районе нашли два свежих пера.
В 1964 г. в смешанном лесу на северо-восточном побережье Чунозера Ю.Г. Гаев
нашел два гнезда бородатых неясытей. Одна пара заняла старое гнездо, в прошлом
году занятое пустельгой, на сосне на высоте 8 м над землей. В день находки, 11. VL
в нем было 4 яйца и самка их насиживала; 18. VI первый наклев; 24. VI в гнезде 3
птенца и 1 яйцо; 13. VII — 4 птенца, частично оперенные, у трех старших разверты-
ваются опахала маховых, но головы еще в пуху; к 24. VII птенцы вылетели, но
находились где-то поблизости, так как сова атаковала Гаева, когда он влезал на
дерево; последний раз неясыть видели около гнезда 29. VII, но 7. IX трех молодых
видели вместе не далее 1 км от него, а 13. IX - одну в том же районе. Другая пара
занимала старое гнездо ястреба-тетеревятника, также на сосне, в 1—2 км от первого.
В день находки, 13. VII, молодые уже вылетели, но одного нашли вместе с взрослой
неясытью в сотне метров от гнезда, а еще двух только слышали. 26. X 1964 г. неясыть
видели над Ленинградской дорогой в 4-5 км к востоку от этих гнезд.
Весной 1980 г. при устье Чуны А. Гилязов нашел кучу перьев погибшей зимой
бородатой неясыти; судя по обгрызенным очинам, съела ее лиса.
15. II 1983 г. на оз. Румельявр филин поймал упитанную взрослую самку борода-
той неясыти с массой тела около 1,6 кг (рис. 45).
9. VI 1983 г. в бору на берегу Чунозера А. Гилязов и О. Семенов-Тян-Шанский
заметили сидящую на гнезде неясыть (рис. 45). Гнездо новое, небольшое (75 см в
диаметре), на развилке сосны в 7 м над землей, вероятно построено самой совой.
Птица сидела крепко, так что с земли ее можно было фотографировать. Вечером
23. VI совы не было в гнезде, а на земле под ним лежали разорванные тела трех едва
начавших оперяться птенцов: сбросил их медведь, следы его когтей остались на коре
сосны. Впоследствии оказалось, что птенцов было пять, два из них остались живы и к
18. VII оставили гнездо. В последний раз неясыть видели в этом районе 11. IX.
Одиночных бородатых неясытей встречали в разных местах, но всегда в бору, в
октябре 1970 и 1973 гг., в сентябре 1978 г., в марте 1982 г., в июне 1986 г., а 9. VIII 1981 г.
ночью на берегу р. Вайкись видели слетка.
В июне 1987 г. О.А. Макарова нашла под деревом 5 больших погадок, по всей
вероятности бородатой неясыти, и до десятка шкурок лесных полевок.
Наблюдения 1964 г. показали, что вывод птенцов бородатой неясыти продолжает-
ся не менее недели (19—26. VI), живут они в гнезде 27—30 дней. В 1983 г. неясыть
гнездилась примерно на неделю раньше: 22. VI 1964 г. совята только выводились из
яиц, а 22. VI 1983 г. масса тела погибших птенцов была около 0,5 кг и они уже
оперялись; выводки оставили гнезда в 1964 г. в одном — к 13. VII, в другом — к
24. VII, в 1983 г. - к 18. VII.
В 1964 г. взрослую самку мы легко отличали от самца не только по большему
росту, но и по травме правого глаза. 13. VI на яйцах сидела самка, подняв хвост
вверх; когда человек полез на дерево, она встала, угрожающе расправила крылья,
распушила оперение, потом взлетела. Самец в это время сидел на соседней сосне; оба
щелкали клювом, шипели, а в общем вели себя сдержанно, так как людей было
двое. Когда Ю. Гаев подходил к гнезду в одиночку, самка на него налетала и
180
Рис. 45. Бородатая неясыть на гнезде, 9. VI 1983 г. Фото А.С. Гилязова
однажды ударила когтями в поясницу, а другой раз в голову; самец также налетал,
но не пытался ударить. При фотографировании и кольцевании птенцов в гнезде
самка садилась на ветку в 2—1,5 м от человека. В 1983 г. мы влезали на дерево
только один раз и неясыти не проявляли агрессии.
Бородатая неясыть считается одной из самых агрессивных 'сов. Микко-
ла сообщает два случая, когда в результате удара этой совы в лицо
Таблица 12. Питание бородатой неясыти летом
Вид добычи 1964 г. экз. 1983 г. экз. 1987 г. экз. Весь ма- териал, %
Млекопитающие Насекомоядные (земле- 3 0,5
ройки) Грызуны (мелкие) Полевка красно-серая 122 40 И 85,5
Полевка рыжая 2 1 — 1,5
Полевка красная — 1 — 0,5
Полевка экономка и 2 8 1 5,5
пашенная• Водяная крыса 1 — — 0,5
Лемминг норвежский — 8 — 4
Хищные (ласка) 1 — — 0,5
Земноводные (травяная 1 — — 0,5
лягушка) Насекомые (жуки)* 2 0 +
Моллюски (наземные 2 — — 0 +
улитки)* Всего объектов: 136 58 12 100%
*Ввиду ничтожного объема жуки и улитки не учтены в последней графе.
181
человек лишился глаза, а еще один сломал ногу, когда неясыть сбила его
с дерева (Mikkola, 1981, с. 103).
Вокализация неясыти разнообразна: как взрослые, так и птенцы с
18-20-дневного возраста щелкают клювами; взрослые, кроме того,
ухают, ’’рычат”, а иногда вскрикивают высоким голосом, подобно
ястребиной сове. Бородатая неясыть активна преимущественно в сумер-
ки, но летом, когда солнце не опускается под горизонт, отлично летает и
охотится при дневном свете.
Летом эта сова питается почти исключительно мелкими млекопита-
ющими. Для изучения ее питания под гнездом 1964 г. собрано 40 погадок
птенцов и 3 остатка добычи, еще 3 погадки найдены в других местах; в
1983 г. у гнезда найдены 23 погадки и 3 желудка птенцов, еще один взят от
жертвы филина. Погадка птенца содержит обычно 3-4 полевки; в одной
погадке взрослой птицы (масса тела 15,3 г.) оказались остатки 6 зверьков.
Сухая масса погадки в среднем 4,8 г (табл. 12).
Врагами неясыти, кроме медведя и филина, можно считать всех
крупных хищных птиц и сов: скорее всего, погибшая неясыть в 1980 г.
была жертвой птицы, а лисица доедала брошенный костяк.
Бородатую неясыть легко отличить от длиннохвостой только спереди,
по концентрическим кольцам на лицевом диске и черной ’’эспаньолке”.
Длиннохвостая неясыть - Strix uralensis Pall.
Местное название: саамы не различают виды неясытей и называ-
ют их одинаково ’’хиппик”.
По материалам Плеске (1887, с. 292), в прошлом веке эта неясыть
встречалась в Лапландии гораздо реже бородатой, а на Кольском полу-
острове ее не находили ни разу. В Лапландском заповеднике до 1941 г.
мы наблюдали только длиннохвостую неясыть, потом соотношение видов
изменилось, и после 1958 г. она встречается гораздо реже бородатой.
Длиннохвостые неясыти были добыты: 6. VI 1934 г. в ельнике, в долине
р. Курка; 2. V 1935 г. - на Охтовараке; 26. III 1939 г. - на берегу Воче-лам-
бины и в ту же зиму близ Мончегорска.
Гнезд этой совы не находили. Взрослых птиц встречали 14. VI 1932 г. в
долине Курки, 24. VI 1939 г. на берегу Чунозера, 19. IV 1940 г. (место не
уточнено), 7. VI 1950 г. в ельнике на берегу озера Ельявр (пара), 9. VI
1940 г. в бору при устье Чуны, 20. VII 1969 г. в ельнике у Нявка-озера,
27. V 1977 г. в бору под Чуна-тундрой, 11. VII 1979 г. и 15. VIII 1982 гг. в
долине Чуны.
Встреча пары в брачное время в 1950 г. - косвенное доказательство
гнездования этого вида. В отличие от бородатой неясыти длиннохвостая,
видимо, предпочитает жить в ельниках.
15. VIII 1982 г. наблюдали попытку нападения длиннохвостой неясыти
на глухаренка, а потом на глухарку. 27. V 1977 г. длиннохвостую неясыть
атаковал дербник, all. VIII 1979 г. за слетевшей совой гнались 12 весни-
чек и 5 гаичек.
Из четырех исследованных желудков один был пуст, в остальных трех
обнаружены одна рыжая полевка, четыре красно-серые и остатки лем-
мингов.
Отличительные признаки этой неясыти - светлая окраска оперения
(особенно лицевого диска) и сравнительно длинный хвост. Размеры двух
182
самок: длина 590 и 590, крыло 365 и 380, хвост 285 и 290 мм, масса тела 800
и 1145 г; масса тела третьей (самец?) 683 г.
Оба вида неясытей - типичные миофаги. Суммируя наблюдения по
обоим видам, находим, что в нашем регионе они встречались на протяже-
нии 31 года из 46 лет наблюдений; из этих лет 10 были годами пика
мелких грызунов, 10 - сравнительно богаты ими, 5 - бедны и 6 - годы
минимума. Прямого соответствия между численностью неясытей и
полевок здесь не видно, но гнезда находили в год, следующий за пиком
полевок (после пиков 1963 и 1982 гг.). Одна неясыть погибла в год мини-
мума полевок (1979/80).
Болотная сова - Asio flammeus Pontopp.
Местное название: саамское - ылшиш.
Ареал болотной совы охватывает почти всю территорию нашей страны,
кроме Крайнего Севера. На Кольском полуострове она встречается
всюду - как в лесной зоне, так и в тундровой, даже на Айновы острова
залетает раз в три года, весной (Татаринкова, 1985, с. 163). В отличие от
остальных наших сов болотная совершает регулярные перелеты, зимуя
вокруг Средиземного моря и в тропической Африке.
В Лапландском заповеднике она появляется около 25 мая (11. V
1978 г. - 6. VI 1969 г. за 10 лет; о ± 7,3 дня, температура + 5,8°), средняя
дата последней встречи 22 сентября (1. IX 1977 г. - 27. X 1938 г. за 9 лет;
о ± 16,6 дней, температура + 5, Г); живет у нас 120 дней.
Болотная сова предпочитает открытые биотипы - болота, луговины и
осоковые заросли по берегам озер и речных проток, лесные гари с редким
древостоем, встречается и на горных тундрах. За 46 лет ее видели на
протяжении 19 сезонов, всего 114 раз; найдено одно гнездо и несколько
выводков, добыты и исследованы 3 экз.
18. VIII 1937 г. в сумерках у саамской избы на берегу Нявка-озера О. Семенов-
Тян-Шанский наблюдал выводок болотных сов и одну добыл. Одиночных видели
также в августе-сентябре на вараке Ельявруайвенч и в устье р. Чуна.
23-25. V 1938 г. в среднем течении Чуны мы наблюдали брачную игру пары сов.
Самец летал с криком ”ту-ту-ту-ту”, похожим на крик мохноногого сыча, но тише,
реже и тоном ниже; иногда на мгновение складывал крылья, падал, а потом продол-
жал горизонтальный полет. Одиночных сов видели также на Нявка-тундре, Купись-
озере, в долинах Ташким-йок и Сылпуая (всего 11 птиц).
В 1978 г. с 11. V по 29. IX зарегистрированы 52 болотные совы, преимущественно в
долинах рек Купись и Вайкись. Пара у оз. Купись, вероятно, гнездилась (совы
налетали на человека); 1—3. VII у Матрениных ламбин наблюдали выводок (6—7 сов);
29. VII на гари под юго-западным склоном Волчьей тундры болотная сова ’’отводила”
от птенцов. Одиночных встречали на Кензисъявреи в долине Чуны, а близ ее устья
нашли остатки болотной совы.
30. VI 1982 г. на болоте при впадении Сылпуая в р. Чуна А. Гилязов спугнул сову с
гнезда, устроенного на кочке, между кустами можжевельника и карликовой
березки; в гнезде было яйцо, сова к нему не вернулась. За сезон отмечены еще
четыре болотные совы, из них одна подобрана на Ленинградской дороге со сломан-
ным крылом в августе.
27. VII 1983 г. в долине Чуны найдены остатки слетка, пойманного лисой или
куницей; за лето 4 встречи в долине р. Купись и на Нявка-озере.
Все случаи гнездования болотной совы и большинство встреч прихо-
дятся на ’’мышиные” годы, однако одиночные особи встречались и в годы
минимума грызунов.
Болотные совы довольно агрессивны: в августе 1937 г. они, как будто
играя, в упор налетали на саамских лаек; в июле 1978 г. одна сова атако-
183
вала двух зимняков, в августе другая гонялась за сизыми чайками. Эти
совы активны как в сумерках, так и днем.
Размеры 3 особей: крыло 315-325, хвост 150, длина одной взрослой
самки 385 мм, масса тела этой самки 314 г, двух истощенных особей 225 и
205 г. Размер яйца 42 х 33 мм.
Мохноногий сыч - Aegolius funereus (L.)
Местное название: саамское - утс-лыеблыкь (=маленький фи-
лин).
Мохноногий сыч - такой же характерный элемент фауны тайги, как
бородатая и длиннохвостая неясыти; сходны и их обширные ареалы.
В лесной части Кольского полуострова сыч обычен, хотя редко попа-
дается на глаза из-за скрытного образа жизни. Плеске упоминает о наход-
ках сыча близ Кандалакши, в истоках р. Нота и на Нотозере; мы добыли
одного в октябре 1930 г. между Колой и Мурмашами, другой был добыт
тогда же на западном склоне Хибин. По-видимому, живет оседло, по
крайней мере попадался нам во все месяцы года - всего 38 визуальных
встреч за 46 лет наблюдений. Услышать брачный крик сыча, напомина-
ющий крик удода, много легче, чем увидеть самую птицу, поэтому люди,
никогда не видавшие сыча, принимают его голос за брачный крик зайца.
Мы слышали его в сумерках или ночью с 20 февраля по 28 мая, реже
осенью, между 26. VIII - 25. IX; один раз сыч кричал около своего гнезда
в июле.
Визуально или на слух сычей наблюдали на протяжении 21 года.
Большинство этих лет были ’’мышиными”, с индексом учета полевок
17-97 на 100 ловушко-суток, только в 6 сезонах снижаясь до 10. Корреля-
ция между индексом учета полевок осенью и числом встреченных за год
сычей не очень велика: г = + 0,46 ± 0,13 (за 35 лет).
Представляют интерес сведения о размножении мохноногого сыча.
2. VIII 1949 г. в бору между реками Чуна и Нявка встречен выводок — три сыча в
одном месте.
5. VII 1974 г. на болоте в том же районе студент Н. Караваев видел самку с
2 слетками.
25. VI 1975 г. в бору на берегу Чунозера в гнездовом ящике для уток (по саам-
ски — ”вуувт”) А. Брагин нашел гнездо сыча с 3 начавшими оперяться птенцами; к
16. VII все птенцы вылетели.
26. V 1982 г. в таком же ”вуувте” на осоковом озерке в бору у берега Чуны
А. Гилязов нашел кладку в 4 яйца; самка насиживала (рис. 46); 2. VI самка из летка
не вылетела, под ней пищали птенцы; 10. VI в гнезде 4 птенца и мертвое яйцо;
старшие птенцы в светлом пуху, четвертый еще голый; 24. VI три птенца в черном
оперении, четвертый в белом пуховом наряде; 27. VI самка в гнезде не сидит; в ночь
на 4. VII последние птенцы оставили гнездо.
В 1983 г. в том же районе А. Гилязов нашел три гнезда сычей: первое 18. V, в
естественном дупле на берегу Чунозера (раньше в нем гнездился гоголь); 9. VI в
нем 5 начинающих оперяться птенцов; 22. VI их масса тела 124, 121, 112, 110, 104 г.;
3. VII все уже вылетели.
Второе гнездо найдено в тот же день 18. V в полукилометре от первого, в ”вуувте”
на краю лесного озерка, в нем 5 яиц; 8. VI в нем 3 почти голых птенца и 2 яйца;
13. VI — 5 совят, все еще слепы; 22. VI их масса 125, 125, 114, 106 и 98 г; 6. VII два
птенца уже вылетели, а 10. VII к ночи вылетел и последний.
Третье гнездо, также в ”вуувте”, на берегу другого озерка в 1,8 км от первого,
обнаружено 19. V, в нем 5 яиц, самка насиживает; 9. VI в нем 5 почти полностью
оперенных птенцов; 22. VI первый уже вылетел, масса тела остальных 165, 157, 157 и
146 г; 29. VI гнездо пусто.
Наблюдения показали, что сычи живут как в сосновых, так и в еловых
лесах, поднимаясь по склонам гор до верхней границы леса. Охотно
184
Рис. 46. Мохноногий сыч, самка, 26. VI 1982 г. Фото О.И. Семенова-Тян-Шанского
занимают искусственные гнездовья для уток (с широким летком); из
шести найденных гнезд только одно находилось в естественном дупле.
Все занятые сычами ’’вуувты” располагались одинаково, на берегу ма-
ленького, до 0,5 га, лесного озерка, заросшего с берегов осокой. Информа-
цию о поведении сычей и хронологии размножения мы получили с трех
гнезд, оборудованных актографами (1975, 1982 гг. и второе гнездо
1983 г.). Эмбрион развивается в яйце не менее 23 сут, а так как инкуба-
ция начинается с откладки первого яйца - продолжительность насижива-
ния (при кладке 5 яиц) составляет не менее 28 сут. Птенцы появляются на
свет почти голыми, прозревают на 13-й день и оставляют гнездо в возрас-
185
Рис. 47. Птенец мохноногого сыча, 6. VII 1983 г. Фото О.И. Семенова-Тян-Шанского
те 29-30 сут. Вылетают совята по старшинству на протяжении 4-7 дней.
Самый ранний вылет наблюдался 22. VI 1983 г., самый поздний - 16. VII
1975 г., средняя дата окончания вылета по 5 гнездам 6 июля, из чего
можно заключить, что кладка начинается около 5-10 мая. Самка прекра-
тила обогревать птенцов в 1975 г. за 15 дней до вылета последнего, в
1982 г. - за 9 дней, в 1983 г. - за 12 дней. Днем мы ни разу не заставали у
гнезда обоих сычей, а ночью однажды слышали поблизости крик самца.
Мохноногий сыч не агрессивен, при осмотре гнезда самка не пыталась
нападать на человека и только беспокойно, молча, перелетала с дерева
на дерево.
186
Таблица 13. Питание мохноногого сыча летом
Вид добычи 1981 г. (экз.) 1982 г. (экз.) 1983 г. (экз.) Случайн. экз. Весь мате- риал, %
Млекопитающие Насекомоядные (зем- леройки ’ — — 2 — 1
Грызуны* (не опреде- ленные) — — — 6 3
Полевка красно- — 17 104 2 57
серая Полевка рыжая — 1 — — 0,5
Полевки лесные* — 3 2 — 2
Полевка пашенная — 5 2 — 3
Полевка экономка — 6 — — 3
Полевки серые* — 1 6 — 3
Лемминг норвеж- ский — 18 8 1 12,5
Птицы (певчие, дятел) 3 — — 3 3
Земноводные (травя- ная лягушка) 21 — — — 10
Насекомые 3 — 2 — 2
Всего объектов: 27 51 126 12 100%
*Ближе не определены.
Мохноногий сыч ведет более ночной образ жизни, чем другие совы.
Его активность в период насиживания яиц и выкармливания птенцов
распределяется по часам суток следующим образом:
Часы суток 09 13 17 21 01 05 09 за 24 ч.
При яйцах 10 2 8 63 12 5 100%
При птенцах 7 6 10 46 21 10 100%
При птенцах взрослые сычи посещали гнездо в среднем в 1975 г. 7,7 раз
в сутки, в 1982 г. - 10,4 раза, в 1983 г. - 5,7 раза. Полагая, что птица
каждый раз приносила корм, находим, что в 1975 и 1982 гг. каждый
птенец получал по 2-3 передачи в сутки, в 1983 г. только по 1-2. Средний
привес птенцов в 1983 г. составлял около 11% в сутки.
При птенцах подстилка гнезда состоит из растоптанных погадок,
смешанных с экскрементами птенцов и гниющими остатками недоеден-
ной добычи, образующих слой до 10 см. Дупло издает резкий запах
аммиака. Возможно, он отпугивает кровососущих насекомых, от которых
птенцы обычно страдают, но непонятно, как они сами дышат? Когда
сычата оперены, один из них обыкновенно сидит в летке (рис. 47).
Пуховые птенцы издают звук, напоминающий чириканье, оперенные,
кроме того, щелкают клювом. Оперение слетков очень темное, почти
черное, глаза ярко-желтые.
Исследованный нами материал по питанию сыча включает подстилку
трех гнезд, недоеденные остатки из разных гнезд, два желудка сычей,
добытых в августе-сентябре 1949 г., несколько погадок и остатки добычи,
в том числе 13 погадок, найденных летом 1981 г. на чердаке лесной
избушки.
187
Мохноногий сыч - типичный миофаг, и только в 1981 и 1985 гг., при
недостатке мелких грызунов, он кормился лягушками, мелкими птичка-
ми и насекомыми. Остатки погибших сычей находили дважды, в 1972 г.
под Нявка-тундрой и в 1985 г. на берегу Чуны.
Размеры двух сычей, добытых осенью 1949 г.: длина самца 253, самки
255 мм, крыло 169 и 187, хвост 104 и 112 мм, масса тела 121 к 147 г. Масса
одного яйца (с погибшим эмбрионом) 10,3 г.
ОТРЯД СТРИЖЕОБРАЗНЫЕ - APODIFORMES
Черный стриж - Apus apus (L.)
В Русской Лапландии, по крайней мере в западной ее части, стриж
встречается регулярно и нередко гнездится. В прошлом веке он доходил
у нас к северу до Нотозера, в Финляндии до оз. Инари, в Норвегии до
Киркенеса (Плеске, 1887, с. 278). В XX в. зарегистрированы залеты стрижа
до о-ва Харлова (июнь, 1935 г.), Святого носа (август, 1922 г.) и даже на
Новую Землю (Птушенко, 1951, с. 624).
В Лапландском заповеднике отмечают в среднем около 50 стрижей в
сезон, или по 11 на одного наблюдателя, с кобеланиями от 2 до 39 и
периодом около 4 лет; при этом в последние 20 лет максимумы приходи-
лись на високосные годы.
Стриж значительную часть дня проводит в воздухе, а потому легче
попадается на глаза, чем лесные птицы; благодаря громкому крику,
напоминающему визг, его легко заметить и на слух. Прилетает стриж
поздно, в Лапландском заповеднике, в среднем за 27 лет, 5 июня (27. V
1968, 1972 гг. - 16. VI 1976 г., о ± 5,8 дней, температура + 8,2°). Нормально
последних стрижей отмечают около 23 августа (8. VIII 1961 г. - 7. IX
1962 г. за 27 лет; о ±7,5, температура + 10,3°), но в отдельные годы их
вполне достоверно наблюдали на месяц позднее, между 19-27. IX (1966,
1969, 1978 и 1985 гг.). Интересно, что так поздно встречали не одиночную,
отсталую или залетную, птицу, а стайку (27. IX 1978 г. над оз. Туарехт-лум-
бол летали 3 стрижа), иногда на протяжении нескольких дней (в 1969 г.
над Чунозером 31. VIII, 6, 8, 14. IX летала стайка, а 21 и 22. IX - одиночный
стриж). Задержку стрижей нельзя объяснить аномально теплой погодой;
напротив, в годы позднего отлета сентябрь был немного холоднее обыч-
ного; в 1966 г. одиночного стрижа видели 19. IX, на следующий день после
образования сплошного снежного покрова.
Подобные случаи задержки отлета стрижей до конца сентября наблю-
дались и в других районах Севера, в частности в долине Печоры (Птушен-
ко, 1951, с. 625); предполагаемая причина этого явления - поздний вывод
птенцов.
Стриж проводит в Лапландском заповеднике в среднем 79 дней,
иногда только 55-70 (1961, 1970, 1975, 1982 гг.), а в годы задержки отлета
больше - 113-116 дней.
Стрижи питаются исключительно насекомыми, пойманными ими в
воздухе. По нашим наблюдениям, они ловят добычу преимущественно
над водой (64% всех встреч), обычно в устье реки или зашейке (место, где
из озера вытекает река), реже просто над озером или рекой. Стрижей
часто наблюдают также в сосновых лесах (24%), гораздо реже над еловым
188
или смешанным лесом (3%), болотом (1,5%) или горной тундрой (0,5%);
довольно часто они летают в городе или возле построек (7% случаев).
Очевидно, они предпочитают насекомых, личинки которых развиваются
в воде (поденки, ручейники, кровососущие двукрылые, возможно и
стрекозы), а в населенных пунктах ловят, вероятно, мух. Стрижи доволь-
но общительны и кормятся иногда стаями до десятка, а иногда и в нес-
колько десятков особей. Распределение стрижей по группам разных размеров
следующее (всего учтено 970 птиц):
Число особей
Вероятность встреч
Распределение особей
1 2 3 4 .5-10' 11-50 51-100
30 25 12 8 14 10 1
6 И 8 7 19 34 15
Большие стаи наблюдаются только
сразу после прилета или перед
отлетом, в августе.
Сведения о размножении скудны. 17. VII 1969 г. в старом бору в долине
р. Чуна. Домнинский и Камалов обнаружили гнездо и видели, как стриж
залетал в дупло на сухой сосне, но сами не смогли до него добраться. В
том же районе 11. VII1970 г. и 5.VIII 1975 г, А. Брагин и А. Гилязов наб-
людали летные выводки стрижей. В начале июня 1977 г. О. и Ю. Пет-
ровские на протяжении недели неоднократно наблюдали стрижа,
залетавшего на чердак кордона на Купись-озере; 15. VI туда
залетала уже пара стрижей, но гнездиться они все же не стали.
В ненастье, особенно при ’’морянке”, сильном северном ветре с резким
похолоданием, дождем или снегопадом, насекомые не летают и стрижам
угрожает голод. По литературным сведениям, птенцы в таких случаях
впадают в анабиоз, а взрослые отлетают на сотни километров, в поисках
мест с более благоприятной погодой, где можно добыть корм; впоследст-
вии они возвращаются к гнездам и продолжают выкармливать птенцов.
Собственных наблюдений по этому вопросу у нас нет. Зимуют наши
стрижи в южной половине Африки.
Изучая питание хищных птиц, единственный раз мы нашли остатки
стрижа (лапку) в погадке ястреба-тетеревятника.
ОТРЯД РАКШЕОБРАЗНЫЕ - CORACIIFORMES
Сизоворонка - Coracias garrulus L.
Эта южная птица, гнездовый ареал которой доходит до южного побе-
режья Балтийского моря, иногда залетает далеко на север, не только в
Лапландию, но даже на Новую Землю (Судиловская, 1951, с. 493). В
октябре 1868 г. одна сизоворонка из пары была добыта у Варангер-фиор-
да, в Финмарке (Плеске, 1887, с. 285). В августе 1950 г. в Лапландском
заповеднике, в долине Суэньлаг-уая, под западным склоном Чуна-тун-
дры студент Ю. Гельцер нашел крыло сизоворонки, ставшей жертвой
хищной птицы.
Удод - Upupa epops L.
Удод гнездится только в южной половине нашей страны, доходя до
Балтийского моря, и в Лапландию только залетает. В середине прошлого
века удоды несколько раз появлялись в сентябре в долине Тана, в
189
Финмарке (Плеске, 1887, с. 286). 8-9 октября 1962 г. залетного удода
наблюдали на Чунозерской усадьбе Лапландского заповедника М. Влади-
мирская и Ю. Гаев. 8-10 сентября 1967 г. одного видели на Большом
Айоновом острове, а 4 октября 1971 г. в г. Кандалакше (Коханов, 1987,
с. 23). Интересно, что все залеты датируются сентябрем - началом октяб-
ря - это то время, когда удоды отлетают на свои зимовки в экваториаль-
ную Африку.
ОТРЯД ДЯТЛООБРАЗНЫЕ - PICIFORMES
Вертишейка - Jynx torquilla L.
Вертишейка широко распространена в лесной полосе Европы и Сибири,
к северу до Белого моря. На Кольском полуострове впервые наблюдалась
3. VI 1940 г. в Лапландском заповеднике: одиночная птица с криком
перелетала по вершинам сосен на Чунозерской усадьбе.
23. VI 1948 г. в центре заповедника, в бору в долине Сылпуая,
Т.В. Кошкина наблюдала вертишейку у ее гнезда в дупле, из которого
слышался писк птенцов.
В 1975 г. вертишейки вторично гнездились в заповеднике. Начиная с
11. V их несколько раз слышали и видели на Чунозерской усадьбе, они
заняли один из скворечников и в конце июля уже носили корм птенцам.
1. VII А.Б. Брагин окольцевал одну из взрослых птиц и 8 птенцов; они
были почти голые, но маховые и рулевые подрастали. Корм приносили и
самец и самка 5 раз в 1,5-2 ч. Птенцы при этом поднимали крик, напоми-
навший стрекотанье кузнечика, взрослая птица для передачи корма
влезала в гнездо, а оттуда выносила очередную капсулу помета. В гнезде
грязно, в подстилке скорлупа яиц, кусочки станиоля и др. Птенцы
вылетели 11 июля, держались поблизости до 13-14. VIII, после чего
пропали из виду. При осмотре пустого гнезда в нем оказались мумии 3
птенцов, из которых один был с кольцом, а два неокольцованные - сле-
довательно, всего было 10 птенцов.
Весной 1981 г. вертишейку неоднократно слышали и видели при устье
р. Верхняя Чуна с 23. V по 1. VI. Крик ее слышали также в 1961 г. у Екост-
ровского пролива на Имандре, в 1976 г. - при устье Верхней Чуны, в
1977 г. - на Чунъявр-вараке, в 1981 г. - в среднем течении реки Нявка и
Чуна, а также на берегу Чунозера, в 1982 г. - в долине Чуны, в 1983 г. - в
лесу под юго-восточным отрогом Чуна-тундры, каждый раз в прилетное
время, между 27. V и 27. VI.
Прилетает вертишейка одновременно с кукушкой, в среднем за 6 лет
29. V (11. V 1975 г. - 13. VI 1977 г.). Время отлета неизвестно.
Черный дятел (желна) - Dryocopus martins (L.)
Местные названия: саамское - кйрыч или чиерычь.
Оседлая птица лесной зоны Европы и Сибири, включая всю лесную
часть Кольского полуострова. Благодаря звонкому, далеко слышному
крику черный дятел самый заметный из всех дятлов, хотя в действитель-
ности он совсем не многочислен, а временами и редок, например в 1977 г.
в Лапландском заповеднике его не встречали ни разу. Численность
колеблется с периодом 5-7 лет; за последние 30 лет минимумы были в
190
Рис. 48. Жировочные дупла желны, 27. VI 1981 г. Фото О.И. Семенова-Тян-Шанского
1958, 1963, 1970, 1975, 1977 и 1984-1987 гг., а максимумы - в 1960-1961,
1966, 1972-1973 и 1978 гг. За время наших наблюдений численность его
сократилась в три раза: в 30-х годах на 1 наблюдателя в год приходилось
4,2 встречи, в 40-х и 50-х - по 2,5, в 60-х - 3,1, в 70-х - 2,0, в 80-х - 1,3.
Черный дятел предпочитает сосновые или смешанные сосново-елово-
березовые леса и гари, вплоть до самых свежих, реже попадается в
еловых и елово-березовых. Встречается на протяжении всего года,
однако в августе-октябре отмечали по 1,0-1,5 птицы в месяц, а в декаб-
ре-феврале только по 0,1-0,2; вероятно, часть особей на зиму откочевы-
191
вает. С марта по июнь встречается чаще, от 0,3 до 0,5 птицы, в июле, во
время выкармливания птенцов, 0,3, а после вылета молодняка встреча-
емость увеличивается втрое, до 1,5 особей в месяц.
Сведения о гнездовании фрагментарны. 18. VII 1941 г. на берегу Чуно-
зера найдено гнездо с птенцами, 30. VII оно было уже пусто. Слетков
наблюдали 22. VI 1940 г. и 29. VI 1941 г. Дупло черного дятла отличается
большим, почти прямоугольным летком, диаметр которого в высоту
больше, чем в ширину (рис. 48).
О питании черного дятла дают представление желудки 6 птиц, добы-
тые в 1933-1941 гг., исследованные Г.А. Новиковым и В.Ю. Фридолиным:
в августе найдены муравьи, личинки, куколки и имаго (жуки, личинки,
иногда имаго); в сентябре и ноябре - муравьи имаго и личинки жуков; в
декабре - только личинки жуков. Черный дятел особенно охотится за
крупными древесными муравьями Camponotus, строящими свои гнезда в
стволах деревьев, пораженных грибами, обычно в прикорневой части
сосен и елей. Добывая их, черный дятел пробивает глубокие дупла в
нижней части ствола, а иногда и в корнях. В мае и июне мы неоднократно
находили свежие ’’жировочные” дупла такого рода. Встречаются и сухие
сосны с десятком отверстий, пробитых черным дятлом: в таких дуплах
нередко гнездятся гоголи, ястребиные совы и стрижи.
Размеры трех черных дятлов: длина 455-480, крыло 242-250, хвост
170-175 мм; средняя масса тела 7 особей 330 г. (самцы 318-359, самки
308-315 г).
Большой пестрый дятел - Dendrocopus major (L.)
Ареал большого пестрого дятла занимает большую часть территории
СССР, кроме Крайнего Севера и безлесных пространств на юге страны. Он
несомненно гнездится и зимует во всей лесной части Кольского полуост-
рова, но его нельзя считать оседлой птицей уже потому, что от времени
до времени вся его популяция полностью исчезает на неопределенное
время, иногда на несколько лет., а потом восстанавливается в результате
перекочевок. Его передвижения отличаются от перелетов и кочевок
других птиц крайней нерегулярностью. За 47 лет наблюдений на протяже-
нии 16 календарных лет мы не зарегистрировали ни одной визуальной
встречи этого дятла (большого пестрого дятла по голосу не всегда можно
отличить от трехпалого, поэтому мы регистрировали его только по
визуальным встречам). Иногда он полностью отсутствовал в течение 2-3
лет (с декабря 1935 г. по май 1938 г.) или даже 5 лет (с января 1963 г. по
июль 1968 г.)7. Эмиграция и иммиграция большого пестрого дятла не
приурочены даже к определенному сезону. Иногда дятлы исчезали, как
только молодняк поднимается на крыло, например в 1981 г. - в июне,
иногда осенью или даже зимой: в 1962 г. в сентябре, в 1972 и 1975 гг. в
октябре, в 1935 и 1959 гг. в ноябре, в 1977 г. в декабре, в 1980 г. в февра-
ле, в 1941 г. в марте, в 1969 г. в апреле. Таким же образом и появлялись
они после длительного отсутствия: в 1938 и 1962 гг. в июне, в 1968 и
1976 гг. в августе, в 1935, 1939 и 1979 гг. в сентябре, в 1969 г. в январе, в
1981 г. в марте, в 1959 г. в апреле, в 1982 г. в мае.
7В некоторые из этих лет попадались следы деятельности дятла (разбитые им
шишки), так что полностью он не исчезал, но был редок.
192
Создается впечатление, что популяция большого пестрого дятла почти
постоянно кочует по лесам, задерживаясь только для гнездования или
там, где достаточно корма для зимовки.
Явной тенденции к сокращению популяции или увеличению числен-
ности этого дятла за последние полвека мы не обнаружили. Число особей,
встреченных одним наблюдателем за год (индекс большого пестрого
дятла) оказалось:
Годы Лет наблю- дений 30-е 10 40-е 5 50-е 4 60-е 10 70-е 10 80-е 8 Всего 47 лет
Индекс дят- 2,4 2,4 1,3 2,4 2,6 3,0 Среднее
ла 2,4
Заметно отклоняются от средней только индексы за 50-е годы, когда
наблюдениями охвачены неполные 4 года; высокий показатель за 80-е
годы объясняется находкой 6 гнезд за 4 года (одно гнездо мы приравни-
вали к 5 птицам - пара взрослых и 3 птенца).
Впервые гнездо большого пестрого дятла найдено в заповеднике 8
июня 1975 г. на 17-летней сосновой гари. Дупло выдолблено в недавно
засохшей сосне, леток на высоте 3,5, обращен на запад; из него слышен
крик птенцов. Молодые вылетели после 14. VI.
В 1981 г. А.С. Гилязов нашел 4 гнезда большого пестрого дятла.
28. VI 1982 г. - в бору на юго-западном берегу Чунозера дупло в
стволе сухой сосны, на высоте 3 м, птенцы громко верещат, взрослые
носят корм (в 4 последних гнездах леток обращен к югу).
30. VI 1982 г. - в бору в долине Чуны, свежее дупло в сухой сосне на
высоте 2,7 м, отверстием на ЮВ; птенцы верещат, под дуйлом на земле
скорлупа 1 яйца.
Приняв во внимание, что большой пестрый дятел насиживает 16 сут, а
птенцы вылетают в возрасте 18-21 день (Harrison, 1975, с. 214), можно
считать, что самка садится на яйца в последней декаде мая, птенцы
выводятся около 10. VI и оставляют гнездо около 30. VI.
Летом большой пестрый дятел кормит птенцов и сам питается насеко-
мыми (муравьями и их коконами, жуками, их личинками, гусеницами
бабочек, лжегусеницами пилильщиков, комарами-долгоножками, наезд-
никами и тлями; Мальчевский, Пукинский, 1983. 1, с. 467), зимой же в
отличие от прочих дятлов питается еловыми и сосновыми семенами.
Добывать их он начинает рано, первые разбитые шишки, иногда еще
зеленые, мы находили обычно между 27. VII и 23. VIII, а в среднем за 6 лет
10 августа. Чтобы достать из шишки семена, дятел вставляет ее основа-
нием в узкую щель, специально для этого выдолбленную в стволе сухой
сосны или ели (если не найдется готовой), ударами клюва отгибает чешуи
и достает из-под них семечки. Использовав одну шишку, он летит за
следующей, отламывает от черешка и, держа в клюве, летит обратно к
своей ’’кузнице”. Таких ’’кузниц” у каждого дятла по нескольку, и под
каждой из них за зиму скопляется множество (иногда до тысячи) брошен-
ных шишек: в учтенных Г.А. Новиковым в 1938 г. 9 ’’кузницах” в среднем
было по 395 шишек, от 98 до 900 (Новиков, 1956, с. 147). В щели ’’кузни-
цы” всегда остается шишка, это означает, что дятел выкидывает исполь-
зованную не раньше, чем принесет новую. Под тремя низко расположен-
ными ’’кузницами” куча брошенных шишек находилась по правую
сторону от работавшего дятла (рис. 49).
13. Зак. 1023
193
Рис. 49. "Отходы производства” большого пестрого дятла, 19. V 1941 г. Фото О.И. Се-
менова-Тян-Шанского
Несмотря на то что питание семенами ставит большого пестрого дятла
в зависимость от урожая еловых и сосновых семян, мы не нашли замет-
ной корреляции между величиной урожая шишек и численностью этой
птицы. Объясняется это, вероятно, тем, что популяция дятла не исполь-
зует всего урожая шишек и птицы концентрируются только в тех местах,
где корм наиболее доступен; иными словами, количество учтенных птиц
зависит не столько от урожая хвойных пород в данном пункте, сколько
от различий в кормовой конъюнктуре на большом пространстве. Дятел
питается семенами хвойных все время, пока лежит снег: до конца апреля
он еще работает на ’’кузницах”. По-видимому, он предпочитает сосновые
шишки еловым, по крайней мере на протяжении 15 зим дятлы кормились
сосновыми семенами, 4 зимы еловыми и 1 зиму как теми, так и другими.
Остатки дятлов, в том числе и большого пестрого, не раз попадались
нам в материалах по питанию ястреба-тетеревятника, кречета и куницы:
последняя, вероятно, ловит их, когда они ночуют в дуплах. З.Х. Аблаева
однажды заметила, что дятел ночевал в ’’вуувте” (искусственной дуп-
лянке для гоголя). Несколько раз мы находили трупы или остатки
больших пестрых дятлов на снегу или на земле: 27.III 1941 г. - труп
самца, слегка поклеванный кукшей или другой птицей, с пустым желуд-
ком; 22. VIII 1949 г. почти свежий труп; 29. V 1969 г. - старый труп самца
на снегу, под ’’кузницей”; 27. V 1980 - перья и подклювье дятла, пойман-
ного хищной птицей. Остатки всех четырех найдены после резкого
сокращения численности дятлов, очевидно в связи с наступившей бес-
кормицей.
Размеры 4 добытых нами самцов: длина 235-266, крыло 136-143, хвост
90-95 мм; масса тела 92,3, 96,4, 97,4 г.
194
Малый пестрый дятел - Dendrocopus minor (L.)
Малый пестрый дятел широко распространен по всей лесной зоне
Палеарктики. В Лапландском заповеднике малый дятел попадается на
протяжении всего года, но раз в 5 реже, чем большой пестрый. На протя-
жении 21 года из 47 лет наблюдений зарегистрировано всего 59 особей,
преимущественно осенью, в сентябре и октябре. Малый пестрый дятел
ведет такой же бродячий образ жизни, как и большой: на протяжении 5,5
лет, с 1959 по 1964 г., его не видели ни разу, зато в 1972-1974 гг. встреча-
ли по 6-7 птиц в год. Гнездование его доказано: 27. VI 1974 г. А.Б. Брагин
нашел гнездо с птенцами в берязняке по левому берегу р. Купись, при
впадении в нее р. Урда. Оно находилось в дупле гнилой березы, в 4 м от
земли. Птенцы уже большие, высовывали головы из летка; их кормили
самка и самец.
Малого пестрого дятла можно узнать уже по голосу, тихому ”пи-пи-
пи-пи”, напоминающему голос вертишейки. Размером он с воробья.
Размеры 2 самцов и 2 самок малых дятлов, добытых в 1938 г.: длина
138-155, крыло 90-98, хвост 58-67 мм, вес 22,0—24,4 г, в среднем 23,0. В
их желудках найдены личинки жуков и двукрылых.
Малый дятел выкармливает птенцов, а летом и сам питается, преиму-
щественно тлями (Мальчевский, Пукинский, 1983, т. 1, с. 474).
Трехпалый дятел - Picoides tridactylus (L.)
Местное название: саамское ’’чайшень” (вероятно, относится и к
двум другим пестрым дятлам).
Трехпалый дятел широко распространен по всей таежной зоне, вклю-
чая и лесную часть Кольского полуострова, где встречается чаще всех
других дятлов: регистрация визуальных встреч дятлов в Лапландском запо-
веднике показала следующее соотношение их численности: трехпалый -
52%, большой пестрый - 29%, малый - 6%, черный - 13%. Трехпалый
дятел наиболее оседлый из всех наших дятлов, встречается постоянно, из
года в год, но зимой несколько реже, чем летом (возможно, по причине
более скрытного образа жизни в снежное время).
Подсчет встреч трехпалых дятлов на одного наблюдателя в год за
полвека обнаружил заметные колебания численности с периодом около
30 лет:
Годы Лет наблю- 30-е 9 40-е 3 50-е 3 60-е 10 70-е 10 80-е 8 Всего
дений 43 года
Индекс 7,1 3,0 2,0 5,0 3,8 2,8 Среднее
4,0
Повышение численности трехпалых дятлов в 30-х и 60-х годах, по всей
вероятности, было следствием обширных пожаров в районе Лапландско-
го заповедника в 1927, 1934 и 1936 гг., а потом в 1960 г.: гари - наиболее
благоприятная стация этого дятла, где он находит обильный корм (рис.
50).
Питание трехпалого дятла изучал Г.А. Новиков, собравший 30 желуд-
ков, содержимое которых было определено энтомологом В.Ю. Фридоли-
ным. Трехпалый дятел питается исключительно насекомыми; зимой в
основном личинками короедов (Ipidae), усачей (Cerambycidae), рогохвос-
тов (Siricidae), стеклянниц (Sesiidae) и других стволовых вредителей;
195
Рис. 50. Трехпалый дятел, 3. IV 1941 г. Фото О.И. Семенова-Тян-Шанского
летом, кроме личинок, ловит жуков, пауков и других, открыто живущих
членистоногих. Личинок он добывает, пробивая клювом кору поражен-
ных деревьев, а иногда отрывая ее пластами. Деревья, поврежденные
огнем, быстро заселяются короедами, усачами и другими вредителями,
почему трехпалый дятел начинает кормиться и гнездиться на гарях уже
на следующее лето после пожара.
В районе заповедника нами учтено 20 гнезд трехпалого дятла, из них
одно, найденное 29. V 1941 г., по-видимому, было с яйцами, а все осталь-
ные были обнаружены по крикам птенцов. 2 гнезда находились на болотах, 5 -
в ельниках, 5 - в смешанном лесу, 5 - в борах, 1 - на свежей гари. Все в
196
дуплах, выдолбленных в стволах сосен (11 случаев) или елей (6 случаев),
как правило, в деревьях засыхающих или со сломанной вершиной и
гнилой сердцевиной. Дупло, по всей вероятности, дятел долбит сам; в
гнезде 1977 г. он продолбил слой крепкой древесины ели со сломанной
вершиной, сделав леток 45 мм в диаметре, а потом вытаскивал по кусоч-
кам гнилую сердцевину. Высота летка варьирует от 1,2 до 10 м от земли,
составляя в среднем 5 м. Отверстие обычно обращено на юг (7 случаев),
реже на восток (3) или юго-восток (1 случай). Большинство гнезд с птен-
цами найдены в третьей декаде июня (10 гнезд); самое раннее - 3. VI, 3 -
во второй декаде июня и 5 - в первой декаде июля: средняя дата всех
находок гнезд с птенцами 24. VI. Вылет птенцов наблюдался 28. VI и дважды
4. VII; слетки вне гнезда наблюдались между 5. VII в 1938 г. и 30. VII в
1985 г. Число птенцов в дупле обычно оставалось неизвестным, но в
гнезде 18. VI 1978 г. были 2-3 птенца, а 28. VI 1978 г. - не менее 3. Вне
гнезда 23. VII 1969 г. встречен выводок из 3 слетков при обоих взрослых,
7. VIII 1973 г. - три, а 30. VII 1985 г. - два слетка, без взрослых. Пока
птенцы в гнезде, их кормят оба родителя, а после вылета они скоро
научаются добывать корм самостоятельно и уже в июле обходятся без
помощи старших.
Размножение происходит в сжатые сроки, в пределах одного месяца.
Принимая, по литературным сведениям, срок инкубации в 14 дней, а
возраст вылетающих птенцов в 21 день, находим ориентировочно средние
даты фаз размножения: кладка яиц с 25. V, начало насиживания 30. V,
вывод птенцов 13. VI, вылет их из гнезд 4. VII.
Остатков трехпалых дятлов, погибших естественной смертью, мы не
находили; в июне 1985 г. в утиной дуплянке нашли остатки одного,
пойманного сычом. Остатки дятлов в материалах по питанию хищников
не удавалось определить до вида, но среди них были и трехпалые.
Размеры 4 самцов: длина 220-240, крыло 124-131, хвост 78-94 мм;
масса тела 31 трехпалого дятла от 53,3 до 79,5 г, средняя масса самца 69,2
(п = 21), самки 64,5 (п = 7), без заметных колебаний по сезонам.
Брачная песнь трехпалого и других дятлов замечательна тем, что
исполняется без участия голосового аппарата: дятел быстро барабанит
клювом по резонирующей щепке на вершине сухого дерева. ’’Барабанную
дробь” дятлов можно слышать уже в феврале, а чаще всего с марта по
май. Летом она раздается уже редко, но можно слышать ее и в сентябре.
Разные виды отличаются по продолжительности и частоте трели, так что
тренированное ухо может определить на слух вид дятла.
ОТРЯД ВОРОБЬИНЫЕ - PASSERIFORMES
СЕМЕЙСТВО ЖАВОРОНКИ - ALAUDIDAE
Лесной жаворонок (юла) - Lullula arbores (L.)
Северная граница ареала лесного жаворонка в Финляндии и Карелии
проходит по 62° с. ш. На западное побережье Белого моря и острова
Кандалакшского заповедника он залетает только изредка (Благоскло-
нов, 1960). В Лапландском заповеднике дважды наблюдались залетные
197
особи: 12. V 1862 г. на Чунозерской усадьбе научный сотрудник Ю. Гаев
наблюдал двух, без уверенности в определении, так как они не пели и не
были добыты; 21. V 1972 г. в сосновом редколесье в низовьях р. Чуна
О. Семенов-Тян-Шанский видел пролетевшую с песней над лесом юлу.
Полевой жаворонок - Alauda arvensis L.
В Мурманской области полевой жаворонок приурочен к сельскохозяй-
ственным угодьям. На полях Хибинской опытной станции поет и, вероят-
но, гнездится, по крайней мере с 1930 г.; регулярно поет также близ г.
Апатиты (совхоз ’’Индустрия”), Мончегорска, Ёны. В прошлом веке, 22.
VIII 1880 г., Ф.Д. Плеске нашел его около г. Колы. На территории заповед-
ника залетных жаворонков наблюдали на Чунозерской усадьбе, Турух-
танном болоте и у протока Меркенч, всего 10 раз, начиная с 1962 г.,
исключительно в пролетное время, не позднее 5 июня.
Прилет жаворонка легко и точно отмечается по песне. Используя
отрывочные наблюдения сотрудников заповедника во всех перечислен-
ных выше пунктах встреч, получаем среднюю дату прилета 13 мая (25. IV
1980 г.- 1. VI 1969 г.; 15 лет).
Рогатый жаворонок (рюм) - Eremophila alpestris (L.)
Для Кольского полуострова Ф.Д. Плеске (1887), а для Лапландского
заповедника М.И. Владимирская (1948) упоминают о рогатом жаворонке
как обычной птице приморских и горных тундр.
В заповеднике до 1956 г. наблюдался в небольшом количестве почти
ежегодно, вероятно и гнездился, хотя ни гнезд, ни слетков не встречали;
после того зарегистрированы только единичные встречи: 7. VII 1973 г. на
Ельнюне, Чуна-тундра, отмечена одна поющая птица, а 25. VII 1985 г.
слышали перекличку рюмов на Поадз-уайвенч, Сальная тундра. В под-
тверждение резкого сокращения популяции за полвека приводим сред-
нее число встреченных за год рогатых жаворонков, по десятилетиям:
Годы 30-е 40-е 50-е 60-е 70-е 80-е
Число птиц в год 13 10 7 4 1 0,3
Сходное уменьшение численности этого вида константировано и на
равнинной тундре северо-востока Кольского полуострова (Михайлов и
др., 1984).
Средняя дата первой встречи рогатого жаворонка за 9 лет наблюдений
- 13 мая (25. IV 1950 г. - 31. V 1969 г.), последней в сезоне, за 11 лет, - 26
сентября (4. IX 1930 г. - 7. X 1941 г.). Пролетные стайки численностью до
двух десятков неоднократно встречали в тундре, на выдувах, на льду озер и
на болотах. В июне-июле парочки жаворонков, вероятно гнездящихся,
несколько раз наблюдали на Чуна-тундре, Ропа-тундре и Сальной тундре.
Размеры 3 самцов: длина 179-190, крыло ПО, хвост - 72-90 мм, масса
тела 34,6-36,5 г; одной самки соответственно - 180, 101 и 65 мм, масса тела
34,3 г.
198
СЕМЕЙСТВО ЛАСТОЧКОВЫЕ - HIRUNDINIDAE
Береговая ласточка - Riparia riparia (L.)
Местное название: саамское - Пяцк-лоодыш (пяцк - обрыв,
лоодыш - уменьшительное от лоньт - птица).
На Кольском полуострове, по литературным данным, гнездовые
колонии береговых ласточек известны по всем крупным рекам (Плеске,
1887; Бианки и др., 1960; Кищинский, 1960). В Лапландском заповеднике
не гнездится за отсутствием подходящих береговых обрывов, но залет-
ных береговушек наблюдали неоднократно: 1. VI 1932 г. - на Верхней
Чуне; 23. VI 1937 г. - трех на Нижней Чуне; 16. VI 1938 г. - на Чунозерской
усадьбе; 7. VII 1983 г. - там же видели пару; 13. VI 1985 г. - над
Чунозером.
Гнездовая колония береговушек впервые появилась в долине р.
Витте, в охранной зоне заповедника, на песчаном откосе карьера, в 1985
г. Все гнездовые норки, 9 жилых и 7 пустых, находились в верхней части
четырехметрового обрыва, обращенного к северу. Средний диаметр
входа 9x10 см (от 4x13 до 6x14 см), глубина - 68 см (от 46 до 130 см; п =
17). 17 июля вокруг летало около десятка ласточек (четыре вылетели из
норок), а в одном из гнезд обнаружены 5 оперенных птенцов с недораз-
витыми еще маховыми и рулевыми; один из них пролетел метров 100 по
наклонной линии (все окольцованы).
Деревенская ласточка (касатка) - Hirindo rustica L.
Распространена в Евразии от Атлантического океана до Тихого и в
Северной Америке. К Северу доходит в Скандинавии - до 7Г, у нас - до
Мурманского побережья (Плеске, 1887; Михайлов и др., 1984). Летом 1977
г. залетная касатка была добыта даже на северной оконечности Новой
Земли, этот экземпляр доставлен в Мурманск и передан в ПИНРО И.В. Бо-
рхиным.
На Чунозерской усадьбе заповедника пара касаток впервые была
замечена 4. VI 1961 г. Я. Янушсом; после того залетных особей, а иногда
пару, неоднократно наблюдали в 1961-1967, 1971-1975 и в 1986 гг. у
домов на Чунозере, а иногда у избушек в долине р. Чуна. Даты залетов от
7. V 1986 г. по 30. VI 1971 г. в среднем за 12 лет - 4 июня. В 1986 г. одна
касатка появилась на Чунозере необычайно рано, когда в лесу еще лежал
снег, и прожила на усадьбе неделю, хватая мух у помойных ям, но исчез-
ла, как только потеплело. 2.VI 1972 г. в устье р. Чуны самец касатки из
пары пел.
Городская ласточка (воронок) - Delichon urbica (L.)
На Кольском полуострове встречается повсеместно по населенным
пунктам, хотя и в незначительном количестве (Плеске, 1887; Владимир-
ская, 1948), а на территорию заповедника только залетает. Впервые пара
городских ласточек была замечена на Чунозерской усадьбе 14. VI 1950 г.
Е.С. Кошкиной; после того их встречали там же в 1958, 1962, 1966, 1967, 1970,
1971, 1973, 1976 и 1984 гг. Объединяя даты первых встреч воронков в
199
заповеднике и ближайших городах, находим, что в среднем они появ-
ляются 1 июня (5. V 1970 г. - 14. VI 1967 г.; 13 лет); средняя дата послед-
ней встречи - 26 августа (18. VIII 1966 г. - 7. IX 1973 г.; 11 лет).
6. X 1950 г. одиночная городская ласточка близко подлетала к Семено-
ву-Тян-Шанскому, плывшему по р. Чуна, - это исключительно поздний
залет, не связанный с отлетом.
Встречи ласточек в заповеднике приурочены к долинам больших рек и
озерам, где летает больше насекомых, чем в лесу или на открытых
участках; обычно залетает одна-две птицы, иногда стайка, до десятка.
В Финляндии известно три места гнездования городских ласточек на
скалах, одно из них в Финской Лапландии, в заповеднике Малла (Jarvi-
nen et. al., 1982). Вполне возможна подобная находка и в Лапландском
заповеднике, тем более, что гнезда воронков находили поблизости от
него. 18. VI 1968 г. в Мончегорске ласточки свили гнездо под коробом
транспорта на комбинате ’’Североникель”, на высоте около 20 м над
землей (А. Брагин). 29. IV 1979 г. в вахтовом поселке лесорубов на р.
Гирвас, под карнизом единственного бревенчатого дома, на высоте 3 м.
А. Брагин и А. Гилязов нашли старую колонию, в которой было более 20
гнезд, большею частью уже разрушенных. 20. VIII 1965 г. в Кандалак-
ше О. Семенов-Тян-Шанский наблюдал гнездовых птенцов.
СЕМЕЙСТВО ТРЯСОГУЗКОВЫЕ - MOTACILLIDAE
Желтая трясогузка - Motacilla flava L.
Ареал этого полиморфного вида охватывает всю Палеарктику, выходя
и за пределы. Из 16 известных подвидов на Кольском полуострове
гнездится свойственная Европе и Сибири М. f. thunbergi Billberg (Влади-
мирская, 1948, с. 229), распространенная в подзонах северной тайги и
лесотундры. На равнинных тундрах желтая трясогузка не гнездится
(Михайлов и др., 1984). Наши желтые трясогузки на зиму отлетают в
Африку, в связи с чем прилетают значительно позднее белых, в среднем
за 43 г. 22 мая, крайние сроки - 11. V 1964 г. и 4. VI 1933 г.; соответственно
рано и отлетают. Первыми появляются одиночные самцы, примерно через
неделю-другую основная масса птиц. В это время можно встретить
стайки до десятка, а иногда и до полусотни желтых трясогузок (6. VI 1978
г. у Райкоряурынча, наблюдение С. Сорокина). Отлет начинается посте-
пенно, в первой половине августа, и происходит менее заметно, чем
прилет. В разные годы между 7. VIII и 17. VIII А. Брагин и А. Гилязов
наблюдали стаю до 70 особей, а отсталые одиночки, чаще молодые,
встречаются и до конца сентября. Последняя встреча за 30 лет приходится
на 23. VIII, крайние сроки - 7. VIII 1967 г. - 24. IX 1975 г.
В заповеднике желтая трясогузка наряду с фифи и луговым конь-
ком - фоновый вид на сфагновых болотах с угнетенными соснами. В
сравнении с другими видами они особенно бросаются в глаза благодаря
яркой окраске оперения и беспокойному поведению на гнездовой терри-
тории. С 1931 г., судя по наблюдениям, численность желтой трясогузки
200
существенно не менялась. Стабильность популяции подтверждают и
маршрутные учеты, проводимые с 1977 г. В начале июня в долине р. Чуна
мы отмечали по 2-4 гнездовые пары на 1 км2, - в долине Ниявки - до 10
пар, а в водораздельном лесу с отдельными пятнами болот - не более
одной пары. В послегнездовой период (первая половина августа), в
водораздельном лесу - по 1-6 особей/км2, а на болотах в долинах рек по
20-60 особей/км2.
В заповеднике желтая трясогузка заселяет кочкарные болота с ерни-
ком и ивняком, заболоченные редколесья по долинам рек и озер, но
избегает горных тундр, березового криволесья и однородных лесных мас-
сивов, не включающих болот. Их излюбленной стацией являются пере-
ходные участки леса к болоту, где растет карликовая березка и подрост
хвойных пород, а нижний ярус образуют багульник, вереск, андромеда,
голубика, морошка, вороника, пушица, осоки и мхи.
Разбивка на пары начинается в конце мая и сопровождается брачными
играми, с погоней в воздухе за самкой иногда нескольких самцов и
спариванием. У самцов желтой трясогузки нет песни как таковой; при
защите территории ее роль, вероятно, выполняют полеты над гнездовым
участком, когда в режиме ’’зависания” самец перелетает с одного дерев-
ца или выворотня на другой сторожевой пункт, издавая своеобразную
’’песню”. При демонстративных полетах его туловище принимает почти
вертикальное положение.
Гнездо строится на сухом месте болота, обычно на кочке, под защитой
кустов ерника, кустарничков или осоки. Оно сплетается из былинок
осоки и пушицы, к внешнему каркасу иногда добавляются тонкие ветки
кустарников, а лоток выстилается более тонкими травинками, часто с
небольшой примесью шерсти лося, оленя, норвежского лемминга или
полевок. Диаметр лотка в начале гнездования 6-7 см, глубина около 5
см, т.е. лоток довольно глубокий. Самая ранняя находка кладки из 4 яиц
- 8. VI 1978 г., но насиженные кладки встречаются вплоть до начала
июля, когда из ранних гнезд птенцы уже вылетели. Самый ранний вылет
наблюдался 1. VII 1938 г. Таким образом, кладка яиц происходит на
протяжении почти всего июня, насиживание - со второй декады июня до
второй декады июля, вывод птенцов - с середины июня.
Полные кладки содержат 4-6 яиц, в среднем 5,1 ( п = 16 ). У гнезда
желтые трясогузки держатся осторожно, самец, как правило, вылетает
навстречу опасности (человек, собака, хищная птица, чайка или кукуш-
ка), с особым сигналом; потом к нему присоединяются самка и другие
особи. Для желтых трясогузок характерна коллективная защита гнезд,
так что несколько соседних пар образуют подобие гнездовой колонии.
Групповое размещение гнезд отчетливо заметно на больших однородных
болотах. Подобное явление мы наблюдаем и в лесотундре Южного Ямала
в июне-июле 1974-1975 гг. (долина р. Хадыта-яха); описано оно также
Мальчевским и Пукинским (1983, т. 2, с. 44).
Самец желтой трясогузки также принимает участие в насиживании и
обогревании птенцов (наблюдение А. Гилязова на Райкоряурынче, 7. VII
1977 г.). Нераспавшиеся выводки при взрослых птицах встречаются до
201
середины августа (16. VIII 1970 г.). Число слетков в выводке от 1 до 6.
Успех размножения желтых трясогузок в значительной мере зависит от
погоды. После снегопада 20-22, VI 1977 г. Г. Катаев и А. Гилязов нашли
два гнезда с брошенными кладками; возле одного из них лежал труп
взрослого самца; через неделю. 28.VI 1977 г., лесники О. и Ю. Петровские
наблюдали, как трясогузки строили гнездо, вероятно, после потери
первой кладки.
Весной, после прилета, желтые трясогузки ищут на проталиных прос-
нувшихся насекомых, на кромке льда или берега ловят вылезающих из
воды веснянок, иногда в компании, не проявляя неприязни к соседям.
Позднее, в гнездовый период, ловят около водоемов различных насеко-
мых и их личинок, летающих или ползающих. В желудках желтых трясо-
гузок находили пауков и насекомых, в частности веснянок, клопов-кру-
жевниц (Tingitidae), листоедов (Donacia), златок (Agrilus), короедов (Ну-
lesininae), двукрылых семейства Cordyluridae, личинок пилильщиков,
орехотворок, муравьев, пчел, ручейников; лишь в одном желудке из
семи найдено 5 семян марьянника (определения Г. Новикова и О. Семе-
нова-Тян-Шанского).
Неоднократно находили мертвых трясогузок: 28. VI 1977 г. и 29. V 1986
г. - взрослых птиц, погибших от бескормицы (Г. Катаев); 8. IX 1977 г.
молодая трясогузка была сбита автомашиной на шоссе; 9. VII 1977 г. в
долине Чуны пойман едва летавший слеток, обреченный на гибель из-за
паразитной инвазии личинками мух Protocalliphora sp. под кожей левого
крыла.
Размеры 6 взрослых самцов: длина 150-182, крыла 78-91, хвост 75-85
мм, масса тела 16,1-18,5 г (в среднем 17,2); одной самки соответствен-
но - 170, 80 и 73 мм, масса тела 18,5 г.
В Мурманской области найдены еще два вида трясогузок, сходных с
желтой:
Желтоголовая трясогузка - Motacilla citreola Pall.
Сибирский вид, проникающий на запад до Британских островов;
изредка гнездится и на Кольском полуострове (Коханов, 1987; Михайлов
и др., 1984). От желтой трясогузки легко отличается желтой окраской
головы, особенно яркой у самца, и серой спиной.
Горная трясогузка - Motacilla cinerea Tunst.
Распространена в горах Западной Европы и на Урале, изредка залетает
и на Кольский полуостров: 31. VII 1931 г. один экземпляр добыт Н.В. Про-
воровым в долине Юкспорйска, в Хибинских горах (Новиков, 1952).
Горная трясогузка отличается от желтой более крупными размерами,
длинным хвостом, голубовато-серой спиной и черным, как у белой
трясогузки, горлом.
202
Белая трясогузка - Motacilla alba L.
Местное название: саамское - йын-пуатынч (=разбивающая лед).
На Кольском полуострове гнездится повсеместно, к северу до Аркти-
ческого побережья.
Из дальних мигрантов белые трясогузки прилетают в Лапландию
одними из первых, как только вскроются устья рек и появятся первые
проталины по берегам, не смущаясь утренними заморозками, которые в
это время достигают - 10е. Средняя за 52 года дата ее прилета - 4 мая (22.
IV 1930 г. - 18. V 1935 г.). С годами сроки ее прилета сдвигаются на более
ранние: в 1928-1941 гг. - 7. V, в 1945-1952 - 5. V, в 1980-1988 - 2. V.
Интересно, что время вскрытия озер подобного сдвига не обнаруживает,
поэтому вероятной причиной более раннего прилета трясогузок может
быть рост народонаселения и возможность найти корм в населенных
пунктах; не исключено, что имело значение и увеличение числа феноло-
гических наблюдателей в заповеднике в последние годы.
Первыми появляются одиночные трясогузки, обычно самцы, иногда
отдельные пары; массовый прилет наступает неделю спустя, в середине
мая, с колебаниями в зависимости от погодных условий. Пролетные
стайки, до нескольких десятков птиц, можно встретить в устьях рек и
ручьев, на порогах и перекатах, где много открытой воды.
Осенью трясогузки сбиваются в стаи в середине августа и держатся по
низовьям рек, в тихих заливах озер, в населенных пунктах. Обычно в
них бывает десяток-другой птиц, иногда до полусотни. Отлет начинается
в конце августа и продолжается до середины сентября, далее миграция
постепенно затухает и в конце сентября - начале октября остаются
только единичные отсталые, обычно молодые, особи.Средняя дата отлета
за 50 лет - 19. IX (9. IX 1930 г. - 5. X 1969 г.). Наряду с перевозчиком
белая трясогузка - самая многочисленная птица на побережьях. Много-
летние наблюдения показали, что численность белой трясогузки доволь-
но постоянна. По учетам, проведенным в последние годы в конце мая -
начале июня в долине р. Чуны, гнездится от 14 до 23 пар на 1 км2 подхо-
дящих стаций. На маленьких ручьях, лесных озерках, в лесу на водо-
разделах она почти не попадается. По осенним (6-21 августа) учетам
численность ее в долинах Чуны и Нявки составляет 20-90 особей на
1км2. Белые трясогузки предпочитают участки рек с протоками, старица-
ми, островками, с невысокими (0,5-1 м) берегами, речные устья, озера с
защищенными от ветра заливами, валунные берега и берега с плавником.
Они гнездятся по горным ручьям до верхнего предела леса, но после
вывода птенцов стягиваются к более кормным стациям, низовьям рек,
устьям ручьев, мелководным прогретым озерам, богатым живностью.
Самцы белых трясогузок занимают участки и начинают петь сразу
после прилета. В это время (13. V - 5. VI) они иногда ожесточенно дерут-
ся: самцы, подобно петухам, некоторое время стоят друг против друга,
вытянув шею с взъерошенными перьями, затем бросаются навстречу и,
сцепившись, нанося удары клювом и крыльями, катаются по земле,
взлетают в воздух, иногда даже падают в воду, но и здесь продолжают
потасовку. Иногда в драке участвуют три самца. Позднее (25. V-18. VI)
203
наблюдаются брачные игры, парные полеты птиц и спаривание. Ухаживая
за самкой, самец принимает особые позы, кружится около нее, опустив
голову, взъерошив перья на шее, распустив крылья и хвост или волоча
одно крыло по земле.
Строительство гнезд наблюдали между 20. V-13. VI; оно занимает
около 6 дней и выполняется самкой; она продолжает таскать материал
для него и после откладки первых яиц. В отличие от желтой трясогузки
гнезда располагаются в самых разнообразных условиях, но всегда близ
воды, не далее 300 м от нее. Из зарегистрированных нами 116 гнезд 39
находились в искусственных гнездовьях, предназначенных для разных
птиц, от синиц до уток, лишь бы они были недалеко от воды; 35 гнезд - в
постройках (под крышей, на балке, под коньком, в пазу и так далее); 9 -
под валежиной в лесу или плавником на берегу; 8 - в углублении
между корней вывороченного дерева; 5 - под камнями; 6 - в нишах
грунта на берегу; 4 - под нависающим дерном; 3 - под кустиком голуби-
ки, багульника или вороники; 3 - в моторе брошенной автомашины или
трактора; 3 - в пустой консервной банке или чайнике и одно - в куче
хвороста. Удобные для гнездования места (строения, выворотни) исполь-
зуются повторно, до трех сезонов подряд. На постройках и деревьях
гнезда строятся на высоте до 8 м, на выворотнях - до 1,5 м от земли.
Гнезда, устроенные в ямках на земле, могут быть свиты только из
травы или с добавлением шерсти; в гнездах, расположенных на поверх-
ности земли, в дуплах или постройках, внешний слой состоит из ягеля,
толстых соломин и мхов, включая и подводный фонтиналис. Обычно
каркас строится из сухих еловых веточек, лишайников, мхов, травы и
пакли, а лоток выстилается тонкими былинками, шерстью млекопитаю-
щих, ивовым пухом (перьев птиц не находили).
Кладка яиц начинается в третьей декаде мая (первая находка - 25. V
1979 г., в отдельных гнездах с 20-22. V); число яиц колеблется от 3 до 7,
в среднем по 63 кладкам 5,4. Самка может оставить гнездо на несколько
дней перед началом кладки или в процессе ее. Насиживание длится
12-13 сут., птенцы оставляют гнездо через 11-13 сут., в зависимости от
величины выводка и стимула беспокойства. Наблюдения А. Брагина 19.
VI 1967 г. и 21. VI 1971 г. показали, что и самец принимает некоторое
участие в насиживании яиц и обогревании птенцов. Большинство их
вылетает из гнезда в третьей декаде июня - начале июля. Часть белых
трясогузок гнездится повторнр, но, судя по встречам поздних кладок, во
втором цикле участвует не более 10% пар. Поздние птенцы вылетают с
конца июля до середины августа.
Белая трясогуска наряду с юрком - самая агрессивная из мелких птиц и
в гнездовый период нападает на многих пернатых, от воробья и зяблика до
чайки, вороны, дербника, ястребиной совы и тетеревятника. У гнезда взрослые
птицы держатся осторожнее, слетают с него скрытно, без демонстраций,
свойственных, например, серым мухоловкам; они отлетают в сторону
низко над землей и подают сигналы беспокойства издали. Поведение
взрослых резко меняется после вылета птенцов, от слетков они энергич-
но ’’отводят”, и в этой демонстрации принимает участие до 6 взрослых
204
птиц (2. VII 1973 г.9 7. VII 1977 г.). Слетки находятся под их присмотром до
10 дней. Со второй половины июля они собираются небольшими стайками
в богатых кормом и безопасных местах.
Основу питания белых трясогузок составляют различные насекомые.
Весной они бегают по кромке льда в поисках веснянок и других беспоз-
воночных, на проталинах и у жилья ловят мух и подбирают отбросы.
Летом, как взрослые, так и молодые ищут корм на прибойной поло-
се озер, по кромке воды на берегах рек и ручьев: ловят и летающих
насекомых, особенно в затишье, а летая низко над водой, подхватывают
насекомых с ее поверхности.
По визуальным наблюдениям и содержимому 6 желудков белые
трясогузки питаются пауками, долгоносиками (Otiorhynchus, Pissodes,
Polydrosus), короедами (Hylesininae), жужелицами, стафилинидами,
веснянками, поденками, комарами (Tipulidae, Chironomidae, Aёdes,
Culex), мухами, сильфидами, наездниками, муравьями, бабочками и их
гусеницами, а при нехватке живого корма едят остатки со стола
человека.
В заповеднике наблюдались случаи разорения гнезд белой трясогузки
куницей, норкой и горностаем, при этом жертвой хищника иногда стано-
вилась наседка; 3 гнезда, построенных в утиных дуплянках, были зало-
жены яйцами гоголя. Десятки птенцов погибли от инвазии их личинками
мух. Отмечены случаи гибели трясогузок на дороге под машиной. Взрос-
лых, а тем более молодых трясогузок ловят все пернатые хищники орни-
тофаги (дербник, ястреб-перепелятник, тетеревятник, ястребиная сова),
но по причине трудности определения остатков птиц в погадках доку-
ментированы лишь единичные находки. 22. VIII 1960 г. О. Семенов-Тян-
Шанский, а 26. VIII 1977 г. А. Гилязов наблюдали нападения дербника на
слетков.
Размеры 4 взрослых самцов: длина 168-198, крыло 90-94, хвост 78-91
мм, масса тела 18,5-22,3 г (среднее 20,5); одной самки соответственно 180,
83 и 88 мм, масса тела 22,0 г.
Лесной конек (щеврица) - Anthus trivialis (L.)
Обычно гнездящаяся перелетная птица, характерна для хвойных и
смешанных лесов. В разреженной северной тайге населяет равномерно
все типы лесов, включая гари и березовое криволесье.
Средняя за 18 лет дата прилета лесного конька 19. V (10. V 1959 г. - 7.
VI 1941 г.). Прилет обычно отмечался по токовой песне, так как по внеш-
нему виду лесного конька едва ли можно отличить от лугового, а в
токовом полете есть характерные отличия.
Численность лесных коньков сравнительно невысока, в смешанных
лесах заповедника она не превышает в июне 1-3 пар/км2, а в июле и
августе - 10 особей/км2.
Гнезда располагаются обычно в лесу, иногда на болоте неподалеку от
опушки (2 случая из 7); при этом конек предпочитает сосновые леса
еловым. Гнездо строится на земле, на невысокой кочке или на бугорочке.
20Ь
Таблица 14. Питание лесного и лугового коньков
(по материалам Г.А. Новикова, с добавлениями, в %)
Состав корма Лесной конек Луговой конек
Пауки Aranei 11 3
Клопы Hemiptera 2 3
Цикадки Jassidae — 25
Тли Aphidoidea 3 —
Жуки Coleoptera 31 19
Слоники Curculionidae (6) (9)
Жужелицы Carabidae (2)
Усачи Cerambycidae (1)
Щелкуны Elateridae (2)
Стафилиниды Staphylinidae (1)
Личинки жуков* (8)
Гусеницы бабочек Lepidoptera 11 3
Перепончатокрылые Hymenoptera 14 28
Наездники Ichneumonidae (5) (15)
Личинки пилиль-
щиков Tentredinidae (2) (3)
Муравьи Formicidae (5) (3)
Двукрылые Diptera 4 3
Насекомые* 17 10
Растительные корма (се-
мена) 7 6
Семена
луговика Deschampsia flexuosa (3)
осок Carex sp. (4)
березы Betula sp. (1)
седмичника Trientalis europaea (1)
марьянника Melampyrun sp. (1)
Всего желудков 19 7
Всего кормовых объек
тов 99 32
*Ближе не определено.
под прикрытием еловой ветки, кустарничков, мха, дерна или камня.
Каркас делается из корешков кустарничков, сухих стеблей злаков, а
лоток выстилается лосиным или оленьим волосом, былинками и стебля-
ми кукушкина льна; измерен один лоток, диаметр его 8, глубина 5 см.
Кладка и насиживание яиц происходят со второй декады июня. В
яйцеводе самки, добытой 10. VI 1938 г., находилось готовое к откладке
яйцо; гнезда с кладками находили с 14. VI по 29. VI, а с птенцами - с 25.
VI по 20. VII. Количество яиц в 5 кладках и птенцов в 3 гнездах равня-
206
лось во всех случаях 5. Птенцы вылетают из гнезд с начала июля. Отлет
лесных коньков происходит в конце августа - начале сентября (самая
поздняя встреча - 27. IX).
Лесной конек использует широкий набор кормов, но находит их
только на поверхности земли: основу его питания составляют насекомые
и пауки, к ним добавляются семена растений (табл. 14).
У взрослого самца, добытого 5. VIII, линька еще не была заметна, у 2
самцов и 1 самки 11-15. VIII линяло оперение туловища.
Размеры 10 самцов: длина 142-171, крыло - 85-95, хвост 62-72 мм,
масса тела 20,5-25,0, в среднем 22,8 г; размеры 4 самок соответственно -
132-167, 88-89 и 56-73 мм, масса тела 15,7-26,2, в среднем 21,6 г.
Луговой конек - Anthus pratensis (L.)
Одна из самых обычных гнездящихся перелетных птиц заповедника,
особенно многочисленная в горной тундре, где гнездится преимущест-
венно в березовом криволесье и в субальписком поясе, на увлажненных
кочковатых тундрах с зарослями ивняков и карликовой березки, отчасти
на травяно-кустарничковых участках, избегая ягельников и каменистых
мест. Во время весенней и осенней миграции их можно встретить в лесу,
по долинам озер и рек.
Средняя за 15 лет дата прилета в Лапландском заповеднике - 15 мая
(7. V 1984 г. - 24. V 1936 г.); дата последней встречи обоих видов коньков
осенью - 13. IX (29. VIII 1982 г. - 2. X 1987 г.; за 45 лет).
Численность луговых коньков, судя по многолетним наблюдениям,
остается на одном уровне, без резких колебаний. По учетам 1978-1987 гг.
в июне, в горной тундре, она составляет 7-11 пар на 1 км2, в болотистой
долине реки - не более 2-4 пар/км2. В сплошном лесу, включая редко-
стойные участки и 20-летние вырубки, этот вид не зарегистрирован. Самцы
луговых коньков начинают петь сразу после прилета и до конца июля.
Весной они токуют даже над озерным льдом. Размер весенних стай не
превышает десятка особей. Сначала они появляются на выдувах и прота-
линах по болотам и южным склонам; поскольку загрязненные дымом участ-
ки тундр и пораженного газами леса вокруг Мончегорска освобождаются
от снега раньше, то первые луговые коньки собираются именно здесь,
но исчезают через неделю-другую, как только стает снег в других
районах.
У самки, добытой 27. V 1938 г., яичники еще не были развиты. 28. V
1981 г. на Сейднотчорре наблюдали брачные игры коньков - в это время
очевидно, они разбиваются на пары и занимают гнездовые территории. 6.
VI 1978 г. на Пельпахке (Нявка-тундра) пара коньков подбирала прош-
логодние листья, причем одна из птиц пыталась отвлечь наблюдателя
от строящегося гнезда, крича и волоча опущенные крылья (других
подобных случаев не наблюдали). Из найденых 35 гнезд 6 располагались
под камнем, 13 - под кочкой, 15 - под кустиком голубики, черники,
карликовой березки и др., 1 - у валежины. Каркас гнезда свивается из
стеблей луговика, овсяницы, белоуса, иногда с добавлением осоки,
вороники, черники, брусники, кусочков ягеля и цетрарии; лоток высти-
207
лается более тонкими стеблями и листьями злаков. Диаметр лотка 5-8 см,
глубина его 4- 6 см.
Кладка яиц начинается в конце первой декады июня, а самая поздняя
полная кладка найдена 19. VII. В июне полные кладки содержат по 4-7
яиц, в среднем - 5,4 ( п = 26 ), в июле по 3-6, среднее - 3,7 ( п = 15 ).
Вторично гнездятся, вероятно, только старые особи и пары, потерявшие
первую кладку. Наличие второго цикла размножения у лугового конька
отмечено на Айновых островах (Коханов, Скокова, 1967) и в Ленинград-
ской области (Мальчевский, Пукинский, 1983, т. 2, с. 64).
Птенцы вылетают из гнезд с начала июля до начала августа (самый
поздний слеток, с недоразвитыми рулевыми и маховыми, встречен 4. VIII
на Ельнюне). Откочевка начинается уже с середины июля (13. VII на
Пельпахке), но пик миграции приходится на конец августа - начало
сентября, когда наблюдаются стаи в сотни птиц. Одиночные луговые
коньки попадаются иногда в лесу до установления снежного покрова в
октябре.
Основу питания лугового конька составляют различные беспозвоноч-
ные с добавлением растительных кормов.
Остатки луговых коньков, пойманных хищными птицами, находили
дважды, 2. VIII 1960 г. на Нявка-тундре, 17. VII 1966 г. - на Нялл-тундре,
а 19. X 1977 г. на Чунозерской усадьбе подобран труп истощенного моло-
дого конька с травмированной лапой.
Размеры 4 взрослых самцов: длина тела 151-155, крыла 78-84, хвоста
58-61 мм, масса тела 14,5-19,5 г, в среднем 17,2 г; размеры 4 самок
соответственно - 132-149, 75-78 и 52-61 мм, масса тела 15,7-18,0, в сред-
нем 17,2 г.
Краснозобый конек - Anthys cervina (Pall.)
Согласно Плеске (1887), встречается по всему Мурманскому побе-
режью, до устья Поноя, а Энвальд наблюдал 17. VII 1883 г. ’’несколько
экземпляров в альпийском поясе Чуни-тундр”:
Нами в Лапландском заповеднике не добыт, но визуально наблюдался
трижды: 27. V 1967 г. Семенов-Тян-Шанский видел пару на Турухтанском
болоте: 1. VI 1977 г. на берегу р. Нявка пара искала корм в нанесенном
водой мусоре; 24. V 1983 г. Гилязов наблюдал на Охтозере в осочнике
пару, по-видимому собиравшую семена. Все эти птицы были доверчивы,
подпускали на 3-5 м.
В брачном наряде краснозобый конек отличается от лугового ржавча-
то-красным зобом, а в остальное время более резкими пестринами на
нижней стороне и надхвостье, а также голосом.
Горный конек - Anthus spinolette (L.)
Распространен по горным системам Европы, Азии и Северной Америки,
а подвид A. s. littoralis Brehm - по морским берегам Британских островов,
Скандинавии и Кольского полуострова (Плеске, 1887, с. 213-214; Коханов
и др., 1967).
208
В Лапландском заповеднике встречался трижды: 14. VII 1938 г. Влади-
мирская добыла взрослого самца на каменной россыпи по северному
отрогу Ыллчорра; 27. VII 1981 г. Гилязов наблюдал одиночного горного
конька с кормом в клюве в сыром кочкарном ивняке на Ельнюнлаге; 3.
VII 1982 г. Брагин нашел на Сейднотлаге (по взлетевшей из-под кустика
вороники самке) гнездо конька с 3 яйцами. Размеры йяца 20,0x14,2
мм, масса 2,05 г (не насижено). Размеры добытого самца: длина 155, крыло
82, хвост 58 мм, масса тела 18,5 г: в желудке остатки насекомых.
Горный конек немного крупнее лугового, верхняя сторона серее,
лапки черно-бурые.
СЕМЕЙСТВО СОРОКОПУТОВЫЕ - LANHDAE
Жулан - Lanius collurio L.
Сорокопут жулан гнездится по всей европейской части Союза, к се-
веру примерно до 64-65°, изредка залетает в Мурманскую область.
19.Х 1964 г. на Чунозерской усадьбе Лапландского заповедника Ю. Га-
ев добыл молодую самку.
2.VI 1978 г. при устье р. Верхняя Чуна Гилязов наблюдал одного сам-
ца; 11.VII 1978 г. Сорокин видел жулана в редком бору по краю болота
близ Витте-озера.
Большой сорокопут - Lanius excubitor L.
Большой сорокопут широко распространен в Европе, Азии и Северной
Америке, причем в Скандинавии и на Кольском полуострове доходит до
северного побережья. Описано около 20 подвидов большого сорокопута,
из них на Кольском полуострове обитает номинальный - L. е. excubitor
(Степанян, 1978). Гнездящиеся у нас сорокопуты отлетают на зиму в За-
падную и Центральную Европу. Средняя за 13 лет дата его прилета в за-
поведник - 16.V (27.1 V 1983 г. - 27.V 1937, 1938 гг.); дата отлета за 14 лет
- 18.IX (3.IX 1950 г. - 5.Х 1934 г.).
Обычно сорокопута встречают в разреженных болотистых елово-бере-
зовых лесах по берегам рек, озер, на вырубках и зарастающих гарях.
Численность низка даже на пригодных стациях: на трансектах длиной до
30 км в начале августа попадается не каждый год, например в 1980 и
1981 гг. его совсем не встречали. Численность повышается в год ’’урожая”
мелких грызунов и следующий за ним сезон: тогда один наблюдатель
отмечает за лето до 10-15 сорокопутов.
Большие сорокопуты приступают к размножению сравнительно рано,
уже 18.V 1978 г. наблюдали на расстоянии 3 км двух поющих самцов,
перелетавших с одной верхушки дерева на другую, выбирая при этом за-
сохшие. У самки, добытой 12.V 1934 г., диаметр самого крупного фолли-
кула превышал 1 см. Самая ранняя встреча летнего выводка - 23.VI
1962 г., а в среднем за 9 лет они появляются 9.VII. Отсюда можно заклю-
чить, что кладка яиц происходит нормально во второй половине мая -
начале июня, а птенцы выводятся в середине июня. Единственное гнездо было
14. Зак. 1023
209
найдено 27.VI 1977 г. в редком елово-березовом лесу на берегу мелководной
Чунозерской ламбины. Оно находилось на высоте 7 м у вершины елки,
прилегая к стволу с юго-западной стороны. Птенцы ’’полухвостики” вы-
летели недавно и сидели на соседних деревьях. Тут же находились и
взрослые сорокопуты, но держались осторожно, лишь изредка подавая сигна-
лы. Каркас гнезда сложен из сухих еловых и сосновых веток, стеблей и корней
вороники, голубики, коры можжевельника, соломин вейника, с вплетенными
снаружи растительными волокнами и ивовым пухом; лоток - из более тонких
стеблей и корешков, травы, перьев куропатки, щура и др., ивового пуха. Раз-
меры гнезда 230x190, высота 108 мм; лотка - 125x111, глубина 46 мм, 9 июля
выводок в составе 4 птенцов и пары взрослых оставался не далее 200-300 м от
гнезда. Число слетков в других семьях варьирует от 1 до 5, в среднем 2,9
(п=17).
Большой сорокопут питается исключительно животными - мелкими
позвоночными и крупными насекомыми. Сорокопутов с добычей в клю-
ве или когтях наблюдали 7 раз, дважды это была птичка, дважды - по-
левка, раз - бабочка пяденица. 17.V 1961 г. сорокопут убил манную че-
четку на точке, где Ю. Гаев ловил птиц для кольцевания, в гнезде соро-
копута 27.VI 1977 г. мы нашли резец полевки, мелкие кости, остатки
шмеля и других насекомых. Во всех шести желудках, исследованных
Г.А. Новиковым, находились остатки насекомых (веснянки, саранчовые,
жуки - усач и щелкуны, ручейники, оса), кроме того, в четырех - пауки,
в двух - остатки полевок и в двух - предположительно лягушки.
В годы высокой численности полевок встречаемость сорокопутов уве-
личивается более чем в 10 раз: такими годами были 1934, 1937, 1938, 1959,
1962, 1970, 1974, 1977, 1978, 1982, 1983. Односторонней тенденции к росту
или сокращению популяции большой сорокопут не обнаруживает: в
30-х годах отмечали в среднем по 5,6 особей в год, в 40-х - 1,4, 50-х -
6,0, 60-х - 5,0, 70-х - 9,4, в 80-х - 2,8.
Размеры (л = 7): длина 240-269 (ср. 250), крыло 113-119 (ср. 117), хвост
111-117 (ср. 114 мм); масса тела взрослой самки 83, трех молодых птиц
65-70 г.
СЕМЕЙСТВО СВИРИСТЕЛЕВЫЕ - BOMBYCILLIDAE
Свиристель - Bombycilla garrulus (L.)
Свиристель встречается по всему Кольскому полуострову, до Мурман-
ского побережья.
По характеру сезонных перелетов свиристель принадлежит к инва-
зионным видам птиц. На территории заповедника он живет с весны до
осени, а на зиму, как правило, отлетает к югу. Численность свиристелей
резко колеблется по годам; вероятно, они так же меняют места гнездо-
вания, как это делает большой пестрый дятел и другие инвазионные
виды. В отдельные зимы (1931/32, 1935/36, 1950/51, 1977/78, 1979/80 и
1982/83) свиристели встречались в заповеднике и зимой. В некоторые
другие годы они появлялись только в пролетное время, а летом практи-
чески отсутствовали (1939,1959, 1969, 1975 и 1983).
210
Свиристели освоили г. Ленинград как место зимовки уже во второй
половине прошлого века. По наблюдениям Д.Н. Кайгородова (1916), не-
обычно рано и в большом количестве они прилетают в зимы с ранними и
обильными снегопадами. Массовые зимовки свиристелей в Ленинграде
наблюдались в 1892/93 г., затем в конце 1956 и 1976 гг. (Мальчевский,
Пукинский, 1983, т. 2, с. 75-76). Свиристель - птица прожорливая, и для
насыщения ему нужен не только хороший запас ягод на деревьях, но и
достаточно длинный световой день, чтобы кормиться; надо думать, не-
прерывная ’’полярная ночь” - одна из главных причин, принуждающих
свиристелей улетать на зиму из северных широт. В послевоенные годы в
озеленении городов Мурманской области первое место прочно заняла ря-
бина, в городских условиях она плодоносит обильнее, чем в лесу, и при-
том ежегодно. Благодаря этому в городах области образовался устой-
чивый кормовой фонд, доступный для птиц и в ’’полярную ночь” благо-
даря искусственному освещению улиц. Впервые 23. XII 1970 г. лесник за-
поведника В. Владимирский наблюдал на рябинах в Кировске большую
стаю свиристелей. В январе 1978 г. в Мурманске О. Семенов-Тян-Шанский
наблюдал несколько стай свиристелей, в то время как А. Гилязов заме-
тил в заповеднике только пару этих птиц. В газете ’’Полярная правда”
З.ХП 1978 г. В. Коханов сообщил о зимовке большой стаи свиристелей в
пос. Мурмаши, а в начале 1981 г. они летели по Мурманску сотенными
стаями. Зимой 1986/87 г. в Мончегорске обитала стая из 56 птиц. Интерес-
но, что, имея обеспеченную подкормку в городах, они стали чаще попа-
даться в последние годы и в заповеднике, где одиночных птиц наблюда-
ли в феврале 1980,1985 и 1986, в марте 1982, 1983 и 1984 гг.
В заповедник свиристели прилетают парами или стайками, в среднем
за 39 лет - 8 мая (2.1 V 1936 г. - 27.V 1934 г.), а осенью последние встречи
датируются 7 октября (4.IX 1936 г. - 22.XI 1986 г.; 38 лет); следовательно,
они проводят на гнездовьях 5 мес. Количество встреченных особей ко-
леблется по годам в 10 раз, больше всего их наблюдалось в 1935-1937, в
1941, 1948-1950, 1963-1968, 1970-1973 и в 1977-1982 гг. (за 1942-1946 и
1952-1957 гг. сведений нет, так как заповедник не функционировал).
В колебаниях численности свиристелей нет явной периодичности, она
может оставаться высокой до 6 лет подряд или низкой - до 4 лет. В то
время как осенне-зимние налеты свиристелей легко и убедительно свя-
зываются с урожайностью ягод рябины, отсутствие их на протяжении
нескольких лет на гнездовых стациях труднее поддается объяснению.
Вероятнее всего, свиристели заселяют территорию неравномерно и вре-
мя от времени меняют места размножения; с другой стороны, возможно,
что они в некоторые годы вообще не гнездятся.
Свиристели занимают гнездовые территории в мае: с 4.V по 6.VI они
встречаются отдельными парами и стайками, в которых пары держатся
уже обособленно. 8.VI 1971 г. О. Семенов-Тян-Шанский наблюдал сценку
ритуального выпрашивания корма самкой. С начала июня птицы чаще
встречаются поодиночке - вероятно, в это время самки начинают наси-
живать. Первые слетки отмечены 1.VII: птенцы хорошо летали, но руле-
вые у них еще недоразвиты, размером менше взрослых, окраска светлее
211
и монотоннее, а хохолок на голове еле выражен. В выводках мы встреча-
ли от 2 до 6 птенцов, в среднем 4,0 (п = 7). Кочующие стаи свиристелей по-
являются со второй половины августа.
Свиристели питаются как растительной, так и животной пищей. В ян-
варе - июне они кормятся почками и соцветиями козьей ивы, березовы-
ми почками, ягодами ”гром-ягоды” (Arctons alpina), толокнянки и воро-
ники, веснянками и пауками. Летом (конец июня - начало августа) при
обилии летающих насекомых свиристели, подобно серой мухоловке,
ловят их на лету: выбрав на берегу наблюдательный пост (камень, дере-
во, корягу), они ловят добычу над водой, зарослью осоки, хвоща и дру-
гих растений короткими бросками. Очевидно, этим кормом они и вы-
кармливают птенцов. В желудках двух птиц, добытых в августе, найде-
ны ягоды вороники, можжевельника, черники наряду с листоедами
(Melosoma lapponum), поденками и комарами (Bibio sp.) (Новиков, 1952).
В октябре (4 желудка) обнаружены семена рябины, ели, вороники, толок-
нянки, лишайник Bryopogon, различные жуки, комары-долгоножки, на-
ездники и другие насекомые, не определенные точнее (Г. Новиков,
О. Семенов-Тян-Шанский).
Размеры 3 взрослых свиристелей: длина 201-204 мм, крыло 110-116,
хвост 61-68 мм; масса тела самцов 59 и 64, самки - 63,3 г; молодого сам-
ца в начале августа - 48 г.
СЕМЕЙСТВО ОЛЯПКОВЫЕ - CINCLIDAE
Оляпка - Cinclus cinclus (L.)
Местное название: саамское - кушк-каргенч (от слов: кушк -
речной порог, кархэ - быстро бегать; ’’бегун по перекатам”).
На Кольском полуострове обычная, хотя и немногочисленная птица,
приуроченная к быстротекущим потокам, почему она более характерна
для западной, гористой, части Русской Лапландии, где живет круглый
год, хотя и не совсем оседло: нуждаясь в открытой воде, по мере замер-
зания ручьев, часть птиц переселяется на зиму на более многоводные,
вытекающие из озер реки. Массовая зимовка оляпок на р. Ниве в
конце XVIII в. привлекала внимание Н. Озерецковского (1804, с. 60). В
Лапландском заповеднике реже всего оляпка попадается на глаза в де-
кабре и январе; только в малоснежные зимы встречаемость оляпок зимой
практически не уменьшается. Далеко от гнездовых мест они не улетают,
иначе трудно объяснить факт моментального появления их на ручьях
вслед за очищением последних от ледяной корки и снежных завалов.
Излюбленные места обитания оляпки летом - небольшие (до несколь-
ких метров шириной), захламленные ветровалом каменистые горные
ручьи, водопады, речные пороги и перекаты. Численность их в гнездовое
время около 0,1-0,2 особи на 1 км - на реках, до 0,3-0,2 - на горных
ручьях.
Пение оляпок слышали в разные годы с 20.11 до 11.VI, чаще в конце
марта - начале апреля: вероятно, в это время они занимают гнездовые
территории; поют они и осенью, но менее интенсивно.
212
Гнезда строятся в конце апреля - начале мая. Гнездо представляет
собой шаровидное сооружение, 16-30 см в длину, 16-25 в ширину, 13-
20 в высоту; толщина стенок после выхода птенцов: боковые 8-12 см,
кровля 3-5, подстилка 4 см; диаметр летка от 6 см в начале гнездования
до 9 см - к вылету птенцов. Вход в гнездо всегда снизу, леток обращен
к водному потоку. Основание, стенки и кровля строятся из различных
мхов, в основном зеленых (Dicranum sp., Pleurozium schreberi, Hylocomium
splendens), водяного мха (Fontinalis sp.) и стеблей осок. Лоток сплетается
из стеблей осок, пушицы, злаков, корней и стеблей кустарничков, а вы-
стилается листьями березы, ивы, брусники, лубом и корой ив, изредка
перьями. Снаружи гнездо камуфлируется свисающими пучками мхов,
ягелей, а вход (окружность летка) укрепляется, а заодно и маскируется
стеблями осок.
Гнездо, найденное 16.V 1983 г. на Среднем Бобровом ручье, было пост-
роено на основе старого, из свежего материала сложена лишь кровля,
летковая часть и, вероятно, лоток. Из найденных 28 гнезд 13 располага-
лись на карнизах каменных стен у берегов ручьев, в 1-2 м над водой;
8 - на камнях посреди потока, на высоте 1-1,5 м; 2 - на стволах ели или
березы, склоненных над потоком, на высоте 1,5-4 м от воды; один - на
развилке растущей у берега березы, на высоте 3,5 м; 4 - на обрыве речно-
го берега, на высоте около 1 м.
7.VII 1986 г. на балке моста через Ельявруай обнаружено недостроен-
ное гнездо, в 1987 г. оляпки достроили его и вывели птенцов. В удобных
местах оляпки гнездятся из года в год,, например на ручье Ельявруай на
одном определенном камне оляпка гнездилась в 1941 г., а потом в 1959—
1961 гг., три года подряд; под водопадом на ручье Мошникова с 1978 по
1986 г. гнездилась ежегодно, а еще раньше, в 1971 г., там были учтены 4
старых гнезда, на разных стадиях разрушения. На Среднем Бобровом
ручье на Каменной стене оляпка гнездилась ежегодно с 1980 по 1987 г.
(рис. 51).
Кладка яиц происходит между 10.V - 8.VI. В кладке 5-6 яиц, в сред-
нем 5,4 (п = 5); форма их удлиненно-овальная, скорлупа белая с розоватым
оттенком; размеры двух яиц 18x25 и 20x26 мм). Гнезда с птенцами нахо-
дили 15.VI - 8.VII, а покинутые выводком - с 7 июля, однако летные вы-
водки попадались и раньше, с 17 июня. Растянутость гнездового периода
на четыре месяца допускает возможность второго цикла размножения в
тот же сезон. Судя по встречам одиночных слетков, ранние выводки рас-
падаются к концу июня. Известны два случая гибели гнезд: одного, с
яйцами, при возврате холодов в июне 1977 г., другого, с выводком,
18.VII 1960 г. из-за гибели взрослых птиц, предположительно пойманных
перепелятником.
Гнездовые птенцы с еще не отросшими маховыми и рулевыми перьями
при опасности (осмотр гнезда) прыгают в воду и ныряют или уплывают
вниз по течению.
Оляпка кормится различными водными беспозвоночными, в основ-
ном личинками веснянок, ручейников, комаров, выискивая их на галеч-
ных отмелях или ныряя на мелководье, на глубине не более 1 м, а чаще
213
Рис. 51. Оляпка у гнезда, 14. VI 1941 г. Фото О.И. Семенова-Тян-Шанского
0,3-0,6 м. Продолжительность нырка от 4 до 10, чаще 5-6 с. Оляпка охот-
но собирает и насекомых на воздухе, с поверхности камня, например
тех же веснянок.
Размеры пяти самцов: длина 180-204, крыло 97-100, хвост 54-59 мм;
масса тела 58,0-70,8, в среднем 65,8 г; размеры одной самки соответствен-
но 192, 86 мм, масса тела 61,5 г.
214
СЕМЕЙСТВО КРАПИВНИКОВЫЕ - TROGLODYTIDAE
Крапивник - Troglodytes troglodytes (L.)
На Кольский полуостров залетает редко. Из шести встреч крапивников
в Лапландском заповеднике в четырех наблюдался поющий самец. Впер-
вые отмечен в 1938 г.: 13.VII А. А. Насимович видел одного в густом и
захламленном ельнике на берегу Тулпъявра, a 6.IX Т. П. Некрасова встре-
тила другого в ельнике по скалистому склону Ельнюна. 6.V 1949 г.
Т.В. Кошкина наблюдала одного в ельнике, в районе Чунозерской усадь-
бы, а 30.VI и 6.VII 1973 г. Н.Н. Скокова - на берегу р. Чуны. Там же, при
устье Среднего Бобрового ручья, в елово-березовом лесу, богатом ветро-
валом, с зарослями ивняка, 21.V 1982 г. А.С. Гилязов заметил очень бес-
покойного самца крапивника: предметом его тревого была забравшаяся
на густую ель норка.
СЕМЕЙСТВО ЗАВИРУШКОВЫЕ -PRUNELLIDAE
Лесная завирушка - Prunella modularis (L.)
Завирушка в Мурманской области доходит до северной границы леса
(Кищинский, 1960). В заповеднике впервые добыта П.С. Мальчевским
22. VIII 1937 г. в ельнике по южному склону Ельнюна (Владимирская,
1948, с. 241). После того одиночные завирушки встречались неоднократно,
хотя и не ежегодно, обычно в пролетное время. Средняя дата первых
встреч за 13 лет - 17 мая (37.V 1984 г. - 3. VI 1985 г.), последних, за 4 го-
да, - 10 сентября (1. IX 1937 г. - 18. IX 1961, 1977 гг.). За 45 лет наблюде-
ний ее встречали в 16 сезонах, всего 38 особей. 3. IX 1962 г. в ельнике под
Ельнюном в мышеловку попалась молодая самка - косвенное доказа-
тельство гнездования у нас завирушки.
Размеры взрослого самца и молодой самки следующие: длина 145 и
152 мм, хвост 69 и 57, крыло самки 71 мм, вес 20,1 и 20,6 г.
СЕМЕЙСТВО ДРОЗДОВЫЕ -TURDIDAE
Зарянка - Erithacus rubecula (L.)
В южной части Кольского полуострова, в районе Кандалакшского за-
лива, зарянка - обыкновенная гнездящаяся птица (Благосклонов, 1960;
Бианки, 1960), севернее гнездится редко и нерегулярно.
В Лапландском заповеднике за 45 лет наблюдений встречалась на про-
тяжении 20 сезонов, всего 44 особи, преимущественно в еловых или сме-
шанных лесах по горным склонам, с частым древостоем и богатым напоч-
венным покровом (долина р. Нявка от Холодного ручья до Падуна, до-
лины Среднего и Нижнего Бобровых ручьев, Второго ручья и Ельявруая,
ельники по берегу Чингильс-явра и склонам Ельнюна Второго и Третье-
го). Первые встречи, в среднем за 13 лет, 13 мая (29. IV 1961 г. - 31. V
215
1982 г.), последние, за 5 лет, около 7 сентября (31. VIII 1966 г. - 11. IX
1967 г.).
6. VIII 1937 г. П.С. Мальчевский наблюдал в долине Второго ручья вы-
водок зарянки, а 27. VIII там же добыл молодую самку; еще 2 молодые
самки попались случайно в капканчики для отлова мелких млекопита-
ющих 8. IX 1937 г. и 30. VIII 1966 г. и одну молодую птицу видели 9. IX
1967 г. на том же месте. Эти находки доказывают, что зарянка в заповед-
нике гнездится. После сильного снегопада 20-22. VI 1977 г. Катаев на-
шел в избушке на оз. Чингльс-явр трупы 2 зарянок, погибших, очевидно,
от голода.
Промеры 2 молодых самок: длина 135-141 мм, крыло 72, хвост 66-60
мм; масса тела 15,1-18,1 г. В их желудках найдены мелкий моллюск,
гусеница бабочки, 3 личинки и остатки нескольких жуков.
Варакушка - Cyanosylvia svecica (L.)
По всей Лапландии варакушка обычная птица. В заповедник приле-
тает обычно в последней декаде мая, в среднем за 39 лет - 24 мая (14.
V 1977,1982 гг. - 3. VI 1950,1966 гг.; о ± 4,7). Отлетает (в среднем за 24 года)
4 сентября (20. VIII 1969 г. - 20. IX 1964 г.; О + 9,3). Средняя температура
на день прилета + 5,5е, на день отлета + 8,4°. Корреляция времени прилета
с температурой мая отчетливая, г = - 0,49 ± 0,13, связь отлета с темпера-
турой августа значительно слабее, г = + 0,28 ± 0,20.
Чаще всего варакушка встречается на стыке березового криволесья
с горной тундрой, в густых ивняках и ерниках по долинам рек, особенно
в лесотундровой северо-западной части заповедника. Отдельные пары
гнездятся по кустарниковым болотам, рединам на заросших гарях и
вырубках. В лесах различных типов, лишайниковой, кустарничковой
и каменистой тундре она не встречается.
Численность варакушки заметно колеблется по годам: за последние
двадцать лет максимумы наблюдались в 1965, 1969, 1976 и 1982, а мини-
мумы - в 1966, 1973, 1979 и 1984 гг.: периодичность колебаний около 6
лет, амплитуда - не менее 4:1. По учетам, проведенным нами в 1977—
1987 гг., численность варакушек в долинах Нявки, Чуны и на Чуна-тундре
(Сейднотчорр, Ельнюнчорр) не превышает 1-2 пар на 1 км2, в долине Ку-
удасйок - до 6 пар, это в мае - первой половине июня, а в конце июля -
первой половине августа соответствено, 5-6 и 23 особи на ту же пло-
щадь. Для сравнения укажем, что в горных березняках Финской Лап-
ландии в июне учтено 9, а на горной тундре - 3 пары/км* (Jarvinen и др.,
1982).
Варакушка начинает петь сразу после прилета. Самец поет обычно си-
дя на верхушке дерева, часто перелетая, но иногда внизу, в кустах, и
так тихо, что кажется, будто до него сотни метров, в действительности же
не более десятка. Варакушка - искусный подражатель, и песня ее, кроме
собственных колен, включает имитацию песен и звуковых сигналов
других птиц, из числа самых обычных, например чирка-свистунка, фифи,
крачки, кукши, белобровика, веснички, юрка и др.
Гнездо свое он помещает в нише кочки, у ствола небольшого дерева,
216
под навесом кустарничков; строится оно из травы, иногда с добавкой
мхов. Гнезда с яйцами мы находили с 15. VI по 24. VII; в полной кладке
4-6 яиц, в среднем 5,4 (п = 5). Птенцы вылупляются начиная с 23. VI,
полные выводки находили с 6. VII, одиночные слетки - с 17. VII, но нерас-
павшиеся семьи встречаются до середины августа.
Судя по содержимому 4 желудков и визуальным наблюдениям, вара-
кушки кормятся преимущественно насекомыми - жуками, в том числе
долгоносиками, муравьями (Formica, Myrmica), гусеницами бабочек, дол-
гоножками (Tipula), мухами, моллюсками, встречаются также единично
семена марьянника, крупа (из отбросов).
Размеры 3 взрослых самцов: длина 132-136, крыло 73-78, хвост 52-
54 мм; масса тела 20,0-23,1 г; двух молодых самок (август) 155, 75-76 и
55-56 мм, масса тела 16,7 и 18,9 г.
В Мурманской области из нескольких подвидов варакушки встречают-
ся только номинальный, С. s. svecica с ржавчатой ’’звездой” на голубом
пятне (у других форм звезда белая или отсутствует).
Синехвостка - Tarsiger cyanurus (Pall.)
Синехвостка - характерная птица темнохвойной сибирской
тайги; ареал ее до начала нашего века простирался от верховьев
Печоры до Тихого океана. Полной неожиданностью явилась наход-
ка студентом ЛГУ П.С. Мальчевским в августе 1937 г. в Лапланд-
ском заповеднике выводка синехвосток, из которого он добыл
3 экз. (двух взрослых и слетка). Место встречи — высокоствольный
ельник на влажной почве с примесью березы, рябины, а в подлеске -
можжевельника, в долине Второго ручья, на юго-восточном склоне
Чуна-тундры (Мальчевский, 1947; Владимирская, 1948, с. 239). Даль-
нейшие поиски синехвосток как здесь, так и в других районах запо-
ведника не дали результатов, зато в последующие годы они были
обнаружены в ряде других мест. С 1949 до 1957 г. залетные сине-
хвостки (самок достоверно не зарегистрировано) найдены в Фин-
ляндии, в 11 пунктах, преимущественно у восточной грани-
цы страны (Helminen, 1958): Хельминен объясняет появление синехвосток
расширением ареала вида путем продления пролетного пути (синехвост-
ка зимует в Китае), обращая внимание на то, что основные сдвиги совпа-
ли с теплыми веснами 1937, 1949 1954 гг. Расширение ареала, подтверж-
дают и позднейшие находки этой птички в Европейской России. В 1938 г.
В.Я. Паровщиков обнаружил ее в юго-западной части Архангельской об-
ласти; в 1960 г. Ю.П. Губарь и А.В. Мошкин (1966) нашли ее обычной в вы-
сокоствольных ельниках Терского побережья Кольского полуострова
и даже определили плотность (2,4 особи/км2). В 1961-1966 гг. В.В. Зимин
наблюдал ее в заповеднике ’’Кивач”, в Карелии, а в 1972-1977 гг. Г.А. Но-
сков и другие люди поймали несколько особей в районе Сивирской губы
Ленинградской области: в обоих пунктах установлено ее гнездование
(Мальчевский, Пукинский, 1983, т. 2, с. 111).
В желудке самца из Лапландского заповедника найдены: жужелица,
листоед (Phytodecta pallida), личинки щелкунов, муха, долгоножки, ли-
217
чинки пилильщиков и гусеницы; у слетка - побег черники, семя березы,
прядь лишайника Bryopogon sp., паук, клоп (Tingitidae), сенокосец (Opili-
ones), гусеница совки.
Размеры самца, самки и слетка: длина 133, 126 и 120 мм, хвост самки 68
мм, птенца 35 мм, вес 13,3, 13,0 и 12,9 г.
Поведением и размерами синехвостка напоминает зарянку; самец ха-
рактеризуется более или менее синей (в зависимости от возраста и сезо-
на) окраской верхней стороны, особенно надхвостья, самка сверху олив-
ково-бурая, нижняя сторона у обоих полов белая с рыжими боками.
Горихвостка-лысушка - Phoenicurus phoenicurus (L.)
На Кольском полуострове обычна всюду, где только есть лес. Средняя
дата появления горихвостки - 11 мая (26. IV 1975 г. - 29. V 1935 г.; о ±
6,8; 44 года), при переходе суточных температур через 2,7°. Отлет проте-
кает незаметно, отчасти по отсутствию транзитного пролета: его удалось
зарегистрировать только за 17 лет, в среднем 1 сентября (25. VIII 1935 г. -
18. IX 1984 г.; а ± 5,4), при температуре + 8,9°. Срок отлета, по-видимому,
не зависит от температуры августа, соответствующий коэффициент кор-
реляции + 0,14 ± 0,24. Интересные данные об осеннем перелете дало коль-
цевание (горихвостки охотно гнездятся в искусственных гнездовьях,
где их систематически кольцуют). Птенец, помеченный 29. VI 1976 г. на
берегу Чунозера, был пойман 11. IX 1977 г. в Швеции, на о-ве Эланд; дру-
гой, помеченный там же 5. VII1961 г., пойман спустя 2 года, 11. IX 1963 г.
в Гельголандской бухте, ФРГ; еще три птенца, окольцованные в 1960,
1961 и 1971 гг.,., обнаружены в сентябре-октябре в год мечения в Польше,
близ Гданьска, и в Италии, у г. Карро и южнее Падуи (45° с.ш., 1. X 1961 г,)
(рис. 52). Вероятно, основная масса горихвосток отлетает в августе, так
как на учетном маршруте в долине р. Чуна длиной 34 км в середине августа
мы их уже не встречали или учитывали единично. Отсталых, обречен-
ных на гибель, горихвосток наблюдали даже после выпадения снега,
11.Х 1973 г. и 25.Х 1980 г.
Горихвостка - типичная лесная птица, предпочитающая спелые насаж-
дения; в разреженных борах она является фоновым видом. Численность
ее существенно не изменилась с 1931 г. В смешанных лесах по берегам
Чунозера и в долине Чуны в начале июня мы учитывали от 2 до 5 гнез-
дящихся пар на 1 км2, в борах по Чуне - 1-2 пары, в березняках с при-
месью ели по берегам реки Чуны - менее 1 пары/км2. В этих стациях из
100 искусственных гнездовий на линии в 10 км горихвостки заселяют не
больше десятка, обычно 2- 6, дуплянок.
Самцы начинают петь в первые же дни после прилета (рис. 53). Интен-
сивность пения нарастает к концу мая - началу июня, в это время они
поют всю ночь; к середине июля они поют уже реже, позднее слышны
песни только отдельных особей (самая поздняя песня - 13. VIII 1966 г.).
Прилетая раньше самок, самцы подыскивают место для гнезда, обычно
дупло, и стаскивают в кучку строительный материал - десятки кусочков
коры, трухи, лишайников. Заготовка гнездового материала носит отчас-
ти и ритуальный характер (практикует ее и мухоловка-пеструшка, также
218
Рис. 52. Места встреч окольцованных в заповеднике горихвосток
гнездящаяся в дуплах). Гнезда строятся в дуплах различного происхож-
дения, как в результате гниения ствола, так и выдолбленных дятлами;
гнездятся они также в берестяных трубах, скворечниках для мелких
птиц и ’’вуувтах” - гнездовых ящиках для уток. В перестойных лесах
заповедника с обилием сушняка достаточный выбор дупел для гнездования.
Высота гнезда (летка) горихвостки колеблется от 0,6 до 10 м, в среднем 3,6 м
(п = 19). Диаметр летка от 3 до 13 х8 см, чаще 5-6 см. Состав гнездового
материала разнообразный, включает сосновую хвою, кору сосны и мож-
жевельника, древесную труху, различные мхи, лишайники, траву; лоток
выстилается шерстью лося, оленя, тонкими стеблями, всегда с добавкой
перьев. Иногда гнездо сделано только из травы и перьев или даже од-
них перьев: 15. VI 1983 г. мы нашли в скворечнике гнездо, сложенное
из перьев косача. Строительство ранних гнезд начинается с 24. V и про-
должается 2-7 дней. Откладка яиц длится с 27. V до 3. VII. Размер кладки
4-8 яиц, в среднем 6,4 (п= 79). 4. VI 1978 г. в долине Чуны найдена нео-
бычная кладка горихвостки, состоявшая из 4 яиц совсем без скорлупы:
из них 3 были раздавлены (вероятно, самой же наседкой).
219
Рис. 53. Самец горихвостки, 14. VI 1980 г. Фото А.С. Гилязова
Птенцы горихвостки вылупляются на 12-й день после откладки послед-
него яйца и покидают гнездо на 10-14 день, в зависимости от размеров
выводка и, следовательно, количества получаемой пищи. Птенцы выле-
тают с конца июня по начало августа, причем основная масса поднимает-
ся на крыло в первой и второй декадах июля.
Корм птенцов и взрослых включает ягоды вороники, черники, а глав-
ным образом состоит из беспозвоночных - пауков, веснянок, короедов
(личинок и имаго), жужелиц, листоедов, мягкотелок, стафилинид, долго-
носиков, щелкунов и их личинок, мух и их личинок, комаров, пилиль-
щиков, наездников, муравьев, бабочек и их гусениц. Насекомых гори-
хвостки ловят как на земле, так и в воздухе, взлетая с земли или с веток.
Основные причины смертности горихвосток в заповеднике - неблаго-
приятная погода и хищники, куница, горностай, норка: проверяя дупла
на своем участке, они ловят самок на гнездах, уничтожают яйца и птен-
цов.
Размеры 4 самцов: длина 141-150 мм, крыло 74-82, хвост 56-65 мм;
масса тела 13,5-16,5 г, в среднем 14,8; 4 самок соответственно - 140-146,
75-79 и 54-62 мм; масса тела 11,1 - 17,3, в среднем 13,9 г.
Луговой чекан - Saxicola rubetra (L.)
Луговой чекан редок и встречается только в юго-западной части
Кольского полуострова.
Б Лапландском заповеднике впервые наблюдался 12. V 1964 г. О. Се-
меновым-Тян-Шанским и Ю. Гаевым; с тех пор зарегистрировано всего
220
25 особей на протяжении 8 сезонов. Дата первой встречи, в среднем за
8 лет, 22 мая (12. V 1964 г. - 7. VI 1979 г.).
В гнездовый период луговой чекан встречается на зарастающих гарях,
в зарослях ерника, ивы, вейника в речной пойме, на болотах с редкими
березками и елями. При всей редкости встреч чекана в заповеднике он
несомненно гнездится: 20. VII 1981 г. в долине р. Чуна у ерниково-богуль-
никового болота мы встретили выводок, пару взрослых птиц и двух слет-
ков с недоразвитыми рулевыми, а 7. VIII 1983 г. на Чунозерской усадьбе
был пойман залетевший на веранду слеток.
Каменка - Oenanthe oenanthe (L.)
Каменка на Кольском полуострове встречается повсеместно.
Средняя дата прилета в Лапландский заповедник - 24 мая (8. V 1968 г. -
6. VI 1972 и 1985 гг.; 41 год), при температуре воздуха +5,6; дата отлета -
2 сентября (20. VIII 1975, 1978 гг. - 15. IX 1958 г.; 26 лет, температура +8,8°).
Каменка прилетает позднее других мелких птиц и отлетает сравнительно рано
по причине большой длины ее пролетных путей.
Гнездится на различных типах тундр - лишайниковых, кустарничко-
вых, злаковых, каменистых - предпочитая лаги, седловины, пересечен-
ные участки; нет ее только на каменных полях на вершинах гор; живет
в лесном поясе, придерживаясь сухих каменистых редин, гарей и выру-
бок, где есть поваленные стволы, камни. Гнездится спорадично, числен-
ность невысока, в июне на горных тундрах 0,5 - 2,0 пар/км2, в лесном по-
ясе не более 0,5 пары, но в отдельные годы локальная численность дости-
гает 8 пар/км2. Численность довольно стабильна, но в 1970,1982 и 1983 гг.
наблюдалось ее повышение в 2-3 раза, вероятно в связи с теплой весной.
Гнездо помещается под выворотнем, обгорелым корнем, камнем,
иногда в ’’синичнике”, на высоте 1,5 м. Каркас гнезда состоит из травя-
нистых стеблей, выстилка - из шерсти копытных и перьев. Откладка
яиц происходит в конце мая - начала июня. В найденных 4 гнездах
было 5, 6, 6 и 7 яиц; они бледно-голубые, сходны по окраске с яйцами му-
холовки-пеструшки. Слетков встречали с первых чисел июля, нераспав-
шиеся выводки самое позднее 25. VIII 1963 г.
По содержанию семи желудков и визуальным наблюдениям за кормеж-
кой пища каменок включает немного ягод брусники и вороники, а в ос-
новном состоит из пауков и насекомых - жуков, в том числе долгоноси-
ков (Otiorhynchus), усачей (Monochamus), щелкунов (Elateridae), перепон-
чатокрылых - наездников, шмелей, ос и муравьев, комаров-долгоножек
и гусениц бабочек.
При возвратах холодов в июне 1977 и 1983 гг. найдено было несколько
погибших от истощения взрослых каменок и наблюдалась откочевка
птиц в долины рек, где они встречались стайками, до десятка. 8. VII
1974 г. в долине Чуны 6 птенцов в гнездах были заражены личинками
мух, причем на голове одного из них было 8 личинок (Н. Караваев).
Размеры двух самцов: длина 132-159, крыло 98-99, хвост 53-60 мм,
масса тела одного 25,5 г; двух самок соответственно - 154-160, 93-95 и
53-58 мм, масса тела 26,1 и 28,1 г.
221
Черный дрозд — Turdus merula L.
В XX в. в Западной Европе черный дрозд постепенно стал обычной го-
родской птицей, что способствует его расселению в северном и восточ-
ном направлениях.
В Лапландском заповеднике впервые наблюдался О.Семеновым-Тян-
Шанским 27.V 1938 г., на Нявка-тундре, всего 5 птиц, без уверенности в
определении; затем 12.V 1964 г. одиночный дрозд замечен и добыт на
Чунозерской усадьбе О.Гаевым; приблизительно в то же время черный
дрозд залетал и на Айновы острова (Коханов, Скокова, 1967). Позднее
черные дрозды в одиночку или парой встречались обычно во время ве-
сенней миграции, что дает возможность определить среднюю дату их при-
лета за 11 лет - 28 апреля (31.III 1979 г. - 22.V 1982 г.); он появляется
раньше других дроздов, иногда даже до начала таяния снега. Летом
встречены 17.VI 1973 г. и 15.VII 1986 г., в последнем случае в березовом
криволесье на южном склоне Чуна-тундры наблюдали слетка с остатками
пуха на голове под опекой взрослой птицы, доказательство нерегуляр-
ного гнездования этого вида в регионе. Осенью отмечено два залета:
10.Х 1966 г. после обильного снегопада на Чунозерской усадьбе появился
черный дрозд; пользуясь подкормкой, он жил на усадьбе до 20.Х, а на
следующее утро на берегу озера нашли только его перья: вероятно, он
замерз (ночью было -12°), а труп расклевали сороки. 28.Х 1969 г. лесники
А.А. Якуничев и И.С. Некрасов видели близ кордона Островское озеро
стайку черных дроздов, вероятно выводок.
Почти всех дроздов встречали возле жилищ, на усадьбе заповедника,
у кордонов и сторожевых изб, раз даже в г. Мончегорске (11-14.IV 1978 г.);
кормились они главным образом у птичьих кормушек или объедками.
Масса тела самца 102 г, самки 91,2 г (длина крыла 129 мм, хвост 109 мм).
Белозобый дрозд - Turdus torquatus L.
Гнездится в горах Европы и Передней Азии, а на Кольский полуостров
только изредка залетает. В середине прошлого века Лильеборг (1852)
’’нашел Turdus torquatus в изобилии около Шурецкой губы на Мурмане и,
вероятно, на основании этого наблюдения и Мела считает его за обитате-
ля Мурманского побережья” (Плеске, 1887, с. 177). Шурецкой губы нет на
современных картах, и авторы позднейших сводок отождествили ее с
Нокуевской губой (Портенко, 1954, с. 238). В 1957 г. А.Кищинский (1960,
с. 122-212) нашел одно гнездо в скалах близ Териберки, a 10.V 1964 г.
залетного самца видели на Айновых островах (Коханов, Скокова, 1967).
8-18.V 1972 г. в Лапландском заповеднике на Чунозерской усадьбе
жил залетный самец белозобого дрозда, он кормился на обтаявшем скло-
не, как все дрозды энергично разбрасывая прелый березовый лист
(рис. 54).
Директор зоопарка г. Галле доктор Р.Брезеке, приехавший в Мурманск
туристом, сообщил нам, что 5.IX 1980 г. в тундре над восточной окраиной
города видел белозобого дрозда.
222
Рис. 54. Белозобый дрозд, 9. V 1972 г. Фото О.И. Семенова-Тян-Шанского
В Финляндии белозобый дрозд редок и встречается только у границы
с Норвегией; в заповеднике Малла в 1979-1980 гг. гнездилось 6 пар, а
общая численность во всей стране оценивалась в 40 пар, не более (Jarvi-
пеп и др., 1982).
Темнозобый дрозд - Turdus ruficollis Pall.
Этот дрозд гнездится в Западной Сибири до верховья Печоры. Заре-
гистрированы залеты этого вида в Архангельскую область (Гладков,
1954), южную Финляндию (Bergman, 1963) и на Кольский полуостров, на
Хибины (Новиков, 1952) и под Мурманск, где один был добыт 15.VIII
1921 г. (Шибанов, 1927).
Рябинник - Turdus pilaris L.
На Кольском полуострове гнездится повсеместно, до Мурманского по-
бережья. В западной части гнездового ареала рябинник зимует, из Во-
сточной отлетает в Западную и Южную Европу. В годы обильного урожая
ягод рябины большие стаи рябинников зимуют даже в Шведской Лаплан-
дии (1945 г.; Карри-Линдал, 1984). При соответствующих условиях воз-
можны зимовки их и у нас: по крайней мере 8.1 1981 г. в Мончегорске
И.Паракецов видел 3 рябинников, а 6.1 V 1981 г. возле усадьбы заповедни-
ка А.Гилязов наблюдал одного; там же одного видели и 25.XI 1963 г.
223
Появляются рябинники, как и другие дрозды, в начале мая, во время
интенсивного таяния снега. Средняя дата прилета за 34 года - 12 мая
(29.IV 1937 г. - 25.V 1980 г.). Прилетные рябинники обычно встречаются
по одиночке, реже стайками по нескольку птиц. Самые поздние даты
прилета возможно объясняются запозданием в регистрации первых
особей в связи с малочисленностью этого дрозда. Отлет лучше выражен,
чем прилет, ввиду сосредоточения дроздов на ягодных участках, где
птицы собираются сотнями. При наличии корма рябинники держатся
вплоть до установления снежного покрова. Средняя дата последних
встреч дроздов (всех видов вместе), в среднем за 46 лет, - 11 октября
(22.1X 1973 г. - 24.Х 1981 г.).
Для рябинников характерны значительные колебания численности по
годам, подобные инвазиям у других растительноядных птиц. За послед-
ние 25 лет (с 1960 г.) рябинники совсем не встречались в заповеднике в
сезон гнездования в 1963, 1965-1967, 1973-1975, 1980, 1983-1984 гг.: в эти
годы их видели только во время весенней и осенней миграции. Колеба-
ния эти нерегулярны; вероятно, они зависят как от урожайности ягодни-
ков, так и от хода весны. Наблюдения заповедника не обнаружили у ря-
бинника тенденции, ни к снижению, ни к повышению численности. Мало-
численность рябинника на гнездовьях в лесотундре северо-восточной
части Кольского полуострова в конце 70-х годов в сравнении с началом
века (Михайлов и др., 1984), вероятно, связана с инвазиями. В сезон гнез-
дования в заповеднике рябинники чаще встречаются в разреженных ель-
никах у верхней границы леса, где черника и вороника плодоносят
лучше, а также в березняках речных долин и на заростающих гарях. Всю-
ду он количественно уступает белобровику и певчему дрозду.
Пение слышно с прилета до конца июня. Строительство гнезд начина-
ется с середины мая. Из 15 известных нам гнезд 10 находились на ветках
елей, 3 - в развилке березового ствола на высоте 1,5-7 м, 2 - на земле,
одно под елкой, другое под камнем. Как и другие дрозды, рябинник
предпочитает гнездиться около воды.
Каркас гнезда строится обычно из стеблей травянистых растений,
иногда с добавлением сухих веток, мхов (Fontinalis, Pleurozium), ягеля;
лоток сложен из тонких былинок и водяного мха. Гнездр всегда укрыто
сверху. Размеры их: наружный диаметр 140-150 мм, внутренний - 90-
120, высота 90-110, глубина лотка 50-90 мм.
Самки начинают нестись с 20.V (1949), в поздних кладках - в начале
июля. Размер кладки 4-8 яиц, в среднем 5,0 (п = 11). Птенцы вылупляют-
ся с середины июня до конца июля. Уже с конца июля выводки стяги-
ваются к долинам рек или на ягодники в березовом криволесье.
В заповеднике описаны три гнездовые колонии рябинников, в каждой
не более 3 гнезд, с расстоянием между ними не более 200 м. Вблизи гнез-
да рябинники беспокоятся, трещат, пикируют на человека и, как прави-
ло, обливают его экскрементами.
По содержимому пяти желудков и визуальным наблюдениям питание
рябинников в гнездовый период смешанное. Из животных кормов преоб-
ладают жуки-жужелицы, долгоносики, личинки щелкунов, водолюбы,
224
встречаются также гусеницы бабочек и муравьи. Растительные корма
состоят из ягод вороники, брусники, черники, коробочек кукушкина
льна, однажды 7. VI найдена хвоя ели. Значение ягод возрастает в конце
лета, с поспеванием черники и брусники, в период подготовки организма
к миграции.
Причины естественной смертности малоизвестны: находили остатки
взрослых птиц, ставших жертвой хищной птицы, а также погибших во
время летних снегопадов.
В 1976 и 1977 гг. найдены гнезда с кладками, брошенными в период
холодов.
Размеры одного самца: длина 255, крыло 146, хвост НО мм, масса тела
97,5 г; двух самок соответственно - 250-267, 140-143 и 100-109 мм, мас-
са тела 96 и 103 г.
Белобровик - Turdus iliacus L.
Местное название: саамское - раштесь (для всех дроздов оди-
наково).
На Кольском полуострове живет всюду, где имеется древесная расти-
тельность. По долинам рек и ручьев, вдоль морского побережья, придер-
живаясь березового криволесья, освоил и тундры на северо-востоке по-
луострова (Шутова и др., 1982; Михайлов и др., 1984).
Прилет белобровика отчетливо выражен и обычно совпадает с появле-
нием первых проталин на склонах, берегах рек и озер. Первыми появля-
ются одиночные особи, через несколько дней подлетает основная масса,
стайками до нескольких десятков птиц, но в теплые весны и белоброви-
ки прилетают дружно. Средняя за 46 лет дата прилета - 8 мая (16.IV 1976 г. -
21.V1945 г), при средней температуре воздуха +2,1° и среднем квадрат-
ном отклонении даты прилета ±7,6. В апреле белобровик прилетает редко
(только в 1950,1960, 1975 и 1976 гг.). Время прилета зависит от температу-
ры апреля, коэффициент корреляции г= -0,55 ± 0,11.
Отлет прослеживается хуже, но как правило, белобровики исчезают до
или с выпадением первого снега. Средняя дата отлета за 42 года 6 октяб-
ря (21 .IX 1966 г. - 22.Х 1987 г.), при температуре +2,4°.
Белобровики поют с момента прилета, но первое время не все самцы
активны. Интенсивность пения постепенно нарастает к концу мая -
первой половине июня и продолжается до середины августа, а иногда
пение слышали и до конца сентября (14.IX 1962 г., 29.IX 1966 г.). Помимо
белобровика, осенью поют, даже в большей мере, веснички; это явление
не имеет отношения к размножению и связано, вероятно, с обучением
видовой песне молодняка. Демонстративная песня белобровиков в запо-
веднике совсем не похожа на песню птиц того же вида на севере Запад-
ной Сибири. Да и в заповеднике, в долине р.Чуна, мы различали, по край-
ней мере, три напева; при этом песня части белобровиков при устье
р.Ташким занимает промежуточное положение между песнями самцов с
устья Чуны и с долины Сылпуая; в то же время здесь встречаются и сам-
цы с другими напевами.
В лесных биотопах белобровик, весничка и юрок фоновые виды. Рас-
15. Зак. 1023 225
пределяется он по лесному поясу неравномерно, высшая плотность
наблюдается в смешанных лесах по долинам рек с развитыми подростом
и кустарничковым покровом, на старых вырубках в елово-березовых
редколесьях с обилием можжевельника, на зарастающих гарях; в общем
белобровик предпочитает те типы лесов, где хорошо развит подлесок.
В смешанных спелых лесах южной части заповедника численность бело-
бровиков составляет в начале июня 1-3, в долинных елово-березовых ле-
сах - 1-6 пар/км2. Послегнездовую численность белобровиков на линей-
ных маршрутах нельзя определить из-за стягивания выводков к густым
зарослям в долинах рек, озер и на вырубках. Локальная плотность в
местах скопления в июле-августе достигает сотни особей на 1 км2.
Строительство гнезд белобровики начинают с середины мая; недостро-
енные находили, самое раннее, с 17.V 1983 г., но часть гнезд строится во
второй декаде мая. В выборе мест для гнездования белобровики непри-
хотливы: из 176 гнезд 43 располагались на деревьях (27 - на елях, 5 - на
соснах, 5 - на березах, 3 - на ивах, 3 - на можжевельнике); 41 - на кор-
нях ветровальных хвойных деревьев; 34 - на земле, 19 - в березовых
кустах; 18 - на торце сломанных хвойных стволов и в берестяных труб-
ках трухлявых берез; 7 - на стволах упавших деревьев; 11 - на прикор-
невых кочках; 2 - на стволах деревьев, нависших над водой; 1 - на тор-
це бревна, прислоненного к стене избы. В пределах каждого типа спосо-
бы укрепления гнезда, его маскировки и защиты от непогоды варьируют.
В выборе мест расположения гнезд белобровики проявляют пластич-
ность. Осенью 1970 г. ураган повалил много деревьев, местами более по-
ловины всего верхнего яруса: после того, в 1975-1980 гг., большая часть
гнезд помещалась на выворотнях. Белобровик гнездится ниже других
дроздов: 48% его гнезд находились ниже 1 м, 20% - на высоте 1-2 м, 19%
- на земле. Наибольшая высота расположения гнезд на торцах стволов -
до 3 м, на деревьях - до 4 м. В начале сезона, пока земля местами еще
покрыта снегом, белобровики помещают гнезда выше, а чем позже они
строятся по ходу весны, тем ниже находятся гнезда (такой же переход
к гнездованию белобровиков на земле мы наблюдали и в 1974 г., в пой-
ме р. Пура, Ямало-Ненецкий автономный округ, но там во время строи-
тельства первых гнезд земля была затоплена половодьем).
Гнездовой материал довольно однотипный, чаще всего гнезда строят-
ся из прошлогодних стеблей травянистых растений, в том числе вейника,
различных осок, к ним добавляются ягели, мхи (Fontinalis, Hylocomium,
Pleurozium), береста, листья; изредка каркас сооружается из сухих веток.
Количество гнездового материала сильно варьирует в зависимости от
расположения гнезда. Особенно громоздки гнезда, построенные на вет-
ках деревьев, корнях, стволах; гораздо меньше материала требуется для
гнезд на земле, в полудуплах, когда жесткий каркас становится не
нужен, так как само расположение гнезда в какой-то мере обеспечивает
его термоизоляцию и механическую прочность. Около 7% гнезд обмазы-
ваются землей (не все наблюдатели проверяли жесткость гнезд, так что
их может быть и больше). Найдено 11 гнезд белобровика, надстроенные
на старое гнездо певчего дрозда или белобровика.
226
Размеры гнезд: наружный диаметр 110-200 мм, внутренний 70-110,
высота 60-216, глубина лотка 40-85 мм.
Откладка яиц происходит со второй декады мая до третьей декады
июля. По расчету наиболее ранняя кладка начата 13.V 1976 г., птенцы вы-
летели 9.VI; самая поздняя - 24.VII, при этом птенцы оставались еще в
гнезде 18.VIII. Весь период кладки более 70 дней. Как известно, часть бе-
лобровиков делает за лето два выводка (Новиков, 1956; Мальчевский,
1959; Кищинский, 1960; и др.), что подтверждается и материалами Лап-
ландского заповедника (Владимирская, 1948). Вторые кладки появляют-
ся у нас в середине июня, но только при благоприятных погодных усло-
виях.
Полная кладка содержит от 2 до 6 яиц, в среднем 4,6 (п = 151). 6 яиц бы-
ло в 22 кладках, 5 - в 72, 4 - в 39, 3 - в 13, 2 - в 5. Продолжительность
инкубации, считая от ©ткладки последнего яйца, 10-13 дней. Птенцы по-
кидают гнездо через 10-14 дней после вылупления, но в случае беспо-
койства некоторые уходят и на 8-9 день. Массовый вылет птенцов пер-
вого цикла наблюдается во второй половине июня, второго цикла - во
второй половине июля - начале августа. Поведение взрослых у гнезда у
разных пар неодинаково, одни своим поведением обнаруживают его, бес-
покойно цикают, щелкают клювом, самые агрессивные пикируют и даже
обливают пометом, как рябинники; другие доверчиво подпускают к гнез-
ду и при осмотре его крутятся рядом; третьи осторожны, при подходе че-
ловека отлетают в сторону, подобно дерябе или певчему дрозду. Разли-
чия в поведении, скорее всего, определяются прежним опытом. Белобро-
вики беспокоятся также при появлении ястребиной совы, вороны, кук-
ши, гадюки. Отгонять врага им помогают другие дрозды, юрки, веснички,
чечетки и белые трясогузки.
Питание (по содержимому 32 желудков и визуальным наблюдениям)
белобровиков, птенцов и взрослых в гнездовой период, смешанное, с
преобладанием животных кормов. Наибольшее значение имеют различ-
ные жуки (долгоносики, жужелицы, щелкуны и их личинки, короеды и
др.), затем пауки, муравьи, личинки пилильщиков, комары, бабочки и
их гусеницы, наездники, веснянки, наконец, улитки. Муравьи в июне
встречены в 63% желудков. Значение растительных кормов увеличивает-
ся весной и осенью. Среди них преобладают ягоды черники, вороники,
брусники, хвоя ели и можжевельника, семена берез, осок, листья
черники.
В заповеднике зарегистрированы случаи гибели взрослых белоброви-
ков от бескормицы при продолжительных снегопадах или дождях с по-
холоданием: в конце июня 1977 г. нашли 10 погибших птиц, в мае 1982г. -
3, в июне 1986 г. - 1. Успех размножения также определяется погодны-
ми условиями. В некоторые годы птенцы несут потери от мелких назем-
ных хищников - горностая, норки и куницы.
Размеры 21 самца и 3 самок: длина самца 192-223, в среднем 209 мм,
крыло 112-120, в среднем 117 мм, хвост 72-96, в среднем 85 мм, масса те-
ла 47,2-71,8, в среднем 60,5 г. Размеры самок соответственно - длина
194-216, хвост 72-84 мм, масса тела 62,6-64,4 г.
227
Певчий дрозд - Turdus philomelos C.L. Brehm
Певчий дрозд прилетает в заповедник несколько позднее белоброви-
ка. Средняя за 42 года дата прилета, отмеченная по первой песне, 11 мая
(1.V 1937 г. - 22.V 1961 г.), при средней суточной температуре +2,7°. Появ-
ляются певчие дрозды незаметно, а проследить их отлет еще труднее,
по причине скрытного образа жизни. Наиболее поздние встречи певчих
дроздов в заповеднике - 19.IX 1970 г. и 28.IX 1978 г.; должно быть, в сред-
нем они отлетают в конце августа - начале сентября.
Гнездовые стации певчего дрозда - разреженные смешанные леса
различных типов: сосняки с примесью ели и березы, елово-березовые ле-
са, ельники с примесью березы или сосны. По склонам они поднимаются
до верхней границы леса, но в березовом криволесье редко встречаются.
Они избегают болотистых редколесий и вообще открытых участков, будь
то гарь, тундра или болото, зато обычны в сплошных массивах леса по
речным долинам и у озер.
По сообщению Владимирской (1948), ’’певчий дрозд многочислен и ши-
роко распространен”. Теперь численность его снизилась и в смешанном
лесу долины Чуны в июне составляет 0,6-2,0 пар/км2; в середине августа,
в период учета боровой дичи, ни слетков, ни взрослых птиц здесь мы
уже не встречали.
Пары распределяются по гнездовым участкам сразу после прилета, и
с этих пор самцы поют до середины июля, в среднем за 31 год до 16.VII
(30.VI 1983 г. - 9.VIII 1964, 1970 гг.). С середины мая до середины июня они
поют регулярно, чаще в вечерние и ночные часы, иногда всю ночь напро-
лет. Песни отдельных особей заметно различаются по длине, за счет
вставки добавочных колен, некоторые из которых подражают голосам
других птиц, например фифи или кукши. В целом же песни певчих дроз-
дов менее индивидуальны, чем песни белобровиков.
Гнезда певчих дроздов, как правило, располагаются на: елях, из 45 об-
наруженных гнезд 38 построены под укрытием еловых веток, 4 - на бере-
зах, из них два открыто, на развилке ствола, 2 - на соснах и одно - на
земле. Чаще всего гнезда находятся на высоте 1-3 м, иногда выше или
ниже; максимум в 11 м от земли. В общем, певчие дрозды строят гнезда вы-
ше, чем белобровики, но занимаемые ими ярусы перекрываются. Случа-
ется, что белобровик занимает старое гнездо певчего дрозда, используя
его как фундамент (4 случая).
Гнездовой материал довольно разнообразен; внешний рыхлый слой
состоит из сухих веточек ели, различных лишайников (Cladonia rangiferi-
na, alpestris, mitis, gracilis; Usnea sp., Bryopogon sp.) и мхов (Sphagnum sp.,
Pleurozium schreberi, Hylocomium splendens, Dicranum sp., Polytrichum sp.,
Fontinalis), стеблей багульника, черники, брусники, хвоща, кусочков ело-
вой и березовой коры. Лоток выстилается спрессованной грибницей дре-
весных грибов и мхами, а верхний край - стеблями луговика и других
злаков. Размеры гнезд; наружный диаметр 150-210 мм, внутренний - 90-
105, высота 100-200, глубина лотка 70-95 мм.
Гнезда с кладками находили с 28.V по 19.VII. Полная кладка содержит
2-6 яиц, в среднем 4,4 (п = 22). У певчих дроздов, как и у белобровиков,
228
два пика размножения. Вылет птенцов из первых кладок, отложенных в
конце мая - начале июня, происходит в конце июня - начале июля, а из позд-
них, вероятно вторых, в конце июля - первой декаде августа. Певчие
дрозды, особенно самцы, осторожны и скрытны. В то время как самка на
гнезде подпускает почти вплотную, а вылетев, беспокойно кружится,
самец молчаливо держится в стороне. Демонстративное поведение, с пи-
кированием и щелканьем клювом, наблюдаются реже, чем у белоброви-
ков, обычно только против зверей и птиц, покушающихся на слетков или
гнездо. Незаметно держатся и выводки.
Питание певчего, как и других дроздов, смешанное и разнообразное.
Первое время после прилета они кормятся ягодами брусники, вороники,
коробочками кукушкина льна (26.V 1938, Г.Новиков). В июне раститель-
ные корма уже не играют большой роли, в желудках встречаются только
единичные листики брусники, ивы и побеги черники. С июля дрозды на-
чинают есть зеленые ягоды вороники. В августе значение растительных
кормов возрастает, они едят в большом количестве плоды и семена
вороники, черники, осок, берез, побеги марьянника, вороники, листья
карликовой березки. Осенью в желудках найдены семена вороники и
шведского дерена. Хвоя ели обнаружена в 7 желудках из 10 (до 17 хвои-
нок, в среднем - 8). Животные корма разнообразнее и в видовом и в ко-
личественном отношении. В 10 желудках встречены: улитка (Patula rude-
rata), паук, неизвестная личинка, тля,‘2 сенокосца, клоп, 2 жужелицы,
3 щелкуна и его личинка, 3 долгоносика (Otiorhynchus dubius), 15 божьих
коровок (Anisocalvia sp.), личинка стафилины, 2 мухи, личинка комара
(Bibio), личинка овода, 2 пилильщика (Cimbex и др.) и его личинка, наезд-
ник, 3 муравья, 6 гусениц бабочек (Argynis sp.; Geometridae). Пища птен-
цов сходная с пищей взрослых, но основу ее составляют различные бес-
позвоночные.
Смертность взрослых птиц и ее причины малоизвестны. Потенциальны-
ми врагами являются различные хищные птицы, например в гнезде
кречета под Ельнюном 1 .VIII 1975 г. найдена обклеванная тушка певчего
дрозда. Гибнут они и от возвратов холодов; так в июне 1977 г. нашли 4
мертвых певчих дроздов, а успех размножения у этого вида в тот год
снизился до 60%. Гнезда дроздов разоряют кукши (наблюдения 27.VII
1976 г., 15.VI 1977 г., 7.VI 1938 г.) и хищники семейства куньих. Много
птенцов гибнет от различных хищников и после вылета из гнезда.
Размеры 2 взрослых самцов: длина 190-192 мм, крыло 110-120, хвост
85-87 мм, масса тела 63,2-79,5 г. Размеры 3 самок соответственно - 186—
214, 95-112 и 76-83 мм, масса тела 75,1-80,5 г.
Деряба - Turdus viscivorus L.
Согласно Плеске (1887, с. 170), деряба свойствен только южной части
нашей области, до 67° с.ш. В Лапландском заповеднике наблюдался с
первых лет его существования, но изредка и не ежегодно; даже осенью
не встречали более 3 птиц одновременно. В 1930-1933, 1936, 1941, 1946—
1964, 1969-1977 и 1984 гг. не попадался ни разу.
Прилетают дерябы к первым проталинам, в первой половине мая (в
229
среднем за 5 лет 13.V), и в это время иногда держатся возле жилищ. Отле-
тают в сентябре - начале октября, самая поздняя встреча 15.Х 1980 г.
Деряба типично лесная птица и у нас распространен преимущественно
по спелым сосновым борам с примесью других пород. Из обнаруженных
в заповеднике 6 гнезд пять находились на соснах и одно на сухой ели.
Гнезда сверху всегда открытые, не замаскированные, помещаются, как
правило, у ствола, на основании толстого сука, однажды в развилке ство-
лов и однажды на развилке ветки в стороне от ствола. Высота над землей
от 4 до 14 м, в среднем 7; таким образом, деряба гнездится выше других
дроздов. Каркас строится на сухих сосновых и еловых веток и эпифитных
лишайников, с вкраплениями ягеля, очевидно для камуфляжа; лоток
выстлан сухой прошлогодней ветошью, стеблями и листьями злаков.
Структура гнезд мягкая, не сцементированная мокрой землей. Размеры:
наружный диаметр 140-200 мм, внутренний 95-105, высота 105-135,
глубина лотка 75 мм. 17.V 1977 г. в долине Чуны нашли строящееся гнез-
до, в виде кома висячих лишайников вперемежку с отдельными сухими
веточками. Сроки размножения, по-видимому, довольно сжатые: в 1977 г.
в четырех гнездах птенцы появились на свет 15, 16, 21 и 25 июня. В трех
кладках было по 4 яйца и в одном - 3. Их размеры: длина 29,0-31,5 мм,
ширина 19,5-20,5 мм. Одно из гнезд птенцы покинули на 14-15-е сутки,
другое - на 13-14-е. В другие годы слетков при взрослых птицах встре-
чали 9-24.VII. Обычно видели по одному птенцу, в одном случае - два
слетка при взрослой птице.
Успех размножения в неблагоприятный по погодным условиям 1977 год
составил 45%; в другие годы он должен быть выше.
По единичным наблюдениям деряба поедал ягоды брусники и личинок
пилильщиков.
. Деряба осторожнее других дроздов, напоминая в этом отношении охот-
ничью дичь. У гнезда или при выводке разные пары ведут себя не одина-
ково: одни держатся в стороне, изредка потрескивая, другие налетают с
трескучими звуками вплотную.
Позыв - грубое трескучее ”трр. . . трр. . . ”; песня медлительная,
свистовая, тоном ниже, чем у певчего дрозда, и без повторяющихся
’’фраз”, напоминает песню черного дрозда. Обычно перелетает по верши-
нам деревьев, не подпуская близко.
СЕМЕЙСТВО СЛАВКОВЫЕ - SYLVHDAE
Камышевка-барсучок - Acrocephalus schoenobaenus (L.)
Сто лет назад в Финской Лапландии барсучок доходил до 68-69°, но
всюду был редок; в Русской - добыт на р. Тулома близ Колы (Плеске,
1887, с. 194). С 1956 г. обнаружен разными лицами в низовьях рек Пялица,
Поноя, Воронья и на Айновых островах, где даже гнездился (Коханов,
Скокова, 1967). В Лапландском заповеднике впервые наблюдался Ю.Гае-
вым 27.VI 1960 г. на р. Чуна, потом встречен в том же районе 28.VII 1972 г.
и с тех пор отмечается ежегодно. Отсутствие наблюдений на протяжении
230
десятков лет невозможно объяснить невнимательностью научных сотруд-
ников; вероятно, как численность популяции, так и граница ее ареала
подвержены колебаниям. С 1972 г. резких колебаний численности не за-
мечено, различия в числе гнездовых пар по годам объясняются обычны-
ми флуктуациями, связанными с погодными условиями весны.
Барсучок прилетает позже всех других певчих птиц, в среднем за 16
лет 13 июня (1.VI 1984 г. - 3.VI 1973 г.), при средней суточной температуре
+9,8°. Интересна эволюция средних сроков прилета - в 70-х годах 18.VI,
в 80-х - 9.VI: создается впечатление, что барсучок в эти годы осваивал
новое место гнездования и искал короткую дорогу к нему. Места встреч
барсучка’в заповеднике ограничены долинами рек Нявка, Чуна и бере-
гами озер Румельявра,Улынчъявра и Чуны в местах, заросших ивняком,
ольхой, ерником, при устьях рек и ручьев, на берегах стариц и протоков.
Мозаичное распределение пар по долинам объясняется недостатком под-
ходящих биотопов. В долине Чуны они постоянно гнездятся в устье Чу-
ны, ее старицы в 8 км от устья и в месте впадения ее притока, Сылпуая,
еще на 14 км выше по течению. В каждом из этих пунктов наблюдали от
одного до четырех поющих самцов в разные годы.
Барсучки поют обычно до середины июля, самую позднюю песню
слышали 13.VIII 1981 г. в устье Чуны. В конце июня - начале июля они
поют интенсивно, иногда всю ночь без перерыва: так, например, 30.VI -
2. VII1973 г. самец пел с 23 ч. до 9 ч. Днем их активность снижается.
Гнезд барсучка в заповеднике не находили, но слетков, от 1 до 3 при
взрослых птицах, встречали 6.VHI 1976 г. - в долине Нявки, 7. VIII 1978 г. -
в долине Чуны, 22.VII 1978 г. - у Окуневого озерка в долине Нявки.
Парочки с агонистическим поведением видели 4.VIII 1982 г. - у протоки
из Улынчъявра и 5. VIII 1982 г. - близ устья Пельйок.
Садовая славка - Sylvia borin (Bodd.)
В окрестностях Кандалакши изредка гнездится (Бианки и др., 1960;
Коханов, 1987, с. 28).
7.VII 1969 г. в ивняке при входе в левобережную старицу р. Чуна О.С
менов-Тян-Шанский наблюдал славку, которую неуверенно определил
как садовую.
Черноголовая славка - Sylvia atricapilla (L.)
15.Х 1975 г. на Чунозерской усадьбе разбилась об окно дома самка или
молодая славка-черноголовка (О. Семенов-Тян-Шанский, А. Брагин) -
единственная встреча этой славки в заповеднике. Несколько раз ее
встречали в г. Кандалакше, а в 1975 г. в Коле (Коханов, 1987, с. 26).
Серая славка - Sylvia communis Lath.
4.VII 1973 г. на северном берегу Чунозера в лиственном лесу по ручью
Куплетского Н.Н. Скокова слышала песню серой славки.
231
Славка-завирушка - Sylvia curruca (L.)
На Кольском полуострове изредка гнездится. 22.VII I960 Ю. Гаев наб-
людал одну на Кировском аэродроме, в кустах под окном. Близ г. Канда-
лакша в 1966-1967 гг. гнездилась (Коханов, 1987, с. 28).
Весничка - Phylloscopus trochilus (L.)
На Кольском полуострове достигает Арктического побережья. В за-
поведнике веснички появляются во время быстрого сокращения снежно-
го покрова. Пролет проходит незаметно. Вначале появляются одиночные
особи, самцы, только пробующие голос (поют не активно и лишь часть
песни). В следующий десяток дней прилетает основная масса особей.
Средняя дата прилета за 42 года - 22 мая (12.V 1964, 1977 гг. - 2.VI
1969 г.), при переходе кривой суточных температур через +5,1°. Весной
редко можно увидеть стайку в десяток птиц (1.VI 1987 г.). Осенние стаи
образуются в конце июля - начале августа, их обычный размер 10-19
птиц, редко больше. Средняя дата последней встречи за 47 лет - 4 сентяб-
ря (19.VIII 1933, 1970 гг. - 17.IX 1948 г.), средняя температура этого
дня +8,4°.
Весничка одна из самых многочисленных птиц заповедника, что облег-
чает точную регистрацию дней прилета и отлета. Дата прилета веснички
отчетливо коррелирует с первым кукованием кукушки (г = +0,55 + 0,11)
и средней температурой мая (г= -0, 61 ± 0,10); среднее квадратическое от-
клонение даты прилета = ±5,1; следовательно, время прилета мало откло-
няется от определенной календарной даты. Осенью весничка ведет себя
иначе. В среднем, одновременно с ее отлетом, на вершинах горных тундр
выпадает первый снег, и это дает основание предполагать, что ее отлет
стимулируется похолоданием воздуха. На деле корреляция между дата-
ми первого снегопада и отлетом веснички близка к нулю, г = +0,16 + 0,15,
при этом амплитуда отклонений дат первого снегопада (о =±13,3)i почти
в два раза больше, чем амплитуда отлета веснички (о = ±7,6). Корреля-
ционная связь между сроками отлета и температурой предшествующего
ему месяца августа примерно вдвое ниже соответствующей корреляции
во время прилета: г = -0,28 + 0,14. Знак минус показывает, что теплый
август не задерживает отлет, а ускоряет его, холодный август, напротив,
задерживает на месте гнездования. Вероятно, объяснение этого парадок-
са в том, что сама по себе температура на отлет не влияет, но теплая по-
года перед началом миграции дает птице возможность раньше накопить
необходимый для перелета энергетический ресурс, что и ускоряет мигра-
цию. Как мы видели, температура для отлета веснички на 3,3° выше, чем
дня прилета, откуда следует ,что улетает она задолго до наступления
неблагоприятных для жизни условий.
Весничка заселяет все типы лесов, поднимаясь по горным склонам до
березового криволесья, встречается и в тундре, в ивняках. Предпочтение
отдает разреженным смешанным лесам, богатым подростом и кустарни-
ком, по берегам озер и рек, по долинам ручьев, а также подросту берез-
няка с примесью хвойных по гарям; реже заселяет однородные сосновые
232
боры и густые ельники, но полностью избегает только открытую горную
тундру. Весничка - самая обычная фоновая птица заповедника, по чис-
ленности уступает только юрку и, в некоторые годы, чечетке. Многолет-
ние наблюдения не обнаружили заметных изменений ее численности.
Плотность гнездовых пар за последние 10 лет в первой половине июня в
смешанных лесах долины Чуны составляет 7-12 пар, в приречных елово-
березовых лесах - 12-21 пар/км2, в сосновых борах, елово-березовых ле-
сах и березовом криволесье - 1-5 пар/км2. В первой половине августа
в прибрежных зарослях ивы, ольхи и березы, куда стягиваются осенние
стаи, локальная численность достигает 800 особей/км2.
Веснички начинают петь через несколько дней после прилета, интен-
сивность пения нарастает к началу мая, снижается к концу июня -
началу июля, но они продолжают петь в течение всего июля. ’’Осеннее
пение” весничек, смысл которого по одной из версий заключается в обу-
чении молодых птиц пению, продолжается до конца августа, иногда да-
же в первой декаде сентября (4JX 1978 г., 5.IX 1968 г., 6JX 1940 г.). Наибо-
лее активно веснички поют утром и вечером, а ночью их не слышно. Во
время затяжного снегопада 20-25 июня 1977 г. веснички продолжали
петь, даже когда другие активные певцы, юрки и дрозды, замолкали.
Брачные полеты и спаривание наблюдались с 4.VI по 2.VII, а гнездо-
строение с 8.VI по 13.VI; готовые, но без яиц гнезда находили 7, 15, 16 и
26 июня. Веснички строят гнезда на земле, обычно на поляне, на кочкар-
ном болоте по опушке, на березовом склоне, как правило на сыроватых
местах. Само гнездо иногда ничем не защищено, не замаскировано и
устроено на ровной земле, но чаще располагается у ствола березы, ивы,
елочки, под кустом можжевельника, березы, рябины, черники, ворони-
ки, голубики, в углублении под кочкой, в норе лемминга, под елкой.
Гнездо представляет собой шарообразный или чуть продолговатый
шалаш размерами 92-120 на 93-100 мм, с диаметром входа 30-50 мм,
таков же и внутренний диаметр. Вход обычно один, но из найденных
нами 76 гнезд одно имело сквозной проход с юго-запада на северо-восток
(птицы пользовались преимущественно последним). Ориентация входа
самая различная, в зависимости от окружающего рельефа. Каркас спле-
тается или из одних стеблей осок и злаков (Deschampsia flexuosa и др.),
или из мхов (Polytrichum sp. Dicranum sp., Pleurozium schreberi, Hylocomium
splendens), или из лесного хвоща (Equisetum silvaticum), или из смеси трав
и мхов. Внутри оно выстилается перьями различных птиц, чаще всего бе-
лыми перьями куропаток, которые в это время линяют, также глухарей,
рябчиков, сов и певчих птиц. Перьями выкладывается вся внутренняя
поверхность ’’шалаша”, включая и свод, но иногда их бывает мало или
совсем нет, и тогда выстилка состоит из одной ветоши. Время кладки
яиц с 10.VI до 16.VII. Полная кладка содержит 3-9 яиц, в среднем 5,7
(п = 71). В отдельных гнездах было:
Число яиц 3 4 5 6 7 8 9
Число кладок 3 9 32 11 13 2 1
Размеры яиц: длина 15-16, ширина 12,8-13 мм, масса 1,1-1,3 г. Птенцы
появляются на 12-14-й день насиживания, вылетают на 14-15-й день, в
233
сроки между 8.VII (1983) до 12.VIII (1981). Из 44 гнезд, судьба которых
была прослежена, 7 разорены наземными хищниками, 2 - полевками и
3 - брошены (из них 2 вследствие снегопада); средний успех размно-
жения 70%.
Часть птенцов погибает после вывода от заражения их личинками па-
разитирующих мух (Trypocalliphora lindneri): 7.VIII 1981 г. в гнезде на бе-
регу Нявки из 4 оперенных птенцов 3 были с личинками (2-5 на каждом);
21.VII 1978 г. в долине Чуны из 3 слетков два не могли летать, а один
взлетел с трудом - из-за поражения тканей личинками.
Количество слетков в выводках, которых взрослые еще кормят, в
среднем 3,3 (п = 11), тогда как вылетает из гнезда в среднем по 5,0 птен-
цов (п = 32). Семьи некоторое время держатся вблизи гнезд, а позднее
стягиваются к густым зарослям кустарников вблизи рек и озер. Кочую-
щие стайки наблюдались самое раннее 31.VII 1979 г. у Ельявра. Слетки
удивительно доверчивы, подпускают вплотную, иногда садятся на руки
или клюют насекомых с рук (18.VII 1976, Курбыш-уай).
Взрослые ведут себя у гнезда различно. Самка вылетает, подпустив
вплотную, а потом перелетает поблизости, мало беспокоясь. Некоторые
самоотверженно нападают, пытаясь клюнуть, или нависают в порхающем
полете - так они поступают по отношению к обычным гнездовым хищни-
кам, кукше, собаке, норке, горностаю. Есть наблюдения, когда веснички
отгоняли перепелятника, неясыть, сорокопута и кукушку; при этом
весничка со скрипучим криком и, распустив крылья, наскакивает на
агрессора. 12.VII 1938 г. Г. Новиков наблюдал близ Чингльс-явра, как пара
весничек ожесточенно атаковала рыжую полевку, залезшую на ель у
гнезда. Непонятны мотивы нападения весничек на перевозчика (3.VII
1978 г., устье Чуны) и сероголовую гаичку (12.VIII 1979 г., долина Чуны).
19.VII 1970 г. А. Брагин стал свидетелем удивительной сцены. У входа
в дом сидел слеток; весничка взяла его за шиворот, пролетела с ношей
около 7-8 м на высоте 2 м и опустила на траву. Птенец при этом трепы-
хал крыльями. Возможно, матка пыталась таким способом удалить его
от опасности. Случай этот, видимо, уникальный, в литературе ничего по-
добного не встречали.
Весничка питается исключительно безпозвоночными. Судя по содер-
жимому 24 желудков, веснички во множестве уничтожают тлей (встре-
чаются в 52% желудков), затем жуков, в том числе различных долгоноси-
ков (в 23%), двукрылых, включая долгоножек, различных комаров и
мух, сетчатокрылых. 16.VIII 1937 г. в одном желудке обнаружено около
100 златоглазок (Chrysops). Часто встречаются пауки, пилильщики, наезд-
ники, но муравьев веснички явно избегают (карточки Г.А. Новикова,
лето 1937 и 1938 гг.). По визуальным наблюдениям, веснички едят бабо-
чек, личинок пилильщиков, тлей, однажды дождевого червя.
Размеры 14 взрослых самцов: длина 113-129 мм, крыло 62-71, хвост
50-58 мм, масса тела 8,1-10,5, в среднем 9,4 г; 2 самок соответственно
124-129, 67-69 и 51-52 мм, масса тела 9,0 и 9,6 г.
Теньковка - Phylloscopus collybita (Vieill.)
Плеске (1887, с. 190) включил теньковку в список птиц Кольского по-
луострова, ссылаясь на Лильеборга и Мела, нашедших ее в юго-западной
части Русской Лапландии.
Характерную песню теньковки в заповеднике слышали с первых же
лет, но довольно редко и не ежегодно. За последние 28 лет наблюдали ее
только в 1959-1961,1964-1966, 1972-1974, 1976 и 1979-1987 гг. Средняя за
21 год дата прилета - 12 мая (ЗОЛУ 1960 г. - 26.V 1938 г.), при средней су-
точной температуре +2,9*. Теньковки поют до середины июля (12.VII
1939 г., 13.VII 1965 г., 10.VII 1983 г.). Осеннее пение слышали только в
1977 г., сЗО.УШ ПО18.1Х.
В отличие от веснички теньковка предпочитает тенистые и сыроватые
высокоствольные ельники с примесью других древесных пород, развитые
преимущественно под южным и восточным склонами Чуна-тундры. Гнезд
не находили, но 24.VII 1937 г. П.С. Мальчевский встретил у побережья
Чунозера летный выводок. 8.VI1964 г. в районе Сейдлухт пели три самца,
во всех остальных случаях пели одиночки.
Таловка - Phylloscopus borealis (Vieill.)
Согласно Плеске (1887), таловка найдена в Лапландии уже в 40-х годах
прошлого века. Встречается она и теперь, но обычно только в юго-восточ-
ной части региона (Пялица; Малышевский, 1962, с. 25). В Лапландском за-
поведнике она попадается значительно реже веснички (Владимирская,
1948, с. 236). Действительно, она, как и теньковка, встречается в заповед-
нике не каждый год и только в отдельные сезоны становится обыкновен-
ной. Так, например, в 1938, 1979, 1982 и 1983 гг. пение таловок можно
было слышать в разных местах в течение всего лета. В 1982 г. в долине
Чуны на протяжении 20 км в июне-июле отмечены 4 поющих самца.
Не встречались таловкив 1930-1936, 1946-1948, 1950-1951, 1959, 1961—
1962, 1966-1975, 1977-1978, 1980 и 1984 гг. За 14 сезонов встреч лишь в
1940 и 1960 гг. таловок замечали в мае, все остальные наблюдения летние.
Возможно, это объясняется ее поздним прилетом в связи с дальностью
миграционного пути. Поют таловки до начала августа (7.VIII 1937 г.,
4.VIII 1983 г.). 2-4.VIII 1937 П. Мальчевский наблюдал летные выводки.
Содержимое 5 желудков состояло из тлей, жуков долгоносиков, мух
(в том числе Scatophaga), цикадок, ручейников, 1 пчелы (Новиков, 1952).
У самцов, добытых 19.VI и 9.VII 1938 г., семенники достигали 6 и 5 мм
в длину.
Размеры 4 самцов: длина 112-135 мм, крыло 68-72, хвост 51-55 мм,
масса тела 10,4-11,0 г; размеры одной самки соответственно 122, 66 и
47 мм, масса тела 9,5 г.
235
СЕМЕЙСТВО КОРОЛЬКОВЫЕ - REGULIDAE
Желтоголовый королек - Regulus regulus (L.)
В заповеднике королек встречается редко и далеко не ежегодно: го-
ды встреч - 1937-1939, 1949, 1965, 1968-1969, 1971, 1973, 1977-1978,
1980, 1982-1983. В зимние месяцы не наблюдался ни разу, самые ранние
встречи в конце апреля - начале мая. Средняя дата прилета за 8 лет -
18 мая (27.1 V 1983 г. - 1.VI 1980 г.); средняя дата из 7 последних встреч -
5 сентября (24.VIII 1938 г. - 27.IX 1965 г.).
Корольки сравнительно часто встречались в 1938, 1971, 1977-1978 и
1983 гг., но даже в эти годы отмечали не более двух-трех десятков осо-
бей. В заповеднике держатся в густых, спелых ельниках с примесью дру-
гих пород, по долинам рек и озер, южным склонам гор. Пение корольков
слышали с прилета до 5.VII 1938 г.; сроки эти косвенно указывают на вре-
мя гнездования. Выводки, от 2-3 до 6 слетков при взрослых птицах,
наблюдались в июле-августе (с 13.VII 1977 г. до 22.VIII 1938 г.). 13.VII
1977 г. в долине Калипсо три слетка из шести сидели кучей на ветке ел-
ки, самка здесь же собирала корм и передавала птенцам, зависая перед
ними в воздухе; при этом все не переставая пищали. Самца поблизости
не было. Осенью иногда корольки держатся в стайках вместе с гаичками.
Как правило, корольки кормятся в верхней части еловых крон.
В желудке одного королька в мае обнаружены паук и жуки. В августе
(3 желудка) корольки питались долгоножками, грибными комариками,
ручейниками, веснянками, орехотворками, сеноедами, гусеницами и
пауками; растительных остатков не обнаружено (Новиков, 1952).
Размеры 4 экз. (d ad, $ ad, d $ j): длина 83-91 мм, крыло 52-59 мм,
хвост 37-45 мм, масса тела 5,5-5,9, в среднем 5,7 г.
СЕМЕЙСТВО МУХОЛОВКИ - MUSCICAPIDAE
Серая мухоловка - Muscicapa striata (Pall.)
На Кольском полуострове северо-восточная граница серой мухоловки
совпадает с границей леса и тундры, но по долинам рек она залетает и в
тундру (Михайлов и др., 1984). (
Серая мухоловка прилетает к нам сравнительно поздно. При теплой
солнечной погоде появление птиц хорошо заметно, но стаями их не
видали никогда. Средняя дата прилета за 30 лет - 2 июня (15.V 1981 г. -
13. VI1951 г.), при средней температуре +7,5°. Отлетают в августе, отдель-
ные особи, чаще молодые, задерживаются до первой декады сентября,
самая поздняя встреча на Чунозерской усадьбе - 26. IX1963 г.
Серая мухоловка одна из обыкновенных птиц лесного пояса. Придер-
живается прогреваемых и защищенных от ветра светлых участков леса у
берегов водоемов, по опушкам леса, избегая густого древостоя. На водо-
разделах, в елово-березовых лесах по горным склонам, в березовом
криволесье не встречается.
Численность довольно резко колеблется по годам. Судя по картотеке,
236
чаще всего встречалась в 1937-1941, 1963, 1966-1969 и 1977-1984, 1986 гг.
По результатам маршрутных учетов в июне 1977-1979 гг. в смешанных
лесах долины Чуны численность держалась на уровне 1 пара/км2, в при-
речном елово-березовом лесу в 1979 г. она не была встречена. В 1980—
1987 гг. в смешанных лесах плотность колебалась от 2 до 14 пар/км2, в
приречном лесу - от 4 до 28 пар/км2. Максимальная численность наблю-
далась в 1984 г.
Серые мухоловки начинают петь сразу после прилета и распределяют-
ся по гнездовым участкам в первые же дни. 30.V 1978 г. на северном бе-
регу Чунозера самец перелетал с одного пня на другой, пел, принимая де-
монстративные позы, самка же следовала за ним; вероятно, эта пара
подыскивала место для гнезда. Брачные игры отмечались до 19.VI, песни -
до конца июня.
Из найденных 38 гнезд 5 располагались в полудуплах на мертвых ство-
лах сосен, 7 - в полудуплах на мертвых березах, 13 - на торцах обломан-
ных стволов, обычно в берестяной ’’чаще”, 3 - на выступе березового,
елового или соснового пня, 3 - на отслоившемся куске бересты или сос-
новой коры, 1 - на развилке березового ствола, 1 - на суку сосны у тол-
стого ствола, 1 - в дупле с летком 6 см, 1 - на бревне избы, 2 - на цветоч-
ном горшке на стене дома. ’’Торцовые” гнезда обычно строятся на конце
бревна, прислоненного к дереву, и нередко прикрыты сверху ветками.
Высота большинства гнезд 1-2 м, 5 - на высоте 2-3 м, одно - в 6 м и
одно на полметра от земли. Чашевидные гнезда строятся снаружи из эпи-
фитных лишайников, травинок, ягеля, мхов, коры ивы, можжевельника,
веток ели, стеблей и корней кустарничков, сосновой и еловой хвои в
различных сочетаниях, причем ягель и мхи скорее служат для маскиров-
ки. Лоток свивается из травинок, шерсти лося, оленя, лошади, с добав-
кой пуха пушицы, перьев, собачьей шерсти, пакли и тряпок. Простейшие
гнезда состоят из травы с камуфляжем из ягеля или мха; наиболее слож-
ные включают почти все перечисленные нами материалы. Нередко серая
мухоловка гнездится на старом месте повторно. Гнезда без яиц или в
процессе стройки находили 7-24 июня, кладка продолжается до первой
декады июля, но однажды П. Мальчевский и Г. Новиков нашли гнездо с
4 яйцами 31.VIII 1937 г. Полная кладка содержит 3-6 яиц, в среднем по
4,2 (п = 30). Птенцы покидают гнезда на 12-14-й день. Слетки при взрос-
лых наблюдались между 2.VII 1980 г. и 19.VIII 1941 г. Серые мухоловки
держатся у гнезда незаметно и подпускают к нему вплотную. Вспугнутые
отлетают скрытно, над самой землей, причась за ствол. У гнезда с птенца-
ми или слетков они беспокоятся больше, даже бросаются на наблюдателя
(10.VII 1980 г.), а 9.VII 1978 г. серая мухоловка атаковала у гнезда кукшу.
Охотятся серые мухоловки, высматривая летящих или ползающих на-
секомых с наблюдательного поста, находящегося на коряге или сушине,
откуда обзор лучше, чем с живого дерева. Содержимое их желудков
включает веснянок, поденок, жуков, в том числе усачей и щелкунов,
мух, комаров, наездников, ос, шмелей, муравьев, сеноедов, ручейников,
бабочек, а также ягоды вороники и брусники.
Поскольку серые мухоловки ловят добычу преимущественно в возду-
237
хе, они больше других насекомоядных птиц страдают от неблагоприятной
погоды. При продолжительном ненастье и похолодании они перебираются
поближе к жилью или водоемам.
Размеры трех взрослых самцов: длина 139-145, крыло 89-90, хвост
62-63 мм, масса тела 15,3-16,8 г; трех самок соответственно - 133-165,
89-90, 60-63 мм, масса тела 14,1-17,4 г.
Мухоловка-пеструшка - Ficedula hypoleuca (Pall.)
Гнездится по всей лесной части Кольского полуострова. Окольцован-
ных в заповеднике птиц отлавливали на пути пролета в Финляндии и
Польше.
Средняя дата прилета за 38 лет - 18 мая (7.V 1949, 1977 гг. - 29.V
1970 г.), при средней температуре воздуха +4,3°. Прилетных стай пестру-
шек не видали ни весной, ни осенью. Отлет происходит в августе, а самые
поздние встречи датируются 13.IX 1977 г. и 14.IX 1982 г.
Мухоловка-пеструшка в Лапландском заповеднике обычная на гнез-
довье, но все же немногочисленная птица. Учеты, начатые в 1977 г.,
показали тенденцию к росту численности с 1981 г. Плотность гнездовых
пар в июне в смешанных лесах по долине Чуны составляла раньше 1-6
пар/км2, в приречном елово-березовом лесу 0-1,4 пар/км2, а в 1982 г.
4 пары/км2.
Любимые стации - спелые смешанные леса (сосна, ель, береза) с раз-
реженным древостоем по долинам рек и озер. Тяга к водоемам, вероятно,
объясняется тем, что береговая зона создает дополнительный источник
кормов насекомоядным птицам. В елово-березовых лесах по речным до-
линам она гнездится реже, еще реже встречается в елово-березовых ле-
сах по склонам, в чистых борах; молодняков на гарях и вырубках раз-
ных лет давности, березового криволесья, а тем более горных тундр
избегает.
Охотно селится в гнездовых ящиках для мелких птиц, их заселенность
в смешанных лесах колеблется от 20 до 50%. В дуплянках, развешанных
в 1966 г. по границе бора с вырубкой, пеструшки ни разу не гнездились,
в то время как горихвостки занимали до 40% их. Интересно, что при на-
личии в северной тайге множества фаутных дуплистых деревьев пест-
рушки предпочитают гнездится в искусственных гнездовьях.
Весной первыми прилетают самцы и при хорошей погоде сразу начи-
нают петь, каждый обычно у выбранного им дупла. В поисках подходяще-
го места для гнезда самец заглядывает во все дупла на избранном им
участке. 25.V 1982 г. А. Брагин наблюдал в долине Калипсо, как самец
пеструшки налетел на сероголовую гаичку, хозяина дупла, садился на
леток, но улетел, вероятно увидев насиживающую самку. 2.VI 1971 г. в
подобном облете участвовала и самка: самец с ’’жиканьем” залетел в
дупло, следом за ним самка; он срезу вылетел и сел рядом, а она пробыла
в гнезде с полминуты, после чего обе птицы улетели (Брагин).
Гнездостроение наблюдается с середины мая до середины июня, редко
позже. По литературным сведениям, гнезда строят только самки. По на-
238
шим наблюдениям, самец, выбрав дупло, заталкивает в него ворох сосно-
вой коры, березовых листьев, бересты еще до подлета самки. 8.VI 1977 г.
в гнездовом ящике у берега Ельлухт обнаружен мертвый самец пеструш-
ки: затаскивая кусок бересты размером 13x4,5 см, он закрыл леток отва-
лившейся частью ветхой стенки дуплянки и оказался в заточении. Рабо-
та самца над гнездом, вероятно, имеет ритуальный характер. В некото-
рых гнездах строительство на этой стадии и прекращается, возможно, от
того, что самец не смог привлечь к нему самку. Для мухоловок-пестру-
шек характерно использование сосновой коры в качестве гнездового ма-
териала, иногда с добавлением бересты, коры и листьев ивы, можжевель-
ника, листьев папоротников. Основа некоторых гнезд состоит из березо-
вых листьев или стеблей злаков и веток. Лоток нередко представляет
просто чашу в ворохе сосновой коры, березовых листьев, или же он спле-
тается из травы, шерсти копытных, но никогда не бывает выстлан перь-
ями. Если пеструшка занимает дупло после другой птицы, то старый гнез-
довый материал хотя бы частично перекрывается новым, особенно в
лотке.
В естественных условиях гнезда чаще строятся в дуплах большого
пестрого или трехпалого дятла (10 случаев из 13), другие во впадине на
месте выпавшего сука (3 случая). Диаметр летка 3-5 см, высота его над
землей 2-8 м. Откладка яиц происходит с 27.V по 30.VI. Кладка содержит
3-8 яиц, а в среднем 5,7 (п = 306). Размеры яиц: длина 16,6-19,4, ширина
12,3-14,6 мм, масса 1,2-1,8 г.
Неблагоприятная погода нередко вынуждает пеструшку сделать пере-
рыв в откладке яиц или насиживании, и это создает условия для появле-
ния смешанных кладок из яиц разных видов дуплогнездников. Пеструш-
ка способна выкормить птенца горихвостки, но в одном случае не выси-
дела птенца из яйца сероголовой гаички. В случае гибели насиживающей
самки самец мухоловки остается возле гнезда (наблюдение Гилязова,
1981): вероятно, он может выкормить птенцов и один, по крайней мере
такой случай имел место в июне 1979 г. в ФРГ (Hellmich, 1981).
Птенцы вылетают с конца июня по 22.VII 1978 г. (самый поздний срок).
Как показали результаты мечения, вторых кладок пеструшек у нас не
бывает. Успех размножения колеблется от 43% в 1977 г. до 89% - в 1980 г.
Основные причины гибели яиц и птенцов - погодные условия, гнездовые
хищники (горностай, рыжая полевка, возможно, дятлы), мухи Trypocal-
liphora. После возврата холодов не раз находили погибших взрослых
птиц, в том числе насиживающих самок в гнездах.
Мухоловка-пеструшка, как и серая, исключительно насекомоядная
птица. Состав корма у взрослых и птенцов совпадает, он включает пау-
ков и их коконы, различных жуков и их личинки, наездников, шмелей,
слепней, мух-журчалок, комаров и муравьев.
В заповеднике окольцованы в гнездах 1569 птенцов, из них 3 самки в
дальнейшем гнездились на расстоянии 0,2, 1,5 и 22 км от места рождения.
Из 155 окольцованных взрослых самок отловлены на гнездах повторно 20
на следующий год - 13, через 2 года - 5, через 3 года - 1, через 4 года - 1.
Наибольший зарегистрированный возраст самки не менее 5 лет. Пять са-
239
мок повторно гнездились в том же дупле, остальные не далее 3 км от мес-
та мечения.
25.V 1977 г. у Чунозера и 8.VI 1978 г. у р. Нявка наблюдались самцы со
светлой окраской верхней стороны (вместо черной): вероятно, это были
молодые годовалые птицы.
Размеры 5 самцов: длина 130-135, крыло 79-79, хвост - 52-54 мм,
масса тела 11,3-13,5, в среднем 11,8 г. Размеры 4 взрослых самок соот-
ветственно - 125-129, 72-76, 42-55 мм, масса тела 12,0 и 14,5 г.
СЕМЕЙСТВО ДЛИННОХВОСТЫЕ СИНИЦЫ - AEGITHALIDAE
Ополовник - Aegithalos caudatus (L.)
По материалам Кандалакшского заповедника, изредка гнездится и
на юге Мурманской области (Бианки и др., 1960).
В Лапландский заповедник ополовники неоднократно залетали
осенью, во время кочевок: в 1949, 1959, 1960, 1962, 1965, и 1984 гг., в раз-
ные сроки между 6 октября и 9 декабря. Разными лицами зарегистриро-
вано в общей сложности около 65 птиц; самая северная встреча в долине
р. Южный Ташким.
Ополовники встречались стайками от 3 до 15 особей, обычно во время
кормежки на березах, ольхах или на снегу. Иногда по соседству с ними,
но обособленно держались гаички. Голос ополовника своеобразный, не
похожий на синичий, напоминает чмоканье.
СЕМЕЙСТВО СИНИЧЬИ - PARIDAE
Пухляк8 - Pams montanus Bald.
Кольский полуостров населяет подвид Parus montanus borealis lonnbergi
Zedl. (Владимирская, 1948, с. 232; Портенко, 1954, с. 116).
По сведениям Ф.Д. Плеске, пухляк встречается по всей Лапландии
гораздо реже сероголовой гаички, приурочен к поясу хвойных лесов и
только во время кочевок появляется на берегах Ледовитого океана
(Плеске, 1887, с. 197-198).
Характер пребывания пухляка в Лапландском заповеднике, в отличие
от более южных широт, различается по годам, в зависимости от кормо-
вых, а возможно, и погодных условий. В некоторые годы пухляки у нас
зимуют, в другие и взрослые и молодые птицы отлетают, в третьи наблю-
дается обратный подлет (с юга?) уже к концу лета. Миграции пухляков,
различные по интенсивности в разные годы, описаны рядом авторов
(очерк Бардина в книге Мальчевского и Пукинского, 1983, с. 273-280).
Подъемы численности наблюдались нами в 1937-1941, 1966-1968, 1972,
8Русские названия синиц из группы гаичек запутаны и не всегда соответствуют их
диагностическим признакам: Бёме, например, называет этот вид "буроголовой
гаичкой”, с характеристикой: ”от черноголовой гаички в природе практически не-
отличима” (Флинт и др., 1968, с. 524). Мы даем название по Иванову (1976).
240
1974-1975, 1977-1981 гг. В Финляндии в 1976 г. по сравнению с 1936 г.
численность пухляка уменьшилась вдвое, что связано с сокращением
его ареала (Jarvinen и др., 1977).
В заповеднике пухляк придерживается смешанных лесов по долинам
больших рек и озер. Пение в разные годы слышали с 1.III по 29.VII,
21.VI 1937 г. П. Мальчевский наблюдал агонистическое поведение пары
пухляков у гнезда.
Выводки из 5-6 слетков встречались между 25.VI (1974) и 28.VII (1980).
В до- и послегнездовые периоды пухляки обычно встречаются поодиноч-
ке или парами, изредка стайками до 8 особей, в среднем по 4 (п = 17).
Иногда пухляки держатся в общей стае с сероголовыми гаичками. В за-
поведнике они гнездятся не каждый год.
Пухляки кормятся, как правило, на деревьях: елях, березах, соснах,
но иногда подбирают семена и с поверхности снега. Характерна привычка
запасать корм и в летнее, и в зимнее время. Добыча прячется в трещины
еловой коры, у основания еловых и сосновых хвоинок, в слоевища
лишайников на елях и березах. Основу питания, в особенности летом,
составляют различные членистоногие, особенно пауки и их коконы, жу-
ки (долгоносики, усачи, короеды) и гусеницы бабочек; встречаются
также мухи и их личинки, комары и их личинки, личинки оводов, му-
равьи, пилильщики, наездники, нематоды. Зимнее питание включает се-
мена ели, берез, в том числе карликовой, можжевельника, брусники,
рябины.
11.VI 1984 г. в долине Чуны в запасах воробьиного сыча в ’’синичнике”
найдена мумия пухляка.
Сероголовая гаичка - Parus cinctus Bodd.
Местные названия: саамское - аввькядзынч или ави-кьядзынч;
относится, вероятно, и к другим синицам.
Одна из немногих оседлых птиц региона. Гнездится предпочтительно в
спелых хвойных, с примесью березы, лесах по долинам крупных рек и
озер; встречаются и на островках леса среди гари, на семенных куртинах
по вырубкам. В гнездовый период избегает чистых сосновых боров и
жердняков, березового криволесья. Во внегнездовое время встречается
практически во всех типах лесов, включая зарастающие сосновым, ело-
вым и березовым молодняком гари и вырубки, болотистые и горные
редколесья.
Сероголовая гаичка - фоновый вид лесного пояса заповедника, в
особенности зимою, и все же ее нельзя назвать многочисленной (рис. 55.).
В смешанных лесах долины Чуны в июне плотность гнездовых пар состав-
ляет 0,5-7, в среднем 3 пары/км2 (1977-1984). В первой половине августа
численность колеблется от 3 до 17 особей/км2, а на участках ельников с
густым подростом локальная численность может достигать 50 особей/км2
(побережья Ельявра, 15.VIII 1979). В первой половине марта численность
составляла от 0-10 до 3-16 особей/км2 на различных участках леса в за-
висимости от густоты и состава древостоя.
16. Зак. 1023 241
Рис. 55. Сероголовая гаичка, 29. VII 1976 г.
Фото О.И. Семенова-Тян-Шанского
Мы не обнаружили у се-
роголовой гаички вековой
тенденции к изменению чи-
сленности, но она сильно
колеблется от года к году.
Годами высокой численнос-
ти в заповеднике были 1938-
1939, 1949, 1962, 1966, 1972,
1976, 1981. Последнее повы-
шение совпадало со сравни-
тельно высоким урожаем
еловых семян; в тот же 1981 г.
повысилась и встречаемость
пухляка, вида, близкого по
экологии. В Финляндии на
основе учетов обнаружено
десятикратное уменьшение
численности гаички к 1976 г. в
сравнении с 1931 г. (Jarvinen
И др., 1977).
Пение нередко слышали
во второй половине марта и
очень редко в августе. По
нашим наблюдениям, в гне-
здовий период гаички не
поют. Мартовское пение,
вероятно связано с распределением птиц по гнездовым участкам.
Сероголовые гаички гнездятся рано, до прилета главных дупло-
гнездников, горихвостки и мухоловки-пеструшки. Ранние сроки
гнездования в сочетании с характерным для северной тайги обилием
дуплистых деревьев и немногочисленностью сероголовых гаичек избав-
ляют их от конкуренции за гнезда с другими птицами. Нам известен один
конфликт из-за гнездовья, но он явно был спровоцирован вмешательст-
вом человека: когда 19.VI 1981 г. на Чунозерской усадьбе уничтожили их
выводок в искусственной дуплянке, воробьи залезли в соседнюю дуп-
лянку, занятую гаичками, ударами клювов убили птенцов и начали
строить в ней собственное гнездо (3. Аблаева).
В природных условиях сероголовые гаички занимают обычно дупла
дятлов, трехпалого или большого пестрого, лишь 3 гнезда из 35 помеща-
лись в полостях на месте выпавшего осинового или березового сука.
Расчистка дупла в трухлявом березовом стволе парой гаичек, наблюдав-
шаяся О. Семеновым-Тян-Шанским 22.V 1934 г., единственный пример
самостоятельного сооружения дупла этим видом. Из найденных нами
гнезд 23 помещались в дуплистых соснах, 6 - в березах, 6 - в осинах, но
ни одного не обнаружено в стволе ели. Объясняется это тем, что и дятлы
предпочитают долбить гладкие, без сучков стволы. Высота летков от зе-
мли от 1 до 8 м: до 2 м - 3 летка, в 2-5 м - 26, в 5-8 м - 6. Диаметр лет-
ка от 23 х 27 до 60 х 60 мм, обычно 30-35 мм; ориентация самая различ-
242
ная, но чаще южная. Гаички заселяют долбленые или дощатые дуплян-
ки, прикрепленные на высоте 1,5 м, выбирая ту, где леток не больше 4 см
в диаметре. В 1966 г. единственный раз гаички гнездились в утиной дуп-
лянке, габариты которой гораздо больше привычных для них (леток
7x8 см, площадь 16 х 16 см, глубина 30 см). Заселенность гаичками
искусственных дуплянок за 10 лет колебалась: в смешанном лесу в доли-
не р. Чуны - от 2 до 14, в среднем 6%, в елово-березовом лесу с примесью
сосны по берегу Чунозера - от 0 до 5, в среднем 2%. Из 24 дуплянок, раз-
вешанных в 1966 г. по границе спелого бора с зарастающей лесосекой,
гаички не гнездились ни в одной. В то же время мухоловка-пеструшка,
плотность которой сравнима с плотностью гаички, занимает в смешанных
лесах до 40% дуплянок, избегая только чистых сосняков.
По результатам мечения, сероголовые гаички живут постоянными
парами. Пара занимает из года в год то же дупло, а если и оставит его, то
селится в соседнем, меняя участок только в случае неуспешного гнездо-
вания.
Гнездостроение наблюдали в разные годы с конца апреля до начала
июня. В начальной стадии его еще лежит глубокий снег, лишь на солнеч-
ной стороне стволов появляются проталины, оголяются береговые
склоны. Дно дупла гаички выстилают трухлявыми волокнами древеси-
ны (до 40% по весу), выше кладется слой зеленого мха толщиной 3-4 см, с
углублением, выстилаемым обычно шерстью леммингов, полевок, реже
зайца, белки, лисы, волка, подпушью и измельченной остью лося или
оленя, смотря по тому, что можно найти поблизости. Состав гнездового
материала довольно стабильный, но иногда слой трухи отсутствует.
Нередко гаички начинают строить 2-3 гнезда одновременно, а потом
бросают лишние, в которых остается кучка зеленого мха; изредка гнездо
бывает брошено на более поздней стадии, с шерстью в подстилке. Иногда
самка продолжает собирать шерсть в процессе кладки и в начале насижи-
вания. Начиная с первых яиц, оставляя гнездо, самка укрывает кладку
ворохом шерсти и только в теплую погоду (выше +10- 15е) оставляет яйца
открытыми.
Первые яйца в разные годы появлялись между 13.V 1983 г. и 22.V
1979 г., в среднем за 9 лет - 18 мая. Сероголовые гаички изредка гнездят-
ся в то же лето повторно; в одной вторичной кладке первое появилось
19.VI. Самец гаички кормит самку не только в процессе насиживания, но
и на стадии кладки. Во время прогулок самка сама добывает корм и
получает добавочный от самца: принимая его, она трепещет крыльями и
издает ’’птенцовые” звуки.
Кладки содержат 4-12 яиц, чаще всего 7-9, а в среднем 7, 3 (п = 60).
Период инкубации с момента откладки последнего яйца составляет
13-16, в среднем 14 дней. Ярвинен для Финской Лапландии определил
время инкубации в 17 дней - результат, сравнимый с нашим, поскольку
гаичка начинает насиживать с одного из последних яиц (Jarvinen, 1978).
Птенцы начинают выходить из гнезда в 11-дневном возрасте, возвра-
щаясь в него на ночь. Вылетают первыми, как правило, наиболее разви-
тые птенцы, вылупившиеся первыми. В среднем птенцы находятся в
243
гнезде 16 дней, максимум - 20. В этом возрасте они начинают самосто-
ятельно добывать корм, получая пищу и от взрослых. Вылет птенцов
наблюдался в разные годы с 8.VI по 15.VII, в самых поздних выводках -
26.VII. Выводки начинают кочевать в августе и держатся вместе до
весны, иногда временно объединяясь с другими гаичками, пухляками,
чечетками и даже клестами. Размеры осенне-зимних стай 13-15 птиц, в
среднем 5,1 (п = 106); наряду со стаями встречаются и пары, и одиночные
гаички.
Успех гнездования гаичек в 1974-1987 гг. колебался по годам от 17 до
94%, в среднем по 67 гнездам - 54%. Резкие колебания успеха размноже-
ния, по нашим наблюдениям, объясняются аномалиями погоды и пресса
хищников. В 1979 и 1982 гг. хищники (в основном куньи) уничтожили
соответственно 55 и 40% яиц и птенцов. В 1977, 1981 и 1982 гг. по погод-
ным условиям погибло 68, 20 и 30% всех кладок и выводков. Взрослые
гаички чаще гибнут зимой: во время весенней проверки дуплянок в них
нередко находили остатки гаичек, их перья или засохшие тушки. Так,
22.V 1984 г. в долине Чуны в ’’синичнике” найдены перья сероголовой
гаички и 2 мумии красно-серых полевок; 11.VI в другой дуплянке, в
1,5 км от первой, - склад из засохших тушек 9 сероголовых гаичек,
1 пухляка и 3 серых полевок. Остатки еще 5 гаичек были найдены в
разных дуплянках в 1980, 1983 и 1984 гг. По всей вероятности, это были
жертвы и запасы воробьиного сыча, так как горностай не может поймать
десяток гаичек в одном месте. Однако в 1979 и 1982 гг. несколько наси-
живающих гаичек были пойманы в гнездах горностаем. Примером смерти
от несчастного случая служит находка трупа гаички лесником В. Станю-
лисом в избушке на Матрениной ламбине 15.Ш 1980 г.: залетев в избу, она
не смогла из нее выбраться.
Гаичка - одна из самых доверчивых птиц нашего региона, часто сама
подлетает или подпускает к себе человека вплотную: 22.Ш 1981 г. на бе-
регу Чунозера одна гаичка из стаи, пролетая, присела на ствол ружья в
руках. Так же доверчивы они у гнезда, залетая в дупло на глазах у наб-
людателя. Когда леток закрывают для отлова, гаичка обычно затаивает-
ся в углу дупла и стремительным броском старается проскочить в любой
просвет при неловком движении. Самка, застигнутая на кладке, иногда с
шипением бросается в закрытый леток, и через слой ткани ощущается
острота ее клюва.
Питается сероголовая гаичка как животной, так и растительной пи-
щей. Последняя преобладает в зимний снежный период (с половины
октября по май) и состоит из семян ели, сосны, берез, ягод можжевель-
ника, семян марьянника, очевидно запасенных с осени. Зимой гаички
сопровождают стаи кормящихся клестов, иногда чечеток, подбирая обро-
ненные ими семена. Зимний животный корм состоит из пауков и их коко-
нов, тлей, жуков, ос, гусениц бабочек. Ос и их личинки гаички добывают
из осиных гнезд, находящихся в дуплах. Гаичек всегда привлекают туши
добытых или павших оленей. В апреле-мае они кормятся возле ручьев и
рек выползающими на снег веснянками. Охотно клюют на птичьих кор-
мушках мясо, сало, мучные изделия.
244
Таблица 15. Размеры синиц (в мм) в районе Лапландского заповедника
Вид Длина Крыло Хвост Масса тела Число экз.
Пухляк, самец 120 63 61 11,0 11
110-137 59-64 57-65 9,2-11,8
самка 112 61 58 10,5 7
109-115 59-63 55-60 9,7-11,9
Сероголовая гаичка, ^5 66 66 12,1 20
самец 119-137 61-70 58-72 9,8-13,7
самка 126 64 62 12,4 5
120-140 63-66 58-67 11,5-13,4
Московка, самцы, самки 105 61 48 9,0 3
95-110 59-63 46-49 7,7-9,8
Большая синица, 150 76 65 19,5 5
самец 137-160 72-79 60-70 18,3-22,7
самка — 75 62 19,0 3
146-147 73-79 60—65 16,6—21,5
Примечание. Здесь в числителе — средняя величина, в знаменателе — предель-
ные.
В зимнем питании гаичек большое значение имеет их привычка запа-
сать корм, когда он в избытке. Заготовка кормов чаще всего наблюдает-
ся в сентябре-октябре, а потом в марте-апреле, в месяцы с достаточно
продолжительным световым днем, но они запасают корм и в другие сезо-
ны, кроме гнездового. Добычу свою они прячут в трещины дерева, под
кору, в основании хвоинок и других подобных местах.
Летом гаички питаются исключительно насекомыми и пауками, ими
же выкармливают и своих птенцов. Из насекомых чаще встречаются гу-
сеницы, куколки и имаго различных бабочек - пядениц (в том числе
осенней, Oporinia autumnata), листоверток, совок, нимфалид; имаго пили-
льщиков, личинки и имаго комаров-долгоножек и различных мух; долго-
носики, божьи коровки, усачи, короеды наездники, муравьи, цикадки,
тли. В общем, жуки гораздо реже встречаются в пище птенцов, чем взрос-
лых. В подстилке гнезда после вылета птенцов всегда можно найти раз-
личных жуков, в том числе долгоносиков, щелкунов, жужелиц, вероят-
но забракованных птенцами из-за их жесткости.
Максимальный возраст самки гаички, установленный при кольцева-
нии, - более 6 лет (табл. 15).
Московка - Parus ater L.
Для южной части Кольского полуострова отмечается как редкий гнез-
дящийся вид (Благосклонов, 1960; Бианки, 1960).
В Лапландском заповеднике в 1935, 1938 и 1939 гг. одиночные москов-
245
ки наблюдались (и добывались) в стаях гаичек в сентябре и с декабря по
февраль; после чего не встречались ни разу. Очевидно, на Кольский полу-
остров московка залетает и на время приживается только в годы ин-
вазий.
Исследовано 3 желудка: 15.IX 1938 г. - около 40 гусениц пяденицы;
20.XII 1938 г. и 23.1 1939 г. - остатки насекомых, паук и оболочки еловых
семян.
Хохлатая синица - Parus cristatus L.
Изредка гнездится у южной границы Мурманской области (о-в Вели-
кий; Коханов, 1987, с. 29). На территорию Лапландского заповедника
только изредка залетает, во время послегнездовой дисперсии молодня-
ка. Впервые одиночную хохлатую синицу видели А. Насимович и М. Вла-
димирская 25.IX 1941 г. на южном берегу Вочеламбины (Владимирская,
1948, с. 233). Позднее В. Федотова наблюдала одиночных птиц на берегах
Чунозера 14.111971 г. и 11.VIII 1973 г.
Большая синица - Parus major L.
Сто лет назад большая синица на Кольском полуострове еще не встре-
чалась (Плеске, 1887, с. 196), а теперь, на правах синантропа, заселила всю
обжитую полосу вдоль железной дороги до побережья Баренцева моря (в
восточной, тундровой части полуострова по-прежнему отсутствует)..
Первая встреча на территории Лапландского заповедника датирует-
ся 5.V 1934 г. До конца 50-х годов наблюдалась не ежегодно, в основном
весной - а апреле-мае и осенью - в сентябре-октябре, обычно по
1-2 особи, редко стайкой, до 5. С 1961 г. стала попадаться и летом. По
материалам картотеки наблюдений большая синица впервые зимовала на
Чунозерской усадьбе в 1966/67 г., а с 1973 г. зимует здесь уже регулярно.
Результаты кольцевания показали, что зимуют не только взрослые пти-
цы, но и сеголетки, вместе с тем часть особей, особенно молодых, мигри-
рует: синица, помеченная птенцом 24.VI 1984 г. на Чунозере, в октябре
обнаружена в г. Ковдоре на расстоянии 95 км к западу.
В заповеднике большие синицы не только зимой, но и в сезон гнездо-
вания держатся у жилья - на усадьбе, у кордонов и сторожевых изб.
Синиц иногда встречали на расстоянии до 4 км от ближней избы (13.V
1982 г. в долине Калипсо), но гнезда - не далее 1 км. Строгая приурочен-
ность к жилью обусловливает малочисленность синиц в заповеднике. В
редкие зимы на усадьбе заповедника доживают до весны 3 пары, а в суро-
вую зиму 1984/85 г., когда в январе морозы достигали -50°, из 7 синиц,
живших здесь в декабре, к марту не осталось ни одной: впоследствии три
из них были найдены мертвыми (взрослая самка и 2 сеголетка). Позднее,
в середине апреля, появилась одна синица, а 27.IV - пара.
Синицы начинают петь уже в декабре-январе, а в конце апреля-мае
они поют наиболее активно. У самца, добытого 5.V 1938 г., семенники
имели длину 6 и 7 мм. Гнезда строятся во второй половине мая - начале
июня на протяжении 3-6 дней. В основном они сложены из зеленого мха,
иногда с добавлением стеблей и корней вороники, а лоток выстилается
246
шерстью леммингов и полевок (случается, вместе с кожей), собаки,
лошади. Толщина гнезда 80-90 см, диаметр лотка 57-70, глубина его
62-70 мм. Как правило, синица пробует строить гнезда в нескольких дуп-
лах, на расстоянии 100-500 м одно от другого.
Мы находили гнезда только в искусственных гнездовьях. По сообще-
нию М.А. Лагутиной, в июне 1975 г. пара синиц гнездилась в колпаке
бездействовавшего уличного светильника, на высоте 6-7 м, в центре
г. Мончегорска.
Величина первых кладок 7-12 яиц, в среднем 9,2 (п = 10), вторых -
5-10, а в среднем 7,4 (п = 5). В кладке яиц возможны перерывы до двух
дней. Продолжительность насиживания, считая с откладки последнего
яйца, - 13-15 дней. Птенцы покидают гнездо на 16- 18-й день. После пер-
вого успешного гнездования, в случае теплого лета, синицы гнездятся
вторично. Вторые кладки достоверно наблюдались в 1984 г., предположи-
тельно также в 1973 и 1981 гг. Птенцы второго выводка вылетают в сере-
дине августа.
Птенцы и кладки часто гибнут от погодных условий или от гнездовых
хищников, поэтому успех размножения невысок, в среднем по И гнез-
дам он составляет 50%, а в 1977 г. падал до 18% (по 4 гнездам).
Во время кладки и насиживания яиц большие синицы осторожны и в
отличие от гаичек при беспокойстве бросают гнездо и принимаются
строить новое, как мы наблюдали в 1977 г. Застигнутая в гнезде самка
ерошит перья и ’’фукает”, резко выдыхая воздух.
Питание большой синицы смешанное. По визуальным наблюдениям и
анализу содержимого 12 желудков, ее корм состоит из семян и плодов
растений, в том числе березы, вороники, черники, и различных насеко-
мых - жуков (короедов, долгоносиков и др.), мух и их личинок, грибных
комариков (Mycetophilidae), комаров, бескрылых тлей, сетчатокрылых
(Hemirobius). Корм птенцов состоит в основном из насекомых. Зимой
доля семян растений возрастает. Для выживания больших синиц зимой
большое значение имеет подкормка у жилищ различными пищевыми
отходами и на птичьих кормушках, где они клюют мясо, сало, хлеб,
кашу. Синицы нередко попадаются в мышиные давилки, наживленные
одним из этих продуктов.
В августе 1985 г. на Чунозерской усадьбе около 20 дней жила пара
синиц необычной расцветки, свойственной среднеазиатским формам
большой синицы: желтый цвет низа заменен белым, спина серо-голубая,
вместо зеленой.
СЕМЕЙСТВО ПОПОЛЗНЕВЫЕ - SITTIDAE
Поползень - Sitta europaea L.
Поползень оседлая птица и в природе запасает на зиму корм, главным
образом орешки лещины и разных древесных пород. За отсутствием на
Кольском полуострове этого пищевого ресурса он не находит здесь не-
обходимых условий для гнездования и акклиматизации, может только
время от времени залетать.
247
Впервые его встретил и сфотографировал 8. III 1977 г. в городском
парке Мончегорска В. Бусуек, а в мае того же года наблюдал на о-ве Хар-
лов Ф. Шкляревич (Коханов, 1987, с. 27). Этот залет явно связан с начав-
шейся осенью 1976 г. инвазией поползня в Финляндию (Sammaliste Lasse,
1978). Потом одиночный поползень жил с 9.Х до 19.XI 1983 г. на Чунозер-
ской усадьбе Лапландского заповедника, питаясь, как и в Мончегорске,
на птичьей кормушке.
2. VI 1984 г. поползня наблюдала в сосновом лесу под Нявчик-тундри-
цей О. Макарова, a 13.IX 1985 г. О. Семенов-Тян-Шанский слышал харак-
терный крик этой птицы в бору в долине Чуны.
СЕМЕЙСТВО ПИЩУХОВЫЕ - CERTHHDAE
Пищуха - Certhia familiaris L.
В лесистой части Кольского полуострова редкий гнездящийся вид. В
Лапландском заповеднике наблюдался с первых лет, но не ежегодно.
Придерживается спелых лесов разных типов, чаще в ельниках, чем в сос-
няках. Встречается обычно поодиночке, иногда в стае гаичек или пухля-
ков. По две птицы вместе наблюдали 27.IX 1932 г. и 13.Х 1939 г., по три -
30. VIII 1966 г., и выводок из нескольких слетков замечен О. Семеновым-
Тян-Шанским 17.VI 1950 г. у Северного ручья: птенцы попрятались в елях
и пищали, взрослые беспокойно лазали по стволам.
С годами пищуха становится в заповеднике все реже, об этом можно
судить по среднему числу отмеченных за год птиц: в 30-х годах - 2,2,
40-х - 2,2,50-х - 1,5, 60-х - 0,4, 70-х - 0,8, 80-х - 0,3.
В 30-х годах были добыты 2 экз., а 8.1 V 1939 г. на снегу у Второго ручья
найдены перья и лапки пищухи, пойманной, вероятно, совой. В желуд-
ках (1.XI и 15.VII 1938 г.) обнаружены комар, остатки жуков, 33 мухи
(Scatophaga), семя можжевельника и еще какие-то семена.
Размеры 2 самцов: длина 127, крыло 64-65, хвост 62 мм, масса тела 8,6
и 8,7 г.
СЕМЕЙСТВО ОВСЯНКОВЫХ - EMBERIZIDAE
Обыкновенная овсянка - Emberiza citrinelia L.
На большей части ареала живет оседло, с Крайнего Севера обычно
отлетает, но может зимовать и на месте, например, у скотных дворов
(Владимирская, 1948, с. 225).
Прилет овсянки хорошо заметен, так как первыми появляются самцы
с песней, и притом около жилья. Средняя за 51 год дата прилета их в за-
поведник 20 апреля (4.IV 1937 г. - 4.V 1973 г.), при средней температуре
воздуха -1,6°. В некоторые годы весной наблюдались стайки в 3-8 овся-
нок. Держатся они у кордонов и на усадьбе - у конюшни, у собачьей
вольеры или конуры и только после схода снега появляются в глубине
заповедника, в борах, на гарях различного возраста и в елово-березовых
лесах долины Чуны, но редко, в силу своей малочисленности: в летние
248
месяцы они регистрировались только в течение 17 сезонов из 45 лет
наблюдений. Стройка гнезда наблюдалась только 1.VI 1938 г. - овсянка
несла тонкую стружку; 9.VI 1937 г. на свежей гари в долине Чуны под сва-
ленной сосенкой найдено гнездо: в нем было 4 яйца и самка насиживала
(Владимирская, 1948, с. 225). Летные выводки встречались в разных
местах в 1938, 1940, 1963, 1964, 1977 и 1978 гг., между 9-29 июля. Овсянки,
гнездившиеся в лесу осенью, не концентрируются у поселков, как вес-
ной, а потому отлет их трудно уловить, так что наблюдения отлета менее
точны. За 29 лет средняя дата последней встречи - 11 октября (23.IX 1973 г. -
3.XI 1977 г.), при средней температуре +1,3°.
По визуальным наблюдениям, с мая по октябрь пища овсянок состоя-
ла из овса, разных круп, семян мокрицы, ягод рябины, семян злаков.
Исследовано 9 желудков, из них два в августе и один в октябре содержа-
ли только растительный корм, в остальных он смешанный. Чаще всего
встречались жуки (долгоносики, стафилины, водолюбы, жужелицы),
затем гусеницы бабочек и личинки пилильщиков; иногда личинки гриб-
ных комариков, стафилинид, однажды муха. Растительный корм состоял из
семян осок, фиалок, звездчатки, вороники и овса; в 7 желудках содер-
жался песок.
Овсянок находили в пище хищников дважды: 24.V 1939 г. у Беличьей
избы на глазах А. Пономарева овсянку схватила хищная птица, a 3.IX
1972 г. в желудке ястребиной совы обнаружена одна лапка.
Размеры 6 взрослых овсянок самцов: длина 157-180, крыло 79-82,
хвост 73-82 мм, масса тела 20,0-34,5 г, в среднем 29,2, размеры 2 самок
соответственно - 167-180, 83-88 и 86-83 мм, масса тела 29,3 и 30,7 г.
Садовая овсянка - Emberiza hortulana L.
Изредка гнездится в южной части Кольского полуострова (Терский
берег, Кандалакша, и поселки вдоль железной дороги, до Апатитов;
Коханов, 1987, с. 31). Севернее, вероятно, только залетает.
20.V 1962 г. на территории Чунозерской усадьбы Лапландского запо-
ведника Ю. Гаев и О. Семенов-Тян-Шанский впервые наблюдали и добы-
ли поющего самца. 14.VII 1962 г. на опушке леса близ поселка Тик-губа
они же слышали песню. Через 3 года, 24.V 1965 г., Ю. Гаев вновь видел на
усадьбе пару садовых овсянок.
Овсянка-крошка - Emberiza pusilla Pall.
В списках птиц Кольского полуострова (Плеске, 1887) и Лапландского
заповедника (Владимирская, 1948) овсянка-крошка не упоминается. В
Хибинах найдена как гнездящийся вид Новиковым (1952). На Терском
берегу обычна на гнездовье (Малышевский, 1962, с. 23; Коханов, 1987,
с. 31).
В Лапландском заповеднике впервые овсянку-крошку обнаружил
Ю. Гаев 25.VI 1962 г., в ельнике между Ельявром и Чингльс-явром: в
разных местах пели 3 самца, одного из которых он добыл. Позднее не-
однократно наблюдалась нами в долинах Чуны, Куудас-йок, района
249
Чунозера и на Румельявре в разные сроки, с 27.V до 8.IX. Весной видели
или поющего самца или пару птиц.
9.VII 1969 г. в долине Второго ручья на поляне в ельнике-черничнике
О. Семенов-Тян-Шанский нашел на земле гнездо с тремя слепыми птен-
цами и одним яйцом; матка слетела с гнезда. 18.VII 1983 г. в горной
тундре на Ельнюне найден труп слетка овсянки-крошки. 6.VII 1986 г. на
берегу Ельлухт 2 слетков кормила взрослая овсянка-крошка.
Овсянка-ремез - Emberiza rustica Pall.
В свое время Ф.Д. Плеске не нашел ремеза на Кольском п-ове и
привел довольно туманные сведения о находках его в Финляндской
Лапландии. Позднее Г. Гебель (1903) наблюдал его в районе Варангер-
фиорда. Для южной части Кольского полуострова отмечен как пролетная
птица (Бианки и др., 1960).
В Лапландском заповеднике ремез впервые встречен 14.V 1937 г.
О.Семеновым-Тян-Шанским в долине Чуны. В 1938 г. в разных местах
было добыто 14 экз., позднее их наблюдали в 1939, 1950, 1956, 1960, 1962 гг., а
начиная с 1964 г. - ежегодно. Нельзя сказать, что численность ремеза за
эти годы увеличилась, так как количество встреченных 2-3 наблюдате-
лями особей не превышает, как и в 30-х годах, нескольких десятков.
Некоторое повышение наблюдалось лишь в 1981-1984 гг.
Средняя дата первой встречи за 16 лет - 19 мая (6.V 1977 г. - 31.V
1970 г.), самой поздней, за 10 лет наблюдений, - 12 сентября (3. IX 1970 г. -
20.1X1971 г.). Чаще всего ремезы попадались в разреженных сырых
елово-березовых лесах, близ ручейка или другого водоема.
За все время найдено 4 гнезда. Одно располагалось на елочке ростом
3-4 м, на ветках у ствола в 0/75 м от земли (Владимирская, 1948, с. 225);
остальные на кочках, под укрытием ветвей карликовой березки или
вороники, или стеблей канареечника (Phalaroides arundinacea), или
внутри березового куста, между стволов. Форма их типичная чашеобраз-
ная, наружный диаметр 120, высота 80, ширина лотка 70, его глубина
50 мм (п = 2). Внешний слой свивается из сухих веток, хвоща, стеблей
канареечника, сухой соломы; лоток выстилается былинками или лоси-
ным волосом. Сравнительно высокое положение гнезд, вероятно, объ-
ясняется тем, что во время стройки земля была покрыта водой или даже
снегом, который в характерных биотопах ремеза поздно стаивает. Само
строительство, по расчету, начинается в середине мая, а кладка яиц - с
25.V по 16.VI. Все четыре кладки были полные и содержали по 5 яиц,
окраска их зеленовато-голубая с неправильной формы темно-серыми пят-
нами разной интенсивности, сгущающимися к тупому концу. Птенцы
вылуплялись между 13-29 июня. Одно из гнезд птенцы покинули на
11-й день. Выводки наблюдались 6.VII 1982 г. у р. Нявка, 9.VII 1938 г. в
долине Чуны, 27.VII 1938 г. у Калзуая, 31.VII 1938 г. между Чунозером и
Охтозером, 11. VIII 1964 г. у речки Роговая.
Питание овсянки-ремеза смешанное, весной и осенью возрастает роль
семян, летом преобладают насекомые. Растительный корм (исследовано
12 желудков) состоял из семян вороники, осок, молинии, седмичника,
250
пушицы, луговика. Животный корм включает пауков, цикадок, тлей,
клопов, жуков (щелкуны, божьи коровки, долгоносики), двукрылых
(долгоножки, комары Aedes и Bibio, мухи), пилильщиков их личинок,
орехотворок, ручейников, гусениц пядениц и листоверток.
Размеры 4 взрослых самцов: длина 127-148, крыло - 76-81 мм, хвост
55-63 мм, масса тела 17,5-20,8, в среднем 19,7 г. Размеры 7 самок: длина
123-146, крыло 70-80, хвост 56-65 мм, масса тела 17,1-19,9, в среднем
18,4 г.
Камышовая овсянка - Emberiza schoeniclus (L.)
На Кольском полуострове распространен до Арктического побережья.
Камышовая овсянка прилетает к нам сравнительно рано, с появлени-
ем первых проталин ча берегах водоемов и у стволов деревьев. Появля-
ется стайками по 5-7 и до 20 птиц. В первых стаях преобладают самцы, позд-
нее через неделю-другую, большинство составляют самки. С их появлением
самцы начинают петь, и скоро пары распределяются по гнездовым уча-
сткам.
Средняя дата прилета за 39 лет - 13 мая (27.IV 1977,1983 гг. - 26.V 1940
г.). Осенняя миграция начинается в середине августа и продолжается в
сентябре, редкие особи задерживаются до октября. Средняя дата послед-
ней встречи за 25 лет - 23 сентября (3.IX 1950 г. - 16.Х 1980 г.).
Любимые гнездовые стации камышовых овсянок - влажные или
заболоченные берега водоемов, заливаемые полой водой, с зарослями
ивняков, карликовой березки, осоки, с редкостойными деревьями. Они
предпочитают берега магистральных рек и больших озер, а у горных
ручьев и речек, у горных озер селятся редко, выбирая опять же заболо-
ченные берега с зарослями кустарников; в глубине леса совсем не встре-
чаются. Хотя камышовая овсянка одна из самых обычных птиц заповед-
ника, многочисленной ее нельзя назвать. Частота встреч отчасти объясня-
ется тем, что и наблюдатели много времени уделяют долинам рек и бере-
гам озер. В середине июня в долине Чуны выше впадения Сылпуая, в при-
речном березняке с примесью ели их плотность в 1979-1984 гг. составля-
ла 14-27 пар/км2. На маршруте в спелом смешанном лесу, захватываю-
щем долину ручейка с озерком, площадью не более 1 га, в 1977 и 1984 гг.
было учтено не более 1 пары/км2.
Камышовые овсянки поют сравнительно короткое время, позже 11. VII
1981 г. их песен уже не слыхали. Кульминация пения приходится на пер-
вую половину июня. В гнездостроении, насиживании и выкармливании
птенцов участвуют • оба партнера. Строительство гнезд начинается в
последней декаде мая, еще по высокой воде. Из найденных 39 гнезд 21
устроены на кочках, под прикрытием кустиков вороники, голубики,
Кассандры, карликовой березки, ивы или в нишах кочек; 12 - на земле,
под кустами березы, черемухи, ивы или под елочкой; 5 - в кустах мож-
жевельника; 1 - на перекрестке стволов ивы на высоте 0,5 м. Из 6 над-
земных гнезд 4 были найдены 26-28.VI 1978 г. на берегу длинного плеса
р. Купись. Постройка гнезд на ветках объясняется тем, что эти овсянки
гнездились на тех участках поймы, откуда вода уходит лишь к вылупле-
нию птенцов.
251
Слои гнезда четко различаются, и каркас состоит из толстых стеблей
осок и злаков, изредка из корней и стеблей кустарничков. Лоток спле-
тается из тонких былинок, оленьего или лосиного волоса, в единичных
случаях из конского, из кукушкина мха и березовых листьев. Для по-
стройки гнезда на ветке предварительно сооружается платформа из со-
ломинок (1.VI 1979 г. на берегу Нявки). Одно из гнезд, найденных в июне
1974 г. на берегу р. Купись, было как бы наколото на ветку можжевель-
ника, конец которой торчал из лотка. Размеры гнезд: внешний диа-
метр 80-88 мм, ширина лотка 52 мм, глубина его 40-50 мм. Откладка яиц
происходит с первых чисел до конца июня. В кладке 4-6 яиц, в среднем
5,4 (п = 42), в том числе с 4 яйцами - 3, с 5 - 21 с 6 - 18 гнезд. Вылупление
птенцов наблюдалось в разные годы между 19.VI и 11.VII. Позднее на-
ступление весны затягивает гнездовый период на неделю-другую. В рай-
оне заповедника камышовые овсянки вторично не гнездятся.
Птенцы уходят с гнезда на 10-12-й день. Со второй половины июля
молодые птицы начинают кочевать. 3-7 VIII, самое позднее 10.VIII 1973 г.
встречались единичные выводки при взрослых птицах.
У гнезда камышовые овсянки довольно доверчивы и подпускают
вплотную, а вспугнутые - активно и упорно ’’отводят”, ковыляя по
земле с распушенным хвостом, сгорбившись и подняв вверх крылья.
Весной камышовые овсянки кормятся на проталинах, на выступающих
из снега и воды кочках, у кромки льда, различными семенами; в том
числе берез, вейника, осок, мятлика, брусники, принесенными водой
личинками ручейников, поденок, выползающими на снег веснянками, а
также летающими насекомыми (16.V 1978 г.). Позднее добавляются другие
корма, например семена марьянника. Судя по анализу 10 желудков,
животный корм состоит из веснянок, ручейников, жуков (долгоносики,
листоеды, короеды), долгоножек, личинок пилильщиков, пауков, кле-
щиков, моллюсков. Известны случаи гибели камышовых овсянок от не-
погоды: 9.IX 1979 г. нашли 2 трупа слетков, 24.VI 1977 г. - взрослой пти-
цы; 15.VII и 5.IX 1977 г. на Чунозерской усадьбе пойманы 2 молодые пти-
цы, неспособные летать, вероятно, из-за зараженности личинками мух.
Размеры 4 взрослых самцов: длина 129-161, крыло 78-80, хвост 47-
73 мм, масса тела 17,1-21,0 г; 4 самок соответственно - 133-148, 67-74,
52-75 мм, масса тела 16,3-19,6 г.
Лапландский подорожник - Calcarius lapponicus (L.)
На кочковатой с мелкими кустарниками тундре, в том числе и на
северо-востоке Кольского полуострова, - фоновый вид; в лесной части
полуострова попадается главным образом на пролете.
В Лапландском заповеднике впервые наблюдался М. Владимирской
24.V 1938 г. в пойменном березовом лесу при впадении ручья Сылпуай в
р. Чуна; хотя птиц была пара и вели они себя территориально, гнездо
найти не удалось (самец добыт в коллекцию). Позднее пролетных подо-
рожников, одиночных или пары, наблюдали на весеннем пролете при
устьях рек, у жилья или в пойменных березняках в 1961, 1966, 1968, 1969,
1971 и 1975 гг. между 3-31 мая, а в среднем за 7 лет - 23.V. Однажды,
252
23.V 1971 г., на усадьбе видели стаю подорожников, a 26.V 1961 г. - оди-
ночного самца в большой стае юрков.
По-видимому, подорожники изредка гнездятся в заповеднике, так как
их встречали в сезон гнездования: 20.VII 1969 г. - самца в березовой
рощице в долине Куудасйок; 11.VIII 1970 г. - три подорожника в бере-
зовом криволесье в долине Рейтлагуая и одна самка на склоне Нявка-
тундры; 9.VI1971 г. токующего в воздухе самца видели на южном склоне
Нявка-тундры у верхней границы леса; 13.VI 1982 г. отмечен самец в ред-
ком бору-скальнике на берегу Ребячьего озера (новая территория запо-
ведника). Собственно на горных тундрах мы подорожников не встречали:
летние встречи приурочены либо к березовому криволесью у границы леса с
тундрой, либо к прибрежным березовым, либо угнетенным хвойным ле-
сам. На осенным пролете не наблюдался.,
Размеры 1 самца: длина 164, крыло 94, хвост 67 мм, масса тела 27,8 г. В
его зобу и желудке найдены семена луговика.
Пуночка - Plectrophenax nivalis (L.)
Местные названия: саамское - пунавка, иногда нюхчь-пытша
(лебяжий хвост; очевидно, по времени прилета и белой окраске).
На Кольском полуострове гнездится по каменистым участкам как гор-
ных, так и равнинных тундр.
Пуночка - один из первых вестников весны. Средняя дата прилета за 53
года - 3 апреля (10.Ш 1986 г. - 18.IV 1945 г.), при средней температуре
воздуха -4,8е и о ±7,3 сут. В конце марта - начале апреля снег едва
начинает таять на южных склонах. В некоторые годы одиночных пуночек
встречали в городах и зимой, например 24.1 1951 г. в Кировске и 15.1
1962 г. - в Апатитах. Птицы держались возле дорог и скотных дворов,
питаясь, как воробьи, зернами из навоза. Вероятно, в 1979/80 г. пуночки
зимовали поблизости, так как отдельные особи появились при потепле-
нии 23.11-2ЛП, затем исчезли и вновь пуночки прилетели уже 7.IV.
Основная масса мигрантов отстает от передовых на 8-12 дней. Размеры
весенних стай колеблются от нескольких особей до нескольких сот, но чаще
всего из 1-2 десятков. Птицы держатся болот, выдувов, населенных
пунктов. Интересно, что, если в 1930-1960 гг. пуночек чаще встречали на
конных дорогах, даже временных ’’зймниках” по льду озер, теперь их
больше видят у шоссе Ленинград-Мурманск. Если на конных дорогах
пуночек интересовал овес и просыпанное зерно, то на автодорогах их
привлекают главным образом семена сорняков, растущих по обочинам.
Количество мигрантов колеблется по годам десятикратно в ту или
другую сторону, в зависимости как от уровня численности популяции,
так и от снежности зимы и быстроты движения весны. Пролет пуночек
идет в два этапа: сначала они живут в тех же биотопах, как и на зимов-
ках, потом, когда горы частично освободятся от снега, занимают гнездо-
вые стации. Осенняя миграция также проходит в два приема: в сентябре
стаи пуночек кочуют по горным тундрам, питаясь семенами злаков, осок,
а главным образом тройчаткой ситника, а когда они скроются под сне-
гом - спускаются на равнины и отлетают уже не вдоль дорог, как весной,
253
а широким фронтом; по этой причине осенний пролет плохо прослежи-
вается.
Крупные стаи, от десятков до сотен птиц, в разные годы встречали с
середины сентября до конца октября.
Средняя дата последних встреч за "37 лет - 24 октября (З.Х 1983 г. -
14.XI 1980, 1987 гг.), при температуре - 1,3е и о ±7,5 дней.
В Лапландском заповеднике пуночка гнездится исключительно в гор-
ной тундре, на высотах 500-1000 м над ур. м., в каменной пустыне со
скудной растительностью в щелях между камнями, состоящей из мха
ракомитриум, исландского лишайника, плауна баранец, тройчатого сит-
ника, ожики изогнутой, вороники, альпийского сердечника и немногих
других видов.
Численность пуночек на гнездовьях ничтожна, можно пройти час-
другой и ни одной не увидеть. По учетам, проведенным в 1976-1984 гг. на
Нявка-тундре и двух участках Чуна-тундры (южном Сейднотчорр-Райкор-
чорр-Ельнюн и северном, Эбрчорр-Аакванюацк), плотность гнездовых пар
во второй половине июня составляла 0,2-1,0 пар/км2. 20.VIII 1975 г. в
районе Эбрчорра за 10,4 ч в тундре отмечено 22 пуночки, что примерно
соответствует 5-6 особям/км2.
Пение пуночек в тундре в разные годы слышали с 30.IV по 13.VII.
Гнездостроение и кладка яиц происходит в первой половине июня. Из
трех найденных гнезд два (19.VI 1965 г. и 22.VI 1966 г.), скрытые под
каменной плитой, были свиты из былинок, выстланы перьями тундряных
куропаток и содержали по 6 яиц (во втором гнезде сильно насиженных);
29.VII 1931 г. А. Васильев нашел на Керка-чорре гнездо с 3 оперенными
птенцами. Слетки при взрослых птицах в разные годы встречались с
13. VII по 8. VIII.
Вблизи гнезда пуночки не проявляют особого беспокойства и подпус-
кают к себе близко, но добраться до гнезда трудно, если оно запрятано в
глубине между камней.
По визуальным наблюдениям в апреле-мае и сентябре-октябре, пита-
ние пуночек состоит из семян тройчатого ситника, овсяницы, луговика,
арктагростиса, осок, карликовой березки, золотой розги, а также семян
ели и веснянок. На дорогах копаются в конском помете, сене, подбирают
овес. Птенцов выкармливают, вероятно, насекомыми, ползающими по
снежникам и поверхности скал.
Размеры 3 самцов: длина 171-180, крыло 108-114, хвост 65-72 мм,
масса тела 33,0-34,2 г; 3 самок соответственно 155-165, 97-111, 60-67 мм,
масса тела 28,8-31,7 г.
СЕМЕЙСТВО ВЬЮРКОВЫЕ - FRINGILLIDAE
Зяблик - Fringilla coelebs L.
Распространен, вероятно, по всей лесной части Кольского полуострова,
но всюду малочислен.
В заповедник зяблики прилетают, в среднем за 46 лет, 30 апреля
(17.IV 1964 - 15.V 1945), при температуре около +0,4°, о = ±6,7 дней. Пер-
254
выми появляются одиночные самцы, позднее стаи до трех десятков, в
которых бывают и самки. Часто зяблики держатся в обществе юрков.
Пролет заканчивается, примерно, к концу мая. За всю весну в заповед-
нике отмечают от нескольких зябликов до 3-4 десятков, в большинст-
ве - возле жилищ. Отлет плохо прослеживается, отчасти из-за малочи-
сленности зябликов, отчасти потому, что осенью им нет нужды прибли-
жаться к жилью. Последних птиц встречали, в среднем за И лет, 21 сен-
тября (9.IX 1977 г. - 4.Х 1987 г.), температура в это время +5,3°, о ±6,6 сут.
В гнездовый период зяблики наблюдаются в заповеднике не каждый
год, лишь с 1976 г. встречи их летом стали более регулярными: возможно,
это признак некоторого увеличения численности.
Поющие самцы или пары зябликов встречаются, как и во время про-
лета, на территории усадьбы или вблизи кордонов и только изредка в
долинах рек и озер. Гнезд зябликов не находили, но встречи слетков
14.VIII 1965 г. и 15.VII 1967 г. на усадьбе заповедника доказывают, что они
здесь гнездятся.
Питание зябликов смешанное; по визуальным наблюдениям и анализу
трех желудков они кормятся различными семенами, в том числе мари,
щавелей, и насекомыми; в частности жуками (щелкуны, усачи) и мухами,
которых часто ловят на стекле.
Известные случаи гибели зябликов: 14.VIII 1965 г. на усадьбе найден
мертвый ’’полухвостик”; 8.Х 1977 г. молодой самец получил контузию от
удара об оконное стекло; 21.IX 1977 г. истощенная молодая самка была
поймана собакой (в брюшной полости много округлых желтых телец
1-1,5 мм в диаметре).
Размеры 2 самцов: длина 149-158, крыло 86-89, хвост 64-69 мм, мас-
са тела 21,7-23,5 г.
Юрок - FringiUa montifringiHa L.
На Кольском полуострове встречается всюду, где только есть древес-
ная растительность, по долинам рек заходит и в тундру.
Юрок прилетает позже зяблика и дроздов. В среднем за 44 года 8 мая
(20.1 V 1965 г. - 22.V 1933 г.), когда в лесу еще лежит снег, при температу-
ре воздуха +2, Г; о ±6,4 дня. В отличие от других мигрантов юрки приле-
тают сразу в массе, но в первом потоке преобладают самцы. Пролет про-
должается до конца мая, самые поздние пролетные стаи отмечены 1.VI
1968 г., 8.VI 1974 г., 11.VI 1969 г. и 17.VI 1982 г. Первые стаи кормятся на
редких проталинах по берегам и склонам, прилетевшие позднее доволь-
но широко расходятся по лесу, придерживаясь речных долин и берегов
озер. Весенние стаи достигают сотен особей, чаще в них по нескольку
десятков птиц. Осенняя миграция хорошо выражена и проходит дружно.
Первые стайки по 1-3 десяткам наблюдаются с начала августа, пик отле-
та приходится на вторую половину августа - в это время можно встре-
тить стаи более 500 юрков. Средняя дата последней встречи за 47 лет
21 сентября (1.IX 1934 г. - 10.Х 1950 г.), при средней температуре +5,3°;
о ±8,6 сут.
255
Юрок самая многочисленная птица заповедника, лишь в годы инвазий
чечетки встречаются чаще юрков. Распространен всюду, где есть деревья,
поднимаясь по склонам до верхней границы леса. Любимая гнездовая
стация юрков в заповеднике - смешанные леса с присутствием ели и
березы; чистых боров, в особенности участков молодняка и жердняка,
они избегают - плотность птиц здесь не превышает нескольких пар на
1км2. Молодые гари, где подрост не превышает 2-3 м, и пояс тундр,
примыкающий к лесу, они осваивают, когда выводки начнут кочевать, с
середины июля. В смешанных лесах по долине Чуны и у Чунозера в
середине июня численность юрков в 1977-1987 гг. колебалась от
9 пар/км2 в 1978 г., до 27 пар в 1983 и 1985 гг.; в приречном елово-березо-
вом лесу в 1979-1987 гг. она составляла 21-29 пар/км* (на северо-западе
Финской Лапландии в заповеднике Малла, в горном березовом лесу в
июне 1979 и 1980 гг. плотность юрков была такая же, 21 пара/км25 Jarvinen,
1982).
Распределение юрков по гнездовым участкам происходит с середины
мая. Самцы начинают петь сразу после прилета, и их ’’вжиканье” можно
слышать до конца июля. В период кульминации пения, в первой полови-
не июня, юрки поют и ночью. Изредка поют и осенью - 4. IX 1938 г. Г. Но-
виков отметил пробу голоса молодыми юрками в стае из двух десятков
птиц.
Гнезда строятся с третьей декады мая до конца июня. Из 19 гнезд 16
помещались на елях (9 - у ствола, 7 - на ветках, на расстоянии до 1 м от
него), 2 - на развилках берез, 1 - на сосне. 10. VII 1964 г. А. Печенюк
нашел у Ельявра гнездо с 6 яйцами на сломе березового ствола на высоте
1,4 м, но не описал его подробнее. Высота гнезд над землей от 1,6 до 10 м,
в среднем 4,1 (л = 14). Строительный материал разнообразный и включает
эпифитные лишайники, ягель, бересту, сфагновый и зеленый мхи, тонкие
веточки, мягкие стебли злаков, растительный пух, шерсть лося и оленя,
перья куропатки, глухаря, клестов, кукш и других птиц; встречаются
собачья шерсть, клочки бумаги, ваты, веревок. Основу каркаса состав-
ляют травы и мхи, в лотке преобладают шерсть и перья. Гнезда с преобла-
данием мхов получаются жесткими и не разваливаются даже брошенные;
гнезда с травяным каркасом рыхлее и сплющиваются к вылету птенцов.
Наружный диаметр гнезд 110-135 мм, высота 70-110, диаметр лотка
54- 69, его глубина 40- 56 мм.
В кладке 3-7 яиц, в среднем 5,4 (п = 14). Длина яиц 20-22, ширина
15-16 мм. Яйца голубого цвета с редким темно-коричневым крапом, от
1-2 мм до мелких точек и редких линий. В одном гнезде юрка насижи-
вание продолжалось 12-13 дней. Из 12 гнезд одно было брошено по
неизвестным причинам, в одном птенцы погибли от дождей, три гнезда
разорены: таким образом, успех размножения у юрков не выше 60%.
Выводки при взрослых птицах в разные годы находили с 30. VI 1987 г. по
4. IX 1976 г. В июле-августе не раз находили мертвых слетков. 26. VII
1977 г. у речки Лылч пойман слеток с опухолью на сгибе крыла, вероятно
след травмы. Погибших взрослых находили реже: 18. VII 1971 г. на р. Чу-
на, в воде, найден труп самца со следами ударов, вероятно клюва: 1. VII
1977 г. после снегопада на берегу Чингльс-явра обнаружена мертвая
самка. К гнездовой смертности можно отнести и три случая выкармлива-
ния юрками кукушенка: 13. VII 1941 г. кормил самец, 13. VIII 1979 г. и
2. VIII 1987 г. - самки.
256
Возле гнезда и слетков, а также на кормежке юрки ведут себя доволь-
но агрессивно и нападают на любую птицу, от шура и белой трясогузки до
кукши, ястребиной совы и тетеревятника. И. VI 1980 г. при осмотре
гнезда самец пикировал на наблюдателя.
Питание юрков летом смешанное, в целом преобладает животный корм,
но в мае и осенью (VIII-X) с уменьшением доступности беспозвоночных
в холодное время доля растительных кормов возрастает. По визуальным
наблюдениям, весной и осенью юрки питаются семенами щавелей, зла-
ков, осок, березы, мокричника, подорожника - наряду с тлями и веснян-
ками. В исследованных Г. Новиковым в 1938 г. 75 желудках в состав
растительного корма входили (в порядке убывания) семена вороники,
черники, брусники, осок, шведского дерена, марьянника, злаков, не
определенных точнее, перловника, березы, живородящего гречишника,
купальницы, а также еловая хвоя, какая-то почка и мох, которые могли
быть случайной примесью; животный корм состоял из жуков (долгоноси-
ки, жужелицы, щелкуны и их личинки, листоеды, усачи, магкотелки,
навозники), гусениц бабочек, пауков, тлей, комаров, мух и их личинок,
перепончатокрылых (наездники, пилильщики, пчелы, осы, муравьи),
ручейников, веснянок, цикадок, клопов и слизней.
Размеры 48 взрослых самцов: длина 130-169, в среднем 148 мм, крыло
80-97, среднее 90 мм, хвост 55-77, в среднем 66 мм, масса тела 19,6-25,0,
в среднем 22,7 г; размеры 12 самок соответственно - 130-159, ср. 144 мм,
83—86, ср. 85 мм, 56- 70, ср. 63 мм, масса тела 17,9-26,5, ср. 22,5 г.
Щегол - Carduelis carduelis (L.)
На Кольский полуостров щегол залетает только изредка. В Лапланд-
ском заповеднике одиночных щеглов наблюдали трижды, почти на одном
месте, на Чунозерской усадьбе и на побережье Ельявра: 7. VII 1963 г.,
25. VI 1971 г. и 15. VII1981 г. (Ю. Гаев, 3. Аблаева). В 1957-1966 гг. щеглов
4 раза видели в Кандалакше (Коханов, 1987, с. 24).
Чиж - Spinus spinus (L.)
В Лапландском заповеднике встречается редко и не ежегодно, но все
же гнездится. Отмечался в 1931, 1937-1939, 1941, 1948-1949, 1961, 1965,
1971-1975,1977 (много), 1979-1981 и 1984-1985 гг., т. е. на протяжении 20
из 44 лет наблюдений; всего зарегистрировано 110-120 особей. Наблю-
даются чижи в основном осенью, что, вероятно, связано с поспеванием
семян березы.
Самое раннее появление чижа в заповеднике - 15. V 1971 и 1977 гг., в
среднем за 5 лет прилетает 21.V (учтены только первые встречи в мае);
самая поздняя встреча - 15. XI 1978 г., а средняя дата последней встречи
за 10 лет (исключая августовские) - 22. IX. Наблюдали их в разреженных
смешанных лесах в долине Чуны и у Чунозера. Годами подъема числен-
ности (по 10-20 птиц за сезон) можно считать 1965, 1974,1977 и 1984.
22. V 1977 г. пара чижей вела себя беспокойно и в клюве одной был
строительный материал. 30. VII 1974 г. и 21. VIII 1984 г. на Чунозерской
усадьбе видели выводки, в первом 4, во втором 5 слетков при взрослой
птице, у которой они выпрашивали корм.
Подобно чечеткам, стайки чижей в 3-5 птиц можно встретить в любой
17. Зак. 1023
257
месяц лета, что может объясняться растянутостью сроков гнездования и
зависимостью его от наличия кормов.
По непосредственным наблюдениям, корм чижей состоит из семян
берез, ели; 15. VII 1977 г. чижи ловили над рекой Чуной насекомых
на лету; в желудке чижа, добытого 24. VII 1939 г., Г. Новиков нашел два
десятка тлей и какие-то семена.
Масса тела взрослого самца 12,4 г, самки 11,8 г.
Обыкновенная чечетка - Acanthis flammea (L.)
Распространена на Кольском полуострове до Арктического побережья.
Современные систематики различают несколько подвидов и форм
чечетки, у нас встречается номинальная (A.f. flammea).
Чечетка - типичный инвазионный вид, характер ее пребывания зависит
от наличия основного корма - семян берез. У нас, как правило, на зиму
остается лишь часть особей, но в годы, богатые кормом, чечетки обычны
и в зимние месяцы. По литературным сведениям, зимой чечетки доле-
тают до Южной Европы, Средней Азии, Японии и Китая. Весенний прилет
мигрантов в заповедник наблюдается в разные гсды в разное время, с
февраля по май. Осенняя миграция начинается в августе, а кульмини-
рует в сентябре - начале октября и заканчивается зимой, в конце нояб-
ря. Миграции лучше заметны в годы с низким урожаем семян, когда
появившиеся птицы долго не задерживаются. В годы, богатые кормом,
картина расплывается за счет местных особей и задержки подлетевших
со стороны чечеток на неопределенное время. Весной и осенью чечетки
собираются иногда сотенными стаями, а скопления их на жировке в
березняках превышают тысячи, но здесь уже трудно выделять отдельные
группировки.
В заповеднике наибольшая встречаемость чечеток наблюдалась в
1932-1933, 1937, 1947, 1959-1960, 1966, 1968, 1971, 1974, 1977, 1980-1981 и
1985-1986 гг. Годы спада численности - 1936, 1940, 1948, 1961-1965, 1976.
Изменения численности чечеток повторяют колебания урожайности
березовых семян. Поскольку семена берез сохраняются в урожайные
годы до следующего июня, высокая численность чечеток может поддер-
живаться за счет семян прошлогоднего урожая.
Длительная депрессия в начале 60-х годов была следствием массового
усыхания берез в результате длительной вспышки размножения осенней
пяденицы в 1963-1965 гг.- (Семенов-Тян-Шанский, 1975). Интересно, что в
период 1963-1981 гг. в Шведской Лапландии наблюдались два пика
чечеток, в 1968 и 1971 гг. и один минимум в 1978 г. (Епетаг и др., 1981),
отмеченные и в Лапландском заповеднике, но за это же время в заповед-
нике прошло 6 максимумов и 5 минимумов. Эта разница, вероятно,
объясняется меньшей стабильностью урожаев березовых семян в наших
пределах в сравнении со Шведской Лапландией. Судя по ежегодным
учетам урожаев древесных семях, проводимым в заповеднике
продуктивность березы все же более устойчива, чем сосны, а тем более
ели. В последние годы заметна тенденция к увеличению численности
чечеток (рис. 56).
Зимой (ноябрь-март) в долине р. Чуна и у берегов Чунозера числен-
ность чечеток достигает 0,6— 9,0 особей/км2, весной (май-июнь) - до 6-18,
осенью (август-сентябрь) - 2,4-34 особей/км2. Локальная плотность в
258
августе может превышать 500 особей/км2. В заповеднике Малла на
северо-западе Финской Лапландии в июне 1979 и 1980 гг. она составляла
9 пар/км2.
Гнездовые стации чечеток разнообразны и включают различные типы
смешанных лесов из ели, сосны и березы. Они предпочитают опушки,
подрост на гарях и вырубках по южным склонам, заросли можжевельни-
ка, ивы, березовые, еловые и сосновые молодняки по долинам рек,
берегам озер, на болотах.
Гнездование и его сроки также зависят от наличия кормов. В годы,
богатые кормом, они начинают гнездиться уже в середине мая, а птенцы
из самых поздних гнезд вылетают в конце августа. Основную массу гнезд
находят в июне-июле. Вполне вероятно, что чечетка гнездится дважды за
сезон, но наши материалы кольцевания этого не подтверждают. Из 36
гнезд 17 построены в кусте можжевельника на высоте 0,2 - 1,0 в среднем
0,5 м; 10 - на елях в 0,5 - 8, в среднем - 2,3 м от земли; 3 - на соснах, на
высоте 0,4 - 1,8 м; 2 - на березах (одно на высоте 2,2 м), по одному в
березовом кусте в 1 м от земли, на карликовой березке и в ивовом кусту,
1 - в полудупле сухого ствола сосны. В общем 79% всех гнезд располага-
лись ниже 1ми12%-в1-3мот земли, остальные на высоте 4,5 и 8 м.
Строительный материал довольно однообразен: внешний слой из
сухих веток и травы, лоток выстилается тонкими травинками, эпифит-
ным лишайником (Bryopogon sp.), растительным пухом, перьями, волоса-
ми копытных, изредка пухом гоголя. Гнезда плотные, крепкие, их
внешний диаметр 9-11 см, внутренний - 4,5 - 7 см, глубина лотка
3,5 - 5 см. На одном можжевеловом кусту на болоте близ Улынчъявра
чечетки гнездились шесть раз, в 1978, 1979, 1980, 1982, 1985 и 1987 гг.,
только обновляя старое гнездо.
259
В 30 кладках в среднем было по 4,5 яиц: в 3 - по 6, в 16 - по 5, в 5 - по
4, в 6- по 3. Птенцы вылупляются на 10-12-й день после откладки
последнего яйца, а вылетают на 9- 14-й день. Насиживает только самка, а
самец ее кормит как во время инкубации, так и в первые дни после
вывода птенцов.
Корм птенцов и взрослых птиц летом смешанный: животный элемент
состоит из личинок пилильщиков, тлей, пауков, куколок мух, долгоносиков,
цикадок; растительный включает семена березы, осок, брусники, вороники и
других растений, почки черники. Семенами березы нового урожая чечетки
кормятся с момента их поспевания в конце июля - начале августа. Дополни-
тельными кормами служат семена ели, сосны, злаков, осок, одуванчика,
крапивы, таволги, щавелей, мокрицы, мари.
Размеры 13 взрослых самцов: длина 112-136, ср. 123 мм, крыло 73-78,
ср. 75 мм, хвост 51-61, ср. 56 мм, масса тела 10,2-16,2, ср. 13,0 г; 11 самок
соответственно 115-142, ср. 126, 72-78, ср. 74,50-63, ср. 57 мм, масса тела
9,9- 14,0, ср. 13,1г.
Тундряная чечетка - Acanthis hormemanni (Holb.)
На Кольском полуострове встречается к северу до Арктического
побережья (устное сообщение В.Д. Коханова).
Чечеток, относимых к этому виду, разные лица наблюдали в заповед-
нике осенью, с 13 по 31. X, и весной, между 6. III - 13. V в отдельные годы.
Тундряные чечетки встречались поодиночке или стайками 4-10 и только
дважды стаей в 30-60 особей. Они отличались светлой окраской; разли-
чий по размерам, голосу или поведению не замечено.
Среди добытых в 1932-1940 гг. и найденных мертвыми в 1974-1985 гг.
41 чечетки не было ни одной тундряной.
Коноплянка - Acanthis cannabina (L.)
Одиночная самка добыта на Чунозерской усадьбе 30. IV 1960 г. Ю.Г. Га-
евым. Встречали коноплянок и в других местах полуострова: 14. V 1963 г.
одну на острове Б. Айнов, а в 1975 и 1977 г. одиночные пары гнездились
на окраине Кандалакши (Коханов, 1987, с. 27).
Чечевица - Carpodacus erythrinus (Pall.)
Чечевица изредка гнездится по южному берегу полуострова, от Чапо-
мы до Кандалакши, и даже на южном побережье Имандры (Коханов, 1987,
с. 30).
В Лапландский заповедник залетала один раз: 6. VIII 1980 г. в ельнике
на берегу Ельявра Д. Карелин видел в стае чечеток одну чечевицу.
Щур - Pinicola enucleator (L.)
На Кольском полуострове распространен к северу и востоку до грани-
цы леса, по долинам рек доходит до побережья Баренцева моря, но на
зиму регулярно отлетает к югу.
В Лапландском заповеднике за 47 лет наблюдений щура только один
раз наблюдали в декабре (5. XII 1934 г.) и дважды - в январе (22.1 1932 г.
и 29. I 1940 г.). Средняя дата прилета за 43 года 2 марта (22. I 1932 г.-
260
22. Ill 1963 г.), при средней температуре - 11,Г и долготе дня около 10 ч.,
однако амплитуда сроков прилета достигает 2мес., соответственно
велика и о = ± 14,7 дней. Появляются щуры постепенно, в некоторые годы
после первой встречи их больше не видят несколько недель. Среди
передовых, как правило, преобладают ярко окрашенные самцы. Основ-
ная часть подлетает с середины марта до начала апреля. Размер весенних
стай не превышает трех десятков, но обычно не более десятка.
Осенние кочевки щуров начинаются в первой половине августа. Отлет
также происходит постепенно и продолжается до конца октября. Дата
последней встречи, в среднем за 42 года, 22 октября (3. X 1975 г. - 5. XII
1934 г.), средняя температура этого дня - 0,9е, долгота дня 9 ч., о дня отлета
± 16,2. В осенних стаях, как и весенних, бывает не более 30 птиц.
После прилета щуры равномерно рассеиваются по территории, их
можно встретить повсеместно в спелых смешанных лесах. В гнездовое и
послегнездовое время щуры чаще встречаются в разреженных елово-бе-
резовых лесах и сосново-еловых с примесью березы; во время кочевок
попадаются и в других типах, включая горные березняки, молодняки на
гарях и вырубках.
Щуры обычны и широко распространены по заповеднику, но в общем
немногочисленны. Плотность их в долине Чуны в июне составляет
0,6-2,1 пары/км2. В августе на трансекте длиной 24 км щуров можно
увидеть не каждый год, в марте же здесь встречается 1-10 особей.
Мелодичное пение щуров слышится сразу после их прилета и продол-
жается до середины июня; осеннее пение, в котором участвуют и моло-
дые птицы, отмечали в разные годы между 12. VIII-10. X.
Гнезда щуров в заповеднике находили в разреженных сосново-еловых
лесах с примесью березы (6) и в сосновом жердняке (1). Они располага-
лись на елях различного возраста (одно на сосне), на ветках у ствола, на
высоте 2-4 м. Все были укрыты от осадков сверху, а одно хорошо замас-
кировано и сбоку. Каркас гнезда состоит из сухихи веточек ели и
березы, стеблей и корней кустарничков максимальной длиной до 56 см.
Средний слой из более тонких концевых веточек березы, ели и кустар-
ничков, а лоток выкладывается из тонких былинок, корешков, изредка
лосиных волос. Гнезда представляют сравнительно непрочную, рыхлую
постройку; в одном из трех было вдвое меньше строительного материала,
чем в других. Размеры: внешний диаметр от 150 х 165 до 220 х 350 мм,
внутренний 68-73, высота 72-110, глубина лотка 30-43 мм.
В гнездах, найденных 6, 13 и 24 июня 1977 г., было 3, 4 и 4 яйца во всех
самки насиживали; в гнездах 11. VI 1964 г. и 9. VI 1986 г. - по 2 яйца
(неполная кладка), судьба их не прослежена; в гнезде 29. VI 1987 г. - 3
птенца (Л. Киренков). Размеры яиц: длина 25-28, ширина 17-18 мм
(п=6). Одно из 7 гнезд найдено брошенным после снегопада, 2 разорены,
предположительно горностаем, в третьем вывелся лишь один птенец, а
три яйца исчезли. Этот птенец на 12-13-й день, достигнув массы тела
38 г, вылезал из гнезда на ветки, затаивался, а на 16-й день его в гнезде
не оказалось. Первая встреча слетка, пойманного руками, 21. VI 1974 г.
Передачу корма взрослыми щурами птенцам наблюдали 16. VII 1964 г. и
6. VIII 1982 г.
Щуры на гнезде и около него доверчиво спокойны, и насиживающая
самка позволяет трогать себя руками, а при осмотре гнезда взрослые
птицы не проявляют заметного беспокойства. Лишь после появления
261
птенца взрослые некоторое время опасались сразу подлетать к гнезду.
Корм гнездовых птенцов (8 проб) смешанный, включает семена брусни-
ки, ели (случайно и еловую хвою), личинок пилильщиков, гусениц, в том
числе пяденицы, пауков, тлей, долгоножек, комаров и моллюсков;
попадаются и кварцевые песчинки.
Судя по анализу желудков и визуальным наблюдениям, в снежный
период, с прилета до схода снега и после выпадения снега до отлета,
щуры в основном питаются женскими цветочными почками ели, а изред-
ка и сосны. Сорванные почки они каким-то образом освобождают от
сухих покровных чешуек, так что в зоб они поступают в виде светло-зе-
леных конических шишечек. В зобу, пищеводе и желудке щура, добыто-
го 13. III 1941 г., найдены 163 цельные шишечки, не считая некоторого
количества измельченных. Щуры, будь они массовой птицей, могли бы
серьезно влиять на семенную продуктивность ели. В апреле, с появле-
нием проталин, щуры^астично кормятся на поверхности земли перези-
мовавшими ягодами брусники и вороники (наблюдения 15. IV 1981 г. и
23. IX 1948 г.). Используют при случае и экзотические корма: 21. IV 1977 г.
подбирали в поселке овес, а 3. V 1975 г. - семена лиственницы в дендро-
логическом питомнике. В бесснежный период они кормятся преимущест-
венно на земле: последние случаи питания еловыми почками отмечены
7. VI 1938 г. и 13. VI1951 г., а первые жировки на вершинах елей - 26. IX
1940 г. и 30. IX 1947 г. Летний рацион включает спорангии кукушкина
льна, ягоды брусники, вороники, толокнянки, можжевельника, семена
осок, марьянника, цветы черники, почки и молодые листья берез; позд-
нее добавляются ягоды черники, семена ожики, а с сентября - березовые
семена и ягоды рябины. У сочных плодов они выбирают мякоть с семена-
ми, отбрасывая кожуру. Иногда щуров наблюдали на водопое (13. X
1960 г.) или поедающих снег (23. IV 1939 г.).
Сведения о смертности скудны: 22. IV 1973 г. у устья Чуны и 1. VI
1981 г. на берегу Чунозера обнаружены остатки щуров, пойманных
хищной птицей. В отличие от насекомоядных птиц щуры, видимо, не
страдают от летних снегопадов.
Размеры 16 взрослых самцов: длина 186-240, в среднем 212 мм, крыло
105-116, ср. 111 мм, хвост 81-99, ср. 89 мм, масса тела 47-60 г, ср. 53,3 г;
16 взрослых самок соответственно 187-227, ср. 208, 102-117, ср. НО,
72-92, ср. 86 мм, масса тела 47-60 г, ср. 53,0 г.
15. VIII 1986 г. на северном берегу Улынчъявр встречены два щура, из
которых один, вероятно самец, был черного цвета, другой - обычного
окраса взрослая самка.
Клест-еловик - Loxia curvirostra L.
Местное название: саамское - Урпша.
Все клесты типичные инвазионные виды. Налеты клестов-еловиков
четко совпадают с хорошим урожаем семян ели и сосны, наоборот, в
неурожайные годы численность их резко уменьшается или они полностью
исчезают.
В Лапландском заповеднике высокая численность еловиков наблюда-
лась в 1935, 1938-1939,1947, 1951, 1958-1959,1961, 1968, 1970-1972, 1974,
1977-1979 и 1987 гг.; низкая численность - в 1933, 1941, : 1949-1950, 1963—
1964, 1969, 1975, 1982 и 1985-1986 гг. Подъем численности не был отмечен
262
Рис. 57. Клесты выщипывали пропитанное солью дерево под окном лесной избушки;
на подоконнике виден пакет с солью. 10. VI 1971 г. Фото О.И. Семенова-Тян-Шан-
ского
в годы хорошего урожая ели 1973 и 1983 гг. и сосны - в 1982-1983 гг.
Вероятно, клесты не иммигрировали к нам в эти годы потому, что не
испытывали затруднений с питанием на местах рождения. Клестов-ело-
виков можно встретить во всех типах хвойных лесов, предпочтительно в
долинах рек и озер.
Материалы по гнездованию клестов-еловиков в Лапландском заповед-
нике, собранные Ю.Г. Гаевым в 1959-1961 гг., опубликованы в работе
Коханова и Гаева (1970), и здесь мы их не переписываем.
Необычайно раннее гнездование клестов (февраль-апрель) объясняет-
ся, тем что они выкармливают птенцов еловыми и сосновыми семенами
и стремятся использовать этот ресурс ”на корню”, прежде чем шишки
раскроются и семена из них начнут выпадать.
Судя по клестам, добытым в 1937-1939 гг., гонады у самцов и самок
начинают увеличиваться в третьей декаде января, достигают максимума
в середине февраля и начинают спадать с конца июля. Строительство
гнезд наблюдалось с 17. П (1939) до 2. IV (1940). Три гнезда, найденные в
1932,1939 и 1940 гг., располагались на елках, одно из них на высоте 2,5 м.
Каркас их состоял из сухих еловых веточек эпифитного лишайника
(Bryopogon sp.) и ягеля, выстилка из того же лишайника, перьев и пакли.
Выводки из 2-5 птенцов отмечались самое раннее 23. IV 1938 г., самое
позднее - 9. VIII 1979 г.
Корм еловиков по анализу 67 желудков, добытых в 1937-1939 гг.,
состоит в основном из семян ели, встречаемость их по сезонам следующая
(в %): весна - 50, лето - 63, осень - 100, зима - 98. Семена сосны обнару-
жены в единичных желудках. В неурожайном 1937 г. дополнительным
кормом служили семена вороники, бодяка, перловника; летом изредка
встречались тли, долгоносики, жужелицы и пауки.
263
Таблица 16. Размеры клестов в районе Лапландского заповедника
Вид клеста Длина, мм Крыло, мм Хвост, мм Вес тела, гр Число экз.
Сосновик, d 176 юз 1 66 54,6 13
148-200 99-107 60-73 51-59
Сосновик, 9 176. 99 64 52,7, 8
152-190 88-106 61-67 49-58
Еловик, d 155 96 61 42,0 31
138-178 91-101 52-65 32-47
Еловик, 9 152 94 60 41,8 22
140-168 90-102 50-66 36-47
Белокрылый 155 94 65 34,0 13
клест, d 139-185 88-105 62-72 30-40
Белокрылый 146 89 59 31,6 5
клест, 9 130-162 85-91 53-63 29-34
Клесты, зимняя диета которых практически состоит из одних семян
хвойных, в отличие от большинства наших птиц остро нуждаются в соли.
В заповеднике во всех лесных избушках держат запас соли, обычно на
подоконнике; \во влажную погоду она размокает и рассол стекает на
наружную сте^У избы. Клесты отдирают клювами просоленные древесные
волокна, освежая таким способом поверхность дерева под окном, так
что это место выглядит светлым пятном на фоне почерневших от непого-
ды старых бревен (рис. 57). Используют они и другой источник соли, снег
пропитанный человеческой мочой. Весной мы не раз наблюдали клестов,
подбиравших рыбьи косточки на местах/ где выливали помои.
Сведения о смертности: 7. II 1941 г. обнаружены остатки клеста-елови-
ка, съеденного куницей; 31. V и 13. VI 1936 г. найдены трупы взрослых
клестов, а 30. VII 1974 г. - слетка, причины смерти которых не опреде-
лены.
Все три вида клестов различаются по размерам, и мы приводим их для
сравнения в одной таблице (табл. 16).
Здесь в числителе среднее значение промера, в знаменателе - мини-
мальное и максимальное.
Клест-сосновик - Loxia pityopsittacus Borkh.
Подобно еловику сосновик живет оседло только в сезон размножения,
а остальной год проводит кочуя по лесам в поисках мест, лучше обеспе-
ченных кормом. Вследствие этого его численность в каждом отдельном
районе резко колеблется по годам, падая иногда до нуля на несколько
лет. За годы работы Лапландского заповедника инвазии клеста-соснови-
ка наблюдались в 1940/41, 1958/59, 1973/74, 1977/78 и 1987 гг. Численность
популяции меняется не только по годам, но и по сезонам: при обильном
урожае шишки, во втором полугодии встречается больше клестов, чем в
264
первом (если предшествующий год был неурожайным). Клесты-соснови-
ки предпочитают водораздельные сосновые боры, сосново-еловые леса,
сосновые редколесья, но могут встречаться и в ельниках (Владимирская,
1948, с. 221; Новиков, 1952).
В общем клесты-сосновики встречаются реже еловиков, об этом
свидетельствует подсчет карточек биосъемки: из 137 добытых в старые
годы клестов на долю еловиков приходится 55%, сосновиков - 26%,
белокрылых - 19%. В годы подъема в смешанном лесу в долине р. Чуна в
июне плотность сосновиков не превышает 0,5— 2,0 пар/км2, в другие годы
на маршрутах длиной более 20 км они .могут и не встретиться. Эффект
обилия клестов (не только сосновиков) создается за счет их заметности,
способности образовать в период кочевок стаи до десятков и сотен птиц и
внезапности их появления.
Клесты-сосновики приступают к размножению в конце января, с
удлинением светового дня. Начало разбивки стай на пары и мелкие
группы в 1939 г. отмечен Г. Новиковым уже 21. I. Клесты-сосновики
могут петь в течение всего года, даже в полярную ночь (3.1 1978 г.), но с
разбивкой на пары интенсивность пения заметно повышается и самцы
иногда дерутся (17. II 1939 г.). Семенники самцов достигают наибольших
размеров к концу февраля, спадают к августу, а в сентябре приходят в
состояние покоя. У самки, добытой 21. III 1941г., яйцевод был увеличен,
265
размер фолликул достигал 8 мм, а 26. III 1941 г. наблюдали совокупление
клестов-сосновиков (О. Семенов-Тян-Шанский).
Слетки при взрослых птицах отмечались в 1935, 1941, 1951, 1973, 1974,
1975, 1982 гг. с 24. IV по 5. VIII. У слетков клестов клюв еще не имеет
перекреста, поэтому они не в состоянии вылущивать семена из шишек
сами и взрослые продолжают их подкармливать целый месяц после
вылета. В выводках встречали от 1 до 5 птенцов. Судя по датам встреч
слетков, сосновики гнездятся вторично в июне-августе.
По содержанию 23 желудков и многочисленным наблюдениям в
природе сосновики в течение всего года питаются семенами ели и сосны,
при этом еловые семена потребляются преимущественно зимой, а сосно-
вые - весной и летом. Изредка в желудках находили семена вороники и
черники. Неоднократно наблюдали, как клесты-сосновики поедали кашу
из собачьих плошек, рыбьи кости, выклевывали известь из печной
трубы, щепки с пропитанных солью досок, подбирали песчинки с земли
или печки, с цементного раствора.
Сведений о смертности мало. Гнезда клестов нередко разоряют кукши;
по крайней мере, из большей части гнезд, обнаруженных Ю. Гаевым
(почти 30), птенцы не вылетели по этой причине. 29. VII 1936 г. в долине
Чуны нашли перья и челюсть сосновика, пойманного хищной птицей;
14. V 1941 г. на усадьбе заповедника подобрали клеста, разбившегося о
провод; 18. IV 1974 г. взрослая самка, а 21. VI 1974 г. самка-сеголетка
подобраны мертвыми на усадьбе.
Сосновик на 28% тяжелее еловика, но различать их в поле довольно
трудно, в бинокль наиболее заметна разница в высоте и массивности
клюва сосновика (рис. 58). Линейные размеры обоих видов перекрывают-
ся, не перекрывается только масса тела.
Белокрылый клест - Loxia leucoptera Gm.
Инвазии всех видов клестов зависят от локализации урожая семян
хвойных пород, поэтому они совпадают во времени, но белокрылый клест
встречается реже других видов и за 45 лет наблюдений был отмечен на
протяжении только 20. Сравнительно часто он встречался в 1935, 1938,
1947, 1965, 1971, 1976, 1978 и 1986/87 гг. Во время миграции белокрылые
часто держатся вместе с другими клестами и легко могут остаться неза-
меченными среди преобладающих в стае еловиков и сосновиков. Бело-
крылому клесту с его сравнительно слабым клювом сосновые семена
становятся доступны, лишь когда шишки сами раскроются, что наступает
летом, с конца мая по август, тогда как сосновики обрабатывают сосно-
вые шишки уже зимой, а еловики - примерно с апреля. Еловые шишки
белокрылые клесты вскрывают с ноября, а другие клесты - с августа.
Сроки созревания и высыпания семян из шишек определяют их доступ-
ность ,для клестов и потому существенно влияют на распределение
клестов между борами и ельниками.
Сроки размножения белокрылых клестов те же, что и у предыдущих
видов, с той разницей, что при хорошем урожае сосны и неурожае ели
белокрылые клесты начинают гнездиться не в феврале, а только в мае-
июне. Подобные колебания сроков размножения в зависимости от дос-
тупности кормов наблюдаются и у других клестов, но в меньшей степени
(Коханов, 1970).
266
Рост семенников у самцов белокрылого клеста замечен с 15. I 1939 г.,
полное их развитие - с 17.П 1939 г. до 5.УШ 1939 г. Хорошо развитые яични-
ки с фолликулами до 3 мм наблюдались с 17. II 1939 г. до 10. VIII 1939 г.
Выводки от 1 до 4 слетков при взрослых птицах встречались в разные
годы с 24. IV 1947 г. по 1. VIII 1938 г.
Пища белокрылых клестов состоит из семян ели и сосны с добавле-
нием плодов других растений, в том числе ягод вороники и коробочек
кукушкина льна; летом часто в небольшом количестве поедаются пауки,
тли, гусеницы бабочек и личинки пилильщиков.
Высота клюва у самцов: белокрылый клест 9,5 мм, еловик 11-11,5,
сосновик 14,5.
Снегирь - Pyrrhula pyrrhula (L.)
Встречается по всему Кольскому полуострову до северной границы
леса, а по речным долинам доходит до морского побережья (Кищинский,
I960), однако редок даже на юге области.
В Лапландском заповеднике снегирь встречается ежегодно, но числен-
ность его заметно колеблется по годам: сравнительно часто он попадался
в 1937-1938, 1965, 1974-1975, 1977, 1980-1981 и 1984 гг. Связать колеба-
ния численности с урожаем семян березы, ели, сосны и различных ягод
нам не удалось.
Миграции снегиря очень нерегулярны, иногда он отлетает к югу уже в
октябре, в другие годы задерживается до середины декабря, а вновь
появляется нормально в марте или первой половине апреля, изредка уже
в феврале. В последние годы снегири, подобно свиристелям, зимуют в
городах Мурманской области - Апатитах, Кировске и Мончегорске: в
Апатитах 8 снегирей прожили всю зиму 1979/80 г. (сообщение Д.М. Шкля-
рова, Лаборатория охраны природы), зимовали они там же в 1985/86 и
1986/87 гг. Средняя за 24 года дата первого появления снегирей в запо-
веднике - 27 марта (1. II 1965 г. - 30. IV 1962 г.), при средней температуре
воздуха -6,2е; средняя дата последней встречи за 29 лет - 16 ноября
(12. X 1975,1977 гг. - 29. XII 1978 г.), при -5,9е. Кочующие снегири держат-
ся стайками до 6-8 птиц, иногда поодиночке в стаях чечеток. Встречают-
ся в лесной зоне повсеместно, но чаще в долинах.
В гнездовой период предпочитают разреженные участки с молодым
подростом и подседом из можжевельника и березы. Сведения о гнездо-
вании фрагментарны. 20. IV 1940 г. А. Назимович наблюдал у берега
Ельлухт в стайке из 5 снегирей драку самцов; 9. VI 1978 г. при устье Чуны
самец передал корм в клюв самки (ритуальное кормление); 25. VI 1938 г.
добыта самка с хорошо развитым яйцеводом, с фолликулами до 8 мм;
9. VIII 1937 г. близ Чунозера наблюдали выводок, а 12. VIII из другого
выводка добыт слеток (П. Мальчевский).
Судя по анализу 8 желудков и визуальным наблюдениям, снегири
питаются плодами и семенами различных растений: ягодами вороники,
черники, брусники, шведского дерена, семенами осок, андромеды,
марьянника, одуванчика, крапивы, щавелей. Зимой имеют большое
значение семена берез, а летом в рацион входит небольшое количество
членистоногих, в частности пауки и долногосики.
Размеры 4 взрослых самцов: длина 145-180 мм, крыло 87-98 мм, хвост
70-74 мм, масса тела 30,0-33,9 г; размеры 3 самок соответственно -
159-172, 86-90, 67-71 мм, масса тела 25,0-34,7 г.
267
Дубонос - Coccothraustes coccothraustes (L.)
В Лапландском заповеднике залетных дубоносов наблюдали дважды:
4. V 1977 г. А. Гилязов заметил в стае зябликов, кормившейся на свалке
на Чунозерской усадьбе, самку дубоноса; 25. V 1986 г. О. Семенов-Тян-
Шанский видел в 2-3 м от себя за окном также самца. Весной 1981 г.
одиночных дубоносов видели в Порьей губе и на о-ве Харлов (Коханов,
1987, с. 24).
СЕМЕЙСТВО ТКАЧИКОВЫЕ - PLOCEIDAE
Домовый воробей - Passer domesticus (L.)
На Кольском полусотрове уже сто лет назад он изредка встречался в
Кандалакше и Коле (Плеске, 1887, с. 230). Массовому расселению воробья
по Мурманской области способствовала постройка в 1915-1916 гг. Мур-
манской железно-дорожной магистрали, в строительстве которой участ-
вовали тысячи людей и широко применялся конный транспорт. К 1955—
1957 гг. воробьи гнездились в Мурманске и Териберке, теперь живут в
поселках по восточному побережью, нет их только в глубинных районах,
например в поселке Краснощелье в апреле 1976 г. мы их не обнаружили.
На территории Лапландского заповедника воробей впервые появился
10. X 1935 г. (Владимирская, 1948, с. 224), через 5 лет после постройки
первого кордона. На опытной станции ВИР в Хибинах уже в 1930 г. мы
застали воробьев, а вслед за организацией в 1931 г. совхоза ’’Индустрия”
(у станции Апатиты) они появились и там, к 1938 г. - во множестве на
Чунозерской усадьбе летом 1937 г. воробьи впервые гнездились и вывели
птенцов, но к зиме исчезли. То же повторялось ежегодно, весной воробьи
появлялись, летом выводили птенцов, а к зиме все исчезали. Когда
заповедник был закрыт, воробьи заселили поселки лесорубов в долине
Чуны и Мавра-губе, но исчезли через год после закрытия лесоучастков. С
1974 г. воробьи постоянно живут и гнездятся на усадьбе заповедника, но
на кордоны и сторожевые избушки они залетают только эпизодически. На
Чунозерской усадьбе доживали до весны от 2 воробьев в 1977 г. до 12 в
1980 г. Должно быть, воробьи, хотя бы частично, переселяются на зиму из
одних населенных пунктов в другие, так как вновь появлялись либо
весной, либо осенью: весной, в среднем за 10 лет, - 9 мая (13. IV 1939 г. -
6. VI 1969 г.), осенью, за 13 лет, - 25 октября (21. IX 1938 г. - 25. XI 1960 г.).
Уменьшение численности зимующих воробьев объясняется большой
смертностью их в суровые зимы, например в 1980/81, 1981/82 и особенно
1984/85 гг., когда морозы достигали -50* и все воробьи вымерли. После
того 17. IV 1985 г. появился один самец, 5. V 1985 г. оказалась уже пара
воробьев, и колония возродилась.
Весеннее возбуждение воробьев в Мончегорске наблюдается уже в
конце января - начале февраля, на усадьбе заповедника между 4. IV
1984г. и 26. IV 1983 г. в разные годы. Стройку гнезд наблюдали самое
раннее 2. IV 1983 г. в Мончегорске, 23. IV 1982 г. - на Чунозере. Из 19
обследованных гнездДббыли устроены под крышами домов, 3 - в дуп-
лянках для мелкйх птиц. Выбирая место для гнезда, воробьи ведут себя
агрессивно. В мае 1981 г. пара воробьев заняла сначала гнездо мухолов-
ки-пеструшки, потом сероголовой гаички, выбросив из дуплянок их яйца
и птенцов.
268
В Мончегорске слетки встречаются со второй декады мая до конца
августа: 17. V 1984 г. у автовокзала видели самца, кормившего 5 слетков,
а 5 VIII 1979 г. видели в гнезде еще пухового птенца. На усадьбе заповед-
ника ранние выводки встречали с середины июня, вторые семьи - с
10. VII. Вылетевших птенцов обслуживает обычно именно самец, вероят-
но, потому, что самка насиживает вторую кладку.
Больше всего воробьев наблюдали в 1980 г., когда в результате разм-
ножения 5-6 пар общая численность их на усадьбе достигла к концу
августа четырех десятков.
Воробьи питаются в основном пищевыми отходами на помойках и у
скотных дворов, регулярно пользуются они и птичьими кормушками.
Входят в их рацион и природные корма - семена сорняков, щавелей,
злаков, крапивы; затем почки ив, очищенные от чешуй, и беспозвоноч-
ные, в частности короеды, долгоносики, личинки пилильщиков, комаров
толкунов, мух.
В южной части Мурманской области гнездится (в Кандалакше с 1975 г.;
Коханов, 1987, с. 34) полевой воробей (Passer montanus (L.)), который
может залететь и в район заповедника.
СЕМЕЙСТВО СКВОРЦЫ - STURNIDAE
Скворец - Sturnus vulgaris L.
Скворец почти такой же синантроп, как и воробей, с той разницей, что
воробьи кормятся пищевыми отходами и фуражом, а скворцу необходи-
ма пашня, на которой он находит червей и личинок насекомых. Поэтому
скворец продвигается на север вместе с земледельческой культурой.
Впервые скворцы появились в районе заповедника еще в 30-х годах:
С.А. Стрелков видел одиночных скворцов весной 1935 г. в Кандалакше, а
29. V 1938 г. - в Мончегорске. 12. VII 1940 г. у домов в Витте-губе (Боль-
шая Имандра) видели 6 птиц (возможно, выводок?). В районе Апатит
скворцы гнездятся, по крайней мере с 1950 г., когда там встречали по
десятку птиц за сезон.
На территории заповедника скворцы впервые появились в конце 50-х
годов, когда он не функционировал и в нем рубили лес. В заповеднике
одиночных наблюдали в 1960, 1961 и 1964 гг., а с 1966 - почти ежегодно,
исключая 1978, 1980 и 1983-1988 гг. Встречали их всегда возле жилья-
на территории усадьбы, у кордонов, поселков лесорубов. Большинство
наблюдений относятся ко времени весеннего пролета, обычно по 1-2
птицы, иногда 3; только в Апатитах и Краснощелье наблюдали до 4 пар
(И. Паракецов).
Средняя дата прилета скворца в районе заповедника за 18 лет - 19
апреля (25. III 1973 г. - 13. V 1966, 1985 гг.), при средней температуре
-1,8е. Позднее июля скворцов не отмечали ни разу. Петь они начинают
сразу после прилета. Несмотря на наличие скворечников, в заповеднике
известны только три случая гнездования:
14. VI1961 г. в поселке бывшего лесоучастка заселенный скворечник; у подросше-
го птенца, взятого из него 24. VI, обнаружено 14 личинок мух, паразитировавших под
кожей (А. Герман);
2. V 1974 г. пара заняла скворечник у кордона Нижняя Чуна; птенцы в нем
появились 1. VI и вылетели к 17. VI;
269
27. IV 1975 г. у того же скворечника появился один скворец, а на следующий день
пара; 12. V они таскали в гнездо сухую траву; 4. VI в гнезде обнаружены 3 мертвых
пуховых птенца (причина их гибели не выяснена; А. Лихачев).
1. VII 1962 г. близ поселка Тик-губа видели стаю около 50 скворцов (объединен-
ные выводки).
СЕМЕЙСТВО ВРАНОВЫЕ - CORVIDAE
Сойка - Carrulus glandarius (L.)
В нашем регионе доходит к северу до южного побережья Белого
моря. В Лапландию только залетает (Плеске, 1887, с. 269).
В Лапландский заповедник сойка залетает нередко, с первых лет его
существования: ее встречали в 1932, 1937-1939, 1947-1950, 1959, 1961—
1967, 1971-1978. 1981 и 1985,1987 гт. Пара соек или одиночная появлялись
обычно осенью (VIII—XI), жили на усадьбе и около нее от 1 до 9 месяцев, а
потом исчезали. В 1961/62,1972/73,1974/75 и 1977/78 гг. - сойки прожили у
нас всю или часть зимы. Интересно, что именно в эти годы сойка в Ленин-
градской области была многочисленна и совершала дальние миграции
типа инвазий (Мальчевский, Пукинский, 1983). Несомненно, что залеты
соек на север связаны с вспышками их численности на юге, в области
основного ареала.
По-видимому, в некоторые годы сойки, прожившие близ жилья до
весны, улетали в лес гнездиться. Так, в 1932 г. И.Г. Эйхфельд видел в
Хибинах выводок соек; в октябре 1961 г. Ю.Г. Гаев на Чунозерской
усадьбе отловил и закольцевал 6 соек, очевидно выводок; 13. X 1971 г.
О.А. Брагина видела там же 5 соек; 7. VI 1977 г. у Второго ручья наблюда-
ли сойку с кормом в клюве; 30. VIII 1976 г. лесник В. Хайретдинов видел
на р.Купись стаю соек, очевидно выводок.
Судя по визуальным наблюдениям, сойки питаются ягодами рябины,
вороники, брусники, семенами березы, полевками, пищевыми отброса-
ми. Желудок сойки, убитой 7. V 1938 г. на Старой базе заповедника, был
наполнен в основном тараканами и их коконами, в нем же найдена
мясная муха, семена вороники, брусники и семена березы.
Из 6 окольцованных на Чунозерской усадьбе осенью 1961 г. соек одна
была поймана вторично там же 15. XII, другая убита 8.XI 1961 г. у Екост-
ровского пролива, в 20 км к востоку от места мечения.
Размеры 2 взрослых самок: длина 335, крыло 179-185, хвост 128-143,
масса тела 146 и 166 г.
Кукша — Perisoreus infaustus (L.)
Местное название: саамское - кузеньк.
Кукша самая оседлая из всех врановых птиц, что связано с ее привыч-
кой запасать корм на зиму. Ее оседлость подтверждают и результаты
кольцевания, проведенного в Лапландском заповеднике в 1959-1960 гг.
Ю. Гаевым. Из двух кукш, окольцованных в сентябре 1959 г., обе отлов-
лены на том же месте, одна - через полгода, вторая - через 13 месяцев;
третья, помеченая на Чунозерской усадьбе сеголетком, 22. IX 1959 г.,
повторно ловилась там же 16. X 1959 г. и 8. III 1960 г.
В заповеднике кукша заселяет различные типы хвойного леса (сосно-
во-еловые, елово-березовые, сосновые боры), в них же и гнездится. В
270
поисках корма, особенно в выводковый период, залетает в горные берез-
няки, зарастающие молодняком гари и вырубки, избегая только горных
тундр и обширных болот.
Хотя кукша принадлежит к числу самых обычных птиц, численность
ее невысока. В 1978-1984 гг. в смешанных лесах долины р. Чуна на
маршруте длиной 34 км в июне плотность ее не превышала 0,6-1,2 особи/
км2, в августе 1,4-1,8 особей/кмг. В 1981 г. кукш на маршруте не встрече-
но. Индивидуальная территория пары кукш определена на острове
Великом в Кандалакшском заповеднике - 0,5 кмг (Коханов, 1982), в
Скандинавии - 1-5 кмг (Blomgren, 1971), в долине Колымы - 1 км2 (Креч-
мар и др., 1978).
Судя по многолетним наблюдениям, численность ее заметно колеблет-
ся. Количество встреч за год в 1938, 1939, 1950, 1960, 1962, 1967, 1971, 1977
и 1978 гг. было в 2-3 раза выше, чем в соседние годы. Подъемы числен-
ности кукш совпадали с вспышками численности полевок в 1938, 1950,
1970 и 1977 гг. Тут связь может быть двоякой: с одной стороны, полевки и
лемминги - объекты ее питания, с другой стороны, куница, часто разо-
ряющая гнезда кукш, при обилии грызунов питается почти исключитель-
но ими и оставляет птиц в покое.
В 1937-1938 гг. встречаемость кукш на одночасовой экскурсии состав-
ляла летом 0,7, зимой 0,1 особь (Новиков, 1956). По нашим наблюдениям
в 1982-1983 гг. соответствующие индексы 0,4-0,6 и 0,1-0,2. Существен-
ных изменений численности кукши за полвека не обнаружено.
В зимнее время кукши в лесу малозаметны, отчасти ввиду спокойной
манеры поведения, отчасти потому, что они концентрируются на корм-
ных местах (у падали, жилья), держась обособленными парами и не
образуя стай.
Кукша гнездится раньше всех других представителей отряда, исклю-
чая только клестов и вброна. Пение ее в разные годы слышали между
5. III 1966 г. и 28. III 1982 г. Вокализация поющей кукши разнообразна и
напоминает одновременно песни дроздов, скворца и бекаса. Поющий
самец тихо передвигается в кроне дерева, самка находится поблизости.
Брачные игры в виде погони друг за другом, ритуального выпрашивания
самкой корма у самца наблюдались в апреле (2. IV 1941 г. - 21. IV 1975 г.).
В те же сроки встречали и птиц с гнездовым материалом (ветка, перо) в
клюве. У самки, добытой 26. IV 1938 г., в яйцеводе оказалось готовое к
откладке яйцо в скорлупе, а в яичнике два желтка 13 и 11 мм.в диаметре;
на брюхе наседное пятно. По процессам кладки и насиживания у нас нет
наблюдений.
Из гнезда, обнаруженного О. Семеновым-Тян-Шанским у подножия
Пыдчельм-варь 10. VI 1940 г., бросились врассыпную 4-5 птенцов, еще
плохо летавших. Гнездо построено на высоте 3,5 м на елке, ростом 5-6 м,
в сыром, заболоченном ельнике на пологом южном склоне; сложено
довольно непрочно из сухих еловых веточек и лишайника (Usnea
plicata). Внешний диаметр гнезда 22 см, лотка, уплощенного птенцами,
13 см, глубина его 2-3 см. Другое гнездо, найденное А. Насимовичем
25. VII 1940 г. в бору с сосново-еловым подростом близ Кислой губы,
находилось на ветке у ствола ели высотой 8 м, с южной стороны в 4,5 м от
земли. Гнездо старое, оставленное выводком; каркас из сухих сосновых
веток, частью с хвоей; лоток сложен из зеленого мха (Pleurozium schrebe-
ri), ягелей (Cladonia alpestris, Cl. rangiferina) и перьев рябчика.
271
Средняя за 25 лет дата встречи первого выводка кукши 10 июня (30. V
1985 г. - 27. VI 1975 г.). Выводки состоят из 1-5, в среднем из 2,8 слетков
(п=52).. Выводки с 5 птенцами встречались лишь дважды. Семьи, скорее
всего, начинают распадаться с установлением снежного покрова, конце
октября. Изредка группы по 3-6 птиц встречаются близ жилищ, где они
могут найти корм, в декабре-январе.
Питание кукши очень разнообразно (Владимирская, 1948; Коханов,
1982). Г.А. Новиков (1956), изучавший питание кукши по содержимому
желудков, нашел мелких грызунов в 41% образцов, пауков - в 43%;
далее - землероек, ос (личинки и куколки), слизней, коробочки кукуш-
кина льна (17%), семена и хвою ели, семена березы, рябины, марьянника;
ягоды Ёороники, черники, брусники, морошки; грибы.
В табл. 17 суммированы материалы по питанию кукши, собранные в
1930-1983 гг. путем прямых наблюдений, с включением 3 желудков,
добытых в старые годы.
Таблица не дает полного представления о рационе кукши, так как в
ней не отражено ее питание в гнездовый период, в частности корм птенцов.
В период размножения кукши живут в лесу, и в их питании большую
роль играют различные беспозвоночные, яйца и птенцы мелких птиц.
Эпифитные лишайники, вероятно, попадают в желудок кукши случайно,
при ловле пауков и насекомых на стволах деревьев.
Характерная особенность питания кукши - запасание кормов на зиму,
наиболее заметное с конца августа до октября. Кукши прячут различный
корм, засовывая в щели, трещины, под кору, подвешивая на ветках
деревьев. Описаны случаи, когда кукши затаскивали рыбьи потроха в
крону. А.В. Кречмар (1978) наблюдал в долине Колымы, как несколько
272
Таблица 17. Питание кукши в бесснежный и снежный периоды
Объекты питания Май-октябрь ’Ноябрь—апрель
число встреч % число встреч %
Отбросы возле жилищ 39 1 1 48 1 30 1 54
В том числе
рыба 18 —
овес 2 2
Падаль 5 6 17 30
В том числе
олень — 12
лось 2 5
тетеревиные 3 —
Мелкие позвоночные 14 17 1 2
В том числе
землеройки 2 —
полевки и лемминги 9 1
воробьиные птицы 1 —
яйца (дрозд?) 2 —
Членистоногие 7 8 4 7
В том числе
пауки и их коконы 2 —
веснянки 2 3
разные насекомые 2 1
имаго
личинки насекомых 1 —
Ягоды и семена 12 15 3 5
Брусника 4 1
Черника 8 —
Вороника — 1
Семена — 1
Грибы 4 5 — —
Эпифитные лишайники 1 1 1 2
Всего: 82 100% 56 100%
кукш за 2-3 дня выбрали и спрятали всех мальков и личинок ручейни-
ков из омутков, образовавшихся на ручейке при понижении его уровня.
Смертность - остатки кукш находили среди жертв различных хищни-
ков, под гнездом перепелятника 24. VII 1937 г., тетеревятника в июне-
июле 1965 г.(2 молодые птицы); кречета в августе 1938 г. (2), в желудке
зимняка (2), куницы (1). В разные годы в марте-мае'трижды находили
остатки кукш, съеденных хищными птицами. 3. III 1974 г. и 6. VIII 1978 г. в
районе усадьбы подобраны мертвые кукши, причина смерти которых не
выяснена. Иногда кукши попадают в капканы, поставленные у падали на
волков, на крючок сушащегося перемета, в рыболовную сеть, привлечен-
ные рыбешкой (всего 5 случаев).
Дважды наблюдали кукш, страдавших от непонятной ’’сонной болез-
ни”. Еще в 30-х годах одну кукшу, спавшую на земле, поймали руками,
взяли в дом и кормили, но она с каждым днем спала все больше и погиб-
ла от истощения. Другой случай произошел 14.Х 1984г.: одна из трех
кукш, привыкших клевать выставленное для синиц сало, повела себя
18. Зак. 1023
273
странно: села на открытую форточку и клевала сало из рук человека,
потом села на землю и уснула под окном. Вскочивший на окно горностай,
а затем пробежавшая мимо собака ее не заметили. Примерно через час
она проснулась и ушла в лес пешком. Она не погибла, так как на следу-
ющий день под окном кормились все три. Возможно, эта кукша отравилась
ею же засушенным мухомором (в заповеднике находили мухоморы,
поклеванные птицами). Способ питания кукши допускает легкую воз-
можность ее отравления и заражения, особенно гельминтами - послед-
ние были найдены в кишечнике и полости тела кукши, добытой 19. VI
1938 г.
Средние размеры 5 самцов: длина 255-299, крыло 135-144, хвост
120-135 мм; 6 самок соответственно - 245-280, 133-139 и 115-134 мм.
Средняя масса тела 64 самцов 88,3 г (75,0-107,6), 48 самок 82,4 г (67,5-
102,5).
Сорока - Pica pica (L.)
На Кольском полуострове встречается преимущественно у населенных
пунктов, питаясь на свалках вместе с воронами всю зиму. Ранней весной
сороки парами подыскивают себе гнездовые участки в лесу, в связи с чем
до 1948 г. они регулярно появлялись на Чунозерской усадьбе в апреле, а
потом исчезали. Начиная с 1948/49 г. сороки иногда зимуют на усадьбе
заповедника или включают ее в свой участок зимовки, наведываясь
время от времени. С этого года весенний прилет удается регистрировать
только на кордонах. Средняя дата перелета сороки с зимовок на гнез-
довья за 22 года - 6 апреля (18. II 1985 г. - 26. IV 1931 г.). В это время они
иногда совершают облет стаей, до 7-12 птиц. 12. IV 1949 г. видели, как
пара сорок перелетала через Чуна-тундру по перевалу Ванчем-нюацкенч.
9. IV 1980 г. близ Апатит сорока летела с веткой для постройки гнезда.
В заповеднике зарегистрированы 7 случаев гнездования:
6. VI 1963 г. близ Нижней Чуны обнаружено гнездо сороки, второе у Чунозерской
усадьбы не локализировано, но 17. VIII здесь появился летный выводок;
В сентябре 1966 г. на усадьбе заповедника держался выводок из двух слетков при
взрослой птице;
26. V 1971 г. в сотне метров от кордона Северного найдено сорочье гнездо на сосне;
30. IV 1972 г. в дендрологическом питомнике у Чунозерской усадьбы замечено
только что отстроенное сорочье гнездо; оно осталось пустым;
9. VIII 1982 г. у нежилого кордона Нявка на ели гнездо с 2 еще нелетными птенца-
ми, а 11. V 1986 г. два гнезда с 4 и 3 яйцами.
Другие наблюдения относятся к г. Мончегорску: 13. V 1980 г. в пали-
саднике при управлении заповедника сороки построили гнездо на иве, на
высоте около 5 м; 14. V 1987 г. в этом же гнезде самка насиживала кладку
из 5 яиц. В зарослях березы, ивы, ольхи вокруг свалки Мончегорска 29. X
1986 г. обнаружены несколько гнезд.
Приводим сведения по зимнему (октябрь-май) питанию сороки: пище-
вые отходы - 29 наблюдений, конский помет - 6, падаль: лось - 6,
олень - 2, собака - 1, рыба - 1 случай.
6. V 1969 г. у Нижней Чуны найдены перья сороки, ставшей жертвой
хищника.
Размеры 2 самок: длина 430-480 мм, крыло 200 мм, хвост 235-245 мм,
масса тела 218-222 г.
274
Кедровка - Nucifraga caryocatactes (L.)
Характерная таежная птица, экологически связанная с кедровой
сосной. Два подвида: толстоклювая кедровка распространена в горных
лесах Центральной Европы и Южной Скандинавии, тонкоклювая кедров-
ка - в таежной зоне Сибири и Приуралья. Поскольку на Кольском полу-
острове кедра нет, кедровка сюда только залетает во время инвазий.
В Сибири и на Урале кедровка зимует и выводит птенцов за счет
сделанных ею запасов кедровых орешков, поэтому ее можно считать
оседлой птицей. В годы неурожая кедровой шишки кедровки кочуют в
поисках пропитания^ перебиваясь случайными кормами, еловыми семе-
нами, насекомыми, но не размножаются и ведут себя как инвазионный
вид. В Лапландский заповедник изредка залетали кедровки, неизвестно
какого подвида.
23. VIII 1968 г. А. Сагайдачный заметил одну кедровку на берегу
оз. Улынчъявр; 24 и 26. IX, возможно, ту же самую кедровку видел
А. Брагин на берегу р. Чуна у Беличьей избы, где она подбирала объедки.
В ту же осень кедровку 8 раз видели на Кандалакшском заливе (Коханов,
1987, с. 24).
9. VI 1976 г. в бору в долине Чуны А. Брагин нашел 3 пера кедровки,
пойманной хищной птицей зимой или весной.
9. III 1981 г. кедровка подлетала к птичьей кормушке на Чунозерской
усадьбе на глазах 3. Аблаевой.
Заметим, что первые встречи совпали с мощной инвазией сибирской
кедровки 1968 г., когда они долетали до Британских островов. Налет
то лоток любой кедровки в европейскую часть СССР осенью 1977, очевид-
но, не коснулся Кольского полуострова.
Галка - Corvus monedula L.
По сообщению агронома В.И. Душечкина, в мае 1940 г. в пос. Хибины
видели залетную галку. По словам лесника М.С. Тютина, еще в 20-х
годах галку видали на свалке на берегу Кольского залива, у Зеленого
мыса (г. Мурманск). Весной 1967 г. одиночную галку видели в районе
Семи островов, а в 1972 г. - пару в пос. Мурмаши (Коханов, 1987, с. 24).
Грач - Corvus frugilegus L.
Изредка залетает и в Мурманскую область. В мае 1940 г. в Хибинах был
застрелен одиночный грач. По словам лесника М.С. Тютина, грачи встре-
чались и раньше, начиная с 1923 г., на восточном берегу Имандры и до
станции Тайбола к северу. В 1982 г. несколько пар грачей гнездились в
Мурманске (Коханов, 1987, с. 27). 27. IV 1986 г. на южном берегу Воче-
ламбины оз. Имандра возле лесной избушки весь день держался грач
(А. Андреев).
Черная ворона - Corvus corone L.
8-10. Ill 1959 г. Ю. Гаев заметил несколько черных ворон в стае серых
на городской свалке у г. Кировска. 3. XI 1959 г. и 6. IV 1960 г. в том же
районе А. Герман и О. Семенов-Тян-Шанский наблюдали до десятка
черных ворон.
275
Серая ворона - Corvus cornix L.
Местное название: саамское - вУрыч.
На Кольском полуострове обычна и многочисленна, но, будучи синант-
ропом, сравнительно редка в глубинных районах, в частности в Лапланд-
ском заповеднике (Владимирская, 1948, с. 217). В связи с ростом народо-
населения области и соответствующим расширением кормовой базы,
численность ворон за последние десятилетия неуклонно растет. До 1950 г.
вороны регулярно отлетали из Мончегорска и Апатит на зиму в полови-
не ноября, возвращаясь в феврале, а в марте они появлялись и в заповед-
нике (Владимирская, 1948).. С 50-х годов они живут в городах оседло и
на свалках можно насчитать до 800 особей в одной стае (6. XI 1968 г.) и
более (X-XI, 1985-1988).
Часть серых ворон, вероятно молодых, откочевывает к югу или юго-за-
паду, как и в прошлые годы. Из 3 ворон, окольцованных птенцами 12. VI
1963 г. в Мавра-губе, 1. XII 1963 г. одна была найдена мертвой в Дании на
острове Глэно на расстоянии 1780 км к юго-западу; другая убита 21. III
1964 г. в Швеции близ Гётеборга в 1464 км к юго-западу. Осенью с 22. VIII
по 21. X в разные годы стаи ворон от 17 до 80 птиц видали у кордонов
Купись, Мавра, Нижняя Чуна и Чунозерской усадьбы. Весенняя миграция
ворон слабее выражена: 30. IV 1975 г. стаю видели на Н. Чуне (А. Лиха-
чев), а 9. IV 1983 г. 12 ворон пролетели над Чунозером к северу (И. Параке-
цов). Средняя дата появления ворон в заповеднике 15 марта (п = 31), а
отлета - 3 ноября (п = 20). В 1971/72, 1973/74, 1976/77 и 1978/79 гг. в районе
Чунозерской усадьбы встречались зимующие особи.
Как отражение подъема численности в окрестных городах численность
ворон в заповеднике с 2 пар/1000 км2 в 30-40-х годах возросла до
13 пар/1000 км2 в 70-х годах и до 20 пар/1000 км2 в 80-х годах, т. е. за 50
лет увеличилась в 10 раз.
В марте-апреле гнездовые пары занимают участки. В апреле 1983 г.
после отстрела поочередно двух самок из пары самец спарился с третьей,
и эта пара вывела потомство. Вероятно, в популяции серых ворон не все
птицы обеспечены гнездовыми участками.
У самки, добытой 6. IV 1983 г., диаметр фолликул достигал 2 мм, 20. IV
1983 г. - 3 мм, 3. V 1981 г. - 5 мм. Строительство гнезд наблюдали 17. IV
1981 г. и 21. IV 1965 г.; насиживающих самок видели 24. IV 1981 г. и 31. V
1978 г., вылупление птенцов - с 7. V 1962 г. Вылупление птенцов в самых
ранних гнездах в Мончегорске происходило в 1978 г. 5-9 V, в поздних, на
территории заповедника в 1983 г. - 17-22. VI. 25 известных нам гнезд
находились поблизости от жилья - поселков, кордонов, часто посеща-
емых лесных избушек. Из них 21 построены на соснах, 2 - на елях, 1 - на
опоре линии электропередачи. Высота гнезд 4-15 м над землей, изредка
выше. Гнезда на соснах, как правило, находятся на ее верхушке или в
верхней части кроны.
Постройка состоит из трех слоев; каркас обычно из сухих веток сосны
и ели, средний слой из тонких гибких свежих березовых веток, сухих
длинных стеблей кустарничков, коры и луба можжевельника и ив, с
добавкой сырого песка, частично проникающего и в подстилку. Лоток
выстилается всяким подручным материалом, осоками, эпифитными
лишайниками, шерстью лося, оленя, собаки, зайца, полевок, паклей,
корой и лубом можжевельника, перьями, тряпками, бумагой. Наружный
276
диаметр гнезд 52 69 см, высота 28 см, диаметр лотка 17-19, глубина его
12 см.
В кладке обычно 5 яиц, от 4 до 7, в среднем 4,8 (п = 21); длина их
40-43 м, ширина 26-29 мм, масса 16-19 г. > Вылет птенцов в заповеднике
наблюдался между 12. VI 1963 г. и 15. VII 1983 г.; в г. Мончегорске самая
ранняя встреча слетков - 2. VI 1978 г. Масса тела птенцов накануне
вылета в среднем 394 г. (290-500, п = 9).
По наблюдениям Н.С. Бойко (1977),ворона насиживает 19-25 дней,
птенцы остаются в гнезде 32-35 дней.
Количество птенцов в гнезде 2-5, в 24 выводках в среднем было по 3
слетка (от 1 до 5). Птенцы, которых взрослые продолжают подкармли-
вать, долго живут в пределах гнездовоо участка; при усадьбе заповедни-
ка нераспавшиеся выводки наблюдали до начала сентября, хотя мигра-
ционные стаи отмечали уже с 22. VIII.
На гнездовом участке серые ворбны отличаются агрессивным поведе-
нием, нередко гоняются за вбронами, дербниками, зимняками, за пус-
тельгой, нападают на собак. Интересно, что позднее, в июне-августе,
вороны сами становятся мишенью других птиц, дербника, крачек, чаек,
кроншнепа, фифи, юрков, белых трясогузок и других мелких птиц,
кладки и птенцы которых страдают от ворбн.
Зимующие ворбны кормятся различными отходами на городских
свалках и животноводческих фермах. На гнездовых участках первое
время также подбирают отбросы возле жилья, кормятся падалью и
остатками добычи хищников, веснянками и леммингами- в годы их
миграций. Летом добавляются разнообразные насекомые, яйца и птенцы. В
заповеднике ворбны разоряли кладки гагары, большого и длинноносого
крохалей, чечеток. Приведем примеры хищнической деятельности ворон:
9. VI 1976 г. на болоте близУлынчъявраза 3 ч наблюдений ворона дважды
подлетала к лебяжьему гнезду, пока оба лебедя кормились вблизи, но
каждый раз один из них успевал отогнать хищницу.
21. V 1978 г. на Мавре-губе пара ворон атаковала бежавшую по льду ондатру
21 VII 1981 г. на разливе речки Нюдовки в Мончегорске 3 вороны
напали на утенка кряквы на глазах у А. Брагина.
В апреле 1978 г. близ Чунозерской усадьбы две вороны напали на
взрослого глухаря. А. Гилязов, подойдя на крики ворон, увидел, как они
поочередно бросались на забившегося под елку глухаря. Ворбны сразу
улетели, глухарь ушел по снегу в токующей позе и взлетел лишь в
300-400 м.
Нам пришлось видеть в Московском зоопарке, как ворона вцепилась в
основание хвоста голубя и тот вырвался, оставив пучок рулевых перьев в клю-
ве вороны. Приведенные примеры показывают, что ворона может играть
роль настоящего хищника, а не только грабителя гнезд. Серая ворона -
единственная птица, которую по правилам охоты в Мурманской области
разрешено уничтожать в течение всего года.
С другой стороны, ворона иногда становится жертвой хищной птицы. В
июне 1975 г. под гнездом кречета на скале Ельнюна мы нашли остатки
двух ворон, взрослой и птенца. 12. VII 1975 г. в устье Мавры был обнару-
жен труп птенца.
Размеры 3 взрослых самцов: крыло 326-335, хвоста 186-194 мм, масса
тела 508-570; 5 самок соответственно - 301-332 (ср. 314,2), 179-189 (ср.
182,8), 370-550 (ср. 447 г.).
277
Ворон - Corvus corax L.
Местное название: саамское - карнесь.
На Кольском полуострове встречается по всей территории до Аркти-
ческого побережья.
В Лапландском заповеднике встречается повсеместно, включая
горные тундры, обширные болота и озера, куда он регулярно залетает в
поисках корма. В иной зимний день случается увидеть или услышать
только ворона - и никаких других птиц. Численность его оценивается в
6-8 пар на 1000 км2 лесной площади. Расстояние между ближайшими
гнездами составляло 10,5, 11 и 13 км. Колебания численности ворона не
зависят от циклов полевок, но, вероятно, связаны с циклом северного
оленя, с которым он связан экологически. Скопления воронов (особенно
зимой) у падали, жертв крупных хищников, на местах отстрела лосей и
оленей, забоя домашних оленей и в районах отела являются временны-
ми; они не дают реального представления о плотности его популяции и
демонстрируют только его удивительную способность концентрироваться
на кормных местах. Так, 11. IV 1941 г. на тундрице Шилд-уайвенч на
тушах двух оленей кормилось 20 воронов (А. Насимович); 2. VI 1966 г. на
Нявка-Тундре на двух пыжиках 17 воронов (А. Брагин); 3. VI 1969 г. у
оленьего стада на Сальных тундрах собрались 42 птицы; на месте отстрела
оленей на р. Толва количество воронов за двое суток 5-7 марта увеличи-
лось с 7 особей до 100 (О. Макарова).
С территории заповедника вороны отлетают на темное время, с конца
ноября до начала января: вероятно, в это время им трудно находить
корм в природе; в этот период они питаются всякими отбросами у насе-
ленных пунктов и животноводческих ферм. Только в 1972-1976 гг.,
когда на озерах Пиренгской системы велся интенсивный промысел
оленей, вороны держались там круглый год.
Игры воронов с кувырканием в воздухе отмечали в разные годы с 4. IX
до 31. V, но частые и активные воздушные игры, с круговыми полетами,
пикированием и пируэтами, наблюдаются только в начале гнездового
периода, с конца февраля до конца марта. 13. III 1979 г. над устьем Чуны в
такой игре участвовали 6 пар воронов; постепенно они поднимались все
выше, пока не стали едва заметными точками в воздухе (М. Заваруев,
А. Гилязов). Подобные игры двух пар воронов наблюдали 27. III 1982 г. на
большой высоте над Куропачьим озером О. Макарова и А. Хохлов.
Гнездятся вороны рано, строительство гнезд в заповеднике наблюдали
с 24.III 1970. Из 17 известных нам гнезд 10 находились в нишах или на
карнизах отвесных скал, 6 - на соснах, 1 - на верхней площадке геодези-
ческого знака. Нередко вороны строят гнездо на месте старого. Гнездо
на уступе отвесной скалы по южному склону Сейднотчорра у ручья Куп-
летского, обнаруженное в 1963 г., находилось позади завесы капавшей
воды: оно повторно перестраивалось после обвала скалы и смыва пост-
ройки водой, но после разорения его, вероятно росомахой, пустовало не
менее четырех лет сряду.
На том же южном склоне Сейднотчорра примерно на 2 км восточнее
находится другая ’’скала воронов”, где вороны три раза гнездились в
разные годы. На обеих скалах одновременно вороны никогда не гнезди-
лись. Подобным образом на отвесной каменной стене в Травяной губе
278
Пиренгского озера в 1960 г. были найдены два гнезда воронов, свежее и
старое. К гнездам на скалах обычно очень трудно добраться.
Гнезда на соснах строятся в верхней части кроны или на макушке, на
высоте до 20 м. Каркас складывается из сухих сосновых сучков, внутрен-
няя выстилка из эпифитных лишайников, оленьей, лосиной, медвежьей
шерсти, березовых листьев и стеблей луговика. Лоток, вначале довольно
глубокий, по мере роста птенцов становится плоским.
29.1 V 1974 г. в гнезде у Куропачьего озера самка еще насиживала.
13.V 1970 г. в гнезде на скале у ручья Куплетского 3 пуховых птенца с
пеньками маховых на крыльях. Самая поздняя встреча птенцов в гнезде
- 14.VI 1966 г., но еще позже, 27.УП 1977 г., А. Белозеров нашел у р. Вайкись
гнездо с двумя птенцами: вероятно, это был случай повторного гнездо-
вания.
Среднее число птенцов в 9 гнездах 3,1, от 2 до 4. Из общего числа 28
птенцов удостоверена гибель 5 (18%): три съедены зверем, вероятно ро-
сомахой, два предположительно снесены ветром с плошадки геодези-
ческой вышки. Средняя за 6 лет дата вылета птенцов из гнезда - 11.VI,
самый ранний, еще плохо летавший выводок встречен 5.VI 1966 г.
Время выкармливания птенцов совпадает с порой интенсивного
таяния снега и схода снежного покрова. В это время появляется на свет
скрытая под снегом падаль, открываются на проталинах ягоды, выбе-
гают на поверхность мелкие грызуны, начинается отел копытных, по-
являются кладки у крупных птиц.
Нераспавшиеся выводки при обеих взрослых птицах встречались до
29.IX 1980 г., самое последнее наблюдение семьи - 28.V 1982 г. Среднее
число слетков в выводке по 7 встречам в июне 3,0, по 27 позднейшим
встречам - 2,0. Если судить по этим цифрам, смертность птенцов в пер-
вые месяцы жизни составляет около 30%. Вероятно, молодые разлетают-
ся после установления снежного покрова, с появлением трудностей в
добыче корма. Из трех птенцов, окольцованных 25.V 1967 г. в гнезде,
через год, 17.VII 1968 г., один обнаружен в Финляндии в ПО км к западу
сравнительно недалеко для ворона.
Питание ворона прослежено главным образом по визуальным наблюде-
ниям, которые мы подразделили на два периода: бесснежный (май-ок-
тябрь) и снежный (декабрь-апрель). Погадки относятся к летнему вре-
мени и характеризуют питание птенцов, так как подобраны в основном
у гнезд (табл. 18).
Зимой ворон питается почти исключительно падалью и остатками до-
бычи хищных зверей - в основном копытными животными. У падали
можно встретить до двух десятков воронов, а в местах забоя или отстре-
ла оленей и лосей - до сотен птиц. В годы массового размножения поле-
вок и леммингов на льду озер и на горных тундрах в феврале-апреле
встречаются остатки леммингов, добытых главным образом воронами и
воронами. Падаль играет большую роль и весной, в гнездовый период.
Рацион ворона летом расширяется за счет яиц, птенцов, снулой рыбы,
насекомых и ягод. Неоднократно наблюдали, как ворон несет в клюве
яйцо крохаля или полевку. Даже ручной вороненок 13.VII 1960 г. пытал-
ся убить птенца дрозда.
279
Таблица 18. Питание ворона
Вид корма Визуальные наблюдения Погадки Всего, %
май—октябрь ноябрь—апрель май—июнь
число встреч % число встреч % число встреч %
Падаль (олень, 72 62 98 78 5 11 61
Мелкая падаль (со- 4 3 16 13 1 2 7
бака, росомаха,, заяц) Полевки, леммин- 1 1 3 2 4 9 3
ги Птицы (куропатки, 5 4 6 5 6 13 6
кулики, крачки) Яйца (крохаля, гагары, глухаря) Рыба (сиг, окунь) 19 И 16 9 — 7 6 16 13 9 6
Лягушки — — — — 3 7 1
Насекомые — — — — 1 2 - 0+
Ягоды (брусника, 1 1 — — 11 25 4
вороника) Семена ситника 1 2 0
Конский помет — — 3 2 — 1
Продукты (хлеб, 2 2 — — — — 1
масло) Мыло 2 2 — — — — 1
Вблизи гнезда с птенцами вороны беспокоятся, не отлетая далеко.
Отмечены случаи нападений ворона на собаку, орлана, тетеревятника,
кречета. На ворона нападали чайки, крачки, ястребиная сова, пустельга,
стрижи, белые трясогузки, чечетки, серые вороны.
Сообразительность ворона общеизвестна. Наши наблюдения подтвер-
ждают сложность и логичность поведения воронов. При осенней охоте на
боровую. дичь разные лица отмечали, что ворон прилетает на лай собаки
или выстрел. 3.VI 1979 г. и 8.VIII 1979 г. на сильно дымящий костер подле-
тали и кружили поблизости в первом случае 2, во втором 6 воронов. При
отстреле оленей после прекращения стрельбы или во время перерыва во-
роны группами по 2-5 моментально накидывались на туши оленей, пы-
тались клевать подраненных.
Зарегистрировано 9 случаев гибели взрослых воронов (не считая упо-
минавшихся выше гнездовых птенцов): два пойманы кречетами, один
неизвестным хищником, один сбит автомашиной, два попались в капка-
ны у падали, три застрелены.
Ворон - самый крупный представитель отряда воробьиных, масса те-
ла взрослой самки в октябре 1500 г, еще 3 особей неизвестного пола -
1200,1236 и 1286 г.
ЗАКЛЮЧЕНИЕ
Во внутренней (горно-лесной) части Русской Лапландии (Кольс-
кий полуостров с прилегающей к нему с запада частью материка) нами
зарегистрированы 192 вида птиц, из них 112 видов гнездятся регулярно,
8 - эпизодически (широконоска, чернозобик, речная крачка, вертишей-
ка , горный конек, синехвостка, сойка и грач), предполагается, но не дока-
зано гнездование еще 7 видов (журавль, филин, воробьиный сыч, длинно-
хвостая неясыть, крапивник, черный дрозд и лапландский подорожник).
Из числа этих 127 видов только 17 регулярно зимуют в нашем регионе.
Для большинства птиц условия зимовки здесь слишком тяжелы. Хотя
сильные морозы (ниже-40°) случаются здесь редко, а средняя темпера-
тура зимних месяцев почти такая же, как и в Средней России, зима про-
должается около полугода и в лесах за это время накапливается масса
снега. Высота снежного покрова в лесах к концу зимы обычно достигает
60-90 см - в борах, 70-120 см - в ельниках (Семенов-Тян-Шанский, 1975,
с 56). Кормовые ресурсы земной поверхности становятся недоступными
под глубоким снегом, и в распоряжении птиц остается только то, что
можно найти в кронах и на стволах деревьев и кустарников, - почки,
побеги, хвоя, семена, а также насекомые под корой и в ее трещинах.
Ограничивает возможность кормодобывания и недостаток дневного
света: солнце полностью отсутствует на широте Мурманска 47 дней, в
Лапландском заповеднике - 24 дня в году, и в эту пору даже сумеречное
освещение продолжается не более 4-6 ч в сутки.
На горных тундрах, где древесные, корма отсутствуют, способна про-
жить зиму только одна птица, тундряная куропатка: пищу она находит
на поверхности земли, разгребая снег на обдуваемых ветром буграх.
Остальные 4 вида тетеревиных птиц радикально решили проблему зим-
него продовольствия путем перестройки на это время режима питания,
переходя на единственно доступные им веточные корма. Специализиро-
ванные энтомофаги, черный и трехпалый дятел, пищуха, а отчасти и си-
ницы как летом, так и зимой достают насекомых на стволах деревьев.
Сходным образом и оляпка зимой находит личинок водных насекомых
на дне не замерзающих из-за быстрого течения рек и ручьев.
Некоторые всеядные птицы запасают корм на зиму, к ним в нашем
регионе относятся кукша, сероголовая гаичка, пухляк, а также воробьи-
ный сыч, а возможно, и ворон. Ряд видов зимует на Крайнем Севере толь-
ко в городах и поселках, питаясь отбросами: одни из них оседлые, посто-
янные синантропы, проникшие сюда недавно, в качестве нахлебников че-
ловека (домовый воробей и голубь); другие гнездятся в лесу и пересе-
ляются в города только на зиму, привлеченные не только доступным кор-
мом, но и возможностью искать его при ярком электрическом освещении
(ворона, сорока и большая синица). В последнее время список зимующих
в городах птиц пополнился свиристелем и снегирем, использующими уро-
281
жай ягод в городских посадках, и ястребом-тетеревятником, который
ловит голубей. Намечается также возможность зимовки кряквы на го-
родских водоемах, в местах сброса теплых вод.
Зимующие на месте составляют около 13% всех гнездящихся у нас
птиц. Большинство видов улетает зимовать на юго-запад, в Западную и
Южную Европу, а некоторые и дальше, в Северную или Экваториальную
Африку. Полярная крачка и некоторые кулики проводят зиму в Южном
полушарии. Несколько видов гнездящихся у нас певчих птиц улетают в
юго-восточном направлении и зимуют в Китае (овсянка-крошка), Японии
(синехвостка) или Индонезии (таловка). В группу регулярно перелетных
птиц входит 85 видов - 67 % всех гнездящихся в регионе.
Особое место занимают птицы, специализированные на использовании
’’дефицитных” кормов, в частности древесных семян, урожаи которых
неустойчивы. При хорошем урожае такие птицы ведут себя как оседлые,
а при неурожае кочуют в поисках мест с удовлетворительным урожаем се-
мян либо перебиваясь случайной пищей. В таких случаях до наступления
хорошего урожая они летом не размножаются. Типичными представите-
лями таких ’’инвазионных” видов являются большой пестрый дятел, все
клесты, чечетка и чиж. В других регионах страны к числу инвазионных ви-
дов относятся кедровка (питается кедровыми орешками), дубонос
(ягодами черемухи), сойка и поползень (орехи, желуди, другие семена
лиственных пород); в голодные годы во время кочевок все перечислен-
ные птицы залетают и на Кольский полуостров.
В той или иной мере дефицитен и корм хищных птиц. Крупные хищники,
охотясь на куропаток и глухарей, зимуют у нас только в годы урожая
боровой дичи (орел, ястреб-тетеревятник, кречет, филин); мелкие, добы-
вающие певчих птиц, как правило, отлетают на зиму вслед за своей до-
бычей (перепелятник, дербник). Из хищников-миофагов одни регулярно,
отлетают на зиму в бесснежные или малоснежные регионы (зимняк, пус-
тельга, болотная сова), другие ведут себя как инвазионные виды, в ’’мы-
шиные” годы живут оседло, а в неурожайные - зимой кочуют, а летом не
гнездятся (ястребиная сова, вероятно, также неясыти и сычи). К группе
зимнекочующих птиц мы относим 18 видов (14 % всех гнездящихся) и
не менее 7 залетных, к числу их относится и белая сова.
65 видов (34 % состава авифауны) появляются у нас только эпизоди-
чески, но не гнездятся. Некоторые из них, черная и белощекая казар-
ки, тулес, краснозобик и куличок-воробей, гнездятся на островах Ледо-
витого океана и на сибирских тундрах и встречаются у нас только в
пролетное время, на пути к гнездовьям или зимовкам. Другие гнездятся
только на периферии Кольского полуострова, на морском побережье
или равнинной тундре, залетая иногда и в глубь страны, одни весной или
летом (баклан, все поморники, бургомистр, травник, кулик-сорока,
камнешарка, морской песочник, белозобый дрозд, горный и краснозо-
бый коньки),другие зимой (кайра, тупик, люрик, гага).
Некоторые птицы залетают в Лапландию с юго-запада или юга, как бы
продолжая весенний пролет за границы своего гнездового ареала (серая
цапля, лысуха, коршун, осоед, горлица, лесной жаворонок, касатка, все
282
славки), другие осенью, в порядке расселения молодняка (ополовник,
хохлатая синица), а возможно, и ошибочного направления отлета (удод).
Залетных гостей с юга около 13, не считая тех, которые при благоприятных
условиях гнездятся (широконоска, чибис, вяхирь, вертишейка).
Влияние человека на авифауну в основном негативное. От прямого
истребления страдают в настоящее время лишь немногие виды (лебедь,
орел, орлан), гораздо большее число - от нарушения биотопов в резуль-
тате деятельности человека; например, от вырубки лесов страдают не
только глухарь и рябчик, но и гназдящиеся в дуплах утки и певчие, а
также хищники, гнездящиеся на старых деревьях. Множество птиц
гибнет от удара о провода и, вероятно, еще больше от загрязнения среды
обитания промышленными отходами. В последние годы резко сократилась
численность большинства певчих птиц, а заодно и экологически связанных
с ними хищников, а также кукушки, но в этом случае, помимо местного
загрязнения, не меньшую роль может играть и загрязнение среды в
бласти зимовок. Наибольший урон понесли дрозды, зяблик, юрок и
другие певчие птицы, зимующие в Европе.
Обнаружено стойкое сокращение некоторых популяций на протяже-
нии последних 50-100 лет: малого веретенника, желны, рогатого жаво-
ронка и рябинника, а с другой стороны - рост численности и расселение
некоторых видов, по-видимому не зависящих от деятельности человека
(золотистая ржанка и камышевка-барсучок). В обоих случаях причины
явления остаются неясными.
Другие птицы распространяются к северу, явно используя появление
новых биотипов и экологических ниш, созданных деятельностью челове-
ка. Развитие сельского хозяйства и появление пахотных полей привлек-
ли на Кольский полуостров чибиса, полевого жаворонка, обыкновенную
и садовую овсянку, скворца и грача. Рост городов не только способство-
вал продвижению на север городской ласточки и озерной чайки, но выз-
вал перестройку экологии и внушительный рост популяций серебристой
и сизой чаек.
В книге приведено много наблюдений по конкурентным отношениям
между хищниками. В основном они проявляются при недостатке пищи
зимой, при этом относительно слабые хищники часто становятся жертвой
более сильных. В первую очередь гибнут обычно совы, но есть и противо-
положный пример, когда кречет стал добычей белой совы.
Лапландским заповедником собран уникальный по длительности ря-
дов материал по срокам прилета, отлета и другим периодическим явле-
ниям у птиц. Не имея возможности привести его полностью, мы ограни-
чились средними и крайними сроками наступления явлений, с указанием
числа лет, а частью также средних квадратических отклонений (сигма)
и нормальной температуры дня наступления явления. Местами приведе-
ны также коэффициенты корреляции между датами явления и средними
температурами соответствующего месяца или сроками соседних по вре-
мени фенологических явлений. Как правило, амплитуда и сигма сроков
прилета по мере движения весны уменьшаются, а для сроков отлета
увеличиваются к концу сезона. Например, сигма прилета щура (начало
283
марта) ± 14,7 дней, лебедя (середина апреля) ± 8,0, белой трясогузки (на-
чало мая) ± 8,6, полярной крачки (конец мая) ± 4,0 дня. При этом у поздно
прилетающих птиц амплитуда и сигма примерно вдвое меньше, чем у со-
седних фито-фенологических или гидрологических явлений. Объясняет-
ся это тем, что время прилета зависит не только от изменчивой темпера-
туры, но и от стабильного изменения условий освещения, вызванных
движением земли вокруг солнца. По этой причине в годы с ранней весной
птицы прилетают при более высокой температуре, чем в случае поздней
весны.
Отлет большинства птиц происходит при температуре на 3-5° выше,
чем прилет. Исключение представляют водоплавающие, пребывание
которых на гнездовьях зависит от наличия открытой воды, а вскрывают-
ся воды при значительно более высокой температуре, чем замерзают. При
относительно низкой температуре отлетают также птицы, способные при
наличии корма зимовать на месте, а именно свиристель, снегирь и
дрозды.
Все птицы без исключения отлетают при гораздо более слабом освеще-
нии, чем прилетают. Весной они летят от коротких дней к длинным;
осенью рано отлетающие птицы летят к более короткому дню, а поздние
мигранты - к более длинному.
Корреляция между сроками прилета и температурой у всех видов от-
рицательная, в пределах - 0,20 - 0,65 (чем теплее, тем раньше прилет).
Отлет поздних птиц обычно задерживается теплой погодой, при этом ко-
эффициент корреляции между температурой октября и отлетом состав-
ляет у гоголя +0,54, у лебедя +0,72. Неожиданно оказалось, что теплая
погода ускоряет отлет ранних мигрантов: корреляция между температу-
рой августа и отлетом фифи равно -0,53, скопы -0,29, веснички -0,22.
Вероятное объяснение этого феномена - птица должна накопить опреде-
ленный энергетический резерв для перелета, а холодная погода этот про-
цесс задерживает.
Неоднозначно влияют на время отлета и условия освещения: юрок от-
летает тем раньше, чем больше было солнечных часов в сентябре (г =
= -0,50), а пуночка тем позже, чем больше солнце светило в октябре (г =
= +0,55) (Семенов-Тян-Шанский, 1978, с. 15). Это можно объяснить тем, что
юрок - перелетная птица и нуждается в запасе жира для миграции, а в
ненастную погоду ему труднее добывать корм; пуночка - кочевник и в
запасах для отлета не нуждается.
Относительно рано отлетают хищные птицы с парящим полетом, кото-
рый требует, как известно, наличия восходящих потоков воздуха от наг-
ретой солнцем земли или даже снежной поверхности, образующих куче-
вые облака. Орлан прилетает 15 апреля при температуре -2,6 и долготе
дня 15,5 ч, отлетает 8 октября, при +3,0° и 10,5-часовом дне; в обоих слу-
чаях еще возможно кучные облака восходящих токов. Прилет скопы
16 мая при+3,8°и? 20-часовом дне, отлет 8 сентября, при 7,7° и 14-часо-
вом дне. Разница температуры в пользу отлета в первом случае 5,6°, во
втором 3,9°. По той же причине рано отлетают и журавли. Хищные птицы,
способные зимовать на месте, пользуются активным полетом.
ЛИТЕРАТУРА
Аблаева З.Х., Семенов-Тян-Шан-
ский О.И. Календарь природы Лап-
ландского заповедника // Сезонная и
разногодичная динамика раститель-
ного покрова в заповедниках РСФСР.
М.: ЦНИЛ Главохоты РСФСР, 1983.
С. 44-58.
Андреев А.В. Адаптация птиц к зимним
условиям Субарктики. М.: Наука,
1980. 175 с.
Андреев А.М. Исторические материалы
о Кольском полуострове, хранящие-
ся в Ленинграде И Сб. материалов по
истории Кольского полуострова в
XVI-XVII вв. Л., 1930. 191 с.
Белко Н.Г. Скопа в Дарвинском запо-
веднике // Хищные птицы и совы в
заповедниках РСФСР. М.: ЦНИЛ
Главохоты РСФСР, 1985. С. 116-130.
Бианки В.В. О гнездовании прибрежных
птиц в районе Кандалакшского зали-
ва // Тр. Кандалакшского заповедни-
ка. Вологда: Кн. изд-во, 1958. Вып. 1.
С. 51-71.
Бианки В.В. Кратка^ история орнитоло-
гических исследовании и состояние
охраны птиц на Белом море // Проб-
лемы изучения и охраны природы
Прибеломорья. Мурманск: Кн. изд-
во, 1987. С. 5-19.
Бианки В.В., Бойко Н.С. К экологии
обыкновенной пустельги на островах
Кандалакшского залива // Хищные
птицы и совы в заповедниках РСФСР.
М.: ЦНИЛ Главохоты РСФСР, 1985.
С. 100-110.
Бианки В.В., Флеров А.И. Список птиц
Кандалакшского залива и его побе-
режий // Тр. Кандалакшского запо-
ведника. Мурманск: Кн. изд-во,
1960. Вып. 2. С. 105-115.
Благосклонов К.Н. Птицы Кандалакш-
ского заповедника и окрестности
биологической Беломорской станции
МГУ // Там же. С. 5-105.
Бойко Н.С. Экология и практическое
значение серой вороны в Мурман-
ской области: Автореф. дис. . . .канд.
биол. наук. Кандалакша, 1977. 16 с.
Брагин А.Б. Водоплавающие птицы
Лапландии, биологические основы и
пути эффективности их размноже-
ния: Автореф. дис. . . . канд. биол.
наук. Петрозаводск, 1974. 21 с.
Владимирская М.И. Птицы Лапландско-
го заповедника // Тр. Лапландского
заповедника. М.: Совмин РСФСР,
1948. Вып. 3. С. 171-245.
Гебель Г.Ф. Материалы к орнитофауне
Лапландии и Соловецких островов //
Тр. Санкт-Петербургского о-ва естест-
воиспытателей. СПб., 1902. Т. 33,
вып. 2. С. 97-137.
Гладков Н.А. Семейство дроздовых //
Птицы Советского Союза. М.: Сов.
наука, 1954. Т. 6. С. 446.
Губарь Ю.П., Мошкин А.В. К расшире-
нию ареала синехвостки (Tarsiger суа-
nurus Pall.) // 4-я Межвузовская зоо-
геогр. конф. 1966 г.: Тез. докл. Одес-
са: Одес. ун-т, 1966. С. 68—69.
Дементьев Г.П. Хищные птицы: Совы //
Птицы Советского Союза. М.: Сов.
наука, 1951. Т. 1. С. 70-420.
Дребенцов В.С., Сурков С.С. Животный
мир Мурманской области. Мур-
манск: Кн. изд-во, 1974. 182 с.
Иванов А.И. Каталог птиц СССР. Л.:
Наука, 1976. 275 с.
Кайгородов Д.Н. Наши зимние пернатые
гости. Пг., 1916.
Карпович В.Н. Кандалакшский заповед-
ник. Мурманск: Кн. Изд-во, 1984.
160 с.
Карпович В.Н. Филин на птичьих база-
рах восточного Мурмана // Хищные
птицы и совы в заповедниках РСФСР.
М.: ЦНИЛ Главохоты РСФСР, 1985.
164 с.
Карри-Линдал К. Птицы над сушей и
над морем. М.: Мысль, 1984. 204 с.
Кириков С.В. Возрастные изменения
жевательной мускулатуры и черепа у
глухарей // Зоол. журн. 1944. Т. 23,
вып. 4. С. 16—25.
Кищинский А.А. К биологии кречета
(Falco gyrfalco gyrfalco) на Кольском
полуострове // Учен. зап. МГУ.
М.: Изд-во МГУ, 1958. Вып. 197.
С. 61-75.
Кищинский А.А. К фауне и экологии
птиц Териберского района Мурман-
ской области // Тр. Кандалакшского
гос. заповедника. Мурманск:
Кн. изд-во, 1960. Вып. 2. С. 122—212.
Коханов В.Д. К экологии кукши в
Мурманской области // Сб. науч,
тр. ЦНИЛ Главохоты РСФСР.
М.: ЦНИЛ Главохоты РСФСР, 1982.
С. 124-137.
Коханов В.Д. О сером журавле в Мур-
манской области // Журавли в СССР.
М.:Наука, 1982. С. 45-48.
Коханов В.Д. Материалы по экологии
дербника и тетеревятника в районе
Кандалакшского залива // Хищные
птицы и совы в заповедниках РСФСР.
М.: ЦНИЛ Главохоты РСФСР, 1985.
С. 100-110.
Коханов В.Д. Обзор изменений, отме;
ченных в орнитофауне Мурманской
области за последнее столетие //
Проблемы изучения и охраны приро-
ды Прибеломорья. Мурманск: Кн.
изд-во, 1987. С. 20—37.
Коханов В.Д., Гаев Ю.Г. Материалы по
экологии клестов в Мурманской
области // Тр. Кандалакшского гос.
285
заповедника. Мурманск: Кн. изд-во,
1970. Вып. 8. С. 236-274.
Коханов В.Д., Скокова Н.Н. Фауна птиц
Айновых островов И Тр. Кандалакш-
ского гос. заповедника. М.: Лесн.
пром-сть, 1967. Вып. 5. С. 185—267.
Красная книга РСФСР: Животные.
М.: Россельхозиздат, 1983. 454 с.
Краснов Ю.В. биологии белой совы на
Восточном Мурмане // Орнитология.
М.: Изд-во МГУ, 1985. Вып. 20.
С. 110-116.
Кречмар А,В., Андреев А.В., Конд-
ратьев А.Я. Экология и распростра-
нение птиц на Северо-Востоке СССР.
М.: Наука, 1978. 196 с.
Курочкин Е.Н., Кошелев А.И. Семей-
ствопастушковых// Птицы СССР:
Курообразные, Журавлеобразные.
Л.:Наука, 1987. С. 335-464.
Малышевский Р.И. Летние наблюдения
над птицами Терского берега Белого
моря Ц Орнитология. М.: Изд-во
МГУ, 1962. Вып. 5. С. 13-27.
Малышевский Р.И. Еще о летней орни-
тофауне Терского берега Белого
моря И Там же. 1963. Вып. 6. С. 475.
Мальчевский А.С. Синехвостка на Коль-
ском полуострове // Природа. 1947.
№ 2. С. 58-59.
Мальчевский А.С. Гнездовая жизнь
певчих птиц. Л.: Изд-во ЛГУ, 1959.
281 с.
Мальчевский А.С., Пукинский Ю.В.
Птицы Ленинградской области.
Л.: Изд-во ЛГУ, 1983. Т. 1. 480 с.;
Т. 2. 504 с.
Меркурьев И.С. Живая речь Кольских
поморов. Мурманск: Кн. изд-во,
1979. 184 с.
Миграция птиц Восточной Европы и
Северной Азии: Гагарообразные —
Аистообразные. М.: Наука, 1978.
270 с.
Михайлов К.Е., Фильчагов А.В. Особен-
ности распространения и расселения
некоторых видов птиц в тундре
Кольского полуострова // Орнитоло-
гия. М.: Изд-во МГУ, 1984. Вып. 19.
С. 22-29.
Насимович А.А. Экология лисицы в
Лапландском заповеднике // Тр.
Лапландского гос. заповедника.
М.: Совмин РСФСР, 1948. Вып. 3.
С. 39-79.
Насимович А.А. Экология лесной куни-
цы // Там же. С. 81-105.
Насимович А.А. Новые данные по эко-
логии росомахи в Лапландском
заповеднике // Там же. С. 107-124.
Новиков Г.А. Экология птиц Хибин-
ских гор // Тр. Зоол. ин-та АН СССР.
Л.: Изд-во АН СССР, 1952 а. Т. 9,
вып. 4. С. 1133-1155.
Новиков Г.А. Материалы по питанию
лесных птиц Кольского полуостро-
ва // 1952 б. Там же. С. 1155-1198.
Новиков Г.А. Еловые леса как среда
обитания и роль в их жизни млеко-
питающих и птиц // Роль животных в
жизни леса. М.: Изд-во МГУ, 1956.
С. 6-165.
Новиков Г.А. Жизнь на снегу и под
снегом. Л.: Наука, 1981. 188 с.
Озерецковский Н. Описание Колы и
Астрахани. СПб., 1804. 131 с.
Плеске Ф.Д. Критический обзор млеко-
питающих и птиц Кольского полу-
острова. СПб.: Императорская АН,
1887. 536 с.
Портенко Л.А. Фауна Анадырского
края: Птицы. Л.: Изд-во Главсевмор-
пути, 1939. Ч. 1. 205 с.
Портенко Л.А. Птицы СССР. М.; Л.: ЗИН
АН СССР, 1954. Ч. 3. 251 с.; Ч. 4. 415 с.
Птушенко Е.С. Длиннокрылые // Птицы
Советского Союза. М.: Сов. наука,
1951. Т. 1. С. 618-645.
Редкие и нуждающиеся в охране живот-
ные и растения Мурманской области.
Мурманск: Кн. изд-во, 1979. 160 с.
Саамско-русский словарь. М., 1985.
567 с.
Семенов-Тян-Шанский О.И. Экология
боровой дичи в Лапландском заповед-
нике // Тр. Лапландского гос. запо-
ведника. М.: СНК РСФСР, 1938.
Вып. 1. С. 217-305.
Семенов-Тян-Шанский О.И. Экология
тетеревиных птиц // Там же. 1959.
Вып. 5. 319 с.
Семенов-Тян-Шанс#ий О.И. Изучение
поведения птиц на гнезде при помо-
щи самописцев // Тр. проблемных й
тематических совещаний при ЗИН.
Л.: Наука, 1960. Вып. 9. С. 279-286.
Семенов-Тян-Шанский О.И. Лапланд-
ский заповедник. Мурманск: Кн.
изд-во, 1975. 244 с.
Семенов-Тян-Шанекий О.И. Индикатор-
ное значение многолетних наблюде-
ний за видами и сообществами //
Биологические методы оценки при-
родной среды. М.: Наука, 1978.
С. 7-28.
Семенов-Тян-Шанский О.И. Звери Мур-
манской области. Мурманск: Кн.
изд-во, 1982. 176 с.
Семенов-Тян-Шанский О.И. Биология
размножения тетеревиных птиц на
Севере. М., 1983. 62 с.
Семенов-Тян-Шанский О.И., Брагин А.Б.
Условия инкубации некоторых вы-
водковых птиц в Субарктике // Бюл.
Моск, о-ва испыт. природы. Отд.
биол. 1969. Т. 74(6). С. 50-66.
Степанян Л.С. Состав и распределение
птиц фауны СССР. Воробьинообраз-
ные. М.: Наука, 1978. 391 с.
Судиловская А.М. Кукушки. Сизово-
ронки. Удоды // Птицы Советского
Союза. М.: Сов. Наука, 1951. Т. 1.
С. 430-464, 486-546.
Татаринкова И.П. О встречах хищных
птиц на Айновых островах // Хищ-
ные птицы и совы в заповедниках
РСФСР. М.: ЦНИЛ Главохоты РСФСР,
1985. С. 162-164.
Татаринкова И.П., Чемякин Р.Г. Допол-
нения к фауне птиц Айновых остро-
вов // Естественная среда и биологи-
286
ческие ресурсы Крайнего Севера.
Л.: Наука, 1975. С. 33-37.
Филъчагов А.В., Черенков А.Е. Числен-
ность водоплавающих птиц на вос-
точном побережье Кольского полу-
острова // Орнитология. М.: МГУ,
1984. Вып. 19. С. 215-216.
Флинт В.Е. Гагарообразные // Птицы
СССР. История изучения. Гагары-
Трубконосные. М.: Наука, 1982.
С. 244-288.
Флинт В.Е., Бёме Р.Л., Костин Ю.В.,
Кузнецов А.А. Птицы СССР. М.: Нау-
ка, 1968. 637 с.
Чернолуский В.В. Заметки о пастьбе и
организации стада у лопарей // Коль-
ский сборник. Л., 1930. С. 23—69.
Шибанов Н.В. К орнитологической
фауне Русской Лапландии // Мему-
ары зоол. отд. О-ва любителей естест-
вознания, антропологии и этнографии.
Л.: АН СССР, 1927. Вып. 3. С. 1-29.
Шкляревич Ф.Н., Краснов Ю.В. К био-
логии лапландского кречета (Falco
gyrfalco J.) на Кольском полуострове //
Экология птиц морских побережий.
М.: ЦНИЛ Главохоты РСФСР, 1980.
С. 17-26.
Шутова Е.В., Бианки В.В», Коханов В.Д.,
Ладыжев Л,С, К экологии дрозда
белобровика на северо-западе
СССР // Сб. научных трудов ЦНИЛ
Главохоты. М.: ЦНИЛ Главохоты
РСФСР, 1982. С. 109-123.
Энгельгард А.П. Русский Север. СПб.,
1897. 258 с.
Юдин К.А.. Фирсова Л.В. Серебристая
чайка // Птицы СССР: Чайковые.
М.: Наука, 1988. С. 126-146.
Bergman У. Mustakaularastas, Turdus ruficollis
strogularis, tuvattu Kupion Haminenlah-
della ja Helsingissa // Ornis fennica. 1965.
Vol. 42. N 2
Blomgren A. Studies of less familiar birds. 162.
Seberian Jay // British Birds. 1971. Vol. 64.
Byrkyedal Ingvar. Time activity budget for
breeding greater golden plovers in Norwe-
gian mountains//Wilson Bull. 1985. Vol. 97.
S. 486-501.
Enemar A., Nystrom B. On grasiskans, Cardue-
lis flammea, bestandsvaxlingar, foda och
hackung ifjallbjorkskog, Sodra Lapland//
Var fagelvard. 1981. Vol. 40, N 6,
S. 409-426.
Evans M.S., Wood N.A., Kear J. Lead shot in
Bewick’s Swans // Wildfowl. 1973. Vol. 24.
S. 56-60.
Grzimek B. Wildes Tier weisser Mann. Brock-
haus Verlag. 1969. 384 s.
Harrison C. A field guide to the nests, eggs and
nestlings of British and European Birds.
Glasgow. 1975. 432 s.
Hellmich J. Zum Futterverhalter eines ”verein-
samten” Trauerschnapper-Mannchens (F.
hypoleuca) am Ende Nestlingszeit // Vogel-
welt. 1981. Vol. 102, N 2. S. 48-51.
Helminen M. Occurence of the Red-flanked
Bluetail (Tarsiger cyanurus) in Finland and
some remarks concerning ets expansion to
the west 11 Ornis fennica. 1958. Vol. 35,
N 3/3 S. 51-64.
Jarvinen A. Talitiaisen poikanen kirijosiepon
esassa // Ornis fennica. 1977. Vol. 54, N 3.
S. 137.
Jarvinen A. Breeding biology of the Siberian
Tit, Parus cinctus, im northern Lapland //
Ornis fennica. 1978. Vol. 55, N 1. S. 24-28.
Jarvinen O. Recent quantitative changes in the
populations of Finnish land birds // Pol.
Ecol. Stud. (PRL). 1977. Vol. 3, N 4.
S. 177-188.
Mikkola N. Der Bartkaus. Wittenberg Luther-
stadt. 1981. 124 s.
Mikkola H., Sulkava S. Mita syo tunturihauk-
ka // Suomen luonto. 1972. V. 31, N 5.
S. 183—185,
Palmer R.S. Handbook of North American
birds. Waterfowl. New Haven — London.
1976. Vol. 2. 521 s. V. 3.560 p.
Palmer R.S. Handbook of North American birds.
Raptors. New Haven — London. 1988. V. 5.
465 s.
Pulliainen E. Behaviour of a Willow Grouse,
Lagopus 1. lagopus, at the nest // Ornis
fennica. 1978. Vol. 55. S. 141-148.
Salomonsen F. Moults and sequence of pluma-
ges in the Rock Ptarmigan (Lagopus mu-
tus) // Vidensk Medd. fra Dansk Natur.
Forcing. 1939. Vol. 103. 491 s.
Sammaliste Lasse. Twenty years of the Finnish
winter bird census // Pol. Ecol. Stud., 1977.
Vol. 3, N 4. S. 199-206.
White C., Cade Y.J. Cliff nesting raptors and
ravens along the Colville river in Arctic
Alaska // The living birds. Cornell Labor, of
ornithology, 10-th annual 1971. Ithaca
New York. 1971. S. 107-150.
СОДЕРЖАНИЕ
Предисловие.................................................................... 3
Отряд Гагары Gaviiformes................................................... 5
Отряд Поганки Podicipediformes............................................. 10
Отряд Веслоногие Pelecaniformes............................................ 10
Отряд Голенастые Ciconiiformes............................................. 10
Отряд Пластинчатоклювые Anseriformes....................................... 11
Отряд Хищные Falconiformes................................................. 55
Отряд Куриные Galliformes.................................................. 90
Отряд Журавлеобразные Gruiformes........................................... 126
Отряд Ржанкообразные Charadriiformes........................................ 127
Подотряд Кулики Charadrii.................................................. 127
Подотряд Чайковые Lari..................................................... 156
Семейство Чайковые Laridae............................................ 157
Семейство Чистиковые Alcidae.......................................... 168
Отряд Голуби Columbiformes.................................................. 169
Отряд Кукушкообразные. Cuculiformes......................................... 170
Отряд Совы Strigiformes..................................................... 172
Отряд Стрижеобразные Apodiformes............................................ 188
Отряд Ракшеобразные Coraciiformes........................................... 189
Отряд Дятлообразные Piciformes.............................................. 190
Отряд Воробьиные Passeriformes.............................................. 197
Семейство Жаворонки Alaudidae......................................... 197
Семейство Ласточковые Hirundinidae.................................... 199
Семейство Трясогузковые Motacillidae.................................. 200
Семейство Сорокопутовые Laniidae...................................... 209
Семейство Свиристелевые Bombycillidae................................. 210
Семейство Оляпковые Cinclidae......................................... 212
Семейство Крапивниковые Troglodytidae................................. 215
Семейство Завирушковые Prunellidae.................................... 215
Семейство Дроздовые Turdidae.......................................... 215
Семейство Славковые Sylviidae......................................... 230
Семейство Корольковые Regulidae....................................... 236
Семейство Мухоловки Muscicapidae..................................... 236
Семейство Длиннохвостые синицы Aegithalidae........................... 240
Семейство Синичьи Paridae............................................. 240
Семейство Поползневые Sittidae........................................ 247
Семейство Пищуховые Certhiidae........................................ 248
Семейство Овсянковые Emberizidae...................................... 248
Семейство Вьюрковые Fringillidae...................................... 254
Семейство Ткачиковые Ploceidae........................................ 268
Семейство Скворцы Sturnidae........................................... 269
Семейство Врановые Corvidae........................................... 270
Заключение
Литература ,
281
285