/























































































































































































Author: Зимин В.Б.
Tags: здравоохранение медицинские науки птицы фауна ссср
ISBN: 5-02-025632-3
Year: 1988




Text
Л
i
В.Б. ЗИМИН
экология
ВОРОБЬИНЫХ
птиц
СЕВЕРО-ЗАПАДА СССР
АКАДЕМИЯ НАУК СССР
КАРЕЛЬСКИЙ ФИЛИАЛ
ИНСТИТУТ БИОЛОГИИ
В. Б. ЗИМИН
ЭКОЛОГИЯ
ВОРОБЬИНЫХ
птиц
СЕВЕРО-ЗАПАДА СССР
я»1
ЛЕНИНГРАД
„НАУКА"
ЛЕНИНГРАДСКОЕ ОТДЕЛЕНИЕ
1988
УДК598.2:591.52 + 591.54 + 591.16 (47 = 16)
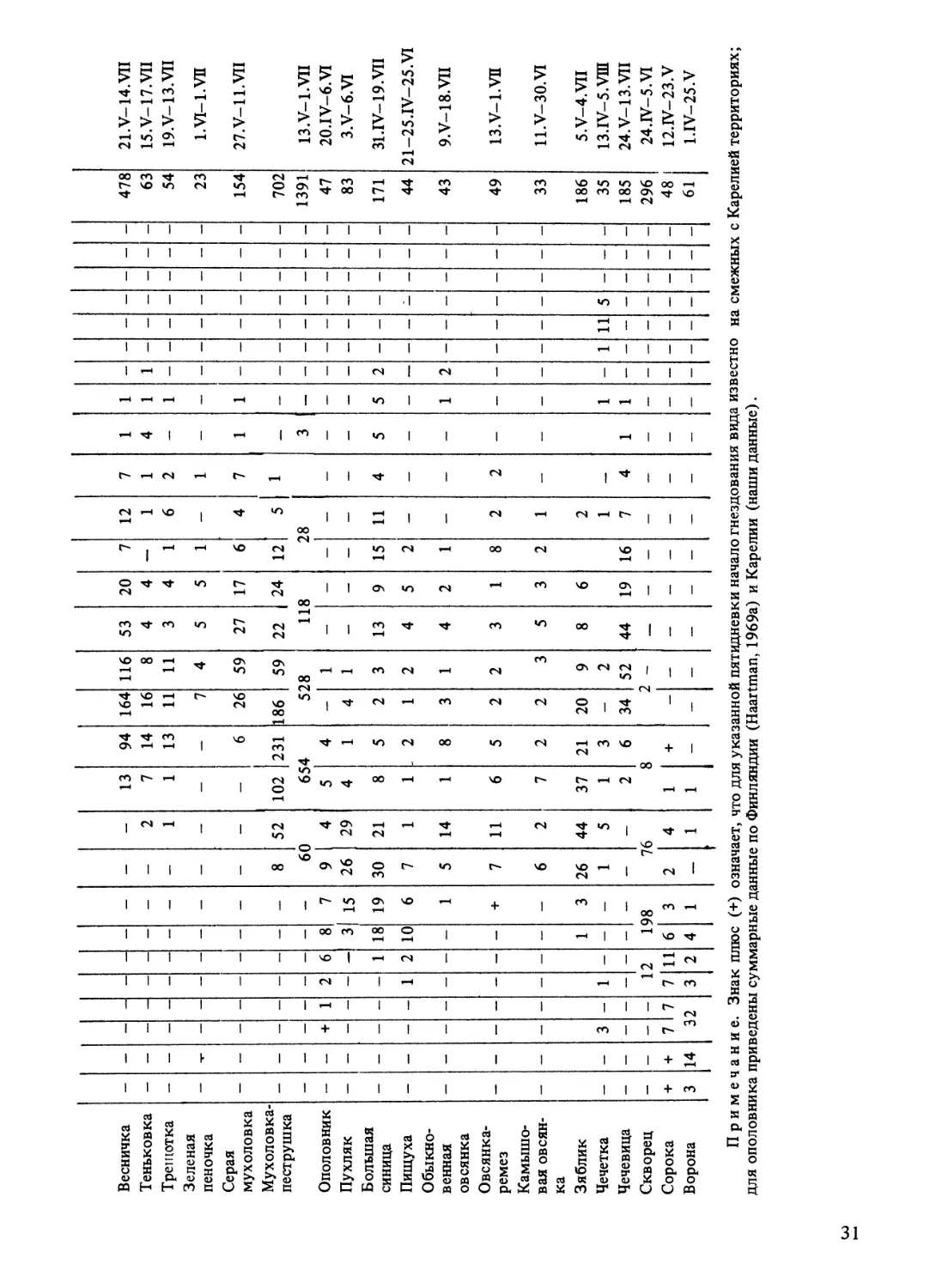
Зимин В. Б. Экология воробьиных птиц Северо-Запада СССР. Л.: Наука. 1988.
В книге анализируются особенности распространения и годовых циклов воробьиных птиц
Северо-Запада СССР, изученных методами постоянного отлова, прижизненного описаниямного-
летнего контроля маркированных популяций. Основные параметры репродуктивного периода и
факторы, определяющие их меж- и внутрипопуляционную изменчивость, рассмотрены в сравни-
тельно-географическом плане.
Предложены гипотезы формирования современных границ распространения, механизма ва-
риабельности величины кладки, путей синхронизации сроков размножения с сезонным развитием
биоценозов, возникновения полициклии и полигинии. Показано, каким образом, используя пози-
тивные и преодолевая негативные воздействия внешних факторов, птицы решают проблему дефи-
цита времени в годовом цикле на севере ареала. Библиогр. 424 назв. Табл. 39. Ил. 47.
ОТВЕТСТВЕННЫЙ РЕДАКТОР
Э.В.ИВ АНТЕР
РЕЦЕНЗЕНТЫ
В. А. БЕРЕСТОВ, В. А. ПАЕВСКИЙ
На обложке: крапивник (Troglodytes troglodytes) с кормом для птенцов (фото Я. А. Пастера).
о 2005020000-563 о_о QQ тт
042 (02)-88 278"88 11
© Издательство „Наука”, 1988 г.
ISBN 5-02-025632-3
ВВЕДЕНИЕ
Популяционная экология — одно из основных направлений современных орнито-
логических исследований, позволяющее решить широкий круг актуальных задач в изу-
чении жизни птиц. В дополнение к традиционным методам популяционная экология,
базируясь на анализе населения птиц достаточно обширных территорий, использует ме-
тоды индивидуального мечения и многолетнего контроля за жизнью маркированных
особей. Отлов и кольцевание птиц позволяют получить более точные данные по их био-
логии, уточнить и выявить многие детали их жизни, обычно ускользающие от внимания
орнитологов, проводящих только визуальные наблюдения.
Основная цель настоящей работы — изучение годовых циклов воробьиных птиц,
обитающих у северных границ ареалов, и выявление адаптаций, позволяющих им суще-
ствовать и поддерживать относительно стабильную численность при увеличивающейся
дальности миграций, кратковременности благоприятных условий и неустойчивости
большинства внешних факторов.
Круг стоявших перед нами задач определялся спецификой географического поло-
жения исследуемой зоны и прежде всего обилием видов, имеющих здесь границы об-
ластей распространения, расселяющихся или периодически предпринимающих попытки
расширения ареалов. Материалом послужили данные, собранные автором в 1957—
1984 гг. на Северо-Западе СССР: на территории Ленинградской обл. (юго-восточное
побережье Ладожского озера), Карельской АССР и в Мурманской обл. (северный архи-
пелаг Белого моря, Кандалакшский заповедник). Стационарные исследования прово-
дились на Белом море, в заповеднике „Кивач”, на Шокшинском (Прионежье) и Ладож-
ском стационарах (д. Гумбарицы и урочище Маячино). В работе частично использованы
также материалы, любезно предоставленные нам Н. В. Лапшиным (о-в Селькьямарьянса-
ари, северо-западная часть Ладожского озера), Т. Ю. Хохловой (о-в Б. Клименецкий,1
Онежское озеро), С. В. Сазоновым (окрестности г. Петрозаводска, заповедник „Кивач”,
с. Кончозеро), Т. А. Рымкевич, Г. А. Яковлевой, В. И. Голованем и В. М. Музаевым
(д. Гумбарицы).
В связи с обилием видов, встречающихся на указанной территории у предела
распространения, важно было определить факторы, препятствующие или способствую-
щие расселению и закреплению вида в определенной широтно-долготной зоне, и выявить
адаптации к жизни на севере.
В северных пограничных областях распространения все популяционные явления
и связи развиваются на фоне гораздо более низкой плотности гнездового населения, чем
в средних и южных частях ареала. Колебания численности свойственны всем видам птиц,
но на севере ее верхний уровень не достигает того рубежа, после которого начинает
четко проявляться зависимость хода различных явлений от плотности населения. Поэ-
тому механизмы регуляции популяционных явлений здесь могут быть иными, чем
в более южных широтах, где к тому же ограничены возможности для проявления дис-
персии. Лесные птицы южных и средних широт обитают в основном в незначительных
В последнее время в печати этот остров стали называть Б. Клим едкий, что неверно, так как
этимологически его название связано с наименованием деревни Клименцы, расположенной
на острове.
3
по площади насаждениях, изолированных друг от друга обширными нелесными место-
обитаниями. В таежной зоне на Северо-Западе СССР, напротив, массивы лесных угодий
занимают большую площадь. Соответственно на севере лесной зоны возможности для
дисперсии особей лесных птиц гораздо шире.
Особая годовая и суточная ритмика освещенности и связанная с этим специфика
суточного баланса энергии местных птиц - характерная черта биолбгии птиц в исследу-
емой зоне, определяющая многие особенности размножения, линьки и миг-
раций.
Многие воробьиные птицы в южных частях ареала ведут оседлый образ жизни,
а проникая на север, становятся мигрантами на ближние и средние расстояния.
Сравнительное изучение биологии таких видов позволяет глубже понять, за счет
каких адаптаций птицы северных популяций справляются с дефицитом вре-
мени, возникающим из-за дополнительных затрат на весенние и осенние миг-
рации.
Таковы наиболее важные особенности исследуемой территории и ее орнитофауны,
определившие как общее направление настоящей работы, так и достаточно широкий
диапазон решаемых в ней конкретных задач.
Анализ любого явления в жизни птиц мы старались по мере возможности про-
водить таким образом, чтобы в итоге была понята его биологическая сущность и можно
было получить ответ на вопрос „почему?”. Отсюда подробности и многократные повто-
рения отдельных фактов и выводов при обсуждении совершенно разных проблем.
Думается, такие повторения неизбежны, и если уж ставить их автору в вину, то в равной
мере можно упрекать в этом и саму природу, в которой все анализируемые явления
прямо или косвенно взаимосвязаны.
При изучении годовых циклов птиц мы по возможности постоянно контролирова-
ли местных маркированных особей в течение всего времени их пребывания на местах
гнездования. Это помогло не только изучить пределы индивидуальной изменчивости,
но и в дополнение к традиционным путям выявить возрастные (годовые) различия
в морфологии, репродуктивных показателях, особенностях линьки и т. п., связав их
с влиянием непостоянных факторов внешней среды, неодинаково воздействующих
на птиц в разные годы.
Низкая численность и рассредоточенность птиц по территории часто не позволяют
выполнить за один сезон достаточное для анализа число наблюдений, поэтому часть
заключений базируется на суммировании данных за весь период исследований.
Если, по мнению автора, такие данные нуждаются в уточнейии, то это специ-
ально оговаривается, а выводы приводятся как предварительные. В тех слу-
чаях, когда это было необходимо, цифровой материал обрабатывался статисти-
чески.
Чтобы не загружать текст, общепринятые положения даны без ссылок на перво-
источник с соответствующими указаниями на широкую известность фактов или
явлений.
В процессе обобщения данных возникали определенные предположения и гипотезы,
недостаточно обоснованные имеющимся материалом. Автор счел возможным опубли-
ковать их, основываясь на логических построениях или литературных сведениях,
полагая, что хотя бы некоторые из высказанных соображений смогут при-
влечь внимание специалистов и побудят их к проверке и развитию начатых
нами исследований. В какой-то мере это определяет и задачи на бу-
дущее.
В сборе полевых материалов в разные годы принимали участие студенты, стажеры,
аспиранты и сотрудники различных учреждений: Калининградского, Одесского, Петро-
заводского, Ленинградского государственных университетов, Карельского педагоги-
ческого института, лаборатории зоологии Института биологии Карельского филиала
АН СССР (КФАН СССР), Зоологического института АН СССР, заповедников „Кивач”,
Кандалакшского и Нижне-Свирского.
4
Большую помощь в работе оказали В. Г. Анненков, А. В. Артемьев, Т. И. Блюмен-
таль, А. Р. Гагинская, В. И. Головань, И. А. Кузьмин, Л. В. Лапшина, Н. В. Лапшин,
В. М. Музаев, Г. А. Носков, И. Б. Савинич, С. В. Сазонов, А. В. Сухов, С. П. Резвый,
А. П. Рипатти, Т. А. Рымкевич, Е. В. Шутенко, Т. Ю. Хохлова, Г. А. Яковлева, М.В. Яков-
лева. В период исследований автор постоянно получал ценные советы и консультации
от профессоров Г. А. Новикова, А. С. Мальчевского и Э. В. Ивантера. Часть работ была
бы неосуществима без поддержки директора Института биологии КФАН СССР проф.
С. Н. Дроздова, директора Института леса КФАН СССР В. И. Ермакова, а также заведу-
ющих лабораториями этих институтов П. И. Данилова и В. И. Шубина. Всем лицам
и коллективам, способствовавшим выполнению данной работы, автор приносит
искреннюю сердечную благодарность.
5
ГЛАВА I
ОБЩИЕ ОСОБЕННОСТИ РАСПРОСТРАНЕНИЯ ПТИЦ
НА СЕВЕРО-ЗАПАДЕ СССР
Наиболее характерной чертой исследуемого региона является обилие растений
и животных, встречающихся здесь у границ своего распространения. Не составляют
исключения и птицы. Так, из 258 видов птиц, зарегистрированных к настоящему вре-
мени на территории Карелии и прилежащих областей, 107 (41.5%) обитают здесь у
северных пределов ареала или даже севернее областей своего постоянного гнездования.
Еще ряд видов имеет на этой территории южные, западные или восточные границы
ареалов.
Только за последние 3 десятилетия в орнитофауне региона произошли значитель-
ные изменения. За это время полностью исчезла с территории Карелии серая куропатка,
в первой половине столетия проникавшая до 65° с. ш. Резко сократилась численность
сапсана и других крупных хищных птиц. Пустельга до 50-х гг. была самой обычной
птицей, но за 2 последующих десятилетия перешла в статус крайне редких; лишь в са-
мое последнее время численность ее постепенно восстанавливается. Буквально на наших
глазах произошло заселение южной части исследуемой территории обыкновенной горли-
цей, черным дроздом, дроздовидной и тростниковой камышевками, дубоносом и др.
Процесс расселения за пределы основных ареалов у различных видов птиц проходит
неодинаково. У одних он носит характер быстро (черный дрозд, чечевица) или постепен-
но (скворец, камышевки) нарастающей экспансии. У других темпы продвижения на
север неравномерны и многолетние периоды относительного обилия сменяются годами
полного или практически полного отсутствия (дубонос, синехвостка, зеленая пеночка).
Для третьих характерны почти ежегодные очень резкие перепады плотности гнездового
населения, никак не связанные с успешностью предыдущего размножения (трещотка).
У ряда видов уже много лет продолжаются эпизодические попытки гнездования неболь-
шого числа особей, не приводящие пока к заметному увеличению и стабилизации числен-
ности (иволга, лесной жаворонок, ястребиная славка, обыкновенный и речной сверчок,
садовая овсянка и др.). Особняком стоит также носящее характер массовых инвазий
ранневесеннее или позднелетнее гнездование чечеток на севере Ленинградской обл.
и юге Карелии.
Обилие птиц, обитающих у границ ареала и предпринимающих более или менее
успешные попытки его расширения, предопределило одно из направлений наших ис-
следований - изучение периферийных зон ареала, демографии и экологии птиц в различ-
ных структурных единицах этих зон и на разных этапах расселения видов. Требовалось
выяснить также, в чем состоит специфика действия факторов внешней среды, ограничи-
вающих распространение столь многих видов птиц и накладывающих отпечаток на такие
особенности их биологии, как фенологически очень раннее начало размножения при его
поздних календарных сроках, большая величина кладки, полицикличность гнездования
и т. п. Возникла необходимость в подробном сравнительном изучении годовых циклов
у аборигенных представителей тайги, а также у птиц, освоивших или пытающихся
заселить ее в настоящее время.
В связи с эпизодичностью формирования временных поселений птиц в периферий-
ных зонах важно было выявить возрастной состав их популяций, характер связей
с территорией, степень гнездового консерватизма. Это, естественно, потребовало орга-
низации массового отлова птиц, их мечения и многолетнего контроля гнездового на-
селения.
6
Широтные изменения в составе растительности
Не останавливаясь на характеристике климата и растительности Северо-Запада
СССР, которая неоднократно публиковалась и автором, и его коллегами (Зимин, Иван-
тер, 1969,i 1974; Ивантер, 1975; Данилов, Туманов, 1976; Зимин, 1976а; Зимин,
Кузьмин, 1980), мы ограничимся лишь анализом наиболее важных особенностей среды,
которые могут влиять на распространение птиц и их биологию. Многие детали связей
птиц с условиями обитания будут специально обсуждаться в следующих главах, поэтому
здесь они представлены в самом общем виде.
В широтном изменении растительного покрова исследуемой территории следует
подчеркнуть следующие основные черты, с которыми может быть связано современное
распространение птиц.
В зоне 60—67° с. ш. в коренных таежных древостоях явно преобладают сосна и
ель, причем степень их доминирования в составе лесообразующих пород быстро увели-
чивается с юга на север при соответствующем сокращении доли лиственных деревьев
и разнообразия кустарников (Яковлев, Воронова, 1959; Раменская, 1960).
Интенсивное использование лесных ресурсов способствовало: 1) смене коренных
таежных древостоев производными типами разновозрастных насаждений со значитель-
ной долей или преобладанием лиственных деревьев, в результате чего обширные тер-
ритории оказались заняты различными типами смешанных лесов, осинников, березня-
ков, сероольшанников с более или менее развитым кустарниковым пологом и под-
ростом хвойных, особенно ели; 2) изменению общего облика ландшафта — лесные мас-
сивы перемежаются обширными открытыми (свежие и необлесившиеся вырубки)
и полуоткрытыми стациями (разнообразные по составу пород, плотности и возрасту
молодняки); 3) возникновению пестроты и мелкоконтурности выделов.
В настоящее время типичные таежные леса представлены на юге региона факти-
чески только лесами 1-й группы, имеющими важное водоохранное значение. На севере
Карелии пока еще сохранились массивы спелых таежных сосновых и еловых лесов,
только местами разреженных рубками или замененных вторичными насаждениями
антропогенного происхождения.
Следует отметить еще одну особенность растительного покрова Северо-Запада
СССР. По неопубликованным данным Ф. С. Яковлева, в коренных таежных лесах этого
региона примерно к середине нашего столетия наметилась отчетливая тенденция к смене
сосны елью. Действительно, и в коренных, и в производных типах сосняков-зелено-
мошников повсеместно заметно формирование елового яруса. Чем бы ни объяснялся
этот процесс, орнитофауна сосняков с развитием подроста ели стала много богаче.
Значительный рост общей численности лесной завирушки, начавшейся в середине теку-
щего столетия, отчасти связан с увеличением обилия ели под пологом смешанных,
лиственных и сосновых лесов. По учетам, выполненным Мерикаллио (Merikallio, 1958)
в первой половине века, и по современным данным (Jarvinen О., 1980), в Финляндии чи-
сленность этого вида возросла примерно в 100 раз.
Расширение сельскохозяйственного производства привело к возникновению об-
ширных пространств пашен и лугов вместо мелких частных угодий такого типа, выру-
бавшихся или выжигавшихся крестьянами внутри лесных массивов, к настоящему вре-
мени заросших кустарниками или лесами с обильным и разнообразным по составу
подлеском и мощным травостоем.
Производные типы леса, особенно лиственные, по своему строению, ярусности,
освещенности, обилию подлеска, характеру фаутности и т. п. имеют много общего
с насаждениями средних широт и широколиственными лесами. Такие стации в первую
очередь осваиваются птицами южных орнитокомплексов при их расселении по таежной
зоне (белоспинный дятел, клинтух, обыкновенная горлица, иволга, лазоревка, соловей,
садовая и черноголовая славки, пересмешка и др.).
Молодняки по своей структуре сходны с кустарниковыми зарослями пойм, овра-
гов и разреженных лесов южных зон. По молодым насаждениям, развивающимся
7
на месте вырубленных лесов, идет расселение к северу козодоя, чечевицы, жулана,
обыкновенной овсянки, всех видов славок, численность которых здесь во много раз
выше, чем в спелых лесах таежного типа, где они иногда заселяют только приопушечный
шлейф по границе с открытыми и полуоткрытыми стациями. Кустообразная поросль,
развивающаяся на заброшенных луговых участках или вдоль осушительных канав
на выпасах и сенокосах, кроме названных видов птиц, привлекает на гнездование садо-
вую и болотную камышевок, коноплянку, местами — садовую овсянку.
Сравнение населения птиц коренных таежных лесов и местообитаний антропоген-
ного происхождения, преобладающих сейчас на большей части территории региона,
отчетливо выявляет процесс „объюжнения” орнитофауны и увеличения численности
многих представителей орнитокомплекса европейских широколиственных лесов,
начавшийся, видимо, еще в прошлом веке (Нейфельдт, 1958; Ивантер, 1962, 19696;
Зимин, Ивантер, 1969, 1974; Зимин, Лапшин, 1975; Зимин, 1976; Данилов и др.,
1979; Зимин, Кузьмин, 1980).
Судя по современному биотопическому размещению птиц, проникших в тайгу
из более южных зон, таежные массивы хвойных лесов до преобразования человеком,
очевидно, долго служили многим птицам своеобразным экологическим барьером на
пути их расселения к северу. Конечно, попытки проникновения через этот барьер навер-
няка имели место, например по пойменным и приозерным лугам с кустарниками. Но
из-за незначительной общей площади таких стаций и их рассеянного распределения
по' территории численность птиц, проникающих в тайгу, была, очевидно, недостаточной
для прочного закрепления вида на новых рубежах. К тому же на небольших луговых
участках, окруженных подступающим вплотную лесом, вегетация растений начинается
позднее, чем на обширных открытых пространствах. Поздние сроки весеннего развития
природы, вероятно, задерживали начало размножения, что при незначительной продол-
жительности благоприятного периода могло служить дополнительным препятствием
для закрепления птиц в северных широтах.
Современное лесное и сельское хозяйство, а также другие виды деятельности
человека существенно сказываются на распределении птиц по территории, обеспечивают
их новыми видами обильных кормов, влияют на выживаемость в критические периоды,
вызывают основательные сдвиги в годовых циклах и т. д. Сорные растения, сопут-
ствующие разведению пропашных и других культур, дают большие урожаи семян,
на которых способно прокормиться несравненно большее число птиц, чем на семенах
травянистых растений в естественных местообитаниях. В предотлетный период на
полях концентрируются очень многие лесные птицы, переходящие на питание раститель-
ными кормами, в основном семенами сорняков. Осенью их поедают жаворонки, конь-
ки, лесные завирушки. В конце августа—сентябре, покидая лесные угодья, выходят
на поля лесные вьюрковые и овсянки. Общая осенняя численность птиц, кормящихся
на сельскохозяйственных угодьях, в сотни, а местами и в тысячи раз выше, чем в лес-
ных стациях и на естественных лугах среди леса (Зимин, 1969а, 19696, 1973).
Семена сорняков — это фактически корма антропогенного происхождения. Во вся-
ком случае, без сельского хозяйства они вряд ли были бы столь обильны. Трудно ска-
зать, могли ли бы птицы в наших широтах, питаясь одними естественными кормами, оста-
ваться на местах гнездования так же долго, как в настоящее время. Многие из них поки-
дают лесные угодья на 1—1.5 мес раньше, чем агроландшафты. Таким образом, вполне
вероятно, что возникновение обильной кормовой базы в местообитаниях, искусственно
созданных человеком, позволило птицам увеличить период пребывания на родине.
Этому способствовало и обеспечение подобными же кормами на всей трассе пролета.
Весной сельскохозяйственные угодья, открытые и полуоткрытые стации по срав-
нению с лесными биотопами на 2—3 недели, а при содействии человека более чем на
месяц раньше освобождаются от снега. Это характерно и для территорий, по которым
идет весенний пролет. Ранний сход снега позволяет птицам обеспечить себя кормом
гораздо раньше, чем внутри лесных массивов. Как будет показано дальше, это опреде-
ляет и более раннее начало гнездования птиц в антропогенном ландшафте.
8
Более ранние прилет и начало размножения вместе с осенней задержкой позволяют
многим видам птиц, приспособившимся к жизни в преобразованной человеком среде,
значительно увеличить период пребывания на родине. Это снимает напряженность годо-
вого цикла и способствует успешному завершению размножения, линьки и подготовки
к миграции.
Таким образом, антропогенное преобразование таежных местообитаний обеспе-
чивает птиц обильными кормами, доступными на всем пространстве ареала и пролет-
ных путях более длительный период, чем в естественных угодьях. Это способствует их
расселению на север и приводит к повышению выживаемости во время предотлетных
скоплений, на местах остановок по трассе миграций, а также и на зимовках. Правда,
практикуемое в современном сельском и лесном хозяйстве широкое применение
пестицидов вызывает обратный эффект.
Для многих синантропных птиц человек стихийно создает богатую и стабильную
кормовую базу. Это всевозможные пищевые отбросы, потерянное при транспортировке
сырье для пищевой промышленности, корма для сельскохозяйственных животных
и т. д. На таких кормах большую часть года существуют многие синантропные птицы -
врановые, сизые голуби, воробьи. Их концентрации на фермах, свалках, в населенных
пунктах, на транспортных магистралях бывают достаточно многочисленными. В послед-
ние десятилетия к питанию пищевыми отходами приспособились и другие птицы. Сизые,
серебристые и озерные чайки с прилета в конце марта и до отлета, затягивающегося
до середины декабря, вместе с врановыми стали постоянными обитателями зверовод-
ческих ферм, где образуют скопления до 2 тыс. особей. Складывается впечатление,
что многие из них, по крайней мере холостые особи, вообще перестают кормиться
естественными кормами. С весны и до осени в этих скоплениях птиц можно увидеть
также скворцов.
Обилие калорийных кормов привело к формированию колоний чаек в необычных
для них местах. Так, в Пряжинском районе в окрестностях зверосовхозов озерные чай-
ки стали гнездиться на прибрежных сфагновых сплавинах озер, а сизые чайки - посреди
болот, на значительном удалении от водоемов. В последние годы в Петрозаводске
и других населенных пунктах все чаще стали вс1речаться озерные чайки, кормящиеся
пищевыми отходами на выгребных ямах или специальных мусорных контейнерах
у жилых домов. Так как обилие и доступность искусственных кормов не зависят
от погоды, то птицы, приспособившиеся к питанию ими, не испытывают затруднений
с выкармливанием потомства. По той же причине, очевидно, улучшается выживаемость
молодых птиц в послегнездовой период и особенно зимой.
Отдельные виды перелетных птиц все чаще стали задерживаться в таежной зоне
на зиму. Это прежде всего грачи и скворцы, практически ежегодно в небольшом коли-
честве зимующие в окрестностях крупных населенных пунктов. Все отмеченные в Каре-
лии случаи зимовки обыкновенных овсянок, зябликов, черных дроздов и зарянок
относятся к населенным пунктам. Да и сойка в условиях тайги вряд ли смогла бы
существовать зимой, если бы не перемещалась на этот период к лесным населенным
пунктам. Очевидно, ворона и сорока также не смогли бы зимовать в наших широтах
только на одних естественных кормах.
Хорошо известно, что на севере нерегулярные массовые зимовки дроздов-рябин-
ников и свиристелей отмечаются в годы обильного плодоношения рябины. Но основные
запасы этого корма сосредоточены здесь отнюдь не в лесах таежного типа, а главным
образом во вторичных лиственных насаждениях, расположенных среди агроландшаф-
та и на относительно более богатых почвах. Запасы ягод рябины внутри коренных
таежных древостоев или вторичных хвойных насаждений настолько малы, что уничтожа-
ются птицами задолго до окончания осенней миграции. В Петрозаводске два зарегистри-
рованных за последние годы случая зимовки зарянок были отмечены в парках и на
кладбище, где птицы кормились интродуцированной здесь красной бузиной и водными
беспозвоночными, которых добывали на порожистых участках рек Лососинки
и Неглинки.
9
Таким образом, все необычные зимовки наших птиц так или иначе связаны с ант-
ропогенными преобразованиями среды. Исключений в этом отношении немного. На
естественных кормах, которые птицы находят на порожистых реках, регулярно зимуют
небольшие группы крякв, отдельные особи чирков-свистунков, гоголей, лебедей-
кликунов. На незамерзающих участках Онежского и Ладожского озер отмечены единич-
ные случаи зимовки больших бакланов, морянок, сизых чаек. На юге Карелии за пос-
ледние 25 лет большие синицы остались зимовать в лесных местообитаниях лишь теплой
зимой 1982—1983 гг. В остальные годы они зимовали только в населенных пунктах.
При расселении за пределы основных ареалов многие виды птиц сначала осваивают
только культурный ландшафт. Щегол, зеленушка, коноплянка, садовая овсянка до сих
пор гнездятся у нас в местообитаниях антропогенного происхождения, а два первых
вида — исключительно в населенных пунктах городского типа.
Итак, антропогенное преобразование таежной зоны существенно изменило состав
и структуру растительных сообществ и ландшафта в целом. Наряду с негативными
последствиями для аборигенов тайги, не сумевших адаптироваться к жизни в новых
условиях, для многих других видов птиц антропогенные изменения среды обитания
имели очень важное положительное значение. Увеличился период благоприятных усло-
вий, необходимых для успешного завершения размножения, линьки и миграций Для
видов, приспособившихся к питанию кормами антропогенного происхождения, это ста-
ло предпосылкой перестройки годового цикла: одним позволило успешно преодолевать
трассу миграций в более поздние сроки осенью и ранние — весной; другим — полностью
или частично исключить миграции. Независимость кормовой базы от внешних воздей-
ствий обеспечила высокую выживаемость птиц в критических ситуациях — при резких
похолоданиях. Все это обусловило, с одной стороны, расширение гнездовых ареалов
в северном направлении, с другой — сдвиг зимовочных областей к северу и сокращение
дальности миграций.
Трудно сказать, были ли наши птицы так же многочисленны на европейских
зимовках в отдаленный исторический период, когда плотность населения человека
была здесь не такой высокой, как сейчас, а степень преобразования коренных природ-
ных ландшафтов — несравненно более низкой. Не исключено, что из-за недостатка при-
годных для жизни стаций многие птицы вынуждены были мигрировать в области с бо-
лее теплым стабильным климатом, а гнездовые ареалы в зоне тайги ограничивались
гораздо более южными широтами.
Скорее всего иным было и соотношение видов. Преимущества, полученные пти-
цами, освоившими местообитания и корма антропогенного происхождения, обеспечива-
ли им лучшую выживаемость и соответствующий рост численности. Не случайно среди
европейских видов доминирующее положение на большей части ареала сейчас занимает
зяблик, 3/4 года обитающий на агроландшафтах. В ближайшее время можно ожидать
подъема численности кряквы, городские, в том числе и зимующие, популяции которой
имеют явную тенденцию к росту. Все успешнее осваивают антропогенный ландшафт
и соответствующие корма сизые и обыкновенные чайки, ставшие наравне с врановыми
обычными обитателями животноводческих ферм, зверосовхозов, городов и т. д.
Сезонность и неустойчивость климата
Характерными чертами климата исследуемого региона, влияющими на распростра-
нение и биолргию птиц, являются резко выраженная сезонность, неустойчивость погоды
в любое время года, незначительная продолжительность вегетационного периода и осо-
бый суточный и сезонный ритм освещенности. Определенное значение имеют для птиц
и некоторые более частные климатические особенности, рассмотренные ниже в этой и
других главах.
Разумеется, такие черты климата,как неустойчивость и сезонность^ той или иной
мере свойственны практически всем природно-климатическим зонам Земли,
10
но в северных широтах они проявляются в наиболее резкой форме. В частности, возвра-
ты холодов с понижением температуры воздуха до О °C и ниже в Северной Карелии,
а тем более в Заполярье, возможны в любое время года, а на юге зоны - в течение
большей части периода массового размножения птиц (табл. 1). Снижение температуры
до 5 °C резко угнетает двигательную активность большинства насекомых, многие
из которых заблаговременно уходят в разнообразные укрытия (Бей-Биенко, 1971).
Это делает их недоступными для многих видов птиц. При похолоданиях, сопровождаю-
щихся обильными снегопадами, кормов лишаются и зерноядные птицы. Даже на юге
Карелии подобные ситуации имеют место вплоть до середины июня.
Похолодания в зависимости от сроков, интенсивности и продолжительности
оказывают на птиц разное воздействие: вызывают массовую обратную миграцию,
перемещение из лесных биотопов в антропогенные местообитания, концентрацию
в населенных пунктах и на побережьях водоемов, переход на питание второстепенными
и нехарактерными кормами, а также прекращение гнездостроения, откладки и инкуба-
ции яиц и более или менее массовую гибель кладок, птенцов и даже взрослых
особей.
Как было показано нами ранее (Зимин и др., 1976), длительные похолодания и
снегопады, приводящие к массовой гибели перелетных птиц, не оказывают заметного
влияния на успешность выкармливания птенцов врановых, хохлатой синицы и пищухи,
адаптированных к зимним условиям и способных извлекать насекомых из укрытий.
Существуют две основные возможности преодоления неожиданно наступающих
похолоданий. Первая — расширение диапазона адаптаций к прямому и косвенному
влиянию низких температур. При холодной погоде преимущество получают виды с бо-
лее совершенным механизмом терморегуляции, адаптированные к низким температу-
рам и наиболее экологически пластичные, обладающие широким спектром питания,
способные переключаться с животной пищи на растительную и, в частности, освоившие
корма антропогенного происхождения. Другой путь приспособления к неустойчивой
погоде и колебаниям в обилии и доступности кормов связан с неодновременностью,
растянутостью начала всех явлений годового цикла птиц в популяции. Весной в ситуа-
циях, сопровождающихся массовой гибелью птиц, неодновременность возвращения
на родину оказывается выгодной — воспроизводство будет осуществляться теми осо-
бями, которые прилетели после похолодания. Общая величина гнездовых потерь при
неодновременном начале размножения также будет ниже, так как внезапные холода
совпадут с критическими стадиями гйездового цикла только у части пар, и т. д.
Растянутость гнездования, равно как и любой другой фазы годового цикла, явля-
ется у птиц и общевидовым, и внутрипопуляционным свойством — одним из проявле-
ний всеобщей разнокачественности признаков. В условиях непостоянства внешней
среды относительная неодновременность прохождения каждой фазы разными особями
приобретает важное значение для поддержания стабильной численности и закрепляется
эволюцией не только у птиц, но и среди других животных, а также у растений. Благо-
даря этому свойству достигается более полное и рациональное использование среды
обитания, а при высокой плотности населения — и более равномерное потребление
пищевых ресурсов, предотвращающее обострение внутривидовой конкуренции. Хотя
положительное значение растянутости особенно заметно в северных широтах, где непо-
стоянство внешних факторов ощущается чаще и в более резкой форме, возможности
для ее проявления с увеличением географической широты быстро сокращаются. Так,
уже в зоне 60—62° с. ш. у некоторых дальних мигрантов период откладки яиц не пре-
вышает 40 дней, и около 80% гнездовых пар в некоторые годы приступают к размноже-
нию в течение 5—10 дней. Но если период массового ьылупления птенцов совпадает
с глубоким и продолжительным похолоданием, то значительная часть их гибнет. В такой
ситуации воспроизводство будет осуществлено птицами, начавшими размножение
в иные, чем основная масса, сроки, а также особями, случайно сохранившими выводок
или успевшими сделать повторную кладку. При строго одновременном начале размно-
жения последствия могли бы оказаться гораздо более тяжелыми.
И
to
Таблица 1
Число лет (%) с заморозками различной интенсивности, отмеченными за соответствующую декаду на юге (Олонец) и (по: Агроклиматический справочник по Карельской АССР, 1959) севере (Лоухи) Карелии
Минималь- Апрель Май Июнь Июль Август Сентябрь Октябрь
ная трмпе- • ратура (°C) 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2 3 1 2
Метеостанция Л оу хи
-5 -3 -1 0 +2 97 97 100 100 100 97 97 100 100 100 74 90 100 100 100 67 83 100 100 100 33 57 100 100 100 7 40 90 100 100 13 60 70 83 7 17 50 3 7 30 3 7 23 13 43 3 15 43 7 15 43 14 17 43 14 35 69 7 14 48 69 83 7 42 69 76 93 38 48 86 86 100 59 83 86 90 100 45 76 90 97 100
Метеостанция Олонец
-5 -3 -1 0 +2 96 100 100 100 100 86 86 96 100 100 35 52 90 100 100 11 46 79 86 97 7 79 86 96 4 29 54 82 11 26 45 74 4 8 30 22 4 - 1 1 1 1 1 4 4 14 4 7 21 7 25 39 54 14 43 54 71 14 29 61 64 96 36 54 71 82 89 39 64 82 86 96
Таблица 2
Годовые вариации сроков некоторых сезонных явлений в Ухте (I) и Петрозаводске (II) (по: Агроклиматический справочник по Карельской АССР, 1959)
Пункт наблюде- ний Дата Амплитуда коле-
Сезонное явление Наиболее ранняя Средняя многолетняя Наиболее поздняя баний (сут)
Окончательный сход снежного покрова Начало пыления серой ольхи Начало зеленения березы Начало цветения черники Начало созревания ягод черники I II I II I II I II I II - 15.04 17.02 30.04 3.04 8.05 26.04 17.05 3.05 28.06 30.06 29.04 6.04 7.05 20.04 25.05 15.05 2.06 25.05 24.07 15.07 18.05 25.04 25.05 10.05 8.06 29.05 18.06 9.06 30.07 35 68 27 38 32 34 33 37 31
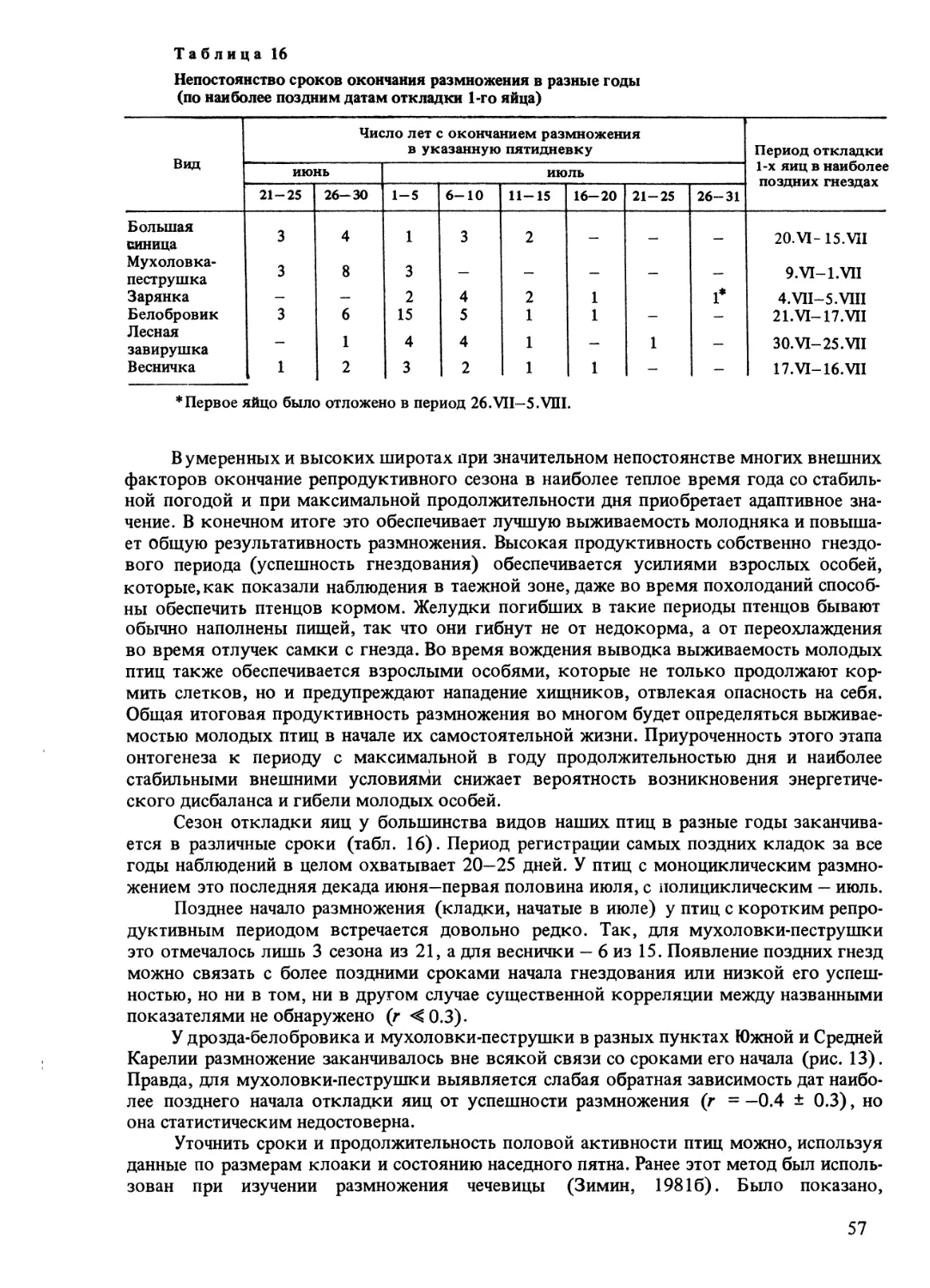
В Субарктике, по данным Данилова (1966), гнездование большинства воробьиных
птиц проходит в очень сжатые сроки — откладка яиц занимает обычно 2-3 декады
(5 только у чечетки). В высоких широтах при массовой гибели птенцов воспроизвод-
ство в популяции осуществляется за счет незначительного числа особей, так как после
потери птгенцов повторное размножение практически невозможно. В этих условиях
становится понятной целесообразность увеличения размеров кладки и выводка у север-
ных птиц: невозможность повторного гнездования особей, потерявших птенцов, в значи-
тельной мере компенсируется тем, что остальные птицы воспитывают крупные выводки.
В наших условиях недостаточно успешное размножение перелетных птиц не всегда
сопровождается сокращением численности гнездового населения вида на следующий
год. Происходит это потому, что динамика численности птиц в периферийных зонах
ареалов осуществляется не только за счет внутренних ресурсов, но и в результате
ежегодного притока птиц извне — дисперсии особей неизвестного происхождения, кото-
рая в разные годы бывает более или менее интенсивной.
Одно из общих следствий неустойчивости климатических факторов — крайняя
неодновременность и неравномерность фенологического развития природы. Сроки
начала отдельных весенних явлений могут различаться по годам более чем на месяц.
Резкие ежегодные различия в характере весны обусловлены состоянием погоды,
в равной мере влияющей на сроки прилета и размножения птиц (табл. 2). Но если
амплитуда колебаний дат прилета у ранних мигрантов (скворец) составляет около
месяца, то сроки начала откладки яиц в целом варьируют меньше. У птиц, обычно при-
ступающих к гнездованию в мае, годовые различия редко превышают две недели, а у на-
чинающих размножение в июне, когда вероятность возврата резких похолоданий сокра-
щается, составляют не более одной декады. В южных и средних широтах сроки начала
гнездования в разные годы менее стабильны, в основном за счет очень ранних дат
откладки яиц в необычно теплые весны (Goodackre, Lack, 1959; Lack, 1966).
В условиях резко выраженной сезонности климата и неустойчивости большинства
факторов внешней среды эволюция годовых циклов птиц (и любых организмов) по-
шла, как известно, по пути строгой приуроченности каждой фазы лишь к определенному
времени года. Но таким образом организм лишь потенциально подготавливается к каж-
дой фазе, а ее реализация определяется уже конкретными условиями изменчивой
среды в местах обитания каждой особи. Поэтому внутри каждого сезона не существует
строго запрограммированных дат начала фазы годового цикла. Эти даты подчинены
колебаниям местных условий вплоть до того, что в особо неблагоприятных ситуациях
отдельные этапы годового цикла могут остаться нереализованными. Примером может
служить отказ птиц от размножения при необычайно холодной погоде в Субарктике
и Арктике (Bertman et al.,1934; Формозов, 1935; Marshall, 1952; Данилов, 1966; Успен-
ский, 1969). При этом сохраняется окологодовая цикличность и реализуется следующая
фаза годового цикла (Marshall, 1960; Дольник, 1964,1975; Farner, 1964; Gwinner, 1971).
При строгой запрограммированности каждой фазы годового цикла существование
вида в изменчивой среде обитания было бы невозможно. Его распространение должно
было бы ограничиваться лишь территориями с наиболее стабильными условиями, а все
попытки расселения за пределы этой зоны заканчивались бы неудачей.
При увеличении дальности миграций у птиц остается очень мало времени на одну
из основных функций — размножение, особенно если учесть, что воспроизводство дол-
жно быть приурочено к периоду благоприятных условий, который укорачивается с уве-
личением географической широты. У птиц возникает необходимость точного определе-
ния сроков и продолжительности каждого активного этапа годового цикла. Мы убежда-
емся в этом на примере дальних мигрантов, у которых и сроки прилета, и даты начала
кладки варьируют по годам гораздо меньше, чем у остальных птиц. Позже всех в нашу
зону прилетает дубровник, сроки прилета и гнездования которого из года в год остают-
ся практически одинаковыми. Так, появление передовых самцов всегда регистрирует-
ся 6—10. VI, формирование гнездового населения завершается за 3—5 сут, откладка
яиц начинается на 3—7 сут после прилета самок (Рымкевич, 1976). Создается
13
Таблица 3
Соотношение времени, необходимого на завершение летних фаз годового цикла при
Вид
Откладка яиц насижива- ние выкармливание птен- цов в гнезде и вы- водке переходный период
Мухоловка-пеструшка 6 12 18 + 12 5-10
Весничка 6 12 12 + 15 5-10
Зарянка (I + II цикл) 6 + 5 12 + 12 (12 + 12) X 2 5-10
Овсянка-ремез (I + II цикл) 6 + 5 12 + 12 (10+10) X 2 5
Дубровник 6 12 9 + 10 —
Чечевица 5 12 12 + 14 —
впечатление строгой датировки сроков прохождения всех фаз годового цикла, которой
на самом деле не существует. Птицы достигают наиболее удаленных от зимовок точек
ареала потенциально полностью готовыми к размножению к моменту стабилизации
погоды. Вероятность возврата холодов ко времени прилета уже низка, и ничто не пре-
пятствует незамедлительному началу размножения. Но дефицит времени на периферии
ареала ощущается остро, поэтому к гнездованию приступают лишь те особи, репродук-
тивное состояние которых еще не выходит за пределы допустимых сроков. Остальные
или остаются холостыми, или частично приступают к размножению, не достигнув ро-
дины, как это случается у нас с вьюрками при затяжных холодных веснах.
Таким образом, у птиц, обитающих в зонах, максимально удаленных от мест
зимовок, и вынужденных большую часть времени затрачивать на миграции и зимовки,
продолжительность и сроки летних фаз годового цикла должны точнее, чем у других,
соответствовать состоянию среды и дефициту времени на местах размножения и трас-
сах миграции. Шансы на участие в воспроизводстве реализуют лишь те особи, у которых
такое соответствие оказывается наиболее полным. Это является причиной, ограничива-
ющей распространение вида, и отчасти объясняет неустойчивость его периферийных
популяций.
Постепенное сокращение благоприятного периода с увеличением географической
широты создает на севере ареалов определенный дефицит времени, необходимого на
реализацию всех фаз годового цикла, приходящихся на весну, лето й осень. Иллюстра-
цией могут служить представленные в табл. 3 данные по продолжительности отдельных
этапов репродуктивного цикла и линьки некоторых видов воробьиных птиц в сравне-
нии с длительностью периода от начала кладки до отлета местных особей в зоне
60-62° с. ш.
Суммарная продолжительность периода, необходимого для размножения, линьки
и подготовки к миграции, рассчитана как сумма времени, требующегося на каждый из
последовательных этапов. Некоторые сложности возникают при определении даты окон-
чания периода, благоприятного для жизни каждого конкретного вида. Очевидно, в ка-
честве такой даты можно принять время отлета местных особей на зимовку.
Четкие критерии „благоприятных условий” трудно определить даже для хорошо
изученных видов птиц. В какой момент отдельные факты среды перестают быть благо-
приятными для вида, если ни один из них не влияет на птиц вне связи с остальными?
Крайне сложно оценить состояние кормовой базы, доступность кормов при разной
погоде и т. д. Ясно, что понижение температуры воздуха сопряжено с увеличением зат-
рат на терморегуляцию и с ухудшением условий кормодобывания, но в зависимости
от продолжительности вынужденного ночного покоя при длительной и более короткой
ночевке суточный баланс энергии организма при одинаковом снижении температуры
будет существенно различаться. У зарянки в Карелии дата начала кладки точно совпада-
ет со средней датой последнего заморозка и переходом среднесуточных температур
примерно через 8 °C. Абсолютно такие же показатели характеризуют среднюю дату
14
классическом разобщении всех этапов (Тн), с реальными затратами его (Тр) у птиц Карелии (сут)
Период от начала гнездо- вания до отлета местных птиц ГР Дефицит времени (Гр “ Гн) на гнездо- вание и линьку
послебрачная линька подготовка к миграции всего
55 5-10 120 25.V-15.VHI 80 -40
55 5 110 l.VI-20.VTII 80 -30
65 5 160 15.V-20.IX 125 -35
60 5 145 15.V-15.IX 120 -25
На зимовке 5 45 10. VI-20. VIII 60 Отсутствует
>> п 5 50 10.VI-20.VIII 60 »♦
начала отлета местных особей. Однако анализ ежегодных дат начала гнездования и отле-
та не подтверждает их связи с названными критериями. К тому же размножение начина-
ется на 18-часовом дне, а отлет — на 12-часовом, т. е. при абсолютно разном балансе
энергии.
По данным табл. 3 очевидно, что уже на 60—62° с. ш. многим видам птиц для за-
вершения гнездования, линьки и подготовки к миграции времени явно недостаточно.
Без дополнительных мер, позволяющих преодолеть данный дефицит, на этих и более
северных широтах птицы существовать не могут. Даже виды, зимующие у нас на местах
гнездования, ощущают недостаток времени с благоприятными условиями. Например,
взрослым пухлякам, ведущим оседлый образ жизни, при очень продолжительной после-
брачной линьке, самые ответственные и энергоемкие стадии, на которых объем сменяю-
щегося оперения максимален, выгоднее пройти до ухудшения условий. В противном
случае напряженность энергетического баланса может оказаться для них чрезмерной.
В процессе детального исследования годовых циклов певчих птиц на Северо-
Западе СССР были выявлены следующие основные пути преодоления дефицита времени,
обеспечивающие успешное завершение всех весенне-осенних фаз годового цикла:
— сокращение периода от прилета до гнездования,
— выпадение отдельных этапов предгнездового поведения,
— совмещение периодов формирования яиц и строительства гнезда,
— совмещение откладки яиц с достраиванием гнезда,
— совмещение откладки яиц с насиживанием,
— совмещение первого и второго циклов размножения,
— совмещение гнездования и линьки у одной особи,
— начало гГостювенильной линьки в более раннем возрасте, особенно припоздаем
размножении,
— сокращение объема сменяющегося оперения,
— совмещение последовательных стадий линьки,
— совмещение окончания линьки с началом миграций,
— полный и частичный перенос линьки на период зимовки,
— формирование промежуточных летних миграций в направлении зимовок.
Сокращение и совмещение последовательных фаз годового цикла и стадий внутри
каждой из них дают местным птицам существенный выигрыш во времени и позволяют
закончить их до наступления неблагоприятных условий.
Специфическое влияние режима суточной освещенности
Еще одна специфическая особенность исследуемой зоны — своеобразная суточная
ритмика освещенности в весенне-летние и зимние месяцы. Зимой на 66° с. ш. минималь-
ная продолжительность дня составляет всего 2 ч 56 мин, на 62° с. ш. — 5 ч 10 мин, летом
на севере исследованной территории солнце в 3-й декаде июня вообще не заходит,
15
а на юге ее остается за горизонтом около 4 ч (рис. 1). Тем не менеее освещенность при
этом сохраняется на высоком уровне, вполне обеспечивающем возможности кормежки
птиц. Белые ночи приходятся на период массового выкармливания птенцов и перехода
сеголетков к самостоятельной жизни.
Летом в суточном энергетическом балансе птиц исследуемой зоны количество
поступающей с пищей энергии значительно превышает чистые затраты на период ночного
покоя. Продолжительность последнего определяется здесь не внешними факторами
(продолжительностью темного времени суток), а только потребностью организма
в отдыхе. Поэтому ночной покой здесь имеет минимальную или близкую к таковой
продолжительность, а при необходимости может прерываться для кратковременной
кормежки и проявления других видов активности. Летом в жизни птиц излишки энер-
гии, образующиеся в суточном балансе, играют очень важную роль. Прежде всего бла-
годаря этому птицы северных популяций получают возможность совмещать
энергоемкие фазы годового цикла (и отдельные стадии внутри них) и смещать гнездова-
ние на более ранние фенологические сроки. Однако это возможно лишь в определенной
широтной зоне. В Субарктике и Арктике этому будут препятствовать более низкие тем-
пературы, в результате чего часть излишков энергии в основном расходуется на термо-
регуляцию. Южнее препятствием к образованию излишков энергии может быть большая
продолжительность периода ночного отдыха. Поэтому отмеченные выше особенности
годовых циклов воробьиных птиц специфичны лишь для южной части зоны белых ночей
с относительно теплым летом. На Северо-Западе СССР это примерно полоса между
59-65° с. ш.
В пределах указанной зоны энергетические потери птенцов воробьиных птиц за
короткий ночной перерыв в их кормлении не успевают достичь летального уровня даже
при не вполне благоприятных условиях кормежки (рис. 2). В результате успешность
выкармливания птенцов на севере связана с величиной выводка в меньшей степени, чем
на юге (подробнее это обсуждается ниже).
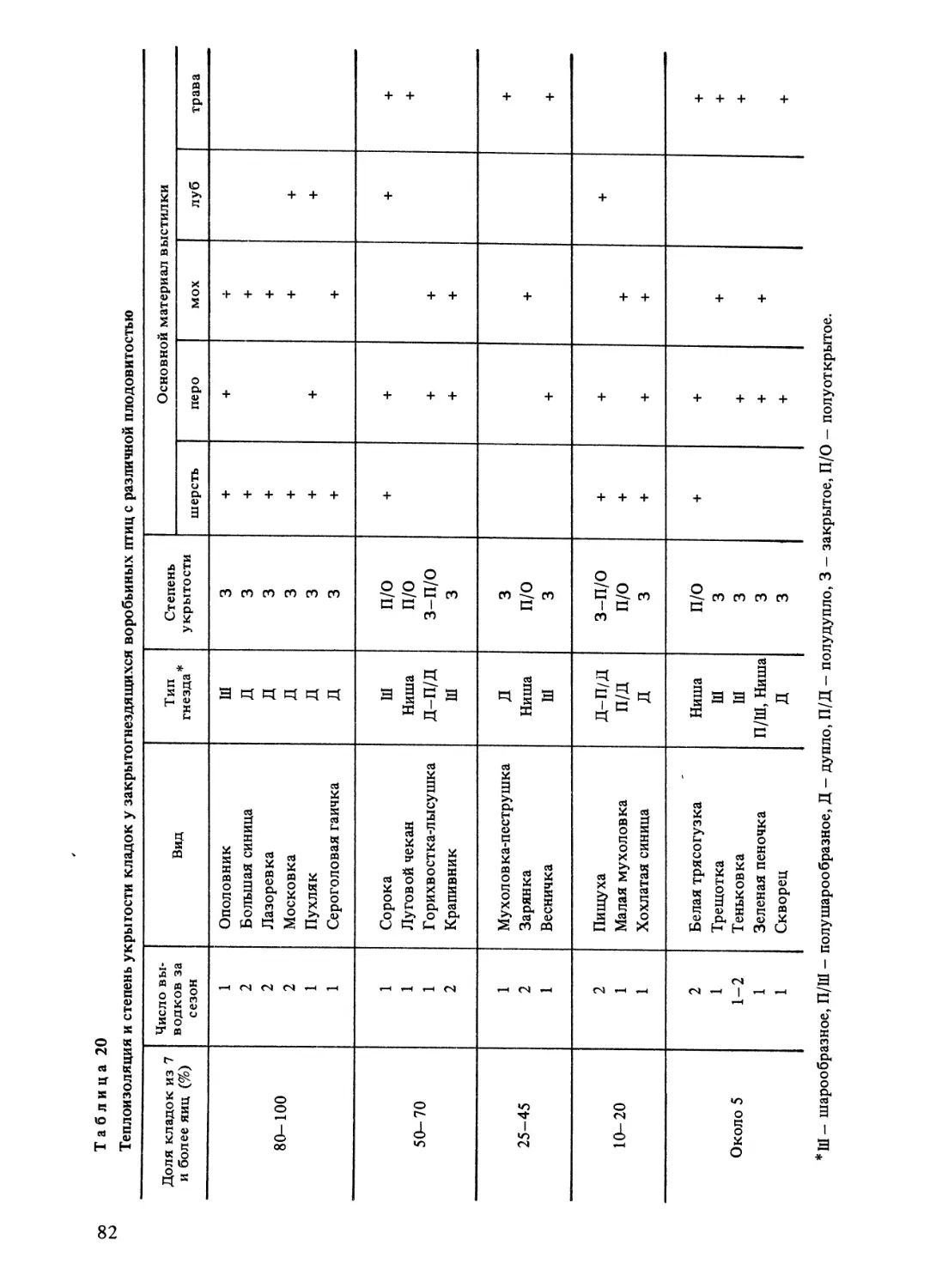
Плотность гнездового населения птиц
Отличительной особенностью исследуемой территории является относительно
невысокий общий и видовой уровень плотности гнездового населения птиц. Новиков
(1960), который проанализировал географическую изменчивости этого показателя,
отметил, что и видовое разнообразие, и численность лесных птиц в период гнездования
закономерно убывают в направлении с юга на север. Результаты учета гнездовой числен-
ности птиц на Северо-Западе (Ивантер, 1962, 19696; Зимин, 1968, 1976, 1978; Зимин,
Ивантер, 1969, 1974; Данилов и др., 1977; Зимин и др., 1978; Зимин, Кузьмин, 1980)
подтверждают выводы Новикова: плотность населения наших птиц находится на более
низком уровне, чем в средних и южных широтах Европы. Особенно низкий уровнь
численности отмечен для периферии видовых ареалов, где отдельные пары или особи
бывают рассеяны по обширной территории, часто расчлененной крупными массивами
стаций, непригодных для жизни птиц данного вида. Во многих случаях это становится
препятствием для встречи полов, приводит к прохолостанию и непостоянству связей
птиц с определенной территорией, а также к формированию высокой подвижности осо-
бей в гнездовой период, направленной на активные поиски партнера по размножению.
Известные механизмы регуляции численности в условиях ее низкого уровня
у наших птиц не проявляются, а зависимые от плотности гнездового населения связи,
как будет показано при обсуждении особенностей размножения, на местном материале
не выявляются. Более важную роль в динамике численности птиц играет здесь измен-
чивость погоды и степени дисперсии особей, родившихся в более южных частях ареалов.
Своеобразие исследуемой территории состоит также в преобладании лесных ланд-
шафтов, расчлененных многочисленными водоемами и относительно небольшими
участками сельхозугодий. Даже болота обычно покрыты редколесьем. На юге лесной
16
Рис. 1. Широтная изменчивость продолжительности дня в течение года (по: Данилевский, 1961).
По оси абсцисс - календарь; по оси ординат - продолжительность дня (ч).
и в лесостепной зоне лесные угодья, напротив, занимают небольшую площадь, основная
часть которой представлена открытыми пространствами агроландшафта. На численности
птиц здесь может сказываться влияние островного эффекта, который иногда проявляет-
ся и в таежной зоне. Так, в Прионежье общая плотность гнездования птиц, населяющих
небольшие (1—2 га) недорубы (приручейные ельники-черничники), окруженные выруб-
ками и молодняками, достигала 800 пар/ км2, что почти в 3 раза превышает соответству-
ющие показатели для крупных массивов еловых лесов.
Другим следствием зональных различий в соотношении лесных и нелесных площа-
дей является разная степень дисперсии молодняка. Ясно, что на юге в лесах островного
типа, разобщенных обширными открытыми пространствами, возможности для проявле-
ния дисперсии молодняка лесных птиц гораздо меньше, чем в таежной зоне. Это может
быть одной из причин известных географических различий в частоте возврата птиц
на места рождения и предыдущего гнездования.
Структура периферийных частей ареала
Периферийные части ареала птиц, в том числе и у северных границ распростране-
ния, имеют довольно сложное строение, которое не остается стабильным и постоянно
изменяется во времени и пространстве. Структура ареала — особый вопрос, не имеющий
прямого отношения к обсуждаемой теме, но поскольку в книге мы постоянно
2 В. Б. Зимин
17
Рис. 2. Принципиальная схема различий в выживае-
мости птенцов воробьиных птиц в больших (Л) и маленьких
(Б) выводках при разной продолжительности ночного
покоя в зоне белых ночей (ON) и в южных широтах (OS).
По оси абсцисс — продолжительность ночи (ч); по оси
ординат - уровень энергетических резервов птенцов. Линия
ВВ — сублетальный уровень энергетических резервов.
используем термины, предложенные ранее для обозна-
чения отдельных структурных единиц (Зимин, 1977),
необходимо привести краткую их расшифровку. Целесообразнее сделать это на примере
происходящей на наших глазах экспансии черного дрозда и других воробьиных птиц.
В середине нашего столетия гнездование черного дрозда было установлено в Юж-
ной Финляндии до 63° с. ш. (Merikallio, 1958) и в Эстонии (Кумари, 1958). В Ленинград-
ской обл. первые сведения о гнездовании этого вида на Карельском перешейке полу-
чены только в 1950 г. (Мальчевский, Пукинский, 1983). Южная Карелия в 50-е гг. отно-
силась к области случайных нерегулярных залетов единичных осо-
бей, которые отмечались в Приладожье и Прионежье (Нейфельдт, 1958).
В начале 70-х гг., когда в Эстонии и Ленинградской обл. происходило быстрое
увеличение численности вида (Йыги, 1967; Мальчевский, Пукинский, 1983), Южная Ка-
релия становится областью регулярных залетов в негнездовое
в р е м я. До 1975 г. черного дрозда ежегодно отмечали в периоды расселения молодых
птиц, а также на осенних и весенних миграциях примерно до 63° с. ш. Область нерегу-
лярных залетов в эти годы переместилась гораздо севернее и захватила
даже острова Баренцева моря (Герасимова и др., 1967; Коханов, Скоко-
ва, 1967).
В 1966 г. в заповеднике „Кивач” и в окрестностях Петрозаводска зарегистрирова-
ны встречи черных дроздов в летнее время, а в 1967 г. около заповедника обнаружена
первая молодая, уже самостоятельная особь. С этого времени Южная Карелия стано-
вится областью летних встреч и непериодического гнездова-
ния черных дроздов.
В связи с различной интенсивностью и темпами расселения птиц встречаемость
их в летний период далеко не всегда совпадает с попытками гнездования. Поэтому
следует разграничивать период (область) летних встречи период
(область) непериодического гнездования. Первый период из-за
низкой численности и рассеянного распределения птиц по территории (в Южной Карелии
это кобчик, глухая кукушка, удод, сизоворонка, зимородок, ястребиная славка и др.)
может длиться десятилетиями.
Гнездование вида на новой территории в течение нескольких лет еще не означает
ее полного освоения. Например, зеленая пеночка в юго-восточном Приладожье гнезди-
лась B40-err.(Paatela,1947). Затем до середины 70-х гг. на территории Карелии ее прак-
тически не было, но с 1976 г. она вновь появилась здесь и в настоящее время как в Юж-
ной Карелии, так и на севере Ленинградской обл. является вполне обычным гнездя-
щимся видом, причем местами численно превосходит теньковку. Тем не менее создает-
ся впечатление, что население зеленых пеночек на местах наших работ ежегодно форми-
руется заново: ни маркированные на гнездах взрослые птицы, ни их птенцы, ни околь-
цованные молодые особи ни разу не обнаружены на месте мечения в последующие годы.
Между тем в зоне постоянного гнездования, например в субвысокогорье Тянь-Шаня,
из 8 окольцованных взрослых самцов и 11 самок в последущие годы найдены на своих
гнездовых участках 4 самца и 2 самки (Ковшарь, 1979). Для обитающей у северных
границ ареала и отличающейся крайне неустойчивой численностью трещотки получены
аналогичные данные — ни одна из птиц, гнездившихся или родившихся на территории
наших стационаров, не вернулась в последующие годы в район первоначаль-
ного мечения.
18
По всей вероятности, периферийные популяции некоторых дальних мигрантов
не способны поддерживать стабильную численность без постоянного притока новых осо-
бей и формируются за счет расселения первогодков из областей постоянного гнездова-
ния вида.
Очевидно, птицы из периферийных частей ареала не успевают своевременно до-
стигнуть мест зимовки. Из-за увеличения дальности миграций и более поздних сроков
их начала в годовых циклах этих птиц постоянно или в отдельные годы создается непре-
одолимый дефицит времени, следствием которого является повышенная ежегодная
смертность особей, размножавшихся или родившихся за пределами областей постоян-
ного гнездования вида. Иного объяснения отсутствию возвратов взрослых особей
на места предыдущего размножения на периферии ареалов мы не имеем. Во всяком слу-
чае, вряд ли виды, для которых доказана высокая степень гнездового консерватизма
в центре ареала, около границ распространения изменяют свое отношение к территории.
Свидетельством формирования периферийных популяций птиц за счет дисперсии
особей из других частей ареалов, помимо данных кольцевания, являются .многочислен-
ные наблюдения за ежегодной встречаемостью ряда видов в исследуемом регионе.
Живущие у пределов ареала длиннохвостая синица и лазоревка уже на 62° 30' с. ш.
гнездятся не каждый год (Зимин, 1968), но в годы с теплой весной проникают до
64—65° с. ш. и даже севернее. Славки — садовая и завирушка — на Кольском полу-
острове гнездятся раз в несколько лет (устное сообщение В. Д. Коханова). Как пра-
вило, это случается в годы их обилия в Карелии.
Одной из причин появления птиц за пределами областей постоянного гнездования
может быть сокращение или исчезновение пригодных для жизни местообитаний. При
современных масштабах и темпах антропогенного преобразования природы это вполне
вероятно, но тогда следует говорить не о естественном расселении вида, а о его вытесне-
нии с исконных территорий. В частности, заселение зимородком Вологодской (Михай-
лов, 1976) и северной части Ленинградской областей (Мальчевский, Пукинский, 1983),
а также залеты этого вида в Карелию (1975, 1977 гг. — юго-восточное Приладожье,
окрестности Петрозаводска), вероятно, связаны с обмелением и загрязнением мелких
рек средней полосы Европы.
Таким образом, одним из этапов освоения птицами новых территорий может быть
довольно длительный период гнездования вида, поддерживаемый за счет ежегодного
обновления особями из области постоянного размножения.
Уже из приведенных примеров ясно, что расселение (или попытки расширения
ареала) у различных видов птиц не всегда идет по предложенной выше схеме. Обычно
вселение вида на новую территорию происходит постепенно и занимает довольно дли-
тельный отрезок времени, но иногда оно приобретает характер массовой инвазии. В по-
следнем случае птицы нередко сразу приступают к гнездованию. Так, малая чайка после
очень длительного перерыва вновь появилась в Свирской губе Ладожского озера
в 1975 г., сформировав колонию из 50 пар. Однако уже в 1979 г. на этом участке гнез-
дилось не менее 200 пар, а чуть севернее, в тростниковых зарослях и на островах около
устья р. Олонки — около 500 пар. В эти же годы произошло заселение малой чайкой
Онежского озера и других водоемов Южной Карелии. Сравнительно быстро проходило
расселение к северу черного дрозда, дубоноса, причем последний двигался на север
достаточно широким фронтом, захватившим Коми АССР (Кочанов, 1983) и северо-за-
пад Сибири (Данилов и др., 1984).
В современных орнитофаунистических сводках намечается тенденция к определе-
нию границ ареалу по крайним точкам гнездовых находок и даже по встречам отдель-
ных особей в внегнездовой период. Любая встреча птиц вдали от известных границ
распространения иногда слишком поспешно объявляется расширением ареала вида.
Но, как показано на примере птиц Северо-Запада СССР, даже факты достаточно мас-
сового гнездования еще не означают полного освоения видом новых территорий и могут
быть расценены лишь как попытка расширения области распространения. Под истинной
экспансией в отличие от попыток расселения следует, на наш взгляд, понимать такое
19
Рис. 3. Расстояние от гнезд белобровика (п = 406)
до опушки спелых насаждений заповедника „Кивач”.
По оси абсцисс — расстояние от опушки (м); по оси
ординат - число известных гнезд (%).
расширение области распространения вида, при ко-
тором его закрепление на новых рубежах по край-
ней мере частично обеспечивается за счет внутрен-
0 50 100 них ресурсов — возврата взрослых и молодых осо-
бей на места размножения и рождения. Это, однако, может быть выявлено лишь путем
длительных стационарных исследований с применением методов контроля местного
населения птиц.
Среди птиц, расселяющихся или предпринимающих попытки расширения ареала
на Северо-Западе и в Скандинавии, обращает на себя внимание преобладание птиц
южного и сибирского орнитокомплексов (Зимин, Лапшин, 1975; Зимин, 1976). На-
правление этих процессов однозначно во всех систематических группах, а доминирование
„южных” и „восточных” видов слишком явно для того, чтобы попытаться объяснить
это только большим видовым разнообразием птиц средних и южных широт. На совре-
менном этапе становления орнитофауны из представителей таежных птиц тенденцию
к расселению в южном направлении демонстрирует лишь белобровик. Но это объясня-
ется скорее всего тем, что белобровик — один из немногих лесных видов таежной зоны,
у которых отмечается явное тяготение к заселению производных типов леса и молодых
насаждений на зарастающих вырубках. Более того, он избегает селиться внутри хвойных
массивов, занимая лишь приопушечную полосу (рис. 3). Освоив в основной части ареа-
ла леса антропогенного происхождения и молодняки на вырубках, по своей конструк-
ции более сходные с южными насаждениями, белобровик адаптировался к жизни в юж-
нотаежных лесах с элементами неморальной флоры, а в последние годы начал гнездить-
ся в зоне широколиственных лесов и лесостепи (Лисецкий, 1981; Шаповал, 1982; Sevcik,
1982). Другие примеры активного расселения северных птиц к югу нам не известны.
Возможно, это связано с отсутствием подходящих местообитаний в более южных зонах.
Так, типично таежные темнохвойные древостои фактически не встречаются уже в Юж-
ной Карелии. Подходящие экологические ниши в южной тайге и южнее заняты родствен-
ными видами птиц с достаточно высоким уровнем численности. Кроме того, птицы
северных широт адаптированы к размножению и линьке на длинных фотопериодах, что
также может препятствовать их расселению в южном направлении. Тем не менее следу-
ет признать, что относительная стабильность южных границ ареалов таежных птиц все же
не имеет удовлетворительного объяснения, а каждый из перечисленных выше пунктов
легко опровергается.
Иногда продолжавшееся десятилетиями расширение ареала внезапно прекращается
и сопровождается отступлением вида с завоеванных позиций. Так, современная волна
экспансии обыкновенной чечевицы, начавшаяся в Европе после 1930 г., является второй
по счету на протяжении двух последних столетий. Первая имела место во второй трети
XVIII-середине XIXв., когда ареал вида продвинулся до верхнего течения Буга и Днес-
тра и южных склонов Татр, Верхней и Нижней Силезии. Но в период 1880—1930 гг.
чечевица полностью исчезла из этой области (Jozefik, 1960). Причины отступления вида
остались невыясненными. Такие колебания границ ареалов, охватывающие десятиле-
тия, возможно, являются следствием многолетних циклов движения численности вида
внутри основной гнездовой области. Если это действительно так, то экспансию можно
рассматривать как один из путей регуляции численности при переуплотнении населения
в зонах регулярного гнездования.
Распространение многих видов птиц на север долгое время сдерживалось отсутст-
вием подходящих для жизни стаций, которые были вкраплены небольшими пятнами
в обширные пространства, занятые коренными таежными древостоями. Их
территориальная разобщенность и незначительная общая площадь препятствовали
20
накоплению численности вида, необходимому как плацдарм для прочного закрепления
на новой территории. Связь низкой численности и спорадичности распространения
птиц с вероятностью их закрепления на новых позициях за пределами областей постоян-
ного гнездования уже рассматривалась выше. Следует указать также, что еще одним
важным следствием низкой численности вида в периферийных зонах ареалов является
слабая вероятность реализации микроэволюционных механизмов приспособления к жиз-
ни в новых условиях. Для возникновения адаптаций к короткому вегетационному пери-
оду на севере и дефициту времени в годовом цикле (увеличение дальности и поздние
сроки миграций) исходного материала (низкая численность, рассеянное распростране-
ние) явно недостаточно. В таких условиях возможны лишь периодические попытки ви-
да выйти за пределы ареала без закрепления на новых рубежах.
Массивы таежных хвойных лесов были экологическим барьером для расселения
к северу птиц широколиственных лесов, многие из которых были отчасти преадаптиро-
ваны к укороченному периоду благоприятных условий и увеличенной дальности мигра-
ций благодаря смещению послебрачной и постювенильной линьки на период зимовки
(чечевица, дубровник), сокращению ее объема (садовая славка, пересмешка, все камы-
шевки, зеленая пеночка,трещотка, таловка, малая и серая мухоловки и др.). В качестве
других преадаптаций, характерных для дальних мигрантов, можно рассматривать также
ускоренные темпы отрастания птенцового оперения и начала постювенильной линьки.
Для нашей зоны это показано на примере овсянок, славок и трясогузок, сравнительное
изучение годовых циклов которых выполнено Кукишем (1976), Рымкевич (1977),
Музаевым (19806).
Антропогенное преобразование таежных лесов способствовало устранению эколо-
гического барьера. Возникновение на месте вырубленных хвойных лесов разновозраст-
ных лиственных и смешанных насаждений создало предпосылки для расселения птиц
европейских широколиственных лесов. Это наглядно проявляется при сравнении соста-
ва орнитофауны и численности птиц в сохранившихся массивах таежных древостоев
и на территориях, преобразованных хозяйственной деятельностью человека. В лесах
таежного типа заповедника „Кивач” общая численность птиц „южного” происхождения
составляла не более 0.4 пар/км2, а в спелых и приспевающих березняках и осинниках,
заменивших вырубленные хвойные древостои, соответственно 10 и 12.4 пар/км2.
Общая плотность населения птиц, относящихся к орнитокомплексу европейских широ-
колиственных лесов, в лесных угодьях заповедника не превышала 3 пар/км2 (без учета
зяблика) или 1.2% от числа гнездящихся птиц, тогда как в окрестных насаждениях
антропогенного происхождения и на зарастающих вырубках этот показатель был в 20—
25 раз выше (Зимин, 1976; Зимин, Кузьмин, 1980).
Вторым важным моментом, предопределившим успешное проникновение „южан”
на север, явилось характерное для трансформированных человеком угодий увеличение
периода благоприятных условий, о чем уже говорилось выше.
Изменение климата
Наряду с антропогенным фактором причиной активного расселения птиц на север
считается общее потепление климата. На северо-западе Европы, как отмечают боль-
шинство климатологов, потепление началось с восьмого десятилетия прошлого века.
В начале нашего столетия оно стало более заметным, а в 20—30 гг. достигло максимума
(Гедеонов, 1973; Будыко, 1974, 1977). Затем наступило некоторое понижение темпе-
ратуры, которое продолжалось до 70-х гг., после чего произошло новое потепление
(Будыко, 1977). При этом в наиболее резкой форме изменения температуры проявля-
лись в высоких широтах.
Потепление климата на севере увеличивает продолжительность вегетационного пе-
риода и улучшает условия для земледелия и существования животных. Менее суровыми
становятся и условия зимовки. Увеличение периода благоприятных условий
21
предоставляет не адаптированным к дефициту времени „южным” видам дополнитель-
ный шанс для своевременного завершения всех летне-осенних фаз годового цикла.
И не случайно именно потеплению климата отдают роль решающего фактора расселения
птиц к северу.
Однако на Северо-Западе СССР далеко не все птицы начали освоение северных
территорий в период преобладания положительных климатических аномалий. Скворец,
например, резко продвинулся на север европейской части СССР в середине нашего
столетия (Полякова и др., 1984; Семенов, 1984). В это же время началась новая волна
экспансии чечевицы и массовое проникновение на север Ленинградской обл. и в Каре-
лию красноголового нырка, обыкновенной горлицы, седого и белоспинного дятлов,
садовой, болотной, тростниковой и дроздовидной камышевок, ястребиной слав-
ки и др. Между тем этот период характеризуется как относительно холодный.
Изменения климата обычно оцениваются по режиму погоды в самые холодные
и наиболее теплые месяцы года. А поскольку абсолютное большинство расселяющихся
птиц — типичные мигранты, для них, очевидно, наиболее важны изменения, происходя-
щие в апреле—сентябре, во время пребывания на местах гнездования. Исчезновение че-
чевицы после первой волны экспансии в Западную Европу произошло в последние де-
сятилетия нрошлого-начале нынешнего века, т. е. в период общего потепления климата
(Jozefik, 1960; Будыко, 1977). Начало второй волны приходилось на самое теплое деся-
тилетие за последние 100 лет — 1931—1940 гг. Затем наступило временное похолодание,
а чечевицы продолжали осваивать все более западные и северные области Европы. В наи-
более удаленных северо-западных частях ареала (Северная Карелия, Кольский полу-
остров, Скандинавия) нарастание численности и периодические попытки гнездования
за пределами областей постоянного размножения начались чуть раньше летнего потеп-
ления климата в этом регионе, хотя наиболее интенсивное расселение отмечено в 1960—
1970 гг. — в период преобладания положительных аномалий летней температуры. Судя
по характеру распространения чечевицы в азиатской части ареала, этот вид может гнез-
диться в лесотундрах и даже в тундрах, так что невысокие летние температуры и ко-
роткий вегетационный период, по-види'мому, не являются для нее существенным пре-
пятствием. В Европе область постоянного гнездования этого вида не выходит за преде-
лы средней подзоны темнохвойной тайги, а периодическое размножение отмечается
лишь в южной полосе северной тайги — на юге Кольского полуострова. Чечевицы, гнез-
дящиеся в азиатской и европейской частях ареала, очевидно, совершают разные по даль-
ности миграции, что может объяснить меньшую степень продвинутости западных попу-
ляций этого вида к северу.
Детальный анализ изменений температуры во внетропической области северного
полушария за все месяцы 1881—1970 гг. выполнен Гедеоновым (1973). Из данных,
приведенных этим автором (табл. 4), следует, что в весенне-осенние месяцы положитель-
ные аномалии температуры воздуха преобладали в Северной Европе в 20-е, 30-е и 60-е гг.
текущего столетия, причем в первые два из этих десятилетий существенным потепле-
нием была охвачена вся Восточная Европа, а в последующем ареал положительных ано-
малий здесь постепенно сокращался. Эти материалы подтверждают вывод, сделанный
выше: на Северо-Западе СССР и в Скандинавии за последние 30 лет расширение границ
ареалов птиц происходило в годы как с относительно более холодными весенне-лет-
ними месяцами (1941—1950), так и с наиболее теплыми (1961 — 1980).
Абсолютные значения аномалий среднемесячных температур вообще невелики
и крайне редко превышают 1—2 °C (Гедеонов, 1973). Однако и это может существенно
влиять на продолжительность вегетационного периода, что важно для закрепления вида
(пусть даже временного) на новых более северных рубежах. Удлинение периода благо-
приятных условий увеличивает шансы птиц на успешное завершение всех фаз годового
цикла, приходящихся на период их пребывания на родине, и может обеспечить лучшую
их выживаемость к следующему репродуктивному сезону. Именно этим, видимо,
объясняется более быстрое нарастание численности расселяющихся птиц в период потеп-
ления климата и их закрепление на новых территориях.
22
Таблица 4
Характеристика аномалий среднемесячных температур воздуха в Северной Европе
(по: Гедеонов, 1973)
Годы Апрель Май Июнь Июль Август Сентябрь
1881-1890 — — — + + —
1891-1900 + — + + +
1901-1910 —+ — + ? — —
1911-1920 + — + + + —
1921-1930 + + + + + +
1931-1940 + + + + + +
1941-1950 + — + — — —
1951-1960 + + + + + —
1961-1970 + + + — — +
Примечание. - в западной части аномалии отрицательные, в восточной - положи-
тельные; - на севере — отрицательные аномалии, на юге - положительные; „?’* - тенденция
не вполне очевидна.
Пролонгация весенней миграции в теплые весны с более или менее стабильным на-
растанием температур способствует иногда достаточно массовому проникновению птиц
за пределы ареала. Тем не менее даже случаи отмеченного здесь гнездования не обяза-
тельно свидетельствуют о расширении ареала вида. В 1975 г. после бурной теплой весны
в Карелии (г. Олонец) отмечена попытка гнездования одной пары кольчатых горлиц
(Носков и др., 1981). Летом того же года 2 птицы этого вида были встречены в г. Кан-
далакше В. Д. Кохановым (устное сообщение). Позднее, вплоть до 1984 г., кольчатые
горлицы в исследуемой зоне не появлялись ,т. е. первые попытки освоения ими север-
ных городов остались безрезультатными.
Для зоны, расположенной севернее 60-й параллели, подобные примеры могут быть
приведены в отношении большого числа видов птиц: малой поганки, серой цапли, боль-
шой выпи, серой утки, кобчика, черной крачки, чегравы, клинтуха, сизоворонки, зимо-
родка, лесного жаворонка, поползня, ястребиной славки, канареечного вьюрка и др.
В последние десятилетия отдельные представители этих видов птиц периодически пред-
принимают небезуспешные попытки гнездования на севере Ленинградской обл. и в Ка-
релии, но закрепиться здесь пока не могут. Так, птицы, однажды гнездившиеся на этой
территории, в последующие годы не возвращались на прежние гнездовые участки даже
в том случае, когда теплая весенняя погода и жаркое лето регистрировались два года
подряд (1983, 1984гг.).
Среди птиц, для которых в последние десятилетия на Северо-Западе СССР отме-
чена экспансия или попытки выхода за границы ареала, можно выделить 3 группы ви-
дов. Первую из них представляют птицы, преадаптированные к короткому северному
лету и дефициту времени. В прошлом их распространение сдерживалось отсутствием
или малой площадью подходящих местообитаний. В основном это птицы, у которых
линька полностью или частично перенесена на зимний период. Заселение таежной зоны
птицами данной группы идет преимущественно по биотопам антропогенного происхож-
дения. Исключение составляют сибирские виды, продвигающиеся в западном и северо-
западном направлениях по коренным таежным лесам (овсянки — крошка и ремез,
большой сорокопут, малая мухоловка, синехвостка, пеночки — зеленая и таловка,
щур). Из птиц восточного происхождения только чечевица, дубровник и садовая камы-
шевка проникли к нам в основном по преобразованным человеком ланд-
шафтам.
Во вторую группу включены виды, у которых в процессе достаточно длительного
существования в зонах с более или менее коротким летом возникли адаптации к дефи-
циту времени и к обитанию в хвойных и смешанных насаждениях (зяблик, певчий
дрозд, мухоловка-пеструшка, хохлатая синица, пищуха и др.).
23
Наконец, к третьей группе относятся птицы, не адаптированные к дефициту време-
ни в годовом цикле. Севернее областей постоянного гнездования во время общего по-
тепления климата из птиц этих видов могут формироваться более или менее многочи-
сленные временные популяции. В периоды многолетних похолоданий и в отдельные хо-
лодные годы они, как правило, исчезают с временно заселенных территорий. В этом слу-
чае закрепления вида на новых рубежах не происходит. В сравнительно теплые 70-е гг.
по Северной Европе прошла волна проникновения в таежную зону дубоноса.
В южной половине Карелии оно было достаточно массовым и сопровождалось
отдельными попытками гнездования и зимовок до 63° с. ш. Но в начале
80-х гг. даже на местах концентрации пролетных птиц - у восточного побе-
режья Ладожского озера — встречаемость дубоноса снизилась до исходного
уровня.
* *
*
Современное „объюжнение” орнитофауны таежной зоны явилось следствием
интенсивного антропогенного преобразования ландшафтов, происходившего на фоне
общего потепления климата внетропических областей северного полушария. Прямое
влияние потепления не следует, видимо, преувеличивать. Вряд ли оно имело бы суще-
ственное значение для распространения птиц без изменения в характере растительности.
Об этом свидетельствует явное преобладание нетаежных элементов фауны в место-
обитаниях антропогенного происхождения, сохраняющееся даже в очень теплые годы,
а также при разных уровнях численности новых для тайги видов. Изменение
температуры — процесс нестабильный: периоды потепления чередуются с более
или менеее продолжительными похолоданиями. При положительных аномалиях
температуры воздуха в таежную зону — в основном по антропогенным ландшаф-
там — могут проникать виды, не адаптированные к короткому вегетационному
периоду. Они образуют временные поселения за пределами областей постоянного
гнездования, но полного закрепления вида на новых территориях не про-
исходит.
При расселении птиц за пределы гнездового ареала возрастает дальность миграций
к местам зимовок и обратно. Все фазы годового цикла при этом сдвигаются на более
поздние календарные сроки. Соответственно, в годовом цикле птиц возникает опреде-
ленный дефицит времени. По нашим данным, в периферийных зонах ареала у мелких
воробьиных он составляет в среднем около 1—1.5 мес. Но, возникая на местах гнездова-
ния, дефицит времени сохраняется и при прохождении трассы миграций. Следовательно,
потепление климата не позволяет виду закрепиться на новых территориях до тех пор,
пока не появится возможность сокращения расхода времени, необходимого для за-
вершения всех фаз годового цикла в благоприятных для жизни вида условиях и обеспе-
чивающего необходимое соотношение ежегодной смертности и плодовитости. Расселе-
ние может дать положительный эффект лишь в том случае, если потеплением будет
охвачена не только периферийная зона ареала, но и вся территория, по которой прохо-
дят пролетные трассы к местам зимовок. Асинхронность в изменении климата этих зон
будет, очевидно, серьезным препятствием для успешного освоения птицами новых тер-
риторий. Если хотя бы одна из фаз годового цикла птиц, пытающихся освоить новые
территории за пределами основного ареала, не будет завершена успешно, закрепления
вида на новых рубежах не произойдет. Дефицит времени и асинхронность в изменениях
климата в областях гнездования и пролета могут привести к тому, что осо-
би, поздно начавшие мигрировать, не успевают своевременно долететь до мест
зимовки.
Вместе с тем периоды потепления климата способствуют росту численности и за-
креплению на новых рубежах птиц, преадаптированных или приобретающих адаптации
24
к укороченному вегетационному периоду у северных пределов их распространения.
Иначе говоря, причины появления птиц за пределами областей постоянного гнездования
для каждой группы видов следует рассматривать дифференцированно.
Северные пределы гнездования птиц в значительной мере определяются соответ-
ствием времени, необходимого для завершения активных фаз годового цикла (размно-
жение, линька, миграции), длительности периода благоприятных для жизни условий
на родине и путях пролета. Поскольку продолжительность этого периода постоянно
варьирует из-за непериодических изменений температуры, периферийные зоны ареала,
численность вида и границы областей его периодического гнездования подвержены зна-
чительным ежегодным или многолетним колебаниям.
25
Г Л А В A II
СРОКИ РАЗМНОЖЕНИЯ
Сезонность размножения
и географическая изменчивость сроков гнездования
Сезонность размножения воробьиных птиц таежной зоны в первую очередь обу-
словлена резко выраженной сезонностью климата и связанного с ним обилия и доступ-
ности кормов. Известные различия в климате отдельных природных зон определяют
и географические вариации в сроках размножения птиц, В целом о географической
изменчивости сроков размножения птиц сложилось вполне определенное мнение, в наи-
более общем виде выраженное в известном правиле Гопкинса о запаздывании
начала размножения на 3—4 сут на каждый градус широты по мере продви-
жения на север.
Данилов (1966), сравнив время гнездования птиц на Южном Ямале и Среднем
Урале, пришел к заключению, что запаздывание размножения у разных видов птиц
в Субарктике происходит не синхронно и что правило Гопкинса имеет частное и огра-
ниченное применение. Не всегда подтверждается оно и в отношении долготного запазды-
вания гнездования в восточном направлении, а также и с увеличением высоты над уров-
нем моря. Последнее подчеркнули Беме (1968) и Ковшарь (1981а), отметив, что более
позднее начало размножения в горах по сравнению с равниной характерно не для всех
видов птиц. Сравнение данных по Ленинградской обл. и Карелии со среднеуральскими
показывает, что на востоке ворона и белобровик приступают к откладке яиц примерно
на 2 декады позднее, рябинник, белая и желтая трясогузки — примерно одновременно,
а каменка и весничка — чуть раньше, чем в западных пунктах (Данилов, 1966; Мальчев-
ский, Пукинский, 1983; наши данные).
При сравнении сроков начала гнездования двух широко распространенных видов
птиц — скворца и полевого воробья — в разных точках их ареалов выяснилось, что
запаздывание в северном направлении идет неравномерно. У полевого воробья более
высокие темпы широтного запаздывания начала откладки яиц намечаются в 2 зонах —
самой южной (до 45° с. ш.) и северной (после 62° с. ш.),но в промежуточной полосе
начальные сроки размножения изменяются медленнее (рис. 4, А), В крайних
зонах начало откладки яиц запаздывает на 4 сут, а в средней — только на 1 сут
Рис. 4. Широтная изменчивость наиболее ранних дат начала откладки яиц у полевого воробья
(А - по: Гагинс1$ая и др., 1981), у скворца (Б).
По оси абсцисс - календарь; по оси ординат - географическая широта (градусы).
26
Рис. 5. Широтная изменчивость средних многолетних дат перехода среднесуточных температур
воздуха через 0, 5 и 10 °C (кривые 1, 2, 3 соответственно) в полосе 30-36° в. д. (по: Основные дан-
ные по климату СССР, 1976).
Обозначения как на рис. 4.
Рис. 6. Теоретические линии регрессии темпов весеннего перехода среднесуточных температур
воздуха через 10 °C в разных широтных зонах в полосе 30—36° в. д. (по: Основные данные по кли-
мату СССР, 1976).
Обозначения как на рис. 4.
Рис. 7. Средняя продолжительность периода со среднесуточными температурами воздуха
10 °C и более в разных широтных зонах полосы 30-36° в. д. (по: Основные данные по климату
СССР, 1976).
По оси абсцисс - продолжительность периода (сут); по оси ординат - географическая широта
(градусы).
на каждый градус широты. У скворца (рис. 4, Б) намечаются 2 подобные зоны: в по-
лосе 44—62° с. ш. запаздывание начала откладки яиц составляет 0.7 сут, а севернее —
4.1 сут на 1° широты.
В пределах более^узкой долготной полосы выявить широтные различия в сроках
начала гнездования ни по одному виду птиц не удалось из-за отсутствия сравнимого
материала. Только в зоне 25—35° в. д. и 56—70° с. ш. это оказалось вполне возможным
для наиболее массовых видов воробьиных птиц. Как выяснилось, у большинства из них
в полосе 56—62—64° с. ш. размножение начинается примерно в одни и те же сроки и так
же, как у скворца и полевого воробья, перелом в темпах запаздывания наступает после
62° с. ш. Так, после этого рубежа откладка яиц начинает более резко смещаться на позд-
ние сроки у сороки, лазоревки, московки, пухляка, длиннохвостой синицы, черного
дрозда, зарянки, обыкновенной овсянки, после 64° — у большой синицы, белобровика,
27
I Рис. 8. Теоретические линии регрессии широтной
™ ^1/=213.6-9.1 х изменчивости продолжительности бесснежного периода
в полосе 30-36° в. д. (по: Основные данные по климату
в СССР, 1976).
Обозначения как на рис. 7.
^\i/=231.5~6.9x
X. веснички, белой и желтой трясогузок, чижа; после
60 - \ \ 66° — у зяблика, вьюрка, чечетки, певчего дрозда,
\и=246.5-5.6х деревенской ласточки. К северу сроки начала
\ гнездования становятся более поздними без рез-
\ ких скачков у полевого жаворонка, рябинника,
\ горихвостки, серой мухоловки, лугового конька
\ (Птушенко, Иноземцев, 1968: Haartman, 1969а; Ма-
50- \ льчевский, Пукинский, 1983; наши данные).
I ( । । 1 । 1 । । 1 Для анализа широтного изменения климати-
160 200 250 ческих показателей в пределах 30—36° в. д. на тер-
ритории СССР использованы среднемесячные температуры воздуха в апреле—июле,
даты перехода среднесуточных температур через 0, 5 и 10 весной и осенью и др.
(рис. 5-8).
Оказалось, что все кривые широтной динамики избранных показателей имеют
более или менее резко выраженный перелом в зоне 60—62° с. ш. В этих же широтах про-
исходит замедление темпов развития весенних фенологических явлений. Так, если су-
дить по датам перехода среднесуточных температур через 0, 5 и 10 °C, то в пределах
45-60° с. ш. они изменяются со скоростью 0.27, 0.44 и 0.44° широты в сутки, а между
60 и 70° с. ш. — соответственно 0.19, 0.21, 0.24° в сутки. Полное исчезновение снежного
покрова на севере также замедляется — 0.21° широты в сутки против 0.36 в полосе
50-60° с. ш. Среднесуточные температуры 5 °C сохраняются в зоне 60—70° с. ш. в те-
чение 90—175 сут, в пределах 45—60° с. ш. — 175—276 сут; период со среднесуточными
температурами 10 °C в тех же широтах соответственно длится 49—125 (18 сут на ст. Пя-
льма Мурманской обл.) и 125—202 сут. Через 15 °C среднесуточная температура уже
около 64° с. ш. переходит не каждый год, а севернее это случается в исключительно
редкие годы.
На рис. 6 дана широтная изменчивость продолжительности периода с температу-
рами 10 °C и более. Величина этого периода быстро убывает к северу, но сокращение
его идет неравномерно — более резко в крайних из сравниваемых зон, т. е. сходно
с кривыми широтной динамики сроков начала откладки яиц у полевого воробья.
Минимальная продолжительность теплого периода в Мурманской обл. составляет всего
19 (ст. Пяльма) и 49 сут (ст. Перевал), а средние многолетние даты наступления перио-
да с температурами 10 °C и более приходятся здесь лишь на последние дни июня—июль.
Ясно, что при таком температурном режиме птицы, особенно насекомоядные, испыты-
вают определенные затруднения при добыче пищи в холодные периоды, приходящиеся
на время выкармливания птенцов и линьки.
Особенности распространения растений на Северо-Западе СССР и размещение гра-
ниц лесорастительных зон (и подзон) также свидетельствуют о переломном характе-
ре широтной полосы 62—66° с. ш., в которой сосредоточены северные пределы ареалов
многих видов деревьев, кустарников и травянистых растений (Яковлев, Воронова,
1959; Раменская, 1960). Между 60 и 63° с. ш. расположены северные границы ареалов
большинства широколиственных пород деревьев и многих кустарников (дуб, клен,
вяз, липа, лещина, черная ольха, татарская и голубая жимолости, калина, волчье лыко,
волчеягодник и др.). Немногим далее на север проникает черемуха. К 65—66° с. ш.
резко сокращается обилие в древостоях осины, серой ольхи, хотя последняя вместе
с березой произрастает и в тундре. В северотаежных лесах примесь этих пород во вто-
ричных древостоях, развивающихся на месте вырубленных хвойных, незначительна и не
идет ни в какое сравнение с их обилием в средней и южной подзонах темнохвойной тайги.
28
во -
52
................
1IV 20IV 10 V 30 V
_J___L
20 VI
AZ o£ <)3 *4 ±5 *6 +7
Рис. 9. Широтная изменчивость начала некоторых феноло-
гических явлений в полосе 30-36° в. д.(по: Фено-
логический ежегодник..., 1965).
1—3 - даты первой весенней встречи: шмелей (У),
дождевых червей (2), рыжих лесных муравьев, вышед-
ших из муравейника (5); 4-6 - даты начала цветения:
козьей ивы (4), черники (5), брусники (6); 7 - дата
зеленения березы. Остальные обозначения как на рис. 4.
Замедление темпов нарастания положительных температур и исчезновения снеж-
ного покрова севернее 60-й параллели соответственно влияют и на скорость широтного
изменения начальных сроков сезонного развития растений и беспозвоночных животных
(рис. 9). Примечательно, что заметное снижение темпов широтной динамики фенологи-
ческих явлений (сут на 1° широты) происходит также около 60—62° с. ш.:
50-60° с. ш. 60-70° с. ш.
Начало зеленения березы 2.4 5.2
Начало цветения ивы 2.2 4.9
черники 2.0 3.2
Первый выход из муравейника
рыжих лесных муравьев 1.4 6.1
Появление дождевых червей 2.3 3.6
Первая встреча шмеля 2.0 6.5
Таким образом, и климатические факторы, и развитие кормовой базы птиц, о тем-
пах которого можно приблизительно судить по приведенным фенологическим данным
для растений и беспозвоночных животных, и связанные со всеми этим факторами сроки
начала размножения птиц изменяются с увеличением географической широты неравно-
мерно, хотя и однозначно. Неоднородные, но взаимосвязанные показатели имеют одина-
ковую тенденцию: в средней полосе их нарастание в меньшей степени зависит от геогра-
фической широты местности, чем в таежной зоне, а заметный перелом в темпах продви-
жения весны на Северо-Западе СССР намечается после 60° с. ш. Следовательно, отмечен-
ное для птиц замедление в сроках начала размножения севернее этой параллели обуслов-
лено аналогичным переломом в темпах развития основных климатических показателей
и весенних фенологических явлений. Приуроченность зоны перелома к исследуемой тер-
ритории во многом определяет специфику ее флоры и фауны — их смены на таежный
тип и обилие границ видовых ареалов. Правда, в современных условиях процесс смены
фауны на таежную в значительной мере сглаживается антропогенным влиянием, спо-
собствующим проникновению в таежную зону многих представителей европейских
широколиственных лесов, о чем говорилось в предыдущей главе.
Фенологические особенности гнездового периода местных птиц
Наши исследования велись в основном на стационарах, которые располо-
жены внутри лесных стаций, удаленных от ближайших населенных пунктов и
29
to
О
Таблица 5
Сроки и динамика откладки яиц у воробьиных птиц Южной Карелии
Название вида Число гнезд с началом откладки яиц в указанную пятидневку (1-6) Всего гнезд Период отклад- ки 1-х яиц в разных гнездах
апрель май июнь июль август
1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4 5 6 1 2 3 4
Береговая ласточка - 7 1 9 4 9 1 1 - 32 22.V-21.VI
Деревен- ская ласточ- 7 9 15 11 10 3 2 — 1 1 2 3 1 2 — — 1 68 29.V-16.VII
ка Белая трясогузка - 2 13 22 22 11 4 5 4 7 2 3 1 1 - 97 9.V-6.VH
Лесной конек - 1 10 22 23 7 3 6 10 9 7 3 - 101 15.V-3.VII
Жулан — 6 12 15 10 4 1 1 1 — 40 29.V-2.VII
Крапивник - 1 5 7 9 3 - - 5 3 4 2 3 5 1 1 - 1 - - ; - 50 15.V-2.VHI
Лесная завирушка - 5 10 19 17 27 12 7 7 13 15 15 9 4 5 1 — 1 - - - - 176 6.V-27.VII
Зарянка — 1 17 34 56 50 28 13 11 19 30 16 14 8 3 2 — — 1 — — — 303 9.V- 5.vm
Горихвост- ка-лысушка - 7 26 36 25 13 8 6 9 7 3 2 - 142 17.V-15.VII
Луговой чекан - 2 6 10 2 2 4 2 1 - 29 18.V-24.VI
Черный дрозд - - - - + - 3 13 10 4 5 5 7 4 5 5 3 3 2 1 1 - 71 1.V-U.VII
Рябинник - 6 62 102 71 22 26 14 14 5 5 2 - 329 1.V-24.VI
Белобро- вик - - — - - 1 16 61 79 71 25 38 31 36 34 36 13 27 23 5 2 - 498 3O.IV-15.VII
Певчий дрозд - - — - - 1 - 22 41 50 50 47 27 17 15 19 21 11 8 3 - 332 30.IV-9.VII
Садовая славка - 4 17 23 43 21 11 14 6 9 2 2 - 152 31.V-18.VII
Черноголо- вая славка - 2 8 3 7 3 3 2 1 - 41 28.V-10.VII
Славка-за- вирушка - 1 9 11 15 14 6 4 5 1 3 - 69 19.V-5.VII
Весничка 13 94 164 116 53 20 7 12 7 1 1 478 21. V-14. VII
Теньковка 2 7 14 16 8 4 4 — 1 1 4 1 1 — 63 15.V-17.VII
Трещотка Зеленая - 1 1 13 11 11 3 4 1 6 2 - 1 - 54 19.V-13.VII
пеночка — — — — — — — — — — — 7 4 5 5 1 — 1 23 i.vi-i.vn
Серая мухоловка - 6 26 59 27 17 6 4 7 1 1 - 154 27.V-11.VII
Мухоловка- пеструшка - 8 52 102 231 186 59 22 24 12 5 1 - 702
60 654 528 118 28 3 — 1391 13.V-1.VII
Ополовник — — + 1 2 6 8 7 9 4 5 4 — 1 — 47 20.IV-6.VI
Пухляк - 3 15 26 29 4 1 4 1 - 83 3.V-6.VI
Большая синица - - - - - 1 18 19 30 21 8 5 2 3 13 9 15 11 4 5 5 2 - 171 31.IV-19.VII
Пищуха Обыкно- - - - - 1 2 10 6 7 1 1 _ 2 1 2 4 5 2 — 44 21-25.IV-25.VI
венная — 1 5 14 1 8 3 1 4 2 1 — — — 1 2 — — — — — — 43 9.V-18.VII
овсянка Овсянка- ремез Камышо- - + 7 11 6 5 2 2 3 1 8 2 2 - 49 13.V-1.VB
вая овсян- — 6 2 7 2 2 3 5 3 2 1 — 33 ll.V-30.VI
ка
Зяблик 1 3 26 44 37 21 20 9 8 6 2 186 5.V-4.VII
Чечетка — — 3 — 1 — — — 1 5 1 3 — 2 1 — 1 — 1 11 5 — — — 35 13.IV-5.Vin
Чечевица — 2 6 34 52 44 19 16 7 4 1 1 — 185 24. V-13. VII
Скворец — — — — 1 2 198 76 3 ) __ 296 24.IV-5.VI
Сорока + + 7 7 7 11 6 3 2 4 1 + — 48 12.IV—23.V
Ворона 3 14 32 3 2 4 1 — 1 1 - - - - - - — - - — - — — — - — - 61 1.IV-25.V
Примечание. Знак плюс (+) означает, что для указанной пятидневки начало гнездования вида известно на смежных с Карелией территориях;
для ополовника приведены суммарные данные по Финляндии (Haartman, 1969а) и Карелии (наши данные).
Рис. 10. Соотношение дат начала откладки яиц у бело-
бровика (1) и певчего дрозда (2) в Южной Карелии.
Горизонтальные линии — общая продолжительность
периода откладки яиц, дополнительно обозначенная край-
ними датами. По оси абсцисс - календарь; по оси ординат -
годы наблюдений.
сельскохозяйственных угодий на 12—20 км. Хорошо
известно, что на культурном ландшафте все фено-
фазы весны смещены на более ранние календарные
сроки. Соответственно и размножение птиц начина-
ется там раньше, что служит основной причиной
некоторых расхождений в датах начала гнездования
птиц, отмеченных нами (табл. 5) и опубликованных
Хаартманом (Haartman, 1969а). Сводка Хаартмана со-
ставлена по фондовой картотеке гнезд и оологическим
коллекциям профессионалов и любителей, работав-
ших преимущественно в ближайших окрестностях
населенных пунктов Финляндии. К тому же за при-
веденный в цитируемой работе 80-летний период
наблюдений необычно теплые весны встречались
чаще, чем за последние 25 лет. Таким образом, отме-
ченные различия имеют скорее всего биотопическую
основу, и им не следует придавать значения географи-
ческих (долготных).
Поскольку годовые различия в сроках размножения отдельных видов птиц ис-
следуемой территории уже рассматривались (Зимин, 1972а, 19726, 1978, 19816; Лап-
шин, 1976; Хохлова, 1976; Хохлова и др., 1983), то мы можем сравнить сроки и дина-
мику размножения за ббльший период, тем более что в последние годы весна была для
наших широт достаточно ранней (табл. 6—8, рис. 10).
В условиях Северо-Запада СССР колебания в начальных датах откладки яиц в раз-
ные годы не столь велики, как на юге Европы. У большинства воробьиных, обычно
приступающих к размножению в первой половине мая, сроки начала откладки яиц
варьируют в пределах 2 (белобровик) или 3 (певчий дрозд и больщая синица) недель
(рис. 10). Приблизительно таков же диапазон колебаний сроков начала откладки яиц
веснички (17 дней) и мухоловки-пеструшки (20), обычно приступающих к гнездова-
нию в конце мая. Однако на фоне общей продолжительности периода откладки яиц
у мухоловки-пеструшки, составляющей для наших широт в среднем 32 дня (18—46,
п = 18), такие 20-дневные различия в сроках начала гнездования уже не кажутся незна-
чительными.
Амплитуда колебаний рассматриваемого показателя для птиц, начинающих гнездо-
вание в июне, составляет около 10 дней (садовая славка, чечевица, серая мухоловка).
Она минимальна у дубровника, у которого и прилет, и начало откладки яиц ежегодно
разнятся не более чем на неделю (откладка яиц начинается с 9—14.VI).
Среди птиц, гнездящихся в таежной зоне, как исключение можно обнаружить
и более значительные смещения в сроках размножения, например у чижа и чечетки, спо-
собных выкармливать птенцов семенами хвойных деревьв. В годы обильного плодоно-
шения ели чижи иногда начинают размножение уже в начале Mapra(Schweppenburg, 1930 и
Bj0mson, Berger, 1966, — цит. по: Eriksson, 1970), а чечетки — по крайней мере в
середине апреля; причем такое раннее гнездование отмечается в иных, чем обычно,
природноклиматических зонах (интерзональное гнездование), на совершенно ином
фотопериоде, температурном режиме и на 1.5—2 мес раньше обычных сроков.
Хотя подобное смещение места и времени гнездования уникально для птиц, оно
тем не менее может вызываться определенными экологическими причинами. Обилие
32
Таблица 6
Изменчивость сроков и хода откладки яиц у зарянки в Южной Карелии
Годы Число кладок, начатых в указанную пятидневку Период от- кладки яиц
май июнь июль
2 3 4 5 6 1 2 3 4 5 6 1 2 3 4
Заповедник . „Кивач”
1961-1967 3 5 3 1 4 - - 2 I2 — I11 | 13.V-13.VII
Шокшинский стационар
1971-1977 - 2 8 8 8 6 1 - 2 - 1 2 2 2 - — 14.V-9.VII
Гумбар* [ЦЫ
1973-1977 - 3 4 11 9 2 3 1 1 2 2 1 12.V-5.VIII
1978 - - 2 9 3 3 - 2 4 3 2 2 2 1 2 18.V-20.VII
Ст ационар „ Маяч] ино”
1979 1 2 6 4 4 1 1 1 4 4 - 1 - 1 - 9.V- 12.VII
1980 - - - 5 4 5 6 2 3 7 5 3 1 - - 24.V-8.VII
1981 - - - 6 8 1 2 - 2 4 2 - 2 - - 22.V-8.VII
1982 6 7 9 12 2 1 2 3 5 1 2 - - - 13.V-3.VII
Таблица 7
Изменчивость сроков и хода откладки яиц у мухоловки-пеструшки в Южной Карелии
Годы Число кладок, начатых в указанную пятидневку Период откладки яиц
май июнь июль
3 4 5 6 1 2 3 4 5 6 1 2
Заповедник „Кивач”
1963 49 1' 1 23.V-29.VI
1964 84 50 16 4 25.V-27.VI
1965 11 и: 3 18 2 1 — 29.V-1.VII
1966 49 3* 7 11 L 2 1 — 26.V-1.VII
1967 89 4' 7 1( ) 2 — — 24.V-28.VI
С ^ационар „Шокшинский”
1972 — — 2 22 8 1 1 1 1 — — — 25.V-24.VI
1973 — — 13 7 4 3 1 — — 1 — — 21.V-27.VI
1974 — — — — 16 8 4 4 — — — — 2.VI-20.VI
1975 — 2 — 4 4 2 — — — — — — 16.V-9.VI
1976 — — 1 6 10 4 3 — — 1 — — 25.V-30.VI
1977 — 2 — 4 4 4 1 1 — — — — 18.V-16.VI
о-в Б. Клименецкий
1975 1 1 22 18 8 2 1 — — — — — 15.V-30.VI
1976 — — 14 21 24 8 2 1 2 — 1 — 23.V-1.VII
1977 — — 6 17 10 6 — — — — — — 20.V-?
Стационар , ,Маячино’
1980 — — — 3 41 6 2 3 1 1 — — 31.V-28.VI'
1981 — — 2 66 6 2 — 2 1 — — — 25. V-24 .VI
1982 — — 2 43 26 7 1 7 5 1 — — 23.V-28.VI
1983 ' 7 47 40 10 2 2 1 1 2 — — — 13.V-25.VI
1984 — 22 42 46 14 2 4 7 — — — — 18.V-18.VI
ЗВ. Б. Зимин
33
Таблица 8
Изменчивость сроков и хода откладки яиц в популяции веснички в Южной Карелии
(по: Лапшин, 1976, с дополнениями)
Годы Число кладок, начатых в указанную пятидневку Период откладки яиц
май июнь июль
4 5 6 1 2 3 4 5 6 12 3 4
Стационар „Шокшинский
2
1970
1971
1972
1973
1974
1975
1976
1977
5
6
7
9
3
7
7
14
4
17
13
1
1
4
3
13
26-31.V-?
3.IV-?
1.VI-?
26.V-17.VI
7.VT-14.VH
26.V-?
25.V-28.VI
26.V-5.VII
1
1
2
3
6
13
37
2
4
7
2
14
9
1
1
1
1
и доступность калорийной пищи, пригодной для выкармливания потомства, иногда
оказывается сигналом к прекращению зимних перемещений и началу гнездования.
В итоге размножение может начаться на фотопериоде (ФП), отличающемся от обычного
для вида почти на 10 ч. В северной тайге чечетка гнездится в июне—июле при круглосуточ-
ном освещении, а на 60° с. ш. — в середине апреля на ФП 14-15 С : 10-9 Т В разных
подзонах тайги птицы начинают гнездиться при разной среднесуточной температуре
воздуха: в северной — в июне при 10 Т, в южной — в апреле при 1—2 °C.
Вынужденный ночной перерыв в кормлении птенцов при зимне-весеннем размно-
жении составляет около 8 ч для чечетки и более 10 ч для чижа. Еще больше этот пере-
рыв у клестов, выкармливающих птенцов в конце февраля—марте, например в Мурман-
ской обл. Очевидно, и у взрослых птиц, и у птенцов должны быть развиты адаптации,
позволяющие им преодолеть столь длинную ночь.
Этому может способствовать замедленное переваривание растительной пищи и ее
более полное усвоение благодаря удлинению тонкого и слепого отделов кишечника.
Вполне вероятно также, что птенцы клестов, чижей и чечеток способны накапливать
определенное количество подкожного жира, который расходуется ночью. Летом подоб-
ное явление мы отметили для чечевицы. — в холодные периоды запасы жира у птенцов
оценивались баллами „средне” и „<средне”. Птенцовый жир накапливается и у других
видов воробьиных, но чаще всего его бывает „мало” или „<мало”. Еще одна важная
форма увеличения энергетических резервов — развитие дополнительных вместилищ
для запасания пищи. В наиболее яркой форме это проявляется у куриных и голубей
(наличие зобов), а у воробьиных встречается реже. В горловых и подъязычных мешках
переносят пищу для птенцов врановые и некотороые вьюрковые (Miller, 1941; French,
1954; Мальчевский, 1959; Ziswiller, 1967; Brooks, 1968; Harrison, 1970). Мальчевский
(1959) считает данную особенность необходимой адаптацией, позволяющей зерноядным
птицам кормить птенцов реже, но каждый раз приносить и передавать им (или самке)
большие порции корма. Действительно, при низкой скорости переваривания раститель-
ной пищи такой путь более целесообразен. У клестов полученная вечером порция корма,
34
очевидно, расходуется по мере освобождения пищеварительного тракта, что приводит
к увеличению периода поступления в организм дополнительных источников энергии.
Вполне вероятно, что именно способность взрослых клестов и их птенцов запасать пищу
впрок в довольно вместительных горловых мешках и определила возможность их гнез-
дования при коротких фотопериодах, тем более в Заполярье, где зимой день, а следова-
тельно и возможность кормежки, не превышает 3—4 ч.
Попутно можно указать, что дивертикул пищевода у некоторых вьюрковых функ-
ционирует не только в гнездовой период, но и зимой. Так, в феврале 1972 г. у двух
щуров, добытых в Карелии перед заходом солнца, горловые мешки были наполнены
почками березы и ольхи, а также личинками, собранными птицами из галлов на иве.
Мешки имели размеры 36 X 2.4 и 40 X 2.7 мм. Их содержимое весило 3.6 и 5.0 г при
массе тела птиц 58.3 и 61.1 г соответственно. У двух других щуров, добытых в том же
месте, но в середине дня, горловые мешки были пустыми, а масса тела составляла
55.2 и 57.6 г. Количество запасенного на ночь корма по массе почти вдвое превышало
содержимое желудка. Запасание корма в „зобу” характерно зимой и для чечеток
(Harvey, Datter, 1961; Brooks, 1968; Андреев, 1980). У чечетки, добытой в декабре, в рас-
ширении пищевода содержалось 0.65 г семян, а в желудке 0.5 г; примерно половину
запасаемого на ночь корма хранят в горловых мешках кедровки (Андреев,
1980).
Согласно Постникову (1983), зимой в ночное время пища может задерживаться
в пищеварительном тракте воробьиных птиц по 18—20 ч, что обеспечивает более полную
ее усвояемость.
Подобные адаптации являются важным условием существования птиц на севере
при коротком зимнем кормовом дне и длительной холодной ночи. Не исключено, что
у клестов, чижей и чечеток подобные адаптации развиты уже в птенцовом возрасте
и это делает возможным их гнездование в столь ранние сроки. Кроме того,
при длительной ночи и низких температурах воздуха подобные приспособления обеспе-
чивают птиц дополнительными источниками энергии, которые могут расходоваться на
оогенез и все виды репродуктивной активности.
Если географические различия в сроках размножения птиц являются следствием
неоднородности среды обитания в пределах ареала вида, то местные вариации (годовые,
сезонные, биотопические, возрастные) прежде всего обусловлены неустойчивостью
большинства внешних факторов, особенно резко проявляющейся в зоне повышенной
циклонической активности, к каковой относится и исследуемая территория.
Скорость развития весенних фенологических явлений варьирует по годам в широ-
ких пределах и отличается неравномерностью, что прежде всего зависит от изменчивости
температуры воздуха, баланса поступающей на Землю и отраженной солнечной энергии,
режима облачности, скорости ветра, глубины и целостности снежного покрова и т. д.
В тесной связи с внешними факторами находятся и даты начала размножения птиц,
ежегодно колеблющиеся в зависимости от темпов развития природы.
Свойственная всем таежным видам птиц пластичность в сроках начала размноже-
ния — существенное эволюционное достижение, обеспечивающее более успешное раз-
множение в неустойчивой среде обитания. Именно отсутствие жестко фиксированных
дат начала размножения позволяет птицам преодолеть непостоянство внешних факто-
ров — погоды и кормовой базы, определяющих баланс энергии организма.
Погода влияет и на состояние кормовой базы, и на величину затрат энергии на тер-
морегуляцию. От обилия потребленной пищи зависит количество поступившей в орга-
низм энергии. Размножение потенциально готовых к нему особей становится возмож-
ным лишь при появлении в организме дополнительных энергетических резервов, необ-
ходимых для быстрого роста гонад, развития наседного пятна, яиц, выводящих путей
и всех дополнительных видов активности, связанных с размножением. Формирование
половых продуктов самки, а также яйцевода и наседного пятна, происходящее на фоне
возрастающих затрат времени и энергии на строительство гнезда, — энергоемкие процес-
сы, требующие поддержания высокого уровня энергетического баланса в течение всего
35
Таблица 9
Запасы подкожного жира (по: Блюменталь, Дольник, 1962) у некоторых птиц,
отловленных на Шокшинском стационаре в период гнездостроения и откладки яиц
(произвольная выборка за 2 сезона)
Вид Число исследованных птиц
всего с баллом жирности всего с жиром
„нет” „>нет” „<мало” „мало” „>мало”
Весничка 83 38 9 16 11 9 45
Мухо- 51 13 7 16 12 3 38
ловка-пе- струшка Зарянка 28 15 8 1 2 2 13
Зяблик 41 20 5 8 4 4 21
периода гнездостроения и откладки яиц. На формирование одного яйца у мелких
воробьиных птиц требуется 4 дня (Lack, 1946), а общая масса кладки у синиц, например,
составляет 100-150% массы тела самки (Perrins, 1970). Собственная масса тела самок
во время откладки яиц увеличивается на 25—30% за счет увеличения размеров яичника
и яйцевода и обводнения тканей наседного пятна.
Для зяблика суммарные затраты продуктивной энергии на гнездо строение и ооге-
нез достигают 5.67 ккал/сут (Дольник и др., 1982). В этой связи становится понятным,
что кормление самок самцами в предгнездовой период имеет у многих видов не только
ритуальное значение, но и возникло как один из путей получения самками необходи-
мого дополнительного питания во время формирования яиц и гнездостроения (Lack,
1966). Энергетический выигрыш получают, очевидно, самки тех видов птиц, у ко-
торых самцы принимают активное участие в гнездостроении или выполняют
основную часть этой работы (крапивник и славки — серая, завирушка, Чер-
ноголовка) .
Еще один путь сокращения затрат времени и энергии на строительные заботы —
использование прошлогодних гнезд с незначительным их ремонтом и заменой выстилки
лотка. Среди воробьиных птиц таежной зоны это характерно лишь для врановых и лас-
точек; дважды отмечено у крапивника.
При высоких энергетических потребностях уже на начальных этапах репродуктив-
ного цикла любое увеличение энергетических расходов будет препятствием для реали-
зации потенциальной готовности к размножению и, наоборот, возможность получения
дополнительной энергии, достаточной для формирования яиц и развития всех видов
активности, будет способствовать началу гнездования.
В светлое время суток повышение расхода энергии может компенсироваться до-
полнительным потреблением пищи, тогда как величина ночных затрат в первую очередь
будет определяться продолжительностью периода покоя и температурой среды. По дан-
ным Дольника (1975), при комнатной температуре птица массой 20 г за ночь в среднем
теряет 0.15 г/ч, а калорический эквивалент ночных потерь массы у мелких воробьиных
варьирует при 0 °C в пределах 5.5-7.5 ккал/г, при 25 °C - 2.2-3.3 ккал/г. В Южной
Карелии в апреле, когда начинают гнездиться чечетки, ночные заморозки могут дости-
гать —25 °C, а в мае —10 °C, что естественно сопровождается повышенными затратами
энергии на терморегуляцию в период ночного покоя, который, кроме того, зависит от
продолжительности темного времени суток. В середине апреля на широте 60° ночь
длится около 8, а в начале мая — почти 6 ч. Если учесть расходы на терморегуляцию,
длительность ночного покоя и необходимость в затратах энергии на оогенез, то ста-
новится понятным, что размножение потенциально готовых к нему птиц может начаться
лишь при оптимальном сочетании фотопериода, внешних температур, скорости ветра
и обилия доступной пищи.
36
Дополнительные источники энергии в период размножения могут быть получены
при накоплении определенного запаса резервов на ночь. Как известно, отложение жира
в качестве источника энергии к моменту начала размножения снижается до минимума,
а жировой обмен сменяется углеводным. Правда, в условиях Карелии воробьиные
птицы сохраняют определенное количество подкожного жира в течение всего периода
размножения (табл. 9), но служит ли он дополнительным источником энергии в этот
период — неизвестно.
Содержание гликогена в организме мелких воробьиных птиц во время размноже-
ния также невелико — его запасы составляют 15—20 мг и быстро расходуются в течение
ночи (Farner et al., 1961; Дольник, 1975). Хотя в годовом цикле гликогена не обнару-
живается заметных пиков его накопления в период размножения, тем не менее гликоген
явно выполняет функцию кратковременного энергетического резерва, по крайней мере
на стадии быстрого роста ооцитов. В этой связи привлекают внимание два обстоятель-
ства. Известно, что у птиц в начале откладки яиц масса печени может увеличиваться
на 60%, но с началом интенсивного роста ооцита быстро снижается (Clavert, 1953). Уста-
новлено также, что в период откладки яиц индекс печени достигает максимума (Шварц,
1953). Добринский (1981) не обнаружил половых различий в годовом цикле этого
показателя, за исключением короткого периода откладки яиц. В это время относитель-
ная масса печени у самцов остается неизменной, а у самок резко увеличивается. С мо-
мента начала насиживания полной кладки эти различия быстро исчезают.
Дольник (1975) на примере мелких воробьиных птиц показал, что уровень гли-
когена достигает максимума к концу дня и быстро расходуется. Это позволяет предпо-
ложить, что с укорочением ночи и увеличением периода кормежки исходный вечер-
ний уровень гликогена может оказаться более высоким, нежели при кратковременной
кормежке и длительной ночи. При одинаковом расходе на формирование яиц в течение
периода постоянного поступления пищи в организм чистое потребление энергетических
резервов при покое во время короткой ночи, вероятно, не успевает снизиться до мини-
мума. Вместимость увеличивающейся в размерах печени самок позволяет в период
откладки яиц накапливать больше гликогена при длительном кормовом дне и компен-
сировать этим затраты не только на образование яиц, но и на другие процессы. Трудно
сказать, влияет ли это на величину кладки, но то, что длинный фотопериод частично
снимает энергетическую напряженность, не вызывает сомнений. Отсюда и фенологиче-
ски более ранние сроки начала размножения птиц на Севере, при меньшем обилии кор-
мов и на более низком уровне внешних температур.
У птиц на период размножения приходится, кроме того, пик годовых циклов про-
теинов, которые, по некоторым данныму могут накапливаться в грудных мышцах.
Температурные пороги начала гнездования
Основными негативными факторами, сдерживающими размножение птиц, являют-
ся низкие температуры воздуха, сильные ветры, недостаточно длинный фотопериод,
дефицит кормов. Низкие температуры воздуха могут сдерживать начало гнездования
как в результате прямого воздействия — через увеличение затрат на терморегуляцию,
так и косвенно, сокращая количество энергии, поступающей в организм с пищей, вслед-
ствие снижения активности беспозвоночных животных, а следовательно, и их доступ-
ности для птиц.
Многие орнитологи, пытаясь объяснить ежегодные вариации в сроках размноже-
ния птиц, выделяют температурные пороги начала откладки яиц. Для этого используют-
ся различные показатели: суммы температур за определенные отрезки времени (от 3—5
дней до 2 мес), минимальные, среднесуточные или среднедекадные температуры, пере-
ход среднесуточных температур через определенные величины и т. д. (Haartman, 1954:
Болотников и др., 1973; Kallander, 1975; Анорова, 1976; Slagsvold, 1976; Hecke, 1979;
Wiehe, 1979; Dhondt, Olaerts, 1980; Хохлова, Головань, 1981; Хохлова и др., 1983). На
37
7Я 2 К Рис. 11. Зависимость сроков начала размножения веснички
~ \ \ от суммы среднесуточных температур за предшествующую пятиднев-
\ \ ку (1) и декаду (2) в разных широтных зонах Карелии в 1975 г.
Q2 - \ \ По оси абсцисс — сумма среднесуточных температур (°C);
Ч \ по оси ординат - географическая широта (градусы).
60\ 1 1—J 1 1 1 L—
и 20 60 100 140 примере некоторых видов птиц можно показать постепенное
снижение температурных порогов начала кладки по направлению к северу. В сводке
данных по полевому воробью (Гагинская и др., 1981) приведены следующие значения
температурных порогов (°C): Астраханская обл. — 17, Чехословакия — 12, Польша —
9.5, Курская обл. - 10, Камское Предуралье - 11-14, Ленинградская обл. - 5-12.
В северных широтах температурные пороги обычно бывают ниже, чем в более южных
частях ареала. Так, при одновременных исследованиях температурным порогом для
начала гнездования мухоловки-пеструшки была среднесуточная температура (°C): 9
приблизительно на 67° с. ш., 7.5 около 68° с. ш., 5 на 69° с. ш. (Jarvinen, Linden,
1980). В Карелии на 62° с. ш. размножение мухоловки-пеструшки начиналось после
того, как минимальные температуры переходили через +3 °C (Хохлова, Головань,
1981). У веснички в северных пунктах наблюдений размножение также начиналось при
более низких температурах (рис. И). Широтные различия в величине температурных
порогов вполне могут быть объяснены спецификой суточного баланса энергии птиц
в зонах с разной продолжительностью темного и светлого времени суток. Следует
учесть, что северные популяции беспозвоночных животных и растений лучше адапти-
рованы к более суровым условиям, а специфика северного фотопериода также оказы-
вает ускоряющее действие на их развитие. Поэтому птицы на севере могут обеспечить
себя достаточным количеством корма при температуре более низкой, чем на юге.
Таким образом, суточный режим освещенности — второй важный фактор, воздей-
ствующий на организм птиц в комплексе с внешними температурами и изменяющий
эффект их влияния на размножение: на длинном световом дне откладка яиц у птиц
на севере может начаться при более низких температурах, тогда как вынужденно дли-
тельная ночевка у птиц в южных широтах увеличивает суммарный расход энергии на
покой даже при более высоких температурах среды.
В пределах одной местности и при одинаковой продолжительности светлого вре-
мени суток отмечаются довольно существенные ежегодные различия в температурных
порогах начала размножения птиц (рис. 11, и 12). Комплексное воздействие на птиц
не ограничено только двумя названными выше факторами. Практически не учитывает-
ся, например, влияние ветра на энергетику птиц и, соответственно, на сроки их размно-
жения. Судя по предварительным данным, сильные ветры приводят у птиц к не менее
(если не более) существенным потерям тепла, чем низкие температуры воздуха. Напри-
мер, чечетки в конце февраля в безветренную погоду при —26 °C проводили на зерно-
вой прикормке немногим более половины светлой части суток и почти столько же вре-
мени кормились менее калорийными естественными кормами. В дни со слабыми
морозами (около 5—6°), но с сильными ветрами (8—10 м/с) они почти не покидали
прикормки и, будучи вспугнуты, возвращались на нее при первой же возможности.
Масса тела меченых особей в такие дни, несмотря на беспрерывную кормежку, снижа-
лась до минимальных значений. Вполне вероятно, что и в другие сезоны года ветровой
режим играет важную роль в энергетике птиц, в частности сказываясь на обилии и до-
ступности беспозвоночных животных. Способы определения температурных порогов
начала размножения выбираются нами произвольно, главным образом с учетом вре-
мени, необходимого на овуляцию. Состояние кормовой базы птиц ко времени, пред-
шествующему их размножению, зависит от очень многих факторов, учесть которые
крайне сложно. Исходная осенняя численность насекомых, условия их зимовки, особен-
ности весеннего развития самих насекомых и объектов их питания, влияние похолада-
ний, глубины снежного покрова и времени его исчезновения — вот далеко не полный
перечень тех факторов, которые определяют состояние кормовой базы насекомоядных
38
140
100 -
- % в
о
60 “о о о • О
Оо
20 - °
I I I I I I I I I I I
13V 17 21 25 29V
12V 20V
о/ *2
Рис. 12. Различия в температурных порогах начала размножения мухоловки-пеструшки (А)
и зарянки (Б) в Южной Карелии (1973-1983 гг.)
1 - сумма среднесуточных температур воздуха за предшествующую пятидневку; 2 - за дека-
ду. По оси абсцисс - календарь; по оси ординат - сумма среднесуточных температур (°C).
птиц. Поэтому какой бы отрезок времени мы ни выбирали для расчета пороговых тем-
ператур начала размножения, на их величине так или иначе должно отразиться состоя-
ние кормовой базы.
Как показали исследования А. Ярвинена (Jarvinen А., 1984), в Финской Лапландии
годовые колебания сроков начала размножения мухоловки-пеструшки коррелируют
с массой тела самок в предгнездовой период: первыми гнездятся самки, у которых
она выше. Большая масса тела птиц независимо от линейных размеров свидетельст-
вует, очевидно, о том, что эти особи не испытывали особых энергетических затруднений
до и после прилета на родину и им не требуется времени на восстановление необходи-
мых для размножения кондиций.
Иногда отмечают совпадение начала пролета и гнездования птиц с фенодатами
развития растений. Однако, как показывают многолетние исследования, даже на одной
территории эта связь подтверждается далеко не всегда. Например, в заповеднике „Ки-
вач” средние многолетние даты начала откладки яиц у белобровика и начала зеленения
березы совпадают, но при этом амплитуда колебания первого показателя составляет
10 сут, а второго — в 2.5 раза больше, и никакой корреляции между ними не обнару-
жено. Недостоверная связь (г =0.73 + 0.48) при совпадении средних многолетних полу-
чена для годовых вариаций дат начала цветения черники и откладки яиц мухоловки-
пеструшки. Подобные связи всегда имеют сугубо местное значение, так как с продвиже-
нием к северу сроки размножения птиц смещаются на более ранние фазы весны.
Кроме годовых различий в датах начала размножения птиц следует отметить также
годовые вариации общей продолжительности периода откладки яиц, которые свойствен-
ны и видам с достаточно длительным репродуктивным сезоном, и дальним мигрантам
(табл. 5-8).
По мере продвижения вида на север общая продолжительность времени откладки
яиц в популяциях сокращается вслед за уменьшением периода благоприятных условий
и смещения начала размножения на более поздние календарные сроки. Так, весничка
в Англии откладывает яйца в течение 97 дней, в Карелии — 58, а в Субарктике, на юж-
ном Ямале - всего за 20 дней (Cramp, 1955; Лапшин, 1976, 1983а; Данилов и др., 1984).
В Англии зарянки гнездятся в любом месяце года, а их массовое размножение растя-
нуто на 4 мес; в Карелии общий период откладки яиц у этих птиц занимает обычно
85 дней, но в некоторые годы длится 44—66 дней.
Наибольшей растянутостью сезона гнездования среди воробьиных птиц отличаются
виды с полициклическим размножением. Так, для обыкновенной чечетки в Южной
Карелии общая продолжительность периода откладки яиц составила 115 дней (13.IV-
5.VHI). Немногим меньше она у чижа, снегиря, коноплянки. С разницей почти в 3 мес
39
15 VII
5vn - : • •+< • •
- *
25VI-
________I__I__1 I I I______|_L
3V 7V 11V 15V
3VII -
25 VI-.*
17VI -
5 15 VII-
e vT
* + 1 5VII - •
25VI ~
9 VI- • , 15VI -
i i i i i i i i i । । i i । i i i
13V 21V 29V 21V 31V 10VI
Рис. 13. Соотношение дат начала (абсцисса) и окончания (ордината) периода откладки яиц
в разные годы у белобровика (А), мухоловки-пеструшки (Б) и веснички (В) в Южной Карелии.
Крест - среднее многолетнее.
могут начинать кладку большие синицы и зарянки, до 2.5 мес — белобровики. У дальних
мигрантов сезон откладки яиц редко превышает 40 дней, а у дубровника завершается
в 2 недели.
Годовые различия в продолжительности периода откладки яиц слабо связаны
с датами ее начала: при опережении обычных сроков гнездование может закончиться
как раньше, так и позже обычного. Даты начала и окончания сезона откладки яиц
оказываются независимыми (рис. 13).
Ожидаемой связи продолжительности периода размножения с его успешностью
не выявляется, так как, во-первых, не все птицы приступают к повтороному размно-
жению, особенно после гибели птенцов в поздние сроки, во-вторых, самки некоторых
видов открытогнездящихся воробьиных исчезают с контролируемых территорий.
Биотопические различия в сроках размножения
Биотопические вариации в сроках размножения птиц, так же как и годовые, отра-
жают способность птиц чутко реагировать на малейшие изменения внешних факторов
и демонстрируют одно из проявлений экологической пластичности вида в неоднородной
среде обитания. В тех случаях, когда биотопические различия достаточно велики, они
становятся одной из причин неодновременности начала гнездования в популяции и уве-
личивают общую растянутость репродуктивного сезона вида.
Наиболее отчетливо биотопические различия проявляются у эвритопных видов
птиц, часть особей которых кроме естественных местообитаний осваивает также агро-
ландшафты и древесно-кустарниковые насаждения внутри населенных пунктов.
Серая ворона в населенных пунктах Карелии и Ленинградской обл. приступает
к откладке яиц почти на 2 недели раньше, чем в естественных местообитаниях - на по-
бережьях крупных озер. Эти различия, очевидно, были бы еще больше, если бы „озер-
ные” вороны были полностью изолированы от антропогенного ландшафта, где они
проводят весь предгнездовой период. При любых похолоданиях весной они покидают
гнездовые участки и перемещаются на дневное время в окрестные населенные пункты,
иногда удаляясь на 10 км и более. Серые вороны и скворцы начинают гнездиться на
морских островах позднее, чем на материке (Tenovuo, 1963; Tenovuo, Lemmetyinen,
1970), а галки в лесных колониях приступают к откладке яиц позже, чем в населенных
пунктах (Antikainen, 1978).
Смещение сроков размножения, также связанное с антропогенным влиянием,
отмечено для уток (Блум и др., 1983). В Латвии хохлатые чернети, приспособившиеся
к питанию искусственными кормами для рыб, гнездятся на рыбоводных прудах много
позднее, чем на эвтрофном озере Энгурес (различия достигают 27 сут). В бедных
40
Таблица 10
Даты установления и разрушения снежного покрова и продолжительность
снежного периода в различных биотопах заповедника „Кивач” в разные годы
(по данным архива заповедника)
Биотоп 1958 1959 1960
Установление снежного покрова
Луг 25.XI 24.XI 20.XI
Ельник 26.XI 27.XI 20.XI
Березняк 25.XI 25.XI 20.XI
Полный сход снега
Луг 5.V 29.IV 15.IV
Ельник 20.V 20.V 20 IV
Березняк 17.V 3.V 20.IV
Продолжительность снежного периода (сут)
Луг 162 158 148
Ельник 176 175 153
Березняк 174 160 153
естественными кормами прудах откладка яиц задерживается до начала весенней под-
кормки рыб, а размножение в целом проходит успешнее, чем на озере.
Роль антропогенной трасформации среды обитания в жизни птиц уже обсуждалась
в предыдущей главе. В данном разделе важно подчеркнуть лишь то, что птицы, приспо-
собившиеся к питанию кормами антропогенного происхождения, менее зависимы
от внешних факторов. По крайней мере важнейший из них — кормовая база — становит-
ся стабильным.
В садах и парках городов раньше гнездятся дрозды и большие синицы (Snow, 1958;
Havlin, 1963; Lack, 1966; Гришанов, 1981). Менее существенные различия обнаружива-
ются при сравнении сроков размножения птиц в разных типах естественных местооби-
таний (Lack, 1966; Stjernberg, 1979; Евдокимов, 1980).
Биотопические различия в микроклимате и фенологии растений длительное время
изучались в заповеднике „Кивач ’’ Насколько существенны такие различия, можно
судить по данным табл. 10. Биотопическая неоднородность в темпах весеннего развития
растительности и энтомофауны служит основной предпосылкой для возникновения
определенных смещений в сроках размножения птиц, населяющих разные биотопы.
В наших широтах размножение многих видов птиц начинается ранней весной,
до появления листвы на деревьях и кустарниках. Концентрация цветущих растений
и привлекаемых ими насекомых в полуоткрытых стациях и разреженных древостоях
отмечается на 2—3 недели раньше, чем, например, внутри массивов еловых лесов.
Отсюда можно было бы ожидать и соответствующего запаздывания начала гнездования
птиц в ельниках, по сравнению с более проницаемыми для солнечных лучей листвен-
ными древостоями. Однако реально наблюдаемые в природе расхождения в датах начала
откладки яиц лесных птиц в разных биотопах обычно не превышают 2—5 сут. Это несо-
ответствие с фенологическим развитием растений обусловлено прежде всего высокой
подвижностью птиц, а также тем, что современные лесонасаждения стали очень неодно-
родными по составу и почти повсеместно отличаются пестротой и мелкоконтурностью
выделов. Хвойные насаждения всюду расчленены вкраплением вырубок, молодняков,
зарастающих кустарниками, заброшенных лесных сенокосов, просеками для линий
электропередач, дорогами, участками вторичных лиственных лесов и т. д. Кольцеванием
установлено постоянное проникновение птиц из одного типа леса в другой в зависи-
мости от состояния погоды. Так, зарянки, отличающиеся достаточно строгой террито-
риальностью, во время похолоданий и утренних заморозков весной покидали уже заня-
тые ими гнездовые участки внутри ельников и вылетали кормиться на освещенные
солнцем участки молодняков. Эти перемещения иногда превышали 0.5 км. Еще
41
в большей мере кормежка вдали от гнезд характерна для врановых, скворцов, дроздов,
чечевиц. В заповеднике „Кивач” скворцы до начала насиживания кладки проводили
в гнездах или около них только ночь, а все светлое время суток кормились на при-
мыкающих к заповеднику полях - в 6—9 км от места гнездования. Значительно ра-
зобщены (до 4—5 км) гнездовые и кормовые стации чечевицы, что позволяет этому
виду осваивать территорию в средне- и северотаежной подзоне темнохвойной тайги
(Зимин, 19816).
Пестрота и сближенность биотопов, различающихся скоростью и степенью продви-
нутости фенологического развития растительности и энтомофауны, а также возмож-
ности использования этих особенностей птицами при их высокой подвижности приводят
к сглаживанию биотопических различий в сроках размножения. В частности, близость
открытых и полуоткрытых стаций позволяет птицам ельников начать размножение рань-
ше, чем это было бы возможно внутри сплошных однородных еловых лесов.
Тем не менее определенная неодновременность начала гнездования у лесных птиц
в разных биотопах все же имеет место, что ранее было показано нами для мухоловки-
пеструшки в заповеднике „Кивач” (Зимин, 19726), большой синицы (Зимин, 1978),
садовой славки (Зимин и др., 1978), чечевицы (Зимин, 19816). Новые материалы,
собранные в различных пунктах Карелии, позволяют еще раз убедиться в том, что
птицы очень чутко реагируют на малейшие отклонения в микроклимате разных типов
леса. Сравнение удобнее выполнить на примере одного наиболее массового эвритопного
вида лесных птиц — мухоловке-пеструшке, легко привлекаемой развеской искусствен-
ных гнездовий даже в такие насаждения, в которых в обычных условиях она не гнез-
дится.
В заповеднике „Кивач ’’мухоловка-пеструшка сначала начинает гнездиться в берез-
няках и осинниках, затем в ельниках с примесью лиственных пород и позднее всего
в сосновых монокультурах. Указанная последовательность сохраняется из года в год,
о чем можно судить по средним датам начала кладки в лиственных, еловых и сосновых
лесах: 1963 г. — 29, 30, 31 мая; 1964 г. — 30, 31 мая и 2 июня; 1965 г. — 3, 6 июня,
и т. д. Гнездовья в заповеднике размещались внутри крупных массивов леса, так что
влияние на птиц соседних биотопов было незначительным. По наблюдениям фенолога
заповедника М. М. Романовской, все фитофенологические даты для лиственных насаж-
дений всегда регистрировались также на несколько дней раньше, чем для хвойных.
Двух- или трехдневное запаздывание в средних датах начала размножения птиц
ежегодно отмечалось в сосново-лиственных молодняках Шокшинского стационара по
сравнению с лиственными лесами на о-ве Б. Клименецкий, расположенном на 0.5° север-
нее. Дополнительное негативное влияние на фенологию островных растений и насеко-
мых оказывало скопление тающих льдов Онежского озера. Тем не менее на острове
птицы всегда гнездились немного раньше, чем на материке в сосново-лиственных молод-
няках. Ранее это. было показано для большой синицы (Зимин, 1978) и может быть про-
иллюстрировано на примере среднегодовых дат начала откладки яиц у мухоловки-
пеструшки, у которой гнездование в спелых и приспевающих лиственных лесах острова
и в шокшинских молодняках в среднем началось соответственно в 1975 г. 28 и 31 мая,
в 1976 г. 3 и 5 июня, в 1977 г. 31 мая и 3 июня.
В юго-восточном Приладожье (урочище Маячино) искусственные гнездовья раз-
мещались в лесах различного типа, в том числе в черноолынанниках. От других листвен-
ных лесов чернооолынанники отличаются сравнительно поздними сроками зеленения
и развертывания листьев. Среднее отклонение от наиболее ранней.даты начала откладки
яиц в каждый год наблюдений составило для мухоловки-пеструшки (сут): в сосняке-
зеленомошнике 5.6, в сосново- и елово-березовых молодняках 6.1, в черноолынанниках
7.3, в сосново-березовых средневозрастных лесах на болоте 7.4 и в ельниках чернично-
травяных 8.5.
Таким образом, основными предпосылками для возникновения внутрипопуля-
ционных биотопических вариаций в сроках размножения эврибионтных видов
птиц являются соответствующие различия в состоянии кормовой базы, которые
42
определяются спецификой видового состава беспозвоночных животных и растений,
динамикой микроклимата и фенологии каждого биотопа. В тех случаях, когда обилие
кормов становится относительно независимым от состояния погоды, размножение
смещается на более ранние (врановые в населенных пунктах, на животноводческих
или звероводческих фермах) или на поздние сроки (утки на рыбоводных прудах).
Возрастные различия в сроках размножения
Возрастные различия в сроках размножения обсуждаются в двух основных аспек-
тах: возраст, в котором птицы приступают к размножению, и различия в датах начала
гнездования старых, ранее уже размножавшихся (ad2) , и впервые участвующих в нем
особей (adi).
Возраст, в котором начинается размножение, — один из показателей продолжитель-
ности жизни птиц в естественных условиях. Ясно, что позднее наступление половозре-
лости должно быть привилегией тех видов птиц, которые обладают меньшей ежегод-
ной смертностью и способны поддерживать стабильный уровень численности за счет
только незначительной части размножающихся особей. Позднее наступление половой
зрелости выгодно для птиц с длительным индивидуальным жизненным циклом и явля-
ется одним из механизмов, препятствующих чрезмерному увеличению численности,
которое в конечном итоге привело бы к перенаселению и уничтожению ограниченных
кормовых ресурсов. Возраст, в котором птицы приступают к размножению, должен
примерно соответствовать средней продолжительности их жизни (Haukioja, Hakala,
1979). Такое соответствие — результат эволюции. Неслучайно поэтому возникновение
новых, чуждых природе, негативных антропогенных факторов (пестициды), к действию
которых многие животные не успевают адаптироваться, в первую очередь оказывается
губительным именно для птиц с низкой плодовитостью, одним из показателей которой
является поздняя половозрел ость.
Среди воробьиных птиц, у большинства из которых продолжительность жизни
едва превышает один год, молодые особи способны к размножению в возрасте около
10—11 мес. При клеточном разведении некоторые из них, в частности зебровые ама-
дины, способны успешно размножаться в возрасте менее 2 мес.
Подавляющее большинство певчих птиц, гнездящихся в таежной зоне, приступает
к размножению в конце 1-го года жизни. Исключений из этого правила немного, и они
свойственны главным образом наиболее крупным представителям отряда. По мере
развития работ по кольцеванию птиц постепенно накапливаются сведения о точном воз-
расте впервые приступающих к размножению особей. Так, у самцов скворца в Голландии
половозрелость наступает в конце второго года жизни, тогда как многие самки начина-
ют гнездиться на год раньше (Kluijver, 1951). Врановые впервые приступают к размно-
жению на 2—3-м году. В наших условиях это установлено для серой вороны (Е. В. Шу-
тенко, устное сообщение) и кукши (Коханов, 1982). В Южной Финляндии не при-
нимают участия в гнездовании около 30 % первогодков мухоловки-пестушки (Haartman,
1956).
В юго-восточном Приладожье годовалые скворцы прилетают на 1—1.5 мес позднее,
чем ad2, и участия в гнездовании не принимают. Эти птицы образуют достаточно круп-
ные стаи (до 300 особей на Олонецкой равнине и до 50 в Северной Карелии), которые
постоянно держатся на сельскохозяйственных угодьях. Годовалые особи не участвуют
в размножении и в Ленинградской обл.,а в Латвии это отмечается лишь как исключение
(Тауринып и др., 1953; Мальчевский, Пукинский, 1983). В центральных областях
РСФСР на долю первогодков приходится 14—30% (Поливанов, 1957; Кошелев, 1978),
а в Черноморском заповеднике — до половины гнездового населения скворцов (Арда-
мацкая и др., 1977). В Северной Польше adi образуют большую часть (66%) популяции
только в год формирования новых колоний, в дальнейшем доля их участия в размноже-
нии сокращается до 31-42% (Gromadzki, 1980).
43
Рис. 14. Динамика развития клоакальных выступов
у самцов чечевицы в возрасте 1 года (1), 2 лет и старше (2).
По оси абсцисс - пятидневки со дня прилета местных
особей; по оси ординат - относительная величина клоакаль-
ных выступов (максимальная величина принята за единицу). За-
штрихованный прямоугольник - период откладки яиц в популя-
ции.
Интересно, что, так же как и у скворцов, размно-
жение годовалых самок чечевиц является нормой,
но молодые самцы, прилетающие на родину позднее ad2, чаще всего довольствуются
ролью гнездовых помощников и реже образуют пары (Зимин, 19816). Судя по состоя-
нию клоакальных выступов, годовалые самцы позднее приходят в состояние готовности
к размножению, чем старые (рис. 14). Они вынуждены селиться в субоптимальных для
вида местообитаниях, и большая часть их остается холостыми. Так как отлов чечевиц
с их обязательным прижизненным описанием производился стационарно, то мы имели
возможность следить за состоянием отдельных особей, многократно ловившихся в те-
чение всего периода их пребывания на местах размножения. Все контролируемые нами
самки чечевиц участвовали в гнездовании. Самцы-первогодки, не нашедшие партнера
для размножения, чаще всего присоединялись к гнездовым парам и выполняли роль
помощников. Такое поведение годовалых самцов чечевиц характерно и для соседних
областей (Risberg, 1970; Нанкинов, 1974; Peiponen, 1974; Stjernberg, 1979). Нанкинов
(1974) под Ленинградом регистрировал у одного гнезда до 5 молодых самцов. По
нашим наблюдениям, на пару гнездящихся особей приходился, как правило, только
один помощник. Возможно, это является следствием неравного соотношения полов.
На Шокшинском стационаре соотношение самцов и самок в разные годы варьировало
от 1.3:1 до 2.2:1. Судя по отловам на зерновых прикормках, ежегодно колебалось
и соотношение возрастных групп среди самцов (Зимин, 19816). Преобладание самцов-
первогодков, характерное для нашего и соседних регионов, связано, вероятно, с их
расположением неподалеку от северо-западных границ ареала, который к тому же
расширялся в годы наблюдений. На первостепенную роль самцов, особенно молодых,
в расселении вида указывал еще Дарвин (1939).
Похоже, что и у белобровиков в Карелии не все годовалые самцы участвуют
в размножении. Во всяком случае, если судить по возрастному составу пар, отловлен-
ных на гнездах в юго-восточном Приладожье и заповеднике „Кивач”, соотношение
ad2 : adi у самок было примерно равным, а среди самцов ad2 явно преобладали (около
75%).
Лишь у немногих видов таежных воробьиных число участвующих в размножении
ad2 превышает или примерно равно adx . Кроме перечисленных выше, это, например,
пухляки, взрослые особи которых отличаются строгим постоянством территории и
состава пар на протяжении всей жизни. На Шокшинском стационаре 75% гнездового
населения пухляков составляли особи в возрасте 2—5 лет. Примерно такое же соотно-
шение возрастов установлено для пухляков и хохлатых синиц в Псковской обл.
(Бардин, 1975а, 19756), а также в юго-восточном Приладожье.
У всех остальных исследованных нами видов воробьиных птиц первогодки состав-
ляли доминирующую часть гнездового населения (табл. 11). При составлении таблицы
мы намеренно использовали лишь данные по отлову птиц непосредственно на гнездах,
хотя располагаем гораздо более многочисленными сведениями по возрастному соста-
ву птиц различных видов, полученными в результате 15-летних стационарных отловов
паутинными сетями, крупногабаритными и другими ловушками. Эти данные лучше
отражают возрастной состав птиц в периоды миграций и летних перемещений, но для
характеристики гнездового населения непригодны. Даже при летних отловах паутин-
ными сетями, установленными в гнездовых стациях, в них постоянно попадаются
посторонние (не местные) особи, перемещающиеся по не вполне понятным пока при-
чинам. Несмотря на то что для некоторых видов птиц до настоящего времени
44
Таблица 11
Возраст партнеров в гнездовых парах некоторых воробьиных птиц
на стационарах „Маячино” (I) и „Шокшинский” (II)
Вид Число исследованных пар Всего особей
всего dad2 9ad2 dad j 9ad2 dad2 9adt dad2 9 adj ad2 adj
d 9 d+9 d 9 d+9
Большая синица, I 29 5 2 11 11 16 7 23 13 22 35
Тот же, II 13 — — 1 12 1 — 1 12 13 25
Пухляк, II 12 7 1 3 1 10 8 18 2 4 6
Мухоловка-пе- струшка, I 163 12 14 49 88 61 26 87 102 137 239
Тот же, II 44 5 8 16 15 21 13 34 23 31 54
Зарянка, I 71 5 3 13 50 18 8 26 53 63 116
Белобровик,I 49 20 6 15 8 35 26 61 14 23 37
Весничка, I [ 89 13 4 29 43 42 17 59 47 72 119
не выявлены четкие возрастные признаки, мы сочли возможным включить в анализ
данные за последние годы по наиболее массовым видам — весничке и мухоловке-
пеструшке, взрослые особи которых в течение 3—4 предыдущих лет контролировались
отловом на гнездах соответственно на 80 и 90—95%. Возраст птиц, окольцованных
в предыдущие годы, определяли как ad2, а всех, впервые гнездящихся на опытных пло-
щадях, считали первогодками.
О преобладании первогодков в гнездовом населении большинства исследованных
видов, кроме прямых данных, позволяет судить степень ежегодного обновления мест-
ного населения птиц, составляющая у разных видов от 60 до 100% (Зимин, Лапшин,
1976).
Благодаря многолетнему кольцеванию птиц на одном месте, мы имеем возмож-
ность установить точный возраст начала первого гнездования. Поскольку таких данных
немного из-за незначительного возврата молодых птиц на места рождения, они приве-
дены полностью (табл. 12). Полученные результаты по всем видам сходны — все они
начинали впервые гнездиться в возрасте 10—12 мес.
Очевидно, у птиц с бициклическим размножением этот показатель может быть
несколько меньшим: родившиеся в конце июля особи начинают размножение в начале-
середине мая (дрозды, зарянка, большая синица, лесная завирушка, крапивник и др.),
т. е. в возрасте около 9.5—10 мес. Еще раньше, на 8.5—9-м месяце жизни, теоретически
возможно начало размножения чечеток: они могут вылупиться во второй половине
июля, а в годы обильного плодоношения семян ели приступить к гнездованию в сере-
дине апреля. Прямых данных, подтверждающих это предположение, у нас нет, но если
Таблица 12
Возраст начала первого размножения у птиц, окольцованных птенцами в Южной Карелии
Вид Пол Сроки Возраст в день откладки первого яйца, (сут)
рождения рачала первого размножения п' lim М
Мухоловка-пестру- d 13.VI-8.VII 15.V-25.VI 12 328-378 347
шка 9 4-24.VI 13.V-21.VI 17 325-379 353
Большая синица d, 9 1. VI-14. VII 28.IV-18.V 5 293-345 328
Весничка d, 9 17.VI-8.VI1 27.V-10.VI 7 336-353 346
Черный дрозд 9 20.V 30.V 1 376
45
10VII
Рис. 15. Соотношение дат начала первого размножения (абсцис-
са) и рождения (ордината) у меченых мухоловок-пеструшек.
30VI ~
2ovj~-*^ * судить по поведению чечеток, содержащихся в неволе
- • «о в • * на естественном фотопериоде Петрозаводска, они могут на-
10VI - • • * чать размножение уже в марте. Во всяком случае, в этом
' , | , i । i i i । I месяце они активно реагируют на предложенный им гнездо-
20 V 1VI 10VI20 VI вой материал, а самцы начинают подкармливать самок.
По нашим данным, птенцы из первых и вторых вывод-
ков большой синицы при полуторамесячной разнице в возрасте начинают гнездиться
одновременно. Это показано и на скворцах (Berthold, 1970). Начало кладки у первогод-
ков мухоловки-пеструшки также не зависит от времени их рождения (рис. 15).
Считается доказанным, что более опытные, уже гнездившиеся ранее самки различ-
ных дуплогнездников — большой синицы (Kluijver, 1951; Лихачев, 1957; Lack, 1957;Рег-
rins, Moos, 1974;Winkel, 1975), мухоловки-пеструшки (Haartman, 1967а; Анорова, 1976;
Лихачев, 1978), скворца (Gromadzki, 1980) — начинают откладку яиц раньше первогод-
ков. Некоторое опережение в средних датах начала гнездования самок ad2 выявлено
также для полевого жаворонка (Delius, 1965) и камышовой овсянки (Haukioja, 1970).
Вместе с тем высказывается мнение и об отсутствии подобных различий. В Южной
Финляндии среди рано гнездящихся самок больших синиц не обнаружено преобладания
птиц старших возрастных групп. Средняя дата откладки первого яйца старыми птицами
(п = 47) была здесь определена как 10.3 мая, а для 18 самок-первогодков — 9.4 мая
(Haartman, 1969а). В Центральной Финляндии средние даты начала кладки у старых
и молодых самок белой трясогузки также оказались одинаковыми, несмотря на более
поздний прилет и вынужденное размещение последних в субоптимальных местообита-
ниях (Leinonen. 1973). В Швеции среди самок больших синиц, получавших в предгнездо-
вой период подкормку „мучными червями”, годовалые особи начали откладку яиц чуть
раньше старых (Kallander, 1974).
Из цитированной литературы следует, что возрастные различия в сроках размноже-
ния выявлены лишь для самок, которым заранее отдается ведущая роль в выборе време-
ни начала кладки; возрасту партнера при этом не придается никакого значения. Это
вряд ли справедливо, поскольку самцы, как показали эекспериментальные работы
Промптова (1956) и исследования других орнитологов (Immelmann, 1971; Хайнд,
1975), могут оказывать существенное стимулирующее влияние на созревание половых
продуктов самок. Даже сам факт присутствия самца ускоряет развитие гонад самки.
Более или менее значительную роль играют при этом, очевидно, и индивидуальные ка-
чества самца, а также определенные преимущества или недостатки занятой им
территории.
У большинства воробьиных птиц решающая роль в выборе территории принадле-
жит самцам. Самцы, занявшие и удерживающие участки в оптимальных для гнездова-
ния местообитаниях, лучше и в более ранние сроки обеспечены кормом. На занятых
ими территориях достаточно надежных укрытий для гнезд, хорошо выражены защитные
условия и т. д. Очевидно, самцы, обладающие лучшей территорией, имеют больше
шансов для привлечения самки, более раннего формирования пары и начала размноже-
ния (примеры биотопических различий в сроках размножения уже приводились). Не ме-
нее важны и индивидуальные качества самца: активность пения, способности к защите
территории от вторжения конкурентов своего или других видов птиц, к обеспечению
самок необходимым количеством корма, к „ухаживанию” и другим формам демон-
стративного поведения.
В том, что самцы могут стимулировать размножение самок, нам довелось убедить-
ся на примере со снегирями. В вольеру к очень активному самцу была подсажена самка,
жившая до этого в неволе около месяца. Обычно только что пойманные птицы даже
при наличии в вольере всех условий для размножения никогда не принимают в нем учас-
тия. Так было и в данном случае: самка не проявляла никаких признаков половой
46
Таблица 13
Возрастные различия в сроках начала размножения самок мухоловки-пеструшки
(Маячино, 1981 — 1983 гг.) ______
Возраст Год n lim M ±т С(%)
adj 1981 62 25.V-24.VI 29.1 мая 0.72 19.6
ad2 1981 12 25.V-1.VI 27.7 мая 0.51 6.4
adj 1982 69 23.V-25.VI 3.0 июня 0.86 20.9
ad2 1982 13 28.V-20.VI 1.1 июня 1.64 18.3
ad. 1983 94 13.V-22.VI 21.7 мая 0.70 31.1
ad2 1983 13 17.V-29.V 20.2 мая 1.17 20.9
активности и на настойчивые ухаживания самца не реагировала. Но после того как
самец начал носить сухие веточки ели и корешки, беспорядчно укладывая их в основа-
ние ветвей елочки, самка также включилась в постройку гнезда и отложила яйца. Труд-
но сказать, возникают ли подобные ситуации в природе среди птиц, у которых самцы
не принимают участия в сборе материала и строительстве гнезд (как и у снегиря), одна-
ко важно подчеркнуть, что в данном случае активные действия самца вынудили самку
начать размножение.
В период формирования пар еще до начала гнездостроения постоянно можно
наблюдать погоню самцов (иногда сразу нескольких) за самками. В отдельных случаях
эти погони заканчиваются попытками копуляции. Такое поведение характерно для
многих воробьиных. Так, в районе Алма-Аты Сема (1969) отмечал попытки копуляции
у скворцов за месяц до начала кладки. Аналогичные данные приводят и другие орнито-
логи: Фетисов (1981) для полевого воробья в Ленинградской обл., Бардин (1975а,
19756) для синиц в Псковской обл.;Лапшин (1978), подробно изучавший предгнездо-
вое поведение веснички в Карелии, пришел к заключению, что такое поведение самца —
необходимое условие для стимулирования самки. Можно предполагать, что самки,
партнеры которых активнее кормят их во время гнездостроения и откладки яиц, полу-
чают больше дополнительной энергии, необходимой для развития яичников, яйцеводов
и наседных пятен; поэтому раннее начало их размножения гораздо вероятнее, чем в па-
рах с менее активными самцами. Вместе с тем такие индивидуальные особенности
самцов, как активность их пения и вокальные данные, по всей вероятности, не имеют
для самок решающего значения при выборе партнера. Они могут образовать пару с сам-
цом, у которого видовая песня имеет значительные дефекты, тогда как активно поющие
самцы с нормальной песней подчас остаются* без самок длительное время или даже
весь сезон (Лапшин, 1978).
Таким образом, несмотря на то что сроки начала откладки яиц в большей степени
зависят от самок, индивидуальные особенности самцов могут прямо или косвенно
влиять на сокращение или увеличение периода между формированием пары и началом
откладки яиц.
Учитывая это, мы попытались проанализировать имеющийся в нашем распоряже-
нии материал в традиционном направлении, т. е. по возрасту самок, и в зависимости
от состава гнездовых пар (табл. 13—15). Величину отклонений от начальной или сред-
ней даты появления первых яиц в гнездах рассчитывали для каждого года отдельно. При
расчете отклонений от средней даты учитывали лишь их абсолютное значение, не при-
нимая во внимание знака опережения ” или запаздывания „+” по отношению к сред-
ней дате. Имеющийся материал не позволяет сделать окончательное заключение о влия-
нии возраста птиц на сроки их размножения, но отражает известную неодновременность
начала гнездования местных особей в любых половых и возрастных группах. В боль-
шинстве случаев различия оказываются статистически недостоверными (р > 0.05).
Даже значимые различия, полученные при анализе с дифференциацией по полу и возрас-
ту одного из партнеров, противоречат данным, отражающим зависимость сроков гнездо-
вания от возрастного состава пар. Так, независимо от возраста партнеров старые самки
47
Таблица 14
Точность совпадения индивидуальных и популяционных сроков размножения
у птиц разного возраста и пола
Пол и воз- раст п Отклонение (сут) от даты
наиболее раннего начала размноже- ния в год наблюдений средней за год
Игл М ± т с (%) Нт М± т с (%)
Большая синица
9ad2 11 0-19 8.0 ± 1.8 70.6 0-13 3.4 ± 1.2 113
9 adj 26 0-22 4.7 ± 0.9 98.3 0-22 2.8 ± 0.7 125.2
d ad 2 24 0-21 5.9 ± 1.1 88.6 0.11 4.0 ± 0.7 82.6
d adj 17 0-25 7.8 ± 1.9 My? 99.6 солов ка-п 0.16 еструшка 4.5 ± 1.2 107.5
9ad2 19 0-21 87. ± 1.4 67.0 1-13 3.8 ± 0.7 78.1
9 adj 44 0-26 9.1 ± 0.9 67.6 0-13 4.0 ± 0.4 71.6
dad2 44 2-29 7.9 ± 0.8 68.7 0-20 4.8 ± 0.5 71.8
d ad. 60 0-25 8.1 ± 0.6 54.5 Зарян 0-16 ка 3.4 ± 0.4 88.2
9 ad2 11 0-22 10.2 ± 2.4 70.0 0-16 4.5 ± 0.9 66.1
9 adj 22 0-33 13.6 ± 2.3 76.9 1-22 8.4 ± 1.4 76.7
d ad2 12 0-12 5.1 ± 1.9 115.1 0-10 3.6 ± 1.0 89.4
dadj 40 0-20 9.8 ± 1.9 60.3 Белобр< 0-12 эвик 5.3 ± 0.5 59.2
9ad2 14 0-13 4.14 ± 1.1 93.6 — — —
9 ad. 11 4-34 7.1 ± 0.9 39.6 — — —
d ad2 15 0-9 4.5 ± 0.8 63.7 — — —
dadj 7 4-10 7.1 ± 1.0 39.4 — — —
Примечание. Здесь и в табл. 15 подчеркнуты достоверные возрастные различия.
Таблица 15
Точность совпадения индивидуальных и популяционных сроков
размножения у птиц при разном возрастном составе гнездовых пар
Состав пар п Отклонение (сут) от даты
наиболее раннего размножения в год наблюдений средней за год
Нт | М±т С (%) Нт | > М ± т | С (%)
Большая синица
9ad2 dad2 9 2-14* 8.2 ±1.3 43.7 1-5 1.9 ±0.5 72.0
9adj dad2 17 0-15 5.2 ± 1.1 84.9 0-11 3.3 ±0.7 92.5
9 adj dadj 17 0-15 4.1 ±1.2 115.3 0-16 3.4+ 1.0 124.8
Мухоловка-пеструшка
9ad2 dad2 13 2-16 8.0— 1.3 57.8 0-14 4.6+ 1.1 79.6
9ad2 dadj 7 3-11 5.6 ±1.2 52.6 1-6 4.3 ±0.8 46.0
9adj dad2 30 2-18 6.8 ±1.0 73.7 0-20 4.9+0.6 72.5
9adj dadj 52 0-16 8.0 ±0.5 48.6 0-16 3.3+0.4 93.1
Весничка
9ad2 dad2 13 0-17 7.3 ±1.5 73.3 1-15 5.8 ±1.3 75.5
9adx dadj 38 0-16 7.9 ± 0.7 52.8 0-12 4.3 ±0.4 59.0
9adj 9ad2 dad2j dadj j 29 0-14 6.4 ± 0.7 56.6 0-14 4.9 + 0.6 70.2
Зарянка
9ad2 dad2 7 0-17 7.0 ±2.8 98.3 1-7 3.9+ 1.1 70.7
9adj dadj 20 2-15 6.1 ±0.8 59.5 0-15 3.8+ 0.6 64.3
9adt 9ad2 dadj dadj j 9 0-10 4.9 ±1.2 72.6 0-10 3.9+ 1.2 87.7
♦Учтены только пары, начавшие откладку яиц в первые 20 дней сезона размножения.
48
и самцы белобровика начинают гнездиться достоверно раньше первогодков (табл. 14).
Логично было бы ожидать, что пары, состоящие из старых птиц, также будут начинать
размножение раньше пар, образованных первогодками. Но на самом деле этого не про-
исходит. Пары из молодых партнеров имеют тенденцию к более раннему началу гнездо-
вания (табл.15).
У больших синиц пары из годовалых особей первые яйца откладывают в среднем
раньше, чем птицы старших возрастных групп (табл. 15), что противоречит общему пра-
вилу. Кроме того, при дифференцированном способе анализа (табл. 14) более раннее
гнездование ad! не подтверждается. Противоречивы и данные для зарянки, у которой
старые самки точнее „определяют” среднюю дату начала откладки яиц, чем годовалые,
но это не подтверждается анализом различий между другими возрастными сочетания-
ми в парах. У веснички намечается противоположная тенденция — наименьшие отклоне-
ния от начальной даты размножения более характерны для пар из партнеров разного
возраста, чем из одновозрастных; расхождения же со средней датой для всех выделен-
ных возрастных групп оказываются практически одинаковыми.
Таким образом, сколько-нибудь существенных возрастных различий в сроках
размножения анализируемых видов птиц на нашем материале не выявлено. Даже тен-
денции к более раннему гнездованию неоднозначны как в пределах одного вида, так
и при межвидовом сравнении. В связи с этим важно еще раз подчеркнуть специфику
возрастного состава птиц, принимающих участие в размножениии на исследуемой тер-
ритории. У большинства воробьиных птиц, населяющих таежную зону, преобладающая
часть гнездового населения представлена первогодками. Это свойственно всем видам
животных с коротким жизненным циклом, и перечисленных выше исключений из этого
правила немного.
Но если более раннее гнездование действительно дает виду преимущества, то по-
чему же оно закреплено лишь за той частью населения, которая составляет меньшин-
ство? С одной стороны, раннее гнездование действительно кажется более выгодным. Оно
может начаться, когда выбранные птицами местообитания не достигли предельного насы-
щения особями того же и других видов. При успешном раннем гнездовании взрослые
особи успевают воспитать потомство до массового вылета молодняка своего и других
видов птиц, т. е. раннее размножение проходит при более низких уровнях численности
конкурентов. Кроме того, хищники и хищничающие животные (обыкновенная белка,
врановые, сорокопуты и др.) не сразу, а по мере увеличения массовости гнездования
приспосабливаются к разорению гнезд (по аналогии с переходом хищников на питание
наиболее массовыми видами кормов). К концу гнездового сезона пресс хищников
вообще возрастает за счет приобретения самостоятельности молодняком хищных и
хищничающих животных и их расселения по территории. К середине лета при относи-
тельной стабилизации температуры на высоком уровне резко возрастает активность
обыкновенной гадюки, от которой страдают в первую очередь мелкие наземногнездя-
щиеся воробьиные, поселяющиеся в полуоткрытых типах местообитаний. Изредка
гадюка нападает также на гнезда кустарниковых птиц (садовая славка, камышевки).
В Приладожье зарегистрированы случаи нападения гадюки на птенцов зяблика, находив-
шихся в гнезде на еловой ветви в 2 м от земли, и даже на птенцов черного дрозда.
В поздних выводках птенцы чаще бывают поражены личинками мух, паразитирую-
щих на птицах, — Protocalliphora sp. и Trypocalliphora sp.
По мнению Лэка (Lack, 1966), раннее размножение дает больше шансов молодым
и старым птицам своевременно завершить линьку и вовремя начать и окончить мигра-
цию к местам зимовки, что обеспечивает лучшую их выживаемость. Правда, Лэк не учи-
тывал при этом синхронизации и ускоренного завершения всех фаз годового цикла птиц
при поздних сроках размножения, что было выявлено сравнительно недавно (Носков,
1969, 1977; Кукиш, 1976; Рымкевич, 1977; Музаев, 19806; Лапшин, 19816; Савинич,
1984). Однако, обладая определенными преимуществами, раннее размножение птиц
в таежной зоне далеко не всегда оказывается самым успешным. Напротив, из-за частого
возврата холодов, — а их вероятность, как было показано в предыдущей главе,
4 В. Б. Зимин
49
максимальна как раз в весенний и предлетний периоды, — раннее гнездование даже
в зоне 60-63° с. ш. нередко сопровождается массовой гибелью птенцов и кладок.
Успешность размножения различных видов птиц подвержена существенным сезонным
колебаниям и асинхронно изменяется в разные годы, что подробнее будет обсуждено
отдельно. Здесь же мы ограничимся только примером с большой синицей. В молод-
няках Шокшинского стационара период откладки яиц у этого вида совпадает с приле-
том самцов вертишейки, которые сразу же начинают активные поиски и расчистку
пригодных для гнездования дупел. В результате многие гнезда больших синиц оказыва-
ются разоренными, причем число погибших кладок положительно коррелирует
(г = 0.84) с численностью вертишейки. Соответственно, успешность раннего гнездова-
ния больших синиц в молодняках оказывается ниже, чем у пар, запаздывающих с нача-
лом кладки.
При неустойчивой весенней погоде в таежной зоне раннее размножение не всегда
является наиболее продуктивным. Следовательно, „личный опыт” старых птиц, то,
чем орнитологи нередко пытаются объяснить возрастные различия в сроках гнездова-
ния, — должен скорее препятствовать раннему началу кладки и способствовать выбору
оптимального для вида времени. Ссылки на большую опытность старых птиц весьма
субъективны, по крайней мере для воробьиных, так как абсолютное большинство
ad2 — это преимущественно двухлетние особи, которые до этого гнездились в большин-
стве случаев только один раз и „оценить” преимущества более раннего или позднего
гнездования, естественно, не могли. Доводы о том, что старые птицы лучше знают свою
территорию, места расположения укрытий и концентрации пищи, тоже достаточно
субъективны. Даже при коротком предгнездовом периоде молодые особи любых видов
при высокой степени подвижности птиц имеют вполне достаточно времени для ознаком-
ления с занятой ими территорией. И коль скоро они дожили до момента начала размно-
жения, то, вероятнее всего, они располагают также и достаточными навыками в отыска-
нии пищи.
Вместе с тем для более раннего гнездования старых птиц существуют вполне объ-
ективные предпосылки. Прежде всего, они первыми возвращаются на родину. Это уста-
новлено для многих видов птиц при массовых отловах крупногабаритными ло-
вушками и паутинными сетями (Носков и др., 1975; Носков и др., 1981; Зимин,
19816, и др.). Более подробно формирование местного населения в наших широтах
было рассмотрено на примере чечевицы (Зимин, 19816) и веснички (Лапшин, 1978;
Лапшин и др., 1981).
Наиболее вероятно, что прилетающие первыми старые самцы занимают террито-
рии, максимально удовлетворяющие требования вида к местам гнездования, и появля-
ющиеся позднее первогодки вынуждены селиться в субоптимальных местообитаниях.
Такое распределение по территории характерно для чечевицы у северо-западных границ
ареала (Risberg, 1970; Peiponen, 1974; Stjernberg, 1979; Зимин, 19816). В сосново-листвен-
ных молодняках 11—17-летнего возраста в обычных условиях большая синица не гнез-
дится, но ее удается привлечь сюда развеской искусственных гнездовий (Зимин и др.,
1976; Зимин, 1978). Первые шесть лет население гнездящихся в молодняках синиц
ежегодно полностью обновлялось только за счет первогодков. Таким образом, ни одна
из окольцованных здесь на гнездах взрослых птиц не возвращалась в молодняки в по-
следующие годы. Для больших синиц шокшинские молодые лесонасаждения по всем
основным показателям относятся к субоптимальным гнездовым стациям (Зимин,
1978). На другом стационаре (о-в Б. Климеке цкий) у больших синиц в стациях с опти-
мальными условиями для гнездования (спелые и приспевающие преимущественно
лиственные древостои) в течение всех лет наблюдений сроки размножения были более
ранними, несмотря на более северное положение и обилие плавающих льдов, снижа-
ющих весенние температуры воздуха.
В центре ареала, в условиях высокой численности гнездящихся птиц, вытеснение
первогодков в субоптимальные местообитания может быть выражено более резко, что,
вероятнее всего, отражается и на задержке начала их размножения. Для скворца,
50
большой синицы и мухоловки-пеструшки это установлено в Окском заповеднике
А. Д. Нумеровым (устное сообщение), а еще раньше было показано для птиц Дарвин-
ского и Приокско-Террасного заповедников (Поливанов, 1957; Лихачев, 1961).
Формирование местного населения птиц, зимующих в наших широтах, тщательно
изучено для пухляка и хохлатой синицы в Псковской обл. (Бардин,1970, 1975а, 19756),
для большой синицы в Ленинградской обл. (Носков, Смирнов, 1981), кукши (Коханов,
1982) и серой вороны (Шутенко, 1983). Старые особи перечисленных видов птиц зиму-
ют на своих гнездовых участках или вблизи от них. Возвращающиеся весной молодые
особи имеют достаточно времени для того, чтобы сформировать гнездовые пары и за-
нять свободные территории. На севере при низкой плотности населения птиц это вполне
возможно. Однако у молодых особей территории будут, очевидно, характеризоваться
менее благоприятными условиями, чем у старых.
На севере, кроме низкой численности популяций, проявлению возрастных разли-
чий в сроках гнездования птиц препятствуют еще два момента. Во-первых, как пока-
зывает сопоставление показателей ежегодной смертности и плодовитости, самые север-
ные периферийные популяции птиц могут существовать только за счет ежегодного
притока иммигрантов извне (Haukioja, 1969; Leinonen, 1973;Solonen, 1979; Jarvinen А.л
1980, 1984). Об этом же свидетельствуют и многолетние данные кольцевания и контро-
ля гнездового населения массовых видов птиц на наших стационарах. Следовательно,
на севере ареала, особенно в зонах спорадического гнездования вида, доминирование
птиц первогодков будет более значительным, что, естественно, снижает вероятность
проявления возможных возрастных различий в особенностях гнездования. Во-вторых,
у птиц северных популяций гнездование начинается более дружно, чем у южных, что
особенно наглядно проявляется в высоких широтах (Irving, 1960; Чемякин, 1982а,
19826). Сроки начала гнездования различных пар здесь настолько сближены, что воз-
растные (и прочие) различия сглаживаются и не могут проявиться в полной мере. Уже
в зоне 60—64° с. ш. до 60, иногда до 80 % гнездового населения многих видов птиц при-
ступает к откладке яиц в течение одной декады или даже пятидневки (рис. 16, 19;
табл. 5—8). Особенно дружно начинается гнездование в годы с затяжной холодной
весной. В нашем регионе 80% кладок певчего дрозда появляются в течение 40, а в Поль-
ше — 70 сут (Dyrcz, 1969). Сходные данные получены и для других птиц. Так, в Англии
период (сут), в течение которого 80% особей в популяции приступают к гнездованию,
намного больше, чем в Карелии.
Карелия Англия (по: Lack, 1946, 1950)
Весничка 20 40
Серая мухоловка 21 45
Зарянка 22 75
Таким образом, у северных пределов распространения вида при укороченном
сезоне размножения, позднем и более дружном его начале вероятность проявления
возрастных различий в сроках гнездования резко сокращается.
Следует отметить, что способ определения возрастных различий в гнездовании
по датам откладки в известных гнездах недостаточно точен. Во многих случаях остается
неясным, действительно ли мы имеем дело с первой попыткой гнездования данной
пары. Особенно много сомнений по этому поводу возникает в отношении второй поло-
вины репродуктивного периода. Уточнить это можно лишь при сплошном индивидуаль-
ном мечении птиц до начала размножения и постоянных контрольных отловах в течение
всего сезона размножения. Это выполнимо лишь для „оседлых” птиц на ограниченной
территории. В то же время для перелетных видов, гнездовое население которых форми-
руется за довольно короткий промежуток времени и не остается постоянным в течение
всего репродуктивного периода, такой путь при современной технике отлова птиц
не гарантирует абсолютно полного контроля.
На наших стационарах население отдельных видов контролировалось на 80—90 %.
Но эти показатели получены путем отлова птиц в течение всего времени пребывания
51
их на родине, а не перед началом гнездового сезона. Более того, при использовании
30—50 паутинных сетей и других способов отлова удавалось более или менее полно
контролировать птиц лишь на площади в 30—40 га. Тем не менее постоянный контроль
местных птиц методом отлова и прижизненного описания позволяет существенно
уточнить ход размножения отдельных птиц и всей популяции в целом и, кроме того,
дает много дополнительных сведений. Так, благодаря применению летних зерновых
прикоромок и модернизированной нами специальной ловушки (Bub, 1966; Зимин,
1973) мы получили возможность контролировать местных чечевиц с момента их приле-
та и до отлета (Зимин, 19816). Некоторые особи ловились практически каждый день
и даже птенцов кормили зерновой смесью из ловушки. Как удалось показать, динамика
развития клоакальных выступов самцов и наседных пятен самок достаточно полно
отражает ход размножения местных чечевиц. Выявленные таким путем возрастные
различия в темпах развития клоакальных выступов подтвердили значительную индиви-
дуальную вариабельность в сроках готовности птиц к размножению, определяющую
растянутость гнездового периода, а также позволили получить другую ценную информа-
цию о различных деталях репродуктивного цикла маркированных особей.
Окончание сезона размножения
Это, пожалуй, наименее изученная часть репродуктивного цикла птиц, редко анали-
зируемая в орнитологической литературе, но заслуживающая самого пристального
внимания. Для ответа на вопрос, чем определяется окончание репродуктивной активно-
сти птиц, необходима всесторонняя оценка накопленных фактов. Определеннее других
высказываются на этот счет авторы экспериментальных исследований годовых циклов,
которые считают, что продолжительность репродуктивного периода птиц генетически
запрограммирована и определяется отсчетом времени от начала фотостимуляции (или
размножения) и что окончание активности гонад связано с наступлением половой реф-
рактерности. Определенная роль иногда отводится и влиянию факторов внешней среды,
контролирующих окончание репродуктивного цикла (Farner, 1967; Дольник, 1975; Но-
викову др., 1982).
При полевых исследованиях, основанных на методах прижизненного изучения
птиц, о времени окончания половой активности птиц приходится судить в основном
по косвенным показателям: по датам откладки яиц, размерам клоакальных выступов
и состоянию наседных пятен, по активности пения, изменению поведения и по началу
линьки. При этом, как правило, остается неясным, сколько времени после откладки
яиц гонады сохраняются в активном состоянии и на какой стадии регресс их становится
необратимым, исключающим возможность повторного размножения. Отстрел птиц
противоречит основным принципам нашей работы, а отрывочные данные, полученные
на случайно погибших при отлове особях, как и сведения о размерах гонад, опубли-
кованные Нейфельдт (1958) и имеющиеся в архиве КФ АН СССР (материалы Л. О. Бе-
лопольского) , позволяют составить по данному вопросу лишь самое общее представ-
ление. Весьма перспективен для прямых наблюдений за активностью гонад метод при-
жизненой лопароскопии (Harrisson, 1978; Burr et al., 1981), но его применение на мелких
воробьиных птицах требует специальной подготовки.
В условиях таежной зоны период окончания кладки у различных видов воробьи-
ных птиц в целом достаточно сильно растянут. Первыми заканчивают гнездование
клесты-еловики, последние кладки у которых появляются 1—15 мая. В эти же сроки
прекращают откладку яиц большинство серых ворон, воронов, кукш, соек и хохлатых
синиц. Лишь отдельные особи этих видов могут приступить к повторному размноже-
нию до последних чисел мая. В первой половине июня к ним присоединяются скворец
и пухляк. Далее число видов, заканчивающих откладку яиц, быстро увеличивается. К
концу июня оно достигает 40%, к 10—15 июля — 75, к 30 июля — 95% всех видов
52
Рис. 16. Географические различия в сроках и продолжительности периода откладки яиц у вес-
нички (А), серой мухоловки (Б), зарянки (В) и певчего дрозда (Г).
1 - Англия (по: Lack, 1946, 1950; Cramp, 1955); 2 - Польша (по: Dyrcz, 1969); 3 - Карелия.
Горизонтальные линии - продолжительность периода, в течение которого 80 % самок приступают
к размножению, символы по краям этих линий соответствуют месту исследования. По оси абсцисс -
календарь; по оси ординат - число исследованных гнезд (%).
воробьиных птиц, гнездящихся в наших широтах. В августе еще приступают к гнездова-
нию единичные пары зарянок, городских и деревенских ласточек, коноплянок, чечеток
и снегирей.
Со второй пятидневки июля общее число видов и особей, еще способных откла-
дывать яйца, сокращается очень резко. Продолжают гнездиться лишь птицы с полицикли-
ческим размножением, единичные особи камышевок и иволга (1 случай в предпослед-
нюю пятидневку июля).
В первой половине июля прекращается также регулярное пение почти всех видов
птиц,токовые полеты и крик кукушек, что является еще одним свидетельством оконча-
ния размножения певчих птиц. К этому же периоду приурочено начало линьки и пролета
дальних мигрантов.
Для июля характерно прогрессирующее сокращение длины дня. За этот месяц
продолжительность светлого времени суток убывает под 66° с. ш. на 3.3 ч, под 62° —
на 2.1 ч. Таким образом, размножение у большинства видов заканчивается на гораздо
более длинном фотопериоде, чем начинается. Казалось бы, можно предположить, что ре-
шающее значение для прекращения активности гонад имеет не какое-то абсолютное,
пороговое значение фотопериода — разное на разных широтах и у различных особей,
а сама тенденция к сокращению длины дня. Но у врановых, хохлатой синицы и пухляка
откладка яиц прекращается на увеличивающемся фотопериоде. У клестов это характер-
но для весеннего размножения, осеннее же завершается на сокращающемся фо-
топериоде.
В широтных различиях сроков гнездования птиц умеренной зоны обращают на
себя внимание несколько особенностей. Общая продолжительность откладки яиц, как
уже указывалось, быстро сокращается по направлению к северу, что связано с укороче-
нием вегетационного периода. Но если начало размножения широко распространенных
видов птиц на севере и юге ареала различается в целом на 2 мес, то его пики оказывают-
ся более сближенными — промежуток между ними в крайних точках зоны умеренного
климата вдвое меньше. Между тем время завершения гнездового сезона для большин-
ства видов оказывается приблизительно одним и тем же. Отсюда следует, что широтные
различия в сроках гнездования и его большая общая растянутость на юге по сравнению
53
Рис. 17. Динамика биомассы основных групп беспозвоночных животных в хвойно-лиственных
молодняках юго-восточного Приладожья.
А - ель; Б - береза; В - почва, подстилка; Г - сосна; Д - травянистые растения. 1 - личин-
ки различных жуков и двукрылых; 2 - олигохеты; 3 - жесткокрылые (имаго); 4 - перепончато-
крылые; 5 - гусеницы чешуекрылых; 6 - двукрылые; 7 - равнокрылые; 8 - пауки; 9 - прочие
беспозвоночные. По оси абсцисс - календарь; по оси ординат - доля от общей биомассы (%).
с севером в значительной мере обусловлены разницей в начальных датах. Всюду в зоне
с умеренным климатом и в Субарктике периодом окончания откладки яиц является
первая половина июля. В этот же период отмечается окончание кладок у подавляющего
большинства видов, нередко значительно различающихся по датам начала гнездования
(рис. 16). Вероятно, именно в это время в среде обитания происходят изме-
нения, резко снижающие половую активность самых разных видов птиц на различ-
ных широтах.
Одним из таких факторов считается уменьшение количества пищи, что, например,
подтверждается прямой связью между ходом откладки яиц в оксфордских популяциях
большой синицы и лазоревки и обилием гусениц пядениц и листоверток (Lack, 1957,
1966). Окончание зимне-весеннего размножения клестов-еловиков также совпадает
с периодом практически полного высыпания семян из шишек. Однако известно и осен-
нее гнездование клестов, которое и начинается, и заканчивается при значительных
запасах пищи. Кроме того, на питании и выкармливании птенцов гусеницами специали-
зируются многие виды певчих птиц с не совпадающими сроками начала и окончания
размножения (хохлатая синица, пухляк, большая синица).
Учет беспозвоночных животных, выполненный в Приладожье, показал, что пики
максимального обилия гусениц некоторых чешуекрылых и ложногусениц пилильщи-
ков разобщены во времени и сменяют друг друга (рис. 17). Согласно многочисленным
исследованиям, птицы легко переходят с одного вида корма на другой, спектр их жертв
очень широк, и они способны быстро переключаться на те виды пищи, которые в дан-
ный момент становятся наиболее массовыми и легкодоступными (Королькова, 1963;
Иноземцев, 1978). В серии работ по питанию гнездовых птенцов и птиц Ленинградской
обл. Прокофьева (1958, 1961, 1963, 1966, 1970, 1979 и др.) на многих видах показала
географические, ежегодные, сезонные биотопические и индивидуальные различия в со-
ставе кормов. В Дарвинском заповеднике гусеницы и имаго чешуекрылых в течение
одного сезона составляли основу пищи, получаемой птенцами скворца, — 55.6% по
массе, а на другой год перешли в разряд второстепенных кормов — 12.4%.
54
Соответственно изменялась и доля приносимых птенцам двукрылых — 8.6 и 45.1%
(Поливанова, 1957).
У чижей зимние выводки выкармливаются семенами хвойных деревьев, обычные
весенние — преимущественно насекомыми, а летние (нормальные вторые) — семенами
травянистых растений и насекомыми. Подобных примеров сезонной изменчивости
питания можно привести достаточно много, но важно отметить, что исчезновение одних
видов корма птицы компенсируют использованием других массовых объектов питания.
Для зерноядных птиц, и в частности для чечевицы, время окончания размножения
вообще приходится на начало массового созревания семян травянистых растений и ягод.
В июле—августе корма вполне достаточно даже в местах повышенной концентра-
ции птиц. Так, в Приладожье в период расселения молодняка и начала миграции общая
численность птиц возрастает в сотни раз по сравнению с любой другой точкой, удален-
ной от направляющих линий (Зимин, 1965; Носков и др., 1975; Резвый, 1976; Резвый,
Савинип. 1978). Об отсутствии дефицита кормов в прибрежной полосе, где происходит
значительная концентрация птиц за счет скопления перемещающихся особей, свидетель-
ствует тот факт, что выживаемость птенцов в расположенных здесь гнездах совершен-
но не отклоняется от видовых норм для июля—августа.
Таким образом, для птиц таежной зоны изменения в обилии пищи не являются
главным фактором, влияюшим на окончание сезона размножения. То же относится
и к температуре воздуха, и к общему состоянию погоды. По крайней мере в северных
широтах июль—середина августа является самым теплым периодом года с наиболее
стабильной погодой (Романов, 1961). Но у большинства птиц начало размножения и
выкармливание гнездовых птенцов проходят в гораздо менее стабильных условиях,
при довольно резких колебаниях температуры воздуха. Таким образом, сезон гнездова-
ния таежных птиц приурочен далеко не к самым лучшим погодным условиям в году.
Подавляющее большинство птиц предпочитают гнездиться раньше, чем наступает самый
теплый и стабильный по состоянию погоды сезон.
К июлю и в течение этого месяца выведенные в нормальные сроки молодые птицы
становятся самостоятельными и многие из них приступают к постювенильной линьке.
Переход к самостоятельному добыванию корма — процесс довольно длительный, а прие-
мы добычи и приобретение навыков отыскания пищи развиваются постепенно (Промп-
тов, 1956; Хайнд, 1975). В связи с этим обилие пищи и ее доступность имеют немало-
важное значение. Чем чаще молодые особи будут сталкиваться с потенциальными объ-
ектами питания, тем быстрее они освоят их активный поиск и приемы добычи. Навыки
распознавания пригодных в пищу насекомых или семян приобретаются методом проб
и ошибок. Так, слетки часто пытаются схватить клювом любые мелкие движущиеся
или колеблющиеся около них предметы (раскачиваемые ветром кусочки бересты,
паутину, перья и другой застрявший в ветвях мусор).
Выведенным в вольерах молодым чечеткам и чижам в качестве дополнительного
корма мы постоянно давали „накошенных” сачком насекомых, среди которых было
немало муравьев, лесных клопов и их личинок. Поначалу молодые птицы постоянно
ошибались и хватали всех разбегающихся насекомых без разбора, но постепенно они
научились отличать муравьев и клопов и в дальнейшем никогда их не трогали. Каждый
новый вид корма вольерный молодняк признавал лишь после того, как им начинали
кормиться взрослые птицы. Все эти моменты специально изучались Промптовым
(1956), который подчеркивал, что становление самостоятельности в кормодобывании —
процесс очень сложный и достаточно длительный.
В природе молодые птицы разных видов становятся самостоятельными в возрасте
1—3 мес. Ясно, что от того, насколько успешно освоят они навыки кормодобывания,
во многом будет зависеть их выживаемость на первых этапах самостоятельной жизни.
Существенную роль в этом играют обилие и доступность пищи, а также состояние
погоды. Ухудшение погоды (похолодания, сильные ветры, длительные осадки) сопро-
вождается, во-первых, увеличением затрат энергии на терморегуляцию и, во-вторых,
уменьшением обилия и доступности пищи (активность насекомых снижается, многие
55
из них заблаговременно уходят в укрытия). Не имеющая достаточного опыта в отыска-
нии пищи и не освоившая всего спектра потенциальных кормов молодая особь в по-
добных ситуациях может погибнуть. Кроме того, на период’становления самостоятель-
ности приходится и начало постювенильной линьки, требующей дополнительной энер-
гии, а следовательно и большего количества пищи.
Поэтому, видимо, не случайно гнездование птиц приурочено к менее теплому
и устойчивому периоду репродуктивного сезона, а переход к самостоятельной жизни
молодых особей и первые ее этапы — к наиболее теплому времени года со стабильными
погодными условиями. Даже при длительных похолоданиях весной взрослые птицы
способны обеспечить птенцов пищей, о чем подробнее говорится ниже. Примечательно,
что все рано заканчивающие гнездование птицы (врановые, хохлатая синица, пищуха,
длиннохвостая синица) имеют и длительные периоды вождения выводков. Иначе го-
воря, наименее экологически стабильную часть репродуктивного сезона молодые
птицы этих видов проводят вместе со взрослыми, а время их перехода к самостоятель-
ности наступает в те же сроки, что и у большинства других птиц, — в период теплой и ус-
тойчивой погоды. Среди наших синиц самые ранние сроки окончания размножения, как
и всюду, имеет хохлатая синица. Птенцы ее после вылета из гнезда получают корм от
родителей в течение более трех недель и становятся самостоятельными в возрасте
примерно 45 дней. У гнездящегося позднее пухляка птенцы переходят к самостоятель-
ности на 10-12 дней раньше — через 15 дней после вылета, в 32—34-дневном
возрасте (Бардин, 1975а), т. е. примерно в то же календарное время, что и хохлатые
синицы.
Такое совпадение в сроках окончания гнездования и перехода молодых особей
к самостоятельности у совершенно разных видов птиц и на всех широтах не может
быть случайным. Для поддержания численности на оптимальном для вида уровне важны
не только успешность выкармливания птенцов в гнезде и выводка, но и послегнездовая
выживаемость молодого поколения. Потери среди молодых птиц в первые месяцы их
жизни во многом зависят от условий, в которых формируются навыки в отыскании
пищи, укрытий от непогоды и хищников. На этот же период приходится энергоемкий
процесс постювенильной линьки — существенная дополнительная нагрузка для молодых
особей, не обладающих достаточным опытом преодоления критических ситуаций. При
похолоданиях, затяжных дождях и сильных ветрах в первую очередь изменяется поведе-
ние насекомых — они становятся малоподвижными, не выходят из укрытий. Трудно
сказать, сможет ли молодая птица в этих условиях обеспечить себя необходимым коли-
чеством корма, доступность которого при низких температурах резко снижается, а она
не успела еще приобрести навыки в отыскании оцепеневших и ушедших в укрытия
насекомых. Кроме того, при холодной и ветреной погоде возрастают затраты энергии
на терморегуляцию. Во время похолодания в конце июля—августе 1977 г. среди погиб-
ших в Приладожье деревенских ласточек были обнаружены лишь сеголетки.
После холодных и ветреных ночей в конце июля—августе в ранние утренние часы
именно молодые особи нередко встречаются „пухлыми”, явно ослабевшими. В таком
состоянии они, очевидно, становятся легкой добычей хищников. Вполне можно допус-
тить, что смертность молодняка, не обладающего еще достаточными навыками в само-
стоятельном отыскании пищи, будет возрастать при нестабильной погоде. Вероятнее
всего, естественный отбор должен был благоприятствовать видам и особям, у которых
критический период перехода молодых птиц к самостоятельной жизни приходился
на время наиболее стабильных в году внешних условий. В умеренных и высоких широ-
тах северного полушария таким периодом является конец июня—первая половина
августа. Кроме устойчивой погоды и самых высоких в году температур воздуха он от-
личается и максимальной продолжительностью светлого времени суток. Последнее
обстоятельство также имеет немаловажное значение для баланса поступающей с пищей
и расходуемой за ночь энергии. На коротком фотопериоде поддержание положительного
баланса было бы для молодых особей гораздо более сложным из-за возрастающего
чистого расхода энергии на длительный ночной покой.
56
Таблица 16
Непостоянство сроков окончания размножения в разные годы
(по наиболее поздним датам откладки 1-го яйца)
Вид Число лет с окончанием размножения в указанную пятидневку Период откладки 1-х яиц в наиболее поздних гнездах
июнь июль
21-25 26-30 1-5 6-10 11-15 16-20 21-25 26-31
Большая синица 3 4 1 3 2 - - - 20.VI-15.VII
Мухоловка- пеструшка 3 8 3 - - - - - 9.VI-1.VII
Зарянка — — 2 4 2 1 1* 4.VII-5.VIII
Белобровик Лесная 3 6 15 5 1 1 — — 21.VI-17.VII
завирушка — 1 4 4 1 — 1 — 30. VI-25. VII
Весничка 1 2 3 2 1 1 — — 17.VI-16.VII
* Первое яйцо было отложено в период 26.VII-5.VIII.
В умеренных и высоких широтах при значительном непостоянстве многих внешних
факторов окончание репродуктивного сезона в наиболее теплое время года со стабиль-
ной погодой и при максимальной продолжительности дня приобретает адаптивное зна-
чение. В конечном итоге это обеспечивает лучшую выживаемость молодняка и повыша-
ет общую результативность размножения. Высокая продуктивность собственно гнездо-
вого периода (успешность гнездования) обеспечивается усилиями взрослых особей,
которые, как показали наблюдения в таежной зоне, даже во время похолоданий способ-
ны обеспечить птенцов кормом. Желудки погибших в такие периоды птенцов бывают
обычно наполнены пищей, так что они гибнут не от недокорма, а от переохлаждения
во время отлучек самки с гнезда. Во время вождения выводка выживаемость молодых
птиц также обеспечивается взрослыми особями, которые не только продолжают кор-
мить слетков, но и предупреждают нападение хищников, отвлекая опасность на себя.
Общая итоговая продуктивность размножения во многом будет определяться выживае-
мостью молодых птиц в начале их самостоятельной жизни. Приуроченность этого этапа
онтогенеза к периоду с максимальной в году продолжительностью дня и наиболее
стабильными внешними условиями снижает вероятность возникновения энергетиче-
ского дисбаланса и гибели молодых особей.
Сезон откладки яиц у большинства видов наших птиц в разные годы заканчива-
ется в различные сроки (табл. 16). Период регистрации самых поздних кладок за все
годы наблюдений в целом охватывает 20—25 дней. У птиц с моноциклическим размно-
жением это последняя декада июня—первая половина июля, с полициклическим — июль.
Позднее начало размножения (кладки, начатые в июле) у птиц с коротким репро-
дуктивным периодом встречается довольно редко. Так, для мухоловки-пеструшки
это отмечалось лишь 3 сезона из 21, а для веснички — 6 из 15. Появление поздних гнезд
можно связать с более поздними сроками начала гнездования или низкой его успеш-
ностью, но ни в том, ни в другом случае существенной корреляции между названными
показателями не обнаружено (г <0.3).
У дрозда-белобровика и мухоловки-пеструшки в разных пунктах Южной и Средней
Карелии размножение заканчивалось вне всякой связи со сроками его начала (рис. 13).
Правда, для мухоловки-пеструшки выявляется слабая обратная зависимость дат наибо-
лее позднего начала откладки яиц от успешности размножения (г = —0.4 ± 0.3), но
она статистическим недостоверна.
Уточнить сроки и продолжительность половой активности птиц можно, используя
данные по размерам клоаки и состоянию наседного пятна. Ранее этот метод был исполь-
зован при изучении размножения чечевицы (Зимин, 19816). Было показано,
57
Рис. 18. Сезонная изменчивость состояния наседного пятна самок (А) и величины клоакаль-
ных выступов самцов (Б) у снегирей.
По оси абсцисс - календарь; по оси ординат - стадии развития наседного пятна (А) и относи-
тельная величина клоакальных выступов (Б).
в частности, что нарастание средних размеров клоакального выступа самцов прекра-
щается после пика откладки яиц, но отдельные особи сохраняют способность к спарива-
нию еще в течение 25—30 сут. Подобные же сведения получены и для снегиря, гнездова-
ние которого в нашем и соседних регионах изучено пока совершенно недостаточно
(рис. 18). Обращает на себя внимание относительно большое число птиц без каких-либо
признаков участия в размножении.Так, среди214 самцов, отловленных в разные годы
с конца апреля до начала августа, 18.2% имели минимальные для вида размеры клоа-
кального выступа, а из 140 самок 15.5% были без наседных пятен. С другой стороны,
клоака отдельных самцов достигала максимальных размеров уже в первых числах мая,
а способность к спариванию сохранялась до первой декады августа включительно. В это
время у отдельных самок наседные пятна находились на I—II стадиях формирования,
т. е. они только приступили к откладке яиц. Это подтверждают и встречи нераспавшихся
выводков снегирей в сентябре (Haartman, 1969b; Мальчевский, Пукинский, 1983; наши
данные).
У птиц с моноциклическим размножением успешное докармливание выводка
в большинстве случаев оказывается сигналом к окончанию активности и быстрому
регрессу гонад. После гибели кладки, птенцов или слетков, если она происходит после
начала линьки, повторного размножения обычно не бывает. Правда, из этого правила
есть исключения, которые подробнее будут рассмотрены в гл. V.
Ряд фактов свидетельствует об относительной независимости времени окончания
половой активности птиц от успешности их размножения. Так, у большого числа видов
воробьиных птиц наших широт линька многих особей начинается независимо от того,
на какой стадии репродуктивного цикла они находятся (гл. V), что указывает на опре-
деленную запрограммированность продолжительности активного состояния гонад
каждой птицы. Это подтверждается и экспериментальным изучением годовых циклов
птиц, содержащихся в неволе (Г. А. Носков, устное сообщение).
Наблюдения в природе свидетельствуют о значительной индивидуальной и ежегод-
ной изменчивости периода активности гонад. Сроки окончания гнездования и начала
линьки меченых особей в разные годы их жизни были неодинаковы. В одних случаях
они совмещали линьку с размножением, тогда как в другие годы, причем в те же самые
календарные сроки, этого не было (гл. V).
Окольцованные на Шокшинском стационаре мухоловки-пеструшки на протя-
жении жизни гнездились в совершенно разные сроки. В 10 случаях самки начинали
откладку яиц на 1—32, в среднем на 13 сут позднее, чем в предшествующем году, а в 5
случаях - на 5—20 (10) сут раньше.2
Самцы в 8 случаях задержались с началом гнездования на 1-19 сут, в 7 случаях
опережали даты предыдущего года на 4—18 сут и дважды гнездились в одни и те же
сроки.
2 Определялась продолжительность репродуктивной активности, а не причина отклонений
в сроках размножения.
58
Таблица 17
Изменчивость дат начала размножения у меченых мухоловок-пеструшек в смежные годы
Пол Воз- раст* Амплитуда колебаний дат в смежные годы (сут) Число особей Величина отклонения (сут) от даты
начала гнездования в популяции средней за сезон
lim м Нт | М | ±т lim | м 1 I ±т
Стационар „Шокшинский” (1972-1977 гг.)
9 а -20...+31 10.8 15 2-19 9.7 1.8 0-9 4.0 0.9
9 а +1 -20...+31 10.8 15 0-35 12.6 2.5 0-24 5.9 1.6
д а 17 0-16 7.2 1.2 0-13 4.0 0.9
6 а +1 — 18..+19 8.9 17 0-19 11.2 1.6 0-15 4.9 1.1
Стационар „Маячино ” (1980- -1984 гг.)
9 а 31 0-26 5.7 0.8 0-22 3.4 0.8
9 а + 1 -34...+9 7.2 31 0-16 5.6 0.6 0-12 3.4 0.4
d а 62 0-33 5.5 0.7 0-22 3.7 0.4
d 1 а +1 -40...+21 9.8 62 0-33 7.7 0.9 0-26 4.5 0.6
* Здесь, в табл. 27 и 39 а - возраст птицы в год мечения, а + 1 - на следующий год.
Для смежных лет амплитуда несовпадений в датах начала откладки яиц у мече-
ных птиц составляла у самок от —20 до +31, у самцов от —18 до +19 сут. В разные
годы в соответствии с ходом весны происходило то опережение дат начала откладки
яиц по сравнению с предыдущим годом, то их запаздывание. При сравнении сезонов
1982 г. (сроки размножения близки к средним) и 1983 г. (самое раннее начало кладки
за последние 25 лет) оказалось, что гнездование 25 из 26 самцов началось в 1983 г.
на 7—40, в среднем на 12 сут раньше, чем в предыдущем. Точно так же все 9 маркиро-
ванных самок в 1983 г. приступили к кладке на 4—34, в среднем на 14 сут раньше, чем
в 1982 г. Таким образом, годовые вариации сроков начала и окончания кладки местных
(т. е. ранее уже гнездившихся здесь) птиц почти полностью соответствуют общей про-
должительности этого периода у мухоловки-пеструшки в Южной и Средней Карелии.
Это означает, что растянутость размножения обусловлена не только иным временем его
начала у первогодков: каждая особь в течение своей жизни может начать кладку и завер-
шить репродуктивный цикл в любое время, свойственное данному виду. Для нее,
следовательно, строго фиксированы лишь крайние сроки возможного начала и оконча-
ния кладки, а не конкретная, строго определенная дата. С увеличением возраста
особи отклонения во времени начала ее гнездования от начальных и средних дат появле-
ния первого яйца в популяции не сокращаются. Иными словами, сколько-нибудь значи-
тельное влияние опыта предыдущего гнездования (возраста) на выбор „лучшего”
для размножения времени по нашим данным для мухоловки-пеструшки не выявляется
(табл. 17). В возрасте а +1 (а — возраст предыдущего размножения) самки гнездились
в среднем на 4, а самцы на 1 день позднее, чем в предыдущем году.
В природе окончание размножения у большинства птиц происходит на фотоперио-
дах, близких к максимальным в году, что подтверждают результаты эксперименталь-
ных исследований — половая рефрактерность у тех видов, которым она свойственна,
наступает при максимальной продолжительности дня (Farner, 1970; Дольник, 1975).
Развитие половой рефрактерности осуществляется, однако, в широком диапазоне
длинных ' фото периодов, и конкретного порогового значения этого фактора, который
служил бы непосредственным сигналом к регрессу гонад, вероятно, не существует.
В этом мы убеждаемся, наблюдая индивидуальные вариации сроков окончания гнездо-
вания птиц в течение одного сезона и их годовые колебания у одних и тех же особей.
Значение ранних и поздних кладок. Если рассмотреть динамику откладки яиц
какого-то массового вида птиц за любой год, нетрудно заметить, что определенная часть
гнездовых пар, даже при дружных сроках начала размножения остальных, уклоняется
59
Рис. 19. Динамика сроков откладки яиц в популяции
мухоловки-пеструшки в годы с дружным началом размно-
жения.
1 - 1965 г. (п = 145); 2 - 1981 г. (п = 79); 3- 1983 г.
(п = 112). По оси абсцисс - календарь; по оси ординат -
доля от общего числа гнезд (%).
в обе стороны от пика массовой кладки. Эти откло-
нения особенно велики у видов с большой продолжи-
тельностью репродуктивного периода, а также в юж-
ных частях ареалов, где в отдельные годы пики вообще сглажены и растянуты во вре-
мени (рис. 16). В Карелии, например, на протяжении 1—2 пятидневок могут начать
откладку яиц до 80% гнездовых пар (мухоловка-пеструшка — 1965, 1983 гг., веснич-
ка — большинство лет наблюдений). Иногда более 80 % пар приступают к размножению
в 6-дневный период (рис. 19).
При полициклии пики начала откладки яиц всегда более сглажены и растянуты.
Например, максимальное число гнездовых пар зарянок, приступивших к гнездованию
в течение одной декады, не превышало 42% (1982 г.), а обычно было меньшим: 33.3
(1979 г.), 34.3 (1978 г.) и 30.1 % по суммарным данным для 303 гнезд. Для белобро-
вика максимум за декаду по всем гнездам (п = 1186) составил 30.9%, а в отдельные
годы - 26.9—37.5% (1972 и 1975 гг.). Даже в годы задержки пика откладки яиц до
3-й декады мая этот показатель не превышал 30 %.
Несмотря на то что в таежной зоне птицы обычно начинают размножение более
дружно, отклонения в сроках откладки яиц у отдельных пар превышают 2-3 недели.
Как правило, гнездование таких птиц начинается и проходит при менее стабильных
условиях среды, особенно ранней весной. В основном это определяется неустойчи-
востью погоды в мае—первой половине июня, а также в августе. В таежной зоне раннее
(до пика) размножение оказывается в отдельные годы полностью или почти полностью
неуспешным. Но иногда даже значительное опережение обычных сроков размножения
вида (чечетка) не сказывается отрицательно на его результатах.
Примеры влияния на птиц весеннего возврата холодов общеизвестны, а для ис-
следуемой территории приводились выше. Позднеиюльские и агустовские похолодания
в южной тайге встречаются реже, но вполне обычны в более высоких широтах. В 1978 г.
прорыв масс холодного арктического воздуха, сопровождавшийся сильными ветрами
и дождем, произошел в конце июля—начале августа, когда в гнездах1 зарянок, дроздов,
теньковок, славок, серых мухоловок, ласточек и др. еще были птенцы. На дроздах —
черном и белобровике, садовой и черногловой славках холодная погода отразилась
в меньшей степени, чем на остальных видах, так как недостаток беспозвоночных они
легко компенсировали ягодами черники. Черникой пытались кормить птенцов также
зарянки и даже теньковки, но ягоды они приносили не раздавленными и маленькие
птенцы не могли их проглотить. В гнездах ласточек, зарянок и мухоловок выводки
в эти дни погибли практически полностью. Для деревенских ласточек, кроме того, была
отмечена гибель слетков и самостоятельно кормящихся сеголетков.
Летние погодные аномалии в отличие от весенних, отмечаемых ежегодно, в южной
тайге наблюдаются раз в 7—8 лет. Однако к северу они значительно учащаются, поэтому
здесь особенно очевиден биологический смысл общей растянутости периода размноже-
ния и индивидуальной неодновременности его начала.
Как было показано выше, птицы, сроки гнездования которых отклоняются от
пика его начала в популяции более чем на 7—10 дней, могут составлять от 10 до 70%
местного гнездового населения.
Лэк (Lack, 1957), а вслед за ним и другие орнитологи, основываясь на данных по
возврату птенцов из ранних и поздних выводков на места рождения, пришли к заключе-
нию, что позднее гнездование, помимо низкой продуктивности, отличается повышенной
смертностью молодых птиц и поэтому элиминируется естественным отбором. Было
60
Рис. 20. Величина возврата молодых весничек, околь-
цованных в разные сроки (Гумбарицы, 1969-1976 гг.).
1 - доля птиц, вернувшихся на место прошлогоднего
кольцевания, где они были мечены в соответствующую
пятидневку (п = 210); 2, 3 - динамика стлова местных (2)
и пролетных сеголетков (и = 9257); 4-5 - период встречае-
мости местных сеголетков, в том числе их массового отлета
(4) (по: Н. В. Лапшин, устное сообщение).По оси абсцисс -
календарь; по оси ординат - доля вернувшихся и отловлен-
ных птиц (%).
предложено и соответствующее объяснение этому: °—©7 •-—•2 о—□ 5 ------4-----5
родившиеся в поздние сроки молодые птицы не успевают вовремя завершить постюве-
нильную линьку, подготовиться к миграции и до наступления неблагоприятных условий
оказаться на местах зимовок. Все эти опасности в равной мере относили и к взрослым
особям, завершающим гнездование в поздние сроки.
При жизни в неустойчивой среде обитания, когда понижение температуры воздуха
и ухудшение условий питания могут наступить в любой момент, неодновременность
начала прилета, гнездования и линьки отдельных особей внутри популяции приобретает
существенное значение для поддержания численности. Успешность гнездования птиц
в течение репродуктивного периода очень непостоянна, а ее колебания в разные годы
асинхронны. Снижение продуктивности размножения из-за непостоянства погоды про-
исходит как в начале и конце гнездового сезона, так и в периоды массового выкарм-
ливания птенцов. В последнем случае воспроизводство популяции осуществляется
в большей мере за счет более раннего и более позднего размножения, особенно когда
восстановление кладок уже невозможно. Кроме того, на севере позднее размножение
чаще проходит при более стабильных условиях и нередко оказывается успешнее, чем
в другие сроки. Часть гнездовых потерь компенсируется повторным размножением,
но на севере при дефиците времени оно никогда не бывает массовым, особенно если
ухудшение условий сопровождалось гибелью уже больших птенцов. В таких случаях
большинство особей моноциклическх видов птиц оказывается неспособным к про-
должению размножения. Так, в Прионежье из-за затяжных дождей, вызвавших мас-
совую гибель птенцов мухоловки-пеструшки (1973, 1976 гг.) и веснички (1976 г.),
к повторному гнездованию приступили лишь единицы. Основной прирост популяции
был достигнут за счет более успешного размножения пар, начавших гнездование чуть
позднее остальных. В итоге общая! продуктивность снизилась не так резко, как это
могло бы произойти при одновременном начале откладки яиц в популяции.
При полициклии неуспешность первого размножения приводит к появлению боль-
шого числа повторных кладок, но это снижает возможность участия птиц в воспитании
второго выводка. В июне 1982 г. в Приладожье похолодание совпало с периодом мас-
сового вылупления птенцов зарянки, зяблика, крапивника и других птиц. Во многих
гнездах выводки погибли полностью. В более выгодном положении оказались самки,
приступившие к откладке яиц на 10—15 сут позднее. Холодная погода совпала у них
с периодом насиживания кладки, а наступившее затем потепление способствовало
благополучному выкармливанию выводков. Среди зарянок, имевших в этом году вто-
рые выводки, явно преобладали птицы, начавшие гнездиться позднее, но успешно
выкормившие первые выводки. К сожалению, в наших широтах величина возврата
птиц, окольцованных птенцами, очень низка (рис. 20), и это является серьезным препят-
ствием для анализа различий в выживаемости птиц из ранних и поздних выводков.
Так, у мухоловки-пеструшки величина возврата на родину оказывается слабо связан-
ной с периодом вылупления птенцов.
Период вылупления до 10.VI 11-20.VI 21-30.VI 1-10.VII > 11.VII
Окольцовано птенцов 441 905 436 61 12
Обнаружено на родине
в следующие годы (%) 1.4 1.5 1.6 1.6 -
61
Средняя дата рождения 42 весничек, вернувшихся на родину, 20.5 июня, а для
всех известных случаев за соответствующие годы наблюдений — 23.2 июня. Для мухо-
ловки-пеструшки эти даты оказались практически одинаковыми — 16.5 и 16.7 июня
соответственно. Даты рождения (21 .V—10.VII, п =8) и кольцевания (25.V— 9.VII, п -14)
птенцов белобровика, вернувшихся на родину или обнаруженных (без случаев гибели)
на зимовках, также охватывают практически весь период их гнездования, за исключе-
нием самых крайних значений. Возвраты птенцов других видов птиц еще малочислен-
нее, в большинстве случаев возвращаются особи, родившиеся во время пиков мас-
сового вылупления вида. При редких возврата^ птенцов наших воробьиных вероятность
их повторного обнаружения во многом будет зависеть от исходного числа особей,
маркированных в разные периоды гнездового сезона. Например, для повторного обна-
ружения хотя бы одного птенца необходимо окольцовывать не 12, как в нашем случае
с мухоловкой-пеструшкой, а по крайней мере 60 птенцов, так как величина возврата
составляет лишь 1.6 %.
Выявленное Лэком (Lack, 1957, 1966) правило о повышенной смертности птиц из
поздних выводков по сравнению с ранними вряд ли можно считать общим для всех
популяций. Если бы высокий уровень смертности особей, родившихся (гнездившихся)
в поздние сроки, проявлялся постоянно и повсеместно, то в итоге это привело бы
к сокращению продолжительности репродуктивного периода, что вряд ли полезно
в неустойчивой среде обитания. Раскрывая механизм элиминации „поздних” особей,
Лэк исходил из того, что на завершение всех послегнездовых фаз годового цикла
они должны затрачивать столько же времени, как и выкормленные в ранние и нормаль-
ные сроки. Между тем подробное изучение годовых циклов воробьиных птиц Прила-
дожья и Прионежья показало, что у особей из поздних гнезд все фазы годового цикла
идут ускоренными темпами за счет их совмещения, а также смещения постювенильной
линьки на более ранний возраст и сокращения ее полноты. В результате „поздние”
особи завершают подготовку к миграции почти одновременно с „ранними”. Таким
образом, несмотря на растянутость гнездового сезона, окончание послегнездовых
явлений годового цикла в популяции синхронизируется.
Трудно сказать, является ли возникновение подобных адаптаций специфической
особенностью птиц северных популяций, ощущающих дефицит времени особенно остро,
или они присущи и птицам других широт. Но сам факт их существования, на наш
взгляд, говорит об отсутствии элиминации молодых птиц позднего размножения. В про-
тивном случае описанные выше адаптации никогда не возникли бы и не были бы закреп-
лены отбором у многих видов наших птиц.
При неустойчивости среды обитания и резко выраженной сезонности условий,
благоприятных для выведения потомства, попытки части особей гнездиться вне периода
массового размножения могут оказаться полезными для вида. Они способствуют расши-
рению диапазона возможных сроков гнездования и более полному использованию
короткого вегетационного периода, характерного для таежной зоны.
Связь сроков размножения птиц
с сезонной цикличностью развития биогеоценозов
В процессе совместной эволюции тесно связанные между собой компоненты при-
родных сообществ выработали однозначные реакции на изменения внешних факторов.
В северных климатических зонах при резко выраженной сезонности годовые циклы
всех растений и животных приобрели однотипную схему, в сроках, продолжитель-
ности и характере протекания последовательных фаз которой решающую роль играют
изменения одних и- тех же первичных факторов среды — света, тепла и влаги, У всех
объектов живой природы - от свободноживущих микроорганизмов до высших
позвоночных животных -- наиболее активные фазы роста, развития, размножения
приурочены к периодам, максимально обеспечивающим эти процессы энергией. Для
62
подавляющего большинства организмов таким периодом является весенне-летний
сезон. Осенью, как правило, происходит сокращение или прекращение различных энер-
гоемких процессов, замедление роста и развития организмов, непосредственно завися-
щих от действия внешних факторов. У многих растений и животных осенний сезон
связан с подготовкой к зимним условиям, накоплением энергетических резервов,
миграциями и т. п. В период минимального поступления солнечной энергии, при низких
температурах среды, у них наступает период относительного покоя, энергозатраты све-
дены до минимума.
Жизнь любых организмов подчинена строгой сезонной цикличности, и в группах
прямо или опосредованно связанных между собой живых компонентов годовая цик-
личность полностью синхронизирована. В условиях неустойчивого климата умеренных
и северных широт такая синхронность может быть достигнута лишь при однозначных
реакциях на изменение внешних факторов. В связи с этим уместно сопоставить основ-
ные закономерности сезонной цикличности воробьиных птиц и нижних звеньев их
трофоценотических цепей — насекомых и растений. Для птиц это более или менее под-
робно рассмотрено выше. Среди разнообразных сведений по экологии насекомых для
нас представляют особый интерес некоторые наиболее общие особенности их биологии,
связанные с сезонностью и неустойчивостью климата умеренных и северных широт
(Данилевский, 1961; Гиляров, 1970; Бей-Биенко, 1971; Воронцов, 1975).
Нижний температурный порог, определяющий минимальную активность и начало
развития лесных членистоногих, варьирует от -1 °C (зимняя пяденица) до +10 °C.
При снижении температуры среды ниже порогового значения они впадают в состояние
холодового оцепенения или депрессии. Примерно’ в тех же температурных пределах
расположена и зона начала вегетации лесных растений. Для большинства из них поро-
говая температура почвы +5 °C, воздуха +10 °C, ниже этих значений вегетация прекра-
щается.
Температура от +15 до +30 °C для многих видов насекомых и растений является
зоной наиболее активной жизнедеятельности, быстрого роста и развития, интенсивного
размножения или цветения. Для полного развития тех и других необходима определен-
ная сумма эффективных температур (произведение разности между средним показате-
лем за каждый день и нижним пороговым значением начала развития на число дней
развития). Сумма эффективных температур видоспецифична как для членистоногих,
так и для растений, а скорость и длительность развития находятся в прямой зависи-
мости от температуры (рис. 21, 22).
Фаза яйца у насекомых, размножающихся в весенне-летний период, длится от
2-3 сут (некоторые мухи) до 2-3 недель (пяденицы, коконопряды, совки, пилильщики
и др.). Темпы развития яиц во многом определяются температурой среды (рис. 22),
режимом влажности и другими факторами. Гусеницы и личинки членистоногих разви-
ваются от 2—3 недель до нескольких лет. Крупные гусеницы пядениц полное развитие
завершают в возрасте 50—60 сут, мелкие — 30—33 сут. Период от начала цветения лес-
ных и луговых трав до появления семян составляет от 2 до 4—5 недель.
Время начала и окончания отдельных фаз развития насекомых и растений в преде-
лах популяций растянуто. Например, выплод гусениц соснового шелкопряда происхо-
дит в течение месяца; лет сосновой пяденицы начинается в мае и продолжается весь
июнь, а при холодной погоде и июль; откладка яиц у сосновой совки обычно продолжа-
ется 3—4 недели (Гусев и др., 1938). Гусеницы сосновой пяденицы встречаются в приро-
де с июня до глубокой осени (Воронцов, 1975). Аналогичные данные можно привести
и для любого вида растений: неодновременность начала вегетации,- цветения и плодоно-
шения обусловлена у них экологическими различиями мест произрастания (почва, свет,
влажность, экспозиция склона, глубина залегания семян и т. д.).
Любое сезонное явление у растений и животных проходит однотипно — по прин-
ципу кривой нормального распределения. Она может идти более круто или полого,
иметь разрывы или несколько вершин, но общая тенденция повышения от минимума
к средней, а затем снижения присуща динамике развития любых сезонных явлений.
63
Рис. 21. Зависимость развития насекомых от температуры (по: Кожанчиков, 1961).
1 - продолжительность развития; 2 - скорость развития; 3 - температурный порог начала
развития. По оси абсцисс - длительность развития (а, сут), скорость развития (б, %); по оси орди-
нат - температуры (°C).
Рис. 22. Зависимость времени развития яиц серой волосистой пяденицы от температуры (по:
Голосова, 1964, - цит. по: Воронцов, 1975).
По оси абсцисс - температура (°C); по оси ординат - длительность развития (сут). Преры-
вистая линия - сумма эффективных температур (°C за сут).
Так же как и у птиц, среди объектов их питания — беспозвоночных животных и
растений — ежегодно сохраняется определенная видовая очередность во времени начала
сезонных явлений. В зависимости от характера весны фенодаты могут ежегодно сме-
щаться в обе строны от средних многолетних, но независимо от этого последователь-
ность начала вегетации различных видов растений, лета насекомых или гнездования
птиц сохраняется.
Относительная разновременность созревания семян различных растений и пиков
обилия насекомых приводит к формированию периода максимального изобилия и раз-
нообразия пищевых ресурсов птиц. В течение этого периода пики биомассы различных
пищевых объектов птиц, сменяя друг друга, образуют как бы один растянутый во вре-
мени пик максимального в году количества пищи. Первоначально он формируется за
счет небольшого числа видов растений и беспозвоночных животных, активные фазы
жизнедеятельности которых приурочены к ранневесеннему периоду с относительно не-
высокими температурами; затем в июне—июле (наиболее теплые месяцы) достигает
максимума, а осенью идет на спад.
Как известно, абсолютное большинство птиц являются полифагами. Это характерно
и для зерноядных, и для насекомоядных птиц, наблюдается и при питании взрослых
особей, и при выкармливании птенцов. Среди обширного набора кормов есть предпо-
читаемые, второстепенные и случайные для птиц виды пищи, причем отсутствие или
снижение обилия одних они легко компенсируют переходом на другие, более много-
численные в данный период кормовые объекты (Прокофьева, 1958, 1961, 1963, 1966,
1979; Мальчевский, 1959; Королькова, 1963; Иноземцев, 1978, и др.). Выше уже при-
водились примеры пластичности птиц в этом отношении.
Относительная полифагия большинства птиц и разновременность пиков обилия
и разнообразия объектов их питания являются предпосылкой для более или менее
успешного выкармливания птенцов на протяжении всего весенне-летнего периода.
Синхронность сезонного развития взаимосвязанных компонентов биоценоза ока-
зывается важным условием их совместного существования. Справедливость гипотезы
(Lack, 1957) о соответствии периодов выкармливания птенцов и максимального обилия
пригодной для этого пищи доказана как самим автором, так и его многочисленными
последователями для многих видов птиц, обитающих в самых разнообразных зонах.
Но, как отмечает Лэк, пик численности гусениц, служащих основным кормом для птен-
цов, наступает много позднее того момента, когда происходит откладка яиц, и не-
посредственным сигналом к началу размножения данный вид корма служить
64
не может. Для этого должны существовать какие-то другие сигнальные факторы, сти-
мулирующие формирование и откладку яиц, тем более что и пики, и начало гнездова-
ния ежегодно варьируют. Чтобы попытаться понять, каким образом птицы определяют
время начала размножения так, чтобы пики вылупления птенцов и обилия пригодной
для них пищи совпадали, необходимо связать воедино ряд общеизвестных фактов.
Как уже говорилось, всем компонентам любого биогеоценоза, объединенным
в трофоценотические цепи, свойственны единая годовая цикличность, обусловленная
резко выраженной в умеренных и северных широтах сезонностью климата. Все сезон-
ные явления в пределах популяции любых организмов всегда развиваются по принци-
пу кривой нормального распределения, причем неодновременность начала каждой
фазы у отдельных особей определяет большую или меньшую растянутость ее во вре-
мени. Разнообразие реакций различных организмов на сезонную изменчивость внешних
факторов (разная пороговая чувствительность) определяет строгую последовательность
начала и очередность пиков любых фенологических явлений в течение года. При этом
адаптации взаимосвязанных организмов к сезонной изменчивости внешних факторов
однозначны и синхронизированы в процессе совместной эволюции, без чего растения
и животные, составляющие трофоценотические цепи, существовать не могут.
В приведенных выше примерах обращает на себя внимание значительное сходство
в продолжительности некоторых фаз развития. У большинства размножающихся весной
насекомых личинки развиваются в яйце 2—3 недели, у цветущих в конце апреля—мае
травянистых растений такой же срок требуется на период от начала цветения до созре-
вания семян. Столько же времени занимает у воробьиных оогенез, откладка и насижива-
ние яиц. На полное развитие гусениц чешуекрылых необходимо при обычном темпера-
турном режиме 30—60 сут, и примерно столько же требуется воробьиным птицам на
гнездовое и послегнездовое развитие до приобретения самостоятельности. Нижние
пороги чувствительности и норма реакции на изменение внешних факторов у взаимо-
связанных организмов примерно одинаковы, что обеспечивает ежегодное совпадение
определенных фаз их фенологического развития.
Десинхронизация годовых циклов в пищевых цепях иногда имеет место, и приме-
ры ее хорошо известны энтомологам. Так, в отдельные годы массовый выход гусениц
дубовой листовертки на несколько дней опережает распускание почек дуба, что приво-
дит к увеличению смертности и снижению численности гусениц (Вороноцов, 1975).
Но из-за неодновременности выплода определенная часть гусениц все же появляется
одновременно с распусканием почек^ так что десинхронизация не бывает полной. Такие
несовпадения в фазах развития взаимосвязанных организмов строго контролируются
естественным отбором и постоянно элиминируются. Кроме того, у многих видов насе-
комых, подобно птицам, развита полифагия, и при частичном несовпадении их циклов
с одним из кормовых растений они переходят на питание другими. Для птиц, у которых
полифагия является нормой пищевых связей, снижение численности отдельных видов
беспозвоночных существенного значения не имеет: недостаток одних видов корма
они легко компенсируют другими (примеры сезонной и ежегодной смены основных
кормов приводились выше). Это важно для сохранения лабильности сроков размноже-
ния птиц и независимости его начала от обилия (или даже наличия) именно тех видов
кормов, которые потом становятся основными для выкармливания птенцов. При сте-
нофагии, напротив, сроки размножения более жестко связаны с годовой цикличностью
пищевых объектов птиц.
В предыдущих разделах было показано, что размножение птиц начинается в мо-
мент, когда фотопериод, состояние погоды и обилие пищи энергетически обеспечивают
нормальное формирование половых продуктов самок, развитие выводящих путей,
наседного пятна, а также всех видов активности, связанных с постройкой гнезда. На
окончательное формирование семенников требуется гораздо меньше дополнительной
энергии, поэтому самцы приходят в состояние готовности к размножению гораздо
раньше самок — на более низких пороговых значениях всех факторов, влияющих на
суточный энергетический баланс. Самки за короткий период продуцируют кладку,
5 В. Б. Зимин
65
общая масса которой равна или на 40—60% превосходит их собственную массу. Кроме
того, в течение всего процесса формирования и откладки яиц у самок остаются увели-
( ченными размеры яйцевода, обводняются ткани наседного пятна. Поэтому масса их тела
в это время возрастает на 20—30%, что требует и соответственного увеличения энергети-
ческих затрат. У большинства воробьиных птиц дополнительной энергетической нагруз-
кой для самок является также строительство гнезда, требующее, по расчетам Дольника
с соавторами (1982), столько же энергии, сколько развитие яиц.
Как бы ни изменялось состояние весенней погоды, биоценотически связанные
растения и животные будут реагировать на это однозначно, а увеличение или уменьше-
ние темпов их развития будет происходить синхронно. У насекомых, например, сроки
появления и пики численности при определенных ежегодных ^вариациях в календарных
датах из года в год повторяются с одной и той же последовательностью и с приблизи-
тельно равными интервалами. Длительность таких интервалов может корректироваться
состоянием погоды и отклоняться на несколько дней от среднего значения, причем
это происходит достаточно синхронно у всех компонентов экосистемы. Поэтому период
массового появления кормов, необходимых для выкармливания птенцов, всегда насту-
пает на определенный срок позже пиков обилия тех видов беспозвоночных, уровень
численности которых обеспечил птицам начало откладки яиц.
Любые значительные изменения внешней среды, происходящие за время насижива-
ния кладки, могут несколько задержать или ускорить развитие насекомых, но вылупле-
ние птенцов произойдет через определенный срок после начала инкубации — независимо
от внешних воздействий. Иными словами, в этот период возможна некоторая десинхро-
низция сроков вылупления птенцов и массового появления пищи, пригодной для их
выкармливания. Абсолютной десинхронизации, однако, не произойдет. Этому будет
препятствовать неодновременность и растянутость всех явлений у любых компонентов
биоценоза. Поэтому если у какой-то части птиц вылупление птенцов придется на начало
появления основных кормов или даже опередит его, то у других совпадет с пиком
обилия, а у третьих — с периодом его снижения. Недостаток основных кормов в любом
случае восполняется второстепенными. Кратковременные возвраты холодов не нару-
шают совпадения пиков вылупления птенцов и обилия их пищи. В то же время длитель-
ные глубокие похолодания одинаково губительны и для птиц, и для объектов их пита-
ния. При повторном размножении птицы переходят на другие виды кормов, которых
в наших условиях всегда вполне хватает. Об этом свидетельствует достаточно хорошая
выживаемость птенцов, нередко даже более высокая, чем в ранние и нормальные сроки
вылупления.
Годовые циклы и особенности распространения птиц
Время, затрачиваемое на каждую из фаз годового цикла — миграции, гнездование,
линьку и зимовку, — у разных видов птиц умеренных широт неодинаково. Соотноше-
ние и продолжительность этих фаз значительно варьируют и в пределах ареала каждого
вида, и индивидуально. Межвидовые различия в распределении времени, приходящегося
на каждую фазу, хорошо известны для так называемых оседлых птиц и мигрантов
на ближние, средние и дальние расстояния, для птиц с моно- и полициклическим типом
размножения, с одной или несколькими линьками в год и т. д.
У птиц, адаптированных к жизни в суровых условиях таежной зимы, отпадает
необходимость в миграциях, что позволяет некоторым из них увеличить продолжитель-
ность других фаз годового цикла — размножения и линьки. В первом случае это реали-
зуется выкармливанием дополнительных выводков — размножение становится поли-
циклическим. Во втором — появляется возможность сделать линьку энергетически ме-
нее напряженной, растянув ее во времени.
У дальних мигрантов годовой цикл проходит более напряженно в связи с тем, что
большую часть года (до 300 сут) они проводят в пути и на зимовках. Время пребывания
66
их на местах размножения сокращается до 2—2.5 мес и занято лишь выведением потом-
ства, тогда как линька целиком переносится на зимовку (чечевица, дубровник). Лишь
немногие дальние мигранты имеют полную послебрачную и частичную постювенильную
смену оперения на родине и при этом осваивают высокие субарктические широты
(весничка). У большинства других мигрантов на дальние расстояния линька на родине
ограничивается лишь сменой контурного оперения.
Нетрудно заметить, что основные различия в годовых циклах воробьиных птиц
умеренных и высоких широт связаны со степенью разобщенности мест их гнездования
и зимовок и сосредоточены на фазе линьки. Именно смена покровов даже среди близ-
кородственных видов птиц достигает максимального разнообразия. Крайний полимор-
физм и неоднозначность проявления адаптаций к данному этапу годового цикла — чис-
ло линек в году, их объем, темпы, приуроченность к разным сезонам года, местам
гнездования или зимовки, существенные географические, меж- и внутрипопуляционные
различия — подчеркивают огромную важность этого процесса для птиц. Как уже указы-
валось, на период смены оперения приходится самый высокий уровень обмена веществ
в году — это самая энергоемкая фаза годового цикла, для успешного завершения кото-
рой необходимы благоприятные условия (Дольник, 1967, 1975). Эти последние, кроме
отсутствия в данный период других энергоемких процессов, определяются обилием
пищи и относительной стабильностью внешних факторов. Следует подчеркнуть, что
именно линька явилась для естественного отбора наиболее благодатным полем деятель-
ности, предоставляющим богатые возможности для разнообразных способов подгонки
фаз годового цикла птиц к сезонности климата в соответствии с биологическими осо-
бенностями каждого вида. Действительно, размножение у абсолютного большинства
видов птиц проходит только на родине, не прерывается, не переносится на другой сезон
или на место зимовки (исключение — полизоналыюе и полисезонное размножение птиц,
потребителей семян хвойных деревьев). Относительно постоянна продолжительность
эмбрионального периода и первых этапов постэмбрионального развития. От дальности
миграций зависит время, затрачиваемое на преодоление пролетных путей, а перенос
зимовок в более северные широты требует возникновения адаптаций к жизни в суро-
вых и неустойчивых условиях. Продолжительность зимовки также определяется стро-
гой сезонностью климата, спецификой питания и кормодобывания вида. Адап-
тации к сезонности климата и связанные с этим особенности биологии и распро-
странения птиц развивались, разумеется, на всех этапах годового цикла, но имен-
но линька оказалась наиболее мобильной, податливой к подобным изменениям
фазой.
Продолжительность различных фаз годового цикла значительно варьирует в преде-
лах ареала — вплоть до того, что отдельные из них, например миграции, могут пол-
ностью или частично исключаться. Известно немало видов, у которых южные популяции
оседлы, а северные совершают регулярные миграции и зимуют на значительном удале-
нии от мест размножения. Зарянка, дрозды, лесная завирушка и другие воробьиные
в условиях мягкого морского климата Англии оседлы, а птицы северных популяций
отлетают на зиму в область Средиземноморья, и только единичные особи иногда пыта-
ются зимовать в южной и средней тайге. Чем дальше от мест зимовок продвинут гнездо-
вой ареал вида, тем больше времени приходится затрачивать на миграции. Даже с учетом
некоторых различий в размещении на зимовках северных и южных популяций чечеви
цам или дубровникам из юго-восточных и северо-западных частей гнездового ареала
приходится преодолевать совершенно разные расстояния во время весенних и осенних
миграций.
Внутрипопуляционные вариации продолжительности отдельных фаз годового
цикла менее значительны. Они возникают за счет половых и возрастных различий в сро-
ках прилета, начала и окончания гнездования, разных темпов и объема линьки в зави-
симости от времени ее начала, различий в продолжительности репродуктивного периода
у птиц с полициклическим гнездованием, выкармливающих один или несколько вывод-
ков, и т. п.
67
Рис. 23. Принципиальная схема формирования периферийных областей ареала и северных
границ распространения птиц.
А - формирование областей постоянного (Ог} йнерегулярного (О2) гнездования: стрелка -
северная граница Ох, крест в позиции III - элиминация особей при несоответствии времени Т сум-
марной продолжительности периода благоприятных условий на местах размножения и трассах
осенней миграции; Б - северные границы распространения птиц, адаптированных и неадапти-
рованных ) к дефициту времени в годовом цикле. По оси абсцисс - календарь; по оси орди-
нат - географическая широта. Остальные объяснения - в тексте.
Чем дальше к северу простирается область гнездования вида, тем короче период
благоприятных условий для размножения и линьки, тем относительно (фенологически)
позднее начинается миграция, которая проходит при менее стабильной погоде и кормо-
вой базе. Любая задержка в завершении очередной фазы влечет за собой запаздывание
последующих этапов годового цикла, реализация которых будет вынужденно осуще-
ствляться при более низких температурах, меньшем обилии пищи и на более короткой
длине дня. Это может привести к тому, что птицы не успеют закончить выкармливание
птенцов, линьку или начать миграцию. Кроме того, очень важно, чтобы благоприятные
условия сохранялись не только во время пребывания на родине, но и по всей трассе
пролета к зимовкам. Это тем более необходимо, что все фазы, приходящиеся на этот
период, требуют повышенных затрат энергии, а следовательно, и постоянного потребле-
ния пищи.
Если на осях координат отложить календарь и географическую широту, то графи-
чески широтная изменчивость продолжительности периода благоприятных для жизни
птиц условий будет выражаться двумя линиями, сходящимися к Северу (рис. 23, Л).
Представленная схема отражает лишь общий принцип широтного сокращения этого
периода, но в целом соответствует географической изменчивости конкретных
68
климатических и фенологических показателей, которые были рассмотрены для тер-
ритории, ограниченной 40—70° с. ш. и 30—36° в. д. (рис. 5—9).
На завершение всех летне-осенних фаз годового цикла птицам требуется опреде-
ленное время Т (рис. 23, А). Оно складывается из затрат времени на гнездование, линь-
ку и миграции, т. е. Т = Тг + Тп + Тм.
По мере увеличения географической широты период гнездования сокращается
в основном за счет более поздних календарных сроков его начала при сохранении вре-
мени окончания. При этом возрастает дальность миграций и соответственно необходи-
мое для этого время Тм.
С расширением ареала вида к северу в зоне II наступает момент, когда Т оказыва-
ется примерно равным по продолжительности периоду благоприятных условий на
местах гнездования и по всей трассе миграций. В данном случае птицы успевают попасть
на места зимовки до наступления неблагоприятных условий. Очевидно, зона II и соот-
ветствует той широте, по которой проходит северная граница области постоянного
гнездования вида, где местное население птиц может поддерживаться за счет внутри-
популяционных ресурсов.
В зоне III размножение, линька и даже начало миграции могут проходить при
благоприятных условиях, но конец миграционной трассы птицы вынуждены будут
преодолевать после наступления неблагоприятных условий, что вряд ли может завер-
шиться успешным попаданием на зимовки. Такие поздние задержки насекомоядных
птиц, мигрантов на дальние и средние расстояния, мы ежегодно регистрируем визуально
при отлове крупногабаритными ловушками и паутинными сетями в конце сентября-
октябре. В этот период температура часто опускается ниже 0 °C и иногда выпадает снег,
а птицы находятся фактически в самом начале миграционного пути. При дефиците
кормов в этот период им вряд ли удастся накопить необходимое для длительного поле-
та количество энергетических резервов.
Случаи, подобные представленнному для зоны III, мы и наблюдаем в периферий-
ных областях ареалов многих видов птиц. Здесь отмечаются лишь попытки вида (у не-
которых птиц достаточно регулярные) расширить ареал. Они могут иметь место и в бо-
лее северных зонах, если этому благоприятствуют, хотя бы временно, внешние условия
(весенняя пролонгация миграции). На завершение гнездования воробьиным требуется
всего 1.5 мес — срок вполне достаточный даже для короткого лета высоких широт.
Но без соответствующих адаптаций к дефициту времени короткого вегетационного
периода северных широт может не хватить даже на завершение линьки. Поэтому на тер-
риториях, расположенных севернее зоны II, местное население птиц при каждой попытке
выхода за пределы основного ареала в большинстве случаев должно формироваться за-
ново из птиц, родившихся южнее. Это мы и наблюдаем в периферийных зонах ареалов
многих воробьиных птиц на Северо-Западе СССР.
Так как период благоприятных условий прежде всего определяется состоянием
погоды, то там, где она неустойчива, его продолжительность может ежеюдно меняться,
причем на всем пространстве от гнездового до зимовочного ареала это происходит чаще
всего асинхронно. Такие изменения могут быть одной из причин пульсации границ
ареала и его периферийных зон. На схеме, во избежание ее усложнения, эти и другие
детали не приводятся. Она отражает лишь основной принцип связи годовых циклов
птиц с их распространением (без учета географической долготы и высоты над уровнем
моря).
Закрепление вида на широтах, расположенных севернее области постоянного
гнездования, возможно лишь после возникновения адаптаций к преодолению дефицита
времени. На схеме, это поясняющей (рис. 23, Б), для упрощения и наглядности весь
отсчет времени дан от начала гнездования (Го) до окончания осенних миграций (Гк).
Соответственно Тх и Т2 — это суммарные затраты времени на гнездование, линьку и
осенние миграции. Линия ВХВ2 обозначает широтное изменение продолжительности
периода благоприятных условий на месте размножения и трассе осенних миграций
(суммарно). OQ — летне-осенние фазы годового цикла на юге ареала, Ох — то же
69
у северной границы области постоянного гнездования птиц, не адаптированных к дефи-
циту времени; Ог — для птиц, адаптированных к дефициту времени. На схеме видно,
что совмещение последовательных фаз годового цикла, даже при увеличении даль-
ности миграций и затрачиваемого на них времени, позволяет птицам, адаптированным
к короткому периоду благоприятных условий, осваивать значительно более северные
широты по сравнению с теми, у которых все фазы годового цикла разобщены по клас-
сической схеме, типичной для средних и южных широт.
Приведенные схемы упрощены и дают лишь самое общее представление о широт-
ных различиях в годовых циклах птиц. На самом деле общая продолжительность пре-
бывания птиц северных популяций на местах размножения сокращается не только
за счет совмещения основных фаз. На севере, например, много короче предгнездовой
период, точнее,— время от прилета до начала гнездования (Новиков, 1944, 1952; Дани-
лов, 1966). Скворцы в Средней Азии появляются на местах гнездования в феврале,
а к откладке яиц приступают только во второй половине апреля (Сема, 1978). В Каре-
лии этот период не превышает 22—30 дней. Предгнездовой период у весничек на Айно-
вых островах Баренцева моря занимает около 2 недель, в южной тайге — 3—3.5 недели,
а в Англии тянется более месяца (Cramp, 1955; Лапшин, 1976; Чемякин, 1982а).
При относительно постоянной для разных зон продолжительности периода гнездо-
вания, откладки яиц, насиживания и выкармливания птенцов птицы северных широт
не получают значительного выигрыша во времени. Правда, отмеченная выше специфика
суточного бюджета энергии, обусловленная особым фотопериодом, позволяет совме-
щать процессы формирования яиц и строительства гнезда. Благодаря этому окончание
постройки гнезда совпадает с откладкой первого яйца и более характерная для южных
широт стадия просушки гнезда выпадает. У некоторых видов, как уже отмечалось,
окончательное оформление выстилки лотка идет одновременно с откладкой яиц (сини-
цы, пеночки, ласточки, крапивник, королек). Но, с другой стороны, обычные на севере
возвраты холодов снижают темпы, а нередко приводят к более или менее длительным
перерывам в гнездостроении и откладке яиц. Линд (Lind, 1960) установил, что в Фин-
ской Лапландии из-за частых отлучек с гнезд во время холодной погоды у деревенских
ласточек удлиняется период инкубации яиц. Для других видов подобной связи, однако,
не выявлено. Ускоренного развития птенцов в условиях круглосуточного полярного
дня, о котором пишут многие авторы, при более строгом подходе не выявляется или
различия оказываются статистически недостоверными. Таким образом, продолжитель-
ность одного гнездового цикла от начала откладки яиц до распадения выводка и на
севере, и на юге ареала оказывается примерно одинаковой.
Принято считать, что полицикличность размножения свойственна лишь птицам
средних и южных широт, а в северных частях ареала невозможна (Lack, 1957; Ковшарь,
1981а, 1982; Ильичев и др., 1982, и др.). Между тем популяционно-экологические
исследования с применением отлова, мечения и контроля местного гнездового населе-
ния птиц на Северо-Западе СССР дали достаточно строгие доказательства двукратного
выведения потомства птицами, для которых в более южных зонах полициклическое
размножение является нормой (см. гл. IV). В отличие от птиц южных широт, у которых
между вылетом птенцов первого выводка и началом новой кладки проходит не менее
двух недель, на севере оба цикла, как правило, совмещены. Так же, как и в горах
(Ковшарь, 1981а), этим достигается значительный выигрыш во времени. Другие адапта-
ции к дефициту времени у птиц с полициклическим размножением, как и у остальных,
касаются в основном линьки — ее совмещения с гнездованием и началом миграций,
меньшего объема, высоких темпов. В результате птицы, обитающие в северных частях
ареалов, успевают завершить все фазы годового цикла, приходящиеся на период их пре-
бывания на родине, на 1 — 1.5 мес быстрее, чем в южных широтах. Например, дубровник
в Южном Приморье встречается с 28 апреля-6 мая до конца августа, а единич-
ные особи регистрируются вплоть до октября (Панов, 1973). У северо-западных
пределов ареала эти овсянки появляются лишь 6—8 июня и отлетают к середи-
не августа.
70
Сокращение и совмещение весенне-осенних фаз годового цикла, разумеется, не
может быть беспредельным. На определенных широтах наступает такая продолжитель-
ность периода благоприятных условий, которая позволяет птицам своевременно попасть
на зимовки при минимальной суммарной затрате времени на успешное завершение
гнездования, линьки и миграций.
Стабильность гнездового населения птиц в любой точке ареала зависит от количе-
ства особей, выживших в течение года и возвратившихся на место предыдущего гнездо-
вания или рождения. Важна не только успешность предыдущего размножения, но и нор-
мальная для вида выживаемость на всех послегнездовых этапах годового цикла. В пери-
ферийных зонах ареала размножение может завершиться успешно, но если любая из
последующих фаз — становление самостоятельности, линька или миграции — хотя бы
частично выйдет за пределы периода благоприятных для вида условий, то результаты
воспроизводства будут сведены до минимума повышенной смертностью. Стабильность
внешних факторов имеет важное значение для выживаемости молодых птиц, начина-
ющих самостоятельную жизнь. Максимальный в году уровень энергозатрат приходит-
ся на период линьки, и ее успешное завершение требует достаточного обилия и доступ-
ности пищи. Накопление энергетических резервов, необходимых для совершения мигра-
ций, также невозможно без устойчивой кормовой базы на всей трассе пролета.
Следовательно, граница области постоянного гнездования вида должна проходить
на таком удалении от мест зимовки, при котором период благоприятных условий оста-
ется чуть большим или равным продолжительности летне-осенних фаз годо-
вого цикла птиц с учетом времени, необходимого на преодоление расстоя-
ния до мест зимовки. Разумеется, это справедливо лишь в том случае, если
распространение вида не ограничивается географическими или экологическими
барьерами.
За указанными пределами расположены все прочие структурные подразделения
периферийных зон ареалов - области нерегулярного и эпизодического гнездования,
летних встреч и пр. (Носков, 1976; Зимин, 1977; Кищинский, 1983). Наличие таких
зон отражает попытки вида расширить область своего распространения, но может быть
связано и с годовыми колебаниями продолжительности периода благоприятных для
птиц условий (ранняя весна и поздняя осень, поздняя весна и ранняя осень). Периоди-
ческое возникновение областей нерегулярного гнездования вида - основная причина
так называемых колебаний или пульсаций границ ареалов. Существование таких коле-
баний уже свидетельствует о неудачных попытках вида закрепиться на новых террито-
риях, о недостаточной адаптированное™ его к местным условиям. Факты гнездования
птиц за пределами основного ареала, пусть даже успешного, еще не означают, что этого
достаточно для поддержания численности вида на новых территориях. Успешное выве-
дение птенцов — это всего лишь одна реализованная фаза годового цикла. При дефиците
времени любой последующий этап может совпасть с наступлением неблагоприятных
условий. Здесь мы имеем частный случай проявления закона Либиха—Блэкмана: выжи-
ваемость птиц в течение года будет ниже нормальной для вида, если хотя бы одна из
послегнездовых фаз годового цикла совпадает с наступлением периода неблагоприят-
ных условий.
По данным массовых отловов в Приладожье, птицы северных популяций мигри-
руют гораздо позднее местных особей. Например, местные веснички исчезают из района
гнездования и рождения в середине августа — при обилии корма, на еще длинном фото-
периоде (15.5 С : 8.5 Т) и при относительно стабильных, близких к максималь-
ным в году среднесуточных температурах воздуха. Но пролет других особей
этого вида продолжается весь сентябрь, а отдельные из них покидают При-
ладожье в начале октября — на фотопериоде 14—9 С: 10—15 Т, при температурах,
быстро приближающихся к отрицательным значениям, и при явном недостатке
кормов.
При экспансии численность вида на новых территориях из года в год достаточно
стабильна или нарастает, и тогда происходит смещение северных границ области
71
постоянного гнездования. В периферийных зонах ареала образуются лишь времен-
ные поселения, судя по всему, ежегодно или с перерывом в несколько лет формиру-
ющиеся заново, а граница области постоянного гнездования сохраняется на прежнем
уровне. Встречи птиц и даже гнездовые находки севернее этих границ отражают лишь
попытки расширения ареала. Так как на новых территориях вид при этом не закрепля-
ется, то периферийные зоны ареалов птиц лучше выделять особо и обязательно в связи
с определенным историческим периодом. Степень освоенности новых территорий опре-
деляют не факты гнездования, а способность вида поддерживать на ней относительно
стабильную численность. Последнее обеспечивается приведением в соответствие уровней
воспроизводства и смертности, что, в свою очередь, достигается приобретением опре-
деленных адаптаций к короткому периоду благоприятных условий.
72
Г Л A В A III
ВЕЛИЧИНА КЛАДКИ И ЕЕ ИЗМЕНЧИВОСТЬ
Повышенный интерес к изучению величины кладки, степени ее постоянства и из-
менчивости внутри отдельных видов и популяций птиц проявляется в большом числе
публикаций по различным аспектам данной проблемы, в частности обширных обзоров,
отражающих разнообразие представлений и взглядов на эволюцию продуктивности
размножения птиц. С одной стороны, это свидетельствует о сложности самой проблемы,
с другой, — указывает на то, что ни одна из гипотез, объясняющих постоянство, изменчи-
вость и эволюцию величины кладки птиц, не является общепринятой.
Наибольшее признание получила концепция Лэка (Lack, 1947), согласно которой
птицы продуцируют столько яиц, сколько птенцов они способны выкормить в конкрет-
ной ситуации. Позднее возникло много новых теорий, в которых проблема величины
кладки решается в связи с энергией существования, расходуемой на репродуктивный
цикл (Cody, 1966), а также с выживаемостью взрослых особей (Williams, 1966), груп-
повым отбором (Wynne-Edwards, 1962), продолжительностью жизни (Haukioja, Hakala,
1979), взаимодействием хищника и жертвы (Ricklefs, 1970), различиями в фенологиче-
ских сроках размножения (Slagsvold, 1975а) и др.
Эти теории подробно рассмотрены на примере американских (Cody, 1966) и евро-
пейских птиц (Klomp, 1970), и нет смысла дублировать их основное содержание. Все
они, касаясь самой общей сущности проблемы в ее наиболее важном филогенетическом
аспекте, обосновывают эволюционную обусловленность величины кладки каждого
вида, но не позволяют понять, что служит внешними и внутренними сигналами к пре-
кращению дальнейшего продуцирования яиц самкой, после того как определенное число
их уже отложено. Известно, - и ниже это будет подтверждено на местном материале, -
что одна и та же особь в течение одного сезона и в разные годы жизни может откла-
дывать совершенно разное число яиц и что величина кладки у различных самок в один
и тот же период и на той же территории также неодинакова. Отсюда следует, что измен-
чивость величины кладки большинства видов птиц, которую Лэк (Lack, 1966) опреде-
лял как важную адаптацию к неоднородности и неустойчивости среды обитания, свой-
ственна не только виду в целом или популяциям, нои каждой конкретной особи. И коль
скоро существуют генетически закрепленные видовые или популяционные пределы
изменчивости размеров кладки, то в процессе эволюции должны были развиться и ме-
ханизмы своевременного прекращения откладки яиц, предотвращающие излишние
затраты энергии и времени на размножение и приводящие к относительному соответ-
ствию количества яиц числу птенцов, которых родители способны воспитать в данных
условиях (по Лэку-Вильямсу).
За последние 30 лет автором и его коллегами в таежной зоне Северо-Запада СССР
было обследовано более 10 тысяч гнезд преимущественно лесных воробьиных птиц.
Так как работа выполнялась стационарно, в большинстве гнезд была точно определена
величина полной кладки, успешность ее насиживания и судьба выводка. С 1970 г. на
Шокшинском стационаре и в Маячино применялся метод постоянного контроля гнездо-
вого населения (отлов взрослых птиц на гнездах и пробных площадях в течение гнездо-
вого и послегнездового периодов, кольцевание птенцов, цветное мечение и т. д.). Это
позволило изучить индивидуальную изменчивость величины кладки и других репро-
дуктивных показателей у маркированных птиц в течение одного сезона и 2-6-летнего
периода их жизни.
73
Внутри- и межвидовая изменчивость величины кладки
Изменчивость величины кладки изучалась преимущественно на территории Южной
Карелии: юго-восточное Приладожье у границ с Ленинградской обл., Прионежье (Шок-
шинский стационар и окрестности Петрозаводска), заповедник „Кивач” и прилежащие
к нему угодья Кондопожского района, острова Заонежья; частично использованы также
материалы по гнездованию птиц, собранные в Калевальском, Муезерском и Беломор-
ском районах Карельской АССР (табл. 18).
Среди северотаежных воробьиных птиц, по величине кладки которых имеется
сравнимый материал, явно преобладают виды, откладывающие не более 6 яиц. К их
числу относятся 37 видов: юла, полевой жаворонок, береговая ласточка, коньки —
луговой, лесной, краснозобый, горный; дрозды — деряба, певчий, черный; соловей,
сверчки — речной и обыкновенный; славки — черноголовая, ястребиная, садовая;
камышевки — дроздовидная, садовая, болотная, тростниковая; овсянки — обыкновен-
ная, садовая, дубровник, ремез, крошка, камышевая; щегол, чиж, чечевица, щур,
клесты — еловик, сосновик, белокрылый; дубонос, иволга, грач, кедровка.
Достаточно разнообразна (24 вида) и II группа птиц, у которых кладки из 7 яиц
и более встречаются сравнительно редко (0.1—10%). К этой группе в указанном регионе
относятся городская и деревенская ласточки; белая и желтая трясогузки, жулан, лес-
ная завирушка, дрозды — рябинник и белобровик; теньковка, пересмешка, славки —
серая и завирушка; камышевка-барсучок, серая и малая мухоловки, хохлатая синица,
зяблик, зеленушка, коноплянка, чечетка, снегирь, скворец, ворон, серая ворона.
В III группе птиц крупные кладки встречаются в 11—50% гнезд. В нее вошли 16
видов: большой сорокопут, крапивник, зарянка, горихвостка, каменка, весничка,
трещотка, таловка, зеленая пеночка, мухоловка-пеструшка, пищуха, вьюрок, домовой
и полевой воробьи, сойка, галка.
IV группу образуют птицы, у которых крупные кладки являются нормой и встре-
чаются более чем в 50% гнезд; всего 9 видов: луговой чекан, королек, ополовник,
сорока, синицы — большая, лазоревка, московка, пухляк, сероголовая гаичка.
Приведенные выше списки птиц и табл. 19 составлены на основании сведений,
известных для Ленинградской обл. (Мальчевский, Пукинский, 1983), Финляндии
(Haartinan, 1969а) и КАССР (наши данные).
Чтобы понять, с чем могут быть связаны существенные межвидовые различия
в величине кладки, мы попытались сравнить выделенные группы видов по дальности
миграций, числу выводкрв за сезон размножения и типу устройства и размещения
гнезд (табл. 19). Как видно из приведенных данных, доля кладок из 7 яиц и более не
имеет сколько-нибудь определенной связи ни с дальностью миграций, ни с числом
выводков за репродуктивный сезон. Но относительное число закрытогнездящихся
видов птиц заметно нарастает от первой группы к четвертой. Так, если в I группе видов
только береговая ласточка имеет гнезда закрытого типа, а остальные размещают их
открыто, то в IV группе, напротив, лишь королек вклинивается в ряд дуплогнездников
и „купольников”. По типу устройства и размещения гнезда и по степени укрытости
кладки королек гораздо ближе к „купольникам”, чем к типичным открытогнездникам.
Верхняя часть лотка в его гнезде сильно стянута внутрь, сама постройка подвешена
под еловой ветвью и закрыта сверху густыми вторичными побегами. Кроме того,
кладка снаружи абсолютно не видна из-за обильной перьевой выстилки, пополняемой
в процессе откладки яиц и даже в первые дни насиживания. Не исключено, что берего-
вая ласточка до недавнего времени гнездилась открыто на уступах береговых обрщвов.
Некоторые исключения из общего правила есть также во II и III группах, но и они впол-
не объяснимы.
Важно отметить, что тенденция к увеличению доли крупных кладок у закрыто-
гнездящихся птиц подтверждается и на внутривидовом уровне. Так, малая мухоловка
в европейской части ареала ведет себя как полудуплогнездник, а нередко строит гнезда
открыто - в основании ветвей деревьев или в кустообразной пристволовой поросли.
74
В Сибири же — это типичный дуплогнездник, один из наиболее массовых видов птиц,
привлекаемых развеской искусственных гнездований. Оказалось, что и средняя величи-
на кладки, и число гнезд, содержащих по 7 яиц и более, у сибирского подвида (по:
Крутовская, 1966) достоверно больше, чем у европейского (рассчитано по: Haartman,
1969а; Мальчевский, Пукинский, 1983; Aleknonis, 1984; наши данные).
Число гнезд lim М ± т Доля круп- ных кладок (%)
Сибирь, заповедник „Столбы” 40 2-9 7.35 ± 0.28 77.5 ± 6.6
Северная Европа 139 3-7 5.14 ± 0.07 5.0 ± 1.8
Лэк (Lack, 1968) разделил европейских птиц на две группы: 1-я — с кладками,
содержащими до 5 яиц включительно, и 2-я — с 6 яйцами и более. Рассчитав среднюю
продолжительность пребывания птенцов в гнезде, он нашел, что у птиц первой группы
она составила 13.5, а у видов второй 16.6 сут. В соответствии со степенью защищенности
величина кладки изменялась от 5.1 до 6.9 яйца. Аналогичные данные, но для американ-
ских видов птиц, получил и Коди (Cody, 1966). Длительный период пребывания птенцов
в гнезде увеличивает риск гибели ог разорения. Но поскольку гнезда дуплогнездников
лучше защищены от хищников, более длительное пребывание птенцов в дуплах позво-
ляет им покидать их уже способными к полету, что несомненно полезно для вида
(Познании, 1946).
Можно было бы предположить, что связь между продолжительностью выкарм-
ливания гнездовых птенцов и величиной кладки развивалась по следующей схеме:
с увеличением числа яиц (а соответственно и птенцов) сокращалось количество получае-
мой птенцами пищи, это приводило к замедлению темпов развития и удлинению перио-
да пребывания птенцов в гнезде. Однако наблюдения за развитием птенцов и экспери-
менты по искусственному увеличению размеров выводка убеждают нас в том, что
темпы развития птенцов у дуплогнездников не зависят от величины выводка. Кроме
того, формирование гнездового наряда у птенцов, отстающих в росте от остальных
(при асинхронном вылуплении масса тела бывает в 2—2.5 раза меньше), точно соответ-
ствует их возрасту. Таким образом, недостаток пищи приводит к резкому замедлению
скорости роста птенцов, но не отражается на темпах развития их оперения. Величина
кладки (выводка) птиц-дуплогнездников не связана с интенсивностью кормления
птенцов их родителями.
Отвлекающая демонстрация — отвод от гнезда, как правило, встречается только
у открытогнездящихся птиц. Для них больше, чем для дуплогнездников, характерно
и прерывистое насиживание (подсиживание) неоконченных кладок. В вольерах при
скученности птиц самки обыкновенной овсянки, зеленушки, снегиря, чижа, щегла
и чечетки всегда начинали плотно насиживать кладки с первого яйца. В природе плот-
ность насиживания возрастает по мере увеличения числа яиц в гнезде. Лишь у немно-
гих таежных видов воробьиных (врановые, клесты, чечетки) самки приступают к плот-
ному насиживанию с откладкой первых яиц, у большинства других это происходит
после появления последнего или предпоследнего яйца. Явление „подсиживания” нео-
конченных кладок подробно изучено на воробьиных птицах и отражено в многочислен-
ных публикациях проф. А. М. Болотникова и его учеников. Одной из функций преры-
вистого насиживания считается и защита неоконченной кладки от хищников. Но, как
показали инструментальные наблюдения, белобровики насиживают неоконченные
кладки половину светлого времени и всю темную часть суток (Tyrvainen, 1969).
У полевого воробья плотность собственно насиживания возрастает с откладкой
каждого яйца: на 2.5% от полных суток после появления первого яйца, на
23%—после третьего и на 80% — после пятого (Болотников, Печерский, 1981).
Чечетки, по наблюдениям в вольерах и в природе, проводят на гнезде боль-
шую часть светлого времени суток и всю ночь уже после снесения пер-
вого яйца.
75
Таблица 18
Изменчивость величины кладки некоторых воробьиных птиц Карелии
Вид Число гнезд с кладкой указанной величины (число яиц) М ± т
2 3 4 5 6 7 8 9 10 И 12 13 14 15 всего
Деревенская ласточка 4 10 30 9 1 54 4.87 ±0.11
Белая трясогузка 3 11 35 2 51 5.71 ± 0.09
Лесной конек 4 22 36 8 70 4.74 ± 0.09
Жулан 2 16 15 33 5.39 ±0.11
Крапивник 3 6 3 3 1 16 6.30 ± 0.50
Лесная завирушка 4 20 101 30 2 157 5.04 ± 0.05
Зарянка 1 9 35 ИЗ 84 6 1 249 6.17 ± 0.05
Горихвостка-лысушка 3 15 34 34 14 100 6.41 ± 0.10
Луговой чекан 8 13 2 23 6.74 ±0.13
Черный дрозд 1 3 32 28 2 /66 4.45 ± 0.10
Рябинник 3 12 81 123 3 222 5.50 ± 0.04
Певчий дрозд 1 12 56 170 7 246 4.69 ± 0.04
Белобровик 3 10 121 443 140 1 718 5.00 ± 0.05
Садовая камышевка 1 2 12 7 22 5.14 ±0.17
Пересмешка 6 7 Г 14 4.64 ± 0.17
Садовая славка 4 9 46 87 2 148 4.50 ± 0.06
Черноголовая славка 1 4 18 1 24 4.92 ± 0.08
Славка-завирушка 1 2 17 32 52 5.54 ± 0.09
Весничка 5 17 49 230 188 5 795 6.12 ± 0.02
Теньковка 3 8 44
Трещотка 2 4 18 31 6
Зеленая пеночка 5 16 1
Серая мухоловка 7 32 97 3
Мухоловка-пеструшка 5 32 177 566 486
Малая мухоловка 1 1 3 5 1
Ополовник
Пухляк 2 1 4
Большая синица 1 - 5
Пищуха 8 18 4
Обыкновенная овсянка 2 4 18 4
Овсянка-ремез 8 18 1
Камышевая овсянка 6 13 8
Зяблик 3 32 95 6 1
Зеленушка 4 5 1
Чечетка 1 2 6 6 3
Коноплянка 1 3 5 6
Чечевица 6 45 84 6
Снегирь 2 6 3
Скворец 4 44 11
Сорока 2 3 2 12 15
Серая ворона 1 12 22 6 1
2 57 5.83 ± 0.09
59 5.66 ± 0.10
22 5.83 ±0.10
139 4.69 ± 0.06
83 11 1360 6.31 ± 0.02
11 5.36 ± 0.35
3 5 1 9 -
6 7 1 25 8.44 ± 0.19
1’4 18 19 33 29 26 5 1 151 10.85 ± 0.15
4 30 5.87 ±0.12
26 4.69 ± 0.18
27 4.74 ± 0.10
27 5.04 ±0.14
137 4.78 ± 0.05
10 5.70 ± 0.22
18 4.44 ± 0.27
15 5.07 ± 0.26
141 4.64 ± 0.05
1 12 5.32 ± 0.32
59 5.12 ± 0.06
13 1 48 6.62 ± 0.20
42 4.86 ± 0.12
Таблица 19
Взаимосвязь плодовитости, типа устройства гнезда и дальности миграции
у воробьиных птиц Северной Европы
Группа видов Доля круп- ных кладок из > 7 яиц (%) Число видов (%)* с
числом вы- водков типом гнезда миграцией
1 2 откры- тый закры- тый дальняя средняя ближняя
I 0 56 44 97 3 55 18 27
II 0.1-10 46 54 70 30 46 21 33
III 11-50 69 31 19 81 44 12 44
IV 51 5 4 1 8 1 4 4
Для IV группы приведено абсолютное значение числа видов.
Прерывистое насиживание в процессе откладки яиц отмечено в таежной зоне для
многих птиц. И хотя на него, по-видимому, расходуется относительно немного энергии
(насиживающая птица определенное время лишена возможности кормиться), оно все
же создает дополнительную энергетическую напряженность в период формирования
яиц — очень энергоемкого процесса. Кроме того, прерывистое насиживание начинается
при не полностью сформированном наседном пятне, не имеющем развитого отека и не
способном нагреть яйца до температур, оптимальных для эмбриогенеза. Правда, как
выясняется, развитие зародыша продвигается на несколько стадий и при субоптималь-
ных температурных режимах насиживания (Пантелеев, Болотников, 1977). Кроме
того, такое насиживание увеличивает асинхронность вылупления птенцов в вы-
водке, которое растягивается обычно до 2, а иногда и до 3—4 сут. Как пра-
вило, это приводит к гибели отстающих в росте птенцов даже в услови-
ях трофического благополучия (например, при избытке корма в вольерах
и клетках).
Несмотря на некоторые негативные последствия, прерывистое насиживание нео-
конченных кладок тем не менее широко распространено среди птиц. Хотя при низких
внешних температурах плотность подсиживания возрастает, последнее очень характер-
но и в летний период при относительно теплой погоде. Следовательно, его нельзя рас-
сматривать только как адаптацию к защите эмбрионов от воздействия низких тем-
ператур.
Одной из важных функций насиживания неоконченных кладок может быть защита
от хищников и конкурентов (растаскивание гнездового материала, борьба за дупла
и т. д.), а также маскировка контрастно выделяющихся, ярко окрашенных яиц. Однако
затрат! времени на насиживание в период такого энергоемкого процесса, как форми-
рование яиц, очевидно, создают повышенную энергетическую напряженность, которую
организм не может выдерживать долго. Таким образом, меньшее число яиц в кладках
открытогнездящихся воробьиных птиц может быть связано с необходимостью дополни-
тельной маскировки и защиты яиц от хищников и других неблагоприятных факторов
среды, что требует увеличения времени нахождения самки на гйезде с неоконченной
кладкой и вызывает повышенную энергетическую напряженность. Как уже указыва-
лось, стремление укрыть кладку наиболее отчетливо проявляется у вольерных птиц.
Становится понятным и отсутствие существенных различий в величине кладки у уток,
гнездящихся открыто и в дуплах: ни те, ни другие не начинают насиживания до отклад-
ки последнего яйца. Каким образом это могло развиться в эволюции птиц? Что было
первичным в возникновении предполагаемой связи* исходно высокая величина кладки
или закрытый способ гнездования? На эти вопросы можно ответить лишь предположи-
тельно. Величина кладки у предков современных дуплогнездников могла быть исходно
78
высокой, а гнездование открытым. При таком размещении гнезд и длительном периоде
откладки вероятность гибели яиц от хищников увеличивалась. Избежать разорения
неоконченных кладок можно было следующими тремя путями.
Во-первых, более ранним началом насиживания. Однако этот способ сохранения
неоконченных кладок не мог поддерживаться естественным отбором, поскольку это
сопровождалось бы значительной разновозрастностью птенцов, неизбежным увеличе-
нием их смертности и сокращением размеров выводка. Иначе говоря, затраты времени
и энергии на продуцирование крупных кладок были бы неоправданными. Вторым
способом компенсации повышенного отхода яиц в плохо укрытых гнездах могло
быть сокращение периода откладки яиц и, следовательно, величины кладки. Однако
параллельно должна была возрасти и ежегодная выживаемость потомства. Но посколь-
ку исходно высокая величина кладки, очевидно, соответствовала и большой ежегодной
смертности, сокращение плодовитости вряд ли могло быть оправданным и поддержан-
ным естественным отбором. Снижение величины кладки могло, правда, компенсиро-
ваться многократным размножением в течение одного года. Такой путь, как известно,
действительно был развит естественным отбором и используется многими современ-
ными видами птиц.
Уязвимость неоконченных кладок определяла лучшую сохранность тех, которые
размещались во всевозможных укрытиях, в гнездах шарообразной формы, где и яйца,
и птенцы были защищены гораздо надежнее, чем на поверхности земли и раститель-
ности. Более высокая успешность размножения при закрытом способе гнездования
обеспечила его закрепление отбором в условиях повышенной величины кладки и вьь
водка.
Большинство орнитологов согласно с тем, что дуплогнездники (воробьиные пти-
цы) первоначально гнездились открыто. Но поскольку у предков наших дуплогнездни-
ков кладка могла быть исходно небольшой, постепенный переход части этих видов
к размещению гнезд в разнообразных нишах и дуплах дал им ряд преимуществ перед
теми, которые сохранили открытый тип гнезда. Лучшая защищенность неоконченных
кладок в укрытых гнездах сняла излишнюю энергетическую напряженность организма
в период откладки яиц и позволила увеличить его продолжительность, а следовательно
и величину кладки.
Среди наиболее плодовитых видов преобладают птицы, зимующие на местах
размножения или неподалеку от них, т. е. в зоне относительно низких температур,
неустойчивой погоды, многоснежья и короткого зимнего дня. Частые погодные анома-
лии (затяжные сильные морозы, тяжелые снегопады, образование наста, ожеледи и т. д.)
при коротком кормовом дне и длительной ночи нередко приводят к значительной
гибели птиц на обширных пространствах. Примеры этого многократно приводились
в отечественной и зарубежной орнитологической литературе, в том числе и для иссле-
дуемого региона. Такие птицы зимуют в зоне потенциально повышенной смертности,
по крайней мере в осенне-весенний период. В зоне относительно неустойчивой погоды
размещены и европейские зимовки птиц.
Кроме величины кладки и числа выводков за год в понятие плодовитости входит
и возраст наступления половой зрелости (Lack, 1957). У абсолютного большинства оби-
тающих в таежной зоне певчих птиц размножение начинается уже на первом
году жизни. Исключений немного — это врановые, самцы скворца и чечевицы
(см. ниже).
Таким образом, уже при поверхностном рассмотрении основных параметров пло-
довитости у певчих птиц тайги выявляются определенные видовые различия в стратегии
размножения, связанные с особенностями пространственного распределения на протя-
жении года.Одним видам для поддержания стабильной численности достаточно однократ-
ного размножения в течение года и воспитания небольшого числа птенцов. Так, высоко-
плодовитая лазоревка на всем пространстве ареала не входит даже в группу содоминан-
тных видов. Зяблик же, не отличающийся высокой плодовитостью, на большей части
гнездовой области количественно доминирует в населении птиц.
79
Если гнездящиеся виды каких-либо более или менее обширных регионов ранжиро-
вать по плодовитости и плотности населения, то положение, занимаемое видом в первом
списке, не будет соответствовать рангу во втором. Более того, оно может оказаться
противоположным, как в приведенных выше примерах. Казалось бы, это легко объяс-
нить разной исходной численностью сравниваемых видов. Однако современные виды
птиц эволюировали под воздействием практически одних и тех же факторов среды
достаточно длительный отрезок времени, так что их современная численность и распре-
деление по территории Земли являются результатом всех форм взаимодействия орга-
низма со средой обитания. И коль скоро виды достигли очень высокой численности
при относительно низком уровне плодовитости, а другие — немногочисленны при ста-
бильно высоком уровне воспроизводства, то это может означать только одно: потом-
ство, оставляемое первыми, оказывается более жизнеспособным и лучше выживает
до следующего сезона размножения.
Если бы это было не так, то виды, обладающие высокой плодовитостью, должны
были бы или повсеместно занять главенствующее положение, или, по крайней мере,
расширить область распространения. На самом же деле и этого не происходит: среди
североевропейских птиц, демонстрирующих на современном этапе отчетливую тенден-
цию к расселению, преобладают, пожалуй, виды с невысокой плодовитостью. Например,
за последние десятилетия на Северо-Западе СССР произошло интенсивное расселение
таких птиц, как черный дрозд, выкармливающий лишь по 3—4 птенца (часть пар делает
это дважды в лето), чечевица (в среднем 3.8 птенца на каждое благополучное гнездо
и 2.9 — на размножавшуюся пару; часть самцов-первогодков не участвуют в размноже-
нии) и зеленая пеночка (5—6 слетков). Не отличаются высокой плодовитостью и другие
новоселы нашей зоны — дубонос, дроздовидная и тростниковая камышевки и многие
неворобьиные птицы.
Как известно, основные различия в численности видов определяются соотноше-
нием их плодовитости и смертности, приведенным в соответствие естественным отбо-
ром (Fisher, 1958, — цит. по: Hussel, 1972;Williams, 1966). Любое нарушение этого баланса
будет отражаться на видовом уровне численности, вызывая его колебания. Смертность
в значительной мере отражает степень адаптированности вида к разнообразным воздей-
ствиям среды обитания внутри области его распространения. Недостаточная адаптиро-
ванность или компенсируется более интенсивным размножением (если оно оказывается
полезным для вида, а не сопровождается увеличением смертности гнездовых птенцов
и не адаптированных к этому родителей), или вызывает исчезновение вида, по крайней
мере из областей, где сила внешних воздействий превышает (пусть даже периодически)
пороги его приспособленности к среде.
Показатели смертности и плодовитости, очевидно, не могут быть одинаковыми
в зонах экологического оптимума и пессимума. Для нормального существования
в пессимальных условиях ответом на неизбежное повышение смертности должно быть
увеличение интенсивности размножения (Fisher, 1958, — цит. по: Hussel, 1972). Если это-
го не произойдет, то вид постепенно исчезнет из зон пессимума при постоянных или
периодических попытках их освоения. В результате дисперсии молодых особей населе-
ние вида будет здесь периодически восстанавливаться и исчезать, пока в результате
микро эволюционных процессов не возникнут адаптации к жизни в данной зоне, позво-
ляющие достичь соответствия плодовитости и смертности. Этому может способствовать
также изменение условий существования (потепление климата, возникновение пригод-
ных для жизни местообитаний у видов, преадаптированных к дефициту времени, и т. д.).
Увеличение затрат на воспроизводство не может быть беспредельным. Во-первых,
оно ограничено способностью взрослых птиц выкормить в данных условиях лишь
определенное число потомков (Lack, 1957), так что увеличение размеров выводка сверх
максимально возможного будет неизбежно сопровождаться повышением смертности
птенцов в гнездовой и послегнездовой периоды (Williams, 1966). Во-вторых, чрезмерные
репродуктивные усилия взрослых особей могут оказаться губительными для них самих.
Судя по возвратам на места размножения, смертность особей, выкормивших много
80
птенцов, оказывается выше, чем у птиц, воспитавших малочисленные выводки (Lack,
1966; Williams, 1966). Но вместе с тем естественный отбор, очевидно, должен был огра-
ничивать и нижние пределы плодовитости. Роль сдерживающего начала могла выполнять
сама малочисленность потомства.
Как известно, в отношении длительности жизни и плодовитости в эволюции живот-
ного мира намечается две стратегии: 1 - увеличение средней продолжительности жизни
особи при низкой интенсивности размножения и 2 — высокая плодовитость при кратко-
временности жизни особей. Эволюция большинства видов воробьиных птиц пошла
по второму пути.
Обращает на себя внимание еще один факт: среди закрытогнездящихся воробь-
иных птиц доля крупных кладок оказывается больше у видов, которые лучше утепляют
гнезда, размещают их в местах, хорошо укрытых от холода и других неблагоприятных
воздействий, используют дополнительные источники тепла — например, более нагретую
землю в норах и земляных нишах (табл. 20).
Среди дуплогнездников, отличающихся наиболее высокой плодовитостью, макси-
мально утепленные гнезда имеют большая синица и лазоревка, откладывающие наиболее
крупные кладки. В выстилке лотка их гнезд преобладает рыхло уложенный, перемешан-
ный с мхом и перьями пух животных (в наших условиях — главным образом зайца-
беляка), известный своими высокими теплоизоляционными свойствами. Кроме того,
оба вида синиц предпочитают гнездиться в дуплах с толстыми стенками, а на время
своего отсутствия прикрывают яйца выстилкой. Менее рыхлая, но также обладающая
хорошими теплозащитными качествами выстилка из шерсти и пуха животных характер-
на для гнезд пухляков, сероголовых гаичек и московок. Выстилки в их гнездах несколь-
ко меньше, чем у двух первых видов, а дупла нередко имеют очень тонкие стенки.
У всех названных видов птиц не менее 80% кладок насчитывают 7 яиц и более. У хохла-
тых синиц основу изолирующего материала составляет мох при относительно неболь-
шом количестве перьев и шерсти. Роль стенки дупла нередко выполняет лишь кора
дерева. К тому же гнездиться они начинают при более низких внешних температурах.
В итоге в гнездах хохлатых синиц выстилка меньше, чем у других видов синиц, защища-
ет яйца от холода, и на долю крупных кладок приходится лишь 10% случаев.
Мох является основным теплоизолятором и в гнездах горихвостки, но она начина-
ет гнездиться на месяц позднее хохлатой синицы. Доля крупных кладок варьирует
у горихвосток, по разным данным, от 35 до 48%.
Скворец и мухоловка-пеструшка используют для выстилки лотка преимуществен-
но тонкие стебли и листья злаков. Скворец утепляет лоток также перьями, но предпочи-
тает гнездиться в довольно обширных дуплах и искусственных гнездовьях, где в отсут-
ствие насиживающей птицы тепло удерживается слабо. По этой причине во время май-
ского похолодания 1965 г. кладки скворцов в скворечниках промерзли и многие яйца
лопнули, тогда как у большой синицы уцелели не только завершенные, но и
неоконченные кладки. Скворец, кроме того, начинает гнездиться на 3—4 недели
раньше мухоловки-пеструшки, при более низких температурах, иногда опуска-
ющихся ниже 0 °C.
У веснички и трещотки основание гнезда размещается в углублении почвы, расчи-
щенном от растений, и большая часть постройки дополнительно защищена моховой
дерновиной или лесной подстилкой. Веснички выстилают лоток большим количеством
перьев, а трещотки — только стеблями и листьями злаков. Перья как основа выстилки
лотка в большом количестве используются также теньковкойли ее гнезда, даже разме-
щаясь на земле, никогда не имеют с ней непосредственного контакта. В лучшем случае
гнездо лежит на лесной подстилке, но чаще значительно приподнято над поверхностью.
Такие гнезда сильнее продуваются ветром, что, по мнению Шилова (1968), может иметь
существенное значение для теплообмена кладки (выводка) с внешней средой. Гнезда
зеленой пеночки фактически не имеют крыши, но размещаются внутри разнообразных
земляных ниш. Правда, для постройки гнезда эти пеночки часто используют ниши
в корневых подошвах ветровала, где земля при похолоданиях быстро остывает.
6 В. Б. Зимин
81
оо
N)
Таблица 20
Теплоизоляция и степень укрытости кладок у закрытогнездящихся воробьиных птиц с различной плодовитостью
Доля кладок из 7 Число вы- Вид Тип Степень Основной материал выстилки
и более яиц (%) водков за сезон гнезда* укрытости шерсть перо мох луб трава
80-100 1 2 2 2 1 1 Ополовник Большая синица Лазоревка Московка Пухляк Сероголовая гаичка Ш д д д д д 3 3 3 3 3 3 + + + + + -г + +
50-70 1 1 1 2 Сорока Луговой чекан Г орихвостка-лысушка Крапивник ш Ниша д-п/д Ш П/О П/О з-п/о 3 + + + + + +
25-45 1 2 1 Мухоловка-пеструшка Зарянка Весничка д Ниша Ш 3 П/О 3 + + +
10-20 2 1 1 Пищуха Малая мухоловка Хохлатая синица Д-П/Д п/д д з-п/о П/О 3 + + + + + +
Около 5 2 1 1-2 1 1 Белая трясогузка Трещотка Теньковка Зеленая пеночка Скворец Ниша Ш Ш П/Ш, Ниша Д П/О 3 3 3 3 + + + + 1 + +
Ш - шарообразное, П/Ш - полушарообразное, Д - дупло, П/Д - полудупло, 3 - закрытое, П/О - полуоткрытое.
По степени теплоизоляции гнезд и по доле крупных кладок виды рода Phylloscopus
образуют ряд: весничка—трещотка—зеленая пеночка—теньковка.
Таким образом, среди близкородственных видов закрытогнездящихся птиц
различия в числе откладываемых яиц соответствуют степени теплоизоляции гнезда,
которая определяется местом его размещения, высотой над землей, особенностями
конструкции, теплопроводностью материалов, температурными условиями, в которых
начинается кладка.
Однако уже среди открытогнездящихся птиц четкой связи теплоизоляционных
свойств гнезд с величиной кладки для родственных видов не выявляется. Например,
среди славок рода Sylvia среднее число яиц максимально у завирушки, строящей наиме-
нее утепленные „прозрачные” гнезда. Следовательно, если такая связь у открытогнездя-
щихся птиц все же существует, она не может реально проявиться из-за более мощного
влияния каких-то других факторов, не учтенные в данном анализе. Шилов (1968)
отмечает, например, что с улучшением теплоизоляции гнезд возникает проблема накоп-
ления в них углекислого газа, которая решается изменением конструкции гнезда или вы-
соты размещения над землей (разная степень вентиляции) и т. д. Выясняется также и
существенное значение для нормального развития эмбрионов режима внутригнездовой
влажности воздуха (Болотников, Калинин, 1974). Важно учитывать и то, что если на
первых этапах размножения в зоне умеренного климата птицам приходится решать
проблему максимального сохранения тепла, то после приобретения птенцами способ-
ности к самостоятельной терморегуляции возникает необходимость преодоления тепло-
вого стресса (Дольник и др., 1982). На севере, при более низких внешних температурах
воздуха, с решением второй задачи птицам справиться легче, и в этом может заключать-
ся одна из предпосылок преобладания более крупных кладок у птиц северных популя-
ций. А так как начало гнездования здесь постоянно сопровождается возвратом холодов
и в целом проходит при более низких температурах, сохранение тепла в период от появ-
ления яиц до достижения птенцами 6—8-дневного возраста является в условиях севера
более существенной проблемой.
Число яиц, откладываемых воробьиными птицами любого вида, даже в пределах
одного региона значительно варьирует (табл. 18). Из обширной орнитологической лите-
ратуры известно о наличии географической, биотопической, сезонной, возрастной и го-
довой изменчивости величины кладки, о связи ее с численностью, высотой над уровнем
моря, обилием пищи, размерами гнездовий и т. д. Уже простое перечисление факторов,
способных влиять на размер кладки, указывает на то, что число яиц, снесенных каждой
самкой в конкретной ситуации, является результатом очень сложных и многообразных
взаимодействий организма и изменчивой среды обитания. Поэтому, изучая влияние
какого-то одного фактора, следует постоянно иметь в виду как возможность воздей-
ствия всего комплекса внешних условий, так и влияние внутренних физиологических
процессов.
В природе вообще невозможно вычленить влияние какого-то одного фактора,
даже если, по нашим представлениям, он занимает положение ведущего. Например,
большинство орнитологов признают решающую роль температуры среды как сигналь-
ного фактора, определяющего начало гнездования потенциально готовых к нему птиц.
Рассчитаны и так называемые температурные пороги начала размножения, которые,
как выясняется, совершенно не одинаковы для одних и тех же видов птиц в разных
частях ареала, где размножение начинается на разных фотопериодах. Они разнятся
и в смежные годы у птиц одной популяции, индивидуальны и непостоянны у одних
и тех же особей в последовательные годы жизни. Эти расхождения вполне объяснимы
недоучетом таких факторов, как фотопериод, влияние ветра (см. выше), состояние
кормовой базы, наличие партнера, масса тела особей в предгнездовой период и т. д.,
в конечном итоге отражающихся и на величине кладки. Можно, конечно, пытаться
решить проблему, используя метод математического моделирования. Но вряд ли на
современном этапе это будет оправданно. Вероятнее всего, для определения значения
каждого из факторов пришлось бы создавать свои модели. Не в состоянии мы оценить
83
количественно, например, кормовую базу насекомоядных птиц, а также качественные
признаки, этологические показатели и т.п.Поэтому, оставляя на будущее такой перспек-
тивный метод.,как моделирование, мы вынуждены ограничиться традиционным анали-
зом собранного материала.
Связь плодовитости с питанием
Пища как один из факторов, влияющих на величину кладки птиц, нередко исполь-
зуется орнитологами для объяснения многих наблюдаемых различий* сезонных, возраст-
ных, биотопических и т. д. Действительно, одним из важнейших показателей пригод-
ности биотопа для птиц считается обилие и доступность кормов. Хорошо известно, что
повышение численности мышевидных грызунов способствует заметному увеличению
числа яиц в кладках птиц миофагов - дневных хищных и сов (Southern, 1955). В Север-
ной Швеции установлено также, что в годы обилия мелких грызунов величина кладки
и выводка у длиннохвостого поморника достоверно возрастает, увеличивается и доля
первогодков, принимающих участие в размножении (Andersson, 1976). Для певчих птиц
убедительных примеров подобной связи крайне мало, что в первую очередь может
объясняться большим разнообразием объектов их питания. Лишь у птиц с более узкой
пищевой специализацией (кедровка в Швеции, — по: Lack, 1957) обнаруживается некото-
рая тенденция к проявлению механизма регуляции величины кладки через обилие пищи.
Правда, зависимость размера кладки кедровки от урожая лесных орехов показана на
малом числе наблюдений и нуждается в подтверждении. Абсолютное большинство
воробьиных птиц — полифаги, и они легко переходят с одного массового вида корма
на другой. Состав их пищи очень разнообразен и быстро изменяется в течение одного
года и даже в разные годы. Примеры вариаций в пищевых связях приводились нами
выше (гл. I). Судя по предварительным данным, полученным на наших стационарах,
такие важные виды кормов, как гусеницы чешуекрылых и ложногусеницы пилиль-
щиков, достигают максимального обилия в разные периоды года, что создает предпо-
сылки для бициклического размножения многих видов птиц в таеж .ой зоне.
Специальные работы, посвященные изучению питания птиц и степени их воздей-
ствия на энтомофауну, показали, что птицы используют лишь незначительную часть
общей биомассы беспозвоночных животных. Таким образом, кормов в таежных лесах
вполне достаточно, и птицы, по крайней мере в северной полосе, гДе их общая числен-
ность невелика, вряд ли испытывают сколько-нибудь существенный недостаток в пище
при благоприятных погодных условиях. Но именно погода на севере является очень
неустойчивым фактором. При похолоданиях активность беспозвоночных животных
резко снижается. Большинство из них заблаговременно уходят в укрытия и становятся
недоступны для многих насекомоядных птиц. В такие периоды дефицит корма ощуща-
ется довольно остро, тем более что низкие температуры и сильный ветер способствуют
росту затрат энергии на теплопродукцию, что само по себе требует дополнительного
потребления пищи.
В 1984 г. во время июньского похолодания и после потепления мы подсчитали
количество насекомых и пауков, встречающихся на одних и тех же соцветиях рябины
при разной температуре воздуха.
Температура (°C) 6.8 9.2 10.6 19.6
Число насекомых и пауков, экз. 3 11 17 204
Число соцветий, экз. 60 100 60 60
Встречаемость беспозвоночных на 1 соцветии, экз. 0.05 0.11 0.28 3.40
При температуре воздуха ниже 10 °C резко сокращается встречаемость активно
перемещающихся насекомых и пауков. Но снижается не сама численность беспозвоноч-
ных, а их доступность для птиц. И именно этот фактор приобретает значение
84
решающего. В холодную погоду даже при высоком уровне биомассы беспозвоночных
животных птицы могут испытывать существенный недостаток в кормах и затрачивать
на их поиски больше времени. Наоборот, снижение запасов пищи будет меньше ощу-
щаться птицами при устойчивой теплой погоде. Конечно, вероятность обнаружения пти-
цей неактивных насекомых должна быть выше при высокой численности последних,
но многолетний опыт полевых наблюдений убеждает нас в том, что даже при недостатке
кормов (например, ранней весной), но при устойчивой теплой погоде птицы легко обес-
печивают себя пищей. Так, например, синицы перестают посещать зерновые прикормки
уже в конце марта, но даже летом, когда обща^ биомасса животных кормов несравнен-
но выше, они вновь появляются у подкормочных пунктов во время внезапных похоло-
даний. В вольерах птицы, подготовленные к размножению, имеют избыточное коли-
чество легкодоступной пищи. Тем не менее величина их кладок бывает не больше обыч-
ной, а чаще даже меньше, чем в естественных условиях. В природе в ситуациях, возника-
ющих при изобилии корма, например у синантропных врановых, кормящихся отходами
в звероводческих хозяйствах, на свалках и в населенных пунктах, средняя величина
кладки, действительно, несколько больше, чем у особей, населяющих территории,
на которых антропогенное влияние не проявляется (устное сообщение Е. В. Шутенко).
Но в первом случае птицы начинают гнездиться на 2—3 недели раньше. И если сопоста-
вить это с сезонной изменчивостью величины кладки, которая, например у ворон, сни-
жается от начала к концу сезона размножения, то не ясно, что же влияет на число откла-
дываемых яиц сильнее — различия в обилии пищи или в датах начала гнездования.
На всех наших стационарах постоянно функционировали зерновые прикормки
из семян подсолнечника, конопли, проса, которые регулярно посещали зяблики, овсян-
ки, снегири, синицы, чечевицы. Последние особенно охотно поедали этот корм в течение
всего периода пребывания на родине, а часто выкармливали им своих птенцов. Есть
основания полагать, что для птиц, гнездившихся неподалеку от мест прикормки, зерно-
вая смесь была основным источником питания в течение всего репродуктивного пери-
ода (многие из них по нескольку раз ежедневно попадались в ловушки, установленные
на прикормках). Обилие и доступность высококалорийных кормов тем не менее никак
не отразились на величине кладки чечевиц, постоянно посещавших места подкормки.
Как в радиусе до 1 км от этих пунктов, так и на более значительном удалении птицы
этого вида откладывали примерно одинаковое количество яиц. Никаких существенных
различий не обнаруживается и при сравнении размеров кладки чечевиц в нашем и смеж-
ных регионах на Северо-Западе СССР и в Скандинавии, где птицы питались только
естественными кормами (Haartman, 1969а; Roostmae, Veroman, 1974; Нанкинов, 1974;
Stjernberg, 1979; Паевский, 19816; Мальчевский, Пукинский, 1983; Aleknonis, 1984,
и др.).
Подкормка больших синиц мучными червями, проводившаяся в течение апреля
в Южной Швеции, вызвала лишь несколько более раннее наступление периода откладки
яиц, а число отложенных яиц не увеличилось (K311ander,1974).
Связь величины кладки с обилием пищи установлена для двух популяций буробо-
кой туайи (Pipio erythrophthalmus) в США (Greenlan, 1978). Но в данном случае сранива-
лись птицы из разных биотопов (дубравы и сосновые леса), имевшие 1—2-недельную
разницу в средних датах начала гнездования. Следовательно, помимо кормового факто-
ра выявленные различия могли быть обусловлены географическими, сезонными и био-
топическими вариациями в размерах кладки. Интересен также эксперимент, поставлен-
ный на сороках в Швеции. Здесь на участках с дополнительными источниками пищи
(весной птиц подкармливали рыбой) сороки приступили к размножению в среднем на
два дня раньше и отложили более крупные яйца и в большем числе, чем на контроль-
ных территориях (Hogstedt, 1981). Сорока — типичный синантропный вид; объекты ее
питания в предгнездовой период — это всевозможные пищевые отходы, всегда в изоби-
лии встречающиеся около жилья человека. Некоторые трудности с добыванием пищи
сороки, очевидно, испытывают лишь во время обильных продолжительных снегопадов
в зимнее время. Весной же, с началом снеготаяния, особых затруднений с добычей
85
пропитания эти птицы не имеют. Даже в периоды затяжных майских похолоданий им уда-
ется выкормить птенцов. Так, в 1965 г. в заповеднике „Кивач” сороки были одними из
немногих птиц, которым холод и снег, пролежавший около 5 дней, не помешали успеш-
но выкормить маленьких птенцов (Зимин, 1968; Зимин и др., 1976). Однако для рас-
сматриваемого вида характерна значительная индивидуальная изменчивость величины
кладки: 48 известных гнезд содержали от 3 до 9 яиц. По нашим данным, уменьшение
майских кладок (6.28 ± 0.41) по сравнению с апрельскими (6.92 ± 0.27) недостоверно,
но даже если оно действительно имеет место, двухдневная разница между датами от-
кладки яиц на опытных и контрольных участках вряд ли могла существенно отразиться
на величине кладки. Было ли дополнительное питание единственным фактором, непо-
средственно повлиявшим на увеличение числа яиц в гнездах сорок, или здесь прояви-
лись какие-то иные связи — осталось невыясненным.
Сомнительно, однако, чтобы регуляция плодовитости через обилие пищи поддер-
живалась естественным отбором у птиц со столь разнообразным и постоянно изменя-
ющимся набором кормов, какой имеют сороки и абсолютное большинство других
представителей отряда воробьиных в умеренной зоне. Размножение птиц начинается
здесь при низких уровнях кормовой базы, когда количество энергии, поступающей
в организм с пищей, превышает ее расход ровно настолько, чтобы обеспечить продуци-
рование яиц. И именно в этот момент, в период относительного дефицита кормов,
а не во время пика, большинство птиц откладывает максимальное число яиц, постепен-
но сокращающееся к концу репродуктивного периода, так что к моменту максималь-
ного обилия и разнообразия кормов величина кладки обычно приближается к своим
минимальным значениям. При строгой зависимости размеров кладки от обилия пищи
сезонное изменение размеров кладки и динамика развития кормовой базы должны
были бы проходить параллельно. Даже если предположить, что о предстоящем после
благополучной зимовки увеличении обилия беспозвоночных птицам сигнализирует
более высокий уровень ранневесенней численности каких-то иных объектов питания,
то в первую очередь это должно отразиться на сроках начала гнездования: излишки
поступающей с пищей энергии должны способствовать компенсации ее затрат на покой
при большей продолжительности ночи, т. е. репродуктивный порог в этом случае будет
пройден в более ранние календарные даты. А так как самые ранние кладки обычно
бывают крупнее, то можно говорить и об опосредованной связи их средних показателей
с обилием пищи.
В условиях неустойчивой весенней погоды в зоне умеренного климата, а тем
более в Субарктике и Арктике, обилие и доступность корма, в частности для насеко-
моядных птиц, очень непостоянны. Поэтому высокая численность насекомых весной
еще не означает, что она обязательно сохраняется на таком же уровне и в более поздние
периоды. Резкие похолодания после массового выплода личинок губительны для насе-
комых и нередко приводят к падению первоначальной численности (Гусев и др., 1938;
Бей-Биенко, 1971). Вряд ли такой изменчивый и непостоянный фактор, как кормовая
база, мог приобрести решающее значение в эволюции величины кладки птиц. Большую
роль играли здесь, по-видимому, общая тенденция сезонной изменчивости кормовой
базы и приобретение большинством видов птиц способности к быстрому переключению
с одного вида корма на другой. В конечном итоге для приведения плодовитости к уров-
ню, необходимому для поддержания стабильной численности вида, наиболее важен
один из интегрированных результатов размножения — число потомков, достигших
самостоятельности Не потому ли именно период развития организма и становления
самостоятельности молодых особей приурочен ко времени максимального обилия
и разнообразия кормов при относительно длинном световом дне и наиболее стабильных
погодных условиях? В этом плане связь пищевого фактора с плодовитостью вполне
закономерна, что неоднократно подчеркивали в своих работах Лэк и его последователи.
Естественный отбор мог благоприятствовать увеличению числа крупных кладок
лишь в том случае, если повышенный уровень кормовой базы на всех этапах
размножения сохранялся в неизменном виде (или нарастал) на протяжении всего
86
репродуктивного периода. Так и происходит, например, в „^мышиные” годы, когда
высокая плотность населения мелких грызунов заметна уже в самом начале весны
и в дальнейшем их обилие постепенно нарастает. Поэтому не случайно именно на мио-
фагах зависимость величины кладки от количества пищи доказана вполне определенно.
Проявления подобной связи можно, очевидно, ожидать и у других птиц-стено-
фагов, но для них мы, к сожалению, не располагаем необходимым сравнитель-
ным материалом.
У большинства других птиц с развитой полифагией такая зависимость не могла
развиться хотя бы потому, что разные этапы их репродуктивного периода проходят
на совершенно различном фоне питания, причем нередко объекты их питания оказыва-
ются не связанными между собой и развиваются в природе независимо друг от друга.
При значительной вариабельности рационов птиц в течение года трудно представить,
чтобы естественный отбор способствовал развитию и закреплению зависимости плодо-
витости птиц-полифагов от уровня пищевых ресурсов в период формирования половых
продуктов и откладки яиц. Это противоречит заключению Лэка (Lack, 1977) о том, что
в годы обилия пищи преимущества получают особи с большой кладкой, а при недостат-
ке кормов - с маленькой. При таком направлении естественного отбора неизбежно
должна была развиться и зависимость размеров кладки от уровня кормовой
базы. Между тем, как показано выше, подобная связь возможна лишь для птиц-
стенофагов.
Географическая изменчивость величины кладки
Этот вопрос неоднократно обсуждался орнитологами с момента опубликования
правила Гопкинса, согласно которому размер кладки птиц увеличивается в направле-
нии к северу и востоку, а также со снижением высоты над уровнем моря (Hopkins,
1938). В дальнейшем это правило получило подтверждение в известной монографии
Лэка (Lack, 1957) и во многих более частных работах по отдельным видам птиц. Было
найдено и объяснение этому явлению: с увеличением продолжительности светлого
времени суток на севере птицы получают возможность выкормить большее число
потомков за счет удлинения кормового дня.
По мере накопления фактического материала правило Гопкинса оказалось не
столь безупречным, каким представлялось сначала. Более того, обработка массовых
данных по размерам кладки некоторых европейских воробьиных птиц на разных широ-
тах и долготах дала совершенно беспорядочное распределение средних величин. Так,
у мухоловки-пеструшки, которая благодаря массовому заселению искусственных гнез-
довий изучена лучше других видов, средняя величина кладки изменяется на протяжении
ареала без каких-либо определенных тенденций (Haartman, 1967b). Однако с учетом вы-
соты над уровнем моря правило Гопкинса начинает проявляться более отчетливо (Berndt,
Winkel, 1967). Тем не менее если даже не вводить поправок на географическую долготу
и высоту над уровнем моря, а ограничиться простым увеличением числа данных по
средней величине кладки в более или менее удаленных по широте пунктах, то нетрудно
заметить две тенденции в географической изменчивости рассматриваемого показателя.
Для мухоловки-пеструшки в южной части ареала характерно постепенное увеличение
размеров кладки в северном направлении, а в полосе 58—69° с. ш. — сокращение числа
откладываемых яиц (рис. 24, А). Следует указать, что на этом рисунке самые северные
точки в основном приведены для территорий, расположенных над уровнем моря в сред-
нем на 200 и более метров выше, чем на юге. Таким образом, отмеченная для северной
части ареала тенденция может иметь лишь региональное значение.
Суммировав собственные и литературные данные по размножению воробьиных
птиц в горных условиях, Ковшарь (1981а, 19816) пришел к заключению, что сроки
размножения и величина кладки изменяются не у всех видов птиц, населяющих высо-
когорья. Сокращение числа откладываемых яиц с увеличением высоты над уровнем
87
Б
о 7 *2
50 Ll_L-L_I 1*1111?!
4.5 5.0 5.5 6.0
Рис. 24. Географическая изменчивость величины кладки
некоторых воробьиных птиц.
А - мухоловка-пеструшка (по: Haartman, 1967b, с дополне-
ниями); Б - полевой воробей (по: Гагинская и др., 1981);
В - белобровик (1) и зяблйк (2). Прерывистые линии - тенден-
ции изменчивости величины кладки. По оси абсцисс - средняя
величина кладки (число яиц); по оси ординат - географическая
широта (градусы).
моря, как правило, происходит параллельно с задержкой сроков начала гнездования.
Оба этих момента важно иметь в виду при изучении географической изменчивости
плодовитости птиц.
Некоторая тенденция к увеличению размера кладки в северном направлении
проявляется у полевого воробья (рис. 24, Б), причем на значении средних показателей
у этого вида могут сказываться также и биотопические различия в сроках гнездования
(естественные и антропогенные местообитания). Шураков с соавторами (1981) ука-
зывают, ссылаясь на различные источники, что в экваториальной зоне в гнездах полево-
го воробья редко бывает более 4 яиц, тогда как в Северной Европе в 28—52% гнезд
содержалось по 6—7 яиц.
Ряд широко распространенных видов птиц демонстрирует относительное постоян-
ство величины кладки в разных частях ареала, что, в частности, отвечено для зяблика
в скандинавских странах (Svensson, 1978). Как выясняется, и на значительно большем
пространстве его ареала различия в средних показателях не столь велики, как у других
воробьиных (рис. 24, В). Размер кладки этого прогрессирующего вида, который в об-
ластях постоянного гнездования почти повсеместно занимает доминирующее положение
по численности, характеризуется, кроме того, и слабой сезонной изменчивостью. Более
95 % самок зяблика имеют кладки, отличающиеся от среднепопуляционной на ± 1 яйцо
(табл. 21). Кроме того, средние размеры кладки и выводки у зябликов мало отлича-
ются друг от друга — степень их соответствия достигает 84%.
Незначительные географические вариации анализируемого показателя характерны
и для чечевицы (табл. 22), у которой отчетливое его снижение отмечено лишь для
горных популяций.
Не обнаруживаются определенные закономерности в широтной изменчивости
величины кладки у скворца (рис. 25) и белобровика (рис. 24, В). У последнего в зоне
20—35° в. д. в целом намечается увеличение данного показателя по направлению к севе-
ру: 4.6—5.1 яйца в Прибалтике, Ленинградской обл., на юге Карелии и Финляндии;
5.2—5.4 — в Северной Финляндии и на островах Баренцева моря (Haartman, 1969а; Tyrva-
inen, 1969; Книстаустас и др., 1981; Шутова и др., 1982; Мальчевский, Пукинский,
1983; наши данные). Однако явно противоречат этому сведения по Псковской обл. —
5.9 яйца (Мешков, Урядова, 1967) и Лапландскому заповеднику — 4.6 яйца (Шутова
и др., 1982). Не укладываются в приведенную схему и данные по Уралу и Западной
88
Таблица 21
Географическая изменчивость величины кладки зяблика
Число гнезд Средняя вели- чина кладки
Место с указанной величиной кладки (число яиц) всего Источник
2 3 4 5 6 7 8
Южная Карелия - 5 35 ИЗ 6 1 - 160 4.74 Наши дан- ные
В том числе:
заповед- ник — 3 9 29 — — 41 4.63 Тот же
„Кивач” Прио- нежье - 1 17 32 3 1 - 54 4.74
Прила- дожье - 1 9 52 3 - - 65 4.88 » ??
Финляндия 4 11 78 208 15 1 1 318 4.71 Haartman, 1969а
Ленинград- ская обл. - 1 65 129 3 1 - 199 4.69 Мальчев- ский, Пукин- ский. 1983
Калинин-
градская обл. 2 68 241 603 41 - - 955 4.64 Дольник и Др., 1982
Литов- ская ССР - 1 18 58 7 - - 84 4.84 Aleknonis, 1984
Всего:
абс. 6 86 437 1111 72 3 1 1716 4.68
% 0.3 5.0 25.5 64.7 4.2 0.3 0.0
Таблица 22
Географическая изменчивость величины кладки чечевицы
Место Средняя величина кладки Источник
Финляндия 5 1 Haartman, 1969а
Южная Финляндия 4.8 Reinikainen, 1939
Южная Финляндия, парки г. Лахти 5.0 Peiponen, 1974
Южная Финляндия, 62.1° с. ш. 5.3 Stjernberg, 1979
Южная Швеция 5.3 Risberg, 1970
Южная Карелия, 60.5-62° с. ш. 4.6 Наши данные
Ленинград и окрестности, парки 4.7 Bozcko, 1974
Ленинградская обл., Старый Петергоф 4.9 Нанкинов, 1974
Эстонская ССР 4.2 По: Srjernberg, 1979
Швеция и Норвегия 4.9 Тот же
ФРГ, ГДР, Польша 4.8 »»
Калининградская обл., п. Рыбачий 4.7 Паевский, 1981
Окрестности г. Уральска, р. Урал 4.8 Левин, 1979
Юг Западной Сибири 4.5 Москвитин,1979
Заилийский Алатау (2-3 тыс.м над ур. м.) 4.1 Ковшарь, 1979
Таласский Алатау, Западный Тянь-Шань 4.0 Губин, 1979
Западный Тянь-Шань 3.6 Третьяков, 1979
89
Рис. 25. Географическая изменчивость величины кладки у скворца. (Источники данных отме-
чены в списке литературы звездочкой).
В кружках - средние размеры кладки; в прямоугольниках - средняя величина кладки:
первой (верхний показатель) и второй (нижний),
Сибири. Левин (1979) определяет среднюю кладку как 5.47, а Москвитин (1979) —
5.19, тогда как на Среднем и Приполярном Урале величина кладки одинакова — 5.3 яй-
ца (Данилов, 1966). Еще далее к востоку, на юго-западе Якутии (60° 30х с. ш.), сред-
няя величина кладки по 20 гнездам составила 5.2 яйца (Гермогенов, 1976).
Изучение географической изменчивости величины кладки птиц усложняется
несоответствием методических подходов, проявлением индивидуальной, сезонной,
годовой и биотопической изменчивости, амплитуда которой в одной местности нередко
перекрывает широтно-долготные различия (Haartman, 1967b). Это полностью подтвер-
ждают и наши данные, обсуждаемые ниже. Кроме того, достоверная зависимость ве-
личины кладки от площади дна синичников и скворечников (различия достигают
2.9 яйца) выявлена для некоторых видов птиц-дуплогнездников (Johansson, 1974; Karls-
son, Nilsson, 1977; Lohrl, 1980). Для привлечения дуплогнездников в отдельных исследо-
вательских центрах используются гнездовья неодинаковых размеров, что может допол-
нительно влиять на получаемые результаты, искажая истинную картину географической
изменчивости величины кладки у птиц данной группы.
Биотопическая изменчивость величины кладки
Хотя в целом биотопические различия в размерах кладки не так велики (обычно
не более 1 яйца), они обнаружены у многих видов птиц, проявляются с достаточным
постоянством и, как правило, однозначны в различных частях ареала.
90
Таблица 23
Биотопическая изменчивость величины кладки мухоловки-пеструшки в Южной Карелии
Число гнезд
Биотоп и м.есто исследований с указанной величиной кладки (число яиц) всего М + т
3 4 5 6 7 8 9
Спелые коренные ельники, „Кивач” - 2 25 97 41 4 3 166 6.14 ± 0.03
Ельник-черничник, Маячино - 1 10 35 25 11 - 82 6.4 ± 0.10
Спелые лишайниковые и зеленомошные сосняки, „Кивач” Зеленомошные сосняки 1 6 49 140 94 9 - 299 6.16 ± 0.05
с подростом ели и подлеском, Маячино — 9 18 59 65 13 - 164 6.33 ± 0.07
Спелые березняки и осинники, „Кивач” - - 6 27 28 1 1 64 6.47 ± 0.10
Спелые лиственные и хвойно-лиственные леса с богатым подлеском, о-в Б. Клименецкий 3 3 И 40 75 30 4 166 6.73 ± 0.08
Березово-сосновые леса на осушенных болотах, Маячино - 2 3 18 28 6 - 57 6.58 ± 0.12
Разреженные чернооль- шанники, Маячино - 1 7 15 13 2 1 39 6.28 ± 0.16
Березово-ольховые
молодняки с подростом ели, „Кивач” — 3 13 22 27 4 - 69 6.23 ± 0.11
Сосновые и
сосново-лиственные — 2 29 71 44 3 2 151 6.15 ± 0.07
молодняки, Прионежье Елово-березовые и сосново-березовые молодняки, Маячино - 1 3 21 18 1 - 44 6.34 ± 0.12
Хорошо известны подобные различия для городских и лесных популяций черного
дрозда (Snow, 1958, и др.). В Англии большие синицы и лазоревки в насаждениях с пре-
обладанием дуба откладывают в среднем на 1 яйцо больше, чем в других местообита-
ниях (Lack, 1966). На севере ареала в березняках, осинниках и сероольшанниках боль-
шая синица также имеет более крупные кладки, чем в ельниках и сосняках (Зимин,
1978). В Швеции гнезда ворон во влажных местообитаниях содержат по 4.6 яйца, а
в сухих — по 3.4 (Loman, 1977). На Северо-Западе СССР птицы этого вида, гнездящиеся
в городах, имеют более крупные кладки по сравнению с естественными местообитания-
ми (устное сообщение Е. В. Шутенко и наши данные). Скворцы на удаленных от побе-
режья островах Ботнического залива откладывают в среднем по 5.65, а на материке —
по 4.89 яйца (Tenovuo, Lemmetyinen, 1970).
У мухоловки-пеструшки связь величины кладки с биотопом однозначно проявля-
ется в разных частях ареала: ФРГ — в березовых лесах ее среднее значение 6.44, а в со-
сновых борах 5.77 (Berndt, Winkel, 1967) ; средняя Карелия - в лиственных древостоях
заповедника „Кивач” 6.47, в хвойных 6.15 (Зимин, 19726); Швеция - в лиственных
насаждениях 6.5, в сосновых 6.2 яйца (Kallander, 1975).
Существование характерных биотопических различий в величине кладки можно
проиллюстрировать нашими данными по мухоловке-пеструшке, легко привлекаемой
на гнездовье даже в такие типы насаждений, где в естественных условиях она не гнез-
дится (табл. 23).
Таблица 24
Биотопическая изменчивость величины кладки у некоторых птиц в Южной Карелии
Биотоп Большая синица * Зарянка* Зяблик Весничка
п М ±т п М ± т п М ±т п М ± т
Сосновые и сосно- во-лиственные 26 10.75 ± 0.24 29 6.21 ± 0.13 54 4.70 ± 0.08 83 6.17 ± 0.08
молодняки Спелые и приспе- вающие леса с преобладанием 29 11.33 ±0.21 59 6.54 ± 0.07 65 4.88 ± 0.06 170 6.34 ± 0.05
сосны и ели Спелые и приспе- вающие листвен- 32 12.75 ±0.18
ные леса
* Учтены только первые кладки.
Спелые и приспевающие лиственные леса, как и насаждения с преобладанием
березы, осины и ольхи, являются основными (оптимальными) гнездовыми стациями
для мухоловки-пеструшки и большой синицы в Карелии. Молодняки, возникающие
на месте вырубленных древостоев и заброшенных лесных сенокосных лугов, обычно
используются большинством дуплогнездников как кормовые стации, но становятся
пригодными для их размножения после развески искусственных гнездовий (Зимин и
др., 1976) и переходят в ранг субоптимальных гнездовых местообитаний. Основные
репродуктивные показатели птиц в оптимальных гнездовых стациях оказываются более
высокими, чем в субоптимальных. Ранее это было показано на примере большой
синицы (Зимин, 1978) и оказалось характерным для величины кладки мухоловки-
пеструшки (достоверные различия подчеркнуты).
Год Оптимальные стации Субоптимальные стации
Число гнезд М± т Число гнезд М± т
1975 54 7.32 ±0.10 12 6.59 ± 0.30
1976 67 6.33 ±0.14 24 6.25 ± 0.22
1977 45 6.49 ±0.15 13 5.92 ± 0.29
Всего 166 6.73 ± 0.08 51 6.25 ±0.13
Биотопические вариации в размерах кладки не везде проявляются одинаково
четко. Так, в Маячино, где гнездовья были развешены многокилометровыми линиями,
часто проходившими по границам между различными типами насаждений, около откры-
тых и полуоткрытых стаций, биотопические различия в величине кладки оказались
статистически недостоверными. Как и всюду на хозяйственно освоенных территориях,
лесонасаждения здесь отличались значительной мелкоконтурностью выделов, пестротой
возрастного и породного состава, плотностью древостоя. При мозаичности условий
и высокой подвижности птиц биотопические различия в сроках начала гнездования
и величине кладки в значительной мере сглаживаются.
Биотопические различия в плодовитости большой синицы, отмеченные нами ранее
(Зимин, 1978), определенно подтверждаются дополнительно собранными данными
(табл. 24). Согласно этим материалам, минимальные кладки у больших синиц встреча-
ются в сосновых и сосново-лиственных молодняках, которые в гнездовое время они
обычно не заселяли, а появились здесь только после развески дуплянок. Помимо этого,
в молодняках для большой синицы характерны более поздние сроки размножения,
а основу гнездового населения составляют птицы-первогодки. Среди 59 особей, гнездив-
шихся в молодняках Шокшинского стационара, 55 (93.2%) были первогодками, при-
чем птицы старших возрастных групп впервые появились здесь лишь после того, как
возраст насаждений достиг 18 лет. В Маячино, где большинство синиц гнездилось
92
в спелых и приспевающих лесах, среди 70 взрослых особей птицы в возрасте 2 лет и
старше составляли 41 % гнездового населения.
Гнезда зарянки в спелых древостоях содержат достоверно больше яиц, чем в мо-
лодняках. Такая же тенденция намечается и для зяблика (табл. 24). Оба вида - типич-
но лесные птицы — начинают осваивать молодые насаждения после того, как деревья
достигают 11 — 13-летнего возраста. Численность этих видов птиц первоначально бывает
невысокой, а основу гнездового населения составляют первогодки. Молодняки являют-
ся для большинства лесных птиц субоптимальными гнездовыми стациями, в противо-
положность весничке, которая именно первые стадии их формирования населяет с доста-
точно высокой плотностью (Зимин, Кузьмин, 1980). Некоторое превышение средней
величины кладок у весничек Приладожья, по сравнению с шокшинскими молодняка-
ми, вероятнее всего связано с различиями в сроках размножения. В годы наблюдений
в Маячино были зарегистрированы самые ранние за все время даты начала гнездования.
Таким образом, наблюдаемые нами биотопические различия в исходной плодови-
тости птиц могут объясняться определенной разницей в сроках начала гнездования и
в возрастном составе гнездового населения вида, что, в свою очередь, обусловливается
экологической спецификой биотопов, динамикой фенологического развития населяю-
щих их насекомых и растений (состоянием кормовой базы), наличием укрытий для
гнезд и выводков, конкурентными взаимоотношениями с представителями своего
и других видов птиц и т. д. Нельзя забывать и о том, что отмеченные биотопические
различия могут определяться как сроками прилета на места размножения и полового
созревания особей разного возраста, так и конкурентоспособностью молодых и старых
птиц при распределении по оптимальным и субоптимальным гнездовым стациям. Ранее
это было продемонстрировано на примере маркированной популяции чечевицы в Каре-
лии (Зимин, 19816).
Годовые колебания размера кладки
Фиксируемые исследователями годовые различия в величине кладки любого вида
воробьиных птиц могут определяться как объективными, так и субъективными причи-
нами. Первые опосредованно связаны с изменчивостью среды обитания в зонах зимовки
птиц, на путях их пролета и местах гнездования. Состояние погоды, ход весенних темпе-
ратур, особенности ветрового режима и снеготаяния определяют темпы и сроки разви-
тия растительности, обилие, разнообразие и доступность кормов, что влияет на время
начала размножения и его динамику в разные годы. Существенные перерывы в размно-
жении птиц наступают при внезапных похолоданиях. Низкие температуры и осадки
уменьшают обилие и доступность кормов, вызывают гибель гнезд и приводят к повтор-
ному гнездованию. Число поздних и восстановленных кладок зависит также от уровня
численности и активности животных — основных разорителей птичьих гнезд. Колеба-
ния численности птиц отражаются на их распределении по биотопам и на возрастном
составе. В годы повышенной плотности населения происходит освоение второстепенных
гнездовых стаций и увеличение доли первогодков, принимающих участие в размножении,
что приводит к снижению средней величины кладки. Однако ежегодные вариации этого
показателя могут иметь и субъективную основу. Более или менее жесткие рамки выбо-
ра изучаемых местообитаний в разные годы в зависимости от решаемых исследователем
задач могут привести к неоднозначности и несравнимости получаемых данных. Это
бывает, например, при развеске новых групп искусственных гнездовий (иные габариты
и биотопы) или смене мест проведения стационарных исследований. Таким образом,
причин, обусловливающих ежегодные различия в величине кладки птиц только в преде-
лах одной местности, достаточно много. Поэтому попытки объяснить наблюдаемые
отклонения влиянием только одного фактора, без комплексной оценки других, могут
привести к ошибочным заключениям. Действительно, организм птиц постоянно, в том
числе и в период откладки яиц, испытывает влияние многообразных, чаще всего
93
Таблица 25
Годовые изменения доли крупных кладок у мухоловки-пеструшки
Год Число гнезд Средняя величина кладки (число яиц) Средняя дата начала откладки яиц
всего с кладкой > 7 яиц
Шокшинский стационар
1971 20 20.0 5.90 5.VI
1972 36 27.8 6.03 31.V
1973 25 40.0 6.32 28.V
1974 28 21.4 5.85 5.VI
1975 12 42 6.59 31.V
1976 24 39.1 6.25 5. VI
1977 13 15 5.92 3.VI
О-в Б. Клименецкий (по: Хохлова, Головань, 1981)
1975 54 85.2 7.32 28.V
1976 67 55.2 6.33 3.VI
1977 45 48.9 6.49 31.V
Стационар , ,Маячино”
1979 14 14 5.69 3.VI
1980 56 26.8 6.00 6.VI
1981 75 40.0 6.41 29.VI
1982 85 48.2 6.33 4.VI
1983 104 73.1 6.79 22.V
взаимосвязанных внешних воздействий. Изменение одного из них так или иначе отра-
жается и на проявлении других, усиливая или подавляя их влияние на организм птиц
и, в частности, на плодовитость. Поэтому, рассматривая возможное влияние какого-то
фактора среды на величину кладки птиц, мы должны постоянно помнить, что его изме-
нение не могло бесследно пройти и для других связанных с ним явлений.
Годовые колебания числа откладываемых яиц в исключительных случаях бывают
2—3-кратными (хищные птицы и совы в „мышиные” и „голодные” годы). Среди воро-
бьиных птиц лишь у большой синицы различия в средней величине кладки в смежные
годы могут достигать 4 яиц (Lack, 1966), а у большинства остальных видов составляют
не более 1 яйца (16—20% при кладке из 5—6 яиц). Собранные нами данные позволяют
проанализировать причины таких колебаний на примере веснички, мухоловки-пеструш-
ки и зарянки.
Фактором, вызывающим годовые вариации средней величины кладки птиц, могут
быть прежде всего разные сроки начала размножения. При этом выявляется правило:
чем раньше начинается гнездование местных особей, тем больше бывает средняя вели-
чина кладки. Это установлено для многих видов воробьиных в различных частях ареала
и проявляется однозначно. Так, в Прионежье и на островах Онежского озера коэффици-
ент корреляции между числом крупных кладок мухоловки-пеструшки (из 7 и более
яиц) и средними датами начала ее гнездования в разные годы оказался равным
—0.69 ± 0.20 (р < 0.05). В приладожских лесах эта связь проявилась еще сильнее —
г = —0.78 ± 0.36 (р < 0.05), что, вероятно, обусловлено более контрастными годовыми
различиями в сроках начала размножения (табл. 25). В целом по Южной Карелии сред-
негодовая величина кладки мухоловки-пеструшки находится в обратной зависимости
от средней даты откладки первого яйца местными птицами. Данные по мухоловке-
пеструшке, собранные за 24 года в одном пункте Юго-Западной Финляндии (Haartman,
1969а), также демонстрируют удовлетворительную отрицательную связь среднегодовой
величины кладки со средней датой начала гнездования 50% самок (рис. 26, А). С запаз-
дыванием этой даты на 1 сут среднегодовая величина кладки сокращается на 0.15
яйца.
94
Таблица 26
Годовая изменчивость величины кладки у веснички в зависимости от сроков размножения
Год Число гнезд Средняя величина кладки (число яиц) Средняя дата начала откладки яиц (июнь) Пятидневка мас- сового начала размножения
всего с кладкой > 7 яиц (%)
1973 22 50 6.41 4 2-я* и 3-я
L974 25 16 6.00 11 4-я и 5-я
1975 17 23 6.12 2 2-я и 3-я
1976 36 25 6.17 8 3-я и 4-я
1977 65 21 6.06 4 3-я
1979 32 25 6.06 3 2-я и 3-я
1980 30 40 6.27 7 3-я и 4-я
1981 37 38 6.19 6 2-я и 3-я
1982 38 58 6.47 7 3-я
1983 51 47 6.25 4 2-я
* Вторая пятидневка - 26-30 мая.
Для веснички средние показатели рассчитаны на сравнительно небольшом материа-
ле и колебались по годам в меньшей степени, чем у предыдущего вида. Поэтому они
не дают четкой и достоверной связи: г - —0.28 (табл. 26). Слабо и также недостоверно
(г = —0.44 ± 0.31) связана доля крупных кладок веснички со сроками массового
начала размножения вида. При сравнительно небольших выборках, использованных
для расчета, существенное влияние на величину средних показателей оказывает число
поздних кладок. У веснички такие кладки, во-первых, ощутимо меньше нормальных,
а во-вторых, их количество значительно колеблется по годам независимо от дат начала
гнездования, его успешности и уровня численности вида. Если поздние кладки, начатые
после 15 июня, исключить из анализа, то средние показатели (дата начала откладки
яиц и размер кладки) оказываются связанными довольно тесно (г = —0.64 ± 0.29 при
р<0.05).
Аналогичная тенденция отмечена для зарянки: в годы с ранним началом гнездова-
ния средняя величина первой кладки составила 6.51 ± 0.07 яйца, а в годы с поздним
размножением — 6.35 ± 0.09 (различия статистически недостоверны,р> 0.05).
Однако, несмотря на относительно небольшой материал, вполне удовлетворитель-
ная отрицательная корреляция получена для величины кладок зарянки, откладка кото-
рых началась до 10.VI, со средней датой начала откладки в разные годы: г =-0.77 ±
±0.28 (р<0.05).
Стьернберг (Stjernberg, 1979), в течение 8 лет изучавший размножение чечевицы
в Южной Финляндии, не обнаружил статистически значимой корреляции между средней
По оси абсцисс — средняя дата начала размножения; по оси ординат - среднегодовая величина
кладки.
Рис. 26. Связь среднегодовой величины кладки мухоловки-пеструшки (А) и чечевицы (Б)
со сроками размножения (А - по: Haartman, 1969а; Б - по: Stjernberg, 1979).
95
величиной ее кладки и датами начала гнездования в разные годы. Однако если не учи-
тывать данных 1968 г., когда было необъяснимо много гнезд с крупными кладками,
то искомая связь проявляется вполне отчетливо (рис. 26, Б).
Увеличение числа откладываемых яиц при смещении начала размножения на более
ранние сроки известно для синиц (Лихачев, 1961, 1967, 1971; Lack, 1966; Louran, Frois-
sart, 1973; Blondel, Isenman, 1979) и мухоловки-пеструшки (Лихачев, 1961, 1967, 1971;
Головань, 1983; Jiirvinen А., 1984).
Следует отметить также, что смещение сроков размножения на более ранние не
у всех видов сопровождается увеличением числа яиц в кладке; например, это не обна-
ружено у скворца в Южной Финляндии (Haartman, 1982). Очевидно, у птиц, размер клад-
ки которых в течение гнездового сезона изменяется от минимума к максимуму и снова
к минимуму, более раннее размножение также не должно сопровождаться увеличением
плодовитости. Характерный пример дает белобровик в Карелии. Лихачев (1967), оцени-
вая результаты многолетних наблюдений в Рязанской обл., обратил внимание на то, что
в годы чрезмерно раннего начала гнездования самые первые кладки мухоловки-пе-
струшки бывают меньше последующих.
Иногда независимо от сроков размножения отмечаются резкие отклонения сред-
негодовых показателей размера кладки, вызванные какими-то другими факторами.
В частности, существует определенная зависимость плодовитости от условий зимовки
птиц. Так, серые куропатки в Англии после мягких малоснежных зим откладывали
в среднем 17.4 яйца, а после суровых — 15.2 (Lack, 1966). Аналогичная связь выявлена
и для наших тетеревиных птиц (Семенов-Тян-Шанский, 1960). В Польше наиболее круп-
ные кладки имели самки полевого воробья, для которых в предгнездовой период была
зарегистрирована высокая жирность (Pinowska, 1979). По мнению автора, такие самки
энергетически были лучше подготовлены к размножению. Ярвинен (Jarvinen А.,1984),
изучавший наиболее северную популяцию мухоловки-пеструшки, обнаружил прямую
связь величины кладки и других репродуктивных показателей маркированных самок
с массой их тела в первые дни после начала плотного насиживания. Эти данные, которые
мы, к сожалению, не можем прокомментировать на собственном материале, заслуживают
самого пристального внимания. Масса тела птиц и ее сезонная изменчивость могут слу-
жить элементарным индикатором энергетического состояния организма, а в репроду-
ктивный период — важным показателем энергетической обеспеченности размножения
особей, имеющих неодинаковые кладки и успешность гнездования.
Сезонная изменчивость величины кладки
За исключением птиц, имеющих постоянную величину кладки (кулики, голуби
и др.), ее размер у большинства видов варьирует в ходе сезона размножения. Измен-
чивость этого показателя для воробьиных птиц является достаточно строгим правилом,
в большей или меньшей степени проявляющимся ежегодно и в любой части ареала вида.
У чибиса (Klomp, 1951) и вяхиря (Lack, 1966) число яиц в гнездах в течение репро-
дуктивного периода остается неизменным. У сов в отдельные годы отмечается постепен-
ное нарастание величины кладки (Lack, 1947). Для полевого воробья также характерно
некоторое увеличение размеров вторых кладок, но причиной этого могут быть „болту-
ны” и „задохлики”, оставшиеся в гнездах после первого размножения и случайно учиты-
ваемые наблюдателями (Фетисов, Корзун, 1979; Фетисов, 1981). Для всех остальных
выделяют 2 основных типа сезонных изменений величины кладки: при первом наблюда-
ется постепенное сокращение ее к концу сезона, при втором сначала происходит увели-
чение числа откладываемых яиц, а после пика размножения — снижение (Klomp, 1970;
Perrins, 1970). К птицам с первым типом изменчивости кладки относят синиц, горихво-
стку, каменку, серую мухоловку, мухоловку-пеструшку, скворца, зяблика, вьюрка;
со вторым — жаворонков, дроздов, зарянку, полевого и городского воробьев,
деревенскую ласточку и др. Первый тип сезонной изменчивости величины кладки
96
Рис. 27. Сезонная изменчивость величины кладки птиц в таежной зоне (цифры в скобках -
число исследованных гнезд).
1 - большая синица (111); 2 - мухоловка-пеструшка (489); 3 - чечевица (236) в Южной
Финляндии (по: Haartman, 1969а); 4 - чечевица в Карелии (141); 5 - горихвостка-лысушка (81);
6 - зарянка (249); 7 - лесная завирушка (124); 8 - славка-завирушка (49); 9 - садовая славка
(129); 10 - лесной конек (109); 11 - серая мухоловка (108); 12 - певчий дрозд (246); 13 - чер-
ный дрозд (66); 14 - весничка (795); 15 - зяблик (160). По оси абсцисс - календарь; по оси
ординат - величина кладки (число яиц).
считается более характерным для птиц с моноциклическим, второй — с полицикличе-
ским размножением (Lack, 1957; Coulson, White, 1961). Однако у большой синицы,
рябинника, скворца и щегла, которые в центре ареала нормально выкармливают по два
выводка, величина кладки сокращается от начала к концу сезона (Klomp, 1970). Напро-
тив, среди птиц с моноциклическим размножением встречаются виды, у которых при
необычно раннем гнездовании самые первые кладки бывают меньше, чем отложенные
в обычные сроки. Для мухоловки-пеструшки это было показано Лихачевым (1967),
а для садовой славки и чечевицы демонстрируется нашими данными по Карелии
(рис. 27).
У зяблика большую часть гнездового периода размер кладки сохраняется практи-
чески на одном уровне и лишь при самых поздних сроках заметно уменьшается (Haart-
man, 1969а; Klomp, 1970; см. также рис. 27). Примерно по такой же схеме изменяется
в нашей зоне размер кладки лесного конька, а в Западной Европе — московки, певчего
дрозда и серой мухоловки (Klomp, 1970).
Преобладание крупных кладок в определенный отрезок времени объясняют дина-
микой кормовой базы птиц (Lack, 1966; Perrins, 1970). Например, у черных дроздов
в Англии период встречаемости наиболее крупных кладок совпадает с временем макси-
мального обилия и доступности их основного корма — дождевых червей (Lack, 1966).
В нашем регионе наибольшее число яиц дрозды (певчий, белобровик и черный) откла-
дывают во второй половине мая. До этого по утрам на почве еще обычны заморозки,
а позднее, в июне, почва начинает подсыхать и дождевые черви уходят в нижние гори-
зонты. По этой причине и в таежной зоне периоды встречаемости наиболее крупных
кладок дроздов и обилия их пищи примерно совпадают. Интересно, что в Англии птенцы
черного дрозда, вылупившиеся из яиц, которые были отложены в такие периоды,
характеризуются наибольшей массой тела (Snow, 1967).
97
7 В. Б. Зимин
30IV 31V
31VII
Рис. 28 Характер сезонной изменчивости величины
кладки у зарянки (Д). веснички (Б) и лугового конька
(В) в разных частях ареала (п - число гнезд).
А-Б: 1 - Дания, Швеция, Эстония, Финляндия,
Норвегия (п = 173), 2 - Англия (А: п- 1091, - по: Lack,
1946; Б: и = 467, - по: Cramp, 1955), 3 - Швейцария, Гер-
мания, Северная и Центральная Франция (/7 = 112), (7, 3 -
по: Lack, 1946), 4 - Карелия {А: п - 249; Б, п- 388, - на-
ши данные); В: 5 - 60-62° с. ш. (п - 32), 6
62 -64° с. ш. (и = 40), 7 - 64-66° с. ш. (п = 30), 8 - 66 -
68° с. ш. (п - 39), 9 - с. ш. (/? = 28) Стрелки
с символом широтной зоны - даты начала откладки яиц
(по: Pulliainen, 1977). По оси абсцисс - календарь; по
оси ординат - средняя величина кладки.
Однако многие виды воробьиных птиц, по крайней мере в северных широтах,
начинают гнездиться при относительном дефиците кормов и при неустойчивой погоде,
отражающейся на доступности пищи. Максимальное разнообразие и стабильность кормо-
вой базы наступают много позднее. Следовательно, для большинства наших воробьиных
птиц сезонные вариации размеров кладки и динамика развития кормовой базы имеют
противоположные направления.
У садовой славки и чечевицы размножение обычно начинается в первой декаде
июня, а иногда задерживается даже до 3-й пятидневки этого месяца. Смещение начала
откладки яиц на конец мая сопровождалось появлением маленьких кладок. И если
раньше (Зимин и др., 1978) мы отмечали постепенное уменьшение величины кладки
садовой славки к концу гнездового периода, то после двух ранних сезонов в последнем
пятилетии схема сезонной изменчивости приняла другой вид (рис. 27) .
У английских весничек (Cramp, 1955) и зарянок (Lack, 1946) размножение начи-
нают особи, откладывающие мало яиц. Позднее величина кладки достигает максимума
и лишь после этого снижается к концу репродуктивного периода. У тех же видов в Ка-
релии, а также у лугового конька в Северной Финляндии (Pulliainen, 1977) самые ранние
кладки бывают и наиболее крупными (рис. 28) .
Таким образом, типы сезонной изменчивости величины кладки, выделенные Клом-
пом-Перринсом, не являются строго видоспецифичными. По крайней мере у некоторых
видов птиц они не одинаковы в различных частях ареала и могут варьировать по годам
в зависимости от сроков размножения.
У трех избранных видов птиц (весничка, зарянка, луговой конек) на юге ареала
на первых этапах репродуктивного периода сначала идет нарастание средней величины
кладки, а потом, к концу сезона, ее уменьшение (II тип сезонной изменчивости по
Кломпу и Перринсу). На севере гнездовых областей величина кладки в течение репро-
дуктивного периода изменяется по I типу: после максимума в начале сезона среднее
число яиц быстро сокращается.
Привлекает также внимание и совпадение календарных периодов встречаемости
наиболее крупных кладок у птиц одного вида, обитающих на разных широтах (рис. 28).
98
Получается, что птицы северных популяций приступают к размножению в период, когда
на юге отмечаются наиболее крупные кладки за сезон. На примере лугового конька
на это явление обратил внимание Пуллиайнен (Pulliainen, 1977), связавший его с геогра-
фической изменчивостью величины кладки.
Анализ особенностей размножения маркированных особей нескольких видов
воробьиных птиц показал, что их плодовитость в наибольшей мере зависит от дат начала
размножения. Независимо от возраста местных особей и их индивидуальных репро-
дуктивных способностей при начале гнездования позже дат, определенных как средние
для популяции, они откладывали меньше яиц, чем в другие годы жизни, и наоборот.
С этих позиций можно попытаться объяснить снижение плодовитости птиц гор-
ных популяций. Особи одного вида, обитающие на той же широте и долготе, но в раз-
ных условиях — в высокогорье и на равнине, весной, очевидно, находятся в состоянии
одинаковой готовности к размножению. Однако на равнине и в предгорьях внешние
условия позволяют реализовать ее раньше, чем в горах, где размножение сдерживается
более поздними сроками фенологического развития природы. В результате птицы гор-
ных популяций приступают к гнездованию в период, когда на равнине уже начинается
закономерное сезонное сокращение числа откладываемых яиц. Тем не менее, как отме-
чает Ковшарь (1981а), в субвысокогорьях Тянь-Шаня сокращение величины кладки
и запаздывание сроков гнездования с увеличением высоты характерно не для всех видов
воробьиных птиц.
Величина кладки и уровень развития репродуктивного состояния
Как можно видеть из вышеприведенных материалов, число откладываемых яиц
у птиц одного вида, популяции и даже в течение жизни каждой особи варьирует в преде-
лах всего видового диапазона изменчивости этого показателя. Но если в географиче-
ских, возрастных, годовых и биотопических вариациях величины кладки мы не обна-
руживаем строгой определенности, то в ее сезонной изменчивости проявляется наиболее
общая и достаточно строгая закономерность. Прежде всего, сезонная изменчивость
размера кладки свойственна абсолютному большинству видов, за исключением неболь-
шой группы птиц, у которых число откладываемых яиц постоянно. В пределах каждого
вида, популяции и у отдельных особей изменчивость этого показателя проявляется
однозначно, одинакова из года в год, а также в любой части ареала. Кроме того, у птиц,
занимающих разное систематическое положение и резко различающихся особенностями
биологии, величина кладки в течение репродуктивного периода изменяется однотипно,
особенно во второй половине гнездового сезона. На фоне значительной неопределен-
ности общей изменчивости этого показателя такая высокая степень общности именно
в сезонной его вариабельности не может не привлечь внимания. Действительно, при всем
многообразии класса птиц для них выделяют всего 2 типа сезонной изменчивости вели-
чины кладки, что обосновано Кломпом и Перринсом (Klomp, 1970; Perrins, 1970).
Оба выделенных типа считались видоспецифичными, и к типу I относили всех
птиц, у которых в течение репродуктивного сезона средняя величина кладки постепен-
но сокращалась, а к типу II — птиц, начинающих размножение с маленьких кладок, раз-
мер которых сначала возрастал, а затем снижался к концу гнездового периода (см. рис.
29, Б\ min I—max—min II).
Обращает на себя внимание тот факт, что из двух кривых, отражающих типы сезон-
ной изменчивости величины кладки, только кривая II типа (рис. 29, Б) соответствует
динамике развития абсолютного большинства сезонных биологических явлений, кото-
рое идет по схеме min I—max—min II. На первый взгляд абсолютно не отвечает этой схе-
ме кривая I типа (рис. 29, А), однако ее можно рассматривать как правую половину
кривой, в которой редуцирована левая, начальная часть (рис. 29, В), В том, что это
действительно так, убеждают случаи появления кладок меньшей, чем обычно, величины
при необычно ранних сроках начала гнездования (рис. 29, Г). Это подтверждается и
99
Рис. 29. Первый (А) и второй (Б) типы сезонной изменчивости величины кладки и трансфор-
мации первого типа во второй при необычно ранних сроках размножения (Г).
В - первый тип изменчивости с нереализованными репродуктивными возможностями (пре-
рывистая линия). Стрелки указывают дату начала откладки яиц. По оси абсцисс - календарь;
по оси ординат - величина кладки.
результатами сравнения географической широтной трансформации типов сезонной из-
менчивости величины кладки у 3 видов воробьиных птиц (рис. 28).
Таким образом, величина кладки птиц в течение года изменяется по тому же
принципу, который типичен и един для сезонной вариабельности любых биологических
процессов. Но в современных климатических условиях не все птицы и не везде могут
полностью реализовать свои потенциальные репродуктивные возможности. В зависимо-
сти от степени редукции начальных этапов развития репродуктивного состояния вели-
чина кладки будет изменяться либо по более редуцированному I типу сезонной вариа-
бельности, либо по II типу, при котором репродуктивные возможности реализуются
гораздо полнее.
Очевидно, единый тип сезонной изменчивости величины кладки птиц был характе-
рен для всех (или большинства) видов в более теплые доледниковые периоды, когда
ведущее значение имела годовая динамика фотопериода, а колебания более высоких
температур не приводили к возникновению столь резко выраженной сезонности кли-
мата, как в настоящее время. Последовавшее затем глобальное похолодание привело
к существенным сдвигам в сроках и динамике развития кормовой базы птиц, сокраще-
нию общей продолжительности гнездового периода и его смещению на самое теплое
время года. При этом у части видов, особенно в южных широтах, редукция репроду-
ктивных возможностей была менее значительной. Во всяком случае, они сохранили,
хотя бы частично, способность начинать размножение на ранних стадиях развития репро-
дуктивного состояния. Но у других видов, особенно там, где благоприятные условия
для размножения наступают поздно, реализация начальных этапов формирования репро-
дуктивного состояния стала невозможной. Кроме того, в связи с развитием более
дальних сезонных миграций годовые циклы птиц усложнились. Возникла необходи-
мость в обязательной реализации весеннего миграционного состояния, предшествую-
щего размножению и препятствующего началу гнездования при наступлении благо-
приятных условий на зимовках и трассах пролета (Дольник, 1975). Так как миграцион-
ное состояние угнетает развитие репродуктивной активности, то у мигрирующих птиц
размножение не может начаться до реализации предшествующей весенней программы
годового цикла. Но после завершения миграций уровень потенциальной готовности птиц
к размножению быстро возрастает, хотя ее реализация может еще длительное время
сдерживаться внешними условиями на местах гнездования. Данные по состоянию гонад
птиц, добытых в зоне наших исследований в первые дни после возвращения на родину,
позволяют предположить, что миграция завершается здесь на низком уровне миграцион-
ного состояния и сравнительно высоком репродуктивном потенциале. Во всяком
случае, гонады самцов веснички в день их появления в Южной Карелии были вполне
развиты (4X5 мм), а при гистологическом обследовании в них обнаружен активный
сперматогенез (стадия образования сперматоцитов), но клоакальный выступ еще не
был развит (Лапшин, 1978). Примерно то же самое характерно и для самцов других
и не только воробьиных птиц. Яичники самок в первые дни после прилета развиты
много слабее, но размеры их быстро увеличиваются непосредственно перед началом
кладки.
100
Создается впечатление, что на севере после прйлета птицы способны очень быстро
перейти к размножению и что сдерживается это только внешними условиями. Доста-
точно интенсивное проявление токового поведения характерно для многих видов
птиц еще на пролете, а у зимующих птиц отмечается даже в разгар зимы (врановые,
дятлы, большая синица, пухляк, пищуха, оляпка и др.). В Петрозаводске мы наблюдали
ворон, пытающихся поправлять свои старые гнезда во время оттепелей в конце февра-
ля-начале марта. Хохлова (1976) на о-ве Б. Клименецкий в Онежском озере отметила
завершение постройки гнезда ополовниками к 8.VI. 1975, а в заповеднике „Кивач”
строящих гнездо птиц этого вида наблюдали в последних числах марта 1985 г. (устное
сообщение Л. Н. Захаровой и М. В. Яковлевой). Выше уже упоминалось о ранних
сроках проявления половой активности у различных вьюрковых птиц, содержащихся
в неволе на естественном фотопериоде.
В этой связи примечательно, что у разных видов птиц прилет в северные части
ареала приходится на тот же календарный период (или даже позднее), в который на юге
начинается размножение.
Все сказанное выше свидетельствует об определенном сходстве в сроках и дина-
мике развития репродуктивного состояния птиц одного вида, обитающего во внетропи-
ческих областях северного полушария. Под репродуктивным состоянием мы понимаем
весь комплекс внутренних процессов, обеспечивающих размножение.
Экспериментально доказано, что интенсивное увеличение массы семенников
у птиц из различных популяций начинается на разных фотопериодах (Ро) и имеет раз-
ную скорость (Дольник, 1975). Оказалось, что у птиц северных популяцийРо характе-
ризуется более высоким значением и повышенной скоростью нарастания массы гонад.
У растений и насекомых генетически закреплено (Данилевский, 1961), что также
вполне допустимо и для птиц (Дольник, 1975). Но строго фиксированная величина
PQ лишает систему контроля годовых циклов определенной гибкости. Так, при глобаль-
ном потеплении климата или других изменениях экологической обстановки благопри-
ятные для размножения условия могут наступить гораздо раньше, чем фотопериод
достигнет значения Ро, но птицы не смо.гут изменить сроки начала размножения.
Кроме Ро, очевидно, должен существовать еще скрытый период медленного
роста гонад, начинающийся у птиц всех популяций примерно в одно время. Такое
единство сроков может быть обусловлено историческими причинами. В теплые доледни-
ковые периоды размножение птиц могло начинаться и заканчиваться одновременно
на всех широтах в период, близкий к весеннему равноденствию, когда различия в длине
дня на разных широтах минимальны. Географические различия в величине Ро в таких
условиях были несущественны. Примечательно в этой связи, что и у современных птиц
на европейских зимовках начало слабого увеличения роста гонад становится заметным
в это же время года.
В зависимости от дальности и продолжительности миграций, развившихся после
глобального похолодания, длительность скрытого периода формирования репродуктив-
ного состояния будет неодинаковой у птиц разных популяций. Пока интенсивность
процессов, обеспечивающих миграции, велика,, развитие репродуктивного состояния
будет угнетено. С началом депрессии миграционного состояния активность процессов,
обслуживающих размножение, возрастает, что внешне выражается в быстром росте
гонад и происходит в зоне с соответствующим Ро. Миграции завершаются, очевидно,
на спаде уровня миграционного состояния от максимальных до нижних пороговых
значений при параллельном увеличении интенсивности процессов, ответственных за
размножение.
Окончание миграции у северных птиц более растянуто и часто сдерживается
неустойчивой погодой, но развитие репродуктивного состояния на этом этапе продолжа-
ется относительно независимо и к моменту прилета достигает достаточно высокого
уровня. Можно сказать, что на север ареала птицы прилетают потенциально более гото-
выми к размножению, чем на юг. Поэтому период от прилета до начала гнездования
на севере короче и составляет иногда лишь считанные дни. На родине уровень
101
A
Рис. 30. Принципиальная схема формирования репродуктивного состояния птиц в разных
широтных зонах ареала от крайних южных (А) до крайних северных (Г).
1-7 - периоды: 1 - фотостимуляция, 2 - начало формирования репродуктивного состояния,
3 - фотопериод Ро, 4 - начало благоприятных для размножения условий, 5 - динамика развития
миграционного состояния, 6 - не реализуемая и 7 - реализуемая стадии репродуктивного состоя-
ния. По оси абсцисс - календарь; по оси ординат - уровень миграционного и репродуктивного со-
стояний.
репродуктивного состояния продолжает повышаться независимо от внешних условий,
но этим достигается только потенциальная готовность птиц к размножению, для реа-
лизации которого необходимо экологическое, этологическое и энергетическое
обеспечение.
Если репродуктивное состояние развивается по принципу сезонной изменчивости
биологических процессов, а его уровень соответствует величине кладки, то этап его
формирования, на котором реализуется размножение, будет определять и весь дальней-
ший ход сезонной изменчивости числа откладываемых яиц (рис. 30). В ситуациях
Б, В, Г период депрессии и окончания миграционного состояния наступает после начала
формирования репродуктивного состояния, и поэтому видовая программа размноже-
ния не может быть реализована целиком, а ее начальные этапы в большей или меньшей
степени редуцируются — сначала угнетены высоким уровнем миграционной активности,
а после перехода через Ро и возвращения на родину сдерживаются внешними
факторами.
Предложенная нами принципиальная схема формирования репродуктивного со-
стояния птиц вполне соответствует рассмотренным выше кривым сезонной изменчи-
вости величины кладки (рис. 29) и удовлетворительно объясняет существование гео-
графических, сезонных, годовых и прочих вариаций этого показателя, его сокращение
в высокогорье.
Следует подчеркнуть, что данная схема отражает лишь самый общий принцип
изменчивости величины кладки, а конкретное число яиц, откладываемых каждой
особью, является результатом очень сложного взаимодействия внутренних процессов
в ее организме с многочисленными изменчивыми факторами внешней среды, зависит
от индивидуальных и популяционных особенностей годовых циклов, успешности
зимовки, общих кондиций в предгнездовой период и т. д.
При поздних сроках гнездования на севере, когда первые гнезда появляются
на спаде репродуктивного потенциала, среднегодовая величина кладки будет меньше
средней многолетней и соответственно меньше, чем в более южных широтах. Это может
объяснить некоторые исключения из правила Гопкинса, в частности для мухоловки-
пеструшки, у которой выше 66° с. ш. размер кладки меньше, чем в зоне 60—64° с. ш.
(Pulliainen, 1977; Брагин, Гилязов, 1984; Jarvinen А.,1984).
102
В центральных и южных частях ареала созревание половых продуктов самок
энергетически вполне может обеспечиваться при устойчиво теплой погоде уже на стадии
перехода репродуктивного состояния через весеннее пороговое значение (min I), т. е.
когда оно еще не достигло максимума. Сочетание физиологических процессов в орга-
низме в этот период будет еще далеко от идеального для размножения (например,
сочетание гормонов). Поэтому величина кладки в данный период не может быть макси-
мальной для вида. А так как число яиц в кладке будет здесь изменяться по II типу
сезонной динамики, то ее среднегодовое значение окажется невысоким. Особи, реали-
зовавшие первую, а у моноциклических видов и единственную попытку размножения,
уже не принимают участия в размножении. Этим объясняется сравнительно редкое
появление крупных кладок и менее высокие значения средних показателей в южных
зонах ареала, и тем самым подтверждается справедливость правила Гопкинса. Не исклю-
чено также, что любая попытка размножения, успешная или нет, каким-то образом
сдерживает дальнейшее развитие репродуктивного состояния и оно не может достичь
максимального уровня.
Репродуктивное состояние птиц южных и северных популяций имеет свои особен-
ности. На юге оно, видимо, поддерживается на более низком уровне в течение более
длительного периода, когда на величине кладки птиц может сказываться так называ-
емая физиологическая усталость. Кроме того, из-за ранних сроков становления репро-
дуктивного состояния к моменту его кульминации появляется большое число повтор-
ных и вторых кладок.
На севере ареала максимальный уровень репродуктивного состояния в целом
может быть выше, но период его действия — гораздо короче. Возможно, с этим связаны
заметные широтные различия в средней величине кладки в кульминационных точках
кривых репродуктивного состояния у популяций, обитающих на разных широтах
(рис. 28). Особенно ярко это выражено у зарянки, но вполне характерно и для веснич-
ки, и для лугового конька.
Сезонную изменчивость величины кладки птиц и ее сокращение к концу гнездо-
вого периода иногда объясняют физиологической усталостью организма. Действительно,
второе и повторное размножение птиц проходит на заметно более слабом эмоциональ-
ном уровне: реже, чем весной, отмечаются драки, заметно ниже интенсивность пения,
проявлений демонстрационного поведения и т. д. С развитием работ по индивидуаль-
ному мечению гнездящихся птиц накапливается все больше точных сведений о вариа-
бельности величины кладки отдельных особей в течение одного или нескольких после-
довательных сезонов размножения. Вопреки априорно сложившемуся мнению о мень-
шей величине повторных и вторых кладок, данные мечения не позволяют утверж-
дать это так однозначно (Haartman, 1969а; Haukioja, 1970; Зимин, 1978; Ковшарь,
1981а, и др.).
Как уже указывалось, ежегодные вариации в числе откладываемых яиц связыва-
ют с особенностями прошедшей зимовки и энергетической подготовленностью отдель-
ных особей к размножению. Показателем последней является масса тела птиц в пред-
гнездовой период. Можно полагать, что особи, которые могут достигать и достаточно
долго поддерживать массу тела на уровне, близком к максимальному для вида (по-
пуляции), отличаются от остальных лучшими способностями удовлетворять энергети-
ческие потребности организма при кормодобывании. В конечном итоге и сроки готов-
ности птиц к размножению, и величина их кладки, и качественные показатели яиц,
и успешность насиживания и выкармливания птенцов прямо или опосредованно свя-
заны с энергетикой взрослых особей, хорошим индикатором которой является масса
тела самок в предгнездовой и гнездовой периоды. Следует помнить, однако, что масса
тела птиц четко коррелирует с размерами, которые могут иметь определенное значение
для конкурентоспособности особи. Наиболее крупные из них скорее всего будут за-
нимать оптимальные для вида местообитания, где, как правило, все репродуктивные
показатели оказываются более высокими. Таким образом, влияние массы тела может
быть и опосредованным.
103
Плотность гнездового населения и величина кладки
В сложном и многообразном комплексе факторов, воздействующих на величину
кладки птиц, важное место нередко отводят плотности населения. Правда, результаты
проведенных в этом направлении исследований на первый взгляд кажутся противоречи-
выми. Лишь для немногих видов воробьиных птиц выявлена более или менее четкая
обратная зависимость величины кладки от плотности населения (Klujiver, 1951; Lack,
1957, 1966). Однако в других, гораздо более многочисленных публикациях, выполнен-
ных на разных видах европейских певчих птиц, констатируется отсутствие подобной
связи. Не подтверждается она и исследованиями, проведенными в Карелии и Скан-
динавии. Практически никаких различий в величине кладки в зависимости от
размеров групп птиц не было получено в небольшой серии опытов по разведению
чечеток в неволе. В одних вольерах птицы содержались группами по 2—4 пары
и вместе с другими вьюрковыми и овсянками, а в других - особи жили отдель-
ными парами. Тем не менее число яиц в гнездах тех и других было примерно
одинаковым. Одна и та же самка чижа в течение одного сезона содержалась
с самцом в отдельной вольере и отложила 3 яйца, а затем при гнездовании в общей
с другими птицами вольере снесла 5 яиц. При экспериментальном изучении влияния
плотности населения на плодовитость скворца было установлено, что в перенасе-
ленных вольерах величина кладки и общее число особей, участвовавших в раз-
множении, были достоверно меньшими (Risser, 1975). Однако при пересчете на
единицу пространства продуктивность размножения при повышенной плотности
населения оказалась втрое больше, что не согласуется с гипотезой ограничения
продуктивности размножения при увеличении численности. Правда, как скворец,
так и чечетка в природе могут гнездиться колониями или достаточно плотными
группами, поэтому „переуплотнение вольерных популяций” может не оказывать
на них существенного влияния.
Увеличением общей численности птиц можно также попытаться объяснить сокра-
щение величины кладки к концу периода гнездования. Действительно, и общая, и видо-
вая численность непрерывно нарастает в течение всего периода размножения и особенно
после вылета и начала расселения молодняка. И если выявленное Лэком (Lack, 1966)
на английских больших синицах сокращение средней величины кладки на 2 яйца про-
исходит уже при двукратном увеличении плотности гнездового населения, то не явля-
ется ли это доказательством связи сезонного сокращения плодовитости с быстрым
нарастанием общей численности птиц после вылета молодняка из гнезд? Ведь обычно
она возрастает в 4—5 раз, а около направляющих линий, где происходит концентрация
расселяющегося молодняка, даже стократно. При этом несомненно учащаются кон-
такты гнездящихся птиц с перемещающимися и резко обостряется трофическая конку-
ренция. Однако если бы это действительно оказывало на гнездящихся птиц какое-то
влияние, то в зонах резко повышенной численности вдоль направляющих линий птицы
должны были бы иметь более мелкие кладки. Общая численность птиц в таких местно-
стях постоянно много выше нормы. Весной длительное время после начала гнездования
продолжают лететь (и останавливаются на кормежку и отдых) птицы более северных
популяций, летом же в массе появляются расселяющиеся молодые, а также сменяющие
места гнездования и мигрирующие взрослые особи. Стационар в Приладожье располо-
жен как раз на такой направляющей линии, благодаря чему мы имеем возможность
сравнить величину кладки гнездящихся здесь птиц с аналогичными данными для других
пунктов Карелии, удаленных от побережий крупных водоемов. Результаты сравнения
не оставляют никаких сомнений в отсутствии влияния высокой численности переме-
щающихся вдоль побережья Ладожского озера взрослых и молодых птиц на величину
кладки местных особей. Точно так же на Куршской косе, где концентрация мигрирую-
щих и расселяющихся птиц еще больше, средняя величина кладки зяблика оказывается
такой же, как в материковых лесах Литовской ССР, — 4.82 и 4.85 яйца соответственно
(Дольник и др., 1982; Aleknonis, 1984). Таким образом, увеличение общей плотности
104
населения вида после вылета молодняка из гнезд не влияет на величину кладки
гнездящихся особей.
Однако на плодовитости птиц может сказываться лишь плотность гнездового
населения. А эта связь может проявиться лишь при высокой степени перенаселенности.
Между тем в таежной зоне, особенно в периферийных частях ареалов многих видов,
численность большинства птиц в гнездовой период слишком низка, чтобы можно
было ожидать ее влияния на размер кладки и другие показатели. Но даже для видов,
гнездовая плотность населения которых местами превышает 200—250 пар/км2, как
например у веснички в Южной Карелии и на севере Ленинградской обл., никакой опре-
деленной связи плотности населения с величиной кладки не выявляется. За последние
12 лет (1972—1983 гг.) в Южной Карелии среднегодовая величина кладки веснички
сохранялась примерно на одном уровне: 6.00—6.47 яйца при двукратных колебаниях
численности. Наиболее вероятными причинами изменения размеров кладки по годам
были разные сроки начала массового размножения вида. Некоторую роль играла также
общая продолжительность периода откладки яиц, с увеличением которой за счет
роста числа повторных кладок меньшей величины средний показатель несколь-
ко снижался. На Шокшинском стационаре при плотности населения весничек
56 пар/км2 средняя величина кладки составила 6.09, а при 116 пар/км2 -
6.20 яйца.
В связи с сукцессией растительности и увеличением возраста насаждений числен-
ность лесных птиц в сосново-лиственных молодняках Шокшинского стационара непре-
рывно нарастала до перехода их в стадию жердняков (Зимин, Кузьмин, 1980). Одно-
временно шел процесс постепенного выселения птиц открытых и полуоткрытых стаций.
Однако ни у тех, ни у других заметных изменений в плодовитости не произошло, хотя,
например, плотность населения зяблика за годы наблюдений возросла здесь с 0 до 64,
а зарянки — с 15 до 68 пар/км2.
Направления широтных различий величины кладки и плотности населения боль-
шинства наших птиц противоположны: к северу численность сокращается, а величина
кладки возрастает. В то же время у представителей северо-таежного орнитокомплекса
направление тенденций одинаково — у чечетки и белобровика, например, и численность,
и плодовитость в северном направлении возрастают (Merikallio, 1958; Haartman, 1969а;
Hilden, 1969; Чемякин, 1982а; Мальчевский, Пукинский, 1983; наши данные).
Как отмечает Лэк (Lack, 1957), нарастание плотности гнездового населения птиц
всегда происходит за счет увеличения доли первогодков, которые, как показано на при-
мере многих видов птиц, откладывают меньше яиц, чем особи старших возрастных
групп.
Итак, нам не удалось найти подтверждения реальности связи величины кладки
птиц с плотностью их гнездового населения. Трудно сказать с уверенностью, могла ли
у воробьиных птиц развиться в процессе эволюции регуляция численности через измене-
ние величины кладки. Не случайно абсолютное большинство воробьиных птиц воспи-
тывает по 3-5 птенцов и лишь немногие способны выкормить более крупные выводки.
Видимо, птицам с коротким жизненным циклом, быстрым эмбриогенезом, постэмбрио-
нальным развитием и ранним становлением половозрелости целесообразнее воспитать
несколько маленьких выводков, чем один крупный. Различные отклонения в величине
кладки от средневидовой нормы (сезонные, годовые, возрастные) у большинства пред-
ставителей отряда воробьиных редко превышают 1 яйцо, что не может оказать столь же
существенного влияния на общий уровень воспроизводства популяции, как например
полициклия размножения.
Регуляция числа выращиваемых молодых через воздействие плотности населения
скорее всего должна была сформироваться у животных с низкой скоростью воспроиз-
водства — медленными темпами развития и поздним наступлением половозрелости.
У многих из них, в частности у крупных представителей класса птиц, полицикличность
размножения невозможна даже теоретически, и один из основных путей изменения
продуктивности их размножения — это увеличение или уменьшение числа однократно
105
выкармливаемых потомков. В этом случае выработка механизма регуляции числен-
ности через изменение величины кладки (выводка) представляется более оправ-
данной.
Влияние возраста птиц на величину кладки
Чтобы не повторять приведенных ранее сведений по влиянию возраста птиц на их
плодовитость, мы ограничимся в данном разделу лишь изложением и оценкой собствен-
ных данных. Их относительно немного, что связано с низкой численностью птиц в зоне
исследований и несовершенством методов отлова открытогнездящихся птиц на гнездах.
Разработке этих методов мы уделяли особое внимание (Зимин, Артемьев, 1981).
При изучении возрастных различий в размножении птиц определенные трудности
возникают также из-за отсутствия для многих массовых видов (пеночки, славки, ряд
овсянок и др.) надежных критериев дифференциации первогодков (adi) и особей стар-
ших возрастных групп (ad2 ). Многолетнее кольцевание и отлов мухоловок-пеструшек
с точно известной датой рождения убедили нас в том, что и для этого вида использова-
ние общепринятых методов определения возраста (Svensson, 1975, 1978; Виноградова и
др., 1976) дает сравнительно большую ошибку. Поэтому во многих случаях мы вынуж-
дены были руководствоваться лишь результатами многолетнего кольцевания местного
населения птиц, которое у веснички и мухоловки-пеструшки контролировалось на
80-95 % (Зимин и др., 1983).
В табл. 27, 28 представлены материалы за последние годы, когда наблюдения за
судьбой гнезд выполнялись особенно тщательно, а отлов взрослых особей на гнездах
производился в абсолютном большинстве случаев. Материал по разным возрастным
группам птиц оказался неравноценным, отражающим явное преобладание первогодков
в гнездовом населении птиц наших стационаров. Только у белобровика доля ad2 в отло-
вах на гнездах была выше. В большинстве случаев выявленные на нашем материале
возрастные отличия в основных параметрах размножения были статистически мало
значимы, что может быть объяснено как их реальной несущественностью, так и недостат-
ком сравниваемых данных.
Лишь двухлетние самки мухоловки-пеструшки откладывали достоверно больше
яиц, чем особи, впервые гнездящиеся в Карелии. Но число птенцов, выкормленных
до вылета из гнезд, в обеих возрастных группах не различалось (табл. 27). Можно было
бы ожидать, что пары мухоловок-пеструшек, состоящие из партнеров в возрасте
Таблица 27
Изменчивость величины кладки и выводка у мухоловки-пеструшки
в зависимости от возраста
Пол и возраст Кладка (число яиц) Выводок (число птенцов)
п М ± т п М ±т
Самка
а 39 6.20 ± 0.12 40 5.12 ±0.12
а +1 43 6.56 ± 0.13 44 5.15 ±0.37
а + 2 14 14
а +5 3 | 6.39 ± 0.28 3 j 5.61 ± 0.43
а +4 1 1
Самец
а 65 6.49 ± 0.10 67 5.66 ± 0.23
а +1 68 6.35 ± 0.13 71 5.45 ± 0.23
а + 2 19 19
а + 3 9 | 6.62 ± 0.18 9 | 5.59 ± 0.37
а + 4 1 1
106
Таблица 28
Зависимость величины кладки и выводка от возраста партнеров
у некоторых воробьиных птиц Карелии
Пол и возраст Кладка (число яиц) Выводок (число птенцов)
n lim M ± m n lim M ± m
Зарянка*
9 ad, 1 41 1 5-8 I 6.61 ± 0.10 50 1-8 1 5.96 ±0.17
9adj | 16 1 6-7 | 6.50 ±0.13 1 22 J 1-7 1 5.68 ±0.37
Большая синица*
9adj 49 9-14 11.16 ± 0.16 66 3-12 8.70 ± 0.24
9ad2 49 10-14 11.70 ± 0.42 14 5-12 8.29 ± 0.58
dadj — — 41 4-12 8.56 ± 0.26
dad2 — — — 36 3-12 8.58 ± 0.40
Весничка
dadj 9adx 92 4-8 6.45 ± 0.06 97 1-7 5.68 t 0.11
dad2 9adj 39 5-7 6.33 ± 0.11 37 3-7 5.62 ± 0.23
dadj 9 ad2 17 5-7 6.29 ± 0.17 14 3-7 5.64 ±0.39
dad2 9ad2 17 5-7 6.59 ±0.18 13 3-7 5.77 ± 0.34
9adx 131 4-8 6.42 ± 0.05 134 1-7 5.68 ± 0.11
9ad2 34 5-8 6.44 ± 0.12 19 3-6 5.70 ± 0.25
Белобровик
dadj 9adt dadj 9ad2') 7 5-6 5.28 ± 0.19 6 3-6 4.33 ± 0.46
dad2 9ad1J 14 4-6 5.07 ± 0.13 14 2-6 4.28 ± 0.32
dad2 9ad/ 18 3-6 4.95 ± 0.21 17 2-6 4.35 ± 0.26
9adt 18 4-6 5.17 ± 0.12 16 2-6 4.44 ± 0.23
9ad2 21 3-6 4.95 ±0.18 21 2-6 4.24 ± 0.24
Мухолов ка-пестру шка
dadt 9adj 100 4-9 6.63 ± 0.09 104 1-8 6.06 ± 0.06
dad2 9adt 14 5-9 6.64 ± 0.30 14 2-8 5.93 ±0.41
dadj 9ad2 31 4-9 6.81 ± 0.16 29 2-8 6.28 ± 0.26
dad2 9ad2 63 4-8 6.65 ± 0.09 62 2-8 6.19 ± 0.14
* Учтены только результаты первого цикла размножения.
ad2, будут иметь максимальное число яиц в кладке и наиболее крупные выводки, но
данные не подтверждают этого (табл. 28).
Очень сложно комментировать материалы для других видов птиц, так как разли-
чия в величине кладки и выводка в гнездах самок и самцов разного возраста чаще
всего были очень малы и противоречивы. Так, самки-первогодки белобровика и зарян-
ки в целом имели несколько более крупные кладки и выводки, чем птицы старших
возрастных групп. Одинаковой оказалась величина кладки и выводка у самцов и самок
весничек, причем как у уже гнездившихся на стационарах в предыдущие годы, так и у
впервые гнездящихся (табл. 28).
Старые самки большой синицы в среднем откладывали больше яиц и воспитывали
более многочисленные выводки, чем первогодки, но данных по старым птицам в наших
сборах было недостаточно, поэтому различия статистически недостоверны. Аналогичные
результаты для этого вида получены и в Юго-Западной Финляндии (Haartman, 1969а).
Старые птицы (и =45) откладывали здесь 10.02 ± 0.17 яйца, а первогодки 9.65 ± 0.39.
Однако в более южных широтах возрастные различия проявляются вполне определенно
(Klujiver, 1951; Lack, 1966). Вероятнее всего, отмеченные расхождения между птицами
южных и северных широт обусловлены тем, что на севере ареала особи разного возраста
начинают гнездиться в одно календарное время (гл. II), а на юге старые самки приступа-
ют к размножению раньше молодых. Как было показано в предыдущей главе, в отдель-
ные годы до 80—90 % гнездового населения некоторых видов наших птиц приступают
к откладке яиц очень дружно — за 1—2 пятидневки. В таких условиях сближенность
107
дат начала размножения старых и молодых особей может служить существенным пре-
пятствием для проявления возможных возрастных различий в плодовитости.
Так как на гнездах в большинстве случаев отлавливлись обе взрослые птицы,
мы получили возможность сравнить репродуктивные показатели гнездовых пар разного
возрастного состава. Однако материалы по мухоловке-пеструшке, весничке и белобро-
вику не позволяют сделать какие-либо определенные заключения — средние величины
кладки и выводка варьировали незначительно и без заметной связи с возрастом партне-
ров по размножению (табл. 28).
Индивидуальная изменчивость величины кладки и выводка у меченых особей
Многолетнее мечение птиц на одной территории и ежегодный контроль местного
гнездового населения птиц наших стационаров наряду с подробным изучением истории
их гнезд позволяют оценить возрастную изменчивость репродуктивных показателей
другим методом - изучением индивидуальной изменчивости величины кладки и вывод-
ка в течение жизни отдельных меченых особей.
У 46 самок мухоловки-пеструшки, гнездование которых было прослежено два
или более года подряд, кладка с увеличением возраста птицы на 1 год в 20 случаях была
больше на 1—2 яйца, в 16 — не изменялась и в 10 — уменьшалась на 1—2 яйца. Различия
в средних показателях оказались статистически мало значимыми (табл. 29). Аналогич-
ные данные получены для маркированных самцов этого вида: увеличение возраста
на 1 год не оказывало никакого влияния на размер кладки.
Выполненный указанным методом анализ данных по весничке также не выявил
существенной связи размеров кладки и выводка с возрастом птиц, хотя слабая тенден-
ция к сокращению средних показателей при увеличении возраста на 1 год оказалась
однозначной у особей обоего пола (табл. 29).
В гл. II подчеркивалось, что даты начала гнездования индивидуально меченых
птиц значительно варьировали в разные годы их жизни и что как у мухоловки-пеструш-
ки, так и у веснички средняя величина кладки быстро сокращается от начала к концу
репродуктивного периода. Сопоставив величины отклонения от дат начала откладки
яиц меченых птиц в смежные годы с разницей в числе отложенных ими яиц (и выкорм-
ленных птенцов), мы обнаружили, что эти показатели взаимосвязаны. У мухоловки-
пеструшки эта связь выражалась отрицательной корреляцией, примерно одинаково
проявляющейся у самцов и самок при анализе данных по величину кладки и выводка
(рис. 31, А-Г).
Таблица 29
Изменение величины кладки и выводка при увеличении возраста меченых птиц на 1 год
Кладка (число яиц) Выводок (число птенцов)
п lim М ± т п lim М ± т
Мухоловка-пеструшка, 9
В год мечения или отлова 44 4-9 1 6.32 ±0.13 43 3-8 5.58 ± 0.20
На следующий год 46 4-9 1 | 6.56 ± 0.14 42 1 4-8 е.98 ± 0.18
Мухоловка-пеструшка, д
В год мечения или отлова 62 I 1 4-9 6.47 ± 0.10 73 1-8 1 5.78 ± 0.13
На следующий год 66 1 4-9 6.42 ± 0.14 93 2-8 6.01 ±0.12
Весничка, 9
В год мечения или отлова 17 1 5-8 6.65 ± 0.21 ! 1 16 1 5-7 6.25 ± 0.20
На следующий год 28 1 5-8 6.43 ± 0.16 1 27 1 2-7 5.61 ± 0.28
Весничка, д
В год мечения или отлова 1.7 5-8 6.65 ± 0.20 18 4-7 I 5.67 ± 0.24
На следующий год 21 5-8 6.33 ± 0.16 22 2-7 5.27± 0.30
108
Е
2-
__।_____1 , I, _______1___.-i—
-30 -10 10' 20
-1 - ...
• •
~2 -
Рис. 31. Соотношение различий в сроках начала откладки яиц и в величине кладки (в вывод-
ке) у меченых особей в смежные годы.
Величина кладки в гнездах с мечеными: А - самками, Б - самцами мухоловки-пеструшки;
Д - самками, Е - самцами веснички; размер выводка мухоловки-пеструшки: В - самок, Г - сам-
цов. По оси абсцисс - запаздывание (+) или опережение (-) сроков откладки первого яйца по срав-
нению с предыдущим годом (сут); по оси ординат - увеличение или сокращение числа отложенных
яиц (выкормленных птенцов).
109
Рис. 32. Примеры индивидуальных колебаний величины кладки меченых мухоловок-
пеструшек (А) и весничек (Б) в разные годы жизни (указаны цифрами).
Кружки, соединенные сплошными линиями, - данные для отдельных особей; прерывистая
линия - случай бигамии (d). По оси абсцисс - календарь; по оси ординат ~ величина кладки (число
яиц).
Аналогичная тенденция выявляется и на небольшом материале по весничке (рис.
31, Д, Е). Иначе говоря, при увеличении возраста особи на 1 год смещение календарных
дат появления первого яйца в гнезде на более поздние по сравнению с предыдущим
сезоном приводит к сокращению числа откладываемых яиц, и наоборот.
Если судить по результатам исследования Аноровой (1976), возраст, с которого
начинается редукция плодовитости мухоловки-пеструшки, составляет 4 года. Посколь-
ку такие старые птицы встречались у нас крайне редко, то возрастные различия в вели-
чине кладки, если они действительно существенны, должны были бы проявиться у боль-
шинства исследованных нами особей. Но, как следует из приведенных выше данных,
одинаковое увеличение возраста птиц (на 1 год) влияло на размер их кладки неодно-
значно. В большей мере на плодовитости птиц сказывалось изменение календарных
(индивидуальных) дат начала гнездования (рис. 31, 32). Однако и эта связь у разных
особей проявлялась неодинаково. Таким образом, годовые вариации величины кладки
у отдельных маркированных птиц зависели не только от анализируемых нами
факторов. В частности, и индивидуальные, и ежегодные колебания плодовитости могли
быть связаны также с различиями условий, в которых каждая особь находилась до
начала размножения (особенности прошедшей зимовки, состояние погоды и кормовой
базы на трассе пролета и в предгнездовой период на родине). Все это могло отразиться
на уровне энергетической готовности к размножению, хорошим показателем которой,
как уже указывалось, могут быть масса тела и величина подкожных жировых
резервов в прегнездовой период. По предварительным данным, подобная связь
намечается для весничек, отловленных на наших стационарах. В 1973 г. у 89 птиц,
исследованных в мае, при средней массе тела 9.14 г средняя величина кладки
была 6.41 яйца,, а в 1976 г. при массе 8.93 г (п = 328) - 6.17. В сравниваемые
годы различались и средние даты откладки первого яйца: в 1973 г. — 4.4. VI,
в 1976 г. - 7.6. VI.
Таким образом, одни и те же самки в различных ситуациях продуцируют разное
количество яиц. В каждом конкретном случае величина кладки является результатом
сложного взаимодействия организма самки с целым комплексом внешних факторов,
оказывающих и положительное, и отрицательное влияние на плодовитость птиц.
ПО
Продуктивность кладок разной величины
На исходную продуктивность размножения птиц - величину кладки кроме наслед-
ственной обусловленности постоянно влияют многочисленные внешние факторы,
воздействующие на организм в течение всего процесса формирования яиц (а вероятнее
всего — и много раньше) и их инкубации. Подобному контролю со стороны внешней
среды подвергается и следующий этап размножения — выкармливание гнездовых
птенцов. О результативности этих фаз мы можем судить вполне объективно по данным
наблюдений за судьбой отдельных гнезд. Однако по числу слетков оцениваются только
начальные этапы размножения, тогда как итог последующих фаз — докармливание вы-
водков, выживаемость молодых птиц в период становления самостоятельности и далее
до начала их гнездования — в подавляющем большинстве случаев объективно оценить не-
возможно. Попытки отдельных исследователей использовать для этих целей данные коль-
цевания и цветного мечения с постоянным визуальным контролем в течение года или
результаты возвратов молодых птиц на места рождения в следующую весну (Lack, 1966;
Williams, 1966, и др.) представляются недостаточно объективными, так как не учитыва-
ют реально существующей дисперсии молрдняка, проявляющейся уже на ранних
этапах онтогенеза. Обоснованные Носковым /(1969, 1981) миграции расселения молод-
няка свойственны многим видам птиц и являются обязательным этапом их онтогенеза.
Иное дело, что проявлению стремления к расселению, как и любому другому биологи-
ческому явлению, свойственна разнокачественность и что на данном этапе разные
особи одного вида (популяции, выводка) оказываются более или менее подвижными.
Но так или иначе, большая часть молодых особей покидает места рождения после распа-
дения выводка, еще до начала или на первых этапах постювенильной линьки. Конечно,
о выживаемости молодняка можно в какой-то мере судить и по тем особям, которые
задерживаются вблизи от гнезд; но, во-первых, еще никто не смог точно определить,
какая часть птиц остается, а какая покидает участок обитания родителей или погибает,
и, во-вторых, судя по отловам крупногабаритными ловушками в Гумбарицах, интенсив-
ность перемещений молодых птиц очень изменчива по годам, причем это непостоянство
оказывается не связанным ни с уровнем численности, ни с успешностью размножения
птиц. Тем не менее миграции расселения молодняка бывают достаточно интенсивными,
ежегодно совершаются примерно в одно время и, если судить по их суточной ритмике,
могут иметь значительную протяженность при разной направленности.
По величине возврата местных молодых птиц на места рождения также нельзя
в полной мере судить о выживаемости в течение первого года жизни, так как современ-
ными методами не удается определить размеры дисперсии, а тем более годовые вариа-
ции этого показателя. Поэтому оценка результатов размножения вынужденно прово-
дится не по его окончательному итогу, каковым является число особей, благополучно
доживших до начала первого гнездового сезона, а только по доступному для объектив-
ной оценки показателю — числу слетков.
При постоянном контроле со стороны внешней среды размер кладки и величина
выводка приводятся в соответствие с особенностями местных условий. Число слетков
на гнездо определяется балансом между влиянием неблагоприятных условий среды
и усилий взрослых птиц на всех предыдущих этапах годового и гнездового циклов.
И если при этом сила влияния хотя бы одного внешнего фактора на одном из этапов
годового цикла окажется непреодолимой для взрослых особей, то результативность
размножения окажется ниже потенциально возможной (или оно будет неуспешным),
т. е. здесь вновь проявляется действие правила Либиха—Блэкмана. Поэтому средние
величины кладки и выводка в обычных условиях, в частности в таежной зоне, не имеют
и не могут иметь абсолютного соответствия. Годовые вариации этих показателей не
были синхронны ни у одного из исследованных нами видов птиц, и между ними не об-
наружено никакой корреляции.
Сила воздействия большинства факторов среды в разные годы или в течение од-
ного сезона размножения не остается постоянной, что является следствием прямого
111
Таблица 30
Продуктивность кладок разной величины у мухоловки-пеструшки в спелых насаждениях
(над чертой) и в сосново-лиственных молодняках (под чертой) Южной Карелии
Число яиц Число птенцов Общая успеш- ность гнездова- ния (%) Доля слетков в популяции (%)
в 1 кладке всего вылупи- лось вылетело из гнезд (%) среднее на 1 гнездо
Q 27 26 85 7.3 81 1.2
У 18 9 4.0 — 1.5
О 224 214 95 7.26 91 11.3
О 8 0 0 — — —
q 917 870 93 6.2 . 88 45.0
287 222 72 3.9 56 29.8
г 708 645 91 5.0 83 32.7
О 420 321 79 3.6 61 47.6
с 195 168 87 3.8 75 8.2
Э 150 121 88 3.5 71 19.8
Л 48 35 80 2.3 58 1.6
16 8 - 1.7 - 1.3
* Абсолютное число птенцов.
или опосредованного влияния неустойчивости внешних условий в зоне наших иссле-
дований. Выше уже приводился пример с зарянкой, которая в необычно теплую весну
1982 г. начала гнездиться раньше известных сроков. Из-за начавшегося в конце мая и
продолжавшегося в течение первой декады июня похолодания результативность раннего
размножения оказалась значительно ниже нормы. Повторное гнездование началось
позднее обычных сроков, что привело к снижению общего уровня воспроизводства, так
как, во-первых, величина повторных кладок и выводков оказалась меньше, чем при нор-
мальных сроках, а во-вторых, из-за позднего окончания выкармливания птенцов боль-
шинство пар не приступило ко второму размножению. В итоге средняя величина кладки
в этом году была выше, чем в другие годы, а размер благополучного выводка —
меньше.
Таким образом, и величина кладки, и размер выводка в значительной мере зави-
сят от того, в каких условиях они формировались. А так как влияние отдельных факто-
ров очень изменчиво, то и соответствия средних показателей этих величин вряд ли сле-
дует ожидать.Не случайно поэтому продуктивность крупных кладок часто оказывается
меньше, чем более мелких, что многими авторами объясняется в основном влиянием
пищевого фактора (Lack, Arn, 1947; Haartman, 1954; Lack, 1957, 1966; Owen,
1960). Иногда, правда, приводятся некоторые уточнения. Так, Джиб (Gibb, 1955) для
большой синицы показал, что при раннем гнездовании смертность птенцов не зависит
от величины выводка, но при позднем — более легкие птенцы из крупных выводков
погибали чаще. Не обнаружено связи успешности выкармливания птенцов с их числом
в гнезде у певчего и черного дроздов, зарянки и горихвостки (Lack, 1966). У мухолов-
ки-белошейки уровень смертности в маленьких выводках оказался выше: гнезда с 5,
6, 7 птенцами дали соответственно 78, 85 и 94% слетков, что объяснялось фенотипиче-
ской адаптированностью величины кладки (Lohrl, 1957).
Результативность гнездования при разной величине кладки и выявление наиболее
продуктивных кладок изучено нами на нескольких видах воробьиных птиц Карелии
(табл. 30, 31). Данные по мухоловке-пеструшке демонстрируют существенные различия
в величине всех показателей для птиц, гнездящихся в оптимальных для вида стациях
и в малоблагоприятных биотопах. Выводки из 3—8 птенцов в нормальных для вида
условиях воспитываются без существенных потерь. Более того, успешность выкармли-
вания выводков из 6—8 птенцов выше, чем меньших. В несвойственных для
112
г=-0.в1±0.16
о
о°
О ООО
---1 L° I
21V 30V 10VI
Рис. 33. Соотношение средней величины кладки со средней датой
начала откладки яиц у мухоловки-пеструшки в Южной Карелии.
„ Л 7.05
По оси абсцисс - средняя дата начала кладки; по оси ординат -
среднегодовая величина кладки (число яиц). 6.75
6.45
мухоловки-пеструшки местообитаниях, в которых до развески в.15
синичников она не гнездилась, успешность выкармливания 5Q5
птенцов снижалась при увеличении размера выводка. Более вы-
ражена и тенденция к росту птенцовой смертности в крупных
выводках, хотя среднее число слетков также связано прямой положительной зависи-
мостью с величиной кладки.
Важным показателем результатов размножения вида представляется также общий
вклад в воспроизводство популяции. Так, в нашей зоне более 3/4 всего молодняка
дают мухоловки-пеструшки, наименее уклоняющиеся от среднепопуляционного значе-
ния величины кладки. Правда, в оптимальных для вида гнездовых стациях основной
прирост достигается за счет кладок из 7, а в субоптимальных — из 6 яиц. В наших усло-
виях мухоловки-пеструшки могут выращивать в среднем наибольшие выводки благо-
даря крупным кладкам, но это, очевидно, требует значительных усилий от взрослых
птиц, и поэтому воспитание крупных выводков не получает широкого распространения.
Кроме того, число кладок из 7 яиц и более тесно коррелирует со средней датой начала
гнездования г = —0.81 ± 0.16 (р< 0.01), а за последние 12 лет, при колебаниях средней
даты начала кладки от 22.V до 6.VI, фактически только в 1983 г. размножение было
необычайно ранним (рис. 33).
Суммируя приведенные данные, можно отметить, что у веснички, садовой славки,
зяблика и чечевицы наиболее часто встречающиеся кладки являются и самыми про-
дуктивными. В более южных зонах это установлено также для большой синицы, ла-
зоревки, скворца, снегиря и стрижа (Lack, 1966; Klomp, 1970). У ряда других видов ве-
личина самых продуктивных кладок чуть превышает наиболее распространенные. Это
характерно для белобровика, мухоловки-пеструшки, зарянки, горихвостки и лесной
завирушки. Южнее это отмечено для певчего и черного дроздов и мухоловки-белошей-
ки (Snow, 1958; Lack, 1966; Klomp, 1970). Только у серой цапли (Owen, 1960) размер наи-
более продуктивного выводка оказался меньше самого обычного в популяции.
Различия между самыми продуктивными и наиболее часто встречающимися в по-
пуляции размерами кладки во всех случаях не превышают 1 яйца, т. е. птицы, делающие
наибольший вклад в воспроизводство, в меньшей степени отклоняются от среднепо-
пуляционных показателей,и их усилия на размножение наиболее сбалансированы с мест-
ными условиями.
Попытки некоторых особей выкормить больше птенцов, чем обычно, ограничива-
ются уже на первых этапах репродуктивного цикла. Большая продолжительность пери-
ода откладки яиц в гнезде увеличивает вероятность их гибели от разорения хищниками.
Следствием преждевременного начала насиживания является увеличивающаяся асин-
хронность вылупления птенцов (потенциально большая в крупных кладках), обычно
приводящая к повышенной смертности потомства в первые дни жизни. Как было
показано выше, смертность птенцов, доживших до 6-8-дневного возраста, в дальней-
шем в выводках разной величины выравнивается. Однако в более южных широтах
при высоких плотностях населения вида и относительно более продолжительном пе-
риоде ночного покоя у ряда видов птиц размер выводка ограничивается повышением
птенцовой смертности. Чаще всего результативность чрезмерно интенсивного размноже-
ния оказывается такой же, как у менее плодовитых особей. Таким образом, увеличение
размеров кладки выше популяционной нормы оказывается непроизводительной тратой
времени и энергии птиц.
В более резкой форме это, очевидно, должно проявляться при рассмотрении сезон-
ной изменчивости продуктивности кладок и выводков разной величины. Мы попыта-
лись провести подобный анализ на мухоловке-пеструшке, у которой в Карелии при
8 В. Б. Зимин
113
Рис. 34. Сезонная изменчивость средней величины
выводка у мухоловки-пеструшки в молодых (1) и спе-
лых (2) насаждениях (3 - недостоверная средняя).
По оси абсцисс - дата откладки первого яйца; по
оси ординат - средняя величина выводка.
относительно коротком репродуктивном периоде
средняя величина выводка сокращается со ско-
ростью 0.6 птенца за каждую декаду (рис. 34).
Результаты сравнения оказались не столь велики,
как ожидалось, и недостоверны (табл. 32). Замет-
на лишь слабая тенденция к снижению продуктив-
ности больших кладок к концу сезона размножения, но в это время они крайне мало-
численны, что можно рассматривать как свидетельство высокой адаптированности
вида к фенологической изменчивости местных условий. Во всяком случае при
Таблица 31
Продуктивность кладок разной величины у некоторых воробьиных птиц Южной Карелии
Величина кладки Число гнезд Общая успешность гнездования (%) Средняя величина выводка Доля слетков в по- пуляции (%)
Зарянка
3 1 — — 0.2
4 9 58.3 3.2 1.8
5 35 76.0 3.8 11.2
6 110 81.5 4.9 45.7
7 78 85.3 5.98 39.2
8 5 57.5 4.6 1.9
9 2 — — —
Весничка
4 3 — — 1.4
5 17 78.8 4.0 9.3
6 64 72.1 5.4 38.7
7 66 77.0 6.4 49.7
8 1 — — 0.8
Лесная завирушка *
3 5 33.0 1.0 1.4
4 13 48.1 2.8 6.9
5 76 68.9 4.0 62.0
6 26 62.8 4.7 27.1
7 2 65 4.5 2.5
Горихвостка-лысушка
5 12 46.6 4.7 12.8
6 23 45.6 5.2 28.9
7 20 67.1 6.3 43.1
8 9 45.8 5.6 15.1
Садовая славка
3 8 70.8 2.8 6.1
4 39 59.8 3.3 28.5
5 55 61.8 4.5 61.0
6 3 — 4.0 4.3
Зяблик
3 3 — 2.5 1.3
4 25 55.0 3.7 13.7
5 92 69.6 44 80.0
6 6 50.0 4.5 4.5
7 1 - 2* 0.5
* Абсолютное число птенцов.
114
10V 30V 20VI 10VII 10V 30V 20VI 10VII 10V 30V 20VI 10V11
Рис. 35. Фенотипическая изменчивость успешности выкармливания выводков разной вели-
чины (цифры у кривых) у мухоловки-пеструшки (А), большой синицы (Б) и веснички (В) в Ка-
релии (без учета разоренных гнезд).
По оси абсцисс - дата откладки первого яйца; по оси ординат - успешность выкармливания
(доля слетков от числа вылупившихся птенцов, %).
Таблица 32
Сезонная изменчивость продуктивности кладок и выводков разной величины
у мухоловки-пеструшки (Маячино, 1984 г.)
Величин кладки и выводка Число слетков (%) в гнездах с началом откладки яиц в указанные периоды
11-20.V 21-31.V 1-10.VI после 11 .VI
Кладка
(число яиц)
8-9 91.5(13) 82.6(18) - (1) —
7 93.6 (40) 86.6(63) 88.8(28) —
6 - (3) 83.3(70) 80.1(41) 70.0(10)
5 78.6(13) 86.2(16) 38.7(13)
4 — - (1) - (2) 63.9 (9)
Выводок
(число птенцов)
8-9 95.4(11) 93.7(14) - (1)
7 97.6(36) 92.6(52) 91.4(25) —
6 -(6) 84.0(66) 83.9(29) - (5)
5 -(2) 91.8(22) 94.8(22) 77.1(7)
4 — - (1) - (1) 78.1(7)
Примечание. В скобках дано число исследованных гнезд.
своевременной откладке 7—8 яиц мухоловки-пеструшки не испытывают особых затруд-
нений с воспитанием такого же количества птенцов.
Не обнаруживается сезонного снижения результативности выкармливания вывод-
ков разной величины и у веснички, у которой более крупные выводки также характер-
ны в основном для двух первых декад после начала кладки (рис. 35).
У большой синицы успешность воспитания выводков из 11 птенцов и менее опре-
деленно снижается к концу сезона. Наиболее часто встречающиеся выводки из 12 птен-
цов даже при повторном и втором гнездовании выкармливаются успешно (рис. 35).
Более крупные выводки встречаются лишь у птиц, начавших откладку яиц до
середины мая, и при ранних сроках дают 93.7 ± 2.0% слетков от числа отло-
женных яиц, а при начале гнездования после 10 мая достоверно меньше —
83.8 ± 4.5%.
В крупных выводках птенцы, как правило, весят меньше, чем в мелких, и с этим
обычно связывают их пониженную выживаемость в послегнездовой период (Gibb,
1955; Lack, 1957, 1966; Williams, 1966; Ricklefs, 1967). Исключением из правила является
лишь мухоловка-пеструшка, у которой в крупных выводках птенцы также весят мень-
ше, но это не влияет на их смертность в гнездовой и послегнездовой периоды (Haartman,
1954, 1967а; Tompa, 1967; см. также табл. 32/. На места рождения возвращается пример-
но половина мухоловок-пеструшек, воспитанных в выводках от 4 до 7 птенцов. Как
было отмечено выше, величина выводка самбк этого вида также не влияла на их выжи-
ваемость в течение года.
115
Вернувшиеся на родину молодые веснички в 7 случаях из 8 были выкормлены
в самых обычных для Карелии выводках — из 5—7 птенцов (1 особь была выкормлена
в выводке из 3 птенцов).
Выводки минимальных для популяции размеров оказываются наименее продук-
тивными не только из-за исходно малого числа потомков. Как явствует из данных
табл. 30—32, общая успешность гнездования и выкармливания птенцов в таких вывод-
как часто оказывается ниже, чем при выращивании более многочисленного потомства,
причем такое иногда случается даже в середине гнездового сезона, хотя, казалось бы,
в это время можно было ожидать очень высокой результативности. Среди особей, отли-
чающихся низкой плодовитостью, преобладают повторно размножающиеся, потерпевшие
неудачу в первой попытке. С другой стороны, выше показано, что повторные кладки да-
леко не всегда бывают меньше предыдущих. Птицы, откладывающие мало яиц, — это
особи, по каким-то причинам задержавшиеся с началом размножения. Не исключено, что
большинство таких птиц начинают гнездиться на спаде репродуктивного состояния,
когда в организме начинается депрессия процессов, ответственных за размножение.
Кроме того, еще одну группу малопродуктивных особей составляют птицы с ис-
ходно низкой репродуктивной активностью и неспособностью к выкармливанию даже
нормального числа птенцов. Это, возможно, энергетически ослабленные, больные или
травмированные птицы. Они в разгар гнездового сезона откладывают мало яиц и не
могут обеспечить пищей всех птенцов даже в маленьких выводках.
У всех рассмотренных нами видов птиц особи, величина кладки которых отлича-
ется от средней на ±1 яйцо, дают абсолютное большинство слетков. Их вклад в воспро-
изводство популяции составил (%): у горихвостки 88, у мухоловки-пеструшки 89—
97, у садовой славки 94, у зарянки и лесной завирушки по 96, у веснички и зяблика
по 98. Следует обратить внимание на то, что у наиболее многочисленных в наших широ-
тах видов — веснички и зяблика — особей, отклоняющихся по величине кладки от видо-
вой нормы, совсем немного. Не исключено, что это может служить особым показателем
прогрессивности вида на данном этапе эволюции и одной из причин сохранения высо-
кого уровня численности на большей части ареала.
Особи, воспитывающие больше или меньше слетков, чем в среднем для популяции
выводке, по крайней мере у рассмотренных нами видов воробьиных птиц в таежной
зоне, не могут оказать существенного влияния на общий уровень воспроизводства.
В связи с этим важно было бы знать, наследуется ли уровень плодовитости. На диких
птицах это до сих пор окончательно не выяснено. По первому впечатлению, время нача-
ла гнездования молодых птиц, окольцованных птенцами, влияло на величину их кладки
больше, чем размеры выв'одков, в которых они были воспитаны. Тем не менее суще-
ствуют и всегда проявляются (в том числе и при одновременном начале размножения)
определенные индивидуальные различия в плодовитости разных особей. Но и они могут
возникать из-за неодинаковой массы тела и разной энергетической готовности птиц
к гнездованию и не обязательно имеют наследственную основу. Известно, однако, что
в крупных выводках масса тела слетков воробьиных птиц бывает меньше, чем в мел-
ких. И если особь в течение всей жизни сохранит массу своего тела на низком уровне,
то величина ее кладки не сможет быть большой. Получается, что потомки плодовитых
родителей не могут иметь высокий репродуктивный потенциал. Соответственно особи
из маленьких выводков, обладая исходно большей массой, должны, согласно Пинов-
ской и Ярвинену (Pinowska, 1979; Jarvinen А., 1984), быть более плодовитыми, чем их
родители. Тогда следующее поколение таких птиц вновь окажется малопродуктивным,
и т. д. При такой смене продуктивных и малопродуктивных поколений могла бы быть
достигнута общая сбалансированность плодовитости в популяции. Но, как было указано
выше, особи, резко уклоняющиеся от средней нормы плодовитости, настолько малочи-
сленны, что не оказывают существенного влияния на общий уровень воспроизводства.
Кроме того, неизвестно, действительно ли особи из крупных выводков сохраняют
малую массу тела на протяжении всей жизни. Все эти вопросы требуют тщательного
исследования. Ясно, однако, что наследуется не конкретная величина кладки, а весь
116
присущий виду диапазон числа откладываемых яиц. Если бы наследование было
жестким, то вряд ли могла бы возникнуть строгая сезонная вариабельность плодови-
тости — достаточно гибкая стратегия размножения в непостоянной среде обитания.
В таких условиях жесткое наследование величины кладки привело бы к непроизводи-
тельным затратам на гнездование и снизило бы общий уровень воспроизводства.
Стратегия плодовитости птиц не была бы столь сбалансированной, какой она стала
благодаря эволюционной поддержке вариабельности величины кладки в пределах
вида, популяций и индивидуальной жизни особи.
Для оценки результативности размножения иногда указывают оптимальный раз-
мер кладки. Но если этот показатель рассматривать как наиболее соответствующий
местным условиям, то он фактически ничего не дает. В любой точке зоны умеренного
климата и севернее условия существования очень изменчивы. Поэтому оптимальные
величины кладки или выводка могут оказаться чрезмерными, например, при существен-
ной задержке начала размножения. В неустойчивой среде обитания мы должны рассчи-
тывать оптимальную величину кладки для каждого биотопа, сезона, года, с учетом
возраста партнеров и т. д. Кроме того, что это значительно усложняет анализ, возникает
необходимость в увеличении объема фактических данных, что в условиях низких плот-
ностей населения птиц не всегда возможно.
Разумеется, и оценку продуктивности кладок разной величины следует проводить
более дифференцированно, что мы и попытались сделать для веснички и мухоловки-
пеструшки (рис. 35, табл. 32). Выявленные различия оказались статистически недосто-
верны, поэтому мы вынуждены обсуждать проблему продуктивности кладок разной
величины по суммарным данным.
Крайне важно определить также назначение и биологический смысл откладки
большого числа яиц, превышающего видовую норму. В какой мере это выгодно для
благополучия вида и как чрезмерная интенсивность воспроизводства отражается на
дальнейшей выживаемости взрослых птиц и их потомства? В поставленных вопросах
заключены основные противоречия в диалектике сложных взаимоотношений продук-
тивности и ежегодной смертности, результат взаимодействия которых в конечном
итоге определяет динамику численности вида, в свою очередь оказывающую влияние
и на плодовитость, и на величину смертности.
Основное противоречие возникает между видом в целом и особью, так как то,
что часто (но не всегда) оказывается полезно для вида, оборачивается гибелью для
особи, затрачивающей чрезмерные усилия на воспроизводство. Материалы массового
кольцевания птиц на гнездах показывают это с достаточной очевидностью (Snow, 1958;
Perrins, 1965; Klujiver, 1966; Williams, 1966; Haartman, 1971; Hussel, 1972; Askenmo,
1979). Правда, подобная связь выявляется далеко не всегда (Klujiver, 1963; Lack, 1966;
Haukioja, 1970; Furrer, 1977; Ricklefs, 1977). Существует, однако, мнение, что невозвра-
щение птиц, выкормивших выводки увеличенных размеров, может быть обусловлено
не только их повышенной смертностью. Они могут изменить свое отношение к террито-
рии, на которой их репродуктивные усилия были чрезмерными (Askenmo, 1979).
Известно также, что повышение плодовитости у многих животных происходит
при снижении плотности их населения — и наоборот. Но при плотности населения, пре-
вышающей определенные нормы, происходит и соответственное увеличение смерт-
ности; таким образом, противоречие между видом (популяцией) и особью решается
в сложном взаимодействии
плодовитость численность смертность
и во многом определяется условиями существования. Повышенная плодовитость особи
может быть и полезна, и вредна для вида в зависимости от уровня численности. Огра-
ничение интенсивности воспроизводства численностью, очевидно, возможно лишь после
какого-то определенного уровня последней, и поэтому в северных зонах ареала птиц
эта связь не проявляется. Однако высокая продуктивность размножения птиц северных
117
популяций может иметь негативное значение на местах зимовки, общих как
с местными, так и с более южными популяциями птиц того же вида.
По-видимому, неодинаковы и затраты энергии на воспитание даже самых круп-
ных выводков. Известно достаточно много примеров успешного выкармливания таких
выводков в наиболее благоприятные сезоны и случаев полной гибели таких же и гораз-
до меньших выводков, например,при очень ранних сроках размножения. При одинако-
вой величине кладки результативность размножения может быть разной в оптималь-
ных и субоптимальных местообитаниях и т. д.
Таким образом, ответ на поставленные выше вопросы не может быть однознач-
ным. Вероятнее всего, для вида полезно иметь особей с повышенной плодовитостью,
не допуская их чрезмерного распространения и доминирования. Механизм такого
ограничения заключается в увеличении смертности таких особей и их потомства (Williams,
1966).
Связь плодовитости, численности и смертности отчетливо проявляется лишь
в оптимальных для вида условиях, где плотность населения его достаточно высока.
В северных периферийных зонах ареала подобная связь имеет свою специфику, с одной
стороны, обусловленную низким уровнем плотности населения, а с другой — предпола-
гаемой повышенной смертностью птиц северных популяций, доказанную пока лишь
для большой синицы (Orell, Ojanen, 1979). Сокращение численности широко распростра-
ненных видов птиц в направлении с юга на север показано Новиковым (1960). Что ка-
сается широтной изменчивости ежегодной смертности, то определенных доказательств
ее увеличения у птиц по мере проникновения к северу пока не получено, за исключе-
нием большой синицы в Финляндии. Тем не менее для повышенной смертности птиц
северных популяций существует целый ряд предпосылок. Часть из них связана с укоро-
ченным вегетационным периодом: при дефиците времени годовой цикл проходит
более напряженно, и энергоемкие фазы — размножение—линька и линька—миграции —
совмещены и проходят при менее благоприятных, неустойчивых, а нередко и экстре-
мальных условиях. Следствием неполной линьки контурного оперения становится
некоторое ухудшение теплоизоляционных свойств покровов, что должно повышать
затраты энергии на терморегуляцию в зимний период. При неполной смене маховых
перьев ухудшаются лётные качества крыла, что вряд ли проходит без последствий при
возрастающей дальности миграций (в д. Гумбарицы таких птиц мы отлавливали глав-
ным образом только осенью).
Другие предпосылки к повышенной смертности птиц северных популяций свя-
заны с увеличением дальности миграций (Lack, 1957). Так, вероятность их гибели на пу-
тях пролета возрастает в связи с большей протяженностью миграционных трасс, на пре-
одоление которых требуется больше времени и энергии. Соответственно увеличивается
вероятность совпадения миграций с периодом внезапных похолоданий. Потенциально
возрастает и возможность отклонений от генерального направления пролетных путей.
Следует подчеркнуть, что зимовки северных птиц также расположены в зонах
с более суровым и неустойчивым климатом. Из-за низкой плотности населения на ме-
стах размножения „северяне” на зимовках могут оказаться менее конкурентоспособ-
ными, чем представители южных популяций, адаптированные к жизни в условиях
высокой численности вида. Кроме того, на весеннем пролете и на местах размножения
северные птицы чаще попадают в критические ситуации в связи с возвратами холодов,
нередко сопровождающихся массовой гибелью местных и пролетных особей.
Разумеется, приобретенные „северянами” адаптации в какой-то мере способствуют
преодолению и дефицита времени, и более суровых условий. Но и адаптации не беспре-
дельны, а частота критических ситуаций, в которые попадают птицы северных популя-
ций, значительно увеличивает вероятность их гибели на всех этапах годового цикла.
Однако до настоящего времени это не получило подтверждения расчетами смертности
различных видов птиц в разных частях ареала. Причина этого, на наш взгляд, заключа-
ется в том, что основные методы определения ежегодной смертности птиц справедливы
лишь для популяций, представляющих собою закрытые саморегулирующиеся системы,
118
все процессы в которых осуществляются только за счет внутренних ресурсов. И если
замкнутость такой системы нарушается эмиграцией или иммиграцией птиц, то
большинство современных методов расчета смертности становятся неприем-
лемыми.
Таким образом, решение вопроса об определении ежегодной смертности птиц и ее
географической изменчивости оказывается в непосредственной связи с проблемой
гнездового консерватизма и дисперсии особей, — проблемой, поднятой и неоднократно
обсуждавшейся Мальчевским (1957, 1968, 1969, 1974а, 19746, 1975, 1977). По мнению
Мальчевского, гнездовой консерватизм и дисперсия — две стороны эволюционного про-
цесса — едины и неотделимы друг от друга. Основная функция первого — поддержание
стабильного генофонда, второго — препятствие инбридингу и удовлетворение стремле-
ния вида к расселению и освоению новых территорий.
Дисперсия — важная сторона эволюции животных, в наиболее яркой форме про-
являющаяся именно в классе птиц, представители которого обладают наибольшей
подвижностью среди животных. Иное дело, что на современном этапе развития орнито-
логии она пока доказывается менее многочисленными фактами, чем консерватизм.
Многолетнее мечение и контроль гнездового населения птиц по понятным причинам
проводятся на ограниченной территории, причем точки массового кольцевания в значи-
тельной мере разобщены. Поэтому данные кольцевания позволяют в большей
мере судить лишь о возврате птиц на места мечения, а расселившиеся за пре-
делы контролируемых территорий птицы обычно исчезают для кольцевателей
бесследно. Для изучения дисперсии нужны принципиально новые методы, а дан-
ные, полученные с помощью кольцевания, единичны и в значительной мере
случайны.
Многолетние исследования на территории, изобилующей периферийными областя-
ми и границами ареалов большого числа видов птиц, кроме прямых данных кольцева-
ния, позволяют привести некоторые косвенные доказательства существенной роли
дисперсии в формировании местного населения птиц, подтверждающие основные идеи
А. С. Мальчевского.
Выше было показано, что население птиц на периферии ареала отличается крайним
непостоянством, что в областях нерегулярного гнездования вида годы относительного
обилия чередуются с годами полного отсутствия птиц, что с этим связаны „колебания
границ ареалов”, что периферийные популяции не могут существовать без ежегодного
притока птиц извне, что, вопреки, традиционному мнению, местное население многих
видов птиц непостоянно даже в течение одного гнездового сезонами т. д. Все это в значи-
тельной мере является следствием постоянно действующей дисперсии особей. Расселе-
ние любого вида за пределы ареала — его расширение и колебания границ распростра-
нения — вообще было бы невозможно, если бы территориальные связи были только
консервативными.
В какой-то мере дисперсия свойственна не только молодым, но и взрослым
особям. Об этом свидетельствуют как данные отловов расселяющихся видов птиц,
так и апериодичность в формировании временных поселений за пределами основного
ареала (старые особи в годы полного отсутствия вида, очевидно, оседают в более юж-
ных зонах).
Постоянное мечение и контроль гнездового населения птиц наших стационаров
дает как прямые, так и косвенные свидетельства ежегодной смены территории отдель-
ными взрослыми птицами. Их высокая подвижность в летний период доказана данными
массовых отловов крупногабаритными ловушками на направляющих линиях (Носков
и др., 1975; Резвый, Савинич, 1978; Лапшин, 1981а; Резвый, 1983). Кроме того, обна-
руживается непостоянство связи с гнездовой территорией в течение сезона размножения
у отдельных особей зарянки, веснички, лесной завирушки и др. У лесной завирушки,
например, состав гнездового населения во второй половине лета обновляется более
чем на треть, причем вновь появившиеся птицы заселяют те же участки леса, на которых
проходило размножение местных (меченых) завирушек.
119
Как отмечалось нами ранее (Зимин, 1981а), в условиях низкой численности
и дисперсного распределения по территории у многих видов певчих птиц в зонах их
спорадического гнездования формируется особый тип территориального поведения -
бродяжничество, функцией которого, очевидно, является активный поиск партнера
по размножению (иволга, болотная камышевка, сверчки и др.).
Наконец, ежегодное практически полное обновление гнездового населения ста-
ционарно контролируемых территорий, достигающее у некоторых видов 60—100%,
также может быть объяснено значительным влиянием дисперсии (Зимин, Лапшин,
1976; Зимин, Кузьмин, 1980).
Прямых доказательств расселения местных молодых птиц у нас мало, так как
в силу определенных обстоятельств размеры контролируемых территорий явно недо-
статочны для определения масштабов дисперсии птиц. О том, что они достаточно велики,
можно судить хотя бы на примере гнездования в ФРГ двух мухоловок-пеструшек,
окольцованных птенцами в Финляндии (Jedicke, 1978) и в заповеднике „Кивач”.
На юго-восточном берегу Ладожского озера с 1968 г. отлавливается ежегодно
по 12-25 тыс. птиц. С 1979 г. параллельно велось кольцевание птиц в урочище Маячино,
в 17 км к северу от первого пункта, на той же направляющей линии, которой является
берег Ладоги. Здесь птиц отлавливают преимущественно паутинными сетями и на гнез-
дах (по 3—8 тыс. в год). В обоих пунктах кольцевали также гнездовых птенцов воро-
бьиных птиц (зябликов, весничек, мухоловок-пеструшек, зарянок и др.). Получены
отдельные сведения по взаимному обмену особями, родившимися в одном пункте,
а гнездившимися — в другом. Для мухоловки-пеструшки, кроме того, установлен
постоянный обмен взрослыми особями. Но подобные данные по каждому из названных
видов пока единичны и недостаточны для обсуждения.
Резюмируя изложенные выше доказательства дисперсии птиц в северных зонах
ареала, можно с достаточной уверенностью заключить, что периферийные популяции
птиц формируются целиком или в значительной мере за счет особей, родившихся где-то
в зонах постоянного гнездования вида. Северные популяции многих воробьиных
вообще не могут существовать без ежегодного притока птиц извне. Таким образом,
по крайней мере в северных частях областей распространения популяции птиц являются
системами, открытыми для эмиграции и иммиграции, без учета которых невозможно
установить реальную величину ежегодной смертности птиц.
120
Г Л А В A IV
МОНО- И ПОЛИЦИКЛИЧЕСКОЕ РАЗМНОЖЕНИЕ
По числу выводков за сезон птицы делятся на две большие группы: виды с моно-
циклическим гнездованием, воспитывающие 1 выводок, и с полициклическим, имею-
щие 2 или более выводков. Внутри второй группы можно выделить птиц с би-, три-
и полициклическим размножением. Считается, что птицы одного вида в зависимости
от возраста, сроков начала кладок, биотопа и общей продолжительности сезона, благо-
приятного для гнездования, могут изменять стратегию размножения, выступая и как
моно-, и как полициклисты. В частности, долгое время бытовало мнение, что у северных
пределов ареала бициклическое гнездование вообще невозможно. Эта точка зрения
поддерживается и в новейших руководствах по птицам.
Знакомство с региональными и крупными фаунистическими сводками, а также
с публикациями по биологии отдельных видов птиц в разных частях ареалов позволяет
убедиться в отсутствии единства взглядов по поводу моно- и полициклии гнездования
птиц. Очень часто заключение о полициклическом размножении дается без достаточных
оснований. На это для западноевропейских популяций воробьиных обратил внимание
Лорль (Lohrl, 1970). Другая крайность — это устоявшееся предубеждение против воз-
можности двукратного размножения вида. Например, абсолютная уверенность в невоз-
можности двукратного размножения обыкновенной пищухи в Карелии привела к тому,
что мы совершенно не обращали внимания на тревожные крики пищух во второй поло-
вине лета, памятуя об аналогичной реакции на человека и у молодых птиц. Но после
того как сначала в первых числах августа был отловлен слеток этого вида, а затем
найдено и гнездо, кладка в котором началась после 15 июня, мы стали метить взрослых
птиц во время весеннего размножения и предприняли специальные поиски их вторых
гнезд. В итоге выяснилось, что практически все гнездовые пары (по крайней мере 80%),
успешно воспитавшие первые выводки, гнездятся у нас вторично, причем в отличие от
других видов вторые гнезда пищухи строят на значительном, до 1 км, удалении от пер-
вых. Обнаружить их бывает нелегко еще и потому, что второй выводок самки, как
правило, выкармливают без самцов и редко тревожатся у гнезд с маленькими птенцами
(во время насиживания они вообще не проявляют признаков беспокойства даже при
появлении человека в непосредственной близости от гнезда).
Основные причины разногласий по вопросу о числе выводков у исследуемого
вида птиц за сезон размножения кроются в несовершенстве методики. До сих пор
отсутствуют надежные способы отлова открытогнездящихся птиц на гнездах, не разра-
ботаны четкие критерии выделения и методы выявления моно- и полициклического
гнездования. Нередко основанием для решения вопроса служат такие показатели,как
длительность репродуктивного периода, продолжительность активного пения и его дина-
мика, наличие двух или более пиков размножения и т. д., а ведь это может быть обу-
словлено не только полициклическим гнездованием. Двух- или трехпиковые кривые
динамики откладки яиц — еще не доказательство, а только повод для более тщательного
исследования числа выводков за сезон.
В связи с необходимостью выделения критериев полициклического размножения
прежде всего важно определить значение постоянно употребляемых терминов „повтор-
ное” и „второе” гнездование.
„Повторными” или восстановленными кладками, как принято, мы считаем случаи
повторной откладки яиц после гибели первоначально отложенных яиц или после потери
121
выводка. Здесь, однако, и возникают весьма существенные трудности. Обычно раз-
множение называют успешным при благополучном вылете из гнезда хотя бы одного
птенца. Между тем в природе может произойти гибель всего выводка до его распаде-
ния. Естественно, в данном случае размножение считать успешным нельзя. Но дело
в том, что, во-первых, мы не всегда знаем об этом и, во-вторых, не учитываем, как
гибель слетков воспринимается их родителями. Судя по всему, потеря выводка в после-
гнездовой период имеет для птиц такое же значение, как и отсутствие успеха на пре-
дыдущих этапах гнездования. И если к этому времени птицы еще сохраняют способ-
ность к размножению, то она и реализуется как повторная кладка. Тем не менее
если судьба первого выводка остается для наблюдателя неизвестной, то такая повтор-
ная кладка будет воспринята им как вторая. Подобный случай был описан для садовой
славки, одну из меченых пар которой обнаружили у нового гнезда после вылета птен-
цов первого выводка, пришедшегося на период дождливой, холодной и ветреной пого-
ды, что, вероятно, послужило причиной гибели слетков (Зимин и др., 1978). Таким об-
разом, может возникнуть видимость доказательства факультативной полицикличности
гнездования, которую фактически следует оценивать лишь как повторное размножение,
связанное с гибелью выводка после вылета из гнезда. Поэтому, кроме обязательной
регистрации фактов двух или более последовательных размножений у меченых особей
в течение одного репродуктивного периода, важным условием прямого, абсолютно
строгого доказательства полициклии должен быть отлов (или достоверная регистра-
ция любым другим способом) достигающих самостоятельности потомков. Получение
подобных доказательств, конечно, сложно, но в ряде случаев совершенно необходимо,
особенно для периферийных популяций и видов с недостоверно выявленной способ-
ностью к полициклии.
Косвенным, но вполне строгим доказательством полициклии может служить так-
же наличие готового к откладке яйца у самок,3 пойманных на гнездах с птенцами или
при докармливании слетков. Разумеется, такие доказательства можно получить лишь
в тех случаях, когда птицы совмещают 2 последовательных цикла размножения. В выяв-
лении полициклии могут помочь также достоверные наблюдения случаев спаривания или
строительства новых гнезд мечеными особями, имеющими гнездовых птенцов или
слетков.
В принципе большинство птиц из отряда воробьиных потенциально способны
к бициклии. Продолжительность активности гонад у птиц, гнездящихся в таежной
зоне, вполне достаточна для этого, а относительный дефицит времени, испытываемый
ими на родине, может быть преодолен совмещением двух смежйых фаз годового
цикла, как это имеет место у многих птиц на севере. Действительно, случаи выкармли-
вания дополнительных выводков изредка выявляются у представителей видов с ти-
пично моно циклическим размножением. Так, у веснички было обнаружено, что одна
из 800 окольцованных на гнездах самок гнездилась дважды за лето (Лапшин, 19816).
Так же у садовых славок: 1 пара из 100 имела в наших условиях второй выводок.
Трижды двукратное выкармливание потомства отмечено для зяблика (п = 96) в Кали-
нинградской обл. (Дольник и др., 1982). Единичные случаи бициклии зарегистрированы
в центре европейской части СССР и для мухоловки-пеструшки (Лихачев, 1955; Аноро-
ва, 1976). Хотя для трех последних видов доказательства бициклии не были абсолютно
строгими (судьба первых выводков после вылета из гнезд не прослежена), можно
допустить, что при благоприятном стечении обстоятельств отдельные особи способны
ко второму размножению. Вместе с тем при нормальной моноциклии реальный прирост
от второго размножения (большей частью случайного и крайне редко встречающегося)
фактически не оказывает никакого влияния на общий баланс воспроизводства попу-
ляции. Среди видов с облигатной полициклией дважды за лето выводят птенцов до 80 %
3 Непосредственно перед откладкой яйца сильно увеличивается клоакальный выступ, прини-
мающий соответствующую форму; кишечник, смещенный за счет увеличения яичника и яйцевода,
рельефно выделяется под кожей брюха.
122
гнездовых пар зарянок и обыкновенных пищух и до 100% больших синиц. По всей
вероятности, моноциклическое размножение большинства видов птиц вполне обеспе-
чивает баланс их плодовитости и ежегодной смертности на уровне, достаточном для
поддержания стабильной численности. Второе размножение, особенно в условиях дефи-
цита времени, создает, видимо, чрезмерную напряженность годового цикла, к которой
птицы с моноциклическим гнездованием недостаточно адаптированы. Неизвестно, как
это отражается на выживаемости взрослых птиц и их потомства, но скорее всего второе
размножение у птиц с моноциклическим гнездованием не поддерживается отбором.
В противном случае оно получило бы более широкое распространение в этой
группе птиц.
Как считает Ковшарь (1981а), строгой грани между птицами сравниваемых групп
проводить не следует, так как, кроме сказанного выше, для каждого вида птиц с поли-
циклическим гнездованием известны географические, биотопические, возрастные
и ежегодные вариации числа выводков. В принципе к увеличению числа выводков по-
тенциально способно большинство воробьиных птиц, но реализуется это только у опре-
деленных видов. Поэтому от облигатной полицикли и, необходимой для
осуществления положительного баланса плодовитости и смертности и поддерживаемой
эволюцией, следует отличать факультативную полицикли ю, не оказыва-
ющую влияния на общий уровень воспроизводства и его баланс с ежегодной смерт-
ностью.
Такой подход, на наш взгляд, поможет устранить разногласия, часто возникаю-
щие при попытках классифицировать птиц по рассматриваемому признаку. В северных
частях видового ареала полициклия размножения может быть факультативной, а в цент-
ре и на юге — облигатной. Но на севере, как было показано выше, периферийные посе-
ления птиц существуют в основном за счет более или менее постоянного притока особей
из южных областей, так что для стабилизации численности характер размножения
имеет здесь меньшее значение, чем в центре ареала.
Кроме средне- и североевропейских птиц, указанных в табл. 33, случаи полицикли-
ческого размножения известны еще для ряда видов, не гнездящихся в наших широтах,
а также для представителей зоны тундры. Среди последних факультативная полициклия
установлена для пуночки на Новой Земле (Успенский,, 1969) и в Гренландии (Tinber-
gen, — цит. по: Haartman, 1969а), для лапландского подорожника на севере Аляски(Си8-
ter, Pitelka, 1977) и предположительно на островах Баренцева моря (Чемякин, 1974),
для рогатого жаворонка и горной чечетки в Северной Норвегии (Schaaning, 1907; L0rens-
kiold, — цит. по: Haartman, 1969а). Кроме того, в Заполярье двукратное выведение по-
томства возможно у белозубого дрозда (Haartman, 1969а) и выявлено также для ряда
других видов (табл. 33).
В зоне 60—64° с. ш. выкармливание двух выводков за лето у воробьиных птиц
абсолютно достоверно установлено для 14 видов, менее строго доказано для 12; с по-
мощью кольцевания подтверждена способность к факультативной полициклии у 5 ви-
дов; по косвенным данным двукратное размножение предполагается или выявляется
еще у 16 видов.
Таким образом, подавляющее большинство представителей отряда воробьиных,
выкармливающих более одного выводка за лето в центре ареала, сохраняют эту спо-
собность на Северо-Западе СССР и в Скандинавских странах, в южной и средней подзо-
нах темнохвойной тайги, а ряд видов — также и в Заполярье. К этому следует добавить,
что далеко не для всех видов птиц, включенных в табл. 33, в центре и на юге областей
их распространения полицикличность размножения можно считать доказанной.
Кроме воробьиных полицикличность размножения установлена или предполага-
ется у представителей еще 8 отрядов европейских птиц. Например, у большой поганки
в ФРГ двукратное размножение не является редкостью, а отдельные самки успевают
сделать за год 3 кладки (Hafke, 1983). В отряде пластинчатоклювых двукратное гнездо-
вание обнаружено у маркированных самок американской кряквы в Северной Дакоте
(Doty, 1975). У наших крякв растянутость периода откладки яиц (вторая декада
123
Таблица 33
Встречаемость полициклического размножения у воробьиных птиц в различных частях ареала
Вид 40—60° с. ш.
Число вывод- ков за год Степень доказан- ности по- лицик- лии Место Источник Число вывод- ков за год
Полевой жаворонок 2-3 + Англия Delius, 1965 2
Лесной жаворонок 2 — СССР Волчанецкий, 1954 2
Береговая ласточка 2 + Англия Cowley, 1983 2
Городская ласточка 2-3 + ФРГ Rheinwald, 1979 2
Деревенская ласточка 2-3 + То же Meller, 1978; Muff, 1979 2
Белая трясогузка 2 - СССР Гладков, 1954a 2
Лесной конек 2-3 4- Бельгия Hecke, 1979 2
Луговой конек 2 + Англия Constant, Eybert, 1980 2
Горный конек 2 - Кавказ Портенко, 1960
Оляпка 2 — Запад УССР Страутман, 1963 2
Крапивник 2 — СССР Судиловская, 1954а 2
Лесная завирушка 2 - Запад УССР Талпош, 1980 2
Зарянка 2-3 + Англия Lack,1946 2
Горихвостка- лысушка 2 — СССР Гладков, 19546 1(2)
Луговой чекан 2 — Англия Fuller, Glue, 1977 1
Черный дрозд 2—4 + То же Snow, 1958 2
Рябинник 2 (-) СССР Гладков, 19546 2
Белобровик 2 (-) Москов- ская обл. Птушенко, Инозем- мцев, 1968 2
Певчий дрозд 2-4 + Северная Ирландия Radford, 1980 2
Деряба 2 (-) СССР Гладков, 19546 2
Тростниковая камышевка 2 (-) ЧССР Ptaci..., 1983а 2
Камышевка-барсучок 2 (-) То же То же 2
Садовая славка 2 (-) СССР Волчанецкий, 1954 1(2)
Серая славка 2 (-) То же То же 1(2)
Славка-завирушка 2 (-) ЧССР Ptaci..., 1983а 1
Черноголовая славка 2 (-) То же То же 1(2)
Весничка 1 (2) Англия Cramp, 1955 1(2)
Теньковка 2 (-) ЧССР Ptaci..., 1983а 2
Желтоголовый королек 2 (-) СССР Судиловская, 19546 2
Серая мухоловка 2-3 (-) То же Портенко, 1960 1(2)
Мухоловка- пеструшка 1(2) Англия Лихачев, 1955; Lack,1966 1
124
60.5-76° с. ш. Наиболее северные пункты регистрации
Степень доказан- ности по- лицик- лии Место Источник Число вывод- ков за год Степень доказан- ности по лицик- лии Место Источник
7 КАССР Наши данные
! Приладожье То же 2 ! Приладожье Наши данные
7 Финляндия Haartman, 1969а
+ Гумбарицы, Наши данные
Финляндия Bergman, 1978
f Гумбарицы Наши данные
1 Гумбарицы, То же 1(2) Баренцево Чемякин, 1980
„Кивач” море
+ Прионежье » а
7 Финляндия Haartman, 1969а 2 + Айновы острова Чемякин, 1974
! Норвегия Mork, 1975
+ „Кивач” Наши данные
! Прионежье КАССР То же 2 + Северная Швеция Swanberg, 1955
! То же »> >>
! »» >> »»
»♦ >1
1 Приладожье »» 2 1 „Кивач” Л. Н. Захарова, устное сообще-
ние
7 „Кивач” »» ) ? 2 7 о. Лодейный, Белое море Наши данные
I КАССР »» 2 + Баренцево море, острова Чемякин, 1974
+ То же 2 7 о-в Лодейный, Белое море Наши данные
? »>
+ Финляндия Raitasuo, 1958 2 + Финляндия Raitasuo, 1958
7 То же Haatrman, 1969a
99 У У To же
99 >> »» »
КАССР Наши данные 1 КАССР Наши данные
Финляндия Haartman, 1969a
КАССР Лапшин, 1981a
+ То же Наши данные
I Швеция Haftorn, 1978
Финляндия Haartman, 1 Северная Haartman,
1969a Финляндия 1969а
КАССР Наши данные 1 То же Jarvinen А., 1984
125
Таблица 33 (продолжение)
Вид 40—60° с. ш.
Число вывод- ков за год Степень доказан- ности по- лицик- лии Место Источник Число вывод- ков за год
Ополовник 2 (-) СССР Волочанецкий, 1954 1
Московка 2-3 (-) ФРГ Zang, 1972 2
Хохлатая синица 2 (-) СССР Воинственский, 1954 1
Большая синица 2 ! Англия Lack,1966 2
Лазоревка 2 f То же Perrins, 1979 2
Пищуха 2 + Англия Flegg, 1973 2
Обыкновенная овсянка Овсянка-крошка Овсянка-ремез 2-3 (-) СССР Портенко, 1960 2-3 2 2
Камышевая овсянка 2 (-) СССР Спангенберг, Суди- ловская, 1954 2
Зяблик 1(2) Калинин- градская обл. Дольник и др., 1982 1
Зеленушка 2-3 (-) ЧССР Ptaci..., 1983а 2
Чиж 2 (-) Москов- ская обл. Птушенко, инозем- цев, 1968 1 + 2*
Щегол 2 (-) СССР Страутман, 1963 2
Коноплянка Чечетка 2 + Англия Eybert, 1980 2 1 + 2*
Снегирь 2 (-) СССР Портенко, 1960 2
Домовой воробей 2-5 (-) ЧССР Ptaci..., 1983а 2
Полевой воробей 3-5 (-) Болгария Шураков и др., 1981 2
Скворец 2 + Южная и Центральная Европа Lack, 1957 1
Примечание. Звездочкой (*) отмечена предполагаемая полизональная полициклия; -
ства не указан.
апреля—середина июля), как и их способность к „усыновлению” чужих птенцов, воз-
можно; также не всегда связана с гибелью самок или потерей первых кладок и вывод-
ков. Я. А. Виксне, например, рассказывал нам о попытке двукратного выведения
потомства у подсадной самки, находившейся на полувольном содержании и имевшей
контакт с дикими самцами. Вполне допустимо, что и в наших условиях какая-то часть
крякв успевает вывести птенцов дважды за лето, особенно если учесть, что птенцы из пер-
вых гнезд могут быть приняты на воспитание в выводки других самок.
Среди куриных возможность двукратного размножения допускается у кеклика
и турача (Иванов, 1953; X. Т. Мустафаев, устное сообщение).
Из журавлеобразных более тщательного исследования заслуживают пастушковые.
Мальчевский и Пукинский (1983) в Старом Петергофе под Ленинградом в 1977 г.
126
60.5-76° с.ш. Наиболее северные пункты регистрации
Степень доказан- ности по- ЛИ ЦИК- ЛИИ Место Источник ] Число вывод- ков за год Степень доказан- ности по- лицик- лии Место Источник
! ! КАССР „Кивач” То же Наши данные > > м 1 2 КАССР Кандалакша Наши данные Бианки, Шуто-
+ Г умбарицы В. И. Головань, 2 ? КАССР ва, 1978 Наши данные
f I ? ? ? Юг КАССР Север Ленин- градской обл. Финляндия КАССР Финляндия КАССР То же »» >> устное сообще- ние Наши данные Т. А. Рымкевич, устное сообщени! Kellomaki et al., 1977 Наши данные Haukioja, 1970 Наши данные То же »> в 2 ? КАССР Наши данные
+ Финляндия Tast, 1970 2 + Финляндия Tast, 1970
? КАССР Наши данные 2 + Баренцево Чем якин, 1974
? ? То же »» а То же >» » » ч» 1 море, острова Мурманская обл. В. Д. Коханов, устное сообще- ние
строго доказанная, „+” - доказанная, „?” - предполагаемая полициклия,,, (-) ” - способ доказатель-
отметили успешное воспитание двух выводков одной изолированной парой камышниц.
Эти же авторы указывают на возможную бициклию гнездования у единичных пар
лысух. В литературе высказывается предположение о бициклии размножения и у других
пастушковых.
В отряде ржанкообразных цветным кольцеванием доказано двукратное гнездова-
ние чибиса в ФРГ (Besser, 1983) : вылупление единственного птенца в первом гнезде про-
изошло здесь 26.IV, а новая кладка была начата по крайней мере 16.V. Правда, судьба
птенца к этому времени осталась неизвестной, а так как молодые чибисы поднимаются
на крыло лишь в возрасте 32—34 сут (Иванова, 1971), то не исключено, что птенец
погиб и новая кладка была лишь повторной. Отловом меченых самок на гнездах дока-
зана бицикличность размножения травника на Черном море (Жмуд. 1983). Судя
127
по растянутости периода тяги и поздним находкам гнезд и выводков, вполне можно
допустить, что дважды в лето в таежной зоне выводят птенцов отдельные самки вальд-
шнепа (Подковыркин, 1972; Мальчевский, Пукинский, 1983; наши наблюдения в Ка-
релии до 64° с. ш.). Пока неясно, как расценить аналогичные данные по бекасу, токо-
вые полеты которого в Ленинградской обл. и в Карелии регистрируются до середины
июля, а гнезда с кладками — с первых чисел мая до середины июля. Одно гнездо най-
дено даже 5 сентября под г. Ломоносов в Ленинградской обл. (Мальчевский, Пу-
кинский, 1983).
У голубей, взрослые особи которых за один раз не могут выкормить более 2 птен-
цов из-за недостаточного количества зобного молочка, эволюция размножения пошла
по пути многократного выведения потомства в течение года. Прямыми наблюдениями
за индивидуально маркированными особями 5-кратное выведение потомства доказано
для сизого полудомашнего голубя в Перми (Ангальт, 1981). В Ленинграде, Петроза-
водске, Кондопоге и других крупных населенных пунктах Северо-Запада СССР гнезда
этого вида можно обнаружить практически в любой месяц года. Для клинтуха в таеж-
ной зоне предполагается бициклия, а для вяхиря — даже трициклия. Размножение обы-
кновенной и кольчатой горлиц, явно полицикличное в южных широтах, на севере —
у границ гнездовых областей — пока изучено крайне недостаточно, и делать какие бы
то ни было заключения на этот счет преждевременно.
Более или менее определенные прямые и косвенные доказательства двукратного
размножения имеются и для некоторых сов. Давно известно, что в Западной Европе
при благоприятных условиях обитания у сипухи, кроме обычных весенних кладок,
бывают также и летние (Schneider, Schneider, 1928, — цит. по: Lack, 1957; Ptaci..., 1983b).
Указания на бициклию размножения приводятся также для ушастой и болотной сов,
обыкновенной неясыти и мохноногого сыча (Иванов, 1953; Пукинский, 1977; Ptaci...,
1983b). В Центральной Финляндии в 1969 г. окольцованная самка мохноногого сыча
была найдена на гнезде со слетками в мае, а 8 июля была повторно обнаружена в 10 км
в новом гнезде с 4 пуховыми птенцами (Kellom’aki et al., 1977).
Среди ракшеобразных воспитание двух выводков за сезон определенно доказано
для зимородка в Окском заповеднике (Нумеров, 1977; Нумеров, Котюков, 1981).
Еще в большей мере полициклическое размножение характерно для птиц южных
зон северного полушария и северных частей южного, но во многих случаях оно также
нуждается в более строгих доказательствах.
Таким образом, факультативная полициклия известна или предполагается для
большого числа видов птиц различных отрядов, представители котбрых обычно огра-
ничиваются воспитанием только одного выводка в год. В принципе двукратное раз-
множение отдельных особей можно в определенных ситуациях ожидать от птиц, у кото-
рых выкармливание одного выводка занимает относительно немного времени, а допол-
нительное гнездование не приводит к резкому увеличению дефицита времени на про-
хождение остальных фаз годового цикла. Поэтому строгой грани между поли- и моно-
циклией, видимо, проводить не следует, тем более что и у видов, для которых полицик-
лия является основной стратегией размножения, далеко не все особи и не везде (даже
в центре ареала) полностью используют свойственные виду репродуктивные возмож-
ности, ограничиваясь выкармливанием только одного выводка. Как будет показано
ниже, число таких пар в разные годы может широко варьировать как в разных точках
ареала, так и в пределах одной местности в зависимости от биотопа, возраста партнеров
ит. д.
Редкая встречаемость факультативной полициклии в пределах вида — свидетель-
ство того, что она не дает ему особых преимуществ, в противном случае она была
бы закреплена в эволюции.
Если для факультативной полициклии обязательным условием является успешное
докармливание слетков первого выводка, то при облигатной второе размножение
нередко начинается еще до окончания первого. В Карелии, например, почти у всех
бициклических видов достоверно установлено совмещение начала второй кладки
128
с выкармливанием гнездовых птенцов (или слетков) в первых гнездах. Подобное
явление широко известно и многократно описывалось в литературе для большой сини-
цы в разных частях ареала. Но рекорд в этом отношении принадлежит зарянке: в юго-
восточном Приладожье вторая кладка у одной из пар началась при 9-дневных птенцах
первого выводка. Следовательно, в подобных случаях второе размножение начинается
независимо от успеха первого.
Моно- и полициклия гнездования — два закрепленных эволюцией направления
репродуктивной стратегии птиц, имеющих своих представителей даже в пределах одного
рода (Motacilla, Phylloscopus, Parus, Emberiza и др.).
Важно понять, почему одним видам для поддержания относительно стабильной
численности достаточно воспитать 1-5 птенцов в год, а другим это приходится делать
дважды или трижды, да еще каждый раз откладывая по 8—14 яиц. Следует учитывать,
что двукратное гнездование требует повышенных энергетических нагрузок на организм,
к тому же птицы в большей степени подвергаются риску при защите гнезда и потомства,
а на севере, кроме того, при дефиците благоприятного для жизни времени у них значи-
тельно возрастает напряженность годового цикла. Естественно было бы ожидать, что
максимальные затраты на воспроизводство связаны с повышенной ежегодной смерт-
ностью птиц, гнездящихся дважды или более в год. Анализ литературных сведений
по смертности европейских видов птиц, суммированных Паевским (1974), не выявил,
однако, никаких существенных различий в ежегодной смертности близкородственных
пар видов с разной стратегией размножения. Не обнаруживаются они и при суммирова-
нии данных по двум группам видов.
У европейских воробьиных птиц с полициклическим гнездованием смерт-
ность составила 54.7% (с колебаниями 34—72), у моноциклических — 53.7%
(33-71).
Если же учесть, что во втором размножении у ряда видов принимает участие
до 60—100% гнездовых пар, то при том же уровне ежегодной смертности итоговый
прирост популяции при полициклии по сравнению с моноциклией должен быть гораздо
выше. Ясно, что излишки воспроизводства должны быть каким-то образом компенсиро-
ваны. Это может быть достигнуто несколькими путями: постоянным ростом числен-
ности; расширением областей гнездования или спектра местообитаний; прохолостанием
части особей, например первогодков; наконец, все-таки повышенной ежегодной смерт-
ностью. Попытаемся рассмотреть каждый из названных аспектов, спираясь на известные
факты.
Численность. Постоянное перепроизводство молодняка в конечном итоге должно
было бы привести к абсолютному доминированию птиц, дающих дополнительные
выводки. На самом деле этого не происходит. Среди птиц таежных и широколиственных
лесов Европы абсолютными доминантами являются зяблик и весничка — виды с редкой
факультативной полициклией и явным преобладанием моноциклии. Аналогичные
результаты дает сравнение численности родственных видов птиц, различающихся осно-
вной стратегией размножения. Так, среди наших пеночек при почти равной величине
кладки плотность населения веснички примерно в 10—20 раз выше, чем у теньковки,
часть пар которой гнездится дважды за сезон. Обратное соотношение обнаруживают
наши овсянки, среди которых моноцикличные дубровник и садовая овсянка оказыва-
ются значительно малочисленное бицикличных обыкновенной, камышезой и овсян-
ки-ремеза. Правда, оба первых вида живут в Карелии у границ своих ареалов,
но и в других зонах совместного обитания садовая овсянка численно уступает
обыкновенной.
Наиболее многочисленным представителем синиц таежной зоны является одно-
кратно размножающийся за сезон пухляк, хотя его родственники - большая синица,
лазоревка и московка — воспитывают за лето вдвое более многочисленное потомство.
В то же время еще один моноциклический вид - хохлатая синица - по степени домини-
рования среди своих родственников занимает в целом лишь третье место, уступая
и пухляку, и большой синице.
9 В. Б. Зимин
129
Конечно, на численность птиц, кроме плодовитости, влияют и многие другие
особенности их биологии и разнообразные связи со средой обитания, но в целом уже
очевидно, что полициклия не приводит к абсолютному доминированию птиц тех видов,
которым она свойственна.
Расширение области распространения и спектра местообитаний также связано
с различными сторонами биологии каждого вида птиц, а не только с размножением.
Тем не менее, если бы полициклии ежегодно приводила к перепроизводству молодняка,
заполнение подходящих для жизни местообитаний привело бы или к освоению новых
стаций, или к регулярному выселению птиц за пределы области постоянного гнездо-
вания вида.
Если сравнить размеры ареалов близкородственных видов евроазиатских
птиц с моно- и полициклическим размножением, то явного преобладания последних
среди широко распространенных видов не обнаруживается. Так, ареал зяблика занимает
гораздо большую площадь, чем у других вьюрковых, обычно размножающихся
дважды или даже трижды в год (чечетка, щегол, зеленушка и др.). Отличается зяблик
и наиболее высокой степенью эврибионтности, в том числе и эвритопности.
Наиболее широко распространенный вид синиц — большая синица. Она повсемест-
но гнездится дважды в год. Тем не менее ареал у пухляка, выкармливающего только
один выводок, гораздо обширнее, чем, например, у лазоревки, которая даже у северных
границ ареала делает дополнительные кладки. К тому же пухляк в целом и более эври-
топен, чем все остальные наши синицы.
Точно так же полициклическая обыкновенная овсянка распространена очень
широко, но дубровник, всюду гнездящийся однократно, освоил более обширные про-
странства Евразии, чем другие азиатские виды, в частности овсянка-крошка с доказан-
ной бициклией гнездования у северо-западных границ ареала (Koivunen et al., 1975).
Среди птиц, демонстрирующих в последние десятилетия на Северо-Западе СССР
интенсивную экспансию или попытки расширения ареала, вйДы с моноциклическим
типом гнездования даже преобладают (гл. I). Таким образом, и экспансия, и расшире-
ние спектра местообитаний не являются прерогативой видов, производящих потомство
более одного раза в год.
Прохолостание части особей также нельзя считать характерной чертой только тех
видов птиц, которые выкармливают дополнительные выводки. Напротив, наиболее
яркие примеры отсутствия первогодков в составе гнездящихся особей (один из косвен-
ных показателей прохолостания) известны для птиц с моноциклическим размножением.
Для Карелии ранее это было показано на примере чечевицы, у которой годовалые
самцы чаще выполняют роль помощников при гнездовых парах, чем образуют их сами
(Зимин, 19816). Как уже упоминалось в гл. I и II, в северных частях ареала не принима-
ют участия в размножении первогодки скворцов и часть белобровиков. На первом
году жизни остаются холостыми отдельные самки мухоловки-пеструшки (Winkel et al.,
1970; Haartman, 1971). Примеры прохолостания молодых особей дают пухляк (Бардин,
19756; наши данные), кукша (Коханоц, 1982) и другие врановые.
Как уже было показано в предыдущих главах, плодовитость годовалых особей,
особенно в южных частях ареалов, оказывается достоверно более низкой — они в целом
воспитывают меньше птенцов и реже участвуют во втором гнездовании. Однако это
характерно не для всех видов птиц с полициклическим размножением и отмечается
не повсеместно.
Таким образом, ни один из перечисленных выше путей не приводит к компенсации
излишков воспроизводства, возникающих в результате выкармливания дополнитель-
ных выводков. Остается предположить, что рассчитываемые показатели ежегодной
смертности птиц не отличаются необходимой точностью. В частности, это может объ-
ясняться недооценкой значения эмиграции и иммиграции при формировании
местного гнездового населения птиц. Иными словами, полицикличность размно-
жения каким-то образом все же должна быть связана с повышенной ежегодной
смертностью.
130
В этой связи обращают на себя внимание характерные различия в размещении
зимовок, дальности, условиях и сроках миграции птиц с разным числом выводков
в год. Из 28 видов воробьиных с доказанной для наших широт бициклией размножения
11 (королек, московка, большая синица, лазоревка, пищуха, крапивник, обыкновен-
ная овсянка, коноплянка, снегирь, домовый и полевой воробьи) зимуют на местах раз-
множения или на незначительном удалении от них. Птицы этой группы за сезон откла-
дывают от 9 до 22, в среднем около 15 яиц.
Группа мигрантов на средние дистанции представлена 12 видами (полевой и лес-
ной жаворонки, лесной конек, белая трясогузка, лесная завирушка, зарянка, дрозды,
камышевая овсянка и теньковка), суммарная величина кладки у которых 9—11,
в среднем 10 яиц за сезон.
Дальних мигрантов среди воробьиных птиц, дважды выводящих птенцов в таеж-
ной зоне, всего 5 видов: городская и деревенская ласточки, тростниковая камышевка
и овсянки — крошка и ремез. За лето птицы этой группы откладывают в среднем около
9 яиц. За исключением тростниковой камышевки, отлет на зимовку остальных птиц
этой группы происходит в необычно поздние для дальних мигрантов сроки. Так, овсян-
ка-ремез проходит трассу пролета на 1.5—2 мес позднее дубровника, хотя оба вида зи-
муют в Юго-Восточной Азии: первый — в Японии и Восточном Китае, второй — в Юго-
Восточном Китае и Индостане (Иванов, 1976). При позднем начале осеннего пролета
ласточки нередко попадают в критические ситуации во время миграции над странами
Западной Европы, о чем свидетельствуют периодически появляющиеся в печати сообще-
ния об их массовой гибели и операциях по спасению птиц.
Хотя доля особей, размножающихся дважды за сезон, более или менее достоверно
установлена лишь для ограниченного числа видов, можно отметить определенную
тенденцию к ее увеличению у птиц, зимующих в более суровых условиях. Так, в Каре-
лии у синиц - большой и лазоревки, у королька, пищухи, крапивника и обыкновен-
ной овсянки дополнительные выводки имеют практически все особи, успешно завер-
шившие первое гнездование (60—100% пар). Среди мигрантов на средние расстояния
этот показатель варьирует от 25—30 (дрозды, теньковка) до 40—80% (белая трясогуз-
ка, зарянка, лесная завирушка). Для дальних мигрантов точных сведений о соотноше-
нии пар с одним и двумя выводками у нас пока нет, но, по предварительным наблюде-
ниям, выше других этот показатель у овсянки-ремеза, а у ласточек он вряд ли превыша-
ет 20-25 %.
Таким образом, большинство видов птиц, дополнительное размножение которых
дает существенный ежегодный прирост популяции, зимует или преодолевает миграцион-
ные трассы в условиях крайне неустойчивой внешней среды. Это, однако, не является
абсолютно строгим правилом, так как в этой же зоне зимуют и птицы, гнездящиеся
обычно только один раз в году (врановые, пухляк, хохлатая и длиннохвостая синицы,
зяблик и др.). Все же зимовочные ареалы большинства других птиц этой группы распо-
ложены в зонах с более стабильными условиями существования. Эта закономерность
вполне отчетливо проявляется и при сравнении родственных видов, различающихся
числом выводков за год, в чем легко убедиться, рассмотрев гнездовые и зимовочные
ареалы некоторых пар видов (рис. 36).
На европейских зимовках резкие похолодания, сопровождающиеся нередко обра-
зованием снежного покрова, кухты, наста и ожеледи, могут внезапно наступить в любое
время осенне-весеннего периода. Иногда они становятся причиной более или менее
массовой гибели зимующих, пролетных и даже вернувшихся на родину птиц.
Там, где гнездовое население большинства воробьиных птиц формируется не
только за счет особей местного происхождения, изменения численности, кроме успеш-
ности размножения и условий зимовки, определяются еще и интенсивностью дисперсии
особей, родившихся в более южных зонах. Размеры дисперсии варьируют по годам,
и каковы бы ни были причины таких вариаций, они в значительной степени влияют
на многолетнюю динамику численности многих видов птиц. Поэтому мы обра-
тились к анализу многолетних колебаний численности европейских воробьиных,
131
M. flava М. alba
Ph. trochilus
Ph.collybita
£. hortulana
E. citrinella
Е. aureola
Рис. 36. Гнездовые и зимовочные
ареалы у близкородственных видов из ро-
дов Motacilla, Phylloscopus и Emberiza, разли-
чающихся по числу выводков за сезон раз-
множения (по: Гладков, 1954а; Птушенко,
1954; Спангенберг, Судиловская, 1954).
1 - область гнездования; 2 - область
круглогодичной встречаемости; 3 — область
зимовок.
Е. rustica
выполненному Свенссоном (Svensson, 1977). В его списке видов, численность которых
подвержена значительным ежегодным колебаниям, явно преобладают птицы с поли-
циклическим типом размножения. Разумеется, есть и исключения, но важно подчерк-
нуть, что депрессии численности птиц (и других животных) с моноциклическим размно-
жением, как правило, бывают более продолжительными и на восстановление поголовья
у них затрачивается несколько лет. Хорошие примеры этому дают наши тете-
ревиные, исходная плотность населения которых, несмотря на большую вели-
чину кладки, иногда после особенно резких снижений достигается через 3—6 лет
(Семенов-Тян-Шанский, 1960; Данилов и др., 1978). Численность пухляка в При-
ладожье после неблагоприятной зимы 1980/81 г. сократилась примерно в 1.5 раза
и по крайней мере до 1984 г. оставалась на низком уровне, хотя плотность
гнездового населения большой синицы уже через год после депрессии оказалась
максимальной за все время наблюдений. На протяжении последних 6 лет плот-
ность населения зарянки в этом районе имела 3 заметных пика и падения. Еще
более резкие колебания численности в смежные годы демонстрируют мелкие
грызуны, белка, заяц-беляк и другие млекопитающие с полициклическим раз-
множением (Ивантер, 1969а, 19696, 1972, 1975, 1978). С другой стороны,
132
Рис. 37. Зависимость числа вторых кладок {абсцисса, %) от плотности гнездового населения
{ордината, пар/км2) у зарянки в Южной Карелии (Л) и у большой синицы в южной Финляндии
{Б) {Б - по: Haartman, 1969а).
млекопитающие с моноциклическим размножением после глубоких депрессий
численности оказываются неспособными к быстрому восстановлению поголовья.
Обращает на себя внимание широкая изменчивость доли птиц, участвующих
в выкармливании дополнительных выводков. Наиболее изучена в этом отношении
большая синица. По данным Лэка (Lack, 1966), в Англии по две кладки имеют в сосно-
вых посадках 20% самок, в дубравах — только 2%. В Голландии вторые выводки дела-
ют в сосновых лесах 27—100, в среднем 68% пар, в лиственных 16-45, в среднем 36%
(Kluijver, 1951). Для Нижней Саксонии доля вторых кладок определена как 90%
(Winkel, 1975), для Дании - 15.9% (Larsen, 1974). В Литовской ССР этот показатель ва-
рьирует по годам от 10 до 71 % (Езерскас, 1960), в Ленинградской обл. по средним мно-
голетним данным составляет около 25% (Носков, Смирнов, 1981), в центре европей-
ской части СССР в отдельные годы достигает 100% (Лихачев, 1957). В Южной Карелии
дважды за лето выводят птенцов в среднем 67 % пар больших синиц, а в отдельные го-
ды — даже до 100 %.
В юго-восточном Приладожье доля дважды гнездящихся зарянок в разные годы
варьировала от 37 до 80%, причем число таких птиц достоверно возрастало в годы
пониженной численности вида (Зимин, 1983; см. также рис. 37, Л). Для большой си-
ницы, численность которой на наших стационарах очень низка, подобной зависимости
не выявлено, но пересчитанные нами многолетние данные по Южной Финляндии (Haart-
man, 1969а) обнаружили удовлетворительную отрицательную корреляцию (г =-0.62)
между числом ежегодно занятых гнездовий и долей самок, выкормивших по два вывод-
ка (рис. 37, Б). Ранее такая же связь была доказана для большой синицы в Голландии
(Kluijver, 1951) и домового крапивника в Америке (Kendeigh, Baldwin, 1937). Если связь
плодовитости и численности у видов с полициклическим размножением действительно
существует, то с учетом разобранных выше обстоятельств причины возникновения по-
лициклии в эволюции птиц (и других животных) становятся достаточно очевидными.
Эта форма репродуктивной стратегии могла возникнуть не как противодействие повы-
шенной смертности вообще, а как результат отбора на способность быстрого восста-
новления численности после ее резких снижений в результате аритмических ухудшений
условий существования, т. е. как способ компенсации неожиданно возрастающей
смертности.
У птиц, обитающих в зоне умеренного климата, подобный механизм мог возник-
нуть прежде всего в связи с жизнью в среде, отличающейся крайней неустойчивостью
климата и аритмией резких отрицательных погодных аномалий. Очевидно, чем севернее
расположены зимовки вида или чем позднее преодолевается трасса пролета у дальних
мигрантов, тем вероятность аномалий больше, тем чаще возникают ситуации, приводя-
щие к увеличению смертности, причем это может происходить с непредсказуемой часто-
той и не обязательно каждый год.
Второй важной причиной возникновения предполагаемого механизма взаимодей-
ствия плодовитости и смертности могла быть относительная стабильность средней
133
величины кладки у абсолютного большинства видов птиц. Исключение в этом плане
составляют лишь некоторые хищные птицы. У остальных величина кладки мало или
совсем не зависит от плотности населения, и даже в тех немногих случаях, когда подоб-
ная зависимость выявлена, уменьшение или увеличение числа откладываемых яиц быва-
ет слишком незначительным, чтобы обеспечить быстрое изменение численности, тем бо-
лее что увеличение размеров выводка нередко сопровождается возрастанием птенцовой
смертности. И наблюдения в природе, и эксперименты с искусственным укрупнением
размеров кладки и выводка убеждают нас в том, что в определенных условиях птицы
способны успешно выкормить лишь ограниченное число птенцов при однократном
размножении. Скорее всего именно поэтому для их эволюции оказалось более прием-
лемым повышение продуктивности гнездования путем многократного выведения по-
томства в течение одного репродуктивного сезона. Наиболее ярко иллюстрирует это
приведенный выше пример с голубями, которые за один раз способны выкормить
только 1—2 птенцов, но благодаря адаптации к полициклии воспитывают до 12 птенцов
за год. В том, что этот путь даже при небольшой величине кладки позволяет обеспечить
очень быстрый рост численности, убеждает история расселения сизого голубя, в считан-
ные годы ставшего одним из самых многочисленных птиц северных городов.
При данном способе поддержания оптимальной численности населения вида ито-
говый уровень общепопуляционной продуктивности может широко варьировать в за-
висимости от соотношения числа особей, гнездящихся один или несколько раз за год.
Основа регуляторного механизма, таким образом, заключается в способности популя-
ций видов с полициклическим размножением реагировать на снижение численности уве-
личением числа птиц, выкармливающих дополнительные выводки. Этим, возможно,
и объясняются существенные различия в доле особей, участвующих в полициклическом
гнездовании в разных частях ареала, а также ежегодные колебания этого показателя
на одной и той же территории.
Не исключено, что биологическая сущность рассматриваемого механизма состоит
не в прямом влиянии численности на количество двукратно размножающихся птиц,
а в ее опосредованном — через изменение возрастного состава — действии. Рост числен-
ности обычно происходит за счет лучшей выживаемости первогодков. При этом соответ-
ственно увеличивается и их доля в составе гнездового населения. Но если дополнитель-
ные выводки воспитываются преимущественно более старыми особями, уже гнездив-
шимися в предыдущие годы, как это показано для черного дрозда (Snow, 1958), то при
повышенной численности снижение их доли в составе гнездового населения должно при-
вести к соответствующему сокращению обилия особей, делающих вторые кладки.
Напротив, после суровой зимовки или неблагоприятных условий во время миграций,
когда выживаемость первогодков оказывается низкой, преобладание „ старых” птиц
в гнездовом населении (при пониженной численности) соответственно выразится
в увеличении доли птиц, гнездящихся дважды за лето.
Кроме численности и возрастного состава, на относительное число особей, участву-
ющих в дополнительном размножении, оказывают влияние и другие факторы. Лэк
(Lack, 1957) считает, например, что биотопические различия этого показателя могут
быть связаны с пищевым фактором: количество гусениц — основного объекта для
выкармливания птенцов больших синиц — быстрее убывает в широколиственных лесах,
чем в хвойных, и поэтому в последних размножение чаще бывает бициклическим.
В Дании и Голландии число вторично размножающихся пар больших синиц связано
обратной зависимостью с величиной первого выводка (Kluijver, 1951; Larsen, 1974). В ка-
кой-то мере это подтверждает гипотезу Лэка (Lack, 1957), согласно которой самки, вы-
кармливающие большие выводки, имеют меньше шансов выжить. Езерскас (1960)
полагает, что обилие вторых выводков большой синицы в разных лесхозах Литвы
находится в прямой зависимости от количества свободных гнездовий к моменту
второго размножения.
Ранее на примере биомассы беспозвоночных животных молодых лесов Прила-
дожья было показано, что в таежной зоне насекомоядные птицы вполне могут
134
обеспечить пищей своих птенцов в течение всего весенне-летнего периода (рис. 17).
Можно допустить, что динамика биомассы насекомых в разные годы имеет свои особен-
ности, но это вряд ли оказывает существенное влияние на число вторых кладок. Во вся-
ком случае, в годы, резко различавшиеся обилием вторых кладок, существенной раз-
ницы в успешности выкармливания птенцов не наблюдалось. Биотопические различия
в доле вторых выводков в таежных лесах также незначительны. Только в молодняках
с преобладанием сосны большие синицы реже выкармливают по 2 выводка, чем в опти-
мальных для гнездования спелых и приспевающих лиственных лесах (Зимин, 1978),
т. е. по сравнению с Англией и Голландией в таежной зоне намечается обратная тенден-
ция. Аналогичные данные получены и для зарянки: в сосново-лиственных молодняках
Прионежья лишь пятая часть контролируемых гнездовых пар выводила птенцов дваж-
ды, а в спелых, преимущественно хвойных древостоях Приладожья — более половины.
Наши данные не подтверждают обратной зависимости между числом вторично
гнездящихся особей и величиной первого выводка. Напротив, средняя величина кладки
и выводка больших синиц, размножавшихся только один раз, была меньше (10.5 ±
±0.4 и 7.8 ± 0.6), чем у бицикличных пар (12.1 ± 0.2 и 9.9 ± 0.5). Очевидно, в Карелии
ко второму размножению чаще приступают особи, отличающиеся более высоким репро-
дуктивным потенциалом. Однако у зарянки подобная связь не выявлена: суммарная
средняя величина кладки и выводка у пар, гнездившихся один и, два раза в год, оказа-
лась практически одинакова.
Вероятность второго гнездования во многом определяется сроками и успеш-
ностью первого. Нами пока не отмечено ни одного случая двукратного размножения
зарянок, потерявших первую кладку или выводок и гнездившихся повторно. Ни одна
из больших синиц не гнездилась дважды, если ее первая кладка была начата в последней
пятидневке мая. Птицы других бициклических видов, потерпевшие неудачу при первом
гнездовании, особенно на последних его этапах, почти всегда ограничивались только
повторным размножением. Многие самки большинства открытогнездящихся видов
птиц в отличие от дуплогнездников после гибели первого гнезда вообще исчезают
с контролируемых территорий, и их дальнейшая судьба остается неизвестной. Вместе
с тем во второй половине репродуктивного периода на наших стационарах появляются
особи, ранее не встречавшиеся на облавливаемых площадях.
Определенную роль в снижении числа вторых кладок могут играть внезапные по-
холодания в период выкармливания птенцов, приводящие к их массовой гибели. После
этого птицы обычно воспитывали лцшь выводки из восстановленных кладок. Аналоги-
чное значение имеют разорение гнезд хищниками и хищничающими животными, а также
конкурентные взаимоотношения, например между вертишейкой и другими обитателя-
ми искусственных гнездовий в молодняках.
135
ГЛАВА V
СОВМЕЩЕНИЕ ГНЕЗДОВАНИЯ И ЛИНЬКИ
В годовом бюджете энергии птиц наибольшие затраты приходятся на периоды
линьки и гнездования. Различные виды расходуют на линьку от 10 до 30, на гнездование
от 25 до 30 и на миграцию 6—7 суточных бюджетов энергии на каждые 1000 км пролет-
ного пути (Дольник, 1975). Увеличение энергетических затрат на линьку, особенно
на такую кратковременную, как у дальних мйгрантов, приводит к сокращению боль-
шинства форм активности вплоть до снижения подвижности и даже потери способности
к полету.
Высокая энергоемкость характерна и для периода гнездования, когда к затратам
энергии на самообеспечение добавляются и разнообразные виды активности, связанные
с родительскими заботами (Дольник, 1975; Дольник и др., 1982). Не случайно поэтому
в годовых циклах большинства видов птиц все энергоемкие фазы бывают четко разоб-
щены во времени.
В процессе изучения годовых циклов воробьиных птиц в исследуемой зоне сначала
было обращено внимание на совмещение линьки птиц с миграцией (Блюменталь и др.,
1963; Блюменталь, Зимин, 1966). Позднее это явление более детально было изучено
на материалах массовых отловов птиц крупногабаритными ловушками в заповеднике
„Кивач” и на юго-восточном побережье Ладожского озера (Зимин, 1968; Носков,
Зимин, 1977; Кукиш, 1976, 1981; Рымкевич, 1977; Музаев, 19806; Лапшин, 19816;
Носков и др., 1981). Затем 'аналогичные данные были получены во время подготовки
к популяционно-экологическим исследованиям, когда у отловленных на гнездах самцов
пухляков и мухоловок-пеструшек, кормящих птенцов, были обнаружены сменяюшиеся
маховые перья.
С освоением и разработкой методов отлова птиц на гнездах (специальные ловуш-
ки) и гнездовых участках (паутинные сети), а также с организацией постоянного конт-
роля местного населения на стационарных пробных площадях совмещенная с гнездова-
нием линька была выявлена у нескольких видов воробьиных и оказалась отнюдь не ред-
ким явлением (Зимин, Лапшин, 1974, 1976).
Первое упоминание о совмещении линьки с размножением у мелких воробьиных
птиц в зоне умеренного климата мы находим в монографии Северцова (1950). Изучая
периодические явления в жизни животных Воронежской губернии, он выделил группу
птиц, у которых линька начинается во время гнездования. К этой группе была отнесена
и большая синица. К сожалению, никаких разъяснений по этому поводу автор не приво-
дит, и невозможно понять, в какой степени эти фазы накладывались друг на друга
и каким путем это было установлено.
Состав видов воробьиных, для которых отмечена совмещенная с гнездованием
линька, полностью не выяснен. Это связано как со спецификой условий, в которых
проводились исследования (лесные угодья), так и с отсутствием простых и достаточно
эффективных методов отлова птиц в гнездовой период. Кроме того, наше внимание
было сконцентрировано в основном на изучении нескольких модельных видов, а осталь-
ные контролировались отловом на гнездах не систематически.
В табл. 34 включены только виды, проходящие полную послебрачную линьку
на родине. Частичная линька также совмещается с гнездованием (садовая славка,
трещотка, садовая камышевка, серая мухоловка и др.), но из-за часто встречающейся
асимметрии в замене оперения на отдельных птерилиях иногда бывает сложно отличить
начало настоящей линьки от восстановления утраченного пера.
136
Таблица 34
Встречаемость линьки, совмещенной с гнездованием, у воробьиных птиц Северо-Запада СССР
Вид Число особей Контролируемый период
всего иссле^ довано с совмещенной линькой
Белая трясогузка 13 1 8-18. VII
Лесной конек 14 — 1O.VI-15.VII
Крапивник 8 — 23.VI-27.VII
Лесная завирушка 81 8 29.V-19.VH-5.Vin
Зарянка, самцы 50 2 17.VH-5.Vin
самки 57 — 17.VII-5.Vni
Черный дрозд 15 — VII-VHI
Певчий дрозд, самцы 8 2 VII-18.VII-VIII
самки 8 — VII-VIII
Белобровик, самцы 22 8 VII-VIII
самки 21 7 VII-VIII
Черноголовая славка 4 — 26.VI-9.VII
Славка-завирушка 20 7 15.VI-24.VI-24.Vn
Весничка, самцы 208 70 16.VI-18.VI-23.Vn
самки 328 18 16.У1-2.УП-З.УП1
Теньковка 34 — 14.VI-22.Vn
Желтоголовый королек — 1
Мухоловка-пеструшка, самцы 384 100 13.VI-18.Vn
самки 404 42 13.VI-19.VI-20.Vn
Большая синица 54 30 19.VI-10.VIII
Пухляк 41 9 1.У1-13.У1-4.УП
Пищуха 25 1 30.V-18.VH-23.VII
Обыкновенная овсянка 16 — 6.VI- 10.VIII
Овсянка-ремез 14 3 6.У1-16.УП-22.УП
Чечетка 10 9 13.VHI- 26.VHI
Зяблик 14 3 15.VI- 4.VII— 20.VII
Коноплянка 2 2 14-16.У1П
Снегирь 8 4 28.VI-27.VII
Примечания. Для зарянки и большой синицы даны результаты отловов после первой
регистрации линьки; самец желтоголового королька отловлен на V стадии линьки с выводком
из 16—18-дневных птенцов; данные о пищухе приведены только для самок, преимущественно при
втором гнездовании. Подчеркнуты даты 1-го отлова линяющей особи.
Следует подчеркнуть, что совмещенной линькой мы называем лишь достоверные
случаи совпадения смены оперения с гнездованием, подтвержденные отло-
вом птиц непосредственно на гнездах. Регистрация особей, линяющих
в гнездовой период, не может служить доказательством такого совмещения, поскольку
такими особями могут быть холостые или потерявшие гнезда и не приступившие к по-
вторному размножению; они, как правило, начинают смену пера намного раньше дру-
гих. В данном случае обсуждается не наложение периодов линьки и гнездования
в масштабах популяции, а непосредственное участие линяющих птиц в размножении.
Детально анализировать видовой состав птиц, совмещающих линьку с гнездова-
нием, пока преждевременно. По многим видам, даже из числа приведенных в табл. 34,
фактический материал слишком мал, а контролируемый период недостаточен. Тем не
менее следует обратить внимание на довольно четко вырисовывающиеся два периода
начала совмещенной линьки: у птиц с моноциклическим размножением - вторая декада
июня, с полициклическим — вторая декада июля. У дальних мигрантов, воспитыва-
ющих только один выводок в год и имеющих короткий период пребывания на родине,
совмещенная линька встречается относительно чаще, чем у птиц с полициклическим
гнездованием. Для последних это вообще мало характерно и отмечается главным
образом при поздних сроках второго размножения (зарянка, лесная завирушка).
Вместе с тем особи, не участвующие во втором гнездовании, приступают к нормальной
замене пера после распадения первых выводков ~ почти на месяц раньше.
137
Среди моноциклических видов привлекает внимание раннее начало линьки у пу-
хляков, кормящих птенцов. В отличие от других представителей этой группы пухляки
фактически не испытывают дефицита времени в связи с тем, что взрослые особи
этого вида живут оседло. Правда, линька у этого вида очень растянута и у отдельных
меченых особей занимала 110—115 дней. Тем выгоднее начать ее как можно раньше,
чтобы наиболее интенсивная смена оперения завершилась при благоприятных условиях:
на более длинном фотопериоде, при относительно более высоких температурах среды,
обилии и доступности пищи — факторах, снимающих излишнюю энергетическую напря-
женность.
Совмещение линьки с гнездованием подробнее изучено у мухоловки-пеструшки
и веснички — дальних мигрантов с относительно коротким периодом пребывания
на родине при моноциклическом размножении и полной послебрачной смене оперения.
Мухоловка-пеструшка прилетает к нам в начале мая и в зависимости от состояния
погоды приступает к откладке яиц в середине мая—1-й пятидневке июня, обычно в 3-й
декаде мая. Вылет птенцов из гнезд чаще всего заканчивается в 1-й декаде июля. Линька
взрослых птиц начинается с середины июня—2-й декады июля. Отлет на зимовки в осно-
вном заканчивается к концу августа. Линяющие самцы ловятся на гнездах уже с 13.VT,
но до конца месяца среди них явно преобладают особи, еще не приступившие к замене
маховых перьев, а после 4.VII — только линяющие. Самки, у которых совмещенная
линька отмечается реже, в целом начинают линять позднее, и многие из них сохраняют
старое оперение даже при очень поздних сроках размножения (табл. 35).
Период пребывания веснички в Карелии также немного превышает 4 мес: прилет
в конце апреля—начале мая, отлет в августе—сентябре. Начало гнездования варьирует
в разные годы от 4-й пятидневки мая до первых чисел июня. На полную послебрачную
линьку требуется 55 дней (Лапшин, 19816). Самцы веснички чаще самок совмещают
линьку с гнездованием и начинают ее на 1-1.5 декады раньше. Начало совмещенной
Таблица 35
Динамика отлова на гнездах самцов (над чертой) и самок
(под чертой) мухоловки-пеструшки с различной стадией линьки (Маячино, 1982-1984 гг.)
Стадия линьки Число особей в Всего
июне июле
10-15 16-20 21-25 26-30 1-5 6-10 11-15 ( 16-20
о* 38 39 47 20 — 146
и 39 55 61 29 192
I 4 5 8 5 2 1 — — 25
1 - 1 3 1 - 2 9
п — 3 6 2 — 1 1 — 13
11 1 1 — - 5 6
III -- 1 6 1 — 2 2 — 12
- - 1 - 4 1 6
IV — — — — — — 1 1 2
V - - - - 1 1 2 1
Всего: 42 48 67 28 4 5 5 1 200
39 56 65 32 1 10 И 1 215
из них 4 9 20 8 2 5 5 1 54
с линь- кой - 1 4 3 — 6 8 1 23
* Здесь и в табл. 36, 37 ноль означает, что линька не началась.
138
линьки приходится на середину июня: 20.VI самец, имеющий вторую стадию линьки,
пойман на гнезде с пятидневными птенцами. Нелиняющие особи регистрировались
у гнезд до 7.VII, и после этой даты все кормящие птенцов самцы находились на I -VI
стадии замены пера (табл. 36).
Первая линная самка веснички (I стадия, птенцам 9 сут) отловлена 2.V1I, но до
20.VII в отловах у гнезд преобладали самки, не приступившие к линьке; последняя такая
особь обнаружена 3.VIII на гнезде с 10-дневными птенцами.
Из данных табл. 35 и 36 можно заключить, что гнездящиеся у нас веснички и му-
холовки-пеструшки способны совмещать выкармливание птенцов с I—V и даже с VI ста-
диями линьки. IV—VI стадии совмещенной линьки отмечены также у самцов овсянки-
ремеза (1 случай), лесной завирушки (3), пухляка (1) и славки-завирушки (1).
Линька гнездящихся птиц чаще всего ограничивается заменой 10—9-го первосте-
пенных маховых перьев (70.4% самцов мухоловки-пеструшки и 71 % весничек). Реже
сменяется и 8-е маховое (22.2 и 17.1% соответственно). На этом совмещеная линька
чаще всего прерывается и лишь при поздних сроках размножения может продвинуться
у самцов до IV—VI стадий: у 7.4% мухоловок-пеструшек и у 11.5 % весничек. У самок
этих видов линька при совмещении с гнездованием в основном также прерывается на
I—ПГ стадиях (22 из 23 мухоловок-пеструшек и 16 из 18 весничек). Ни II, ни III стадии
линьки никогда не бывают строго дифференцированы. II стадия всегда проходит одно-
временно с I (если линька не прекращается после выпадения 10-го махового пера),
а III почти всегда совмещена с серединой или окончанием периода дорастания 9—10-го
маховых.
Объем совмещенной линьки, как правило, бывает незначительным, и одновремен-
ное выпадение и рост одного-двух маховых перьев не может существенно ухудшить
лётные качества крыла. В результате линяющие во время гнездования птицы сохраняют
способность к активным перемещениям в поисках пищи.
Многие самцы с совмещенной линькой, продолжающейся после окончания III-IV
стадий, по-видимому, прекращают кормить птенцов. Выше уже упоминалось об этом
на примере разных видов воробьиных птиц нашей зоны. При позднем гнездовании это
явленйе особенно характерно как раз для веснички и мухоловки-пеструшки. Пока
ни разу не отмечены у гнезд со вторыми выводками и самцы пищухи. У самцов линька,
очевидно, начиналась почти сразу после завершения самками вторых кладок, поскольку
в конце августа некоторые самцы отлавливались на своих гнездовых участках на X—XI
стадиях линьки. Случаи, когда интенсивно линяющие самки прекращали бы выкармли-
вать птенцов, нам не известны.
Точно определить время начала совмещенной линьки пока не удается: во-первых,
не установлено, каковы темпы линьки при разном объеме сменяющегося оперения и
на разных этапах гнездового цикла; во-вторых, перья на правом и левом крыльях
часто выпадают неодновременно - с разницей в 1-2 стадии. Кроме того, 2-3 последова-
тельные стадии бывают нередко совмещены. Поэтому о скорости совмещенной линьки
можно судить лишь весьма приблизительно, в частности по отдельным немногочислен-
ным данным для мухоловки-пеструшки. Самец, пойманный 13 июня на гнезде с 2-днев-
ными птенцами, находился на I стадии линьки, а 25 июня — на III. Таким образом, на две
стадии ушло 13 дней. У другого самца I стадия линьки обнаружена 10 июля на одном
крыле, а 12 июля — на обоих. Еще один самец, отловленный при 1—2-дневных птенцах,
14 июня был на I, а 23 июня — на II стадии линьки, т. е. на одну стадию ему понадоби-
лось 10 дней. 28 июня насиживающая кладку самка еще не линяла, 8 июля при 7-днев-
ных птенцах у нее была I, а 13 июля — II стадия линьки, т. е. прохождение одной ста-
дии заняло 6 дней. Самка, выкармливающая гнездовых птенцов, с 26 июня по 8 июля
продвинулась в смене маховых перьев с I до III стадии (2 стадии за 13 дней). Еще у од-
ной повторно гнездившейся самки 28 июня при откладке второго яйца на одном крыле
отсутствовали 9-е и 10-е маховые (асимметричная II стадия), а 25 июля она имела уже
IV стадию линьки (10-е и 9-е — новые, полностью отросшие перья; в основании 8-го
махового сохранились остатки чехлика, 7-е имело половину нормальной длины).
139
Таблица 36
Динамика отлова на гнездах самцов (над чертой)
и самок (под чертой) веснички с различной стадией линьки
Стадия линьки Число особей в Всего
июне июле августе
16-20 21-25 26-30 1-5 6-10 11-15 16-20 21-25 26-31 1-5
п 7 36 64 17 14 — — — — 138
и 20 66 100 59 41 тг 310
т 1 7 9 4 1 1 — 23
1 — — 3 — — — Т" 4
тт 3 4 4 8 7 1 — — — 27
11 — — — — — — т 8
III 1 1 3 4 1 — 2 — 12
— — 1 — 1 — •*— 1 4
IV — — — — 3 1 — “Г - — 4_t 1
V — — — — — — 2 “Г — — 2 1
VI — — — — — — 2 — — — 2
Всего: 10 42 76 37 32 4 5 2 — 208
20 66 100 63 41 12 10 10 327
из них 3 6 12 20 18 4 5 2 — — 70
с линькой — — 4 — 1 3 7 Т” 18
Получается, что на прохождение одной стадии совмещенной с гнездованием линьки
мухоловке-пеструшке требуется 6—10 дней. Однако у отдельных особей процесс смены
маховых перьев затормаживается — на одной и той же стадии они могут оставаться
более двух недель. В целом мухоловки-пеструшки, сменившие за время совмещенной
линьки только 9-е и 10-е маховые, получают выигрыш во времени по меньшей мере
в 6 дней по сравнению с особями, приступившими к смене оперения при вождении
слетков, и около трех недель против птиц, начавших линять после распадения выводка.
Периоды, в которые возможно начало совмещенной линьки у разных особей
мухоловок-пеструшек и весничек в Южной Карелии, представлены на рис. 38
и в табл. 35—36. Обращает на себя внимание, что и самки, и самцы этих видов при
разных сроках начала линьки приступают к ней достаточно дружно: около 70% в тече-
ние двухнедельного периода. К первой декаде июля линяющими оказываются практи-
чески все особи, за исключением отдельных самок весничек, у которых регенерация
оперения задерживается до 4-й пятидневки этого месяца.
Связь совмещенной линьки с датами начала размножения прослеживается уже
на основании изложенных выше данных — при поздних сроках выкармливания птенцов
линяющие особи встречаются чаще, а стадии линьки у них обычно выше. Поскольку да-
ты откладки первого яйца в контролируемых гнездах нам известны, можно попытаться
выяснить, какой будет доля птиц, совмещающих линьку и гнездование, при разных
сроках начала размножения (табл. 37). Общее число таких особей быстро нарастает в те-
чение репродуктивного сезона. Линяющие самцы начинают преобладать в отловах уже
в середине гнездового периода, хотя совмещать обе фазы годового цикла могут также
птицы, приступившие к размножению в ранние сроки. Исключением являются лишь сам-
ки веснички, начавшие откладку яиц после 16 июня. Но и при самых поздних сроках
начала гнездования многие самки веснички и мухоловки-пеструшки остаются
в старом наряде до окончания выкармливания гнездовых птенцов. Для самцов
140
100
Рис. 38. Динамика встречаемости мухоловок-пеструшек и весничек, совмещающих гнездова-
ние и линьку
1 - мухоловки-пеструшки; 2 - веснички. По оси абсцисс - календарь; по оси ординат -
доля линяющих особей среди отловленных на гнездах за каждую пятидневку (%).
мухоловки-пеструшки, в гнездах которых откладка яиц начиналась после 10.VI, совме-
щение линьки с гнездованием становится обязательным. Для самцов веснички такой
рубеж наступает на пятидневку позднее. В связи с этим важно выяснить, на какой
фазе гнездования особи может начаться смена оперения. Приблизительно зная темпы
линьки при ее совмещении с размножением, мы рассчитали возможные сроки ее на-
чала и получили следующее распределение (по числу особей) .
Таблица 37
Связь совмещенной с гнездованием линьки со сроками
начала откладки яиц (над чертой - линяющие особи, под чертой - не линяющие)
Вид и пол Число птиц, у которых размножение началось в
мае июне июле
11-15 16-20 21-25 26-31 1-5 6-10 11-15 16-20 21-25 26-30 1-5 6-10
Мухоловка- пеструшка 3 3 24 8 6 3 7 1
самцы 30 48 54 14 2 1
самки Весничка 1 36 3 42 1 1 74 10 3 26 15 65 1 9 20 2 3 13 6 7 7 2 22 3 1 — —
самцы 2 50 29 2 3 1 7 3 2
самки "8? 94~ 26~ 10 6 4 2 2
141
Весничка
Мухоловка-пеструшка
самцы самки самцы самки
Откладка яиц 1 (?) 2 1 2
Насиживание кладки 19 12 24 14
Выкармливание птенцов 50 4 29 7
Разумеется, эти данные следует рассматривать как предварительные. Тем не менее
они позволяют с достаточной определенностью констатировать, что большинство самцов
веснички (около 70%) приступают к линьке после того, как их самки начинают наси-
живание кладок, а остальные — при выкармливании гнездовых птенцов. Только один
самец веснички, видимо, начал линьку в период откладки яиц его партнершей: 20.VII
при десятидневных птенцах у него была VI стадия смены маховых перьев (первое
яйцо появилось в гнезде 25.VI).
Две трети самок веснички приступают к совмещенной линьке во время насижива-
ния кладки. Одна из них в день вылупления птенцов находилась на IV—V стадии линьки
и, вероятно, могла начать ее или одновременно с откладкой яиц, или до этого. Еще
одна самка, видимо, прекратила линьку на время откладки яиц: в середине инкубацион-
ного периода 9-е и 10-е маховые были у нее полностью отросшими, а на 8-м еще сохра-
нялся длинный роговой чехлик при 4/5 нормальных размеров пера.
Самцы мухоловки-пеструшки в основном также начинают линять во время
выкармливания птенцов в гнездах (54%) или на предыдущей стадии гнездования
(46%). Самки чаще совмещают начало линьки с насиживанием яиц и вдвое реже —
с выкармливанием птенцов. В двух случаях линька самок совпала с откладкой яиц.
Об одном из них упоминалось выше, а в другом - после потери гнезда с кладкой,
начатой 21.V, самка приступила к повторному размножению и 16—19.VI отложила 4 яй-
ца в новое гнездо; 26.VI у нее была уже III стадия линьки, на которой она оставалась
по крайней мере до 8.VII. За 7—11 дней линька у нее, конечно, не могла про-
двинуться на три стадии и наверняка началась еще до 16.VI, а затем, при нача-
ле нового размножения, была прервана, хотя заменившиеся перья продолжа-
ли расти.
Успешность гнездования при совмещенной линьке
Можно ожидать, что линяющие птицы из-за дополнительных расходов энергии
на линьку и ухудшения лётных качеств крыла будут отличаться пониженной успешно-
стью гнездования. Сравнение данных по гнездам мухоловок-пеструшек с различным
сочетанием линяющих и не линяющих партнеров показывает, что совмещение
двух смежных фаз годового цикла не снижало успешность выкармливания
птенцов.
Самец и самка не линяют линяют Линяет только самка Линяет самка, самец не кормит птенцов
Число вылупившихся птенцов 806 80 422 93
Успешность выкармлива- ния птенцов (%) 94.5 ± 0.8 95.0 ± 2.4 9 6.2 ±0.9 78.5 ±4.3
Лишь при отсутствии самца линяющие самки хуже справлялись с воспитанием
птенцов.
142
A
-------О 1983
&------------о III 1939.
Б
£--------ОI 1982
-------О
х--------О Л 1984
А—О7Л 1982
---------О
л--------С 1984
JK-------OZ 1982
а--------°Л
-------О 1984
< 1982
—ОII-III 1983
▲ А с 7 1973 1974
£~" о III 1975
А о 1976
А он 1980
-JSZ । 1 cl 1 1981
10V 31VI 20 VI 10VII
1982
1983
А/ $2 оЗ
_1__
10V
О 1983
—ОЛ 1984
ОТ 1982
ОН 1984
г--------о 1982
I
31V 20 VI 10VH
Рис. 39. Состояние оперения самцов мухо-
ловки-пеструшки (А) и веснички (Б), отлов-
ленных на гнездах в разные годы их жизни.
1-3 - даты: 1 - откладки первого яйца,
2 - отлова до начала линьки, 3 - отлова на раз-
ных стадиях линьки. Римскими цифрами обозна-
чены стадии линьки; арабскими - годы отлова.
По оси абсцисс - календарь.
Аналогичные данные получены и для веснички: нелиняющие самец и самка вы-
кормили 94.5 ± 0.9% птенцов (п = 601), а в тех случаях, когда к смене оперения при-
ступил один из партнеров или оба, — 93.3 ± 1.4% птенцов (и = 313).
Не подтверждается и предположение о том, что при совмещенной с гнездованием
линьке птицы испытывают дополнительную нагрузку и Поэтому хуже выживают в по-
слегнездовой период. Величина возврата особей, совмещавших две фазы годового
цикла, и птиц, линявших после вылета птенцов из гнезда, оказалась практически равной.
Линяли после
гнездования
Совмещали линьку
и гнездование
Мухоловка-пеструшка,
самцы:
окольцовано (особей)
обнаружено на следую-
щий год (%)
самки:
окольцовано (особей)
обнаружено на следую-
щий год (%)
Весничка, самцы:
окольцовано (особей)
обнаружено на следую-
щий год (%)
222
43.2 ± 3.3
309
20.4 ± 2.3
102
9.8 ± 2.8
43
48.8 ± 7.6
23
26.1 ± 9.1
46
8.7 ±4.1
Является ли совмещение линьки с гнездованием приви-
легией отдельных особей? Многолетнее кольцевание и контроль местного
гнездового населения птиц позволяет ответить на этот вопрос отрицательно (рис. 39).
Правда, из-за годовых различий в сроках начала откладки яиц и контрольных отловов
143
Рис. 40. Примеры непостоянства совмещения гнездования и линьки у меченых самцов мухо-
ловки-пеструшки в разные годы их жизни (каждая особь обозначена отдельным символом).
По оси абсцисс - дата контрольного отлова; по оси ординат - стадии линьки (0 - линьки нет).
Рис. 41. Связь стадии линьки, совмещенной с гнездованием (абсцисса), с величиной df - d0
(ордината), где d0 - дата начала совмещенной линьки в популяции, df - дата контрольного отло-
ва особи.
на гнездах комментировать полученные данные довольно сложно. Тем не менее ясно, что
совмещенная линька не является каким-то особым признаком, присущим только отдель-
ным особям: в течение последовательных сезонов размножения одни и те же меченые
птицы начинали смену оперения как во время гнездования, так и после его завершения.
Не существует также и строго запрограммированных индивидуальных дат начала
линьки: ежегодные различия достигали у отдельных особей двух недель и более.
На рис. 40 показаны годовые изменения связи дат отлова меченых птиц с состоя-
нием их оперения. Примечательно, что большинство мухоловок-пеструшек, не линяв-
ших во время гнездования, отлавливалось в период до 20.VI (8 из 11), а все линя-
ющие — после 19.VI. Выше, правда, было показано, что и после 20.VI некоторая часть
самцов не совмещает линьку с гнездованием (табл. 35). По наклону прямых на рис. 40
можно судить также о тенденции к увеличению вероятности совмещения линьки с гнез-
дованием при поздних сроках размножения особи.
Частота совмещения линьки с гнездованием, а также степень ее продвинутости
(стадия линьки) у самцов мухоловки-пеструшки оказывается положительно связан-
ной с величиной dt — do, тде dt — дата отлова исследуемой особи, a d0 — дата наиболее
раннего начала совмещенной линьки (г =0.52 £ 0.16) при р> 0.99 (рис. 41).
Так как у мухоловки-пеструшки совмещенная линька чаще отмечалась при позд-
них сроках размножения, была предпринята попытка определить ее корреляцию с датами
начала гнездования меченых особей в разные годы. Для приведения последних к обще-
му знаменателю определялась разность между конкретными датами начала откладки
яиц и наиболее ранним сроком откладки первого яйца в Южной Карелии. Корреляция
данного показателя со стадией совмещенной линьки (от 0 до V) оказалась достаточно
сильной и достоверной (г =0.745 ± 0.126 прир> 0.99; рис. 42).
Таким образом, совмещение линьки с гнездованием может произойти у любого
самца мухоловки-пеструшки в течение жизни, если по каким-то причинам его размноже-
ние в данный год начнется в поздние сроки. Действительно, доля птиц, у которых обе
фазы годового цикла проходят одновременно, в годы с поздним началом размножения
(по средней дате откладки первого яйца) возрастает. Соответствующие коэффициенты
корреляции: для самцов 0.74, для самок 0.51.
У веснички годовые различия в относительном обилии особей, совмещающих
линьку с гнездованием, оказались недостоверны. Но определенная тенденция к увеличе-
нию числа таких особей в годы с поздним началом и более растянутым размножением
намечается и для веснички, и для других видов птиц. В числе последних особенно хара-
ктерны птицы, выкармливающие по 2 выводка за год.
144
Рис. 42. Зависимость стадии линьки, совмещенной с гнез-
дованием, от даты начала откладки яиц в гнездах маркирован-
ных самцов мухоловки-пеструшки в разные годы их
жизни.
По оси абсцисс - стадии линьки; по оси ординат - 30
d\- d0 (сут), где di - дата откладки первого яйца в данный
год, d0 — наиболее ранняя дата откладки первого яйца
в популяции за все годы наблюдений.
20
Таким образом, наиболее энергоемкие фазы
годового цикла — гнездование и линька, считавшиеся м
несовместимыми, у целого ряда мелких воробьиных
птиц таежной зоны могут проходить одновременно.
Соотношение особей, линька которых накладывается
или разобщена с гнездованием, неодинаково для раз-
ных видов и подвержено внутривидовым, годовым
и сезонным колебаниям. Доля особей с совмещенной линькой в пределах изученных
нами видов возрастает в годы с поздним и растянутым гнездованием, а также к концу
сезона. Способность к совмещению двух фаз годового цикла не индивидуальна: у одних
и тех же особей в течение жизни она проявляется лишь в годы с более поздним началом
размножения.
При совмещенной линьке чаще всего сменяются 10—8-е первостепенные маховые
перья (I—III стадии), тогда как три последующие стадии у особей, не закончивших вы-
кармливание гнездовых птенцов, встречаются много реже. На этих и более поздних ста-
диях смены пера большинство самцов прекращают кормить птенцов. При указанных
объемах сменяющегося оперения лётные качества крыла оказываются вполне достато-
чными для сохранения высокой подвижности. Успешность выкармливания птенцов
при этом не снижается.
Энергетические затраты на совмещенную линьку, очевидно, не являются для птиц
чрезмерными, во всяком случае величина их возврата на места предыдущего гнездова-
ния приблизительно такая же, как у особей, линявших после завершения гнездования.
У самцов совмещенная линька чаще всего начинается при выкармливании гнездо-
вых птенцов, у самок — при насиживании кладок. Достоверно выявлены отдельные слу-
чаи откладки яиц линяющими самками мухоловки-пеструшки. Предполагается также,
что начальные этапы линьки могут быть прерваны на время нового — вероятнее всего,
повторного — размножения. Совмещенная линька может быть прервана также в про-
цессе гнездового цикла, причем темпы замены и роста пера, как правило, оказываются
в этом случае более низкими, чем при обычной — разобщенной с гнездованием -
линьке.
Совмещение двух энергоемких фаз годового цикла требует увеличения энергети-
ческих затрат, что оказывается возможным лишь при отсутствии дефицита кормов,
при стабильной кормовой базе и при наличии каких-то специфических условий, обеспе-
чивающих восполнение дополнительных затрат на линьку. Возможно также, что расход
энергии на совмещенную линьку не столь значителен, как при разобщенной с гнездова-
нием смене оперения.
Сезон, в котором чаще всего отмечается совмещение линьки с гнездованием в зо-
не наших исследований, характеризуется прежде всего максимальной в году продолжи-
тельностью светлого времени суток, наиболее стабильными погодными условиями,
самыми высокими в году положительными температурами, обилием, разнообразием
и постоянной доступностью кормов. К экологическим предпосылкам возникновения
совмещенной линьки следует, кроме того, отнести и общую невысокую плотность насе-
ления птиц в таежной зоне, т. е. отсутствие существенной конкуренции за пищу.
Специфика условий, благоприятствующих совмещению гнездования и линьки
в исследуемой зоне, несомненно определяется особенностями местного фотопериода —
72 10 В. Б. Зимин
145
практически круглосуточной освещенностью во время гнездования и совмещенной
линьки. Кормовой день — точнее, возможность добыть необходимое дополнительное ко-
личество пищи — сохраняется здесь круглосуточно. Ночной покой в условиях белых
ночей ограничивается не столько внешними факторами, сколько потребностями птиц
в отдыхе, которые могут быть близки к минимально допустимым. Кроме того, ночной
покой здесь может быть при необходимости прерван для кратковременной кормежки.
Снижение до минимума чистого расхода энергии в покое является существенным
преимуществом птиц северных популяций для получения необходимого дополнитель-
ного количества энергии. Это также может быть одной из предпосылок для возникно-
вения совмещенной линьки. Сказанное объясняет заметное преобладание особей, совме-
щающих линьку и гнездование, именно в период максимальной длины дня. И, видимо,
не случайно такое совмещение редко встречается при более поздних сроках размноже-
ния, например у з^янки, лесной завирушки и дроздов. Вполне вероятно, что в меньшей
степени оно развито и в условиях более высоких широт, где из-за низких температур
энергия в большей мере расходуется на восполнение теплопотерь и ее излишков не
образуется.
Как было показано выше, объем линьки, совмещенной с гнездованием, невелик
(сменяются 1—2, реже 4—6 маховых); смена оперения растянута во времени и проходит
с меньшей скоростью, чем обычно. Контурное оперение начинает обновляться в конце
IV и VI стадий, т. е. до этого момента целостность покровного оперения, а следователь-
но, и его теплоизоляционные свойства сохраняются на высоком уровне. Дополнитель-
ные затраты на терморегуляцию при совмещенной линьке не требуются.
При более продвинутой линьке резко увеличивается общий объем выпадающего
и одновременно растущего пера, что особенно заметно после начала линьки контурных
перьев крупных птерилий. При этом, естественно, должны возрастать и общие затраты
энергии на линьку. Совмещенная линька достигает таких стадий крайне редко. Чаще
это происходит у самиов, приступающих к регенерации оперения раньше самок. Боль-
шинство из них в подобных ситуациях прекращают кормление птенцов, и их линька
после этого становится фактически разобщенной с гнездованием.
Таким образом, наиболее важными особенностями совмещенной линьки явля-
ются: незначительнй объем сменяющегося оперения, низкая скорость роста пера, сохра-
нение нормальных теплоизоляционных свойств покровов и лётных способностей. Судя
по всему, такая линька не требует сколько-нибудь значительного увеличения расходов
энергии. Кроме того, дополнительные затраты могут компенсироваться указанными
выше преимуществами, которые получают птицы в исследуемой нами зоне.
Если дальние мигранты расходуют на линьку около 30 суточных бюджетов энер-
гии (Дольник, 1975), то с учетом очевидной неравномерности ее распределения на
средние и конечные стадии, включающие одновременную замену маховых, рулевых и
покровных перьев, должна приходиться большая часть общих затрат. Первые пять ста-
дий нормальной линьки — до начала смены рулевых и контурных перьев, — видимо,
вообще идут при низких энергетических затратах. Кроме того, при совмещенной линьке
по сравнению с обычной на рост первых пяти маховых расходуется примерно вдвое
больше времени. Таким образом, суточное потребление энергии на рост пера не должно
существенно влиять на общий энергетический баланс организма.
Для совмещения гнездования и линьки в зоне наших работ существуют вполне
определенные морфологические, экологические и физиологические предпосылки.
Механизм совмещения линьки с размножением до настоящего времени не выяв-
лен. Нам известны лишь внешние проявления процесса, среди которых наиболее приме-
чательны приуроченность его ко второй половине репродуктивного периода и способ-
ность отдельных особей прерывать начатую линьку или даже совмещать ее с откладкой
яиц. Следует также иметь в виду, что в зоне наших исследований и севернее из-за дефи-
цита времени, приходящегося на благоприятный период года, фазы гнездования и линь-
ки у большинства видов сближены в гораздо большей степени, чем на юге ареалов,
и почти всегда перекрываются. Кроме того, при значительно более поздних сроках
146
начала гнездования на севере его окончание приходится примерно на то же календар-
ное время, что и на юге. Сроки линьки воробьиных в разных частях ареала различаются
в гораздо меньшей степени, чем время гнездования. Поэтому вынужденно позднее
начало размножения на севере частично накладывается на период потенциально возмож-
ного начала линьки.
Совмещенная линька, вероятнее всего, возникает в переходный период между
окончанием активности гонад и подготовкой организма к смене покровов. В наиболее
общем виде процесс ее развития можно представить следующим образом. Сближены
и перекрываются около своих пороговых значений циклы гормонов, ответственных за
размножение и линьку. При возникновении внешних стимулов к гнездованию актив-
ность половых гормонов (если она еще не перешла нижней пороговой границы) может
временно повышаться. На период спаривания и откладки яиц соответственно заторма-
живается уже начавшееся развитие активности гормонов линьки. После завершения
кладки циклы гормонов продолжают развиваться в своих первоначальных направле-
ниях. Таким образом, в околопороговой области эти циклы, видимо, еще обратимы.
Если при сближенности двух фаз годового цикла никаких стимулов не возникает,
гнездование и линька оказываются разобщенными, и наоборот. Это отчасти объясняет
внутрипопуляционные различия в совмещении или разобщении двух фаз годового
цикла.
Кроме того, существуют, видимо, индивидуальные различия в продолжитель-
ности отдельных фаз годового цикла, в частности половой активности. Иначе трудно
объяснить, почему при размножении в одинаковые сроки совмещенная линька отмеча-
ется только у части особей. Вероятнее всего, годовые циклы гормонов размножения
и линьки индивидуальны. Об этом свидетельствует, например, разновременность начала
гнездования и линьки в популяции. Следовательно, у разных особей в одной местности
циклы гормонов будут иметь не вполне одинаковые продолжительность, сроки и сте-
пень сближенности. Во многом это будет, видимо, зависеть от того, насколько успешно
и в какие сроки прошли все предшествующие этапы годового цикла особи, как было
реализовано ее размножение. На меченых птицах установлено, например, что прохоло-
стание сопровождается более ранним началом линьки (пухляк, мухоловка-пеструшка).
Точно так же зарянки и лесные завирушки, не имеющие вторых кладок, приступают
к смене оперения на месяц раньше, чем их сородичи, обремененные воспитанием допол-
нительных выводков. Не исключено, что разнокачественность любых внутрипопуля-
ционных явлений хотя бы отчасти является следствием дисперсии особей, приводящей
к разнородности состава популяции и ее формированию из птиц, родившихся не только
вблизи пункта исследований.
Все эти предположения требуют дальнейшего подтверждения. Но для того чтобы
понять происходящие в организме птиц изменения в динамике, необходимы методы
прижизненного исследования отдельных особей в течение годового цикла. Для мелких
воробьиных птиц такие методы для применения в полевых условиях пока не разработа-
ны. Некоторые перспективы открывает известный метод лопароскопии. С его освое-
нием можно будет, по крайней мере, изучить основные нюансы в динамике развития
и состояния гонад особей, имеющих совмещенную и разобщенную с гнездованием
линьку.
В последнее время выясняется, что совмещение линьки с гнездованием характерно
не только для северных периферийных популяций воробьиных птиц. Оно обнаружено
у птиц, обитающих в более южных зонах (Ковшарь, 1977, 1981а; Губин, Ковшарь,
1982; Губин, 1983). С. П. Резвый и И. Б. Савинич (устное сообщение) обнаружили
линяющих на гнездах воробьиных птиц в лесостепной дубраве „Лес на Ворскле” (Белго-
родская обл.), где период благоприятных условий достаточно продолжителен. В субвы-
сокогорьях, где гнездование птиц начинается позднее, чем на равнине, птицы так же,
как и на севере, испытывают определенный дефицит времени. В этих условиях совмеще-
ние гнездования и линьки дает определенный выигрыш во времени и сокращает сум-
марные затраты его на преодоление довольно длительных фаз годового цикла. Кроме
147
того, это позволяет птицам завершить обе фазы в наиболее благоприятный период года.
На севере и в горах совмещение гнездования и линьки, а также линьки и миграции
позволяет птицам начать миграцию до наступления неблагоприятных условий: при отно-
сительно стабильных положительных температурах, большей продолжительности дня
и изобилии кормов — факторах, имеющих первостепенное значение для поддержания
положительного баланса энергии без излишнего напряжения.
В заключение следует еще раз напомнить, что совмещение гнездования и линьки
к настоящему времени более подробно исследовано только на двух видах воробьиных
птиц и в одной географической точке. Более широкое и углубленное изучение этого
явления позволит уточнить сделанные в данной главе обобщения, которые пока сле-
дует рассматривать лишь как предварительные.
148
ГЛАВА VI
ФОРМЫ БРАЧНЫХ ОТНОШЕНИЙ И РОЛЬ ПОЛИГИНЙИ
В РАЗМНОЖЕНИИ ТАЕЖНЫХ ВОРОБЬИНЫХ ПТИЦ
Наиболее распространенной формой брачных отношений у воробьиных птиц
исследуемой области является моногамия. Обычно при моногамии семья формируется
только на время размножения, но иногда распадается до его завершения, например
после разорения гнезда. В этом случае судьба семейной связи оказывается различной.
В большинстве случаев пара распадается и самка покидает территорию первоначально
выбранного самца. Иногда сменяет индивидуальный участок и самец, как это отмеча-
лось выше для зарянки. Реже после потери кладки или выводка гнездовая пара сохра-
няется и строит новое гнездо на прежнем или ближайшем свободном участке.
Иногда партнеры сохраняют верность друг другу на протяжении ряда лет, хотя
в целом для мелких воробьиных это не характерно. За исключением пухляка, хохла-
той синицы и врановых, маркированные на наших стационарах лесные воробьиные
птицы обычно каждый год размножались с новыми партнерами даже в тех случаях,
когда оба возращались на место предыдущего гнездования.
Как отмечалось в предыдущей главе, при интенсивной совмещенной линьке самцы
веснички, мухоловки-пеструшки, зяблика и пищухи прекращают кормить птенцов.
Это также означает распадение пары, хотя такие самцы обычно остаются на своих
гнездовых участках и сигналами тревоги оповещают выводок об опасности. Тем не ме-
нее вряд ли это свидетельствует о сохранении какой-то связи с семьей, поскольку
многие линяющие птицы издают крики беспокойства и после распадения выводка.
У чечевицы при позднем размножении самцы также покидают выводки, включаясь
в миграцию на зимовки. Начиная со второй декады июля количество их быстро сокра-
щается, а в первых числах августа из Карелии исчезают все „старые” (ad2) самцы. У пи-
щух при втором гнездовании ни в одном из 14 контролируемых гнезд самцы не корми-
ли птенцов и не подлетали к гнездам даже на тревожные крики самок. Благодаря тому
что при первом гнездовании все Взрослые особи были окольцованы, мы по данным
повторных отловов можем утверждать, что большинство самцов оставались на своих
участках, но с гнездами и вторыми выводками прямого контакта не имели.
Необычны брачные отношения у теньковки, самцы которой, как правило, не кор-
мят птенцов, хотя прекрасно знают место расположения гнезда, подлетают к нему,
сопровождают самку и выводок. В течение всего периода размножения самцы теньков-
ки, судя по активности пения и состоянию клоакального выступа, вполне способны
образовать полигамную семью. Однако, несмотря на специально предпринятые поиски,
полигиния у теньковки пока установлена только на одном из 50 обследованных гнездо-
вых участков (Н. В. Лапшин, устное сообщение).
В целом брачные связи таежных воробьиных птиц недостаточно прочны, и по раз-
ным причинам гнездовая пара может распасться на любой стадии репродуктивного ци-
кла. При относительно продолжительном сохранении половой активности птиц непроч-
ность семейных уз может служить одной из предпосылок к развитию так называемой
последовательной или разновременной полигамии.
Последнее время в связи с расширением работ по индивидуальному мечению слу-
чаи полигамии выявляются у многих видов птиц, в том числе и в районе наших иссле-
дований. Например, подробные материалы по факультативной полигинии у веснички
уже опубликованы (Лапшин, 1975, 19836). Готовятся к печати и соответствующие
данные по мухоловке-пеструшке, собранные за последние годы А. В. Артемьевым.
Частично опубликованы сведения о полигинии у славок (Музаев, 1980а, 19806,
149
1981), дубровника (Рымкевич, 1977), лесной завирушки (Яковлева, 1983). Отдельные
случаи образования полигиничных семей выявлены нами у белой трясогузки, крапив-
ника, горихвостки, трещотки, обыкновенной овсянки. Детальный обзор полигинии
у воробьиных птиц выполнен Хаартманом (Haartman, 1969b) по европейским видам и
Ориансом (Orians, 1969, 1978) по американским.
По гипотезе Вернера—Орианса, в основе полигинии лежит способность самок
оценивать качество территории самцов и дифференцировать перспективность партнера
для размножения. Если качество занятой самцом территории настолько высоко, что
привлекает двух разных самок, то, очевидно, такие территории полнее соответствуют
требованиям птиц к гнездовым стациям и будут поэтому чаще заселяться птицами, чем
пустовать (Verner, Willson, 1966; Orians, 1969).
Полигиния у мухоловки-пеструшки в Карелии изучалась на птицах, селящихся
в искусственных гнездовьях, расположенных несколькими изолированными линиями.
Ежегодно отловом на гнездах контролировалось около 90% взрослых особей (Зимин
и др., 1983). В разные годы бигиния была выявлена у 4.8-10, в среднем у 6.5 % самцов
мухоловки-пеструшки (и = 352).
При этом только в одном из 42 синичников, в разные годы занимаемых бигам-
ными самцами, полигиния была отмечена дважды за 6 лет. Обычно же каждый год ее
регистрировали на новых участках. Тем не менее в целом заметна определенная тенден-
ция к образованию полигиничных семей в одних и тех же, но достаточно больших груп-
пах гнездовий (20—30 синичников на 1—1.5-километровой линии). Эти материалы
не дают убедительного подтверждения гипотезе Вернера—Орианса. Можно отметить
лишь, что суммарная заселенность дуплянок, самцы в которых хотя бы один раз были
бигамными, оказалась достоверно выше, чем в остальных гнездовьях, занятых только
моногамными семьями (54.3 ± 4.8 и 34.3 ± 2.3% соответственно). Очевидно, у бигам-
ных самцов более привлекательны или сами гнездовья, или участки, на которых они
размещались. Этот вывод не вызывал бы никаких сомнений, если бы не одно обстоя-
тельство, на которое следует обратить внимание. У мухоловки-пеструшки, веснички,
лесной завирушки гнезда самок, имеющих общего самца, бывают не только удалены
друг от друга на значительные расстояния, но нередко между ними расположены гнез-
довые участки других пар птиц того же вида. На рис. 43 представлена одна из подобных
ситуаций. В данном случае самки, имеющие общего самца, отложили первые яйца
с интервалом в один день. В гнездах соседних пар кладка началась в те же сроки. Самец,
у которого первая самка гнездилась в синичнике № 47, на пути к своему второму
гнезду (№ 69) пересекал гнездовые участки 4 моногамных семей мухоловок-пеструшек,
пролетая около 1500 м. Хорошо известно, что самцы у этого вида являются политер-
риториальными особями. Кроме того, установлено, что одну и ту же дуплянку в период
гнездостроения, откладки и инкубации яиц могут посещать многие самцы. Так, у одной
из них за неделю было отловлено 7 разных самцов. Следовательно, вполне возможно,
что на участки с особо привлекательными гнездовьями могут претендовать несколько
самцов, в том чирле и уже имеющие самок. Самка № 69 могла бы стать партнершей
любого из соседних самцов — № 67 и № 73, которые наверняка чаще появлялись около
синичника № 69, расположенного в непосредственной близости от их собственных гнезд.
Во всяком случае территории, на которых размещались все названные дуплянки,
качественно были примерно одинаковы (синичники № 67 и № 73 заселялись мухолов-
ками честрушками по 3 сезона, а № 69 — только 2 раза за 6 сезонов). Синичник № 69
фактически находился на территории самца № 67, и тем не менее самка предпочла
„дальнего” самца. Таким образом, качество территории в данном случае не имело реша-
ющего значения.
На Шокшинском стационаре и в Маячино, где места размещения всех найденных
гнезд были точно картированы, мы имели возможность проследить в течение ряда лет
за распределением гнезд бигамных самцов веснички. Численность этого вида достигала
на пробных площадях 200 пар/км2, бигамия в разные годы отмечалась для 5—17%
контролируемых гнезд (Лапшин, 19836). Если вторых самок привлекает главным
150
52 60 61 67 69 73
В 0 0 0 0 * 0 0 00000»« 00 000*0В000«
26V 25V 27V 27V 26V 27V 29V
I 1500м I
7 *2 o3
Рис. 43. Пример одновременной политерриториальной полигинии у мухоловки-пеструшки
(Маячино, 1981 г.).
1-3 - гнездовья: 1 - полигиничного самца, 2 - занятые моногамными семьями, 3 - свобод-
ные. Верхние цифры - порядковые номера гнездовий; нижние - даты откладки первого яйца.
образом качество занятой самцами территории, то можно ожидать, что полигиния
в разные годы будет регистрироваться на одних и тех же участках. Однако за 5 лет
наблюдений лишь на двух гнездовых участках полигиния была отмечена в течение
2 и 3 лет подряд, тогда как на остальных 19 участках регистрировалась по одному
разу. Следовательно, территории самцов, которым удается привлечь вторых самок,
не обладают какими-то особыми достоинствами.
Как было показано нами ранее, перевод сосново-лиственных насаждений в сосно-
вые монокультуры при любом способе устранения лиственных деревьев приводил
к резкому сокращению численности весничек (Зимин, Кузьмин, 1980). С восстановле-
нием поросли лиственных деревьев плотность населения этого вида постепенно нараста-
ла (Зимин, 1983). Однако в момент депрессии численности и в годы ее подъема полиги-
ния у веснички на осветленных участках молодняков была обнаружена только один
раз. Правда, у мухоловки-пеструшки, гнездовое население которой также уменьшалось
после проведения ухода за сосновыми культурами, соотношение полигамных и моно-
гамных самцов на опытных и контрольных площадях было одинаковым. Результаты
этого сравнения также противоречивы. Явное ухудшение условий гнездования вызвало
у веснички уменьшение числа случаев полигинии, а на мухоловку-пеструшку никакого
влияния не оказало.
При ежегодных вариациях заселенности искусственных гнездовий мухоловкой-
пеструшкой от 10 до 32% и доли бигамных самцов от 0 до 23% (по отдельным уча-
сткам развески гнездовий) никакой корреляции между этими показателями не обнару-
жено (г = 0.075). Для веснички же при колебаниях плотности населения на отдельных
пробных площадях в разные годы от 22 до 200 пар/км2 и доли бигамных самцов от О
до 20% отмечена слабая и недостоверная связь этих показателей (г =0.39). Полигиния
у этого вида регистрировалась лишь на участках с численностью от 68 до 150 пар/км2.
На примере индиговой овсянки в штате Индиана (США) было показано, что часто-
та образования полигиничных семейных групп возрастала в годы с более асинхронным
формированием местного гнездового населения (Carey, Nolan, 1979). Мы пытались вы-
явить подобную связь на наших видах птиц. В качестве соответствующего показателя
синхронности бралась продолжительность периода, за который в 80 % гнезд была начата
кладка. У мухоловки-пеструшки в Маячино этот показатель варьировал от 5 до 16 дней,
у веснички — от 12 до 24, но никакой связи его с частотой встречаемости бигамных
самцов не обнаружено (рис. 44, Л). Другим показателем синхронности прохождения
отдельных фаз годового цикла местными птицами может служить общая продолжитель-
ность периода откладки яиц (по датам появления первого яйца в разных гнездах).
У мухоловки-пеструшки, по наблюдениям 1979—1984 гг., этот период занимал от 23 до
44 дней, у веснички (1973—1983 гг.) — от 21 до 58 дней. Связь растянутости гнездового
периода с долей бигамных самцов и в этом случае остается пока недоказанной (рис. 44,
Б). К тому же она оказалась неоднозначной: для мухоловки-пеструшки - положитель-
ная, а для веснички — отрицательная.
Для понимания сущности полигинии важно определить период ее возникновения
в течение репродуктивного сезона. У мухоловки-пеструшки вторые самки бигамных
самцов приступали к размножению во 2—7-й пятидневках, в среднем через 14 (3—32)
151
25Y-
20
15
10
5
^=[0.11]
10
о
4Z? -
30 -
20 -
0 2
6
. о
1^046+0.33]
гг= [0.5+ ±0.+2] °1
J____I__I___I_
+ б 8 10 Л2
Рис. 44. Частота встречаемости полигинии у веснички (I) и мухоловки-пеструшки (2) в зави-
симости от продолжительности пика массовой откладки яиц (А) и общего периода откладки яиц
у местных птиц (Б)
По оси абсцисс - доля бигамных самцов в популяции (%) ; по оси ординат'. А - период, в тече-
ние которого 80 % самок приступили к откладке яиц (сут), Б - продолжительность периода отклад-
ки яиц в популяции (сут).
дней после начала размножения в популяции, когда в 66.5 % (45—83) гнезд моногам-
ных семей были начатые или оконченные кладки. Соответствующие показатели для ве-
снички почти такие же: 2—5-я пятидневка, 12(5—20) дней, 78.3% (47—84) гнезд. Разли-
чия в фенологии гнездования моногамных и полигиничных семей веснички и мухолов-
ки-пеструшки наглядно представлены на рис. 45. Кроме запаздывания начала размноже-
ния, обращает на себя внимание тот факт, что кладки вторых самок в основном появля-
ются после пика откладки яиц в популяции. Лишь в редких случаях вторые самки обо-
их видов успевают найти самца и начать кладку в период массового начала гнездования,
т. е. одновременно с большинством моногамных пар. После гибели гнезд меченые самки
мухоловки-пеструшки ведут себя неодинаково. Одни из них остаются вблизи гнезд
с тем же самцом, другие меняют территорию и партнера, третьи становятся вторыми
самками самцов, уже имеющих гнезда, четвертые, несмотря на поздние сроки размно-
жения, образуют нормальные моногамные семьи. Таким образом, птицы, оказывающие-
ся в роли вторых самок, не являются какими-то неполноценными особями. Моногам-
ное размножение среди контролируемых нами весничек и мухолов ок-пеструшек еже-
годно отмечается уже после завершения откладки яиц всеми вторыми самками в поли-
гиничных группах (рис. 45).
Более низкая продуктивность размножения вторых самок бигамных самцов
по сравнению с первыми считается правилом. Но если сопоставить при этом известную
сезонную изменчивость величины кладки и выводка с более поздними сроками размно-
жения вторых самок, то пониженная продуктивность их размножения оказывается
вполне закономерной и не связанной с низкими репродуктивными возможностями
таких птиц, тем более что в моногамных семьях при 1—4-недельном запаздывании
начала кладки число откладываемых яиц и выкормленных птенцов тоже резко сокра-
щается. За непродолжительный гнездовой период величина кладки у веснички в Каре-
лии уменьшается на 1.4, а у мухоловки-пеструшки — почти на 2 яйца. В целом
же сравнение основных репродуктивных показателей у самок, имеющих раз-
ный статус в полигиничных семьях, не выявило существенных различий (рис. 46;
табл. 38).
Таким образом, несмотря на более низкий ранг, занимаемый вторыми самками
в полигиничных семьях, для воспроизводства популяции они имеют такое же значение,
как и первые самки бигамных самцов. Различия возникают в основном в результате
более позднего размножения вторых самок, что может быть обусловлено более позд-
ним прилетом на места гнездования, задержкой с развитием репродуктивного состоя-
ния, сменой места размножения из-за гибели самца, разорения гнезда, беспокойства
хищниками или конкурентами и тому подобными причинами. Хорошо известно, что на
первых этапах размножения связь самок с гнездом очень непрочна. При малейшем бес-
покойстве они легко оставляют построенные гнезда, начатые и даже полные кладки.
152
В. Б. Зимин
Таблица 38
Основные репродуктивные показатели первых и вторых самок в полигиничных семейных группах мухоловки-пеструшки и веснички
Статус самки Дата откладки 1-го яйца Величина кладки (число яиц) Число слетков на 1 гнездо Величина благополучного выводка (число птенцов)
lim | М д м, м, А4 м.
Первая I I 16.V-4.VI 1 25V 1 25 v 1 I 6.65 ±0.15 Мухоловка-пеструшк 1 6.65 ±0.15 I 5.92 ±0.31 1 :а I 5.92 ±0.31 I [ 6.16 ± 0.20 I 6.16 ± 0.20
Вторая 21.V-24 .VI 2.VI 1 29.VI 1 1 6.39 ±0.18 | 6.58 ±0.18 4.77 ± 0.43 | 5.21 ±0.51 | 5.39 ± 0.28 1 1 5.82 ± 0.31
Первая | 21JV-9.VI I 31.V 1 3LV 1 1 6.63 ±0.16 I Весничка I 6.63 ±0.16 I 4.56 ±0.46 I 4.56 ± 0.46 I 5.00 ± 0.37 1 5.00 ± 0.37
Вторая 1-18.VI 8.VI 6.VI 6.11 ±0.15 6.23 ±0.17 4.41 ± 0.53 4.13 ± 0.77 5.39 ± 0.33 5.64 ± 0.47
Примечание. — средняя для всего периода, — средняя для общего периода откладки яиц первыми и вторыми самками.
Таблица 39
Возраст самок в полигиничных группах веснички и мухоловки-пеструшки
Статус самки Число птиц в указанных возрастных группах
а adf а + 1,ad2 а + 2, ad3 а + 3, ad4 а + 4, ad5
Первая 1 36 Весничка (по: Лапшин, 1 3 1 19836, с дополнениями) 1 1 1 1 1
Вторая J 34 1 1 2 1 5 — — 1 1
Первая 1 21 Мухоловка-пеструшка 1111
Вторая 19 1 2 j 1 | 1 ~ -
Рис. 45. Различия в сроках и динамике размножения при моно- (1) и полигамии (2) у веснич-
ки (А) и мухоловки-пеструшки (Б).
По оси абсцисс - порядковый номер пятидневки от начала откладки яиц в популяции; по оси
ординат - число начатых кладок (%).
Рис. 46. Соотношение величины кладки первых и вторых самок в полигиничных семьях
мухоловки-пеструшки (А) и веснички (Б) при разных интервалах между сроками откладки пер-
вого яйца.
По оси абсцисс - разница в числе яиц, отложенных первыми и вторыми самками; по оси
ординат - различия в датах откладки первого яйца (сут).
Нередко птицы бросают гнезда по вине наблюдателей, регулярно осматривающих гнез-
да или выполняющих другие работы на пробных площадях.
Среди поздно приступающих к гнездованию птиц, как правило, преобладают
особи, потерпевшие неудачу при первом размножении. После гибели гнезд маркирован-
ные самки веснички, как и многих других видов открытогнездящихся птиц, исчезали
с контролируемых территорий и ни разу за все годы исследований на них повторно
не отмечались. Между тем окольцованные самки мухоловки-пеструшки после разоре-
ния первых гнезд нередко снова гнездились в непосредственной близости от них или
смещались на расстояние до 20 км.
О происхождении и дальнейшей судьбе первых и вторых самок в полигиничных
группах известно немного. Четыре самки мухоловки-пеструшки родились в 1—15 км
от места будущего гнездования с бигамными самцами. На положении вторых они ока-
зались в возрасте 1—2 лет и два раза — трехлетними, соответственно выкормив по 4, 6,
7 и 0 птенцов. После участия в полигиничном размножении ни одещ из них в следующие
годы на контролируемой территории не обнаружена.
После успешного размножения 2 первые и 2 вторые самки возвратились на место
гнездования (3 — на следующий год и 1 — в течение двух сезонов), когда они образо-
вывали нормальные моногамные семьи. Все эти самки были членами полигиничных се-
мей в год их первого обнаружения на стационаре, а еще одна — в возрасте ,/z + 7”.
Самка веснички, окольцованная птенцом, в возрасте 3 и 4 лет гнездилась в статусе
„первая” с двумя разными бигамными самцами. Еще 9 самок после гнездования в поли-
гиничных семейных группах в следующие годы на контролируемой территории не были
встречены.
Среди самок, имеющих ранг „вторая”, обычно отмечают преобладание первогод-
ков (Martin, 1974; Crawford, 1977, и др.). Однако наши материалы по мухоловке-
пеструшке и весничке не позволяют утверждать это с полной определенностью
(табл. 39). Первогодки среди местного населения этих птиц вообще составляют боль-
шинство, а в положении вторых самок особи 2—4-летнего возраста встречались одина-
ково редко.
Размножение 18 самцов мухоловки-пеструшки и 6 самцов веснички с достоверно
выявленной полигинией удалось проследить на протяжении 2—4 лет (рис. 47). В течение
этого периода все самцы становились бигамными только по одному разу, хотя в подав-
ляющем большинстве случаев занимали одни и те же территории, качество которых
вряд ли сколько-нибудь значительно изменялось в разные годы. При низкой
154
Рис. 47. Индивидуальная изменчивость числа слетков, выкормленных маркированными
самцами мухоловки-пеструшки (А) и веснички (Б) в разные годы жизни при полигамии (1) и мо-
ногамии (2).
По оси абсцисс - возраст птиц (годы); по оси ординат — число выкормленных слетков.
заселенности гнездовий на участке любого самца мухоловки-пеструшки всегда были
свободные синичники. Как видно из приведенных данных, самцы становились полига-
мами в любом возрасте, но относительно чаще — в возрасте „л + 7”.
Вышеупомянутые самцы веснички и мухоловки-пеструшки в течение контроли-
руемого периода их жизни вели себя и как моногамы, и как полигамы, т. е. не обладали
какими-то генетически закрепленными или приобретенными качествами, особенно
привлекательными для самок.
Важно подчеркнуть, что в отличие от самок многие самцы в следующие годы после
бигамного размножения благополучно возвращались на родину, т. е. участие в выкарм-
ливании 2 выводков не оказало отрицательного влияния на их выживаемость.
У мухоловки-пеструшки для бигамных самцов в целом характерны ранние сроки
начала гнездования (с первыми самками). Способность к раннему размножению такие
самцы демонстрировали и в другие годы их жизни, когда они имели моногамные семьи.
Однако каких-то исключительных способностей в выкармливании птенцов бигамные
самцы не проявляют. В годы, когда они были моногамами, средняя величина успешно
выкормленного ими выЁодка была 5.73 ± 0.32, т. е. почти такая же, как среднепопуля-
ционная — 5.65 ± 0.05 (р > 0.05). Последний показатель рассчитан по величине всех
выводков в популяции, в том числе* и поздних, с небольшим числом птенцов. Если же
определить его только для периода, в который регистрируется гнездование бигамных
семей, то он окажется достоверно выше: 5.97 ± 0.08 (р< 0.05).
Обычно считается, что полигиничные самцы оставляют в целом более многочислен-
ное потомство (Price, Bock, 1973; Carey, Nolan, 1979; Balfour, Cadburry, 1979, и др.). Изу-
ченные в Карелии птицы также не являются исключением в этом отношении. По данным
Лапшина (19836), на каждого полигиничного самца веснички приходилось от 0 до 13
слетков, в среднем 8.2 ± 0.7, что почти вдвое больше среднепопуляционной нормы.
Соответствующие показатели для мухоловки-пеструшки следующие: при бигамии
от 4 до 14 слетков (10.77 ± 0.52), при моногамии в среднем 6.65 ± 0.07.
Многие исследователи отмечают также, что при одновременной полигинии самец
обычно активно кормит только один из выводков. При ухудшении условий, например
во время дождливой погоды или при похолоданиях, бигамные самцы дроздовидной
камышевки кормили только один выводок (Dyrcz, 1977). У рисовых трупиалов самец
кормит птенцов первой самки, а вторая выкармливает выводок одна, причем частота
кормления у второй самки гораздо выше, поэтому птенцы обоих выводков получают
примерно одинаковое количество пищи (Martin, 1974).
Истинная роль бигамного самца в выкармливании двух выводков весьма неясна.
Нередко его участие во втором размножении, особенно при поздних сроках, фактически
сводится лишь к оплодотворению самки. Поздние выводки вторые самки веснички и
мухоловки-пеструшки очень* часто выкармливают без самцов. При одновременной
155
полигинии в нормальные сроки размножения не все самцы веснички в одинаковой
мере кормят птенцов обоих выводков. При последовательной полигинии сначала отдает-
ся предпочтение первому выводку, а после его вылета самцы переключаются на воспи-
тание второго (Лапшин, 19836). Следовательно, ни один из выводков не воспитывает-
ся самцом полностью. Аналогичное распределение родительских забот наблюдается
и в полигиничных семьях мухоловки-пеструшки. Таким образом, при полигинии основ-
ную роль в воспитании вторых выводков играют самки, а помощь самцов часто ока-
зывается малосущественной. Тем не менее успешность размножения вторых самок
оказывается почти такой же, как у первых. Так, общая успешность размножения у вес-
нички составляла для первых самок 69.5 ± 4%, для вторых 73.9 ± 4.1 %; у мухоловки-
пеструшки 89.0 ± 2.4 и 82.1 ± 3.1 % соответственно.
Относительно способности самок самостоятельно выкормить выводок и о ее зна-
чении как важной предпосылки для развития полигинии существует две точки зрения.
По одной из них, полигиния может развиться только в условиях трофического благо-
получия (Lack, 1978), по другой - ступенью к полигинии служит постоянный недостаток
корма, поэтому самке выгоднее использовать его одной, чем делить с самцом (Willis et
al., 1978). Последнее показано на примере мухоловки Pipromorpha macconnelli в Брази-
лии. Самцы этого вида после спаривания покидают территорию самки и могут найти
других, с которыми также не остаются. Родительские заботы у этого вида целиком
возложены на самок.
Однако успех размножения зависит не только от обилия или недостатка корма.
При низких температурах среды возникает необходимость в более плотном насижива-
нии кладок и дополнительном обогреве маленьких птенцов. Для самки-одиночки
это создает очень напряженный режим с дефицитом времени на сбор корма для себя
и птенцов. При температурах, превышающих оптимальные, также возрастают затраты
времени и энергии на предупреждение перегрева кладки и птенцов. Интересная работа
в этом плане выполнена на саванной овсянке, в южных популяциях которой полигиния
встречается часто, а в условиях тундры не обнаружена (Weatherhead, 1979). Работая на
Аляске, автор выяснил, что без участия самцов, которых искусственно устраняли, сам-
ки не в состоянии справиться с выкармливанием птенцов.
Хаартман (Haartman, 1969b), проанализировавший распространение полигинии
среди европейских воробьиных птиц, обратил внимание на то, что наиболее часто она
встречается среди дуплогнездников и „купольников”, и связал это с лучшей защищен-
ностью выводка и меньшими потерями тепла в гнездах закрытого типа. У многих видов
воробьиных и других птиц самки во время насиживания часть необходимого корма
получают от самцов, что позволяет им проводить на гнезде больше времени. Кроме
более плотного режима насиживания, это способствует лучшей защищенности кладки
от хищников. Окраска яиц часто резко контрастирует с окружающим фоном, из-за чего
в открытых гнездах при отсутствии самки яйца легче обнаружить. Самцы многих видов
воробьиных птиц не кормят самок в период инкубации яиц и не обогревают птенцов.
Такие виды есть как среди закрыто-, так и среди открытогнездящихся птиц, и их пред-
ставители одинаково хорошо адаптированы к подобному разделению родительских за-
бот. Не получая дополнительного корма от самцов, самки успешно справляются с инку-
бацией яиц, обогревом и выкармливанием маленьких птенцов. Самцы теньковок не кор-
мят ни самок, ни птенцов. Птенцы первых выводков иногда вылупляются у них на дека-
ду раньше, чем у веснички, нередко во время похолоданий. Кроме того, гнезда теньков-
ки чаще всего приподняты над поверхностью, поэтому, согласно Шилову (1968), темпе-
ратурный режим в них должен быть более зависим от внешних воздействий, чем в назем-
ных гнездах, и на стабилизацию внутригнездовой температуры необходимы дополни-
тельные затраты времени и энергии самки. Тем не менее самки теньковки без помощи
самцов выкармливают в среднем столько же птенцов, сколько и другие виды пеночек,
у которых самцы активно участвуют в воспитании потомства.
Предпосылкой к полигинии является также освобождение самцов от участия
в посторойке гнезд, инкубации яиц и кормлении самок. Лишь у немногих видов
156
воробьиных птиц в таежной зоне самцы существенно обременены подобными заботами.
В строительстве гнезда не участвуют самцы жаворонков, большинства синиц, пищух,
пеночек, камышевок, сверчков, завирушек, трясогузок, свиристелей, овсянок, вьюрко-
вых. У врановых самцы выполняют роль подносчиков гнездового материала, у пухляка
и хохлатой синицы гнезда в совместно выдолбленных дуплах строят только самки.
Еще до прилета самок самцы славок делают несколько гнездовых „набросов” (садовая
славка) или каркасов (славка-завирушка, черноголовая славка), которые, однако, не
всегда используются самками. И лишь у немногих наших воробьиных постройкой гнезда
заняты оба партнера (ласточки, оляпка, длиннохвостая синица, королек, сорокопуты,
скворец, воробьи). В зоне наших исследований только у крапивника роль самки в гнез-
достроении ограничена выбором одного из 6—8 построенных самцом гнезд и выстилкой
лотка небольшим количеством перьев. Кстати, постройка „лишних” гнезд самцом кра-
пивника, как полагают, может быть одним из средств привлечения нескольких самок.
При одновременной полигинии связь бигамных самцов с новой самкой может
формироваться у веснички и мухоловки-пеструшки в период строительства гнезда,
которым самцы не заняты.
Хотя сведения об участии самцов воробьиных птиц в насиживании кладки и обог-
реве птенцов часто очень противоречивы, два наиболее важных обобщения все же можно
сделать. Во-первых, у большинства видов самцы не имеют наседных пятен. К этой груп-
пе принадлежат и весничка, и мухоловка-пеструшка. Во-вторых, принимающие участие
в насиживании самцы как при наличии наседных пятен — ласточки, скворцы, некоторые
славки рода Sylvia (Parkes, 1953; Мальчевский, 1959; Lloyd, 1965; Люлеева, 1967; Ефре-
мов, Паевский, 1973), так и не имеющие их — желтоголовый королек, жулан, овсянка-
ремез, дубровник, в целом проводят на гнезде гораздо меньше времени, чем их самки.
Известно также, что родительский вклад отдельных самцов в воспитание птенцов
оказывается очень разным и на всех этапах гнездовой жизни варьирует в достаточно
широких пределах.
Пища, которой самцы подкармливают самок на начальных этапах гнездования,
является дополнительным источником энергии, необходимой для гнездостроения и фор-
мирования половых продуктов (Lack, 1966). Видимо, не случайно в зоне умеренного
климата такое поведение в большей мере свойственно птицам, приступающим к гнездо-
вании? в ранневесеннее время, в условиях относительно низких температур, более ко-
роткого дня и при дефиците кормов.Тем не менее, пожалуй, лишь у клестов самцы спо-
собны полностью обеспечить пищей практически не слетающих с гнезда самок (Коханов,
Гаев, 1970). У всех прочих воробьиных птиц таежной зоны насиживающие самки вынуж-
дены периодически кормиться сами, так как приносимой самцами пищи им не хватает.
При одновременной полигинии самцы мухоловки-пеструшки кормят только
одну (обычно первую) самку и приносят ей до 100 порций корма в день. Самки длин-
нохвостых синиц в период инкубации получают корм каждые 15—20 мин. Правосудов
(1983) провел хронометрирование кормления самцами самок пухляка и сероголовой
гаички и пришел к заключению, что доля корма партнера в суточном рационе самок
в период откладки яиц составила у обоих видов лишь по 0.4 ± 0.1 %, а при насижива-
нии 2.3 ± 0.4 у пухляка и 2.4 ± 0.3 % у сероголовой гаички. В целом во время насижива-
ния самки обоих видов проводят в поисках корма 5—6 ч, т. е. почти треть светлого
времени суток. По-видимому, полученные Правосудовым результаты все же несколько
занижают роль самцов. О доле корма, переданного самке, автор судил по числу кор-
млений, но масса пищевого комка по понятным причинам осталась не выясненной.
И если при самостоятельной кормежке самка пухляка склевывает корм с интервалом
0.12 мин, а самец кормит ее через 3.5 мин, то, видимо, за это время он может собрать
достаточное число пищевых объектов.
Даже в тех случаях, когда кормление насиживающей самки является обязатель-
ным актом, у самцов остается достаточно времени для других видов активности.
Интенсивность кормления самок у самцов в значительной мере индивидуальна, как
и участие в выкармливании птенцов. У мухоловок-пеструшек, например, отдельные
самцы при моногамии кормят самок лишь 2—3 раза в час.
Таким образом, в периоды постройки гнезд, откладки яиц и их инкубации самцы
практически всех видов птиц расходуют на это незначительную часть времени и его
им вполне достаточно для проявления всех других видов активности, в том числе и для
установления связи со второй самкой. Следовательно, незначительная занятость самцов
родительскими заботами также может быть одной из предпосылок для возникновения
полигинии.
У самцов некоторых видов птиц после начала насиживания кладки проявление
всех видов брачной активности заметно снижается. Судить об этом можно хотя бы по
резкому сокращению активности пения (Мальчевский, 1959). Подобное поведение
характерно для дроздов, зарянки, лесной завирушки, пищухи, королька и синиц. Оно
может быть существенным препятствием для возникновения полигинии. По наблюде-
ниям Лапшина (1978, 19836), самцы веснички после того, как их самки приступают
к насиживанию кладки, ведут себя так же, как в предгнездовой период, т. е. как холо-
стые особи. Частота исполнения песен при отсутствии визуального контакта с самкой
достигает 67 песен за 10 мин (при контакте — 12—16 песен). Практически не снижается
активность пения и у самцов мухоловки-пеструшки. После того как самки приступают
к насиживанию кладки, они поют так же интенсивно, причем не только около своего
гнезда, но и на свободных территориях, на удалении до 2 км. По данным отловов пау-
тинными сетями и визуальной регистрации маркированных самцов на Шокшинском
стационаре, где контролировалась территория немногим более 40 га, самцы мухоловки-
пеструшки в мае повторно находились в среднем в 220 м от гнезда (0—750 м),
в период с 1 по 20 июня — в 165 м, а позднее, до начала вождения выводка, — в 120 м.
Проявление высокого уровня половой активности у самцов после завершения
самкой откладки яиц, очевидно, вводит в заблуждение других самок, по разным причи-
нам еще не имеющих партнера или потерпевших неудачу в начале гнездования, и они
вполне могут воспринимать таких самцов как холостых. И коль скоро эти самцы и за-
нятые ими участки уже пользовались успехом у первых самок, то, очевидно, они имеют
такие же шансы на привлечение новой самки. Возможно, этим объясняется одновремен-
ное наличие холостых и бигамных самцов в одной и той же местности. Причины прохо-
лостания отдельных самцов до сих пор остаются неизвестными и малопонятными.
Холостыми, например, могут оставаться самцы с Нормальной видовой песней и высокой
активностью пения, тогда как особи с дефективной песней, живущие на соседней тер-
ритории, успешно образуют пару. Нередко не участвуют в размножении активные сам-
цы, занимающие заведомо пригодную территорию, на которой в предыдущем и в после-
дующие годы отмечалось успешное гнездование других особей. Не всегда гаран-
тировано образование семьи даже в оптимальных для гнездования вида стаци-
ях, и, напротив, в субоптимальных местообитаниях пары формируются вполне успешно.
Сосуществование таких явлений, как прохолостание части особей и полигиния,
отмечается всюду, для многих видов птиц с достаточным постоянством и скорее всего
не случайно. Это наводит на мысль о том, что полигиния могла возникнуть в эволюции
птиц как один из механизмов репродуктивной изоляции особей, по каким-то пока
не вполне ясным причинам не соответствующих требованиям, предъявляемым
к партнеру по размножению. Не исключено, что особей, остающихся холостыми, отлича-
ют какие-то особенности внешних признаков или поведения, отклоняющиеся от популя-
ционных (видовых) норм. Тогда прохолостание таких особей, их исключение из воспро-
изводства может оказаться полезным для вида, так как препятствует развитию чуждых
популяции (виду) признаков и их проникновению в генофонд. В таком случае поли-
гамию можно рассматривать как барьер, способствующий сохранению чистоты генофон-
да и целостности вида.
Полигиния у веснички и мухоловки-пеструшки была как поли-, так и монотерри-
ториальной, а также гаремной (одновременной)и последовательной (разновременной).
У веснички в 45 % случаев самец спаривался со второй самкой до завершения кладки
в первом гнезде, в 47% — во время насиживания яиц первой самкой и в 8% - после
158
гибели выводка. В большинстве случаев полигиния была монотерриториальной, причем
самки не проявляли агрессивности друг к другу (Лапшин, 19836).
У мухоловки-пеструшки соотношение моно- и политерриториальной полигинии
было несколько иным (число случаев).
Монотерриториальная Политерриториальная
Одновременная 10 4
Разновременная 8 4
Здесь обращает на себя внимание заметное преобладание монотерриториальной
полигинии, что в наших условиях может быть связано с общей невысокой численностью
и слабой заселенностью искусственных гнездовий, когда свободные синичники имелись
на участке каждого самца. Таким образом, политерриториальность самцов мухоловки-
пеструшки хотя и важна для возникновения полигинии, но строго обязательным усло-
вием все же не является.
Определенным препятствием для образования полигиничных семейных групп
птиц могут быть агрессивные взаимоотношения между самками. Среди таежных птиц
в наиболее яркой форме это проявляется у горихвостки и обыкновенной овсянки
(Haartman, 1969b). Гем не менее факультативная полигиния известна и для этих видов,
у которых, по мнению Хаартмана, самцы легко находят пути, устраняющие данную
помеху. У веснички, как уже указывалось, первые и вторые самки бигамных самцов
вполне терпимо относятся друг к другу. В значительной мере это справедливо и для
мухоловки-пеструшки. Если бы у этого вида самки активно препятствовали проникно-
вению на занятую территорию других самок того же вида, то политерриториальная по-
лигиния должна была бы чаще возникать в период откладки яиц, чем во время насижи-
вания, когда первая самка большую часть суток находится внутри дупла. Однако на
самом деле роль первых самок в предотвращении полигинии их партнеров весьма сом-
нительна: и моно- и политерриториальная полигиния возникают на любом этапе гнездо-
вания первой самки (число случаев).
Монотерриториальная Политерриториальная
«Откладка яиц 8 4
Инкубация яиц 8 3
Выкармливание птенцов 1 1
В отличие от веснички самцы мухоловки-пеструшки способны спариваться со вто-
рой самкой даже во время выкармливания птенцов первого выводка. В одном случае
первое яйцо во втором гнезде появилось при четырехдневных птенцах первого вывод-
ка, в другом — при восьмидневных птенцах. Известен также случай, когда самец обра-
зовал новую пару (!) после гибели выводка. У веснички подобные случаи отмечены
для 8 % бигамных самцов (Лапшин, 19836).
Возникает вопрос: является ли это полигинией, как иногда считают? Поскольку
самец последовательно спаривается с двумя самками, то на первый взгляд это типичная
полигиния. Но если его вторая самка до встречи с ним также имела и потеряла гнездо,
то и она последовательно спаривалась с двумя самцами, а это уже полиандрия. В этой и
подобной ситуациях не соблюдено еще одно условие полигамии — в момент возникно-
вения новой связи один из партнеров уже должен иметь семью. В приведенном примере
образуется лишь гнездовая пара, а не полигамная группа. Единственное отличие от
нормальной моногамной пары лишь в том, что оба партнера (или один) размножаются
повторно в данный сезон.
Теоретически возможно (но у птиц с моноциклическим размножением нами пока
не обнаружено), что самец после успешного докармливания слетков вновь образует
пару с новой партнершей, например потерявшей гнездо. Изредка такое случается у птиц
с бициклическим гнездованием и отмечено нами для зарянки, большой синицы, лесной
завирушки и дроздов. Но в данном случае также заново образуется п а р а, а не много-
159
членная семья. Поэтому подобные брачные связи не следует называть полигамией,
хотя и можно рассматривать как определенный шаг к ее возникновению.
В приведенных примерах выявляется определенная связь между полициклией
размножения и полигинией. Спаривание самца, успешно выкормившего один выводок,
с новой партнершей в принципе можно рассматривать как одну из разновидностей
полициклического размножения или как шаг к его развитию. При облигатном поли-
циклическом гнездовании оба члена пары выводят птенцов дважды в год (или более),
а при полигинии - один самец с двумя самками. Один и тот же результат - два выводка
в год — достигается разными путями. При полициклии это осуществляется двумя
особями при последовательном размножении в течение длительного гнездового сезона,
при полигамии — тремя (или более) особями, но за гораздо более короткий про-
межуток времени.
Первый путь могут использовать птицы с относительно продолжительным гнездо-
вым сезоном, второй — те, у которых период размножения вынужденно укорочен
из-за большей растянутости других фаз годового цикла, таких как линька или миграции.
Полигиния, пожалуй, является одним из реальных путей повышения общего
уровня воспроизводства в популяциях дальних мигрантов, в годовых циклах которых
гнездовой сезон занимает незначительную часть времени. Полицикличность размно-
жения для них недоступна, так как она создавала бы чрезмерную напряженность годо-
вого цикла и значительный дефицит времени на линьку и миграции. Не для всех видов
приемлемо и увеличение размера кладки, требующее повышенных затрат энергии (и
времени) на воспитание больших выводков, в итоге приводящее к росту смертности
производителей и потомства. Важно обратить внимание на то, что среди всех лесных
певчих птиц таежной зоны полигиния наиболее развита именно у анализируемых нами
видов — веснички и мухоловки-пеструшки, являющихся дальними мигрантами и име-
ющих короткий период размножения. Это не означает, однако, что в процессе эволюции
птиц данный путь повышения продуктивности размножения был строго разграничен.
Среди птиц с продолжительным сезоном размножения также есть виды, использующие
для повышения продуктивности размножения и полициклию, и полигамию. В частности,
к таким птицам относятся крапивник, белая трясогузка, обыкновенная овсянка, гори-
хво стк а-л ысушка.
Итак, полигамия возникла и поддерживается естественным отбором как один из
путей оптимального использования популяционных ресурсов, как особая стратегия
размножения, обеспечивающая более высокий уровень воспроизводства. Не исключено
также, что отбор благоприятствует полигамии и как средству, которое не допускает
участия в размножении некоторых особей и предупреждает в связи с этим проявление
каких-то негативных последствий, нарушающих целостность вида и изменяющих его ге-
нофонд.
важнейшей предпосылкой к возникновению полигинии, по крайней мере у иссле-
дованных нами видов птиц, является способность самок самостоятельно, без помощи
самца построить гнездо, успешно завершить насиживание кладки и выкармливание
выводка. Возникновению полигинии способствует также относительная непрочность
семейных и территориальных связей, особенно характерная для начальных этапов гнез-
дований, и способность самок к неоднократному восстановлению брошенных или утра-
ченных кладок не с одним, а с разными самцами и на новых территориях. Кроме того,
существенное значение для развития полигинии имеет сохранение высокой половой
активности у самцов после завершения самками откладки яиц и при относительной
или абсолютной необремененности заботами о постройке гнезд, инкубации яиц, кормле-
нии самки и обогреве птенцов. В результате этого самки, по разным причинам задержи-
вающиеся с началом размножения, могут воспринимать самцов как холостых. Нако-
нец, для развития последовательной разновременной полигинии особенно важны отно-
сительная растянутость гнездового периода и сохранение высокого уровня половой
активности в течение более или менее длительного периода.
160
Г Л А В A VII
ПРЕОДОЛЕНИЕ НЕГАТИВНЫХ И ИСПОЛЬЗОВАНИЕ ПОЗИТИВНЫХ
ФАКТОРОВ СРЕДЫ ПТИЦАМИ, ОБИТАЮЩИМИ НА ПЕРИФЕРИИ АРЕАЛА
В СЕВЕРО-ЗАПАДНОЙ ЧАСТИ СССР
Суммируя основные выводы из предыдущих разделов книги, необходимо оценить
в целом суть проблемы „жизнь на пределе возможного” и определить важнейшие пути
преодоления негативных и использования позитивных внешних воздействий, обеспечи-
вающие существование птиц в пессимальных областях ареала.
Специфика суточного режима освещенности и его влияние на птиц. Известно,
что суточный энергетический баланс животных определяется как соотношение энергии,
поступающей в организм с пищей, и энергии, расходуемой на поддержание жизнедея-
тельности и всех видов активности. Ночью энергетические резервы только расхо-
дуются — и, очевидно, тем полнее, чем длиннее период ночного сна. На юге сон дневных
птиц жестко ограничен темным временем суток, а у „северян” — только внутренними
потребностями, которые могут соответствовать минимальной необходимости и быть
максимально кратковременными. К тому же на севере, в зоне белых ночей, птицы
способны прерывать ночной покой для непродолжительной кормежки, обеспечивающей
дополнительное поступление энергии с пищей. Суточный энергетический баланс север-
ных птин даже при возрастании затрат энергии на терморегуляцию из-за относительно
более низких температур при низком уровне чистого расхода энергии на покой способ-
ствует образованию излишков — резервов, которые дополнительно могут использовать-
ся на размножение, линьку или миграции.
Размножение птиц может начаться лишь при положительном энергетическом балан-
се, когда суммарное количество поступающей в организм энергии, кроме обычных
затрат, расходуется на формирование половых продуктов и на все виды активности,
связанные с гнездованием. На севере это осуществляется при более низких уровнях
темйературных порогов и на более ранних фенофазах весны, т. е. при меньшем обилии
и разнообразии кормов. Недостаток пищи и повышенные затраты на терморегуляцию
компенсируются низким суммарном расходом энергетических резервов на покой.
В результате гнездование птиц в северных широтах начинается при более глубоком
негативном влиянии большинства факторов внешней среды, за исключением фотопери-
ода, оказывающего положительное воздействие. Этого достаточно для того, чтобы
размножение птиц на севере начиналось в более ранние фенологические (но не календар-
ные) сроки.
В выводках больших размеров вероятность повышения птенцовой смертности
возрастает: недостаточная обеспеченность пищей определяет более низкий вечерний
уровень энергетических резервов птенцов, расход которых может зависеть от продолжи-
тельности ночного покоя и превышать запасы. На севере родители начинают кормить
птенцов раньше, чем уровнь энергетических резервов приблизится к летальным или
сублетальным значениям. С этим, очевидно, связано отсутствие существенных различий
в смертности птенцов в больших и маленьких выводках у птиц исследуемой зоны.
Поэтому не исключено, что способность птиц северных популяций воспитывать крупные
выводки имела определенное положительное влияние на эволюцию величины кладки.
Во всяком случае это можно рассматривать как одну из предпосылок к увеличению
кладки в северных широтах.
Если у птиц, выкармливающих более одного выводка в год, в южных и средних
широтах оба цикла размножения, как правило, строго разобщены во времени, то на севе-
ре их ареалов откладка яиц во вторые гнезда нередко начинается еще при
выкармливании гнездовых птенцов. Так, у одной пары меченых зарянок в Южной
161
Карелии новая кладка была начата при 9-дневных птенцах. Ясно, что подобное совме-
щение возможно лишь в условиях трофического благополучия и при соответству-
ющем энергетическом обеспечении: три энергоемких процесса — кормление птенцов,
постройка нового гнезда и формирование яиц второй кладки - проходят одно-
временно. Увеличенный расход энергии в подобных случаях компенсируется,
вероятно, более низким уровнем использования резервов за время сна и их
постоянным пополнением при активной кормежке. Важным условием для совме-
щения двух циклов размножения является также способность самцов самостоя-
тельно докормить выводок после того, как самки приступают к насиживанию
вторых кладок.
Спецификой суточного баланса энергии птиц северных популяций можно объ-
яснить явление совмещений: гнездостроения с формированием половых продуктов,
выводящих путей и наседных пятен; окончания постройки гнезда с откладкой яиц;
гнездования с линькой; линьки с миграцией расселения молодняка и осенней мигра-
цией, и т. д.
В июле—начале августа, т. е. в период окончания размножения и начала смены опе-
рения, вдоль направляющих линий формируется поток перемещающихся воробьиных
птиц. По своему характеру и направленности эти перемещения отличаются от типичных
миграций фактически лишь сроками и состоянием оперения участвующих в них особей.
Молодые птицы летят в ювенильном наряде или на начальных этапах его замены, взрос-
лые - до начала или на I—V стадиях послебрачной линьки. Функция таких летних пере-
мещений до конца пока не ясна. Отсутствуют и фактические данные о дальности летних
миграций (от большого числа окольцованных птиц мы не имеем возвратов), но расчеты
по косвенным показателям позволяют предположить, что они приобретают значение
промежуточных перелетов. И если это действительно так, то участвующие в летних
миграциях особи должны получать ощутимый выигрыш во времени, так как еще до за-
вершения линьки оказываются значительно ближе к местам зимовки, чем остальные.
Следует отметить, однако, что в летних перемещениях участвуют далеко не все местные
особи, и вполне вероятно, что в данном случае мы имеем дело с начальным этапом
формирования новых адаптаций, направленных на преодоление дефицита вре-
мени и более рациональное использование среды обитания на всем пространстве
ареала.
Птицы, сравнительно недавно освоившие исследуемую зону (черный дрозд,
коноплянка, зеленушка, трещотка, малая чайка и др.), приступают к гнездованию
раньше, чем их родственники, давно обитающие в таежной зоне или являющиеся ее
аборигенами. Возможно, „южане” адаптированы к более напряженному суточному
балансу энергии и началу размножения на коротких фотопериодах, когда преиму-
щества жизни при повышенных температурах среды теряются из-за больших затрат
энергии на вынужденно длительный ночной покой. В северных широтах, где такие затра-
ты минимальны, южные по происхождению птицы, видимо, раньше достигают того поро-
га. после которого могут быть реализованы потенциальные репродуктивные возмож-
ности, — их Ро (по: Дольник, 1975) сохраняется после расселения на более низком
уровне, чем у аборигенов тайги. Энергетическое обеспечение более раннего начала
размножения „южан” достигается за счет сокращения затрат на период покоя. Поэтому
при одинаковых внешних условиях они приступают к откладке яиц раньше та-
ежных птиц.
Поскольку часть населения многих видов воробьиных птиц в периферийных зонах
ареала ежегодно формируется за счет дисперсии особей, родившихся в более южных
зонах, то можно допустить, что они способны приступать к размножению чуть панаше
птиц местного происхождения. Это создает одну из предпосылок для разновременности
начала гнездования птиц, обитающих в одной местности.
Таким образом, многие специфические черты биологии воробьиных птиц таежного
Северо-Запада СССР являются следствием особого суточного баланса энергии, в свою
очередь обусловленного круглосуточной освещенностью и коротким периодом ночного
162
покоя в весенне-летний сезон. Количество энергии, поступающей с пищей за день,
превышает ее расход во время ночного покоя, что дает птицам определенные преиму-
щества, позволяющие мобилизовать потенциальные возможности организма на более
полное использование позитивных и преодоление негативных воздействий среды
обитания.
Способность к началу гнездования в более ранние фенологические сроки и совме-
щение энергоемких фаз годового цикла имеют адаптивное значение для жизни птиц
на севере, позволяя им завершить размножение и линьку, а затем начать отлет на зи-
мовку до наступления неблагоприятных условий на родине. В конечном итоге это спо-
собствует преодолению более или менее существенного дефицита времени в напряжен-
ном годовом цикле с поздними календарными сроками гнездования, коротким летом
и увеличенной дальностью и продолжительностью миграций.
Неустойчивость факторов внешней среды. Все явления годового цикла птиц,
равно как и других животных и растений, адаптированы лишь к общей тенденции
в фенологическом развитии природы, но конкретных адаптаций к непостоянству пого-
ды не существует. Такие адаптации возникают в эволюции, но на надорганизменном,
популяционном уровне, причем они, как известно, оказались однозначными и у живот-
ных, и у растений, совместно эволюировавших под влиянием одних и тех же факторов
среды. Преодоление неустойчивости климата и связанных с этим внешних воздействий
развивалось по пути совершенствования внутривидовой экологической пластичности
и неоднородности индивидуальных и популяционных реакций. Все сезонные явления
в пределах местного населения птиц любой ограниченной территории отличаются
неодновременностью начала у разных особей и как следствие этого — растянутостью.
Это способствует более полному использованию возможностей среды обитания,
а в условиях неустойчивости внешних факторов приобретает значение популяционной
адаптации к непредсказуемым изменениям погоды, позволяющей сократить общепо-
пуляционные потери. Например, при неодновременном прилете местных птиц на родину
под губительное влияние внезапных и продолжительных снегопадов и холодов попадает
только какая-то часть, а не все особи. Точно так же наиболее тяжелы последствия возвра-
та холодов для маленьких неоперенных птенцов, но в те же дни и в таких же биотопах
погода не оказывает влияния на успешность инкубации яиц и выкармливания птенцов,
уже способных к самостоятельной терморегуляции. Потери в воспроизводстве в данном
случае бывают лишь частичными, а не полными, какими они могли бы стать при одно-
временном начале размножения в популяции. В этом заключается наиболее важное
значение растянутости биологических явлений и, в частности, гнездования птиц.
При высокой вероятности возврата холодов на севере раннее начало гнездования
не всегда бывает успешным, а развитие птенцов из поздних и вторых выводков нередко
проходит при более благоприятных условиях, но благодаря синхронизации годовых
циклов в популяции они успевают перелинять и начать отлет на зимовку практически
одновременно с молодыми птицами из ранних выводков. Вероятнее всего, именно
поэтому, а также из-за кратковременности всего периода размножения, лучшая выжива-
емость ранних выводков не подтвердилась. Кроме того, у птиц на севере даже ранние
выводки появляются в период, соответствующий времени вылета поздних выводков
на юге.
Поскольку биоценотически связанные животные и растения эволюировали под
влиянием одних и тех же факторов среды, то на современном этапе их эволюции мы
всюду наблюдаем достаточно строгую синхронность в их фенологическом развитии.
В природе это достигается однозначностью реакций птиц, растений и беспозвоночных
животных на изменения погоды. Поэтому нет ничего удивительного в том, что пики
массового вылупления птенцов и обилия подходящих для их выкармливания насеко-
мых совпадают. Начало вегетации и цветения растений, выхода из диапаузы или выпло-i
да насекомых — опылителей или потребителей растений, откладка яиц птицами — кон-
сументами обеих предыдущих групп происходят почти одновременно. У взаимосвязан-
ных компонентов биоценоза в разные годы в зависимости от хода весны этот период
163
одинаково смещается на более ранние или поздние сроки, развитие начинается в одном
диапазоне температур, при одинаковом суммарном количестве поступившей за весну
тепловой энергии солнца и т. д. Темпы развития взаимосвязанных организмов также
оказываются примерно сходными, так что все наиболее ответственные фазы их годо-
вых циклов приходятся на одинаковые календарные и фенологические периоды года.
Пики фенофаз у различных видов растений и животных ежегодно чередуются
в определенной последовательности. Вариации в сроках и степени сближенности пиков
однозначно регулируются состоянием погоды. Размножение птиц-полифагов может на-
чинаться на пике обилия одних кормов, а выкармливание птенцов — совершенно иных.
Но последовательность появления пиков в природе в разные годы будет смещаться
синхронно, а промежутки между ними будут примерно одинаковыми. Поэтому мас-
совое вылупление птенцов и появление кормов, которыми птицы их выкармливают,
наступают практически одновременно. Определенная десинхронизация может возник-
нуть лишь в тех случаях, когда похолодание приходится на период массового насижива-
ния кладок у птиц. Тогда эмбриогенез птиц идет обычными темпами, а рост и развитие
насекомых или растений сдерживаются низкими внешними температурами. Однако
полной десинхронизации никогда не происходит. Этому препятствует эволюционно
поддерживаемая всеобщая растянутость любых фаз у объектов питания птиц и всех
явлений годового цикла птиц, а также экологическая пластичность видов. При миниму-
ме основных кормов птицы способны выкормить птенцов второстепенными, о чем
свидетельствует известная сезонная и ежегодная изменчивость в составе приносимой
птенцам пищи.
Весенние возвраты холодов в северных широтах часто существенно влияют
на численность приступающих к размножению птиц и общий уровень воспроизводства.
Кроме роста затрат на терморегуляцию, основная причина гибели птиц заключается
в снижении доступности кормов и увеличении времени на поиски оцепеневших, мало-
подвижных насекомых, многие из которых заблаговременно уходят в разнообразные
укрытия. Поэтому для птиц, адаптированных к добыванию беспозвоночных из всевоз-
можных убежищ (дятлы, синицы, пищухи, корольки), негативное влияние холодной
и ветреной погоды бывает менее существенным. У остальных обогреву птенцов не уде-
ляется должного внимания из-за возрастающих потерь времени на поиски пищи. Желуд-
ки птенцов, погибших в такие периоды, как правило, бывают наполнены непереварен-
ными остатками пищи. Иначе говоря, несмотря на холодную погоду, родители все же
снабжают птенцов пищей, но отсутствие обогрева приводит к переохлаждению тела
птенцов (терморегуляция еще не совершенная), причем обмен веществ, видимо, снижа-
ется настолько, что получаемая пища не может перевариваться и усваиваться. Характер-
но, что в тех же биотопах и в те же дни оперенные большие птенцы с более развитой
терморегуляцией выживают нормально, т. е. пищей они вполне обеспечены. Об этом
же свидетельствуют и различия в выживаемости птенцов зарянки в зависимости от вы-
соты размещения гнезд. При типичном для этого вида способе размещения гнезд
в укрытиях (земляные ниши под корнями, в кочках) тепла, излучаемого стенками
укрытий, оказалось достаточно для того, чтобы за время длительного отсутствия самок
при холодной погоде маленькие птенцы не переохлаждались до состояния гипотермии,
потери двигательной активности и способности к усвоению пищи, что при прочих рав-
ных условиях было характерно для птенцов в приподнятых над землей гнездах. Таким
образом, снижение доступности пищи при весенних возвратах холодов является не пря-
мой, а косвенной причиной повышения птенцовой смертности.
Неустойчивость урожая семян хвойных деревьев и обусловленные этим уникаль-
ные черты биологии потребителей данного корма — клестов — общеизвестны. Биология
и особенно территориальные связи этих птиц изучены между тем еще недостаточно.
По-видимому, они могут полнее использовать возможности среды обитания. Факты
осеннего размножения клестов при урожае семян ели указывают на вероятность дву-
кратного размножения их в течение календарного года (Коханов, Гаев, 1970; Мальчев-
ский, Пукинский, 1983).
164
Выясняется также, что и другие потребители семян хвойных деревьев — чижи и
чечетки могут размножаться в позднезимний и ранневесенний периоды на значитель-
ных расстояниях от своих основных гнездовых областей, в совершенно иных природно-
климатических зонах. Это происходит в тех случаях, когда на зимовках или путях
пролета чижи и чечетки попадают на территории с обильным урожаем семян ели. Подоб-
ное усложнение годового цикла и полизональное размножение, прерываемое проме-
жуточными миграциям#, оказывается выгодным для птиц — потребителей семян хвой-
ных деревьев, позволяет им максимально использовать неустойчивую и неравномерно
распределенную по ареалу кормовую базу, пригодное для размножения время и репро-
дуктивные возможности, значительно повышая общий уровень воспроизводства.
Чередование периодов высокой подвижности и размножения должно иметь уни-
кальное физиологическое обеспечение, изучение которого заслуживает самого присталь-
ного внимания.
Пути обеспечения стабильной численности гнездового населения. Относительная
стабильность численности любого вида птиц достигается приведением в соответствие
уровней ежегодной смертности и плодовистости. Как известно, этот баланс может
поддерживаться или увеличением плодовитости, или снижением ежегодной смертности.
Действительно, для сохранения оптимального уровня численности одним видам птиц
достаточно однократного размножения в течение года и выкармливания 1—4 птенцов,
а другим не хватает даже 2—4-кратного увеличения числа потомков или выращивания
нескольких выводков за сезон.
Увеличение плодовитости достигается в отряде воробьиных птиц разными спосо-
бами: 1) размножением в возрасте около одного года, что при короткой средней про-
должительности жизни становится необходимостью; 2) увеличением числа потомков
при моноциклическом размножении (в нашей зоне — сероголовая гаичка, пухляк,
ополовник); 3) увеличением числа выводков при обычной для воробьиных птиц вели-
чине кладки (большинство видов с полициклией гнездования); 4) 2-3-кратным воспи-
танием больших выводков (большая синица, лазоревка, московка, желтоголовый
королек, крапивник); 5) полизональной полициклией размножения (чиж, чечетка,
возможно, клесты); 6) полигамией, которую можно рассматривать как частный случай
полициклии, цри которой два или более выводка выкармливает один из партнеров
(мухоловка-пеструшка, весничка и др.); 7) использованием полигинии при полицикли-
ческом гнездовании (обыкновенная овсянка, большая синица).
Реальных путей снижения смертности при жизни в неустойчивой среде обитания
не так много. Более высокую выживаемость можно ожидать при переносе зимовок
в области с более устойчивым теплым климатом. Но это связано с увеличением затрат
времени и энергии на миграции и соответствущим сокращением продолжительности
остальных фаз годового цикла. Не случайно поэтому ни для одного вида птиц не извест-
но стремления к увеличению дальности миграций в современных условиях. Напротив,
у многих из них в Северной Европе отмечается обратная тенденция — стремление зи-
мовать вблизи от мест размножения (кряква, грач, скворец, зяблик, черный дрозд
и др. в зоне 60—64° с. ш.).
Одним из путей преодоления неустойчивости кормовой базы и климата может
быть совершенствование уже имеющихся и приобретение новых адаптаций, позволя-
ющих легче переносить периодическое исчезновение кормов и снижение температуры
воздуха. Но такие адаптации не беспредельны, а период их становления весьма продол-
жителен. Здесь получают определенные преимущества те виды птиц, которые освоили
трансформированные человеком местообитания и новые виды соответствующих кор-
мов — от семян кормовых растений и сопутствующих им сорняков до отходов и отбро-
сов, на которых также развивается комплекс их потребителей — беспозвоночных и дру-
гих животных. В отличие от естественных эти корма обильны, доступны и, что особен-
но важно, многие из них стабильно встречаются в течение всего года и независимо от
состояния погоды. Можно с достаточной долей уверенности утверждать, что, если бы не
антропогенное преобразование среды обитания, границы современных гнездовых
165
областей и мест зимовки многих видов птиц были бы расположены много южнее, чем
в настоящее время, а общий уровень численности был бы гораздо ниже.
В северных частях ареалов, по крайней мере в пределах изученной нами зоны,
воробьиные птицы реализуют все возможные пути увеличения плодовитости. Не всегда
используется, пожалуй, лишь один из них — размножение первогодков (скворец, чече-
вица). Скворец пока остается и единственным видом среди воробьиных птиц, для
которого в отличие от более южных зон не доказано двукратное размножение. Все
остальные виды с более или менее достоверно установленной для средних широт поли-
циклией гнездования у нас, а иногда и в Заполярье, выводят птенцов дважды за сезон,
причем доля птиц, воспитывающих дополнительные выводки, бывает у нас даже выше,
чем в южных частях ареала (до 80—100% пар больших синиц, зарянок, пищух). Скла-
дывается впечатление, что для некоторых видов птиц в северных частях ареала воспи-
тание двух выводков является обязательным условием для закрепления на данной тер-
ритории. Это может быть связано с большей, чем у птиц южных популяций, напряжен-
ностью годовых циклов, более суровыми условиями зимовки, более дальними мигра-
циями, проходящими при менее благоприятных условиях.
Максимальное число потомков за сезон оставляют синицы и корольки — виды,
зимующие на местах размножения или неподалеку от них, т. е. в наиболее суровых усло-
виях. В целом абсолютное большинство видов птиц с достоверно выявленной поли-
цикличностью размножения зимуют в более северных широтах, чем их менее плодови-
тые родственники.
В эволюции птиц сформировались две основные стратегии, обеспечивающие отно-
сительную стабильность численности и приводящие в соответствие соотношение пло-
довитости и смертности. Одна из них характеризуется более высокой выживаемостью
и обеспечивается зимовкой птиц в стабильно благоприятных условиях. Это требует уве-
личения дальности миграций и соответственно сокращает период пребывания птиц на
родине, время на размножение и линьку. В связи с этим гнездование уже не может быть
полициклическим, а линька — растянутой, т. е. менее энергетически напряженной.
Не случайно у многих дальних мигрантов послебрачная и постювенильная линьки на ро-
дине бывают лишь частичными или вообще переносятся на период зимовки. Плодови-
тость птиц этой группы, как правило, невысока.
Лишь немногие дальние мигранты способны увеличить время пребывания на ро-
дине и успевают вывести птенцов дважды (ласточки, овсянки - ремез и крошка).
У первых это достигается благодаря высокой скорости миграции, но из-за ее поздних
сроков они нередко попадают в критические ситуации на европейских трассах пролета.
О местах зимовки названных видов овсянок точных сведений пока нет. Но судя по
литературе по крайней мере овсянка-ремез заканчивает пролет по азиатской части СССР
уже в декабре, т. е. при низких температурах и многоснежье.
Другая стратегия свойственна птицам, зимующим в относительно суровых и край-
не неустойчивых условиях зоны умеренного климата. Области их размножения и зимов-
ки нередко перекрываются. Неустойчивость климата, холодная погода с тяжелыми
снегопадами при коротком зимнем кормовом дне и длительной ночевке нередко приво-
дят к массовой гибели птиц. Миграции у них кратковременны или вообще не выражены.
Продолжительность гнездового периода возрастает, что позволяет выкормить дополни-
тельные выводки и иметь растянутую, менее энергетически напряженную линьку. За
немногими исключениями высокая плодовитость достигается увеличением размеров
кладки или числа выводков либо тем и другим вместе.
Средняя для популяции величина кладки изменяется по годам слишком незначи-
тельно, чтобы существенно влиять на общий уровень воспроизводства. К тому же
величина кладки и выводка далеко не всегда изменяются синхронно — увеличение раз-
меров выводка нередко (особенно на юге ареала) сопровождается ростом птенцовой
смертности и сокращением числа слетков. У исследованных видов птиц в зоне наших
работ ежегодные вариации величины кладки не обнаруживают связи с плотностью
гнездового населения, хотя примеры обратной зависимости между этими показателями
166
известны (большая синица в Англии и Голландии). Не исключено, правда, что выявлен-
ная для этого вида зависимость могла быть опосредованной: пики численности форми-
руются в основном за счет первогодков, которые, по свидетельству ряда исследовате-
лей, менее плодовиты.
Трудно представить, чтобы у воробьиных птиц эволюция поддерживала развитие
механизма регуляции численности лишь через изменение величины кладки, которая
в значительной мере зависит от сроков начала размножения, т. е. фактически от состоя-
ния весенней погоды, отличающейся крайней неустойчивостью. К тому же у воробьиных
птиц годовые колебания величины кладки, выражающиеся обычно в десятых долях,
вряд ли могут существенно повлиять на общий уровень воспроизводства, тем более что
успешность размножения при непостоянстве погоды в северных широтах в большей ме-
ре зависит от совпадения периода массового вылупления птенцов с благоприятными
или неблагоприятными условиями. И, видимо, вполне закономерно, что после глубоких
депрессий численность птиц с моноциклическим гнездованием даже при большой вели-
чине кладки (пухляк, тетеревиные) восстанавливается медленно — в течение несколь-
ких лет.
Годы депрессии численности у птиц с полициклическим размножением (зарянка,
большая синица, королек и др.) быстро сменяются годами подъема. Одновременно
изменяется и доля птиц, размножающихся дважды за лето. Оба показателя — плотность
населения и относительное число гнездовых пар, воспитывающих дополнительные
выводки, — связаны обратной зависимостью. Следовательно, на снижение плотности
гнездового населения птицы этой группы реагируют увеличением числа особей, гнездя-
щихся дважды за сезон. Поскольку во втором гнездовании у зарянок, больших синиц
и пищух принимают участие до 80—100% пар, то суммарное увеличение поголовья
становится весьма существенным — число выкормленных в двух выводках птенцов
в 1.6—1.8 раза больше, чем при однократном гнездовании.
Такой способ регуляции численности эффективен не только и даже не столько
при повышенной смертности вообще, сколько при постоянных ее колебаниях, что обу-
словлено существованием птиц в неустойчивой среде обитания. Если бы полициклия
была простым средством компенсации повышенной смертности, не зависящим от уровня
численности, то в условиях неустойчивости большинства внешних факторов такой
путь был бы менее рационален. Глубже и продолжительнее были бы периоды депрессии
численности, а после ее восстановления ничем не сдерживаемое полициклическое раз-
множение приводило бы к перенаселенности. В принципе такой путь вполне возможен,
но только в среде, отличающейся более или менее строгой стабильностью условий.
Там, где внешне факторы то увеличивают, то уменьшают смертность без строгой перио-
дичности, более целесообразна регуляция плодовитости в зависимости от плотности
населения птиц.
Механизм этой зависимости сходен, вероятно, с механизмом, определяющим
указанную выше связь между величиной кладки и численностью: увеличение последней
достигается главным образом посредством лучшей выживаемости первогодков в годы
с благоприятными условиями существования. Преобладание среди гнездового населе-
ния годовалых особей, относительно реже принимающих участие в воспитании дополни-
тельных выводков, отражается на доле бициклических пар в популяции.
Изучение кормовой базы птиц, выполненное в Приладожье, выявило, что пики
численности различных групп беспозвоночных животных и урожаи растительных кор-
мов, служащих пищей птицам и их птенцам, сменяют друг друга в течение всего периода
размножения и линьки, так что на этих фазах годового цикла птицы в таежной зоне
вполне обеспечены кормами. Это обстоятельство, а также климатическая обстановка,
определенное сокращение затрат времени на гнездование и линьку, синхронизация
индивидуальных годовых циклов у особей из первых и вторых выводков являются
важными предпосылками и экологической основой для развития полицикличе-
ского гнездования в зоне наших исследований, где оно долгое время считалось
невозможным.
167
Динамика численности многих видов птиц, обитающих у северных границ ареала,
подвержена значительным ежегодным колебаниям. Одна из важнейших причин такого
непостоянства — это влияние дисперсии особей из областей постоянного гнездования.
Интенсивность этого процесса также изменчива и, в свою очередь, определяется общим
уровнем численности вида внутри основного ареала, условиями прошедшей зимовки,
состоянием погоды в период весенней миграции (степенью ее пролонгации), соотноше
нием ежегодных различий в развитии и реализации миграционного и репродуктивного
состояний.
Колебания численности птиц в периферийных зонах ареала определенно связаны
с неустойчивостью погоды, нередко приводящей к более или менее массовой гибели
на местах зимовки, трассах миграций и на родине. Особенно губительны весенние
возвраты холодов, сопровождающиеся продолжительными тяжелыми снегопадами
на обширных территориях. Неустойчивость погоды и кормовой базы отрицательно
влияют также на успешность размножения птиц и общий уровень воспроизводства.
Запаздывание начала размножения в годы с затяжной холодной весной и раннее насту-
пление осенних холодов увеличивают напряженность годового цикла и дефицит вре-
мени, что снижает шансы птиц на выполнение всей годовой программы. Вероятность ее
успешной реализации снижается также из-за асинхронности в состоянии погоды на
родине и трассах пролета к зимовкам.
Вместе с тем ежегодные колебания численности птиц периферийных популяций
происходят без сколько-нибудь существенной связи с основными демографическими
параметрами за предшествующий сезон (плотность населения, величина кладки и вы-
водка, успешность гнездования, общий уровень воспроизводства).
Единственным показателем плодовитости, зависимым от плотности гнездового
населения, оказалось относительное число пар, делающих по два выводка за сезон.
Но механизм этой связи до конца еще не ясен, а ее проявление обнаружено пока только
у двух видов птиц наших широт.
Между величиной кладки и плотностью населения ни для одного вида птиц, иссле-
дованных на нашей и соседних территориях, никакой зависимости не выявлено.
При низком уровне численности птиц в пограничных зонах ареала степень насыще-
ния подходящих для размножения местообитаний далека от предельной, поэтому
в таежной зоне птицы имеют неограниченные возможности для рассредоточения Нередко
здесь заселяются только биотопы с оптимальными условиями для гнездования, и лишь
при существенном увеличении численности птицы осваивают и некоторые из субопти
мальных местообитаний.
Богатые возможности для рассредоточения по территории служат одним из основ'
ных препятствий для проявления популяционных механизмов регуляции численности
в исследуемой зоне. Даже максимальные концентрации местных птиц на гнездовье,
видимо, не достигают в таежной зоне тех уровней плотности населения, после которых
она начинает оказывать влияние на плодовитость как ограничивающий фактор. В свою
очередь, отсутствие сдерживающего влияния плотности населения птиц на их плодови
тость позволяет птицам северных популяций поддерживать величину кладки на макси-
мальном видовом уровне.
В исследуемой зоне не подтвердилось и влияние плодовитости на плотность населе
ния. Основной причиной этого является непостоянство состава населения периферий
ных популяций и их ежегодное практически полное обновление за счет птиц, родив
шихся в более южных областях. Точные места рождения особей, образующих поселения
за пределами основного ареала, пока неизвестны. Решение этой задачи, равно как и изу
чение дисперсии и территориальных связей птиц, может быть успешным лишь после
значительного увеличения числа кольцующих центров или разработки принципиально
новых методов массового мечения птиц на обширных территориях. Одним из возмож
ных направлений поиска таких методов может быть самомечение птиц какими-io
химическими маркерами. Это должны быть не токсичные вещества, образующие
стойкие соединения в растущем пере, но распадающиеся в природе за 1--L5 мео
168
Вводить в среду обитания их можно специальными прикормками или авиарассеиванием,
т. е. они должны легко проникать в трофоценотические цепи птиц, закрепляться в пере
птиц во время линьки и определяться на спектрофотограммах при сжигании неболь-
шого количества перьев. При современном уровне развития химии эта задача представ-
ляется вполне разрешимой (аналоги таких маркеров уже есть, но это токсичные
содержащие ртуть соединения), а ее решение открывает перед орнитологами широкие
перспективы.
12 В. Б. Зимин
169
ЛИТЕРАТУРА
Агроклиматический справочник по Карельской АСССР. Л., 1959.184 с.
Ангальт В. 3. Размножение сизого голубя в условиях большого города // Экологоя и охрана
птиц. Тез. докл. 8-й Всесоюз. орнитол. конф. Кишинев, 1981. С. 8.
Андреев А. В. Адаптации птиц к зимним условиям Субарктики. М., 1980. 175 с.
* Анорова Н. С. Размножение популяции мухоловки-пеструшки в зависимости от возраста
птиц // Орнитология. М., 1976. Вып. 12. С. 77-86.
* Анорова Н. С. Фактор возраста в плодовитости диких и домашних птиц // Орнитология. М.,
1979. Вып. 14. С. 150-157.
* Ардамацкая Т. Б., Приклонский С. Г., Прокопчук Л. Н., Хохлова Н. А. Изменчивость сроков
гнездового цикла и плодовитости скворца обыкновенного // Тез. докл. 7-й Всесоюз. орнитол. конф.
Киев, 1977. Ч. 1.С. 194-195.
* Артемьев Ю. Т., Попов В. А. Дроздовые // Птицы Волжско-Камского края. М., 1977.
С. 70-94.
Бардин А. В. Территория обитания и структура синичьих стай // Материалы 7-й Прибалт,
орнитол. конф. Рига, 1970. Т. 1. С. 21-24.
Бардин А. В. Сравнительное изучение жизненных циклов некоторых представителей рода
Parus (Paridae, Aves): Автореф. дис. ... канд. биол. наук. Л., 1975а. 24 с.
Бардин А. В. Территориальное поведение скандинавского подвида буроголовой гаички ( Parus
montanus borealis Selis-Longchamps) // Вестник ЛГУ. Биология. Л., 19756. № 9. С. 24-34.
Бей-Биенко Г. Я. Сбщая энтомология. М., 1971. 479 с.
Беме Р. Л. Размножение птиц в горах и на равнине // Орнитология. М., 1968. Вып. 9. С. 27—48.
Бианки В. В., Шутова Е. В. К экологии большой синицы в Мурманской области // Бюл.
МОИП. Отд. биол. М., 1978. Т. 83, № 2. С. 63-69.
* Бианки В. В., Шутова Е. В. Использование искусственных гнездовий для привлечения птиц
на северном архипелаге (Кандалакшский залив) // Проблемы охраны природы в бассейне Белого
моря. Мурманск, 1984. С. 42-50.
Блюменталь Т. И., Дольник В. Р. Оценка энергетических показателей птиц в полевых усло-
виях //Орнитология. М., 1962. Вып. 4. С. 394—407.
Блюменталь Т. И., Зимин В. Б. Широтные географические различия в особенностях размноже-
ния и линьки у трех видов воробьиных птиц в Прибалтике // Материалы 6-й Прибалт, орнитол.
конф. Вильнюс, 1966. С. 15-17.
Блюменталь Т. И., Дольник В. Р., Зимин В. Б., Носков Г. А., Урядова Л. П., Шильдмахер Г.
Географические различия в сроках и соотношении послебрачной линьки, предмиграционного отложе-
ния жира и начала осенней миграции у зяблика // Тез. докл. 5-й Прибалт, орнитол. конф. Тарту,
1963. С. 19-21.
Блум П., Меднис А., Бауманис Я. Различие в выживаемости молодняка хохлатой чернети на
эвтрофном озере и рыбоводных прудах // Тез. докл» 11-й Прибалт, орнитол. конф. Таллин, 1983.
С. 58-60.
* Бойко Н. Я., Шкарин В. С., Маркс Л. П. К биологии размножения скворца на юге Западной
Сибири // Тез. докл. 7-й Всесоюз. орнитол. конф. Киев, 1977. Ч. 1. С. 22-23.
Болотников А. М., Калинин С. С. Температура и относительная влажность в гнезде обыкновен-
ной чайки // Сб. статей по орнитологии. Пермь, 1974. С. 10-15. (Учен. зап. Пермск. пед. ин-та;
Т.122).
* Болотников А. М., Печерский А. С. Насиживание, инкубация // Полевой воробей (характе-
ристика вида на пространстве ареала). Л., 1981. С. 157—161.
♦ Болотников А. М., Фуфаев А. А. К фенологии гнездовой жизни некоторых воробьиных птиц
Камского Предуралья // Сб. статей по орнитологии. Пермь, 1974. С. 6-9. (Учен. зап. Пермск. пед.
ин-та, Т. 122).
Звездочкой* отмечены работы, использованные как источники данных при составлении
рис. 4, 24, 25.
170
* Болотников А. М., Пантелеев М. Ф., Каменский Ю. Н. Основные параметры размножения
обыкновенного скворца в Камском Предуралье // Материалы 3-го Всесоюз. совещ. „Вид и его про-
дуктивность в ареале”. Вильнюс, 1980. С. 85- 86.
* Болотников А. М., Шураков А. И., Шкарин В. С. К экологии размножения дроздов в Пред-
уралье // Сб. статей по орнитологии. Пермь, 1973. С. 29—34. (Учен. зап. Пермск. пед. ин-та; Т. 113).
* Брагин А. Б., Гилязов А. С. Результаты привлечения в искусственные гнездовья птиц таеж-
ной зоны Кольского полуострова // Проблемы охраны природы в бассейне Белого моря. Мурманск,
1984. С. 42-50.
Будыко М. И. Изменения климата. Л., 1974. 280 с.
Будыко М. И. Современное изменение климата. Л., 1977. 48 с.
* Букина Т. Г. Некоторые особенности биологии певчего дрозда и белобровика в У АССР //
Фауна и экология животных УАССР и прилежащих районов. Ижевск, 1981. С. 13-20.
* Васильчук А. Н. Опыт привлечения птиц в искусственные гнездовья // Тр. по лесному опыт-
ному делу в России. Петроград, 1915. 117 с.
Виноградова Н. В., Дольник В. Р., Ефремов В. Д., Паевский В. А. Определение пола и возраста
воробьиных птиц фауны СССР: Справочник. М., 1976. 191 с.
Воинственский М. А. Семейство Синицевые. М., 1954. С. 726 -772. (Птицы Советского
Союза; Т. 5).
* Воинственский М. А., Боярчук В. П. Материалы по гнездованию скворца в Черноморском
заповеднике // Тез. докл. 7-й Всесоюз. орнитол. конф. Киев, 1977. Ч. 1. С. 218-220.
Волчанецкий И. Б. Семейство Славковые. М., 1954. С. 330-368. (Птицы Советского Со-
юза; Т. 6).
Воронцов А. И. Лесная энтомология. Изд. 3-е. М., 1975. 367 с.
* Гагинская А. Р., Фетисов С. А., Шураков А. И., Болотников А. М.» Нанкинов Д. Н. Сроки
репродуктивного периода. Число циклов размножения // Полевой воробей (характеристика вида
на пространстве ареала). Л., 1981. С. 133—139.
Гедеонов А. Д. Изменение температуры воздуха на северном полушарии за 90 лет. Л.,
1973. 146 с.
Герасимова Т. Д., Горчаковская Н. Н., Карпович В. Н., Рахилин В. К. Дополнения к фауне птиц
Айновых островов // Тр. Кандалакш. гос. заповедника. М., 1967. Вып. 5. С. 339-344.
Гермогенов Н. И. Материалы по распространению и экологии дроздов певчего и белобровика
в юго-западной Якутии // Тез. докл. 7-го симп. Биологические проблемы севера. Зоология. Петроза-
водск, 1976. С. 81—83.
Гиляров М. С. Закономерности приспособления членистоногих к жизни на суше. М.,
1970. 276 с.
Гладков Н. А. Семейство Трясогузковые. М., 1954а. С. 594—691. (Птицы Советского
Союза; Т. 5).
Гладков Н. А. Семейство Дроздовые. М., 19546. С. 399—621. (Птицы Советского Союза; Т. 6).
*Головань В. И. О географических особенностях размножения мухоловки-пеструшки // Мате-
риалы 9-й Прибалт, орнитол. конф. Вильнюс, 1976. С. 72—75.
* Головань В. И. Сравнительная экология европейских видов мухоловок (биология размноже-
ния, линька и территориальное поведение) : Автореф. дне. ... канд. биол. наук. Л., 1982. 24 с.
Головань В. И. О характере брачных связей у мухоловки-пеструшки // Тез. докл. 11-й При-
балт. орнитол. конф. Таллин, 1983. С. 76—78.
Гришанов Г. В. Особенности биологии гнездования черного дрозда (Turdus merula L.) в ланд-
шафтах с различной степенью окультуривания на территории Калининградской области // Тез. докл.
10-й Прибалт, орнитол. конф. Рига, 1981. Ч. 2. С. 47-48.
* Губин Б. И. Величина кладки некоторых воробьиных птиц в Таласском Алатау (Западный
Тянь-Шань) // Тез. Всесоюз. конф. „ Экология гнездования птиц и методы ее изучения”. Самарканд,
1979. С. 55-57.
Губин Б. И. О совмещении сроков размножения и линьки у воробьиных птиц в долине сред-
него течения Урала // Тез. докл. 2-й Сибир. орнитол. конф. „Птицы Сибири”. Горно-Алтайск, 1983.
С. 168-170.
Губин Б. И., Ковшарь А. Ф. О совмещении линьки и размножения у воробьиных птиц // Тез.
докл. 18-го Между нар. орнитол. к он гр. М., 1982. С. 161-162.
Гусев В. И., Полу боя ринов И. И., Римский-Корсаков М. Н., Шиперович В. Я., Яцентков-
ский А. В. Лесная энтомология. Изд. 2-е Л., 1938.448 с.
Данилевский А. С. Фотопериодизм и сезонное развитие насекомых. Л., 1961. 24 3 с.
* Данилов Н. Н. Размножение мухоловки-пеструшки на Среднем Урале // Тез. докл. 2-й Все-
союз. орнитол. конф. М., 1959. Т. 2. С. 62-63.
Данилов Н. Н. Пути приспособления наземных позвоночных к условиям существования
в Субарктике. Т. 2: Птицы. Свердловск, 1966. С. 1—148. (Тр. Ин-та биологии Уральск, гос. ун-та;
Вып. 56).
Данилов Н. Н., Рыжановский В. Н., Рябицев В. К. Птицы Ямала. М., 1984. 134 с.
Данилов П. И., Туманов И. Л. Куньи Северо-Запада СССР. Л., 1976. 255 с.
171
Данилов По И», Зимин В. Б., Ивантер Т. В., Лапшин Н. В., Марковский В. А., Анненков В. Г.
Фаунистический обзор наземных позвоночных // Биологические ресурсы района Костомукши, пути
освоения и охраны. Петрозаводск, 1977. С. 109-127.
Данилов II. И., Зимин В. Б., Ивантер Э. В., Анненков В. Г. Материалы по численности тетереви-
ных птиц в Карелии // Фауна и экология птиц и млекопитающих таежного Северо-Запада СССР.
Петрозаводск, 1978. С. 54-67.
Данилов П. И., Зимин В. Б., Ивантер Э. В. Изменение зоогеографической структруры и пути
формирования современной фауны наземных позвоночных Карелии // Тез. докл. 7-й Всесоюз. зоогео-
граф. конф. М., 1979. С. 20-23.
Дарвин Ч. Происхождение видов // Соч. Т. HI. М.; Л., 1939. 839 с.
Добринский Л. Н. Динамика морфо-физиологических особенностей птиц. М., 1981. 124 с.
Дольник В. Р. О механизме фотопериодического контроля эндогенного ритма половой цик-
личности птиц // Зоол. журн. 1964. Т. 43. С. 720-733.
Дольник В. Р. Годовые циклы биоэнергетических приспособлений к условиям существования
у 16 видов Passeriformes // Миграции птиц Прибалтики. Л., 1967. С. 115-163.
Дольник В. Р. Миграционное состояние птиц. М., 1975. 399 с.
Дольник В. Р., Виноградова Н. В., Гаврилов В. М., Дольник Т. В., Ильина Т. А., Люлеева Д. С.,
Паевский В. А., Соколов Л. В., Шумаков М. Е., Яблонкевич М. Л. Популяционная экология зяблика.
Л., 1982. 302 с. (Тр. Зоол. ин-та; Т. 90).
Евдокимов В. Д. О величине кладки и размерах яиц рябинника в разных биотопах // Гнездо-
вая жизнь птиц (Межвузовский сборник научных турдов). Пермь, 1980. С. 80-82.
* Езерскас Л. И. О биологии птиц-дуплогнездников семейства синицевых и мухоловковых
в Литовской ССР // Труды 4-й Прибалт, орнитол. конф. Рига, 1960. С. 32-33.
Ефремов В. Д., Паевский В. А. Поведение насиживания и наседные пятна самцов у пяти видов
птиц рода Sylvia // Зоол. журн. 1973. Т. 52. С. 721-727.
Жмуд М. Е.О двукратной откладке яиц травником (Tringa totanus) в течение одного сезона //
Бюл. МОИП. Отд. биол. 1983. Т. 88, вып. 2. С. 15-17.
Зимин В. Б. Пролет птиц на восточном берегу Ладожского озера осенью 1962 года // Сообщ.
Прибалт, комис. по изучению миграций птиц. Тарту, 1965. № 3. С. 28-50
Зимин В. Б. Экология птиц Южной Карелии: Автореф. дис.... канд. биол. наук. Л., 1968. 16 с.
Зимин В. Б. Значение фаутности древостоя для гнездования птиц // Тр. заповедника „Кивач”.
Петрозаводск, 1969а. Вып. 1. С. 103-114.
Зимин В. Б. Особенности размещения гнезд дуплогнездников в лесах заповедника „Кивач” //
Вопросы экологии животных. Петрозаводск, 19696. С. 104-109.
Зимин В. Б.Особенности гнездования обыкновенного крапивника (Troglodytes troglodytes L.)
в Карелии // Зоол. журн. 1972а. Т. 51, вып. 5. С. 770-772.
Зимин В. Б. Экология размножения мухоловки-пеструшки в Южной Карелии // Экология.
19726. №5. С. 23-29.
Зимин В. Б. Осенние миграции птиц в заповеднике „Кивач” // Тр. гос. заповедника „Кивач”.
Петрозаводск, 1973. Вып. 2. С. 64-125.
Зимин В. Б. Орнитофауна хвойно-лиственных молодняков Южной Карелии // Экология птиц
и млекопитающих Северо-Запада СССР. Петрозаводск, 1976. С. 38-44.
Зимин В. Б. Материалы по гнездованию большой синицы (Parus major L.) в Карелии // Фауна и
экология птиц и млекопитающих таежного Северо-Запада СССР. Петрозаводск, 1978. С. 17-31.
Зимин В. Б. Возрастная и сезонная изменчивость размеров у некоторых воробьиных птиц //
Экология птиц Приладожья. Л., 1981а. С. 151-161. (Тр. Биол. НИИ ЛГУ; № 32).
Зимин В. Б. К экологии обыкновенной чечевицы (Carpodacus erythrinus Pall.) в Карелии // Эко-
логия наземных позвоночных Северо-Запада СССР. Петрозаводск, 19816. С. 13-31.
Зимин В. Б. О полицикличности размножения птиц // Тез. докл. 11-й Прибалт, орнитол. конф.
Таллин, 1983. С. 52-68. ч
Зимин В. Б., Артемьев А. В. О методах отлова открытогнездящихся дендрофильных птиц //
Экология наземных позвоночных Северо-Запада СССР. Петрозаводск, 1981. С. 32-37.
Зимин В. Б., Ивантер Э. В. Фаунистический обзор наземных позвоночных заповедника „Ки-
вач” // Тр. заповедника „Кивач”. Петрозаводск, 1969. Вып 1. С. 22-64.
Зимин В. Б., Ивантер Э. В. Птицы. Петрозаводск, 1974. 256 с.
Зимин В. Б., Кузьмин И. А. Экологические последствия применения гербицидов в лесном
хозяйстве. Л., 1980.175 с.
Зимин В. Б., Лапшин Н. В. Совмещение сроков размножения и линьки у некоторых воробь-
иных птиц Южной Карелии // Тез. докл. Науч. конф, биологов Карелии, посвященной 250-летию
Академии наук СССР. Зоология, паразитология, физиология и биохимия животных. Петрозаводск,
1974. С. 21-22.
Зимин В. Б., Лапшин Н. В. Экологические русла расселения птиц в Карелии // Актуальные воп-
росы зоогеографии. Кишинев. 1975. С. 94.
Зимин В. Б., Лапшин Н. В. Результаты отлова и мечения птиц в гнездовой период // Материалы
9-й Прибалт, орнитол. конф. Вильнюс. 1976. С. 96—102.
172
Зимин В. Б., Лапшин Н. В., Хохлова Т. Ю. Влияние весеннего возврата холодов на птиц Каре-
лии // Тез. докл. 7-го симп. „Биологические проблемы севера”. Зоология. Петрозаводск, 1976^
С. 127-129
Зимин В. Б., Лапшин Н. В., Хохлова Т. Ю. Биология размножения садовой славки в Карелии //
Фауна и экология птиц и млекопитающих таежного Северо-Запада СССР. Петрозаводск, 1978.
С. 5- 16.
Зимин В. Б., Лапшин И. В., Артемьев А. В. Эффективность различных методов контроля
возврата птиц на места кольцевания // Тез. докл. 11-й Прибалт, орнитол. конф. Таллин, 1983.
С.91-93.
Зубцовский Н. Е. Эффективность размножения птиц в Ильменском заповеднике // Эколо-
гия. 1981. №2. С. 94-96.
Иванов А. И. Птицы СССР. Л., 1953.
Иванов А. И. Каталог птиц СССР. Л., 1976. 276 с.
Иванова Н. С. Сравнительное изучение размножения и постэмбрионального развития куликов
(Limicola, Charadriiformes): Автореф. дис. ... канд. биол. наук. Л., 1971. 18 с.
Ивантер Э. В. Птицы заповедника „Кивач” // Орнитология. М., 1962. Вып. 5. С. 68-85.
Ивантер Э. В. Заяц-беляк в Карелии // Вопросы экологии животных. Петрозаводск, 1969а.
С. 137-156.
Ивантер Э. В. Птицы северо-восточной Карелии // Вопросы экологии животных. Петрозаводск,
19696. С. 93-103.
Ивантер Э. В. К экологии белки в Карелии // Учен. зап. Петрозавод. гос. ун-та. Биол. и сельхоз.
науки. Петрозаводск, 1972. Т. 19, вып. 5. С. 139—160.
Ивантер Э. В. Популяционная экология мелких млекопитающих таежного Северо-Запада
СССР. Л., 1975.244 с.
Ивантер Э. В. Возрастная структура популяции грызунов и ее адаптивное значение (на приме-
ре рыжей полевки) // Фауна и экология птиц и млекопитающих таежного Северо-Запада СССР.
Петрозаводск, 1978. С. 93-119.
Ильичев В. Д., Карташев Н. Н., Шилов И. А. Общая орнитология. М., 1982. 464 с.
Иноземцев А. А. Роль насекомоядных птиц в лесных биоценозах. Л., 1978. 264 с.
Йыги А. И. Миграция эстонских дроздов по данным кольцевания // Сообщ. Прибалт, комис.
по изучению миграций птиц. Таллин, 1967. № 4. С. 136-149.
Кищинский А. А. О структруе и динамике областей гнезования птиц на Севере // Исследования
по морфологии и биологии птиц. Л., 1983. С. 47-57. (Тр. Зоол. ин-та; Т. 116).
* Книстаустас А., Люткус А., Бараускас Р. Основные фенологические и гнездовые показатели
дроздов в Литве // Тез. докл. 10-й Прибалт, орнитол. конф. Рига, 1981. Ч. 2. С. 70-75.
Ковшарь А. Ф. Совмещение сроков размножения и линьки у певчих птиц в условиях субвысо-
когорья Тянь-Шаня // Тез. докл. 7-й Всесоюз. орнитол. конф. Киев, 1977. Ч. 2. С. 19-22.
Ковшарь А. ,Ф. Певчие птицы в субвысокогорье Тянь-Шаня (очерки летней жизни фоновых
видов). Алма-Ата, 1979. 312 с.
Ковшарь А. Ф. Особенности размножения птиц в субвысокогорье (на материалах
Passeriformes в Тянь-Шане). Алма-Ата, 1981а. 260 с.
Ковшарь А. Ф. Полициклия и гипсоморфный эффект в размножении горных птиц // Экология
и охрана птиц. Тез. докл. 8-й Всесоюз. орнитол. конф. Кишинев. 19816. С. 108.
Ковшарь А. Ф. Полициклия и гипсоморфный эффект в размножении горных птиц // Тез. докл.
18-го междунар. орнитол, конгр. М., 1982. С. 27—28.
Кожанчиков И. В. Методы исследования экологии насекомых. М., 1961. 286 с.
Королькова Г. Е. Влияние птиц на численность вредных насекомых. М., 1963.126 с.
* Костин Ю. В. Птицы Крыма. М., 1983. 241 с.
* Котов А. А. Экология скворца в городе Новокузнецке // Бюл. МОИП. Отд. биол. М., 1977.
Т. 82, № 1.С. 18-22.
Коханов В. Д. Ареал гнездования некоторых птиц в районе Кандалакшского залива Белого
моря // Природа и хозяйство Севера. Мурманск, 1969. Вып. 1. С. 216-219.
Коханов В. Д. К экологии кукши в Мурманской области // Экология и морфология птиц на
крайнем Северо-Западе СССР. М., 1982. С. 124-137.
Коханов В. Д., Гаев Ю. Г. Материалы по экологии клестов в Мурманской области // Гр. Канда-
лакша гос. заповедника. Мурманск, 1970. Вып. 8. С. 236-274.
Коханов В. Д., Скокова Н. Н. Фауна птиц Айновых островов // Тр. Кандалакш. гос. заповед-
ника. М., 1967. Вып. 5. С. 185-267.
Кочанов С. К. Структура населения птиц г. Сыктывкара // Структура населения птиц европей-
ского Северо-Востока СССР. Сыктывкар, 1983. С. 50-56.
* Кошелев Н. Т. Особенности биологии продуктивной и непродуктивной частей популяции
скворцов по данным отлова // Доклады МОИП. Зоол. и ботан. М., 1978. С. 70-71.
Крутовская Е. В. Материалы к экологии птиц искусственных гнездовий заповедника „Стол-
бы” // Тр. гос. заповедника „Столбы”. Красноярск, 1966. Вып. 5. С. 232-267.
Кукиш А. И. Сравнительная характеристика годовых циклов двух вигов рода Motacilla-
Автореф. дис. ... канд. биол. наук. Л., 1976.17 с.
173
Кукиш А. И. О соотношении сроков миграции и линьки у белой трясогузки (Motacilla alba L.) //
Экология птиц Приладожья. Л., 1981. С. 144-151. (Тр. Биол. НИИ ЛГУ; № 32).
Кумари Э. В. Основные черты новейшей динамики распространения фауны птиц Восточной
Прибалтики // Ornitoloog. kogumik. Тарту, 1958. № 1. С. 7-20.
Лапшин Н. В. К вопросу о полигинии у некоторых воробьиных птиц Южной Карелии // Тез.
докл. 2-й Всесоюз. конф, по вопросам сравнительной морфологии и экологии животных. М., 1975.
С. 168-169.
* Лапшин Н. В. Биология гнездования пеночки-веснички в Южной Карелии // Экология птиц
и млекопитающих Северо-Запада СССР. Петрозаводск, 1976. С. 28—38.
Лапшин Н. В. Поведение пеночки-веснички в предгнездовой период // Фауна и экология птиц
и млекопитающих таежного Северо-Запада СССР. Петрозаводск, 1978. С. 32-39.
Лапшин Н. В. Особенности территориального поведения веснички на Северо-Западе СССР //
Экология и охрана птиц. Тез. докл. 8-й Всесоюз. орнитол. конф. Кишинев, 1981а. С. 132.
Лапшин Н. В. Соотношение циклов размножения и линьки у веснички на Северо-Западе
РСФСР // Тез. докл. 10-й Прибалт, орнитол. конф. Рига, 19816. Ч. 2. С. 89-91.
Лапшин Н. В. Материалы по размножению четырех видов пеночек на севере Ленинградской
области и в Южной Карелии // Тез. докл. 11-й Прибалт, орнитол. конф. Таллин, 1983а. С. 126—128.
Лапшин Н. В. Факультативная полигиния у веснички, Phylloscopus trochilus (L.) в условиях та-
ежного Северо-Запада РСФСР // Фауна и экология птиц и млекопитающих Северо-Запада СССР. Пет-
розаводск, 19836. С. 34-41.
Лапшин Н. В., Большаков К. В., Резвый С. П. Весенняя миграция пеночки-веснички (Phyllosco-
pus trochilus L.)// Экология птиц Приладожья. Л., 1981. С. 86-100. (Тр. Биол. НИИ ЛГУ; № 32).
* Левин А. С. О величине кладок лесных воробьиных птиц в пойме реки Урал // Тез. Всесоюз.
конф. „Экология гнездования птиц и методы ее изучения”. Самарканд, 1979. С. 129—130.
* Левин А. С. О биологии дрозда-белобровика в долине среднего течения р. Урал //
Изв. АН КазССР. 1980. № 3. С. 45-49.
Лисецкий А. С. Новые сведения о распространении птиц на северо-востоке Украины // Эколо-
гия и охрана птиц. Тез. докл. 8-й Всесоюз. орнитол. конф. Кишинев, 1981. С. 139.
* Лихачев Г. Н. Мухоловка-пеструшка (Muscicapa hypoleuca Pall.) и ее связ^ с гнездовой терри-
торией // Тр. Бюро кольцевания. М., 1955. Вып. 8. С. 123—156.
* Лихачев Г. Н. Дополнительные данные по характеру размножения большой синицы в искус-
ственных гнездовьях // Тр. Приокско-Террасного гос. заповедника. М., 1957. Вып. 1. С. 248-265.
* Лихачев Г. Н. Материалы по биологии птиц, гнездящихся в искусственных гнездовьях //
Тр. Приокско-Террасного гос. заповедника. М., 1961. Вып. 4. С. 82-146.
Лихачев Г. Н. О величине кладки некоторых птиц в центре европейской части СССР // Орнито-
логия. М., 1967. Вып. 8. С. 165-174.
Лихачев Г. Н. Материалы по динамике численности птиц, гнездящихся в искусственных гнез-
довьях на юге Подмосковья // Тр. Приокско-Террасного гос. заповедника. М., 1971. Вып. 5.
С. 66-110.
* Лихачев Г. Н. Размножение и численность мухоловки-пеструшки на юге Московской
области // Сб. тр. Зоол. музея МГУ. М., 1978. Т. 17. С. 119—140. »
Люлеева Д. С. О наседном пятне у воробьиных птиц // Экология млекопитающих и птиц. М.,
1967.С. 301-308.
♦ Люняк М. Численность гнездовой популяции и продуктивность выводков скворца в Варша-
ве // Материалы 6-й Всесоюз. орнитол. конф. М., 1974. Ч. 2. С. 335—336.
Мальчевский А. С. Явление гнездового консерватизма у воробьиных птиц // Вести. ЛГУ. Био-
логия. 1957. № 9. С. 58-70.
* Мальчевский А. С. Гнездовая жизнь певчих птиц. Л., 1959. 282 с.
Мальчевский А. С. О консервативном и дисперсном типах эволюции популяций птиц // Зоол.
журн. 1968. Т. 47, № 6. С. 833-842.
Мальчевский А. С. Дисперсия особей и эволюция видов и популяций у птиц // Орнитология
в СССР. Ашхабад, 1969. Кн. 1. С. 111-124.
Мальчевский А. С. Отношение животных к территории как фактор эволюции (на примере
птиц) // Вести. ЛГУ. Биология. 1974а. № 3. С. 5-15.
Мальчевский А. С. Степень постоянства и подвижности территориальных связей птиц как
фактор эволюции // Материалы 6-й Всесоюз. орнитол. конф. М., 19746. С. 37-39.
Мальчевский А. С. О путях и методах изучения популяционной структуры вида у птиц // Ис-
следование продуктивности вида в ареале. Вильнюс, 1975. С. 77-86.
Мальчевский А. С. Об эволюционном значении подвижности территориальных связей птиц
в гнездовой области // Адаптивные особенности и эволюция птиц. М., 1977. С. 72-77.
* Мальчевский А. С., Пукинский Ю. Б. Птицы Ленинградской области и сопредельных терри-
торий. Л., 1983. Т. 2. 504 с.
* Мешков М. М., Урядова Л. П. Материалы по гнездованию птиц в Псковской области // Тр. 5-й
Прибалт, орнитол. конф. Таллин, 1967. С. 66-75.
Михайлов В. Н. Гнездование зимородка в Белозерском районе Вологодской области // Орни-
тология. М., 1976. Вып. 12 С. 239.
174
* Москвитин С. С. О величине кладки и размерах яиц у некоторых воробьиных// Тез. Всесоюз.
конф. „Экология гнездования птиц и методы ее изучения”. Самарканд, 1979. С. 145 -146.
Музаев М. М. Явление бигамии у серой славки (Sylvia communis Lath.) // Биолог науки.М.,
1980а. №12. С. 38-42.
Музаев В. М. Сравнительная экология, территориальное поведение и годовые циклы некото-
рых представителей рода Sylvia: Автореф. дис. ... канд. биол. наук. Л., 19806. 22 с.
Музаев В. М. Особенности территориального поведения бигамных самцов серой славки // Эко-
логия и охрана птиц. Тез. докл. 8-й Всесоюз. орнитол. конф. Кишинев, 1981. С. 158- 159.
Музаев В. М. К экологии садовой славки в Ленинградской области // Тез. докл. 11 -й Прибалт,
орнитол. конф. Таллин, 1983. С. 140- 142.
Нанкинов Д. Н. Об экологии обыкновенной чечевицы Ленинградской области // Материалы
6-й Всесоюз. орнитол. конф. М., 1974. Ч. 2. С. 92-94.
* Нанкинов Д. Н О степени растянутости сроков размножения птиц// Acta omithol. Warszawa,
1978. Vol. 16, № 10. P. 295-308.
Нейфельдт И. А. Об орнитофауне Южной Карелии // Тр. Зоол. ин-та АН СССР. Л., 1958. Т. 25.
С. 183-254.
Новиков Б. Г., Руднева Л. М., Булдакова А. Н., Иванова Л. С., Гармагина С. Н., Данилова О. В.
Роль гипоталамо-гипофизарной системы в годичном цикле линьки и функции гонад // Тез. докл.
18-го Междунар. орнитол. конгр. М., 1982. С. 50-51.
Новиков Г. А. Особенности гнездовой жизни птиц на Кольском полуострове // Природа. 1944.
№1. С. 78-80.
Новиков Г. А. Экология птиц Хибинских гор (Кольский полуостров) // Тр. Зоол. ин-та
АН СССР. Л., 1952. Т. 9./вып. 4. С. 1155-1198.
Новиков Г. А. Географическая изменчивость плотности населения лесных птиц в европейской
части СССР и сопредельных странах // Зоол. журн. 1960. Т. 39, № 3. С. 433 -447.
Носков Г. А. О географической и сезонной изменчивости линьки у некоторых воробьиных
птиц // Механизмы биологических процессов. Л., 1969. С. 62.
Носков Г. А. Некоторые особенности вида у птиц // Материалы 2-го Всесоюз. совещ. „ Вид
и его продуктивность в ареале” Вильнюс, 1976. С. 94-96.
Носков Г. А. Линька зяблика (Fringilla coelebs). 2. Фотопериодическая регуляция и место в го-
довом цикле // Зоол. журн. 1977. Т. 56., № 11. С. 1676-1686.
Носков Г. А. Сравнительная характеристика территориального поведения воробьиных птиц //
Экология и охрана птиц. Тез. докл. 8-й Всесоюз. орнитол. конф. Кишинев, 1981. С. 164-165.
Носков Г. А., Зимин В. Б. Основные итоги работ Ладожского орнитологического стациона-
ра // Тез. докл. 2-й Всесоюз. орнитол. конф, по миграциям птиц. Алма-Ата, 1977. Ч. 1. С. 111-113.
* Носков Г. А., Смирнов О. П. Территориальное поведение и миграции большой синицы (Parus
m. major L.)// Экология птиц Приладожья. Л., 1981. С. 100-130. (Тр. Биол. НИИ ЛГУ; № 32).
Носков Г. А., Зимин В. Б., Резвый С. П. Миграции птиц на Ладожском озере // Сообщ. Прибалт,
комис. по изучению миграций птиц. Таллин, 1975. № 8. С. 3-50.
Носков Г. А., Зимин В. Б., Резвый С. П., Рымкевич Т. А., Лапшин Н. В., Головань В. И. Птицы
Ладожского орнитологического стационара и его окрестностей // Экология пгиц Приладожья. Л.,
1981. С. 3-86. (Тр. Биол. НИИ ЛГУ; № 32).
Нумеров А. Д. К биологии зимородка в Окском заповеднике // Тез. докл. 7-й Всесоюз.
орнитол. конф. Киев, 1977. Ч. 1. С. 292—293.
Нумеров А. Д., Котюков Ю. В. Материалы по численности, продуктивности и структуре попу-
ляции зимородка // Тез. докл. 10-й Прибалт, орнитол. конф. Рига, 1981. Т. 2. С. 151 -153.
Основные данные по климату СССР: Справочник. Обнинск, 1976. 391 с.
Паевский В. А. Продолжительность жизни и ежегодная смертность птиц // Исследования по
биологии птиц. Л., 1974. С. 142-185. (Тр. Зоол. ин-та АН СССР; Т. 55).
Паевский В. А. Демографические показатели популяции обыкновенной чечевицы на Куршской
косе // Тез. докл. 10-й Прибалт, орнитол. конф. Рига, 19816. Т. 2. С. 153-155.
Панов Е. Н. Птицы южного Приморья (Фауна, биология, поведение). Новосибирск,
1973.412 с.
Пантелеев М. Ф„, Болотников А. М. Температурные условия раннего онтогенеза птиц-дупло-
гнездников // Гнездовая жизнь птиц. Пермь, 1977. С. 37-41.
Подковыркин Б. А. Позднее гнездование вальдшнепа // Охота и охотничье хозяйство. М.,
1972. №4. С. 43.
Познании Л. П. Постом бринальный рост некоторых видов лесных птиц в связи с особеннос-
тями их экологии // Докл. АН СССР. 1946. Т. 54, № 3. С. 285-288.
* Поливанов В. М. Местные популяции у птиц и степень их постоянства // Тр. Дарвин, гос.
заповедника, Вологда, 1957. Вып. 4. С. 79-155.
Поливанова Н. Н. Питание птенцов некоторых видов полезных насекомоядных птиц в Дарвин-
ском заповденике // Тр. Дарвин, гос. заповедника. Вологда, 1957. Вып. 4 С. 157-244.
Полякова А. Д., Бианки В. В., Зимин В. Б., Естафьев А. А., Семенов-Тян-Шанский О. И.
Расширение ареала обыкновенного скворца на севере европейской части СССР// Гр. Окск. гос. запо-
ведника. М., к 934. Вып. 15. С. 158—169.
175
Портенки Л. А, Птицы СССР. М.; Л., 1960. Т. 4. 416 с
Постников С. Н. Проблемы экологической энергетики птиц в высоких широтах // Тез. докл.
2-й Сибир. орнитол. конф. „Птицы Сибири”. Горно-Алтайск, 1983. С. 120-122.
Правосуден В. В. Эффективность кормления самцами самок сероголовой гаички и пухляка
в гнездовой период // Тез. докл. 11-й Прибалт, орнитол. конф Таллин, 1983. С. 166-169.
Прокофьева И. В. О питании гнездовых птенцов некоторых видов лесных птиц // Учен. зап.
Ленингр. гос. пед. ин-та им. А. И. Герцена. Л., 1958. Т. 143, № 7. С. 49-66.
Прокофьева И. В. О роли различных беспозвоночных и позвоночных животных в питании
птенцов некоторых лесных птиц // Докл. АН СССР. Сер. биол. 1961. Т. 136, № 2. С. 497-499.
Прокофьева И. В. О некоторых особенностях питания мелких лесных птиц // Учен. зап.
Ленингр. гос. пед. ин^га им. А. И. Герцена. Л., 1963. Т. 230, № 9. С. 33-56.
Прокофьева И. В. О питании и хозяйственном значении мухоловок (Muscicapidae) в гнездовой
период // Зоол. журн. 1966. Т. 45, № 8. С. 1210-1215.
Прокофьева И. В. О некоторых особенностях питания птиц в разных биотопах (по наблюде-
ниям в Ленинградской области) // 12-е Герценовские чтения: Естествознание. Л., 1970. С. 114-117.
Прокофьева И. В., Драгункина Н. С., Огородникова Л. И. О географической изменчивости пи-
тания белой трясогузки // Тез. докл. 7-й Всесоюз. зоогеограф, конф. М., 1979. С. 303-305.
Промптов А. Н. Очерки по проблеме биологической адаптации поведения воробьиных птиц.
М.; Л., 1956.311 с.
Птушенко Е. С. Семейство Славковые. М., 1954. С. 146-330. (Птицы Советского Союза; Т. 6).
Птушенко Е. С., Иноземцев А. А. Биологическое и хозяйственное значение птиц Московской
области и сопредельных территорий. М., 1968.462 с.
Пукинский Ю. Б. Жизнь сов. Л., 1977. 240 с.
Раменская М. Л. Определитель высших растений Карелии. Петрозаводск, 1960.485 с.
Резвый С. П Летне-осенние миграции длиннохвостой синицы на юго-восточном берегу Ладож-
ского озера в 1968-1975 гг. // Материалы 9-й Прибалт, орнитол. конф. Вильнюс, 1976. С. 221-226.
Резвый С. П. Летние миграции зарянки (Erithacus rubecula L.) на северо-востоке Ленинградской
области // Сообщ. Прибалт, орнитол. комис. по изучению миграций птиц. Тарту, 1983. № 14. С.'’0-84.
Резвый С. И., Савинич И. Б. О летних миграциях зарянки на юго-восточном берегу Ладож-
ского озера // Тез. докл. 2-й Всесоюз. конф, по миграциям птиц. Алма-Ата, 1978. Ч. 2. С. 131-132.
Романов А. А. О климате Карелии. Петрозаводск, 1961.140 с.
Рымкевич Т. А. Овсянка-дубровник в Ленинградской области // Материалы 9-й Прибалт,
орнитол. конф. Вильнюс, 1976. С. 229-232.
Рымкевич То А. Сравнительное изучение годовых циклов некоторых представителей рода
Emberiza: Автореф. дис.... канд. биол. наук. Л., 1977. 23 с.
Савинич И. Б. Формирование оперения зарянки (Erithacus rubecula L.) на первом году жизни и
фотопериодическая регуляция постювенальной линьки //Вести.ЛГУ. 1984. № 9. Биология, вып. 2.
С. 27-31.
Сдобников В. М. О причинах перерывов в гнездовании у арктических птиц // Зоол. журн.
1971. Т. 50, вып. 5. С. 734-740.
Северцов Но А. Периодические явления в жизни зверей, птиц и гад Воронежской губернии.
М., 1950. 308 с
♦ Сема А. М. О количестве кладок у туркестанского скворца // Орнитология в СССР. Ашха-
бад, 1969. Кн 2. С. 574-576.
* Сема А. М. Биология обыкновенного скворца на юго-востоке Казахстана // Тр. Ин-та зоол.
АН КазССР. Алма-Ата, 1978. Т. 38. С. 120-126.
Семенов Б Т. Изменения северной границы распространения скворца в Архангельской об-
ласти за 60-летний период // Тр. Окск. гос. заповедника. М., 1984* Вып. 15. С. 170-182.
Семенов-Тян-Шанский О. И. Экология тетеревиных птиц // Тр Лапланд. гос. заповедника. М.,
1960. Вып. 5. С. 1-318.
♦ Скрылева Л. Ф. Некоторые данные о размножении обыкновенного скворца в Тамбовской
области // Гнездовая жизнь птиц. Пермь, 1981. С. 132-135.
* Смогоржевская Л. И„ Смогоржевский Л. А., Прокопчук Л« Н. Рост птенцов обыкновенного
скворца // Тез. докл< 7-й Всесоюз. орнитол. конф. Киев, 1977. Ч. 1. С. 325-326.
Спангенберг Е. IL, Судиловская А. М. Род Овсянка. М., 1954. С. 376-510. (Птицы Советского
Союза; Т. 5).
Страутман Ф. И. Птицы западных областей УССР. Львов, 1963. Т 2. 182 с.
Судиловская А. М. Год Крапивник. М., 1954а. С. 661—670. (Птицы Советского Союза; Т. 6).
Судиловская А. М. Род Королек. М., 19546. С. 126-141. (Птицы Советского Союза; Т. 6).
* Сухов А. В., Кабанен О. А. Закономерности роста и развития птенцов мухоловки-пеструшки
в Южной Карелии // Экология наземных позвоночных Северо-Запада СССР. Петрозаводск, 1981.
С. 63-69.
Талпош В. С. Европейская лесная завирушка (Prunella modularis L.) на западе Украины // Вести,
зоол. Киев, 1980. № 5. С. 67- 73.
♦ Таращук В. И. Птицы полезащитных насаждений степной зоны УССР и возможность исполы
зования их для борьбы с вредителями. Киев, 1953.124 с.
176
Тауриньш Э. Я., Вилкс К. А., Михельсон X. А. Сезонное размещение и миграции некоторых
видов птиц по данным кольцевания орнитологической станции АН Латвийской ССР // Перелеты птиц
в европейской части СССР. Рига, 1953. С. 57-102.
Третьяков Г. П. Материалы по биологии красношапочного вьюрка и обыкновенной чечевицы
в высокогорьях Западного Тянь-Шаня // Тез. Всесоюз. конф. „Экология гнездования птиц и методы
ее изучения”. Самарканд, 1979. С. 212-213.
Успенский С. М. Жизнь в высоких широтах на примере птиц. М., 1969. 463 с.
Фенологический ежегодник за 1961 год. Европейская территория СССР. Л., 1965. 158 с.
♦Фетисов С. А. Биология полевого воробья (Passer montanus L.) в репродуктивный период:
Автореф. дис.... канд. биол. наук. Л., 1981. 22 с.
* Фетисов С. А., Корзун А. П. О величине кладки полевого воробья в условиях Ленинградской
области // Тез. Всесоюз. конф. „Экология гнездования птиц и методы ее изучения”. Самарканд,
1979. С. 218-219.
Формозов А. Н. Колебания численности промысловых животных. М.; Л., 1935. 108 с.
* Хазиева А. Н., Болотников А. М., Каменский Ю. Н„ Никольская В. И. Материалы о гнездя-
щихся птицах правобережья реки Камы Добрянского района Тамбовской области // Гнездовая
жизнь птиц. Пермь, 1975. С. 117-127. (Учен. зап. Пермск. гос. пед. ин-та; Т. 146).
Хайнд Р. Поведение животных. М., 1975. 855 с. (Пер. с англ.).
Хохлова Т. Ю. Фенология прилета и размножения птиц в условиях ранней весны 1975 года //
Экология птиц и млекопитающих Северо-Запада СССР. Петрозаводск, 1976. С. 38-44.
♦ Хохлова Т. Ю. Соотношение сроков вылупления и линьки у трех видов дроздов в Южной
Карелии // Тез. докл. 11-й Прибалт, орнитол. конф. Таллин, 1983. С. 213-215.
* Хохлова Т. Ю., Захарова Л. С. К характеристике двух циклов размножения дрозда-белобро-
вика // Тез. докл. 11-й Прибалт, орнитол. конф. Таллин, 1983. С. 215-218.
* Хохлова Т. Ю., Головань В. И. К биологии размножения мухоловки-пеструшки в Южной
Карелии // Экология наземных позвоночных Северо-Запада СССР. Петрозаводск, 1981. С. 11-29.
* Хохлова Т. Ю., Захарова Л. С., Зимин В. Б. Сроки и динамика явлений годового цикла у дро-
зда-белобровика в Карелии // Фауна и экология птиц и млекопитающих Северо-Запада СССР. Петро-
заводск, 1983. С. 11-29.
♦ Чемякин Р. Г. Сроки гнездования воробьиных на Айновых островах // Материалы 6-й Все-
союз. орнитол. конф. М., 1974. Ч. 2. С. 149-151.
* Чемякин Р. Г. Интересные случаи в гнездовании воробьиных на Айновых островах // Эколо-
гия птиц морских побережий. М., 1980. С. 94-95.
Чемякин Р. Г. Изменчивость сроков весеннего пролета и размножения воробьиных птиц на
Айновых островах // Экология и морфология птиц на крайнем Северо-Западе СССР. М., 1982а.
С. 137-141. .
♦Чемякин Р. Г. О влиянии погоды на начало размножения лугового конька (Anthus pratensis
L.) // Экология и морфология птиц на крайнем Северо-Западе СССР. М., 19826. С. 142-146.
Шаповал А. П. Гнездовые находки желтоголовой трясогузки и дрозда-белобровика в Полтав-
ской области // Вести, зоол. Киев, 1982. №4. С. 76-77.
Шварц С. С. Опыт экологического анализа некоторых морфо-физиологических признаков
наземных позвоночных: Автореф. дис. ... докт. биол. наук. М., 1953. 20 с.
Шилов И. А. Регуляция теплообмена у птиц (эколого-физиологический очерк). М., 1968. 252 с.
♦ Шураков А. И., Болотников А. М., Печерский А. С., Елсуков С. В. Величина кладки // Поле-
вой воробей (Характеристика вида на пространстве ареала). Л., 1981. С. 153-161.
Шутенко Е. В. О летне-осеннем территориальном поведении молодых ворон в лесном биото-
пе // Тез. докл. 11-й Прибалт, орнитол. конф. Таллин, 1983. С. 231-234.
♦ Шутова Е. В. О сроках начала гнездования лесных насекомоядных птиц в Мурманской об-
ласти // Тез. докл. 11-й Прибалт, орнитол. конф. Таллин, 1983. С. 234-236.
* Шутова Е. В., Бианки В. В., Коханов В. Д., Ладыжев Л. С. К экологии дрозда-белобровика
(Turdus iliacus L.) на Сейеро-Западе СССР // Экология и морфология птиц на крайнем Северо-Западе
СССР. М., 1982. С. 109-123.
Яковлев Ф. С., Воронова В. С. Типы лесов Карелии и их природное районирование. Петро-
заводск, 1959.192 с.
Яковлева М. В. К биологии лесной завирушки, Prunella modularis (L.), на юго-восточном побе-
режье Ладожского озера // Фауна и экология птиц и млекопитающих Северо-Запада СССР. Петро-
заводск, 1983. С. 52-57.
♦ Aleknonis A. Giriu Giesmininkai. Vilnus, 1984. 174 s.
Andersson M. Clutch size in the Longtailed Skua, Stercorarius longicaudus: some field experiments //
Ibis. 1976. Vol. 118, N4. P. 586-588.
Antikainen E. The breeding adaptation of the Jackdaw Corvus monedula L. in Finnland // Savonia.
1978. N 2. P. 1-45.
♦Askenmo C. Reproductive effort and return rate of male Pied Flycatcher // Amer. Natur. 1979. Vol.
114, N 5. P. 748-753.
Balfour E., Cadburry C. J. Polygyny, spacing and sex ratio among hens Harriers Circus cyaneus in
Orkney, Scotland // Ornis Scand. 1979. Vol. 10, N 1. P. 133-141.
177
Bergman G. Auswerfen von Zweitbruteiern und Beeintrachtigung des Brutresultat beider Mehlschwal-
be (Delichon urbica) im regnerischen Sommer 1977 // Mem. Soc. Fauna et Flora Fen. 1978. Vol.
54, P. 29-30.
Berndt R., Winkel W. Die Gelegegrosse des Trauerschnappers (Ficedula hypoleuca) im Beziehung zu
Ort, Zeit, Biotop und Alter // Vogelwelt. 1967. Bd 88, N 4/5. S. 97-136.
Berthold P. Zur Jahresperiodik von Staren (Sturnus vulgaris) aus Friih-and Spatbruten // Vogelwelt.
1970. Bd91,N3.S. 88-95.
Bertman G., Lack D., Roberts B. Notes on East Greenlands Birds with discussion of the periodical
Nonbreeding among Arctic birds // Ibis. 1934. Vol. 4, N 4.
Besser H. J. Schachtelbrutbeim Kiebitz (Vanellus vanellus) // Charadrius. 1983. Vol. 19, N 1. P. 11.
Blondel J., Isenman P. Insularite et demographic des mesanges du genre Parus (Aves) // C. r. Acad,
sci., Ser. D. 1979. Vol. 289, N 2. P. 161-164.
Bozcko S. I. The ecology of the nesting period in the Scarlet Grosbeak, Carpodacus erythrinus Pall.,
in the parks of Leningrad // Acta ornithol. 1974. Vol. 14, N 3. P. 39-57.
Brooks W. S. Comparative adaptations of the Alaskan redpolls to the Arctic environment // Wilson.
Bull. 1968. Vol. 80, N 3. P. 253-280.
Bub H. Vogelfang und Vogelberingung. Wittenberg Lutherstadt. 1966. 100 s. (Die Neue Brehm-Buche-
rei; T. 1).
Burr E. W., Huchzermeyer F. W., Riley A. E. Laparoscopic examination to determine sex in monomor-
phic avian species // J. of the South African Veterinary Association. 1981. Vol. 52, N 1. P. 45-47.
Carey M., Nolan V. J. Population dynamics of indigo bunting and die evolution of avian polygyny //
Evolution. 1979. Vol. 33, N 4. P. 1 180-1192.
Clavert J. La biochimie de V ovogenese // Arch, neerl. zool. 1953. T. 10, N 1. P. 30-46.
Cody M. L. A general theory of Clutch size // Evolution. 1966. Vol. 20, N 1. P. 174-184.
Constant P., Eybert M. Chr. Donnees sur la biologie de la reproduction du Pipit farlous, Anthus
pratensis L., dans les landes bretones // Nos Oiseaux. 1980. T. 35, N 381. P. 349-360.
Coulson J. C., White E. An analyses of the factors influencing the clutch-size of the Kittiwake //
Proc. Zool. Soc. London. 1961. Vol. 45, N 1. P. 205-213.
Cowley E. Multi-brooding and mate infidelity in the Sand Martin //Bird Study. 1983. Vol.
30,Nl.P. 1-7.
* Cramp S. The breeding of the Willow Warbler // Brit. Birds. 1955. Vol. 48, N 2 (3). P. 121-135.
Crawford R. Breeding biology of year-old and older females Red-winged and Yellow-headed black-
birds // Wilson. Bull. 1977. Vol. 89, N 1. P. 73-80.
* Creutz G. Die brutbiologie des Tranecfliegenschnapper (Muscicapa h. hypoleuca Pall.) // Ber. Ver.
Schles Ornithol. 1943. Bd 28. S. 1-10.
* Creutz G. Der Trauerschnapper (Muscicapa hypoleuca Pall.) eine populationstudie // J. Ornithol.
1955. Bd 96. S. 241-326.
Custer T. W.? Pitelka F. A. Demographic features of Lapland Longspur population near Barrow,
Alaska // Auk. 1977. Vol. 94, N 3. P. 505-525.
Delius J. D. A population study of Skylark, Alauda arvensis // Ibis. 1965. Vol. 107, N 4. P. 466-492.
Dhondt A. A., Olaerts G. Ringing dates of first broods of the Great Tit in difference parts of Belgium
and in relation to spring temperature// Gerfaut. 1980. Vol. 70, Nl.P. 113-118. '
Doty H. A. Renesting and second broods of wild Mallard // Wilson. Bull. 1975. Vol. 87, N 1. P. 115.
* Dunnet G. M. The breeding of Starling, Sturnus vulgaris, in relation to its food supply // Ibis. 1955.
Vol. 97, N 4. P. 619-662.
* Dyrcz A. The ecology of the Song Trush (Turdus philomelos Brehm) and Blackbird (Turdus merula
L.) during the breeding season in area of their joint occurence // Ecol. Polska. 1969. T. 17, N 39. Ser. A.
S. 735-793.
Dyrcz A. Polygyny and breeding success among Great Reed Warbler Acrocephalus arundinaceus at
Milicz, Poland // Ibis. 1977. Vol. 119. N 1. P. 73-77.
Eriksson K. The authumn migration and wintering ecology of the Siskin, Carduelis spinus /f Ornis
fenn. 1970. Vol. 47, N 2. P. 52-68.
Eybert M. Chr. Dynamique de la reproduction de la linotte melodieuse (Acanthis cannabina) sur une
Lande Bretonne // Bull. ecol. 1980. T. 11, N.3-4. S. 543-548.
Earner D. S. Fotoperiodical control of reproductive cycles in birds // Amer. Scientist. 1964. Vol.
52 P. 137-156.
Farner D. S. The control of avian reproductive cycles // Proc. XIV Intern. Ornithol. Congr. Copenga-
gen, 1967. P. 107-133.
Farner D. S. Predictive functions in the control of annual cycles // Environment. Res. 1970. Vol. 3.
P. 119-131.
Farner D. S., Oksche A., Kamemoto F. I., King J. R., Cheyney H. E. A comparison of the effect
of long daily photoperiods on the pattern of energy storage in migratory and non-migratory finches // Comp.
Biochern. Physiol. 1961. Vol. 2, N 2. P. 125-142.
Flegg J, J. A study of Tieecreepers // Bird Study. 1973. Vol. 20, N 4. P. 281—302.
* Flux J. E. C., Flux M. M. Population dynamics and age structure of Starling (Sturnus vulgaris) in
New Zealand // N. Z. J. Ecol. 1981. Vol. 4. P. 65-72.
178
French N. R. Notes on breeding activities and gular sacks in the Pine Grosbeak // Condor. 1954.
Vol. 74. P. 83-85.
Fuller R. J., Glue D. E. The breeding biology of the Stonechat and Whinchat // Bird Study. 1977.
Vol. 24, N 4. P. 215-228.
Furrer R. Beringung, Zugverhalten und Mortalitat schweizericher Wacholdedrosseln Turdus pilaris:
eine Ringfundanalyse // Ornithol. Beobachter. 1977. Bd 74, N 2. S. 37-53.
Gibb J. A. Feeding rates of Great Tits // Brit. Birds. 1955. Vol. 48, N 1. P. 49-58.
* Goodackre M. J., Lack D. Early breeding in 1957 // Brit. Birds. 1959. Vol. 52, N 3. P. 74-82.
Greenlan J. S. The relation of breeding schedule and clutch size to food supply in the Rufous-sided
Towhee // Condor. 1978. Vol. 80, N 1. P. 24-33. v
* Gromadzki M. Reproduction of the Starling, Sturnus vulgaris, in Zulavy Wislane, North Poland //
Acta ornithol. 1980. T. 17, N 16. S. 195-224.
Gwinner E. A. A comparative study of circannual rhythms in warblers // Biochronometry Washing-
ton Nat. Acad. Sci. 1971. P. 405-427.
* Haartman L. von. Der Trauerfliegenschnapper. III. Die Nahrungsbiologie // Acta Zool. Fenn. 1954.
N 83. P. 1-96.
* Haatrman L. von. The life of the Pied Flycatcher (Ficedula hypoleuca) // Proc. XI Intern. Ornithol.
Congr. Copenhagen, 1956. P. 267-272.
Haartman L. von. Clutch-size in the Pied Flycatcher // Proc. XIV Intern. Ornithol. Congr. Oxford;
Edinburg, 1967a. P. 155-164.
* Haartman L. von. Geographical variation in clutch-size uf the Pied Flycatcher // Ornis fenn. 1967b.
Vol. 44, N 4. P. 89-98.
Haartman L. von. The nesting habits of Finnish Birds. 1. Passeriformes // Comment. Biolog. Soc.
Sci. Fenn. 1969a. Vol. 32. P. 1-187.
Haartman L. von. Nest-site and evolution of polygamy in European passerine biids // Ornis fenn.
1969b. Vol. 46, N l.P. 1-12.
Haartman L. von. Population dynamics // Avian Biology. New York; London, 1971. Vol. 1.
P. 391-459.
Haartman L. von. Two modes of clutch-size determination in passerine birds // Misc. Repts Gama-
shina Inst. Ornithol. 1982. Vol. 14, N 2-3. P. 214-219.
Hachfeld B. Brutbiologie und Ethologie der Klappergrasmiicke (Sylvia curruca L.) // Voliere. 1982.
Vol. 5, N l.P. 1-9.
Hafke J. Die Bestandsentwicklund des Haubentauchers (Podiceps cristatus) an einem neubesiedelten
Gewiisser // Charadrius. 1983. Bd 19, N 4. S. 205-219.
Haftorn S. Cooperation between the male and female Goldcrest, Regulus regulus, when rearing over-
lapping double broods // Ornis Seand. 1978. Vol. 12, P. 124-129.
Harrisson C. J. The crop of the Chaffinch, Fringilla coelebs // Bull. Brith. Ornithol. Club. 1970. Vol.
90, N l.P. 15-16.
Harriston G. J. Endoscopic examination of avian gonadal tissues // Veterinary Medicine Small Animal
Clinicial. 1978. Vol. 73, N 4. P. 479-484.
Harvey J., Datter E. Esophagial diverticula in Redpoll, Acanthis flammea // Auk. 1961. Vol. 78, N 4.
P. 528-531.
Haukioja E. Mortality rates of some Finnish Passerines // Ornis fenn. 1969. Vol. 46, N 4. P. 171-178.
Haukioja E. Clutch size of the Reed Bunting, Emberiza schoeniclus // Ornis fenn. 1970. Vol. 47, N 3.
P. 107- 135.
Haukioja E., Hakala T. On the relationship between avian clutch size and life span // Ornis fenn.
1979. Vol. 56, N73.P-45-55.
Havlin J. Reproduction in the Blackbird (Turdus merula L) Zool. listy. 1963. Roc. 12, Cis. 4.
C. 1-18, 195-216.
Hecke P. von. Zur Brutbiologie des Baumpiepers (Anthus t. trivialis): Legeperiode, Gelegegrosse,
Bruterfolg // J. Ornithol. 1979. Bd 120, N 1. S. 12-29.
Hilden O.Uber Vorcommen und Brutbiologie des Birkenzeisigs (Carduelis flammea) in Finnish-Lapland
im Sommer 1968 // Ornis fenn. 1969. Vol. 46, N 3. P. 93-112.
Hopkins A. D. Oioclimatics, a science of life and climate relations // U. S. Depart. Agr., Misc., Publ.
1938. 280 p.
Hussel D. J. Factors affecting clutch size in Arctic passerines // Ecol. Monogr. 1972. Vol. 42, N 3.
P. 317-364.
Hogstedt G. Effect of additional food on reproductive success in the Magpie (Pica pica) //J. Anim.
Ecol. 1981. Vol. 50, N 1. P. 219-229.
Immelmann K. Ecological aspects of periodic reproduction// Avian Biology. New York; London.
1971. Vol. l.P. 341-389.
Irving L. Birds of Anaktuvuk Pass, Kobuk and old Crow. A study in Arctic adaptation. Washington,
1960. 409 p. (U. S. Nat. Mus. Bull., 217).
*Jarvinen A. Population dynamics in the Pied Flycatcher, Ficedula hypoleuca, at subarctic Kilpisjarvi,
Finnish Lapland // Ornis fenn. 1980.. Vol. 57, N l.P. 17-25.
179
♦Jarvinen A. The breeding ecology of hole-nesting passerines in extreme northern condition //
Kilpisjarvi Notes. 1984. Vol. 8. P. 1-12.
♦Jarvinen A., Linden H. Timing of breeding and clutch size in Pied Flycatcher, Ficedula hypoleuca,
in Finnish Lapland // Ornis fenn. 1980. Vol. 57, N 3. P. 112-116.
Jarvinen O. Ecological zoogeography of Northern European bird communites. Helsinki, 1980. 108 p.
Jedicke E. Finnischer Trauerschnapper brutet in Nordhessen // Vogelk. Hefte. 1978. N 4. S. 123-124.
Johansson H. Kullstorlek och hackningframgang hos vissa holkhackande smafaglar i centrale
Sverige // Fauna och Flora. 1974. Bd 59, N 6. P. 212-218.
Jozefik M. Modyfikacja poludniowo-zachodnej granicy zasegu Erythrina e. erythrina (Pall.) na przes-
trzeni dwoch ostatnich stuleci // Acta Ornithol. 1960. Vol. 5. P. 307-324.
Kallander H. Advancement of laying of Great Tits by provision of food // Ibis. 1974. Vol. 116,
N3.P. 365-367.
♦ Kallander H. Breeding data for Pied Flycatcher, Ficedula hypoleuca, in southermost Sweden //
Ornis fenn. 1975. Vol. 52, N 3. P. 97-102.
Karlsson J., Nillsson S. G. The influence of nest-box area on clutch size in some hole-nesting passeri-
nes // Ibis. 1977. Vol. 119, N 2. P. 207-211.
Kellomaki E., Heinonen E., Tiainen H. Helmipollo pesinyt kahdesti samana kesana // Ornis fenn. 1977.
Vol. 54, N 3. P. 134-135.
Kendeigh S. C., Baldwin S. P. Factors affecting yearly abundance of passerines birds // Ecol. Monogr.
1937. Vol. 7. P. 91-123.
Klomp H. Over de achteruitgang van de Kievit (Vanellus vanellus) in Nederland en gegevens over het
legmechanisme en het eiproductie // Ardea. 1951. Vol. 39.
Klomp H. The determination of clutch-size in birds. A review // Ardea. 1970. Vol. 58, N 1-2.
P. 1-124.
♦Kluijver H. N. Waarnemingen over de levenswijze van den Spreenw (Sturnus vulgaris L.) met behulp
van geringde individuen // Ardea. 1935. Vol. 24, N 2. P. 133-166.
Kluijver H. N. The population ecology of Great Tit, Parus m. major L. // Ardea. 1951. Vol. 39, N 1.
P. 1-135.
Kluijver H. N. The determination of reproductive rate in Paridae // Proc. XIII Intern. Ornithol. congr.
Jthaca, 1963. P. 706-716.
Kluijver H. N. Regulation of a bird population // Ostrich. 1966. Suppl. 6. P. 389-396.
Koivunen P., Nyholm E. S., Sulkava S. Occurence and breeding of Little Bunting, Emberiza pusilla
in Kuusamo (NE Finland) // Ornis fenn. 1975. Vol. 52, N 3. P. 85-96.
♦Korpimaki E. Breeding biology of the Starling, Sturnus vulgaris, in western Finland // Ornis fenn.
1978. Vol. 55, N 3. P. 93-104.
Lack D. Clutch and brood size in the Robin // Brit. Birds. 1946. Vol. 39. P. 98-109, 130-135.
Lack D. The significance of clutch-size. Pts 1-2 // Ibis. 1947. Vol. 89, N 3. P. 302-352.
Lack D. Breeding seasons of European birds // Ibis. 1950. Vol. 92, N 2. P. 288-316.
* (Lack D.) Лэк Д. Численность животных и ее регуляция в природе. М., 1957. 403 с.
Lack D. Population studies of birds. Oxford, 1966. 341 p.
Lack D. Ecological adaptation in birds. London; Methuen, 1968. 409 p.
Lack D. Nesting conditions as a factor controlling breeding time in birds // Patternal Behavior Birds.
Stroudsburg, 1977. P. 31-36.
Lack D. The significance of the pair-bond and sexual selection in birds // Reading Sociobiol. San-Fran-
cisco, 1978. P. 328-341.
Lack D., Arn A. Die Bedeutung der Gelegegrosse beim Alpensegler // Ornithol. Beobachter. 1947.
Jg 44. P. 186-210.
Larsen V. H. En analyse af yngleresultafer hos mejser Paridae i Danmark // Dan. ornithol. foren.
tidsskr. 1974. Bd 68, N 1-2. S. 49-62.
Leinonen M. Comparisons between the breeding biology of year-old and older females of the White
Wagtail, Motacilla alba, in Central Finland // Ornis fenn. 1973. Vol. 50, N 3. P. 126-133.
Lind E. A. Zur Ethologie und Okologie der Mehlschwalbe, Delichon u. urbica (L.) // A. Zool. Soc.
„Vanamo”. 1960. Vol. 21. P. 1-123.
♦Lloyd J. A. Seasonal development in the incubation patch in Starling // Condor. 1965. Vol. 67,
N l.P. 67-72.
Loman J. Factors affecting clutch and brood size in the Crow, Corvus cornix// Oikos, 1977. Vol. 29,
N2.P. 294-301.
Louran H., Froissart Y. Statut de la Mesange alpestre dans la region a Briancon (Hautes-Alpes) //
Nos ois^ux. 1973. T. 32, N 349. P. 73-82.
♦Lohrl H. Populationsokologische Untersuchungen beim Halsbandschnapper (Ficedula albicollis) //
Bonn. Zool. Beitr. 1957. Bd 8. S. 130-177.
Lohrl H. Nachweis und Problematik von Zweitbruten // Vogelwarte. 1970. Bd 91, N 6. S. 223-230.
Lohrl H. Weitere Versuche zur Frage „Brutraum und Gelegegrojte” bei der kohlmeise, Parus major //
J. Ornithol. 1980. Vol. 121, N 4. P. 403-405.
Marshall A. J. Non-breeding among arctic birds // Ibis. 1952. Vol. 94, N 3 P. 310-333.
Marshall A. J. (Edit) Biology and comparative physiology of birds. New York; London, 1960.
180
Martin S. G. Adaptation for polygynous breeding in the Bobolink, Dolichonyx orizivorus // Amer.
Zool. 1974. Vol. 14, N 1. P. 109-119.
Meller W. Zur Brutbiologie der Rauchschwalbe (Hirundo rustica) in dem Dorf Edertal-Anraff //
Vogelk. Hefte. 1978. N 4. S. 62-82.
Merikallio E. Finnish birds. Their distribution and numbers. Helsinki, 1958. 181 p
*Meidel O. Life history of the Pied Flycatcher and Redstart in Norvegian mountain area // Nytt.
Magas. Zool. 1961. Vol. 10. P. 5-48.
Miller Ch. The Buccal food-carrying pouches of the Rosy Finch // Condor. 1941. Vol. 43. P. 72-73.
Mork K.Bigamy-og tokulli same sesSng-pavist hos fossekali // Sterna. 1975. Vol. 14, N 3. P. 131-134.
Muff J. Hoher Bruterfolg bei Rauchschwalben // Ornithol. Beobachter. 1979. Jg 76, N 2. S. 135-136.
Orell M., Ojanen M. Mortality rates of the Great Tit, Parus major in northern population // Ardea.
1979. Vol. 67, N 3-4. P. 130-133.
Orians G. M. On the evolution of mating system in birds and mammals // Amer. Natur. 1969. Vol.
103, N 934. P. 589-603.
Orians G. M. On the evolution of mating system in birds and mammals // Reading SociobioL San-Fran-
cisco, 1978. P. 115-132.
Owen D. F. The nesting success of the Heron, Ardea cinerea, in relation to the availability of food //
Proc. Zool. Soc. London. 1960. Vol. 133. P. 597-617.
Paatela J. Laatokan itarannikon linnustosta // Ornis. fenn. 1947. Vol. 24, N 3-4. P. 93-105.
Parkes K. S. The incubation patch in males of suborder Tyranni // Condor. 1953. Vol. 55, N 3. P.
218-219.
Peiponen V. A. Food and breeding of Scarlet rosefinch (Carpodacus e. erythrinus Pall.) in southern
Finland // Ann. Zool. Fenn. 1974. Vol. 11. N ?. P. 155-165.
Perrins С. M. Population fluctuation and clutch size in Great Tit, Parus major L // J. Anim. Ecol.
1965. Vol. 11, N 3. P. 601-647.
Perrins С. M. The timing of bird’s breeding seasons // Ibis. 1970. Vol. 112, N 2. P. 242- 255.
Perrins С. M. British Tits. London, 1979. 304 p.
Perrins С. M., Moos D. Survival of young Great Tit in relation to age female parent // Ibis. 1974.
Vol. 116, N 2. P. 220-224.
Pinowska B. The effect of energy and building resources of females on the production of House
Sparrow (Passer domesticus L.) population // Ecol. Polska. 1979. T. 27, N 3. S. 363-396.
Price F. E.. Воск С. E. Polygyny in Dipper // Condor. 1973. Vol. 75, N 4. P. 457.
Ptaci — Aves / Edit. Balat F. // Fauna CSSR. Praha, 1983a. 704 s.
Ptaci - Aves / Edit. Balat F. // Fauna CSSR. Praha, 1983b. 709-1034 s.
Pulliainen E. Habitat selection and clutch-size in the Meadow pipit, Anthus pratensis, in Finland,
especially in Lapland //Aquila. Ser. zool. 1977. Vol. 17, N 1. P. 1-16.
Radford D. J. Pair of Song Trushes with four broods in season // Bird Study. 1980. Vol. 27, N 2.
P. 121.
Raitasuo K. Havaintoa rytikerttusen (Acrocephalus scirpaceus) ja ruokokerttusen (Ac schoenobaenus)
pesimisvaiheesta // Ornis fenn. 1958. Vol. 35, N 1. P. 18-28.
Reinikainen A. Zur Brutbiologie des Karmingimpels// Ornis fenn. 1939. Vol. 16. N 1. P. 73-95.
Rheinwald G. Brutbiologie der Mehlchwalbe (Delichon urbica) im Bereich der Voreifel//Vogelwelt.
1979. Bdl00,N3.S. 85-107.
Ricklefs R. E. Relative growth, body constituents and energy content of nestling barn swallows
and blackbirds // Auk. 1967. Vol. 84, N 4. P. 560-570.
Ricklefs R. E. Clutch size in birds: Outcome of opposing predator and prey adaptations // Science.
1970. Vol. 168, N 3931. P. 599-600.
Ricklefs R. E. On the evolution of reproductive strategies in birds: reproductive effort // Amer.
Natur. 1977. Vol. Ill, N 979. P. 453-478.
Risberg E. L. Immigration of Scarlet Grosbeak, Carpodacus erythrinus, into Sweden and studies
of its breeding biology // Var Fagelvarld. 1970. Vol. 29, N 2. P. 77-89.
Risser A. Experimental modification of reproductive performance by density in captive Starlings //
Condor. 1975. Vol. 77, N 2. P. 125-132.
*Roostm‘aeL., Veroman H. Eesti Laulu-linnud. Tallin, 1974. 296 s.
*Schneider W. Der Star. Wittenberg Lutherstadt. 1972. 128 s (Die Neue Brehm-Biicherei, N 248).
Slagsvold T. Breeding time of birds in relation to latitude // Norw. J. Zool. 1975a. Vol. 23, N 2.
P. 213-218.
*Slagsvold T. Hypotheses on breeding time and clutch-size in birds // Norw. J. Zool. 1975b. Vol.
23, N 2.P. 219-222.
*Slagsvold T. Annual and geographical variations in the time of breeding of the Great Tit, Parus ma-
jor, and Pied Flycatcher, Ficedula hypoleuca, in relation to environmental phenology and spring tempera-
ture // Ornis Scand. 1976. Vol. 7. P. 127-145.
Snow D. W. The breeding of Blackbird Turdus merula at Oxford // Ibis. 1958. Vol. 100, N 2. P. 1 -30.
Snow D. W. How many eggs Blackbird lay? // Birds. 1967. Vol. 1, N 9. P. 181-183.
181
Solonen T. Population dynamics of the Garden Warbler, Sylvia borin, in southern Finland // Ornis
fenn. 1979. Vol. 56, N 1. P. 3- 12.
Southern H. N. Problems of Arctic bird life // Evolution, 1955. Vol. 9, N 1.
Stjernberg T. Breeding biology and population dynamics of the Scarlet Rosefinch Carpodacus erythri-
nus // Acta Zool. Fenn. 1979. N 157. P. 1-89.
Svensson S. Identification quid to European passerines / 2th ed. Stokholm, 1975. 184 p.
Svensson S. Population trends of common birds in Sweden // Pol. Ecol. Stud. (RRL). 1977. Vol. 3,
N 4. P. 207-213.
Svensson S. Identification quid to European passerines / 3th ed. Stockholm, 1978. 312 p.
Swanberg P. O. Om dubls kullar i fjallen norr om polcirkeln // Van F^gelvarld. 1955. Vol. 14, N 2.
P. 89-96.^
Sevcik I. Hnizdeni cvrcaly (Turdus iliacus L. 1766) na Trebonsku // Sb. Tiehiesk muz. Cesk. Bud^jov.
Prir. vedy. 1982. Roc. 22, Cis. 1. C. 89-96.
Tast J. Group nesting and breeding season of the Linnet, Carduelis cannabina, in Finland // Ornis
fenn. 1970. Vol. 47, N 2. P. 69-73.
Tenovuo R. Zur Brutzeitlichen Bioiogie der Nebelkrahe (Corvus corone cornix L.) im ausseren Schna-
lenhof Siidwestfinnlands // Ann. Zool. Soc.<£Vanamo>?. 1963. Vol. 25. P. 1—147.
* Tenovuo R., Lemmetyinen R. On the breeding ecology of the Starling, Sturnus vulagaris, in the
archipelago oi south-western Finland // Ornis fenn. 1970. Vol. 47, N 4. P. 159-166.
* Tompa F. S. Reproductive success in relation to breeding density in Pied Flycatcher, Ficedula hypo-
leuca (Pallas) // Acta Zool. Fenn. 1967. N 118. P. 1-28.
* Tyrvainen H. The breeding biology of the Redwing (Turdus iliacus L.) // Ann. Zool. Fenn. 1969.
Vol. 6, N 1. P. 1-46.
Verner J., Willson M. F. The influence of habitats on mating systems of North Americani Passerine
birds // Ecology. 1966. Vol. 47, N 1. P. 143-147.
Weatherhead P. J. Ecological correlates of monogamy in tundra-breeding Savannah Sparrows // Auk.
1979. Vol. 96, N 2. P. 391-401.
Wiehe H. Brutbiologische untersuchungen an Gelbspottern (Hippolais icterina) // Ornithol. Mitt.
1979. Bd 31, N 7. S. 151-155.
Williams G. C. Natural selection, the cost of reproduction and a refinement of Lack’s principle //
Amer. Natur. 1966. Vol. 100. P. 687-690.
Willis E. O., Wechsler D., Oniki Y. On behavior and nesting of McConnell’s flycatcher (Pipromorpha
macconelli): does female rejection lead to male promiscuity? // Auk. 1978. Vol. 95, N l.P. 1-8.
Winkel W. Vergleichend-brutbiologische Untersuchungen an fiinf Meisen-Arten (Parus spp.) in einem
niedersa’chsischen Aufforstungsgebiet mit Japanischer Larche, Larix leptolepis // Vogelwelt. 1975. Bd 96,
N 3. S. 104-114.
* Winkel W. Nachweis von Irigynie beim Trauerschnapper (Ficedula hypoleuca) // Vogelwelt. 1979.
Bd 100, N 4. S. 156-159.
*Winkel W., Richter D., Berndt R. Uber Beziehungen zwischen Farbtyp und Lebensalter mannlicher
Trauerschnapper (Ficedula hypoleuca)// Vogelwelt. 1970. Bd 91, N 4. S. 161-170.
Wynne-Edwards V. C. Animal dispersion in relation to social behavior.Edinburg; London, 1962. 653 p.
Zang H. ijber Zweit- und Drittbruten der Tannenmeise (Parus ater) // Vogelwelt. 1972. Bd 93, N 5.
S. 180-182.
Ziswiller V. Der Verdauungstrakt kornerfressender Singvbgel als taxonomischer Merkmalskomplex //
Rev. suisse zool. 1967. Bd 74, N 3. S. 620-628.
182
ОГЛАВЛЕНИЕ
Стр.
Введение............................... ............................................ 3
Глава I. Общие особенности распространения птиц на Северо-Западе СССР................... 6
Широтные изменения в составе растительности....................................... 7
Сезонность и неустойчивость климата............................................... ю
Специфическое влияние режима суточной освещенности............................... 15
Плотность гнездового населения птиц............................................ 16
Структура периферийных частей ареала........................................... 17
Изменение климата................................................................ 21
Глава II. Сроки размножения.......................................................... 26
Сезонность размножения и географическая изменчивость сроков гнездования . , ,.... 26
Фенологические особенности гнездового периода местных птиц....................... 29
Температурные пороги начала гнездования........................................ 37
Биотопические различия в сроках размножения................................. . 40
Возрастные различия в сроках размножения ...................................... 43
Окончание сезона размножения..................................................... 52
Связь сроков размножения птиц с сезонной цикличностью развития биогеоценозов... 62
Годовые циклы и особенности распространения птиц................................ 66
Глава III. Величина кладки и ее изменчивость......................................... 73
Внутри- и межвидовая изменчивость величины кладки............................... 74
Связь плодовитости с питанием . ................................................ 84
Географическая изменчивость величины кладки...................................... 87
Биотопическая изменчивость величины кладки....................................... 90
Годовые колебания размера кладки................................................. 93
Сезонная изменчивость величины кладки........................................ 96
Величина кладки и уровень развития репродуктивного состояния..................... 99
Плотность гнездового населения и величина кладки............................... 104
Влияние возраста птиц на величину кладки........................................ 106
Индивидуальная изменчивость величины кладки и выводка у меченых особей.......... . 108
Продуктивность кладок разной величины........................................... 111
Глава IV. Моно-и полициклическое размножение......................................... 121
Г л а в а V. Совмещение гнездования и линьки..................................... . 136
Успешность гнездования при совмещенной линьке................................... 142
Глава VI. Формы брачных отношений и роль полигинии в размножении таежных воробьиных
птиц........................................................................ 149
Глава VII. Преодоление негативных и использование позитивных факторов среды тицами,
обитающими на периферии ареала в северо-западной части СССР..................... 161
Литература.......................................................................... 170
183
НАУЧНОЕ ИЗДАНИЕ
Владимир Борисович Зимин
ЭКОЛОГИЯ ВОРОБЬИНЫХ ПТИЦ СЕВЕРО-ЗАПАДА СССР
Утверждено к печати
Институтом биологии Карельского филиала Академии наук СССР
Редактор издательства Л. В. Славошевская
Художник Г. В. Смирнов
Технический редактор Т, В. Шаврина
Корректор О. И. Буркова
ИБ №21522
Набор выполнен в издательстве на наборно-печатающем автомате. Подписано к печати 29.02.88.
М- 38061. Форма 70 X 100 Бумага офсетная № 1. Печать офсетная. Усл. печ. л. 14.95. Усл. кр.-от.
15.27. Уч.-изд.л. 16.24. Тираж 1100. Тип. зак. № 2100. Цена 2 р. 40 к.
Ордена Трудового Красного Знамени
издательство „Наука”. Ленинградское отделение
199034, Ленинград, В-34, Менделеевская лин., 1.
Ордена Трудового Красного Знамени
Первая типография издательства „Наука”.
199034, Ленинград, В-34, 9 линия, 12.
184
2 р. 40 к.
Издательство НАУКА»
Ленинградское отделение