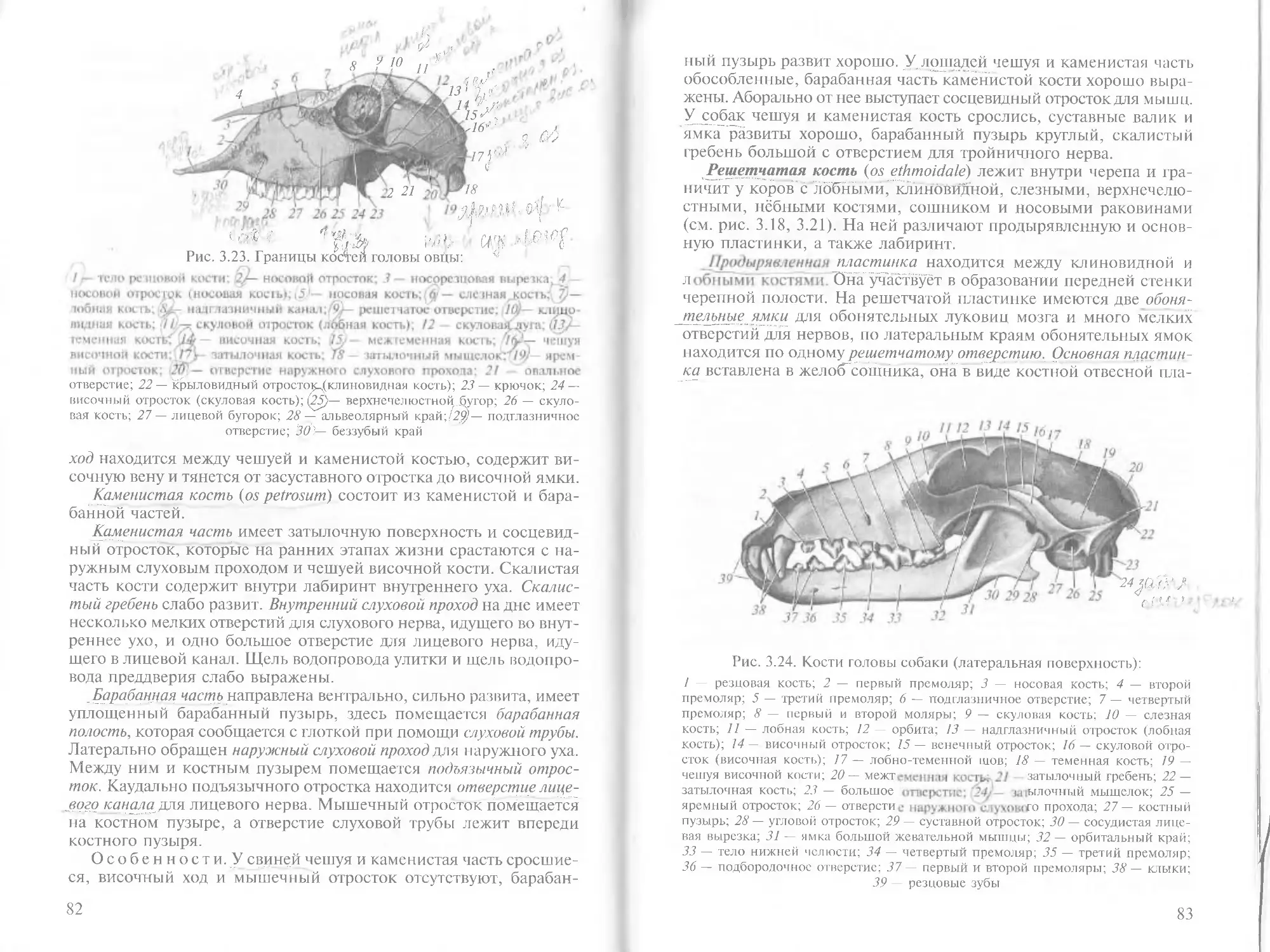
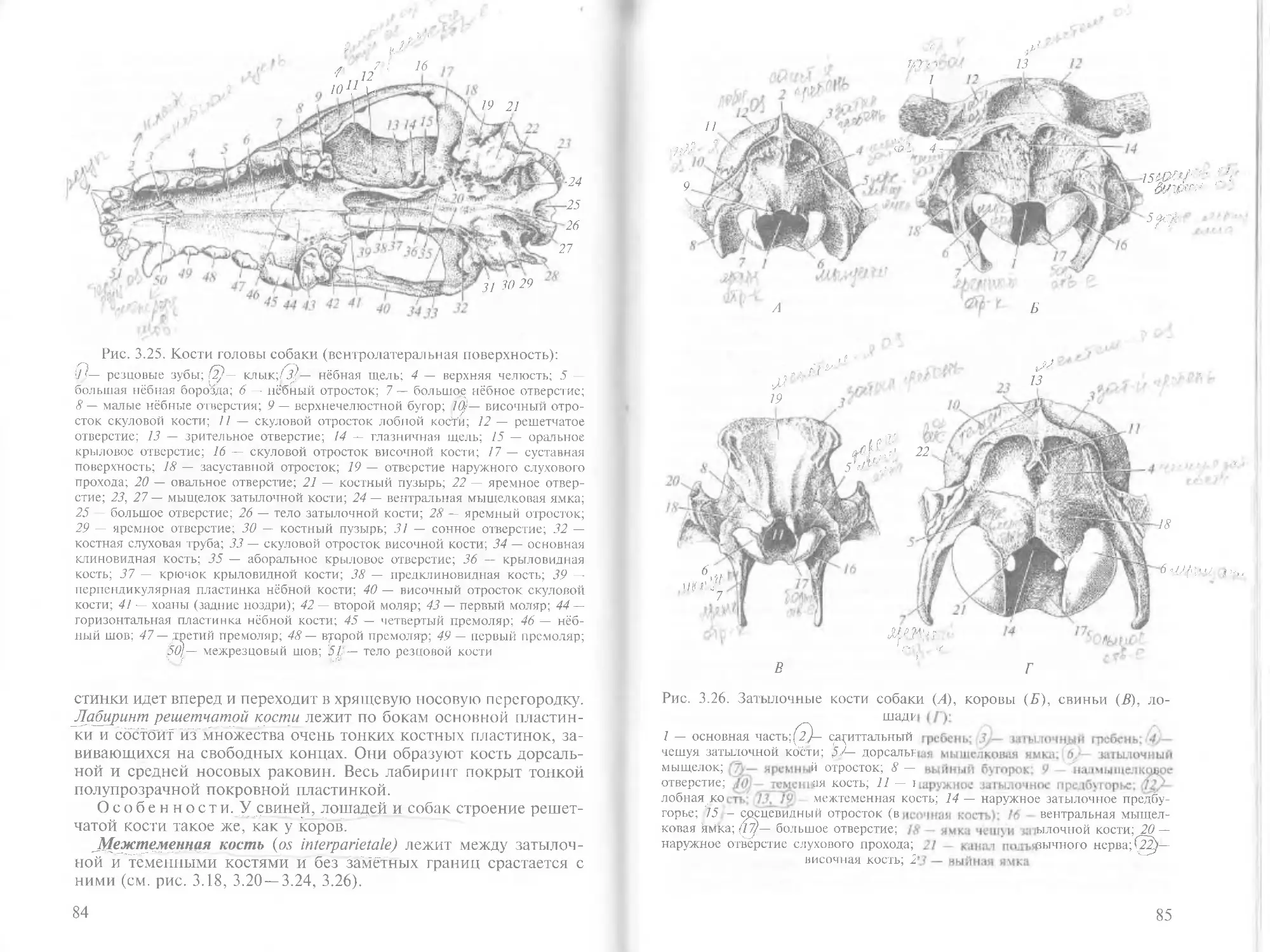
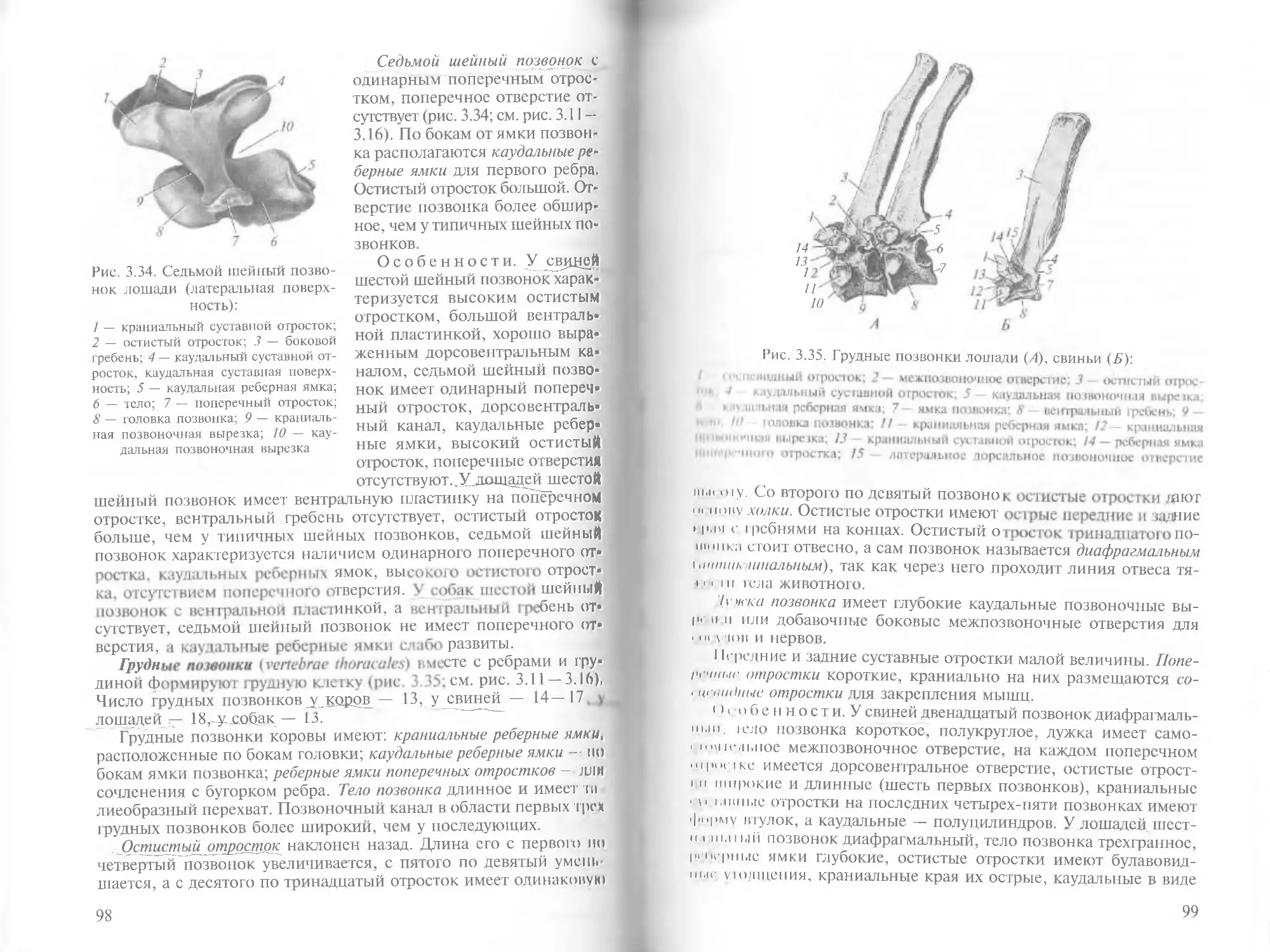
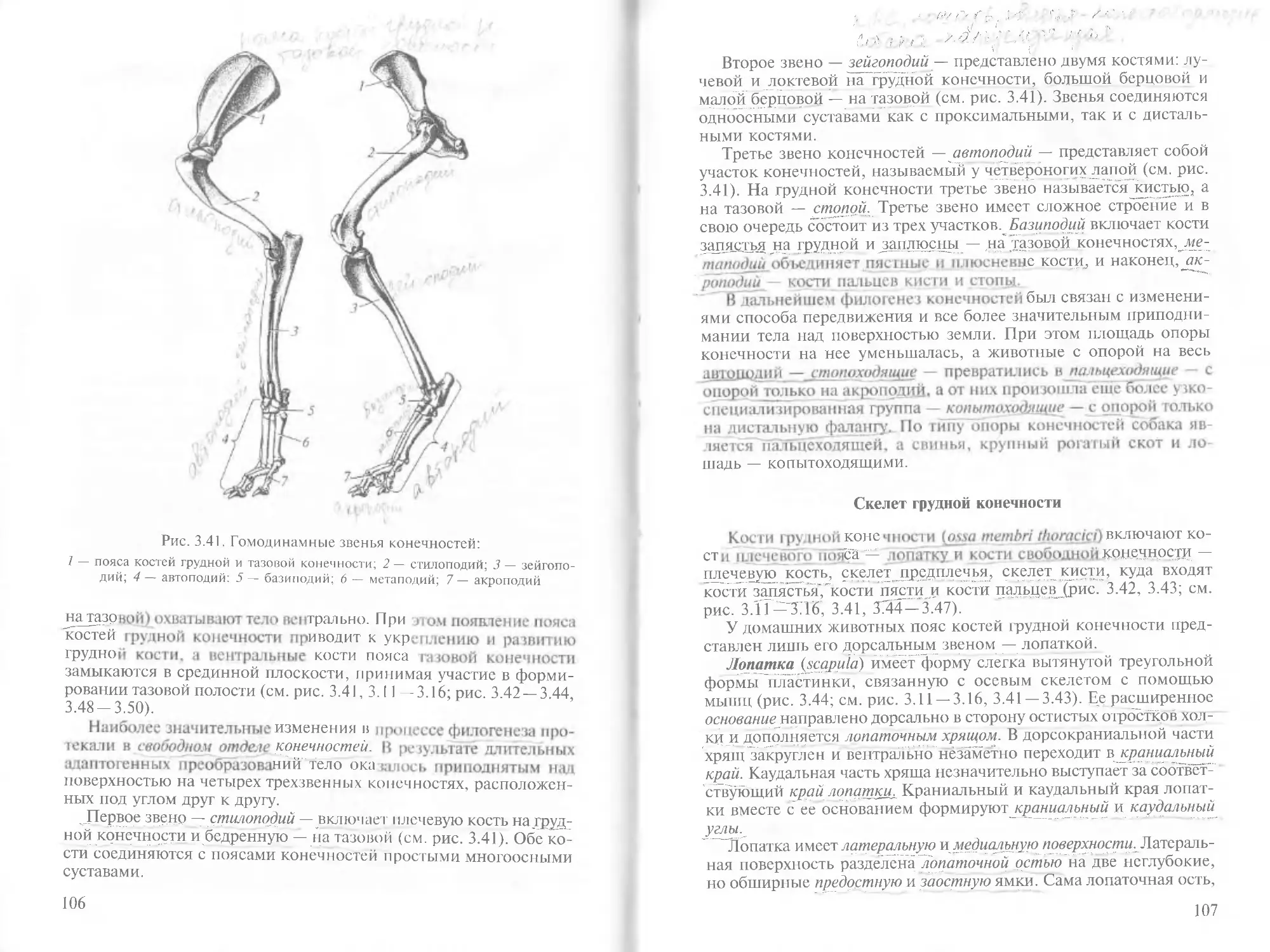
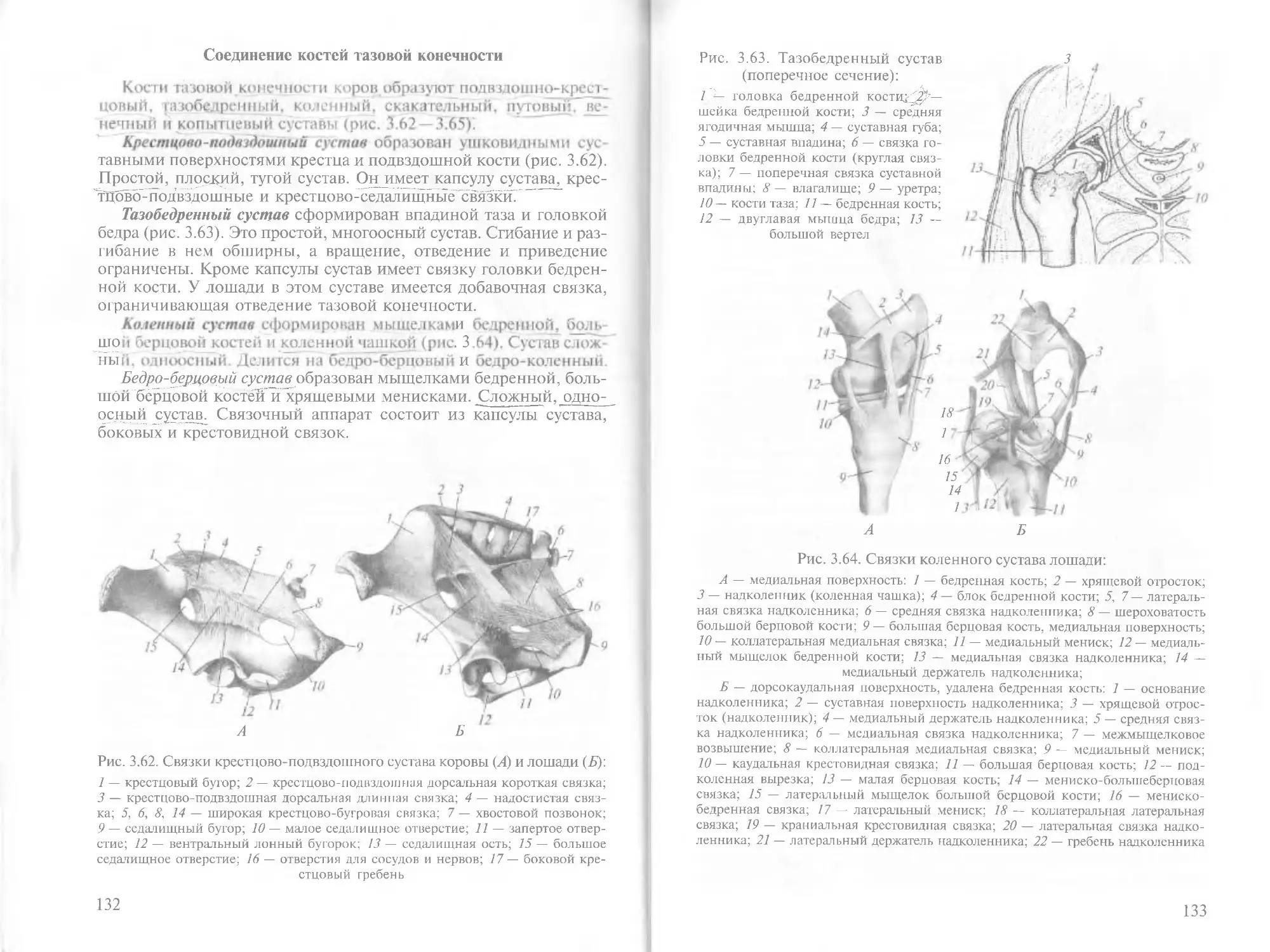
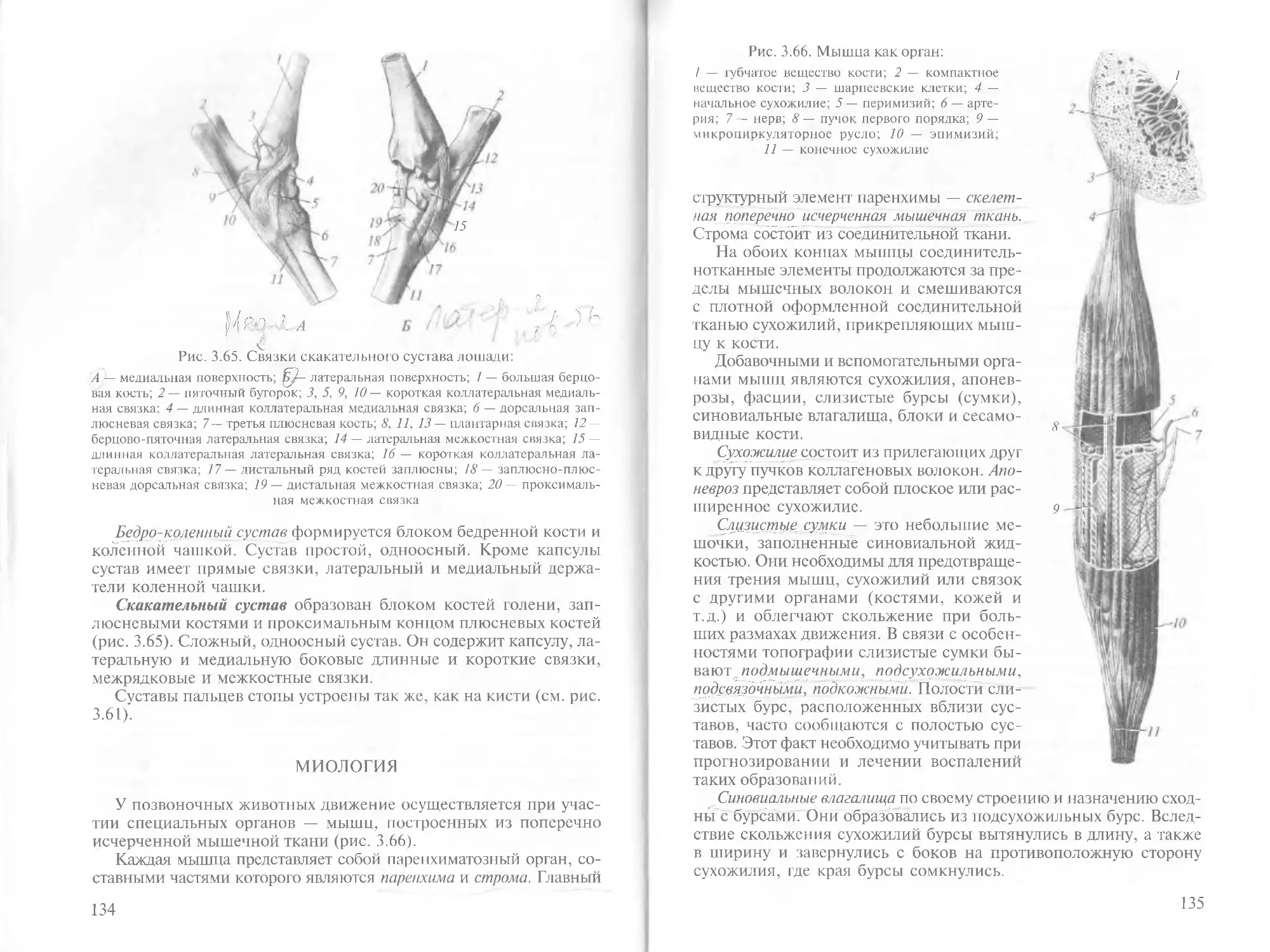
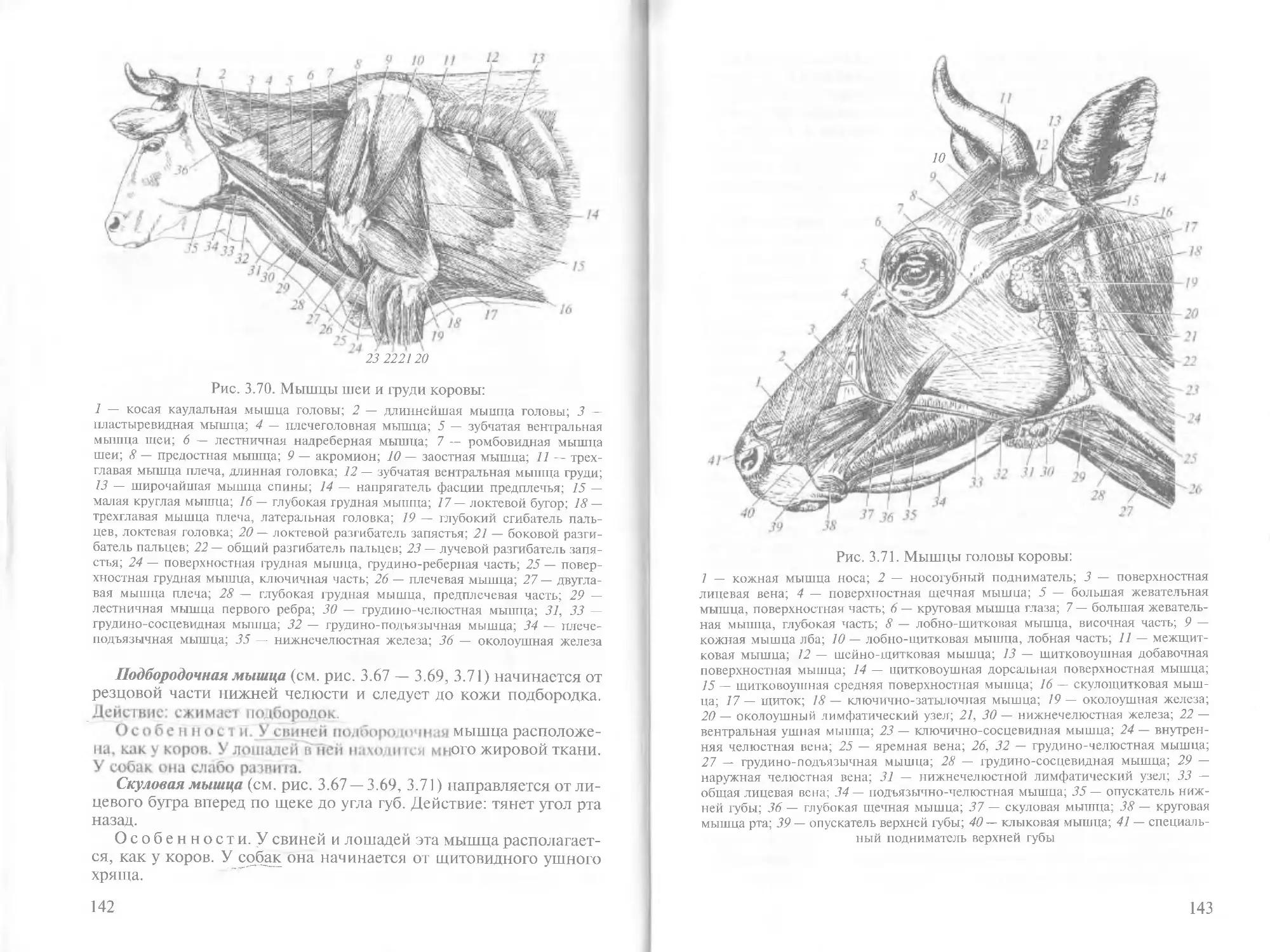
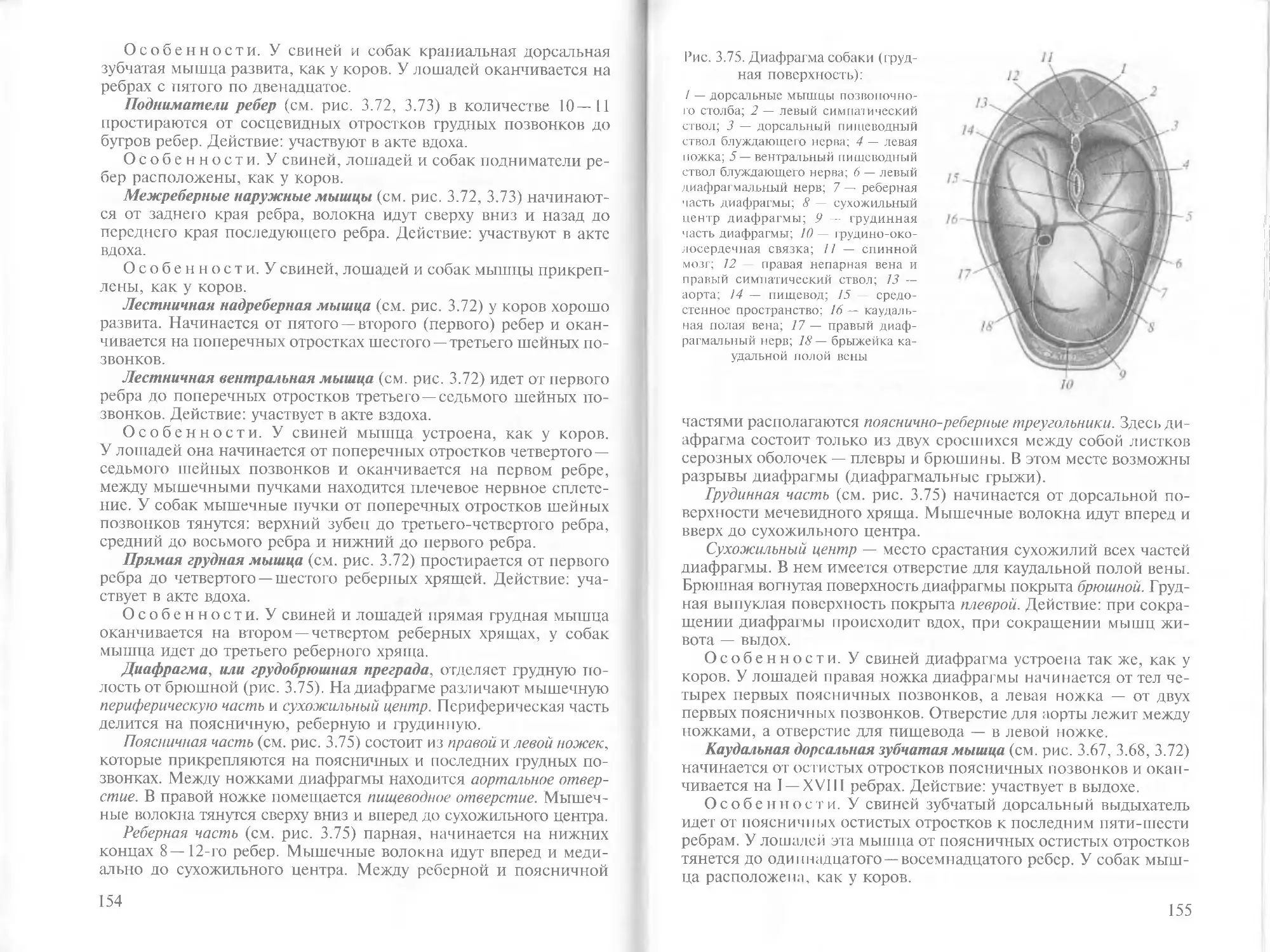
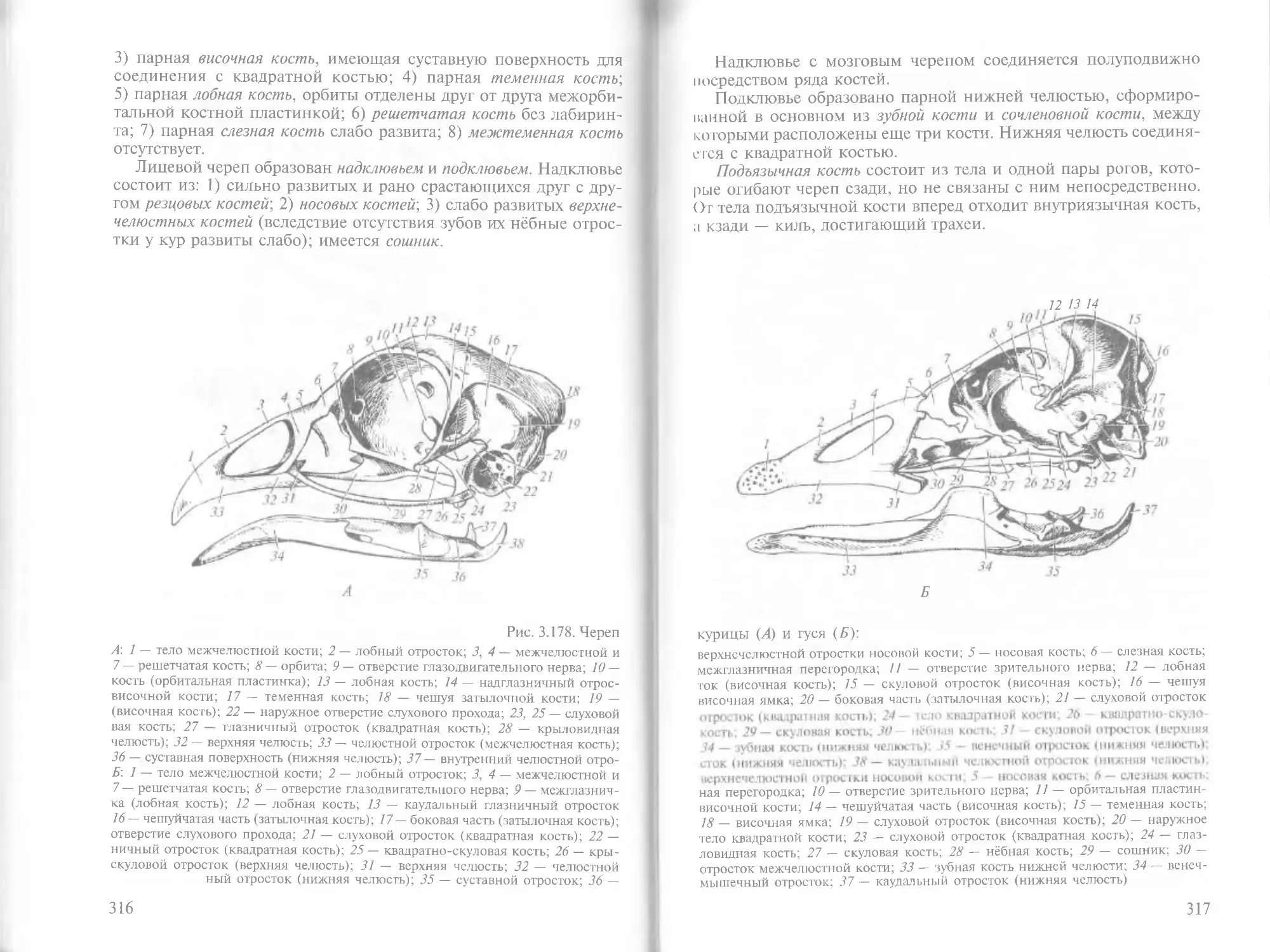
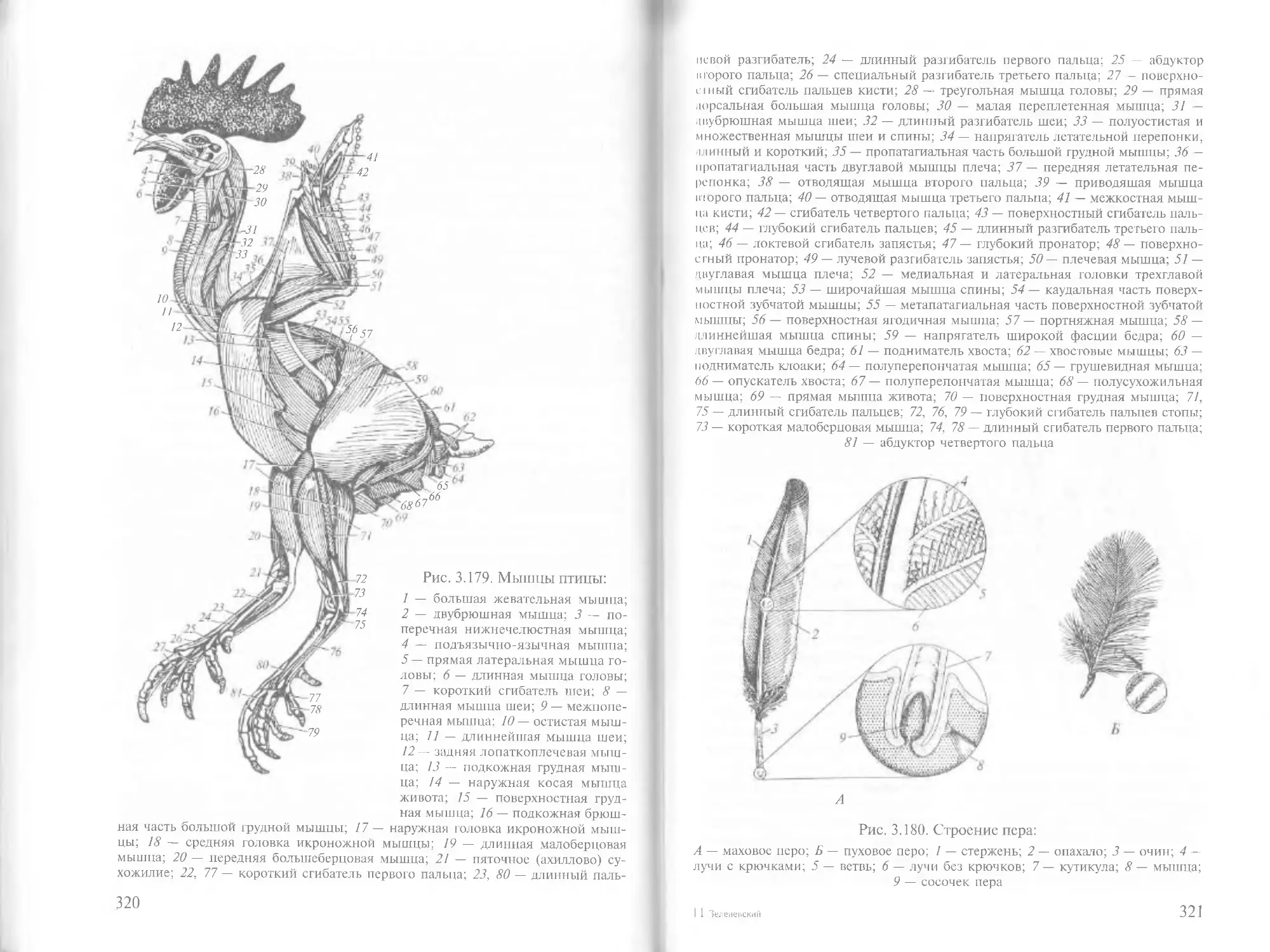
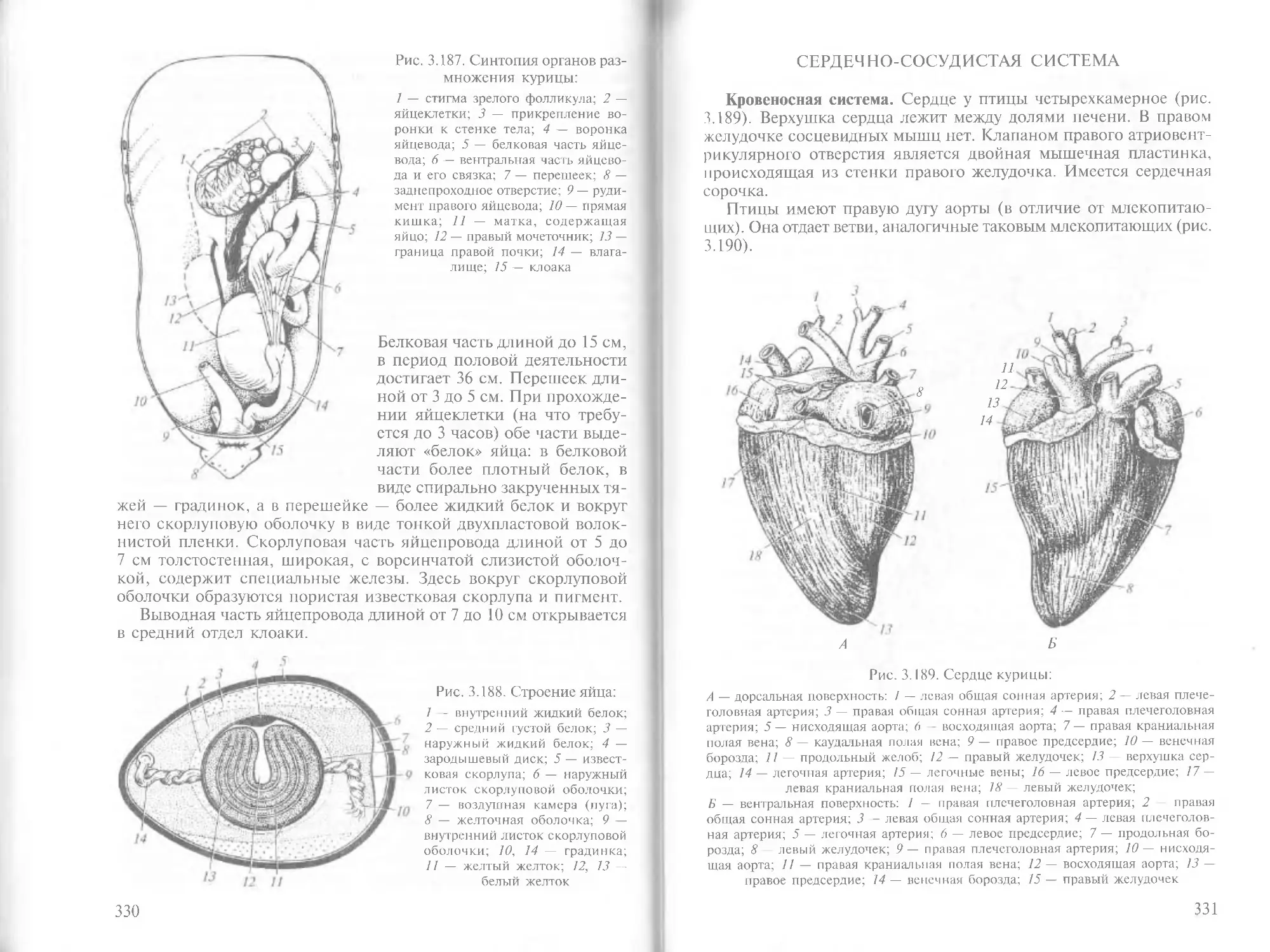
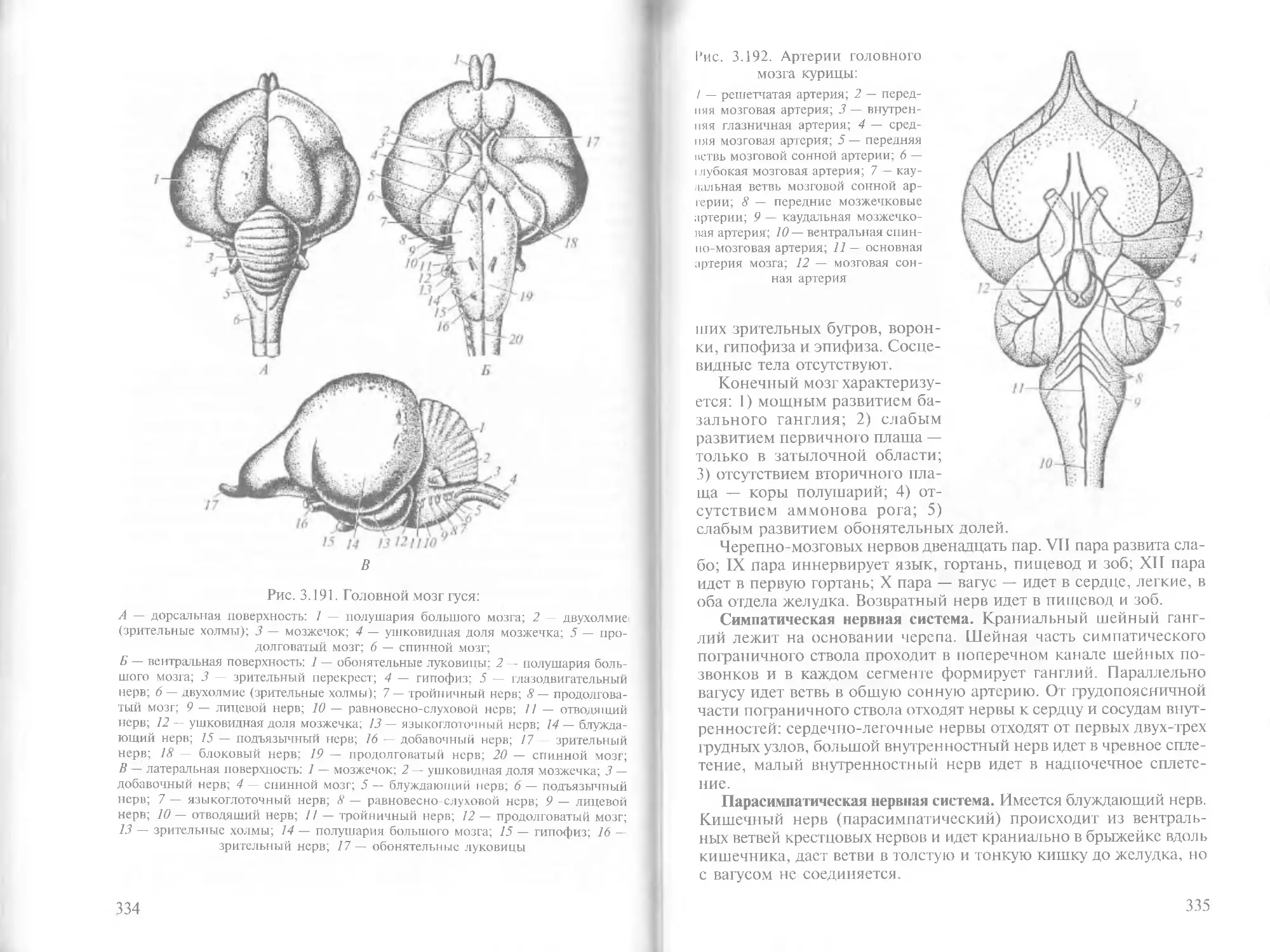
/




















































































































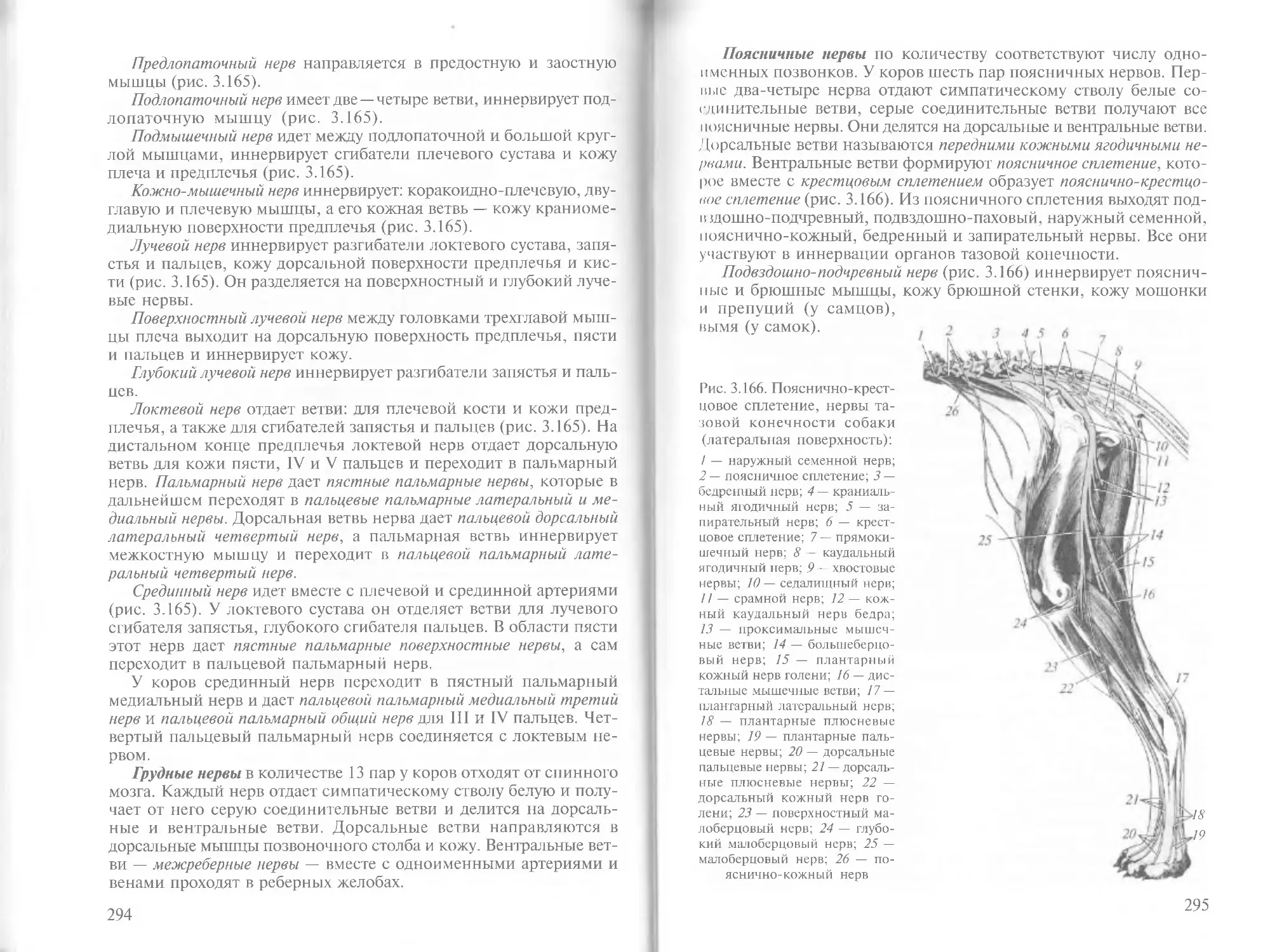

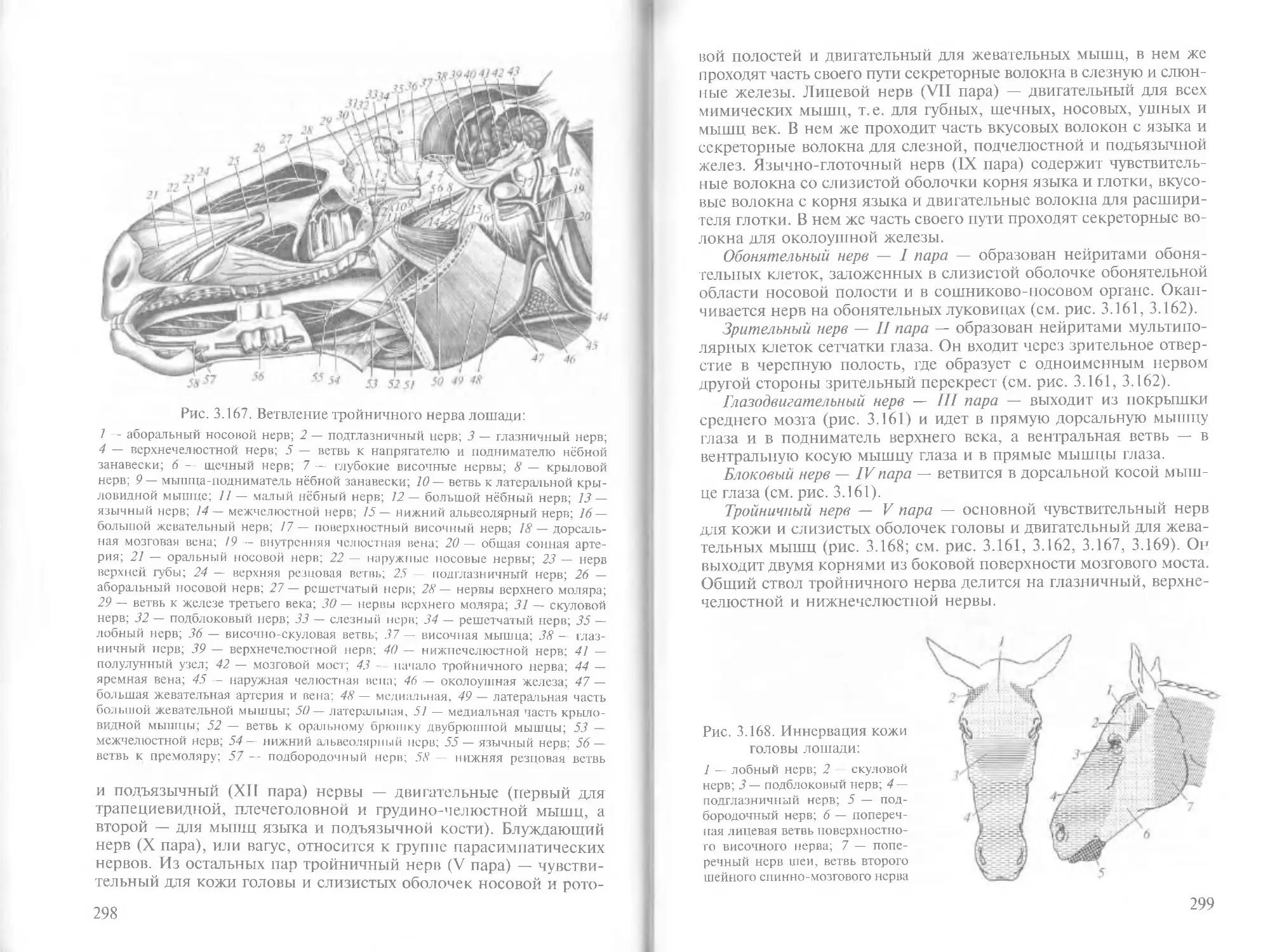




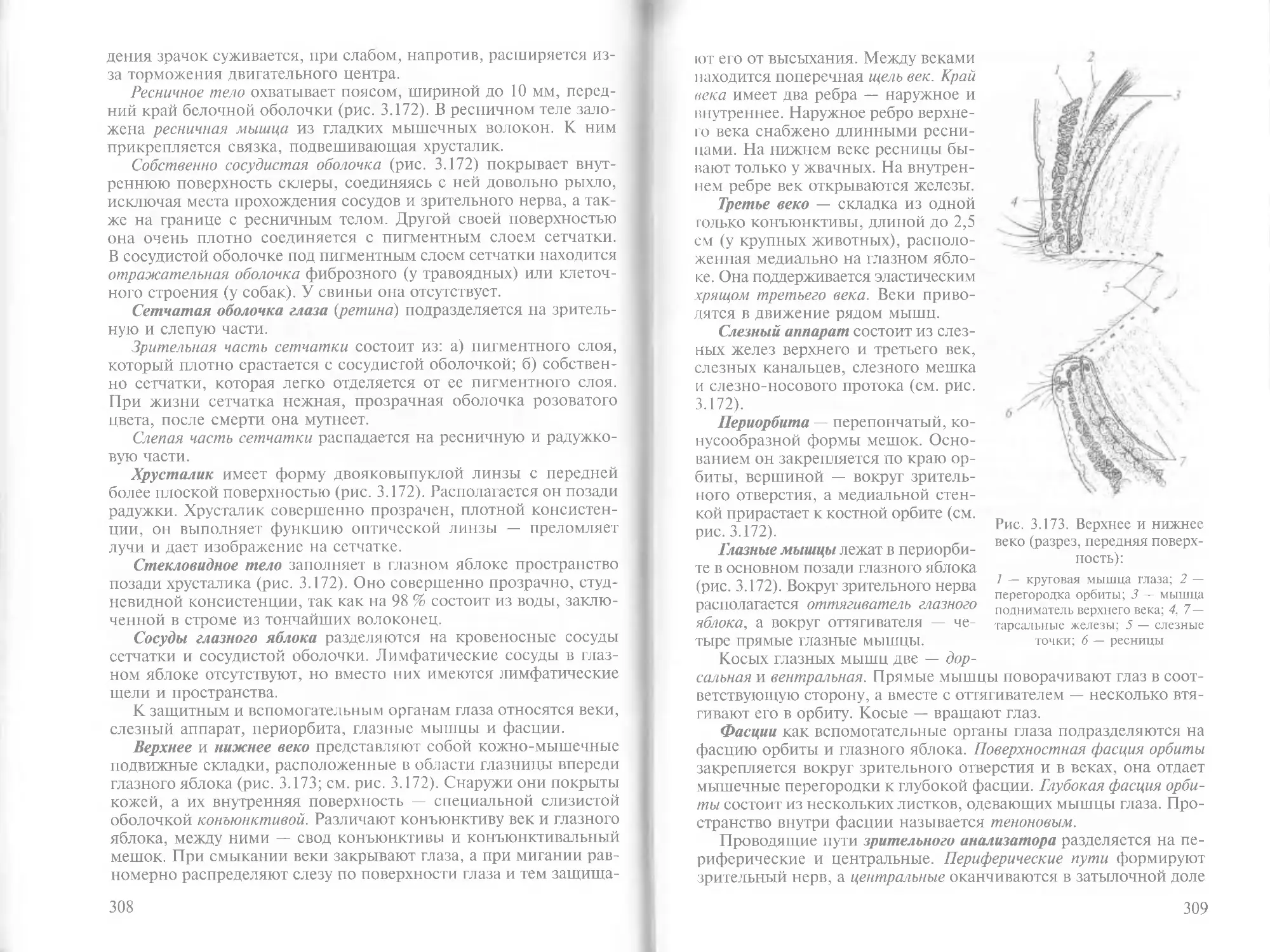


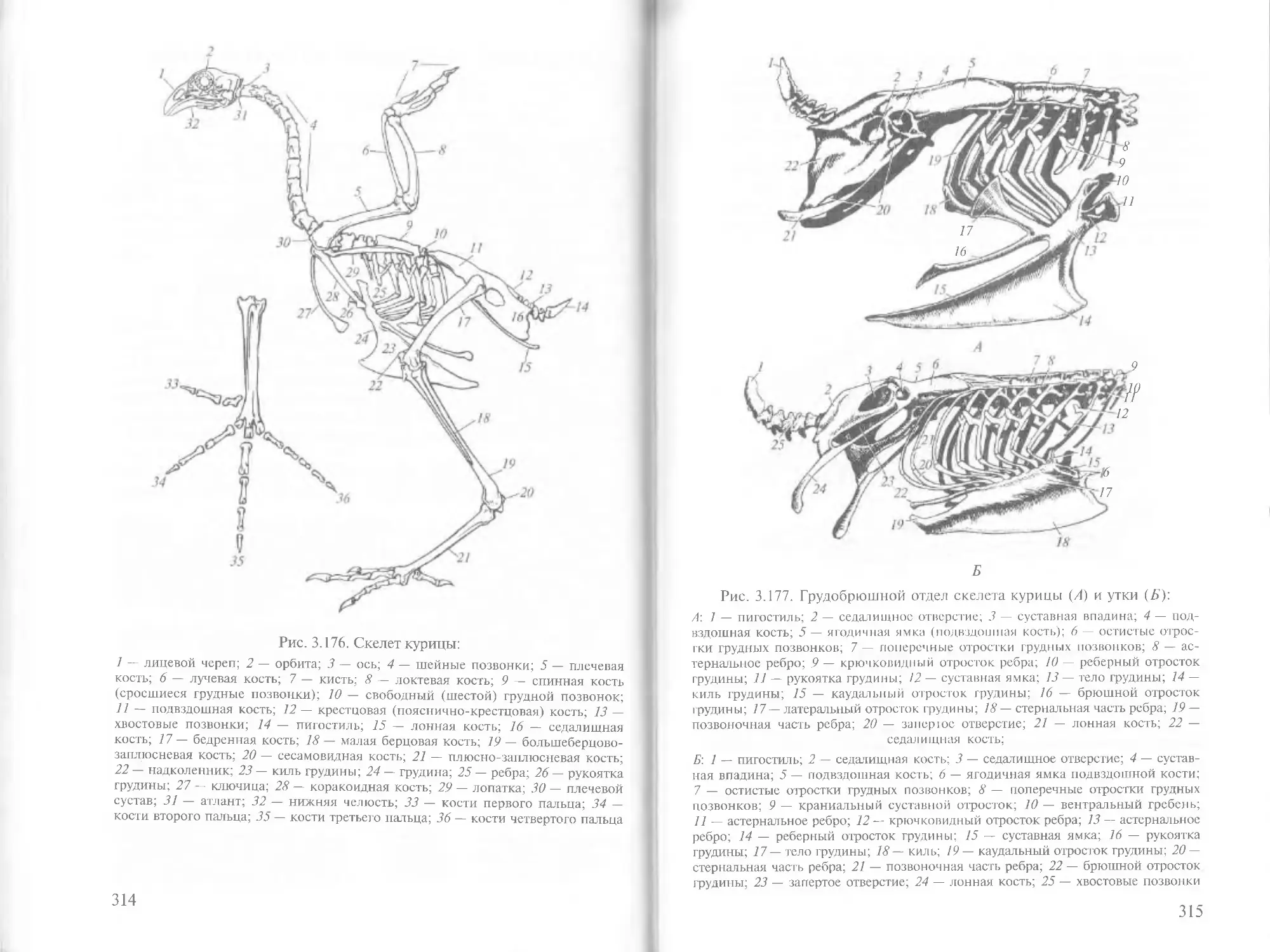



































































Author: Зеленевский Н.В.
Tags: морфология животных органология животных анатомия животных зоология физиология животных ветеринария
ISBN: 978-5-7695-5760-6
Year: 2009




Text
Н. В.Зеленевсний
А. П. Васильев
Л. К. Логинова
АНАТОМИЯ
И ФИЗИОЛОГИЯ
ЖИВОТНЫХ
2-е издание
СЕЛЬСКОЕ ХОЗЯЙСТВО
СРЕДНЕЕ ПРОФЕССИОНАЛЬНОЕ ОБРАЗОВАНИЕ
Н. В.ЗЕЛЕНЕВСКИЙ, А. П. ВАСИЛЬЕВ, Л. К. ЛОГИНОВА
АНАТОМИЯ
И ФИЗИОЛОГИЯ
ЖИВОТНЫХ
Допущено
Министерством образования Российской Федерации
в качестве учебника для студентов образовательных учреждений
среднего профессионального образования
2-е издание, исправленное
ACADEMA
Москва
Издательский центр "Академия»
2009
УДК 591.4:591.1(075.32)
ББК 28.66:28.673я723
3-48
Рецензенты:
зав. кафедрой ветеринарии Санкт-Петербургского аграрного университета,
д-р ветеринарных наук, проф. Г. И. Кожурина;
зам. директора по учебной работе, преподаватель ветеринарных дисциплин
Волоколамского аграрного техникума «Холмогорка» Е. Н. Букарева
Зеленевский Н. В.
3-48 Анатомия и физиология животных : учебник для студ.
образоват. учреждений сред. проф. образования / Н. В. Зеле-
невский, А.П. Васильев, Л.К.Логинова. — 2-е изд., испр. —
М. : Издательский центр «Академия», 2009. — 464 с.
ISBN 978-5-7695-5760-6
В учебнике изложены сведения по цитологии, гистологии, анатомии
и физиологии животных с учетом последних достижений отечественной
и зарубежной науки. Приведены русские и латинские термины, соответ-
ствующие четвертой редакции Международной номенклатуры (Москва,
2003).
Для студентов образовательных учреждений среднего профессиональ-
ного образования, обучающихся по специальностям «Ветеринария» и «Зоо-
техния».
УДК 591.4:591.1(075.32)
ББК 28.66:28.673я723
Оригинал-макет данного издания является собственностью
Издательского центра «Академия», воспроизведение его любым способом
без согласия правообладателя запрещается
© Зеленевский Н.В., Васильев А. П., Логинова Л. К., 2005
© Образовательно-издательский центр «Академия», 2005
ISBN 978-5-7695-5760-6 © Оформление. Издательский центр «Академия», 2005
ПРЕДИСЛОВИЕ
Сельскохозяйственное производство — наиболее обширная и
важная отрасль народного хозяйства, обеспечивающая промыш-
ленность сырьем, а людей пищей. Населению такой огромной стра-
ны, как Российская Федерация, для удовлетворения своих нужд
и поддержания здоровья нужно много полноценных продуктов
питания.
В настоящее время достаточно остро стоит вопрос не только о
повышении полноценности и количества продуктов питания, ка-
чества животного сырья, но и об их безопасности. Нарушение эко-
ногии, вызванное антропогенным воздействием на окружающую
среду, химизация сельскохозяйственного и промышленного про-
изводства, все более широкое и подчас неконтролируемое выра-
щивание и использование генетически измененных сельскохозяй-
ственных культур ставят перед учеными, медицинскими и вете-
ринарными работниками, практическими животноводами задачи
по гарантированию экологической и химической чистоты того,
что используется в пищу людям и на корм животным. Кроме того,
ветеринарные специалисты должны со своей стороны нести от-
ветственность за успешную профилактику и лечение уже извест-
ных и ряда вновь появляющихся заболеваний, многие из которых
свойственны как животным, так и человеку.
Решить поставленные задачи смогут молодые специалисты, не
только отличающиеся широкой эрудицией и высоким професси-
онализмом, но и адаптированные к новым рыночным условиям
груда в сельском хозяйстве. В связи с этим активное освоение и
развитие научных знаний о строении и физиологии животных, о
питательных веществах, необходимых им для нормальной жизне-
деятельности и продуктивности, а также для поддержания здоро-
вья, позволяют разрабатывать более рациональные методы разве-
дения, кормления и содержания животных, проводить целена-
правленную селекционную работу, зоотехнические и ветеринар-
но-профилактические мероприятия.
Для того чтобы постичь живой организм, научиться управлять
ими помогать ему, нужно в первую очередь разобраться в самом
гпавном — как этот живой организм устроен и как он функцио-
нирует. Необходимая для этого информация собрана в таких био-
чогических дисциплинах, как морфология, физиология и биохи-
мия.
3
Предлагаемый учебник ставит своей задачей по возможности
подробно познакомить студентов с первыми двумя из них.
Учащимся нужно помнить, что, знакомясь с двумя частными
сторонами весьма сложной и многосторонней организации живо-
го организма, нельзя упускать из виду главный закон природы —
признание этого организма единым и неделимым целым.
ГЛАВА 1
МОРФОЛОГИЯ И ФИЗИОЛОГИЯ
(ЗНАЧЕНИЕ, РАЗВИТИЕ, СВЯЗЬ
С ДРУГИМИ БИОЛОГИЧЕСКИМИ НАУКАМИ)
Морфология (morphe — форма, logos — учение) — это наука о
происхождении, развитии, формах и строении живого организма.
Современная морфология животных включает три главные дис-
циплины: цитологию, гистологию и анатомию, позволяющие про-
водить изучение и описание строения тела животного во всей его
полноте на разных уровнях расчлененности.
Цитология (cytos — клетка, logos — учение) — наука о наибо-
лее мелкой частице живой материи — клетке: ее значении, стро-
ении и функционировании. Учение о клетке и ее жизнедеятельно-
сти известно в биологии под названием клеточной теории, нача-
ло которой относится к первым исследованиям английского фи-
зика Роберта Гука, проведенным с помощью созданного им в
1665 г. микроскопа (рис. 1.1). Клеточная теория, сформулирован-
ная в 1839 г. немецким биологом Т. Шванном, наряду с теорией
Рис. 1.1. Микроскоп Роберта Гука (А) и сделанный им рисунок микро-
скопического строения пробки (£)
эволюции Ч. Дарвина и теорией гена является одной из осново-
полагающих теорий современной биологии.
На определенном этапе накопления информации из цитологии
выделилась в самостоятельный раздел эмбриология (embryon — за-
родыш, logos — учение). Она изучает индивидуальное развитие
живого организма от момента слияния половых клеток до рожде-
ния и вместе с возрастной анатомией, исследующей превращения
организма после рождения, определяет полную картину онтогене-
за (ontos — существо, genesis — развитие), т.е. комплекс сложных
преобразований организма от зарождения до естественной смерти.
«Зернышки» индивидуальных программ развития, или онтоге-
неза, представителей определенного вида животных от простей-
ших форм до современных, прослеженные на протяжении огром-
ного числа поколений, последовательно сменяющих друг друга,
составляют мозаику исторического {эволюционного) развития дан-
ного вида животных, или филогенез (phylon — племя, род, genesis —
развитие). Филогенез предусматривает сведение воедино данных
палеонтологии (palaios — древний, ontos — существо), т.е. учения
об ископаемых животных, сравнительной анатомии, гистологии
и эмбриологии.
Гистология (histos — ткань, logos — учение) как морфологи-
ческая наука изучает ткани (эпителиальные, опорно-трофические,
мышечные и нервную), т. е. особые сложные комплексы клеток и
неклеточного вещества, отличающиеся общностью происхожде-
ния, строения и функции.
Цитология и гистология относятся к микроанатомии (mikros —
малый), так как изучение отдельных клеток возможно только при
значительном увеличении и после специальной обработки. Для
изучения внутреннего устройства клеток используют световой,
электронный, сканирующий электронный и другие микроскопы,
обеспечивающие многократное увеличение рассматриваемого
объекта.
Анатомия (anatome — разрез, рассечение) — одна из самых
древних биологических наук. Она изучает закономерности внешних
форм и внутреннее устройство здорового, не измененного болез-
нью организма в целом, а также отдельных его органов, систем
органов и отдельных его частей. Обычно объектом исследования
анатома служат трупы животных, но сведения, получаемые на
мертвых телах, дают право и возможность судить о закономерно-
стях строения живого организма.
Основным приемом работы анатома-исследователя является
последовательное расчленение скальпелем более крупного и слож-
ного комплекса структур на более мелкие и простые, т.е. «анато-
мирование», или «препарирование». При этом проводится под-
робное описание увиденного невооруженным глазом или с помо-
щью лупы (описательная анатомия).
6
По мере расширения и углубления данных о строении орга-
низма в составе анатомии оформились в самостоятельные разде-
лы: 1) системная анатомия, описывающая строение систем ор-
ганов — костной, мышечной, внутренних органов, кровеносной,
нервной; 2) топографическая анатомия, содержащая сведения о
естественной форме органов, их взаимной связи и расположении
в теле; 3) рентгеноанатомия, дающая возможность изучать и опи-
сывать скелет, форму и расположение внутренних органов и сосу-
дов на живом животном; 4) пластическая анатомия, изучающая
внешние формы тела животного с определением соотношения или
пропорций отдельных его частей; 5) сравнительная анатомия,
выявляющая общие черты и различия в строении и форме сход-
ных органов и организмов животных разных классов от низших
до высших; она стала надежной теоретической основой для оп-
ределения и правильного понимания общих закономерностей эво-
люции животных в их индивидуальном (онтогенез) и истори-
ческом (филогенез) развитии; 6) возрастная анатомия, объеди-
няющая сведения об изменениях размеров и строения органов на
протяжении всей жизни отдельной особи от момента рождения
до смерти.
Физиология (physis — рождение, происхождение, logos — уче-
ние) — наука о процессах жизнедеятельности организма, а так-
же механизмах, обеспечивающих взаимодействие отдельных ча-
стей организма и функционирование его как единой целостной
и самонастраивающейся системы в условиях обмена веществ и
энергии с окружающей средой. Физиология является второй по
объему и значимости после морфологии биологической наукой.
На всем пути развития физиологии сложилась ее тесная взаимо-
связь с морфологическими науками, при которой функция и
форма бессмысленны одна без другой. Область интересов физио-
логии включает раскрытие и изучение тонких механизмов таких
актов, как дыхание, питание, движение, поддержание опреде-
ленного уровня обмена веществ и др. Важная сторона физиоло-
гических исследований — выявление качественных различий в
функционировании организмов животных, находящихся на раз-
ных ступенях эволюционной лестницы, и организмов одного
вида, живущих в разных условиях обитания (сравнительная фи-
зиология).
В настоящее время в физиологии принято выделять несколько
направлений исследования: общее, видовое, возрастное, эволю-
ционное и т. д. Сравнительно новой областью физиологии являет-
ся экологическая физиология, рассматривающая потенции, харак-
тер и скорость функциональных преобразований при адаптации
животного организма к условиям постоянно ухудшающейся окру-
жающей среды и разведении животных в экологически неблаго-
получных регионах.
7
Основной метод физиологических исследований — экспери-
мент (опыт), т.е. воздействие на организм животного посредством
специальных приборов и установок, дающих возможность стиму-
лировать или, наоборот, подавлять те или иные функции и впос-
ледствии наблюдать и регистрировать возникающие ответные ре-
акции (например, изменение двигательной активности, секретор-
ной деятельности и т.п.).
Глубокое проникновение анатомии, гистологии и физиологии
в онто- и филогенез, в закономерности строения и функциони-
рования органов, тканей и клеток дает возможность биологичес-
ким дисциплинам внести существенный вклад в познание живого
мира и определение картины развития жизни на Земле.
Эти науки имеют и вполне ощутимое прикладное использова-
ние. Не вызывает сомнения значимость анатомии и физиологии
для ветеринарного и зоотехнического образования. Осознанное и
кропотливое изучение всех аспектов строения и функционирова-
ния организма животного является необходимой базой для фор-
мирования врачебного мышления, дает почву для более глубоко-
го понимания всех клинических дисциплин, решающих пробле-
мы профилактики и лечения животных. Без знания топографичес-
кой анатомии нельзя стать хорошим хирургом или терапевтом. Без
понимания тонкого устройства и «профессиональной деятельно-
сти» органов размножения, а также знания всех сторон эмбрио-
логии нет акушера-гинеколога. Недостаток подготовки по нормаль-
ной анатомии, гистологии и физиологии делает ветеринарного
специалиста безоружным в патологической анатомии и патологи-
ческой физиологии при определении тех или иных изменений в
функциональной деятельности организма и в органах, поражен-
ных болезнью.
Краткие сведения по истории морфологии и физиологии
сельскохозяйственных животных
Анатомия как наука о строении тела человека и животного является
одной из древнейших биологических наук. За много миллионов лет она
прошла путь от примитивных наскальных рисунков первобытных охот-
ников до изучения молекулярного строения клетки.
Развитие анатомии происходило неравномерно и тесно увязывалось
как с историей развития общества, так и с появлением и развитием
наук, обслуживающих это общество, в том числе естественных и осо-
бенно медицины. Точное время возникновения анатомии определить прак-
тически невозможно. Вполне логично допустить, что первые анатоми-
ческие «уроки» о наличии, размещении и о жизненной важности орга-
нов получали древние люди в первобытном обществе более чем за 20 тыс.
лет до н. э.
Накопленные в первобытном обществе отрывочные сведения о строе-
нии тела и основных проявлениях жизнедеятёл ьности животных значитель-
8
но расширились и начали систематизироваться с переходом к скотоводству
в развитых рабовладельческих государствах Древнего Востока — Китае,
Индии, Египте и др. О высоком уровне этих знаний свидетельствуют в пер-
вую очередь успехи, достигнутые в Древнем Египте в связи с культом баль-
замирования трупов и изготовления мумий человека и животных.
Особое развитие анатомия получила в античной Греции. Большин-
ство ученых Древней Греции отличались широтой кругозора, богатством
и разнообразием интересов. Преодолевая влияние религиозных воззре-
ний о божественной предопределенности мира и человека в нем, живя в
условиях строгого запрета на вскрытие трупов людей, Гиппократ (460 —
377 гг. до н. э.), Аристотель (384 — 322 гг. до н.э.), Демокрит и другие
ученые занимались рассечением и изучением трупов животных, чем
существенно продвинули учение о строении тела животных. Выдаю-
щимся энциклопедистом своего времени был римский врач Клавдий
Гален (131 — 201 гг. до н.э.).
В конце первого и начале второго тысячелетия нашей эры центром
научной мысли становится Восток. История сохранила нам труд вели-
чайшего мыслителя и врача Абу Али Хусейн Ибн-Абдаллах Ибн-Сины
(980—1037), известного больше под именем Авиценны, — «Канон вра-
чебной науки», в котором он обобщил имевшиеся на тот момент сведе-
ния по диагностике и лечению основных заболеваний. Значительное ме-
сто в этой книге уделено анатомии.
В эпоху феодализма (V—XV вв.) в идеологии европейских стран уста-
новилось господство богословия, в первую очередь католицизма, а ос-
новой экономики стало натуральное хозяйство. Это привело к упадку
всех прогрессивных наук. И анатомические сведения, хотя и продолжали
понемногу накапливаться, не систематизировались. Новый прогрессив-
ный период в развитии человечества, названный эпохой Возрождения,
начался в конце XV в. В этот период активно развиваются все виды искус-
ства и науки. Открываются медицинские школы и первые университеты.
Пышный расцвет пытливой мысли захватил и анатомию. Перевод ее
на научную основу связан с двумя главными именами — Леонардо да
Винчи (1452—1519) и Андреем Везалием (1514—1564). Итальянский ху-
дожник Леонардо да Винчи — человек фантастической одаренности и
многогранности интересов — явился первым иллюстратором анатомии,
чьи рисунки отличались реалистичностью. Эти рисунки послужили осно-
вой для развития пластической анатомии. Профессор Падуанского уни-
верситета Андрей Везалий, врач, мыслитель и художник, прославился
своим монументальным 8-томным трудом «О строении человеческого
тела», в котором выявлены существенные отличия в строении органов
человека и животных. Везалий по праву считается «отцом» описательной
и сравнительной анатомии. Для работы с трупами и публичного их вскры-
тия в 1594 г. в Падуе и в 1637 г. в Болонье были созданы анатомические
театры. Извлеченные органы стали подвергать особой обработке, позво-
ляющей сохранять их длительное время. Голландский анатом Р. Рюиш
(1638—1731) усовершенствовал метод бальзамирования и создал боль-
шую коллекцию анатомических препаратов, в том числе новорожденных
уродов. Часть этой коллекции, купленная Петром I, хранится до сих пор
в Кунсткамере Санкт-Петербурга.
9
Развитие сравнительной анатомии связано с именами К. Линнея
(1707-1778), П.С.Палласа (1741-1811), Ж.Кювье (1769-1832). Пос-
ледний считается основателем палеонтологии. Несмотря на все достиже-
ния биологической науки, до середины XIX в. большинство получаемых
результатов интерпретировалось исходя из представления, что весь мир
создан Богом однажды и все в нем неизменно, в том числе животные и
человек. Изменение этих воззрений и возникновение эволюционной ана-
томии во многом подготовили Ж.-Б. Ламарк, изложивший свои взгляды
в книге «Философия зоологии», и особенно английский ученый-есте-
ствоиспытатель Ч.Дарвин (1809—1882), который считал, что животные
появились на Земле естественным путем в виде очень простых форм и в
течение тысячелетий постепенно изменялись и совершенствовались до
современных форм.
Таким образом, в XVII —XIX вв. анатомия получила научную направ-
ленность и оформилась в самостоятельную науку, которая дифференци-
ровалась на ряд более конкретных дисциплин. В это же время было поло-
жено начало физиологии, эмбриологии и микроскопической анатомии.
Развитие анатомии домашних животных в России первоначально было
связано с практическими потребностями скотоводства и коневодства,
которыми в XV—XVII вв. занимались монахи в монастырях, превратив
их в прибыльную отрасль. В большинстве своем «коневые» и «скотские»
лекари были практикующими самоучками. Только с переходом к капи-
тализму во второй половине XVIII в. в связи со значительным оживлени-
ем промышленности, сельского хозяйства и торговли увеличилась по-
требность в научных знаниях.
В 70-х гг. XVIII в. на медицинских факультетах Петербургского, Мос-
ковского, Киевского, Харьковского и Казанского университетов начали
готовить первых ветеринарных врачей (скотоврачей). Руководство «Крат-
кое начертание анатомии домашних животных», написанное прозекто-
ром кафедры анатомии при Московском университете И. С. Андреевским
(1759 — 1808), является первым российским учебником по анатомии жи-
вотных, а дата его выхода в свет (1804) считается официальной датой
рождения в России анатомии домашних животных как науки. Наиболее
выдающимся представителем ветеринарной науки XIX в. был академик
ветеринарного отделения Петербургской медико-хирургической акаде-
мии В.И.Всеволодов (1790—1863). Этот энциклопедически образован-
ный человек, основоположник отечественной зоотехнии, создал двух-
томное руководство «Анатомия домашних животных», в котором строе-
ние тела излагалось в связи с функцией и развитием. Его последователя-
ми стали Н.П.Чирвинский (1848—1920) и П.Ф.Лесгафт (1837—1909),
которые отстаивали мнение о формирующем влиянии среды на орга-
низм, в частности на скелет. Они стали основателями функциональной
анатомии. Хирург Н.И.Пирогов (1810—1881) заложил основы топогра-
фической анатомии.
В 1873 г. в Казани, Харькове и Варшаве были открыты самостоятельные
высшие учебные ветеринарные заведения, где изучение анатомии заня-
ло важное место в подготовке грамотных ветеринарных специалистов.
На формирование основных научных направлений в анатомии боль-
шое влияние оказывали и продолжают оказывать ветеринарные школы,
10
например Казанская, откуда вышли корифеи отечественной анатомии
(Д. М. Автократов, А. Ф. Климов, А. Н. Акаевский, Н. А. Васнецов,
Н. В. Михайлов, Ю.Ф. Юдичев), заложившие прочную научную базу для
понимания и изучения многих сторон нервной системы животных. Про-
фессор А. Ф. Климов (1878 —1940) сумел создать достаточно большой кол-
лектив Московской школы анатомов. Его ученики (профессора Б. В. Бо-
городский, С. М. Смиренский, П. А. Глаголев и др.) с успехом продол-
жили дело своего учителя. Долгое время кафедру анатомии при Москов-
ской ветеринарной академии возглавляла профессор И. В. Хрусталева,
передавшая эстафету достойным своим ученикам Н. А. Слесаренко и
Б. В. Криштофоровой. Много лет Московская школа анатомов изучает
вопросы сравнительной морфофизиологии органов движения, а также
закономерности и возможные пределы адаптации этих органов к новым
экологическим условиям содержания домашних животных. Профессор
Д.М.Автократов (1868—1953) с коллегами и учениками (П.П.Виногра-
довым, В. Н.Жедёновым, Г. М.Удовиным и др.) длительное время изу-
чали особенности органов дыхания животных. Академик Б. А. Домбров-
ский, а затем его ученик академик В.Г. Касьяненко основали Киевскую
школу сравнительных и эволюционных морфологов, из которой вышли
Г.А.Гиммельрейх, П.М. Мажуга, С.Ф.Манзий, С. К. Рудик. Огромный
вклад в изучение анатомии сосудистой и нервной систем животных вне-
сли и продолжают вносить ученые Ленинградской (Санкт-Петербургс-
кой) анатомической школы (профессора П.Я. Янушкевич, Н.Ф.Богда-
шев, М. И.Лебедев, М.А. Соколова, М. И. Климентов, А.А. Малышев,
Н. В. Зеленевский и др.). Активно работают по изучению сравнительной
анатомии пуховых коз на кафедре анатомии Оренбургского сельскохо-
зяйственного института, возглавляемой профессором В. В.Дегтярёвым.
Подробные исследования пушных зверей клеточного содержания прово-
дятся на кафедре анатомии Омского ветеринарного института во главе с
профессором Г.А.Хониным.
Краткая история физиологии на всем пути ее развития неразрывно
связана с морфологией. Общепринятым днем рождения физиологии как
науки считается 1628 г., когда Вильям Гарвей (1578—1657) опублико-
вал итог своих многолетних исследований — «Анатомическое исследова-
ние о движении сердца и крови животных». В ней приведены сведения о
движении крови по двум замкнутым на сердце кругам кровообращения
(большому и малому). Толчком для этого исследования послужило от-
крытие в 1553 г. итальянским ученым Коломбо и испанским физиологом
Сервето малого (легочного) круга кровообращения, а дополнил и как
бы завершил теорию кровообращения М. Мальпиги, который в 1661 г.
обнаружил при помощи микроскопа кровеносные капилляры, связыва-
ющие артериальные и венозные сосуды в единый ансамбль. Очень долго
физиологические наблюдения были лишь сопровождением анатомичес-
ких исследований, к тому же господство в идеологии религиозных воз-
зрений о непознаваемости божественным образом возникшего организ-
ма (витализм) значительно тормозило правильное понимание физиоло-
гической информации.
Только в XVIII —XIX вв. в связи с развитием других естественных
наук физиология постепенно начинает занимать достойное место. Во
11
многих странах Европы (Франция, Германия, Англия) открываются
первые физиологические школы, в которых объединяются передовые
биологические, в том числе и физиологические, умы — И. Мюллер
(1801 — 1858), Г. Гельмгольц (1821 — 1894), Р.Гейденгайн (1834—1897),
Ф. Мажанди (1783 — 1855) и др.
Революционный поворот в физиологии произошел в связи с откры-
тием итальянским физиком и естествоиспытателем Л. Гальвани «живот-
ного электричества» — явления, которое легло в основу электрофизио-
логии.
Особые заслуги в изучении биоэлектричества принадлежат Э.Дюбуа-
Реймону, Л. Герману, Э.Пфлюгеру, Ю. Бернштейну. Последний указал
на электрохимическую природу нервных импульсов и сформулировал
мембранную теорию их возникновения.
Развитие физиологии в России, как и анатомии, начинается в местах
активного скотоводства и коневодства при монастырях. Первые попытки
более подробного изучения физиологии животных были предприняты
на скотоврачебных кафедрах, открытых в 1807 г. на медицинских факуль-
тетах при Московском, Петербургском, Казанском и Харьковском уни-
верситетах, однако достойное место как наука физиология заняла по-
зднее, в специализированных ветеринарных институтах.
Первым учебником по физиологии в России стало трехтомное изда-
ние «Физиология для руководства своих слушателей», написанное про-
фессором Московского университета А. М. Филомафитским в 1836 г.
Приоритетную роль российские ученые сыграли в развитии рефлектор-
ной теории. Работы И. М. Сеченова и И. П. Павлова — «отцов-основате-
лей» отечественной физиологии, знает весь мир. Стараниями их сорат-
ников и учеников (Б. В. Вериги, М. Н. Шатерникова, А.А. Лихачева,
Л.А. Орбели, А. Д. Сперанского, К. М. Быкова, Н. А. Рожанского и др.)
было всесторонне разработано учение о рефлекторной природе всех про-
цессов, происходящих в организме, введено понятие условного рефлек-
са как основы поведения животных и психики человека (учение о выс-
шей нервной деятельности животных и человека).
В книге И. М. Сеченова «Рефлексы головного мозга» отражен результат
его многолетнего труда по изучению деятельности головного мозга как
материального субстрата сознания, а также открыто явление торможе-
ния в ЦНС («сеченовское торможение»). В. М. Бехтерев внес существен-
ный вклад в изучение функций головного мозга, выявил роль подкор-
ковых структур в формировании эмоциональных и двигательных реак-
ций. А. А. Ухтомский создал учение о доминанте как ведущем принципе
работы головного мозга. Л.А. Орбели и А.В.Тонков предложили идею
об адаптационно-трофическом влиянии вегетативной нервной систе-
мы. Л.А.Орбели основал принципиально новое направление в физио-
логии — эволюционную физиологию.
Изучение деятельности висцеральных органов, основы которого были
заложены И. П. Павловым (Нобелевская премия за работы в области
физиологии пищеварения), продолжили его ученики и последователи
(Г. П. Зеленый, А.А. Кудрявцев, Э.П. Кокорин, В. И. Георгиевский,
Б.Д.Кальницкий, Р.Н. Ордынцев и др.). В 1960 г. был основан Всесоюз-
ный (ныне: Всероссийский) НИИ физиологии, биохимии и питания
12
сельскохозяйственных животных, ставший ведущим центром в разра-
ботке основ жизнедеятельности и кормления животных.
Контрольные вопросы
1. Что изучает анатомия и каковы методы анатомических исследова-
ний?
2. Назовите основные вопросы, изучаемые физиологией, и методы
физиологических исследований.
3. Дайте определение понятий «эволюция», «филогенез» и «онтоге-
нез».
4. Почему ветеринарному врачу нужны подробные знания по анато-
мии и физиологии животных?
ГЛАВА 2
ЦИТОЛОГИЯ, гистология
И ЭМБРИОЛОГИЯ
цитология
Клетка. В основе строения и жизнедеятельности всех живых
организмов лежит элементарная частица живой материи — клетка.
Она представляет собой открытую саморегулирующуюся систе-
му, в которой материя выработала способность к постоянному
избирательному обмену веществом и энергией с окружающей сре-
дой. Клетки способны к росту, размножению и реагируют на вне-
шние воздействия. В них хранится генетический код, т. е. програм-
ма передачи наследственных признаков от родителей детям. Вне
пределов клетки жизнь невозможна.
Живой организм может состоять из одной клетки (Protozoa —
простейшие) или представлять собой сложноорганизованное со-
общество огромного числа клеток (Metazoa — многоклеточные).
В многоклеточном организме животных клетки теряют свою са-
мостоятельность и в зависимости от потребностей целого орга-
низма, закрепленных в генетической программе, специализиру-
ются по «местоположению» и по выполняемой функции. Форми-
руются огромные клеточные «кланы», связанные общностью про-
исхождения, строения и выполняемой работой, т.е. ткани (кост-
ная, мышечные, нервная и др.), из которых строятся более круп-
ные структуры — органы. Взаимная связь и взаимная зависимость
между огромным числом различных по строению и функции кле-
ток, неклеточных образований и образованных ими тканей и ор-
ганов обеспечивают морфологическую и физиологическую цело-
стность организма.
Формы клеток многообразны (рис. 2.1) и тесно связаны с вы-
полняемой функцией: пластинки, призмы, шары, веретена. Клетки
могут иметь отростки, хвостики, могут образовывать временные
ложные выросты, благодаря которым приобретают способность
передвигаться.
Размер клеток животных значительно меньше растительных
клеток и колеблется от 0,5 до 200 мкм. Самыми мелкими клетками
в организме животных и человека являются клетки крови (эрит-
роциты — 7 мкм, малые лимфоциты — 4 — 6 мкм), а самыми
большими — яйцеклетки (120 — 200 мкм). Средние размеры сома-
14
Рис. 2.1. Различные формы клеток (схема):
/ — призматические клетки каемчатого эпителия тонкой кишки; 2 — кубичес-
кий эпителий канальцев нефрона; 3 — плоские клетки мезотелия брюшины; 4,
Л' - округлые клетки крови; 6 — бокаловидная клетка эпителия толстой киш-
ки; 7 — веретенообразная гладкая мышечная клетка; 9 — хвостатая клетка (спер-
мин); 10 — многоядерная клетка (мегакариоцит); 11 — отростчатая клетка
(нейрон)
гических клеток — от 20 до 30 мкм, в то время как нервная клет-
ка, имеющая микроскопическое тело, вместе с отростками мо-
жет достигать до 1 м у человека и 1,5 — 2 м у лошади.
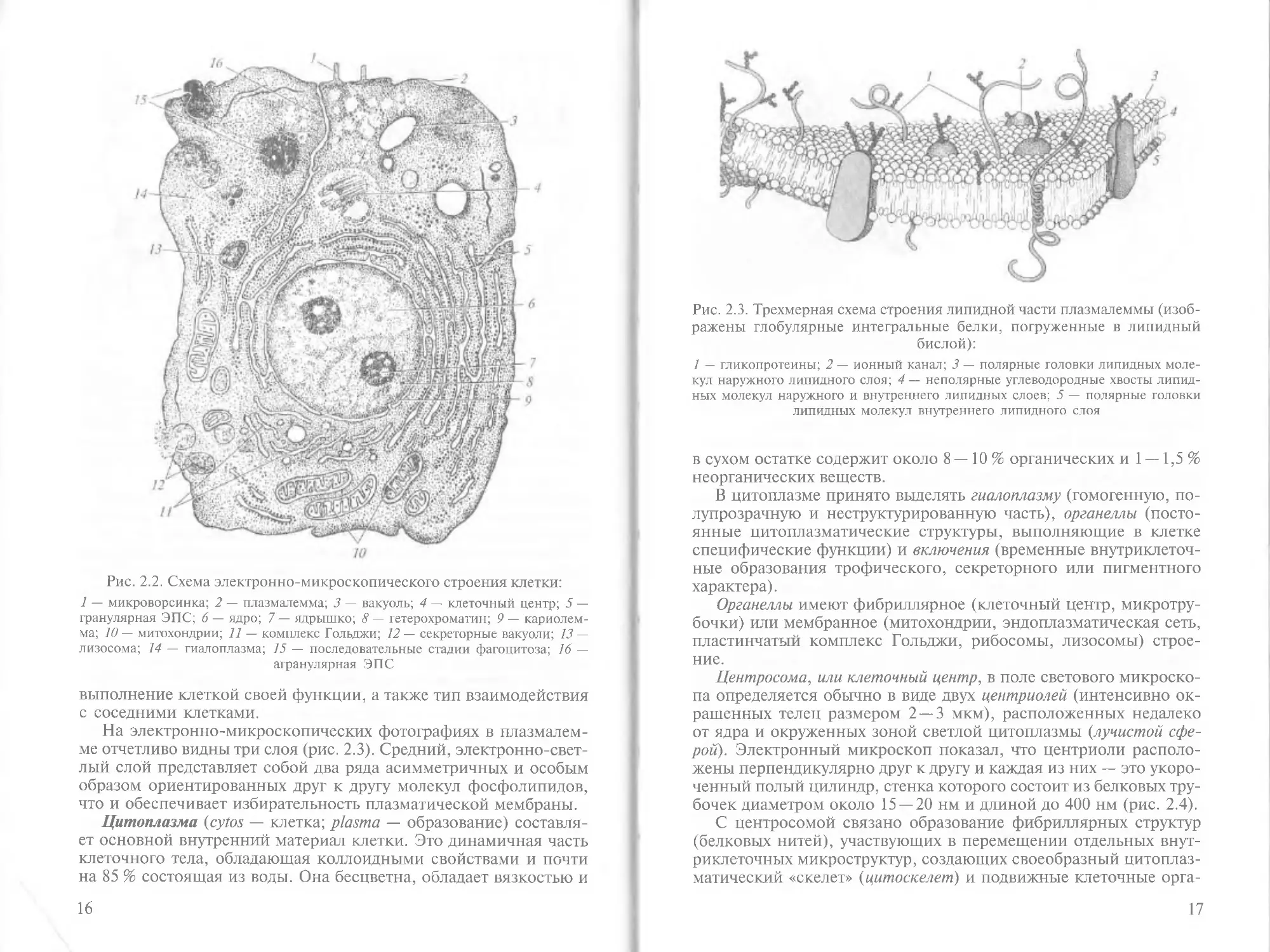
Строение клетки. Клетка отличается значительной сложностью
с троения и в то же самое время поражает воображение своей упо-
рядоченностью (рис. 2.2). Все жизненные процессы в клетках, а
также выполняемая ими функция обеспечиваются более мелки-
м и, чем сама клетка, субклеточными структурами, составляющи-
ми ее рабочие органы. В клетке различают плазмалемму, цито-
плазму и ядро.
Плазмалемма, или цитолемма (cytos — клетка, lemma — огра-
ничение), — это особым образом устроенная наружная граница
клетки, выделяющая ее из окружающего пространства в качестве
самостоятельной единицы. Плазмалемма по своим физико-хими-
ческим свойствам является полупроницаемой «пленкой», поэто-
му именуется также биологической мембраной (membrana — кожи-
ца, оболочка, пленка). Она обладает высокой прочностью, элас-
тичностью и уникальным свойством — избирательностью. Кле-
точная оболочка играет роль барьера, препятствующего свобод-
ному перемещению химических веществ и воды в клетку и обрат-
но, определяя тем самым химический состав клетки, характер
обмена между ней и окружающей ее средой, обеспечивающий
15
ю
Рис. 2.2. Схема электронно-микроскопического строения клетки:
1 — микроворсинка; 2 — плазмалемма; 3 — вакуоль; 4 — клеточный центр; 5 —
гранулярная ЭПС; 6— ядро; 7— ядрышко; 8 — гетерохроматин; 9 — кариолем-
ма; 10 — митохондрии; 11 — комплекс Гольджи; 12 — секреторные вакуоли; 13 —
лизосома; 14 — гиалоплазма; 75 — последовательные стадии фагоцитоза; 16 —
агранулярная ЭПС
выполнение клеткой своей функции, а также тип взаимодействия
с соседними клетками.
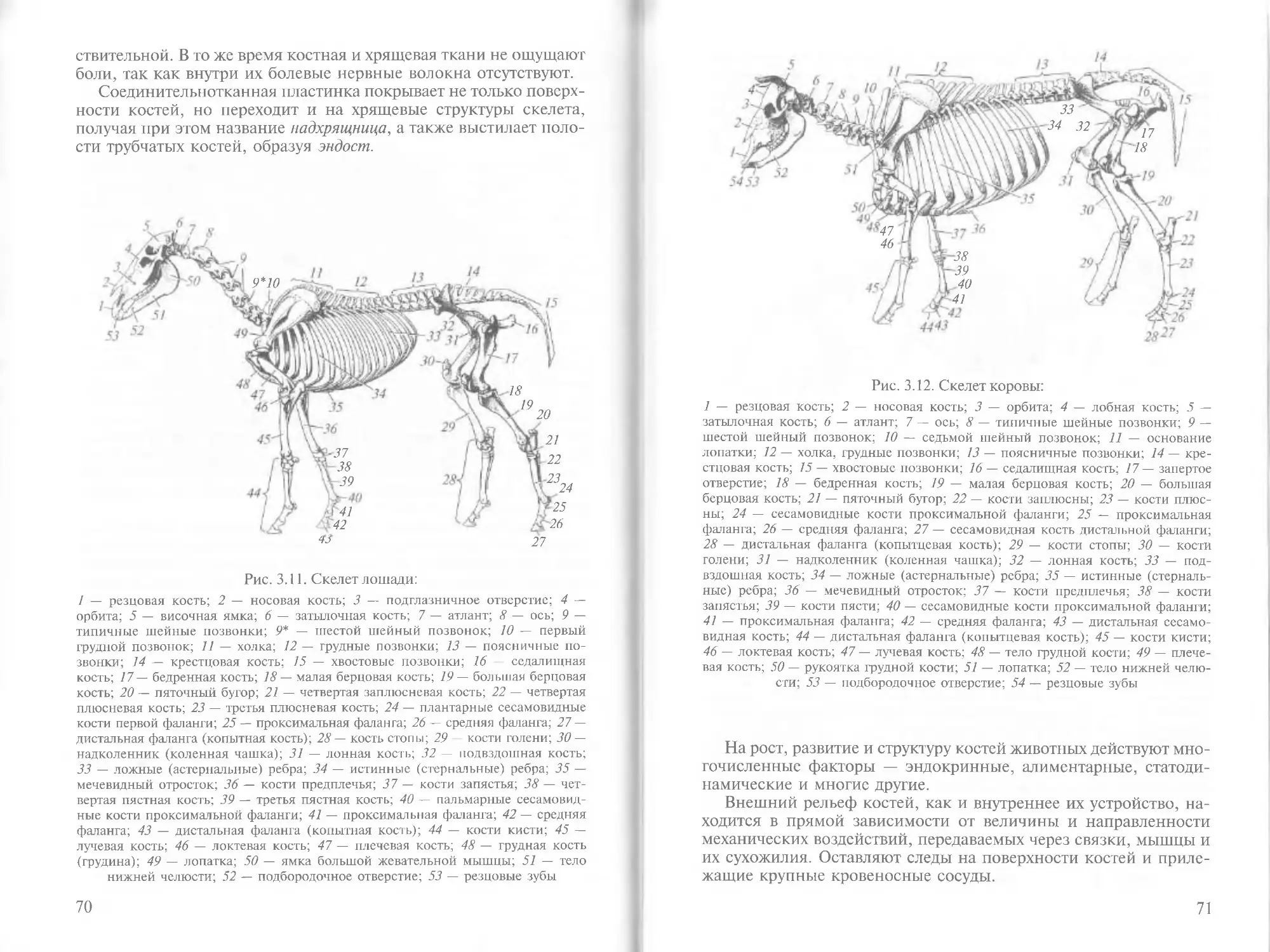
На электронно-микроскопических фотографиях в плазмалем-
ме отчетливо видны три слоя (рис. 2.3). Средний, электронно-свет-
лый слой представляет собой два ряда асимметричных и особым
образом ориентированных друг к другу молекул фосфолипидов,
что и обеспечивает избирательность плазматической мембраны.
Цитоплазма (cytos — клетка; plasma — образование) составля-
ет основной внутренний материал клетки. Это динамичная часть
клеточного тела, обладающая коллоидными свойствами и почти
на 85 % состоящая из воды. Она бесцветна, обладает вязкостью и
16
2
Рис. 2.3. Трехмерная схема строения липидной части плазмалеммы (изоб-
ражены глобулярные интегральные белки, погруженные в липидный
бислой):
7 — гликопротеины; 2 — ионный канал; 3 — полярные головки липидных моле-
кул наружного липидного слоя; 4 — неполярные углеводородные хвосты липид-
ных молекул наружного и внутреннего липидных слоев; 5 — полярные головки
липидных молекул внутреннего липидного слоя
в сухом остатке содержит около 8 —10 % органических и 1 — 1,5 %
неорганических веществ.
В цитоплазме принято выделять гиалоплазму (гомогенную, по-
лупрозрачную и неструктурированную часть), органеллы (посто-
янные цитоплазматические структуры, выполняющие в клетке
специфические функции) и включения (временные внутриклеточ-
ные образования трофического, секреторного или пигментного
характера).
Органеллы имеют фибриллярное (клеточный центр, микротру-
бочки) или мембранное (митохондрии, эндоплазматическая сеть,
пластинчатый комплекс Гольджи, рибосомы, лизосомы) строе-
ние.
Центросома, или клеточный центр, в поле светового микроско-
па определяется обычно в виде двух центриолей (интенсивно ок-
рашенных телец размером 2 — 3 мкм), расположенных недалеко
от ядра и окруженных зоной светлой цитоплазмы {лучистой сфе-
рой). Электронный микроскоп показал, что центриоли располо-
жены перпендикулярно друг к другу и каждая из них — это укоро-
ченный полый цилиндр, стенка которого состоит из белковых тру-
бочек диаметром около 15 — 20 нм и длиной до 400 нм (рис. 2.4).
С центросомой связано образование фибриллярных структур
(белковых нитей), участвующих в перемещении отдельных внут-
риклеточных микроструктур, создающих своеобразный цитоплаз-
матический «скелет» {цитоскелет) и подвижные клеточные орга-
17
Рис. 2.4. Клеточный центр (центросома):
А — ув. х 280: 1 — ядро клетки; 2 — центриоли; 3 — центросфера; Б — электрон-
но-микроскопическая реконструкция центриоли: / — микротрубочки; 2 —
мостики; 3 — сателлиты
неллы {реснички, микроворсинки, хвостики), а также образующих
веретено при делении клеток.
Митохондрии {mitos — нить, chondros — зерно) являются энер-
гетическими станциями клетки. При световой микроскопии они
имеют вид палочек диаметром 0,5—1 мкм и длиной 0,5 — 7 мкм.
На электронно-микроскопической реконструкции (рис. 2.5) вид-
но, что митохондрия представляет собой полую структуру, за-
полненную зернистым электронно-светлым веществом {матрик-
сом) и окруженную стенкой из двух не связанных между собой
биологических мембран.
Эндоплазматическая {endo — внутренний), или цитоплазмати-
ческая, сеть (здесь и дальше сокращенно ЭПС) — неотъемлемая
часть общей метаболической системы клетки. Структуры ЭПС вы-
являются только при электронной микроскопии и представлены
сложной системой складок двойных биологических мембран тол-
щиной 5 —6 нм (рис. 2.6). По строению и характеру синтетической
деятельности мембраны ЭПС условно разделяют на шероховатые
{гранулярные) и гладкие {агранулярные). Гранулярная ЭПС распола-
гается ближе к ядру и несет на себе мелкие электронно-плотные
гранулы (рибосомы), на которых происходит синтез белков. Осо-
Рис. 2 5. Электронно-микроско-
пическая реконструкция мито-
хондрии:
1 — криста; 2, 3 — внутренняя и
наружная мембраны; 4 — матрикс
18
Рис. 2.6. Электронно-микроскопи-
ческая реконструкция эндоплазма-
тической сети в нервной клетке:
I — цитоплазматические мембраны;
2 — анастомозы между цистернами;
3 — плоские цистерны; 4 — окно; 5 —
трубкообразная цистерна
Рис. 2.7. Расположение комплек-
са Гольджи:
/I — в клетке спинно-мозгового ган-
глия кошки; Б — в клетке экзокрин-
ной части поджелудочной железы
аксолотля; / — комплекс Гольджи;
2 — гранулы секрета; 3 — ядро
бенного развития она достигает в клетках, выделяющих белковые
секреты (ферменты, гормоны). Агранулярная ЭПС не связана с
рибосомами и обеспечивает синтез длинноцепочных жирных кис-
лот и липоидов.
Пластинчатый комплекс (комплекс Гольджи) играет в клетке
роль распределительной и выделительной системы и тесно связан
с ЭПС. Он расположен в околоядерном пространстве и в поле
светового микроскопа имеет вид тонких параллельных нитей с
петельками на концах (рис. 2.7). Электронно-микроскопическая ре-
конструкция отчетливо показывает мембранное строение комп-
лекса Гольджи (рис. 2.8). Последний наиболее развит в секретиру-
ющих клетках различных желез, так как именно в нем накаплива-
ются, а возможно и синтезируются химические вещества (секре-
ты).
Рибосомы — ультрамикроскопические (диаметр 0,01 — 0,015
мкм) электронно-непрозрачные образования, состоящие из двух
разных по длине молекул ДНК, связанных с белками (нуклео-
протеиды) и свернутых в виде двух (большой и малой) субъеди-
ниц (рис. 2.9). Рибосомы образуются в ядре при активном участии
ядрышка, а затем выходят в цитоплазму и располагаются на мем-
бранах гранулярной ЭПС и кариолемме. Они являются центрами
синтеза белковых молекул.
Лизосомы (Hseo — растворяю, soma — тело) — мелкие (0,3 —
0,5 мкм) сферические пузырьки, окруженные мембраной и види-
19
Рис. 2.8. Электронно-микроскопическая реконструкция комплекса Голь-
джи:
1 — цистерна; 2 — цитоплазматические мембраны; 3 — вакуоли; 4 — пузырьки
мые только с помощью электронного микроскопа. Они содержат
особые ферменты, способные «переваривать» как вещества, по-
ступившие в клетку извне, так и конечные продукты метаболизма
самой клетки, а также отработавшие или поврежденные внутри-
клеточные структуры.
Цитоплазматические включения по химическому составу и фи-
зиологическому значению объединяются в три основные группы:
1) трофические (trophe — пища) — капли жира, глыбки гликоге-
на, зерна белка; 2) секреторные (капли пищеварительных секре-
тов и глыбки гормонов); 3) пигментные (меланин, липохромы).
Ядро клетки (nucleus — орех) — центр управления жизнедея-
тельностью клетки, без которого цитоплазма не может нормаль-
Рис. 2.9. Рибосома:
1 — малая, 2 — большая субъеди-
ницы
но функционировать. Руководящая
роль ядра в клетке заключается в
выполнении двух основных функ-
ций: 1) ядро хранит генетическую
информацию клетки, закодиро-
ванную в ДНК; 2) ядро реализует
исполнение генетической про-
граммы построения и функциони-
рования цитоплазмы клетки, кон-
тролируя все обменные процессы.
В ядре принято различать (рис. 2.10):
оболочку (кариолемму), ядерное
вещество (кариоплазму), хроматин
и микротельце (ядрышко). В пери-
20
Рис. 2.10. Электронная микрограм-
ма ядра:
1 — внутренняя, 2 — наружная мемб-
раны кариолеммы; 3 — перинуклеар-
ное пространство; 4 — связь перинук-
леарного пространства с цистернами
ЭПС; 5 — ядерная пора; 6 — карио-
плазма; 7 — эухроматин; 8 — ядрыш-
ко; 9 — гетерохроматин
од деления клетки ядро претер-
певает сильные изменения и ис-
чезает, восстанавливаясь только
в дочерних клетках.
Элементарный химический
состав живой клетки. Определя-
ет характер ее биохимических и
физиологических процессов,
целиком зависит от элементов,
которые присутствуют в земной
коре. Из 90 природных химичес-
ких элементов, обнаруженных в
клетке, с очень большим отры-
вом (около 95 %) преобладают
всего четыре элемента — О, С,
Н и N (макроэлементы), боль-
шинство элементов (Р, К, S, С1,
Na, Mg, Fe и др.) встречаются
в небольшом количестве и со-
ставляют чуть меньше 4 % (мик-
роэлементы). Остальные элемен-
ты (Си, I, F, Аи и др.) присут-
ствуют в клетках в минимальных
количествах, измеряющихся тысячными долями процента (улът-
раэлементы).
Одним из самых востребованных клеткой неорганических веществ
является вода. Она составляет 75 % от сырой массы клетки и са-
мым активным образом участвует во всех процессах ее жизнедея-
тельности. Вода как наиболее приемлемый растворитель для боль-
шинства внутри- и внеклеточных веществ формирует ту среду, в
которой осуществляется вся молекулярная жизнь клетки. Потеря
клеткой более 20 % воды приводит к ее гибели.
Молекулярные основы биохимии и физиологии клетки. Основ-
ными отличительными свойствами живой материи являются об-
мен веществ, избирательность в поглощении веществ из окру-
жающей среды, раздражимость и способность к росту и размно-
жению.
Поглощая вещества из внешнего мира, живой организм по-
чти неузнаваемо меняет их структуру и придает им совершенно
новые свойства. Эти вещества названы органическими, или био-
логическими.
Органические вещества клетки имеют общий план строения и
состоят из углеродного «каркаса», к которому подсоединяются
фосфатные, карбоксильные и другие химические группы. По та-
кому принципу в клетке синтезируется огромное число разнооб-
разных протяженных органических молекул (макромолекул, или
21
полимеров). К основным органическим соединениям, определяю-
щим все аспекты жизнедеятельности клетки и жизнедеятельности
всего организма, относятся: белки, жиры и углеводы.
Белки являются самой многочисленной группой органических
соединений и составляют почти половину сухого клеточного ос-
татка. Это строительный компонент для цитоплазмы, ядра и всех
клеточных мембран, основа для веретена, ресничек, жгутиков и
многих сократимых структур клетки. Белки входят в состав фер-
ментов и гормонов и обеспечивают этим тонко сбалансирован-
ный метаболизм на уровне клетки и всего организма.
Жиры, или липиды {lipos — жир), по химическому составу — это
сложные эфиры глицерина и жирных кислот. Они нерастворимы в
воде, обладают высокой энергетической емкостью (9,5 ккал/г энер-
гии). Утилизация жиров в клетке сопровождается не только выде-
лением энергии, но и образованием воды, которая также может
использоваться клетками. Из жиров, поступающих с пищей, каж-
дый организм вырабатывает жиры, свойственные его виду (ове-
чий, свиной, рыбий и т.д.).
Углеводы присутствуют в клетках как источник энергии (энер-
гетическая емкость углеводов — 4,5 ккал/г), которая образуется в
результате утилизации их посредством внутриклеточного дыхания
и переводится в митохондриях в химическую энергию молекул
АТФ. Используются углеводы и для синтеза многих биологически
активных веществ (нуклеиновых кислот, различных ферментов).
По молекулярному строению углеводы подразделяются на мо-
носахариды (глюкоза, фруктоза) и полисахариды (гликоген в жи-
вотных и крахмал в растительных клетках). Углеводы вступают в
связь с молекулами других веществ: с белками {гликопротеиды) и
между собой {мукополисахариды).
Нуклеиновые кислоты (НК) были впервые открыты в ядре
(nucleus). Различают два типа НК: дезоксирибонуклеиновую (ДНК)
и рибонуклеиновую (РНК). Обе кислоты по своей структурной орга-
низации — это линейные молекулярные цепи, образованные по-
лимеризацией очень похожих по своему строению мономерных
звеньев {мононуклеотидов), расположенных в строго определен-
ном порядке. Типизация НК проводится по двум параметрам их
нуклеотидов: по углеводу, входящему в их молекулу (в ДНК —
дезоксирибоза, а в РНК — рибоза), и по набору азотистых основа-
ний (в ДНК — аденин (А), гуанин (Г), цитозин (Ц) и тимин (Т),
а в РНК — урацил (У), А, Г и Ц). Но, несмотря на очень неболь-
шие химические отличия, НК значительно различаются по свой-
ствам и биологическим функциям.
ДНК расположена в ядерном веществе клетки и входит в со-
став генетического материала {хромосомы). Она отличается очень
большим линейным размером своей молекулы, имеет вид двойной
цилиндрической спирали и содержит генетический код, т. е. наслед-
22
Рис. 2.11. Схема молекулы ДНК:
А — двухспиральное строение ДНК с обозначением участка спаривания азотистых
оснований; Б — линейные размеры одного витка двойной спирали ДНК; / — глу-
бокая; 2 — мелкая бороздки в молекуле ДНК; стрелками показано направ-
ление спирализации отдельных цепей ДНК
ственную информацию о строении и развитии, передаваемую в
дочерние клетки и «записанную» посредством разнообразия в чере-
довании пар азотистых оснований по длине молекулы ДНК (рис. 2.11).
Передача генетической «записи» в дочерние клетки в полном объе-
ме обеспечивается способностью молекул ДНК изготавливать аб-
солютно идентичные свои копии (редубликация, или удвоение).
Такой процесс протекает как обязательный при делении клеток.
Вне времени деления (интерфаза) двойные спирали молекулы
ДНК используются для «считывания» части своей информации с
одной из цепей и транскрипции (синтеза) из свободных мононук-
леотидов молекул РНК.
РНК в живых клетках присутствует в виде трех типов: рибосо-
мальная (рРНК), информационная (иРНК), или матричная (мРНК),
и транспортная (тРНК). Вместе все типы РНК являются «инстру-
ментом» при ДНК, посредством которого закодированная в ДНК
23
генетическая информация переводится «на язык» определенной пос-
ледовательности аминокислотных остатков в процессе синтеза бел-
ка и передается в качестве «руководства к действию» в каждую клетку
организма. Таким образом, создается молекулярный механизм, бла-
годаря которому осуществляется генетический контроль над про-
цессами синтеза белков — самых важных клеточных веществ.
Аденозинтрифосфорная кислота (АТФ) — это основное энерге-
тическое соединение клетки во всех живых организмах. АТФ обра-
зуется в митохондриях и накапливает в своих молекулярных мак-
роэргических фосфатных связях около 7 ккал химической энер-
гии. Энергия, освобождающаяся при окислении молекулы АТФ,
обеспечивает протекание абсолютного большинства энергозатрат-
ных реакций клеточного метаболизма.
Транспорт веществ через оболочку клетки. Осуществляется в двух
основных направлениях: окружающая среда — клетка и клетка —
окружающая среда, является важной стороной жизнедеятельнос-
ти клетки. При этом первое направление обслуживается процесса-
ми, не требующими затраты энергии (пассивный транспорт), а
второе направление предусматривает использование энергии (ак-
тивный транспорт) (рис. 2.12).
Пассивный транспорт обеспечивает в основном избирательное
поступление веществ внутрь клетки и осуществляется тремя ос-
новными путями: 1) простой диффузией, т. е. прямым проникно-
вением в клетку по градиенту (показателю) концентрации мел-
ких ионов, воды и водорастворимых веществ через каналы или
поры; 2) фильтрацией жирорастворимых неэлектролитов посред-
Транспортируемая Канадообра- Белок-
Простая Облегченная Активный
диффузия диффузия транспорт
I______________________I
Пассивный транспорт
Рис. 2.12. Схема транспорта веществ через липидный бислой плазмалеммы
24
Рис. 2.13. Схема пиноцитоза с лизосомами:
1, 2 — крупные и мелкие частицы в околоклеточной среде; 3, 4 — захват частиц
путем пиноцитоза (микропиноцитозные пузырьки); 5, 6 — возникновение лизо-
сом и образование их ферментативного содержимого; 7 — слияние лизосом и
микропиноцитозных вакуолей; 8, 9 — расщепление содержимого вакуолей; 10 —
выделение продуктов расщепления
ством растворения их в липидной части плазмалеммы и прямого
проникновения в цитоплазму клетки; 3) облегченной диффузией —
проникновением небольших молекул сахаров, аминокислот и
некоторых ионов при помощи специализированных транспорт-
ных белков-переносчиков.
Активный трансмембранный транспорт отличается тем, что мо-
лекулы веществ перемещаются через мембрану против гради-
ента их концентрации с использованием высокомолекулярных
белков-переносчиков и химической энергии АТФ. Таким спосо-
бом в клетку проникают некоторые сахара и аминокислоты, а из
клеток выводятся продукты метаболизма, заряженные частицы
(ионы) и секретируемые клетками вещества (секреты).
Особым вариантом активного транспорта является эндоцитоз —
захват частиц вещества, ионов и крупных молекул клеточной мем-
браной. Существует два способа захвата частиц вещества из меж-
клеточного пространства: фагоцитоз (phagein — поедаю) — захват
твердых веществ, и пиноцитоз (pynein — пью) — захват капелек
растворенного вещества (рис. 2.13). Процесс фагоцитоза играет важ-
ную роль в реакции организма на инфекцию, обеспечивая захват
и разрушение болезнетворных микробов.
Обмен веществ, или метаболизм, — одно из основных прояв-
лений жизнедеятельности клетки. В клетку из околоклеточной среды
постоянно поступают химические вещества, внутри клетки про-
исходят их сложные преобразования (для поддержания химичес-
25
кого статуса цитоплазмы, обновления внутриклеточных структур —
мембран, органелл и др., и образования секретов), а в околокле-
точную среду клетка выделяет продукты своей жизнедеятельнос-
ти и секреты. Благодаря множеству разнообразных химических ре-
акций синтеза из простых веществ новых, более сложных {процес-
сы ассимиляции), и распада сложных веществ на простые {процес-
сы диссимиляции), существующих в единстве своей противополож-
ности, обеспечиваются все жизненные проявления клетки. Наи-
более важным из синтетических процессов клеточного метаболизма
является синтез белка.
Преобладание ассимиляции над диссимиляцией — непремен-
ное условие для обеспечения роста, т.е. накопления клеточного
материала и увеличения размеров клетки.
Дифференциация клеток — это процесс постепенного и неук-
лонного расхождения клеток в ходе эмбрионального развития в
структурном и функциональном отношении от первичного нача-
ла (зиготы). Этот сложный комплекс внутриклеточных преобразо-
ваний обусловлен генетически, т. е. для каждого типа клеток су-
ществует свой временной механизм начала и конца (наступление
зрелости) дифференциального развития.
Раздражимость — важная и неотъемлемая часть жизненных
свойств клетки. Естественным ее проявлением являются волнооб-
разные колебания клеточной мембраны, перемещение по цито-
плазме ядра, митохондрий, транспортных пузырьков и вакуолей,
а также способность к движению самой клетки и целенаправлен-
ное усиление или ослабление метаболических процессов в ответ
на изменения околоклеточной среды (химического состава, pH,
температуры и т.п.).
Жизненный цикл любой клетки принято делить на два периода:
интерфазу (период между двумя клеточными делениями, когда
клетка увеличивается в размере, происходит синтез различных
органических веществ, РНК, удвоение ДНК) и деление.
Деление клеток. Биологическая способность к самовоспроиз-
водству — одно из уникальных свойств живой материи. Этот ко-
роткий период занимает 10 % жизненного времени клетки, но
является самым ярким моментом в ее жизни. Он обеспечивает по-
явление новых молодых клеток, способных заменить в тканях и
органах старые, больные и погибшие клетки. Передача в дочерние
клетки генетической информации обеспечивается особыми ген-
содержащими структурами — хромосомами.
Хромосомы в делящихся клетках представляют собой компакт-
ные хроматиновые тела, напоминающие клюшки для гольфа с
двумя неравными частями {плечами), соединенными тонкой пер-
вичной перетяжкой {центромерой, или кинетохором) для прикреп-
ления к нитям веретена деления (рис. 2.14). Многочисленные нити
молекул ДНК {хромонемы), погруженные в своеобразный орган-
26
но-минеральный матрикс, содержат генетичес-
кий код. В процессе подготовки к делению клет-
ки хромосомы удваиваются {хроматиды) за счет
редублицикации (удвоения) входящей в них мо-
лекулы ДНК. Установлено, что для каждого вида
животных форма и число хромосом строго спе-
цифичны {кариотип). Например, у лошади и
коровы 60 хромосом, у свиньи 40, у челове-
ка 46. У млекопитающих все хромосомы разде-
лены на одинаковые по виду пары {гомологич-
ные хромосомы), кроме двух {половых) хромо-
сом, имеющих разный вид и обозначаемых как
Хи Y.
Существует три способа деления клеток:
1) прямое деление {амитоз), при котором клет-
ка просто и быстро делится путем перетяжки
ядрышка, ядра и цитоплазмы, но без гарантии
равномерного распределения генетического
материала между дочерними клетками (так де-
лятся одноклеточные организмы, а также клет-
ки поврежденных тканей); 2) непрямое деле-
ние {митоз); 3) редукционное деление {мей-
оз), характерное только для половых клеток
(яйцеклетки и сперматозоида); в результате ре-
дукционного деления на раннем этапе жизнен-
ного цикла половых клеток число их хромосом
Рис. 2.14. Внешний
вид и схема внут-
реннего строения
хромосомы:
1 — пелликула; 2 —
хроматида; 3 — центр
организации ядрыш-
ка (вторичная пере-
тяжка); 4 — матрикс;
5 — центромера (пер-
вичная перетяжка);
6— двойная спираль
ДНК; 7 — спутник
уменьшается вдвое.
Митоз, или кариокинез {karyon — ядро, kinesis — движение),
является достаточно сложным универсальным механизмом деле-
ния, благодаря которому родительская клетка способна поровну
распределить генетический материал между дочерними клетками.
Он состоит из четырех фаз: профазы, метафазы, анафазы и
телофазы (рис. 2.15). Наиболее заметные трансформации при ми-
тозе протекают в ядре. В течение профазы (наиболее длинной фазы
митоза) проявляются хромосомы, ядро постепенно теряет обо-
лочку и ядрышко, а карио- и цитоплазма объединяются в миксо-
плазму, в которой между центриолями протягиваются нити вере-
тена. В метафазе разбросанные по всей клетке хромосомы подтя-
гиваются нитями веретена в центральную зону миксоплазмы и
выстраиваются в виде тонкой экваториальной пластинки {мате-
ринской звезды), при этом каждая разделяется на две хромонемы
{дочерние хромосомы). Анафаза — самая короткая, но наиболее зре-
лищная фаза митоза, в течение которой дочерние хромосомы ак-
тивно перемещаются к полюсам клетки и формируют там тонкие
пластинки, или дочерние звезды. Этот процесс сопровождается за-
метным волнообразным движением миксоплазмы. В течение тело-
27
Рис. 2.15. Схема последовательных стадий митоза.
Профаза: 1 — формирование хромосом в ядре делящейся клетки (стадия плотно-
го клубка) и начало расхождения удвоенных центриолей с образованием микро-
трубочек веретена деления; 2 — активная спирализация хромосом (стадия рых-
лого клубка) и завершение формирования веретена деления; 3 — разрушение
ядерной оболочки и прикрепление хромомеров хромосом к нитям веретена де-
ления в гиалоплазме. Метафаза: 4 — раздвоение хромосом на хромонемы и рас-
положение их в виде экваториальной пластинки. Анафаза: 5 — расхождение хро-
монем (дочерних хромосом) к полюсам делящейся клетки. Телофаза: 6 — рыхлые
клубки дочерних хромосом, вокруг которых формируется ядерная оболочка; 7 —
разделение цитоплазмы материнской клетки между двумя дочерними клетками
28
фазы на каждом полюсе делящейся клетки дочерние хромосомы
преобразуются в хроматиновые нити, восстанавливаются ядерные
оболочки и ядрышки и образуются дочерние ядра, растворяются
короткие нити веретена и по центру клетки воздвигается перего-
родка, которая обеспечивает быстрое и полное разделение ци-
топлазмы (цитокинез) материнской клетки на две дочерние.
Контрольные вопросы
1. Какие внутриклеточные «помощники» обеспечивают все жизнен-
ные процессы в клетке?
2. Какую роль играет в клетке ядро?
3. Дайте определение понятия «метаболизм». Какая структура и каким
образом регулирует поступление веществ в клетку?
4. Какое значение для жизнедеятельности клетки имеют белки, жиры
и углеводы?
5. Откуда клетка берет энергию?
6. Какую роль играют нуклеиновые кислоты в синтезе белков и пере-
даче генетической информации?
7. Опишите строение хромосомы.
8. Какие существуют способы деления клеток; в чем состоят их разли-
чия и значение?
ГИСТОЛОГИЯ
Ткани — это эволюционно сложившаяся система гистологи-
ческих элементов (клеток и неклеточных компонентов), объеди-
ненных общностью развития, строения, обмена веществ, специ-
ализированная на выполнении определенной функции. Ткани как
неотъемлемая часть многоклеточного организма создают второй,
после клеток, структурный уровень его организации. Ткани при-
нято делить на эпителиальные, опорно-трофические, мышечные
и нервную.
Эпителиальные ткани, или эпителии (epithelium). Располагают-
ся, как правило, на границе между внутренней средой организма
и внешней, т. е. окружающей его средой. Выделяют (отграничива-
ют) живой объект как самостоятельно существующую единицу в
окружающем пространстве. В зависимости от расположения и ис-
пытываемых механических и иных воздействий эпителии преоб-
разуются, и их принято делить на несколько типов (схема 2.1). Но
несмотря на многообразие вариантов, эпителиальные ткани сбли-
жают важные особенности: 1) они построены из одного или не-
скольких слоев высокоспециализированных клеток; 2) обладают
полярной дифференциацией (неодинаковым строением глубоких
и наружных слоев в многослойных эпителиях и верхнего (апи-
кального) и нижнего (базального) концов клеток в однослойном
29
Схема 2.1
эпителии; 3) в эпителиальных тканях нет сосудов, а питание их
осуществляется диффузным образом; 4) между клетками нет меж-
клеточных пространств и они плотно сомкнуты посредством плаз-
матических выростов (интердигитация)', 5) эпителии укреплены
на особой тонкой соединительнотканной пластинке (базальной
мембране), благодаря которой прочно удерживаются на подлежа-
щей ткани.
Однослойные эпителии расположены в относительно защищен-
ных местах и характеризуются тем, что состоят из одного ряда
(слоя) клеток. Однослойные эпителии подразделяются на одно-
рядные и многорядный призматический мерцательный.
В однорядных эпителиях все клетки имеют одинаковую высоту и
отличаются только местоположением и формой. Среди них разли-
чают призматический, кубический, плоский эпителий (рис. 2.16).
Однослойный призматический эпителий покрывает слизистые
оболочки конечных выводных протоков экзокринных желез, мо-
чеотводящих канальцев мозгового слоя почки, семяпроводов.
Однослойный однорядный призматический железистый эпителий
располагается на поверхности железистой части слизистой обо-
лочки желудка и постоянно выделяет защитную слизь; впячива-
ясь в собственный слой слизистой оболочки, он участвует в обра-
зовании пищеварительных желез.
Однослойный призматический каёмчатый эпителий в целом по-
строен аналогично типичному призматическому эпителию, но
на апикальном, обращенном в просвет кишечника конце каёмча-
тых клеток выступает до 3000 ультрамикроскопических цитоплаз-
матических выростов (микроворсинок), увеличивающих клеточную
поверхность в 30 раз и видимых в световой микроскоп как темная
полоска (щеточная кайма). Они обеспечивают очень важную фун-
30
Рис. 2.16. Различные виды однослойного эпителия:
А — каёмчатый; Б — кубический; В — плоский; 1 — эпителиальные клетки; 2 —
подэпителиальная соединительная ткань
кцию — всасывание из просвета кишки простых химических ве-
ществ, образовавшихся в результате переваривания пищи в же-
лудке и кишечнике.
Однослойный кубический эпителий встречается в извитых каналь-
цах почечных нефронов, а также в секреторных отделах многих
экзокринных желез (молочных, слюнных, потовых) и желчном
протоке. Помимо покровной и защитной функции кубические
клетки в извитых канальцах нефронов выполняют еще и важную
обменную функцию, обеспечивая реабсорбцию (обратное всасы-
вание) воды и некоторых веществ из просвета канальцев в кровь.
Как особый вариант однослойного кубического эпителия мож-
но выделить однослойный железистый эпителий. Клетки этого эпи-
телия обладают способностью к образованию и выделению осо-
бых химических веществ (секретов) и располагаются одиночно
(бокаловидные клетки) или создают специализированные секре-
тирующие отделы в экзокринных, а также некоторых эндокрин-
ных железах (кора надпочечников, щитовидная железа, эндок-
ринные островки поджелудочной железы).
Однослойный плоский эпителий (мезотелий) выстилает сероз-
ные оболочки: плевру, покрывающую стенки и органы грудной
полости; брюшину, покрывающую стенки и органы брюшной по-
лости; наружную поверхность сердца (эпикард); внутреннюю
поверхность перикарда. Он состоит из одного слоя плоских плот-
но сомкнутых клеток, покрытых многочисленными короткими
микроворсинками (диаметром 0,1 мкм и длиной 1 — 3 мкм), кото-
рые обеспечивают скользкую прослойку, облегчающую смеще-
31
Рис. 2.17. Электронная микро-
фотография мерцательного эпи-
телия из выстилки слизистой
оболочки бронха. Поля реснит-
чатых клеток с длинными (7)
и короткими (2) ресничками;
(ув.х 1500)
ние отдельных органов в полостях
тела по отношению к стенкам и
другим органам. Клетки мезотелия
выделяют небольшое количество се-
розной жидкости, которая увлаж-
няет серозные покровы.
Однослойный многорядный мерца-
тельный эпителий покрывает слизи-
стую оболочку дыхательных путей,
яйцеводов и (в отдельные периоды)
эндометрий. Он построен из трех
типов (камбиальных, мерцательных
и бокаловидных) плотно сомкну-
тых клеток различной высоты и
формы. Многорядность проявляет-
ся в расположении ядер на разной
высоте. Отличительной особеннос-
тью мерцательной клетки является
наличие на ее апикальном конце
многочисленных (более 250) под-
вижных ресничек (рис. 2.17), пост-
роенных из особых белковых микротрубочек. Среди мерцательных
клеток встречаются отдельные железистые (бокаловидные) клет-
ки, выделяющие вязкую жидкость (слизь), которая служит для
защиты слизистой оболочки от высыхания.
Многослойные эпителии встречаются, как правило, в местах,
подвергающихся значительным, в первую очередь механическим,
воздействиям. Обычно в многослойном эпителии выделяют три
главных слоя: глубокий (ростковый), средний, или основной (не-
сколько рядов шиповатых клеток), и наружный (несколько рядов
клеток-пластинок). Отличительной особенностью многослойного
эпителия является постоянное перемещение клеток из глубокого
к наружному слою, слущивание клеток-пластинок и быстрое об-
новление эпителиальной поверхности при ее повреждении.
Существует три типа многослойных эпителиев: многослойный
плоский неороговевающий, многослойный плоский ороговеваю-
щий и переходный. Такое деление определено местом их располо-
жения, а разница в строении касается главным образом наружно-
го эпителиального слоя.
Многослойный плоский неороговевающий эпителий встречается
наиболее часто. Им выстланы преимущественно слизистые обо-
лочки некоторых органов пищеварения (до желудка), мочеиспус-
кательного канала и влагалища у самок, а также наружная повер-
хность роговицы. Название этот эпителий получил за плоскую фор-
му клеток наружного слоя, цитоплазма которых частично запол-
нена кератогиалином.
32
Рис. 2.18. Многослойный плоский ороговевающий эпителий (схема):
1 — базальный слой; 2 — шиповатый слой; 3 — зернистый слой; 4 — блестящий
слой; 5 — слой роговых чешуек
Многослойный плоский ороговевающий эпителий составляет глав-
ным образом наружный слой кожи, покрывающей тело животно-
го (эпидермис), и отличается повышенной прочностью и способ-
ностью к активному восстановлению (регенерации) при повреж-
дении. Наружный слой эпидермиса самый широкий; он состоит
из мертвых клеток (роговые чешуйки), тела которых сплошь запол-
нены кератином. Постепенное накопление в шиповатых клетках
кератина привело к образованию двух тонких, но хорошо замет-
ных дополнительных слоя: зернистого и блестящего, более выра-
женных в эпидермисе безволосых участков кожи, особенно в мя-
кишах (рис. 2.18). Преобладание процесса слущивания над процес-
сом восполнения снижает защитную функцию эпидермиса и мо-
жет привести к образованию кожных язв.
Переходный эпителий покрывает слизистую оболочку мочевы-
водящих путей (почечных лоханок, мочеточников, мочевого пу-
зыря, мочеполового канала у самцов). Он способен сильно растя-
гиваться и спадаться без образования сколько-нибудь заметных
складок. Построен переходный эпителий из трех слоев: глубокого,
среднего и наружного, различающихся по морфофизиологичес-
ким свойствам клеток. Способность клеток переходного эпителия
менять свою форму придает ему значительную потенцию на рас-
тяжение (это важно для мочевого пузыря) и делает поверхность,
обращенную в просвет органа, очень гладкой (это способствует
быстрому продвижению мочи по мочеточникам).
2 Зелепевский
33
Опорно-трофические ткани. Объединяют большую группу тка-
ней, обеспечивающих внутренние потребности организма (схе-
ма 2.2). Биологическое значение этих тканей связано с выполне-
нием трех главных функций: трофической (питающей), защитной
и опорной. Все опорно-трофические ткани объединяет два суще-
ственных момента: 1) они развиваются из одного источника —
мезенхимы; 2) в своем строении используют как клетки, так и
неклеточное (межклеточное) вещество. Среди опорно-трофичес-
ких тканей различают: кровь и соединительные ткани (собствен-
но соединительную, хрящевую и костную).
Мезенхима (mesenchyma) на ранних этапах эмбрионального раз-
вития является основной тканью зародыша и представляет собой
сетевидное клеточное образование, погруженное в гелеобразное меж-
клеточное вещество, в котором каждая клетка имеет собственную
оболочку. К моменту рождения мезенхимные клетки дифференци-
руются в полноценные клетки всех опорно-трофических тканей.
Эндотелий {endothelium) — это особый вариант однослойной
выстилки внутренней поверхности стенки всех кровеносных и
лимфатических сосудов, а также полостей сердца. Эндотелиаль-
ные клетки развиваются на ранних этапах эмбриогенеза из мезен-
химы и не только защищают подлежащие ткани, но и активно
участвуют в обмене веществ между кровью и окружающей сосуды
соединительной тканью.
Схема 2.2
34
Кровь (sanguis) как ткань диф-
ференцируется на ранних этапах
развития зародыша одной из пер-
вых и отличается по своему строе-
нию набором разнообразных кле-
ток (форменных элементов) и
жидким межклеточным веществом
(плазмой). У плода кровь первона-
чально образуется, как и другие
опорно-трофические ткани, из ме-
зенхимы, а после рождения обнов-
ление клеточных элементов идет за
счет ретикулярной ткани. Среднее
количество крови у животных со-
ставляет 7 — 9 % от массы тела.
Плазма крови составляет 58 —
65 % от ее общего объема. Это свет-
ло-желтая жидкость, содержащая
до 93 % воды и 7 % органических
и неорганических веществ. Среди
органических веществ особое зна-
чение имеют фибриноген, играю-
щий основную роль в процессах
свертывания крови, а также небел-
ковые азотистые вещества (амми-
ак, мочевина), определяющие во
многом физические свойства плаз-
мы. Углеводы, жиры, белки, кис-
лород, воду, витамины и соли
Рис. 2.19. Кровь крупного рогато-
го скота:
1 — эритроцит; 2, 6 — эозинофиль-
ный гранулоцит; 3 — средний лим-
фоцит; 4 — кровяные пластинки; 5,
9 — нейтрофильные гранулоциты;
7 — базофильный гранулоцит; 8 —
малый лимфоцит; 10 — моноцит;
И — большой лимфоцит
клетки получают из плазмы арте-
риальной крови, которая в виде
тканевой жидкости проникает че-
рез стенку капилляров. В нее же
выделяются все продукты жизне-
деятельности клеток.
Форменные элементы крови под-
разделяются на красные и белые
кровяные клетки (рис. 2.19). Эрит-
роциты, или красные кровяные
клетки (erythros — красный), — это безъядерные в зрелом состоя-
нии клетки, имеющие форму двояковогнутого (на месте бывшего
ядра) диска диаметром 4,5 — 7 мкм и толщиной около 2 мкм,
обеспечивающие перенос О2 и СО2. Количество эритроцитов в
крови млекопитающих измеряется несколькими миллионами в
1 мм3, что придает крови красный цвет и делает ее непрозрачной.
Среди клеток крови эритроциты являются долгожителями: в токе
35
крови они циркулируют НО —120 дней, после чего утилизируют-
ся в селезенке.
Лейкоциты, или белые кровяные клетки {leikos — бесцветный), —
это более крупные, чем эритроциты, округлые подвижные клет-
ки с различным по форме ядром и набором клеточных органелл.
Лейкоциты обеспечивают защиту организма животных от микро-
бов и токсинов либо посредством фагоцитоза, либо за счет выде-
ления особых веществ, нейтрализующих бактериальные яды. Ко-
личество лейкоцитов у млекопитающих значительно уступает ко-
личеству эритроцитов и измеряется тысячами в 1 мм3 крови. Лей-
коциты принято разделять на две группы: зернистые (гранулоци-
ты) и незернистые (агранулоциты). Количественное соотношение
разных типов лейкоцитов сравнительно постоянно и обычно в
общем анализе крови определяется как лейкограмма. Лейкоциты
развиваются из общего источника — стволовой клетки. Моноциты
и все гранулоциты образуются в костном мозге, а лимфоциты — в
специальных лимфоидных органах (лимфатических узлах, селе-
зенке, миндалинах, тимусе).
Зернистые лейкоциты (гранулоциты) имеют обычно лопастное
или сегментированное ядро, а в цитоплазме их выявляются плот-
ные частицы — гранулы или зерна. По характеру их окраски гра-
нулоциты подразделяются на: базофильные {базофилы}, эозино-
фильные {эозинофилы) и нейтрофильные {нейтрофилы) лейкоци-
ты.
Незернистые лейкоциты не содержат в цитоплазме сколько-
нибудь заметной зернистости и представлены лимфоцитами и
моноцитами.
Лимфоциты — это наиболее распространенные из лейкоцитов
клетки. Они являются активными фагами (макрофагами) и в оча-
ге микробного поражения приходят на помощь нейтрофилам (мик-
рофагам), а также участвуют в формировании клеточного и гумо-
рального иммунитета.
Моноциты — наиболее крупные клетки крови (от 12 до 20 мкм),
в условиях нормы они составляют 3 — 5 % от общего числа лейко-
цитов. Как и лимфоциты, моноциты имеют базофильную цито-
плазму и сравнительно большое рыхлое ядро. Выселяясь из сосу-
дов в очаги поражения на поздних стадиях воспаления, моноциты
могут превращаться в макрофаги. Они обладают и бактерицидной
способностью.
Кровяные пластинки, или тромбоциты, — мелкие, прозрачные,
слабобазофильные образования, являющиеся частями цитоплаз-
мы огромных клеток, расположенных в красном костном мозге
(мегакариоцитов). Размер тромбоцитов обычно не превышает 2 —
4 мкм при толщине 0,5 —0,7 мкм, а содержание в крови — 200 —
300 тыс. в 1 мм3. Кровяные пластинки участвуют в процессе свер-
тывания крови и остановке кровотечений.
36
Лимфа (lympha — чистая влага) вместе с плазмой крови и тка-
невой жидкостью составляет внутреннюю среду организма. Она
представляет собой прозрачную светло-желтую жидкость. Как и
кровь, лимфа относится к особому виду опорно-трофической ткани
и состоит из жидкой части (плазмы) и форменных (клеточных)
элементов. Плазма лимфы образуется из тканевой жидкости при
попадании последней в лимфатические капилляры и содержит про-
дукты жизнедеятельности клеток, чем объясняется разный состав
лимфы в разных частях тела. Лимфа, оттекающая от кишечника и
обогащенная жиром, имеет молочно-белый цвет и получает на-
звание — хилюс, или млечный сок. Образовавшаяся в капиллярах
лимфа оттекает по лимфатическим сосудам к соответствующим
данному органу или части тела лимфатическим узлам, выполня-
ющим барьерно-фильтрационную и иммунную функции. В них
лимфа очищается, обезвреживается и обогащается лимфоцита-
ми, составляющими 95 — 97% всего клеточного состава лимфы.
В конечном итоге лимфа собирается со всего тела в крупные лим-
фатические протоки, впадающие в краниальную полую вену.
Соединительные ткани отличаются хорошо развитым межкле-
точным веществом и подразделяются на собственно соединитель-
ные, хрящевую и костную ткани.
Собственно соединительные ткани выполняет трофическую,
опорную, защитную, а также и заместительную (участвуют в про-
цессах регенерации любых поврежденных тканей и заживления
ран) функции. Это обеспечивает им значительную востребован-
ность в организме. В зависимости от клеточного состава и характе-
ра организации межклеточного вещества собственно соединитель-
ные ткани, в свою очередь, принято делить на специальные (ре-
тикулярную и жировую) и волокнистые (рыхлую и оформлен-
ную).
Ретикулярная ткань сохраняет наибольшее сходство с мезен-
химой, имеет аналогичное сетевидное строение и состоит из свя-
занных в виде перекладин ретикулярных клеток и межклеточного
вещества, пронизанного большим числом ретикулиновых (арги-
рофильных) волокон. Ретикулярные клетки способны при необ-
ходимости отделяться и преобразовываться либо в макрофаги, либо
в кроветворные клетки, что и определяет биологическое значе-
ние ретикулярной ткани как защитной и кроветворной. Ретику-
лярная ткань сравнительно широко распространена в организме.
Она составляет основу красного костного мозга, лимфатических
узлов, селезенки, тимуса, миндалин, в небольших количествах
присутствует в печени, слизистых оболочках трубчатых органов, в
стенках кровеносных и лимфатических сосудов.
Жировая ткань выполняет трофическую и защитную функции.
Основу этой ткани составляют дифференцировавшиеся ретику-
лярные клетки, способные накапливать и отдавать жир, — жиро-
37
Рис. 2.20. Рыхлая неоформленная
соединительная ткань:
7 — эластическое волокно; 2 — кол-
лагеновые волокна; 3 — фибробласт;
4 — лимфоцит; 5 — гистиоцит
вые клетки. По цвету жира и характеру его расположения в цито-
плазме выявлены два типа жировых клеток: 1) белые жировые клет-
ки, из которых составлена основная масса жира; 2) бурые жиро-
вые клетки, встречающиеся более редко, жир в которых собран в
мелкие капли и слегка окрашен в коричневатый цвет. Жировые
клетки прослоены элементами рыхлой неоформленной соедини-
тельной ткани, по которой проходит много кровеносных сосудов.
Волокнистая рыхлая неоформленная соединительная ткань
(рис. 2.20) встречается практически во всех органах как сопровож-
дающая сосуды и нервы, а также образует различной толщины
прослойки: подслизистый слой в стенке трубчатых органов, меж-
дольковые (в железах), является основой для подкожного слоя (клет-
чатки) и жировой ткани.
Основную и постоянную массу клеток этой ткани составляют:
фибробласты (камбиальные клетки), фиброциты (дифференциро-
ванная форма фибробластов) и гистиоциты (активно мигрирую-
щие макрофаги, или блуждающие клетки). Непостоянно и в раз-
ных количествах в соединительной ткани присутствуют: ретику-
лярные, жировые, пигментные, тучные и плазматические клет-
ки, а также тканевые макрофаги (лимфоциты, выселившиеся из
кровеносных сосудов).
Волокнистые элементы межклеточного вещества соединитель-
ной ткани представлены в основном коллагеновыми, эластичес-
кими и аргирофильными (ретикулиновыми) волокнами. Коллаге-
новые волокна состоят из фибриллярного белка коллагена, имеют
плоскую, цилиндрическую и спиральную форму и толщину от 20
до 400 нм. Коллагеновые волокна не растягиваются, имеют попе-
речную исчерченность и обладают значительной прочностью. Эла-
стические волокна имеют гладкую поверхность, состоят из белка
эластина, поддаются растяжению и лежат одиночно в виде длин-
ных тонких и прямых нитей. Ретикулиновые (аргирофильные) во-
локна — это очень нежные, активно ветвящиеся волокна, имею-
щие поперечную исчерченность и выявляемые гистологически с
использованием серебра.
38
Аморфное (бесструктурное) межклеточное вещество — это по-
лупрозрачная, не видимая при световой микроскопии, вязкая мас-
са, продуцируемая главным образом фибробластами. Оно связы-
вает большое количество воды, активно участвует в обменных
процессах и способствует отделению волокнистых белков.
Волокнистая плотная неоформленная соединительная ткань вы-
полняет главным образом биомеханическую функцию. Из нее по-
строены основная часть среднего слоя кожи (дермы), а также
фиброзные капсулы и строма органов, наружные покровы (ад-
вентиции) трубчатых органов шеи и тазовой полости, основа бры-
жеек и сальников, мозговые оболочки. Плотная неоформленная
ткань построена аналогично рыхлой неоформленной, но отлича-
ется бедным клеточным составом (в основном фибробласты и фиб-
роциты) и значительным преобладанием в межклеточном веще-
стве волокнистых элементов (в основном коллагеновых и неболь-
шого числа эластических волокон) над аморфной массой. По сво-
ему ходу коллагеновые волокна активно обмениваются пучками
своих фибрилл и связываются тонкими волокнистыми связями
(рис. 2.21), образуя вместе с эластическими волокнами единый
функциональный биомеханический ансамбль.
Волокнистая плотная оформленная соединительная ткань пост-
роена преимущественно из хорошо развитых волокон, располо-
женных в небольшом количестве аморфного вещества. В зависимо-
сти от характера преобладающих волокон эта ткань подразделяет-
ся на два типа: плотную оформленную коллагеновую и плотную
оформленную эластическую ткани.
Плотная оформленная коллагеновая
ткань встречается там, где имеет-
ся повышенная механическая на-
грузка определенной направленно-
сти (сухожилия, фасции, апонев-
розы, капсулы и связки в суставах).
Эта ткань отличается значительной
прочностью. Ее межклеточное ве-
щество представлено в основном
толстыми пучками (I,II, III и т.д.
порядка) коллагеновых волокон,
ориентированных вдоль длинной
оси связки или сухожилия. Фиброб-
ласты и фиброциты зажаты между
пучками в виде своеобразных вер-
тикальных колонок. В плотной офор-
мленной эластической ткани преоб-
ладают эластические волокна, об-
разующие густые резиноподобные
сети, узкие ячеи которых растяну-
Основные волокна (7) связаны меж-
ду собой системой тонких связоч-
ных волокон (2). Сканирующая элек-
тронная микроскопия (ув. х 4500)
39
ты вдоль линий силовых деформирующих нагрузок и содержат
небольшое количество фиброцитов и аморфного вещества, а так-
же отдельные тонкие коллагеновые волокна. Из такой ткани по-
строены главным образом эластические желтые связки позвоноч-
ного столба (выйная, межостистые, межпоперечные, междуго-
вые).
Хрящевая ткань (или хрящ) построена из крупных с повышен-
ной упругостью клеток (хондробластов и хондроцитов) и плотного
межклеточного вещества, состоящего из волокон и аморфной ча-
сти. Эта ткань является чисто опорной. По своему строению хрящ
подразделяется на три типа: гиалиновый, эластический и волок-
нистый. Снаружи все типы хрящей покрыты надхрящницей (пери-
хондром). Кровеносных сосудов в хрящевой ткани нет, а питание
их осуществляется диффузно из надхрящницы.
Гиалиновый (стекловидный) хрящ является наиболее распрост-
раненным видом хрящевой ткани. Из него построены реберные и
суставные хрящи, носовая перегородка и все хрящевые включе-
ния в стенках органов дыхания. Он составляет основную массу
хрящевого скелета в эмбриональный период. Гиалиновый хрящ
состоит из хондроцитов и гомогенного межклеточного вещества.
Макроскопически он имеет очень ровную и гладкую поверхность
и похож на бело-голубое стекло. Использование специальных ме-
тодов обработки показало наличие в межклеточном веществе топ-
ких коллагеновых волокон, придающих гиалиновому хрящу по-
вышенную упругость и прочность. Хондроциты в наружных (мо-
лодых) участках хряща лежат одиночно, а в более глубоких (зре-
лых) участках — в виде групп похожих клеток, заключенных в
особые капсулы из молодого хря-
ща (рис. 2.22).
Эластический хрящ построен в
основном аналогично гиалиново-
му хрящу, но содержит меньше
изогенных групп, в его межклеточ-
ном веществе преобладают элас-
тические волокна, придающие
хрящу желтый цвет и формирую-
щие тонкие, разные по густоте
сети. Он образует ушные ракови-
ны, хрящевые части слуховой тру-
бы и наружного слухового прохо-
Рис. 2.22. Гиалиновый хрящ:
1 — надхрящница; 2 — зона моло-
дых хрящевых клеток (хондроблас-
тов); 3 — хрящевое основное веще-
ство; 4 — группы зрелых хрящевых
клеток (хондроцитов)
да, а также надгортанник, рожко-
вые и голосовые отростки черпа-
ловидных хрящей гортани.
Волокнистый, или коллагеново-
волокнистый, хрящ отличается зна-
чительной прочностью на разрыв
40
обусловленной наличием в межклеточном веществе толстых пуч-
ков параллельно идущих коллагеновых волокон. Из волокнистого
хряща построены связка головки бедренной кости, хрящевой диск
в височно-челюстном и мениски в коленном суставах, волокнис-
тое кольцо в межпозвоночных хрящах, а также лонная часть тазо-
вого сращения; встречается он и в местах прикрепления связок и
сухожилий к костям.
Костная ткань отличается повышенной прочностью и одно-
временно упругостью. Она выполняет защитную, опорную и, в
определенной мере, амортизационную функции, депонирует не-
органические вещества и активно участвует в минеральном об-
мене. Из нее построены все кости скелета, а также сердечные и
половочленные кости некоторых животных. Костная ткань со-
стоит из небольших отростчатых костных клеток (остеоцитов) и
межклеточного вещества, включающего аморфную массу (оссео-
мукоид), большое количество оссеиновых (коллагеноподобных)
и немного эластических волокон. Твердость межклеточному ве-
ществу придают минеральные соли, главным образом фосфаты
и карбонаты Са и Mg, масса которых в костях составляет более
97 % от общего содержания в организме. Минеральные вещества
постоянно выводятся из костей в кровь и используются для нужд
организма животных, что требует постоянного их пополнения с
кормом.
Снаружи кости покрыты надкостницей, или периостом (peri —
вокруг, около, os — кость), которая помимо соединительной тка-
ни, чувствительных нервов и питающих кость сосудов содержит
два вида клеток: остеобласты, или костеобразователи (камбиаль-
ные клетки), и остеокласты, или костеразрушители. Эти клетки
выселяются в костную ткань и в зависимости от испытываемых
ею в данный момент нагрузок обеспечивают постоянную каче-
ственную и количественную перестройку костей, а также их сра-
щивание после перелома.
По характеру распределения волокон в межклеточном веще-
стве принято различать два типа костной ткани: грубоволокнис-
тую и тонковолокнистую (пластинчатую). Грубоволокнистая кост-
ная ткань используется для формирования скелета на ранних ста-
диях эмбриогенеза и к моменту рождения плода путем преобразо-
вания и упорядоченности всех гистологических элементов пре-
вращается в тонковолокнистую, или пластинчатую, костную ткань.
В тонковолокнистой, или пластинчатой, костной ткани пучки ос-
сеиновых волокон, склеенные оссеомукоидом и пропитанные ми-
неральными солями, располагаются параллельно и формируют
прочные и упругие на изгиб структуры — костные пластинки тол-
щиной 5 — 7 мкм. Внутри они пронизаны тонкими канальцами,
заполненными тканевой питающей жидкостью и соединяющими
особые костные полости, в которых располагаются остеоциты.
41
Рис. 2.23. Перпендикулярный срез малой берцовой кости собаки:
1 — центральный канал остеона; 2 — циркулярные костные пластинки остеона;
3 — вставочные пластинки. Сканирующая электронная микроскопия (ув. х 300)
В костях пластинки располагаются вдоль линий силовых нагрузок,
формируя губчатое и компактное костное вещество. Структурной
единицей, значительно повышающей прочность компактного ко-
стного вещества, является остеон, или гаверсова система, — осо-
бое многослойное образование, развернутое вдоль главной оси
кости и состоящее из цилиндрических костных пластинок, встав-
ленных одна в другую, с внутренним (гаверсовым) каналом (рис.
2.23). Пространство между остеонами плотно заполнено дугооб-
разно изогнутыми (вставочными) костными пластинками, кото-
рые являются остатками прежде существовавших остеонов. Эти
пластинки пронизаны идущими от надкостницы поперечными
каналами, составляющими с гаверсовыми каналами единую сис-
тему для распространения внутри костей сосудов и нервов. Снару-
жи оба типа костного вещества покрыты продольно идущими об-
щими (генеральными) костными пластинками. По характеру внут-
рикостной организации кости в скелете подразделяются на пять
типов: губчатые, пластинчатые (плоские), пневматизированные (со-
держащие внутри полости или пазухи, заполненные воздухом),
трубчатые (имеющие два губчатых конца — эпифиза и полую труб-
чатую середину — диафиз, стенка которого построена из компак-
тного костного вещества) и смешанные, имеющие губчатые и пла-
стинчатые части.
Мышечные ткани. Обладают способностью к активному укоро-
чению (сокращению) и осуществляют все двигательные процес-
сы в организме животных и человека. По происхождению и мор-
фофизиологическим характеристикам различают три типа мышеч-
ной ткани: поперечно исчерченную (скелетную), гладкую (орган-
ную) и специальную (сердечную).
42
Поперечно исчерченная (скелетная) мышечная ткань развива-
ется из мезодермы (среднего зародышевого листка), отличается
способностью к быстрым и сильным сокращениям и в своей ра-
боте подчиняется воле животного и человека (произвольная мус-
кулатура). Из этой ткани построены все скелетные мышцы, мыш-
цы глазного яблока и ушной раковины, а также язык и мышеч-
ные оболочки глотки и шейной части пищевода. Иерархические
уровни строения скелетной мышцы показаны на рис. 2.24. Струк-
турными единицами этой ткани являются многоядерные прото-
плазматические симпласты (поперечно исчерченные мышечные во-
локна) диаметром около 0,1 мм и длиной до 10—12 см, объеди-
ненные в различные по размеру пучки посредством прослоек из
рыхлой соединительной ткани (эндо- и перимизиев), в которых
. _ Актиновый филамент
-Миозиновые филаменты
Рис. 2.24. Иерархические уровни организации скелетной мышцы
43
проходят сосуды и нервы. Под сарколеммой волокон располага-
ются многочисленные удлиненные ядра, а в саркоплазме распо-
ложено несколько десятков основных структур сократительного
аппарата — поперечно исчерченных миофибрилл. На электронно-мик-
роскопическом уровне видно, что они состоят из тонких сократи-
тельных миозиновых и актиновых филаментов.
Неисчерченная гладкая мышечная ткань развивается из мезен-
химы и отличается медленными ритмичными сокращениями. Она
относится к непроизвольной, т.е. не подчиняющейся воле, мус-
кулатуре. Из нее построены мышечные оболочки практически всех
трубчатых органов и средняя оболочка (медия) кровеносных и
лимфатических сосудов; встречается она и в некоторых паренхи-
матозных органах (органная, или внутренностная, мускулатура).
Структурными единицами этой ткани являются одноядерные ве-
ретенообразные мышечные клетки (гладкие мышечные волокна)
длиной от 40 до 300 мкм (в беременной матке до 500 мкм), объе-
диненные в пучки разного размера. Ядро в гладких мышечных во-
локнах лежит в их средней расширенной части, а саркоплазма
содержит большое число тонких сократительных элементов (мио-
филаментов), не имеющих сколько-нибудь заметных поперечных
полосок.
Сердечная мышечная ткань встречается только в стенке серд-
ца, образуя ее среднюю оболочку — миокард. Миокард способен
к длительной работе без значительного перерыва (неутомляемая
мускулатура) и отличается ритмичными непроизвольными со-
кращениями, скорость и сила которых больше, чем у гладкой,
но несколько меньше, чем у поперечно исчерченной мышечной
ткани. Структурными единицами миокарда являются прямоуголь-
ные одноядерные мышечные клетки — кардиомиоциты. В отличие
от клеток других мышечных тканей миофибриллы в кардиомио-
цитах, имеющие слабо выраженную поперечную исчерченность,
располагаются не в центре, а по периферии клетки под сарко-
леммой. Удлиненное небольшое ядро лежит, как правило, в цен-
тре клетки. Кардиомиоциты соединены посредством вставочных
пластинок в тяжи, или трабекулы, идущие в разных направлени-
ях или образующие слои. Во многих местах между клеточными
тяжами образуются анастомозы, что создает иллюзию симпласта
и способствует одновременному их сокращению (рис. 2.25). По-
Рис. 2.25. Сердечные мышечные во-
локна (миокард):
1 — кардиомиоциты; 2 — вставочные
пластинки
44
мимо рабочих кардиомиоцитов,
обеспечивающих сокращение и
расслабление, в миокарде есть и
проводящие возбуждение (атипич-
ные) кардиомиоциты (волокна
Пуркинье), которые обеспечивают
синхронность в работе мускулату-
ры предсердий и желудочков; они
отличаются незначительным чис-
лом миофибрилл и более светлы-
ми ядром и саркоплазмой.
Нервная ткань. Является своеоб-
разной и высокодифференциро-
ванной тканью. Особое свойство
нервной ткани — возбудимость, т. е.
способность отвечать на раздраже-
ние в виде специфической реак-
ции — образовании нервного импуль-
са, и быстро проводить нервный
импульс на значительные рассто-
яния — свойство проводимости.
Такая специфика нервной ткани
определила функции построенной
из нее нервной системы: 1) осу-
ществление интеграции — взаимо-
связи тканей и органов в организ-
ме; 2) поддержание гомеостаза —
структурного и физико-химиче-
ского постоянства в организме;
3) обеспечение адаптации — при-
спосабливаемое™ организма к по-
стоянно изменяющимся условиям
внешней среды. В нервной системе
можно легко выделить две состав-
ляющие: главную морфофункциональную единицу — нервную
клетку (нейрон) с отростками, а также вспомогательный аппарат,
представленный клетками нейроглии. Нейроны и основная часть
нейроглии (макроглия) развиваются из утолщения эктодермаль-
ного зародышевого листка (нервной пластинки), а незначительная
часть нейроглии (микроглия) происходит из мезенхимы.
Нейрон состоит из довольно большого тела (нейросомы) и от-
ростков (дендритов и аксона) (рис. 2.26).
Тело нейрона, как правило, имеет многоугольную (звездчатую)
форму. В нем находятся светлое крупное ядро с 1 — 2 ядрышками и
основные органеллы: большое число митохондрий, комплекс Голь-
джи и хорошо развитая гранулярная ЭПС с крупными рибо-
Рис. 2.26. Мотонейрон спинного
мозга (схема):
1 — тело; 2 — дендриты; 3 — аксон;
4 — перехват Ранвье; 5 — шваннов-
ская клетка; а — синапсы; б — ак-
сонный холмик; в — терминальные
окончания аксона; стрелками пока-
заны направления передачи сигналов
45
сомальными гранулами, выявляющаяся при обычной микроско-
пии как тигроидное вещество, или глыбки Ниеля. Здесь же можно
увидеть густую сеть из тонких белковых нитей (нейрофиламентов
и нейрофибрилл), распространяющихся в отростки и выполняю-
щих биомеханическую функцию.
Среди отростков, отходящих от тела нейрона, различают два
типа: дендриты (dendron — дерево) — это обычно ветвящиеся от-
ростки и аксон (ахоп — ось), или нейрит, — это прямой, неветвя-
щийся и выходящий их аксонного холмика тела отросток. По ден-
дритам к телу нейрона приходят афферентные (восходящие, или
центростремительные) нервные импульсы, а по аксону от тела
нейрона к клеткам других тканей или другим нейронам идут эф-
ферентные (нисходящие, или центробежные) нервные импульсы.
Нейрон окружен оболочкой {неврилеммой), которая с тела пере-
ходит на отростки {безмякотное, или безмиелиновое, волокно) и
может дополняться окружением из специальных клеток макро-
глии {мякотные, или миелиновые, волокна). Местом передачи не-
рвных импульсов от нейрона к мышечной или железистой клет-
ке, а также от нейрона к нейрону является особое терминальное
{концевое) образование — синапс.
По функциям различают чувствительные, двигательные и про-
межуточные нейроны. Чувствительные (афферентные, или рецеп-
торные) нейроны на концах своих дендритов образуют специаль-
ные терминалии (рецепторы). Последние имеют разное строение;
они воспринимают внешние или внутренние раздражения, пере-
водят их в нервный импульс и передают в спинной или головной
мозг. Двигательные (эфферентные) нейроны передают нервный
импульс от спинного и головного мозга к мышцам или железам,
т.е. органам-эффекторам (органам-исполнителям). Промежуточ-
ные (ассоциативные, или вставочные) нейроны передают нервный
импульс от чувствительных к двигательным нейронам. Объедине-
ние этих нейронов напрямую или через промежуточный нейрон
при передаче нервного импульса является основным средством
обеспечения работы нервной системы {рефлекторная дуга).
Клетки нейроглии весьма многообразны и встречаются как в
сером, так и в белом мозговом веществе. Они выполняют опор-
ную, трофическую и защитную функции. Среди глиальных клеток
принято различать: макроглию (эпендимоглиоциты, астроглиоци-
ты, олигодендроглиоциты) и микроглию (микроглиоциты).
Контрольные вопросы
1. Что такое ткани? Какие типы тканей встречаются в животном орга-
низме?
2. Как построены и какое значение для организма имеют эпителиаль-
ные ткани?
46
3. Опишите строение крови как ткани.
4. Что определяет способность мышечных тканей к сокращению и какое
это имеет значение для мышечной работы?
5. Что делает кость прочной и как эту прочность сохранить и увели-
чить?
6. Что является основной структурно-функциональной единицей не-
рвной ткани?
ЭМБРИОЛОГИЯ
В ходе длительного пути исторического развития у млекопита-
ющих выработался особый, половой, путь размножения. Он харак-
теризуется четырьмя основными особенностями: 1) половым ди-
морфизмом между самцами и самками, т. е. различным строением
полового аппарата и половых клеток (гамет); 2) внутренним опло-
дотворением, при котором половые клетки самца вводятся в по-
ловые органы самки; 3) внутриутробным развитием плода, т. е.
достаточно длительным созреванием (вынашиванием) его внутри
материнского организма; 4) последующим после рождения вскар-
мливанием детеныша материнским молоком (отсюда название клас-
са — млекопитающие).
Полевой диморфизм. Особое значение для полового размноже-
ния имеют высокоспециализированные (половые) клетки гаме-
ты — спермий у самцов и яйцеклетка у самок, отличающиеся от
обычных соматических клеток одним набором хромосом. Биоло-
гическое значение гамет заключается в том, что, сливаясь при
оплодотворении, они образуют зиготу — уникальную структуру,
из которой и развивается новый живой организм того же вида,
что и родители.
Спермий (спермиоцит, или спермоцит) представляет собой
микроскопическую клетку, длина которой у сельскохозяйствен-
ных животных находится в пределах 35 — 78 мкм. Она отличается
своеобразным строением. У каждого зрелого спермия принято раз-
личать три основные части: овально-уплощенную головку, где раз-
мещается ядро клетки и комплекс Гольджи, формирующий осо-
бое тельце, заполненное ферментами (акросому); суженную
часть — шейку, где располагается центросома, дающая начало наи-
более длинной части (70 % от общей длины) — хвостику, движе-
нием которого спермий перемещается. Форма и размер отдельных
частей спермия, особенно головки, строго специфичны для каж-
дого вида животных.
Яйцеклетка (овоцит, или ооцит) является самой крупной клет-
кой в организме животных. Зрелая яйцеклетка имеет круглое ядро
с крупным ядрышком и отложения в цитоплазме желтка (трофи-
ческих включений жировой, белковой и углеводной природы,
предназначенных для питания зародыша на ранних этапах его раз-
47
Рис. 2.27. Схема спермио- и оогенеза (овогенеза):
А — аутосомы; Хи Y — половые хромосомы (отцовские — черные, материнские —
белые); 1 — спермиогонии; 2— овогонии; 3 — спермиоциты первого порядка; 4 —
овоциты первого порядка; 5 — образование тетрад; 6 — спермиоциты второго
порядка; 7 — овоциты второго порядка; 8 — редукционные (полярные) тельца
1-го и 2-го порядков; 9 — спермиды; 10 — спермин; 11 — зрелая яйцеклетка
(с гаплоидным набором хромосом)
вития). Она окружена двумя оболочками: первичной, состоящей из
внутреннего (желточного) слоя и тонкой плазмалеммы, и вто-
ричной {прозрачной оболочки), образованной фолликулярными клет-
ками. Средний диаметр яйцеклетки у коровы — до 180 мкм, у
собаки и кошки — 120—150 мкм. Яйцеклетка не способна к само-
стоятельному движению.
Развитие половых клеток (гаметогенез) в организме самца и
самки определяется соответственно как спермиогенез и овогенез
(рис. 2.27).
Спермиогенез протекает в извитых канальцах семенника. При
этом одновременно в половых органах самца развиваются милли-
арды половых клеток и каждая проходит четыре периода сложных
преобразований: размножение, рост, созревание, формирование.
Размножение — 7 — 9 генераций митотического деления спермио-
гоний, имеющих диплоидный (двойной) набор хромосом и рас-
положенных под оболочкой извитого канальца. Рост — ассимиля-
48
ция питательных веществ, увеличение в размере и превращение в
более крупные клетки — спермиоциты I порядка. Период созрева-
ния — последовательно протекающие деления — мейоз, или ре-
дукционное деление, и следующий за ним митоз, или уравни-
тельное деление; в результате получаются сначала два спермиоци-
та II порядка, а затем четыре равноценных спермиды с одним на-
бором хромосом (гаплоидные клетки). Располагаясь почти у про-
света канальца, спермиоциты II порядка погружаются передним
концом (головкой) в особые вспомогательные сертолиевы клетки
и подвергаются значительным морфологическим преобразовани-
ям (этап формирования)', уменьшается объем цитоплазмы, переме-
щаются ядро и органеллы; в результате всех преобразований из
каждой спермиды получается клетка особой формы — спермий.
Зрелые спермин выталкиваются в просвет извитых канальцев
семенника. Образование спермиев в организме самца начинается
в период полового созревания и продолжается достаточно долго.
Спермиогенез нарушается у самцов (особенно у производителей)
при плохом содержании, а также при неполноценном или недо-
статочном питании.
Овогенез в отличие от спермиогенеза начинается задолго до
рождения самки и протекает с разной интенсивностью в три эта-
па: размножение, рост и созревание. Кроме того, весь цикл раз-
вития яйцеклетки проходит внутри специального защитного об-
разования — фолликула. Этап размножения и малого роста у яйце-
клеток начинается в ранний эмбриональный период развития са-
мой самки, когда среди мелких клеток, активно проникающих
внутрь закладки яичника из наружной ее части, образуются от-
дельные более крупные — первичные половые клетки, или ового-
нии. Сначала они многократно делятся митозом, создавая запас
«зародышей» половых клеток, во много раз превышающий по-
тенциальные возможности деторождения у самок, но к моменту
рождения самки овогонии теряют способность к делению и пере-
ходят к активной ассимиляции питательных веществ, существен-
но увеличивая свой размер и преобразуясь в овоцит I порядка. После
рождения самки рост половых клеток приостанавливается вплоть
до момента полового созревания, после чего они вновь начинают
расти (период большого роста), а в цитоплазме овоцита I порядка
начинает накапливаться желток. За весь период роста овоцит I
порядка увеличивается в размере по сравнению с овогонией в
сотни раз.
На всем протяжении двух описанных периодов параллельно с
развитием яйцеклеток развиваются и фолликулярные клетки. Они
возникают в эмбриональной закладке яичника одновременно с
овогониями. Вокруг овоцитов I порядка фолликулярные клетки
формируют рыхлое обрамление (первичные, или примордиальные,
фолликулы) — начало формирования фолликулярной зоны яич-
49
2
Рис. 2.28. Схематическое изображение оплодотворения яйцеклетки и дроб-
ления зиготы:
1 — граафов пузырек (пузырчатый фолликул); 2 — яичник; 3 — воронка яйцево-
да; 4 — лопнувший фолликул; 5 — яйцеклетка; 6 — фолликулярные клетки;
7 — оплодотворение в ампуле; 8 — спермин; 9 — зигота; 10 — редукционные
(полярные) клетки; 11 — рог матки; 12 — двухбластомерная стадия; 13 — четы-
рехбластомерная стадия; 14 — восьмибластомерная стадия; 75 — морула; 16 —
зародышевый пузырь; 17 — слизистая оболочка матки; 18 — железы матки
ника. После рождения в период большого роста фолликулярные
клетки размножаются и образуют вокруг половой клетки сначала
многослойную стенку вторичного фолликула, а затем путем рас-
слоения — граафов пузырек, заполненный фолликулярной жид-
костью. Непосредственно вокруг самой половой клетки группиру-
ются «приближенные» фолликулярные клетки {лучистый венец),
прикрепляющие ее к стенке фолликула {яйценосный бугорок), обес-
печивающие питанием и создающие вокруг нее вторичную {блес-
тящую) оболочку.
Период созревания яйцеклетка проходит уже в яйцеводе (рис.
2.28), куда она попадает после разрыва граафова пузырька {овуля-
ции), побуждаемого нервной системой, и выпадения вместе с лу-
чистым венцом на стадии овоцита I порядка с поверхности яич-
ника в воронку яйцевода. Этот период, как и в спермиогенезе,
заключается в двух быстро следующих друг за другом делениях.
Сначала редукционным делением (мейозом) из овоцита I поряд-
ка образуются большой и малый {первое редукционное тельце) ово-
циты II порядка, имеющие по одному набору хромосом. Затем обе
клетки делятся обычным митотическим делением, в результате
которого из большого овоцита II порядка получается одна боль-
шая клетка — зрелая яйцеклетка и небольшое клеточное тело —
50
второе редукционное тельце, а из малого овоцита 11 порядка (пер-
вое редукционное тельце) — два редукционных тельца. Таким об-
разом, из одного овоцита 1 порядка получаются одна зрелая и
способная к оплодотворению яйцеклетка и три мелких побочных
и не способных к оплодотворению клетки (редукционных, или
полярных, тельца). Овогенез в отличие от спермиогенеза сопря-
жен у самки с определенной периодизацией половой жизни (по-
ловыми циклами), отличающимися у разных видов животных своим
ритмом и продолжительностью.
В спермиогониях и овогониях содержатся по две гетерохромо-
сомы (непарные), определяющие пол потомства. При этом в спер-
миогониях они неодинаковые (Хи У), а в овогониях — одинако-
вые (Хи X). После редукционного деления в каждой из половых
клеток сохраняется только одна гетерохромосома. Значит, при оп-
лодотворении яйцеклетка, имеющая Х-непарную хромосому, мо-
жет получить либо Y-, либо Х-непарную хромосому спермия.
В первом случае родится самец, а во втором — самка.
Оплодотворение у млекопитающих, в том числе у сельскохо-
зяйственных животных, происходит, как правило, в яйцеводе. Оно
состоит в слиянии спермия с яйцеклеткой и осуществляется пос-
ле процесса осеменения, т. е. введения спермиев во влагалище (у
рогатого скота) или матку (у лошади, свиньи и собаки) и само-
стоятельного продвижения спермиев к месту оплодотворения. На
продвижение по половым путям самки спермин затрачивают от
одного (у собак и лошадей) до пяти-шести часов (у крупного и
мелкого рогатого скота). При этом из нескольких миллиардов вве-
денных спермиев до яйцевода доходят не более нескольких сотен
тысяч, и на первой стадии оплодотворения они атакуют каждую
из овулировавших яйцеклеток (рис. 2.29), разрушая ферментами
окружающий ее лучистый венец. Внутрь яйцеклетки для осуще-
Рис. 2.29. Яйцеклетка, атакованная спермиями:
А — ув. х 580; Б - ув. х 2900
51
ствления оплодотворения проникает обычно только одна половая
клетка самца (рис. 2.30), после чего наружный, прилежащий к
плазмалемме слой цитоплазмы яйцеклетки уплотняется и образу-
ется дополнительная оболочка (желточная, или оболочка оплодот-
ворения), исключающая проникновение в эту яйцеклетку других
спермиев.
После этого начинается непродолжительный период подготов-
ки к оплодотворению, в ходе которого в ядрах обеих половых кле-
ток уплотняется хроматиновая субстанция, в цитоплазме яйце-
клетки происходят активные процессы сложного перемещения и
зонирования, а спермий теряет хвостик и шейку. Затем осуществ-
ляются слияние ядерного материала спермия и яйцеклетки (про-
нуклеуса самца и пронуклеуса самки) и объединение их гаплоид-
ных хромосомных наборов (сингамия), т.е. совершается процесс
оплодотворения. В результате образуется особая клетка (зигота) с
новыми свойствами, отличающими ее и от спермия, и от яйцек-
летки, с общим ядром, содержащим двойной набор хромосом,
объединивших наследственные качества отца и матери (ядро дроб-
ления). Редукционные тельца помещаются на анимальном полюсе
зиготы и постепенно утилизируются.
Индивидуальное развитие (онтогенез) сельскохозяйственных
животных. Развитие представителей плацентарных млекопитающих
складывается из двух основных периодов: внутриутробного, или
пренатального, и внеутробного, или постнатального.
Внутриутробное развитие детеныша происходит внутри тела
матери и охватывает временной период от возникновения зиготы
до рождения плода. Он включает весь комплекс генетически обус-
ловленных преобразований различных участков клеточного тела
зиготы с постепенным формированием клеток, тканей и органов
зародыша, а также комплекса внезародышевых структур, обеспе-
чивающих защиту и питание зародыша на всех этапах внутриут-
робного развития. Особенно сложные и быстро протекающие про-
цессы прогрессивного изменения зародышевого материала осу-
ществляются в самом начале внутриутробного развития зароды-
ша, поэтому в нем принято выделять два периода: эмбриональ-
Рис. 2.30. Ранняя стадия развития брюшной по-
лости (схема):
1 — нервная трубка; 2 — миотом; 3 — склеротом; 4 —
закладка мочеполовых органов; 5 — хорда; 6 — ки-
шечник; а — висцеральный и б — париетальный ли-
стки серозной оболочки; в — брыжейка; г — перито-
неальная полость
52
ный (от оплодотворения до образования плаценты) и плодный
(далее до рождения).
Эмбриональный, или зародышевый, период характеризуется про-
цессами постоянного размножения, перемещения и дифферен-
циации клеток зародыша, протекающими в определенной после-
довательности. Он начинается с дробления зиготы, которое насту-
пает у большинства сельскохозяйственных животных на 2-е сутки
после оплодотворения, и длится у крупного рогатого скота обыч-
но 6 — 7 суток (у овец — 3 — 5 суток, у свиньи — 3 — 4 суток) по
мере продвижения зиготы по яйцеводу к верхней части рога мат-
ки за счет волнообразного движения их стенок.
Дробление зиготы имеет генетическую обусловленность и про-
исходит посредством деления, в результате которого она активно
распадается на быстро увеличивающееся число мелких клеток —
бластомеров. Последние при этом не расходятся, а постепенно
образуют сначала округлый зародышевый комочек (морулу), а за-
тем однослойный пузырек (бластулу, или бластоцисту), в кото-
ром вследствие неравномерно протекающего дробления форми-
руются два полюса — зародышевый узелок, или эмбриопласт, и
трофобласт. Снаружи пузырек покрыт сохранившейся со времен
яйцеклетки прозрачной оболочкой, а внутри имеет небольшую
полость (первичная полость зародыша, или бластоцель). У круп-
ного рогатого скота к 9-м суткам он достигает размера 1 мм.
В течение последующих 1 — 2 суток бластоциста, до этого сво-
бодно перемещавшаяся в роге матки, теряет блестящую оболочку
и погружается (имплантируется) в слизистую оболочку матки,
выделяющую особый секрет (маточное молочко), который благо-
даря ворсинкам трофобласта используется для питания зародыша
вплоть до образования плаценты, т. е. до конца зародышевого пе-
риода. Таким образом устанавливается первоначальная связь мате-
ри и зародыша.
Активизация размножения и преобразования бластомеров в
зародышевом узелке после имплантации зародыша приводит к
образованию зародышевого диска с более плотной средней час-
тью (зародышевый щиток), где смещением клеток образуются пер-
вичная полоска и первичный (гензеновский) узелок.
По мере количественного роста происходит и качественное
преобразование зародышевого материала. Клеточная масса заро-
дыша начинает расщепляться сначала на два зародышевых лист-
ка — эктодерму (наружный листок) и энтодерму (внутренний ли-
сток). К 20-м суткам у крупного рогатого скота и 12-м суткам у
овцы путем сложного клеточного перемещения их зародышевой
полоски между экто- и энтодермой появляется третий (средний)
зародышевый листок — мезодерма, а из гензеновского узелка на-
чинает образовываться хорда. Эти первые организационные груп-
пы клеток эмбриона отличаются генетически обусловленной внут-
53
ренней специализацией и в дальнейшем значительно расходятся
как по морфологической организации, так и по биологическому
значению. На протяжении раннего эмбрионального периода каж-
дый из зародышевых листков проходит свой путь морфогенети-
ческих преобразований. Плоские клетки энтодермы, активно раз-
растаясь вниз вдоль трофобласта, формируют первичный желточ-
ный мешок, куда заключается находящаяся в бластоцеле жидкость,
временно, до образования плаценты, питающая зародыш. Из эк-
тодермального листка образуются эпидермис кожи и нервная пла-
стинка, которая преобразуется в нервную трубку и даст начало
всем составляющим нервной системы.
Наиболее сложные процессы структурирования происходят в
материале мезодермы. Первоначально она развивается по бокам
от осевых органов зародыша (хорды и нервной трубки) в виде
двух растянутых вдоль зародышевой полоски мешков между экто-
и эндодермой. С 20—22-х суток у крупного рогатого скота и с 16 —
17-х суток у овец оба мезодермальных мешка разделяются на две
части: верхнюю и нижнюю.
Верхняя часть постепенно, начиная от головного конца заро-
дыша, сегментируется на отдельные участки (сомиты) и дает на-
чало почти всем основным элементам стенки тела животного, обус-
ловив общие принципы его построения: двустороннюю (билате-
ральную) симметрию и частичную метамерию. Каждый мезодер-
мальный сомит подразделяется натри части (см. рис. 2.30): 1) дер-
матом — наружная часть, из которой образуется основа кожи;
2) склеротом лежит под дерматомом и дает начало всем хрящам и
костям скелета; 3) миотом — внутренняя часть, зачаток скелет-
ной мускулатуры.
Нижняя часть мезодермальных мешков (спланхнотом) состоит
из двух клеточных пластинок, или листков: пристенного (парие-
тального) и органного (висцерального), свободное пространство
между которыми именуется вторичной полостью тела (целомом)
и впоследствии полностью разделяется специальной перегород-
кой (диафрагмой) на грудную и брюшную полости, где появля-
ются закладки соответственных органов. Оба мезодермальных ли-
стка образуют в названных полостях серозные листки двух оболо-
чек — плевры и брюшины, покрывающих изнутри стенки полос-
тей и одевающих расположенные в них органы. Кроме того, из
материала висцерального мезодермального листка, или спланх-
ноплевры, образуются гладкие мышцы и соединительнотканные
элементы внутренних органов, а из париетального мезодермаль-
ного листка, или соматоплевры, образуются мышцы конечностей.
Промежутки между всеми зародышевыми листками заполняет пер-
вичная соединительная ткань — мезенхима.
Очень важным моментом раннего эмбриогенеза является раз-
деление зародышевых листков на две части: 1) зародышевую, из
54
материала которой постепенно сформируется тело зародыша;
2) внезародышевую, из которой создадутся временные структуры
для защиты и питания развивающегося организма. Процесс раз-
деления всех зародышевых листков на зародышевую и внезароды-
шевую части начинается у крупного рогатого скота с 20-х суток, а
у овцы — с 16-х суток и осуществляется посредством парной ту-
ловищной складки, образующейся в центре эмбриональной зак-
ладки по бокам от зародышевой полоски. В результате зародыш
округляется и вбирает в себя центральную (зародышевую) часть
экто-, мезо- и энтодермы, т.е. центральную часть всей эмбрио-
нальной закладки, приподнимается над остальной (внезародыше-
вой) частью зародышевых листков, составляющей боковые части
эмбриональной закладки, и постепенно от них обособляется.
С появлением плаценты эта связь сохраняется только посредством
пупочного канатика. Клетки зародышевой части зародышевых ли-
стков начинают формировать зачатки кожи, костей, мышц и внут-
ренних органов.
К внезародышевым органам (плодным оболочкам), временно
появляющимся во внутриутробном периоде развития млекопита-
ющих животных, относятся образующиеся на ранних этапах эмб-
риогенеза: амнион, желточный мешок, аллантоис и хорион
(рис. 2.31). В конце эмбрионального периода формируется основ-
ной питающий плод орган — плацента. Амнион (амниотическая
полость) образуется вокруг зародыша коровы к 19-м суткам из
парной амниотической складки внезародышевых частей эктодер-
мы и париетального листка мезодермы. Он имеет вид шатра и за-
полнен амниотической жидкостью. Эта жидкость в разные перио-
ды внутриутробного развития имеет разный химический состав и
создает вокруг зародыша среду, в которой он, подвешенный на
пупочном канатике, свободно «плавает» вплоть до момента свое-
Рис. 2.31. Временные внезародышевые органы эмбриона свиньи (30 мм):
1 — амнион; 2 — амниотическая полость; 3 — внезародышевая полость; 4 —
полость аллантоиса; 5 — плодная часть плаценты; 6 — пупочный канатик; 7 —
желточный мешок
55
го рождения. Желточный мешок, заполненный питающей жидко-
стью, образуется у овец к 13-м суткам из боковых (внезародыше-
вых) частей энтодермы и висцерального листка спланхнотома ме-
зодермы. У плацентарных животных желточный мешок в допече-
ночный период вплоть до 27 — 29-х суток становится первым кро-
ветворным органом, чему способствует и сеть сосудов, развиваю-
щихся в его стенке. Полость первичной кишки первоначально свя-
зана с полостью желточного мешка желточным каналом, от кото-
рого по мере развития зародыша остается лишь тонкий желточ-
ный стебелек, входящий в состав пупочного канатика. К концу
эмбрионального периода с формированием плаценты желточный
мешок обычно редуцируется.
Хорион развивается несколько позже других плодных оболочек
на базе трофобласта и является сложноустроенной наружной за-
родышевой оболочкой. Он окружает всю эмбриональную закладку
(плодный пузырь) и формируется из эктодермы, расположенной
снаружи и образующей крупные ворсинки, благодаря которым
он устанавливает связь со стенкой матки и участвует в образова-
нии плаценты. Изнутри хорион подслоен париетальным листком
спланхнотома мезодермы, дающим начало кровеносным сосудам.
Между хорионом, желточным мешком и аллантоисом располага-
ется внезародышевая полость (внезародышевый целом).
Аллантоис представляет собой вырост стенки задней части киш-
ки. Он состоит из энтодермы и подслаивающего и укрепляющего
его висцерального листка спланхнотома мезодермы, из материала
которого развивается широкая сеть кровеносных сосудов. Аллан-
тоис закладывается у крупного рогатого скота к 19-м суткам, у
овцы — к 17-м суткам и очень сильно разрастается, особенно в
плодный период. При этом он касается изнутри хориона и с ним
частично срастается (аллантохорион), участвуя в образовании «дет-
ской» части плаценты, так как сам хорион с зародышем непос-
редственно не связан, и связь плаценты и сосудов плода устанав-
ливается через сосуды аллантохориона. Частично аллантоис срас-
тается и с амнионом, образуя совмещенную часть стенки — ал-
лантоамнион. В аллантоисном мешке скапливается коричневатая
экскреторная жидкость (моча) зародыша (до 5 л перед рождени-
ем теленка). Во время родов амниотическая и аллантоисная жид-
кости помогают осуществлять давление плодного пузыря на орга-
ны самки, помогая открывать родовые пути, а после разрыва плод-
ных оболочек способствуют более легкому продвижению плода
наружу. Выходящий вслед за рожденным детенышем разорванный
плодный пузырь именуется последом.
Образование плаценты завершает комплекс сложных морфо-
генетических преобразований зародыша и его окружения, со-
вершающихся на ранних этапах эмбриогенеза. Эти преобразова-
ния характерны для живородящих (плацентарных) млекопитаю-
56
щих и преследуют цель обеспечить очень раннее и полноценное
питание зародыша за счет крови матери. Она формируется в кон-
це эмбрионального периода и первой половине плодного перио-
да (у крупного рогатого скота с 27-х до 50-х суток) и состоит из
двух частей: материнской и плодной, или «детской». Последняя
соединяется с зародышем посредством пупочного канатика, ос-
нову которого составляет тонкий желточный стебелек и толстый,
заполненный студневидной массой стебель аллантоиса, в кото-
ром проходят крупные пупочные артерия и вена. Плацента обра-
зуется как сложное двустороннее объединение участков слизис-
той оболочки матки и аллантохориона зародыша, обеспечиваю-
щее тесную связь между материнским организмом и зародышем
в период внутриутробного развития. Через плаценту путем изби-
рательной фильтрации (плацентарный барьер) осуществляется
передача зародышу из материнской крови кислорода, питатель-
ных веществ, воды, солей, витаминов и гормонов для полно-
ценного развития и роста, а из крови плода — продуктов обмена
и углекислого газа. Особенностью трансплацентарных обменов
является то, что на всем протяжении развития плода кровь мате-
ри не смешивается с кровью плода, между ними всегда суще-
ствует клеточно-тканевая преграда. При формировании плацен-
тарной связи специальные выросты (ворсинки) хориона входят
в особые углубления (ямки, или крипты) на поверхности слизи-
стой оболочки матки. При этом у различных видов животных ха-
рактер расположения ворсинок хориона и глубина проникнове-
ния их в слизистую оболочку матки различны. Для сельскохозяй-
ственных и домашних животных характерно несколько типов
плацент: диффузная (у кобылы, свиньи, верблюдицы), котиле-
донная (у жвачных), кольцевидная (у хищных), дисковидная (у
приматов и грызунов).
Развитие тела зародыша. Начинается с образования зародыше-
вой полоски и появления трех зародышевых листков — эктодер-
мы, мезодермы и энтодермы, каждый из которых разделяется на
зародышевую и внезародышевую части. Все структуры тела заро-
дыша развиваются путем сложного перемещения и дифференци-
ации отдельных участков зародышевых частей каждого из зароды-
шевых листков. К производным эктодермы у взрослого животного
относятся: эпидермис кожи, волосы, потовые и молочные желе-
зы, многослойные плоские эпителии внутренних органов, хрус-
талик глаза, зубная эмаль, а также сетчатка глаза, пигментные
клетки и практически все структуры нервной системы. Из мезо-
дермы образуются хорда, соединительные ткани, в том числе кровь,
кости, хрящи и мышцы, серозные оболочки. Энтодерма образует
выстилки пищеварительных и дыхательных путей, печень, под-
желудочную железу, эндокринные железы (щитовидную, пара-
щитовидные и др.).
57
Особой активностью этих процессов отличаются поздний эмб-
риональный и ранний плодный периоды развития зародыша, со-
впадающие по времени с формированием плаценты. К концу эм-
брионального периода зародыш имеет уже вполне проявленное
вытянутое и полностью сегментированное тело, имеющее харак-
терный изгиб более широкого переднего и более узкого заднего
концов. Видны находящиеся в стадии раннего органогенеза учас-
тки головного и спинного мозга, крупных туловищных магист-
ральных сосудов, а в брюшной полости — основного набора внут-
ренних органов.
В поздний плодный период внутриутробного развития процес-
сы роста все активнее преобладают над процессами дифференци-
ровки органов и тканей. Начинается становление собственных
функций органов, подготовка их к участию в процессе рождения
и раннего (приспособительного) периода постнатальной жизни.
Естественно, интенсивность этих процессов находится в тесной
связи с продолжительностью беременности: она тем выше, чем
короче беременность.
Развитие плода внутри организма матери является весьма про-
грессивным приспособлением, обеспечившим плацентарным мле-
копитающим возможность появления на свет более крупных и
высокоорганизованных детенышей, многие из которых способны
следовать за матерью почти сразу же после рождения.
Контрольные вопросы
1. Какие черты отличают процесс воспроизводства плацентарных мле-
копитающих от других животных?
2. Где и как развиваются спермий и яйцеклетка?
3. Как происходит оплодотворение и в чем его биологическое значе-
ние?
4. Как формируется плацента?
5. Опишите процессы, происходящие в эмбриональный период раз-
вития зародыша.
6. Чем характеризуется плодный период внутриутробного развития
сельскохозяйственных животных?
ГЛАВА 3
АНАТОМИЯ
ОБЩИЕ ТЕРМИНЫ, УПОТРЕБЛЯЕМЫЕ
ПРИ ОПИСАНИИ СТРОЕНИЯ ТЕЛА ЖИВОТНОГО
Для точного описания строения органов, их частей, располо-
жения и взаимосвязи с другими частями тела и органами приме-
няют специальные анатомические термины. Прежде всего услов-
но тело животного разделяют рядом плоскостей на части: одни из
них, продольные, идут вдоль тела животного, а другие — поперек.
Продольная плоскость, проведенная по оси симметрии и разде-
ляющая тело на две симметричные половины, правую и левую,
называется срединной, или медианной. Она проходит от рта до кон-
чика хвоста вдоль позвоночного столба. Продольные плоскости,
проведенные параллельно срединной, называются сагиттальны-
ми (рис. 3.1). Они рассекают тело животного на правые и левые
Рис. 3.1. Плоскости на теле лошади:
А — медианная; В — поперечная; С — горизонтальная
59
Рис. 3.2. Плоскости, поверхности и направления на теле собаки.
Плоскости: А — медианная; В — горизонтальная; С — поперечная. Направления:
1 — краниальное, 2 — каудальное, 3 — дорсальное, 4 — вентральное, 5, 7 —
проксимальное, 6, 8 — дистальное, 9 — ростральное, 10 — аборальное. Поверх-
ности: 11, 13 — дорсальная, 12 — пальмарная, 14 — плантарная
участки. Определение положения органа, его частей или сторон
по направлению к этим плоскостям принято обозначать термина-
ми: латеральный {lateralis), или боковой, направленный в боко-
вую (правую или левую) сторону от срединной плоскости, и ме-
диальный (medialis), или средний, направленный в сторону к сре-
динной плоскости (рис. 3.2). К продольным относятся также гори-
зонтальные плоскости (см. рис. 3.1). Они проводятся вдоль тела пер-
пендикулярно сагиттальным плоскостям и разделяют его на спин-
ные и брюшные участки.
Положение органов и их частей относительно друг друга и по
отношению к этим плоскостям принято определять терминами
дорсальный (dorsalis), или спинной, направленный в сторону спи-
ны, и вентральный (ventralis), или брюшной, направленный в сто-
рону живота. Этими терминами обозначают все тело животного,
за исключением свободных конечностей (см. рис. 3.2).
Поперечные плоскости (см. рис. 3.1) проходят поперек тела жи-
вотного и разделяют его на ряд сегментов. Положение органов и
их частей относительно друг друга и по направлению к этим плос-
60
костям определяют терминами: на туловище — краниальный
(cranialis), или черепной, направленный в сторону черепа, и кау-
дальный (caudalis), или хвостовой, направленный в сторону хвос-
та (см. рис. 3.2); на голове — ростральный (rostralis), направленный
в сторону рта, и аборальный (aboralis), или противоротовой, на-
правленный в противоположную сторону, т. е. в сторону шеи. В пре-
делах головы, особенно в дорсальной ее части (носовая полость,
головной мозг), употребляются термины тех же направлений: на-
зальный (nasalis), или носовой, направленный в сторону верхуш-
ки носа, и каудальный, направленный в сторону хвоста (см. рис. 3.2).
Если же на теле животного или на отдельных органах необхо-
димо описать косо лежащие поверхности, углы, отростки и т.п.,
то можно использовать сложные термины, составленные из вы-
шеуказанных, например дорсолатеральный, вентролатеральный,
вентромедиальный, дорсомедиальный и т.д.
При описании органов свободных отделов грудной и тазовой
конечностей соответственно до кисти и стопы применяются тер-
мины краниальный, каудальный, латеральный и медиальный.
На кисти различают спинку {dorsum manus) и ладонь (palma
manus). Отсюда и вся передняя поверхность кисти получает назва-
ние дорсальной (спинковой), а противоположная ей задняя име-
нуется как палъмарная, противоспинковая (см. рис. 3.2).
На стопе различают спинку (dorsumpedis) и подошву (plantapedis).
Вся передняя поверхность стопы называется дорсальной (спинко-
вой), а противоположная — плантарной (противоспинковой). На-
правления по длине оси свободных конечностей определяют тер-
минами: проксимальный (proximalis), т. е. ближайший к телу конец
конечности или какое-либо ближайшее к телу ее звено, и дис-
тальный (distalis), т.е. дальний конец. Кроме того, у многопалых
животных при описании структур в области кисти и стопы ис-
пользуются термины: осевой (axialis) и неосевой (abaxialis).
ПОНЯТИЕ О ЧАСТЯХ И ОБЛАСТЯХ ТЕЛА ЖИВОТНОГО
Тело животного принято подразделять на стволовую часть и
конечности.
Стволовая часть. В стволовой части различают голову, шею,
туловище и хвост (рис. 3.3; 3,4; 3,5). Голова (caput) делится на моз-
говой и лицевой отделы (рис. 3.6). Мозговой отдел для более деталь-
ной ориентировки подразделяют на области: затылочную, темен-
ную, лобную, на области ушной раковины, век, височную. Лице-
вой отдел подразделяют на носовую область, которая в свою оче-
редь подразделяется на спинку носа и боковую область носа, под-
глазничную область, области ноздрей, верхней губы, нижней губы,
подбородка. На боковых поверхностях лицевого отдела выделяют
61
Рис. 3.3. Области тела лошади:
1 — область головы; 2 — дорсальная область шеи; 3 — область холки (область
между лопатками); 4 — область лопатки; 5 — область спины; 6 — реберная об-
ласть; 7 — область голодной ямки; 8 — область поясницы; 9 — область маклока;
10 — область крестца; 11, 17 — область бедра; 12 — область хвоста; 13 — ягодич-
ная область; 14 — вертлужная область; 15 — область седалищного бугра; 16 —
хвост; 18 — область коленной чашки; 19 — область голени; 20 — область заплюс-
ны; 21 — область плюсны; 22 — область пальца стопы; 23 — паховая область;
24 — лонная область; 25 — средняя брюшная область; 26 — пупочная область;
27 — левая подреберная область; 28 — область мечевидного хряща; 29 — грудин-
ная область; 30 — область локтя; 31 — область предплечья; 32 — область запяс-
тья; 33 — область пясти; 34 — область пальца кисти; 35 — область плеча; 36 —
область плечевого сустава; 37 — вентральная шейная область
щечную область, область большой жевательной мышцы, подчелю-
стную область (рис. 3.6).
На шее (cervix, s. collum) принято выделять верхнюю выйную
область с ее выйным краем и боковые шейные области. Все они
лежат над телами шейных позвонков. Вдоль тел шейных позвон-
ков широкой полосой лежит область плечеголовной мышцы. Вен-
трально от нее расположена шейная вентральная область с ее гор-
танной областью впереди и трахеальной областью сзади (см. рис. 3.3,
3.4, 3.5, 3.6).
62
Рис. 3.4. Области тела коровы:
1 — область головы; 2 — дорсальная область шеи; 3 — область холки (область
между лопатками); 4 — область лопатки; 5 — область спины; 6 — реберная об-
ласть; 7 — левая подреберная область; 8 — область поясницы; 9 — область голод-
ной ямки; 10 — область маклока; 11 — область крестца; 12 — область хвоста; 13 —
ягодичная область; 14 — область седалищного бугра; 75 — вертлужная область;
16 — область бедра; 17 — область коленной чашки; 18 — область голени; 19 —
область заплюсны; 20 — область плюсны; 21 — область пальца стопы; 22 — пахо-
вая область; 23 — лонная область; 24 — средняя брюшная область; 25 — пупоч-
ная область; 26 — область мечевидного хряща; 27 — грудинная область; 28 —
область локтя; 29 — область пальца кисти; 30 — область пясти; 31 — область
запястья; 32 — область предплечья; 33 — предгрудинная область; 34 — область
плеча; 35 — область плечевого сустава; 36 — вентральная шейная область
Туловище (tcuneus) включает спинно-грудной, пояснично-брюш-
ной и крестцово-ягодичный отделы (см. рис. 3.3; 3.4; 3.5).
Спинно-грудной отдел разделяется на область холки и спинную
область. Грудь с поверхности подразделяется на грудную боковую
область, обширную нижнюю грудинную область и предгрудин-
ную область. На боковой грудной области различают реберную
область и область плечевого пояса и плеча, принадлежащие уже
грудной конечности (см. рис. 3.3; 3.4; 3.5; 3.7).
Пояснично-брюшной отдел охватывает поясничную область и
обширную брюшную область (брюхо, живот). Эта область двумя
поперечными линиями, из которых одна проходит на уровне са-
мой выпуклой части последнего ребра, а вторая — на уровне мак-
лока, делится на три участка. Передний участок, от первой попе-
речной линии до контура реберной дуги, дает область мечевидно-
го хряща (см. рис. 3.3; 3.4; 3.5; 3.7).
63
Рис. 3.5. Области тела собаки (левая латеральная поверхность):
1 — область лицевого черепа; 2 — область мозгового черепа; 3 — дорсальная
область шеи; 4— область лопатки; 5 — область спины; 6 — реберная область; 7 —
область поясницы; 8 — область крупа; 9 — область бедра; 10 — область голени;
11 — стопа; 12 — лонная область; 13 — средняя брюшная область; 14 — область
мечевидного хряща; 75 — грудинная область; 16 — кисть; 17 — область предпле-
чья; 18 — область плеча; 19 — предгрудинная область; 20 — латеральная область
шеи; 21 — вентральная область шеи
Средний участок разграничивается на правую и левую под-
вздошные области, примыкающие сверху к пояснице. Место у
поясницы впереди маклока принято называть голодной ямкой.
Позади области мечевидного хряща располагается пупочная об-
ласть. Продолжаясь назад, правая и левая подвздошные области
переходят в правую и левую паховые области, а пупочная — в
лонную (срамную) область (см. рис. 3.3; 3.4; 3.5).
Крестцово-ягодичный отдел, продолжающий поясницу, подраз-
деляется на крестцовую область, которая кзади переходит в хвост,
а сбоку — в ягодичную область (см. рис. 3.3; 3.4; 3.5).
Конечности. Грудная конечность примыкает к спинно-грудному
отделу туловища. Здесь принято выделять область лопатки, кото-
рая в свою очередь подразделяется на область лопаточного хряща,
примыкающую к холке, предостную и заостную области. Впереди,
на границе области плечевого пояса и области плеча, выступает
плечевой сустав. Область плеча служит главным образом местом
расположения трехглавой мышцы плеча и отделена сзади ясно
выраженной локтевой линией. Далее на конечности выделяют пред-
64
Рис. 3.6. Области головы собаки (левая латеральная поверхность):
/ — область ноздрей; 2 — верхнечелюстная область; 3 — латеральная область
носа; 4 — щечная область; 5 — подглазничная область; 6 — орбитальная область;
7 — лобная область; 8 — височная область; 9 — скуловая область; 10 — область
ушной раковины; 11 — область височно-нижнечелюстного сустава; 12 — область
околоушной железы; 13 — латеральная область шеи; 14 — дорсальная область
шеи; 15 — вентральная область шеи; 16 — область гортани; 17 — область глотки;
18 — область большой жевательной мышцы; 19 — нижнечелюстная область;
20 — подбородочная область; 21 — области верхней и нижней губы
Рис. 3.7. Области тела собаки
(вентральная поверхность):
1 — предгрудинная область: 2,
33 — область плеча; 3, 32 — об-
ласть предплечья; 4, 31 — область
локтевого сустава; 5, 30 — область
локтя; 6, 28 — реберная область;
7 — грудинная область; 8, 27 —
область подреберья; 9 — левая
подвздошная область; 10 — ле-
вая паховая область; 11, 17— об-
ласть бедра; 12, 21 — медиальная
область колена; 13, 20 — область
коленного сустава; 14, 19 — об-
ласть голени; 15, 18 — подколен-
ная область; 16 — область корня
хвоста; 22 — срамная (лонная)
область; 23 — правая паховая
область; 24 — пупочная область;
25 — правая подвздошная об-
ласть; 26 — область мечевидно-
го хряща; 29 — подмышечная
область
3 Зеленевский
65
Рис. 3.8. Области промежности кобеля:
1 — область хвоста; 2 — седалищно-прямокишечная ямка; 3 — кожная область
ануса; 4, 13 — области седалищных бугров; 5 — седалищная дуга; 6 — корень
полового члена; 7 — мошонка; 8 — шов мошонки; 9 — область бедра; 10 — тело
полового члена; 11 — ножка полового члена; 12 — мочеполовая область; 14 —
область ануса; 15 — анус; 16 — ягодичная область
Рис. 3.9. Области промежности суки:
1 — область хвоста; 2 — ягодичная область; 3 — анус; 4 — преддверие; 5, 10 —
области седалищных бугров; 6 — вульва; 7, 8 — область бедра; 9 — мочеполовая
область; 11 — анальная область; 12 — седалищно-прямокишечная ямка
66
плечье и кисть. В состав последней входят запястье, пясть и паль-
цы (см. рис. 3.3; 3.4; 3.5).
Тазовая конечность тазовым поясом примыкает к крестцово-
ягодичной области туловища. Здесь находится ягодичная область,
ограниченная снизу ломаной линией, проходящей от маклока к
тазобедренному суставу, а от него — до седалищного бугра. В сум-
ме правая и левая ягодичные и крестцовые области называются
крупом. Задний участок под начальной частью хвоста называется
анальной областью. Под ним до половых губ у самок или до мо-
шонки у самцов лежит область промежности (см. рис. 3.3; 3.4; 3.5;
3.8; 3.9).
От ягодичной области до коленного сустава располагается бед-
ро с областью коленной чашки. Впереди и несколько выше нее ле-
жит коленная складка. Ниже бедра располагаются голень и далее
стопа. В последней различают заплюсну, плюсну и пальцы (см. рис.
3.3; 3.4; 3.5).
Контрольные вопросы
1. Перечислите основные плоскости и направления на теле живот-
ного.
2. Какие анатомические области выделяют на голове и шеи животно-
го?
3. Какие анатомические области выделяют на грудной и тазовой ко-
нечности животного?
4. Какие анатомические области выделяют на туловище животного?
5. Перечислите направления и поверхности на грудной и тазовой ко-
нечности животного.
ЛОКОМОТОРНЫЙ АППАРАТ
Характерным свойством животного организма является локо-
моция (движение). Она осуществляется с помощью костей, свя-
зок и мышц. Локомоция составляет предмет исследования трех
направлений анатомии: остеологии (учение о костях), артроло-
гии, или синдесмологии (учение о соединениях костей), и мио-
логии (учение о мышцах).
ОСТЕОЛОГИЯ
Кости (ossw, ед. число — os). Располагаясь внутри тела, кости
выполняют функцию рычагов для прикрепления и приложения
действия скелетной мускулатуры, а также служат вместительным
депо минеральных и органических веществ, необходимых орга-
низму, и местом размещения красного костного мозга.
67
Рис. 3.10. Кость как орган:
1 — костные пластинки; 2 — внутрикостные сосуды; 3 — остеон; 4 — надкостни-
ца; 5, 6 — артерия и вена; 7 — компактное вещество; 8 — губчатое вещество
Кость построена из костной ткани и покрыта тонким слоем
соединительной ткани, образующей надкостницу. Основу кост-
ной ткани образуют костные клетки — остеоциты и костные пла-
стинки толщиной 3 — 7 мкм, состоящие из параллельно идущих
коллагеновых волокон, пропитанных солями извести и замуро-
ванных в особое плотное бесструктурное вещество — матрикс (рис.
3.10). Последний состоит из воды (50 %), органических (около 28 %)
и неорганических, или минеральных (около 22 %), веществ. Орга-
нические соединения и вода придают кости эластичность, а ми-
неральные — твердость.
Химический состав костей испытывает значительные колеба-
ния в зависимости от возраста, условий питания и физиологичес-
кого состояния организма. Кости молодых животных за счет боль-
шого количества влаги и органических веществ отличаются повы-
шенной эластичностью. С возрастом они теряют влагу и органи-
ческие компоненты, становясь более ломкими.
68
Кости построены из плотного компактного и рыхлого губча-
того веществ. Губчатое вещество пористое и состоит из тонких
костных пластинок — перекладин, взаимно переплетающихся под
различными углами соответственно направлению действующих
на кость сил. Они образуют ячейки, заполненные костным моз-
гом.
Компактное вещество плотное и имеет сложную архитекто-
нику, структурно-функциональной единицей которой являет-
ся остеон, или гаверсова система. Остеон представляет собой
комплекс большого числа костных пластинок. За счет волокни-
стого строения пластинки свернуты в трубочки разного диа-
метра и вставлены одна в другую. Трубочки плотно сомкнуты, и
между ними расположены слоями костные клетки, отростки
которых проникают в соседние костные пластинки и связыва-
ют их.
Особую прочность остеону придает то, что коллагеновые во-
локна в соседних пластинках идут по взаимно перпендикулярным
направлениям. Внутри каждого остеона имеется канал для про-
хождения кровеносных сосудов и вазомоторных нервов. Компакт-
ное вещество костей построено из многих остеонов, ориентиро-
ванных в основном вдоль длинной оси кости. Между ними, связы-
вая остеоны, располагаются так называемые вставочные пластин-
ки, имеющие дугообразную форму. Снаружи компактное веще-
ство костей покрыто несколькими слоями прямых продольных об-
щих, как бы упаковывающих, костных пластинок, над которыми
располагается надкостница.
Надкостница (периост) — пластинка соединительной ткани,
образованная снаружи коллагеновыми волокнами (волокнистый
слой надкостницы), а внутри особыми клетками — остеокласта-
ми (костеобразователями) и остеобластами (костеразрушителя-
ми). Наружный волокнистый слой является покровным, защит-
ным, а внутренний (клеточный) — костеобразующим, остеоген-
ным. За счет этого слоя надкостницы кость растет в толщину. При
переломах костей именно надкостница образует новую молодую
кость (костную мозоль) для сращения костных осколков.
Надкостница участвует в перестройке костей и в течение жиз-
ни животного в соответствии с изменяющимися условиями дей-
ствия на кость различных сил. Усиление мышечной нагрузки на
кости способствует укреплению костной ткани за счет увеличе-
ния числа остеонов и изменения их взаимного расположения. На-
против, при уменьшении действия мышц кости становятся тонь-
ше и мягче.
Надкостница густо пронизана кровеносными и лимфатичес-
кими сосудами, проникающими по остеонным каналам внутрь
кости и осуществляющими ее питание. Много в надкостнице и
нервных окончаний болевых рецепторов, что делает ее весьма чув-
69
ствительной. В то же время костная и хрящевая ткани не ощущают
боли, так как внутри их болевые нервные волокна отсутствуют.
Соединительнотканная пластинка покрывает не только поверх-
ности костей, но переходит и на хрящевые структуры скелета,
получая при этом название надхрящница, а также выстилает поло-
сти трубчатых костей, образуя эндост.
Рис. 3.11. Скелет лошади:
1 — резцовая кость; 2 — носовая кость; 3 — подглазничное отверстие; 4 —
орбита; 5 — височная ямка; 6 — затылочная кость; 7 — атлант; 8 — ось; 9 —
типичные шейные позвонки; 9* — шестой шейный позвонок; 10 — первый
грудной позвонок; 11 — холка; 12 — грудные позвонки; 13 — поясничные по-
звонки; 14 — крестцовая кость; 75 — хвостовые позвонки; 16 — седалищная
кость; 17 — бедренная кость; 18 — малая берцовая кость; 19 — большая берцовая
кость; 20 — пяточный бугор; 21 — четвертая заплюсневая кость; 22 — четвертая
плюсневая кость; 23 — третья плюсневая кость; 24 — плантарные сесамовидные
кости первой фаланги; 25 — проксимальная фаланга; 26 - средняя фаланга; 27 —
дистальная фаланга (копытная кость); 28 — кость стопы; 29 — кости голени; 30 —
надколенник (коленная чашка); 31 — лонная кость; 32 — подвздошная кость;
33 — ложные (астернальные) ребра; 34 — истинные (стернальные) ребра; 35 —
мечевидный отросток; 36 — кости предплечья; 37 — кости запястья; 38 — чет-
вертая пястная кость; 39 — третья пястная кость; 40 — пальмарные сесамовид-
ные кости проксимальной фаланги; 41 — проксимальная фаланга; 42 — средняя
фаланга; 43 — дистальная фаланга (копытная кость); 44 — кости кисти; 45 —
лучевая кость; 46 — локтевая кость; 47 — плечевая кость; 48 — грудная кость
(грудина); 49 — лопатка; 50 — ямка большой жевательной мышцы; 57 — тело
нижней челюсти; 52 — подбородочное отверстие; 53 — резцовые зубы
70
Рис. 3.12. Скелет коровы:
1 — резцовая кость; 2 — носовая кость; 3 — орбита; 4 — лобная кость; 5 —
затылочная кость; 6 — атлант; 7 - ось; 8 — типичные шейные позвонки; 9 —
шестой шейный позвонок; 10 — седьмой шейный позвонок; 11 — основание
лопатки; 12 — холка, грудные позвонки; 13 — поясничные позвонки; 14 — кре-
стцовая кость; 15 — хвостовые позвонки; 16 — седалищная кость; 17— запертое
отверстие; 18 — бедренная кость; 19 — малая берцовая кость; 20 — большая
берцовая кость; 21 — пяточный бугор; 22 — кости заплюсны; 23 — кости плюс-
ны; 24 — сесамовидные кости проксимальной фаланги; 25 — проксимальная
фаланга; 26 — средняя фаланга; 27 — сесамовидная кость дистальной фаланги;
28 — дистальная фаланга (копытцевая кость); 29 — кости стопы; 30 — кости
голени; 31 — надколенник (коленная чашка); 32 — лонная кость; 33 — под-
вздошная кость; 34 — ложные (астернальные) ребра; 35 — истинные (стерналь-
ные) ребра; 36 — мечевидный отросток; 37 — кости предплечья; 38 — кости
запястья; 39 — кости пясти; 40 — сесамовидные кости проксимальной фаланги;
41 — проксимальная фаланга; 42 — средняя фаланга; 43 — дистальная сесамо-
видная кость; 44 — дистальная фаланга (копытцевая кость); 45 — кости кисти;
46 — локтевая кость; 47 — лучевая кость; 48 — тело грудной кости; 49 — плече-
вая кость; 50 — рукоятка грудной кости; 51 — лопатка; 52 — тело нижней челю-
сти; 53 — подбородочное отверстие; 54 — резцовые зубы
На рост, развитие и структуру костей животных действуют мно-
гочисленные факторы — эндокринные, алиментарные, статоди-
намические и многие другие.
Внешний рельеф костей, как и внутреннее их устройство, на-
ходится в прямой зависимости от величины и направленности
механических воздействий, передаваемых через связки, мышцы и
их сухожилия. Оставляют следы на поверхности костей и приле-
жащие крупные кровеносные сосуды.
71
Выросты на костях в зависимости от формы именуются: отро-
сток, бугор, бугорок, ость, головка, блок, гребень, мыщелок,
вертел, шероховатость. Углубления называются: ямка, полость,
плоское вдавливание, желоб (борозда), щель, отверстие, канал,
вырезка. Некоторые отростки в процессе эмбрионального разви-
тия имеют собственные точки окостенения и получают название
«апофиз».
Рис. 3.13. Скелет овцы:
1 — резцовая кость; 2 — подглазничное отверстие верхней челюсти; 3 — носовая
кость; 4 — слезная кость; 5 — орбита; 6 — затылочная кость; 7 — атлант; 8 — ось;
9 — типичные шейные позвонки; 10 — шестой шейный позвонок; 11 — седьмой
шейный позвонок; 12 — лопаточный хрящ; 13 — холка, грудные позвонки; 14 —
поясничные позвонки; 15 — крестцовая кость; 16 — хвостовые позвонки; 17 —
седалищная кость; 18 — бедренная кость; 19 — малая берцовая кость; 20 —
большая берцовая кость; 21 — пяточный бугор; 22 — кости заплюсны; 23 -
кости плюсны; 24 — сесамовидные кости проксимальной фаланги; 25 — прокси-
мальная фаланга; 26 — средняя фаланга; 27 — сесамовидная кость дистальной
фаланги; 28 - дистальная фаланга (копытцевая кость); 29 — кости стопы; 30 —
большая берцовая кость; 31 — шероховатость большой берцовой кости; 32 —
надколенник (коленная чашка); 33 — тазобедренный сустав; 34 — подвздошная
кость; 35 — ложные (астернальные) ребра; 36 истинные (стернальные) ребра;
37 — мечевидный отросток; 38 — локтевая кость; 39 — лучевая кость; 40 — кости
заплюсны; 41 — кости плюсны; 42 — сесамовидные кости проксимальной фа-
ланги; 43 — проксимальная фаланга; 44 — средняя фаланга; 45 — сесамовидная
кость дистальной фаланги; 46 — дистальная фаланга; 47 — кости кисти; 48 —
кости предплечья; 49 — плечевая кость; 50 — лопатка; 57 — ветвь нижней
челюсти; 52 — тело нижней челюсти; 53 — подбородочное отверстие; 54 —
резцовые зубы
72
Рис. 3.14. Скелет козы:
/ — резцовая кость; 2 — носовая кость; 3 - орбита; 4 — лобная кость; 5 —
теменная кость; 6 — атлант; 7 — ось; 8 — типичные шейные позвонки; 9 —
шестой шейный позвонок; 10 — седьмой шейный позвонок; 11 — лопаточный
хрящ; 12 — грудные позвонки; 13 — поясничные позвонки; 14 — крестцовая
кость; 15 — хвостовые позвонки; 16 — седалищная кость; 17 — запертое отвер-
стие; 18 — бедренная кость; 19 — большая берцовая кость; 20 — кости заплюсны;
21 — кости плюсны; 22 — проксимальные сесамовидные кости; 23 — прокси-
мальная фаланга; 24 — средняя фаланга; 25 — дистальная сесамовидная кость;
26 — дистальная фаланга; 27 — кости стопы; 28 — малая берцовая кость; 29,
30 — тазобедренный сустав; 31 — подвздошная кость; 32 — ложные (астерналь-
ные) ребра; 33 — истинные (стернальные) ребра; 34 — локтевая кость; 35 —
лучевая кость; 36 — кости заплюсны; 37 — кости плюсны; 38 - проксимальные
сесамовидные кости; 39 — проксимальная фаланга; 40 — средняя фаланга; 41 —
дистальная сесамовидная кость; 42 — дистальная фаланга; 43 — кости кисти;
44 — кости предплечья; 45 — плечевая кость; 46 — тело грудной кости; 47 —
рукоятка грудной кости; 48 — лопатка; 49 - нижняя челюсть; 50 — резцовые
зубы
Кости подвижно соединяются между собой суставами, а мыш-
цы, присоединяющиеся к частям, шероховатостям или углубле-
ниям на них, обеспечивают функцию движения.
Кости и их соединения (сращения и суставы) образуют скелет.
Скелет (skeleton — высушенный). Выполняет опорную функ-
цию и формирует полости тела, являющиеся вместилищем внут-
ренних органов, центральной нервной системы, некоторых желез
и органов чувств.
Скелет, согласно его историческому и индивидуальному раз-
витию, принято делить на осевой и периферический. В состав осе-
73
вого скелета входят кости головы (скелет головы, или череп), по-
звоночный столб, ребра и грудная кость (грудина); периферичес-
кий скелет включает кости грудных и тазовых конечностей. Наи-
более древней частью осевого скелета является позвоночный столб.
Он состоит из шейных, грудных, поясничных, крестцовых и хво-
стовых позвонков. Ребра и грудная кость образовались несколько
позднее.
Позвоночный столб построен по принципу двусторонней сим-
метрии и имеет выраженные черты сегментальное™. Сегменты
представляют повторяющиеся участки, имеющие сходное строе-
ние и расположенные один за другим. В грудной части позвоноч-
Рис. 3.15. Скелет свиньи:
/ — резцовая кость; 2 — носовая кость; 3 — подглазничное отверстие; 4 —
слезная кость; 5 - орбита; 6 — височная ямка; 7 — затылочная кость; 8 —
атлант; 9 — ось; 10— типичные шейные позвонки; 11 — шестой шейный позво-
нок; /2 — седьмой шейный позвонок; 13 — грудные позвонки; 14 -- поясничные
позвонки; 15 — крестцовая кость; 16 — хвостовые позвонки; 17 — седалищная
кость; 18 — бедренная кость; 19 — большая берцовая кость; 20 — ма лая берцовая
кость; 21 — пяточный бугор; 22 — кости заплюсны; 23 — кости плюсны; 24 —
сесамовидные кости проксимальной фаланги; 25— проксимальная фаланга; 26 —
средняя фаланга; 27— дистальная фаланга; 28— кости стопы; 29— кости голе-
ни; 30— надколенник (коленная чашка); 31 — лонная косп>; 32 — подвздошная
кость; 33 — ложные (астернальные) ребра; 34 — реберная avia; 35 — истинные
(стернальные) ребра; 36 — грудная кость; 37 — локшпои бугор; 38 — кости
предплечья; 39 — кости запястья; 40 — кости пясти; 4! сесамовидная кость
проксимальной фаланги; 42 — проксимальная фаланга; 43 средняя фаланга;
44 — дистальная фаланга; 45 — кости кисти; 46 — лучевая кость; 47 — локтевая
кость; 48 — плечевая кость; 49 — рукоятка грудной кости; 50 — первое ребро;
51 — скуловая дуга; 52 — ветвь нижней челюсти; 53 — тело нижней челюсти;
54 — подбородочное отверстие; 55 — резцовые зубы
74
Рис. 3.16. Скелет собаки:
1 — резцовые зубы; 2 — резцовая кость; 3 — верхняя челюсть; 4 — подглазнич-
ное отверстие; 5 — орбита; 6 — височная ямка; 7 — затылочная кость; 8 —
атлант; 9 — ось; 10 — типичные шейные позвонки; 11 — шестой шейный позво-
нок; 12 — седьмой шейный позвонок; 13 — лопатка; 14— грудные позвонки; 15 —
поясничные позвонки; 16 — крестцовая кость; 17 — хвостовые позвонки; 18 —
запертое отверстие; 19 — седалищная кость; 20 — сесамовидные кости; 21 —
малая берцовая кость; 22 — большая берцовая кость; 23 — пяточный бугор; 24 —
кости заплюсны; 25 — кости плюсны; 26 — сесамовидные кости проксимальной
фаланги; 27 — проксимальная фаланга; 28 — средняя фаланга; 29 — дистальная
фаланга; 30 — кости стопы; 31 — кости голени; 32 — надколенник (коленная
чашка); 33 — бедренная кость; 34 — лонная кость; 35 — подвздошная кость; 36 —
ложные (астернальные) ребра; 37 — реберная дуга; 38 — истинные (стерналь-
ные) ребра; 39 — грудная кость; 40 — локтевой бугор; 41 — кости предплечья;
42 — добавочная кость запястья; 43 — кости запястья; 44 — кости пясти; 45 —
проксимальная фаланга; 46 — средняя фаланга; 47 — дистальная фаланга; 48 —
кости кисти; 49 — лучевая кость; 50 — локтевая кость; 51 — плечевая кость; 52 —
рукоятка грудной кости; 53— барабанный пузырь; 54— угловой отросток; 55 —
ямка большой жевательной мышцы; 56 — тело нижней челюсти; 57 — подборо-
дочное отверстие; 58 — резцовая часть нижней челюсти
кого столба каждый из сегментов является полным и представлен
позвонком, двумя ребрами и сегментом грудины. Общий вид ске-
летов лошади, коровы, овцы, козы, свиньи, собаки показан на
(рис. 3.11 — 3.40).
Череп — сильно специализированная часть осевого скелета (рис.
3.11-3.40).
75
Скелет головы
Кости головы формируют ротовую, цосовукти черепную поло-
сти, являющиеся вместилищем начальных отделов органов пище-
варения и дыхания, головного мозга и некоторых анализаторов.
В соответствии с историческим и индивидуальным развитием ске-
лет головы — череп — подразделяют на два отдела: мозговой и
лицевой череп. Скелет головы (череп) коровы, лошади, овцы,
свиньи, собаки показан на рис. 3.17 — 3.29, а также на схемах об-
щего скелета (см. рис. 3.11 —3.16).
Мозговой череп
Мозговой череп (cranium cerebrate) образуется затылочной, кли-
новидной, височными, решётчатой, межтеменной, теменными,
лобными, крыловидными костями и сошником (рис. 3.17 — 3.27).
Затылочная кость (os occipitale) находится в аборальной обла-
сти черепа (см. рис. 3.11 — 3.26). Она состоит из основной, двух
боковых частей и чешуи затылка. Чешуя затылка сливается с те-
менными и межтеменной костями.
Рис. 3.17. Границы костей головы коровы:
1 — резцовая кость; 2 — носо-резцовая вырезка; 3 — носовая кость; -7, 6 — слезная
кость; 5. 9— лобная кость; 7— глазничное крыло клиновидной кости; 8 — скуло-
вой отросток (лобная кость); 10 — зрительное отверстие; // — роговой отросток
(лобная кость); 12 — теменная кость; 13 — чешуя височной кости; 14 — затылоч-
ный мыщелок; 75 — яремный отросток (затылочная кость); 16 — отверстие наруж-
ного слухового прохода; 17 — барабанная часть; 18 — скуловой отросток (височ-
ная кость); 19 — височный отросток (скуловая кость); 20 — крыловидный отрос-
ток (клиновидная кость); 21 — лобный отросток (скуловая кость); 22 — перпенди-
кулярная пластинка (нёбная кость); 23 — скуловая кость; 24 — лицевой бугор; 25 —
верхняя челюсть; цб)— подглазничное отверстие; 27 — беззубый край
76
Рис. 3.18. Череп коровы (сагиттальный разрез):
1 — тело резцовой кости; 2 — носовой отросток; 3 — верхняя челюсть; 4, 5 —
латеральный и медиальный носовые отростки; 6 — носовая кость; 7 — кость
вентральной носовой раковины; 8 — кость дорсальной носовой раковины; 9 —
сошник; 10 — кость средней носовой раковины; 11 — лобная кость; 12, 19 —
лобная пазуха; 13 — решетчатый лабиринт; 14 — петуший гребень; 15 — обо-
нятельная ямка; 16 — желоб перекреста; 17 — глазнично-клиновидный гребень;
18 — роговой отросток; 20 — каменистая часть; 21 — чешуя затылка; 22 — внут-
реннее затылочное предбугорье; 23 — отверстие височного канала; 24. 25 —
отверстия мыщелкового канала; 26 — затылочный мыщелок; 27 — подъязычный
канал; 28 — яремный отросток; 29 — яремное отверстие; 30 — каменисто-бази-
лярная щель; 31 — отверстие внутреннего слухового прохода; 32, 34 — мышечные
отростки; 33 — спинка седла; 35 — овальное отверстие; 36 — глазнично-круглое
отверстие; 37 — предклиновидная кость; 38 — крыловидная кость; 39 — пазуха
предклиновидной кости; 40 — перпендикулярная пластинка; 41 — клинонёбное
отверстие; 42 — хоана; 43 — вход в верхнечелюстную пазуху; 44 — третий премо-
ляр; 45 — нёбная пазуха; 46 — беззубый край; 47 — нёбный отросток
Основная часть затылочной кости лежит в основании черепа и
срастается с клиновидной костью. На наружной поверхности за-
тылочной кости, на месте соединения ее с клиновидной костью,
находится парный мышечный бугорок для прикрепления мышц го-
ловы. Латеральные острые края основной части затылочной кости
образуют рваное отверстие для сосудов и нервов. На мозговой по-
верхности тела затылочной кости слабо заметны вдавливание про-
долговатого мозга и вдавливание моста. Задний конец тела заты-
лочной кости ограничивает нижнюю часть большого отверстия.
^Боковые части затылочной кости ограничивают большое от-
верстие и имеют затылочные мыщелки для сочленения с атлантом..
Внизу они отделены друг от друга межмыщелковой вырезкой. Выше
мыщелка расположена надмыщелковая ямка, а ниже — подмы-
щелковая ямка с двумя-тремя подъязычными отверстиями подъ-
77
Рис. 3.19. Череп коровы (вентральная поверхность):
1,2— затылочныелребень и мыщелок; 3, 24 — центральная мыщелковая ямка; 4 —
лобная кость; 5,\6i — яремный и роговой отростки; 7 — яремное отверстие; 8 —
височная кость; 9 — мышечный бугорок; К), 13 крыловидны» iребень и кость;
11 - надглазничный канал; 12 — решетчатое отверстие; 14 — крючок; Z5 — верх-
нечелюстной бугор; 16 — хоана (задняя ноздря); /7 малые нёбные отверстия;
18 — большое нёбное отверстие; 19^- нёбный шов; 20 нёбный отросток (резцо-
вая кость); 21 — резцовая щель; 22— большое отверстое; 23 — подъязычный канал;
25 — шиловидный отросток; 26 — засуставное отверстие; 27 - отверстие наружного
слухового прохода; 28— засуставной отросток; 29 — сусшвной бугорок; 30— оваль-
ное отверстие; 31 — глазнично-круглое отверстие; 32 — трительное отверстие; 33 —
слезный пузырь; 34 — крыло сошника; 35 — клинонёбное отверстие; 36 — верхне-
челюстное отверстие; 37 каудальное нёбное отверстие; 38 — нёбный отросток
(верхняя челюсть); 39 — диастема (беззубый край); 40 — нёбная щель
78
14 15 /6
Рис. 3.20. Череп лошади (латеральная поверхность):
7 — гсло рентной кости 2— носовой отросток; носовая кость; 4— носо-
резшИпй вырезка; 5. — птшлазничние отверстие; ~7— скуловая кость;!&^— слез-
ная кость; орбита; лобная кость; 77 — надглазничное отверстие; 12 —
скуловой отросток лобной кости; 13 — венечный отросток; 14 — чешуя височ-
ной кости; 75 — наружный сагиттальный гребень; 16 — отверстия-выпускники;
77- наружный затылочный гребень; 18 — затылочная кость; 19 — забарабанный
отросток; 20 — сосцевидный отросток; 21 — большое отверстие; 22 — яремный
отросток; 23 — основная часть (затылочная кость); 24 — наружное слуховое от-
верстие; 25 — мыщелковый отросток; 26 — нижнечелюстная вырезка; 27 — круг-
лое отверстие; 28— ростральное крыловое отверстие, 29 .-перпендикулярная
пластинка (нёбная кость/ Зч верхнечелюстной tiyiop, 31 — альвеол ярим и
край:.?/— ямка большой жевательной иишпы; '^3— угол нижней чслюсти;^^—
лицевая сосудистая вырезка; — альвеолярный край; 36 - щечная цоверх-
ность; 37 — беззубый кра№ бу— полбородочноелпиерстне. 39 — клыки. 4 >
резцовая часть; f4l‘— рещовыс зубы; 43 — лицевой гребень
язычного канала. Л агеральнес мыщелка находится яремный отрос-
ток для мышц. Между наружной и внутренней костными плас-
тинками лежит воздушная затылочная пазуха.
О с о б е нн о ст и. У овец чешуя затылочной кости имеет те-
менную и затылочную поверхности, затылочный гребень слабый,
затылочная пазуха отсутствует. У свиней чешуя высокая, выемча-
тая, треугольная, расширяется вверху. Яремные отростки прямые
и длинные. Подъязычное отверстие одно. У лошадей на затылоч-
ной кости имеются теменная, собственно затылочная и основная
поверхности. У собак теменная поверхность затылочной кости вкли-
нивается вперед. Затылочный гребень хорошо развит.
Клиновидная кость (os sphenoidale) коровы лежит вентрально и
соединяется с решетчатой костью, сошником, крыловидными,
нёбными, лобными, височными и затылочной костями (см. рис.
3.17 — 3.25). На ней различают тело, височные и глазничные кры-
лья. Тело клиновидной кости состоит из двух частей: передней —
79
Рис. 3.21. Череп лошади (медианный разрез):
! JJ— резцовые зубы; 2 ic.io резиопой кости ре шпкый канал; 4 — нёбный
Отросток; 5 — носовой отросток; 6 — верхняя челюсть, 7 — носо-резцовая вы-
резка; 8 — носовая кость; 9 — дорсальный носовой ход; 10 — кость вентральной
носовой раковины; 11 средн и it носовой ход; 12 — кость дорсальной носовой
раковины; 13 лобная пазуха; 341— решетчатый лабиринт; /,5 — клиновидный
клюв; /6 — орбитальное крыло; /7 чешуйчатая часть (височная кость»; /У
каменистая часть (височная кость); 19 — височная крыло, 20 — лобная кость, 21 —
теменная кость; 22t межтеменная кость; 23 — выйный бугорок; 24 — чешуя
затылка; 25 — подъязычное отверстие; 26 — большое отверстие; 27 — яремный
отросток; 28 — основная часть (затылочная кость); 29 — разорванное отверстие;
30 — основная клиновидная кость; 31 — борозда глазничного нерва; 32 — глаз-
ничная щель; 33 - клиновидная пазуха; 34 — крыловидная кость; 35 - нижне-
челюстное отверстие;/За — хоаны ?7 крючок; 38 — вентральный носовой ход;
39 горизонтальная Пластинка небной косой 40 нёбный отросток 1верхттяя
челюсть); 41 ямка крыловидной мышцы: 42) — угол нижней челюсти; 431 —
1ИИСВЛЯ сосудистая пыреися; 44 — язычная поверхность. 45 регионал часть
(нижняя челюсть); 46 — диастема (беззубый край); 47 — клыки
пресфеноида и задней — базисфеноида, Ростральные — глазнич-
ные крылья клиновидной кости, каудальные — височные крылья при-
легают к подвисочной ямке. На наружной поверхности тела кли-
новидной кости ростровентрально выступает крыловидный отрос-
ток', его передний край дает слабый крыловидный гребень.
С внутренней поверхности тела клиновидной кости рострально
выступает клюв для соединения с решетчатой и лобной костями.
Каудально 01 клюва находится поперечный желоб перекреста для
зрительного нерва Из него в орбиту проходит зрительное отвер-
стие Каудальное перекреста лежат турецкое седло и спинка седла.
Ямка седла называется гипофизарной. По бокам ее расположен боль-
шой желоб, который ведет в круглоглазничнре отверстие. От клюва
80
вниз тянется костная пластинка, разделяющая пазухи клиновид-
ной к,ости. В основании височного крыла находится овальное от-
верстие.
Особенности. У свиней круглоглазничное отверстие боль-
шое, овальнЬё~бтсутствует, спинка седла высокая. У лошадей сильно
развиты глазничные крылья. У основания крыловидных отростков
находится большой крыловой канал. У собаки глазничные крылья
меньше височных. Глазничное и круглое отверстия самостоятель-
ны.
Височная кость (as teniporaleljpac положе на у коровы с боков
мозгового черепа и граничит с затылочной, теменными, лобны-
ми. скуловыми, клиновидной н верхнечелюстыми костями (рис.
127; см. рис. 3.14 — 3.25). Она состоит из чешуйчатой части и каме-
нистой кости, которые обычно срастаются. Чешуя височной кости
снаружи имеет скуловой отросток Он отделяет височную ямку от
подвисочной, а также принимает участие в образовании скуловой
дуги. На скуловом отростке расположен суставной аппарат, состо-
ящий из суставного валика, суставной ямки и засуставного отрос-
тка. Внутренняя поверхность чешуи почти вся шовная. Височный
Рис. 3.22. Границы костей головы свиньи:
1 — резцовая кость; 2 — носовая кость; 3 — верхняя челюсть; 4 — скуловая
кость; 5 — слезная кость; 6 — слезное отверстие; 7 — орбита; 8 — надглазничный
канал; 9 — лобная кость; 10 — глазничное крыло (клиновидная кость); 11 —
теменная кость, височная ямка; 12 — чешуйчатая часть (височная кость); 13 —
выйная линия; 14 — отверстие наружного слухового прохода; 15 — барабанная
часть (височная кость); 16 — затылочный мыщелок; 17 — скуловой отросток
височной кости; 18 — яремный отросток; 19 — барабанный пузырь; 20 — скуло-
вая дуга; 21 — крыловидный отросток (клиновидная кость); 22 — крючок; 23 —
крыловидный отросток (нёбная кость); 24 — верхнечелюстной бугор; 25 — лице-
вой гребень; 26 — подглазничное отверстие; 27 — первый премоляр; 28 — клык;
29 — третий резцовый зуб; 30 — второй резцовый зуб; 31 — первый резцовый
зуб; 32 — хоботковая кость
81
I - тело резцовой кости: 2/— носовой отросток; 3 — носорсзиопая вырезка^4
носовой отросток (носовая кость*. |5'- носовая кость;: ф — слсчнаяj<ocn>; 1/—
зобнпи кость; надглазничный канал;решетчатое отверстие; 10) k-ii.iuo
пидиця кость; 7/^ скуловой отросток (лобная кость); 12 скуловая^уга; ЦЗ/~
теменная кость. 14 — височная кость; Лу — межтеменная кость; 'ly-г чешуя
височной кости;.затылочная Кость; TS затылочный мыщелок: ।19/— ярем-
ный (чросзок; 26 — oiiiepiTHC наружного сзухового прохода: 21 опальное
отверстие; 22 — крыловидный отросто^клиновидная кость); 23 — крючок; 24 —
височный отросток (скуловая кость);(25)— верхнечелюстной.бугор; 26 — скуло-
вая кость; 27 — лицевой бугорок; 28 — альвеолярный край;/^' — подглазничное
отверстие; 30 — беззубый край
ход находится между чешуей и каменистой костью, содержит ви-
сочную вену и тянется от засуставного отростка до височной ямки.
Каменистая кость (os petrosum) состоит из каменистой и бара-
банной частей.
Каменистая часть имеет затылочную поверхность и сосцевид-
ный отросток, которые на ранних этапах жизни срастаются с на-
ружным слуховым проходом и чешуей височной кости. Скалистая
часть кости содержит внутри лабиринт внутреннего уха. Скалис-
тый гребень слабо развит. Внутренний слуховой проход на дне имеет
несколько мелких отверстий для слухового нерва, идущего во внут-
реннее ухо, и одно большое отверстие для лицевого нерва, иду-
щего в лицевой канал. Щель водопровода улитки и щель водопро-
вода преддверия слабо выражены.
Барабанная часть направлена вентрально, сильно развита, имеет
уплощенный барабанный пузырь, здесь помещается барабанная
полость, которая сообщается с глоткой при помощи слуховой трубы.
Латерально обращен наружный слуховой проход для наружного уха.
Между ним и костным пузырем помещается подъязычный отрос-
ток. Каудально подъязычного отростка находится отверстие лице-
вого канала для лицевого нерва. Мышечный отросток помещается
на костном пузыре, а отверстие слуховой трубы лежит впереди
костного пузыря.
Особенности. У свиней чешуя и каменистая часть сросшие-
ся, височный ход и мышечный отросток отсутствуют, барабан-
82
ный пузырь развит хорошо. У лошадей чешуя и каменистая часть
обособленные, барабанная часть каменистой кости хорошо выра-
жены. Аборально от нее выступает сосцевидный отросток для мышц.
У собак чешуя и каменистая кость срослись, суставные валик и
ямка развиты хорошо, барабанный пузырь круглый, скалистый
гребень большой с отверстием для тройничного нерва.
Решетчатая кость (os ethmoidale) лежит внутри черепа и гра-
ничит у коров с лобными, клиновидной, слезными, верхнечелю-
стными, нёбными костями, сошником и носовыми раковинами
(см. рис. 3.18, 3.21). На ней различают продырявленную и основ-
ную пластинки, а также лабиринт.
JЦюдырявленния пластинка находится между клиновидной и
лобными костями. Она участвует в образовании передней стенки
черепной полости. На решетчатой пластинке имеются две обоня-
тельные ямки для обонятельных луковиц мозга и много мелких
отверстий для нервов, по латеральным краям обонятельных ямок
находится по одному решетчатому отверстию. Основная пластин-
ка вставлена в желоб сошника, она в виде костной отвесной пла-
Рис. 3.24. Кости головы собаки (латеральная поверхность):
I — резцовая кость; 2 — первый премоляр; 3 — носовая кость; 4 — второй
премоляр; 5 — третий премоляр; 6 — подглазничное отверстие; 7 — четвертый
премоляр; 8 — первый и второй моляры; 9 — скуловая кость; 10 — слезная
кость; 11 — лобная кость; 12 — орбита; 13 — надглазничный отросток (лобная
кость); 14 - височный отросток; 15 — венечный отросток; 16— скуловой отро-
сток (височная кость); 17 — лобно-теменной шов; 18 — теменная кость; 19 —
чешуя височной кости; 20 — межтеменная Косты- 21 затылочный гребень; 22 —
затылочная кость; 23 — большое отверстие; 24/ затылочный мыщелок; 25 —
яремный отросток; 26 — отверстие наружного слухового прохода; 27 — костный
пузырь; 28 — угловой отросток; 29 — суставной отросток; 30 — сосудистая лице-
вая вырезка; 31 — ямка большой жевательной мышцы; 32 — орбитальный край;
33 — тело нижней челюсти; 34 — четвертый премоляр; 35 — третий премоляр;
36 — подбородочное отверстие; 37 — первый и второй премоляры; 38 — клыки;
39 — резцовые зубы
83
Рис. 3.25. Кости головы собаки (вентролатеральная поверхность):
резцовые зубы; \2)— клык;/5?— нёбная щель; 4 — верхняя челюсть; 5 —
большая нёбная борозда; 6 — небный отросток; 7 — большое нёбное отверстие;
8 — малые нёбные отверстия; 9 — верхнечелюстной бугор; 10}— височный отро-
сток скуловой кости; 11 — скуловой отросток лобной кости; 12 — решетчатое
отверстие; 13 — зрительное отверстие; 14 — глазничная щель; 15 — оральное
крыловое отверстие; 16 — скуловой отросток височной кости; 17 — суставная
поверхность; 18 — засуставной отросток; 19 — отверстие наружного слухового
прохода. 20 — овальное отверстие; 21 — костный пузырь; 22 яремное отвер-
стие; 23. 27 — мыщелок затылочной кости; 24 — вентральная мыщелковая ямка;
25 — большое отверстие; 26 — тело затылочной кости; 28 — яремный отросток;
29 — яремное отверстие; 30 — костный пузырь; 31 — сонное отверстие; 32 —
костная слуховая труба; 33 — скуловой отросток височной кости; 34 — основная
клиновидная кость; 35 — аборальное крыловое отверстие; 36 — крыловидная
кость; 37 — крючок крыловидной кости; 38 — предклиновидная кость; 39 —
перпендикулярная пластинка нёбной кости; 40 — височный отросток скуловой
кости; 41 — хоаны (задние ноздри); 42 — второй моляр; 43 — первый моляр; 44 —
горизонтальная пластинка нёбной кости; 45 — четвертый премоляр; 46 — нёб-
ный шов; 47 — .третий премоляр; 48— второй премоляр; 49 — первый премоляр;
50; — межрезцовый шов; 51 — тело резцовой кости
стинки идет вперед и переходит в хрящевую носовую перегородку.
Лабиринт решетчатой кости лежит по бокам основной пластин-
ки и состоит из множества очень тонких костных пластинок, за-
вивающихся на свободных концах. Они образуют кость дорсаль-
ной и средней носовых раковин. Весь лабиринт покрыт тонкой
полупрозрачной покровной пластинкой.
Особенности. У свиней, лошадей и собак строение решет-
чатой кости такое же, как у коров.
Межтеменная кость (os interparietale) лежит между затылоч-
ной и теменными костями и без заметных границ срастается с
ними (см. рис. 3.18, 3.20 — 3.24, 3.26).
84
Рис. 3.26. Затылочные кости собаки (Л), коровы (Б), свиньи (В), ло-
шади (г):
1 — основная часть; fS)- сагиттальный гребень; .? - шн ючный гребень
чешуя затылочной кости; дорсальная мыщелковая ямка. О - кныючпый
мыщелок; (ТУ— яремный отросток; 8 — выйный бугорок; У — надмыщелковое
отверстие; $0)— теменная кость; 11 — наружное затылочное предбугорье;
лобная кость /Л_Т9 межтеменная кость; 14 — наружное затылочное предбу-
горье; 15 — сосцевидный отросток (височная кость): 16 - вентральная мыщел-
ковая Ямка; (17]— большое отверстие; 18 — ямка чешуи шылочной кости; 20 —
наружное отверстие слухового прохода; 21 — канал подъязычного нерва;(2^—
височная кость; 2'3 — выйная ямка
85
Рис. 3.27. Барабанная часть височ-
ной кости лошади:
/У— мышечный отросток: 2 пыре:<ка
грпнничного Hernia. 3 каменисто-ба-
рабанная шель;№ височный ход; 5
огнерстне наружного слухового прохо-
да, И чешуя височной кости; 7 —
сосневилный отросток. А' желоб ка-
ЩМЛЪНОЙ артерии мозговых оболочек;
Ч‘ uni юншшо-сосцевидное отверстие;
10 — ИШТОИИЛН1.П1 отросток; /7/— ба-
рабанный пузырь
Особенности. Усвиней
межтеменная кость еще у пло-
дов срастается с затылочной и
теменными костями. Внутренне-
го затылочного предбугорья нет.
У лошадей межтеменная кость
снаружи имеет сагиттальный
гребень, а внутри — внутреннее
затылочное предбугорье. Кост-
ный намет образуется скалисты-
ми гребнями и краями внутрен-
него татылочного предбугорья.
У собак межтеменная кость сра-
стается с затылочными костями
и внутри образует большое внут-
реннее затылочное предбугорье.
Теменные кости (ossa
parietalia) находятся у корон "и
затылочной и височных облас-
тях (см. рис. 3.11 — 3.17, 3.20 —
3.24). Чешуя затылочной кости,
межтеменная и теменные кости
срослись в одну подковообраз-
ную кость. В ней имеется пазу-
ха, которая сообщается с лоб-
ной пазухой. Внутреннее заты-
лочное предбугорье отсутствует.
Особенности. У свиней теменные кости массивные, между
их наружной и внутренней пластинками находятся теменные па-
зухи. У лошадей теменные кости имеют выпуклую наружную по-
верхность с "выступающим теменным гребнем. У собак сагитталь-
ный гребень теменных костей хорошо развит.
Лобные кости (ossa frontalid) граничат у коров с носовыми,
слезными, решетчатыми,"межтеменной, теменными, височны-
ми, нёбными и верхнечелюстными костями (см. рис. 3.11 —3.26).
На наружной поверхности лобных костей выделяют три части:
лобную, глазничную и височную. От лобной кости в латеральном
направлении выступает скуловой отросток, направляющийся к ску-
ловой кости. На орбитальной поверхности глазничного отростка
имеются две ямки — слезная для слезной железы и блоковая для
блока склеры. У основания глазничного отростка в середине же-
лоба находятся два надглазничных отверстия. На нижнем крае глаз-
ничной части лежит решетчатое отверстие. Каудальные края лоб-
ных костей образуют поперечные валики, от концов которых от-
ходят роговые отростки. Носовые отростки вытянуты вперед. Внут-
ренняя поверхность лобных костей поперечным гребнем делится
86
па две части: мозговую и носовую. В лобных костях помещается об-
ширная лобная пазуха. Она разделена костной перегородкой на
левую и правую.
Особенности. У свиней лобная пазуха обширная, решетча-
тое отверстие находится на лобной кости. У лошадей глазничный
отросток соединен со скуловой дугой, на нем находятся слезная и
блоковая ямки и надглазничное отверстие. У собак глазничные
отростки не доходят до скуловой дуги.
Крыловидная кость (os pterygoideum) парная, граничит у коров
с нёбной,клиновидной костями и сошником (см. рис. 3.17 — 3.19,
3.21 — 3.25). Оральный конец кости образует крючок.
Особенност и.У свиней каудальный конец кости расширен
У лошадей эти кости тонкие,'лентовидные с большим крючком.
У соба^Трьыовидные кости квадратной формы, вогнутые, с тре-
угольным крючком.
Сошник (уотег) — непарная кость с расширенным аборальным
концом — граничит у коров с клиновидной, крыловидной, верх-
нечелюстной и резцовой костями (см. рис. 3.18, 3.19, 3.21, 3.25).
Сбоку на сошнике находятся крылья, между которыми лежит же-
лоб носовой перегородки. Сошник изогнут продольно, сошнико-
вая вырезка отсутствует.
Особенности. У свиней сошник сзади узок, снизу желобо-
ват. У лошадей он аборально имеет большие крылья с вырезкой и
соединен с нёбными, верхнечелюстными и резцовыми костями.
У собак сошник с маленькой вырезкой.
Лицевой череп
^Лицевой череп (сгш/ш/л viscerate) является остовом ротовой и
носовой полостей. В его образовании участвуют нижняя челюсть,
верхняя челюсть, резцовая, носовая, скуловая, слезная, небная
кости, кость дорсальной и кость вентральной носовых раковин,
подъязычная и хоботковая кости (см. рис. 3.11 — 3.29).
Нижняя челюсть (mandibula) сочленяется у коровы со скуло-
вым отростком височной кости (см. рис. 3.11—3.16, 3.20, 3.21,
3.24, 3.28). На ней различают тело, на котором располагаются зуб-
ные альвеолы и челюстные ветви. Граница между ними проходит
по лицевой сосудистой вырезке. На теле нижней челюсти выделя-
ются резцовая и коренная части. На резцовой части находятся две
поверхности: наружная выпуклая подбородочная и внутренняя вог-
нутая язычная. На альвеолярном крае, где сходятся обе поверхно-
сти, располагаются четыре зубные альвеолы. Альвеолярный край пе-
реходит в беззубый край. На коренной части тела различают внут-
реннюю язычную поверхность со слабой челюстно-подъязычной
линией для межчелюстной мышцы и щечную поверхность. Верх-
ний альвеолярный и нижний края закруглены. Суженная ростраль-
87
Рис. 3.28. Нижние челюсти собаки (Л), коровы (Б), свиньи (В),
лошади (Г) (дорсолатеральная поверхность):
1 — резцовые зубы; 2 — клык; 3 — резцовая кость; 4 — предкоренные зубы; 5 —
коренные зубы; 6 — нижнечелюстное отверстие; 7 — ямка крыловидной мыш-
цы; 8 — tK-нс'шын отросток: — иижнечелюснш мцюэка; (0)— мышелконый
отросток; II — шейка Нкжшф челюсти; 12 — имка большой жтТОЛЫЮ* мыш-
цы; 13 — Vi юной отросток: /•#•' Двцамя сосудистая вырезки; /5 — шло нижней
челюсти; /б — резцовая часгь;^/7. /Л/- подбородочные отиерстие и поиерхноегь
ная область нижнечелюстной кости называется резцовой частью.
На ней сбоку находится подбородочное отверстие.
Ветви нижнечелюстной кости тянутся от челюстного угла до
суставного отростка. На латеральной поверхности ветвей имеется
ямка большой жевательной мышцы, а на медиальной — крыловид-
ная ямка. На краниальном крае ветвей вверху выступает мышечный
отросток для височной мышцы. На каудальном крае вверху поме-
щается вогнутый суставной отросток. Он отделен челюстной вы-
резкой от мышечного отростка. В крыловой ямке лежит нижнече-
люстное отверстие, ведущее в нижнечелюстной канал. Этот канал
заканчивается подбородочным отверстием.
Особенности. У свиней нижние челюсти сращены в одну
кость, на теле кости имеется несколько подбородочных отвер-
стий. У лошадей нижние челюсти рано срастаются в одну кость,
88
челюстной угол утолщен и закруглеп. У собак ямка жевательной
мышцы глубокая, беззубого края нет, подбородочных отверстий
несколько.
Верхняя челюсть (maxilla) парная, граничит с носовыми, слез-
ными, скуловыми, лобными, нёбными, резцовыми костями, со-
шником, вентральной носовой раковиной (см. рис. 3.11 — 3.25). На
ней различают лицевую, носовую и нёбную поверхности. На лице-
вой поверхности против первого коренного зуба находится под-
глазничное отверстие. Им заканчивается подглазничный канал, а
начинается он в клинонёбной ямке верхнечелюстным отверстием.
Каудально на верхней челюсти выступает височный отросток, аль-
веолярный край кости позади каудальной альвеолы переходит
в верхнечелюстной бугор. Против третьего коренного зуба находит-
ся лицевой бугор. Вентрально на верхнечелюстной кости лежат аль-
веолярный и беззубый края. Дорсально на ней выступает носо-
вой отросток. Носовая поверхность верхней челюсти ограничивает
носовую полость. На ней дорсально находится слезный желоб, ниже
желоба располагается раковинный гребень, а вентрально — носовой
гребень и желоб сошниково-носового органа. Нёбная поверхность
верхнечелюстной кости обращена в ротовую полость. Ее образует
нёбный отросток. Между костными пластинками верхнечелюст-
ной кости имеется верхнечелюстная пазуха, которая сообщается
большим отверстием с носовой полостью. Часть пазухи заходит в
нёбный отросток и называется нёбной пазухой.
Особенности. У свиней лицевая поверхность верхнечелюс-
тной кости вогнута, подглазничное отверстие лежит против тре-
тьего коренного зуба, верхнечелюстные пазухи малы. У лошадей
на верхней челюсти имеется скуловой гребень, челюстной бугор
большой. _У собак носовой отросток доходит до лобных костей,
имеется верхнечелюстная пазуха.
Резцовая кость (os incisivum) парная, граничит у коров с верх-
нечелюстной носовой костями и сошником (см. рис. 3.11 — 3.25).
Тело резцовой кости имеет вид утолщенной пластинки. На самой
кости различают губную, нёбную и медиальную поверхности. Пе-
редний край резцовой кости не имеет луночек для зубов и назы-
вается беззубым. Между медиальными поверхностями находится
резцовая щель. От тела каудально простирается носовой отросток,
рядом с ним — нёбный отросток. Между этими отростками имеет-
ся нёбная щель.
Особенности. У свиней тело резцовых костей массивное с
тремя луночками для рсзпов.У лошадей тело резцовой кости мас-
сивное, с изогнутым резцовымГканалом, с тремя луночками для
резцов и половиной луночки для клыка (у самцов), носовые от-
ростки большие. У собак тело резцовой кости массивное, нёбная
щель хорошо видна у длинномордых, хуже — у короткомордых
собак.
89
Носовая кость (os nasale) парная^ граничит у коров с лобны-
ми, слезными, верхнечелюстными и резцовыми костями (см. рис.
3.11 — 3.25). Носовые кости соединены плоским швом: наружно —
между собой, внутри — с дорсальными носовыми раковинами и
с носовой перегородкой. Передний конец носовых костей двой-
ной свободный, их наружная поверхность выпуклая, внутренняя —
вогнутая, имеет два гребня: носовой — для перегородки и раковин-
ный — для прикрепления дорсальной раковины. Между латераль-
ным краем носовой кости и смежными с ней костями — слезной,
верхнечелюстной и резцовой — образуются щели.
Особенност и. У свиней носовые кости не соединены со слез-
ными костями, носовой отросток заострен. У лошадей носовые
кости имеют округлое основание, в толще кости находится носо-
вая пазуха, раковинный гребень большой. У собак кости не соеди-
нены со слезными, их передняя часть сильно расширена и вырез-
кой разделена на два отростка: ла1ср;ыьный и медиальный.
Скуловая кость (<« гууотайсит) парная., граничит у коров с
височными, слезными КОСТЯМИ и верхней челюстью (см. рис. 3.11 —
3.25). Она участвует в образовании скуловой дуги, глазницы и че-
люстной пазухи. Основание скуловой кости обращено ростраль-
но, а височный и лобный отростки — каудально. На наружной по-
верхности скуловой кости различают лицевую, глазничную и же-
вательную площадки. Ее внутренняя поверхность образует часть
верхнечелюстной пазухи.
Особенности. У свиней скуловые кости имеют два отрост-
ка, глазничная и жевательная поверхности закруглены. У лошадей_
эти кости с одним височным отростком, жевательная поверхность
шероховата. Удгобак указанные кости имеют основание из двух
корней, глазничная поверхность и лобный отросток малы.
Слезная кость (os lacrimale) парная? граничит у коров с лоб-
ными, носовыми, скуловыми костями и верхней челюстью (см.
рис. 3.11 — 3.25). Внутри к слезным костям прилежат дорсальные
носовые раковины. На их наружной поверхности различают лице-
вую и глазничную части, разделенные глазничным краем. На лице-
вой части имеется задний слезный отросток. На глазничной части
находится ямка слезного мешка, а в ней отверстие слезно-носового
канала. Здесь же образуется два тонкостенных слезных пузыря', ла-
теральный — больший, медиальный — меньший. Полости пузы-
рей сообщаются с верхнечелюстными пазухами. Внутренняя по-
верхность слезной кости является стенкой пазухи.
Особенност и. У свиней лицевая часть слезной кости уча-
ствует в образовании клыковой ямки, слезный бугорок в виде
гребня. У лощадей-слезные кости содержат оральный и абораль-
ный слезные бугорки, отверстие слезно-носового канала ворон-
кообразное. У собакони не имеют лицевой части, не доходят до
носовых костей, но граничат с нёбными костями.
90
Нёбная кость (os palaiinum) парная; укоров нёбные кости об-
разуют хоань1'(заднйё ноздрйТ «граничат с верхней челюстью,
лобными, решетчатой, клиновидной, крыловидной костями, вен-
тральной носовой раковиной и сошником (см. рис. 3.18 — 3.19, 3.21,
3.25). На нёбной кости различают горизонтальную и перпендику-
лярную пластинки. Горизонтальная пластинка занимает одну треть
твердого нёба. Она имеет воздушную пазуху, нёбную и носовую
поверхности, носовой гребень. Ростральный и латеральный края
небной кости снабжены швами для верхней челюсти. Перпендику-
лярная пластинка имеет ровную медиальную и шовную латераль-
ную поверхности. Последняя с клиновидной костью и верхней
челюстью образует, клинонёбную ямку. В ней находятся три отвер-
стия; верхнечелюстное — ведет в подглазничный канал, клинонёб-
ное — проникает в носовую полость и каудальное небное — следует
в небный канал. К крыловидной кос in npivieiaci крыловидный
отросток нёбной кости для прикрепления крыловидной мышцы.
Нёбный канал образован только нёбной костью.
Особенност и. У свиней горизонтальная пластинка нёбной
кости треугольной формы, носовой гребень хорошо развит, кры-
ловидный отросток большой. У лошадей горизонтальная пластин-
ка узкая, нёбный канал образован нёбной и верхнечелюстной
костями. У собак нёбные кости имеют большую горизонтальную
пластинку с несколькими отверстиями в нёбный канал, перпен-
дикулярная пластинка велика и соединена со слезной костью,
воздушных пазух нет.
Кость дорсальной носовой раковины (os conchae nasalis dorsalis)
у коров меньше вентральной (см. рис. 3.18, 3.21). Она представляет
собой нежную продырявленную костную пластинку, согнутую в
трубку веретенообразной формы. Ростральная часть закручивается
в вентральном направлении на полтора оборота, а каудальная часть
не закручивается. Спиральный отдел носовой раковины соединен
узкой щелью со средним носовым ходом, а полость каудальной
части дорсальной носовой раковины сообщается с лобными и
верхнечелюстными пазухами. Дорсальная носовая раковина лате-
рально прикреплена к раковинному гребню носовой кости, а або-
рально — к пластинке решетчатой кости. С поверхности она по-
крыта слизистой оболочкой.
Особенности. У свиней дорсальная носовая раковина уст-
роена так же, как у коров..У лошадей она одинаковых размеров с
вентральной носовой раковиной. У собак эта кость образует вто-
ричные разветвления в виде трубочек.
Кость вентральной носовой раковины (os conchae nasalis ventralis)
у коров прикреплена к гребню верхнечелюстной кости (см. рис.
3.18, 3.21). Рострально ее часть закручивается спирально на пол-
тора оборота вверх и на полтора оборота вниз, широко сообщаясь
со средним носовым ходом. Аборальная часть вентральной рако-
91
вины представляет одну полость, которая сообщается с лобными
и верхнечелюстными пазухами.
Особенности. /свиней строение вентральной раковины та-
кое же, как у коров. У лошадей раковина делает полтора оборота
только вверх. У собак строение раковины в основном такое же,
как у коров, но образуются вторичные разветвления.
Подъязычная кость {подъязычный аппарат) {os hyoideunr,
apparatus hyoideus) непарная, соединяется у коров с каменистой
костью и служит основанием для корня языка и гортани (рис. 3.29).
На подъязычной кости имеются тело, большие и малые рога. Тело
(базигиоид) — это небольшая поперечная пластинка, от которой
краниально выступает округлый язычный отросток. Каудально от
Рис. 3.29. Подъязычные кости собаки (А), коровы (Б), свинья (В), лоша-
ди (Г) (дорсолатеральная поверхность):
7 — тимпаногиоид (проксимальный членик); 2— стилогиоид (средний членик);
3— эпигиоид (дистальный членик); 4 — кератогиоид (малый рог); 5 — базигио-
ид (тело); 6 — тирогиоид (большой рог); 7 — язычный отросток; 8 — мышеч-
ный отросток
92
концов тела идут большие рога — тирогиоид — для соединения с
гортанью. От тела дорсально тянутся малые рога — кератогиоид. К
ним от каменистой кости спускаются ветви, на которых различа-
ют три членика: проксимальный — тимпаногиоид.—хрящевой, со-
единяется с височной костью; средний — стилогиоид — длинный,
плоский, с мышечным отростком; дистальный — эпигиоид — со-
единяется с малыми рогами.
Особенности. Усвиней подъязычная кость имеет сильно
развитое тело, язычный отросток и суставы отсутствуют. У лоша-
д,ей тело подъязычной кости с длинным острым язычным отрос-
тком. Средний членик длинный, пластинчатый, с малым мышеч-
ным отростком. У собак средний и дистальный членики развиты
одинаково хорошо, язычного отростка нет, большие и малые рога
хорошо развиты.
Череп в целом
Череп имеет форму неправильной четырехугольной пирамиды
(см. рис. 3.11 —3.25). На его дорсальной поверхности различают лоб-
ную и носовую ооласш. Первая из них гладкая, обширная и имеет
надглазничное отверстие. На венгральной поверхности черепа на-
ходятся область основания черепа, область хоан и нёбная область.
В области мозгового черепа лежит височная ямка, глазница и кли-
нонёбная ямка. Височная ямка тянется от скуловой дуги до темен-
ного валика лобной кости. Она соединяется с глазницей, клино-
нёбной и подвисочной ямками.^Глазница — это вместилище для
глаза. Кольцо глазницы образовано лобными, слезными, скуло-
выми костями. В нее открываются решетчатое, зрительное, круг-
ло-глазничное, надглазничное отверстия. На слезной кости лежат
отверстие слезно-носового канала и мышечная ямка. На глазнич-
ном отростке лобной кости видна ямка слезной железы. Клино-
нёбная ямка имеет следующие отверстия: верхнечелюстное, кли-
нонёбное и каудальное нёбное.
Полости черепа подразделяются на мозговую, носовую и рото-
вую. Мозговая полость ограничена семью костями, образующими
свод и основание.
^Носовая полость ограничена: вверху — лобными и носовыми
костями, внизу — нёбными отростками верхней челюсти, резцо-
вых и нёбных костей, латерально — верхней челюстью, слезной,
скуловой, резцовой костями и носовыми раковинами, медиаль-
но — сошником, вертикальной пластинкой решетчатой кости и
хрящевой носовой перегородкой.
Ротовая^полость образуется резцовыми костями, верхней и
нижней челюстями.
Околоносовые пазухи располагаются между наружной и внут-
ренней пластинками костей мозгового и лицевого черепа. Верхне-
93
челюстная пазуха находится между пластинками верхнечелюст-
ной кости. Слезная пазуха образована слезной костью и имеет два
сообщающихся между собой пузыря: латеральный, больший, и
медиальный, меньший. Лобная пазуха сформирована лобной кос-
тью. Она имеет большие размеры, передняя часть пазухи находит-
ся между глазницами, а задняя — примыкает к затылочной кости
и переходит в роговые отростки, с возрастом объем лобных пазух
увеличивается. Носовая пазуха лежит в носовой кости (часто от-
сутствует). Клинонёбная пазуха образована пластинками нёбной и
клиновидной костей.
Позвоночный столб
Позвоночный столб (columna vertebralis) состоит из позвонков.
Позвонок (vertebrae) имеет тело, головку и ямку (см. рис. 3.30 —
3.38, 3.11 —3.16). Тело — самая массивная часть позвонка. На его
краниальном конце имеется выпуклость — головка позвонка, на
каудальном конце — ямка позвонка. На вентральной поверхности
тела находится вентральный гребень. Дорсально от тела вытягива-
ется дужка позвонка. Между дужкой и телом позвонка образуется
отверстие позвонка. Все отверстия позвонков, соединяясь, фор-
мируют позвоночный канал. У основания краниального края дужки
находится краниальная позвоночная вырезка, а у основания кау-
дального края — каудальная позвоночная вырезка. Вырезки двух со-
седних позвонков формируют межпозвоночное отверстие. По кра-
ям дужек выступают краниальные и каудальные суставные отрост-
ки. По бокам от тела позвонка отходят поперечные отростки. От
середины дужки поднимается дорсально остистый отросток.
Шейные позвонки (vertebrae cervicales) по своему строению нео-
динаковые (см. рис. 3.11 —3.16, 3.31 — 3.34). Наиболее сходны меж-
ду собой лишь средние (третий, четвертый и пятый) позвонки.
Остальные, особенно второй и первый, имеют выраженные осо-
Рис. 3.30. Части позвонка:
7 — остистый отросток; 2, 72 — дужка позвонка;
3 — каудальный суставной отросток; 4 — доба-
вочный отросток; 5 — каудальная позвоночная
вырезка; 6 — поперечный отросток; 7 — сосце-
видный отросток; 8 — тело позвонка; 9 — вент-
ральный гребень; 10 — головка позвонка; 77 —
краниальная позвоночная вырезка; 13 — отвер-
стие позвонка; 14 — краниальный суставной от-
росток
94
Рис. 3.31. Атланты собаки (А), свиньи (Б), лошади (В), коровы (Г):
1 — боковое позвоночное отверстие; 2 — крыловое отверстие; 3 — крыловая вы-
резка; 4 — дорсальная дужка; 5 — дорсальный бугорок; 6 — крыло атланта; 7 —
поперечное отверстие; 8 — каудальная суставная поверхность; 9 — вентральная
дужка; 10 — вентральный бугорок; 11 — позвоночное отверстие; 12 — ямка зуба
бенности в строении. У всех домашних животных по семь шейных
позвонков.
Первый шейный позвонок — атлант (atlas) сильно отличается
от остальных шейных позвонков (рис. 3.31, см. рис. 3.11 — 3.16). На
нем выделяют дорсальную и вентральную дужки. На дорсальной
дужке имеется дорсальный бугорок, на вентральной дужке — вент-
ральный бугорок — для прикрепления мышц, обеспечивающих сги-
бание и разгибание головы. Поперечные отростки образовали мас-
сивные прямые крылья атланта. Под крыльями находятся крыло-
вые ямки. На крыльях атланта имеются два парных отверстия: кры-
ловое отверстие открывается в крыловую ямку, межпозвоночное
отверстие сообщается с отверстием позвонка. На краниальном
конце крыльев атланта располагаются краниальные суставные ямки
яйцевидной формы для сочленения с мыщелками затылочной
кости. На каудальном конце крыльев выступают каудальные сус-
тавные поверхности для соединения со вторым шейным позвон-
ком. Поперечное отверстие отсутствует.
Особенности. У^виней атлант имеет массивные крылья,
дорсальный и вентральный бугорки большие, краниальное кры-
ловое и межпозвоночное отверстия хорошо развиты, а попереч-
ное отверстие лежит на заднем крае крыла атланта^ У лошадей
атлант имеет широкие тонкие пластинчатые крылья с утолшен-
95
ними краями, загнутыми вниз, крыловые ямки глубокие, в кры-
ле расположены межпозвоночное, крыловое и поперечное отвер-
стия.. У собак крылья атланта горизонтальные, тонкие, длинные,
прямые, крыловая ямка плохо выражена, крыловое отверстие на
переднем крае крыла образует вырезку, поперечное отверстие
хорошо выражено.
Второй шейный позвонок — ось, или эпистрофей {axis, s.
epistropheus), на переднем конце длинного тела вместо головки
позвонка имеет зуб с суставной поверхностью для сочленения с
атлантом (рис. 3.32; см. рис. 3.11 — 3.16). Его дорсальный гребень слегка
понижается в краниальном направлении, задний край дорсаль-
ного гребня более массивный. Каудальные суставные отростки
начинаются от дуг. Краниальное межпозвоночное отверстие круглое.
Поперечный отросток одинарный, имеет небольшое поперечное
отверстие.
Особенност и..У свиней цилиндрический зуб оси с перехва-
том у основания, гребень позвонка высокий, толстый, постепен-
но возвышающийся в каудальном направлении, поперечный от-
росток мал, поперечное отверстие большое. У лошадей ось с длин-
Рис. 3.32. Эпистрофеи собаки (А), свиньи (/>'), коровы (5), лошади (Г):
1 — остистый отросток (гребень эпистрофея); 2 — каудальный суставной отросток;
3 — каудальная позвоночная вырезка; 4 — ямка позвонка; 5 — поперечный отрос-
ток; 6 — вентральный гребень; 7 — тело; 8 — поперечное отверстие; 9 — кра-
ниальная суставная поверхность; 10 — зуб; II — боковое позвоночное отверстие;
12 — краниальная позвоночная вырезка
96
ным телом, конусовидным зу-
бом, дорсальный гребень по-
томка большой, разветвляется
и переходит в каудальные сус-
тавные отростки. У собак ось
имеет сильно выдвинутый впе-
ред тонкий гребень позвонка,
поперечное отверстие хорошо
развито.
Средние шейные позвонки
одинаковые по своему строению
и называются типичными (рис.
3.33; см. рис. 3.11 — 3.16). У них
короткое тело, а головка и ямка
позвонка хорошо развиты. Ости-
стые отростки наклонены впе-
ред, на конце имеются утолще-
ния. На поперечных отростках
находятся краниальная и кау-
дальная ветви, у основания по-
перечных отростков расположе-
но поперечное отверстие. Крани-
альные суставные отростки мас-
сивные, с обширными плоски-
Рис. 3.33. Типичный шейный позво-
нок лошади (латеральная поверх-
ность):
7 — краниальный суставной отросток;
2 — дорсальный бугорок; 3 — боковой
гребень; 4 — каудальный суставной
отросток, каудальная суставная повер-
хность; 5 — каудальная позвоночная
вырезка; 6— поперечное отверстие; 7 —
ямка позвонка; 8 — вентральный гре-
бень; 9—тело; 10 — поперечно-ребер-
ный отросток; 11 — поперечный отро-
сток; 12 — реберный отросток; 13 —
краниальная позвоночная вырезка;
14 — головка позвонка
ми суставными поверхностями
вверху, такие же поверхности имеются и на каудальных суставных
отростках, но они расположены внизу. Между отростками тянут-
ся соединительные гребни.
Особенности. У свиней типичные шейные позвонки име-
ют короткое тело, длинные остистые и поперечные отростки,
вентральные гребни отсутствуют, вентральные пластинки попе-
речных отростков заходят одна за другую как черепица, в осно-
вании поперечных отростков лежит дорсовентральный канал, кра-
ниальные, каудальные суставные отростки и поперечные отвер-
стия развиты хорошо. У лошадей типичные шейные позвонки
имеют длинное тело, крутую головку и глубокую ямку, остис-
тые отростки развиты слабо, краниальные и каудальные сустав-
ные отростки массивные и соединены гребнями. У собак на кау-
дальных суставных отростках хорошо развиты добавочные’бтрос-
тки.
Шестой и седьмой шейные позвонки отличаются по строению
от типичных шейных позвонков (см. рис. 3.11 — 3.16). Шестой шей-
ный позвонок имеет массивную вентральную пластинку, попереч-
ное отверстие у него большое, вентральный гребень отсутствует,
остистый отросток развит сильнее, чем у остальных шейных по-
звонков.
4 Зеленонский
97
Седьмой шейный позвонок с
одинарным поперечным отрос-
тком, поперечное отверстие от-
сутствует (рис. 3.34; см. рис. 3.11 —
3.16). По бокам от ямки позвон-
ка располагаются каудальные ре-
берные ямки для первого ребра.
Остистый отросток большой. От-
верстие позвонка более обшир-
ное, чем у типичных шейных по-
звонков.
Рис. 3.34. Седьмой шейный позво-
нок лошади (латеральная поверх-
ность):
1 — краниальный суставной отросток;
2 — остистый отросток; 3 — боковой
гребень; 4 — каудальный суставной от-
росток, каудальная суставная поверх-
ность; 5 — каудальная реберная ямка;
6 — тело; 7 — поперечный отросток;
8 — головка позвонка; 9 — краниаль-
ная позвоночная вырезка; 10 — кау-
дальная позвоночная вырезка
Особенности. У свиней
шестой шейный позвонок харак-
теризуется высоким остистым
отростком, большой вентраль-
ной пластинкой, хорошо выра-
женным дорсовентральным ка-
налом, седьмой шейный позво-
нок имеет одинарный попереч-
ный отросток, дорсовентраль-
ный канал, каудальные ребер-
ные ямки, высокий остистый
отросток, поперечные отверстия
отсутствуют. .У.лошадей шестой
шейный позвонок имеет вентральную пластинку на поперечном
отростке, вентральный гребень отсутствует, остистый отросток
больше, чем у типичных шейных позвонков, седьмой шейный
позвонок характеризуется наличием одинарного поперечного от-
ростка. каудальных реберных ямок, высокою остистого отрост-
ка. отсутствием поперечного отверстия. У собак шестой шейный
позвонок с вентральной пластинкой, а вентральный [ребень от-
сутствует, седьмой шейный позвонок не имеет поперечного от-
верстия, а каудальные реберные ямки слабо развиты.
Грудные позвонки (vertebrae thoracalcs} вместе с ребрами и гру-
диной Формируют грудную клетку (рис. 3.35; см. рис. 3.11 — 3.16),
Число грудных позвонков у коров — 13, у свиней — 14—17. \
лошадей .— IX, у собак — 13.
Грудные позвонки коровы имеют: краниальные реберные ямки,
расположенные по бокам головки; каудальные реберные ямки по
бокам ямки позвонка; реберные ямки поперечных отростков — дли
сочленения с бугорком ребра. Тело позвонка длинное и имее т тп
лиеобразный перехват. Позвоночный канал в области первых грех
грудных позвонков более широкий, чем у последующих.
... Остистый отросток наклонен назад. Длина его с первого по
четвертый позвонок увеличивается, с пятого по девятый умень-
шается, а с десятого по тринадцатый отросток имеет одинаковую
98
Рис. 3.35. Грудные позвонки лошади (Л), свиньи (Б);
I < •ц.исшшшй огросток; 2 - межпозвоночное отверстие; .7 — остистый отрос
1 каудальный суставной отросток; 5 каудальная по(ноночпля вырезка.
► п игп.ная реберная ямка; 7 ямка позвонка; & центральный |ребень; V -
• • । • IH головка позвонка: // краниальная реберная ямка; 12 краниальная
......... вырезка; 13 краниальный суставной отросток; 14 — реберная ямка
......... и 'iiioin отростка; 15 латеральное дорсальное позвоночное отверстие
т.н «ну. Со второго по девятый позвонок остистые отростки дают
..иву холки. Остистые отростки имеют ост рые передние и задние
трап с гребнями на концах. Остистый отросток тринадцатогопо-
..... стоит отвесно, а сам позвонок называется диафрагмальным
ыптт шпальным), так как через него проходит линия отвеса тя-
.1 'ч in тела животного.
1\>кка позвонка имеет глубокие каудальные позвоночные вы-
Iи и и или добавочные боковые межпозвоночные отверстия для
। in \ к>в и нервов.
I (средние и задние суставные отростки малой величины. Попе-
речные отростки короткие, краниально на них размещаются со-
. н< видные отростки для закрепления мышц.
<) ( обенности. У свиней двенадцатый позвонок диафрагмаль-
|||.in. leno позвонка короткое, полукруглое, дужка имеет само-
। шиleiiuioe межпозвоночное отверстие, на каждом поперечном
>и|ич1кс имеется дорсовентральное отверстие, остистые отроет -
। и широкие и длинные (шесть первых позвонков), краниальные
• м i.iiiiibie отростки на последних четырех-пяти позвонках имеют
Jnipmv вгулок, а каудальные — полуцилиндров. У лошадей шест-
и.। ни п.1Й позвонок диафрагмальный, тело позвонка трехгранное,
|п1ч рц|,1с ямки глубокие, остистые отростки имеют булавовид-
ные уюдщения, краниальные края их острые, каудальные в виде
99
1 — остистый отросток; 2 — попе-
речный отросток; 3 — краниаль-
ный суставной отросток; 4 — каудальный суставной отросток; 5 — позвоночное
отверстие; 6— краниальная позвоночная вырезка; 7— каудальная позвоночная
вырезка: 8 — вентральный гребень; 9 — головка позвонка; 10 — ямка позвонка;
11 - суставная поверхность поперечного отростка; 12 — суставная поверхность
каудального суставного отростка
шероховатых площадок, реберная фасетка поперечных отростков
на последних позвонках сливается с краниальной реберной ям-
кой.У собак рдинадцатый позвонок диафрагмальный, гребни на
телах позвонков отсутствуют, остистые отростки округлые, с пер-
вого по девятый дугообразно изогнуты вверх, краниальные сус-
тавные отростки на последних четырех позвонках в виде верти-
кальных пластинок с сосцевидными отростками; на последних
грудных позвонках каудально выступают добавочные отростки.
Поясничные позвонки (vertebrae lumbales) характеризуются на-
личием длинных, плоских, лентовидных поперечных и хорошо
развитых суставных отростков (рис. 3.36; см. рис. 3.11 — 3.16). У ко-
ров шесть поясничных позвонков, у свиней — шесть—восемь, у
лошадей — шесть, у собак — семь.
У коров тела позвонков длинные, с талиеобразным перехва-
том. Поперечные отростки с острыми неровными краями и загну-
100
ты вперед. Остистые отростки стоят отвесно, одинаковы по высо-
те и ширине. Краниальные суставные отростки образуют полуци-
линдрические втулки, а каудальные — такие же блоки.
Особенност и. У свиней_поясничные позвонки устроены так
же, как у коров, но имеют дорсовентральное отверстие в основа-
нии поперечного отростка (иногда вырезку), краниальные сус-
тавные отростки втулкообразные, каудальные — валиковидные.
У лошадей тела первых позвонков трехгранные, с гребнем для
диафрагмы; тела последних позвонков — овальные, остистые от-
ростки наклонены вперед, высота их в два раза больше ширины,
суставные отростки имеют плоскую форму. У собак остистые от-
ростки наклонены вперед, поперечные отростки обращены впе-
ред, вниз и латерально, суставные поверхности находятся в сред-
ней плоскости, под каудальными суставными отростками имеют-
ся добавочные отростки.
Крестцовые позвонки (vertebrae sacrales) слились в одну крест-
цовую кость — os sacrum (рис. 3.37; см. рис. 3.11 —3.16). У коров
крестцовую кость образуют пять позвонков, у свиней — четыре, у
лошадей — пять, у собак — три.
Рис. 3.37. Крестцовые кости собаки (А), свиньи (Б), коровы (5), лоша-
ди (Г):
1 — основание крестцовой кости; 2 — мыс; 3 — крыло крестцовой кости; 4 —
тазовая поверхность; 5 — ушковидная поверхность; 6 — краниальный суставной
отросток; 7— суставная поверхность; 8 — срединный крестцовый гребень; 9 —
промежуточный крестцовый гребень; 10 — латеральный крестцовый гребень;
11 — каудальный суставной отросток; 12— междужковые пространства
101
У крестцовой кости остистые отростки слились в крестцовый
грепснь. междужковые отверстия отсутствуют. Позвоночные вы-
резки образую! четыре пары дорсальных крестцовых отверстий
Поперечные отростки слились в острые и зазубреш!ые боковые
части. Первые два поперечных отростка образуют четырехуголь-
ные массивные крылья крестцовой кости. На них имеются ушко-
видные поверхности для сочленения с тазом.
Передний край крыла крестцовой кости гладкий и тупой, боко-
вой и задний края — заостренные. Вентральная поверхность крест-
цовой кости вогнутая, на ней видны поперечные линии — места сли-
яния тел позвонков. Здесь же проходят сосудистый желоб и четыре
пары вентральных крестцовых отверстий. Головка первого крестцо-
вого позвонка выпуклая; выступая вниз, она образует мыс.
Крестцовый канал суживается в каудальном направлении.
Широкий край крестцовой кости направлен вперед, боковые ее
края острые. Крылья крестцовой кости образуют правый и левый
углы. Они соединяются с крыльями подвздошной кости, а задний
угол соединен с хвостовыми позвонками.
Особенности. У свиней крылья крестцовой кости сильно
повернуты в сторону сагиттальной плоскости, остистые отростки
отсутствуют, междужковые отверстия большие, вентральных кре-
стцовых отверстий три пары, краниальные суставные отростки
имеют вид полуцилиндрических втулок, боковые части крестцо-
вой кости гладкие и округленные. У лошадей остистые отростки с
булавовидными утолщениями, наклоненьГназад и не сливаются
между собой, дорсальных и вентральных крестцовых отверстий
по четыре пары, боковые части крестцовой кости массивные, ее
крылья треугольной формы и лежат горизонтально, на краниаль-
ном крае крыла имеется суставная поверхность для сочленения с
поперечным отростком поясничного позвонка, краниальные сус-
тавные отростки лежат в сагиттальной плоскости. Уцобак крылья
крестцовой кости расположены в сагиттальной плоскости, остис-
тые отростки сливаются в гребень, вентральных и дорсальных от-
верстий по две пары, краниальные суставные отростки направле-
ны, как у лошади.
Число хвостовых позвонков (vertebrae coccygeae) у животных
сильно колеблется: у коров — 18 — 20, у свиней — 20 —25,.у ло-
шадсй — 18 — 20. v ,с.оба к — 20 — 23.
-^У коров краниальные суставные отростки хвостовых позвон-
ков хорошо развиты только на первых пяти-шести позвонках, но
они не имеют суставных поверхностей, поперечные отростки име-
ют вид широких, загнутых вниз пластинок, на восьмом—десятом
позвонках эти отростки отсутствуют, на вентральной стороне вто-
рого—тринадцатого позвонков хорошо развиты гемальные отро-
стки, между которыми проходит артерия (рис. 3.38; см. рис. 3.11 —
3.16).
102
Особенности. У свиней
первые четыре-пять позвонков
имеют краниальные и каудаль-
ные суставные отростки с сус-
тавными поверхностями, попе-
речные отростки, начиная с пя-
того-шестого позвонка, слабо
развиты. У лошадей краниальные
суставные отростки имеют вид
бугорков, каудальные отсутству-
ют, гемальные дуги, начиная с
четвертого позвонка, исчезли,
поперечные отростки массив-
ные, но с восьмого позвонка
редуцируются. У..собак на пер-
Рис. 3.38. Хвостовые позвонки со-
баки:
1 — краниальный суставной отросток;
2 — головка позвонка; 3 — попереч-
ный отросток; 4 — остистый отросток;
5 — отверстие позвонка; 6 — гемаль-
ная дужка
вых четырех-пяти позвонках кра-
ниальные и каудальные суставные отростки сочленяются между
собой, поперечные отростки направлены назад и в стороны, на
пятом—десятом позвонках находятся гемальные отростки.
Ребра и грудная кость
Ребра (costae) состоят из двух сросшихся частей: верхней ре-
берной кости и нижней — реберного хряща (рис. 3.39: см. рис. 3.11 -
3.16). У корон имеется 13 пар ребер, у свиней — 14— 17, у лоша-
дей 18. у собак — 13. Различают истинные и ложные ребра. Ис-
тинные ребра соединяются хрящами непосредственно с грудиной.
Ложные ребра не доходят до грудины, они соединяются с лежа-
щими впереди реберными хрящами, которые вместе с последним
ребром образуют реберную дугу. У коров истинных ребер восемь
пар, ложных — пять.
На ребре различают концы — позвоночный, обращенный к
позвонку, и грудинный, или стернальный, направленный в сто-
рону грудины. На позвоночном конце находится головка ребра с
двумя выпуклыми суставными поверхностями для сочленения с
телами двух смежных позвонков, в стороне or головки выступает
бугорок ребра с суставной поверхностью, сочленяющейся С ребер-
ной суставной поверхностью поперечного отростка позвонка.
Ниже головки имеется шейка ребра. Позади бугорка ясно выс-
тупает угол ребра для прикрепления мышц. Тело ребра широкое, а
межреберное пространство узкое. На теле находится две поверх-
ности: наружная — выпуклая, внутренняя - вогнутая. Передний
край тела ребра заостренный, задний — притупленный.
Снаружи у переднего края ребра лежит мышечный желоб для
прикрепления межреберных мышц. Вдоль заднего края на внут-
103
Рис. 3.39. Ребра:
А — лошадь, первое ребро; Б — собака; В — свинья; Г — корова; Д — лошадь,
18-е ребро; / — головка ребра; 2 — шейка ребра; 3 — бугорок ребра; 4 — угол
ребра; 5 — тело ребра
реннеи поверхности ребра помещается сосудистый желоб для раз-
мещения сосудов и нервов. У коров на грудных концах ребер от
второго до десятою имеются суставы для сочленения с реберны-
ми хряшами Грудинные концы ребер шире тела. Длина ребер от
первого до седьмого увеличивается, от седьмого до десятого не
изменяется, а от десятого до тринадцатого уменьшается.
Реберные хрящи от второго до десятого имеют вверху сустав-
ные поверхности для сочленения с ребром, ширина реберных
хрящей уменьшается в каудальном направлении, длина реберных
хрящей от первого до десятого ребра увеличивается, а от десятого
до тринадцатого уменьшается.
Особенности. У свиней истинных ребер шесть— восемь пар,
ложных — восемь-девять. Мышечные желоба хорошо выражены.
На стернальных концах от второго до V ребра имеются суставные
поверхности. У лошадей истинных ребер восемь пар. Длина их уве-
личивается от первого до десятого, а от десятого до восемнадца-
того ребра уменьшается. У собак истинных ребер девять пар, сус-
тавы бугорка и головки последних двух-трех ребер сливаются.
Грудная кость, или грудина (sternum), состоит у коров из семи
костных сегментов, на ней различают рукоятку, тело и мечевид-
ный отросток. Рукоятка грудины — первый сегмент трехгранной
формы, слегка приподнята вверх, имеет две реберные ямки для
первой пары реберных хрящей Тело грудины сплюснуто сверху вниз
104
Рис. 3.40. Грудные кости собаки
(Л), свиньи (Б), овцы (S), коро-
вы (Г), лошади (Д):
1 — рукоятка грудины; 2 — тело
грудины; 5 мечевидный отросток;
4 — мечевидный хрящ
и соединяется с рукояткой грудины суставом. На боковых сторо-
нах тела грудины лежит шесть пар суставных ямок для реберных
хрящей. Мечевидный отросток — каудальная часть грудины — имеет
вид широкой хрящевой пластинки (рис. 3.40; см. рис. 3.11 — 3.16).
Особенности. У свиней тело грудины состоит из шести сег-
ментов, рукоятка массивная и лежит горизонтально, тело груди-
ны сплюснуто дорсовентрально, мечевидный хрящ хорошо выра-
жен. У лошадей грудина сильно сжата с боков, рукоятка грудины
спереди дополняется хрящевым соколком, а каудально переходит
в тело. У собак тело грудной кости состоит из восьми цилиндри-
ческих сегментов, мечевидный хрящ с небольшой округлостью
на конце.
Периферический скелет
Периферический ckc.'ici представлен костями грудных и таю-.,
вых конечностей 3.41—3.56, см. рис. 3.11— 3 16). Они появля-
ются у млекопитающих н результате длительного филогенеза пу-
тем преобразования плавников предков костистых рыб в сторону
укрепления поясов конечностей около осевого скелета и приоб-
ретения возможности приподнять тело над поверхностью земли
благодаря двум парам трехчленных конечностей.
Пояса костей грудной и тазовой конечности оказались постро-
енными из трех костей: по одной кости (лопатка на грудной ко-
нечности_и.додвздошнаяна талоном) присоединяются’ к осево-
му скелету с дорсальной поверхности. ио две кости (коракоилпая
кость и ключ и ца на грудной конечности, лонная и седалищная —
105
Рис. 3.41. Гомодинамные звенья конечностей:
7 — пояса костей грудной и тазовой конечности; 2 — стилоподий; 3 — зейгопо-
дий; 4— автоподий: 5— базиподий; 6 — метаподий; 7— акроподий
на тазовой»охватывают тело вентрально. При ном поянтение пояса
костей [рудной конечности приводит к укреплению и развитию
грудной кости, а вентральные кости пояса газовой конечности
замыкаются в срединной плоскости, принимая участие в форми-
ровании тазовой полости (см. рис. 3.41,3.11 —3.16; рис. 3.42 — 3.44,
3.48-3.50).
Наиболее значительные изменения в процессе филогенеза про-
текали в свободном отделе конечностей. В роультате длительных
алаптогенных преобразований тело оказалось приподнятым нал
поверхностью на четырех трехзвенных конечностях, расположен-
ных под углом друг к другу.
Первое звено — стилоподий — включает плечевую кость нагрудг
ной конечности и бедренную — на тазовой (см. рис. 3.41). Обе ко-
сти соединяются с поясами конечностей простыми многоосными
суставами.
106
№ Л ':' ;z '"
Второе звено — зейгоподий — представлено двумя костями: лу-
чевой и локтевой на'грудной конечности, большой берцовой и
малой берцовой — на тазовой (см. рис. 3.41). Звенья соединяются
одноосными суставами как с проксимальными, так и с дисталь-
ными костями.
Третье звено конечностей — автоподий — представляет собой
участок конечностей, называемый у четвероногих лапой (см. рис.
3.41). На грудной конечности третье звено называется кистью, а
на тазовой — стопой. Третье звено имеет сложное строение и в
свою очередь состоит из трех участков. Базиподий включает кости
запястья на грудной и заплюсны — на тазовой конечностях,jue-
тапидий объединяет пясшыс и иноеневые кости, и наконец, ак-
роподий — кости пальцев кисти и стопы.
В ia.ii пейшем филогенез конечностей был связан с изменени-
ями способа передвижения и все более значительным приподни-
мании тела над поверхностью земли. При этом площадь опоры
конечности на нее уменьшалась, а животные с опорой на весь
автоцодий — стопоходящие — превратились » пальцеходящие — с
опорой только на акроподий. а от НИХ пром юшла'ёшё более у жа-
спе циализирова иная группа — копытоходящие — с опорой только
на дистальную фалангу. По niiTy опоры конечностей собака яв-
ляется пайьцеходяшей, а свинья, крупный рогатый скот и ло-
шадь — копытоходящими.
Скелет грудной конечности
Кости i рудной конечное in (oyvu membri thoracic!) включают ко-
сти плечевою пояса — лопатку и кости свободной конечности —
плечевую кость, скелет предплечья, скелет кисти, куда входят
кости запястья, кости пясти и кости пальцев (рис. 3.42, 3.43; см.
рис. 3.113.16, 3.41, 3.44- 3.47).
У домашних животных пояс костей грудной конечности пред-
ставлен лишь его дорсальным звеном — лопаткой.
Лопатка (scapula) имеет форму слегка вытянутой треугольной
формы пластинки, связанную с осевым скелетом с помощью
мышц (рис. 3.44; см. рис. 3,11 — 3.16, 3.41 — 3.43). Нерасширенное
основание направлено дорсально в сторону остистых отростков хол-
ки и дополняется лопаточным хрящом. В дорсокраниальной части
хрящ закруглен и вентрально незаметно переходит в краниальный
край. Каудальная часть хряща незначительно выступает за соответ-
ствующий край лопатки. Краниальный и каудальный края лопат-
ки вместе с ее основанием формируют краниальный и каудальный
углы.
Л опатка имеет латеральную и медиальную поверхности.Латераль-
ная поверхность разделена лопаточной остью на две неглубокие,
но обширные предостную и заостную ямки. Сама лопаточная ость,
107
Рис. 3.42. Кости грудной конечности
коровы (медиальная поверхность):
/краниальный угол; (2/— основание
лопатки; ср каудальный угол;^— кау-
дальный край. _> дпейм лопатки; ^—
плечевой СТстад; Z*- ПОЗадисктввНоЛ
бугорок; 70— локтевой бугор, W— .кис-
тевой отросток; Iff проксимальная
межкостная шелъ предплечья; 7р—лок-
тевая гость; 72,— шиловидный отрос-
ток; /.? добавочная кость «пястья;
14 — пальмарные сссамовддные кости
проксимальной фаланги; 15.— пальмар-
ные сездмовилпые кости дистальной фа-
ланги; 1б) подлопаточная ямка;, IV
краниальный край; Д’/>акромион:Т/5-
налсустакной бугорок. большой бу-
горок (плечевая кость). малыйДуу-
горок:5^- гребень малого бугорка;’2.1'
тело плечевой гости; 24: мслнадСНын
надмыщелок. 2> - тело лучевой кости;
•2ft'— >1С1налы1ый шиловидный отрос
ток: 27'— лучевая кость «пястья. 28/-
вторая г третья кость «пястья. 29 - тре-
тья + четвертая пястная кос1ь;ЗЙ— блок
головки «ретьей пястной кости,
проксимальные фалпнгн: 52 средние
фаланги, Лу- дистальные фаланги (ко-
1< ? пыгиевые кости)
опускаясь вентрально, постепен-
но сходит на нет, не достигая су-
ставного угла. Дистальный учас-
ток ости выступает в виде акро-
миона, а в средней части ее об-
наружен вается шероховатое утол-
щение — бугор ости. Предостная ямка по площади значительно (в
два-три раза) уступает аналогичному показателю заостной ямки.
Медиальная поверхность лопатки формирует обширную и не-
глубокую подлопаточную ямку пля закрепления одноименной мыш-
цы. В дорсальном направлении ямка ограничена зубчатой линией.
Дорсальнее от последней лежит зубчатая поверхность, наиболее
сильно развитая ближе к краниальному углу лопаткйГ
Проксимальная половина краниального края лопатки выпук-
лая. В дистальном направлении краниальным край лопатки посте-
пенно приближается к ее ости, формируя вырезку лопатки. Дис-
тальнее последней располагается надсуетавной бугорок для при-
крепления двуглавой мышиы плеча _ и г.ной поверхности
108
Рис. 3.43. Кости грудной конечности
лошади (латеральная поверхность):
1 — краниальный угол; 2 — основание ло-
патки; 3 — каудальный угол. •/ каудаль-
ный край; ! • ! яМКЯ шейка
лопатки; 7— позадисуспшной бугорок;;^—
головка плечевой коспг. 9 тело плече-
вой кости;(70/— локтевая ямка; //- лок-
тевой бугор; 12 — проксимальная межкост-
ная щель предплечья; 13 — латеральный
шиловидный отросток; 14 — добавочная
кость запястья; 15 — четвертая пястная
кость, 16 пальмарные проксимальные
якамоцинвв kociii; 12/— краниальный
i.n.iif. 7.Y — предостная ямка//у~ ость ло-
патки. 20 - надсуставной бугорок; 21 —
большой бугорок; 22 — дельтовидная ше-
роховатость; 23 — латеральный мыщелок;
24 — головка лучевой кости; 25 — лучевая
кость; 26 — проксимальный ряд костей за-
пястья; 21 — дистальный ряд костей запя-
стья; 28 — третья пястная кость; 29 — про-
ксимальная фаланга; 130'— средняя фалан-
га; 31)- дистальная фаланга
бугра располагается кордкоидный от-
Лосток — место прикрепления клю-
вовито-плечевой мышцы. Каудаль-
ный кран лопатки ровный. в дис-
тальной трети кости он приближа-
ется к ости лопатки и вместе с вы-
резкой лопатки формирует шеицуло-
патки.
~Угол лопатки, направленный в
сторону свободного отдела конечно-
сти, сильно утолщен и служит мес-
том соединения с плечевой костью.
Он называется. суставным углам. На нем располагается пологая
округлая суставная впадина для образования лопаточно-плечево-
го сустава.
Особенност и._У свиней лопатка широкая и короткая, бугор
ости большой, шейка и лопаточный хрящ хорошо выражены.
У лошадей лопатка длинная, предостная ямка занимает одну треть
109
Рис. 3.44. Лопатки кошки (Л). собаки (Б), свиньи (В), коровы (Г). лоша-
ди (Д) (латеральная поверхность);
/' — основание лопатки; краниальный угол; 3 каудальный усол: 4
мешкая ямка; 5 каудальный край; 6,1 шейка лопатки;. 7 потадисуставной
бугорок; d щалкрючковидный отросток; 9 — бугор оши допаткн. 10. предр-
ешай ямка; // краниальный край; /2- ость лопатки; /3 акромион. 14
наасустивной бугорок; 7.у— суставная впадина
лопатки, акромион мал. У собак краниальный и каудальный углы
лопатки закруглены, акромион большой, бугор ости отсутствует.
Плечевая кость (os humeri, х. brachii) мощная трубчатая кость,
важный рычаг при движении (рис. 3.45; см. рис. 3.11 —3.16, 3 41 —
3.43). На ней выделяют средний участок диафиз (тело) и два кон-
ца, или проксимальный и дистальный эпифизы. Первый из них на-
правлен в сторону лопатки и обрагует с ней плечевой сустав, а
irropon - в сторону костей предплечья, формируя с ними локте-
вой сустав.
Проксимальный конец несет головку плечевой кости. Поверх-
ность ее, покрытая гиалиновым хрящом, значительно больше со-
110
Рис. 3.45. Плечевые кости кошки (А), собаки (Б), свиньи (В), лошади (Г),
коровы (Д) (краниолатеральная поверхность):
/ — головка плечевой кости, £ большой бугорок; поверхность эаостной
muuiiiu: •£> межбугорковый желоб; Д - шероховатость малой круглой мышцы:
6 дельтовидная шероховатость; 7^- гребень плечевой кости; А/- налмышедко-
вое отверстие, Чу тучевая ямка. W— гребень лагери.иного надмышелка.
блок плечевой копи. 12 латеральный надмыщелок; IJ. малый бугорок;
средний бугорок
111
пленяющейся с ней суставной впадиной лопатки, что обеспечи-
вает значительный размах в этом суставе при движении. Головка
переходит в тело, формируя шейку плечевой кости. Вокруг головки
кости располагаются возвышения и шероховатости для закрепле-
ния мышц и связок.
С дорсолатеральной поверхности располагается большой буго-
рок. с дорсоме шальной поверхности — малый бугорок. бугорки
разделены межбугорковым желобом лля прохождения сухожилии
двуглавой мыпшы плеча. С ллеральной поверхности большого
бугорка, у его основания, располагается шероховатость для заве-
тной мышцы. Дистальнее оз последней проходит гребень большого
йг. о/гг <,•. шканчмваюшиися г!< ттовндной шероховатосщьюГ
С медиальной поверхности от малого”бу(ра плечевой кости
опускается гребень малого_дугврка,заканчивающийся большой круг-
лой шероховатостью. Ниже шероховатости находится сосудистое
отверстие. Дистальный эпифиз образует поперечно поставленный
блок плечевой кости, имеющий почт посередине желоб с синови
ильной ямкой. Дорсально блок ограничен венечной ямкой, а при
переходе на каудальную поверхность кости ^локтевой ямкий^\а-
герально локтевузо ямку ограничиваез латеральный (расгибатель-
ный) надмыщелок, ас медиальной поверхности — медиальный (сги-
бательный) надмыщелок.
О с о б е н н о с т и._У свиней тело плечевой кост и сжато с боков,
дельтовидная шероховатость слабо выражена, суставной блок го-
ризонтальный. У лошадей плечевая кость имеет пологую головку,
вперед от нее выступают латеральный, средний и медиальный
бугорки и два желоба, дельтовидная шероховатость большая, ве-
нечная и локтевая ямки хорошо выражены. У собак плечевая кость
имеет не сильно загнутый латеральный бугорок, дельтовидная
шероховатость мала, дистальный блок поставлен косо.
Скелет предплечья (skeleton anebraMi) представлен двумя срос-
шимися костями — лУЧсвбй и локтевой (рис. 3.46; см. рис. 3.11 —
3.16, 3.4| -3.43).
^Лучевая кость (луч) — os radii_ (radius) — представляет собой
слабо июгнутую |рубчзгую копь. уплощенную краниокаудально.
Она развита сильнее, чем локтевая, и опускается отвесно от лок-
тевого сустава. Луч имеет проксимальный и дистальный эпифизы,
между которыми заключено тело — диафиз.
Проксима наши эпифиз — гаывка_луча несет на себе неглубо-
кую обширную ямку головки, отграниченную суставной окружно-
стью. По переднему краю, обрамляющему суставную поверхность,
проходит венечный отросток, который при максимально согну-
том локтевом суставе входит в одноименную ямку плечевой кости.
Латерально и медиально на проксимальном эпифизе находятся
связочные бугры. Головка луча дистально отделена от диафиза слабо
выраженной шейкой луча. Ниже шейки на дорсомедиальной по-
112
Рис. 3.46. Кости предплечья кошки (Л), собаки (5). свиньи (в>. коровы (/').
лошади (//> (латеральная поверхность):
I юкгевой бугор:: 2; локтевой игросток;^— крючковидный отростчк;/^-
блоконаи вырезка; 5 медиальный венечный отросток; ф- латеральный венеч-
ный отросток;/7 — ямка головки лучсвойхостм; К шероховатость лучевой
кости; 9 - латеральный связочным бугорк/Ф— ирокенма тьная^межкостная шель
предплечья; II/ гело лудсвои кости; 72И- локтевая, кость: IJ?— латеральный
шиловидный отросток. блок лучевоп кости: (Зл медиальный шиловид-
ный отросток
верхности располагается шероховатость лучевой кости, на кото-
рой оканчивается двуглавая мышца плеча.
Тело лучевой кости слегка искривлено по длине с выпуклостью
краниально? ОтюТтесколько сплюснуто краниокаудально, в связи
с чем на нем различают краниальную и каудальную поверхности,
закругленный латеральный и медиа юный края.
JucrtHLibHbiu лиифил — блок лучевой кости — несет на себе не-
ровную суставную поверхность запястья, отграниченную от тела
кости изрезанным поперечным гребнем Латеральный и медиаль-
ный части блока несколько выдаются дистально и вместе с руди-
ментом локтевой кости формируют соответствующие шиловидные
отростки,
Локтеваркость (os ulnae) длиннее лучевой кости, на дисталь-
ном конце имеет латеральный грифелевидный отросток. На про-
ксимальном конце локтевой кости выступает массивный локте-
вой отросток, на конце которого помещается раздвоенный лок-
тевой бугор. Здесь же вперед выпячивается клювовидный отросток.
113
а ниже него находится полулунная вырезка — для сочленения с
плечевой костью. Тело лок тевой кости трехгранное. Срастаясь не-
подвижно с лучевой костью, оно образует проксимальную и дис-
тальную межкостные щели.
Особен пос г и. У свиней локтевая кость хорошо развита и
неподвижно соединена связками с лучевой костью. У лошадей
лучевая шероховатость обширная, проксимальная межкостная щель
разви та, а дис тальная отсутствует. У собак лучевая и локтевая ко-
сти хорошо развиты и подвижно соединяются между собой.
Скелет кисти {skeleton manus) включает кости запястья, пясти_
и кости пальцев (см. рис. 3.11 — 3.16, 3.41 — 3.43, 3.47).
Кости запястья {ossa carpi) располагаются у коровы в дв^ ряда
(рис. 3.47; см. рис. 3.11 3.16, 3.41—3.43). В проксимальномряду_
находятся четыре кости; jyw для кость запястья - прямоеiо и,нои
формы, paciio.iai аекя медиально; промежуточная кость запяс-
тья — клиновидной формы; локтевая кость запястья — распола-
гается латерально, неправильной многоугольной формы; добавоч-
Рис. 3.47. Видовые особенности костей кисти собаки {А), свиньи {Б),
коровы {В), лошади (Г):
7 —лучевая кость; 2 юктевая коси. 7 — лучевая кость запястья; .4)— проме-
жуточная кость запястья; У— Локтева» кость запястья; .6 — добавочная кость
запястья; 7 - мерная кость запястья. V птопии мчть запястья; 9 - четвдпая
кость запястья; 10 — третья кость запястья. II первая пястная кость; /2 —
вторая пястная кость; 13 — пятая пястная кость. 14 — четвертая пястная кость.
'15 — третья пястная кость; /р проксимдзы1.1Я <|>тз.пип; 17 — средняя фаланга;
' 18' шетальная фа таит т
114
пая кость запястья — имеет округленную форму, соединяется толь-
ко с локтевой костью запястья. В дистальном ряду первая кость
запястья отсутствует; вторая и третья кости запястья срослись в
одну кость; четвертая кость запястья хорошо разни га.
Особе ННОСТИ..У свиней в проксимальном ряду запястья,
как и у коров, четыре кости, в дистальном ряду запястья v них
лежат первая, вторая, третья и четвертая запястные кости.; У ло-
шадей в проксимальном ряду четыре кости, а в дистальном
первая, вторая, третья и четвертая 'запястные кости._У_собак в
проксимальном ряду только три косги, гак как .лучевая и проме-
жуточная кости запястья слились в одну промежуточно-лучевую
кость, в дистальном ряду имеются четыре запястные кости: пер-
вая, вторая, третья и четвертая (см. рис. 3.11 — 3.16, 3.41 — 3.43,
3.47).
Кости пясти (ossa metacarpi} состоят у коровы из третьей и
четвертой сросшихся"между собой костей. На латеральной сторо-
не костей находи гея рудиментарная пятая пястная кость. Первая
и вторая пяс тные кости у них отсутствуют. На проксимальном конце
костей пясти находится плоская суставная поверхность. На дор-
сальной поверхности выступает пястная шероховатость. Тело пяс-
тной кости с пальмарной стороны плоское, а с дорсальной —
округлое с продольным сосудистым желобом. Дистальный конец
ее расширен и имеет^двр суставных блока. Каждый блок разделен
сагиттальным гребнем. Суставные поверхности, прилегающие к
межблоконой нырезке. всегда меньше красных. Медиальная крае-
вая поверхность блока шире, чем латеральная. На боковых повер-
хностях блоков находятся связочные ямки.
Особенности. У сдиней четыре пястных кости: вторая, тре-
тья, четвертая и пятая, вторая и пятая пястные кости короче и
менее развиты, чем остальные. У лошадей третья пястная кость
является главной опорной, вторая и четвертая пястные кости ру-
диментарные и называются грифелевпдными. У собак первая пяс
гная кость развита слабо. Остальные пястные кости развиты хоро-
шо (см. рис. 3.11 —3.16. 3.41 —3.43, 3.47).
Кости пальцев (ossa digitorum) состоят из грех фаланг. У коровы
функционируют только третий и четвертый пальцы. Каждый из
них имеет по три фаланги и сесамовидные кости. Проксимальная^
фаланга — короткая трубчатая трехгранная кость. которая имеет
на проксимальном расширенном конце углубленную суставную
поверхность с сагиттальным желобом, а но боковым поверхнос-
тям — свя зочные бугры. Тело ее суживается к дистальному концу,
дорсальная и боковая поверхности тела слегка выпуклые, а паль-
марная и межпальцевая — более плоские. Дистальный суставный
блок разделен сагиттальным желобом на две части. Позади про-
ксимального конца кости располагаются две сесамовидные кости
проксимальной фаланги.
115
Средняя фаланга — трубчатая короткая кость. На проксималь-
нохТ'конце кости находятся суставная ямка и сагиттальный гре-
бень, а на дистальном конце — суставной блок с сагиттальным
желобом.
Дистальная фаланга имеет вид трехгранной пирамиды с вер-
шиной, обращенной вперед. На ней различают проксимальную
(суставную), стенную и подошвенную поверхности Суставная
, ясность разделена сагиттальным гребнем на две части Стен
пая поверхность пронизана отверстиями и шероховата. Подошвен-
ная поверхность состоит из сгибательной и опорной площадок.
Спереди выступает разгибательный отросток для прикрепления
разгибателя пальца Сзади каждой листал иной фаланги располага-
ется по одной сесамовидной (челночной) кости.
О со б е н н о с т и. У свиней хорошо развиты четыре пальца: тре-
тий и четвертый, опирающиеся всегда, а второй и пятый висячие
пальцы опираются только при движении по мягкому грунту, у всех
пальцев имеется по три фаланги, которые внешне похожи на фа-
ланги коровы. У лошадей развит только третий палец, проксималь-
ная фаланга на верхнем конце имеет углубленную суставную по-
верхность с сагиттальным желобом, а на боковых поверхностях по-
мещаются связочные бугры. На пальмарной поверхности тела нахо-
дятся шероховатая треугольная площадка для связок, дистальный
блок с сагиттальной бороздой, средняя фаланга в виде трубчатой
кости, ширина которой больше высоты. На проксимальном конце
фаланги находятся суставная ямка с сагиттальным гребнем, дис-
тальный конец ее образует блок для третьей фаланги, дистальная
фаланга — короткая копытная кость, имеет суставную, стенную и
подошвенную поверхности. Подошвенная поверхность вогнута ку-
полом и состоит из передней опорной и задней сгибательной час-
тей. На этой поверхности различают сосудистый желоб, цодошвец-
ные отверстия и подошвенный канал, проксимальный край с раз-
гибательным отростком, подошвенный край полукруглой формы.
У собак первый палеи слабо развив и состоит из двух фаланг, jbto^
poii, третий. четвертый к пятый па1Ы1Ы хорошо развиты и состоят^
из трех фалаш (см. рис 3.11 - 3.16. 3.41 — 3.43. 3.47).
Скелет тазовой конечности
Кости, тазовой конечности (ossa membri pelvini) включают пояс
тазовой конечности, состоящий и i трех парных костей, — под-
вздошной, лонной и седалищной, объединенных в правую и левую
безымянные (тазовые) кости, а также свободный отдел конечнос-
ти, к которому относятся бедренная кость, скелет голени (боль-
шой берцовой и малой берцовой костей) и скелет стопы, состо-
ящий из костей заплюсны, костей плюсны и костей пальцев (рис.
3.48-3.53; см. рис. 3.11-3.16, 3.41).
116
Рис. 3.48. Кости тазовой конечности
лошади (латеральная поверхность):
/ гюдвмошпый гребень;^— крест-
цовый бугор; .? большая седалищная
вьцх жя —седалищная ость;боль-
шой исрчсл; 4 малая седалищная вы-
рс!кл, 7 седалищный бугор;за-
пертое отверстие;, У2- ягодичная повер-
\носп.. 10 — третий вертел; (77/— над-
мыщелковая ^ямка; (72/ — латеральный
мыте .ют, — межмыщелковое воз-
вышение: 14 малая берцовая кость;
13 пяточный бугор; ,</б9— пяточная
кость. /7- четвертая кбеть заплюсны;
- четвертая плюсневая кость; 19 —
Плантарные сееаМОШШШе кости про-
ксимл/1.мои фаланги. 26 — д.чгеральный
подвздошный бугор, миклок; 21)— тело
подвздошной крс.ти; 22 — тшйвка бед-
ренной кости. '27)- краниальная часть
............... 24 — тело бедрен-
ной кости, 25,— коленная чашка (над-
коленник): 2(у- блок бедренной кос-
ти; 27 — шероховатость большой бер-
цовой кости; 2А' — тело большой бер-
цовой кости 26 латеральная лодыж-
ка; 30 — таранная кость; 31 — цент-
ральная кость кнытосны; 32 — третья
тип поуны; 33)— третья плюсне-
ШШ ко, и.; мевка плюсневой,ко-
сти 25 проксимальная фаланга; 3$ —
средняя <|>.т тинга; J0— копыто, дисталь-
ная "фаланга
Правая и левая безымянные
кости вместе с крестцовой костью
образуют таз, служащий остовом
тазовой полости и связью свобод-
ной конечности с туловищем.
Подвздошная кость (os ilium)
треугольной формы, имеет крыло
и тело. Дорсально она соединяет-
ся с крестцом, а нижним концом срастается с седалищной и лон-
ной костями (рис 3.49. 3.50, см. рис. 3.11 — 3.16, 3.41, 3.48).
Крыло тн)тдоитой кости соединяется с крестцом, а тело под-
вздошной кости нижним концом срастается с седалищной и лон-
ной костями. Ягодичная поверхность подвздошной кости вогнутая,
на ней находится ягодичная линия. Внутренняя поверхность кости
выпуклая, на ней имеется ушковидная суставная площадка для
крестцовой кости.
117
Рис. 3.49. Тазовые кости собаки (А), коровы (Б), свиньи (В), лошади (Г)
(дорсальная поверхность):
1 — подвздошный гребец!.. .’ ттеральный подвиошиый бугор, маклок^—
ягодичная поверхность; 4 - ягодичная линия; 5 - крестовый бугор; ,6/- тело
подвздошной кости; 7.— большая седалищная вырезка; 8 — подвздошно-лонное
писнис; 9 — ipe6cni. лонной кости; ЛЯ - КрШСМЛШМ пствт^лонной кости;
II модальная ветвь лонной кости: /2. --суставная ин.ijhii.i.''.;’ седалищная
осп.. !4 - малая седалищная вырезка; 15 — седалищный osiop; — тело седа-
ИИЦИОЙ mh.hi. /7 запертое огверыие; /Л седалищная дуга
На подвздошной кости различают краниальный, медиальный
и латеральный края. Краниальный край называется подвздошным
гребнем. Медиальный край образует большую седалищную вырезку
для выхода сосудов и нервов. Латеральный край выемчатый. На
подвздошной кости имеются медиальный, латеральный и каудаль-
118
ный углы. Медиальный угол, или крестцовый бугор, лежит против
первого остистого отростка крестца. Латеральный угол называется
маклоковым бугром. Каудальный, или суставной, угол участвует в
образовании суставной впадины.
Седалищная кость (os ischii) расположена каудально (см. рис.
3.11—3.16, 3.41, 3.48 — 3.50). Тело ее имеет вентральную выпук-
лую и дорсальную вогнутую поверхности. Краниально тело кости
ограничивает часть запертого отверстия, а каудально образует се-
далищную дугу и седалищный бугор для прикрепления мышц. Лате-
ральный край седалищной кости формирует малую седалищную
вырезку, а медиальный край ее участвует в образовании седалищ-
Рис. 3.50. Тазовые кости собаки (А), свиньи (Б), коровы (В), лошади (Г)
(латеральная поверхность):
I латеральный птвиоишый бугор; 2/— подвздошный гребень. X—крестцо-
вый бугор; болында седалищная вырезка.^,— седалищная ость. 6 мадам
ссмлншная вырезка. 7 — хчыалищньти бугор. .V хг.-к» подвздошной кости; У —
ягодичная поверхность. 761 - ягодичная пгния; Н тело седалищной кости;
12 — запертое отверстие: /5 каудальная ветвь донной кости; ' Ц лонная
кость; /5 — суставная впадина; /б мышечная ямка
119
ного шва. Вперед выступают две ветви: шовная ветвь участвует в
создании тазового шва, впадинная ветвь — в образовании сустав-
ной впадины.
Лонная кость {os pubis') лежит краниально (см. рис. 3.11 — 3.16,
3.41, 3.48 — 3.50) и образует лонное сращение, которое вместе с
седалищным составляет тазовое сращение. Лонная кость имеет две
ветви. Впадинная ветвь срастается в суставной впадине с седалищ-
ной и подвздошной копями. Краниально она имеет лонный гре-
бень и гребешковый бугор. Шовная ветвь расположена продольно.
Обе ветви ограничивают яшертоё отверстие. Впереди на копи
находится лонный бугор, массивный у самцов. Суставная впадина
лежит на середине длины безымянной кости, где срастаются между
собой три кости. Медиально от суставной впадины заметна вырез-
ка впадины. Выше впадины выступает седалищная ость.
Особенности.„У.свиней направление осей таза параллель-
ное, седалищная ость высокая, крыло подвздошной кости боль-
шое, повернуто в сторону сагиттальной плоскости, подвздошный
гребень выпуклый. У лошадей оси безымянных костей сближают-
ся сзади, маклоковый бугор в виде толстого гребня, подвздош-
ный гребень вогнутый. У собак оси безымянных костей сходятся
впереди, подвздошная кость сочленяется с крестцом почти в са-
гиттальной плоскости, седалищный бугор в виде утолщенных ва-
ликовидных пластин.
Бедренная кость (os femoris) — самая крупная трубчатая кость
коровы (рис. 3.51’;см. рис. 3.11 — 3.16, 3.41,3.48). Она расположена
косо от таза вниз и вперед. На проксимальном конце медиально
выступает головка бедренной кости. На ней имеется маленькая ямка
головки для прикрепления связки. Латерально от головки выступа-
ет большой вертел. Под основанием вертела находится вертлужная,
ямка. На теле бедренной кости ниже головки лежит малый вертел
Ниже большого вертела размещается слабо выраженный третий
вертел.
В дистальной трети тела бедренной кости лежит плоская над-
мыщелковая ямка.* На дистальном конце с плантарной стороны
находятся мыщелки: латеральный и медиальный для сочленения с
костями голени. Они разделены межмыщелковой ямкой. Спереди
находится суставной блок для коленной чашки, он состоит из ла-
терального малого и медиального большого блоковых гребней и
блокового желоба. Над блоком находится неглубокая ямка для ко-
ленной чашки. Сбоку дистального конца латерального гребня по-
мещается разгибательная ямка.
Коленная, чашка {надколенник) (patella) — кость клиновидной
формы, суставной поверхностью присоединяется к бедренной
кости (см. рис. 3.11 — 3.16, 3.48).
Особенности.Усвиней головка и шейка бедренной кости
хорошо выражены, вертелы устроены так же, как у коровы, ко-
120
Рис. 3.51. Бедренные кости собаки (Л), свиньи (£), коровы (В), лошади
{Г) (каудальная поверхность):
/ — большой вертел; 2 к- шейка бедренной коеги.^Л- головка бедренной кости;
4 ямка головки белрендой кости .5 вырезка галошей бедренной кости;/6,4
ягодичная, поверхность: 7J вертлужная ямка, У.'— малый ncpie.i, 9 третий
мтел;,^^ латеральная губ.ц 7$— тело бедренной кости. /Д— медиальная губа;
1лу— питающее ртверстие; 14,— ншмыителкошн бугорок: /5/— надмыщелковая
шероховатость; /й) — надмыщелковая ямка, /77—ла шпальный мышелок. /4
межмыщелковая ямка; j /9,— медиальный мышелок; 2ft1 блок бедренной кости
ленная чашка сжата с боков._У лошади имеется глубокая вырезка
головки бедренной кости, большой вертел отделен от среднего,
третий вертел сильно развит, коленная чашка сжата краниокау-
дально. У собак большой вертел ниже головки, вместо надмыщел-
ковой ямки имеется шероховатость, над мыщелками с каудаль-
ной поверхности имеются суставные поверхности для сесамовид-
ных костей, коленная чашка овальной формы.
Скелет голени {skeleton cruris) представлены двумя костями —
большой’мГмалой берцовыми (рис. 3.52; см. рис. 3.11 — 3.16, 3.41,
3.48).
Большая берцовая кость {tibia) длинная, трубчатая. На прокси-
мальном конце ее находятся два мыщелка — латеральный имеди-.
альный. Между мышечками имеется межмыщелковое возвышение.
Мыщелки с <а.ш ра:иеле}П4_по<^квкрОырезкш7. Спереди мыще.н
ков имением шероховатое утолщение и массивный гребень, загиу-
гыи мтеральнб Между гребнем и латеральным мыщелком нахо-
дится мышечный желоб. На латеральном мыщелке выступает не-
121
Рис. 3.52. Кости голени собаки (Л), свиньи (Б), коровы (В), лошади (Г)
(краниолатеральная поверхность):
Л межмыщелковое возвышение:'^)- медиальный мыщелок/J латеральный
мыщелок: / шероховатость большой берцовой кости; 5)— разгибательный
желоб; Л — макая берцовая кость: 7 межкостная щель: 8 тело большой
берцовой кости. у медиальная лодыжка: W— латеральная лодыжка; // —
лодыжковая кость
большой малоберцовый отросток (рудимент малой берцовой кос-
ти). Тело большеберцовой кости трехгранной формы, имеет лате-
ральную, медиальную и плантарную поверхности. На дистальном
конце большой берцовой кости имеется прямо поставленный блок.
Боковые части блока называются медиальная и латеральная ло-
дыжки. На латеральной лодыжке находится лодыжковая кость —
нижний конец рудиментарной малой берцовой кости. Малая бер-
цовая кость (fibula) как самостоятельная кость у коров отсутст-
вует.
Особенности.^£свщщей на большой берцовой кости мы-
шечная вырезка небольшая, малая берцовая кость хорошо разви-
та, межкостное пространство выражено по всей длине кости.
У лошадей малая берцовая кость имеет головку, а тело истончает-
ся й сходит на нет к середине голени. У собака тело большой бер-
цовой кости изогнуто, малая берцовая кость в виде тонкой кост-
ной пластинки с утолщенными концами внизу прикасается плот-
но к большой берцовой кости.
122
Скелет стопы (skeleton pedis) состоит из костей заплюсны,
костей плюсны и костей пальцев.
Кости заплюсны (ossa tarsi) у коровы состоят из шести костей,
расположенных в три ряда (рис. 3.53; см. рис. 3.11 —3.16, 3.41.3.48).
В проксимальном ряду находятся таранная и пяточная кости^/д^
ранная кость лежит медиально, снабжена тремя блоками — про-
ксимальным, дистальным и плантарным. Проксимальныйблок пред-
назначен для большеберцовой кости, дистальный — для централь-
ной кости, плантарный — для пяточной кости; латёрально к та-
ранной кости присоединяется лодыжковая кость. Пяточная кость
находится латерально и соединяется с таранной, центротарсаль-
иой и лодыжковой костями. Туло пяточной кости массивное. Вверх
от нею тянется пяточный отросток, на конце которого находится
пяточный бугор для ахиллова сухожилия. Вниз направлен_сус/пов-
ной отросток для центральной кости, а медиально выступает ejep-
жатель таранной кости, его плантарная поверхность служит бло-
ком для мышц.
В центральном ряду находится центральная кость, которая сли-
лась с четвертой заплюсневой костью в одну центротарсальную
кость.
В дистальном ряду лежат три заплюсневые кости, из них наи-
более крупные третья и четвертая. Первая заплюсневая кость ма-
ленькая, цилиндрической формы, соединяется с центральной и
третьей плюсневой костями. Вторая и третья заплюсневые кости
Рис. 3.53. Скелетотопия костей заплюсны и плюсны собаки (Л), свиньи (Б),
коровы (/Ml лошади (Г) (дорсальная поверхность):
'ДО-пяточная кость. — шрапная кость./.? — четвертая крсть заплюсны;-4 —
центральная коси, ктплюсны. 5. - третья кость иплихтны; О' вторая костьзап-
люсны;^- первая кость заплюсны; А’— третья ЛЛЮСНСвая кость; fy— четвертая
плюсневая кость; fiO)— вторая плюсневая кость, 11 - пятая плюсневая кость; '72'—
первая плюсневая кость
123
вытянутые, слегка изогнутые, короткие, соединяются с централь-
ной и третьей плюсневой костями. Четвертая заплюсневая кость
влилась в центротарсальную кость.
Особенности. У свинец таранная и пяточная кости устрое-
ны так же, как у коров. У лошадей таранная кость имеет винтовой
блок, пяточный отросток длинный, держатель таранной кости с
четырьмя фасетками, четвертая заплюсневая кость кубовидной
формы. У собак таранная кость дистально несет головку, цент-
ральная заплюсневая кость вверху вогнутая.
Кости плюсны [ossa melatamij у коров сходны с костями пяс-
ти: третья и четвертая плюсневые кости слились в одну трубча-
тую кость (см. рис. 3.1 1 — 3.16, 3 41, 3.48, 3.53).
Особенности. У свиней развиты вторая, третья, четвертая
и пятая плюсневые кости, из которых третья и четвертая развиты
сильнее, а вторая и пятая — слабее.^У лошади кости плюсны по-
добны костям пясти. У собак развиты вторая, третья, четвертая и
пятая плюсневые кости; наиболее длинные третья и четвертая
плюсневые кости, они имеют сзади проксимальные отростки, на
дистальных валиках есть гребни только на плантарной поверхности.
Кости пальцев тазовой конечности (ossa digitorum) коров пост-
роены так же, как на грудной конечности, но проксимальная фа-
ланга тазовой конечности длиннее и более сдавлена с боков, чем
на грудной, средняя фаланга длиннее и уже, чем на грудной,
дистальная фаланга длиннее и сдавлена с боков (см. рис. 3.11 —
3.16, 3.41-3.43, 3.47, 3.48).
Ос о бе н н ости. У свиней и лошадей проксимальная, сред-
няя и дистальная фаланги тазовой конечности длиннее и уже та-
ковых грудной конечности. У собак кости пальцев тазовой конеч-
ности устроены так же, как и грудной.
АРТРОЛОГИЯ
Скелет представляет собой систему рычагов движения и опо-
ры; одновременно он формирует полости и является защитой для
мягких тканей тела. В связи с этим в организме возникает необхо-
димость в разных способах соединения частей скелета между со-
бой (рис. 3.54 — 3.65).
С анатомической точки зрения, все соединения костей можно
подразделить на прерывные и непрерывные.
Прерывный (синовиальный) тип соединения костей или сустав —
articulatio (от греч. arthroon — сустав) — характеризуется наличи-
ем между соединяющимися костями шелевидной суставной поло-
сти (рис. 3.54, 3.63).
Большинство суставов имеет вид более или менее выраженного
угла, в котором различают вершину, костные лучи и угол сустава.
124
Вершина сустава обычно хорошо проявлена либо в виде круп-
ного бугра, либо отдельной косточки (коленная чашка). Имеются
суставы и без ясно выраженной вершины (височно-нижнечелюс-
гной).
Костные лучи участвуют в образовании сустава; угол сустава —
пространство между костными лучами. В суставах с ясно выражен-
ной вершиной костные лучи при разгибании стремятся достичь
угла 180°.
По анатомическому строеттю суставы характеризуются нали-
чием: 1) суставных поверхностей; 2) суставной капсулы: 3) сус-
тавной полости.
Суставные поверхности расположены на эпифизах костей и
представляют собой различные по форме и размеру гладкие пло-
щадки, окруженные и отделенные от кости краевыми гребешками.
Они могут покрывать головки, ямки, отростки, блоки, мыщелки
и впадины соединяющихся костей. Для суставных поверхностей
соединяющихся костей, не соответствующих друг другу, в суставе
имеются диски, или мениски, способствующие выравниванию ио-
не рхносги.
Суставные поверхности покрыты гиалиновым хрящом. Он не
имеет налхряшнииы. лишен волокон, полупрозрачен и по внеш-
нему виду напоминает голубоватое стекло. В хряще отсутствуют
кровеносные и лимфатические сосуды, а питание его осуществ-
ляется диффузно'йз синовиальной жидкости. Хрящевое покрытие
имеет разную толщину в зависимости от испытываемых нагрузок:
в среднем 0,5— 1,2 мм.
Суставная капсула прикрепляется по краю суставной поверх-
ности сочленяющихся костей, прочно срастается с надкостни-
цей. образуя ымкнучую шеленилную суставную полость, запол-
ненную небольшим количеством суставной жидкости — синовии.
Капсула сосюп i ш шух оболочек — наружной фиброзной, являю-
щейся переходом фиброзного слоя надкостницы с одной кости
на другую, и внутренней. Внутренняя оболочка капсулы — сино-
виальная, Она выстлана специальными клетками, вырабатываю-
щими синовиальную жидкость. На отдельных суставах капсула
может образовывать выпячивания или вывороты, иногда сооб-
щающиеся с полостями слизистых бурс и (или) синовиальных
влагалищ.
Суставные поверхности костей крайне разнообразны по фор-
ме, что полностью зависит от характера движения в суставе и
одновременно определяет его. В связи с этим суставы делятся на
пять типов: одноосные, двухосные, многоосные, комбинированные и
скользящие^
Суета в, в образовании которого принимают участие две кос-
ти, называется простым в отличие от сложного сустава, в кото-
ром между двумя костями располагаются внутрисуставные ме-
125
Рис. 3.54. Схема строения сустава:
/ - полость сустава; 2 — синовиальная оболочка кап-
сулы сустава; 3 — суставной хрящ; 4 — фиброзная обо-
лочка капсулы сустава; 5— надкостница; 6— компак-
тное вещество кости
писки, или диски, или они образуются тремя и более костями
(рис. 3.54).
В суставах возможны различные виды движения, однако в це-
юм они следующие: сгибание, разгибание, отведение (аддукция),
приведение (абдукция), вращение (супинация — вращение нару-
жу, пронация — вращение внутрь), кружение.
Непрерывный тип соединений костей - синиртроз (synartrvsis)
характеризуется тем, что между соединяющимися костями в ка-
честве их связи располагается мышечная, эластическая, соедини-
тельная, хрящевая или костная ткани (рис. 3.55, 3.56).
Соединение костей осевого скелета
Тела позвонков соединяются между собой межпозвоночными
волокнистыми хрящевыми лисками и продольными дорсальной и
вентральной связками. Межпозвоночные диски располагаются меж-
ду головками и ямками соседних позвонков; с поверхности они
образуют волокнистое кольцо, в центре лежит пульпозное ядро
(рис. 3.55).
Дуги позвонков и поперечные отростки по звонков соединяются
между собой посредством соответственных связок. Междуговые
связки и межпоперечные связки находятся между дугами и понс-
126
Рис. 3.56. Выйная связка коровы:
I канатик иыйион свяжи; — п син: 3 ось; '! ' пластинка тайной связки;
5] — капюшоннця часть выйной свяжи; §}— межостистая свяжи.;'7/— первое
ребро. 8 ~ вентральная пластинка (пксгав шейный полюйок)
речными отростками и состоят из эластической ткани (см. рис.
3.55).
Суставные отростки позвонков соединяются между собой сус-
тавными капсулами, которые прикрепляются к краям суставных
поверхностей, а остистые отростки — выйной и межостистыми
связками.
Выйная связка, самая крупная, построена из эластической тка-
ни: она состоит из столбиковой и пластинчатой частей (рис. 3.56).
Столбиковая часть начинается на наружном затылочном пред-
бугорьеТпрохбдйгв виде парного тяжа над шейными позвонками
и оканчивается на первом грудном позвонке. Часть выйной связ-
ки, соединяющая остистые отростки грудных позвонков, называ-
ют^надостиспюй связкой. Она начинается от первого грудного по-
звонка и заканчивается на крестновых буграх подвздошной кости.
Под этой связкой лежат три слизистые сумки: первая — над дугой
атланта, вторая — над осью, третья — над вторым грудным по-
звонком.
Пластинчатая чде/пь имеет вид широкой парной пластинки;
она начинается от гребня оси и остистых отростков третьего —
шестого шейных позвонков и оканчивается на столбиковой части.
Первый и второй шейные позвонки соединены атлантозаты-
лочным и атлантоаксиальным суставами.
Атлантозатылочный сустав образован мыщелками затылоч-
ной кости и краниальными суставными поверхностями атланта.
Сустав простой, двуосный. Движения — в виде сгибания и разги-
бания оси, а повороты вправо и влево — вокруг отвесной оси.
В суставе имеются капсула сустава, дорсальная и вентральная ат-
лантозатылочные мембраны и боковые связки.
127
Рис. 3.57. Реберно позвонковые су-
станы:
/Л дорсальная про лажная связка; Q-
краниальный суставной отросток: А:—
связка бугодка ребра; ч/— связка шей-
ки ребра; .у — связка головки ребра;
О— вентральная продольная связка;
'7/ сосдиннтслыиы связка ребер: 8
тело позвонка; ч) - головка ребра
Атлантоаксиальный сустав
образован зубом оси и атлантом.
Сустав простой, одноосный, ко-
ловратный. В суставе, кроме кап-
сулы сустава, имеются связки: ме-
жостистая, междуговая, внут-
ренняя зубовидная.
Ребра с позвонками соедине-
ны посредством двух суставов
(рис. 3.57). Один из них — сустав
реберной головки, второй — сус-
тав бугорка ребра. В суставе име-
ются капсула сустава и четыре
специальные связки: радиальная
связка головки ребра~соедмнй
тельная связка реберных головок,
связка реберного бугра, связка ре-
берной шейки.
Реберные хрящи с грудиной
образуют грудино-реберный сус-
тав, который имеет капсулу су-
става и грудино-реберную связку.
Рукоятка грудной кости со-
единяется с телом грудной кос-
ти суставом рукоятки. Этот сус-
тав имеет капсулу сустава и две
специальные связки: внутреннюю
и наружную специальные грудин-
ные связки. Сегменты грудной кости соединяются между собой син-
хондрозом. ' "
Кости черепа соединяются между собой швами. Нижняя челюсть
соединяется с чешуей височной кости височно-нижнечелюстным
суставом. Между суставными поверхностями вставлен суставной
диск, поэтому сустав сложный. Движения: сгибание, разгибание,
отведение вправо и влево. Связочный аппарат сустава состоит из
капсулы, боковых и задних связок.
Соединение костей грудной конечности
Кости грудной конечности сочленяются между собой посред-
ством суставов. На грудной конечности коров различают следую-
щие суставы: плечевой, локтевой, запястный, путовый, венеч-
ный, копытный.
Плечевой сустав образован ямкой лопатки и головкой плече-
вой кости (рис. 3.58). Сустав простой, многоосный. Движения: сги-
128
Рис. 3.58. Плечевой сустав лошади:
I — акромион; 2 — предостная ямка;
3 — надсуставной бугорок; 4 — шейка
лопатки; 5 — позади суставной буго-
рок; 6 — капсула сустава; 7 — боль-
шой бугорок; 8 — гребень большого
бугорка; 9 — плечевая кость
Рис. 3.59. Локтевой сустав собаки
(латеральная поверхность):
Л/— плечевая кость; капсула
сустава; 3 — локтевой отросток; 5 —
локтевая коллатеральная латеральная
связка (лучевая часть); 6/1— лучевая
кость; ( Т-1— локтевая коллатеральная
латеральная свя >ка (локтевая часть);
$ — межкостная мембрана; — лок-
тевая кость
бание, разгибание, приведение, отведение, пронация, супина-
ция. В суставе имеется только капсула сустава.
Локтевой сустав образован блоком плечевой кости, головкой
лучевой кости и локтевым отростком (рис. 3.59). Сустав простой,
одноосный. Движения: сгибание и разгибание. Сустав имеет кап-
сулу, боковые, поперечные и межкостную связки.
Запястный сустав образован дистальным блоком предплечья,
костями запястья и проксимальным концом пястных костей (рис.
3.60). Это сложный одноосный сустав. Связочный аппарат его пред-
ставлен капсулой, латеральной и медиальной боковыми длин-
ной и короткой связками, межкостными и межрядовыми связ-
ками.
Путовый сустав образован дистальным суставным блоком пя-
стной кости и проксимальной фалангой третьего (четвертого) паль-
ца и их сесамовидными костями (рис. 3.61). Сустав простой, одно-
осный; движения: сгибание и разгибание. Кроме капсулы сустава
имеются боковые, межпальцевые и сесамовидные связки.
Венечный сустав образован блоком путовой и проксимальным
концом венечной кости (см. рис. 3.61). Сустав имеет капсулу, бо-
ковые и пальмарные связки.
Копытцевый сустав образован венечной, копытцевой и чел-
ночной костями. Кроме капсулы сустав имеет боковые и челноч-
ные связки (см. рис. 3.61).
5 3,-(сцепекн,
129
Рис. 3.60. Запястный сустав коровы (А — дорсолатеральная поверхность;
Б — медиопальмарная поверхность) и лошади (В — схема, сечение; Г —
дорсолатеральная поверхность):
1 — лучевая кость; 2 — локтевая кость; 3, 14 — коллатеральная латеральная связка
запястья; 4 — межзапястная дорсолатеральная связка; 5 — пятая пястная кость; 6 —
межкостная мембрана; 7— третья + четвертая пястная кость; 8, 17, 20 — колла-
теральная медиальная запястная связка; 9 межзапястная дорсальная; 10— меж-
костная дорсальная проксимальная медиальная связка; 11 — дорсальная луче-
запястная связка; 12 — запястно-пястная дорсальная связка; 13 — межкостная
дистальная связка; 75 — пальмарная запястно-пястная латеральная связка; 16 —
пальмарная запястно-пястная медиальная связка; 18 — межрядовая пальмарная
связка; 19 — запястно-лучевая пальмарная связка; 21, 24 — капсула запястного
сустава; 22 — дистальный ряд костей запястья; 23 — пястная кость; 25, 27 — по-
лость сустава; 26 — проксимальный ряд костей запястья; 28 — межкостная дор-
сальная дистальная связка; 29 — запястно-пястная дорсальная латеральная
связка; 30— запястно-пястная дорсальная медиальная связка; 31 — межкостная
дорсальная проксимальная латеральная связка
130
Рис. 3.61. Суставы пальца лошади:
'4' — латеральная поверхность://J— грифельная костьц2, 12 — третья межко-
сткйя мышца; 3 — проксимальная сесамовидная кость; (4у- боковая связка сеса-
мовидной кости; 5 — косая связка сесамовидных костей; 6, 7, 14 — путово-
всисчная связка; 8 - латеральная путово-венечная связка. 9 — сижка поддон
кости и мякишного хряща, 10 ~ связка копытной кости и мякишного хряпы; // *•
третья пястная кость; !у- боковая связка сустава проксимальной фаланги. 75
проксимальная фаланта; /6 — боковая связка сустава средней фаланги; 17 -
связка средней фалант и мякишного хряща. /Л боков ля смэка дистальной
фаланги;
/» — пальмарная поверхность: 1 суставная поверхность гретьей пястной
кости; 2. 3 — грифельные кости; 4 грезья межкостная дяышна;^ л — ветви
третьей межкостной мышцы на юрсальную поверхность.' 77- мсжсссамовилная
связка: 8 — боковая связка сесамовидной кости; 91- прямая связка сесамовид-
ных костей; 10 — косая связка сесамовидной кости: II пугодо-венечная связ-
ка; 12. 13 латеральная путово-венечная связка; >14,’ сухожилие глубокого
сгибателя пальца; /5 — капсула сустава дистальной фаланги; челночная
кость; 17 — сгибательная поверхность дистальной фаланги, /X дисальнпя фа-
< Тани», пальмарная поверхность.
IB)— сагиттальный разрез: / — связка цуговой кости и мякишного хряща; 2,
7— мякишный хрящ; 3, 4, 5 косые н прямая связки сесамовидных костей; 6 —
сухожилие общего разгибателя пальца; 8 — блок; 9 — дистальная фаланга; 10 —
челночная кость; 11 — сухожилие глубокого сгибателя пальца; 12, 13 — паль-
марная связка мякишного хряща и дистальной фаланги
131
Соединение костей тазовой конечности
Кости тазовой конечности коров образуют подвздошно-крест-
новый, тазобедренный, коленный, скакательный, пугоный. ве-
нечный и копытневый суставы (рис. 3.62 — 3.65).
Крестцово-подвздошный суета» образован ушковидными сус-
тавными поверхностями крестца и подвздошной кости (рис. 3.62).
Простой, плоский, тугой сустав. Он имеет капсулу сустава, крес-
тцбво’-подвздошные и крестцово-седалищные связки. ~
Тазобедренный сустав сформирован впадиной таза и головкой
бедра (рис. 3.63). Это простой, многоосный сустав. Сгибание и раз-
гибание в нем обширны, а вращение, отведение и приведение
ограничены. Кроме капсулы сустав имеет связку головки бедрен-
ной кости. У лошади в этом суставе имеется добавочная связка,
ограничивающая отведение тазовой конечности.
Коленный сустав сформирован мыщелками бедренной, боль
шой берцовой костей и кошенной чашкой (рис. 3 .64). Сустав слож-
ный, одноосный . К'.ннся на белро-бериовый и бедро-коленный
Бедро-берцовый сустав образован мыщелками бедренной, боль-
шой берцовой костей и хрящевыми менисками. Сложный, одно-
осный сустав. Связочный аппарат состоит из капсулы сустава,
боковых и крестовидной связок.
Рис. 3.62. Связки крестцово-подвздошного сустава коровы (Л) и лошади (5):
1 — крестцовый бугор; 2 — крестцово-подвздошная дорсальная короткая связка;
3 — крестцово-подвздошная дорсальная длинная связка; 4 — надостистая связ-
ка; 5, 6, 8, 14 — широкая крестцово-бугровая связка; 7 — хвостовой позвонок;
9 — седалищный бугор; 10 — малое седалищное отверстие; 11 — запертое отвер-
стие; 12 — вентральный лонный бугорок; 13 — седалищная ость; 15 — большое
седалищное отверстие; 16— отверстия для сосудов и нервов; 17— боковой кре-
стцовый гребень
132
Рис. 3.63. Тазобедренный сустав
(поперечное сечение):
/ — головка бедренной кости;-2” —
шейка бедренной кости; 3 — средняя
ягодичная мышца; 4 — суставная губа;
5 — суставная впадина; 6 — связка го-
ловки бедренной кости (круглая связ-
ка); 7 — поперечная связка суставной
впадины; 8 — влагалище; 9 — уретра;
10 — кости таза; 11 — бедренная кость;
12 — двуглавая мышца бедра; 13 —
большой вертел
Рис. 3.64. Связки коленного сустава лошади:
А — медиальная поверхность: 1 — бедренная кость; 2 — хрящевой отросток;
3 — надколенник (коленная чашка); 4 — блок бедренной кости; 5, 7 — латераль-
ная связка надколенника; 6 — средняя связка надколенника; 8 — шероховатость
большой берцовой кости; 9— большая берцовая кость, медиальная поверхность;
10 — коллатеральная медиальная связка; 11 — медиальный мениск; 12 — медиаль-
ный мыщелок бедренной кости; 13 — медиальная связка надколенника; 14 —
медиальный держатель надколенника;
Б — дорсокаудальная поверхность, удалена бедренная кость: 1 — основание
надколенника; 2 — суставная поверхность надколенника; 3 — хрящевой отрос-
ток (надколенник); 4 — медиальный держатель надколенника; 5 — средняя связ-
ка надколенника; 6 — медиальная связка надколенника; 7 — межмыщелковое
возвышение; 8 — коллатеральная медиальная связка; 9 — медиальный мениск;
10 — каудальная крестовидная связка; 11 — большая берцовая кость; 12 — под-
коленная вырезка; 13 — малая берцовая кость; 14 — мениско-большеберцовая
связка; 15 — латеральный мыщелок большой берцовой кости; 16 — мениско-
бедренная связка; 17 — латеральный мениск; 18 — коллатеральная латеральная
связка; 19 — краниальная крестовидная связка; 20 — латеральная связка надко-
ленника; 21 — латеральный держатель надколенника; 22 — гребень надколенника
133
Рис. 3.65. Связки скакательного сустава лошади:
А — медиальная поверхность; латеральная поверхность; / — большая берцо-
вая кость; 2 — пяточный бугорок; 3. 5. 9, 10 — короткая коллатеральная медиаль-
ная связка; 4 — длинная коллатеральная медиальная связка; 6 — дорсальная зап-
люсневая связка; 7 — третья плюсневая кость; 8, И, 13— плантарная связка; 12 —
берцово-пяточная латеральная связка; 14 — латеральная межкостная связка; 75 —
длинная коллатеральная латеральная связка; 16 — короткая коллатеральная ла-
теральная связка; 17 — дистальный ряд костей заплюсны; 18 — заплюсно-плюс-
невая дорсальная связка; 19 — дистальная межкостная связка; 20 — проксималь-
ная межкостная связка
Бедро-коленный сустав формируется блоком бедренной кости и
коленной чашкой. Сустав простой, одноосный. Кроме капсулы
сустав имеет прямые связки, латеральный и медиальный держа-
тели коленной чашки.
Скакательный сустав образован блоком костей голени, зап-
люсневыми костями и проксимальным концом плюсневых костей
(рис. 3.65). Сложный, одноосный сустав. Он содержит капсулу, ла-
теральную и медиальную боковые длинные и короткие связки,
межрядковые и межкостные связки.
Суставы пальцев стопы устроены так же, как на кисти (см. рис.
3.61).
МИОЛОГИЯ
У позвоночных животных движение осуществляется при учас-
тии специальных органов — мышц, построенных из поперечно
исчерченной мышечной ткани (рис. 3.66).
Каждая мышца представляет собой паренхиматозный орган, со-
ставными частями которого являются паренхима и строма. Главный
134
Рис. 3.66. Мышца как орган:
/ — губчатое вещество кости; 2 — компактное
вещество кости; 3 — шарпеевские клетки; 4 —
начальное сухожилие; 5 — перимизий; 6 — арте-
рия; 7 - нерв; 8 — пучок первого порядка; 9 —
микроциркуляторное русло; 10 — эпимизий;
11 — конечное сухожилие
структурный элемент паренхимы — скелет-
ная поперечно исчерченная мышечная ткань.
Строма состоит из соединительной ткани.
На обоих концах мышцы соединитель-
нотканные элементы продолжаются за пре-
делы мышечных волокон и смешиваются
с плотной оформленной соединительной
тканью сухожилий, прикрепляющих мыш-
цу к кости.
Добавочными и вспомогательными орга-
нами мышц являются сухожилия, апонев-
розы, фасции, слизистые бурсы (сумки),
синовиальные влагалища, блоки и сесамо-
видные кости.
Сухожилие состоит из прилегающих друг
к другу пучков коллагеновых волокон. Апо-
невроз представляет собой плоское или рас-
ширенное сухожилие.
Слизистые сумки — это небольшие ме-
шочки, заполненные синовиальной жид-
костью. Они необходимы для предотвраще-
ния трения мышц, сухожилий или связок
с другими органами (костями, кожей и
т.д.) и облегчают скольжение при боль-
ших размахах движения. В связи с особен-
ностями топографии слизистые сумки бы-
вают _ подмышечными, подсухожильными,
подсвязочными, подкожными. Полости сли-
зистых бурс, расположенных вблизи сус-
тавов, часто сообщаются с полостью сус-
тавов. Этот факт необходимо учитывать при
прогнозировании и лечении воспалений
таких образований.
Синовиальные влагалища по своему строению и назначению сход-
ны с бурсами. Они образовались из подсухожильных бурс. Вслед-
ствие скольжения сухожилий бурсы вытянулись в длину, а также
в ширину и завернулись с боков на противоположную сторону
сухожилия, где края бурсы сомкнулись.
135
Фасции — соединительнотканная прослойка, окружающая от-
дельные мышцы (специальные фасции), группы мышц (глубокие
фасции) или все тело (поверхностные фасции).
Сесамовидные кости располагаются в области кисти и стопы
соответственно на пальмарной и плантарной поверхности. Они
уменьшают трение и располагаются под сухожилиями.
Блок — участок кости, покрытый гиалиновым хрящом. Блоки
располагаются там, где сухожилие мышцы перебрасывается через
кость, меняя направление (блок плечевой кости, блок пяточного
бугра).
Мышцы и фасции туловища
Желтая брюшная фасция покрывает наружную косую мышцу
живота и грудную клетку. Она начинается от ребер и грудной ко-
сти и тянется назад до таза и глубокой фасции бедра. Фасция со-
держит много эластической ткани и поэтому имеет желтый цвет.
Действие: поддерживает внутренние органы.
Особенности.,)/ свиней хорошо развита только грудобрюш-
ная фасция. Удошадей на груди и животе развита поверхностная
грудобрюшная фасция, большая кожная мышца простирается от
глубокой грудной мышцы до коленной складки. У собак развиты
грудобрюшная и кожная шейные мышцы.
Трапециевидная мышца (рис. 3.67, см. рис. 3.69) располагается
поверхностно и состоит у коров из двух частей: шейной и спин-
ной. Шейная часть начинается пластинчатым сухожилием от стол-
биковой части выйной связки на протяжении от первого (второ-
го) шейного до третьего грудного позвонка и оканчивается на
ости лопатки. Спинная часть берет начало от надостистой связки
на уровне от третьего до десятого грудного позвонка и оканчива-
ется на заднем крае верхней трети ости лопатки.
Особенности. У свиней трапециевидная мышца начинается
от всех шейных и десяти грудных позвонков. У лошадей шейная
часть начинается от выйной связки со второго шейного по третий
грудной позвонок, а спинная часть — с третьего по одиннадца-
тый грудной позвонок. У собак шейная часть указанной мышцы
сильнее развита.
Плечеатлантная мышца (см. рис. 3.67, рис. 3.69) простирается
от крыла атланта и поперечного отростка оси до фасции лопатки.
Действие: тянет лопатку вперед.
Особенности. У свиней эта мышца располагается, как у ко-
ров. У лошадей ее пет.У собак она сильно развита.
Ромбовидная мышца (рис. 3.68, см. рис. 3.70) располагается
под трапециевидной мышцей и делится на шейную и спинную
части.
136
Рис. 3.67. Мышцы коровы:
/ — круговая мышца глаза; 2 — подкожная мышца лба; 3 — большая жевательная
мышца; 4 — щиток; 5 — околоушная железа; 6— вентральная ушная мышца; 7 —
ключично-затылочная мышца; 8- плечепоперечная мышца; 9— 1рапсциевидная
мышца, шейная часть; 10 — трехглавая мышца плеча, длинная головка; //, 13 —
широчайшая мышца спины; 12 — трапециевидная мышца, грудная часть; 14 —
пояснично-грудная фасция; 75 — зубчатая каудальная дорсальная мышца; 16 —
косая наружная мышца живота; 17 — маклок; 18, 19 — средняя ягодичная мышца;
20 — поверхностная ягодичная мышца; 27 — крестцово-хвостовая латеральная
мышца; 22 - хвостовая латеральная мышца; 23 — межпоперечная хвостовая мыш-
ца; 24 — крестцово-хвостовая дорсальная мышца; 25 — полусухожильная мышца;
26 — каудальная часть двуглавой мышцы бедра; 27 — краниальнаая часть двугла-
вой мышцы бедра; 28 — длинная малоберцовая мышца; 29 — пяточное сухожи-
лие; 30 — глубокий сгибатель пальцев стопы; 31 — сухожилие специального разги-
бателя четвертого пальца; 32 — средняя межкостная мышца; 33, 34 — сухожилие
длинного разгибателя пальцев; 35 — специальный разгибатель четвертого пальца;
36 — третья малоберцовая мышца + длинный разгибатель пальцев; 37 — напряга-
тель широкой фасции бедра; 38 — апоневроз косой наружной мышцы живота;
39 — наружная косая мышца живота; 40 — зубчатая вентральная мышца груди;
41 — глубокая грудная мышца; 42 — локтевой сгибатель запястья; 43 — лучевой
сгибатель запястья; 44 — поверхностный сгибатель пальцев; 45 — средняя межко-
стная мышца; 46 — добавочная кость запястья; 47 — сухожилие глубокого сгиба-
теля пальцев; 48 — специальный разгибатель четвертого пальца; 49 — сухожилие
общего разгибателя пальцев; 50 — сухожилие специального разгибателя третьего
пальца; 57 — длинный абдуктор большого пальца; 52 — лучевой разгибатель запя-
стья; 53 — трехглавая мышца плеча, латеральная головка; 54 - поверхностная
грудная мышца, ключичная часть; 55 — ключично-плечевая мышца; 56 — ярем-
ная вена; 57 — грудино-сосцевидная мышца; 58 — грудино-челюстная мышца; 59 —
ключично-сосцевидная мышца; 60 — грудино-подъязычная мышца; 61 — наруж-
ная челюстная вена; 62— нижнечелюстная железа; 63 — опускатель нижней губы;
64— щечная мышца; 65 — скуловая мышца; 66 — опускатель верхней губы; 67 —
специальный подниматель верхней губы и клыковая мышца; 68 — кожная мышца
носа; 69 — носогубный подниматель; 70 — опускатель нижнего века
137
4
Рис. 3.68. Мышцы лошади:
1 — клыковая мышца; 2 — носогубным подниматель; 3 — околоушная железа; 4 —
вентральная ушная мышца; 5— ключично-затылочная мышца; 6— пластыревид-
ная мышца; 7 — плечепоперечцая мышца; 8 — трапециевидная мышца, шейная
часть; 9 — зубчатая вентральная мышца шеи; 10— трапециевидная мышца, груд-
ная часть; 11 — широчайшая мышца спины; 12 — зубчатая каудальная дорсальная
мышца; 13 — средняя ягодичная мышца; 14 — напрягатель широкой фасции бед-
ра; 15 — поверхностная ягодичная мышца; 16 — полусухожильная мышца; 17,
18 — двуглавая мышца бедра; 19 — икроножная мышца, латеральная головка;
20 — длинный разгибатель пальца; 21 — коленная складка; 22 — косая наружная
мышца живота; 23 — глубокая грудная мышца; 24 — зубчатая вентральная мышца
груди; 25 — дельтовидная мышца; 26 — трехглавая мышца голени; 27 — локтевой
разгибатель запястья; 28 — общий разгибатель пальцев; 29 — лучевой разгибатель
запястья; 30 — трехглавая мышца плеча, латеральная головка; 31 — плечевая мышца;
32 — предостная мышца; 33 — плечеголовная мышца; 34 — наружная яремная
вена; 35 — грудиноголовная мышца; 36 — большая жевательная мышца; 37 —
скуловая мышца; 38 — щечная мышца; 39 — круговая мышца рта
Особенности. У свиней ромбовидная мышца имеет три ча-
сти: головную, шейную и спинную. У собак ромбовидная мышца
имеет такое же строение, как у свиней.
Плечеголовная мышца (см. рис. 3.67 — 3.72) состоит из трех час-
тей: ключично-затылочной, ключично-сосцевидной и ключично-
плечевой.
138
Ключично-затылочная мышца начинается на затылочной кости
и на выйной связке.
Ключично-сосцевидная мышца берет начало на сосцевидном от-
ростке височной кости и на нижней челюсти. Обе они оканчива-
ются на ключичной полоске. От последней к плечу простирается
ключично-плечевая мышца.
Особенност и.,У свиней плечеголовная мышца располагает-
ся, как у коров. У лошадей она одной частью начинается на со-
сцевидном отростке и затылочном гребне, а другой — на попе-
речных отростках второго — пятого шейных позвонков. Плечего-
ловная мышца прикрывает длинную мышцу головы, межпопе-
речные мышцы, плечеподъязычную мышцу, лестничную мышцу
и предлопаточную часть глубокой грудной мышцы.
Широчайшая мышца спины (см. рис. 3.67 — 3.69) начинается от
нояснично-спинной фасции и надостистой связки на уровне от
первого поясничного до четвертого грудного позвонка и оканчи-
вается у всех животных на гребне малого бугра. Действие: тянет
плечевой сустав назад.
ОсобенносткУ свиней широчайшая мышца спины начи-
нается на пояснично-спинной фасции и на последних ребрах.
У лошадей эта мышца располагается в основном так же, как у
коров. У собак расположение мышцы такое же, как у свиней.
Вентральная зубчатая мышца (рис. 3.69; см. рис. 3.70, 3.72) имеет
веерообразную форму и состоит из двух частей — шейной и груд-
ной. Шейная часть начинается от поперечных отростков шейных
позвонков с третьего по седьмой и оканчивается на зубчатой по-
верхности лопатки. Грудная часть берет начало от грудинных кон-
цов ребер с первого по девятое и оканчивается на зубчатой по-
верхности лопатки. Действие: шейная часть тянет лопатку вперед,
а грудная — назад.
Особенности. У свиней шейная часть начинается на попе-
речных отростках всеЗГшеиных позвонков, а грудная — на ребрах
с первого по восьмое. У лошадей и собак эта мышца начинается и
оканчивается, как у коров.
Грудная поверхностная мышца (см. рис. 3.67 — 3.70, 3.72) состо-
ит из двух частей — плечевой и предплечевой. Плечевая часть на-
чинается от рукоятки грудной кости и оканчивается на гребне
большого бугра. Предплечевая часть простирается от тела грудной
кости со второго по шестой реберный хрящ до фасции предпле-
чья. Действие: приводит конечность.
Особенности. У свиней мышца начинается и оканчивает-
ся, как у коров. У лошадей плечевая часть развита в виде коротко-
го и почти округлого брюшка, предплечевая часть более широкая
и плоская. У собак имеется только плечевая часть.
Грудная глубокая мышца (см. рис. 3.67 — 3.70, 3.72) имеет раз-
витую только плечевую часть, которая начинается от грудной ко-
139
сти и реберных хрящей и оканчивается на гребне большого бугра.
Действие: приводит конечность.
Особенност и. У свиней грудная глубокая мышца начинает-
ся от всей грудины, оканчивается плечевая часть мышцы на боль-
Рис. 3.69. Мышцы свиньи:
7 — ключично-сосцевидная мышца; 2 — грудино-головная мышца; 3 — ключич-
но-затылочная мышца; 4 — плечепоперечная мышца; 5 — ключичная полоска;
6 — трапециевидная мышца, шейная часть; 7 — предостная мышца; 8 — дельто-
видная мышца; 9 — трапециевидная мышца, грудная часть; 10 — напрягатель
фасции предплечья; 11+36 — трехглавая мышца плеча, длинная головка; 12 —
широчайшая мышца спины; 13 — зубчатая дорсальная каудальная мышца; 14 —
подвздошно-реберная мышца; 15 — средняя ягодичная мышца; 16 + 17— поверх-
ностная ягодичная мышца; 16 — поверхностная ягодичная мышца, краниальная
часть; 17 — поверхностная ягодичная мышца, средняя часть; 18 — крестцово-
хвостовая дорсальная мышца; 19 — крестцово-хвостовая вентральная мышца;
20 — полуперепончатая мышца; 21 — полусухожильная мышца; 22 — поверхно-
стная ягодичная мышца, каудальная часть; 23 — напрягатель широкой фасции
бедра; 24+25 — двуглавая мышца бедра: 24 — двуглавая мышца бедра, краниаль-
ная часть, 25 — двуглавая мышца бедра, каудальная часть; 26 — длинная мало-
берцовая мышца; 27 — специальный разгибатель пятого пальца; 28 — икронож-
ная мышца; 29 — сухожилие специального разгибателя четвертого пальца; 30 —
третья малоберцовая мышца; 31 — передняя большеберцовая мышца; 32 — апо-
невроз; 33 — наружная косая мышца живота; 34 — зубчатая вентральная мышца
груди; 35 — глубокая грудная мышца; 37 — трехглавая мышца плеча, латераль-
ная головка; 38 — локтевой разгибатель запястья; 39 — поверхностный сгибатель
пальцев; 40 — общий разгибатель пальцев; 41 — длинный абдуктор большого
пальца; 42 — специальный разгибатель четвертого пальца; 43 — лучевой разгиба-
тель запястья; 44 — плечевая мышца; 45 — ключично-плечевая мышца; 46 —
грудино-подъязычная мышца; 47 — вентральная ушная мышца; 48 — околоуш-
ная железа; 49 — большая жевательная мышца; 50 — скуловая мышца; 57 —
щечная мышца; 52 — опускатель нижней губы; 53 — круговая мышца рта; 54 —
клыковая мышца; 55 — носогубный подниматель; 56 — специальный под-
ниматель верхней губы; 57 — опускатель хоботка
140
том бугре, предлопаточная — на фасции предостной мышцы.
У лошадей плечевая часть простирается от мечевидного хряща гру-
дины до малого и большого бугра. Предлопаточная часть мышцы
начинается от первых четырех реберных хрящей и оканчивается
впереди лопатки, вплетаясь в фасцию предостной мышцы._У со-
бак имеется только плечевая часть.
Подключичная мышца (см. рис. 3.69) имеет шнуровидную фор-
му. Начинается от рукоятки грудной кости, направляется к плече-
головной мышце и оканчивается в области ключичной попереч-
ной сухожильной полоски. Действие: поворачивает шею вправо и
влево.
Особенност и. У свиней и собак подключичная мышца от-
сутствует. У лошадей она слилась с грудино-головной мышцей.
Мышцы и фасции головы
Фасции головы подразделяются на поверхностную и глубокую.
Поверхностная фасция головы в отдельных местах образует са-
мостоятельные фасции: околоушно-жевательная фасция покрыва-
ет околоушную железу и жевательную мышцу; поверхностная ви-
сочная фасция располагается поверх височной мышцы; лобная фас-
ция лежит в области лба; щечная фасция находится в области щек;
подчелюстная фасция покрывает подкожную мышцу губ.
Глубокая фасция головы в отдельных участках имеет свои назва-
ния: щечно-глоточная фасция покрывает массетер и щечные мыш-
цы; глубокая височная фасция срастается с височной мышцей.
Мышцы головы делятся на две группы: лицевые (мимические у
человека) и жевательные.
Лицевые мышцы. Располагаются в основном в области естествен-
ных отверстий головы (рот, нос, глаз) и выполняют функции
сжимателей (констрикторов) или расширителей (дилататоров).
Круговая мышца рта (см. рис. 3.67 — 3.69, 3.71) слабо развита у
коров. Она лежит в основе верхней и нижней губы. Действие: зак-
рывает ротовое отверстие.
Особенности. У свиней круговая мышца развита слабо.
У лошадей она хорошо выражена..У собак эта мышца в верхней
губе развита сильнее, чем в нижней.
Резцовые мышцы (рис. 3.70; см. рис. 3.67 — 3.69) делятся на вер-
хние и нижние. Верхняя резцовая мышца выражена слабее, чем
нижняя резцовая. Начинается она от тела резцовой кости и тянется
до круговой мышцы рта.
Нижняя резцовая простирается от резцовой части нижней че-
люсти до края нижней губы. Действие: прижимает губы.
Особенност и. У свиней и лошадей резцовые мышцы хоро-
шо выражены. У собак они~слабо развиты.
141
Рис. 3.70. Мышцы шеи и груди коровы:
1 — косая каудальная мышца головы; 2 — длиннейшая мышца головы; 3 —
пластыревидная мышца; 4 — плечеголовная мышца; 5 — зубчатая вентральная
мышца шеи; 6 — лестничная надреберная мышца; 7 — ромбовидная мышца
шеи; 8 — предостная мышца; 9 — акромион; 10 — заостная мышца; 11 — трех-
главая мышца плеча, длинная головка; 12 — зубчатая вентральная мышца груди;
13 — широчайшая мышца спины; 14 — напрягатель фасции предплечья; 75 —
малая круглая мышца; 16 — глубокая грудная мышца; 17 — локтевой бугор; 18 —
трехглавая мышца плеча, латеральная головка; 19 — глубокий сгибатель паль-
цев, локтевая головка; 20 — локтевой разгибатель запястья; 21 — боковой разги-
батель пальцев; 22 — общий разгибатель пальцев; 23 — лучевой разгибатель запя-
стья; 24 — поверхностная грудная мышца, грудино-реберная часть; 25 — повер-
хностная грудная мышца, ключичная часть; 26 — плечевая мышца; 27 — двугла-
вая мышца плеча; 28 — глубокая грудная мышца, предплечевая часть; 29 —
лестничная мышца первого ребра; 30 — грудино-челюстная мышца; 31, 33 —
грудино-сосцевидная мышца; 32 — грудино-подъязычная мышца; 34 — плече -
подъязычная мышца; 35 — нижнечелюстная железа; 36 — околоушная железа
Подбородочная мышца (см. рис. 3.67 — 3.69, 3.71) начинается от
резцовой части нижней челюсти и следует до кожи подбородка.
Действие: сжимает подбородок.
Особенное! и. У свиней подбородочная мышца расположе-
на, каку коров. У лошадей иней находи гей много жировой ткани.
У собак она слабо развита.
Скуловая мышца (см. рис. 3.67 — 3.69, 3.71) направляется от ли-
цевого бугра вперед по щеке до угла губ. Действие: тянет угол рта
назад.
Особенности. У свиней и лошадей эта мышца располагает-
ся, как у коров. У собак она начинается от щитовидного ушного
хряща.
142
Рис. 3.71. Мышцы головы коровы:
1 — кожная мышца носа; 2 — носогубный подниматель; 3 — поверхностная
лицевая вена; 4 — поверхностная щечная мышца; 5 — большая жевательная
мышца, поверхностная часть; 6 — круговая мышца глаза; 7 — большая жеватель-
ная мышца, глубокая часть; 8 — лобно-щитковая мышца, височная часть; 9 —
кожная мышца лба; 10 — лобно-щитковая мышца, лобная часть; 11 — межщит-
ковая мышца; 12 — шейно-щитковая мышца; 13 — щитковоушная добавочная
поверхностная мышца; 14 — щитковоушная дорсальная поверхностная мышца;
15 — щитковоушная средняя поверхностная мышца; 16 — скулощитковая мыш-
ца; 17 — щиток; 18 — ключично-затылочная мышца; 19 — околоушная железа;
20 — околоушный лимфатический узел; 21, 30 — нижнечелюстная железа; 22 —
вентральная ушная мышца; 23 — ключично-сосцевидная мышца; 24 — внутрен-
няя челюстная вена; 25 — яремная вена; 26, 32 — грудино-челюстная мышца;
27 — грудино-подъязычная мышца; 28 — грудино-сосцевидная мышца; 29 —
наружная челюстная вена; 31 — нижнечелюстной лимфатический узел; 33 —
общая лицевая вена; 34 — подъязычно-челюстная мышца; 35 — опускатель ниж-
ней губы; 36 — глубокая щечная мышца; 37 — скуловая мышца; 38 — круговая
мышца рта; 39 — опускатель верхней губы; 40— клыковая мышца; 41 — специаль-
ный подниматель верхней губы
143
Носогубный подниматель (см. рис. 3.67 — 3.69, 3.71) состоит из
поверхностной и глубокой частей и мышцы нижнего века. Повер-
хностная часть от подкожной мышцы лба и носа направляется до
латерального крыла носа и верхней губы. Глубокая часть лежит
латерально, начинается там же, но оканчивается в области угла
рта под клыковой мышцей.
Мышца нижнего века (рис. 3.70; см. рис. 3.67 — 3.69, 3.71) про-
стирается от скуловой кости до круговой мышцы нижнего века.
Действие: поднимает губу и нос.
Особенности. У свиней носогубной подниматель идет от
носовой кости косо к верхней губе. У лошадей и собак он при-
крепляется, как у коров.
Клыковая мышца (см. рис. 3.67 — 3.69, 3.71) начинается от ли-
цевого бугра и оканчивается на верхней губе и латеральном крыле
носа. Действие: поднимает верхнюю губу.
Особенности. У свиней клыковая мышца берет начало в
ямке слезной, скуловой и верхнечелюстной костей и оканчивает-
ся у края отверстия носа. У лошадей клыковая мышца от передне-
го конца лицевого гребня идет, расширяясь, до латерального крыла
носа и круговой мышцы рта. У собак она начинается около под-
глазничного отверстия и оканчивается в верхней губе и латераль-
ном крыле носа.
Щечная мышца (см. рис. 3.67 — 3.69, 3.71) заложена в толще щеки
и состоит из поверхностной и глубокой частей. Поверхностная
часть — перистая мышца, у которой посередине проходит сухо-
жильная полоска. От этой полоски мышечные пучки тянутся вверх
и вперед до верхней челюсти, а в нижней половине они идут вниз
и вперед до нижней челюсти. Глубокая часть начинается от верхней
и нижней челюсти. Обе части соединяются вместе и оканчиваются
в области угла рта. Действие: прижимает щеку к зубам.
Особенности. У свиней и лошадей щечная мышца устрое-
на, как у коров. У собак она слабо развита.
Апикальный расширитель носа начинается от резцовой кости и
следует до кожи медиальных углов носа. Мышца разделяется на
два расширителя носа — медиальный и латеральный.
Медиальный расширитель носа направляется от боковой хря-
щевой стенки носа до медиального крыла носа.
Латеральный расширитель носа начинается от носового отрос-
тка резцовой кости и тянется до латерального крыла носа и якоря.
Действие: расширяют носовое отверстие.
Особенности. У свиней носовые мыш цы не выражены. У ло-
шадей выражены очень хорошо поперечная и боковая мышцы носа.
Поперечная мышца носа лежит на крыловидных хрящах носа,
простирается от пластинки и рожка правого крыловидного хряща
до пластинки и рожка левого крыловидного хряща. Действие: под-
нимает медиальное крыло носа.
144
Боковая мышца носа состоит из четырех маленьких расширите-
ией носа, которые следуют от крыловидного хряща до кожи лате-
рального крыла носа. Действие: все четыре мышцы расширяют
вход в носовую полость.
У собак носовые мышцы выражены слабо.
Жевательные мышцы хотя и немногочисленны, но отличаются
большой мощностью.
Большая жевательная мышца (см. рис. 3.67 — 3.69, 3.71) начи-
нается на лицевом бугре и скуловой дуге и разделяется на лате-
ральный и медиальный слои. Латеральный слой идет веером впе-
ред и вниз до ветви и коренной части тела нижней челюсти. Ме-
диальный слой идет косо вниз и назад до жевательной ямки. Дей-
ствие: сжимает челюсти.
Особенности. У свиней и лошадей жевательная мышца при-
крепляется, как у коров. У собак она плотно связана с височной
мышцей.
Крыловидная мышца состоит из двух слоев: медиального и ла-
терального. Медиальный слой начинается от крыловидной и нёб-
ной костей и оканчивается в крыловой ямке. Латеральный слой
идет от крыловидного отростка до крыловой ямки. Действие: сжи-
мает челюсти.
О с о б е н н о сти. У свиней и лошадей эта мышца развита силь-
нее, чем у коров. У собак ее брюшко разделено на два слоя не так
резко, как у коров.
Височная мышца (см. рис. 3.67 — 3.69, 3.71) направляется от
височной ямки к мышечному отростку нижней челюсти. Мышца
многоперистая. Действие: сжимает челюсти.
Особенности. У свиней, лошадей и собак височная мышца
сильно развита.
Двубрюшная мышца (см. рис. 3.67 — 3.69, 3.71) простирается от
яремного отростка до нижнего края нижней челюсти. Действие:
разжимает челюсти.
Особенности. У свиней мышца не разделяется на два брюшка.
У лошадей она делится на медиальную и латеральную части. Ме-
диальная часть имеет два брюшка, начинается она от яремного
отростка и скоро переходит в сухожилие. После прободения ко-
нечного сухожилия подъязычная мышца в дальнейшем формиру-
ет новое мышечное брюшко, которое оканчивается в области ли-
цевой сосудистой вырезки. Латеральная часть простирается от ярем-
ного отростка до угла нижней челюсти. У собак двубрюшная мышца
сильно выражена, но деление ее на два брюшка отсутствует.
Дорсальные мышцы позвоночного столба
Пластыревидная мышца головы и шеи (рис. 3.72; см. рис. 3.68) у
коров развита слабо, начинается от остистых отростков первых
145
г
Рис. 3.72. Мышцы туловища лошади (поверхностный пласт):
1 — краниальная косая мышца головы; 2— ключично-сосцевидная мышца; 3, 8 —
длиннейшая мышца головы; 4 — каудальная косая мышца головы; 5, 6, 12 —
пластыре видная мышца; 7 — полуостистая мышца головы; 9 — длиннейшая мышца
атланта; 10, 13 — длиннейшая мышца шеи; 11 — подвздошно-реберная мышца
шеи; 14 — капюшон выйной связки; 15 — остистая мышца спины; 16 — зубчатая
дорсальная краниальная мышца; 17 — подвздошно-реберная мышца спины; 18 —
зубчатая дорсальная каудальная мышца; 19 — длиннейшая мышца спины; 20, 33,
37 — наружная косая мышца живота; 21 — маклок; 22, 25 — полусухожильная
мышца; 23 — крестцово-хвостовая дорсальная медиальная мышца; 24 — крест-
цово-хвостовая дорсальная латеральная мышца; 26 — широкая крестцово-бугро-
вая связка; 27 — подвздошно-реберная мышца; 28— паховый канал; 29 — медиаль-
ная ножка; 30 — вход в брюшную полость; 31 — паховая связка; 32, 34 — внут-
ренняя косая мышца живота; 35, 40 — внутренние межреберные мышцы; 36 —
прямая мышца живота; 38 — зубчатая вентральная мышца груди; 39 — прямая
мышца груди; 41 — подмышечная вена; 42 — подмышечная артерия; 43 — по-
верхностный шейный лимфатический узел; 44 — трахея; 45, 51 — общая сонная
артерия; 46 — длинная мышца шеи; 47 — яремная вена; 48 — плече-подъязыч-
ная мышца; 49 — грудино-нижнечелюстная мышца; 50 — ключично-поперечная
мышца; 52 — длинная мышца головы; 53 — затылочная вена; 54 — внутренняя
челюстная вена; 55 — яремно-нижнечелюстная мышца
трех-четырех грудных позвонков и поперечно-остистой фасции и
оканчивается на крыле атланта, на поперечном отростке оси и
затылочной кости. Действие: разгибает шею.
Особенности. У свиней мышца оканчивается тремя ветвя-
ми: на затылочной кости, сосцевидном отростке и атланте. У ло-
шадей она оканчивается шейной частью на поперечных отростках
пятого, четвертого, третьего шейных позвонков, а головной — на
затылочном гребне и сосцевидном отростке. У собак пластыре-
видная мышца тянется от первых четырех-пяти грудных позвон-
ков до затылочного гребня.
146
Длиннейшая мышца спины (рис. 3.73; см. рис. 3.72) начинается
от подвздошной кости и от остистых отростков поясничных и пос-
ледних грудных позвонков. Волокна мышцы идут вперед и вниз в
виде латеральных и медиальных зубцов. Латеральными зубцами
мышца достигает поперечных отростков поясничных позвонков
всех ребер и поперечного отростка последнего шейного позвон-
ка, а медиальными — сосцевидных отростков поясничных и груд-
ных позвонков. Действие: разгибает позвоночный столб.
Особенности. У свиней эта мышца простирается от под-
вздошной кости до пятого шейного позвонка. У лошадей она на-
правляется так же, как у коров, но сзади в области поясницы
имеет углубление для прикрепления средней ягодичной мышцы.
У собак указанная мышца оканчивается медиальными зубцами на
добавочных отростках грудных позвонков — с тринадцатого по
восьмой.
Длиннейшая мышца шеи (см. рис. 3.72, 3.73) направляется от
поперечно-остистой фасции и поперечных отростков первых груд-
ных позвонков до поперечных отростков последних четырех шей-
ных позвонков. Действие: разгибает шею.
Особенности. У свиней, лошадей и собак длиннейшая мыш-
ца шеи устроена, как у коров.
Рис. 3.73. Мышцы туловища коровы (глубокий пласт):
1 — прямая дорсальная большая мышца головы; 2 — косая каудальная мышца
головы; 5, 5 — множественная мышца шеи; 4 — пластинка выйной связки; 6 —
капюшонная бурса; 7 - остистая мышца спины; 8, 12 — подниматели ребер; 9 —
полуостистая мышца спины; 10 — поперечно-остистая мышца; 11 — остисто-
поперечная мышца; 13, 15 — межпоперечная мышца поясницы; 14, 24 — мно-
жественная мышца спины; 16 — средняя ягодичная мышца; 17 — маклок; 18 —
напрягатель широкой фасции бедра; 19 — поперечная мышца живота; 20, 21 —
внутренние межреберные мышцы; 22 — наружные межреберные мышцы; 23 —
полуостистая мышпа шеи; 25, 28, 29 — межпоперечные мышцы шеи; 26 — су-
хожилие остистой и нолуостистой мышцы шеи; 27, 30 — длинная мышца шеи;
31 прямая вентральная мышца головы; 32— краниальная косая мышца головы
147
Длиннейшая мышца головы и атланта (см. рис. 3.72, 3.73) начи-
нается от суставных отростков первого грудного и последних четы-
рех шейных позвонков и оканчивается на крыле атланта и сосце-
видном отростке. Действие: разгибает шею и поднимает голову.
Особенности. У свиней мышца начинается от первых груд-
ных позвонков и оканчивается на затылочной кости и на атланте.
У лошадей она начинается и оканчивается, как у коров. У собак
мышца начинается от поперечных отростков пятого—третьего шей-
ных позвонков и оканчивается на крыле атланта.
Подвздошно-реберная мышца (см. рис. 3.72, 3.73) начинается от
гребня подвздошной кости, поперечных отростков поясничных
позвонков и ребер. Оканчивается в трех точках: поясничная часть —
на последнем ребре, грудная — на углах ребер, шейная — на по-
перечных отростках трех последних шейных позвонков. Действие:
разгибает поясницу, спину и шею.
Особенности. У свиней подвздошно-реберная мышца име-
ет тонкую поясничную часть и хорошо развитую спинную; шей-
ная часть простирается до атланта. У лошадей эта мышца имеет
такое же строение, как у коров. У собак поясничная часть начина-
ется от подвздошной кости и оканчивается на поперечных отрос-
тках поясничных позвонков, спинная часть сильно развита.
Остистая и полуостистая мышцы спины и шеи (см. рис. 3.72,
3.73) в области поясницы срастаются с длиннейшей мышцей спи-
ны, а в области первого поясничного позвонка они обособляются
от длиннейшей мышцы спины и оканчиваются на остистых отро-
стках последних четырех-пяти шейных позвонков. Действие: раз-
гибают поясницу, спину и шею.
Особенности. У свиней остистая мышца идет до шестого
(пятого) шейного позвонка. У лошадей она постепенно формиру-
ет самостоятельное мышечное брюшко, направляющееся к ости-
стым отросткам холки. У собак остистая мышца спины и шеи окан-
чивается на остистых отростках грудных и последних двух шейных
позвонков.
Полуостистая мышца головы (см. рис. 3.72, 3.73) начинается от
основания остистых отростков первых восьми-девяти грудных и
последних шейных позвонков, а оканчивается на затылочной ко-
сти. Действие: разгибает шею и поднимает голову.
Особенности. У свиней полуостистая мышца состоит из
брюшной и большой комплексной части. Двубрюшная мышца ле-
жит дорсомедиально, а большая комплексная — вентролатераль-
но. Обе части начинаются от остистых отростков первых грудных
позвонков и оканчиваются на затылочной кости. У лошадей полу-
остистая мышца головы располагается, как у коров. У собак она
имеет такое же строение, как у свиней.
Множественная мышца спины (см. рис. 3.73) начинается от со-
сцевидных отростков поясничных позвонков и поперечных отро-
148
стков грудных позвонков, а оканчивается на остистых отростках,
пропуская два—пять сегментов. Действие: разгибает поясницу, спи-
ну и шею.
Особенноети. У свиней множественная мышца спины ле-
жит между поперечными и остистыми отростками. У лошадей она
имеет такое же строение, как у коров. У собак множественная
мышца спины и шеи сильно развита.
Межпоперечные мышцы (см. рис. 3.73) сильнее выражены на
шее, слабее на пояснице. Действие: изгибает шею и поясницу впра-
во или влево.
Особенности. У свиней и собак межпоперечные мышцы хо-
рошо развиты. У лошадей они подразделяются на дорсальные, вен-
тральные и средние межпоперечные мышцы и хорошо развиты
только на шее.
Межостистые мышцы (см. рис. 3.73) лежат между остистыми
отростками. Действие: разгибают позвоночный столб.
Особенности. У свиней и собак межостистые мышцы хоро-
шо развиты. У лошадей они превратились в межостистые связки.
Косая головная каудальная мышца (см. рис. 3.72, 3.73) прости-
рается от крыла атланта до дорсального гребня оси. Действие: по-
ворачивает крыло атланта.
Особенности. У свиней, лошадей и собак мышца устроена,
как у коров.
Косая головная краниальная мышца (см. рис. 3.72, 3.73) при-
крепляется одним концом к яремному отростку, а другим — к
крылу атланта. Действие: поворачивает голову.
Особенности. У свиней и лошадей устроена, как у коров.
У собак она лежит между сосцевидным отростком и крылом ат-
ланта.
Большая дорсальная прямая мышца головы (см. рис. 3.72, 3.73)
идет от затылочной кости до дорсального гребня оси. Действие:
поднимает голову.
Особенности. У свиней, лошадей и собак эта мышца рас-
положена, как у коров.
Малая дорсальная прямая мышца головы лежит под предыду-
щей мышцей, начинается от затылочной кости и оканчивается на
дорсальном бугре атланта. Действие: поднимает голову.
Особенности. У свиней, лошадей и собак эта мышца ле-
жит, как у коров.
Длинный и короткий подниматели хвоста (см. рис. 3.72) хоро-
шо развиты, направляются от остистых отростков до сосцевид-
ных.
Действие: поднимают хвост.
Особенности.У свиней сухожилия поднимателей хвоста спи-
ралевидно перекручены. У лошадей и собак они прикрепляются,
как у коров.
149
Межпоперечные мышцы хвоста лежат между поперечными от-
ростками хвостовых позвонков. Действие: поворачивают хвост.
Особенности. У свиней межпоперечные мышцы хвоста слабо
развиты. У лошадей и собак они хорошо выражены.
Вентральные мышцы позвоночного столба
Длинная мышца шеи (см. рис. 3.73) лежит на вентральной повер-
хности шейного и грудного отделов позвоночника. Грудная часть
начинается от тел первых шести грудных позвонков и заканчивает-
ся на поперечном отростке седьмого шейного позвонка. Шейная
часть берет начало от поперечно-реберных отростков и тел после-
дних пяти шейных позвонков, а оканчивается на гребне шейных
позвонков и вентральном бугре атланта. Действие: сгибает шею.
Особенности. У свиней, лошадей и собак мышца прикреп-
ляется, как у коров.
Длинная мышца головы (см. рис. 3.73) следует от поперечных
отростков пятого — второго шейных позвонков до мышечного бу-
горка затылочной кости. Действие: сгибает шею и голову.
Особенности. У свиней, лошадей и собак она располагает-
ся, как у коров.
Прямая вентральная мышца головы направляется от мышечно-
го бугорка затылочной кости до вентрального бугорка атланта.
Действие: опускает голову.
Особенности. У свиней, лошадей и собак мышца устроена,
как у коров.
Прямая латеральная мышца головы от яремного отростка идет
до вентральной дуги атланта. Действие: опускает голову.
Особенности. У свиней, лошадей и собак мышца лежит так
же, как у коров.
Грудино-головная мышца (см. рис. 3.67 — 3.72) состоит из двух
частей: грудино-сосцевидной и грудино-челюстной мышц. Груди-
но-сосцевидная мышца тянется от рукоятки грудной кости до со-
сцевидного отростка. Грудино-челюстная мышца идет от рукоятки
грудины до нижней челюсти. Действие: опускает голову и шею.
Особенности. У свиней и собак мышца представлена только
грудино-сосцевидной мышцей. У лошадей устроена, как у коров.
Плечеподъязычная мышца (см. рис. 3.67 — 3.72) начинается от
поперечных отростков третьего, четвертого (пятого) шейных по-
звонков, оканчивается на подъязычной кости. Действие: оттяги-
вает назад подъязычную кость.
Особенности. У свиней и лошадей плечеподъязычная мышца
устроена, как у коров. У собак она отсутствует.
Грудино-щитовидная мышца (см. рис. 3.67 — 3.72) простирается
от рукоятки грудной кости до щитовидного хряща. Действие: от-
тягивает гортань назад.
150
Особенности. У свиней грудино-щитовидная мышца начи-
нается от первого ребра. У лошадей она построена, как у коров.
У собак грудино-щитовидная мышца начинается вместе с груди-
но-подъязычной мышцей на рукоятке грудины.
Грудино-подъязычная мышца (см. рис. 3.67 — 3.72) начинается
от рукоятки грудины, оканчивается на теле подъязычной кости.
Действие: оттягивает подъязычную кость назад.
Особенности. У свиней грудино-подъязычная мышца на-
чинается от первого ребра. У лошадей и собак она расположена,
как у коров.
Квадратная поясничная мышца лежит под поперечными отро-
стками поясничных позвонков. Она простирается от последних ре-
бер до крыла крестца. Действие: сгибает поясницу.
Особенности. У свиней, лошадей и собак мышца устроена,
как у коров.
Поясничная малая мышца направляется от вентральной повер-
хности тел поясничных и последних грудных позвонков к пояс-
ничному бугорку таза. Действие: сгибает поясницу.
Особенности. У свиней, лошадей и собак мышца прикреп-
ляется, как у коров.
Подвздошно-поясничная мышца (см. рис. 3.67, 3.69, 3.72, 3.73)
состоит из двух самостоятельных мышц: большой поясничной и
подвздошной. Большая поясничная мышца направляется от тел по-
ясничных и последних грудных позвонков к малому вертелу. Под-
вздошная мышца состоит из двух головок: латеральная головка на-
чинается от бугра подвздошной кости, а медиальная от крестцо-
вой кости и тела подвздошной кости. Обе головки оканчиваются
на малом вертеле бедренной кости. Действие: сгибает поясницу и
тазобедренный сустав.
Особенности. У свиней, лошадей и собак мышца устроена,
как у коров.
Длинный опускателъ хвоста начинается на вентральной повер-
хности крестца и поперечных отростков первых хвостовых позвон-
ков и оканчивается на вентральной поверхности поперечных от-
ростков и тел позвонков. Действие: опускает хвост.
Особенности. У свиней опускатель хвоста развит слабо.
У лошадей и собак он располагается, как у коров.
Короткий опускатель хвоста берет начало от вентральной по-
верхности крестца и первых семи (восьми) хвостовых позвонков
и оканчивается сухожилиями на телах позвонков. Действие: опус-
кает хвост.
Особенности. У свиней опускатель хвоста развит слабо.
У лошадей и собак он прикрепляется, как у коров.
Хвостовая мышца начинается от седалищной ости таза, окан-
чивается на поперечных отростках первых трех (четырех) хвосто-
вых позвонков. Действие: опускает хвост.
151
Особенности. У свиней, лошадей и собак хвостовая мышца
располагается, как у коров.
Мышцы брюшной стенки
Мышцы брюшной стенки поддерживают внутренние органы и
выполняют ряд других функций.
Косая наружная мышца живота (рис. 3.74; см. рис. 3.67 — 3.69,
3.72 — 3.73) начинается от ребер с пятого по тринадцатое, ее мы-
шечные пучки идут косо сверху вниз и назад и оканчиваются ши-
роким апоневрозом, который образует брюшную, тазовую и бед-
ренную пластинки.
Брюшная пластинка самая широкая, срастается с одноимен-
ной пластинкой другой стороны по белой линии живота. Тазовая
пластинка оканчивается на паховой связке. Бедренная пластинка
заканчивается на медиальной поверхности широкой бедренной
фасции. Между брюшной и тазовой пластинками у самцов образу-
ется наружное паховое кольцо довольно больших размеров, вытя-
нутой веретеновидной формы. От наружного пахового кольца на-
чинается паховый канал конусовидной формы, в котором нахо-
дится семенной канатик. Паховый канал расположен между на-
ружной и внутренней косыми мышцами живота. Снаружи он по-
Рис. 3.74. Мышцы брюшной стенки лошади:
1 — наружная косая мышца живота; 2 — внутренняя косая мышца живота; 3 —
поперечная мышца живота; 4 — прямая мышца живота
152
крыт желтой брюшной фасцией. Заканчивается канал внутренним
паховым кольцом небольших размеров (толщиной с карандаш),
которое ограничено задним краем внутренней косой мышцы жи-
вота и паховой связкой.
Особенности. У свиней, лошадей мышца устроена, как у
быков. У собак наружное паховое кольцо хорошо выражено не
только у самцов, но и у самок.
Косая внутренняя мышца живота (см. рис. 3.72 — 3.74) начина-
ется от маклока и от поперечных отростков поясничных позвон-
ков и оканчивается на последних ребрах и белой линии живота.
Мышечные пучки идут сверху, вниз и вперед.
Особенности. У свиней, лошадей и собак она расположе-
на, как у коров.
Поперечная мышца живота (см. рис. 3.73, 3.74) берет начало от
поперечных отростков поясничных позвонков и от последних ре-
берных хрящей и следует до белой линии живота. Мышечные пуч-
ки направлены сверху вниз. Медиально эта мышца прикрыта внут-
рибрюшной поперечной фасцией и брюшиной.
Особенности. У свиней, лошадей и собак поперечная мышца
живота прикрепляется, как у коров.
Прямая мышца живота (см. рис. 3.72, 3.74) начинается от ниж-
них концов четвертого—девятого реберных хрящей и грудной ко-
сти и оканчивается на лонном гребне. Мышечные пучки идут спе-
реди назад справа и слева вдоль белой линии живота и имеют пять
сухожильных перемычек. Мышца заключена в сухожильное влага-
лище, образованное апоневрозами наружной и внутренней косых
мышц живота (наружная пластинка), поперечной мышцы и внут-
рибрюшной фасции (внутренняя пластинка). В области второй или
третьей сухожильной перемычки у коров располагается молоч-
ный колодец для прохождения подкожной брюшной вены.
Действие: все мышцы живота не только поддерживают внут-
ренности, но принимают участие в акте выдоха и помогают опо-
рожнению брюшных органов при дефекации, мочеиспускании, а
у самок участвуют в родах.
Мышцы грудной стенки
Мышцы грудной стенки, сокращаясь, или расширяют груд-
ную полость, или, наоборот, суживают ее. Расширение грудной
полости сопровождается вдохом, а сужение — выдохом. Мышцы,
обеспечивающие вдох, называются инспираторами (вдыхателями),
а вызывающие выдох — экспираторами (выдыхателями).
Дорсальная краниальная зубчатая мышца (см. рис. 3.72) у коров
развита слабо. Начинается от остистых отростков первого — пятого
грудных позвонков, идет сверху вниз и назад до пятого — восьмо-
го ребер. Действие: участвует в акте вдоха.
153
Особенности. У свиней и собак краниальная дорсальная
зубчатая мышца развита, как у коров. У лошадей оканчивается на
ребрах с пятого по двенадцатое.
Подниматели ребер (см. рис. 3.72, 3.73) в количестве 10—11
простираются от сосцевидных отростков грудных позвонков до
бугров ребер. Действие: участвуют в акте вдоха.
Особенности. У свиней, лошадей и собак подниматели ре-
бер расположены, как у коров.
Межреберные наружные мышцы (см. рис. 3.72, 3.73) начинают-
ся от заднего края ребра, волокна идут сверху вниз и назад до
переднего края последующего ребра. Действие: участвуют в акте
вдоха.
Особенности. У свиней, лошадей и собак мышцы прикреп-
лены, как у коров.
Лестничная надреберная мышца (см. рис. 3.72) у коров хорошо
развита. Начинается от пятого — второго (первого) ребер и окан-
чивается на поперечных отростках шестого — третьего шейных по-
звонков.
Лестничная вентральная мышца (см. рис. 3.72) идет от первого
ребра до поперечных отростков третьего — седьмого шейных по-
звонков. Действие: участвует в акте вздоха.
Особенности. У свиней мышца устроена, как у коров.
У лошадей она начинается от поперечных отростков четвертого —
седьмого шейных позвонков и оканчивается на первом ребре,
между мышечными пучками находится плечевое нервное сплете-
ние. У собак мышечные пучки от поперечных отростков шейных
позвонков тянутся: верхний зубец до третьего-четвертого ребра,
средний до восьмого ребра и нижний до первого ребра.
Прямая грудная мышца (см. рис. 3.72) простирается от первого
ребра до четвертого — шестого реберных хрящей. Действие: уча-
ствует в акте вдоха.
Особенности. У свиней и лошадей прямая грудная мышца
оканчивается на втором — четвертом реберных хрящах, у собак
мышца идет до третьего реберного хряща.
Диафрагма, или грудобрюшная преграда, отделяет грудную по-
лость от брюшной (рис. 3.75). На диафрагме различают мышечную
периферическую часть и сухожильный центр. Периферическая часть
делится на поясничную, реберную и грудинную.
Поясничная часть (см. рис. 3.75) состоит из правой vt левой ножек,
которые прикрепляются на поясничных и последних грудных по-
звонках. Между ножками диафрагмы находится аортальное отвер-
стие. В правой ножке помещается пищеводное отверстие. Мышеч-
ные волокна тянутся сверху вниз и вперед до сухожильного центра.
Реберная часть (см. рис. 3.75) парная, начинается на нижних
концах 8—12-го ребер. Мышечные волокна идут вперед и меди-
ально до сухожильного центра. Между реберной и поясничной
154
Рис. 3.75. Диафрагма собаки (( руд-
ная поверхность):
7 — дорсальные мышцы позвоночно-
го столба; 2 — левый симпатический
ствол; 3 — дорсальный пищеводный
ствол блуждающего нерва; 4 — левая
ножка; 5— вентральный пищеводный
ствол блуждающего нерва; 6 — левый
диафрагмальный нерв; 7 — реберная
часть диафрагмы; 8 — сухожильный
центр диафрагмы; 9 — грудинная
часть диафрагмы; 10 — грудино-око-
лосердечная связка; 7 7 — спинной
мозг; 12 — правая непарная вена и
правый симпатический ствол; 13 —
аорта; 14 — пищевод; 15 — средо-
стенное пространство; 16— каудаль-
ная полая вена; 17 — правый диаф-
рагмальный нерв; 18 — брыжейка ка-
удальной полой вены
частями располагаются пояснично-реберные треугольники. Здесь ди-
афрагма состоит только из двух сросшихся между собой листков
серозных оболочек — плевры и брюшины. В этом месте возможны
разрывы диафрагмы (диафрагмальные грыжи).
Грудинная часть (см. рис. 3.75) начинается от дорсальной по-
верхности мечевидного хряща. Мышечные волокна идут вперед и
вверх до сухожильного центра.
Сухожильный центр — место срастания сухожилий всех частей
диафрагмы. В нем имеется отверстие для каудальной полой вены.
Брюшная вогнутая поверхность диафрагмы покрыта брюшной. Груд-
ная выпуклая поверхность покрыта плеврой. Действие: при сокра-
щении диафрагмы происходит вдох, при сокращении мышц жи-
вота — выдох.
Особенности. У свиней диафрагма устроена так же, как у
коров. У лошадей правая ножка диафрагмы начинается от тел че-
тырех первых поясничных позвонков, а левая ножка — от двух
первых поясничных позвонков. Отверстие для аорты лежит между
ножками, а отверстие для пищевода — в левой ножке.
Каудальная дорсальная зубчатая мышца (см. рис. 3.67, 3.68, 3.72)
начинается от ос тистых отростков поясничных позвонков и окан-
чивается на I —XVIII ребрах. Действие: участвует в выдохе.
Особеннос ти. У свиней зубчатый дорсальный выдыхатель
идет от поясничных остистых отростков к последним пяти-шести
ребрам. У лошадей эта мышца от поясничных остистых отростков
тянется до одиннадцатого —восемнадцатого ребер. У собак мыш-
ца расположена, как у коров.
155
Межреберные внутренние мышцы (см. рис. 3.72, 3.73) начина-
ются от заднего края ребер, мышечные пучки идут косо снизу,
назад и вверх до переднего края следующего ребра. Они распола-
гаются и между реберными хрящами. Действие: выдыхатели.
Особенности. У свиней, лошадей и собак межреберные
мышцы расположены, как у коров.
Пояснично-реберная мышца (см. рис. 3.72, 3.73) простирается
от поперечных отростков первого —третьего поясничных позвон-
ков до последнего ребра. Действие: выдыхатель.
Особенности. У свиней и лошадей мышца прикрепляется,
как у коров. У собак она отсутствует.
Поперечная грудная мышца берет начало от дорсальной повер-
хности грудной кости и оканчивается на внутренней поверхности
второго — восьмого реберных хрящей. Действие: выдыхатель.
Особенности. У свиней, лошадей и собак поперечная груд-
ная мышца расположена, как у коров.
Фасции и мышцы грудной конечности
Фасции грудной конечности делятся на поверхностную и глу-
бокую.
Поверхностная фасция одевает лопатку и плечо с латеральной
стороны.
Глубокая фасция в различных участках получает специальные
названия: подлопаточная фасция покрывает подлопаточную и пре-
достную мышцы; лопатко-плечевая фасция располагается с лате-
ральной стороны в области плечевого пояса; фасция предплечья
сильно развита на дорсальной и латеральной поверхностях пред-
плечья. Глубокая фасция доходит до запястья и формирует попе-
речную связку запястья, которая укрепляет все сгибатели. Она фор-
мирует кольцевые связки сухожилий в области путового сустава и
соединяется с напрягателем фасции предплечья.
Мышцы грудной конечности включают мышцы плечевого по-
яса — присоединяющие грудную конечность к туловищу (изложе-
ны для удобства в начале раздела), мышцы плечевого сустава,
мышцы локтевого сустава, мышцы запястного сустава и суставов
пальцев. На тазовой конечности выделяют мышцы тазобедренно-
го, мышцы коленного суставов, мышцы заплюсневого сустава и
суставов пальцев.
Мышцы плечевого сустава представлены разгибателями, сги-
бателями, а также приводящими и отводящими мышцами.
Мышцы-разгибатели плечевого сустава. Предостная мышца (рис.
3.76, 3.77; см. рис. 3.67 — 3.70) начинается от всей поверхности
предостной ямки лопатки, от ее краниального края и от крани-
альной поверхности ости лопатки. Хорошо развитое мышечное
156
Рис. 3.76. Мышцы грудной конеч-
ности лошади (латеральная поверх-
ность):
/ — ОСНОП.1НИС лопатки, 2 большая
круглая мышца; 3 — йосгная мышца,
дсльтонндная мьшша: ? - rpcxi ш-
иая мышца плеча, липшая юлоика: 6-'-
। ре\глаяяя_мышиа гшеча, латеральная
штопка: 7 - юктения головка глубо-
кою diiliii.c |я ппыга; 8 — локтевой
разгибатель запястья;.9 — боковой раз-
гибатель пальца; 10 —-^латеральная гри-
фелевидная кость; (11),14 — сухожилие
поверхностного сгибателя пальца; 12,
15 - сухожилие глубокого сгибателя
папьиа; 13 третья межкостная мыш-
ца; /6'—прелестная мышца;'/Л- глу-
бокая грудная мыши», 1грсллопаточная
часть: 18 — таосгноя мыший. 19)- ми-
лая круглая мышиа: 2Q .туманам
мышца плеча; 21/ плечевая мышца;
22/— лучевой ра ггибагелъ запястья, 232-
общий разгибатель пальпа:)2-/ — длин-
ный абдуктор большого пальца; 25 —
сухожилие лучевого разгибателя запя-
стья; 26 — сухожилие общего разгиба-
теля пальца; 27 — сухожилие бокового
разгибателя пальца; 28 — ветвь третьей
межкостной мышцы к сухожилию об-
щего разгибателя пальца
брюшко полностью заполняет
предостную ямку, дистально на-
легает над краниальным краем
лопатки, опускается вниз и над
лопаточным бугром разделяется
на две ножки, пропуская сухожи-
лие двуглавой мышцы плеча.
Оканчивается мышца на большом
и малом бугорках плечевой кости.
Действие: разгибает плечевой су-
став.
Особенности. У свиней, лошадей и собак предостная мыш-
ца расположена, как у коров.
Клювовидно-плечевая мышца (см. рис. 3.77) проходит по меди-
альной поверхности плечевого сустава. Начинается длинным уз-
ким уплощенным сухожилием от коракоидного отростка лопат-
ки, проходит нал медиальной поверхностью сухожилия подлопа-
точной мышцы. Оканчивается округлым сухожилием на медиаль-
157
3.67 — 3.69, 3.76) делится
Рис. 3.77. Мышцы грудной конечности
овцы (медиальная поверхность):
1 — ромбовидная мышца шеи; 2 — лопаточ-
ный хрящ; 3 — зубчатая вентральная мышца
груди; 4 — широчайшая мышца спины; 5 —
большая круглая мышца; 6 — напрягатель
фасции предплечья; 7 — трехглавая мышца
плеча, длинная головка; 8— трехглавая мыш-
ца плеча, медиальная головка; 9 — локтевая
головка глубокого сгибателя пальцев; 10 —
локтевой сгибатель запястья; 11 — лучевой
сгибатель запястья; 12, 13, 16 — поверхност-
ный сгибатель пальцев; 14— сухожилие глу-
бокого сгибателя пальцев; 15 — средняя меж-
костная мышца; 17 - зубчатая вентральная
мышца шеи; 18+19+21 — подлопаточная
мышца; 20 — предостная мышца; 22 — глу-
бокая грудная мышца; 23 — двуглавая мыш-
ца плеча; 24 — коракоидно-плечевая мыш-
ца; 25 — поверхностная грудная мышца; 26 —
плечевая мышца; 27 — сухожильный тяж;
28 — круглый пронатор; 29 — длинный аб-
дуктор большого пальца; 30 — сухожилие
специального разгибателя третьего пальца;
31 — ветвь к сухожилию общего разгибателя
пальцев
ной поверхности проксимальной ча-
сти плечевой кости вблизи большой
круглой шероховатости.
Особенности. У свиней предо-
стная мышца сильно развита. У ло-
шадей и собак она заканчивается на
большом и малом буграх плечевой
кости.
Мышцы-сгибатели плечевого сус-
тава. Дельтовидная мышца (см. рис.
акромиальную и лопаточную части.
Обе части оканчиваются на дельтовидной шероховатости. Действие:
сгибает плечевой сустав.
Особенности. У свиней и собак дельтовидная мышца силь-
но развита. У лошадей развита только ее лопаточная часть.
Малая круглая мышца (см. рис. 3.70) лежит под дельтовидной
мышцей, простирается от нижней части заднего края лопатки до
локтевой линии. Действие: сгибает плечевой сустав.
Особенности. У свиней, лошадей и собак эта мышца рас-
положена, как у коров.
158
Большая круглая мышца (см. рис. 3.77) лежит позади подлопа-
точной мышцы. Она начинается от каудального угла лопатки и
оканчивается на гребне малого бугра. Срастается с широчайшей
мышцей спины. Действие: сгибает плечевой сустав.
Особенности. У свиней, лошадей и собак эта мышца при-
крепляется, как у коров.
Приводящие и отводящие мышцы плечевого сустава. Подлопа-
точная мышца (см. рис. 3.77) начинается от подлопаточной ямки
и оканчивается на медиальном мышечном буре плечевой кости.
Действие: приводит плечевой сустав.
Особенности. У свиней, лошадей и собак имеет точки при-
крепления, как у коров.
Заветная мышца (см. рис. 3.67 — 3.70, 3.76) берет начало от за-
остной ямки и оканчивается двумя ветвями: мясисто на латераль-
ном мышечном бугре плечевой кости и сухожильно на латераль-
ной поверхности большого бугра, здесь имеет слизистую сумку.
Действие: отводит плечевой сустав.
Особенности. У свиней, лошадей и собак заостная мышца
сильно выражена.
Мышцы локтевого сустава представлены разгибателями и сги-
бателями.
Разгибатели локтевого сустава. Трехглавая мышца плеча (см. рис.
3.67 — 3.70, 3.76, 3.77) занимает пространство между лопаткой и
локтевым отростком. Она состоит из трех головок: длинной, лате-
ральной и медиальной. Действие: разгибает локтевой сустав.
Особенности. У свиней и лошадей она устроена, как у ко-
ров. У собак имеется еще добавочная головка, которая начинается
от шейки плечевой кости и оканчивается на локтевом бугре.
Локтевая мышца направляется от локтевой ямки к локтевому
отростку. Действие: разгибает локтевой сустав.
Особенности. У свиней, лошадей и собак локтевая мышца
прикрепляется, как у коров.
Напрягатель фасции предплечья (см. рис. 3.77) идет от каудаль-
ного края лопатки и апоневроза широчайшей мышцы спины к
локтевому бугру, а частью — к фасции предплечья. Действие: раз-
гибает локтевой сустав и напрягает фасцию предплечья.
Особенности. У свиней, лошадей и собак эта мышца рас-
положена, как у коров.
Сгибатели локтевого сустава. Двуглавая мышца плеча (см. рис.
3.70, 3.76, 3.77) начинается от надсуставного бугорка лопатки.
Проксимальное сухожилие ее перебрасывается через блок плече-
вой кости, а оканчивается мышца на шероховатости лучевой ко-
сти. На плечевом блоке мышца имеет слизистую сумку. Действие:
сгибает локтевой сустав.
Особенности.У свиней и собак двуглавая мышца плеча рас-
полагается, как у коров. У лошадей — это округлая, пронизанная
159
от начала до конца многочисленными сухожильными пучками
мышца.
Плечевая мышца (см. рис. 3.77) простирается от шейки плече-
вой кости до лучевой шероховатости. Действие: сгибает локтевой
сустав.
Особенности. У свиней, лошадей и собак эта мышца при-
крепляется, как у коров.
Мышцы запястного сустава представлены разгибателями, сги-
бателями, супинаторами и пронаторами.
Разгибатели запястного сустава. Лучевой разгибатель запястья
(рис. 3.78; см. рис. 3.67 — 3.70, 3.76) начинается от разгибательного
надмыщелка плечевой кости и прикрепляется к пястной шерохо-
ватости. На дорсальной поверхности запястья сухожилие окутыва-
ется синовиальным влагалищем. Действие: разгибает запястный
сустав.
Рис. 3.78. Сравнительная анатомия мышц кисти собаки (Л), свиньи (Б),
коровы (Б), лошади (Л):
1 — лучевой разгибатель запястья; 2 — плечелучевая мышца; 3 — обший разгиба-
тель пальцев; 4 — специальный разгибатель третьего пальца; 5 — боковой разги-
батель пальцев; 6 — локтевой разгибатель запястья; 7— длинный абдуктор боль-
шого пальца; 8 — локтевой сгибатель запястья; 9 — глубокий сгибатель пальцев,
локтевая головка; 10 — поверхностный сгибатель пальцев; 11, 12— межкостные
мышцы
160
Особенности. У свиней лучевой разгибатель запястья окан-
чивается на третьей пястной кости. У лошадей к его сухожилию
подходит сильно развитый сухожильный тяж от двуглавой мыш-
цы плеча.
Длинный абдуктор большого пальца (см. рис. 3.67—3.70, 3.76—
3.78) начинается от латерального края лучевой кости в ее средней
и нижней трети, идет косо по краниальной поверхности предпле-
чья через лучевой разгибатель запястья и оканчивается на меди-
альной шероховатости проксимального конца пястных костей. Здесь
его сухожилие имеет синовиальное влагалище. Действие: разгиба-
ет запястный сустав.
Особенности. У свиней и лошадей оканчивается на второй
пястной кости, у собак — на первой пястной кости.
Сгибатели запястного сустава. Локтевой разгибатель запястья
(см. рис. 3.67, 3.77) берет начало от разгибательного надмыщелка
плечевой кости. Заканчивается мышца на основании пятой пяст-
ной кости и на добавочной кости запястья. Действие: сгибает за-
пястный сустав.
Особенности. У свиней локтевой разгибатель оканчивается
на добавочной кости запястья и пятой пястной кости; у лошадей —
на добавочной кости и четвертой пястной кости; у собак — на
пятой пястной кости.
Локтевой сгибатель запястья (см. рис. 3.77, 3.78) начинается
от сгибательного надмыщелка плечевой кости и от локтевого от-
ростка и оканчивается на добавочной кости запястья. Действие:
сгибает запястный сустав.
Особенности. У свиней, лошадей и собак локтевой сгиба-
тель устроен, как у коров.
Лучевой сгибатель запястья (см. рис. 3.77) простирается от сги-
бательного надмыщелка плечевой кости до медиальной поверх-
ности пястных костей. На запястье имеет синовиальное влагалище.
Действие: сгибает запястный сустав.
Особенности. У свиней лучевой сгибатель оканчивается на
третьей пястной кости. У лошадей — на второй, у собак — на
второй и третьей пястных костях.
Супинаторы запястного сустава. Плечелучевая мышца (см. рис.
3.78) у собак слабо развита, начинается от разгибательного над-
мыщелка и оканчивается на медиальном крае лучевой кости. Дей-
ствие: супинирует предплечье.
Особенное г и. У коров, свиней и лошадей эта мышца от-
сутствует.
Короткий супинатор развит у собак, идет от разгибательного
надмыщелка и о г латеральной боковой связки до медиального края
лучевой кости. Действие: супинирует предплечье.
О с о б е н н о с т и. У свиней эта мышца слабо развита. У коров и
лошадей — отсутствует.
О Зеленевский
161
Пронаторы запястного сустава. Круглый пронатор у собак раз-
вит сравнительно сильно, идет от сгибательного надмыщелка к
медиальному краю лучевой кости. Действие: пронирует предп-
лечье.
Особенности. У свиней круглый пронатор развит слабо, у
лошадей в виде сухожильного тяжа.
Квадратный пронатор имеется только у собаки, лежит в попе-
речном направлении между лучевой и локтевой костями, запол-
няя все межкостные пространства предплечья. Действие: прони-
рует предплечье (поворачивает внутрь).
Мышцы суставов пальцев кисти представлены разгибателями
и сгибателями.
Разгибатели суставов пальцев кисти. Общий разгибатель паль-
цев (см. рис. 3.67 — 3.70, 3.76, 3.78) начинается на разгибательном
надмыщелке плечевой кости двумя ветвями и оканчивается на
разгибательных отростках копытцевых костей. На запястье сухо-
жилие имеет синовиальное влагалище, а на путовом суставе каж-
дое сухожилие имеет слизистую сумку. Действие: разгибает путо-
вый, венечный и копытцевый суставы.
Особенности. У свиней общий разгибатель пальцев распа-
дается на три мышечных брюшка. У лошадей он оканчивается на
разгибательном отростке дистальной фаланги. У собак мышца имеет
четыре брюшка.
Боковой разгибатель пальцев (см. рис. 3.67 — 3.69, 3.76, 3.78)
начинается от латеральной боковой связки локтевого сустава и
оканчивается на средней фаланге четвертого пальца. Действие: раз-
гибает путовый, венечный суставы.
Особенности. У свиней боковой разгибатель пальца распа-
дается на специальный разгибатель четвертого пальца и специ-
альный разгибатель пятого пальца. У лошадей он оканчивается на
проксимальном конце проксимальной фаланги. У собак боковой
разгибатель пальца имеет два брюшка.
Специальный разгибатель второго пальца у коров отсутствует.
Особенности. У свиней и лошадей специальный разгиба-
тель второго пальца идет от локтевой кости к общему разгибателю
пальцев. У собак он оканчивается двумя ветвями.
Специальный разгибатель третьего пальца (см. рис. 3.78) начи-
нается от разгибательного надмыщелка и идет до разгибательного
отростка копытцевой кости третьего пальца. Действие: разгибает
суставы третьего пальца.
Особенности. У свиней и лошадей разгибатель третьего паль-
ца слился с общим разгибателем пальца. У собак он слился с бо-
ковым разгибателем пальца.
Сгибатели суставов пальцев кисти. Поверхностный сгибатель
пальцев (см. рис. 3.77) начинается от сгибательного надмыщелка и
оканчивается на вторых фалангах третьего и четвертого пальцев.
162
На запястье пальмарно имеет сухожильное влагалище. Между кон-
цевыми ветвями сухожилия поверхностного сгибателя пальцев на
поверхность выходит сухожилие глубокого сгибателя пальцев. Дей-
ствие: сгибает путовый и венечный суставы.
Особенности. У свиней сухожилие поверхностного брюшка
тянется двумя ветвями до средней фаланги четвертого пальца, а
глубокое сухожилие — до второй фаланги третьего пальца. У ло-
шадей она тянется к связочным буграм венечной кости. У собак
сухожилие делится на четыре ветви.
Глубокий сгибатель пальцев (см. рис. 3.77) начинается тремя го-
ловками: плечевой — от сгибательного надмыщелка (тройная);
лучевой — от лучевой кости; локтевой — от локтевой кости. Сухо-
жилие, общее для трех головок, на пальмарной поверхности за-
пястья имеет синовиальное влагалище общее с поверхностным
сгибателем пальцев. Над суставами первых фаланг сухожилие рас-
щепляется на две ветви, которые прободают соответствующее су-
хожилие поверхностного сгибателя пальца и оканчиваются на паль-
марной поверхности копытцевых костей третьего и четвертого
пальцев. Здесь они имеют синовиальное влагалище и подблоко-
вую слизистую сумку. Действие: сгибает путовый, венечный и ко-
пытцевый суставы.
Особенности. У свиней и собак плечевая головка распада-
ется на два брюшка. У лошадей глубокий сгибатель пальцев имеет
три головки.
Короткие мышцы пальцев. Представлены следующими флексо-
рами, абдукторами и аддукторами.
Межкостная средняя мышца мясистая у молодых животных,
сухожильная у взрослых. Начинается от пальмарной поверхности
проксимального конца пястных костей. В дистальной трети пясти
делится на три главные ветви. Латеральная и медиальная ветви
идут к сесамовидным костям, а затем направляются на дорсаль-
ную поверхность к сухожилиям специальных пальцевых разгиба-
телей.
Действие: сгибает путовый сустав.
Особенности. У свиней третья и четвертая межкостные мыш-
цы начинаются от третьей и четвертой пястных костей.
Короткий сгибатель второго пальца простирается от медиаль-
ного края сухожилия глубокого сгибателя пальцев до проксималь-
ной фаланги второго пальца. Действие: сгибает суставы второго
пальца.
Особенности. Имеется только у свиней и собак.
Короткий сгибатель пятого пальца начинается от добавочной
кости запястья, оканчивается на проксимальной фаланге пятого
пальца.
Действие: сгибает сустав проксимальной фаланги.
Особенности. Имеется только у свиней и собак.
163
Отводящая мышца второго пальца начинается от третьей и вто-
рой пястных костей и оканчивается на проксимальной фаланге
второго пальца. Действие: отводит второй палец.
Особенности. Встречается только у свиней.
Отводящая мышца пятого пальца у свиней идет от третьей и
четвертой пястных костей к проксимальной фаланге пальца. Дей-
ствие: отводит пятый палец.
Особенности. У коров и лошадей эта мышца отсутствует.
У собак берет начало от добавочной кости запястья и оканчи-
вается на латеральной сесамовидной кости проксимальной фа-
ланги.
Приводящая мышца второго пальца начинается от проксималь-
ных концов третьей и четвертой пястных костей и оканчивается
на проксимальной фаланге второго пальца. Действие: приводит
второй палец.
Особенности. Развит только у свиней и собак.
Приводящая мышца пятого пальца берет начало от проксималь-
ного конца третьей и четвертой пястных костей и заканчивается
на проксимальной фаланге пятого пальца. Действие: приводит
пятый палец.
Особенности. Развита только у свиней и собак.
Короткий сгибатель первого пальца начинается на пальмарной
поверхности запястья и оканчивается на сесамовидных костях
проксимальной фаланги первого пальца. Действие: сгибает пер-
вый палец.
Особенности. Развит только у собак.
Отводящая мышца первого пальца простирается от пальмарной
поверхности запястья и сухожилия поверхностного сгибателя паль-
цев до медиальной поверхности проксимальной фаланги первого
пальца. Действие: отводит первый палец.
Особенности. Развита только у собак.
Приводящая мышца первого пальца идет от пальмарной повер-
хности запястья до латеральной поверхности проксимальной фа-
ланги первого пальца. Действие: приводит первый палец.
Особенности. Мышца развита только у собак.
Ладонная длинная мышца начинается от глубокого сгибателя
пальцев в нижней трети предплечья и оканчивается тонкими су-
хожилиями в ветвях поверхностного сгибателя третьего и четвер-
того пальцев. Действие: сгибает запястный и первый фаланговый
сустав третьего и четвертого пальцев.
Особенности. Развита только у собак.
Ладонная короткая мышца берет начало от сухожилия поверх-
ностного сгибателя пальца в области запястья и оканчивается на
первом фаланговом суставе пятого пальца. Действие: сгибает сус-
тавы первых фаланг четвертого и пятого пальцев.
Особенности. Имеется только у собак.
164
Фасции и мышцы тазовой конечности
Фасции тазовой конечности. Бывают поверхностными и глубо-
кими.
Поверхностная фасция лежит под кожей. Глубокая фасция полу-
чает специальные названия: ягодичная фасция — покрывает груп-
пу ягодичных мышц, широкая бедренная фасция — охватывает все
мышцы бедра и колена и переходит в фасцию голени. Последняя
является продолжением широкой фасции бедра, она хорошо вы-
ражена на голени и распространяется на плюсну.
Общее распределение мышц тазовой конечности по звеньям
тазовой конечности соответствует схеме, существующей на груд-
Рис. 3.79. Мышцы тазовой конечно-
сти лошади (латеральная поверх-
ность):
1 — средняя ягодичная мышца; 2 —
поверхностная ягодичная мышца; 3, 6,
7 — двуглавая мышца бедра; 4 — полу-
перепончатая мышца; 5 — полусухо-
жильная мышца; S — латеральная го-
ловка икроножной мышцы; 9 — пяточ-
ная мышца; 10 — длинный сгибатель
большого пальца; 11 — сухожилие трех-
главой мышцы голени; 12 — латераль-
ный разгибатель пальца стопы; 13 — пя-
точный бугор; 14, 18, 19 — сухожилие
поверхностного сгибателя пальца; 15,
20 — сухожилие глубокого сгибателя
пальца; 16 — четвертая плюсневая кость;
17 — третья межкостная мышца; 21 —
маклок; 22 — напрягатель широкой фас-
ции; 23 — прямая мышца бедра; 24 —
широкая латеральная мышца; 25 — фас-
ция голени; 26 — длинный разгибатель
пальца; 27 — третья малоберцовая мыш-
ца; 28 — передняя большеберцовая
мышца; 29 — проксимальный удержи-
ватель сухожилий; 30 — короткий раз-
гибатель пальца; 31 — дистальный удер-
живатель сухожилий; 32 — сухожилие
длинного разгибателя пальца; 33 — тре-
тья плюсневая кость; 34 — ветвь к длин-
ному разгибателю пальца
165
ной конечности. Однако важно заметить, что углы суставов тазо-
вой конечности (тазобедренный, коленный и скакательный), при-
способленные к толканию туловища вперед, направлены верши-
нами в противоположную сторону по сравнению с соответствую-
щими углами суставов грудной конечности, адаптированными к
восприятию толчков сзади.
Мышцы тазобедренного сустава по выполняемой функции де-
лятся на разгибатели, сгибатели, супинаторы и аддукторы.
Разгибатели тазобедренного сустава. Поверхностная ягодичная
мышца (рис. 3.79; см. рис. 3.67 — 3.69) у коров отсутствует, ее ме-
диальная часть вошла в состав двуглавой мышцы, а латеральная
— в состав напрягателя широкой фасции бедра. Действие: разги-
бает тазобедренный сустав.
Особенности. У свиней поверхностная ягодичная мышца
срастается, как у коров. У лошадей она начинается от ягодичной
фасции, маклока и крестцовой кости. У собак мышца медиальной
частью начинается от крестцовой кости и первых хвостовых по-
звонков.
Средняя ягодичная мышца (см. рис. 3.67 — 3.69, 3.79) начинает-
ся от ягодичной поверхности маклока и крестцового бугра, от
дорсальной крестцово-подвздошной связки и оканчивается на
большом вертеле. Действие: разгибает тазобедренный сустав.
Особенности. У свиней и собак средняя ягодичная мышца
развита, как у коров. У лошадей она очень массивная.
Глубокая ягодичная мышца (см. рис. 3.67 — 3.69, 3.79) начинает-
ся от седалищной ости и тянется до большого вертела. Действие:
разгибает тазобедренный сустав.
Особенности. У свиней и собак глубокая ягодичная мышца
начинается и оканчивается, как у коров. У лошадей она оканчива-
ется на среднем вертеле.
Заднебедренная группа разгибателей действует на тазобедрен-
ный, коленный и скакательный суставы. В эту группу входят четы-
ре мышцы: двуглавая мышца бедра, полусухожильная и полупе-
репончатая мышцы, квадратная мышца бедра.
Двуглавая мышца бедра (см. рис. 3.67 — 3.69, 3.79) имеет две
головки. Позвоночная головка слилась с ягодичной мышцей и на-
зывается ягодично-двуглавой мышцей. Она начинается от остис-
тых отростков крестца и от крестцово-седалищной связки. Седа-
лищная головка начинается от седалищной кости. Брюшко ягодич-
но-двуглавой мышцы передним краем сливается с напрягателем
широкой фасции бедра. Оканчивается мышца на боковой прямой
связке, на гребне большой берцовой кости, на пяточном бугре.
Действие: разгибает тазобедренный сустав, сгибает коленный и
разгибает скакательный суставы.
Особенности. У свиней ягодично-двуглавая мышца устрое-
на, как у коров. У лошадей двуглавая мышца бедра позвоночной
166
головкой не сливается с поверхностной ягодичной мышцей. У собак
она начинается одной головкой на крестцово-седалищной связ-
ке, а другой — на седалищном бугре.
Полусухожильная мышца (рис. 3.80; см. рис. 3.67 — 3.69, 3.79)
начинается от седалищной кости, оканчивается на медиальной
стороне гребня большой берцовой кости и пяточном бугре. Дей-
ствие: разгибает тазобедренный и скакательный суставы, сгибает
коленный сустав.
Особенности. У свиней и собак полусухожильная мышца,
как у коров. У лошадей она начинается от последних крестцовых
позвонков, крестцово-седалищной связки и седалищного бугра и
оканчивается, как у коров.
Полуперепончатая мышца (см. рис. 3.79, 3.80) начинается от
седалищной кости и тянется до медиальных мыщелков бедренной
Рис. 3.80. Мышцы тазовой конеч-
ности коровы (медиальная по-
верхность):
I — малая поясничная мышца; 2 —
медиальная подвздошная мышца;
3 — средняя ягодичная мышца; 4 —
поверхностная ягодичная мышца;
5 — хвостовая мышца; 6 — подни-
матель хвоста; 7 — внутренняя за-
пирательная мышца; 8 — полупере-
пончатая мышца; 9, 26 — стройная
мышца; 10 - полусухожильная мыш-
ца; 11 — икроножная мышца, ме-
диальная головка; 12 — икронож-
ная мышца, латеральная головка;
13 — длинный сгибатель пальцев
стопы; 14 — длинный сгибатель
большого пальца стопы; 15 — сухо-
жилие поверхностного сгибателя
пальцев; 16 — сухожилие глубокого
сгибателя пальцев; 17 — средняя
межкостная мышца; 18 — ветвь к
длинному разгибателю пальцев; 19 —
большая поясничная мышца; 20 —
паховая связка; 21 — напрягатель
широкой фасции; 22 — прямая
мышца бедра; 23 — портняжная
мышца; 24 — широкая медиальная
мышца; 25 — прямая связка надко-
ленника; 27 — третья малоберцовая
мышца; 28— проксимальный удер-
живатель сухожилий; 29 — дисталь-
ный удерживатель сухожилий; 30 —
короткий разгибатель пальцев; 31 —
сухожилие длинного разгибателя
пальцев
167
и большеберцовой костей. Действие: разгибает тазобедренный и
сгибает коленный суставы.
Особенности. У свиней и собак полуперепончатая мышца
располагается в основном, как у коров. У лошадей она начинается
от крестцово-седалищной связки и от вентральной поверхности
седалищного бугра.
Квадратная мышца бедра берет начало от вентральной повер-
хности седалищной кости и оканчивается на каудальной поверх-
ности бедренной кости. Действие: разгибает тазобедренный сус-
тав.
Особенности. У свиней, лошадей и собак эта мышца в ос-
новном сходна с таковой у коров.
Сгибатели тазобедренного сустава. Напрягатель широкой фас-
ции бедра (см. рис. 3.67 — 3.69, 3.80) простирается от маклока до
широкой бедренной фасции. Действие: сгибает тазобедренный су-
став и натягивает широкую бедренную фасцию.
Особенности. У свиней напрягатель широкой фасции бедра
располагается, как у коров. У лошадей задний край этой мышцы
тесно срастается с передней частью поверхностной ягодичной
мышцы. У собак эта мышца имеет две части.
Портняжная мышца (см. рис. 3.80) начинается от сухожилия
малой поясничной мышцы и от столбиковой части подвздошной
кости, оканчивается на коленной чашке. Действие: сгибает тазо-
бедренный сустав и приводит конечность в действие.
Особенности. У свиней и лошадей эта мышца начинается и
оканчивается, как у коров. У собак она двойная.
Приводящие мышцы тазобедренного сустава. Стройная мышца
(см. рис. 3.80) начинается от тазового сращения и тянется до ме-
диальной боковой связки бедро-берцового сустава. Действие: при-
водит конечность в действие.
Особенности. У свиней, лошадей и собак эта мышца такая
же, как у коров.
Приводящая мышца (см. рис. 3.80) берет начало от вентральной
поверхности седалищной кости и оканчивается на медиальной и
плантарной поверхности бедренной кости. Действие: приводит
конечность в действие.
Особенности. У свиней, лошадей и собак эта мышца рас-
полагается так же, как у коров.
Вращатели тазобедренного сустава. Наружная запирающая мыш-
ца (см. рис. 3.80) начинается от запертого отверстия и тянется су-
хожилием до вертлужной ямки. Действие: супинирует (вращает
наружу) тазобедренный сустав.
Особенности. У свиней, лошадей и собак мышца сходна с
таковой у коров.
Внутренняя запирающая мышца (см. рис. 3.80) идет от внутрен-
ней поверхности подвздошной кости, проходит через малую се-
168
далищную вырезку и оканчивается в вертлужной ямке. Действие:
супинирует тазобедренный сустав.
Особенности. У свиней, лошадей и собак имеет такое же
расположение, как у коров.
Двойничные мышцы (см. рис. 3.80) отходят от малой седалищ-
ной вырезки и тянутся до вертлужной ямки. Лежат между обеими
запирающими мышцами. Действие: супинируют тазобедренный су-
став.
Особенности. У свиней, лошадей и собак начинаются и
оканчиваются, как у коров.
Мышцы коленного сустава по выполняемой функции подраз-
деляются на сгибатели и разгибатели.
Разгибатели и сгибатели коленного сустава. Четырехглавая
мышца бедра (см. рис. 3.79, 3.80) имеет прямую, латеральную,
медиальную и промежуточную головки. Прямая головка начинает-
ся выше суставной впадины таза; латеральная головка — от лате-
ральной поверхности бедренной кости; медиальная головка — от
медиальной поверхности бедренной кости; промежуточная голов-
ка — от дорсальной поверхности бедренной кости. Все головки,
сливаясь между собой, оканчиваются на коленной чашке. Дей-
ствие: разгибает коленный сустав.
Особенности. У свиней, лошадей и собак мышца устроена,
как у коров.
Подколенная мышца прикрыта икроножной мышцей. Начина-
ется она сухожильно от латерального мыщелка бедренной кости,
постепенно расширяясь, направляется косо внутрь и оканчивает-
ся на плантарной шероховатой поверхности большеберцовой ко-
сти.
Действие: сгибает коленный сустав.
Особенности. У свиней, лошадей и собак эта мышца уст-
роена, как у коров.
Мышцы скакательного сустава подразделяются на разгибатели
и сгибатели.
Разгибатели скакательного сустава. Трехглавая мышца голени
(см. рис. 3.67 — 3.69, 3.79 — 3.81) состоит из двух мышц: икронож-
ной и пяточной.
Икроножная мышца начинается двумя головками по бокам над-
блоковой ямки бедренной кости. На половине длины большебер-
цовой кости мышца переходит в крепкое пяточное (ахиллово) су-
хожилие и прикрепляется к пяточному бугру.
Пяточная мышца берет начало от латерального мыщелка боль-
шеберцовой кости и тянется до пяточного бугра. Действие: разги-
бает скакательный сустав.
Особенности. У свиней, как у коров, имеются две хорошо
развитые мышцы. У лошадей пяточная мышца слабо развита. У собак
трехглавая мышца голени имеет только две головки.
169
Сгибатели скакательного сустава. Большеберцовая передняя
мышца (см. рис. 3.67 — 3.69, 3.79 — 3.81) берет начало от прокси-
мального конца большеберцовой кости и направляется до про-
ксимального конца плюсневых костей. Действие: сгибает скака-
тельный сустав.
Особенности. У свиней мышца тянется от гребня больше-
берцовой кости до второй плюсневой кости. У собак большебер-
цовая передняя мышца начинается на латеральном мыщелке и
гребне большеберцовой кости.
Малоберцовая третья мышца (см. рис. 3.67 — 3.69, 3.79 — 3.81)
начинается от разгибательной ямки бедренной кости, оканчива-
ется на проксимальном конце плюсневых костей. Действие: сгиба-
ет скакательный сустав.
Особенности. У свиней эта мышца располагается, как у ко-
ров. У лошадей она превратилась в крепкий сухожильный тяж.
У собак она отсутствует.
Большеберцовая задняя мышца (см. рис. 3.80) у коров сливается
с глубоким пальцевым сгибателем. Действие: сгибает скакатель-
ный сустав.
Особенности. У свиней и лошадей большеберцовая задняя
мышца устроена, как у коров. У собак она прикрыта брюшком
длинного пальцевого сгибателя и начинается на верхнем конце
малоберцовой кости. Оканчивается мышца на медиальной боко-
вой связке скакательного сустава.
Малоберцовая длинная мышца (см. рис. 3.67 — 3.69, 3.79 — 3.81)
начинается от латерального мыщелка большеберцовой кости и
оканчивается на первой предплюсневой кости. Действие: сгибает
скакательный сустав.
Особенности. У свиней эта мышца располагается, как у ко-
ров. У лошадей она отсутствует. У собак мышца начинается от ла-
теральной связки коленного сустава и малоберцовой кости.
Малоберцовая короткая мышца (см. рис. 3.67 — 3.69, 3.79 — 3.81)
имеется не у всех животных. Действие: сгибает скакательный сустав.
Особенности. У свиней и лошадей эта мышца, как и у
коров, отсутствует. У собак она начинается от нижней половины
малоберцовой кости, оканчивается на верхнем конце плюсне-
вой кости.
Мышцы пальцев по выполняемой функции подразделяются на
сгибатели и разгибатели.
Разгибатели пальцев стопы. Длинный разгибатель пальцев (рис.
3.81; см. рис. 3.67 — 3.69, 3.79 — 380) начинается от разгибательной
ямки бедренной кости, в верхней трети голени превращается в
два сухожилия, которые в области скакательного сустава имеют
синовиальные влагалища. Оканчивается мышца на разгибатель-
ных отростках копытцевых костей третьего четвертого пальцев.
Действие: разгибает суставы пальцев.
170
Рис. 3.81. Сравнительная анатомия мышц стопы собаки (А), свиньи (Б),
коровы (5), лошади (Г):
1 — передняя большеберцовая мышца; 2 — третья малоберцовая мышца; 3 —
длинная малоберцовая мышца; 4 — длинный сгибатель большого пальца; 5 —
длинный разгибатель пальцев; 6 — поверхностный сгибатель пальцев; 7— икро-
ножная мышца; 8 — пяточная мышца; 9 — боковой разгибатель пальцев; 10 —
короткая малоберцовая мышца; 11 — короткий разгибатель пальцев; 12 — меж-
костные мышцы; 13 — специальный разгибатель четвертого пальца; 14 — специ-
альный разгибатель пятого пальца; 15 — сухожилия длинного разгибателя паль-
цев; 16 — сухожильная ветвь к длинному разгибателю пальцев; 17 — короткий
разгибатель пальцев; 18 — средняя межкостная мышца; 19 — сухожильная ветвь
к длинному разгибателю пальцев; 20 — сухожилие третьей малоберцовой мыш-
цы; 21 — короткий разгибатель пальца; 22 — сухожилие длинного разгибателя
пальца; 23 — третья межкостная мышца; 24 — сухожильная ветвь к длинному
разгибателю пальца; 25 — пяточное (ахиллово) сухожилие; 26 — проксималь-
ный удерживатель сухожилий; 27 — дистальный удерживатель сухожилий
Особенности. У лошадей сухожилие в области плюсны сли-
вается с коротким разгибателем и сухожилием бокового разгиба-
теля пальцев, идет по дорсальной поверхности пальца и оканчи-
вается на разгибательном отростке копытной кости. У собак мыш-
ца оканчивается на дистальной фаланге всех пальцев.
171
Боковой разгибатель пальцев (см. рис. 3.67 — 3.69, 3.79 — 3.81)
начинается на латеральной связке коленного сустава и оканчива-
ется на второй фаланге четвертого пальца. Действие: разгибает
суставы проксимальной и средней фаланг.
Особенности. У лошадей сухожилие этой мышцы на дор-
сальной поверхности плюсны сливается с сухожилием длинного
разгибателя пальца. У собак боковой разгибатель пальца начина-
ется под головкой малоберцовой кости и на пятой плюсневой кости
сливается с ветвью сухожилия длинного пальцевого разгибателя,
идущей к пятому пальцу.
Короткий разгибатель пальцев (см. рис. 3.67 — 3.69, 3.79 — 3.81)
начинается на дорсальной поверхности предплюсны и оканчива-
ется в сухожилии длинного разгибателя пальцев на уровне верх-
ней трети плюсны. Действие: разгибает суставы пальцев.
Особенности. У свиней эта мышца начинается на заплюсне
и образует четыре брюшка. У лошадей короткий разгибатель паль-
ца начинается от дорсальной связки скакательного сустава и окан-
чивается в сухожилии длинного разгибателя пальца. У собак он
начинается на верхних концах плюсневых костей.
Сгибатели пальцев стопы. Поверхностный сгибатель пальцев (см.
рис. 3.67 — 3.69, 3.79 — 3.81) начинается в надблоковой ямке бед-
ренной кости, перекручивается с сухожилием икроножной мыш-
цы так, что из глубины выходит на поверхность и проходит по
вершине пяточного бугра. Здесь сухожилие расширяется, закреп-
ляется на обеих сторонах пяточного бугра короткими крепкими
пучками, а затем следует вниз по плантарной поверхности стопы.
Над пяточным бугром под сухожилием имеется слизистая сумка.
Затем в области плюсны сухожилие разделяется на две ветви. Каж-
дая ветвь разделяется на две части, между которыми проходит
сухожилие глубокого сгибателя пальцев, и оканчивается на верх-
нем конце средней фаланги третьего и четвертого пальцев. На се-
самовидных костях имеется синовиальное влагалище. Действие:
сгибает суставы проксимальной и средней фаланг третьего и чет-
вертого пальцев.
Особенности. У свиней поверхностный сгибатель пальцев
занимает такое же положение, как у коров. У лошадей его брюш-
ко содержит мало мышечной и много фиброзной ткани. У собак
он оканчивается на вторых фалангах всех пальцев.
Глубокий сгибатель пальцев (см. рис. 3.67 — 3.69, 3.79—3.81) име-
ет три головки: латеральную поверхностную, латеральную глубо-
кую и медиальную. Все головки сливаются в одно сухожилие, ко-
торое проходит медиальнее пяточного отростка. Здесь оно имеет
синовиальное влагалище. Сухожилие расщепляется над прокси-
мальной фалангой на две ветви для третьего и четвертого пальцев.
Каждая ветвь выходит из глубины на поверхность и оканчивается
на дистальной фаланге третьего и четвертого пальцев. Здесь под
172
сухожилиями имеются подблоковые слизистые сумки. Действие: сги-
бает все суставы пальцев.
Особенности. У лошадей и свиней глубокий сгибатель паль-
цев имеет три головки. У собак мышца имеет только две головки.
Межкостные мышцы начинаются в области заплюсны и име-
ют глубокую и поверхностную пластинки. Действие: сгибают сус-
тав проксимальные фаланги и фиксируют сесамовидные кости.
Особенности. У свиней, лошадей и собак они устроены,
как на грудной конечности.
Квадратная мышца подошвы характерна лишь для собак. Дей-
ствие: сгибает сустав проксимальной фаланги и напрягает сухо-
жилие глубокого сгибателя пальца.
Особенности. У коров, свиней и лошадей эта мышца от-
сутствует. У собак она начинается от дистального конца пяточной
кости, от латеральной боковой связки скакательного сустава и
оканчивается на пятой плюсневой кости.
Контрольные вопросы
1. Какие системы объединяет локомоторный аппарат?
2. Какие кости входят в состав осевого скелета?
3. Какие кости входят в состав периферического скелета?
4. Перечислите суставы грудной и тазовой конечности.
5. На какие морфофункциональные группы делятся мышцы головы?
6. Какие морфофункциональные группы мышц характерны для суста-
вов грудной и тазовой конечностей животных?
КОЖНЫЙ ПОКРОВ
Общий покров (кожа) [integumentum commune (cutis)] — это на-
ружная оболочка тела животного, покрытая волосами и непос-
редственно соприкасающаяся с внешней средой (рис. 3.82; см. рис.
3.83 — 3.88). Она состоит из эпидермиса, основы кожи и подкож-
ного слоя.
Эпидермис (рис. 3.82) образован многослойным плоским эпи-
телием .ДДТосновы кожи он отделяется основной мембраной. На ней
располагается производящий слой эпидермиса. Клетки этого слоя,
размножаясь, перемешаются на поверхность, стареют, высыха-
ют, становятся более плоскими, постепенно подвергаются орого-
вению и в конечном итоге превращаются в чешуйчатые пластин-
ки, предохраняющие кожу от механических повреждений и вы-
сыхания. В процессе отпадения чешуек кожа очищается от грязи,
паразитов, микробов и различных посторонних тел. В эпидермисе
залегают пигментные клетки, кровеносных сосудов нет, питание
эпидермиса происходит осмотическим путем.
173
Основа кожи (рис. 3.82) состоит из поверхностного сосочково-
го и глубокого сетчатого слоев. Она построена из плотной нео-
формленной соединительной ткани. В ней располагаются сальные
и потовые железы, артерии, вены, лимфатические сосуды, ка-
пилляры, нервные волокна, концевые чувствительные аппараты —
рецепторы, корни волос и гладкие мышечные пучки. Толщина
основы кожи определяет толщину всей кожи. Самая толстая осно-
ва кожи у коров, самая тонкая — у овец и коз. У коров толщина
основы кожи достигает 3 — 7 мм (и даже 13 мм), у овец и коз —
0,5 — 3 мм. У старых животных основа кожи толще, чем у молодых.
У самцов кожа толще, чем у самок. На спине основа кожи толще,
чем на животе. На латеральной поверхности конечностей кожа
толще, чем на медиальной.
Подкожный слой состоит из рыхлой соединительной ткани и
прикрепляет основу кожи к фасциям и мышцам. Иногда в нем
Рис. 3.82. Кожа:
7, 8, 10 — сальная железа; 2 — сеть лимфатических капилляров; 3 — луковица
волоса; 4 — корень волоса; 5 — стержень волоса; 6 — сеть кровеносных капилля-
ров сосочкового слоя основы кожи; 7 — новый (сменяющийся) волос; 9 — ста-
рый (сменяемый) волос; 11 — наружное корневое влагалище; 12 — внутреннее
корневое влагалище; 13 — кутикула волоса; 14 — мозговой слой волоса; 15 —
корковый слой волоса; 16 — эпидермис; 17 — сосочковый; 18 — сетчатый слой
основы кожи (дермы); 19 — лимфатический сосуд; 20 — артерия; 21 — вена; 22 —
нерв; 23 — подкожный слой; 24 — мышца-подниматель волоса; 25, 28 — кровя-
ной синус; 26 — синусовый волос; 27 — волосяной сосочек; 29 — подкожный
слой; 30, 31 — потовая железа; 32 — волосяная сумка
174
против бугров или отростков образуются слизистые сумки (на-
пример, над локтевым бугром и над маклоком).
В подкожном слое имеются различные соединительнотканные
клетки, в том числе и жировые. Последние у хорошо упитанных
животных формируют подкожный жировой слой. Подкожный жир
служит запасом питательных веществ и предохраняет животных
от переохлаждения.
Особенности. У коров кожа под грудиной образует боль-
шую складку — подгрудок. У свиней кожа достигает наибольшей
толщины на вентральной поверхности шеи и в области груди. Здесь
она у самцов образует так называемый щит. Жир откладывается в
подкожном слое в течение всего года.
Производные кожного покрова. Волос имеет вид плотной, но
гибкой и эластичной нити. Наружный слой волоса составляет кути-
кула. Под ней находится толстый корковый слой. Последний содер-
жит высокие клетки с пигментом, который и определяет цвет во-
лос. В центре волоса располагается сердцевина из округлых, много-
угольных и кубических клеток.
Волосы в различных участках кожи имеют неодинаковую дли-
ну и толщину. По строению они подразделяются на три вида:
покровные, длинные и синуозные. У лошадей покровные волосы
короткие, синуозные — хорошо развиты и растут на веках, гу-
бач. щеках, подбородке. Длинные волосы формируютзшд^.лдр-
стовые волосы, щетки (позади их юных сч с танов), чему (В обла^
ни лба). '
Кроме волос кпроишодным кожного покрова оi нося (см же-
лезы_кожи, молочные же.теия (вымя), мякиши, копыт. копыт-
ца, когти и рога.
Молочные железы (рис. 3.83, 3.84) имеют сложное трубчато-
альвеолярное строение. У свиней и собак молочные железы пред-
ставляют собой отдельные доли, которые все вместе образуют
множественное вымя. У коров и кобыл оно образовалось от слия-
ния двух пар долей в одно компактное вымя, располагающееся на
брюшной стенке между бедрами.
Молрч назыжелеза состоит из тела и сосков. Каждая доля молоч-
ной жей&Зыимёет паренхиму'тгбстов. Паренхима железы состоит
из долек. В образовании их участвуют молочные альвеолы, молочные
трубочки, молочные каналы и молочные протоки. Последние откры-
ваются в молочную цистерну. В молочных альвеолах и молочных
трубочках секретируется молоко, а во время усиленной лактации
оно может секретироваться и в молочных каналах. Обычно молоч-
ные каналы и молочные ходы служат для выведения молока в
молочную цистерну.
От молочной цистерны к верхушке соска тянется сосковый ка-
нал, который открывается на конце соска отверстием соскового
канала (см. рис. 3.84). В основе выходного отверстия соскового ка-
175
Рис. 3.83. Вымя коровы (поперечный
разрез):
7.7— наружная косая мышца живота; 2 —
Внутренняя косая мышца живота; 3 —
лимфатический коллектор; 4 — наруж-
ная срамная цена; 5 наружная срам-
ная артерия; г, прямаздышца живо-
та; V/—белая шипи. V.брюшина;
Iff латеральная iubujiiiikj полк-рдн-
ваюшего аппарата: II средняя пддр-
гинка поддерживающего аппарата: •'12
сосковый канал; 133— молочная нПс-
Рис. 3.84. Молочная цистерна
_ (продольный разрез):
/. 2, 3 — молочные пазухи малом
'ной цистерны: 4~ отверстие молоч-
ного хала; 5 — сосковый проток; б/ -
отверстие соскового протока. 7—-ве-
нозное сплетение стенки соска;(Я—
венозное сплетение основания соска
терна
нала лежит сфинктер соска. Снаружи сосок покрыт безволосой
кожей. Между кожей соска и слизистой оболочкой соскового ка-
нала залегают эластическая и мышечная ткани.
Остов железы состоит из соединительной ткани с жировыми
прослойками. От крупных перегородок отходят более мелкие пуч-
ки соединительной ткани. Окутывая молочные альвеолы, молоч-
ные трубочки и молочные каналы, они поддерживают их. По со-
единительнотканным прослойкам проходят артерии, вены, ка-
пилляры, лимфатические сосуды и нервы.
Особенности. У коровы молочная железа, или вымя, хоро-
шо развита и сое гои 1 из двух пар долей, имеет полушаровидную
или чашеобразную форму (см. рис. 3.83, 3.84). Правая половина
вымени отделяется от левой срединной бороздой. На каждой по-
176
ловине вымени имеется два соска. Соски цилиндрической формы,
в длину достигают 7—10 см и содержат по одному выходному
отверстию. Передняя и задняя доли вымени сращены друг с дру-
гом. В паренхиме каждой половины вымени имеется две системы
молочных трубок с самостоятельным выходом каждой системы на
своем соске. На уровне основания соска находится круговая складка.
Она разделяет молочную цистерну на верхнюю железистую часть
и нижнюю сосковую. Вымя покрыто тонкой кожей с редкими во-
лосками, а у беспородных маломолочных коров оно сильно об-
росшее волосами. Кожа на задней поверхности вымени образует
две складки, которые тянутся от половой щели вниз и в стороны
к боковым поверхностям вымени. Между ними располагается мо-
лочное зеркало. Под кожей находится поверхностная фасция выме-
ни, под ней простирается глубокая фасция вымени. Она отходит от
желтой фасции живота, на середине вымени отдает два эласти-
ческих листка, идущих от белой линии живота к основанию вы-
мени. Эти листки образуют перегородку вымени, разделяющую его
на правую и левую половины, и в то же время являются подвеши-
вающей связкой вымени.
У свины (множествен ное вымя расположено по вентральной стен-
ке гела от грудины до лонных костей и состоит из пяти восьми
парных отдельных молочных холмов с короткими сосками. На каж-
дом соске открывается два-три сосковых канала. Молочные цистер-
ны в виде маленьких расширений находятся в основании сосков.
Рис. 3.85. Копытная стенка:
/ — основа кожи копытной каймы;/2 — основа кожи копытного венчика; За-
роет трубчатого рога копытной стенки; 4 —/трубчатый рог копытной стенки; (5 —
листочки основы кожи копытной стенки; 'б/— наружный слой копытной стенки
177
Рис. 3.86. Копыто (подошвенная
поверхность):
1 — подошвенный край роговой
стенки (зацепной участок); 2 — бе-
лая линия; 3 — роговая подошва;
4 — заворотная часть роговой стен-
ки; 5 — пяточный угол; 6 — рого-
вой слой подушки мякиша; 7 —
ножка стрелки; 8— роговая стрел-
ка мякиша
У лошади вымя состоит из двух __
половин. В каждой половине находит-
ся по две доли с двумя сосковыми_
каналами на одном соске. Сфинкте-
ры сосковых каналов хорошо разви-
ты. Передняя и задняя доли в каж-
дой половине вымени срослись. Внут-
ри половины вымени молочные си-
стемы трубок не сообщаются между
собой. Молочные цистерны хорошо
развиты в каждой доле вымени. Вымя
укрепляется хорошо выраженной
поддерживающей связкой. Кожа вы-
мени нежная с тонкими волосками.
У собак множественное вымя расг__
полагается вдоль белой линии живо-
та и состоит из 4 — 6 парныхмолоч-'
ных холмов. Кожа сосков содержит
нежные волоски. На сосках открыва-
ются от 6 до 20 молочных холоп. Мо-
лочные цистерны отсутствуют.’
Мякиши — это подушковидное
утолщение кожи задней поверхности
лап. Мякиши состоят из трех слоев:
эпидермиса, основы кожи и подкож-
ного слоя, У лошадей пальцевой .мя-
киш глубоко вклинивается в рогйную__
подошву и выполняет функцию пру-
жинящего аппарата копыта (рис. 3.85,
3.86). Он состоит из подушки, стрелки и мякишных хрящей.
Запястный,.щмсщдщстречается у собак и лошадей. У последних
он расположен в области дистального конца предплечья с меди-
альной стороны в виде каштана. У собак он находится в области
добавочной кости запястья.
Зап иосневый мякиш ра твит только у лошадей на каудомедиаль-
ной поверхности заплюсны (каштан).
Копыто (ungula) лошади состоит из роговой капсулы, основы
кожи и подкожного слоя (рис. 3.85, 3.86).
Роговая капсудн.^оцыта^, или роговой башмак, является произ-
водным эпидермиса. Она поставлена вертикально, укорочена, силь-
но расширена книзу и опирается не только подошвой, но и стен-
кой. На роговой капсуле копыта находятся следующие четыре ча-
стиц копытная кайма, копытный венчик, копытная стенка и ко-
пытная подошва.
Подкожный слой копыта хорошо развит только в области ко-
пытной каймы и копытного венчика.
178
Рис. 3.87. Копыт-
це:
/ — основа кожи
копытцевого вен-
чика; 2 — листоч-
ки основы кожи
копытной стенки;
3 — венчиковый
желоб; 4 — боко-
вая стенка копыт-
ца; 5 — роговые
листочки
Рис. 3.88. Рог коровы:
7, 6 — роговая капсула; 2 — роговой отросток лобной ко-
сти; 3 — периост; 4 — основа кожи рога; 5 — сосочки
основы кожи и трубочки рога; 7 — лобная пазуха
Копытце {ungula) коров и свиней состоит из копытцевой кай-
мы, копытцевого венчика, копытцевой стенки и копытцевой по-
дошвы (рис. 3.87).
Коготь (unguiculus'l состоит из трех частей: когтевого валика,
когтевой стенки и когтевой подошвы.
Рога (сотиа\ состоят из эпидермиса и основы кожи (рис. 3.88).
Эпидермис образует роговую капсулу. Основа кожи рога плотно
срастается с надкостницей рогового отростка лобной кости, она
очень богата кровеносными сосудами. На роге различают корень,
тело и верхушку.
Корень рога — самая тонкая его часть, которая находится на
месте перехода' рога в кожу. Тело рога продолжается от корня до
верхушки и является самой обширной и массивной частью. Вер-
хушка рога^- заостренный свободный конец рога.
На рогах часто встречаются поперечные кольца, которые зави-
сят от различной скорости роста роговой капсулы. У коров на по-
перечном разрезе рога по своей форме приближаются к кругу.
Контрольные вопросы
1. Перечислите слои и производные кожи.
2. Укажите части копыта и копытца.
179
3. Перечислите части молочной железы. Вспомните особенности стро-
ения молочной железы у разных видов сельскохозяйственных животных.
4. Из чего состоит паренхима молочной железы?
СПЛАНХНОЛОГИЯ
Спланхнология (splanchnologia) — раздел анатомии, изучаю-
щий внутренности (splanchne, s. viscera) (см. рис. 3.89 — 3.134). Внут-
ренности осуществляют жизненно важные функции организма, в
связи с чем подразделяются на системы: пищеварительную и ды-
хательную. объединенные н науку Himepaioeuio. Органы мочевы-
деления и половой аппарат изучает нефрогонология.
Пищеварительная система, или аппарат пищеварения, состоит
из четырех отделов: головной, передней, средней и задней ки-
шок, каждая из которых в свою очередь состоит из подотделов.
Головную кишку подразделяют на ротовую полость и глотку; в
состав передней кишки входят пищевод и желудок; средняя киш-
ка включает в себя двенадцатиперстную, тощую и подвздошную
кишки; задняя кишка состоит из слепой, ободочной и прямой
кишок. Последняя оканчивается заднепроходным отверстием —
анусом (см. рис. 3.89 — 3.109).
Аппарат дьрцания включает носовую полость, глотку, гортань,
трахею и легкие. Последние являются основным органом, в кото-
ром происходит газообмен между внешней средой и кровью (см.
рис. 3.110 — 3.115).
Мочеполовой аппарат включает в себя органы мочеотделения,
состоящие из почек, мочеточников, мочевого пузыря и мочеис-
пускательного канала, и органы размножения. Органы размножения
самцов состоят из семенников с их придатками, семенникового
мешка, семяпровода, придаточных половых желез, мочеполового
канала и полового члена с препуцием. Органы размножения самок,
включают яичники, маточные трубы, матку, влагалище, преддве-
рие влагалища и наружные половые органы (см. рис. 3.116 — 3.134).
Внутренние органы располагаются определенным образом и в
определенной последовательности в естественных полостях тела.
I рудная и брюшная полос i n выстланы серозной оболочкой и по-
лому называются серозными полостями. Кроме того, в организме
имею гея серозная околосердечная и тазовая полости. Последняя не
выстлана серозной оболочкой.
Грудная полость возникает у животных в результате разделения
диафрагмой в начале индивидуального развития единой полости
тела. Она занимает часть грудной клетки, образованной грудными
позвонками, ребрами и грудной костью.
Грудная полость изнутри выстлана врутригрудной фасцией_н_
тонкой серозной оболочкой — плеврой. Последнюю'Принято де-
лить на два листка — париетальный и висцеральный. Первый из них
180
покрывает стенки грудной полости, в связи с чем делится на ре-
ерную и диафрагмальную плевру. Висцеральный листок формирует
срединную перегородку грудной полости — средостение. Между
листками средостения лежат аорта, трахея, с проходящими по
ней нервами, пищевод, полые вены, лимфатические сосуды и
узлы, сердце с околосердечной сумкой. Часть средостенной плев-
ры, покрывающей околосердечную сумку, называется перикарди-
альной плеврой. Средостенные листки переходят на бронхи и лег-
кое, покрывают их и называются ^легочной плеврой. Между парие-
тальным и висцеральным листками плевры заключена шслсвид-
пая парная плевральная полость, содержащая небольшое количё-
ство серозной жидкости. Последняя значительно уменьшает тре-
ние между серозными листками при дыхании. Париетальная плев-
ра в отличие от висцеральной содержит большое количество бо-
левых рецепторов, чем объясняется возникновение болей при
плевр игах. (см. рис. 3.114).
Брюшная полость достигает значительных размеров, так как в
ней располагается основная часть .органов пищеварения, некото-
рые органы мочеотделения и размножения. Дорсально она огра-
ничена крышей, сформированной последними iрудными, пояс-
ничными и первыми крестцовыми позвонками. Латерально и вен-
трально стенку брюшной полости формируют мышцы живот, а
краниально Гранину се определяе, диафрагма. В норме брюшная
полость у животных имеет яйцевидную форму: тупой конец се
направлен кранновентрально и ограничен куполом диафрагмы, а
несколько суженный заостренный конец - каудодорсально и пе-
реходи г в газовую полость. Однако ее форма и объем могут значи-
тельно изменяться в зависимости от наполнения органов пищева-
рения, а также от периода развития плодов у беременных самок.
Внутренняя поверхность брюшной полости выстлана попереч-
нойурасцией. С которой прочно срастается серозная оболочкабрюш-
||| полости, получившая здесь нагвание_й/до<ммд<^__Как и сероз-
ной оболочкой грудной полости, она дели гея на ДМ hicikb. Пари-
етальный листок выстилает стенку брюшной полости и каудаль-
ную поверхность диафрагмы. Поднимаясь дорсально к позвоноч-
ному столбу, правый и левый листки объединяются п^цгереходя
на внутренние органы, образуют брыжейку, Между ее удвоенным
листком располагаются внутренние органы брюшной полости, в
связи с шум он получает название висцеральный. Последний обра-
зует наружную серозную оболочку на органах брюшной полости.
Между париетальным и висцеральным листками серозной оболоч-
ки образуется щелевидное пространство^- перитонеальная полость,
гаполненная ис|х>зно>1.жЦДкостыо£см. рис. 3.98).
Серозная оболочка брюшной полости, переходя от одного орга-
на на другой, формирует связки. Между печенью и малой кривиз-
ной желудка серозная оболочка образует малый еалытк. Серозная
181
оболочка желудка, опускаясь с его большой кривизны, формиру-
ет удвоенный листок. Последний по вентральной брюшной стенке
направляется каудально и у входа в тазовую полость заворачива-
ется дорсально, меняя направление на краниальное, и достигает
желудка. Таким образом, формируется большой сальник и сальни-
ковый мешок.
Тазовая полость в большинстве своем не выстлана серозной
оболочкой. Все щелевидные пространства между органами выпол-
нены рыхлой соединительной тканью и представляют собой тазо-
вое забрюшинное пространство. Стенки ее образуют кости тазово-
го пояса, крестцово-седалищные связки и мышцы. Вход в тазовую
полость ограничен лонными и подвздошными костями, а выход
вентрально ограничен седалищной .'ivioii. .кпершн.но крестцо-
во-бу|ровыми связками и дорсально — крестцовой коегьк» и пер-
выми хвостовыми позвонками.
ПИЩЕВАРИТЕЛЬНЫЙ АППАРАТ
Пищеварительный аппарат (apparatusdigestorius) представляет
комплекс внутренних органов, обесценивающих захватывание,
механическую обработку, продвижение, переваривание и всасы-
вание пищи и воды.
Органы пищеварения расположены в основном в брюшной
полости, частично — в грудной и тазовой полостях, а также в
области шеи и головы. Схематично их можно представить в виде
сквозной трубки, простирающейся через весь организм от пере-
днего конца тела к заднему (см. рис. 3.89). :
Рис 3.89. Схема расположения органон пишспарсния собаки:
/ ротовая подоегь. 2— Гхьчьшие слюнные желсия. J глотка; •/ пищевод;
'.V - желудок; б печень; 7 лвена.пытнперешая кишка; 8: — поджелудочная
Жслета. Ю сотая кишка, переходящая в подвздошную кишку: 10 — слепня
кишка: 11. 12. !.< — ободочная кишка. 14 прямая кишка; 15 пнус
182
Ротовая полость
Ротовая полость (сауит oris) делится на преддверие рта и соб-
ственно ротовую полость (рис. 3.90) Преддверие рта представляет
собой шелевилное пространство между губами и щеками.с одной
сюроны, зубами и деснами друг о it Собственно ротовая по-
меть ограничена спереди центовыми зубами, сзади — мягким
нёбом, с боков корегщымн зубами, сверху — твердым нёбом и
снизу — дном собственно роговой полости. Вход в ротовую по-
лость — ротовую щель — находится между верхней и нижней губа-
ми. а между краем мягкого нёба и корнем языка располагается
' ' -О.» '
Рис. 3.90. Ротовая полость лошади:
/ — круговая мышца губ: 2у верхняя губа; .? верхняя резцовая мышца. 4.
сухожилие специального подниматели верхней губы: 3 репювый сосочек; (у—
нёбное сплетение, 7 — носовая кость; .V хрящ перегородки носа; 9 — венозное
.плетение слизистой оболочки перегородки носа; Ю твердое нёбо; II. 13
головной moj[; /4 лобная пазуха; 14— сигиттальный синус; /.у. мёжтемен-
ная кость; 16 - канатик вышюп связки: /? - мозолистое тело; /Л> — средний
мозг, /9— Мозжечок; .2^/— продолговатый мам. 21 клиновидная пазуха; &)—
прямая дорсальная большая мышца головы; 2.£> дорсальная дуга атланта; 24>.—
спинкой маа^.25— вснпуцьная дуга anairraTzd здщнтан мышца галопы; f}
пищевод; 28 - трахея. 29'- трахеальные хрящи; 1?02— груди но-поип>язычная
мып1п.т;^ГТ своя глотки: ’ хоаны, ?.Г,- глоточное отверстие слуховой
|рубы;, .Н — боковой гортанный карман. >’5 голосовой нросток. лй. 37 *-
надгортанник. т’л' подъязычно надюргаинач мышиз; тгпе по.’п.я тычннв
кости; 40 — нижнечелюстной лимфатический узел; 4! — челюстно-подъязычная
мышца; 42 — подбородочно-подъязычная мышца; 43 — подбородочно-язычная
мышца; 44— нижняя челюсть; 45— подбородок; 46— нижняя резцовая мышца;
47 — нижняя губа; 48 — мягкое нёбо; 49 — тело языка; 50 — верхушка языка;
51 — резцовые зубы; 52 — черпаловидный хрящ; 53 — кольцевидный хрящ
183
выход из ротовой полости — зев. В ротовую полость открываются
протоки слюнных желез.
Губы (labia) — это кожно-мышечные складки, ограничиваю-
щие ротовую щель (рис. 3.90; см. рис. 3.110). Различают верхнюю и
нижнюю губы. На боковой поверхности головы, где края губ сли-
ваются между собой, образуются правая и левая спайки губ, оги-
бающие соответствующий угол рта. У' коров губы короткие,.тол-
стые, малоподвижные. Верхняя гу^а примыкает к носовым отвер-
стиям и прикрепляется к резцовой кости.
Кожа верхней губы вдоль губного края лишена волос и образует
носогубное зеркало. На нем заметны отверстия выводных протоков
носогубных желез. Они выделяют секрет, увлажняющий зеркальце.
Нижняя губа прикреплена к резцовой части нижней челюсти.
Губные железы располагаются в подслизистом слое углов губ и
открываются многочисленными отверстиями в преддверие рта.
Нижняя губа переходит в подбородок. На слизистой оболочке вер-
хней губы имеются конусовидные сосочки.
Особ е нн о сти._У..свиней губы малоподвижные, короткие и
слабоволосатые, нижняя губа кпереди заострена, ротовая щель
довольно обширна, у клыков губы не смыкаются, верхняя губа
переходит в хоботок, губные железы развиты слабо. У лошадей_
губы длинные, тонкие, мягкие, очень подвижные и весьма чув-
ствительные, нижняя губа переходит в ясно выраженный подбо-
родок с тонкой кожей и осязательными волосами. У собак губы
менее подвижны, ротовая щель большой величины, угол рта силь-
но свисает вниз, губные железы выражены слабо.
Щеки (Ьисса) — это кожно-мышечные складки, формирую-
щие боковые мягкие стенки ротовой полости (см. рис. 3.110). Щека
простирается от угла рта до крыловидно-челюстной складки слизи-
стой оболочки. Снаружи щеки покрыты кожей, а внутри выстла-
ны слизистой оболочкой желто-розового цвета с черными участ-
ками. На слизистой оболочке щеки на уровне четвертого верхнего
коренного зуба имеется низкий слюнный сосочек. В нем открывает-
ся устье протока околоушной железы. Щеки вместе с Коренными
зубами формируют защечное преддверие. Щечные железы рдсийдаг
гаются в толще щеки в три ряда и подразделяются \\адарсальные.^
средние и вентральные. Слизистая оболочка щёк снабжена много-
численными щечными сосочками (до 1,0 см длиной).
Особенное ти. У свиней щеки i юкрьп ы изнутри гладкой сли-
зистой оболочкой, щечные железы располагаются в два ряда.
У лошади.слизистая оболочка щек гладкая, слюнный сосочек от-
крывается на уровне третьего верхнего коренного зуба, имеются
дорсальные и вентральные щечные железы. У собак щеки корот-
кие, слизистая оболочка гладкая, местами пигментированная.
Десны (gingiva) представляют собой складки слизистой обо-
лочки, покрывающие челюсти около основания зубов (см. рис. 3.90,
184
3.92). Позади последних зубов десны переходят в крыловидно-челю-
стную складку.
Язык (Jingua) — массивный, весьма подвижный мышечный
орган, который служит для захватывания корма, определения его
вкуса, перемешивания и продвижения его из полости рта в глот-
ку (см. рис. 3.90, 3.91, 3.111). Корень языка тянется от начала горта-
ни до последнего коренного зуба; он лежит глубоко в ротовой
полости и прикреплен к подъязычной кости, Между корнем языка
и нёбно-язычной дужкой на кадодой стороне находится по нёбной
миндалине.
Тело языка относительно толстое, размещается между корен-
ными зубами, на поперечном сечении имеет почти трехгранную
форму. Нижняя грань тела языка связана с дном ротовой полости.
Правая и левая его грани округлые, они отделяют боковые повер-
хности языка от его спинки. Слизистая оболочка вентральной по-
верхности образует уздечку языка. На его спинке выделяется по-
душка языка.
Верхушка языка прикасается к резцовым зубам.. Она имеет пра-
вый и левый края и дне поверхности: дорсальную и вентральную.
Дорсальная поверхность. шеромнштая, вентро и,ная гладкая.
Спинка языка усеяна сосочками разнообразной величины й
формы. Нитевидные — грубые, ороговевшие, крупные, наклоне-
Рис 3.91. Язык собаки (Л), кошки (Л). г пиньи (5), коровы (/’), лошади (.7):
/ — нёбная мин.ы.шна^?.' срединная борозшкЗ нитевидные сосочки; 4 —
листовидныйi сосочек. 5' - пазуха мин шдины. ь надгортанник. 7 псиная
миндалина; Л; in ыкопнлиыс сосочки. 9 подушка языка; 10 — ямка языка:
III- грибовидные сосочки
185
ны назад. Конусовидные сосочки располагаются в основном на кор-
не языка.
Кроме указанных сосочков, выполняющих механическую фун-
кцию, на спинке языка имеются грибовидные. валиковидные и ли
стовидные вкусовые сосочки. Лист овили ые сосочки > коров отсуг-
ствуют. В толще слизистой оболочки корня языка и по его краям
заложены язычные железы.
На корне языка в слизистой оболочке находятся многочислен^,
ные язычные лимфатические фолликулы, формирующие язычную
миндалину.
Основу языка составляет язычная мышца,^состоящая из. про-
дольных, поперечных и перпендикулярных мышечных пучковГ
В вентральной части перегородки языка слабо развит язычный хрящ.
Движение языка обеспечиваются мышцами языка и подъязыч-
ной кости.
Особенности. У свиньи язык относительно узкий, ните-
видные сосочки языка_МЯ1 кие и гонкие, грибовидные сосочки
ясно видны на спинке языка, валиковидные сосочки расположен
ны на корне языка по одному с каждой стороны, листовидные
сосочки слабо развиты. У лошади яиак длинный, на его спинке”
имеются два крупных валиковидных сосочка, по бокам корня языка
находятся два листовидных сосочка, а каудальные их — две нёб-
ные миндалины, грибовидные сосочки находятся на спинке и по
краям языка. У собаки язык широкий, плоский, с заостренными
краями, имеется язычный хрящ — лисса,
Твердое небо {palatum durum) образовано резцовыми, верхнече-
люстными и небными костями (рис. 3.92; см. рис. 3.90, 3.111). Сни-
зу кости покрыты слизистой оболочкой. На твердом нёбе_имеется__
небньш шов. Он проходи! по срединной линии твердою неба, а
поперек него располагаются нёбные валики. У корон ИХ 15—20,
причем передние валики высокие, с зазубренными краями, тая-
ние — меньшей высоты и более гладкие, ближе к глотке они со-
всем исчезают. Впереди первых валиков за зубной пластинкой на-
ходится небольшой резцовый сосочек. Справа и слева от него двумя
отверстиями открывается носонёбный канал.
Нёбная занавеска, или мягкое нёбо (palatum molle)^ представляет
собой складку слизистой оболочки, спускающуюся от твердого нёба
назад (см. рис. 3.90, 3.92, 3.11 1). Задний конец мягкого нёба называ-
ется нёбной дужкой. Между нёбной дужкой и корнем языка нахо-
дится зев. Мышечную основу мягкого нёба составляют: нёбная мыш-
ца, подниматель и напрягатель нёбной занавески.
Ротовая поверхность нёбной занавески покрыта слизистой обо-
лочкой, выстланной многослойным плоским эпителием, имеет
много слизистых желез. Глоточная поверхность нёбной занавески
тоже покрыта слизистой оболочкой, которая выстлана мерцатель-
ным эпителием и содержит большое число отверстий серозных
186
Рис. 3.92. Твердое и мягкое нёбо собаки (вентральная поверхность):
мягкое небо (небная зшзавескаЦ,£)— резцовый соазчек; j)— клык: 4,—
премоляры: с первого по четвертый; Л- лицевая артерия; У- лицевая вена: 7-—
центральный щечный нерв: К — первый и второй моляры; W проток околоуш-
ной железы; 1U — вентральный щечный нерп; И'— нижнечелюстная же лета:
12 внутренняя челюстная вена; 132— заглоточный латеральный лимфатиче-
ский узел; 14 — хоаны (эадцис ноздри); (/У— наружная сонная артерия; /б)—
грудино-галовная мыиша^/7 нёбный шов, /V/ нёбные валики; J4)— щека;
20 нижняя челюсть; 21' большая жевательная мышца; 22) - двубрюш-
ная мышца;.2?)— глоточная миндалина
желез. В слизистой оболочке мягкого нёба находится непарная нёб-
ная миндалина. Между нёбно-язычной дугой и корнем языка слева
и справа располагаются парные нёбные миндалины.
Дно ротовой полости (Jundus cavi oris) располагается, с одной
стороны, между боковыми поверхностями языка и деснами ко-
ренных зубов, с другой — под верхушкой языка и резцовой час-
тью нижней челюсти. На дне заметен ряд больших сосочков, меж-
ду которыми открываются маленькие отверстия протоков подъя-
зычной железы. Под верхушкой языка выступает подъязычная (го-
лодная) бородавка. В ней открываются протоки нижнечелюстной и
подъязычной желез.
Особенности. У свиньи твердое нёбо имеет извилистый
нёбный шов^20 —25 гладких нёбных валиков. Мягкое нёбо корот-
кое, толстое и поставлено почти горизонтально. Голодной боро-
187
давки нет.У лошади твердое нёбо имеет 16—18 пар валиков со
швами. Мягкое нёбо длинное, задним концом доходит до основа-
ния надгортанника. У собак твердое нёбо сильно расширяется в
аборальном направлении. Нёбный шов хорошо виден.
Зубы (dentes) размещаются в зубных луночках — альвеолах че-
люстей (рис. 3.93; см. рис. 3.92, 3.94). На продольном распиле в
центре зуба находится зубная полость, которую заполняет зубная
^мякоть” или пульпа зуба. В нижнем конце корня зуба имеется не-
большое отверстие для сосудов и нервов, идущих к пульпе. Основ-
ным веществом зуба является дентин. Он содержит 70 S0 % ми-
неральных веществ и 10—30% коллагена, В области коронки ден-
тин снаружи покрыт слоем змали. Эмаль — самое твердое веще-
ство. состоящее на 95 — 98% из неорганических веществ. В облас-
ти корня зуба дентин покрыт зубным цементом, сходным но стро-
ению с костной гкамью. Он содержит 60 — 70% минеральных ве-
ществ. К зубам подходят кровеносные сосуды и нервы от подглаз~
ничных и нижнечелюстных артерий и нервов.
Зубы по функции, строению и расположению делятся на резцы,
клыки и коренные. Последние, в свою очередь, подразделяются на
премоляры и моляры. РезЩ)/ обо-
значаются буквой !, клыки — С,
премоляры I’. моляры —
Рис. 3.93. Продольный разрез корот-
кокоронкового зуба (резцовый зуб
собаки):
(?¥- эмаль;(2)— пульпа;(з)— десна; @—
цемент; (^—отверстие верхуннц, зуба,
кровеносные сосуды, нерв^б, 7 — ден-
тин; 8 — периодонт
Зубы бывают двух поколе-
ний — молочные и постоянные.
Различают: молочные резцы — Id,
молочные клыки — Cd и молочные
премоляры — Pd. На каждом зубе
имеются коронка, шейкам корень.
Корова имеет восемь резцов
на нижней челюсти, по четыре
зуба с каждой стороны, четвер-
тая пара резцов — это видоиз-
мененные клыки. Верхняя че-
люсть не имеет резцов, на их
месте находится роговая зубная
пластинка.
Постоянныя. рез.цав.ые_зубы
имеют форму слегка изогнутой
лопатки с заостренным краем,
коронка резца резко отличается
от округлого корня шейкой. Кор-
ни неглубоко сидят в зубных аль-
веолах, губная поверхность ко-
ронки слегка выпуклая, языч-
ная — вогнутая. Молочные резцы
отличаются от постоянных лишь
188
Рис. 3.94. Длиннокоронковый зуб (резцовый зуб лошади):
А — сагиттальный распил зуба; Б — зуб со стороны язычной поверхности; В
зацеп joiii.i.nt различною жвраста; 1 — чашка зуба; гребни min: 3 —
язычнзи iiom-pxihiL11. 7 пвиент; 5, 14 — пульпа; л КСДОНЬ г.б.,. покрытый
цементом; 7 - коронка пба; 8 — шейка зуба; 9 — корень зуба, h) — ipo года;
// пян> .Гт, 12 — девять лет; 13 — старше IH iei
. Л
меньшей величиной, по мере стирания свободный край резцов
становится тупым, клыки отсутствуют.
Даренные зубы в количестве 24 относятся к типу лунчатых зу-
бов. премоляры имеют"молочных предшественников, которые в
два раза тоньше моляров, особенно мал третий премоляр. Жева-
тельная поверхность премоляров разделена полукруглым высту-
пом, в язычную и щечную стороны от выступа находится по од-
ной луночке. Моляры не имеют молочных предшественников, они
объемистее премоляров, их трущиеся поверхности двумя высту-
пами поделены на четыре луночки. Формула зубов у коров имеет
следующий вид: молочных — Id 0/4, Cd0/0, Pd3/3 = 20 зубов;
постоянных — I 0/4, С 0/0, Р 3/3, М 3/3=32 зуба.
Особенности. У свиней насчитывается 28 молочных зубов,
из них 12 резцовых, 4 клыка и 12 коренных зубов. Постоянные
клыки у самок небольшого размера; _у_самцов они необычайно
массивны, глубоко сидят в луночках и длительно растут. Постоян-
ные коренные зубы представлены четырьмя премолярами и тремя
молярами. Формула постоянных зубов у свиней имеет следующий
вид: I 3/3, С 1/1, Р 4/4, М 3/3.
У лошадей формула постоянных зубов имеет вид: I 3/3, С 1/1,
Р 3/3, М 3/3 (рис. 3.94). Постоянные резцы в виде изогнутых клинь-
ев. Коронка зубов без выраженной шейки переходит в корень. Клы-
ки отделены беззубым краем от резцов и коренных зубов. Они име-
ют изогнутую конусовидную форму. У кобыл клыки часто отсут-
ствуют. Коренные зубьцю шали .ииннокорои ковыс. складчатъ1е_
У собак шейка зубов хорошо выражена, также ясно выделяется
поясок основания коронки, на нем возвышаются от двух до четы-
189
рех зубов. У собак 32 молочных зуба, формула зубов имеет следу-
ющий вид: Id 3/3, Cd 1/1, Pd 4/4. Постоянных зубов у них 42,
зубная формула: 1 3/3, С 1/1, Р 4/47~NT2/3.
Околоушная железа (glandula parotis) размещается у коровы по-
зади ветви нижней челюсти (рис. 3.95; см. рис. 3.92). В нижнем, более
узком, участке она частично прикрыта большой жевательной мыш-
цей, а в среднем участке сама прикрывает нижнечелюстную железу.
В сторону уха она шире и толще, лежит впереди основания ушной
раковины. Околоушная железа имеет плотную консистенцию и буро-
красный цвет. Ее наружная поверхность покрыта подкожной мыш-
цей головы. Внутренняя поверхность железы прикасается с нижне-
челюстной железой, жевательной мышцей, лимфатическим узлом,
Рис. 3.95. Слюнные железы коровы (4), лошади (Б), свиньи (В), собаки (Г):
О — щечная дорсальная; (2.щечная вентральная железа; 3 — околоуш-
ная железа; /4'— нижнечелюстная железа; [5J— подъязычная железа; \61— орби-
тальная железа
190
суставом и скуловой дугой. Передний край железы слегка вогну-
тый, задний расширен и округлен, а нижний заострен. Проток око-
лоушной железы лежи ! на медиальной стороне ветви нижней че-
люсти, отсюда он переходит через-лицевую сосудистую вырезку,
поднимается дорсально и открывается в щеке на уровне четверто-
го верхнего коренного зуба низким слюнным сосочком.
Особенности. У свиньи околоушная железа лежит позади
нижней челюсти и имеет треугольную форму. У лошади околоуш-
ная железа располагается под ушной раковиной. Околоушной про-
ток проходит в лицевой сосудистой вырезке и прободает щеку на
уровне третьего верхнего коренного зуба. У собак околоушная
железа треугольной формы и охватывает верхним участком осно-
вание ушной раковины. Проток железы направляется поперек боль-
шой жевательной мышцы и открывается на уровне третьего верх-
него коренного зуба.
Нижнечелюстная железа (glandula mandibutaris) задневерхним
концом лежит под крылом атланта, а передненижним находится
на уровне тела подъязычной кости (см. рис. 3.95). Задний край же-
лезы выпуклый. Проток ее направляется вперед и открывается в
подъязычной бородавке.
Особенност и. У свиней железа небольшая, округлой фор-
мы, дольчатого строения, прикрыта околоушной железой. Про-
ток ее открывается около уздечки языка. У лошадей железа в виде
вытянутого и изогнутого тела протянулась от ямки атланта до тела
подъязычной кости. Проток железы тянется до подъязычной бо-
родавки. У собак железа округлая, ее дольки плотно соединены
друг с другом, лежит она у угла нижней челюсти, проток ее на-
правляется к подъязычной бородавке.
Подъязычная железа (glandula sublingualis) двойная (см. рис. 3.95).
Подъязычная короткопротоковая железа расположена под слизис-
той оболочкой дна ротовой полости. Ее малые выводные протоки
открываются на слизистой оболочке. Подъязычная длиннопротоко-
вая железа достигает впереди подбородочного угла. Ее длинный
проток открывается на подъязычной бородавке.
Особенности. Усвиньиполъязычная железа двойная. У ло-
шадей подъязычная железа только одна — короткопротоковая. Она
тянется от третьего коренного зуба до подбородочного угла, име-
ет около 30 малых протоков, которые открываются на слизистой
оболочке.^У-СОбак^подъязычная железа двойная; имеется орбиталь-
ная железа.
Глотка
Глотка (pharynx) — это полый конусовидный мышечный орган,
соединяющий ротовую полость с пищеводом, а носовую — с гор-
191
Г
Рис. 3.%. Схема строения ыотки:
Cj)— ВОСОМЙ ||<1,н1ст«.: S ротонах пи.1лн. mhimx небо ^)— носоглотка;^)—
корень^ ника, ff— гортань; у/ — ротоглотка: 5 — нёбно-глоточная дуга;
л нишеволу li'i 1П.1С1ИНКЗ Ko.ihiieHH/iHoio хрята;/7у—трахея
танью (рис. 3.96; см. рис. 3.90, 3.92, 3.111). В глотке имеются две
части — дыхательная и пищеварительная. Границей между ними
служит нёбно-глоточная дуга. Вершина глотки обращена к пище-
воду, а основание — к хоанам. Глотка коров короткая и широкая,
состоит из трех оболочек: слизистой, мышечной и соединитель-
нотканной.
Слишстая оболочка в дыхагсльной части покрыта мерцатель-
ным эпителием, а в iiHiiiuHapitTc.ii.iioii м hoi ослоп пым плоским.
В се толте находятся ели шетые желе чч и шмфатичеекие фоллику-
лы, которые в сумме формируют непарную глоточную миндалину.
Мышечная оболочка глотки представлена тремя парными сужи-
вателями (констрикторами): краниальным, средним и каудаль-
ным, и одним парным расширителем.
Адвентиция глотки является внешней оболочкой, состоящей
из рыхлой соединительной ткани.
Особенное! и .У свиней глотка имеет такое же строение,
как у коров. У лошади глотка сильно вытянута спереди назад. Вхо-
ды в слуховые грубы представ лены в виде щелей длиной до 4 —
5 см. Они ведут в воздухоносные мешки. Воздухоносный мешок пред-
ставляет собой выпячивание слизистой оболочки'слуховс>й'тру-
бы, емкость его 300 — 500 мл; он располагается сзади и латераль-
ное глоткшУсобак крыша глотки около хоан имеет щелевидные
косо поставленныё~входы в слуховые трубы.
Пищевод
Пищевод (esophagus) — длинная трубка, соединяющая глотку
с желудком (см. рис. 3.89, 3.90, 3.92, 3.96, 3.98 — 3.102). Он подраз-
деляется на шейную, грудную и брюшную части. Слизистая обо-
192
дочка пищевода покрыта многослойным плоским эпителием и
собрана в продольные складки. В начальной части пищевода она
содержит слизистые железы. Мышечная оболочка пищевода по-
строена из поперечно исчерченной мышечной ткани. Слои мышц
взаимно пронизывают друг друга, как дранки плетеной корзин-
ки. Мышечная оболочка на каудальном конце пищевода включа-
ет наружные продольные и внутренние кольцевые мышечные
пучки. Шейная часть пищевода снаружи покрыла оболочкой ИЗ
рыхлой соедтительной ткани, грудная и брюшная час i n еероз-
нойЪ5олочкой. Шейная часть пищеводаЛСЖИТ выше трахеи. В ниж-
ней трети шеи пищевод слегка свешивается на левую сторону,
затем снова поднимается на трахею и проникает в грудную по-
лость. Грудная часть пищевода находится дорсально от трахеи
между листиками средостения. На уровне десятого ребра он про-
никает через диафрагму в брюшную полость и впадает в пред-
дверие рубца.
Особенност и.ЗДсвиней шейная часть пищевода относитель-
но короткая, железы слизистой оболочки находятся только в шей-
ной части пищевода. У лошадей начальная часть пищевода лежит
между правым и левым воздухоносными мешками; на уровне три-
надцатого ребра он переходит через диафрагму в брюшную по-
лость.,У собак пищевод имеет пищеводное преддверие с мелкими
продольнымй"складками, железы рассеяны по всему пищеводу.
Желудок
Желудок и дистально расположенные органы пищеварения
сосредоточены в основном в брюшной полости. Для более точно-
го описания их расположения брюшную полость принято делить
на ряд областей. При этом вначале она разделяется на три отдела
рис. 3.97). Первый и| них расно unaeioi ОТ иннррлi мы до попереч7
НОЙ плоскости, проведенной по выпуклому краю последнего реб-
ра Это — эпигастрии. Второй простирается от указанной выше
плоскости до второй поперечной плоскости, проведенной каса-
тельно краниального края маклоков. Он получил название мезога-
стрий (см. рис. 3.97). Третий занимает часть брюшной полостгГбт
указанной выше второй сегментальной плоскости до входа в тазо-
вую полость, ограниченный телом подвздошной и краниальной
ветвей лонной костей. Он называется гипогастрий (см. рис. 3.97).
Эпигастральный отдел брюшной полости, в свою очередь, де-
лится ТГа'три области? Дополнительная плоскость, проведенная
касательно правой и левой реберных дуг, отсекает от него вент-
рально расположенную[ область мечевидного хряща. Оставшаяся часть
срединной плоскостью делится на правое и левое подреберье
(см. рис. 3.97).
I Зеленевский 193
Мезогастральный отдел двумя сагиттальными плоскостями, про-
веденными касательно поперечных отростков поясничных по-
звонков, делится на три части. Латерально от них располагаются
правая и левая подвздошные области^ А оставшийся больший сред-
ний участок делится фронтальной плоскостью на верхнюю пояс^
ничную (почечную) область и нижнюю пупочную область (рис. 3.97)..
^ гипогастральный отдел правая и левая подвздошные области
переходят как правая и левая паховые области, поясничная об-
ласть переходит в тазовую полость, а пупочная область — в лон-
ную (срамную) область (рис. 3.98; см. рис. 3.97).
Различают желудки однокамерные Jy лошади, свиньи и соба-
ки) и многокамерные (у жвачных). В зависимости от характера сли-
зистой оболочки желудки бывают нишеводного (безжелешетые).
ральную (Б) поверхности, передний (В) и средний (Г) отделы:
линия прикрепления диафрагмы; 2 — поперечная плоскость, касательная
реберной дуги; 3 — поперечная плоскость, касательная маклоков; 4 — граница с
тазовой полостью; 5 — левая паховая область; 6 — левая подвздошная область; 7 —
область мечевидного хряща; 8 — реберная дуга; 9 — левое подреберье;
Б.1 — линия прикрепления диафрагмы; 2 — левое подреберье; 3 — реберная
дуга; 4 — поперечная плоскость, касательная реберной дуги; 5 — левая под-
вздошная область; 6 — поперечная плоскость, касательная маклоков; 7 — левая
паховая область; 8 — лонная область; 9 - правая паховая область; 10 — правая
подвздошная область; 11 — пупочная область; 12 — область мечевидного хряща;
13 — правое подреберье,
В, Г. 7 — линия прикр. । । leaee подреберье; правое
подреберье;!41- область мечевидного хряппг.А”— поясничная обзжн.; <> — пупоч-
ная область; у/— левая подвздошная область: У правая подвздошная область
194
Рис. 3.98. Схема строения серозной оболочки брюшной полости (попереч-
ное сечение брюшной полости):
I ОСТИСТЫЙ отросток ПОЯСНИЧНОГО IIOJBOHKJ.A?,’— JlopcA'IMIUe МЫШЦЫ ПОЛИ!'
ночного сщлба:,.}/— каудальная полажена; < М,*— почки; 5/— связки печени и
мглудкэ: б,— поджелудочная железа.(л*- печсночно-двенадиатиперст|иш связка,
двенадцатиперстная кишка, У печень, [О' вход в сальниковый мешок.
II - желудок; 12 сальниковый мешокьИЦ — белая линия ж и ища ; 14 — порта;
15 - кожа: ./7?- фэсиня;^. мышцы; v’V- последнее ребро, 20)— Поперечная
брюшная фасиня: 21. 23. 24.— висцеральный листок серозной оболочки брюш-
ной полости — брюшины;, ZJz париетальный листок серозной оболочки брюш-
ной полости — брюшины; 2$ -тощая кишка; $ — большой сальник; 27; —
селезенка; — прямая мышца живота
кишечного {железистные) и пищеводно-кишечного {смешанные) ти-
пов. -- -
Однокамерный желудок {ventriculus, s. gaster) — это мешкооб-
разное расширение пищеварительной трубки позади диафрагмы
(рис. 3.99; см. рис. 3.89, 3.103, 3.104, 3.106).
Желудок лошади однокамерный, пищеводно-кишечного типа^
В нем различают выпуклую большую кривизну и вогнутую малую
кривизну. В желудке выделяют пристенную поверхность, обращен-
ную к диафрагме, и висцеральную, граничащую с внутренностями.
Кроме того, различают вход пищевода в желудок, или кардия^
выход из желудка в двенадцатиперстную кишку, или пилорус, рас-
ширенную часть — дно желудка. Желудок разделяют на три части:
левую — кардиальную, граничащую с кардиа, правую — пилори-
ческую. прилегающую к пилорусу, и дойную, или фундальную.
расположенную между кардиальной и пилорической частями. От
малой кривизны внутрь желудка вдастся глубокая угловая складка,.
являющаяся границей между его кардиальной и пилорическои
195
Рис. 3.99. Желудок лошади (про-
дольный разрез):
(7/— слепой мешок; ’2)— пищевдд;
Лу— двенадцатиперстная кишка;(4Р-
пилорус; X — пилорическдя часть
желудка;/^- дно желудка; 7 склад-
чатый край
частями. Кардиальная часть желудка лошади хорошо выражена и
имеет .V('///fT\_. Jl jiJi. LopiLliL'ci<oii 4ac rn желудка двумя б<>ро на-
ми выде.тяется пилорическая полость.
Стенка желудка суетой г из слизистой, мышечной и серозной
оболочек^
~ Слизистая.оболочкр в области слепого мешка образует складча-1
тый край, являющийся границей между безжелезистой и желези- I
стой частями желудка. В безжелезистой части слизистая оболочка
беловатого цвета, прочная и грубая, выстлана многослойным плос-
ким эпителием. В железистой части желудка она нежная, розова-
того цвета, покрыта цилиндрическим железистым эпителием,
содержит лимфатические фолликулы. Поверхность слизистой обо-
лочки неровная, усеяна небольшими желудочными ямочками, в
которые открывается желудочные железы. В зависимости от зоны
желудка различают железы трех типов: кардиальные, пилорические
и донные.
Мышечная оболочки состоит из (.никои мышечной ткани и обра-
зует три слоя: продольный, кольцевой и косой. Продольный слой
лежит на большой и малой кривизне и на пилорическом мешке.
Кольцевой слой сильно развит в пилорической части, в кардиальной
он отсутствует. В складках пилорического мешка он формирует два
сжимателя пилоруса. Косой слой находится только в кардиальном
отделе и подразделяется на наружные косые пучки и на внутренние
косые пучки, образующие очень прочный кардиальный сфинктер.
Серозная оболочка покрывает желудок снаружи. Она образует
большой и малый сальники, а также связки (см. рис. 3.98).
Желудок .loiUiUH pacnt) i.ii к л в основном в левом iioapeoe-
^рьд лишь небольшой участок ею пилорической части захопп в~
правое подреберье. Он прилежит к брюшной стенке вверху слева
против четырнадцатого-пятнадцатого межреберного промежутка,
а внизу против девятого —одиннадцатого ребра на линии плече-
вого сустава. Большая кривизна желудка обращена влево и назад.
Особенност и..У свиньижслудок пищеводно-кишечного типа.
В его кардиальной части имеется слепое выпячивание — диверти-
196
кул .желуОка.Усобаки^желудок объемистый кишечного типа. Стро-
ение серозной оболочки и расположение желудка такие же, как у
СВИНЬИ^-.
Многокамерный желудок
Многокамерный желудок коровы состоит из четырех камер:
рубца, сетки, книжки и сычуга. Ближе к пищеводу лежит рубец,
за ним"следует сетка, затем книжка и сычуг (рис. 3.100; см. рис.
3.101-3.102, 3.105).
Объем камер у взрослых животных постоянен. У коров самая
большая камера — рубец, за ним идет книжка, а затем сычуг.
Объем книжки и сычуга часто одинаковый. Самая малая камера —
сетка. Стенка каждой камеры желудка состоит из слизистой, мы-
шечной и серозной оболочек.
Рубец (rumen) — начальная камера желудка (см. рис. 3.100 —
3.102, 3.105). Он занимает всю левую половину брюшной полос-
ти, а вентральным краем заходит даже в ее правую половину. Ру-
Г. -,0 f0
' У
Рис. 3.100. Желудок овцы (девая сторона):
пищевод; 2 — преддверие рубца; книжка; 4 — листочки книжки; 5 —
дорсальный поперечный тяж. Л i ni.i и.и ы рубцовая борозда; 7 — правый
продольный тяж; 8 — дорсальный край рубца; 9 — дорсальный рубцовый мешок;
10 — каудодорсальная поперечная борозда; 11 — каудодорсальный слепой ме-
шок; 12 — каудальная рубцовая борозда; 13 — каудовентральный слепой мешок;
14 — каудовентральная поперечная борозда; 15 — левая продольная борозда; 16 -
вентральный руононый мешок; /7 — i не нал ижрт гретая киши; 18 - спираль-
ные складки сычуга. 19 — паруса СЫЧУПК 20 — сычу|, 21 — рубцово-сетковое
oTBcpc ine: 2? ВЫХОД из пишенсна и жетулок. 23 - рубиово-сетксиШЙ желоб;
24 — правам rvw рубионо-сеткпшно желоба; 25;-- сетка; 2b — ячейки сетки;
27 лно книжки. 28 книжково-сычужное отверстие
197
бец слегка сжат с боковых сторон и несколько вытянут по длине
тела. На нем различают левую и правую поверхности, верхний и
нижний края, передний и задний концы. ^Левая, или париеталь-
ная, поверхность обращена к брюшной стенке; правая, или висце-
ральная, поверхность прилегает к внутренностям. Верхний край об-
ратен вверх и влево, нижний край направлен вниз и вправо. Пере-
дний конец соприкасается с диафрагмой, задний коней обращен
назад..
Рубец делится продольными правым и левым желобами на дор-
сальный и вентральный рубцовые мешки. Мешки спереди и сзади
отграничены друг от друга глубокими бороздами, образующими
снаружи краниальный и каудальный рубцовые желоба, а в полости
рубца — краниальный и каудальный тяжи.
На каудальном конце рубца имеются каудодорсальный и каудо-
вентральный слепые мешки. Границами этих мешков являются сна-
ружи — каудодорсальный и каудовентральный венечные желоба, в
полости рубца — каудодорсалъные и каудовентральные тяжи. Кра-
ниальный конец рубца, куда впадает пищевод, называется пред-
дверием рубца. Оно отграничивается от дорсального мешка снару-
жи краниодорсальным венечным желобом, а внутри рубцасоот-
ветствующим тяжом.
Слизистая оболочка рубца безжелезистая, покрыта многослой-
ным плоским эпителием. На ней возвышается множество сосоч-
ков, имеющих различную форму и величину (до 10 мм у коров).
Они и придают шероховатость внутренней поверхности рубца.
Сосочки отсутствуют только на тяжах, поэтому последние явля-
ются гладкими и более светлыми.
Мышечная оболочка рубца состоит из гладкой мышечной тка-
ни, которая формирует продольный, кольцевой и косой слои. Она
очень толстая, и сокращения ее очень сильные.
Серозная оболочка рубца покрывает всю его поверхность, в
области правого и левого продольных желобов она переходит в
большой сальник. Рубец прикрепляется рыхлой соединительной
тканью к левой ножке диафрагмы и к поясничным мышцам.
Сетка (reiiculum):- небольшой округлый мешок (см. рис. 3.100—
3.102, 3.105). Она является продолжением преддверия рубца, от
которого сетка отделяется с вентральной стороны рубцово-сетко-
вым тяжом. С рубцом сетка сообщается широким отверстием, с
книжкой — щелевидным. Кроме того, она связана с пищеводом
посредством рубцово-сеткового желоба. Берегами желоба служат
его губы, между которыми лежит дно желоба. Рубцово-сетковый
желоб в виде спирально изогнутых складок слизистой оболочки
идет от пищевода к книжке. В толще его губ заложены гладкие
мышечные пучки, сокращение которых вызывает смыкание губ
желоба и превращение его в трубку; в этом случае жидкий корм
может попадать из пищевода прямо в книжку. Дно желоба содер-
198
жит поперечный слой гладких мышечных пучков, при сокраще-
нии которых наступает размыкание губ, и грубый корм свалива-
ется в рубец или сетку.
Стенка сетки построена из слизистой, мышечной и серозной
оболочек. Слизистая оболочка сетки образует пластинчатые складки
(10—12 мм), соединяющиеся друг с другом в четырех-, пятн-
или шестисторонние ячейки, напоминающие собой пчелиные соты.
На дне ячеек находятся более низкие складочки слизистой обо-
лочки, отграничивающие более мелкие ячейки второго порядка.
Вся слизистая оболочка усеяна мелкими ороговевшими сосочка-
ми. Вблизи рубца и пищеводного желоба ячейки у коров посте-
пенно понижаются, а затем совсем исчезают.
Мышечная и серозная оболочки сетки построены, как в рубце.
Сетка располагается в области мечевидного хряща на вентраль-
ной брюшной стенке и может быть пропущена сразу за мечевид-
ным отростком грудины.
Книжка (omasumV\\'Avc\ округлую форму (рис. 3.101; см. рис.
3.100, З.Т02, 3.1(b), С одной стороны она служит непосредствен-
ным продолжением сетки, а с другой — переходит в сычуг. Сли-
зистая оболочка книжки имеет тонкие пластинчатые складки, по-
лучившие название листочков, которые по величине бывают: боль-
шие, средние, малые и самые малые. Больших листочков насчиты-
вается 12 — 14. По сторонам и краям листочков находятся соСочки.
Внутри листочки состоят из продольных и поперечных мышечных
пучков, которые при сокращении удлиняют или укорачивают ли-
сточки. Пространство между двумя большими листочками назы-
вается межлисточковой нишей. В середине ниши помещаются один
средний листочек, два малых и четыре самых малых.
Дно книжки с внутренней стороны имеет вид короткого жело-
ба, идущего из сетки в сычуг. Желоб отграничен по сторонам дву-
Рис. 3.101. Желудок теленка (пра-
вая сторона):
/ — правая продольная борозда; 2 —
дорсальный рубцовый мешок; 3 —
рубец; 4, 8 — книжка; 5 — преддве-
рие рубца; 6 — пищевод; 7— сетка;
9 — сычуг; 10 — дно сычуга; 11 —
выход из желудка в двенадцатиперст-
ную кишку; 12 — двенадцатиперстная
кишка; 13 — вентральный рубцовый
мешок; 14 — каудовентральная попе-
речная борозда; 15 — каудовентраль-
ный слепой мешок; 16 — каудальная
рубцовая борозда; /7—каудодорсаль-
ный слепой мешок; 18 — каудодор-
сальная поперечная борозда
199
мя валиками с высокими сосочками. Дно его собрано в очень ма-
ленькие продольные складочки. Перед отверстием в сычуг гладкая
мышечная ткань в толще дна книжки формирует сфинктер. На
границе между книжкой и сычугом по сторонам отверстия распо-
лагаются паруса книжки.
Между желобом дна и свободными краями листочков остается
канал книжки, который сообщается с нишами. Книжка распола-
гается в правом подреберье на уровне нижней трети восьмого-
девятого ребер.
Сычуг (abomasum) вытянут в длину в виде изогнутой груши
(рис 1102: см рис 3 100, 3.101, 3.105). Сычуг в правом подреберье
Рис. 3.102. Топография желудка коровы:
А — желудок за левой стенкой; Б — желудок за левой стенкой, вскрыты сетка,
книжка и сычу); В — жедудок за правой стенкой: Г — желудок за правой стен-
кой. удален кишечник; Ч — пищевод; 2; — селезенка; 3. 7 — дорсальный, 4. 10 —
вентральный рубцовый ме11юк;(3~/6, 2/ сетка; 6 — преддверие рубца; 8. 23 —
каулопцрсильный слепой мешок; Ч 22 — кауловситральный слепой мешок; //
сычуг. 12 сетка. 13 рубио во-се i копий желоб; 14 большой сальник. покры-
ваютпий кишечник: 15 — двепад шпниерс гния кишка; 17 — книжка. прикрытия
большим сальником; /А —сычуг. /9 — пи дорическая ч: кть сычуга: 20 - вентраль-
ный кран печени
200
п в области мечевидного хряща, пилорическая часть обращена
назад и загибается вверх. Суживающаяся часть сычуга — пилорус —
переходит в двенадцатиперстную кишку. В нем различают вент-
ральную выпуклую большую кривизну и дорсальную вогнутую
малую кривизну. Стенка сычуга состоит из слизистой, мышечной
и серозной оболочек.
Слизистаа_оболочка сычуга выстлана однослойным цилиндри-
ческим эпителием и "содержит кардиальные, фундальные и пило-
рические желудочные железы. Она гладкая, нежная, мягкая, со-
брана в 12—14 спиральных складок, увеличивающих поверхность
секреции желудочного сока.
Мышечная оболочка сычуга состоит из наружного продоль-
ного и внутреннего-кольцевого слоев. От конца малой кривизны
сычуга в просвет пилоруса выступает полукольцевой попереч-
ный валик, который и служит своеобразным сфинктером пило-
руса.
Серозная оболочка снаружи окутывает весь сычуг. От висце-
ральной поверхности печени отделяется складка брюшины —
малый сальник, он переходит на сычуг и двенадцатиперстную
кишку.
Тонкая кишка
Тонкая кишка (intestinum tenue) характеризуется большой дли-
ной (у кОрбв около 60 м), относительно узким просветом и свое-
образным расположением (см. рис. 3.89, 3.102 — 3.106).
Слизистая оболочка тонкой кишки покрыта каемчатым элите-
лйёмТГсобрангГв складки различных направлений, которые уве-
личивают секреторную и всасывательную поверхность. Различают
продольные складки, поперечные, или кольцевые, и сетевидные. Кроме
того, имеются микроскопические складки, или кишечные ворсин-
ки. В толще слизистой оболочки залегают железы двух видов: об-
щекишечные и дуоденальные. Общекишечные железы относятся к
простым трубчатым железам и распространены по всему кишеч-
нику. Их устья открываются на поверхности тонкого отдела киш-
ки в промежутках между ворсинками. Дуоденальные железы тоже
имеют трубчатое строение и располагаются у коров на протяже-
нии 6 —9 м длины кишки, начиная от пилорического сфинктера
сычуга. Железы выделяют кишечный сок для переваривания бел-
ков, жиров и углеводов. На поверхности слизистой оболочки слегка
выпячиваются солитарные фолликулы величиной с булавочную
головку. Многочисленные скопления фолликулов получили на-
звание пейеровых бляшек.
Мышечная оболочка имеет поверхностный продольный и более
глубокий кольцевой слои. Она обеспечивает движение кишечника
201
в двух направлениях: спереди назад — перистальтика, сзади впе-
ред — антиперистальтика.
Серозная оболочка образует брыжейку. В состав тонкой кишки
входят три отдела: двенадцатиперстный, тощий и подвздошный.
Двенадцатиперстная кишка (intestinum duodenum) достигает
длины от 90 до 120 см (рис. 3.103; см. рис. 3.89, 3.102, 3.104 —
3.106); начинается от пилоруса и направляется вперед и вверх до
печени. В правом подреберье против ворот печени эта кишка обра-
зует S-образный изгиб. Затем она поднимается косо вверх и назад,
принимает протоки печени и поджелудочной железы и подходит к
правой почке. Здесь она поворачивает назад, образуя первый пово-
рот, а дальше направляется горизонтально в сторону таза, дохо-
дит до подвздошной кости, поворачивает налево, где образует
второй поворот, а затем поворачивает вперед, образуя третий по-
ворот и, направляясь по горизонтали вперед, вновь достигает пе-
чени. Здесь она делает двенадцатиперстно-тощий поворот и без
резкой границы переходит в тощую кишку. Ворсинки находятся
на сетевидных складках слизистой оболочки кишки, а лимфати-
Рис. 3.103. Синтопия органов пище-
варения брюшной полости собаки:
1 — ампула прямой кишки; 2 — обо-
дочно-двенадцатиперстная связка; 3 —
нисходящая ободочная кишка; 4 — вос-
ходящая ободочная кишка; 5 — попе-
речная ободочная кишка; 6, 8— нисхо-
дящая часть двенадцатиперстной киш-
ки; 7 — желудок; 9 — тощая кишка;
10 — тощекишечные лимфатические
узлы; 11 — подвздошная кишка; 12 —
каудальная часть двенадцатиперстной
кишки; 13 — слепая кишка
слизистой оболочки.
Двенадцатиперстная кишка
соединяется с желудком, пече-
нью, почками, слепой и ободоч-
ной кишками при помощи связок.
Тощая кишка (jntestium jeju-
num) образует гирлянду завит-
ков (рис. 3.104; см. рис. 3.89, 3.102,
3.103, 3.105, 3.106). Ворсинки по-
мешаются на складках слизистой
оболочки. Лимфатические узел-
ки и пейеровы бляшки имеются
в большом количестве. Кишка
расположена в правом подребе-
рье, в подвздошной и паховой
областях. Она огибает и прикры-
вает лабиринт ободочной киш-
ки. Тощая кишка подвешена на
корне брыжейки таким образом,
что между листками брыжейки
ближе к ее корню оказывается
заключенным весь спиральный
диск ободочной кишки, с кото-
рого брыжейка в виде своеобраз-
ной бахромы переходит на пет-
ли тощей кишки.
Подвздошная кишка (intes-
tinum ileum) начинается от пос-
202
Рис. 3.104. Синтопия органов пищеварения брюшной полости лошади:
/ — прямая кишка; 2 — нисходящая ободочная кишка; 3 — восходящая часть
двенадцатиперстной кишки; 4 - тазовый изгиб; 5— поперечная ободочная кишка;
6 — левое дорсальное положение ободочной кишки; 7 — межободочная связка;
8 — левое вентральное положение ободочной кишки; 9 — диафрагмальный из-
гиб ободочной кишки; 10 — нисходящая часть ободочной кишки; 11 — желудок;
12 — грудинный изгиб ободочной кишки; 13 — правое вентральное положение
ободочной кишки; 14 — верхушка слепой кишки; 15 — тощая кишка; 16 —
брыжейка тощей кишки; 17 — слепо-ободочная связка; 18 — подвздошная киш-
ка; 19 тело слепой кишки; 20 — основание слепой кишки; 21 — поперечная
часть двенадцатиперстной кишки; 22 — правое дорсальное положение ободоч-
ной кишки
леднего завитка тощей кишки, располагается в правом подвздо-
хе, между слепой и концевой извилинами ободочной кишки (см.
рис. 3.89, 3.102 — 3.106). Она впадает в толстую кишку на границе
слепой и ободочной кишок. Выходное отверстие подвздошной кишки
находится на уровне четвертого поясничного позвонка. В стенке
выходного отверстия образуется заслонка подвздошной кишки.
Особенности. У свиней длина тонкой кишки в среднем равна
20 м. Двенадцатиперстная кишка (40 — 90 см) подвешена на ко-
роткой брыжейке. Желчный проток и проток поджелудочной же-
лезы открываются в разных местах кишки. У лошади двенадцати-
перстная кишка длиной до 1 м. Ее начальная часть прилегает к
печени и образует на ней S-образный изгиб. На месте впадения
желчного протока и протока поджелудочной железы слизистая
оболочка образует дивертикул двенадцатиперстной кишки. Тощая
кишка подвешена на длинной брыжейке (до 50 см) переднего
корня. У собаки двенадцатиперстная кишка подвешена на длин-
ной брыжейке. Желчный проток и проток поджелудочной железы
впадают раздельно на одном общем сосочке.
203
Толстая кишка
Толстая кишка (jntestinum crassum) состоит из слепого, ободоч-
ного и прямого отделов (см. рис. 3.89, 3.103— 107).
Слепая кишка (intestinum cecum) достигает у коров длины 30 —
70 см, гладкая, цилиндрической формы, большого диаметра (рис.
3.105). Она лежит в верхней четверти правой половины брюшной
полости. Начинается слепая кишка от подвздошной кишки на уров-
не середины поясницы, она слегка искривлена и верхушкой об-
ращена назад. На ней различают тело и верхушку. На месте соеди-
нения слепой кишки с ободочной впадает подвздошная кишка и
образует подвздошно-слепо-ободочное отверстие, которое закрыто
сфинктером подвздошной кишки. Слепо-ободочное отверстие сфин-
ктера не имеет. Стенка слепой кишки построена из слизистой,
мышечной и серозной оболочек. Слизистая оболочка не имеет вор-
синок. Лимфатические фолликулы встречаются часто, пейеровы
бляшки редко. Трубочки обще-
кишечных желез длинные. Мы-
шечная и серозная оболочки
слепой кишки построены, как
и в тонкой кишке.
Особенности. У свиней
слепая кишка относительно ко-
роткая, но большого диаметра
(рис. 3.106). Она несет на себе три
ленты и три ряда кармашков, а
простирается от середины пояс-
ницы до ее конца. Тело кишки
лежит рядом с задним концом
левой почки, а верхушка опус-
Рис. 3.105. Синтопия органов пище-
варения брюшной полости коровы:
1 — ампула прямой кишки; 2 — попе-
речная часть двенадцатиперстной киш-
ки; 3 — ободочно-двенадцатиперстная
связка; 4 — восходящая часть двенад-
цатиперстной кишки; 5 — нисходящая
ободочная кишка; 6, 11 — тощая киш-
ка; 7 — рубец; 8 — сигмовидная изви-
лина; 9 — пищевод; 10 — сетка; 12 —
брыжейка тощей кишки; 13 — центро-
стремительные, 14 — центробежные из-
вилины ободочной кишки; 15 — цент-
ральный изгиб ободочной кишки; 16 —
подвздошно-слепокишечная связка;
17 — слепая кишка; 18 — проксималь-
ная петля ободочной кишки; 19 — дис-
тальная петля ободочной кишки
кается вентрально, назад и впра-
во. У лошадей слепая кишка
большая (130 см) и имеет фор-
му буквы С (см. рис. 3.104, 3.107).
На ней различают головку, тело
и верхушку. Продольный мышеч-
ный слой стенки кишки образу-
ет ленты — тении, а продоль-
ный — кармашки. У собак сле-
пая кишка образует 2 — 3 изги-
ба, подвешена на очень корот-
кой брыжейке между вторым и
четвертым поясничными по-
звонками (см. рис. 3.103).
Ободочная кишка (intestinum
colon) достигает у коров длины
204
6 — 9 м и разделяется на три от-
резка: 1) начальную петлю;
2) спиральный лабиринт; 3) ко-
нечную петлю (см. рис. 3.105).
Начальная петля — это пря-
мое продолжение слепой киш-
ки; границей между ними явля-
ется входное отверстие под-
вздошной кишки в слепую. От
него начальная петля направля-
ется вперед, под правой почкой
она круто поворачивает назад и
идет над своим начальным уча-
стком в сторону таза, где пере-
ходит на левую сторону, делает
около слепой кишки еще один
поворот, идет вперед и на уров-
не третьего поясничного по-
звонка переходит в спиральный
лабиринт.
Спиральный лабиринт нахо-
дится на правой стенке рубца и
образует спирали двух порядков:
центрипетальные и центрифу-
гальные. Центрипеталъные изви-
лины кишки в 1,5 —2,0 оборота
идут в лабиринте к центру дис-
ка по часовой стрелке. В центре
диска кишка образует централь-
ный изгиб, после чего делает со-
Рис. 3.106. Синтопия органов пище-
варения брюшной полости свиньи:
1 — прямая кишка; 2 — нисходящая
ободочная кишка; 3 — восходящая часть
двенадцатиперстной кишки; 4 — попе-
речная ободочная кишка; 5 — нисхо-
дящая часть двенадцатиперстной киш-
ки; 6 — желудок; 7 — тощая кишка; 8 —
брыжейка тощей кишки; 9 — центро-
стремительные извилины; 10 — цент-
ральный изгиб ободочной кишки; 11 —
центробежные извилины ободочной
кишки; 12 — подвздошная кишка; 13 —
подвздошно-слепокишечная связка; 14 —
слепая кишка; 15 — поперечная часть
двенадцатиперстной кишки
ответствующее число центрифугальных оборотов, идущих против
часовой стрелки, достигает начальной петли и переходит в ко-
нечную петлю.
Конечная петля меньшего диаметра, чем начальная петля. Она
направляется в сторону таза, затем делает поворот под позвоноч-
ником вперед и переходит без границ в прямую кишку. Толстая
кишка находится в правой половине брюшной полости.
Особенности. У свиней ободочная кишка формирует лаби-
ринт в виде конуса (рис. 3.106), который широким основанием
обращен дорсально в область поясницы и левой подвздошной
области, а вершиной направлен в лонную область. От широкого
основания конуса к центру его вершины направляется центрипе-
тальная спираль. Она образует периферию конуса. Начальная пет-
ля кишки большого диаметра и имеет две ленты и два ряда кар-
машков. От центра она переходит в центрифугальную спираль,
следующую обратно к основанию конуса и заключенную внутри
205
Рис. 3.107. Топография слепой
кишки лошади:
/ — основание слепой кишки; 2 —
верхушка слепой кишки; 3 — тело
слепой кишки; 4 — восемнадцатое
ребро
него. Диаметр этой части петли
меньший, и она не имеет ни лент,
ни кармашков. Конечная петля пе-
реходит в прямую кишку.
У лошадей ободочная кишка
разделяется на большую и малую
ободочные кишки (рис. 3.107).
Большая ободочная кишка начи-
нается от слепой кишки, направ-
ляется вперед по правой стороне
брюшной полости и образует вен-
тральное правое положение.
Вблизи диафрагмы большая
ободочная кишка поворачивает
справа налево и образует грудин-
ное положение. Затем она направ-
ляется по левой стороне брюшной
полости назад до входа в тазовую полость и образует вентральное
левое положение. У входа в тазовую полость большая ободочная
кишка поворачивает вверх и назад, образуя тазовый изгиб. От из-
гиба ободочная кишка следует обратно по пройденному уже пути.
Располагаясь над вентральным положением, она образует дорсаль-
ное левое положение, диафрагмальное положение и дорсальное пра-
вое положение. Последнее в области поясницы сильно суживается
и переходит в малую ободочную кишку. Большая ободочная киш-
ка имеет четыре ленты и четыре ряда кармашков.
Малая ободочная кишка является продолжением правого дор-
сального положения большой ободочной кишки. Она имеет не-
большой диаметр, одинаковый на всем протяжении кишки, две
ленты и два ряда кармашков. Кишка висит на длинной брыжейке
и без видимых границ переходит в прямую кишку.
У собак ободочная кишка состоит из трех частей (см. рис. 3.89,
3.103): 1) правой восходящей части, идущей медиально от две-
надцатиперстной кишки вперед до желудка; 2) короткой попе-
речной части; 3) левой нисходящей части, направляющейся на-
зад под левой почкой и формирующей в поясничной области по-
логую извилину.
Прямая кишка (intestinum rectum) проходит в тазовой полости
под позвоночником (см. рис. 3.89, 3.103 — 3.106). Перед анусом про-
дольный слой мышечной оболочки с правой и левой сторон фор-
мирует мышцу прямой кишки и хвоста, которая направляется вверх
и назад к первым хвостовым позвонкам. Слизистая оболочка пря-
мой кишки и ануса собрана в продольные складки. Она лишена
ворсинок, но имеет общекишечные железы и много бокаловид-
ных клеток, секрет которых придает слизистой оболочке скольз-
кость.
206
Особенности. У свиней прямая кишка заканчивается ану-
сом. Она обычно окружена жировой тканью, имеется ампула пря-
мой кишки (см. рис. 3.106). Довольно сильно развита пейерова бляш-
ка (до 7 см). Лимфатические узелки многочисленны.
У лошадей прямая кишка образует ампулообразное расшире-
ние (см. рис. 3.104). Серозная оболочка имеется только в передней
части, а у четвертого-пятого крестцового позвонка кишка снару-
жи покрыта рыхлой соединительной тканью. Мышца прямой киш-
ки и хвоста развита хорошо.
У собак прямая кишка образует ампулообразное расширение
(см. рис. 3.89, 3.103). Сбоку от ануса располагаются небольшие же-
лезистые мешочки, параанальные синусы. Они лежат между под-
нимателями ануса и его наружным сфинктером. Секрет парааналь-
ных желез придает собакам специфический запах.
Задний проход (anus) формируется каудальным концом пря-
мой кишки (см. рис. 3.89). Слизистая оболочка ануса выстлана мно-
гослойным плоским эпителием. Мышечная оболочка образует
сфинктер. В глубине он состоит из гладкой мышечной ткани —
внутренний сфинктер ануса, а на поверхности — из поперечно-
исчерченной мышцы — наружный сфинктер ануса.
Анус прикрепляется к первым хвостовым позвонкам и к тазу
мышцей прямой кишки и хвоста, поднимателем ануса, а также
подвешивающей связкой ануса.
Особенности. У свиней анус устроен в основном так же,
как у коров.
У лошадей кожа ануса содержит сальные и потовые железы;
подслизистая рыхлая соединительная ткань, подвешивающая связ-
ка, внутренний и наружный сфинктеры хорошо развиты.
Печень
Печень (hepar) — самая крупная пищеварительная железа (рис.
3.108; см. рис. 3.89). Она состоит из большого числа печеночных
клеток, которые формируют печеночные балки и печеночные
дольки величиной 1380 микрон. Печень коровы массивная, вы-
пукло-вогнутой формы, буро-красного цвета; она лежит в правом
подреберье, имеет желчный пузырь и слабо выраженную дольча-
тость. На печени различают дорсальный тупой край и вентраль-
ный острый край. Печень имеет выпуклую диафрагмальную поверх-
ность, примыкающую к диафрагме, и вогнутую висцеральную по-
верхность, обращенную к желудку и кишечнику. На остром крае
печени имеются вырезки. Они делят печень на левую, правую и
среднюю доли.
Левая доля находится влево от круглой связки. Правая доля рас-
полагается вправо от желчного пузыря. Средняя доля лежит между
207
Рис. 3.108. Видовые особенности строения печени лошади (А),
коровы (Б), свиньи (В), собаки (Г):
1 — каудальная полая вена; 2 — воротная вена; 3 — хвостатый отросток; 4 —
ворота печени; 5 — правая доля; 6 — правая латеральная доля; 7— правая меди-
альная доля; 8 — квадратная доля; 9 — желчный пузырь; 10 — желчный проток;
11 — левая доля; 12 — левая медиальная доля; 13 — левая латеральная доля
левой и правой долями. Желчный пузырь помещается на висцераль-
ной поверхности печени, между ее правой и средней долями. Он
сильно свешивается за пределы вентрального края печени.
В центре органа лежат ворота печени — место входа в печень
кровеносных сосудов и нервов. Они делят среднюю долю печени
на нижнюю квадратную долю и верхнюю хвостатую долю. На хво-
стовой доле находятся два отростка: сосцевидный — направлен в
левую сторону, хвостатый — сильно нависает сзади на правую
долю печени. На правой доле и хвостатом отростке имеется почеч-
ное вдавливание. На тупом крае печени между ее левой и хвостатой
долями находится пищеводная вырезка. Между правой и хвостатой
долями печени имеется борозда, в которой помещается каудаль-
ная полая вена, плотно прилегающая к печени.
От желчного пузыря отходит пузырный проток, который вместе
с печеночным протоком формирует желчный проток. Последний
впадает в двенадцатиперстную кишку. На месте впадения протока
в кишку (50 — 70 см от пилоруса) выступает сосок.
Печень снаружи покрыта серозной оболочкой и укрепляется в
своем положении специальными связками. Она располагается в
208
правом подреберье, простираясь от плоскости шестого-седьмого
ребра до второго-третьего поясничного позвонка.
Особенности. У свиней левая и правая доли печени глубо-
кими вырезками разделяются на левые и правые латеральные и
медиальные доли. Хвостатая доля сильно выступает своим хвоста-
тым отростком в правую сторону и по нему проходит задняя по-
лая вена. На поверхности печени хорошо видны дольки. У лоша-
дей печень буро-красного цвета. Желчного пузыря нет, дольча-
тость и вырезки печени выражены хорошо. У собак печень боль-
шая с глубокими вырезками, разделяющими ее на семь долей.
Поджелудочная железа
Поджелудочная железа (pancreas) — дольчатая сложная труб-
чато-альвеолярная железа желто-бурого цвета с розовым оттен-
ком (см. рис. 3.89, 3.109). В ней различают слабо выраженную и
обращенную к печени среднюю долю, или головку, правую долю и
левую долю, или хвост железы. Головка и доли лежат вправо от
средней сагиттальной плоскости; они простираются от двенадца-
того ребра до второго — четвертого поясничного позвонка. Правая
доля направлена назад, лежит под правой почкой и соприкасает-
ся с двенадцатиперстной и нисходящей петлей ободочной кишки.
Короткая левая доля находится между рубцом и ножкой диафраг-
мы и граничит с селезенкой. Поджелудочная железа окружает во-
ротную вену.
Выводной проток поджелудочной железы выходит из конца пра-
вой доли и открывается в двенадцатиперстную кишку обособлен-
но от желчного протока на удалении 30 — 40 см от него и на рас-
Рис. 3.109. Поджелудочная железа свиньи с дорсальной (Л) и вентраль-
ной (Б) поверхности:
], 2, 3 — левая, правая, промежуточная доли соответственно; 4 — почечное
вдавливание; 5 — вдавливание ободочной кишки
209
стоянии около 80 — 110 см от пилоруса на уровне IV поясничного
позвонка.
Особенности. У свиней поджелудочная железа желтоватого
цвета. Она состоит из головки, левой и правой (хвост железы)
долей. Выводной проток открывается на расстоянии 15-25 см от
пилоруса. У лошадей поджелудочная железа имеет розовато-се-
рый цвет, находится позади печени между желудком и двенадца-
типерстной кишкой в правом и левом подреберьях. Выводных про-
токов два. У собак поджелудочная железа имеет вид узкой и длин-
ной ленты. Выводных протоков бывает один-два, или даже три.
Контрольные вопросы
1. Перечислите органы пищеварения по ходу пищи.
2. Сколько зубов у коровы, лошади, свиньи и собаки?
3. Какие особенности строения желудка у лошади, свиньи и собаки?
4. Укажите камеры многокамерного желудка коровы.
5. Как устроена ободочная кишка у лошади, коровы и свиньи?
ОРГАНЫ ДЫХАНИЯ
К органам дыхания относятся нос с носовой полостью, глот-
ка, гортань, трахея и легкие (см. рис. 3.110 — 3.115).
Нос
Hoc (nasus) — начальный отдел дыхательного аппарата (рис.
3.110; см. рис. 3.90, 3.111). Он содержит обширную носовую по-
лость. Различают спинку, боковые стенки, корень и верхушку носа.
Спинка носа является сводом носовой полости. Она постепенно
переходит в боковые стенки носа. Корень носа лежит на границе
носа с лобной областью и является продолжением его спинки и
боковых стенок. Верхушка носа находится над верхней губой. В ней
помещаются входные отверстия в носовую полость — ноздри.
У коров они сравнительно небольшие, широко расставленные,
овальные с удлиненным дорсолатеральным углом в виде неболь-
шого крыловидного желоба. Ноздри по окружности отграничены
толстыми, малоподвижными крыльями носа. Между ноздрями кож-
ный покров формирует носогубное зеркало
Носовая полость расположена в лицевой части головы коров и
служит для проведения воздуха, определения запаха, обогрева-
ния, увлажнения и очищения воздуха. Остовом носовой полости
служат носовые, резцовые, верхнечелюстные, лобные, решетча-
тые, нёбные кости, сошник, носовые раковины, хрящевая носо-
вая перегородка и хрящи носа. Внутри остов носовой полости по-
крыт слизистой оболочкой.
210
Рис. 3.110. Нос коровы (А), лошади (Б), козы (В), овцы (Г), свиньи (Д),
собаки (£)
Носовая полость разделена носовой перегородкой на правую и
левую половины, из которых каждая имеет входное (ноздря) и
выходное (хоана) отверстия, крышу, дно, латеральную и меди-
альную стенки. Носовая перегородка представляет собой вертикаль-
ную пластинку решетчатой кости, передняя, неокостеневшая часть
этой пластинки, разделяющей носовую полость, называется хря-
щевой носовой перегородкой (см. рис. 3.90).
Внутренняя поверхность стенки носовой полости в переднем
ее отделе покрыта кожей. Эта часть полости называется преддвери-
ем носа. На границе преддверия носа и собственно носовой поло-
стью открывается отверстие слезно-носового канала. Собственно
носовая полость выстлана слизистой оболочкой. Последняя розо-
вого цвета, образует много складок, они формируют на боковых
стенках носовой полости дорсальные и вентральные раковины, а
в аборальном отделе полости — лабиринт решетчатой кости, где
цвет слизистой оболочки желтоватый.
Дорсальная носовая раковина длинная и узкая, ее слизистая обо-
лочка впереди переходит в прямую складку. Назад дорсальная но-
совая раковина простирается до продырявленной пластинки ре-
шетчатой кости. Вентральная носовая раковина широкая, ее слизи-
стая оболочка впереди в вверх образует крыловую складку. Або-
ральный конец вентральной раковины сходит на нет в области
хоан. Носовые раковины разделяют носовую полость на дорсаль-
ный, средний, вентральный и общий носовые ходы (рис. 3.111).
Дорсальный носовой ход узкий, расположен между крышей но-
совой полости и дорсальной носовой раковиной и ведет в обоня-
тельную область лабиринта решетчатой кости (рис. 3.111).
211
Средний носовой ход расположен между дорсальной и вентраль-
ной носовыми раковинами, задний его конец делится на два хода,
из среднего носового хода воздух проникает в челюстную, нёб-
ную, лобную пазухи, а также и в полости верхней и нижней но-
совых раковин (рис. 3.111).
Вентральный носовой ход широкий, расположен между вент-
ральной носовой раковиной и дном носовой полости. Вблизи хоан
он соединяется с аналогичным ходом другой стороны и ведет в
узкие, но длинные хоаны. Через носонёбный канал вентральный
носовой ход сообщается с ротовой полостью. В носонёбный канал
открывается сошниково-носовой орган (рис. 3.111).
Общий носовой ход' находится между носовой перегородкой и
носовыми раковинами. Он сообщается со всеми предыдущими
ходами. Слизистая оболочка краниальной дыхательдой части но-
12
Рис. 3.111. Сагиттальный разрез головы свиньи (ноёовая перегородка уда-
лена):
1.— хоботковая кость;! 2, 6,— вентральная носовая раковина; 3 — дорсальный
носовой ход; :.4 — дорсальна^я носовая ракопинард— средний носовой ход; (/.—
вентральный носовой хол; 8 - торная пазуха; -9 — средняя носовая раковдна;
10 — мбирюп рсшспмой косые '//. лобная кость; (7^/— головной мозг; (13)—
затылочная кость. 14 мозжечок; /5 клиновидная кость; основная йаСть
(затылочная кость); 77 — глотка: 7' — надгортанник. /9 — полъязычно-надгор-
танная мышца; ipx.nuio псипяаычная мышца. 21. 24 мяжос небо. 22-
тело подъязычной кис i i. . «ев, 25 ||илйородо’1!н^полъязьг1ная мыший; 26'—
тело языка; 27 — подборейоч!го-язычная мышца; '28]— нижняя челюсть; 29 —
подъязычный карман; 30 — твердое нёбо; 31 — преддверие рта; 32 — нижняя
губа; 33 — верхняя губа; 34 — костное нёбо; 35 — хоботок
212
Рис. 3.112. Проекция верхнечелюстной пазухи лошади в возрасте:
А — одного месяца; Б — одного года; В — четырех лет; Г — 12 лет
совой полости покрыта мерцательным эпителием и имеет розо-
вый цвет, а в каудальной обонятельной части она покрыта обоня-
тельным эпителием и имеет желтоватый оттенок. Подслизистый
слой носовой перегородки и вентральной стенки носа имеет сети
сосудов (кавернозное тело). Латеральные носовые железы у коров
отсутствуют (рис. 3.111).
Особенное! и. У свинец носовая полость относительно ут
к ля. длинная, верхушка носа обра 1ует хоботок. У лошади спинка,
боковые частя, верхушка и кореньноса хорошо выражены Нозд-
ри большие и по бокам ограничены крыльями носа. Боковая стен-
ка носа между носовыми и резцовыми костями образует мягкий
нос. На ней имеется носовой дивертикул длиной до 5 — 7 см. Вход в
дивертикул находится в дорсальной части ноздри. В вентральном
углу носа расположено отверстие слезно-носового канала. У собак
нос отличается подвижностью переднего участка.
Околоносовые пазухи (sinus paranasales) — воздухоносные поло-
сти, расположенные в основном в области лица между костными
пластинками. Внутренняя полость их выстлана слизистой оболоч-
кой. Наиболее сильно у животных развиты верхнечелюстная (гай-
морова) пазуха, лобная и клинонёбная пазухи (рис. 3.112).
Гортань
Гортань (larynx) — хрящевой непарный полый орган (рис. 3.113;
см. рис. 3.90, 3.96, 3.111). Она служит для проведения вдыхаемого и
выдыхаемого воздуха и издания звука. Остов гортани составляют
пять хрящей: кольцевидный, щитовидный, два черпаловидных и
213
Рис. 3.113. Гортань собаки (Л), свиньи (5), коровы (В), лошади (Г) (ме-
дианный разрез):
/ г- надгортанник, 2)— черпаловидно-иадгодцшная складка; J рожковый отро-
сток; 4 — кольцевидный хрящ. пластинка; 5.— трахеальные хрящи;.6 — кольце-
видный хрящ, дуга; 7 голосовая складка; Л желудочек гортани. У дтитовлл-
пый х ряш; 10 тело подъятычной кости. //— подъязычно-надгортанная мышца
надгортанный. Все хрящи гортани соединены между собой связ-
ками, а между кольцевидным и черпаловидными хрящами имеет-
ся сустав с суставной капсулой.
Кольцевидный хрящ состоит из дужки и дорсально лежащей пла-
стинки. Пластинка направляется к дужке под тупым углом и посе-
редине имеет мышечный гребень (см. рис. 3.90, 3.96, 3.111, 3.113).
Щитовидный хрящ формирует вентральную и боковые стенки
гортани. На нем различают четырехугольные боковые пластинки,
которые вентрально соединены в длинное тело щитовидного хря-
ща. На заднем конце тела хряща расположено незначительное гор-
танное возвышение, а по краям тела лежат небольшие ростральная
и аборальная вырезки (см. рис. 3.90, 3.96, 3.111, 3.113).
Черпаловидный хрящ парный, имеет вид неправильной треуголь-
ной пирамидки. Передневерхний угол его завернут крючком вверх,
состоит из эластического хряща и называется рожковым хрящом.
Вентральный угол этого хряща хорошо развит и называется голо-
совым отростком. Верхнезадний угол черпаловидного хряща при-
крепляется связкой к пластинке кольцевидного хряща (см. рис. 3.90,
3.96, 3.111, 3.113).
214
Надгортанный хрящ построен из эластического хряща, имеет
овальную форму. Он закрепляется на мембране щитовидного хря-
ща и подъязычной кости (см. рис. 3.90, 3.96, 3.111, 3.113).
Внутри гортань выстлана слизистой оболочкой. Между надгор-
танным и черпаловидными хрящами слизистая оболочка форми-
рует сильно развитые черпаловидно-надгортанные складки. На бо-
ковых стенках гортани, между голосовыми отростками черпало-
видных хрящей и телом щитовидного хряща формируются голосо-
вые складки, основу которых составляют голосовые связки и мышцы.
Голосовые складки располагаются по отношению к дну гортани
под прямым углом, а вместе с черпаловидными хрящами форми-
руют широкую голосовую щель.
Голосовые складки делят полость гортани на преддверие гор-
тани, голосовую щель и каудально расположенную собственную
полость гортани.
Преддверие гортани находится впереди голосовых складок, го-
лосовая щель располагается между черпаловидными хрящами и
голосовыми складками, полость гортани размещается позади го-
лосовых складок. Слизистая оболочка преддверия гортани и голо-
совых складок выстлана плоским многослойным эпителием, а ка-
удальной полости гортани — мерцательным.
На наружной стороне гортанных хрящей располагаются мыш-
цы гортани. Они делятся на расширители и суживатели.
Особенности. У свиней гортань длинная, надгортанный
хрящ широкий. У лошади дужка кольцевидного хряща расположе-
на под прямым’углом к пластинке^У собак кольцевидный хрящ с
обширной дорсальной пластинкой и широкой дужкой. Щитовид-
ный хрящ относительно короткий.
Трахея
Трахея {trachea) имеет форму трубки, в стенках которой зало-
жены хряЩевйе’кольца (см. рис. 3.90, 3.96, 3.113). У коров таких
колец 48 — 50. Наличие в ее стенке хрящевых колец позволяет тра-
хее быть постоянно раскрытой. Трахея начинается от гортани, тя-
нется вдоль всей шеи и в области первых ребер входит в грудную
полость. Она лежит под шейными позвонками и пищеводом или
рядом с ним. Вентрально трахея прикрыта грудино-щитовидны-
ми, грудино-подъязычными и грудино-головными мышцами. На
уровне четвертого-пятого межреберного пространства имеется
бифуркация трахеи. До бифуркации она отдает в верхушку правого
легкого трахейный {эпартериалъный) бронх. Просвет трахеи имеет
в среднем 6 — 7 см в высоту и 4 — 5 см в ширину.
Слизистая оболочка трахеи выстлана мерцательным эпители-
ем и содержит трахеальные железы.
215
Особенности. У свиней трахея цилиндрической формы, хря-
щевых колеи в ней. 32 —36. Конны колец заходят один за другой.
У лошади в трахее 4S - 60 хрящевых колец; ее длина достигает 1 м.
Бифуркация трахеи находится на уровне пятого-шестого межре-
берного пространства. У собак трахея содержит 42 — 46 хрящевых
колеи.
Легкие
Легкие (pulmones) парный орган дыхания, в котором проис-
ходит газообмен Между вдыхаемым воздухом и кровью.
Правое легкое и левое легкое включают проводящие пути — брон-
хи и их разветвления, и отделы, где непосредственно присходит
газообмен между кровью и воздухом — альвеолярные ходы, аль-
веолярные мешки и альвеолы. Оба легких располагаются вместе с
сердцем в грудной полости. Они покрыты серозной оболочкой —
плеврой и имеют форму конуса (рис. 3.114). Основание конуса об-
ращено каудально к выпуклой поверхности диафрагмы, вершина
конуса направлена краниально. 11а каждом лет ком различают: вог-
нутую — диафрагмальную поверхность, выпуклую реберную по-
верхность и средостенную поверхность. 11а средостенной поверхно-
сти находится углубление — норота легкого. Через них в легкие
входят главный бронх, легочная артерия, нервы и выходят легоч-
Рис. 3.114. Схема строения серозной
оболочки грудной полости (попе-
речное сечение грудной полости):
I — остистый отросток грузного позвон-
ка; 2 — дорсальные мышцы позвоноч-
ного столба; J спинной мозг; 4. 23
плевральная полость; 5 — аорги, 6 —
средостение; 7 — изопект. 8 — брон-
хи; 9— трахея; 70—легкие; // —пред-
сердие: 12 — межжелудочковая перего-
родка; 13 желудочек сердца; 14— ми-
окард; 15 — грудино-перикардиальная
связка; /б- кожа, 17 фас пня. 18-
рсбро; 19 — мышны туловища; 20 —
ннутригрудная фасшгя; 21 — париеталь-
ный листок серозной оболочки ре-
берная плевра; 22 — висцеральный ли-
сток серизной оболочки — легочная
плевра; 24 перикардиальная плевра;
25 фиброзный листок сердечной со-
рочки, 26 париетальный листок се-
розного перикарда; 27 - перикарлихдь-
ная полос зь. 28 - висцеральный лис-
ток серозного нерп карда зпнкард
216
Рис. 3.115. Бронхиальное древо собаки:
/ трахея (дыхательное горло); 2 кра- '\
ииальная доля левого легкого; бифур-
кация трахеи;; 4 корень легкого; 5 — сред-
няя доля левого легкого; 6 — каудальная
юля левого легкого; ~ краниальная доля
правого jo мио; л* — средняя воля правого
lei'Koin; О — главный бронх. 10 - кау.ылг.
пая доля правого легкого; 77 — сегменталь-
ные бронхи; 72 !— добавочная доля
)
ные вены. Бронх вместе с сосудами и нервами образует корень
легкого. На легком различают тупой дорсальный и острый вент-
ральный края. Вырезками левое легкое делится на переднюю кра-
ниальную, среднюю и каудальную доли. Подобными вырезками пра-
вое легкое подразделяется на верхушечную краниальную, верхушеч-
ную каудальную, среднюю, каудальную и добавочную доли. Добавоч-
ная доля лежит с медиальной стороны и отделяется от правого
легкого каудальной полой веной (рис. 3.115).
Главный бронх в каждом легком разветвляется на крупные бронхи.
От крупных бронхов отделяются средние бронхи. От средних — мел-
кие бронхи. От мелких бронхов отходят бронхиолы, которые вступа-
ют в легочные дольки. В результате разветвления главного бронха на
все более и более мелкие бронхи внутри легких формируется брон-
хиальное древо. Дольковый бронх ветвится в дольке на дыхательные
бронхиолы, которые оканчиваются альвеолярными ходами, стенки
последних образуют альвеолярные мешки и альвеолы (легочные пу-
зырьки). У коров насчитывают до 3 млрд альвеол; дыхательную
площадь считают ранной 300 м2. Параллельно с бронхами в каж-
дом легком ветвится легочная артерия, она распадается на капил-
лярные сети, оплетающие альвеолы. От капиллярных сетей начи-
наются легочные вены.
Стенка бронхов состоит из хрящевых колец и пластинок и снаб-
жена гладкими мышцами, внутри бронхи выстланы слизистой
оболочкой, покрытой мерцательным эпителием и содержащей
железы. Альвеолярные ходы, альвеолярные мешки и альвеолы со-
стоят из эластической ткани и покрыты респираторным эпите-
лием.
На легком сильно развит сетчатый рисунок. Это обусловлива-
ется сильным развитием соединительнотканных прослоек между
дольками. Правое легкое больше левого.
Особенности. У свиней на правом легком краниальная доля
па переднюю и заднюю не делится. Левое легкое имеет краниаль-
ную, среднюю и каудальную доли. На поверхности легкого сетча-
217
тый рисунок выражен, но слабее, чем у коров. У лошади дольча-
тость легкого слабо выражена. Острый край легкого имеет поло-
гую сердечную вырезку, она подразделяет каждое легкое на кра-
ниальную и каудальную доли. У правого легкого есть добавочная
доля. собак сетчатый рисунок не заметен, доли легкого отделя-
ются глубокими междолевыми вырезками, достигающими основ-
ных бронхов.
Контрольные вопросы
1. Перечислите органы дыхания.
2. Укажите хрящи гортани.
3. Сколько долей легких у лошади, кровы, свиньи и собаки?
ОРГАНЫ МОЧЕОТДЕЛЕНИЯ
К органам мочеотделения относятся: почки, мочеточники,
мочевой пузырь и мочеиспускательный канал (см. рис. 3.116 — 3.120,
3.123, 3.127).
Почки
Почки Irenes) — это парный орган красно-бурого цвета (рис.
3.116 — 3.118; см. рис. 3.120, 3.125). У коров почки бороздчатые мно-
гососочковые.
На почках различают два конца — краниальный и каудальный,
два края — латеральный и медиальный и две поверхности — дор-
сальную и вентральную. Каудальный конец почки шире и толще
краниального, ее латеральный край выпуклый, опущен вниз, а
медиальный прямой или слегка вогнутый, направленный дорсаль-
но. На медиальном, вогнутом крае почки располагается почечная
ямка. С поверхности почки покрыты фиброзной капсулой. Поверх
фиброзной капсулы лежит жировая капсула.
На продольном разрезе почки различают три зоны: мочеотде-
лительную, мочеотводящую и пограничную. Мочеотделительная
зона находится по периферии почки, окрашена в темно-красный
цвет. Мочеотводящая зона помещается в центре почки, имеет бо-
лее светлую окраску. Пограничная зона простирается между двумя
предыдущими зонами, имеет интенсивно красную окраску.
В пограничной зоне расположены дуговые артерии и вены —
ветви междолевых артерий и вен, берущие начало от почечных
артерии и вены. От дуговых артерий в мочеотделительную зону
отходят междольковые артерии. На их боковых ветвях образуются
многочисленные сосудистые клубочки.
Сосудистые клубочки располагаются в чашеобразных углубле-
ниях почечных капсул, вместе с которыми они и формируют по-
218
Рис. 3.116. Скелетотопия почек коровы (А), свиньи (Б) (вентральная по-
верхность):
/ — правый надпочечник; 2 — правая почка: 3 — почечная артерия и вена; 4 —
левый надпочечник; 5 — левая почка; 6— мочеточник; 7 — брюшная аорта; 8 —
каудальная полая вена. Th XII —Th XIV — положение грудных позвонков; L I —
L V — положение поясничных позвонков
Рис. 3.117. Скелетотопия почек собаки (Л), овцы (5) (вентральная поверх-
ность):
/ — правый надпочечник; 2 — правая почка; 3 — почечная артерия и вена; 4 —
левый надпочечник; 5 — левая почка; 6 — мочеточник; 7 — брюшная аорта; 8 —
каудальная полая вена. Th XII —Th XIII — положение грудных позвонков;
L I —L VI — положение поясничных позвонков
219
Th XVI
Th XVII
Th XVIII
LI r
LII
L III л 1
L IV
Рис. 3.118. Скелетотопия по-
чек лошади (вентральная по-
верхность):
) правый надпочечник; 2 —
правая почка; 3 — почечная ар-
терия и вена; 4 — левый надпо-
чечник; 5 — левая почка; 6 —
мочеточник; 7 — брюшная аор-
та; 8 — каудальная полая вена
Th XVI —Th XVIH — положе-
ние грудных позвонков; L I —
L IV — положение поясничных
позвонков
чечные тельца. От почечных телец начинаются извитые мочеотде-
лительные канальцы, в целом представляющие мочеотделитель-
ную зону почек. Извитые канальцы переходят в прямые канальцы,
в корковой зоне из них складываются мозговые лучи. Прямые ка-
нальцы направляются в мочеотводящую зону, придавая ей ради-
альную исчерченность. Здесь они соединяются в собирательные
канальцы, открывающиеся на поверхности почечного сосочка. Все
сосочки вершинами обращены в почечную ямку и свисают в обо-
собленные почечные чашечки, имеющиеся на концах разветвлен-
ных стеблей мочеточника. Стебельки мочеточника в пределах по-
чечной ямки соединяются в два крупных концевых хода. После-
дние сливаются друг с другом и впадают в мочеточник. Число со-
сочков варьирует (от 18 до 22).
Правая почка располагается в поясничной области от ХП реб-
ра до второго-третьего поясничного позвонка. Дорсально она при-
лежит к большой поясничной мышце и правой ножке диафраг-
мы, ее передний конец находится в почечном углублении печени.
Вентрально она граничит с поджелудочной железой, а также со
слепой и ободочной кишками.
Левая почка подвешена на собственной брыжейке и перемеща-
ется вправо и влево на уровне второго — пятого поясничного по-
звонка. От длительного давления рубца на латеральной поверхности
почки, ближе к ее переднему концу, имеется рубцовое углубление.
Особенности. J/ свиней почки гладкие многососочковые,
бобовидной формы. У лошадей почки гладкие однососочковые. Ле-
вая почка продолговатая, бобовидная, а правая — сердцевидная.
У собак почки гладкие однососочковые.
Мочеточники
Мочеточники (иге teres) выходят из почечной ямки и направ-
ляются назад к тазу (см. рис. 3.116 —3.120, 3.123, 3.125). У самцов
220
мочеточники лежат в мочеполовой складке брюшины, на дор-
сальной стенке мочевого пузыря они сближаются и вблизи шей-
ки впадают в его полость. У самок мочеточники помещаются в
широкой маточной связке, подходят к мочевому пузырю сбоку
матки.
Слизистая оболочка мочеточников выстлана многослойным
переходным эпителием и содержит слизистые мочеточниковые
железы. Мышечная оболочка построена из двух продольных слоев
и одного циркулярного слоя гладких мышечных волокон. Снару-
жи мочеточники покрыты соединительной тканью — адвентици-
ей и серозной оболочкой.
Мочевой пузырь
Мочевой пузырь (vesica urinaria) имеет грушевидную форму, в
ненаполненном состоянии размером приблизительно с кулак (рис.
3.119; см. рис. 3.120, 3.123, 3.125). Он лежит в тазовой полости над
лонными костями, под мочеполовой складкой (у самцов) или
под маткой и влагалищем (у самок).
На мочевом пузыре различают тело пузыря, суженную, направ-
ленную каудально шейку пузыря, округлую, обращенную в брюш-
ную полость верхушку пузыря. Стенка мочевого пузыря состоит из
слизистой, мышечной и серозной оболочек.
Слизистая оболочка пузыря довольно толстая, выстлана пере-
ходным эпителием, в опорожненном пузыре она собрана в мно-
гочисленные складки. Мышечная оболочка образует три слоя глад-
ких мышечных волокон: наружный продольный, средний кольцевой
Рис. 3.119. Соединение мочеточника и мочевого пузыря:
Л — умеренное наполнение мочевого пузыря: 5?— максимальное наполнение
мочевого пузыря: С— мочеточникJ2 — полость мочевого пузыря; /3 )— стенка
мочевого пузыря;1,'4)— шейка мочевого пузыря
221
и внутренний продольный. На шейке пузыря мышечные пучки фор-
мируют сфинктер пузыря.
Серозная оболочка покрывает спереди вершину и тело мочевого
пузыря.
Мочеиспускательный канал
Мочеиспускательный канал (urethra) начинается от шейки мо-
чевого пузыря, тянется назад и открывается: у самок на вент-
ральной стенке влагалища, образуя на границе между влагали-
щем и мочеполовым преддверием дивертикул уретры, у самцов —
на головке полового члена. Стенка мочеиспускательного канала
состоит из слизистой, мышечной и соединительнотканной обо-
лочек (см. рис. 3.119, 3.120, 3.124-3.127, 3.129, 3.130, 3.132-
3.134).
ОРГАНЫ РАЗМНОЖЕНИЯ САМЦОВ
К органам размножения самцов относятся: семенники, при-
датки семенников, семенной канатик, семяпровод, семеннико-
вый мешок, мочеполовой канал, придаточные половые железы,
половой член и препуций (рис. 3.120 — 3.122; см. рис. 3.123 — 3.126).
Семенники (testes) — парные половые железы, в которых выра-
батываются спермин и половые гормоны (рис. 3.120 — 3.122, см.
рис. 3.125, 3.126). У быков они крупные, имеют форму удлинен-
ных вертикальных эллипсоидов. На каждом семеннике различают
два конца — головчатый и хвостатый, два края — придатковый и
свободный и две поверхности — латеральную и медиальную.
Остов семенника состоит из фиброзной белочной оболочки. Она
врастает в семенник со стороны головки придатка, глубоко вне-
дряется в толщу органа и образует средостение семенника толщи-
ной 2 — 8 мм. От средостения к белочной оболочке отходят пере-
городки семенника, они разделяют семенник на множество ка-
мер, в которых заложены извитые семенные канальцы, составля-
ющие паренхиму семенника. Длина канальца достигает 75 см при
диаметре 0,1 —0,2 мм. Извитые канальцы переходят в прямые се-
менные канальцы, последние располагаются в средостении семен-
ника и формируют в нем семенниковую сеть. Паренхима семен-
ника у быка желтоватого цвета. Трабекулы слабо развиты. Снару-
жи семенник покрыт специальной влагалищной оболочкой. Эта обо-
лочка, кроме семенника, покрывает и его придаток, а затем пе-
реходит в брыжейку семенника, при помощи которой семенник
прикрепляется к общей влагалищной оболочке семенникового
мешка.
222
Рис. 3.120. Схема строения и взаимосвязи органов мочеотделения
и репродуктивных органов самца:
7 - мочеточ1П1к; 2 предстательная железа; 3 — прямая кишка; 4, 77,7- семя-
провод Д — мышна-огтягнвагель пенис:,; кавернозное тетйГпёниса; 7^—
придаток семенника; Д' — семенник; ^ 7- семенной кан.чтнк; 10 - раковый канал;
12 — препуций; 13' — полость препуция; 14 кость пениса: '7.5/ — луковица
головки полового члена;(1д\— моченой пузырь: /7 почка
Особенност и. У хряков семенники эллипсоидальной фор-
мы, продольная ось их направлена косо вверх и назад, цвет па-
ренхимы семенников сероватый. У жеребцов семенники имеют
эллипсоидальную, слегка сдавленную с бокоФформу. Перегород-
ки, извитые и прямые канальцы, семенниковая сеть и гайморово
тело хорошо заметны. У кобелей семенники имеют округло-эл-
липсоидную форму. Длинная ось их идет дорсокаудально. Средо-
стение хорошо развито.
Придаток семенника (epididymis) у быка расположен вдоль при-
даткового края семенника. На нем различают головку, тело и
хвост. Головка придатка лежит дорсально, она образуется несколь-
кими выносящими канальцами, которые впадают в проток придат-
ка. Каждый выносящий каналец вместе с покрывающими его обо-
лочками имеет коническую форму и составляет маленькую дольку
придатка. Тело придатка образуется сильно извивающейся частью
протока придатка. Хвост придатка соединен с семенником специ-
альной связкой семенника, а с общей влагалищной оболочкой и
мошонкой — паховой связкой.
Снаружи придаток семенника покрыт специальной влагалищ-
ной оболочкой, переходящей на него с семенника. На латеральной
поверхности между придатком и краем семенника эта оболочка
формирует синус придатка (см. рис. 3.120 — 3.122, 3.125, 3.126).
223
Рис. 3.121. Семенниковый мешок
и его оболочки:
1 — кожа Моин, 1>.1' _мышечно-
эластичес км ооо io* ка О/.—ня|
ная семенная фасция. !. 10 — общая
влагалищная оболочка: 5. .? — семя-
нронол; л «. 14- специальная влага-
лищная оболочка: 7 — мышпа-под-
ниматель семенника; 12 — перегород-
ка мошонку; 13 — хвост придатка;
15 — семяпровод
Семенной канатик (funiculus
spermaticus) у быка длинный,
округлый, основанием прикреп-
лен к головчатому концу семен-
ника и к головке придатка, а вер-
шиной оканчивается в области
брюшного пахового кольца. В кра-
ниальной части он содержит внут-
реннюю семенную артерию, не-
рвы, вены и лимфатические со-
суды. Здесь вены многократно
анастомозируют друг с другом и
вместе с артериями формируют
«и удистое сплетение. Внутренний
подниматель семенника развит
слабо. Все эти части окутываются
собственной влагалищной обо-
лочкой. На медиальной поверхно-
сти семенного канатика проходит
семяпровод. В области внутренне-
го пахового кольца семенной ка-
натик «рассыпается» (см. рис.
3.120-3.122, 3.125, 3.126).
Семяпровод (ductus deferens)
выходит из хвоста придатка и
является прямым продолжением
канала придатка семенника (см.
рис. 3.120-3.123, 3.125, 3.126).
В составе семенного канатика он
направляется через паховый канал в брюшную полость.
Вблизи шейки мочевого пузыря семяпровод соединяется с про-
током пузырьковидной железы и переходит в семяизвергающий
канал. Стенка конечной части семяпровода, лежащей над моче-
вым пузырем, утолщена за счет скопления в ней желез. Этот уча-
сток семяпровода называется ампулой семяпровода. Правый и ле-
вый семяизвергающие каналы открываются в мочеиспускатель-
ный канал, который после этого называется мочеполовым каналом.
Семяпровод у быка незначительного диаметра и тонкостенный.
Железистая часть его достигает в длину 12 см и в толщину 1,5 см.
Семенниковый мешок (saccus testicularis) состоит из мошонки,
наружного поднимателя семенника и общей влагалищной обо-
лочки (см. рис. 3.120 — 3.123, 3.125, 3.126). Мошонка у быков распо-
лагается впереди краниального края лонных костей. Несколько
выше семенника выделяется шейка мошонки. Мошонка состоит из
тесно сросшихся кожи и мышечно-эластической оболочки. Кожа
мошонки розоватого цвета, иногда пигментирована. Мышечно-эла-
224
стическая оболочка является прямым продолжением желтой фас-
ции живота. За счет названной оболочки образуется перегородка
мошонки. В каждой из них заключен семенник вместе с придатком
п семенным канатиком.
Общая влагалищная оболочка состоит из двух прочно сросшихся
между собой листков — наружного фиброзного и внутреннего се-
розного. Семенник покрыт висцеральным листком брюшины —
специальной влагалищной оболочкой. Между общей и специальной
влагалищными оболочками имеется влагалищная полость, в кото-
рой находится небольшое количество серозной жидкости. Влага-
лищная полость сообщается с перитонеальной полостью через
влагалищный канал.
Особенности.,У хряков мо-
шонка расположена недалеко от зад-
непроходного отверстия, семенники
располагаются в ней косо, кожа мо-
шонки покрыта редкими вол осам и.З7.
жеребцов мошонка располагается у
края лонных костей, под мышечно-
пластической оболочкой находится
наружный подниматель семенника. У
кобелей мошонка расположена вбли-
ш заднепроходного отверстия.
Мочеполовой канал (canalis urogeni-
Kili.s.s. urethra masculina) служит для
выведения мочи и спермы (см. рис.
3.120, 3.124—3.126). Он подразделяется
на тазовую и удовую части.
Тазовая часть мочеполового кана- ’
на у быка длиной 10—12 см. Она ле-
жит на лонных и седалищных костях.
Здесь в просвет мочеполового кана-
ла открываются протоки предста-
тельных и луковичных желез. Сюда
же впадают семяпроводы и пузырь-
ковидные железы.
Удовая часть мочеполового канала
расположена на вентральной повер-
хности полового члена. Она полнос-
тью окружена белочной оболочкой
11ещеристых тел полового члена. Мо-
чеполовой канал состоит из трех обо-
лочек — слизистой, сосудистой и
мышечной.
Слизистая оболочка собрана в про-
дольные складки, покрыта много-
8 Зеленевский
Рис. 3.122. Семенник на про-
дольном разрезе:
/ - лаюнилпос венозное сплете-
ние; 2 — семявыносяшля прото-
ка; 4 проток придатка; 4)- тело
придатка; 5- хиост придатка: Дг-
извитые семенные канальцы; ?)—
специальная влаташшпая и белом
нам ободочки; Ж — головка при-
датка; V — выносящие канальцы
семенника.. 10 сеть семенника
// средостение семенники; /2
прямые семенные канальцы
225
Рис. 3.123. Придаточные половые железы жеребца (А), быка (Б), хря-
ка (В), кобеля (Г):
1)— мочеточник; 2 — семявыносящий проток; 3 — мочевой пузырь; 4)~ ампула
семявыносящего протока; '5 — пузырьковидная железа; .6 — предстательная же-
леза;['7 — уретра;, б1/— бельбоуретральная железа;;^— корень пениса
слойным переходным эпителием. Сосудистая оболочка, или кавер-
нозное тело, в тазовой части уретры развита слабее, чем в удовой.
Остов его построен из соединительной ткани, пронизанной мно-
гочисленными эластическими и гладкими мышечными волокна-
ми. В остов заложено густое венозное сплетение с кавернами, вы-
стланными эндотелием. При эрекции кавернозное тело набухает,
просвет канала начинает зиять, благодаря чему создается свобод-
ный проход для спермы.
Мышечная оболочка мочеполового канала образована толстым
слоем преимущественно поперечной счерченных мышечных воло-
кон. Мышечные волокна в тазовой части уретры формируют моче-
половую мышцу, в удовой части — луковично-кавернозную.
Придаточные половые железы открываются в тазовую часть
мочеполового канала. К ним относятся пузырьковидная, предста-
тельная и луковичная железы (рис. 3.123; см. рис. 3.120, 3.125, 3.126).
Пузырьковидная железа (glandula vesicularis) парная, имеет буг-
ристую поверхность. Она размещается сбоку и дорсально от шей-
ки мочевого пузыря. Снаружи железа покрыта толстой фиброз-
ной капсулой с мышечными волокнами. От капсулы в толщу
железы отходят трабекулы, разделяющие ее на дольки. Главные
выводные протоки пузырьковидных желез открываются в моче-
половой канал вместе с семяпроводами (см. рис. 3.120, 3.123, 3.125,
3.126).
Особенности. У хряков пузырьковидные железы большие,
имеют дольчатое строение, цвет бледно-розовый. Снаружи желе-
226
ii.i покрыты тонкой фиброзной капсулой, выводные протоки сли-
паются в общий проток, который открывается на семенном хол-
мике. У жеребцов пузырьковидные железы имеют вид гладких ме-
шочков грушевидной формы. Стенка мешочка построена из сли-
шстой, мышечной и серозной оболочек. Выводной проток каж-
дой железы сливается с семяпроводом в общий семяизвергающий
проток, открывающийся на семенном холмике. У кобелей пузырь-
ковидные железы отсутствуют.
Предстательная железа (prostata) состоит у быков из неболь-
шого тела и рассеянной части. Тело предстательной железы распо-
лагается поперек шейки мочевого пузыря, а рассеянная часть
железы лежит между слизистой и мышечной оболочками уретры.
Дорсальный участок ее толще (10—12 мм), чем вентральный (2 мм).
Железу хорошо видно на поперечном разрезе. Протоки предста-
тельной железы открываются в мочеполовой канал.
Особенности. У хряков тело предстательной железы дости-
1ает в ширину 2,0 —2,5 см. Оно лежит дорсально над шейкой мо-
чевого пузыря и прикрыто пузырьковидными железами, рассеян-
ная часть железы крупная, У жеребцов предстательная железа рас-
положена над шейкой мочевого пузыря. Она состоит из правой и
левой долей и среднего участка — перешейка. Железа каждой сто-
роны открывается 16—18 выводными протоками. У кобел ей пред-
стательная железа относительно большая, желтоватого оттенка,
плотная и дольчатая. Медиальной бороздой она разделена на две
доли. Рассеянная часть железы слабо развита (см. рис. 3.120, 3.123,
3.125, 3.126).
Луковичная железа (glandula bulbourethralis) парная, у быка не-
большая (2,8 —1,8 см), снаружи покрыта толстым слоем фиброз-
ной ткани и луковично-кавернозной мышцей. Каждая железа имеет
по одному протоку. Протоки открываются в мочеполовой канал,
их отверстия прикрыты складками слизистой оболочки (рис. 3.120,
3.123, 3.125, 3.126).
Особенности. У хряков луковичные железы очень большие
(12 см), плотные, бугристые, лежат дорсолатерально от уретры. У
жеребцов луковичные железы прикрыты луковично-кавернозной
мышцей, они овальной формы, длиной около 4 см, каждая желе-
за открывается 5 — 8 протоками. У кобелей луковичная железа от-
сутствует.
Половой член (уд) (penis) (рис. 3.124; см. рис. 3.120, 3.123 — 3.126)
быка цилиндрической формы, длинный и тонкий. Он состоит из
пещеристых тел и удовой части мочеиспускательного канала. Пе-
щеристые тела развиты у быков слабо. Поэтому половой член у
них при эрекции увеличивается в объеме незначительно. Различа-
ют три пещеристых тела: два — собственно полового члена и
одно — мочеиспускательного канала. Наружным слоем пещерис-
того тела является белочная оболочка, состоящая из фиброзной
227
Рис. 3. ] 24. Поперечное сечение пениса жеребца (А — дистальнее корня;
Б — в средней части; В — у головки):
/ — белочная оболочка кавернозных тел; 2 —перегородка пениса; 3 — каверноз-
ное тело; 4 — губчатое тело пениса; 5 — луковично-губчатая мышца; 6 — оттяги-
ватель пениса; 7 — уретра; 8 — дорсальный отросток головки пениса
ткани. Она в толще полового члена формирует срединную перего-
родку, от которой отходят в разные стороны многочисленные тра-
бекулы — перегородки.
На половом члене различают корень, тело и головку. Корень
полового члена образуется двумя ножками, которые прикрепля-
ются к седалищным буграм. Каждая ножка покрыта седалищно-
кавернозной мышцей. Эта мышца, сокращаясь, препятствует отто-
ку венозной крови из пещеристых тел при эрекции. Корень поло-
вого члена двумя подвешивающими связками прикрепляется к шву
седалищных костей.
Тело полового члена тонкое, длинное, цилиндрической фор-
мы. Позади мошонки на половом члене имеется S-образный изгиб.
Во время эрекции этот изгиб выпрямляется. К каудальному концу
изгиба прикрепляется ретрактор полового члена. Головка полово-
го члена заострена и закручена влево. На головке полового члена
различают отросток мочеполового канала, шейку головки, колпачок
головки и венчик головки. Колпачок имеет розовый цвет и гладкую
поверхность.
Особенности. У хряка половой член длиной до 40 — 45 см,
имеет S-образный изгиб,"который лежит краниальнее мошонки,
головка члена спиралеобразно закручена, наружное отверстие мо-
чеполового канала лежит рядом с заостренным концом полового
члена, ретрактор полового члена тянется от третьего-четвертого
сегмента крестца до вентрального колена S-образного изгиба, у
жеребцов половой член сильно развит. Ножки образуют корень
228
полового члена, они начинаются от седалищных бугров и при-
крыты седалищно-кавернозной мышцей. Тело полового члена мас-
сивное, перегородкой разделяется на две симметричные полови-
ны, белочная оболочка толстая, на вентральной стороне полово-
го члена формирует продольный желоб для мочеполового канала.
У кобелей член в каудальном участке имеет два хорошо развитых
пещеристых тела. В краниальный участок полового члена, или го-
ловку, заложена кость полового члена, достигающая в длину 8 —
10 см.
Препуций (praeputium) представляет собой кожную складку,
состоящую из двух листков: париетального и висцерального (рис.
3.125, 3.126; см. рис. 3.120, 3.126).^У быков препуций длинный (35 —
40 см) и узкий (до 3 см). Отверстие препуция лежит на 4 — 5 см
каудальнее пупка. Снаружи оно окружено длинными волосами.
Внутренний (париетальный) листок препуция выстлан многослой-
ным плоским эпителием и заключает в себе трубчатые железы,
висцеральный листок окрашен в розовый цвет и лишен желез. Лим-
фатические узелки располагаются в каудальной части препуция.
Препуций быков имеет две препуциальные мышцы — краниаль-
ную и каудальную, которые тянут его начальную часть вперед или
назад.
Особенности. У хряков препуций открывается узким пре-
пуциальным отверстием, в краниальной части его дорсальной стен-
ки имеется круглое отверстие, ведущее в слепой мешок (до 12 см
шириной) — дивертикул препуция. У^жсребцов различают наруж-
ный и внутренний препуций, каждый из них состоит из двух —
поверхностного и глубокого —
листков и имеет два — наружное
и внутреннее — препуциальных
отверстия. У кобелей препуций
формирует полый кожный чехол
Рис. 3.125. Органы размножения
быка:
/ бу-lьбоуре гралышя железа: 2у—
предстательная же.кча; .? ^мпуда
семявыносящего протока; / пу-
(ырышвилная железа; 5 мочеточ-
ник b — семявыносящий проток. 7 —
мочевой путырь v седалищно-ка-
вернозная мыпны ч— оттягиватсль
пениса.-/0 сигмовидной изгиб пе-
ниса; II — семенник; 12 ~ иритагок
семенника; /2 - головки пениса
229
Рис. 3.126. Органы размножения
хряка:
/ — правая подвздошная кость; '2 —
придаток семенника;. 3 — мошонка; $)—
семявыносящий проток; 5 — сигмовид-
ный изгиб полового члена; пенис;
! 7’— препуций; ‘8,— дивертикул препу-
ция; '9 — мочевой пузырь; 10)— пузырь-
ковидная железа, скрывающая под собой
предстательную железу; 11 — бульбо-
уретральная железа
вокруг краниальной части чле-
на. Наружным листком препуция
является кожа, внутренний ли-
сток тонкий, розового цвета,
лишен желез и содержит боль-
шое число лимфатических узел-
ков.
ОРГАНЫ РАЗМНОЖЕНИЯ САМОК
К органам размножения самок относятся: яичники, маточные
трубы, матка, влагалище, преддверие влагалища, половые губы и
клитор (рис. 3.127 — 3.134).
Рис. 3.127. Схема строения и взаимосвязи органов мочеотделения
и репродуктивных органов самки:
11— яичник;^— мочеточник: з)— шейка матки; 4 — влагалище;(5/— преддверие
влагалища; б'— клитор; 7.— мочеиспускательный канал; 8 — мочевой пузырь; 91—
рог матки; 10 — маточная труба; !11 V почка
230
I iiiiyipcHifHft яичниковый нерп; 2— яичниковая вена; 3 — яичниковая арте-
рия. 4 лимфатический сосуд; 5 брюшина, б1- мозговая (сосудистая* зона
яичника; 7 — зачатковый эпителий; Ц белейшая оболочка; 9 пузырьковид-
ный фолликул (граафов пузырек); 10 — полость граафова пулырькл. заполнен-
ная фолликулярной жидкостью; 1 i — лучистый пенен. /2 первичная оболоч-
ка; 13 — ядро яйцеклетки, _Н яйценосный бугорок; 15 — интерстициальные
клетки; /6яйцеклетка; 17>— атретическнй фолликул; /<?— зпоричный фолли-
кул; 19— неркичные фолликулы; 2rf— желтое тело; 21 - брыжейка
Яичники (ovarium) — парные половые органы, в которых обра-
зуются яйцеклетки и половые гормоны (см. рис. 3.127 — 3.130,
3.132 — 3.134). Специальных выводных протоков яичники не име-
ют. Половые клетки выделяются периодически, когда разрывают-
ся созревшие в яичниках фолликулы.
У короту яичники овально-удлиненной формы, сравнительно
небольших размеров, располагаются в области поясницы за поч-
ками. Снаружи они покрыты зачатковым эпителием. На разрезе
яичника видны фолликулярная и сосудистая зоны. Фолликулярная.
зона лежит более поверхностно и содержит много фолликулов, в
которых развиваются яйцеклетки. Сосудистая зона представляет
собой соединительнотканный остов с многочисленными сосуда-
ми и нервами.
Особенност и. У свиней яичник относительно большой (до
5 см в длину), округлый^ слегка вытянутой формы, бугристый,
закреплен на яичниковой брыжейке и специальной яичниковой
связке. У кобыл яичник большой (5 — 8 см), бобовидной формы,
231
у молодых кобыл он больше, чем у старых. Свободный край яич-
ника вогнут, на нем имеется овуляционная ямка. У сук яичник
имеет удлиненную (2 см) и слегка сдавленную с боков эллипсо-
идную форму. Брыжейка яичника и яичниковая связка образуют
бурсу яичника.
Маточная труба {tuba uteri па) — тонкая, извитая трубка, слу-
жащая в основном для'выведения зрелых яйцеклеток, образую-
щихся в яичнике (см. рис. 3.127, 3.129, 3.130, 3.132 — 3.134). Длина
ее у коров незначительная — от 25 до 30 см.
На маточной трубе различают два конца — яичниковый и ма-
точный. Яичниковый конец расширен, имеет вид воронки. Края
воронки называются бахромкой маточной трубы. Часть бахромки,
соединенную с краниальным краем яичника, называют бахром-
кой яичника. В суженной части воронки помещается брюшное от-
верстие маточной трубы. Этим отверстием маточная труба откры-
вается в перитонеальную полость.
Маточный конец маточной трубы открывается маточным от-
верстием в полость рога матки. Стенка маточной трубы состоит из
слизистой, мышечной и серозной оболочек. Серозная оболочка
образует брыжейку яйцевода.
Матка (uterus) — толстостенный мышечный орган, в котором
развивается зародыш (см. рис. 3.127, 3.129, 3.130, 3.132 — 3.134).
У коров матка двурогого типа, в ней различают рога, тело и шейку.
Рогц.матки начинаются от ее тела, идут вперед и вниз, слегка
отклоняясь в латеральную сторону; они спирально изогнуты в
дорсальном направлении и по форме напоминают рога барана.
В задней своей части рога соединяются межроговыми связками.
Рис. 3.129. Типы маток:
А — двойная (кролик); Б — днхро.аи с перегородкой (ларрОКШШшнс); В —
Двурепя Гкч licpvlopo.lKH (Ko6bii.ii. 1 — ip.Ki.ia (человек) ,/ — воронка маточ-
ной трувы; - мвго’ишя груба: 3 — poi матки. 4 гсЛО НГГКИ;/5Э“ шейка
матки: 6 - влагалище; 7 — преддверие
232
Рис. 3.130. Репродуктивные органы
коровы (дорсальная поверхность,
стенка вскрыта):
1 — яичник;2 — миомная груба. 2 —
норовка маючиой грубы. J — рог мат-
ки. 4 межроговая связка; 5'- перего-
родка; 6 -- стенка тела матки; 7 1— тело
матки; А шейка матки; 0— влагалищ-
ная порция шейки; 10 — свод влагали-
ща^/Л)— влагалище;. Q2)— положение
девственной плевы; .13 — наружное от-
верстие уретры и ихрл в подуретраль-
ный дивертикул;' 14 L большая железа
преддверия и отверстие ее протока; /5 V
премверие цзагалища; So'— головка
клитора., 17 - правая губа вульвы
Рис. 3.131. Развитие карункулов матки коровы:
1 — карункулы в небеременной матке; 2 — карункул в матке двухнедельной
стельности; 3 — карункул в матке шестимесячной стельности; 4 — карункул
матки стельной коровы и плодные оболочки; 5 — разрез плаценты коровы; 6 —
плацента овцы
233
Рис. 3.132. Репродуктивные орга-
ны кобылы (дорсальная поверх-
ность):
1 — яичник; 2 — маточная труба; 3 —
рог матки; 4 — тело матки; 5 — шей-
ка матки; 6 — свод влагалища; 7 —
влагалищная порция шейки; 8— вла-
галище; 9 — преддверие влагалища;
10 — вульва; 11 — правая губа; 12 —
головка клитора; 13 — стенка пред-
дверия; 14 — правая губа; 15 — пре-
пуций клитора; 16 — головка клитора
Мышечная оболочка (myomet
Тело матки снаружи достигает
в длину 10—15 см, но внутри на
протяжении примерно 10 см де-
лится продольной перегородкой
пополам, в связи с чем неразде-
ленная часть тела матки не пре-
вышает у коров 5 — 6 см.
Шейка матки сравнительно
длинная (7 — 10 см), толстостен-
ная; она небольшой частью вда-
ется в полость влагалища, обра-
зуя влагалищную долю (порцию)
матки. В шейке матки имеется ше-
ечный канал с двумя отверстиями.
Наружное маточное отверстие
открывается во влагалище, а внут-
реннее маточное отверстие в по-
лость матки.
Стенка матки состоит из трех
оболочек: внутренней — слизис-
той, средней — мышечной и на-
ружной — серозной. Слизистая
оболочка — endometrium — выст-
лана цилиндрическим эпителием.
В ней имеются многочисленные
маточные железы. Секретом маточ-
ных желез первое время питается
зародыш. У коров на слизистой
оболочке рогов и тела матки выс-
тупают четыре ряда возвыше-
ний — маточных карункулов.
чт) состоит из внутреннего слоя
циркулярных и наружного слоя продольных мышечных пучков
гладких мышечных волокон. В области шейки матки циркулярные
пучки формируют сфинктер.
Серозная оболочка (perimetrium) окружает матку снаружи, с бо-
ковых поверхностей матки серозная оболочка переходит в птиро-
кую маточную связку — маточную брыжейку. Брыжейка удержива-
ет матку в брюшной полости, в ней проходят артерии, вены, не-
рвы.
Особенности. У кобыл матка двурогая, рога направлены
вперед и завершаются тупыми округленными концами, каждый
рог имеет форму пологой дуги с выпуклым краем, обращенным
вентрально. У свиней матка двурогая, рога матки весьма длинные
(до 200 см) и узкие, образуют многочисленные петли, целиком
лежат в брюшной полости, подвешены на маточной брыжейке,
234
тело матки длиной до 5 см и лежит
между рогами и шейкой. Оно прибли-
зительно в 3 раза короче шейки, шей-
ка матки длиной до 15—18 см пред-
ставляет суженную часть матки, не-
заметно переходит во влагалище. На
слизистой оболочке шейки матки
имеются многочисленные (14 — 20)
складки. Канал шейки волнообразный
и крепко замкнутый. У сук матка дву-
рогая, отличается длинными, тонки-
ми и прямыми рогами, расходящи-
мися в виде цифры V, шейка матки
толстостенная, короткая, вдается во
влагалище.
Влагалище (vagina) — непарный
орган в виде мышечной трубки (см.
рис. 3.127, 3.129, 3.130, 3.132-3.134).
Длина его у коров 22 — 28 см. Стенка
влагалища состоит из слизистой обо-
лочки, мышечного слоя и серозной
оболочки или соединительнотканной
адвентиции.
Слизистая оболочка выстлана мно-
гослойным плоским эпителием и со-
брана в глубокие продольные складки.
Мышечный слой состоит из внутрен-
них циркулярных и наружных продоль-
ных пучков гладких мышечных воло-
кон. Серозной оболочкой покрыта
лишь краниальная часть влагалища,
вся остальная его часть - соединитель-
нотканной адвснтицией.
Преддверие влагалища (vestibulum
vaginae) находится в тазовой полос-
ти под прямой кишкой (см. рис. 3.127,
Рис. 3.133. Репродуктивные
органы свиньи (дорсальная
поверхность, стенка вскрыта):
/, 10 — рог матки; 2 — тело мат-
ки; 3 — шейка матки; 4 — канал
шейки матки; 5 — наружное
отверстие матки; 6 — влагалище;
7 — наружное отверстие уретры;
8 — преддверие влагалища; 9 —
головка клитора; 11 — широкая
маточная связка; 12 — яичник;
13 орыжеик.1 маточной трубы;
14 — маточная груба; 15 — мо-
чевой пузырь; 16 левая губа
вульвы
3.129, 3.130, 3.132-3.134).
Оно ограничено от влагалища поперечной складкой — девствен-
ной плевой. На вертикальной стенке преддверия ясно заметно вы-
пячивание, в котором помещается отверстие мочеиспускательно-
го канала, под выпячиванием находится слепой мешок — дивер-
тикул мочеиспускательного канала.
Половые губы (labia vulvae) состоят из кожи, сжимателя вуль-
вы и слизистой оболочки (см. рис. 3.127, 3.130, 3.132 — 3.134). Они
образуют дорсальный и вертикальный углы. Дорсальный угол за-
кругленный, вертикальный — заостренный. От вертикального угла
у коров свисает пучок длинных волос.
235
Рис. 3.134. Репродуктивные органы
суки (дорсальная поверхность):
1 — яичниковая бурса; 2 — маточная
труба; 3 — вход в яичниковую бурсу; 4 —
рог матки; 5 — широкая маточная связ-
ка; 6 — тело матки; 7 — влагалищная
порция шейки; 8 — адвентициальная
оболочка влагалища; 9 — мышечная обо-
лочка влагалища; 10 — слизистая обо-
лочка влагалища; 11 — полость влагали-
ща; 12 — наружное отверстие уретры;
13 — железы преддверия; 14 — преддве-
рие влагалища; 75 — головка клитора
Клитор (clitoris) состоит из пе-
щеристого тела, он начинается от
седалищных бугров двумя ножками,
прикрытыми седалищно-каверноз-
ной мышцей (см. рис. 3.127, 3.130,
3.132 — 3.134). Сливаясь вместе, нож-
ки формируют тело клитора дли-
ной до 12 см, которое заканчива-
ется головкой клитора. Она прикрыта
препуцием клитора.
АНГИОЛОГИЯ
Ahiио.'Ю!пя (angiologia) раздел анатомии, изучающий строе-
ние сердечно-сосудистой системы. Последняя включает кровенос-
ную и лимфатическую системы, условно объединяемые органами
кроветворения (рис. 3.135 — 3.155).
КРОВЕНОСНАЯ СИСТЕМА
Кровь (haema, s. sanguis) — основная функционирующая и со-
ставная часть кровеносной системы. Она представляет собой жид-
кую ткань, состоящую из плазмы и клеточных элементов. В плазме
растворены питательные и подлежащие выведению из организма
вещества. К клеткам крови относятся красные кровяные тельца —
эритроциты, переносчики кислорода. Белые кровяные клетки —
лейкоциты — имеют разнообразное строение и выполняют защит-
ную функцию.
Кровь может выполнять свои функции только при условии дви-
жения ее в кровеносных сосудах по двум кругам: по большому
(системному) и малому (легочному).
236
Большой круг кровообращения — это путь крови из левого желу-
дочка сердца в его правое предсердие (рис. 3.135). Он начинается
из левого желудочка сердца самым крупным артериальным сосу-
дом аортой. От нее ко всем органам отходят артерии. Отток веноз-
ной крови от органов и тканей происходит по краниальной и
каудальной полым венам. Последние впадают в правое предсер-
дие, где и заканчивается большой круг кровообращения. Необхо-
димо помнить, что от всех органов пищеварения, расположенных
в брюшной полости (исключая каудальную часть прямой кишки),
отток венозной крови осуществляется в воротную вену печени.
Отток крови из печени происходит по печеночным венам, от-
крывающимся в каудальную полую вену (подробнее см. на с. 262).
Из правого предсердия кровь поступает в правый желудочек
сердца, откуда начинается малый круг кровообращения — путь
крови из правого желудочка в левое предсердие через капилляры
легких.
Рис. 3.135. Круги кровообращения:
/ правое предсердие; 2 правый желудочек; •’ — левое предсердие; 4 - левый
желудочек: 5 - дуга аорты л легочный ствол; 7 каузальная полая ясна. 8 —
краниальная полая вена; v — яремная вена, 10 - подмышечная артерия; II
сосуды легких; 12 — по .мышечная вена. 13 сосуды органов грудной конечно
сти; /4 — сосуды органов головы. 15 — общая сонная артерия; /й плече-
головной ствол; /7 легочные вены. IS печеночная артерия; 14 - печеноч-
ная вена; 20 воротная вена печени: 21 — чревная артерия. 22 грудная аорта.
23 клудатьная полая вена; 24 левая желудочная артерия; 25 перикард
(сердечная сорочка); 2b |руднно-перикарлн.ыьная снягка; 27 - сосуды печени;
28 — сосуды желудка; 2*1 сосуды селезенки; 30 селе геномная артерия; 31
брыжеечная артерия. 32 — почечная артерия. 33 почечная вена. 34 брюш
ная аорта; 35 — наружная подвяюшная артерия; 36 кишка. 37 — внутренняя
полнхюшная вена. 38 наружная полиглотная вена: 34 внутренняя под
вхюшнпя артерия; 40 — сосуды органов таловой полости и таговой стенки. 41
сосуды органов тазовой конечности
237
Малый круг кровообращения (рис. 3.135) начинается из правого
желудочка стволом легочных артерий. Последний над сердцем де-
лится на правую и левую легочные артерии, вступающие через
ворота легких в его ткани. В капиллярах легких происходит газооб-
мен, а насыщенная кислородом кровь по легочным венам возвра-
щается в левое предсердие. Из левого предсердия кровь поступает
в левый желудочек, т. е. к началу большого круга кровообраще-
ния.
Сердце
Сердце (cor, kardia) — центральный орган сердечно-сосудис-
той системы (рис. 3.136 — 3.140), располагается в грудной полости
и заключено в перикард (околосердечную сумку, сердечную со-
рочку).
Перикард представляет собой прочный соединительнотканный
мешок, в котором помещается сердце (рис. 3.136; см. рис. 3.114). Он
состоит из двух серозных и заключенного между ними фиброзно-
го листков. Наружный серозный листок представляет собой учас-
ток средостенной плевры и называется перикардиальной плеврой.
Последняя спускается на грудную кость,
формируя грудино-перикардиальную связ-
ку, а затем без видимых границ перехо-
дит на грудную стенку в виде реберной
плевры.
Внутренняя серозная выстилка око-
лосердечной сумки образуется специаль-
ной серозной оболочкой, которая фор-
мирует околосердечную серозную по-
лость и называется серозным перикардом.
На нем различают два листка: висцераль-
ный покрывает стенку сердца, образуя
его наружную оболочку — эпикард. Он
поднимается по сердцу дорсально и у
его основания переходит в париеталь-
ный листок. Последний выстилает внут-
реннюю поверхность сердечной сороч-
ки, замыкается вентрально, формируя
щелевидную полость перикарда, запол-
ненную небольшим количеством сероз-
ной жидкости.
Между двумя серозными листками сер-
дечной сорочки со стороны грудной кос-
ти проникает внугригрудная фасция, об-
разуя перикардиальный фиброзный листок.
Рис. 3.136. Схема строения
сердечной сумки:
/ — средостенная плевра; 2 —
фиброзный листок перикар-
да; 3 — париетальный листок
серозного перикарда; 4 — пе-
рикардиальная полость; 5 —
висцеральный листок сероз-
ного перикарда; 6 — сердце;
7— грудино-перикардиальная
связка; 8 реберная плевра
238
Рис. 3.137. Сердце лошади:
4 — левая поверхность: I — общим плсчс-нмонной ствол: 2 линия прикрепле-
ния перикарда;.? - артериальная связка. 4 - дуга аорты; 5 легочный ствол; л —
правая легочная артерия, 7— легочные вены; 8 — венечная борозла. 9 — левое
предсердие, ушко предсердия: К) — левый желудочек; II верхушка сердца;
12 - паракональная межжелудочковая борозда, ветвь левой венечной артерии,
большая сердечная пена. А? правый желудочек; 14 — субэпикардиальный жир
в венечной борозде; 15 — ушко правого предсердия;
Б — правая поверхность: /, 4 — легочные вены; 2, 3 — левая и правая легочные
артерии; 6 — правая непарная вена; 7 — общий плече-головной ствол; 8 —
краниальная ползя вена; 9 — правое предсердие 10 - линия прикрепления
перикарда; 11 правый же-луцочек; верчмпка сердца; 13 — ясшаи Жвлудр
чек; 14 — субсинуозная межжелудочковая борозда, ветвь правой венечной арте-
рии, средняя сердечная вена. 15 — большая сердечная вена; 16 — каудальная
полая вена; 17 - левое предсердие
Сердце — это темно-красный полый, конусовидный, мышеч-
ный орган, приводящий в движение кровь и лимфу (рис. 3.137 —
3.140). На нем различают основание и верхушку, левую и правую
поверхности, передний и задний края.
У основания сердца проходит венечная борозда, которая охва-
тывает его кольцом. В ней находятся правая и левая венечные арте-
рии. От венечной борозды по правой и левой повсрхност и в сторо-
ну верхушки тянутся правая и левая продольные борозды, а на зад-
нем крае сердца тянется добавочная продольная борозда (рис. 3.137).
Стенки сердца состоят из эндокарда, миокарда и эпикарда.
Эндокард — внутренняя оболочка сердца — состоит из соеди-
нительной ткани и покрыта эндотелием. Миокард — средняя обо-
лочка — состоит из сердечной мышечной ткани, мышечные пуч-
ки предсердий и желудочков не смешиваются, а отделяются фиб-
розными кольцами. Различная толщина стенок предсердий и же-
239
лудочков зависит от толщины миокарда. Эпикард — наружная се-
розная оболочка сердца (рис. 3.138).
Сердце перегородками делится на четыре полости. Две из них
расположены у основания сердца — дорсально от венечной бо-
розды и две вентрально. Первые две полости — правое и левое
предсердия, а вторые — правый и левый желудочки (см. рис. 3.137,
Рис. 3.138. Сердце лошади (вскрыты
правое предсердие и правый желу-
дочек):
1 — овальная ямка; 2 — межвенозный
бугорок; 3 — устье правой непарной
вены; 4— краниальная полая вена; 5 —
пограничный гребень, пограничная бо-
розда; 6 — эндокард правого предсер-
дия; 7 — трабекулы ушка; 8 — гребеш-
ковые мышцы; 9 — правое предсердно-
желудочковое отверстие; 10— правая ве-
нечная артерия; 11 — угловая створка
грехш ворчагого клапана; /2 — сухо-
жильная струна; 13 — передняя септаль-
ная сосочковая мышца; !4 — трабеку-
ла; 15 — задняя септальная сосочковая
мышца; /6 — верхушка сердца; /7—тра-
бекулы; 18 — правая поперечная сер-
дечная мышца; /9 — перегородковая
створка трехстворчатого клапана; 20 —
сухожильные струны; 22 — пристенная
створка трехстворчатого клапана; 23 -
правая венечная артерия; 24 — венеч-
ный синус (устье большой сердечной
вены); 25 — каудальная полая вена;
26 — межжелудочковая перегородка
3.138). Правое предсердие со-
единяется с правым желудоч-
ком, а левое предсердие — с
левым желудочком. Сообщения
правой половины сердца с ле-
вой не существует. Только в пе-
риод внутриутробного развития
оба предсердия сообщаются
между собой через овальное
отверстие, которое вскоре пос-
ле рождения зарастает, остав-
ляя после себя след в виде
овальной ямки, заметной на пе-
регородке правого предсердия.
Предсердия располагаются на
основании сердца и отграниче-
ны от желудочков венечной бо-
роздой (см. рис. 3.137, 3.138).
Каждое предсердие имеет тон-
кие стенки и образует слепое
выпячивание — правое и левое
сердечное ушко. Под эндокардом
предсердий за счет миокарда
образуются гребешковые мышцы.
Правое предсердие в верхней
стенке имеет отверстие пере-
дней и задней полых вен. Первое
из них находится в передневер-
хней стенке предсердия, а вто-
рое — на противоположной сто-
роне. Кроме того, в правом пред-
сердии находятся отверстия
малых сердечных вен, большой
сердечной вены, а также отвер-
стия левой непарной вены. Часть
предсердия, располагающаяся
между устьями полых вен, об-
разует мясистое валикообразное
утолщение — межвенозный бу-
горок. Правое предсердие и пра-
240
вый желудочек сообщаются че-
рез обширное правое атриовен-
трикулярное отверстие.
Левое предсердие в дорсальной
стенке имеет четыре —семь от-
верстий для легочных вен, кото-
рые открываются в три лакуны:
правую, левую и центральную,
несущие насыщенную кислоро-
дом кровь из легких. Выходное
отверстие из него — левое атрио-
вентрикулярное — ведет в левый
желудочек.
Желудочки занимают боль-
Рис. 3.139. Клапаны сердца лошади
при диастоле (удалены легочный
ствол, дуга аорты, оба предсердия):
1 — правая венечная артерия; 2— пра-
вый атриовентрикулярный (трехствор-
чатый) клапан; 3 — правое предсерд-
но-желудочковое отверстие: 4— венеч-
ный синус 5 — клапан аорты (полу-
лунный); 6— левый атриовентрикуляр-
ный (двухстворчатый, митральный)
клапан; 7 — левая венечная артерия;
8 — клапан легочного ствола (полулун-
ный)
шую часть сердца и лежат вент-
рально от венечной борозды (см.
рис. 3.137, 3.138). Внутренняя по-
верхность их покрыта эндокар-
дом, под которым миокард об-
разует мышечные перекладины,
поперечные и сосочковые мышцы.
К концам сосочковых мышц
прикрепляются сухожильные
струны, идущие к концам ство-
рок атриовентрикулярных кла-
панов. Снаружи стенки желудоч-
ков покрыты эпикардом.
Правый желудочек направлен
вперед и вправо (см. рис. 3.138). Стенка его значительно толще
стенки предсердий. Внутри желудочка расположены три сосочко-
вые мышцы. Отверстие, ведущее из правого предсердия в желудо-
чек — правое атриовентрикулярное, окружено фиброзным кольцом и
прикрыто трехстворчатым клапаном. Створки последнего явля-
ются складками эндокарда. Основание створок прикрепляется к
фиброзному кольцу, а свободные края обращены в желудочек и
сухожильными струнами прикреплены к сосочковым мышцам.
Поэтому при сокращении желудочка, захлопнувшись, они не могут
выворачиваться в сторону предсердия. В правом желудочке нахо-
дится также артериальное отверстие, которое ведет в легочную ар-
терию. Оно также окружено фиброзным кольцом. Его прикрывает
полулунный клапан легочной артерии, состоящий из трех створок.
Выпуклая поверхность створки обращена в желудочек, а вогну-
тая — в сторону сосуда. Свободные края створок имеют узелки,
способствующие лучшему закрыванию отверстия. Полулунный кла-
пан закрывается при расслаблении желудочка и препятствует воз-
вращению крови (рис. 3.139).
241
Левый желудочек лежит сзади и слева. Стенка его в 2,5 раза толще
стенки правого желудочка. На ее внутренней поверхности имеются
две сосочковые мышцы. Из левого предсердия в правый желудочек
ведет левое атриовентрикулярное отверстие. Оно прикрыто двухствор-
чатым клапаном, состоящим из двух створок с сухожильными стру-
нами (рис. 3.139). Аортальное отверстие ведет в аорту, оно прикры-
то полулунным клапаном аорты. В основе атриовентрикулярного от-
верстия находится фиброзное кольцо, в котором лежат две сердечные
косточки. Из них правая длиной 5 — 6 см, имеет вид изогнутой пла-
стинки треугольной формы, служит местом прикрепления правой
задней полулунной створки. К левой косточке, длиной 2 — 3 см,
прикрепляется левая задняя полулунная створка.
Сердце помещается в средостении от третьего до шестого реб-
ра. На 5/7 оно сдвинуто влево и между третьим и четвертым реб-
рами прилежит к левой грудной стенке. Основание сердца лежит
под шестым грудным позвонком. Верхушка сердца находится про-
тив пятого реберного хряща, не достигая грудины на 2 см и впе-
реди диафрагмы на 2 —5 см.
Синовентрикулярная (проводящая, нервномышечная) система сер-
дца обеспечивает согласованную работу камер сердца (см. рис. 4.2).
Она образована атипичными кардиомиоцитами. В ней различают:
два узла — синоаурикулярный и атриовентрикулярный, атрио-
вентрикулярный пучок и волокна Пуркинье.
Синоаурикулярный (синусный) узел бледно-розового цвета, ле-
жит в пограничной борозде, расположенной между передней по-
лой веной и правым сердечным ушком. Форма его серповидная,
длина 3,5 см, ширина 1 см и толщина 3 мм.
Атриовентрикулярный (предсердный) узел лежит вентрально от
овальной ямки близ устья большой сердечной вены. Краниаль-
ным концом узел переходит в атриовентрикулярный пучок.
Атриовентрикулярный пучок имеет вид тяжа длиной 0,9— 1,2 см,
шириной 5 — 7 мм. На перегородке желудочков пучок делится на
правую тонкую и левую толстую ножки. Правая ножка в составе
правых поперечных мышц сердца переходит на его стенку, левая
ножка по левым поперечным мышцам сердца подходит к основа-
нию сосочковых мышц. В дальнейшем они делятся до волокон
Пуркинье.
Симпатические нервы сердца происходят из шейно-грудного
(звездчатого) узла, а от блуждающего нерва отходят парасимпати-
ческие нервы (рис. 3.140).
Сосуды сердца представлены правой и левой венечными артерия-
ми, большой, средней и малыми сердечными венами.
Особенности. У свиней сердце с притупленной верхушкой.
Овальное отверстие в 20 % случаев не зарастает. В аортальном фиб-
розном кольце находится один сердечный хрящ. У лошадей верхуш-
ка сердца более тупая, чем у коров. В аортальном фиброзном коль-
242
6
5
Рис. 3.140. Иннервации сердца собаки:
7 — позвоночный нерв; 2 — шейно-грудной (звездчатый) ганглий; 3 — шейно-
грудной каудодорсальный сердечный нерв; 4 — третий грудной ганглий (симпа-
тический); 5 — грудные сердечные нервы; 6 седьмой грудной ганглий (симпа-
тический); 7— блуждающие краниальный и каудальный сердечные нервы; 8 —
левый возвратный гортанный нерв; 9 — левое предсердие; 10 — левый желудо-
чек; /7 — артериальный конус (правый желудочек); 12 — позвоночные сердеч-
ные нервы; 13 — шейно-грудной кау до вентральный сердечный нерв; 14 — сред-
ний шейный ганглий (симпатический); 15 — вагосимпатический ствол (блужда-
ющий нерв + симпатический ствол); 16 — симпатический ствол; 17 — подклю-
чичная петля
це от одного до трех сердечных хрящей, у старых животных они
могут окостеневать. У собак сердце более округлое и с более при-
тупленной верхушкой. При устье задней полой вены, на границе
с устьем большой сердечной вены, располагается клапан задней
полой вены.
Артерии и вены малого круга кровообращения
Малый круг кровообращения начинается из правого желудоч-
ка легочным стволом (см. рис. 3.135). Легочный ствол выходит из
артериального конуса и сначала направляется каудодорсально.
У корня легкого он делится на две ветви для правого и левого
легкого. Каждая из них располагается вентролатерально от ма! ис-
грального бронха и полностью повторяет его ветвления. Гемомик-
243
роциркуляторное русло образует сеть вокруг легочных альвеол,
где и происходит газообмен.
Отток артериальной крови от легких осуществляют легочные
вены. Впадают они в левое предсердие, формируя три лакуны:
правую, левую и центральную.
Артерии большого круга кровообращения
Большой круг кровообращения начинается самым крупным арте-
риальным сосудом — аортой (см. рис. 3.135, 3.141). Последняя вы-
ходит из левого желудочка и сразу же над полулунным клапаном
образует незначительное расширение — синус аорты.
Дуга аорты (см. рис. 3.135, 3.141, 3.144) располагается справа
от легочной артерии и поднимается дорсально к пятому (шесто-
му) грудному позвонку, а затем переходит в грудную аорту. В об-
Рис. 3.141. Схема расположения кровеносных сосудов свиньи:
7 — наружная челюстная артерия; 2 — наружная сонная артерия; 3 — глубокая
шейная артерия; 4 — поперечная артерия шеи; 5 — плечеголовная артерия; 6 —
легочный стол; 7 — дуга аорты; 8 — общая сонная артерия; 9 — яремная вена;
10 — подмышечная артерия; 11 — подмышечная вена; 12 — краниальная полая
вена; 13 — сосуды грудной конечности; 14 — внутренние грудные артерия и
вена; 15 — сердце; 16 — печень; 17 — желудок; 18 — каудальная полая вена; 19 —
печеночная вена; 20 — воротная вена печени; 21 — селезенка; 22 — кишечник;
23 — чревная артерия; 24 — селезеночная артерия; 25 - брыжеечная вена; 26 —
почечная вена; 27 — брыжеечная артерия; 28 — межреберные артерии и вены; 29,
30 — брюшная аорта; 31 — каудальная полая вена; 32 — окружная подвздошная
глубокая артерия; 33 — наружная подвздошная артерия; 34 — средние крестцо-
вые артерия и вена; 35 — внутренняя подвздошная артерия и вена; 36 — над-
чревно-срамной ствол; 37, 38 — артерии и вены тазовой конечности
244
нети створок полулунного клапана от луковицы аорты отходят
правая и левая венечные артерии. Помимо венечных артерий от дуги
порты отходит крупный сосуд — общий плечеголовной ствол.
Общий плечеголовной ствол направляется от дуги аорты крани-
ипыю (см. рис. 3.135, 3.141). Разветвляясь наряд сосудов, он снаб-
жает кровью переднюю часть грудной стенки, шею, голову и груд-
ные конечности. На уровне второго грудного позвонка от общего
плечеголовного ствола отходит в левую сторону левая подключич-
ная артерия, а продолжающийся сосуд называется плечеголовной
артерией. Плечеголовная артерия, отдав общий ствол общих сонных
артерий, переходит в правую подключичную артерию. Каждая под-
киючичная артерия отделяет: общий реберно-шейный ствол, внут-
реннюю грудную артерию, плечешейный ствол и наружную груд-
ную артерию. После ответвления от подключичной артерии на-
ша иных сосудов продолжающийся ствол называется подмышеч-
ной артерией. Последняя снабжает кровью грудную конечность.
Общий реберно-шейный ствол (см. рис. 3.141) отдает переднюю
межреберную, поперечную шейную, глубокую шейную артерии
и переходит в позвоночную артерию, передняя межреберная арте-
рия идет назад вдоль четырех позвонков и отдает вторую, третью,
четвертую (пятую) межреберные артерии для спинного мозга, меж-
реберных мышц, разгибателей спины, поперечная артерия шеи
впереди первого ребра идет к холке и питает ее мышцы, глубокая
шейная артерия лежит впереди первого грудного позвонка, пита-
ет разгибатели шеи и головы.
Позвоночная артерия идет в межпозвоночном канале и отдает в
каждом сегменте спинно-мозговую, дорсальные и вентральные
мышечные ветви (см. рис. 3.141).
Внутренняя грудная артерия идет вниз и назад по грудной кос-
ти до седьмого-восьмого реберного хряща (см. рис. 3.141). Она пи-
тает тимус, средостение, поперечную грудную мышцу, сердеч-
ную сорочку, грудные мышцы и кожу, межреберные мышцы, ди-
афрагму и сердечную сорочку. Мышечно-диафрагмальная артерия
питает реберную часть диафрагмы, поперечную брюшную мышцу
и анастомозирует с межреберными артериями. Краниальная над-
чревная артерия идет назад по брюшной стенке и анастомозирует
с каудальной надчревной артерией.
Плечешейный ствол идет в плечеголовную, лестничную, плече-
подъязычную и поверхностную грудную мышцы.
Наружная грудная артерия тонкая, огибает первое ребро и пи-
тает глубокую грудную мышцу (см. рис. 3.141).
Артерии головы
Общая сонная артерия в области затылочного сустава делится
па наружную и внутреннюю сонные артерии (рис. 3.142). От общей
245
16
Рис. 3.142. Скелетотопия артерий головы лошади (поверхностный пласт):
I — резцовая артерия; 2 — артерия преддверия носа; 3 — верхняя губная арте-
рия; 4 — латеральная, 5 — дорсальная артерии носа; 6 - подглазничная артерия;
7— артерия угла глаза; 8 — латеральная артерия нижнего века; 9 — надглазнич-
ная артерия; 10 — поверхностная височная артерия; 11 — поперечная артерия
лица; /2 — общий ствол поверхностной височной и поперечной артерии лица;
13 — щечная артерия; 14 — артерия большой жевательной мышцы; 75 — лицевая
артерия; 16 — каудальная артерия мозговых оболочек; 17 — общий ствол лате-
ральной и средней ушной артерий; 18 — затылочная ветвь; 19 — нисходящая
ветвь; 20 — каудальная артерия мозговых оболочек; 21 — затылочная артерия;
22 — внутренняя сонная артерия; 23 — глубокая ушная артерия; 24 — большая
ушная артерия; 25 — ветвь околоушной железы; 26 — восходящая глоточная
артерия; 27— левая общая сонная артерия; 28— краниальная щитовидная арте-
рия; 29 — средняя артерия нижнечелюстной железы; 30 — наружная сон-
ная артерия; 31 — лицевая сосудистая вырезка; 32 — артерия нижней губы;
33 -- подбородочная артерия
сонной артерии, кроме мышечных, пищеводных и трахеальных
ветвей, отходят следующие артерии: краниальная щитовидная,
глоточная, гортанная и внутренняя сонная.
Краниальная щитовидная артерия (рис. 3.142) отдает ветвь к глот-
ке глоточной артерии, а сама переходит в гортанную артерию. Гло-
точная артерия питает мышцы глотки. Гортанная артерия питает
мышцы гортани. Внутренняя сонная артерия имеется только у те-
лят. Она проходит через переднюю часть рваного отверстия в че-
репную полость, где и участвует в образовании чудесной сети.
У взрослых животных внутренняя сонная артерия редуцирована.
Наружная сонная артерия является продолжением общей сонной
артерии (рис. 3.142). Она отделяет следующие артерии: затылочную,
наружную челюстную, большую ушную, жевательную, поверхност-
ную височную. После ответвления названных артерий наружная сон-
ная артерия переходит во внутреннюю челюстную артерию.
246
Затылочная артерия слабо развита, от нее ответвляются следу-
ющие ветви: восходящая нёбная артерия, нисходящая ветвь, сред-
няя артерия мозговой оболочки, затылочная ветвь и мыщелковая
артерия. Восходящая нёбная артерия идет в стенку глотки и в нёб-
ную занавеску, нисходящая ветвь очень тонкая, в крыловом от-
верстии атланта она анастомозирует с позвоночной артерией, сред-
няя артерия мозговой оболочки тонкая, проходит через рваное от-
верстие, затылочная ветвь идет в короткие разгибатели головы,
мыщелковая артерия направляется через подъязычное отверстие в
черепную полость для образования чудесной сети.
Наружная челюстная артерия развита слабо, отделяет язычную
артерию, ряд мышечных ветвей и переходит в лицевую артерию.
От лицевой артерии отходят верхняя и нижняя губные артерии.
Большая ушная артерия достигает ушной раковины (рис. 3.142).
От нее отделяются артерия лицевого канала (питает среднее ухо),
ютеральная ушная ветвь (питает кожу ушной раковины), глубо-
кая ушная артерия (питает кожу внутренней поверхности ушной
раковины), общий ствол средних ушных ветвей (питает кожу уш-
ной раковины).
Жевательная артерия (рис. 3.142) питает жевательную мышцу
и околоушную железу.
Поверхностная височная артерия направляется в височную об-
ласть (рис. 3.142). Она отдает поперечную лицевую, добавочную
мозговой оболочки, поверхностную слезную и общий ствол для
артерий рога. Поперечная лицевая артерия питает жевательные
мышцы; добавочная артерия мозговой оболочки идет через височ-
ный канал и анастомозирует с мыщелковой артерией; поверхнос-
тная слезная артерия дает ветви к верхнему и нижнему веку, к
нобной мышце и слезной железе; общий ствол для артерий рога
целится на латеральную и медиальную ветви.
Верхнечелюстная артерия является продолжением наружной
.'онной артерии после ответвления от нее поверхностной височ-
ной артерии (рис. 3.142). Она направляется в клинонёбную ямку.
От внутренней челюстной артерии отходят следующие артерии:
нижняя зубная, глубокие височные, щечная, ветви для чудесной
мозговой сети, наружная глазничная, нижнего века, подглазнич-
ная, клинонёбная, большая нёбная.
Нижняя зубная артерия следует в нижнечелюстной канал, вы-
ходит как подбородочная артерия и направляется в нижнюю губу,
в нижнечелюстном канале она отдает веточки для всех коренных
зубов и десен нижней челюсти.
Глубокие височные артерии направляются в височную мышцу.
Щечная артерия разветвляется в жевательной и щечной мыш-
цах, от нее отделяется артерия глазничного жира.
Ветви для чудесной мозговой сети вступают в черепную полость
через овальное и глазнично-круглое отверстия.
247
Наружная глазничная артерия образует чудесную сеть, которая
лежит на зрительном нерве. Из чудесной сети выходят: ветвь глаз-
ного яблока, слезная глубокая артерия, ветви для глазных мышц,
Артерия нижнего века сильно развита, начинается общим ство-
лом из подглазничной артерии и выходит из орбиты по вырезке
на слезной кости. В орбите она отдает: мышечные ветви, артерию
третьего века и назальную артерию нижнего века. По выходе из
орбиты от артерии нижнего века отделяются: артерия угла глаза и
дорсальная носовая артерия.
Подглазничная артерия (рис. 3.142) питает верхние зубы и по
выходе из подглазничного канала дает латеральную носовую арте-
рию для мышц и кожи лица.
Клинонёбная артерия вступает в носовую полость и образует
сплетение.
Большая нёбная артерия отдает малую нёбную артерию для нёб-
ной занавески, а сама переходит на твердое нёбо, где одной вет-
вью направляется в зубную пластинку, а другой проникает через
нёбную щель в носовую полость.
Артерии грудной конечности
Подмышечная артерия является продолжением подключичной
артерии. Она огибает первое ребро, лежит на медиальной поверх-
ности плечевого сустава (рис. 3.143; см. рис. 3.141). У коров подмы-
шечная артерия впереди сустава отдает акромиальную артерию, а
позади сустава делится на подлопаточную и плечевую артерии. Ак-
ромиальная артерия идет дорсально и питает предостную и подло-
паточную мышцы, капсулу сустава и плечевую кость.
Особенности. У свиней и собак акромиальная артерия от-
сутствует, подмышечная артерия делится на подлопаточную и пле-
чевую артерии. У лошадей ветвление подмышечной артерии такое
же, как у коров.
Подлопаточная артерия лежит между подлопаточной и боль-
шой круглой мышцами (рис. 3.143). На своем пути подлопаточная
артерия отдает окружную латеральную, грудоспинную, окружную
лопаточную артерии и мышечные ветви.
Окружная плечевая латеральная артерия сильно развита, она
отделяет нисходящую ветвь для низания трехглавой мышцы, на-
прягателя фасции предплечья, плечевой мышцы.
Грудоспинная артерия (рис. 3.143) идет назад и питает большую
круглую, широчайшую спинную, кожную туловищную мышцы.
Окружная лопаточная артерия питает подлопаточную, заост-
ную, дельтовидную, предостную мышцы и отдает питающую ар-
терию лопатки.
Мышечные ветви питают подлопаточную, большую круглую и
трехглавую мышцы.
248
I’iic. 3.143. Артерии грудной конечно-
сти лошади:
1.4— подлопаточная артерия; 2 — мы-
шечные ветви; 3 — грудоспинная артерия;
- плечевая артерия; 6 — глубокая арте-
рия плеча; 7 — возвратная межкостная ар-
нрия; 8 — срединная артерия; 9 — колла-
кральная локтевая артерия; 10 — латераль-
ная глубокая пальмарная пястная артерия;
II. 12 — поверхностная пальмарная пяст-
ная артерия; 13 — медиальная дорсальная
пястная артерия; 14 — вторая общая паль-
марная пальцевая артерия; 15 — медиаль-
ная пальцевая артерия; 16— венечная ар-
к-рия; 17 — окружная лопаточная артерия;
18 — акромиальная артерия; 19 — каудаль-
ная окружная плечевая артерия; 20— под-
мышечная артерия; 21 — наружная груд-
ная артерия; 22 — краниальная окружная
п лечевая артерия; 23 — артерия двуглавой
мышцы плеча; 24 — коллатеральная лок-
ц'вая артерия; 25 — коллатеральная луче-
вая артерия; 26 — мышечная ветвь к раз-
шбателям запястья; 27 — общая межкост-
ная артерия; 28— артерия пальмарной сети
запястья
Плечевая артерия (рис. 3.143) спускается косо по медиальной
поверхности плеча и локтевого сустава, отдает общую межкост-
ную артерию и переходит в срединную артерию. Она отдает следу-
ющие артерии:
окружная плечевая медиальная артерия выходит впереди плече-
вой кости и анастомозирует с окружной плечевой латеральной
артерией, питает клювовидно-плечевую, глубокую грудную и дву-
главую мышцы;
глубокая плечевая артерия тонкая, питает трехглавую мышцу
плеча (рис. 3.143);
артерия двуглавой мышцы тонкая, начинается в области дис-
тальной трети плеча и васкуляризирует одноименную мышцу;
коллатеральная локтевая артерия (рис. 3.143) идет медиально
от локтевого отростка и питает трехглавую мышцу, напрягатель
фасции, поверхностную грудную мышцу и кожу предплечья;
249
коллатеральная лучевая артерия (рис. 3.143) идет на сгибатель-
ную поверхность локтевого сустава, питает капсулу локтевого су-
става, плечевую мышцу, лучевой разгибатель запястья, общий
пальцевой разгибатель, длинный абдуктор большого пальца, кожу
и дорсальную сеть запястья;
возвратная локтевая артерия слабо развита у коров, питает
разгибатели запястья и пальцев (рис. 3.143);
общая межкостная артерия ответвляется от плечевой артерии,
через проксимальную межкостную щель выходит на латеральную
поверхность предплечья и отдает возвратную межкостную арте-
рию, дорсальную межкостную артерию, мышечные ветви.
После ответвления этих артерий и ветвей общая межкостная
артерия продолжается как пальмарная межкостная артерия. Пос-
ледняя располагается в межкостном желобе, дает крупную дор-
сальную ветвь в дорсальную сеть запястья и в кожу пясти. Ее более
тонкая пальмарная ветвь идет через дистальную щель на пальмар-
ную поверхность запястья и анастомозирует с локтевой и средин-
но-лучевой артериями. Затем она спускается как латеральная паль-
марная пястная (глубокая) артерия и вливается в четвертую об-
щую пальмарную пальцевую артерию.
Срединная артерия является продолжением плечевой артерии
после ответвления от нее общей межкостной артерии, она спус-
кается на пясть, отдает срединно-лучевую артерию и делится на
вторую, третью и четвертую пальмарные поверхностные пястные
артерии, которые на дистальном конце пясти переходят во вто-
рую, третью и четвертую общие пальмарные пальцевые артерии
(рис. 3.143).
Вторая общая пальмарная пальцевая артерия после соединения
со второй пястной пальмарной глубокой артерией отдает соедини-
тельную ветвь к четвертой общей пальмарной пальцевой артерии
и образует глубокую пальмарную дугу.
Далее вторая общая пальмарная пальцевая артерия переходит
в специальную медиальную пальмарную третью пальцевую арте-
рию.
Четвертая общая пальмарная пальцевая артерия сливается с чет-
вертой пальмарной глубокой пястной артерией, принимает соеди-
нительную ветвь от второй пальцевой артерии, а сама переходит в
специальную латеральную четвертую пальмарную пальцевую арте-
рию.
Третья общая пальмарная пальцевая артерия идет между глав-
ными пальцами, отдает общий ствол для обеих мякишных арте-
рий главных пальцев, затем ответвляет пальмарные ветви для пер-
вых фаланг и общий ствол для дорсальных пальцевых артерий и
делится на латеральную третью специальную пальмарную пальце-
вую артерию и медиальную четвертую специальную пальмарную
пальцевую артерию, которые идут в копытцсвые кости (рис. 3.143).
250
Срединно-лучевая артерия ответвляется от срединной артерии
на середине предплечья, отдает проксимальную пястную пробода-
ющую артерию, которая в свою очередь дает начало средней паль-
марной пястной глубокой артерии. Последняя вливается в глубокую
пальмарную дугу, анастомозирует с пальмарной ветвью межкост-
ной артерии и выходит через проксимальный межкостный канал
пясти на ее дорсальную поверхность, где соединяется с третьей
дорсальной пястной артерией. Отдав прободающую артерию, сре-
цинно-лучевая артерия идет на пясти как вторая пальмарная пяс-
тная глубокая артерия.
Дорсальная сеть запястья образуется ветвями срединно-луче-
вой, локтевой и обеими межкостными артериями. Из сети выхо-
дит дорсальная третья пястная артерия, которая получает под-
крепление от срединной и срединно-лучевой артерий, а между
пальцами — и от третьей пальмарной общей пальцевой артерии.
Она дает специальные дорсальные пальцевые артерии — латераль-
ную третью и медиальную четвертую.
Артерии грудной и брюшной аорты
Грудная аорта тянется под телами грудных позвонков, проходит
через аортальное отверстие диафрагмы в брюшную полость и там
। (азывается брюшной аортой (рис. 3.144). От грудной аорты у коров
отходят межреберные, бронхиальная и пищеводная артерии.
Рис. 3.144. Ветви грудной и брюшной аорты:
/, 25 — левая подключичная артерия; 2 — правая и левая общие сонные арте-
рии; 3 — общий ствол сонных артерий; 4 — правая подключичная артерия; 5 —
реберно-шейный ствол; 6 — правые дорсальные межреберные артерии; 7— пра-
вые поясничные артерии; 8 — внутренняя семенная артерия; 9 — правая окруж-
ная подвздошная глубокая артерия; 10 — правая наружная подвздошная арте-
рия; 11 — правая внутренняя подвздошная артерия; 12 — средняя крестцовая
артерия; 13 — бедренная артерия; 14 — левая наружная подвздошная артерия;
15 — левая окружная подвздошная глубокая артерия; 16 — каудальные брыжееч-
ные артерии; 17 — левые поясничные артерии; 18 — краниальная брыжеечная
артерия; 19 — чревная артерия; 20 — диафрагмальная артерия; 21 — левые дор-
сальные межреберные артерии; 22 — пищеводная и бронхиальная артерии;
23 — дуга аорты; 24 — правая и левая венечные артерии
251
Рис. 3.145. Схема ветвления чревной артерии свиньи (А), собаки (А),
лошади (В):
1 — чревная артерия; 2 — селезеночная артерия; 3 — селезенка; 4 — левая желу-
дочная артерия; 5 — печеночная артерия; 6 — правая желудочная артерия; 7 —
правая, 8— левая желудочно-сальниковые артерии; 9 — ветви к поджелудочной
железе; 10 — желудочные вс>ви; // — селезеночные ветви; 12 — желудочно-
двенадцатиперстная артерия; 13 — желудок; 14 — желудочные ветви
Межреберные артерии (рис. 3.144) парные, выходят из аорты в
количестве 10 пар. От них отделяются дорсальные и спинно-мозго-
вые ветви. Первые из них снабжают кровью разгибатели спины и
кожу, а вторые входят через межпозвоночные отверстия в позвоноч-
ный канал, анастомозируют между собой, питая спинной мозг.
Бронхиальная артерия отходит от грудной аорты в области 5-го
грудного позвонка, снабжает кровью легкие (рис. 3.144).
Пищеводная артерия разветвляется в пищеводе.
Брюшная аорта (рис. 3.144) является продолжением грудной
аорты позади диафрагмы. От нее отходят каудальные диафрагмаль-
ные, чревная, краниальная и каудальная брыжеечные, почечные,
внутренние семенные и поясничные артерии.
Каудальная диафрагмальная артерия парная, ответвляется у ко-
ров от аорты, чревной или поясничной артерии и направляется к
ножкам диафрагмы.
252
Чревная артерия (рис. 3.145; см. рис. 3.144) непарная, длиной до
18 см, начинается под первым поясничным позвонком и идет на
правую поверхность рубца. От чревной артерии отходят печеноч-
ная, селезеночная, левая рубцовая и левая желудочная артерии.
Печеночная артерия (рис. 3.145) отдает ряд сосудов, в том числе:
1) ветви для поджелудочной железы; 2) ветвь для желчного пузы-
ря; 3) правую желудочную артерию, которая направляется в пило-
рическую часть сычуга и на начальную часть двенадцатиперстной
кишки; 4) желудочно-двенадцатиперстную артерию. Последняя
является непосредственным продолжением печеночной артерии,
она делится на правую желудочно-сальниковую артерию, идущую
по большой кривизне сычуга, и краниальную поджелудочно-две-
надцатиперстную артерию, направляющуюся на начальную часть
двенадцатиперстной кишки и в поджелудочную железу.
Селезеночная артерия (рис. 3.145) отделяет толстую правую руб-
цовую артерию, а сама направляется в селезенку. Правая рубцовая
артерия идет по правой про-
дольной борозде рубца и выхо-
дит на его левую поверхность.
Левая рубцовая артерия про-
ходит в левой продольной бо-
розде рубца и отдает: 1) ветвь
для преддверия рубца; 2) арте-
рию сетки; 3) каудальную ди-
афрагмальную артерию.
Левая желудочная артерия
(рис. 3.145) является продолже-
нием чревной артерии. Она идет
справа между рубцом и сеткой
и, достигнув книжки, отдает на
большую кривизну сычуга левую
желудочно-сальниковую арте-
рию, а сама на малой кривизне
сычуга анастомозирует с правой
желудочной артерией.
Краниальная брыжеечная ар-
терия (рис. 3.146; см. рис. 3.144)
непарная крупная, охватывает
в виде дуги весь лабиринт обо-
дочной кишки. От нее отделя-
ются следующие сосуды: кау-
дальная поджелудочно-двенадца-
типерстная, средняя ободочная,
подвздошно-слепо-ободочная ар-
терии, коллатеральный ствол и
артерии тощей кишки.
Рис. 3.146. Схема васкуляризации
кишечника собаки:
/ — пилорическая часть желудка; 2 —
s-образный изгиб двенадцатиперстной
кишки; 3 — нисходящее. 4 — попереч-
ное, 5 — восходящее колено двенадца-
типерстной кишки; 6 — начало тощей
кишки; 7 — тощая кишка; 8 — под-
вздошная кишка; 9 — слепая кишка;
10 — восходящее, 11 — поперечное,
12 — нисходящее колено ободочной
кишки; 13 — прямая кишка; 14 — кау-
дальная прямокишечная артерия; 15 —
каудальная брыжеечная артерия; 16 —
подвздошно-ободочная артерия; 17 —
краниальная брыжеечная артерия; 18 —
правая ободочно-кишечная артерия;
19 — артерии тошей кишки
253
Рис. 3.147. Кровоснабжение
репродуктивных органов су-
ки (дорсальная стенка влага-
лища вскрыта):
/ — воронка яичника; 2 — яич-
ник; 3, 13 — артерия яичника;
4 — широкая маточная связка;
5, 10 — влагалищная артерия;
6 — влагалище; 7— преддверие
влагалища; 8 — вульва; 9 — ка-
удальная ветвь влагалищной ар-
терии; И — маточная артерия;
12 — маточная ветвь яичнико-
вой артерии; 14 — рог матки;
15 — тело матки
Каудальная брыжеечная артерия (см. рис. 3.144) непарная, вы-
ходит из аорты напротив четвертого поясничного позвонка. Она
делится на левую ободочную артерию и краниальную прямокишеч-
ную артерию.
Почечная артерия (см. рис. 3.144) парная, начинается от аорты
и идет в почки.
Внутренняя семенная артерия парная (рис. 3.147; см. рис. 3.144),
у быков она идет в семенники, а у коров делится на яичниковую
ветвь и краниальную маточную артерию: для рога матки и анасто-
моза со средней маточной артерией.
Поясничные артерии выходят из аорты в количестве 5 — 6 пар
(см. рис. 3.144). Каждая поясничная артерия отделяет дорсальную
ветвь в разгибатели спины, вентральную ветвь в поясничные мыш-
цы и спинно-мозговую ветвь в мозговые оболочки.
Артерии таза и тазовой конечности
К органам тазовой полости и к тазовым конечностям кровь
поступает из брюшной аорты по средней крестцовой, внутренней
и наружной подвздошным артериям (см. рис. 3.144, 3.148).
Средняя крестцовая артерия (см. рис. 3.144, 3.148) является про-
должением брюшной аорты на крестец после ответвления от нее
наружной и внутренней подвздошных артерий. Она сильно разви-
254
та у коров. От нее отходят четыре пары крестцовых латеральных
артерий и парная хвостовая латеральная артерия.
Крестцовые латеральные артерии снабжают кровью мышцы и
кожу области крестца и хвоста и отдают в канал крестцовой кости
спинно-мозговые ветви для спинного мозга и его оболочек.
Хвостовая латеральная артерия (см. рис. 3.144, 3.148) парная,
делится на дорсальную и вентральную хвостовые артерии.
Средняя крестцовая артерия от первого хвостового позвонка
идет как хвостовая артерия (см. рис. 3.144, 3.148).
Внутренняя подвздошная артерия (см. рис. 3.144, 3.148) отходит
от брюшной аорты под шестым поясничным позвонком и направ-
ляется каудально под крестцово-седалищной связкой. От нее от-
ходят пупочная, подвздошно-поясничная, краниальная ягодич-
ная и каудальная маточная артерии, запирательные ветви и внут-
ренняя срамная артерия.
Пупочная артерия (см. рис. 3.144, 3.148) остается проходимой
для крови только в верхней части до мочевого пузыря, в котором
и разветвляется как краниальная пузырная артерия. Из начальной
части пупочной артерии у быка отходит тонкая артерия семяпро-
вода, а у коровы — очень толстая средняя маточная артерия. Пос-
ледняя анастомозирует с краниальной и каудальной маточными
артериями, особенно сильно она увеличивается во время бере-
менности.
Подвздошно-поясничная артерия слабо развита, проходит по
медиальной поверхности подвздошной кости, снабжает кровью
поясничные и ягодичные мышцы, напрягатель фасции бедра (рис.
3.148).
Краниальная ягодичная артерия отходит обычно несколькими
ветвями на уровне дорсального края крыла подвздошной кости.
Каудальная маточная артерия коров сильно развита, питает
матку, влагалище и мочевой пузырь, анастомозирует со средней
маточной и с каудальной прямокишечной артериями (рис. 3.148).
У быков на ее месте имеются веточки для мочевого пузыря и до-
бавочных половых желез.
Запертые ветви развиты очень слабо, питают запирательные
мышцы.
Внутренняя срамная артерия коров развита слабо, питает толь-
ко клитор (рис. 3.148). У быков внутренняя срамная артерия хоро-
шо развита, она отдает ряд ветвей, в том числе: каудальную пря-
мокишечную артерию, бульбоуретральную артерию, глубокие удо-
вые артерии, артерию промежности. Внутренняя срамная артерия
у быков после ответвления от нее названных сосудов переходит в
дорсальную удовую артерию.
Наружная подвздошная артерия отделяется от брюшной аорты
под пятым поясничным позвонком, идет вдоль тела подвздошной
кости и проникает в бедренный канал (рис. 3.148). Она еще до
255
Рис. 3.148. Артерии тазовой
конечности коровы (ме-
диальная поверхность):
J — правая внутренняя под-
вздошная артерия; 2 — пу-
почная артерия; 3 — средняя
крестцовая артерия; 4 — под-
вздошно-поясничная арте-
рия; 5 — краниальная ягодич-
ная артерия; 6 — артерия хво-
стовой мышцы; 7 — каудаль-
ная ягодичная артерия; 8 —
внутренняя срамная артерия; 9, 11 — запертые ветви; 10 — мочеполовая артерия;
12 — глубокая бедренная артерия; 13 — надчревно-срамной ствол; 14 — наруж-
ная срамная артерия; 15 — каудальная надчревная артерия; 16 — окружная бед-
ренная медиальная артерия; 17 — каудальная бедренная артерия; 18 — каудаль-
ная, 19 — краниальная большеберцовая артерия; 20 — медиальная заплюсневая
артерия; 21 — анастомотическая ветвь к дорсальной сети заплюсны; 22 — пробо-
дающая заплюсневая артерия; 23 — медиальная поверхностная плантарная плюс-
невая артерия; 24 — дистальная прободающая артерия; 25, 26 — неосевая третья
собственная пальцевая артерия; 27 — брюшная аорта; 28 — внутренняя семенная
артерия; 29— каудальная брыжеечная артерия; 30— правая наружная подвздошная
артерия; 31 — окружная подвздошная глубокая артерия; 32 — мышечная ветвь;
33 — латеральная окружная бедренная артерия; 34 — анастомотическая ветвь;
35 — бедренная артерия; 36 — артерия сафена; 37 — подколенная артерия; 38 —
дорсальная сеть заплюсны; 39 — третья дорсальная плюсневая артерия; 40 —
дорсальная общая пальцевая артерия
256
входа в бедренный канал отдает окружную глубокую подвздош-
ную и глубокую бедренную артерии.
Окружная глубокая подвздошная артерия развита очень сильно.
Опа направляется к маклоку и отдает краниальную и каудальную
ветви. Первая питает поясничные и брюшные мышцы, а вторая —
коленную складку и молочную железу.
Глубокая бедренная артерия идет каудовентрально по направле-
нию к бедру (рис. 3.148). Она отделяет надчревно-срамной ствол,
ia 11 ирательную ветвь и медиальную окружную бедренную арте-
рию.
Надчревно-срамной ствол идет вперед и отделяет каудальную
надчревную артерию для мышц брюшной стенки, наружную срам-
ную артерию, а у быков, кроме того, наружную семенную арте-
рию для препуция и мошонки. Наружная срамная артерия у коров
крупная, питает вымя и называется выменной артерией (рис. 3.148).
Запертая ветвь направляется в запирательные мышцы.
Медиальная окружная бедренная артерия снабжает кровью при-
водящую, квадратную и двуглавую мышцы бедра (рис. 3.148).
Бедренная артерия (рис. 3.148) является продолжением наруж-
ной подвздошной артерии после отделения от нее глубокой бед-
ренной артерии. Она находится в бедренном канале, пересекает
бедренную кость с медиальной стороны. От бедренной артерии
отходят краниальная бедренная артерия, окружная латеральная
бедренная артерия, артерия сафена, мышечные ветви, каудаль-
ная бедренная артерия. После ответвления перечисленных арте-
рий она переходит в подколенную артерию.
Краниальная бедренная артерия питает четырехглавую мышцу
бедра.
Окружная латеральная бедренная артерия питает разгибатели
коленного сустава.
Артерия сафена (медиальная подкожная артерия бедра и голени)
тянется медиально по ахиллову сухожилию и отдает латеральную
и медиальную лодыжковые артерии, а сама медиально от пяточ-
ной кости делится на медиальную и латеральную плантарные ар-
терии (рис. 3.148). Плантарные артерии формируют проксималь-
ную и дистальную плантарную дуги. Из медиальной плантарной
артерии происходят вторая и третья общие плантарные пальце-
вые артерии, а из латеральной плантарной артерии выходит чет-
вертая общая плантарная пальцевая артерия. Вторая и четвертая
общие пальцевые артерии отдают специальные плантарные паль-
цевые артерии для внешних поверхностей III и IV главных паль-
цев и для II и V рудиментарных пальцев. От третьей общей план-
тарной пальцевой артерии берут начало специальные плантарные
пальцевые артерии, латеральная для III пальца и медиальная для
IV пальца. Они анастомозируют с дорсальными пальцевыми арте-
риями.
9 Зелеисвский
257
Мышечные ветви хорошо развиты, питают мышцы бедра.
Каудальная бедренная артерия идет каудально, дает восходящую
и нисходящую ветви для длинных разгибателей тазобедренного
сустава, анастомозирует с возвратной большеберцовой артерией
и артерией сафена.
Подколенная артерия является продолжением бедренной арте-
рии, идет между мыщелками бедренной кости и головками икро-
ножной мышцы. Под латеральным мыщелком большеберцовой
кости она отдает тонкую заднюю большеберцовую артерию, а сама
переходит в переднюю большеберцовую артерию.
Передняя большеберцовая артерия тонкая и короткая (рис. 3.148).
Она отдает малоберцовую артерию для разгибателей пальцев, пи-
тающую артерию для большеберцовой кости, плантарную ветвь
для дорсальной заплюсневой артериальной сети. После ответвле-
ния названных сосудов передняя большеберцовая артерия пере-
ходит в дорсальную артерию стопы.
Дорсальная артерия стопы (рис. 3.148) дает прободающую зап-
люсневую артерию, а от нее ответвляется средняя плюсневая план-
тарная артерия, вливающаяся в прободающую плюсневую артерию.
Продолжение дорсальной артерии стопы на плюсну называется
плюсневой дорсальной третьей артерией в дорсальном сосудистом
желобе плюсны. На дистальном конце плюсны плюсневая дорсаль-
ная третья артерия отдает прободающую плюсневую артерию для
формирования дистальной плантарной дуги и затем переходит в
общую дорсальную третью пальцевую артерию, которая делится на
две дорсальные специальные пальцевые артерии, анастомозирую-
щие со специальными плантарными пальцевыми артериями на
обращенных друг к другу поверхностях III и IV пальцев.
Вены большого круга кровообращения
Вены головы и шеи представлены краниальной полой веной,
парными наружными и внутренними яремными и челюстными
венами (рис. 3.149, 3.150).
Краниальная полая вена — основная венозная магистраль, по
которой отводится кровь от головы, шеи, грудных конечностей и
грудных стенок в правое предсердие (рис. 3.149). Она образуется
наружными и внутренними яремными венами, формирующими
правую и левую общую яремную вену, выносящую кровь из головы,
и подмышечными венами, выносящими кровь из грудных конеч-
ностей. Краниальная полая вена тянется от первого ребра до пра-
вого предсердия. В нее впадают внутренняя грудная, позвоночная
и реберно-шейная вены.
Внутренняя грудная вена собирает кровь из грудных мышц и
принимает подкожную брюшную вену. Подкожная брюшная вена
258
/
Рис. 3.149. Вены собаки:
/ — поверхностная височная вена; 2 — анастомотическая ветвь; 3 — глубокая
лицевая вена; 4 — дорсальная вена носа; 5 — вена верхней, 6 — вена нижней
। убы; 7 — язычная вена; 8 — наружная челюстная вена; 9 — наружная яремная
вена; 10 — внутренняя яремная вена; 11 — подкожная вена плеча; 12 — подкож-
ная вена предплечья; 13 — дорсальная вена пясти; 14 — дорсальные пальцевые
пены; 15 — пальмарные пальцевые вены; 16 — плечевая вена; 17 — внутренняя
। рудная вена; 18 — печеночная вена; 19 — краниальная надчревная вена; 20 —
чревная вена; 21 — латеральная вена сафена; 22 — краниальная большеберцовая
пена; 23 — дорсальные вены плюсны; 24 — дорсальные вены пальцев стопы; 25 —
плантарные вены пальцев стопы; 26 — плантарные вены плюсны; 27 — медиаль-
ная вена сафена; 28 — бедренная вена; 29 — внутренняя срамная вена; 30 —
наружная, 31 — внутренняя подвздошная вена; 32 — вены тощей кишки; 33 —
поротная вена печени; 34— каудальная полая вена; 35— межреберная вена; 36 —
правая непарная вена; 37 — краниальная полая вена; 38 — венозное сплетение
спинного мозга; 39 — верхнечелюстная вена
прободает брюшные мышцы против восьмого реберного хряща и
образует там отверстие, называемое молочным колодцем. Подкож-
ная брюшная вена выносит кровь из вымени, и поэтому называ-
ется молочной веной. Часто вена имеет несколько коллатералей
(рис. 3.149, 3.150).
Позвоночная вена собирает кровь из спинного мозга и мышц
шеи (рис. 3.149).
Реберно-шейная вена образуется от слияния трех вен — попе-
речной шейной, передней межреберной и глубокой шейной.
Наружная яремная вена толстая, начинается верхнечелюстной
и наружной челюстной венами, лежит в яремном желобе, при-
259
нимает восходящую шейную вену и подкожную вену плеча (рис.
3.149).
Внутренняя яремная вена тонкая, идет рядом с общей сонной
артерией. Она образуется в области глотки затылочной, гортан-
ной и щитовидной венами.
Затылочная вена несет кровь из базальной системы синусов и
анастомозирует с глубокой шейной веной.
Гортанная вена отводит кровь из гортани.
Щитовидная вена собирает кровь из щитовидной железы, мышц
глотки и гортани.
Наружная челюстная вена начинается из поверхностной лице-
вой вены. В нее впадают язычная вена, подъязычная вена, мус-
кульные ветви, вена нижнечелюстной железы.
Поверхностная лицевая вена образуется дорсальной носовой
веной (двойной) и веной угла глаза, в которую впадает лобная
вена, выходящая через надглазничный канал из орбиты. В лице-
вую вену впадают верхняя и нижняя губные вены.
Верхнечелюстная вена начинается в клинонёбной ямке. В нее
впадают: вена нижнего века, глубокие височные вены, большая
ушная вена, вена большой жевательной мышцы, вена околоуш-
ной железы, вентральная мозговая вена, щечная и височная вены.
Вены грудной конечности образуют две венозные магистрали:
глубокую и подкожную (рис. 3.149).
Глубокая венозная магистраль начинается из общей пальмарной
пальцевой вены, образованной специальными пальмарными паль-
цевыми венами — медиальной IV пальца и латеральной III пальца.
Общая пальмарная пальцевая вена на середине пясти анастомо-
зирует с медиальной пальмарной пястной веной и переходит в
срединную вену. Последняя принимает срединно-лучевую вену,
анастомозирующую в области предплечья с подкожной веной плеча.
Срединная вена, приняв в себя общую межкостную вену, перехо-
дит в плечевую вену. Плечевая вена принимает в себя следующие
вены: коллатеральную лучевую, коллатеральную локтевую, глу-
бокую плечевую, окружную плечевую медиальную, грудоспин-
ную. Далее плечевая вена сливается с подлопаточной веной и пе-
реходит в подмышечную вену, которая соединяется с общим ство-
лом яремных вен. В подмышечную вену впадают две вены: наруж-
ная грудная и акромиальная.
Подкожная венозная магистраль начинается от неосевых паль-
цевых вен — латеральной IV пальца и медиальной III пальца. Эти
вены образуют пальмарную глубокую венозную дугу, из которой
выходят медиальная и латеральная пальмарные пястные вены.
Последние переходят в подкожную вену предплечья, которая ана-
стомозирует со срединно-лучевой веной, принимает добавочную
подкожную вену и затем продолжается как подкожная вена плеча.
Добавочная подкожная вена начинается из внутренних дорсаль-
260
ных пальцевых вен — латеральной III пальца — и медиальной IV
пальца, которые переходят в дорсальную третью пястную вену,
анастомозирующую с общими пальмарными пальцевыми венами
и с латеральной пальмарной пястной веной. Дорсальная третья
пястная вена переходит в добавочную подкожную вену.
Каудальная полая вена собирает кровь из органов тазовой и
брюшной полостей, их стенок и тазовых конечностей (рис. 3.149,
3.150). Она является продолжением средней крестцовой вены, после
того как к ней присоединяется под пятым поясничным позвон-
ком правая и левая общие подвздошные вены, которые произошли
от слияния наружных и внутренних подвздошных вен. Каудальная
полая вена идет вперед справа от брюшной аорты, спускается к
отверстию сухожильного центра диафрагмы, где получает две (три)
диафрагмальные вены. В каудальную полую вену впадают пояс-
ничные вены, внутренние семенные вены, почечные вены, пече-
ночные вены и диафрагмальные вены.
Средняя крестцовая вена (рис. 3.149) собирает кровь из хвосто-
вой, двух латеральных хвостовых и крестцовых латеральных вен.
Внутренняя подвздошная вена принимает кровь из слабо разви-
той подвздошно-поясничной вены, краниальной ягодичной вены,
Рис. 3.150. Краниальная и каудальная полые вены коровы:
1 — аорта; 2 — каудальная полая вена; 3 — срединные крестцовые артерия и
вена; 4 — внутренняя срамная артерия и вена; 5 — вентральные промежностные
артерия и вена; 6 — наружные подвздошные артерия и вена; 7 — артерия и вена
вымени (наружная срамная артерия и вена); 8 — каудальные надчревные арте-
рия и вена; 9 — краниальные надчревные артерия и вена; 10 — внутренняя
грудная артерия; И — краниальная полая вена
261
слабо развитых запирательных ветвей, пупочной вены, сильно
развитой каудальной маточной вены, внутренней срамной и кау-
дальной ягодичной вен.
Вены тазовой конечности образуют три венозные магистрали:
глубокую, подкожную латеральную и медиальную.
Глубокая венозная магистраль сформирована следующими ве-
нами: прободающей заплюсневой, средней дорсальной плюсне-
вой, передней большеберцовой, подколенной, бедренной и на-
ружной подвздошной.
Подкожная венозная латеральная магистраль толще медиаль-
ной. В ее состав входят пальцевая дорсальная третья латеральная
вена и пальцевая дорсальная четвертая медиальная вена. Эти вены
формируют пальцевую дорсальную общую вену, которая прокси-
мально переходит в плюсневую дорсальную латеральную вену. Пос-
ледняя дает дорсальную ветвь, которая сливается с плантарной
ветвью латеральной подкожной вены. Плантарная ветвь латераль-
ной подкожной вены формируется за счет пальцевых плантарных
латеральной и медиальной вен. Названные вены проксимально
образуют плюсневые плантарные глубокие латеральную и меди-
альную вены. На середине голени дорсальная и плантарная ветви
соединяются в латеральную подкожную вену стопы и голени, или
малую вену сафена, которая впадает в глубокую бедренную вену
или в подколенную вену.
Подкожная венозная медиальная магистраль начинается из паль-
цевой плантарной венозной дуги двумя плюсневыми плантарны-
ми поверхностными латеральной и медиальной венами. Эти вены
при слиянии образуют лодыжковую вену, переходящую прокси-
мально в медиальную подкожную вену стопы и голени, или боль-
шую вену сафена.
Наружная подвздошная вена (рис. 3.149) принимает глубокую
бедренную вену и окружную подвздошную глубокую вену. Глубо-
кая бедренная вена сформирована малой веной сафена, окруж-
ной бедренной медиальной веной, подкожной брюшной веной и
наружной срамной веной.
Наружная срамная вена у коров отводит кровь из вымени и
сливается с краниальной и каудальной венами вымени. Крани-
альная вена вымени отводит кровь из паренхимы железы и веноз-
ного сплетения сосков. В наружную срамную вену вливаются кра-
ниальная и каудальная вены основания вымени, они анастомози-
руют с подкожной брюшной и внутренней срамной венами.
Бедренная вена собирает кровь из многих вен. В нее вливаются
каудальная бедренная вена, большая вена сафена, окружная бед-
ренная медиальная вена и краниальная бедренная вена.
Воротная вена печени (рис. 3.149) образуется у коров желудоч-
но-двенадцатиперстной веной, которая в свою очередь образует-
ся из краниальной поджелудочно-двенадцатиперстной вены и
262
общего ствола правой желудочной и правой желудочно-сальни-
ковой вены. Затем в воротную вену вливается левая желудочно-
селезеночная вена и общая брыжеечная вена. Общая брыжеечная
вена образуется от слияния следующих вен: большой вены подже-
лудочной железы, каудальной поджелудочно-двенадцатиперстной
вены, средней ободочной вены, левой ободочной вены, подвздош-
по-слепо-ободочного и кишечного стволов.
ЛИМФАТИЧЕСКАЯ СИСТЕМА
Лимфология — раздел анатомии, изучающий лимфатическую
систему.
Лимфатическая система (systema lymphaticum) состоит из внут-
риорганного и внеорганного лимфатического сосудистого русла,
объединяющего приносящие и выносящие лимфатические сосу-
ды, регионарные лимфатические узлы и лимфатические протоки
(рис. 3.151 — 3.155).
Рис. 3.151. Схема строения лим-
фатической системы млекопита-
ющего (вид сверху):
1 — наружная и внутренняя ярем-
ные вены; 2 — краниальная полая
вена; 3 — лимфатические узлы голо-
вы и шеи; 4 — лимфатические узлы
грудной конечности; 5 — грудной
проток; 6 — лимфатические узлы ор-
ганов грудной полости; 7 — пояснич-
ная цистерна; 8 — лимфатические
узлы органов брюшной полости; 9 —
лимфатические узлы поясничной об-
ласти и почек; 10 — тазовые лимфа-
тические узлы; 11 — лимфатические
узлы тазовой конечности
263
Лимфатические сосуды
По лимфатическим сосудам движется лимфа. В ее состав вхо-
дит плазма лимфы и форменные элементы. Клеточные элементы
лимфы представлены главным образом лимфоцитами. Плазма лим-
фы сходна с плазмой крови, но отличается от нее тем, что со-
держит часть продуктов обмена веществ тех органов, из которых
она оттекает.
В лимфу, оттекающую от кишечника, всасывается жир, поэто-
му она принимает молочный вид и называется хилюсом, а лимфа-
тические сосуды кишечника — млечными сосудами.
Лимфатические капилляры по строению напоминают кровенос-
ные, а лимфатические сосуды — вены. Лимфатические сосуды обыч-
но идут в тканях и органах вместе с артерией и спутницей веной.
Лимфатические сосуды, за исключением самых мелких внут-
риорганных, содержат двухстворчатые клапаны. Последние рас-
полагаются так часто, что разделяют лимфатический сосуд на от-
дельные клапанные сегменты — лимфангионы, являющиеся их
структурно-функциональными единицами. Стенка каждого лимфан-
гиона способна сокращаться, продвигая лимфу к центру.
Лимфа, оттекающая от органов и тканей, проходит через лим-
фатические узлы. Лимфатические сосуды, несущие лимфу к лим-
фатическому узлу, называются приносящими. Они проникают в лим-
фатический узел по всей поверхности, образуя его «корни», и
имеют меньший диаметр по сравнению с выносящими лимфати-
ческими сосудами, выходящими из лимфатического узла только
через его ворота.
Главные лимфатические сосуды
Поясничная цистерна имеет вид продолговато-овального ме-
шочка, расположенного на уровне первого — четвертого пояснич-
ных позвонков правее и выше аорты. В ее каудальный конец впа-
дают правый и левый поясничные стволы, отводящие лимфу от ор-
ганов тазовой полости и тазовой конечности. В среднюю часть ци-
стерны с вентральной поверхности открывается кишечный ствол,
отводящий лимфу от тонкой и толстой кишки. В переднюю часть
цистерны впадает чревный лимфатический ствол, дренирующий
желудок, селезенку и печень (рис. 3.151).
Грудной проток выносит лимфу из 3/4 тела. Он начинается из
переднего конца поясничной цистерны, тянется вперед правее и
выше аорты. На уровне третьего-четвертого грудного позвонка
проток переходит в прекардиальную (левую) часть. Здесь он обра-
зует S-образный изгиб и после ампулообразного расширения,
находящегося у первого ребра, впадает в краниальную полую вену
(рис. 3.151).
264
Правый и левый трахеальные стволы формируются выносящи-
ми лимфатическими сосудами краниальных лимфатических узлов
и тянутся каудально вдоль дорсолатеральной поверхности трахеи.
У первого ребра они открываются в наружные яремные вены (рис.
3.151).
Правый лимфатический проток имеет длину около 3 см при
диаметре 3 — 6 мм. Он образуется выносящими сосудами правого
поверхностного шейного лимфатического узла, а открывается в
правую подключичную вену (рис. 3.151).
Лимфатические узлы
Лимфатические узлы имеют постоянную топографию и при-
нимают лимфу только от определенных органов, в связи с чем
получают название — регионарный лимфатический узел.
Лимфа, оттекающая от органов, проходит, как правило, не
один, а несколько (до 14) последовательно расположенных лим-
фатических узлов, поэтому последним приписывается еще и по-
рядковый номер. По форме лимфатические узлы бывают бобовид-
ной формы, реже округлые, овальные, или сильно вытянутые, а
иногда плоские и неправильной формы.
Лимфатические узлы выполняют функцию биологического и
механического фильтра. Как в любом паренхиматозном органе, в
нем выделяют строму и паренхиму. В состав первой входит капсула
лимфатического узла, состоящая, в основном, из плотной соеди-
нительной ткани, эластических волокон и гладких миоцитов. От
нее внутрь узла отходят перекладины — трабекулы.
Паренхима лимфатического узла состоит из ретикулярной тка-
ни с большим количеством ретикулоэндотелиальных клеток и
клеток белой крови. За счет этой ткани формируются фолликулы
и мозговые тяжи. Фолликулы прилежат к капсуле и формируют
корковое вещество узла. Мозговые тяжи образуют центральное пе-
реплетение, или мозговое вещество узла. Фолликулы отделены от
капсулы и трабекул узкими пространствами — лимфатическими
синусами. По лимфатическим сосудам лимфа доставляется в крае-
вой синус, затем в корковый и промежуточный и, наконец, в
мозговой. По ним лимфа продвигается к воротам узла и выходит
из них по выносящим сосудам. Синусы пронизаны волокнами ре-
тикулярной ткани, в ячейках которой часто встречаются лимфо-
циты и макрофаги.
У домашних и сельскохозяйственных животных, за исключе-
нием свиньи, корковое вещество лимфатического узла располага-
ется по его периферии, а мозговое — в центре. При этом принося-
щие лимфатические сосуды впадают в лимфатический узел по
всей его поверхности, а выносящие выходят через его ворота. Для
265
лимфатических узлов свиньи характерна обратная картина: моз-
говое вещество узла располагается по его периферии, а корко-
вое — по центру; приносящие лимфатические сосуды входят в
узел через его ворота, а выносящие — по всей его поверхности.
Лимфатические узлы по происхождению своих «корней» раз-
деляются на внутренностные, мышечные, кожные, кожно-мы-
шечные и мышечно-внутренностные. Внутренностные лимфати-
ческие узлы располагаются над внутренними органами и выносят
из них лимфу. Мышечные лимфатические узлы лежат в опреде-
ленных наиболее подвижных частях тела: на границе головы и
шеи, при входе в грудную полость, в области суставов. Кожные
лимфатические узлы имеются только в области коленной склад-
ки, а в остальных частях тела встречаются кожно-мышечные, или
кожно-мышечно-внутренностные узлы.
На голове располагаются околоушный, нижнечелюстной и два
заглоточных лимфатических узла.
Околоушный лимфатический узел (рис. 3.152) длиной 6 — 9 см у
коров, 3 — 6 см у свиней, 2 — 7 см у лошадей, 2,5 см у собак лежит
ниже челюстного сустава под околоушной слюнной железой. Со-
бирает лимфу из органов и тканей головы. Отток лимфы идет в
заглоточные латеральные узлы у коров и свиней, в узлы воздухо-
носного мешка у лошадей и в медиальные заглоточные узлы у
собак.
Нижнечелюстной лимфатический узел (рис. 3.152) длиной 3,0 —
4,5 смукоров, 3,0 —3,5 смусвиней, 0,2 — 3,5 смулошадей, 1,0—3,5
см у собак находится в межчелюстном пространстве позади лицевой
сосудистой вырезки, у собак позади углового отростка. Собирает
лимфу из ротовой и носовой полостей, языка, слюнных желез. От-
ток лимфы совершается в заглоточный латеральный узел у коров,
свиней и лошадей, в заглоточный медиальный узел у собак.
Заглоточный медиальный лимфатический узел длиной 3 — 6 см у
коров, 0,2 —2,0 см у свиней, 0,3 — 4 см у лошадей, 1,5 —8,0 см у
собак располагается между глоткой и сгибателями головы. Соби-
рает лимфу из ротоглотки, носовой полости, слюнных желез, гор-
тани и нижней челюсти. Отток лимфы происходит в заглоточный
латеральный узел у коров, в трахеальный ствол у свиней и собак,
в передние шейные узлы у лошадей.
Заглоточный латеральный лимфатический узел длиной 4 —5 см
у коров, 0,2 —0,9 см у свиней, 0,3— 1,5 см у лошадей, 0,2 —0,4 см
у собак собирает лимфу из ротовой полости, нижней челюсти,
ушной раковины, слюнных желез, лимфатических узлов головы,
мышц и костей шеи. Отток лимфы идет в трахеальный проток у
коров, в поверхностные шейные узлы у свиней, в передние шей-
ные узлы у лошадей и собак.
На шее встречаются поверхностные, глубокие шейные и ре-
берно-шейные лимфатические узлы.
266
Рис. 3.152. Поверхностные лимфатические узлы лошади:
1 — подподвздошный лимфатический узел; 2 — лимфатические сосуды в повер-
хностные паховые лимфатические узлы; 3 — подколенный лимфатический узел;
4 — локтевой лимфатический узел; 5 — подмышечный лимфатический узел; 6 —
поверхностный шейный лимфатический узел; 7 — нижнечелюстной лимфати-
ческий узел; 8 — околоушный лимфатический узел
Поверхностный шейный лимфатический узел (рис. 3.152) длиной
7 — 9 см у коров (4—5 см вентральные и дорсальные у свиней),
1,5 —3,0 см у лошадей, 2,7 —3,5 см у собак помещается впереди
предостной мышцы выше плечевого сустава под плечеголовной и
трапециевидной мышцами. Собирает лимфу из кожи, мышц и
костей шеи, грудной конечности, подгрудка и грудной клетки.
Отток лимфы идет справа в трахеальный проток, а слева — в груд-
ной проток.
Глубокие шейные лимфатические узлы длиной 0,5 —3,0 см у ко-
ров подразделяются на передние, средние и задние. Располагают-
ся на дорсальной поверхности трахеи у коров, свиней и лошадей,
а у собак часто отсутствуют. Собирают лимфу из глотки, гортани,
трахеи, вентральных мышц шеи, реберно-шейных и подмышеч-
ных узлов. Отток лимфы происходит слева в трахеальный и груд-
ной проток у коров, в грудной проток у свиней, справа — в пра-
вый лимфатический проток.
267
Реберно-шейный лимфатический узел длиной 1,5 —3,0 см встре-
чается только у коров, находится сбоку от пищевода и трахеи
впереди и медиально от первого ребра. Собирает лимфу из мышц
шеи, плечевого пояса и грудной стенки, плевры, средостенных
узлов. Отток лимфы справа идет в трахеальный проток, слева — в
грудной проток.
На грудной конечности имеются подмышечный и локтевой лим-
фатические узлы.
Подмышечный лимфатический узел длиной 2,0 —3,5 см у коров,
4,0 —7,0 см у лошадей, 3,0 —5,0 см у собак находится позади и
медиальное плечевого сустава. Собирает лимфу из мышц, суста-
вов и кожи грудной конечности. Отток лимфы происходит в под-
мышечный узел первого ребра у коров, в задние глубокие шей-
ные узлы у лошадей, в грудной проток у собак.
Подмышечный лимфатический узел первого ребра длиной 1,5 см
у коров, 2,0 —3,5 см у свиней слит с глубоким шейным узлом у
лошадей, отсутствует у собак, лежит между глубокой грудной
мышцей и плечевым суставом против первого ребра. Собирает
лимфу из мышц шеи, грудной клетки и грудной конечности. От-
ток лимфы совершается справа в трахеальный проток, слева в
грудной проток у коров, в трахеальный ствол у свиней.
Локтевой лимфатический узел (рис. 3.152) длиной 4 — 5 см име-
ется только у лошадей и помещается медиально от локтевого сус-
тава. Собирает лимфу из мышц, суставов и костей грудной конеч-
ности. Отток лимфы идет в подмышечный узел.
На тазовой конечности располагаются следующие лимфати-
ческие узлы.
Подколенный лимфатический узел (рис. 3.152) длиной 3 — 4,5 см
у коров, 0,5 — 2 см поверхностный и глубокий у свиней, 3 — 5 см
у лошадей, 5 см у собак помещается на головках икроножной
мышцы. Собирает лимфу из кожи, костей, связок, мышц и сухо-
жилий стопы и голени. Отток лимфы происходит: в глубокий па-
ховый, крестцовый латеральный узел — у коров, в подвздошные
медиальные, глубокие паховые и крестцовые латеральные узлы —
у свиней, в глубокие паховые узлы — у лошадей, в медиальные
подвздошные узлы — у собак.
Надколенный лимфатический узел длиной 6—11 см у коров,
5,5 см у свиней, 6—10 см у лошадей (отсутствует у собак) поме-
щается под кожей впереди напрягателя широкой фасции бедра.
Собирает лимфу из кожи брюшных и грудных стенок, таза, бедра
и голени. Отток лимфы совершается в глубокий паховый узел у
коров, в подвздошные латеральные и медиальные узлы у свиней
и лошадей.
Поверхностные паховые лимфатические узлы (рис. 3.152) дли-
ной 3 — 6 см у быка лежат сбоку пениса впереди и сзади семенно-
го канатика. Собирают лимфу из мошонки, препуция, пениса,
268
кожи бедра и голени. Отток лимфы идет в глубокий паховый узел
v быка и жеребца.
У самок этот узел называется надвыменным. Длина узла равна
(> —10 см у коров, 5 — 7 см у свиней, 10—14 см у кобыл, 1 — 2 см
у сук. Он лежит под кожей сзади над основанием вымени у коров,
свиней и кобыл, а у сук впереди лонных костей. Собирает лимфу
из молочной железы. Отток лимфы происходит в глубокие пахо-
вые и медиальные подвздошные узлы.
Глубокий паховый лимфатический узел длиной 3,5 —9,5 см у ко-
ров, 1 — 3 см у свиней, 8—12 см у лошадей располагается в бед-
ренном канале между бедренной и глубокой бедренной артерия-
ми. У собак он отсутствует. Узел собирает лимфу из кожи, мышц и
костей тазовой конечности, брюшных стенок и мочеполовых ор-
ганов. Отток лимфы идет в медиальные подвздошные узлы у ко-
ров и лошадей, в поясничную цистерну у свиней.
На грудной стенке и органах грудной полости различают следу-
ющие лимфатические узлы.
Межреберные лимфатические узлы (рис. 3.153) дайной 0,3— 1,5 см
у коров (нет у свиней), 0,3 —0,6 см у лошадей, 0,2 —0,7 см у
собак помещаются около реберных головок в межреберных про-
странствах. Собирают лимфу из костей и мышц грудной клетки,
спины, плеча, плевры и диафрагмы. Отток лимфы происходит в
Рис. 3.153. Лимфатические узлы грудной полости коровы:
1 — краниальный средостенный лимфатический узел; 2 — левый трахеобронхиаль-
ный лимфатический узел; 3 — средние средостенные лимфатические узлы; 4 —
лимфатический узел первого ребра; 5 — грудной проток; 6, 7 — межреберные
лимфатические узлы; 8 — каудальные средостенные лимфатические узлы; 9 —
каудальный грудинный лимфатический узел; 10— краниальные грудинные лим-
фатические узлы; 11 — наружная яремная вена
269
дорсальные средостенные узлы и грудной проток у коров, лоша-
дей и собак.
Грудинный лимфатический узел (рис. 3.153) длиной 1,5 —2,5 см
у коров, есть у свиней и собак. Располагается на грудине у первого
ребра. Собирает лимфу из мышц грудной клетки, диафрагмы, сер-
дечной сорочки, легких, средостенных нижних узлов. Отток лим-
фы ведется в передний грудинный узел и в грудной проток у ко-
ров, свиней и собак.
Дорсальные средостенные узлы (рис. 3.153) длиной 1,0—3,5 см у
коров, 0,2 —0,4 см у свиней, 0,3 —0,5 см у лошадей (нет у собак)
лежат между аортой и грудными позвонками. Собирают лимфу из
мышц грудной стенки, средостения, легких. Отток лимфы проис-
ходит в грудной проток.
Вентральные средостенные лимфатические узлы (рис. 3.153) дли-
ной 1 — 3 см у коров, у других животных отсутствуют. Находятся в
средостении между диафрагмой и сердечной сорочкой. Собирают
лимфу из плевры, сердечной сорочки и диафрагмы. Отток лимфы
идет в грудинный узел.
Передние средостенные лимфатические узлы (рис. 3.153) длиной
2 — 7 см у коров, 0,1 — 0,5 см у свиней, 0,1 — 0,8 см у лошадей, 3 —
4 см у собак лежат в прекардиальном средостении. Собирают лим-
фу из грудных и брюшных стенок, молочных желез, трахеи, пи-
щевода, сердца, аорты, вилочковой железы, бронхиальных и меж-
реберных узлов. Отток лимфы совершается в грудной проток.
Средние средостенные лимфатические узлы длиной 0,5 — 5 см у
коров, 0,2—0,4 см у свиней, 0,2 —6,0 см у лошадей (нет у собак)
собирают лимфу из трахеи, пищевода, легких, плевры. Отток лим-
фы ведется в грудной проток.
Задние средостенные лимфатические узлы длиной 10—15 см у
коров, 0,1 —0,7 у свиней, 0,1 —2,5 см у лошадей (нет у собак)
собирают лимфу из трахеи, пищевода, легких, печени и селезен-
ки. Отток лимфы происходит в грудной проток.
Бронхиальные (бифуркационные) лимфатические узлы (рис. 3.153)
подразделяются на правый, левый, дорсальный и эпартериальный,
лежат в области бифуркации трахеи у всех животных. Собирают лим-
фу из легких, бронхов, пищевода, сердца и легочных узлов. Отток
лимфы идет в передние средостенные узлы и грудной проток.
Легочные лимфатические узлы длиной 0,5 — 1,5 ему коров, 0,3 —
1,0 см у лошадей, 0,1 —0,4 см у собак лежат на бронхах и легких.
Собирают лимфу из легких. Отток лимфы идет в бифуркационные
узлы.
Лимфатический узел сердечной сорочки (рис. 3.153) длиной 1,5 см
имеется только у коров, лежит под дугой аорты. Собирает лимфу из
сердечной сорочки. Отток лимфы происходит в грудной проток.
На брюшной и тазовой стенках имеются следующие лимфати-
ческие узлы.
270
Поясничные лимфатические узлы длиной 0,5 — 5 см у коров, 0,3 —
0,5 см у свиней, 0,3 —3,5 см у лошадей, 0,2 —0,4 см у собак распо-
пагаются: слева — дорсально от аорты, справа — дорсально от
каудальной полой вены, иногда встречаются у межпозвоночных
отверстий.
Собирают лимфу из спинных и поясничных мышц, брюшных
стенок, почек и надпочечников. Отток лимфы идет в поясничную
цистерну.
Медиальные подвздошные лимфатические узлы длиной 0,5 — 5 см
у коров, 0,5 —2,0 см у свиней, 0,2 —5,5 см у лошадей, 0,5 —6,0 см
у собак лежат впереди корня окружной подвздошной глубокой
артерии. Собирают лимфу из поясницы, таза, бедра, мочеполо-
вых органов, надколенных, латеральных подвздошных, глубоких
паховых, тазовых и крестцовых лимфатических узлов. Отток лим-
фы идет в поясничный ствол у коров и лошадей, в поясничную
। шстерну — у свиней и собак.
Подподвздошные лимфатические узлы длиной 1 — 2,5 см у коров,
0,3 —0,8 см у свиней, 0,2 —3,5 см у лошадей (нет у собак) лежат
под брюшиной ниже маклока между передней и задней ветвями
глубокой окружной подвздошной артерии. Собирают лимфу из
мышц таза, живота, тазовых конечностей. Отток лимфы идет в
тазовый ствол у коров, поясничную цистерну — у свиней, пояс-
ничные узлы и поясничный ствол — у лошадей.
Тазовые лимфатические узлы длиной 0,5 —4,5 см у коров, 0,2 —
0,5 см у свиней, 0,3 — 2,0 см у лошадей, часто отсутствуют у собак.
Помещаются между корнями правой и левой внутренних подвздош-
ных артерий. Собирают лимфу из органов тазовой полости и бедра.
Отток лимфы идет в медиальные подвздошные узлы.
Крестцовые лимфатические узлы длиной 0,4—4,5 см у коров,
0,2 —0,5 см у лошадей, часто отсутствуют у собак. Разделяются на
латеральные и медиальные. Латеральные лежат на крестцово-се-
далищной связке около большой седалищной вырезки, а меди-
альные — под верхним концом крестцово-седалищной связки
(иногда отсутствуют). Латеральные собирают лимфу из стенок та-
зовой полости, а медиальные — из мочеполовых органов. Отток
лимфы идет в тазовые узлы.
Седалищные узлы длиной 2,5 — 3,5 см имеются только у коров,
располагаются снаружи на крестцово-седалищной связке около
малой седалищной вырезки. Собирают лимфу из таза, тазобедрен-
ного сустава, прямой кишки, мочеполовых органов. Отток лимфы
происходит в тазовые узлы.
В органах брюшной и тазовой полости есть следующие лимфа-
тические узлы. Чревные лимфатические узлы (рис. 3.154) длиной
1 — 2 см у коров, 0,3 — 0,8 см у свиней, 0,2 —2,5 см у лошадей,
0,4—0,9 см у собак лежат у корня чревной артерии. Собирают
лимфу из диафрагмы, брюшины, надпочечников, печеночных,
271
селезеночных, желудочных и сальниковых узлов. Отток лимфы про
исходит по чревному стволу в поясничную цистерну.
Желудочные лимфатические узлы (рис. 3.154, 3.155) длиной 0,5 --
4 см у коров, свинеи они встречаются по одиночке и группами,
у лошадей группой 15 — 35 узлов, у собак часто отсутствуют.
У коров на рубце находятся правые и левые рубцовые узлы, на
сетке и книжке — по 3 — 4 лимфатических узла, на сычуге име-
ются дорсальные и вентральные сычужные узлы. Собирают лим-
фу из рубца, сетки, книжки, сычуга, селезенки, двенадцати-
перстной кишки. Отток лимфы происходит в поясничную цис-
терну.
Печеночные лимфатические узлы (рис. 3.154, 3.155) длиной 1 —
7 см у коров, 0,4—9,0 см у лошадей, 0,2 —3,0 см у собак лежат в
воротах печени. Собирают лимфу из печени, поджелудочной же-
лезы, двенадцатиперстной кишки и сычужных лимфатических
узлов. Отток лимфы идет в кишечный ствол.
Селезеночные лимфатические узлы (рис. 3.154) длиной 0,5 —
1 см у свиней, 0,2 — 7 см у лошадей, 0,2 — 2 см у собак лежат в
Рис. 3.154. Лимфатические узлы органов брюшной полости лошади:
1 — селезеночные лимфатические узлы; 2 — сальниковые лимфатические узлы;
3 — желудочные лимфатические узлы; 4 — чревная артерия; 5 — чревные лим-
фатические узлы; 6 — портальные лимфатические узлы; 7 — чревный ствол; 8 —
тощекишечные лимфатические узлы; 9 — краниальная брыжеечная артерия; 10 —
поясничная UHciepua; 11 — ободочнокишечныи ствад; !2 - добавочные пра-
вые ободочно-кишечные лимфатические уыы, 13 — кишечный стам; /4 —лим-
фатические узлы ча.юй ободочной кишки. 1> поясничный ствол; /о - кау-
дальная брыжеечная артерия, 7 7—каудальные брыжеечные лимфатически;
18 — оболочно-кишечные лимфатические узлы; /9 — латеральные слепокишеч-
ные лимфатические узлы; 20 — поджелудочно-дуоденальные лимфатические узлы;
21 — селезеночные лимфатические узлы; 22 — грудной проток
272
воротах селезенки; у коров от-
сутствуют. Собирают лимфу из
желудка, селезенки, сальнико-
вых узлов. Отток лимфы идет в
чревные узлы.
Передние брыжеечные узлы
(рис. 3.154) лежат около пере-
дней брыжеечной артерии у ко-
ров, свиней, лошадей и собак.
Собирают лимфу из двенадца-
типерстной, ободочной кишок,
аорты, надпочечников, из лим-
фатических узлов тонкой и тол-
стой кишки. Отток лимфы идет
в поясничную цистерну.
Лимфатические узлы тощей
кишки (рис. 3.154; см. рис. 3.151)
длиной 0,5 — 15 см у коров, 0,8 —
10 см у свиней, 0,3 — 6 см у ло-
шадей, 0,6 — 3 см у собак, ле-
жат в брыжейке тощей кишки.
Собирают лимфу из тощей киш-
Рис. 3.155. Лимфатические узлы ко-
ровы:
1 — подподвздошный лимфатический
узел; 2 — седалищный лимфатический
узел; 3 — поверхностный паховый (над-
выменный) лимфатический узел
ки. Отток лимфы идет в поясничную цистерну.
Лимфатические узлы слепой кишки (рис. 3.154) собирают лимфу
из слепой и подвздошной кишок. Отток лимфы происходит в ки-
шечный ствол и поясничную цистерну.
Лимфатические узлы ободочной кишки (рис. 3.154) лежат в бры-
жейке ободочной кишки. Собирают лимфу из ободочной кишки.
Отток лимфы происходит в поясничную цистерну.
Задние брыжеечные лимфатические узлы (рис. 3.154) лежат в
брыжейке около задней брыжеечной артерии у всех животных.
Собирают лимфу из ободочной и прямой кишки. Отток лимфы
идет в поясничную цистерну.
Лимфатические узлы прямой кишки лежат на верхней стенке
прямой кишки у всех животных. Собирают лимфу из прямой киш-
ки. Отток лимфы идет в поясничную цистерну.
Заднепроходные лимфатические узлы лежат выше ануса под кожей
у всех животных. Собирают лимфу из ануса и промежности, хвоста,
вульвы. Отток лимфы происходит в медиальные подвздошные узлы.
Почечные лимфатические узлы (см. рис. 3.151) лежат в воротах
почки у всех животных. Собирают лимфу из почек. Отток лимфы
идет в поясничную цистерну.
Яичниковый лимфатический узел (см. рис. 3.154) маленький, име-
ется только у лошадей, лежит в связке яичника.
Маточные лимфатические узлы хорошо развиты у свиней, ле-
жат в широкой маточной связке.
273
Органы иммуногенеза и кроветворения
Система органов иммуногенеза и кроветворения обеспечивает
поддержание постоянства морфологического состава крови, им-
мунного гомеостаза, барьерную и защитную функции. К ним от-
носятся красный костный мозг, тимус, лимфатические узлы, се-
лезенка, а также лимфоидные узелки в пищеварительной и дру-
гих системах. В основе почти всех их лежит ретикулярная ткань,
создающая микросреду для размножения и дифференцировки
клеток. Красный костный мозг и тимус относят к центральным
органам иммуногенеза и кроветворения в связи с тем, что в них
происходит антигеннезависимое генетически запрограммирован-
ное размножение лимфоцитов. Остальные органы, в которых про-
исходит развитие клеток крови, являются периферическими, так
как в них происходит антигензависимое развитие и дифференци-
ровка лимфоцитов, мигрирующих сюда из центральных органов.
Красный костный мозг находится в губчатом веществе эпифи-
зов трубчатых костей, позвонков, ребрах и в грудной кости. В нем
миелоидная ткань продуцирует все виды клеток белой крови, эрит-
роциты, кровяные пластинки, здесь также проходят начальные
стадии развития популяции Т- и В-лимфоцитов. Строма органа
состоит из ретикулярной ткани, в которой находятся свободные
клетки крови на разных стадиях формирования. Дифференциро-
ванные клетки поступают в просвет синусоидных капилляров. Этому
способствует то, что стенки этих микрососудов тонкие и имеют
миграционные поры в эндотелии.
Ткань, в которой развиваются и созревают клетки крови, рас-
полагается очажками вокруг кровеносных сосудов. Наиболее ин-
тенсивно кроветворение происходит вблизи эндоста.
Желтый костный мозг находится в диафизах трубчатых костей
и содержит преимущественно жировые клетки, содержащие пиг-
менты типа липохромов. Небольшое количество жировых клеток
постоянно встречается и в красном костном мозге.
Тимус является центральным органом иммуногенеза, хорошо
развит только у молодых животных и состоит из двух шейных и
непарной грудной долей. Шейные части лежат по бокам трахеи, а
грудная находится в предсердечном средостении. Снаружи тимус
покрыт коллагенизированной капсулой. Последняя проникает
внутрь органа, разделяя его на дольки. Междольковая соедини-
тельная ткань выполняет функцию стромы: в ней проходят крове-
носные и лимфатические сосуды, а также нервы. В каждой дольке
различают периферическую зону — корковое вещество и цент-
ральную — мозговое вещество. В мозговом веществе тимуса име-
ются тельца Гассаля.
После наступления половой зрелости наблюдается прогресси-
рующее уменьшение долек и замещение их жировой тканью. Так,
274
к 2 — 3 годам жизни собаки наблюдается значительная редукция
шейных долей тимуса.
Тимус является центральным органом, контролирующим все
иммунные реакции и лимфоцитопоэз. В отличие от других лимфо-
идных органов, он состоит не только из ретикулярной и лимфо-
идной ткани. Более важное значение здесь имеет эпителий, обра-
|ующий в дольках органа сеточку.
Дольки тимуса не полностью отделены друг от друга соеди-
нительной тканью. В каждой дольке выделяется корковое и моз-
говое вещество. Ретикулоэпителиальная сеточка коры густо за-
полнена малыми лимфоцитами (тимоцитами), которые активно
размножаются и далее мигрируют в мозговое вещество дольки.
Здесь их количество значительно меньше. По мере продвижения
। имоцитов из коры в мозговое вещество происходит их диффе-
ренцировка и проникновение в синусоидные кровеносные ка-
пилляры.
Эпителиальные клетки долек, отмирая, образуют в мозговом
веществе компактные островки — тимусные тельца Гассаля.
Помимо Т-лимфоцитов тимус выбрасывает в кровь биологичес-
ки активные гормоноподобные вещества, которые содействуют
созреванию Т-лимфоцитов в тимусе и периферических органах.
Селезенка представляет собой плоский, серповидной формы
орган сине-красного или сине-фиолетового цвета с поверхности
и интенсивного красно-коричневого цвета на разрезе, довольно
мягкой консистенции. Она выполняет разнообразные функции и
кроме лимфоцитопоэтической роли и биофильтра крови участву-
ет в реутилизации отработавших эритроцитов и тромбоцитов. Кроме
того, она является мощным депо крови: в ней может задержи-
ваться до 15 % всей крови организма.
На селезенке различают вентральный расширенный конец,
или основание, и дорсальный суженный конец, или верхушку. Пе-
редний край селезенки, вогнутый и острый, несет на себе вы-
резку, а задний — выпуклый и тупой. Вдоль вогнутого края на
висцеральной поверхности (обращенной к внутренним органам)
проходит желоб: в нем лежат и через него входят в селезенку
кровеносные сосуды и нервы. Вследствие этого желоб называет-
ся воротами селезенки. В области ворот селезенка толще. Узкой
полосой, лежащей впереди ворот, селезенка прилежит к боль-
шой кривизне желудка, а остальной, значительно большей час-
тью примыкает к большому сальнику и граничит с поджелудоч-
ной железой.
Снаружи селезенка покрыта серозной оболочкой, образован-
ной листком большого сальника. Она плотно прилежит к капсуле
и переходит на окружающие органы, образуя связки.
Селезенка состоит из стромы и паренхимы. Последняя называ-
ется еще селезеночной пульпой. Строма образована капсулой, одева-
275
ющей селезенку с поверхности. Она построена из плотной соеди-
нительной ткани с примесью эластических и гладкомышечных
волокон.
От внутренней поверхности капсулы отходят многочисленные
перегородки — трабекулы. Анастомозируя друг с другом, они фор-
мируют остов селезенки в виде губки. По элементам стромы орга-
на проходят кровеносные и лимфатические сосуды, а также не-
рвы.
Толщина капсулы составляет около 50 мкм, а тканевой состав
трабекул сходен с капсулой.
Межтрабекулярная ткань — пульпа — разделяется на красную и
белую пульпу. В красной пульпе в венозных синусах депонируется
кровь, а белую составляет ткань лимфоидных узелков.
Особенности. У овец селезенка плоская, округло-треуголь-
ной формы, красно-коричневого цвета. Верхнепередний угол се-
лезенки на висцеральной поверхности имеет ворота. У свиней се-
лезенка вытянутая, длинная, с суженными концами, на попе-
речном разрезе треугольная. На висцеральной поверхности селе-
зенки виден продольный гребень, вдоль которого находятся во-
рота селезенки.
Цвет селезенки ярко-красный, консистенция довольно плот-
ная. На разрезе ясно выступают фолликулы. Длина селезенки 38 —
45 см, ширина 5 — 8 см, масса 0,1 — 0,3 %. Лежит она в левом под-
реберье на большой кривизне желудка.
У лошадей селезенка плоская, серповидной формы, сине-крас-
ного или сине-фиолетового цвета с поверхности, а на разрезе
красного цвета, мягкой консистенции. На селезенке различают
дорсальный расширенный конец, или основание, и суженный
конец, обращенный вниз, или верхушку. Передний край селезен-
ки вогнутый и острый, а задний край выпуклый и тупой. Висце-
ральной поверхностью селезенка прилегает к большой кривизне
желудка, париетальной—к диафрагме. Основание ее лежит против
2 — 3 последних ребер и первого поясничного позвонка, а вер-
хушка — против середины девятого —одиннадцатого ребер. Се-
розная оболочка покрывает капсулу селезенки. Длина селезенки
30 — 55 см, ширина 17 — 25 см. Абсолютная масса достигает 0,5 —
1,5 кг, или 0,2 —0,4% от массы тела.
У собак селезенка плоская, неправильной треугольной фор-
мы, вытянутая сверху вниз. Нижний конец селезенки расширен, а
верхний сужен. Задний край прямой, а передний вогнут. Середина
ее более тонкая и узкая. Цвет селезенки красный с синеватым
оттенком, консистенция плотная. Лежит селезенка в левом подре-
берье. Задний и верхний ее концы находятся против второго —
четвертого поясничного позвонка, а передний и нижний концы —
против седьмого—десятого реберно-хрящевого соединения. Мас-
са селезенки колеблется от 0,08 до 0,4 % от массы тела.
276
ОРГАНЫ ВНУТРЕННЕЙ СЕКРЕЦИИ
К железам внутренней секреции относятся органы, ткани и
। руппы клеток, выделяющие в кровь гормоны — биологически ак-
цизные вещества, регулирующие обмен веществ, функции орга-
низма, его рост и развитие.
Все эндокринные железы имеют ряд общих морфологических
признаков строения: 1) их секреторные клетки имеют развитый
синтетический и секреторный аппарат; 2) они пронизаны плот-
ной сетью капилляров; 3) у них отсутствуют выводные протоки.
В этой единой системе выделяются центральные и периферичес-
кие органы. К центральным регуляторным образованиям эндок-
ринной системы относятся: нейросекреторные ядра гипоталаму-
са, гипофиз и эпифиз. Периферические эндокринные железы вклю-
чают щитовидную и паращитовидную железы, надпочечники с
их корковым и мозговым веществом. Кроме того, эндокринная
функция присуща гонадам (семенник, яичник), поджелудочной
железе, плаценте, почкам, сердцу, слюнным железам и другим
органам.
Гипофиз (питуитарная железа) — этот комплексный эндокрин-
ный орган бобовидной формы лежит в турецком седле клиновид-
ной кости, соединяется с гипоталамусом и разделяется на адено-
гипофиз и нейрогипофиз. В аденогипофизе, в свою очередь, выделя-
ют инфундибулярную (ворон новую), промежуточную и дистальную
части. Между промежуточной и дистальной частями располагает-
ся полость гипофиза.
Аденогипофиз (передняя доля гипофиза) выделяет шесть гор-
монов. Четыре из них (адренокортикотропный, тиротропный, фол-
шкулостимулирующий и лютеонизирующий) регулируют деятель-
ность других желез внутренней секреции — надпочечников, щи-
товидной железы, половых желез. Два гормона аденогипофиза —
соматотропный и лютеотропный — находятся под двойным гипо-
таламическим контролем — стимулирующим и тормозным.
Соматотропный гормон (соматотропин, гормон роста СТГ)
представляет собой полипептид, структура которого различна у
Рис. 3.156. Гипофиз лошади (А), коровы (А), свиньи (В), собаки (Г):
/ — карман третьего мозгового желудочка; 2 — нейрогипофиз; 3 — промежуточ-
ная часть; 4 — аденогипофиз
277
разных видов животных. Его недостаток у молодых животных при
водит к замедлению роста тела или полной его остановке. Нс
нормальность роста — карликовость, гигантизм, акромегалия
(утолщение костей и рост мягких тканей отдельных частей тела у
взрослых животных) обусловлены также дисфункцией аденоги
пофиза.
Пролактин (лактогенный гормон, лютеотропный гормон) — по
липептид, сходный по строению с СТГ. Он стимулирует проли
ферацию тканей молочной железы и секрецию молока, является
одним из пусковых гормонов лактопоэза. Другие эффекты про-
лактина заключаются в проявлении родительского инстинкта.
Тиротропный гормон (ТТГ) представляет собой гликопротеид,
основная функция которого — стимуляция роста щитовидной
железы (увеличение размеров и числа фолликулярных клеток). ТТ Г
влияет на скорость поглощения йода из крови, включение йода в
состав тироидных гормонов и освобождение гормонов из железы.
Адренокортикотропный гормон (АКТЕ) — это полипептид, ос-
новная функция которого — стимуляция роста коры надпочечни-
ков (пучковой и сетчатой зон), регуляция синтеза и секреции
кортикостероидных гормонов.
Гонадотропные гормоны — фолликулостимулирующий (ФСГ) и
лютеинизирующий гормон (ЛГ) являются глюкопротеидами и нс
обладают половой специфичностью: они вырабатываются у сам-
цов и самок и оказывают на них в принципе аналогичное влияние.
Основная роль этих гормонов заключается в контроле за развити-
ем и нормальным функционированием гонад. У молодых живот-
ных после удаления гипофиза нс происходит полового развития,
а у половозрелых происходит атрофия гонад и наружных половых
органов.
Основными гормонами нейрогипофиза являются окситоцин и
вазопрессин. Они образуются в ядрах гипоталамуса и в виде гранул
секрета аксональным током поступают в заднюю долю гипофиза.
Основная функция вазопрессина (антидиуретического гормона —
АДГ) — регуляция водного баланса.
Окситоцин — гормон, стимулирующий сокращение гладких
миоцитов матки, а также миоэпи гслиальных клеток альвеол и
протоков молочных желез. Естественным стимулом для выработ-
ки и выделения окситоцина является раздражение сосков мо-
лочной железы при сосании, а также растяжение матки и влага-
лища.
Эпифиз (шишковидная железа). Располагается между передней
парой холмов пластинки четверохолмия среднего мозга, имеет
форму шишки с длиной 2,1 мм и шириной 0,5 мм с заостренной
верхушкой. Вырабатываемые клетками эпифиза биологически ак-
тивные вещества оказывают стимулирующее действие на функ-
цию половой системы, а регуляторные пептиды (их вырабатыва-
278
i-1 ся около 40) — на ряд важнейших функций организма, в том
числе на минеральный обмен (повышается уровень калия в
крови).
Щитовидная железа. Лежит позади щитовидного хряща горта-
ни на первых трахейных кольцах и состоит из двух миндалевидной
формы долей, объединенных перешейком. Снаружи покрыта со-
единительнотканной капсулой, от которой внутрь органа прони-
кают прослойки — трабекулы. Последние делят орган на отдель-
ные дольки, содержащие паренхиму.
Железа секретирует два гормона — L-тироксин и L-трийодти-
ронин, обладающие двумя основными эффектами: метаболичес-
ким и ростовым. Попав с током крови, гормоны увеличивают ско-
рость обмена веществ животного.
Парафолликулярными клетками щитовидной железы выраба-
। ывается гормон кальцитонин (КТ). Объектами действия КТ явля-
ются кости, почки, возможно, кишечник.
Паращитовидные железы. Топографически связаны со щитовид-
ной железой, но в отношении эмбриогенеза, строения и функ-
ции являются самостоятельными органами. Они представлены дву-
мя парами (внутренней и наружной) небольших желез, тесно
прилежат к поверхности щитовидной железы и имеют с после-
дней общую иннервацию и кровоснабжение. Гормон паращито-
видных желез — паратгормон (ПТГ) — повышает уровень каль-
ция в крови за счет его потери в костной ткани. Тирокальцитонин
же, наоборот, тормозит резорбцию кости.
Надпочечники (предпочечники). Эти парные инкреторные орга-
ны состоят из двух самостоятельных гормонопродуцирующих же-
лез, формирующих в органе кору и мозговое вещество. Железы
лежат краниально от почек и снаружи покрыты соединительно-
тканной капсулой. Корковые эндокриноциты образуют три зоны:
клубочковую, пучковую и сетчатую.
Клубочковая зона представлена короткими столбами плоских
клеток. В ней вырабатывается минералокортикоидный гормон —
альдостерон. Он контролирует баланс электролитов в организме
(задерживает натрий, хлориды, воду и способствует усиленному
выделению калия, фосфора и кальция) и кислотно-щелочное рав-
новесие.
Пучковая зона занимав! значительный объем в коре. Ее клетки
синтезируют глюкокортикоидные гормоны. Из большого числа кор-
тикостероидов два обладают наибольшей физиологической актив-
ностью и постоянно определяются в крови — это кортизол и кор-
тикостерон. Они участвуют в регуляции: 1) обмена жиров, белков
и углеводов; 2) реакций организма на стресс-факторы (травма,
инфекция и пр.); 3) водно-электролитного обмена; 4) механизма
стабилизации артериального давления и 5) воспалительной реак-
ции, сопровождающейся снижением в периферической крови
279
лимфоцитов и эозинофилов. Кроме того, глюкокортикоиды уско-
ряют глюконеогенез (образование глюкозы за счет белков) и от-
ложение гликогена в печени и миокарде.
Клетки сетчатой зоны вырабатывают гормоны, по химическо-
му составу и физиологическому действию напоминающие поло-
вые гормоны.
Мозговое вещество надпочечника относится к адреналовой сис-
теме. Клетки его секретируют гормон норадреналин и адреналин.
Действие этих веществ в основном аналогично: они повышают
частоту сокращений и возбудимость сердца, повышают печеноч-
ный кровоток, систолическое давление и периферическое веноз-
ное давление, расслабляют мускулатуру бронхов, усиливают ле-
гочную вентиляцию, стимулируют углеводный обмен.
jj „ НЕВРОЛОГИЯ
Неврология — раздел анатомии, изучающий строение нервной
системы.
Нервная система (systema nervosum) координирует работу всех
органов внутри организма и согласовывает их деятельность в
зависимости от меняющихся условий внешней среды. Ее при-
нято подразделять топографически на центральную и перифери-
ческую, а функционально — на соматическую, висцеральную (па-
расимпатическую) и сосудистую (симпатическую) (рис. 3.157 —
3.171).
К центральной нервной системе (ЦНС) относится спинной и
головной мозг. К периферической нервной системе относятся все
нервы, "
Рис. 3.157. Типы нейронов:
А — мульти полярный, Б — биполярный; В — псевдоуниполярный нейрон; 1 —
дендриты; 2 — тела нервных клеток; 3 — аксоны
280
Нервная система состоит из трех структурных элементов — ней-
ронов, нейроглии и мезоглии. Нейрон является основной структур-
но-функциональной единицей нервной системы. Тела нейронов
называют нервными клетками (рис. 3.157).
ЦЕНТРАЛЬНАЯ НЕРВНАЯ СИСТЕМА
ЦНС функционально и топографически разделяется на спин-
ной мозг, лежащий в позвоночном канале, и головной мозг, по-
мещающийся в черепной полости (рис. 3.158 — 3.164). Она постро-
ена из серого и белого мозгового вещества. Серое мозговое веще-
ство состоит из тел нервных клеток и их отростков, а белое мозго-
вое вещество только из отростков нервных клеток, образующих
центральные проводящие пути.
Серое мозговое вещество располагается в центре мозга вокруг
мозговых полостей — центрального спинно-мозгового канала и
желудочков головного мозга. В спинном мозге оно макроскопи-
чески воспринимается как сплошная масса, в то время как в го-
ловном мозге разбито на огромное число ядер. Другая особенность
строения головного мозга млекопитающих заключается в том, что
серое мозговое вещество залегает и на его периферии, образуя
кору большого мозга и мозжечка. Коре большого мозга функцио-
нально подчинены все глубже лежащие ядра серого мозгового ве-
щества — подкорковые ядра.
Кроме коры полушарий и подкорковых ядер в головном и спин-
ном мозге имеется сетчатая формация. Она сформирована сетью
нервных волокон и нервных клеток разнообразной величины и
формы.
Спинной мозг
^Спинной мозг {medulla spinalis) в виде шнура лежит в позво-
ночном канале (рис. 3.158, 3.159). Передний конец его без замет-
ных границ переходит в продолговатый мозг. Задний конец, начи-
ная с пояснично-крестцовой части спинного мозга, суживается и
образует мозговой конус, который в дальнейшем iiepe\(uiri_jw<o«-
цевую нить (р\к. 3.159). На спинном мозге имеются шейное и по-
яснично-крестцовое утолщение как места концентрации клеток и
волокон нервов грудных и тазовых конечностей.
Через каждое межпозвоночное отверстие из спинного мозга
отходят спинно-мозговые нервы. Они в его передних частях распола-
гаются под прямым углом к продольной оси, а в поясничной ча-
сти отходят в косом направлении и придают мозговому конусу
вид конского хвоста.
281
Рис. 3.158. Поперечное сечение спинного мозга:
Л/— .татеральный канатик; 2 - серая спайка, У центральный канал; 4/.— дор-
сальная боротда: 5. - дорсальный канатик; 6/— дорсальный рог; 7/- нити дор-
сального корешка; 8 — нейтральный рог; 9,— НИТИ неНТВЫЫЮГО кнрсшка;
центра 1М1ЫЙ канатик; 1Ц иси|ра.и.ная щель. /? ос гая сп.|ика
Снизу спинного мозга находится срединная щель, в которой
помещается спинно-мозговая вентральная артерия. Сверху спинно-
го мозга располагаются дорсальная срединная борозда и латераль^
ные дорсальные борозды для дорсальных чувствительных корешков.
Латерально от вентральной срединной щели лежат латеральные
вентральные борозды для вентральных двигательных корешков.
На поперечном разрезе видно, что спинной мозг состоит из
белого мозгового вещества, расположенного по периферии, и
серого мозгового вещества, занимающего центральную часть спин-
ного мозга и имеющего вид буквы «Н».
Рис. 3.159. Положение спинного мозга в спинно-мозговом канале
кошки (А), собаки (/>), свиньи (5), коровы (Г), лошади (Д)
282
Серое мозговое вещество вдается в белое мозговое вещество в
виде столбов (рогов) дорсальных чувствительных и вентральных
двигательных, соединенных между собой серой спайкой, в центре
которой располагается центральный спинно-мозговой канал
(рис. 3.158).
Каждый спинно-мозговой нерв начинается двумя корешками —
вентральным и дорсальным. В области межпозвоночного отверстия
вентральный и дорсальный корешки соединяются в смешанный
нерв. На дорсальном корешке лежит спинно-мозговой узел. Каждый
смешанный нерв делится на дорсальную и вентральную ветви для
иннервации мышц и кожи и присоединяет ветви от пограничного
симпатического ствола.
Белое мозговое вещество сильно развито в передних отделах спин-
ного мозга. Оно столбами серого мозгового вещества разделено на
гри пары мозговых канатиков — дорсальные, вентральные и лате-
ральные. Вентральные канатики соединены между собой вентраль-
ной белой спайкой.
Масса спинного мозга у коров равна 260 г. Спинной мозг пита-
ют вентральная спинно-мозговая артерия, дорсальные спинно-моз-
говые правая и левая артерии. В каждом межпозвоночном отвер-
стии эти артерии получают ветви от позвоночных, межреберных,
поясничных и крестцовых артерий. Спинно-мозговые вены идут вдоль
вентральных корешков и отводят кровь в венозные сплетения и
парный позвоночный вентральный синус, а из него — в сегментные
вены туловища.
Особенности. У свиней спинной мозг весит 45 — 70 г, у ло-
шади — 250 —300, у собакц.. —13 г. У лошадей мозговой конус
доходит до 2 —3-го крестцового, а у собак до 6 —7-го пояснично-
го позвонка.
Мозговые оболочки
Оболочки спинного мозга (meninges) разделяются на: твердую,
паутинную и мягкую (рис. 3.160).
Твердая-оболочка (наружная) построена из плотной соедини-
тельной ткани, очень прочная, бедна кровеносными сосудами,
покрывает спинной мозг и его нервы, прикрепляется по краям
межпозвоночных отверстий. Она состоит из внутренней и наруж-
ной пластинок. Первая из них прочно срастается с надкостницей,
а внутренняя поверхность второй выстлана эндотелием.
Между указанными пластинками твердой оболочки располага-
ется эпидуральное пространство, заполненное рыхлой соединитель-
ной тканью.
Паутинная оболочка (средняя) нежная и бессосудистая. Обе
поверхности ее выстланы эндотелием. Паутинная оболочка, по-
283
крывая спинной мозг, не заходит в его щели и борозды. Между
твердой и паутинной оболочками находится узкое щелевидное
субдуральное пространство, заполненное лимфой. Паутинная обо-
лочка соединяется с твердой оболочкой кровеносными сосудами,
зубовидными связками мягкой оболочки, нервами, а также под-
вешивающими связками, которые располагаются впереди первой
пары шейных нервов.
Мягкая оболочка (внутренняя) очень тонкая, но плотная, она
срастается со спинным мозгом, заходя во все его извилины. Сна-
ружи мягкая оболочка покрыта эндотелием. Между паутинной и
мягкой оболочками находится подпаутинное пространство, запол-
ненное спинно-мозговой жидкостью. На боковых поверхностях спин-
ного мозга мягкая оболочка формирует правую и левую боковые
связки. От этих связок между сегментами к твердой оболочке отхо-
дят зубовидные связки.
Оболочки головного мозга являются продолжением оболочек
спинного мозга и также подразделяются на твердую, паутинную
и мягкую (рис. 3.160).
. Твердая оболочка головного мозга.поверхностная. Она состоит из
двух .пластинок. Наружная из них срастается с надкюстнгГПЯГче-
репных костей/Между ней и внутренней пластинкой располага-
ются безмышечные вены, образующие две системы венозных со-
судов — дорсальную и вентральную.
Дорсальная система синусов складывается из сагиттального,
прямого, поперечных, дорсальных каменистых и затылочных си-
нусов.
Вентральная система синусов состоит из циркулярного и бази-
лярного синусов.
Рис. 3.160. Оболочки головно-
го мозга:
1 — твердая оболочка головного
мозга; 2 — кости черепа; 3 —
грануляции паутинной оболочки;
4 — сагиттальный синус; 5 — ко-
жа; 6 — эмиссарная вена; 7 — па-
утинная оболочка; 8 — субарах-
ноидальное пространство; 9 — бе-
лое мозговое вещество; 10 — серп
большого мозга; И — серое моз-
говое вещество
284
От твердой оболочки головного мозга в субдуральное простран-
ство отходят серп мозга и перепончатый мозжечковый намет.
Паутинная оболочка головного мозга состоит из соединитель-
ной ткани, на извилинах мозга она срастается с мягкой мозговой
оболочкой, подпаутинное пространство встречается только в об-
ласти щелей и борозд. В субдуральное пространство, заполненное
лимфой, отходят ворсинки паутинной оболочки.
Мягкая оболочка головного мозга тесно срастается с мозгом,
проникает во все его щели и борозды, а также в боковые, третий
и четвертый мозговые желудочки. Здесь она участвует в формиро-
вании сосудистых покрышек, в которых находятся сосудистые спле-
тения. Между ней и паутинной оболочкой располагается подпау-
тинное пространство. В нем находится спинно-мозговая жидкость.
Последняя выделяется клетками сосудистых сплетений мозга и
мозгового вещества. Движется жидкость в подпаутинном простран-
стве краниально, а в центральном канале каудально. Отток ее осу-
ществляется в венозную и лимфатическую системы.
Головной мозг
Головной мозг (encephalon} подразделяется на большой и ром-
бовидный. Большой моя (cerebrum} делится на конечный,
меж\ (очный и срс иши. а ромбовидный моя (rhombencephahn) на
laaiiiiii и нродо.иовагый (рис 3 161 3 164)
Конечный мозг
Конечный мозг {telencephalon) состоит из правого и левого по-
лушарий большого мозга (рис. 3.161 — 3.163). В каждом полуша-
рии различают плащ, обонятельный мозг, полосатые тела и боко-
вые мозговые желудочки. Границей между плащом и обонятель-
ным мозгом на нижней мозговой поверхности является базальная
пограничная борозда. Полосатое тело помещается над обонятель-
ным мозгом.
Плащ состоит из серого и белого мозгового вещества. Серое
мозговое вещество находится снаружи и составляет кору большого
мозга. Она покрыта извилинами и щелями. На плаще различают лоб-
ную, височную, теменную, затылочную и обонятельную доли.
Белое мозговое вещество находится под корой плаща (рис. 3.161 —
3.163). Оно формирует проводящие пути.
Обонятельный мозг помещается в передненижней части боль-
шого мозга (рис. 3.161 — 3.163). К нему относятся обонятельные
луковицы, обонятельные тракты и извилины, обонятельные тре-
угольники, грушевидные доли, аммоновы рога, свод и полосатое
тело.
285
Рис. 3.161 Базальная поверхность и артерии головного мозга лошади:
/•— продольная борозда; 12-— полость луковицы, медиальная обонятельная
ишнлинп; •/ - артерия мозолистого тела, 5 краниальная артерия мозговых обо-
лочек; 6} оральная соединительная ветм,.. 7 средняя мо новая артерия,.4
передняя apiepitn сосудистого сплсшгия, внутренняя сонная apieptia, 10/
аборальная соединительная ветвь; II - глубокая мозговая артерия; 12 — Перс
зияя артерш мозжечка; 7J, базальная мозговая артерия; 14 задняя мозговая
артерия; '/5 — продолговатый мои; !<г щгдьязычный нерв; 77 — базальная
мозговая артерия: 1Ц — подъязычный нерв. 14 — зобаммный нерв; 2(> — яздео-
глоточный нерк-2/. блуждающий нерв, 22 равновесно-слуховой лгерв; 2J -
лииенои нерв; £?4‘- тройничный нерв;'25‘ нижнечелюстной нерв; 26 — отводя-
щий нерв; ,27. — г>щзодвигательный нерв; 2S'— верхнечелюстной нерв,
блоковый нерв. 10,— шззничный нерв; 21 глазодвнгательшй нерв. 22
зрительный перекрест; 32,- латератьный обонятельный тракт: 24 — зрительный
иерв;,55— общий обонятельный трякг;<7$— обонятельная борозда; ^7/— обоня-
тельная луковица
286
Рис. 3.162. Головной мозг собаки (латеральная поверхность):
/2- обонятельная луковица; 2 . обонятельная борозда; 5/ сильвиева борозда;
•/ — супрасильвиеаа борозда; 5 — крестовидная борозда; б,.— маргинальная бо-
розда;-7— маргинальная и тимина; л — жгосилышенз борозда;, Ч 1.1 «ь жто-
сильвиева извилина. 10 сильвиева извилина; II зктомаргннальная борозда;
/2 эктомарпталыгая извилина; ./4 супрасидьвиева извилина; червя
чок; 16 — полушарие мозжечка; 17 - кло'Ю^Ц/^)—подъязычный^ерв; ДО)—
1<><).‘ночпыи нерв. 20 блуждающий нерв. ,2//— языкоглоточный нерв; 22 —
|рсд1нерно-ули1>.оныи нерв: 2.? — лицевой нерв; 24 — отводящий нерв;(2?,) —
тройничный нерв; -26)— мост; /27j— грушевидная доля; $8у— зрительный нерв;
(2УЬ— общий обонятельный тракт
Обонятельная луковица (рис. 3.161—3.163) —парное образова-
ние, которое помещается в ямках решетчатой кости. Верхнемеди-
альный участок луковицы состоит из серого мозгового вещества,
а нижнелатеральный — из белого. Внутри луковицы находится по-
лость — желудочек обонятельной луковицы. В луковицу вступают
многочисленные обонятельные нити от обонятельных клеток сли-
зистой оболочки носовой полости, которые в сумме образуют обо-
нятельный нерв
Обонятельные тракты и извилины соединены проводящими
тлями с корой большого мозга и содержат ядра серого мозгового
вещества. Различают общий, медиальный и латеральный обоня-
тельные тракты (рис. 3.161).
Обонятельный треугольник построен из серого мозгового веще-
ства. Он ограничен обонятельными трактами.
Грушевидные доли помещаются позади латерального обоня-
тельного тракта и обонятельного треугольника, латеральнее но-
жек большого мозга (рис. 3.161). Задний участок бокового моз-
287
гового желудочка распространяется в грушевидную долю и об-
разует в ней полость. Здесь находится нижний конец аммонова
рога.
Аймонов рог (гиппокамп) является парной складкой коры мозга
в ббластиФрушевидной доли (см. рис. 3.164). Он своей верхней ча-
стью образует дно бокового мозгового желудочка позади хвоста-
того ядра. Между аммоновым рогом и хвостатым ядром находится
сосудистое сплетение бокового мозгового желудочка.
Свод образован проводящими пучками между аммоновыми
рогами и сосцевидным телом промежуточного мозга (рис. 3.163).
На своде различают желобоватый листок, кайму аммонова рога,
ножки, столбы и тело свода, комиссуру аммоновых рогов.
Полосатое тело состоит из хвостатого, чечевицеобразного и
миндалевидного "ядер, ограды, внутренней и наружной капсул.
Промежуточный мозг
Промежуточный мозг (diencephalon) находится между полоса-
тыми телами конечного мозга и средним мозгом (рис. 3.161 —
3.164). Выше промежуточного мозга располагается сосудистая по-
крышка третьего мозгового желудочка и аммоновы рога. Проме-
жуточный мозг состоит из таламуса (основная часть), эпиталаму-
са и гипоталамуса.
Таламус (рис. 3.163) состоит из зрительных бугров и третьего
мозгового желудочка.
Третий мозговой желудочек (рис. 3.163) имеет кольцевидную
формул Он п(жёщаётсОгежду зрительными буграми и сообщается
через мозговой водопровод с четвертым мозговым желудочком, а
около назальной спайки мозга — с правым и левым боковыми
мозговыми желудочками через межжелудочковое отверстие.
Эпиталамус (рис. 3.163) состоит из сосудистой покрышки тре-
тьего мозгового желудочка, эпифиза и парного узла уздечки.
Сосудистая покрышка образует позади валика мозолистого тела
надэпифизарный выступ. Эпифиз — железа внутренней секреции,
грушевидной формы, помещается в ямке между зрительными буг-
рами и четверохолмием. По краям ямки зрительных бугров видны
узкие белые мозговые полоски зрительных бугров. На них распо-
лагается парный узел уздечки. Последний затем переходит в уздеч-
ку. На уздечке и укреплен эпифиз.
^Гипоталамус (см. рис. 3.161, 3.163) располагается ниже зри-
тельных бугров, образует стенку третьего мозгового желудочка. В
гипоталамусе различают серый бугор, гипофиз и сосцевидное
тело.
Серый бугор находится сзади перекрестка зрительных нервов,
между ножками большого мозга и служит вегетативным цент-
ром.
288
Рис. 3.163. Головной мозг собаки (сагиттальный разрез):
I — задняя комиссура; 2 — ножка большого мозга: ? волопрснюл среппего
• '•i.i. ростральный парус; 5- передня комиссура. ЛА межжс,|у.|очкоь<к-
• i перстне; JA колсни-мозолисгого тцла; 8,— кора головного MOjr.i;tJ>x- мозоли
•к тело. J0 — енол;tii — т.сшмус; /<— эпиталамус;. 15. — эпифиз;-^ поясная
(тороид. 15 налик мозолистого тела, 1а каудальные холмы: 17- кора .мог
«.емка; 1л)— древо жизни. 79 — четвертый мозцпюИ желудочек: /(> — мост: 2L —
ростральные холмы; 22 — сосцсшиное тело; gA гипофиз; 24' воронка. 2$.'
срый бугор: 26 зрительный перекрест: 27) питмус; 2а обонятельная
луковица
Гипофиз (см. рис. 3.156, 3.161, 3.163) имеет вид округлого тела,
нескольйгГсдавленного сверху вниз. Он имеет небольшую цент-
ральную полость, которая сообщается с третьим мозговым желу-
дочком. Гипофиз — железа внутренней секреции, которая выде-
ляет несколько очень важных гормонов. Он подразделяется на три
части: мозговую, промежуточную и железистую.
(tn ueeudjioeme.io находится позади серого бугра. Оно через свод
связывается С обонятельным мозгом, со зрительными буграми и
сетчатой субстанцией.
Средний мозг
Средний мозг (mesencephalon) состоит из ножек большого моз-
га. пластинки четверохолмия й покрышки, или чепца (рис. 3.161 —
3.164). Полость среднего мозга превратилась в мозговой (сильвиев)
водопроводов стенках водопровода находится центральное серое
мозгово^вёше ст во.
10 Зелсневский
289
/Л и
Рис. 3.164. Головной мозг собаки (поперечное сечение между средним и
промежуточным мозгом):
/ грстий желудочек мозга: 2 — бокраои желудочек мозга; .? — полушарие
большого мозга. 4/ мозолистое тело:(5 свод; /> - латеральное коленчатое
ядро; 7. /?'— фитсльный трак г; 10)— медиальное коленчатое ядро; 11 черная
субстанция. 12 ножка мозга. 11 средняя ножка мозжечка; 14 - ядро ножки;
15 межножковое ядро; 16 кмулальная спайка. 17 — ко.юироно.1 среднего
мозга; IX — красное ядро, боковой желудочек мозга. 20 внутренняя
капсула;.’/ ядра таламуса;'22 нипюкамп
Ножки большого мозга в виде двух толстых валиков выпячива-
ются на нижней поверхности головного мозга между зрительны-
ми трактами и мостом (рис. 3.161 — 3.164). Они состоят из прово-
дящих путей, соединяющих кору и зрительные бугры со средним,
ромбовидным и спинным мозгом.
Пластинка четверохолмия лежит в верхней части среднего моз-
га. Она состоит из двух передних и задних холмов. Передние холмы
являются зрительным центром, а задние — слуховым.
Покрышка ножек, или чепец (см. рис. 3.163) помещается в цен-
тре среднего мозга между ножками большого мозга и четверохол-
мием. Он состоит из белого мозгового вещества, в котором зало-
жены парные ядра серого вещества. Наиболее крупными из них
являются: красное, ядро глазодвигательного нерва, парасимпати-
ческие ядра, ядро блокового нерва, ядро пятой пары черепных
нервов.
290
Ромбовидный мозг
Ромбовидный мозг (rhombencephalon) состоит из продолгова-
|ого и заднего мозга (см. рис. 3.161 — 3.163).
Продолговатый мозг тянется от мо нового моста назад и без
ш меткой границы переходи! в спинной моя (см. рис. 3 161 —3.163).
< нижней стороны мозга помешается вентральная срединная щель.
11 о обе стороны от срединной щели проходят боковые борозды.
Между последними накояятсяргирамиды продолгдвапцзго..мозга. Из
передней части продолговатого мозга латерально от пирамид вы-
ходит отводящий нерв. Латерально от перекрестка пирамид выхо-
лит подъязычный нерв. Из боковых частей продолговатого мозга
выходят добавочный, блуждающий я язычно-глоточный нервы.
Задний мозг состоит из мозгового моста, мозжечка и чётверто-
1о мозгового желудочка (см. рис. 3.161 — 3.163).
Мозговой мост помещается на переднем конце продолговатого
мозга в виде поперечного валика, который спереди граничит со
средним мозгом (см. рис. 3.161 —3.163). Боковые концы моста тя-
нутся к мозжечку как боковые ножки мозжечка. Мост и ножки
образованы проводящими путями. Из боковых частей моста начи-
нается тройничный нерв. Позади моста в поперечном направлении
помещается трапециевидное тело в виде узкого и низкого валика.
От боковых концов трапециевидного тела выходят равновесно-слу-
ховой и лицевой нервы.
Мозжечок имеет вид шаровидного образования, расположен-
ного наддтрбдолговатым мозгом (см. рис. 3.161 — 3.163). Двумя бо-
ковыми продольными бороздами он разделяется на червячок и две
боковые доли. Серое мозговое вещество образует кору мозжечка и,
кроме того, ядра в белом мозговом веществе, расположенном цен-
трально. На поверхности червячка имеются многочисленные, по-
перечные борозды и щели.
Четвертый мозговой желудочек (см. рис. 3.161 — 3.163) имеет
свод, состоящий йз червячка и мозговых парусов, и дно, которое
называется ромбовидной ямкой.
Особенности. Абсолютная масса головного мозгц_у_коров
составляет 410 — 550 г^у^винсй — 96—145, у лошади — 372 —
570, у_собак — 46—138 г.
ПЕРИФЕРИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
Периферическая нервная система (systema nervosum peripheri-
сит) включает черепные и спинно-мозговые нервы, сплетения,
ганглии и нервные окончания, заложенные в органах и тканях
(рис. 3.165 — 3.171). В составе периферической нервной системы при-
нято выделять три части: соматическую — сегментальную, связы-
291
вающую центры со скелетной мускулатурой и кожей, симпати-
ческую (сосудистую) — связанную с гладкой мускулатурой сосу-
дов, парасимпатическую (висцеральную) — связанную с железами
внутренней секреции и гладкой мускулатурой внутренних орга-
нов.
Нерв образуется пучками нервных волокон, т. е. отростками
нейронов, и способен проводить электические импульсы. Он име-
ет вид белого тяжа разнообразной формы — то более округлого,
то более плоского. Толщина его колеблется от микроскопических
размеров до нескольких сантиметров.
Соматическая часть периферической нервной системы вклю-
чает спинно-мозговые и черепные нервы (см. рис. 3.157, 3.161). Спин-
но-мозговые нервы образованы отростками нервных клеток, тела
которых лежат в спинном мозге. Черепные нервы (рис. 3.161; 3.167)
входят в состав соматической части периферической нервной си-
стемы, но отходят они от головного мозга.
Спинно-мозговые нервы
Спинно-мозговые нервы отходят от спинного мозга и в соот-
ветствии с отделами позвоночного столба делятся на шейные (С),
грудные (Th), поясничные (L), крестцовые (S) и хвостовые (Сс).
Число грудных, поясничных и крестцовых нервов соответствует
числу позвонков соответствующего отдела позвоночного столба,
шейных нервов больше на одну пару (восемь пар).
Шейные нервы в количестве восьми пар выходят через межпоз-
воночных отверстий шейных позвонков. Из отдельных вентраль-
ных ветвей шейных нервов образуются диафрагмальный, дорсаль-
ный лопаточный и надключичный нервы. Вентральные ветви трех
(четырех) последних шейных и первых двух грудных спинно-моз-
говых нервов образуют плечевое сплетение.
Плечевое сплетение лежит ниже лестничной мышцы и меди-
альное лопатки (рис. 3.165). Из него выходят грудные, предлопа-
точный, подлопаточный, подмышечный, кожно-мышечный, лу-
чевой, локтевой и срединный нервы. Все они участвуют в иннер-
вации грудной конечности.
Грудные нервы подразделяются на передние и задние (рис. 3.165).
Передние грудные нервы иннервируют тремя-четырьмя ветвями груд-
ные мышцы впереди плечевого сустава. Задние грудные нервы под-
разделяются на длинный, верхний, боковой и нижний грудные
нервы. Длинный грудной нерв иннервирует грудную часть нижней
зубчатой мышцы. Верхний грудной нерв иннервирует широчайшую
мышцу спины. Боковой грудной нерв иннервирует кожу латераль-
ной грудной стенки. Нижний грудной нерв иннервирует глубокую
грудную мышцу и кожу.
292
Рис. 3.165. Нервы, образующие
плечевое сплетение лошади
(медиальная поверхность):
/ — четвертый спинно-мозговой
нерв; 2 — вентральная ветвь
пятого спинно-мозгового нерва;
3, 4 — дорсальные ветви шесто-
го спинно-мозгового нерва; 5 —
плечевое сплетение; 6 — первый
грудной спинно-мозговой нерв;
7 — грудной вентральный нерв;
8 — грудной латеральный нерв:
9 — грудной дорсальный нерв;
10 — диафрагмальный нерв;
11 — локтевой нерв; 12 — сре-
динный нерв; 13 — лучевая кость;
14— пальмарный пястный нерв;
15 — медиальный пальмарный
пальцевой нерв; 16, 19 — кра-
ниальный грудной нерв; 17 —
предлопаточный нерв; 18— мы-
шечно-кожный нерв; 20, 21 —
лучевой нерв; 22 — мышечно-
кожный нерв; 23 — медиальный
дорсальный пальцевой нерв
293
Предлопаточный нерв направляется в предостную и заостную
мышцы (рис. 3.165).
Подлопаточный нерв имеет две—четыре ветви, иннервирует под-
лопаточную мышцу (рис. 3.165).
Подмышечный нерв идет между подлопаточной и большой круг-
лой мышцами, иннервирует сгибатели плечевого сустава и кожу
плеча и предплечья (рис. 3.165).
Кожно-мышечный нерв иннервирует: коракоидно-плечевую, дву-
главую и плечевую мышцы, а его кожная ветвь — кожу краниоме-
диальную поверхности предплечья (рис. 3.165).
Лучевой нерв иннервирует разгибатели локтевого сустава, запя-
стья и пальцев, кожу дорсальной поверхности предплечья и кис-
ти (рис. 3.165). Он разделяется на поверхностный и глубокий луче-
вые нервы.
Поверхностный лучевой нерв между головками трехглавой мыш-
цы плеча выходит на дорсальную поверхность предплечья, пясти
и пальцев и иннервирует кожу.
Глубокий лучевой нерв иннервирует разгибатели запястья и паль-
цев.
Локтевой нерв отдает ветви: для плечевой кости и кожи пред-
плечья, а также для сгибателей запястья и пальцев (рис. 3.165). На
дистальном конце предплечья локтевой нерв отдает дорсальную
ветвь для кожи пясти, IV и V пальцев и переходит в пальмарный
нерв. Пальмарный нерв дает пястные пальмарные нервы, которые в
дальнейшем переходят в пальцевые пальмарные латеральный и ме-
диальный нервы. Дорсальная ветвь нерва дает пальцевой дорсальный
латеральный четвертый нерв, а пальмарная ветвь иннервирует
межкостную мышцу и переходит в пальцевой пальмарный лате-
ральный четвертый нерв.
Срединный нерв идет вместе с плечевой и срединной артериями
(рис. 3.165). У локтевого сустава он отделяет ветви для лучевого
сгибателя запястья, глубокого сгибателя пальцев. В области пясти
этот нерв дает пястные пальмарные поверхностные нервы, а сам
переходит в пальцевой пальмарный нерв.
У коров срединный нерв переходит в пястный пальмарный
медиальный нерв и дает пальцевой пальмарный медиальный третий
нерв и пальцевой пальмарный общий нерв для III и ГУ пальцев. Чет-
вертый пальцевый пальмарный нерв соединяется с локтевым не-
рвом.
Грудные нервы в количестве 13 пар у коров отходят от спинного
мозга. Каждый нерв отдает симпатическому стволу белую и полу-
чает от него серую соединительные ветви и делится на дорсаль-
ные и вентральные ветви. Дорсальные ветви направляются в
дорсальные мышцы позвоночного столба и кожу. Вентральные вет-
ви — межреберные нервы — вместе с одноименными артериями и
венами проходят в реберных желобах.
294
Поясничные нервы по количеству соответствуют числу одно-
именных позвонков. У коров шесть пар поясничных нервов. Пер-
вые два-четыре нерва отдают симпатическому стволу белые со-
единительные ветви, серые соединительные ветви получают все
। юясничные нервы. Они делятся на дорсальные и вентральные ветви.
Дорсальные ветви называются передними кожными ягодичными не-
рвами. Вентральные ветви формируют поясничное сплетение, кото-
рое вместе с крестцовым сплетением образует пояснично-крестцо-
вое сплетение (рис. 3.166). Из поясничного сплетения выходят под-
вздошно-подчревный, подвздошно-паховый, наружный семенной,
иояснично-кожный, бедренный и запирательный нервы. Все они
участвуют в иннервации органов тазовой конечности.
Подвздошно-подчревный нерв (рис. 3.166) иннервирует пояснич-
ные и брюшные мышцы, кожу брюшной стенки, кожу мошонки
и препуций (у самцов),
вымя (у самок).
Рис. 3.166. Пояснично-крест-
цовое сплетение, нервы та-
зовой конечности собаки
(латеральная поверхность):
/ — наружный семенной нерв;
2 — поясничное сплетение; 3 —
бедренный нерв; 4 — краниаль-
ный ягодичный нерв; 5 — за-
пирательный нерв; 6 — крест-
цовое сплетение; 7 — прямоки-
шечный нерв; 8 — каудальный
ягодичный нерв; 9 — хвостовые
нервы; 10 — седалищный нерв;
11 — срамной нерв; 12 — кож-
ный каудальный нерв бедра;
13 — проксимальные мышеч-
ные ветви; 14 — большеберцо-
вый нерв; 15 — плантарный
кожный нерв голени; 16 — дис-
тальные мышечные ветви; 17 —
плантарный латеральный нерв;
18 — плантарные плюсневые
нервы; 19 — плантарные паль-
цевые нервы; 20 — дорсальные
пальцевые нервы; 21 — дорсаль-
ные плюсневые нервы; 22 —
дорсальный кожный нерв го-
лени; 23 — поверхностный ма-
лоберцовый нерв; 24 — глубо-
кий малоберцовый нерв; 25 —
малоберцовый нерв; 26 — по-
яснично-кожный нерв
295
Подвздошно-паховый нерв (рис. 3.166) иннервирует большую по-
ясничную, квадратную поясничную, брюшные мышцы, кожу бед-
ра, кожу мошонки и препуций (у самцов), вымя (у самок).
Наружный семенной нерв иннервирует подвздошно-поясничную,
квадратную поясничную, брюшные мышцы, кожу бедра, вымя (у
самок), мошонку и препуций (у самцов).
Пояснично-кожный нерв иннервирует: мышечная ветвь — боль-
шую поясничную мышцу, кожная ветвь под названием кожного
латерального нерва бедра — кожу коленного сустава.
Бедренный нерв (рис. 3.166) выходит между медиальной голов-
кой подвздошной мышцы и портняжной мышцей, проходит в
бедренном канале, делится на две ветви и иннервирует: мышеч-
ная ветвь — подвздошную и четырехглавую мышцы, подкожная
ветвь под названием нерв сафенус кожу голени и стопы.
Запирательный нерв иннервирует запирательные мышцы.
Крестцовые нервы выходят через дорсальные и вентральные
крестцовые отверстия и присоединяют серые соединительные ветви
симпатического ствола. Дорсальные ветви направляются в разги-
батели тазобедренного сустава и в кожу как кожные ягодичные сред-
ние нервы (рис. 3.166). Вентральные ветви формируют крестцовое
сплетение, из которого выходят передний и задний ягодичные,
каудальный кожный, срамной, задний прямокишечный, седалищ-
ный нервы.
Передний ягодичный нерв иннервирует ягодичные мышцы. Зад-
ний ягодичный нерв иннервирует двуглавую и ягодичные мышцы.
Каудальные кожные нервы бедра выходят на поверхность бедра
позади двуглавой мышцы и под названием кожных ягодичных зад-
них нервов иннервирует кожу бедра.
Срамной нерв (рис. 3.166) идет через седалищную дугу рядом со
срамной артерией. У самцов он переходит на половой член как
дорсальный удовый нерв и иннервирует пенис. От срамного нерва
отходят средний нерв прямой кишки и нерв промежности для кожи
половой щели, ануса и вымени у самок, кожи промежности и
мошонки у самцов.
Задний прямокишечный нерв иннервирует прямую кишку, анус,
а у самок и половые губы.
Седалищный нерв иннервирует всю тазовую конечность (рис.
3.166). Он выходит через большую седалишную вырезку таза, про-
ходит между большим вертелом и седалищным бугром. Позади
тазобедренного сустава седалищный нерв делится на большебер-
цовый и малоберцовый нервы, которые идут вместе до коленного
сустава. В области таза седалищный нерв дает ветви для глубокой
ягодичной, внутренней запирательной, двойничных и квадрат-
ной бедренной мышц.
Большеберцовый нерв (рис. 3.166) отдает в области тазобедрен-
ного сустава проксимальные мышечные ветви для двуглавой, полу-
296
сухожильной и полуперепончатой мышц, на середине бедра —
плантарный кожный нерв голени, который идет рядом с латераль-
ной подкожной веной стопы и голени и иннервирует кожу голени
и плюсны, против коленного сустава — дистальные мышечные ветви
для разгибателей предплюсневого сустава и сгибателей пальцев,
на середине голени — медиальный кожный нерв голени для кожи
голени и плюсны. Выше предплюсны большеберцовый нерв де-
лится на латеральный и медиальный плантарные нервы. Плантарные
нервы делятся на плюсневые плантарные нервы, которые в даль-
нейшем переходят в пальцевые нервы.
У коров плантарный медиальный нерв отдает пальцевой плантар-
ный IIIмедиальный нерв и общий пальцевой плантарный нерв для III
и IV пальцев. Плантарный латеральный нерв переходит в пальцевой
плантарный латеральный IV нерв.
Малоберцовый нерв (рис. 3.166) находится в области бедра под
двуглавой мышцей, а на голени — впереди большеберцовой кос-
ти рядом с передней большеберцовой артерией и веной. На уров-
не коленного сустава малоберцовый нерв дает дорсальный кожный
нерв голени. У латерального мыщелка большеберцовой кости он
делится на поверхностный и глубокий малоберцовые нервы. По-
верхностный малоберцовый нерв направляется в сторону пальцев
между боковым и длинным разгибателями пальцев. Глубокий мало-
берцовый нерв идет вместе с большеберцовой передней артерией и
веной и иннервирует дорсальные мышцы голени. На плюсне он
дает плюсневые дорсальные нервы.
У коров поверхностный малоберцовый нерв отдает пальцевые
дорсальные латеральный IVи медиальный IIIнервы и общий пальце-
вой нерв для III и IVпальца. Глубокий малоберцовый нерв присо-
единяется к общему пальцевому нерву.
Хвостовые нервы выходят в числе пяти-шести пар позади соот-
ветствующих хвостовых позвонков. Дорсальные ветви хвостовых
нервов соединяются в продольный дорсальный нерв хвоста, а вен-
тральные — в продольный вентральный хвостовой нерв для мышц
и кожи хвоста.
Черепные нервы
Черепных (головных) нервов у позвоночных животных насчи-
тывается XII пар (рис. 3.167 — 3.169; см. рис. 3.161, 3.162). Они вы-
полняют различные функции. Обонятельный (I пара), зритель-
ный (II пара) и равновесно-слуховой нерв (VIII пара) — чув-
ствительные и служат проводящими путями анализаторов — обо-
нятельного, зрительного и равновесно-слухового. Глазодвигатель-
ный (III пара), блоковый (IV пара) и отводящий (VI пара) нервы
являются двигательными для глазных мышц. Добавочный (XI пара)
297
Рис. 3.167. Ветвление тройничного нерва лошади:
1 — аборальный носовой нерв; 2 — подглазничный нерв; 3 — глазничный нерв;
4 — верхнечелюстной нерв; 5 — ветвь к напрягателю и поднимателю нёбной
занавески; 6 — щечный нерв; 7 — глубокие височные нервы; 8 — крыловой
нерв; 9 — мышца-подниматель нёбной занавески; 10 — ветвь к латеральной кры-
ловидной мышце; 11 — малый нёбный нерв; 12 — большой нёбный нерв; 13 —
язычный нерв; 14— межчелюстной нерв; 15 — нижний альвеолярный нерв; 16 —
большой жевательный нерв; /7 — поверхностный височный нерв; 18 — дорсаль-
ная мозговая вена; 19 — внутренняя челюстная вена; 20 — общая сонная арте-
рия; 21 — оральный носовой нерв; 22 — наружные носовые нервы; 23 — нерв
верхней губы; 24 — верхняя резцовая ветвь; 25 — подглазничный нерв; 26 —
аборальный носовой нерв; 27 -- решетчатый нерв; 28 — нервы верхнего моляра;
29 — ветвь к железе третьего века; 30 — нервы верхнего моляра; 31 — скуловой
нерв; 32 — подблоковый нерв; 33 — слезный нерв; 34 — решетчатый нерв; 35 —
лобный нерв; 36 — височно-скуловая ветвь; 37 — височная мышца; 38 — глаз-
ничный нерв; 39 — верхнечелюстной нерв; 40 — нижнечелюстной нерв; 41 —
полулунный узел; 42 — мозговой мост; 43 - начало тройничного нерва; 44 —
яремная вена; 45 — наружная челюстная вена; 46 — околоушная железа; 47 —
большая жевательная артерия и вена; 48 - медиальная, 49 — латеральная часть
большой жевательной мышцы; 50 — латеральная, 57 — медиальная часть крыло-
видной мышцы; 52 — ветвь к оральному брюшку двубрюшной мышцы; 53 —
межчелюстной нерв; 54— нижний альвеолярный нерв; 55 — язычный нерв; 56 —
ветвь к премоляру; 57 — подбородочный нерв; 58 — нижняя резцовая ветвь
и подъязычный (XII пара) нервы — двигательные (первый для
трапециевидной, плечеголовной и грудино-челюстной мышц, а
второй — для мышц языка и подъязычной кости). Блуждающий
нерв (X пара), или вагус, относится к группе парасимпатических
нервов. Из остальных пар тройничный нерв (V пара) — чувстви-
тельный для кожи головы и слизистых оболочек носовой и рото-
298
вой полостей и двигательный для жевательных мышц, в нем же
проходят часть своего пути секреторные волокна в слезную и слюн-
ные железы. Лицевой нерв (VII пара) — двигательный для всех
мимических мышц, т.е. для губных, щечных, носовых, ушных и
мышц век. В нем же проходит часть вкусовых волокон с языка и
секреторные волокна для слезной, подчелюстной и подъязычной
желез. Язычно-глоточный нерв (IX пара) содержит чувствитель-
ные волокна со слизистой оболочки корня языка и глотки, вкусо-
вые волокна с корня языка и двигательные волокна для расшири-
теля глотки. В нем же часть своего пути проходят секреторные во-
локна для околоушной железы.
Обонятельный нерв — I пара — образован нейритами обоня-
тельных клеток, заложенных в слизистой оболочке обонятельной
области носовой полости и в сошниково-носовом органе. Окан-
чивается нерв на обонятельных луковицах (см. рис. 3.161, 3.162).
Зрительный нерв — II пара — образован нейритами мультипо-
лярных клеток сетчатки глаза. Он входит через зрительное отвер-
стие в черепную полость, где образует с одноименным нервом
другой стороны зрительный перекрест (см. рис. 3.161, 3.162).
Глазодвигательный нерв — III пара — выходит из покрышки
среднего мозга (рис. 3.161) и идет в прямую дорсальную мышцу
глаза и в подниматель верхнего века, а вентральная ветвь — в
вентральную косую мышцу глаза и в прямые мышцы глаза.
Блоковый нерв — IV пара — ветвится в дорсальной косой мыш-
це глаза (см. рис. 3.161).
Тройничный нерв — V пара — основной чувствительный нерв
для кожи и слизистых оболочек головы и двигательный для жева-
тельных мышц (рис. 3.168; см. рис. 3.161, 3.162, 3.167, 3.169). Он
выходит двумя корнями из боковой поверхности мозгового моста.
Общий ствол тройничного нерва делится на глазничный, верхне-
челюстной и нижнечелюстной нервы.
Рис. 3.168. Иннервация кожи
головы лошади:
1 — лобный нерв; 2 — скуловой
нерв; 3 — подблоковый нерв; 4
подглазничный нерв; 5 — под-
бородочный нерв; 6 — попереч-
ная лицевая ветвь поверхностно-
го височного нерва; 7 — попе-
речный нерв шеи, ветвь второго
шейного спинно-мозгового нерва
299
Глазничный нерв выходит из черепной полости через глазную
щель и делится на четыре нерва: слезный, лобный, носоресничный и
подблоковый (см. рис. 3.167, 3.168, 3.169).
Верхнечелюстной нерв является нервом общей чувствительнос-
ти для области верхней челюсти. Он выходит в глазницу через
круглое отверстие и делится на скуловой, подглазничный и клино-
нёбный нервы (рис. 3.169; см. рис. 3.167, 3.168).
Нижнечелюстной нерв является нервом общей чувствительнос-
ти для нижней челюсти, височной области и двигательным не-
рвом для жевательных мышц (см. рис. 3.167, 3.168, 3.169). Нерв
выходит из черепной полости через рваное отверстие и отдает три
мышечных нерва в большую жевательную, глубокую височную и
крыловую мышцы, а также поверхностный височный нерв для кожи,
щечный нерв для слизистой оболочки щеки, язычный нерв для сли-
зистой оболочки языка, межчелюстной мышечный нерв. Далее он
переходит в альвеолярный нерв нижней челюсти.
Отводящий нерв — VI пара — иннервирует оттягиватель глазно-
го яблока и глазную прямую латеральную мышцу (см. рис. 3.161).
Лицевой нерв — VII пара — двигательный для мимической мус-
кулатуры, содержит вкусовые и парасимпатические секреторные
Рис. 3.169. Иннервация слизистых оболочек головы лошади:
1 — внутренняя носовая ветвь подглазничного нерва; 2 — каудальный носовой
нерв; 3 — решетчатый нерв; 4 — подблоковый нерв; 5 — лобный нерв; 6 —
языкоглоточный нерв; 7— блуждающий нерв; 8, 10 — язычный нерв; 9 — малый
нёбный нерв; 11 — большой нёбный нерв; 12 — щечный нерв; 13 — подбородоч-
ный нерв
300
волокна (см. рис. 3.161, 3.162). В лицевом канале на нерве лежит
коленчатый узел, принадлежащий барабанной струне.
По выходе из отверстия лицевой нерв идет по наружной по-
верхности большой жевательной мышцы, соединяется с вентраль-
ной ветвью поверхностного височного нерва и отдает: каудальный
ушной нерв, внутренний ушной нерв, нерв двубрюшной мышцы, веко-
ушной нерв, шейную ветвь, дорсальный щечный нерв, вентральный
щечный нерв. Последние два из них иннервируют мимическую муску-
латуру.
Равновесно-слуховой нерв — VIII пара — чувствительный, обра-
зован нейритами улиткового и вестибулярного ганглиев внутрен-
него уха. Он вступает в продолговатый мозг впереди лицевого не-
рва (см. рис. 3.161, 3.162).
Язычно-глоточный нерв — IX пара — является нервом обшей
чувствительности для корня языка, нёбной занавески и глотки,
вкусовым нервом для корня языка и двигательным нервом для
расширителя глотки (см. рис. 3.161, 3.162).
Блуждающий нерв Хпара — описан ниже (см. рис. 3.161, 3.162,
3.171).
Добавочный нерв — XI пара — идет в плечеголовную, трапецие-
видную и в грудино-челюстную мышцу (см. рис. 3.161, 3.162).
Подъязычный нерв — XII пара — двигательный нерв для мышц
языка и подъязычной кости (см. рис. 3.161, 3.162).
АВТОНОМНАЯ НЕРВНАЯ СИСТЕМА
Автономная (вегетативная) нервная система (systema nervosum
autonomicum) представляет собой специализированную часть еди-
ной нервной системы, осуществляющую иннервацию сердечно-
сосудистой системы и внутренних органов, в том числе желез
внутренней и внешней секреции Она подразделяется на симпа-
тическую и nupai имнатическую части (рис 3. 170, 3.171). Для пер-
вой характерен медиа юр норадрена тн. ня второй — ацетил-
холин.
Симпатическая нервная система
Симпатическая (сосудистая) часть вегетативной нервной сис-
темы (рис. 3.170) осуществляет иннервацию мышечной оболочки
кровеносных и лимфатических сосудов. Она состоит из центров и
периферической части.
Центры симпатической системы лежат в промежуточно-лате-
ральных ядрах боковых рогов серого мозгового вещества спинного
мозга в области от первого грудного до четвертого поясничного
301
Рис. 3.170. Схема строения симпатической не-
рвной системы (вентральная поверхность):
1 — центры симпатической нервной системы в бо-
ковых рогах спинного мозга с первого грудного по
третий поясничный сегменты; 2 — краниальный
шейный ганглий; 3 — вагосимпатический ствол; 4 —
позвоночный нерв; 5 — шейно-грудной (звездча-
тый) узел; 6, 7 — подключичная петля; 8 — сим-
патический ствол; 9 — соединительные ветви; 10 —
большой чревный нерв; 11 — чревный ганглий; 12 —
краниальный брыжеечный ганглий; 13, 14 — ма-
лые чревные нервы; 15 — каудальный брыжеечный
ганглий; 16 — подчревные нервы
сегментов. Периферическая часть включа-
ет: 1) белые и серые соединительные вет-
ви; 2) парный симпатический ствол с па-
равертебральными (околопозвоночными)
нервными узлами; 3) превертебральные
(предпозвоночные) нервные узлы; 4) не-
рвы и сплетения сосудов.
Белые соединительные ветви выходят из
промежуточно-латеральных ядер серого
мозгового вещества спинного мозга в со-
ставе вентральных корешков спинно-моз-
говых нервов. На выходе из межпозвоноч-
ных отверстий они образуют преганглионар-
ные мякотные волокна, оканчивающиеся в
околопозвоночных ганглиях этого же сег-
мента, или же направляются краниально
(каудально) до предпозвоночного узла.
В результате формируются правый и левый
симпатические стволы. Из околопозвоночных грудных, поясничных,
крестцовых и хвостовых узлов к спинно-мозговым нервам отходят
серые соединительные ветви, иннервирующие кровеносные и лим-
фатические сосуды.
Преганглионарные мякотные волокна, выходящие из первых
четырех-пяти сегментов грудной части спинного мозга, направ-
ляются краниально и оканчиваются в шейно-грудном, в среднем
шейном или же в краниальном шейном узлах, формируя шейную
часть симпатического ствола.
Часть преганглионарных мякотных нервных волокон, выхо-
дящих в области VI —XIII грудных сегментов, обособляются от
симпатического ствола в виде большого и малого внутренностных
нервов. Последние оканчиваются в предпозвоночных чревном и
302
краниальном брыжеечном узлах, лежащих на одноименных арте-
риях. Эти узлы находятся в солнечном сплетении, в образовании
которого принимают участие правый и левый блуждающие не-
рвы.
Преганглионарные мякотные нервные волокна, выходящие из
поясничных сегментов спинного мозга, дают начало подчревному
нерву, направляющемуся через каудальный брыжеечный ганглий
и подчревное сплетение сосудов органов тазовой полости.
Парный симпатический ствол лежит на вентролатеральной
поверхности грудных, поясничных, крестцовых и первых хвосто-
вых позвонков и состоит из преганглионарных белых соедини-
тельных ветвей и околопозвоночных симпатических узлов. После-
дние, как правило парные, располагаются в каждом сегменте, и
только в области третьего (четвертого) хвостового позвонка обра-
зуется непарный хвостовой узел. От каждого узла к спинно-мозго-
вому нерву отходят серые соединительные ветви. Последние следу-
ют в составе спинно-мозговых нервов, достигая мышечной обо-
лочки стенки кровеносных и лимфатических сосудов, проникают
в гладкие мышечные волокна и железы кожи.
Симпатический ствол располагается в области шеи вместе с
блуждающим нервом на латеральной поверхности трахеи, дости-
гая основания черепа. В области груди, поясницы, крестца и хво-
ста он лежит на телах позвонков и подразделяется на головную,
шейную, грудную, поясничную и крестцово-хвостовую части.
Головная часть симпатического ствола представлена краниаль-
ным шейным ганглием (рис. 3.170). Последний, имеющий непра-
вильную продолговато-овальную форму, лежит вблизи тела заты-
лочной кости и затылочной артерии. Из ганглия выходят постган-
глионарные ветви, имеющие специальные названия и иннерви-
рующие сосуды головы.
Шейная часть симпатического ствола простирается от крани-
ального шейного до шейно-грудного ганглиев и прилежит к блуж-
дающему нерву, образуя с ним вагосимпатический ствол (рис. 3.170).
Последний проходит по дорсомедиальной поверхности трахеи
рядом с общей сонной артерией. При входе в грудную полость
блуждающий нерв продолжается каудально по трахее, а симпати-
ческий нерв поднимается к головке первого ребра и вступает в
шейно-грудной (звездчатый) ганглий.
Шейно-грудной {звездчатый) узел лежит на уровне шейки ребер
в первом межреберном прост ранстве у дорсального края длинной
мышцы шеи. Из него выходят позвоночный нерв, сердечные ветви,
серые соединительные ветви.
Грудная часть симпатического ствола простирается от звезд-
чатого узла до диафрагмы. Она лежит на вентролатеральной по-
верхности тел позвонков, прикрытая внутригрудной фасцией и
плеврой (рис. 3.170). У каждой реберной головки на симпатичес-
303
ком стволе располагается грудной ганглий. Из последних к каждо-
му грудному спинно-мозговому нерву отходят серые соединитель-
ные ветви.
От грудных ганглиев отходят: ветвь в аортальное сплетение, ветвь
в легочное сплетение, большой и малый внутренностные нервы к
сосудам органов брюшной полости.
Поясничная и крестцово-хвостовая части симпатического ство-
ла тонкие и в области поясницы прикрыты вентральными мыш-
цами позвоночного столба (рис. 3.170). В области третьего крест-
цового позвонка они делятся на латеральную и медиальную вет-
ви, затем на уровне третьего (четвертого) хвостового позвонка
сливаются в непарный хвостовой ганглий. Белые соединительные
ветви отходят только от трех (четырех) первых поясничных не-
рвов, в то время как серые соединительные ветви, напротив,
идут ко всем поясничным, крестцовым и хвостовым спинно-
мозговым нервам.
Парасимпатическая нервная система
Парасимпатическая (висцеральная) часть автономной нервной
системы (рис. 3.171) иннервируе! i какую мускулатуру органон и
железы. Она состоит и 1 пре- и постганглионарных нервных воло-
кон, а также ядер, сосредоточенных в среднем и продолговатом
мозге, а также в крестцовой части спинного мозга. Ядра парасим-
патической системы лежат на уровне среднего мозга, продолгова-
того мозга и в крестцовом отделе спинного мозга.
От ядер среднего мозга отходят парасимпатические нервные
волокна, идущие в сфинктер зрачка и ресничную мышцу (рис.
3.171).
Парасимпатический отдел продолговатого мозга содержит сле-
зо- и слюноотделительные ядра, а также двигательные и секре-
торные ядра для внутренностей (рис. 3.171). Пре- и постганглио-
нарные парасимпатические волокна образуют слезоотделительный,
ростральный и каудальный слюноотделительные пути, а также
вагус — блуждающий нерв
Блуждающий нерв (см. рис. 3.161, 3.162, 3.171) по функции сме-
шанный: чувствительные волокна в нем идут от слизистой обо-
лочки пищеварительного тракта (начиная с глотки) и дыхатель-
ного тракта (начиная с гортани) к клеткам проксимального и
дистального ганглиев; двигательные соматические волокна — к
поперечноисчерченной мускулатуре глотки и гортани; двигатель-
ные парасимпатические волокна — к гладкой мускулатуре пище-
вода. желудка, тонкой и отчасти толстой кишки, а также трахеи
и бронхов, миокарда; секреторные волокна — к железам желуд-
ка, кишечника и бронхов.
304
Рис. 3.171. Схема строения парасимпатической
нервной системы (вентральная поверхность):
/ — парасимпатическое ядро глазодвигательного
нерва (средний мозг); 2 — ростральное и медиаль-
ное парасимпатическое ядро продолговатого мозга;
I — дорсальное ядро блуждающего нерва (продол-
। оватый мозг); 4 — блуждающий нерв; 5 — возврат-
ный гортанный нерв; 6 — парасимпатические вет-
ви к легким и сердцу; 7 — вентральный пищевод-
ный ствол блуждающего нерва; 8 — дорсальный пи-
щеводный ствол блуждающего нерва; 9, 10 — пара-
симпатические ветви к органам брюшной полости;
/1 парасимпатические ядра крестцового отдела
спинного мозга; 12 — тазовые нервы
Блуждающий нерв следует вдоль трахеи
в грудную полость вместе с шейной час-
тью симпатического ствола как вагосимпа-
тический ствол, сопровождая по дорсоме-
диальной поверхности общую сонную ар-
терию.
У входа в грудную полость вагус отде-
ляется от симпатического ствола и по пи-
щеводу вступает в грудную, а затем и в
брюшную полости.
По ходу блуждающий нерв делится на
шейную, грудную и брюшную части. Груд-
ная часть вагуса отдает ветви в каудальное
трахеальное сплетение и в интрамуральные
сплетения пищевода и сердца.
В брюшной полости от дорсального ство-
ла вагуса отходит толстая ветвь в крани-
альный брыжеечный узел солнечного спле-
тения, из которого выходят ветви в интрамуральные сплетения
тонкой и толстой кишки.
В стенках органов желудочно-кишечного тракта между продоль-
ным и циркулярным слоями гладких миоцитов располагается меж-
мышечное сплетение, а в подслизистом слое — подслизистое спле-
тение.
Кроме того, существует еще и подсерозное сплетение. Все они
относятся к постганглионарному парасимпатическому сплетению,
соединяются друг с другом многочисленными анастомозами и
содержат большое число ганглиозных клеток.
Крестцовая часть парасимпатической нервной системы содер-
жит двигательные и секреторные волокна для органов тазовой
полости (рис. 3.171).
305
АНАЛИЗАТОРЫ
Анализатор (anahsator), по И. П. Павлову, — это «сложный не-
рвный механизм, начинающийся наружным воспринимающим
аппаратом и кончающийся в мозге». Анализаторы, являющиеся
обязательной частью нервной системы, состоят из: а) рецепто-
ров; б) периферических проводящих путей; в) мозгового отдела
с подкорковыми и корковыми центрами, соединенными цент-
ральными проводящими путями (рис. 3.172 — 3.175).
ЗРИТЕЛЬНЫЙ АНАЛИЗАТОР
Зрительный анализатор состоит из: 1) глазного яблока, в кото-
ром заключен рецепторный аппарат анализатора — сетчатка глаза;
2) защитных и вспомогательных органов глаза; 3) проводящих пу-
тей анализатора; 4) подкорковых и корковых центров анализаторов.
Глазное яблоко {bulbus oculi) имеет шарообразную, сплющен-
ную спереди назад форму, с передней выпуклой и задней не-
сколько уплощенной поверхностью (рис. 3.172).
Самое крупное глазное яблоко (по отношению к массе тела)
имеет кошка, за ней следуют собака, овца, лошадь, приматы,
корова, свинья, бык.
Угол между зрительными осями у собаки составляет 92°, у сви-
ньи — 118°, у коровы — 119°, у овцы — 134°, у лошади — 137°;
угол между осями глазниц у собаки — 79°, у свиньи — 85°,
у коровы — 94°, у лошади — 115°, у овцы — 129°.
В глазном яблоке рассматривают: I) оболочки глаза (считая сна-
ружи внутрь) — фиброзную, сосудистую и сетчатую; 2) свето-
преломляющие среды — хрусталик, стекловидное тело, внутриглаз-
ную жидкость; 3) сосуды и нервы.
Фиброзная оболочка глаза {наружная) разделяется на белоч-
ную оболочку и роговицу (рис. 3.172).
Белочная оболочка {склера) глаза занимает около 4/5 всей по-
верхности глазного яблока (рис. 3.172). Она непрозрачная, плот-
ная и бедна сосудами. В заднем латеровентральном квадранте ее
находится продырявленная пластинка склеры. Через ее отверстия из
глазного яблока выходит зрительный нерв.
Роговица занимает по площади около 1/5 всей поверхности глаз-
ного яблока (рис. 3.172). Она совершенно прозрачная, очень плот-
ная и имеет толщину до 0,6 —0,7 мм в центре (по краям толще).
Роговица, за исключением краевой зоны, лишена сосудов, но
богата безмякотными нервами.
Сосудистая оболочка глаза {средняя) состоит из радужной обо-
лочки, ресничного тела и собственно сосудистой оболочки (рис.
3.172).
306
Рис. 3.172. Схема строения глазного яблока:
/ — венозное сплетение склеры; 2 — задняя глазная камера; 3 — ресничное тело;
4 — центральное поле сетчатки; 5 — зрительный сосок; 6 — хрусталиковая связ-
ка и лимфатические щели; 7 - тарсальная железа; 8 — хрусталиковая сумка; 9 —
роговица; 10 — передняя глазная камера; 11 - хрусталик; 12 — радужная обо-
лочка; 13 — нижнее веко; 14 — ресничная мышца; 15 — конъюнктивальный
мешок; 16 — мышца-опускатель нижнего века; /7 — поверхностная фасция; 18 —
белочная оболочка; /9 — собственно сосудистая оболочка; 20 — пигментный
слой сетчатки; 21 — собственно сетчатка; 22 — прямая вентральная мышца; 23 —
мышца-оттягиватель глазного яблока; 24 — теноново пространство; 25, 26 —
зрительный нерв; 27 — влагалище зрительного нерва; 28 — нейроэпителий; 29 —
мышца-наружный подниматель верхнего века; 30 — мышца-внутренний под-
ниматель верхнего века; 31 — слезная железа; 32 — дорсальная прямая мышца;
33 — экстраорбитальное жировое тело; 34 — глубокая фасция; 35 — ганглий
зрительного нерва; 36 — ганглий сетчатки
Радужная оболочка (радужка) располагается позади роговицы,
в центре она имеет отверстие — зрачок (рис. 3.172). Радужная обо-
лочка содержит пигментные клетки, которые обусловливают «цвет»
глаз. Циркулярные гладкие мышечные волокна радужной оболоч-
ки формируют сфинктер зрачка. При сильном свете в силу возбуж-
307
дения зрачок суживается, при слабом, напротив, расширяется из-
за торможения двигательного центра.
Ресничное тело охватывает поясом, шириной до 10 мм, перед-
ний край белочной оболочки (рис. 3.172). В ресничном теле зало-
жена ресничная мышца из гладких мышечных волокон. К ним
прикрепляется связка, подвешивающая хрусталик.
Собственно сосудистая оболочка (рис. 3.172) покрывает внут-
реннюю поверхность склеры, соединяясь с ней довольно рыхло,
исключая места прохождения сосудов и зрительного нерва, а так-
же на границе с ресничным телом. Другой своей поверхностью
она очень плотно соединяется с пигментным слоем сетчатки.
В сосудистой оболочке под пигментным слоем сетчатки находится
отражательная оболочка фиброзного (у травоядных) или клеточ-
ного строения (у собак). У свиньи она отсутствует.
Сетчатая оболочка глаза (ретина) подразделяется на зритель-
ную и слепую части.
Зрительная часть сетчатки состоит из: а) пигментного слоя,
который плотно срастается с сосудистой оболочкой; б) собствен-
но сетчатки, которая легко отделяется от ее пигментного слоя.
При жизни сетчатка нежная, прозрачная оболочка розоватого
цвета, после смерти она мутнеет.
Слепая часть сетчатки распадается на ресничную и радужко-
вую части.
Хрусталик имеет форму двояковыпуклой линзы с передней
более плоской поверхностью (рис. 3.172). Располагается он позади
радужки. Хрусталик совершенно прозрачен, плотной консистен-
ции, он выполняет функцию оптической линзы — преломляет
лучи и дает изображение на сетчатке.
Стекловидное тело заполняет в глазном яблоке пространство
позади хрусталика (рис. 3.172). Оно совершенно прозрачно, студ-
невидной консистенции, так как на 98 % состоит из воды, заклю-
ченной в строме из тончайших волоконец.
Сосуды глазного яблока разделяются на кровеносные сосуды
сетчатки и сосудистой оболочки. Лимфатические сосуды в глаз-
ном яблоке отсутствуют, но вместо них имеются лимфатические
щели и пространства.
К защитным и вспомогательным органам глаза относятся веки,
слезный аппарат, периорбита, глазные мышцы и фасции.
Верхнее и нижнее веко представляют собой кожно-мышечные
подвижные складки, расположенные в области глазницы впереди
глазного яблока (рис. 3.173; см. рис. 3.172). Снаружи они покрыты
кожей, а их внутренняя поверхность — специальной слизистой
оболочкой конъюнктивой. Различают конъюнктиву век и глазного
яблока, между ними — свод конъюнктивы и конъюнктивальный
мешок. При смыкании веки закрывают глаза, а при мигании рав-
номерно распределяют слезу по поверхности глаза и тем защища-
308
ют его от высыхания. Между веками
находится поперечная щель век. Край
века имеет два ребра — наружное и
внутреннее. Наружное ребро верхне-
го века снабжено длинными ресни-
цами. На нижнем веке ресницы бы-
вают только у жвачных. На внутрен-
нем ребре век открываются железы.
Третье веко — складка из одной
только конъюнктивы, длиной до 2,5
см (у крупных животных), располо-
женная медиально на глазном ябло-
ке. Она поддерживается эластическим
хрящом третьего века. Веки приво-
дятся в движение рядом мышц.
Слезный аппарат состоит из слез-
ных желез верхнего и третьего век,
слезных канальцев, слезного мешка
и слезно-носового протока (см. рис.
3.172).
Периорбита — перепончатый, ко-
нусообразной формы мешок. Осно-
ванием он закрепляется по краю ор-
биты, вершиной — вокруг зритель-
ного отверстия, а медиальной стен-
кой прирастает к костной орбите (см.
рис. 3.172).
Глазные мышцы лежат в периорби-
те в основном позади глазного яблока
(рис. 3.172). Вокруг зрительного нерва
располагается оттягиватель глазного
яблока, а вокруг оттягивателя — че-
тыре прямые глазные мышцы.
Косых глазных мышц две — дор-
сальная и вентральная. Прямые мышцы поворачивают глаз в соот-
ветствующую сторону, а вместе с оттягивателем — несколько втя-
гивают его в орбиту. Косые — вращают глаз.
Фасции как вспомогательные органы глаза подразделяются на
фасцию орбиты и глазного яблока. Поверхностная фасция орбиты
закрепляется вокруг зрительного отверстия и в веках, она отдает
мышечные перегородки к глубокой фасции. Глубокая фасция орби-
ты состоит из нескольких листков, одевающих мышцы глаза. Про-
странство внутри фасции называется теноновым.
Проводящие пути зрительного анализатора разделяется на пе-
риферические и центральные. Периферические пути формируют
зрительный нерв, а центральные оканчиваются в затылочной доле
Рис. 3.173. Верхнее и нижнее
веко (разрез, передняя поверх-
ность):
1 — круговая мышца глаза; 2 —
перегородка орбиты; 3 — мышца
подниматель верхнего века; 4. 7 —
тарсальные железы; 5 — слезные
точки; 6 — ресницы
309
коры полушарий большого мозга (высший центр зрительного ана-
лизатора).
СТАТОАКУСТИЧЕСКИЙ АНАЛИЗАТОР
Статоакустический (равновесный и слуховой) анализатор со-
стоит из: 1) рецепторного отдела, представленного наружным, сред-
ним и внутренним ухом (внутреннее ухо обслуживает равновесный
и слуховой анализаторы, а среднее и наружное ухо — исключи-
тельно слуховой); 2) проводящих путей анализатора; 3) подкорко-
вых и корковых центров анализаторов (рис. 3.174, 3.175).
Наружное ухо состоит из ушной раковины, двигательного ап-
парата ушной раковины и наружного слухового прохода (рис. 3.174).
Все они являются вспомогательными органами акустического (слу-
хового) рецептора.
Наружный слуховой проход имеет костную основу, которая сна-
ружи дополняется кольцевидным хрящом с медиальной щелью, за-
тянутой эластической перепонкой (рис. 3.174). Внутреннее отвер-
стие слухового прохода обращено в полость среднего уха, оно ог-
раничено косо поставленным барабанным кольцом, затянутым ба-
рабанной перепонкой. К наружному отверстию слухового прохода
прикрепляется ушная раковина.
Ушная раковина — это кожная складка воронкообразной формы
(рис. 3.174). Суженным концом она укреплена на наружном слухо-
вом проходе, а косо срезанным основанием направлена дистально.
Наружная поверхность раковины образует спинку раковины, а внут-
ренняя поверхность — ладьевидную ямку. Последняя переходит в
ямку завитковой части, а затем — и в наружный слуховой проход.
Основание ушной раковины покоится на жировой подушке, что
обеспечивает подвижность раковины, неодинаковую у разных живот-
ных. Мышцы ушной раковины многочисленны и сильно развиты.
Среднее ухо состоит из барабанной перепонки и четырех слухо-
вых косточек с их мышцами и связками, размещающимися в ба-
рабанной полости (рис. 3.174). Все они являются вспомогатель-
ными органами акустического рецептора.
Барабанная полость находится в барабанной части каменистой
кости. На медиальной ее стенке расположены: окно преддверия,
закрытое стремечком, и окно улитки, закрытое внутренней бара-
банной перепонкой, между ними — мыс. На передней стенке по-
лости расположено отверстие, ведущее в слуховую трубу. В дор-
сальной стенке проходит канал лицевого нерва, а латеральную
стенку образует барабанная перепонка.
Барабанная перепонка построена из соединительнотканных ра-
диальных и циркулярных волоконец, изнутри она покрыта слизи-
стой оболочкой с однослойным плоским эпителием, а снаружи —
плоским многослойным эпителием.
310
Рис. 3.174. Анатомия наружного, среднего и внутреннего уха (сегмен-
тальный разрез, схема):
/, 24 — клетки равновесного пятна; 2 — полукружные каналы; 3 — овальный
мешочек (маточка, утрикулюс); 4 — стремечко; 5 — чечевицеобразная косточка;
ti — наковальня; 7 — молоточек; 8 — хрящ ушной раковины; 9 — наружный
слуховой проход; 10 — барабанная перепонка; 11 — мышца-напрягатель бара-
банной перепонки; 12 — улитка; 13 — слуховая труба; 14 — мышечный отросток;
15 — спиральный ганглий; 16 — лестница преддверия; 17 — кортиев орган; 18
канал улитки; 19 — барабанная лестница; 20 — круглый мешочек (саккулюс);
21 — внутренний слуховой проход; 22 — равпоиссно-слуховой нерв; 23 — эндо-
лимфатический проток и мешочек
Слуховых косточек четыре: молоточек, наковальня, чечевицеоб-
разная косточка и стремечко (рис. 3.174).
Слуховая труба проходит вдоль мышечного отростка каменис-
той кости до полости глотки. Она способствует выравниванию
давления воздуха внутри барабанной полости (рис. 3.174).
Воздухоносный мешок лошадей образован выпячиванием слизи-
стой оболочки слуховой трубы.
Внутреннее ухо состоит из костного и перепончатого лабирин-
тов (рис. 3.174, 3.175). Костный лабиринт расположен в скалистой
части каменистой кости. В нем различают три отдела — преддве-
рие, полукружные костные каналы и костную улитку.
Преддверие — шаровидная полость диаметром до 5 мм (рис. 3.174,
3.175). В медиальной ее стенке находится перфорированное дно
311
Рис. 3.175. Перепончатый лабиринт:
7 — полукружные каналы; 2 — ампулы;
3 — утрикулюс (маточка); 4 — эндо-
лимфатический проток; 5, 8 — маку-
лы; 6 — саккулюс (круглый мешочек);
7 — канал улитки
внутреннего слухового прохода,
через которое проходит слухо-
вой нерв. В латеральной стенке
находится окно преддверия, за-
крытое стремечком со стороны
барабанной полости. В каудаль-
ной стенке расположены четыре
отверстия трех костных полу-
кружных каналов. В назальной
стенке начинается канал костной
улитки, а назовентрально от не-
го — водопровод преддверия, на-
правляющийся на медиальную
поверхность каменистой кости.
Три костных полукружных
канала лежат дорсокаудально от
преддверия, из них латераль-
ный — в горизонтальной плос-
кости, дорсальный — в сагит-
тальной плоскости и каудаль-
ный — в сегментальной плос-
кости.
Костная улитка лежит назо-
вентрально от преддверия. В ней
различают ось улитки и спираль-
ный канал, который образует вокруг оси от 1,5 до 5 завитков (у
собаки 3 завитка, у свиньи 4, у жвачных 3,5 и лошади 2,5). Верши-
на оси улитки, или купол спирального канала, обращена латераль-
но к мысу на стенке барабанной полости. На оси лежит спиральная
пластинка, она заканчивается под куполом улитки крючком. В ос-
новании спиральной пластинки лежит спиральный ганглий улитки.
Спиральная пластинка подразделяет спиральный канал улитки на
два отдела: лестницу преддверия и барабанную лестницу. Обе лест-
ницы под куполом переходят одна в другую.
Перепончатый лабиринт (рис. 3.175) состоит из овального ме-
шочка с тремя перепончатыми полукружными каналами, круглого
мешочка с перепончатым каналом улитки и эндолимфатического
протока. Овальный мешочек с полукружными каналами и круглый
мешочек относятся к вестибулярному (равновесному) аппарату, а
улитка с Кортиевым органом — к акустическому (слуховому).
Проводящие пути слухового анализатора доводят слуховые им-
пульсы до высшего слухового центра — височных долей коры боль-
шого мозга.
Проводящие пути вестибулярного анализатора через зритель-
ные бугры доводят импульсы до корковых центров в височных
долях коры большого мозга.
312
ОСОБЕННОСТИ СТРОЕНИЯ ДОМАШНИХ ПТИЦ
СКЕЛЕТ
Скелет туловища. Число шейных позвонков у кур 13 — 14, у
у гок 14— 15, у гусей 17 — 18 (а у лебедя 23 — 25). Остистые отрост-
ки слабо развиты или отсутствуют, на поперечных отростках име-
ются рудименты ребер, концы их направлены каудально (рис. 3.176).
Гела позвонков соединяются суставами с хрящевыми прослойка-
ми. Суставные поверхности седловидные, что обеспечивает под-
вижность шеи в сагиттальной и фронтальной плоскостях (т. е. впра-
во-влево и вверх—вниз).
Атлант кольцевидный, маленький, с ямкой для мыщелка за-
тылочной кости. Эпистрофей имеет хорошо выраженный зуб.
Грудных позвонков у кур и индеек 7, у уток и гусей 9. Первый
грудной позвонок соединяется со вторым так же, как и шейные,
а второй—пятый позвонки сращены друг с другом, причем ости-
стые и поперечные отростки образуют гребни. Имеются вентраль-
ные остистые отростки: шестой грудной позвонок у кур соединя-
ется подвижно, а седьмой позвонок срастается с поясничными и
крестцовыми позвонками в пояснично-крестцовую кость из 14 —
16 позвонков.
Хвостовых позвонков у кур 5, у уток и гусей 4 — 6. Кроме того,
и меется копчик, состоящий из 4 — 5 сросшихся последних хвосто-
вых позвонков.
Первые два (три) ребра астернальные, остальные — стерналь-
ные (рис. 3.176, 3.177). Каждое стернальное ребро разделяется на
позвоночный (вертебральный) и грудинный (стернальный) кос-
тные участки. Каждое ребро соединяется с соответствующим по-
звонком головкой и бугорком и друг с другом особыми крючко-
видными отростками.
Грудная клетка в целом имеет конусовидную форму — основа-
ние конуса направлено каудально (рис. 3.176, 3.177).
Грудная кость сильно развита, на вентральной поверхности не-
сет гребень грудины (киль). Задний край грудины у кур имеет парную
глубокую вырезку, вследствие чего образуются средний, боковой и
реберный отростки грудины (рис. 3.176, 3.177). На боковых отростках
грудины имеются ямки для ребер, а у краниального края — сустав-
ные поверхности для сочленения с коракоидными костями. У гусей
вместо вырезок на грудной кости имеются замкнутые отверстия.
Скелет головы — череп. Череп у кур сравнительно небольшой
(рис. 3.178; см. рис. 3.176). Отдельные кости очень рано срастаются,
а швы между ними исчезают. В мозговом черепе имеются следую-
щие кости: 1) затылочная кость, характерная наличием лишь од-
ного мыщелка для соединения с атлантом; 2) клиновидная кость',
313
Рис. 3.176. Скелет курицы:
1 — лицевой череп; 2 — орбита; 3 — ось; 4 — шейные позвонки; 5 — плечевая
кость; 6 — лучевая кость; 7 — кисть; 8 — локтевая кость; 9 — спинная кость
(сросшиеся грудные позвонки); 10 — свободный (шестой) грудной позвонок;
11 — подвздошная кость; 12 — крестцовая (пояснично-крестцовая) кость; 13 —
хвостовые позвонки; 14 — пигостиль; 75 — лонная кость; 16 — седалищная
кость; 17 — бедренная кость; 18 — малая берцовая кость; 19 — болыиеберцово-
заплюсневая кость; 20 — сесамовидная кость; 21 — плюсно-заплюсневая кость;
22 — надколенник; 23 — киль грудины; 24 — грудина; 25 — ребра; 26 — рукоятка
грудины; 27 — ключица; 28 — коракоидная кость; 29 — лопатка; 30 — плечевой
сустав; 31 — атлант; 32 — нижняя челюсть; 33 — кости первого пальца; 34 —
кости второго пальца; 35 — кости третьего пальца; 36 — кости четвертого пальца
314
Б
Рис. 3.177. Грудобрюшной отдел скелета курицы (А) и утки (Б):
А: 1 — пигостиль; 2 — седалищное отверстие; 3 — суставная впадина; 4 — под-
вздошная кость; 5 — ягодичная ямка (подвздошная кость); 6 — остистые отрос-
тки грудных позвонков; 7 — поперечные отростки грудных позвонков; 8 — ас-
тернальное ребро; 9 — крючковидный отросток ребра; 10 — реберный отросток
грудины; 11 — рукоятка грудины; Г2 — суставная ямка; 13 — тело грудины; 14 —
киль грудины; 75 — каудальный отросток грудины; 16 — брюшной отросток
грудины; 77—латеральный отросток грудины; 18 — стернальная часть ребра; 19 —
позвоночная часть ребра; 20 — запсрюс отверстие; 21 — лонная кость; 22 —
седалищная кость;
Б: 1 — пигостиль; 2 — седалищная кость; 3 — седалищное отверстие; 4 — сустав-
ная впадина; 5 — подвздошная кость; 6 — ягодичная ямка подвздошной кости;
7 — остистые отростки грудных позвонков; 8 — поперечные отростки грудных
позвонков; 9 — краниальный суставной отросток; 10 — вентральный гребень;
77 — астернальное ребро; 12 — крючковидный отросток ребра; 13 — астернальное
ребро; 14 — реберный отросток грудины; 75 — суставная ямка; 16 — рукоятка
грудины; 77 — тело грудины; 18 — киль; 19 — каудальный отросток грудины; 20 —
стернальная часть ребра; 21 — позвоночная часть ребра; 22 — брюшной отросток
грудины; 23 — запертое отверстие; 24 — лонная кость; 25 — хвостовые позвонки
315
3) парная височная кость, имеющая суставную поверхность для
соединения с квадратной костью; 4) парная теменная кость;
5) парная лобная кость, орбиты отделены друг от друга межорби-
тальной костной пластинкой; 6) решетчатая кость без лабирин-
та; 7) парная слезная кость слабо развита; 8) межтеменная кость
отсутствует.
Лицевой череп образован надклювьем и подклювьем. Надклювье
состоит из: 1) сильно развитых и рано срастающихся друг с дру-
гом резцовых костей; 2) носовых костей; 3) слабо развитых верхне-
челюстных костей (вследствие отсутствия зубов их нёбные отрос-
тки у кур развиты слабо); имеется сошник.
Рис. 3.178. Череп
А: 1 — тело межчелюстной кости; 2 — лобный отросток; 3, 4 — межчелюстной и
7— решетчатая кость; 8 — орбита; 9— отверстие глазодвигательного нерва; 10 —
кость (орбитальная пластинка); 13 — лобная кость; 14 — надглазничный отрос-
височной кости; 17 — теменная кость; 18 — чешуя затылочной кости; 19 —
(височная кость); 22 — наружное отверстие слухового прохода; 23, 25 — слуховой
вая кость; 27 — глазничный отросток (квадратная кость); 28 — крыловидная
челюсть); 32 — верхняя челюсть; 33 — челюстной отросток (межчелюстная кость);
36 — суставная поверхность (нижняя челюсть); 37 — внутренний челюстной отро-
Б: 1 — тело межчелюстной кости; 2 — лобный отросток; 3, 4 — межчелюстной и
7— решетчатая кость; 8 — отверстие глазодвигательного нерва; 9— межглазнич-
ка (лобная кость); 12 — лобная кость; 13 — каудальный глазничный отросток
16— чешуйчатая часть (затылочная кость); 17 — боковая часть (затылочная кость);
отверстие слухового прохода; 21 — слуховой отросток (квадратная кость); 22 —
ничный отросток (квадратная кость); 25 — квадратно-скуловая кость; 26 — кры-
скуловой отросток (верхняя челюсть); 31 — верхняя челюсть; 32 — челюстной
ный отросток (нижняя челюсть); 35 — суставной отросток; 36 —
316
Надклювье с мозговым черепом соединяется полуподвижно
посредством ряда костей.
Подклювье образовано парной нижней челюстью, сформиро-
ванной в основном из зубной кости и сочленовной кости, между
которыми расположены еще три кости. Нижняя челюсть соединя-
ется с квадратной костью.
Подъязычная кость состоит из тела и одной пары рогов, кото-
рые огибают череп сзади, но не связаны с ним непосредственно.
От тела подъязычной кости вперед отходит внутриязычная кость,
а кзади — киль, достигающий трахеи.
курицы (А) и гуся (/>):
верхнечелюстной отростки носовой кости; 5 — носовая кость; 6 — слезная кость;
межглазничная перегородка; II — отверстие зрительного нерва; 12 — лобная
ток (височная кость); 15 — скуловой отросток (височная кость); 16 — чешуя
височная ямка; 20 — боковая часть (затылочная кость); 21 — слуховой отросток
(Просим (квадратная кость); 24 — тело квадратной кости; 26 кжщратноскуло
кость. 29 — скуловая кость. 30 небная кисть; .?/ скуловой отросток (верхняя
<4 — зубная кость (нижняя челюсть); >5 — венечный отросток (нижняя челюсть):
сток (нижняя челюсть): .W — кау ы льныи челюстной отросток (нижняя челюсть),
верхнечелюстной oi ростки носовой кости, 5 - носовая кость: 6— слезная кость,
ная перегородка; 10 — отверстие зрительного нерва; 11 — орбитальная пластин-
височной кости; 14 — чешуйчатая часть (височная кость); 15 — теменная кость;
18 — височная ямка; 19 — слуховой отросток (височная кость); 20 — наружное
тело квадратной кости; 23 — слуховой отросток (квадратная кость); 24 — глаз-
ловидная кость; 27 — скуловая кость; 28 — нёбная кость; 29 — сошник; 30 —
отросток межчелюстной кости; 33 — зубная кость нижней челюсти; 34 — венеч-
мышечный отросток; 37 — каудальный отросток (нижняя челюсть)
317
Скелет конечностей (см. рис. 3.176). Плечевой пояс представлен
тремя костями: лопатка — в виде узкой, костной пластинки, со-
членяется с плечевой костью и, кроме того, с ключицей и кора-
коидной костями. Коракоидная кость самая мощная, она соединя-
ется с грудной костью тугим суставом. Ключица дистальным кон-
цом соединяется с одноименной костью другой стороны, образуя
дужку или вилку (у кур дужка имеет плоский отросток).
Плечевая кость хорошо развита. Локтевая кость развита силь-
нее лучевой, локтевой отросток небольшой. Межкостное простран-
ство обширное.
В запястном суставе только две кости: запястная лучевая кость
срастается с промежуточной, а запястная локтевая — с добавоч-
ной. Дистальные кости запястья срослись с пястными костями.
Из запястно-пястных костей наиболее сильно развита третья,
слабее — четвертая, обе кости своими концами сращены друг с
другом; вторая кость очень короткая, прирастает к третьей кости.
Кости пальцев редуцированы: более сильно развит третий па-
лец с двумя фалангами, второй и четвертый пальцы состоят толь-
ко из одной фаланги.
Тазовый пояс состоит из подвздошной, седалищной и лонной кос-
тей. Правая и левая кости таза срастаются с пояснично-крестцо-
вой костью, но вентрально таз широко открыт. Лонная кость лен-
тообразная лежит вдоль седалищной кости. Имеется запертое
отверстие. Бедренная кость короче костей голени. Большая берцовая
кость большая и длинная, дистально сращена с проксимальными
костями заплюсны.
Малая берцовая кость редуцирована у кур и сращена с большой
берцовой. Она соединяется с латеральным мыщелком большой бер-
цовой кости. Кости заплюсны как таковые отсутствуют: кости про-
ксимального ряда срастаются с большой берцовой костью, а кости
дистального ряда — с костями плюсны. Заплюсно-плюсневая кость —
цевка — длинная, образована сросшимися второй—четвертой плюс-
невыми костями и дистальным рядом костей заплюсны. Дисталь-
ный конец цевки несет четыре блока для пальцев. Первый палец
направлен назад, состоит из двух фаланг, второй палец состоит из
трех фаланг, третий — из четырех, четвертый — из пяти.
МЫШЕЧНАЯ СИСТЕМА
У хорошо летающих птиц скелетные мышцы темно-красные, у
плохо или совсем нелетающих мышцы более бледные. В сухожили-
ях часто встречаются окостеневшие участки (рис. 3.179).
Кожные мышцы хорошо развиты, они оказывают влияние на
перья, особенно на маховые и рулевые. Кроме того, они напряга-
ют летательную перепонку крыла.
318
Лицевые мышцы отсутствуют. Жевательные мышцы сильно раз-
виты и более разнообразные (действуют на квадратную кость).
На туловище особенно хорошо развиты мышцы, действующие
на хвостовой отдел. Брюшные мышцы развиты слабо. На грудной
клетке мышцы обеспечивают опускание грудной кости и увели-
чение грудной клетки в глубину. Диафрагма слабо развита.
Мышцы крыла многочисленны, особенно сильно развиты груд-
ные мышцы, составляющие до 45 % массы всей мускулатуры.
Мышцы тазовой конечности также многочисленны, особенно в
области бедра. Интересен механизм сгибания пальцев при сгиба-
нии коленного сустава, что обеспечивает возможность без затра-
ты мышечной силы сидеть и даже спать на сучках. Механизм обус-
ловлен наличием сухожилия, ответвляющегося от стройной мыш-
цы. Это сухожилие проходит по дорсальной поверхности колен-
ной чашки и через латеральную сторону коленного сустава на
каудальную поверхность голени в поверхностный сгибатель паль-
цев. На своем пути сухожилие прикрепляется к малоберцовой ко-
сти. Таким образом, при сгибании коленного сустава автомати-
чески сгибаются пальцы.
КОЖНЫЙ ПОКРОВ птиц
Эпидермис кожного покрова в местах, не покрытых перьями,
сильно развит: на клюве, когтях, мякишах пальцев, чешуях ног и
шпорах петуха (рис. 3.180). В таких местах основа кожи имеет хоро-
шо выраженный сосочковый слой. В основе кожи отсутствуют по-
товые и сальные железы и мало сосудов, за исключением гребня
и бородки, где имеется подобие кавернозных тел. Подкожный слой
тонкий, но всюду выражен. Над последними крестцовыми позвон-
ками находится копчиковая железа. Ее секрет служит для смазы-
вания перьев, поэтому у водоплавающих эта железа развита наи-
более сильно.
Для птиц характерно наличие перьев, которые облегчают ее
полет и сохраняют постоянство температуры тела. На развитом
пере отмечают: стержень, опахало (бородку). В стержне имеется
очин и стебель. Очин — свободная от опахала часть пера, содер-
жит сердцевину, или дужку, помещается в перьевой сумке кожи.
Стебель находится в опахале, от него отходят лучи, снабженные
крючками.
Различают контурные, или покровные, перья, пуховые перья с
опахалом без крючков; они более мелкие, лежат под покровными
перьями и обеспечивают сохранение тепла. Покровные перья раз-
виваются в определенных участках кожного покрова, а проме-
жутки между ними остаются без перьев. Маховые перья на крыльях
и рулевые перья на хвосте.
319
Рис. 3.179. Мышцы птицы:
/ — большая жевательная мышца;
2 — двубрюшная мышца; 3 — по-
перечная нижнечелюстная мышца;
4 — подъязычно-язычная мышца;
5— прямая латеральная мышца го-
ловы; 6 — длинная мышца головы;
7 — короткий сгибатель шеи; 8 —
длинная мышца шеи; 9— межпопе-
речная мышца; 10— остистая мыш-
ца; 11 — длиннейшая мышца шеи;
12 — задняя лопаткоплечевая мыш-
ца; 13 — подкожная грудная мыш-
ца; 14 — наружная косая мышца
ная часть большой грудной мышцы;
живота; 15 — поверхностная груд-
ная мышца; 16 — подкожная брюш-
17 — наружная головка икроножной мыш-
цы; 18 — средняя головка икроножной мышцы; 19 — длинная малоберцовая
мышца; 20 — передняя большеберцовая мышца; 2/ — пяточное (ахиллово) су-
хожилие; 22, 77 — короткий сгибатель первого пальца; 23, 80 — длинный паль-
320
иевой разгибатель; 24 — длинный разгибатель первого пальца; 25 — абдуктор
второго пальца; 26 — специальный разгибатель третьего пальца; 27 — поверхно-
с1ный сгибатель пальцев кисти; 28 — треугольная мышца головы; 29 — прямая
дорсальная большая мышца головы; 30 — малая переплетенная мышца; 31 —
двубрюшная мышца шеи; 32 — длинный разгибатель шеи; 33 — полуостистая и
множественная мышцы шеи и спины; 34 — напрягатель летательной перепонки,
длинный и короткий; 35 — пропатагиальная часть большой грудной мышцы; 36 —
нропатагиальная часть двуглавой мышцы плеча; 37 — передняя летательная пе-
репонка; 38 — отводящая мышца второго пальца; 39 — приводящая мышца
второго пальца; 40 — отводящая мышца третьего пальца; 41 — межкостная мыш-
ца кисти; 42 — сгибатель четвертого пальца; 43 — поверхностный сгибатель паль-
цев; 44 — глубокий сгибатель пальцев; 45 — длинный разгибатель третьего паль-
ца; 46 — локтевой сгибатель запястья; 47 — глубокий пронатор; 48 — поверхно-
стный пронатор; 49 — лучевой разгибатель запястья; 50— плечевая мышца; 51 —
двуглавая мышца плеча; 52 — медиальная и латеральная головки трехглавой
мышцы плеча; 53 — широчайшая мышца спины; 54 — каудальная часть поверх-
ностной зубчатой мышцы; 55 — метапатагиальная часть поверхностной зубчатой
мышцы; 56 — поверхностная ягодичная мышца; 57 — портняжная мышца; 58 —
длиннейшая мышца спины; 59 — напрягатель широкой фасции бедра; 60 —
двуглавая мышца бедра; 61 — подниматель хвоста; 62 — хвостовые мышцы; 63 —
нодниматель клоаки; 64 — полуперепончатая мышца; 65 — грушевидная мышца;
66 — опускатель хвоста; 67 — полуперепончатая мышца; 68 — полусухожильная
мышца; 69 — прямая мышца живота; 70 — поверхностная грудная мышца; 7/,
75 — длинный сгибатель пальцев; 72, 76, 79 — глубокий сгибатель пальцев стопы;
73 — короткая малоберцовая мышца; 74, 78 — длинный сгибатель первого пальца;
81 — абдуктор четвертого пальца
Рис. 3.180. Строение пера:
А — маховое перо; Б — пуховое перо; / — стержень; 2 — опахало; 3 — очин; 4 —
лучи с крючками; 5 — ветвь; 6 — лучи без крючков; 7 — кутикула; 8 — мышца;
9 — сосочек пера
I 1 Зелененекий
321
ПИЩЕВАРИТЕЛЬНЫЙ АППАРАТ
Ротоглотка (рис. 3.181). Так как нёбная занавеска у птицы от-
сутствует, рот и глотка представляют собой одно целое. Отсут-
ствуют зубы, десны, губы и щеки. Челюсти покрыты роговым чех-
лом — клювом, который подразделяется на надклювье и подклю-
вье. У кур и индеек клюв твердый, конусообразный.
В твердом нёбе по средней линии находится нёбная щель, ко-
торая в области глотки становится более широкой, — хоаны. По
обе стороны от нёбной щели располагаются сосочки в пять рядов.
В переднем участке твердого нёба открываются протоки челюст-
ных желез, на боковых краях нёба — протоки латеральных нёбных
желез, а в заднем участке нёба — протоки медиальных нёбных
желез. В глоточной части ротоглотки имеется общее отверстие слу-
ховых труб, а с боков от него — отверстия глоточных желез и
глоточные миндалины.
Рис. 3.181. Ротоглотка курицы:
1 — медиальный и латеральный нёб-
ные валики; 2 — отверстия слюнных
желез; 3 — хоаны; 4 — отверстия нёб-
ных желез; 5 — глоточное отверстие
слуховых труб; 6 — вход в гортань; 7 —
механические сосочки; 8 — гортанное
возвышение; 9 — пищевод; 10 — трахея
У уток и гусей клюв удлинен-
ный, уплощенный, относитель-
но мягкий, покрыт восковицей,
по краям входа в ротоглотку
имеются поперечные пластин-
ки, содержащие осязательные
тельца. Нёбная щель отсутству-
ет, хоаны имеются, сосочки на
нёбе конусовидные, рассеян-
ные.
Язык по форме соответству-
ет клюву, в его корне распола-
гается внутриязычная кость.
Слизистая оболочка языка име-
ет толстый роговой слой, сосоч-
ки языка у кур расположены
поперек основания языка, а у
уток и гусей — по боковым кра-
ям языка, параллельно средин-
ному желобу языка. Вкусовые
сосочки у птиц отсутствуют.
У кур имеются передние и зад-
ние нижнечелюстные железы и
железы угла рта.
Пищевод (рис. 3.181). Грани-
ца между ротоглоткой и пище-
водом образована глоточными
сосочками. На пищеводе имеет-
ся зоб, наиболее сильно разви-
тый у зерноядных птиц, у кото-
322
Рис. 3.182. Желудок курицы:
I — наружная поверхность; Б — вскрытый желудок; 1 — пищевод; 2 — двенад-
цатиперстная кишка; 3 — краниальный слепой мешок; 4 — каудальный слепой
мешок; 5 — стенка железистого желудка; 6 — полость железистого желудка; 7 —
пилорическое отверстие; 8 — стенка мышечного желудка; 9 — полость мышеч-
ного желудка
рых он расположен с правой стороны, перед входом в грудную
клетку. В слизистой оболочке зоба имеются слизистые железы и
। устая капиллярная сеть. У уток и гусей зоб веретенообразной
формы. В каудальной части пищевода у кур и уток находится пи-
щеводная миндалина.
Желудок (рис. 3.182). Желудок у птиц состоит из двух частей —
железистой и мышечной. Железистая часть имеет веретенообраз-
ную форму, стенки ее утолщены, более сильно развит циркуляр-
ный мышечный слой. Слизистая оболочка содержит железы, со-
ответствующие железам дна желудка млекопитающих животных.
У кур выводные протоки желез открываются на небольших желе-
зистых возвышениях. Железистый желудок лежит между двумя
долями печени.
Мышечная часть желудка особенно сильно развита у зернояд-
ных птиц, имеет округлую сдавленную форму. Мышечный желу-
док состоит из гладкой мышечной ткани с синеватым оттенком,
формирующей две боковые мышцы, спаянные сухожильными зер-
калами, расположенными на боковых поверхностях. У петухов
боковые мышцы располагаются на желудке дорсально и вентраль-
но, а у кур — справа и слева. На переднем и заднем концах желуд-
323
ка имеются тонкостенные промежуточные мышцы. Слизистая обо-
лочка мышечного желудка складчатая, содержит железы, выделя-
ющие секрет, который превращается в твердую кератиноидную
кутикулу. Она обеспечивает раздробление и перетирание корма.
В краниальный конец мышечного желудка входит железистый
желудок и выходит двенадцатиперстная кишка. Сальник у птиц
отсутствует.
Тонкая кишка (рис. 3.183, 3.184). Тонкая кишка разделяется на
двенадцатиперстную, тощую и подвздошную. Двенадцатиперст-
ная кишка образует петлю, в которой залегает поджелудочная
железа. Петля огибает справа желудок и достигает таза. Тощая кишка
лежит между воздухоносными мешками, а подвздошная между
двумя слепыми кишками. На тонкой кишке у гусей встречается
остаток желточного мешка в виде дивертикула.
Печень у птиц крупная, состоит из двух долей, на правой доле
имеется желчный пузырь. Левый печеночный проток идет непо-
средственно в конец двенадцатиперстной кишки, а правый идет в
желчный пузырь, из которого уже выходит пузырный проток.
Поджелудочная железа имеет у кур три протока, а у уток и
гусей — два протока, которые открываются самостоятельно в ко-
нец двенадцатиперстной кишки.
Рис. 3.183. Изолированный
кишечник с детализацией
подвздошно-слепокишеч-
ного соединения курицы:
1 — подвздошная кишка; 2 —
слепая кишка, правая и ле-
вая; 3 — тощая кишка; 4 -
место впадения желчного про-
тока и протока поджелудоч-
ной железы; 5 — пилорус; 6,
7 — дорсальная и вентраль-
ная доли поджелудочной же-
лезы; 8 — подвздошно-ки-
шечная миндалина; 9 — изгиб
двенадцатиперстной кишки;
Ю — тошая кишка; 11 — ди-
вертикул тощей кишки; 12 —
подвздошная кишка; 13 —
подвздошно-слепокишечная
связка; 14, 15 — правая и левая
слепая кишка; ]6 — прямая
кишка; 17 — клоака; 18 —
заднепроходное отверстие
324
Рис. 3.184. Васкуляриза-
ция кишечника курицы:
/ — брыжеечно-хвостовая
вена; 2 — яйцевод; 3 — нис-
ходящая аорта; 4 — желе-
1истый желудок; 5 — поч-
ка; 6 — яичник; 7 — чрев-
ная артерия; 8 — краниаль-
ная брыжеечная артерия;
9 — селезенка; 10 — ворот-
ная вена; 11 — печень; 12 —
петля двенадцатиперстной
кишки с поджелудочной
железой; 13, 14, 15 — то-
щая кишка; 16 — подвздош-
но-слепокишечная связка;
/7 — подвздошная кишка;
18 — слепая кишка; 19 —
мышечный желудок; 20 —
клоака; 21 — прямая кишка
Рис. 3.185. Медианный разрез
клоаки:
/ — копчиковая железа; 2 —
сосочек копчиковой железы; 3 —
хвостовое перо; 4 — сосочек се-
мяпровода; 5 — бурса клоаки
(фабрициева сумка); 6 — кожа;
7 — дорсальная проктодеумная
железа; 8 — заднепроходное от-
верстие; 9 — проктодеум (зад-
ний отдел клоаки); 10 — уро-
деу.мно-проктодеумная складка;
/1 — уродеум (средний отдел
клоаки); 12— положение отвер-
стия яйцевода (только слева);
13 — копродеумная складка;
!4 — отверстие мочеточника;
15 — копродеум (передний от-
дел клоаки); /6 — прямая киш-
ка; 17 — пигостиль
325
Толстая кишка (рис. 3.183, 3.184). Тонкая кишка состоит из двух
слепых кишок и прямой кишки, слизистая оболочка ее образует
ворсинки. Ободочная кишка отсутствует. Слепые кишки своими
концами направлены вперед. В слизистой оболочке начальной ча-
сти слепых кишок находятся миндалины слепой кишки. Прямая
кишка представляет короткую трубку, которая кзади заканчива-
ется клоакой.
Клоака (рис. 3.185). Двумя поперечными кольцеобразными склад-
ками клоака подразделяется на три отдела — краниальный, сред-
ний и конечный. В средний отдел открываются мочеточники и
выводящие пути половых органов. Конечный отдел клоаки откры-
вается наружу поперечным анальным отверстием. В конечный от-
дел клоаки у молодых птиц открывается бурса, в стенках которой
имеются лимфатические фолликулы. Бурса редуцируется с наступ-
лением половой зрелости.
ОРГАНЫ ДЫХАНИЯ
Носовая полость. Ноздри округлые или овальные, у кур имеет-
ся носовой клапан, а вокруг ноздрей — венчик коротких щетин-
кообразных перьев. Края ноздрей покрыты восковицей.
У гусей и уток ноздри лежат впереди носовой перегородки и
сообщаются друг с другом.
В носовой полости имеются три парных носовых раковины,
основой которых служат хрящи. Лабиринт решетчатой кости от-
сутствует. Вблизи медиального угла глаза в лобной кости лежат
носовые железы, выводные протоки которых открываются в но-
совую полость.
Гортань. Расположена на дне ротоглотки, вход в нее окружен
сосочками слизистой оболочки. Остов гортани образован кольце-
видным хрящом сложного строения и черпаловидными хрящами.
Кольцевидный хрящ состоит из вентральной вогнутой пластинки,
рано окостеневающей, с ней соединяются боковые пластинки,
которые скрепляются дорсальной пластинкой. С последней соеди-
няются черпаловидные хрящи, которые состоят из вентрального
костного и дорсального хрящевого участков. Надгортанник отсут-
ствует, вместо него имеется лишь поперечная складка слизистой
оболочки впереди входа в гортань. Слизистая оболочка гортани
покрыта мерцательным эпителием. Голосовые складки отсутствуют.
Звукообразование обеспечивается певчей гортанью. Остов гортани
подвижен и управляется четырьмя парными мышцами.
Трахея. Трахея длинная, на поперечном сечении округлая или
поперечно-овальная. Остов образован кольцевидными хрящами,
окостеневающими у старых гусей и уток. У места ветвления трахеи
на бронхи находится нижняя, или певчая, гортань. Она состоит
326
из барабана, мостика с полулунной мембраной и барабанных пе-
репонок в виде эластичных складок.
Барабан состоит из утолщенных колец трахеи, которые у гусей
сливаются друг с другом. У селезней с левой стороны развит осо-
бый костный барабанный пузырь (резонатор). Мостик лежит в месте
деления трахеи на бронхи, он или хрящевой, или окостеневший.
Слизистая оболочка на мостике образует полулунную складку.
Внутренние барабанные перепонки соединяют мостик с медиаль-
ной стенкой бронхов, а наружные барабанные перепонки соеди-
няют латеральные стенки бронхов с барабаном. Барабанные пере-
понки заменяют голосовые складки гортани и участвуют в звуко-
образовании. У певчих птиц имеются специальные мышцы, на-
прягающие или расслабляющие мембраны.
Легкие. Легкие птиц построены своеобразно. Они расположены
под позвоночником и заходят в углубления между позвоночными
отделами ребер. Каудально легкие прилежат к рудиментарной ди-
афрагме. Они простираются от первого ребра до почек. В каждое
легкое вступает главный бронх, который за пределами заднего
края легкого расширяется в обширный брюшной воздухоносный
мешок. От главного бронха отходят дорсально и вентрально вто-
ричные бронхи: внутренние — энтобронхи и наружные — экто-
бронхи. Все вторичные бронхи внутри легкого соединены мелкими
бронхами — парабронхами, от которых уже отходят выпячивания,
формирующие респираторные участки легкого. Эктобронхи уча-
ствуют в образовании воздухоносных мешков, которые являются
выпячиваниями слизистой оболочки, покрытой серозной оболоч-
кой с сосудами. От воздухоносных мешков происходят ответвле-
ния, проникающие в кости (исключая череп), от этого последние
становятся пневматизированными (кости таза, плечевые и бед-
ренные кости, позвонки: шейные, грудные, поясничные и крес-
тцовые; ребра).
Воздухоносных мешков девять, и лишь один из них непарный —
межключичный. Он происходит из вентральных ключичных эк-
тобронхов и образует подмышечные дивертикулы. Парные шей-
ные мешки происходят из вентральных шейных эктобронхов, ле-
жат над трахеей и пищеводом. Краниальные грудные мешки про-
исходят из каудовентральных эктобронхов. Они лежат под легки-
ми до последнего ребра. Каудальные грудные мешки происходят
от главного бронха. Они прилежат к печени, желудку, кишечнику
и к брюшным мешкам. От задних грудных и брюшных мешков
отходят мешковые бронхи, которые ветвятся в толще легких и
соединяются с их респираторными участками.
Воздухоносные мешки являются запасными резервуарами вды-
хаемого воздуха, обеспечивающими поступление свежего воздуха
даже при выдохе (через мешковые бронхи). Подмышечные дивер-
тикулы при полете играют роль мехов, заменяющих движения
327
грудной клетки, которая является опорой для мышц, обеспечи-
вающих полет. Воздухоносные мешки выполняют и другие функ-
ции, в частности предохраняют тело от перегревания.
ОРГАНЫ МОЧЕОТДЕЛЕНИЯ
Почки (рис. 3.186). Почки у птиц лежат вентрально от пояснич-
но-крестцового отдела позвоночника и подвздошной кости. Они
простираются от конца легких до прямой кишки. Почки темно-
красного цвета, мягкой консистенции, проходящими по ним со-
Рис. 3.186. Кровеносные сосуды и нервы почек (вентральная поверхность):
1 — ветвь второго порядка мочеточника; 2 — ветвь первого порядка мочеточни-
ка; 3 — краниальная брыжеечная артерия; 4 — чревная артерия; 5 — аорта; 6 —
каудальная полая вена; 7 — краниальная почечная артерия; 8 — слияние общих
подвздошных вен; 9 — анастомоз к позвоночному венозному синусу; 10 — кра-
ниальная почечная вена; 11 — общая подвздошная вена; 12 — краниальная во-
ротная вена почки; 13 — воротный клапан; 14 - каудальная воротная вена поч-
ки; 15 — каудальная почечная вена; 16 — медиальная почечная артерия; 17 —
каудальная почечная артерия; 18 — каудальная брыжеечная вена; 19 — внутрен-
няя подвздошная вена; 20 — мочеточник; 21 — седалищная вена; 22 — седалищ-
ная артерия; 23 — седалищный нерв; 24 — наружная подвздошная артерия; 25 —
наружная подвздошная вена; 26 — бедренный нерв; I, II, III — соответственно
краниальный, средний и каудальный участки почки
328
(удами они подразделяются на переднюю, среднюю и заднюю
/юли. Внутреннее строение почек примитивное: мочеотделитель-
ные канальцы, соединяясь, переходят прямо в мочеточник.
Мочеточники. У птиц мочеточники проходят по медиальному
краю почек и открываются в средний отдел клоаки: у самцов —
медиально от семяпроводов: у самок — справа от яйцепровода.
Мочевого пузыря нет.
ОРГАНЫ РАЗМНОЖЕНИЯ
Половые органы самцов. Семенники имеют бобовидную или
яйцевидную форму, лежат краниовентрально от почек, на корот-
кой брыжейке. Левый семенник более крупный. В период половой
деятельности семенники становятся крупнее. На медиальном крае
семенника выходят семявыносящие канальцы и впадают в неболь-
шой канал придатка семенника. Канал придатка переходит в се-
мяпровод, который сильно извивается и открывается в сосочке
средней части клоаки.
У селезней и гусаков имеется половой член из фиброзной тка-
ни, образующей кавернозное тело. На поверхности полового чле-
на проходит спирально семенной желоб, который при эрекции
превращается в канал. Придаточных половых желез у птицы нет.
Половые органы самок (рис. 3.187, 3.188). У птиц функциониру-
ет только левый яичник. Правый яичник хотя и закладывается, но
вскоре редуцируется. Яичник подвешен на брыжейке под перед-
ней долей левой почки. Глубокий слой яичника образует сосудис-
тую зону. Поверхностный слой представлен разнообразной вели-
чины фолликулами, которые подвешены на выростах серозной
оболочки, богатой сосудами. Фолликулы содержат яйцевые клет-
ки, окруженные желтком. Вместе с фолликулами яичник имеет
гроздевидную форму. На оболочках фолликулов заметны дугооб-
разные белые линии. По белой линии происходит разрыв оболоч-
ки, при выделении созревших яйцеклеток, после чего остаются
полые чашки, впоследствии подвергающиеся редукции. На яйце-
вой клетке, богатой питательным желтком, имеется зародыше-
вый диск с клеточным ядром. При любом положении яйца заро-
дышевый диск всплывает кверху. После оплодотворения в диске
начинается процесс дробления яйцеклетки. Питательный желток
по своему составу и цвету подразделяется на белый и желтый.
Яйцепровод имеет слизистую, мышечную и серозную оболоч-
ки, подвешен на брыжейке и образует три поперечных колена. По
строению и функции яйцепровод может быть подразделен на пять
участков: воронку, белковую часть, перешеек, скорлуповую и
выводную части. Воронка длиной от 2 до 4 см, начинается широ-
ким отверстием. В ней происходит оплодотворение яйцеклетки.
329
Рис. 3.187. Синтопия органов раз-
множения курицы:
1 — стигма зрелого фолликула; 2 —
яйцеклетки; 3 — прикрепление во-
ронки к стенке тела; 4 — воронка
яйцевода; 5 — белковая часть яйце-
вода; 6 — вентральная часть яйцево-
да и его связка; 7 — перешеек; 8 —
заднепроходное отверстие; 9 — руди-
мент правого яйцевода; 10 — прямая
кишка; 11 — матка, содержащая
яйцо; 12 — правый мочеточник; 13 —
граница правой почки; 14 — влага-
лище; 15 — клоака
Белковая часть длиной до 15 см,
в период половой деятельности
достигает 36 см. Перешеек дли-
ной от 3 до 5 см. При прохожде-
нии яйцеклетки (на что требу-
ется до 3 часов) обе части выде-
ляют «белок» яйца: в белковой
части более плотный белок, в
виде спирально закрученных тя-
жей — градинок, а в перешейке — более жидкий белок и вокруг
него скорлуповую оболочку в виде тонкой двухпластовой волок-
нистой пленки. Скорлуповая часть яйцепровода длиной от 5 до
7 см толстостенная, широкая, с ворсинчатой слизистой оболоч-
кой, содержит специальные железы. Здесь вокруг скорлуповой
оболочки образуются пористая известковая скорлупа и пигмент.
Выводная часть яйцепровода длиной от 7 до 10 см открывается
в средний отдел клоаки.
Рис. 3.188. Строение яйца:
1 — внутренний жидкий белок;
2 - средний густой белок; 3 —
наружный жидкий белок; 4 —
зародышевый диск; 5 — извест-
ковая скорлупа; 6 — наружный
листок скорлуповой оболочки;
7 — воздушная камера (пуга);
8 — желточная оболочка; 9 —
внутренний листок скорлуповой
оболочки; 10, 14 — градинка;
11 — желтый желток; 12, 13 —
белый желток
330
СЕРДЕЧНО-СОСУДИСТАЯ СИСТЕМА
Кровеносная система. Сердце у птицы четырехкамерное (рис.
3.189). Верхушка сердца лежит между долями печени. В правом
желудочке сосцевидных мышц нет. Клапаном правого атриовент-
рикулярного отверстия является двойная мышечная пластинка,
происходящая из стенки правого желудочка. Имеется сердечная
сорочка.
Птицы имеют правую дугу аорты (в отличие от млекопитаю-
щих). Она отдает ветви, аналогичные таковым млекопитающих (рис.
3.190).
Рис. 3.189. Сердце курицы:
А — дорсальная поверхность: / — левая общая сонная артерия; 2 — левая плече-
головная артерия; 3 — правая общая сонная артерия; 4 — правая плечеголовная
артерия; 5 — нисходящая аорта; 6 — восходящая аорта; 7 — правая краниальная
полая вена; 8 — каудальная полая вена; 9 — правое предсердие; 10 — венечная
борозда; 7 7 — продольный желоб; 12 — правый желудочек; 13 — верхушка сер-
дца; 14 — легочная артерия; 15 — легочные вены; 16 — левое предсердие; 17 —
левая краниальная полая вена; 18 — левый желудочек;
Б — вентральная поверхность; 1 — правая плечеголовная артерия; 2 — правая
общая сонная артерия; 3 — левая общая сонная артерия; 4 — левая плечеголов-
ная артерия; 5 — легочная артерия; 6 — левое предсердие; 7 — продольная бо-
розда; 8 — левый желудочек; 9 — правая плечеголовная артерия; 10 — нисходя-
щая аорта; II — правая краниальная полая вена; 12 — восходящая аорта; 13 —
правое предсердие; 14 — венечная борозда; 75 — правый желудочек
331
332
Рис. 3.190. Артерии курицы:
/ - нёбная артерия; 2 — лицевая артерия; 3 — артерия подъязычной кости; 4 —
ыгылочная артерия; 5 — внутренняя сонная артерия; 6 — артерия, сопровожда-
ющая вагус; 7 — позвоночная артерия; 8 — акромиальная артерия; 9— подклю-
чичная артерия; 10 — глубокая плечевая артерия; И — окружная краниальная
ппечевая артерия; 12 — мышечные ветви лучевой артерии; 13 — подъязычная
артерия; 14 — левая обшая сонная артерия; 15 — пальцевые ветви локтевой
артерии; 16 — локтевая артерия; 17 — лучевая артерия; 18 — мышечные ветви;
/9 — глубокая плечевая артерия; 20 — нисходящая аорта; 21 — окружная бедрен-
ная артерия; 22 — бедренная артерия; 23 — седалищная артерия; 24 — мышеч-
ные ветви; 25 — срамная артерия; 26 — тазовая артерия; 27 — глубокая бедрен-
ная артерия; 28 - питающая артерия; 29, 30 — каудальная бедренная артерия;
?/ — каудальная большеберцовая артерия; 32 — латеральная большеберцовая
артерия; 33 — грудино-ключичная артерия; 34 — ключичная артерия; 35 — гру-
аипная артерия; 36 — медиальная большеберцовая артерия; 37 — краниальная
большеберцовая артерия; 38 — общая дорсальная плюсневая артерия
Брюшная аорта в области таза отдает правую и левую наружные
подвздошные артерии и далее делится на правую и левую седалищ-
ные артерии, являющиеся основными магистралями для тазовых
конечностей. Имеется средняя крестцовая артерия. Краниальных
полых вен две; они являются продолжением яремных и подклю-
чичных вен. Каудальная полая вена принимает печеночные вены,
непарную вену брюшной стенки и обе общие подвздошные вены.
Воротных вен печени две: левая выносит кровь из селезенки и же-
лудка, правая — из кишечника и находится в связи с хвостовой
веной. Наружные и внутренние подвздошные вены отводят кровь в
большую почечную вену, отходящую от хвостовой вены.
Лимфатическая система. Лимфатические узлы встречаются толь-
ко у водоплавающих — в области входа в грудную клетку, у конца
яремных вен и в области поясницы между краниальными конца-
ми почек (в лимфатических образованиях— миндалинах). Лимфа-
тических грудных протоков два. Имеются лимфатические сосуды.
Селезенка небольшая, лежит на правой стороне желудка, име-
ет округлую форму.
НЕРВНАЯ СИСТЕМА
Спинной мозг. Имеет шейное и более сильно развитое пояснич-
ное утолщение.
Головной мозг (рис. 3.191, 3.192). Состоит из продолговатого
мозга, мозжечка, который имеет сильно развитый червячок и слабо
развитые клочки. Полушарий мозжечка нет.
Средний мозг состоит из чепца и очень сильно развитого дву-
холмия. Мост отсутствует. Промежуточный мозг состоит из неболь-
333
в
Рис. 3.191. Головной мозг гуся:
А — дорсальная поверхность: / — полушария большого мозга; 2 — двухолмие;
(зрительные холмы); 3 — мозжечок; 4 — ушковидная доля мозжечка; 5 — про-
долговатый мозг; 6 — спинной мозг;
Б — вентральная поверхность: 1 — обонятельные луковицы; 2 — полушария боль-
шого мозга; .? — зрительный перекрест; 4 — гипофиз; 5 — глазодвигательный
нерв; 6 двухолмие (зрительные холмы); 7 — тройничный нерв; 8— продолгова-
тый мозг; 9 — лицевой нерв; 10 — равновесно-слуховой нерв; 77 — отводящий
нерв; 12 — ушковидная доля мозжечка; 13 — языкоглоточный нерв; 14 — блужда-
ющий нерв; 75 — подъязычный нерв; 16 — добавочный нерв; 17 — зрительный
нерв; 18 — блоковый нерв; 19 — продолговатый нерв; 20 — спинной мозг;
В — латеральная поверхность: 7 — мозжечок; 2 — ушковидная доля мозжечка; 3 —
добавочный нерв; 4 — спинной мозг; 5— блуждающий нерв; 6— подъязычный
нерв; 7— языкоглоточный нерв; 8 — равновесно-слуховой нерв; 9 — лицевой
нерв; 10 — отводящий нерв; 11 — тройничный нерв; 12 — продолговатый мозг;
13 — зрительные холмы; 14 — полушария большого мозга; 15 — гипофиз; 16 —
зрительный нерв; 17 — обонятельные луковицы
334
Рис. 3.192. Артерии головного
мозга курицы:
/ — решетчатая артерия; 2 — перед-
няя мозговая артерия; 3 — внутрен-
няя глазничная артерия; 4 — сред-
няя мозговая артерия; 5— передняя
ветвь мозговой сонной артерии; 6 —
। лубокая мозговая артерия; 7— кау-
дальная ветвь мозговой сонной ар-
терии; 8 — передние мозжечковые
артерии; 9 — каудальная мозжечко-
вая артерия; 10—вентральная спин-
но-мозговая артерия; 11 — основная
артерия мозга; 12 — мозговая сон-
ная артерия
ших зрительных бугров, ворон-
ки, гипофиза и эпифиза. Сосце-
видные тела отсутствуют.
Конечный мозг характеризу-
ется: 1) мощным развитием ба-
зального ганглия; 2) слабым
развитием первичного плаща —
только в затылочной области;
3) отсутствием вторичного пла-
ща — коры полушарий; 4) от-
сутствием аммонова рога; 5)
слабым развитием обонятельных долей.
Черепно-мозговых нервов двенадцать пар. VII пара развита сла-
бо; IX пара иннервирует язык, гортань, пищевод и зоб; XII пара
идет в первую гортань; X пара — вагус — идет в сердце, легкие, в
оба отдела желудка. Возвратный нерв идет в пищевод и зоб.
Симпатическая нервная система. Краниальный шейный ганг-
лий лежит на основании черепа. Шейная часть симпатического
пограничного ствола проходит в поперечном канале шейных по-
звонков и в каждом сегменте формирует ганглий. Параллельно
вагусу идет ветвь в общую сонную артерию. От грудопоясничной
части пограничного ствола отходят нервы к сердцу и сосудам внут-
ренностей: сердечно-легочные нервы отходят от первых двух-трех
грудных узлов, большой внутренностный нерв идет в чревное спле-
тение, малый внутренностный нерв идет в надпочечное сплете-
ние.
Парасимпатическая нервная система. Имеется блуждающий нерв.
Кишечный нерв (парасимпатический) происходит из вентраль-
ных ветвей крестцовых нервов и идет краниально в брыжейке вдоль
кишечника, дает ветви в толстую и тонкую кишку до желудка, но
с вагусом не соединяется.
335
Рис. 3.193. Продольный разрез глаз-
ного яблока курицы:
7 — склеральная пластинка; 2 — ра-
дужная оболочка; 3 — роговица; 4 —
сетчатка; 5— сосудистая оболочка; 6 —
склера; 7 — зрительный нерв; 8 — гре-
бень; 9 — центральная ямка
Рис. 3.194 . Внутреннее ухо курицы:
1 — ампула полукружного канала; 2 — преддверие; 3 — глазничный канал; 4 —
канал сонной артерии; 5 — наружный слуховой проход; 6 — канал блуждающего
нерва; 7 — улитка; 8 — полукружные каналы
336
АНАЛИЗАТОРЫ
Органы зрения. В глазном яблоке тапетум отсутствует (рис. 3.193).
1рачок округлый. Сфинктер зрачка представлен поперечноисчер-
ченной мышечной тканью. В толще стекловидного тела имеется
। ребень, который проходит от входа зрительного нерва до капсу-
лы хрусталика.
Нижнее веко хорошо развито и подвижнее верхнего. Сильно
развито третье веко. Мигательная перепонка без хряща, она мо-
жет с помощью мышц закрывать всю переднюю часть глазного
яблока. Слезная железа развита слабо.
Органы слуха. В органе слуха наружное ухо с хрящевой ракови-
ной отсутствует. Имеется лишь наружный слуховой проход, до-
вольно широкий, но короткий.
Слуховая косточка только одна — столбик (рис. 3.194), соеди-
няющий барабанную перепонку с овальным окном внутреннего
уха. С помощью мышцы, действующей на столбик, возможно на-
пряжение барабанной перепонки. Улитка лабиринта внутреннего
уха слабо развита.
ГЛАВА 4
ФИЗИОЛОГИЯ
СИСТЕМА КРОВИ
Кровь — жидкая ткань, циркулирующая в кровеносной систе-
ме животных, разновидность соединительной ткани. Она пред-
ставляет собой непрозрачную, вязкую жидкость солоноватого вкуса
и своеобразного запаха. В артериях кровь ярко-красная, в венах —
вишневого цвета. Красный цвет крови обеспечивает поглощение
фиолетовой и ультрафиолетовой части солнечного спектра, что
способствует усвоению организмом солнечной энергии. Она со-
стоит из жидкой части — плазмы и взвешенных в ней форменных
элементов: эритроцитов, лейкоцитов и тромбоцитов. Если свежую
кровь с добавлением лимоннокислого натрия центрифугировать,
то она расслоится. На дно пробирки осядут более тяжелые эритро-
циты, над ними образуется небольшой слой лейкоцитов и тром-
боцитов, а наверху останется полупрозрачная, желтоватого цвета
жидкость — плазма. Объем плазмы больше (58 — 65 %), чем объем
форменных элементов (35 — 42 %).
Кровь вместе с лимфой и тканевой жидкостью образуют внут-
реннюю среду организма.
Для нормального функционирования организма необходимо от-
носительное динамическое постоянство состава и физико-хими-
ческих свойств внутренней среды, называемое гомеостазом. Он
является результатом сложных регуляторных взаимоотношений на
всех структурных уровнях организма. Гомеостаз обеспечивается ней-
рогуморальными, гормональными, барьерными и выделительны-
ми механизмами.
Сохраняя постоянство состава кровь, тем не менее, отражает
происходящие в организме изменения как в норме, так и в пато-
логии. Поэтому в ветеринарии широко используются гематологи-
ческие анализы.
Кровь вместе с органами, в которых происходит образование и
разрушение форменных элементов (костный мозг, печень — в эм-
бриональный период, тимус, селезенка, лимфатические узлы,
миндалины и лимфоидные образования), объединяют в единую
систему крови, деятельность которой регулируется нейрогумораль-
ными механизмами.
338
ФУНКЦИИ КРОВИ
Циркулируя по замкнутой системе сосудов, кровь выполняет
важнейшие физиологические функции.
Транспортная функция — кровь переносит питательные веще-
ства: продукты расщепления белков, углеводов, липидов, а также
витамины, гормоны, минеральные соли и воду от системы орга-
нов пищеварения к клеткам организма. Из крови питательные и
другие вещества поступают в тканевую жидкость. Эти вещества
нужны клеткам в качестве строительного и энергетического мате-
риала.
Выделительная функция — из клеток организма в тканевую
жидкость, а из нее в кровь и лимфу поступают продукты клеточ-
ного метаболизма, основная часть которых переносится кровью к
органам выделения.
Дыхательная функция — кровь переносит О2 от легких к тка-
ням, а образующийся в них СО2 (диоксид углерода) к легким,
откуда он удаляется при выдохе. В капиллярах легких кровь отдает
СО2, который выдыхается, и поглощает О2. В переносе О2 основ-
ную роль выполняет гемоглобин, в переносе СО2 — соли, раство-
ренные в плазме крови.
Регуляторная функция — через кровь осуществляется гумораль-
ная регуляция деятельности органов и систем организма за счет
поступающих в кровь биологически активных веществ.
Защитная функция — кровь защищает организм от губитель-
ного действия микробов, вирусов и их токсинов, а также чуже-
родных организму веществ. Эта функция осуществляется за счет
бактерицидных свойств плазмы крови, фагоцитарной активности
лейкоцитов и лимфоцитов, ответственных за тканевый и клеточ-
ный иммунитет.
Терморегуляторная функция — кровь выполняет эту функцию
благодаря непрерывной циркуляции и высокой удельной тепло-
емкости. Образующееся в организме тепло аккумулируется кро-
вью и равномерно распределяется по всему организму.
Все функции крови направлены на поддержание гомеостаза,
т.е. относительного постоянства состава внутренней среды.
У животных в состоянии покоя примерно половина всей крови
циркулирует в кровеносных сосудах (55 — 60 %), а другая часть (40—
45 %) депонируется в селезенке (16 %), печени (15 — 20 %) и коже
(10%). Соотношение между циркулирующей и депонированной
кровью меняется в зависимости от функционального состояния
организма. Когда организм испытывает недостаток О2, часть де-
понированной крови рефлекторно поступает в общий кровоток.
Депонированная кровь содержит больше форменных элементов,
чем кровь, циркулирующая в сосудах. Обе фракции крови нахо-
дятся в динамическом равновесии.
339
Количество крови зависит от вида животного, пола, породы,
хозяйственного использования, кормления, времени года. У круп-
ного рогатого скота и овец оно составляет в среднем 8 % массы
тела, у свиней — 7, у лошадей — 9,8 (у спортивных лошадей 14—
15, а у тяжеловозов 7 — 8 %).
Чем интенсивнее процессы обмена веществ в организме и чем
выше потребность в О2, тем больше крови у животного. Резкое
уменьшение количества крови может привести животное к ги-
бели.
ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА КРОВИ
Вязкость и плотность крови. У животных вязкость и плотность
крови в 3 — 6 раз больше вязкости и плотности воды, что зависит
от количества эритроцитов, лейкоцитов и белкового состава пла-
змы.
Плотность крови равна 1,035 — 1,056, плазмы — 1,02—1,03,
эритроцитов — 1,08—1,09. Относительная плотность лейкоцитов
и кровяных пластинок ниже, чем эритроцитов, поэтому при цен-
трифугировании они образуют слой над эритроцитами.
Увеличивают вязкость крови белки, особенно фибриноген.
В капиллярах вязкость крови уменьшается. Вязкость крови влияет
на гемодинамику и величину кровяного давления.
Минеральный состав и белки плазмы крови. В плазме и фор-
менных элементах находятся: минеральные соли, белки, глюко-
за, мочевина и др. Они создают осмотическое давление — силу,
которая вызывает движение растворителя через полупроницае-
мую мембрану из менее концентрированного раствора в более
концентрированный раствор. В плазме крови содержится около
0,9 % минеральных солей. Они находятся в виде катионов и ани-
онов, и осмотическое давление, равное 7,6 атм., в основном
создается за их счет. Осмотическое давление обеспечивает в орга-
низме обмен воды между кровью и тканями. Постоянство осмо-
тического давления крови, лимфы и тканевой жидкости имеют
значение для функции клеток. Мембраны, в том числе и клеток
крови, полупроницаемы. Если из плазмы крови теряется много
воды и концентрация солей в ней повышается, то вода из эрит-
роцитов в силу законов осмоса начинает поступать в плазму че-
рез их полупроницаемую мембрану, и эритроциты сморщивают-
ся. При понижении концентрации солей в плазме вода из нее
поступает в эритроциты, и они набухают. При очень низком ос-
мотическом давлении оболочки эритроцитов разрушаются и со-
держимое их поступает в плазму. В стенках кровеносных сосудов,
в тканях, в гипоталамусе имеются специальные осморецепторы.
Раздражение их вызывает рефлекторное изменение деятельно-
340
ci и выделительных органов, и они удаляют избыток воды или
солей, поступивших в кровь.
Осмотическое давление создают и коллоиды — белки плазмы,
поэтому кровь обладает также и коллоидным давлением. Коллоид-
ное давление называется также онкотическим. Оно составляет ме-
нее 1 % от осмотического давления. Из общего осмотического дав-
нения на долю белков приходится 1/20— 1/30 атм, т. е. 30 — 35 мм
рт. ст. Несмотря на малую величину, онкотическое давление игра-
ет важную роль в обмене воды между кровью и тканями. Это та
сила, которая удерживает воду внутри сосудов и способствует пе-
реходу ее из тканевой жидкости в кровь. Коллоидно-осмотическое
давление складывается из осмотического и онкотического. При
уменьшении количества белков в плазме крови происходят отеки
тканей.
Реакция крови и буферные системы. Кровь животных имеет сла-
бощелочную реакцию. Активная реакция крови обусловлена
концентрацией водородных (Н+) и гидроксильных (ОН ) ионов.
Для характеристики реакции крови пользуются водородным по-
казателем, обозначаемым pH. Кровь животных имеет pH 7,35 —
7,55. Эта величина сохраняется на относительно постоянном уров-
не, несмотря на непрерывное поступление кислых и щелочных
продуктов обмена веществ. Сдвиг pH на 0,3 —0,4 может вызвать
смерть животного.
Поддержание pH на оптимальном уровне обеспечивается бу-
ферными системами крови и деятельностью выделительных орга-
нов, удаляющих избытки кислот и щелочей. В крови имеются сле-
дующие буферные системы: гемоглобиновая, карбонатная, фос-
фатная и система белков плазмы крови.
Гемоглобиновая буферная система — самая мощная система, так
как примерно 75 % буферов крови составляет гемоглобин. Он при-
соединяет как кислоты, так и щелочи.
Белки плазмы крови обладают амфотерными свойствами: с
кислотами вступают в реакцию как основания, с основаниями —
как кислоты, благодаря чему участвуют в поддержании pH на от-
носительно постоянном уровне.
Карбонатная буферная система состоит из угольной кислоты
(Н2СО3) и ее солей — бикарбонатов натрия и калия (NaHCO3 и
КНСО3).
Когда в процессе обмена веществ в кровь поступают кислоты,
они нейтрализуются бикарбонатами. На долю карбонатной системы
приходится относительно небольшая часть буферных веществ кро-
ви, но ее роль в организме значительна, так как с деятельностью
этой системы связано выведение СО2 легкими, что обеспечивает
почти мгновенное восстановление нормальной реакции крови.
Фосфатная буферная система образована натриевыми солями
фосфорной кислоты: NaH2PO4 и Na2HPO4. Первое соединение
341
слабо диссоциирует и ведет себя как слабая кислота, второе —
имеет свойства слабой щелочи. Вследствие небольшой концентра-
ции фосфатов в крови роль этой системы менее значительна.
В крови имеется определенное и довольно постоянное соотно-
шение между кислотными и щелочными компонентами, его на-
зывают кислотно-щелочным равновесием.
Бикарбонаты плазмы образуют щелочной резерв крови. Благо-
даря ему трудно вызвать сдвиг реакции крови в кислую сторону.
Например, кормление коров кислым силосом, напряженная мы-
шечная работа лошадей уменьшают щелочной резерв, и при по-
ступлении большого количества кислот в кровь наступает ацидоз.
Скармливание большого количества щелочных продуктов или за-
держка их в организме вызывают алкалоз.
В цельной крови основная буферная способность (более 75 %)
обеспечивается гемоглобином, в плазме крови — карбонатной си-
стемой. Принцип действия буферных систем основан на замене силь-
ной кислоты слабой, при диссоциации второй образуется меньше
ионов Н+ и, следовательно, pH плазмы снижается в меньшей сте-
пени. Молочная кислота, образующаяся в организме, более силь-
ная, чем угольная. Поэтому она нейтрализуется бикарбонатом и
замещается угольной кислотой. Свободная угольная кислота спо-
собна связывать и ОН -ионы с образованием ионов бикарбоната.
Сущность действия фосфатной системы основана на диссоциации
двузамещенного фосфата натрия с образованием двух ионов на-
трия и ионов вторичного фосфата. Последние связывают протоны
и дают первичный фосфат, который, в свою очередь, может дис-
социировать на ион водорода и анион вторичного фосфата.
Организм надежно защищен от сдвига реакции в кислую сто-
рону, важная роль в этом принадлежит карбонатной буферной
системе, которая обеспечивает до 20 % буферной емкости всей
крови и основную часть буферной емкости плазмы.
Запас бикарбонатов плазмы называют щелочным резервом кро-
ви. Щелочной резерв крови во много раз превышает кислотный.
ПЛАЗМА И ФОРМЕННЫЕ ЭЛЕМЕНТЫ КРОВИ
Состав плазмы крови. Плазма крови — это сложная биологи-
ческая среда, тесно связанная с тканевой жидкостью организма.
Плазма представляет собой полупрозрачную жидкость желтовато-
го цвета с вязкостью 1,7 —2,2, относительной плотностью 1,030 —
1,035. В плазме крови содержится 90 — 92 % воды и 8 — 10 % сухих
веществ.
В плазме крови растворены также О2, СО2 и N. Растворы, осмо-
тическое давление которых равно плазме, называются изотониче-
скими. Растворы с меньшим осмотическим давлением называются
342
гипотоническими, с большим — гипертоническими. Отклонение ос-
мотического давления плазмы от нормальных величин отражает-
ся на структуре и функции клеточных элементов крови. Это следу-
ет учитывать при внутривенном введении животным питательных
веществ или лечебных препаратов в большом количестве жидко-
сти.
Основную часть сухого вещества плазмы составляют белки,
липиды, углеводы, минеральные и другие биологически актив-
ные вещества. Их количество равняется 8 —10 %. Белки представ-
лены главным образом альбуминами и глобулинами. Соотношение
альбуминов и глобулинов в плазме крови называют белковым ко-
эффициентом. У свиней, овец, коз, собак, кроликов и человека
он больше единицы, а у лошадей, крупного рогатого скота мень-
ше единицы. Считают, что от величины белкового коэффициента
«висит скорость оседания эритроцитов. Она повышается при уве-
личении количества глобулинов.
Альбумины и фибриноген образуются в печени, глобулины,
кроме печени, еще и в костном мозгу, селезенке, лимфатических
узлах.
Белки плазмы являются резервом для построения тканевых
белков, выполняют функцию переносчиков биологически актив-
ных веществ — гормонов, витаминов, пигментов, метаболитов,
микроэлементов. Принимают участие в регуляции кислотно-ще-
лочного равновесия (pH) крови. Из гамма-глобулинов образуют-
ся антитела, которые создают иммунитет в организме и защища-
ют его от бактерий и вирусов.
Форменные элементы крови. Выделяют три группы форменных
элементов крови: эритроциты, лейкоциты и тромбоциты. Общий
объем форменных элементов, главным образом эритроцитов, в
100 объемах крови называют показателем гематокрита, который
выражается в процентах (40 — 45 %).
Эритроцит покрыт белково-липидной оболочкой, внутри имеет
сетчатое строение, ячейки которого заполнены гемоглобином.
Количество эритроцитов в крови определяют под микроскопом с
помощью счетных камер или электронных приборов целлоскопов.
В 1 мм3 крови у животных разных видов содержится неодинаковое
количество эритроцитов.
Функции эритроцитов', переносят О2 от легких к тканям, уча-
ствуют в транспортировании СО2 от тканей к легким, на своей
поверхности транспортируют аминокислоты, гормоны, витами-
ны и различные продукты обмена веществ.
При изменении осмотического давления плазмы крови, по-
ступлении в кровь химических веществ или при физическом воз-
действии эритроциты разрушаются. Разрушение оболочки эрит-
роцитов и выход из них гемоглобина называется гемолизом. Разли-
чают осмотический, химический, физический гемолиз.
343
Способность эритроцитов сохранять целостность своей струк-.
туры при изменении осмотического давления называется осмоти-J
ческой стойкостью, или резистентностью.
Эритроциты образуются и созревают в красном костном мозге.
В процессе созревания они теряют ядро и после этого поступают в
кровь. Юные клетки эритроцитов, непосредственные предшествен-
ники зрелых эритроцитов, получили название ретикулоцитов.
У взрослого животного они составляют 5 — 10 % всех эритроцитов
крови.
Эритроциты у лошади разрушаются и заменяются новыми в
среднем через 100 дней, у крупного рогатого скота — 120— 160, у,
овцы — 130, у северного оленя — 95, у кролика — через 45 —
60 дней. Эритроциты разрушаются в печени и селезенке. За сутки
обновляется в среднем 0,8 — 1 % эритроцитов. Однако скорость эрит-
ропоэза может резко возрастать при кровопотерях, недостатке О2,
патологическом укорочении длительности жизни эритроцитов. При
этом возрастает содержание в плазме особого стимулирующего
вещества — эритропоэтина. По химической структуре это глюко-
протеин, который образуется при расщеплении одного из глобу-
линов плазмы под воздействием ферментов почек.
Роль гемоглобина. В эритроцитах находится сложное химическое
соединение — гемоглобин. Он состоит из двух частей: белка глоби-
на и четырех молекул гема. Молекула гема, содержащая атом двух-
валентного железа, обладает способностью присоединять и
отдавать О2. В капиллярах легких гемоглобин присоединяет О2 и
становится оксигемоглобином (НЬО2), а в капиллярах тканей от-
дает О2 и превращается в восстановленный гемоглобин. Артери-
альная кровь, содержащая оксигемоглобин, имеет ярко-алый цвет.
Венозная кровь, в которой содержится восстановленный гемо-
глобин, темно-вишневая. Восстановленным или редуцированным
называют гемоглобин (НЬ), отдавший О2.
Различают две разновидности гемоглобина: тип А у взрослых
животных и тип F (фетальный) у плода. Фетальный гемоглобин
обладает более высоким сродством к О2, чем гемоглобин взрос-
лых, что создает оптимальные условия для перехода О2 из крови
матери в кровь плода. Окисление F-гемоглобина происходит за счет
гемоглобина А матери, который находится в кровеносной системе
плаценты, по другую сторону трансплацентарного барьера.
При действии на гемоглобин сильных окислителей (бертолето-
ва соль, нитробензол и др.) железо окисляется и переходит из
двухвалентного в трехвалентное, а гемоглобин превращается в
метгемоглобин (MtHb) и приобретает коричневую окраску. В слу-
чае образования в крови большого количества метгемоглобина О2
тканям не отдается и наступает смерть от удушья.
Гемоглобин присоединяет также угарный газ (СО), причем в
150 — 200 раз активнее, чем кислород, при этом образуется кар-
344
i«оксигемоглобин (НЬСО). Это соединение очень прочное, и ге-
моглобин перестает быть переносчиком О2. Поэтому примесь в
воздухе даже 0,1 % угарного газа ведет к тому, что 80 % гемогло-
бина связывается с ним, а это опасно для жизни: через 30 — 60 мин
Развиваются тяжелые последствия кислородного голодания (рво-
ia, потеря сознания). При концентрации угарного газа в количе-
стве 1 % через несколько минут наступает смерть.
Гемоглобин и его производные определяют с помощью спект-
роскопа, рассматривая разведенную гемолизированную кровь.
В скелетных и сердечной мышцах находится мышечный гемогло-
бин (миоглобин). По строению он сходен с гемоглобином. Миогло-
бин способен присоединять больше О2, что имеет важное значе-
ние для снабжения О2 сокращающихся мышц.
Количество гемоглобина в крови зависит от вида животного,
возраста, пола, породы, его кормления, а также от высоты над
уровнем моря. У новорожденных животных эритроцитов и гемо-
глобина больше, чем у взрослых, у самцов больше, чем у самок.
У овец и коров, находящихся на горных пастбищах, содержание
гемоглобина повышено. Количество гемоглобина в крови опреде-
ляют с помощью специального прибора — гемоглобинометра
(рис. 4.1) и выражают в грамм-процентах (г %) или в граммах на
литр крови (г/л). В среднем у животных в крови содержится гемог-
лобина от 9 до 17 г; у крупного рогатого скота — 100— 130 г/л, у
свиней — 100—120, у лошадей — 90—150 г/л.
Недостаток гемоглобина является причиной анемии. Под этим
гермином понимают снижение способности крови переносить О2.
При анемии уменьшается либо число эритроцитов, либо содер-
жание в них гемоглобина (а иногда — и то и другое).
Гемолиз — это разрушение оболочки эритроцитов и выход из
них гемоглобина. Гемолиз может быть химический, при воздей-
Рис. 4.1. Гемоглобинометр ГС-3:
I — корпус прибора; 2 — градуиро-
ванная пробирка для исследованной
крови; 3 — стеклянная палочка; 4 —
пипетка для дистиллированной воды;
5 — пробирки, запаянные со стандарт-
ным раствором солянокислого гемати-
на; 6 — капиллярная пипетка для взя-
тия крови
345
ствии химических веществ, растворяющих жиры (эфира, хлоро-
форма, сапонинов, змеиного яда) и нарушающих структуру или
целостность мембраны, и физический, который подразделяют на
механический (при сильном встряхивании), температурный (под
действием высокой и низкой температуры), лучевой (под дей-
ствием рентгеновских или ультрафиолетовых лучей).
В гипотоническом растворе эритроциты поглощают воду, на-
бухают, приобретают сферическую форму, в результате их обо-
лочка разрывается и гемоглобин выходит в среду. Этот процесс
называется осмотическим гемолизом. В гипертоническом растворе
эритроциты, наоборот, теряют воду и сморщиваются. Гемолиз на-
чинается при концентрации раствора ниже 0,55 % Nad.
Скорость оседания эритроцитов (СОЭ). Плотность эритроцитов
выше, чем плазмы (1,095 и 1,030 соответственно), поэтому при
отстаивании в пробирке крови, предохраненной от свертывания,
эритроциты медленно оседают на дно. Скорость оседания эритро-
цитов зависит от вида животных, возраста, физиологического
состояния. Так, у взрослого крупного рогатого скота СОЭ состав-
ляет 0,7 мм, у птиц — 4,0, у свиней — 8,0, у лошадей — 64 мм за
первый час.
Величина СОЭ зависит от числа эритроцитов, их размера и
особенно от белкового состава плазмы. Во время беременности,
при инфекционных болезнях, воспалительных процессах СОЭ
повышена. Усиленная мышечная работа замедляет эту реакцию.
Для определения СОЭ кровь смешивают с раствором лимонно-
кислого натрия и набирают в стеклянную трубочку с миллимет-
ровыми делениями. Через некоторое время отсчитывают высоту
верхнего прозрачного слоя.
Лейкоциты — это белые кровяные клетки, размеры их дости-
гают 20 мкм. Они имеют цитоплазму и ядро. Их подразделяют на
две большие группы: зернистые (гранулоциты) и незернистые (аг-
ранулоциты). В цитоплазме зернистых лейкоцитов содержатся зер-
нышки (гранулы), в цитоплазме незернистых лейкоцитов грану-
лы отсутствуют.
Зернистые лейкоциты — в зависимости от окраски гранул раз-
личают эозинофильные, базофильные и нейтрофильные. Эти клет-
ки, а также моноциты образуются и дифференцируются в крас-
ном костном мозгу. К незернистым, формам относятся лимфоциты
и моноциты.
Лимфоциты имеют крупное ядро, окруженное узким поясом
цитоплазмы. В зависимости от размера различают большие, сред-
ние и малые лимфоциты. Общее количество лейкоцитов зависит
от физиологического состояния животного. Процентное соотно-
шение различных форм лейкоцитов называется лейкограммой. Уве-
личение количества лейкоцитов называют лейкоцитозом, а умень-
шение — лейкопенией. Лимфоциты составляют большую часть бе-
346
пых кровяных клеток: у крупного рогатого скота — 50 — 60 % всех
лейкоцитов, у свиней 45 — 60, у овен 55 — 65, у коз — 40 — 50, у
кроликов — 50 — 65, у кур 45 — 65 %. Этим видам животных при-
сущ так называемый лимфоцитарный профиль крови. У лошадей и
плотоядных — нейтрофильный профиль крови. Лейкоциты способ-
ны к амебовидному движению, они проходят через стенки ка-
пилляров. Все виды лейкоцитов играют важную роль в защитных
реакциях организма, но каждый вид осуществляет это особым спо-
собом. Поглощение и переваривание лейкоцитами микробов, от-
мерших клеток организма и всяких чужеродных белков и других
веществ, попадающих в организм, называют фагоцитозом. Явле-
ние фагоцитоза открыл И. И. Мечников.
Нейтрофилы {микрофаги) фагоцитируют бактерии и продукты
распада тканей и разрушают их ферментами. Они оказывают так-
же противовирусное действие, вырабатывая особый белок — ин-
терферон.
Базофилы синтезируют противосвертывающее вещество — ге-
парин, а также гистамин, участвующий в воспалительных реак-
циях по месту внедрения микробов.
Эозинофилы участвуют в обезвреживании токсинов белкового
происхождения. Содержат фермент гистаминазу, разрушающий
гистамин и снижающий местную воспалительную реакцию.
Моноциты — самые большие клетки крови, обладают хорошо
выраженной фагоцитарной и бактерицидной активностью. Они
фагоцитируют микробы, погибшие лейкоциты, поврежденные
клетки тканей, очищая очаг воспаления.
Лимфоциты участвуют в выработке антител и образовании им-
мунитета — невосприимчивости к болезням. Они также ответствен-
ны за реакции на введение чужеродного белка и отторжение чу-
жеродных тканей при пересадке органов. Образуются в лимфати-
ческих узлах, миндалинах, пейеровых бляшках кишечника, селе-
зенке, тимусе, фабрициевой сумке (у птиц). Среди них выделяют
Т-лимфоциты (на их долю приходится 40 — 70% всех лимфоци-
тов) и В-лимфоциты (они составляют 20 — 30% циркулирующих
лимфоцитов).
Тромбоциты — это маленькие плоские безъядерные тельца
овальной или веретенообразной формы. Время пребывания их в
кровотоке 8—10 суток. У животных их содержится от 220 до 400 тыс.
в 1 мм3 крови. При повреждении кровеносных сосудов тромбоци-
ты разрушаются. При этом из них выходит ряд веществ (тромбо-
цитарные факторы), участвующих в остановке кровотечения и
свертывании крови.
Стволовые клетки крови (СКК). Под стволовой кроветворной
клеткой крови понимается родоначальная клетка, способная к
развитию в различные виды зрелых клеток. СКК относительно
редко делятся, в среднем 1 раз за 10 суток, поэтому они более
347
радиорезистентны, чем их потомки. С возрастом общее число СКК
не меняется. Циркулирующие в периферической крови СКК со-
ставляют 0,1 % общего количества клеток крови. Они сходны по
виду с лимфоцитами, их диаметр равен 8 — 10 мкм.
Процесс развития стволовых клеток в зрелые форменные эле-
менты крови складывается из пролиферации, дифференцировки
и созревания.
СВЕРТЫВАНИЕ КРОВИ
Пока кровь течет по неповрежденным сосудам, она остается
жидкой. Но стоит поранить сосуд, как довольно быстро образует-
ся кровяной сгусток (тромб), он закрывает рану и кровотечение
останавливается. В основе свертывания крови лежит физико-хи-
мический процесс превращения растворенного в плазме крови
белка фибриногена в нерастворимой фибрин. Фибрин выпадает в
виде тонких нитей. Нити фибрина образуют густую мелкоячеис-
тую сеть, в которой задерживаются форменные элементы. Свер-
тывание крови происходит в три фазы.
Первая фаза — образование тромбопластина. В результате сопри-
косновения крови с краями раны или инородной поверхностью
разрушаются тромбоциты. Из них выделяются тромбоцитарные фак-
торы; они взаимодействуют с факторами свертывания плазмы и
ионами кальция, в результате чего образуется тромбопластин.
Вторая фаза — превращение протромбина в тромбин. Протром-
бин вырабатывается в печени и постоянно находится в крови. При
воздействии тромбопластина с участием ионов кальция, факто-
ров плазмы и тромбоцитов протромбин переходит в тромбин.
Третья фаза — образование из растворенного в плазме фибри-
ногена нерастворимого фибрина.
Известно наследственное заболевание людей — гемофилия, при
котором резко понижена свертываемость крови. Подобное заболе-
вание, связанное с нарушением синтеза одного из факторов плаз-
мы, встречается также у собак и свиней.
Регуляция свертывания крови находится под непосредственным
контролем центральной нервной системы.
Скорость свертывания крови является видовым признаком. Так,
кровь крупного рогатого скота свертывается в течение 7 — 9 мин,
лошади — 10—12, свиньи — 3 — 4, птицы — 2 — 3 мин. У беремен-
ных животных она свертывается быстрее. Сильно замедляется свер-
тывание крови у крупного рогатого скота после поедания гнию-
щего сена или силоса из медового клевера — донника.
Противосвертывающая система предохраняет внутрисосудис-
тое свертывание крови и участвует в растворении образовавшихся
тромбов.
348
ГРУППЫ КРОВИ
В основе определения групп крови лежит явление агглютина-
ции (склеивание), открытое в начале XX в. Группы крови опреде-
ляются по наличию в эритроцитах специфических антигенов —
агглютиногенов, а в плазме — специфических антител к ним —
агглютининов. Впервые в эритроцитах крови людей К. Ландштей-
нер и Я. Янский обнаружили два вещества белковой природы,
которые они назвали агглютиногенами (склеиваемыми вещества-
ми), а в плазме — два агглютинина (склеивающие вещества). Агг-
лютиногены обозначаются буквами латинского алфавита А и В,
агглютинины — буквами греческого алфавита а (альфа) и р (бета).
Агглютинин а склеивает эритроциты, содержащие агглютино-
ген Л, агглютинин р склеивает эритроциты с агглютиногеном В, в
результате чего наступает агглютинация эритроцитов и их разру-
шение — гемолиз. Свойство эритроцитов склеиваться при действии
на них плазмы или сыворотки крови другого человека послужило
основой для разделения крови всех людей на четыре группы.
Кровь людей I группы не содержит агглютиногенов, поэтому
ее можно переливать человеку с любой другой группой крови. Кровь
же IV группы нельзя переливать людям с кровью других групп,
так как она содержит оба агглютиногена — А и В. Человек, даю-
щий свою кровь для переливания, называется донором, а получа-
ющий эту кровь — реципиентом. Переливание крови донора, со-
держащей агглютинины а и [3, к реципиенту с кровью, содержа-
щей агглютиногены А и В, неопасно, так как агглютинины в кро-
ви реципиента быстро разбавляются его кровью, и низкая кон-
центрация агглютининов не вызывает склеивания эритроцитов и
их гемолиз.
В эритроцитах был открыт еще один агглютиноген. Его впервые
обнаружили у обезьян (Macacus rhesus') и назвали резус-фактор.
Если кровь человека, содержащую резус-фактор (резус-положи-
тельную), перелить человеку, не имеющему его (резус-отрица-
тельному), то у последнего образуются специфические антигены.
Повторное переливание такому человеку крови, содержащей ре-
зус-фактор, вызывает агглютинацию эритроцитов и тяжелые ос-
ложнения. У сельскохозяйственных животных резус-фактор обна-
ружен только в крови лошадей.
Агглютиногены эритроцитов и резус-фактор образуются в пе-
риод эмбрионального развития, поэтому группы крови не меня-
ются в течение всей жизни.
В эритроцитах сельскохозяйственных животных обнаружено
большое количество антигенных факторов. У крупного рогатого
скота изучено 100 антигенных факторов, объединенных в 12 сис-
тем. У свиней выявлено 14, у овец — 7, у кур — 14 систем групп
крови. У лошадей открыто 10 агглютиногенов.
349
Кровь животных независимо от групповой принадлежности
несовместима с кровью человека.
КРОВЕТВОРЕНИЕ
Кроветворение — одна из наиболее рано формирующихся функ-
ций организма. Процесс образования, развития и созревания фор-
менных элементов крови носит название гемопоэза. Форменные
элементы крови живут недолго, поэтому непрерывно образуются
новые и разрушаются старые. Относительное постоянство их и не-
прерывное обновление за счет стволовых полипотентных клеток
крови осуществляется в органах кроветворения и иммуногенеза,
и представляет собой физиологическую регенерацию крови. Глав-
ным органом кроветворения, в котором образуются эритроциты,
тромбоциты и лейкоциты, является красный костный мозг. Лим-
фоциты формируются также в лимфоузлах и тимусе. У плода орга-
нами кроветворения служат костный мозг, печень и селезенка.
Все форменные элементы крови образуются из одной стволо-
вой клетки костного мозга. Разрушение эритроцитов и лейкоци-
тов происходит в печени и селезенке.
Кроветворение связано с обменом веществ. Образование эрит-
роцитов происходит при участии витамина В12 и фолиевой кисло-
ты, а также микроэлементов — железа, кобальта, меди. Кроветво-
рение регулируется нервной системой, железами внутренней сек-
реции и гуморальными факторами.
Влияние ЦИС на кроветворение осуществляется через вегета-
тивную нервную систему. Симпатическая нервная система стиму-
лирует кроветворение, а парасимпатическая — угнетает.
Гормоны гипофиза (кортикотропин и соматотропин) и про-
дукты распада тканей, возникающие при их повреждении и вос-
палении, способствуют увеличению образования лейкоцитов.
Среди гуморальных факторов большую роль играют эритропо-
этины. Они образуются в почках и усиливают образование эритро-
цитов. При кислородном голодании в результате кровопотери,
разрушении эритроцитов при отравлении, пребывании высоко в
горах количество эритропоэтинов увеличивается.
ЛИМФАТИЧЕСКАЯ СИСТЕМА.
ЛИМФА И ТКАНЕВАЯ ЖИДКОСТЬ
В организме наряду с системой кровеносных сосудов имеется
система лимфатических сосудов, по которым возвращается в кровь
тканевая жидкость. Тканевая жидкость, поступившая в лимфати-
ческие сосуды, называется лимфой.
350
Тканевая жидкость представляет собой среду, которая окружа-
ет все клетки организма. Из нее клетки получают необходимые
питательные вещества, гормоны, витамины, минеральные веще-
ства, Н2О, О2 и выделяют в эту жидкость СО2 и другие продукты
внутриклеточного обмена веществ. Важнейшая функция лимфы —
возврат белков из тканевых пространств в кровь, участие в пере-
распределении воды в организме, молокообразовании, пищева-
рении и обмене веществ.
Лимфа — полупрозрачная, слегка желтоватая жидкость щелоч-
ной реакции (pH 7,5 —9,0) с удельным весом 1,023—1,025, за-
полняющая лимфатические сосуды. Она состоит из лимфоплазмы
и форменных элементов. Химический состав ее близок к составу
плазмы крови, но отличается более низким содержанием белка
(60% от количества в плазме крови), меньшей вязкостью, более
низким коллоидно-осмотическим давлением.
В лимфе содержатся белки, преобладают альбумины, есть им-
муноглобулины, фибриноген и протромбин, не белковые азотис-
тые вещества, глюкоза, соли, ферменты, гормоны, витамины и
антитела. Вследствие низкого содержания белков плотность и вяз-
кость лимфы меньше, чем плазмы.
Лимфа, оттекающая от кишечника, молочно-белого цвета,
непрозрачная в связи с тем, что в ней содержатся капельки эмуль-
гированного жира. Лимфа, оттекающая от печени, содержит мно-
го белков. Количество лимфоцитов в лимфе после прохождения
через лимфоузлы увеличивается и в грудном протоке составляет
5 — 20 тыс. в 1 мм3. Это обусловлено тем, что лимфоциты образу-
ются в лимфоузлах и с током лимфы уносятся в кровь. Кроме
лимфоцитов, в лимфе имеется небольшое количество моноци-
тов, нейтрофилов, базофилов и эозинофилов. В лимфе нет тром-
боцитов, но она способна свертываться, так как содержит фиб-
риноген и ряд факторов свертывания. Однако защитных белков в
лимфе меньше, чем в крови, поэтому она является хорошей сре-
дой для размножения и распространения возбудителей инфекции
и опухолевых клеток. Около 90 % форменных элементов лимфы
составляют лимфоциты. Из общего количества лимфоцитов, цир-
кулирующих в крови и лимфе, примерно 75 % приходится на долю
Т-лимфоцитов, 15 % — на долю В-лимфоцитов и 10 % — на клет-
ки, не относящиеся ни к той, ни к другой группе (ноль-клетки,
или К-клетки).
Лимфатические капилляры называют корнями лимфатической
системы, в них и образуется лимфа. Интенсивность образования
тканевой жидкости и лимфы зависит от проницаемости капилля-
ров. Поэтому те вещества, которые увеличивают проницаемость
капилляров, называются лимфогонными. Белки крови, повышая
онкотическое давление в капиллярах, препятствуют лимфообра-
зованию.
351
Скорость движения лимфы в десятки раз ниже, чем крови. Дав-
ление лимфы в мелких лимфатических сосудах составляет 8—10 мм
водяного столба. Вся лимфа в конечном итоге поступает в крани-
альную полую вену.
Контрольные вопросы
1. Расскажите о внутренней среде организма.
2. Что такое гомеостаз?
3. Назовите основные функции крови.
4. Состав и форменные элементы крови, их значение.
5. Опишите процесс свертывания крови.
6. Назовите группы крови у домашних животных.
7. Расскажите о составе, свойствах и значении лимфы и тканевой жид-
кости.
8. Назовите органы кроветворения.
ФИЗИОЛОГИЯ ИММУННОЙ СИСТЕМЫ
ИММУНИТЕТ, ЕГО ЗНАЧЕНИЕ. ИММУННАЯ СИСТЕМА
Иммунная система обеспечивает защиту организма от инфек-
ций, а также способность организма защищать себя от генетически
чужеродных тел и веществ, сохранять генетический гомеостаз.
Иммунитет рассматривают как способность организма различать
«свое» и «не свое», сохранять свою биологическую индивидуальность.
Морфологически иммунная система представляет собой сово-
купность всех лимфоидных органов и скоплений лимфоидных кле-
ток тела. Связь между ними осуществляется через кровеносную и
лимфатическую системы. Главной клеточной формой иммунной
системы является лимфоцит.
Лимфоидные органы условно делятся на центральные и пери-
ферические. К центральным (первичным) органам иммунной сис-
темы относят тимус, фабрициеву бурсу у птиц и костный мозг.
К периферическим (вторичным) относят селезенку, лимфатиче-
ские узлы, лимфоидную ткань пищеварительного тракта {миндали-
ны глотки, пейеровы бляшки и солитарные фолликулы кишечника), а
также лимфоидную ткань органов дыхания. Известно, что органы
дыхания и пищеварительный тракт служат главными «входными
воротами» для антигенов, поэтому содержащиеся там многочис-
ленные лимфатические фолликулы сходны по строению с тако-
выми селезенки и лимфатических узлов.
В центральных лимфоидных органах образуются исходные ство-
ловые клетки, осуществляется пролиферация и первичная диф-
ференцировка иммунокомпетентных (ответственных за иммуни-
тет) клеток — лимфоцитов. В периферических лимфоидных орга-
352
нах происходит созревание лимфоцитов, их пролиферация в от-
вет на антигенную стимуляцию.
Первичные лимфоидные органы. В красном костном мозге и пе-
чени (у плодов) находятся стволовые клетки, дающие начало всем
типам клеток крови. Некоторые из стволовых клеток, запрограм-
мированные как лимфоцитарные, мигрируют с током крови в
тимус, где размножаются и дифференцируются в лимфоциты
(Т-лимфоциты, или тимусзависимые). Другая группа стволовых
клеток поселяется и дифференцируется в фабрициевой бурсе
птиц — дивертикуле клоаки (В-лимфоциты, или бурсозависи-
мые). У млекопитающих эквивалента фабрициевой бурсы не най-
дено. Предполагают, что ее функцию выполняет либо сама крове-
творная ткань костного мозга, либо лимфатические пейеровы
бляшки, расположенные в стенке тонкой кишки. С наступлением
половой зрелости тимус и фабрициева бурса уменьшаются в раз-
мерах и затем подвергаются инволюции.
Вторичные лимфоидные органы. Часть лимфоцитов, прошедших
«курс обучения» в тимусе и фабрициевой бурсе и обладающих
полной иммунокомпетентностью, переносится (еще в эмбриональ-
ный период) в периферические лимфоидные органы: лимфати-
ческие узлы, селезенку, миндалины, лимфоэпителиальные обра-
зования в слизистой желудочно-кишечного тракта, дыхательных
и мочеполовых путей. В лимфатических фолликулах этих образова-
ний различают тимусзависимые и тимуснезависимые зоны. В пер-
вых селятся 'Т-лимфоциты, во вторых — ^-лимфоциты.
В лимфатических узлах тимуснезависимой зоной является кор-
тикальный слой, расположенный ближе к поверхности; в его фол-
ликулах находятся главным образом В-лимфоциты. Паракортикаль-
ный слой, прилегающий к медуллярным синусам, составляет ти-
мусзависимый слой, содержащий Т-клетки. Однако резкой грани-
цы между зонами нет, поскольку иммунный ответ требует, как
правило, взаимодействия между Т- и В-лимфоцитами. В лимфати-
ческих узлах происходит антигензависимая пролиферация (кло-
нирование) и дифференцировка Т- и В-лимфоцитов и образова-
ние клеток памяти.
В селезенке, выполняющей роль фильтра для крови, обе зоны
находятся по соседству в белой пульпе. Непосредственно вдоль
артерий расположена тимусзависимая зона, кнаружи от нее ти-
муснезависимая зона.
Селезенка участвует в защите организма и иммунологических
реакциях за счет того, что эндотелиальные клетки способны за-
хватывать чужеродные частицы и электроотрицательные коллои-
ды. А в связи с тем, что она состоит из ретикулярной и лимфоид-
ной тканей, выполняет функции кроветворения.
Антигены, их характеристика. В современной иммунологии ан-
тигенами называют иммуногены и гаптены, которые, активируя
I 2 Зеленевский
353
иммунокомпетентные клетки, вызывают образование иммуногло-
булинов и развитие многих других иммунологических (защитных)
процессов. Иммуногены представляют собой высокомолекулярные
соединения, а низкомолекулярные, только реагирующие с анти-
телами, называют гаптенами. Способность стимулировать антите -
лообразование гаптены приобретают при введении их в комплек-
се с другими компонентами, например белками.
Антитела, их структура, основные функции. Антитела — имму-
ноглобулины разных классов, образующиеся в организме под воз-
действием антигена и обладающие специфическим сродством к
нему. Антитела обладают функцией защиты организма от микро-
организмов и генетически чужеродных элементов. Они образуют-
ся в организме в результате инфицирования (естественная имму-
низация) или вакцинации (искусственная иммунизация), инъек-
ции чужеродного белка, пересадки тканей от других организмов.
Антитела содержатся в гамма-глобулиновой фракции сыворот-
ки крови и в лимфе (циркулирующие антитела), в молозиве и
секретах (секретирующие антитела), на поверхности клеток, на-
пример эпителия слизистых оболочек (связанные с мембраной
антитела), спинно-мозговой жидкости, воспалительных выпотах.
По функции антитела условно подразделяют на нейтрализую-
щие, лизирующие и коагулирующие. К нейтрализирующим анти-
телам относят антитоксины, вируснейтрализующие антитела и
антиферменты; к лизирующим — бактериолизины, гемолизины,
цитолизин; к коагулирующим — преципитины и агглютинины.
Основными продуцентами антител в организме являются плаз-
матические клетки, образующиеся из лимфоцитов и ретикулоцитов.
В крови сельскохозяйственных животных (крупного рогатого
скота, свиней, овец, коз и лошадей) обнаружено три класса им-
муноглобулинов: IgG, IgA, IgM. В молозиве животных содержатся
все три класса иммуноглобулинов, но преимущественно IgG, в
молоке преобладают IgA и IgM. У новорожденных животных анти-
тела в крови отсутствуют, впервые они получают их с молозивом
матери. Собственный синтез антител начинается у телят с двухне-
дельного возраста, но происходит очень медленно, что связано с
функциональной незрелостью лимфоидной ткани.
ВИДЫ ИММУНИТЕТА
По происхождению различают естественный, или врожденный,
и приобретенный иммунитет.
Естественный иммунитет — видовой признак, присущий оп-
ределенному виду животных и передающийся по наследству. Он
определяет видовую невоспримчивость к тому или иному заболе-
ванию.
354
Приобретенный иммунитет возникает у животного вследствие
естественного переболевания (естественно приобретенный) или
в результате искусственной иммунизации (искусственно приоб-
ретенный).
Приобретенный иммунитет бывает активный и пассивный. Ак-
тивный иммунитет вырабатывается либо после перенесенной ин-
фекции, либо в результате вакцинации. В обоих случаях в организ-
ме вырабатываются антитела против возбудителя. Активный им-
мунитет может быть весьма напряженным и длительным. Пассив-
ный, или сывороточный, иммунитет создается введением в орга-
низм иммунной сыворотки, содержащей готовые антитела. Он
наступает через несколько часов после введения препарата, но
проявляется непродолжительно (10—14 суток, реже до 3 недель).
Разновидностью пассивного иммунитета является колостраль-
ный (молозивный) иммунитет, возникающий у новорожденных
организмов при передаче им готовых антител (иммуноглобули-
нов) с молозивом от матери.
ИСПОЛЬЗОВАНИЕ ИММУНИТЕТА В ЖИВОТНОВОДСТВЕ
И ВЕТЕРИНАРИИ
Деятельность иммунной системы направлена на поддержание
генетического гомеостаза организма, поэтому стимуляция меха-
низмов специфической и неспецифической защиты способствует
сохранению функциональной целостности организма, повыше-
нию его устойчивости к неблагоприятным факторам среды.
Контрольные вопросы
1. Расскажите об иммунитете и его значении.
2. Опишите типы лимфоцитов, роль тимуса.
3. Дайте характеристику антигенам и антителам.
4. Назовите виды иммунитета.
КРОВО- И ЛИМФООБРАЩЕНИЕ
СИСТЕМА ОРГАНОВ КРОВООБРАЩЕНИЯ
Непрерывное движение крови по кровеносным сосудам осу-
ществляется сердцем — центральным органом сердечно-сосудис-
той системы. В передвижении крови по организму активное учас-
тие принимают кровеносные сосуды.
У млекопитающих сердце четырехкамерное с полностью изо-
лированными потоками венозной и артериальной крови. Кровь
движется по двум кругам — большому и малому (см. гл. 3).
355
Сердце
Основу сердца составляет сердечная мышца — миокард, пост-
роенная из сердечной поперечно исчерченной мышечной ткани.
Анатомическое строение сердца изложено в разделе «Анатомия
сердечно-сосудистой системы».
Физиологические свойства сердечной мышцы. Организм всегда
приспосабливает ритм работы сердца к характеру выполняемой
работы. Например, у рысаков в процессе бега частота сокращений
сердца достигает 200 и более ударов в минуту, что превосходит
исходный уровень в 4 —5 раз. У коров в период отела она может
повышаться до 110 ударов. Такой широкий диапазон работы серд-
ца объясняется физиологическими свойствами сердечной мышцы.
Сердечная мышца обладает свойствами автоматии, возбудимос-
ти, проводимости, сократимости и рефрактерное™ (рис. 4.2).
Автоматизм сердца — способность сердца ритмически сокра-
щаться без внешних раздражений, под влиянием импульсов, воз-
никающих в нем самом. Ритмичность работы сердца обеспечивает
Рис. 4.2. Проводящая система
сердца лошади:
1 — каудальная полая вена; 2 —
краниальная полая вена; 3 —
синусно-предсердный узел; 4 —
предсердно-желудочковый узел;
5 — предсердно-желудочковый
пучок; 6 — левая ножка и ее раз-
ветвления до волокон Пуркинье;
7 — левая септомаргинальная
трабекула; 8 — правая септомар-
гинальная трабекула; 9 — пра-
вая ножка и ее разветвления до
волокон Пуркине
особая система, проводящая возбуж-
дение. Эта система состоит из синус-
ного узла, атриовентрикулярного узла
и атриовентрикулярного пучка, а об-
разуется из атипичных кардиомиоци-
тов. Синусный узел расположен в по-
граничной борозде, между устьем пе-
редней полой вены и правым ушком,
атриовентрикулярный узел — на пе-
регородке предсердий. Атриовентри-
кулярный пучок проходит от атриовен-
трикулярного узла через фиброзное
кольцо на перегородку желудочков и
делится на две ножки, которые пере-
ходят на боковые стенки желудочков.
Ножки разветвляются на особые мы-
шечные волокна, называемые волок-
нами Пуркинье, которые обильно вет-
вятся под эндотелием желудочков.
Синусный узел является центром ав-
томатии. Частота сокращений сердца
определяется частотой возбуждений,
возникающих в данном узле, которой
поэтому называют водителем сердеч-
ного ритма первого порядка. Разные
отделы сердца проявляют различный
автоматизм. Автоматия атриовентри-
кулярного узла выражена в меньшей
степени. Это центр автоматии второго порядка. Далее по ходу про-
водящей системы автоматия продолжает убывать. В случае пораже-
ния проводящей системы ритм сердца сильно замедляется, воз-
никают аритмии.
Возбуждение сердечной мышцы. При действии на сердце раз-
дражителей в сердечной мышце возникает возбуждение и сокра-
щение. В ответ на раздражение пороговой силы сердце отвечает
сокращением максимальной силы (закон «все или ничего»). При
дальнейшем усилении раздражения сила сокращений не меняется.
Во время возбуждения она не воспринимает новые раздражения.
Такое состояние невозбудимости называют абсолютной рефрак-
терностъю, она длится столько же времени, сколько сокращается
мышца. По окончании абсолютной рефрактерности возбудимость
постепенно возвращается к норме; это период относительнойреф-
рактерности, он совпадает с расслаблением сердечной мышцы.
За периодом относительной рефрактерное™ наблюдается повы-
шенная возбудимость — это период экзальтации.
Сократимость сердечной мышцы. Возбужденная сердечная мыш-
ца сокращается. Она отвечает сокращением только на одиночный
импульс возбуждения, на серию частых импульсов не реагирует.
Установлено, что сила сокращения сердечной мышцы прямо про-
порциональна длине мышечных волокон перед началом сокраще-
ния. Чем больше растягиваются волокна во время диастолы, тем
больше сила сокращения. Эта особенность получила название «за-
кон сердца». Поставщиком энергии для сокращения сердца слу-
жат макроэргические фосфорсодержащие соединения — адено-
зинтрифосфат и креатинфосфат.
Сердечный цикл. Сердце работает ритмично — в две фазы. Со-
кращение сердца называется систолой, расслабление — диасто-
лой. Систола и диастола составляют сердечный цикл. В первую фазу
сокращаются предсердия, желудочки находятся в состоянии рас-
слабления. Во вторую фазу происходит сокращение желудочков и
расслабление предсердий. За второй фазой следует общая диасто-
ла предсердий и желудочков. Желудочки и предсердия заполня-
ются кровью, поступающей из вен. При систоле давление в пред-
сердиях повышается до 10 — 30 мм ртутного столба, в правом же-
лудочке — до 70 мм, а в левом — до 180 — 200 мм.
Длительность сердечного цикла зависит от частоты сокраще-
ний сердца. При 60 сокращениях в минуту (например, у коров)
продолжительность одного сердечного цикла составляет 1 с (60:60).
Из этого времени примерно 0,1 с длится систола предсердий, а
диастола — около 0,9 с. Систола желудочков равна примерно 0,3 с,
а диастола — 0,7 с. Таким образом, в течение одного сердечного
цикла систола предсердий и желудочков длится 0,4 с, а на общую
диастолу сердца приходится 0,6 с. Такой длительный отдых обес-
печивает восстановление работоспособности сердца и дает ему воз-
357
356
можность беспрерывно работать без утомления. Учащение работы
сердца происходит в основном за счет укорочения диастолы, дли-
тельность систолы при этом уменьшается незначительно.
Частота сердечных сокращений у животных разных видов раз-
личная: у лошадей — 32 — 42, верблюдов — 32 — 52, крупного ро-
гатого скота и свиньи — 60 — 80, собак — 70—120, кроликов —
120—140, кур — до 300 сокращений в минуту. Чем крупнее жи-
вотное, тем медленнее сокращается его сердце.
Тоны сердца. Это звуковые явления, которыми сопровождается
работа сердца. При выслушивании сердца посредством фонендос-
копа слышны два звука, или тона: систолический — низкий и
протяжный и диастолический — высокий и короткий. Систоли-
ческий тон возникает при систоле и обусловлен сокращением мышц
сердца, колебанием атриовентрикулярных клапанов и прикреп-
ленных к ним сухожильных нитей. Диастолический тон слышен в
начале диастолы и образуется при захлопывании полулунных кла-
панов. По изменениям тонов и по возникающим шумам опреде-
ляют расстройства сердечной деятельности.
Систолический и минутный объем кровотока. Количество кро-
ви, выбрасываемой желудочками сердца в минуту, называется
минутным объемом кровотока. Минутный объем сердца у лошади
равен 20 — 30 л, коров — 35 л, овец — 4 л, собак — до 1,5 л.
Разделив минутный объем на число сокращений сердца в минуту,
можно вычислить систолический объем кровотока. У лошадей он
Рис. 4.3. Регистрация биопотенциалов сердца:
А — электрокардиограмма: 1 — схема ЭКГ; 2, 3 — ЭКГ во втором отведении у
лошади и коровы; Б — векторкардиограмма лошади: Р — возбуждение предсер-
дий; комплекс зубцов QRS — возбуждение желудочков в момент их сокращения;
Т — конечная фаза возбуждения желудочков
Рис. 4.4. Схема ин-
нервации сердца:
1 — кора больших по-
лушарий головного моз-
га; 2 — продолговатый
мозг; 3, 4 — симпати-
ческие центры и волок-
на (соответственно пре-
и постганглионарные);
5 — шейные симпати-
ческие узлы; 6 — во-
локна блуждающего
нерва (пре- и постган-
глионарные); 7—про-
межуточный мозг
составляет 850 мл, у крупного рогатого ско-
та — 580, овец — 55, собак массой 10 кг —
14 мл. При напряженной работе систоличес-
кий и минутный объемы сердца резко увели-
чиваются.
Биоэлектрические явления в сердце. Потен-
циалы действия, возникающие в сердце при
прохождении волны возбуждения, распрост-
раняются по всему телу и могут быть зарегис-
трированы с помощью электродов, приложен-
ных к поверхности тела (область груди, ко-
нечности). Метод регистрации потенциалов сер-
дца называется электрокардиографией (ЭКГ),
а прибор, с помощью которого их регистри-
руют, — электрокардиографом.
Электрокардиограмма (ЭКГ) животных
состоит из отдельных зубцов и интервалов
между ними, обозначаемых буквами Р, Q, R,
S, Т. Небольшой зубец Р отражает возбужде-
ние предсердий, комплекс зубцов Q, R, S,
наибольший по амплитуде, характеризует
процесс возбуждения желудочков в момент их
систолы. Зубец Тсвязан с восстановительны-
ми процессами в миокарде после его возбуж-
дения. Метод электрокардиографии позволя-
ет судить о последовательности распростра-
нения возбуждения по сердцу; его можно ис-
пользовать при постановке диагноза наруше-
ний работы сердца (рис. 4.3).
Регуляция работы сердца. Сердце относит-
ся к числу автоматически функционирующих
органов, но его деятельность должна соответ-
ствовать меняющимся потребностям организма в кровоснабже-
нии. Это осуществляется за счет нервной и гуморальной регуляции
сердечной деятельности.
Нервная регуляция. Работа сердца регулируется нервными им-
пульсами, поступающими к нему из центральной нервной систе-
мы по блуждающим и симпатическим нервам (рис. 4.4). Блуждаю-
щие нервы доходят до синусного и атриовентрикулярного узлов,
симпатические нервы направляются непосредственно к мышеч-
ным волокнам сердца. Блуждающие нервы тормозят сердечную
деятельность: замедляют частоту сокращений, ослабляют силу со-
кращений, понижают возбудимость и проводимость сердечной
мышцы. Симпатические нервы, наоборот, учащают и усиливают
сокращения сердца, увеличивают возбудимость и проводимость
сердечной мышцы.
358
359
Центральная нервная система обеспечивает рефлекторную регу-
ляцию функций сердца посредством центров, лежащих в продолго-
ватом и спинном мозге, коре больших полушарий, гипоталамусе.
Важную роль в регуляции сердца играют рецепторы, находя-
щиеся в дуге аорты, в области разветвления сонных артерий (ка-
ротидный синус), в устье полых вен. Расположенные здесь прес-
сорецепторы и хеморецепторы образуют сосудистые рефлексоген-
ные зоны. Раздражение рецепторов дуги аорты и каротидного сину-
са при повышении кровяного давления вызывает рефлекторное
ослабление и урежение сердечных сокращений. Повышение дав-
ления в устье полых вен раздражает расположенные здесь рецеп-
торы и рефлекторно усиливает и учащает сокращения сердца.
Гуморальная регуляция. Хеморецепторы сосудистых рефлексо-
генных зон возбуждаются при изменении состава омывающей их
крови: увеличении СО2, уменьшении О2. Рецепторы есть также в
миокарде и эндокарде, их раздражение изменяет работу сердца.
Для нормальной работы сердца имеют большое значение элек-
тролиты: калий и кальций влияют на автоматию сердца и его со-
кратительные свойства. При избытке ионов калия сердце сокра-
щается реже и слабее и даже может остановиться в диастоле. Ионы
кальция учащают и усиливают сокращения сердца.
Движение крови по сосудам
Движение крови по кровеносным сосудам происходит по зако-
нам гидродинамики. Течение жидкости обеспечивается разностью
давления: жидкость продвигается от большего давления к меньшему.
Давление крови в сосудах создается работой сердца: при каждой си-
столе в аорту и легочный ствол нагнетается определенное количе-
ство крови. Часть энергии давления крови, создаваемой работой сер-
дца, идет на передвижение крови по сосудам, а большая ее часть
расходуется на устранение возникающего при этом сопротивления.
Движение крови по венам имеет ряд особенностей. Вены из-за
небольшой толщины их мышечного слоя легко растягиваются и
сдавливаются. Движение крови в венах происходит из-за разности
давления в мелких и крупных венах. Так, давление в мелких венах
составляет 10 — 20 мм рт. ст., а в крупных венах, находящихся в
грудной полости, — оно отрицательное. Однако эта разность дав-
лений невелика, и потому кровотоку в венах способствует ряд
дополнительных факторов. В венах имеются клапаны, пропускаю-
щие кровь только по направлению к сердцу. Тонкостенные вены
лежат между мышцами или рядом с ними. При сокращении по-
следних кровь выжимается из вен по направлению к сердцу. Дви-
жению крови по венам способствует присасывающее действие
грудной полости, особенно во время вдоха.
360
Скорость кровотока. В различных сосудах скорость кровотока
неодинаковая, что связано с их суммарным диаметром. Различают
объемную и линейную скорость кровотока. Объемной скоростью
кровотока называют количество крови, протекающее в единицу
времени через поперечное сечение сосудов. Объемная скорость во
всех участках кровеносных сосудов неодинаковая. Линейной скоро-
стью кровотока называют скорость движения частиц крови вдоль
сосуда в секунду. Она обратно пропорциональна суммарной ши-
рине сосудов, по которым течет кровь.
Кровь, как и любая жидкость, течет быстрее в том отделе со-
судистой системы, где общий просвет сосудов наиболее узкий; по
мере расширения общего просвета скорость кровотока замедляет-
ся. Если сравнивать суммы просветов различных отделов крове-
носной системы, то наиболее узкой частью сосудистой системы
является аорта. Например, суммарный диаметр капилляров в 600 —
800 раз больше диаметра аорты. Вследствие этого в аорте кровь
течет быстрее, чем в капиллярах. Скорость течения крови в аорте
во время систолы равна 500 — 600 мм в секунду. В артериях она
уменьшается до 250 мм, в капиллярах — до 0,5 мм в секунду.
Артериальный пульс. При каждой систоле желудочки сердца
выбрасывают в аорту определенное количество крови, которая
растягивает ее стенки. Во время диастолы растянутые стенки воз-
вращаются в исходное положение. Это растяжение и спадение сте-
нок аорты вызывает ее ритмические колебания, которые переда-
ются по стенкам артерий. Такие растяжения и спадения стенок
аорты соответствуют частоте сокращений сердца. Ритмические
колебания стенок артерий называют артериальным пульсом. Пульс
можно прощупать у лошади и коровы на лицевой или хвостовой
артерии, у мелких животных — на бедренной артерии. Пульс от-
ражает особенности работы сердца и состояние самих кровенос-
ных сосудов, поэтому его показатели имеют большое клиниче-
ское значение в оценке функционального состояния сердечно-
сосудистой системы.
Венный пульс наблюдается в крупных, расположенных близко
к сердцу, венах (полые и яремные). В мелких и средних венах пуль-
сация их стенки отсутствует. Кровь по венам течет непрерывно,
но она поступает в сердце только во время диастолы. При систоле
предсердий и желудочков задержка крови в венах повышает там
давление, а при диастоле давление снижается. Эти колебания сте-
нок в крупных венах и называют венным пульсом. Венный пульс
имеет диагностическое значение при некоторых болезнях сердца.
Давление крови. В кровеносных сосудах кровь находится под
определенным давлением. Величина кровяного давления неоди-
накова в разных участках сосудистой системы и служит одним из
показателей функционального состояния организма. Оно макси-
мально в аорте и крупных артериях, снижается в мелких артериях
361
и венах и падает ниже атмосферного в полых венах. Различают
артериальное, капиллярное и венозное давление.
По мере удаления от сердца давление крови падает. Так, в аор-
те давление равно 200 — 240 мм рт. ст., в артериях — 120—130, в
артериолах — около 70, в капиллярах до 40 мм, а в крупных венах
ниже атмосферного на 2 — 5 мм рт. ст.
В малом круге кровообращения давление крови в 5 — 6 раз ниже,
чем в большом круге.
Давление крови меняется в зависимости от фазы сердечного
цикла. Подъем давления крови во время систолы желудочков на-
зывают максимальным, или систолическим, снижение давления во
время диастолы называют диастолическим, или минимальным. Раз-
ность между систолическим и диастолическим давлением называ-
ют пульсовым давлением. У крупного рогатого скота систолическое
давление в хвостовой артерии равно 110—140 мм рт. ст., диасто-
лическое — 35 — 50, а пульсовое — 75 — 90 мм рт. ст. Давление
крови зависит от вида, пола, возраста, породы животного, состо-
яния нервной системы.
Давление крови измеряют прямым или косвенным методом. При
прямом методе в артерию вводят полую иглу, соединенную с ма-
нометром. Этот метод требует хирургического вмешательства и
применяется редко, для массовых исследований он непригоден.
Косвенный метод основан на измерении той величины давления,
которая необходима для сдавливания артерии и прекращения в
ней кровотока. С этой целью у крупного рогатого скота на репицу
хвоста накладывают резиновую манжету, соединенную с резино-
вой грушей и ртутным или пружинным манометром. У мелких
животных манжету накладывают на область бедра.
Регуляция кровообращения. Оптимальные условия кровоснаб-
жения органов и тканей в соответствии с их физиологическим
состоянием обеспечиваются нервной и гуморальной регуляцией ра-
боты сердца, изменением просвета кровеносных сосудов и коли-
чества циркулирующей крови.
Изменение просвета кровеносных сосудов осуществляется имею-
щимися в них гладкими мышцами, которые могут длительно нахо-
диться в состоянии сокращения без признаков утомления. Такое со-
стояние гладких мышц называется тонусом. Увеличение тонуса мышц
приводит к сужению сосудов, ослабление — к их расширению.
На тонус мышц кровеносных сосудов, а следовательно, и на
просвет сосудов главное влияние оказывает нервная система. Со-
суды иннервируются двоякого рода нервами: сосудосуживающими
и сосудорасширяющими. Сосудосуживающие нервы принадлежат
симпатическому отделу нервной системы. Они вызывают сужение
кровеносных сосудов кожи, органов брюшной полости, легких и
других органов. Лишь сосуды сердца и головного мозга под влия-
нием симпатических нервов расширяются.
362
Сосудорасширяющими являются парасимпатические нервы. Их
волокна имеются во многих смешанных нервах, а также содер-
жатся в составе дорсальных корешков спинного мозга.
Сосудодвигательный центр находится в продолговатом мозгу и
состоит из двух отделов: сосудосуживающего (прессорного) и со-
судорасширяющего (депрессорного).
Рефлекторные влияния на сосуды. Сосудодвигательный центр
находится в состоянии постоянного возбуждения, что обеспечи-
вает тонус сосудистой системы. Тонус сосудодвигательного цент-
ра зависит от импульсов, поступающих от рецепторов, располо-
женных в некоторых сосудистых областях, а также от влияния
гуморальных раздражителей. В дуге аорты, в области разветвления
сонных артерий (каротидный синус), и в устье полых вен имеют-
ся рецепторы, которые возбуждаются при изменении давления
крови. Их называют прессорецепторами, а места их расположения —
сосудистыми рефлексогенными зонами.
Гуморальная регуляция. Некоторые химические вещества, дей-
ствуя непосредственно на стенки сосудов, вызывают сужение или
расширение сосудов. Гормоны надпочечников адреналин и норад-
реналин обусловливают сужение артерий и артериол кожи, орга-
нов брюшной полости и легких, а сосуды сердца и головного мозга
они расширяют. Гормон задней доли гипофиза вазопрессин вызы-
вает сужение артериол и капилляров. В почках образуется веще-
ство ренин; поступая в кровь, он вызывает сужение сосудов.
К сосудосуживающим веществам относятся также серотонин; он
образуется в мозгу, в слизистой оболочке кишечника, а также
при разрушении тромбоцитов.
К сосудорасширяющим веществам относятся ацетилхолин, ги-
стамин, простагландины, аденозинтрифосфорная, молочная и уголь-
ная кислоты.
Кровоснабжение сердца, мозга, легких, печени, селезенки, а
также почек и поджелудочной железы осуществляется по конце-
вому типу ветвления артерий. Преимущество концевых артерий
заключается в том, что по ним кровь поступает в ткани органа
под большим и одинаковым давлением. Крупным же недостатком
их является то, что в случае закупорки такого сосуда (тромбом)
соответствующий участок органа не получает крови, что немину-
емо ведет его к омертвлению.
Кровообращение в венечных сосудах сердца происходит преиму-
щественно во время диастолы. Во время систолы сердечная мыш-
ца сдавливает расположенные в ней сосуды, поэтому кровоток в
них ослабевает. Сердце получает до 10% крови, выбрасываемой
при систоле левым желудочком.
Кровоснабжение головного мозга происходит через внутренние
сонные артерии. Проникая к головному мозгу через рваные отвер-
стия, они образуют вокруг гипофиза артериальное кольцо. Пуль-
363
сация протекающей крови в нем гасится. Кровь от мозга оттекает
в вены, образующие дорсальную и вентральную системы сину-
сов, расположенных в твердой мозговой оболочке.
Циркуляция крови в легких обеспечивается как малым (через ле-
гочные артерии), так и большим (через бронхиальные артерии)
кругом кровообращения. Но газообмен между венозной кровью и
воздухом, поступающим в легкие, осуществляется только за счет
малого круга. В легочной артерии лошадей кровяное давление колеб-
лется в пределах 30 — 60 мм рт. ст., у крупного рогатого скота —
40 — 80 мм рт. ст., у собак систолическое давление в легочной ар-
терии равно 40 мм рт. ст., диастолическое — 10 мм рт. ст. Во время
вдоха кровяное давление сильно понижается, а при выдохе повы-
шается.
В печень кровь поступает из печеночной артерии и из воротной
вены, причем через воротную вену поступает в 2 — 3 раза больше
крови, чем через печеночную артерию. Воротная вена печени рас-
падается на сеть капилляров, которые, объединяясь, образуют
печеночные вены. Такое образование получило название «чудес-
ная венозная сеть» печени. Значение чудесной сети заключается в
замедлении тока крови в ней, что обеспечивает более тесный кон-
такт крови с печеночными клетками и освобождение крови от
ядовитых продуктов обмена.
Кровообращение в селезенке связано с особенностями строения
ее капилляров. На концевых веточках капилляров селезеньей рас-
положены кисточки, которые заканчиваются слепыми расшире-
ниями с отверстиями. Через эти отверстия кровь переходит в пуль-
пу, а оттуда в синусы, имеющие отверстия в стенках. Поэтому
селезенка, как губка, может впитывать кровь и служит ее депо.
Кровь селезенки содержит больше эритроцитов и на 15 % больше
гемоглобина, чем кровь, циркулирующая в сосудах.
Особенностью почечного кровообращения является наличие со-
судистых клубочков, представляющих собой сплетение артерий и
капилляров. Причем приносящие и выносящие к сосудистому клу-
бочку кровь сосуды являются артериями. Такое образование полу-
чило название «чудесная артериальная сеть». Давление крови в при-
носящих к сосудистому клубочку артериях определяется кровя-
ным давлением в аорте, а в выносящих сосудах — значительно
меньшим диаметром этих сосудов.
ЛИМФООБРАЩЕНИЕ
Лимфатическая система обеспечивает отток тканевой жидко-
сти от органов и тканей. Тканевая жидкость, всосавшаяся в лим-
фатические сосуды, называется лимфой. Образование лимфы про-
исходит в результате фильтрации жидкой части крови через стен-
364
ки капилляров. Фильтрация обусловлена разницей гидростатиче-
ского и онкотического давления в кровеносных капиллярах и тка-
невой жидкости. Увеличение гидростатического давления крови в
капиллярах способствует образованию лимфы, а повышение он-
котического давления — препятствует. Фильтрация жидкости из
крови происходит в артериальном конце капилляра, а возвращается
она в кровь в венозном конце. В венозных концах капилляров и в
венулах гидростатическое давление крови составляет 15 — 20 мм
рт. ст., а онкотическое давление остается таким же — около
25 мм рт. ст. (поскольку белки не проходят в тканевую жидкость).
Поэтому часть тканевой жидкости, в основном низкомолекуляр-
ные вещества, всасывается в венозные капилляры и мелкие вену-
лы. Тканевая жидкость образуется непрерывно, но количество ее
зависит от уровня обмена веществ в данном органе. Стенки ка-
пилляров обладают активной избирательной способностью, про-
пуская одни вещества и не пропуская другие. Лимфатическая сис-
тема начинается в виде замкнутых слепых меш-
ков сетью лимфатических капилляров, кото-
рые образуют анастомозы между собой. В эти
капилляры поступает тканевая жидкость.
Лимфатические капилляры образуют лим-
фатические сосуды, в которых имеются клапа-
ны, пропускающие лимфу по направлению к
сердцу. Вся оттекающая по ним лимфа собира-
ется в поясничную цистерну, грудной лимфа-
тический проток, в правый лимфатический
ствол, трахеальные протоки и по ним посту-
пает в краниальную полую вену. Лимфатиче-
ская система выполняет дренажную функцию,
отводя в кровь тканевую жидкость (рис. 4.5).
По ходу лимфатических сосудов имеется
большое количество лимфоузлов. Лимфа про-
ходит через лимфоузлы, где обогащается лим-
фоцитами. Эти узлы служат одновременно и
фильтром, в котором задерживаются и разру-
шаются попавшие в лимфу бактерии и ино-
родные тела.
Передвижение лимфы обеспечивает ряд
факторов: разность давления в лимфатических
капиллярах и грудном протоке, присасываю-
щее действие грудной полости во время вдо-
ха; сокращение мышц тела, ритмические со-
кращения стенок лимфатических сосудов и
капсулы лимфатических узлов. Лимфа движется
непрерывно, но очень медленно. Скорость дви-
жения ее составляет 270 — 300 мм в минуту.
Рис. 4.5. Схема взаи-
моотношений кро-
веносных и лимфа-
тических капилля-
ров:
/ — кровеносный ка-
пилляр; а — б — пере-
ход артериального ка-
пилляра в венозный;
2 — лимфатический
капилляр; 3 — клетки;
4 — волокна соеди-
нительной ткани
365
Давление лимфы в лимфатических сосудах равно 8—10 мм, а в
грудном протоке — 4 мм вод. ст.
Лимфатические сосуды иннервируются симпатическим отде-
лом нервной системой, ее возбуждение вызывает их сокращение.
Скорость течения лимфы изменяется рефлекторно под влиянием
болевых раздражений, а также при раздражении рецепторов ка-
ротидных синусов.
Количество лимфы в организме в среднем составляет 50 мл/кг
массы тела.
Контрольные вопросы
1. Что такое круги кровообращения?
2. Назовите свойства сердечной мышцы.
3. Что такое сердечный цикл?
4. Опишите тоны сердца.
5. Расскажите о проводящей системе сердца.
6. Что такое артериальный и венозный пульс?
7. Как измеряется давление крови?
8. Расскажите об особенностях движения венозной крови и лимфы.
ДЫХАНИЕ
Дыхание — совокупность процессов, обеспечивающих обмен
О2 и СО2 между внешней средой и тканями организма. Комплекс
органов, участвующих в процессе дыхания, называют системой
органов дыхания. В основе дыхания лежат тканевые окислительно-
восстановительные процессы. Дыхание животных состоит из сле-
дующих процессов: внешнее дыхание — обмен газов между внеш-
ней средой и альвеолами легких; транспортировка газов кровью —
перенос кровью О2 от легких к тканям и СО2 от тканей к легким;
клеточное дыхание — потребление О клетками и выделение ими
СО2.
Внешнее дыхание. Осуществляется в результате ритмических дви-
жений грудной клетки, что обеспечивает поддержание постоян-
ства напряжения диоксида углерода (СОД, концентрации водо-
родных ионов (Н+) и напряжения кислорода (О2) в артериальной
крови.
Механизм вдоха и выдоха. При увеличении объема грудной клет-
ки обогащенный кислородом воздух поступает в легкие — проис-
ходит вдох, при уменьшении — выдох воздуха с большим содер-
жанием СО2. При спокойном вдохе сокращаются мышцы-инспи-
раторы, все ребра приподнимаются вверх и частично смещаются
вперед, объем грудной клетки увеличивается в поперечном и про-
дольном направлении. Расширению грудной клетки спереди назад
способствует и сокращение диафрагмы. Она давит на органы брюш-
366
ной полости, оттесняя их назад. Брюшная стенка при этом выпя-
чивается, по ее движению можно определить частоту дыхания. За
расширением грудной клетки следует расширение легких, кото-
рые легко растягиваются вследствие своей эластичности. Давле-
ние в легких становится ниже атмосферного, и воздух через воз-
духоносные пути входит в легкие.
После прекращения вдоха наступает выдох. Он осуществляется
обычно пассивно вследствие расслабления мышц-инспираторов
и частичном сокращении мышц-экспираторов. Г рудная клетка при
этом возвращается в исходное положение. Расслабляются и мыш-
цы диафрагмы, оттесненные при вдохе диафрагмой, органы брюш-
ной полости подаются вперед, диафрагма становится более вы-
пуклой. Объем грудной клетки уменьшается, она сдавливает лег-
кие и выжимает из них воздух — происходит выдох. При усилен-
ном выдохе активно сокращаются все мышцы-выдыхатели и мыш-
цы брюшного пресса.
Давление в грудной полости отрицательное. Образование отри-
цательного давления в полости обусловлено особенностями эмб-
рионального развития.
Отрицательное давление в плевральной полости обеспечивает
расширение легких. Если нарушить целостность грудной стенки
так, чтобы воздух вошел в плевральную полость, то атмосферное
давление на наружную и внутреннюю поверхность легких сравня-
ется, легкие сразу же спадутся и не будут следовать за дыхатель-
ными движениями грудной клетки. Это явление называют пневмо-
тораксом. При двустороннем пневмотораксе дыхание становится
невозможным, животное погибает от удушья.
Типы и частота дыхания. Различают три типа дыхания: ребер-
ное, или грудное, брюшное, или диафрагмальное, и грудобрюш-
ное. Грудное дыхание совершается главным образом сокращением
межреберных мышц; брюшное — за счет сокращения диафрагмы.
Когда при дыхании в равной степени участвуют диафрагма, меж-
реберные и брюшные мышцы, оно относится к грудобрюшному
типу. Первый тип дыхания наблюдается преимущественно у собак.
У сельскохозяйственных животных преобладает грудобрюшной тип
дыхания, но при беременности он заменяется на грудной.
Частота дыхания в минуту у животных имеет видовые особен-
ности. Частоту дыхания определяют по движению грудной или
брюшной стенки, а у лошадей и кролика — по движению крыль-
ев носа.
Жизненная емкость легких. В покое животные вдыхают и выды-
хают определенный объем воздуха. Этот объем называют дыха-
тельным воздухом. У овец и крупных собак он составляет в сред-
нем 0,3 —0,5, у лошадей — 5 — 6 л. После спокойного вдоха жи-
вотные могут вдохнуть дополнительно еще некоторый объем воз-
духа. Данный объем называют дополнительным воздухом, у овец
367
и собак он равен 0,5— 1 л, у лошади — 10—12 л. Воздух, который
животное может еще выдохнуть после спокойного выдоха, назы-
вают резервным. Его объем приблизительно равен дополнитель-
ному. Сумма объемов дыхательного, дополнительного и резервно-
го составляет жизненную емкость легких. У собак она равна 1,5 —
3 л, у лошадей — 26 — 30, у крупного рогатого скота — 30 —35 л
воздуха.
После максимального выдоха в легких у животных остается еще
некоторое количество воздуха. Его называют остаточным возду-
хом. У лошади он составляет около 12 л. Остаточный воздух входит
в легкие при первом вдохе новорожденного животного, и легкие
никогда уже не освобождаются от него, даже после смерти. Жиз-
ненная емкость легких и остаточный воздух составляет общую
емкость легких.
Легочная вентиляция. Циркуляция воздуха при вдохе и выдохе
называется легочной вентиляцией. Умножая объем отдельного вдоха
на число дыханий в минуту, можно рассчитать минутный объем
вентиляции легких. У лошади минутный объем в покое равен 40 —
60 л, при движении — 100—120 л, а при очень напряженной
работе может дойти до 500 л.
Газообмен происходит только в альвеолах. В них поступает только
70 % вдыхаемого воздуха, а 30 % его остается в дыхательных пу-
тях.
Воздух, заполняющий альвеолы, называют альвеолярным; в
основном он состоит из остаточного и резервного, поэтому его
объем значительно больше вдыхаемого воздуха. В результате этого
при каждом вдохе и выдохе альвеолярный воздух заменяется не
полностью, а частично, примерно на 1/5 —1/6 его объема.
Газообмен в легких и тканях. В легких О2 переходит из альвео-
лярного воздуха в кровь, а СО2 из крови поступает в легкие. Это
происходит за счет разницы парциального давления О2 и СО2 в
альвеолярном воздухе и крови. Парциальным давлением называ-
ется та часть общего давления, которая приходится на долю каж-
дого газа в газовой смеси.
Диоксид углерода в венозной крови имеет напряжение 47 мм
рт. ст., а в альвеолярном воздухе его парциальное давление 40 мм
рт. ст., вследствие чего СО2 будет выходить из крови в альвеоляр-
ный воздух. В тканях кровь отдает О2 и поглощает СО2. Клетки
усиленно поглощают кислород, поэтому его напряжение в клет-
ках очень низкое и может быть даже равно нулю. В притекающей
артериальной крови напряжение О2 100—105 мм рт. ст., и О2 пу-
тем диффузии переходит в клетки. Напряжение СО2 в клетках
может достигать 60 мм рт. ст., а в притекающей к тканям артери-
альной крови — 40 мм рт. ст., вследствие этого СО2 переходит в
тканевую жидкость, из нее в кровь, которая становится веноз-
ной.
368
Перенос газов кровью. Кровь переносит О2 от легких к тканям и
СО2 от тканей к легким. В крови эти газы могут находиться в двух
состояниях: физически растворенном и химически связанном.
Основной переносчик О2 — гемоглобин крови. Он вступает в
соединение с О2, образуя оксигемоглобин', 1 г гемоглобина может
связать 1,34 мл кислорода.
В альвеолярном воздухе парциальное давление кислорода 100—
110 мм рт. ст., при этих условиях 97 % гемоглобина связывается с
О2. В виде оксигемоглобина он кровью приносится к тканям, где
оксигемоглобин отдает тканям О2 и превращается в восстанов-
ленный гемоглобин.
В венозной крови содержится 55 — 58 об. % СО2, из этого коли-
чества около 3 об. % растворено в плазме крови, а остальная его
часть находится в химически связанном виде. Большое количество
СО2 переносится кровью в виде угольной кислоты (Н2СО3) и ее
солей: бикарбонатов калия и натрия (КНСО3 и NaCO3). Некото-
рое количество СО2 связано с гемоглобином, образуя соедине-
ние карбогемоглобин.
Венозная кровь поступает в легкие. Здесь в капиллярах бикар-
бонаты калия и натрия распадаются, высвобождая угольную кис-
лоту. Под влиянием фермента карбоангидразы угольная кислота
расщепляется на воду и углекислый газ, который выделяется лег-
кими в атмосферу.
Клеточное дыхание. Использование О2 клетками и образова-
ние СО2 в результате окисления углеводов, жиров и белков на-
зывается тканевым дыханием. При окислении питательных ве-
ществ в клетках не происходит прямого присоединения к ним О2.
Их окисление начинается с отнятия водорода от окисляемых ве-
ществ и передачи его через ряд реакций О2, в результате чего
образуются вода и СО2. Эта реакция происходит при участии же-
лезосодержащих дыхательных ферментов. Кроме того, в восстано-
вительно-окислительных процессах в клетках принимает участие
аскорбиновая кислота (витамин С). Реакция окисления, которая
происходит в организме, протекает при температуре тела и в
водной среде.
Регуляция дыхания. Дыхание животных меняется в зависимос-
ти от их состояния. Оно спокойное и редкое в состоянии покоя и
учащается и углубляется при физических нагрузках (рис. 4.6).
Рефлекторная регуляция. Дыхание регулируется дыхательным
центром, расположенным в ретикулярной формации продолгова-
того мозга, в области дна четвертого мозгового желудочка. Он яв-
ляется парным образованием и состоит из двух отделов: центра
вдоха и центра выдоха. Дыхательный центр находится постоянно в
состоянии активности. Изменение дыхания происходит рефлек-
торно. Автоматизм дыхательного центра регулируется нервными
импульсами, приходящими к нему от рецепторов самих легких,
369
Рис. 4.6. Схема регуляции дыхания:
/ — кора больших полушарий; 2 —
дыхательный центр продолговатого
мозга; 3 — центр диафрагмального
нерва; 4 — центры межреберных не-
рвов в спинном мозге; 5 — межре-
берные нервы; 6 — диафрагмальный
нерв; 7 — диафрагма; 8 — легкие; 9 —
волокна блуждающего нерва, несу-
щие импульсы от рецепторов легких;
10 — пути импульсов от кожных и
других рецепторов
сосудистых рефлексогенных зон, дыхательных мышц. Вдох реф-
лекторно вызывает выдох, а выдох, в свою очередь, рефлекторно
стимулирует вдох.
Гуморальная регуляция. На состояние дыхательного центра ока-
зывает влияние химический состав крови. Накопление угольной
кислоты в крови новорожденного, когда прерывается связь плода
с организмом матери, является причиной его первого вдоха.
Частота и глубина дыхания зависят от интенсивности обмена
веществ. Дыхание при мышечной работе становится не только более
частым, но и более глубоким. Вентиляция легких увеличивается.
Мышечная работа сопровождается также усилением кровообра-
щения и повышением содержания гемоглобина в крови. Вслед-
ствие увеличения обмена веществ при мышечной работе возрас-
тает образование СО2 и молочной кислоты в мышцах и поступле-
ние их в кровь. Такое состояние называется кислородной задолжен-
ностью. Окисление образовавшейся молочной кислоты заверша-
ется уже после окончания работы.
Особенности дыхания у птиц. Дыхание у птиц отличается от
дыхания млекопитающих животных, что обусловлено особеннос-
тью строения их дыхательного аппарата (рис. 4.7). У птиц в акте
вдоха участвует сильно развитая грудная клетка. Реберная стенка,
смещаясь назад и вниз, увеличивает объем грудной полости. В лег-
кие засасывается воздух, который проникает и в воздухоносные
мешки. При выдохе сжимаются грудная полость и воздухоносные
мешки (под воздействием грудных и брюшных мышц), и воздух
из них проходит через легкие в обратном направлении.
Таким образом, воздух через легкие проходит дважды — при вдо-
хе и при выдохе. За один акт вдоха и выдоха дважды извлекается О2
370
Рис. 4.7. Схема расположения лег-
ких и воздухоносных мешков птиц:
/ — межключичный мешок; 2 — под-
мышечные дивертикулы; 3 — шей-
ные мешки; 4 — ход в плечевую кость;
5 — краниальные грудные мешки;
6 — легкие; 7 — каудальные груд-
ные мешки; 8 — брюшные мешки;
а — преддверие главного бронха с
отверстиями во вторичные бронхи;
б — медиальные края легких; в — эк-
тобронх в брюшные мешки; г, д —
мешковые бронхи
из воздуха и дважды происходит отдача СО2. Важное значение воз-
духоносные мешки имеют в процессах терморегуляции, спо-
собствуя охлаждению тела и внутренних органов. Во время полета
грудная клетка неподвижна и воздух засасывается воздухоносны-
ми мешками при взмахах крыльев.
Частота дыхания у разных видов птиц неодинакова (в 1 мин):
куры — 20 — 40; утки — 50 — 75; гуси — 15 — 25; индюшки — 12 —
14; голуби — 40 — 60.
Контрольные вопросы
1. Каково значение дыхания для организма?
2. Опишите механизм вдоха и выдоха.
3. Расскажите о составе вдыхаемого и выдыхаемого воздуха.
4. Какова жизненная емкость легких?
5. Как происходит обмен газов между альвеолярным воздухом и кро-
вью?
6. Что такое регуляция дыхания?
7. Каковы особенности дыхания у птиц?
ПИЩЕВАРЕНИЕ
Пищеварение — это совокупность механических, физико-хи-
мических и биологических процессов, обеспечивающих расщеп-
ление поступающих с кормом сложных питательных веществ на
относительно простые соединения, которые могут быть ассими-
лированы организмом. Большинство белков, жиров и углеводов
371
являются высокомолекулярными соединениями, которые без пред-
варительной подготовки не могут всасываться из пищеваритель-
ного канала в кровь и лимфу, а следовательно, быть усвоены клет-
ками и тканями организма. В пищеварительном канале белки рас-
падаются на аминокислоты, жиры — на глицерин и жирные
кислоты, углеводы — на поли- или моносахариды. Хотя общие
принципы пищеварения одинаковы для всех видов животных,
структура и форма отделов их пищеварительного тракта существен-
но различаются, что и обусловлено характером питания.
Органы пищеварения выполняют следующие основные функ-
ции-. секреторную, двигательную, всасывательную, выделитель-
ную, эндокринную, защитную и рецепторную.
Секреторная функция состоит в выделении пищеварительны-
ми железами в пищеварительный канал соков: слюны, желудоч-
ного, поджелудочного, кишечного соков, слизи и желчи. Двига-
тельная функция осуществляется жевательной и мимической мус-
кулатурой ротовой полости, мускулатурой органов пищеварения
и обеспечивает прием корма, его перемешивание и продвижение
по пищеварительному каналу. Всасывательная функция выполня-
ется слизистой оболочкой пищеварительного канала и обеспечи-
вает поступление воды и расщепленных питательных веществ до
простых соединений в кровь и лимфу. Выделительная функция со-
стоит в том, что органы пищеварения выделяют из организма не-
которые продукты обмена (желчные пигменты, мочевину и др.),
минеральные и случайно попавшие в организм вещества. Эндок-
ринная функция заключается в синтезе и выделении биологиче-
ски активных веществ и гормонов. Защитная — защита организма
от вредных агентов (бактерицидное, бактериостатическое и де-
зинтоксикационное действие). Рецепторная — осуществление не-
рвных связей. В пищеварительном тракте находятся рецепторные
поля многих рефлекторных дуг висцеральных систем и соматиче-
ских рефлексов.
Основные типы пищеварения. Различают три основных типа
пищеварения: внутриклеточное, внеклеточное и мембранное. У мле-
копитающих внутриклеточное пищеварение свойственно только
лейкоцитам — фагоцитам крови.
У высших млекопитающих основными типами пищеварения
являются полостное и пристеночное.
Роль ферментов в пищеварении. Ферментативная система пи-
щеварительного тракта включает: а) ферменты пищеварительных
секретов, выделяемых внутристенными или застенными пищева-
рительтными железами; б) ферменты, образуемые микроорганиз-
мами пищеварительного тракта; в) ферменты, содержащиеся в
растительных кормах.
Ферменты являются биологическими катализаторами белковой
природы, которые осуществляют переваривание питательных ве-
372
Рис. 4.8. Схема операции изолированного желудочка по И. П. Павлову:
А — схема разреза желудка; Б — изолированный желудочек, образованный в
результате операции; 1 — линия разреза; 2 — нервы, идущие к изолированному
желудочку; 3 — участок желудка, из которого образуется изолированный желу-
дочек; 4 — полость большого желудочка; 5 — полость изолированного желудоч-
ка; 6 — брюшная стенка; 7 — пробка; 8 — трубка (фистула) для вытекания сока
ществ. Ферменты пищеварительных соков подразделяются на три
группы: протеазы — расщепляют белки (протеины), липазы —
расщепляют липиды и амилазы — расщепляют углеводы.
Методы изучения функций органов пищеварения. Наиболее со-
вершенным и объективным методом исследования функции пи-
щеварительных органов считается павловский метод, дополнен-
ный современными методиками.
Н.П.Павлов разработал метод хронических опытов, с помо-
щью которых удалось установить основные закономерности в де-
ятельности органов пищеварения (рис. 4.8). Были разработаны ме-
тодики выведения протоков слюнных желез, поджелудочной же-
лезы, создания изолированного желудочка. Павловские методики
с некоторыми усовершенствованиями применяются и сейчас при
изучении пищеварения у животных.
ПИЩЕВАРЕНИЕ В РОТОВОЙ ПОЛОСТИ
Пищеварение в полости рта состоит из трех этапов: приема
корма, собственно ротового пищеварения, глотания.
Прием корма. Животные разных видов принимают корм и пьют
воду по-разному. Это зависит от подвижности и формы губ, стро-
ения языка, формы верхней и нижней челюстей, от строения зу-
бов. Лошадь, овца, коза имеют подвижные губы и при приеме
зерна захватывают его губами, траву подрезают резцами и при
помощи языка направляют в ротовую полость. У коров губы ко-
роткие и менее подвижны; корм захватывается языком. Свиньи из
корыта захватывают корм зубами и языком. Плотоядные живот-
373
ные захватывают корм резцами и клыками. Животные пьют воду,
насасывая ее через небольшую щель у середины губ после их смы-
кания. Плотоядные лакают воду и жидкую пищу языком.
Слюна и слюноотделение. Слюна — это продукт секреции трех
пар застенных слюнных желез: околоушных, подъязычных, ниж-
нечелюстных и пристенных: губных, щечных, язычных, которые
открываются в ротовую полость многочисленными протоками. Для
изучения деятельности слюнных желез И. П. Павлов разработал
на собаках методику наложения хронических фистул протоков
слюнных желез.
В состав слюны входят: вода, органические вещества (белок —
муцин и др.), неорганические вещества (хлориды, сульфаты, кар-
бонаты натрия, калия, кальция).
Слюна — вязкая жидкость слабощелочной реакции с плотнос-
тью 1,002— 1,012: содержит 99 — 99,4 % воды и 0,6— 1 % сухих ве-
ществ; pH слюны жвачных 8,1 —8,3, лошади и свиньи — 7,2 —7,5.
В слюне свиньи имеются ферменты птиалин и мальтаза. Птиалин
расщепляет полисахарид, крахмал до дисахарида мальтозы, а маль-
таза превращает мальтозу в глюкозу. В слюне лошади, жвачных и
плотоядных этих ферментов нет.
Слюна, смачивая корм, облегчает процесс жевания. Посред-
ством муцина она склеивает и обволакивает пищевой ком, облег-
чает его проглатывание. Своими щелочными основаниями слюна
нейтрализует кислоты желудка.
Слюноотделение у животных различных видов. У лошади слюно-
отделение происходит при приеме корма и его пережевывании. Она
жует корм попеременно то на одной, то на другой стороне, поэто-
му и слюна выделяется больше на той стороне, на которой проис-
ходит жевание. В течение суток у нее выделяется до 40 л слюны.
У свиньи слюна отделяется периодически, т.е. при приеме корма и
его пережевывании; за сутки выделяется до 15 л слюны. У жвачных
околоушные железы выделяют слюну постоянно. Подчелюстные и
подъязычные железы выделяют слюну периодически — при при-
еме корма и жвачке. У крупного рогатого скота за сутки выделяется
90— 190 л, у овец — 6 — 10 л слюны. В слюне жвачных органические
вещества составляют 0,3 %, неорганические — 0,7 %, ее концент-
рация способствует нормализации биотических процессов в пред-
желудках. Обильное количество слюны, поступающей в рубец, ней-
трализует кислоты, образующиеся при брожении клетчатки. У со-
бак слюна выделяется только при приеме корма.
Регуляция слюноотделения. Слюноотделение происходит реф-
лекторно. Когда в ротовую полость попадает корм, он раздражает
вкусовые рецепторы. Возбуждения от рецепторов по центростре-
мительным нервам передаются в продолговатый мозг, где нахо-
дится центр слюноотделения, а оттуда — по центробежным не-
рвам к слюнным железам. В центробежных нервах имеются волок-
374
ua вегетативной нервной системы. Парасимпатические нервы вы-
зывают образование в большом количестве жидкой слюны, сим-
патические — небольшое количество густой слюны.
Глотание. Глотание — сложнорефлекторный акт. Пережеванный
и смоченный слюной корм превращается в скользкий пищевой
ком, движением языка и щек он направляется в глотку. Корм,
раздражая слизистую корня языка и глотки, вызывает рефлектор-
ное сокращение мышц глотки и мягкого нёба, а корень языка
прижимает надгортанник к гортани, поэтому при глотании ком
не попадает в верхние дыхательные пути, а проглатывается. Даль-
нейшее продвижение пищевого кома осуществляется перисталь-
тическими движениями пищевода. Центр глотания находится в
продолговатом мозге.
ПИЩЕВАРЕНИЕ В ЖЕЛУДКЕ
В однокамерном желудке корм подвергается химической обра-
ботке желудочным соком, перемешивается и продвигается в ки-
шечник вследствие сокращений мышц желудка.
Состав желудочного сока. Желудочный сок — бесцветная, про-
зрачная жидкость кислой реакции, содержащая органические и
неорганические вещества. Он выделяется железами слизистой обо-
лочки желудка — главными, добавочными и обкладочными. Клетки
главных желез образуют ферменты, добавочных — слизь, обкла-
дочных — соляную кислоту.
Чистый желудочный сок собаки — это бесцветная прозрачная
жидкость кислой реакции, плотностью 1,002— 1,006. Желудочный
сок состоит из воды, органических веществ (ферменты, амино-
кислоты), минеральных солей и соляной кислоты. В желудочном
соке содержатся ферменты: пепсин, химозин (сычужный фермент),
желатиназа и липаза. Пепсин выделяется в неактивной форме в
виде пепсиногена, а затем активируеася соляной кислотой; он рас-
щепляет белки до альбумоз и пептонов, а у взрослых животных
створаживает молоко. Химозин, или сычужный фермент, содер-
жится в основном у молодняка, действует на белок молока —
казеиноген, превращая его в казеин, в результате чего происхо-
дит свертывание молока. Желатиназа действует на белок соедини-
тельной ткани — желатин, разжижая его. Липаза расщепляет жиры
•за глицерин и жирные кислоты, однако она действует только на
эмульгированный жир.
Регуляция желудочного сокоотделения. Желудочный сок выде-
ляется в две фазы: рефлекторную и гуморальную.
Рефлекторная фаза — когда корм возбуждает рецепторы рото-
той полости, от которых импульс по афферентным нервам пере-
дается в центр пищеварения, расположенный в продол!оватом
375
мозге. Далее по эфферентным волокнам блуждающего нерва воз-
буждение передается ганглиозным клеткам, расположенным в
стенке желудка.
Гуморальная фаза — когда выделение желудочного сока проис-
ходит под влиянием химических раздражителей.
Эвакуация из желудка начинается тогда, когда оно становится
полужидким или жидким. Скорость эвакуации зависит от степени
наполнения двенадцатиперстной кишки: при растяжении ее хи-
мусом эвакуация временно задерживается. Блуждающий нерв ус-
коряет, а симпатический тормозит эвакуацию.
Пищеварение в желудке жвачных. У жвачных в преджелудках —
рубце, сетке и книжке — корм подвергается механическим и био-
логическим воздействиям. За счет энергичных сокращений рубца
и сетки корм хорошо перетирается и размельчается. Растительные
ферменты частично расщепляют белки, жиры и особенно углево-
ды. В рубце происходит брожение с образованием пропионовой,
уксусной и масляной кислот. Эти летучие жирные кислоты всасы-
ваются в кровь и служат исходным материалом для образования
гликогена, откладываемого в печени.
В рубце жвачных имеется огромное количество анаэробных мик-
роорганизмов'. бактерий, инфузорий и грибков. Общее количество
бактерий может достигать в 1 г рубцового содержимого до 100
млрд, а инфузорий — до 1 млн. Все симбионтные бактерии —
анаэробы. В рубце переваривается до 50 % сухого вещества рацио-
на. Температура в рубце поддерживается в пределах 38 — 42 °C. Под
действием целлюлозолитических бактерий в преджелудках жвач-
ных расщепляется до 60 — 70% переваримой клетчатки. Бактерии
расщепляют клетчатку до дисахаридов и моносахаридов, которые
подвергаются в рубце различным видам брожения.
Микроорганизмы рубца синтезируют витамины группы В и
витамин К. Поэтому взрослые животные при сбалансированном
кормлении не нуждаются в наличии этих витаминов в рационе.
Однако молодняк, у которого рубец еще не функционирует, дол-
жен получать указанные витамины с кормом.
Значение микроорганизмов состоит не только в том, что они
расщепляют корм в рубце. За счет микроорганизмов жвачные по-
лучают за сутки около 100 г полноценного белка.
В процессе жизнедеятельности микроорганизмов в рубце обра-
зуются газы: углекислый газ, метан, азот, водород, сероводород.
Они используются в реакциях, в результате которых образуется ряд
ценных питательных веществ. У крупного рогатого скота за сутки
может образоваться до 1000 л газов в зависимости от вида корма.
Наибольшее количество газов образуется при скармливании зеле-
ных сочных кормов. Избыток газов рубца, не используемый микро-
организмами, в основном удаляется при отрыжке, частично вса-
сывается в кровь и выделяется через легкие при дыхании.
376
Сетка является органом сортировки и отрыгивания корма. Че-
рез сетку проходит только измельченная разжиженная масса, а
грубые частицы остаются в рубце для дальнейшего переварива-
ния.
Между листочками книжки задерживаются недостаточно из-
мельченные частицы корма, прошедшего через сетку. При сокра-
щении книжки происходит дальнейшее измельчение частиц кор-
ма. В книжке всасываются летучие жирные кислоты и вода.
Моторика преджелудков. В преджелудках корм многократно пе-
ремешивается вследствие периодических сокращений рубца, сет-
ки, книжки и продвигается в сычуг. Сокращение рубца можно
исследовать, прощупывая его рукой в области левой голодной ямки.
В норме рубец сокращается 2 — 4 раз в 2 мин.
Сокращения преджелудков согласованы между собой и проис-
ходят в определенной последовательности: вначале сокращается
сетка, затем преддверие рубца, дорсальный и вентральный меш-
ки рубца. Во время сокращения дорсального мешка отрыгиваются
газы, а при сокращении вентрального мешка и слепых выступов
находящаяся в них жидкость выдавливается и омывает содержи-
мое других отделов.
Сетка сокращается в два приема каждые 30 — 60 с: сначала напо-
ловину, затем она слегка расслабляется, после чего сокращается
полностью. При сокращении сетки грубые крупные частицы содер-
жимого выталкиваются обратно в рубец, а измельченная и полу-
жидкая масса поступает в книжку, а затем в сычуг. Во время сокра -
щения сетки расширяется сычуг, в нем создается отрицательное
давление и жидкая масса насасывается из книжки в сычуг. Грубые
частицы поступают в межлисточковые пространства книжки, при
сокращении которой измельчаются. Книжка сокращается в попе-
речном и продольном направлениях. Сокращается она медленно:
обычно на два сокращения сетки приходится одно сокращение
книжки. В книжке интенсивно всасывается вода, растворенные в
ней органические вещества и минеральные соли, а уплотненное
содержимое переходит в сычуг. В сычуге оно снова разбавляется, но
уже желудочным соком, содержащим соляную кислоту.
Функция рубцово-сеткового желоба. У молодняка жвачных мо-
локо, минуя рубец и сетку, переходит из пищевода в книжку по
так называемому рубцово-сетковому желобу, образованному склад-
ками слизистой оболочки. Этот желоб начинается у конца пище-
вода (кардиального отверстия) и заканчивается у отверстия меж-
ду сеткой и книжкой. Сюда же открывается и отверстие в сычуг.
Края желоба могут рефлекторно смыкаться и образовывать труб-
ку, по которой жидкость попадает непосредственно в сычуг. Усло-
вием для рефлекторного смыкания желоба является акт сосания.
Рецепторы, раздражение которых вызывает этот рефлекс, нахо-
дятся в слизистой оболочке языка, губ и глотки. Если края желоба
377
смыкаются недостаточно, а жидкости поглощено много, она по-
ступает в преджелудки. Рефлекс смыкания краев желоба выражен
с первого дня жизни теленка и до 2 — 2,5 мес. К 6 —8-му мес этот
рефлекс постепенно теряет свое значение.
Пищеварение в сычуге. Сычуг является истинным желудком. В нем
выделяется сычужный сок, его pH 1,5 —2,5. В сычужном соке со-
держатся ферменты: пепсин, химозин и липаза. Секреция сы-
чужного сока происходит непрерывно, так как в сычуг постоянно
поступает рубцовое содержимое. Прием корма усиливает секре-
цию желез сычуга. За сутки у коров образуется 40 — 80 л сычужно-
го сока.
ПИЩЕВАРЕНИЕ В ТОНКОЙ КИШКЕ
Кормовые массы, частично переваренные в желудке, поступа-
ют в тонкую кишку. Здесь они смачиваются поджелудочным и ки-
шечным соками и желчью.
Поджелудочный сок вырабатывается поджелудочной железой
и через проток поступает в двенадцатиперстную кишку. У крупно-
го рогатого скота за сутки его выделяется 6 —7 л, у свиней — 8 л.
Поджелудочный сок — прозрачная бесцветная жидкость щелоч-
ной реакции, pH 7,2 —8,0; он содержит ферменты, расщепляю-
щие белки, углеводы и липиды.
Методы изучения секреции поджелудочного сока. Секрецию под-
желудочного сока изучают в острых и хронических опытах. При
острых опытах собаке под наркозом в проток поджелудочной же-
лезы вводят канюлю, соединенную с регистратором. Хронические
опыты на собаках разработал И. П. Павлов. Вырезают небольшой
участок двенадцатиперстной кишки вместе с впадающим в него
протоком поджелудочной железы. С обоих концов изолированный
участок зашивают и вставляют в него фистульную трубку. Сшива-
ют концы перерезанной кишки и вставляют в нее фистульную
трубку. Обе фистульные трубки выводят наружу и соединяют меж-
ду собой резиновой трубкой (рис. 4.9). Во время опыта снимают
резиновую трубку с фистульной трубки, вставленной в изолиро-
ванный участок кишки, и собирают поджелудочный сок.
Желчь. Секрет печени — желчь поступает в полость двенадца-
типерстной кишки. Образование и выделение желчи изучают в
хронических опытах, накладывая фистулы на желчный пузырь и
желчный проток. В состав желчи входят вода, соли желчных кис-
лот (гликохолевая и таурохолевая), желчные пигменты: билиру-
бин и биливердин, а также холестерин, лецитин, мочевина, мо-
чевая кислота и другие вещества.
Желчь эмульгирует жир, образуя мельчайшие капельки, что
облегчает действие липазы на жир. Желчь способствует нейтрали-
378
(ации кислого содержимого, поступающе-
го в кишечник из желудка, усиливает дви-
жения кишечника.
Желчеобразование является не только
секреторным, но и экскреторным процес-
сом, в результате которого выводятся из
организма желчные пигменты, холестерин.
мочевина, пуриновые основания и др.
Печень вырабатывает желчь непрерыв-
но. Она накапливается в желчном пузыре
и отсюда поступает в кишечник в период
пищеварения. У лошади, верблюда и оле-
ня желчного пузыря нет, его функцию
выполняют желчные ходы, имеющие боль-
шие размеры.
Блуждающие нервы усиливают выделе-
ние желчи, а симпатические — тормозят.
В сутки выделяется желчи у лошади 6 — 7 л,
у крупного рогатого скота — 7 — 9, у овец
и коз — 1 — 1,5, у свиней — 2,4 — 4 л. Ко-
личество и качество желчи зависит от со-
става рациона.
Состав кишечного сока. Для изучения
секреции кишечного сока вырезают учас-
ток кишки. Один конец отрезка зашивают
Рис. 4.9. Схема оператив-
ного выведения прото-
ка поджелудочной же-
лезы:
/ — фистульная трубка в
двенадцатиперстной киш-
ке; 2 — фистульная труб-
ка в изолированном отрез-
ке двенадцатиперстной ки-
шки; 5 — резиновая труб-
ка, которой соединяются
фистульные трубки вне пе-
риодов получения сока;
4 — проток поджелудоч-
ной железы; 5— брюшная
стенка; 6— поджелудочная
железа; 7 — место соеди-
нения перерезанной киш-
ки
наглухо, другой выводят наружу и вшива-
ют в кожу. Концы перерезанного кишечника сшивают для восста-
новления целости пищеварительного тракта (рис. 4.10). По другой
методике в кожу вшивают оба конца кишечного отрезка. А. Д. Си-
нещеков предложил изучать кишечное пищеварение при помощи
методики внешних анастомозов. По этой методике кишку перере-
зают, концы наглухо зашивают и в оба участка кишки вставляют
фистульные трубки, которые выводят наружу и соединяют между
собой резиновой трубкой. Образуется обходной пут ь — внешний
анастомоз — для перехода содержимою из вышележащего отдела
в нижележащий.
Методика внешних анастомозов позволяет’ расчленять кишеч-
ник на отдельные части и изучать процессы пищеварения в каж-
дом отделе.
Кишечный сок — бесцветная, слегка мутноватая жидкость
щелочной реакции (pH 8,2 — 8,7). В нем содержатся ферменты, дей-
ствующие на промежуточные продукты расщепления белков и
углеводов. Смесь пептидаз (полипептидазы и дипептидазы) рас-
щепляет полипептиды и дипептиды до аминокислот. Дисахариды
до моносахаридов расщепляются ферментами мальтазой, инвер-
тазой и лактазой. В небольшом количестве имеются амилаза, ли-
379
Рис. 4.10. Схема наложения внешних анастомозов на кишечник:
1 — желудок; 2 — тонкая кишка; 3 — толстая кишка; 4, 5 — кишечные анастомозы
паза и нуклеаза. В кишечном соке содержится также энтерокиназа,
превращающая трипсиноген в трипсин, и щелочная фосфатаза,
обеспечивающая процессы фосфорилирования углеводов и ами-
нокислот и их всасывание.
Полостное и пристеночное пищеварение. В результате перевари-
вания питательных веществ корма и смешивания его с пищевари-
тельными соками содержимое тонкой кишки приобретает вид
однородной жидкой массы, которую называют химусом. Общее
количество химуса составляет у лошадей 190 л, у крупного рога-
того скота — 150, у свиней — 50, у овец — 15 — 20 л.
В тонкой кишке расщепляется значительная часть полипепти-
дов, полисахаридов, жиров, нуклеиновых кислот. Крупные час-
тицы питательных веществ расщепля-
ются в полости кишечника до более
мелких — полостное пищеварение. По-
следние соприкасаются со слизистой
оболочкой кишечника, где также идет
процесс переваривания питательных ве-
ществ. Он называется пристеночным, или
мембранным, пищеварением.
Пристеночному пищеварению спо-
собствует строение слизистой оболочки
тонкой кишки (рис. 4.11). На поверхнос-
ти ворсинок имеется огромное количе-
Рис. 4.11. Схема взаимоотношения поло-
стного (/) и пристеночного (//) пище-
варения в полости кишки:
1 — микроворсинки; 2 — ферменты (на по-
верхности микроворсинок они строго ори-
ентированы); 3 - микроб (по своим разме-
рам он не может проникнуть в поры щеточ-
ной каймы); 4 — пищевые вещества на раз-
ных стадиях расщепления
380
сгво микроворсинок. Между ними есть небольшие пространства, в
которых находятся ферменты, фиксированные на мембране клеток
микроворсинок. В результате движений кишечника происходит не-
прерывное перемешивание химуса и его соприкосновение с мик-
роворсинками. Пищевые частицы, размеры которых меньше рас-
сеяния между микроворсинками, поступают в пространства меж-
ду ними и здесь подвергаются пристеночному перевариванию.
У жвачных животных пристеночное пищеварение происходит
не только в кишечнике, но и в преджелудках под влиянием фер-
ментов, выделяемых бактериями.
Нервная регуляция движения кишечника осуществляется блуж-
дающими и симпатическими нервами. Блуждающие нервы усилива-
ют движения, а симпатические тормозят.
ПИЩЕВАРЕНИЕ В ТОЛСТОЙ КИШКЕ
Невсосавшаяся часть химуса из тонкой кишки переходит в тол-
стую кишку. Этот переход у жвачных регулируется особым клапа-
ном, а у лошади, свиньи и собаки — сфинктером. Сфинктер и
клапан рефлекторно периодически открываются в результате раз-
дражения рецепторов вышележащего отдела кишки и химус посту-
пает небольшими порциями. Если толстая кишка переполнена, сфин-
ктер плотно закрывается и задерживает химус в тонкой кишке.
Сок толстой кишки содержит в основном слизь и небольшое
количество слабо активных ферментов. Пищеварение здесь проис-
ходит преимущественно за счет ферментов, принесенных с химу-
сом из тонкой кишки, а также под влиянием бактерий. В толстой
кишке находится огромное количество бактерий (до 15 млрд в 1 г
содержимого), которые разрушают клетчатку, сбраживают угле-
воды, разлагают белки и жир. В результате образуются летучие
жирные и другие кислоты, различные газы: сероводород, метан,
углекислый газ, а при гниении белков — ядовитые продукты: фе-
нол, крезол, индол, скатол.
Всасывание. Этот физиологический процесс происходит в ре-
зультате активной деятельности клеток слизистой оболочки ки-
шечника. В клетках во время всасывания усиливается обмен ве-
ществ.
Всасывание углеводов. Углеводы всасываются в основном в ки-
шечнике в виде моносахаридов - ииокозы, галактозы, фруктозы.
Различные моносахариды усваиваю'1ся с различной скоростью,
быстрее всасываются глюкоза и галактоза.
Всасывание белков. Белки всасываются в кишечнике после их
расщепления до аминокислот. Аминокислоты всасываются актив-
но, т.е. с затратой энергии. Кроме аминокислот, в тонкой кишке
могут всасываться низкомолекулярные полипептиды и дипепти-
.381
ды. Некоторые белки частично всасываются без расщепления, это
происходит в основном у новорожденных животных. Например, у
них без изменения всасываются глобулины молозива, и в резуль-
тате этого организм получает готовые иммунные тела.
Всасывание жиров. В пищеварительном тракте расщепляется
примерно 30 —45 % всего количества жира, поступающего с кор-
мом. Поэтому всасывание жира происходит как в виде продуктов
его расщепления — глицерина и жирных кислот, — так и в виде
нерасщепленного эмульгированного жира. Глицерин хорошо ра-
створяется в воде и поэтому легко всасывается. Жирные кислоты
вступают в соединение с желчными кислотами и образуют комп-
лексное соединение, которое растворяется в воде и поэтому вса-
сывается. Всосавшийся жир в основном поступает в лимфатиче-
ские сосуды и лишь в небольшой части — в кровеносные.
Всасывание воды и минеральных веществ. Вода всасывается во
всех отделах пищеварительного тракта, но основное ее всасыва-
ние происходит в тонкой и толстой кишках. Вместе с водой вса-
сываются и растворенные в ней минеральные вещества.
Формирование кала и дефекация. В дистальном отделе толстой
кишки в результате всасывания воды содержимое кишечника сгу-
щается и начинается формирование кала. Он состоит из неперева-
ренных остатков корма, отмерших клеток слизистой кишечника,
минеральных веществ, холестерина. Более 30 % сухого вещества
кала составляют отмершие микроорганизмы и некоторая часть
живых. Крупный рогатый скот за сутки выделяет до 40 кг, ло-
шадь — до 17, овца — около 3 кг кала. Каловые массы накаплива-
ются в заднем отделе толстой кишки.
Выход из прямой кишки закрыт двумя сфинктерами: внутрен-
ним, состоящим из гладкой мускулатуры, и наружным — из по-
перечноисчерченной. При дефекации сфинктеры открываются.
Дефекация — выбрасывание кала через анальное отверстие — яв-
ляется сложнорефлекторным актом. Она происходит при участии
центральной нервной системы в результате безусловных и услов-
ных рефлексов. Центр дефекации находится в поясничной части
спинного мозга. Акту дефекации помогает сокращение диафраг-
мы и мышц брюшной стенки.
ПИЩЕВАРЕНИЕ У ДОМАШНЕЙ ПТИЦЫ
В ротовой полости птицы корм не задерживается, быстро про-
глатывается и поступает в зоб. У птиц есть небольшие слюнные
железы. Слюны выделяется мало. Она содержит фермент птиалин
и слизь, которая облегчает проглатывание корма. У кур, индеек и
других зерноядных птиц хорошо развит зоб. У гусей и уток вместо
зоба имеется веретенообразное расширение пищевода.
382
В зобу корм смешивается со слизью, выделяемой зобными же-
лезами. Здесь перевариваются углеводы, белки и жиры под влия-
нием ферментов растительных кормов и микроорганизмов. Мяг-
кий корм быстро переходит в желудок, твердые зерновые кор-
ма — медленнее.
Желудок птиц состоит из двух отделов — железистого и мышеч-
ного. Корм в железистом отделе желудка не задерживается, а про-
ходит в мышечный отдел. Слизистая оболочка его выделяет кол-
лоидный секрет, который превращается в роговую пленку — ку-
тикулу. Она предохраняет стенки желудка от повреждений. Птица
заглатывает мелкие камешки и другие твердые предметы, кото-
рые способствуют перетиранию корма при сокращении желудка.
В мышечном отделе желудка происходит и переваривание корма
соком, выделяемым железистым отделом желудка. Этот сок выде-
ляется постоянно, имея сложнорефлекторную и гуморальную фазы
секреции. При показе птице корма или если она видит другую
птицу, клюющую корм, секреция сока у нее усиливается. Гумо-
ральная фаза секреции желудочных желез связана с поступлени-
ем в кровь продуктов переваривания белка.
Сок железистого желудка содержит соляную кисло ту и фер-
мент пепсиноген, который переходит в пепсин под влиянием соля-
ной кислоты. Соляная кислота расщепляет белки до пептонов.
Пищеварение в кишечнике. У птиц кишечник по отношению к
длине тела короче, чем у млекопитающих. Корм проходит через
кишечник быстро. В тонкой кишке происходит переваривание бел-
ков, жиров и углеводов. В двенадцатиперстную кишку поступает
поджелудочный сок щелочной реакции, содержащий те же фер-
менты, что и у млекопитающих. Печень у птиц большая, желчи
образуется и выделяется много. Кишечные железы выделяют сек-
рет, ферменты которого в пищеварении имеют небольшое значе-
ние. В желудочно-кишечном тракте у птиц нет ферментов для пе-
реваривания клетчатки; она частично переваривается микроорга-
низмами в слепой кишке. В кишечнике птиц корм перемешивает-
ся посредством маятникообразных движений и ритмичсского сег-
ментирования.
Пищеварение и всасывание у птиц весьма интенсивное. В пи-
щеварительном тракте у кур корм находи гея менее 4, а у цып-
лят — 2 — 3 ч. У птиц толстая кишка заканчивается расширенным
отделом — клоакой. Каловые массы выделяются вместе с мочой,
они полужидкие. На поверхности кала образуется белая пленка из
кристаллов мочевины.
Контрольные вопросы
1. Расскажите о сущности пищеварения и основных функциях орга-
нов пищеварения.
2. Что такое полостное и пристеночное пищеварение?
383
3. Каковы особенности приема корма и пищеварение в ротовой поло
сти у разных животных?
4. Каковы особенности слюноотделения у животных?
5. Перечислите состав и свойства слюны и желудочного сока.
6. Опишите пищеварение в однокамерном желудке.
7. Расскажите о пищеварении в многокамерном желудке.
8. Охарактеризуйте моторику преджелудков и функцию рубцово-сег
кового желоба.
9. Каковы особенности пищеварения в тонкой кишке?
10. Перечислите особенности пищеварения в толстой кишке.
11. Что такое жвачный процесс?
12. Каковы особенности пищеварения у птиц?
ОБМЕН ВЕЩЕСТВ И ЭНЕРГИИ
Обмен веществ и энергии — единый процесс. Источником энер-
гии являются углеводы, жиры и белки. Благодаря обмену веществ,
происходит расщепление и синтез молекул, входящих в состав
клеток, образование, разрушение и обновление клеточных струк-
тур и межклеточного вещества. Например, синтез одной молеку-
лы белка в клетке длится всего 3 — 4 секунды.
ОБМЕН ВЕЩЕСТВ
Обмен веществ принято подразделять на общий, межуточный
и основной, а также на углеводный, белковый, жировой, мине-
ральный и водный.
Общий обмен — это обмен веществ и энергии между внешней
средой и организмом при обычных условиях жизни.
Межуточный обмен — это внутриклеточный обмен: совокупность
химических превращений веществ в клетках тканях и органах.
Основной обмен — это энергетические затраты организма в строго
определенных условиях (относительный покой, постоянная темпе-
ратура, очищенный кишечник).
Обмен веществ складывается из двух взаимосвязанных и одно-
временно протекающих в организме процессов — ассимиляции и
диссимиляции, или анаболизма и катаболизма.
Ассимиляция — это процесс усвоения организмом питательных
веществ, поступающих из внешней среды.
Диссимиляция — это процесс распада сложных органических
веществ организма на более простые химические соединения; со-
провождается освобождением энергии и образованием конечных
продуктов метаболизма.
Ассимиляция и диссимиляция неразрывно связаны между со-
бой и составляют единый процесс обмена веществ и энергии. Все
384
реакции обмена осуществляются, прежде всего, на клеточном
\ ровне и регулируются ферментами. В основе автоматической ре-
|уляции обмена веществ лежит принцип обратной связи, когда
концентрация вещества определяет направленность химических
процессов.
Часть энергии используется для построения новых клеток, рас-
ходуется в процессе их жизнедеятельности, например для сокра-
щения мышц, а часть ее освобождается в виде тепла.
При превращениях углеводов, жиров и белков формируются
особые химические соединения, накапливающие запас энергии —
макроэрги. В организме роль макроэргов выполняют в основном
различные фосфорные соединения, главным образом АТФ — аде-
иозинтрифосфорная кислота. При отщеплении одного остатка фос-
форной кислоты АТФ превращается в АДФ — аденозиндифос-
форную кислоту с выделением большого количества энергии,
используемой в процессе жизнедеятельности. В АТФ концентри-
руется 60 —70 % энергии. АТФ рассматривается как универсальный
посредник, обеспечивающий перенос химической энергии от пи-
тательных веществ к метаболическим процессам, требующим ее
затрат.
Обмен углеводов. Углеводы — основной источник энергии в
организме: при окислении 1 г углеводов выделяется 4,1 ккал теп-
ла. Некоторые углеводы, соединяясь с белками и липидами, обра-
зуют структурные компоненты клеток. Углеводы содержатся в ра-
стительных кормах в виде полисахаридов (глюкоза, фруктоза). Они
всасываются из кишечника в виде глюкозы. Глюкоза расходуется в
организме для энергетических целей, откладывается в печени и
мышцах в форме гликогена, а в жировых депо превращается в
жир. Гликоген и жнр являются запасным энергетическим матери-
алом.
Уменьшение содержания глюкозы в крови ниже нормы назы-
вается гипогликемией, а увеличение — гипергликемией. При ги-
погликемии появляется мышечная слабость, понижается темпе-
ратура тела, нарушается деятельность центральной нервной сис-
темы, возникают судороги, и животные могут погибнуть. Гипер-
гликемия может возникать после приема корма, богатого глюко-
зой и сахарозой. Избыточное количество глюкозы в крови выво-
дится почками, появление ее в моче называется глюкозурией.
Расщепление углеводов в организме с освобождением энергии
может происходить как без участия О2 — анаэробное расщепление,
так и с его участием — аэробное расщепление.
При анаэробном расщеплении углеводов образуется молочная
кислота, которая затем при участии О2 окисляется до воды и СО2,
либо снова превращается в гликоген. Важнейшим процессом окис-
ления углеводов в тканях животных является их аэробное расщеп-
ление, при котором конечными продуктами являются СО2 и Н2О.
I 3 ?е .ц? цс >.ц,и
385
При этом полностью освобождается заключенная в углеводах энс.р
гия, которая в основном накапливается в АТФ. Гормоны подже
лудочной железы — инсулин и глюкагон регулируют окисление,
глюкозы в тканях, синтез гликогена в печени и мышцах.
Обмен белков. Белки, или протеины, — сложные высокомоле
кулярные органические соединения, построенные из аминокис
лот. Белки в обмене веществ занимают особое место, они являют
ся главной составной частью живого вещества и материальной
основой процессов жизнедеятельности.
В состав белков входят С, О2, Н, N, иногда S, Р, Fe. Молекула
белка состоит из десятков и сотен аминокислот. Структура белко-
вых молекул животных специфична и свойственна только данно-
му животному. В пищеварительном тракте белки расщепляются до
аминокислот и при этом лишаются специфических свойств. Из
аминокислот, принесенных кровью к клеткам, синтезируются
белки, свойственные уже данному животному.
Аминокислоты, идущие на построение белков организма, не-
равноценны. Одни из них являются заменимыми, другие — неза-
менимыми. К заменимым относятся те аминокислоты, которые мо-
гут синтезироваться в организме из других аминокислот. Незаме-
нимыми называются такие кислоты, которые не синтезируются в
организме. К ним относятся: валин, изолейцин, лейцин, лизин,
метионин, треонин, триптофан, фенилаланин. Если этих амино-
кислот нет в корме, то в организме нарушается обмен веществ,
синтез белков, некоторых гормонов и т.д. Животное постепенно
худеет и, в конце концов, погибает.
Центральная нервная система регулирует обмен белков через
железы внутренней секреции: щитовидные, половые, надпочеч-
ники (см. раздел «Железы внутренней секреции»).
Биологическая ценность белков. Белки и корма, содержащие
все незаменимые аминокислоты, называются полноценными. К ним
относятся животные белки (молоко, мясо, яйца). В большинстве
растительных белков (рожь, пшеница, овес, кукуруза, горох)
некоторые незаменимые аминокислоты отсутствуют или находят-
ся в очень малом количестве. Такие белки не обеспечивают всех
потребностей животного организма, и они называются неполно-
ценными. Поэтому при составлении рациона для животных и птиц
необходимо учитывать аминокислотный состав кормов.
Обмен липидов. Липиды — это общее название для жира и жи-
роподобных веществ.
Жиры состоят из одной молекулы глицерина и трех молекул
жирной кислоты. У разного вида животных состав жира, точка его
плавления, содержание различных жирных кислот неодинаковы.
Жиры имеют большое значение в организме. Они входят в состав
клеток (цитоплазма, ядро, клеточные мембраны), являясь их струк-
турной частью.
386
Жир служит основным источником энергии в организме. При
окислении 1 г жира выделяется 9,3 ккал тепла. С жирами в орга-
низм поступают растворимые в них витамины A, D, Е, К.
Жиры в организме животных составляют 10 —20 % живой мас-
сы, а при откорме — 30% и более.
Жиры могут образовываться ИЗ углеводов и белков. Однако жиры
корма нельзя заменять полностью углеводами и белками, так как
такие жирные кислоты, как линолевая, линоленовая и арахидо-
новая, в организме не синтезируются. При их недостатке у живот-
ных нарушается половая функция, снижается эластичность сте-
нок кровеносных сосудов, нарушается обмен жиров.
Регуляция жирового обмена осуществляется центральной не-
рвной системой и железами внутренней секреции. Центры регуля-
ции находятся в гипоталамусе, они оказываю г свое влияние на
жировой обмен через вегетативную нервную систему. Симпати-
ческие нервы усиливают распад, а парасимпатические — синтез
жира. Деятельность гипоталамуса контролируется корой больших
полушарий.
Взаимосвязь обмена белков, углеводов и жиров. Обмен белков,
углеводов и жиров имеет специфические особенности, но наряду
с этим существуют и общие закономерности. В процессе обмена
белков, углеводов и жиров образуется пировиноградная кислота,
которая является общим продуктом их обмена. Эта кислота может
служить продуктом для синтеза углеводов и жиров.
В процессе обмена из аминокислот образуются углеводы и жиры,
из углеводов — жиры, а из жира — углеводы. В процессе обмена
белков, углеводов и жиров образуется энергия: 60 — 70% ее на-
капливается в аденозинтрифосфорной кислоте (АТФ). 30 40 %
превращается в тепловую энергию, которая выделяется из орга-
низма во внешнюю среду в процессе теплоотдачи.
Обмен воды и электролитов. Все биохимические реакции в орга-
низме идут в водных растворах. Вода обеспечивает основу внутри-
клеточного обмена. Внутри клеток заключено 71 % всех водных
запасов организма. Внеклеточная вода находится в крови, лимфе,
спинно-мозговой жидкост и и составляет 10%, а в межклеточном
пространстве — 19 %. Вода в организме находится в виде солевых
растворов, что обусловливает тесную связь водного обмена с об-
меном минеральных веществ. После исключения воды из рациона
животных они погибают через несколько суток. Вода и минераль-
ные соли создают внутреннюю среду организма, являясь состав-
ной частью плазмы, лимфы и тканевой жидкости. Они участвуют
в поддержании осмотического давления и реакции крови.
Обмен воды тесно связан с обменом электролитов. Для биоло-
гических мембран (оболочка клеток, стенки капилляров) харак-
терна полупроницаемость, т.с. они проницаемы для воды и не-
проницаемы для крупных моллекул. При повышении осмотиче-
387
ского давления вода легко проникает через этот участок и проис-
ходит выравнивание концентраций осмотически активных веществ.
Вода участвует в регуляции температуры тела; испаряясь, она ох-
лаждает тело и предохраняет его от перегрева.
Потребность в воде у животных неодинаковая, она зависит также
и от вида корма. При сухих кормах воды потребляется больше. На
каждый 1 кг сухого вещества корма корова потребляет 4 —6 л
воды, лошадь и овца — 2 — 3, свинья — 7 —8 л. Регуляция водного
обмена происходит центральной нервной системой и железами
внутренней секреции.
Минеральный обмен. Роль минеральных веществ в организме
разнообразна. С ними связаны перенос газов и секреция пищева-
рительных желез. Они составляют основу костной ткани, участву-
ют в процессах обмена веществ, поддерживают кислотно-щелоч-
ное равновесие, создают осмотическое давление, возбудимость
нервной и мышечной тканей. Они входят в состав гемоглобина,
сложных белков — металлопротеидов, содержащих атомы метал-
лов (Fe, Mg, Си, Zn, Со, Мп и др.).
Минеральные вещества обеспечивают процессы роста, размно-
жения, поддержания физиологического равновесия и продуктив-
ности животных, поскольку они участвуют во всех жизненных про-
цессах организма: дыхании, работе сердца и мышц, деятельности
нервной системы и др. В минеральных веществах особенно нужда-
ются молодые, растущие и высокопродуктивные животные. В орга-
низм они поступают с пищей и водой.
Химические элементы, содержащиеся в организме в значитель-
ных количествах, называются макроэлементами, другие — в не-
значительных количествах — называются микроэлементами. К мак-
роэлементам относят Na, К, Cl, Са, Р, Fe, Mg, S.
Натрий и калий. Ионы натрия и калия оказывают влияние на
возбудимость нервной системы, на сердечную деятельность. В ос-
новном благодаря NaCl поддерживается нормальное осмотическое
давление крови. Он необходим для процессов роста. Калий уча-
ствует в транспорте СО2 кровью. Натрия в растительных продуктах
мало, поэтому травоядные должны получать достаточное количе-
ство NaCl, однако избыточное поступление Na тоже вредно, осо-
бенно для птицы и свиней. Нормальная деятельность организма
возможна при соотношении Na: К = 1 : 2. Любые отклонения при-
водят к нарушению деятельности сердца, кишечника, мышечной
и нервной ткани.
Кальций вместе с фосфором составляет основную массу кост-
ной ткани. В основном (99%) он находится в костях в виде фос-
форных и углекислых солей. Кальций, помимо чисто механиче-
ской функции, необходим для многих процессов жизнедеятель-
ности. Так, Са участвует в процессах свертывания крови, стимули-
рует сердечную деятельность, влияет на проницаемость клеточ-
388
ной мембраны для натрия и калия, участвует в процессе мышеч-
ного сокращения. Кальций понижает возбудимость нервной сис-
1емы, поэтому при его недостатке в крови у животных наступают
судороги. Особо нуждаются в Са молодняк и лактирующие живот-
ные, выделяющие с молоком много его соединений. Кальций есть
ко всех кормах, но в грубых кормах его больше.
Фосфор. Обмен Р тесно связан с обменом Са. Соотношение Са
и Р в рационе должно быть примерно 2 или 1,5:1. Кальций и фос-
фор составляют в теле животных 65 —70 % всех минеральных со-
единений. Фосфор необходим для нормального межуточного об-
мена. Соли фосфорной кислоты входят в состав всех клеток и меж-
клеточных жидкостей, они имеются в различных белках, липидах
и участвуют в процессах их обмена. Фосфор является важнейшей
частью нуклеиновых кислот, входит в состав аденозинтрифосфор-
ной кислоты и креатинфосфата, в которых аккумулируется энер-
гия, образующаяся при обмене веществ. Фосфор — активный ка-
тализатор и стимулятор обменных процессов в организме.
Сера входит в состав белков, аминокислот, гормона инсулина,
витаминов В] (тиамина) и биотина. Особую роль она играет в
формировании шерстного покрова. Соединения серы в организме
участвуют в детоксикации, связывая ядовитые вещества — фено-
лы, индоксилы и др. продукты обмена. Сера поступает в организм
вместе с белками корма, выводится с мочой, калом и потом у
овец.
Хлор — важнейший анион в составе жидкостей организма. Ани-
оны С1 — непременные участники процессов возбуждения ЦНС.
Он участвует в образовании соляной кислоты желудочного сока.
Участвует в транспорте СО2 кровью, в водном обмене.
Железо входит в состав гемоглобина, миоглобина (мышечного
гемоглобина), ферментов, участвующих в тканевом дыхании.
В организме железо находится в соединении с белками и отклады-
вается в печени, селезенке и слизистой оболочке кишечника. При
недостатке железа нарушается образование эритроцитов, что при-
водит к возникновению у животных анемии. Это наблюдается у
молодняка в подсосный период, особенно у поросят, так как в
молоке железа очень мало. 11оэ1ому препараты железа необходимо
давать молодняку в виде подкормки. Потребность в железе у взрос-
лых животных покрывается гем количеством, которое имеется в
кормах. Излишний Са конкурирует с железом, а низкая кислот-
ность желудочного сока снижает усвояемость Fe. Дефицит вита-
мина А и В нарушает процесс всасывания Fe.
Магний — 60 % его находится в костях в виде фосфорнокисло-
го магния, 20% в мышцах в соединении с белками. Остальные
20% находятся в других тканях, больше всего его обнаружено в
печени. Магний участвует в процессе мышечного сокращения,
активирует выработку антител организмом, входит в систему,
389
обеспечивающую, естественную резистентность организма к раз-
личным возбудителям.
К микроэлементам относят Со, J, Си, Мп, Zn, F, Br, Sr и др.
Они принимают участие в росте и развитии животных, способ-
ствуют устойчивости к различным заболеваниям, повышают пло-
довитость и продуктивность.
Кобальт требуется для синтеза витамина В|2, в состав которо-
го он входит. В организм животных поступает с кормом, отклады-
вается главным образом в печени, поджелудочной железе, мыш-
цах. Он необходим для образования эритроцитов и гемоглобина,
для внутриутробного развития плода. Кобальт стимулирует рост
молодняка, повышает молочную и шерстную продуктивность жи-
вотных, улучшает качество спермы. При недостатке кобальта у жи-
вотных развивается анемия, возникают заболевания (лизуха и др.).
Йод является важнейшим компонентом гормона щитовидной
железы — тироксина, роль которого в организме исключительно
велика. Недостаток йода нарушает функцию щитовидной железы,
молодняк рождается слабым, нежизнеспособным. У взрослых жи-
вотных при недостатке йода снижаются продуктивность и плодо-
витость животных. Йод поступает в организм с кормами и водой.
Медь — один из незаменимых микроэлементов для организма.
Она находится в мышцах, костях и печени. В крови медь содер-
жится в эритроцитах и лейкоцитах. Она входит в состав некоторых
ферментов. Главное биологическое значение ее состоит в стиму-
ляции тканевого дыхания, процессов кроветворения и синтеза ге-
моглобина. При недостатке меди у животных нарушается функция
нервной, мышечной и кровеносной систем. У крупного рогатого
скота снижается молочная продуктивность, воспроизводительная
способность, развивается анемия.
Цинк обнаружен во всех органах и тканях, но наибольшее его
количество находится в скелетных мышцах, а также в гипофизе,
половых железах, печени и сперме. Он является составной частью
фермента карбоангидразы, участвующего в процессах дыхания.
Недостаток Zn задерживает рост, нарушает процессы размноже-
ния, рост шерсти, приводит к развитию рахита и остеопороза.
Избыток цинка вызывает у животных тяжелые отравления.
Марганец содержится во всех органах и тканях животных, но
его больше в печени, костях, почках. Он входит в состав некото-
рых ферментов, участвует в окислительно-восстановительных про-
цессах. При дефиците Мп у животных замедляется рост скелета,
нарушаются функция нервной системы, равновесие, животные
не способны к размножению. Избыток марганца также неблаго-
приятно влияет на организм: происходит задержка роста, нару-
шается эмаль зубов, изменения в костях напоминают рахит.
Фтор практически весь в организме входит в состав твердых
тканей (кости, зубы) и спермы. При его дефиците у животных
390
наблюдают задержку роста, снижение плодовитости и продолжи-
тельности жизни, кариес зубов.
Стронций содержится во всех органах и тканях животных, боль-
ше его в костях и зубах. Отсутствие стронция вызывает кариес
зубов, а избыток — стронциевый рахит.
Регуляция минерального обмена. Минеральный обмен тесно свя-
зан с водным обменом. Регуляция минерального обмена осуще-
ствляется гипоталамусом и железами внутренней секреции —
щитовидной, паращитовидной, гипофизом, надпочечниками.
Витамины и их роль в обмене веществ. Витаминами называют
особую группу низкомолекулярных, биологически активных орга-
нических соединений, обеспечивающих нормальные биохимиче-
ские и физиологические процессы в организме.
Витамины открыл в 1881 г. русский ученый Н. И. Лунин, а на-
звание им предложил в 1912 г. польский ученый К. Функ. В насто-
ящее время известно более 30 витаминов, установлена их хими-
ческая структура. Многие витамины входят в состав ферментов,
поэтому жизнь без них невозможна. Некоторые витамины образу-
ются в организме животных из провитаминов, другие — в желу-
дочно-кишечном тракте синтезируются микроорганизмами.
По физико-химическим свойствам витамины делят на две груп-
пы: жирорастворимые и водорастворимые.
Жирорастворимые витамины. К их числу относят: витамин А
(ретинол), витамин D (кальциферол), витамин Е (токоферол),
витамин К (нафтохинон).
Ретинол образуется в организме животных из растительного
пигмента каротина, который является провитамином А. Из каро-
тина в стенке тонкой кишки образуется ретинол. Он участвует в
процессах обмена веществ, поддерживает нормальное состояние
эпителия пищеварительного тракта, дыхательных, мочеполовых
путей, кожного покрова, глаз. При его недостатке эти клетки оро-
говевают. Ретинол участвует в процессах зрения, из него образу-
ется в темноте зрительный пигмент родопсин.
Кальциферол объединяет целую группу витаминов (D2, D3, D4,
Ds, D6). Для животных имеют значение витамины D2 и D3. В орга-
низме витамин D3 образуется из эргостерина под влиянием уль-
трафиолетовых лучей. Витамин D2 синтезируется в высушенной
на солнце траве. Наиболее богат витаминами группы D рыбий жир;
они содержатся также в молоке, сливочном масле, яичном желт-
ке. Витамины группы D регулируют в организме обмен кальция и
фосфора. С фосфорно-кальциевым обменом самым тесным обра-
зом связаны процессы роста и развития. Когда в организме не
хватает витаминов D, минерализация костной ткани нарушается,
останавливаются процессы ее образования и регенерации. При не-
достатке витаминов D у молодняка развивается рахит, а у взрос-
лых — остеомаляция.
391
Токоферол (витамины группы Е) представлен тремя видами,
которые участвуют в обмене жиров, белков, углеводов, способ-
ствуют усвоению витамина А, процессам размножения и разви-
тия зародыша у самок. Витамины Е содержатся в зеленых кормах,
зародышах семян злаков, молоке, масле (коровьем и раститель-
ном), мясе, яйцах. При недостатке этого витамина нарушается
образование спермиев, у самок гибнет плод.
Филлохинон (витамин К) представлен тремя витаминами. Они
содержатся в зеленых частях растений, а у животных участвуют в
образовании протромбина, необходимого для свертывания крови.
У взрослых животных витамины К синтезируются микроорганиз-
мами желудочно-кишечного тракта, а при их недостатке у живот-
ных появляются кровоизлияния в мышцах, кишечнике. Особенно
чувствительны к недостатку витамина К птицы.
Водорастворимые витамины. К ним относится большая группа
витаминов В, витамин С (аскорбиновая кислота), витамин Р (цит-
рин).
Тиамин (витамин Bt) содержится в зернах злаковых, горохе, дрож-
жах. У жвачных и лошади он синтезируется в желудочно-кишечном
тракте и играет важную роль в процессах обмена веществ. Тиамин
образует активную часть ферментов, участвующих в углеводном об-
мене, влияет на обмен ацетилхолина. При его недостатке нарушает-
ся нервная проводимость. Кроме того, в результате накопления не-
доокисленных продуктов обмена углеводов возникают воспаление
нервной системы, судороги, параличи, расстройство движений.
Рибофлавин (витамин В2) содержится в зеленых кормах, дрож-
жах, печени, почках, молоке, яйцах. Рибофлавин необходим для
синтеза ферментов, участвующих в процессах обмена белков и
углеводов, для процессов цветового зрения, для синтеза гемогло-
бина, функции нервной системы и половых желез. От недостатка
рибофлавина чаще страдают свиньи и птицы. У них ухудшается
аппетит, воспаляется слизистая оболочка пищеварительного трак-
та, появляется понос. У крупного рогатого скота рибофлавин син-
тезируется в желудочно-кишечном тракте.
Пантотеновая кислота (витамин В3) широко представлен в
растительных и животных тканях. Наиболее богаты ею печень, яич-
ный желток, почки, надпочечники, сердце, арахис, горох, дрож-
жи, а также зеленые растения и злаки. Она синтезируется микро-
флорой желудочно-кишечного тракта. Пантотеновая кислота яв-
ляется составной частью фермента, который участвует в процес-
сах обмена углеводов, жиров и белков, необходима для синтеза
ацетилхолина, нормальной функции надпочечников. Дефицит ее
у птиц проявляется в виде массовых параличей, а у свиней разви-
ваются дерматит и язвенный колит.
Холин (витамин В4) содержится в зеленых листьях, хлебных
злаках, жмыхах, дрожжах, печени, рыбной и мясной муке, со-
392
евых бобах, арахисе, капусте. Он необходим для образования ме-
диатора ацетилхолина, для предупреждения жирового перерож-
дения печени. Холин принимает участие в процессах роста, улуч-
шает сопротивляемость организма к инфекционным болезням.
Никотиновая кислота (витамин В5, или витамин РР) — анти-
пеллагрический витамин, содержится в молоке, мясе, яйцах, сыре,
бобах, семенах кунжута и подсолнечника, цельном зерне и пивных
дрожжах, пшеничных отрубях, пшенице, ячмене. Синтезируется в
пищеварительном тракте животных, если животные получают бел-
ки, содержащие аминокислоту триптофан. При недостатке витами-
на РР у животных, особенно у свиней и птиц, возникает тяжелое
заболевание пеллагра, которая протекает с явлениями нарушения
функции коры больших полушарий и поражениями кожи.
Пиридоксин (витамин В6) — содержится в печени, мясе, рыбе,
молоке, зернах хлебных злаков, бобовых, жмыхах, картофеле, дрож-
жах. Он участвует в процессе обмена белков, являясь активной ча-
стью ферментов, влияет на процессы кроветворения. При недо-
статке пиридоксина у свиней и птиц появляются дерматиты, ане-
мия, судороги, параличи. У молодняка происходит задержка роста.
Фолиевая кислота (витамин В9) содержится в зеленых листьях
растений, цветной капусте, хлебных злаках, сое, грибах, дрож-
жах, печени. Она входит в состав ферментов, обеспечивающих
эритропоэз, деление и дифференцировку лейкоцитов, предупреж-
дает жировое перерождение печени. При недостатке фолиевой
кислоты у цыплят, индюшат, поросят развивается анемия, задер-
живается рост.
Биотин (витамин Н) содержится в печени, почках, молоке,
зернах хлебных злаков, овощах, дрожжах, частично синтезирует-
ся кишечной микрофлорой. При участии биотина совместно с АТФ
происходят реакции присоединения СО2 к органическим кисло-
там (реакция карбоксилирования). Снижает уровень глюкозы в
крови.
Цианкобаламин (витамин В12) синтезируется в кишечнике у
моногастральных и в рубце у жвачных. Поступая в кровь накапли-
вается в печени, почках и селезенке. В его состав входит кобальт и
цианогруппы. Цианкобаламин участвует в синтезе нуклеиновых
кислот, холина. Он стимулирует синтез белков. Витамин необхо-
дим для образования эритроцитов и гемоглобина. Авитаминоз
может наблюдаться у свиней, птиц и собак. При этом нарушается
белковый обмен, возникают анемия и расстройства функции не-
рвной системы.
Парааминобензойная кислота (витамин Hj) содержится в про-
дуктах растительного и животного происхождения, особенно бо-
гаты ею дрожжи и печень. Она способствует синтезу РНК и ДНК,
входит в состав фолиевой кислоты. При отсутствии этого витами-
на задерживается рост волос и наступает их поседение.
393
Пангамовая кислота (витамин В15) усиливает кислородный
обмен в клетках и тканях, предупреждает жировое перерождение
печени. Содержится в растениях, тканях животных, дрожжах.
Аскорбиновая кислота (витамин С) содержится в плодах ши-
повника, ягодах черной смородины, в помидорах, цитрусовых,
капусте, картофеле, зеленой траве, хвое сосны, листьях березы,
липы, хрене, петрушке и др. растениях. Витамин С синтезируется у
всех животных, за исключением человека, обезьян и морских сви-
нок. Он необходим для синтеза ряда гормонов, ферментов, уча-
ствует в углеводном обмене, обеспечивает нормальную проницае-
мость капилляров, ускоряет заживление ран, повышает сопротив-
ляемость организма к различным инфекциям и неблагоприятным
воздействиям внешней среды, стимулирует образование антител.
Цитрин (витамин Р) содержится вместе с аскорбиновой кис-
лотой в растительных продуктах. В организме он повышает проч-
ность капилляров, нормализует их проницаемость. Витамин Р про-
являет активность только в присутствии аскорбиновой кислоты и
способствует более экономному ее использованию в организме.
Антивитамины. Соединения, которые химически похожи на тот
или иной витамин, но по своему действию обладают противопо-
ложными свойствами, называются антивитаминами. Открыты они
у тиамина, пиридоксина, фолиевой кислоты, биотина и др. Ме-
ханизм действия антивитаминов заключается в конкурентных от-
ношениях с витаминами при образовании ферментов. В ряде слу-
чаев свойства антивитаминов используются в лечебных целях.
ОБМЕН ЭНЕРГИИ
В результате сложных превращений в процессе диссимиляции
потенциальная энергия питательных веществ частично расходует-
ся на процессы ассимиляции, на механическую работу (сокраще-
ние сердца, скелетных мышц и т.п.), электрическую энергию, но
в большей своей части превращается в тепловую энергию. Уста-
новлено, что жиры, белки и углеводы при окислении в организ-
ме дают определенное количество тепла: 1 г жира — 9,3 ккал; 1 г
белка — 4,1 ккал; 1 г углеводов — 4,1 ккал.
Регуляция обмена энергии. Ведущая роль в регуляции обмена
энергии принадлежит коре головного мозга. На обмен энергии
влияют гипоталамус, в котором находятся центры вегетативной
нервной системы: симпатическая нервная система усиливает об-
мен энергии. Гипофиз, щитовидная железа, надпочечники тоже
влияют на обмен энергии; гормон щитовидной железы — тирок-
син, надпочечников — адреналин усиливают его.
Методы изучения обмена энергии. Количество энергии, выде-
ляемой организмом, определяют методами прямой и непрямой
394
калометрии. Прямая калометрия производится с помощью специ-
альных аппаратов — калориметрических камер. В практике широ-
ко применяется непрямая калометрия — метод измерения энергии
по выделенному углекислому газу и потребленному кислороду.
У животных измеряют количество выдохнутого воздуха за опреде-
ленное время, содержание в нем СО2 и О2 и вычисляют дыхатель-
ный коэффициент.
Дыхательным коэффициентом называется объемное отношение
выдохнутого СО2 к потребленному О2. При окислении углеводов
дыхательный коэффициент равен 1; белков — 0,8; жиров — 0,7.
Для вычисления образовавшейся энергии учитывают количество
потребленного О2 или выдохнутого СО2, так как потребление 1 л
О2 или выделение 1 л СО2 соответствует образованию определен-
ного количества тепла.
Контрольные вопросы
1. Охарактеризуйте значение обмена веществ в животном организме.
2. Какова роль белков, жиров и углеводов в обмене веществ?
3. Опишите обмен воды в организме и его регуляцию.
4. Расскажите о минеральном обмене.
5. Какова роль витаминов в организме?
6. Что такое регуляция обмена веществ и энергии?
ТЕПЛОРЕГУЛЯЦИЯ
Поддержание температурного гомеостаза в организме высших
животных осуществляется благодаря деятельности сложного фи-
зиологического механизма, регулирующего теплопродукцию и теп-
лоотдачу. Теплопродукция — процесс химический, а теплоотда-
ча — физический.
Для каждого вида теплокровных животных характерна опреде-
ленная температура тела. Жизнь теплокровных животных возмож-
на в сравнительно узких температурных пределах — от 37 до 42 °C.
Гибель их наступает при снижении температуры ниже 24 °C и подъе-
ме ее выше 44 °C. Из внутренних органов самая высокая темпера-
тура в печени и сердце.
Образование тепла в организме сопровождается его отдачей.
Организм теряет столько тепла, сколько его в нем образуется. Тепло
в теле животных не задерживается, иначе они погибли бы в тече-
ние нескольких часов.
Химическая теплорегуляция. Тепло в теле животных образуется
в результате окисления питательных веществ до конечных про-
дуктов их распада. Примерно 2/3 тепла, образующегося в организ-
ме, приходится на мышцы. В них образование тепла происходит
даже тогда, когда животные находятся в полном покое, так как
395
мышцы сохраняют определенный тонус. Много тепла образуется в
печени, пищеварительном тракте, во время приема корма, при
пережевывании жвачки.
Физическая теплорегуляция. Организм непрерывно выделяет
тепло. Это происходит в основном через поверхность тела, лишь
небольшая часть тепла выделяется с выдыхаемым воздухом, ка-
лом и мочой. Любое нагретое тело теряет тепло, если температура
окружающего воздуха ниже температуры тела. Такой способ отда-
чи тепла называется теплопроводностью. Путем теплопроводнос-
ти животные отдают значительную часть тепла в холодное время
года. Движение воздуха усиливает теплопроводность. Тепло удаля-
ется с поверхности тела животных путем излучения невидимых
инфракрасных лучей. Животные с хорошо развитыми потовыми
железами (лошадь, крупный рогатый скот и овна) отдают тепло с
потом, который в основном состоит из воды, обладающей высо-
кой теплоемкостью. При испарении 1 мл воды теряется примерно
0,58 ккал. В зависимости от количества образующегося тепла и тем-
пературы окружающей среды выделяется различное количество
пота. У собак потовые железы развиты слабо, теплоотдача у них
происходит путем испарения слюны с поверхности языка, из ро-
товой полости, через дыхательные пути.
Коже принадлежит основная роль в теплоотдаче. В результате
раздражения кожи холодом рефлекторно сокращаются подкож-
ные мышцы, поднимаются волосы на теле. Между взъерошенны-
ми волосами образуется воздушная прослойка, и организм задер-
живает тепло. При этом выделение пота на время прекращается.
К зиме животные обрастают густой шерстью, у них увеличивается
отложение жира в подкожной клетчатке. После весенней линьки
шерсть и мех становятся реже, что увеличивает отдачу тепла с
поверхности кожи.
Регуляция температуры тела производится корой больших по-
лушарий, гипоталамусом и железами внутренней секреции. В ги-
поталамусе имеются центры теплорегуляции. Возбуждение центра
теплорегуляции происходит рефлекторным и гуморальным путем.
Контрольные вопросы
1 Перечислите видовые особенности температуры тела у животных.
2. Назовите виды теплорегуляции.
3. Что такое регуляция температуры тела?
ВЫДЕЛЕНИЕ
Заключительным этапом обмена веществ является выведение
из организма продуктов диссимиляции. Живые клетки организма
постоянно выделяют продукты своей жизнедеятельности. Выведе-
396
ние их из организма является обязательным условием жизни, иначе
наступает самоотравление и гибель животного. Выведение продук-
тов жизнедеятельности называется экскрецией, а органы, их уда-
ляющие, называются экскреторными, или выделительными. К ним
относятся почки, потовые железы, легкие, кишечник, слизистые
оболочки. Через легкие выводятся из организма диоксид углерода
(СО2), вода и некоторые летучие вещества. Кишечник выделяет
некоторые соли в составе кала, непереваренные части корма. По-
товые железы выводят воду, соли, некоторые органические ве-
щества.
Основное значение в выделительных процессах принадлежит
почкам, которые выполняют следующие функции: 1) удаляют из
организма конечные продукты белкового обмена: аммиак, моче-
вину, мочевую кислоту, креатинин и др.; 2) поддерживают нор-
мальное содержание в организме воды, минеральных веществ,
глюкозы и других веществ; 3) регулируют pH крови; 4) сохраня-
ют постоянное осмотическое давление; 5) выводят из организма
некоторые вещества, поступившие извне (лекарственные веще-
ства, краски и др.); 6) выполняют важную инкреторную функ-
цию, образуя биологически активные вещества, регулирующие
артериальное давление.
Состав мочи. Вся циркулирующая кровь проходит через почки
за несколько минут. У свиньи массой 90 — 100 кг в течение суток
через почки протекает до 1,5 тыс. л крови. Такой интенсивный
кровоток обеспечивает интенсивный процесс образования мочи.
Ее образование происходит в нефроне. Кровь в почках проходит
через две капиллярные сети: капилляры сосудистого клубочка и
капилляры стенок извитых канальцев. В почке крупного рогатого
скота 8 млн нефронов, овец — 1 млн, у свиней — 1,5 млн, соба-
ки — 816 тыс., а у человека около 2 млн.
Животные выделяют за сутки значительное количество мочи:
крупный рогатый скот — 6 — 12 л, лошади — 3 — 4,5, свиньи —
2 — 4, овцы — 1—1,5, собаки — 0,5—1, кошки — 0,1 —0,2 л. Моча
у лошади мутная, так как содержит много мелких кристаллов
СаСО3. У крупного и мелкого рогатого скота и свиней моча про-
зрачная. Реакция мочи у травоядных щелочная, у свиней бывает и
кислой, что зависит от состава кормов. Плотность мочи колеблет-
ся от 1,018 до 1,040. Составные части мочи те же, что и в плазме
крови, за исключением белков, жиров и глюкозы, которых нет в
нормальной моче.
Образование мочи. Образование мочи идет в две фазы. Первая
фаза — фильтрационная осуществляется в сосудистом клубочке.
Из плазмы крови через стенку кровеносных сосудов клубочков
отфильтровываются в капсулу клубочка под влиянием высокого
давления большое количество воды, минеральные вещества и все
растворимые органические вещества (аминокислоты, глюкоза и
397
др.). Белки не могут пройти через стенку капилляров в полость
капсулы. Такой ультрафильтрат жидкой части крови, не содержа-
щий белков, получил название первичной мочи. Из каждых 10 л
крови, протекающей через капилляры клубочка, отфильтровыва-
ется около 1 л первичной мочи.
Вторая фаза — реабсорбционная, или обратного всасывания.
Безбелковый ультрафильтрат плазмы крови при прохождении по
системе почечных канальцев подвергается значительному изме-
нению и превращается в конечный продукт — конечную мочу.
В системе канальцев происходит обратное всасывание большого
количества воды, а также продуктов межуточного обмена (глюко-
за, аминокислоты и др.); в моче остаются только типичные ее
составные части в обычной для нее концетрации. Для образования
1 л конечной мочи необходимо, чтобы через канальцы почек про-
шло 90 л первичной мочи.
В извитых канальцах также идут процессы синтеза. В них из бен-
зойной кислоты и гликоколя образуется гиппуровая кислота. Много
ее синтезируется у травоядных животных.
Выведение мочи. Почка работает непрерывно, а моча выводит-
ся из организма периодически. Образующаяся моча поступает в
почечную лоханку, из нее по мочеточникам — в мочевой пузырь.
Обратное поступление мочи из мочевого пузыря в мочеточники
невозможно. В месте выхода мочеиспускательного канала имеются
внутренний и наружный сфинктеры. Во время наполнения пузы-
ря мочой сфинктеры сокращены, при мочеиспускании они рас-
слабляются. Мочеиспускание осуществляется рефлекторно. Сфин-
ктеры и мочевой пузырь иннервируются симпатическими и пара-
симпатическими нервами.
Контрольные вопросы
1. Охарактеризуйте выделительные органы.
2. Опишите механизм образования мочи.
3. Расскажите о нефроне и его строении.
4. Как регулируются функции почек?
ФИЗИОЛОГИЯ кожи
Кожа защищает организм от внешних неблагоприятных воз-
действий (от механических повреждений, проникновения микро-
организмов, световых и тепловых воздействий, от излишнего ис-
парения воды). Она функционирует как выделительный орган, депо
крови, воды и солей. В ней имеется много различных рецепторов,
поэтому она является важнейшим анализатором и мощным реф-
лексогенным полем.
398
В коже идут процессы синтеза и распада белков, углеводов и
других органических и неорганических веществ.
Температура кожи обычно на 6 — 8° ниже температуры тела. Цвет
кожи и волос зависит от наличия пигментов гемосидерина (крас-
ный пигмент) и меланина (черный пигмент). Пигментация кожи
играет защитную роль, предохраняя организм от повреждающего
действия ультрафиолетовых лучей.
Секреторная функция кожи связана с отделением пота, кож-
ного сала, а у самок и молока. Выделение пота имеет значение
। лавным образом для процессов теплорегуляции. Потовые железы
участвуют в водно-солевом обмене, способствуя постоянству ос-
мотического давления.
Потовые железы иннервируются симпатическими нервами.
Центры потоотделения находятся в спинном, продолговатом и
промежуточном мозгу. Потоотделительные центры возбуждаются
рефлекторно и гуморально. Стимулятором потоотделения являет-
ся температура окружающей среды. Потоотделение усиливается так-
же и тогда, когда центры омываются кровью, имеющей повы-
шенную температуру.
Секреция кожного сала. Сальные железы расположены вблизи
волос, их протоки открываются в волосяной мешок. Они образу-
ют кожное сало, которое состоит из ненасыщенных жиров и хо-
лестерина. Под влиянием кислот пота кожное сало разлагается с
образованием летучих жирных кислот со специфическим запахом.
Сальные железы иннервируются симпатическими нервами. Кож-
ное сало защищает эпидермис от высыхания и образования тре-
щин, а кожу и волосы — от проникновения воды, делая кожу
мягкой и эластичной. У плода толстая жировая смазка кожи пре-
пятствует пропитыванию тела жидкостью амниона.
Смесь кожного сала и пота называется жиропотом, который
предохраняет волос от намокания и загрязнения, а у овец обеспе-
чивает образование руна. Количество жиропота зависит от породы
и условий кормления животных, от климата. Жиропота больше у
тонкорунных овец. У них количество жиропота составляет от 7 до
30 % к общей массе шерсти.
Волосяной покров животных. Волосы располагаются на всей
поверхности тела животных. Густота и длина волос зависят от вида,
породы, условий содержания, кормления, возраста животных,
сезона года, климата и т.д. У лошадей на 1 см2 кожи находится в
среднем 700 волос, у романовских овец — до 5000, у мериносо-
вых — до 8000, у кроликов породы шиншилла — от 6000 до 12 000.
Волосы у молодых животных растут интенсивнее, чем у старых.
Замена волосяного покрова называется линькой.
Линька бывает постоянной, сезонной и возрастной. При посто-
янной линьке частичное выпадение волос происходит на протя-
жении всего года. Постоянная линька наблюдается у тонкорунных
399
овец. Длинные волосы гривы и хвоста лошадей держатся в течение
5 — 6 лет, смена их происходит в течение всего года по мере ста-
рения волосяных луковиц.
При сезонной линьке волосяной покров меняется полностью
за сравнительно короткое время. У лошадей и крупного рогатого
скота линька бывает 2 раза в год — весной и осенью. Этим они
предохраняют себя от перегревания летом, а зимой — от переох-
лаждения.
Примером возрастной линьки, не зависящей от сезона года,
может служить линька жеребят и телят в 5 — 7-месячном возрасте.
У жеребят только после этой линьки устанавливается постоянная
окраска волосяного покрова (масть).
Контрольные вопросы
I. Каковы основные функции кожи?
2. Расскажите о значении потоотделения для животного.
3. Каково значение сальных желез, жиропот?
4. Перечислите виды линьки.
5. Что такое руно?
ВНУТРЕННЯЯ СЕКРЕЦИЯ
ОБЩАЯ ХАРАКТЕРИСТИКА
ЖЕЛЕЗ ВНУТРЕННЕЙ СЕКРЕЦИИ
Железами внутренней секреции называют органы или группы
клеток, которые способны вырабатывать специфические физио-
логически активные вещества — гормоны, регулирующие различ-
ные функции организма. Гормоны выделяются непосредственно в
кровь или лимфу. Железы внутренней секреции небольшой вели-
чины, имеют богатое кровоснабжение и иннервируются вегета-
тивным отделом нервной системы.
Методы изучения функций желез внутренней секреции. Один из
наиболее старых методов — метод экстирпации — оперативного
удаления какой-либо железы с последующим изучением измене-
ний, которые произойдут в организме в результате такой опера-
ции. Вместо оперативного удаления можно выключать функцию
желез химическими веществами. Например, функцию щитовид-
ной железы можно выключить метилтиурацилом, бета-клетки ос-
тровков Лангерганса поджелудочной железы — аллоксаном. При-
меняется также метод пересадки желез из одного участка тела в
другой у одного и того же животного или от одного животного
другому.
400
Свойства гормонов. По химическому строению гормоны явля-
ются белками, полипептидами, пептидами, йодированными ами-
нокислотами, стероидами, катехоламинами и обладают высокой
биологической активностью. Основное назначение их — участие в
>уморальной регуляции обмена веществ. Они существенно влияют
на рост, дифференцировку, половое созревание, размножение и
другие процессы.
Независимо от химической природы и функции гормоны име-
ют ряд общих свойств: специфичность, высокую биологическую
активность, дистантность действия, высокую проницаемость,
быструю обновляемость и отсутствие видовой специфичности. При
этом каждый гормон оказывает влияние на определенные, чув-
ствительные к нему органы, т.е. органы-мишени.
Механизм действия гормонов. Одни гормоны изменяют прони-
цаемость клеточных мембран отдельных внутриклеточных струк-
тур и внутриклеточные ферментные системы. Другие проявляют
свое действие на уровне генетического аппарата клетки. Прони-
кая в ядро клетки, они влияют на синтез информационной РНК
и таким путем на синтез белка и ферментов. Гормоны могут ока-
зывать свое влияние на органы и ткани путем воздействия на нер-
вную систему, как на нервные центры, так и на рецепторы. Дей-
ствуя на специальные рецепторы, они рефлекторно меняют функ-
цию нервных центров.
Нервная регуляция желез внутренней секреции. В регуляции фун-
кции желез внутренней секреции участвуют кора больших полу-
шарий, промежуточный мозг и другие отделы головного мозга. На
некоторые железы внутренней секреции (мозговой слой надпо-
«Нагрузка»
401
Рис. 4.12. Схема регуляции
функций желез внутренней
секреции:
1 — кора больших полушарий;
2 — внешние раздражители; 3 —
гипоталамус; 4, 5 — обратная
связь периферических желез внут-
ренней секреции, гипофиза и ги-
поталамуса
1 4 Зсзепеиекий
чечников, паращитовидные железы, поджелудочная железа) цен-
тральная нервная система оказывает свое влияние через симпати-
ческие и парасимпатические нервы. Однако на ряд желез внутрен-
ней секреции центральная нервная система влияет через гипо-
физ, функцию которого регулирует гипоталамус (рис. 4.12).
Гипоталамус и гипофиз образуют единую гипоталамо-гипофи-
зарную систему, обеспечивающую переключение нервных импуль-
сов на эндокринную регуляцию функций. В гипоталамусе имеются
скопления нервных клеток, которые не только проводят нервный
импульс, но и выделяют специфические вещества, получившие
название нейросекретов. Одним из путей регуляции функции же-
лез внутренней секреции является система обратной связи пери-
ферической железы внутренней секреции, гипофиза и гипоталамуса.
Система обратной связи состоит в том, что при увеличении коли-
чества гормона какой-либо периферической железы происходит
торможение образования нейросекретов в гипоталамусе, тропных
гормонов в гипофизе и наоборот.
ЖЕЛЕЗЫ ВНУТРЕННЕЙ СЕКРЕЦИИ
Гипофиз. Передняя доля гипофиза выделяет шесть гормонов:
соматотропин (СТГ, гормон роста), тиротропин (ТТГ), корти-
котропин (КТГ), фоллитропин (ФСГ), лютропин (ЛГ), пролактин
(ЛТГ, лактотропный гормон). Все они, за исключением сомато-
тропина, влияют на деятельность других желез внутренней секре-
ции и называются тропными гормонами (см. раздел «Органы внут-
ренней секреции»).
Щитовидная железа. В щитовидной железе образуются три гор-
мона — тироксин (преобладает), трийодтиронин и тиреокальци-
тонин. Гормоны щитовидной железы образуются из аминокисло-
ты тирозина и йода (см. раздел «Органы внутренней секреции»).
Паращитовидные железы. Регулируют концентрацию кальция
и фосфора в крови, стимулируют активность остеокластов — кле-
ток, разрушающих кости (см. раздел «Органы внутренней секре-
ции»).
Надпочечники. Состоят из двух самостоятельных желез: коры и
мозгового вещества. Корковый слой надпочечников выделяет три
группы стероидных гормонов: глюкокортикоиды, минералокорти-
коиды и половые гормоны (см. раздел «Органы внутренней секре-
ции»).
Поджелудочная железа. Железа имеет двойную секрецию: вне-
шнюю (выделяет пищеварительные ферменты) и внутреннюю.
Внутрисекреторную функцию выполняют панкреатические ост-
ровки (Лангерганса). Они занимают от 2 до 10% массы органа,
которые состоят из а- и р-клеток. P-Клетки выделяют инсулин,
402
состоящий из двух пептидных цепей. Он регулирует углеводный,
жировой и белковый обмен. Под влиянием инсулина происходит
процесс окисления глюкозы в клетках, превращение глюкозы в
печени и мышцах в гликоген, а в жировой ткани — в жир.
Половые железы. Стероидные половые гормоны подразделяют-
ся на мужские {андрогены) — тестостерон, андростерон и женские
(эстрогены) — эстрадиол, эстрон и эстриол. Они образуются из
холестерина в семенниках и яичниках. Тестостерон стимулирует
рост и развитие органов размножения и вторичных половых при-
знаков, а также вызывает половое поведение самцов — влечение
к самкам.
Женские половые железы — яичники — синтезируют эстроге-
ны и прогестерон. Эстрогены возбуждают центральную нервную
систему, вызывая у самок состояние половой охоты, обеспечивая
этим спаривание с самцом.
Прогестерон синтезируется в желтом теле, которое образуется
на месте разорвавшегося фолликула. Прогестерон способствует
закреплению оплодотворенной яйцеклетки в роге матки, бере-
менности, развитию железистой ткани в молочной железе. Он тор-
мозит проявление охоты, мышцы матки делает нечувствительны-
ми к окситоцину.
Плацента. Плацента обладает эндокринной функцией и выра-
батывает плацентарные эстрогены, прогестерон, релаксин и пла-
центарный гонадотропин. Прогестерон обеспечивает беременность
вплоть до родов. Плацентарный гонадотропин по своему действию
сходен с гонадотропными гормонами гипофиза, он предохраняет
от самопроизвольных абортов, так как стимулирует синтез проге-
стерона. У лошадей, начиная с 40-го по 120-й день жеребости, в
матке вырабатывается гонадотропный гормон, называемый гона-
дотропином сыворотки жеребой кобылы — СЖК.
Тимус. Гормоны тимуса — тимопоэтин, тимозин стимулирует
развитие Т-лимфоцитов. Тимус как центральный орган иммуни-
тета одновременно является эндокринным органом и способству-
ет выработке антител.
Эпифиз. Его гормоны тормозят преждевременное половое со-
зревание, а также играют существенную роль в работе механизма
«биологических часов»: регулируют периодичность функций орга-
низма в разное время суток. Гормоны эпифиза повышают функ-
циональную активность иммунной системы.
Применение гормонов и гормональных препаратов в животновод-
стве и ветеринарии. В арсенал ветеринарного врача входит большое
количество гормональных препаратов. Гормоны и гормональные
препараты могут быть использованы в целях увеличения поголо-
вья скота, повышения мясной, молочной и шерстной его про-
дуктивности. Например, они могут применяться при трансплан-
тации эмбрионов для улучшения молочного стада, для синхрони-
403
зации половых циклов у самок при искусственном осеменении.
При использовании гормональных препаратов можно добиваться
многоплодия. Окситоцин используют для улучшения сокращений
матки при родах, для отделения последа при его задержании; он
облегчает выведение мумифицированных плодов. В этих же целях
применяют и питуитрин — комплексный препарат, содержащий
гормоны окситоцин и вазопрессин. Получают питуитрин путем
экстракции из задней доли гипофиза.
Применение бетазина и йодистого калия улучшает внутриут-
робное развитие плодов, повышает резистентность и сохранение
новорожденных телят. Гормоны используются для улучшения и
ускорения откорма скота: например, эстрогены способствуют ус-
воению корма и стимуляции роста животных.
Контрольные вопросы
1. Дайте общую характеристику желез внутренней секреции.
2. Назовите механизм действия гормонов.
3. Перечислите железы внутренней секреции.
4. Назовите железы двойной секреции, их функциональное значение.
5. Как используют гормоны в животноводстве и ветеринарии?
РАЗМНОЖЕНИЕ
Половая и физиологическая зрелость самцов и самок. Способ-
ность к размножению и передаче своих качеств потомству обеспе-
чивает преемственность жизни на нашей планете и сохранение
определенной численности видов животных. При ослаблении этой
способности или с ее прекращением виды животных обречены на
вымирание.
Размножение возможно с наступлением половой зрелости,
когда начинают функционировать органы размножения. В яични-
ках самок периодически развиваются фолликулы, в которых со-
зревают яйцеклетки, а у самцов в семенниках развиваются спер-
мин. У половозрелых животных усиливается выделение половых
гормонов и появляется особая форма поведения — половые реф-
лексы. У самок возникают течка и охота. Самец способен оплодот-
ворить самку, а самка может стать беременной и вынашивать плод.
Половая зрелость крупного рогатого скота наступает в 8 —
12 мес, овец и коз — в 7 —8, свиней — в 5 — 8, лошадей — в 16 —
18 мес, т.е. значительно раньше, чем заканчивается физиологи-
ческое созревание всего организма. Физиологическая зрелость те-
лок наступает в 16 — 18 мес, овец — в 12—13, свиней — в 8 —
11 мес, кобыл — в 3 — 4 года. По достижении такого возраста их
можно спаривать. В эти же сроки начинают использовать и сам-
цов-производителей.
404
Физиология органов размножения самцов. К органам размноже-
ния самцов относятся: семенники с придатками, семяпроводы,
придаточные половые железы, мочеполовой канал и совокупи-
тельный орган — половой член. В извитых семенных канальцах
развиваются спермин, этот процесс называется сперматогенезом
(рис. 4.13). Сперма состоит из двух частей: спермиев и плазмы,
являющейся смесью секретов придаточных половых желез. Спер-
мин состоит из головки, шейки, тела и хвоста (рис. 4.14). В головке
находится ядро, а шейка, тело и хвост функционируют как двига-
тельный аппарат спермия. Спермии образуются непрерывно, а вы-
водятся наружу только при спаривании. До этого они хранятся в
придатке семенника.
Сперма быка и барана нейтральная (pH 7,0) или спабокислая
(pH 6,7 —6,9); сперма хряка и жеребца — щелочная (pH 7,2 —7,6).
Спермии гибнут при нарушении осмотического давления в окру-
жающей их среде, а также ее избыточной кислотности или ще-
лочности.
В тазовую часть мочеполового канала открываются протоки при-
даточных половых желез: пузырьковидных, предстательной, лу-
ковичных и мелких уретральных. У хищных животных имеется
только предстательная железа. При совокуплении (половом акте)
секреты придаточных половых желез выделяются в определенной
последовательности. Они разбавляют сперму во много раз, обра-
зуя эякулят, и способствуют продвижению спермиев в половых
путях самки.
Рис. 4.13. Схема спермиогенеза и оогенеза:
А — участок стенки извитого канальца; Б — сперматогенез; В — оогенсц / —
сперматогоний; 2 — сперматоцит I порядка; 3 — сперматоцит II порядка; 4 —
сперматиды; 5 — спермии; 6 — оноюнии; 7 — ооцит 1 порядка; У — ооцит
II порядка; 9— зрелая яйцеклетка; а, b — первое и второе направительные тельца
405
Рис. 4.14. Строение спермиев быка
(А), жеребца (Б), барана (5), хря-
ка (Г); ультрамикроскопическое
строение головки и связующего
отдела (Д):
7 — головка; 2 — связующий отдел;
3 — хвостик; 4 — акросома; 5 — ядро;
6 — проксимальная и 7 — дистальная
центриоли; 8 — осевая нить; 9 — ми-
тохондриальная спираль; 10 — цент-
ральное кольцо; 11 — толстое волокно
Спермин выводятся из хвоста придатка семенника в семяпровод
при половом акте. В канале придатка спермин накапливаются в боль-
ших количествах (20 — 40 млрд у быка). Здесь они претерпевают
дальнейшие морфофункциональные изменения (дозревают) в те-
чение 8 — 20 дней. В кислой бескислородной среде канала придатка
спермин впадают в состояние, подобное анабиозу. Оплодотворяю-
щую способность они сохраняют в придатках семенника до 2 — 3
мес. Порция спермы, выделенная самцом за одну садку, называет-
ся эякулятом, а процесс семяизвержения называется эякуляцией.
Объем эякулята и концентрация в нем спермиев у нормальных
самцов млекопитающих разных видов различны. Промежуточная
ткань семенника вырабатывает половые гормоны.
Физиология органов размножения самок. К органам размноже-
ния самок относятся: яичники, яйцепроводы, матка, влагалище,
мочеполовое преддверие и наружные половые органы — вульва
(половые губы, клитор).
Яичники — парная половая железа самки и железа внутренней
секреции, в которой развиваются яйцеклетки и синтезируются по-
ловые гормоны. Наружный слой яичника с расположенными в нем
фолликулами, находящимися нт разной стадии развития или атри-
зии (гибели), и желтых тел называется корковым слоем. Во внут-
реннем, мозговом слое яичника разветвляются нервы и сосуды.
К моменту рождения в яичниках самок содержатся все оогонии,
из которых впоследствии будут развиваться яйцеклетки. Общее число
оогониев в одном яичнике составляет: у коров — около 140 тыс., у
свиней — 120, у кур — 60—150 тыс. В дальнейшем этот запас не
пополняется. Однако полной зрелости в период размножения жи-
406
вотного достигают лишь некоторые из них. Например, у коровы
за всю продуктивную жизнь созревают не более 300 фолликулов,
т.е. по 1 — 2 на каждый половой цикл, у овец в зависимости от
породы — от 1 до 4 фолликулов; у свиней в обоих яичниках одно-
временно созревает 15 — 20 фолликулов. Остальные фолликулы на
разных стадиях развития подвергаются дегенерации и атрезии.
Яйцеклетка в фолликуле проходит определенные стадии рос-
та, развития и созревания (см. раздел «Эмбриология»), После со-
зревания фолликула (граафова пузырька) происходит овуляция.
На месте разорвавшегося фолликула развивается желтое тело, ко-
торое является временной железой внутренней секреции. Если сам-
ка оплодотворилась, оно сохраняется в течение всей беременнос-
ти. Если оплодотворение не наступило, то, начиная с 10—15-го
дня после овуляции, желтое тело постепенно рассасывается, а в
яичниках развиваются новые фолликулы.
Половой цикл —1 периодически повторяющиеся у половозрелых
самок морфофизиологические процессы, связанные с размноже-
нием. Половой цикл у животных делят на две основные стадии:
одна стадия — течка и половая охота, другая — межтечковый пе-
риод, или половой покой.
Продолжительность полового цикла и его стадий неодинако-
вая у разных видов животных. У кобыл и свиней половой цикл
составляет 20 — 21 день, у коров — 19 —21, у овец — 17 дней.
Течка — процесс выделения слизи из половых органов как след-
ствие морфологических изменений полового аппарата. Во время
течки происходит периодическое сокращение и расслабление
мышц рогов, тела и шейки матки.
Половая охота. Под влиянием эстрогенов повышается возбуди-
мость нервной системы. Самки становятся беспокойными, отка-
зываются от корма. У лактирующих коров уменьшаются удои. Про-
должительность течки и охоты у разных видов животных неодина-
ковая. У коров охота продолжается 10 — 20 ч, овец — 24 — 40, сви-
ней — 48 —72 ч, у кобыл — 5 — 7 дней.
Время овуляции у разных видов животных также неодинаковое.
У коров овуляция обычно происходит через 7— 15 ч после окон-
чания охоты. У овец она наступает через 31—32 ч, у свиней —
через 25 — 40 ч от начала охоты. Овуляция у кобыл происходит за
24 —48 ч до окончания охоты. Во время половой охоты самка до-
пускает самца к спариванию.
У коров, овец и свиней течка и половая охота могут повто-
ряться в течение всего года, но у отдельных видов и пород живот-
ных проявляется сезонность размножения, унаследованная ими
от диких предков. В связи с этим и сроки спаривания — половой
сезон — приходятся на разное время года.
Регуляция полового цикла. Ведущую роль в проявлении функ-
ции размножения выполняет центральная нервная система. Цент-
407
ры этой регуляции расположены в гипоталамусе. В нем образуются
нейросекреты — фоллиберин и люлиберин, они стимулируют
выделение в передней доле гипофиза гонадотропных гормонов фол-
литропина и лютропина. Фоллитропин вызывает рост и развитие
фолликулов, лютропин — разрыв фолликулов. Под влиянием лют-
ропина в развивающихся фолликулах образуется большое количе-
ство эстрогенных гормонов, они вызывают течку и половую охоту
(половую доминанту).
После овуляции на месте разорвавшегося фолликула образует-
ся желтое тело, оно выделяет гормон прогестерон, тормозящий
образование в гипоталамусе люлиберина; в гипофизе уменьшает-
ся выделение лютропина, что снижает выработку в яичниках эст-
рогенных гормонов. Прогестерон также активирует нервные цент-
ры, связанные с материнством. В результате течка и охота прекра-
щаются — наступает стадия полового покоя; при оплодотворении
самки этот покой называется стадией материнской доминанты.
Спаривание. Оплодотворение. Типы осеменения. Спаривание —
сложнорефлекторный процесс, состоящий из ряда половых реф-
лексов: приближения, обнимательного, эрекции, совокупитель-
ного, эякуляции. В их осуществлении участвуют зрительный, обо-
нятельный, кожный и другие анализаторы.
При спаривании у коров, овец и коз сперма попадает во влага-
лище (влагалищный тип осеменения), а у кобыл и свиней — в мат-
ку (маточный тип осеменения). В результате сокращений мышц
Рис. 4.15. Схема строения яйца (А) и развития зиготы (£>'):
/ — стадия двух бластомеров: 11 — стадия четырех бластомеров; III — стадия
морулы; /И — стадия бластоцисты / — фолликулярные клетки яйценосного бу-
горка; 2 — клетки лучистого венца; 3 - прозрачная оболочка; 4 — околожелточ-
ное пространство; 5 — желточная оболочка; 6 — цитоплазма; 7 — ядро; 8 —
прозрачная оболочка яйцеклетки: 9 — бластомеры; 10 — трофобласт; 11 — эмб-
риобласт; Р — полярные тельца
408
спермии быстро оказываются в яйцепроводе, где могут встретить
яйцеклетку (рис. 4.15).
Спермии у коров достигают воронки яйцепровода в течение
2 — 4 мин, у овец — 6, у свиней — через 10—15 мин, у лошадей
они достигают вершины рогов в первые же секунды после спари-
вания. В половых органах коров и овец продолжительность жизни
спермиев 36 — 48 ч, свиней и кобыл — 24 — 48 ч.
У сельскохозяйственных животных широко применяется искус-
ственное осеменение — введение спермы самке без спаривания с
самцом. Искусственное осеменение дает возможность улучшить
племенные качества животных, используя ценных в породном
отношении самцов-производителей (рис. 4.16).
Оплодотворение — процесс проникновения спермиев в цито-
плазму яйца, слияние ядер спермиев и яйца и образование зиго-
ты (см. раздел «Эмбриология»).
Беременность. Состояние самки от оплодотворения до рожде-
ния плода называется беременностью. Для кобыл применяют тер-
мин жеребость, для коров — стельность (рис. 4.17), овец — суяг-
ность, у свиней — супоросность. С наступлением беременности у
самок изменяется функционирование многих органов. Самки ста-
новятся более спокойными, у них улучшается аппетит. В первой
половине беременности они лучше усваивают корм, делаются более
упитанными, однако к концу беременности некоторые из них
худеют. В период беременности изменяется функция некоторых
Рис. 4.16. Схема оплодотворе-
ния и перемещения эмбрио-
на (по Пичману—Кроллиш):
1 — яичник; 2 — желтое тело’;
3 — зачатковый эпителий; 7 -
первичный, 5 — вторичный, 6 -
пузырчатый фолликулы (i раафои
пузырек); 7 — яйценосный бу-
горок; 8 — покрышка фоллику-
ла; 9 — выход яйцеклетки и т
яичника (овуляция); 10 — ворон-
ка яйцепровода; // — яйцепро-
вод; 12 — спермии у яйцеклет-
ки; 13 - оплодотворение; 14 —
эмбрион на стадии двух блас-
томеров, 15 - четырех бласто-
меров; 16 — морула; 17 — плод-
ный пузырск (бластоциста); 18—
рог матки
409
5-
6
Рис. 4.17. Детская и материнская плаценты коровы (по Г.А.Шмидту):
А — плодный пузырь с плодом в возрасте 4 мес; Б — карункул слизистой обо-
лочки матки коровы; й — пупочный канатик; 1 — котилидоны; 2 — пупочные
apiepnn и иена; 3 — пупочный канатик; 4 — крипты карункулов; 5 — брюшная
сгснка; 6 — остаюк желточного пузыря; 7— мочевой проток (урахус)
желез внутренней секреции. Так, в гипофизе больше образуется
соматотропина; желтые тела и плацента вырабатывают больше про-
гестерона. В яичниках самок прекращается рост фолликулов. Во
время беременности в печени накапливается гликоген. Усилива-
ется деятельность сердца, почек, учащается дыхание.
Продолжительность беременности у животных разных видов
неодинаковая (см. раздел «Эмбриология»),
Размножение домашней птицы. Семенники у самцов птиц рас-
положены в брюшной полости. Придаточные половые железы от-
сутствуют. Петух вводит семя, прижимая свою клоаку к клоаке
самки.
Объем эякулята у петуха примерно 0,8 мл, количество сперми-
ев в I мл эякулята составляет 5 — 7 млрд. Спермии длительное
время сохраняются в яйцеводе кур, не теряя оплодотворяющей
способости. Например, куры несут оплодотворенные яйца еще в
течение 20 суток после отсадки самцов, у индеек и гусынь этот
срок более продолжителен. У кур, постоянно находящихся с пету-
хами, все отделы яйцевода наполнены спермиями. Свет стимули-
рует деятельность семенников. После спаривания первые опло-
410
дотворенные яйца курица несет лишь на iprши inn. Uiihuh в
пик яйцекладки напоминает кисть винограда, и ш м « и в р i im ч
от 500 до 3500 ооцитов на разных стадиях pa пип ня
Развиваясь и накапливая желток, яйцеклетки щк npain.> и и
большие шары — яичные желтки 3,5 —4 см и днамснн Hi Mi.p.i < >
вание желтка таких размеров требуется около II /пнш llu нчш р\
ности желтка расположен зародышевый диск (coin пинии ininci.
летка) в виде белого пятнышка 1—2 мм в диаметре < <ир> niniiti
желток после разрыва фолликула проскальзываем и iuipo.ni \ ниш.
вода. Последний, перисталически сокращаясь, iipoiiiuii ас । и .
к клоаке. В процессе движения желток вращается и шн н ш........
покрывается белком, который выделяется железами iiiiiiiiiha ।
В яйцеводе образуется около 40 — 50 % белка, остапынЩ <|к,1рмпр\
ется в матке, являющейся расширенной частью aitiiciuia.i 11> •
образуется белок, подскорлупная оболочка и известковая оПп п>ч
ка яйца — скорлупа. Из матки яйцо переходит в клоаку и выпи
дится путем сокращения мышц наружу.
Продолжительность эмбрионального развития при iiiikMaipii
вании: цыплята выводятся на 21-й день, утята, гусята, ин/iunti.i
та — на 28-й день.
Размножение пушных зверей. Важной биологической ocotlriin<«
стью хищных пушных зверей, сохранившейся в условиях их i и
точного разведения, является сезонность размножения. Пргш ы
вители семейств куньих и псовых моноэстричны и принося! п<>
томство 1 раз в год, тогда как грызуны — нутрии и шиншин.пы
полиэстричны и размножаются круглый год.
Спаривание (гон) клеточных норок, песцов, лисиц, енот
видных собак происходит 1 раз в году — в конце зимы и ранней
весной, у соболей — летом, у хорька фуро — с третьей декады
марта до середины августа. У гибридных хорьков, полученных <н
прямого скрещивания фуро с черным хорьком — тхорзофреток,
которые отличаются большой плодовитостью, можно получит ь по
2 приплода в год. Первый сезон гона у них начинается обычно в
конце марта, второй в конце июня—июля.
Беременность у хищных пушных зверей имеет также свои осо
бенности.
У клеточных песцов и лисиц продолжительность беременное i и
варьирует от 50 до 52 сут, у клеточных норок — от 40 до /<.
енотовидных собак в условиях разведения в неволе — от 58 до (i-l
у хорьков фуро — от 40 до 42 сут, соболей — от 7 до 8 мег
У соболей и норок удлинение сроков беременности свяыно <
эмбриональной диапаузой, во время которой развитие >м(1|пи>нн
замедлено. Период же интенсивного роста плода у норок < < < ып
ляет около 30 сут, у клеточных соболей 30 — 35 сут.
У нутрии продолжительность беременности колебден и m I '
до 137, у клеточной шиншиллы от 106 до 111 сут.
411
Сезон щенения хищных довольно растянут. У клеточных норок
он охватывает конец апреля и середину мая, у песцов — апрель-
июнь, соболей — конец марта—апрель, енотовидной собаки —
начало апреля, лисиц — конец марта—апрель. Хищные пушные
звери относятся к незрелорождающимся млекопитающим. Дете-
ныши рождаются слепыми, беззубыми, с плотно замкнутыми
слуховыми проходами. Они большеголовы, с удлиненным .телом
и относительно короткими конечностями и хвостом. Новорож-
денные покрыты эмбриональным пухом.
Контрольные вопросы
I. Перечислите критерии половой и физиологической зрелости сам-
цов и самок.
2. Охарактеризуйте строение спермия, состав спермы, эякулят.
3. Какова функция придатка семенника и придаточных половых же-
лез?
4. Половой цикл и его стадии.
5. Каков механизм оплодотворения?
6. Расскажите о продолжительности беременности у разных видов жи-
вотных.
7. Охарактеризуйте функцию плаценты в развитии плода.
ЛАКТАЦИЯ
Лактацией называется процесс образования, накопления и
выведения молока из молочной железы во время сосания или до-
ения. Длительность лактации у животных различная. У коров она
длится 10 мес, у буйволиц и коз — 7 —8, у овец — 4 — 6, у сви-
ней — 2, у кобыл — 5—11, у верблюдиц — 16—18 мес.
Рост и развитие молочной железы. Молочные железы (вымя)
начинают функционировать, когда достигнут определенного раз-
вития. Они наиболее сильно развиты у коров. Сильный рост и окон-
чательное развитие вымени наступают в процессе первой бере-
менности. К четвертому месяцу стельности увеличивается объем
железистой ткани, развивающиеся протоки и альвеолы вытесня-
ют жировую ткань. В это время увеличивается количество крове-
носных сосудов и нервных волокон. Перед родами происходит раз-
витие протоков и альвеол, быстро формируется железистая ткань.
После отела наступает период наибольшей активности молочной
железы, альвеолы становятся крупными, а количество соедини-
тельной ткани в вымени уменьшается. Однако общее развитие
вымени происходит еще в течение нескольких лет лактации.
Через десять месяцев лактации удои значительно уменьшаются
и коров прекращают доить до нового отела. После перевода жи-
вотных на сухостой происходит инволюция вымени: альвеолы
412
вымени уменьшаются в размерах, железистая ткань частично за-
меняется жировой, часть мелких протоков атрофируется.
Молоко. Молозиво. Молоко имеет сложный химический состав.
Оно состоит из воды, белка, жира, углеводов, минеральных ве-
ществ. У каждого вида животных молоко имеет строго определен-
ный состав.
Молоко по биологической ценности превосходит все продук-
ты питания, имеющиеся в природе. Оно является естественной
пищей новорожденных. Чем быстрее растет новорожденный, тем
калорийнее молоко и богаче оно белками и минеральными веще-
ствами, очень необходимыми для построения скелета. В молоке
имеются соли неорганических и органических кислот. Соотноше-
ние в молоке кальция к фосфору 1,2: 1, что способствует хороше-
му усвоению организмом кальция.
Молоко первых 7—10 дней лактации называется молозивом,
оно отличается от обычного молока и выполняет в организме две
важные функции — питательную и защитную. Молозиво желтова-
то-белого цвета, солоноватое на вкус, с особым запахом, при
нагревании свертывается. В нем много альбуминов и глобулинов,
которые лучше усваиваются, чем казеин. В молозиве содержатся
соли магния, вызывающие послабляющее действие и блаюприят-
но влияющие на перистальтику кишечника. Молозиво обеспечи-
вает постепенный переход в питании новорожденного от крови
матери к молоку.
Процесс молокообразования. Структурно-функциональной
единицей молочной железы является альвеола. Молоко образу-
ется в эпителиальных секреторных клетках альвеол и протоков
из составных частей крови. Для образования 1 л молока необхо-
димо, чтобы через вымя прошло около 500 л крови. Витамины,
минеральные вещества переходят в молоко из крови без изме-
нений.
В образовании молока различаю! четыре тина секреции. В мо-
лозивный период наблюдается апокриновый тин, при котором
отторгается верхушечная часто секреторной клетки вместе с об-
разовавшимся секретом. В разгар лактации секреция молока идет
мерокриновым типом. Образующиеся капельки жира и другие со-
ставные части молока перемещаются в верхушечную часть клет-
ки и отсюда просачиваются через ее мембрану без повреждения
клетки. При леммокриновом типе капли секрета уносят на себе
частицы плазматической мембраны. Считают, что этот тип сек-
реции происходит одновременно с мерокриновым и выделяется
только жир. В стадию инволюции вымени наблюдается голокри-
новый тип, при котором происходит преобразование всей клет-
ки в секрет молока.
Молоко образуется непрерывно, заполняя вначале альвеолы и
протоки, а затем перемещается в цистерны.
413
Рис. 4.18. Схема регуляции молоко-
отдач и:
1 — чувствительные рецепторы соска;
2 — афферентный путь; 3 — спинной
мозг; 4 — эфферентный путь; 5 — ги-
поталамус; 6 — кора больших полуша-
рий; 7— гипофиз; 8— окситоцин; 9 —
звездчатые миоэпителиальные клетки;
10 — сосок
Регуляцию процесса образо-
вания молока осуществляют не-
рвная система, железы внутрен-
ней секреции и гипоталамуса,
в которых располагается центр
лактации. (
Рефлекс молокоотдачи. В реф-
лексе молокоотдачи различают
две фазы (рис. 4.18). Первая
фаза — нервная', при раздраже-
нии рецепторов вымени во вре-
мя сосания или доения импуль-
сы передаются в центры пояснич-
но-крестцового отделов спинного
мозга, а оттуда к сфинктерам
сосков; сфинктеры раскрывают-
ся и выделяется цистернальная
порция молока. К этому момен-
ту в цистерне содержится около
25 % удоя, но при правильной
подготовке (массаж, обмывание
вымени) в цистерне скапливается (85 — 97%) молока.
Вторая фаза — нейрогуморальная'. импульсы от рецепторов вы-
мени передаются в гипоталамус, в котором образуется гормон
окситоцин, поступающий в заднюю долю гипофиза. Окситоцин
из гипофиза выделяется в кровь, с нею приносится к вымени, где
вызывает сокращение звездчатых миоэпителиальных клеток аль-
веол, в результате чего сокращаются альвеолы и выводится альве-
олярная порция молока.
Физиология доения. Доение является сложным процессом. Все
приемы, связанные с доением: подход к животному, массаж вы-
мени, его обтирание и пр. — должны проводиться в определенной
последовательности. Это способствует образованию и закрепле-
нию у коров условных рефлексов, связанных с рефлексом моло-
коотдачи.
Машинное доение — наиболее целесообразный способ получе-
ния молока. При ручном доении одновременно выдаивают две
четверти, а доильной машиной сразу все четыре. При выдаивании
поочередно одной четверти за другой из четверти, выдоенной
последней, получают меньше молока и с пониженной жирнос-
тью. Это происходит потому, что молоко из нее частично перехо-
дит в протоки и альвеолы. Выделение и действие окситоцина длится
4 — 7 мин. С учетом этого доение должно быть закончено в период
действия окситоцина на миоэпителий альвеол. Поэтому коров
нужно доить быстро и энергично, так как рефлекс молокоотдачи
длится лишь 5 — 7 мин.
414
Контрольные вопросы
1. Расскажите о лактации, ее продолжительности у разных видов жи-
вотных.
2. Какова структурная и функциональная единица молочной железы?
3. В чем отличие молока от молозива?
4. Каков механизм образования молока?
5. Перечислите преимущества машинного доения коров.
ФИЗИОЛОГИЯ МЫШЦ И НЕРВОВ
Согласно современным представлениям, нервная и мышечная
ткани могут находиться в трех основных состояниях — физиоло-
гическом покое, возбуждении и торможении.
Физиологический покой — это такое состояние, когда ткань или
орган не проявляет признаков присущей им деятельности.
Возбуждение — деятельное состояние живой ткани, в которое
она приходит под влиянием раздражения.
Торможение — такое состояние, когда деятельность ткани или
органа ослабляется или полностью прекращается.
Раздражители. При раздражении мышца приходит в состояние
возбуждения, которое распространяется по мышце и вызывает ее
сокращение. По биологическому действию раздражители бывают
адекватные и неадекватные. Адекватный — это такой раздражи-
тель, к которому данный орган или ткань приспособились в про-
цессе эволюции. Для мышцы адекватным раздражителем является
нервный импульс. Неадекватными — будут такие раздражители,
действию которых ткань или орган в естественных условиях обыч-
но не подвергаются. Так, сокращение мышцы можно вызвать ме-
ханическим раздражением (укол, кислота, ток) и т.д.
Все раздражители по своей силе делят на пороговые, подпоро-
говые и сверхпороговые. Пороговыми называют минимальные раз-
дражители, которые могут вызвать возбуждение. Цодпороговые —
это раздражители, сила которых меньше пороговой, сверхпорого-
вые — раздражители более сильные, чем пороговые.
Биоэлектрические явления. Возникновение и распространение
возбуждения связано с изменением электрического заряда на по-
верхности клеточной мембраны и внутри клетки (рис. 4.19).
Потенциал покоя. Мембрана нервной или мышечной клетки в
состоянии покоя с наружной стороны заряжена положительно, а
с внутренней — отрицательно. Разность зарядов между наружной
и внутренней поверхностью клеточной мембраны в состоянии
физиологического покоя клетки называется потенциалом покоя,
или мембранным потенциалом.
Потенциал действия. Под влиянием раздражения проницае-
мость клеточной мембраны для ионов натрия повышается в согни
415
Рис. 4.19.Схема потенциалов покоя (А), действия (Б) и механизмы их
возникновения (В, Г)\
1 — деполяризация (уменьшение потенциала покоя под влиянием раздражите-
ля); 2 — перезарядка мембраны; 3 — восстановление первоначального заряда;
4 — потенциал действия; 5 — потенциал покоя; Р — направление раздражения;
а — мембрана; b — цитоплазма; прямые стрелки указывают направление движе-
ния ионов К+ и \а в сторону их наименьшей концентрации; стрелка с кружка-
ми — ионный натрий-калиевый насос; Еь — порог возбудимости
раз. Происходит перезарядка клеточной мембраны: внутренняя ее
сторона заряжается положительно, а наружная — отрицательно.
Затем значительно повышается проницаемость мембраны для ионов
калия, они начинают быстро перемещаться из клеток во внекле-
точную жидкость и вновь восстанавливают первоначальный заряд
мембраны. Пикообразное колебание потенциала, возникающее в
результате кратковременной перезарядки мембраны и последую-
щего восстановления ее заряда, называется потенциалом действия.
Такие колебания потенциала продолжаются 1 — 5 мс.
Проведение возбуждения. Круговой ток, проходя через участок
покоя, раздражает его и вызывает появление потенциала действия
(рис. 4.20). Когда в одном из участков нервной клетки, нервного
или мышечного волокна возникает возбуждение и появляется по-
тенциал действия, между возбужденным и соседним невозбуж-
денным участком возникает разность потенциалов. Эта разность
потенциалов является причиной появления круговых (локальных)
токов. Внутри клетки ток идет от возбужденного участка к участку
покоя, по внешней стороне — от участка покоя к возбужденному.
В участке, который ранее был возбужден, происходит восста-
новление потенциала покоя. Данный процесс многократно по-
вторяется и обусловливает распространение импульсов возбужде-
416
ния. Скорость проведения возбуждения
по скелетным мышцам 12 —15 м/с, глад-
ким мышцам — 1 — 18 см/с, в безмя-
котных нервах — 0,5 — 3 м/с.
Основные свойства живой ткани. Лю-
бая живая клетка обладает свойствами
раздражимости, возбудимости и лабиль-
ности (функциональной подвижности).
Раздражимость. С раздражимостью
Рис. 4.20. Схема распрост-
ранения возбуждения:
связаны все процессы роста и размно-
жения клеток и тканей. На разнообраз-
ные воздействия внешней или внутрен-
ней среды — раздражения протоплазма
живой клетки отвечает специфической
реакцией, при которой происходят уси-
ление или ослабление обмена веществ,
количественное и качественное его из-
менение. Эта реакция на раздражение
получила название раздражимости, или
А — волокна мышц и безмя-
котных нервных волокон; Б —
мякотное нервное волокно;
/ — мембрана; 2 — волокно;
а, Ь, с — перехваты Ранвье;
заштрихованы возбужденные
участки; светлые — участки
покоя; стрелками показано
реактивности.
Возбудимость — свойство нервной
движение круговых токов
или мышечной ткани отвечать на действие раздражителя возбуж-
дением. Для измерения величины возбудимости определяют порог
возбудимости и хронаксию. Порогом возбудимости называется наи-
меньшая сила раздражителя, способная вызвать возбуждение; его
выражают в вольтах постоянного тока.
Лабильность — скорость, с которой в ткани возникает и успе-
вает закончиться полный период отдельного импульса возбужде-
ния. Мера лабильности — это максимальное число импульсов воз-
буждения за 1 с в ответ на такое же максимальное число раздра-
жений. Высокой лабильностью обладают мякотные нервы, у без-
мякотных нервов она низкая. Так, максимальный ритм возбужде-
ний мякотного нервного волокна 500, безмякоз noi о — 200. Мак-
симальный ритм возбуждений скелетной мышцы 200 импульсов в
1 с, а гладкой мышцы — в десятки раз меньше.
При очень частых раздражениях сокращения мышцы умень-
шаются или даже совсем прекращаются. Такая частота называет-
ся пессимальной, или пессимумом. Пессимум возникает вследствие
того, что возбуждение езце не закончилось и ткань находится в
состоянии абсолютной или относительной рефрактерное™, а на
нее действует новое раздражение. Частые раздражения, превы-
шающие меру лабильности, вызывают не возбуждение, а тормо-
жение.
Учение Н. Е. Введенского о парабиозе устанавливает связь меж-
ду возбуждением и торможением. Возбуждение и торможение —
это различные реакции ткани на раздражение, исход которого
417
зависит от лабильности. При высокой лабильности возникает воз-
буждение, снижение лабильности вызывает торможение.
ФИЗИОЛОГИЯ мышц
(
Скелетные мыищи, входящие в состав опорно-двигательного
аппарата позвоночных животных, состоят из группы мышечных
пучков, каждый из них составлен из тысяч мышечных волокон,
которые представляют собой клетки цилиндрической формы дли-
ной до 12 см и диаметром 10—100 мкм. Каждое волокно окружено
оболочкой сарколеммой и содержит тонкие нити — миофибрил-
лы (рис. 4.21). Поперечные мембраны делят каждую миофибриллу
на отдельные участки — саркомеры. Сократимым веществом мы-
шечного волокна являются миофибриллы, состоящие из множе-
ства (около 2500) тонких и толстых белковых нитей — протофиб-
рилл. Толстые протофибриллы образованы из белка миозина, тон-
кие — из актина. Нити актина прикреплены к мембране саркоме-
ра; они образуют светлые участки миофибриллы. В темных участ-
ках находятся нити миозина. Актиновые нити частично входят сво-
ими концами в промежутки между миозиновыми нитями. Нити
актина и миозина соединяются между собой многочисленными
поперечными мостиками, которые образованы скрученными в
спираль отростками — мостиками миозиновой нити. Чередование
нитей в миофибрилле обусловливает ее поперечную исчерченность.
Гладкие мышцы, образующие стен-
Рис. 4.21. Схема чередова-
ния нитей в миофибрилле:
А — электронно-микроско-
пическая фотография мио-
фибриллы; Б — строение мио-
фибриллы; Z — поперечная
мембрана, разделяющая ми-
офибриллу на отдельные уча-
стки — саркомеры; 7 — ми-
озиновая нить. 2 — актино-
вая нить
ки внутренних органов, состоят из кле-
ток различной формы длиной от 30 до
500 мкм и диаметром 2—10 мкм; у кле-
ток имеются боковые отростки, благо-
даря которым клетки гладких мышц
группируются в длинные пучки. Они, в
свою очередь, при помощи тяжей соеди-
няются друг с другом, обеспечивая дея-
тельность мышцы как единой системы.
Сократительным аппаратом гладких
мышц являются миофибриллы, в кото-
рых содержатся тонкие актиновые нити
с прикрепленными к ним короткими от-
ростками миозиновых нитей, называе-
мых димерами.
Свойства скелетных мышц. Проведе-
ние возбуждения в мышцах происходит
изолированно, т. е. потенциалы действия
не переходят с одного мышечного во-
локна на другое. Возбудимость скелетных
418
мышц меньше возбудимости нервов. Нервное волокно оканчива-
ется в середине мышечного волокна, поэтому возбуждение рас-
пространяется в обе стороны со скоростью 4—15 м/с.
Скелетная мышца является упругим телом. Если к мышце под-
весить груз, то она растягивается, это свойство называется растя-
жимостью. Эластичностью мышцы называется возвращение мыш-
цы к первоначальной своей длине после растяжения, когда будет
удален груз. Пластичностью мышцы называется свойство сохра-
нять удлиненную форму после удаления груза, вызвавшего ее ра-
стяжение. В скелетных мышцах лучше выражено свойство эластич-
ности, чем пластичности.
Виды мышечных сокращений. При нанесении на мышцу оди-
ночного раздражения она отвечает одиночным сокращением. За-
писывая это сокращение на двигающейся ленте какого-либо са-
мописца (например, в простейшем случае на кимограф), можно
отметить три периода: латентный — период от раздражения до
начала сокращения, период сокращения и период расслабления.
У икроножной мышцы лягушки одиночное сокращение длится
0,1 с, у млекопитающих — 0,04 —0,1 с. В латентный период в мыш-
це происходят процессы, при которых освобождается энергия для
мышечного сокращения.
Если к мышце поступает несколько частых импульсов возбуж-
дения, наступает длительное сокращение мышцы, которое назы-
вается тетаническим, или тетанусом. В зависимости от частоты
возбуждений тетанус будет зубчатым или гладким (рис. 4.22).
Зубчатый тетанус наблюдается при такой частоте импульсов
возбуждений, когда каждый импульс действует на мышцу в тот
момент, когда она уже начинает расслабляться. Если же импульсы
возбуждения настолько частые, что они воздействуют на мышцу
до начала ее расслабления, то получатся длительные непрерыв-
ные сокращения мышцы — гладкий тетанус.
Рис. 4.22. Сокращение скелетной мышцы при различной частоте раздра
жения:
А — одиночное сокращение в ответ на одно раздражение; Б — зубчатый тетанус
(до 15 раздражений в I с); В — гладкий тетанус (свыше 25 раздражений); М -
механограмма — запись сокращения мышц; Р — частота раздражений; / -- фаза
сокращения; 2 — фаза расслабления
419
Механизм мышечного сокращения (теория скольжения). В со-
стоянии покоя тонкие нити актина лежат частично своими кон-
цами в промежутках между толстыми миозиновыми нитями. При
возбуждении мышцы под влиянием потенциала действия нити ак-
тина сдвигаются в промежутке между нитями миозина, т. е. сколь-
зят навстречу друг другу, вызывая сокращение миофибдэилл
(рис. 4.23). Считают, что скольжение происходит в результате уко-
рочения мостиков — боковых отростков миозиновых нитей под
влиянием ионов кальция. Укорачиваясь, мостики тянут актино-
вую нить навстречу миозиновой.
Работа и сила мышц. При сокращении мышца укорачивается и
тем самым совершает работу. Она будет наибольшей при средних
нагрузках и среднем ритме сокращений.
Силу мышцы определяют по максимальному сокращению ее
при поднятии максимального груза. Наибольшей силой обладают
мышцы с косыми волокнами, имеющими перистое строение.
Утомление мышц. В процессе сокращений мышцы утомляются,
при этом понижаются их возбудимость, лабильность и величина
сокращения. В утомленной мышце понижается содержание глико-
гена и накапливаются продукты обмена. В опытах на нервно-мы-
шечном препарате Н. Е. Введенский установил, что прежде всего
утомляются синапсы в связи с их низкой лабильностью. В целом
организме утомление в первую очередь наступает в нервных цен-
трах коры больших полушарий. И. М. Сеченов доказал, что быст-
рое восстановление работоспособности утомленных мышц насту-
пает не при полном их покое, а при работе других, до этого не
сокращавшихся мышц. Импульсы от вновь вовлеченных в работу
мышц повышают возбудимость нервных центров, возбуждение
одних нервных центров снижает и даже снимает утомление других
центров. Утомление зависит от состояния симпатической нервной
системы и желез внутренней секреции: утомленная мышца вновь
А
Рис. 4.23. Схема расслабленной (А) и сократившейся (Б) миофибриллы
420
начинает сокращаться при раздражении симпатического нерва или
введении адреналина, усиливающих обмен. Тренировка мышц
позволяет отсрочить наступление утомления. При ней увеличива-
ется объем мышц в результате утолщения мышечных волокон,
повышается содержание гликогена, АТФ и креатинфосфата, ус-
коряются восстановительные процессы.
Тонус мышц. Тонус скелетных мышц играет важную роль для
поддержания определенного положения тела в пространстве и
деятельности двигательного аппарата. Скелетные мышцы в покое
расслабляются не полностью, а находятся в некотором напряже-
нии, т.е. тонусе. Он обусловлен поступлением к мышцам редких
нервных импульсов, возбуждающих мышечные волокна не одно-
временно, а поочередно.
Гладкие мышцы обладают способностью длительно находить-
ся в тонусе. Длительные тонические сокращения этих мышц осо-
бенно отчетливо выражены в сфинктерах полых органов, в стен-
ках кровеносных сосудов.
ФИЗИОЛОГИЯ НЕРВНОГО ВОЛОКНА
Нерв состоит из множества нервных волокон, которые, объе-
диняясь в пучки, образуют нервы. Однако возбуждение по каждо-
му нервному волокну проводится изолированно, не переходя на
соседние.
Функциональной единицей нервной системы является нейрон,
который представляет собой нервную клетку с отростками. Отрос-
тки нервных клеток образуют нервные волокна; они являются
проводниками нервных импульсов. Различают мякотные (покры-
тые миелиновой оболочкой) и безмякотные нервные волокна.
Свойства нервных волокон. Нервные волокна обладают высо-
кой возбудимостью, у мякотных волокон опа выше, чем у бсзмя-
котных. Лабильность нервных волокон наивысшая. Мякотные во-
локна могут воспроизводить до 500 импульсов в I с. У безмякот-
ных волокон лабильность очень низкая.
Обмен веществ в нерве очень незначителен. Энергия образует-
ся на 1 г нерва примерно в миллион раз меньше, чем на 1 г мыш-
цы. Нерв практически неутомляем, что обусловлено его низким
обменом веществ и высокой лабильностью. Это создает благопри-
ятные условия для выполнения нервом основной функции — про-
ведения импульсов возбуждения (нервных импульсов).
После перерезки нервного волокна его периферический от-
резок, отделенный от тела нейрона, дегенерирует. Это свиде-
тельствует о том, что нервная клетка является трофическим цен-
тром для нервных волокон, которые не могут существовать вне
связи с клеткой.
421
Рис. 4.24. Схема нервно-мышечного синапса:
I — неврилемма, переходящая в наружный слой сарколеммы; 2 — осевой ци-
линдр; 3 — митохондрии; 4 — синаптические пузырьки; 5 — миофибрилла,
состоящая из протофибрилл; 6— синаптическое пространство (щель); 7— пост-
синаптическая мембрана; 8 — пресинаптическая мембрана; 9 — сарколемма
Перерезанные концы центрального отрезка могут регенериро-
вать, они образуют колбы роста — утолщения, которые растут в
направлении периферического отрезка. Врастание нервных воло-
кон из центрального отрезка в периферический начинается уже
через 2 — 3 дня после перерезки. Регенерация нервных волокон
происходит очень медленно (0,3— 1 мм в сутки). Например, в мыш-
цах начальные признаки восстановления функций регенерирую-
щего нерва обнаруживают примерно через 1,5 мес после перерез-
ки. Полная регенерация нервов происходит в течение многих ме-
сяцев и даже лет.
Синаптическая передача возбуждения. Передача возбуждения с
нервного волокна на мышечное происходит через специализиро-
ванное структурное образование, обеспечивающее контакт аксо-
на двигательного нейрона с мышечным волокном — нервно-мы-
шечный синапс (рис. 4.24). Он состоит из трех основных элементов:
пресинаптической мембраны, синаптического пространства
(щели) и постсинаптической мембраны.
Контрольные вопросы
1. Что такое физиологический покой, возбуждение и торможение?
2. Назовите биоэлектрические явления в возбудимых тканях.
422
3. Охарактеризуйте потенциал покоя и действия, проведение возбуж-
дения по нервному и мышечному волокну.
4. Что такое раздражимость, возбудимость, лабильность?
5. Какова теория мышечного сокращения?
6. Опишите утомление мышц.
7. Каковы свойства нервных волокон?
8. Что такое синаптическая передача возбуждения?
ФИЗИОЛОГИЯ ЦЕНТРАЛЬНОЙ
И ПЕРИФЕРИЧЕСКОЙ НЕРВНОЙ СИСТЕМЫ
Центральная нервная система осуществляет следующие жиз-
ненно важные функции:
- анализирует поступающие раздражения из внешней и внут-
ренней среды и формирует ответные приспособительные реакции;
- интегрирует механизмы управления на всех уровнях, органи-
зует и обеспечивает согласованную, гармоничную деятельность
органов;
-является материальным субстратом психических процессов —
ощущений, восприятий, эмоций, памяти, навыков и прочих,
лежащих в основе сложных форм поведения животных; эта фун-
кция осуществляется корой головного мозга и подкорковыми цен-
трами.
Координирующую роль ЦНС осуществляет через перифериче-
ские проводники — соматические и вегетативные нервы при уча-
стии органов гуморальной регуляции — желез внутренней секре-
ции и сенсорных органов. Основными функциональными элемен-
тами ЦНС являются нейроны', в организме млекопитающих их со-
держится примерно 50 млрд, из них лишь небольшая часть распо-
ложена на периферических участках тела.
Нейроны составляют 10—15% оз общего числа клеточных эле-
ментов ЦНС, основную большую ее часть занимают клетки нейро-
глии, выполняющие трофическую, опорную и защитную функции.
Основной формой деятельности центральной нервной систе-
мы является рефлекс. Рефлексом называется реакция организма на
раздражение рецепторов, осуществляемая при участии централь-
ной нервной системы. Возбуждение проходит по определенному
пути — рефлекторной дуге, которая состоит из рецептора, аффе-
рентного нейрона, промежуточного нейрона, эфферентного ней-
рона и эффектора (рис. 4.25). В организме рефлекторную дугу обра-
зует множество нейронов, расположенных как в спинном мозге,
так и в различных отделах головного мозга, включая и кору боль-
ших полушарий. В настоящее время учение о рефлексе дополнено
понятием об обратной связи, поэтому говорят не только о реф-
лекторной дуге, но и о рефлекторном кольце.
423
Рис. 4.25. Схема рефлек-
торной дуги с обратной
связью:
1 — рецептор; 2 — аффе-
рентный нейрон; 3 — про-
межуточный нейрон; 4 —
эфферентный нейрон; 5 —
эффектор (мышца); 6 —
проприорецептор (мышеч-
ное веретено); 7 — нейрон
обратной связи; стрелки —
направление прохождения
импульса
Контакты между нейронами в ЦИС
обеспечивают межнейронные синапсы. Ак-
сон одного нейрона может оканчиваться
на теле, дендрите или (реже) аксоне дру-
гого нейрона, образуя синапс. По функции
различают возбуждающие и тормозящие си-
напсы. Возбуждающие синапсы находятся в
основном на дендритах, а тормозящие —
на клетках нейрона. Они отличаются друг
от друга строением пре- и постсинаптичес-
ких мембран и шириной синаптической
щели. Химическим передатчиком возбуж-
дения (медиатором) в возбуждающих си-
напсах является ацетилхолин, в тормозя-
щих синапсах — аминокислота глицин и
гамма-аминомасляная кислота.
Нервные центры и их свойства. Не-
рвным центром называется группа ней-
ронов в центральной нервной системе,
связанная с осуществлением определен-
ных рефлекторных действий. Нервным
центрам свойствен ряд физиологических особенностей.
Одностороннее проведение от рецепторного нейрона через проме-
жуточный нейрон к эфферентному нейрону определяется свойства-
ми синапсов, которые проводят импульсы от аксона к дендриту.
Суммация импульсов в нервных центрах может быть во времени
и пространстве. Например, слабое раздражение лапки лягушки не
вызывает рефлекса сгибания. Если же нанести несколько частых
слабых раздражений, то лягушка сгибает лапку.
Иррадиация возбуждения', сильное возбуждение одного центра
вызывает возбуждение и других центров.
Конвергенция возбуждения (или схождение) обусловлена тем,
что в центральной нервной системе афферентных путей в 4 — 5 раз
больше, чем эфферентных. В результате этого к одному эфферент-
ному нейрону возбуждение поступает по многим путям.
Последействие в нервных центрах проявляется в том, что раз-
дражение рецепторов уже закончено, а ответная реакция еще
продолжается.
Пластичность нервных центров — перестройка функции не-
рвных центров, если рабочий орган, с которым связан данный
центр, заменить другим.
Инертность нервных центров — способность длительно сохра-
нять следы возбуждения.
Доминанта — временное, стойкое возбуждение центра, кото-
рый занимает в данный момент господствующее положение в цен-
тральной нервной системе.
424
Доминантный центр способен накапливать, усиливать свое
возбуждение за счет импульсов, адресованных другим центрам,
перехватывая эти импульсы.
Нервный центр находится в состоянии незначительного посто-
янного возбуждения, или тонуса. Например, тонус двигательных
центров поддерживают непрерывные потоки импульсов, идущие
от чувствительных нервных окончаний, заложенных в самих мыш-
цах. Слабое возбуждение, идущее от нервных центров к мышцам,
поддерживает также их тонус.
Обмен веществ в нервных центрах по сравнению с нервным
волокном высок в состоянии покоя, а при возбуждении увеличи-
вается в 3 — 4 раза. Поэтому центры весьма чувствительны к недо-
статку кислорода и глюкозы. Вследствие очень высокого уровня
обмена веществ нервные центры быстро утомляются, это связано
с нарушением проведения возбуждения в межнейронных синап-
сах.
Торможение в центральной нервной системе — это процесс
ослабления или прекращения какой-либо деятельности.
Двигательные рефлексы можно затормозить, если в центрах
встретятся возбуждения, идущие от двух рецептивных полей. Так,
рефлекс отдергивания лапки на раздражение ее слабым раство-
ром серной кислоты тормозится при сильном сжимании пинце-
том другой лапки. Торможение наблюдается при наложении за-
крутки на губу лошади или щипцов на носовую перегородку быка:
сильное болевое раздражение тормозит двигательные реакции
животных.
Координация деятельности нервных центров происходит в ре-
зультате взаимодействия процессов возбуждения и торможения.
ФИЗИОЛОГИЯ ГОЛОВНОГО МОЗГА
Головной мозг — самый важный отдел центральной нервной
системы. Его подразделяют на ромбовидный и большой мозг. Ром-
бовидный мозг включает продолговатый мозг и задний мозг, пред-
ставленный мозжечком и мостом. В состав большого мозга входят
средний, промежуточный и конечный мозг.
Продолговатый мозг. Содержит жизненно важные рефлектор-
ные центры. Белое вещество продолговатого мозга состоит из длин-
ных и коротких волокон. Длинные входят в состав восходящих и
нисходящих проводящих путей, короткие — осуществляют связи с
мостом и мозжечком.
Серое мозговое вещество в нем образует скопление соматиче-
ских и вегетативных ядер. Одни из них являются промежуточными
ядрами, расположенными по ходу основных проводящих путей,
425
другие служат центрами V—ХП пар черепно-мозговых нервов,
третьи представляют собой чрезвычайно важные нервные цент-
ры: дыхания, сосудодвигательный, сердечной деятельности, по-
тоотделения, жевания, глотания, сосания, секреции пищевари-
тельных желез; центры ряда защитных рефлексов: чихания, каш-
ля, моргания, слезоотделения, рвоты.
Мост мозга — выполняет проводниковую функцию, связывая
вышележащие и нижележащие отделы центральной нервной сис-
темы между собой и мозжечком, а мозжечок — с корой больших
полушарий мозга. В нем находится центр пневмотаксиса, участву-
ющий в регуляции дыхания.
Мозжечок является органом координации движения. Он связан
афферентными и эфферентными путями с другими отделами цен-
тральной нервной системы. К нему идут импульсы от спинного и
продолговатого мозга, из четверохолмия. Мозжечок получает аф-
ферентные импульсы по каналам обратной связи от всех рецепто-
ров, раздражение которых происходит во время движения тела.
К нему приходят импульсы от зрительных, слуховых и тактиль-
ных рецепторов.
Средний мозг относится к стволу головного мозга. Он располо-
жен между мозговым мостом и промежуточным мозгом. В нем раз-
личают ножки большого мозга, лежащие в основании среднего
мозга, пластинки четверохолмия (крыша), расположенные дор-
сально, и покрышки ножек с красным ядром, парасимпатичес-
ким ядром Якубовича и черной субстанцией. В передних холмах
четверохолмия находятся подкорковые центры зрительных, а в
задних холмах — слуховых ориентировочных рефлексов. При их
участии осуществляется поворот головы в сторону зрительных или
звуковых раздражителей. В покрышке лежат ядра глазодвигатель-
ного, блокового, часть ядра тройничного нервов. Красное ядро
регулирует тонус мышц, влияя на центр мышечного тонуса про-
долговатого мозга. Блоковый и глазодвигательный нервы подхо-
дят к мышцам глазного яблока. Черная субстанция связана с под-
корковыми ядрами и участвует в регуляции жевания, глотания,
сложных тонких двигательных актов, эмоционального поведения,
а также ряда вегетативных функций (дыхания, тонуса сосудов).
Промежуточный мозг. Состоит из таламуса, гипоталамуса и эпи-
таламуса.
Таламус (зрительный бугор} — самая большая часть промежу-
точного мозга. В зрительных буграх находится большое число ядер
(см. гл. 3).
Большое число нервных путей связывают таламус со всеми
отделами головного и спинного мозга. Через таламус проходят все
афферентные импульсы, направляющиеся в кору больших полу-
шарий. В таламусе происходит первоначальный анализ и синтез
афферентных импульсов.
426
Гипоталамус — одно из древнейших образований головного
мозга. Он связан со всеми отделами центральной нервной систе-
мы. В нем находятся высшие центры вегетативной нервной систе-
мы — центры, регулирующие все виды обмена веществ: белково-
го, липидного, углеводного, водно-солевого; имеются центры теп-
лорегуляции. Гипоталамус находится в тесной морфологической
и функциональной связи с гипофизом, образуя гипоталамо-гипо-
физарную систему. В ядрах гипоталамуса образуются нейросекре-
ты, которые по аксонам нейросекреторных клеток поступают в
кровеносные сосуды и кровью приносятся в переднюю и сред-
нюю доли гипофиза. Гипоталамус через гипофиз влияет на функ-
цию почти всех желез внутренней секреции. Он принимает учас-
тие в эмоциональных и поведенческих реакциях животных.
Эпиталамус состоит из сосудистой покрышки третьего мозго-
вого желудочка, эпифиза и парного узла уздечки. Эпифиз — желе-
за внутренней секреции грушевидной формы. Имеются сведения,
что эпифиз функционирует только у молодых животных, в даль-
нейшем он подвергается инволюции. Предполагают, что эпифиз
участвует в регуляции компасной ориентации животных. Он тесно
связан с гипоталамо-гипофизарной системой регуляции обмен-
но-вегетативных функций и приспособительных реакций орга-
низма.
Базальные, или подкорковые, ядра представляют скопления кле-
ток, расположенных в толще белого вещества больших полуша-
рий головного мозга. К ним относят полосатое тело, бледное ядро,
ограду и миндалевидное тело. Они участвуют в регуляции движе-
ний, в проявлении инстинктов — врожденных форм поведения
животных.
Серое мозговое вещество плаща формирует кору больших по-
лушарий мозга — высший отдел центральной нервной системы.
В коре располагаются отделы всех анализаторов и двигательные
центры всех органов тела.
Ретикулярная формация. В центральной нервной системе име-
ются скопления крупных нервных клеток, окруженные многочис-
ленными волокнами, идущими в различных направлениях и об-
разующими подобие сети. Она получила название ретикулярная
(сетчатая) формация. Нейроны ретикулярной формации распола-
гаются в промежуточном, среднем и продолговатом мозге, а так-
же в шейной части спинного мозга. В ретикулярной формации
насчитывают более 40 ядер. Они получают импульсы от всех ре-
цепторов по коллатералям различных афферентных путей и по-
стоянно находятся в активном состоянии. Эти нейроны очень чув-
ствительны также к различным химическим веществам — гормо-
нам и продуктам обмена веществ. Кроме восходящих, ретикуляр-
ная формация получает также импульсы от коры больших полу-
шарий. Ретикулярная формация имеет большое значение для ре-
427
гуляции возбудимости и тонуса всех отделов центральной нервной
системы. В ней различают нисходящий и восходящий отделы. Нис-
ходящий отдел оказывает как активирующее, так и тормозящее
влияние на рефлекторную деятельность спинного мозга. Изменяя
деятельность вегетативной нервной системы, этот отдел влияет
также на функцию внутренних органов. Восходящий отдел оказы-
вает активирующее воздействие на кору больших полушарий. С его
активностью связано протекание в коре процессов возбуждения и
торможения, сна и бодрствования, образование условных реф-
лексов. Будучи связанной с лимбической системой (см. ниже),
ретикулярная формация участвует в проявлении различных эмо-
ций, таких, как ярость, страх, удовольствие и т.д.
Лимбическая система. В состав лимбической системы входят
нервные структуры: поясная извилина, гиппокамп, миндалевид-
ное ядро, извилина гиппокампа, свод, расположенные на меди-
альной поверхности больших полушарий. Она принимает учас-
тие в регуляции процессов обмена веществ, эндокринных функ-
ций, способствует поддержанию гомеостаза, влияет на сердеч-
но-сосудистую систему, дыхание, функцию желудочно-кишечного
тракта.
В лимбической системе, гипоталамусе и ретикулярной форма-
ции расположены центры, управляющие эмоциями (ярость, страх,
тоска, удовлетворение, радость, успокоение и т.д.). С лимбиче-
ской системой связано пищевое и половое поведение животных.
ФИЗИОЛОГИЯ СПИННОГО МОЗГА
Спинной мозг физиологически самый древний отдел централь-
ной нервной системы. Здесь замыкаются большое число рефлек-
торных дуг как соматических, так и вегетативных рефлексов. В их
осуществлении и координации большую роль играют межнейрон-
ные взаимоотношения.
В шейном отделе лежат центры диафрагмальных нервов, нару-
шение этих центров приводит к расстройству дыхания. В грудной
части спинного мозга находятся центры мышц, приводящих в
движение передние конечности и туловище; в поясничной облас-
ти — центры мышц задних конечностей. В спинном мозге имеются
центры симпатических и парасимпатических нервов: симпатиче-
ских — в грудо-поясничном отделе, парасимпатических — в кре-
стцовой части спинного мозга. Все центры спинного мозга нахо-
дятся под контролем центров головного мозга.
Важной функцией спинного мозга является проведение им-
пульсов в высшие центры головного мозга от периферических
рецепторов и обратно — к исполнительным органам. Проводящие
пути его подразделяются на восходящие и нисходящие. Одни из них
428
короткие и объединяют между собой соседние сегменты спинно-
го мозга; другие — длинные, соединяющие спинной мозг с раз-
личными отделами головного мозга. Импульсы от рецепторов кожи,
мышц, сухожилий, связок, внутренних органов, кровеносных
сосудов по восходящим путям проводятся в продолговатый, сред-
ний, промежуточный мозг и мозжечок. Далее от промежуточного
мозга по специальным путям импульсы поступают в кору боль-
ших полушарий. Нисходящие пути проводят импульсы от пира-
мидных клеток двигательной области коры больших полушарий к
эфферентным нейронам спинного мозга. Часть восходящих и нис-
ходящих путей перекрещиваются на уровне продолговатого и спин-
ного мозга, поэтому каждая половина головного мозга получает
импульсы от рецепторов как своей, так и противоположной сто-
роны тела, а сигналы от высших двигательных центров коры боль-
ших полушарий головного мозга поступают на противоположную
сторону тела.
ФИЗИОЛОГИЯ ВЕГЕТАТИВНОЙ НЕРВНОЙ СИСТЕМЫ
Вегетативная нервная система регулирует деятельность внут-
ренних органов (кровообращения, пищеварения, мочеполовой
системы и т.д.) и обмен веществ. Все эти процессы называют ве-
гетативными. Вегетативная нервная система имеет структурные и
функциональные отличия от соматической.
Центробежный (эфферентный, двигательный) путь вегетатив-
ной нервной системы состоит из двух нейронов и прерывается в
ганглиях (нервныхузлах). Волокна первого центробежного нейрона
вегетативной нервной системы после выхода из центральной не-
рвной системы заканчиваются на нейроне вегетативного ганглия -
второго нейрона центрального пути, волокна которого доходят
непосредственно до органа.
Волокна, идущие из центральной нервной системы до ганг-
лия, называют преганглионарными, а волокна, идущие от ганг-
лиев к органу, — постганглионарными. Вегетативные волокна очень
тонкие, менее возбудимые, возбуждение по ним распространяет-
ся медленно.
Высшие центры вегетативной нервной системы расположены
в гипоталамусе и полосатом теле. Вегетативный отдел нервной
системы состоит из симпатической и парасимпатической нервной
систем.
Симпатическая нервная система. Центры симпатической нервной
системы лежат в грудопоясничном отделе спинного мозга. Пре-
ганглионарные симпатические нервные волокна заканчиваются в
ганглиях, одни из которых располагаются вблизи позвоночного
столба и образуют цепочку (пограничный симпатический ствол).
429
Часть ганглиев располагается на некотором удалении от позво-
ночного столба. В окончаниях преганглионарных волокон выделя-
ется медиатор ацетилхолин, а в большинстве постганглионарных —
норадреналин, и только в окончаниях нервов, подходящих к пото-
вым железам, выделяется ацетилхолин. Симпатическая или сосу-
дистая нервная система иннервирует всю сердечно-сосудистую си-
стему.
Раздражение симпатической нервной системы вызывает: уча-
щение и усиление сокращений сердца, сужение всех периферичес-
ких кровеносных сосудов и внутренних органов (сосуды сердца и
мозга расширяются), торможение, моторики и секреции желудоч-
но-кишечного тракта, усиливает процессы диссимиляции, повы-
шает выделение адреналина надпочечниками, расширяет зрачки.
Парасимпатическая нервная система. Центры парасимпатиче-
ской нервной системы находятся в среднем и продолговатом
мозге, а также в крестцовом отделе спинного мозга. Аксоны пер-
вого нейрона, выходя из центральной нервной системы, окан-
чиваются на нейронах ганглиев (второй нейрон), которые рас-
положены внутри иннервируемых органов или вблизи от них. Ме-
диатором парасимпатической нервной системы является ацетил-
холин. При раздражении парасимпатических нервов тормозится
работа сердца, усиливается секреция и моторика желудочно-ки-
шечного тракта, увеличивается диурез, суживаются зрачки, по-
вышается ассимиляция и т.д. Оба отдела обеспечивают поддер-
жание постоянства внутренней среды организма. Их деятельность
регулируют кора больших полушарий, ретикулярная формация,
гипоталамус и мозжечок. В коре больших полушарий имеются
области, связанные нисходящими путями с ретикулярной фор-
мацией и гипоталамусом.
Трофическая функция нервной системы. Нервная система регу-
лирует обмен веществ в тканях и органах. Учение о трофической
функции нервной системы обосновал И. П. Павлов. Он указывал,
что каждый орган находится под тройным нервным контролем:
нервов функциональных, вызывающих его деятельность; нервов
сосудистых, регулирующих просвет сосудов и доставку по ним
питательных веществ кровью; нервов собственно трофических,
определяющих точный размер использования питательных веществ
каждым органом и тканью.
Установлено, что трофическое влияние на органы и ткани не-
рвная система оказывает как через вегетативные (в основном),
так и через соматические нервы. Ведущее значение в регуляции
трофических процессов в организме принадлежит коре больших
полушарий, которая воздействует на эти процессы через подкор-
ковые ядра (полосатое тело), ретикулярную формацию и гипота-
ламус. Опыты на животных показали, что при нарушении функ-
ции коры наступают трофические расстройства.
430
Контрольные вопросы
1. В чем состоит координирующая и интегрирующая роль ЦНС?
2. Что является основной формой деятельности ЦНС?
3. Каково значение возбуждающих и тормозящих синапсов?
4. Перечислите свойства нервных центров.
5. Охарактеризуйте кору большого мозга и ее значение.
6. Опишите проводящие пуги головного и спинного мозга.
7. Каково значение ретикулярной формации в возбудимости отделов
ЦНС?
8. Расскажите о значении лимбической системы в регуляции процес-
сов жизнедеятельности.
9. Каковы функциональные особенности симпатического и парасим-
патического отделов нервной системы?
10. Что такое трофическая функция нервной системы?
ВЫСШАЯ НЕРВНАЯ ДЕЯТЕЛЬНОСТЬ
Строение и методы исследования функций коры больших полу-
шарий головного мозга. Большие полушария головного мозга явля-
ются высшим отделом центральной нервной системы. В ней распо-
ложены сенсорные, моторные и ассоциативные зоны. Сенсорные
зоны — это корковые центры анализаторов: слухового, зрительно-
го, тактильного, обонятельного и др. Зрительная зона находится в
затылочных долях больших полушарий, слуховая — в височных,
обонятельная — в аммоновых рогах, зона тактильной чувствитель-
ности (осязания) — в области центральной извилины.
В моторной зоне сосредоточены двигательные центры к скелет-
ным мышцам, сюда же поступают и анализируются импульсы от
рецепторов, заложенных в толще мышц, сухожилиях и суставах.
Их локализация имеет видовую специфичность.
Ассоциативные, или вторичные сенсорные, зоны окружают пер-
вичные сенсорные зоны полосой в I — 5 см. На них конвергируют
афферентные пути, идущие от различных рецепторных систем.
Функцию коры больших полушарий изучают различными ме-
тодами:
- метод наблюдения за поведением животного в различных ус-
ловиях и при различных состояниях;
- метод раздражения коры, для этого у животного под нарко-
зом обнажают большие полушария и, раздражая электрическим
током их отдельные участки, наблюдают за сокращением мышц и
функциями других органов;
- метод удаления коры ил и отдельных ее участков дает возмож-
ность выяснить функции разных зон и коры в целом;
- запись биотоков коры больших полушарий (электроэнцефало-
графия), в том числе в условиях свободного поведения животного.
431
В настоящее время разработана методика записи биотоков отдель-
ных нервных клеток и их волокон;
- метод условных рефлексов. Величайшей заслугой И. П. Павло-
ва является создание метода условных рефлексов при исследова-
ния коры больших полушарий. Он объективно отражает основной
принцип деятельности головного мозга, направленный на осуще-
ствление взаимодействия организма с внешней средой.
Условный рефлекс. Изучая процессы пищеварения, И. П. Пав-
лов обратил внимание на то, что у собаки слюноотделение начи-
нается еще до того, как пища попала в рот. Достаточно появиться
лицу, которое кормит собаку, или внести пустую посуду, в кото-
рой она получает корм, как у собаки начинается обильное слю-
ноотделение. Это явление до Павлова называлось «психическая
секреция». И. П. Павлов научно проанализировал его и сделал
вывод, что «психическая секреция» является рефлексом, но не-
обычным, а возникающим как временная связь организма с пред-
метами внешней среды. Эти рефлексы Павлов назвал условными
рефлексами. Другие рефлексы, известные и изученные раньше,
он назвал безусловными. И. П. Павлов сделал вывод, что условные
рефлексы — это основная форма деятельности коры больших по-
лушарий и высшая нервная деятельность животных, их поведение
являются взаимодействием условных и безусловных рефлексов.
Безусловные рефлексы. Эти рефлексы возникают при раздра-
жении определенных рецепторов, и для каждого безусловного реф-
лекса имеется своя постоянная рефлекторная дуга. Условные реф-
лексы могут образоваться на любые раздражения любого рецепто-
ра и не имеют постоянных рефлекторных дуг: один и тот же ус-
ловный рефлекс, например слюноотделительный, можно выра-
ботать на свет лампочки, на звук звонка, на прикосновение к
коже и т.д. Безусловные рефлексы осуществляются и при отсут-
ствии коры больших полушарий, они результат деятельности низ-
ших отделов центральной нервной системы. Условные рефлексы
являются функцией коры больших полушарий, после ее удаления
исчезают и вновь не образуются.
Отличие условных рефлексов от безусловных. Все безусловные
рефлексы врожденные; так, новорожденный теленок начинает со-
сать, вылупившийся из яйца цыпленок — клевать. Условные реф-
лексы — приобретенные, вырабатываются животными в течение
всей жизни. Безусловные рефлексы — постоянные, условные —
временные, непостоянные. Безусловные рефлексы являются ви-
довыми, т.е. свойственными всем представителям данного вида;
например, все ежи при опасности свертываются в клубок. Услов-
ные рефлексы индивидуальны, так одно животное реагирует на
одну кличку, другое — на другую.
Процесс образования условного рефлекса. Для образования ус-
ловного рефлекса необходимо, чтобы условный раздражитель (сиг-
432
нал) начал действовать на несколько секунд раньше безусловно-
го раздражителя и в течение некоторого времени сопровождал
его (рис. 4.26). Например, для образования у собаки условного реф-
лекса на звонок он должен начать звонить на 5 с раньше, чем
собака получит корм, и некоторое время сопровождая еду. В даль-
нейшем по мере повторного применения условного и безусловно-
го раздражителей условный рефлекс становится прочным.
Биологическое значение условных рефлексов. Условные рефлек-
сы присущи всем видам животных и человеку. С помощью услов-
ных рефлексов животные приобретают опыт, который помогает
им в добывании корма, в защите от вредных влияний. Свой жиз-
ненный опыт, всю совокупность своих условно-рефлекторных свя-
зей старшее поколение передает молодому поколению при помо-
щи подражательных условных рефлексов. В обычных условиях су-
ществования у животных вообще не бывает чистых безусловных
рефлексов. Каждый безусловный рефлекс обрастает условными
рефлексами различной сложности. Рефлекторные акты, являющи-
еся результатом слияния условных и безусловных рефлексов, на-
зывают сложнорефлекторными.
Анализ и синтез в коре больших полушарий. Высшая нервная
деятельность животного проявля-
ется в форме анализа и синтеза
явлений внешнего мира. Рецеп-
торы избирательно улавливают
раздражения, исходящие из внеш-
ней и внутренней среды. Пери-
ферический конец анализатора
(рецепторы) производит элемен-
тарный анализ раздражений, а
корковый конец — тончайший их
анализ, с помощью которого и
обеспечивается приспособление
животного к условиям своего су-
ществования. Кора больших по-
лушарий производит не только
анализ, но и синтез поступающих
сигналов. Последний проявляет-
ся в связывании, обобщении,
объединении возбуждений, воз-
никающих в различных участках
коры. Образование условного
рефлекса, т. е. формирование вре-
менной связи между двумя воз-
бужденными участками коры,
есть форма проявления синтети-
ческой деятельности коры.
15 Зеленсвский
Рис. 4.26. Схема рефлекторной
дуги безусловного и условного
рефлексов:
1 — эфферентные пути от вкусовых
рецепторов; 2 — центр слюноотделе-
ния продолговатого мозга; 3 — эффе-
рентные пути к слюнной железе; 4 —
пищевой центр коры; 5 — афферент-
ные пути от слуховых рецепторов; 6 —
слуховой центр коры; 7— временная
связь между слуховым и пищевым
центрами коры; стрелки показывают
направление движения импульсов воз-
буждения
433
Одним из проявлений процессов синтеза в коре больших по-
лушарий является образование динамического стереотипа при
действии на животное комплекса условных раздражителей, пред-
лагаемых в строго определенной последовательности. Много-
кратно повторяясь изо дня в день, такие стереотипные комп-
лексы раздражителей приводят к выработке у животных систе-
мы условных рефлексов, которые связаны между собой. При
образовавшемся динамическом стереотипе условный сигнал вос-
принимается не как отдельный изолированный раздражитель,
а как элемент (часть) определенной системы сигналов, нахо-
дящийся в связи с предыдущими и последующими раздражите-
лями. У животных в течение жизни вырабатывается сложная
система условных рефлексов — динамический стереотип — тес-
но связанных друг с другом. Примером может служить система
условных рефлексов на доильный процесс, на раздачу кормов
и т.д.
Типы нервной системы. В основу деления животных по типам
нервной деятельности И. П. Павлов взял свойства нервных про-
цессов — возбуждения и торможения: силу, уравновешенность и
подвижность. Сила нервных процессов определяется работоспо-
собностью клеток коры больших полушарий. Уравновешенность ха-
рактеризуется соотношением между силой возбуждения и тормо-
жения. Иногда эти процессы развиты одинаково, иногда же один
процесс (чаще возбуждение) преобладает над торможением. Под-
вижность нервных процессов обусловлена тем, как быстро в клет-
ках коры процесс возбуждения сменяется торможением, и нао-
борот.
Классификация типов высшей нервной деятельности по И. П. Па-
влову совпадаете классификацией темпераментов по Гиппократу.
У животных сильного, уравновешенного, подвижного типа услов-
ные рефлексы вырабатываются быстро и прочно удерживаются;
животные быстро приспосабливаются к изменениям окружающей
среды. Это идеальный тип животных для хозяйств.
Животные сильного, уравновешенного, инертного типа спокой-
ные, могут выдержать воздействие сильных раздражителей, но они
трудно приспосабливаются к быстро меняющимся условиям сре-
ды обитания.
У животных сильного, неуравновешенного типа возбуждение пре-
обладает над торможением. У них трудно вырабатываются тор-
мозные условные рефлексы и дифференцировки, особенно тон-
кие. При напряженной нервной деятельности у животных возни-
кают неврозы — срывы нервной деятельности.
Слабый тип характеризуется слабостью процессов возбуждения
и торможения. Это боязливые, всего избегающие животные. Ус-
ловные рефлексы вырабатываются с трудом. Этот тип мало приго-
ден для хозяйства.
434
Установлена определенная взаимосвязь типа высшей нервной
деятельности и продуктивности животных. У коров сильного, урав-
новешенного и подвижного типа наблюдается высокий устойчи-
вый уровень лактации. Они хорошо приспосабливаются к услови-
ям содержания и кормления. Коровы с неуравновешенной нервной
системой редко обладают устойчивой лактацией, обычно она у
них снижена. У коров слабого типа низкий уровень лактации, удои
сильно колеблются от разных причин.
Лошади сильного, уравновешенного, подвижного типа обла-
дают высокой работоспособностью при всех видах использования,
особенно в спорте.
Свиноматки сильного, уравновешенного, подвижного типа
спокойны, мало реагируют на посторонние раздражители. Они
многоплодны, их поросята имеют наибольшую массу тела, хоро-
шо растут.
Сон и гипноз. Сон является важнейшей физиологической по-
требностью животного организма. Лишение сна животные пере-
носят намного труднее, чем отсутствие пищи. Щенки погибают от
бессоницы на 4—5 сутки. Длительная работа нервных клеток коры
больших полушарий ведет к их утомлению и истощению, вслед-
ствие чего они переходят в торможение. И. П.Павлов рассматри-
вал сон как внутреннее торможение, распространившееся на всю
массу полушарий и на лежащие ниже отделы головного мозга. Во
время сна основной обмен понижается на 13%, пульс на 20%,
кровяное давление падает на 10 % и более, расширяются мелкие
сосуды мозга, замедляется дыхание. Во сне тонус мышц резко
понижен, скелетная мускулатура расслаблена, условные рефлек-
сы заторможены. Кора перестает воспринимать раздражения. В на-
ступлении сна участвуют ретикулярная формация, таламус и ги-
поталамус, в последнем находится центр спа, который при ак-
тивном состоянии коры заторможен. Эксперимсшально ycianoB-
лены два типа спа: медленный и бышрый. В течение ночи 4 5 раз
медленный сон сменяется быстрым сном. Быстрый сои длится 3—
4 мин, затем снова на 50 S0 мин насгупасч медленный сон. У
лошадей, крупных и мелких жвачных, свиней продолжительность
сна 5 — 7 час в сутки. Сон животных периодический и многократ-
ный, в сутки они сня т 7 —S раз. Все животные спя т лежа, а лоша-
ди могут спать стоя.
При гипнозе, как и при сне, кора больших полушарий нахо-
дится в состоянии торможения. Механизм гипноза такой же, что
и при сне. Различия заключаются в том, что при гипнозе процесс
торможения вызывается искусственно и распространяется только
на отдельные участки коры.
Первая и вторая сигнальная система. И. П. Павлов указывал, что
и для животных, и для человека общими являются анализ и син-
тез непосредственных конкретных раздражителей внешней и внут-
435
ренней среды. Такую деятельность мозга он назвал первой сиг-
нальной системой действительности. У человека в отличие от все-
го животного мира особое значение приобрело слово — речь. Сло-
во составило вторую сигнальную систему действительности, яв-
ляясь сигналом первых сигналов. Словами человек обозначает все
то, что он воспринимает при помощи своих рецепторов. Времен-
ные связи в коре больших полушарий человека образуются под
влиянием не только непосредственных раздражений, но и под
влиянием слов, произносимых, слышимых и видимых. Слово как
сигнал сигналов дает возможность отвлечься от конкретных пред-
метов и явлений. Сигнальное значение слова связано не с простым
звукосочетанием, а с его смысловым содержанием, понятием, ко-
торое в него вложено. Вторая сигнальная система неразрывно свя-
зана с социальной жизнью человека. Словесная сигнализация, речь,
язык являются средством общения людей. Слово возникло у людей
в процессе коллективного труда. Вне общества, без общения с дру-
гими людьми вторая сигнальная система не развивается.
Контрольные вопросы
1. Какова роль И.М.Сеченова и И.П.Павлова в изучении физиологии
коры большого мозга?
2. В чем отличие условных рефлексов от безусловных?
3. Опишите типы высшей нервной деятельности и их связь с продук-
тивностью животных.
ЭТОЛОГИЯ
Врожденное поведение — инстинкт. Этология — это наука о
поведении животных. Ее название происходит от греческого слова
«этос», что значит привычка, обычай, поведение. Этология изу-
чает биологические основы поведения животных, которое рас-
сматривают как проявление внешней преимущественно двигатель-
ной активности, обеспечивающей их приспособление к условиям
существования. В основе поведения животного лежит состояние и
деятельность нервной и гуморальной систем. Люди еще в глубо-
кой древности изучали поведение животных, чтобы удачнее охо-
титься. Одомашнивание животных тоже требовало знаний поведе-
ния животных. Этология как наука получила развитие с конца
XIX столетия, когда возникло два направления исследования по-
ведения животных: изучение физиологических механизмов услов-
но-рефлекторной деятельности, лежащей в основе поведения
(И. П. Павлов) и анализ инстинктивного поведения животных. Это-
логия в последнее время приобретает большое прикладное значе-
ние. Знание поведения животных дает возможность найти более
экономичные способы их содержания.
436
Для изучения поведения животных применяют различные ме-
тоды. Широко используется метод наблюдения за поведением жи-
вотного в естественных условиях. При этом подробно отмечают
все формы поведения и ведут хронометраж, как долго животные
передвигаются, стоят, лежат, принимают корм и т.д.
В этологии применяются также различные экспериментальные
методы. Например, выращивание детенышей изолированно от
родителей дает возможность выяснять, что в поведении передает-
ся по наследству, а что приобретается в процессе жизни. Метод
обходного пути — животному для достижения цели дают возмож-
ность обойти одну или несколько преград. Метод лабиринтов по-
зволяет изучать сообразительность животных в поиске выхода из
лабиринта. Метод вживления электродов в различные структуры
мозга позволяет изучать, какие отделы мозга отвечают за то или
иное поведение.
Приобретенное поведение на основе обучения. Различают роди-
тельское, половое, пищевое, оборонительное, исследовательское
поведение и т.д. В основе поведения животных лежит инстинкт —
системы сложных цепных безусловных рефлексов, при которых
конец одного рефлекса вызывает начало другого. В процессе жиз-
ни на инстинктивное поведение наслаивается масса условных реф-
лексов.
При изучении поведения животных большое значение имеет
выяснение вопроса, как происходит обучение животных. Обуче-
ние — это процесс приобретения жизненного опыта. В основе
многочисленных видов обучения лежит выработка условных реф-
лексов различной сложности. Наиболее часто обучение происхо-
дит способом подражения. Так, молодняк всех видов животных,
подражая взрослым животным, приобретает их опыт. Или, па-
пример, при погрузке лошадей в вагоны некоторые из них со-
противляются и не входят в вагон. Если па глазах такой лошади
ввести другую лошадь, то вслед за ней идет и сопротивлявшаяся.
Метод проб и ошибок — ч тобы достичь цели, животное соверша-
ет различные поисковые реакции. Когда одна из поисковых ре-
акций приводит к цели, то в аналогичной ситуации животное
всегда использует ее.
Большое значение в поведении животных имеют ориентиро-
вочные реакции. При вводе в новое помещение, появлении незна-
комых людей, предметов животные осматриваются, прислушива-
ются, принюхиваются, обнюхивают землю, предметы. В соответ-
ствии с данными ориентировки животные ведут себя определен-
ным образом. Ориентировочные реакции особое место занимают
в поведении молодых животных. Исследовательская активность,
подражание движениям матери и других животных помогает мо-
лодому животному совершенствовать свои приемы добывания кор-
ма и другие необходимые для жизни действия.
437
Важное значение имеет групповое поведение животных. Поведе-
ние каждой особи в группах определяется стадной иерархией —
порядком подчинения одних особей другим по строго определен-
ному ранговому месту в группе. Такая организация способствует
диким животным выжить в борьбе за существование. В настоящее
время условия жизни сельскохозяйственных животных измени-
лись, но поведение их при групповом содержании определяется
стадной иерархией, во главе стада всегда стоит лидирующее жи-
вотное — вожак. Законы стадной иерархии необходимо учитывать
при формировании стада животных, проведении их перегруппи-
ровок.
Поведение сельскохозяйственных животных в условиях промыш-
ленных комплексов. Содержание животных и производственные
процессы нужно организовывать с учетом особенностей поведе-
ния того или иного вида, добиваться выработки у них различных
условных рефлексов, формирующихся уже с первых дней жизни.
Так, у поросят можно выработать условный рефлекс на сосание
определенного соска свиноматки. В овцеводстве в отаре овец обычно
вожаками стада держат коз, так как у них быстро вырабатываются
условные рефлексы на словесные команды. В коневодстве табун-
щики для управления табуном вырабатывают у лошадей условные
рефлексы на словесные приказания и свистки.
В настоящее время организовано много крупных птицефабрик,
свиноводческих и молочных комплексов. В них существенно изме-
нены условия содержания животных, что сказывается на их пове-
дении. В промышленном животноводстве в результате большой
плотности размещения животных, насыщения комплексов раз-
нообразной техникой резко увеличена физиологическая нагрузка
на организм животных, и в первую очередь на нервную систему.
Некоторые животные, особенно неуравновешенного, а также сла-
бого типов нервной системы не выдерживают такой нагрузки.
Продуктивность падает и их приходится выбраковывать. Для про-
мышленного животноводства необходимо отбирать животных силь-
ного уравновешенного типа нервной системы, которые обладают
быстрой адаптацией к таким условиям существования, устойчи-
вы к стрессовым состояниям и заболеваниям. Все это следует учи-
тывать в селекционно-племенной работе и разводить животных с
нужными качествами нервной системы.
Контрольные вопросы
1. Какая наука изучает поведение животных?
2. Опишите инстинкты врожденные и приобретенные.
3. Расскажите о методах изучения поведения животных.
4. Каково поведение животных в условиях промышленных комплексов?
5. Как надо использовать знания науки о поведении животных для
организации их содержания?
438
ФИЗИОЛОГИЯ ОРГАНОВ ЧУВСТВ
На животное постоянно влияют множество раздражителей: свет
различной силы и спектра, всевозможные звуки, запахи, колеба-
ния температуры, давления, прикосновения. Разнообразнейшие
воздействия внешнего мира воспринимаются органами чувств,
благодаря которым и осуществляется связь организма с окружаю-
щей его средой.
Во всех частях организма имеются окончания чувствительных
нервов. Их называют рецепторами. Различают экстерорецепторы,
воспринимающие раздражения из внешней среды, и интероре-
цепторы — сигналы из внутренней среды организма. Раздражение
рецепторов теплом, холодом, светом, звуком, давлением, хими-
ческими веществами и другими раздражителями вызывает появ-
ление в них нервных импульсов, которые поступают в мозг и пе-
редают соответствующую информацию.
В процессе эволюции у животных чувствительные нервные окон-
чания специализировались и объединялись в комплексы. Так об-
разовались органы зрения, слуха, обоняния и другие органы чувств.
Органы чувств воспринимают бесчисленное количество сигналов,
идущих из внешней и внутренней среды организма и передающих
информацию о них в центральную нервную систему. Это помогае г
животному приспосабливаться к изменениям окружающей среды
той или иной поведенческой реакцией.
Большое значение для дальнейшего развития физиологии ор-
ганов чувств имело учение И. П. Павлова об анализаторах. И. П. Пав-
лов анализатором называл единую систему, состоящую из трех
функционально связанных между собой частей: рецептора — пе-
риферической части анализатора, проводниковой части и цент-
ральной, находящейся в определенном месте коры больших по-
лушарий. Рецепторы специализированы для восприятия опреде-
ленного раздражителя. Проводниковая часть передает возбужде-
ние от рецепторов в кору больших полушарий центр анализа-
тора, здесь происходит тончайший анализ поступившего возбуж-
дения, которое воспринимается как ощущение.
ЗРИТЕЛЬНЫЙ АНАЛИЗАТОР
При помощи зрения животные воспринимают форму и вели-
чину предметов, их освещенность, определяют расстояние до них,
перемещение в пространстве; некоторые виды животных различа-
ют цвет предметов.
Оптическая система глаза. Состоит из роговицы, передней и зад-
ней камер глаза, заполненных внутриглазной жидкостью, хруста-
лика и стекловидного тела. Хрусталик похож на двояковыпуклое
439
увеличительное стекло. По краю всей окружности хрусталика к
нему прикреплена связка, называемая хрусталиковой. Хрусталик
заключен в капсулу и посредством хрусталиковой связки прикреп-
лен к ресничной мышце. При сокращении ресничной мышцы из-
меняются выпуклость хрусталика и соответственно его преломля-
ющая сила. Пройдя через оптическую систему глаза, световой луч
попадает на сетчатую оболочку. На последней появляется умень-
шенное, обратное изображение предмета. В нормальном глазу в
состоянии покоя ресничные мышцы расслаблены, связки хруста-
лика натянуты, хрусталик имеет небольшую выпуклость и лучи от
удаленных предметов фокусируются на сетчатке. Предметы, нахо-
дящиеся вдали, хорошо видны, а близкие — кажутся расплывча-
тыми. При рассматривании близких предметов сокращается рес-
ничная мышца, связка хрусталика расслабляется и хрусталик де-
лается более выпуклым, увеличивается его преломляющая сила.
Поэтому близкие предметы будут видны хорошо, а удаленные —
плохо, они кажутся расплывчатыми. Свойство глаза видеть хоро-
шо предметы, находящиеся вблизи и вдали, называется аккомо-
Рис. 4.27. Схема строения сетчатки
глаза:
7 — пигментный слой; 2 — слой пало-
чек (а) и колбочек (6); 3 — биполяр-
ные нейронные; 4 — горизонтальные
(звездчатые), 5 — амакриновые клет-
ки, соединяющие по горизонтали би-
полярные и ганглиозные клетки; 6 —
ганглиозные клетки: 7— волокна зри-
тельного нерва
дацией. Аккомодация осуществля-
ется за счет изменения кривиз-
ны хрусталика. Качество зрения
изменяется в зависимости от воз-
раста животного. У старых живот-
ных хрусталик теряет свою элас-
тичность, и при рассматривании
близких предметов его выпук-
лость не увеличивается. При этом
состоянии развивается дально-
зоркость, при которой хорошо
видны удаленные предметы и
плохо близкие. Дальнозоркость
развивается и в том случае, ког-
да глазное яблоко укорочено.
Иногда глазное яблоко удли-
нено, лучи от дальних предме-
тов фокусируются перед сетчат-
кой, и видны плохо дальние
предметы; такое явление назы-
вается близорукостью. Дально-
зоркость и близорукость часто
наблюдаются у лошадей; наибо-
лее близоруки овцы.
Световоспринимающие струк-
туры глаза. Сетчатка глаза состо-
ит из нескольких слоев, толщи-
на ее 0,1 —0,2 мм. Наружный
440
слой состоит из пигментных клеток, содержащих пигмент фус-
цин; он поглощает свет и препятствует его рассеиванию, при силь-
ном освещении зерна пигментных клеток перемещаются и засло-
няют от яркого света палочки и колбочки (рис. 4.27). Затем идет
слой палочек и колбочек, они являются зрительными рецептора-
ми — фоторецепторами. Фоторецепторы сетчатки содержат свето-
чувствительные вещества: палочки — родопсин, или зрительный пур-
пур (красного цвета), колбочки — йодопсин (фиолетового цвета).
На свету родопсин распадается на ретинен (альдегид витамина
А) и белок опсин. В темноте происходит восстановление родопси-
на, для этого необходим витамин А, который содержится в пиг-
ментном слое.
Палочки в 1000 раз чувствительнее к свету, чем колбочки. По-
этому при слабом освещении функционируют только палочки. Они
расположены в основном по периферии сетчатки, и поэтому в
сумерки лучше видны предметы, расположенные по сторонам. При
ярком свете восстановление родопсина не поспевает за его распа-
дом, и основными световоспринимающими рецепторами становятся
колбочки. Таким образом, палочки являются аппаратом сумереч-
ного зрения, а колбочки — дневного. У животных, ведущих днев-
ной образ жизни, в сетчатке преобладают колбочки (куры, голу-
би), а у ночных животных (летучая мышь, сова, крот, кошка,
еж) — палочки. Однако кошки прекрасно видят днем, а содержа-
щиеся в неволе ежи легко приспосабливаются к дневному образу
жизни.
Проводниковый отдел зрительного анализатора. От фоторецеп-
торов возбуждение через отростки передается второму нейрону —
биполярным клеткам, а те передают возбуждение третьему ней-
рону — ганглиозным клеткам. Аксоны ганглиозных клеток образу-
ют волокна зрительного нерва, в нем содержи гея до Х00 ()()() воло-
кон. Одна биполярная клетка кон тактирует с несколькими палоч-
ками. Каждая колбочка соединена только с одной биполярной клет-
кой. Поэтому импульсы ог рядом находящихся палочек сливают-
ся, а от двух рядом находящихся колбочек передаются отдельно.
При рассматривании задней стенки глазного яблока (т.е. глаз-
ного дна) с помощью специального вогнутого зеркала — офталь-
москопа, можно увидеть участок, от которого расходятся крове-
носные сосуды и выходит зри тельный нерв. Этот участок называ-
ют слепым пятном, поскольку он не содержит нейроэпителия с
палочками и колбочками. Приблизительно в центре сетчатки нахо-
дится центральная ямка — это место наилучшего видения. В ней
имеются одни только колбочки.
Участок вокруг центральной ямки окрашен в желтый цвет и
называется желтым пятном.
Волокна зрительного нерва, отходящие от сетчатки, на базаль-
ной поверхности мозга перекрещиваются. После зрительного пе-
441
рекреста зрительные нервы обоих глаз переходят в зрительные
тракты, в составе которых имеются прямые проводящие пути из
латеральных отделов сетчаток глазных яблок и перекрещенные пути
из медиальных отделов сетчаток.
Таким образом, каждый зрительный тракт содержит волокна
из обоих глаз. Этим достигается лучшее качество зрения (стерео-
скопичность). Волокна зрительных трактов оканчиваются в трех
первичных (подкорковых) зрительных центрах: а) латеральных
коленчатых телах; б) в каудальных ядрах зрительных бугров; в) в
назальных холмах четверохолмия. Из перечисленных первичных
подкорковых центров происходят четвертые нейроны, образую-
щие центральные проводящие пути зрительного анализатора. Из
латерального коленчатого тела (и из каудальных ядер зрительных
бугров) четвертые нейроны передают импульсы в корковые зри-
тельные центры затылочной доли коры полушарий. Из назальных
холмов четверохолмия четвертые нейроны формируют трактус тек-
тоспиналис, по которому импульсы передаются: а) на моторные
клетки вентральных столбов шейногрудной части спинного мозга
(эти клетки представляют собой нейроны, через которые осуще-
ствляются рефлекторные движения головы и шеи) и б) на клет-
ки ядер третьего, четвертого и шестого двигательных нервов глаз-
ных мышц. Назальными холмами четверохолмия при участии ней-
ронов, заложенных в парасимпатическом ядре Якубовича и в рес-
ничном узле, управляются также рефлекторные сокращения сфин-
ктера зрачка и ресничного тела.
Мышечный аппарат глаза. Имеет большое значение для нор-
мального процесса зрения.
Глаз все время находятся в движении в результате сокращения
мышц глазного яблока. Глазные мышцы не только устанавливают
глаз на наилучшее видение, но и помогают определить направле-
ние, оценивать расстояние и величину предмета в результате би-
нокулярного зрения. Ресничные мышцы обеспечивают аккомода-
цию глаза. В радужной оболочке имеются кольцевые и радиальные
мышцы. При ярком освещении зрачок суживается в результате
сокращения кольцевых мышц, и на сетчатку поступает меньше
световых лучей. В темноте происходит расширение зрачка вслед-
ствие сокращения радиальных мышц. Этот процесс носит назва-
ние адаптация глаза к силе света.
Защитный аппарат глаза. У млекопитающих животных глаз за-
щищен веками: верхним, нижним и рудеминтированным треть-
им, которые рефлекторно закрываются при раздражении рого-
вицы. По краям век расположены железы, выделяющие глазную
смазку, которая при мигании расплывается по глазному яблоку и
предохраняет его от высыхания и предотвращает скатывание сле-
зы через край века. Слезный аппарат состоит из слезных желез
верхнего и третьего века, слезных канальцев, слезного мешка и
442
слезного протока. Железы выделяют слезы, которые увлажняют и
очищают конъюнктиву и роговицу глаза. В слезах содержится фер-
мент лизоцим, обладающий бактерицидным действием.
Роговица, хрусталик и стекловидное тело не имеют кровенос-
ных сосудов. К клеткам этих тканей питательные вещества посту-
пают из внутриглазной жидкости, заполняющей переднюю и зад-
нюю камеры глаза. В радужной оболочке и ресничном теле, бога-
тых кровеносными сосудами, питательные вещества из крови пе-
реходят в камеры глаза. Но через стенки сосудов проникают лишь
те вещества, которые входят в состав водянистой влаги, а состав
ее отличается от состава крови. Например, аскорбиновой кислоты
в водянистой влаге в несколько раз больше, чем в крови, белков
же нет совсем. Данное свойство стенок кровеносных сосудов глаза
пропускать одни вещества и задерживать другие называется гема-
тоофтальмическим, или глазным, барьером.
РАВНОВЕСНО-СЛУХОВОЙ АНАЛИЗАТОР
Слуховой анализатор. Воспринимает звуковые колебания и дает
ощущение звука. Наружное ухо улавливает звуки. При звуковых
раздражениях животное настораживает уши, поворачивая их в
сторону звука. Барабанная пере-
понка передает колебания через
систему слуховых косточек сред-
него уха к овальному окошку, а
от него — жидкости внутренне-
го уха. Внутреннее ухо состоит из
преддверия, полукружных кана-
лов и улитки, представляющий
собой костный спирально зак-
рученный канал. Улитка распо-
ложена в скалистой части каме-
нистой кости (рис. 4.28). 11о всей
длине почти до конца улитки
костный канал разделен рейс-
неровой и основной мембрана-
ми на три канала.
Основная мембрана сос тоит из
соединител ьн от ка н н ы х вол о ко н,
имеющих поперечное направле-
ние. Эти волокна короткие у ос-
нования улитки и удлиняются к
ее вершине. Внутри среднего ка-
нала на основной мембране рас-
положен звуковоспринимающий
Рис. 4.28. Поперечный разрез улитки:
/ — лестница преддверия; 2 — рейс-
всрова мембрана; 3 — костная стенка
улитки; 4 — спиральная связка; 5 —
канал перепончатой улитки; 6 — кор-
тиева мембрана; 7 — основная мемб-
рана и кортиев орган; 8— барабанная
лестница; 9 — спиральная пластинка;
10 — спиральный ганглий; 11 — от-
ходящие от ганглия нервные волокна;
12 — ось улитки
443
Рис. 4.29. Кортиев орган:
/ — эпителий; 2 — желобок лимба; 3 — кортиева мембрана; 4 — рецепторные
волосковые клетки; 5 — клетки Гензена; 6 — основная мембрана; 7 - опорные
клетки; 8 — спиральный сосуд; 9 — клетки столбов; 10 — спиральная пластинка;
11 — спиральный ганглий; 12 — лимб
аппарат — кортиев орган, который содержит рецепторные волос-
ковые клетки (рис. 4.29).
Звуковые колебания, переданные на мембрану овального окна,
вызывают колебания жидкости в верхнем и нижнем каналах улит-
ки и достигают круглого окошка. Если бы не было круглого окош-
ка, то колебания жидкости были бы невозможны, так как жид-
кость несжимаема.
Современная теория слуха. Длительное время широким призна-
нием пользовалась резонансная теория Г. Гельмгольца. Согласно
ей, при высоких звуках колеблются короткие волокна основной
мембраны у основания улитки, при низких — длинные у верши-
ны улитки. В настоящее время эта теория пересмотрена и предло-
жена новая. В улитке колеблется столб жидкости определенной
длины. Чем выше звук, т.е. чем больше частота колебаний, тем
меньше дл ина колеблющегося столба жидкости в каналах улитки
и тем ближе к основанию улитки и овальному окну расположено
место максимальной амплитуды колебания. При действии звуков
низкой частоты длина колеблющегося столба жидкости увеличи-
вается, а место максимальной амплитуды колебаний отодвигает-
ся в сторону вершины улитки. При колебании жидкости в каналах
улитки колеблются не отдельные волокна основной мембраны, а
определенной величины ее участки. В результате этого возбужда-
ется разное количество рецепторных волосковых клеток кортиева
органа, расположенных на основной мембране. Чем ниже звук
444
(меньше частота колебаний), тем больше рецепторных клеток бу-
дет возбуждаться. Таким образом, каждой высоте тона соответ-
ствуют определенная длина участка основной мембраны, вовле-
ченного в колебание, и определенное число возбужденных ре-
цепторных клеток.
Слуховая чувствительность у разных видов животных неодина-
ковая. Так, собака воспринимает до 80 000 Гц в секунду, лошади и
крупный рогатый скот воспринимают более высокую частоту, чем
человек (человек — до 20 000 Гц). Животные могут определять на-
правление, откуда доносится звук, в результате слушания дву-
мя ушами.
Равновесный (вестибулярный) анализатор. Дает информацию
о положении и движении тела в пространстве. Рецепторы этого
анализатора находятся в лабиринте внутреннего уха: преддверии
и полукружных каналах. Рецепторные клетки соединяются с би-
полярными клетками, отростки которых образуют волокна вес-
тибулярной ветви слухового нерва. Импульсы по нему передают-
ся в продолговатый мозг, в ядро Дейтерса и другие вестибуляр-
ные ядра, от них к мозжечку, а также в височную область коры
больших полушарий, где находится центр анализатора. В пред-
дверии имеются два мешочка, в них расположены рецепторные
клетки, на волосках которых лежат отолиты (рис. 4.30). Они пред-
ставляют собой войлокоподобную студенистую массу, в которой
находится большое количество микроскопических кристалликов
аррагонита (соли углекислого и фосфорнокислого кальция). Раз-
дражителями рецепторных клеток служат наклоны головы, тряс-
ка, ускорение или замедление прямолинейного движения тела.
При этом отолиты или давят, или растягивают волоски и возни-
кает возбуждение.
Рис. 4.30. Схема строения отолитового аппарата:
1 — нервные волокна вестибулярного нерва; 2— опорные клетки; 3 рецептор-
ные клетки; 4 — волоски рецепторных клеток; 5 — желатинозная масса; 6 —
отолитовая мембрана; 7 — отолиты
445
Рис. 4.31. Схема обонятель-
ного эпителия носовой по-
лости:
/ — обонятельная клетка; 2 —
обонятельные волоски и мик-
роворсинки; 3 — опорная клет-
ка; 4 — базальная клетка; 5 —
обонятельные нити
Перепончатые полукружные капа
лы представляют собой три тонкие ду
гообразные трубки, заполненные эн
долимфой, они расположены в грех
взаимно перпендикулярных плоско
стях.
В ампуловидных расширениях капа
лов на небольшом выступе (гребешке)
находятся рецепторные клетки с но
лосками. Рецепторные клетки раздра
жаются в результате смещения эндо
лимфы при угловом ускорении и за
медлении движений. При сильном раз
дражении рецепторов полукружных
каналов возникает рефлекторное из-
менение деятельности сердца, дыха
ния, потоотделения, иногда рвота
(морская болезнь у человека).
ОБОНЯТЕЛЬНЫЙ И ВКУСОВОЙ АНАЛИЗАТОРЫ
Обонятельный анализатор, воспринимающий запахи, является
древнейшим.
Рецепторы обоняния (обонятельные клетки) находятся в носо-
вой полости, в области верхнего носового входа и в задней верх-
ней части носовой перегородки. На поверхности обонятельных
клеток множество микроскопических ресничек, увеличивающих
поверхность соприкосновения с пахучими веществами. От обоим
тельных клеток идут аксоны, образуя обонятельные нити, часть
которых оканчивается в обонятельной луковице, а часть — в ам
меновых рогах (гиппокампе). Эти образования относятся к обоня
тельному мозгу, входящему в состав древней коры (рис. 4.31).
Механизм обоняния. Пахучие вещества проникают к обонятель
ным рецепторам при вдыхании воздуха через нос и при еде. От
пахучих тел непрерывно отделяются молекулы, они очень лету
чие и специфичны для каждого вещества. Молекулы пахучего вс
щества взаимодействуют с обонятельными рецепторами и, адсор
бируясь на их мембране, вызывают возбуждение рецепторов. Me
ханизм этого взаимодействия изучен недостаточно. Существует две
теории запахов: химическая и физическая. Согласно химической
теории, молекула пахучего вещества раздражает рецептор в том
случае, если ее форма строения соответствует углублению на по
верхности мембраны рецептора (по аналогии соответствия между
ключом и замком). Физическая теория объясняет причину запаха
тем, что молекулы пахучих веществ обладают электромагнитны
446
ми колебаниями, частота которых у разных веществ различна. Эти
колебания и улавливаются рецепторами.
Классификация запахов точно не разработана. По одной из
классификаций считают, что имеется семь основных запахов: кам-
фарный, мускусный, цветочный, мятный, эфирный, острый и
гнилостный. Все остальные запахи возникают при их смешивании
в определенной пропорции.
Большинство животных обладают хорошо развитым обоняни-
ем, их называют макросматиками. Слабо развито обоняние у птиц,
китов, обезьян и человека — это микросматики. Особенно хоро-
шо развито обоняние у собак: она может определить присутствие
одной молекулы пахучего вещества в 1 л воздуха. Служебные со-
баки различают индивидуальные запахи следов людей и других
животных среди множества всевозможных запахов. У сельскохо-
зяйственных животных обоняние тоже развито хорошо. Так, коро-
вы различают запахи трав, они ощущают запах раствора аммиака
в концентрации 1 : 100 000.
Среди пахучих веществ особое сигнальное значение имеют па-
хучие вещества, выделяемые животными — феромоны. Они слу-
жат средством общения особей одного вида, с помощью их мар-
кируется занимаемая территория. Обоняние играет важную роль в
половых инстинктах. Запахи, выделяемые самкой во время течки,
возбуждают самца. Животные посредством обоняния отыскивают
корм, обнаруживают других животных в таких условиях, когда
это невозможно сделать с помощью зрения или слуха.
Вкусовой анализатор дает информацию о характере веществ,
входящих в состав корма и поступающих в ротовую полость. Ре-
Рис. 4.32. Вкусовые сосочки языка:
I грибовидные; Б — желобоватые; В — листовидные; Г — вкусовая луковица;
/ поры луковицы; 2 — вкусовая клетка; 3 — опорные клетки; 4 — разветвле-
ния нервною волокна вокруг вкусовой клетки; 5 — нервные волокна
447
цепторы — вкусовые луковицы находятся во вкусовых сосочках сли-
зистой оболочки языка, нёба и в глотке (рис. 4.32). Вкусовые луко-
вицы имеют овальную форму и состоят из веретенообразных вку-
совых клеток с микроворсинками и опорных клеток цилиндри-
ческой формы. В них со слюной проникают химические вещества,
которые возбуждают микроворсинки вкусовых клеток. Животные раз-
личают сладкие, горькие, кислые, соленые вещества и их комби-
нации. Импульсы от вкусовых клеток передаются по нервам в про-
долговатый мозг, а оттуда в таламус и кору больших полушарий.
Кроме вкусовых луковиц, в слизистой оболочке ротовой поло-
сти и на языке имеются рецепторы, воспринимающие прикосно-
вение, боль и температурные раздражения. Вкусовой анализатор
играет важную роль в деятельности аппарата пищеварения. Вкусо-
вой анализатор хорошо развит у травоядных животных. На паст-
бище животные поедают не все травы подряд, а выборочно.
У хищных животных вкусовой анализатор развит хуже.
КОЖНЫЙ АНАЛИЗАТОР
Кожный анализатор ощущает прикосновение, давление, боль,
тепло и холод. Каждое из этих ощущений возникает вследствие
раздражений, которые воспринимаются специальными рецепто-
рами (рис. 4.33).
Рецепторы отличаются друг от друга строением и глубиной рас-
положения в толще кожи. Разные виды рецепторов имеют также
различные проводящие пути. От спинного мозга идут восходящие
чувствительные пути до таламуса, а от него в теменную область
коры, где находится центр кожного анализатора.
Тактильные рецепторы представляют собой специальные об-
разования — осязательные тельца Мейснера, меркелевы диски и
пачиниевы тельца. Они воспринимают прикосновение и давление. Так-
Рис. 4.33. Рецепторы кожи:
1 — пачиниево тельце; 2 — мейснерово тельце; 3 — нервное сплетение вокруг
луковицы; 4 — свободные нервные окончания; 5 — тельца Меркеля; 6 — колба
Краузе; 7 — тельца Фатера—Пачини
448
тильную чувствительность повышают волосы кожи — слеш
ные осязательные волоски — вибриссы, расположенные на
Раздражитель вызывает ощущение прикосновения или давления и
том случае, когда он деформирует поверхность кожи. Различные
участки ее обладают неодинаковой тактильной чувствительнос-
тью, так как рецепторы распределены неравномерно. Тактильные
рецепторы быстро адаптируются, поэтому ощущается не само при-
косновение и давление, а только их изменение.
Болевые ощущения возникают при раздражении специальных
рецепторов в виде свободных нервных окончаний. Чувство боли
возникает при действии самых различных раздражителей: меха-
нических, температурных, химических, электрических. Ощуще-
ние боли сопровождается повышенным выделением в кровь адре-
налина, увеличением количества сахара в крови, возрастанием
силы и частоты сердечных сокращений, повышением кровяного
давления и т.д. Чувство боли появляется при значительно боль-
шой силе раздражителя. Ощущение боли вызывает у животных
защитные рефлексы, которые предохраняют их от вредных воз-
действий окружающей среды.
Кожный анализатор имеет большое значение в жизни живот-
ного. Благодаря большой поверхности и наружному расположе-
нию кожи ее рецепторам принадлежит важная роль в восприятии
внешнего мира.
ДВИГАТЕЛЬНЫЙ АНАЛИЗАТОР
Двигательный анализатор, или мышечно-суставное чувство,
дает представление о положении тела в пространстве и его от-
дельных частей, остепени сокращения мышц, натяжении связок.
С помощью этого анализатора животное знает о положении свое-
го тела и без зрительного контроля при ыкрытых ишзах. Это
происходит потому, что в мышцах, сухожилиях, связках и па по-
верхности суставов находятся рецепторы, которые называются
проприорецепторами'. в мышцах эго мышечное веретено, в сухожи-
лиях — тельца Гольджи. Проприорецепторы возбуждаются при мы-
шечных сокращениях, они не способны к адаптации. Центр дви-
гательного анализатора находится в теменной области коры боль-
ших полушарий. Проводниковый отдел этого анализатора переда-
ет импульсы также в другие центры головного мозга: продолгова-
тый и средний мозг, мозжечок.
Каждый анализатор имеет центры в определенной области коры
больших полушарий. Эти центры между собой взаимосвязаны.
В естественных условиях одновременно раздражается несколько
анализаторов, например, слуховой и зрительный, вкусовой и обо-
нятельный и т.д. Двигательный анализатор взаимодействует с дру-
449
гими анализаторами, например с анализаторами положения тела
в пространстве, слуховым, зрительным. Возбуждение одних ана-
лизаторов влияет на другие, поэтому при освещении звуки ка-
жутся более громкими, чем в темноте; ультразвуки повышают
остроту зрения. В результате взаимодействия анализаторов осуще-
ствляются сложные акты поведения животных.
Контрольные вопросы
1. Опишите строение анализатора по И.П.Павлову.
2. Назовите общие свойства анализатора.
3. Что такое зрительный анализатор, механизм адаптации и аккомо-
дации глаза?
4. Какова оптическая система глаза?
5. Расскажите о рецепторном аппарате зрительного анализатора.
6. Слуховой анализатор, его аккомодационный и рецепторный аппарат.
7. Объясните механизм функционирования равновесного анализатора.
8. Каков механизм восприятия запахов?
9. Что такое вкусовой анализатор?
10. Охарактеризуйте двигательный анализатор.
11. Перечислите виды кожной чувствительности.
ФИЗИОЛОГИЯ АДАПТАЦИИ
Адаптация — приспособление организмов к жизни посредством
свойств, обеспечивающих их выживание и размножение в усло-
виях меняющейся внешней среды. Под физиологической адапта-
цией понимают совокупность морфофизиологических процессов
в организме, лежащих в основе его приспособления к конкрет-
ным условиям внешней среды. Адаптация повышает устойчивость
организма к перепадам температуры воздуха, недостатку кисло-
рода, воды, освещения, различным видам корма и многим дру-
гим факторам, связанным с экологическими условиями обита-
ния и поведения разных видов животных.
Изменение условий среды обитания естественно вызывает фун-
кциональные сдвиги в организме, которые при определенных ус-
ловиях могут вызывать болезненные состояния животного.
По эколого-генетической классификации физиологическую адап-
тацию подразделяют на видовую, или унаследованную, и индивиду-
альную, или приобретенную. Так, одни виды животных приспособи-
лись к холодному климату, другие — к умеренному, третьи — к
высокогорным условиям, а некоторые — к тропическому климату.
Критериями адаптации служат реакции сердечно-сосудистой и
дыхательной систем, картина крови, функция желудочно-кишеч-
ного тракта, состояние водного обмена, температура тела. Адап-
тация лучше проявляется у молодняка.
450
Механизмы адаптации. В процессе адаптации организм живот-
ных реагирует как единое целое с участием всех его органов и
систем при ведущей роли центральной нервной системы. Но пер-
вым с различными воздействиями окружающей среды на орга-
низм соприкасается рецепторный аппарат анализаторов. Анализа-
торы, реагируя на силу раздражителя, передают сигналы в соот-
ветствующие подкорковые и корковые центры. Ответные реакции
имеют, прежде всего, защитно-приспособительный характер и
направлены на адаптацию организма к новым условиям. Установ-
лено исключительно важное значение в адаптации организма сим-
патической нервной системы (Л. А. Орбели). С помощью ее проис-
ходит мобилизация энергетических ресурсов, стимулируется фун-
кция сердечно-сосудистой системы, усиливается работоспособ-
ность мышц, активизируются иммунологические процессы. В раз-
витии общей адаптации организма большое значение имеет гипо-
физарно-надпочечниковая система. Совокупность реакций организ-
ма в ответ на возбуждение этой системы называется адаптацион-
ным синдромом, или стрессом.
Стресс развивается в три стадии. Первая стадия — «реакция
тревоги» — характеризуется активацией надпочечников и выбро-
сом в кровь катехоламинов и глюкокортикоидов, что способству-
ет повышению устойчивости организма к неблагоприятным фак-
торам.
Во вторую стадию — «стадию резистентности» — повышается
устойчивость организма к ряду чрезвычайных раздражителей. Уве-
личивается количество циркулирующей крови, возрастает арте-
риальное давление, усиливается глюконеогенез. Глюкокортикои-
ды активизируют иммунологические процессы защиты, образо-
вание антител, фагоцитоз и др.
Третья стадия — «стадия истощения» — возникает тогда, ког-
да действие стрессоров продолжается. Надпочечники, особенно
их корковая часть, истощаются и в этот период может наступить
гибель организма.
Адаптация животных к внешней температуре, газовой среде, ос-
вещению и другим условиям содержания. Для каждого возрастного
периода имеется свой температурный оптимум. Например, для
новорожденных телят в первые дни жизни он равен 16—18°С, от
года до двух лет 3 — 5 °C, а для продуктивных коров ниже О °C.
У телят температурный гомеостаз удерживается преимуществен-
но за счет физической терморегуляции. Поросята до 10-дневного
возраста в условиях как высоких, так и низких температур не спо-
собны поддерживать температурный гомеостаз вследствие слабо-
развитых механизмов терморегуляции. Только к месячному возра-
сту терморегуляция у них достигает уровня взрослых животных.
В поддержании температурного гомеостаза у жвачных живот-
ных важную роль играют преджелудки, особенно рубец, который
45!
служит температурной рефлексогенной зоной. Температура в руб-
це постоянно высока, что связано с высоким уровнем обменных
реакций, которые сглаживают влияние окружающей среды.
Животные некоторых видов приспособились к недостатку воды:
например, верблюды и курдючные овцы имеют хорошо развитые
жировые ткани. В горбах верблюдов хранится до 100 кг жира, ко-
торый в условиях водного голодания животных, окисляясь, мо-
жет выделить до 40 —50 л воды. Верблюды могут обходиться без
питья в течение 10—12 дней. При ограниченном приеме воды тем-
пература тела у них может повышаться на 5 °C и более без види-
мых нарушений функций организма. Лошади, крупный рогатый
скот и некурдючные овцы при таком повышении температуры
погибают. При питании сочной растительностью верблюд может
совершенно не пить воды. Летом на сухом подножном корме вер-
блюд подходит к воде не чаще одного раза в четыре дня. В самое
жаркое время года он может находиться на безводной диете около
семи дней, но теряя при этом почти четверть массы.
Животным, адаптированным к жаркому климату, свойственна
более светлая окраска шерсти при пигментированной коже. Круп-
ный рогатый скот с довольно длинной шерстью хуже переносит
длительное пребывание на солнце, чем животные с короткой
шерстью.
Животные в определенной степени адаптируются и к низким
температурам. При минусовой температуре воздуха у молодняка
наблюдается мышечная дрожь — своеобразный согревательный
механизм, и усиливается рост шерсти. Сужение сосудов кожи мо-
жет снизить потери тепла до 70 %.
Адаптация к пониженному содержанию кислорода у животных
продолжается в течение 20 — 25 дней. В этом процессе выделяют
три стадии: первая связана с усилением легочной вентиляции,
увеличением кровотока, повышением активности ферментов кро-
ви, изменением кислотно-щелочного равновесия. Вторая стадия
проявляется увеличением содержания гемоглобина и активнос-
тью тканевых ферментов. В третьей отмечают усиление анаэробно-
го гликолиза, снижение потребления кислорода и повышение об-
шей резистентности организма. Происходит перестройка обмен-
ных процессов на более низкий уровень. Легче и быстрее на тка-
невый тип адаптации переходит молодняк.
Адаптация животных в промышленных комплексах. У животных,
в том числе и лактируюших коров, в искусственно созданных ус-
ловиях микроклимата сохраняется обычное физиологическое со-
стояние, но частота дыхания, работа сердца, температура и кро-
вяное давление находятся на верхних границах нормы.
Скученное размещение животных не обеспечивает физиологи-
чески необходимую для них двигательную активность. Гиподина-
мия и высокий уровень ненормированного кормления создают
452
условия для ожирения коров, что служит предрасполагающим
фактором в развитии кетоза, яловости и другой патологии, что
указывает на неполную физиологическую адаптацию.
Для поддержания нормальных физиологических функций и
профилактики нарушений обмена веществ в помещении для ко-
ров и нетелей рекомендуется поддерживать следующий микро-
климат: температура воздуха зимой 5 — 16 °C, летом не выше 25 °C,
влажность воздуха 70 — 85 %, содержание двуокиси углерода не
более 0,25 об %, аммиака 0,01 — 0,02 мл/л, сероводорода 0,005 —
0,01 мг/л. Кроме того, лактирующие и стельные сухостойные ко-
ровы должны постоянно иметь активный моцион.
Контрольные вопросы
1. Что такое процесс адаптации?
2. Каковы механизмы адаптации?
3. В чем заключается адаптация животных в промышленных комплек-
сах?
4. Перечислите возрастные границы адаптации.
Таблица П.З
Гематологические показатели у животных
Показатели Крупный рогатый скот Овцы Свиньи Лошади Кролики Пушные звери Птицы
Количество эритроцитов, млн в 1 мм3 5-7,5 7,5-12,5 6-7.5 6-9 5-7,5 8,5-11 2,5-4,5
Количество лейкоцитов, тыс. в 1 мм3 6-10 6-11 8-16 7-12 5,5-9 4-10 20-40
Количество тромбоцитов, тыс. в мм3 450 350 210 350 190 300 50
Содержание гемоглобина, г % 9-12 7-11 9-11 8-14 10-12 12-17 8-13
СОЭ(по Неводову), мин:
15 0,15 0,2 1 35 0 0,5 0,5
30 0,35 0,4 з 54 0,3 0,9 2,0
45 0,50 0,6 5 58 0,9 1,7 3,5
60 0,70 0,6 8 64 1,5 2,5 4,0
Лейкограмма, %:
нейтофилы 31 44 48 58.5 43 48 -
эозинофилы 6,5 7,5 2.0 4,0 2,0 4,5 -
базофилы 1.0 0,5 0,5 0,5 1,0 0,5 2,0
лимфоциты 57 45 45 34 52 45 54
МОНОЦИТЫ 4,5 3,0 4,5 3,0 2,0 2.0 6,0
Скорость свертывания крови, мин 7-9 4-5 3-4 10-12 5-6 4-5 2-3
Таблица П.4
Парциальное давление кислорода и углекислого газа в альвеолярном
воздухе и их напряжение в крови и тканях при 37 С
Исследуемая среда Парциальное давление, мм рт.ст. Общее давление, мм рт. ст.
Н2О со2 О2 n2
Вдыхаемый воздух 6 0,3 159 595 760
Выдыхаемый воздух 47 32 116 565 760
Альвеолярный воздух 47 40 100-110 573 760
Артериальная кровь 47 40 100-105 573 755
Смешанная венозная кровь 47 47 40 573 706
Тканевая жидкость 47 60 20 573 700
Таблица П.5
Ректальная температура у различных видов животных
Животные Температура, °C Животные Температура, °C
Коровы 37,5-39,0 Буйволы 37,0-38,5
Лошади 37,5-38,5 Верблюды 37,5-38,5
Свиньи 38,0-40,0 Олени 38,0-38,5
Овцы 38,5-40,0 Кролики 38,5-39,5
Козы 38,5-40,0 Норки 39,5-40,5
Собаки 37,5-39,5 Куры 40,5-42,5
Кошки 38,0-39,5 Утки 41,0-43,0
456
Таблица П.6
Состав мочи и плазмы крови
Состав Моча, % Плазма крови, % Во сколько раз данного вещества больше в моче, чем в плазме крови
Вода 93-95 90-93 Одинаково
Белки Г 7-9
Глюкоза 0,1 -
Мочевина 2,00 0,03 70
Мочевая кислота 0,05 0,002 25
Na 0,35 0,32 Одинаково
К 0,15 0,02 7
Mg 0,04 0,001 40
Са 0,006 0,0025 2,4
С1 0,6 0,37 1,6
РО4 0,27 0,009 30
SO4 0,18 0,002 90
Таблица П.7
Средние показатели спермы
Животные Объем эякулята, мл Общее число спермиев во всем эякуляте, млрд Концентрация спермиев в 1 мл, млрд
Быки 4-5 4-10 1,0-1,5
Жеребцы 50-120 6-15 0,1-0,25
Бараны 1-2 2-10 2,0-4,0
Хряки 150-300 20-80 0,1-0,2
457
Таблица П.8
Продолжительность беременности у животных, дни
Животные в среднем Колебания Животные В среднем Колебания
Лошади 340 307-412 Собаки 62 59-65
Ослы 380 360-390 Кошки 58 55-60
Коровы 285 240-310 Лисицы 51 51-54
Верблюды 365 335-395 Норки 42 37-80
Северные олени 225 195-243 Соболи 250-300
Свиньи 114 110-140 Кролики 30 28-33
Овцы и козы 150 140-160
Таблица П.9
Химический состав молока различных животных, %
Животные Сухие вещества Жир Белок Молочный сахар Мине- ральные вещества
Коровы 13,0 3,7 з,з 4,8 0,7
Козы 13,4 4,3 3,6 4,5 0,85
Овцы 18,5 7,2 5,7 4,6 0,9
Буйволицы 17,5 7,7 4,2 4,7 0,8
Верблюдицы одногорбые 13,0 4,5 3,6 4,9 0,7
Верблюдицы двугорбые 15,0 5,4 3,8 5,0 0,7
Самки яка 17,8 6,8 5,0 5,0 0,9
Кобылицы 10,7 1,8 2,1 6,4 0,35
Ослицы 9,9 1,4 1,9 6,2 0,5
Свиньи 17,4 5,9 6,2 4,0 1,1
Собаки 21,1 8,6 7,1 4,1 1,3
Крольчихи 30,5 10,5 15,5 2,0 1,0
Самки северного оленя 33,8 18,7 10,0 3,6 1,4
458
Таблица П.10
Средний состав молока и молозива коровы, %
Вещества Молоко Молози во первого дня Вещества Молоко Молозиво первого дня
Вода 87,5 75,42 Молочный сахар 4,7 3,31
Белок з,з 15,08 Минераль- ные соли 0,7 1,2
Жир 3,8 5,4
Таблица П.11
Классификация типов высшей нервной деятельности
Типы высшей нервной деятельности, по И. П. Павлову Классификация темпераментов человека, по Гиппократу
по силе по уравновешенности по подвижности
Сильный У равнове шейный Подвижный Сангвиник
Сильный Уравновешенный Инертный Флегматик
Сильный Н еура вновешенный - Холерик
Слабый Меланхолик
ОГЛАВЛЕНИЕ
Предисловие................................................ 3
Глава 1. Морфология и физиология (значение, развитие,
связь с другими биологическими науками)..................... 5
Глава 2. Цитология, гистология и эмбриология...............14
Цитология..........................................14
Гистология.........................................29
Эмбриология........................................47
Глава 3. Анатомия..........................................59
Общие термины, употребляемые при описании
строения тела животного....................................59
Понятие о частях и областях тела животного.................61
Локомоторный аппарат.......................................67
I. Остеология........................................67
II. Артрология......................................124
III. Миология.......................................134
Кожный покров..............................................173
Спланхнология..............................................180
Пищеварительный аппарат............................182
Органы дыхания....................................210
Органы мочеотделения..............................218
Органы размножения самцов.......................222
Органы размножения самок........................230
Ангиология ................................................236
Кровеносная система...............................236
Лимфатическая система.............................263
Органы внутренней секреции.................................277
Неврология.................................................280
Центральная нервная система.......................281
Периферическая нервная система.....................291
Автономная нервная система.........................301
Анализаторы ...............................................306
Зрительный анализатор..............................306
Статоакустический анализатор ......................310
Особенности строения домашних птиц........................313
Скелет............................................313
Мышечная система..................................318
Кожный покров птиц................................319
Пищеварительный аппарат...........................322
460
Органы дыхания...........
Органы мочеотделения....... OS
Органы размножения........................ . 129
Сердечно-сосудистая еиезема ...................... И|
Нервная система........................... И '
Анализаторы..................................... И /
Глава 4. Физиология....................................... ИК
Система крови............................................338
Функции крови.....................................339
Физико-химические свойства крови..................340
Плазма и форменные элементы крови.........342
Свертывание крови.................................348
Группы крови......................................349
Кроветворение ....................................350
Лимфатическая система. Лимфа и тканевая жидкость...350
Физиология иммунной системы...............................352
Иммунитет, его значение. Иммунная система.........352
Виды иммунитета ................................ 354
Использование иммунитета в жпвозиоводстве
и ветеринарии............ .. .. .................355
Крово- и лимфообращение................................. 355
Система органов кровобращения........... ... 155
Лимфообращение .................................. ?64
Дыхание ............................................... '<>(>
Пищеварение................................................371
Пищеварение в ротовой полости ....................373
Пищеварение в желудке.............................375
Пищеварение в тонкой кишке........................378
Пищеварение в толстой кишке.......................381
Пищеварение у домашней птицы......................382
Обмен веществ и энергии.................................. 384
Обмен веществ.....................................384
Обмен энергии.....................................394
Теплорегуляция............................................395
Выделение.................................................396
Физиология кожи...........................................398
Внутренняя секреция.......................................400
Общая характеристика желез внутренней секреции....400
Железы внутренней секреции .......................402
Размножение...............................................404
Лактация .................................................412
Физиология мышц и нервов..................................415
Физиология мышц................................. 418
Физиология нервного волокна.......................421
Физиология центральной и периферической нервной системы.... 423
Физиология головного мозга........................425
Физиология спинного мозга.........................428
Физиология вегетативной нервной системы...........429
461
Высшая нервная деятельность................................431
Этология.................................................. 436
Физиологии органон чувств..................................439
Зрительный анализатор.............................439
Равновесно-слуховой анализатор....................443
Обонятельный и вкусовой анализаторы...............446
Кожный анализатор.................................448
Двигательный анализатор...........................449
Физиология адаптации.......................................450
Приложение.................................................454
Учебное издание
Зеленевский Николай Вячеславович,
Васильев Александр Петрович,
Логинова Людмила Константиновна
Анатомия и физиология животных
Учебник
Редактор Г.Г.Есакова
Технические редакторы О. С. Александрова, Е.Ф.Коржуева
Компьютерная верстка: Е. Ю. Матвеева
Корректоры В. А. Жилкина, Г. Н. Петрова, Н. В. Савельева
Изд № 102107184. Подписано в печать 22.09.08. Формат 60x90/16.
Гарнитура «Таймс». Бумага офсетная № 1. Печать офсетная. Усл. печ. л. 29,0.
Тираж 1000 экз. Заказ № 27430.
Издательский центр «Академия», www.academia-moskow.ru
Санитарно-эпидемиологическое заключение № 77.99.02.953.Д.004796.07.04 от 20.07.2004.
117342, Москва, ул. Бутлерова, 17-Б, к. 360. Тел./факс: (495)330-1092, 334-8337.
Отпечатано в соответствии с качеством предоставленных издательством
электронных носителей в ОАО «Саратовский полиграфкомбинат».
410004, г. Саратов, ул. Чернышевского, 59. www.sarpk.ru
АНАТОМИЯ
И ФИЗИОЛОГИЯ
ЖИВОТНЫХ
Издательский центр
«Академия»
www. academia-moscow. ru