


















































































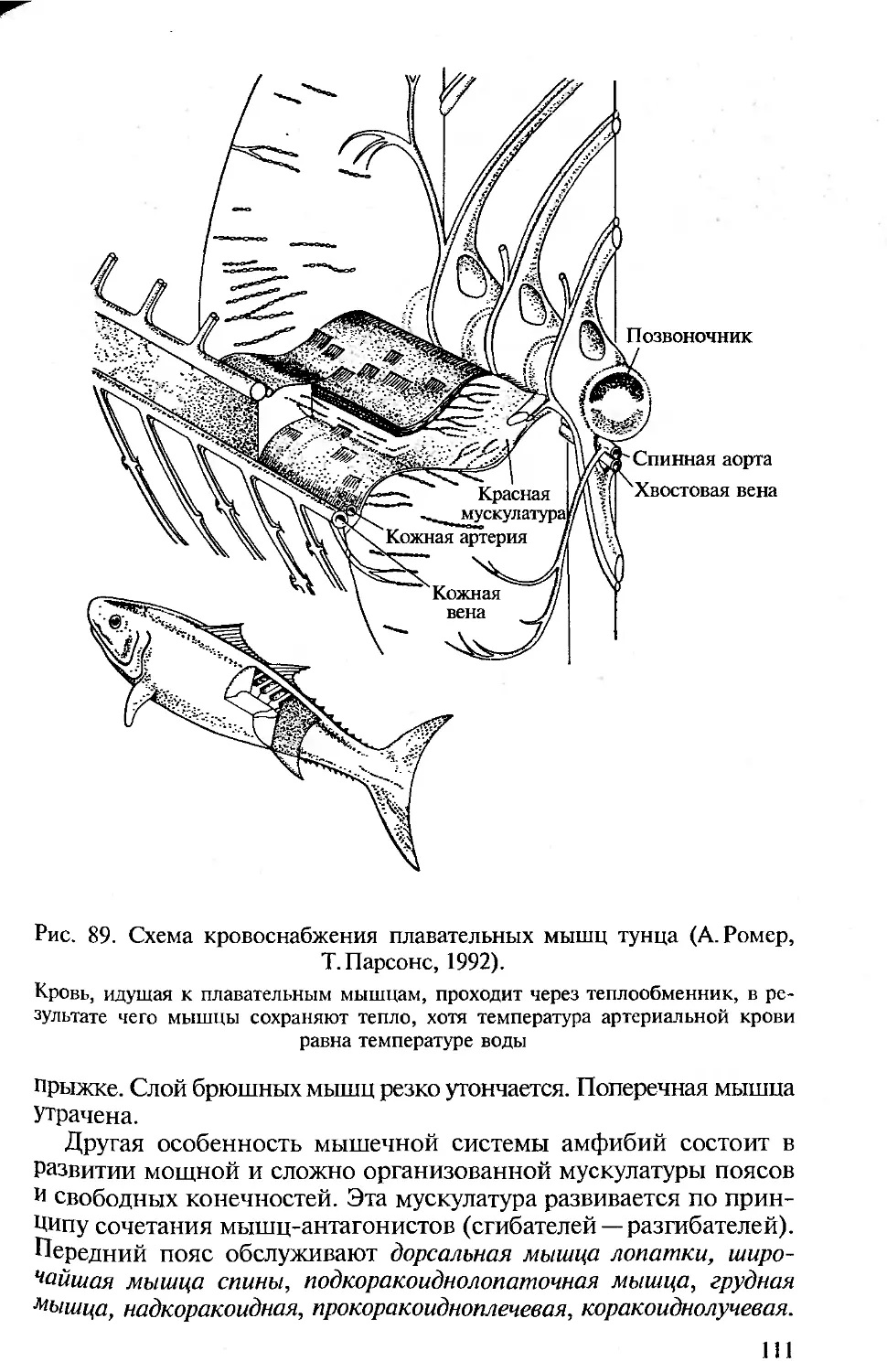
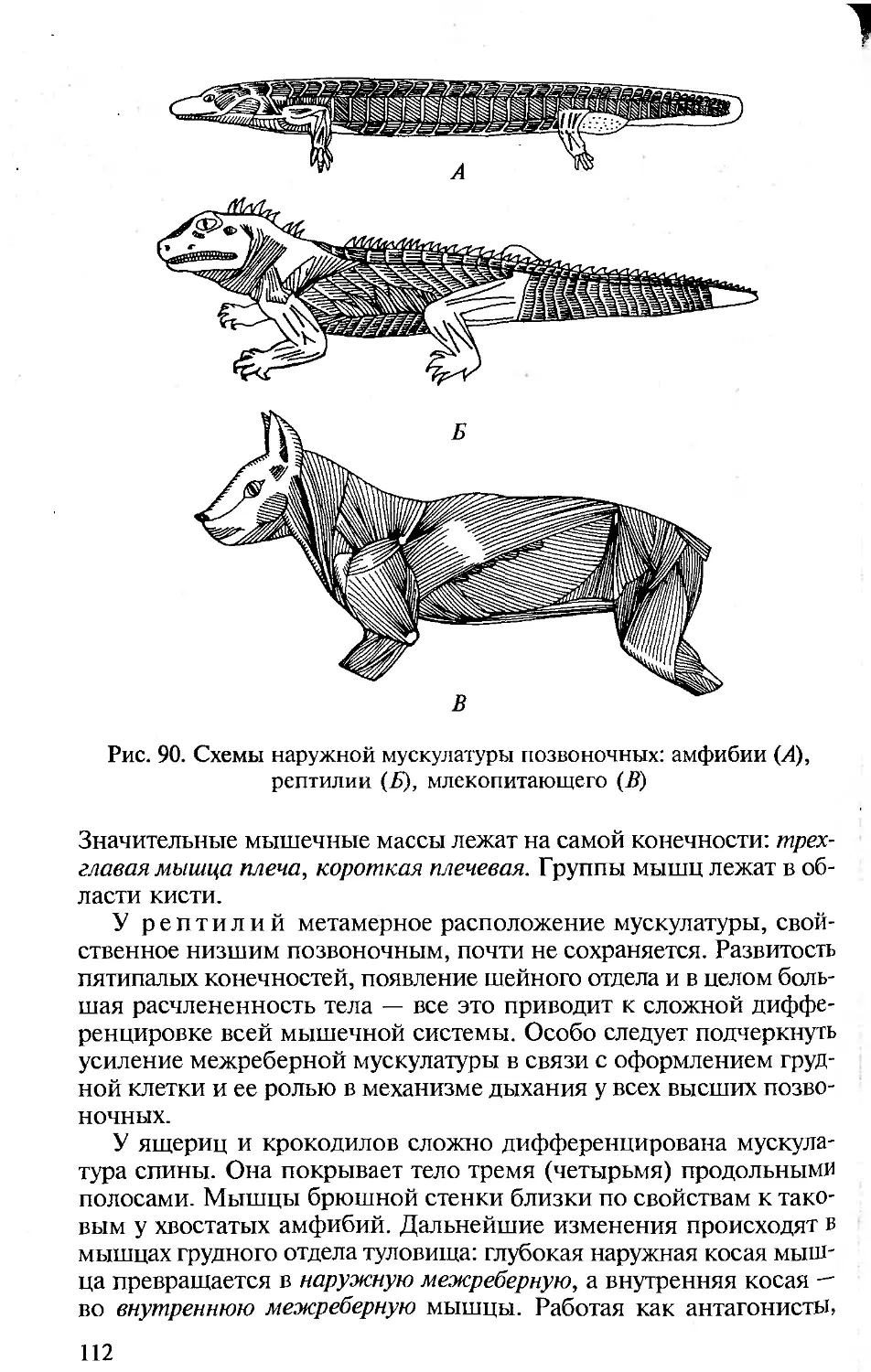







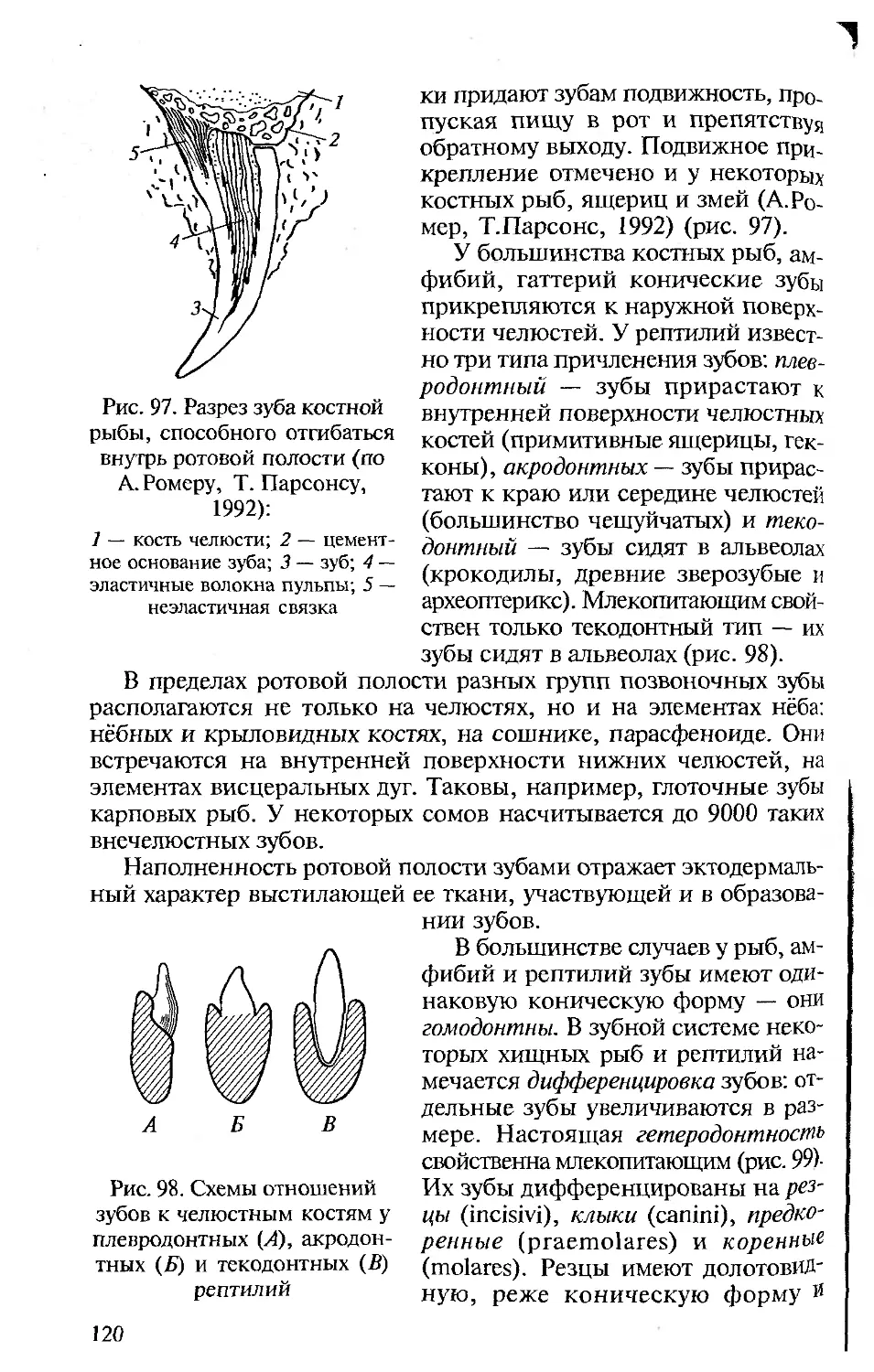



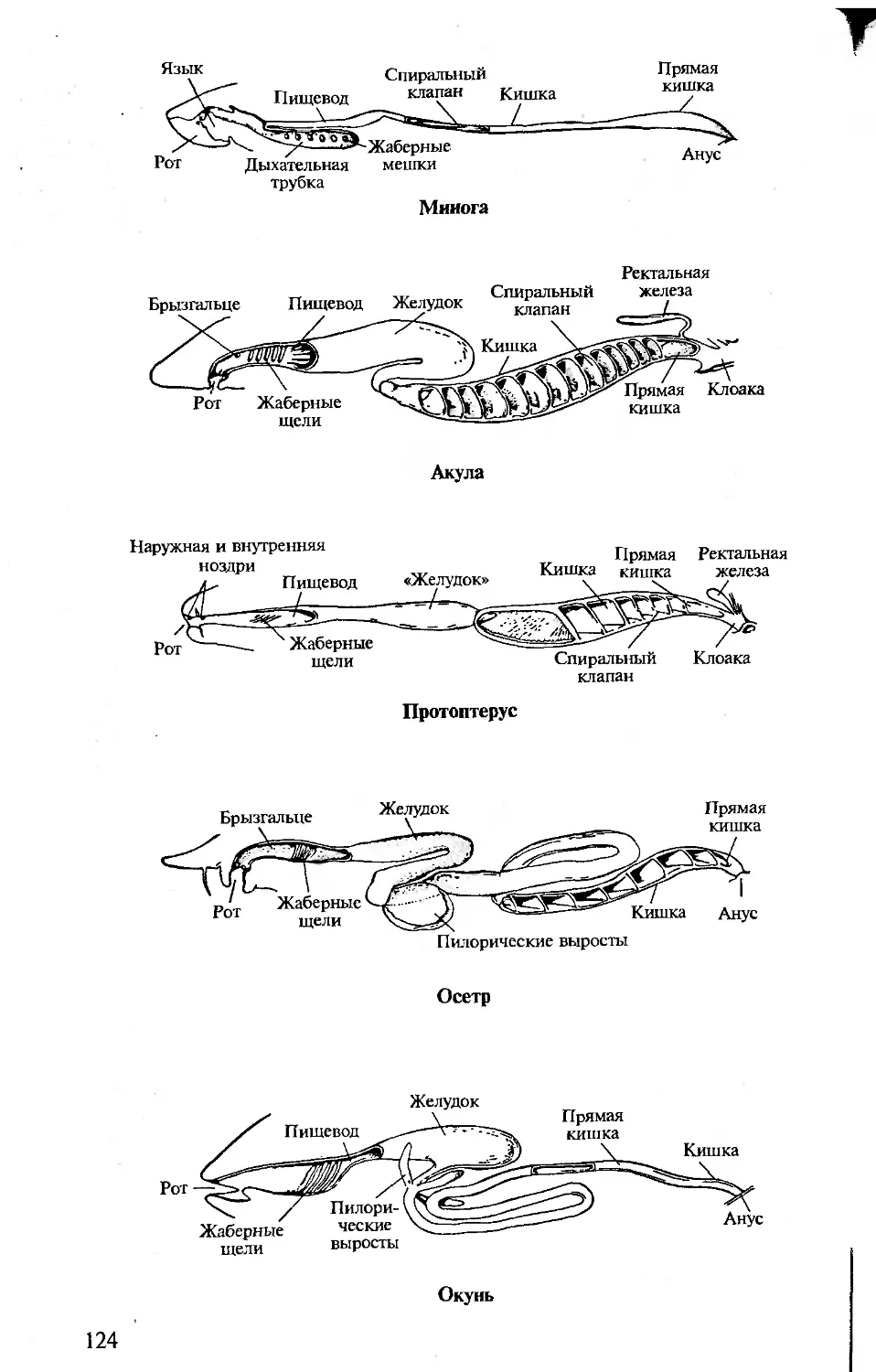














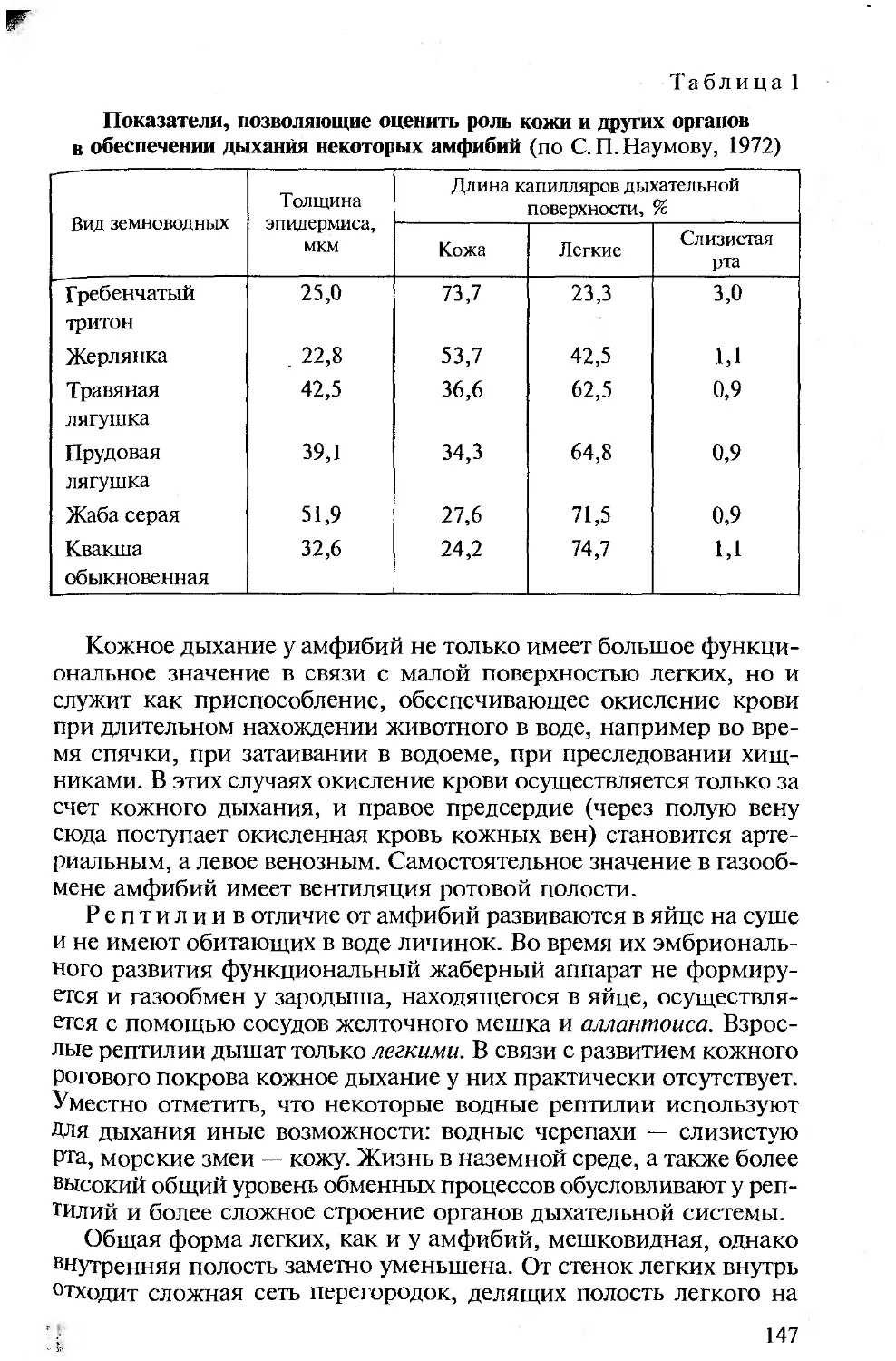
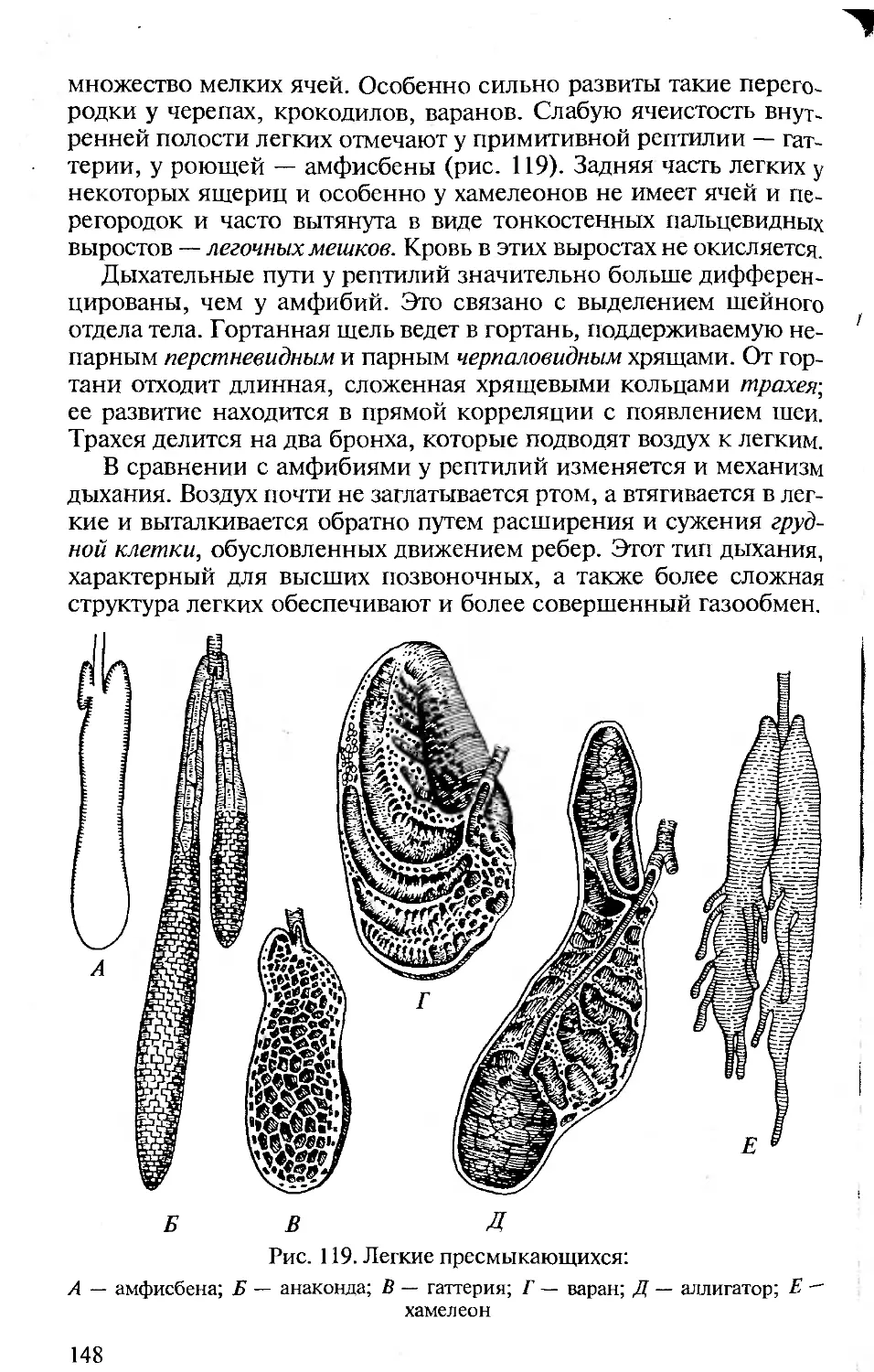


















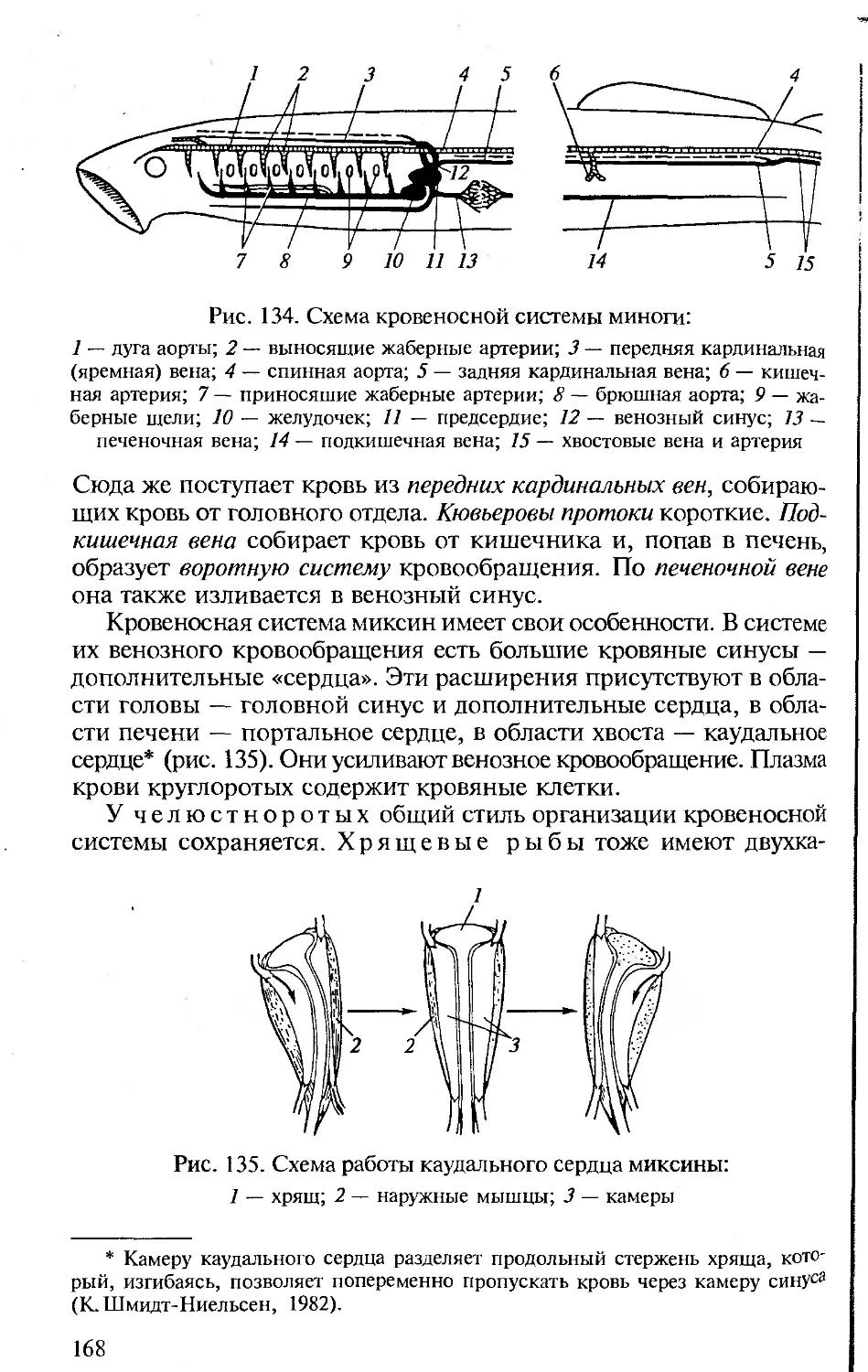
































































































Author: Константинов В.М. Шаталова С.П.
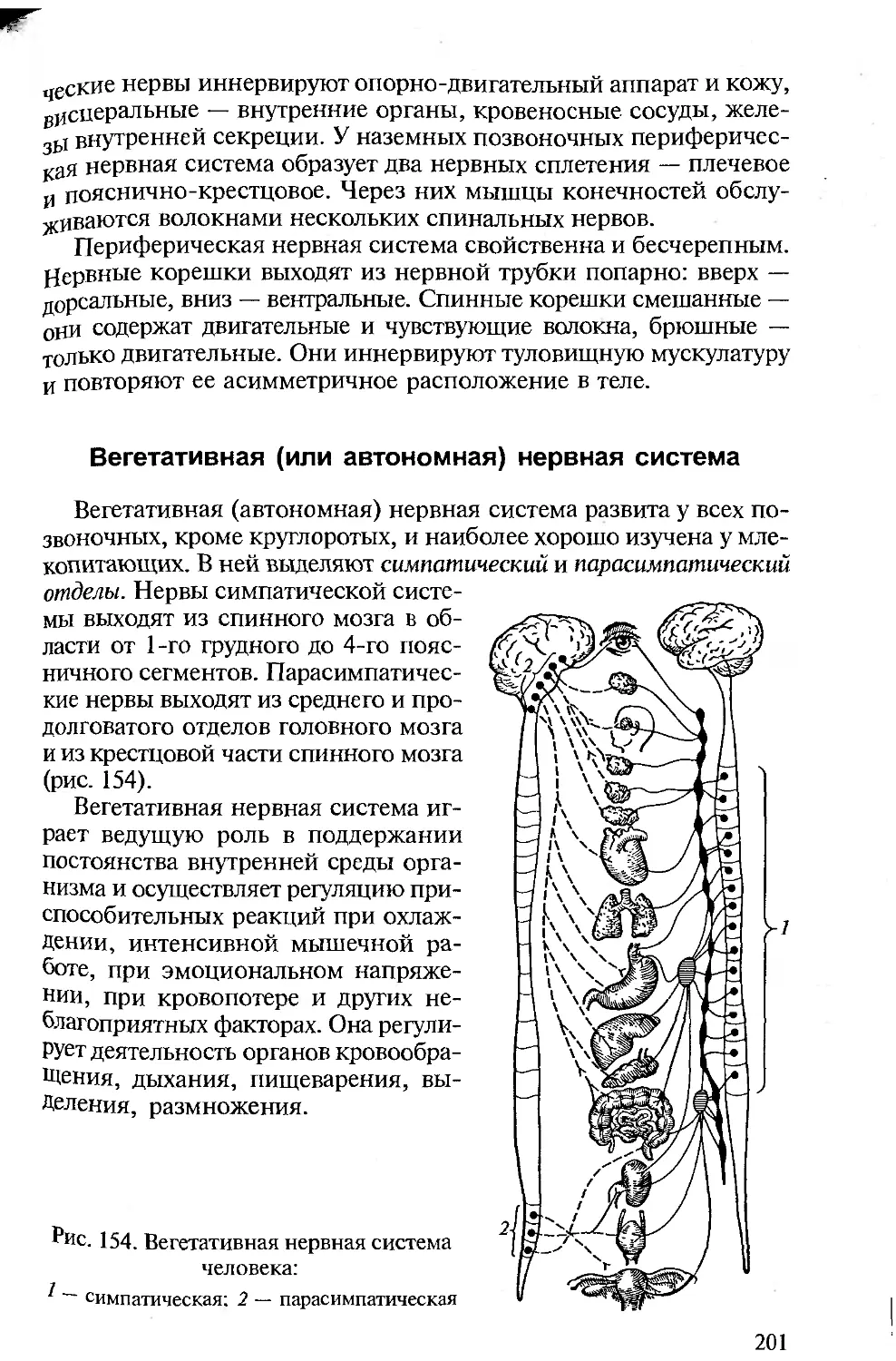
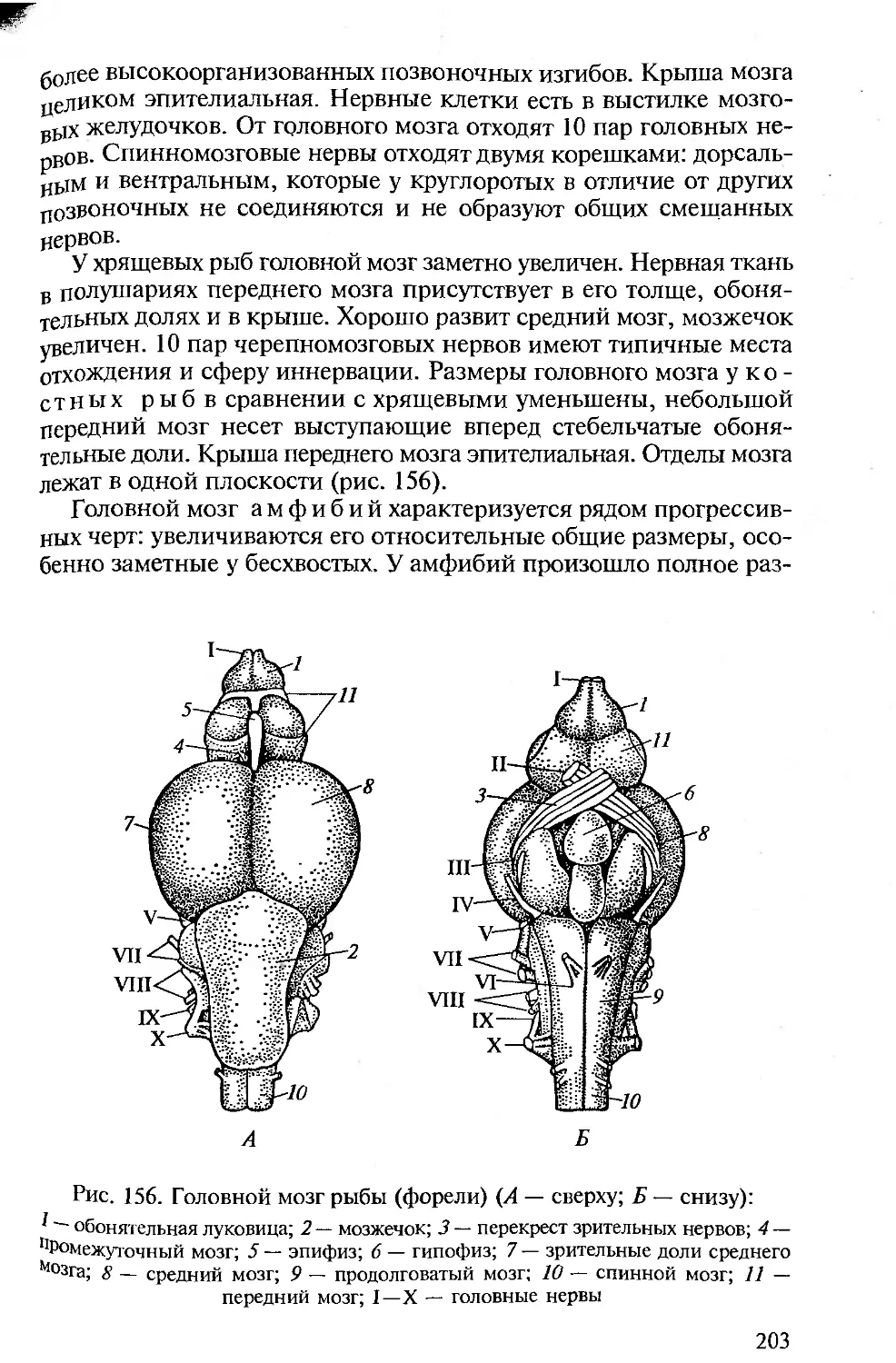
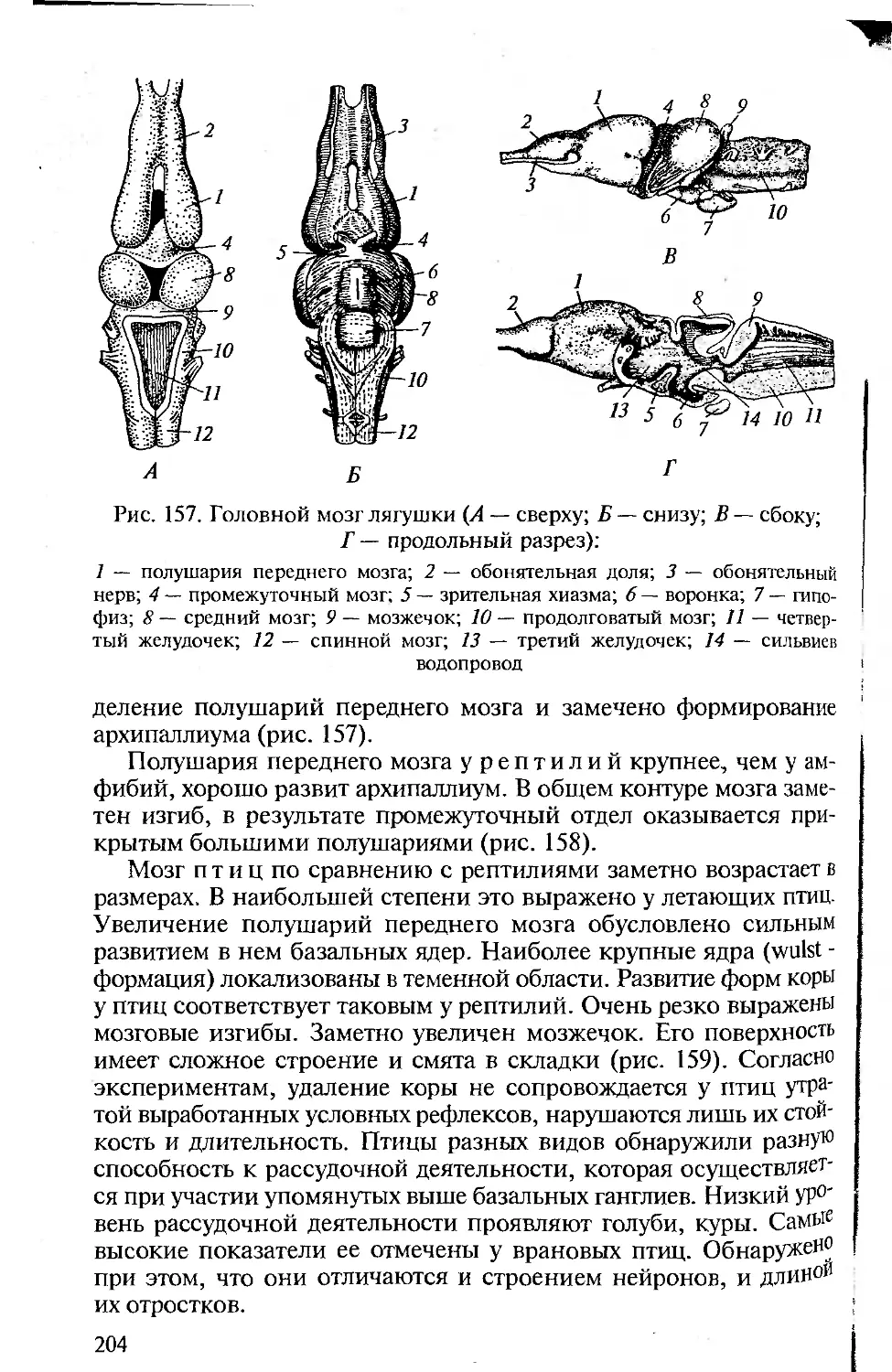
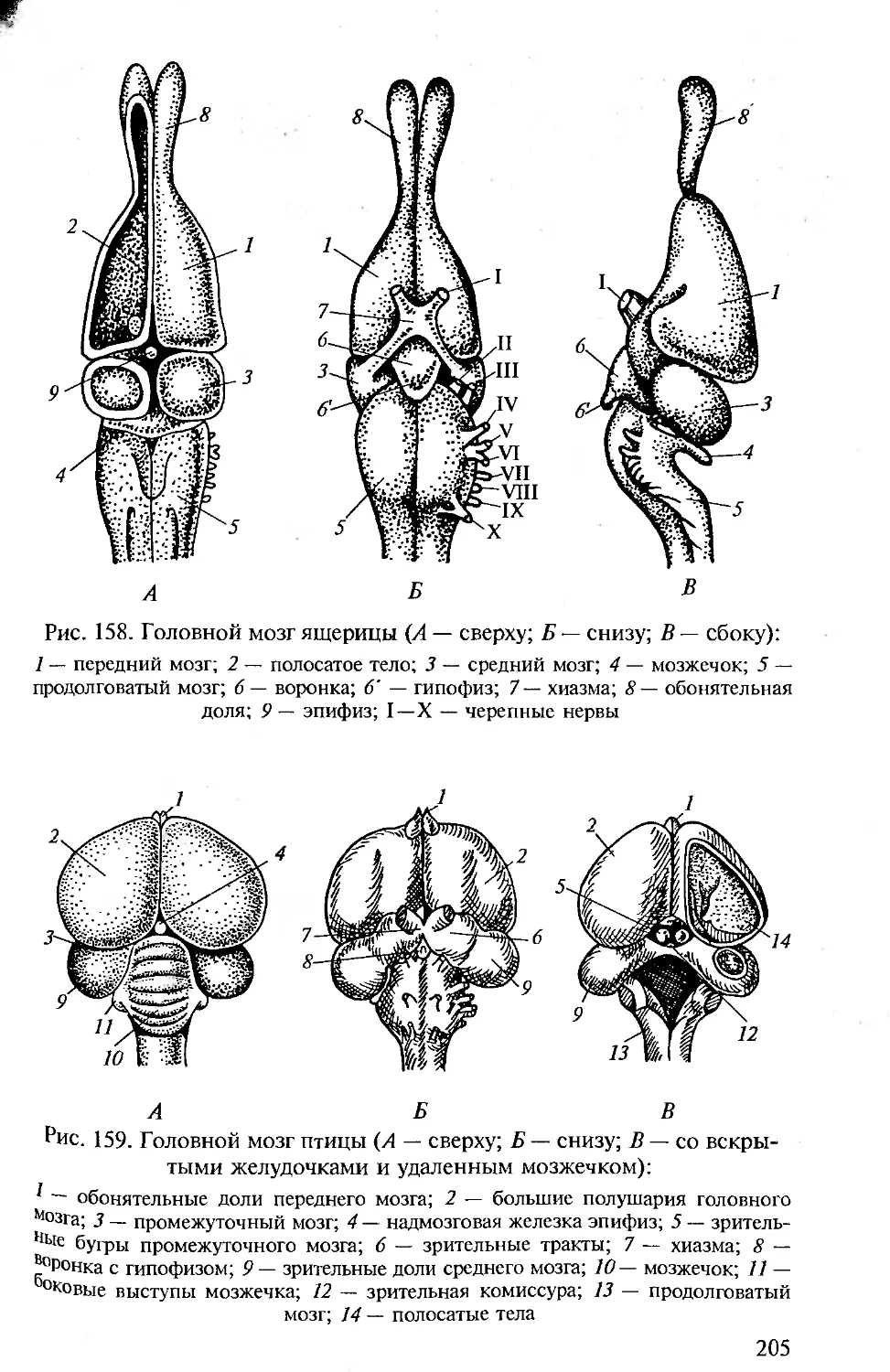
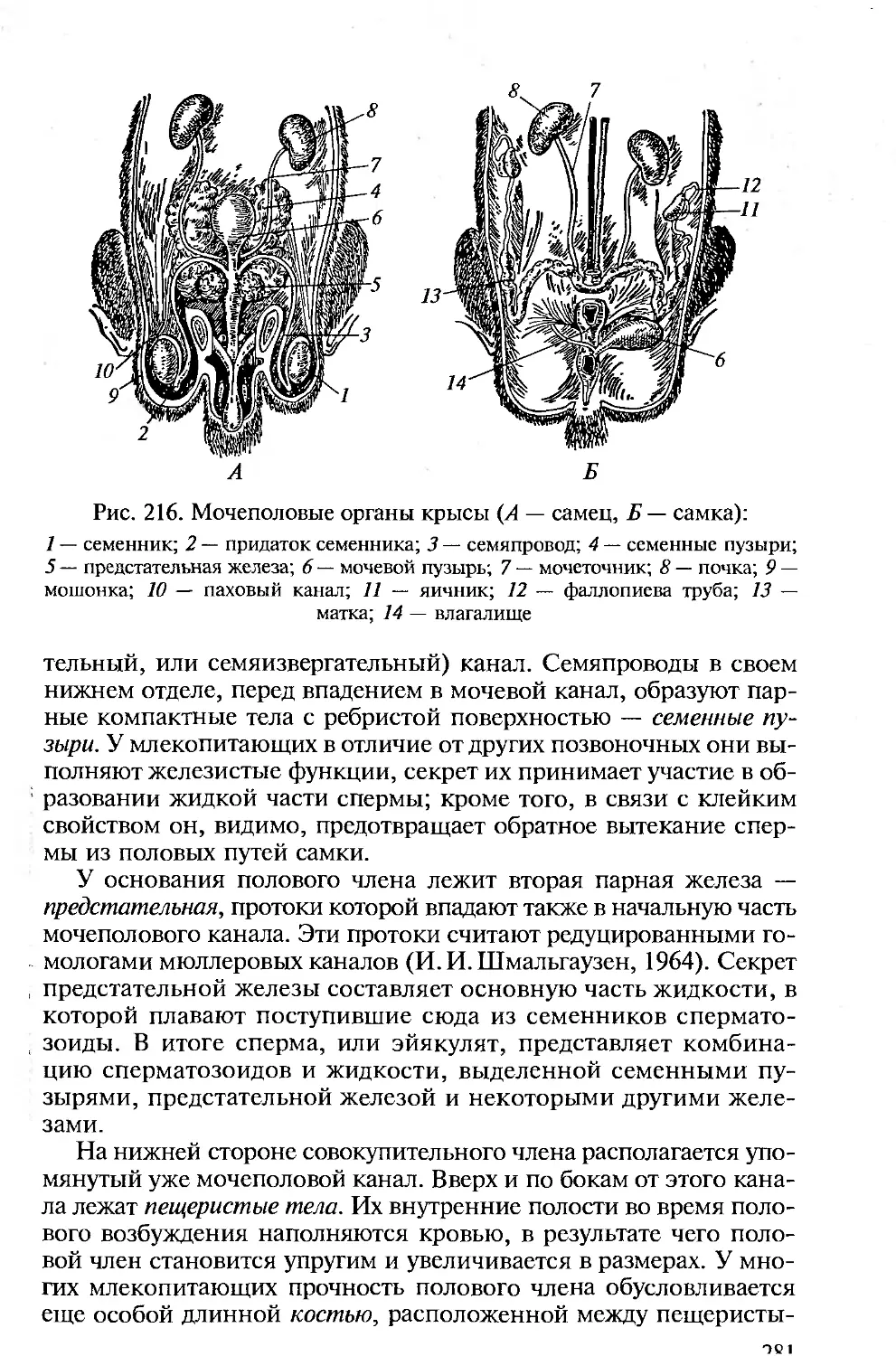
Tags: морфология животных органология животных анатомия животных зоология систематика животных специальные зоологические науки биология анатомия животные морфология
ISBN: 5-7695-1770-0
Year: 2005




Высшее профессиональное образование
В.М. Константинов
С.П. Шаталова
СРАВНИТЕЛЬНАЯ
АНАТОМИЯ
ПОЗВОНОЧНЫХ
ЖИВОТНЫХ
ф
х
ю
о
и
О
С
ф
о
X
10
ф
О
Педагогические
специальности
-
К
пЫСШЕЕ ПРОФЕССИОНАЛЬНОЕ ОБРАЗОВАНИЕ
I .
-А
В. И. КОНСТАНТИНОВ, С. П. ШАТАЛОВА
СРАВНИТЕЛЬНАЯ
АНАТОМИЯ
ПОЗВОНОЧНЫХ
ЖИВОТНЫХ
Допущено
Учебно-методическим объединением по специальностям
педагогического образования в качестве учебного пособия
для студентов высших учебных заведений, обучающихся
по специальности 032400 «Биология»
библио г.. ; А
Омского
у н и верите та
Москва
ACADEMA
2005
УДК 591.4 (075.8)
ББК 28.693.3я73
К64
Рецензенты:
д-р биол. наук, проф. А. Г. Резанов (зав. кафедрой биологии Московского
городского педагогического университета);
канд. биол. наук, доц. А. О. Шубин (Московский педагогический
государственный университет)
Константинов В. М.
К64 Сравнительная анатомия позвоночных животных: Учеб,
пособие для студ. высш. пед. учеб, заведений / В. М. Константи-
нов, С. П. Шаталова. — М.: Издательский центр «Академия»,
2005. - 304 с.
ISBN 5-7695-1770-0
Пособие отражает современные научные достижения в функциональ-
ной морфологии позвоночных животных и последовательно рассматрива-
ет: кожные покровы и их производные, опорно-двигательную, пищева-
рительную, дыхательную, кровеносную, нервную системы и органы чувств.
В книге показана связь строения и функций. Учебный материал располо-
жен по эволюционному принципу: от примитивных бесчелюстных до
высших позвоночных — птиц и млекопитающих.
Для студентов высших педагогических учебных заведений, обучаю-
щихся по специальности «Биология».
УДК 591.4 (075.8)
ББК 28.693.3я73
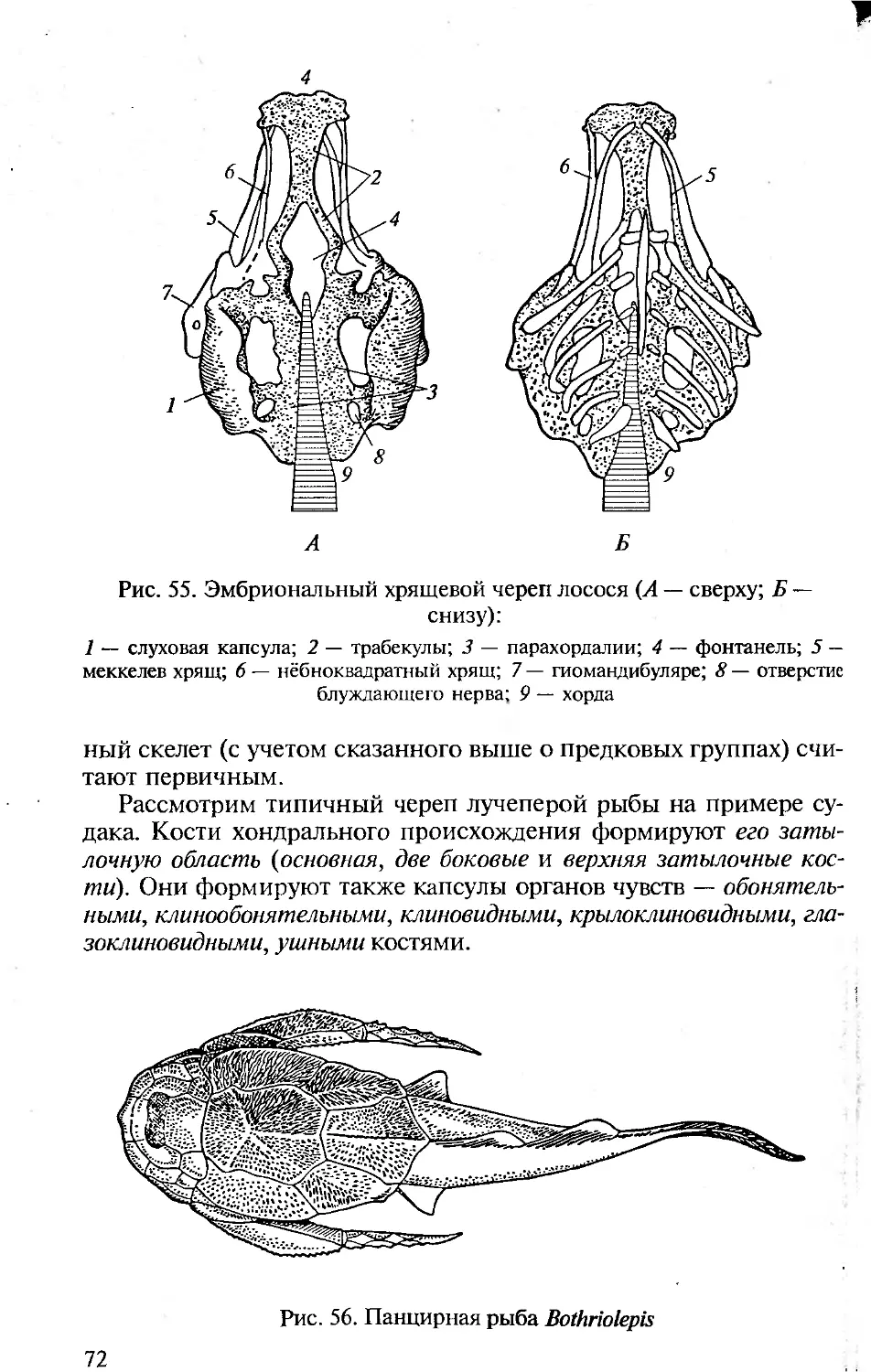
ISBN 5-7695-1770-0
© Константинов В.М., Шаталова С П., 2005
© Издательский центр «Академия», 2005/
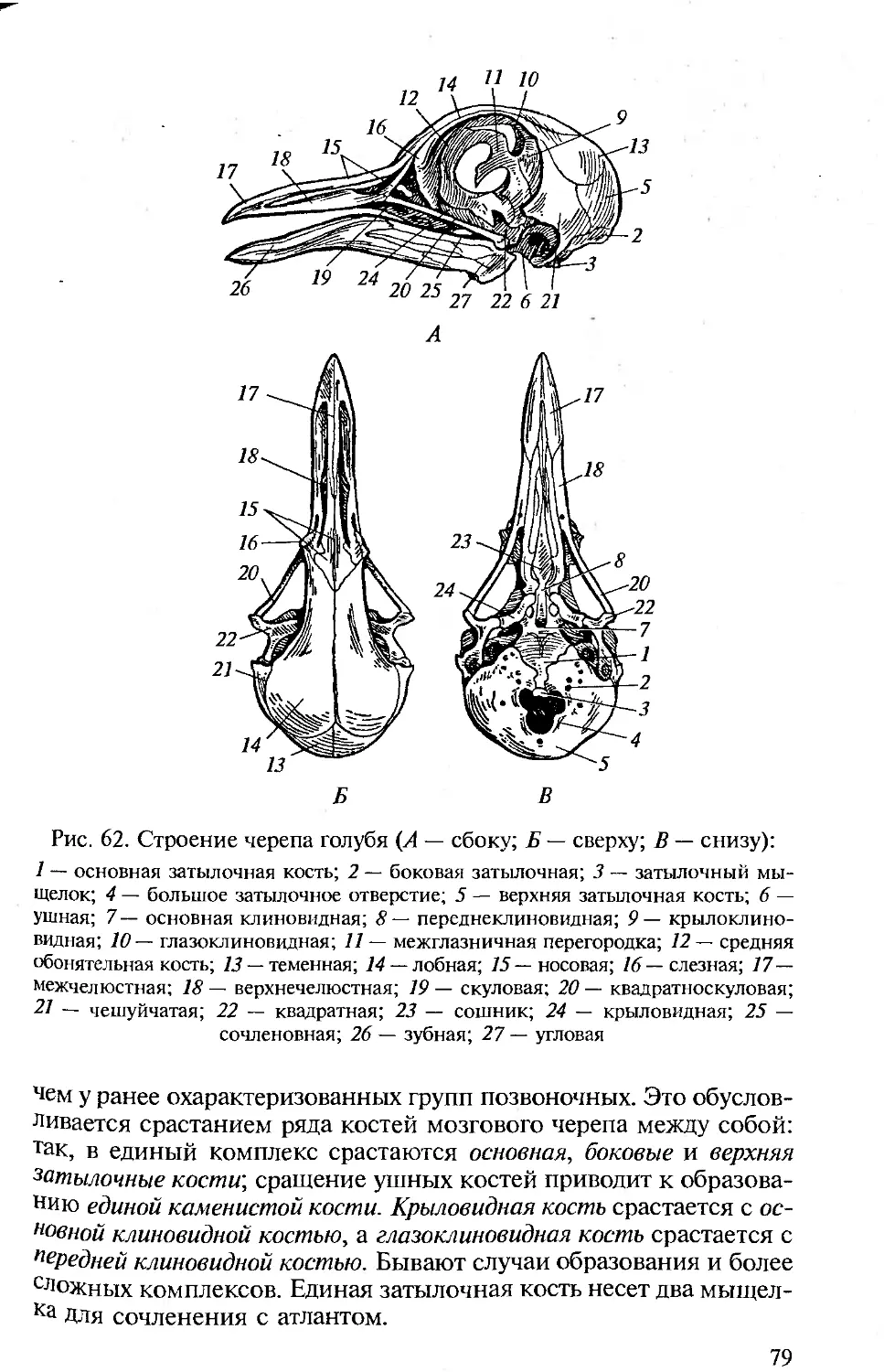
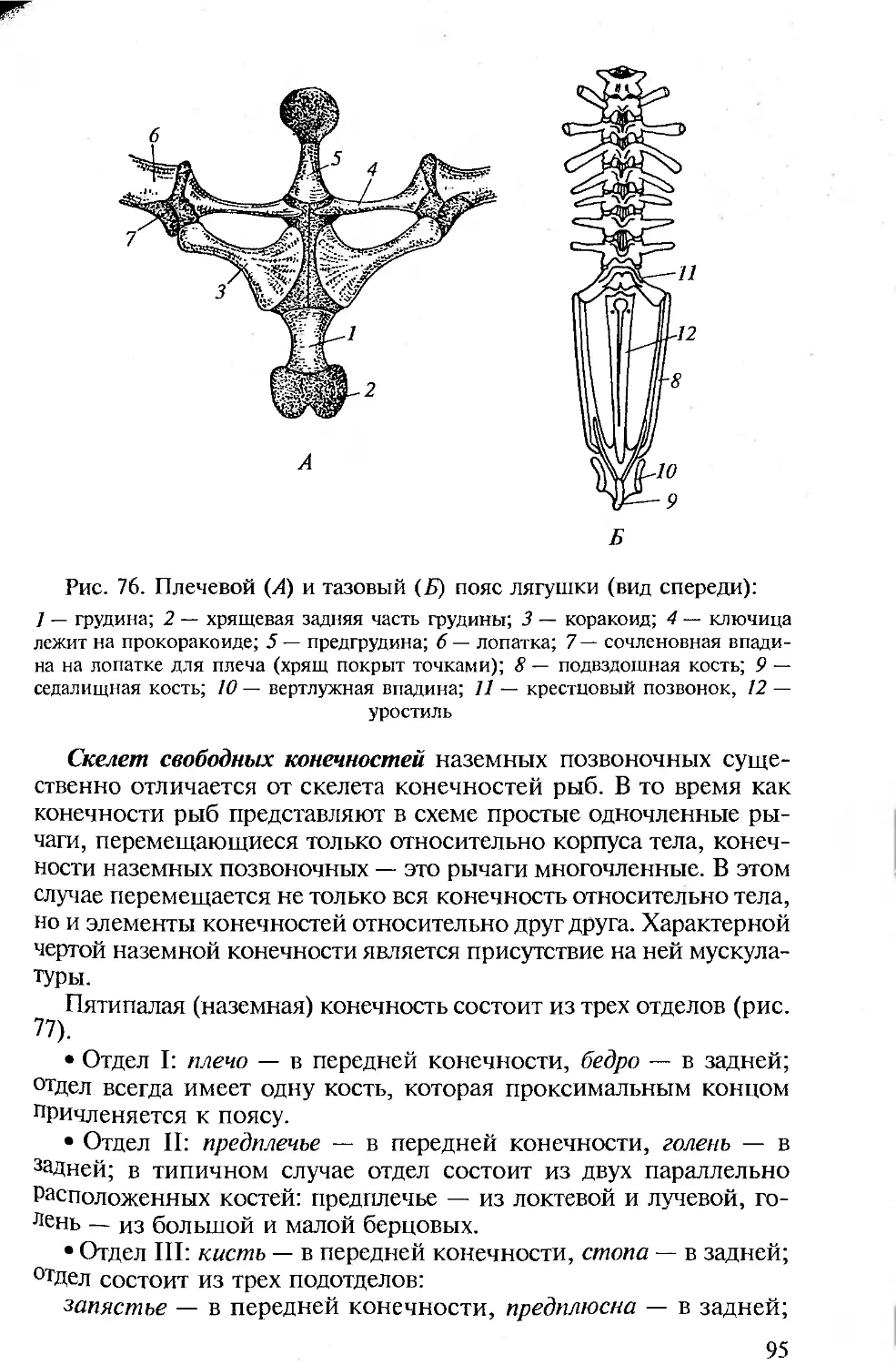
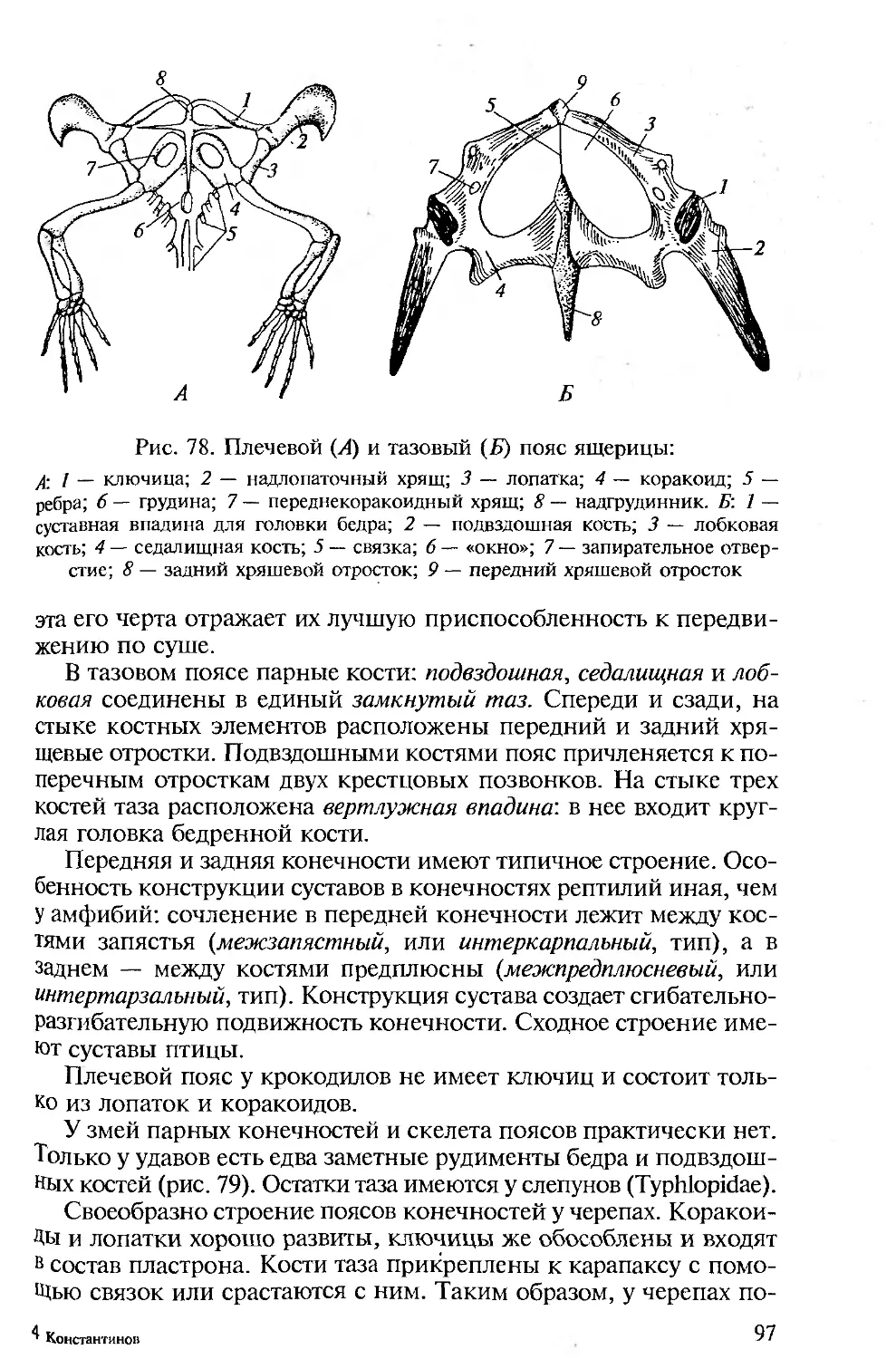
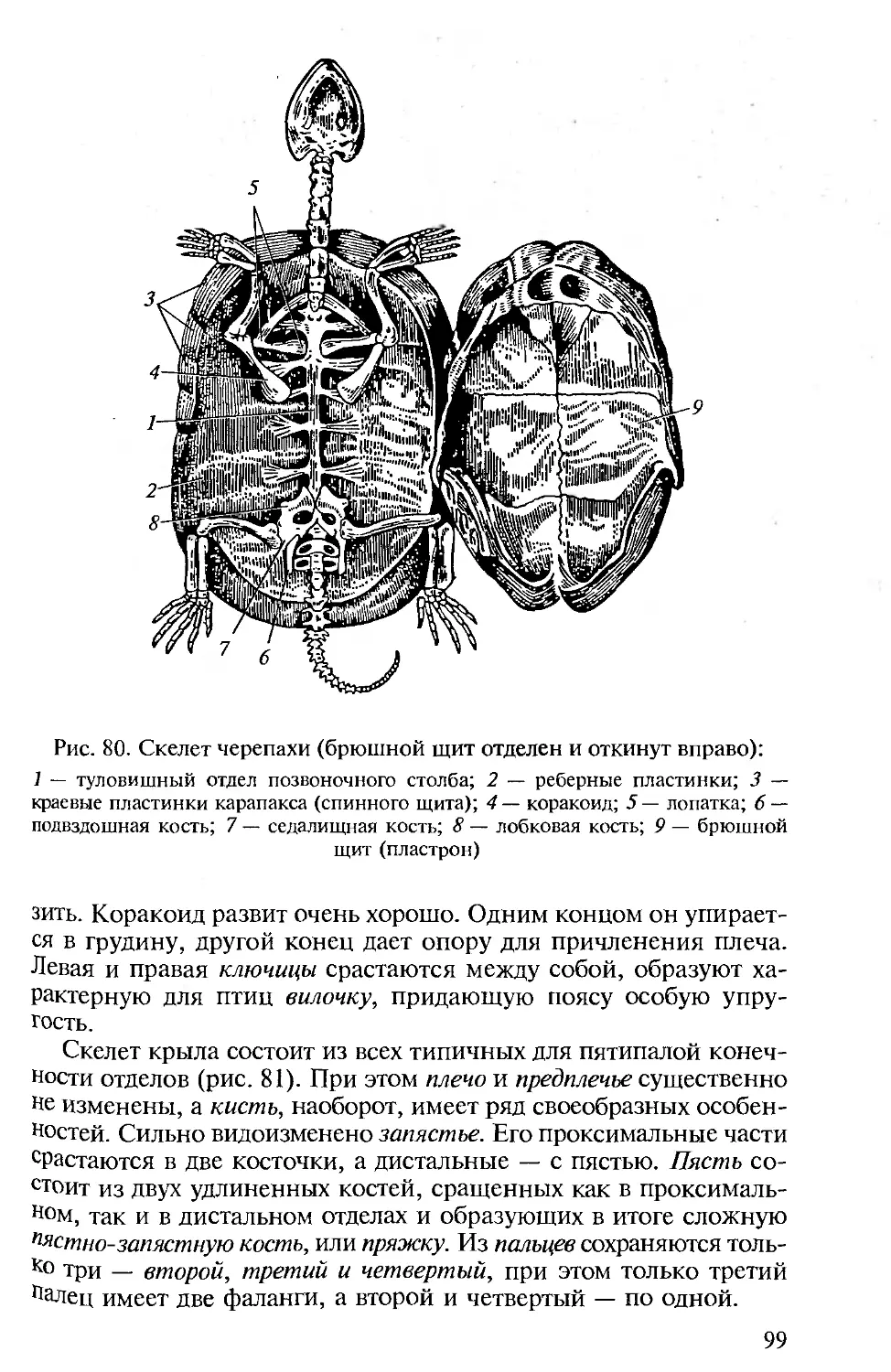
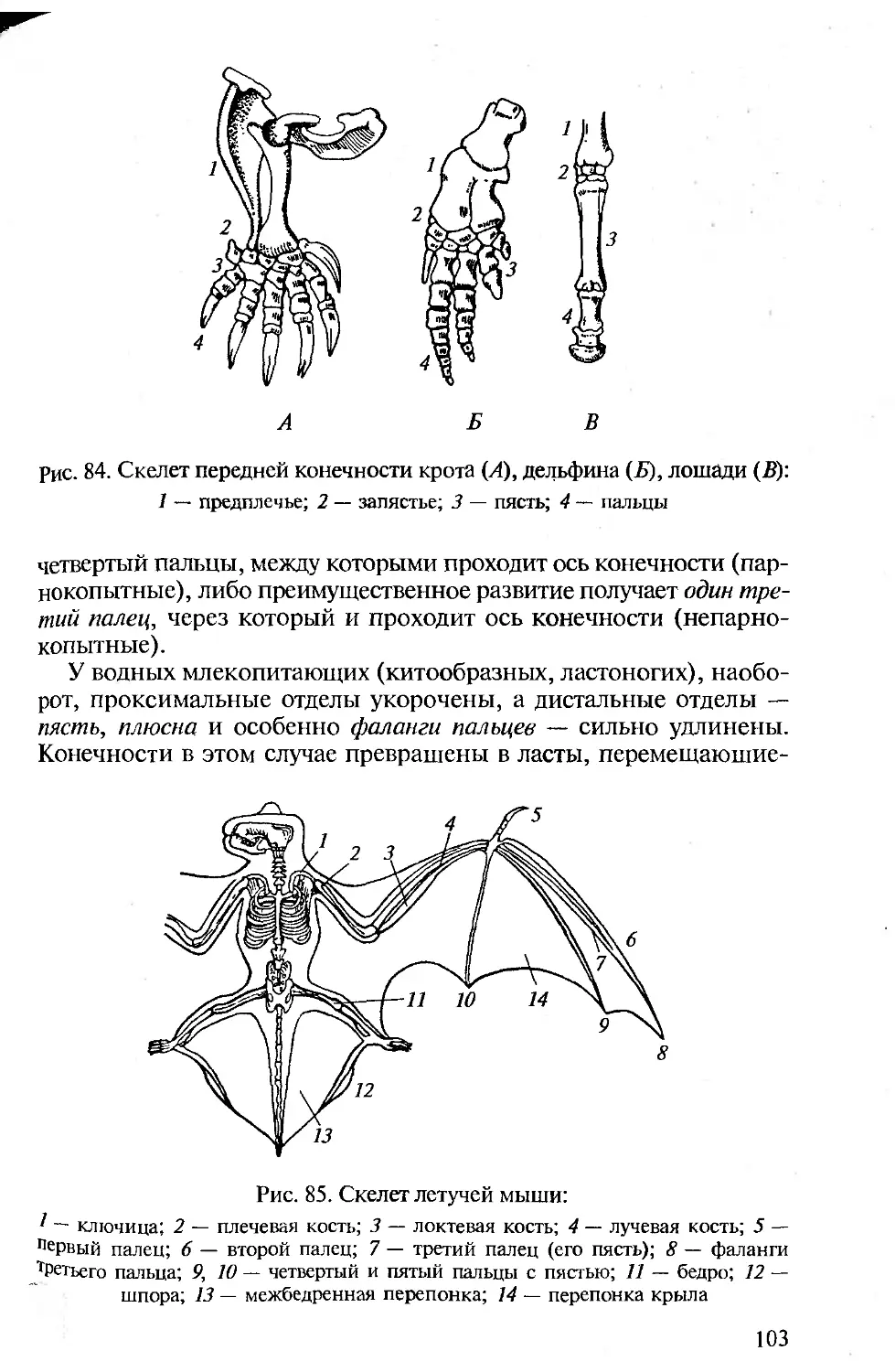
ПРЕДИСЛОВИЕ
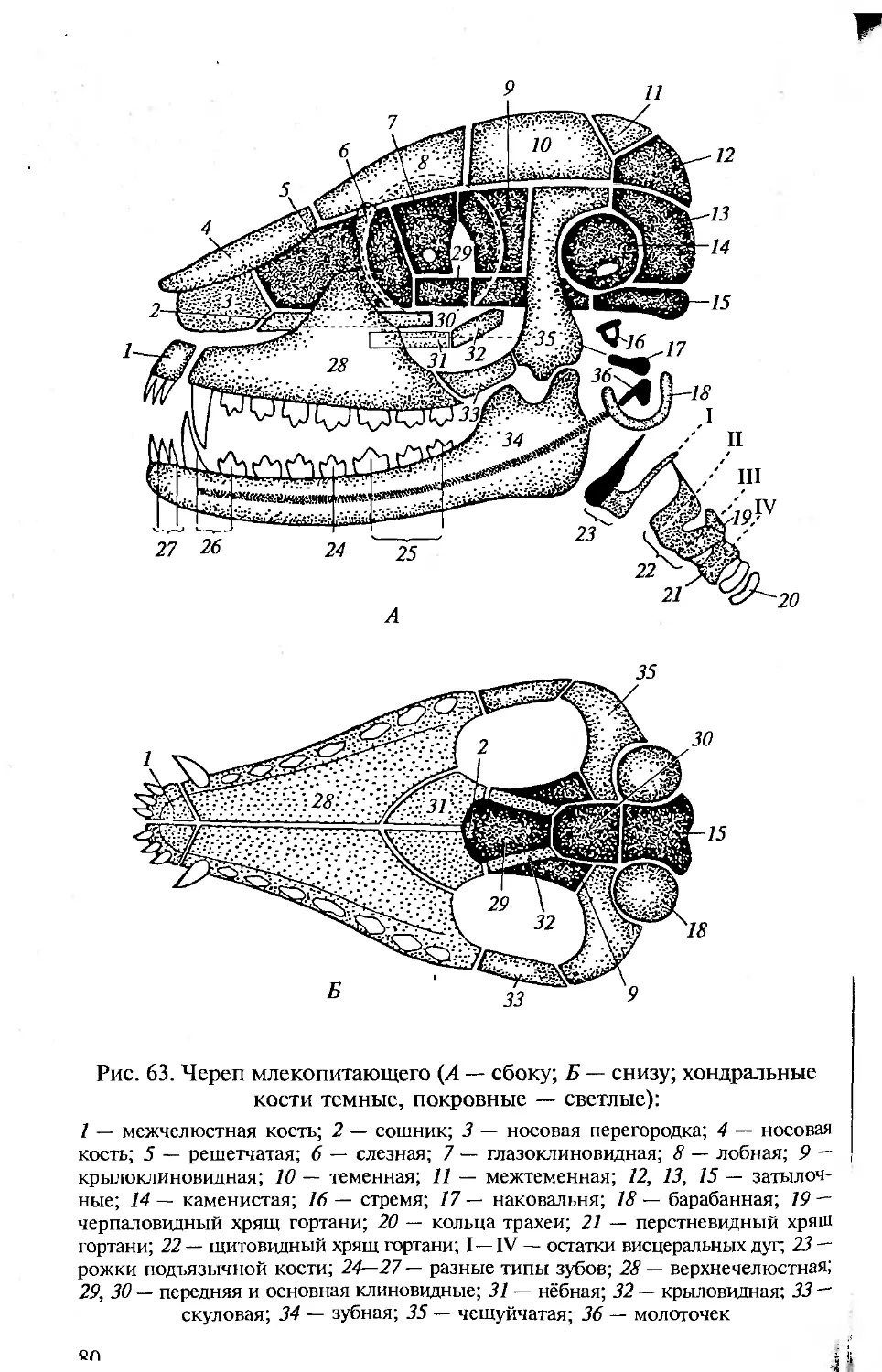
Настоящая книга посвящена сравнительной анатомии позво-
ночных животных. Книги, рассматривающие строение животных
в сравнительно-эволюционном плане, выходят редко. Так, широко
известное руководство для студентов биофаков университетов «Ос-
новы сравнительной анатомии позвоночных» И. И. Шмальгаузена
последний раз было опубликовано около 40 лет назад (4-е изд. —
1964 г.). Двухтомная «Анатомия позвоночных» А. Ромера и Т. Пар-
сонса, которой пользуются отечественные зоологи, была переве-
дена на русский язык с шестого американского издания (1986) и
издана в России в 1992 г.
В 1998 г. вышла в свет книга профессора кафедры зоологии по-
звоночных (Московский государственный университет им. М.В. Ло-
моносова) Ф. Я.Дзержинского «Сравнительная анатомия позво-
ночных животных». Для педагогических же вузов, где в отличие от
университетов эта дисциплина преподается факультативно сту-
дентам старших курсов биологических факультетов и слушателям
ФПК (с 1970-х годов), общие руководства, посвященные сравни-
тельной анатомии позвоночных, ранее не издавались. Исключе-
ние составляют опубликованные в последнее время небольшие
по объему книги И. X. Шаровой с соавторами (2002) «Сравнитель-
ная анатомия и эволюция животных», В. Н.Яценко и др. «Анато-
мо-морфологические особенности кровеносной системы живот-
ных» (2001), Л. Ф. Скрылевой и др. «Анатомо-морфологические осо-
бенности скелета хордовых животных» (2002), которые рассмат-
ривают эволюционные изменения в системах органов беспозвоноч-
ных и позвоночных животных. Очень полезными стали также книги
Н. Г.Андреевой и Д. К. Обухова «Эволюционная морфология не-
рвной системы позвоночных» (1999), С. В. Савельева «Сравнитель-
ная анатомия нервной системы позвоночных» (2001), в которых
эволюция нервной системы рассматривается на широком фоне
эволюции позвоночных. В настоящем издании авторы постарались
Учесть позиции всех упомянутых ученых.
Еще одним источником знаний, использованных авторами при
подготовке данного пособия, послужили несколько поколений
Учебников по зоологии позвоночных для студентов биофаков уни-
Верситетов и педагогических вузов, в частности книги С. И. Огне-
Ва (1945), Б.С.Матвеева и др. (1966), Н.П.Наумова и Н.Н.Карта-
3
шева (1979), С. П. Наумова (1983). Фундаментальным руководство^
для детального изучения морфофункциональной организации
позвоночных служит трехтомная «Практическая зоотомия позво-
ночных» (Н.Н. Гуртовой и др., 1976, 1978, 1992).
Хочется предостеречь читателя от поверхностного подхода к
изучению сравнительной анатомии. Перечитывание даже очень
хороших учебников не может заменить сравнительного изучения
реальных объектов. Как только слова начинают подтверждаться
словами и изображениями, так заканчивается естественно-науч-
ный подход. «Виртуальный анализ приводит к виртуальным ре-
зультатам» (С. В. Савельев, 2001). Это особенно нужно осознавать
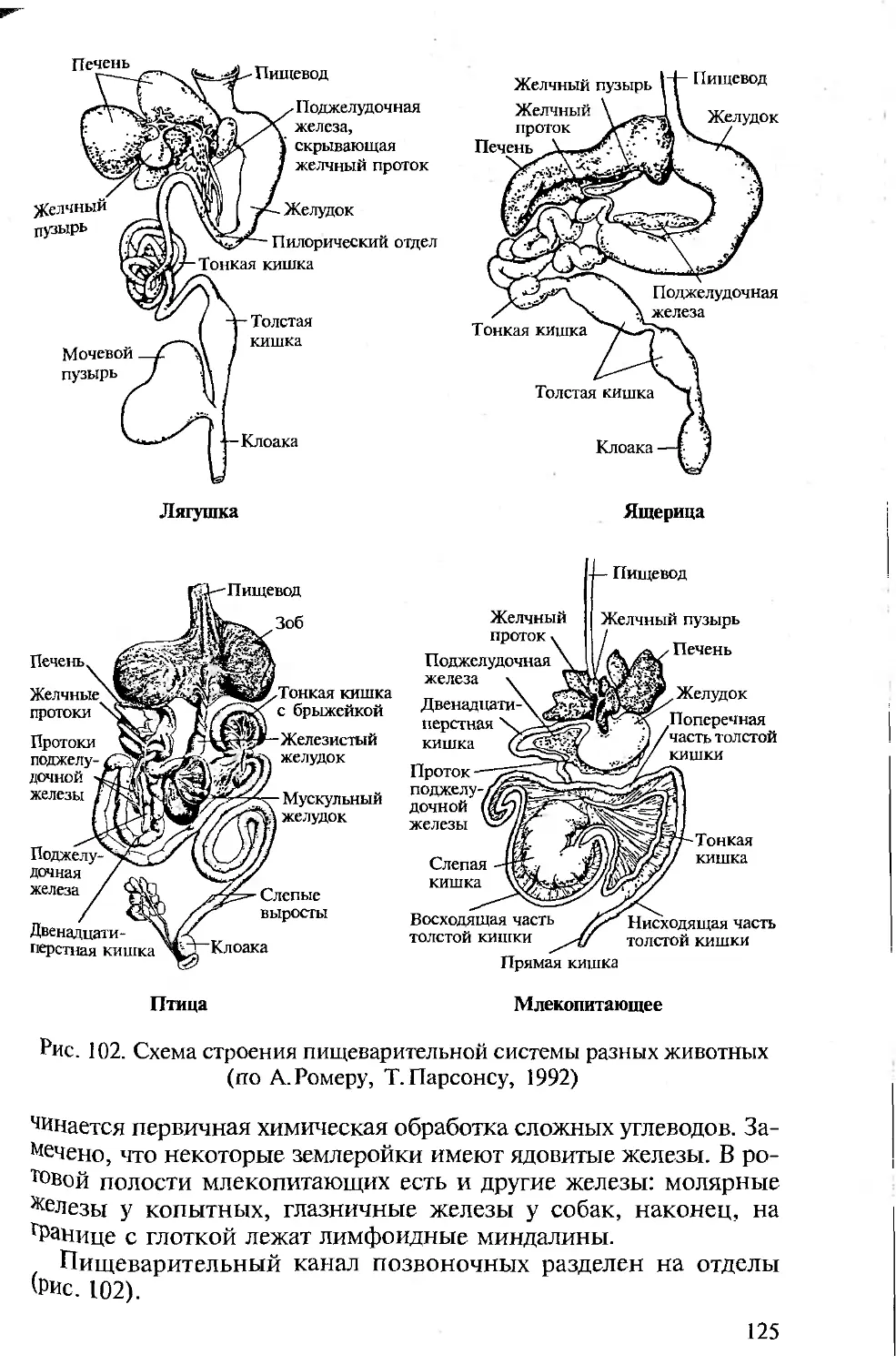
в современных условиях, когда материальная база педвузов резко
ухудшилась, а изучение анатомии животных проводится лишь на
отдельных модельных объектах, которые с невероятными трудно-
стями могут достать сами преподаватели. Это, однако, не меняет
той принципиально важной в биологии позиции: правильность
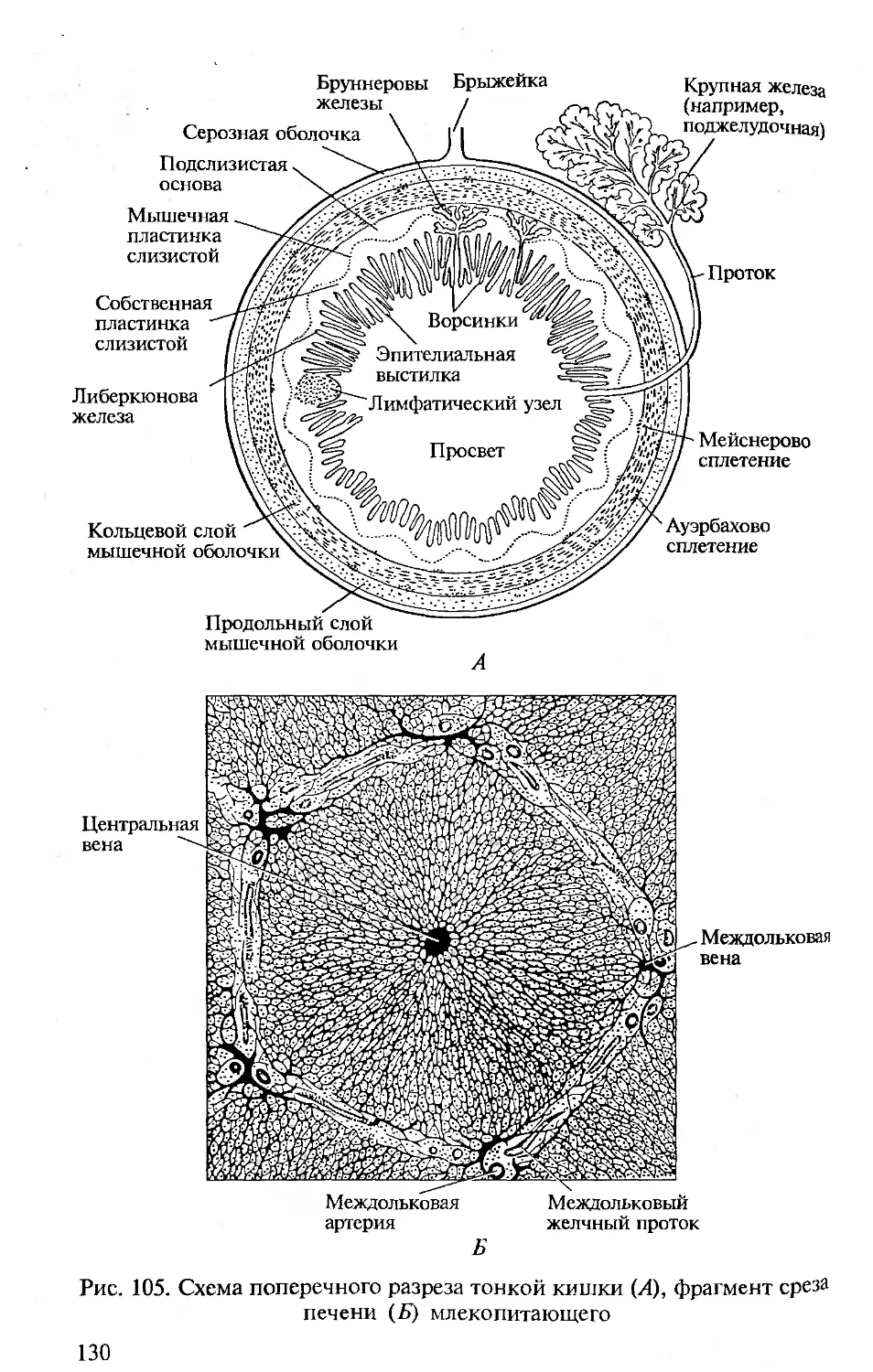
теоретических положений может проверяться только на практике.
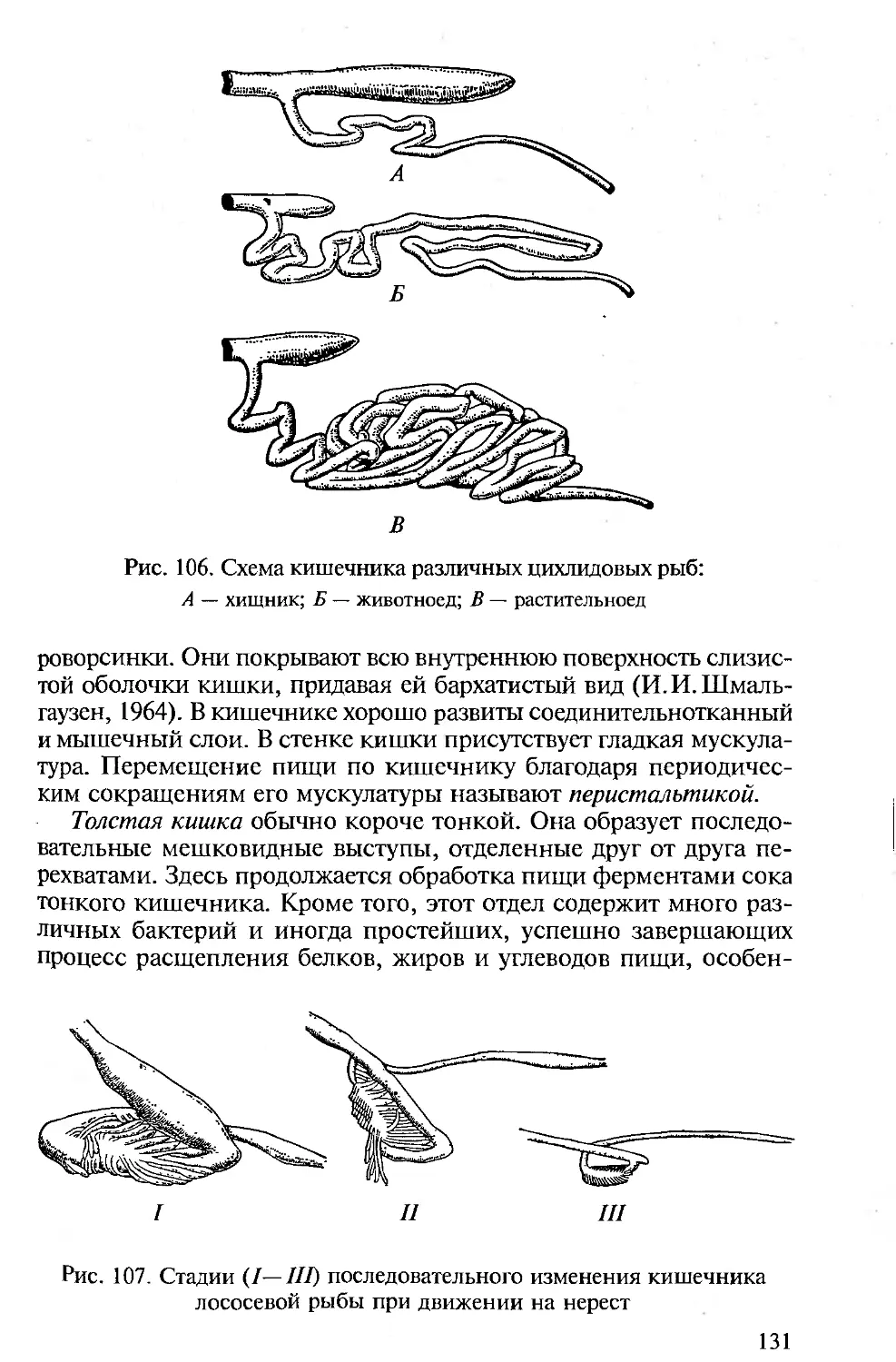
Существенно, что во всех вузовских и школьных зоологических
курсах использованы современные сравнительно-анатомические
знания, которые служат основой для построения эволюционных
теорий.
Есть еще одна причина важности сравнительной анатомии
животных (включая человека) — это убежденность многих уче-
ных и даже биологов других специальностей в научной завер-
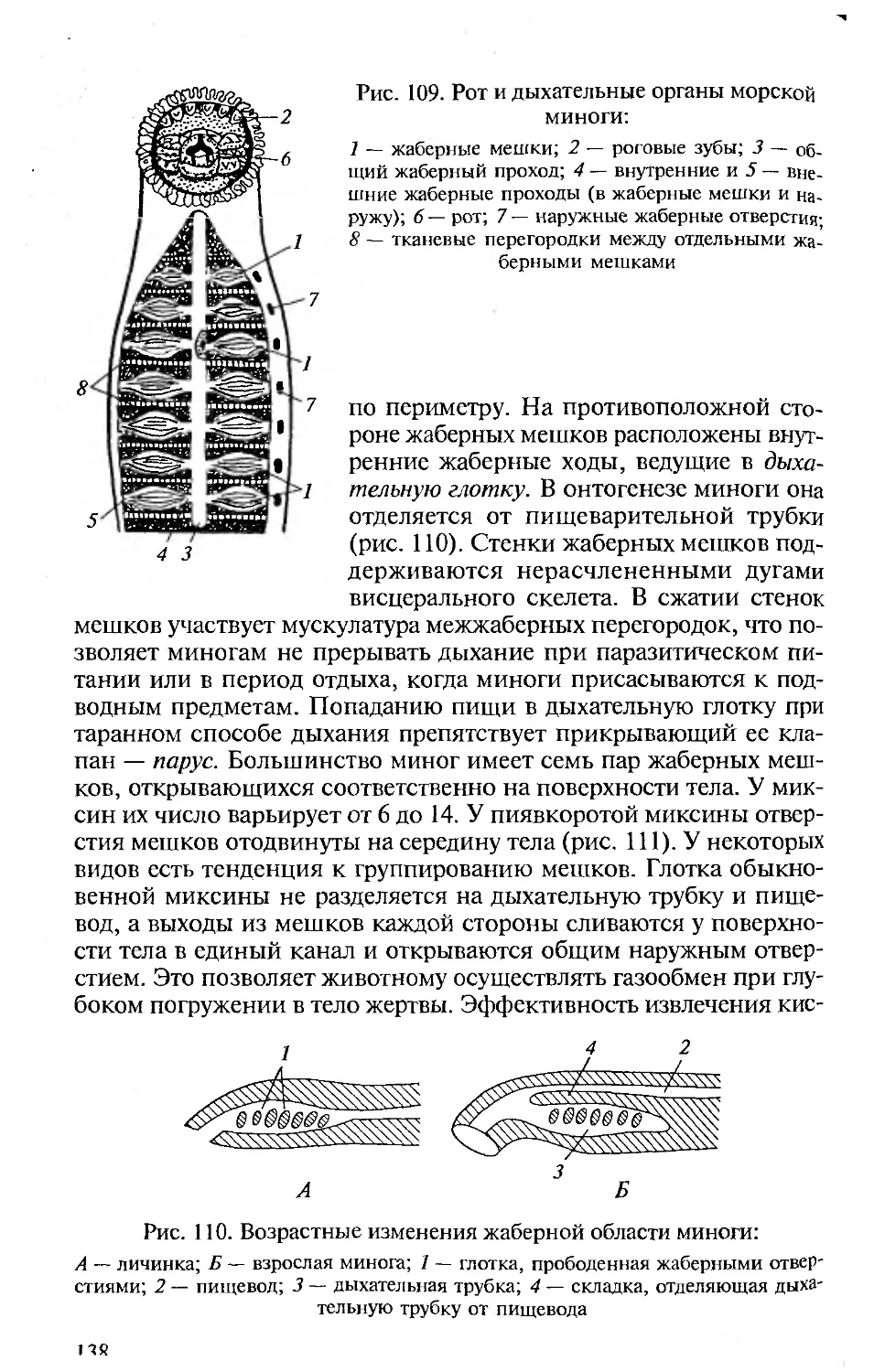
шенности, полной изученности морфологии, что приводит, как
правило, к минимальному поступлению новых фактов и искус-
ственному занижению общественного интереса к данной облас-
ти знаний. Следствием этого часто являются теоретические пост-
роения, основанные на каком-нибудь одном (иногда весьма со-
мнительном) методе или на комбинировании явно устаревших
сведений, что упрощает сравнительную анатомию, часто доводя
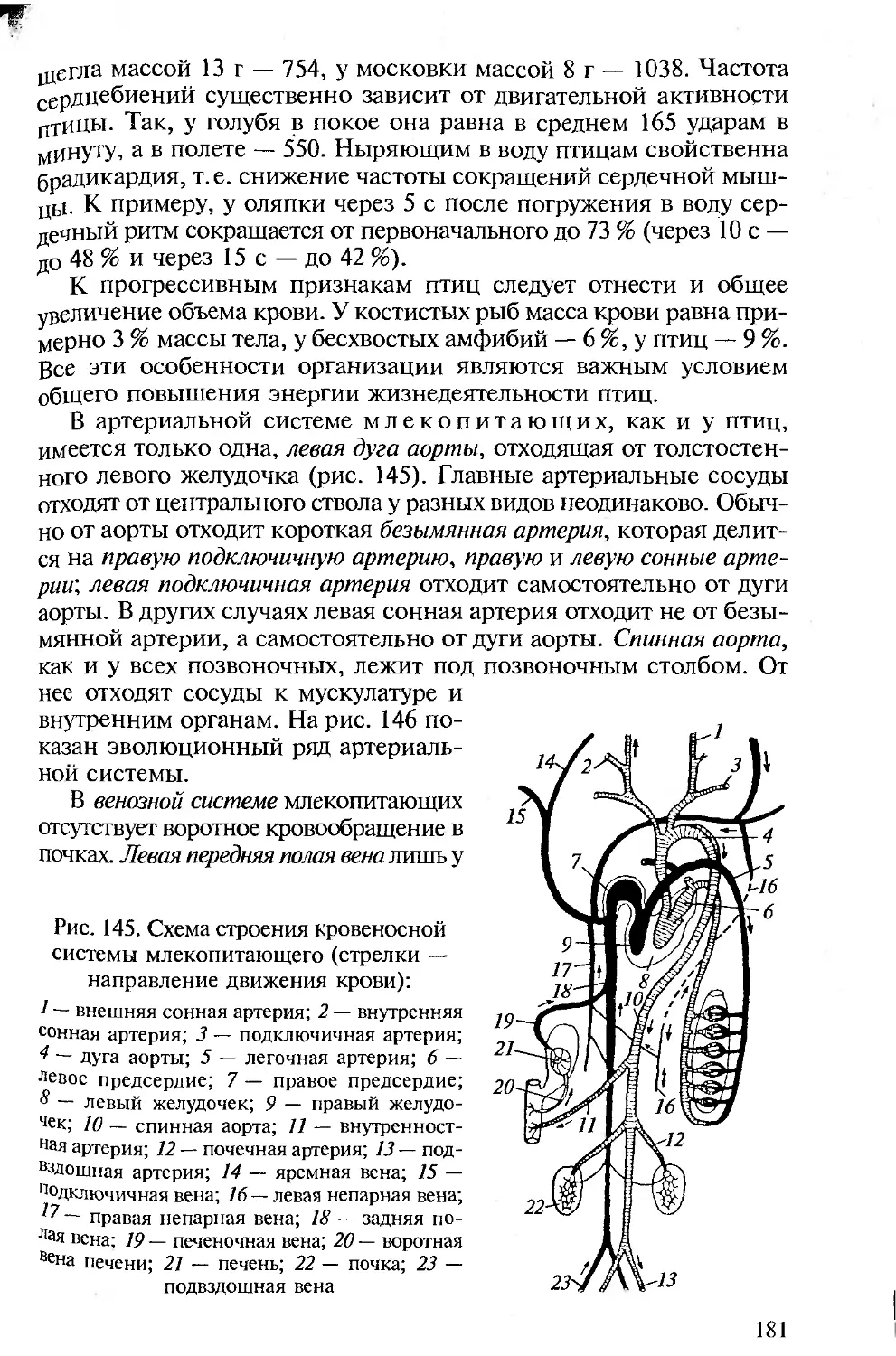
ее до абсурда.
Основная цель сравнительной анатомии — практическое срав-
нительное изучение эволюционных изменений систем органов у
систематически разных групп животных. Именно здесь важны те-
оретические построения, хорошо подтвержденные проверенны-
ми фактами. В этом случае очень полезны рисунки, схемы и дру-
гие изображения, позволяющие лучше запомнить большой и слож-
ный материал. Однако читатель должен избежать чисто книжного
восприятия сравнительной анатомии животных. При написании
этого пособия, как и других подобных книг, авторы опирались на
многолетний собственный опыт преподавания зоологии позво-
ночных студентам педвузов, чтения лекций на факультете повы-
шения квалификации вузовских преподавателей. Материал книги
подан под определенным углом зрения, изложен с определенных
позиций и в известной степени субъективно. Очень хотелось, что-
4
v изучающих сравнительную анатомию животных, в том числе
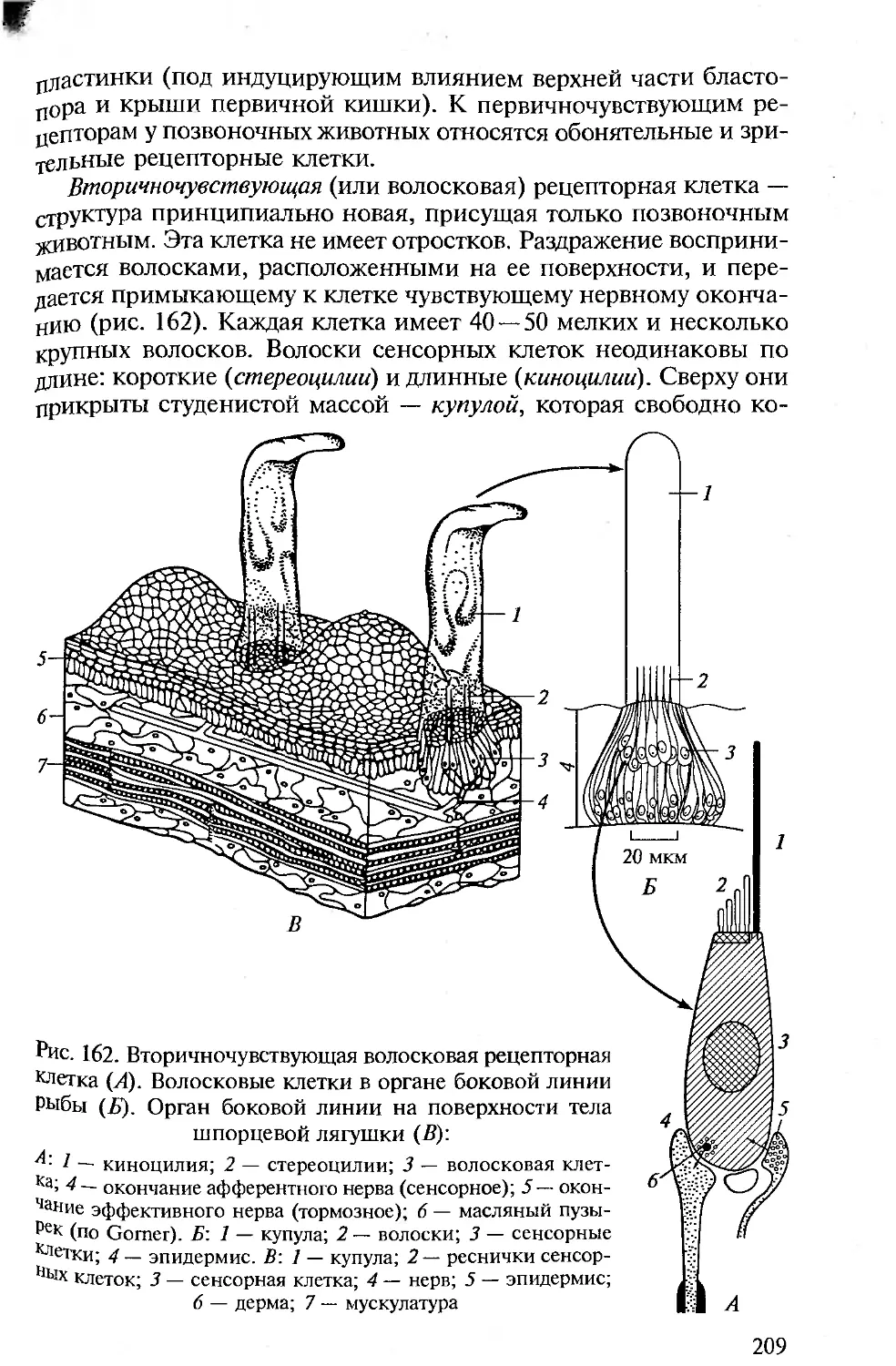
использованием настоящей книги, сложились собственные пред-
С явления об интересных объектах и фактах, важных для изучения
С подтверждения уже сложившихся теорий или опровержения их.
И Т Парсонс в предисловии к «Анатомии позвоночных» (1992)
приводит определенные требования к подобного рода изданиям.
Эти требования А. Ромер перечислил во вступлении к новому из-
данию названной книги. Они и сейчас не утратили своего значе-
ния. С некоторыми изменениями уместно напомнить о них здесь.
• Необходимость соблюдения подлинно сравнительного подхода в
описании систем органов в последовательных рядах позвоночных
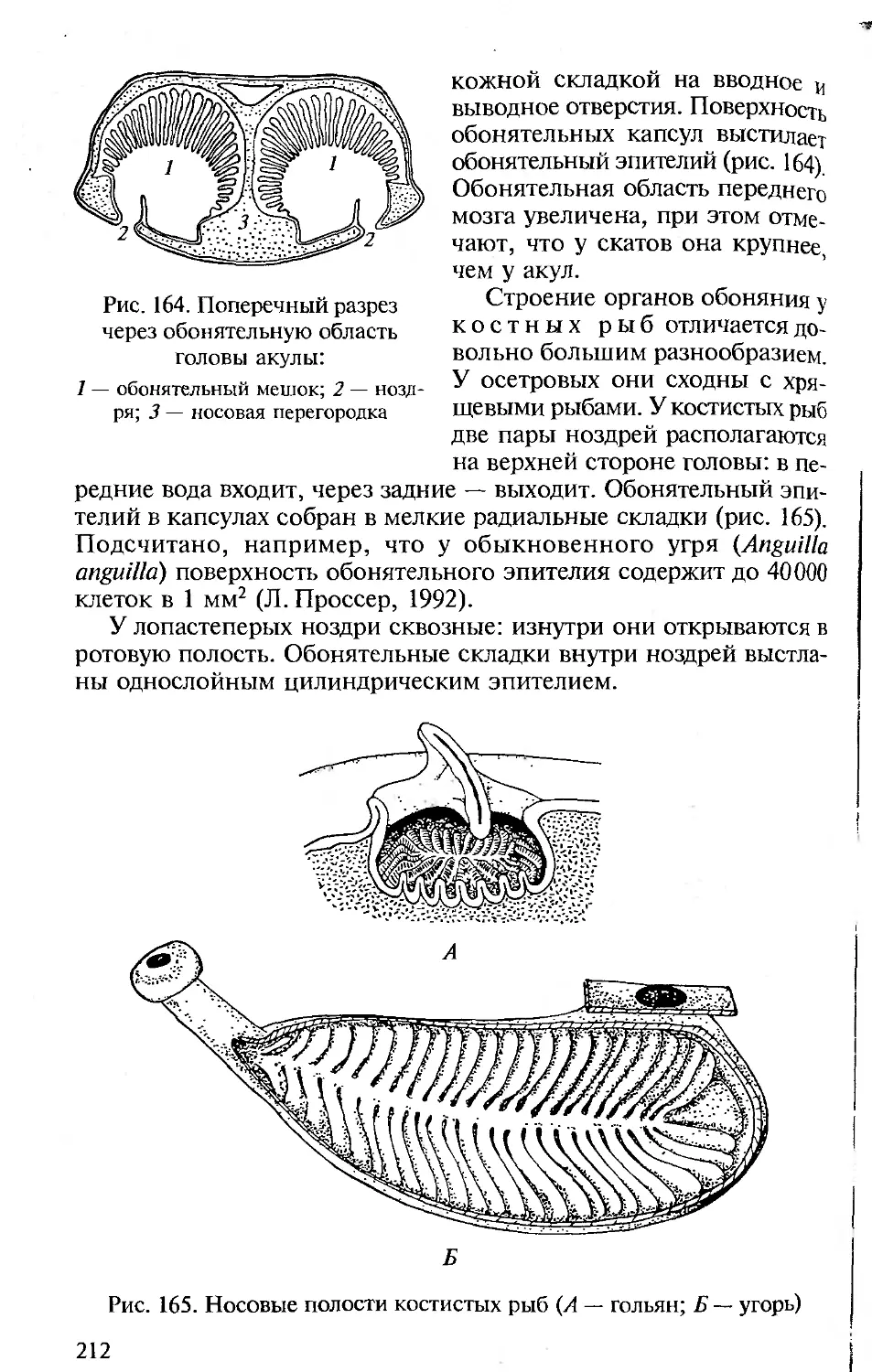
животных по мере их эволюционного совершенствования. При этом
мы согласны с А.Ромером, что не следует злоупотреблять сравне-
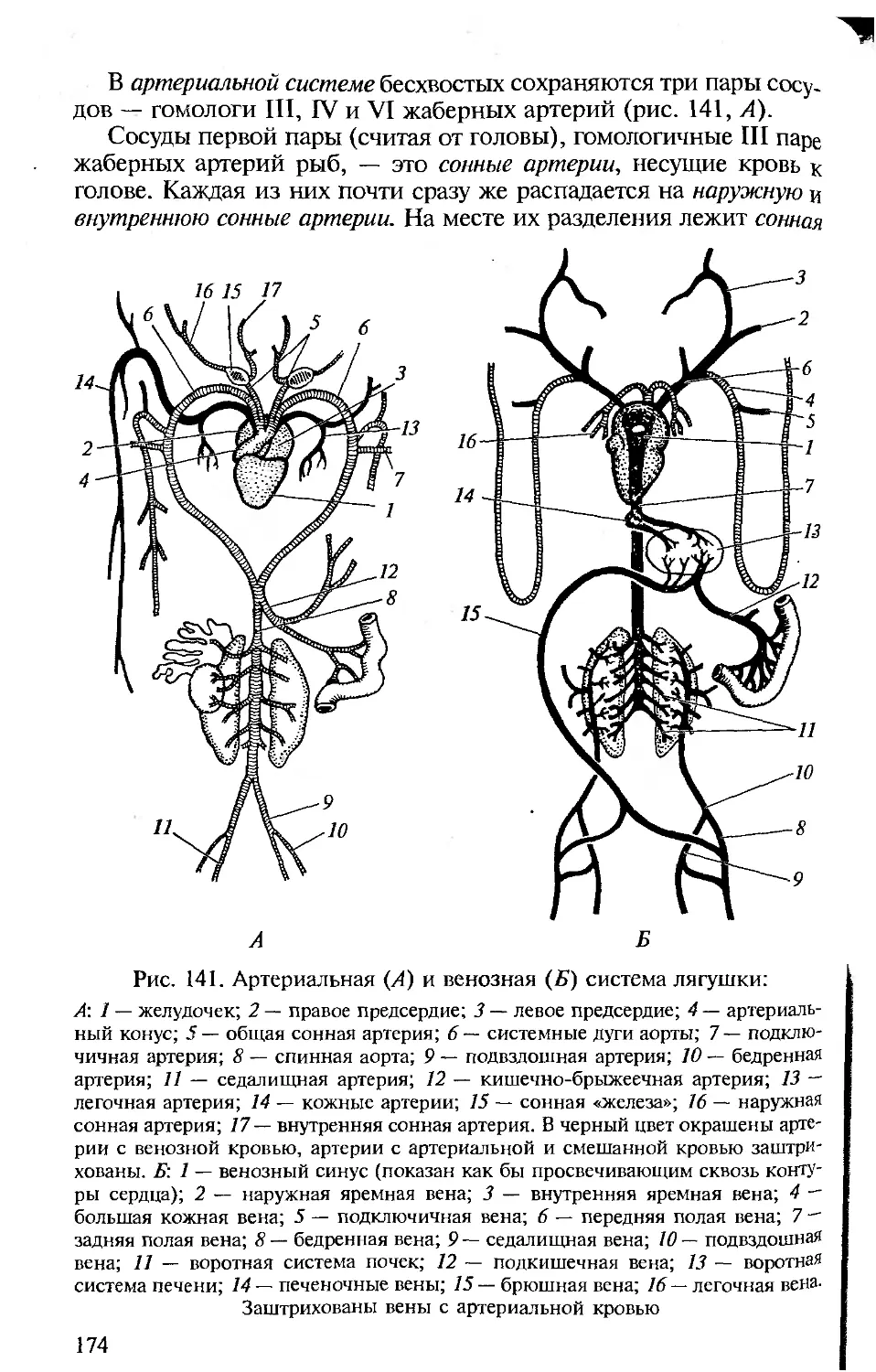
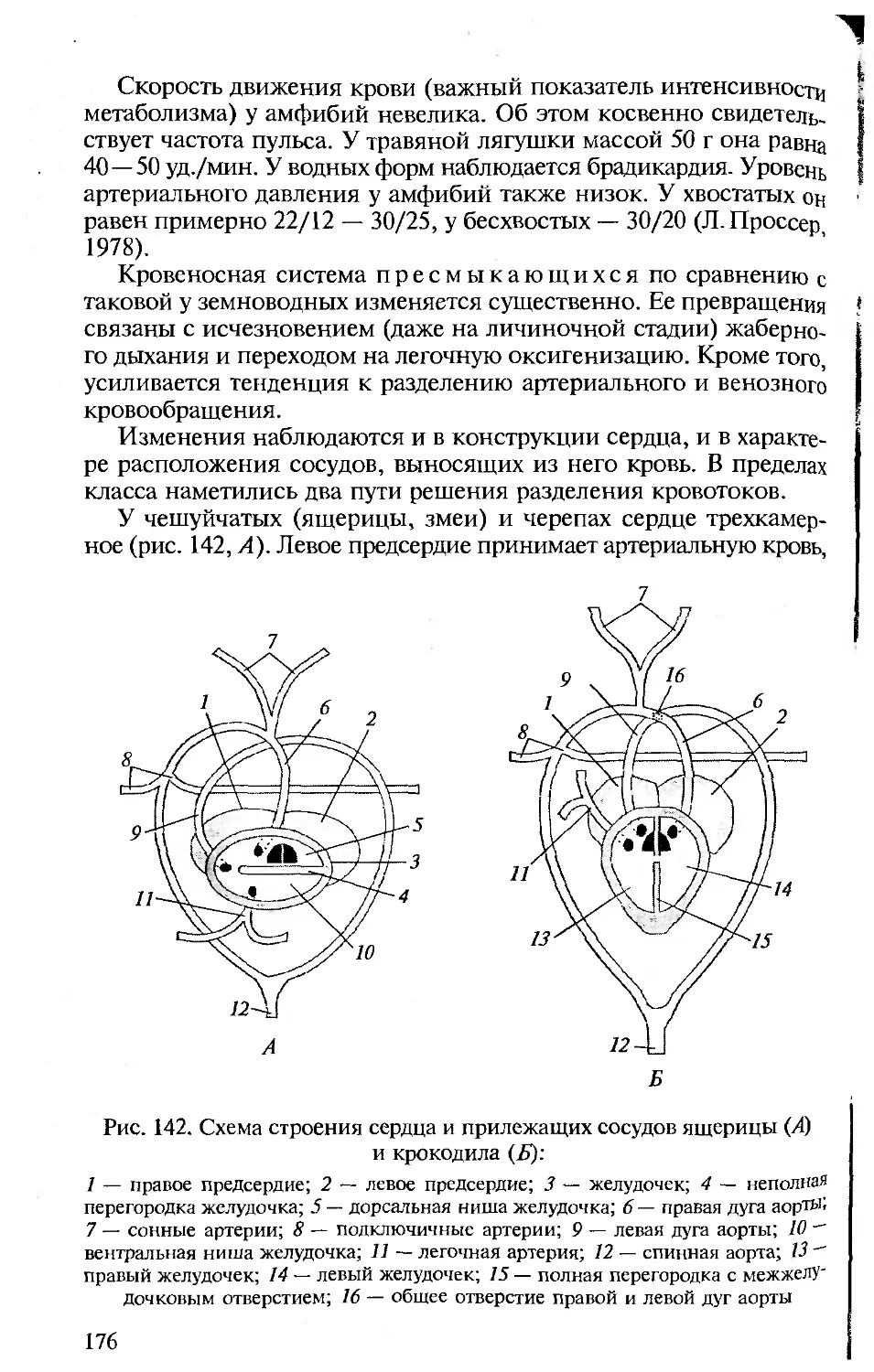
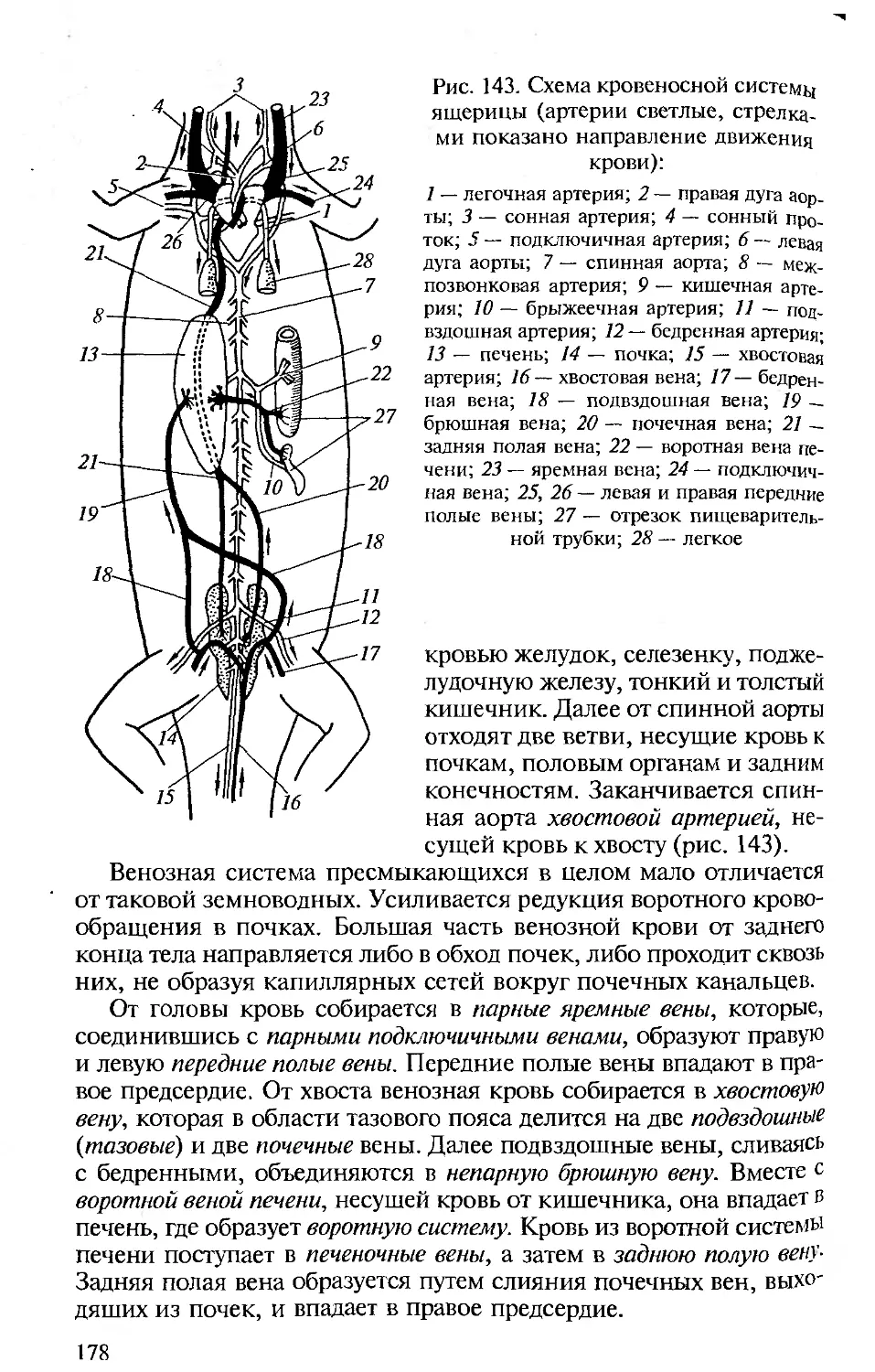
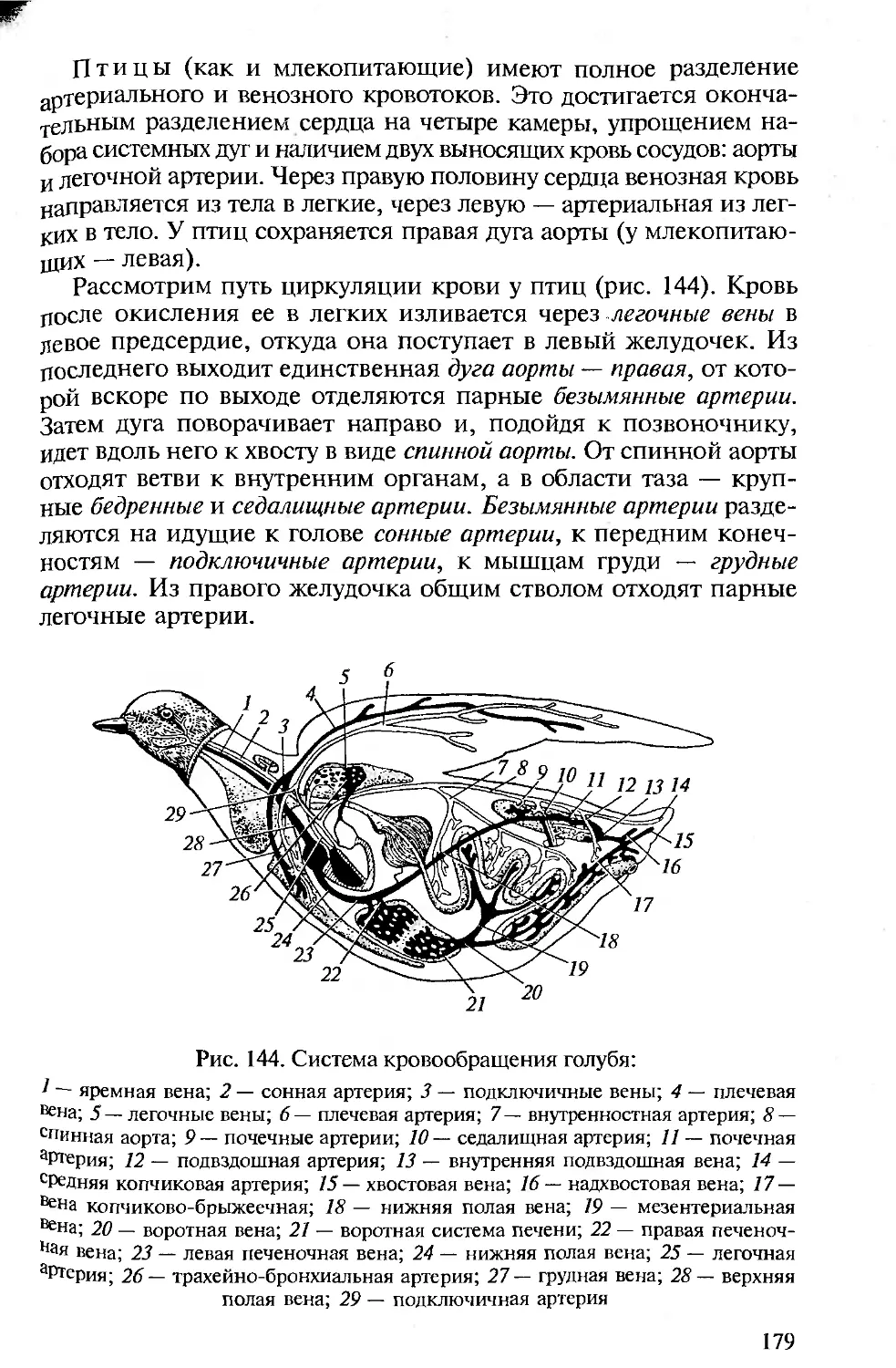
ниями с человеком. Анатомия и физиология человека подробно
рассматриваются в соответствующих курсах, изучаемых на биоло-
гических факультетах университетов и педвузов, и упоминание о
тех или иных чертах организации человека вполне достаточно для
создания общей картины эволюции позвоночных и места челове-
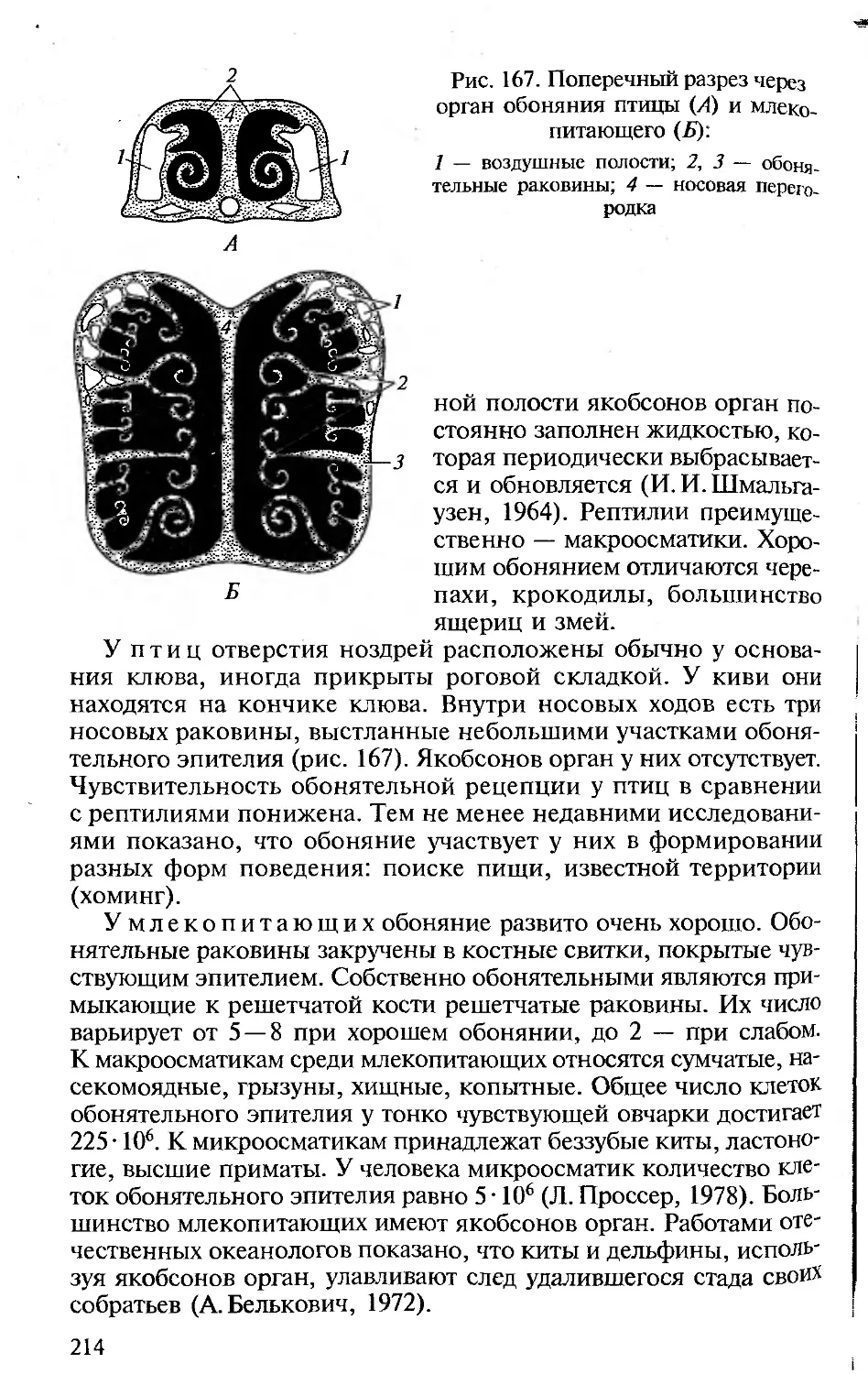
ка в ней.
• Сохранение функционального подхода при описании морфоло-
гических структур. Такой подход соответствует требованиям ко всем
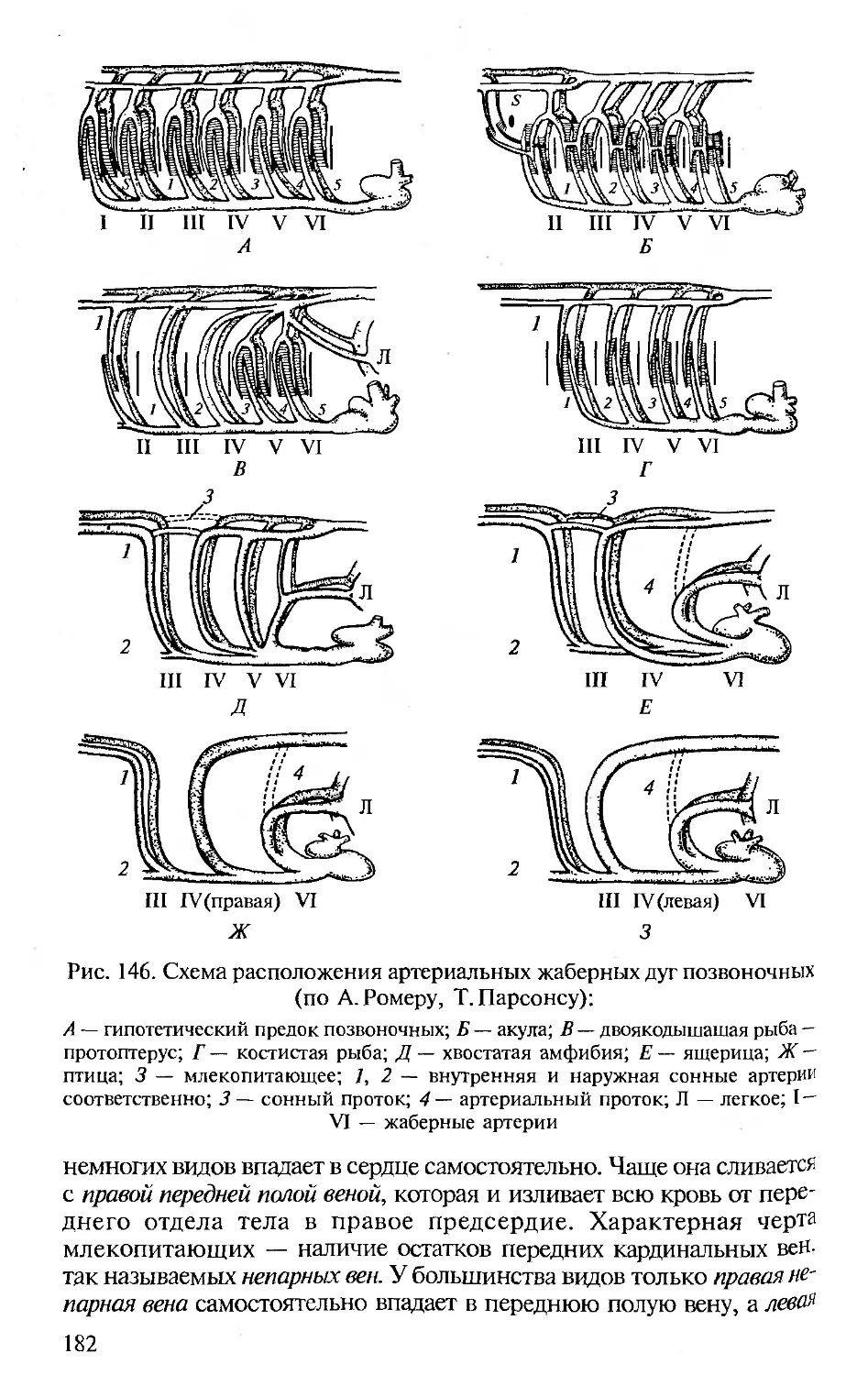
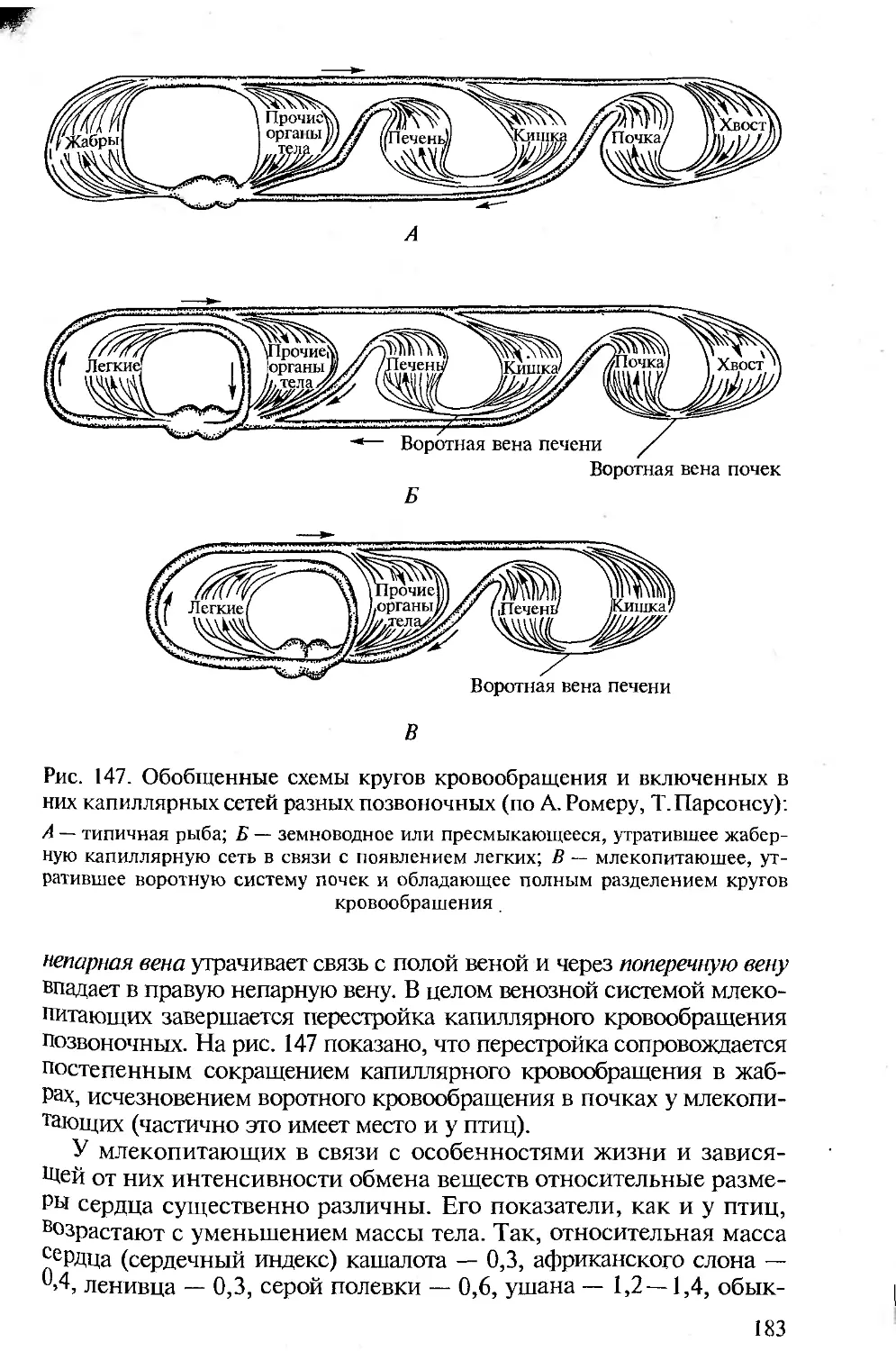
отечественным изданиям по анатомии, поэтому во всех разделах
настоящей книги принципиальным является рассмотрение строе-
ния органов и их систем у позвоночных в связи с выполняемыми
ими функциями.
• Должное внимание к эмбриональному развитию. Рассмотрение
эмбрионального развития разных групп животных важно для ус-
тановления гомологии, на которой основана вся сравнительная
анатомия. При изучении строения тела любого позвоночного сле-
дует иметь в виду, что взрослый организм — одна из многих пос-
ледовательных стадий его развития. Именно поэтому в первом раз-
деле предлагаемой вашему вниманию книги рассматриваются осо-
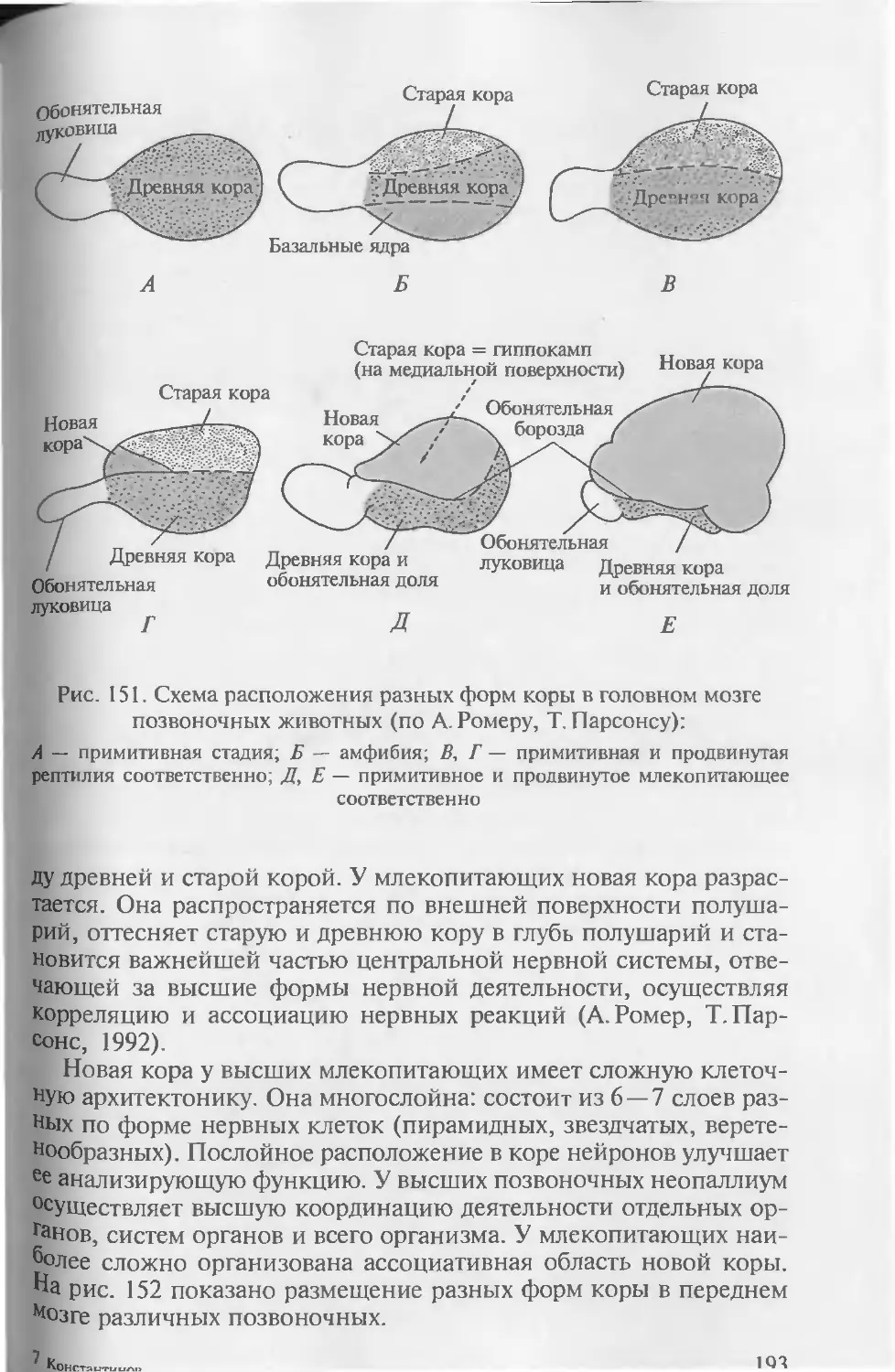
бенности развития хордовых, а при описании систем органов ав-
торы постоянно опираются на то, как происходит их формирова-
ние в онтогенезе.
• Привлечение к анализу гистологических данных. При рассмотре-
нии морфологии покровов, органов опорно-двигательной, пи-
щеварительной, дыхательной, выделительной, репродуктивной и
Других систем для понимания особенностей их функционирова-
ния необходимо знание клеточного строения их органов.
• Надлежащий палеонтологический фон. Использование сведе-
ний по историческому развитию разных групп позвоночных важно
Для понимания их эволюции и современных филогенетических
взаимоотношений между ними, поэтому в качестве обобщаю-
щей главы рассматриваются происхождение и эволюция хордо-
вых.
• Хорошие иллюстрации. Каким бы обширным ни был лабора-
торный практикум, изучение строения всегда проводится на оп-
ределенных «типичных представителях» каждой систематической
группы, на определенных «модельных видах», на основе знаний,
по которым формируются представления о группе в целом. Для
демонстрации предлагается общая (типичная) схема (рисунок,
модель) их строения. Поэтому в подобного рода изданиях столь
важны иллюстрации.
Авторы настоящего пособия, имея опыт анатомического вскры-
тия и изучения морфологических особенностей позвоночных жи-
вотных, старались учесть перечисленные требования при изложе-
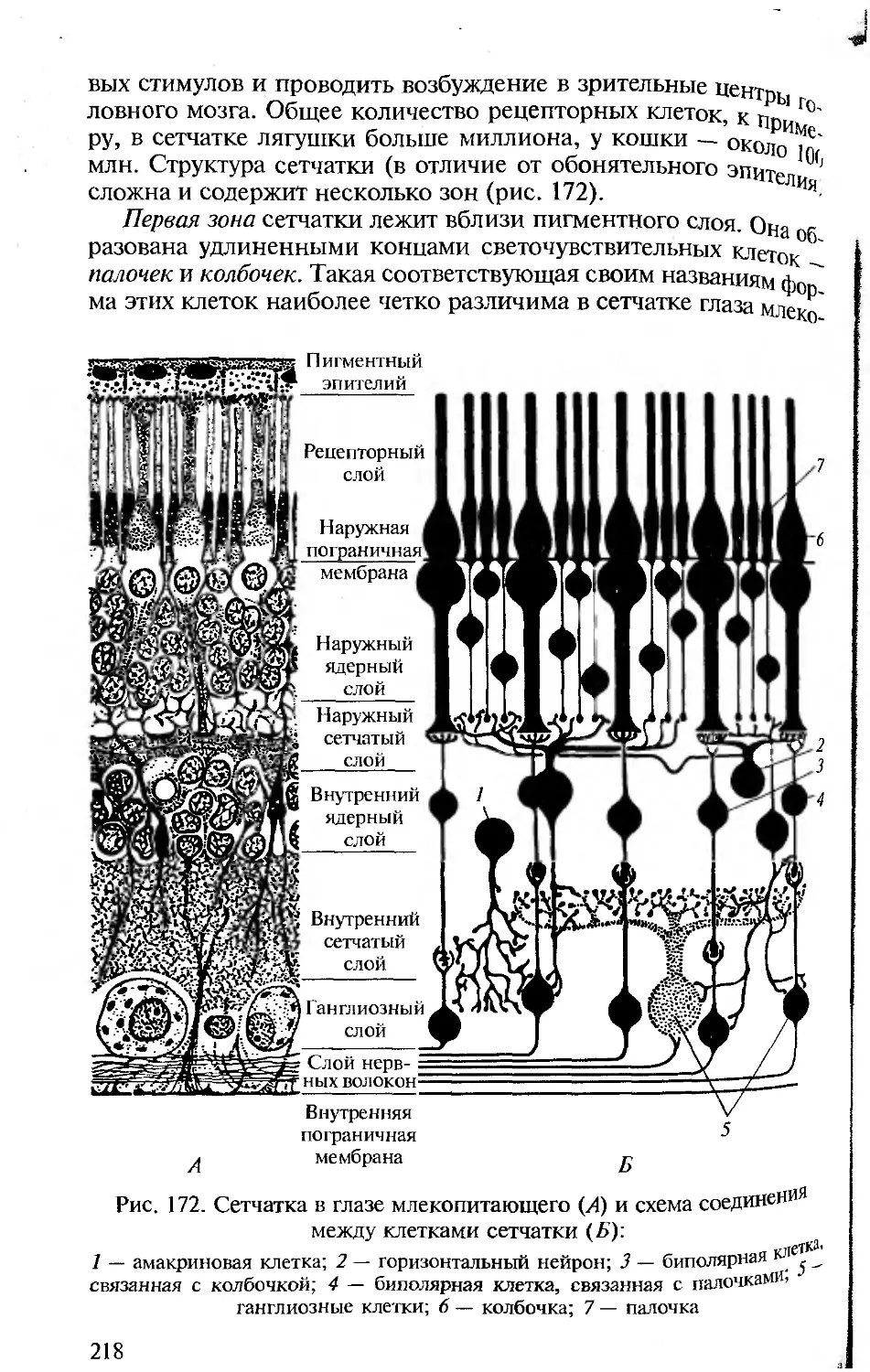
нии конкретных разделов. В книгу включены многие традицион-
ные иллюстрации, взятые из изданных ранее учебников и руко-
водств по сравнительной анатомии позвоночных. В тех случаях,
когда использовались оригинальные рисунки, приведены соот-
ветствующие ссылки.
Есть еще одна причина, побудившая авторов к написанию дан-
ной книги. Относительно недавно нами подготовлено второе из-
дание учебника для педвузов «Зоология позвоночных» («Акаде-
мия», 2004), в основу которого в свое время был положен учеб-
ник профессора С. П. Наумова (4-е изд. — 1983 г.). За сравнительно
небольшой отрезок времени (2000—2003), прошедший между под-
готовкой первого и второго изданий, удалось существенно попол-
нить учебник новыми научными сведениями, включив новые срав-
нительные материалы, затрагивающие принципиальные позиции
зоологической науки: происхождение птиц, начальные этапы эво-
люции позвоночных, в том числе роль и место щитковых и пан-
цирных рыб в эволюции современных бесчелюстных и челюстно-
ротых, охрану животного мира. Эти и другие сведения вошли так-
же в уже изданный «Лабораторный практикум по зоологии позво-
ночных» («Академия», 2004). При написании названных книг по-
стоянно ощущалась необходимость в более последовательном срав-
нительно-анатомическом подходе, лежащем в основе зоологичес-
ких и эволюционных курсов, изучаемых на биологических факуль-
тетах педагогических вузов. Предлагаемая книга полезна студен-
там-биологам при обобщении знаний на завершающих этапах обу-
чения, а также аспирантам биологических специальностей.
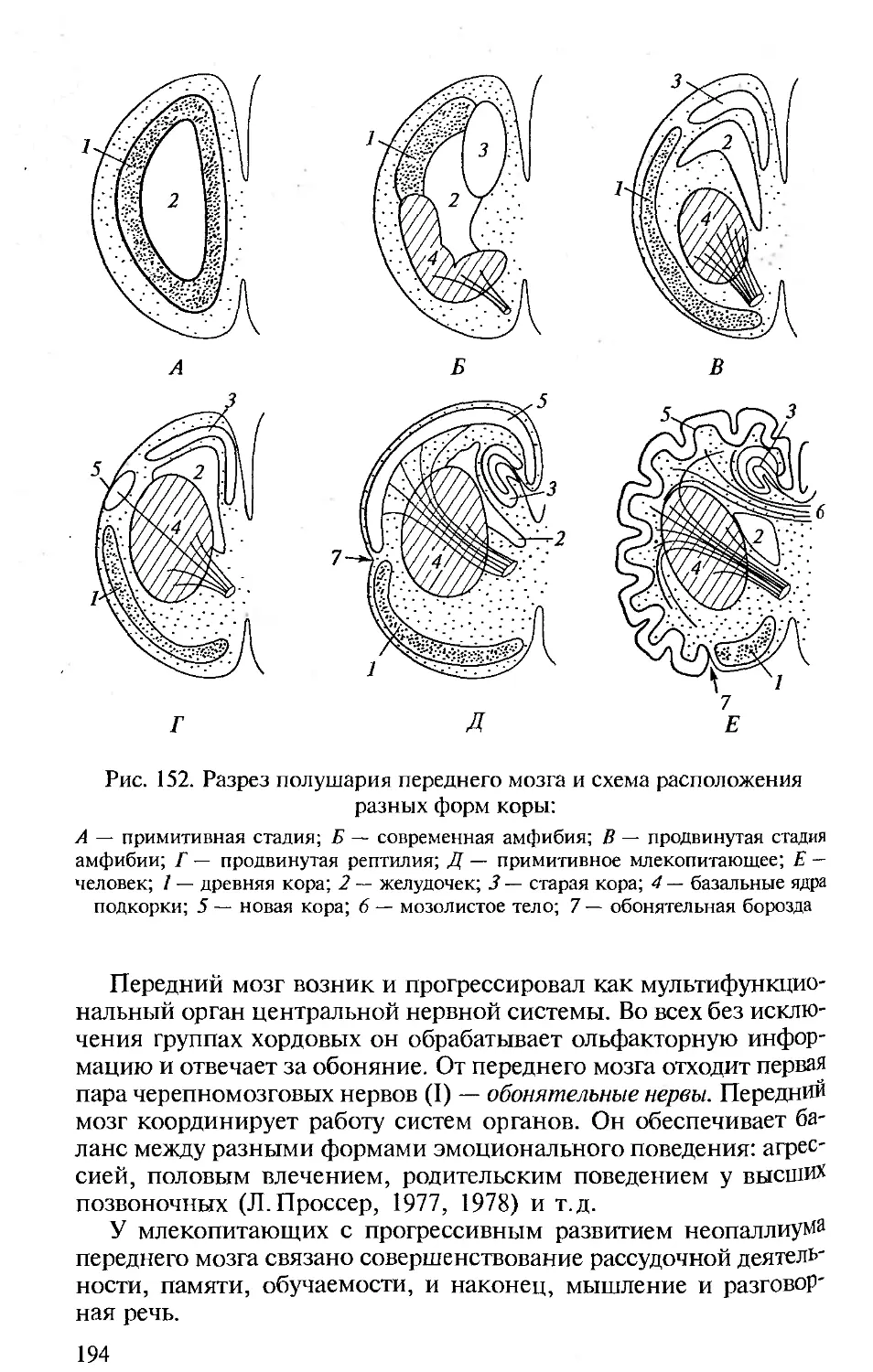
ВВЕДЕНИЕ
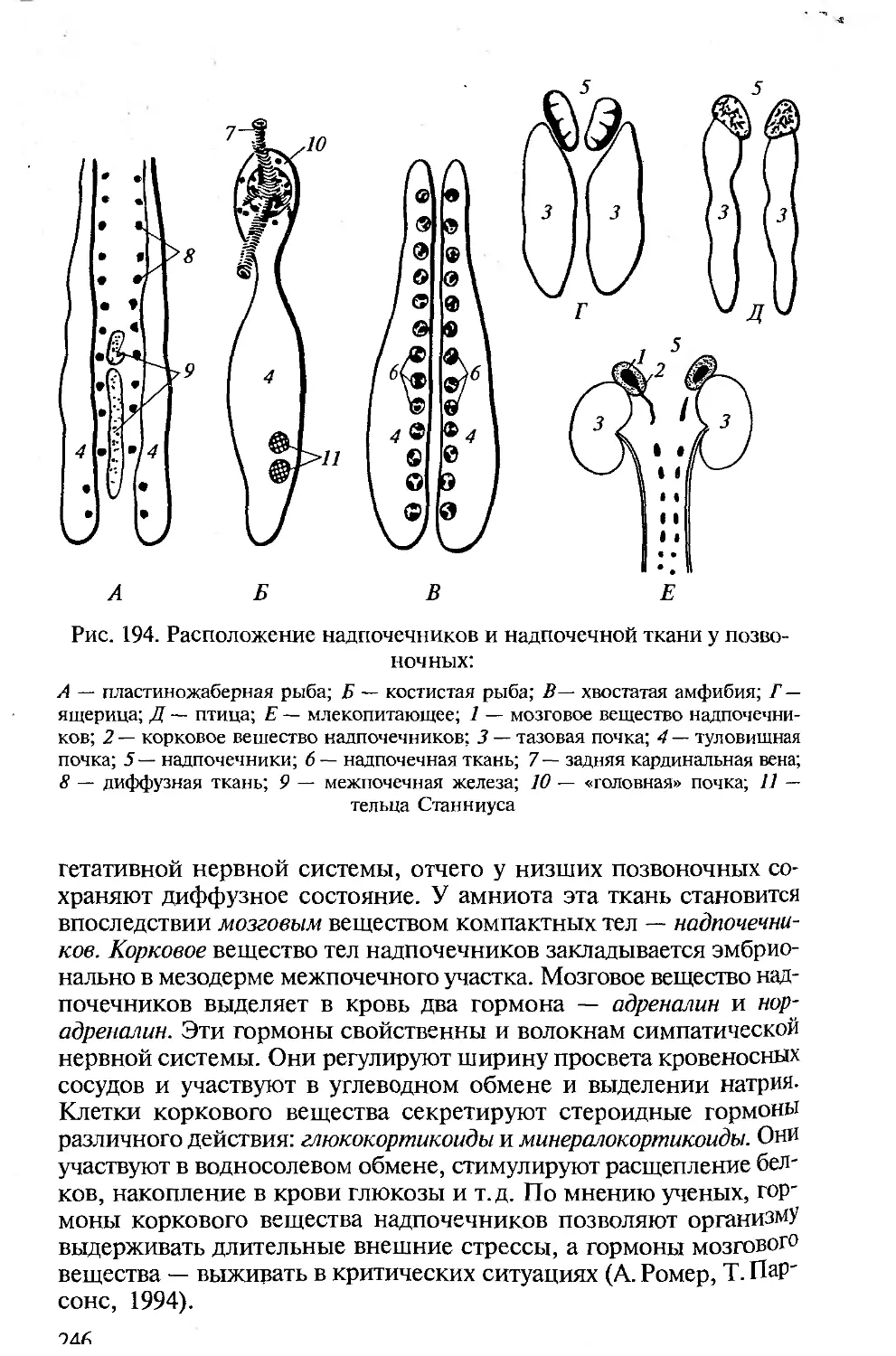
Особенности морфофизиологической организации животных,
принадлежащих к типу Хордовые, большую часть которых состав-
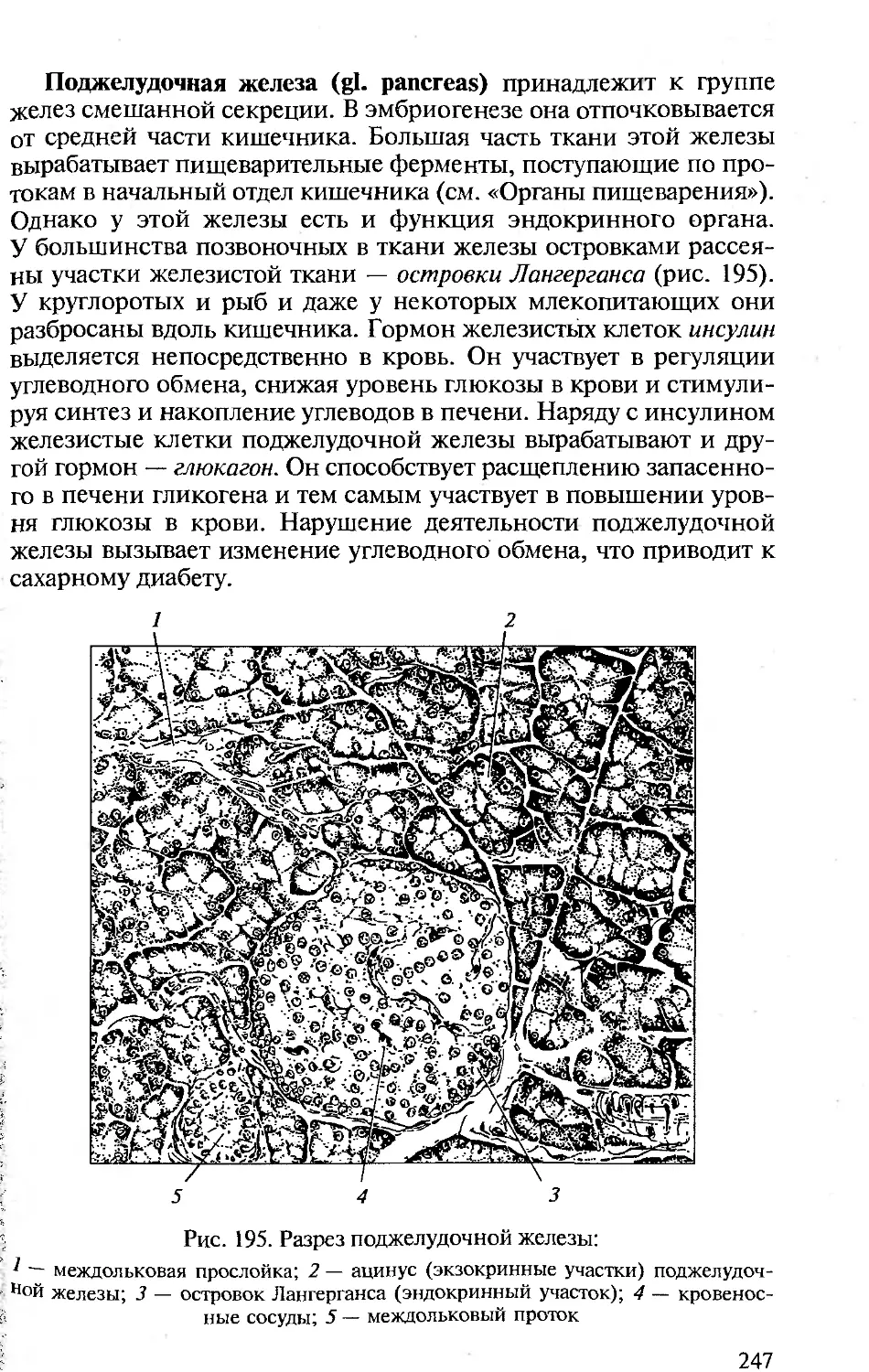
ляют позвоночные (отсюда традиционное название «Сравнитель-
ная анатомия позвоночных»), представляют большой интерес для
биологов с различных позиций.
Эволюция хордовых складывалась в общей картине историчес-
кого развития царства Животные, со многими типами которого
они имеют общие черты: вторичноротость, целомичность, дву-
стороннюю симметрию тела, метамерию. В свою очередь, ориги-
нальность хордовых в отличие от представителей других типов
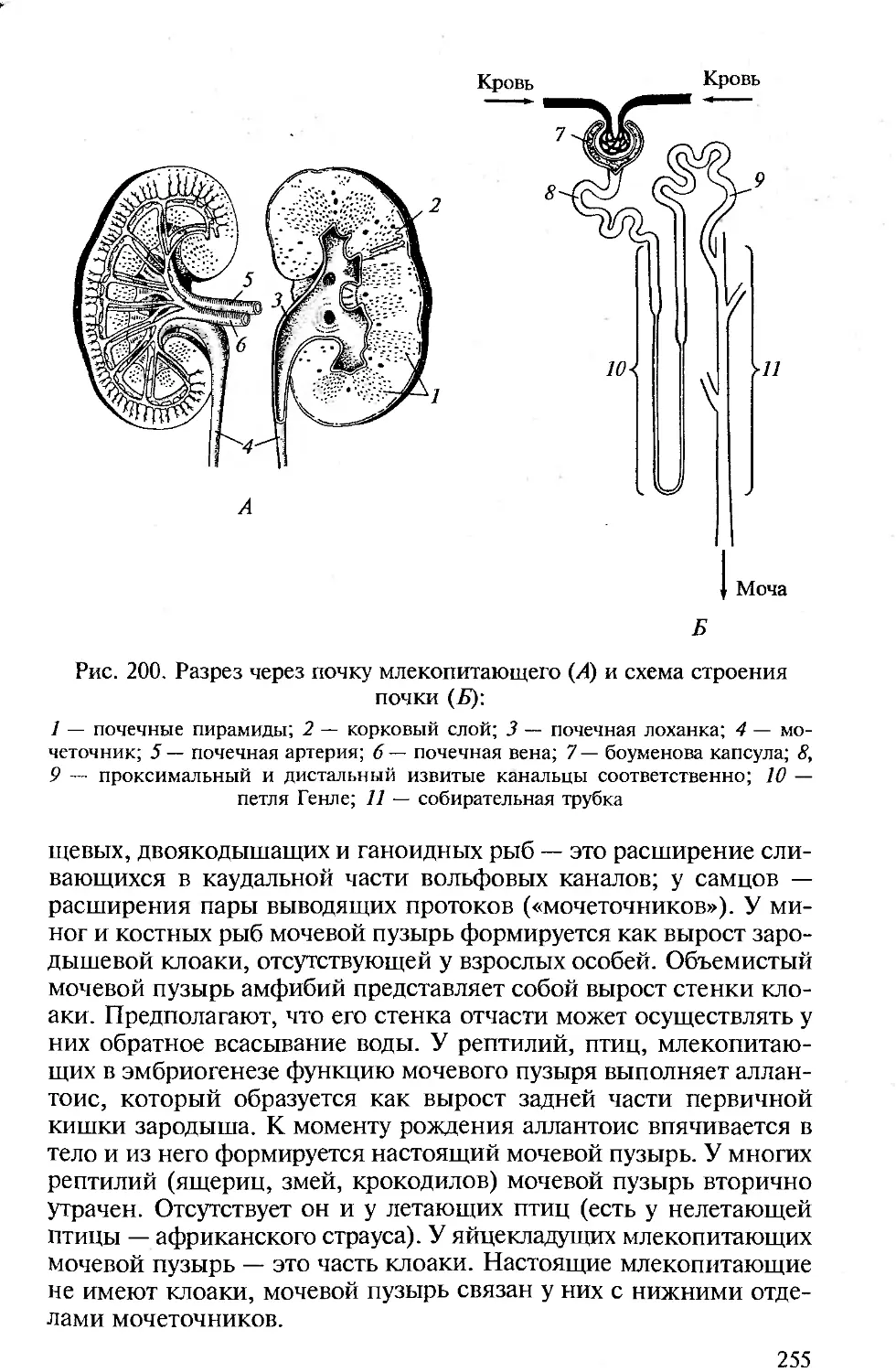
животных проявляется в своеобразии их анатомо-морфологичес-
кой организации: наличии внутреннего скелета в виде хорды или
позвоночника, центральной нервной системы в виде дорсально
расположенной по отношению к хорде нервной трубки, висце-
ральных щелей в глоточном отделе кишечника, изначальная фун-
кция которых — фильтрация пищи и дыхание*.
Известно, что в поступательной морфологической эволюции
хордовых каждый исторический этап сопровождался чередовани-
ем крупных прогрессивных изменений (ароморфозов) с много-
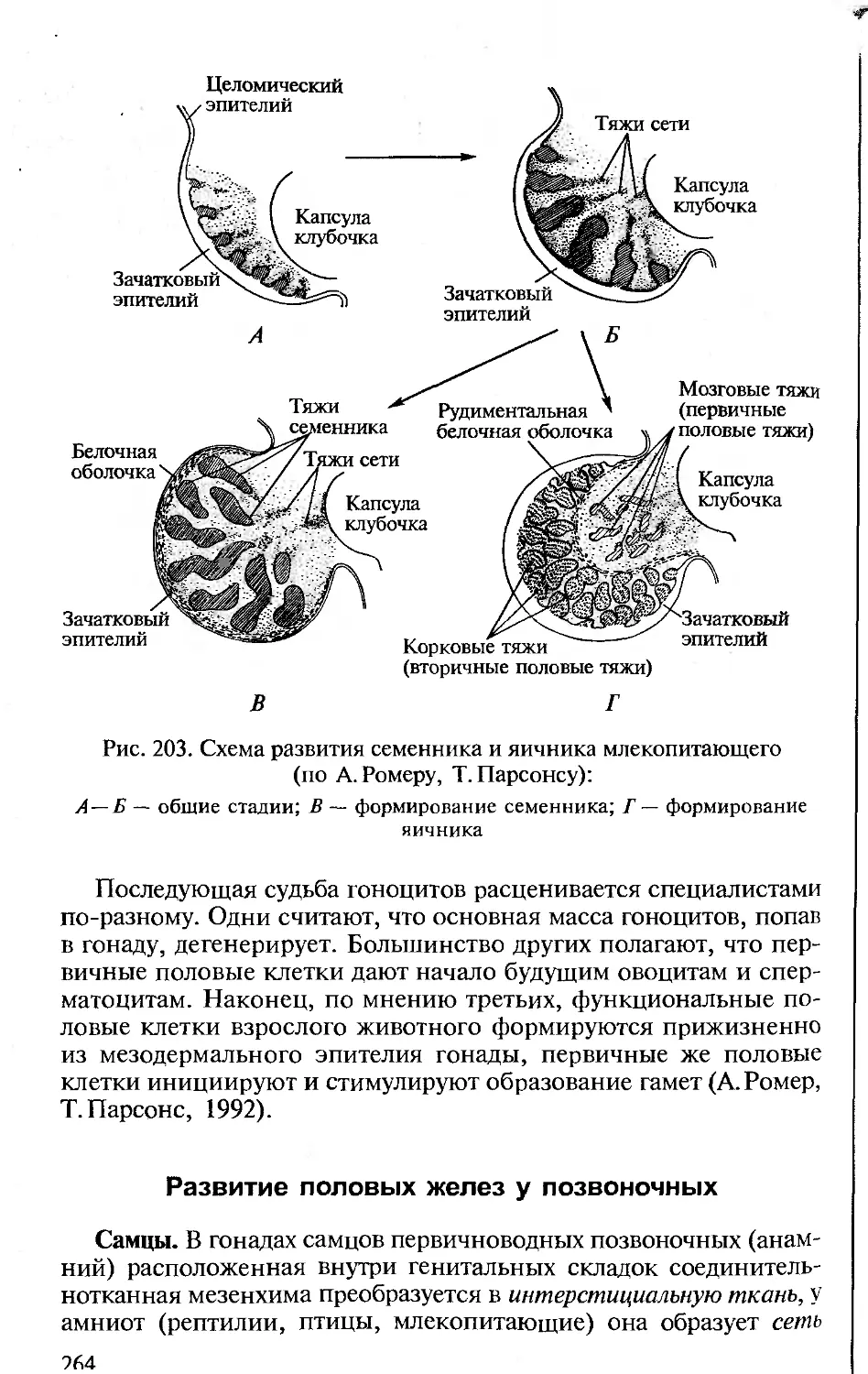
образными частными приспособлениями (алломорфозами). Это
принципиально важно учитывать при рассмотрении конкретных
групп животных.
Морфоэкологическая эволюция сопровождалась в этом типе
освоением животными разных сред обитания, в том числе пере-
ходом водных позвоночных к жизни в наземно-воздушной среде.
Изначальный морфологический тип хордовых оформился в вод-
ной среде. Современные первично-водные хордовые (бесчереп-
ные, круглоротые, рыбы амфибии) во многом сохранили исход-
ный план строения. Освоение наземно-воздушной среды обита-
ния принципиально изменило ракурсы морфофизиологической
организации представителей крупных систематических таксонов
(классов) подтипа Позвоночные. Водная среда была освоена пред-
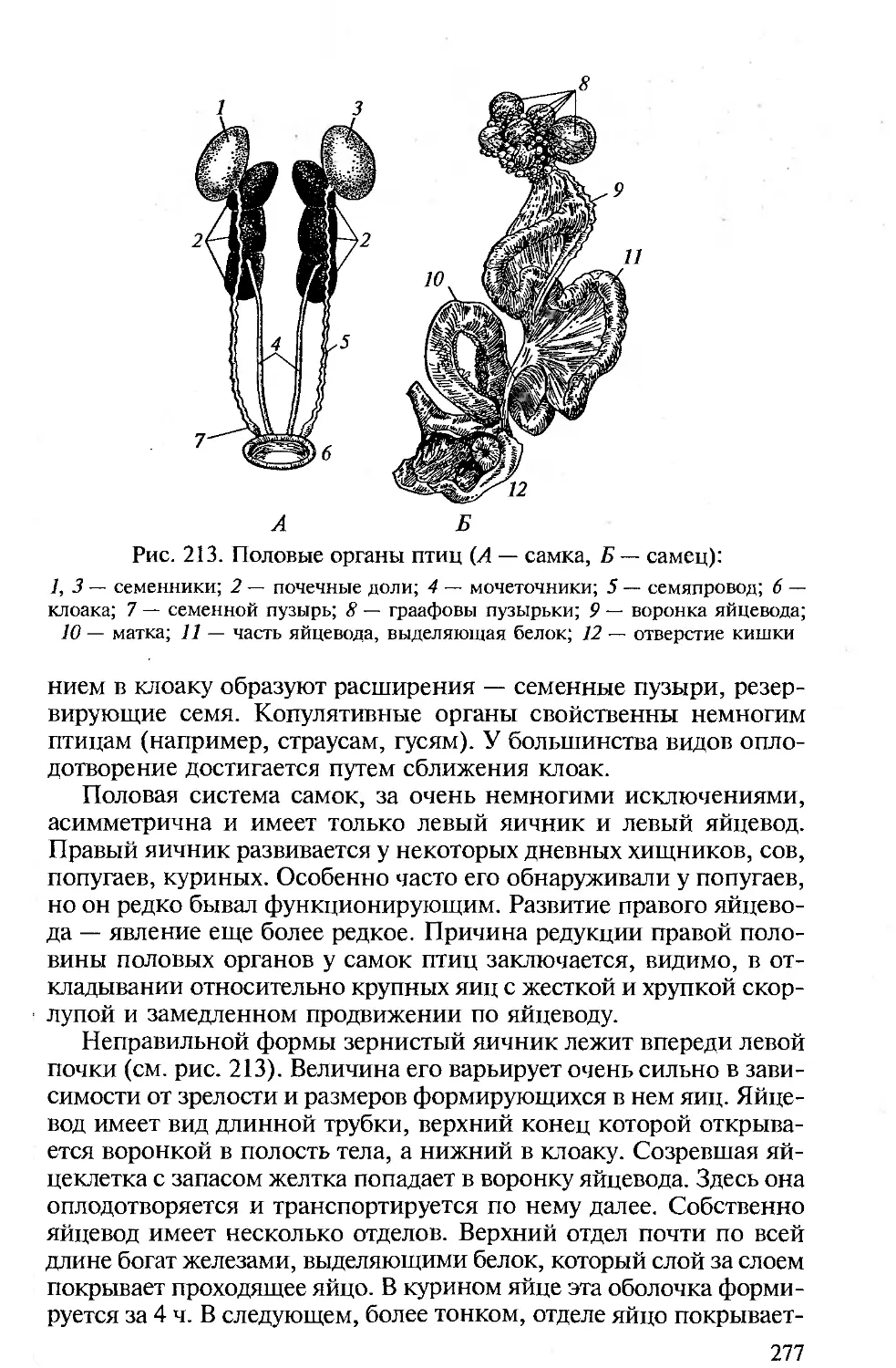
ставителями различных групп наземных позвоночных вторично,
но уже на ином, более высоком уровне организации.
* Некоторые черты хордовых свойственны и относительно маловыразитель-
ному типу Полухордовые (Hemichordata).
7
Особенности морфофизиологической организации позволили
хордовым достичь наивысшего уровня развития живых существ —
появлению человека, способного к высшим формам рассудочной
деятельности: абстрактному мышлению, развитому интеллекту,
осознанному поведению и т.д. Познание морфологической эво-
люции позвоночных приближает к пониманию эволюции, орга-
низации и особенностей биологической природы человека.
Остановимся подробнее на важнейших аспектах общей орга-
низации хордовых.
• Вторичноротость — достаточно раннее свойство животных,
оформившееся у червей. Кроме хордовых она присуща членисто-
ногим иглокожим, погонофорам, полухордовым. При их эмбрио-
нальном развитии рот оформляется (прорывом) на конце тела,
противоположном бластопору, а первичный рот становится аналь-
ным отверстием пищеварительной системы. Известно, что у всех
вторичноротых параллельно развивается и ряд других оригиналь-
ных признаков: в их теле в виде парных выпячиваний первичной
кишки энтодермально закладывается целом. Эти выросты дают
начало мезодерме. Параллельно формируется замкнутая кровенос-
ная система.
• Хордовые — вторичнополостные (целомические) животные. Это
свойство филогенетически объединяет их с кольчатыми червями,
плеченогими, членистоногими, погонофорами, иглокожими, полу-
хордовыми. Вторичная полость тела закладывается в эмбриогенезе
как парные выросты первичной кишки. Она образует простран-
ство между стенкой тела и внутренними органами. Полость выст-
лана серозной оболочкой и содержит целомическую жидкость.
Изначальная (первичная) функция целома — опорная: у части
беспозвоночных животных вторичная полость служит гидростати-
ческим скелетом. У большинства других животных, в том числе и
у хордовых, целом поддерживает биохимическое постоянство внут-
ренней среды организма. Выросты целома проникают в ткани и
органы тела животного и снабжают их питательными веществами
и кислородом. С целомом связаны образование и функционирова-
ние кровеносной и выделительной систем.
У низших позвоночных пространство целома поделено на пе-
редний отдел — околосердечную (грудную) полость, в которой ле-
жит сердце, и задний — брюшную полость, где располагаются все
внутренние органы. Околосердечную полость ограничивает пери-
кард — соединительнотканный мешок, образующий околосердеч-
ную сумку. Брюшную полость выстилает брюшная (серозная) обо-
лочка. Она тоже образована соединительной тканью, покрытой с
поверхности эпителием. К спинной стенке брюшной полости по-
средством спинной связки — брыжейки — подвешен кишечник.
У млекопитающих от брюшной полости отделяется грудная.
Поперечной перегородкой служит диафрагма. В центральной части
„фрагмэ образована сухожилиями, по периферии располагает-
'ДН мышечный слой. В виде соединительнотканной складки диаф-
Сдгма присутствует уже у некоторых рептилий.
Р В грудной полости находятся сердце и легкие. Сердце заключе-
но в околосердечную сумку. По бокам его, ближе к спине, распо-
ложены легочные полости, окружающие легкие. Они выстланы
плевральной оболочкой, отчего их называют плевральными. Че-
ез грудную полость проходят пищевод и крупные кровеносные
сосуды. Все остальные органы лежат в брюшной полости. Пери-
кард, плевра, брюшина — это разные варианты (по местоположе-
нию) серозной оболочки. Чаще они состоят из двух листков, между
которыми находится пространство, заполненное целомической
(серозной) жидкостью.
Целомическая полость у ланцетника в связи с образованием '
вдоль туловища обширной атриальной полости оттеснена к спине.
Участки целома окружают гонады и участки пищеварительной
трубки и др. Через выделительные трубочки она сообщается с внеш-
ней средой (атриальной полостью).
У пластинчатожаберных и некоторых костных рыб присутству-
ют короткие парные каналы (брюшные поры), лежащие в облас-
ти анального отверстия, или клоаки. Через них из целомической
полости выводится избыток жидкости. У самок всех позвоночных
в целом открываются воронки мюллеровых каналов (яйцеводов),
выносящие наружу продукты размножения.
• Двусторонняя {билатеральная) симметрия тела — свойство
многих типов животных впервые оформляется у плоских червей.
Исторически она развивается в связи с усилением двигательной
активности животных и обособлением у них переднего конца тела.
Приобретение двусторонней симметрии было важным историчес-
ким шагом в морфологической эволюции животных, значитель-
но расширившим возможности последующих экологических адап-
таций. У позвоночных животных разных классов двусторонняя
симметрия закладывается в раннем эмбриогенезе разновременно:
У головохордовых, осетровых рыб и земноводных — до начала
Дробления яйца, у хрящевых рыб, рептилий, птиц, млекопитаю-
щих — в конце периода дробления.
Активное движение сопровождалось у животных и развитием
метамерии. У высших многоклеточных она связана с совершен-
ствованием механизмов движения (например, волнообразного
изгибания тела при плавании и ползании), с упорядочением внут-
ренней организации, интенсификацией деятельности различных
органов. У хордовых метамерия непременно закладывается в ран-
нем онтогенезе. Она явственно выражена во взрослом состоянии у
низших форм. У высокоорганизованных позвоночных метамерия
сменяется олигомеризацией, а в расположении внутренних орга-
нов «маскируется» при специализации.
Кроме черт, объединяющих хордовых с животными других ти-
пов, они имеют и собственные оригинальные особенности мор-
фофункциональной организации.
• Все хордовые обладают внутренним осевым скелетом. Его ос-
новной элемент — хорда закладывается энтомезодермально и ок-
ружена соединительной тканью. У позвоночных эта ткань стано-
вится основой позвоночника и мозгового черепа. Внутренний ске-
лет составляет опору для скелетной мускулатуры. Сложенную та-
ким образом комплексную осевую структуру тела хордовых назы-
вают иногда миохордом. У низших хордовых с ней связано разви-
тие локомоции: основным органом поступательного движения тела
животного в воде служит хвостовой отдел тела. С появлением мно-
горычаговых парных конечностей, обеспечивающих многообраз-
ное движение в воде и устойчивое перемещение тела в наземно-
воздушной среде, ось тела, образованная позвоночником, служит
опорой для них.
• Центральная нервная система организована в нервную трубку ~
это еще одна оригинальная черта хордовых. В отличие от хорды
нервная трубка закладывается эктодермально. Внутренняя полость
нервной трубки — невроцелъ заполнена жидкостью, омывающей
мозг как изнутри, так и снаружи. Колебание жидкости обеспечива-
ет в разных отделах центральной нервной системы активное пита-
ние нервной ткани и удаление продуктов метаболизма.
• Следующая оригинальная черта типа — наличие в переднем
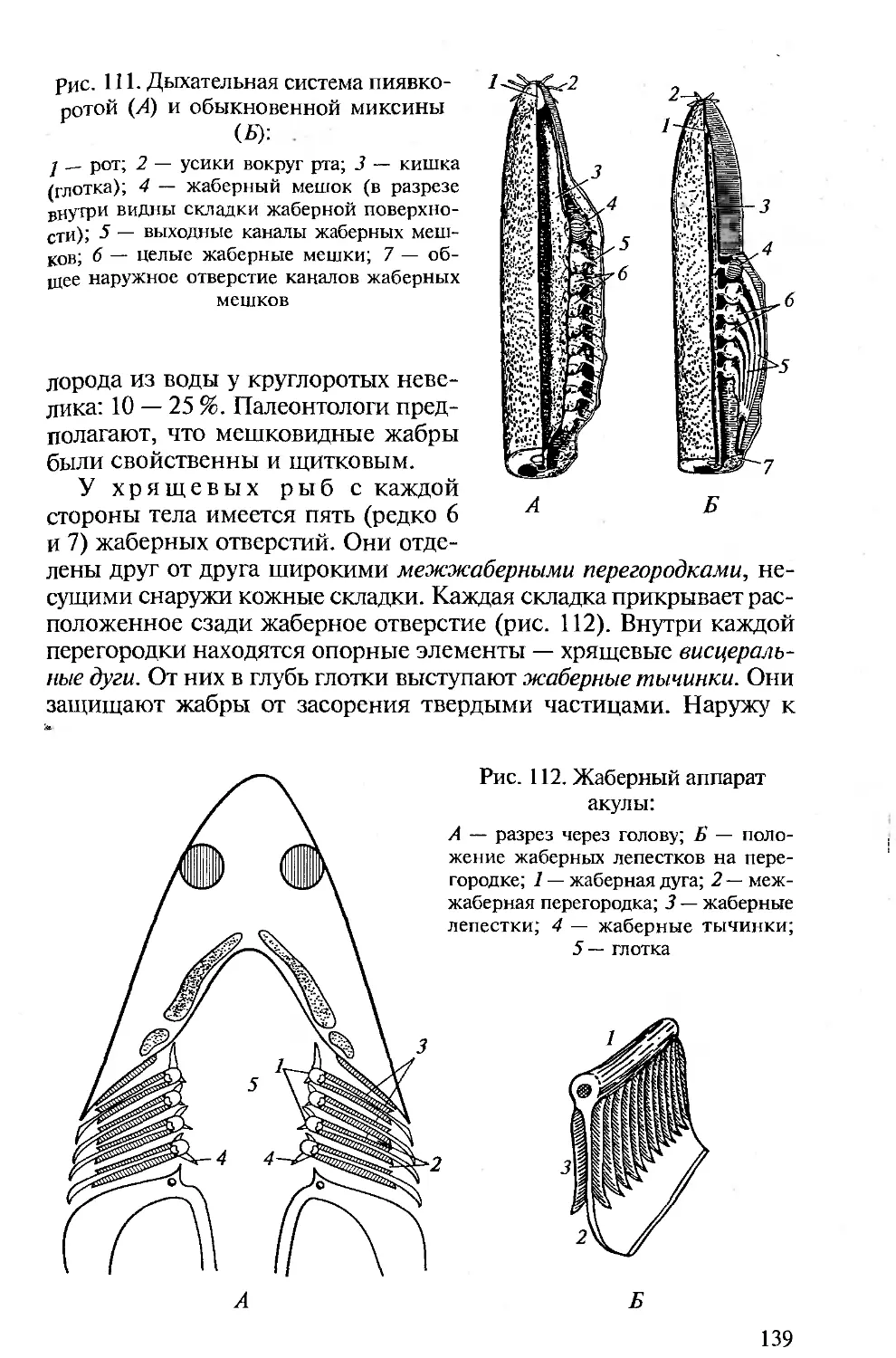
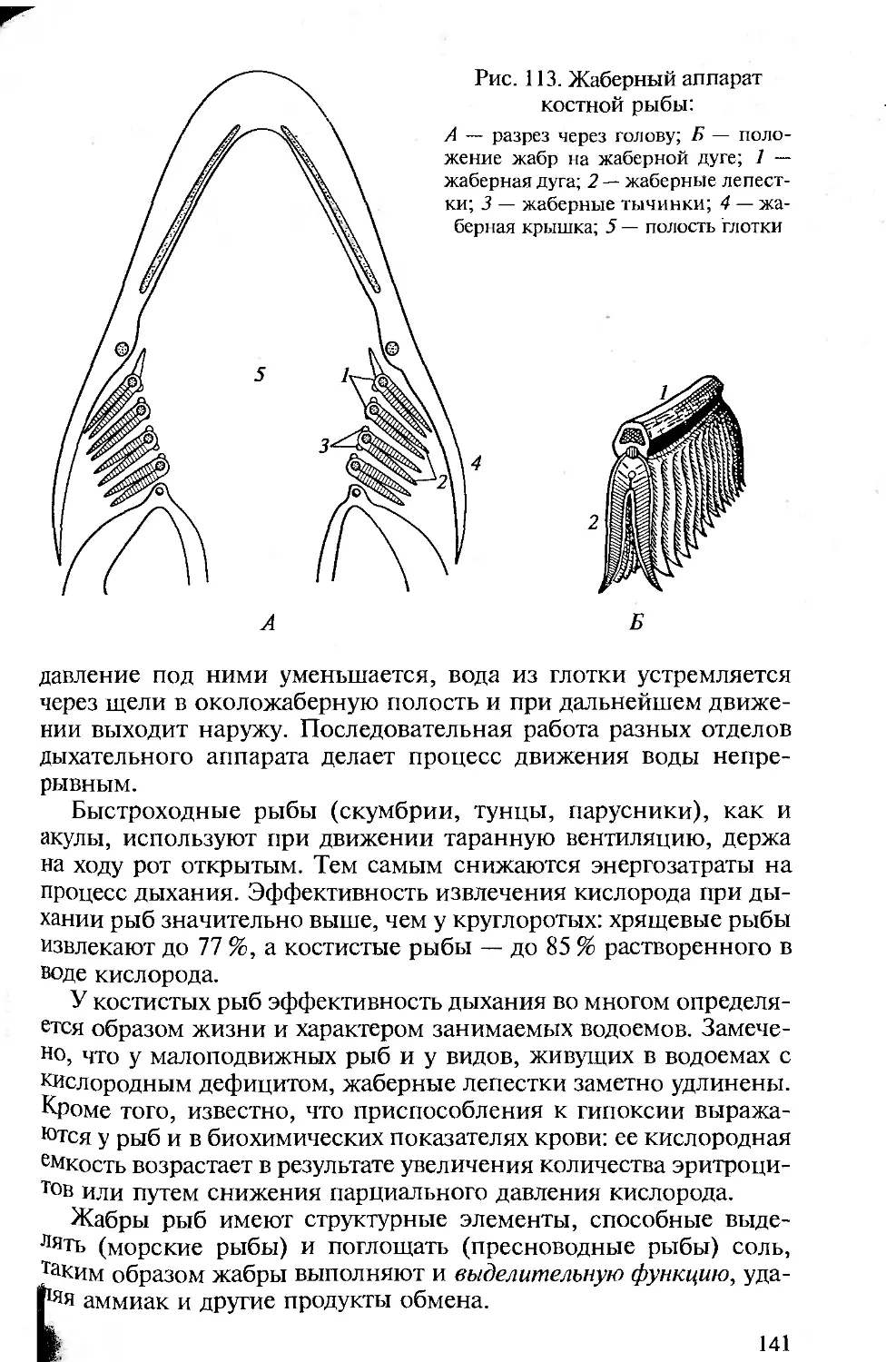
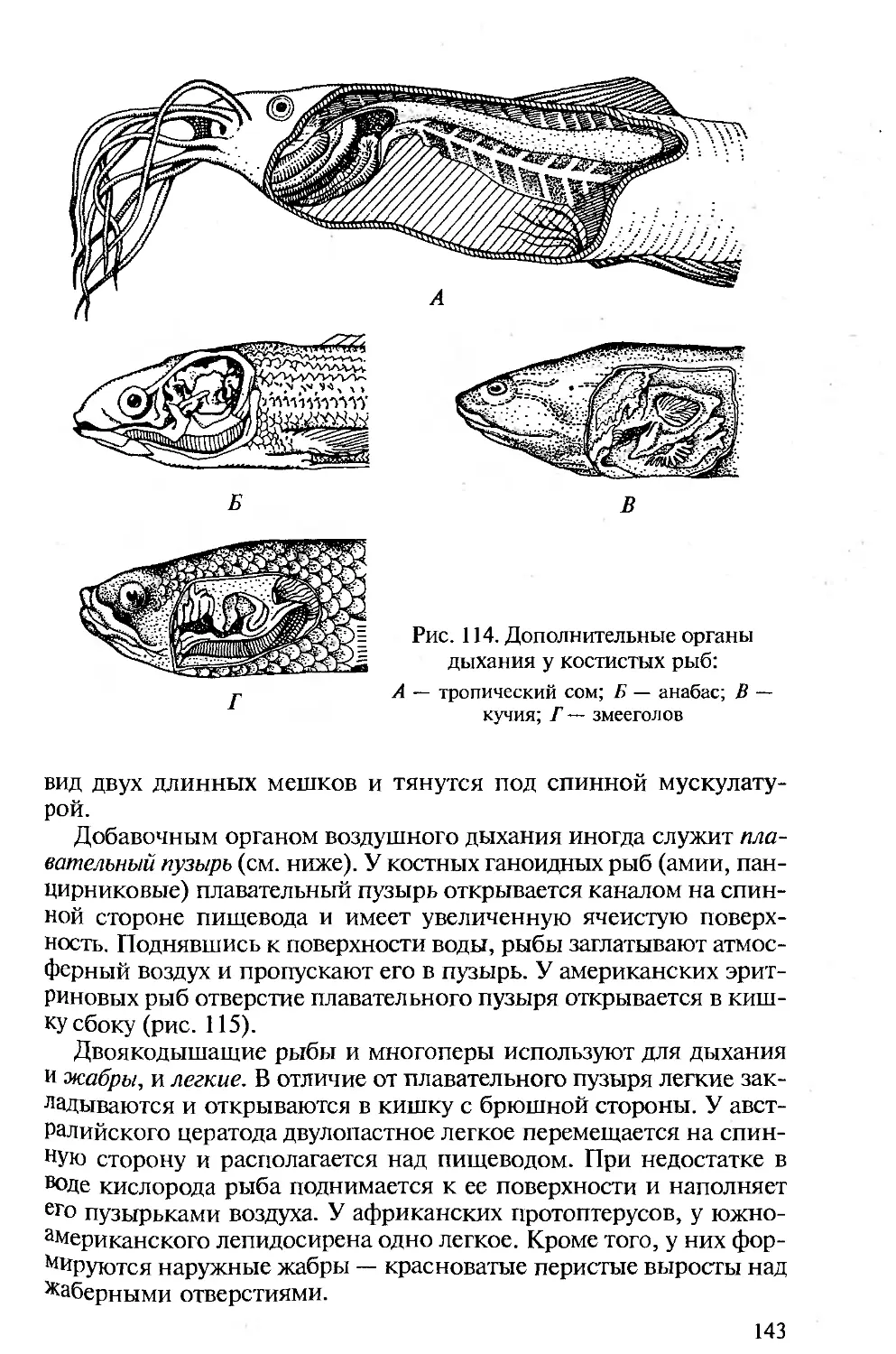
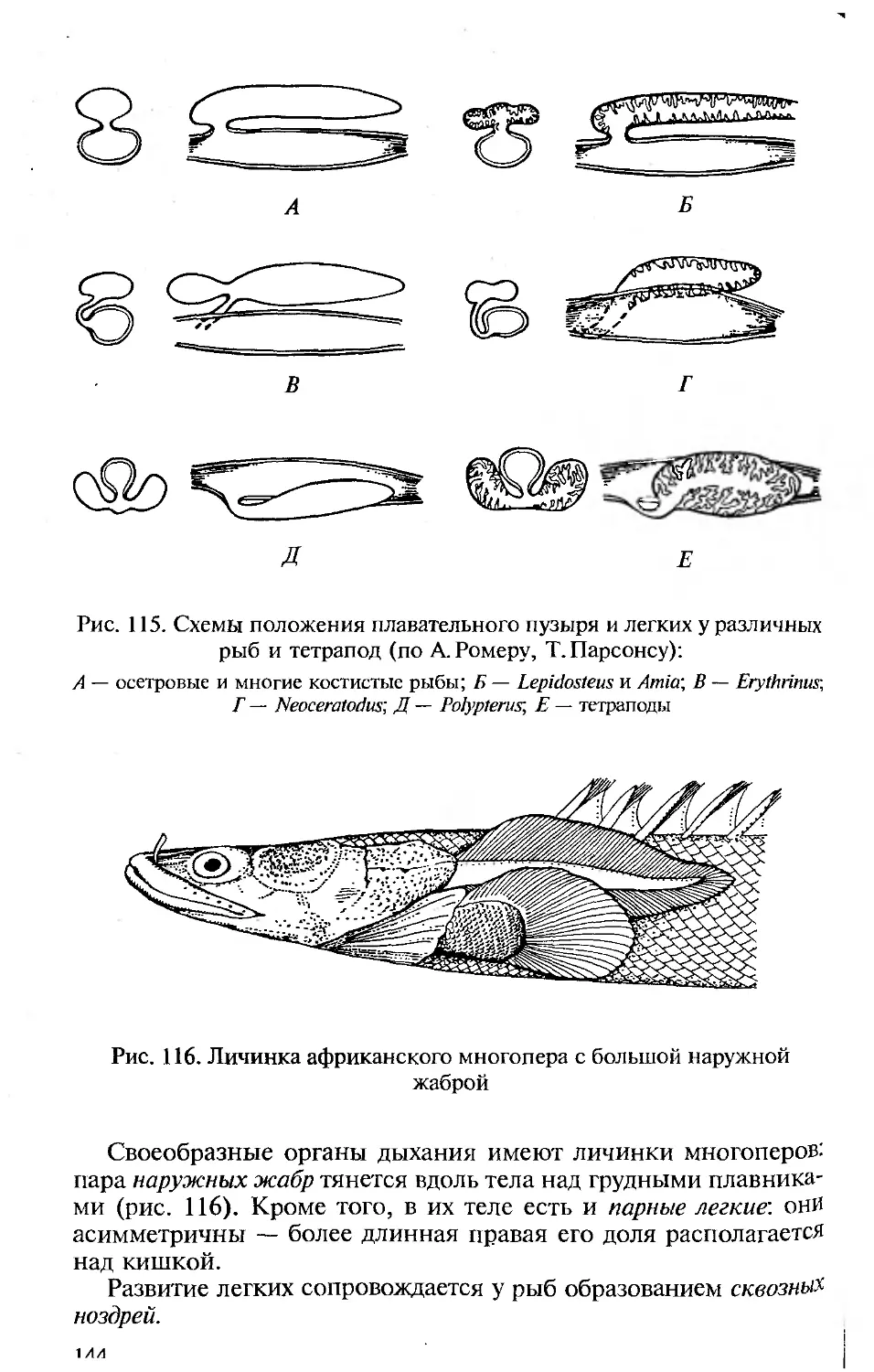
отделе пищеварительной трубки, в глотке, висцеральных щелей. Они
закладываются эктоэнтодермалъно и, будучи сквозными, участвуют
в отфильтровывании пищи и выведении воды, омывающей стен-
ки межжаберных перегородок. Последнее подготовило их участие
в дыхании хордовых. В процессе эволюции органы дыхания совер-
шенствуются. У первичноводных ими являются энто- или экто-
дермальные жаберные лепестки. У наземных позвоночных органы
дыхания — легкие — образуются как энтодермальные выпячива-
ния брюшной стороны пищеварительной трубки.
Таким образом, имея единый план строения (или всего лишь
два — для первичноводных и наземных) организма, четкие об-
щие черты морфофизиологической организации, хорошо пред-
ставленные в палеонтологической летописи, хордовые традици-
онно используются для общебиологических заключений и обоб-
щений. Так, эволюционная теория построена в основном на срав-
нительно-морфологических сведениях о хордовых животных.
ОСОБЕННОСТИ РАЗВИТИЯ ХОРДОВЫХ*
Взрослый организм — завершающая стадия длительного инди-
видуального развития любого хордового животного. Он формиру-
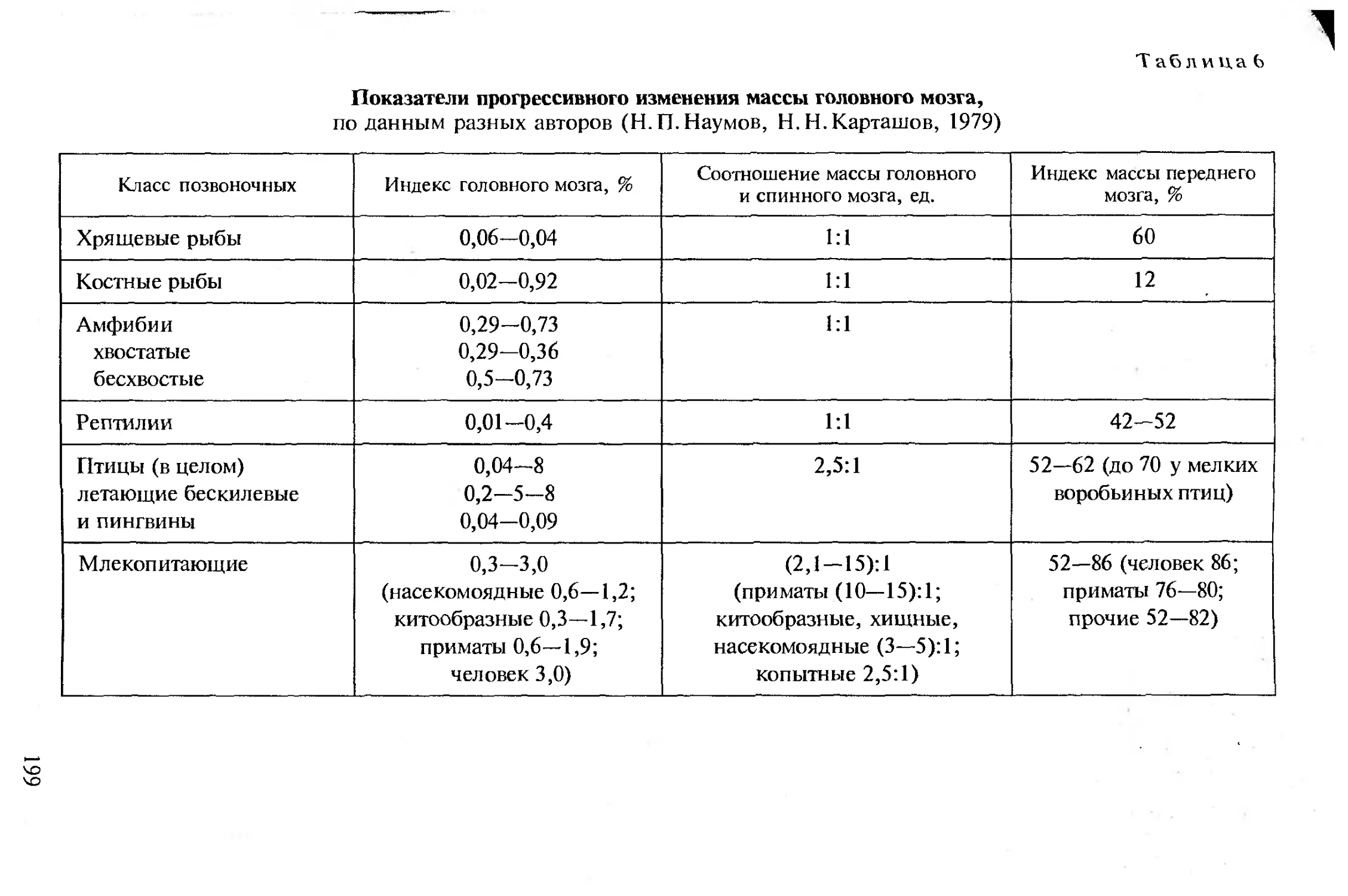
ется в процессе последовательно протекающих стадий онтогене-
за включающем эмбриональное и постэмбриональное развитие.
’ Как известно, половые клетки формируются и созревают в го-
надах (см. раздел «Размножение»). Овоциты (ооциты) хордовых
чрезвычайно разнообразны по величине. К примеру, бедное жел-
тком яйцо обыкновенной полевки имеет в диаметре 50 мкм. Наи-
более крупное яйцо у африканского страуса. Оно богато желтком
и достигает (без белочной оболочки) в диаметре 80 мм. Овоциты
имеют, как правило, сферическую форму, содержат ядро, про-
зрачную цитоплазму и желток с запасом питательных веществ,
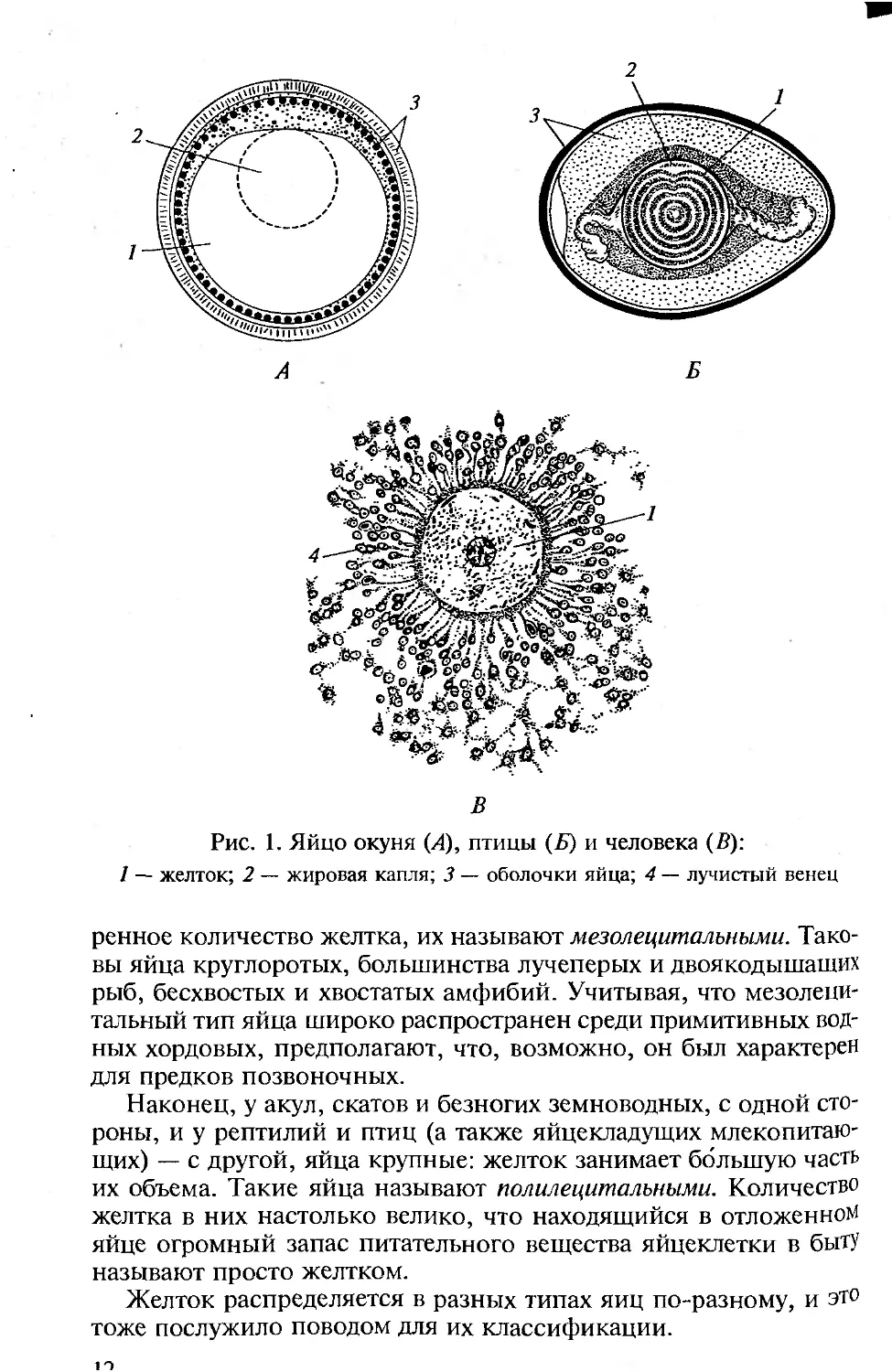
служащий резервом пищи для развивающегося зародыша. На рис. 1
показаны яйца разных позвоночных. Величина яйца не зависит от
размеров животного, но обычно обратно пропорциональна пло-
довитости. Такая корреляция отсутствует у тех позвоночных, за-
родыши которых развиваются под покровом материнского орга-
низма, например у плацентарных млекопитающих.
Зрелая яйцеклетка после выхода из яичника готова к последу-
ющему развитию. У абсолютного большинства видов хордовых
эмбриональное развитие начинается после оплодотворения — сли-
яния с яйцеклеткой сперматозоида, при котором объединяются
их гаплоидные наборы хромосом. С этого времени оплодотворен-
ную яйцеклетку — зиготу — называют яйцом. Некоторым видам
рыб и рептилий свойственна партеногенетическая форма размно-
жения.
~ Количество желтка, наполняющего вышедшую из яичника
яйцеклетку, сильно варьирует. Это обстоятельство влияет на раз-
меры яйца и тип его последующего дробления. У одних живот-
ных, в частности у ланцетников и плацентарных млекопитаю-
щих, яйца содержат мало желтка. Такие яйца называют олиголеци-
талъными. Яйца другого типа несколько крупнее и содержат уме-
Рассматриваются особенности эмбрионального развития основных групп
РДовых, последовательность образования у них тканей и органов. Материал
агается в несколько упрощенном и схематическом виде, вполне достаточном
я Формирования представлений у студентов-биологов.
11
2
В
Рис. 1. Яйцо окуня (Л), птицы (Б) и человека (В):
1 — желток; 2 — жировая капля; 3 — оболочки яйца; 4 — лучистый венец
ренное количество желтка, их называют мезолециталъными. Тако-
вы яйца круглоротых, большинства лучеперых и двоякодышащих
рыб, бесхвостых и хвостатых амфибий. Учитывая, что мезолеци-
тальный тип яйца широко распространен среди примитивных вод-
ных хордовых, предполагают, что, возможно, он был характерен
для предков позвоночных.
Наконец, у акул, скатов и безногих земноводных, с одной сто-
роны, и у рептилий и птиц (а также яйцекладущих млекопитаю-
щих) — с другой, яйца крупные: желток занимает большую часть
их объема. Такие яйца называют полилециталъными. Количество
желтка в них настолько велико, что находящийся в отложенном
яйце огромный запас питательного вещества яйцеклетки в быту
называют просто желтком.
Желток распределяется в разных типах яиц по-разному, и это
тоже послужило поводом для их классификации.
1 о
Яйиа, в которых желток распределен довольно равномерно,
например, в олиголецитальных, называют изолециталъными.
'пмезо- и полилециталъных яйцах более обильный желток чаще со-
едоточен в одной половине яйца: например, в отложенных в
о икринках рыб и амфибий желток, как правило, сосредото-
чен в их нижней половине. Такие яйца называют телолециталъны-
juu Концентрация желтка в одном полушарии свидетельствует о
наличии в яйце полярности: на верхней его части находится ани-
мальный полюс, на нижней — вегетативный.
Проследим последовательность этапов раннего онтогенеза в
разных типах яиц, приняв в качестве модели эмбриональное раз-
витие ланцетника. Согласно предложенной выше типологии,
яйцо ланцетника — классический пример олиго- и изолециталъ-
ного яйца.
Рассмотрим основные эмбриональные стадии, последователь-
но сменяющие друг друга в индивидуальном развитии ланцетни-
ка: 1) дробление и образование бластулы', 2) гаструляцию и заклад-
ку зародышевых листков', 3) образование нервной трубки и мезодер-
мальных структур', 4) органогенез.
Процесс дробления яйца, приводящий к образованию бласту-
лы, протекает у ланцетника в полном соответствии с классичес-
кой схемой, известной и для беспозвоночных животных. Первая
борозда деления проходит меридионально от полюса к полюсу и
приводит к образованию двух одинаковых по размерам клеток —
Рис. 2. Дробление и образование бластулы в олиголецитальном яйце
ланцетника (анимальный пояс яйца находится сверху):
Результат первого дробления; Б — образование 4 бластомеров в результате
второго деления; В — результат третьего деления; Г,Д— результаты последующих
РОбдений; Е — бластула в разрезе; бластоцель окружена одним слоем клеток
бластомеров (рис. 2). Эти клетки примыкают одна к другой, кая<- ;
дая имеет шарообразную форму. Такая форма клеток сохраняется
и при последующих делениях. У ланцетника двусторонняя сим-
метрия закладывается еще до оплодотворения, и две клетки, воз-
никающие в результате первого деления при нормальном разви-
тии, образуют в дальнейшем правую и левую половины тела. Вто-
рая борозда деления располагается параллельно первой.
Третья борозда дробления яйца перпендикулярна двум пред-
шествующим. Она проходит почти по экватору яйца, разделяя
каждый из четырех уже имеющихся бластомеров на верхнюю и
нижнюю половины, в результате чего образуется зародыш из 8
бластомеров. В образовавшемся скоплении клеток развивается цен-
тральная полость бластоцель, которая при дальнейшем дробле-
нии расширяется.
Олиголецитальное яйцо ланцетника содержит мало желтка. Его
концентрация несколько возрастает к вегетативному полюсу. Клет-
ки делятся не по общему центру, а равномерно делят лишь ци-
топлазму. Относительно инертный желток реже участвует в дроб-
лении. Поэтому третье деление, приводящее к образованию 8 бла-
стомеров, проходит не строго по экватору яйца, а несколько
выше его середины. В результате клетки верхней группы (ани-
мальный полюс) чуть мельче нижних и соответственно содержа-
ние желтка в клетках нижней части (вегетативный полюс) не-
много больше.
Следующие деления проходят в вертикальном направлении:
каждый бластомер делится надвое, и возникает стадия зародыша,
состоящая из 16 клеток. Затем в анимальном и вегетативном полу-
шариях происходят горизонтальные деления с образованием 32-
клеточного зародыша с четырьмя неравными по размерам коль-
цами клеток. Процесс деления проходит в геометрической про-
грессии, и вслед за стадией зародыша из 32 клеток число их быс-
тро возрастает до 64, 128 и т. д. За короткое время исходная одно-
клеточная зигота дает начало нескольким сотням клеток, образу-
ющих однослойную стенку вокруг центральной полости. На этой
стадии (бластула) клетки в области анимального полюса мельче,
а на вегетативном полюсе они крупнее и богаче желтком. В сред-
ней части зародыша находятся клетки, промежуточные по разме-
ру. Таким образом, уже на стадии бластулы намечается дифферен-
цировка клеток зародыша на разные группы, судьба которых бу-
дет реализована впоследствии в соответствии с их функциональ-
ной направленностью.
Следующий важный этап эмбрионального развития — процесс
гаструляции — преобразование сферы бластулы в двухслойную
структуру. С началом гаструляции клетки вегетативного полюса в
области губ бластопора начинают вворачиваться внутрь. Наруж-
ный слой клеток двухслойного зародыша ланцетника образует
14
Рис. 3. Эмбриональное развитие ланцет-
ника (по И. И. Шмальгаузену, 1964).
Продольные разрезы сделаны на стадиях блас-
тулы (Л), гаструляции (Б, В, Г), образования
мезодермы, хорды и нервной системы (Д):
1 — анимальный полюс; 2 — вегетативный
полюс; 3 — гастральная полость; 4 — гастро-
пор; 5 — нервный канал; б — нервно-кишеч-
ный канал; 7 — невропор; 8 — складка мезо-
дермы; 9 — целомические мешки; 10 — хорда
эктодерму, или наружный зародышевый листок, соответствующий
будущей коже. Позднее в поверхностном эктодермальном слое
выделяется особый участок — нейральная эктодерма (модулярная
пластинка), из которой впоследствии образуется нервная трубка
(рис. 3).
Внутренние клетки гаструлы составляют энтодерму, или внут-
ренний зародышевый листок. Энтодерма выстилает первичную кишку
зародыша. Возникшее в результате инвагинации бластулы отвер-
стие, ведущее в кишку, называют бластопором, а его края — губа-
ми — спинной, брюшной, боковыми. Оставшиеся снаружи бога-
тые желтком бластомеры образуют желточную пробку. Впослед-
ствии бластопор становится анальным отверстием.
В процессе гаструляции закладывается третий зародышевый ли-
сток — мезодерма. Она ложится двумя полосками по сторонам
первичной кишки. В дальнейшем из мезодермы сформируется боль-
шая часть внутренних органов хордового животного.
Закладка мезодермы трактуется в специальной научной лите-
ратуре неоднозначно. Согласно И. И. Шмальгаузену (1964), на спин-
ной стороне первичной кишки появляется продольное впячива-
ние. В последующем этот желобок отшнуровывается и превраща-
ется в хорду (рис. 4). Примерно в это же время по бокам от зачатка
хорды появляются два ряда симметрично расположенных выпя-
чиваний кишки. По мере разрастания они отшнуровываются от
нее и образуют парный ряд метамерно расположенных целомичес-
Ких мешков — зачатков мезодермы. В процессе развития каждый
Целомический мешок делится на два отдела: верхний — сомит и
нижний — боковую пластинку. Полости сомитов не сливаются между
собой и в последующем исчезают. Полости же боковых пласти-
15
5
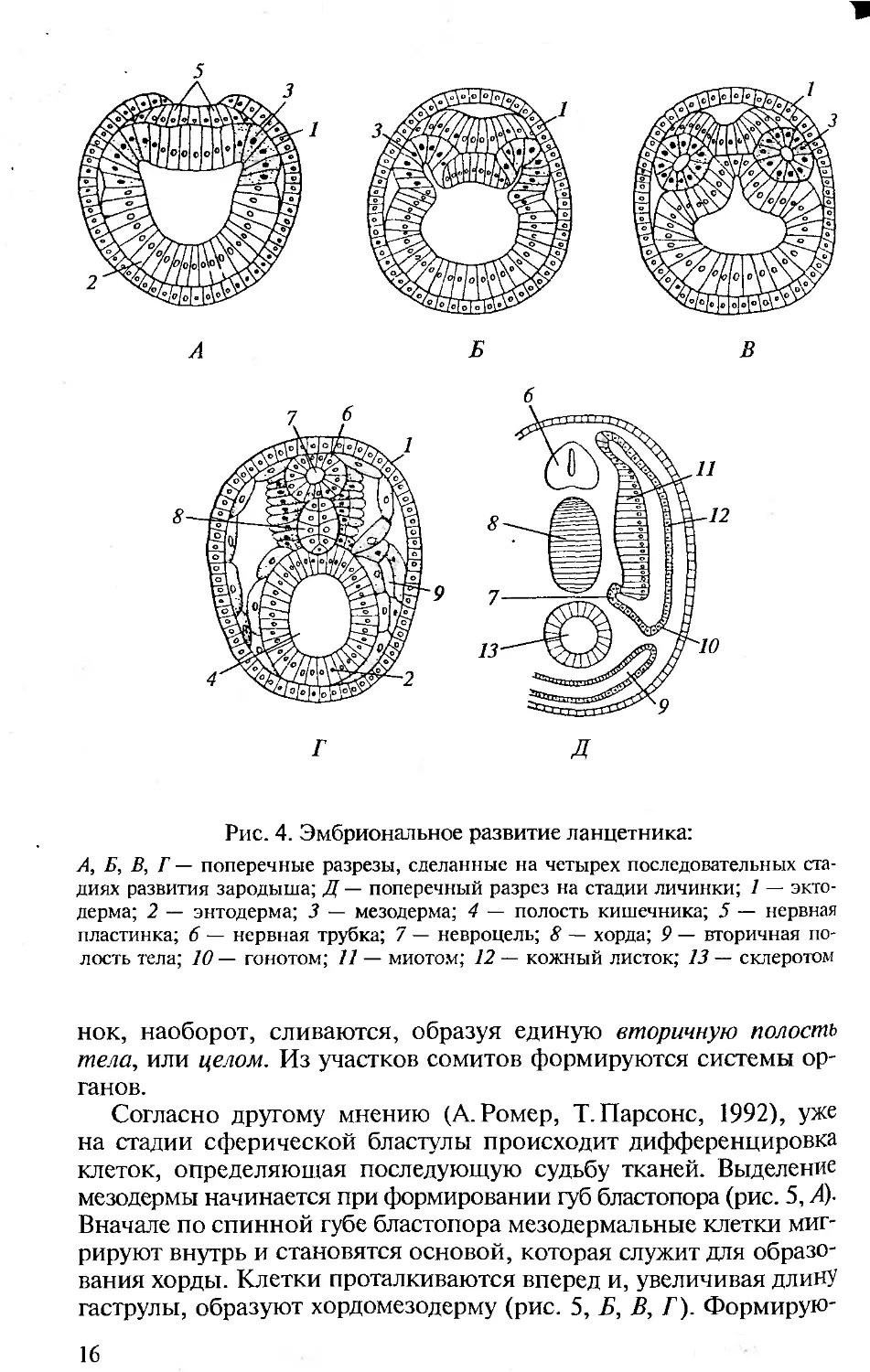
Рис. 4. Эмбриональное развитие ланцетника:
А, Б, В, Г — поперечные разрезы, сделанные на четырех последовательных ста-
диях развития зародыша; Д — поперечный разрез на стадии личинки; 1 — экто-
дерма; 2 — энтодерма; 3 — мезодерма; 4 — полость кишечника; 5 — нервная
пластинка; 6 — нервная трубка; 7 — невроцель; 8 — хорда; 9 — вторичная по-
лость тела; 10 — гонотом; 11 — миотом; 12 — кожный листок; 13 — склеротом
нок, наоборот, сливаются, образуя единую вторичную полость
тела, или целом. Из участков сомитов формируются системы ор-
ганов.
Согласно другому мнению (А. Ромер, Т. Парсонс, 1992), уже
на стадии сферической бластулы происходит дифференцировка
клеток, определяющая последующую судьбу тканей. Выделение
мезодермы начинается при формировании губ бластопора (рис. 5, А)-
Вначале по спинной губе бластопора мезодермальные клетки миг-
рируют внутрь и становятся основой, которая служит для образо-
вания хорды. Клетки проталкиваются вперед и, увеличивая длину
гаструлы, образуют хордомезодерму (рис. 5, Б, В, Г). Формирую-
16
6
1
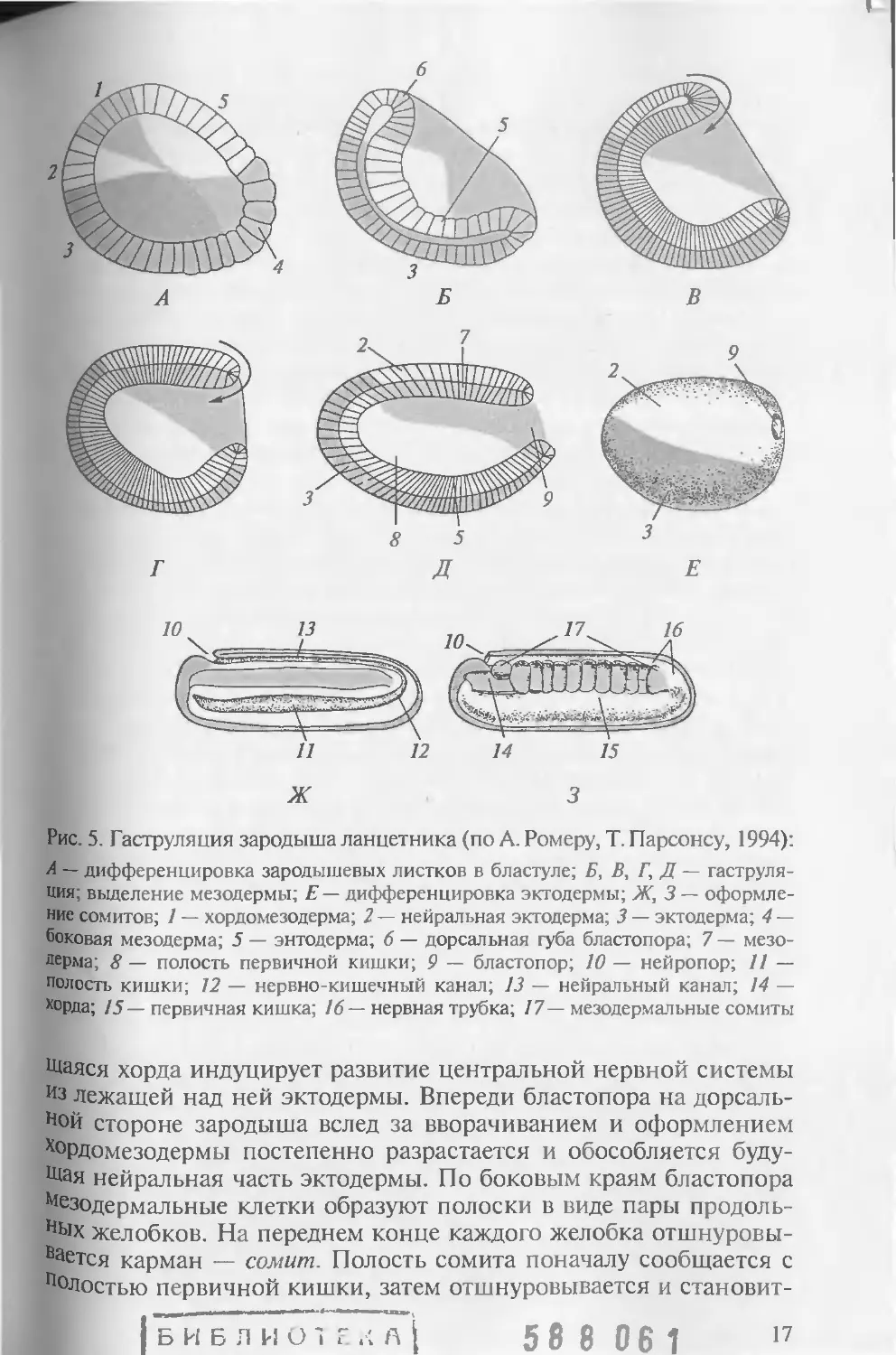
Рис. 5. Гаструляпия зародыша ланцетника (по А. Ромеру, Т. Парсонсу, 1994):
А — дифференцировка зародышевых листков в бластуле; Б, В, Г, Д — гаструля-
ция; выделение мезодермы; Е— дифференцировка эктодермы; Ж, 3 — оформле-
ние сомитов; 1 — хордомезодерма; 2 — нейральная эктодерма; 3 — эктодерма; 4 —
боковая мезодерма; 5 — энтодерма; 6 — дорсальная губа бластопора; 7 — мезо-
дерма; 8 — полость первичной кишки; 9 — бластопор; 10 — нейропор; 11 —
полость кишки; 12 — нервно-кишечный канал; 13 — нейральный канал; 14 —
хорда; 15— первичная кишка; 16— нервная трубка; 17— мезодермальные сомиты
Щаяся хорда индуцирует развитие центральной нервной системы
Из лежащей над ней эктодермы. Впереди бластопора на дорсаль-
ной стороне зародыша вслед за вворачиванием и оформлением
ХоРДомезодермы постепенно разрастается и обособляется буду-
щая нейральная часть эктодермы. По боковым краям бластопора
Мезодермальные клетки образуют полоски в виде пары продоль-
ных желобков. На переднем конце каждого желобка отшнуровы-
Вается карман — сомит. Полость сомита поначалу сообщается с
Полостью первичной кишки, затем отшнуровывается и становит-
| Б И Б Л И О 1 Г .< А [ 5 8 8 0 6 1 17
ся частью целома. Пары сомитов отпочковываются назад по все^
длине туловища. Стенки сомитов дифференцируются впоследствии
в мезодермальные ткани (см. рис. 4). Уместно подчеркнуть, что
такой тип закладки мезодермы у ланцетника в виде карманов на,
поминает аналогичный процесс, наблюдаемый у полухордовых и
иглокожих, и служит серьезным аргументом в пользу доказательств
общности их происхождения.
Таким образом, зародыш ланцетника по окончании гастру.
ляции имеет форму вытянутого сфероида с расположенным сза-
ди единственным ведущим внутрь отверстием — бластопором
(рис. 5, Ж, 3). Снаружи переднюю поверхность зародыша покры-
вает кожная эктодерма; дорсально располагается медулярная пла-
стинка (нейральная эктодерма). Изнутри мешок гаструлы выстлан
энтодермой. Вдоль крыши первичной кишки располагается хор-
домезодерма, ограниченная с боков мезодермальными структу-
рами в виде сомитов.
После завершения гаструляции, сопровождающейся переме-
щением клеток и вычленением основных тканей зародыша — кож-
ной эктодермы, энтодермы, нейральной эктодермы, хордомезо-
дермы и боковой мезодермы, начинается процесс формирования
органов. Ранний этап закладки органов происходит у зародыша
ланцетника на стадии нейрулы.
Как сказано выше, потенциальная нервная трубка занимает
у ланцетника обширный овальный участок на спинной и зад-
ней поверхности гаструлы. По обе стороны этого участка, на
границе между будущей кожной и нейральной эктодермой воз-
никает складка ткани. Образование этой складки сопровожда-
ется у ланцетника выделением двух типов эктодермы. Края кож-
ной эктодермы с правой и левой сторон нарастают на нейраль-
ную область и в конечном итоге сходятся, покрывая спинную
поверхность тела сплошным слоем клеток. Скрытые под этим
покровом боковые края нейроэктодермального листка постепен-
но заворачиваются на дорсальную сторону и, соединяясь, обра-
зуют круглую нервную трубку. Процесс смыкания трубки начи-
нается у ланцетника в средней (по длине) части зародыша и
продолжается оттуда в обоих направлениях, вперед и назад. Пе-
редний конец будущей нервной трубки некоторое время остает-
ся открытым, а его отверстие называется невропором. На заднем
конце зародыша складки эктодермы смыкаются над бластопо-
ром. По завершении образования нервной трубки бластопор не
открывается на поверхности зародыша, а сообщается с задним
концом нервной трубки. В результате первичная кишка оказыва-
ется связанной с поверхностью зародыша через нервный канал
и невропор. Канал, связывающий кишку с нервной трубкой,
называют нервно-кишечным (canalis neuroentericus). При дальней-
шем развитии, с оформлением хвостового отдела тела, этот
18
закрывается, а нервная трубка и кишка обособляются
прут от друга-
ДИ В итоге сомиты становятся основой формирования будущих
ней и внутренних органов: 1) нижняя внутренняя часть соми-
тК ___ скдеротом — дает начало клеткам, образующим соедини-
Тедьнотканное влагалище хорды и нервной трубки, опорные лучи
Т плавниках и, видимо, миосепты; 2) часть сомита, прилегающая
к хорде, миотом, формирует туловищную мускулатуру; 3) на-
ужная часть сомита, кожный листок, образует соединительно-
тканную часть кожи, т.е. кутис (см. рис. 4, Д).
Из боковой пластинки развиваются брюшина, брыжейки (в
которых в виде продольных каналов возникают основные крове-
носные сосуды), мускулатура кишечника и поперечная мышца.
Нефридиальные канальцы появляются в виде пальцевидных вы-
пячиваний стенок вторичной полости тела. Гонады формируются
из гонотома — выпячивания той части стенок полости тела, кото-
рая соответствует месту подразделения сомита и боковой плас-
тинки.
Рот образуется путем выпячивания энтодермы первичной кишки
на конце, противоположном бластопору (первичному рту), и
встречного впячивания эктодермы. В месте встречи этих образова-
ний возникает прорыв. У ланцетника ротовое отверстие заклады-
вается асимметрично на нижней левой стороне зародыша. Асим-
метрично и своеобразно развиваются и жаберные щели. Первона-
чально жаберные щели (в числе 14) возникают на брюшной сто-
роне тела, а затем перемещаются на правую сторону. Здесь появ-
ляется еще один ряд щелей (в числе 8), расположенных выше от
упомянутых ранее 14. Затем нижний ряд перемещается на брюш-
ную сторону и лишь после этого на левую сторону. В это время
число щелей сокращается с 14 до 8. В последующем онтогенезе
число жаберных отверстий у ланцетника с обеих сторон резко
возрастает. Рот передвигается на брюшную сторону тела. На зад-
нем конце тела зарастает нервно-кишечный канал.
Атриальная полость возникает первоначально на нижней по-
верхности тела в виде желобка. Образующие этот желобок мета-
плевральные складки, расположенные по бокам, растут навстре-
чу друг другу и смыкаются. Так образуется полость, которая от-
крывается наружу лишь в задней своей части, где складки расхо-
дятся, образуя отверстие — атриопор. Личиночное развитие лан-
цетников длится около трех месяцев.
Раннее эмбриональное развитие животных с другими типами
яиц принципиально сходно с ланцетником, но отличается от него
некоторыми собственными чертами.
Яйцо лягушки — пример типичного мезолецитального яйца'.
гранулярный желток рассеян в нем по всему объему с несколько
°льщей концентрацией в вегетативной части.
19
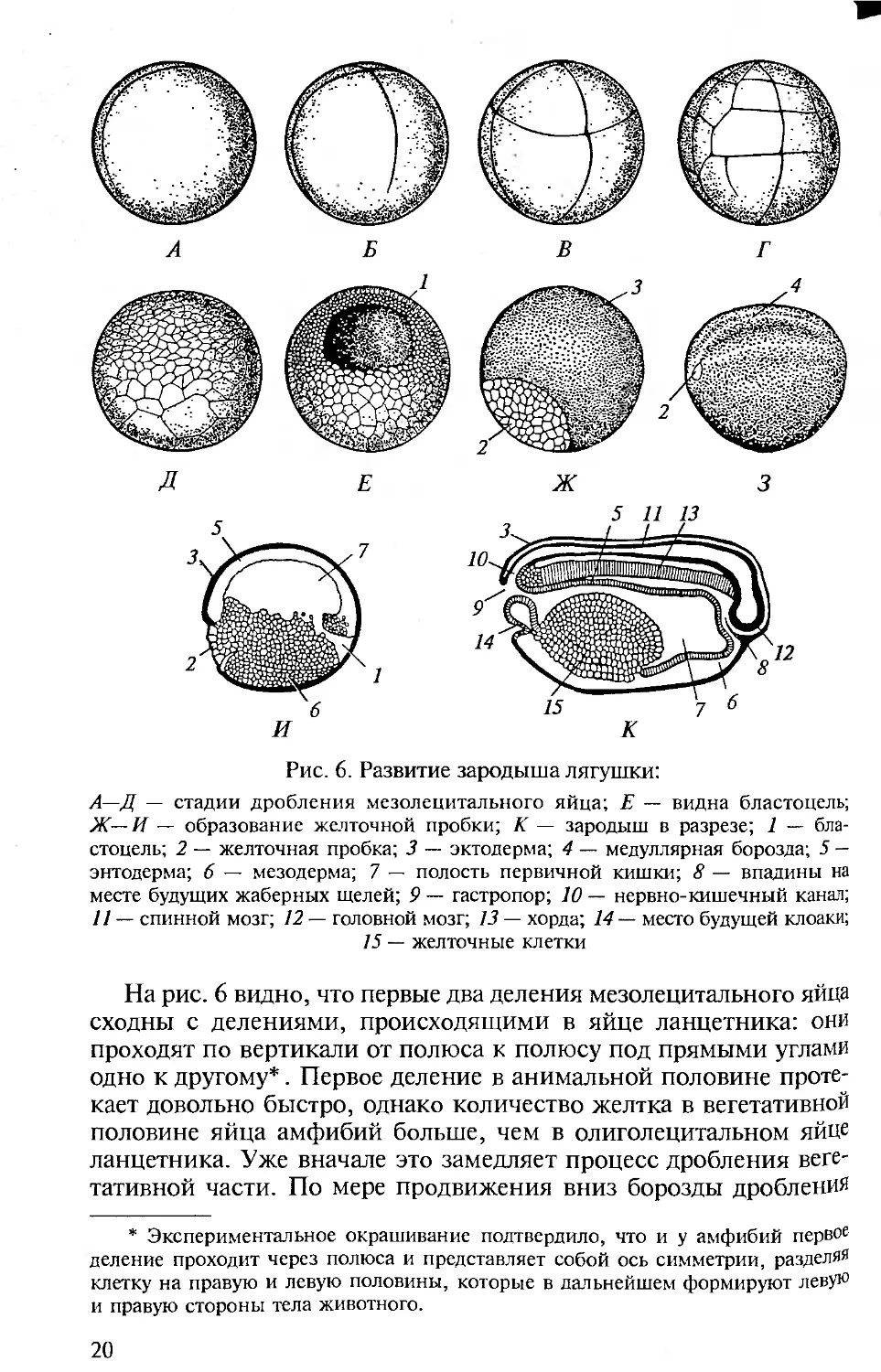
Рис. 6. Развитие зародыша лягушки:
А—Д — стадии дробления мезолецитального яйца; Е — видна бластоцель;
Ж—И — образование желточной пробки; К — зародыш в разрезе; 1 — бла-
стоцель; 2 — желточная пробка; 3 — эктодерма; 4 — медуллярная борозда; 5 —
энтодерма; 6 — мезодерма; 7 — полость первичной кишки; 8 — впадины на
месте будущих жаберных щелей; 9 — гастропор; 10 — нервно-кишечный канал;
11 — спинной мозг; 12 — головной мозг; 13 — хорда; 14 — место будущей клоаки;
15 — желточные клетки
На рис. 6 видно, что первые два деления мезолецитального яйца
сходны с делениями, происходящими в яйце ланцетника: они
проходят по вертикали от полюса к полюсу под прямыми углами
одно к другому*. Первое деление в анимальной половине проте-
кает довольно быстро, однако количество желтка в вегетативной
половине яйца амфибий больше, чем в олиголецитальном яйце
ланцетника. Уже вначале это замедляет процесс дробления веге-
тативной части. По мере продвижения вниз борозды дробления
* Экспериментальное окрашивание подтвердило, что и у амфибий первое
деление проходит через полюса и представляет собой ось симметрии, разделяя
клетку на правую и левую половины, которые в дальнейшем формируют левую
и правую стороны тела животного.
20
тпечают сопротивление со стороны инертного желтка, и про-
вС с деления замедляется. Поэтому второе деление начинается еще
Це того, как завершилось первое. Эта тенденция к задержке деле-
ния клеток в нижней половине яйца сохраняется на всем протя-
» периода дробления. В результате дробления яйца амфибий
бразуется бластула, сходная с бластулой ланцетника. Однако пе-
еполненная желтком цитоплазма у вегетативного полюса яйца
делится медленно, и крупные, богатые желтком клетки частично
заполняют внутреннюю область. Поэтому у амфибий бластоцель
не так обширна, как у ланцетника. Дробление яйца при образова-
ний бластулы в этом случае называют голобластическим типом.
Обилие богатых желтком клеток, заполняющих большую часть
бластоцели, видимо, изменило у амфибий и процесс гаструля-
ции. На поверхности бластулы, в месте, соответствующем верх-
ней (дорсальной) губе бластопора ланцетника, образуется бороз-
дка. В нее снаружи последовательно устремляются клетки — зачат-
ки хорды и мезодермы. Одновременно участок спинной губы бла-
стопора нарастает сверху на желток, и в конечном итоге на по-
верхности зародыша остается только небольшая желточная проб-
ка, закрывающая уменьшенный бластопор.
Из крыши первичной кишки, как и у ланцетника, вычленяют-
ся хорда и боковые скопления мезодермы. Хорда постепенно рас-
полагается по средней линии спины зародыша, а элементы мезо-
дермы выворачиваются наружу. В закладке мезодермы нет карма-
нов: ее клетки, разрастаясь между экто- и энтодермой, образуют
непрерывный слой по обе стороны хорды. Целомические полости
возникают в ней позднее.
Д Е
Рис. 7. Развитие из мезолецитального яйца хвостатой амфибии —
протея:
А —-
поздняя гаструла; Б — формирование нервных валиков; В — головной мозг
ВЬ1стУпает над желточным мешком; Г — оформление туловищного и хвостового
отделов; Д, Е — появление наружных жабр, глаз, конечностей
21
Из энтодермы развиваются выстилка кишки и ее производ- -
ные, в том числе легкие, глоточные железы, печень, поджелу-
дочная железа. Мезодерма становится основой скелета, мышц
соединительной ткани, кровеносной, выделительной и репродук-
тивной систем.
Большая часть эктодермы становится основой покровов тела. Как
и у ланцетника, в ней вычленяется нейральная эктодерма — осно-
ва будущей нервной трубки. Развитие нервной трубки идет под ин-
дуцирующим влиянием хорды, однако ее дифференцировка про-
текает иначе. Со спинной стороны в эктодерме выпячиваются вверх
складки — нервные валики (рис. 7). Они растут навстречу друг другу.
В каждой складке кожная и нейральная эктодермы разделяются и
попарно объединяются, так что развитие кожи и нервной трубки
завершается одновременно. В головном отделе трубки в виде утол-
щений зародышевой эктодермы — плакод — закладываются эле-
менты будущих отделов нервной системы и органов чувств.
К перечисленным особенностям дробления и развития эмбри-
онов из олиго- и мезолецитальных яиц следует добавить, что у
амфибий и большинства костистых рыб количество желтка, кото-
рый расходует зародыш в раннем онтогенезе, ограничено, и мо-
лодь при вылуплении имеет небольшие размеры. Молодые особи
заметно отличаются от взрослых и внешним видом, и образом
жизни: они непременно живут в воде, потребляют иную пищу, у
них развиваются временные органы, соответствующие потребно-
стям их возрастного состояния. В процессе индивидуального раз-
вития у них возникает личиночная стадия, а переход ко взрослому
состоянию происходит путем метаморфоза, в котором важную роль
играют гормоны щитовидной железы. Хорошо известными при-
мерами личиночных стадий служат личинки акуловых и костис-
тых рыб, головастики лягушек и жаб (рис. 8). Личинки хвостатых
амфибий живут в воде и дышат жабрами. Большинство их видов
после метаморфоза теряет жабры и переходит к полуназемному
образу жизни (рис. 9), однако у некоторых форм метаморфоз про-
исходит лишь частично или вообще не происходит (американс-
кий и европейский протей, тигровая амбистома). Личинка тигро-
вой амбистомы, аксолотль, хорошо известен как аквариумное ла-
бораторное животное. Личиночную стадию имеют многие костис-
тые рыбы. Особенно резко отличаются от взрослой формы про-
зрачные листовидные личинки европейского угря (рис. 10), оби-
тающие в открытом океане, о связи которых со взрослыми фор-
мами долгое время даже не подозревали. Личиночная стадия есть у
круглоротых. Например, взрослая морская минога — это крупное
хищное животное, а ее маленькая личинка — пескоройка — ведет
малоподвижный образ жизни на дне, питаясь микроскопически-
ми организмами, которых она отфильтровывает из ила. Особен-
ности морфофизиологической организации сближают ее с личин-
22
Рис. 8. Стадии постэмбрионального развития остромордой лягушки
(по А. Г. Банникову, 1960):
1 икринки; 2 — личинки в момент вылупления; 3 и 4 — увеличение наружных
Жабр и разрастание плавниковой складки; 4а — тот же головастик снизу (видна
присоска); 46 — наружная жабра; 5 — развитие жаберной крышки, прикрывшей
НаРУЖные жабры; 5а — тот же головастик сверху (виден прорвавшийся рот); 6 —
явление конечностей; 6а — ротовой аппарат головастика; 7— полностью сфор-
мованные конечности; 8 — прорыв наружу передних конечностей, начало
резорбции хвоста; 9 — стадия выхода на сушу
74
Рис. 9. Стадии постэмбрионального развития обыкновенного тритона
(по А. Г. Банникову, 1960):
1 — икринки; 2 — личинка в момент вылупления; 2а — она же, вид сверху; 3 —
прорыв рта, начало ветвления наружных жабр; 4 — полное развитие наружных
жабр, расчленение передних конечностей; 4а — наружные жабры; 5 — заверШ®'
ние расчленения задних конечностей; 6 — начало редукции наружных жабр И
плавниковой складки; 7 — стадия выхода на сушу
24
Рис. 10. Метаморфоз европейского угря и миграции его личинок
(в кружочках — средняя длина личинок, мм):
1 — только что вышедшая личинка; 2 — годовалая личинка; 3 — двухгодовалая
личинка; 4 — личинка перед началом преврашения; 5 — стеклянный угорек; 6 —
взрослый угорь
кой ланцетника. Такое проявление рекапитуляции свидетельству-
ет об эволюционном родстве этих групп животных.
Яйцахрящевых рыб (акул, скатов), безногих ам-
фибий, рептилий, птиц и низших млекопитаю-
щих полилецитального типа содержат большое количество желт-
ка. Для таких яиц в отличие от яиц амфибий характерен мероблас-
тический тип дробления. На анимальном полюсе клетки, содер-
жащие цитоплазму, ограничены небольшой областью, где они
покрывают желток как бы шапочкой. В дроблении и образовании
бластулы участвует только этот участок клеток с прозрачной ци-
топлазмой, а масса желтка на вегетативном полюсе почти не дро-
бится.
Первая и вторая борозды дробления в полилецитальном яйце
акУЛовых рыб, как и в яйцах других типов, проходят через центр
ВДимального полюса. На рис. 11 видно, что эти борозды делят не
Все яйцо, а только клетки цитоплазматической «шапочки», не
затРагивая желток. Следующая «экваториальная» борозда дробле-
ния также проходит выше середины яйца, отделяя клетки, окру-
жающие полюс, от периферических клеток. Дальнейшее деление
Р°Должается до тех пор, пока все цитоплазматические клетки
25
Рис. 11. Дробление полилецитального яйца (акулы, ящерицы, птицы):
А—Г— образование «шапочки»: Д — зародыш в разрезе
в области анимального полюса не будут разделены. В результате
образуется плоская структура, которая соответствует бластуле. Она
получила название бластодермы. Напомним, что в олиголециталь-
ном яйце ланцетника стенка бластулы состоит из одного ряда
клеток, а в мезолецитальном яйце амфибий стенка бластулы го-
раздо толще и состоит из нескольких слоев клеток. В полилеци-
тальном яйце вся стенка бластулы представляет собой толстый
слой клеток в форме плоского диска, слегка приподнятого над
лежащим под ним желтком. Как сказано выше, его края, сопри-
касающиеся с желтком, образованы клетками, которые в яйцах,
менее богатых желтком, находятся в вегетативном полушарии.
Представляет интерес то, что, несмотря на небольшие разме-
ры, яйца костистых рыб отчасти относятся к полилецитальному
типу, и большая часть их с прозрачной цитоплазмой образует ша-
почку над плотной массой желтка. Однако по типу дробления и
эмбриональному характеру развития они сходны с более крупны-
ми мезолецитальными яйцами амфибий.
Характер перемещения клеток во время гаструляции в полиле-
цитальном яйце, естественно, иной, чем в других типах яиц, по-
тому что в этом случае вместо сферической бластулы имеется уп-
лощенная бластодерма.
Дифференцировка клеток на поверхности бластодермы — по-
тенциальных зародышевых листков — сходна с их закладкой на
поверхности яйца амфибий. На рис. 12 показана последователь-
ность выделения тканей в результате гаструляции зародыша. Эк-
тодермальные клетки вворачиваются внутрь и на поверхности
желтка образуют тяж. Хордомезодерма и мезодерма отделяются от
энтодермы, занимая свое положение между двумя зародышевыми
листками.
Зародыш акулы в результате описанных процессов раннего
эмбриогенеза достигает строения, характерного для гаструлы ам-
26
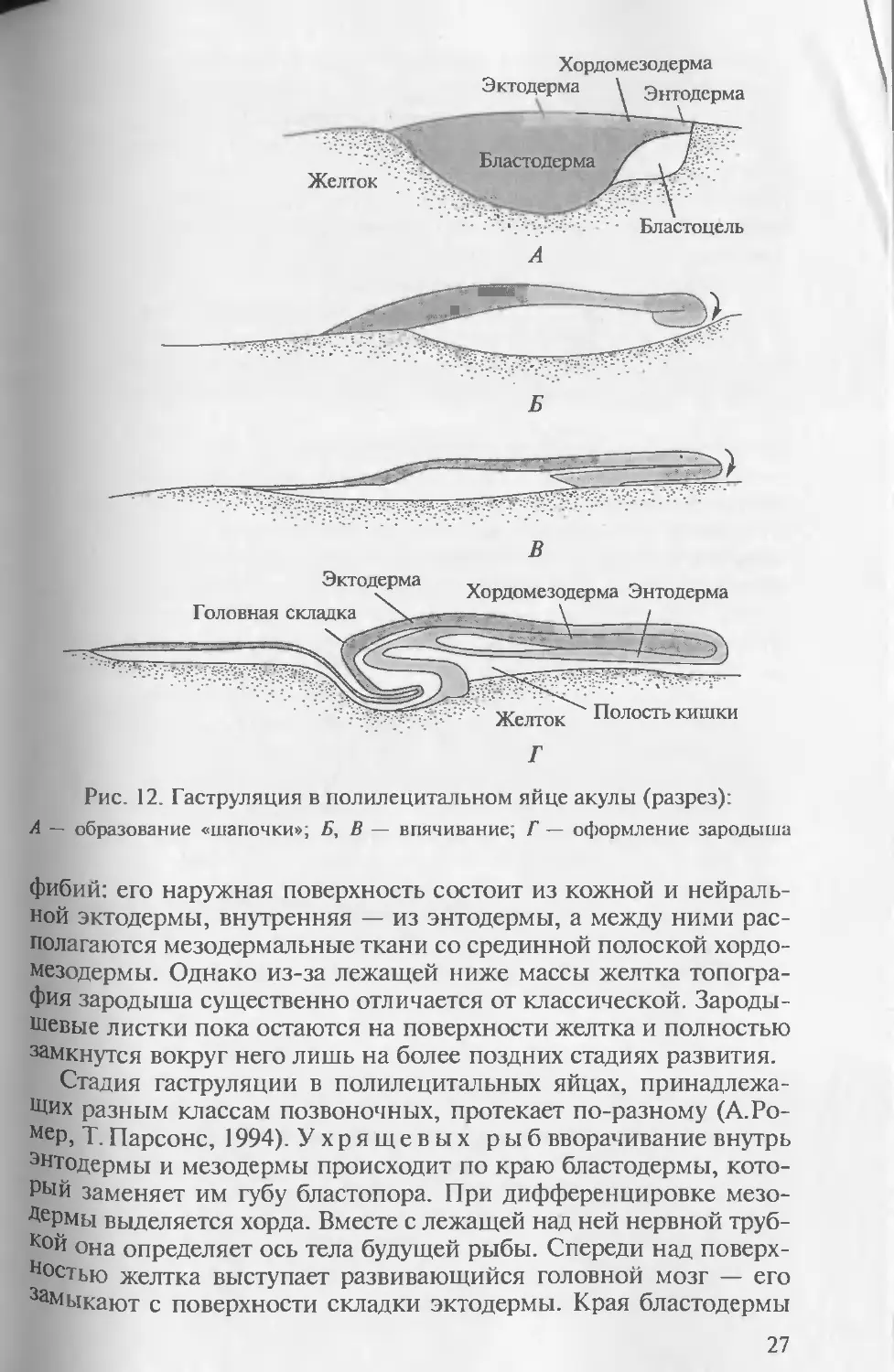
Рис. 12. Гаструляция в полилецитальном яйце акулы (разрез):
А — образование «шапочки»; Б, В — впячивание; Г — оформление зародыша
фибий: его наружная поверхность состоит из кожной и нейраль-
ной эктодермы, внутренняя — из энтодермы, а между ними рас-
полагаются мезодермальные ткани со срединной полоской хордо-
мезодермы. Однако из-за лежащей ниже массы желтка топогра-
фия зародыша существенно отличается от классической. Зароды-
шевые листки пока остаются на поверхности желтка и полностью
замкнутся вокруг него лишь на более поздних стадиях развития.
Стадия гаструляции в полилецитальных яйцах, принадлежа-
щих разным классам позвоночных, протекает по-разному (А.Ро-
МеР, Т. Парсонс, 1994). Ухрящевых рыб вворачивание внутрь
энтодермы и мезодермы происходит по краю бластодермы, кото-
рый заменяет им губу бластопора. При дифференцировке мезо-
Дермы выделяется хорда. Вместе с лежащей над ней нервной труб-
кой она определяет ось тела будущей рыбы. Спереди над поверх-
ностью желтка выступает развивающийся головной мозг — его
3аМыкают с поверхности складки эктодермы. Края бластодермы
27
нарастают на желток, в результате чего он оказывается полнос-
тью заключенным в желточный мешок, полость которого пред-
ставляет собой по существу внезародышевую часть полости киш-
ки* . В прилегающем слое мезодермы образуются кровеносные со-
суды, обеспечивающие питание и дыхание зародыша. Желток в
процессе роста и развития постепенно расходуется; желточный
мешок истощается и в конечном итоге рассасывается. На рис. 13
показано поэтапное формирование зародыша акулы.
У самок живородящих пластиножаберных образуются дополни-
тельные структуры, с помощью которых развивающиеся в матке
мальки получают питательные вещества от тела матери. Зародыше-
вым приспособлением у некоторых акул являются пучки наружных
жабр. Стадию личинки молодые особи проходят в яйце или в утро-
бе матери. Малек акулы в момент появления на свет представляет
собой маленькую копию взрослой особи.
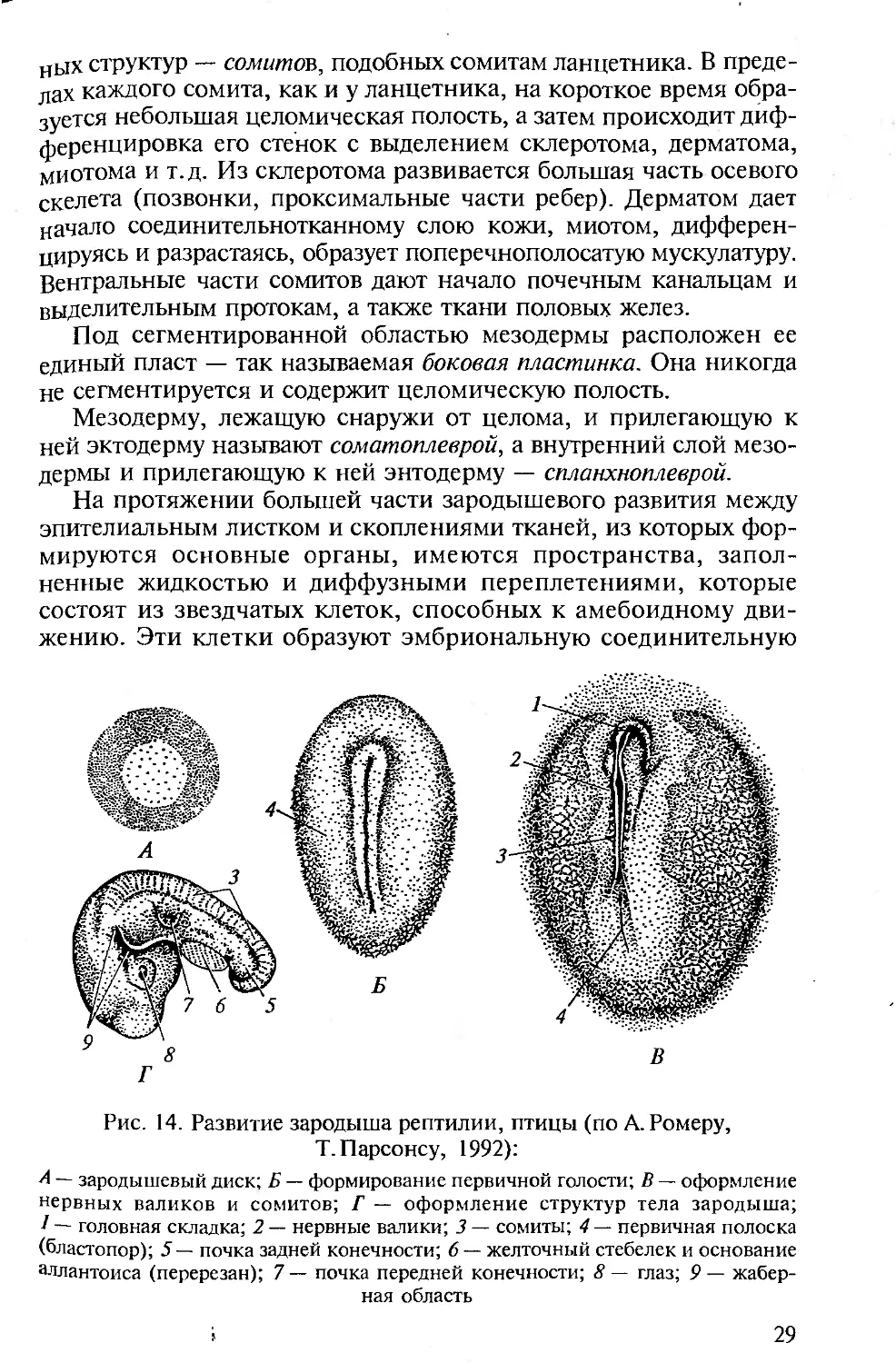
У рептилий и птиц мезодерма и энтодерма формируются
только в центральной части бластодермы (рис. 14). Края же зароды-
шевого диска служат для образования эктодермы. По средней линии
дифференцируется хорда. Мезодерма вблизи хорды утолщается и
подразделяется на каждой стороне на продольный ряд метамер-
Рис. 13. Развитие детеныша из полилецитального яйца акулы:
А — диск бластодермы при гаструляции; Б, В — разрастание диска, выделение
нервных валиков; Г — зародыш с желточным мешком; Д — малек акулы с жел-
точным мешком
* У зародышей костистых рыб желточный мешок не имеет энтодермальной
выстилки и отделен от первичной кишки.
28
ных структур — сомитов, подобных сомитам ланцетника. В преде-
дах каждого сомита, как и у ланцетника, на короткое время обра-
зуется небольшая целомическая полость, а затем происходит диф-
ференцировка его стенок с выделением склеротома, дерматома,
миотома и т.д. Из склеротома развивается большая часть осевого
скелета (позвонки, проксимальные части ребер). Дерматом дает
начало соединительнотканному слою кожи, миотом, дифферен-
цируясь и разрастаясь, образует поперечнополосатую мускулатуру.
Вентральные части сомитов дают начало почечным канальцам и
выделительным протокам, а также ткани половых желез.
Под сегментированной областью мезодермы расположен ее
единый пласт — так называемая боковая пластинка. Она никогда
не сегментируется и содержит целомическую полость.
Мезодерму, лежащую снаружи от целома, и прилегающую к
ней эктодерму называют соматоплеврой, а внутренний слой мезо-
дермы и прилегающую к ней энтодерму — спланхноплеврой.
На протяжении большей части зародышевого развития между
эпителиальным листком и скоплениями тканей, из которых фор-
мируются основные органы, имеются пространства, запол-
ненные жидкостью и диффузными переплетениями, которые
состоят из звездчатых клеток, способных к амебоидному дви-
жению. Эти клетки образуют эмбриональную соединительную
Рис. 14. Развитие зародыша рептилии, птицы (по А. Ромеру,
Т. Парсонсу, 1992):
А — зародышевый диск; Б — формирование первичной голости; В — оформление
нервных валиков и сомитов; Г — оформление структур тела зародыша;
— головная складка; 2 — нервные валики; 3 — сомиты; 4 — первичная полоска
(бластопор); 5 — почка задней конечности; 6 — желточный стебелек и основание
аллантоиса (перерезан); 7 — почка передней конечности; 8 — глаз; 9 — жабер-
ная область
29
ткань — мезенхиму. Мезенхима имеет в основном мезодермальное
происхождение, так как возникает за счет разрастания клеток
сомитов и боковой пластинки. В некоторых случаях начало мезен-
химоподобному материалу дают экто- и энтодерма.
Хотя мезенхима выполняет функцию эмбриональной соеди-
нительной ткани, ее активность не ограничивается такой, в сущ-
ности, пассивной и временной ролью. Мезенхима обладает очень
большой пластичностью. Она участвует в образовании соедини-
тельной ткани скелета, дает начало большой части кровеносной
системы — кровеносным сосудам и форменным элементам крови.
Из мезенхимы происходит большая часть мускулатуры: все глад-
кие мышцы, специализированная сердечная мышца и даже зна-
чительная часть поперечнополосатых мышц.
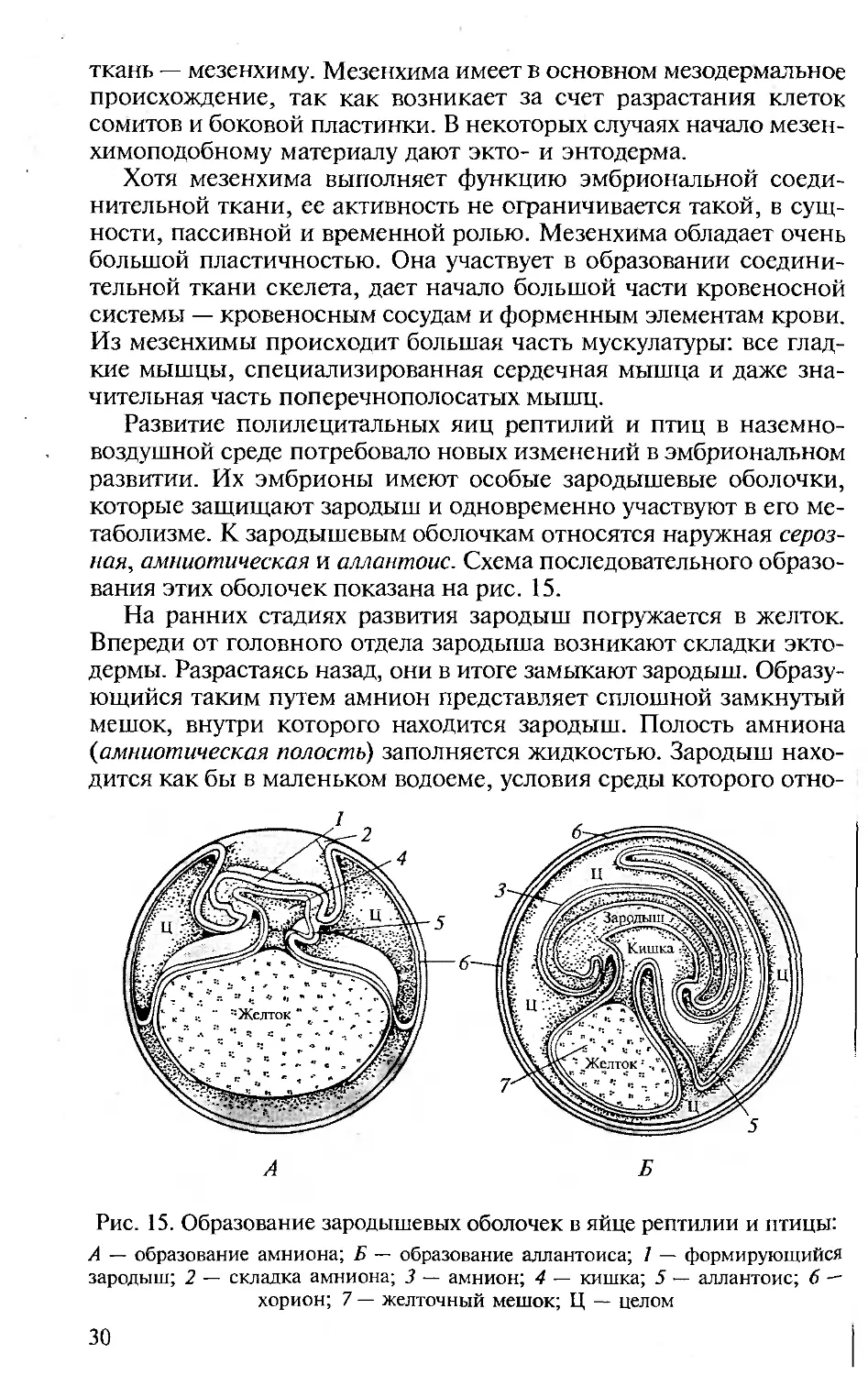
Развитие полилецитальных яиц рептилий и птиц в наземно-
воздушной среде потребовало новых изменений в эмбриональном
развитии. Их эмбрионы имеют особые зародышевые оболочки,
которые защищают зародыш и одновременно участвуют в его ме-
таболизме. К зародышевым оболочкам относятся наружная сероз-
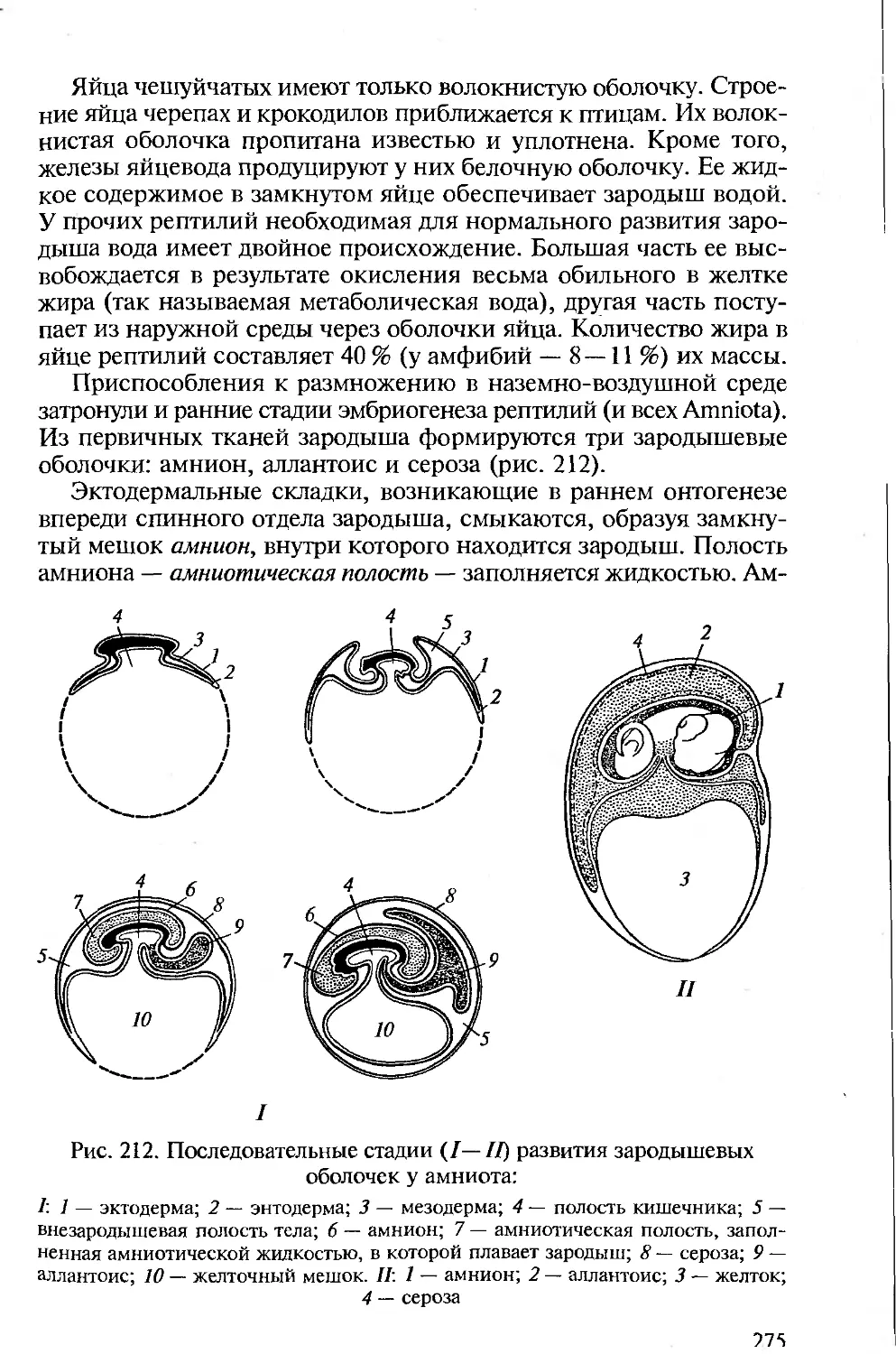
ная, амниотическая и аллантоис. Схема последовательного образо-
вания этих оболочек показана на рис. 15.
На ранних стадиях развития зародыш погружается в желток.
Впереди от головного отдела зародыша возникают складки экто-
дермы. Разрастаясь назад, они в итоге замыкают зародыш. Образу-
ющийся таким путем амнион представляет сплошной замкнутый
мешок, внутри которого находится зародыш. Полость амниона
{амниотическая полость) заполняется жидкостью. Зародыш нахо-
дится как бы в маленьком водоеме, условия среды которого отно-
Рис. 15. Образование зародышевых оболочек в яйце рептилии и птицы:
А — образование амниона; Б — образование аллантоиса; 1 — формирующийся
зародыш; 2 — складка амниона; 3 — амнион; 4 — кишка; 5 — аллантоис; б —
хорион; 7 — желточный мешок; Ц — целом
30
сительно постоянны. Роль амниона заключается еще и в том, что
он предохраняет зародыш от соприкосновения с плотными яйце-
выми оболочками.
Яйцевые оболочки наземных позвоночных усложняют процес-
сы метаболизма в яйце. Поэтому естественно возникает вопрос:
как дышит зародыш и как выводятся наружу его продукты обмена?
Первоначальным приспособлением для дыхания может служить
густая сеть кровеносных сосудов, оплетающих желток, — желточ-
ный круг кровообращения. В дальнейшем эта сеть постепенно за-
мещается вновь образующейся сетью сосудов, развивающихся в
аллантоисе. Аллантоис возникает как мешковидный вырост брюш-
ной стенки задней кишки и по мере развития приобретает вид
большого пузыря, лежащего между желточным мешком и сероз-
ной оболочкой. К концу развития аллантоис полностью окружает
тело зародыша и желточный мешок. Наружная часть аллантоиса у
некоторых рептилий срастается с серозой, образуя хорион. Кро-
веносные сосуды аллантоиса переносят кислород от поверхности
к зародышу и освобождают его от углекислоты.
Аллантоис одновременно является и зародышевым мочевым
пузырем. При этом он не только пассивно принимает и накапли-
вает мочу, но и забирает из нее излишнюю воду, которая через
сеть кровеносных сосудов возвращается в тело зародыша. Часть
конечных продуктов обмена веществ зародыша выводится наружу
в газообразном состоянии. Это обусловлено тем, что основным
энергетическим материалом в яйце рептилии и птицы является
жир, окисляющийся до воды и двуокиси углерода. Освобождение
аллантоиса от продуктов выделения происходит при рождении.
У некоторых ящериц и змей хорион и аллантоис близко со-
прикасаются, образуя хориоаллантоидную плаценту, желток при
этом редуцируется. Эти черты характерны для млекопитающих.
Ранние стадии эмбрионального развития млекопитающих
специализированы и не похожи на соответствующие стадии раз-
вития других позвоночных. Это обусловлено тем, что все млеко-
питающие вынашивают развивающихся детенышей в собствен-
ном теле. Питательные вещества, необходимые для развития заро-
дыша, вначале выделяются стенками матки, затем поступают к
зародышу из кровотока матери через плаценту. Исключение со-
ставляют представители подкласса Первозвери, яйца которых мало
чем отличаются от покрытых пергаментными оболочками яиц
Рептилий. Плацента образуется путем срастания части зародыше-
вых оболочек (серозы и аллантоиса) и ворсинок стенки матки.
Яйцеклетка оплодотворяется в верхней части яйцевода, затем за-
родыш опускается в матку и через несколько дней имплантирует-
ся в ее стенку. Своеобразие ранних стадий эмбрионального разви-
тия млекопитающих объясняется необходимостью быстрого раз-
вития плаценты.
31
У млекопитающих, как и у ланцетников, олиголецитальные яйца.
Они имеют очень маленькие размеры и практически лишены жел-
тка. Подобно яйцам ланцетников они претерпевают полное дроб-
ление, которое вскоре приводит к образованию нескольких де-
сятков клеток (рис. 16). У многих млекопитающих дифференци-
ровка тканей начинается уже на стадии бластулы. Перед гаструля-
цией около зародышевого диска формируются полости, сравни-
мые с амниотической и желточной. В зародыше выделяется плот-
ное скопление клеток — внутренняя клеточная масса, и располо-
женный вне этой массы обширный шар из клеток — трофобласт,
который в дальнейшем становится частью хориона. Трофобласт
развивается быстро и при наличии соответствующих условий всту-
пает в контакт с тканями матки. Трофобласт никогда не встреча-
ется ни у взрослых особей, ни у зародышей низших позвоночных
и служит прекрасным примером эмбриональной адаптации (цено-
генезом). Бластопор представлен у млекопитающих специфичес-
кой структурой — первичной полоской.
Расположенная между амниотической полостью и полостью
желточного мешка бластодерма соединяется с трофобластом сте-
бельком из мезодермальных клеток. Вырост первичной кишки вдоль
этого стебелька разрастается в аллантоидный мешок, который
располагается под трофобластом. Энтодермальную выстилку жел-
точного мешка и аллантоиса укрепляют мезодермальные ткани,
в которых развиваются кровеносные сосуды. Мезодерма укрепляет
и эктодерму, образуя вместе с ней прочный амнион. Одновремен-
но она одевает трофобласт изнутри, образуя хорион. В итоге в ин-
Рис. 16. Дробление олиголецитального яйца млекопитающего:
А—Г— фазы дробления, сходные с ланцетником; Д — формирование внутрен-
ней клеточной массы — трофобласта
32
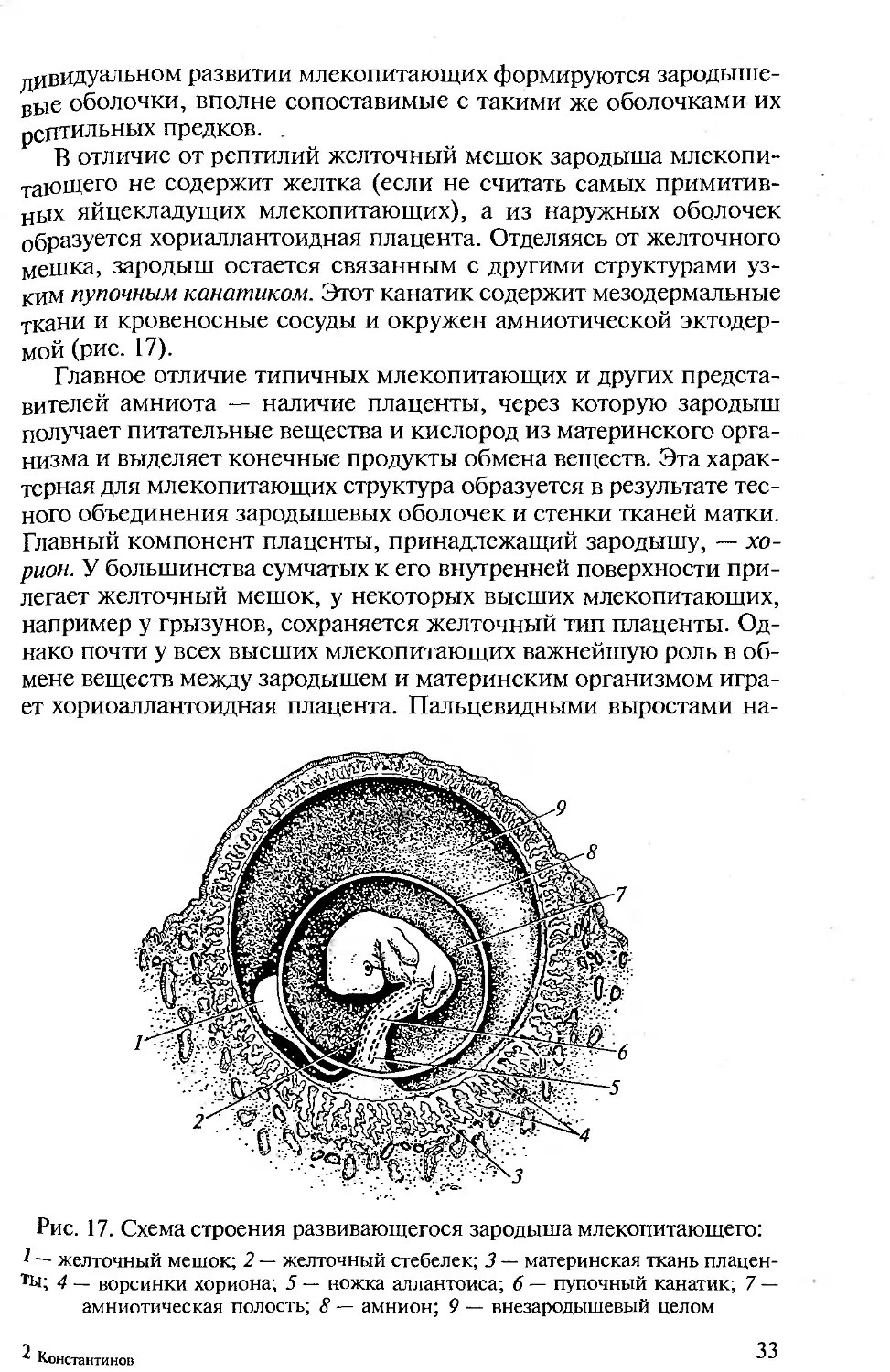
дивидуальном развитии млекопитающих формируются зародыше-
вые оболочки, вполне сопоставимые с такими же оболочками их
рептильных предков. .
В отличие от рептилий желточный мешок зародыша млекопи-
тающего не содержит желтка (если не считать самых примитив-
ных яйцекладущих млекопитающих), а из наружных оболочек
образуется хориаллантоидная плацента. Отделяясь от желточного
мешка, зародыш остается связанным с другими структурами уз-
ким пупочным канатиком. Этот канатик содержит мезодермальные
ткани и кровеносные сосуды и окружен амниотической эктодер-
мой (рис. 17).
Главное отличие типичных млекопитающих и других предста-
вителей амниота — наличие плаценты, через которую зародыш
получает питательные вещества и кислород из материнского орга-
низма и выделяет конечные продукты обмена веществ. Эта харак-
терная для млекопитающих структура образуется в результате тес-
ного объединения зародышевых оболочек и стенки тканей матки.
Главный компонент плаценты, принадлежащий зародышу, — хо-
рион. У большинства сумчатых к его внутренней поверхности при-
легает желточный мешок, у некоторых высших млекопитающих,
например у грызунов, сохраняется желточный тип плаценты. Од-
нако почти у всех высших млекопитающих важнейшую роль в об-
мене веществ между зародышем и материнским организмом игра-
ет хориоаллантоидная плацента. Пальцевидными выростами на-
Рис. 17. Схема строения развивающегося зародыша млекопитающего:
— желточный мешок; 2 — желточный стебелек; 3 — материнская ткань плацен-
ТЫ; 4 — ворсинки хориона; 5 — ножка аллантоиса; 6 — пупочный канатик; 7 —
амниотическая полость; 8 — амнион; 9 — внезародышевый целом
2 Константинов
33
ружной поверхности хорион врастает в ткани стенки матки.
выросты — ворсинки — различаются по величине, расположен^
и способу соединения со стенками матки.
Плацента обильно снабжена кровеносными сосудами матери и
зародыша. Между ними непрерывно происходит обмен веществ
однако кровеносные русла матери и зародыша никогда не вступи
ют в непосредственный контакт. Они всегда разделены мембрана,
ми, через которые проходят небольшие молекулы, но не могут
проникнуть ни белки крови, ни ее форменные элементы. В разньц
группах млекопитающих образуются плаценты различных типов
(см. разд. «Репродуктивная система»).
Зародышевые листки, возникая в процессе раннего эмбриоге-
неза хордовых (позвоночных) животных, дают начало тканям й
органам будущих взрослых организмов.
Общая кожная эктодерма формирует поверхностный эпители-
альный слой (эпидермис) кожи, покрывающий тело снаружи и
заходящий внутрь на обоих концах пищеварительного тракта,
выстилая ротовую полость и область клоаки и прямой кишки.
Производными эпидермиса являются роговые образования. Ос-
новные из них следующие: роговые чешуи рептилий и роговой
панцирь черепах, перья, роговые щитки и чехлы на клювах птиц,
волосы, колючки, ногти, когти, роговые чешуи на хвосте многих
грызунов, роговой панцирь броненосцев и панголинов, рога и
копыта зверей. Кроме этого эктодерма образует кожные железы
(млечные, потовые, сальные и их производные — пахучие), сен-
сорный эпителий полости носа и внутреннего уха, хрусталик гла-
за, эмалевый слой плакоидных чешуй и зубную эмаль.
Нейральная часть эктодермы участвует в организации почти всей
нервной системы и сетчатки глаза. Нервный гребень дает сенсор-
ные ганглии нервной системы и некоторые ненейральные струк-
туры.
Энтодерма выстилает большую часть пищеварительного тракта
(эндотелий) и связанных с ним органов (печени, поджелудочной
железы); большую часть эндокринных желез (тимуса, паращито-
видных желез, щитовидной железы); большую часть жаберных
лепестков низших позвоночных и образует выстилку дыхательной
системы высших позвоночных, дышащих легкими, концевые уча-
стки мочевых и половых путей.
Мезодерма служит основой для развития соединительной тка-
ни и большей части скелета, мышц, сосудистой системы, значи-
тельной части мочеполовой системы, выстилки целомических
полостей, брыжейки, хорды.
* * *
Таким образом, в эмбриональном развитии зародыши хорД0'
вых проходят последовательно несколько стадий: бластула, гасТ'
34 I
нейрула, которые рассматриваются на классической моде-
рн отражающей общие черты раннего онтогенеза всего типа —
лИ’оа3витии ланцетника. Закладка органов у представителей раз-
На х систематических групп позвоночных происходит сходно из
нь* х и тех же зачаточных листков: эктодермы, энтодермы и ме-
° дермы. Различия в продолжительности и завершенности основ-
ах стадий развития зародыша у представителей различных сто-
матических групп зависят от обилия запасных питательных ве-
ществ (желтка) в оплодотворенной яйцеклетке. Эти различия воз-
растают в эволюционно прогрессивных группах в связи с увели-
чением эмбриональных адаптаций и формированием дефинитив-
ных признаков животных на завершающих стадиях эмбриональ-
ного развития.
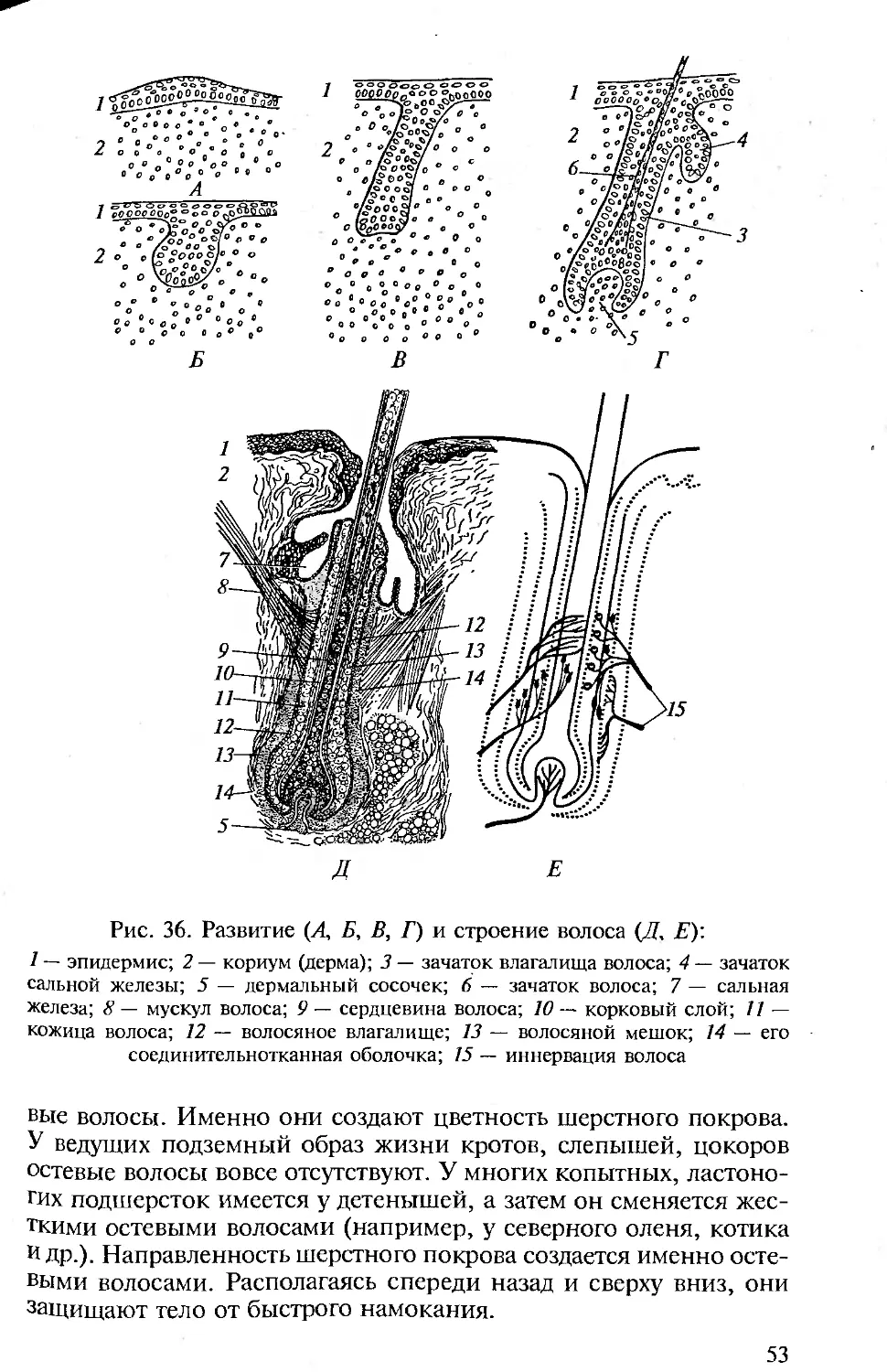
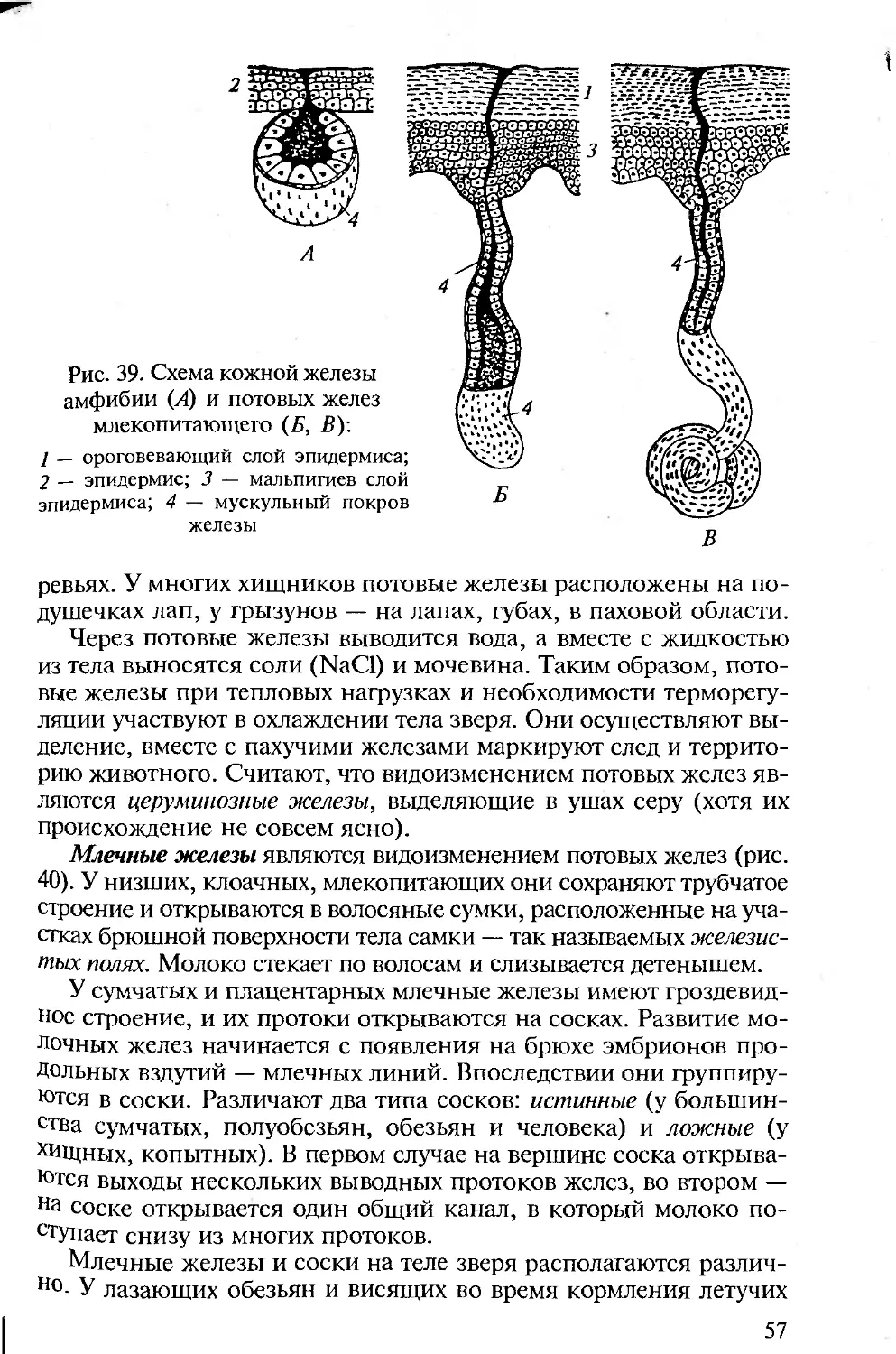
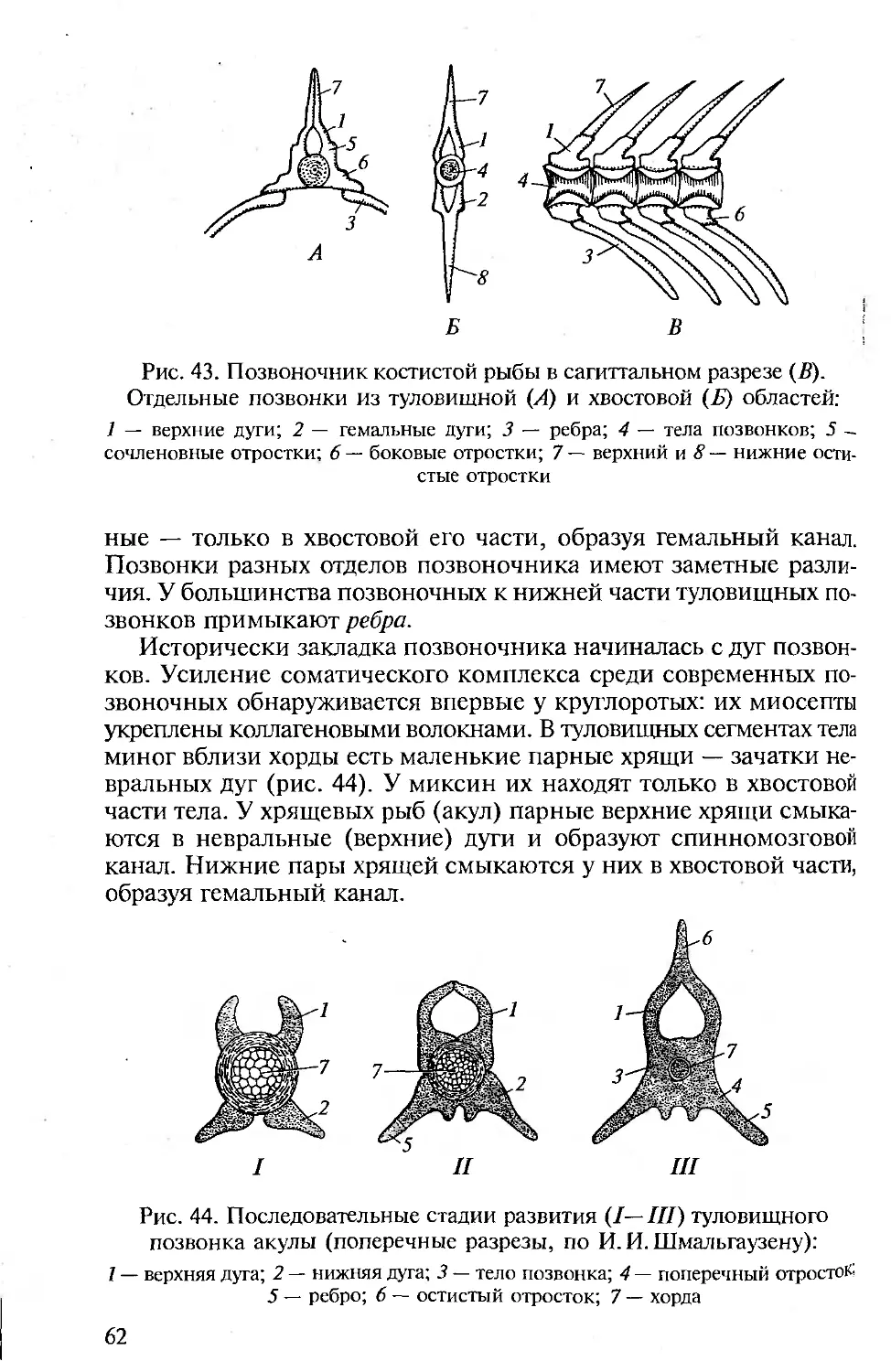
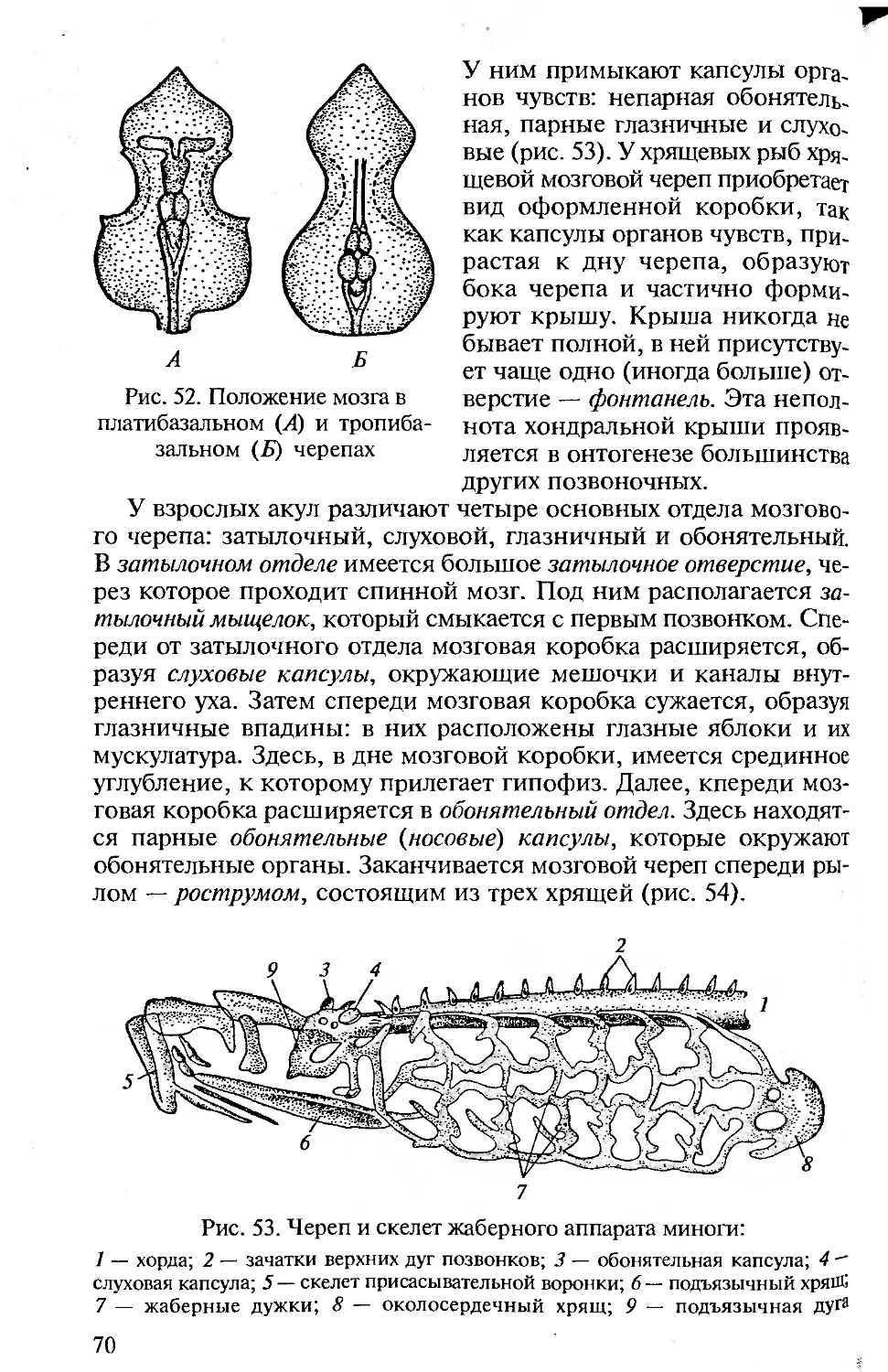
КОЖА И ЕЕ ПРОИЗВОДНЫЕ
Покровы тела хордовых животных — система многофункцио-
нальная. Они ограничивают тело животного и вместе с мускулату-
рой придают ему форму и удерживают внутренние органы. По-
кровы защищают тело от механических повреждений, препятству-
ют проникновению в организм животного бактерий, микроорга-
низмов, химических веществ. Покровы участвуют в регуляции вод-
ного баланса, защищая тело и от потери воды, и от чрезвычайно-
го ею насыщения. Благодаря наличию специализированных струк-
тур, желез и обильной гиперемии кровеносными сосудами кожа
участвует в дыхании и выведении конечных продуктов.
Кожа и ее производные пигментированы. Окраска тела также
многофункциональна: она делает одних животных незаметными,
других — демонстративными. Пигменты кожи защищают тело жи-
вотного от избыточного проникновения ультрафиолетовых лучей.
Кожа снабжена нервными окончаниями и специализирован-
ными рецепторными образованиями, которые осуществляют сен-
сорную функцию при общении животного со средой. Видимо, не-
случайно большинство органов чувств эмбрионально возникает
из специализированных участков наружных покровов зародышей.
Производные кожи принимают участие также в формировании
органов передвижения (перепонки на лапах, между конечностя-
ми и телом, маховые и рулевые перья), служат для хватания, борь-
бы, защиты (роговые рога, щитки, острые выросты).
Строение кожи. Кожа позвоночных (хордовых) двухслойная: она
состоит из наружного эпидермиса и внутреннего кутиса.
Между эпидермисом и следующим, соединительнотканным,
слоем кожи — кутисом — лежит базальная мембрана, образо-
ванная чаще эпителиальной, но отчасти и соединительной тка-
нью.
Эпидермис представлен цилиндрическими клетками и проис-
ходит из эктодермы зародыша. У ланцетника он образован одним
слоем цилиндрических клеток, у личинки клетки эпидермиса
имеют реснички. Бокаловидные клетки выделяют на поверхность
эпидермиса тонкую пленку — кутикулу. Структурность кутиса
выражена у ланцетников слабо.
У оболочников эпидермис выделяет на поверхность тела сту-
денистую оболочку, которая в результате проникновения в нее
36
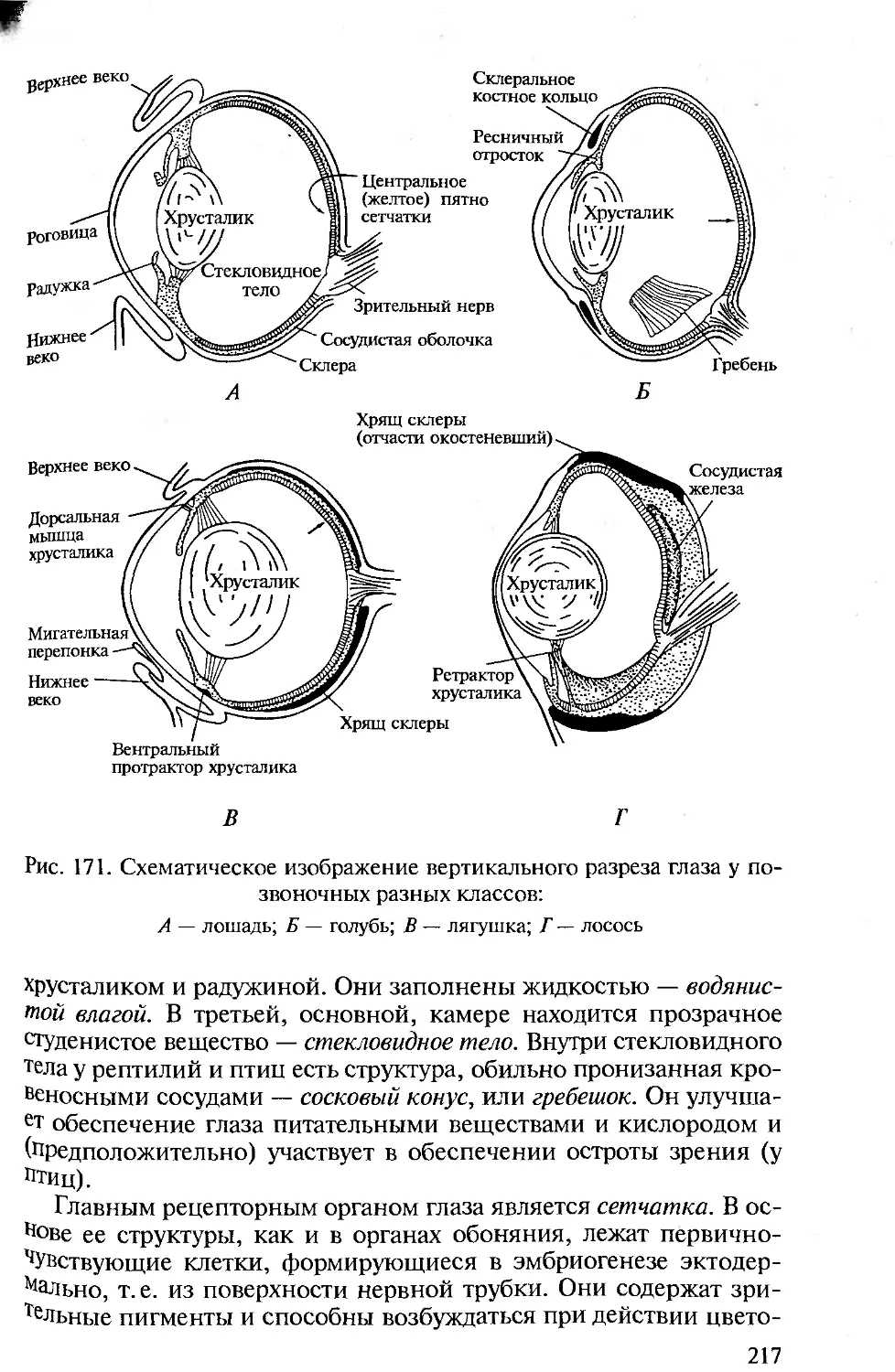
А Б
Рис. 18. Схематический разрез через кожу ланцетника (Л) и миноги (Б):
/ — эпидермис; 2 — кутис; 3 — кутикула; 4, 5 — железистые клетки; 6 — базаль-
ная мембрана
снизу мезодермальных элементов становится плотной опорной
тканью. В состав такого экзоскелета оболочников входит целлюло-
за (Л. Проссер, 1977).
Эпидермис у позвоночных — всегда многослойная ткань, не
имеющая кутикулы. У низших позвоночных (круглоротых, рыб,
амфибий) эпидермис отграничен от подстилающей его соеди-
нительной ткани базальной мембраной — бесструктурной пере-
понкой, которая выделяется со-
вместно эпидермисом и кутисом
(рис. 18, 19).
У рыб и личинок амфибий
верхний слой эпидермиса (как и
у личинок ланцетника) образует
на поверхности тонкую пористую
каемку, которую считают остат-
ком общего мерцательного покро-
ва предковых форм (И. И. Шмаль-
гаузен, 1964). У современных
взрослых амфибий под слоем эпи-
дермиса обильно ветвятся крове-
носные капилляры, через них осу-
ществляется кожное дыхание (га-
зообмен между кровью и окружа-
ющей средой). Эпидермис богат
Многоклеточными железами. Их
Рис. 19. Схематический разрез
через кожу рыбы:
1 — эпидермис; 2 — кутис; 3 — ба-
зальный слой; 4, 5, 6 — железистые
клетки; 7 — нервы; 8 — сосуды
секрет создает на поверхности
Кожи жидкостную пленку, кото-
рая препятствует иссушению по-
кровов и делает возможным га-
зообмен.
37
Рис. 20. Строение кожи млекопитающего:
1 — эпидермис; 2 — кориум (дерма, или кутис); 3 — подкожная жировая клет-
чатка; 4 — дермальные сосочки; 5 — нервные окончания; б — капилляры; 7 —
сальная железа; 8 — мускул, двигающий волос; 9 — кровеносные сосуды; 10 —
корень волоса; 11 — потовая железа
Эпидермис в коже наземных позвоночных (рептилий, птиц,
млекопитающих) — ткань многослойная и достаточно сложная
(рис. 20). Его нижний базальный мальпигиев слой пожизненно ос-
тается деятельным и продуцирует к поверхности новые слои
клеток, поэтому поверхностные нарушения эпидермиса у них легко
восстанавливаются. Верхний слой эпидермиса — роговой — состо-
ит обычно из клеток уплощенной формы. У наземных позвоноч-
ных (в том числе и у некоторых амфибий) в них накапливается
водонепроницаемый белок (кератин или кератогиалин). При из-
нашивании и повреждении верхние клетки слущиваются и заме-
няются новыми, лежащими ниже. Кровеносных сосудов эпидер-
мис не имеет.
Эпидермис дает начало различным роговым производным в виде
роговых чешуй и щитков, перьев, волос, копыт, когтей, ногтей,
полых рогов, перепонок (см. ниже). В эпидермисе развиваются и
разнообразные кожные железы (см. ниже). Наконец, клетки глубо-
ких слоев эпидермиса нередко содержат красящие вещества. Вмес-
те с пигментами кутиса они обусловливают окраску тела живот-
ного.
Кутис — внутренний волокнистый слой кожи позвоночных.
Его называют также кориум, дерма, собственно кожа. В эмбриоге-
незе кутис развивается из мезодермального зачатка — кожного
листка сомита. Слой кутиса (дермы), как правило, толстый и со-
38
ставляет основную часть кожи. В этом слое плотно переплетаются
волокна соединительной ткани. Дерма придает коже прочность и
эластичность. В кутисе развиваются разнообразные окостенения:
чешуя рыб, покровные окостенения скелета, костные рога ко-
пытных, костные пластины и панцири некоторых рептилий и мле-
копитающих. У млекопитающих в кутис погружены основания
кожных желез, волосяные сумки, у птиц — перьевые фолликулы.
В наружной части кутиса располагаются нервные окончания
тепловых и осязательных рецепторов. Здесь ветвится сложная сеть
кровеносных и лимфатических сосудов.
В наружной же части кутиса и его производных (как и в нижних
слоях эпидермиса) лежат многообразные пигментные клетки —
хроматофоры. Среди них различают меланофоры, липофоры, иридо-
циты. Меланофоры содержат меланин и придают телу коричне-
вую окраску, липофоры содержат каротиноиды и создают крас-
но-желтые тона окраски, иридоциты содержат кристаллы гуани-
на. Они отражают свет и способны менять эффект другого пиг-
мента, придавая животному серебристый блеск.
Пигментные клетки обладают подвижностью и способны пе-
реползать в межклеточные пространства. Одновременно в них мо-
жет меняться положение пигментных зерен. Этим объясняется
способность некоторых животных менять окраску (камбалы, скор-
пены, амфибии, ящерицы, хамелеоны) (рис. 21). Перемещение
клеток стимулируется подходящими к хроматофорам веточками
нервов.
Более глубокая часть кутиса имеет рыхлую структуру и служит
основным местом образования жировой ткани — подкожной жи-
ровой клетчатки. Здесь накапливается жир — термоизоляционный
и питательный материал тела животного. Большой толщины этот
слой достигает у морских млекопитающих — китов и дельфинов.
Рис. 21. Изменение окраски камбалы в зависимости от цвета субстрата
(по Н.П.Наумову, Н.Н.Карташову, 1979)
39
Например, подкожный слой жира у крупного синего кита дости-
гает 36 см на спине и 20 —25 см на боках, а общая масса подкож-
ного жира при массе кита более 100 т составляет 15—20 т. Жир
существенно предохраняет тело от излишней потери тепла, улуч-
шает плавучесть и восполняет недостаток питания при сезонных
голодовках.
У некоторых млекопитающих на отдельных участках тела кожа
чрезвычайно тонкая, например у зайцев на спине, у сонь и мы-
шей на хвосте. Это приводит к частичной аутотомии ее участков с
последующей их регенерацией.
Производные кориума — чешуя рыб, покровные окостенения
скелета, костные пластины рептилий, костные рога копытных.
Остановимся на них подробнее.
Тела древних примитивных челюстноротых, стоявших у исто-
ков возникновения позвоночных животных (щитковые, панцир-
ные рыбы), были заключены в костный панцирь. Основу их наруж-
ного скелета составляли покровные кости. Бугорки и гребни этих
костей были покрыты эмалью.
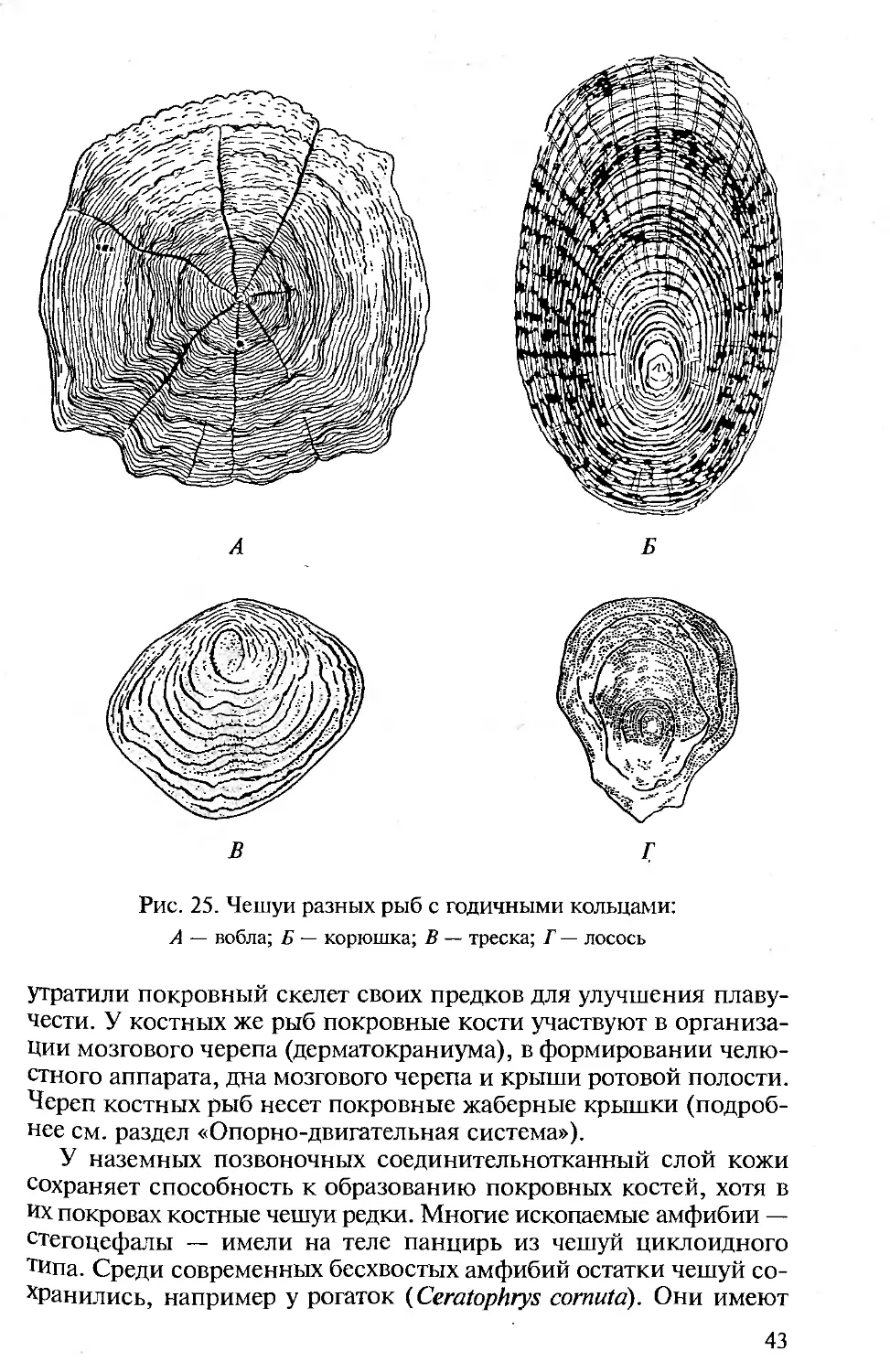
Тело современных рыб покрыто костной чешуей, различной по
химическому составу и морфологическим особенностям.
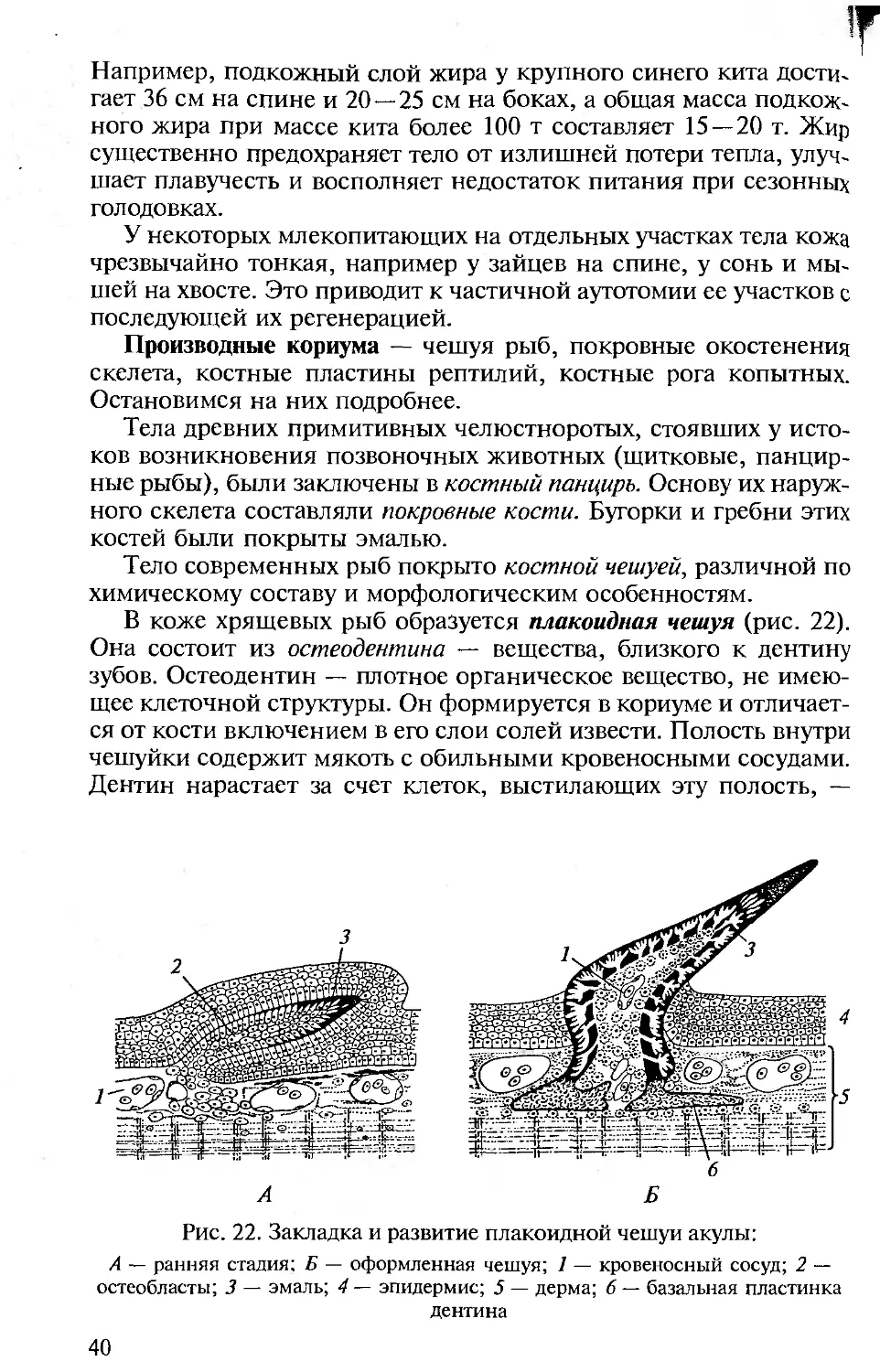
В коже хрящевых рыб образуется плакоидная чешуя (рис. 22).
Она состоит из остеодентина — вещества, близкого к дентину
зубов. Остеодентин — плотное органическое вещество, не имею-
щее клеточной структуры. Он формируется в кориуме и отличает-
ся от кости включением в его слои солей извести. Полость внутри
чешуйки содержит мякоть с обильными кровеносными сосудами.
Дентин нарастает за счет клеток, выстилающих эту полость, —
Рис. 22. Закладка и развитие плакоидной чешуи акулы:
А — ранняя стадия; Б — оформленная чешуя; 1 — кровеносный сосуд; 2 —
остеобласты; 3 — эмаль; 4 — эпидермис; 5 — дерма; б — базальная пластинка
дентина
40
склеробластов. Снаружи зубец чешуи покрыт тонким слоем эмали
(витродентином), производным клеток эпидермиса.
1 На челюстях акул плакоидная чешуя укрупняется и выполняет
функцию зубов (кожные зубы). Изношенные зубы сбрасываются и
замешаются новыми. На поверхности тела некоторых скатов от-
дельные чешуи разрастаются в шиповатые структуры. В спинных
плавниках некоторых акул (например, колючих) есть увеличен-
ные шипы, покрытые эмалью.
Тела относительно небольшой группы современных костных
рыб (панцирников, многоперов) покрывает ганоидная чешуя. Это
вошло в их систематическое название— ганоидные рыбы. Ганоид-
ная чешуя — производное только мезодермального слоя кожи. Ее
нижние слои строятся из настоящей костной ткани (покровное
окостенение), а поверхность формируется из остеодентина, свой-
ственного плакоидной чешуе. В ганоидной чешуе остеодентин ле-
жит слоями, нарастая одновременно снизу и сверху. Верхний слой
называют ганоин, нижний — изопедин (рис. 23). Чешуя нарастает
одновременно нижним и верхним слоями. Наиболее демонстра-
тивна такая чешуя у современной панцирной щуки, тело которой
покрыто твердыми ромбическими пластинками.
Ганоидная чешуя была свойственна древним двоякодышащим
и кистеперым, но у них она имела свои особенности. Основу че-
шуи составлял космин — комплексная ткань, включавшая срос-
шиеся плакоидные чешуи с системой ветвящихся в них каналь-
цев, заполненных мякотью (космоидная чешуя). Чешуя современ-
ного целаканта — латимерии — отличается от покрова ее предше-
ственников. Она имеет четыре слоя: поверхностный — эмалепо-
добный, с зубчиками и порами, губчатокостный, костногубча-
тый и слой плотных костных пластинок. У современных двояко-
Рис. 23. Образование (А—В) и строение (Г) ганоидной чешуи:
I — поверхность чешуи; 2 — слой ганоина; 3 — канальцы в слое космина; 4 —
слои изопедина
41
6
Рис. 24. Кожа костистой рыбы (А) и костные чешуи (Б)
(по Н.П.Наумову, Н.Н.Карташову, 1979):
1 — эпидермис; 2 — кориум; 3 — базальный слой эпидермиса; 4 — костная
чешуя; 5 — чувствующая концевая почка; б — слизеотделительная одноклеточ-
ная железа; 7 — колбовидная одноклеточная железа
дышащих рыб эмалевый слой и дентин утрачены и чешуя приоб-
рела волокнистую структуру (А. Ромер, Т. Парсонс, 1992).
В ганоидной чешуе разных родов отряда многоперообразных
рыб и древних палеонисцид обнаружено сочетание нижних кост-
ных слоев с космином и ганоином.
У ганоидной — ильной рыбы (или амии) ганоидный слой утон-
чается и чешуя почти полностью костная. Эта рыба как бы про-
кладывает переход к наиболее распространенному типу покровов
костных рыб — костной чешуе.
У современных костистых рыб чешуя костная (рис. 24). Чешуй-
ки формируются из мезодермальных склеробластов. Они лежат в
кожных карманах, черепицеобразно налегая друг на друга. Кост-
ные чешуйки могут быть округлыми {циклоидные) или зазубрен-
ными (ктеноидные). Ряды бляшек на теле осетровых рыб и мелкие
чешуйки на их теле — это тоже костная чешуя.
Костная и ганоидная чешуя — образования постоянные и не
сменяются. На протяжении жизни рыбы чешуи увеличиваются
в размерах. Скорость их нарастания неодинакова в разные сезо-
ны, что послужило разработке методики определения возраста
рыбы по шлифам чешуи путем подсчета нарастающих в ней сло-
ев (рис. 25).
Кроме чешуи в кутисе (кориуме) рыб образуются покровные
окостенения скелета. Как уже сказано выше, тела многих древних
первичноводных позвоночных имели панцирь из покровных кос-
тей. У многих щитковых это были разнообразные крупные и мел-
кие пластины. У панцирных рыб можно различить пластины груд-
ной области, которые приблизительно соответствуют элементам
плечевого пояса современных, впереди от которых был располо-
жен щит, покрывавший голову. Предполагают, что хрящевые рыбы
42
Рис. 25. Чешуи разных рыб с годичными кольцами:
А — вобла; Б — корюшка; В — треска; Г — лосось
утратили покровный скелет своих предков для улучшения плаву-
чести. У костных же рыб покровные кости участвуют в организа-
ции мозгового черепа (дерматокраниума), в формировании челю-
стного аппарата, дна мозгового черепа и крыши ротовой полости.
Череп костных рыб несет покровные жаберные крышки (подроб-
нее см. раздел «Опорно-двигательная система»),
У наземных позвоночных соединительнотканный слой кожи
сохраняет способность к образованию покровных костей, хотя в
Их покровах костные чешуи редки. Многие ископаемые амфибии —
стегоцефалы — имели на теле панцирь из чешуй циклоидного
типа. Среди современных бесхвостых амфибий остатки чешуй со-
хранились, например у рогаток (Ceratophrys cornuta). Они имеют
43
вид лежащих глубоко в коже маленьких костных пластинок. Кост-
ные чешуйки есть у безногих.
У гаттерий и крокодилов сохраняются так называемые брюш-
ные ребра (гастралии). Их считают остатками брюшного панциря
стегоцефалов. Возможно именно они приняли участие и в образо-
вании пластрона у черепах. У некоторых рептилий новообразова-
ния кожного скелета есть и на спинной стороне тела: костные
пластинки под роговыми чешуями у крокодилов, некоторых яще-
риц (гекконов, сцинков).
Панцирь черепах заслуживает особого внимания. Костный пан-
цирь современных черепах является уникальным наружным ске-
летом среди всех наземных позвоночных. У большинства видов
под роговыми щитками лежит прочный костный панцирь. Он со-
стоит из выпуклого овального спинного щита — карапакса и плос-
кой брюшной пластины — пластрона. По бокам обе части соеди-
нены костными перемычками. Для головы, конечностей и хвоста
панцирь имеет переднее и заднее отверстия.
Карапакс включает три набора пластин. Его края оформлены
краевыми пластинами; по средней линии спереди назад пролега-
ет ряд почти квадратных невральных или позвоночных пластин,
большинство из которых прирастает к расположенным под ними
позвонкам (рис. 26, А). Между краевыми и невральными пласти-
нами лежат восемь вытянутых поперечно-плевральных, или ре-
берных, пластин, сросшихся с лежащими под ними ребрами. Эле-
менты карапакса плотно сращены, швы между ними с возрастом
утрачиваются. Границы поверхностных роговых щитков в основ-
ном не совпадают с костными пластинами, а лишь отпечатыва-
ются на поверхности костного ка-
рапакса. Как правило, границы меж-
ду роговыми щитками карапакса и
пластрона чередуются со швами
между костными пластинами, что
придает большую прочность конст-
рукции панциря в целом.
Пластрон типичных черепах со-
стоит из четырех парных пластин и
Рис. 26. Щитки панциря болотной
черепахи:
А — карапакс; Б — пластрон; / — плевраль-
ные шитки; 2 — краевые; 3 — надхвостовые;
4 — позвоночные; 5 — горловые; б — груд-
ные; 7 — брюшные; 8 — плечевые; 9 —
бедренные; 10 — заднепроходные; 11 —
шейные
44
переднего срединного элемента (рис. 26, Б). У примитивных чере-
пах бывают дополнительные парные пластины. Из трех пластин
переднего края пластрона две являются ключицами и одна — меж-
^лючичной вставкой кожных элементов плечевого пояса. Осталь-
ные элементы пластрона и все пластины карапакса черепах, по-
видимому, являются новообразованиями, хотя предполагают, что
в состав пластрона вошли покровные брюшные ребра (А. Ромер,
Т. Парсонс, 1992).
Похожие костные пластины свойственны млекопитающим. Так,
ими покрыто тело современных броненосцев. Известно, что в ис-
торическом прошлом сплошной костный панцирь покрывал спи-
ну близких к ним глиптодонтов. У вымерших ленивцев кожа была
укреплена участками покровных костей.
Наконец, следует упомянуть о рогах копытных — костных вы-
ростах на лобных костях. Парные костные выросты образуются
при участии особого кожного окостенения (oscornu), которое
прирастает к мозговому черепу. Как правило, костные рога отра-
стают у самцов оленей. К примеру, длина рогов у пятнистого
оленя может достигать 93 см. Рога у обоих полов свойственны
северному оленю. Костные рога вследствие разрушения костной
ткани их основания периодически сбрасываются. К периоду се-
зонной половой активности рога вырастают вновь, иногда ус-
ложняющейся формы (рис. 27). Число отростков на рогах измен-
чиво у разных видов: у пятнистого оленя их в норме 4, может
быть 6, марала — 6 — 7.
У полорогих копытных костные выросты на лбу невелики и
покрыты постоянными толстыми роговыми чехлами. Уникальным
исключением является вилорог Antilocapra americana, теряющий
роговой чехол ежегодно. У жирафа костные рога одеты мягкой
кожей. У оленей кожа с волосами покрывает только молодые рога
(панты), а затем она отмирает и отпадает.
Рис. 27. Схемы рогов разных типов:
— настоящий рог коровы, овцы; Б — олений рог, покрытый кожей; В — оле-
ний рог без кожи; Г — рог вилорога; Д — рог жирафа; Е — рог носорога
45
Роговые производные эпидермиса — приобретения наземных
позвоночных животных. Их основное назначение — предохране-
ние тела от иссушения и защита от повреждений. Эмбрионально
роговая чешуя закладывается как выпячивание эпидермиса с ме-
зодермальным сосочком внутри. Такое сочетание тканей сохраня-
ется и у взрослых животных.
Утолщения рогового слоя есть на теле жаб в виде бородавок или
участков сплошного покрова. На теле рептилий образуется сплош-
ной покров из роговых чешуй или щитков.
Поверхность тела гекконов, агам, хамелеонов, гаттерий по-
крывает примитивная бугорчатая роговая чешуя. На теле настоя-
щих ящериц и змей чешуя расположена черепицеобразно или ря-
дами (рис. 28, 29). Голова змей и многих ящериц покрыта упоря-
доченными рядами роговых щитков. Их расположение учитывает-
ся при систематической характеристике видов (рис. 30). Видоизме-
ненные роговые щитки на хвосте гремучей змеи образуют «погре-
мушку» (рис. 31). Роговой покров подвержен смене: у змей, оттор-
гаясь вначале в области челюстей, он сползает с тела полностью,
«чулком», у ящерицы отделяется участками. У некоторых репти-
лий под роговыми чешуями
слоя кожи.
есть окостенения мезодермального
Рис. 28. Схематические разрезы
чешуи рептилий:
А — яшерицы; Б — змеи; В — крокодила;
1 — роговая чешуя; 2 — эпидермис; 3 —
дерма; 4 — костная основа роговой
чешуи
Рис. 29. Продольный разрез через кожу ящерицы:
1 — эпидермис; 2 — собственно кожа; 3 — роговой слой; 4 — пигментные клет-
ки; 5 — кожные окостенения
46
16
Рис. 30. Расположение щитков на голове ящерицы сверху (А) и сбоку (Б):
1 — межчелюстной щиток; 2 — передние и задние носовые щитки; 3 — лобно-
носовой щиток; 4 — предлобные щитки; 5 — лобный щиток; 6 — надглазничные
щитки; 7 — лобно-теменные щитки; 8 — теменные щитки; 9 — межтеменной
щиток (с отверстием для теменного глаза); 10— затылочный щиток; 11 — пере-
дний скуловой щиток; 12 — задний скуловой щиток; 13 — верхнересничные
щитки; 14 — ушной щиток; 15 — верхнегубные щитки; 16 — подглазничные
щитки; 17 — центрально-височный щиток; 18 — подбородочный щиток; 19 —
нижнегубные щитки; 20 — нижнечелюстные щитки; 21 — задневисочные щитки
Роговая чешуя покрывает черепицеобразно и тело ящерооб-
разных млекопитающих — панголинов. Тело крокодилов, панцирь
черепах и броненосцев покрыто с поверхности роговыми щитка-
ми. Щитки и чешуйки сохраняются на ногах птиц, на хвостах не-
которых млекопитающих (крысы, бобры и др.).
Видоизменениями рогового покрова следует считать роговые
чехлы на челюстях — клювы. Они свойственны черепахам, некото-
рым вымершим рептилиям, однопроходным млекопитающим и,
наконец, всем птицам. Многообразие форм клювов птиц отража-
ет специфику их кормодобывающей де-
ятельности (рис. 32).
Роговые структуры располагаются на
конечностях — когти, ногти, роговые пе-
репонки и складки на пальцах птиц, по-
дошвенные подушечки у млекопитающих,
кожные гребешки на лапах приматов,
копыта на концах пальцев копытных
млекопитающих (рис. 33). Любопытным
роговым производным являются рога у
полорогих копытных млекопитающих:
быков, козлов, баранов, антилоп. Пря-
мые или изогнутые, иногда закручен-
ные, эти роговые чехлы покрывают ко-
стные выросты лобных костей черепа.
Они нарастают на протяжении всей
жизни и не отторгаются.
Рис. 31. «Погремушка»
на хвосте гремучей змеи
47
Рис. 32. Клювы птиц:
1 — зеленоклювого тукана; 2 — колпицы; 3 — большеклювого попугая; 4 —
тупика; 5 — водореза; 6 — большого пестрого дятла; 7 — фламинго; 8 — большо-
го кроншнепа; 9 — утки; 10 — шилоклювки; 11 — ибиса; 12 — клеста; 13 — марабу;
14 — козодоя-лягушкорта
48
Рис. 33. Продольные разрезы концевых фаланг разных млекопитающих:
Л — коготь хищника; Б — копыто лошади; В — ноготь примата; Г — ноготь
человека
К роговым производным эпидермиса относятся также перьевой
покров птиц и шерстный {волосяной) покров млекопитающих.
Перьевой покров. Тело птицы покрыто перьями. Развитие перьев
в онтогенезе птиц указывает на их тесную генетическую связь с
чешуями рептилий. Как и зачаток роговой чешуи, зачаток пера
возникает в виде бугорка соединительнотканного слоя кожи, по-
крытого снаружи эпидермисом (рис. 34). По мере роста бугорок
Рис. 34. Схема развития пера:
А~В — продольные разрезы через зачатки пера разного возраста; Г — эмбрио-
нальное перо в разрезе; Д — стереограмма развивающегося контурного пера;
1 ~ эпидермис; 2 — кутис; 3 — сосочек пера; 4 — зачаток стержня; 5 — зачатки
бородок. После сбрасывания наружной кожицы бородки опахала освобождаются
и раздвигаются в обе стороны по линии, указанной стрелкой
49
отклоняется назад, а его основание опускается в глубь кожи, об-
разуя влагалище будущего пера и его сосочек, обильно снабжае-
мый кровью, через который происходит питание растущего пера.
Эктодермальная часть зачатка, разрастаясь, дифференцируется на
продольное утолщение — будущий стержень и два продольных
киля этого утолщения, которые позднее распадаются на бородки
опахала. Весь зачаток покрыт снаружи тонким роговым чехликом,
разрушающимся по мере оформления пера. После удаления чех-
лика опахала освобождаются, левая и правая его половинки раз-
двигаются в стороны. Кроме эмбриональной закладки на генети-
ческую близость пера к чешуе рептилии указывает и то обстоя-
тельство, что при глубоких повреждениях участков кожи на теле
птиц вместо перьев отрастает чешуя.
Перья у птиц дифференцированы на контурные, пуховые, соб-
ственно пух, нитевидные, щетинки. Они располагаются на теле не
равномерно, а участками — птерилиями. Такое их распределение
не ограничивает свободу движения птицы.
Опахало контурного пера сложено удлиненными бородками
1-го порядка (рис. 35). Каждая из них несет короткие бородки 2-го
порядка. В сомкнутое опахало они объединяются крючочками. Птица
поддерживает сомкнутость опахал, особенно маховых и рулевых
перьев, восстанавливая их целостность клювом. Бородки пуховых
перьев рассучены и крючков не имеют. Опрятный вид и здоровое
состояние перьевого покрова поддерживается нанесением на его
поверхность секрета копчиковой железы. Содержащийся в ее вы-
делениях витамин А способствует хорошему состоянию покровов.
Перья регулярно сменяются. В онтогенезе птицы наиболее рано
происходит смена ювенильного пуха на более зрелое оперение.
У взрослых птиц оперение сменяется регулярно. В году у многих
птиц бывает не одна, а две или три линьки. У многих птиц линька
связана со свойственным птицам сезонным полиморфизмом и при-
обретением брачного наряда. В этом случае меняется обычно не
все оперение, а только определенные его участки. Характер линь-
ки у птиц различен. Хищники и насекомоядные виды, ловящие
добычу в воздухе, линяют постепенно, не теряя способности к
полету. Куриные птицы, обитающие в лесах, кустарниковых и тра-
вянистых зарослях, линяют быстрее. Во время линьки они с тру-
дом могут подняться в воздух, поэтому держатся, затаиваясь в
укромных местах. Тетерева и глухари на короткий период вовсе
теряют способность к полету. Очень своеобразно линяют утки, гуси,
лебеди, чистики, поганки, гагары и большинство пастушковых.
Маховые перья у них выпадают почти одновременно, и птицы на
долгое время лишаются возможности летать. Гуси, некоторые утки,
лебеди в это время собираются в глухих, труднодоступных местах
по берегам рек, озер и морей, концентрируясь там иногда по многу
тысяч особей.
50
Рис. 35. Контурное перо птицы (А) и схема
строения опахала (Б):
1 — стержень; 2 — наружное опахало; 3 — внут-
реннее опахало; 4 — ствол; 5 — очин; 6 — отвер-
стие очина; 7 — дужка пера; 8 — бородка; 9 —
бородочки; 10 — крючочки
При линьке у ряда видов сменяется не только масса оперения,
но и его структура. Так, в летнем оперении чижа насчитывается
примерно 1500 контурных и пуховых перьев, в зимнем оперении
число пуховых увеличивается до 2100 — 2400. У одного из видов
синиц летом насчитывается до 1100, а зимой 1700 перьев. У белой
куропатки длина контурных перьев зимой равна в среднем 5,4 см,
а летом 3, 8 см; пуховая часть их соответственно равна 1,8 и 1,4 см;
Побочный ствол — 3,7 и 2,5 мм (А. В. Михеев, 1960).
Перьевой покров — образование многофункциональное. Его
окраска скрывает птицу от взора хищника или, наоборот, демон-
стративна в половых играх. Структурно и механически (распуша-
ясь) он участвует в терморегуляции. И наконец, главное — перь-
евой покров обеспечивает птице возможность движения в воздухе.
51
Край крыла образован маховыми перьями: первостепенные махо-
вые прикрепляются к кисти, второстепенные — к предплечью,
третьестепенные — к плечу (см. рис. 81). Стабильное количество
10—12 первостепенных маховых является систематическим при-
знаком отряда. Несущую плоскость хвоста образуют рулевые пе-
рья. Их число тоже регламентировано.
Шерстный (волосяной) покров — характерная черта облика мле-
копитающих. Волосы — тоже роговое производное эпидермального
слоя кожи. Однако развитие волоса в эмбриогенезе начинается иначе,
чем чешуи и пера. Плотный участок эпидермальных клеток приоб-
ретает цилиндрическую форму и не выпячивается наружу, а на-
оборот, погружается в мезодермальную часть кожи. Крайние клет-
ки участвуют в образовании влагалища волоса (фолликула) и саль-
ных желез, а расположенные по центру превращаются в зачаток
самого волоса. Клетки зачатка волоса ороговевают (рис. 36, Д, Е).
В структуре развитого волоса по длине можно различить высту-
пающий над кожей ствол и сидящий в коже корень. Ствол состоит
из сердцевины, коркового слоя и кожицы. Сердцевина представляет
собой пористую ткань, между клетками которой находится воз-
дух; именно эта часть волоса сообщает ему малую теплопровод-
ность. Корковый слой, наоборот, очень плотный и придает воло-
су прочность. От механических и химических воздействий волос
защищен снаружи тонкой кожицей. Ствол волоса является мерт-
вым роговым образованием, не способным к росту и изменению
формы. Корень волоса в своей верхней части имеет цилиндричес-
кую форму и является прямым продолжением ствола. В нижней
части корень расширяется и заканчивается колбообразным взду-
тием — луковицей волоса, которую, как колпачком, охватывает
вырост кутиса — волосяной сосочек. Кровеносные сосуды, входя-
щие в этот сосочек, обеспечивают жизнедеятельность клеток лу-
ковицы волоса. Формирование и нарастание волоса идет за счет
размножения и видоизменения клеток луковицы (рис. 36, Д, Г).
В волосяной мешок (влагалище волоса) открываются протоки
сальных желез, секрет которых смазывает волос, что придает ему
большую прочность и водоупорность. К нижней части волосяного
мешка прикрепляются мышечные волокна, сокращения которых
обусловливают движение мешка и сидящего в нем волоса. Это дви-
жение и вызывает «ощетинивание» зверя. Обычно волосы сидят в
коже не перпендикулярно ее поверхности, а вершины их направ-
лены в какую-либо сторону, так что волосяной покров более или
менее прилегает к коже.
Шерстный покров состоит из волос различного типа. Основ-
ные из них — это пуховые волосы, или подшерсток, остевые воло-
сы, или ость, чувствующие волосы, или вибриссы.
Основу шерстного покрова у большинства видов млекопитаю-
щих составляет подшерсток. Между пуховыми сидят жесткие осте-
52
д
Рис. 36. Развитие (Л, Б, В, Г) и строение волоса (Д £):
1 — эпидермис; 2 — кориум (дерма); 3 — зачаток влагалища волоса; 4 — зачаток
сальной железы; 5 — дермальный сосочек; б — зачаток волоса; 7 — сальная
железа; 8 — мускул волоса; 9 — сердцевина волоса; 10 — корковый слой; 11 —
кожица волоса; 12 — волосяное влагалище; 13 — волосяной мешок; 14 — его
соединительнотканная оболочка; 15 — иннервация волоса
вые волосы. Именно они создают цветность шерстного покрова.
V ведущих подземный образ жизни кротов, слепышей, цокоров
остевые волосы вовсе отсутствуют. У многих копытных, ластоно-
гих подшерсток имеется у детенышей, а затем он сменяется жес-
ткими остевыми волосами (например, у северного оленя, котика
и др.). Направленность шерстного покрова создается именно осте-
выми волосами. Располагаясь спереди назад и сверху вниз, они
защищают тело от быстрого намокания.
53
Шерстный покров периодически меняется: звери линяют. Бел-
ки, песцы, лисицы, кроты сменяют мех дважды в год. Другие же
виды линяют только один раз в год: начав менять шерстный по-
кров весной, они полноценно обрастают новой шерстью к осени
(суслики). У человека волосы растут всю жизнь, прирастая в день
в среднем на 0,35 мм.
Шерстный покров, как и перья, образование полифункцио-
нальное. Большинство видов млекопитающих окрашено невзрач-
но, это скрывает их в местах обитания. В редких случаях окрас и
длина шерстного покрова отражает половой диморфизм (у неко-
торых хищных, приматов). Велико значение шерстного покрова
в термоизоляции тела. У северных видов его густота и высота
существенно меняются по сезонам. Исследования показали, что
у обыкновенной белки на 1 см2 задней части спины летом быва-
ет в среднем 4200 волос, а зимой 8100, у зайца-беляка соответ-
ственно 8000 и 14 700. Длина волос тоже изменчива: например, у
тех же обыкновенной белки и зайца-беляка к зиме они удлиня-
ются в 1,5 — 2 раза. У тропических животных в силу малой разни-
цы в температурных условиях зимой и летом таких резких изме-
нений шерстного покрова не бывает.
Особую категорию волос представляют вибриссы: очень длин-
ные жесткие волосы, выполняющие осязательную функцию. Они
сидят чаще на голове (так называемые «усы»), на нижней части
шеи, на груди, а у некоторых лазающих древесных форм (напри-
мер, у белки) и на брюхе. В основании волосяного мешка чувству-
ющего волоса и в его стенках располагаются мякотные нервные
волокна, воспринимающие соприкосновение стержня вибриссы
с посторонними предметами. Вибриссы снабжены поперечнопо-
лосатой мускулатурой и иннервируются на голове ветвями трой-
ничного нерва.
Видоизменениями волос являются щетина, иглы. Роговым про-
изводным эпидермиса у китообразных являются пластины кито-
вого уса. Например, в ротовой полости гренландского кита выра-
стает до 350—400 роговых пластин разной величины. Наибольшие
пластины имеют длину 4,5 м. Внутренние расщепленные края,
смыкаясь друг с другом в плотное сито, обеспечивают фильтра-
цию пищи.
Кожные железы в эпидермисе позвоночных многочисленны и
разнообразны по своим функциям.
Распространенным типом желез у первично-водных позвоноч-
ных — миног, миксин, хрящевых и костных рыб — являются
многочисленные бокаловидные железистые клетки, выделяющие
на поверхность тела слизь (см. рис. 18, 19). Она уменьшает трение
тела при плавании, очищает его поверхность, коагулирует ино-
родные частицы, содержит бактерицидные вещества и обеззара-
живает тело; создавая водную пленку, она участвует в кожном
54
Рис. 37. Светящиеся органы костистой рыбы
(по А. Ромеру, Т. Парсонсу, 2002):
А — орган рыбы Cyclothone в разрезе; Б — размещение органов на теле рыбы
Myctophunr, 1 — пигментный слой; 2 — рефлектор; 3 — железистая ткань; 4 —
линза; 5 — проток железы; 6 — эпидермис
дыхании животного. У миксин эпидермис почти на половину
толщины состоит из железистых клеток. При содержании мик-
син в сосуде обильные слизистые выделения кожных желез быс-
тро превращают воду в слизь. Бокаловидные клетки есть и в эпи-
дермисе ланцетника.
Модификацией слизистых желез считают светящиеся органы
некоторых глубоководных рыб (фотофоры): свечение возникает в
результате сложных процессов окисления в железистой ткани.
У других рыб свечение обусловлено находящимися в органе спе-
цифическими светящимися бактериями (рис. 37).
Одноклеточные железистые клетки закладываются в эпидер-
мисе и у личинок земноводных, однако у взрослых амфибий кож-
ные железы всегда многоклеточные (рис. 38). Участок эпидермиса
погружается в соединительную ткань, и железа приобретает ша-
ровидную форму. Эти железы поддерживают кожу амфибии во
влажном состоянии, что важно, в частности, для газообмена че-
рез водную пленку. Слизь обеззараживает покровы, так как содер-
жит бактерицидное вещество лизоцим.
У некоторых амфибий кожные железы видоизменяются в ядо-
витые, выделяющие зернистый секрет, и защищают их от врагов.
Значительные скопления таких желез — паротид образуются час-
то по бокам головы жаб, саламандр. Их секрет может быть чрезвы-
чайно ядовит (например, яд кураре, содержащийся в выделениях
жабы аги). Многие ядовитые земноводные ярко окрашены в крас-
Hbie, желтые, синие тона. Таким способом они предупреждают о
собственной ядовитости своих врагов.
Ядовитые железы встречаются и в коже рыб. У некоторых хря-
Щевых (скатов, акул), у отдельных костистых рыб они локализо-
55
Рис. 38. Разрез через кожу саламандры:
1 — выступающий наружу секрет железы; 2 — пигментный слой; 3 — слизистые
кожные железы; 4 — ядовитая кожная железа; 5 — перерезанные кровеносные
сосуды; 6 — эпидермис; 7 — волокнистый слой кожи
ваны вблизи шипов хвоста, жаберной крышки, у оснований плав-
ников. Таковы, к примеру, скаты-хвостоколы.
Сухая кожа рептилий лишена желез, однако у крокодилов,
черепах есть пахучие железы. Они располагаются по бокам нижней
челюсти, на спине, в области клоаки и используются, видимо,
при внутривидовых взаимоотношениях. Зачатки таких желез есть у
змей. В связи с железами всегда обсуждаются бедренные поры
ящериц, выделяющие вязкий секрет, напоминающий массу оро-
говевших клеток. Их секреция усиливается в сезон размножения.
Эти выделения остаются малоизученными.
У птиц, как и у рептилий, кожа сухая, безжелезистая. Един-
ственная парная копчиковая железа располагается над корнем хво-
ста и открывается парой отверстий на коже. Эта железа имеет слож-
ное трубчатое строение и выделяет маслянистый секрет. Птица с
помощью клюва смазывает оперение, что придает ему водонеп-
роницаемость и снабжает витамином А.
Млекопитающие обилием кожных желез близки к амфибиям.
По форме различают трубчатые и альвеолярные железы, по функ-
циям выделяют потовые, млечные, сальные, пахучие.
Потовые железы имеют вид длинных трубок, иногда мешко-
видных, чаще закрученных в клубок (рис. 39). Они открываются на
поверхности кожи или в волосяные сумки. Секрет потовой желе-
зы — жидкий или вязкий, слизистый, часто сильно пахнущий —
выделяется железистыми клетками ее стенки. Пот чаще бесцвет-
ный, но у бегемота он красноватый, у африканской антилопы
голубой, у большого кенгуру пот подкрашивает шерсть в рыжий
цвет, у лошади он содержит белок, создающий на теле пену.
Лазающим по деревьям обезьянам секрет, выделяемый на го-
лых участках конечностей, позволяет лучше удерживаться на де-
56
Рис. 39. Схема кожной железы
амфибии (Л) и потовых желез
млекопитающего (Б, В):
1 — ороговевающий слой эпидермиса;
2 — эпидермис; 3 — мальпигиев слой
эпидермиса; 4 — мускульный покров
железы
ревьях. У многих хищников потовые железы расположены на по-
душечках лап, у грызунов — на лапах, губах, в паховой области.
Через потовые железы выводится вода, а вместе с жидкостью
из тела выносятся соли (NaCl) и мочевина. Таким образом, пото-
вые железы при тепловых нагрузках и необходимости терморегу-
ляции участвуют в охлаждении тела зверя. Они осуществляют вы-
деление, вместе с пахучими железами маркируют след и террито-
рию животного. Считают, что видоизменением потовых желез яв-
ляются церуминозные железы, выделяющие в ушах серу (хотя их
происхождение не совсем ясно).
Млечные железы являются видоизменением потовых желез (рис.
40). У низших, клоачных, млекопитающих они сохраняют трубчатое
строение и открываются в волосяные сумки, расположенные на уча-
стках брюшной поверхности тела самки — так называемых железис-
тых полях. Молоко стекает по волосам и слизывается детенышем.
У сумчатых и плацентарных млечные железы имеют гроздевид-
ное строение, и их протоки открываются на сосках. Развитие мо-
лочных желез начинается с появления на брюхе эмбрионов про-
дольных вздутий — млечных линий. Впоследствии они группиру-
ются в соски. Различают два типа сосков: истинные (у большин-
ства сумчатых, полуобезьян, обезьян и человека) и ложные (у
хищных, копытных). В первом случае на вершине соска открыва-
ются выходы нескольких выводных протоков желез, во втором —
на соске открывается один общий канал, в который молоко по-
купает снизу из многих протоков.
Млечные железы и соски на теле зверя располагаются различ-
Но- У лазающих обезьян и висящих во время кормления летучих
57
Рис. 40. Развитие потовой (Л) и млечной (£) железы млекопитающего:
1 — эпидермис; 2 — кориум
мышей имеется одна пара сосков на груди, у бегающих копытных
соски приурочены только к паховой области. У насекомоядных и
хищных соски располагаются двумя рядами по всей нижней по-
верхности туловища. Число сосков состоит в прямой связи с разо-
вой плодовитостью самок и в некоторой степени соответствует
числу одновременно рождаемых детенышей. Минимальное число
сосков (два) характерно для обезьян, овец, коз, слонов и др. У ви-
дов с многочисленным потомством — грызунов, насекомоядных,
многих хищных, у свиней, некоторых сумчатых количество со-
сков варьирует от 10 до 24 и располагаются они на брюхе в два
длинных ряда. Любопытно расположены соски у нутрии. Они раз-
двинуты по бокам тела так, что самка может кормить, лежа в
грязи.
Продукт млечных желез — молоко, питательная жидкость, со-
держащая казеин, лактозу, жиры, соли. Насыщенность молока
питательными веществами у разных млекопитающих может суще-
ственно различаться. В качестве примера ниже приведен химичес-
кий состав молока коровы и обыкновенного дельфина (%). Обра-1
Животное Жиры Белки Лактоза Вода
Корова 3 3,0 8,0 86,0
Дельфин 51,2 Н,5 1,6 35,3
щает на себя внимание насыщенность молока дельфина жирами И
белками:
Сальные железы — альвеолярные. Они имеют гроздевидную
форму и открываются почти всегда в волосяную сумку. Жирный
секрет этих желез смазывает волос и поверхностный слой эпидер'
58
миса кожи, предохраняя их от намокания и снашивания. Сальные
железы отсутствуют у китообразных.
Пахучие железы — видоизмененные потовые или сальные же-
лезы, иногда образующие комбинацию тех и других. Пахучие же-
лезы располагаются в разных участках тела, часто в анальной и
паховой областях. Из их числа укажем на анальные железы куньих,
секрет которых имеет очень резкий запах и служит в основном для
защиты от преследующих зверя врагов. Особенно сильно эти же-
лезы развиты у американских скунсов, или вонючек {Mephitis),
способных выпрыскивать большие порции резко, пахнущих выде-
лений на значительное расстояние. Мускусные железы имеются у
кабарги, выхухоли, бобра, ондатры. Известны подглазничная же-
леза оленей, хвостовая — северного оленя, тарзальная — черно-
хвостатого оленя, спинная — даманов, средобрюшная — песчанок,
подбородочная — кроликов и т.д. (В. Е. Соколов, 1975).
В целом значение кожных желез в теле млекопитающих чрез-
вычайно велико. Кроме чисто физиологических свойств, они уча-
ствуют в разных сторонах этологии животных: в распознавании
особей своего вида, в опознании врага и жертвы, половом пове-
дении, мечении территории, наконец, в защите от врагов.
* * *
Таким образом, наружные кожные покровы хордовых, образу-
ющиеся из эктодермы и энтодермы, полифункциональны. Вместе
со скелетом и мышцами кожа придает определенную форму те-
лам животных. Она защищает организм животного от механичес-
ких повреждений, от вредных веществ и от болезнетворных мик-
роорганизмов, участвует в газовом и водном балансе организмов,
выведении продуктов метаболизма и терморегуляции. Кожа вы-
полняет сенсорную функцию. Окраска покровов имеет защитное
и сигнальное значение.
Примитивные покровы ланцетника состоят из одного слоя кле-
ток эпидермиса и тонкого слоя кутиса. В процессе эволюции эпи-
дермис становится многослойным. Его наружный ороговевающий
слой периодически сменяется. Известны многочисленные произ-
водные эпидермиса — роговые щитки и панцири рептилий, пе-
рья птиц, шерсть, рога и копыта зверей; производные толстого
слоя кутиса — многочисленные накладные костные образования.
У млекопитающих в кутисе формируется значительный жировой
слой, служащий запасом питательных веществ и надежной защи-
той от низких температур.
В процессе эволюции возрастает специализация кожных желез:
вначале это слизистые и увлажняющие покровы, в дальнейшем —
ядовитые, жировые, потовые, млечные, пахучие.
ОПОРНО-ДВИГАТЕЛЬНАЯ СИСТЕМА
Одна из ведущих черт морфофункциональной организации хор-
довых — непременное формирование внутреннего скелета. Он зак-
ладывается на ранних этапах их онто- и филогенеза в виде спинной
струны, или хорды.
На более поздних стадиях (онто- и филогенеза) основу осевого
скелета позвоночных животных составляют позвоночник и мозго-
вой череп. Производные соединительной ткани образованы хря-
щем или костью, в которых накапливаются соли кальция. Хорда
же сохраняется в относительно редуцированном состоянии. В со-
став опорно-двигательной системы кроме внутреннего скелета
входит и соматическая мускулатура.
Хорда, позвоночник
Хорда представляет собой нечленистый веретеновидный тяж,
имеющий внутри относительно мягкое студенистое содержи-
мое, образованное сильно вакуолизированными клетками. С по-
Рис. 41. Участок поперечного
разреза через хорду личинки
миноги:
1 — ткань хорды; 2 — волокнис-
тая оболочка; 3 — наружная эла-
стичная оболочка; 4 — соедини-
тельная ткань
верхности она ограничена волокни-
стой оболочкой, наружной мемб-
раной и сложенными кольцевыми
волокнами. Они придают спинной
струне эластичность и способству-
ют легкому ее изгибанию (рис. 41).
В раннем эмбриогенезе хорда зак-
ладывается энтодермально как вы-
рост первичной кишки (или спин-
ной губы бластопора). Продвигаясь
вперед, она размещается под не-
рвной трубкой.
Эта осевая структура сложилась
исторически на ранних этапах фило-
генеза типа хордовых. Как пожизнен-
ная основа осевого скелета хорда со-
хранилась у современных бесчереп-
ных, круглоротых, цельноголовых
хрящевых рыб, двоякодышащих рыб,
60
у кистеперой рыбы (латимерии)
и хряшевьи ганоидов (осетровых
й веслоносов)* (рис. 42).
Эмбрионально хорда заклады-
вается у всех позвоночных. Ее пе-
редний конец доходит до гипо-
физарной воронки, а задний —
до основания мясистой лопасти
хвоста. У взрослых форм хорда
или сохраняется пожизненно (см.
выше), или присутствует в реду-
цированном виде, замещаясь те-
лами позвонков. У низших позво-
ночных хорда имеет четковидную
Рис. 42. Хорда и позвоночник
осетровой рыбы:
1 — хорда; 2 — верхние дуги; 3 —
нижние дуги; 4 — вставочные хря-
щи; 5 — остистые отростки
форму, утончаясь внутри тел позвонков и расширяясь между их
соседними телами. У большинства тетрапод (включая человека)
во взрослом состоянии хорда представлена только студенистым
содержимым хрящевых межпозвонковых дисков.
Осевой скелет позвоночных сложен главным образом скелето-
генной тканью, закладывающейся в раннем эмбриогенезе в скле-
ротомах сомитов. С простейшим выражением скелетогенной ме-
зенхимы мы встречаемся у ланцетника, у которого соединитель-
нотканный чехол покрывает с поверхности хорду и нервную трубку
и в виде миосепт проникает между мышечными сегментами. Внут-
ренние скелетные образования складываются в соматический ком-
плекс.
В теле позвоночного хорду замещает позвоночник, образуя его
основную ось. Таким образом, эластичность и гибкость первичной
оси — хорды — усиливаются прочностью возникающих позже ске-
летных структур. Кроме того, элементы позвоночника, разраста-
ясь от хорды вверх, образуют невральный канал и защищают спин-
ной мозг, а снизу создают защиту крупным кровеносным сосудам
в виде гемального канала.
Позвоночник в отличие от хорды — структура, более прочная
и сегментированная. Он состоит из позвонков и подразделяется
на отделы. У рыб выделяют туловищный и хвостовой отделы. У на-
земных позвоночных туловищная часть дифференцируется далее
на шейный, грудной, поясничный и крестцовый.
Структурной единицей осевого скелета является позвонок (рис. 43).
Число позвонков, как правило, соответствует числу эмбриональ-
но закладывающихся сомитов. В оформленном виде позвонок имеет
^ело и две дуги: верхнюю (невральную) и нижнюю (гемальную).
Невральные дуги сомкнуты по всей длине позвоночника, гемаль-
* У оболочников хорда закладывается в эмбриогенезе, а затем редуцируется.
61
Рис. 43. Позвоночник костистой рыбы в сагиттальном разрезе (В).
Отдельные позвонки из туловищной (Л) и хвостовой (Б) областей:
1 — верхние дуги; 2 — ге.мальные дуги; 3 — ребра; 4 — тела позвонков; 5 ~
сочленовные отростки; 6 — боковые отростки; 7 — верхний и 8 — нижние ости-
стые отростки
ные — только в хвостовой его части, образуя гемальный канал.
Позвонки разных отделов позвоночника имеют заметные разли-
чия. У большинства позвоночных к нижней части туловищных по-
звонков примыкают ребра.
Исторически закладка позвоночника начиналась с дуг позвон-
ков. Усиление соматического комплекса среди современных по-
звоночных обнаруживается впервые у круглоротых: их миосепты
укреплены коллагеновыми волокнами. В туловищных сегментах тела
миног вблизи хорды есть маленькие парные хрящи — зачатки не-
вральных дуг (рис. 44). У миксин их находят только в хвостовой
части тела. У хрящевых рыб (акул) парные верхние хрящи смыка-
ются в невральные (верхние) дуги и образуют спинномозговой
канал. Нижние пары хрящей смыкаются у них в хвостовой части,
образуя гемальный канал.
Рис. 44. Последовательные стадии развития (1—Ш) туловищного
позвонка акулы (поперечные разрезы, по И. И. Шмальгаузену):
Z — верхняя дуга; 2 — нижняя дуга; 3 — тело позвонка; 4 — поперечный отростой
5 — ребро; 6 — остистый отросток; 7 — хорда
62
рис. 45. Различные типы позвонков у позвоночных (схемы сагиттальных
разрезов; передние концы обращены влево):
— примитивный амфицельный; Б — опистоцельный; В — процельный; Г —
ацельный (предшественник платицельного)
Тела позвонков по форме весьма разнообразны. У круглоротых роль
осевого скелета выполняет хорда и тела позвонков отсутствуют.
У акул эти тела имеют вид двояковогнутых коротких цилинд-
ров (амфицельные) (рис. 45). Они образованы концентрическими
слоями хряща вокруг хорды внутри ее волокнистой оболочки. Клет-
ки скелетогенной мезенхимы проникают через поры в наружной
мембране хорды и вызывают обызвествление волокнистой обо-
лочки. Так формируется одна — внутренняя — часть тела позвонка.
Другая часть образуется путем разрастания оснований верхних и
нижних дуг.
У двоякодышащих рыб, латимерий и осетровых тела позвонков
образованы только хрящевыми основаниями дуг, поэтому хорда со-
храняется как основная ось тела в виде толстого тяжа без перехватов.
Тела позвонков костистых рыб, тоже амфицельные по форме,
образованы костной тканью, заместившей хрящ. Между собой по-
звонки соединяются через студенистую подушку — остаток хорды.
Среди тетрапод амфицельные позвонки есть у примитивных
хвостатых амфибий, некоторых ящериц, у гаттерий.
У большинства наземных позвоночных и некоторых рыб между
позвонками развивается в виде дисков замещающий хорду хрящ.
Внутри дисков заключено желеобразное ядро — остаток хорды.
Суставная щель между позвонками ориентирована по-разному.
В опистоцелъных позвонках она выпуклая, причем выпуклость на-
правлена вперед (высшие хвостатые амфибии, панцирная щука),
в процельных позвонках (бесхвостые амфибии, высшие рептилии)
выпуклость обращена назад. Тела гетероцельных позвонков птиц
имеют сложную седловидную форму. Наконец, платицельные по-
звонки млекопитающих имеют плоские сочленяющиеся поверх-
ности. Их межпозвонковые диски состоят в основном из кольце-
видного волокнистого хряща, придавая позвонкам взаимную под-
вижность и гибкость.
В туловищной части позвоночника у всех костистых рыб и на-
земных позвоночных развиваются ребра (см. рис. 48). Туловищные
позвонки костистых рыб несут ребра от затылочной кости черепа
4о основания хвоста.
63
Различают верхние и нижние ребра. Их структура и эмбриональ-
ная закладка неоднозначно объясняется специалистами. Согласно
одним авторам (А. Ромер, Т. Парсонс, 1992), верхние ребра раз-
виваются из мезенхимы склеротомов, расположенной рядом с
телами позвонков, а нижние — из мезодермы боковой пластин-
ки. Их считают гомологами гемальных дуг. По мнению других
(Ф.Я. Дзержинский, 1998), нижние ребра формируются в эмбрио-
генезе из хряща вблизи тел позвонков и растут центробежно от
позвоночника к периферии. Верхние же не имеют хрящевого пред-
шественника. Они формируются в покровных тканях и растут от
периферии к центру (центростремительно).
К позвонку ребро прикрепляется головкой. У наземных позво-
ночных головка раздвоена. Один выступ — истинная головка —
прикреплен к нижней дуге или непосредственно к телу позвонка,
другой — бугорок — соединен суставом с основанием верхней
дуги (поперечным отростком).
Круглоротые ребер не имеют. Считают, что короткие ребра акул
принадлежат к нижнему ряду. У многоперов и большинства кос-
тистых рыб есть и верхние, и нижние ребра. Они укрепляют мус-
кулатуру, ограничивающую полость тела. Некоторые костистые
рыбы имеют только нижние ребра. Ребра тетрапод, вероятно, со-
ответствуют верхним ребрам рыб (А. Ромер, Т. Парсонс, 1992). По-
лагают, что у предков тетрапод ребра присутствовали на всех по-
звонках от головы до основания хвоста.
У современных наземных позвоночных ребра развиты по-раз-
ному. Шейные ребра у рептилий укорочены, у птиц и млекопита-
ющих они прирастают к позвонкам. Ребра грудно-
«gb го отдела у современных бесхвостых амфибий силь-
но редуцированы и даже могут отсутствовать вов-
се- У хвостатых они не достигают грудины и раз-
двоены на дистальном конце (рис. 46). У безногих
ребра развиты хорошо.
.sin Ы У большинства настоящих наземных животных
ребра утолщенные и дугообразно изогнутые. Они
обеспечивают опору туловища на плечевой пояс.
У рептилий, птиц и млекопитающих грудные реб-
Ра связаны с грудиной и образуют с ней грудную
V |М\клетку. Грудная клетка защищает внутренние орга-
'If ны и участвует в механизме легочного дыхания.
Рис. 46. Позвоночник бесхвостой амфибии:
/ — шейный позвонок; 2 — поперечные отростки туловииг
ных позвонков; 3 — поперечные отростки хвостовой облас-
ти; 4 — ребра; 5 — крестцовый позвонок; 6 — уростиль
64
у рептилий (ящериц, крокодилов) груд-
ные ребра обычно имеют два сегмента —
проксимальный (верхний) окостеневший
н дистальный (грудинный) хрящевой. Под-
вижный сустав между этими сегментами
позволяет грудной клетке расширяться и
сжиматься. У змей грудная клетка отсутству-
ет вторично. Грудные ребра у черепах (8
пар) прочно сращены с прилегающими к
ним сверху пластинами карапакса.
Грудные ребра крокодилов, клювоголо-
вых и птиц имеют крючковидные отрост-
ки для прикрепления мышц (рис. 47). Это
создает (особенно птицам) высокую эф-
фективность расширения грудной клетки
при вдохе.
У млекопитающих 10 пар истинных ре-
бер соединены с грудиной посредством ре-
берных хрящей. Лежащие за ними 2—3 пары
«ложных ребер» с грудиной не связаны.
Поперечные отростки поясничных позвонков закладываются
эмбрионально как ребра. В хвостовом отделе ребра отсутствуют.
Отделы позвоночника. Как сказано выше, в позвоночнике рыб
выделяют два отдела — туловищный и хвостовой (рис. 48). Голова и
Рис. 47. Грудные ребра
птиц:
1 — лопатка; 2 — позво-
ночник; 3 — ребра; 4 —
грудина; 5 — киль; 6 —
коракоид; 7 — ключица;
8 — плечо
туловище у них взаимно неподвижны, что достигается соответ-
ствующим соединением позвоночника и мозгового черепа. Назем -
Рис. 48. Скелет костистой рыбы (окуня):
— кости черепа; 2 — основные элементы спинного плавника; 3, 4 — лучи
спинного плавника; 5 — уростиль; 6 — хвостовые позвонки; 7 — основные эле-
менты анального плавника; 8 — туловищные позвонки; 9 — ребра с придатками;
Ю — кости и лучи брюшного плавника; 11 — кости и лучи грудного плавника;
12 — жаберная крышка; 13 — верхняя и нижняя челюсти
Константинов
65
I
Рис. 49. Отделы позвоночника млекопитающего:
1 — шейный; 2 — грудной; 3 — поясничный; 4 — крестцовый; 5 — хвостовой
ный образ жизни потребовал большой подвижности головы и уси-
ления опоры парным конечностям. В этой связи у наземных позво-
ночных в туловищной области выделяются отделы: шейный, груд-
ной, поясничный, крестцовый (рис. 49).
Шейный отдел. Впервые один шейный позвонок выделяется у
амфибий. Двумя затылочными мыщелками на него опирается за-
тылочный отдел черепа. Их сочленение создает небольшую под-
вижность головы вверх-вниз.
У рептилий, птиц и млекопитающих число шейных позвонков
возрастает. У птиц длина шеи меняется заметно, количество по-
звонков варьирует от 11 до 22. Большинство млекопитающих имеет
постоянное число шейных позвонков — 7. Длина шеи у них опреде-
ляется величиной самих позвонков. Такая стабильность шейного
отдела у зверей — следствие большой механической нагрузки на
этот отдел позвоночника (тяжелая голова, опора плечевого пояса
через мышцы), которая и обусловила скоординированность мор-
фогенеза ее конструкции (Ф.Я. Дзержинский, 1998). Есть и исклю-
чения. У ленивцев число шейных позвонков варьирует от 6 до 10, у
ламантина их 6. Их голову поддерживает сильная мускулатура. Под-
вижность головы очень важна для хищников, что обусловлено
способом добывания пищи. Наоборот, у грызунов, норников и
особенно у землероев шейные позвонки укорочены, а подвиж-
ность шеи ограничена. Наиболее мощный шейный отдел с боль-
шими остистыми отростками характерен для копытных.
Подвижность головы улучшается также благодаря выделению в
шейном отделе у настоящих наземных позвоночных двух первых'
позвонков: атланта и эпистрофея (рис. 50). Невральная дуга ат-
ланта образует кольцо с суставной поверхностью, а его тело при-
растает в виде отростка к телу эпистрофея. У рептилий и птицс
атлантом сочленяется суставом один затылочный мыщелок чере-
па, что обеспечивает высокую вращательную подвижность голо-
вы. У птиц наблюдается угол поворота головы, наибольший среди
позвоночных (у сов, например, до 270 °). У млекопитающих и птиП
66
Рис. 50. Атлант (/) и эпистрофей (II) гуся (А) и носорога (Б):
1 — сочленовная ямка для мыщелка черепа; 2 — зубовидный отросток
1
на теле эпистрофея имеется зубовидный отросток. При эмбрио-
нальной закладке он возникает за счет прирастания тела атланта
к телу эпистрофея. При двух затылочных мыщелках у зверей через
зубовидный отросток проходит ось вращения.
Грудной отдел позвоночника у хвостатых амфибий не отделя-
ется от поясничного и имеет ребра. У бесхвостых они отсутствуют.
Число пояснично-грудных позвонков у рептилий 22. Все они не-
сут ребра, но только ребра первых пяти из них присоединены к
грудине. Грудные позвонки у птиц срастаются в единую кость.
У млекопитающих грудной отдел включает чаще 12—15 по-
звонков, однако возможны отклонения. К примеру, самое малое
количество позвонков — 9 — обнаружено у одного из видов бро-
неносцев и клюворылого кита. Максимальное число грудных по-
звонков — 24 — у ленивца (р. Choloepus).
К передним грудным позвонкам (обычно к семи) у зверей при-
членяются ребра, соединенные с грудиной (истинные ребра). Ос-
тальные грудные позвонки несут ребра, не доходящие до грудины
(ложные ребра). Грудина представляет сегментированную костную
пластинку, заканчивающуюся удлиненным хрящом — мечевидным
отростком. Расширенный передний сегмент носит название руко-
ятки грудины. У летучих мышей и у зверей, конечности которых
хорошо приспособлены для рытья (кроты), грудина теряет ясно
выраженную сегментацию и несет киль, служащий, как и у птиц,
ВДя прикрепления сильных грудных мышц.
Позвонки поясничного отдела рептилий имеют ребра. У птиц,
срастаясь, они входят в состав сложного крестца. У млекопитаю-
щих этот отдел позвоночника имеет рудиментарные ребра и со-
стоит из 2 — 9 позвонков.
67
Крестцовый отдел впервые выделяется у амфибий в виде од.
ного. позвонка с короткими ребрами, к которым причленяютсц
кости тазового пояса. Рептилии имеют 2 крестцовых позвонка,
птицы и млекопитающие — от 3 до 5 (чаще 4). У птиц они сраста-
ются и входят в состав сложного крестца. У млекопитающих срас-
таются в крестцовую кость. При этом только первые два позвонка
истинно крестцовые, а остальные — прирастающие к крестцу хво-
стовые позвонки. У хищных зверей 3 крестцовых позвонка, у ут-
коноса, как и у рептилий, — 2.
Хвостовой отдел. У бесхвостых амфибий хвостовые позвонки
сливаются в единую стержневую кость — уростиль. Хвостатые и
безногие амфибии, рептилии, многие млекопитающие имеют в
хвостовом отделе от трех (у гиббона) до нескольких десятков по-
звонков (49 — у длиннохвостого ящера). Концевые позвонки в
хвосте птиц сливаются в единую копчиковую кость — пигостиль —
место прикрепления рулевых перьев.
Хвостовой отдел позвоночника некоторых современных яще-
риц, гаттерий, некоторых древних рептилий имеет свои особен-
ности. По середине тела отдельных хвостовых позвонков есть нео-
костеневающая прослойка, разделяющая каждый позвонок на
переднюю и заднюю части. По этой прослойке позвонки могут
разрываться, и хвост отбрасывается усилием окружающих мышц
(аутотомия), но впоследствии он регенерирует. Отрыв хвоста за-
мечен и у некоторых хвостатых амфибий, однако у них разлом
происходит у основания хвоста между соседними позвонками.
Мозговой череп
Мозговой, или осевой, череп (cranium) позвоночных животных
относят к осевому скелету. Мозговой череп возникает в связи с
развитием головного мозга и органов чувств как их вместилище и
защитное образование.
В эмбриогенезе эта часть осевого скелета закладывается разны-
ми путями.
Первичная структура мозгового черепа наиболее демонстра-
тивна при его эмбриональной закладке у хрящевых рыб. По бокам
хорды под передним концом нервной трубки образуются парные
хрящи — парахордалии, трабекулы и затылочные хрящи. Они со-
ставляют основу дна черепа. С боков и спереди к дну примыкают
хрящевые капсулы органов чувств — обонятельные, глазные й
слуховые (рис. 51).
Отделы мозгового черепа формируются путем смешанного со-
четания элементов. Парахордалии и затылочные хрящи заклады-
ваются из ткани, общей с позвоночником, — мезенхимы склеро-
томов, трабекулы и капсулы органов чувств — из скелетогенной
68
Рис. 51. Расположение хрящевых закладок
мозгового черепа акулы:
у __ трабекулы; 2 — парахордалии; 3 — слуховая
капсула; 4 — глазница; 5 — обонятельная капсула;
6 — хорда; 7 — закладка позвонков
ткани кожного происхождения, которую
продуцирует нервный гребень (Ф. Я. Дзер-
жинский, 1998).
Закладка мозгового черепа издавна
привлекала внимание специалистов. «Про-
блема головы» известна с трудов И. Гёте,
Л.Окена, Р. Оуэна (конец XVIII —начало
XIX в.) (Я.Д.Бляхер, 1970). В обособлении
и развитии мозгового черепа они уловили
сходство с закладкой позвоночника. В ос-
нову дна черепа были положены сначала 6, затем 3 первых по-
звонка. Более поздние эмбриологические исследования подтверди-
ли сегментарную закладку первичного черепа. Как факт, подтверж-
дающий близость черепа позвоночнику, приводится возможная го-
мология черепномозговых нервов всех позвоночных спинномозго-
вым корешкам круглоротых (Ф. Я. Дзержинский, 1998).
В процессе поступательной эволюции позвоночных структура
мозгового черепа претерпела серьезные изменения разного плана.
Основу скелетной ткани первичного мозгового черепа составил
хрящ, дополнявшийся покровными и хондральными окостене-
ниями. В итоге мозговой отдел вошел составной частью в общий
комплекс черепа, включающий и висцеральный скелет. У высших
позвоночных в черепе различают его мозговой и лицевой отделы.
У позвоночных выделяют два типа строения мозгового черепа:
платибазапъный и тропибазалъный. Платибазальный череп имеет
широкое основание. Глазницы широко раздвинуты, мозговая по-
лость продолжается между ними. Такой тип черепа характерен для
хрящевых рыб, круглоротых, амфибий. Настоящие наземные по-
звоночные имеют тропибазальный тип черепа: глазницы в нем
сближены и часто разделены лишь межглазничной перегородкой.
Мозговая полость при этом оттеснена назад. О различии в органи-
зации двух типов мозгового черепа можно судить по расположе-
нию в них головного мозга (рис. 52).
Рассмотрим особенности строения мозгового черепа последо-
Вательно по группам животных.
М круглоротых и хрящевых рыб дно черепа закла-
дывается в виде парных хрящей — парахордалий и трабекул.
69
Рис. 52. Положение мозга в
платибазальном (А) и тропиба-
зальном (Б) черепах
У ним примыкают капсулы орга-
нов чувств: непарная обонятель-
ная, парные глазничные и слухо-
вые (рис. 53). У хрящевых рыб хря-
щевой мозговой череп приобретает
вид оформленной коробки, так
как капсулы органов чувств, при-
растая к дну черепа, образуют
бока черепа и частично форми-
руют крышу. Крыша никогда не
бывает полной, в ней присутству-
ет чаще одно (иногда больше) от-
верстие — фонтанелъ. Эта непол-
нота хондральной крыши прояв-
ляется в онтогенезе большинства
других позвоночных.
У взрослых акул различают четыре основных отдела мозгово-
го черепа: затылочный, слуховой, глазничный и обонятельный.
В затылочном отделе имеется большое затылочное отверстие, че-
рез которое проходит спинной мозг. Под ним располагается за-
тылочный мыщелок, который смыкается с первым позвонком. Спе-
реди от затылочного отдела мозговая коробка расширяется, об-
разуя слуховые капсулы, окружающие мешочки и каналы внут-
реннего уха. Затем спереди мозговая коробка сужается, образуя
глазничные впадины: в них расположены глазные яблоки и их
мускулатура. Здесь, в дне мозговой коробки, имеется срединное
углубление, к которому прилегает гипофиз. Далее, кпереди моз-
говая коробка расширяется в обонятельный отдел. Здесь находят-
ся парные обонятельные {носовые) капсулы, которые окружают
обонятельные органы. Заканчивается мозговой череп спереди ры-
лом — рострумом, состоящим из трех хрящей (рис. 54).
Рис. 53. Череп и скелет жаберного аппарата миноги:
1 — хорда; 2 — зачатки верхних дуг позвонков; 3 — обонятельная капсула; 4 "
слуховая капсула; 5 — скелет присасывательной воронки; 6 — подъязычный хряш;
7 — жаберные дужки; 8 — околосердечный хрящ; 9 — подъязычная дуг8
70
в мозговой коробке имеются многочисленные отверстия, через
которые проходят черепные нервы и кровеносные сосуды. В сре-
динное углубление крыши мозговой коробки открываются Кана-
ды из полостей внутреннего уха. Снизу в области коробки распо-
дагаются отверстия для сонных артерий, снабжающих кровью
мозг, а сбоку другие отверстия для артериальных и венозных со-
судов. Через хордальный канал в дне мозговой коробки у взрослых
акул и зародышей других рыб вперед до гипофиза тянется хорда.
У эволюционно прогрессивных костных рыб и у наземных по-
звоночных мозговой череп эмбрионально всегда развивается как
хрящевой, подобно акулам (рис. 55). У осетровых рыб при удале-
нии покровных (кожных) костей череп оказывается хрящевым.
Считают, что в историческом прошлом мозговой череп позво-
ночных был покрыт панцирем из покровных костей. К примеру, у
щитковых голову покрывали костные щитки — крупные у разно-
щитковых (Heterostraca), мелкие у бесщитковых (Anaspida) и те-
лодонтов (Thelodontia). Цельный костный панцирь покрывал го-
лову костнощитковых (Osteostraca). Костные пластины покрывали
голову панцирных рыб (рис. 56). Акантоды имели на поверхности
головы мелкую мозаику ганоидной чешуи. Что касается хрящевых
рыб, то они, видимо, утратили костный покров вторично ради
облегчения тела.
Мозговой череп костных рыб слагается из костей разного
происхождения. Одни — хондральные — замещают хрящ в первич-
ном хрящевом черепе, другие — покровные, формируются в со-
единительнотканном слое кожи. Исторически покровный кост-
Рис. 54. Череп акулы:
1 ~ носовая капсула; 2 — носовой вырост (рострум); 3 — орбита; 4 — затылочная
Капсула; 5 — нёбноквадратный хрящ (верхняя челюсть); 6 — меккелев хрящ
(нижняя челюсть); 7 — губные хрящи; 8 — подвесок (гиомандибуляре); 9 —
гиоид; 10 — жаберные дуги
71
Рис. 55. Эмбриональный хрящевой череп лосося (А — сверху; Б —
снизу):
1 — слуховая капсула; 2 — трабекулы; 3 — парахордалии; 4 — фонтанель; 5 —
меккелев хрящ; 6 — нёбноквадратный хрящ; 7 — гиомандибуляре; 8 — отверстие
блуждающего нерва: 9 — хорда
ный скелет (с учетом сказанного выше о предковых группах) счи-
тают первичным.
Рассмотрим типичный череп лучеперой рыбы на примере су-
дака. Кости хондрального происхождения формируют его заты-
лочную область (основная, две боковые и верхняя затылочные кос-
ти). Они формируют также капсулы органов чувств — обонятель-
ными, клинообонятелъными, клиновидными, крылоклиновидными, гла-
зоклиновидными, ушными костями.
Рис. 56. Панцирная рыба Bothriolepis
72
Покровные кости — носовые, лобные, теменные— закрывают с
поверхности крышу черепа. Они укрепляют дно мозгового черепа —
парасфеноид и сошник. Сбоку около глаза из мелких покровных
косточек формируется окологлазничное кольцо. Ряд костей имеет
смешанную природу: предлобная (или боковая обонятельная), зад-
нелобная (или клиновидноушная), надвисочная (крыловидноуш-
цая). Схема размещения покровных и хондральных окостенений в
черепе костистой рыбы показана на рис. 57.
В ранней эволюции костных рыб, например древних рипидис-
тий, двоякодышащих рыб и палеонисков, в крыше черепа была
заметна тенденция к олигомеризации — замене множества мелких
мозаичных костных элементов крупными костями.
У современных хрящевых ганоидов (осетровых рыб) голова по-
крыта панцирем из покровных щитков, близких к «жучкам», ря-
дами покрывающими тело. С возрастом эти щитки сливаются и
укрупняются (рис. 58).
30 25 26 27 28
Рис. 57. Схема строения черепа костистой рыбы (жаберная крышка и
окологлазничное кольцо удалены, хрящевые кости обозначены
точками):
1 ~ нижняя затылочная кость; 2 — боковая затылочная; 3 — верхняя затылочная;
4 ~~ ушные; 5 — основная клиновидная; 6 — крылоклиновидная; 7 — глазок-
линовидная; 8 — межобонятельная; 9 — боковая обонятельная; 10 — теменная;
— лобная; 12 — носовая; 13 — парасфеноид; 14 — сошник; 15 — нёбная; 16 —
Квадратная; 17 — крыловидные; 18 — межчелюстная; 19 — верхнечелюстная; 20 —
с°членовная; 21 — зубная; 22 — угловая; 23 — гиомандибуляре; 24 — симплекти-
кум; 25—29 — I—V жаберные дуги; 30 — гиоид
73
Рис. 58. Крыша черепа осетра:
1 — ноздри; 2 — носовые кости; 3 — лобные; 4 — глазница; 5 — жаберная
крышка; 6 — теменные кости; 7 — брызгальце; 8 — каналы боковой линии
Заслуживает внимания особенность, отмеченная уже в черепе
древних кистеперых рыб — рипидистий. Мозговая коробка была
разделена у них на две части. Передняя часть была прочно связана
с нёбными и челюстными структурами и включала клиновидный
и обонятельный отделы вплоть до гипофизарной ямки. Задняя часть
объединяла слуховой и затылочный отделы мозговой коробки. Обе
части были подвижно сочленены через связку и мышцу друг с
другом и, кроме того, имели гибкое соединение с помощью хор-
ды, которая проходила в переднюю часть черепа. Такую подвиж-
ность костей черепа называли кинетизмом. Кинетизм обеспечивал
хишным репидистиям большую силу сжатия челюстей. Известно,
что и у наиболее древних амфибий мозговая коробка, по крайней
мере частично, разделялась на два отдела и сохраняла широкий
хордальный канал (А. Ромер, Т. Парсонс, 1992). Как будет рас-
смотрено далее, кинетизм характерен для черепа современных
змей, птиц, однако у них он проявляется в основном в лицевой
части черепа.
У современных амфибий заметна тенденция к относительному
расширению и уплошению мозговой коробки и редукции ее око-
стенений. Мозговой череп амфибийвзначительной части оста-
ется хрящевым, что обусловлено слабым развитием как хондраль-
ных, так и накладных окостенений. У современных амфибий в
сравнении с ранними тетраполами сильнее выражена редукция
крыши черепа. В широком и плоском черепе современных бесхво-
стых и хвостатых амфибий сохранилась лишь малая доля покров-
ной крыши предковых форм (рис. 59).
В мозговой коробке амфибий развиваются следующие хондраль-
ные кости. В затылочной области есть две боковые затылочные кос-
ти. На них заметны два затылочных мыщелка. Места, соответствуй
ющие основной и верхней затылочным костям рыб, у амфибий
остаются хрящевыми. В области слуховой капсулы формируется
ЗА
19
Рис. 59. Схема строения черепа земноводного (А — сбоку; Б — снизу;
хрящевые элементы показаны точками, хондральные кости — черным
цветом, покровные — светлым):
] — межчелюстная кость; 2 — сошник; 3 — носовая полость; 4 — носовая кость;
5— клинообонятельная (поясок); 6 — лобная; 7— теменная; 8 — переднеушная;
9— чешуйчатая; 10 — боковая затылочная; 11 — стремячко; 12— квадратная;
13 — сочленовная; 14 — гиоид; 15 — подъязычная пластинка; 16 — угловая; 17 —
зубная; 18 — нёбная; 19 — верхнечелюстная; 20 — квадратно-скуловая; 21 —
крыловидная; 22 — парасфеноид; 23 — хоаны
одна небольшая переднеушная кость, большая часть капсулы оста-
ется хрящевой. В передней части глазницы мозговой череп сужает-
ся. Здесь у бесхвостых амфибий развивается клинообонятелъная
кость-, у хвостатых эта кость парная. Обонятельная капсула остает-
ся хрящевой.
Покровных костей также немного. Крышу черепа составляют
теменные и лобные кости, которые у бесхвостых срастаются в
лобно-теменные кости. Кпереди от них находятся носовые кос-
ти, у безногих они срастаются с предчелюстными костями. По
бокам задней части черепа расположены чешуйчатые кости, осо-
бенно сильно развитые у безногих. Дно черепа подстилает боль-
шой крестообразный парасфеноид, а впереди от него лежат пар-
ные сошниковые кости. Снизу на черепе видны отверстия для
выхода нервов (зрительного и тройничного). На поверхности
крыши слабо заметными желобками проступают трубчатые ходы
боковой линии. У древних тетрапод поверхность крыши была
структурирована гребнями и ямками, к которым прикрепля-
лась кожа.
Общей особенностью мозгового черепа рептилий является
Почти полное окостенение первичного хрящевого черепа и разви-
тие большого числа кожных костей, формирующих крышу, бока
75
и дно черепа. Рассмотрим скелет мозгового черепа рептилий на
примере крупной ящерицы — варана (рис. 60).
Затылочная область мозгового черепа состоит их четырех кос-
тей хондрального происхождения: верхнезатылочной, основной за-
тылочной и двух боковых затылочных. Все эти кости окаймляют
затылочное отверстие, книзу от которого лежит единственный за-
тылочный мыщелок. В его образовании участвуют обе боковые за-
тылочные и основная затылочная кости. Спереди от основной за-
Рис. 60. Череп ящерицы (А — сбоку; Б — сверху; В — снизу):
1 — теменные кости (Г — отверстие для теменного глаза); 2 — лобные; 3 —
предлобные; 4— надглазничные; 5— заглазничные; 6— носовые; 7— верхнече-
люстные; 8 — межчелюстные; 9 — сошники; 10 — нёбные; 11 — хоаны; 12 —
крыловидные кости (12' — зубы на крыловидных костях); 13 — квадратные; 14 —
поперечные; 15 — основная затылочная; 16 — затылочный мыщелок; 17 — ос-
новная клиновидная; 18 — остаток парасфеноида; 19 — скуловая; 20 — слезная;
21 — столбчатая (надкрыловидная); 22 — чешуйчатая; 23 — надвисочная; 24 —
зубная; 25 — сочленовная; 26— угловая; 27 — надугловая; 28 — венечная кость
76
2
2
Рис. 61. Схемы расположения височных ям в черепах разных типов
рептилий:
/ — анапсидный тип (котилозавры, черепахи); Б — синапсидный тип (зверопо-
добные рептилии); В — эвриапсидный тип (вымершие плезиозавры и др.); Г —
диапсидный тип (клювоголовые; архозавры, а также ящерицы и змеи, утратив-
шие одну или обе височные дуги); 1 — скуловая кость; 2 — теменная; 3 — заглаз-
ничная; 4 — чешуйчатая
тылочной кости лежит основная клиновидная кость, принимаю-
щая участие в образовании дна черепа. У некоторых рептилий —
змей, крокодилов — есть дополнительные боковые клиновидные
кости. Парасфеноид мал и расположен у переднего конца основ-
ной клиновидной кости. Сошниковые кости редуцированы.
В области слуховых капсул развиваются три ушные кости. Одна
из них — переднеушная — пожизненно остается самостоятельной,
две другие срастаются: заднеушная — с боковой затылочной кос-
тью, а верхнеушная — с верхнезатылочной костью.
В обонятельной области хондральные окостенения не развива-
ются и она остается хрящевой.
Крыша черепа образована парными накладными костями: но-
совыми, предлобными, лобными, заднелобными, теменными и не-
парной межтеменной. Последняя имеет в середине отверстие для
теменного органа. Это отверстие есть у ящериц и гаттерий, однако
в черепе змей, черепах и крокодилов оно отсутствует.
Бока черепа слагают многочисленные накладные кости: непар-
ная межчелюстная, парные верхнечелюстные, надглазничные, ску-
ловые, чешуйчатые. Под крышей черепа, ниже височной кости, у
ящериц расположена большая впадина — боковая височная яма.
Многие важные изменения в крыше черепа рептилий связаны
с развитием височных впадин (окон, ям). Для ранних рептилий
(котилозавров, черепах) характерна сплошная крыша без впадин.
Этот примитивный тип называли анапсидным* (рис. 61). Черепа
* Апсидой называют дугу, окаймляющую впадину.
77
современных черепах в сравнении с древними изменились: у многих
видов теменные кости часто образуют глубокие выемки. Иногда
вся височная область превращается во впадину, и элементы кры-
ши от глазницы до заднего края черепа отсутствуют.
У большинства рептилий в процессе эволюции в области вис-
ка и щеки развились одна или две впадины. Этот признак сыг-
рал большую роль в классификации древних рептилий. Нижняя
височная впадина свойственна синапсидным (звероподобные),
верхняя — параноидным и эвриапсидным (ихтиозавры, плезио-
завры). Наконец, в черепе диапсидных образовались две впади-
ны: одна в верхней части крыши, другая — низко, в области
щеки. Горизонтальную костную дугу между ними образуют заг-
лазничная и чешуйчатая кости. Диапсидными были многочис-
ленные динозавры, широко распространенные в мезозойскую
эру. От ранних диапсидных рептилий произошли чешуйчатые (в
частности, ящерицы). Считают, что современные виды ниж-
нюю впадину утратили и у них сохранилась только одна боко-
вая височная впадина.
Мозговой череп птиц по общей схеме строения весьма бли-
зок к черепу рептилий (рис. 62). Затылочный отдел оформлен обыч-
ными четырьмя затылочными костями {основной, двумя боковыми
и верхней). Затылочный мыщелок, как и у рептилий, один. Слухо-
вая капсула закладывается, как у рептилий, из трех ушных кос-
тей, которые у взрослых птиц сливаются в одну.
Дно черепа представлено основной клиновидной и переднеклино-
видной костями, а также нёбными и крыловидными костями. Крышу
черепа формируют парные носовые, лобные, теменные и чешуйча-
тые кости.
В связи с тонкостью костей, ранним и прочным их срастанием
череп птицы значительно облегчен. Швы между костями зараста-
ют рано и незаметны. Объем мозговой коробки и размеры глазниц
существенно увеличены. Глазницы как бы сжали щечный отдел
исходно диапсидного черепа, а перекладины позади глазниц и
между двумя височными впадинами исчезли. Увеличение мозго-
вой коробки произошло за счет разрастания парных элементов
крыши — лобных, теменных, чешуйчатых и комплекса боковых
клиновидных костей.
Череп млекопитающих (рис. 63) также характеризуется от-
носительно более крупной в сравнении с рептилиями мозговой
коробкой, что связано, как и у птиц, с большими размерами
головного мозга. Швы между костными комплексами у млекопи-
тающих зарастают сравнительно поздно, что обеспечивает воз-
можность увеличения объема головного мозга по мере роста жи-
вотного. У молодых зверей мозговая коробка по сравнению с ли-
цевой частью обычно развита относительно сильнее, чем у взрос-
лых. Число отдельных костей в черепе млекопитающего меньше.
78
14
11 10
12
Рис. 62. Строение черепа голубя (А — сбоку; Б — сверху; В — снизу):
1 — основная затылочная кость; 2 — боковая затылочная; 3 — затылочный мы-
щелок; 4 — большое затылочное отверстие; 5 — верхняя затылочная кость; 6 —
ушная; 7— основная клиновидная; 8 — переднеклиновидная; 9 — крылоклино-
видная; 10 — глазоклиновидная; 11 — межглазничная перегородка; 12 — средняя
обонятельная кость; 13 — теменная; 14 — лобная; 75—носовая; 16— слезная; 77—
межчелюстная; 18 — верхнечелюстная; 19 — скуловая; 20 — квадратноскуловая;
27 — чешуйчатая; 22 — квадратная; 23 — сошник; 24 — крыловидная; 25 —
сочленовная; 26 — зубная; 27 — угловая
чем у ранее охарактеризованных групп позвоночных. Это обуслов-
ливается срастанием ряда костей мозгового черепа между собой:
так, в единый комплекс срастаются основная, боковые и верхняя
затылочные кости', сращение ушных костей приводит к образова-
нию единой каменистой кости. Крыловидная кость срастается с ос-
новной клиновидной костью, а глазоклиновидная кость срастается с
вредней клиновидной костью. Бывают случаи образования и более
сложных комплексов. Единая затылочная кость несет два мыщел-
Ка Для сочленения с атлантом.
79
Рис. 63. Череп млекопитающего (А — сбоку; Б — снизу; хондральные
кости темные, покровные — светлые):
1 — межчелюстная кость; 2 — сошник; 3 — носовая перегородка; 4 — носовая
кость; 5 — решетчатая; 6 — слезная; 7 — глазоклиновидная; 8 — лобная; 9 —
крылоклиновидная; 10 — теменная; 11 — межтеменная; 12, 13, 15 — затылоч-
ные; 14— каменистая; 16 — стремя; 17 — наковальня; 18 — барабанная; 19 —
черпаловидный хрящ гортани; 20 — кольца трахеи; 21 — перстневидный хряш
гортани; 22 — щитовидный хрящ гортани; I—IV — остатки висцеральных дуг; 23 —
рожки подъязычной кости; 24—27— разные типы зубов; 28 — верхнечелюстная;
29, 30 — передняя и основная клиновидные; 31 — нёбная; 32 — крыловидная; 33 —
скуловая; 34 — зубная; 35 — чешуйчатая; 36 — молоточек
on
1 !‘
Крыша черепа образована парными носовыми, лобными, те-
менными и непарной межтеменной костями. Такое строение кры-
щй характерно, к примеру, для черепа собаки. Весьма изменчиво
в черепе различных млекопитающих расположение ноздрей. У форм
с гибким рылом (тапиры) или с хоботом (слоны, сирены) на-
ружное отверстие ноздрей бывает сдвинуто по крыше черепа на-
зад- У китообразных такое «дыхало» находится на вершине чере-
па, так как носовая область сместилась назад, а затылочные кости
продвинулись вперед.
Бока черепной коробки образованы чешуйчатыми костями, от
них наружу и вперед отходят скуловые отростки. Последние со-
единяются со скуловой костью, которая, в свою очередь, спереди
сочленена со скуловым отростком верхнечелюстной кости. В итоге
образуется весьма характерная для млекопитающих скуло-
вая дуга.
Парасфеноид в дне мозгового черепа отсутствует. Дно образу-
ют по-разному сочетающиеся основная клиновидная, крылокли-
новидная и крыловидная кости. В отдельных случаях они срастают-
ся в единую клиновидную кость. Переднеклиновидные и глазокли-
новидные кости тоже могут образовывать перед ней единую струк-
туру. У приматов, грызунов и хищников под ними развивается
срединная обонятельная кость, разделяющая две носовые полос-
ти. В пределах носовых полостей развиваются костные или хря-
щевые завитки, либо раковины. Они увеличивают поверхность
обонятельных ходов.
Слуховые капсулы окостеневают несколькими центрами и в
конечном счете в дне черепа формируется каменистая кость.
Кроме того, здесь располагается свойственная только млекопи-
тающим покровная барабанная кость (bula timpany). Она окру-
жает среднее ухо. В слуховой барабан входит наружный слухо-
вой проход.
Весьма примечательна изменчивость общего облика черепа раз-
ных млекопитающих. К примеру, у мелких млекопитающих его
Дорсальная поверхность выглядит вздутой из-за относительно боль-
шого мозга. У крупных видов на облике мозговой коробки отража-
ется развитие височных и шейных мышц, в результате чего на
Поверхности черепа образуются гребни (например, сагиттальный
гребень у хищных, выйный гребень у копытных). Высокая куполооб-
разная крыша черепа слона объясняется в основном не увеличе-
нием мозга, а развитием широкой поверхности для прикрепле-
ния шейных мышц и связок, поддерживающих тяжелую голову с
бивнями и хоботом.
Поверхность черепа сверху и снизу несет множество отверстий —
’пест выхода нервов, кровеносных сосудов, каналов (в том числе
ПоДглазничный канал, слезно-носовой канал, резцовое отверстие
ит.д.)_
81
Висцеральный череп
Висцеральный череп* (скелет) (splanchnocranium) филогене-
тически формировался независимо от мозгового черепа. Первона-
чально у ранних позвоночных висцеральный скелет был представ-
лен многочисленным рядом однообразных опорных элементов
висцеральных дуг, которые располагались в передней части ки-
шечника между жаберными щелями и служили для поддержания
дыхательного аппарата. На последующих этапах эволюции позво-
ночных число дуг сократилось. Сокрашение сопровождалось зна-
чительными преобразованиями висцеральных дуг, сыгравшими
большую роль в морфологической эволюции скелета головы у
позвоночных животных. Элементы передних висцеральных дуг вош-
ли в состав дна мозгового черепа (передние парахордалии).
В эмбриогенезе дуги висцерального скелета образуются из ме-
зенхимы эктодермального происхождения (мезоэктодермы). Эта
ткань образуется из клеток нервного гребня, участвует в образо-
вании жаберных дуг и предположительно (А. Ромер, Т. Парсонс,
1992) покровных костей и некоторой части осевого скелета (см.
«Мозговой череп»).
У круглоротых элементы передних висцеральных дут пре-
образуются в скелет языка, а задние образуют нерасчлененную
решетку, окружающую жаберную область и соединяющуюся с моз-
говым черепом. Вертикальные 9 пар дуг соединены в ней рядами
горизонтальных хрящей (см. рис. 53). Возможно, скелет висцераль-
ных дуг щитковых был слит со скелетом мозгового черепа, у дру-
гих животных он был похож на скелет современных миног. Неко-
торые элементы висцерального скелета у миног специализирова-
ны для поддержания мускулатуры специфического рогового орга-
на скребущего языка, который используется при паразитическом
питании. Сзади к висцеральному скелету примыкает околосердеч-
ная капсула.
Дифференцировка висцерального скелета начинается у рыб
(рис. 64). Прежде всего исчезают связывающие дуги горизонталь-
ные ряды хрящей. Приобретенная дугами свобода и воплощается
в их последующем преобразовании. Жаберный скелет состоит из
членистых дуг в стенке глотки, последовательно расположенных
между жаберными щелями. У акул две первые дуги висцерально-
го скелета преобразуются в губные хрящи. Наиболее серьезным
преобразованием стало превращение третьей пары дуг в челюст-
ную дугу.
Третья висцеральная дуга представлена с каждой стороны па- i
рой хрящей. Верхний — нёбноквадратный — хрящ выполняет У
* Название висцеральный получено по названию передней части пищеварИ'
тельной трубки (внутренности).
ST li
Рис. 64. Преобразование двух висцеральных дуг (III и IV) в ряду позво-
ночных:
А — акула; Б — костистая рыба; В — амфибия; Г — пресмыкающееся; Д — терио-
донт; Е — млекопитающее; 1 — нёбноквадратный хрящ; 2 — меккелев хрящ; 3 —
квадратная кость (у млекопитающих — наковальня); 4 — нёбная; 5 — крыловид-
ные кости; 6 — сочленовная кость — молоточек млекопитающих; 7 — зубная
кость; S — угловая кость; 9 — гиомандибулярный хрящ — слуховая косточка
(стремечко) наземных позвоночных; 10 — чешуйчатая кость; 11 — гиоид
хрящевых рыб функцию верхней челюсти. Нижняя половина че-
люстной дуги — нижняя челюсть — представлена также парным
меккелевым хрящом. У животных с костным скелетом верхняя по-
ловина первичной челюстной дуги, т.е. кости, замещающие нёб-
ноквадратный хрящ, теряют функцию верхней челюсти (перехо-
дящую к двум парам новых накладных костей) и входят в состав
дна черепа.
Челюстной сустав устойчиво сохраняет свое положение между
верхним и нижним висцеральными элементами во всех классах
позвоночных, кроме млекопитающих (см. ниже).
За челюстной располагается четвертая висцеральная дуга —
подъязычная {гиоидная) дуга, специализированная на поддержа-
нии челюстей и укреплении дна ротовой полости. Важным эле-
ментом подъязычной дуги является парный гиоид, или рожково-
подъязычный элемент. Сзади гиоид сочленен с подвеском, или гио-
мандибуляре, который, в свою очередь, неподвижно прикреплен
к слуховому отделу мозгового черепа. Снизу парные подвески при-
креплены связками к челюстям, прочно соединяя их с мозговой
Коробкой. Такая конструкция прикрепления челюстей к черепу
Называется гиостилической (рис. 65). У некоторых акул возможно и
Двоякое причленение челюстей к черепу — через гиомандибуляре
и прямое сочленение верхней челюсти с передней частью мозго-
II
83
1
Рис. 65. Типы причленения висцеральных дуг к мозговому черепу:
1 — амфистилический череп древней акулы; // — амфистилический череп кост-
ного ганоида; III — гиостилический череп современной акулы; IV — аутостили-
ческий череп наземного позвоночного (лягушка); 1 — нёбноквадратный хрящ;
2, 3 — выросты нёбноквадратного хряща; 4 — меккелев хрящ; 5 — нижняя
челюсть; 6 — гиомандибуляре; 7 — гиоид
вой коробки. Такую конструкцию называют амфистилической. Ко-
личество жаберных дуг у хрящевых рыб 5 — 7.
У костных рыб в связи с окостенением скелета висцеральный
череп претерпевает более заметные изменения, чем мозговой (см.
рис. 57).
Верхняя часть челюстной дуги, гомологичная нёбноквадратно-
му хрящу, замещена в передней части смешанной по происхож-
дению нёбной костью, в серединной — тремя крыловидными кос-
тями, из которых две кожного и одна хондрального происхожде-
ния; в задней части она замещена хондральной квадратной кос-
тью. Система указанных костей у костных рыб полностью теряет
функцию верхней челюсти и в большей мере формирует дно че-
репа. Роль верхней челюсти выполняют парные кости кожного про-
исхождения: верхнечелюстная и предчелюстная.
Нижняя челюсть представлена большой зубной костью кожно-
го происхождения, покрывающей меккелев хрящ, угловой костью,
образующей нижнезадний угол челюсти, и единственной хонд-
ральной костью — сочленовной, которая сочленяется с квадратной
костью.
Подъязычная дуга представлена в основе теми же элементами,
что и у хрящевых рыб, т.е. парными гиомандибуляре, гиоидами и
непарной копулой. Все эти элементы костные, хондрального про-
исхождения. К заднему краю нижней части подъязычной дуги при-
членяется ряд длинных косточек — лучей жаберной перепонки,
подстилающих глотку.
Жаберные дуги такого же строения, как и у хрящевых рыб,
при этом последняя (пятая) дуга сильно редуцирована. Новопрй'
обретением костных рыб является жаберная крышка, представ-
ленная четырьмя плоскими накладными костями {крышка, пред-
крышка, подкрышка, межкрышка).
Висцеральный череп амфибийв сравнении с рыбами изме-
нен значительно.
84
Личинки амфибий (и некоторые взрослые) имеют жабры, со-
ответственно у них развиваются жаберные дуги. Количество жа-
берных дуг уменьшается до четырех; часто они не имеют расчле-
нения на элементы и являются цельными.
У амфибий, прошедших метаморфоз, висцеральные дуги пре-
образуются в скелетные элементы, характерные для наземных
позвоночных. Они входят в состав подъязычного аппарата, служа-
щего опорой языку, в состав скелета гортани и кольцевидных хря-
щей трахеи, а также участвуют в совершенствовании слухового
аппарата. Подъязычный аппарат располагается у лягушки в дне
ротовой полости между ветвями нижней челюсти и представляет
собой хрящевую пластину с рожками. Ее передние рожки, загиба-
ясь вверх, охватывают кишечную трубку и прикрепляются к слу-
ховым капсулам (см. рис. 59).
Функцию верхних челюстей у земноводных выполняет, как и у
костистых рыб, костная дуга, состоящая из покровных межчелю-
стных (предчелюстных), верхнечелюстных и квадратных костей.
Элементы первичного челюстного комплекса, образованного
нёбноквадратным хрящом, принимают участие в формировании
дна мозгового черепа: нёбная кость прилегает к сошнику, а квад-
ратная кость — к чешуйчатой кости.
В нижней челюсти меккелев хрящ прикрыт снаружи зубной и
угловой костями.
Череп амфибий аутостилический: челюстная дуга прирастает
непосредственно к мозговому черепу (см. рис. 65). Подъязычная
дуга не принимает участия в прикреплении челюстного аппарата
к черепу. Верхний элемент этой дуги — гиомандибуляре — пре-
вращен в маленькую кость — стремя, которая, как подвесок у
рыб, верхним концом упирается в слуховую капсулу. Она лежит в
преобразованной из брызгальца полости и выполняет роль слухо-
вой косточки.
Висцеральный скелет рептилий (ящериц) в значительной сте-
пени близок к амфибиям. Верхние челюсти образованы межчелю-
стными и верхнечелюстными костями, несущими зубы.
Нёбноквадратный хрящ подвергается дальнейшим превраще-
ниям. Хондральная квадратная кость своим верхним отделом свя-
зана с мозговой коробкой; снизу к ней причленяется нижняя че-
люсть. Остальная часть нёбноквадратного хряща покрывается кож-
ными костями — в передней части нёбными, а в серединной —
крыловидными. Все эти кости участвуют в укреплении дна черепа.
Появляются новые покровные поперечные кости, соединяющие
Крыловидные кости с верхнечелюстными, а у ящериц и гаттерий
еЩе и хондральные столбчатые кости, соединяющие крыловид-
ные кости с теменными.
Нижняя челюсть, гомологичная меккелеву хрящу, состоит из
Хондральной сочленовной кости, которая причленяется к квадрат-
85
ной, и ряда накладных костей, облегающих меккелев хрящ: зуб-
ной, угловой, надугловой, венечной, пластинчатой. Верхний отдел
подъязычной дуги, гомологичный гиомандибуляре, представлен
(как и у земноводных) слуховой косточкой — стременем. Осталь-
ные элементы висцерального скелета образуют подъязычный ап-
парат, состоящий из хрящевой пластинки (гомолог копулы) д
трех пар рожков (гомологи гиоидов и жаберных дуг).
Весьма своеобразно строение черепа у змей. Для них харак-
терно подвижное соединение элементов челюстного аппарата и
связанных с ними костей: нёбных, крыловидных и чешуйчатых.
Эта особенность позволяет змее заглатывать пищу целиком, даже
довольно крупных животных. Механизм заглатывания достигает-
ся попеременным движением левой и правой половин нижней
челюсти. При наблюдении за кормящейся змеей создается впе-
чатление, что она как бы наползает на свою добычу. У ядовитых
змей подвижность костей облегчает перемещение ядовитых зу-
бов (рис. 66).
У крокодилов зубы расположены на межчелюстных, верхнече-
люстных и зубных костях и сидят в альвеолах, как у млекопитаю-
щих. В связи с разрастанием нёбных отростков межчелюстных и
верхнечелюстных, а также нёбных костей у крокодилов развивает-
ся костный помост — вторичное костное нёбо. Оно разделяет по-
лость рта на верхний отдел, носоглоточный ход, и нижний отдел,
собственно ротовую полость (рис. 67). Таким образом, внутренние
отверстия ноздрей (хоаны) открываются не непосредственно в ро-
товую полость, а в ее заднюю часть, поэтому пища, находящаяся
во рту, не мешает поступлению воздуха к органам дыхания.
Рис. 66. Череп гремучей змеи с закрытой (А) и раскрытой (Б) пастью:
1 — ядовитый зуб; 2 — верхнечелюстная кость; 3 — поперечная кость; 4 "
крыловидная кость; 5 — чешуйчатая кость; 6 — квадратная кость
86
рис. 67. Вторичное костное нёбо у кроко-
дила:
j межчелюстные кости; 2 — верхнечелюст-
нЫе кости; 3 — нёбные кости; 4 — крыловидные
кости; 5 — хоаны
У черепах нёбо прирастает к мозго-
вому черепу. Квадратная кость массив-
ная и на большом протяжении контак-
тирует с ушным отделом, повышая
прочность задней части черепа.
В висцеральном черепе птиц мно-
го общего с черепом рептилий. Верх-
няя челюсть включает парные межче-
люстные и верхнечелюстные кости. К
последним причленяются палочковид-
ные скуловые и квадратноскуловые ко-
сти, которые, в свою очередь, соеди-
няются с квадратными костями. В итоге образуется весьма ха-
рактерная для птиц нижняя височная дуга, отграничивающая снизу
глазницу и височную яму. Нижняя челюсть состоит из гомоло-
гичной меккелеву хрящу сочленовной кости и из кожного проис-
хождения зубной, пластинчатой, угловой и венечной костей. Вмес-
те с верхней челюстью она образует характерный для птиц клюв.
Челюсти, как правило, сильно вытянуты, их кости срастают-
ся между собой. Возникает весьма совершенный, прочный хва-
тательный аппарат (см. рис. 62).
Слуховая косточка, стремячко, одна, как и у рептилий, и ей
гомологична.
Костный подъязычный аппарат имеет вид удлиненной плас-
тинки с очень длинными рожками, гомологичными первой паре
жаберных дуг. Подъязычная пластинка участвует у птиц в движе-
нии языка при захвате пищи, а также при звукообразовании. Спе-
циалисты отмечают, что, например, у токующего глухаря, во вре-
мя второго колена призывной песни, когда птица глубоко втяги-
вает язык в гортань, загнутые рожки сдвигают набухшую кровью
кожу и соединительнотканную пленку на голове, отчего глухарь
временно глохнет (Н. Н. Руковский, 2001) (рис. 68).
Строение висцерального черепа млекопитающихв сравне-
нии с рептилиями существенно меняется. Верхние челюсти состоят
из парных межчелюстных и верхнечелюстных костей. Для всех мле-
копитающих характерно развитие вторичного костного нёба, обра-
зованного нёбными отростками межчелюстных и верхнечелюстных
Костей и нёбными костями. В связи с его образованием хоаны от-
87
Рис. 68. Подъязычный аппарат глухаря (А) и его положение относи-
тельно слухового прохода (Б) (по Н. Н. Руковскому, 2003):
1 — язык; 2 — ороговевшие шипики языка; 3 — костная основа языка; 4 —
передние рожки; 5 — задние рожки; а — обычное положение языка; б — язык
втянут в глотку
крываются не между верхнечелюстными костями, как у других на-
земных позвоночных, а как у крокодилов и черепах — позади нёб-
ных костей. Такое строение нёба предотвращает закупорку хоан (т.е.
перерыв дыхания) в то время, когда пищевой комок задерживает-
ся в ротовой полости при обработке пиши во рту и при глотании.
Крыловидные кости уменьшаются и прирастают к дну черепа.
Нижняя челюсть образована только парными зубными костя-
ми, которые несут два отростка — венечный и угловой. Нижние
челюсти причленяются непосредственно к чешуйчатым костям.
Наиболее серьезным преобразованием становится превращение
сочленовной кости в слуховую косточку — молоточек, а квад-
ратной — в наковальню. Вместе со стременем они лежат в полос-
ти среднего уха и участвуют в звукопередаче. Наружная стенка
последней, а также часть наружного слухового прохода окру-
жены барабанной костью, гомологичной, по-видимому, угло-
вой кости нижней челюсти прочих позвоночных. Таким обра-
зом, у млекопитающих наблюдается дальнейшее превращение
части висцерального аппарата в слуховой аппарат среднего уха.
Остатки подъязычного аппарата располагаются у млекопитаю-
щих в нижней части глотки и входят в состав части гортани. Фун-
кциональное значение скелета подъязычного аппарата опреде-
ляется его ролью в движениях языка, связанных с захватывани-
ем, обработкой в ротовой полости и проглатыванием пиши, с
88
движениями глотки и гортани во время глотания, при воспроиз-
реДении звуков и т.д.
Изменчивость подъязычного аппарата в эволюционном ряду
позвоночных показано на рис. 69.
Ранее уже было упомянуто о подвижности частей черепа по
отношению друг друга у рипидистных рыб — кинетизме.
Свойство кинетизма признано также для лицевого скелета реп-
тилий и птиц. Обстоятельное описание этого явления было дано
ф.Я. Дзержинским (1998). Под кинетизмом он понимает подвиж-
ность верхних челюстей относительно затылка и дна черепа (за-
тылочных, ушных и клиновидных костей).
Предполагают, что такой подвижностью обладали стегоцефа-
лы и копториноморфные котилозавры. Подвижность верхней че-
люсти позволяла ускорять встречное движение челюстей, мани-
пулировать с добычей, амортизировать толчки и рывки добычи
и т.д. Кинетизм в черепе создается разными механизмами. В кры-
ше черепа зона подвижности проходит на границе покровных
костей с затылочной областью. Такая особенность наблюдается,
например, у гаттерии: покровный скелет головы вместе с верх-
ними челюстями может поворачиваться вверх и вниз около по-
перечной оси, расположенной на заднем крае крыши черепа.
Уящерицв крыше черепа есть еще одна гибкая зона: между
теменной и лобной костями. В таком черепе рыло вместе с лобны-
ми костями задирается относительно теменной области вверх, она
же, в свою очередь, опускается относительно затылочной облас-
ти. Подвижность приобретают и кости висцерального черепа: квад-
ратная кость вместе с чешуйчатой и переднеушной поворачивает-
ся нижним концом вперед, увлекая костное нёбо.
Упомянутую ранее продольную подвижность половинок челю-
стей в черепе змей Ф. Я. Дзержинский (1998) назвал гиперкине-
тизмом. Такая подвижность, усиленная наличием эластичных свя-
зок, используется при заглатывании добычи. Половинки челюст-
ного аппарата работают поочередно: с помощью одной из них
Рис. 69. Изменение подъязычного аппарата в ряду позвоночных:
бесхвостая амфибия; Б — рептилия; В — эму; Г — летучая мышь; 1 —
тело подъязычного аппарата; 2 — шиловидный отросток
89
Рис. 70. Схема черепа ворона, иллюстри
руюшая механизм кинетизма (подвижно,
ста) надклювья (по Ф.Я. Дзержинском^
1998).
Кружками показаны подвижные сочленения
/ — гибкая зона надклювья; 2 — соединение
надклювья с нёбным аппаратом; 3 — челюс-
тной сустав; 4 — квадратно-черепной сустав
змея втягивает добычу в глубь рта, а другая половинка в это время
раскрывается и выносится вперед для следующего захвата. У ядо-
витых змей вынесением нёба вперед обеспечивается поворот ка-
чающейся верхнечелюстной кости для приведения сидящего на
ней ядовитого зуба в рабочее положение (см. рис. 66).
Кинетизм характерен для черепа всех без исключения птиц,
при этом разные виды имеют свои особенности. Для относитель-
но архаичных групп птиц (тинаму, бескилевых, некоторых ржан-
кообразных и ибисовых) характерна примитивная форма кине-
тизма — ринхокинетизм (от лат. названия клюва — rhynchos). У них
гибкая зона крыши располагается не в задней части черепа, а
впереди и локализуется в пределах надклювья. Иногда она захва-
тывает предчелюстную кость и предчелюстные отростки носовых
костей. Носовая перегородка в этом месте прерывается. Высокий
уровень специализации при такой форме кинетизма отмечен у
вальдшнепа. Гибкая зона располагается у него на границе конеч-
ной трети клюва и позволяет раскрыть его при погружении в по-
чву на всю длину.
Для большинства птиц характерна другая форма кинетизма —
прокинетизм. Гибкая зона в этом случае образована почти исклю-
чительно носовыми костями вблизи основания верхней челюсти.
Межглазничная перегородка в этом месте отделена от носовой
перегородки широким окном — так называемой черепнолицевой
щелью. Кинетизм такого типа позволяет (в отличие от ранее
рассмотренного варианта) лучше сжимать клюв на всем его протя-
жении. Представление о подвижности клюва птицы дает рис. 70.
В черепе млекопитающих кинетизм не выражен.
Скелет парных конечностей и их поясов
Скелет парных конечностей у первичноводных позвоночных
имеет следующие элементы: базалии и радиалии. У наземных пере-
дняя конечность представляет многочленный рычаг. Опорой пар'
ным конечностям служат плечевой и тазовый пояса.
Эмбриональное развитие парных конечностей начинается с
выпячивания эктодермы. Внутрь выпячивания из париетального
90
листка боковой пластинки выселяются клетки скелетогенной ме-
зенхимы. Вслед за ними сюда врастают мышечные зачатки в виде
почек миотомов сомитов. В пределах выростов обособляются дор-
сальные и вентральные пласты мускулатуры (рис. 71). У рыб сгу-
щения скелетогенной мезенхимы образуют зачатки радиалий. Ба-
залии формируются как утолщенные членики радиалий.
Предполагают, что парные плавники исторически возникли в
результате обособления парных брюшных складок кожи с набо-
ром скелетных образований и пластами управляющей ими муску-
латуры, подобно непарным плавникам на теле. Складки посте-
пенно дифференцировались, в результате чего выделились две пары
плавников: грудные и брюшные. У древних акулоподобных рыб —
акантодий — между грудными и брюшными плавниками сохра-
нялось еще несколько пар (до 7) промежуточных плавничков
(рис. 72). Первично парные плавники служили горизонтальными
стабилизирующими килями, предотвращавшими вращение тела
вокруг оси и килевую качку. При этом основание примитивных
плавников было широким.
Ископаемые панцирные рыбы имели грудные и брюшные плав-
ники с длинным костным шипом в основе.
У бесчелюстных (щитковых и круглоротых) парные плавники
как таковые отсутствуют. Однако палеонтологами замечено, что
если среди разнощитковых нет и намека на парные выросты, то у
некоторых костнощитковых в передней части туловища обнару-
жены ластоподобные образования. У бесщитковых некоторые фор-
мы имели стабилизирующие боковые складки.
Парные плавники современных рыб имеют относительно уз-
кие основания, что предполагает достаточно широкую амплитуду
их движений.
Кроме собственно конечностей в теле животного располагают-
ся опорные скелетные элементы — пояса конечностей. У водных
позвоночных пояс служит опорой для конечностей, у наземных —
посредником, передающим на конечность всю массу тела. Пред-
полагают, что у панцирных рыб внутри тела присутствовал насто-
Рис. 71. Эмбриональные компоненты парной
конечности на схеме поперечного разреза
зародыша (по Ф.Я. Дзержинскому, 1998):
I ~~ эктодерма; 2 — нервная трубка; 3 — миотом соми-
та; 4 — дерматом; 5 — мускульная почка миотома; 6 —
Члавниковая складка; 7 — выселение скелетогенной
Мезенхимы; 8— париетальный листок боковой пластин-
ки; 9 — целом; 10 — хорда
91
Рис. 72. Акантодия (Climatius)
ящий пояс, а боковые стенки головогрудного панциря служили
дополнительным покровным плечевым поясом (А.Ромер,
Т. Парсонс, 1992). В теле хрящевых рыб располагаются только
элементы внутреннего плечевого пояса — сочлененные между со-
бой лопаточный и коракоидный отделы. Сливаясь снизу, обе поло-
вины образуют дугообразную конструкцию. Пояс лежит свободно
в туловищной мускулатуре и к осевому скелету не прикреплен.
К стыкам лопаточного и коракоидного отделов прикрепляются
парные плавники. У акул они лежат в горизонтальной плоскости
и включают три базалии, множество радиалий и эластоидиновые
нити мягкой лопасти плавника. Грудные плавники скатов вторич-
но изменены в широкие боковые складки (рис. 73).
У акул две половины тазового пояса сращены в единый эле-
мент. В структуру брюшных плавников включены две базалии, много
радиалий и эластоидиновые нити.
Укостных рыб пояс передних конечностей частично или пол-
ностью костный. Он состоит из маленьких, хондрального проис-
хождения коракоидов и «лопаток». К указанным хондральным ко-
стям, составляющим первичный пояс, присоединяются кожные
кости «вторичного» пояса. Основная из них — большая серповид-
ная клейтрум. Посредством более мелких костей она сочленяется
с мозговой частью черепа. Скелет грудных плавников не имеет
базалий и состоит из костных радиалий и костных лучей, причле-
Рис. 73. Скелет поясов и парных конечностей самца акулы:
А — плечевой пояс и грудные плавники: 1 — коракоидный отдел; 2 — лопаточ-
ный отдел; 3, 4, 5 — базалии; 6 — радиалии; 7 — эластоидиновые нити. Б "
пояс задних конечностей и брюшные плавники: 1 — пояс; 2, 3, 4— базалии; 5"
радиалии; 6 — эластоидиновые нити
92
5
Рис. 74. Грудной пояс костистой рыбы судака.
Первичный пояс: 1 — лопатка; 2 — коракоид. Вторичный пояс: 3 — клейтрум; 4 —
супраклейтрум; 5 — заднетеменная кость; б — подклейтрум. Скелет плавника: 7—
радиалии; 8 — лепидотрихии (кожные костные лучи)
няющихся непосредственно к поясу (рис. 74). Пояс задних конечно-
стей лежит в толще мускулатуры и представлен непарной вытя-
нутой пластинкой. Скелет брюшных плавников содержит только
кожные костные лучи. Таким образом, видно, что скелет парных
конечностей у костистых рыб в сравнении со скелетом парных
конечностей хрящевых рыб упрощен за счет утраты в обеих парах
плавников базалий, а в задней паре и радиалий.
Структура скелета плавников древних кистеперых рыб — осте-
олепиформных рипидистий стала прототипом конечности назем-
ного типа (рис. 75, А). Своеобразное расположение радиалий по
отношению к базалиям (унисериальный тип плавника) способ-
ствовало концентрации мускулатуры на конечности. В этом улав-
ливается сходство в расположении скелетных элементов* конеч-
ностей древней кистеперой рыбы и стегоцефала (рис. 75, Б).
Пояса конечностей наземных позвоночных значитель-
но изменяются и в связи с увеличением размеров конечностей, и
в результате распределения массы мускулатуры, начинающейся
на поясе. В скелете земноводных пояс передних конечностей —
плечевой — тоже имеет вид дуги, обращенной вершиной к брюш-
ной поверхности животного (рис. 76), однако в нем появляются
новые элементы. Верхняя (спинная часть) представлена лопат-
ой, которая заканчивается широким надлопаточным хрящом.
* Расположение скелетных элементов в плавниках других лопастеперых —
Двоякодышащих рыб и поролепиформных рипидистий было иным (бисериаль-
Чый тип плавника). Конечности этого типа эволюционировали лишь в направле-
нии углубления узкой специализации.
93
Рис. 75. Плечевой пояс и плавник древней кистеперой рыбы (А),
тазовый пояс и плавники неоцератода (Б):
1 — пояс; 2 — базалии; 3 — радиалии
Нижняя (брюшная) часть состоит из коракоида и лежащего впе-
реди него прокоракоида. Три элемента пояса сходятся в точке при-
членения плечевой кости и формируют сочленовную ямку. Впере-
ди от места соединения левого и правого коракоидов находится
предгрудина, а сзади — грудина.
Обе эти кости заканчиваются хрящами. У бесхвостых амфибий
между предгрудиной и лопаткой параллельно прокоракоиду рас-
положена тонкая палочковидная ключица. Плечевой пояс лежит
свободно в толще мускулатуры.
Пояс задних конечностей — тазовый — образован тремя пар-
ными элементами, сходящимися в области вертлужной впадины,
которую они и образуют. Длинные подвздошные кости своими про-
ксимальными концами прикреплены к поперечным отросткам
крестцового позвонка. Направленный вперед и вниз лобковый эле-
мент пояса у лягушек остается хрящевым. Сзади него расположи'
на седалищная кость. Такое расположение элементов тазового по-
яса характерно для всех наземных позвоночных.
94
Б
Рис. 76. Плечевой (А) и тазовый (Б) пояс лягушки (вид спереди):
1 — грудина; 2 — хрящевая задняя часть грудины; 3 — коракоид; 4 — ключица
лежит на прокоракоиде; 5 — предгрудина; б — лопатка; 7— сочленовная впади-
на на лопатке для плеча (хрящ покрыт точками); 8 — подвздошная кость; 9 —
седалищная кость; 10 — вертлужная впадина; 11 — крестцовый позвонок, 12 —
уростиль
Скелет свободных конечностей наземных позвоночных суще-
ственно отличается от скелета конечностей рыб. В то время как
конечности рыб представляют в схеме простые одночленные ры-
чаги, перемещающиеся только относительно корпуса тела, конеч-
ности наземных позвоночных — это рычаги многочленные. В этом
случае перемещается не только вся конечность относительно тела,
но и элементы конечностей относительно друг друга. Характерной
чертой наземной конечности является присутствие на ней мускула-
туры.
Пятипалая (наземная) конечность состоит из трех отделов (рис.
77).
• Отдел I: плечо — в передней конечности, бедро — в задней;
отдел всегда имеет одну кость, которая проксимальным концом
причленяется к поясу.
• Отдел И: предплечье — в передней конечности, голень — в
задней; в типичном случае отдел состоит из двух параллельно
Расположенных костей: предплечье — из локтевой и лучевой, го-
лень — из большой и малой берцовых.
• Отдел III: кисть — в передней конечности, стопа — в задней;
Отдел состоит из трех подотделов:
запястье — в передней конечности, предплюсна — в задней;
95
Рис. 77. Схема строения парных конеч.
ностей наземных позвоночных:
А — передняя конечность; Б — задняя ко.
нечность; I — плечо —бедро; И — предпЛе.
чье — голень; III — кисть—стопа; 1 — цЛс
чевая кость; 2 — локтевая кость; 3 — луче.
вая кость; 4 — запястье; 5 — пясть; 6 — фа
ланги пальцев; 7 — бедренная кость; g _
большая берцовая кость; 9 — малая берц0.
вая кость; 10 — предплюсна; 11 — плюсна-
12 — коленная чашечка
подотдел представлен в типичном случае 9—10 мелкими кос-
точками, расположенными в три ряда;
пясть — в передней конечности, плюсна — в задней; подот-
дел в типичном случае состоит из 5 удлиненных костей, распо-
ложенных в один ряд веерообразно, от запястья или предплюс-
ны;
фаланги пальцев представляют продолжение пясти или плюсны
и состоят из пяти (или меньше) рядов, по нескольку косточек в
каждом.
У хвостатых амфибий скелет конечностей почти полностью
соответствует приведенной схеме. У лягушек наблюдаются неко-
торые изменения, в частности оба элемента предплечья и голени
срастаются в одну кость. Срастаются между собой также большая
часть костей запястья и предплюсны. Перед первым пальцем зад-
ней конечности есть рудимент добавочного пальца. Эти особенно-
сти имеют вторичный характер и связаны с приспособлением бес-
хвостых земноводных к передвижению прыжками.
Особенность конструкции суставов в конечностях амфибий —
их сочленение между кистью и предплечьем (кистевой), между
стопой и голенью (голеностопный) — характерна также для мле-
копитающих.
У рептилий, в частности у ящериц и крокодилов, плече-
вой пояс включает в основном те же элементы, что и у амфибий
(рис. 78). Брюшная его часть представлена коракоидами, несущими
сочленовные ямки для причленения плечевых костей. Спинная часть
пояса состоит из лопатки и надлопаточного хряща. Коракоид свя-
зан с грудиной и обладает подвижностью. Впереди от него распо-
ложена ключица. Характерно наличие крестообразного надгрудин-
ника, лежащего перед грудиной и связывающего ее с ключицами-
В целом плечевой пояс рептилий более прочен, чем у амфибий, и
96
8
9
Рис. 78. Плечевой (А) и тазовый (Б) пояс ящерицы:
А: 1 — ключица; 2 — надлопаточный хрящ; 3 — лопатка; 4 — коракоид; 5 —
ребра; б — грудина; 7 — переднекоракоидный хрящ; 8 — надгрудинник. Б\ 1 —
суставная впадина для головки бедра; 2 — подвздошная кость; 3 — лобковая
кость; 4 — седалищная кость; 5 — связка; 6 — «окно»; 7 — запирательное отвер-
стие; 8 — задний хрящевой отросток; 9 — передний хряшевой отросток
эта его черта отражает их лучшую приспособленность к передви-
жению по суше.
В тазовом поясе парные кости: подвздошная, седалищная и лоб-
ковая соединены в единый замкнутый таз. Спереди и сзади, на
стыке костных элементов расположены передний и задний хря-
щевые отростки. Подвздошными костями пояс причленяется к по-
перечным отросткам двух крестцовых позвонков. На стыке трех
костей таза расположена вертлужная впадина', в нее входит круг-
лая головка бедренной кости.
Передняя и задняя конечности имеют типичное строение. Осо-
бенность конструкции суставов в конечностях рептилий иная, чем
У амфибий: сочленение в передней конечности лежит между кос-
тями запястья {межзапястный, или интеркарпалъный, тип), а в
заднем — между костями предплюсны (межпредплюсневый, или
интертарзальный, тип). Конструкция сустава создает сгибательно-
разгибательную подвижность конечности. Сходное строение име-
ют суставы птицы.
Плечевой пояс у крокодилов не имеет ключиц и состоит толь-
ко из лопаток и коракоидов.
У змей парных конечностей и скелета поясов практически нет.
Только у удавов есть едва заметные рудименты бедра и подвздош-
ных костей (рис. 79). Остатки таза имеются у слепунов (Typhlopidae).
Своеобразно строение поясов конечностей у черепах. Коракои-
ды ц лопатки хорошо развиты, ключицы же обособлены и входят
в состав пластрона. Кости таза прикреплены к карапаксу с помо-
щью связок или срастаются с ним. Таким образом, у черепах по-
Константинов 97
Рис. 79. Скелет пояса
задних конечностей удава.
Видны рудименты подвздош-
ной кости (7) и бедра (2)
яса конечностей срастаются с панцц.
рем, что отражается на их двигатель,
ной способности (рис. 80).
Уместно вспомнить, что в эволю-
ции динозавров в свое время опреде-
лились два направления, связанные
с конструкцией таза. У ящеротазовых
динозавров (Aurischia) лобковая и се-
далищная кости приобрели стержне-
видную форму, и таз был трехлуче-
вым (например, у тираннозавра). Кон-
струкция таза современных крокоди-
лов напоминает тираннозавра. То же
демонстрируют птицы. У птицетазо-
вых динозавров (Ornithischia) тонкий
стержень лобковой кости был повер-
нут назад и лежал параллельно седа-
лищной кости.
Несколько слов о локомоции. У реп-
тилий (и хвостатых амфибий) конеч-
ности расставлены в стороны от тела. Животные перемещаются,
прижимая тело к земле, используя одновременно возможности
хвоста. Такая позиция требует значительного расхода энергии на
поддержание тела и движение. В истории наземных позвоночных
соотношение тела и конечностей решалось различными путями.
Экономичность движения была достигнута перемещением обеих
пар конечностей под тело и изменением ориентации локтевого и
коленного суставов.
Попытка приподнять тело над землей при четырехногой ло-
комоции свойственна некоторым крокодилам, черепахам, яще-
рицам. Известно, что некоторые из них могут быстро бегать.
Ящерица-василиск, например, встает на задние конечности при
беге.
Бипедальная локомоция, при которой задние ноги удлинены,
а колени вывернуты вперед под тело, известна уже у некоторых
динозавров.Позднее это стало основой движения в наземной сре-
де и привело у птиц и некоторых млекопитающих к бипедальной
(двуногой) локомоции. Другой путь — утрата конечностей, как
это свойственно змеям (и безногим амфибиям).
У п т и ц скелет передних конечностей (пояс и свободная ко-
нечность) имеет черты, преемственные от рептилий, но в связи
с приспособлением к полету отличается рядом особенностей. Пояс
включает лопатки, коракоиды (вороньи кости) и ключицы, кото-
рые своими проксимальными концами формируют площадки для
причленения плечевых костей. Лопатка длинная, саблевидно изог-
нутая, лежит на ребрах, по которым она может свободно сколь-
98
Рис. 80. Скелет черепахи (брюшной щит отделен и откинут вправо):
1 — туловищный отдел позвоночного столба; 2 — реберные пластинки; 3 —
краевые пластинки карапакса (спинного щита); 4 — коракоид; 5 — лопатка; 6 —
подвздошная кость; 7 — седалищная кость; 8 — лобковая кость; 9 — брюшной
щит (пластрон)
зить. Коракоид развит очень хорошо. Одним концом он упирает-
ся в грудину, другой конец дает опору для причленения плеча.
Левая и правая ключицы срастаются между собой, образуют ха-
рактерную для птиц вилочку, придающую поясу особую упру-
гость.
Скелет крыла состоит из всех типичных для пятипалой конеч-
ности отделов (рис. 81). При этом плечо и предплечье существенно
не изменены, а кисть, наоборот, имеет ряд своеобразных особен-
ностей. Сильно видоизменено запястье. Его проксимальные части
срастаются в две косточки, а дистальные — с пястью. Пясть со-
стоит из двух удлиненных костей, сращенных как в проксималь-
ном, так и в дистальном отделах и образующих в итоге сложную
пястно-запястную кость, или пряжку. Из пальцев сохраняются толь-
ко три — второй, третий и четвертый, при этом только третий
Палец имеет две фаланги, а второй и четвертый — по одной.
99
Рис. 81. Скелет крыла птицы и расположение маховых перьев:
1 — кисть (первостепенные); 2 — предплечье (второстепенные); 3 — плечо (тре-
тьестепенные)
В связи с наличием особых мускулов, сухожилий и кожистых
перепонок кости крыла соединены между собой так, что все кры-
ло не может быть вытянуто по прямой, и его части расположены
под некоторым углом друг к другу. Отделы конечности могут дви-
гаться только в одном направлении — в плоскости крыла, скла-
дывая и расправляя его.
Скелет задних конечностей (пояс и свободная конечность)
птиц также имеет ряд особенностей, связанных с тем, что при
движении по земле птица переносит на них всю тяжесть тела.
Прочность осевого корпуса тела определяется срастанием боль-
ших подвздошных костей по всей их длине со сложным крест-
цом. С подвздошными костями срастаются также весьма крупные
седалищные кости. Лобковые кости, наоборот, малы; в виде тон-
ких палочек они присоединены к наружному краю костей седа-
лищных. Как и у птицетазовых динозавров (Ornithischia), лобко-
вая кость у современных птиц отклонена назад, по всей вероят-
ности конвергентно. Ее передний отросток развит слабо. Все три
кости таза участвуют в образовании вертлужной впадины. В отли-
чие от других наземных позвоночных (рептилий, млекопитаю-
щих) у птиц отсутствует лобковый симфиз — сочленение тазо-
вых костей внизу: две половины тазового пояса широко расстав-
лены, что связано с откладыванием крупных яиц, покрытых твер-
дой скорлупой, и необходимостью в широком тазовом выход-
ном отверстии. Такой тип таза называют открытым. Закрытый
таз имеют только африканские страусы.
Бедро типичного строения. Голень состоит из двух типичных
костей, однако полностью развивается только большая берцовая
кость, малая берцовая кость рудиментарна и прирастает к первой-
100
рис. 82. Скелет задней конечности голубя (Л)
и часть задней конечности его птенца (Б):
j __ бедро; 2 — большая берцовая кость; 3 — реду-
цированная малая берцовая кость; 4 — цевка; 5 —
чашечка; 6 — большая берцовая кость; 7 — сросши-
еся проксимальные элементы предплюсны, прира-
стающие позднее к большой берцовой кости; 8 —
сросшиеся дистальные элементы предплюсны, при-
растающие позднее к основаниям плюсневых ко-
стей; 9 — срастающиеся кости плюсны
к дистальной части голени прирастает
проксимальный ряд косточек предплюсны.
Срастание здесь столь полное, что у взрос-
лой птицы швы утрачиваются полностью.
Лежащий вслед за голенью отдел конеч-
ности носит название цевки. У взрослой
птицы он состоит из одной длинной кос-
ти, однако эмбрионально эта кость возни-
кает в результате срастания костей плюс-
ны и нижнего (дистального) ряда костей
предплюсны (рис. 82). В итоге подвижный
сустав у птиц (как и у пресмыкающихся)
расположен между двумя рядами костей
предплюсны, а не между предплюсной и
голенью, как у амфибий. Это сочленение
именуется межпредплюсневым, или интер-
тарзалъным.
Пальцев у птиц чаще бывает 4, реже 3
и лишь у африканского страуса их 2.
А
У млекопитающих плечевой пояс сравнительно прост. Его
основу составляет лопатка, к которой прирастает рудиментарный
коракоид. Только у однопроходных коракоид и прокоракоид суще-
ствуют в виде самостоятельных костей. Ключица имеется у тех мле-
копитающих, передние конечности которых совершают разнооб-
разные сложные движения: наличие ключицы требуется для бо-
лее прочного причленения плечевой кости и упрочнения всего
Плечевого пояса. Таковы, например, обезьяны, кроты, летучие
мыши. Наоборот, у видов, которые перемешают передние ко-
нечности только или преимущественно в плоскости, параллель-
ной главной оси тела (бег, ходьба), ключицы рудиментарны или
отсутствуют. Таковы копытные, хищные, грызуны, зайцеобраз-
ные.
В тазовом поясе млекопитающих произошли серьезные изме-
нения, связанные с перемещением конечностей под тело и пре-
образованием обслуживающей их мускулатуры. Тазовый пояс со-
101
стоит из трех типичных для наземных позвоночных парных
тей: подвздошных, седалищных и лобковых. У многих видов эти кос,
ти срастаются в одну безымянную кость. Подвздошные кости на-
правлены вперед, а лобковые и седалищные назад.
У массивных копытных (лошадей, быков) и у слонов с кругк
ными ягодичными мышцами подвздошные кости сильно расши-
ряются. На границе лобковой и седалищной костей лежит запира-
тельное отверстие. Таз замкнутый. У самок выходное отверстие часто
бывает шире, чем у самцов. У насекомоядных — кротов и земле-
роек — кости таза не соединяются и он остается открытым.
У однопроходных и сумчатых имеется пара сумчатых костей
которые тянутся вперед от лобковых и поддерживают брюшную
стенку.
Скелет парных конечностей сохраняет у млекопитающих все
основные черты строения типичной пятипалой конечности: в пе-
редней конечности — плечо, предплечье (локтевая и лучевая кос-
ти), кисть (запястье, пясть, фаланги пальцев), в задней — бедро,
голень (большая и малая берцовые кости), стопа (предплюсна,
плюсна и фаланги пальцев).
Однако в связи с разнообразием условий существования и ха-
рактером использования конечностей детали их строения у раз-
ных групп различны. У наземных форм значительно удлинены про-
ксимальные отделы. К примеру, у быстро бегающих зверей пред-
плюсна, плюсна, запястье и пясть располагаются более или менее
отвесно, и эти животные опираются только на пальцы. Таковы,
например, собаки (рис. 83). У наиболее совершенных бегунов, ко-
пытных, сокращается число пальцев. Крайние пальцы редуциру-
ются, и животные ступают либо на одинаково развитые третий и
Рис. 83. Задние конечности пальце- и стопоходящих млекопитающих.
Элементы стопы затемнены:
А — обезьяна павиан; Б — собака; В — лама
102
Рис. 84. Скелет передней конечности крота (А), дельфина (Б), лошади (В):
1 — предплечье; 2 — запястье; 3 — пясть; 4 — пальцы
четвертый пальцы, между которыми проходит ось конечности (пар-
нокопытные), либо преимущественное развитие получает один тре-
тий палец, через который и проходит ось конечности (непарно-
копытные).
У водных млекопитающих (китообразных, ластоногих), наобо-
рот, проксимальные отделы укорочены, а дистальные отделы —
пясть, плюсна и особенно фаланги пальцев — сильно удлинены.
Конечности в этом случае превращены в ласты, перемещаюшие-
Рис. 85. Скелет летучей мыши:
— ключица; 2 — плечевая кость; 3 — локтевая кость; 4 — лучевая кость; 5 —
Первый палец; 6 — второй палец; 7 — третий палец (его пясть); 8 — фаланги
Третьего пальца; 9, 10 — четвертый и пятый пальцы с пястью; 11 — бедро; 12 —
шпора; 13 — межбедренная перепонка; 14 — перепонка крыла
103
ся относительно тела в основном как единое целое (рис. 84). Пере-
мещение же отделов конечностей относительно друг друга разви-
то сравнительно слабо. У летучих мышей только первый палец
передних конечностей развит нормально, остальные пальцы ц
плюсна сильно удлинены; между ними расположена кожистая пе-
репонка, образующая основную часть поверхности крыла (рис. 85).
Скелет непарных плавников
Этот скелет формируется на спинной части тела рыб над верх-
ними остистыми отростками, а в хвостовой — под гемальными ду-
гами (см. рис. 48). Верхние и нижние остистые отростки хвостовых
позвонков могут сами непосредственно участвовать в поддержании
хвостового плавника. В непарных спинных и анальном плавниках опор-
ными элементами обычно служат радиальные лучи из хряща и ко-
сти, так называемые птеригофоры. Они могут причленяться к
невральным или гемальным элементам позвоночного столба, но
часто бывают отделены от них мышечным промежутком. У многих
акул и химер, у некоторых костистых рыб непарные плавники не-
сут спереди шипы, которые работают как водорезы. Шипы присут-
ствовали и на непарных плавниках ископаемых рыб — акантодий.
Спинных плавников бывает обычно один или два (редко три).
В прошлом это имело место у щитковых, у панцирных рыб, ныне
характерно для хрящевых и костных. Среди ранних костных рыб
лучеперые имели исходно один спинной плавник, а ранние лопа-
степерые — два. Спинные плавники могут редуцироваться или
сливаться с хвостовым, как у современных угрей, мурен, церато-
дов, а могут быть раздроблены на множество мелких, как у мно-
гопера.
Позади анального или клоакального отверстия почти всегда
находится анальный плавник. Он может утрачиваться у придонных
рыб или сливаться с хвостовым плавником.
Хвостовой плавник состоит из плотной кожной перепонки, под-
держиваемой мощными плавниковыми лучами. Основными ске-
летными опорными элементами в нем являются невральные и
гемальные дуги конечных позвонков. Иногда в плавнике развива-
ются дополнительные радиальные элементы.
Различают несколько типов хвостовых плавников: протоцер-
кальный, гетероцеркальный, гипоцеркальный, дифицеркалъный и го-
моцеркальный (рис. 86). Протоцеркальный тип плавника характе-
рен для бесчерепных и круглоротых. Плавниковая складка равно-
мерно охватывает хвостовую ось позвоночника. Гетероцеркальный
тип плавника характерен для акул. Дистальный конец позвоноч-
ного столба поворачивает вверх, а свободная часть плавниковой
складки развивается ниже этой оси. Некоторые из древнейших
104
позвоночных, например щитковые, имели хвосты перевернутого
геТероцеркального (гипоцеркального) типа. Позвоночник откло-
нялся в них вниз, а не-вверх. Такой тип плавника соответствовал
придонному образу жизни. Дифицеркальный тип плавника харак-
теризуется тем, что позвоночный столб тянется по прямой назад
до конца тела, и плавник симметрично развит и над, и под по-
звоночником. Таковы ныне живущие двоякодышащие, многопе-
ру. Гомоцеркальный тип плавника характерен для большинства
костистых рыб. Снаружи он симметричен, однако при вскрытии
видно, что последние позвонки несколько отклоняются вверх, а
нижняя лопасть плавника поддерживается увеличенными нижни-
ми остистыми отростками.
Гетероцеркальный тип хвостового плавника характерен для
современных акул. Гетероцеркальный плавник может обеспечи-
вать наклонную силу тяги (вперед и вверх) и тем самым противо-
действовать силе тяжести. У скатов произошла редукция плавни-
Рис. 86. Типы (А— Д) и формы (£) хвостовых плавников:
~~ протоцеркальный (ланцетник, минога); Б — гетероцеркальный (акулы, осет-
Р°°бразные); В — гомоцеркальный (большинство костистых рыб); Г — дифицер-
Кальный; Д — гипоцеркальный (щитковые); 1 — полулунная (тунец); 2 — виль-
чатая (сельдь); 3 — выемчатая (лосось); 4 — усеченная (треска); 5 — округ-
лая (налим); 6 — заостренная (бельдюга)
105
ковой складки до жгутообразного состояния: их плавание совер.
шается главным образом с помощью волнообразных движение
увеличенных грудных плавников. Хорошо развитый гетероцеркаль-
ный плавник имели палеозойские палеонисциды; он был выра_
жен у мезозойских ганоидных рыб. Гетероцеркальную форму со.
хранили осетры и веслоносы, у современных панцирников конец
позвоночника заметно укорочен.
Ранние двоякодышащие рыбы имели типичный гетероцеркаль-
ный хвостовой плавник, однако, согласно палеонтологической
летописи, анальный и спинной плавники удлинились в направ-
лении хвостового, а позвоночная ось слегка укоротилась, в ре-
зультате у современных видов хвостовой плавник превратился в
дифицеркальный. Гетероцеркальный плавник имели и примитив-
ные девонские кистеперые рыбы, но выпрямление конца позво-
ночного столба привело у них к характерному трехлопастному хво-
стовому плавнику дифицеркального типа.
Образование симметричных лопастей хвостового плавника го-
моцеркального и дифицеркального типов характерно в основном
для костных рыб, имеющих либо легкие, либо плавательный пу-
зырь.
У наземных позвоночных (за исключением самых ранних древ-
них амфибий) непарные плавники, изначально присущие рыбам,
были полностью утрачены. Головастик лягушки, тритон, протей,
аксолотль и некоторые другие хвостатые амфибии хорошо плава-
ют с помощью дорсовентрально расширенного хвостового плав-
ника и совершенно не имеют внутренних скелетных образований.
К водному образу жизни вернулись многие настоящие наземные
позвоночные (амниота). Некоторые из них, например черепахи,
вымершие плезиозавры, пингвины и тюлени, для передвижения
в воде воспользовались не хвостом, а конечностями. Среди млеко-
питающих вновь приспособились к плаванию и воспользовались
хвостовыми плавниками для движения китообразные и сирене-
вые. У них лопасти хвостового плавника ориентированы горизон-
тально и имеют волокнистую основу. Наиболее близко к образо-
ванию заново хвостового плавника рыбьего типа подошли в про-
шлом ихтиозавры. Позвоночная ось входит у этих животных в ниж-
нюю лопасть хвостового плавника. У ихтиозавров и китообразных
появились и спинные плавники, но они тоже лишены скелетной
основы.
* * *
Таким образом, внутренний скелет хордовых энто- и мезодер-
мального происхождения поддерживает тело, служит для укреп-
ления скелетных мышц, участвует в движении, обеспечивает за-
щиту внутренних органов. Прогрессивные изменения связаны с
заменой в осевом скелете единого нечлененного упругого тяжа —
106
ордЫ сначала хрящевым, а затем костным позвоночником, его
сЛедовательной дифференцировкой на отделы и появлением
розгового черепа. В висцеральном скелете в эволюционном ряду
позвоночных висцеральные дуги редуцируются, развиваются че-
дюсти, формируются слуховые косточки. Мозговой и висцераль-
ный скелет объединяются в единое морфофункциональное обра-
зование — череп с лицевой и мозговой частями. Скелет органов
движения у первичноводных хордовых — первоначально непар-
ных плавников — прогрессивно развивается в связи со специали-
зацией к условиям обитания, затем дополняется парными плав-
никами, используемыми как стабилизаторы и рули при движе-
нии и поворотах. Появление мускулатуры на унисериальных пар-
ных плавниках, использовавшихся в качестве опоры при движе-
нии по грунту дна, послужило основой для формирования пяти-
палой конечности наземного типа.
МЫШЕЧНАЯ СИСТЕМА
Роль мускулатуры в теле позвоночных животных многоплано-
вая. Будучи связанной со скелетом, она прежде всего обеспечива-
ет двигательную активность животного. Кроме того, в различных
каналах тела — пищеварительной трубке, кровеносных сосудах,
каналах выделительной и репродуктивной систем и др. — муску-
латура способствует транспортировке веществ к органам. Сердеч-
ная мышца участвует в поддержании непрерывного тока крови.
В покровах мускулатура обеспечивает работу отдельных кожных
элементов (кожных желез, волосяного и перьевого покровов). Осо-
бая роль принадлежит мышечной системе в создании тепла в теле
позвоночного животного.
В мускулатуре энергия химических связей превращается не толь-
ко в механическую, но и в тепловую. У теплокровных животных
скелетная мускулатура поддерживает температуру тела на высо-
ком и постоянном уровне. Мышечная ткань может создавать элек-
трические поля. Деятельность мышечной системы регулируется ней-
рогуморальной системой, питание осуществляется потоками кро-
ви и лимфы.
Мускулатура представляет собой совокупность сократимых эле-
ментов мышечной ткани, объединенных обычно в мышцы, свя-
занные между собой соединительной тканью.
В теле хордового животного различают париетальную {скелет-
ную) мускулатуру, составляющую основу мускулатуры туловища,
в основном осевую, и мускулатуры конечностей, и висцеральную —
мускулатуру внутренних органов.
Париетальная мускулатура образована поперечнополосатой
мышечной тканью. Она формируется эмбрионально из миотомов
сомитов.
В состав париетальной мускулатуры у первичноводных входит
мускулатура туловища и хвоста, а у наземных — кроме названно-
го, мускулатура шеи и конечностей. Вместе они образуют осевую
мускулатуру.
При обособлении головного отдела тела осевая мускулатура дает
начало глазной и подъязычной мышцам. Из специальных мускуль-
ных почек миотомов сомитов развивается мускулатура парных И
непарных конечностей: плавников рыб и конечностей наземного
типа.
108
Париетальная мышечная система
у первичновод ных животных осевая мускулатура служит
идя передвижения в воде и горизонтальном изгибании тела. У лан-
цетника она сегментирована и представлена мышечными сегмен-
тами миомерами, разделенными поперечными соединительноткан-
ными миосептами. Число миомеров соответствует числу заклады-
вающихся эмбрионально миотомов (рис. 87).
Миомеры левой и правой сторон тела ланцетника расположе-
ны асимметрично. Миосепты ориентированы косо, их угол на-
правлен к переднему концу тела — это обеспечивает свободу со-
кращениям мышечных сегментов. Мускулатура круглоротых
мало отличается от мускулатуры ланцетников, что соответствует
относительно слабой их подвижности.
У р ы б активное плавание повлекло за собой серьезные пере-
стройки осевой мускулатуры. Миомеры представляют собой гир-
лянды вложенных друг в друга конусов (рис. 88). Миосепты имеют
многократные изгибы и укрепляются окостенениями (мускуль-
ными косточками), особенно хорошо развитыми у костистых рыб.
Горизонтальная миосепта в области боковой линии отделяет бо-
лее мощную спинную часть туловишной мускулатуры с продоль-
ным направлением волокон от брюшной стороны с косо ориен-
тированными волокнами. У пелагических костистых рыб к основ-
ной массе белой мускулатуры обычно прилегает красная мускуль-
ная полоска (Ф.Я.Дзержинский, 1998). Расчеты показали, что у
тунцов кровь, идущая к плавательным мышцам, проходит через
теплообменник красной мускулатуры, в результате чего мышцы
сохраняют тепло (К. Шмидт-Ниельсен, 1982). У хищных рыб от
затылка назад распространяется длиннейшая мышца спины, уча-
ствующая в некотором задирании головы перед схватыванием до-
бычи (рис. 89).
Мускулатура парных плавников (или первичная мускулатура
конечностей) включает мышцу, поднимающую и отводящую плав-
ник и отпускающую и приводящую его в нормальное положение.
Рис. 87. Мускулатура ланцетника:
1 — миомеры; 2 — миосепты
109
1 2 б
Рис. 88. Мускулатура круглоротых и рыб: миноги (Л), лосося (Б),
щуки (В), схема миомера акулы (Г):
1 — миомеры; 2 — миосепты; 3 — длиннейшая мышца спины; 4 — наружная
косая мышца живота; 5 — внутренняя косая мышца живота; 6 — горизонтальная
перегородка
Отдельные мышечные волокна могут выдвигать плавник вперед
или оттягивать назад.
Мышечная система амфибий заметно отличается от рыб
рядом важных особенностей, связанных с движением животных с
помощью конечностей наземного типа. Во-первых, развивается
мощная и сложно организованная мускулатура на свободных ко-
нечностях, а не в корпусе тела. В связи с более сложными движе-
ниями характерная для рыб метамерия нарушена и осевая муску-
латура значительно более дифференцирована. В пределах класса
амфибий структура туловищной мускулатуры различна. У хвоста-
тых слабо выражена метамерия миомеров. Единая дорсальная мыш-
ца туловища сменяется парными — длиннейшей мышцей спины И
подвздошно-поясничной мышцей. Брюшная стенка уплотняется за счет
лежащих в четыре слоя мышечных масс, которые создают подо-
бие кожно-мускульного мешка. Среди них выделяют наружные
косые мышцы (поверхностную и глубокую), внутреннюю косую,
поперечную и прямую мышцу живота (рис. 90). У бесхвостых изме-
нения глубже. В редуцированной хвостовой области у них развива-
ется парная мощная подвздошно-копчиковая мышца, контролиру-
ющая подвижность длинных подвздошных костей, участвующих В
110
Рис. 89. Схема кровоснабжения плавательных мышц тунца (А. Ромер,
Т. Парсонс, 1992).
Кровь, идущая к плавательным мышцам, проходит через теплообменник, в ре-
зультате чего мышцы сохраняют тепло, хотя температура артериальной крови
равна температуре воды
прыжке. Слой брюшных мышц резко утончается. Поперечная мышца
Утрачена.
Другая особенность мышечной системы амфибий состоит в
развитии мощной и сложно организованной мускулатуры поясов
и свободных конечностей. Эта мускулатура развивается по прин-
ципу сочетания мышц-антагонистов (сгибателей — разгибателей).
Передний пояс обслуживают дорсальная мышца лопатки, широ-
чайшая мышца спины, подкоракоиднолопаточная мышца, грудная
мышца, надкоракоидная, прокоракоидноплечевая, коракоиднолучевая.
111
Рис. 90. Схемы наружной мускулатуры позвоночных: амфибии (Л),
рептилии (Б), млекопитающего (В)
Значительные мышечные массы лежат на самой конечности: трех-
главая мышца плеча, короткая плечевая. Группы мышц лежат в об-
ласти кисти.
У рептилий метамерное расположение мускулатуры, свой-
ственное низшим позвоночным, почти не сохраняется. Развитость
пятипалых конечностей, появление шейного отдела и в целом боль-
шая расчлененность тела — все это приводит к сложной диффе-
ренцировке всей мышечной системы. Особо следует подчеркнуть
усиление межреберной мускулатуры в связи с оформлением груд-
ной клетки и ее ролью в механизме дыхания у всех высших позво-
ночных.
У ящериц и крокодилов сложно дифференцирована мускула-
тура спины. Она покрывает тело тремя (четырьмя) продольными
полосами. Мышцы брюшной стенки близки по свойствам к тако-
вым у хвостатых амфибий. Дальнейшие изменения происходят в
мышцах грудного отдела туловища: глубокая наружная косая мыш-
ца превращается в наружную межреберную, а внутренняя косая —
во внутреннюю межреберную мышцы. Работая как антагонисты,
112
дТЙ мышцы участвуют в дыхательных движениях грудной клетки,
способствуя вдоху и выдоху.
Своеобразно распределение мышц в туловище черепахи. Попе-
оечная мышца, участвующая в сжимании внутренностей, в том
<щсле легких, облегчает выдох, а наружная косая мышца образует
Р паховой области панциря вогнутую стенку. При сокращении она
уплощается и участвует во вдохе.
В передних конечностях рептилий наблюдается дальнейшая
дифференцировка мышц и отрабатываются механизмы их сухо-
жильного прикрепления к костям. Возникает двуглавая мышца плеча.
У п т и ц мускулатура отличается рядом особенностей, связан-
ных с характерными чертами образа жизни. Их мышечная система
еше более дифференцирована, даже по сравнению с рептилия-
ми, что обусловлено значительно более сложными видами дви-
жения птиц — полет, хождение (в т.ч. с подпрыгиванием), лаза-
ние, плавание и т.д. Наиболее крупные мышцы, приводящие в
движение конечности, располагаются на туловище, а к самим
конечностям направлены сухожилия.
В связи с огромной работой, выполняемой крыльями, основ-
ная масса мускулатуры располагается в области груди и прикреп-
ляется к килю грудины. Массивные грудные мышцы достигают
20 % общей массы птицы и служат для опускания крыльев. Лежа-
щие под ними и несколько меньшие по размеру подключичные
мышцы поднимают крылья.
Очень сложную мускулатуру имеют задние конечности (до 35
мышц). Большой интерес представляет имеющаяся у некоторых
древесных птиц обходящая мышца. Она начинается на тазовых
костях, тянется вдоль бедра, далее в виде тяжа перекидывается
через колено и затем соединяется со сгибателем пальцев. Птица,
севшая на присаду, сгибает колени и этим движением натягива-
ет обходящую мышцу, а следовательно, и сгибатель пальцев, в
результате чего пальцы сжимаются и плотно обхватывают ветку.
Чем ниже приседает птица, тем сильнее натягивается обходя-
щая мышца и тем сильнее птица пальцами обхватывает ветку.
Это особенно важно во сне, когда птица автоматически удержи-
вается на ветке.
У других видов птиц (например, воробьиных) механизм авто-
матического сгибания пальцев иной. Он обусловлен действием
Мышцы, называемой глубоким сгибателем пальцев. Ее сухожилия,
идущие к концам пальцев, имеют сильно шероховатую нижнюю
Поверхность и двигаются во влагалищах, внутренняя поверхность
Которых несет поперечные ребра. Когда птица садится на ветку и
обхватывает ее пальцами, шероховатая поверхность сухожилий под
тяжестью птицы прижимается к влагалищу и закрепляется на его
Ребрышках. В итоге пальцы фиксируются в согнутом положении и
Плотно обхватывают ветку без участия мышц (рис. 91).
ИЗ
Рис. 91. Схема расположения костей плюсны и сгибающего их сухожи-
лия в ноге птицы:
1 — нога вытянута; 2 — нога согнута; 3 — схема закрепляющего приспособления;
а — ребрышки сухожильной сумки; б — поперечные выросты на сухожилии; п —
пяточное сочленение; х — закрепляющее приспособление
Мышечная система млекопитающих сильно дифферен-
цирована и отличается большим числом разнообразно располо-
женных мускулов (см. рис. 90). Характерно наличие куполообраз-
ной мышцы диафрагмы, отделяющей брюшную полость от груд-
ной. Ее основная роль заключается в изменении объема грудной
полости в связи с актом дыхания. Значительное развитие получает
подкожная мускулатура, которая прикрепляется к коже и приво-
дит в движение те или иные ее участки. У ежей и ящеров она
обусловливает возможность свертывания тела в клубок. Поднятие
игл у ежей и дикобразов, «ощетинивание» зверей и движение чув-
ствующих волос — вибрис, подергивание кожи у лошадей также
вызываются действием этой мускулатуры.
В целом подкожная мускулатура сложна и своеобразна. У рыб с
кожей связана поверхность скелетных мышц. Заметно обособле-
ние подкожных мышц у змей: к каждой из крупных брюшных
чешуй змеи прикрепляется мощная подкожная мышца, что об-
легчает движение животного при цепля-
Рис. 92. Жевательная
мускулатура у грызуна
(белка)
нии за грунт.
У млекопитающих подкожная мускула-
тура комбинирована. Значительную ее
часть в переднем конце тела образует под-
кожная мышца туловища, происходящая
из грудной мышцы, обслуживающей ко-
нечности. Одновременно сжиматель шеи,
образуя кольцо поверхностных волокон
вокруг шеи, является частью висцераль-
ной мускулатуры. Этот подкожный пласт,
давая пучки от шеи на череп и щеки, со-
114
„местно с другими участвует в формировании мускулатуры лица
(рис. 92).
Висцеральная мышечная система дает начало гладкой мускула-
туре внутренних органов. Она имеет иное, чем париетальная мус-
кулатура, происхождение. Начало мускулатуре кишечной трубки
дает висцеральный листок боковой пластинки. Из других участков
мезодермы формируется мускулатура подкожная и кожных же-
лез, стенок кровеносных сосудов, выводных протоков выдели-
тельной и репродуктивной систем. Висцеральная мускулатура в
основном гладкая. Исключения составляют мышца сердца и мус-
кулатура глотки, сложенные поперечнополосатой тканью. В ходе
эволюции значительным изменениям подверглась висцеральная
мускулатура, связанная с глоткой и челюстями. У млекопитаю-
щих ее перестройки привели к формированию сложных мимичес-
ких мышц.
Особое место в мышечной системе занимает сердечная мышца.
Сердце развивается в эмбриогенезе из первичного парного зачат-
ка кровеносного сосуда, расположенного под глоткой (брюшной
аорты). Из его стенок образуется эндокард, а из прилегающего
листка брюшины — миокард и эпикард. Мускулатура сердца (мио-
кард) работает в автономном режиме. Сокращения сердечной
мышцы вызываются периодически появляющимися в ней элект-
рическими импульсами возбуждений, которые возникают в клет-
ках проводящей системы сердца. Энергия сокращения зависит от
степени растяжения мышечных волокон. В эволюции позвоночных
сердечная мышца усложняется. В том числе на внутренней повер-
хности желудочков образуются сосочковые мышцы: от них тянут-
ся сухожильные тяжи к свободным краям клапанов, препятствуя
выворачиванию створок клапанов в сторону предсердий, что обес-
печивает движение крови в одном направлении.
* * *
Таким образом, мускулатура обеспечивает двигательную актив-
ность позвоночного животного и работу его внутренних органов.
Сердечная мышца поддерживает непрерывный поток крови.
ОРГАНЫ ПИЩЕВАРЕНИЯ
Хордовые, как и иглокожие, — животные вторичноротые: их
ротовое отверстие при эмбриогенезе прорывается на конце, про-
тивоположном бластопору. Сквозной пищеварительный канал фор-
мируется из эмбриональной первичной кишки, т. е. закладывается
энтодермально. Со стороны ротового и анального отверстий на-
встречу энтодерме незначительно впячивается эктодерма, и энто-
дермальный эпителий пищеварительного тракта в этих местах не-
заметно переходит в эктодермальный. По существу эпителий ро-
товой полости представляет собой эктодерму, а следующий от-
дел, глотка, преимущественно выстлана энтодермой. То же каса-
ется клоаки и анального отверстия.
Общая организация пищеварительной трубки бесчерепных
проста. Пищевые частицы с током воды поступают в нее через
предротовую воронку, а затем рот. Глотка объемна и прободена
многочисленными жаберными щелями. По средней линии на
брюшной стороне глотки тянется желобок, выстланный желези-
стыми и мерцательными клетками, — эндостилъ. Железистые клет-
ки обволакивают пищевые частицы слизью, а движением ресничек
они увлекаются в обратном направлении. У ротового отверстия эн-
достиль раздваивается и переходит в наджаберную борозду, по ко-
торой пищевые частицы перемещаются в кишечник (рис. 93, 94).
Рис. 93. Продольный разрез тела ланцетника (схема):
1 — предротовое отверстие, окруженное щупальцами; 2 — хвостовой плавник; 3 "
спинной плавник; 4 — метаплевральная складка; 5 — атриальное отверстие; 6"
хорда; 7 — миомер (показан только в хвостовой области); 8 — миосепта; 9 "
нервная трубка; 10 — парус; 11 — жаберные щели; 12 — кишка; 13 — печеноч-
ный вырост кишечника; 14 — околожаберная полость; 15 — эндостиль; 16 "
половые железы
116
рлс. 94. Поперечный разрез ланцетника в
области глотки:
у __ хорда; 2 — нервная трубка; 3 — миосепта;
4 — миомер; 5 — целом; 6 — околожаберная
полость; 7 — эндостиль; 8 — полость глотки; 9 —
жаберное отверстие; 10 — межжаберная перего-
подка; И — нефридий; 12 — печеночный вы-
F рост кишечника; 13 — половая железа
Кишечник простой, на отделы не под-
разделяется. Сразу за глоткой от киш-
ки вперед отходит полый слепой вы-
рост (печеночный), гомологичный по
возникновению и функциям печени
позвоночных*. Переваривание пиши
7 6
происходит за счет ферментов, выделяемых железами в стенках
кишки и печеночного выроста. По стенке выроста пиша переме-
щается дальше в кишечник. Непереваренные остатки пищи удаля-
ются через анальное отверстие, расположенное у основания хво-
стового плавника.
Прогрессивное усложнение кишечного тракта в ряду позво-
ночных идет по пути его удлинения и дифференцировки на отде-
лы. Функционально это обеспечивает лучшую переработку и усво-
Рис. 95. Схема строения позвоночного:
— хорда; 2 — спинной мозг; 3 — головной мозг; 4 — жаберные щели; 5 —
сеРдце; 6 — легкое; 7 — головная почка, или пронефрос; 8 — туловищная поч-
ка, или мезонефрос; 9 — тазовая почка, или метанефрос; 10 — половая железа;
— желудок; 12 — кишка; 13 — поджелудочная железа; 14 — печень; 15 —
селезенка; 16 — мочевой пузырь; 17 — клоака
* Этот вырост закладывается эмбрионально у всех хордовых, в онтогенезе
п°звоночных он превращается в печень.
117
ение пищи, реализацию ее энергетических и строительных воз-
можностей. Пищеварительный тракт позвоночных — система слож-
ная. В нем выделяют следующие основные отделы: ротовую по-
лость, глотку, пищевод, желудок, кишечник (рис. 95).
Ротовая полость
Ротовая полость служит для принятия пищи. Она выстлана
многослойным эктодермальным эпителием. Снаружи рот ограни-
чивают кожные складки — губы, образованные эпидермисом и
соединительной тканью. Не во всех классах они выражены четко.
У миног кольцевой край кожи образует присоску с сосочками, у
миксин рот окружен щупальцами. У рыб, амфибий, рептилий зубы
и челюсти прикрываются слабо выраженными кожными складка-
ми. У черепах челюсти имеют роговые чехлы. У птиц губы превра-
щаются в клюв (надклювье и подклювье). Наконец, только у мле-
копитающих благодаря расположенной в их толще мускулатуре
губы становятся мясистыми и подвижными. Разрастание губных
складок приводит у них к формированию дополнительной полос-
ти между губами и зубами — преддверия рта. С боков ее ограничи-
вают щеки. У некоторых грызунов и обезьян Старого Света за ще-
ками образуются защечные мешки, пригодные для временного со-
хранения пищи, чаще всего при ее транспортировке.
У рыб и амфибий ротовую полость сверху ограничивает дно
мозгового черепа. У легочнодышащих амфибий и рыб (двоякоды-
шащие) в ней видны внутренние отверстия ноздрей — хоаны.
У рептилий и птиц на верхних челюстных костях появляются вда-
ющиеся внутрь ротовой полости гребневидные выросты. У кроко-
дилов и млекопитающих сошник, нёбные кости и нёбные отрос-
тки верхнечелюстных костей образуют вторичное костное нёбо.
Вместе с мягкой нёбной завесой оно отделяет ротовую полость и
ротоглотку от носоглотки.
Ротовая полость выстлана изнутри многослойным эпителием.
У амфибий он имеет реснички и густую сеть капилляров.
Зубы. На нёбе у костных рыб, амфибий и некоторых рептилий
можно увидеть нёбные зубы. Копытные и хищные млекопитаю-
щие имеют на нёбе ороговевшие поперечные гребни, обеспечи-
вающие удержание и перетирание грубой пищи. У усатых китов на
верхних челюстях из этих гребней сформировались мощные рого-
вые пластины китового уса, фильтрующие планктон.
Практически у всех позвоночных, кроме круглоротых, ротовая
полость имеет костные зубы. В некоторых случаях при пищевой
специализации у млекопитающих они недоразвиваются и даже
утрачиваются (неполнозубые, трубкозубы и др.). Зубы удержива-
ют пищу во рту, помогают откусывать, а у млекопитающих —
118
размельчать и перетирать ее. У низших хордовых и бесчелюстных
Настоящих зубов нет. Твердые зубы предротовой воронки миног,
помогающие разрушать покровы жертв, образованы ороговевшей
эпидермальной тканью. Кусательный аппарат вымерших панцир-
ных рыб был образован режущими краями костных челюстей.
Острые зазубренные, одно-, а чаще трехвершинные зубы свой-
ственны акулам — это видоизмененная плакоидная чешуя. Они
развиваются в мягких тканях на внутренней поверхности челюс-
тей: каждый зуб постепенно сдвигается на край челюсти и по мере
старения отпадает, заменяясь другим (рис. 96).
Настоящие конические зубы свойственны костным рыбам, хотя
у многих лучеперых на верхних челюстях они исчезают. У австра-
лийского неоцератода челюсти несут систему пологих радиальных
гребней. Исследования показали, что эмбрионально и филогени-
чески каждый гребень построен из сросшегося ряда изначально
обособленных конических зубов.
Зубы образуются в покровной соединительной ткани, окружа-
ющей челюсти. Толщу зуба слагает плотный дентин — вещество,
близкое по химическому составу к кости и имеющее мезодермаль-
ное происхождение. Внутри зуба сохраняется полость — пульпа,
заполненная мягкой тканью; она содержит питающие зуб крове-
носные сосуды и нервы. Дентин снаружи покрыт эмалью — слоем
блестящего твердого вещества, богатого известью. Эмаль форми-
руется клетками эктодермы. У большинства зубы плотно прирас-
тают к челюстям. У млекопитающих и крокодилов зубы сидят в
ячейках (альвеолах) и имеют один или несколько корней.
Зубы прикрепляются к челюстям губчатым костным цементом.
У акул зубы прикреплены к хрящевым челюстям связками. Связ-
Рис. 96. Разрез челюсти акулы, показывающий последовательные
стадии (/— VII) развития зуба (по А. Ромеру, Т. Парсонсу, 1992):
1 ~~ эмаль; 2 — дентин; 3 — эпидермис ротовой полости; 4 — плакоидная чешуя;
•' ~~ дермальный сосочек ранней зубной закладки; 6 — слой амелобластов; 7 —
сЛой одонтобластов; 8 — нижнечелюстной хрящ; 9 — кориум; 10 — эпидермис
119
ки придают зубам подвижность, про-
пуская пищу в рот и препятствуя
обратному выходу. Подвижное при-
крепление отмечено и у некоторых
костных рыб, ящериц и змей (А.Ро-
мер, Т.Парсонс, 1992) (рис. 97).
У большинства костных рыб, ам-
фибий, гаттерий конические зубы
прикрепляются к наружной поверх-
ности челюстей. У рептилий извест-
но три типа причленения зубов: плев-
родонтный — зубы прирастают к
внутренней поверхности челюстных
костей (примитивные ящерицы, гек-
коны), акродонтных — зубы прирас-
тают к краю или середине челюстей
(большинство чешуйчатых) и теко-
донтный — зубы сидят в альвеолах
(крокодилы, древние зверозубые и
археоптерикс). Млекопитающим свой-
ствен только текодонтный тип — их
Рис. 97. Разрез зуба костной
рыбы, способного отгибаться
внутрь ротовой полости (по
А. Ромеру, Т. Парсонсу,
1992):
1 — кость челюсти; 2 — цемент-
ное основание зуба; 3 — зуб; 4 —
эластичные волокна пульпы; 5 —
неэластичная связка
Рис. 98. Схемы отношений
зубов к челюстным костям у
плевродонтных (А), акродон-
тных (/>) и текодонтных (В)
рептилий
зубы сидят в альвеолах (рис. 98).
В пределах ротовой полости разных групп позвоночных зубы
располагаются не только на челюстях, но и на элементах нёба:
нёбных и крыловидных костях, на сошнике, парасфеноиде. Они
встречаются на внутренней поверхности нижних челюстей, на
элементах висцеральных дуг. Таковы, например, глоточные зубы
карповых рыб. У некоторых сомов насчитывается до 9000 таких
внечелюстных зубов.
Наполненность ротовой полости зубами отражает эктодермаль-
ный характер выстилающей ее ткани, участвующей и в образова-
нии зубов.
В большинстве случаев у рыб, ам-
фибий и рептилий зубы имеют оди-
наковую коническую форму — они
гомодонтны. В зубной системе неко-
торых хищных рыб и рептилий на-
мечается дифференцировка зубов: от-
дельные зубы увеличиваются в раз-
мере. Настоящая гетеродонтностпь
свойственна млекопитающим (рис. 99)-
Их зубы дифференцированы на рез-
цы (incisivi), клыки (canini), предко-
ренные (praemolares) и коренные
(molares). Резцы имеют долотовИД-
ную, реже коническую форму 14
120
Рис. 99. Зубы собаки сбоку (Л) и снизу (Б) (по А.Видерсгейму):
1 — резцы; 2 — клыки; 3 — ложнокоренные; 4 — коренные зубы
1
предназначены для откусывания пищи. Их количество 2 — 3 в каж-
дой половине челюсти. Клыки — зубы одиночные, с длинным
корнем, остроконечной формы. Хорошо развитые у хищников,
они используются при нападении на добычу. Предкоренные име-
ют расширенную коронку, а коренные — сложную бугорчатую ее
поверхность. Они используются при жевании и измельчении пищи.
Есть правило написания зубной формулы млекопитающего в виде
дроби: в числителе записывается число зубов в половине верхней
челюсти, в знаменателе — в половине нижней, в последователь-
ности U С Pm М. Зубная формула отражает пищевую специализа-
цию систематических групп млекопитающих и учитывается как
систематический признак при их определении и классификации.
3.1.3.3 3.1.3.3
Например, зубная формула лошадей — j j j j; коров — qq J
Ю23 3142 ,
грызунов — хищных — (рис. 100).
У всех позвоночных зубы подвержены смене, кроме клыков
Моржей, бивней слонов, нарвалов, которые растут всю жизнь.
У рыб, амфибий, рептилий замещение зубов происходит волна-
ми. Относительно недавно расшифрован ее механизм. Зубы эмб-
рионально закладываются путем выпячивания эктодермы вдоль
всей челюсти и образования зубной пластинки. В ней развиваются
Сдельные зубные зачатки. Впоследствии зубная пластина редуци-
РУется, а ткани с зубными зачатками сохраняются в глубине че-
ДЮсти, часть зачатков остается как бы спрятанными в ней. Сменя-
121
Рис. 100. Схема строения зубного ряда у некоторых млекопитающих:
А — насекомоядные (землеройка); Б — хищные (лисица); В — непарнокопытные
(лошадь); Г — зайцеобразные (заяц)
ющие друг друга зубы отпочковываются от зачатков и постепенно
перемещаются к поверхности (см. рис. 96).
У большинства млекопитающих смена сопровождается заме-
ной части молочных зубов постоянными зубами. У отдельных от-
рядов млекопитающих сменяют друг друга 2 — 3 ряда зубов, у хо-
ботных стершиеся коренные зубы заменяются новыми, нараста-
ющими сзади. Такая смена происходит 5 — 7 раз.
Язык. На дне ротовой полости располагается язык. Его скелет-
ная часть образована подъязычным аппаратом. У рыб этим скеле-
том поддерживается складка слизистой оболочки без мускулату-
ры. У наземных позвоночных язык — подвижный орган, снабжен-
ный мускулатурой. Язык служит для захватывания, удержания и
перемещения пищи во рту, для ощущения ее вкуса, объема, кон-
систенции. У рептилий язык служит органом, доставляющим па-
хучие вещества к якобсонову органу. Он участвует в глотании,
питье, некоторым помогает собирать пищу (хамелеон, муравьед,
лягушки). Форма языка очень изменчива и отражает пищевую спе-
циализацию животного, например у птиц. У бесхвостых амфибий
язык прикреплен обычно спереди к стыку нижних челюстей, его
свободный конец откинут назад к глотке. Есть группа амфибий,
лишенная языка, — безъязычные лягушки.
У высших позвоночных язык закладывается в нижней части
глотки вблизи щитовидной железы. Формируются корень и тело
языка. Рожки подъязычного аппарата служат местом прикрепле-
ния мышц, движущих языком: его выдвигает подбородочно-языч-
ная мышца и втягивает подъязычно-язычная мышца. Собственная
мускулатура языка обособляется в онтогенезе от подъязычной
мышцы. Мускулатура языка иннервируется подъязычным нервом
(XII) и ветвями тройничного нерва (V).
У птиц язык спереди заострен и в толще лишен мускулатуры-
Он отличается сильным развитием на поверхности рогового слоя-
У дятлов, колибри длинный язык выдвигается с помощью подъя-
зычного аппарата (рис. 101). Его задние рожки огибают череп сза-
122
рис. 101. Механизм втягивания (Л) и
выбрасывания (Б) языка дятла:
] — язык; 2 — рожки подъязычного
аппарата
2
дИ и ложатся сверху, доходя до
ноздрей. У многих насекомоядных
птиц язык снабжен шипиками.
Мясистый язык свойствен попу-
гаям.
Язык млекопитающих очень
подвижен. Он снабжен мускулату-
рой, чрезвычайно богат железами
и несет многочисленные вкусовые
сосочки.
Железы ротовой полости. При питании первичноводных по-
звоночных вода смачивает пищу и облегчает ее проглатывание,
поэтому у них во рту пищеварительные железы отсутствуют. Во
рту у рыб есть отдельные секретирующие клетки, у миног есть
железа, секрет которой препятствует свертыванию крови жертвы.
Настоящие слюнные железы существуют в ротовой полости лишь
у наземных позвоночных.
Секрет слюнных желез смачивает слизистую оболочку ротовой
полости и попавшую в рот пищу. У амфибий есть крупная трубча-
тая межносовая (или межчелюстная) железа, нёбные железы и мел-
кие железы на языке. Слюна у большинства амфибий лишь смачи-
вает пишу, в ней нет ферментов. У рептилий есть еще подъязычные,
губные, зубные. У ядовитых змей и ящериц-ядозубов пара задних
верхнезубных желез преобразуется в ядовитый аппарат, располо-
женный в основании пары соответствующих зубов. Яд использует-
ся ими для обездвиживания добычи, в редких случаях как сред-
ство защиты. Птипы, добывающие корм в воде, слюнных желез
почти не имеют. В отличие от них зерноядным птицам секрет слюн-
ных желез крайне необходим, так как их пища содержит крахмал
и клетчатку. Некоторые птицы используют слюну при постройке
гнезд (стрижи-саланганы и др.).
У большинства амфибий и рептилий секрет слюнных желез не
содержит химически активных веществ. Лишь у некоторых бесхво-
стых земноводных, некоторых рептилий и у птиц в слюне обнару-
жен фермент птиалин, участвующий в переваривании крахмала.
Наибольшего развития слюнные железы достигают у млекопи-
тающих. У них есть во рту крупные железы: подъязычная, заднея-
зычная, подчелюстная, околоушная и многочисленные мелкие
слизистые железы, разбросанные по его поверхности. Во рту на-
123
Спиральный
Пищевод клапан Кишка
Язык
Дыхательная
трубка
Прямая
кишка
Анус
Жаберные
мешки
Минога
Акула
Наружная и внутренняя
ноздри
Пищевод
Кишка
«Желудок»
Прямая
кишка
Ректальная
железа
Клоака
Жаберные
щели
Спиральный
клапан
Протоптерус
Осетр
Окунь
124
Печень
iiL' Пищевод
ЭКелчный
пузырь
Мочевой
пузырь
Желудок
—Клоака
Толстая
кишка
Поджелудочная
железа,
скрывающая
желчный проток
Лягушка
Пилорический отдел
Тонкая кишка
Поджелу
дочная
железа
Пищевод
Печень
Желчные
протоки
Протоки
поджелу-
дочной
железы
Зоб
Клоака
Тонкая кишка
с брыжейкой
Слепые
выросты
•Железистый
желудок
Мускульный
желудок
Двенадцати-
перстная кишка
Птица
Желчный
проток
Поджелудочная
железа
- — Пищевод
Желчный пузырь
Слепая
кишка
Проток
поджелу-
дочной
железы
Печень
Прямая кишка
Тонкая
кишка
Желудок
Поперечная
часть толстой
кишки
Двенадцати-
перстная
кишка
Восходящая часть
толстой кишки
Нисходящая часть
толстой кишки
Млекопитающее
Рис. 102. Схема строения пищеварительной системы разных животных
(по А. Ромеру, Т. Парсонсу, 1992)
чинается первичная химическая обработка сложных углеводов. За-
мечено, что некоторые землеройки имеют ядовитые железы. В ро-
товой полости млекопитающих есть и другие железы: молярные
Железы у копытных, глазничные железы у собак, наконец, на
гРанице с глоткой лежат лимфоидные миндалины.
Пищеварительный канал позвоночных разделен на отделы
ФИс. 102).
125
Глотка и пищевод
Глотка — это короткий участок пищеварительного тракта, рас-
положенный между ротовой полостью и пищеводом. Ее стенки
выстланы многослойным энтодермальным эпителием и снабже-
ны поперечнополосатой мускулатурой. У наземных позвоночных
животных сверху в глотку открываются евстахиевы трубы—гомо-
логи IV жаберной щели, сообщающиеся с полостями среднего
уха, снизу — гортанная щель ведет в легочный аппарат. Глоточ-
ный отдел пищеварительной трубки играет важную роль в эмбри-
огенезе позвоночных: здесь развивается дыхательный аппарат, а
также железы внутренней секреции — щитовидная, паращито-
видная, зобная и аденогипофиз. Мускулатура глотки участвует в
акте глотания и проталкивании пищи в пищевод.
Пищевод — отдел собственно кишечной трубки, служащий для
проведения пищи. У рыб, амфибий, чешуйчатых рептилий он
выстлан многослойным эпителием, клетки которого часто имеют
реснички и активно транспортируют пищевой комок. Иногда внут-
ренний слой эпителия пищевода ороговевает. На внутренней его
поверхности есть продольные складки, способствующие его рас-
тяжению. У многих птиц пищевод имеет расширение — зоб, где
пища резервируется и увлажняется. У голубей во время насижива-
ния и выкармливания птенцов стенка зоба утолщается, клетки
его стенок испытывают частичное жировое перерождение и выде-
ляют полужидкий (творожистый) продукт, которым птицы вы-
кармливают птенцов.
Желудок
Желудок — мешковидное, мускулистое расширение кишечно-
го тракта, где пища накапливается и начинается ее химическая
обработка, главным образом белков, чаще всего пепсином в кис-
лой среде. Благодаря мощной мышечной ткани пища размягчает-
ся и раздавливается, подготавливаясь к химической обработке.
Форма желудка весьма изменчива. У рыб с удлиненным телом он
вытянут в длину, имеет мешковидную форму или У-образно изог-
нут. В мешковидном желудке млекопитающих передняя стенка
желудка вогнута и образует малую кривизну, задняя, выпуклая,
образует большую кривизну. Стенка желудка выстлана слизистой
оболочкой, собранной в сетевидные складки.
Внутри желудка выделяют зоны по типу выстилающего их эпи-
телия. Наиболее многообразны эти зоны у млекопитающих. Часть
желудка, ближайшая к пищеводу, — пищеводная — выстлана ли-
шенным желез многослойным эндотелием, и ее по существу счи-
тают продолжением пищевода. Далее к ней примыкает кардиал^
126
^ая зона, по-настоящему выраженная у млекопитающих. Она вы-
стлана однослойным цилиндрическим эпителием и имеет трубча-
тые железы, выделяющие слизистый секрет. Следующая область
желудка — донная. Ее многочисленные трубчатые железы выраба-
тывают пищеварительные ферменты: пепсиноген и прохимозин,
воздействующие на белки, липазу, расщепляющую жиры. Здесь
же выделяется соляная кислота, активирующая пищеварительные
ферменты желудочного сока.
Область желудка, примыкающую к кишке, называют пилори-
ческой. Ее завершает кольцевая складка — пилорический клапан.
В пилорической области пища частично нейтрализуется перед ее
переходом в щелочную среду кишечника.
Разделение на зоны неодинаково в желудках животных различ-
ных систематических групп и даже разных видов позвоночных жи-
вотных. На рис. 103 приведена сравнительная схема расположения
зон в желудках различных животных. В подковообразно загнутом
желудке акуловых рыб различают части: кардиальную, донную и
пилорическую. Загнутая часть желудка костистых рыб иногда снаб-
жена мешком с донной зоной. В его пилорической части располо-
жены слепые выросты — пилорические придатки. Они увеличивают
пищеварительную поверхность и нейтрализуют среду.
У амфибий, ящериц, змей желудок прямой, слегка расширен-
ный в мешок. У крокодилов за большим мешковидным и муску-
листым железистым отделом следует пилорическая часть. У птиц
желудок четко дифференцирован на железистый и мускульный.
Рис. ЮЗ. Расположение зон в желудках разных позвоночных животных:
— акула; Б — костистая рыба; В — тритон; Г — черепаха; Д — птица; Е —
Человек; Ж — заяц; 3 — суслик; И — даман; К — дельфин; Л — кенгуру; М —
Корова; 1 — пищеводная; 2 — кардиальная; 3 — донная; 4 — пилорическая
127
Рис. 104. Желудок коровы: размещение в теле (А), строение и схема
перемещения пищи (Б):
1 — пищевод; 2 — рубец; 3 — сетка; 4 — книжка; 5 — сычуг; 6 — тонкая кишка
В железистом желудке донные железы распределены в виде кар-
машков. Мускульный желудок имеет толстые мышечные стенки,
а внутри выстлан твердой рогоподобной оболочкой с бугорчаты-
ми стенками. Эта область вместе с заглатываемыми камешками
участвует в перетирании пиши. Наибольшего развития мускуль-
ный желудок достигает у зерноядных птиц. У хищных птиц здесь
формируются погадки и отрыгиваются по мере накопления.
Чрезвычайно изменчивы желудки у млекопитающих. Напри-
мер, у однопроходных, кенгуру, даманов, у некоторых мышей
значительная часть желудка является пищеводной: она выстлана,
как пищевод, многослойным плоским эндотелием и не имеет желез.
Желудок жвачных заслуживает специального описания (рис. 104).
У быков, баранов, козлов, оленей желудок имеет четыре камеры:
рубец, сетку, книжку, сычуг. Проглоченная пища попадает сначала в
мешковидный рубец и сетку. Здесь она увлажняется, разминается
и под влиянием бактерий сбраживается. У верблюдов в рубце и
сетке есть водяные клетки, где надолго задерживается запас воды
(верблюд единожды может выпить количество воды, равное од-
ной трети его веса). Затем животное отрыгивает жвачку в рот для
дополнительного пережевывания. Обработанная вновь пиша спус-
кается по глубокой складке сетки в книжку. Ее стенка характери-
зуется множеством складок. Следующий отдел — сычуг — имеет
три типа эпителия, характерных для отделов желудка других мле-
копитающих: кардиальный, донный и пилорический, и является
настоящим переваривающим пищу отделом, т.е. собственно же-
лудком.
В многокамерном желудке китообразных разрастаются области
донных и пилорических желез. Желудок хищных — однокамер-
ный, но в нем хорошо развиты кардиальные, донные и пилори-
ческие железы. То же свойственно и человеку.
128
Кишечник
Кишечник считают- главным отделом пищеварительной систе-
мы. Здесь пища проходит основную химическую обработку (пере-
варивание) и всасывается через кишечную стенку в кровь. Функ-
ционально кишечник подразделен на отделы: тонкий и толстый.
Передний отдел тонкой кишки у высших позвоночных называют
двенадцатиперстной кишкой, по названию соответствующего от-
дела в кишечнике человека. Задний отдел толстой кишки состав-
ляет прямую кишку. На границе тонкой и толстой кишок у амниот
лежит объемистая слепая кишка.
Внутренняя поверхность тонкого кишечника выстлана эндоте-
лием (рис. 105). В нем рассеяны бокаловидные клетки, выделяю-
щие слизь, и трубчатые пищеварительные железы. В области две-
надцатиперстной кишки лежат бруннеровы железы, близкие к пи-
лорическим железам желудка. В средней части многочисленны мел-
кие либеркюновы железы, вырабатывающие желудочно-кишечные
гормоны, которые стимулируют деятельность поджелудочной же-
лезы, желчного пузыря, печени и создают щелочную среду в ки-
шечнике. Кроме собственно кишечных желез в просвет начала тон-
кого кишечника открываются протоки поджелудочной железы и
желчных протоков печени. Кишечный сок способствует окончатель-
ному расщеплению всех видов органической пищи.
Поверхностный эпителий кишки имеет микроворсинки, осу-
ществляющие всасывание расщепленных пищевых веществ. Через
их стенки из кишечника в кровь и лимфатические сосуды прони-
кают вода, соли, простые сахара, жирные кислоты, аминокисло-
ты, витамины, кишечные гормоны. Всасывающая поверхность
эпителия кишки в эволюционном ряду позвоночных развивается
разными путями. У одних в кишечнике есть большая складка эпи-
телия и соединительной ткани, делающая в полости кишки кру-
тые обороты — так называемый спиральный клапан. Эти обороты
напоминают вложенные друг в друга конуса. Спиральный клапан
существовал у панцирных рыб, он есть у акул, скатов и химер,
Двоякодышащих и костных ганоидов, миног. Костистые рыбы спи-
рального клапана не имеют, однако у них по сравнению с хряще-
выми рыбами кишечник заметно удлиняется, причем особенно
заметно у растительноядных рыб (рис. 106). Длина его в среднем в
1,5 раза превышает длину тела рыбы. Кроме того, у многих кости-
стых рыб есть система многочисленных слепых отростков (до 900)
На границе с желудком: их называют пилорическими выростами.
Они увеличивают всасывающую поверхность кишечника (рис. 107).
Важную роль в усилении всасывающей способности кишечни-
ка играют многочисленные складки, перегородки слизистой обо-
лочки кишечника. Они есть у амфибий и рептилий. Высшим позво-
ночным птицам и млекопитающим свойственны в кишечнике мак-
Константинов 129
Кольцевой слой
мышечной оболочки
Мышечная
пластинка
слизистой
Собственная
пластинка
слизистой
Либеркюнова
железа
Бруннеровы Брыжейка
железы
Серозная оболочка
Подслизистая
основа
Проток
Ворсинки
Просвет
Мейснерово
сплетение
Продольный слой
мышечной оболочки
Ауэрбахово
сплетение
Эпителиальная
выстилка
Лимфатический узел
Крупная железа
(например,
поджелудочная)
А
Рис. 105. Схема поперечного разреза тонкой кишки (А), фрагмент среза
печени (Б) млекопитающего
130
Рис. 106. Схема кишечника различных цихлидовых рыб:
А — хищник; Б — животноед; В — растительноед
роворсинки. Они покрывают всю внутреннюю поверхность слизис-
той оболочки кишки, придавая ей бархатистый вид (И.И.Шмаль-
гаузен, 1964). В кишечнике хорошо развиты соединительнотканный
и мышечный слои. В стенке кишки присутствует гладкая мускула-
тура. Перемещение пищи по кишечнику благодаря периодичес-
ким сокращениям его мускулатуры называют перистальтикой.
Толстая кишка обычно короче тонкой. Она образует последо-
вательные мешковидные выступы, отделенные друг от друга пе-
рехватами. Здесь продолжается обработка пищи ферментами сока
тонкого кишечника. Кроме того, этот отдел содержит много раз-
личных бактерий и иногда простейших, успешно завершающих
процесс расщепления белков, жиров и углеводов пищи, особен-
Рис. 107. Стадии (/— ПГ) последовательного изменения кишечника
лососевой рыбы при движении на нерест
131
но трудно перевариваемой растительной клетчатки — целлюлозы.
Стенки толстой кишки всасывают воду, а в ее просвете формиру-
ются фекальные массы.
На границе тонкого и толстого кишечника у всех животных
группы амниота лежит слепой вырост — слепая кишка. Наиболь-
ших размеров она достигает у травоядных млекопитающих. В ее
стенках находится много лимфатических узлов. В химусе слепой
кишки содержится множество бактерий и простейших. У многих
грызунов, полуобезьян, обезьян (и у человека) слепая кишка силь-
но редуцирована и представлена червеобразным отростком — ап-
пендиксом, который считают рудиментом этого отдела кишечника.
Учитывая обилие лимфатических узлов в его стенках, можно по-
лагать, что он играет роль лимфатической железы.
Задний отдел толстой кишки — прямая кишка — снабжена
не только гладкой, но и поперечнополосатой мускулатурой, об-
разующей кольцеобразные мышцы-замыкатели — сфинктеры.
Прямая кишка наиболее четко выделяется у млекопитающих.
Она завершается у них анальным отверстием. У пластинчатожа-
берных и двоякодышащих рыб, у амфибий, рептилий, птиц,
однопроходных млекопитающих толстый кишечник завершает-
ся клоакой. Клоака — это комплексный орган, который имеет в
эмбриогенезе двойное происхождение. Ее главная часть у заро-
дышей всех позвоночных образована расширением заднего конца
энтодермальной кишечной трубки. С ней объединяется возни-
кающее в виде впячивания углубление эктодермы. Полагают,
что именно эктодерма играет ведущую роль в образовании кло-
аки и ее производных — мочевого пузыря, наружных половых
выростов. В клоаку открываются протоки половых желез и моче-
точники* .
У птиц в дорсальной части клоаки развивается лимфоидный
орган — фабрициева сумка.
Пищеварительные железы
Различные выросты пищеварительной трубки дают начало мно-
гим специфическим органам: пищеварительным железам (слюн-
ным, печени и поджелудочный железе), железам внутренней сек-
реции (щитовидной, паращитовидной, зобной), органам дыхания,
плавательному пузырю рыб, селезенке.
Как сказано выше, в тонкий кишечник открываются пищева-
рительные железы.
* В отдельных случаях в клоаку объединяются половые и выделительные
протоки: у самки латимерии, костных ганоидов. Частью клоаки у некоторых ко-
стистых рыб является мочеполовой синус.
132
Поджелудочная железа эмбрионально развивается из несколь-
выростов (в виде небольших почек) среднего отдела кишеч-
дика. Чаще всего образуются один спинной и два брюшных выро-
ста, которые затем объединяются вместе. Поджелудочная железа
имеет один или несколько протоков, открывающихся в началь-
ный отдел тонкой кишки.
У птиц и млекопитающих (а также акул, амфибий и репти-
лий) железа располагается обычно в первой петле кишечника,
у миног и двоякодышащих рыб она скрыта в стенках кишечника,
у некоторых костистых рыб и миксин примыкает к ткани печени
или рассеяна вдоль кишечника.
Поджелудочная железа имеет сложное альвеолярное строение.
Ее отдельные дольки (ацинусы) вырабатывают секрет, поступа-
ющий сначала в мелкие, а затем в крупные протоки. Выделения
поджелудочной железы содержат важные пищеварительные фер-
менты (трипсиноген, эректазу, диастазу, липазу и др.) (около
10) для переваривания трех основных составных частей органи-
ческой пищи: белков, углеводов и жиров.
Часть поджелудочной железы выполняет эндокринную функ-
цию (см. рис. 195).
Печень развивается в эмбриогенезе как направленный вперед
вырост брюшной стенки кишечника. У низших хордовых (ланцет-
ники и личинки миноги-пескоройки) печеночный вырост выст-
лан железистым эпителием и участвует в усвоении пищи (и регу-
ляции венозного кровообращения).
В теле эмбрионов высших позвоночных животных на начальных
этапах развития печень представляет собой сложную трубчатую же-
лезу. За счет многочисленных анастомозов она приобретает сетчатую
структуру. Печень взрослых особей состоит из множества печеноч-
ных клеток эктодермального происхождения (см. рис. 105, Б). У взрос-
лых амниот эти клетки имеют многогранную форму и собраны в
множество мелких долек, пронизанных сетью мелких желчных ка-
пилляров. Они собирают поступающие из долек секрет печени —
желчь — в один или несколько печеночных протоков. Желчь накап-
ливается в желчном пузыре и по общему желчному протоку (иног-
да вместе с протоком поджелудочной железы) поступает в началь-
ный отдел тонкой кишки*. Печень имеет две или три лопасти, ко-
торые у млекопитающих могут быть поделены на дольки.
Выделяемая печенью желчь имеет щелочную реакцию. Она ней-
трализует желудочный сок, эмульгирует жиры, активизирует ли-
ПазУ, ферменты поджелудочной железы, способствует расщепле-
нию крупных молекул жирных кислот и возбуждает перистальти-
ку Кишечника. В состав желчи входят продукты расщепления жи-
* В отдельных случаях желчный пузырь может отсутствовать (минога, голубь,
КРЫса и т.д.), в других случаях желчных протоков может быть несколько.
133
ров и белковых соединений, желчные кислоты, желчные пигмен-
ты (билирубин, биливердин), образующиеся из гемоглобина оч-
мерших и переработанных в печени эритроцитов.
Печень — орган многофункциональный. Для жизнедеятельнос-
ти собственных клеток печень снабжается артериальной кровью
через печеночную артерию. Однако объем венозной крови, проте-
кающей через воротную систему кровообращения печени, намного
больше ее артериального кровоснабжения. Через воротную систе-
му в печень притекает кровь от кишечника. Клетки печени отби-
рают из крови питательные вещества для запасания и химической
переработки, очищают кровь от ядов, освобождают ее от отмер-
ших кровяных клеток.
Печень участвует в запасании углеводов. Углеводы откладыва-
ются в печени в виде гликогена (животного крахмала). По мере
необходимости они вновь поступают в кровь. У некоторых рыб —
акуловые, тресковые — в печени запасается жидкий жир. Клетки
печени синтезируют белки, превращают белки и жиры в углево-
ды. Они преображают конечные продукты азотистого обмена (ам-
миак) в мочевину и мочевую кислоту, поступающие затем в поч-
ки и выделяемые ими наружу.
* * *
Таким образом, возникающая энтодермально нечлененная
пищеварительная трубка функционирует у взрослых ланцетников.
В процессе эволюции происходит последовательная ее дифферен-
цировка и подразделение на специализированные отделы. В пере-
днем отделе выделяются ротовая полость, пищевод, желудок;
кишечник подразделяется на тонкую (ее передний отдел — две-
надцатиперстную) и толстую кишку. Сравнительно рано возника-
ют зубы, сначала только удерживающие и отделяющие пищевые
порции, затем измельчающие и перетирающие пищу. Наиболее
совершенная дифференцированная альвеолярная зубная система
свойственна млекопитающим. В ротовой полости появляется пред-
дверие рта, пищевые пути в глотке отделяются от дыхательных.
Слюнные железы, возникшие впервые у амфибий, только смачи-
вали пищу. При дальнейшей специализации развивается несколь-
ко слюнных желез, выделяющих разные ферменты, в том числе
расщепляющие сложные сахара. У рептилий слюнные железы пре-
образуются в ядовитые. Пищевод развивается в связи с выделени-
ем шейного отдела тела. У зерноядных и хищных птиц появляется
зоб, а желудок подразделяется на железистый и мускульный. Наи-
большей дифференцировки достигает желудок у жвачных парно-
копытных, из которых только последний отдел является истин-
ным гомологом желудка других зверей, а остальные отделы пред-
ставляют собой видоизменения пищевода. Непременным услови-
ем успешной работы кишечника является его длина. Вначале
134
замедления движения и лучшего усвоения пищи служит спираль-
ный клапан у круглоротых и хрящевых рыб, затем многократно
возросшая длина тонкого кишечника у птиц и млекопитающих.
Увеличение поверхности кишечника достигается и слепыми вы-
ростами, которые возникали неоднократно у разных системати-
ческих групп. У рыб их бывает много (3 — 22), у растительноядных
млекопитающих присутствует слепая кишка. Долгое время в эво-
люционном ряду (до первозверей) сохраняется последний отдел
толстой кишки — клоака. Пищеварительные железы возникают
как выпячивания первичной кишки. В процессе эволюции возрас-
тает специализация пищеварительных ферментов: они активно
действуют на определенные вещества при определенном уровне
кислотности и в узком температурном интервале.
ОРГАНЫ ДЫХАНИЯ
Органы дыхания осуществляют газообмен между организмом
животного и окружающей средой. Из внешней среды животное
поглощает атмосферный (или растворенный в воде) кислород и
выделяет в эту среду углекислый газ, образовавшийся в теле в
результате окислительных процессов метаболизма.
Органы дыхания имеют и дополнительные функции: у наземных
позвоночных они участвуют в терморегуляции (полипное), у неко-
торых рептилий, птиц и млекопитающих — в голосообразовании, у
рыб в выделении аммиака и других конечных продуктов белкового
метаболизма, легкие амфибий и рептилий используются в различ-
ных поведенческих реакциях (например, при раздувании тела).
Хордовым свойственны органы дыхания двух типов: у первич-
новодных — жабры, у наземных — легкие. У многих существенное
значение имеет кожное дыхание.
Органы дыхания закладываются в онтогенезе позвоночных в
переднем отделе пищеварительной трубки — в глотке. На ранних
стадиях эмбриогенеза в энтодерме глотки выпячиваются карма-
нообразные выросты. Одновременно в наружных покровах появ-
ляются встречные эктодермальные впячивания. Прорывающиеся
в результате их смыкания сквозные висцеральные щели имеют сме-
шанное энто-эктодермальное происхождение. Висцеральные щели
закладываются эмбрионально у всех позвоночных, однако у на-
земных в ходе онтогенеза они зарастают и исчезают. У всех сохра-
няется только одна висцеральная щель: она участвует в усложне-
нии органов слуха.
У первичноводных позвоночных функцию газообмена в сквоз-
ных щелях осуществляют жаберные лепестки. У бесчелюстных и
челюстноротых жаберные лепестки имеют разное происхождение,
что стало, видимо, важным признаком, подтверждающим их об-
щее происхождение и раннюю дивергенцию. Они закладываются
или в виде меридиональных складок эктодермальной стенки (щит-
ковые, круглоротые), или в виде складок слизистой эктодермы
(все рыбы, личинки амфибий, некоторые взрослые амфибии)-
У некоторых рыб, а также у личинок амфибий развиваются соот-
ветственно эктодермальные наружные перистые жабры.
Легкие — органы дыхания наземных позвоночных возникают
как выпячивания дна глотки позади жаберного аппарата, т. е. име-
136
jot эктодермальное происхождение. Они перемещаются в даль-
нейшем в полости тела на спинную сторону, сохранив вход в глотку
с брюшной стороны.
Рассмотрим подробнее изменения дыхательного аппарата в эво-
люционном ряду хордовых.
Дыхание водных позвоночных
Жаберный аппарат
Глотка ланцетника с обеих сторон прободена многочис-
ленными (несколько десятков пар) косо расположенными жа-
берными щелями. В межжаберных перегородках, не ветвясь, прохо-
дят тонкостенные жаберные артерии. Ток воды, перемещаясь из
глотки в околожаберную полость, омывает эти артерии и осуще-
ствляет газообмен. У оболочников вода проходит из глотки через
многочисленные мелкие отверстия — стигмы, как и у ланцетни-
ка, изливаясь в околожаберную полость (рис. 108). Газообмен у
оболочников происходит в многочисленных мелких сосудах, оп-
летающих глотку.
У круглоротых узкие округлые наружные жаберные отвер-
стия ведут в обширные линзообразные жаберные мешки, стенки
которых покрыты многочислен-
ными складками эктодермально-
го жаберного эпителия, прони-
занными обильной сетью кро-
веносных капилляров (рис. 109).
У личинок миног жаберные ле-
пестки представлены полужабра-
ми, расположенными на передней
и задней поверхностях соседних
мешков. У взрослых жаберные ле-
пестки опоясывают каждый мешок
Рис. 108. Вскрытая одиночная асци-
дия:
1 — ротовой сифон; 2 — клоакальный си-
Фон; 3 — туника; 4 — полость глотки; 5 —
Жаберные щели; 6 — эндостиль; 7 — спин-
ная борозда; 8 — околожаберная полость;
стенка околожаберной полости; 10 —
Желудок; 11 — анальное отверстие; 12 —
СеМенник; 13 — яичник; 14 — протоки по-
ловых желез; 15 — околосердечная сумка;
16 — сердце; 17 — нервный узел
137
4 3
Рис. 109. Рот и дыхательные органы морской
миноги:
1 — жаберные мешки; 2 — роговые зубы; 3 — об-
щий жаберный проход; 4 — внутренние и 5 — вне-
шние жаберные проходы (в жаберные мешки и на-
ружу); 6 — рот; 7 — наружные жаберные отверстия;
8 — тканевые перегородки между отдельными жа-
берными мешками
по периметру. На противоположной сто-
роне жаберных мешков расположены внут-
ренние жаберные ходы, ведущие в дыха-
тельную глотку. В онтогенезе миноги она
отделяется от пищеварительной трубки
(рис. ПО). Стенки жаберных мешков под-
держиваются нерасчлененными дугами
висцерального скелета. В сжатии стенок
мешков участвует мускулатура межжаберных перегородок, что по-
зволяет миногам не прерывать дыхание при паразитическом пи-
тании или в период отдыха, когда миноги присасываются к под-
водным предметам. Попаданию пищи в дыхательную глотку при
таранном способе дыхания препятствует прикрывающий ее кла-
пан — парус. Большинство миног имеет семь пар жаберных меш-
ков, открывающихся соответственно на поверхности тела. У мик-
син их число варьирует от 6 до 14. У пиявкоротой миксины отвер-
стия мешков отодвинуты на середину тела (рис. 111). У некоторых
видов есть тенденция к группированию мешков. Глотка обыкно-
венной миксины не разделяется на дыхательную трубку и пище-
вод, а выходы из мешков каждой стороны сливаются у поверхно-
сти тела в единый канал и открываются общим наружным отвер-
стием. Это позволяет животному осуществлять газообмен при глу-
боком погружении в тело жертвы. Эффективность извлечения кис-
1
Рис. ПО. Возрастные изменения жаберной области миноги:
А — личинка; Б — взрослая минога; 1 — глотка, прободенная жаберными отвер-
стиями; 2 — пищевод; 3 — дыхательная трубка; 4 — складка, отделяющая дыха-
тельную трубку от пищевода
1 TR
рис. Ill- Дыхательная система пиявко-
ротой (А) и обыкновенной миксины
(Б)--
у — рот; 2 — усики вокруг рта; 3 — кишка
(глотка); 4 — жаберный мешок (в разрезе
внутри видны складки жаберной поверхно-
сти); 5 — выходные каналы жаберных меш-
ков; 6 — целые жаберные мешки; 7 — об-
щее наружное отверстие каналов жаберных
мешков
А
лорода из воды у круглоротых неве-
лика: 10 — 25 %. Палеонтологи пред-
полагают, что мешковидные жабры
были свойственны и щитковым.
У хрящевых рыб с каждой
стороны тела имеется пять (редко 6
и 7) жаберных отверстий. Они отде-
лены друг от друга широкими межжаберными перегородками, не-
сущими снаружи кожные складки. Каждая складка прикрывает рас-
положенное сзади жаберное отверстие (рис. 112). Внутри каждой
перегородки находятся опорные элементы — хрящевые висцераль-
ные дуги. От них в глубь глотки выступают жаберные тычинки. Они
защищают жабры от засорения твердыми частицами. Наружу к
Рис. 112. Жаберный аппарат
акулы:
А — разрез через голову; Б — поло-
жение жаберных лепестков на пере-
городке; 1 — жаберная дуга; 2 — меж-
жаберная перегородка; 3 — жаберные
лепестки; 4 — жаберные тычинки;
5 — глотка
А Б
139
поверхности тела от дуг направлены жаберные лучи, поддержива-
ющие жабры.
Передние и задние стенки жаберных щелей выстланы слизис-
той оболочкой, образующей пластинчатые выросты с бахромой
жаберных лепестков — это полужабры. В эктодермальных жаберных
лепестках ветвятся приносящие жаберные артерии, образуя ка-
пиллярную сеть, здесь происходит газообмен. Движение потоков
крови и воды в жаберных отверстиях происходит встречно по прин-
ципу противотока. Такое взаимонаправленное движение жидко-
стей позволяет извлекать из воды максимальное количество кис-
лорода. Хрящевым рыбам свойственна таранная вентиляция. В этом
случае вода активно пропускается через открытый рот к жабер-
ным щелям при стремительном движении рыбы вперед. В спокой-
ном состоянии рыбы движение воды создается мускулатурой ро-
товой полости и глотки. Таким способом осуществляется насосная
вентиляция: при открытой ротовой щели и закрытых наружных
жаберных отверстиях глотка расширяется, и рыба через рот наби-
рает воду. Затем рот закрывается, глотка сжимается и выталкивает
воду через висцеральные щели наружу.
Между челюстной и подъязычной дугами у хрящевых рыб рас-
положены парные каналы — брызгальца — видоизмененные жа-
берные щели. Брызгальца служат каналами, пропускающими воду
в глотку для дыхания во время питания рыбы, что особенно важ-
но для видов, питающихся вблизи дна. На задней поверхности
каждого брызгальца есть ложная жабра. Проходящая через нее
артерия участвует в снабжении кислородом глаз и головного моз-
га. У быстроходных акул брызгальца малы, а у медлительных ви-
дов и у обитающих вблизи дна скатов они заметно увеличены.
В отличие от пластинчатожаберных у цельноголовых хрящевых
рыб все жабры прикрыты одной цельной складкой кожи, похо-
жей на жаберную крышку костистых рыб.
В дыхательном аппарате костных рыбв отличие от хряще-
вых межжаберные перегородки редуцированы, а у лучеперых ут-
рачены вовсе. Жаберные лепестки сидят непосредственно на жа-
берных дужках — так из полужабр хрящевых рыб образуются пары
полных жабр костных рыб. Весь жаберный аппарат прикрыт кост-
ной жаберной крышкой с мягкой складкой по краю. У большин-
ства лучеперых на внутренней поверхности жаберной крышки раз-
вивается дополнительная «ложная» жабра. Между жаберной крыш-
кой и жабрами располагается околожаберная, или оперкулярная,
полость (рис. 113).
В механизме насосного дыхания у костных рыб участвует мус-
кулатура ротовой полости, глотки и жаберной крышки. При ра-
стяжении ротовой полости вода всасывается через рот в глотку,
при этом жаберные крышки и их оторочки прижимаются к телу-
Когда рот закрыт, а жаберные крышки начинают отодвигаться,
140
Рис. 113. Жаберный аппарат
костной рыбы:
А — разрез через голову; Б — поло-
жение жабр на жаберной дуге; 1 —
жаберная дуга; 2 — жаберные лепест-
ки; 3 — жаберные тычинки; 4 — жа-
берная крышка; 5 — полость глотки
давление под ними уменьшается, вода из глотки устремляется
через щели в околожаберную полость и при дальнейшем движе-
нии выходит наружу. Последовательная работа разных отделов
дыхательного аппарата делает процесс движения воды непре-
рывным.
Быстроходные рыбы (скумбрии, тунцы, парусники), как и
акулы, используют при движении таранную вентиляцию, держа
на ходу рот открытым. Тем самым снижаются энергозатраты на
процесс дыхания. Эффективность извлечения кислорода при ды-
хании рыб значительно выше, чем у круглоротых: хрящевые рыбы
извлекают до 77 %, а костистые рыбы — до 85 % растворенного в
воде кислорода.
У костистых рыб эффективность дыхания во многом определя-
ется образом жизни и характером занимаемых водоемов. Замече-
но, что у малоподвижных рыб и у видов, живущих в водоемах с
кислородным дефицитом, жаберные лепестки заметно удлинены.
Кроме того, известно, что приспособления к гипоксии выража-
ются у рыб и в биохимических показателях крови: ее кислородная
емкость возрастает в результате увеличения количества эритроци-
тов или путем снижения парциального давления кислорода.
Жабры рыб имеют структурные элементы, способные выде-
лять (морские рыбы) и поглощать (пресноводные рыбы) соль,
ТаКим образом жабры выполняют и выделительную функцию, уда-
ряя аммиак и другие продукты обмена.
141
Дополнительные органы дыхания водных позвоночных. У голо,
кожих рыб (угри, сомы, прыгуны и др.) в водном газообмене
участвует гиперемированная кровеносными сосудами кожа. Мно-
гим сомовым, в том числе вьюнам, свойственно кишечное дыха-
ние. Поднявшись к поверхности воды, рыба заглатывает атмос-
ферный воздух и пропускает его вдоль всего кишечного канала,
обильно снабженного капиллярами. Броняковые сомы при дыха-
нии используют кишечник, пропуская в него проглоченные с по-
верхности воды пузырьки воздуха. Будучи изолированными от по-
верхности воды, они испытывают удушье. Рыбы сем. Слитножа-
берниковые имеют недоразвитые жабры и дышат атмосферным
воздухом благодаря обильной кровеносной сети в глотке и ки-
шечнике. Они могут жить в болоте, выходить на рисовые поля, а
при пересыхании водоемов зарываться в ил. Прыгуны набирают
резерв воды в околожаберные полости и могут долго охотиться на
поверхности водоема, передвигаясь по растениям. При необходи-
мости они еще опускают в воду хвост и восполняют дефицит кис-
лорода кожным дыханием.
Специальные приспособления для дыхания атмосферным воз-
духом есть у лабиринтовых рыб (Anabantidae). В наджаберной об-
ласти их глотки от первой жаберной дуги разрастается сложная
система костных пластинок — лабиринт (рис. 114). Покрывающая
их слизистая оболочка обильно пронизана кровеносными сосу-
дами.
Рыбы заглатывают воздух с поверхности воды и в результате
этого могут держаться либо в обедненных кислородом водоемах,
либо вне воды, используя для дыхания захваченный резерв возду-
ха. Весьма своеобразно, что обогащенная кислородом в наджабер-
ной области кровь поступает у них в кардинальные вены, в ре-
зультате этого через их сердце протекает смешанная кровь. Похо-
жие образования есть у азиатских змееголовов (Channa, или
Ophiocephalus). Они постоянно заглатывают воздух в наджаберный
орган, расположенный в виде пластинчатых отростков первой
жаберной дуги и подвеска, обильно пронизанный кровеносными
сосудами.
Весьма своеобразны некоторые тропические сомы, имеющие раз-
личные приспособления к кислородному дефициту в воде. У кларие-
вых сомов (Clariidae), обитателей пресных водоемов Юго-Восточ-
ной Азии, от жаберной полости тоже отходит древовидно развет-
вленный парный наджаберный орган с густой сетью кровеносных
сосудов. Эти рыбы либо попеременно пользуются жабрами и столь
своеобразными «легкими», либо используют его при переполза-
нии между водоемами (за это они получили название «шагают®
сомы»).
У мешкожаберных сомов (Heteropneustes fossilis) дополнитель-
ные органы дыхания отходят от жаберной полости. Они имеЮт
142
Рис. 114. Дополнительные органы
дыхания у костистых рыб:
А — тропический сом; Б — анабас; В —
кучия; Г — змееголов
вид двух длинных мешков и тянутся под спинной мускулату-
рой.
Добавочным органом воздушного дыхания иногда служит пла-
вательный пузырь (см. ниже). У костных ганоидных рыб (амии, пан-
Цирниковые) плавательный пузырь открывается каналом на спин-
ной стороне пищевода и имеет увеличенную ячеистую поверх-
ность. Поднявшись к поверхности воды, рыбы заглатывают атмос-
ферный воздух и пропускают его в пузырь. У американских эрит-
риновых рыб отверстие плавательного пузыря открывается в киш-
ку сбоку (рис. 115).
Двоякодышащие рыбы и многоперы используют для дыхания
и жабры, и легкие. В отличие от плавательного пузыря легкие зак-
ладываются и открываются в кишку с брюшной стороны. У авст-
ралийского цератода двулопастное легкое перемещается на спин-
ную сторону и располагается над пищеводом. При недостатке в
воде кислорода рыба поднимается к ее поверхности и наполняет
его пузырьками воздуха. У африканских протоптерусов, у южно-
аМериканского лепидосирена одно легкое. Кроме того, у них фор-
мируются наружные жабры — красноватые перистые выросты над
Жаберными отверстиями.
143
Рис. 115. Схемы положения плавательного пузыря и легких у различных
рыб и тетрапод (по А. Ромеру, Т. Парсонсу):
А — осетровые и многие костистые рыбы; Б — Lepidosteus и Amia; В — Erythrinus-,
Г — Neoceratodus", Д — Polypterus-, Е — тетраподы
Рис. 116. Личинка африканского многопера с большой наружной
жаброй
Своеобразные органы дыхания имеют личинки многоперов:
пара наружных жабр тянется вдоль тела над грудными плавника-
ми (рис. 116). Кроме того, в их теле есть и парные легкие, они
асимметричны — более длинная правая его доля располагается
над кишкой.
Развитие легких сопровождается у рыб образованием сквозных
ноздрей.
1 ЛЛ
Дыхание наземных позвоночных
Органы дыхания у амфибий характеризуются переходным
состоянием — сочетанием черт, типичных для водных и наземных
позвоночных. У личинок хвостатых амфибий прорываются три пары
жаберных щелей. Над ними развиваются перистые красноватые
выросты — эктодермальные наружные жабры (рис. 117). Это состо-
яние, характерное для водных личинок всех амфибий, у некото-
рых хвостатых сохраняется пожизненно, и стало их систематичес-
ким признаком (протеи, сирены, аксолотли амбистом). Большин-
ство взрослых хвостатых амфибий используют для дыхания легкие,
кожу, слизистую поверхность рта. Воздух в легкие поступает через
сквозные ноздри, ротоглоточную полость, обозначившуюся гор-
тань. У некоторых хвостатых амфибий легкие вторично утрачены
(безлегочные саламандры). С этим связаны и регрессивные изме-
нения в кровеносной системе (двухкамерное сердце и один круг
кровообращения) и усиление ротового и кожного дыхания.
У личинок бесхвостых амфибий — головастиков — образуются
четыре пары жаберных щелей. На ранней стадии постнатального
онтогенеза около них заметны наружные жабры. Их развитие у
головастиков связывают с дефицитом кислорода в стоячей воде,
где развиваются личинки большинства бесхвостых амфибий. В та-
ких условиях увеличение дыхательной поверхности улучшает воз-
можность использования ограниченных запасов кислорода в сто-
ячей воде, что особенно важно для молодых растущих организмов.
В дальнейшем по бокам головы отрастают оперкулярные складки,
которые прикрывают жаберные щели. Часто под этими временно
возникающими кожными образованиями — крышечками бывают
скрыты и передние конечности.
Во взрослом состоянии бесхвостые амфибии дышат легкими и
через кожу. Легкие представляют парные мешки с тонкими слабо-
ячеистыми стенками. Дыхательная поверхность легких в отличие
от жабр рыб располагается внутри тела животного и непосред-
ственно с водой не соприкасается. Газообмен между кровью и
внешней средой осуществляется через водную пленку, покрыва-
ющую легочный эпителий изнутри. Дыхательные пути у бесхвос-
Рис. 117. Наружные жабры личинки хвостатой амфибии — аксолотля
145
Ноздри
Рис. 118. Последовательные этапы (I—IV) дыхательного акта лягушки
тых оформлены слабо. Воздух через ноздри, ротоглоточную по-
лость проходит в легкие через гортанную щель. Механизм легочно-
го дыхания в связи с отсутствием грудной клетки своеобразен
(рис. 118). Роль насоса выполняет дно ротоглоточной полости, ко-
торое то опускается (воздух через открытые ноздри всасывается),
то поднимается (воздух при закрытых ноздрях проталкивается в
легкие). В этой связи заслуживает внимания структура головы ам-
фибий: их череп при небольшой высоте мозговой части чрезвы-
чайно широк за счет значительного разведения ветвей челюстей.
В этом случае эффективность легочного дыхания будет тем боль-
шей, чем шире расстояние между ветвями нижней челюсти (на-
пример, у жаб, рогаток).
Легкие амфибий примитивны, их дыхательная поверхность мала.
У лягушек она составляет 0,25 м2/кг. Недостаточность легочного
дыхания компенсируется у них кожным дыханием. Значение кож-
ного дыхания чрезвычайно велико. Влажность кожи постоянно
поддерживается выделениями слизистых желез. Соотношение по-
верхности легких и кожи у многих амфибий составляет 2:3 (табл. 11-
Значение кожи в дыхательном процессе особенно заметно воз-
растает при водном образе жизни (тритоны, жерлянки, зеленые
лягушки). Например, у прудовой лягушки через кожу поступает
51 % кислорода, употребляемого для окисления крови, и 49 % его
поступает через легкие. Еще большее значение кожное дыхание
имеет для выведения из тела амфибии углекислоты: 86 % СО2 вы-
деляется через кожу и лишь 14 % через легкие. Как сказано выше,
некоторые амфибии (например, саламандры р. Plethodontidae) вов-
се не имеют легких и дышат только через кожу.
146
Таблица 1
Показатели, позволяющие оценить роль кожи и других органов
в обеспечении дыхания некоторых амфибий (по С. П. Наумову, 1972)
Вид земноводных Толщина эпидермиса, мкм Длина капилляров дыхательной поверхности, %
Кожа Легкие Слизистая рта
Гребенчатый 25,0 73,7 23,3 3,0
тритон Жерлянка . 22,8 53,7 42,5 1,1
Травяная лягушка 42,5 36,6 62,5 0,9
Прудовая лягушка 39,1 34,3 64,8 0,9
Жаба серая 51,9 27,6 71,5 0,9
Квакша обыкновенная 32,6 24,2 74,7 1,1
Кожное дыхание у амфибий не только имеет большое функци-
ональное значение в связи с малой поверхностью легких, но и
служит как приспособление, обеспечивающее окисление крови
при длительном нахождении животного в воде, например во вре-
мя спячки, при затаивании в водоеме, при преследовании хищ-
никами. В этих случаях окисление крови осуществляется только за
счет кожного дыхания, и правое предсердие (через полую вену
сюда поступает окисленная кровь кожных вен) становится арте-
риальным, а левое венозным. Самостоятельное значение в газооб-
мене амфибий имеет вентиляция ротовой полости.
Рептилиив отличие от амфибий развиваются в яйце на суше
и не имеют обитающих в воде личинок. Во время их эмбриональ-
ного развития функциональный жаберный аппарат не формиру-
ется и газообмен у зародыша, находящегося в яйце, осуществля-
ется с помощью сосудов желточного мешка и аллантоиса. Взрос-
лые рептилии дышат только легкими. В связи с развитием кожного
рогового покрова кожное дыхание у них практически отсутствует.
Уместно отметить, что некоторые водные рептилии используют
Для дыхания иные возможности: водные черепахи — слизистую
Рта, морские змеи — кожу. Жизнь в наземной среде, а также более
высокий общий уровень обменных процессов обусловливают у реп-
тилий и более сложное строение органов дыхательной системы.
Общая форма легких, как и у амфибий, мешковидная, однако
внутренняя полость заметно уменьшена. От стенок легких внутрь
отходит сложная сеть перегородок, делящих полость легкого на
147
множество мелких ячей. Особенно сильно развиты такие перего-
родки у черепах, крокодилов, варанов. Слабую ячеистость внут-
ренней полости легких отмечают у примитивной рептилии — гат-
терии, у роющей — амфисбены (рис. 119). Задняя часть легких у
некоторых ящериц и особенно у хамелеонов не имеет ячей и пе-
регородок и часто вытянута в виде тонкостенных пальцевидных
выростов — легочных мешков. Кровь в этих выростах не окисляется.
Дыхательные пути у рептилий значительно больше дифферен-
цированы, чем у амфибий. Это связано с выделением шейного
отдела тела. Гортанная щель ведет в гортань, поддерживаемую не-
парным перстневидным и парным черпаловидным хрящами. От гор-
тани отходит длинная, сложенная хрящевыми кольцами трахея-,
ее развитие находится в прямой корреляции с появлением шеи.
Трахея делится на два бронха, которые подводят воздух к легким.
В сравнении с амфибиями у рептилий изменяется и механизм
дыхания. Воздух почти не заглатывается ртом, а втягивается в лег-
кие и выталкивается обратно путем расширения и сужения груд-
ной клетки, обусловленных движением ребер. Этот тип дыхания,
характерный для высших позвоночных, а также более сложная
структура легких обеспечивают и более совершенный газообмен.
Рис. 119. Легкие пресмыкающихся:
А — амфисбена; Б — анаконда; В — гаттерия; Г — варан; Д — аллигатор; Е
хамелеон
148
у крокодилов в полости тела есть складка, напоминающая ди-
афрагму млекопитающих, которая может участвовать в акте дыха-
щ1Я. Затягивание воздуха частично сохраняется у черепах. У них ребра
неподвижно прирастают к панцирю. Эти рептилии используют для
вдоха и выдоха движения парных конечностей и их поясов.
Пресмыкающиеся — пойкилотермные животные и частота вен-
тиляции легких имеет приспособительное значение. Она участвует
в терморегуляции тела животного и меняется в зависимости от
температуры окружающей среды. При длительном нахождении на
солнце тело рептилии нагревается и быстро остывает при значи-
тельном падении температуры окружающего воздуха. В терморегу-
ляцию включаются разные механизмы. К примеру, у некоторых
пустынных ящериц при подъеме температуры воздуха увеличива-
ется частота дыхательных движений от 26 до 37 в минуту (напри-
мер, у заборных игуан рода Sceloporus). У некоторых пустынных
черепах (например, Testudo sulcata) при значительном повыше-
нии температуры воздуха резко возрастает отделение слюны: вы-
текая изо рта, она смачивает нижнюю часть головы, шею и ко-
нечности. Испарение слюны с поверхности кожи существенно
повышает теплоотдачу. Реакцией на повышение (и понижение)
температуры является у рептилий оцепенение, впадение в спячку.
Однако эти приспособления имеют временное значение и не мо-
гут обеспечить этой группе наземных позвоночных постоянство
температуры тела. Неслучайно рас-
пространение рептилий ограничено
жаркими климатическими поясами.
Органы дыхания птиц сложны
и крайне своеобразны. Они, с одной
стороны, имеют черты приспособле-
ния к жизни в наземной среде, с
другой — специализированы в связи
с движением птиц в воздухе. Дыха-
тельная система птиц включает гор-
тань, трахею, систему бронхов, лег-
кие, воздушные мешки (рис. 120).
Рис. 120. Схема дыхательной системы
птицы, вид с брюшной стороны:
— трахея; 2 — легкое; 3 — шейный мешок;
— межключичный мешок; 5, 6, 7 и 8 —
выросты межключичного мешка; 9 — перел-
Негрудной мешок; 10 — заднегрудной ме-
шок; 11 — брюшной воздушный мешок
149
Рис. 121. Нижняя, или певчая,
гортань голубя:
А — вид спереди; Б — вид сбоку; ]
бронхотрахейные мешки; 2 — нару^.
ная голосовая перепонка; 3 — после,
днее кольцо трахеи; 4 — первое коль-
цо бронха
Верхнюю часть трахеи, как и у рептилий, образует гортань,
поддерживаемая непарным перстневидным и парными черпало-
видными хрящами. У птиц эта гортань известна под названием
верхней. Она пропускает воздух и не участвует в голосообразова-
нии. Функцию голосового аппарата выполняет так называемая
нижняя гортань — орган, свойственный только птицам (рис. 121).
Она располагается в месте разделения трахеи на два бронха и пред-
ставляет собой расширение, поддерживаемое костными кольца-
ми. Здесь внутрь полости трахеи сверху, от ее наружных стенок,
вдаются наружные голосовые перепонки, а снизу, уже от места де-
ления трахеи на бронхи, вдаются внутренние голосовые перепонки.
Голосовые перепонки благодаря сокращению специальных пев-
чих мышц могут менять свое положение и форму, с чем связано
разнообразие издаваемых птицами звуков.
Легкие птиц — это плотные губчатые тела, расположенные в
спинной части грудной клетки. Первичные бронхи, войдя в легкие,
многократно дихотомически ветвятся на вторичные бронхи, распо-
ложенные в легких со спинной и брюшной сторон. Ветви вторич-
ных бронхов соединяются между собой в легких тонкими каналами —
третичными бронхами, или парабронхами (рис. 122, Б). Их стенки
имеют множество слепых кармашков — бронхиол и микробронхов,
вокруг которых ветвятся капилляры кровеносных сосудов (рис. 123).
Трахея и бронхи образованы хрящевыми кольцами, препятствую-
щими изменению их просвета. Внутренние стенки парабронхов
имеют гладкую мускулатуру, регулирующую их диаметр. Централь-
ные бронхи и их ветви пронизывают легкие насквозь, выходят за
пределы собственно легких и впадают в огромные тонкостенные
воздушные мешки, объем которых во много раз превосходит объем
легких. Воздушные мешки располагаются между различными внут-
ренними органами, а их ответвления проникают между мышцами
под кожу и заходят в пневматичные кости (рис. 122, А).
С образованием воздушных мешков связано у птиц сложное
подразделение полости тела. Всего мешков 9: два шейных тянутся
150
Рис. 122. Схема расположения воздушных мешков (Л) и дыхательных
путей (Б) в теле птицы (по А. Ромеру, Т. Парсонсу):
j — ключичный мешок; 2 — шейный мешок; 3 — левое легкое; 4 — главный
бронх, идущий через легкое в задние воздушные мешки; 5 — бронхи, возвраща-
ющие воздух из задних мешков к легкому; 6 — брюшной мешок; 7 — задний
грудной мешок; 8 — передний грудной мешок; 9 — сердце; 10 — парабронхи
по бокам вдоль шеи, один межключичный находится под вилоч-
кой; две пары грудных располагаются в грудной области и пара
очень крупных брюшных тянется назад между внутренними орга-
нами. Стенка воздушного мешка, тонкая и гладкая, самостоятель-
но в газообмене не участвует. Мешки играют большую роль в вен-
тиляции легких, определяя механизм дыхания птицы.
Рис. 123. Ультраструктура легкого птицы. Видны два парабронха
с кармашками и губчатой тканью
151
Цикл I
Цикл II
Рис. 124. Схема движения воздуха через дыхательную систему птицы
(по К. Шмидт-Ниельсену, 1982):
А, Б — вдох (I—II); В, Г — выдох (I, II)
Дыхательный процесс у птиц происходит благодаря сокраще-
нию мускулатуры грудной клетки, работающей по принципу на-
соса: ребра движутся вперед и вверх, а грудина — вперед и вниз.
Сами легкие сжимаются и расширяются слабо, резко меняется
объем воздушных мешков. На изменении их объема отражается
изменение общего объема целомической полости тела. Воздух про-
ходит через систему органов дыхания, как и у всех амниот, при
чередовании вдохов и выдохов. Главная роль в вентиляции легких ।
принадлежит задним воздушным мешкам. Как показали экспери-
ментальные исследования, при вдохе большая часть воздуха про-
ходит по главным бронхам в задние — грудные и брюшные воз-
душные мешки. Более мелкие возвратные бронхи соединяют эти
мешки с легкими. На первом выдохе воздух перемещается по ним
в легкие, где в парабронхах совершается газообмен. При последу-
ющем вдохе воздух перемещается в передние группы воздушных
мешков, а затем, на втором выдохе, покидает дыхательную сис-
тему* (рис. 124).
Таким образом, воздух, участвующий в газообмене, циркулирует
в дыхательной системе птиц однонаправленно: из задних воздушных
мешков через легкие в передние воздушные мешки.
Газообмен в легких осуществляется по принципу противотока'.
направление движения воздуха по парабронхам противоположно
* Речь идет о радиоактивно меченной порции кислорода (|7О), направлен-
ной в дыхательную систему птиц экспериментально. В норме воздух перемещает-
ся в дыхательной системе птиц непрерывно.
iso
движению крови по оплетаю-
щим их капиллярам (рис. 125).
При таком расположении пара-
бронхов и сосудов обеспечива-
ется высокая эффективность из-
влечения кровью кислорода из
воздуха. Экспериментальные ис-
следования показали, что высо-
кий уровень насыщения крови
кислородом сохраняется и у
птиц, пребывающих в разрежен-
ном пространстве на больших
высотах. Воробей активен на
Рис. 125. Система противотока при
дыхании птицы (по К. Шмидт -
Ниельсену, 1982)
высоте 8600 м, а равная ему по массе домовая мышь испытывает
состояние гипоксии (К. Шмидт-Ниельсен, 1976).
Движения крыльев при полете углубляют дыхание птицы, так
как могут влиять на расширение и сжатие грудной клетки и всей
целомической полости, но не определяют частоту ее дыхательных
движений.
Не менее важна роль воздушных мешков в терморегуляции: во
время полета они предохраняют тело птицы от перегревания, так
как воздух обтекает практически все внутренние органы и муску-
латуру. Потери тепла у птиц в полете в сравнении с состоянием
покоя увеличиваются примерно в 5 раз (Л. Проссер, 1977). Ниже
приведены данные о сравнительном объеме органов дыхательной
системы птиц и млекопитающих (табл. 2). Объем воздушных меш-
ков, в которых отсутствует дыхательный эпителий, в 3—4 раза и
более превышает объем легких.
Наряду с воздушными мешками важное значение в терморегу-
ляции имеют верхние дыхательные пути — гортань, трахея, бронхи.
Установлено, что при повышении температуры окружающей сре-
ды дыхание птицы резко учащается и становится весьма поверх-
ностным. Одновременно в полости рта и глотке происходит очень
Таблица 2
Объем органов дыхательной системы птицы и млекопитающего
(по К. Шмидт-Ниельсену, 1982)
Орган Объем, мл
Птица Млекопитающее
Легкие 29,6 53,5
Трахея 3,7 0,9
Воздушные мешки 127,5 —
Общий объем дыхательной 160,8 54,4
системы
153
сильное расширение кровеносных сосудов, которым сопровожда-
ется усиленная отдача тепла из организма птицы (полипное).
Частота дыхательных движений птицы зависит и от ее физио-
логического состояния, и от ее размеров. К примеру, голубь в
состоянии покоя совершает 26 дыхательных движений в минуту
при ходьбе — 77, в полете — 400, утка в покое — 30—43, мелкая
воробьиная птица — 90—100. Приведенные показатели отражают
состояние метаболизма этих птиц и коррелируют с правилом по-
верхности теплокровного животного. Кожа как орган дыхания у
птиц, как правило, не обсуждается.
У млекопитающих, как и у птиц, легкие являются по
существу единственным органом дыхания. Роль кожи в их газооб-
мене мала: только около 1 % кислорода поступает у наземных зве-
рей через кожные кровеносные сосуды. Суммарная дыхательная
поверхность легких у млекопитающих в 50—100 раз превышает
поверхность кожи.
В дыхательных путях млекопитающих происходит дальнейшее
усложнение гортани по сравнению с рептилиями (рис. 126). В ее
основании лежит кольцеобразный перстневидный хрящ. Над ним
по бокам спинной стороны гортани находятся парные черпаловид-
ные хрящи. Передняя и боковые стенки гортани образованы свой-
ственным только млекопитающим щитовидным хрящом. К пере-
днему краю щитовидного хряща примыкает тонкий лепесткооб-
разный надгортанник. Между перстневидным и щитовидным хря-
щами расположены небольшие мешковидные полости — желу-
дочки гортани. Голосовые связки в виде парных складок слизис-
той оболочки гортани лежат между щитовидным и черпаловид-
ными хрящами. Расположенные за гортанью трахея и бронхи сло-
жены хрящевыми кольцами. Бронхи дихотомически ветвятся в
легких на большое число мелких трубочек. Самые мелкие из них —
бронхиолы — заканчиваются пузырьками — альвеолами, имеющи-
ми ячеистое строение (рис. 127).
Рис. 126. Гортань кролика
спереди (Л) и сзади (Б):
1 — надгортанник; 2 — щитовидный
хрящ; 3 — перстневидный хрящ; 4 —
трахея; 5 — сантонинов хрящ; 6 —
черпаловидный хрящ
В стенке альвеолы ветвятся кро-
веносные сосуды (рис. 128).
Такая структура легких млеко-
питающих как бы продолжает ге-
неральную эволюционную линию
усложнения органов дыхания, по-
лучившую начало у амфибий и реп-
тилий. Число альвеол огромно и
отражает образ жизни животного:
у крупных подвижных хищников их
300 — 500 млн, у малоподвижных
ленивцев — около 6 млн. Альвеолы
образуют огромную поверхность
для газообмена. Площадь дыхатедь-
154
Рис. 127. Схема строения легкого
млекопитающего:
1 — альвеолярный ход; 2 — бронх; 3 —
бронхиола; 4 — конечные пузырьки с
альвеолами
Рис. 128. Схема строения
легочных пузырьков млекопи-
тающего.
На левом осталась только сеть ка-
пиллярных сосудов, на правом —
вскрытый пузырек без сосудов: 1 —
бронх; 2 — артерия; 3 — вена
ной поверхности альвеол (см2) при расчете на 1 г массы тела зверя:
у ленивца — 6, домашней кошки— 28, домовой мыши — 54, летучей
мыши — 100. Общая поверхность альвеол у человека 90 м2.
Механизм дыхания обусловлен у млекопитающих изменением
объема грудной клетки, возникающим в результате сокращения
межреберной мускулатуры {грудное дыхание) и особой, куполооб-
разной выдающейся в грудную полость мышцы — диафрагмы (ди-
афрагмальное дыхание). Число дыхательных движений коррелирует
с размерами тела животного, что связано, как и у птиц, с раз-
личной интенсивностью обмена веществ. Так, у лошади оно рав-
но 8 — 16 в минуту, у человека — 15 — 20, у крысы — 100—150, у
мыши — около 200.
Вентиляция легких, кроме участия в газообмене, и у млекопи-
тающих имеет существенное значение для теплорегуляции. Это осо-
бенно важно для видов с относительно слабо развитыми потовы-
ми железами. Охлаждение тела при его перегревании в значитель-
ной мере достигается усиленным испарением воды, пары кото-
рой выводятся вместе с воздухом, выдыхаемым легкими (поли-
пное). Например, собака в покое выдыхает 2 л воздуха в 1 минуту.
При беге или при резком повышении температуры окружающей
сРеды количество выдыхаемого воздуха доходит до 50 —75 л в ми-
нуту, а количество испаряемой воды может составить 200 см3 за
Час. В отличие от птиц в дыхательной системе млекопитающих нет
воздушных мешков и ее общий объем значительно меньше (см.
табл. 2).
155
Плавательный пузырь
В теле костных рыб присутствует особый орган — плавательный
пузырь. Это новообразование возникает у них параллельно с обра-
зованием костного скелета. Роль плавательного пузыря неодноз-
начна. У большинства лучеперых этот орган создает подъемную
силу, необходимую для поддержания нейтральной плавучести в
разных горизонтах воды. Обширная полость, наполненная смесью
газов, по составу близкой к атмосферному воздуху, уменьшает
удельный вес их тела и служит своеобразным поплавком. У при-
донных рыб — многочисленных видов камбал, не нуждающихся в
удержании тела в толще воды, этот орган вторично отсутствует.
Уместно отметить, что акуловым рыбам, не имеющим плаватель-
ного пузыря, в удержании тела помогают горизонтальные парные
плавники, значительное количество жира в тканях тела и посто-
янное поступательное движение.
Плавательный пузырь расположен обычно в верхней части по-
лости тела под позвоночником и имеет овальную форму с одной
или двумя камерами. Этот орган формируется в онтогенезе рыбы
как вырост дорсальной стороны пищеварительной трубки (рис. 129).
У открытопузырных рыб пожизненно сохраняется его связь с пи-
щеводом. Таковы сельдеобразные, лососеобразные, карпообраз-
ные и др. У многих рыб, называемых закрытопузырными, связь
пузыря с пищеводом в ходе онтогенеза утрачивается (трескооб-
разные, щукообразные и др.). У открытопузырных плавательный
пузырь может наполняться смесью газов при заглатывании ими
воздуха с поверхности воды. Однако эксперименты показали, что
у большинства рыб наполнение и освобождение плавательного
пузыря совершается его собственными внутренними механизма-
ми, основную роль в которых играет кровеносная система. В стен-
ке передней части плавательного пузыря есть газовая железа, или
«красное тело» {corpus rubrum). Кровь поступает в газовую железу
через множественное ветвление кровеносных сосудов («чудесное
Рис. 129. Схемы продольных разрезов плавательного пузыря разных
костистых рыб;
А — открытопузырный тип; Б, В — закрытопузырный тип; I — кишечная трубка;
2 — передняя камера; 3 — задняя камера (поглощение газа); 4 — красное тело
(выделение газа); 5 — овал (поглощение газа)
156
сплетение») и, передав в пузырь газы, покидает ее. Во время сек-
реции газа сосуды, где происходит его поглощение, закрыты и
тОк крови в них приостановлен. Подсчитано, что сосудистая сеть в
газовой железе обыкновенного угря содержит около 200000 ка-
пилляров общей длиной 800 м (Л. Проссер, 1977). Удаление газов
из пузыря происходит в противоположном его участке — в задней
камере двухкамерного пузыря или сверху задней части однока-
мерного (рис. 130).
Механизм перехода газов из кровеносных капилляров в пузырь
и обратно пока до конца не расшифрован. Известно, в частности,
что кислород диффундирует благодаря секреции в газовой железе
молочной кислоты. У глубоководных рыб связывание и резерви-
рование кислорода внутри пузыря осуществляется, возможно, ли-
пидными соединениями (Л. Проссер, 1977).
Соотношение газов, наполняющих плавательный пузырь, весь-
ма изменчиво. К примеру, у обыкновенной трески и некоторых
других закрытопузырных рыб пузырь наполнен смесью газов кис-
лорода, азота и углекислоты в соотношении соответственно 63,1;
28,1 и 8,8%. У лососевой рыбы сига (Coregomis albus) плаватель-
ный пузырь на 95 % наполнен азотом. Нередки случаи сочетания
азота и аргона.
У некоторых рыб павательный пузырь используется как допол-
нительный орган дыхания. Таковы самая большая в мире пресно-
водная рыба арапайма (Arapaima), обитающая в реках Южной
Америки, некоторые примитивные костные рыбы, жительницы
пресных водоемов Северной Америки: виды панцирников
(Lepisosteus) и ильная рыба (Amia calva).
Участие плавательного пузыря в дыхательном процессе пре-
сноводных двудышащих рыб вызвало дискуссию о возможной
J Рис. 130. Схема кровоснабжения плавательного пузыря рыбы
1 (по А. Ромеру, Т. Парсонсу, 1992)
157
морфологической близости легких и плавательного пузыря
И. И. Шмальгаузеном (1964) было убедительно показано, что лег-
кие у многоперов и двоякодышащих рыб закладываются с брющ_
ной стороны тела и в процессе онтогенеза перемещаются на спин-
ную сторону, возможно принимая на себя функцию плавательно-
го пузыря. Позднее были высказаны предположения о возможном
перемещении у морских костистых рыб одного легкого на спин-
ную сторону, утрате им дыхательной функции и превращении в
плавательный пузырь.
***
Таким образом, специализированные органы дыхания возни-
кают экто-энтодермально как идущие навстречу выпячивания гло-
точного отдела первичной кишки и впячивания эктодермы, т.е.
жабры у водных позвоночных экто-эктодермального происхожде-
ния. Эволюционные изменения жабр направлены на увеличение
их дыхательной поверхности и большую защиту. Легкие позвоноч-
ных формируются как боковые карманы задней стенки жаберной
полости. Сначала они представляют малоячеистые мешки, затем
ячеистость возрастает. Параллельная эволюция органов дыхания
птиц и млекопитающих привела у птиц к окислению крови в стен-
ках парабронхов, а у млекопитающих в альвеолах. Эффективность
окисления крови у птиц выше, чем у зверей, за счет постоянного
движения воздуха через легкие (парабронхи) из задних воздуш-
ных мешков в передние. Насосный механизм дыхания у млекопи-
тающих за счет работы межреберных мышц и диафрагмы приво-
дит к частичному обмену воздуха в альвеолах, и окисление крови
у них менее эффективно, чем у птиц.
ОРГАНЫ КРОВООБРАЩЕНИЯ
Кровеносная система хордовых замкнутая, непрерывающаяся.
Она состоит из сообщающихся между собой кровеносных сосу-
дов, повторяя стиль организации этой системы у кольчатых чер-
вей. Однако у хордовых меняется направление движения крови:
по брюшному сосуду она движется к переднему концу тела, а по
спинному — назад, от головы к хвосту.
Кровеносная система имеет мезодермальное происхождение.
Она закладывается эмбрионально во внутренних листках боко-
вых пластинок. Из зародышевой соединительной ткани энтоме-
зенхимы формируются кровеносные сосуды и форменные эле-
менты крови.
Значение кровеносной системы трудно переоценить. Она осу-
ществляет доставку к органам, тканям, клеткам организма пита-
тельных веществ и кислорода, гормонов и ферментов, удаляет из
них углекислоту и продукты обмена. Клетки крови обладают спо-
собностью к внутриклеточному пищеварению и участвуют в за-
щите организма от инфекций. Капиллярная сеть в покровах уча-
ствует в терморегуляции. Прогрессивная эволюция кровеносной
системы — важное условие поступательного развития всех позво-
ночных.
Рассмотрим различные характеристики кровеносной системы.
Кровь
Сосуды кровеносной системы наполнены кровью. Это жидкая
соединительная ткань, удерживаемая в кровеносном русле стен-
ками сосудов. Жидкая часть крови — плазма. Ее основу составляет
тканевая жидкость, проникающая в сосуды из межклеточного
пространства. Однако плазма отличается от тканевой жидкости.
Большую часть сухой массы плазмы составляют белки крови: аль-
бумины, глобулины и фибриноген. В силу структуры молекул эти
белки не покидают кровяного русла и присутствуют в плазме по-
стоянно.
Альбумины повышают осмотическое давление крови по срав-
нению с тканевой жидкостью. Глобулины осуществляют защиту
°Рганизма от болезней, так как нейтрализуют микроорганизмы и
159
посторонние химические вещества. Фибриноген участвует в свер.
тывании крови при повреждениях, сосудов. Малые концентрации
имеют ферменты и гормоны, переносимые кровью между соот
ветствующими органами.
В состав плазмы входят также соли, глюкоза, аминокислоты,,,
также продукты распада: бикарбонат, мочевина, мочевая кислота
Значительная часть этих составляющих плазмы свободно переме
щается в тканях через мембраны сосудов. Однако в норме кровь
позвоночных непосредственно с тканевой жидкостью нигде ц
соприкасается.
В плазме крови во взвешенном состоянии находятся кровяные
клетки различных типов: эритроциты (красные кровяные тель-
ца), лейкоциты (белые кровяные тельца) и тромбоциты. Эритро-
циты по форме — небольшие плотные диски. Их цвет обусловлен
красящим веществом — белком гемоглобином, осуществляющим
дыхательную функцию крови. Молекула гемоглобина включает
ионы железа и способна связывать и переносить кислород. В эво-
люционном ряду позвоночных количество эритроцитов и их фор-
ма, а следовательно, и содержание гемоглобина в крови — вели-
чины весьма изменчивые.
У млекопитающих кислородная емкость крови заметно возрас-
тает в связи с безъядерностью их эритроцитов. Вместе с тем лан-
цетники и некоторые холоднокровные рыбы гемоглобина вовсе
не имеют: существенную роль в газообмене у них играют хими-
ческие и физические свойства плазмы крови. В крови оболочников
кислород связывают многочисленные специфические клетки —
ваннадоциты, содержащие ваннадий и серную кислоту. Известна
их химическая специфика. Она содержит большой набор неорга-
нических элементов: никель, медь, марганец, железо, кальций,
алюминий, натрий, кремний, хром, титан.
Лейкоциты — бесцветные подвижные амебообразные клетки,
содержащие ядро. Они участвуют в уничтожении попавших в тело
микроорганизмов, а также в иммунных и аллергических реакциях.
Их число среди кровяных телец невелико: у амниот — менее 1 %,
у анамний (рыб) — до 10 % и более. Лейкоциты — весьма разно-
родная группа клеток. Среди них наиболее многочисленны грану-
лоциты и лимфоциты. Функция гранулоцитов (или зернистых лей-
коцитов) — захват и переваривание бактерий. Лимфоцитам (или
незернистым лейкоцитам) принадлежит ведущая роль в иммун-
ных реакциях.
Тромбоциты — это мелкие плоские клетки. У большинства по-
звоночных тромбоциты имеют веретенообразную форму. Они иг*
рают важную роль в свертывании крови при нарушении целост-
ности сосудов.
У млекопитающих тромбоциты представлены кровеносными пла-
стинками.
i
14Л
Кроветворение
В раннем эмбриогенезе рыб, амфибий, рептилий и птиц клет-
ки крови формируются в основании желточного мешка в виде
скоплений, называемых кровяными островками. У млекопитающих
они образуются в области хориона. По мере развития кроветвор-
ные органы локализуются в разных отделах тела эмбриона. У хря-
щевых и костных рыб, амфибий, рептилий и птиц важным источ-
ником клеток крови становятся почки, у млекопитающих — пе-
чень и селезенка (А. Ромер, Т. Парсонс, 1994).
В постэмбриональном состоянии животных клетки крови обра-
зуются в разных органах: почках, гонадах, печени, тимусе, селе-
зенке, костном мозге, части кишечника, лимфатических узлах и
т.д-*
Важным органом кроветворения у наземных позвоночных стал
красный костный мозг, заполнивший полости трубчатых костей.
У рептилий и птиц в нем прижизненно образуются все типы кро-
вяных клеток. У млекопитающих он продуцирует главным образом
эритроциты и некоторые формы лейкоцитов (гранулоциты и мо-
ноциты).
Специализированными структурами кроветворения, свойствен-
ными главным образом млекопитающим, являются лимфатичес-
кие узлы. Они содержат кроветворную ткань и продуцируют лим-
фоциты. Эти макрофаги уничтожают в теле животного бактерии и
прочие инородные элементы. При обилии этих элементов лимфа-
тические узлы увеличиваются в размерах.
Важным органом кроветворения, накопления и разрушения
клеток крови у позвоночных является селезенка. В эмбриогенезе
она развивается как область зародышевой кишечной трубки и уча-
ствует в кроветворении у зародыша. У круглоротых и двоякодыша-
щих рыб селезенка не образует компактного органа: ее ткань раз-
бросана по кишечнику. Селезенка у остальных позвоночных — это
оформленный компактный орган, расположенный обычно рядом
с желудком и окруженный соединительнотканной капсулой. Внутри
селезенки соединительная ткань образует ячеистую (ретикуляр-
ную) сеть с белой и красной пульпой внутри (рис. 131). Белая
Пульпа содержит скопления лимфоцитов, красная — преимуще-
* Согласно А. Ромеру, Т. Парсонсу (1994), у миног, хрящевых и большинства
костных рыб, амфибий существенным источником клеток крови в течение всей
«изни являются почки и селезенка. У хрящевых рыб лейкоциты образуются в
гонадах. У осетровых кроветворная ткань окружает сердце. У многих позвоночных
Кроветворение идет в стенках ротовой полости и глотки. У акул в пищеводе есть
ткань, образующая лимфоциты. У многих животных лимфоциты образуются в
стенках тонкой кишки (пейеровы бляшки), у человека — в аппендиксе. У птиц
Местом образования лимфоцитов является нижняя часть кишки — фабрициева
сумка.
® Константинов 161
Рис. 131. Схема участка селезенки млекопитающего:
1 — капилляр; 2 — венозный синусоид; 3 — капсула; 4 — трабекула; 5 — красная
пульпа; 6 — фолликулярная артерия; 7 — артерия пульпы; 8 — артерия; 9 — вена
трабекулы; 10 — белая пульпа; 11 — артерия покидает трабекулу; 12 — артерия
трабекулы
ственно эритроциты и гранулоциты. В селезенке клетки крови об-
разуются, хранятся и здесь же они разрушаются с помощью круп-
ных лимфоцитов. Кровь в селезенку поступает через артерию и
отводится веной.
Таблица 3
Показатели массы крови к массе тела у позвоночных разных классов
(Л. Проссер, 1978)
Класс позвоночных Относительная масса крови, % Класс позвоночных Относительная масса крови,
Хрящевые рыбы 2-7 Рептилии 4-15
Костные рыбы 2-7 Птицы 6-10
Амфибии 3-9 Млекопитающие 6-13 _
162
Важным органом кроветворения и в эмбриональном, и постэм-
бриональном состоянии позвоночных животных является зобная
железа (тимус). Кроветворные клетки этого органа развиваются из
мезенхимы, мигрирующей из других кроветворных структур. Зоб-
ная железа интенсивно продуцирует лимфоциты и участвует в ста-
новлении иммунитета развивающегося организма. По достижении
половой зрелости орган дегенерирует (у млекопитающих исчезает
полностью).
В эволюционном ряду позвоночных общая масса крови по от-
ношению к массе тела прогрессивно возрастает (табл. 3).
Органы кровеносной системы
Органами кровеносной системы являются сосуды — артерии,
вены и капилляры, и главный двигатель крови по сосудам — сердце.
Сосуды. Стенка кровеносного сосуда (в обобщенной схеме)
имеет три оболочки: внутреннюю, среднюю и наружную. Изнутри
сосуд выстлан слоем эндотелия. Клетки этого слоя обладают высо-
кой митотической активностью и способствуют регенерации и
восстановлению кровеносного сосуда при его повреждении. В круп-
ных сосудах эндотелий подстилается эластичной внутренней ба-
зальной мембраной.
Средняя оболочка — мышечная, содержит преимущественно
гладкую мышечную ткань и нервные волокна. В крупных сосудах
снаружи ее покрывает наружная эластичная мембрана.
По артериям кровь течет под большим давлением, создавае-
мым сокращениями сердца. Просвет артерий на всем протяжении
остается постоянным и в них отсутствуют клапаны. Стенки вен в
сравнении с артериями имеют более простое строение. Большин-
ство вен не имеет внутренней эластичной мембраны, подстилаю-
щей эндотелий. Мышечный слой развит слабо или вовсе отсутству-
ет. Крупная вена может иметь мешковидное расширение — синус,
например, перед впадением ее в предсердие. Обратному току крови
в венах препятствуют клапаны. На рис. 132 продемонстрированы в
сравнении схемы строения двух сосудов: артерии и вены.
Капилляры — мельчайшие сосуды в артериальной и венозной
системах. Чаще их стенки имеют только слой эндотелия. Он обес-
печивает избирательную проницаемость сосудов. Диаметр самых
Мелких капилляров достаточен для прохождения одного эритро-
цита. Капилляры проникают между клетками тканей и через их
стенки идет активная диффузия веществ и растворенных в крови
газов — О2 и СО2. В типичном случае в своей проксимальной (на-
чальной) части капилляр отдает кислород и питательные веще-
ства наружу, а в дистальной (конечной) принимает от тканей
Двуокись углерода и конечные продукты обмена.
163
1
2
3
1
Рис. 132. Поперечный разрез через артерию (А) и вену (Б):
1 — эндотелий (часть внутренней оболочки); 2 — наружная оболочка; 3 — ка-
пилляр; 4 — жировая клетка; 5 — средняя оболочка (мышечный слой)
Капиллярные сети обычно соединяют артериальные сосуды с
венозными. Однако капиллярные разветвления могут возникать и
по ходу сосудов. Например, жаберное кровообращение рыб, круг-
лоротых, личинок амфибий осуществляют ветвящиеся на капил-
ляры артерии. Артериальные клубочки в мальпигиевых тельцах
почек образованы приносящими и выносящими артериолами.
В механизме работы плавательного пузыря костных рыб участвует
чудесное сплетение (или «красное тело») — система мелких ка-
пилляров артериолы. В лицевых ямках ямкоголовых змей, в дис-
тальных отделах конечностей длинноногих птиц, в ластах водных
млекопитающих, в хвостах млекопитающих артериолами созда-
ются так называемые противоточные системы (у человека — коп-
чиковый клубочек) (А. Ромер, Т. Парсонс, 1994).
Разветвления венозных сосудов (венул) образуют так называе-
мые воротные системы почек, печени, гипофиза, надпочечников.
Сердце. Движение крови по сосудам происходит благодаря со-
кращениям мускульного органа — сердца (исключение составля-
ют бесчерепные, у которых движение крови создается сокраще-
ниями брюшного артериального ствола и артериальных расшире-
ний жаберных сосудов — «жаберные сердца»). В целом же сосуды к
автономной пульсации не способны*.
В самом раннем онтогенезе позвоночного животного первые
кровеносные сосуды закладываются в виде пары вен в нижней
стенке эмбриональной кишки. Далее они сливаются в единый со-
* У взрослой асцидии единственный сосуд, попеременно пульсируя, на-
правляет кровь то к дыхательной части глотки, то изливая ее в лакуны.
164
суд — подкишечную вену. Из ее переднего участка развиваются
брюшная аорта и сердце, остальная часть дает начало печеночной
Бене и воротной вене печени.
Сердце формируется как ряд расширений задней части брюш-
ной аорты, которая свертывается в изогнутую петлю. Передний
отдел дает начало желудочку, задний предсердию. Предсердие
Принимает кровь, желудочек направляет ее по кровеносным со-
судам. В наиболее простом выражении — у рыб — сердце представ-
ляет собой трубку из четырех последовательно расположенных
камер: венозный синус принимает венозную кровь и передает ее
в тонкостенное и растяжимое предсердие. Желудочек, основная
мускулистая часть сердца, принимает кровь из предсердия и вы-
талкивает ее в начало брюшной аорты (в артериальный конус,
или в артериальную луковицу). Клапан, расположенный между
предсердием и желудочком, препятствует обратному току крови.
Число камер в сердце животных разных классов последовательно
прогрессивно изменяется: 2—3—4. Соответственно меняются и от-
носительные размеры сердца (табл. 4).
Структура стенки сердца по организации близка к крупному
кровеносному сосуду, но имеет и свои существенные особеннос-
ти. Тонкая внутренняя оболочка сердца — эндокард соответствует
эндотелию сосуда. Наружная оболочка — эпикард образована со-
единительной тканью и покрыта с поверхности целомическим
эпителием.
Основную массу сердца образует средний слой — миокард, со-
стоящий из поперечнополосатой мускулатуры и соединительной
ткани. Сокращение сердца позвоночного не подчинено волевым
импульсам, оно работает в автономном режиме. Импульсы воз-
буждения возникают ритмично в нервных узлах, расположенных
в толще миокарда. Между камерами сердца в местах входа и выхо-
да центральных кровеносных сосудов расположены образованные
эндокардом клапаны сердца (например, у млекопитающих четыре
клапана: трехстворчатый, двухстворчатый, или митральный, и два
полулунных). Кроме того, в полости желудочков есть тяжи соеди-
Таблица4
Сердечный индекс у позвоночных разных классов
Класс позвоночных —— Сердечный индекс (масса сердца/масса тела, %) Класс позвоночных Сердечный индекс (масса сердца/масса тела, %)
^Рящевые рыбы 0,3 Рептилии 2,1
Костные рыбы 0,6 Птицы 2,5
Амфибии 1,0 Млекопитающие 1,7
165
нительной или мышечной ткани, предохраняющие их от чрез-
мерного расширения при большом давлении крови, и сухожиль-
ные тяжи, ограничивающие вдавливание клапанов в закрываемое
отверстие.
Стенка сердца снабжается кровью через сеть коронарных сосу-
дов. Кровь поступает к сердцу от спинной аорты. Сердце окружено
околосердечной сумкой, отграничивающей часть целома (пери-
кардиальную полость).
В кровеносном русле позвоночных (хордовых) выделяют сис-
тему артериальную — несущую кровь от сердца, и систему веноз-
ную — возвращающую кровь к сердцу.
В процессе усложнения позвоночных структура органов крово-
обращения изменяется: вместо одного круга обращения, несуще-
го смешанную кровь, образуются два, что позволило в конечном
итоге полностью отделить богатую кислородом артериальную кровь
от обедненной венозной крови. Переход сопровождался постепен-
ной редукцией артериальных жаберных дуг и увеличением числа
камер в сердце (см. рис. 146).
Особенности организации кровообращения
в эволюционном ряду хордовых
Филогенетические преобразования кровеносной системы в эво-
люционном ряду хордовых представляют собой одно из интерес-
нейших направлений морфофизиологической эволюции хордовых.
В артериальной системе ланцетника под глоткой тянется
сократимая брюшная аорта (рис. 133). Венозная кровь движется по
ней к головному концу и поступает в парные жаберные артерии,
Рис. 133. Схема основных кровеносных сосудов ланцетника:
I — передние кардинальные вены; 2 — кювьеровы протоки; 3 — задние карДИ'
нальные вены; 4 — спинная аорта; 5— воротная вена печеночного выроста; б''
печеночный вырост; 7 — печеночная вена; 8 — брюшная аорта
166
проходящие в перегородках между жаберными щелями. Жаберные
артерии (несколько десятков) тонкостенны и не образуют в пере-
городках капиллярных, разветвлений: газообмен осуществляется
через их стенки. Из жаберных артерий кровь изливается в корни
спинной аорты. На уровне заднего конца глотки они сливаются и
образуют основной артериальный ствол — спинную аорту. Распо-
лагаясь под хордой, она тянется вдоль тела до его заднего конца.
На своем пути спинная аорта распадается на множество мелких
артериальных сосудов, снабжающих кровью внутренние органы.
Вперед от корней аорты отходят сонные артерии к головному от-
делу-
Обедненная кислородом и насыщенная углекислым газом ве-
нозная кровь собирается от переднего конца тела в парные пере-
дние кардинальные вены, несущие ее назад. От заднего конца тела
кровь выносится вперед парными задними кардинальными венами.
Позади глотки передние и задние кардинальные вены попарно
сливаются, образуя два (левый и правый) широких тонкостенных
кювьеровых протока. Из них кровь изливается в венозный синус, от-
куда и берет начало брюшная аорта (см. выше). От внутренних
органов венозная кровь собирается в подкишечную вену. Достигнув
печеночного выроста, она распадается в нем на сеть венозных
капилляров, образуя воротную систему печеночного выроста. Далее
по печеночной вене эта кровь тоже попадает в венозный синус.
Движение крови по сосудам обусловлено у ланцетника пульса-
цией брюшной аорты и нижних частей жаберных сосудов («жа-
берные сердца»). Сердце отсутствует. Газообмен осуществляют же-
лезосодержащие вещества плазмы крови.
У оболочников кровеносная система незамкнутая, лакунарная.
Сердце имеет вид сосуда. Пульсируя, оно гонит кровь поперемен-
но к жабрам и органам тела, в которых она изливается в лакуны.
У круглоротых общий план строения кровеносной систе-
мы близок к ланцетнику (рис. 134). Однако у них есть сердце, со-
стоящее их двух камер — предсердия и желудочка. Оно расположе-
но в начале брюшной аорты и способствует усилению кровотока.
К предсердию примыкает тонкостенный венозный синус, прини-
мающий кровь из всех венозных сосудов. От брюшной аорты отхо-
дят 7 — 15 пар (у разных видов) приносящих жаберных артерий.
В жаберных лепестках они образуют капиллярную сеть для газооб-
мена, причем каждая, раздваиваясь, обслуживает половины со-
седних жаберных мешков. Выносящие жаберные артерии впадают
в непарный корень аорты, который продолжается назад в основ-
ной артериальный ствол — спинную аорту, расположенную под
ХоРдой. Вперед от корня аорты отходят сонные артерии. Кровь по
спинной аорте поступает ко всем органам тела.
Венозная кровь от почек, гонад и мускулатуры тела собирается
в Парные задние кардинальные вены, впадающие в венозный синус.
167
Рис. 134. Схема кровеносной системы миноги:
1 — дуга аорты; 2 — выносящие жаберные артерии; 3 — передняя кардинальная
(яремная) вена; 4 — спинная аорта; 5 — задняя кардинальная вена; 6 — кишеч-
ная артерия; 7 — приносящие жаберные артерии; S — брюшная аорта; 9 — жа-
берные щели; 10 — желудочек; 11 — предсердие; 12 — венозный синус; 13 —
печеночная вена; 14 — подкишечная вена; 15 — хвостовые вена и артерия
Сюда же поступает кровь из передних кардинальных вен, собираю-
щих кровь от головного отдела. Кювьеровы протоки короткие. Под-
кишечная вена собирает кровь от кишечника и, попав в печень,
образует воротную систему кровообращения. По печеночной вене
она также изливается в венозный синус.
Кровеносная система миксин имеет свои особенности. В системе
их венозного кровообращения есть большие кровяные синусы —
дополнительные «сердца». Эти расширения присутствуют в обла-
сти головы — головной синус и дополнительные сердца, в обла-
сти печени — портальное сердце, в области хвоста — каудальное
сердце* (рис. 135). Они усиливают венозное кровообращение. Плазма
крови круглоротых содержит кровяные клетки.
У челюстноротых общий стиль организации кровеносной
системы сохраняется. Хрящевые рыбы тоже имеют двухка-
Рис. 135. Схема работы каудального сердца миксины:
1 — хрящ; 2 — наружные мышцы; 3 — камеры
* Камеру каудального сердца разделяет продольный стержень хряща, кото-
рый, изгибаясь, позволяет попеременно пропускать кровь через камеру синуса
(К. Шмидт-Ниельсен, 1982).
168
мерное сердце. К предсердию прилегает широкий тонкостенный
венозный синус, принимающий венозную кровь. К основанию же-
лудочка прилегает артериальный конус. Наличие в нем поперечно-
полосатой мускулатуры свидетельствует о его принадлежности к
желудочку (рис. 136, А). Внутри конуса есть полулунный клапан.
Внешне артериальным конусом начинается брюшная аорта. От нее
к жабрам отходят приносящие жаберные артерии. В эмбриогенезе
спереди назад закладываются 7 пар жаберных артерий (артериаль-
ных дуг). Капиллярные сети начинают функционировать после про-
рывания жаберных отверстий. При этом первая пара (I) у эмбри-
она закладывается целиком, а у взрослой акулы сохраняется лишь
ее выносящая часть. Она несет кровь от ложной жабры, располо-
женной в переднем крае брызгальца, и участвует в кровоснабже-
нии лицевой части головы, глаза и головного мозга. Отходящие от
первой пары короткие наружные сонные артерии несут кровь к ниж-
ней челюсти. Остальные пять пар (II —VI) жаберных артерий омы-
вают жаберный аппарат. Каждая приносящая жаберная артерия об-
служивает у хрящевых рыб лепестки соседних жаберных отверстий.
Выносящие жаберные артерии впадают в парные продольные
сосуды — корни аорты, которые, сливаясь, образуют основной
артериальный ствол — спинную аорту. Она лежит под позвоноч-
ником и снабжает кровью внутренние органы. Вперед от корней
аорты отходят внутренние сонные артерии. По ходу спинной аорты
отходят сосуды к передним и задним конечностям (подключич-
ные артерии) (рис. 136, Б).
Венозная кровь собирается от головы в парные боковые вены.
Они вливаются в парные передние кардинальные вены. От туловища
кровь собирается в парные задние кардинальные вены. На уровне
сердца они попарно сливаются, образуя кювъеровы протоки, впа-
дающие в венозный синус. Кардинальные вены в почках прерыва-
ются и образуют воротную систему кровообращения. Венозный
сосуд, войдя в почку, разветвляется и образует сеть капилляров,
окружающую почечные канальцы. Затем кровь вновь собирается в
кардинальные вены и направляется к кювьеровым протокам. От
кишечника кровь поступает в подкишечную вену, которая образует
в печени воротную систему кровообращения. Из печени кровь по
печеночной вене изливается в венозный синус. От туловищной мус-
кулатуры и конечностей кровь собирают парные брюшные вены,
впадающие в кювьеровы протоки.
В полости тела хрящевых рыб появляется селезенка (она при-
мыкает к нижней части желудка). Это депо крови и одновременно
Кроветворный орган, где образуются эритроциты, лейкоциты,
тромбоциты. Кроме селезенки клетки крови образуются в печени
и верхушечных частях почек.
Кровеносная система костных рыб отличается от хрящевых
в Деталях. Одновременно наблюдается расхождение и в разных си-
169
Рис. 136. Кровеносная система
акулы:
А — схема движения крови в сердце;
Б — схема расположения основных
сосудов
Височная (стапедиальная) артерия
1яя кардинальная вена
Спинная аорта
\ Общая
\ кардинальная вена
Чревная артерия
Выносящая артерия брызгальпа
Выносящие жаберные артерии
Задняя кардинальная вена
Боковая вена головы
Внутренняя сонная артерия
Глазничный синус
Печень
Подвздошная артерия
Подвздошная вена
Нижний яремный синус
Брюшная аорта
Артериальный конус
Предсердие
Желудочек |
Венозная пазуха
Селезенка
Селезеночно-желудочная артерия
Почка Воротная вена почки
Хвостовая артерия
Хвостовая вена
Кишка
Боковая брюшная вена
Передняя брыжеечная артерия
Поджелудочная железа
Желудок
Воротная вена печени
Печеночная вена
Подключичная артерия
Подключичная вена
схематических группах костных рыб. У кистеперых, двоякодыша-
щих и ганоидных рыб (осетровых, многоперов) сохраняется ар-
териальный конус с полулунным клапаном. У большинства луче-
перых брюшная аорта в самом начале имеет вздутие — артериаль-
ную луковицу. Внешне она похожа на артериальный конус, но ее
стенка содержит не поперечнополосатую, а гладкую мускулатуру,
и она не способна к самостоятельному пульсированию. Число при-
носящих жаберных артерий равно четырем (III—VI). Пройдя жаб-
ры, кровь поступает в корни спинной аорты. Сонные артерии несут
ее к голове, непарная спинная аорта снабжает кровью внутрен-
ние органы и конечности (рис. 137, А, Б).
4
19
Рис. 137. Схема кровеносной
. системы костистой рыбы:
А — схема расположения основных со-
судов: 1 — предсердие; 2 — желудо-
чек; 3 — луковица аорты; 4 — брюш-
ная аорта; 5 — приносящие жаберные
артерии; б— выносящие жаберные ар-
терии; 7 — корни аорты; 8 — спинная
аорта; 9— сонные артерии; 10— под-
ключичные артерии; 11 — хвостовая
вена; 12 — правая задняя кардиналь-
ная вена; 13 — левая задняя карди-
нальная вена; 14 — воротная система
левой почки; 75 — подкишечная вена;
16 — воротная система печени; 17 —
кювьеров проток; 18— печеночная вена;
19 — передние кардинальные вены;
Б — схема движения крови в сердце
Б
171
Для венозной системы большинства видов рыб характерна не-
прерывность правой задней кардинальной вены: только левая карди-
нальная вена образует в соответствующей почке воротную систему
кровообращения. При парном слиянии кардинальных вен образу-
ются кювьеровы протоки.
Кровеносная система двоякодышащих рыбв связи с ча-
стичной редукцией жабр и развитием легочного дыхания имеет свои
специфические особенности: часть жаберных артерий проходит че-
рез жаберную область, не образуя капилляров. Кровь поступает в
легкие по сосудам, отходящим от последней пары жаберных арте-
рий и, видимо, гомологичным легочным артериям. От легких специ-
альные сосуды несут кровь к сердцу: их можно считать гомологами
легочных вен (рис. 138). Небольшая перегородка частично делит пред-
сердие на левую и правую половины: в левую часть поступает кровь
из легочных вен, в правую — вся венозная кровь из тела. На выходе
из сердца складка в артериальном конусе частично разделяет их по-
токи. Изменяются задние кардинальные вены: левая сохраняется, а
правая расщепляется. В результате ее расщепления возникает новый
крупный венозный ствол — задняя полая вена. Она берет начало от
почек, проходит через печень и несет кровь к сердцу. Значительно
увеличивается венозный синус. Брюшные вены отсутствуют.
В кровеносной системе амфибий усиливаются изменения, на-
метившиеся у двоякодышащих рыб. При этом у хвостатых сохра-
няются черты примитивности, а бес-
хвостые демонстрируют следующий
шаг преобразований.
У хвостатых амфибий предсердия
частично разделены. Сохраняется си-
стема артериальных жаберных арте-
рий (жаберных дуг) «рыбьего типа»
(рис. 139). Четыре пары этих артерий
(III—VI) у взрослых амфибий пред-
ставлены непрерывными сосудами,
не имеющими капиллярной сети. По-
ток артериальной крови несколько
перераспределяется: III пара артерий
направляет кровь к голове во внут-
ренние сонные артерии; IV—VI жа-
Рис. 138. Схема артериального кровооб-
ращения двоякодышащей рыбы:
1—4 (I —IV) — пары жаберных артерий; 5^
спинная аорта; 6 — брюшная аорта; 7 — ле'
точная артерия; 8 — легочная вена
172
Рис. 139. Артериальные дуги
хвостатой амфибии;
1 — сонные артерии (I артери-
альная дуга); 2 — системные дуги
(И артериальная дуга); 3 — III ар-
териальная дуга; 4 — легочные
артерии (IV артериальная дуга);
5— сонные протоки; 6 — боталов
проток
Рис. 140. Схема вскрытого сердца
лягушки:
1 — правое предсердие; 2 — левое пред-
сердие; 3 — желудочек; 4 — клапаны, зак-
рывающие общее отверстие, ведущее из
обоих предсердий в желудочек; 5 —- арте-
риальный конус; 6 — общий артериаль-
ный ствол; 7 — кожно-легочная артерия;
8 — дуга аорты; 9 — общая сонная арте-
рия; 10— сонная «железа»; 11 — спираль-
ный клапан артериального конуса
берные артерии несут ее назад в корни спинной аорты. У хвоста-
тых и безногих сохраняется связь сонных артерий с корнями аор-
ты в виде сонных протоков. Тенденцию к редукции имеет V пара
жаберных артерий. Легкие снабжаются кровью через VI пару жа-
берных артерий, от которых отходят легочные артерии. У хвоста-
тых и безногих сохраняется их связь в виде боталовых протоков с
системными дугами аорты.
У более высокоорганизованных бесхвостых амфибий (лягуш-
ки) сердце трехкамерное (рис. 140). Полная перегородка делит пред-
сердие на левое и правое. Они сообщаются с желудочком общим
отверстием со сложным клапаном. С правым, более объемистым
Предсердием сообщается венозный синус (пазуха), принимающий
венозную кровь от всего тела. В левое предсердие поступает кровь
только из легких. Желудочек имеет толстую мышечную стенку. Его
внутренняя поверхность несет многочисленные карманы, способ-
ствующие частичному разделению поступившей в него крови,
брюшная аорта редуцируется до короткого канала. При сердечном
сокращении кровь из желудочка выталкивается в артериальный
к°нус. Закрученная складка (спиральный клапан) разделяет по-
ступающие сюда потоки крови по трем парам артерий.
173
В артериальной системе бесхвостых сохраняются три пары сосу-
дов — гомологи III, IV и VI жаберных артерий (рис. 141, А).
Сосуды первой пары (считая от головы), гомологичные III паре
жаберных артерий рыб, — это сонные артерии, несущие кровь к
голове. Каждая из них почти сразу же распадается на наружную и
внутреннюю сонные артерии. На месте их разделения лежит сонная
Рис. 141. Артериальная (Л) и венозная (Б) система лягушки:
А: 1 — желудочек; 2 — правое предсердие; 3 — левое предсердие; 4 — артериаль-
ный конус; 5 — общая сонная артерия; 6 — системные дуги аорты; 7 — подклю-
чичная артерия; 8 — спинная аорта; 9 — подвздошная артерия; 10 — бедренная
артерия; 11 — седалищная артерия; 12 — кишечно-брыжеечная артерия; 13 —
легочная артерия; 14 — кожные артерии; 15 — сонная «железа»; 16 — наружная
сонная артерия; 17 — внутренняя сонная артерия. В черный цвет окрашены арте-
рии с венозной кровью, артерии с артериальной и смешанной кровью заштри-
хованы. Б: 1 — венозный синус (показан как бы просвечивающим сквозь конту-
ры сердца); 2 — наружная яремная вена; 3 — внутренняя яремная вена; 4 —
большая кожная вена; 5 — подключичная вена; 6 — передняя полая вена; 7 —
задняя полая вена; 8 — бедренная вена; 9 — седалищная вена; 10 — подвздошная
вена; 11 — воротная система почек; 12 — подкишечная вена; 13 — воротная
система печени; 14 — печеночные вены; 15 — брюшная вена; 16 — легочная вена-
Заштрихованы вены с артериальной кровью
174
«железа», регулирующая, по-видимому, давление крови в сонных
артериях.
По сосудам второй пары (гомологичной IV паре жаберных ар-
терий рыб) — системным дугам аорты — кровь направляется к
внутренним органам и передним конечностям. Системные дуги
огибают сердце соответственно с левой и правой стороны и сли-
ваются под позвоночником в общий ствол — спинную аорту. От
спинной аорты кровь по соответствующим артериям поступает к
внутренним органам и задним конечностям.
Сосуды, гомологичные V паре жаберных артерий рыб, у ля-
гушки отсутствуют. Поэтому третья пара артериальных сосудов
гомологична VI паре жаберных артерий рыб — это легочные арте-
рии. По ним кровь направляется в легкие. От каждой легочной ар-
терии отходит крупная кожная артерия, по которой кровь прите-
кает к коже для обеспечения кожного дыхания, поэтому третью
пару сосудов иногда называют кожно-легочными артериями.
Полного разделения потоков артериальной и венозной крови у
амфибий нет. Поступающая к органам тела кровь содержит боль-
шой процент углекислоты. Это несомненно отражается на уровне
метаболизма амфибий и их сезонном поведении.
Венозная система низших амфибий, как и артериальная, по-
хожа на венозную систему двоякодышащих рыб. Хвостовая вена
делится на две воротные вены почек. Из них кровь поступает в
непарную заднюю полую вену и в парные задние кардинальные вены.
Последние на уровне сердца сливаются с парными яремными, под-
ключичными и кожными венами и образуют кювъеровы протоки,
изливающие кровь в венозный синус. От кишечника кровь собира-
ется по подкишечной и брюшной венам. Сливаясь, они образуют
воротную вену печени. Из печени кровь поступает в заднюю полую
вену по печеночной вене.
У бесхвостых амфибий кардинальные вены не сохраняются, и
вся кровь туловищной области, пройдя воротную систему по-
чек, собирается в заднюю полую вену и вливается в венозный синус
(рис. 141, Б). Брюшная и подкишечная вены образуют в печени
воротную систему кровообращения. В связи с отсутствием карди-
нальных вен у бесхвостых отсутствуют и кювьеровы протоки. Ярем-
ные вены, объединяясь с подключичными, образуют в этом случае
парные передние полые вены, впадающие в венозный синус. В пе-
редние полые вены впадают и кожные вены соответствующей
стороны, которые несут не венозную, а артериальную кровь.
Таким образом, венозная кровь, поступающая в правое пред-
сердие, имеет довольно значительную примесь содержащей кис-
лород крови, которая поступает в передние полые вены из кож-
йьгх вен.
У земноводных появляется новый кроветворный орган — крас-
ный костный мозг трубчатых костей.
175
Скорость движения крови (важный показатель интенсивности
метаболизма) у амфибий невелика. Об этом косвенно свидетель-
ствует частота пульса. У травяной лягушки массой 50 г она равна
40—50 уд./мин. У водных форм наблюдается брадикардия. Уровень
артериального давления у амфибий также низок. У хвостатых он
равен примерно 22/12 — 30/25, у бесхвостых — 30/20 (Л. Проссер
1978).
Кровеносная система пресмыкающихся по сравнению с
таковой у земноводных изменяется существенно. Ее превращения
связаны с исчезновением (даже на личиночной стадии) жаберно-
го дыхания и переходом на легочную оксигенизацию. Кроме того,
усиливается тенденция к разделению артериального и венозного
кровообращения.
Изменения наблюдаются и в конструкции сердца, и в характе-
ре расположения сосудов, выносящих из него кровь. В пределах
класса наметились два пути решения разделения кровотоков.
У чешуйчатых (ящерицы, змеи) и черепах сердце трехкамер-
ное (рис. 142, А). Левое предсердие принимает артериальную кровь,
Рис. 142. Схема строения сердца и прилежащих сосудов ящерицы (Я)
и крокодила (Б):
1 — правое предсердие; 2 — левое предсердие; 3 — желудочек; 4 — неполная
перегородка желудочка; 5 — дорсальная ниша желудочка; 6 — правая дуга аорты;
7 — сонные артерии; 8 — подключичные артерии; 9 — левая дуга аорты; 10 —
вентральная ниша желудочка; 11 — легочная артерия; 12 — спинная аорта; 13 "
правый желудочек; 14 — левый желудочек; 75 — полная перегородка с межжелУ'
дочковым отверстием; 16 — общее отверстие правой и левой дуг аорты
176
правое — венозную. Венозный синус, известный в прежних клас-
сах, сливается (поглощается) с правым предсердием. В желудочке
имеется неполная перегородка. Она прикреплена к левой части
желудочка и располагается горизонтально. Перегородка делит по-
лость желудочка на верхнюю (дорсальную) и нижнюю (вентраль-
ную) камеры.
Предсердия открываются в желудочек самостоятельными сбли-
женными отверстиями, снабженными полулунными клапанами.
Кровь из обоих предсердий поступает в верхнюю (дорсальную)
камеру. Артериальный конус отсутствует. Из желудочка выходят
самостоятельно три артериальных ствола. Их отверстия располага-
ются вокруг свободного края перегородки. Несколько ниже, на
вентральной стенке желудочка находится отверстие легочной ар-
терии, правее — отверстие левой дуги аорты, выше, на дорсаль-
ной стенке желудочка — отверстие правой дуги аорты.
При такой конструкции сердца в щелевидном пространстве
вокруг неполной перегородки желудочка устанавливается гради-
ент кислорода крови. После сокращения предсердий артериаль-
ная кровь из левого предсердия оказывается в дорсальной нише
желудочка и вытесняет венозную кровь, излившуюся из правого
желудочка, в вентральную нишу. В правой части желудочка оказы-
вается смешанная кровь. При сокращении желудочка каждая пор-
ция крови устремляется к ближайшему отверстию: артериальная
кровь из дорсальной ниши — в правую дугу аорты, венозная кровь
из вентральной ниши — в легочную артерию, смешанная кровь
из правой части желудочка — в левую дугу аорты. Таким образом,
артериальный и венозный кровотоки оказываются в значитель-
ной мере разделенными. От правой дуги берут начало две сонные
и две подключичные артерии.
У крокодилов потоки крови разделяются иначе (рис. 142, Б).
Вертикальная перегородка делит желудочек на левую и правую
половины практически полностью. В межжелудочковой перегородке
есть отверстие (паницциево отверстие), допускающее частичное
смешение крови в желудочке*. От правого желудочка отходят два
сосуда: по легочной артерии венозная кровь поступает в органы
Дыхания. Другой сосуд — левая дуга аорты объединяется с правой
дугой аорты, несущей артериальную кровь из левого желудочка в
общий артериальный ствол — спинную аорту со смешанной кро-
вью.
У всех пресмыкающихся от начала спинной аорты отходит ки-
шечно-брыжеечная артерия. Она делится на брыжеечную и кишеч-
ную артерии, которые образуют густую сеть сосудов, питающих
* Согласно другой точке зрения, камеры желудочка у крокодилов изолиро-
ваны полностью. Паницциево отверстие находится в месте соприкосновения ос-
нований двух дуг аорты (С.Левушкин, И. Шилов, 1994).
177
Рис. 143. Схема кровеносной системы
ящерицы (артерии светлые, стрелка-
ми показано направление движения
крови):
1 — легочная артерия; 2 — правая дуга аор.
ты; 3 — сонная артерия; 4 — сонный про-
ток; 5 — подключичная артерия; 6 — левая
дуга аорты; 7 — спинная аорта; 8 — меж-
позвонковая артерия; 9 — кишечная арте-
рия; 10 — брыжеечная артерия; 11 — под-
вздошная артерия; 12 — бедренная артерия;
13 — печень; 14 — почка; 15 — хвостовая
артерия; 16 — хвостовая вена; 17— бедрен-
ная вена; 18 — подвздошная вена; 19 —
брюшная вена; 20 — почечная вена; 21 —
задняя полая вена; 22 — воротная вена пе-
чени; 23 — яремная вена; 24 — подключич-
ная вена; 25, 26 — левая и правая передние
полые вены; 27 — отрезок пищеваритель-
ной трубки; 28 — легкое
кровью желудок, селезенку, подже-
лудочную железу, тонкий и толстый
кишечник. Далее от спинной аорты
отходят две ветви, несущие кровь к
почкам, половым органам и задним
конечностям. Заканчивается спин-
ная аорта хвостовой артерией, не-
сущей кровь к хвосту (рис. 143).
Венозная система пресмыкающихся в целом мало отличается
от таковой земноводных. Усиливается редукция воротного крово-
обращения в почках. Большая часть венозной крови от заднего
конца тела направляется либо в обход почек, либо проходит сквозь
них, не образуя капиллярных сетей вокруг почечных канальцев.
От головы кровь собирается в парные яремные вены, которые,
соединившись с парными подключичными венами, образуют правую
и левую передние полые вены. Передние полые вены впадают в пра-
вое предсердие. От хвоста венозная кровь собирается в хвостовую
вену, которая в области тазового пояса делится на две подвздошные
(тазовые) и две почечные вены. Далее подвздошные вены, сливаясь
с бедренными, объединяются в непарную брюшную вену. Вместе с
воротной веной печени, несущей кровь от кишечника, она впадает в
печень, где образует воротную систему. Кровь из воротной системы
печени поступает в печеночные вены, а затем в заднюю полую вену-
Задняя полая вена образуется путем слияния почечных вен, выхо-
дящих из почек, и впадает в правое предсердие.
178
Птицы (как и млекопитающие) имеют полное разделение
артериального и венозного кровотоков. Это достигается оконча-
тельным разделением сердца на четыре камеры, упрощением на-
бора системных дуг и наличием двух выносящих кровь сосудов: аорты
и легочной артерии. Через правую половину сердца венозная кровь
направляется из тела в легкие, через левую — артериальная из лег-
ких в тело. У птиц сохраняется правая дуга аорты (у млекопитаю-
щих — левая).
Рассмотрим путь циркуляции крови у птиц (рис. 144). Кровь
после окисления ее в легких изливается через легочные вены в
левое предсердие, откуда она поступает в левый желудочек. Из
последнего выходит единственная дуга аорты — правая, от кото-
рой вскоре по выходе отделяются парные безымянные артерии.
Затем дуга поворачивает направо и, подойдя к позвоночнику,
идет вдоль него к хвосту в виде спинной аорты. От спинной аорты
отходят ветви к внутренним органам, а в области таза — круп-
ные бедренные и седалищные артерии. Безымянные артерии разде-
ляются на идущие к голове сонные артерии, к передним конеч-
ностям — подключичные артерии, к мышцам груди — грудные
артерии. Из правого желудочка общим стволом отходят парные
легочные артерии.
Рис. 144. Система кровообращения голубя:
1 — яремная вена; 2 — сонная артерия; 3 — подключичные вены; 4 — плечевая
,1ена; 5—легочные вены; б— плечевая артерия; 7— внутренностная артерия; 8 —
спинная аорта; 9— почечные артерии; 10— седалищная артерия; 11 — почечная
артерия; 12 — подвздошная артерия; 13 — внутренняя подвздошная вена; 14 —
средняя копчиковая артерия; 15 — хвостовая вена; 16 — надхвостовая вена; 17 —
,1ена копчиково-брыжеечная; 18 — нижняя полая вена; 19 — мезентериальная
,1ена; 20 — воротная вена; 21 — воротная система печени; 22 — правая печеноч-
ная вена; 23 — левая печеночная вена; 24 — нижняя полая вена; 25 — легочная
аРтерия; 26 — трахейно-бронхиальная артерия; 27 — грудная вена; 28 — верхняя
полая вена; 29 — подключичная артерия
179
Таким образом, у птиц в отличие от рептилий из сердца выхо-
дят только два артериальных ствола: правая дуга аорты и ствол
легочных артерий. Левой дуги аорты, отходящей у рептилий от
правой части желудочка (а у крокодилов — от самостоятельного
правого желудочка) и несущей, следовательно, венозную кровь,
которая изливается в спинную аорту, у взрослых птиц нет. Однако
первоначально у зародыша закладываются обе дуги аорты, и его
артериальная система в общих чертах сходна с таковой у крокоди-
лов. В последующем левая дуга исчезает, и этим устраняется сме-
шивание в спинной аорте артериальной и венозной крови.
Венозная система не имеет столь же принципиальных особен-
ностей. По небольшим хвостовым венам кровь изливается в две
воротные вены почек. Однако не вся кровь поступает в капилляр-
ную сеть воротной системы, и часть ее проходит через вещество
почек по специальным сосудам насквозь. По выходе из почек эти
сосуды, слившись с бедренными венами, образуют парные подвздош-
ные вены, а слияние подвздошных вен приводит к образованию
основного венозного ствола туловищной области — задней полой
вены. От внутренних органов кровь собирается в копчиково-брыже-
ечную вену, а от брюшной стенки тела — в надкишечную вену. Обе
вены сливаются в воротную вену печени. По выходе из печени
кровь по печеночным венам изливается в заднюю полую вену. Та-
ким образом, в заднюю полую вену собирается вся венозная кровь
от большей части туловищной области. Эта вена впадает в правое
предсердие.
У птиц в сравнении с другими позвоночными заметно возраста-
ют относительные размеры сердца, что, безусловно, связано с
высокой интенсивностью обмена веществ, особенно во время по-
лета. Так, сердечный индекс у утки равен 0,6, а у кролика всего 0,2.
Масса сердца у мелких птиц относительно большая по сравне-
нию с крупными, что связано с более интенсивным обменом ве-
ществ (в свою очередь, это обусловлено тем, что у мелких живот-
ных менее выгодное соотношение между объемом тела и его по-
верхностью, отдающей тепло). Например, у снегиря при массе
тела 23 г относительная масса сердца равна 1,3 %, у чечетки мас-
сой 13 г — 1,6 %, а у синицы-московки массой 8 г — 1,8 %.
Прослеживается также зависимость между относительной мас-
сой сердца и энергичностью движений. У хороших летунов соко-
ла-чеглока и пустельги сердечный индекс равен 1,2 %, а у плохо-
го летуна — сороки — всего около 0,9 %.
Работа сердца у птиц более энергична, чем у низкоорганизо-
ванных наземных позвоночных. У травяной лягушки число сокрЗ'
щений сердца равно 40 — 50 уд./мин, у снегиря — в среднем 730
уд./мин. Хорошо видна зависимость частоты сокращений сердца и
от размеров тела. У голубя массой 250 г число сокращений серди8
в минуту равно в среднем 248, у зеленушки массой 22 г — 697, У
180
щегла массой 13 г — 754, у московки массой 8 г - 1038. Частота
сердцебиений существенно зависит от двигательной активности
[-[типы. Так, у голубя в покое она равна в среднем 165 ударам в
минуту, а в полете — 550. Ныряющим в воду птицам свойственна
брадикардия, т. е. снижение частоты сокращений сердечной мыш-
цы. К примеру, у оляпки через 5 с после погружения в воду сер-
дечный ритм сокращается от первоначального до 73 % (через 10с —
до 48 % и через 15 с — до 42 %).
К прогрессивным признакам птиц следует отнести и общее
увеличение объема крови. У костистых рыб масса крови равна при-
мерно 3 % массы тела, у бесхвостых амфибий — 6 %, у птиц — 9 %.
Все эти особенности организации являются важным условием
общего повышения энергии жизнедеятельности птиц.
В артериальной системе млекопитающих, как и у птиц,
имеется только одна, левая дуга аорты, отходящая от толстостен-
ного левого желудочка (рис. 145). Главные артериальные сосуды
отходят от центрального ствола у разных видов неодинаково. Обыч-
но от аорты отходит короткая безымянная артерия, которая делит-
ся на правую подключичную артерию, правую и левую сонные арте-
рии’, левая подключичная артерия отходит самостоятельно от дуги
аорты. В других случаях левая сонная артерия отходит не от безы-
мянной артерии, а самостоятельно от дуги аорты. Спинная аорта,
как и у всех позвоночных, лежит под
нее отходят сосуды к мускулатуре и
внутренним органам. На рис. 146 по-
казан эволюционный ряд артериаль-
ной системы.
В венозной системе млекопитающих
отсутствует воротное кровообращение в
почках. Левая передняя полая вена лишь у
Рис. 145. Схема строения кровеносной
системы млекопитающего (стрелки —
направление движения крови):
1 — внешняя сонная артерия; 2 — внутренняя
сонная артерия; 3 — подключичная артерия;
4 — дуга аорты; 5 — легочная артерия; б —
левое предсердие; 7 — правое предсердие;
— левый желудочек; 9 — правый желудо-
чек; 10 — спинная аорта; 11 — внутренност-
ная артерия; 12 — почечная артерия; 13 — под-
вздошная артерия; 14 — яремная вена; 15 —
Подключичная вена; 16— левая непарная вена;
17 — правая непарная вена; 18 — задняя по-
лая вена; 19 — печеночная вена; 20 — воротная
вена печени; 21 — печень; 22 — почка; 23 —
подвздошная вена
позвоночным столбом.
От
20
19
21
9
17
18
181
I II III IV V VI
A
III rv V VI
г
Рис. 146. Схема расположения артериальных жаберных дуг позвоночных
(по А. Ромеру, Т. Парсонсу):
А — гипотетический предок позвоночных; Б — акула; В — двоякодышащая рыба —
протоптерус; Г — костистая рыба; Д — хвостатая амфибия; Е — ящерица; Ж —
птица; 3 — млекопитающее; 1,2— внутренняя и наружная сонные артерии
соответственно; 3 — сонный проток; 4— артериальный проток; Л — легкое; I—
VI — жаберные артерии
немногих видов впадает в сердце самостоятельно. Чаще она сливается
с правой передней полой веной, которая и изливает всю кровь от пере-
днего отдела тела в правое предсердие. Характерная черта
млекопитающих — наличие остатков передних кардинальных веН-
так называемых непарных вен. У большинства видов только правая не-
парная вена самостоятельно впадает в переднюю полую вену, а левая
182
в
Рис. 147. Обобщенные схемы кругов кровообращения и включенных в
них капиллярных сетей разных позвоночных (по А. Ромеру, Т. Парсонсу):
А — типичная рыба; Б — земноводное или пресмыкающееся, утратившее жабер-
ную капиллярную сеть в связи с появлением легких; В — млекопитающее, ут-
ратившее воротную систему почек и обладающее полным разделением кругов
кровообращения
непарная вена утрачивает связь с полой веной и через поперечную вену
впадает в правую непарную вену. В целом венозной системой млеко-
питающих завершается перестройка капиллярного кровообращения
Позвоночных. На рис. 147 показано, что перестройка сопровождается
постепенным сокращением капиллярного кровообращения в жаб-
Рах, исчезновением воротного кровообращения в почках у млекопи-
тающих (частично это имеет место и у птиц).
У млекопитающих в связи с особенностями жизни и завися-
щей от них интенсивности обмена веществ относительные разме-
ры сердца существенно различны. Его показатели, как и у птиц,
возрастают с уменьшением массы тела. Так, относительная масса
Сердца (сердечный индекс) кашалота — 0,3, африканского слона —
"4, ленивца — 0,3, серой полевки — 0,6, ушана — 1,2 —1,4, обык-
183
Таблица s
Частота пульса разных млекопитающих
Вид животного Масса тела, г Частота пульса, уд./мин
Мышь 0,025 600
Собака 6,500 120
Овца 50,000 70-80
Бык 500000 43
новенной землеройки — 1,4. Приведенный ряд показателей отра-
жает одновременно и зависимость интенсивности обмена веществ
от величины поверхности тела, отдающей тепло. Ту же зависимость
отражает и частота сокращений сердца (частота пульса) (табл. 5).
Интенсивность частоты сердцебиения имеет и иное приспосо-
бительное значение. Учащение пульса (тахикардия) наблюдается
при усилении работы мышечной системы. Наиболее быстро кровь
продвигается в аорте.
Своеобразные приспособления возникают при водном образе
жизни зверей, когда возможность атмосферного дыхания перио-
дически прерывается. С одной стороны, это выражается в резком
увеличении количества связывающего кислород глобина в мыш-
цах {миоглобина) — около 50 % всего глобина организма. Кроме
того, у зверей, способных погружаться в воду на длительное вре-
мя, отключается периферическое кровообращение и сохраняется
стабильным кровоснабжение мозга и сердца. У них снижена чув-
ствительность дыхательного центра мозга к накоплению углекис-
лоты в крови.
С другой стороны, у водных и полуводных млекопитающих при
погружении в воду уменьшается число сердцебиений: возникает
так называемая брадикардия. Она приводит к замедленному крово-
току и более полному использованию кислорода крови. Напри-
мер, у тюленя {Phoca vitulina), находящегося в погруженном со-
стоянии, число сокращений сердца в минуту равно 1280. Через
11 с после погружения оно уменьшается до 60, через 27 с — до 35
и далее устанавливается на уровне 30 в течение всего времени
нахождения зверя под водой. Антарктический тюлень Уэддола, на-
пример, опускается на глубину до 600 м и способен провести под
водой до 43 мин.
Прогрессивные особенности в кровообращении у млекопитаю-
щих заключаются не только в строении и функционировании орга-
нов сосудистой системы. У них увеличивается общее количество кро-
ви и повышается ее кислородная емкость. Это связано с больший
числом эритроцитов, их безъядерностью и увеличением в них ко-
личества гемоглобина. У низших позвоночных (рыбы, амфибий)
содержание гемоглобина в среднем 5 —10 г, а у млекопитающих ''
184
jO—15 г на 100 см3 крови (X.С. Коштоянц, 1950). Важным показа-
телем кровообращения является уровень кровяного давления.
Лимфатическая система
Наряду с собственно кровеносной системой у позвоночных есть
другая, связанная с ней сосудистая система — лимфатическая. Она
состоит из лимфатических сосудов и лимфатических желез. Лимфа-
тическая система в отличие от кровеносной незамкнутая. Только
крупные сосуды имеют самостоятельные стенки, в то время как
их разветвления располагаются в межклеточных пространствах
различных органов. Лимфатические сосуды содержат бесцветную
жидкость — лимфу, в которой плавают лимфоциты — белые кровя-
ные тельца, образующиеся в лимфатических железах. Движение
лимфы создается сокращением стенок некоторых участков круп-
ных сосудов (так называемых лимфатических сердец) и периоди-
чески повторяющимся давлением на сосуды различных органов.
Лимфатическая система служит посредницей обмена веществ
между кровью и тканями тела животного. Она регулирует осмоти-
ческое давление в тканях. Кроме того, образующиеся в лимфати-
ческих узлах и железах белые кровяные клетки — лимфоциты унич-
тожают болезнетворные организмы, выполняя в теле защитную
роль, участвуя в иммунологических реакциях.
* * *
Таким образом, возникающая мезодермально замкнутая кро-
веносная система хордовых доставляет питательные вещества,
кислород и гормоны органам и тканям, транспортирует вредные
продукты метаболизма к специализированным органам: углекис-
лый газ — к легким, вредные продукты белкового обмена — к
печени и почкам. Кровь защищает организм животных от инфек-
ций. Единым планом строения обладает кровеносная система пер-
вичноводных хордовых — один круг кровообращения и окисле-
ние крови в жабрах. Движение крови у ланцетника происходит
благодаря сокращениям оснований жаберных артерий, а у после-
дующих в эволюционном ряду групп движение крови обеспечива-
ется работой сердца. Важным условием поступательного развития
Позвоночных является эволюция кровеносной системы у назем-
ных позвоночных с двумя кругами кровообращения. Она направ-
лена на все более полное разделение потоков артериальной и ве-
крови, в том числе и путем усложнения строения сердца,
разделение потоков крови в организме достигнуто при
Параллельной независимой эволюции птиц и млекопитающих.
Эволюция крови шла путем увеличения числа форменных эле-
ментов и большей их специализации.
Нозной
Полное
ТЕРМОРЕГУЛЯЦИЯ
Известно, что температура тела животных зависит от уровня их
метаболизма и сама влияет на скорость обменных процессов, обус-
ловливая многие формы жизнедеятельности организмов.
Среди позвоночных различают две физиологические группы:
пойкилотермные и гомойотер.мные животные.
Пойкилотермные (или холоднокровные) животные не имеют
постоянной температуры тела. Она меняется в широких пределах в
зависимости от изменений температуры окружающей среды. К этой
группе принадлежат круглоротые, рыбы, амфибии и рептилии.
Обычно их температура тела на 1 — 2 °C превышает температуру
окружающей среды. Уровень метаболизма пойкилотермных живот-
ных невысок. Соответственно температура окружающей среды (воды
или воздуха) выступает ведущим природным фактором, опреде-
ляющим физиологическое состояние, поведение, экологию ви-
дов, а также ограничивающим их географическое и вертикальное
распространение и в воде, и на суше.
Отдельные поведенческие или физиологические адаптации
позволяют им повысить (или понизить) теплопродукцию тела
разными путями. Например, тела быстроходных рыб тунцов, скум-
брий, акул разогреваются при усиленной мышечной работе во
время плавания; температура тела при этом повышается на 10—
15 °C и может превысить температуру окружающей воды. Своеоб-
разные теплообменники расположены у тунцов непосредственно
под кожей в виде темных красных мышц, обильно снабженных
кровеносными сосудами (К. Шмидт-Ниельсен, 1982) (см. рис. 89)-
Рептилии используют гелиотермию при переваривании пищи или
в период размножения. Улавливать солнечное тепло им помогают
темная окраска и умение распластывать тело и конечности на суб-
страте. Температура тела у них может возрасти в десятки раз по
сравнению с температурой окружающей среды. Важный элемент
жизни пойкилотермных животных — этологическая форма термо-
регуляции. Сезонные и суточные понижения температуры среды
сопровождаются у них впадением в спячку, оцепенением, изме-
нением ритма суточной активности. При перегреве тела кроме
поведенческих реакций (зарывание в песок, уход в укрытие, вле-
зание на кусты и т.д.) они используют тепловую одышку (поли-
пноэ), благодаря которой ящерицы и крокодилы, например, Д®1'
186
тельное время удерживают температуру тела на 2 — 5 °C ниже тем-
пературы окружающей среды. Черепахи для охлаждения исполь-
зуют обильную слюну.
Гомойотермные животные. Тела гомойотермных животных об-
ладают способностью поддерживать постоянную температуру тела,
благоприятную для их обменных процессов и комфортного состо-
яния. Этой уникальной способностью среди позвоночных облада-
ют птицы и млекопитающие. Они имеют высокий уровень энерге-
тического обмена и характеризуются сложными механизмами тер-
морегуляции. Различают химическую и физическую терморегуляцию.
Эффект химической терморегуляции состоит в освобождении теп-
лопродукции при питании и дыхании животного и рефлекторном
повышении эндогенного теплообразования при понижении тем-
пературы окружающей среды или других явлениях. Богатые энерги-
ей вещества в теле позвоночных животных запасаются в «депо» —
печени, полостных сальниках, подкожной жировой клетчатке и
скелетной мускулатуре. Подкожный жир толстым слоем лежит в
коже китов, тюленей, белых медведей. Высвобождение тепла при
тонусе покоящейся мускулатуры — основной источник теплооб-
разования в организме.
Механизмы физической терморегуляции контролируют тепло-
отдачу. Поднятие и распушение, сезонная густота волосяного и
перьевого покровов, так же как и сужение кожных кровеносных
сосудов, уменьшают теплоотдачу. Наоборот, расширение сосудов,
уменьшение густоты покровов способствуют охлаждению тела. Теп-
лоотдача заметно усиливается при вентиляции ротовой полости и
верхних дыхательных путей (полипноэ), так как испарение жид-
кости с их поверхностей сопровождается использованием эндо-
генного тепла. При повышении температуры среды или усилен-
ной мышечной работе у животных учащается дыхание (параллельно
учащается сердцебиение).
У некоторых млекопитающих с разреженным волосяным по-
кровом в удалении избыточного тепла путем испарения жидкости
Участвуют потовые железы, расположенные в разных участках тела.
Соотношение уровней теплопродукции и теплоотдачи у гомой-
отермных животных контролируется центром терморегуляции, рас-
положенным в гипоталамусе. Центр получает информацию от теп-
ловых и холодовых терморецепторов кожи и осуществляет кор-
рекцию температуры тела. Дополнительную информацию о темпе-
ратуре сообщает центру артериальная кровь, поступающая к го-
ловному мозгу по сонным артериям. Раздражение холодовых тер-
морецепторов сопровождается увеличением теплопродукции за счет
Интенсификации обмена веществ, появления холодовой дрожи,
сУл<ения поверхностных кровеносных сосудов. Раздражение тепло-
Нувствительных нейронов, наоборот, сопровождается снижением
МЬ1щечного ТОНуеа, расширением кровеносных сосудов и т.д.
187
Работа центра терморегуляции гипоталамуса находится у мле-
копитающих под контролем коры больших полушарий, у птиц
эта ответственность принадлежит ядрам подкорки. Присущие по-
звоночным животным формы теплообмена со средой являются
одним из важнейших механизмов поддержания их гомеостаза. Они
обеспечивают им способность к выживанию в различных природ-
ных условиях, определяют границы расселения и формы поведе-
ния.
* * *
Таким образом, температура тела позвоночных животных оп-
ределяется уровнем обменных процессов и совершенством термо-
регуляции. Большинство позвоночных пойкилотермны. Темпера-
тура тела у них повышается при усиленной мышечной работе.
У наземных пойкилотермных позвоночных отмечена поведенчес-
кая терморегуляция. Эволюционно прогрессивные изменения в ряду
позвоночных привели к устойчивой гомойотермности птиц и мле-
копитающих и выработке совершенных механизмов терморегуля-
ции.
НЕРВНАЯ СИСТЕМА
В теле позвоночного есть две системы, координирующие и ин-
тегрирующие работу всех систем органов и обеспечивающие гар-
моничную деятельность целого организма: нервная и гуморальная.
Центральная нервная система хордовых отличается от таковой
у представителей других типов животного царства. Она имеет вид
нервной трубки, расположенной дорсально (со спинной стороны
тела). В онтогенезе животного нервная трубка закладывается путем
впячивания эктодермы внутрь тела эмбриона (рис. 148). Эктодер-
мальная закладка предопределила, согласно И. И. Шмальгаузену
(1964), чувствующее назначение нервной трубки. У низших пред-
ставителей типа — бесчерепных — это выразилось в ее светочув-
ствительности (наличии глазков Гессе) и способности восприни-
мать химические раздражители; у позвоночных — свидетельство
тому эмбриональная закладка ряда органов чувств как выростов
головного мозга. В процессе длительной эволюции позвоночных
центральная нервная система сохранила чувствующие функции и
одновременно стала системой, координирующей работу всех ор-
ганов тела.
Характерная черта нервной трубки — наличие внутри нее не-
вроцеля, т. е. полости, содержащей
ных невроцель сообщается с
внешней средой посредством от-
верстия в головной части тела —
невропора и с кишкой посред-
ством нервно-кишечного канала.
У взрослых ланцетников на мес-
те заросшего невропора сохра-
няется чувствующий орган — обо-
нятельная ямка. У взрослых позво-
ночных полость невроцеля при
зарастании нервно-кишечного
Канала становится замкнутой.
Изнутри и снаружи нервную
трубку омывает спинномозговая
Жидкость. Она создает вокруг не-
рвного ствола защитный слой и
вЫполняет транспортную функ-
жидкость. У личинок бесчереп-
Рис. 148. Последовательные
стадии развития (закладки)
центральной нервной системы
(А-ДУ
1 — эпидермис; 2 — ганглионарная
пластинка
189
цию. Через эту жидкость нервные клетки обеспечиваются питали,
ем и освобождаются от продуктов метаболизма. Источником спщ^
номозговой жидкости служит плазма крови, проникающая из спде.
тений кровеносных сосудов в сосудистой оболочке, выстилаю,
щей полости невроцеля и мозговых желудочков. Отток жидкости
происходит через венозную систему твердой мозговой оболочки
Колебательные движения спинномозговой жидкости обусловле-
ны изменениями положения тела, дыхательными движениями
пульсацией сердца. В головном отделе через жидкость поддержи-
вается внутричерепное давление.
Нервная трубка у низших хордовых практически не дифферен-
цирована* . У позвоночных в ней четко выделяются два отдела: го-
ловной (cephalon) и спинной (medulla sphinalis) мозг, образующие
центральную нервную систему. От центральной нервной системы к
мышцам и органам тела отходят нервы. Они составляют перифери-
ческую нервную систему. Наряду с центральной позвоночные имеют
вегетативную (симпатическую и парасимпатическую) нервную сис-
тему, которая осуществляет иннервацию всех внутренних органов,
сосудов, поровых желез и обеспечивает целостное реагирование
организма на внешние и внутренние воздействия.
Головной мозг
Головной мозг позвоночного закладывается эмбрионально в виде
трех мозговых пузырей (рис. 149). Впоследствии в результате их диф-
ференцировки формируются пять отделов головного мозга. При
поперечном делении первого мозгового пузыря выделяются пере-
дний мозг (telencephalon) и промежуточный (diencephalon). Из вто-
рого пузыря формируется средний мозг (mesencephalon). Третий
мозговой пузырь продольно разделяется на продолговатый мозг
(myencephalon) и мозжечок (metencephalon или cerebellum). Сред-
ний и продолговатый отделы составляют ствол головного мозга,
в нем сохраняются признаки метамерности (посегментное распо-
ложение ядер и отхождение черепномозговых нервов), характер-
ные для спинного мозга. Рис. 150 дает представление об оформле-
нии отделов головного мозга в онтогенезе позвоночного.
Невроцель внутри отделов головного мозга образует четыре
связанные между собой полости — мозговые желудочки. Два боко-
вых желудочка расположены симметрично в переднем мозге, один
в промежуточном и один в продолговатом мозге. Головной мозг
покрыт оболочками. У рыб — это однослойная первичная соедини-
тельнотканная мембрана. У наземных позвоночных она диффереН-
* Она закладывается в онтогенезе асцидий. Однако по мере взросления осо-
бей уменьшается до одного узла.
190
I
рис- 149. Схемы развития отделов
головного мозга позвоночного
животного:
. __ стадия трех пузырей (/, 2, .?); Б —
оТделЫ мозга: I — передний; 2 — проме-
жуточный; 3 — средний; 4 — мозжечок;
у — продолговатый; 6 — глазной бокал
Мозжечок
IMES
I ENCEPHALON
I
Обонятельный
стебелек
1 Таламус
Обонятельная
луковица ।
Концевая пластинка
Глазной стебелек
| Гипофиз ।
MES- ।
ENCEPHALbN
(MET-
Полушарие
конечного мозга!
(парное)^..*
TELENCEPHALON DIENCEPHALON
'ENCEPHALON MYENCEPHALON
Тектум ’1
(Мост) ।
(Продолговатый мозг
Кора, или плащ
Парафиз
Мозжечок
Теменной орган
щфиз
I Сосудистое сплетение
к-ЗТектум
Ьазальныел-;
1 Продолговатый мозг
Зрительный перекрест
Таламус
Гипоталаму<1
Обонятельная Сос»1исто
луковица сплетение
Рис. 150. Схемы, отражающие последовательность расположения
основных отделов и структур головного мозга позвоночного животного
(по А. Ромеру, Т. Парсонсу):
А — внешняя дифференцировка; Б — продольный разрез
Боковой
желудочек
Чйруется на твердую и мягкую оболочки, в свою очередь, у мле-
копитающих из мягкой формируются паутинная и сосудистая. Про-
странства между оболочками заполнены жидкостью.
191
Передний, или конечный, мозг (telencephalon)
Передний мозг у большинства позвоночных представлен дву^
полушариями, в эволюционном ряду позвоночных их разделецц
усиливается. Спереди отдела обособляются обонятельные д0Л11
Внутри переднего мозга невроцель образует два боковых желудок
ка, заполненных жидкостью.
Структурно масса переднего мозга характеризируется нали-
чием серого и белого вещества. Серое вещество образовано тела-
ми нейронов. Скопления серого вещества в толще переднего
мозга в виде базальных ядер образуют полосатое тело. В поверх,
ностных областях мозга изнутри и снаружи серое вещество обра-
зует кору полушарий головного мозга. Значительную часть передне-
го мозга составляет белое вещество, образованное длинными
отростками нервных клеток (аксонами), покрытыми миэлино-
выми оболочками. Базальные ядра переднего мозга вместе с яд-
рами промежуточного мозга противопоставляются коре в каче-
стве подкорковых образований. Базальные ядра переднего мозга —
структуры, преемственные в ряду позвоночных. Специалисты
улавливают их явную гомологию, однако мнения ученых на этот
счет не едины.
В эволюционном ряду позвоночных проявляются три формы
коры переднего мозга: 1) древняя — палеокортекс (палеопаллиум);
2) старая — архикортекс (архипаллиум); 3) новая — неокортекс
(неопаллиум) (рис. 151).
Изначально у примитивных позвоночных масса серого веще-
ства обнаруживается в обонятельных долях переднего мозга, тон-
ким слоем она выстилает полости мозговых желудочков. Эта поло-
са нервной ткани получила название древней коры (палеопаллиум,
палеокортекс). Область древней коры остается участком обоня-
тельных долей на всех стадиях эволюции позвоночных (А. Ромер,
Т. Парсонс, 1992).
Дорсальнее и ближе к центральной оси мозга развивается ста-
рая кора (архипаллиум, архикортекс). Эволюционно она заклады-
вается позднее древней коры. У низших позвоночных (рыбы, зем-
новодные) она также содержит обонятельные нейроны и связана
с базальными ядрами. Позднее у рептилий она выделяется в само-
стоятельную область, расположенную над промежуточным моз-
гом, именуемую гиппокамп. Предполагают, что эта область вместе
с отделами промежуточного мозга играет существенную роль в
поддержании постоянства внутренней среды организма, участву-
ет в координации функций размножения, эмоциональном пове-
дении, в процессах обучения и сохранения памяти у позвоноч-
ных.
Новая кора (неопаллиум, неокортекс) впервые обнаруживает-
ся у некоторых рептилий. Поначалу она занимает положение меХ'
192
Обонятельная
Старая кора
А Б В
Новая
Старая кора
Древняя кора Древняя кора
Обонятельная обонятельная
луковица
кора
Старая кора = гиппокамп
(на медиальной поверхнос
Новая
кора
и
доля
луковица Древняя кора
и обонятельная доля
Рис. 151. Схема расположения разных форм коры в головном мозге
позвоночных животных (по А. Ромеру, Т. Парсонсу):
А — примитивная стадия; Б — амфибия; В, Г — примитивная и продвинутая
рептилия соответственно; Д, Е — примитивное и продвинутое млекопитающее
соответственно
ДУ древней и старой корой. У млекопитающих новая кора разрас-
тается. Она распространяется по внешней поверхности полуша-
рий, оттесняет старую и древнюю кору в глубь полушарий и ста-
новится важнейшей частью центральной нервной системы, отве-
чающей за высшие формы нервной деятельности, осуществляя
корреляцию и ассоциацию нервных реакций (А. Ромер, Т. Пар-
сонс, 1992).
Новая кора у высших млекопитающих имеет сложную клеточ-
ную архитектонику. Она многослойна: состоит из 6 —7 слоев раз-
ных по форме нервных клеток (пирамидных, звездчатых, верете-
нообразных). Послойное расположение в коре нейронов улучшает
ее анализирующую функцию. У высших позвоночных неопаллиум
осуществляет высшую координацию деятельности отдельных ор-
ганов, систем органов и всего организма. У млекопитающих наи-
более сложно организована ассоциативная область новой коры.
На рис. 152 показано размещение разных форм коры в переднем
Мозге различных позвоночных.
КоНСТамтиили 1Q4
Рис. 152. Разрез полушария переднего мозга и схема расположения
разных форм коры:
А — примитивная стадия; Б — современная амфибия; В — продвинутая стадия
амфибии; Г — продвинутая рептилия; Д — примитивное млекопитающее; Е —
человек; I — древняя кора; 2 — желудочек; 3 — старая кора; 4 — базальные ядра
подкорки; 5 — новая кора; 6 — мозолистое тело; 7 — обонятельная борозда
Передний мозг возник и прогрессировал как мультифункцио-
нальный орган центральной нервной системы. Во всех без исклю-
чения группах хордовых он обрабатывает ольфакторную инфор-
мацию и отвечает за обоняние. От переднего мозга отходит первая
пара черепномозговых нервов (I) — обонятельные нервы. Передний
мозг координирует работу систем органов. Он обеспечивает ба-
ланс между разными формами эмоционального поведения: агрес-
сией, половым влечением, родительским поведением у высших
позвоночных (Л.Проссер, 1977, 1978) и т.д.
У млекопитающих с прогрессивным развитием неопаллиум3
переднего мозга связано совершенствование рассудочной деятель-
ности, памяти, обучаемости, и наконец, мышление и разговор'
ная речь.
194
Промежуточный мозг (diencephalon)
В эмбриогенезе промежуточный мозг образуется из задней час-
ти первого мозгового пузыря. В промежуточном мозге находится
третий мозговой желудочек. Спереди и сверху промежуточный мозг
граничит с передним, а снизу и сзади — со средним мозгом. Не-
рвное вещество промежуточного мозга состоит из множества свя-
занных между собой и другими отделами головного мозга ядер
(скоплений нейронов).
Центральную область промежуточного мозга — таламус обра-
зуют зрительные бугры. Здесь происходит переработка зрительной
информации и передача ее в высшие зрительные центры (у хря-
щевых рыб, земноводных и рептилий — в крышу среднего мозга,
у млекопитающих — в кору больших полушарий). Крышу проме-
жуточного мозга образует надбугорная область — эпиталамус (см.
рис. 150, Б). Как мозговой центр, он не имеет большого самосто-
ятельного значения: здесь расположен так называемый габенуляр-
ный ганглий, переключающий нервное возбуждение от переднего
мозга в его стволовую часть.
Нижнюю подбугорную область промежуточного мозга образу-
ет гипоталамус. Филогенетически это древний отдел промежуточ-
ного мозга. Он известен у всех позвоночных и наивысшего разви-
тия достигает у птиц и млекопитающих. Во всех классах позвоноч-
ных животных гипоталамус является важным интегративным цен-
тром, координирующим деятельность всех систем внутренних ор-
ганов. Структурность и функциональная ответственность гипота-
ламуса в эволюционном ряду позвоночных возрастает, при этом
усиливается дробность его морфологических структур. В ядра ги-
поталамуса поступают сигналы от органов обоняния и вкуса. Его
центры осуществляют регуляцию водно-солевого обмена, влияют
на функцию почек и осмотическую проницаемость жабр у рыб,
регулируют температуру тела, кровяное давление, частоту сер-
дечных сокращений и дыхания, деятельность кишечника, влияют
на сон, чувство голода и сытости и т.д. Гипоталамус участвует в
поддержании гомеостаза, в регуляции размножения, в реализа-
ции защитно-приспособительных реакций животных. Вместе с
гипофизом он образует единый гипоталамо-гипофизарный комп-
лекс и осуществляет связь двух регуляторных систем: нервной и
гуморальной (см. ниже).
В промежуточном мозге внешне сверху и снизу заметны выпя-
чивания. У зародышей большинства позвоночных спереди проме-
жуточного мозга отрастает вверх тонкостенный мешок — парафиз.
Как провизорный (зародышевый) орган он участвует в снабжении
м°зга гликогеном (А. Ромер, Т. Парсонс, 1992). Далее сверху разви-
Ьаются два выроста эпиталамуса: передний (париетальный) — те-
менной орган и задний (пинеальный) — эпифиз. Оба эти органа пред-
195
ставляют собой непарные стебельчатые глазоподобные структуры
В эволюционном ряду позвоночных они (вместе или попеременно)
проявляют себя как светочувствительные органы (непарный темен-
ной глаз), а эпифиз еще и как железа внутренней секреции.
Боковые выпячивания промежуточного мозга образуют глаз-
ные пузыри (парные глаза). В них закладываются специализирован-
ные корковые структуры мозга — сетчатка и пигментная оболоч-
ка глаз. Внутренние части этих выпячиваний превращаются в зри-
тельные нервы (II пара черепномозговых нервов), идущие к сет-
чатке глаза. В передней части дна промежуточного мозга они пере-
крещиваются, образуя зрительный перекрест, или хиазму*.
От гипоталамуса вниз отрастает важная мозговая железа — ней-
рогипофиз.
Средний мозг (mesencephalon)
В эмбриогенезе средний мозг образуется из второго мозгового
пузыря. Вместе с продолговатым мозгом он составляет ствол го-
ловного мозга и сохраняет следы сегментированности. Невроцель
не образует здесь объемного желудочка, а проходит через средний
мозг узким каналом, который называют сильвиев водопровод.
Толща среднего мозга сформирована волокнистыми пучками
белого вещества («ножки большого мозга»). Верхняя поверхность
среднего мозга заметно приподнята: серое вещество, заключен-
ное в толще белого, образует здесь над полостью невроцеля мощ-
ное утолщение — крышу, или тектум. У большинства позвоноч-
ных тектум представлен двухолмием, у млекопитающих — четыре-
холмием. Утолщения серого вещества в боковых стенках среднего
мозга называют тегментум. В тектуме заканчиваются волокна зри-
тельного тракта, идущие от зрительных бугров промежуточного
мозга к расположенным здесь зрительным центрам. У млекопита-
ющих зрительной областью остаются передние бугры, а задние
играют роль центра, воспринимающего слуховые сигналы.
У низших позвоночных в среднем мозге локализованы высоко-
развитые ассоциативные механизмы, управляющие адаптивным
поведением и участвующие вместе с мозжечком в координации
движений и поддержании мышечного тонуса. Так, у лягушки со-
хранение стволовой части мозга при поврежденных полушариях
обеспечивает ей вполне нормальное поведение. У млекопитающих
деятельность среднего мозга подчиняется полушариям переднего
мозга (А. Ромер, Т. Парсонс, 1992).
От среднего мозга берут начало две пары черепномозговых не-
рвов: глазодвигательные (III пара) и блоковые (IV пара).
* Парные и непарные глаза подробнее обсуждаются в разделе, посвященном
органам чувств, гипофиз и эпифиз — в разд. «Эндокринные железы».
196
Мозжечок (metencephalon или cerebellum)
Мозжечок в эмбриогенезе позвоночных образуется из верхней
части третьего мозгового пузыря. Внешне он заметно возвышается
над мозговым стволом на границе среднего и продолговатого моз-
ра. Размеры и форма мозжечка у позвоночных весьма изменчивы.
Он слабо развит у круглоротых и амфибий, незначителен у репти-
лий, больших размеров достигает у рыб. Наиболее сложная струк-
тура и возросшие размеры мозжечка свойственны птицам и мле-
копитающим. У рыб толща мозжечка образована белым веществом
с включением ядер серого вещества. У амфибий и рептилий серое
вещество появляется в его поверхностной части (корковая струк-
тура). У птиц и млекопитающих серое вещество разрастается и
образует поверхностную сложную извилистую слоистую структу-
ру. Клетки мозжечка осуществляют тонкий обмен сенсорными
сигналами. В коре мозжечка есть специализированные клетки Пур-
кине: их дендиты образуют густо ветвящиеся «деревца». Они соби-
рают информацию с обширных участков коры мозжечка и посы-
лают ее в центральное ядро отдела.
Морфологически в мозжечке выделяют тело и боковые ушки
(или клочки). Считают, что у рыб именно ушки обеспечивают
равновесие. Тело — средняя и основная область мозжечка у всех
позвоночных.
Получая информацию от внутреннего уха и органов боковой
линии у низших позвоночных, мозжечок участвует в ориентации
тела в пространстве и поддержании его равновесия. У всех позво-
ночных он получает информацию от сухожилий и скелетных мышц
и участвует в регуляции мышечного тонуса и координации дви-
жений. Через мозжечок проходят моторные сигналы, идущие от
головного мозга к спинному.
Продолговатый мозг (myencephalon)
Продолговатый мозг образуется в онтогенезе позвоночного из
нижней части третьего мозгового пузыря. Вместе со средним моз-
гом он составляет ствол головного мозга, имеет следы сегменти-
рованности и плавно переходит в спинной мозг. Здесь лежат ядра
большинства (V—XII) черепномозговых нервов.
В верхней части отдела расположен четвертый мозговой желу-
дочек. Он имеет утолщенные дно и стенки, а сверху прикрыт тон-
ким эпителием с включением сосудистого сплетения и примы-
кающей миэлиновой оболочки мозга. Полость четвертого желудоч-
Ка — ромбовидная ямка — продолжается непосредственно в цент-
ральный канал спинного мозга.
Значительная часть продолговатого мозга — белое вещество. Се-
Рое вещество выстилает дно и стенки полости желудочка, образуя
Яцра.
197
В толще продолговатого мозга лежат центры, регулирующИе
работу значительной части систем органов (дыхательный, пище.
варительный, сердечный, первичный слуховой, вкусовой и пр.)
и ядра 6—8 пар черепномозговых нервов (V — тройничный, VI J
отводящий, VII — лицевой, VIII — слуховой, IX — языкоглоточ-
ный, X — блуждающий)*. У млекопитающих имеется еще две пары-
XI — добавочный, идущий к мускулатуре головы и плечевого по-
яса; XII — подъязычный, иннервирующий мускулатуру языка
подъязычного аппарата и шеи.
Спинной мозг (medulla sphinalis)
Спинной мозг — филогенетически наиболее древняя часть цен-
тральной нервной системы позвоночных. Он расположен в позво-
ночном канале, сохраняет вид нервной трубки и имеет выражен-
ную сегментированность. Внутри спинного мозга сохраняется за-
полненная жидкостью полость — центральный канал (невроцель).
Спинномозговая жидкость омывает спинной мозг изнутри и сна-
ружи. Поверхность мозга покрыта тремя оболочками — твердой,
мягкой и паутинной.
В разрезе спинной мозг имеет овальную (или округлую) форму
с глубокими бороздами сверху и снизу. В каудальном направлении
(к хвосту) он сужается и заканчивается на уровне задних пояс-
ничных позвонков.
На поперечном сечении мозга (в разрезе) видно, что серое
вещество расположено в центре, окружая невроцель в виде бабоч-
ки. Оно образовано телами нейронов. Серое вещество окружено
белым веществом — это проходящие вверх и вниз миелинизиро-
ванные нервные волокна (отростки нейронов). Восходящие и нис-
ходящие проводящие пути спинного мозга соединяют все основ-
ные отделы центральной нервной системы в единую систему. Спин-
ной мозг осуществляет элементарные двигательные и чувствую-
щие реакции. Выходящие из спинного мозга передние (вентраль-
ные) корешки содержат двигательные волокна. Они иннервируют
соматическую мускулатуру, а также участвуют в иннервации глад-
кой мускулатуры внутренних органов. Задние (дорсальные) ко-
решки спинного мозга содержат чувствующие волокна, передаю-
щие информацию в мозг от кожи, суставов и внутренних органов.
* V — тройничный нерв — иннервирует кожу головы, лицевую мускулатура
зубы; VI — отводящий — мышцы глаза, мигательную перепонку; VII — лицевой —
у рыб орган боковой линии, у млекопитающих — мимическую мускулатуру; VIII -
слуховой — связывает органы слуха и равновесия с головным мозгом; IX "
языкоглоточный — иннервирует мышцы глотки; X — блуждающий — иннерви-
рует мышцы жаберных дуг, плавательный пузырь, органы боковой линии, лег-
кие, сердце, кишечник.
198
Табли ПаЬ
Показатели прогрессивного изменения массы головного мозга,
по данным разных авторов (Н.П.Наумов, Н.Н.Карташов, 1979)
Класс позвоночных Индекс головного мозга, % Соотношение массы головного и спинного мозга, ед. Индекс массы переднего мозга, %
Хрящевые рыбы 0,06-0,04 1:1 60
Костные рыбы 0,02-0,92 1:1 12
Амфибии хвостатые бесхвостые 0,29-0,73 0,29-0,36 0,5-0,73 1:1
Рептилии 0,01-0,4 1:1 42-52
Птицы (в целом) летающие бескилевые и пингвины 0,04-8 0,2-5-8 0,04-0,09 2,5:1 52—62 (до 70 у мелких воробьиных птиц)
Млекопитающие 0,3-3,0 (насекомоядные 0,6—1,2; китообразные 0,3—1,7; приматы 0,6—1,9; человек 3,0) (2,1-15):! (приматы (10—15): 1; китообразные, хищные, насекомоядные (3—5):1; копытные 2,5:1) 52—86 (человек 86; приматы 76—80; прочие 52—82)
За пределами спинного мозга передние и задние корешки объе-
диняются в единые спинномозговые нервы.
Пары спинномозговых нервов отходят от спинного мозга стрр.
го периодично, посегментно. Информация с чувствующего на дви-
гательный нейрон передается в сером веществе мозга через вста-
вочный рон, образуя рефлекторную дугу. Деятельность спинного
мозга находится под контролем высших отделов головного мозга.
У наземных позвоночных на уровне поясов конечностей в спин-
ном мозге образуются плечевое и поясничное утолщения.
В табл. 6 приведены показатели изменения мозга в эволюцион-
ном ряду позвоночных.
Периферическая нервная система
Головной и спинной мозг вместе образуют центральную не-
рвную систему. От них отходят многочисленные пары нервов —
10 — 12 пар черепномозговых и несколько десятков (у человека 31)
пар спинномозговых нервов (рис. 153). Как сказано выше, нервные
волокна выходят из мозга и входят в него попарно: входящие ко-
решки — афферентные, или чувствующие, выходящие — эффе-
рентные, или двигательные.
Парные нервы, иннервирующие все органы и ткани организ-
ма, образуют периферическую нервную систему. В ее состав вхо-
дят соматические и висцеральные {вегетативные} нервы. Сомати-
Рис. 153. Головные нервы акулы (схема):
I — X, XII — пары головных нервов; 1—5— жаберные щели; 6 — брызгальне; 7—
передний мозг; 8 — эпифиз; 9 — средний мозг; 10 — мозжечок; 11 — продолго-
ватый мозг; 12 — подъязычный нерв; 13 — кишечный нерв; 14 — подъязычная
ветвь лицевого нерва; 15 — боковой нерв
700
ческие нервы иннервируют опорно-двигательный аппарат и кожу,
генеральные — внутренние органы, кровеносные сосуды, желе-
зы внутренней секреции. У наземных позвоночных периферичес-
кая нервная система образует два нервных сплетения — плечевое
й пояснично-крестцовое. Через них мышцы конечностей обслу-
живаются волокнами нескольких спинальных нервов.
Периферическая нервная система свойственна и бесчерепным.
Нервные корешки выходят из нервной трубки попарно: вверх —
дорсальные, вниз — вентральные. Спинные корешки смешанные —
они содержат двигательные и чувствующие волокна, брюшные —
только двигательные. Они иннервируют туловищную мускулатуру
и повторяют ее асимметричное расположение в теле.
Вегетативная (или автономная) нервная система
Вегетативная (автономная) нервная система развита у всех по-
звоночных, кроме круглоротых, и наиболее хорошо изучена у мле-
копитающих. В ней выделяют симпатический и парасимпатический
отделы. Нервы симпатической систе-
мы выходят из спинного мозга в об-
ласти от 1-го грудного до 4-го пояс-
ничного сегментов. Парасимпатичес-
кие нервы выходят из среднего и про-
долговатого отделов головного мозга
и из крестцовой части спинного мозга
(рис. 154).
Вегетативная нервная система иг-
рает ведущую роль в поддержании
постоянства внутренней среды орга-
низма и осуществляет регуляцию при-
способительных реакций при охлаж-
дении, интенсивной мышечной ра-
боте, при эмоциональном напряже-
нии, при кровопотере и других не-
благоприятных факторах. Она регули-
рует деятельность органов кровообра-
щения, дыхания, пищеварения, вы-
деления, размножения.
Рис. 154. Вегетативная нервная система
человека:
~ симпатическая; 2 — парасимпатическая
201
Симпатические и парасимпатические центры находятся под
контролем гипоталамической области промежуточного мозга ц
коры больших полушарий.
На большинство органов симпатическая и парасимпатическая
системы, будучи антагонистами, оказывают противоположное
воздействие. Как правило, симпатическая система осуществляет
активизирующее действие. Она обеспечивает мобилизацию, под-
готовку органа к усиленной деятельности. Парасимпатическая си-
стема способствует расслабляющему или спокойному состоянию.
На рис. 154 показано расположение соответствующих волокон в
теле человека.
При эмоциональных проявлениях под влиянием вегетативной
нервной системы усиливается выделение в кровеносную систему
гормонов желез внутренней секреции: надпочечников, гипофи-
за, щитовидной железы.
Особенности строения нервной системы различных
классов позвоночных
Рассмотрим кратко строение нервной системы в эволюцион-
ном ряду позвоночных животных. В табл. 4 приведены некоторые
показатели прогрессивного изменения мозга в эволюционном ряду.
У круглоротых головной мозг имеет четыре типичных для
позвоночных отдела: передний, промежуточный, средний и про-
долговатый мозг (рис. 155). Мозжечок не оформлен. Размеры мозга
относительно малы. Все отделы
расположены в одной плоскости,
т.е. не образуют типичных для
Рис. 155. Головной мозг миноги
(А — сверху; Б — снизу):
1 — полушария переднего мозга; 2 —
обонятельные доли; 3 — обонятельный
нерв; 4 — промежуточный мозг; 5 и 6 —
правый и левый габенулярные ганглии;
7 — теменной орган, прикрывающий
эпифиз; 8 — зрительный нерв; 9 — моз-
говая воронка; 10 — зрительные доли;
11 — отверстие в крыше среднего моз-
га; 12 — дно среднего мозга; 13 — niaJ
зодвигательный нерв; 14 — тройничный
нерв; 75 — слуховой нерв; 16 — продол-
говатый мозг; 77— ромбоидальная ямка,
18 — зачаточный мозжечок
202
более высокоорганизованных позвоночных изгибов. Крыша мозга
целиком эпителиальная. Нервные клетки есть в выстилке мозго-
рЬ1Х желудочков. От головного мозга отходят 10 пар головных не-
рвов. Спинномозговые нервы отходят двумя корешками: дорсаль-
цЬ1М и вентральным, которые у круглоротых в отличие от других
позвоночных не соединяются и не образуют общих смешанных
нервов.
У хрящевых рыб головной мозг заметно увеличен. Нервная ткань
Б полушариях переднего мозга присутствует в его толще, обоня-
тельных долях и в крыше. Хорошо развит средний мозг, мозжечок
увеличен. 10 пар черепномозговых нервов имеют типичные места
отхождения и сферу иннервации. Размеры головного мозга у к о -
стных рыб в сравнении с хрящевыми уменьшены, небольшой
передний мозг несет выступающие вперед стебельчатые обоня-
тельные доли. Крыша переднего мозга эпителиальная. Отделы мозга
лежат в одной плоскости (рис. 156).
Головной мозг амфибий характеризуется рядом прогрессив-
ных черт: увеличиваются его относительные общие размеры, осо-
бенно заметные у бесхвостых. У амфибий произошло полное раз-
Рис. 156. Головной мозг рыбы (форели) (А — сверху; Б — снизу):
1 ~~ обонятельная луковица; 2 — мозжечок; 3 — перекрест зрительных нервов; 4 —
Промежуточный мозг; 5 — эпифиз; 6 — гипофиз; 7 — зрительные доли среднего
м°зга; 8 — средний мозг; 9 — продолговатый мозг; 10 — спинной мозг; 11 —
передний мозг; I—X — головные нервы
203
Рис. 157. Головной мозг лягушки (А — сверху; Б — снизу; В — сбоку;
Г — продольный разрез):
1 — полушария переднего мозга; 2 — обонятельная доля; 3 — обонятельный
нерв; 4 — промежуточный мозг; 5 — зрительная хиазма; 6 — воронка; 7 — гипо-
физ; 8 — средний мозг; 9 — мозжечок; 10 — продолговатый мозг; 11 — четвер-
тый желудочек; 12 — спинной мозг; 13 — третий желудочек; 14 — сильвиев
водопровод
деление полушарий переднего мозга и замечено формирование
архипаллиума (рис. 157).
Полушария переднего мозга урептилий крупнее, чем у ам-
фибий, хорошо развит архипаллиум. В общем контуре мозга заме-
тен изгиб, в результате промежуточный отдел оказывается при-
крытым большими полушариями (рис. 158).
Мозг п т и ц по сравнению с рептилиями заметно возрастает в
размерах. В наибольшей степени это выражено у летающих птиц.
Увеличение полушарий переднего мозга обусловлено сильным
развитием в нем базальных ядер. Наиболее крупные ядра (wulst -
формация) локализованы в теменной области. Развитие форм коры
у птиц соответствует таковым у рептилий. Очень резко выражены
мозговые изгибы. Заметно увеличен мозжечок. Его поверхность
имеет сложное строение и смята в складки (рис. 159). Согласно
экспериментам, удаление коры не сопровождается у птиц утра'
той выработанных условных рефлексов, нарушаются лишь их стой-
кость и длительность. Птицы разных видов обнаружили разную
способность к рассудочной деятельности, которая осуществляет-
ся при участии упомянутых выше базальных ганглиев. Низкий УР0'
вень рассудочной деятельности проявляют голуби, куры. СамЫе
высокие показатели ее отмечены у врановых птиц. Обнаружен®
при этом, что они отличаются и строением нейронов, и длиной
их отростков.
204
Рис. 158. Головной мозг ящерицы (А — сверху; Б — снизу; В — сбоку):
1 — передний мозг; 2 — полосатое тело; 3 — средний мозг; 4 — мозжечок; 5 —
продолговатый мозг; 6 — воронка; 6' — гипофиз; 7 — хиазма; 8 — обонятельная
доля; 9 — эпифиз; I—X — черепные нервы
А Б В
рис. 159. Головной мозг птицы (А — сверху; Б — снизу; В — со вскры-
тыми желудочками и удаленным мозжечком);
— обонятельные доли переднего мозга; 2 — большие полушария головного
Иозга; 3 — промежуточный мозг; 4 — надмозговая железка эпифиз; 5 — зритель-
ные бугры промежуточного мозга; 6 — зрительные тракты; 7 — хиазма; 8 —
®°Р°нка с гипофизом; 9 — зрительные доли среднего мозга; 10— мозжечок; 11 —
°Ковые выступы мозжечка; 12 — зрительная комиссура; 13 — продолговатый
мозг; 14 — полосатые тела
205
Рис. 160. Мозг кролика (А — сверху.
Б — снизу; В — сбоку; Г— продояС
ный разрез):
1 — большие полушария; 2 — обоця.
тельные доли; 3 — зрительный нерв;
эпифиз; 5 — средний мозг; 6 — мозже-
чок; 7 — продолговатый мозг; 8 — ги-
пофиз; 9 — варолиев мост; 10 — мозго-
вая воронка; 11 — мозолистое тело
В головном мозге млекопи-
тающих все названные выше
отделы оформлены наиболее
четко (рис. 160). Передний мозг
характеризуется развитием вто-
ричного мозгового свода — но-
вой коры — неопаллиума. В но-
вой коре нейроны организова-
ны в проекционные и ассоциа-
тивные поля. Они, с одной сто-
роны, анализируют различную
сенсорную информацию, а с дру-
гой — осуществляют сложный
интегрированный ответ на комплекс внешних и внутренних воз-
действий. В новой коре расположены центры, связанные с высшей
нервной деятельностью. В пределах класса млекопитающих повер-
хность неопаллиума прогрессивно возрастает. Наивысшего разме-
ра она достигает у китообразных и приматов. Кора полушарий
переднего мозга имеет извилины и борозды. В промежуточном мозге
усилена функциональная дифференцировка его отделов, наибо-
лее выраженная в области гипоталамуса. В среднем мозге форми-
руется четверохолмие, в мозжечке хорошо заметно выделение от-
делов и расширение поверхностной коры. От головного мозга от-
ходят 12 пар черепномозговых нервов.
Подводя итог описанию особенностей нервной системы по-
звоночных, следует отметить, что ее эволюция сопровождалась
рядом последовательно протекавших явлений, важнейшие из них:
— дифференцировка нервной трубки на головной и спинной
мозг;
— морфологическое и функциональное разделение головного
мозга на 5 отделов;
— выделение в переднем мозге нервного вещества в виде ба-
зальных ганглиев и коры;
206
— - последовательное нарастание форм коры переднего мозга:
Падеопаллиума, архипаллиума, неопаллиума;
— функциональное подчинение всех форм нервной деятельно-
сти новой коре больших полушарий (неопаллиум);
— прогрессивное нарастание массы мозга, массы головного
мозга, массы переднего мозга;
— усиление дифференцировки и функциональной ответствен-
ности гипоталамуса, гипофиза и других отделов мозга;
— развитие вегетативной нервной системы.
* * *
Таким образом, своеобразная трубчатая центральная нервная
система хордовых, эктодермальная по происхождению, располо-
жена дорсально по отношению к осевому скелету. Она отвечает за
согласованную деятельность систем органов и всего организма.
Исходно у хордовых (ланцетника) она представляет собой еди-
ную нервную трубку, метамерность которой обозначается отходя-
щими от нее чувствительными и двигательными нервами. Эволю-
ция центральной нервной системы у позвоночных связана с вы-
делением головного мозга и дифференцировкой его на пять отде-
лов, отвечающих за деятельность определенных органов, и про-
грессивное развитие в них нервных центров — анализаторов, со-
вершенствованием корковых центров, отвечающих за сложное
поведение животных. Наибольшего развития достигает передний
мозг, в котором последовательно выделяются древнейшая, древ-
няя и новая кора. Самый крупный мозг (относительно массы тела)
у хищных зверей, дельфинов и приматов. У последних многослой-
ная кора переднего мозга образует многочисленные борозды и
складки, увеличивающие ее поверхность.
ОРГАНЫ ЧУВСТВ
Рис. 161. Первичночув-
ствующая рецептор-
ная клетка:
7 — периферический от-
росток; 2 — центральный
отросток
К чувствующей системе принадлежат органы обоняния, зре-
ния, слуха, вкуса, осязания и некоторые другие. Через органы
чувств животные получают информацию о состоянии и объектах
окружающей их среды: они оценивают свойства среды, использу-
ют поступающую информацию при поисках пищи, в половых кон-
тактах, при обнаружении и избегании врагов.
Рецепторы органов чувств распознают механические воздей-
ствия, химические раздражители, а также тепловую, световую и
электромагнитную энергию.
К механическим раздражителям относятся вибрация и прикос-
новение, давление, сила тяжести, инерция, наконец, звук и его
отражение. Химические раздражители воспринимаются через вкус
и запах. Наконец, животные воспринимают из внешней среды
электрическую, магнитную, тепловую и
световую энергию. Они чувствительны к
теплу и холоду, к свету, ультрафиолетово-
му и инфракрасному излучениям.
Органы чувств животных своим проис-
хождением и функционально связаны с
центральной нервной системой.
Структурные особенности органов чувств
позвоночных животных основываются на
двух типах рецепторных клеток: первично-
и вторичночувствующих. Строение этих
клеток различно. Первичночувствующая ре-
цепторная клетка имеет два отростка — пе-
риферический и центральный (рис. 161). Пе-
риферический отросток воспринимает раз-
дражение, а по центральному отростку оно
передается в центральную нервную систе-
му (анализатор). Первичночувствующие ре-
цепторные клетки считаются филогениче-
ски более древней группой, имеющейся уже
у беспозвоночных животных. У позвоноч-
ных животных клетки этого типа заклады-
ваются в раннем эмбриогенезе в эктодерме
одновременно с оформлением нервной
208
пластинки (под индуцирующим влиянием верхней части бласто-
пора и крыши первичной кишки). К первичночувствующим ре-
цепторам у позвоночных животных относятся обонятельные и зри-
тельные рецепторные клетки.
Вторичночувствующая (или волосковая) рецепторная клетка —
структура принципиально новая, присущая только позвоночным
лшвотным. Эта клетка не имеет отростков. Раздражение восприни-
мается волосками, расположенными на ее поверхности, и пере-
дается примыкающему к клетке чувствующему нервному оконча-
нию (рис. 162). Каждая клетка имеет 40 — 50 мелких и несколько
крупных волосков. Волоски сенсорных клеток неодинаковы по
длине: короткие (стереоцилии) и длинные (киноцилии). Сверху они
прикрыты студенистой массой — купулой, которая свободно ко-
20 мкм
Рис. 162. Вторичночувствующая волосковая рецепторная
клетка (А). Волосковые клетки в органе боковой линии
Рыбы (/>'). Орган боковой линии на поверхности тела
шпорцевой лягушки (В):
А'- I — киноцилия; 2 — стереоцилии; 3 — волосковая клет-
ка; 4~ окончание афферентного нерва (сенсорное); 5— окон-
чание эффективного нерва (тормозное); 6 — масляный пузы-
Рек (по Gomer). Б'. 1 — купула; 2 — волоски; 3 — сенсорные
Клетки; 4 — эпидермис. В: 1 — купула; 2— реснички сенсор-
ных клеток; 3 — сенсорная клетка; 4 — нерв; 5 — эпидермис;
6 — дерма; 7 — мускулатура
209
лышется окружающей водой. В основании волосковых клеток есть
масляные пузырьки, обеспечивающие возможность химической
передачи нервных импульсов с клеток на нервные окончания
(Л. Проссер, т. 2, 1987).
Вторичночувствующие клетки закладываются в эмбриогенезе
тоже в эктодерме, но по времени позднее рассмотренных ранее
первичночувствующих, под индуцирующим влиянием уже офор.
мившейся нервной трубки.
Волосковые клетки в соответствующих органах чувств окруже-
ны жидкостью: пресная или морская вода для боковой линии рыб
и личинок амфибий, жидкость перепончатого лабиринта (пери-
лимфа, эндолимфа), жидкая среда ротовой полости.
Вторичночувствующие клетки входят в состав сейсмосенсор-
ной системы (боковой линии круглоротых, рыб, личинок амфи-
бий, некоторых взрослых земноводных), вестибулярного и слухо-
вого аппарата и органа вкуса.
Первичночувствующая рецепция
Органы обоняния
Рис. 163. Поверхность обоня-
тельного эпителия:
1 — опорная клетка; 2 — обоня-
тельная клетка; 3 — слизистая
оболочка носа; 4 — нервное во-
локно
Обонятельная (запаховая) рецепция — важная чувствующая
система всех позвоночных. Пользуясь органами обоняния, живот-
ные разыскивают пищу, обнаруживают врагов, оценивают химизм
среды. Это важное средство внутривидовой коммуникации: оно
проявляется при взаимодействии полов, родителей и детей, рав-
но- и разновозрастных особей популяции. Запаховый след, запа-
ховые метки животные оставляют в виде выделений специфиче-
ских кожных желез, выделений из пищеварительного тракта, мо-
чеиспускательных и половых путей.
Органы обоняния в эмбриогенезе
закладываются в виде парных утолще-
ний эктодермы на переднем конце тела
зародыша. При погружении в глубь по-
кровов образуются обонятельные ямки,
а вслед за ними — мешки, выстлан-
ные цилиндрическими клетками обо-
нятельного эпителия (рис. 163). Пери-
ферический отросток обонятельной
клетки выходит на поверхность в слой
слизи, покрывающей обонятельный
эпителий. Здесь он заканчивается бу-
лавовидным утолщением, несущи*1
пучок длинных обонятельных волос-
ков, контактирующих с молекулами
пахучих веществ.
ип
В передней стенке переднего мозга навстречу утолщениям эк-
зодермы развиваются парные выпячивания — обонятельные доли,
изнутри в них проникают полости боковых желудочков. Форма
обонятельных долей изменчива у разных видов животных: «сидя-
чде» на мозге, вытянутые в длину, «сидящие» на стебельках и т.д.
Параллельно с обонятельными долями мозга формируется скелет
обонятельных капсул, которые в дальнейшем входят в состав моз-
гового черепа.
Органы обоняния иннервируются I парой черепномозговых
нервов. Особенность обонятельного нерва состоит в том, что он
слагается из центральных отростков первичночувствующих кле-
ток обонятельного эпителия. Войдя в обонятельные доли перед-
него мозга, эти отростки вступают в связь с дендритами других
нервных клеток. Так идет передача ольфакторной (обонятельной)
информации: сама обонятельная клетка является и рецептором, и
проводником нервного возбуждения. Такой принцип организации
наблюдается и в соответствующих органах беспозвоночных жи-
вотных и свидетельствует об эволюционной древности чувства
обоняния. У позвоночных животных с органами обоняния связана
древняя кора переднего мозга.
Органы обоняния в эволюционном ряду хордовых заметно ус-
ложняются. Поверхность обонятельного эпителия прогрессивно
разрастается. Это сопровождается образованием многочисленных
меридианальных складок внутри обонятельных капсул и измене-
нием объема обонятельных долей переднего мозга.
Пороговая чувствительность обонятельных клеток очень высо-
кая. Для стимуляции одной клетки достаточно энергии, развивае-
мой 1 — 4 молекулами пахучего вещества (Э.Хадорн, Р. Венер, 1989).
По уровню чувствительности обонятельного аппарата к восприя-
тию запахов среди позвоночных различают макро- и микроосма-
тиков. Первые имеют высокочувствительное обоняние, у вторых
его чувствительность понижена, и в ориентации им существенно
помогают другие органы чувств.
У бесчерепных обонятельный эпителий сосредоточен в об-
ласти дорсальной непарной обонятельной ямки (ямка Келлике-
Ра), в том месте, где невроцель сообщался с поверхностью тела у
их личинок.
У круглоротых обонятельный мешок тоже непарный. У мик-
син он открывается непарной ноздрей в верхней части рыла, у
миног — на верхней поверхности головного отдела. Снизу непар-
ный срединный мешок связан с гипофизарной трубкой. Расшире-
ние и сжатие этой трубки способствуют движению воды вблизи
°бонятельного эпителия. У миксин эта трубка открывается в глот-
КУ и участвует в акте дыхания при паразитическом питании.
У хрящевых рыб пара несквозных ноздрей расположена
На нижней поверхности рыла. Каждая ноздря частично разделена
211
Рис. 164. Поперечный разрез
через обонятельную область
головы акулы:
1 — обонятельный мешок; 2 — нозд-
ря; 3 — носовая перегородка
кожной складкой на вводное и
выводное отверстия. Поверхность
обонятельных капсул выстилает
обонятельный эпителий (рис. 164)
Обонятельная область переднего
мозга увеличена, при этом отме-
чают, что у скатов она крупнее,
чем у акул.
Строение органов обоняния у
костных рыб отличается до-
вольно большим разнообразием.
У осетровых они сходны с хря-
щевыми рыбами. У костистых рыб
две пары ноздрей располагаются
на верхней стороне головы: в пе-
редние вода входит, через задние — выходит. Обонятельный эпи-
телий в капсулах собран в мелкие радиальные складки (рис. 165).
Подсчитано, например, что у обыкновенного угря (Anguilla
anguilla) поверхность обонятельного эпителия содержит до 40000
клеток в 1 мм2 (Л. Проссер, 1992).
У лопастеперых ноздри сквозные: изнутри они открываются в
ротовую полость. Обонятельные складки внутри ноздрей выстла-
ны однослойным цилиндрическим эпителием.
Б
Рис. 165. Носовые полости костистых рыб (А — гольян; Б — угорь)
212
Все четвероногие имеют сквозные ноздри, пропускающие ат-
мосферный воздух в ротоглоточную полость. Среди обонятельных
клеток размещены специальные железы, выделяющие внутрь хо-
дов водянистый секрет и жироподобные вещества. В результате
запаховые раздражители поглощаются пленкой жидкости, покры-
вающей обонятельный эпителий. Предполагают, что увлажнению
способствует и поступление жидкости от глаз через слезоносовые
протоки (возможно, это пара бывших задних ноздрей костных рыб)
(А. Ромер, Т. Парсонс, 1992).
У личинок амфибий обонятельный эпителий представлен,
как и у рыб, радиальными складками. У взрослых амфибий в обо-
нятельных капсулах появляются окостенения — обонятельные ра-
ковины, а эпителий приобретает вид закрученных выростов. У без-
ногих есть своеобразные обонятельные щупальца, выпячивающи-
еся из ноздрей. Считается, что среди амфибий обоняние лучше
развито у хвостатых.
Сквозные ноздри у амфибий полифункциональны: кроме обо-
няния они пропускают воздух к легким. Полость обонятельного
мешка подразделяется у них на две части: внутреннюю, собствен-
но обонятельную, выстланную обонятельным эпителием, и на-
ружную — респираторную, выстланную покровным эпителием.
У амфибий впервые намечается обособление от обонятельной кап-
сулы специализированного участка обонятельного эпителия —
якобсонова органа. У хвостатых этот орган выражен нечетко, у про-
чих же амфибий он организован в слепой мешок, открывающий-
ся в основную носовую полость. Якобсонов орган информирует о
запахе пищи, находящейся во рту, и иннервируется самостоя-
тельной ветвью обонятельного нерва.
У рептилий происходит даль-
нейшая дифференцировка сквозных
ноздрей на преддверие, средний и
задний отделы (рис. 166). Верхняя об-
ласть носовой камеры имеет не-
сколько выростов — раковин, вы-
стланных обонятельным эпителием.
У крокодилов и некоторых черепах
Носовая камера вместе с носоглоточ-
ным ходом отделены от рта вторич-
ным костным нёбом.
Якобсонов орган хорошо развит
У ящериц и змей; он парный и отде-
лен от сквозных ноздрей. Химическая
информация улавливается кончиком
Раздвоенного языка и переносится
6 слизь чувствующего эпителия это-
го органа. В отличие от обонятель-
Рис. 166. Дифференцировка
обонятельного мешка у
ящерицы:
1 — наружная ноздря; 2 — пред-
дверье; 3 — респираторный отдел;
4 — носоглоточный канал; 5 —
хоана (внутренняя ноздря); 6 —
обонятельный отдел: 7 — якобсо-
нов орган
213
A
Рис. 167. Поперечный разрез через
орган обоняния птицы (А) и млеко-
питающего (Б):
1 — воздушные полости; 2, 3 — обоня-
тельные раковины; 4 — носовая перего-
родка
ной полости якобсонов орган по-
стоянно заполнен жидкостью, ко-
торая периодически выбрасывает-
ся и обновляется (И. И. Шмальга-
узен, 1964). Рептилии преимуще-
ственно — макроосматики. Хоро-
шим обонянием отличаются чере-
пахи, крокодилы, большинство
ящериц и змей.
У п т и ц отверстия ноздрей расположены обычно у основа-
ния клюва, иногда прикрыты роговой складкой. У киви они
находятся на кончике клюва. Внутри носовых ходов есть три
носовых раковины, выстланные небольшими участками обоня-
тельного эпителия (рис. 167). Якобсонов орган у них отсутствует.
Чувствительность обонятельной рецепции у птиц в сравнении
с рептилиями понижена. Тем не менее недавними исследовани-
ями показано, что обоняние участвует у них в формировании
разных форм поведения: поиске пищи, известной территории
(хоминг).
Умлекопитающих обоняние развито очень хорошо. Обо-
нятельные раковины закручены в костные свитки, покрытые чув-
ствующим эпителием. Собственно обонятельными являются при-
мыкающие к решетчатой кости решетчатые раковины. Их число
варьирует от 5 —8 при хорошем обонянии, до 2 — при слабом.
К макроосматикам среди млекопитающих относятся сумчатые, на-
секомоядные, грызуны, хищные, копытные. Общее число клеток
обонятельного эпителия у тонко чувствующей овчарки достигает
225 • 106. К микроосматикам принадлежат беззубые киты, ластоно-
гие, высшие приматы. У человека микроосматик количество кле-
ток обонятельного эпителия равно 5 • 106 (Л. Проссер, 1978). Боль-
шинство млекопитающих имеют якобсонов орган. Работами оте-
чественных океанологов показано, что киты и дельфины, исполь-
зуя якобсонов орган, улавливают след удалившегося стада своих
собратьев (А. Белькович, 1972).
214
Органы зрения
Восприятие света — еще одно древнее свойство нервной труб-
ки у хордовых. У бесчерепных светочувствительностью обладают
глазки Гессе, лежащие в невроцели* (рис. 168). Позвоночные име-
ют специализированные светочувствительные органы: парные и
непарные глаза. Они воспринимают высокочастотные волны кос-
мических излучений.
Парные глаза закладываются в эмбриогенезе животного позднее
органов обоняния, после замыкания нервной пластины в трубку.
От нервной трубки на уровне таламуса промежуточного мозга
выпячиваются наружу округлые глазные пузыри. Их соединения с
мозгом — глазные стебельки — постепенно сужаются и оттесня-
ются на брюшную сторону нервной трубки. Из них впоследствии
оформляются зрительные нервы (рис. 169).
Наружная стенка каждого глазного пузыря впячивается и фор-
мируется двухслойный глазной бокал. Наружный тонкий слой кле-
ток бокала превращается в пигментную оболочку камерного глаза.
Утолщенный внутренний слой становится его рецепторной обо-
лочкой — сетчаткой, содержащей чувствительные, нервные и
опорные элементы. У взрослых животных сосудистая и пигмент-
ная оболочки и сетчатка срастаются (рис. 170). Снаружи глазной
бокал обрастает мезодермой. Из нее выделяются 2 оболочки: внут-
ренняя — сосудистая и поверхностная — склера.
Парные глаза имеют сложное строение. Кроме названных оболо-
чек в структуру глаза входят хрусталик, сосудистая оболочка, радуж-
ка, ресничное тело, склера, роговица, стек-
ловидное тело. Структура глаза разных
позвоночных показана на рис. 171.
Хрусталик закладывается и обособ-
ляется в эктодерме зародыша напро-
тив глазного пузыря. Замкнувшись в
пузырек, он отшнуровывается от эк-
тодермы и погружается в полость бо-
кала.
Сосудистая оболочка примыкает к
пигментной. Она обильно пронизана
кровеносными сосудами, пигментиро-
вана и поглощает большую часть све-
та, попадающего на нее сквозь сетчат-
ку • В этой оболочке есть светоотража-
ющие устройства, они помогают кон-
центрировать лучи на сетчатке в сла-
Рис. 168. Светочувствитель-
ные клетки в нервной
трубке ланцетника:
1 — невроцель; 2 — нервные
клетки; 3 — пигментная клетка
* Каждый глазок состоит из 2 клеток — светочувствительной и пигментной.
1аИцетник воспринимает контраст света и тени.
215
6-
Рис. 169. Схема последовательных стадий развития глаза (А—Г)
(по И. И. Шмальгаузену, 1947):
1 — промежуточный мозг; 2 — глазной пузырь; 3 — ножка глазного пузыря; 4 —
хрусталик; 5 — сетчатка; 6 — пигментный слой; 7 — сосудистая оболочка; 8 —
склера; 9 — роговица
бо освещенной среде и хорошо заметны по свечению глаз у ноч-
ных млекопитающих, рептилий и птиц, у глубоководных рыб.
Спереди глаза сосудистая оболочка вместе с утончающейся
пигментной образуют окрашенную радужную оболочку (радужку).
Будучи пигментированной, она обусловливает цвет глаз. Радужи-
на ограничивает зрачок и содержит мышечные волокна. Концент-
рически и радиально расположенные мышечные волокна способ-
ны сужать и расширять зрачок, регулируя размеры пучка света,
пропускаемого в глаз. Спереди склера покрыта конъюнктивой.
Вблизи хрусталика расположено ресничное тело. Мускулатура
этого тела участвует в его аккомодации.
С поверхности глаз окружен склерой. У круглоротых и млеко-
питающих она состоит из плотной соединительной ткани, у дру-
гих — укреплена хрящом и даже костной тканью. Спереди глаза
склера утончается и превращается в прозрачную роговицу. Форма
роговицы изменчива: у рыб она плоская, у наземных и вторично-
водных сферически выпуклая. Вместе с хрусталиком роговица уча-
ствует в преломлении потока света, поступающего в глаз.
Полость глазного яблока имеет внутри три камеры-, передняя
располагается между роговицей и хрусталиком, средняя — между
716
Рис. 170. Схематический разрез глаза,
иллюстрирующий расположение форми-
рующих его слоев в эмбриогенезе (по
А. Ромеру, Т. Парсонсу):
1 — склера; 2 — сосудистая оболочка; 3 "
сетчатка (2 слоя); 4 — хрусталик; 5 — рогови-
ца; 6 — стекловидное тело
протрактор хрусталика
Рис. 171. Схематическое изображение вертикального разреза глаза у по-
звоночных разных классов:
А — лошадь; Б — голубь; В — лягушка; Г — лосось
хрусталиком и радужиной. Они заполнены жидкостью — водянис-
той влагой. В третьей, основной, камере находится прозрачное
студенистое вещество — стекловидное тело. Внутри стекловидного
тела у рептилий и птиц есть структура, обильно пронизанная кро-
веносными сосудами — сосковый конус, или гребешок. Он улучша-
ет обеспечение глаза питательными веществами и кислородом и
(предположительно) участвует в обеспечении остроты зрения (у
Птиц).
Главным рецепторным органом глаза является сетчатка. В ос-
нове ее структуры, как и в органах обоняния, лежат первично-
Нувствующие клетки, формирующиеся в эмбриогенезе эктодер-
^ально, т. е. из поверхности нервной трубки. Они содержат зри-
тельные пигменты и способны возбуждаться при действии цвето-
217
вых стимулов и проводить возбуждение в зрительные пентрь
ловного мозга. Общее количество рецепторных клеток, к rin г°'
ру, в сетчатке лягушки больше миллиона, у кошки — около Т
млн. Структура сетчатки (в отличие от обонятельного эпите ™
сложна и содержит несколько зон (рис. 172). Ия
Первая зона сетчатки лежит вблизи пигментного слоя. Она
разована удлиненными концами светочувствительных клеток
палочек и колбочек. Такая соответствующая своим названиям <Ьо"
ма этих клеток наиболее четко различима в сетчатке глаза млек
А
Пигментный
эпителий
” Внутренний
7 сетчатый
слой
Внутренняя
пограничная
мембрана
Слой нерв-
ных волокон
Внутренний
ядерный
слой
Ганглиозный
слой
Рецепторный
слой
Наружная
пограничная
мембрана
Наружный
ядерный
слой
Наружный
сетчатый
слой
Рис. 172. Сетчатка в глазе млекопитающего (А) и схема соединения
между клетками сетчатки (Б):
1 — амакриновая клетка; 2 — горизонтальный нейрон; 3 — биполярная кле^
связанная с колбочкой; 4 — биполярная клетка, связанная с палочками. •
ганглиозные клетки; 6 — колбочка; 7 — палочка
218
аЮщего. Ядра палочек и колбочек слегка сдвинуты в глубь сет-
и образуют ядерную зону. Вторая зона включает биполярные
с короткими отростками. Они принимают возбуждение от
^очек и колбочек и передают его клеткам третьего слоя.
^рпетъя зона содержит нейроны с длинными отростками. Эти
остки образуют волокна зрительного нерва, по ним возбужде-
е передается в промежуточный мозг. То место, где длинные от-
мостки нервных клеток отходят от глаза и начинается зрительный
Р в не содержит чувствительных клеток: это слепое пятно сет-
атки. У млекопитающих зрительная информация проходит через
названные зоны сетчатки, через таламическую часть промежуточ-
ного мозга и передается коре переднего мозга, в которой сосредо-
точены основные зрительные зоны.
Своеобразие сетчатки позвоночных животных состоит в ее ин-
вертированное™ (перевернутости рецепторного аппарата): палочки
и колбочки размешаются не со стороны поступления в глаз света, а
в глубине оболочки. От нее отсчитываются слои сетчатки. Прежде
чем достичь рецепторного слоя, свет должен пройти через всю тол-
щу клеток сетчатки. Происхождение и функциональный эффект та-
кой структуры сетчатки пока до конца не расшифрованы.
Расположение чувствующих клеток — палочек и колбочек — в
сетчатке имеет некоторую упорядоченность. Колбочки сконцент-
рированы в центральном поле — желтом пятне. Это место соот-
ветствует наибольшей остроте зрения. Палочки группируются боль-
ше по периферии сетчатки — поэтому боковое зрение нечеткое и
черно-белое. В сетчатке глаза лягушки желтого пятна нет, палочки
и колбочки распределены по ней относительно равномерно. Во-
локна зрительного нерва отличаются у них разнокачественностью.
В их составе есть элементы, реагирующие на мелкие темные дви-
жущиеся объекты, вне зависимости от освещенности (К. Шмидт-
Ниельсен, 1982).
Палочки и колбочки функционально различны. Палочки — ре-
цепторы, реагирующие на слабое освещение, они передают изоб-
ражение в черно-белых тонах. Палочки преобладают в сетчатке у
ночных наземных животных, у акул и глубоководных костных рыб.
Колбочки чувствительны к интенсивному освещению, они хоро-
шо передают тонкие детали изображения и обеспечивают цветное
3Рение. Колбочковое, цветное, зрение имеют костные рыбы, пти-
Цы и дневные пресмыкающиеся. Значительной части млекопита-
Ших, ведущих сумеречный, ночной или подземно-роющий об-
3 Жизни, цветное зрение не свойственно. Известно, что цвет-
зрением среди млекопитающих обладают приматы.
с в °снове цветного зрения лежит неодинаковая спектральная чув-
ительность колбочек к свету. Например, у приматов известно
Ст w типа колбочек, максимум чувствительности которых соответ-
Ует синей, желтой и зеленой областям спектра. Это обусловле-
219
но наличием и количественным сочетанием в них зрительные
пигментов. У позвоночных в колбочках сочетаются главным обра,
зом два пигмента родопсин и порфиропсин (Л. Проссер, 1992). р(к
допсин, или зрительный пурпур, свойствен разным типам жи-
вотных (в том числе и хордовым). Под действием света он распа-
дается на ретиналь (форма витамина А) и опсин — белок. Эти
производные родопсина довольно вариабельны. Их различная спек-
тральная чувствительность и лежит в основе цветного зрения.
Порфиропсин известен только у позвоночных. Он отличается
от родопсина присутствием хроматофора. Например, порфироп-
син обнаружен в сетчатке пресноводных рыб, амфибий. Исследо-
вания показали, что у них соотношение родопсина и порфироп-
сина в сетчатке находится под непосредственным контролем сре-
ды. Например, у проходных рыб при переходе из соленой воды в
пресную происходит смена пигмента. У личинок хвостатых и бес-
хвостых амфибий при метаморфозе порфиропсин сменяется ро-
допсином и т.д., у жаб присутствует только родопсин. У человека
родопсин расходуется днем и восстанавливается во время сна
(Л. Проссер, 1978).
При переходе в течение суток от дневного зрения к сумеречно-
му исчезает чувствительность к восприятию цветов — это явление
называют цветовой слепотой. При цветовой слепоте наблюдается
так называемая ретиномоторная реакция сетчатки: палочки и кол-
бочки несколько меняют свое местоположение в сетчатке.
Поток света, попадающий в глаз животного, проходит через
систему фокусировки, позволяющей получать более или менее чет-
кое изображение предмета. В этом участвуют роговица, хрусталик и
стекловидное тело.
Рыбы имеют плоскую роговицу, слабо преломляющую свето-
вые лучи. У наземных она приобретает выпуклую сферическую
форму и участвует в коррекции изображения. Водные млекопита-
ющие сохраняют выпуклую (наземную) форму роговицы. Опре-
деленную оптическую роль играет стекловидное тело.
Наибольшая ответственность за передачу изображения принад-
лежит хрусталику. Форма его у позвоночных изменчива. Сфери-
ческая, округлая форма хрусталика и приближенность его к рого-
вице создает у рыб эффект максимального увеличения и четкого
изображения предмета на сетчатке только при близком рассмот-
рении предмета: рыбы — «близоруки». У наземных позвоночных
хрусталик уплощен, имеет вид линзы и отодвинут от роговицы.
Для получения четкого изображения имеет значение изменение
формы хрусталика — аккомодация.
Механизмы аккомодации в глазах позвоночных довольно раз-
нообразны. В этом принимают участие мезодермальные мыш®1
ресничного тела и эктодермальные мышцы радужки. У круглоро-
тых, хрящевых и костных рыб, у личинок амфибий аккомодация
220
достигается перемещением хрусталика внутри глазного яблока,
у рептилий, птиц и млекопитающих приспособление к четкому
рдению происходит за счет перемещения и изменения формы
хрусталика. При рассмотрении удаленных предметов хрусталик
уддощается, при ближнем зрении он округляется. Это осуществ-
ляется мышцей ресничного тела. Замечено, что спокойное со-
стояние хрусталика у разных позвоночных может решить различ-
ные задачи: в одних случаях проблему близкого зрения, в других —
дальнего.
К глазу позвоночного примыкают вспомогательные структуры',
веки, мышцы, слезные железы и пр.
У большинства рыб веки практически отсутствуют. В отдельных
случаях около глаз слегка разрастаются складки кожи, у некото-
рых акул могут быть верхнее и даже нижнее веки. У наземных
позвоночных глаза всегда прикрыты веками. Они защищают рого-
вицу от попадания соринок, пыли и от иссушения. У амфибий,
рептилий и птиц сильнее развито нижнее веко, у млекопитающих
и крокодилов — верхнее. Под подвижными веками у амфибий,
рептилий, птиц есть третье веко — мигательная перепонка, при-
крывающая глаз спереди назад. У змей веки срастаются, они дос-
таточно тонкие, отчего роговица оказывается прикрытой прозрач-
ной кожей. При линьке кожный покров отделяется от поверхнос-
ти глаз, что вызывает нарушение зрения. Считают, что после линь-
ки зрение у змей становится особенно острым.
Глазное яблоко приводят в движение шесть глазных мышц. Они
иннервируются ветвями III, IV и VI пар черепномозговых нервов.
У амфибий, некоторых рептилий и млекопитающих есть допол-
нительные мышцы: они втягивают глазные яблоки внутрь глазной
впадины и поднимают верхние веки.
Важное приобретение наземных позвоночных — слезные желе-
зы: они располагаются в заднем наружном углу глаза. Выделяемая
ими жидкость смачивает и промывает роговицу. В переднем углу
глаза лежит гардерова железа, секретирующая маслянистую жид-
кость. Гардеровы железы хорошо развиты у земноводных, пре-
смыкающихся и птиц. Млекопитающим они, как правило, не свой-
ственны, исключая китообразных (и ластоногих), у которых сек-
рет этих желез предохраняет глаза в воде, а над водой увеличивает
Дальнозоркость, по принципу контактной линзы.
У наземных позвоночных в переднем углу глаза есть носослез-
ный проток. Он выводит избыток слезной жидкости в носовую
полость, увлажняя слизистую носа. У морских черепах и некото-
рых морских птиц через слезные железы выделяется гипертони-
ческий раствор соли, что позволяет им использовать для питья
Морскую воду и освобождаться от лишних солей.
Парные глаза у позвоночных заметно отличаются разнообрази-
еЫ. Глазное яблоко может быть очень большим, равным по объе-
221
му, к примеру, у хищных птиц, всему мозгу, и маленьким, чаще
сферическим, или вытянутым. Пресноводную тропическую рыбку
Anableps из Южной Америки называют четырехглазкой: в каждое
глазном яблоке есть две роговицы и два хрусталика, поэтому оца
может видеть одновременно в воде и над ее поверхностью.
Отсутствие освещения приводит к редукции и упрощению глаз
Это свойственно глубоководным рыбам и некоторым норным
животным, живущим под землей, — кротам, слепышам, слепу-
шонкам, гоферам, землеройкам, слепозмеям.
Большинство животных обладают монокулярным боковым зре-
нием, но у некоторых, например птиц, возникает частичное или
полное бинокулярное зрение, при этом глаза сближаются на лице-
вом диске. Полное бинокулярное зрение позволяет охватывать
большее пространство. Например, человек охватывает перед со-
бой бинокулярным зрением 150° пространства, воробьиная птица —
лишь 30°, сова — 60°.
Непарные глаза. Непарные глаза развиваются как два полых
выроста эпиталамуса — крыши промежуточного мозга (рис. 173).
Отшнуровавшись, они превращаются в два пузырька, соединен-
ных с мозгом посредством тонких ножек: передний (париеталь-
ный, или парапинеальный) называют теменным органом, задний
(пинеальный) — эпифизом. Оба органа могут иметь фоторецеп-
торные клетки с неинвертированной сетчаткой (Л. Проссер, 1992)
и в эволюционном ряду позвоночных сильно изменчивы. Свето-
чувствительная функция присуща теменному органу, например у
рыб, хотя факты показывают, что у круглоротых и бесхвостых
Рис. 173. Теменной глаз ящерицы и смежные с ним структуры:
1 — кожа; 2 — крышка черепа; 3 — эпифиз; 4 — средний мозг; 5 — парафиз; 6
теменной глаз
222
а>1фибий она принадлежит эпифизу. Большое разнообразие в стро-
ении теменного органа обнаружено у рептилий. Например, те-
менной орган гаттерии имеет вид пузырчатого глаза. Его передняя
стенка утолщена и образует хрусталик. Задняя стенка пигментиро-
вана и в отличие от парного глаза имеет многослойную неинвер-
тИрованную ретину: светочувствительные клетки обращены в по-
лость глазного яблока (И. И. Шмальгаузен, 1964). У некоторых бес-
хвостых амфибий и ящериц на уровне промежуточного мозга, на
коже или под ней, есть светочувствительное пятно (М.М. Куре-
пина, 1972). Во всех случаях непарный глаз воспринимает уровень
освещенности, а не образ предмета. Предполагают, что теменной
«третий глаз» присутствовал у древних позвоночных: щитковых,
панцирных, кистеперых, двоякодышащих рыб, стегоцефалов и
даже ранних зверообразных рептилий (И. И. Шмальгаузен, 1964).
Эпифиз, утратив зрительную функцию, превращается в желе-
зистое образование — шишковидную железу. Удаление этого орга-
на у птиц подавляет годовой ритм миграционного беспокойства
(Э.Хадорн, Р. Венер, 1989). У млекопитающих шишковидная же-
леза участвует в регуляции деятельности половых желез и форми-
ровании вторичных половых признаков.
Вторичночувствующая рецепция
Орган боковой линии (сейсмосенсорная система)
Этой системой обладают только первичноводные позвоночные
(Anamnia): круглоротые, рыбы, живущие в воде амфибии и их
личинки.
Органы боковой линии представляют собой кожные каналы,
расположенные по бокам тела и на голове животного. У круглоро-
тых, низших примитивных рыб и химер, у амфибий они лежат на
поверхности кожи или погружаются в неглубокие ямки. У боль-
шинства рыб каналы погружены глубоко в кожу, прикрыты че-
шуей и вдоль всего канала имеют множество выходов сквозь от-
верстия в чешуйках (рис. 174). Главные каналы боковой линии рас-
полагаются по бокам вдоль тела. На голове рыбы боковая линия
образует сложную сеть. И. И. Шмальгаузен (1964) называет пять
пар каналов в области головы и уточняет их иннервацию: 1) над-
глазничный — проходит над глазом на верхнюю сторону рыла и
иннервируется глазничной ветвью лицевого нерва; 2) подглазнич-
ный — проходит под глазом вперед на нижнюю сторону рыла и
иннервируется щечной ветвью лицевого нерва; 3) подъязычно-че-
люстной — проходит в области предкрышки (и подвеска) на ниж-
Нюю челюсть и иннервируется подъязычно-челюстной ветвью
Лицевого нерва; 4) главный боковой — переходит в боковую ли-
223
1
2 3
2
Рис. 174. Схема органа боковой линии костистой рыбы (А, Б)
и амфибии (В):
А, Б: 1 — канал; 2 — наружные отверстия канала; 3 — чешуя; 4 — рецепторы
боковой линии; 5 — боковая ветвь блуждающего нерва; 6 — ответвление нерва,
идущее к органу боковой линии. В: 1 — опорные клетки; 2 — чувствующие клет-
ки; 3 — нерв
нию вдоль тела и иннервируется боковой ветвью блуждающего
нерва и отчасти веточкой языкоглоточного нерва; 5) заднетемен-
ной — соединяет главные боковые каналы обеих сторон в заты-
лочной области и тоже иннервируется ветвью блуждающего нерва
(рис. 175).
В прошлом боковая линия была свойственна щитковым, при-
митивным рыбам, ранним амфибиям (рис. 176).
Рис. 175. Каналы боковой линии (светлые) и их иннервация на голов6
трески
224
Рис. 176. Расположение каналов боковой линии на голове первичновод-
ных позвоночных (по А. Ромеру, Т. Парсонсу, 1992):
А, В — щитковые; Б — панцирная рыба; Г~ двоякодышащая рыба; Д — кистепе-
рая рыба; Е — лабиринтодонт
В онтогенезе позвоночного боковая линия закладывается в уча-
стке эктодермы зародыша, расположенном вблизи нервной плас-
тинки. В ней образуются утолщенные участки, так называемые
чувствующие холмики. Каждый такой участок имеет два рода кле-
ток: опорные — длинные цилиндрические, идущие от поверхнос-
ти до основания, и сенсорные — клетки грушевидной формы. Они
занимают центральное положение в участке и не доходят до его
основания. Сенсорные клетки на свободном конце имеют плот-
ную щеточку волосков, а у основания оплетены нервными окон-
чаниями. Скопления чувствующих клеток называются невромас-
тами.
Органы боковой линии воспринимают легкие движения и ко-
лебания воды большой частоты, в основном вблизи источника
колебаний. Экспериментальное удаление чувствующих клеток в
области головы рыб приводит к потере способности восприни-
мать волнения воды, исходящие от пищевого источника (Л. Прос-
сер, 1987).
Видимо, информация, поступающая от боковой линии, по-
могает рыбе ориентироваться в воде, разыскивать пищу, избе-
гать препятствия, ускользать от врагов и т.д. Клетки боковой ли-
нии участвуют в электрорецепции (см. ниже). Среди амфибий боко-
вая линия свойственна живущим в воде личинкам бесхвостых и
Хвостатых и немногим взрослым хвостатым. Так как амфибии тон-
кокожи и не имеют в покровах чешуй, каналы боковой линии
°ткрыты. У большинства бесхвостых при смене образа жизни в
°Нтогенезе она утрачивается. У вторичноводных в группе Amniota
(У рептилий, птиц, млекопитающих) боковая линия не развива-
ется.
Константинов
225
Органы равновесия и органы слуха
В эволюции позвоночных органы равновесия и органы слуха
претерпевает серьезные изменения. В наиболее оформленном виде
эти органы свойственны птицам и млекопитающим. Органы слуха
включают следующие три отдела: внутреннее, среднее и нарух.
ное ухо.
Внутреннее ухо (или перепончатый лабиринт) наиболее древ-
няя часть этой системы. Типографически оно располагается в слу-
ховой капсуле мозгового черепа. С поверхности перепончатый ла-
биринт омывается жидкостью — перилимфой, поступающей из
лимфатических полостей черепа через перилимфатический про-
ток.
Капсула внутреннего уха у позвоночных содержит кроме ре-
цепторов слуха рецепторы гравитации (вестибулярный аппарат).
Вестибулярный аппарат. Предполагают, что внутреннее ухо про-
исходит из специализированной области боковой линии. Оно зак-
ладывается в эмбриогенезе в виде пары утолщений (плакод) в
эктодерме зародыша по обеим сторонам нервной трубки на уров-
не продолговатого мозга. Погружаясь в подлежащую мезенхиму,
плакоды преобразуются в пару слуховых пузырьков (рис. 177). Не-
которое время пузырьки сообщаются с поверхностью эмбриона
эндолимфатическими протоками, а затем обособляются. В каждом
слуховом пузырьке первоначально оформляется поле рецептор-
ных клеток — macula communis. Далее слуховой пузырек делится
VII
Рис. 177. Последовательные стадии (/— VII) развития внутреннего уха
позвоночных животных (по И. И. Шмальгаузену, 1964):
1 — овальный мешочек; 2—4— полукружные каналы; 5— круглый мешочек; 6"
эндолимфатический канал; 7 — полый выступ круглого мешочка — лагена; 8 -
слуховые чувствительные поля; 9 — оттолиты
226
дерехватом на два отдела: верхний — овальный мешочек (утрику-
rjyc) и нижний — круглый (саккулус). Оба мешочка вместе назы-
вают преддверием. От овального мешочка вверх отходят взаимно
перпендикулярные полукружные каналы — два у бесчелюстных и
три У всех челюстноротых: один горизонтальный и два вертикаль-
ных-
Нижняя часть круглого мешочка слегка выпячивается, образуя
дагену. В обоих мешочках и в лагене оформляются рецепторные
области — пятна, или макулы: макула утрикулуса (macula utriculi),
макула саккулуса (macula sacculi), макула лагены (macula lagenae).
В ампулах полукружных каналов рецепторные участки оформля-
ются в виде гребешков {крист) — по одному в каждом канале.
Мешочки и каналы заполнены жидкостью — эндолимфой. Внут-
ри мозгового черепа челюстноротых есть эндолимфатический ме-
шок, поставляющий эту жидкость по каналу. Волоски рецепторных
клеток на всех участках погружены в студенистые купулы. Движе-
ние эндолимфы вызывает колебание купул.
В эндолимфе мешочков, полукружных каналов и лагены взве-
шены кристаллические включения карбоната кальция — крупные
(рттолиты) и мелкие {оттоконии). Форма оттолитов видоспеци-
фична и позволяет специалистам, например у рыб, определить их
систематическую принадлежность.
Чувство равновесия — ранняя исходная функция внутреннего
уха. Ее и осуществляют вторичночувствующие волосковые клетки
полукружных каналов. Многочисленными исследованиями пока-
зано, что рецепторы внутреннего уха фиксируют положение тела
и головы по отношению к гравитационному полю Земли. Сенсор-
ные волоски, воспринимая перемещение эндолимфы и сдвиг от-
толитов, регистрируют наклоны головы и линейные ускорения.
Информацию о поворотах головы и тела в трех пространственных
измерениях и дают полукружные каналы.
В прогрессивной эволюции челюстноротых от рыб до высших
млекопитающих строение вестибулярного аппарата принципиально
не изменяется. Возможно, это объясняется постоянством характе-
ристик гравитационного поля Земли, стабильных по направле-
нию и величине и независимых от среды обитания животных.
Орган слуха. У рыб специализированного слухового рецептора
во внутреннем ухе нет. У рыб, земноводных и пресмыкающихся
отсутствует и звукоорганизующая система — внешнее ухо. Звук
°Ни воспринимают всей поверхностью тела. Органами, восприни-
мающими колебания среды, у рыб и личинок земноводных следу-
ет считать внутреннее ухо, боковую линию и клетки кожного ана-
лизатора. Слуховую функцию во внутреннем ухе выполняют от-
дельные волосковые клетки круглого мешочка и лагены — это
Рецептор давления. Они улавливают звуки, отдаленные на боль-
шие расстояния. Орган боковой линии — рецептор смещения —
227
улавливает звуки вблизи их источника. Интенсивные звуки может
воспринимать кожный анализатор. Обнаружено, что хрящевые рыбы
воспринимают поверхностью тела звуковые колебания низких ча-
стот.
У костных рыб в восприятии звуков участвуют система
полостей в черепе и плавательный пузырь. Например, у сельде-
образных звуковые колебания с поверхности тела передает труб-
чатый отросток плавательного пузыря, соприкасающийся с пе-
репончатым лабиринтом (рис. 178). У карпообразных механизм
иной: плавательный пузырь связан у них с перепончатым лаби-
ринтом системой косточек — производных отростков передних
позвонков {веберов аппарат). Полагают, что они действуют по-
добно слуховым косточкам млекопитающих (А. Ромер, Т. Пар-
сонс, 1992).
Звукоизлучение в воде имеет разную природу. Это внешние
звуки, связанные с течением, приливами и отливами, ветром,
землетрясениями, с биологической жизнью обитателей водоемов,
с индустриальной деятельностью людей. Важными раздражите-
лями являются звуки, издаваемые самими рыбами. Это разнооб-
разные варианты механических звуков, возникающих с участи-
ем стридуляционных (инструментальных) механизмов (И.Яма-
лова, 1970). Специалисты выделяют несколько типов таких звуков:
звуки, связанные с движением жаберных крышек, сочленений
скелета, трением лучей плавников, срывом водяных вихрей с
поверхности тела и т.д. Диапазон
Б В
частот этих звуков от 20 Гц до
5 кГц. Такие звуки возникают при
питании рыб, захвате и перети-
рании пищи. Другие звуки изда-
ются при участии плавательного
пузыря. Они воспроизводятся
импульсивно и напоминают се-
рии ударов, барабанный бой,
Рис. 178. Связь плавательного
пузыря с внутренним ухом:
А — сельдь атлантическая; Б — треска,
В — карповые; 1 — выросты плаватель-
ного пузыря; 2 — внутреннее ухо; 7 -
головной мозг; 4 — косточки веберов
аппарата; 5 — передние позвонки; 6 "
общий эндолимфатический проток
228
молоточные удары, хрюканье. Эти звуки реализуются в диапазо-
не 50 Гп — 2,5 кГц и т.д.*
В наземной среде условия передачи звуков иные. Скорость
распространения звуковой волны в воздухе меньше, чем в воде:
330 м/с. Звуки быстро рассеиваются и затухают. В воздушной среде
механические звуки и голоса, столь распространенные у рыб, ста-
новятся малоэффективными. Поэтому системы звуковосприятия
и звукоизлучения у наземных позвоночных совершенствовались в
процессе эволюции.
У амфибий рецепторный аппарат внутреннего уха приобре-
тает новообразования — сосочки. На границе между утрикулусом
и саккулусом располагается амфибиальный сосочек. Это оригиналь-
ная черта амфибий, несвойственная прочим наземным позвоноч-
ным. При его участии воспринимаются звуки в довольно узком
диапазоне: 200 — 940 Гц. Полагают, что амфибиальный сосочек уча-
ствует у них и в системе водного восприятия звуков (И. И. Шмальга-
узен, 1964). Другой сосочек, базиллярный, располагается в саккулу-
се вблизи лагены. Он воспринимает звуки в диапазоне 3 — 4 Гц —
1 кГц. Передача звуков на рецепторный аппарат внутреннего уха
осуществляется у амфибий двумя путями: водным (через лимфа-
тические мешки, венозную систему и эндолимфу внутреннего уха)
и воздушным (через среднее ухо).
Во внутреннем ухе настоящих наземных позвоночных (амнио-
та) важную роль для восприятия звуков в воздушной среде играет
расположенная в слуховой капсуле (и выходящая за ее пределы в
полость черепа) перилимфатическая цистерна — мешочек с одно-
именным протоком. Проток отделен от базиллярного сосочка под-
стилающей базиллярной мембраной. Это образование считают важ-
нейшим компонентом звуковоспринимаюшего аппарата внутрен-
него уха всех амниота (А. Ромер, Т. Парсонс, 1994).
Наземным позвоночным свойственны и иные формы совер-
шенствования звуковоспринимающей системы. У них образуется
новый отдел — среднее ухо (рис. 179). В этот орган преобразуются:
IV жаберная щель — брызгальце и участок IV висцеральной дуги —
подвесок (гиомандибуляре). Снаружи полость среднего уха огра-
ничена тонкой мембраной — барабанной перепонкой. Она образо-
вана листком соединительной ткани и прикрыта с поверхности
кожей (у амфибий, многих ящериц). Через широкое отверстие или
* Вода как среда для проведения звуков имеет ряд особенностей. Скорость
Распространения звука в воде в силу ее большой плотности и малой сжимаемо-
сти в 4,5 раза больше, чем в воздухе, и составляет 1440 м/с. Она меняется в
зависимости от температуры воды и ее плотности. При понижении температуры
скорость звука уменьшается, при увеличении плотности, например с глубиной,
возрастает. Начиная с глубины 1220 м, где температура воды остается практичес-
ки постоянной, скорость звука меняется за счет изменения давления.
229
10
Рис. 179. Схема развития среднего уха в ряду позвоночных:
А — хрящевые рыбы; Б — земноводные и пресмыкающиеся; В — млекопитаю-
щие; 1 — гиомандибулярный хрящ; 2 — брызгальце; 3 — гиоид; 4 — слуховая
капсула черепа; 5 — ротовая полость; 6 — столбик (стремя); 7 — барабанная пере-
понка; 8 — среднее ухо; 9 — евстахиева труба; 10 — стремечко; II — наковальня;
12 — молоточек; 13 — наружный слуховой проход; 14 — барабанная кость
через канал (евстахиева труба) эта полость соединяется с глоткой.
Гиомандибуляре теряет роль подвеска челюстей и превращается в
столбчатую косточку — стремечко (stapes, или columella). Расши-
ренный в виде подошвы конец стремечка упирается в овальное
окно слуховой капсулы и передает колебания барабанной пере-
понки на систему: перилимфатический проток — базилярная мем-
брана — лагена. Таким путем звук из воздушной среды передается
на жидкость внутреннего уха (рис. 180).
Описанная выше система свойственна большинству бесхвос-
тых земноводных. Хвостатые амфибии, находясь постоянно в воде,
не имеют среднего уха и не пользуются системой звуковосприя-
тия в наземной среде. Безногие вторично его утратили. Эти амфи-
бии не используют акустические раздражители в воздушной среде.
Рис. 180. Поперечный разрез че-
рез голову лягушки в области уха
(схема):
1 — черепная коробка; 2 — про-
долговатый мозг; 3 — слуховой нерв;
4 — полукружные каналы; 5 — по-
лость среднего уха; 6 — евстафиева
труба; 7 — глотка; 8 — стремя; 9 —
барабанная перепонка
230
вторичную утрату среднего уха связывают с недостаточностью его
защитных и акустических свойств в воде и почве, в результате
чего барабанная перепонка исчезает, а полость редуцируется
(Б. Д. Васильев, 1975).
Хвостатые амфибии в большинстве не имеют голоса. Однако
некоторые саламандры пищат, амфиумы и сирены свистят, тихо-
океанская амбистома {Dieamptodon ensatu) лает, Megalobatrachus
громко мычит. Амбистомовые имеют голосовые связки и подают
звуки при выдохе. Безлегочные саламандры издают звуки при со-
кращении объема ротоглоточной полости.
Разнообразно звукообразование у бесхвостых. Они излучают
довольно широкий диапазон звуков от 100 Гц до 12 — 15 кГц. Ор-
ганом звукоизлучения у них служит гортанная щель. Интенсив-
ность звука усиливается за счет голосовых мешков (резонаторов).
Отсутствие голосовых мешков сочетается иногда с полной безго-
лосостью, как, например, хвостатая лягушка гладконог {Ascaphus
truel), или со способностью издавать звуки под водой {Pipa). Вос-
производство звуков имеет у бесхвостых земноводных важное био-
логическое значение при коммуникации между особями во время
брачных игр, криков агрессии, испуга, защиты.
В органе слуха у многих рептилий оформились три отдела:
внутреннее, среднее и наружное ухо. Внутреннее ухо по организа-
ции близко к амфибиям, за исключением отсутствующего амфи-
биального сосочка. В круглом мешочке (саккулусе) заметен вы-
рост, получивший название — кохлеарный канал (или улитка).
В нем концентрируются волосковые клетки базиллярного сосочка
и лагены. Структура внутреннего уха в разных систематических и
экологических группах рептилий изменчива (Б. В. Васильев, 1975).
Наиболее примитивный рецепторный аппарат обнаружен у агам,
игуан, веретениц. Более специализирован он у гекконов, чешуе -
ногов. Диапазон звуковых частот, воспринимаемый ящерицами, —
100 Гц — 6 кГц. У черепах рецепторный аппарат упрощен. Их диа-
пазон — 20 Гц — 4 кГц, а область лучшей слышимости сужена до
80 — 450 Гц. Рецепторный аппарат крокодилов совершеннее: их
волосковые клетки отличаются разнообразием форм, а диапазон
их слышимости — 100 Гц — 5 кГц (Manley, 1981).
Среднее ухо хорошо развито у крокодилов, черепах и несколь-
ких семейств ящериц (Gecconidae, Xantuscidae, Condylidae,
Lacertidae). Его положение на голове в сравнении с амфибиями
Несколько смещено вниз. Через евстахиеву трубу барабанная по-
лость сообщается с глоткой. В целом стремечко повышает чувстви-
тельность слуха у рептилий в широком диапазоне частот.
У гаттерий и змей среднее ухо полностью редуцировано. Ис-
чезновение его, как правило, связано с ползающим и роющим
образом жизни. Обитание в водной среде к редукции этой части
Уха обычно не приводит (И.Ямалова, 1970).
231
У многих рептилий звукоорганизующей системой становится
наружный слуховой проход. Он защищает барабанную перепонку в
нем поддерживаются постоянная температура и давление. Развщ
тость этого канала меняется у рептилий в широких пределах. Наи-
более хорошо наружный слуховой проход оформлен у крокодилов
и некоторых гекконов. Кольцевые мышцы позволяют сужать и
перекрывать наружное отверстие. Слуховой проход хорошо обо-
значен у ящериц, укрывающихся в тесных убежищах, под камня-
ми, в узких щелях — сцинки, золотистая мабуя, полосатый голо-
глаз. У ящериц, пользующихся просторными убежищами среди
крупных камней, в трещинах скал, между корнями деревьев, слу-
ховой проход или слегка обозначен, или неразвит вовсе (агамы
скальные ящерицы). У змей он отсутствует.
Способы звукообразования у рептилий довольно разнообраз-
ны. Настоящим голосом обладают крокодилы, гекконы, хамелео-
ны. Многие рептилии шипят, резко выдыхая воздух из легких (че-
репахи, змеи, ящерицы). Аризонский аспид, змея-носорог, аф-
риканская гадюка издают довольно громкие звуки, приводя в дви-
жение мускулатуру клоаки и стенки брюшной полости.
У рептилий встречаются стридуляционные механизмы. К ним
относятся быстрые движения покрытого крупными чешуями хво-
ста у сцинкового геккона, погремушки на хвосте гремучих змей.
Считают, что в целом у рептилий акустическая сигнализация
слаба, хотя многие могут издавать предупреждающие шумовые
сигналы. Крики гекконов имеют, по-видимому, привлекающее и
территориальное значение. У крокодилов есть призывные, пре-
дупреждающие и территориальные сигналы частотой 0,8 — 3,4 кГц
(Л.Проссер, 1977 — 1978).
В органе слуха птиц хорошо оформлены все три отдела уха:
внутреннее, среднее и наружное. Во внутреннем ухе, как и у реп-
тилий, рецепторным органом является базиллярный сосочек. Он
располагается в выросте саккулуса — улитке, имеющей вид слабо
изогнутой трубки (0,5 оборота). На ее дистальном конце располо-
жена лагена. Волосковые клетки у птиц многообразны по форме.
Они представлены тремя модификациями (высокие, низкие, ско-
шенные) и отличаются строением, типографией и иннервацией
(И.Ямалова, 1977, 1978, 1979, 1980). В улитке увеличены размеры
овального и круглого окон.
Общий план строения среднего уха птиц и рептилий сходен:
барабанная перепонка, барабанная полость, стремечко, евстахи-
ева труба. Через стремечко сохраняется однокосточковая передача
колебаний от барабанной перепонки к внутреннему уху. По мор-
фологическим особенностям специалисты выделяют у птиц не-
сколько типов среднего уха: клипеолярный (совы), шатровый (во-
робьиные), поршневой (многие группы) и др. (В. Д. Ильичев и др-,
1972).
737
Наружное ухо оформлено у птиц в короткий и сложный по
рельефу наружный слуховой проход с расположенными у входа
ложными складками (дперкулум и птерилозис), валиками и ушными
п1Перлиями. Выделяют несколько морфофункциональных типов
наружного уха: 1) рупорный (совы, луни, козодои), обладающий
высокой акустической чувствительностью к пространственному
анализу звуков низкой интенсивности; 2) рефракторный (воробь-
иные птицы), имеющий широкий диапазон акустических возмож-
ностей; 3) водозащитный, сформировавшийся у плавающих птиц
в связи с необходимостью защищать барабанную перепонку от
воды; у этих птиц (Gaviformes, Padicipediiformes, Alciformes) нет
кожных складок, наружное отверстие уменьшено и прикрыто пе-
рьями; 4) универсальный тип, характерный для Columbiformes,
Galliformes, Gruiformes, — полагают, что слабое развитие звуко-
вого общения не требует у этих птиц чуткого специализированно-
го слуха, отверстие мало, кожные складки отсутствуют, валики
выражены плохо (В. Д. Ильичев, 1972).
Птицы воспринимают звуки в очень широком диапазоне час-
тот: от 120 Гц до 15 кГц. Отдельные виды способны к эхолокации.
Например, обитающий в пещерах Южной Америки козодой (гуа-
харо) при полетах в темноте подземелий издает ультразвуки с
частотой 7,3 кГц. Используя ультразвуки в мутной воде, охотятся
пингвины, ориентируются в тумане кроншнепы.
Для воспроизведения звуков птицы используют настоящий (ды-
хательный) голос и инструментальный (стридуляционный). Насто-
ящий голос образуется на выдохе при участии голосовых связок
нижней гортани и резонирующих полостей тела (верхняя гортань,
пищевод, трахея, воздушные мешки). Издавая дыхательные зву-
ки, птица может варьировать их параметры и управлять ими. По-
этому в акустической ориентации птиц именно дыхательный го-
лос играет ведущую роль и служит основным источником разно-
образных и сложных сигналов общения. Инструментальные звуки
воспроизводятся оперением, клювом, лапами (В.Д. Ильичев, 1972).
Выполняя те же функции, что и настоящий голос, они расширя-
ют возможности общения за счет дополнительных каналов связи,
заменяя, дополняя и дублируя настоящий голос.
Млекопитающие имеют в органе слуха три хорошо раз-
витых отдела (рис. 181).
Во внутреннем ухе улитка спирально закручена в 1 — 5 оборо-
тов и полностью изолирована от вестибулярного аппарата, у од-
нопроходных она имеет 0,5 оборота и напоминает улитку у птиц.
Оригинальной чертой млекопитающих считают постоянное число
Рецепторных клеток и одинаковый характер их размещения, не-
зависимо от специализации животного (И.Ямалова, 1977).
Заметно преобразуется среднее ухо. В передаче звуков с бара-
банной перепонки на овальное окно внутреннего уха участвуют
733
Рис. 181. Схема органа слуха
млекопитающего:
1 — наружный слуховой проход; 2 —
эндолимфатический канал; 3 — круглое
окно; 4 — наковальня; 5 — молоточек;
6 — барабанная перепонка; 7 — слухо-
вой нерв; 8— евстахиева труба; 9— нерв
улитки; 10 — стремя
три слуховые косточки — мОло~
точек, наковальня и стремечк.о
гомологи соответственно квад-
ратной, сочленовной костей и
подвеска. Считается, что систе-
ма косточек эффективнее пре-
дохраняет внутреннее ухо от на-
грузок при быстрых изменениях
интенсивности звуков. Нару-
шения в системе косточек не
ведут к полной потере слуха.
В этом случае звуковые колеба-
ния к внутреннему уху поступа-
ют через прилежащие кости че-
репа.
У видов, обладающих эхоло-
кацией и воспринимающих уль-
тразвуки (землеройки, летучие
мыши, некоторые грызуны, зу-
батые киты), между молоточком
и наковальней появляется дополнительный рычаг, а молоточек
жестко присоединяется к барабанной перепонке. Это повышает
жесткость системы и улучшает восприятие и проведение специ-
фических сигналов.
Наружное ухо у наземных млекопитающих включает слуховой
проход и ушную раковину, форма и подвижность которой сильно
варьируют. Животное «настораживает» уши благодаря мускулату-
ре, обслуживающей ушные раковины. Наружное ухо — орган мно-
гофункциональный: с его помощью животное улавливает звуки,
осуществляет терморегуляцию, демонстрирует эмоциональное
состояние. У полуводных (например, ластоногих) ушные ракови-
ны укорачиваются, а специальные кожные выросты при нахож-
дении зверя под водой закрывают ушной проход. У настоящих
тюленей раковина редуцируется, а слуховой проход имеет коле-
нообразный изгиб. У дельфинов ушная раковина редуцирована, а
слуховой проход замкнут и заполнен воздухом, однако это не
мешает им хорошо слышать. Исследования показали, что дель-
фин определяет направление сигнала с точностью до 1,5°, а при
отсутствии помех — до 0,5° (П. Белькович, 1970).
Звуковая коммуникация важна в жизни млекопитающих, хотя
имеет меньшее значение, чем у птиц. Им свойственны лай, вой,
писк, рев, мяуканье, скуление и др. Эти звуки отражают эмоцио-
нальное состояние животного, но в малой степени видоспеии-
фичны. Отдельные группы зверей способны к эхолокации. Напри'
мер, большой бурый кожан (Eptesicus fuscus) издает звуковые сиг-
налы с частотой 2500 — 50000 Гц. Высокочастотные звуки, изДа'
234
|раемые летучими мышами и воспринимаемые чутким ухом, по-
Зроляют им успешно охотиться в темноте. Их ушная раковина при
зтом приобретает сложную архитектонику. Дельфины используют
локационную систему при охоте в мутной воде или ночью, при
обнаружении предметов.
Рекорд дальности в звуковой коммуникации млекопитающих
поставлен горбатыми китами: их песни могут восприниматься
другими китами с расстояния в несколько десятков километров,
у китообразных большую роль в восприятии звуков играют спе-
циализированные структуры головы. Эхолокацией пользуются так-
же землеройки. Охотясь в подстилке, они издают импульсные зву-
ки с частотой до 50000 Гц и более.
Орган вкуса
Рис. 182. Вкусовая почка
ротовой полости млеко-
питающего:
1 — опорная клетка; 2 — вку-
совая пора; 3 — рецепторная
клетка; 4 — вкусовое нервное
волокно; 5 — соединительная
ткань; 6 — многослойный
эпителий
Орган вкуса тоже принадлежит к системе вторичночувствую-
щей рецепции. Рецепторами вкуса служат вкусовые луковицы (или
почки). Они состоят из длинных цилиндрических клеток с тонким
и коротким чувствующим волоском. Между сенсорными лежат
длинные опорные клетки. Снизу во вкусовую почку проникают
нервные волокна, которые и оплетают чувствующие клетки свои-
ми разветвлениями (рис. 182).
Вкусовые почки располагаются главным образом в ротовой по-
лости и глотке. Однако у костных рыб они распространены и на
внешней поверхности тела в покровах встречаются на жаберных
дугах, на плавниках, а особенно на губах и усиках. Например,
осетры получают вкусовую информацию с нижней поверхности
рыла, американские сомики-кошки
(Amiurus nembulosus) — от клеток, раз-
бросанных по телу (рис. 183). Обнаруже-
но, что у другого сомика — пятнистого,
или канального, (Yctalurus punctatus) —
Хеморецепторы на усиках весьма чув-
ствительны к малым концентрациям раз-
личных аминокислот (К. Шмидт-Ниель-
сен, 1982). Вкусовые почки у амфибий
встречаются в коже. У рептилий и птиц
они находятся в глотке, редко на языке.
В основном на языке вкусовые почки кон-
центрируются у млекопитающих. У гры-
зунов, хищных и копытных млекопита-
ющих на дорсальной поверхности языка
Присутствуют три вида вкусовых сосоч-
ков, несущих почки: грибовидные, валь-
к°ватые и листовидные (И. И. Шмальга-
Узен, 1964, 3. В.Любимова, 1980).
235
Рис. 183. Вкусовые почки на теле сома (А — сбоку; Б — сверху; В —
снизу)
Вкусовые сенсорные клетки каждые несколько дней замеща-
ются новыми (Л. Проссер, 1977—1978). У большинства позвоноч-
ных вкусовые почки иннервируются ветвями лицевого (VII), язы-
коглоточного (IX) и блуждающего (X) нервов. У млекопитающих
вкусовыми нервами являются ветвь лицевого нерва — барабанная
струна и язычная часть языкоглоточного нерва.
Позвоночные различают четыре основных вкусовых качества:
горькое, соленое, кислое, сладкое. Рецепция вкуса происходит
путем распознавания молекул и ионов в жидкой среде: кислот —
как кислое, хлоридов — как соленое, ощущение горького вызы-
вается алкалоидами, ощущение сладкого — сахарами и спиртами.
Органы осязания
Органы осязания принадлежат у позвоночных к специфиче-
ским чувствующим системам. Они не имеют первично- или вто-
ричночувствующих клеток. Их воспринимающим аппаратом явля-
ются свободные нервные окончания, которые разбросаны по всей
коже, имеются также в кишечнике и других органах.
В покровах осязательную функцию несут группы клеток эпи-
дермиса, оплетенные сетью нервных волокон, — осязательные
пятна, или осязательные тельца. В эволюционном ряду позвоноч-
ных существует большое разнообразие этих телец. В разных систе-
матических группах описаны тельца Фатер — Пачини, восприни-
мающие давление и растяжение, колбы Краузе — холод, тедьиа
Гербста, осязательные тельца Мейссенера, тельца Меркеля, ам-
пулы Лоренцини на голове хрящевых рыб и др. В органах осязания
236
различают тактильные осязательные тельца, или механорецепто-
ры, и терморецепторы.
Механическая чувствительность. В плавниках хрящевых рыб
осязательные тельца реагируют на синусоидальное механическое
раздражение. Убесхвостых амфибий органы осязания раз-
бросаны по всему телу и представляют собой быстроадаптирую-
щиеся и высокочувствительные органы (Л. Проссер, 1977).
У чешуйчатых рептилий осязательные тельца распола-
гаются на заднем крае каждой чешуйки и прикрыты тонким сло-
ем эпидермиса. Кожные рецепторы змей воспринимают вибра-
цию субстрата. В отличие от амфибий тактильный аппарат репти-
лий — медленно адаптирующаяся система (Л. Проссер, 1977).
У птиц осязательную чувствительность раньше ограничива-
ли восковицей и поверхностно языка (И. И. Шмальгаузен, 1964).
Однако сейчас значительно расширилось представление о чув-
ствующих способностях участков кожи, расположенных в основа-
нии перьевого покрова. К примеру, у утки основания пуховых
перьев образуют рецептивные поля. Показано, что и нитевидные
перья имеют осязательную функцию, сигнализируя о нарушени-
ях перьевого покрова.
У млекопитающих осязательные тельца обнаружены на
участках кожи, лишенных волос, и во влагалищах разных типов
волос. Кожа сигнализирует о боли, прикосновении, тепле, холоде.
Разные тактильные рецепторы отличаются у зверей тонкостью и
скоростью адаптации.
Исследованиями показано, например, что у кролика и кошки
тактильные осязательные тельца (механорецепторы) волос под-
шерстка имеют низкие пороги возбуждения и быстро адаптируются
к раздражителю, а рецепторы остевых
волос имеют высокие пороги возбужде-
ния и адаптируются медленно. В подушеч-
ках лап кошки обнаружено три типа ме-
ханорецепторов (А. Ромер, Т. Парсонс,
1992). Наиболее совершенную осяза-
тельную систему представляют вибрис-
сы млекопитающих (рис. 184). Их воло-
сяные сумки иннервируются ветвями
тройничного нерва и снабжены попе-
речнополосатой мускулатурой. Чаще
они расположены на голове в виде усов
и бровей. Вибриссы значительно рас-
ширяют возможности животного в вос-
приятии окружающей среды. Они реа-
гируют не только на прикосновения,
Но и на колебание воздуха. У норников
вибриссы разбросаны по всему телу.
Рис. 184. Схема расположе-
ния чувствующих волос
(вибрисс) на морде кроли-
ка. Очерчена «осязательная
зона»
237
Тактильные рецепторы, кроме кожи, присутствуют у позво-
ночных в скелетных мышцах (рецепторы мышечного веретена ц
сухожилия создают «мышечное чувство»). Они передают в цент-
ральную нервную систему сигналы о степени сокращения или
растяжения мышц, информируют о положении частей тела в про-
странстве. Механическая рецепция есть на языке, в кишечнике, в
кровеносных сосудах и других органах.
Другие виды кожной чувствительности. Наряду с тактильными
рецепторами в коже позвоночных находятся терморецепторы. Счи-
тают, что у акул и скатов эту функцию выполняют лежащие на
поверхности головы ампулы Лоренцини.
У ямкоголовых змей, охотящихся на теплокровных живот-
ных, на переднем конце головы есть углубления — лицевые ямки
(рис. 185). Наружная камера каждой ямки имеет на дне пленку с
подстилающей воздушной подушкой (внутренней камерой). Эти
органы воспринимают инфракрасные излучения теплокровных жи-
вотных с длиной волны 0,5 — 2 мкм. Полагают, что лицевые ямки
чувствительны к легкому повышению температуры. Чувствитель-
ность их достигает в темноте до 0,001 °C. У удавов эти органы рас-
полагаются в верхней части головы. Иннервируются лицевые ямки
ветвями тройничного нерва.
Терморецепторы в коже млекопитающих в большинстве случа-
ев представлены свободными (голыми) нервными окончаниями,
некоторые из них инкапсулированы. Тепло и холод воспринима-
ются в коже разными рецепторами. Замечено, что холодовые ре-
цепторы лежат ближе к поверхности кожи и более многочислен-
ны (Л. Проссер. Т. II, 1978).
Наконец, среди органов чувств есть органы специфической рецеп-
ции, в том числе генерирующие и воспринимающие электричес-
2
3
Рис. 185. Термолокатор на голове
гремучей змеи:
1 — наружная камера; 2 — перепон-
ка; 3 — внутренняя камера
кий ток (электрорецепция), вос-
принимающие магнитное поле
Земли. Эти виды рецепции изуче-
ны значительно меньше. Ограни-
чимся кратким их описанием.
У относительно небольшого
числа морских и пресноводных
рыб, таких, как скаты рода Torpe-
do, южно-американский электри-
ческий угорь (Elextrophorus), аф-
риканский электрический сом,
пресноводные тропические рыбы
рода Gymnotus, развиваются спе-
циальные органы, способные
генерировать электрический ток
(рис. 186). Электрические органы
участвуют в обездвиживании ДО'
238
Рис. 186. Расположение электрических органов у электрических рыб
(по Гриффину и Новику):
А — электрический скат; Б — морская лисица; В — электрический угорь; Г —
электрический сом; Д — слонорыл; Е — гимнарх; Ж — звездочет. Выделенные
участки на контурном изображении каждой рыбы представляют собой проекции
электрических органов
бычи, защите от врагов. Считается, что у них в электрические
органы преобразуются или поперечнополосатые мышцы, или не-
рвномышечные комплексы (рис. 187). В результате образуются мно-
гочисленные пластины — электроплаксы, между которыми воз-
никают разности потенциалов:
200 В, электрический угорь —
свыше 500 В. У африканского
сома электрическая ткань немы-
шечного происхождения распо-
лагается под кожей на всем теле
от головы до хвоста (А. Ромер,
Т. Парсонс, 1994).
У низковольтных рыб наибо-
лее часто волны, излучаемые
электрическими органами, со-
здают вблизи тела электричес-
кое поле. Кожными рецептора-
ми являются ампулы Лоренци-
Ии (рис. 188). Приближающиеся
объекты возмущают это поле,
Что и улавливается рецептора-
ми боковой линии хозяина. Это
свойство особенно важно для
скат генерирует напряжение до
Рис. 187. Схема строения электри-
ческого органа ската:
I — IV — электрические нервы; I — элек-
трические доли головного мозга; 2 —
электрические столбики, сложенные
пластинами (электроплаксами)
239
рыб, живущих в темной и мут-
ной воде, таких, например, как
пресноводные костистые рыбы
Южной Америки и Африки —
нильский гимнарх (Gymnarchus
niloticus) и др. Хрящевые рыбы
чаще хищники, неспособные
сами подавать электрические
Рис. 188. Ампулы Лоренцини сигналы, весьма чувствительны
на голове акулы к слабым электрическим полям,
возникающим при обычной
мышечной активности тела жертвы.
Предполагают, что электрическая чувствительность была свой-
ственна древним первичноводным позвоночным. Она сохранилась
у современных миног и пластинчатожаберных. Предполагают, что
чувствительность к электрическим полям отражается на биологи-
ческих свойствах животных: например, она обусловливает нави-
гацию угрей в океане (Л. Проссер, 1978).
Магнитная рецепция — малоизученная область. Участок чере-
па с магнитными свойствами обнаружен у обыкновенного дель-
фина. У птиц областью, воспринимающей магнитное поле Зем-
ли, считают эпифиз промежуточного мозга. Птицы используют
этот механизм при навигации.
К специфическим органам можно отнести и инфундибулярный
орган рыб. Он образован группой первичночувствующих клеток в
стенке сосудистого мешка промежуточного мозга. Высказано пред-
положение, что этот орган, воспринимая давление в желудочках
мозга, рефлекторно регулирует глубину погружения животного в
толщу воды (И. И. Шмальгаузен, 1964).
* * *
Таким образом, у позвоночных функционируют разнообраз-
ные органы чувств, обеспечивающие им возможность восприни-
мать раздражение из окружающей среды и посредством цент-
ральной нервной системы адекватно на них реагировать. Органы
чувств — важное средство их внутри- и межвидовой коммуника-
ции.
ЭНДОКРИННЫЕ ЖЕЛЕЗЫ
Железы внутренней секреции (эндокринные) не имеют спе-
циальных выводных протоков в органы. Секреты эндокринных
желез (гормоны) поступают в кровеносное русло через капилляр-
ную сеть и с током крови разносятся по всему телу. Они регулиру-
ют обмен веществ, влияют на развитие и рост органов и всего
организма в целом, корректируют сезонное состояние организма
и вместе с нервной системой осуществляют нейрогуморальную
регуляцию функций всех органов и систем в организме.
Для большинства эндокринных желез выделение гормонов —
единственная их специфическая функция (щитовидная, паращи-
товидная, зобная, надпочечники, гипофиз), для других — выде-
ление гормонов — их дополнительная функция наряду с другой
основной (поджелудочная железа, половые железы).
Железы внутренней секреции топографически располагаются
в разных участках тела животного. Щитовидная, паращитовидная
и зобная железы закладываются в онтогенезе как парные выпячи-
вания глотки и концентрируются в передней части тела (рис. 189).
Рис. 189. Расположение и развитие эндокринных желез в области
глотки:
4 — акула; Б — хвостатая амфибия; В — ящерица; Г — курица; Д — млекопита-
ющее; 1 — преджаберный карман; 2— 6 — жаберные карманы; 7 — щитовидная
*елеза; 8 — зобная железа (тимус); 9 — паращитовидная железа; 10 — постбран-
Хиальные тельца (функция неизвестна); II — эпителиальные тельца; 12 — щель
для эмбриональной глотки
9 Константино»
241
Щитовидная железа (glandula thyreoidea) закладывается эмб-
рионально как карманообразное выпячивание брюшной стен-
ки глотки. У круглоротых ткань железы разбросана вдоль глот-
ки, у рыб и некоторых рептилий это единый компактный орган
у акул она лежит между ветвями нижней челюсти впереди брюцы
ной аорты; у костистых рыб охватывает брюшную аорту в области
передних жаберных дуг; у земноводных расположена в области
подъязычного аппарата; у рептилий — под трахеей; у птиц — в
основании бронхов; у млекопитающих — в области гортанных
хрящей (рис. 190). У амфибий, птиц и млекопитающих железа под-
разделена на две симметричные доли.
В микроструктуре железы присутствует много мелких округлых
фолликулов и развита богатая сеть кровеносных сосудов (рис. 191).
Полости фолликулов содержат в коллоидном состоянии белок ти-
реоглобулин. Его молекула включает иод, отчего это вещество на-
зывают иодтиреоглобулин. В результате гидролиза этого белка в кровь
выделяются гормоны тироксин и трииодтиронин. Они участвуют в
регуляции общего тканевого и энергетического обменов. У птиц и
млекопитающих эти гормоны участвуют в поддержании постоян-
ной высокой температуры тела, у амфибий они контролируют
метаморфоз. Ослабление (гипофункция) деятельности щитовид-
ной железы замедляет обмен веществ, задерживает рост, наруша-
ет терморегуляцию. Усиленная деятельность (гиперфункция) же-
лезы приводит к тяжелому заболеванию — базедовой болезни,
нарушающей работу репродуктивной, кровеносной систем и др.
В
Рис. 190. Положение щитовидной железы и тимуса у ящерицы (А),
птицы (Б), млекопитающего (В):
1 — щитовидная железа; 2 — зобная железа (тимус); 3 — эпителиальные тельца;
4 — постбранхиальные тельца; 5 — гортань; 6 — трахея; 7 — сердце; 8 — ДУ1"®
аорты; 9 — сонная артерия
Рис. 191. Гистологическое строение щитовидной и паращитовидной
желез крысы:
I — кровеносный сосуд; 2 — фолликулы щитовидной железы; 3 — мышца; 4 —
паращитовидная железа; 5 — соединительнотканная прослойка между фолли-
кулами
Эндокринная активность щитовидной железы контролируется ти-
реотропным гормоном гипофиза.
Предполагают, что щитовидная железа гомологична эндости-
лю низших хордовых. У бесчерепных ткань эндостиля продуцирует
тирозин и трииодтироксин. То же наблюдается при метаморфозе
личинок круглоротых (С. И. Левушкин, И. А. Шилов, 1994).
Зобная железа (вилочковая, или тимус) (gl.thymus) возникает в
виде выпячиваний жаберных карманов спинной стороны глотки.
У рыб зачатки этой железы появляются вблизи жаберных меш-
ков, и она остается в области жаберного аппарата. У наземных
позвоночных тимус развивается из меньшего числа зачатков и рас-
полагается по бокам шеи. В онтогенезе позвоночного животного
эта железа непостоянна: она растет до наступления половой зре-
лости, а затем постепенно уменьшается в размерах и даже вовсе
Исчезает (у млекопитающих).
Секрет зобной железы влияет на формирование и функцию
Половых желез, стимулирует рост организма и обмен веществ в
Нем, участвует в становлении иммунных реакций. Однако главная
243
функция этого органа — кроветворение в раннем онтогенезе. Ти-
мус интенсивно продуцирует лимфоциты в начале постэмбрио-
нальной жизни. Впоследствии эта функция переходит к селезен-
ке, лимфатическим узлам, костному мозгу.
Паращитовидная железа (gLparathyreoidea) в эмбриогенезе зак-
ладывается с брюшной стороны глотки одновременно со щито-
видной железой в виде нескольких долек. Структурно она отлича-
ется от щитовидной железы характером расположения клеток —
они вытянуты в виде тяжей (см. рис. 191). Эту железу в разное вре-
мя выделяли как эпителиальные тельца, околощитовидные желе-
зы, пост- или ультимобранхиальные тельца. До настоящего вре-
мени эта железа довольно плохо изучена. Известно, что она не
оформлена у рыб и хорошо развита у змей и млекопитающих. Гор-
мон паращитовидной железы — белок паратгормон — регулирует
концентрацию кальция и фосфора в тканях тела животного и ва-
жен для поддержания химического состава костной ткани (А. Ро-
мер, Т. Парсонс, 1994).
Гипофиз (hypophysis cerebri) представляет собой небольшую ком-
коватую железу, прилегающую к нижней части промежуточного
мозга (к воронке гипоталамуса)*.
Гипофиз — сложный орган.
В эмбриогенезе животного он фор-
мируется комплексно (рис. 192).
Снизу промежуточного мозга, в
области гипоталамуса выпячива-
ется полая яйцевидная воронка.
Навстречу ей от первичной рото-
вой полости выпячивается вверх
энтодермальный гипофизарный
Рис. 192. Схемы эмбрионального раз-
вития гипофиза млекопитающего (Я)
и строения сформировавшегося гипо-
физа (Б):
А: а — выпячивание воронки промежу-
точного мозга; 1—6— изменение гипо-
физарного кармана.
Б: 1 — нейрогипофиз; 2 — центральная
полость; 3 — промежуточная часть; 4 ~~
дистальная часть; 5 — бугорная часть; 6 —
третий желудочек
* Предполагают, что передний конец нервной трубки ланцетника содержит
секреторные клетки, близкие по организации к первичной системе гипоталамУ'
са. Их секрет стимулирует работу репродуктивной системы.
244
карман. Во взрослом состоянии жи-
вотного он отшнуровывается от
этой полости. Из нервной ткани во-
ронки формируется задняя доля ги-
пофиза — нейрогипофиз. Внутри него
есть полость — отрог третьего моз-
гового желудочка. В свою очередь,
из гипофизарного кармана форми-
руется большая и более гормональ-
но значимая область — передняя
доля гипофиза — железистый, или
аденогипофиз (рис. 193).
Рис. 193. Схема строения
Экспериментально показано, гипофиза рыбы:
ЧТО гормоны, накапливающиеся В у—?—разные области аденоги-
нейрогипофизе, вырабатываются пофиза; 4 — нейрогипофиз
ядрами гипоталамуса. Тела клеток
гипоталамуса обладают секреторной активностью, а их аксоны
передают гормоны в нейрогипофиз. Наиболее четко, как компак-
тное тело, нейрогипофиз оформлен у млекопитающих. Гормоны
нейрогипофиза — вазопрессин и окситоцин. Вазопрессин, или АДГ
(антидиуретический гормон), стимулирует реабсорбцию воды в
почках и регулирует водный баланс тела животного. Окситоцин
влияет на процессы, связанные с размножением: он стимулирует
сокращение мускулатуры матки и выделение молока.
Аденогипофиз состоит из железистых клеток и дифференци-
рован на три части: внешнюю — дистальную часть; примыкаю-
щую к нейрогипофизу промежуточную часть и участок, охватыва-
ющий муфтой стебелек воронки, — бугорную часть. Железистые
клетки содержатся в дистальной и промежуточной частях адено-
гипофиза. Бугорная часть не имеет секреторных клеток. Аденоги-
пофиз вырабатывает большинство гормонов гипофиза: гормон ро-
ста стимулирует рост скелета и мускулатуры, регулирует обмен
жиров и углеводов; пролактин стимулирует у млекопитающих сек-
рецию молока, у голубей — птичьего «зобного молочка»; гона-
дотропные гормоны регулируют функции яичников и семенни-
ков; тиреотропный (ТТГ) и аденокортикотропный (АКТГ) гормо-
ны стимулируют работу щитовидной железы и надпочечников. Де-
ятельность аденогипофиза регулируется со стороны мозга через
воротную систему кровообращения, расположенную на его гра-
нице с нейрогипофизом.
Надпочечные железы (gl. suprarenalis). Парные надпочечные
Железы находятся у рептилий, птиц, млекопитающих обычно у-
переднего верхнего края почки. У рыб и амфибий их участки раз-
бросаны вдоль почек (рис. 194).
Эти железы имеют сложное происхождение. Они возникают в
эмбриогенезе как производные зачатков симпатических узлов ве-
245
Рис. 194. Расположение надпочечников и надпочечной ткани у позво-
ночных:
А — пластиножаберная рыба; Б — костистая рыба; В— хвостатая амфибия; Г —
ящерица; Д — птица; Е — млекопитающее; 1 — мозговое вещество надпочечни-
ков; 2 — корковое вещество надпочечников; 3 — тазовая почка; 4 — туловищная
почка; 5— надпочечники; 6 — надпочечная ткань; 7— задняя кардинальная вена;
8 — диффузная ткань; 9 — межпочечная железа; 10 — «головная» почка; 11 —
тельца Станниуса
гетативной нервной системы, отчего у низших позвоночных со-
храняют диффузное состояние. У амниота эта ткань становится
впоследствии мозговым веществом компактных тел — надпочечни-
ков. Корковое вещество тел надпочечников закладывается эмбрио-
нально в мезодерме межпочечного участка. Мозговое вещество над-
почечников выделяет в кровь два гормона — адреналин и нор-
адреналин. Эти гормоны свойственны и волокнам симпатической
нервной системы. Они регулируют ширину просвета кровеносных
сосудов и участвуют в углеводном обмене и выделении натрия.
Клетки коркового вещества секретируют стероидные гормоны
различного действия: глюкокортикоиды и минералокортикоиды. Они
участвуют в водносолевом обмене, стимулируют расщепление бел-
ков, накопление в крови глюкозы и т. д. По мнению ученых, гор-
моны коркового вещества надпочечников позволяют организму
выдерживать длительные внешние стрессы, а гормоны мозгового
вещества — выживать в критических ситуациях (А. Ромер, Т. Пар-
сонс, 1994).
Поджелудочная железа (gl. pancreas) принадлежит к группе
желез смешанной секреции. В эмбриогенезе она отпочковывается
от средней части кишечника. Большая часть ткани этой железы
вырабатывает пищеварительные ферменты, поступающие по про-
токам в начальный отдел кишечника (см. «Органы пищеварения»).
Однако у этой железы есть и функция эндокринного органа.
У большинства позвоночных в ткани железы островками рассея-
ны участки железистой ткани — островки Лангерганса (рис. 195).
У круглоротых и рыб и даже у некоторых млекопитающих они
разбросаны вдоль кишечника. Гормон железистых клеток инсулин
выделяется непосредственно в кровь. Он участвует в регуляции
углеводного обмена, снижая уровень глюкозы в крови и стимули-
руя синтез и накопление углеводов в печени. Наряду с инсулином
железистые клетки поджелудочной железы вырабатывают и дру-
гой гормон — глюкагон. Он способствует расщеплению запасенно-
го в печени гликогена и тем самым участвует в повышении уров-
ня глюкозы в крови. Нарушение деятельности поджелудочной
железы вызывает изменение углеводного обмена, что приводит к
сахарному диабету.
Рис. 195. Разрез поджелудочной железы:
1 — междольковая прослойка; 2 — ацинус (экзокринные участки) поджелудоч-
ной железы; 3 — островок Лангерганса (эндокринный участок); 4 — кровенос-
ные сосуды; 5 — междольковый проток
247
Половые железы (gonadas). Секреторная функция свойственна
также половым железам — гонадам. Эндокринная деятельность этих
желез играет основную роль в формировании вторичных половых
признаков, проявлениях полового диморфизма, в окраске тела
рыб, амфибий, рептилий, характере сезонного оперения птиц ц
волосяного покрова у млекопитающих, участвует в регуляции
процессов, связанных с размножением.
Ведущие половые гормоны — это вырабатываемый в семенни-
ках тестостерон, в яичниках — эстрадиол, прогестерон. Гонады
вырабатывают гормоны в значительной мере под влиянием гона-
дотропных гормонов аденогипофиза.
Гормоны половых желез по химической структуре — это сте-
роидные гормоны, сходные с гормонами коры надпочечников. Их
сходство объясняют общностью эмбриональной закладки этих ор-
ганов из мезодермы брюшной полости.
В теле позвоночных животных гормоны выделяются не только
специфическими эндокринными железами. Показано, что гормон
печени соматомедин регулирует действие гормона роста, выделя-
емого аденогипофизом. Гормон стенки желудка гастрин стимули-
рует секрецию желудочного сока. Наконец, ряд гормонов выраба-
тывается клетками двенадцатиперстной кишки: секретин — сти-
мулирует секрецию пищеварительного сока поджелудочной же-
лезы; холецистокинин стимулирует выделение желчи из желчного
пузыря печени; энтерогастрон тормозит желудочную секрецию
(К. Шмидт-Ниельсен, 1982).
* * *
В целом железы внутренней секреции вместе с нервной систе-
мой осуществляют нейрогуморальную регуляцию всех функций
живого организма. Механизмы гуморальной регуляции протекают
по времени значительно дольше, чем нервные, но отличаются
многофункциональностью и разнонаправленностью.
ОРГАНЫ ВЫДЕЛЕНИЯ
Тело животного в процессе жизнедеятельности освобождается
от конечных продуктов обмена. Пищеварительная система выво-
дит непереваренные органические остатки пищи, через органы
дыхания удаляется углекислый газ. Органы выделения служат для
выведения избытка воды и растворенных в ней солей, а также
токсичных биологически активных веществ, в том числе и азоти-
стых соединений, образовавшихся в процессе белкового метабо-
лизма или поступивших с пищей. У низших водных позвоночных
функцию выделения частично выполняют жабры и покровы тела,
у рептилий и птиц — слезные железы, у млекопитающих — пото-
вые железы. Кроме выведения воды органам выделения принадле-
жит важнейшая роль в поддержании постоянства химического
состава жидкостей внутренней среды — необходимого условия
эффективной деятельности всех органов и систем. В свою очередь,
постоянство внутренней среды (гомеостаз) обеспечивает организму
животного свободу и независимость существования в непрерывно
меняющихся условиях среды.
Наряду с выделительной функцией почки регулируют артери-
альное давление, активизируют действие витамина D, участвую-
щего в кальциевом обмене и т. д.
Эволюция органов выделения. Структурной единицей органов
выделения многих беспозвоночных животных являются нефридии —
канальцы. В простейшем случае у кольчатых червей они сообща-
ются одним концом с полостью тела — целомом, а другим — с
внешней средой. Организация и функционирование выделитель-
ной системы у наиболее примитивных хордовых — ланцетников —
близка к кольчатым червям (рис. 196). Нефридиальные канальцы
расположены у них метамерно в жаберной области, вблизи жа-
берных щелей. Расширенный конец каждого канальца (нефростом
с клетками соленоцитами) открывается многочисленными отвер-
стиями в целом, а выводное отверстие — в околожаберную по-
лость. Выводимые продукты азотистого обмена у ланцетника —
аммиак и мочевина.
Некоторым хордовым — оболочникам, пресмыкающимся, как
и многим беспозвоночным (нематодам, ракообразным, паукооб-
разным, многоножкам, насекомым), свойственна выделительная
система накопительного типа. Например, у оболочников, веду-
749
Рис. 196. Нефридии ланцетника:
А — целый каналец с многими нефр0_
стомами и соленоцитами; Б — часть
стенки выделительного канальца с си-
дящими на ней соленоцитами; 7 — вер-
хний конец жаберной щели; 2 — отвер-
стие почечного канальца в околожа-
берную полость
щих одиночный образ жизни, на внутренней поверхности мантии
образуются мелкие вздутия — почечные пузырьки. Это место по-
жизненного накопления мочевой кислоты, так как во внешнюю
среду она не выделяется (у колониальных форм оболочников че-
рез жаберные отверстия выводится аммиак). У многих рептилий
конечные продукты обмена накапливаются в покровах и сбрасы-
ваются при линьке.
Выделительная система позвоночных организована в специаль-
ные органы — почки. Они лежат в полости тела и снабжены выво-
дящими протоками.
В эмбриогенезе почки формируются в мезодерме зародыша:
нефрогенная ткань образуется на границе сомита и боковой пла-
стинки. Там же закладываются и органы размножения. В ходе онто-
и филогенеза происходит параллельное развитие, взаимосвязь и
взаимообслуживание выделительной и репродуктивной систем,
поэтому особенности их организации и функционирования в раз-
ных группах позвоночных рассматриваются вместе.
В филогенезе и онтогенезе позвоночных прослеживается по-
следовательная смена трех типов выделительных органов: пред-
почка, или головная, почка — пронефрос (pronephros), первичная,
или туловищная, почка — мезонефрос (mesonephros) и вторичная,
или тазовая, почка — метанефрос (metanephros) (рис. 197).
У зародышей низших позвоночных (круглоротых,
рыб, амфибий) в начале эмбрионального развития из нефроген-
ной ткани закладывается система выделительных канальцев, кото-
рые, как и у взрослого ланцетника, открываются воронками —
нефростомами — в полость тела и служат для непосредственного
транспорта продуктов выделения во внешнюю среду. Вблизи во-
ронок у них обнаружены микроскопические вздутия со сплетени-
ем кровеносных сосудов — сосудистые клубочки. Они участвуют в
250
Рис. 197. Схема последовательного расположения
генераций почек:
у — спинная аорта; 2 — внешний мальпигиев клубо-
чек; 3 — воронки канальцев головной и туловищной
почек; 4 — мальпигиев клубочек в боуменовой капсу-
ле; 5— задняя кишка; 6— мюллеров канал; 7— голов-
ная почка; 8 — туловищная почка; 9 — тазовая почка;
10 — проток туловищной почки (вольфов канал); 11 —
вторичный мочеточник; 12 — клоака
транспорте продуктов выделения путем их
фильтрации из крови. Противоположные кон-
цы выделительных трубочек постепенно сли-
ваются в общий выводящий проток. Такой тип
выделительного органа носит название голов-
ной почки, или предпочки (pronephros). Ее вы-
водной проток называют пронефрическим ка-
налом. Пронефрос можно назвать почкой лишь
условно. Головная почка никогда не бывает
компактным органом, а представляет собой
совокупность канальцев. Важность ее обязатель-
ной закладки состоит именно в непременном
формировании пронефрического канала, пред-
шественника вольфова и мюллерова каналов.
У взрослых круглоротых (миног) головная почка со-
храняется, а пронефрические воронки сообщаются у них с око-
лосердечной полостью. У большинства взрослых рыбиам-
ф и б и й личиночная головная почка исчезает*.
На последующих стадиях онтогенеза у низших позвоноч-
ных позади пронефрических (головных) закладываются компак-
тные — первичные, или туловищные (мезонефрические), почки. Они
и становятся настоящими органами выделения взрослых круг-
лоротых, рыб и амфибий. Частично в мезонефрических
Почках сохраняется связь с полостью тела через нефростомы. Од-
новременно их выделительные канальцы получают продукты, под-
лежащие выведению из организма, непосредственно из крови.
Кровеносные сосуды в почках образуют сложные сплетения —
сосудистые клубочки. Они располагаются в воронковидных вырос-
тах выделительных канальцев — боуменовых капсулах. Сочетание
* У некоторых рыб и у миксин она сохраняется в течение всей жизни в виде
скопления лимфоидной ткани (А.Ромер, Т. Парсонс, 1994).
251
сосудистого клубка и боуменовой капсулы называют мальпигие-
вым тельцем (рис. 198). Диаметр артерии, входящей в капсулу
больше диаметра выходящей артерии. За счет возникающей при
этом разницы давлений кровь фильтруется через стенки капилля-
ров в боуменову капсулу и образует первичную мочу. Строение не-
фронов у разных видов рыб и амфибий отличается. У одних более
примитивных наряду с мальпигиевым тельцем сохраняется не-
фростома, а через нее связь с целомом. У других канальцы начи-
наются боуменовыми капсулами и выделение идет только из кро-
ви.
При завершении формирования туловищной почки у зароды-
шей рыб и амфибий пронефрическая почка редуцируется, а про-
нефрический канал расщепляется на два. С одним их них продол-
жают сообщаться выделительные канальцы мезонефрической поч-
ки. Он функционирует как мочеточник и именуется вольфовым
каналом. Развитие другой части пронефрического канала зависит
от половой принадлежности. У самцов она исчезает, у самок же
она функционирует как яйцевод и именуется мюллеровым каналом.
Таким образом, выводной канал, исходно принадлежавший сис-
теме выделительной, становится у самок функциональным орга-
ном системы репродуктивной, а пронефрическая воронка стано-
вится приемником яйцеклетки.
Рис. 198. Схема строения нефронов туловищной почки:
А — с нефростомом; Б — без нефростома; В — кровеносные сосуды; 1 — боуме
нова капсула; 2 — сосудистый клубочек; 3 — мальпигиево тельце; 4 — нефрос
том; 5 — почечный каналец; 6 — собирательная трубочка
252
Представления об участии вольфова канала в выведении про-
дуктов выделения у особей мужского пола в различных система-
тических группах позвоночных в последнее время значительно
расширились (рис. 199). У самцов круглоротых вольфовы каналы
обслуживают только выделительную систему и с гонадами не свя-
заны. У хрящевых рыб иначе: у мужских особей всех акул и скатов
через вольфов канал выводятся в клоаку только продукты раз-
множения. Мочу из каждой почки собирает веер тонких трубочек,
объединяющийся на краю почки в общий канал — дополнитель-
ный выделительный проток — мочеточник (А. Ромер, Т. Парсонс,
1994). У самцов двоякодышащих и некоторых ганоидных рыб —
протоптеров, лепидосиренов, многоперов — вольфов канал слу-
жит мочеточником, а в заднем нижнем своем конце принимает и
продукты размножения. У самцов костистых рыб вольфов канал
принадлежит только выделительной системе.
По-разному выводится моча и у самцов амфибий. Например, у
жабы-повитухи и обыкновенного тритона мочу из почки выводит
дополнительный выводной проток — собственно мочеточник, а
вольфов канал служит семяпроводом. Однако у самцов других ви-
дов хвостатых и бесхвостых амфибий вольфов канал выводит и
мочу, и половые продукты. У самок пути репродуктивной и выде-
лительной систем всегда разделены.
Из сказанного выше следует, что ведущей линией эволюции
мезонефрической почки в разных группах первичноводных позво-
ночных стала концентрация выделительной функции в ее нижней
каудальной части. Эта особенность, видимо, реализовалась в даль-
нейшем в закладке следующего типа почек — вторичных, или тазо-
А Б В Г Д
Рис. 199. Соотношение выделительных и половых протоков у самцов
позвоночных:
А — круглоротые; Б — осетровые, панцирники; В — акулы, хвостатые амфибии;
Г — костистые рыбы; Д — рептилии (амниота); / — семенник; 2 — почка; 3 —
вольфов канал; 4 — семенной канал; 5 — канал-семяпровод; б — почка
253
вых (metanephros). Вторичные почки характерны для всех Anmiota —
настоящих наземных позвоночных.
У зародышей рептилий, птиц и млекопитаю-
щих из нефрогенной ткани последовательно формируются три типа
почек. Предпочка {пронефрос) выводит мочу через пронефрический
канал из тела зародыша в самом раннем онтогенезе. Первичная
почка {мезонефрос) — это орган выделения, функционирующий
на протяжении их позднего зародышевого и раннего постнаталь-
ного состояния. Во взрослом состоянии мезонефрические почки
практически исчезают. Их передняя часть сохраняется только у
самцов в виде придатков семенников, а вольфовы каналы превра-
щаются в семяпроводы.
Тазовые {метанефрические) почки формируются в результате
сильного разрастания почечной ткани в каудальном отделе туло-
вища, где они превращаются в парные компактные утолщенные
тела. В поясничном отделе они вдаются в брюшную полость. Мочу
из метанефрической почки выводит самостоятельный канал —
мочеточник. В онтогенезе он отделяется от вольфова канала.
У рептилий тазовые почки состоят из множества мелких
долек. Отходящие от каждой дольки почечные канальцы — соби-
рательные трубочки — выносят мочу в многочисленные ветви мо-
четочников, которые сливаются в единый канал. Количество со-
бирательных канальцев в почках разных видов ящериц колеблется
от 3 до 30 тыс.
У птиц почки крупные: на их долю приходится около 2 %
массы тела. Они распадаются на крупные и более мелкие дольки.
Собирательные канальцы в почках птиц еще более многочислен-
ны, чем у рептилий: например, у домашней курицы их число
достигает 20 тыс.
Почки млекопитающих — это компактные тела бобовид-
ной или дольчатой формы. Дольчатая форма почек свойственна ко-
пытным, хищным, слонам, китам. Число долек в почках крупных
китов доходит до нескольких тысяч. Каждая долька сообщается с
мочеточником самостоятельным канальцем. Мочеточник образует
расширение — почечную лоханку. Она подразделена в почке на уг-
лубления — почечные чашечки. К каждой чашечке из ткани почки
сходятся собирательные трубочки, образуя почечные пирамиды.
На рис. 200 показано строение бобовидной почки человека. На
срезе четко различимы два слоя. Наружный корковый слой включа-
ет почечные {мальпигиевы) тельца и отходящие от них извитые
канальцы. Внутренний мозговой слой содержит почечные пирамиды
и состоит из собирательных трубочек (канальцев). Количество ка-
нальцев в почках у человека исчисляется несколькими миллиона-
ми (к примеру, у мыши их около 20 тыс.).
Еще один орган выделительной системы — мочевой пузырь. Он
по-разному закладывается у позвоночных. Например, у самок хря-
254
Моча
Рис. 200. Разрез через почку млекопитающего (Л) и схема строения
почки (Б):
1 — почечные пирамиды; 2 — корковый слой; 3 — почечная лоханка; 4 — мо-
четочник; 5 — почечная артерия; 6 — почечная вена; 7 — боуменова капсула; 8,
9 — проксимальный и дистальный извитые канальцы соответственно; 10 —
петля Генле; 11 — собирательная трубка
щевых, двоякодышащих и ганоидных рыб — это расширение сли-
вающихся в каудальной части вольфовых каналов; у самцов —
расширения пары выводящих протоков («мочеточников»). У ми-
ног и костных рыб мочевой пузырь формируется как вырост заро-
дышевой клоаки, отсутствующей у взрослых особей. Объемистый
мочевой пузырь амфибий представляет собой вырост стенки кло-
аки. Предполагают, что его стенка отчасти может осуществлять у
них обратное всасывание воды. У рептилий, птиц, млекопитаю-
щих в эмбриогенезе функцию мочевого пузыря выполняет аллан-
тоис, который образуется как вырост задней части первичной
кишки зародыша. К моменту рождения аллантоис впячивается в
тело и из него формируется настоящий мочевой пузырь. У многих
рептилий (ящериц, змей, крокодилов) мочевой пузырь вторично
утрачен. Отсутствует он и у летающих птиц (есть у нелетающей
птицы — африканского страуса). У яйцекладущих млекопитающих
мочевой пузырь — это часть клоаки. Настоящие млекопитающие
не имеют клоаки, мочевой пузырь связан у них с нижними отде-
лами мочеточников.
255
Механизм образования и выделения мочи. Основная работающая
структура почки позвоночных, как сказано выше, нефрон (у низ-
ших — нефридиальная трубочка), или почечный каналец. В про-
стейшем случае — в пронефрической почке — он выделяет про-
дукты обмена непосредственно из полости тела. В мезонефричес-
кой (туловищной) и метанефрической (тазовой) почках силой
кровяного давления из клубочков в капсулы фильтруется плазма
крови с растворенными в ней солями, сахарами, витаминами,
аминокислотами {первичная моча).
Клубочковая фильтрация обеспечивает интенсивное выведение
воды из тела. Например, пресноводные рыбы и круглоротые вы-
деляют в сутки более 300 мл/кг первичной слабоконцентрирован-
ной мочи. У млекопитающих скорость клубочковой фильтрации в
10—100 раз выше.
Кровоснабжение почки осуществляет почечная артерия. Почеч-
ные канальцы оплетены снаружи кровеносными сосудами. У мор-
ских и проходных рыб, у земноводных, пресмыкающихся (и не-
которых птиц) в оплетении почечных канальцев участвуют и ве-
нозные сосуды воротной системы почек (ренопортальные вены), что
создает условия для быстрого выведения продуктов обмена у жи-
вотных с относительно невысоким уровнем минутного объема
сердца и интенсивности артериального снабжения почки. Кровь
от почек оттекает у низших позвоночных по почечным венам в
кардинальные вены, у высших позвоночных — в заднюю полую
вену. На рис. 201 показано кровоснабжение нефрона в туловищ-
ной почке при наличии воротного кровообращения. Почечная ар-
териола, выйдя из боуменовой капсулы, объединяется с веноз-
ным капилляром, отделившимся от воротной вены почки, и оп-
Рис. 201. Кровоснабжение нефрона в туловищной почке:
1 — сосуд от почечной артерии; 2 — боуменова капсула; 3 — клубочек; 4 —
извитой каналец; 5 — сосуд от воротной вены почки; б — сосуд к почечной вене
Рис. 202. Кровоснабжение нефрона
в тазовой почке:
1 — почечная артерия; 2 — почечная вена; 3 -
боуменова капсула; 4 — сосудистый клубочек
5 — проксимальный отдел канальца; 6 — ди
стальный отдел канальца; 7 — петля Генле; 8 -
собирательный каналец
Почечная лоханка
летает нефрон. У большинства птиц и
у всех млекопитающих такой тип во-
ротной системы отсутствует и каналь-
цы оплетены только артериальными
сосудами (рис. 202). Через стенку по-
чечного канальца осуществляется
обратное всасывание (реабсорбция).
В кровеносную сеть из первичной мочи
уходят вещества, нежелательные для
потери: вода, сахара, аминокислоты.
Кроме того, стенка почечного каналь-
ца обладает секретирующей функци-
ей. Ее клетки дополнительно извлекают из крови и выделяют в
него вещества, подлежащие удалению. Таким способом в просвет
канальца поступает мочевина (или мочевая кислота). Образовавша-
яся на выходе из почки вторичная моча выводится из организма.
Она представляет собой концентрированный раствор солей и мо-
чевины (мочевой кислоты).
Система органов выделения необходима и для поддержания
постоянства внутренней среды организма, которое осуществляет-
ся путем водно-солевого обмена.
Водно-солевой обмен. Постоянство осмотической концентрации,
ионного состава, кислотно-щелочного равновесия и объема жид-
костей внутренней среды регулируется механизмами водно-соле-
вого обмена. Выяснено, что проницаемость кожи, жабр и почек
по отношению к воде и растворенным в ней солям регулируется
различными гормонами гипофиза (Л. Проссер, 1977).
Организация водно-солевого обмена в разных классах и эколо-
гических группах позвоночных имеет свою специфику. Для живот-
ных, перешедших к жизни в пресных водоемах и на суше, а также
У мигрировавших из пресных вод снова в море или ставших после
Длительного обитания на суше вторичноводными, механизмы ос-
морегуляции решаются в соответствии с требованиями среды.
Пресноводные виды рыб живут в гипотонической сре-
де. Осмотическое давление в их тканях выше осмотического давле-
ния Окружающей пресной воды, поэтому вода по осмотическому
257
градиенту проникает в их тело через кожу, жабры, эпителий ро-
товой полости, слизистую кишечника. Чтобы не допускать обвод-
нения тела, почки активно выводят избыток воды наружу: пример-
но 300 мл на 1 кг веса в сутки. При этом потери солей почками
компенсируются притоком их с пищей, а также через солевые клет-
ки, находящиеся в жабрах, которые способны к абсорбции одно-
валентных ионов (Na+ и СГ). Отдельные виды пресноводных рыб
способны осваивать осолоненные водоемы благодаря взаимодей-
ствию почек и абсорбционной активности жаберного эпителия. Икра
пресноводных рыб тоже гиперосмотична по отношению к среде,
поэтому в течение первых же часов после откладывания в воду обо-
лочки икринок быстро набухают и теряют проницаемость. Пресно-
водные миноги имеют осморегуляцию, сходную с описанной выше.
У первичноводных морских обитателей механиз-
мы стабилизации жидкостной внутренней среды сложились по-
своему. Ткани тела миксин имеют концентрацию жидкостей, сход-
ную с морской водой.
Морские костистые рыбы живут в гипертоническом
растворе, извлекают из него кислород и используют его для питья.
Предположительно, тела морских рыб должны были бы терять
воду, однако этому препятствует ряд функциональных особенно-
стей. В их почках уменьшено число сосудистых клубочков (агломе-
рулярные почки). Кроме того, вода из нефронов активно реабсор-
бируется в кровь, что позволяет значительно снизить количество
выделяемой воды (до 20 мл на 1 кг массы в сутки). Избыток солей
у морских костистых рыб выводится с экскрементами, выделяет-
ся почками и солевыми клетками жаберного эпителия. В отличие
от пресноводных рыб меняется функциональная направленность
работы этих клеток: абсорбционная активность сменяется удале-
нием из тела рыбы одновалентных ионов (К. Шмидт-Ниельсен,
1982).
Продукты азотистого обмена у костных рыб активно выводятся
через жабры (аммиак) и почки (мочевина).
Осморегуляция морских хрящевых рыб складывается по-иному.
Концентрация солей в их теле несколько выше, чем в окружаю-
щей воде, однако осмотическое давление внутренней среды орга-
низма оказывается почти равным внешнему, так как кровь и тка-
невая жидкость хрящевых рыб содержат большое количество со-
лей. Естественным компонентом жидкостей тела акуловых рыб яв-
ляется мочевина. Ее содержание в крови в 100 раз выше, чем У
млекопитающих. Мочевина удерживается в тканях тела акуловых
рыб вследствие непроницаемости для нее кожи и эпителия жа-
берных лепестков. Из организма она удаляется вместе с избытком
воды и солей через почки в составе вторичной мочи. Строение
нефронов и оплетающих их кровеносных капилляров позволяет
эффективно реабсорбировать мочевину, выравнивая за счет этого
258
осмотическое давление в теле хрящевых рыб с морской водой.
Таким образом, накопление в крови мочевины позволило хряще-
вым рыбам решать проблему обитания в соленой воде иначе, чем
морским костистым рыбам. Накопление мочевины в теле отмече-
но также у современных кистеперых и двоякодышащих рыб.
Дополнительным органом выведения солей из организма хря-
щевых рыб и кистеперой рыбы — латимерии является ректальная
железа. Ее выделения изоосмотичны по отношению к крови и со-
держат значительное количество хлорида натрия.
У первичноводных древних бесчелюстных (щит-
ковых, панцирных рыб) и у современных хрящевых и костных
рыб проблему содержания воды в организме ограничили (наряду
с механизмами выделительной системы) мощные костные по-
кровы в виде панцирей или чешуи.
Механизм водно-солевого обмена у земноводных в целом
близок к тому, который описан выше для пресноводных рыб. Ам-
фибии, как и рыбы, воду почти не пьют. Она практически бес-
препятственно поступает через покровы («лягушка пьет воду ко-
жей»). Избыток воды и растворенную в ней мочевину амфибии
выводят через почки. Темпы клубочковой фильтрации у них дос-
таточно высоки: IO-— 25 мл на 1 кг массы в час. Земноводные вы-
деляют очень жидкую мочу со слабой концентрацией солей. Моча
в теле некоторых лягушек составляет до 30 % общей массы тела.
Препятствием к потере солей у амфибий служит реабсорбция ионов
в почечных канальцах: более 90 % солей, поступивших в первич-
ную мочу, возвращается (реабсорбируется) обратно в кровенос-
ное русло. Реабсорбция мочевины осуществляется стенкой моче-
вого пузыря. У пустынных видов вода может накапливаться в мо-
чевом пузыре.
Однако среди земноводных есть и исключения. Например, в
коже южноамериканских филломедуз (Phyllomedusae) имеются
специфические железы, выделяющие на поверхность восковид-
ный секрет. Это делает кожу водонепроницаемой. Лягушки р. Хи-
романтис (Chiromantls) выделяют через почки не мочевину, а
мочевую кислоту. Наряду с уменьшением испарения с покровов
это сближает их с рептилиями. Обитающая в мангровых зарослях
морского побережья в Юго-Восточной Азии крабоядная лягушка
{Rana cancrivora) в поисках пищи плавает в морской воде. Осмо-
тической потере воды препятствует у нее накопление в жидкостях
тела мочевины, как у хрящевых рыб.
У хвостатых амфибий сохраняется фильтрация жидкости из
Полости тела через нефростомы: например, у тритонов таким пу-
тем выводится до 22 % всего объема мочи (С. И. Левушкин,
И. А. Шилов, 1994).
Переход позвоночных к наземному образу жизни сопровож-
дался приобретением плотных покровов с многослойным эпидер-
мисом, однако исследования показали, что они не всегда эффек_
тивно защищают тело от испарения воды. Значительно более важ-
ным оказалось приобретение выделительной системы функцио-
нально нового типа — тазовых почек. В них уменьшается способ-
ность клубочков к первичной фильтрации плазмы и усиливается
реабсорбция воды в почечных канальцах. Например, у рептилий
структура сосудистого клубка упрощается, число капиллярных
петель сокращается до 2 — 3. Но при этом усиливается извилис-
тость почечных канальцев. В результате этих преобразований до
95 % первичного фильтрата возвращается обратно.
У всех настоящих наземных позвоночных продукты азотистого
обмена выделяются через почки. Урептилий и птиц продук-
том выделения является мочевая кислота. Такой тип выделения
сложился в условиях дефицита воды. Мочевая кислота имеет ка-
шицеобразную структуру, что значительно уменьшает объем вы-
водимой воды. Осаждению кристаллов мочевой кислоты в выде-
лительных путях препятствует большое количество слизи, выде-
ляемой клетками почечных канальцев и мочеточников*. Выдели-
тельные системы птиц и рептилий в значительной степени близ-
ки. Почечные канальцы разнообразны: с короткой петлей, с длин-
ной петлей. Мочевая кислота попадает через мочеточники в клоа-
ку и вместе с фекальными массами выбрасывается наружу. Высох-
шие экскременты некоторых морских птиц (гуано) содержат до
25 % мочевой кислоты.
Предполагают, что мочекислотный тип выделения развился в
связи с размножением путем откладки яиц с системой яйцевых и
зародышевых оболочек: накопление в аллантоисе токсичного ам-
миака в виде мочевины вызывало бы отравление организма. Мо-
чевая кислота, выпадающая в осадок, для него менее токсична.
Экспериментально показано, что при развитии птенца в яйце до
середины инкубационного периода образуются аммиак и мочеви-
на, а затем усиливается отложение мочевой кислоты: например, в
курином яйце к концу зародышевого развития количество выде-
ляемого азота достигает 40 мг в день: 23 % выделяется в виде ам-
миака и мочевины, 77 % — в виде мочевой кислоты.
Наземные позвоночные: рептилии, птицы и млекопитающие
производят гиперосмотическую мочу, т. е. с большей концентра-
цией солей и органических веществ, чем жидкости тела. У птиц
концентрация солей в моче в 2 раза, а у млекопитающих в 25 раз
выше их концентрации в крови. Механизм образования концент-
рированной мочи основан на реабсорбции воды из канальцевой
жидкости. Это оказалось возможным в результате того, что выде-
лительные канальцы у птиц и млекопитающих резко удлиняются
* Мочевая кислота выделяется также некоторыми амфибиями родов Chiromantus,
Phyllomedusae (К.Шмидт-Ниельсен, 1982).
260
за счет образования петель Генле, образующих мозговой слой почки
(в целом у птиц их меньше). У рыб, амфибий и рептилий этих
петель нет, поэтому они не могут вырабатывать мочу более высо-
кой концентрации, чем плазма крови. У птиц и млекопитающих,
обитающих в аридных зонах, есть дополнительные механизмы удер-
жания воды в организме, — это и откладывание в запас жировой
ткани, и увеличение количества длиннопетельных почечных ка-
нальцев, и усиление всасывания в них воды, и, наконец, способ-
ность к длительному воздержанию без воды (торможение «чувства
жажды»).
Выведение солей большинством птиц осуществляется через
почки. Однако есть и другие способы. Так, у обитателей морс-
ких побережий (бакланов, пеликанов, альбатросов, некоторых
чаек, чистиков) в черепе над глазницами хорошо развиты со-
левые носовые железы — их соленый секрет каплями стекает с
клюва. Солевые железы специализированы на выведении ионов
натрия и хлора. Такие же железы выводят соли из тела кроко-
дилов, черепах и некоторых других рептилий («крокодиловы
слезы»).
Моча, выводимая почками млекопитающих, содержит
соли и растворимую в воде мочевину. В их почках максимального
развития достигает мозговое вещество, содержащее длиннопе-
тельные канальцы. Замечено, что у человека при нарушениях
водно-солевого обмена возможно отложение мочевой кислоты в
почках в виде камней и накопление ее в суставах. Вода и некото-
рые соли выводятся у млекопитающих также с потом через по-
кровы тела.
В целом механизмы поддержания постоянства внутренней сре-
ды организма (гомеостаз) у птиц и млекопитающих выражены
наиболее совершенно. Они обеспечивают слабую изменчивость
объема крови и других внеклеточных жидкостей, концентрации в
них осмотически активных веществ (изоосмия), pH крови и со-
става в ней белков, углеводов и липидов. В относительно узких
пределах меняется и температура их тела (изотермия). Дополни-
тельные механизмы обеспечивают устойчивость в состоянии и
свойствах жидкостей, окружающих жизненно важные органы —
головной и спинной мозг, сердце, легкие, глаза и т.д., регулиру-
ют артериальное давление. Все это способствует расширению при-
способительных возможностей птиц и зверей к жизни в меняю-
щихся условиях внешней среды.
* * *
Таким образом, мезодермально развившаяся система органов
выделения удаляет из организма вредные продукты белкового об-
мена: аммиак, мочевину, мочевую кислоту и обеспечивают вод-
но-солевой баланс. Исходной структурной единицей выделитель-
261
ной системы являются нефридиальные канальцы, явственно офор-
мленные у ланцетника. Они составляют основу наиболее прими-
тивной почки позвоночных — предпочки. В эволюционном ряду
позвоночных в выделительной системе последовательно сменя-
ются пронефрическая, головная почка (предпочка), первичная,
или туловищная (мезонефрическая), и наконец, вторичная, или
тазовая (метанефрическая) почка. При этом уменьшается роль
нефростом, выделяющих продукты обмена из целомических по-
лостей. У настоящих наземных позвоночных (амниота) фильтра-
ция мочи из крови полностью происходит в мальпигиевых тель-
цах. Возрастает роль извитых канальцев, обеспечивающих реаб-
сорбцию воды и полезных веществ из первичной мочи и секре-
цию продуктов азотистого обмена, что окончательно формирует
вторичную мочу. У первичноводных позвоночных в моче из про-
дуктов метаболизма преобладает аммиак, у амфибий и млекопи-
тающих — мочевина, у рептилий и птиц — мочевая кислота, от-
ражающие возрастающую необходимость экономии воды в телах
представителей указанных групп.
РЕПРОДУКТИВНАЯ СИСТЕМА
Половое размножение — характерная черта хордовых. Иные
формы репродукции (партеногенез, гиногенез) редки.
Система органов размножения включает половые железы, или
гонады, половые протоки, совокупительные органы и сопутствую-
щие железы.
В раннем эмбриогенезе хордового животного гонады заклады-
ваются как выпячивания участков стенок полости тела, соответ-
ствующих гонотому, — месту подразделения сомитов и боковой
пластинки. Парные продольные утолщения — половые, или гени-
тальные, складки — со временем погружаются в полость тела и
оказываются подвешенными на брыжейке, располагаясь медиаль-
нее (по бокам) почек. С поверхности эти складки образованы утол-
щенным энтодермальным эпителием, а изнутри имеют пласт со-
единительной ткани. Поначалу вытянутые, они впоследствии уко-
рачиваются и приобретают компактную форму.
В начале эмбриогенеза гонады некоторое время пребывают в
индифферентной стадии и не проявляют принадлежности к полу.
Разделение по полу оформляется по ходу эмбрионального разви-
тия. К концу эмбриогенеза гонада оформляется как яичник или
семенник. Семенники по форме, как правило, более компактные
образования, чем яичники.
Первичные половые клетки гоноциты закладываются у эмбри-
онов рано и вне генитальных складок. На стадии дифференциров-
ки зародышевых листков они обнаруживаются в выстилке энто-
дермы первичной кишки и желточного мешка и гистологически
непохожи на обычные клетки пищеварительного тракта. Затем либо
с кровотоком, либо путем амебоидных движений они проникают
в целомический эпителий генитальных складок. На следующем
этапе целомический эпителий с запасом полученных клеток вра-
стает в соединительнотканную строму складок будущей гонады в
виде многоклеточных шнуров — первичных половых тяжей. Их пос-
ледующая судьба различна у разных полов. У самцов первичные
половые тяжи разбиваются на фолликулы и становятся репродук-
тивной тканью (рис. 203), у самок же первичные половые тяжи
дегенерируют. Репродуктивной тканью у них становятся вторич-
ные половые тяжи, врастающие в тело гонады из зачаткового эпи-
телия.
263
(вторичные половые тяжи)
Рис. 203. Схема развития семенника и яичника млекопитающего
(по А. Ромеру, Т. Парсонсу):
А—Б — общие стадии; Б — формирование семенника; Г — формирование
яичника
Последующая судьба гоноцитов расценивается специалистами
по-разному. Одни считают, что основная масса гоноцитов, попав
в гонаду, дегенерирует. Большинство других полагают, что пер-
вичные половые клетки дают начало будущим овоцитам и спер-
матоцитам. Наконец, по мнению третьих, функциональные по-
ловые клетки взрослого животного формируются прижизненно
из мезодермального эпителия гонады, первичные же половые
клетки инициируют и стимулируют образование гамет (А. Ромер,
Т. Парсонс, 1992).
Развитие половых желез у позвоночных
Самцы. В гонадах самцов первичноводных позвоночных (анам-
ний) расположенная внутри генитальных складок соединитель-
нотканная мезенхима преобразуется в интерстициальную ткань, у
амниот (рептилии, птицы, млекопитающие) она образует сеть
764
семенника (rete testis). Оформившаяся в первичных половых тяжах
толщи семенника репродуктивная ткань разделяется на многочис-
ленные пустотелые структуры. У рыб и амфибий это семенные пу-
зырьки, а у амниот — семенные канальцы. Эти образования выстла-
ны спермиогенным эпителием, в сезон размножения он выраба-
тывает половые клетки и содержит опорные клетки {клетки Сер-
толи) (рис. 204).
Семенные пузырьки низших позвоночных напоминают фолли-
кулы яичника. К сезону размножения в части пузырьков созрева-
ют половые клетки: высвободив сперматозоиды, они рассасыва-
ются. Вслед затем активизируется другая часть пузырьков, спер-
матогенез в которых был замедлен, и продуцируется следующая
порция половых клеток.
Семенные канальцы высших позвоночных в отличие от семен-
ных пузырьков низших образования постоянные. В сезон размноже-
ния они регулярно производят сперматозоиды и в течение половой
жизни особи меняются нерезко, затухая и активизируясь соответ-
ственно природным сезонам. Первичный зачатковый эпителий,
покрывающий изначально поверхность семенника, рассасывается
и замещается плотной соединительнотканной белочной оболочкой.
Во взрослом состоянии у большинства самцов позвоночных го-
нады сохраняют свое первичное положение в верхней части цело-
мической полости. У млекопитающих же в ходе онтогенеза положе-
ние семенников в теле существенно изменяется. В дне брюшной
Рис. 204. Фрагмент канальца семенника млекопитающего (в разрезе):
1 — кровеносный сосуд; 2 — созревающие сперматозоиды; 3 — интерстициаль-
ная клетка; 4 — клетка Сертоли
265
полости образуется пара карманов, выпячивающихся наружу в виде
мошонки. Они представляют собой часть целома и выстланы изнут-
ри целомическим эпителием. Семенники сдвигаются в карманы мо-
шонки вместе с семяпроводами и кровеносными сосудами. У неко-
торых видов после окончания сезона размножения семенники мо-
гут втягиваться через паховый канал обратно в брюшную полость.
Перемещение семенников за пределы тела связано с необхо-
димостью поддержания более низкой температуры при спермато-
генезе, так как при высокой температуре тела животного созрева-
ние половых клеток замедляется.
Выведение половых продуктов из гонад у большинства самцов
позвоночных связано с каналами выделительной системы — воль-
фовыми каналами (см. рис. 199, 206). Нижняя часть вольфовых ка-
налов у многих расширяется в семенные пузыри, резервирующие
семя. В нижних частях семяпроводов формируются железы, секре-
ты которых разжижают семенную жидкость и стимулируют под-
вижность сперматозоидов.
Самки. Зачаток гонады, превращающейся в яичник, развивает-
ся по иной, чем семенник, программе. Как было сказано выше,
первичные половые тяжи, углубившись в строму гонады, дегене-
рируют. Снаружи из зачаткового эпителия в тело гонады врастают
вторичные половые тяжи. Содержащиеся в них половые клетки
многократно делятся и превращаются в дефинитивные (зароды-
шевые, первичные) овоциты.
Вокруг каждого из овоцитов концентрируются клетки фолли-
кулярного эпителия, обеспечивающие питание растущей поло-
вой клетки. Ранние фолликулы с одним слоем фолликулярного
эпителия называют первичными, или примордиальными. Стенка фол-
ликула образована соединительной тканью. С наступлением зре-
лости овоцит и фолликул увеличиваются в размерах. В них заметно
возрастает число слоев фолликулярного эпителия. В период репро-
дуктивной активности фолликулы перемещаются к поверхности
гонады. Созревшие фолликулы — граафовы пузырьки разрывают-
ся и овоциты выпадают в целомическую полость (рис. 205). У боль-
шинства позвоночных содержимое фолликула впоследствии рас-
сасывается, однако у всех млекопитающих (и некоторых акул) на
месте разорвавшихся фолликулов формируются желтые тела (corpus
luteum), производящие гормон прогестерон.
Число одновременно созревающих овоцитов в яичнике у на-
земных позвоночных невелико (1 —20), тогда как у амфибий их
сотни и даже тысячи, а у лучеперых рыб — сотни тысяч и милли-
оны. К примеру, самка трески выметывает за один сезон размно-
жения 4 млн икринок.
Известно, что нередко число созревающих фолликулов суШе'
ственно превышает число овулирующих впоследствии яйцеклеток.
Значительная их часть превращается в атретические фолликулы,
Белочная
оболочка
Оболочка
фолликула
Первичный
фолликул
Граафов
пузырек
Растущий
фолликул
Белое
тело
Созревающий
фолликул
Атретический
фолликул
фолликул
Желтое тело
Мозговое
вещество
Зачатковый
эпителий
Интерстициальная
ткань
—-Разорвавшийся
Дегенерирующее
желтое тело
Рис. 205. Схема разреза яичника млекопитающего
(по А.Ромеру, Т.Парсонсу, 1994)
и их материал расходуется на дозревание других (внутригонадная
овофагия).
Яичники у большинства позвоночных образования парные. В по-
кое они часто имеют небольшие размеры и овальную форму, а в
Период половой активности становятся крупными и имеют не-
правильную форму. Половые продукты выводятся у самок через
парные яйцеводы (мюллеровы каналы), возникающие эмбриональ-
но при расщеплении пронефрических каналов. Яичник и яйцевод
непосредственно не соединяются. Передний конец каждого яйце-
вода расположен вблизи соответствующего яичника и образует
ресничную воронку с гофрированным краем, открывающуюся в
целом. Овоциты, выделившиеся при овуляции из яичника, улав-
ливаются воронкой и транспортируются по яйцеводу. У многих
костистых рыб яичник полностью изолирован от целома: икрин-
ки выпадают во внутреннюю полость яичника и по протоку, об-
разованному поверхностной тканью гонады, транспортируются во
внешнюю среду.
Мюллеров канал в ряду позвоночных усложняется. В наиболее
оформленном виде в нем различают собственно яйцевод (фалло-
пиеву трубу), маточный отдел и влагалище, образованное сраста-
нием половых путей. В эпителии разных отделов яйцевода у репти-
лий и птиц присутствуют железы: одни секретируют белок, дру-
гие — материал волокнистых и скорлуповых оболочек (есть желе-
зы и у хрящевых рыб).
Механизмы становления пола у некоторых позвоночных живот-
ных сбалансированы нечетко, и гонады долгое время пребывают в
состоянии выбора превращения в семенник или яичник. В этих слу-
чаях в гонаде присутствуют и женские, и мужские гаметы. Напри-
мер, у миксин в разных частях одной гонады закладываются оба
типа гамет, в ходе же полового созревания усиливается преобла-
дание одного из них, и животное обретает пол. Некоторые особи
остаются стерильными интрасексами в течение всей жизни (А. Ро-
мер, Т. Парсонс, 1992). Размножающиеся гермафродиты известны
в нескольких семействах морских костистых рыб, например у ка-
менных окуней (род Serranus), смаригдовых (Centracanthidae), до-
рад (Chrysophys). Половые железы функционируют у них попере-
менно то как семенники, то как яичники, и самооплодотворение
исключено. У морского окуня разные части гонады могут одно-
временно выделять яйца и сперматозоиды. Интересно проявляет-
ся гермафродитизм у морских карасей (Sparidae). Например, у оби-
тающего в Средиземном море (иногда появляется в Черном море)
красного пагела (Pagellus erythrinus) часть самок и часть самцов с
возрастом меняют свой пол на противоположный. Сходное явление
свойственно некоторым губановым рыбам (сем. Labridae), например
обитающему в Черном море морскому юнкеру (Coris julis). Иногда гер-
мафродитные особи встречаются у трески, макрели, сельдей.
Некоторые отклонения в становлении пола зафиксированы У
амфибий. Например, у самцов обыкновенной жабы, кроме се-
менников, есть орган, похожий на яичник, с не вполне понят-
ной функцией (биддеров орган). Молодые самцы травяной лягуш-
ки проявляют особенности самок, а генетические самки в старо-
сти способны вырабатывать сперму (А. Ромер, Т. Парсонс, 1994)-
268
Изменения в репродуктивной системе в эволюционном
ряду хордовых
У ланцетника сходные по внешнему виду семенники и
яичники в виде парного ряда округлых тел (около 25 пар) распо-
ложены в жаберном отделе целома. Созревшие половые продукты
через разрывы стенок тела и специальные временно возникаю-
щие половые протоки выводятся в атриальную полость, а из нее
через атриопор наружу. Оплодотворение происходит в воде.
У круглоротых половые железы непарные и тоже не име-
ют специальных протоков. Половые продукты через разрывы сте-
нок гонады выпадают в полость тела, откуда через временные от-
верстия попадают в мочеполовой синус и далее через канал моче-
полового сосочка наружу. Оплодотворение у круглоротых внешнее.
У миксин на поверхности яиц обнаружена тонкая скорлупа. На
одном участке яйца (микропиле) она утончена и не препятствует
оплодотворению (А. Ромер, Т. Парсонс, 1994).
У большинства самок хрящевых рыб яичники парные. Пар-
ные яйцеводы (мюллеровы каналы) открываются в полость тела в
непосредственной близости от гонад. В верхнем отделе яйцеводов
расположены скорлуповые железы. Здесь яйцо покрывается белком
и роговой оболочкой. Расширенные нижние отделы яйцеводов
открываются в клоаку. Половые и мочевые пути у самок на всем
протяжении разделены (рис. 206).
В полости тела самцов лежат парные семенники; правый в боль-
шинстве случаев крупнее левого, а иногда они могут сливаться.
Системой канальцев семенники связаны с передними отделами
мезонефрических почек. Канальцы проходят через вещество по-
чек и впадают в вольфовы каналы, которые у самцов хрящевых
рыб служат семяпроводами. Передние отделы почек не имеют
выделительной функции (это свойство задних отделов) и служат
придатками семенников.
В нижнем отделе семяпроводы расширены и образуют тонко-
стенные семенные пузыри. Оба семяпровода (вольфовы каналы)
впадают в мочеполовой синус, который открывается в клоаку.
Оплодотворение у большинства хрящевых рыб внутреннее. Ис-
пользуя пальцевидные выросты брюшных плавников, самец вво-
дит сперматозоиды в клоаку самки. Оплодотворение происходит в
верхней части яйцевода до того момента, как яйцеклетка опус-
тится до скорлуповой железы. У многих акул и скатов наблюдает-
ся живорождение. При этом плотная оболочка на поверхности яйца
не образуется, эмбрион развивается в нижнем расширенном от-
деле яйцевода («яйцевой капсуле»).
Похожие органы размножения имеют двоякодышащие
Рыб ы. У многоперов и ильной рыбы короткие яйцеводы широ-
кой воронкой открываются в полость тела и тесно прилегают к
269
270
Рис. 206. Схемы мочеполовой системы позвоночных.
Самцы: А — акулы, амфибии; Б — рептилии; В — млекопитающие. Самки; Г —
акулы, амфибии; Д — рептилии; Е — млекопитающие. 1 — головная почка; 2 —
семенник; 3 — выносящие протоки; 4 — туловищная почка; 5 — мочевой пу-
зырь; 6 — прямая кишка; 7 — клоака; 8 — проток мезонефроса; 9 — воронка
яйцевода; 10 — яичник; 11 — мюллеров канал; 12 — рудиментарный мюллеров
канал у самца; 13 — придаток семенника (преобразованная туловищная почка);
14 — семяпровод; 15 — тазовая почка; 16 — вторичный мочеточник; 17 — ру-
диментарная туловищная почка самок амниот; 18 — яйцеклетка; 19 — яйцевод;
20 — белковая оболочка, выделяемая железами стенки яйцевода; 21 — матка; 22 —
влагалище; 23 — семенной пузырек; 24 — предстательная железа; 25 — анус; 26—
пенис; 27 — мочеполовой синус; 28 — эмбрион в матке, окруженный эмбрио-
нальными оболочками; 29 — промежность
яичнику. Канальцы семенников проникают через нижнюю часть
почки в вольфов канал (рис. 207).
Большинство костистых рыб (а также панцирная щука из
рыб ганоидных) имеют иное строение органов размножения. Оно
связано с особенностями, определяющими стиль размножения
рыб: большая плодовитость, наружное оплодотворение, откла-
дывание мелкой икры в тонкой студенистой оболочке. Половые
продукты никогда не попадают в целом. У самок нет мюллеровых
каналов, а у самцов семенники не связаны канальцами с почками
и вольфовы каналы выполняют только выделительную функцию.
Парные семенники и яичники имеют внутри свободные полости,
куда и выпадают половые клетки. Полости открываются на моче-
половом сосочке. Возможны отклонения: у самцов осетровых ка-
нальцы пронизывают почти всю ткань почки и впадают в вольфов
канал. У лосососевых рыб воронка яйцевода примыкает снизу к
цельному яичнику (см. рис. 207).
Икра оплодотворяется, как правило, вне тела матери. У отно-
сительно немногих видов наблюдается внутреннее оплодотворе-
ние и яйцеживорождение. Таковы, например, американские суб-
тропические зубастые карповые (сем. Cyprinodontidae). В случае жи-
ворождения оплодотворение икринок и развитие мальков проис-
ходит в полости яичника. Количество потомков при живорожде-
нии заметно уменьшено. Один из наиболее известных живородя-
щих видов — гамбузия. Современная кистеперая рыба латимерия
тоже является яйцеживородящей и производит малое число дете-
нышей, в разное время в ее теле находили от 5 до 19 мальков.
Парные семенники у амфибий, как и у акул, не имеют са-
мостоятельных выводных путей (рис. 208). Семявыносящие канальцы
проходят через передний отдел почки и впадают в вольфов ка-
нал, который выносит половые продукты в клоаку. Как и у акул,
каждый вольфов канал у самцов образует расширение — семен-
ной пузырек, временно резервирующий семя. Копулятивных ор-
Рис. 207. Мочеполовая система рыб (по А. Ромеру, Т. Парсонсу, 1994):
А, Б, В — самцы акулы, южноамериканской двоякодышащей рыбы (Lepidosirien)
и костистой рыбы соответственно; Г, Д, Е — самки акулы, лосося и костистой
рыбы соответственно; 1 — передняя часть почки акулы, сравнимая с придатком
семенника; 2 — мочеточник; 3 — клоака; 4 — гонада; 5 — вольфов канал; 6 —
семяпровод; 7 — половое отверстие у костистых рыб; 8 — выделительное отвер-
стие у костистых рыб; 9 — яйцевая капсула, или «матка», у акул; 10 — яйцевод;
11 — скорлуповая железа; 12 — воронка яйцевода
ганов у подавляющего большинства амфибий нет. Над семенника-
ми лежат образования неправильной формы желтого цвета — жи-
ровые тела. Они служат для питания семенников и развивающих-
ся в них сперматозоидов. Величина жировых тел меняется по сезо-
нам: осенью они крупные, весной, во время интенсивного спер-
матогенеза, их вещество энергично расходуется и размеры затем
резко сокращаются.
Яичники у амфибий парные; над ними также лежат жировые
тела. Созревшие яйца выпадают из гонад в полость тела и отсюда
272
10
Рис. 208. Мочеполовая система
самца лягушки:
1 — почка; 2 — мочеточник; 3 — по-
лость клоаки; 4 — мочеполовое отвер-
стие; 5 — мочевой пузырь; 6 — отвер-
стие мочевого пузыря; 7 — семенник;
8 — семявыносящие каналы; 9 — се-
менной пузырек; 10 — жировое тело;
11 — надпочечник
Рис. 209. Мочеполовая система
самки лягушки:
1 — воронка яйцевода; 2, 6 — яйце-
вод; 3 — жировое тело; 4 — надпочеч-
ник; 5 — почка; 7 — вольфов канал;
8 — яйцевая капсула; 9 — выходы в
клоаку; 10 — яичник
поступают в воронкообразные расширения парных яйцеводов
(мюллеровых каналов). Яйцеводы — длинные, сильно извитые труб-
ки, их задний конец открывается в клоаку (рис. 209).
Большинству амфибий, как и костистым рыбам, свойственны
наружное оплодотворение и многочисленные половые продукты.
Немногие хвостатые и бесхвостые амфибии живородящие. У них
вынашивание детенышей происходит, как и у акул, в яйцевых
капсулах — специальных расширениях нижней части яйцеводов.
У рептилий (рис. 210) половые железы лежат в полости тела
по бокам позвоночника. К семенникам примыкает придаток — ос-
таток верхней части мезонефрической почки. Канальцы придатка
впадают в вольфов канал, служащий у самцов рептилий (и у ос-
тальных высших позвоночных) семяпроводом. Все рептилии, кроме
гаттерии, имеют совокупительные органы. У ящериц и змей это
парные выросты задней стенки клоаки, которые в период воз-
буждения выворачиваются наружу. У крокодилов и черепах копу-
лятивный орган непарный, также представляющий вырост стен-
ки клоаки.
У самок в качестве яйцеводов функционируют парные мюлле-
ровы каналы — тонкостенные трубки, открывающиеся воронка-
ми в полость тела, а другим концом — в клоаку. Яйцо оплодотво-
10 Клыгтяытынпп 0’7'2
Рис. 210. Наружные
копулятивные мешки
самца ящерицы
ряется на воронке яйцевода. У черепах и
крокодилов в эпителии средних отделов
яйцеводов имеются железы, выделяющие
белковую оболочку яйца. Железы, фор-
мирующие волокнистую или пропитан-
ную известью скорлуповую оболочку,
расположены в нижней части яйцеводов.
В размножении рептилий много осо-
бенностей, свидетельствующих о приспо-
соблении к наземному образу жизни. Оп-
лодотворение всегда внутреннее. Разви-
тие происходит без превращений: отсут-
ствует личиночная стадия. Основные при-
способления к размножению вне воды
заключаются в особенностях строения
яйца и развития зародыша. В сравнении
с амфибиями у рептилий яйца крупные. Они увеличиваются за
счет обогащения их желтком, а у черепах и крокодилов к тому же
за счет появления «белка» — белочной яйцевой оболочки. Боль-
шой запас питательного вещества в яйце создает возможность
прямого (без превращения) развития эмбриона. Большинство реп-
тилий откладывают яйца на суше, у некоторых видов наблюдают-
ся живорождение и яйнеживорождение. В связи с откладыванием
яиц в условиях наземной среды у рептилий возникает ряд при-
способлений, защищающих их от иссушения, механического по-
вреждения и обеспечивающих развивающемуся эмбриону возмож-
ность газообмена, снабжения его водой и хотя бы частичного вы-
ведения продуктов обмена. Это прежде всего оболочки, покрыва-
ющих яйцо (волокнистая и белочная) (рис. 211). Они образуются
при прохождении оплодотворенного яйца по яйцеводу и проду-
цируются разными отделами его стенки.
Рис. 211. Схема образования яйцевых оболочек в половых путях самки
степной черепахи:
1 — яйцеклетка; 2 — белковая оболочка; 3 — волокнистая оболочка; 4 — скорлу-
повая оболочка
Э7/!
Яйца чешуйчатых имеют только волокнистую оболочку. Строе-
ние яйца черепах и крокодилов приближается к птицам. Их волок-
нистая оболочка пропитана известью и уплотнена. Кроме того,
железы яйцевода продуцируют у них белочную оболочку. Ее жид-
кое содержимое в замкнутом яйце обеспечивает зародыш водой.
У прочих рептилий необходимая для нормального развития заро-
дыша вода имеет двойное происхождение. Большая часть ее выс-
вобождается в результате окисления весьма обильного в желтке
жира (так называемая метаболическая вода), другая часть посту-
пает из наружной среды через оболочки яйца. Количество жира в
яйце рептилий составляет 40 % (у амфибий — 8 — 11 %) их массы.
Приспособления к размножению в наземно-воздушной среде
затронули и ранние стадии эмбриогенеза рептилий (и всех Amniota).
Из первичных тканей зародыша формируются три зародышевые
оболочки: амнион, аллантоис и сероза (рис. 212).
Эктодермальные складки, возникающие в раннем онтогенезе
впереди спинного отдела зародыша, смыкаются, образуя замкну-
тый мешок амнион, внутри которого находится зародыш. Полость
амниона — амниотическая полость — заполняется жидкостью. Ам-
Рис. 212. Последовательные стадии (Z— II) развития зародышевых
оболочек у амниота:
1.1 — эктодерма; 2 — энтодерма; 3 — мезодерма; 4 — полость кишечника; 5 —
внезародышевая полость тела; 6 — амнион; 7 — амниотическая полость, запол-
ненная амниотической жидкостью, в которой плавает зародыш; 8 — сероза; 9 —
аллантоис; 10 — желточный мешок. II: 1 — амнион; 2 — аллантоис; 3 — желток;
4 — сероза
775
ниотическая оболочка создает постоянство среды и предохраняет
зародыш от соприкосновения с плотными оболочками яйца. Из
наружных частей эктодермальных складок образуется сероза, за-
щищающая зародыш от механических воздействий, проникнове-
ния болезнетворных микроорганизмов.
Еще одна зародышевая оболочка — аллантоис — возникает как
энтодермальный мешковидный вырост брюшной стенки задней
кишки. По мере развития аллантоис приобретает вид большого
пузыря, лежащего между желточным мешком и серозой. К концу
эмбрионального развития аллантоис почти полностью окружает
тело зародыша и желточный мешок.
Стенка аллантоиса имеет богатую сеть кровеносных сосудов.
Аллантоис выполняет одновременно две функции: он является
органом дыхания и зародышевым мочевым пузырем. При этом
аллантоис не только пассивно принимает мочу, но воспринимает
из нее лишнюю воду, которая через сеть кровеносных сосудов
возвращается в тело зародыша. Кроме того, через стенку алланто-
иса частично осуществляется газообмен зародыша. Первоначаль-
ным же приспособлением для дыхания зародыша служит густая
сеть кровеносных сосудов, оплетающих желток, — желточный круг
кровообращения.
Наряду с откладыванием яиц у некоторых рептилий существу-
ет яйцеживорождение и живорождение. В простейшем случае оп-
лодотворенные яйца задерживаются в половых путях самки и про-
ходят здесь все стадии развития. Детеныши выклевываются из яиц
тотчас же после откладывания их во внешнюю среду. Так бывает у
гадюки (Vipera bems), живородящей ящерицы {Lacerta vivipara) и
веретеницы (Anguisfragilis). Яйцеживорождение свойственно не-
которым удавам (песчаному удавчику), морским змеям, ужам.
Немногим видам рептилий свойственно настоящее живорож-
дение. У сцинков, например, кровеносные сосуды желточного
мешка эмбриона тесно сближены с сосудами маточного отдела
яйцевода (желточная плацента), и питание эмбриона идет в зна-
чительной степени за счет организма матери.
Есть сведения, что у некоторых змей и ящериц формируется
настоящая аллантоисная плацента, при которой ворсинки хорио-
аллантоиса внедряются в слизистую выстилку половых путей сам-
ки, как это свойственно плаценте млекопитающих.
У птиц парные семенники бобовидной формы расположены
над верхними долями почек по бокам позвоночника (рис. 213). Их
величина существенно меняется по сезонам, увеличиваясь к сезо-
ну размножения в 1000—1500 раз. К внутренним краям семенни-
ков присоединяются слабо выраженные придатки, гомологичные
передним отделам туловищных почек эмбриона. От придатков от-
ходят семяпроводы, которые тянутся параллельно мочеточникам
и впадают в клоаку. У некоторых видов семяпроводы перед впаде-
276
8
Рис. 213. Половые органы птиц (А — самка, Б — самец):
1, 3 — семенники; 2 — почечные доли; 4 — мочеточники; 5 — семяпровод; 6 —
клоака; 7 — семенной пузырь; 8 — граафовы пузырьки; 9 — воронка яйцевода;
10 — матка; 11 — часть яйцевода, выделяющая белок; 12 — отверстие кишки
нием в клоаку образуют расширения — семенные пузыри, резер-
вирующие семя. Копулятивные органы свойственны немногим
птицам (например, страусам, гусям). У большинства видов опло-
дотворение достигается путем сближения клоак.
Половая система самок, за очень немногими исключениями,
асимметрична и имеет только левый яичник и левый яйцевод.
Правый яичник развивается у некоторых дневных хищников, сов,
попугаев, куриных. Особенно часто его обнаруживали у попугаев,
но он редко бывал функционирующим. Развитие правого яйцево-
да — явление еще более редкое. Причина редукции правой поло-
вины половых органов у самок птиц заключается, видимо, в от-
кладывании относительно крупных яиц с жесткой и хрупкой скор-
лупой и замедленном продвижении по яйцеводу.
Неправильной формы зернистый яичник лежит впереди левой
почки (см. рис. 213). Величина его варьирует очень сильно в зави-
симости от зрелости и размеров формирующихся в нем яиц. Яйце-
вод имеет вид длинной трубки, верхний конец которой открыва-
ется воронкой в полость тела, а нижний в клоаку. Созревшая яй-
цеклетка с запасом желтка попадает в воронку яйцевода. Здесь она
оплодотворяется и транспортируется по нему далее. Собственно
яйцевод имеет несколько отделов. Верхний отдел почти по всей
длине богат железами, выделяющими белок, который слой за слоем
покрывает проходящее яйцо. В курином яйце эта оболочка форми-
руется за 4 ч. В следующем, более тонком, отделе яйцо покрывает-
277
ся двумя волокнистыми пергаментообразными подскорлуповыми
оболочками. На выходе из этого отдела яйцо задерживается. Его
оболочки, набирая воду, набухают. Далее следует маточный от-
дел, богатый железами, образующими известковую скорлупу яйца
и окрашивающими ее пигментами. Куриное яйцо задерживается
здесь в течение 15 —16 ч. С поверхности скорлупу покрывает тон-
кая кутикулярная надскорлуповая оболочка. Она имеет бактери-
цидное значение, предотвращая проникновение через поры скор-
лупы болезнетворных бактерий. Последний отдел яйцевода, «вла-
галище», короток, обладает значительной мускулатурой; из него
яйцо выходит в клоаку и далее наружу.
Весь период прохождения яйца по яйцеводу составляет у кури-
цы около суток, у голубя — 41 ч.
Строение яйца птицы. Структура яйца птицы (рис. 214) отчасти
сходна с яйцом рептилий. Собственно яйцеклеткой является лишь
то, что в обиходе называют «желтком». Остальные образования
(например, «белок») представляют его яйцевые оболочки, про-
дуцируемые, как сказано выше, не яичником, а яйцеводом.
На анимальном полюсе желтка находится небольшой зародыше-
вый диск. В неоплодотворенном яйце — это овоцит, состоящий из
ядра и цитоплазмы, в оплодотворенном — зародыш на стадии по-
здней бластулы или гаструлы. Такой стадии развития зародыш дос-
тигает во время движения по яйцеводу. Развитие зародыша продол-
жается с началом насиживания. Остальная часть желтка состоит из
мельчайших пузырьков — желтого желтка, прослоенного яйце-
вой плазмой, или светлым желтком. Слои светлого и желтого жел-
тка располагаются концентрически. Снаружи весь желток одет тон-
кой оболочкой. Желток является запасом питательных веществ и
воды, которые расходуются раз-
вивающимся эмбрионом и служат
материалом для построения его
тела. В составе желтка обращает
внимание обилие жира и воды. Ку-
риный желток, например, содер-
жит (%): воды — 50, жиров — 23,
протеинов — 16, липидов — 11.
Поэтому при развитии зародышей
яйца птиц не поглощают воду из
наружной среды, подобно яйцам
чешуйчатых рептилий, так как
весь запас воды, необходимый для
развития, находится в самом яйце,
в частности в желтке и белке. Пи-
тательные вещества сосредоточе-
ны вблизи нижнего вегетативно-
го полюса яйца.
2
Рис. 214. Строение яйца птицы:
1 — халаза; 2 — скорлупа; 3 — воз-
душная камера; 4 — наружная под-
скорлуповая оболочка; 5 — жидкий
белок; 6 — плотный белок; 7 — заро-
дышевый диск; 8 — светлый желток;
9 — темный желток
278
Белок представляет собой яйцевую оболочку полужидкой кон-
систенции. Белок неоднороден, основная его масса — жидкий бе-
лок. Он служит важным источником получения эмбрионом воды.
В составе куриного белка вода составляет 87 %, протеины — 12 %.
На остальные вещества приходится около 1 %. Желток не плавает
свободно в толще белка, а подвешен на уплотненных скрученных
белковых шнурах — халазах, что предохраняет его от толчков. Кроме
того, зародышевый диск благодаря халазам всегда оказывается
наверху. Это имеет большое значение при инкубации, так как обес-
печивает лучший его обогрев.
Над белком расположены две тонкие подскорлуповые оболочки.
Они образованы сетью органических волокон. Волокнистый слой
наружной оболочки связан с внутренней поверхностью скорлупы
через маммиллярные бугорки — центры кристаллизации кальцита
при формировании скорлуповой оболочки. Вскоре после откла-
дывания яйца подскорлуповые оболочки на тупом конце яйца
расходятся и образуют воздушную камеру, обеспечивающую воз-
можность изменения объема яйца при перемене температуры и
газообмен зародыша.
Скорлуповая оболочка. Из всех яйцевых оболочек наиболее сложно
устроена скорлупа (рис. 215). Связано это с тем, что через нее
происходит газообмен развивающегося зародыша, к которому бес-
препятственно должен поступать кислород и свободно выходить
S
2,0
1,8
1,6
1,4
1,2
1,0
0,8
0,6
0,4
0,2
0
Рис. 215. Строение скорлупы яйца птицы:
7 — африканский страус; 2 — нанду; 3 — лебедь;
4 — гусь; 5 — киви; 6 — курина; 7 — тинаму; 8 —
зарянки; 9 — колибри
7 2 3 4 5 6 7 8 9
э-то
углекислый газ. Одновременно скорлупа защищает его от механи-
ческих повреждений и потери воды. Скорлупа куриного яйца (мас-
сой 60 г) весит 5 г. Она состоит в основном из карбоната кальция,
представленного колонками (столбиками) кристалликов кальци-
та, и небольшого количества органического вещества. Между кри-
сталлическими колонками находятся мелкие поры, которые за-
кладываются при формировании скорлупы в яйцеводе. Коничес-
кое устье каждой поры постепенно сужается и переходит в ци-
линдрический канал диаметром в несколько микрометров, про-
низывающий всю толщину скорлупы. Снаружи устья пор не вид-
ны: они частично прикрыты секретированным бактерицидным ве-
ществом и кутикулой. У каждого вида птиц число пор в скорлупе
постоянно и пропорционально размерам яйца. К примеру, в скор-
лупе куриного яйца их 10000, общей площадью сечения около
2 мм. При возрастании массы яйца и толщины скорлупы число
пор увеличивается. Несколько большее число пор имеет скорлупа
закрытогнездящихся птиц, например, их больше в скорлупе яйца
береговой ласточки, гнездящейся в норах, по сравнению со скор-
лупой яйца городской ласточки, гнездящейся открыто. В течение
всего времени инкубации яйцо теряет за счет испарения воды
через поры около 15 % своей массы.
Во время развития эмбриона скорлупа частично расходуется
на построение его скелета. У курицы, в частности, масса извест-
ковых элементов скорлупы за время насиживания снижается по-
чти на 8 %. Скорлупа предохраняет яйцо от растекания, механи-
ческих повреждений, проникновения бактерий и не препятствует
газообмену зародыша.
Снаружи скорлупа покрыта тонкой надскорлуповой мембраной,
или кутикулой, состоящей из органического вещества и имею-
щей трещиноватую поверхность, обеспечивающую успешный га-
зообмен, равномерное испарение воды и защиту от болезнетвор-
ных микроорганизмов.
У млекопитающих половые железы самца — семенники —
имеют характерную овальную форму (рис. 216). У однопроход-
ных, некоторых насекомоядных и неполнозубых, у слонов и ки-
тообразных они в течение всей жизни находятся в полости тела.
У большинства других животных семенники первоначально рас-
полагаются в полости тела, но по мере полового созревания они
опускаются вниз и попадают в мошонку — расположенный сна-
ружи мешочек, сообщающийся с полостью тела паховым кана-
лом. К семенникам прилегают вытянутые по их оси зернистые
тела — придатки семенников. Они представляют собой клубок сильно
извитых семявыносящих канальцев семенников, гомологичных пе-
редним отделам туловищных почек. От придатков семенников от-
ходят гомологичные вольфовым каналам парные семяпроводы,
впадающие у корня полового члена в мочеполовой (мочеиспуска-
8.
7
Рис. 216. Мочеполовые органы крысы (А — самец, Б — самка):
1 — семенник; 2 — придаток семенника; 3 — семяпровод; 4 — семенные пузыри;
5 — предстательная железа; 6 — мочевой пузырь; 7 — мочеточник; 8 — почка; 9 —
мошонка; 10 — паховый канал; 11 — яичник; 12 — фаллопиева труба; 13 —
матка; 14 — влагалище
тельный, или семяизвергательный) канал. Семяпроводы в своем
нижнем отделе, перед впадением в мочевой канал, образуют пар-
ные компактные тела с ребристой поверхностью — семенные пу-
зыри. У млекопитающих в отличие от других позвоночных они вы-
полняют железистые функции, секрет их принимает участие в об-
: разовании жидкой части спермы; кроме того, в связи с клейким
свойством он, видимо, предотвращает обратное вытекание спер-
мы из половых путей самки.
У основания полового члена лежит вторая парная железа —
предстательная, протоки которой впадают также в начальную часть
мочеполового канала. Эти протоки считают редуцированными го-
мологами мюллеровых каналов (И. И. Шмальгаузен, 1964). Секрет
, предстательной железы составляет основную часть жидкости, в
которой плавают поступившие сюда из семенников спермато-
, зоиды. В итоге сперма, или эйякулят, представляет комбина-
цию сперматозоидов и жидкости, выделенной семенными пу-
зырями, предстательной железой и некоторыми другими желе-
зами.
На нижней стороне совокупительного члена располагается упо-
мянутый уже мочеполовой канал. Вверх и по бокам от этого кана-
ла лежат пещеристые тела. Их внутренние полости во время поло-
вого возбуждения наполняются кровью, в результате чего поло-
вой член становится упругим и увеличивается в размерах. У мно-
гих млекопитающих прочность полового члена обусловливается
еще особой длинной костью, расположенной между пещеристы-
OQ 1
ми телами. Таковы хищные, ластоногие, многие грызуны, неко-
торые летучие мыши. По бокам мочеполового канала расположе-
ны парные куперовы железы, имеющие с поверхности мускуль-
ную оболочку. Выделения этих желез участвуют в активизации
сперматозоидов.
Парные яичники млекопитающих всегда лежат в полости тела
и брыжейками прикреплены к спинной стороне брюшной полос-
ти. Парные яйцеводы, гомологичные мюллеровым каналам, от-
крываются передними концами в полость тела в непосредствен-
ной близости от яичников, образуя широкие воронки. Верхние
извитые отделы яйцеводов носят название фаллопиевы трубы. Да-
лее идут расширенные отделы — матки, которые открываются в
непарный у большинства зверей отдел — влагалище. Последнее
переходит в короткий мочеполовой канал, в который, кроме вла-
галища, открывается мочеиспускательный канал. На брюшной
стороне мочевого канала располагается небольшой вырост — кли-
тор, обладающий пещеристыми телами и соответствующий пе-
нису самца. У некоторых видов в клиторе имеется кость. В нижнюю
часть влагалища открываются бартолиниевые железы, выделяю-
щие слизистый секрет.
Строение женских половых путей существенно различается у
разных групп млекопитающих. Так, у однопроходных яйцеводы на
всем протяжении парные и дифференцированы только на фалло-
пиевы трубы и матки, которые самостоятельными отверстиями
Рис. 217. Схема половых путей у разных млекопитающих:
А — клоачное; Б — низшее сумчатое (опоссум); В — высшее сумчатое (кенгуру);
Г — плацентарное млекопитаюшее с двойной маткой; Д — то же, с двурогой
маткой; £ — то же, с простой маткой; 1 — фаллопиева труба; 2 — матка; 3 —
влагалище; 4 — мочеполовой синус; 5 — мочевой пузырь; 6 — клоака; 7 — пря-
мая кишка
открываются в мочеполовой синус. У сумчатых обособляется вла-
галище, но часто оно остается парным. Соответственно бывают
раздвоены совокупительные органы самцов. У плацентарных вла-
галище всегда непарное, а верхние отделы яйцеводов сохраняют
парный характер. Матка в простейшем случае парная, ее левый и
правый отделы открываются во влагалище самостоятельными от-
верстиями. Такую матку называют двойной', она свойственна мно-
гим грызунам, некоторым неполнозубым. Матки могут быть со-
единены только в нижнем отделе — двуразделъная матка некото-
рых грызунов, летучих мышей, хищников. Слияние значительной
части левой и правой маток приводит к образованию двурогой матки
хищных, китообразных, копытных. Наконец, у приматов, полу-
обезьян, некоторых летучих мышей матка непарная — простая, и
парными остаются только верхние отделы яйцеводов — фаллопи-
евы трубы. Разные варианты строения матки млекопитающих пред-
ставлены на рис. 217.
Плацента. В матке млекопитающих во время развития эмбриона
формируется характерное для них образование, известное под на-
званием детского места, или плаценты. Она отсутствует только у
однопроходных. Плацента возникает путем срастания наружной
стенки аллантоиса с серозой, в результате чего формируется губ-
чатое образование — хориоаллантоис. Хорион образует выросты —
ворсинки, которые соединяются или срастаются с разрыхленным
участком эпителия матки. Здесь оформляются и близко соприка-
саются кровеносные сосуды матери и развивающегося зародыша.
Сосуды эмбриона и матери часто сплетаются (но не сливаются),
и таким образом устанавливается связь между кровяными русла-
ми эмбриона и матери. Через кровеносную сеть обеспечиваются
Рис. 218. Плаценты млекопитающих (А — сумчатое, Б — плацентарное):
1 — амнион; 2 — сероза; 3 — зародыш; 4 — аллантоис; 5 — амниотическая
полость; 6 — полость кишки; 7 — ворсинки хориона
283
газообмен в теле зародыша, его питание и удаление конечных
продукта обмена.
Плацента сумчатых млекопитающих примитивна. Хорион не
образует ворсинок и сохраняет, как и у яйцекладущих, лишь кон-
такт кровеносных сосудов матки с сосудами желточного мешка
(так называемая «желточная плацента»). У высших плацентарных
животных хорион всегда образует выросты — ворсинки, соединя-
ющиеся со стенками матки (рис. 218). Характер расположения вор-
синок различен у разных групп зверей. Основываясь на этом, вы-
деляют несколько типов плаценты: диффузную, когда ворсинки
распределяются равномерно по хориону (китообразные, многие
копытные, полуобезьяны); дольчатую, когда ворсинки собраны в
группы, распределенные по всей поверхности хориона (большин-
ство жвачных); дискоидалъную, когда ворсинки располагаются на
ограниченном, имеющем вид диска участке хориона (насекомо-
ядные, грызуны, обезьяны), и др.
Степень связанности детской и материнской частей плаценты
различна. В одних случаях эта связь непрочная, и при родах слизи-
стая матки не отрывается. В других — связь более прочная, и при
родах соответствующая часть слизистой оболочки матки отпадает
и выводится наружу как послед. В первом случае плацента называ-
ется неотпадающей, во втором — отпадающей. На местах прикреп-
ления эмбрионов в матке сохраняются темные пятна.
* **
Таким образом, для позвоночных животных характерно поло-
вое размножение. Яичники самок и семенники самцов закладыва-
ются сходно в мезодерме. Примитивное наружное оплодотворе-
ние свойственно ланцетнику: из сходных по внешнему виду гонад
по временным протокам в воду у самок выделяются яйцеклетки,
у самцов — сперматозоиды, где после оплодотворения происхо-
дит развитие зародышей. Такой способ размножения присущ круг-
лоротым, большинству рыб, амфибиям. В этом случае выделяется
огромное количество половых продуктов, так как эффективность
размножения очень мала из-за отсутствия заботы родителей о по-
томстве. Внутреннее оплодотворение возникало у позвоночных в
разных систематических группах. Эволюционная прогрессивность
внутреннего оплодотворения особенно ярко проявляется при раз-
витии зародыша в теле матери. Высоких показателей эффективно-
сти размножения достигли птицы и млекопитающие: одни — пу-
тем совершенствования яйцекладности, другие — улучшения кон-
тактов детского и материнского организмов при живорождении.
Важнейшую роль играет забота о потомстве: защита, обогрев,
выкармливание и его обучение.
ПРОИСХОЖДЕНИЕ и эволюция позвоночных
О происхождении и ранней эволюции хордовых можно лишь
строить обшие гипотезы, так как ископаемые остатки, позволяю-
щие достоверно судить об их связях с другими, в том числе с
более примитивными типами животных, отсутствуют. Общий план
строения и особенности развития (вторичноротость и вторично-
полостность) сближают их с иглокожими, полухордовыми, по-
гонофорами.
Согласно наиболее распространенной сейчас точке зрения,
предками хордовых были целомические червеобразные животные,
перешедшие к малоподвижному или сидячему (прикрепленному)
образу жизни. Эволюция привела к уменьшению числа сегментов
в их теле и образованию вторичного рта. Питались эти животные
пассивно, отфильтровывая из воды пищевые частицы. Эти обита-
тели морского дна дали начало нескольким современным типам
животных. Среди них, вероятно, были иглокожие, которые, об-
разовав водно-сосудистую, амбулакральную систему и сложный
аппарат захвата пищи, приспособились к движению по разным
субстратам и стали активно питаться малоподвижными животны-
ми. Это обеспечило им биологический успех: иглокожие процве-
тают во многих морских донных биоценозах, не имея серьезных
конкурентов.
Другая ветвь привела к обособлению примитивных хордовых
животных. Видимо, в самом начале эволюции от нее отделилась
своеобразная немногочисленная группа животных — Полухордовые
(Hemichordata), которой теперь придают ранг типа.
Становление типа Хордовые (Chordata) шло, видимо, в до-
кембрии. Одна из гипотез предполагает, что формы, близкие к
примитивным оболочникам (Tunicata), обладавшие плавающей
личинкой, перешли к неотении, утратив прикрепленную стадию.
Примитивные бесчерепные (Acrania primitiva), возникшие от та-
ких форм, возможно, стали прямыми предками современных лан-
цетников. В ходе дальнейшей эволюции они дали начало подтипу
Позвоночные (Vertebrata).
Позвоночные животные в ранней эволюции разделились на две
крупные ветви: Бесчелюстные (Agnatha) и Челюстноротые
(Gnathostomata). Бесчелюстные были широко представлены в на-
чале палеозоя различными Щитковыми (Ostracodermii) — медли-
285
тельными существами с голой или покрытой щитками кожей,
сосущим детрит ртом и непарным обонятельным органом. Они не
имели челюстей и парных конечностей. Гипоцеркальный хвосто-
вой плавник свидетельствовал об обитании близ дна. Среди щитко-
вых выделяют несколько подклассов: Телодонты (Thelodontia) —
небольшие беспанцирные формы, обитавшие в прибрежных уча-
стках морей и в морских лиманах. Развитием костного панциря на
поверхности тела отличались Разнощитковые (Heterostraca). Они
тоже были придонными животными, проводившими большую
часть жизни в морских лиманах и в дельтах рек. Согласно предпо-
ложениям специалистов, они обладали электрическими органа-
ми, а при достижении максимальных размеров приобретали на-
ружный костный панцирь. Костные щитки покрывали тело Кост-
нощитковых (Osteostraca). Были среди них и Бесщитковые
(Anaspida).
Современные представители бесчелюстных Круглоротые
(Cyclostomata) произошли, вероятно, от нектонных бесщитко-
вых подкласса Anaspida, а может быть, и телодонтов, часть кото-
рых использовала тела погибших панцирных бесчелюстных в ка-
честве источников пищи и временных укрытий. Возникшие таким
образом Миноги (подкласс Petromyzones) перешли к хищниче-
ству и эктопаразитизму в солоноватых и пресных водоемах, а
Миксины (подкласс Myxini) — к бентосоядности и своеобразно-
му хищничеству в океане. Разделение этих двух ветвей круглоро-
тых произошло, видимо, до начала карбона. Круглоротые — пер-
вый класс позвоночных, у которых сформированы отделы голов-
ного мозга, характерные для остальных позвоночных животных.
Значительные эволюционные преимущества перед бесчелюст-
ными приобрели ранние Челюстноротые (Protognathostomata), у
которых развились парные плавники, а первые висцеральные дуги
были преобразованы в челюстной аппарат, служащий для актив-
ного захватывания пищи. Способность к активному питанию и
быстрому маневренному движению повысили жизнедеятельность
челюстноротых, позволили им широко распространиться в вод-
ной среде, сформировав самый многочисленный по числу видов
и особей и разнообразный по жизненным формам надкласс Рыбы
(Pisces).
Исторически сравнительно рано среди рыб оформились два
класса: ныне полностью вымерший класс Панцирные рыбы
(Placodermi) и сохранившийся до настоящего времени класс Хря-
щевые рыбы (Chondrichthyes). Появившись в начале девонского
периода, хрящевые рыбы сохранили основные особенности орга-
низации своих предковых форм.
Немногочисленные ныне среди хрящевых рыб Цельноголовые,
или Химеровые, в особенностях организации сочетают черты пла-
стинчатожаберных и панцирных рыб, занимая как бы промежу-
точное положение между ними. В мелу цельноголовые обитали в
прибрежных мелководных участках морей, однако в дальнейшем
вследствие развития и распространения костных рыб были вытес-
нены на большие глубины.
Крупнейшая группа хрящевых рыб Пластинчатожаберные (под-
класс Elasmobranchii) поначалу, в палеозое, были многочислен-
ными обитателями солоноватых вод морских лиманов, в кайно-
зое, за единичными исключениями, они стали обитателями мо-
рей. Среди современных пластинчатожаберных выделяют надот-
ряды Акулы и Скаты. В настоящее время сохранились (по-видимо-
му, весьма консервативные) реликты палеозоя — плащеносные
акулы, известны с юры морские ангелы и гребнезубые акулы, с
мела — пилоносы, пилы-рыбы, разнозубые акулы. Прогрессив-
ную группу пластинчатожаберных рыб составляют ламноидные,
кархарины и некоторые скаты. Именно они играют сейчас важ-
ную роль в экосистемах морей, занимая вершины трофических
пирамид. Небезынтересно, что скатообразные формы не раз воз-
никали в длительной эволюции хрящевых рыб.
Современные Хрящевые рыбы — процветающая группа, засе-
лившая обширные районы Мирового океана и некоторые пре-
сные водоемы. Она насчитывает около 50 семейств, несколько сот
видов акул, скатов и химер.
Своеобразны обитатели солоноватых и пресных водоемов пер-
вой половины палеозоя — Челюстножаберные рыбы, или аканто-
дии (класс Acanthodii — сравнительно быстро вымершие). Разные
авторы относят их то к истокам хрящевых, то к началу становле-
ния класса Костные рыбы (Osteichthyes). Этот класс самый много-
численный среди рыб. Исторически очень рано, возможно, на
уровне общих предковых форм произошло разделение класса Ко-
стные рыбы на подклассы Лучеперые (Actinopterygii) и Лопасте-
перые (Sarcopterygii). Древние лучеперые рыбы принадлежали над-
отряду Палеониски (Palaeonisci). Они очень широко дивергирова-
ли уже в палеозое. От одного из стволов палеонисков серией вет-
вей возникли Ганоидные рыбы (надотряд Holostei), заселявшие в
мезозое солоноватые и пресные воды. Ныне они представлены
двумя группами реликтов мелового периода — панцирными щу-
ками и ильными рыбами пресных водоемов Северной и Цент-
ральной Америки. Полагают, что в результате эволюционной пе-
рестройки (возможно, нескольких ветвей) в группе панцирнико-
вых сформировался надотряд Костистые рыбы (Teleostei). Появив-
шись в морских лиманах палеозоя, они широко распространились
в раннем мелу в морях и пресных водоемах, а впоследствии, в
кайнозое, стали господствовать во всех водоемах планеты.
По-видимому, от палеонисков возникли хрящевые ганоиды
(надотряд Chondrostei), в среднем мелу они уже разделились на
осетровых рыб и веслоносов.
987
Лопастеперые отличались от Лучеперых мясистыми конечнос-
тями (мышцы располагались на самих парных конечностях) и
возможностью использовать органы дыхания для потребления
кислорода воздуха, так как имели легкие и сквозные ноздри. Они
развивались в палеозое двумя самостоятельными параллельными
линиями, воплотившимися в подклассы Двоякодышащие (Dipneusto-
morpha) и Кистеперые (Crossopterygii) рыбы.
Кистеперые рыбы, имевшие унисериальные парные плавни-
ки, были неоднородной группой хищников солоноватых и пре-
сных водоемов палеозоя. Одна их ветвь, известная под общим на-
званием остеолепиформные рипидистии (Rhipidistia), дала нача-
ло наземным, или четвероногим (надкласс Tetrapoda), позвоноч-
ным. Представители другой ветви — целаканты — в девоне, еще
не успев приобрести все общие для кистеперых черты организа-
ции, переместились жить в океан, освоив относительно глубокие
морские воды; здесь они очень медленно изменялись и дожили до
наших дней в качестве единственного «живого ископаемого» —
латимерии (Latimeria chalumnae).
Двоякодышащие рыбы проигрывали кистеперым в организа-
ции парных конечностей: основой структуры их парных плав-
ников был бисериальный тип, располагавший к специализации.
У протоптерусов конечности превратились в усики. Современные
двоякодышащие рыбы (надотряд Dipnoi) сохранились в пресных
водоемах Африки, Австралии и Южной Америки. Полагают, что
они отделились от общих с кистеперыми предковых форм еще в
палеозое. Две современные ветви двоякодышащих рыб — австра-
лийская и африкано-южноамериканская — известны, согласно
палеонтологической летописи, с конца палеозоя. Такие поведен-
ческие адаптации, как зарывание в грунт, образование ильного
«кокона» при высыхании водоемов, впадение в спячку, видимо,
сформировались у них в среднем и позднем девоне.
По современным представлениям, первые наземные позвоноч-
ные произошли от остеолепиформных рипидистий (Rhipidistia).
Полагают, что климатические условия в девонском периоде были
очень изменчивы. Это способствовало адаптивной радиации ри-
пидистий. Одни активно осваивали пресные водоемы, другие пе-
реходили к жизни в морских лагунах, которые периодически пе-
ресыхали. Приспосабливаясь к полуводному образу жизни, они
приобретали свойства, послужившие в дальнейшем предпосыл-
ками к выходу на сушу. Приобретение адаптаций к полуназемно-
му существованию происходило среди различных групп рипидис-
тий, и часть «четвероногих рыб», или «полуамфибий», так и оста-
лись слепыми ветвями эволюции. Наиболее реальными переход-
ными к амфибиям формами считают ихтиостегид. Они положили
начало крупной группе амфибий — стегоцефалам, которые также
были неоднородны. Среди них выделяются лабиринтодонты, ле-
поспондилы и эмболомеры. Эволюционная судьба двух первых
групп реализовалась в последующем многообразии класса Амфи-
бии. Потомки третьей группы, эмболомеров, по-настоящему всту-
пили на сушу (И.И.Шмальгаузен, 1964).
Современные амфибии, составляющие надотряд Lissamphibia,
представлены тремя отрядами: Хвостатые (Urodela), Бесхвостые
(Anura) и Безногие (Apoda). Они сформировались в палеозое от
сильно различающихся предковых групп. Считается, что бесхвос-
тые амфибии стали развиваться обособленно от хвостатых и без-
ногих более 350 млн лет назад.
В позднем периоде палеозоя, в перми, с объединением Гонд-
ваны и Лавразии в огромный сверхматерик Пангею, климат на
нем стал континентальным и более засушливым. В таких условиях
влаголюбивые стегоцефалы постепенно вымирали. Им на смену
пришли рептилии, лучше приспосабливавшиеся к наземным био-
ценозам. Среди рептилий сложились две разные эволюционные
ветви: одна из них — зауропсидная — включает классы Рептилии
(Reptilia) и Птицы (Aves), а другая, терапсидная, последователь-
но выразилась в классах Зверообразные рептилии (Theromorpha)
и Млекопитающие (Mammalia).
Наиболее ранними рептилиями считают котилозавров. Их от-
личал анапсидный (безвпадинный) тип черепа. Среди современ-
ных рептилий прямыми потомками анапсидных котилозавров
могут рассматриваться черепахи, во многом особняком стоящие
среди других рептилий. Из современных видов только пресно-
водные черепахи мелких водоемов сохранили образ жизни и уро-
вень организации своих далеких предков. Мягкокожистые чере-
пахи мало изменились за последние 100 млн лет, сохранив мно-
гие связи со средой, установившиеся у Них на рубеже раннего
мела. Предки морских черепах отделились от общего для черепах
ствола почти 120 млн лет назад, а бокошейные — еще раньше,
почти с триаса.
Другие группы рептилий имели в черепе впадины. В соответ-
ствии с их расположением и развивавшейся активно мускулатурой
выделены группы синапсидных, диапсидных, парапсидных и эв-
риапсидных. Две последние ветви, давшие начало ихтиозаврам и
плезиозаврам, оказались тупиковыми ветвями эволюции рептилий.
От мелких, подвижных насекомоядных котилозавров ведет свое
начало группа диапсидных рептилий. Среди них — сохранившаяся
с начала мезозоя до нашего времени в Новой Зеландии реликто-
вая форма из отряда Клювоголовые (Rhynchocephalia) — гатте-
рия, или туатара. По всей вероятности, она обитает в малоизме-
нившихся за этот долгий период условиях небольших островов.
Широкая адаптивная радиация современных ящериц (отряд
Lacertilia) происходила в основном с середины мезозоя. Нынеш-
ние представители гекконовых, агамовых, сцинковых, настоящих
TRQ
ящериц, варановых и других семейств развивались, по-видимо-
му, параллельно от общих предковых форм рептилий.
Змеи (Serpentes), филогенетически самый молодой отряд со-
временных рептилий, в меловом периоде прошли стадию роющих
предков, имевших конечности. Свидетельством наличия конечно-
стей у предков змей являются их рудименты у некоторых удавов,
сохранившиеся в рудиментарном состоянии мышцы, управляю-
щие конечностями, и соответствующие нейронные образования в
спинном мозге. В группе безногих ящериц (отряд Amphisboenia)
утрата конечностей и переход к роющему образу жизни также
вторичен по отношению к их активным бегающим предкам.
Большую группу диапсидных рептилий составили архозавры (под-
класс Archosauria). В мезозое некоторые представители этой крупной
группы (типичные представители — динозавры) отличались слож-
ным поведением и, возможно, некоторые из них приобрели даже
уровень теплокровности (динозавры, летающие ящеры Ptero-
sauria). Единственные современные представители архозавров —
крокодилы (отряд Crocodilia) отличаются сравнительно высоким
уровнем организации. Наиболее примитивные среди них аллигато-
ры и кайманы. Среди динозавров, видимо, возникли птицы.
Происхождение птиц трактуют по-разному. Согласно одним
воззрениям, предками птиц были ранние архозавры, в частности
текодонты (отряд Thecodontia). Другое предположение допускает
происхождение птиц от группы мелких динозавров из отряда
Saurischia. Они обладали бипедией (бегали на двух ногах), имели
увеличенный мозг и некоторые другие общие с птицами черты.
Полагают, что в эволюции позвоночных было несколько попыток
выйти в воздушную среду, и некоторые из них оказались тупико-
выми (например, хорошо известный всем Archeopteryx). В послед-
нее время широко обсуждается вопрос о существовании одной из
возможных переходных форм от рептилий к птицам. Эта форма,
названная Protoavis, старше археоптерикса более чем на 75 млн
лет, обладала большим набором «птичьих» черт, чем археопте-
рикс. Расхождение основных стволов птиц на килегрудых и бески-
левых свершилось очень рано — в мелу, когда шло интенсивное
формирование, над отряд ов и отрядов этого класса.
Полагают, что представители из отрядов казуаров, тинаму,
сорных кур и сейчас живут в условиях, близких к тем, в которых
жили их предки в меловом периоде мезозоя.
Особенности и детали становления класса Млекопитающих
сегодня известны в большей степени, чем какого-либо другого
класса позвоночных. Их эволюция имеет длительную историю, и
формирование зверей большую часть времени происходило на фоне
небывалого расцвета рептилий. Млекопитающие оформились сре-
ди синапсидных рептилий: их череп имел нижнюю впадину. В по-
следовательной эволюции они прошли стадии Звероподобных
эап
(Theromorpha), затем — Зверозубых (Theriodontia). Таким обра-
зом, два главных ствола класса Млекопитающие (Mammalia) не-
сомненно выделились еще на «рептильной» стадии и ныне пред-
ставлены подклассами Первозвери (Prototheria) и Настоящие Звери
(Theria).
Еще в перми основную массу наземных четвероногих представ-
ляли потомки котилозавров — зверообразные рептилии. В течение
всей перми и большей части триаса в этой группе многократно
появлялись и исчезали растительноядные, хищные и всеядные
виды, образуя сменяющие друг друга волны адаптивной радиа-
ции. Среди них были как крупные (трех-, четырехметровые), так
и мелкие (10-сантиметровые) животные совершенно разных жиз-
ненных форм. В перми зверообразные процветали и господствова-
ли во всех экосистемах суши. Они вымерли лишь в юре, уступив
место архозаврам, в первую очередь динозаврам. Без преувеличе-
ния пермский период можно назвать эрой зверообразных.
Среди зверообразных выделялась одна группа рептилий — Зве-
розубые, или Териодонты, представители которой стали прямы-
ми предками млекопитающих. Появились они очень рано (как и
все зверообразные) — в конце карбона и во времени предшество-
вали большинству завропсидных рептилий. Зверозубые имели мно-
жество примитивных признаков, к которым можно отнести: ам-
фицельные позвонки, малый размер головного мозга, подвиж-
ные ребра, которые располагались и в шейном, и в поясничном
отделах, богатую железами кожу. Эти животные во многом похо-
дили на своих предков котилозавров, сохраняя в то же время «ам-
фибийные» признаки. С другой стороны, зверозубые уже на заре
возникновения обладали определенными «маммальными» черта-
ми. Зубы сидели в альвеолах и были дифференцированы. У многих
было вторичное костное нёбо. Нижняя челюсть ранних рептилий
имела разное сочленение с мозговым черепом — через сочленов-
ную и квадратную кости (как у большинства позвоночных, кроме
млекопитающих) и через зубную и чешуйчатую кости (как у мле-
копитающих). В результате в нижней челюсти зубная кость увели-
чилась, а сочленовная уменьшилась. Демаркационной линией
между настоящими рептилиями и млекопитающими палеозооло-
гии считают исчезновение в нижней челюсти всех элементов,
кроме зубной кости. Конечности у зверозубых располагались под
туловищем, а не по бокам, как у других первых наземных позво-
ночных.
В перми Зверозубые были представлены десятками специали-
зированных групп, обитавших по всей Пангее. Среди них преоб-
ладали хищные формы, например такие, как четырехметровая
иностранцевия, открытая В.П.Амалицким и названная в честь
профессора Александра Иностранцева, или трехметровые иван-
тозавры. Растительноядных было меньше, среди них очень круп-
291
ные клыкастые дицинодонты и листрозавры (всего найдены сот-
ни видов зверозубых). Все эти специализированные формы не могли
дать начало млекопитающим и вымерли уже в триасе. Для нас
наибольший интерес среди зверозубых представляют Цинодонты
(Собакозубые). Мелкие неспециализированные формы Цинодон-
тов по своей организации крайне близки к примитивным млеко-
питающим. В частности, у этих рептилий были чувствительные
волоски (вибриссы), а возможно, и настоящий шерстный покров.
Некоторые ученые допускают наличие примитивной терморегу-
ляции и повышенной температуры тела у поздних цинодонтов, в
частности у циногнат. На протяжении триаса большинство зверо-
образных вымирает, а их экологические ниши занимают более
прогрессивные архозавры, которые господствовали на Земле в
течение всего мезозоя. Примитивные млекопитающие в течение
всего мезозоя жили бок о бок с динозаврами и занимали подчи-
ненное положение, не играя заметной роли в биосфере Земли.
Таким образом, прямые предки млекопитающих — зверозубые —
возникли очень рано, имели множество архаичных признаков и в
буквальном смысле были еще полуамфибиями — полурептилия-
ми. Именно этим объясняется множество «амфибийных» черт со-
временных млекопитающих. В то же время в конце своего суще-
ствования они приобрели ряд признаков, которые в дальнейшем
помогли их потомкам занять господствующее положение во всех
экосистемах Земли.
В верхнем триасе млекопитающие уже представлены двумя вет-
вями — многобугорчатыми (Multituberculata) и трехбугорчатыми
(Trituberculata — Tribotheria).
Многобугорчатые млекопитающие (Multituberculata) были спе-
циализированными растительноядными животными, с хорошо
развитыми резцами и без клыков. Размеры их были мелкие, с крысу
или сурка. Специализированность многобугорчатых не позволила
им выделить новые группы млекопитающих, и они вымерли в
начале кайнозоя. Интересно, что зубы многобугорчатых крайне
похожи на зубы зародыша утконоса, что дает основание утверж-
дать их филогенетическую близость. Видимо, ранние многобугор-
чатые дали ветвь, которая позже привела к однопроходным (прав-
да, последних находят лишь в отложениях плейстоцена).
Трехбугорчатые были мелкими зверьками и питались в основ-
ном насекомыми и мелкими позвоночными. Их зубной ряд был
сплошным и менее специализированным, чем у многобугорчатых.
Головной мозг был значительно больше, чем у их предков терио-
донтов, особенно хорошо развиты большие полушария переднего
мозга. В меловом периоде от трехбугорчатых произошли настоящие
звери — сумчатые и плацентарные. Обе эти ветви возникли при-
близительно в одно время, по крайней мере плацентарные не по-
зднее сумчатых и эволюционировали они параллельно. Десятки
292
миллионов лет в мелу настоящие звери развивались достаточно мед-
ленно на фоне подавляющего господства архозавров. По мере выми-
рания рептилий роль млекопитающих в экосистемах заметно возра-
стала, и в начале кайнозоя уже произошла их широкая адаптивная
радиация. В это время млекопитающие освоили все среды жизни: не
только наземную, но и воздушную, водную, подземную. Такое раз-
витие в эволюционном масштабе вполне можно назвать бурным.
Сумчатые и плацентарные представляют собой две ветви мле-
копитающих, отошедшие от общей предковой группы триботе-
рий (Tribotheria) в среднем мелу. Возможно, консервативными
реликтами того времени среди сумчатых сейчас являются амери-
канские автохтонные опоссумовые. Исходя из современных нахо-
док, родиной сумчатых следует считать Северное полушарие, где
они долгое время были широко распространены и активно рассе-
лились на юг (до сих пор они сохранились в Америке). Большое
разнообразие сумчатых Австралии объясняется ранним отделени-
ем ее от других материков в меловом периоде, когда плацентар-
ные еще не были так широко распространены, как сумчатые. Поз-
же благодаря более эффективному размножению плацентарные
заняли господствующее положение на других материках, а сумча-
тые везде были вытеснены и, кроме Австралии, сохранились только
в Америке. Многие вопросы, связанные с путями расселения сум-
чатых, до сих пор остаются дискуссионными.
Наиболее архаичные плацентарные относились к ныне вымер-
шим представителям отряда Protheutheria, которые жили с конца
раннего мела до палеогена. От ранних форм Protheutheria отдели-
лись предки современных неполнозубых и насекомоядных, раз-
вивающиеся далее тремя параллельными ветвями: тенрековые,
ежеобразные и землеройкообразные. В конце мела от Protheutheria
отделились ветви, в дальнейшем давшие первых приматов, руко-
крылых и шерстокрылых. Группы протеутерий, давшие начало гры-
зунам, уже в то время стояли далеко от форм, которые были пред-
ками современных зайцеобразных.
Одной из важных групп плацентарных, возникших от проуте-
рий, были древние копытные (отряд Condylarthra), сформировав-
шиеся в позднем мелу. Используя растительные корма, как кон-
сументы первого порядка, они резко повысили свою численность,
широко распространились по суше и дали широкую адаптивную
радиацию. Полагают, что из группы древних копытных развились
такие ветви плацентарных, как отряды Трубкозубые, Даманы,
Сирены, Хоботные, Парнокопытные, Мозоленогие, Непарноко-
пытные, Китообразные и некоторые другие. При этом даже в пре-
делах одного отряда разные группы иногда возникали от разных
предковых форм, как, например, зубатые и усатые киты.
Становление крупнейшего отряда плацентарных — Хищные
млекопитающие (Carnivora) — шло непосредственно от протеу-
293
терий или от очень ранних, древних копытных, от группы, тяго-
тевшей к плотоядности. Среди хищных наиболее близки к своим
палеогеновым предкам по общей морфофизиологической орга-
низации представители семейств Виверровые и Псовые, предста-
вители же семейства Кошачьи наиболее продвинуты и специали-
зированы по сравнению со своими предковыми формами. Древ-
ние хищные, приспособившись к жизни в воде, дали начало группе
ластоногих, среди которых ушастые тюлени, видимо, наименее
изменились по сравнению с ранними формами ластоногих, жив-
шими на рубеже палеогена и неогена.
* * *
Таким образом, возникнув в меловом периоде, плацентарные
уже 50 — 55 млн лет назад были представлены практически всеми
современными отрядами, а в олигоцене (26 — 38 млн лет назад)
разнообразие зверей достигло максимума и в дальнейшем лишь
сокращалось. Из 32 известных отрядов зверей 14 отрядов вымерло.
Итак, эволюция основных групп позвоночных во многих слу-
чаях шла независимыми, параллельными ветвями, крайне нерав-
номерно по темпу и времени. Это существенно отразилось на их
морфофизиологической организации.
ЗАКЛЮЧЕНИЕ
Существование типа Хордовые (Chordata) было обосновано
известным русским зоологом А.О. Ковалевским (1840—1901), ус-
тановившим принципиальное сходство в организации и развитии
бесчерепных (Acrania) и оболочников (Tunicata) с позвоночны-
ми (Vertebrata) животными. Название типа Chordata было предло-
жено Беллом в 1878 г. Современная систематика типа Хордовые
включает следующие его подразделения:
— подтип Бесчерепные (Acrania)
— класс Головохордовые (Cephalochordata)
— подтип Личиночно-хордовые (Urochordata), или Оболочни-
ки (Tunicata)
— класс Асцидии (Acidiae)
— класс Сальпы (Salpae)
— класс Аппендикулярии (Appendicularia)
— подтип Черепные (Craniata), или Позвоночные (Vertebrata)
— надкласс Бесчелюстные (Agnatha)
— класс Круглоротые (Cyclostomata)
— надкласс Рыбы (Pisces)
— класс Хрящевые рыбы (Chondrichthyes)
— класс Костные рыбы (Osteichthyes)
— надкласс Четвероногие (Tetrapoda)
— класс Амфибии (Amphibia)
— класс Рептилии (Reptilia)
— класс Птицы (Aves)
— класс Млекопитающие (Mammalia).
Кроме того, в пределах подтипа Позвоночные (Черепные)
принято выделять две группы:
А. Первичноводные (Anamnia) — включают классы Круглоро-
гые, Хрящевые рыбы, Костные рыбы, Амфибии.
Б. Наземные (Amniota) — включают классы Рептилии, Птицы
и Млекопитающие.
Как было сказано во Введении к настоящему изданию, тип
Кордовые объединяет животных, характеризующихся единством
морфологической организации. Анализ данных сравнительной ана-
томии и морфологии, эмбриологии и палеонтологии позволяет
/видеть родственные связи между систематическими группами типа,
"енеральный путь становления типа Хордовые определил следу-
ющие признаки: внутренний осевой скелет (хорда), трубчатый
тип центральной нервной системы, висцеральные щели в пище-
варительной трубке, замкнутая кровеносная система.
Каждый этап эволюции типа, характеризовавшийся возник-
ноЬением очередного класса, сохраняя генеральные черты орга-
низации типа, сопровождался своими крупными преобразова-
ниями. В последовательной морфофизиологической эволюции хор-
довых сочеталось чередование крупных прогрессивных измене-
ний с многообразием на каждом этапе эволюции частных при-
способлений.
Становление подтипа Позвоночные (Черепные) сопровожда-
лось приобретением заместившего хорду хрящевого и костного
осевого скелета в виде позвоночника и мозгового черепа, образо-
вавшегося вновь, — висцерального скелета; вычленением в не-
рвной трубке головного мозга, в кровеносной системе сердца; об-
разованием в дыхательной системе жаберных лепестков и в выде-
лительной системе почек с гломерулярными структурами извле-
чения мочи.
Совершенствованию жизни в воде способствовало образова-
ние парных конечностей — плавников и парных челюстей. Эти
черты полностью оформились у представителей надкласса Рыбы.
В механизме дыхания существенную роль сыграло приобретение
костными рыбами жаберной крышки.
Внедрение в наземно-воздушную среду обитания оказалось
возможным путем смены органов движения — хвостового плав-
ника и одночленных парных плавников — на наземный пятипа-
лый тип конечности, а органов дыхания на легкие. Впервые это
сочетание реализовалось в классе Амфибий, представители кото-
рого во многом сохранили черты своих водных предков.
Последующая прогрессивная эволюция типа связана с освое-
нием экологического разнообразия условий наземно-воздушной
среды обитания. Этому прежде всего сопутствовала принципиаль-
ная перестройка характера размножения. Наружное оплодотворе-
ние полностью сменилось внутренним; яйца приобрели большой
запас питательных веществ и защитные яйцевые оболочки. Диф-
ференцировка тканей эмбрионов в раннем онтогенезе сопровож-
далась образованием зародышевых оболочек. Массовыми способа-
ми размножения стали: яйцекладность, доведенная до совершен-
ства у птиц, и живорождение, выкармливание детенышей моло-
ком у млекопитающих с последующей заботой о потомстве, ха-
рактерной для представителей классов птиц и млекопитающих.
Вершиной морфологических приспособлений в размножении стало
образование плаценты.
Значительная изменчивость показателей экологических факто-
ров в наземно-воздушной среде обитания (колебания температу-
ры, влажности, инсоляции, плотности среды и т.д.) способство-
г
вала интенсификации обменных процессов у животных. Это со-
провождалось перестройками в системах органов. Образование 3-
и 4-камерного сердца привело к разделению артериального и ве-
нозного потоков крови в двух кругах кровообращения. В органах
дыхания мешковидные легкие сменились трубчатыми (у птиц) и
альвеолярными (у млекопитающих), что увеличило поверхность
соприкосновения крови с воздухом и естественно отразилось на
интенсивности метаболизма. Образование грудной клетки (и диа-
фрагмы у млекопитающих) активно изменило механизм венти-
ляции легких. Формирование в органах выделения тазовых почек
сопровождалось усилением механизма реабсорбции воды, позво-
лившим максимально экономить ее в организме. Изменение струк-
туры покровов также способствовало уменьшению и регуляции
потери жидкости телом животного.
Крупные эволюционные изменения происходили также в не-
рвной и гуморальной системах. Разделение нервной трубки на го-
ловной и спинной мозг, формирование пяти отделов головного
мозга, выделение в переднем мозге нервных структур — коры и
подкорки, совершенствование периферической и образование
вегетативной нервной системы и, наконец, совершенствование
органов чувств — все названные черты приняли участие в усиле-
нии нервно-чувствующей деятельности, повышении ее ответствен-
ности за работу органов тела, в усложнении поведенческих актов
и формировании у высших позвоночных элементарной рассудоч-
ной деятельности.
Замечательным достижением морфофизиологической органи-
зации хордовых следует считать теплокровность птиц и млекопи-
тающих со сложными механизмами терморегуляции. Интенсив-
ное теплообразование и удержание температуры тела на постоян-
ном уровне — это результат интеграции функциональных прояв-
лений всех систем органов.
Сейчас известно, что млекопитающие воспринимают объем-
ность и размерность пространства, оценивают направление по-
ступающего сигнала, способны к счету, экстраполяции инфор-
мации, к обучению. Высокий уровень нервно-чувствующей дея-
тельности проявляется в особенностях образа жизни в поведении,
организует внутривидовые отношения, структуру вида и весь спо-
соб его существования.
Большое внимание в современной оценке эволюционных пре-
образований хордовых отводят морфоэкологической эволюции их
основных систематических групп. Наряду с поступательной эво-
люцией в пределах всего типа (как и всего животного царства)
конвергентно оформились жизненные формы, или адаптивные
типы.
Исторический процесс привел не только к заметным крупным
изменениям морфофизиологической организации групп, но и к
797
увеличению их видового богатства и экологического разнообра-
зия. Хордовые распространены практически во всех климатиче-
ских поясах земного шара. Они не только разделены на первично-
водных и первичноназемных. В пределах водной и наземно-воз-
душной оболочек Земли ими освоены все многочисленные вари-
ации местообитаний. Естественно, экологическая радиация в пре-
делах классов привела к значительным изменениям анатомомор-
фологического облика и особенностей физиологии входящих в
них видов.
Экологическая радиация первичноводных позвоночных позво-
лила им освоить пресную и соленую воду. В любом водоеме они
заселяют его различные «этажи», распределяясь по ярусам обита-
ния. Не менее масштабна экологическая радиация наземных по-
звоночных. Они освоили сушу всех природных зон и климатиче-
ских поясов. Мир позвоночных особенно богат и разнообразен в
тропических лесах и саваннах и беден в зонах безводных пустынь и
вечных снегов. Местами обитания позвоночных стали все этажи
наземно-воздушной среды: поверхность почвы, травянистый и
древесный ярусы. Они внедрились в почву и освоили воздух. Мно-
гие освоили переходные местообитания — полуводные, полуна-
земные. Часть наземных вторично вернулась в водную среду.
* * *
Многообразие морфоэкологических адаптаций хордовых — са-
мостоятельная область исследований. В настоящем издании мы
выборочно прикоснулись лишь к некоторым характеристикам.
Хотелось бы еще раз напомнить читателю о необходимости уг-
лубленного изучения сравнительной анатомии и морфологии. Пе-
речитывание даже очень хороших учебников не может заменить
знакомства с реальными объектами, которые надо изучать в срав-
нении.
СПИСОК ЛИТЕРАТУРЫ
Андреева Н. Г., Обухов Д. К. Эволюционная морфология нервной систе-
мы позвоночных. — СПб.: Изд-во «Лань», 1999.
Гуртовой Н. И, Матвеев Б. С., Дзержинский Ф.Я. Практическая зоото-
мия позвоночных. — Т. 1. Низшие хордовые, Бесчерепные, Рыбы. — М.:
Высшая школа, 1976; — Т. 2. Земноводные, Пресмыкающиеся, 1978; — Т. 3.
Птицы, Млекопитающие, 1992.
Дзержинский Ф.Я. Сравнительная анатомия позвоночных животных. —
М.: Изд-во МГУ; «ЧеРо», 1998.
Константинов В. М., Наумов С. П., Шаталова С. П. Зоология позвоноч-
ных. — М.: Изд-во «Академия», 2003.
Кэрролл Р. Палеонтология и эволюция позвоночных: в 3 т. — М.: Мир.
Т. 1 - 1992; Т.2-3 - 1993.
Левушкин С. И., Шилов И. А. Общая зоология. — М.: Высшая школа,
1994.
Мануйлова Н.А. Гистология с основами эмбриологии. — М.: Просве-
щение, 1973.
Наумов Н. П., Карташев Н. Н. Зоология позвоночных: в 2 т. — М.: Выс-
шая школа, 1979.
Наумов С.П. Зоология позвоночных. — М.: Просвещение, 1973.
Огнев С. И. Зоология позвоночных. — М.: Советская наука, 1945.
Проссер Л. (ред.) Сравнительная физиология животных: в 3 т. — М.:
Мир, 1977-1978.
Ромер А., Парсонс Т. Анатомия позвоночных: в 2 т. — М.: Мир, 1992,
1994.
Савельев С. В. Сравнительная анатомия нервной системы позвоноч-
ных. — М.: Гэотар-Медицина, 2001.
Скрылева Л. Ф., Яценко В.Н., Микляева М.А. Анатомо-морфологические
особенности скелета хордовых животных. — Мичуринск, 2002.
Токин Б.Н. Общая эмбриология. — М.: Высшая школа, 1970.
Хадорн Э., Венер Р. Общая зоология. — М.: Мир, 1989.
Шарова И.Х., Макаров КВ. Сравнительная анатомия и эволюция жи-
вотных. — М.: Энас, 2003.
Шмальгаузен И. И. Основы сравнительной анатомии позвоночных жи-
вотных. — М.: Советская наука, 1964.
Шмидт-Ниельсен К. Физиология животных. Приспособление и среда:
в 2 т. - М.: Мир, 1982.
СОДЕРЖАНИЕ
Предисловие................................................3
Введение...................................................7
Особенности развития хордовых.............................11
Кожа и ее производные.....................................36
Опорно-двигательная система...............................60
Хорда, позвоночник.....................................60
Мозговой череп.........................................68
Висцеральный череп.....................................82
Скелет парных конечностей и его поясов.................90
Скелет непарных плавников.............................104
Мышечная система.........................................108
Органы пищеварения.......................................116
Ротовая полость.......................................118
Глотка и пищевод......................................126
Желудок...............................................126
Кишечник..............................................129
Пищеварительные железы................................132
Органы дыхания...........................................136
Дыхание водных позвоночных............................137
Жаберный аппарат...................................137
Дыхание наземных позвоночных..........................145
Плавательный пузырь...................................156
Органы кровообращения....................................159
Кровь.................................................159
Кроветворение.........................................161
Органы кровеносной системы............................163
Особенности организации кровообращения
в эволюционном ряду хордовых..........................1^6
Лимфатическая система.................................185
Терморегуляция...........................................186
Нервная система..........................................189
Головной мозг.........................................196
Передний, или конечный, мозг.......................192
Промежуточный мозг.................................195
Средний мозг.......................................196
Мозжечок...........................................1^7
300
Продолговатый мозг....................................197
Спинной мозг........................................ 198
Периферическая нервная система..........................200
Вегетативная (или автономная) нервная система...........201
Особенности строения нервной системы различных классов
позвоночных.............................................202
Органы чувств..............................................208
Первичночувствующая рецепция............................210
Органы обоняния.......................................210
Органы зрения............................;............215
Вторичночувствующая рецепция............................223
Орган боковой линии (сейсмосенсорная система).........223
Органы равновесия и органы слуха......................226
Орган вкуса...........................................235
Органы осязания.......................................236
Эндокринные железы.........................................241
Органы выделения...........................................249
Репродуктивная система.....................................263
Развитие половых желез у позвоночных....................264
Изменения в репродуктивной системе в эволюционном ряду
хордовых................................................269
Происхождение и эволюция позвоночных.......................285
Заключение.................................................295
Список литературы..........................................299
Учебное издание
Константинов Владимир Михайлович,
Шаталова Светлана Петровна
Сравнительная анатомия позвоночных животных
Учебное пособие
Редактор Г. Г. Есакова
Технический редактор Н. И. Горбачева
Компьютерная верстка: В.А.Крыжко
Корректоры В. А. Жилкина, В. В. Кожуткина
Диапозитивы предоставлены. издательством
Изд. № А-310-1. Подписано в печать 25.10.2004. Формат 60 х 90/16.
Бумага тип. № 2. Гарнитура «Таймс». Печать офсетная. Усл. печ. л. 19,0.
Тираж 5100 экз. Заказ №14045.
Лицензия ИД № 02025 от 13.06.2000. Издательский центр «Академия».
Санитарно-эпидемиологическое заключение № 77.99.02.953.Д.0034796.07.04 от 20.07.2004.
117342, Москва, ул. Бутлерова, 17-Б, к. 328. Тел./факс: (095) 330-1092, 334-8337.
Отпечатано на Саратовском полиграфическом комбинате.
410004, г. Саратов, ул. Чернышевского, 59.
academa
Книги Издательского центра
«АКАДЕМИЯ»
можно приобрести
В розницу:
• Выставка-продажа литературы издательства
(Москва, ул. Черняховского, 9, здоние Института развития
профессионального образовония). Тел./факс: (095) 152-1878
• Книжный клуб «Олимпийский» (Москва, Олимпийский пр-т, 16,
5-й этаж, место 20; 3-й этож, место 166)
• Книжная ярмарка но Тульской (Москво, Варшавское шоссе, 9,
могозин-склод «Марко»)
• Московский дом книги (Москво, ул. Новый Арбот, 8)
• Дом педагогической книги (Москва, ул. Б. Дмитровко, 7/5; ул. Кузнецкий мает, 4)
• Торговый дом «Библио-Глобус» (Москво, ул. Мясницкая, 6)
• Дом технической книги (Москва, Ленинский пр-т, 40)
• Дом медицинской книги (Москва, Комсомольский пр-т, 25)
• Могозин «Библиосфера» (Москво, ул. Марксистская, 9)
• Сеть магазинов «Новый книжный» (Москва, Сухаревская пл., 12;
Волгоградский пр-т, 78)
Оптом:
• Москво, ул. Бутлерова, 17-Б, 3-й этож, к. 328 (здание ГУП «Книгоэкспорт»).
Тел./факс: (095)334-7873, 330-1092, 334-8337.
E-moil: sales@acodemia-moscow.ru
• Москва, Автомобильный пр-д, д. 10 (территория ГУП «Таганское»).
Тел./факс: (095) 975-8927, 975-8928. E-mail: sales@academia-moscow.ru
• Санкт-Петербург, ноб. Обводного конала, д. 211-213, литер «В».
Тел./фокс: (812) 259-6229, 251-9253. E-mail: fspbocad@comset.net
(оптово-розничная торговля)
СТР «АКАДЕМ И Я »
УВАЖАЕМЫЕ ЧИТАТЕЛИ!
-^1 ИЗДАТЕЛЬСКИЙ ЦЕНТР
ACADEMA «АКАДЕМИЯ»
ПРЕДЛАГАЕТ ВАШЕМУ ВНИМАНИЮ
СЛЕДУЮЩИЕ КНИГИ:
5₽ss:
Г
Г
►
L
*
I
[
с
Г
t
к...—..
В.М. КОНСТАНТИНОВ, С. П. НАУМОВ, С.П. ШАТАЛОВА
ЗООЛОГИЯ позвоночных
Объем 464 с.
При написании учебника авторы учитывали современные тен-
денции развития зоологической науки и опирались на данные,
утвердившиеся в ней за последние 20 лет. В соответствии с дос-
тижениями экологии и эволюционного учения особое внимание
уделено важнейшим сведениям о практическом значении позво-
ночных животных, рациональном использовании и охране живот-
ного мира. Учтена специфика применения зоологических знаний
выпускниками педвузов в их преподавательской деятельности.
Для студентов биологических факультетов педагогических ву-
зов, может быть полезен студентам биофаков университетов, учи-
телям биологии.
В. М. КОНСТАНТИНОВ, С. П. ШАТАЛОВА, В. Г. БАБЕНКО и др.
ЛАБОРАТОРНЫЙ ПРАКТИКУМ ПО ЗООЛОГИИ
ПОЗВОНОЧНЫХ
ПОД РЕД, В.М.КОНСТАНТИНОВА
Объем 272 с.
В книге основное внимание уделяется работам, рассматрива-
ющим внешнее и внутреннее строение типичных представителей
каждого класса хордовых животных; занятиям по систематике клас-
сов; семинарам по происхождению и эволюции рыб, земноводных,
пресмыкающихся, птиц и млекопитающих. Практикум снабжен
рекомендациями для студентов по выполнению лабораторных
работ.
Для студентов биологических специальностей педагогических
вузов.