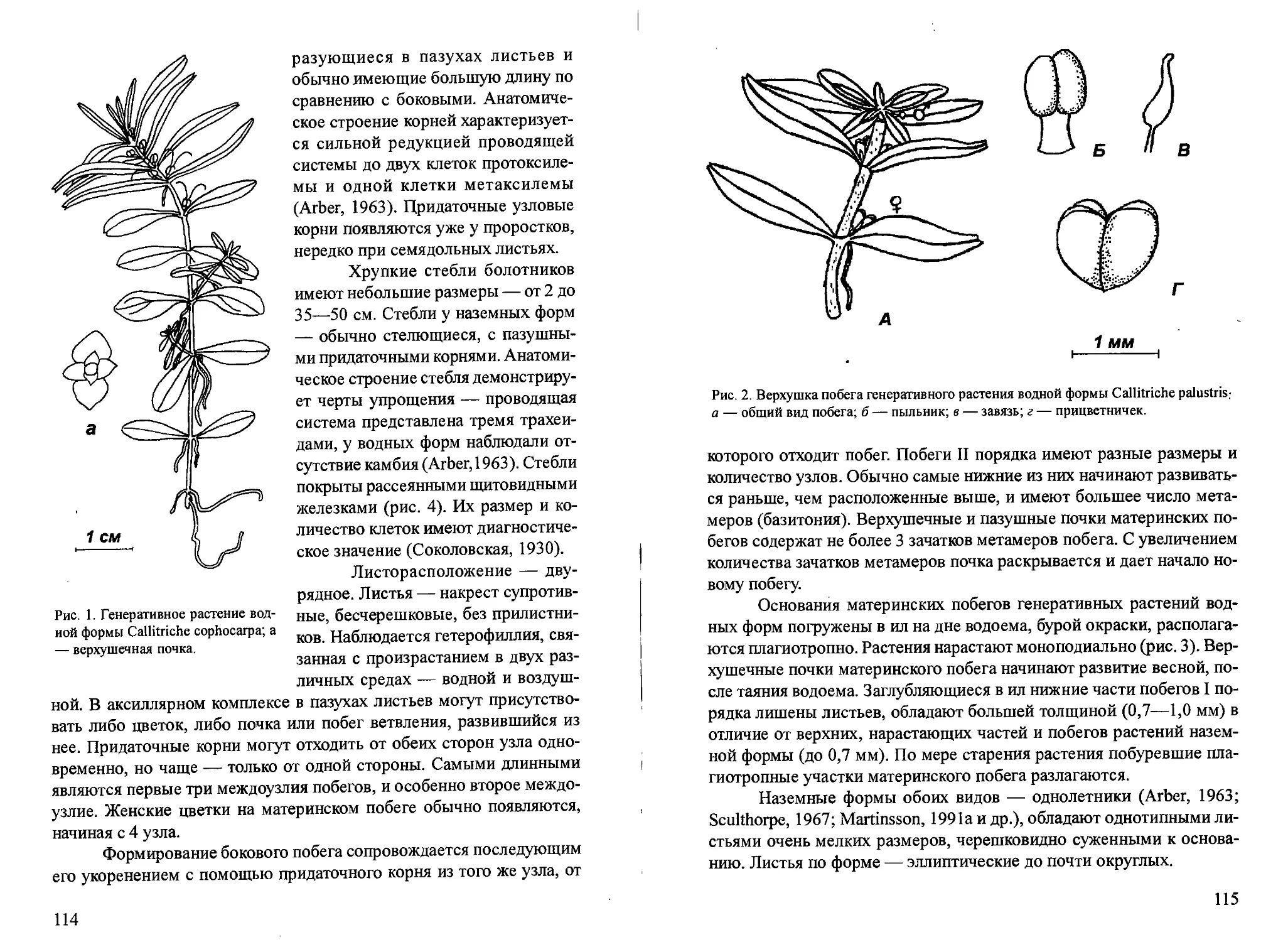
/





















































































































Author: Тихомиров В.Н. Павлов В.Н.
Tags: география растений фитогеография флора географическое распространение растений
ISBN: 5-86871-012-6
Year: 1997




Text
БИОЛОГИЧЕСКАЯ ФЛОРА
МОСКОВСКОЙ ОБЛАСТИ
Выпуск тринадцатый
Москва
1997
БИОЛОГИЧЕСКАЯ ФЛОРА
МОСКОВСКОЙ
ОБЛАСТИ
Под редакцией
профессора В.Н. Павлова,
члена-корреспондента РАН В.Н. Тихомирова
Выпуск тринадцатый
Москва
1997
УДК 581.9 (471.311)
Биологическая флора Московской области. Вып. 13. Под ред.
В.Н.Павлова, В. Н. Тихомирова. — М.: Изд-во Полиэкс, 1997. — 237 с.
ISBN 5-86871-012-6
В книге обобщены итоги оригинальных наблюдений, экспери-
ментальных работ и литературные данные по систематике, географи-
ческому распространению, морфологии, онтогенезу, экологии, консор-
тивным связям, фитоценологии, хозяйственному значению и охране
видов растений, произрастающих в Московской области. Охаракте-
ризованы как широко распространенные лесные и луговые виды и бо-
лее редкие виды растений.
Для студентов, аспирантов, преподавателей высших учебных за-
ведений и широкого круга специалистов-биологов, работников сель-
ского и лесного хозяйства, деятелей охраны природы.
Редакторы выпуска М.Г. Вахрамеева, Н.Г. Уланова
Рецензенты: доктор географических наук О.Л. Лисс,
кандидат биологических наук Л.С. Исаева-Петрова
Печатается по постановлению Редакционно-издательского Совета
Московского университета
Biological Flora of the Moscow Region. Partl3. Moscow: Poliex ,1997.—
237 p.
Chief editors - Prof. V.N. Pavlov, Prof. V.N. Tikhomirov
Edited by M.G. Vakhrameeva, N.G. Ulanova
Издание осуществлено при поддержке Российского фонда фун-
даментальных исследований по проекту А& 95-04-286506, ГНТП
«Биологическое разнообразие» и гранта государственной
поддержки ведущих научных школ № 96-15-97965
077(02) - 97 — заказное Кафедра геоботаники
МГУ, 1997
ISBN 5-85549-148-Х
ПРЕДИСЛОВИЕ
Начиная с 1974 г., опубликовано 13 выпусков сборника «Биоло-
гическая флора Московской области», в которых охарактеризовано око-
ло 260 видов растений из 50 семейств. В настоящем выпуске прини-
мали участие сотрудники Московского государственного университе-
та и Московского педагогического государственного университета,
Тверского государственного университета, Центрально-Лесного
государственного заповедника и других учреждений. Растения опи-
саны в традиционном для данного издания плане: номенклатура и сис-
тематическое положение, географическое распространение, морфоло-
гическое описание, онтогенез, способы размножения и распростране-
ния, сезонное развитие, экология, фитоценология, консортивные связи,
биологическая продуктивность, хозяйственное значение. Во всех стать-
ях раздел «номенклатура» написан чл.-корр. РАН В.Н. Тихомировым.
В разделе «экология», как и в предыдущих выпусках, использо-
ваны экологические шкалы Л.Г. Раменского (Раменский и др., 1956),
Г. Элленберга (Ellenberg, 1974) и Э. Ландольта (Landolt, 1977). В тексте
приведены только фамилии авторов экологических шкал без ссылки
на источники. Краткий перевод последних двух шкал опубликован в
6-м выпуске «Биологической флоры Московской области» (1980).
Авторы обработок таксонов во «Флорах» и «Определителях»
приведены в номенклатурных цитатах, в списке литературы даны толь-
ко названия источников. Во всех статьях сборника для обозначения
возрастных состояний растений применяются следующие условные
обозначения: р— всходы, j_ проростки, jm— имматурные, v — вир-
гинильные, g — генеративные, s сенильные.
Основную работу по комплектации и редактированию выпуска
осуществляли сотрудники кафедры геоботаники Московского государ-
ственного университета — проф. В.Н. Павлов, с.н.с. М.Г. Вахрамеева,
доц. Ю.Е. Алексеев, с.н.с. Н.Г. Уланова, а также с.н.с. Института
лесоведения РАН А.А. Маслов.
3
E.B. Maepoduee
РОГОЗ УЗКОЛИСТНЫЙ
Номенклатура, систематическое положение,
внутривидовая систематика
Typha angustifolia L. 1753, Sp. PL: 971; Кауфман, 1866, Моск.
фл.,изд. 1:591; Цингер, 1885, Сборник свед.: 404; Шмальгаузен, 1897,
Фл. Средн, и Южн, России, 2: 531; Петунников, 1901, Крит, обзор, 3:
132; Сырейщиков, 1906, Илл. фл. Моск, губ., 1: 68; он же, 1914, цит.
соч., 4: 4; он же, 1927, Опред. раст. Моск, губ.: 45; Б. Федченко, 1934,
Фл. СССР, 1: 215; Назаров, 1940, в Маевский, Фл., изд. 7: 62; Побе-
димова, 1964, в Маевский, Фл., изд. 9: 694; В. Тихомиров, 1966, Оп-
ред. раст. Моск, обл.: 54; он же, 1986, Опред. раст. Мещеры, 1: 62;
Леонова, 1979, Фл. европ. части СССР, 4: 329; C.D.K. Cook, 1980, Fl.
Europ., 5: 275; Черепанов, 1995, Сосуд, раст. России: 949. — Рогоз
узколистный (семейство Typhaceae — Рогозовые).
' Среди исследователей Typha angustifolia L. нет единой точки
зрения на объем этого таксона, но можно говорить о тенденции к его
принятию sensu latiore. Р. Graebner (1900) рассматривает Т. australis
Schum. et Thonn в качестве подвида Т. angustifolia s.L, а европейские и
американские образцы Т. angustifolia относит к типовому подвиду Т.
euangustifolia Graebner. По мнению этого автора, Т. australis не при-
надлежит к родству Т. domingensis Pers.: Р. Graebner (1900) приводит
под этим названием лишь африканские низкорослые рогозы с особым
строением мужских цветков. Существуют попытки выделения в пре-
делах Т. angustifolia внутривидовых таксонов. ТакР. Graebner (1900),
не осуществляя необходимых номенклатурных комбинаций, предла-
гает принять ряд разновидностей, описанных М. Кронфельдом
(Kronfeld, 1889) в ранге форм, но до выхода в свет монографии иногда
принимавшихся в ранге самостоятельных видов. 1) Var. media Kronfeld
— растение от 1,5 до 3 м высотой, листья плоские, их ширина 3—5
мм, мужской и женский початки по длине одинаковые. Эта разновид-
ность отмечена в ряде стран Западной Европы, в Крыму, где собира-
лась еще Д. Сырейщиковым и нами в окрестностях Красноармейско-
4
го района Волгограда. 2) Var. inaequalis Kronfeld — мужской поча-
ток значительно длиннее женского. Она отмечена в Западной Европе,
но ее распространение в целом не ясно. 3) Var. sonderi Kronfeld—муж-
ской и женский початки сближены. Наш анализ образцов Т. angustifolia
s.l. из ряда юго-восточных районов России показывает, что формы
со сближенными початками встречаются там достаточно часто. Раз-
новидность приводится для Западной Европы (Kronfeld, 1889).
Из Германии известна f. uechtritzii — с длинным (60—80 см)
неопадающим кроющим листом мужского соцветия (Kronfeld, 1889).
Из Северной Америки описана разновидность var. elongata Wiegand.
По мнению F. Segadas-Viana (1951), она отличается от типичных
представителей Т. angustifolia очень плотным парциальным кустом,
состоящим из значительного числа вертикальных побегов.
Из Австралии и Полинезии известен Т. brownii Kunth, также
считающийся разновидностью Т. angustifolia s.l. (Kronfeld, 1889) или
иногда принимавшейся в ранге proles последнего вида (например,
Graebner, 1900). Он отличается от типичных представителей вида
сближенными початками, зубчатым краем рыльца, особым строени-
ем прицветников женских цветков и волосков при основании муж-
ских цветков (Graebner, 1900).
Ряд вопросов внутривидовой систематики Т. angustifolia затра-
гивается в недавнем малодоступном исследовании А.Н. Красновой
(1993). Описанный Е.Г. Победимовой (1949) Т. foveolata является
осенней формой Т. angustifolia (Белавская, Леонова, 1975). Мы соглас-
ны, наконец, с мнением В.П. Ткачик (1993), что отнесение Т. pontica
Klokov fil. et A. Krasnova к родству T. angustifolia требует дополни-
тельных доказательств.
В качестве возможных гибридов Т. angustifolia и Т. latifolia ино-
гда описывались внутривидовые таксоны Т. latifolia s.l. (Kronfeld, 1899).
Попытка синонимизации Т. elata Boreau, который обычно принимает-
ся в ранге одного из внутривидовых таксонов Т. latifolia (Ascherson,
Graebner, 1898), с гибридом Т. latifolia х Т. angustifolia (Cook, 1980),
требует критического обсуждения, но можно утверждать, что Т.
elata Boreau морфологически отличен от известных гибридов Т. latifolia
х Т. angustifolia первого поколения.
Показано, что гибрид широколистного и узколистного рогозов
(Т. х glauca Godron) частично фертилен, а гибрид Т. domingensis Pers
s.l. х Т. angustifolia (Т. х geze Rothmaler) фертилен полностью (Smith,
1967). L.C. Marsch (1962, цит. no: Smith, 1967) показал, что первый из
5
отмеченных гибридов (Т. х glauca Godron) может быть получен только
при опылении растений Т. angustifolia пыльцой Т. latifolia, а для вто-
рого гибрида выбор материнского вида не существенен. Отношение
Т. х glauca Godron к Т. kuzmichovii A. Krasnova неясно. Известен гиб-
рид Т. schuttleworthii х Т. angustifolia, установленный К. Haussknecht
(1888) и называемый Т. х bavarica Graebner (1900). Показана возмож-
ность интрогрессии Т. х glauca с родительскими видами (Lee, 1975).
Интрогрессия гибрида Т. angustifolia х Т. domingensis с каждым из ро-
дителей предположена (Smith, 1967), но экспериментально не показа-
на.
Основное число хромосом Т. angustifolia равно 15 (Roscoe, 1927).
Известный из Австралийского флористического царства Т. mulleri
Rohr. (Rohrbach, 1869), принимавшийся в ранге subspecies Т. angusti-
folia (Graebner Р., 1900), является тетраплоидом последнего (Roscoe,
1927). Тетраплоиды Т. angustifolia указываются отечественными авто-
рами (Флора европ. части СССР, 1979), отмечены в США (Heiser,
Whitaker, 1948), но их таксономический ранг не обсуждался.
Географическое распространение
> Рогоз узколистный произрастает преимущественно в теплых и
умеренных зонах, доходя на севере почти до полярного круга. Он
растет в Европе, Западной и в ряде районов Восточной Сибири (глав-
ным образом по Ангаре и ее притокам), почти по всей Средней Азии,
на Кавказе, в Средиземноморье, Иране, Монголии, Китае, Японии,
на Дальнем Востоке, в Северной Америке, Африке (Северная Афри-
ка и территория Капского флористического царства), Австралии и
Новой Зеландии, на Канарских островах. В.Н. Голубев (1996), а так-
же Н.И. Рубцов и Л. А. Привалова (1961) относят ареал рогоза узколи-
стного к голарктическому типу. О. Sendtner (1854,1860, цит. по: Ми-
хайлова, 1940) отмечает, что верхней высотной границей для
Т. angustifolia в южной Баварии является 532 м. В других горных сис-
темах он встречается на высотах от 1500 до 1700 метров над уров-
нем моря. В Калифорнии популяции вида обитают на высотах 1525—
1830 м (Smith, 1967).
В Московской области рогоз узколистный относительно редок.
6
Морфологическое описание
Крупный длиннокорневищный многолетник высотой до 4,5 м.
От узлов и междоузлий корневища, а также от основания вертикаль-
ного побега отходят многочисленные придаточные корни двух типов:
укороченные (3—7 см длиной), обычно формирующие корни второго
порядка, и удлиненные (до 30 см), обычно без корней второго поряд-
ка. Диаметр корней — около 1 мм (Михайлова, 1940). Высказыва-
лось предположение (Loew, 1908), что придаточные корни первого
типа служат для обеспечения минерального питания растения, а кор-
ни второго типа — для его успешного закрепление в субстрате. По
нашим данным, число придаточных корней каждого типа сильно за-
висит от экологических условий конкретного местообитания.
Корневая система — короткая и глубокая, отмечено образова-
ние корневых клубней (Голубев, 1996). Корневища формируются из
диагеотропных побегов часто одинаковых по диаметру на всем протя-
жении, иногда с развитой аэренхимой, розоватые, желтоватые или зе-
леноватые, разветвленные (Loew, 1908; Grace, Wetzel, 1982а), до 0,5—
1(1,5) м длиной, с многочисленными быстро отмирающими чешуе-
видными листьями или их о статками. Диаметр корневищ—3 и более
см (Смиренский, 1950). Корневище обычно состоит из 10—12 мета-
меров до основания его дуги, которая выносит на поверхность почвы
вертикальные побеги, в основании которых оно может несколько
утолщаться (Schnizlein, 1845). В пазухах нижних чешуевидных ли-
стьев вертикального побега и на укороченных междоузлиях дуги кор-
невища часто образуются немногочисленные боковые побеги, форми-
рующие зоны кущения, и иногда значительно утолщенные в основа-
нии.
Структура надземных побегов рогоза узколистного позволяет
отнести его к полурозеточным растениям (Голубев, 1996). Срединные
листья на генеративном побеге расположены по двум ортостихам в
числе от 8 до 12 и обычно по длине превышают ось соцветия. Они
состоят из пластинки и прямого, в сечении округлого, голого влага-
лища, иногда слегка ребристого от многочисленных продольных жи-
лок, развитых на всем влагалище, включая его пленчатые края. Корот-
кие поперечные жилки есть всегда, но заметны хуже. Влагалище не-
замкнутое, часто плотно охватывает ось побега или влагалища ниже-
лежащих срединных листьев. В пределах пленчатого края влагалища
можно выделить две зоны—светлую узкую, проходящую украявла-
7
галища, и более широкую (обычно светло-коричневую) область меж-
ду краем и центральной (обычно зеленой) частью влагалища. Под дей-
ствием воды влагалища нижних срединных листьев снаружи покры-
ваются особыми пленчатыми структурами, природа которых не ясна.
Язычка нет, ушки всегда хорошо развиты.
Эпидермис клеток внутренней стороны влагалища в изобилии
выделяет слизь, функциональное значение которой не вполне ясно.
Показано (Михайлова, 1940), что в отсутствие слизи, вызванном, на-
пример, поражением рогоза узколистного некоторыми паразитами, пре-
вращающими слизь в пенистую массу, пластинки срединных листьев
засыхают. У Т. latifolia слизь выступает в роли смазки, предохраняю-
щей листья от неблагоприятного воздействия трения при ветре
(Dickerman, Wetzel, 1985). А.А. Смиренский (1950) высказывает пред-
положение, что слизь способна защищать растение от неблагоприят-
ного воздействия растворенных в воде солей.
Влагалищно-пластиночные сочленения седловидные. Пластинка
листа длинная (часто до 3 м и более), гладкая, на конце закругленная,
с остроконечием, линейно продолжает влагалище, с многочислен-
ными хорошо развитыми параллельными жилками. Края пластинки
никогда не бывают острыми. На поперечном срезе пластинки хоро-
шо заметны многочисленные воздухоносные полости (от 10 до 18),
нередко сплошь заполненные аэренхимой, отчего пластинка обычно
оказывается слегка утолщенной. Толщина пластинки неодинакова на
всем ее протяжении — ближе к остроконечию пластинка становится
тоньше. Форма поперечного среза пластинки разнообразна — от поч-
ти двояковыпуклой до дугообразной, реже почти полукруглой.
Ширина листовой пластинки у побегов, растущих из толщи воды все-
гда больше, чем у сухопутных и прибрежных побегов. Пластинка
может спирально изгибаться, делая до полутора оборотов вокруг
своей оси, снижая тем самым поверхность сопротивления ветру (Ми-
хайлова, 1940).
По нашим данным, последняя четверть пластинки срединного
листа во второй половине лета всегда хотя бы немного поникает. С ниж-
ней стороны пластинки киль не выражен, а цвет пластинки варьирует,
хотя и постоянен в пределах одной популяции и может быть зеленым
либо темно-зеленым. Иногда развивается тонкий слой сизоватого на-
лета.
Считается, что ширина пластинки срединного листа является
диагностическим признаком Т. angustifolia и в многочисленных оп-
8
ределителях и обработках Typha часто приводится верхний предел ее
изменчивости, равный 10 мм. По нашим данным, у юго-восточных
представителей Т. angustifolia s.l. этот предел может быть намного вы-
ше: ширина пластинки, хотя и очень редко, может достигать 20 мм,
несколько чаще она равняется 13—18 мм, а ее значения 8—12 мм нуж-
но считать обычными. Л.Н. Михайлова (1940) также отмечает, что ши-
рина листа у образцов рогоза узколистного из Астраханского заповед-
ника может достигать 13 мм. По нашим данным, этот показатель на
вегетативных побегах рогоза узколистного всегда несколько больше,
чем на побегах репродуктивных. Л.Н. Михайлова (1940) отмечает, что
самые нижние из срединных листьев генеративного побега всегда уже
и тоньше прочих срединных листьев.
Растение однодомное. Соцветие состоит из женского (нижне-
го) и мужского (верхнего) початков, хотя иногда соцветия Typha рас-
сматривают как один початок, поделенный на мужскую и женскую час-
ти (Celakovsky, 1885; Тахтаджян, 1966). Длина мужского початка обыч-
но составляет 10—30 см, женского — от 10 до 40 и более см, но, как
правило, оба початка, расположенные на одном побеге имеют примерно
одинаковую длину. Величина промежутка между початками раз-
лична — иногда початки соприкасаются друг с другом или расстав-
лены на расстояние до 0,5 см, чаще всего они расставлены на рас-
стояние около 4 см, изредка — до 30 см. Диаметр женского початка
в среднем равняется 1,9 см, но варьирует в пределах от 0,75 до
3,4 см.
Выяснению деталей строения соцветий, особенностей разви-
тия и цветорасположения у рогозов посвящены оригинальные ис-
следования L. Celakovsky (1885), М. Kronfeld(1889)HD. Muller-Doblies
(1970). Частично результаты L. Celakovsky (1885), как и ряд дан-
ных относительно ранних стадий развития женских соцветий Т. angusti-
folia, приводит К. Goebel (1931). Сведения о цвете молодых женских
початков различаются: указывается, что молодые початки коричневые
(Smith, 1967) или зеленые (Михайлова, 1940). Диаметр генеративно-
го побега всегда больше в основании, к вершине он сужается, дуго-
образно изгибается, его поверхность иногда становиться ребристой,
что дает повод некоторым авторам называть его «остью» (Горбик,
1988). Этим же автором приводятся данные относительно диаметра
оси женского початка у его основания и верхушки: в среднем, это
3,2±0,4 и 2,3±0,2 (у нижневолжских рогозов эти показатели обычно
намного выше). При этом длина початка изменяется в пределах от
9
152,8±14,9 до 281,3±27,9 мм. Ось женского початка, иногда называе-
мая цветоложем (Горбик, 1988), усажена многочисленными ступенча-
тыми выростами (нередко называемых «цветоножками»), на каждом
из которых расположены несколько женских цветков. Линейные раз-
меры выростов и их форма считаются систематическими признаками.
Цветки однополые. В мужских цветках тычиночная нить обыч-
но разветвляется и несет 3 пыльника. Пыльник состоит из 4 микро-
спорангиев, структура стенок которых описана В.А. Поддубной-Ар-
нольди (1982), имеет дистальный надсвязник и вскрывается двумя ще-
лями. Вскоре после цветения пыльники вместе с тычиночными нитя-
ми опадают. По всей оси мужского початка и в основании каждой ты-
чиночной нити обычно расположено несколько (2—4) уплощенных
нитевидных выростов («волосков»), иногда называемых прицветни-
ками (Morong, 1888). Их природа не ясна. У типичных представите-
лей вида они примерно одинаковы по ширине на всем протяжении и
нередко разделяются надвое в верхней части. У Т. angustifolia они обыч-
но коричневого цвета, короче тычинок и хотя бы частично долго со-
храняются после цветения на оси мужского початка, хорошо заметны
на сухом материале даже при небольшом увеличении. Пыльца состо-
ит из одиночных зерен.
Рогоз узколистный имеет 4 типа женских цветков: плодущие,
стерильные, карподии (Kronfeld, 1889; Белавская, Леонова, 1975) и
рудиментарные в виде пучка волосков (Krattinger, 1975). Завязь пло-
дущих цветков верхняя. «Околоцветник» состоит из многочисленных
волосков, отходящих от основания ножки завязи и от самой ножки.
Длина околоцветных волосков примерно равна длине прицветника,
но оба последних значительно короче рылец. Прицветник развит у всех
цветков, кроме рудиментарных. Он очень вариабелен: J.B. Geze (1912)
насчитывает до 15 его типов. Стерильные женские цветки встречают-
ся нечасто и отличаются от плодущих длиной прицветников, которая
превосходит длину рыльца. Карподии, иногда неверно называемые сте-
рильными цветками (Morong, 1888), — обратноконические, на вер-
хушке с заострением или без, развиты в большем числе, белые либо
коричневатые. Карподий обычно примерно равен по длине околоцвет-
ным волоскам.
Завязь на ножке, с 1 висячим семезачатком(Тахтаджян, 1966).
После оплодотворения ножка завязи вытягивается, образуя так назы-
ваемый «гинофор». Его длина обычно составляет 6—9 мм, но он (как
и столбик) может быть скручен в силу механической деформации (Ле-
10
онова, 1976). Околоплодник прозрачный или матовый, пленчатый, на-
верху переходит в столбик, и далее, в линейное рыльце, которое часто
опадает. По нашим наблюдениям, оно сохраняется нередко у части
цветков данного початка, что согласуется с данными Л.Н. Михайло-
вой (1940).
Единственное семя заключено внутри перикарпия, который не
срастается с семенной кожурой, разрываясь через 0,3—72 часа по-
сле того, как плод попал в воду. На поперечном срезе в перикарпии
выделяются тонкостенный экзокарпий и эндокарпий с утолщенными
оболочками. По вопросу о структуре пленчатой семенной кожуры ро-
гоза узколистного имеются противоречивые данные (Schnizlein, 1845;
Сравнительная анатомия семян, 1985). Анатомическое строение по-
перечного среза плода различных представителей Typha и его систе-
матическое значение обсуждаются Р. Rohrbach (1869) и Т. Morong
(1888). Против микропиллярной части семени плоды имеют две кры-
шечки: внутреннюю, образованную интегументами семени, и внеш-
нюю, образованную околоплодником (Сравнительная анатомия семян,
1985). Длина семени составляет в среднем около 1 мм (Marsch, 1962,
цит. по: Grace, Harrison, 1986). Она варьирует в пределах 0,9—1,2 мм,
при этом распределение семян по длине мало отличается от нормаль-
ного.
После того, как плод отрывается от оси женского початка, око-
лоцветные волоски сильно отгибаются назад. Верхние концы волос-
ков не утолщены, либо слегка утолщены (Победимова, 1949), но все-
гда иначе, чем волоски рогозов из группы Т. minima s.l. Они содержат
различное количество темного пигмента, распределенного внутри по-
лого волоска более или менее равномерно, или же в виде нескольких
пятен различного размера и формы (Smith, 1967). Прицветник обычно
долго сохраняется, при определенном навыке хорошо заметен на по-
верхности зрелого початка, часто почти прозрачный в основании, но
на верхушке всегда темнее рыльца и обычно уже его (Smith, 1967).
Нередко прицветник отваливается вскоре после отделения плода от
оси женского початка.
Онтогенез и сезонный ритм развития
Данные относительно всхожести и особенностей прорастания
семян расходятся у разных авторов. Отмечалось, что прорастание се-
мян подземное (Сравнительная анатомия семян, 1985) и подводное
(Coops, van der Velde, 1995) происходит во второй половине марта или
11
в апреле (Kronfeld, 1889). Продолжительность прорастания зависит
от температуры и оптимальна при 14—16°С (Kronfeld, 1889), хотя А.А.
Смиренский (1950) отмечает, что семена рогоза вообще не прораста-
ют, если температура воды ниже 14°. Всхожесть семян обычно состав-
ляет, как и у Т. latifolia, от 20 до 30% (Kronfeld, 1889). По другим,
уточненным данным, при оптимальной температуре в 24° семена Т.
angustifolia достигали 50% всхожести в течение трех дней неза-
висимо от предшествовавшей обработки, но при снижении темпера-
туры до 18° 50% семян прорастало только через 12 дней. Повыше-
ние температуры с 24 до 30° никак не сказывалось на интенсивности
прорастания семян Т. angustifolia (McNaughton, 1966). Было показа-
но, что при температурах прорастания ниже 24° можно добиться 3 0%
всхожести семян только после их предварительной обработки холо-
дом (Горбик, 1988).
Рогоз узколистный может иметь в почве долговременный банк
жизнеспособных семян (Grace, Harrison, 1986). В наших опытах даже
незрелые (!) сухие семена рогоза оказались способны к успешному
прорастанию: они сохраняют всхожесть после 16,5 и более месяцев
хранения в гербарии, но при этом всхожесть снижается не менее чем в
два раза. Удалось прорастить семена рогоза, взятые из образцов грун-
та со дна озера после семилетнего пересыхания последнего, при этом
грунт озера был «осолонцован» (Воронов, 1948).
Развитие проростков зависит от химического состава воды и
содержания взвесей (Sharma, Gopal, 1978), а также от гидрологиче-
ского режима местообитания: семена рогоза узколистного могут про-
растать на глубине до 40 см, но лучше всего прорастают на глубине в
1—4 см: именно при этих глубинах структура донного грунта, опреде-
ляемая размерами его частиц, оптимальна для развития проростков
(Keddy, Constabel, 1986). Для песчаных грунтов указываются и другие
значения оптимума глубины (Keddy, Ellis, 1985).
По нашим данным, проростки рогоза узколистного чрезвы-
чайно похожи на проростки Т. latifolia, морфология которых изучена
хорошо (Velenovsky, 1907; Серебряков, 1952). При прорастании семе-
ни приподнимается микропиллярная крышечка, вытягивается гипоко-
тиль, а за ним (базальная) часть семядоли, которая частично и произ-
вольным образом разрывает семенные оболочки. Длина семядоли в
несколько раз больше продольной длины семени, в дальнейшем семя-
доля нередко оказывается сильно изогнутой или даже перекручен-ной
вокруг продольной оси проростка. Через свернутое семядольную щель
12
ченной вокруг продольной оси проростка. Через свернутое семядоль-
ную щель пробивается почечка (рис.1, в). Обычно это происходит на
третий день после начала прорастания.
Корневая система проростка первоначально представлена ко-
ротким главным корнем и кольцом боковых корней, расположенных
на утолщении главного корня. «Зачаток» главного корня дифференци-
руется практически синхронно с гипокотилем и отделен от него не-
большим кольцеобразным утолщени-
ем. Первоначально он очень мал по
размеру (рис. 1, а, б). Уже на вто-
рые—третьи сутки после начала
прорастания главный корень значи-
тельно вытягивается в длину и на
нем формируются многочисленные
прозрачные или молочно-белые бо-
ковые корешки, активно врастающие
в субстрат. Гипокотиль становится
почти прозрачным (ранее он был бе-
ловатым или бело-зеленым), отчетли-
во отличается от семядоли и ста-
новится короче ее (по времени это
совпадает с формированием первого
листочка проростка). Длина гипоко-
тиля у 10-дневного проростка состав-
ляет 0,7 мм, ширина листочка — око-
ло 0,3 мм, а длина — до 10 мм, но ча-
ще 7—7,5 мм. Придаточные корни
Рис. 1. Проростки Typha angustifolia:
А — через 10 часов после начала про-
растания, Б —через сутки, В — через
8 дней, Г — через 12 дней.
не выражены и появляются на гра-
нице гипокотиля и влагалища семядоли позднее (рис.1, г).
В погруженных условиях проростки рогоза узколистного
всегда формируют наиболее длинные листья (Coops, van der
Velde, 1995). По данным Е. Warming (1897, цит.: по Loew, 1908),
семена растения способны прорастать непосредственно в пе-
рикарпии, разрывая последний, что хорошо видно из приве-
денного у Е. Loew (1908) рисунка. По нашим данным, кото-
рые согласуются с результатами Р. Rohrbach (1869), это не так
— в воде семена всегда прорастали только после того, как
выпадали из перикарпия. Пересадка проростков узколистного
рогоза на различные глубины показала, что в отсутствии про-
13
до 115 см (Grace, Wetzel, 1981b; Sharma, Gopal, 1978).
Ювенильное состояние характерно для растений первого года
жизни. Первый лист развивается через 1—1,5 месяца (рис. 2, а), а че-
рез 4 месяца растение имеет уже 3—4 листа (рис. 2, б, в). У одно-
годичного растения корневая система мочковатая. Из его диагеотроп-
ного побега формируется тонкое корневище со стеблеродными кор-
нями, первоначально отходящими от его узлов (рис. 2, г, д). Верху-
шечная почка корневища трогается в рост осенью текущего года и
образует розетку из 5—6 метамеров. Образование побега третьего по-
рядка из пазушных почек дуги начинается о сенью того же года или
весной следующего. Как отмечает S.J. McNaughton (1966), пробужде-
ние пазушных почек происходит в марте и обычно в течение 1—3
дней. Особенности роста листьев Typha изучены на примере Т. latifo-
lia (Дудинский, Бажутина, 1976). Ими в частности показано, что лист
Typha имеет единственную интеркалярную зону роста, влагалище за-
кладывается очень рано и его зачатки видны уже у второго листового
примордия.
Растение второго года имеет симподиальное корневище и мо-
жет формировать в основании вертикального побега зону кущения,
образованную, по крайней мере, одним небольшим побегом п поряд-
ка, растущим апогеотропно, хотя, согласно Е. Warming (1897), первый
вертикальный побег начинает развиваться в зоне кущения уже у юве-
нильного растения. Двулетнее растение узколистного рогоза можно
отнести к виргинильным. Листья виргинильных растений могут дос-
тигать до 1 м длины (Warming, 1897, цит. по: Михайлова, 1940).
Побеги рогоза узколистного должны быть охарактеризованы
как полициклические; генеративные побеги обычно возникают лишь
к третьему году существования парциального куста или еще позд-
нее (Михайлова, 1940). Между началом роста репродуктивного побе-
га и началом его цветения проходит от 40 до 43 дней (McNaughton,
1966). В мае популяции рогоза узколистного нередко состоят исклю-
чительно из нецветущих побегов, но уже в июне доля репродуктив-
ных побегов составляет 55—60%. Почти такое же соотношение побе-
гов можно наблюдать в июле. В августе доля репродуктивных побегов
составляет 50% или ниже, в сентябре — более 85% (Grace, Wetzel,
1982в). По данным Л.Н. Михайловой (1940), большинство клонов
Т. angustifolia никогда не дает генеративных побегов, а если и дает,
то их доля не превышает 30%. S.J. McNaughton (1966) наблюдал в зре-
лой популяции генеративные побеги у 76—98% парциальных кустов.
14
Рис. 2. Ювенильные растения Typha angustifolia ' Л —через месяц после
начала прорастания, Б—В — дальнейший рост, Г— начало формирования
диагеотропного корневища, Ц — со сформированными корневищами.
Как показали предварительные наблюдения, генеративные побеги ро-
гоза одревесневают и к сентябрю нередко становятся ломкими.
Побеги третьего года несут в основании фрагменты прошлогод-
них срединных листьев в виде немногочисленных продольных воло-
кон, нередко совершенно разрушающихся и плохо заметных, в зоне их
15
кущения имеется, по крайней мере, один апогеотропный побег ц
порядка (Grace, Wetzel, 1982а), от основания которого может отходить,
по крайней мере, еще один апогеотропный побег ш порядка (Schnizlen,
1845,1846). Таким образом, в парциальных кустах Т. angustifolia обыч-
но оказывается не менее 2 или 3 вертикальных побегов, а с учетом
того, что в зоне кущения рогоза узколистного нередко возникает не-
сколько новых диагеотропных побегов, образующих побеги ni поряд-
ка, старый парциальный куст рогоза может состоять и из большего
числа вертикальных побегов, например, из 4, 5 или более (Schnizlen,
1845). Но нередко растения рогоза узколистного образуют парциаль-
ные кусты еще более сложной структуры: по нашим данным, у взрос-
лых особей десятилетний парциальный куст имеет побеги ix поряд-
ка.
Переход растения в сенильный период сопровождается парти-
куляцией клонов и практически не изучен. Продолжительность жизни
корневищ неизвестна.
Способы размножения и распространения
Рогоз узколистный, видимо, следует отнести к облигатным ане-
мофилам, ибо возможность гидрофилии у ряда представителей Typha
не получила экспериментального подтверждения. Рогоз обычно зацве-
тает во второй половине мая или в первой половине июня (Grace,
Wetzel, 1982b), или в апреле—мае и обычно раньше, чем Т. latifolia и
Т. domingensis s.l. Конкретные сроки цветения зависят от географиче-
ского положения данной популяции. Период цветения продолжается
от 28—30 (McNaughton, 1966; Krattinger, 1975) до 45—50 дней
(McNaughton, 1975).
Для вида характерна протогиния (Белавская, 1994). Мужское
соцветие производит до 170 миллионов пыльцевых зерен (Krattinger,
1975). При сильном ветре пыльца способна переноситься на расстоя-
ние до 1 км. Наибольшее количество жизнеспособной пыльцы появ-
ляется к концу первой недели с момента начала цветения, созревание
пыльцы происходит на 1—2 дня раньше, чем раскрывание пыльни-
ков. Пыльца жизнеспособна в температурном диапазоне от —20 до
+60°. Большее количество пыльцы оседает в верхней части женского
початка, но количество пыльцы, достигшей основания женского по-
чатка у Т. angustifolia выше, чем в тех же условиях у Т. latifolia
(Krattinger, 1975). Между клонами этого вида существует факульта-
16
тивное перекрестное опыление, выраженное сильнее, чем у Т. latifolia
(Krattinger, 1975). Объяснить это можно тем, что пыльца Т. angustifolia
легче, чем пыльца Т. latifolia, она успешнее переносится ветром, а также
тем, что мужской початок Т. angustifolia обычно приподнят над жен-
ским (Krattinger, 1975).
Число опыленных женских цветков составляет 58%, из них оп-
лодотворено обычно 37%. Апомиксис не обнаружен (Поддубная-Ар-
нольди, 1982). На 1 см оси женского початка приходится в среднем 96
женских цветков (Горбик, 1984, 1988). Данные о плотности цветков в
мужском початке отсутствуют.
Семенная продуктивность Т. angustifolia изучена В.П. Горбик
(1984) на примере популяций рогоза узколистного ряда украинских
водохранилищ, причем была разработана подробная методика изме-
рения семенной продуктивности рогоза. Предложена простая форму-
ла для вычисления потенциальной семенной продуктивности (ПСП)
единичного початка: ПСП = p.L.K [(D + d)/2], где L — длина фрагмен-
та генеративного побега, несущего женский початок, D — диаметр
генеративного побега в основании початка, d — его же диаметр в верх-
ней части початка, К — число плодущих женских цветков на 1 см по-
верхности оси женского початка. Результаты расчетов и наблюдений
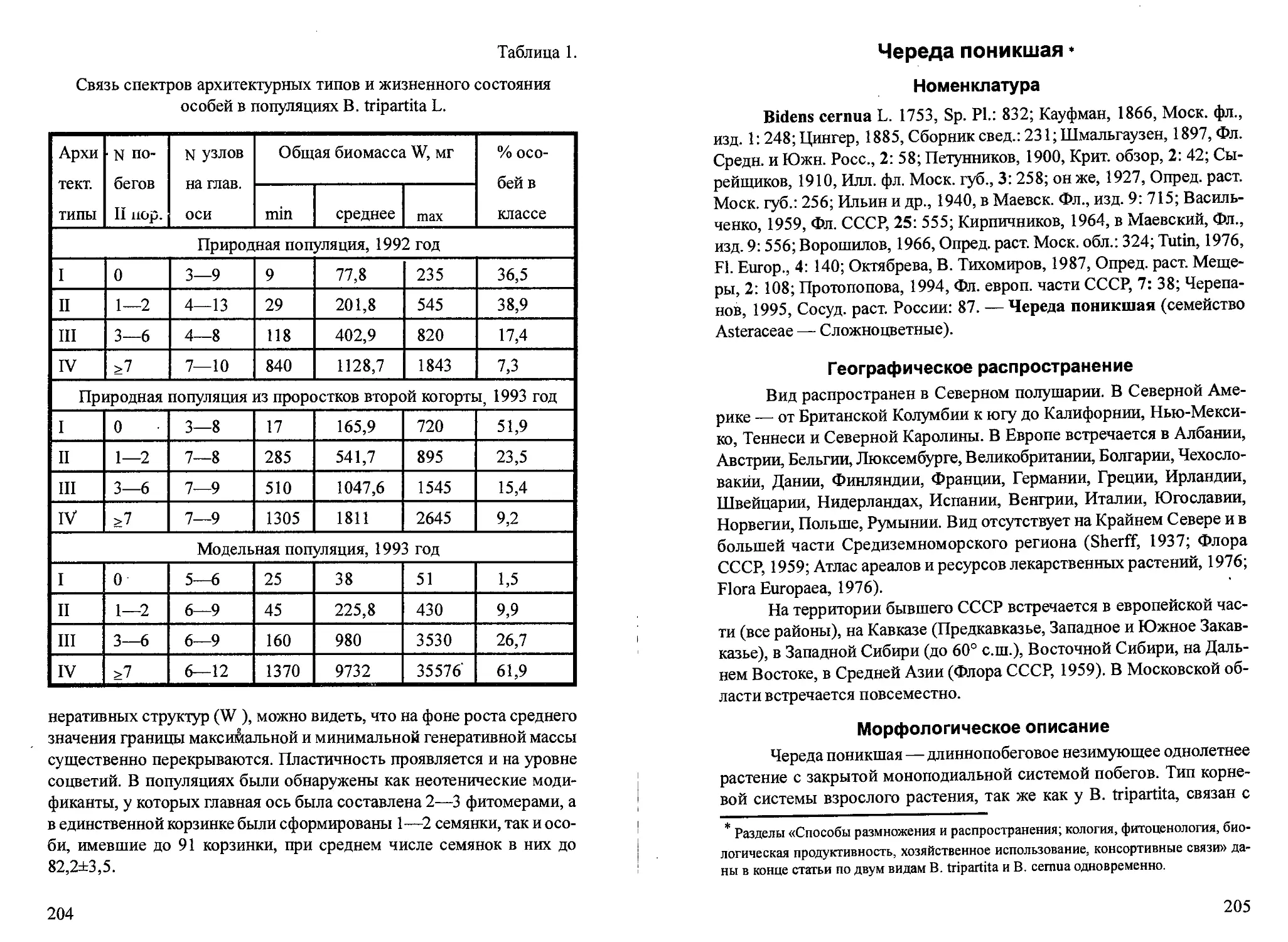
В.П. Горбик приводятся в таблице 1.
Таблица 1.
Потенциальная (ПСП) и фактическая семенная продуктивность (ФСП),
доля семенификации (ДС) и численность семян рогоза узколистного
некоторых водохранилищ Украины (по: Горбик, 1988)
Водохра- нилище ПСП, тыс. шт. ДС, % ФСП, тыс. шт. Численность, тыс. шт.\м2
Киевское 162,3±11,5 63,0±4,0 102,7±9,5 595,9±123,5
Каневское 184,0±16,1 46,8±2,6 90,3±13,0 338,8±256,7
Каховское 288,3±37,2 39,6±2,8 110,6±11,6 543,9±134,4
Показана прямая положительная зависимость среднего веса еди-
ничного семени от процента генеративных побегов в данном место-
обитании (McNaughton, 1966).
Плод рогоза узколистного имеет приспособления для эффек-
17
тивного переноса ветром (анемохории). Однако значение этого спо-
соба расселения рогоза нельзя переоценивать. В течение пяти поле-
вых сезонов мы ни разу не наблюдали массового лета «летучек» рого-
за узколистного и никогда не видели, чтобы у существенной части ге-
неративных побегов в данной популяции ось женского соцветия была
свободна от зрелых плодиков. Значительное число женских початков
со зрелыми плодами вообще остается целыми вплоть до поздней
осени (или даже зимы), когда эффективный лет плодиков уже затруд-
нен, например, ввиду намокания початков. Иногда структура зрелого
женского початка не нарушается без активного воздействия птиц
(Смиренский, 1950) или (косвенно) ветра. Особенно плохо разруша-
ются женские соцветия с большим числом карподиев, но и после
разрушения початка значительная часть плодов по-прежнему остает-
ся на его оси, а освободившееся плодики нередко падают на суб-
страт большими агрегатами и не покидают пределов местообитания,
что совпадает с наблюдениями, сделанными J.B. Grace и J.S. Harrison
(1986).
Роль гидрохории в распространении рогоза узколистного мож-
но оценить лишь условно. При попадании в воду волоски околоцвет-
ника нередко слипаются, а плоды распространяются по поверхности
воды или погружаются в воду из-за гигроскопичности волосков
(Kjattinger, 1975). Благодаря своей острой форме семена рогоза могут
застревать в чешуе рыб и таким образом выноситься за пределы ме-
стообитания. Данные о распространении рогоза узколистного стад-
ными животными и птицами носят характер предположений (Михай-
лова, 1940). Мирмекохория не отмечалась.
Хотя семенная продуктивность рогоза узколистного доволь-
но высока, однако увеличение числа особей в новых популяциях за
счет семенного возобновления не очень эффективно (McNaughton,
1966). В пользу последнего вывода косвенно свидетельствуют и дан-
ные о том, что популяции Typha (в т.ч. и Т. angustifolia) генетически
относительно однородны (Lee, 1975), т. е. возникают преимущест-
венно за счет вегетативного размножения очень небольшого числа ин-
дивидуумов и могут рассматриваться в качестве клонов. Сопоставле-
ние числа вновь образованных и погибших вертикальных побегов Т.
angustifolia привело J.B. Grace и R.G. Wetzel (1982а) к выводу о том,
что увеличение числа дочерних рамет не всегда точно отражает дей-
ствительный рост популяции — вегетативное воспроизводство боль-
ше необходимо для воспроизводства отмерших побегов, чем для соб-
18
ственно размножения.
Рост популяций Т. angustifolia происходит преимущественно за
счет образования нормально развитых вегетативных побегов, его тем-
пы невелики. Этот вывод хорошо согласуется с отсутствием обратной
зависимости между продукцией корневищ и продукцией листьев, яв-
ляющейся косвенным показателем степени развития вертикальных по-
бегов. Зависимость между двумя названными показателями у Т.
angustifolia близка к прямой, тогда как у Т. latifolia имеет обратный
характер (McNaughton, 1975). Средняя длина горизонтальных побе-
гов одного растения рогоза узколистного составляет 130 см (Aario,
1933), чаще всего вегетативные побеги возникают на глубинах от 50
до 100 см (Grace, Wetzel, 1982b).
Экология
Рогоз узколистный — воздушно-водный гидрофит, растущий в
прибрежной зоне озер, рек, прудов и стариц, а также на низинных бо-
лотах. Н. Ellenberg с соавторами (1991) относят его к растениям часто
затопляемых почв (10-я ступень шкалы увлажнения). Корневища обыч-
но размещены в донном грунте на глубине 8(12)—29 см, хотя нередко
(особенно на илистых грунтах) они оказываются непосредственно на
поверхности дна водоема (Смиренский, 1950). У вертикальных побе-
гов нижняя (обыкновенно, меньшая) часть находится в воде, а боль-
шая —1 в воздушной среде.
Глубина слоя воды в местообитаниях рогоза не превышает
300 см, но, как правило, составляет не менее 15 см (Grace, Wetzel,
1981b), по крайней мере, весной (наши наблюдения). На песчаных суб-
стратах проростки Т. angustifolia одинаково хорошо развивались на
глубинах от 5 до 10 см (Keddy, Ellis, 1985). Уровень воды является
одним из факторов, влияющих на зацветание рогоза узколистного
(Graebner et al., 1919). Доля генеративных побегов растения может за-
висеть от глубины водоема: при глубине 30—60 см она составляет 50%,
при 60—90 см — 70%, при 90—120 см — около 40%.
Максимальное количество генеративных побегов (до 100%) раз-
вивается при глубине водоема 80 см (Grace, Wetzel, 1981b; 1982а, b).
По другим данным, число генеративных побегов может быть макси-
мальным на небольших глубинах и равномерно уменьшаться по мере
увеличения глубины местообитания: на 1 м2 дна при глубине 20 см в
среднем размещалось 8, при 50 см — 6, а при 70 см — 2 генеративных
побега (Горбик, 1988). Цветение рогоза лучше всего происходит на
19
прибрежных мелководьях, а на более глубоких участках водоема «об-
ширные заросли рогоза могут быть полностью лишены цветущих по-
бегов» (Смиренский, 1950).
Плотность распределения корней и корневищ в субстрате зави-
сит от уровня воды в местообитании. Такие показатели как вес корне-
вищ, число дочерних диагеотропных побегов, длина корневищ и их
биомасса максимальны при глубине воды 100 см, а минимальны при
глубине 50 см. Длина листовой пластинки срединного листа и ее вес
зависят от уровня воды в данном местообитании и от времени года:
оба показателя максимальны в августе при глубине 80 см и минималь-
ны в мае при глубине 15 см (Grace, Wetzel, 1982а).
Рогоз узколистный плохо переносит затенение (Grace, Wetzel,
1981а) и относится к гелиофитам (Голубев, 1996). По экологическим
шкалам Элленберга он занимает промежуточное положение между по-
лусветовыми и световыми растениями (8-я ступень световой шкалы).
Число дочерних рамет клона и общая длина дочерних корневищ пря-
мо пропорциональны проценту полного солнечного света (ППС) в со-
ответствующем местообитании. Затенение, однако, положительно
влияет на линейные размеры срединных листьев: длина листа рогоза,
а также отношение длины листа к его весу, оказываются максималь-
ными при ППС равном 24%, а при ППС 100% оба показателя мини-
мальны. Средний вес рамет данной популяции также прямо зависит
от процента ППС. Таким образом, прирост биомассы клонов рогоза
при полном освещении происходит в основном за счет прироста его
подземной части (Grace, Wetzel, 1981а).
Рогоз узколистный — растение со средней потребностью в теп-
ле (Loew, 1908), по шкалам Элленберга — умеренно теплолюбив (7-я
ступень). Он хорошо растет как на прогреваемых мелководьях, так и в
холодных водоемах при температуре 14—16° С. Высокая температура
воды (в среднем 24°) во время паводков благоприятно сказывается на
его вегетации. В горных источниках рогоз не развивается, если темпе-
ратура воды в них ниже 9—12° (Михайлова, 1940). В центральной Ка-
наде рогоз узколистный распространен в тех местах, где зимой темпе-
ратура не опускается ниже —13° (Grace, Harrison, 1986). Обычно ро-
гоз начинает развиваться при температуре воды 10—12° (Белавская,
1994).'
Рогоз узколистный растет на умеренно-кислых и нейтральных
почвах (pH от 4,1 до 7,7), при содержании кальция в них от 500 до
10 000 ppm (Segadas-Vianna, 1951). Нормальный рост наблюдается и
20
на слабощелочных грунтах (pH 8,3), при этом содержание калия в почве
составляет 348 ppm, фосфора 340 ppm, натрия 440 ppm, а доля азота
составляет 0,038% (McNaughton, 1966). По шкалам Элленберга, Т.
angustifolia растет в основном на нейтральных, но также на слабо кис-
лых и основных субстратах, предпочитая почвы, богатые азотом (7-я
ступень кислотности почвы и богатства азотом).
Растение предпочитает илистые и песчано-илистые субстраты
(Белавская, 1994; Краснова, 1988), но растет также на различных гли-
нистых и торфянистых грунтах (при этом толщина торфа под рогозом
обычно не превышает 1 м). Встречается на песчаных аллювиальных
наносах, островах, отмелях и антропогенно нарушенных субстратах
— вдоль насыпей железных дорог, по карьерам, ямам, обочинам до-
рог. Процент песка в верхнем почвенном горизонте стандартного био-
топа узколистного рогоза составляет 4%, ила — 21%, глины — 75%
(McNaughton, 1966). Высокое содержание ила в местообитаниях ро-
гоза может быть результатом вторичного накопления в зарослях. От-
мечалось, что семена рогоза чаще всего прорастают на илистых грун-
тах (Loew, 1908). В рыхлых грунтах рогоз укрепляется плохо и может
отрываться при сильном ветре (Warming, 1897, цит. по: Михайлова,
1940).
Рогоз узколистный предпочитает пресную и слабосоленую во-
ду. В пределах лесной зоны он обычно поселяется в водоемах с мяг-
кой, бедной известью водой (Белавская, Леонова, 1975). Вид спосо-
бен занимать песчаные отмели в море далеко от побережья, но высо-
кая концентрация солей для него губительна (Михайлова, 1940). В.Н.
Голубев (1996) относит Т. angustifolia к гликофитам. По нашим наблю-
дениям, рогоз узколистный зачастую заселяет влажные западины на
солончаках (иногда совместно с Т. laxmannii Lepechin), но при этом
длина его вертикальных побегов может сильно снижаться. Он также
встречается по берегам солончаковых озер, в мангровых зарослях, в
умеренно засоленных каналах оросительных систем. Нередко Т. an-
gustifolia предпочитает слабосоленые воды пресным, а высокая семен-
ная продуктивность способствует успешному «поиску» и заселению
новых соленых водоемов, как правило, свободных от Т. latifolia
(McNaughton, 1975).
Рогоз узколистный хорошо переносит паводки, длящиеся до 3
месяцев, такие паводки могут благоприятно сказываться на угнетен-
ных и пораженных вредителями растениях (Михайлова, 1940). Одна-
ко вид, видимо, не выносит длительного многолетнего повышения
21
уровня воды. Растение не способно возобновляется после длительно-
го (например, двухлетнего) пересыхания местообитаний, но хорошо
восстанавливается после кратковременного затопления пересохших
биотопов. В качестве одной из причин уменьшения численности
Т. angustifolia в течение 25 лет в окрестностях Берлина указывается
снижение уровня воды в исследованных местообитаниях (Sukopp,
Markstein, 1989).
Действие волн на популяции рогоза узколистного происходит
как непосредственно, так и опосредовано, путем изменения механи-
ческого состава грунта и его минерального состава (Coops et al., 1991).
Обычно вид растет в местах, защищенных от прямого действия волн,
а опыты с выращиванием растений рогоза в культуре на грунтах, взя-
тых из местообитаний, защищенных от волнобоя, а также подвержен-
ных ему постоянно и спорадически показали, что рогоз узколистный
лучше развивается на грунтах первого типа. Зимой влияние волн на
популяции рогоза узколистного уменьшается из-за отсутствия живых
надземных частей растений, и их выживаемость зависит от стабиль-
ности поверхности почвы.
По нашим наблюдениям, регулярные осенние и весенние пожа-
ры, в местообитаниях с низким, непостоянным уровнем воды и значи-
тельным участием Phragmites australis, не влияли заметно на величину
и состояние зарослей рогоза узколистного. Пожары благоприятно дей-
ствуют на заросли рогоза, уничтожая оставшиеся побеги прошлого
года (Михайлова, 1940).
Выпас скота угнетает заросли рогоза узколистного. При вы-
кашивании рогоза ниже уровня воды его заросли практически не
возобновляются и растения погибают — очевидно, вследствие попа-
дания воды в воздухоносные ткани. Зарастание участка в таких случа-
ях происходит только за счет невыкошенной части зарослей и завер-
шается лишь к 4-му году после скашивания. При скашивании выше
уровня воды побеги в первый год отрастают медленно и большинство
их гибнет, а в последующие годы новые побеги отличаются от естест-
венных зарослей низким ростом, и более высокой плотностью (Гор-
бик, 1988).
Рогоз узколистный предпочитает более или менее глубокие
участки водоемов. Существует «дифференцировка» пространственных
ниш Т. latifolia и Т. angustifolia в местах их совместного произраста-
ния: рогоз узколистный предпочитает глубоководные участки водо-
емов и обычно не растет там, где уровень воды меньше указанного в
22
начале раздела, а Т. latifolia доминирует на мелководьях, обычно не
встречаясь на глубинах более 80 см (Grace, Wetzel, 1982а, 1981b). «Рас-
хождение ниш» возникает в результате конкуренции между двумя ви-
дами рогоза: рогоз широколистный имеет большую поверхность ли-
стьев и затеняет растения рогоза узколистного, с другой стороны, на
больших глубинах энергетически выгоднее формировать листья с мень-
шей поверхностью (Grace, Wetzel, 1981b, 1982а), что дает преимуще-
ство Т. angustifolia.
Конкуренция между Т. latifolia и Т. angustifolia, которая оцени-
валась через изменение плотности их рамет, была однозначно показа-
на S.E.B. Weisner (1993). Многолетние стационарные полевые наблю-
дения за естественными популяциями рогоза в одном из эвтрофных
озер Швеции, а также наблюдения за парциальными кустами рогоза
узколистного, пересаженными непосредственно в однородные попу-
ляции Т. latifolia, показали, что первый вид угнетает рогоз широколи-
стный на всем диапазоне глубин, начиная от 0,25 м.
Наиболее сомкнутые монодоминантные сообщества рогоз уз-
колистный образует на песчаных и супесчаных, слегка заиленных грун-
тах при глубине воды в 30—50 см (Белавская, Леонова, 1975). Проро-
стки Т. latifolia и Т. angustifolia успешно развиваются на больших глу-
бинах, чем проростки ряда широко распространенных прибрежных и
водных макрофитов: Iris pseudacorus, Phalaris arundinacea, Phragmites
australis и Scirpus lacustris, что позволяет растениям рогоза занимать
более глубокие прибрежные участки водоемов по сравнению с назван-
ными видами (Coops, Van der Velde, 1995).
В.Н. Голубев (1996) относит рогоз узколистный к летнезеленым
растениям. Отмирание семенных особей в популяциях Т. angustifolia
обычно не превышает 31%, при этом популяции способны полностью
самовозобновляться преимущественно за счет образования новых ве-
гетативных побегов (Grace, Wetzel, 1982а).
Полевые эксперименты в солонцеватых местообитаниях ро-
гоза узколистного (США) показали, что отмершие и отмирающие по-
беги растений этого вида обеспечивают необходимую для его покоя-
щихся корневищ аэрацию: так, срезание отмерших вертикальных по-
бегов или их сгибание до уровня земли, проделанные осенью текуще-
го года в ряде популяций, снижали концентрацию кислорода в тканях
вертикальных побегов рогоза, что отрицательно сказывалось на его
продуктивности, а также снижало общее число генеративных побегов
в следующим году. Степень выраженности каждого из названных
23
эффектов находилась в обратной зависимости от времени срезания или
сгиба. Срезание отмирающих вертикальных побегов рогоза способст-
вовало также уменьшению числа листовых пластинок, появляющихся
в следующем году, а сгиб увеличивал их количество (Jordan et al., 1990).
Газообмен в корневищах рогоза узколистного после скашивания вер-
тикальных побегов ниже уровня воды происходит на основе эф-
фективного гликолитического окисления сахаров в горизонтальных
побегах, благодаря чему в анаэробных условиях его корневища начи-
нали продуцировать этанол и практически не погибали (Sale, Wetzel,
1983).
Фитоценология
Рогоз узколистный — доминант-коннектор 4 яруса (высоких
трав), часто формирует моно доминантные сообщества (Быков, 1962).
Образует формацию Typheta angustifoliae, обладающую наибольшим
разнообразием и сложностью среди группы рогозовых формаций и
широко распространенную в лесостепной, степной и пустынных зо-
нах. Чаще остальных встречается Typhetum purum, представляющая
собой густые чистые заросли на «открытой воде» (Быков, 1962; Крас-
нова, 1988). Проективное покрытие рогоза узколистного в тех сооб-
ществах, где он доминирует, варьирует от 15 -20 до 100%, но чаще боль-
ше 40%, при этом угнетенные заросли имеют покрытие в 5—7(10)%.
На 1 м2 приходится от 6 до 60 вертикальных побегов, но при выкаши-
вании рогоза выше уровня воды число побегов на единицу площади
резко возрастает (Смиренский, 1950; Горбик, 1988; Segadas-Viana,
1951).
Вид входит в число диагностических для класса Phragmitetalia
W. Koch, 1926 и союза Phragmition communis W. Koch, 1926. Описаны
ассоциации Typhetum angustifoliae Pignatti и Sparganio erecti —
Typhetum angustifoliae Golub et Losev (1988) (Korotkov et al., 1991).
Существуют попытки оценить эколого-ценотические стратегии
Т. angustifolia. S.J. McNaughton (1975) склонен рассматривать его как
преимущественного «К-стратега», что может считаться доводом в поль-
зу его виолентности в условиях доминирования или содоминирова-
ния. Рогоз узколистный оказывается первым поселенцем на только что
появившихся мелях и отмелях рек, озер и водохранилищ и в дальней-
шем обычно не вытесняется другими гидрофитами (Белавская, Лео-
нова, 1975).
24
Консортивные связи
Наибольший вред листьям рогоза наносит совка Arsilonche
albovenosa (Lepidoptera), которая способна съедать их целиком. Пла-
стинки листьев рогоза узколистного поражаются также гусеницами
Lymnaecia phrargitella Staint (Tineidae, Lepidoptera), которые обитают
в их полостях (Claasen, 1921). Ткани стебля разрушаются гусеницами
радужницы Donacia cinerea (Lepidoptera), а влагалища срединных ли-
стьев поражаются клопом Ichnodemus caspius (Myodochidae), который
вспенивает выделяемую здесь слизь, способствуя ее скорому высыха-
нию (Михайлова, 1940). Вертикальные побеги рогоза узколистного по-
ражает тля Aphis persicae Sulz. (Wilson, Vickery, 1918; цит. no: Claasen,
1921), а также бабочка Bellura obliqua (Lepidoptera, Noctuidae) (Penko,
Pratt, 1986). Оставшиеся женские початки предыдущего года весной
оказываются зараженными Ischnorrhynchus resedae Panz. (Lygaeidae,
Hemiptera), причем молодые нимфы могут развиваться в сухих семе-
нах, а нимфы старших возрастов способны переползать на початки,
сформированные в текущем году (Claasen, 1921). В зарослях водных
макрофитов (Typha angustifolia и Phragmites australis) в массе развива-
ются комары-звонцы (Glyptotendipes viridis, Endochironomus albipennis,
Pentapedilum sordens, Cricotopus sylvestris agg.) (Dvorak, 1996). На по-
чатках рогоза узколистного растет целый ряд эпифитных водорослей
(Nakano, 1985).
Молодые побеги рогоза — едва ли не основной корм водяной
крысы, ондатры и нутрии; рогоз поедается бобрами, выхухолью (го-
ризонтальные побеги), лосями (молодые вертикальные побеги) (Рас-
тит. ресурсы России..., 1997).
В дельтах южных рек, в степях Казахстана и юга Русской рав-
нины заросли рогоза служат укрытием для животных (преимущест-
венно кабанов) от хищников, но в отличие от тростника, защитная роль
рогозовых зарослей, видимо, исчерпывается летним и осенним перио-
дами. Они служат укрытием лягушкам и ужам, местом охоты много-
численных чаек, которые нередко гнездятся в обширных зарослях ро-
гоза. По нашим данным, здесь гнездятся выпь, различные утиные и
куликовые. В южной Калифорнии заросли служат местом гнездовок
Quiscalus major, причем последняя использует для устройства гнезд
как молодые побеги, так и побеги прошлого года.
Известно, что рыбы лини (Tinea tinea) светлое время суток час-
то проводят в зарослях рогоза узколистного. Поскольку именно этот
25
рогоз способен формировать относительно глубоководные популяции,
а малая плотность парциальных кустов позволяет линю проникать в
заросли достаточно глубоко (Perrow et al., 1996).
В ризосфере рогоза развиваются бактерии, относящиеся к трем
основным функциональным группам азотфиксаторов, причем, все они
встречаются и вне пределов корневой системы рогоза. В основном это
— грамм-отрицательные микроорганизмы, нередко демонстрирующие
облигатный оксидативный метаболизм. В отсутствии нитратов в ризо-
сфере Typha angustifolia доминируют фиксаторы диоксида азота, кото-
рые в присутствии нитрата, но в строго бескислородных условиях, раз-
виваются здесь совместно с денитрифицирующими псевдомонадами
(Brunel et al., 1992).
Микориза в подземной сфере растений рогоза узколистного не
образуется (Grace, Harrison, 1986), сведения о поражаемости ржавчин-
ными грибами отсутствуют.
Биологическая продуктивность
Урожайность рогоза узколистного сильно зависит от географи-
ческого положения популяций, состава фитоценозов, времени прове-
дения наблюдений, а также от многочисленных абиотических факто-
ров (например, годичных изменений солености грунта). Влияние сред-
них дневной и ночной температур, а также фотопериода на урожай-
ность рогоза узколистного имеет сложный характер (McNaughton,
1966). В силу изложенного, оценки биологической продуктивности,
приводимые разными авторами, не сопоставимы между собой, к тому
же далеко не всегда указываются конкретные сроки проведения на-
блюдений.
Некоторые оценки урожайности Т. angustifolia в различных ре-
гионах России и сопредельных государств приводятся в таблице 2, в
ряде случаев указаны среднее значение по нескольким выборкам, чис-
ло которых приводится в скобках. Отмечалось также, что корневище
и корни составляют от 45 до 60 % от веса свежей биомассы данного
клона (Sculthorpe, 1967).
С увеличением толщины водного слоя увеличивается процент
дочерних корневищ, а биомасса листьев родительских вертикальных
побегов снижается. Доля биомассы соцветий максимальна при слое
воды в 50 см, а минимальна — при глубине 100 см. Чистый вес боль-
шинства вегетативных вертикальных побегов составляет летом 2—15
г, а репродуктивных — 16—45 г (Grace, Wetzel, 1982а, b).
26
Таблица 2.
Биомасса надземных побегов рогоза узколистного
из разных регионов (г\м2).
Источник Район Биомасса
сырая сухая
Смиренский, 1950 Курганская лесо- степь (4) 5750 960
Вампилов, 1950' дельта Селенги (4) 4400 1400
Михайлова, 1940 дельта Волги 350—700
Катанская, 1960 Амударья 1500
Быков, 1960 дельта Сырдарьи 520
Горбик, 1988 Украина 1228±328
* По: Смиренский, 1950.
Хозяйственное значение
Представители рода Typha известны человечеству с глубокой
древности: они упоминаются уже в книге Иова (VIII, 11). Рогоз узко-
листный (кивух, коловей, куга, мячка, тырлик, цветуха, чакан, «камыш»)
издревле используется в ткацком деле. Слово «рогожа» — название
грубой плетеной упаковочной ткани из узких мочальных лент — про-
исходит именно от слова «рогоз», «рогозник», который в старину слу-
жил основным источником сырья для изготовления рогожи (Черных,
1994) и до сих пор используется для производства грубых тканей (меш-
ковины, дорожек и пр.).
Волокно рогоза, обработанное щелочью, может быть длинным
(от 300 до 1000 мм) или куделообразным (от 50 до 300 мм), оно имеет
преимущество перед волокном пеньки, льна и кенафа в том, что у него
отсутствует костра, оно гигроскопично и легко окрашивается в любой
цвет стандартными красителями для хлопчатобумажных и шерстяных
тканей. Выход волокна составляет до 35—40%, но оно довольно хруп-
кое, что, впрочем, делает его ценным сырьем для производства подби-
вочных материалов. Очесы и отходы чаканного волокна пригодны для
изготовления войлока (Михайлова, 1940).
27
Настой подземной части растений Т. angustifolia используют в
качестве наружного ранозаживляющего и кровоостанавливающего
средства, примочки из отвара корневищ применяются в Южной Амери-
ке для лечении опухолей (Растит, ресурсы России..., 1997). Пыльца
применяется в Китае в качестве антисклеротического средства, а в со-
ставе многокомпонентного сбора — для лечения уретритов и цисти-
тов.
Подземная часть рогоза узколистного пригодна для получения
крахмала, в измельченном виде она использовалась в качестве добав-
ки к ржаной и пшеничной муке, а также как самостоятельное сырье
для выпечки хлеба, приготовления кваса и киселей. Считается диети-
ческим продуктом для больных сахарным диабетом, а в жареном из-
мельченном виде является суррогатом кофе. Сырое корневище рогоза
содержит 60% воды, 6—8% белка, 15% крахмала, до 8% сырой «клет-
чатки», 0,3% жиров и 2,7% золы (Смиренский, 1950). В сухих генера-
тивных побегах Т. angustifolia содержание крахмала достигает 58% и
более (Новикова, 1940).
Молодые вертикальные побеги в свежем, маринованном, соле-
ном виде используются в качестве салатов, а также могут добавляться
в супы (Grace, Harrison, 1986); будучи сваренными, они могут заме-
нить спаржу, жареные используются как приправы к мясным и рыб-
щ.1м блюдам. Семена рогоза съедобны и могут использоваться в кон-
дитерском деле. В.Н. Голубев (1996) относит рогоз узколистный к ви-
таминоносным растениям
Высокое содержание в листьях калийных солей щавелевой ки-
слоты (Михайлова, 1940) позволяло использовать их в качестве сырья
для производства поташа (отсюда сербское название рогоза — «по-
ташник»). Женские соцветия употребляли в качестве заменителя ва-
ты, а также набивочного и термоизоляционного материала (в качестве
источника сырья для производства рогозита — заменителя натураль-
ной пробки в холодильной промышленности). В XVIII веке волоски
использовались в качестве примеси к козьему пуху при изготовлении
фетра для шляп (Михайлова, 1940). Волоски околоцветника женских
цветков раньше использовались для изготовления искусственного шел-
ка, кинопленки, а также получения особой целлюлозы для производ-
ства нитроглицерина (Федченко, 1927).
Вертикальные побеги рогоза используются в качестве топлива,
подвязочного материала в садоводстве, подстилки для скота, а также
для приготовления низкосортной бумаги и картона, для плетения ма-
28
тов, корзин, сумок, циновок, шляп, веревок, шпагатов. После обра-
ботки битумом надземные побеги можно использовать как кровель-
ный материал. В бондарном деле побегами пробивают и прокладыва-
ют швы в бочках и кадках (Михайлова, 1940).
Все растение пригодно для силосования, а также в качестве па-
стбищного для свиней, охотно поедающих корневища. Хороший био-
фильтр для очистки бытовых и промышленных стоков, нефтяных за-
грязнений. Помимо удовлетворительного осаждения органических
взвесей из сточных вод, рогоз узколистный преимущественно аккуму-
лирует из них азот, а не фосфор (Ansola et al., 1995). В последнее вре-
мя предлагается использовать биомассу рогоза в качестве источника
«самовозобновляющегося» сырья для производства энергии, а также
для изготовления химических реагентов.
Высокоэффективное испарение воды растениями рогоза узко-
листного наряду со способностью его зарослей осаждать значитель-
ные количества ила, при определенных условиях приводит к загнива-
нию заросших рогозом биотопов (Grace, Harrison, 1986). Все рогозы
создают серьезные проблемы при орошении полей, поскольку способ-
ствуют быстрому зарастанию соответствующих систем и сооружений.
29
Т.Ю. Минаева
ШЕЙХЦЕРИЯ БОЛОТНАЯ
Номенклатура, систематическое положение,
внутривидовая систематика
Scheuchzeria palustris L. 1753, Sp. Pl.: 338; Кауфман, 1866,
Моск, фл., изд. 1: 488; Цингер, 1885, Сборник свед.: 412:'Шмаль-
гаузен, 1897, Фл. Средн, и Южн. Росс., 2: 531; Петунников, 1901,
Крит, обзор, 3: 66; Сырейщиков, 1906, Илл. фл. Моск, губ., 1: 84;
он же, 1914, цит. соч., 4: 10; он же, 1927, Опред. раст. Моск, губ.: 50;
Б. Федченко, 1934, Фл. СССР, 1:278; Назаров, 1940, в Маевский, Фл.,
изд. 7: 72; Юзепчук, 1964, в Маевский, Фл., изд. 9: 642; В. Тихомиров,
1966, Опред. раст. Моск, обл.: 60; Цвелев, 1979, Фл. европ. части
СССР, 4: 171; Walters, 1980, FI. Europ., 5: 6; Новиков, 1986, Опред.
раст. Мещеры, 1: 69; Черепанов, 1995, Сосуд, раст. России: 913.
— Шейхцерия болотная (семейство Scheuchzeriaceae—Шейхцерие-
вые).
' Syn.: S. paniculata Gilib. 1798 Exerc. Phyt. II. 502; Papillaria
palustris Dulac 1867, Fl. Hautes-Pyr.: 45. Относится к монотипному ро-
ду Scheuchzeria L., названому в честь Иоганна Якоба Шейхцера (Johann
Jakob Scheuchzer) 1672—1733, швейцарского профессора ботаники, и
к монотипному семейству Scheuchzeriaceae Rudolphi 1830.
В Северной Америке встречается подвид — subsp. americana
(Fem.) Hult., который отличается большим размером побега и семян, а
также наличием столбика, более загнутым носиком плодиков (листо-
вок) и их более темным, почти черным цветом (Hooker, 1840; Britton,
1909; Femald, 1923; Clarkson, 1961; Henry, 1963). В Японии описаны
экземпляры с сидячим, как у европейских экземпляров, рыльцем и
крупными, как у американского подвида, плодиками.
Географическое распространение
Шейхцерия болотная имеет почти циркумполярный голаркти-
ческий ареал (Meusel, Jflger, Weinert, 1965). На севере граница рас-
пространения этого вида проходит южнее северной границы леса. На
30
юге доходит до степной зоны (Лавренко, 1936), где встречается
на торфяных сфагновых болотах. В Евразии ареал вида охватывает на
востоке Камчатку, Сахалин, Северную Японию, Приамурье, захваты-
вает Северную Монголию. Вид широко распространен в Сибири и
Восточной Европе, встречается спорадически в средней части
Западной Европы, где приурочен к субальпийскому поясу гор. На
севере в Европе ареал включает Лапландию до 68°28' с.ш., Шотлан-
дию, Ирландию, острова Соловецкого архипелага, юго-западную часть
Кольского полуострова. Южная граница в Евразии проходит по линии
Абакан, Тобольск, Южный Урал, Средняя Волга, верховья Северного
Донца, среднее течение Днепра, верховья Ю. Буга, Днестра и Прута
(Андр1енко, 1975), в Украинских и Южных Карпатах, Южных
Альпах. Изолированные местообитания отмечены также на Кавказе
(Кимеридзе, 1964) и в Центральных Пиренеях (Tallis, Birks, 1965).
Ареал североамериканской разновидности на севере доходит до
Ньюфаундленда, Манитобы, Вашингтона. Южная граница ареала фраг-
ментирована и проходит по штатам Нью-Джерси, Пенсильвания,
Огайо, Иллинойс, Айова, Небраска, Нью-Мексико, Калифорния
(Femald, 1923; Markgraf, 1981). Современный ареал вида значительно
уступает его древнему ареалу (Sledge, 1949; Tallis, Birks, 1965).
Согласно точки зрения И.Д. Богдановской-Гиенэф (1946), шейхцерия
болотная отнесена к элементу флоры четвертичных болот, осно-
ву которого составляют «...потомки термофильных безусловно
гидрофильных видов. Эти виды не приобрели значительной холодо-
стойкости, вследствие чего их ареалы не простираются далеко на
север... Эволюция видов этой группы в неогене шла в сторону повы-
шения холодостойкости... и уменьшения евтрофности». Мощное
развитие аэренхимы и адаптация метаболизма к анаэробным услови-
ям указывает на исходную приуроченность шейхцерии болотной к за-
стойно-водным местообитаниям. Это же подтверждается массовыми
находками остатков шейхцерии в древних сапропелях совместно с
семенами Brasenia (Дохтуровский, 1935). В связи с этим в период суб-
атлантического похолодания климата ареал шейхцерии значительно
сократился на севере (Тихомиров, 1941), в Альпах опустились ниже
границы ее высотного распространения (Tallis, Birks, 1965), а южная
граница спорадического распространения на равнинах достигла степ-
ной зоны, где до настоящего времени вид обнаруживается на неболь-
ших сфагновых болотах (Андреенко, 1975). По мнению И.Д. Богда-
новской-Гиенэф (1949), именно в этот период из-за слишком низкой
31
температуры питающих низинные болота грунтовых вод, происходит
вытеснение шейхцерии из евтрофных местообитаний, олиготрофиза-
ция местообитаний и установление постоянной связи с гигрофильны-
ми сфагновыми мхами.
J.H. Tallis и H.J. Birks (1965) показывают максимум распростра-
нения шейхцерии болотной в Европе именно в субатлантических тор-
фах на территории Германии, Франции и Британии. В ледниковый пе-
риод для Западной Европы находки шейхцерии в торфах показаны толь-
ко лишь в Нидерландах. Эта точка считается данными авторами ис-
точником расселения шейхцерии по Западной Европе в атлантический
период. В настоящий момент шейхцерия болотная, вид, широко рас-
пространенный в Европе в атлантический период, практически отсут-
ствует на западе континента не только в связи с похолоданием клима-
та, но и в результате уничтожения торфяных болот. Последняя причи-
на объясняет сокращение ареала шейхцерии в Северной Германии
(Dierssen, Mierward, 1982), Шотландии (Scartch, 1911; Sledge, 1949),
Ирландии (Moore, 1955).
В Московской области шейхцерия болотная произрастает пре-
имущественно на переходных мезотрофных болотах, по берегам за-
растающих сфагновыми сплавинами озер, на верховых сосново-сфаг-
новых болотах, где приурочена к мезотрофным водотокам. В районе
Клинско-Дмитровской гряды обычна на верховых болотах с грядово-
мочажинным комплексом, где типичным местообитанием шейхцерии
болотной являются мочажины различной степени увлажнения: от сфаг-
новых ковров до обводненных мочажин.
Морфологическое описание
Многолетний травянистый длиннокорневищный полицентриче-
ский поликарпик. Геофит. Корневая система придаточная. Корневищ-
ные побеги шейхцерии — ярко белые, сочные. Во внешней коре кор-
невища под эпидермисом выделяются три слоя мелких клеток с утол-
щенными стенками, во внутренней коре — крупные воздухоносные
ходы (Metsflvaino, 1931). Корни ветвятся до уровня III порядка,
образуя короткие 1—5 см корешки, снабженные корневыми волоска-
ми. Междоузлия плагиотропной части корневища покрыты продолго-
ватыми пленчатыми чешуевидными листьями, реже, в сильно обвод-
ненных местообитаниях — настоящими листьями. Ортотропная
часть корневищного побега покрыта настоящими листьями с расши-
32
ренными влагалищами или их остатками. Расширенные основания
трубчатых листьев плотно охватывают передний, приподнимающий-
ся конец основного побега, несущего в пазухах листьев почки возоб-
новления, а затем надземные побеги. У шейхцерии в подземной сфере
корневищные побеги всегда связывают несколько (3—5 или более) над-
земных побегов. Корневище шейхцерии, нарастая в длину на перед-
нем конце в среднем на 25—30 см в год, постепенно отмирает в части,
удаленной от надземных побегов (Солоневич, 1956).
Надземные побеги растения — от желто-зеленого на сфагно-
вых коврах, до темно-зеленого в обводненных местообитаниях цвета,
голые, в первой половине лета — блестящие. Вегетативные побеги
20—30(40) см высотой с 5—7 очередными листьями. Последние вла-
галищные листья — с линейной вальковатой пластиной, имеющей апи-
кальную пору. У основания внутри листового влагалища имеется гус-
той пазушный ряд волосков. Устьица на стебле и листьях тетрацитные
(Тахтаджян, 1987). В конце лета на вегетативном надземном побеге
выделяются следующие типы листьев: 1—2 коротких (3—6 см)
листьев и один длинный лист (15—17 см) с широким пленчатым вла-
галищем (к концу лета все они отмирают); 1—2 цилиндрических лис-
та (18—22 см) без выраженного влагалища, также не зимующие;
1—2 цилиндрических листа (0,2—1,5 см) без выраженного влагали-
ща, которые станут первыми листьями надземного побега следующе-
го года. Длина междоузлий вегетативного побега варьирует от 0,1 до
0,9 см. Надземный генеративный моноциклический побег несет
обычно 2—3 коротких (5—8 см) листа и 2 средних (10—14 см) листа
с широким пленчатым влагалищем. Эти листья прикреплены к осно-
ванию генеративного побега и разделены междоузлиями 0,1—0,3 см
длиной. Кроме того, генеративный побег несет один, реже два стебле-
вых листа с узким влагалищем 5—7 см длиной (от нижних листьев
они отделены междоузлием 5—6 см) и 4—5 брактей длиной 1—2 см
(рис. 1).
Соцветие — терминальная кисть, с короткими гладкими ветвя-
ми в пазухах расширенных во влагалища брактей (рис. 2, а). Цветки
на цветоножках 15—25 мм длиной, с брактеями (кроме верхушечно-
го), обоеполые, актиноморфные, трехчленные (рис. 2, б). Околоцвет-
ник зеленоватый, из 6 свободных, расположенных в два круга ланцет-
ных листочков, 3 мм длиной и 1 мм шириной, сохраняющихся сухими
при плодах; доли внутреннего круга несколько уже наружных.
33
1 CM
12 3 4 5 6 7 8 10 11 12 13 14 16 16
Рис. I. Строение надземных побегов Scheuchzeria palustris L. (рис. О.В.Че-
редниченко). 1 — междоузлия вегетативного побега; 2—4 — листья веге-
тативного побега с выраженным влагалищем; 5—7 — цилиндрические ли-
стья вегетативного побега; 8 — развивающийся лист следующего года; 9
— верхушка листа с выраженной апикальной порой; 10 — генеративный
побег; 11 — нижний лист генеративного побега; 12, 13 — стеблевые ли-
стья генеративного побега; 14—16 — брактеи.
Тычинок—6, свободных, в двух кругах, тычиночные нити длин-
ные, тонкие, в конце цветения волосовидные. Пыльники 1 мм длиной
(рис. 2, б), прикрепленные у основания, при цветении удлиняются, с
выдающимся заостренным связником, вначале прямые, затем повис-
лые, вскрываются продольно, экстрорзные. Микроспорангия—4, мик-
роспорогенез сукцессивный, микроспоры соединены в диады. Зрелые
пыльцевые зерна—двух-трехклеточные, безапертурные с сетчатой экзи-
ной (Поддубная-Арнольди, 1982; Moore et al., 1991).
Гинецей из 3—6 плодолистиков, хотя описаны случаи форми-
рования гинецея из 13 плодолистиков (Cerbmesco-Jitariu, 1966). По
Е. Eber (1934), гинецей формируется из 4—5 плодолистиков, 3 из ко-
торых развиваются равномерно. U. Posiuszny (1983) настаивает на из-
начальной трехчленности гинецея. Нами обнаружено от 1 до 5
плодиков в многолистовке сформированных плодов. Гинецей апокарп-
34
Рис. 2. Генеративные органы
Scheuchzeria palustris L.: а — гене-
ративный побег (по: Karsten, 1895);
б — цветок, в — соплодие (по: Фло-
ра европ. части СССР, т. 4, 1979).
ный,однако Е. Eber(1934)nG. eerbmesco-Jitariu( 1966) показано сра-
стание плодолистиков между собой в основании гинецея вентральны-
ми частями и с тканью оси цветка. Плодолистики монолокулярные, с
не полностью сросшимися краями (Posiuszny, 1983), с двумя семяза-
чатками. Рыльце сидячее, расположе-
но в верхней части плодолистика
вдоль дорсальной жилки, низбегаю-
щее, экстрорзное, покрытое сосочка-
ми. У американской разновидности
рыльце на тонком столбике, форми-
рующем изогнутый носик 0,5—1 мм
длиной (Femald, 1923).
Плод — сухая многолистовка.
Плодики—яйцевидные, 6—8 мм дли-
ной, со слегка оттянутым носиком
(рис. 2, в). Вначале плодики желто-зе-
леные, по созревании—сухие и взду-
тые, содержат по 1—2 семени, рас-
крываются с внутренней стороны. Се-
мена —продолговатой реже элипсои-
дальной формы, довольно крупные (4
мм длиной и 1,8 мм шириной), свет-
ло- и темно-коричневого цвета, глад-
кие. Семенная кожура — кожистая,
плотная, толстая, образована в основном наружным интегументом
(Плиско, 1985). Семена со слаборазвитым (8—10 слоев клеток) эн-
доспермом, содержащим крахмал и алейрон, без перисперма и с круп-
ным зародышем. Зародыш заполняет все семя, дифференцированный,
односемядольный, хлорофиллоносный, содержит крахмал и масло,
подвесок из одной крупной пузыревидной клетки, гаусторий не най-
ден (Поддубная-Арнольди, 1982). Хромосомное число 2n=22 (Manton,
1949).
Онтогенез
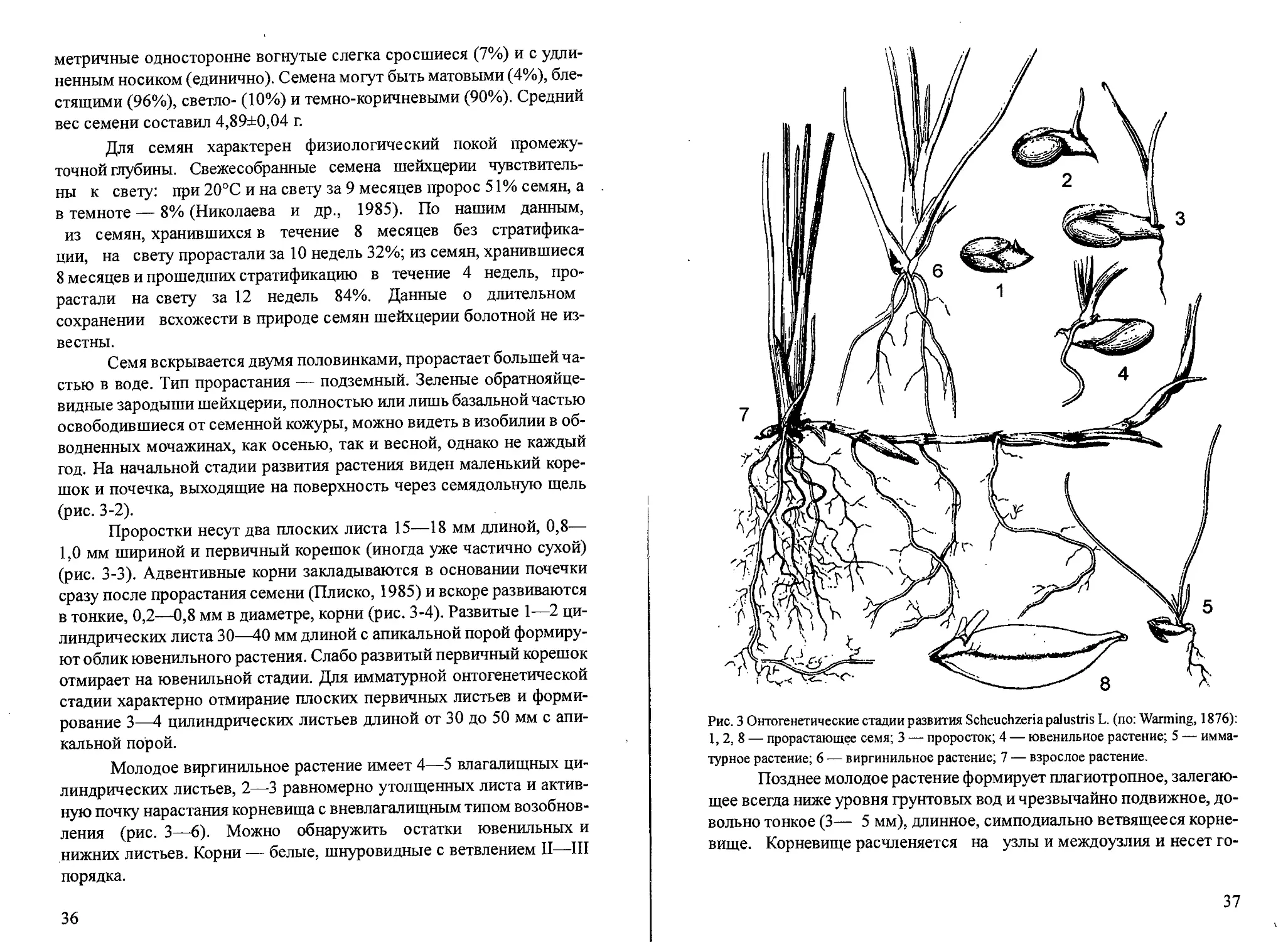
По нашим наблюдениям, растение имеет несколько типов се-
мян: продолговатые без рубца (63% из 1015 проанализированных се-
мян), эллипсоидальные с рубцом (15%), эллипсоидальные двояковог-
нутые с килем на месте семенного шва (12%) и без него (3%), асим-
35
метричные односторонне вогнутые слегка сросшиеся (7%) и с удли-
ненным носиком (единично). Семена мотут быть матовыми (4%), бле-
стящими (96%), светло- (10%) и темно-коричневыми (90%). Средний
вес семени составил 4,89±0,04 г.
Для семян характерен физиологический покой промежу-
точной глубины. Свежесобранные семена шейхцерии чувствитель-
ны к свету: при 20°С и на свету за 9 месяцев пророс 51% семян, а
в темноте — 8% (Николаева и др., 1985). По нашим данным,
из семян, хранившихся в течение 8 месяцев без стратифика-
ции, на свету прорастали за 10 недель 32%; из семян, хранившиеся
8 месяцев и прошедших стратификацию в течение 4 недель, про-
растали на свету за 12 недель 84%. Данные о длительном
сохранении всхожести в природе семян шейхцерии болотной не из-
вестны.
Семя вскрывается двумя половинками, прорастает большей ча-
стью в воде. Тип прорастания — подземный. Зеленые обратнояйце-
видные зародыши шейхцерии, полностью или лишь базальной частью
освободившиеся от семенной кожуры, можно видеть в изобилии в об-
водненных мочажинах, как осенью, так и весной, однако не каждый
год. На начальной стадии развития растения виден маленький коре-
шок и почечка, выходящие на поверхность через семядольную щель
(рис. 3-2).
Проростки несут два плоских листа 15—18 мм длиной, 0,8—
1,0 мм шириной и первичный корешок (иногда уже частично сухой)
(рис. 3-3). Адвентивные корни закладываются в основании почечки
сразу после прорастания семени (Плиско, 1985) и вскоре развиваются
в тонкие, 0,2—0,8 мм в диаметре, корни (рис. 3-4). Развитые 1—2 ци-
линдрических листа 30—40 мм длиной с апикальной порой формиру-
ют облик ювенильного растения. Слабо развитый первичный корешок
отмирает на ювенильной стадии. Для имматурной онтогенетической
стадии характерно отмирание плоских первичных листьев и форми-
рование 3—4 цилиндрических листьев длиной от 30 до 50 мм с апи-
кальной порой.
Молодое виргинильное растение имеет 4—5 влагалищных ци-
линдрических листьев, 2—3 равномерно утолщенных листа и актив-
ную почку нарастания корневища с вневлагалищным типом возобнов-
ления (рис. 3—6). Можно обнаружить остатки ювенильных и
нижних листьев. Корни — белые, шнуровидные с ветвлением II—III
порядка.
36
Рис. 3 Онтогенетические стадии развития Scheuchzeriapalustris L. (по: Warming, 1876):
1, 2, 8 — прорастающее семя; 3 — проросток; 4 — ювенильное растение; 5 — имма-
турное растение; 6 — виргинильное растение; 7 — взрослое растение.
Позднее молодое растение формирует плагиотропное, залегаю-
щее всегда ниже уровня грунтовых вод и чрезвычайно подвижное, до-
вольно тонкое (3— 5 мм), длинное, симподиально ветвящееся корне-
вище. Корневище расчленяется на узлы и междоузлия и несет го-
37
ризонтально простирающиеся надузловые корни, которые чуть толще
(0,6—1,0 мм) корней на первичном побеге семенного происхождения.
Корневище образует вегетативные полициклические и генеративные
моноциклические надземные побеги или «раметы», которые и рассмат-
риваются нами как взрослые растения и счетные единицы при популя-
ционных исследованиях (рис. 3-7). Скорость партикуляции варьирует
в связи с условиями местопроизрастания. Наиболее долго связь между
побегами сохраняется в обводненных местообитаниях.
Сезонный ритм развития
Шейхцерия болотная — условно зимнезеленое растение: моло-
дые надземные побеги следующего года начинают формироваться осе-
нью, оставаясь в течение предзимья и зимы, как правило, погребенны-
ми в сфагнум, побеги растут ортотропно и приобретают зеленую окра-
ску. На старых надземных вегетативных побегах среди отмерших ли-
стьев под поверхностью сфагнума сохраняются 2—3 формирующихся
листа. Подснежный весенний рост шейхцерии не наблюдался, что объ-
ясняется длительным промерзанием мочажин — основных местооби-
таний шейхцерии на верховых болотах. По среднемноголетним дан-
ным на болоте Старосельский мох (Тверская обл.) 20 марта мощность
дьда в мочажинах составляет 28,3±3,1 см (п=9) при глубине снежного
покрова 19,5±5,2 см (п=9).
Появление первых надземных побегов на поверхности проис-
ходит в первой декаде мая в Московской области (Серебряков, Галиц-
кая, 1951) и на 1—2 декаду позднее в более северных районах (табл.
1). Для этих сроков показаны довольно высокие значения суммы эф-
фективных температур воздуха (70—100°С), однако ограничением для
вегетации является низкая температура воды, которая остается ниже
порога 10°С в среднем до достижения суммы эффективных темпера-
тур воздуха 200°С. Интенсивный рост побегов приходится на начало
июня, максимум вегетации при замедлении ростовых процессов — на
конец июня. К этому времени на вегетативных побегах формируется
3—4 листа. В конце июля — начале августа формируются 5-ый и ко-
роткие 6—7-ой листья. На генеративных побегах до начала цветения
образуется 1—3 листа и брактеи по числу веточек соцветия за исклю-
чением терминальной. Динамика нарастания и отмирания листьев
шейхцерии болотной для южной Швеции представлена на рис. 4 по
данным I. Васкйиз (1985).
38
Рис. 4. Скорость роста в длину семи последовательных листьев (I—7) одного побега
Scheuchzeria palustris в течение двух вегетационных сезонов (по: Backflus, 1985). По
оси ординат: скорость роста (мм/день), по оси абсцисс: месяцы наблюдений.
Бутонизация и цветение в средней полосе приходится на конец
мая — первую декаду июня (табл. 1). Довольно короткие сроки между
началом роста надземного генеративного побега и массовым цветени-
ем объясняются формированием генеративных органов в почках во-
зобновления в год предшествующий цветению (Серебряков, Галиц-
кая, 1951). Цветение в средней полосе длится 3—6 дней, в Карелии —
7±1 дней (Юдина, Максимова, 1993). Завязи достаточно быстро раз-
виваются, к концу июня формируются зеленые плоды. Они достигают
размеров зрелых плодов ко второй декаде июля.
Массовое созревание плодов отмечается в конце августа, за чем
следует быстрое растрескивание листовок и опадение семян. Некото-
рые листовки опадают при отмирании надземных побегов закрыты-
ми. Фаза отмирания надземных побегов начинается с усыхания верху-
шек листьев в зависимости от мест обитания: в первой декаде июля в
мочажинах и в первой декаде августа в проточных обводненных ме-
стообитаниях, где жизненность этого вида значительно выше. Полно-
му отмиранию надземных частей побегов предшествуют устойчивые
ночные заморозки, обычно наступающие в средней полосе России в
последней декаде сентября. При анализе характера прохождения фе-
нофаз шейхцерии болотной в связи с климатическими параметрами
выявляется значимая зависимость от температурного режима (Юди-
на, Максимова, 1993).
39
Среднемноголетние показатели сезонного развития шейхцерии
Дата наступления фенофаз (М±т)
Фенофаза Карелия1’ п=15 Швеция2’ Тверская область3’ п=9
Появление новых вегетативных побеговмая 25,05±12 2 декада 12,05±2,3
Массовая вегетация 3,06±2 25,06±3,6
Первое пожелтение верхушек листьев 27,07±4 3 декада августа 7,07±6,4
Полное отмирание надзе- мных вегетативных побегов 1 декада октября 2,10±2,1
Осеннее появление новых вегетативных побегов 28,07±10,4
Появление новых генера- тивных побегов 1,06±1,5
Начало бутонизации 3,06±2 5,06±2,6
Массовое цветение 18,06±2 2 декада 11,06±1,4 июня
Первые зеленые плоды 23,06±2 3 декада 27506±5э1 июня
Первые зрелые плоды 10,09±3 2,08±6,7
Начало опадения плодов 17,09±4 2 декада 1,09±8,7 сентября
Полное отмирание генера- тивных побегов 4,10±4 21,09±6,1
*’ Юдина, Максимова, 1993;2) Backftus, 1985; 4 собственные
Способы размножения и распространения
Шейхцерия болотная как длиннокорневищный многолетник
размножается в основном вегетативно, образуя также достаточное ко-
личество семян. Генеративные побеги — однодомные с обоеполыми
цветками. Апомиксис не обнаружен, описана полиэмбриония
40
болотной в различных пунктах Таблица 1.
Сумма эффективных температур (М±т)
Московская область4’ Карелия л=15 Швеция Тверская область п=9
1 декада мая 81±3 72 121,6±17
1 декада июня 183±12 487,7±36
2 декада августа 686±42 860 611,2±71
1 декада октября 1070 1318,6±13
3 декада июля 789,1±115
1 декада мая 240,8±12
3 декада мая 260±20 281,7±24
1 декада июня 302±23 190 326,8±15
3 декада июня 324±25 260 507,8±58
3 декада августа 1039±40 914±99
1 декада сентября 1097±44 1010 1235,2±106
1 декада октября 1155±52 1300,8±59
данные;4) Серебряков, Галицкая, 1951.
(Поддубная-Арнольди, 1982). В основном в многолистовке насчиты-
вается 3 плодика, в каждом из которых развивается по 2 семени. Одна-
ко эти показатели значительно варьируют (табл.2), что определяет
сложную структуру семенной продуктивности вида. Так, доля плоди-
ков с неразвитыми семязачатками, с одним и двумя семенами - при-
близительно одинакова, тогда как число плодиков в многолистовке-
более устойчивый показатель.
41
Таблица 2.
Показатели семенной продуктивности’ Scheuchzeria palustris L.
(п побегов = 76; n плодиков = 954)
Число плодиков в многолистовке 2,99±0,07
Число семян в плодике 0,92±0,3
Число многолистовок на побег 4,69±0,07
Число семян в многолистовке 2,88±0,08
Число семян на побег 12,12±0,73
Численность генеративных побегов, шт./м2 5,2±0,09
Урожайность, шт./м2 68,3±9,1
’ Среднее ± ошибка средней.
Таблица 3.
Естественные повреждения семян Scheuchzeria palustris L. (n = 1015)
Тип повреждения %
Физические повреждения 36,27
Неразвитые семязачатки 3,05
Сухие семена 3,55
Повреждения фитофагами 1,08
Повреждения плесневыми грибами 0,49
Урожайность семян, по В.В. Мазингу (1955), составляет 30—
50 семян на 1 м2. Для зрелых семян шейхцерии болотной характерна
высокая лабораторная всхожесть. Данных о полевой всхожести семян
этого вида нет. По нашим данным, семена шейхцерии болотной в зна-
чительной мере подвержены различным естественным повреяедени-
ям, и количество нормальных семян не превышает 55% (табл. 3).
Отмечаются случаи распространения семян шейхцерии болот-
ной как на различные расстояния с помощью воды, и на относительно
большие расстояния — способом эндозоохории (Мазинг, 1955). Спо-
собность семян к гидрохории обеспечивается за счет воздухоносной
полости в верхней части семени между зародышем и семенной ко-
журой (Плиско, 1985). Семена имеют низкий коэффициент парус-
42
ности (9 см2/г по Мазингу, 1955) и большой вес (по нашим данным,
вес 1000 семян — 5,08 г, по данным Мазинга — 6,7 г). Это делает
невозможным распространение семян ветром. Птицы и мелкие мле-
копитающие изредка поедают уже опавшие семена. Чрезвычайно ред-
ко встречаются следы погрызов лосей, которые, вероятно, поедают це-
ликом генеративные побеги с плодами, что, однако не подтверждено
исследованием экскрементов.
Данные о всходах шейхцерии болотной чрезвычайно редки.
Н.Г. Солоневич (1956) отмечает, что не обнаружила ни одного всхода
шейхцерии на протяжении всей работы с этим видом. В.В. Мазинг
(1955) регистрировал проростки шейхцерии в отдельных случаях на
сфагновом покрове в мочажинах и редко — на ослабленном сфагно-
вом покрове в местах экскрементов птиц. По нашим данным, доста-
точно большое число проростков при низкой их выживаемости может
быть обнаружено в обводненных мочажинах.
Вегетативное размножение осуществляется путем разрастания
плагиотропных корневищ, захвата пространства и старческой парти-
куляции. Длина междоузлий корневищ составляет 1,11±0,05 см (п =
194) на плагиотропных и 1,25±0,05 см (п = 346) на ортотропных уча-
стках. В мочажинах со сфагновым покровом и низким уровнем стоя-
ния грунтовых вод отмечается одна зона кущения и, соответственно,
один надземный побег на каждые 8 междоузлий плагиотропной части
корневища, одна живая почка на 5 междоузлий и одно ответвление
корневища на 4,5 междоузлий той же части корневища. Для сильно
обводненных мочажин эти показатели составляют 9,2, 5,5 и 8 междо-
узлий соответственно. Чередуя ортотропный и плагиотропный рост
корневищ, шейхцерия болотная поддерживает численность популяций
и остается доминантам и основным торфообразователем в широком
спектре сообществ верховых и переходных болот.
Экология
Шейхцерия болотная — облигатный гелофит, и этим ограничен
спектр экологических условий, в которых вид формирует жизнеспо-
собные популяции. Болотные местообитания вида большей частью не
затенены, что и позволило Г. Элленбергу (1991) отнести шейхцерию
болотную к 9-ой (максимальной) ступени по шкале освещенности. Экс-
периментальные работы, проясняющие истинное отношение шейхце-
рии болотной к фактору освещенности, на настоящий момент не из-
вестны.
43
Отмеченное многими авторами происхождение шейхцерии бо-
лотной от группы теплолюбивых гигрофитов, определяет требователь-
ность этого вида к фактору температуры. Приуроченность шейхцерии
болотной в своем распространении к областям с умеренным клима-
том или невысоким горным областям определяется климатическими
ограничениями на распространение торфяных сфагновых верховых и
переходных болот с омбротрофным или смешанным типом питания и
легко прогреваемым верхним горизонтом торфяной залежи. В связи с
этим, на южных пределах своего распространения, как в Евразии, так
и в Америке, вид приурочен к высоким гипсометрическим отметкам,
не превышая при этом умеренных климатических условий, и не спус-
кается в долину, где болотные местообитания представлены лишь ни-
зинными болотами грунтового питания без сфагнового покрова. На
севере в своем распространении вид также ограничен умеренными
условиями лесной зоны. Относительно позднее начало вегетации и
отмирание надземных побегов с началом осенних заморозков также
свидетельствует о теплолюбивости этого вида. В микроэкологических
исследованиях шейхцерия болотная показывает высокую степень кор-
реляции фактора температуры субстрата и жизненности популяций.
Оптимальные для шейхцерии болотной среднемесячные температу-
ры в июле—августе верхних горизонтов субстрата составляют 13—
15°С.
В отношении условий питания многими исследователями шейх-
церия болотная относится к эвритопным видам. В.Д. Лопатин (1983)
определяет по предложенным им шкалам спектр условий произраста-
ния вида от олиготрофных до мезоэвтрофных. По отношению к азоту
для шейхцерии болотной характерны условия между бедными азотом
почвами и испытывающими дефицит азота (3-я ступень шкалы Эл-
ленберга). По шкале богатства и засоленности почв Л.Г. Раменского
(1956) шейхцерия болотная встречается от особо бедных почв, вклю-
чая торф (1—3-я ступени), до небогатых мезотрофных почв (7-я сту-
пень).
Прямые измерения некоторых факторов богатства субстрата в
местообитаниях шейхцерии болотной А.П. Соколом (1981) в предгорь-
ях Северного Урала показали следующие результаты: зольность тор-
фа= 1,4—12,1%,содержаниеК 0 = 25,0—145,0мг/100г;Nлегко гид-
ролизуемый = 0,04—0,2%; содержание Р О = 2,5—35,0 мг/100 г; со-
держание Са = 105,7—120,4 мг-экв/100 г;2содержание Mg = 18,6—28,4
мг-экв/100 г; содержание F2O3 = 35,1—52,4 мг/100 г.
44
D.J. Bellamy (1962) приводит данные по содержанию кальция
— 0,02—6,1 мг-экв/1 г, анионов — 0,3—0,8 мг-экв/1 г и катионов —
0,3—0,8 мг-экв/1 г. Приведенные данные демонстрируют широкие пре-
делы варьирования показателей богатства субстрата в местообитани-
ях шейхцерии болотной.
Достаточно индифферентное отношение проявляет шейхцерия
болотная и к реакции почвы. По данным, сведенным J.H. Tallis и
H.J. Birks (1965) по европейским местообитаниям шейхцерии болот-
ной, значения pH субстрата варьируют от 3,2 до 7,5. А.П. Сокол (1981)
для болот северного Урала приводит значения pH = 2,8—4,5. Мень-
шее варьирование значений pH показано для Карелии: 3,92—4,76 (Ло-
патин, 1956) и Тверской области: 3,79—4,06 (наши данные). По шкале
реакции почвы Элленберга шейхцерии болотной соответствует 3-я сту-
пень, к которой отнесены растения-индикаторы кислых и нейтральных
почв.
Для шейхцерии болотной значимым фактором является увлаж-
нение. Г. Элленберг (1991) отно сит вид к индикаторам влажных плохо
аэрируемых, часто затопляемых почв (9-ая ступень), а в шкалах Ра-
менского уровню увлажнения местообитаний шейхцерии соответст-
вуют ступени 89—100 (от лугово-болотных до болотных почв). Одна-
ко микроэкологические исследования показали, что многие парамет-
ры жизненности популяций шейхцерии болотной коррелируют с по-
казателем уровня стояния болотных вод и их проточности. Проточ-
ность определяется по содержанию кислорода в воде, которое в мес-
тах обитания шейхцерии колеблется от2,5 до 5,7 мг/л, или по коэффи-
циенту фильтрации, который изменяется здесь от 1,17 до 5,6 см/сек
(Лебедева, 1957).
Фитоценология
Относительная эвритопность шейхцерии болотной обеспечи-
вает виду в пределах достаточно узкого спектра условий верховых и
переходных болрт широкую синэкологическую амплитуду и пред-
ставленность в различных растительных ассоциациях. В континенталь-
ных условиях, в олиготрофных мало обводненных мочажинах с доми-
нированием в моховом покрове Sphagnum fallax (Klinggr.) Klinggr. *
и Sph. balticum (Russ.) C. Jens., а в Прибалтике—Sph. papillosum Lindb.,
шейхцерия болотная служит основным ценозообразователем.
* Названия сфагновых мхов даны по: Домбровская, Шляков (1967).
45
При этом жизненность вида невелика: высота побегов не превышает
20 см, цвет побегов — желто-зеленый, урожайность семян низкая.
Условия произрастания, по нашим данным (Тверская область),
в среднем за июль—август имеют следующие характеристики: уро-
вень грунтовых вод (УГВ) — 5 см, содержание кислорода в воде —
3,16 мг/л, pH = 3,99, общая электрическая проводимость раствора
(ОВП) = 66,6, температура воды —+11,4°С. Вследствие бедности этих
сообществ по сравнению с классической ассоциацией Caricetun limosae
(Dierssen, Reichelt, 1988), мы, в соответствии с точкой зрения М.С. Боч
(1986), относим эти сообщества к ассоциации Cuspidato-
Scheuchzerietum palustris (Тх. 1937) PRSG et Tx. 1958, субассоциации
C-Sch. palustris sphagnetosum fallacis.
В сообществах сильно обводненных мочажин и водотоков из
той же ассоциации с разреженным, ввиду высокого уровня стояния
воды, моховым покровом из Sph. cuspidatum Hoffin. и Sph. majus (Russ.)
C. Jens, уровень прогреваемости воды, высокая степень аэрации и сле-
дующее за этим увеличение трофности обеспечивают условия, близ-
кие к синэкологическому оптимуму шейхцерии болотной. Условия сре-
ды для Тверской области в среднем за июль—август следующие:
УГВ > 0 см, содержание кислорода в воде—5,67 мг/л, pH = 4,06, ОВП
= 89,8, температура воды — +15,6°С.. Побеги шейхцерии — темно-
зеленого цвета и достигают высоты 30 см; отмечены высокая вызре-
ваемость семян и наличие всходов.
В сообществах мезоэвтрофных сфагновых ковров переходных
болот среднего уровня увлажнения с доминированием Sph. fallax и Sph.
flexuosum Dozy et Mo Ik. (из ассоциации Caricetun limosae Paul 1910 ex
Osv. 1923 emend. Di. 1982) шейхцерия болотная произрастает вместе с
Carex limosa L., Menyanthes trifoliata L., Drosera anglica Huds., Carex
rostrata Stokes, Rhynchospora alba (L.) Vahl, Equisetum fluviatile L. Здесь
шейхцерия болотная не проявляет ожидаемой высокой жизненности
популяций, что мы связываем с конкурентным влиянием других ви-
дов.
Плоские гряды и низкие кочки из ассоциации Chamaedaphne-
Sphagnetum magellanici характеризуются для Тверской области в сред-
нем за июль—август УГВ = —15 см, содержанием кислорода в воде
2,88 мг/л, pH = 3,79, ОВП = 68,6 и температурой воды+10,3°С. Шейх-
церия болотная «залезает» на гряды случайно и практически не обра-
зует генеративных побегов. Жизненность ценопопуляций в этих усло-
виях низкая.
46
Крайне неблагоприятными для произрастания сосудистых рас-
тений являются условия в сообществах на деградированном сфагно-
вом покрове в местах выхода болотных газов, вымерзания или вымо-
кания растительности. Непроницаемая корка из живых и отмерших
печеночников Cladopodiella fluitans (Nees) Buch * и Gymnocolea inflata
(Huds.) Dum. создает условия низкой аэрации, теплопроводности и
трофности: УГВ.= —4 см, содержание кислорода в воде — 2,56 мг/л,
pH = 3,92, ОВП = 58,6 и температура воды — +12,6°С. Однако и здесь
численность побегов шейхцерии болотной достаточно высока при
очень низких морфометрических показателях.
Для всех приведенных сообществ показана специальными ис-
следованиями сходная возрастная структура и численность ценопопу-
ляций шейхцерии болотной. Различия проявлялись в значениях над-
земной продукции, урожайности семян и некоторых морфометриче-
ских показателей. Этот факт, наряду с широкой синэкологической ам-
плитудой вида и выраженной монодоминантностью в большинстве це-
нозов, свидетельствует о реализации черт патиентности в жизненной
стратегии шейхцерии болотной. Активность, с которой шейхцерия ос-
ваивает бесконечно изменяющуюся поверхность болота, не удивитель-
на, если принять во внимание предполагаемый (Богдановская-Гиэнэф,
1949) исключительно древний возраст этого вида и разнообразие ус-
ловий, им пережитых.
Биологическая продуктивность
Биологическая продуктивность шейхцерии болотной во мно-
гом определяет интенсивность процесса торфонакопления на верхо-
вых и переходных болотах. При этом наиболее значимую роль играет
подземная фитомасса, процессы накопления которой все еще мало изу-
чены. По данным анализов торфяных монолитов, в олиготрофных мо-
чажинах корни шейхцерии болотной составляют 31%, корневища —
58% от общей подземной фитомассы всех видов, составляющей 58,5
ц/га (Елина и др., 1984).
Характер сезонной динамики формирования фитомассы опре-
деляется тем, что в первой половине вегетационного сезона активно
нарастают исключительно вегетативные части побегов. Во второй по-
ловине лета вегетативный рост практически прекращается (рис. 4),
* Названия печеночников даны по: Константинова, Потемкин, Шляков (1992).
47
Рис. 5. Структура надзем-
ной фитомассы Scheuchze-
ria palustris в июне и авгу-
сте в сообществах олиго-
трофных мочажин (Твер-
ская _ область).
Обозначения: 1 — почки
возобновления, 2 — междо-
узлия, 3 — листья, 4 —
стебли и соцветия генера-
тивных побегов. Отрица-
тельным значениям соот-
ветствует мертвая фитомас-
са прироста текущего года.
начинаются активные
процессы отмирания листьев, тогда как вклад генеративных органов
становится значимым. Так, по нашим данным, в сообществах олиго-
трофных мочажин Тверской области в конце вегетационного сезона
доля генеративных побегов в фитомассе резко возрастает (рис. 5).
Таблица 4.
Фитомасса надземных побегов Scheuchzeria palustris L., ц/га
Регион Олиготрофные мочажины Мезотрофные ковры Мезоэвтрофные топи
Карелия11 2,60 0,36 —
Швеция4 3,55 0,45 —
Белоруссия3’ 5,55 0,26 0,92
Тверская область4’ 4,14 0,57 —
° Елина и др., 1984; 2’ Backeus, 1985; 3) Валетов, 1989;4) наши
данные.
В зависимости от условий местообитания у шейхцерии болот-
ной значительно изменяются такие характеристики как интенсивность
прироста и отмирания фитомассы, морфометрия листьев (длина, ши-
рина, площадь, удельный вес), вклад генеративных и вегетативных ор-
ганов в формирование фитомассы. Наряду с вероятной различной чис-
ленностью ценопопуляций, эти факторы определяют различия в пред-
48
ставленных значениях фитомассы в некоторых регионах и типах со-
обществ (табл. 4). Таким образом, шейхцерия болотная не имеет вы-
раженного пика продуктивности и, несмотря на монодоминантность
сообществ, не формирует высокопродуктивных ценозов.
Консортивные связи и хозяйственное значение
Шейхцерия болотная практически не поедается позвоночными
животными, быть может, в связи с присутствием в тканях цианогенно-
го гликозида триглохинина (Ruijgrok, 1974). В основаниях цилиндри-
ческих листьев часто обнаруживаются личинки Coleoptera. Личинки
из той же группы часто выедают содержимое семян шейхцерии бо-
лотной еще в плодах, оставляя на семенной кожуре характерное от-
верстие.
Шейхцерия болотная является основным торфообразователем
верховых и переходных болот. Как растительное сырье не использует-
ся.
49
М.Г. Вахрамеева, Т.И. Варлыгина,
А.Е. Баталов, И.А. Тимченко,
Т.И. Богомолова
РОД ДРЕМЛИК
В роде Дремлик (Epipactis Zinn) — около 25 видов, распростра-
ненных в умеренном поясе Евразии, в Африке и Северной Америке. К
роду относятся многолетние травянистые корневищные растения с со-
цветием в виде прямой рыхлой кисти, состоящей из довольно круп-
ных поникающих зеленоватых, пурпуровых, реже беловато-желтова-
тых цветков. Околоцветник — простой зигоморфный из 6 свободных
расходящихся листочков, расположенных в 2 круга. Средний листо-
чек внутреннего круга — губа не имеет шпорца, разделена посереди-
не глубокой вырезкой на две доли: заднюю (гипохилий) — чашевидно
вогнутую, выделяющую нектар, и переднюю (эпихилий) — почти пло-
скую, отогнутую книзу, при основании с бугорками или гребешками.
Нить единственной сохранившейся тычинки срастается со столбиком
в короткую колонку. Рыльце — почти квадратное с широким клюви-
ком. Пыльник—сердцевидный, порошковидная пыльца собрана в дву-
раЗдельные поллинии, связанные маленькой железкой. Завязь — ниж-
няя, плод — коробочка.
В Московской области встречаются 3 вида рода Дремлик, два
из которых — Epipactis helleborine (L.) Crantz и E. palustris (L.) Crantz
распространены довольно широко, а один — E. atrorubens (Hoffin. ex
Bemh.) Schult, очень редок, но нельзя исключить, что могут быть об-
наружены его новые местонахождения.
Ключ для определения видов, произрастающих
в Московской области
1. Корневище, с удлиненными междоузлиями, нижние листья снизу
голые, 8—15 см длиной, продолговатые; гипохилий слегка вогну-
тый с двумя тупыми боковыми лопастями, розовато-белый с фио-
летовыми жилками, посередине с оранжевыми бородавочками;
эпихилий — белый с розовыми жилками, при основании с двумя
выдающимися гребешками, отделен от гипохилия узкой перемыч-
кой. Epipactis palustris — Дремлик болотный.
50
— Корневище укороченное, нижние листья 4—9(15) см длиной, ши-
рокоовальные или широко ланцетные, снизу по жилкам шерохо-
ватые; гипохилий глубоко вогнутый, без боковых лопастей, отде-
лен от эпихилия широкой перемычкой; эпихилий при основании
с двумя бугорками..................................... 2.
2. Цветки красновато-пурпуровые с запахом ванили, бугорки у осно-
вания эпихилия морщинистые; листочки околоцветника собраны
колокольчато, нижние листья продолговатые или широко ланцет-
ные, острые; стебель в верхней половине буроватофиолетовый;
ось соцветия, цветоножки и завязь густоопушенные. Е. atrorubens
— Д. темно-красный.
— Цветки зеленоватые или зеленовато-пурпуровые, без запаха, бу-
горки у основания эпихилия гладкие, листочки околоцветника про-
стертые; нижние листья чаще широкоовальные или яйцевидные;
стебель зеленый, ось соцветия, цветоножки и завязь более или
менее гладкие. Е. helleborine— Д. широколистный.
Дремлик болотный
Номенклатура, систематическое положение,
внутривидовая систематика
Epipactis palustris (L.) Crantz, 1769, Strip. Austr. ed. 2, 2: 462;
Кауфман, 1866, Моск, фл., изд. 1: 476; Цингер, 1885, Сборник свед.:
422; Шмальгаузен, 1897, Фл. Средн, и Южн. Росс., 2: 451; Петунни-
ков, 1901, Крит, обзор, 3: 55; Сырейщиков, 1906, Илл. фл. Моск, губ.,
1: 265; он же, 1914, цит. соч., 4: 49; он же, 1927, Опред. раст. Моск,
губ.: 97; Невский, 1935, Фл. СССР, 4: 623; Назаров, 1940, в Маевский,
Фл., изд. 7: 243; Голицын, 1964, в Маевский, Фл., изд. 9: 678; Сквор-
цов, 1966, Опред. раст. Моск, обл.: 122; Смольянинова, 1976, Фл. ев-
роп. части СССР, 2: 26; D.M. Moore, 1980, Fl. Europ., 5: 327; Новиков,
1986, Опред. раст. Мещеры, 1: 134; Черепанов, 1995, Сосуд, раст. Рос-
сии: 619. — Serapias helleborine var. palustris L.
1753, Sp. PL: 950. — Дремлик болотный (семейство Orchidaceae —
Орхидные).
Дремлик болотный относительно мало изменчив. Известны его
формы: f. ochroleuca Baria (с желто-зелеными листочками околоцвет-
ника), f. ericetorum (низкорослые растения до 10 см высотой с немно-
51
гими цветками), f. silvatica (высокое растение с широкими листьями и
удлиненным рыхлым соцветием), кроме того, выделяют f. palustris, f.
ampla, f. parviflora, f. longibracteata, f. submerse (Hegi, 1909; Prochazka,
Velisek, 1983; Fuller, 1986). Этим формам обычно не придают высоко-
го таксономического ранга, нередко, по крайне мере некоторые из них
рассматриваются как экологические формы. По мнению ряда иссле-
дователей, дремлик болотный ближе других дремликов к роду
Cephalanthera (Summerhayes, 1951).
Дремлики менее склонны к гибридизации, чем ятрышники и
пальчатокоренники, это особенно относится к Epipactis palustris. Из-
вестен лишь единственный (крайне редко встречающийся) гибрид это-
го вида с Е. atrorubens (Fuller, 1986). Он, в частности, указан для Вин-
ницкой области на Украине (Флора европ. части СССР, 1976).
Географическое распространение
Дремлик болотный распространен на большей части Европы
от Британских островов и Скандинавии (отсутствует на Крайнем Се-
вере) до Испании, Средиземноморья (кроме островов), Балканского
полуострова и Крыма. Встречается в Северной Африке. В Азии его
ареал охватывает Малую Азию, Кавказ, Центральную Азию, Сибирь,
Внешнюю Монголию, Гималаи (Флора СССР, 1936; Ziegenspeck, 1936;
Summerhayes, 1951; Флора европ. части СССР, 1976; Flora Europaea,
1980; Fuller, 1986; Губанов, 1996).
В России встречается в европейской части (в пределах всей лес-
ной зоны), в Западной и Восточной Сибири, доходя до оз. Байкал (Фло-
ра СССР, 1935; Флора европ. части СССР, 1976; Флора Сибири, 1987;
Опр. сосуд, раст. центра европ. России, 1995).
В Московской области встречается изредка по всей территории,
в южных районах реже, иногда образует значительные скопления (Во-
рошилов и др., 1966).
Морфологическое описание
Многолетнее травянистое длиннокорневищное растение. Кор-
невище с длинными междоузлиями, подземное, с чешуевидными (ни-
зовыми) листьями, в пазухах которых находятся почки. От каждого
узла отходят по 2—4 придаточных корня. На приподнимающейся час-
ти корневища (в основании ортотропных побегов) развиваются 1—3
низовых влагалищных листа (Ziegenspeck, 1936; Summerhayes, 1951;
52
Смирнова, 1990). Стебель—20—50 см высотой, несущий 3—5(6) нор-
мально развитых (срединных) листьев, в нижней части голый, в верх-
ней — немного опушенный. Листья — очередные, продолговатые или
продолговато-ланцетные, заостренные, голые; нижние—8—15(30) см
длиной, 1,5—2,5(4)см шириной, длиннее междоузлий; верхние — бо-
лее мелкие, ланцетные. Самый верхний лист — еще меньше, с корот-
ким влагалищем, Е.С. Смирнова (1990) называет его листовидной брак-
теей.
Соцветие — редковатая кисть, 6—15(20) см длиной, из 6—
15(24) поникающих цветков. Прицветники—ланцетные, короче цвет-
ков, лишь самые нижние равны им. Цветки — довольно крупные: бо-
лее декоративны, чем у других видов рода. Околоцветник образован 6
свободными листочками. Листочки наружного круга—ланцетные или
овально-ланцетные, 8—12 мм длиной, буровато-зеленоватые, с неяс-
ными грязновато-фиолетовыми полосками с внутренней стороны, за-
остренные, с тремя жилками, снаружи мелко рассеянно опушенные.
Листочки внутреннего круга немного короче наружных, ланцетные или
узкоовальные, тупые, голые, с тремя жилками, беловатые, при основа-
нии с нерезкими полосками. Губа — 10—12 мм длиной, равна или
длиннее наружных листочков, разделена глубокой вырезкой на две до-
ли: заднюю (гипохилий) и переднюю (эпихилий). Гипохилий — слег-
ка вогнутый в виде желоба, выделяющий со дна нектар, треугольный,
с двумя короткими, тупыми, вперед обращенными лопастями,
4,5—5,5 мм длиной; снаружи розовато-белый, внутри с розова-
то-фиолетовыми жилками и оранжевыми бородавочками. Эпихи-
лий— плоский, широкоовальный, белый с розовыми жилками,
5,5—6,5 мм длиной, 5—6 мм шириной, тупой, на конце немного вы-
емчатый, по краю волнисто-городчатый, при основании с двумя вы-
дающимися гребешками. Колонка—прямая, 4—5 мм длиной, рыльце
— почти квадратное с широким клювиком; пыльник — сердцевид-
ный.
Поллинии — двураздельные, грушевидные. Завязь — прямая,
опушенная. Плод—поникающая коробочка с многочисленными очень
мелкими семенами (Hegi, 1909; Флора СССР, 1935; Fuller, 1986; Смир-
нова, 1990).
Сведения о диплоидном наборе хромосом разноречивы: 2п =
40 (Fuller, 1986), 40 и 24 (Флора европ. части СССР, 1976); 40, 44, 46,
48 (Flora Europaea, 1980).
53
Онтогенез
Рис. 1. Растения Epipactis palustris
семенного происхождения; пк —
сохранившийся протокорм.
Об онтогенезе дремлика болотного известно немного. Семена
начинают прорастать в первую же осень при участии грибов, и к весне
образуется протокорм (Ziegenspeck,
1936). В асимбиотических условиях
в лабораторных опытах семена дрем-
лика не прорастали (Arczewska, 1993).
В природных условиях через 2—3 го-
да после прорастания семян образу-
ются мясистые придаточные корни, а
затем и первый надземный побег, не-
сущий зеленые листья (Summerhayes,
1951). Общая продолжительность
жизни в условиях питомника—не ме-
нее 20 лет (Работнов, 1947).
В природных условиях найти особи
семенного происхождения удается
очень редко. Летом 1997 г. нам удалось
их обнаружить на низинном болоте в
Вологодской области (рис. 1). Это бы-
ли ювенильные и имматурные расте-
ния, еще сохранившие протокорм и рез-
ко отличающиеся по размерам от осо-
бей вегетативного происхождения, рас-
тущих рядом (высота ювенильного рас-
тения — 5,8 см, имматурного — 7,5 см).
В.Г. Собко и О.Н. Нефедова (1983) проследили онтогенез осо-
бей вегетативного происхождения в природе и культуре. Дочерние кор-
невища дремлика болотного образуются из пазушных почек чешуе-
видных листьев материнского корневища, они значительно тоньше ма-
теринского корневища. Место соединения дочернего и материнского
корневища быстро отмирает, и дочерние растения переходят к са-
мостоятельному существованию. В природных условиях однолетние
особи вегетативного происхождения имеют 1—2 влагалищных (низо-
вых) листа и 1—2 зеленых (срединных) ланцетно-заостренных
листа, у двулетних особей — 3—4 зеленых листа. Виргинильный
период длится 5—6 лет, на 6—7-й год появляются генеративные побе-
ги, которые после цветения и плодоношения отмирают. Остается
54
V
Рис. 2. Возрастные состояния Epipactis palustris семенного происхождения.
лишь почка возобновления, заложенная в основании генеративного по-
бега. На следующий год из этой почки, как правило, образуется веге-
тативный побег, а генеративный будет сформирован лишь через не-
сколько лет, в связи с чем цветение и плодоношение таких особей ве-
гетативного происхождения наблюдается не ежегодно.
В Московской области мы наблюдали подобные особи вегета-
тивного происхождения (рис. 2), некоторые их морфологические па-
раметры приведены в таблице 1.
Таблица 1.
Морфометрические показатели дремлика болотного в Московской обл.
Показатель Онтогенетическое состояние
j im V g
Число листьев 1—2 3—4 4-5(6) (5)6—7
Длина нижнего листа, см 18—26 22—27 23—30 21—31
Ширина нижнего листа, см 0,5—1,2 0,7—1,3 1,0—1,5 1,2—2,0
Число цветков — — — 3—24
Длина цветоноса, см — — — 44—83
55
В условиях культуры развитие дремлика болотного происходит
значительно быстрее (Собко, Нефедова, 1983). Дочерние особи веге-
тативного происхождения зацветают уже на второй год, при этом цве-
тут и плодоносят ежегодно, а размеры их — значительно крупнее, чем
в природе (табл. 2).
Таблица 2.
Морфометрические показатели генеративных растений дремлика
болотного в природных условиях и культуре (Собко, Нефедова, 1983)
Показатель В природе В культуре
Высота стебля, см 34—41 45—50
Число листьев 7—8 8—12
Длина срединных листьев, см 10—12 12—14
Ширина листьев, см 2—3 8—9,5
Число цветков в соцветии 8—10 15—30
Число плодов 2—5 9—15
Число корней на одном метамере корневища 1—3 7—9
Способы размножения и сезонное развитие
Дремлик болотный размножается как семенным, так и вегета-
тивным путем, причем вегетативное размножение сопровождается глу-
боким омоложением (Тимченко, 1996). В.Н. Голубев (1984) относит
дремлик болотный к типичным летнезеленым растениям. В Москов-
ской области дремлик зацветает в конце июня, цветение длится до кон-
ца июля — начала августа, плодоносит в августе—сентябре. Сроки
цветения и плодоношения в зависимости от погодных условий могут
сдвигаться на 2—3 недели. Плодоношение обычно интенсивное, сред-
ний процент плодообразования в разных частях ареала колеблется в
пределах 65—85% (Ziegenspeck, 1936; Summerhayes, 1951).
На Украине среднее число цветков на генеративном побеге со-
ставляет 9,4—11,6, среднее число плодов — 6,4—9,9, в типичных
условиях произрастания процент плодообразования составляет
68—87%, но в нарушенных фитоценозах эти показатели заметно сни-
жаются. В Германии, по данным О. Wildhaber (1970), у всех видов
56
дремликов в каждом плоде содержится около 4500 семян, из которых
80% — жизнеспособны. В Московской области, по нашим данным, в
ненарушенных местообитаниях среднее число цветков в соцветии
составляет около 10, максимальное число — 41, процент плодообра-
зования — около 85.
Экология
Дремлик болотный — светолюбивое растение, лишь изредка
встречается при небольшом затенении (4-я ступень шкалы Ландольта,
2-я ступень шкалы Элленберга), гелиосциофит по В .Н. Голубеву (1984).
В тенистых местообитаниях имеет тонкий стебель, тонкие широкие
листья, зеленоватые цветки. Гигрофит, встречается, главным образом,
на сырых почвах (выдерживает периодическое затопление), избегает
сухих почв (4-я ступень шкалы Ландольта, 8-я ступень шкалы Эллен-
берга). В условиях Крыма ведет себя как мезофит (Голубев, 1984). На
осушенных болотах может сохраняться, но приобретает карликовые
размеры (Summerhayes, 1951). К богатству почв (особенно азотом) не-
требователен, может расти на бедных и очень бедных почвах (2-е сту-
пени шкал Ландольта и Элленберга), к содержанию гумуса довольно
безразличен. На богатых почвах отмечено более интенсивное ветвле-
ние корневищ (Summerhayes, 1951) и быстрое расширение занятой тер-
ритории. Растет в основном на нейтральных и щелочных почвах (4-я
ступень шкалы Ландольта, 8-я ступень шкалы Элленберга), избегает
очень кислых почв. В Польше встречается на почвах с pH 6,5—7,4
(Sarosiek et al., 1990; Kaminski et al., 1990), в Чехословакии указан на
почвах от кислых до нейтральных, преимущественно с pH 6,5—8,0
(Prochazka, Velisek, 1983). Ряд авторов относит этот вид к кальцефи-
лам (Muller, 1974; Sundermann, 1975). Обычно встречается на тонко-
структурных (глинистых, торфянистых) почвах (5-я ступень шкалы
Ландольта).
Вид растет как на равнинах, так и в горах, поднимаясь до 1500
м над уровнем моря (Summerhayes, 1951).
Фитоценология
Дремлик болотный растет преимущественно на известковых
низинных осоковых болотах, реже — в заболоченных лесах, зарослях
кустарников, на заболоченных лугах (Флора СССР, 1935; Ziegenspeck,
1936; Флора европ. части СССР, 1976; Flora Europaea, 1980;
57
Флора Сибири, 1987; Опр. сосуд, раст. центра европ. России, 1995).
Переходных болот со сфагновыми мхами, где повышенная кислотность
субстрата, он обычно избегает (Ziegenspeck, 1936).
Отмечают, что в условиях Средней Европы оптимальные для
себя условия Epipactis palustris находит в фитоценозах с присутствием
Carex davalliana (класс Scheuchzerio-Caricetea fuscae, порядок Caricion
davallianae) (Prochazka, Velisek, 1983; Sarosiek et al., 1990; Kaminski et
al., 1990). В Германии (Fuller, 1986) дремлик болотный — характер-
ный вид союза Eriophorion latifolii Br. Bl. et Tx., часто растет на низин-
ных болотах вместе с Dactylorhiza incarnata, Pinguicula vulgaris,
Ophioglossum vulgatum, образуя скопления до нескольких сотен экзем-
пляров. Встречается он также в осоково-гипновых сообществах на по-
бережье и в прибрежных дюнах Северного моря (Fuller, 1986). На Ук-
раине (Тимченко, 1996) вид растет преимущественно на лесных мезо-
трофных и безлесных болотах, а также на торфянистых лугах, а В.Г.
Собко (1989) отмечает его привязанность к гипновым и осоково-сфаг-
новым группировкам.
В России фитоценотический ареал дремлика болотного также
обширен. В Тверской области в районе Вышневолоцко-Новоторжско-
го вала (Дементьева, 1990), где наблюдается повышенное видовое бо-
гатство орхидных, дремлик болотный встречается (помимо заболочен-
ных лугов) в необычных для него фитоценозах — по опушкам сосно-
во-березовых перелесков, на сухих лугах с доминированием клевера
горного, в зарослях ольхи и ив, в весьма широком диапазоне влажно-
сти почв. В этом районе дремлик болотный образует крупные ценопо-
пуляции с высокой плотностью — 27—70 особей на м2, в том числе —
до 40 генеративных растений. Высота генеративных растений в этих
условиях — около 50 см (от42,6±1,28 до 50,7±1,29). В ценопопуляци-
ях, как правило, преобладают молодые особи.
При изучении возрастного состава популяций этого и других
видов дремликов в качестве фитоценотической счетной единицы (Це-
нопопуляции растений, 1976) использовали надземный побег, посколь-
ку определить целостность особей без их выкапывания невозможно.
В Московской области дремлик болотный встречается в типич-
ных для него условиях местообитания — на низинных болотах,
заболоченных лугах, в зарослях кустарников. Как правило, образует
многочисленные нормальные популяции с надежным самоподдержа-
нием (преимущественно вегетативным путем) и высокой плотностью
(табл. 3). Изученные нами ценопопуляции дремлика болотного в раз-
58
ных частях ареала (Вологодская область, Ставропольский край) нахо-
дились также в типичных условиях местообитания — чаще на низин-
ных болотах и заболоченных лугах. Для сравнения приведены литера-
турные сведения по Украине (Тимченко, 1993) и Польше (Sarosiek et
al., 1990; Kaminski et al., 1990).
Дремлик болотный может встречаться во вторичных местооби-
таниях, осваивая новые территории (Prochazka, Velisek, 1983; Вахра-
меева, Варлыгина, 1996; Вахрамеева и др., 1997), однако при осуше-
нии и загрязнении территории может полностью исчезнуть (Nilsson,
1979; Тимченко, 1994). Занятую территорию вид удерживает стойко,
поэтому иногда может расти в необычных условиях, например, на месте
бывших болот (Summerhayes, 1951). При незначительном антропоген-
ном воздействии (сенокошение, слабая рекреация) нередко образует
крупные популяции (тысячи особей) со значительным участием моло-
дых особей (Тимченко, 1994).
Таблица 3.
Возрастная структура (%) и плотность ценопопуляций (особей на м2)
дремлика болотного в различных частях ареала
Местонахождение Онтогенетическое состояние Плот- ность
j im V g
Московская обл. 13 39 26 22 24
Вологодская обл. 7 32 49 12 81
Ставропольский край 9 12 62 17 12
Житомирская обл. (Тимченко, 1993) 9 31,3 42,9 16,8 23,3
Львовская обл. (Тимченко, 1993) 0,5 18,4 58,0 23,1 4,24
Польша (Sarosiek et al., 1990) 7,5 31,7 55,9 8,9 —
Польша (Kaminski et al., 1990) 0,6 26,9 66,5 6,0 —
Примечание: счетная единица — условная особь (преимущественно
вегетативного происхождения).
59
Возрастная структура ценопопуляций вида в разных частях ареа-
ла показывает сходную в общих чертах картину (табл. 3). Возрастной
спектр — чаще правосторонний (большая часть ценопопуляции пред-
ставлена взрослыми, вегетативными и генеративными) особями. Лишь
очень редко ювенильные растения составляют более или менее значи-
тельную часть ценопопуляции (8—13%), а нередко молодые растения,
даже и в благоприятных условиях, практически отсутствуют (менее
1%). Плотность ценопопуляций колеблется в значительных пределах
от 2 до 38 особей на м2.
Проведенный на Украине по методике Ю. А. Злобина (1989) ана-
лиз виталитетной структуры ценопопуляций дремликов показал, что
индикаторными признаками для этой цели могут служить площадь и
ширина листа, высота растения (Тимченко, 1996). Среди популяций
дремлика болотного преобладают дигрессивные, а процветающие по-
пуляции встречаются редко и характерны для условий слабого воздей-
ствия антропогенного фактора.
Консортивные связи
Энтомофил, привлекающий насекомых нектаром. Цветки до рас-
крывания и после оплодотворения повисают, а к моменту опыления
зайимают вертикальное положение так, что губа представляет собой
удобную посадочную площадку (Summerhayes, 1951). Строение цвет-
ка высоко специализировано и приспособлено к перекрестному опы-
лению (Фегри, Пейл, 1982). Нижняя губа производит очень простые
пассивные движения под действием тяжести опылителя. Дремлик бо-
лотный имеет относительно высокую колонку, пыльник его — удли-
ненный, поллинии — также длинные и узкие, прочные только в перед-
ней части, а в задней — более рыхлые и непрочные. Когда насекомое
прилетает на цветок, оно приземляется на гибкий подвижный эпихи-
лий, который под грузом склоняется вниз, создавая насекомому опре-
деленное неудобство. К тому же нектар вырабатывается только на дне
гипохилия, что вынуждает насекомое переместиться выше и достиг-
нуть такого места, где оно может своим коротким хоботком достать
нектар. При этом насекомое не только разрывает тонкий эпидермис,
но и одновременно приподнимает пыльник, в результате чего полли-
ний приклеивается к голове насекомого и при дальнейшем полете пе-
реносится на рыльце других цветков (Fuller, 1986).
60
Дремлик болотный посещается многими насекомыми, но об ос-
новных опылителях мнения разных авторов расходятся. L. Nilsson
(1978) указывает, что в Швеции цветки посещаются пчелами, мухами,
бабочками, муравьями и пауками, но наибольшее количество пыльцы
переносят осы семейства Eumenidae, которые лучше других приспо-
соблены к взаимодействию с колебательным механизмом губы цвет-
ка. В основном опыляют цветки самцы одиночных ос. Особенности
строения их первого брюшного сегмента, вероятно, способствовали
коэволюции колебательно-вращательного механизма цветка (Nilsson,
1980). По мнению V. Summerhayes (1951), основным опылителем это-
го вида является медоносная пчела (Apis mellifera). Н. Ziegenspeck
(1936) среди постоянных посетителей цветков дремлика болотного от-
мечает медоносных пчел, реже — ос (Grabo brevis). F. Prochazka и V.
Velisek (1983) называют вероятными опылителями мух и медоносную
пчелу.
Следует заметить, что многие исследователи признают большое
значение самоопыления в жизни дремлика болотного (Summerhayes,
1951; Wiefelsputz, 1970; Fuller,1986 и др.). Возможность самоопыле-
ния определяется положением пыльника и слабостью поллиниев. Ко-
гда основная часть поллиния уносится опылителем, в пыльнике обыч-
но остаются единичные тетрады пыльцы, которые затем дуновением
ветра или в результате сотрясения цветка падающими дождевыми ка-
плями попадают на рыльце (Fuller, 1986).
В отличие от других видов рода, у дремлика болотного грибная
инфекция корней очень слабая, но на бедных почвах и при низкой ос-
вещенности степень микотрофности возрастает. Интенсивное мико-
ризообразование наблюдается также на очень сырых почвах с низким
содержанием гумуса (Ziegenspeck, 1936; Summerhayes, 1951).
Хозяйственное значение и охрана вида
Хозяйственного значения вид не имеет. Из дремликов этот вид
наиболее декоративен, он легко поддается культивированию (Собко,
Нефедова, 1983; Собко, 1989). Разводится в ботанических садах Мо-
сквы, а также на Украине — в Киеве, Харькове. А.А. Алехин (1992)
относит его к категории видов, очень перспективных для интродукции.
Дремлик болотный — вид с сокращающейся численностью, что
связано главным образом с антропогенным воздействием (особенно
загрязнением и осушением). Так, в Англии 50 лет назад этот вид был
61
обычным, а ныне полностью исчез (Nilsson, 1979). Дремлик болот-
ный быстро сокращает свое распространение и на Украине (Андриен-
ко, 1980), где ранее встречался широко (например, в окрестностях Кие-
ва), однако местами все еще сохранились крупные популяции (Андри-
енко, 1987). В настоящее время известно 100 местонахождений этого
вида в пределах Украины, но часть из них, возможно, могла исчезнуть
в результате мелиорации (Тимченко, 1994).
Дремлик болотный внесен в списки охраняемых растений в Мо-
сковской области и других регионах России (Татарстан, Ямало-Ненец-
кий национальный округ, Владимирская, Тамбовская обл.), а также в
Красную книгу Украины (1980).
Дремлик широколистный
Номенклатура, систематическое положение,
внутривидовая систематика
Epipactis helleborine (L.) Crantz, 1769, Stirp. Austr. ed. 2, 2: 467;
Скворцов, 1966, Опред. раст. Моск, обл.: 123; Смольянинова, 1'976,
Фл. европ. части СССР, 2: 26; D.M. Moore, 1980, Fl. Europ., 5: 327;
Новиков,. 1986, Опред. раст. Мещеры, 1:134; Черепанов, 1995, Сосуд,
раст. России: 619. — Serapiashelleborine L. 1753, Sp. Pl.: 949, p.p., quoad
a latifolia L. — Epipactis latifolia (L.) All. 1785, Fl. Pedem., 2: 151;
Кауфман, 1866, Моск, фл., изд. 1: 476; Цингер, 1885, Сборник свед.:
423; Шмальгаузен, 1897, Фл. Средн, и Южн. Росс., 2: 452; Петун-
ников, 1901, Крит, обзор, 3: 55; Сырейщиков, 1906, Илл. фл. Моск,
губ., 1: 264; он же, 1914, цит. соч., 4: 48; он же, 1927, Опред. раст.
Моск, туб.: 96; Невский, 1935, Фл. СССР, 4: 624; Назаров, 1940, в
Маевский, Фл., изд. 7: 244; Голицын, 1964, в Маевский, Фл., изд. 9:
679 — Дремлик широколистный (Семейство Orchidaceae — Орхид-
ные).
Наиболее полиморфный вид рода Epipactis, внутри которого вы-
делено множество подвидов, разновидностей, отличающихся по раз-
мерам и форме листовой пластинки, форме и окраске цветков. Неко-
торых из них иногда возводят в ранг вида (Rhichards, Porter, 1982;
Tyteca, Dufrem, 1994). E. Peitz (1970, цит. no: Ftiller, 1986) упоминает
около 40 форм, однако, ряд авторов (Senghas, 1970, и др.) считают, что
многие из них не имеют таксономического значения и их, вероятно,
следует считать экологическими формами.
62
F. Prochazka и V. Veh'sek (1983) выделяют на территории Чехо-
словакии 2 подвида (ssp. helleborine и ssp. viridis), 6 разновидностей,
15 форм. Для территории Украины Е.И. Бордзиловский (Флора УРСР,
1950) отмечает 6 разновидностей, встречающихся в основном в за-
падных областях. С.В. Голицын (в Определителе... Маевского, 1964)
указывает две разновидности: var. platyphylla (высокий толстый сте-
бель, часто с фиолетовым оттенком, листья почти равны междоузли-
ям, кисть всегда удлиненная, густая, основание эпихилия с треуголь-
ным бугорком) и var. viridiflora (стебель короче и тоньше, листья зна-
чительно длиннее междоузлий, продолговато-яйцевидные, кисть бо-
лее редкая и короткая, с менее опушенной осью, бугорок у основания
эпихилия малозаметен, листочки околоцветника зеленые, иногда с ро-
зовым оттенком). Те же разновидности на территории Западной Евро-
пы отмечает Н. Ziegenspeck (1936).
Epipactis helleborine образует внутриродовые гибриды с Epipactis
atrorubens, Е. leptochila, Е. microphylla, Е. purpurea, а также межродо-
вой гибрид с Cephalanthera damasonium (Senghas, 1970; Fuller, 1986;
Potiicek, 1990).
Географическое распространение
Наиболее широко распространенный вид рода дремлик, встре-
чается в Европе (от Скандинавии и Британских островов до Среди-
земноморья и Урала), в значительной части Азии (Малая Азия, Иран,
Средняя Азия, Сибирь, Монголия, Гималаи), а также в Северной Аф-
рике (Флора СССР, 1935; Ziegenspeck, 1936; Summerhayes, 1951; Фло-
ра европ. частиСССР, 1976; Flora Europaea, 1980; Флора Сибири, 1987;
Губанов, 1996).
Дремлик широколистный — один из немногих видов орхидных
Старого Света, натурализовавшихся в Северной Америке — США и
Канаде (Sunders, 1978; Catling, 1983; Brunton, 1986 и др.).
В России распространен в европейской части от Карелии до
Волжско-Донского и Заволжского районов, а в Азиатской части — в
Западной и Восточной Сибири до северо-восточного побережья Бай-
кала и Якутии (Флора СССР, 1935; Флора европ. части СССР, 1976;
Флора Сибири, 1987; Опред. сосуд, растений центра европ. России,
1995). В Московской области встречается нередко во всех районах (Во-
рошилов и др., 1966), но крупных скоплений не образует.
63
Морфологическое описание
Многолетнее корневищное травянистое растение, с побегами,
междоузлия которых разновелики (Смирнова, 1990). По И.Г. Серебря-
кову (1962,1964), дремлик широколистный (так же, как и дремлик тем-
но-красный) следует относить к классу наземных травянистых поли-
карпических растений с ассимилирующими побегами несуккулентно-
го типа, подклассу корневищных многолетников. Корневище — гипо-
геогенное, симподиально формирующееся, укороченное (в понимании
Е.Л. Любарского, 1963), плагиотропное. По И.В. Татаренко (1996), жиз-
ненная форма дремлика широколистного (и дремлика темно-красно-
го) — короткокорневищная.
Почки возобновления закладываются в пазухах чешуевидных
листьев при основании монокарпического побега. Наиболее развитой
оказывается верхняя по положению почка, несущая зачаток генера-
тивного побега. Из нижележащей почки (редко двух) формируются
вегетативные побеги неполного цикла развития. Направление роста
корневища — поступательное (тип «А» по Н. Rasmussen, 1986), побе-
ги возобновления, развивающиеся из верхней по положению почки,
ежегодно растут в одном направлении (Татаренко, 1996). Глубина за-
легания корневища зависит от характера субстрата и поддерживается
за счет контрактильной деятельности корней. Корневище сформиро-
вано серией из двух первых междоузлий, сохраняющихся от каждого
годичного прироста (Татаренко, 1996), на первом из них располага-
ются спящие почки и придаточные корни, на втором — почки возоб-
новления. Каждый годичный прирост корневища взрослого растения
имеет от 2 до 6 корней. Корни растут несколько лет, достигая мине-
рального горизонта почвы (длина корней в среднем 15—17 , иногда
до 60 см). Одновременно у взрослых особей функционируют около
20—25 корней. От корневища обычно отходит 1 побег (реже 2), но
иногда — до 5—6 побегов.
Стебель — прямой, крепкий, 25—80(100) см высотой, внизу —
голый, вверху—рассеянно опушенный, иногда пурпуровый в нижней
части. Зеленые (срединные) листья в числе 4—10(12), спирально рас-
положенные, эллиптически-ланцетные, тупые или немного заострен-
ные, 5—10(12) см длиной, 3—5,5(8) см шириной. Нижние листья —
широкоовальные, яйцевидные, переходящие во влагалище, верхние —
яйцевидно-ланцетные, не образующие влагалища.
Соцветие — прямая терминальная кисть 10—40 см длиной, со-
стоящая из 6—25(50), иногда до 100 цветков (Summerhayes, 1951).
64
Прицветники — ланцетные, зеленые, нижние — превышающие цве-
ток. Цветки — без запаха, варьируют по окраске от бледных желтова-
то-зеленых до зеленовато-вишневых. Листочки околоцветника — рас-
простертые, овальные или овально-ланцетные, заостренные, зелено-
ватые, 1—1,3 см длиной, внутренние немного короче — до 1,1 см,
эллиптически-овальные, бледно-зеленые, в нижней половине чаще бо-
лее или менее окрашенные в розовато-фиолетовый цвет. Губа — 9—
11 мм длиной. Гипохилий 5—5,5 мм длиной и шириной, округлый,
полушаровидно-мешковидный, чашевидно-вогнутый, внутри—голый,
красновато-темнобурый, снаружи — зеленоватый, по краям — зеле-
новато-беловатый. Эпихилий — 5—5,5 мм длиной и шириной, широ-
ко-сердцевидно-яйцевидный, зеленовато-бледнофиолетый, немного за-
остренный, при основании с двумя гладкими или почти гладкими бу-
горками. Колонка — 3—3,5 мм длиной, с широким носиком, полли-
нии двухраздельные, с маленькой железкой.
Завязь — голая или слабо опушенная, вместе с цветоножкой
7—9 мм длиной, постепенно сужена к основанию. Плод — коробочка,
повислая, овальная, нередко шестигранная, раскрывается двумя ще-
лями. Семена — мелкие бледно-желтые, 1,2—1,4 мм длиной (Флора
СССР, 1935; Ziegenspeck, 1936; Summerhayes, 1951; Флора европ. час-
ти СССР, 1976; Смирнова, 1990; Татаренко, 1996).
Диплоидный набор хромосом 2п = 36, 38, 40, 44 (Флора европ.
части СССР, 1976; Флора Сибири, 1987; Flora Europaea, 1980).
(Этногенез, особенности размножения,
ритм сезонного развития
Сведений об онтогенезе мало. Семена прорастают с помощью
грибов, образуя протокорм (микоризом), который находится на значи-
тельной глубине, что затрудняет его изучение (Ziegenspeck, 1936). Опы-
ты с другим представителем рода, Е. gigantea, показали, что незрелые
семена прорастают лучше, чем зрелые. Обработка семян ауксином и
этрелом не дала положительного эффекта (Arditti et al., 1981). Прото-
корм интенсивно инфицируется грибом, каждый год образуя новые
междоузлия, несущие корни и чешуевидные листья, в пазухах
которых заложены спящие почки. Отмечено, что протокорм до 7 лет
растет моноподиально, а позже — симподиально. На 9-й год после
прорастания семени развивается первый надземный олиственный по-
бег. Корневище начинает расти более интенсивно в горизон-
65
тальном направлении. В 10—11 лет растение зацветает (Ziegenspeck,
1936). К этому возрасту отмирают корни, образовавшиеся первыми (в
неблагоприятных условиях это происходит через 3 года).
По сведениями V. Summerhayes (1951), от момента прораста-
ния семени до первого цветения происходит не менее 8 лет. После
этого растение может цвести ежегодно много лет без перерыва. Внеш-
ний вид дремлика широколистного в различных возрастных состоя-
ниях показан на рис. 3, некоторые морфологические показатели этих
состояний приведены в таблице 4. Мы наблюдали за экземпляром дрем-
лика широколиственного в Подмосковье в березняке орляково-ланды-
шевом, который цвел каждый год в течение 24 лет, однако, в послед-
ние годы образовывал меньше цветков и плодов. Таким образом, мож-
но предположить, что жизненный цикл дремлика широколистного мо-
жет длиться 30—35 лет (или более).
Таблица 4.
Некоторые морфометрические показатели дремлика
широколистного в Московской области
Показатель Онтогенетическое состояние
j im V g
Число листьев 1—2 3 4 4-6(7)
Длина листа, см 5—6 6—8 7—10,5 7,5—12
Ширина листа, см 1—1,5 1,8—2,9 2,1—3,9 3,7—5,5
Высота побега, см 6—11 10—21 20—32 38—72
Число цветков — — — 4—32
Дремлик широколистный размножается преимущественно се-
менным путем. Процент плодообразования в различных частях
ареала вида сильно колеблется: в Московской области — 65%,
в Ставропольском крае (Тебердинский заповедник) — 36(0—100)%,
на Украине 2—91%. Однако, ювенильные растения встречаются ред-
ко, в большинстве местонахождений даже в благоприятных условиях
они составляют не более 14% от численности популяции (Тимченко,
1993). В.Г. Собко (1989) отмечает, что семенное размножение этого
вида на Украине чаще всего происходит под пологом дуба обыкновен-
ного. В.В. Назаров (1995) для дремлика широколистного установил
в Крыму следующие показатели: реальная семенная продуктив-
66
s
Рис. 3. Возрастные состояния Epipactis helleborine.
ность цветка — 3 284 (потенциальная 3 518), реальная семенная про-
дуктивность особи — 26 604 (потенциальная — 60 792).
По данным А.Е. Баталова, в Архангельской обл. среднее число
семян в одном плоде у растений этого вида составило 3 024±1 756,
причем в пределах соцветия этот показатель резко различается: в верх-
ней части 1156±170, в средней 2764±665, в нижней 5152±689 семян в
одном плоде.
В Московской области молодые семенные растения встречают-
ся также очень редко, но длительное пребывание во взрослом (осо-
бенно генеративном) состоянии обеспечивает устойчивое существо-
вание популяции на занятой территории. Дремлик широколистный, по
нашим наблюдениям, может пребывать в состоянии вторичного покоя
в течение многих лет. Так, например, в березняке ландышево-орляко-
во-разнотравном (Одинцовский р-н Московской области) часть осо-
бей вегетирует и цветет ежегодно, а некоторые пребывают в состоя-
нии вторичного покоя 2—7 (возможно, и более) лет.
67
Вегетативное размножение дремлика широколистного может
осуществляться путем «деления» и отмирания старого участка корне-
вища при образовании двух и более надземных побегов, но так как
расстояние между этими побегами незначительное, то расходятся вновь
возникшие особи в стороны очень медленно (Summerhayes, 1951). По
нашим наблюдениям, число особей, имеющих более одного побега, в
разных частях ареала невысоко (не более 5%), а расстояние между
побегами составляет от 0,5 до 1,2 см. В.Г. Собко (1990) отмечает воз-
можность вегетативного размножения этого вида в культуре делением
корневища на 2—3 части.
Дремлик широколистный — один из самых поздноцветущих
видов наших орхидных. В Московской области вегетировать начина-
ет лишь в начале июня, зацветает во второй половине — конце июля,
иногда даже в начале августа, цветение продолжается 2—3 (иногда 4)
недели. В зависимости от погодных условий разница в сроках созре-
вания плодов может доходить до 1 месяца. Например, в 1995 г. в по-
следних числах июля семена уже начали высыпаться, а в 1996 г. 26
августа все плоды еще были закрыты. Заканчивается вегетация этого
вида в Московской области в конце сентября. Сроки цветения дремли-
ка широколистного в других частях ареала близки к вышеуказанным
— в Германии (Ziegenspeck, 1936) конец июня—августа (в тени — до
сентября), в Англии (Summerhayes, 1951) — июль — конец авгу-
ста.
К концу вегетации побег будущего года в почке полностью сфор-
мирован. И.В. Татаренко (личное сообщение), рассматривая в октяб-
ре 1996 г. почки генеративных особей дремлика широколистного, об-
наружила, что в почке сформированы зеленые листья, прицветники и
цветки, а также заложены почки, из которых разовьются побеги в 1998
и 1999 гг, то есть внутрипочечное развитие побега происходит в тече-
ние трех лет.
Экология
Среди дремликов, произрастающих в России, широколистный
отличается наибольшей экологической амплитудой (Вахрамеева и др.,
1994, Тимченко, 1996). Теневынослив, чаще растет в полутени, хотя
может встречаться и на открытых местах, например, на лесных поля-
нах, вырубках, по обочинам дорог (3-я ступень шкалы Ландольта и
шкалы Элленберга). Обычно растет на участках с увлажнением
68
от свежего до влажного, индикатор среднего увлажнения (3-я ступень
шкалы Ландольта, 5-я ступень шкалы Элленберга). Иногда может рас-
ти на сухих склонах благодаря глубокому проникновению корней в
почву (Summerhayes, 1951). В условиях Крыма — ксеромезофит (Го-
лубев, 1984).
По отношению к кислотности почв сходен с другими видами
дремликов — в основном растет на нейтральных и щелочных почвах,
иногда — на слабо кислых, но никогда на очень кислых (4-я ступень
шкалы Ландольта, 7-я ступень шкалы Элленберга). В Польше встре-
чается на почвах с pH 5,4 (Krukowski et al., 1993), в Чехословакии —
на почвах с pH 6,3—8,0 (Prochazka, Vellsek, 1983). Предпочитает поч-
вы, богатые гумусом (4-я ступень шкалы Ландольта), средне богатые
другими питательными веществами (2-я ступень шкалы Ландольта, 5-
я ступень шкалы Элленберга), но может встречаться и на бедных поч-
вах. Растет чаще на тонкоструктурных почвах (5-я ступень шкалы Лан-
дольта), но отмечен также и на каменистых и песчаных почвах
(Summerhayes, 1951). Поднимается в горы до 13 00 м над уровнем моря
(Prochazka, Vellsek, 1983).
В Московской области чаще встречается в осветленных лесах,
по обочинам дорог, на лесных полянах (ближе к стене леса), изредка и
в глубокой тени (но в таком случае растения развиты слабо и не пло-
доносят), чаще — на дерново-подзолистых среднеувлажненных поч-
вах с pH 4,7—6,5.
Фитоценология
Дремлик широколистный встречается в широколиственных ле-
сах (буковых, дубовых, грабовых), мелколиственных (березовых, ре-
же — осиновых), смешанных, изредка в хвойных (еловых, пихтовых,
сосновых), а также в зарослях кустарников, на лугах (Флора СССР,
1935; Summerhayes, 1951; Флора европ. части СССР, 1976; Fuller, 1986).
В Германии—характерный вид сообществ Fagetalia (Oberdorfer, 1970),
но также растет в пихтовых лесах, в лесах лесостепной зоны, на при-
брежных лугах Северного моря, где встречается низкорослая форма
этого вида (Fuller, 1986). В Крыму растет преимущественно в лесах из
дуба пушистого, на Карпатах — в буковых лесах (Собко, 1989).
В Эстонии дремлик широколистный встречен нами в коротко-
ножковых березняках с можжевельником, в Калининградской области
— в сосновых и смешанных лесах, в Вологодской области — в бе-
69
резняках, осинниках, ольшаниках, в заболоченных сосняках с бере-
зой, на опушках и обочинах лесных дорог. На Северном Кавказе (Те-
бердинский заповедник, Ставропольский край) — в буковых, сосно-
вых и пихтовых лесах. Чаще растет небольшими группами или оди-
ночными растениями, но иногда может образовать и значительные ско-
пления. Так F. Fuller (1986) описал крупную популяцию в ельнике-
черничнике, где на площади ЗООХЗОО м обитало более 800 растений,
причем окраска цветков сильно варьировала от бело-зеленоватого до
грязно-красного.
Дремлик широколистный образует ценопопуляции с возрастны-
ми спектрами двух типов: большей частью (около 80%) — неполно-
членными (отсутствуют преимущественно ювенильные и имматурные
особи или только первые), реже (около 20%) — полночленными (таб-
лица 5). Неполночленные ценопопуляции найдены нами в Московской,
Вологодской, Тверской областях, подобные популяции описаны на Ук-
раине (Тимченко, 1996) и в Польше (Krukowski et al., 1993). В полно-
членных ценопопуляциях, которые встречаются в разных частях
ареала и на территориях различного хозяйственного использова-
ния, численность молодых особей может быть значительной —
до 35% ювенильных и до 45% имматурных растений. В большинстве
случаев возрастной спектр ценопопуляций — правосторонний с мак-
симумом на группе взрослых вегетативных, реже — генеративных осо-
бей.
Анализ виталитетной структуры ценопопуляций (по Ю.А. Зло-
бину, 1989), произрастающих на Украине (Тимченко, 1996) показал,
что украинские ценопопуляции дремлика широколистного — преиму-
щественно дигрессивные (75%), процветающие и равновесные соста-
вили 25%. В тенистых грабовых лесах даже на охраняемых террито-
риях ценопопуляции дигрессивны, что объясняется низким уровнем
освещенности. Процветающие ценопопуляции чаще встречаются в
более светлых лесах, нередко — при слабом антропогенном наруше-
нии.
Дремлик широколистный относится к числу немногих европей-
ских орхидных, которые не только хорошо приживаются во вторич-
ных местообитаниях — в посадках деревьев, на вырубках, обочинах
шоссейных и железных дорог (Вахрамеева и др., 1996; Вахрамеева и
др., 1997), но и натурализуются за пределами естественного ареала в
США и Канаде (Catling, 1983; Brunton, 1986 и др.), образуя там до-
вольно значительные скопления.
70
Таблица 5.
Возрастные спектры (%) и плотность (особей на м2) ценопопуляций
дремлика широколистного в различных местонахождениях
Район исследования Онтогенетическое состояние Плот-
j im V g ность
Московская обл. 3 8 18 71 —
Московская обл. 0 2 11 29 0,1
Вологодская обл. 2 3 16 79 2,5(6)
Ставропольский край 5 7 14 74 4,9(8)
Крым 0 0 23 77 0,14
Крым 4 31 54 11 —
Хмельницкая обл. 11,8 35,3 47 5,9 1,3
Крым 12,7 27,3 29,1 30,9 0,24
Польша 1 13 9 77 —
В 1979 г. нами была обнаружена крупная популяция дремлика
широколистного в Москве на Воробьевых горах. Позже в 1996 г. дрем-
лик широколистный был найден в дендрарии Ботанического сада МГУ
и на территории Института металлургии РАН. Все они возникли на
месте бывших пахотных земель, где при строительстве в конце 40-х и
50-х гг. был полностью нарушен почвенный покров, примерно тогда
же были пересажены из леса липы (Tilia cordata). Самая крупная по-
пуляция дремлика широколистного насчитывает свыше двух тысяч осо-
бей. Возрастной спектр этих популяций — правосторонний, с макси-
мумом в генеративной группе особей (67—93%). Вегетативные особи
составляют 7—33% (в том числе ювенильные и имматурные — в сум-
ме не более 10%, а чаще — менее 5%). Средняя плотность ценопопу-
ляций — от 0,5 до 5,4 особей на м2. Интересно отметить, что за про-
шедшие годы популяция на Воробьевых горах заметно расширила за-
нятую территорию, переселившись через проезжую часть улицы в по-
садки березы. Поддержание этой популяции и расширение террито-
рии происходит в основном семенным путем.
71
Консортивные связи
Дремлик широколистный — энтомофил, привлекает насекомых
нектаром, стекающим по губе цветка (Ziegenspeck, 1936). Опыляется
преимущественно осами. Цветки посещают также шмели и мухи-жур-
чалки, но последние вряд ли осуществляют опыление. По мнению V.
Summerhayes (1951), шмели с их крупной головой не могут проник-
нуть внутрь цветка, а журчалки слишком малы, чтобы суметь достать
нектар. Nilsson (1979) считает, что не только осы, но шмели и пчелы
успешно участвуют в опылении этого вида, причем нектар цветка мо-
жет оказывать на насекомое наркотический эффект. Натурализовав-
шиеся в Канаде дремлики нашли «своего» опылителя — осу Vespula
vulgaris (Judd, 1972;Dowden, 1975). Хотя вид — преимущественно ал-
ло гамный (Назаров, 1995), ряд исследователей указывает на наличие
у него и автогамии (Ziegenspeck, 1936; Kunkele, 1969,1970; Nieschalk,
Nieschalk, 1970 и др.).
Дремлик широколистный — микотроф, но интенсивность ми-
коризообразования весьма изменчива в зависимости от возрастного
состояния особи и качества субстрата, на котором растет растение
(Ziegenspeck, 1936; Summerhayes, 1951). Более микотрофны молодые
особи. Во всех возрастных состояниях микоризообразование в сырых
местообитаниях более интенсивно. В хвойных лесах на умеренно ув-
лажненных глинистых почвах корни располагаются поверхностно на
границе с подстилкой и практически лишены микоризы (Ziegenspeck,
1936). Взрослые растения с хорошо развитыми листовой и корневой
системами, как правило, автотрофны.
И.В. Татаренко (личное сообщение), просмотрела корни осо-
бей дремлика, собранного на Воробьевых горах в октябре, и обнару-
жила наличие микоризы в корнях всех возрастных групп, причем у
генеративных растений интенсивность микоризообразования оказалась
выше, чем у других возрастных групп. В корнях взрослых растений
были отмечены участки, заново инфицированные грибом. Эти сведе-
ния противоречат данным, изложенным выше. Вероятно, микотроф-
ность или автотрофность взрослых особей в большой степени зави-
сит от условий произрастания и может сильно изменяться в течение
сезона.
Хозяйственное значение и охрана вида
В подробной сводке L. Lawler (1984) указывается, что дремлик
широколистный был завезен в Северную Америку как средство
от подагры, а впоследствии натурализовался и даже, по мнению
72
A. Hawkes (1943, 1944), стал сорным видом. Есть также сведения, что
дремлик широколистный использовали в качестве слабительного сред-
ства (Planchon, Collin, 1895, цит. по: Lawler, 1984) и при артрите (Maout,
Decarius, 1976). Сведения об употреблении этого вида в нашей народ-
ной медицине не были обнаружены.
Дремлик широколистный успешно приживается в культуре (Соб-
ко, 1989), образуя через 2—3 года куртину из 5—7 особей (побегов).
А. А. Алехин (1992), занимаясь интродукцией этого вида в Ботаниче-
ском саду Харьковского университета, оценивает его как перспектив-
ный для интродукции. В Ботаническом саду МГУ взрослые особи этого
вида выпадали через 1—4 года после посадки. Однако под пологом
деревьев образовался его самосев, из которого в условиях культуры
сформировалась устойчивая популяция.
Дремлик широколистный внесен в Списки охраняемых видов
растений в Московской области и других регионах России (Татарстан,
Владимирская обл.), а также в Красную книгу Украины (1980).
Дремлик темно-красный
Номенклатура, систематическое положение,
внутривидовая систематика
Epipactis atrorubens (Hoffin.) Bess. 1809, Prim. Fl. Galic. 2: 220;
Скворцов, 1966, Опред. раст. Моск, обл.: 122; Черепанов, 1973, Свод
дополн. и измен.: 357; он же, 1995, Сосуд, раст. России: 619; Смолья-
нинова, 1976, Фл. европ. части СССР, 2:26; D.M. Moore, 1980, Fl. Europ.,
5: 328. — Serapias atrorubens Hoffin. 1804, Deutschl. Fl. ed. 2, 2, 1: 182.
— Epipactis rubiginosa Crantz, 1769, Stirp. Austr., 6:467; Невский, 1935,
Фл. СССР, 4:626. — Дремлик темно-красный (семейство Orchidaceae
— Орхидные).
Вид довольно изменчив. Помимо типового, описан подвид subsp.
borbasii (Soo) Soo, эндемичный для венгерских песчаных степей.
F. Prochazka с V. Velisek (1983) выделяют разновидности —
var. atrorubens (типичная форма с высотой более 15 см, шириной ли-
стьев более 1,5 см и многоцветковым соцветием) и var. radnensis (вы-
сокогорная форма, менее 15 см высотой, с узкими листьями и мало-
цветковым соцветием). В свою очередь, var. atrorubens представлен
рядом форм (иногда им придают более высокий таксономический
ранг), различающихся размерами листьев и прицветников, окраской
73
околоцветника, особенностями строения губы (f. atrorubens, f. latifolia,
f. longibracteata, f. majoriflora, f. laeviconica и др.). Ряд авторов (Борд-
зиловский, 1950; Prochazka, Velisek, 1983; Fuller, 1986; Собко, 1989)
отмечают, что помимо растений с типичными темно-красными цвет-
ками, встречаются формы с другой окраской околоцветника:
f. viridiflora Sanio — с желтовато-зеленой, f. pallens Beckhaus — с зе-
леновато-белой, f. lutescens Coss, et Germ. — co светло-желтой. По-
следняя форма отмечена в Эстонии (Schmeidt, 1996). J. Gelbrecht и G.
Hamel (1987) в окр. Бранденбурга (Германия) в сухих сосновых лесах
обнаружили триплоидную форму дремлика темно-красного, отличаю-
щуюся большими размерами, окраской листьев и фенологией (зацве-
тает на несколько недель позже основной формы).
Epipactis atrorubens способен образовать межвидовые гибриды,
чаще всего — с Е. helleborine, реже — с Е. microphylla, Е. palustris;
существует и межродовой гибрид с Cephalanthera damasonium (Борд-
зиловский, 1950; Senghas, 1970; Prochazka, Velisek, 1983; Fuller, 1986;
Potucek, 1990).
Географическое распространение
Евразиатский вид, ареал которого идет на восток и юг не так
далеко, как у Epipactis helleborine. В Европе распространен широко от
Британских островов и Скандинавии до Испании, Италии, Балканско-
го полуострова и Крыма. На севере достигает 71°07' с.ш., встречаясь
вдоль побережья океана, где особенно ощутимо смягчающее влияние
Гольфстрима (Alm, 1992). Отсутствует на побережье Атлантического
океана и на юге Средиземноморской области. Ареал охватывает и зна-
чительную часть Азии—Малую Азию, север Ирана, Кавказ, Централь-
ную Азию, Сибирь (Ziegenspeck, 1936; Summerhayes, 1951; Meusel et
al., 1965; Флора европ. части СССР, 1976; Flora Europaea, 1980; Fuller,
1986; Собко, 1989).
В России встречается в европейской части и в Сибири. Ареал
вида простирается от Кольского полуострова до Нижне-Донского и
Заволжского районов, далее переходит через Урал в азиатскую часть
России, где вид встречается в южной части Западной Сибири (Тюмен-
ская, Курганская, Омская и Новосибирская области). На севере
России отмечен в центральной части Кольского полуострова (Хиби-
ны, Ловозерские горы) и у Адахского ущелья на р. Усе, но нигде не
достигает пределов тундровой зоны, за исключением местонахожде-
74
ния близ г. Никель в Мурманской области (Флора СССР, 1935; Флора
европ. части СССР, 1976; Флора Северо-востока европ. части СССР,
1976; Флора Сибири, 1987; Похилько, 1993).
В Московской области найден в 1927 г. на территории Можай-
ского района (по р. Озерне), но нельзя исключить, что он может быть
обнаружен и в других пунктах.
Морфологическое описание
Многолетнее травянистое корневищное растение с побегами,
междоузлия которых (как и у дремлика широколистного) — разной
длины (Смирнова, 1990). Корневая система образована придаточны-
ми корнями, число которых у мощных экземпляров может достигать
40—50 и даже 100 (Ziegenspeck, 1936). Длина корней, по нашим на-
блюдениям, в зависимости от условий местообитания варьирует от
10—15 см (на рыхлых субстратах — дерново-подзолистых, дерново-
карбонатных почвах) до 25 см и более (на каменистых обнажениях).
Генеративный побег — 25—60(100) см высотой. Стебель в верх-
ней части — коротко густо опушенный и обычно окрашенный в крас-
но-фиолетовый цвет. Листья — в числе 7—9(12), жесткие и слегка
шероховатые, овальные или овально-ланцетные, заостренные, длин-
нее междоузлий, 4—8 см длиной и 1,5—4 см шириной.
Соцветие — прямая однобокая кисть 7—20 см длиной и более,
с густо опушенной осью. Цветки — в числе 5—23(30) на скрученных
цветоножках, темно-пурпурные или красновато-пурпурные, с запахом
ванили. Прицветники — ланцетные, нижние — немного длиннее
цветков. Наружные листочки околоцветника — яйцевидные, заост-
ренные, с тремя жилками, снаружи — рассеянно мелко опушен-
ные, 6—7 мм длиной, 3 мм шириной. Листочки внутреннего круга
— эллиптические, немного короче; все листочки околоцветника, кро-
ме губы, — сходящиеся, собранные колокольчато. Губа — без шпор-
ца, 5,5—6,5 мм длиной; гипохилий — чашевидно - вогнутый,
2,5—3 мм длиной и примерно столько же шириной, с широким перед-
ним входом. Эпихилий — сердцевидно-почковидный, 3—3,5 мм дли-
ной, 4—5 мм шириной, с коротеньким остроконечием и мелко тупо
зазубренными краями; при основании — с двумя морщинисто-зубча-
тыми буграми.
Колонка — прямая, короткая, до 2,5 мм длиной; завязь и цвето-
ножка — густо опушенные (Флора СССР, 1935; Ziegenspeck, 1936;
75
Summerhayes, 1951; Маевский, 1964; Флора европ. части СССР, 1976;
Fuller, 1986; Prochazka, Vellsek, 1983). Плоды — коробочки, от почти
шарообразных до грушевидных, слегка повисающих и повернутых в
одну сторону. Плодоножки — короткие, около '/2 длины коробочки
(Fuller, 1986).
Диплоидный набор хромосом 2п = 40 (Love , Love , 1944, 1961;
Hagerup, 1944; Kliphuis, 1963; Tanaka, Kamemoto, 1974, 1984; Флора
европ. части СССР, 1976; Fuller, 1986, Флора Сибири, 1987). Отмече-
ны хромосомные расы с 2п = 60, а также анеуплоиды: 2п = 40 + (0—
7В) (Flora Europaea, 1980); 2n = 40 + (0—12В), 60 + 2В (Prochazka,
Vellsek, 1983).
Онтогенез и сезонное развитие
Онтогенез вида к настоящему времени изучен недостаточно.
Сведений о прорастании семян дремлика темно-красного мало. При
отсутствии симбиотрофного компонента не удалось добиться прорас-
тания, как при грунтовых посадках, так и в лаборатории (Похилько,
1993). По Н. Ziegenspeck (1936), семена дремлика темно-красного про-
растают после перезимовки весной, к осени образуется протокорм (ми-
коризом) с точкой роста на одном из полюсов. В течение второго года
развиваются 2 междоузлия микоризома с двумя чешуевидными листь-
ями и 1—2 придаточных корня, а к осени закладывается почка, содер-
жащая зачаток будущего надземного побега. Первый зеленый лист по-
является на третий год жизни, и растение переходит от микосимбио-
трофного к смешанному способу питания. Моноподиальный способ
нарастания сменяется симподиальным.
По данным А.А. Похилько (1993) и И.В. Блиновой (1995), под-
земное развитие в условиях Мурманской области может длиться доль-
ше — до 5 лет. В дальнейшем увеличивается число междоузлий и кор-
ней на корневище. На 4-й год растение образует два, на 5-й год — три
зеленых листа. В каком возрасте дремлик темно-красный зацветает—
точно не известно (Ziegenspeck, 1936). Может цвести несколько лет
подряд или с перерывами. По нашим данным, продолжительность боль-
шого жизненного цикла этого вида не менее 25—30 лет. На основе
изучения дремлика темно-красного в условиях Архангельской облас-
ти нами были выделены критерии онтогенетических (возрастных) со-
стояний (рис. 4).
Ювенильные растения, высотой 3,9±0,25 см, с 1—2 листьями,
длина которых 1,9 см, ширина — 0,6 см. Корневище короткое (0,5—
76
см) и тонкое (2—2,5 мм), с 3—5 придаточными корнями, залегает
на глубине от 3 до 4,5 см (здесь и далее глубина залегания корневища
указана для растений из сосняка разнотравного, сформированного на
среднесуглинистой дерново-карбонатной почве).
Имматурные растения, высотой 6,3±0,42 см, с 3—4 листьями
до 4 см длиной и 1,5 см шириной. Корневище достигает 1,5 см в дли-
ну, 3—4 мм в диаметре; число придаточных корней увеличивается до
6—11. Глубина залегания корневища—до 7 см. Заглубление корневи-
ща, обусловленное контрактильной деятельностью корней, продолжа-
ется в течение всей жизни растения.
Среди взрослых вегетативных растений мы выделяем две груп-
пы — слабо развитые v и мощно развитые v . Растения группы v
имеют высоту 10—12 см, с 3—4 листьями, длина которых превышает
3,9 см, а ширина—1,5 см. Корневище длиной 1,7—2,3 см, диаметром
4—5,5 мм, залегающее на глубине 7,4—8 см, несет от 15 до 22 корней.
У v2 растений число листьев достигает 5—7, побег — до 22 см высо-
той. Длина корневища — 2,4—3,6 см, диаметр — 5,5—7 мм. Глубина
залегания корневища — 9—12 см, придаточных корней — 14—35. У
растений этого возрастного состояния отмечено ветвление корневи-
ща. Помимо верхней по положению пазушной почки, за счет которой
осуществляется симподиальное нарастание корневища, закладываются
1—2 почки, несущие зачатки побегов, и имеющие после отрастания
черты строения v или im растений.
Общая длительность виргинильного периода, по нашим наблю-
дениям, составляет 8—16 лет; из них в состоянии протокорма расте-
ние пребывает 3—5 лет, в ювенильном состоянии— 1—3 года. Имма-
турная стадия длится 2—3 года, во взрослом вегетативном состоянии,
в зависимости от условий существования, растение находится от 2 до
5 лет. В среднем, растения приступают к цветению в возрасте 10—12
лет.
Среди генеративных растений может быть выделено 3 группы:
молодые g средневозрастные g2 и старые g3 особи. Молодые генера-
тивные растения имеют высоту до 30 см и 4—5 листьев. Корневище,
залегающее на глубине 9—12 см, имеет длину 2,7—3,5 см, диаметр —
5,5—8,5 мм; число придаточных корней — 14—29. Корневище, как
правило, ветвится и, кроме генеративного, может нести 1—2 вегета-
тивных побега. Соцветие длиной 3,5—9,2 см, из 5—10 цветков.
Взрослые генеративные особи достигают 50 см в высоту, име-
ют до 7 нормально развитых фотосинтезирующих листьев. Соцветие —
77
Рис. 4. Возрастные состояния Epipactis atrorubens (на рисунках корневищ изобра-
жены не все корни).
10—20 см длиной, с 10—23 цветками. От корневища может отходить
несколько генеративных и вегетативных побегов. В результате отми-
райия старых частей корневища происходит партикуляция, не сопро-
вождающаяся омоложением. Длина корневища может достигать 5 см,
диаметр — 8—9,5 мм, глубина залегания — 10—15 см. Число прида-
точных корней — 23—43.
У старых генеративных растений корневища более короткие (до
1,7—2,6 см), диаметром 4,5—6,5 мм, число придаточных корней —
11—19. Корневище продолжает погружаться в почву и располагается
на глубине 12—17 см, ветвление его ослабевает или прекращается.
Генеративный побег высотой 18—25 см несет от 3 до 5 нормально
развитых листьев. Соцветие — 5,5—8 см длиной с 5—7 цветками.
Длительность генеративного состояния окончательно выяснить
не удалось. По-видимому, она составляет 10 и более лет. Перерывы в
цветении, как правило, длятся не более 2—3 лет. Различить в полевых
условиях молодые и старые генеративные растения без их выкапыва-
ния невозможно, поэтому при популяционных исследованиях все ге-
неративные особи рассматриваются как единая группа, либо в ее
пределах выделяют слабо развитые g , g3 и мощно развитые g2 расте-
ния.
78
Рис. 4 (продолжение).
Сенильные растения (s) внешне отчасти сходны с ювенильны-
ми или имматурными. Они имеют 2—3 листа длиной 1,6—2,4 см, ши-
риной 0,5—0,8 см. Корневище, в отличие от молодых особей, — ко-
роткое (0,3—0,5 см) и более толстое (5,1—5,5 мм). Придаточных
79
корней — 2—4, часто темно-коричневых, со следами загнивания. Глу-
бина залегания корней — 12—15 см. Почки возобновления отсутству-
ют, либо имеется одна почка, несущая зачаток побега сенильного ти-
па.
По ритму сезонного развития дремлик относится к весенне-лет-
не-осеннезеленым растениям с периодом зимнего покоя (Татаренко,
1996). В.Н. Голубев (1984), в условиях Крыма, относит этот вид к эфе-
мероидам, отрастающим весной. Сроки наступления отдельных фаз
различаются в разных частях ареала. Так, по многолетним наблюде-
ниям в Жигулевском заповеднике (Саксонов, Терентьева, 1992), нача-
ло весеннего отрастания дремлика отмечается, в среднем, 13 мая. На-
чало цветения — 6—24 июня, массовое цветение — с 20 июня по 3
июля, созревание семян — с 4 июля по 22 августа. Отмирание надзем-
ных побегов происходит с 7 сентября по 27 октября. В Мурманской
области вегетация начинается в конце мая — начале июня, начало бу-
тонизации — во второй декаде июня, цветение наступает во второй-
третьей декадах июля. Плоды созревают через 25—30 и более дней
после начала цветения. Зрелые семена образуются в конце августа.
Отмирание надземных побегов заканчивается обычно не позднее 20
сентября (Похилько, 1993). В Германии вид зацветает первым из дрем-
ликов, иногда — в начале, но чаще — в середине июня, и цветет до
июля (Fuller, 1986).
Нами исследованы некоторые особенности развития монокар-
пического побега дремлика темно-красного в условиях Архангельской
области. Почка возобновления первого года развития, представленная
меристематическим бугорком, закладывается в период конца бутони-
зации — начала цветения (начало — середина июля). Длина почки —
0,25 мм, диаметр — 0,20 мм. К концу лета формируется 1—2 листо-
вых примордия, почка увеличивается до 0,5—0,6 мм в длину. В тече-
ние следующего года продолжается образование листовых приморди-
ев, их число достигает 7—10(15). В пазухах второго, третьего или чет-
вертого примордиев также начинается образование почек возобнов-
ления, причем зачаток генеративного побега несет только верхняя по
положению почка.
К концу второго года развития длина почки возобновления дос-
тигает 7,5—9,0 мм, ширина — 4,0—5,5 мм. На третий год с самого
начала вегетации в почке активно закладываются зачатки цветков на
побеге будущего года. Длина почки достигает 12—16 мм при ширине
4—5 мм. К концу лета эти размеры увеличиваются до 20—21 и
80
5—6 мм соответственно. Вегетация начинается, обычно, в конце мая,
цветение — в первой—второй декадах июля, плодоношение — с конца
июля до конца августа, отмирание надземной части—во второй—третьей
декадах сентября. Цикл развития монокарпического побега в условиях
Архангельской области, таким образом, длится более 3 лет.
Способы размножения и распространения
Дремлик темно-красный размножается как семенным, так и ве-
гетативным путем. Опыление — аллогамное, сведений о самоопыле-
нии в природе нет. Плодоношение происходит регулярно во всех час-
тях ареала, в том числе на севере (в Мурманской и Архангельской об-
ластях). Процент образования плодов довольно высок — до 93%
(в среднем — от 50 до 75%). Значительно (до 36%) снижено плодооб-
разование в условиях сильного затенения (ольшаник разнотравный),
где, очевидно, затруднено опыление. Данные о потенциальной семен-
ной продуктивности отсутствуют. Реальная семенная продуктивность
изучалась нами в 1995—1996 годах в различных фитоценозах Архан-
гельской области. Максимальное отмеченное число семян на коробочку
— 2823. В то же время, по данным А.А. Похилько (1993), в Мурман-
ской области средняя семенная продуктивность достигает 3 500 и да-
же 4 000 семян на один плод.
Вегетативное размножение играет намного меньшую роль, чем
у длиннокорневищных видов, например Е. palustris. Ветвление корне-
вища начинается при переходе растения в генеративное состояние, хотя
у некоторых, наиболее мощно развитых, особей может осуществлять-
ся и раньше (в виргинильном периоде). У средневозрастных генера-
тивных растений происходит партикуляция, приводящая к образова-
нию клонов. А.А. Похилько (1993) отмечает образование на корнях
придаточных почек, дающих начало побегам, зацветающим на третий
год. V. Summerhayes (1951) пишет о возможности (в редких случаях)
вегетативного размножения дремлика темно-красного с помощью по-
чек, образующихся на корнях. Возникшие таким способом растения
могут оказаться на некотором расстоянии от материнского. При изу-
чении живого и фиксированного материала из Архангельской облас-
ти, а также гербарных образцов из различных частей ареала, нами не
было обнаружено ни одного случая наличия корневых отпрысков. Об-
разование почек на корнях дремлика темно-красного, видимо, носит
факультативный характер, как и у ряда других видов орхидных (Irmish,
1853; Вахрамеева и др., 1996; Татаренко, 1996).
81
Таблица 6.
Семенная продуктивность дремлика темно-красного
в различных условиях местообитания
Фитоценоз Число цветков на побег % пло- до обра- зования Число семян в плоде Число плодов на побег Число семян на побег
Сосняк разнотравный* И,5 66,9 1524±68 7,72 11765
Вырубка на месте сосняка* 10,9 61,58 1413±79 6,71 9481
Ольшаник разнотравный* 12,9 36,29 1073±64 4,68 5022
Сосняк с мож- жевельником* 11,7 51,13 1356±62 5,99 8122
Гипсовое обнажение" 9,5 46,35 1087±73 4,40 4783
Обочина автодороги" 9,8 50,11 1006±68 4,91 4934
Березовое криволесье*** 12,8 76,5 4100 9,80 —
Примечание: * Каргопольский р-н Архангельской обл., "Пинеж-
ский р-н Архангельской обл. (данные авторов), ‘“Мурманская обл. (по:
Похилько, 1993).
Экология
По экологическим особенностям дремлик темно-красный бли-
зок к дремлику широколистному, но, в отличие от него, тяготеет к бо-
лее сухим и светлым местообитаниям (Собко, 1989; Вахрамеева и др.,
1994; Татаренко, 1996). Обычно встречается под пологом леса или по
опушкам, в «окнах» древесной растительности, в полутени, при осве-
щенности более 10% (6-я ступень шкалы Элленберга, 3-я ступень шка-
лы Ландольта), нередко при полном освещении (Summerhayes, 1951),
изредка при сильном затенении; в последнем случае проявляют-
ся признаки угнетения — снижается степень образования пло-
дов и семенная продуктивность, плотность популяций. В.Н. Го-
лубев (1984) относит дремлик темно-красный к гелиосциофи-
82
там. При резком изменении условий освещения способен переходить
в состояние вторичного покоя (Summerhayes, 1951).
Дремлик темно-красный предпочитает сухие или средне сухие
почвы, избегая очень сухих (3-я ступень шкалы Элленберга, 2-я сту-
пень шкалы Ландольта). Е. Lando It (1977) указывает на приурочен-
ность дремлика к глинистым или торфянистым плохо аэрируемым поч-
вам (5-я ступень шкалы), но, по нашим наблюдениям, гораздо чаще
вид встречается на хорошо аэрируемых субстратах, от мелкопесчаных
до грубых по механическому составу, иногда на скальных обнажени-
ях. Кальцефил, часто произрастает на выходах гипсов, известняков
или доломитов (Summerhayes, 1951; Bringer, 1961). Предпочитает
нейтральные или щелочные почвы (8-я ступень шкалы Элленберга,
4-я ступень шкалы Ландольта), избегает кислых почв. По дан-
ным F. Prochazka и V. Velisek (1983), встречается в диапазоне
pH от 7,5 до 9,0. Почвы, на которых произрастает дремлик темно-
красный, обычно бедны питательными веществами, особенно мине-
ральным азотом (1-я ступень шкалы Элленберга, 2-я ступень шкалы
Ландольта).
По мнению А.А. Похилько (1993), дремлик темно-красный —
гемикриптофит. По нашим данным — геофит. Почки возобновления
находятся в почве на глубине от 4 до 20 см (в зависимости от структу-
ры и механических свойств субстрата). В условиях Архангельской об-
ласти заморозками не повреждается. Растет как на равнинах, так и в
горах, где достигает субальпийского пояса, поднимаясь до 2000 м
(Ziegenspeck, 1936; Summerhayes, 1951; Prochazka, Velisek, 1983).
Фитоценология
Дремлик темно-красный — лесной вид, встречается в листвен-
ных (дубовых, буковых, грабовых) и хвойных (преимущественно со-
сновых, лиственничных, реже еловых) и смешанных лесах, по их опуш-
кам, в зарослях кустарников, на морских дюнах, на известняковых,
мергелевых и алебастровых обнажениях, у подножия скал, на песча-
ных осыпях, по щебнистым бичевникам (Флора СССР, 1935;
Ziegenspeck, 1936; Summerhayes, 1951; Флора европ. части СССР, 1976;
Опр. сосуд, раст. центра европ. России, 1995).
На севере Германии, в странах Прибалтики, Калининградской
области России дремлик темно-красный нередко встречается на дюнах
Балтийского моря, где часто представлен очень крупными (до
83
1 м высотой) экземплярами (Ziegenspeck, 1936; Summerhayes, 1951;
Андронова и др., 1980; Талла, 1980; Fuller, 1986; Schmeidt, 1996). На
Украине дремлик темно-красный произрастает в сосново-дубовых, ду-
бово-грабовых лесах совместно с такими орхидными, как Platanthera
bifolia, Listera ovata, Epipactis helleborine, Neottia nidus-avis (Прядко,
1987). В Молдавии — редок, может быть обнаружен в буковых дубра-
вах, в тенистых, умеренно влажных местах (Киртока, Истратий, 1987).
В Жигулевском заповеднике (Саксонов, Терентьева, 1992) встречает-
ся в сложных борах (со вторым ярусом из клена и березы), сосняках с
березой, остепненных борах, реже — в нагорных дубравах и дубово-
липово-кленовых лесах, но нигде не образует больших скоплений. На
южном Урале отмечен в хвойных и смешанных лесах, на сырых лугах
(Кучеров, Мулдашев и др., 1987).
На севере ареала иногда произрастает в сырых смешанных ле-
сах, пихтово-сфагновых и папоротниковых ельниках, на прибрежных
закустаренных «лужках» с покровом из ягодных кустарничков, овся-
ницы овечьей, перловника, пиона (Флора северо-востока европ. части
СССР, 1976). В Мурманской области встречается на каменистых рос-
сыпях в субальпийских березняках Хибин (Мишкин, 1953). По дан-
ным А. А. Похилько (1993), популяции дремлика обитают в ценозах,
общее проективное покрытие которых колеблется от 70 до 95%. От-
дельные особи могут встречаться у больших камней или среди них,
где проективное покрытие растений составляет всего 10—20%.
В Архангельской области (бассейны рек Пинеги и Сотки), на
гипсовых и известняковых обнажениях, дремлик темно-красный встре-
чен в составе реликтовых комплексов, совместно с арктоальпийскими
видами (Юдин, 1963). По нашим данным, из этой группы наиболее
часто сопутствуют ему Dryas octopetala, D. punctata, Arctous alpina,
Saussurrea alpina. Довольно крупные скопления дремлика отмечены в
сосняке можжевельниковом, по лесным опушками, а также на скаль-
ных участках. В Каргопольском районе Архангельской области, на тер-
ритории с неглубоким залеганием известняков, дремлик темно-крас-
ный входит в состав травяного яруса специфических сосняков разно-
травных. В ассоциациях этого типа общее проективное покрытие
травяного яруса обычно не превышает 60—70%, а мохово-лишайни-
кового яруса — 85—90%. Встречается дремлик и на открытых
участках — вырубках лесов, сохраняя в этих условиях способность к
семенному и вегетативному размножению. На богатых почвах,
в густом травостое, дремлик испытывает сильное воздействие со
84
стороны более конкурентно-способных видов, что проявляется в сни-
жении плотности популяций и семенной продуктивности и, как след-
ствие, способности к удерживанию занятой территории.
Таблица 7.
Возрастные спектры (%) и плотность (особей на м2) ценопопуляций
дремлика темно-красного в разных условиях местообитания
Фитоценоз Онтогенетические группы Плот- ность
j im V g s
Сосняк разнотравный 8,3 15,0 29,3 45,1 2,3 4,43
Вырубка на месте сосняка 9,3 5,5 29,6 48,2 7,4 2,16
Ольшаник разнотравный 3,6 3,6 32,1 53,6 7,1 1,87
Сосняк с мож- жевельником 6,3 10,7 33,0 46,4 3,6 2,80
Скальный участок 6,4 6,4 25,5 59,1 2,7 2,75
Обочина автодороги 1,6 4,7 34,4 59,4 0 1,28
Исследованные популяции относятся к нормальным полночлен-
ным (таблица 7). В возрастном спектре преобладают генеративные и
взрослые вегетативные особи (76—84%). Доля молодых растений не-
велика — до 24%, что объясняется меньшей длительностью этих
стадий по сравнению с взрослыми, более высокой элиминацией осо-
бей на ранних этапах развития и активным разрастанием и вегетатив-
ным размножением генеративных экземпляров, в результате чего из
почек возобновления развиваются побеги с признаками взрослых рас-
тений.
В поясе березового криволесья на Кольском полуострове, на кру-
тых крупнокаменистых осыпях с общим проективным покрытием 80—
95%, популяции дремлика темно-красного — неполночленные. Воз-
растные спектры — правосторонние. Ювенильные особи отсутст-
85
вуют, доля имматурных растений не превышает 4,9%, а генеративные
особи составляют более 50% (Похилько, 1993).
Дремлик темно-красный способен выдерживать умеренную ан-
тропогенную нагрузку, может расти во вторичных местообитаниях
(Summerhayes, 1951; Fuller, 1986; Вахрамеева, Варлыгина, 1996; Вах-
рамеева и др., 1997). Например, в охранной зоне Пинежского заповед-
ника дремлик активно заселяет обочины автодороги Пинега—Архан-
гельск, прочно удерживает занятую территорию и активно размножа-
ется. По нашим данным, дремлик способен выдерживать умеренную
рекреационную нагрузку — сбор в букеты, вытаптывание. При резких
изменениях условий обитания переходит в состояние вторичного по-
коя (Summerhayes, 1951; Татаренко, 1996).
Консортивные связи
Дремлик темно-красный—энтомофил (Ziegenspeck, 1936; Sum-
merhayes, 1951; Nilsson, 1979), цветки содержат нектар и имеют запах
ванили (у мощных растений запах значительно сильнее), привлекаю-
щий насекомых (Schmeidt, 1996). И.В. Татаренко (1996) определяет
характер опыления дремлика как слабо специализированную энтомо-
филию, т.е. наряду с преобладающими опылителями, его осуществля-
ют й некоторые другие группы насекомых.
F. Fuller (1986) указывает, что у дремлика темно-красного
имеются довольно значительные различия в величине цветков,
причем околоцветник может быть более или менее открытым или же
колокольчато сходящимся. Открытые цветки предпочитают пчелы,
а колокольчатые — шмели. Прилетевшее насекомое находит на
эпихилии надежную опору, однако, для эффективного опыления
более важной оказывается позиция насекомого на гипохилии. Это
обеспечивает строение краев гипохилия, утолщающихся в передней
части так, что в месте перехода к эпихилию образуется очень узкое
отверстие, через которое протискивается насекомое, корректируя свое
положение. Хоботок опылителя достигает нектара, скрытого в
углублении («чаше») гипохилия. При этом устанавливается необ-
ходимое расстояние ото лба насекомого до липкой поверхности рыль-
ца, к которой приклеиваются принесенные поллинии. Пчела, приле-
тевшая без поллиниев, должна при проникновении в гипохилий разо-
рвать железку, после чего комочки пыльцы приклеиваются к ее голо-
ве. Обычно в пределах соцветия пчела посещает только один цветок,
86
редко 2 или 3, затем она летит на другие соцветия. Быстрая смена по-
сещаемых цветков обеспечивает эффективное опыление в пределах
популяции.
На ранних этапах развития дремлик темно-красный — обли-
гатный микосимбиотроф. Во взрослом состоянии интенсивность ми-
коризной инфекции — от слабой до средней (Татаренко, 1996). Расте-
ния, особенно во взрослом состоянии, мало зависят от грибов
(Summerhaues, 1951; Nilsson, 1979). Н. Ziegenspeck (1936) отмечает
четкую сезонность инфицирования и переваривания мицелия грибов
в корнях дремлика темно-красного. Изредка встречаются бесхлоро-
филльные особи, существующие исключительно за счет микосимбио-
трофии (Prochazka, Velisek, 1983; Schmeidt, 1996). Данных о таксоно-
мической принадлежности и видоспецифичности микосимбионта
дремлика.темно-красного нет. По-видимому, он относится к формаль-
ному роду Rhizoctonia (Burgeff, 1936).
Хозяйственное значение и охрана
Хозяйственного значения вид не имеет. Высоко декоративен, но
широкого распространения в культуре не получил. Сведения об ус-
пешности его интродукции противоречивы. По данным В.Г. Собко
(1989), в первичной культуре вид хорошо размножается вегетативно и
дает самосев. В Ботаническом саду Уральского научного центра РАН
(Екатеринбург) вид успешно цветет и плодоносит. В то же время пере-
садка взрослых растений в питомник Полярно-альпийского ботаниче-
ского сада (Похилько, 1993) не дала положительных результатов. В
большинстве случаев растения выпадали через 3—4 года, чаще всего,
оставаясь в вегетативном состоянии. В некоторых случаях удается по-
лучить молодые растения, подсевая семена к взрослым экземплярам
(Prochazka, Velisek, 1983).
Численность вида сокращается в результате, главным образом,
вырубки лесов и рекреации. Она резко снизилась в ряде регионов —
Латвии (Талла, 1980), Литве (Балявичене, 1980), на Украине (Загуль-
ский, 1987; Косых, 1987), Калининградской области в России (Андро-
нова и др., 1980). Вид включен в списки охраняемых растений Мос-
ковской области и других регионов России (Татарстан, Ямало-Ненец-
кий национальный округ, Владимирская, Ульяновская обл. и др.), а
также в Красные книги Украины (1980), Латвии (1985) и Белоруссии
(1993).
87
М.А. Гуленкова, С.К. Пятунина
ДРЕМА БЕЛАЯ
Номенклатура
Melandrium album (Mill.) Garcke, 1858, Deutchl. FL, 4 Aufl.: 55;
Сырейщиков, 1907, Илл. фл. Моск, губ., 2: 105; он же, цит. соч., 4: 65;
он же, 1927, Опред. раст. Моск, обл.: 123; Шишкин, 1936, Фл. СССР,
6: 725; он же, 1940, в Маевский, Фл., изд. 7: 345; он же, 1964, цит. соч.,
изд. 9:229; Черепанов, 1995, Сосуд, раст. России: 323. — Lychnis alba
Mill. 1768, Gard. Diet. ed. 8, № 4; Шмальгаузен, 1895, Фл. Средн, и
Южн. Росс., 1: 145. — L. pratensis auct.: Кауфман, 1866, Моск. фл. изд.
1: 86. — Melandrium pratense Rohl. 1812, Deutschl. FL, 2 Aufl., 1: 274;
Цингер, 1885, Сборник свед.: 99. — Silene alba (Mill.) E.H.L. Krause,
1901, in Sturm, Deutschl. FL, 2 Aufl., 5: 98; Chater a. Walters, 1964, FL
Europ., 1: 174; В. Тихомиров, 1966, Опред. раст. Моск, обл.: 157; он
же, 1986, Опред. раст. Мещеры, 1: 169. — Дрема белая (семейство
Caryophyllaceae — Гвоздичные).
Географическое распространение
Melandrium album имеет евразийский ареал, в пределах которо-
го довольно обычна на лугах, полях, около канав, на огородах, в садах
и посевах яровых. Распространена почти по всей европейской части,
за исключением Арктики, на Кавказе, в Западной и Восточной Сиби-
ри (за исключением альпийских областей). В Средней Азии имеет ара-
ло-каспийский ареал (Флора СССР, 1938). За пределами евразиатско-
го континента распространена в Канаде и Северной Африке (Baker,
1947). В Московской области это — довольно обычное расте-
ние, хотя в северной части области встречается реже (Ворошилов и
ДР, 1966).
Морфология
Melandrium album—двудомное многолетнее травянистое расте-
ние. В начале онтогенеза — стержнекорневое, а затем — укорочен-
но - корневищное с мочковатой корневой системой и с симподиально
88
возобновляющимися полурозеточными ортотропными побегами, ко-
торые развиваются по моно-, ди-, и полициклическому типу. Дрема
белая — гемикриптофит, но проявляет себя и как протогемикрипто-
фит (Baker, 1947; Ellenberg, 1974).
Стебель на поперечном срезе — округлый, в узлах слегка утол-
щен, опушен простыми и железистыми волосками. Стебель у женских
особей имеет больший диаметр, и более развитую механическую ткань,
залегающую под корой, что повышает его прочность и упругость. Это,
вероятно, связано с баллистическим способом распространения се-
мян. Кроме того, женские растения испытывают большую механиче-
скую нагрузку на стебель во время плодоношения. Интересно отме-
тить и различие в проводящей системе мужских и женских особей.
Как показали наши исследования, у мужских экземпляров М. album на
всем протяжении стебля сохраняется пучковое строение. Тогда как у
женских особей только в верхней его части — проводящая система в
виде пучков, а на большей части стебля ксилема и флоэма образуют
сплошное кольцо. На уровне узла (однопучкового, однолакунного) раз-
личий обнаружено не было.
Листорасположение — супротивное. Листья — простые, без
прилистников, сидячие, покрыты простыми кроющими и железисты-
ми волосками. Их пластинка — цельнокрайняя, овально-продолгова-
тая, верхушка — острая, основание — клиновидное, жилкование —
перистое.
Сравнительный морфологический анализ побегов мужских и
женских особей позволил выявить некоторые различия в их организа-
ции. Женские растения, произрастающие в тех же условиях что и муж-
ские, имеют более мощные побеги: их длина в 1,5 раза выше, а число
листьев на особь в 2 раза больше по сравнению с мужскими. Суммар-
ная площадь пластинок листьев на удлиненной части побега свиде-
тельствует о большей фотосинтезирующей поверхности у женских осо-
бей, что биологически оправдано и объясняется большими затратами
энергии на формирование плодов и семян.
Цветки — однополые, актиноморфные, с двойным околоцвет-
ником. Чашечка — пятичленная, сростнолистная, опушенная просты-
ми кроющими и железистыми волосками, часто — слегка окрашена
антоцианом. У тычиночных цветков она — трубчато-колокольчатая, у
пестичных—широкояйцевидная. Венчик—белый, пятичленный, раз-
дельнолепестный, отгиб лепестка — двулопастной переходит в удли-
ненный ноготок. Зев окаймлен привенчиком до 2 мм высотой.
89
Периметр чашечки женского цветка в 1,5 раза больше, чем у
мужского цветка. По мере развития плода, размеры чашечки увеличи-
ваются, ее жилки становятся более мощными, возрастает число ана-
стомозов между ними. Усиливается наряду с защитной и трофическая
функция чашечки (обеспечение растущего плода питательными веще-
ствами). Анализ поперечных срезов через чашечку показал, что у жен-
ских растений наблюдается большее число слоев хлорофиллоносных
клеток, которые располагаются более компактно, в отличие от чашеч-
ки мужского цветка; форма клеток у мужских и женских цветков одно-
типна.
Гинецей — ценокарпный, пятичленный, завязь — верхняя.
А.Г. Девятовым (1988) описаны цветки с различными отклонениями
развития: с дополнительным плодолистиком, развитым так же, как и
все остальные, с расщеплением плодолистика, с появлением добавоч-
ных плодолистиков на колонке. Иногда меняется число тычинок или
лепестков, например, был описан четырехчленный цветок Melandrium
album с 8-членным гинецеем.
Особенностью гинецея дремы белой является неодинаковая
структура завязи на разных ее уровнях. В нижней части гинецей —
синкарпный, выше — лизикарпный и еще выше — паракарпный. Со-
ответственно плацентация в основании завязи — центрально-уголко-
вая, выше (на большем протяжении) — центрально-ложно-осевая, а в
самой верхней части завязи — париетальная.
Механизмы, приводящие к возникновению одногнездной завя-
зи, были описаны G. Bocquet (1960). Плодолистики в цветках Meland-
rium album возникают как аппендикулярные образования, т.е. занима-
ют боковое положение на апексе цветоложа. Вначале закладывается 5
бугорков, которые, разрастаясь, сливаются краями, образуя кольцо. От
места слияния бугорков в центростремительном направлении продол-
жается рост плодолистиков, что приводит к образованию септ и воз-
никновению пяти гнезд с центрально-уголковой плацентацией. В са-
мой верхней расширенной части формирующегося пестика септ
не образуется, и завязь остается одногнездной с париетальной
плацентацией. При формировании завязи внешняя сторона пло-
долистиков растет быстрее, чем внутренняя и септы. Впоследствии
за счет лизиса перегородок в средней части завязи протяженность
одногнездного участка увеличивается. Следы разрушения септ
обнаруживаются по сохраняющимся их остаткам на внутренней сто-
роне плодолистиков, а затем и перикарпия. Возникающая при этом в
90
центре завязи колонка образована также остатками плодолистиков; на
ней — сформированные семязачатки.
Процессы, происходящие при развитии завязи, подтверждают,
что у М. album разрушение перегородок — активный и закономерный
процесс (Девятов, 1992). Об этом свидетельствует расположение об-
туратов в колонке: у М. album они имеют вид сплошных широких по-
лос между рядами семязачатков.
Андроцей представлен 10 тычинками в 2 круга на длинных ты-
чиночных нитях. Пыльники в бутонах имеют почти те же размеры,
что и во взрослом цветке. Пыльники первого круга созревают раньше,
второго — позднее. Разновременное созревание и раскрытие пыльни-
ков продлевает сроки использования пыльцы. Мужские особи зацве-
тают раньше и продолжают цветение дольше, чем женские. В жен-
ских цветках имеется дисковидное утолщение в основании завязи (ру-
димент андроцея), которое выполняет функцию нектарника (Карта-
шова, 1965).
В центре мужского цветка находится рудимент гинецея (Baker,
1947). Наши наблюдения показали, что его размеры у разных особей
неодинаковы. В одних цветках рудиментарный пестик вытягивается
до ’/ длины тычиночной нити, в других он едва заметен. Между ними
возможны все переходы. На продольных срезах мужских цветков
«пестик» хорошо заметен. Внешнее гистологическое сходство между
ним и нектарником в женском цветке позволило сделать вывод
о его секретирующей функции (Brantjes, 1976). Однако Н.Н. Кар-
ташова (1965) считает, что мужские цветки дремы белой этой функ-
ции лишены.
В природных популяциях дремы белой в Ульяновской области,
отмечено три структурных типа цветков (Крискович, Старшова, 1994):
мужские, женские и частично андростерильные. Описаны и четыре
половых типа особей: 1) мужские; 2) женские; 3) мужские и частично
андростерильные; 4) частично андростерильные. Случаев реверсии
мужских цветков к обоеполости не обнаружено. Соотношения полно-
ценных мужских и женских цветков почти равны (1,1:1,0), а на долю
частично андростерильных приходится 0,4. Поскольку частично анд-
ростерильные цветки являются существенными донорами пыльцы,
флоральное соотношение несколько сдвинуто в пользу мужских цвет-
ков, а опылительный потенциал больше 1—1,5.
Плод — многосемянная, яйцевидная, лизикарпная коробочка,
при созревании вскрывается наверху десятью отверстиями в резуль-
91
тате растрескивания околоплодника. Место вскрывания определяется
нижележащими слоями перикарпия, которые подвергаются структур-
ным изменениям, а эпидермальные и субэпидермальные слои не из-
меняются (Девятов, 1992). Около каждого отверстия имеется по зуб-
чику. При формировании плода происходит склерификация околоплод-
ника. Раньше начинается и заканчивается одревеснение в верхней части
формирующейся коробочки, тогда как нижняя ее часть, оставаясь па-
ренхимной, по мере созревания заметно разрастается и увеличивает-
ся в диаметре. Поэтому в процессе морфогенеза коробочки ее форма
изменяется от цилиндрической до конусовидной. Как указывает Н.
Baker (1947), «сгибание» зубцов коробочки во время влажной погоды
и прикрывание ими отверстий сохраняет семена сухими, препятству-
ет их слипанию.
Цветки дремы белой собраны в сложный дихазий. Цветет рас-
тение с июня по август. Длительность цветения определяется после-
довательным развертыванием цветков соцветия, а также разновремен-
ным отрастанием генеративных побегов в пределах одной особи. По-
следнее обусловливает наличие на одном экземпляре одновременно
бутонизирующих, цветущих и плодоносящих побегов.
Число хромосом 2п=24 (Хромосомные числа..., 1969). Их на-
бор состоит из 22 аутосом и 2 половых хромосом: XX —- женские осо-
би: XY — мужские, причем половые хромосомы отличаются друг от
друга размерами (У-хромосома — больше) и особенностями строе-
ния (Сидорский 1983).
Способы размножения и распространения
Melandrium album — растение энтомофильное. Опыляется ноч-
ными бабочками. Размножение и расселение осуществляется в приро-
де в основном семенами. Дрема белая — растение баллист. Созрева-
ние и раскрытие коробочек происходит через 4—5 недель после опы-
ления. К моменту обсеменения коробочка занимает вертикальное по-
ложение, благодаря карпотропическому изгибу плодоножки. Как пра-
вило, у растений баллистов чашечка играет барьерную роль, но у дан-
ного вида эта функция не выражена, к моменту созревания плода
чашечка подвергается деструкции. Семена высыпаются из коробочки
и разбрасываются под влиянием механического воздействия на побе-
ги. На полях они могут распространяться вместе с семенами клевера
или злаковых культур (Baker, 1947). Возможно также распростране-
92
ние семян птицами. Новые места обитания осваиваются благодаря се-
менному размножению. Вегетативное размножение — процесс мед-
ленный и может иметь значение только в агрофитоценозах (Baker,
1947).
Онтогенез и сезонное развитие
Семена — округло-почковидные, симметричные, более широ-
кие у спинки, черные, с восковидным налетом. Боковые их стороны
— более или менее выпуклые, в местах перехода от боковых сторон к
брюшной стороне видны неглубокие впадинки. Спинка — выпуклая
или слегка уплощена, без ложбинки. Семенная кожура близ микропи-
ле незначительно углублена, поверхность семян — отчетливо бугор-
чатая, с хорошо выраженной кутикулой. Размеры семян — 1,2—1,5
мм (длина), 0,8—1,0 мм (ширина). Средняя толщина экзотесты —
0,04—0,15 мм (Федосеев, 1986).
После созревания семена переходят в состояние покоя «дремо-
ты», которое продолжается меньше года. При благоприятных услови-
ях (температуре около 25°, глубине залегания не глубже 4 см) семена
прорастают в любое время года (Baker, 1947).
Прорастание семян—надземное, гипокотилярное. Окончатель-
ная длина гипокотиля зависит от глубины залегания семян в почве и
освещенности. Чем глубже залегание семян и чем меньше света, тем
длиннее подсемядольное колено. Семенная кожура разрывается при
интеркалярном разрастании гипокотиля. Семядоли — овальные или
яйцевидные, после образования первой пары листьев они вскоре отми-
рают.
К концу первого года жизни формируется укороченный побег.
Длительность виргинильного периода определяется условиями про-
израстания. В густом травостое и при большой задернованности пре-
бывание в розеточном состоянии может продлиться несколько лет (2—
3 года). Чаще верхушечная почка укороченного побега I порядка пере-
ходит к формированию генеративного побега. Но возможно перевер-
шинивание, и к цветению переходят побеги следующих порядков. Раз-
витие их идет по типу моноциклических (яровых или озимых), ди-, и
полициклических. Имеются сведения, что растение может'вести себя
и как однолетник и как многолетник (Baker, 1947).
93
Экология, фитоценология
Melandrium album — «любитель солнца» (Baker, 1947), относи-
тельная освещенность во все периоды вегетации должно быть не ни-
же 40% (8-я ступень шкалы Элленберга). В условиях сильного затене-
ния особи остаются в вегетативном состоянии. Лучший рост наблю-
дается в условиях среднего увлажнения субстрата. Будучи мезофитом
(4-я ступень шкалы Элленберга), растение, однако, хорошо переносит
невысокую атмосферную влажность и проявляет устойчивость к вет-
РУ
Согласно Н. Baker (1947), дрема произрастает на разных по хи-
мическому и механическому составу почвах («индифферентна» к суб-
страту по Элленбергу). Ее можно встретить на скалах (южная Анг-
лия), меловых отложениях и почвах бедных кальцием, в частности,
песчаных. Дрема положительно реагирует на азотное питание (7-я сту-
пень шкалы Элленберга). Встречается как на слабокислых, так и на
щелочных почвах (pH в пределах 6,1—8,2). Качество субстрата и сте-
пень освещенности сказывается на внешнем облике растения. На поч-
вах, богатых минеральным и органическим питанием при хорошем
водообеспечении и в условиях благоприятного освещения побеги дре-
мы белой достигают более 1 м длины, их листья — крупные, а при
богатстве почв доступным азотом — темно-зеленые. Напротив, на ска-
лах произрастают растения невысокие, с узкими густоопушенными
листьями. Карликовые формы дремы с недоразвитыми цветками опи-
саны на переувлажненных субстратах; у этих растений отмечается на-
копление в клетках антоциана.
В разных природных популяциях соотношение между мужски-
ми и женскими растениями неодинаково. Чаще преобладают женские
особи (Джапаридзе, 1963 и др.), что, согласно В.Н. Наугольных и Т.Н.
Бурковой (1951), может объясняться более высокой толерантностью
первых уже с ранних этапов онтогенеза.
По нашим данным, в одних и тех же местообитаниях мужские
особи характеризуются меньшими размерами листьев, большим
числом устьиц на единицу площади и большей водоудерживающей
способностью. Это позволяет сделать вывод о повышенной засухоус-
тойчивости мужских растений по сравнению с женскими. Эту точку
зрения разделяют В.Г. Александров (1962) и Н.В. Первухина (1970).
Напротив, В.Н. Наугольных (1945), Ф.Ф. Лейсле и Н.А. Макарова
(1950) считают, что более ксерофильны женские особи. Доказатель-
94
ством последнего, согласно Лейсле и Макаровой, является тот факт,
что в поставленном ими эксперименте при снижении влажности поч-
вы выпадало меньше женских растений, чем мужских. Повышение об-
водненности субстрата (от 40 до 60% от общей влагоемкости) способ-
ствовало увеличению числа как мужских, так и женских особей. Даль-
нейшее повышение влажности (до 80%) стимулировало увеличение
числа только мужских растений.
На соотношение полов в потомстве влияет «возраст» мужских
и женских цветков. Исследования чешских ученых (Procopova, 1964)
показали, что в потомстве, полученном от опыления женских цветков
в 2-дневном (после развертывания бутона) возрасте, достоверно по-
вышен процент женских растений. В потомстве от опыления женских
цветков с возрастом больше двух дней (3—5) доля женских растений
падала. Возраст мужских цветков не оказал никакого влияния на соот-
ношение полов в потомстве.
Отмечено изменение половой дифференциации под влиянием
повышенной радиации. В результате наших наблюдений в 1986 году
(авария на Чернобыльской АЭС) в Московской области (Истринский
р-н, Павловская слобода) обнаружено появление у дремы белой дву-
полых цветков в результате развития гинецея в мужских цветках. В
последующие годы (1987—88) размеры гинецея у взятых под наблю-
дение особей уменьшались, и вновь появлялись побеги только с муж-
скими цветками.
Довольно обычна дрема белая на пашнях среди злаков и разно-
травья, иногда на землях, оставленных под пар; на пустынных местах,
пастбищах, насыпях, окраине лесов, по краю леса и вдоль лесных до-
рог. Вид никогда не формирует крупных популяций, и обычно встре-
чается в виде единичных растений или небольших групп.
Консортивные связи
Дрема белая охотно поедается домашними животными (лошадь-
ми, овцами), кролики ее не едят (Baker, 1947).
Возможно заражение грибом Fusarium, который вызывает гние-
ние семян и угнетает развитие сеянцев (Baker, 1947). Гриб Ustilago
violacae стимулирует развитие тычинок в пестичных цветках или их
стерилизацию, а также поражает почки и проростки дремы. После за-
ражения грибом рост и развитие растения замедляется.
Растение опыляется ночными бабочками, в том числе Nadena
Bickuris Hutu. Согласно N.B. Brantjes (1976), при посещении бабоч-
95
кой женских цветков она откладывает яйца возле завязи. В мужских
цветках откладка яиц тоже возможна, но, тем не менее, бабочки пред-
почитают женские особи. Последнее вполне объяснимо, т.к. первые
стадии жизни личинка проводит внутри семени под защитой кожуры.
На последних стадиях развития личинка покидает семя. В результате
проведенных N.B. Brantjes экспериментов по «распознаванию» бабоч-
кой цветка, выявлено, что большое значение в восприятии растения
насекомым имеет запах. Запах мужских и женских цветков сущест-
венно отличается: число составляющих компонентов летучего веще-
ства и интенсивность запаха у женских цветков гораздо больше. Хо-
боток бабочки покрыт хеморецепторами, что, видимо, помогает ей рас-
познавать цветки. Интересно отметить, что перед откладкой яиц ба-
бочка всегда питается нектаром. Автором подсчитано, что во время
попеременного посещения женских и мужских цветков, бабочки пе-
реносят 316 пыльцевых зерен за визит, максимально — 605, мини-
мально — 37.
Сложившиеся в данном случае отношения между растением и
насекомым можно, видимо, охарактеризовать как своеобразную фор-
му мутуализма с элементами паразитизма со стороны насекомого.
Хозяйственное значение и охрана вида
. Сведений о хозяйственном Значении дремы белой нет. В связи с
малой декоративностью вид не подвергается прямому уничтожению.
96
Р.П. Барыкина, Н.В. Чубатова
КУПАЛЬНИЦА ЕВРОПЕЙСКАЯ
Номенклатура
Trollius europaeus L. 1753, Sp. Pl.: 556; Кауфман, 1866, Моск,
фл., изд. 1: 32; Цингер, 1885, Сборник свед.: 44: Шмальгаузен, 1895,
Фл. Средн, и Южн. Росс., 1: 24; Сырейщиков, 1907, Илл. фл. Моск,
губ., 2:-137; он же, 1927, Опред. раст. Моск, губ.: 131; Шипчинский,
1937, Фл. СССР, 7: 45; Невский, 1940, в Маевский, Фл., изд. 7: 358;
Маценко, 1964, в Маевский, Фл., изд. 9: 72; Tutin, 1964, Fl. Europ., 1:
210; Ворошилов, 1966, Опред. раст. Моск, обл.: 164; Новиков, 1986,
Опред. раст. Мещеры, 1: 176; Черепанов, 1995, Сосуд, раст. России:
844. — Купальница европейская (семейство Ranunculaceae—Люти-
ковые).
Внутривидовая изменчивость вида выражена слабо. Только на
крайнем северо-востоке области распространения (северный Урал)
встречается var. aperta Per. с полуоткрытыми и открытыми цветками, с
ярко-желтыми или почти оранжевыми чашелистиками (Рычин, 1948).
Кроме научного названия вида «купальница европейская» известны
его другие названия: авдотка, золотые бубенцы и купава.
Географическое распространение
Купальница европейская — евро-сибирский вид. В Европе его
ареал охватывает Скандинавию, Среднюю и Атлантическую Европу,
Средиземноморье, Балкано-Малоазиатский район и проходит через
всю Центральную и Восточную Европу, включая Прибалтику, Бело-
руссию, Украину (KpoMv Крыма), европейскую Россию, за исключе-
нием Нижневолжского района. Встречается на севере Западной Сиби-
ри (Флора СССР, 1937; Растительные ресурсы СССР, 1984; Флора Си-
бири, 1987). В Московской области нередко встречается во всех бота-
нико-географических районах (Ворошилов и др., 1966).
97
Морфологическое описание
Купальница европейская — кистекорневой короткокорневищ-
ный гемикриптофит (Рычин, 1948; Голубев, 1962; Рысина, 1973;
Ellenberg, 1974; Гуланян, 1976; Landolt, 1987). Побеги — ортотроп-
ные, полурозеточные, дициклические, монокарпические, нередко вет-
вящиеся, в основании одеты остатками прошлогодних листьев, высо-
той 15—90 см, с одним терминальным цветком, реже несколькими
цветками, собранными в цимозные соцветия. Прикорневые листья —
длинночерешковые (до 65 см), пальчато 5-раздельные, с ромбически-
ми, остро зубчато-надрезанными долями. Стеблевые листья генера-
тивных побегов — в числе 3—7, нижние из них — на черешках, верх-
ние — сидячие с более мелкими пластинками. Цветоножки — 3—15
см длиной, при плодах сильно удлиняющиеся (до 30 см), продолгова-
то-бороздчатые.
Цветки — слегка душистые, крупные, до 5 см в диаметре,
шаровидные, ациклические, актиноморфные. Цветоложе — выпуклое.
Околоцветник — двойной. Чашелистики — светло-желтые, более
или менее округлые, в числе 7—15 (до 20), сильно вогнутые,
черепитчато налегающие один на другой, скрывающие все остальные
части цветка. Лепестки - нектарники — оранжевые, около 7 мм
длщюй, короче чашелистиков и тычинок, узкие, почти линейные, с
лопатчато - расширенной тупой верхушкой и нектарной ямкой
на внутренней стороне на высоте около 2 мм от основания.
Лепесток чуть расширен вокруг нектарной ямки и сужен в ос-
новании. Цветок — протерандрический. Тычинки — многочислен-
ные с длинными нитями, неподвижными продолговатыми пыль-
никами. Созревание тычинок, как и пестиков, центростремитель-
ное.
Пыльцевые зерна — 3-бороздные, эллипсоидальные или сфе-
роидальные, с полюса — округлые, слабо 3-лопастные, с экватора —
от узкоэллиптических до округлых, 19(17—20) мкм длиной, 16(13—
20) мкм в диаметре. Поверхность пыльцевого зерна — сглаженная,
струйчатая. Ультраструктура поверхности перфорированная; микро-
отверстия, которые чаще встречаются в области апертур, — округлые,
размером 0,1—1 мкм. Борозды — складковидные, узкие, длинные, края
борозды ровные, концы тупые, мембрана крупнобугристая, бородав-
чатая. Экзина — тонкая, 0,7—1 мкм. Внешний край покрова — волни-
стый, складчатый (Савицкий, 1982).
98
Гинецей — апокарпный, пестиков много. Семязачатки — ана-
тропные, с двумя интегументами. Плод — почти шаровидная, ацикли-
ческая, голая многолистовка (Каден, 1965). Плодик — многосемянная
листовка до 10 мм длиной с коротким, около 1 мм, загнутым к брюш-
ному шву стилодием. Семена — обратнояйцевидные с острым высо-
ким килем по всей длине, 1,3—2 мм длиной и 0,7—1,2 мм в диаметре
(Кац и др., 1965), черно-бурые, гладкие, блестящие (Флора СССР, 1937).
Вес 100 семян— около 70 мг.
Диплоидное число хромосом равно 16 (Langlet, 1927; Жукова,
1961; Числа хромосом..., 1993), указывают также числа 32
(Doroczewska, 1964), 18 (Delay, 1947, цит. по: Хромосомные числа...,
1969), 24 (Lundegardh, 1909, цит. по: Хромосомные числа..., 1969).
Онтогенез и сезонное развитие
Семена купальницы европейской, имеющие маленький (0,5 мм
длиной) двусемядольный зародыш, окруженный эндоспермом, харак-
теризуются замедленным прорастанием (Kinzel, 1913) и сохраняют
всхожесть в течение 5—6 лет (Рысина, 1973). Промораживание значи-
тельно ускоряет прорастание. В природе всходы появляются весной в
апреле.
Прорастание — надземное котиледонарное, начинается с появ-
ления коричневого главного корня, который обгоняет в росте гипоко-
тиль. У корневой шейки развивается кольцо густых бурых волосков. В
начале мая, удлиняющиеся (до 1 см) свободные черешки семядолей
поднимают над поверхностью почвы овальные светло-зеленые пла-
стинки, 4—6 мм длиной и 2 мм шириной, выполняющие функцию пер-
вых фотосинтезирующих органов проростка (рис., а).
При прорастании почечки развивается укороченный побег с
единственным ассимилирующим листом. Этот лист имеет округлую в
очертании трехлопастную пластинку, 5—8 мм в диаметре, с неравны-
ми зубцами по краю (рис., б). В его пазухе закладывается боковая поч-
ка, остающаяся, как правило, в последующие годы спящей. По наблю-
дениям В.Н. Голубева (1956а), при благоприятных условиях число и
мощность листьев у всходов может возрастать. С развертыванием фо-
тосинтезирующих семядолей и листьев на побеге развиваются прида-
точные узловые корни. В середине лета главный корень начинает от-
мирать. У растений, обитающих на хорошо аэрируемой почве, глав-
ный корень сохраняется более длительное время (Голубев, 19566).
99
1 CM
Возрастные состояния Trollius europaeus. Обозначения: A — всход, Б — ювениль-
ное, В — генеративное растение в осенний период, Г — генеративное растение в
весенний период, Ц — цветочная почка в продольном сечении (октябрь); с —
семядоли, чл — чешуевидный лист, л — ассимилирующий стеблевой лист, рл —
розеточный лист, опб — отмерший генеративный побег, цп — цветочная почка, пв
— вегетативная почка возобновления, б — бутон, кщ — эпигеогенное корневище,
гк — главный корень, пк — придаточный корень.
100
Терминальная почка побега, защищенная кроющей чешуей
и имеющая на конусе нарастания 3—4 листовых зачатка, благодаря
контрактильности придаточных корней, заглубляется в почву. Весной
следующего года она образует розеточный побег с одним чешуевид-
ным, быстро разрушающимся листом, и двумя-тремя ассимилирую-
щими листьями, имеющими тройчатораздельные пластинки. У боль-
шинства растений к этому времени полностью отмирают как главный
корень, так и гипокотиль. В течение лета на узлах годичного прироста
главного побега развиваются новые слабо разветвленные придаточ-
ные корни, в пазухах всех листьев закладываются почки. Базальные
участки главной оси, продолжая погружаться в почву за счет активно-
го сокращения придаточных корней, дают начало эпигеогенному
корневищу, диаметр которого у двулетних растений не превышает
1 мм.
На протяжении последующих 5—6 лет главный побег остается
розеточным моноподиальным, в пределах годичных приростов наблю-
дается смена первых чешуевидных листьев последующими ассими-
лирующими. Увеличиваются размеры отдельных компонентов элемен-
тарных метамеров побега (листьев, почек, междоузлий) и растения
в целом. Число ассимилирующих листьев возрастает до 3—4, увели-
чивается число спящих почек и придаточных корней, возрастает сте-
пень расчлененности пластинок (они становятся трех-пятираздельны-
ми).
У взрослых вегетативных особей в листовой розетке насчиты-
вается 3—4 пальчато-рассеченных листа, черешки которых, по дан-
ным В.Н. Голубева (1962), достигают длины 65 см. Базальные участки
эпигеогенного корневища постепенно отмирают, так что у взрослых
растений оно включает лишь 2—4 последних годичных прироста
и обычно не превышает 3,5 см. По мере увеличения числа и размеров
ежегодно образующихся листьев, корневище постепенно утолщается
до 5—12 мм. От корневища отходят около 140 придаточных корней,
достигающих глубины 60 см и простирающихся горизонтально на рас-
стояние 75 см (Голубев, 1962), образуя густое сплетение полусфери-
ческой формы. Придаточные корни, темно-каштановые, тонкие
(в основании 1,2 мм в диаметре), сохраняют первичное строение и
обильно ветвятся по всей длине до III порядка. Корни II порядка дос-
тигают 5 см длины при диаметре 0,5 мм, III порядка — 5—10 мм дли-
ны, 0,2—0,3 мм в диаметре (Linkola, Tiirikka, 1936; Рысин, Рысина,
1987).
101
С конца виргинильного периода наблюдается ветвление корне-
вища в результате раскрытия 1—3 пазушных почек (рис., в). Боковые
побеги — розеточные. Скученное размещение их определяется незна-
чительными годичными приростами корневища.
В природных условиях генеративный период развития наступа-
ет у купальницы на 7—8-й год жизни (Работнов, 1951), а в культуре —
на 2—3-й год (Голубев, 1956а; Евтюхова, 1968). Генеративные побеги
— дициклические, реже моно- или полициклические, полурозеточные,
чаще с одним прикорневым листом, заканчивающиеся терминальным
цветком (рис., г). Иногда формируются моно- или плейохазиальные
соцветия. В культуре, по данным Г.П. Рысиной (1973), у наиболее мощ-
ных экземпляров образуется большое число (более 10) генеративных
побегов. Не исключено, что в данном случае речь идет о нескольких
особях — клоне, образовавшемся из материнской особи вегетативным
путем.
После плодоношения генеративные побеги отмирают до базаль-
ных укороченных междоузлий. Весной параллельно с ними раскрыва-
ются почки возобновления. В течение лета они образуют розеточный
побег с 4—5 ассимилирующими листьями, достигающими максималь-
ных размеров в июле (ширина пластинки иногда составляет 22 см).
При развитии монокарпического побега по дициклическому типу в
терминальной почке вегетативного побега в октябре—ноябре уже
полностью сформирован цветоносный побег второго года, включая
цветки.
Просмотр гербарного материала выявил случаи вторичного цве-
тения купальницы европейской в конце августа после сенокошения в
результате пробуждения терминальной почки вегетативного побега без
периода зимнего покоя.
Купальница европейская — длительно вегетирующее, летнезе-
леное растение с поздневесенне-раннелетним ритмом цветения. В Под-
московье, как в природных ценозах, так и в культуре рост генератив-
ных побегов начинается в конце апреля — начале мая, цвете-
ние начинается с середины мая и продолжается до середины июня (Ин-
тродукция лекарственных..., 1965),. Длительность цветения одного
цветка — 7—10 дней. Плоды созревают с конца мая по август, мас-
совое плодоношение приходится на середину июля. Период диссеми-
нации сильно р астянут и длится до конца августа. Генеративные по-
беги отмирают в начале сентября. Розетка зеленых листьев укорочен-
ного вегетативного побега, которая образуется из почки возобновле-
на
ния следующего порядка, функционирует с конца мая до конца октяб-
ря. В терминальной почке такого вегетативного побега с середины июля
начинается заложение метамеров цветоносного побега второго года,
включая зачатки одной—трех почечных чешуй (рис., в), зеленых ли-
стьев и цветков (рис., д). Дифференциация элементов цветков продол-
жается и в зимнее время; завершается она в середине мая (Серебря-
ков, 1947). В пазухе почечной чешуи закладывается боковой монокар-
пический побег III порядка, дальнейшее развитие которого протекает
также по дициклическому типу. В условиях лесостепи, по данным В.Н.
Голубева (1965), фенофазы укорочены во времени и сдвинуты на бо-
лее ранние сроки.
Продолжительность большого жизненного цикла растения оп-
ределить трудно в связи с постоянным отмиранием базальных участ-
ков корневища. Исходя из того, что виргинильный период у травяни-
стых многолетних лесных растений составляет, согласно И.Г. Сереб-
рякову (1952), одну десятую всей их жизни, можно предположить, что
максимальный возраст купальницы европейской достигает 70 и более
лет.
Способы размножения и распространения
Размножается купальница в основном семенами. Семенная про-
дуктивность заметно меняется в онтогенезе. Она возрастает с перехо-
дом растения в средневозрастное генеративное состояние, что обуслов-
лено увеличением как числа монокарпических побегов и цветков на
отдельном побеге, так и числа формирующихся в цветке плодолисти-
ков. У взрослых особей может сформироваться 1—2, очень редко — 3
генеративных побега, несущих 1—3, редко 4, цветка (табл. 1).
В терминальном цветке, раскрывающемся первым, закладыва-
ется наибольшее число плодолистиков (до 70), в нижерасположенных
пазушных цветках число их обычно уменьшается примерно на чет-
верть. В каждом плодолистике формируется от 12 до 17 семяпочек (в
среднем 14). Таким образом, потенциальная семенная одного цветка
достигает 1 000, а продуктивность однопобегового генеративного рас-
тения составляет около 2500 семян.
Реальная семенная продуктивность растения может резко от-
личаться от потенциальной, что в значительной степени зависит
от условий местообитания и погодных условий года. В терминальном
цветке, по нашим подсчетам, вызревает обычно не более 52 листо-
103
вок, несущих от 10 до 15 семян. Продуктивность однопобегового рас-
тения составляет не более 1100 семян, а всего клона, в котором иногда
насчитывается до 15 особей, — 14500 семян.
Таблица 1.
Семенная продуктивность купальницы европейской в разных
фитоценозах (Одинцовский р-н Московской области)
Фитоценоз Число цветков на побеге Число плодолистиков Число семян на 1 генер. побег
терминаль ный цветок боковой цветок
Луг 1 1,7±0,5 40,3±6,0 32,О±8,3 766±356
2,1±0,8 29,1±13,0 28,4±7,7 649±349
Обозначения: 1 —лугразнотравно-манжетково-купальницевый
вдоль железнодорожной насыпи; 2—луг крапивно-таволговый по дни-
щу оврага.
В Московской области, по нашим наблюдениям, купальница ев-
ропейская в разных фитоценозах продуцирует до 1100 семян на 1 ге-
неративный побег (табл. 1) и до 6000—30000 семян на м2. Максималь-
ный урожай семян отмечен в разнотравно-манжетково-купальницевом
луговом сообществе.
Купальница европейская — баллист. Листовки ее в момент об-
семенения занимают вертикальное положение и наиболее широко рас-
крываются вверху. Книзу щель суживается и часто не доходит до ос-
нования плодика. Отдельные листовки прижаты друг к другу и обра-
щены швами к центру плода. Такое расположение плодиков препятст-
вует свободному высыпанию семян. Зрелые семена рассеиваются при
раскачивании стебля на расстояние, равное его высоте или несколько
большее, до 150—170 см (Левина, 1957). Иногда отмечается эндозо-
охория. Семена поедают птицы и распространяют их с экскрементами
(Жизнь растений, 1980).
В ходе онтогенеза, как отмечалось ранее, в результате разруше-
ния базальных участков корневища происходит дезинтеграция гене-
ративной особи на несколько дочерних. В состав клона наряду с гене-
ративными могут входить и вегетирующие растения. Однако, клони-
рование — видимое увеличение числа особей, хотя и сопровож-
104
дающееся нередко их частичным омоложением, нельзя считать эффек-
тивным способом вегетативного размножения купальницы европей-
ской, так как в возникающих клонах дочерние растения располагают-
ся скученно и, видимо, существуют непродолжительное время. В свя-
зи с этим, купальницу европейскую следует рассматривать, как слабо
подвижный вегетативный малолетник. Роль вегетативного размноже-
ния по сравнению с семенным в сохранении вида невелика. Распро-
странение купальницы европейской осуществляется исключительно
семенами и с высокой эффективностью, тогда как поддержание кон-
кретных популяций происходит преимущественно благодаря клони-
рованию.
В культуре купальницу размножают делением клона и корневи-
ща, а также семенами. Лучшие результаты дает посев под зиму; в этом
случае всходы появляются весной (Евтюхова, 1968).
Экология
Купальница европейская—бореальный (Тихомиров, 1941) или
умеренно-бореальный вид (2-я ступень шкалы Ландольта, 3-я ступень
шкалы Элленберга), встречающийся почти повсеместно, за исключе-
нием областей с континентальным климатом (3-я ступень шкалы Лан-
дольта, 5-я ступень шкалы Элленберга). Она относится к светолюби-
вым видам, но способна выдерживать 50% затенение (4-я ступень шка-
лы Ландольта, 9-я ступень шкалы Элленберга); при сильном затене-
нии не цветет (Рысина, 1973).
Купальница европейская — мезофит (Ellenberg, 1974) или ме-
зосемигигрофит (Цыганов, 1976). Массово и обильно произрастает
на небогатых почвах (7—9-я ступени шкалы Раменского, 1956) подзо-
листого, дерново-подзолистого, подзолисто-глеевого и торфянистого
типов легкого механического состава, нередко супесчаных и песча-
ных (5-я ступень шкалы Ландольта). Характерна слабо кислая,
нейтральная или слабо щелочная реакция почвы с pH 4,5—7,5 (3-я
ступень шкалы Ландольта, 7-я ступень шкалы Элленберга). Чаще встре-
чается в условиях влажно-лугового увлажнения (65—75-я ступени
шкалы Раменского, 7-я ступень шкалы Элленберга). В умеренных и
малых количествах встречается на довольно богатых (10-я ступень
шкалы Раменского) и бедных торфянистых почвах (4—6-я ступени
шкалы Раменского) в условиях от сухолугового до сыролугового ув-
лажнения (56—88-я ступени шкалы Раменского). Предпочитает поч-
105
вы, богатые гумусом (4-я ступень шкалы Ландольта). По данным Л.Г.
Раменского с соавторами (1956), лишь единичные экземпляры, испы-
тывающие заметное угнетение, отмечаются на бедных почвах (до 4-
ой ступени). Ограничивающая ступень в сторону усиления бедности
почвы не установлена, однако, как отмечает Ландольт, купальница ев-
ропейская не встречается на очень бедных почвах по содержанию ми-
нерального азота (3-я ступень шкалы Ландольта, 6-я ступень шкалы
Элленберга), избегает также очень кислых и сухих почв.
Купальница европейская произрастает в широком экологиче-
ском диапазоне, но в неблагоприятных условиях испытывает замет-
ное угнетение. Не выносит длительного затопления, но успешно пе-
реносит временное затопление полыми водами. Отрицательно реаги-
рует на выпас и сенокошение (Работнов, 1951).
Фитоценология и биологическая продуктивность
Купальница европейская произрастает в разреженных смешан-
ных и лиственных лесах, на лесных полянах, лугах, в степных колках,
зарослях кустарников, моховых и ивняковых тундрах, по берегам рек,
ручьев и стариц (Флора СССР, 1937; Растительные ресурсы СССР, 1984;
Ядовитые растения..., 1950).
В Московской области купальница европейская также является
весьма обычным растением и обладает широкой фитоценотической
амплитудой. Часто она образует обширные популяции на опушках и
полянах в различных типах леса — в ельниках простых и сложных, в
смешанных, широколиственных и мелколиственных сыроватых лесах,
сероолыпаниках, осинниках, ивняках. В изобилии встречается на за-
болоченных и сырых лугах, а также в разных нарушенных местооби-
таниях — в канавах по обочинам грунтовых и шоссейных дорог, на
олуговевших участках в полосе отчуждения вдоль железнодорожных
насыпей, изредка — на торфяниках. В некоторых фитоценозах купаль-
ница европейская достигает высокого обилия и выступает в качестве
доминанта, в момент массового цветения образует аспект. Наиболее
частыми спутниками ее в луговых фитоценозах являются различные
виды Alchemilla, Geum rivale, Anthoxantum odoratum, Deschampsia
caespitosa, на заболоченной луговине — Filipendula ulmaria, Polygonum
bistorta, Urtica dioica, Geranium palustre, Lysimachia vulgaris, на лесных
полянах и опушках Convallaria majalis, Melampyrum nemorosum,
Potentilla erecta, Stellaria holostea.
106
Наблюдения, проведенные нами в Одинцовском районе Мос-
ковской области в 3 фитоценозах показали (табл. 2), что участие ку-
пальницы в травостое может быть очень велико. Ее проективное по-
крытие нередко достигает 60%. Плотность ценопопуляций может варь-
ировать в значительных пределах. Молодые ценопопуляции составля-
ли, как правило, одиночные растения, зрелые — более или менее об-
ширные клоны из двух—семи дочерних особей. Число особей в клоне
в ходе онтогенеза в целом увеличивается, так что плотность ценопо-
пуляции постепенно растет. В исследованных нами ценопопуляциях
плотность варьировала от 9 до 45 особей на м2.
В возрастном спектре всех трех исследованных ценопопуляций
преобладали генеративные и взрослые вегетативные растения как се-
менного, так и вегетативного происхождения. Молодые (ювенильные
и имматурные) особи составляли менее 10%. При этом соотношение
между числом генеративных и взрослых вегетативных растений часто
зависело от условий произрастания, особенно от освещенности. На
разнотравно-манжетково-купальницевом лугу генеративные особи со-
ставляли 63%, а на крапивно-таволговом лугу по днищу оврага — 70%,
плотность популяции здесь относительно невысока — 9 клонов (28
особей) на 1 м2. На лугу высота генеративных растений колебалась от
24 до 84 см, общее число прикорневых листьев особей одного клона
— от 2 до 46, а на лугу по днищу оврага соответственно — 60—103
см, 2—30 (табл. 2).
Таблица 2.
Биоморфологические показатели генеративных особей купальницы
европейской в разных фитоценозах Одинцовского р-на
Фито- ценоз 1 Прикорневые листья Генеративный побег
ЧИСЛО в клоне 14,0±11,0 длина че- решка, см 26,5±7,7 ширина пла- стинки, мм 119,7±21,8 число на м2 32 высота, см 66,5±13,3
2 12,2±9,3 31,7±7,8 136,0±28,0 25 77,9±10,3
3 4,7±2,9 35,0±7,4 120,0±35,0 6 70,0±11,0
Обозначения фитоценозов (здесь и в табл. 3): 1 — луг разно-
травно-манжетково-купальницевый вдоль железнодорожной насыпи;
2—луг крапивно-таволговый по днищу оврага; 3 — разнотравно-шуч-
ково-душистоколосковый луг с ивовым подростом по днищу балки.
107
Биологическая продуктивность купальницы в разных фитоце-
нозах тесно сопряжена как с биометрическими характеристиками рас-
тений (табл. 2), так и с плотностью популяций (табл. 3). В исследован-
ных ценозах она колебалась от 280 г/м2 сырого веса (разнотравно-ман-
жетково-купальницевый луг) до 724 г/м2 (крапивно-таволговый луг).
Выход воздушно-сухой массы изменяется от 15 до 25%.
Таблица 3.
Плотность популяции и биологическая продуктивность купальницы
европейской в разных фитоценозах (Одинцовский р-н)
Фитоце- ноз Число клонов на м2 Число генер. особей Надземная био- масса клона, г Продуктивность популяции, г/м2
сырая сухая сырая сухая
1 27 32 36,7±48 6,9±6 724 134
2 9 25 39,4±26 9,9±6 280 70
Консортивные связи
Сведений о консортивных связях купальницы европейской в ли-
тературе крайне мало. Энтомофил. Элемент аттрактации закрытого
типа (Фегри, Пейл, 1982). Последовательное раскрывание цветков
в соцветии обеспечивает эффективное перекрестное опыление. Цвет-
ки купальницы, имеющие яркую окраску околоцветника, исто-
чающие тонкий запах, выделяющие нектар и обильную пыльцу, при-
влекают насекомых - опылителей, в основном пчел и жуков, ко-
торые добираются до нектара, раздвигая чашелистики (Нейштадт,
1963). X. Кюглер (Kiigler, 1970) в числе опылителей называет также
муху Chiastochaeta trollii, которая откладывает яйца на завязи.
Вылупившиеся личинки, видимо, прогрызают завязь и в течение 10
дней развиваются внутри нее, используя в качестве пищи ткани семя-
почек. В непогоду маленькие насекомые забираются в цветок, находя
в нем временное жилище, что также способствует опылению (Ней-
штадт, 1963).
Иногда отмечается эндозоохория. Установлено, что северный
олень в арктических областях поедает плоды и семена купальницы
европейской и с экскрементами распространяет их (Жизнь растений,
108
1980). В то же время купальница европейская, из-за присутствия в ли-
стьях и стеблях ядовитых веществ, обычно животными не поедается
(Раменский, 1956).
Корни купальницы европейской, по данным А.Е. Катенина
(1962), лишены всасывающих волосков. Поглощению воды способст-
вуют гифы гриба, которые, проходя по межклетникам, проникают в
клетки двух внутренних слоев первичной коры корня. В последнем
слое на границе с эндодермой происходит переваривание гиф. Эндо-
трофная микориза — везикулярного типа, образованная фикомицета-
ми.
Хозяйственное значение
Все части растения вследствие значительного содержания ал-
калоидов (до 0,05%) ядовиты. В корнях и корневищах присутствует
алкалоид магнофлорин, в надземном побеге — магнофлорин и холин,
а также флавоноиды. Цветки содержат флавоноиды и каротиноиды,
семена — алкалоиды и жирное масло (18,4—32,0%). Настой травы,
обладающий антибактериальными и желчегонными свойствами (Са-
цыперова, 1948), используется при отеках, заболеваниях желудка и пе-
чени, а также наружно — при фурункулезах и опухолях. Настой цвет-
ков рекомендуется при цинге и болезнях печени. В ветеринарии рас-
тение применяется при лечении вымени у коров (Растительные ресур-
сы СССР, 1984). Цветки содержат краситель, пригодный для окраши-
вания хлопчатобумажных тканей в желтый цвет (Стоянов, Китанов,
1960; Нейштадт, 1963). Хороший нектаронос (Глухов, 1955).
Купальница европейская, благодаря ярко окрашенным крупным
цветкам, относится к числу признанных декоративных растений. Ее
культивирование началось в садах Европы в XVI —XVII веках (Евтю-
хова, 1968; Малышева, 1972; Жизнь растений, 1980). Отмечается ма-
лая изменчивость вида в культуре. Известны всего две разновидно-
сти: одна — характеризуется бледно-желтыми цветками, другая — бо-
лее крупными размерами. Выведены гибридные формы между купаль-
ницей европейской и азиатскими видами рода, в частности, купальни-
цей азиатской и купальницей алтайской, отличающиеся величиной и
окраской цветков. Используется для групповых и одиночных посадок,
а также на срезку (Декоративное садоводство, 1949).
109
О.А. Горбачевская
РОД БОЛОТНИК
Систематика и номенклатура'
Семейство Callitrichaceae (болотниковые или красоволосковые)
является монотипным с единственным родом Callitriche (болотник).
При разном понимании объема таксонов в мировой флоре насчитыва-
ют от 17 (Mabberley, 1989; Тахтаджян, 1989) до 25 (Willis, 1988) и 50
видов (Martinsson, 1991 в). В России встречается 6—7 видов рода
Callitriche (Кречетович, 1949; Цвелев, 1975; Черепанов, 1995). В Мос-
ковской области — 3 вида.
Систематика рассматриваемой группы имеет сложную историю,
и дискуссии по некоторым вопросам уходят глубоко в долиннеевские
времена. С одной стороны, это связано с трудностью установления
места рода Callitriche в системе: не случайны применявшиеся к его
представителям названия Lenticuba Haller, Stellaria Bauhin, Corisper-
mum L., как не случайно и доныне твердо не установившееся пред-
ставление о родственных связях семейства Callitrichaceae. С другой
стороны, и это в нашем случае более существенно, большие трудно-
сти в систематике на уровне видов создает широкая амплитуда измен-
чивости, прежде всего экологической, которая свойственна земновод-
ным видам рода.
К. Линней (Linnaeus, 1753) принял род Callitriche как монотип-
ный с единственным видом С. palustris L. и тремя разновидностями в
его пределах. Несколько позже он выделил из состава этого вида
С. hermaphroditica L. (Linnaeus, 1755а) и С. vema (Linnaeus, 17556).
Впоследствии был описан ряд новых видов, разновидностей и форм
(Hoppe, 1792; Schultz, 1806; Kutzing, 1832, и др.), причем споры об их
таксономическом ранге никогда не прекращались. В отечественной ли-
тературе первым, по-видимому, попытался разобраться в систематике
группы и сопоставить между собой описанные к тому времени таксо-
ны F. Ruprecht (1860), не признавший описанные Кютцингом виды,
но, в конечном счете, не столько прояснивший, сколько запутавший
таксономическую и номенклатурную ситуацию.
*
Раздел написан В.Н. Тихомировым и О.А. Горбачевской.
НО
Большое значение для формирования современных представ-
лений о таксономии Callitriche имели появившиеся в 60-х годах XIX
века монографии J.E. Lebel (1863) и особенно F. Hegelmaier (1864),
которые, по-видимому, остались неизвестны Н.Н. Кауфману (1866).
Кауфман в своей «Московской флоре» «... шел по стопам Рупрехта»
(Петунников, 1900), рассматривая в пределах С. palustris L. многочис-
ленные разновидности, выделяемые преимущественно по признакам
вегетативной сферы — форме розеточных и подводных листьев, их
жилкованию, характеру верхушки и т.д. Эта традиция в русской фло-
ристической литературе сохранялась долго (см., напр., Цингер, 1885;
Шмальгаузен, 1895; Сырейщиков, 1907, 1927 и др.). В пяти первых
изданиях «Флоры средней России» П.Ф. Маевского (1892,1895,1902,
1912, 1917) отображена та же самая точка зрения, и только
В.А. Петров (Маевский, 1933) в шестом издании этой сводки
впервые совершенно правильно охарактеризовал существенные
признаки встречающихся в нашей флоре видов — С. autumnalis
L. (= С. hermaphroditica L.), С. polymorpha Loennr. (= С. cophocarpa
Sendtner) и С. vema L. emend. Loennr. (= C. palustris L.). В настоя-
щее время такая трактовка «устоялась» и получила широкое при-
знание. Сказанное избавляет нас от необходимости приводить обшир-
ную синонимику с рассмотрением старой литературы и множества
внутривидовых таксонов, позволяя ограничиться лишь самыми необ-
ходимыми цитатами.
Все три названных выше вида: Callitriche cophocarpa Sendtner
(болотник короткоплодный), С. palustris L. (болотник болотный) и
С. hermaphroditica L. (болотник обоеполый) встречаются в Московской
области. Два из них — С. cophocarpa и С. palustris относятся к типовой
секции Callitriche и носят название «водяная звездочка», что связано с
наличием розетки плавающих супротивных листьев на верхушке по-
бега. Они существуют в трех жизненных формах — наземной, водной
с плавающими на поверхности розетками и подводной, полностью по-
груженной в толщу воды.
Третий вид, С. hermaphroditica L., однолетник из секции Pseudo-
callitriche, резко отличается от двух этих видов. Растения полностью
погружены в воду и не образуют наземной жизненной формы. Поэто-
му в данной статье С. hermaphroditica мы не рассматриваем.
Callitriche cophocarpa Sendtner, 1854, Veg. Siidbayems: 773;
idem, 1864, in Hegelm. Monogr. Gatt. Callitriche: 55; Скворцов, 1966,
111
Опред. раст. Моск, обл.: 228; Цвелев, 1966, Новости сист. высш, раст.,
3: 296; Schotsman, 1972, Fl. Europ., 3: 125; Цвелев, 1975, Новости сист.
высш, раст., 12: 238; он же, 1978, Фл. европ. части СССР, 3: 212; Нови-
ков, 1987, Опред. раст. Мещеры, 2: 9; Черепанов, 1995, Сосуд, раст.
России: 292. — С. polymorpha Loennr. 1867, Rot. Notis. (Lund), 1867:4;
Петров, 1933, в Маевский, Фл., изд. 6:468; Б. Федченко, 1940, в Маев-
ский, Фл., изд. 7: 502; Кречетович, 1949, Фл. СССР, 14:498; Проханов,
1964, в Маевский, Фл., изд. 9: 365. — С. palustris auct. pl. Ross. Med.
p.p., non L. — Болотник короткоплодный (семейство Callitrichaceae
— Болотниковые).
Callitriche palustris L. 1753, Sp. Pl.: 969, s. str.; Проханов, 1964,
в Маевский, Фл., изд. 9: 365; Скворцов, 1960, Опред. раст. Моск, обл.:
228; Schotsman, 1972, Fl. Europ., 3: 125; Цвелев, 1975, Новости сист.
высш, раст., 12: 238; он же, 1978, Фл. европ. части СССР, 3:212; Нови-
ков, 1987, Опред. раст. Мещеры, 2: 9; Черепанов, 1995, Сосуд, раст.
России: 292. — С. vema L. 1755, Fl. Succ., ed. 2, 1: 2; Петров, 1933, в
Маевский, Фл., изд. 6:468; Б. Федченко, 1940, в Маевский, Фл., изд. 7:
502; Кречетович, 1949, Фл. СССР, 14: 496. — Болотник болотный
(семейство Callitrichaceae — Болотниковые).
Диплоидное число хромосом С. cophocarpa 2n=10, С. palustris
— 2n=20 (Schotsman, 1967, цит. по: Martinsson, 19916). Растения обо-
их видов внешне очень похожи. Генеративное растение Callitriche
cophocarpa показано на рис. 1, С. palustris — на рисунке 2. Ниже при-
водится ключ для определения видов.
1. Прицветнички обычно длиннее половины плода. Стилодии б.м.
отклоненные в стороны, долго сохраняются при плодах (рис.
3, в). Эремы (части плода) — овальные. Кили эремов — едва
крылатые или бескрылые, с жесткими ребрами, или без них. В
розетке наземной формы — от 10 до 22 овальных листьев. Фор-
ма и размеры листьев срединной формации сильно варьируют
от овальных до линейных. Длина листьев срединной формации
у наземной формы — обычно 3—14 мм, водной —4—16 мм,
ширина соответственно — 1,5—6,0 мм и 1,0—8,5 мм.
Плавающие розетки у водных форм вогнутые. На листьях желез-
ки диаметром 0,1—0,2 мм, содержат 8 клеток (рис. 4, б). С.
cophocarpa Sendtner. — Б. короткоплодный.
112
— Прицветнички обычно короче половины плода. Эремы широко
обратнояйцевидные (рис. 2, в). Стилодии — прямые, опадающие
при плодах. Кили эремов всегда узкокрылатые в верхней части. В
розетке наземной формы от 4 до 10 линейных листьев. Листья
срединной формации длиной обычно 8—17 мм у наземных и 8—
40 мм у водных форм, шириной соответственно — 1,5—4,5 мм и
1,0—4,0 мм. Плавающие розетки у водных форм — плоские. Же-
лезки на листьях имеют диаметр 0,05—0,07 мм и содержат 16
клеток (рис. 4, а). С. palustris L. — Б. болотный.
Географическое распространение
Виды рода Callitriche встречаются по всему земному шару, ис-
ключая Южную Африку, некоторые пустыни, высокогорья, а также се-
вер тундровой зоны. Наземная и водная формы болотника коротко-
плодного и болотника болотного широко распространены в районах
умеренного климата. На территории бывшего СССР ареал болотника
короткоплодного охватывает всю европейскую часть (кроме крайнего
юго-востока), западное Закавказье, Западную Сибирь и часть районов
Восточной Сибири. Вне бывшего СССР вид встречается в Скандина-
вии, Средней и Атлантической Европе, в Средиземноморье (Белавская,
1994; Martinsson, 1991а, б).
Ареал бблотника болотного обширнее. Он включает европей-
скую часть бывшего СССР, Кавказ, Сибирь и Дальний Восток, Казах-
стан, юго-восток Средней Азии. Ареал вне бывшего СССР частично
совпадает с ареалом Callitriche cophocarpa, но кроме этого занимает
также юго-восточную Азию, Малую Азию, Балканский полуостров,
Северную Америку и Австралию (Белавская, 1994).
На территории Московской области болотники широко распро-
странены во влажных местообитаниях Мещерской низменности, в сы-
рых лесах севера области и даже в черте города Москвы (по материа-
лам гербария). Болотник болотный в Московской области встречается
реже, чем болотник короткоплодный (Щербаков, 1990).
Морфологическое описание
Корневая система у наземной и водных форм имеет слабо раз-
витый главный корень, который отмирает рано -— в стадии проростка.
Боковые корни обычно тоже слабо развиты, и основную роль в пита-
нии и закреплении растения в почве играют корни придаточные, об-
113
Рис. 1. Генеративное растение вод-
ной формы Callitriche cophocarpa; а
— верхушечная почка.
разующиеся в пазухах листьев и
обычно имеющие большую длину по
сравнению с боковыми. Анатомиче-
ское строение корней характеризует-
ся сильной редукцией проводящей
системы до двух клеток протоксиле-
мы и одной клетки метаксилемы
(Arber, 1963). Придаточные узловые
корни появляются уже у проростков,
нередко при семядольных листьях.
Хрупкие стебли болотников
имеют небольшие размеры — от 2 до
35—50 см. Стебли у наземных форм
— обычно стелющиеся, с пазушны-
ми придаточными корнями. Анатоми-
ческое строение стебля демонстриру-
ет черты упрощения — проводящая
система представлена тремя трахеи-
дами, у водных форм наблюдали от-
сутствие камбия (Arber, 1963). Стебли
покрыты рассеянными щитовидными
железками (рис. 4). Их размер и ко-
личество клеток имеют диагностиче-
ское значение (Соколовская, 1930).
Листорасположение — дву-
рядное. Листья — накрест супротив-
ные, бесчерешковые, без прилистни-
ков. Наблюдается гетерофиллия, свя-
занная с произрастанием в двух раз-
личных средах — водной и воздуш-
ной. В аксиллярном комплексе в пазухах листьев могут присутство-
вать либо цветок, либо почка или побег ветвления, развившийся из
нее. Придаточные корни могут отходить от обеих сторон узла одно-
временно, но чаще — только от одной стороны. Самыми длинными
являются первые три междоузлия побегов, и особенно второе междо-
узлие. Женские цветки на материнском побеге обычно появляются,
начиная с 4 узла.
Формирование бокового побега сопровождается последующим
его укоренением с помощью придаточного корня из того же узла, от
114
Рис. 2. Верхушка побега генеративного растения водной формы Callitriche palustris:
а — общий вид побега; б — пыльник; в — завязь; г — прицветничек.
которого отходит побег. Побеги II порядка имеют разные размеры и
количество узлов. Обычно самые нижние из них начинают развивать-
ся раньше, чем расположенные выше, и имеют большее число мета-
меров (базитония). Верхушечные и пазушные почки материнских по-
бегов содержат не более 3 зачатков метамеров побега. С увеличением
количества зачатков метамеров почка раскрывается и дает начало но-
вому побегу.
Основания материнских побегов генеративных растений вод-
ных форм погружены в ил на дне водоема, бурой окраски, располага-
ются плагиотропно. Растения нарастают моноподиально (рис. 3). Вер-
хушечные почки материнского побега начинают развитие весной, по-
сле таяния водоема. Заглубляющиеся в ил нижние части побегов I по-
рядка лишены листьев, обладают большей толщиной (0,7—1,0 мм) в
отличие от верхних, нарастающих частей и побегов растений назем-
ной формы (до 0,7 мм). По мере старения растения побуревшие пла-
гиотропные участки материнского побега разлагаются.
Наземные формы обоих видов — однолетники (Arber, 1963;
Sculthorpe, 1967; Martinsson, 1991а и др.), обладают однотипными ли-
стьями очень мелких размеров, черешковидно суженными к основа-
нию. Листья по форме — эллиптические до почти округлых.
115
л
Рис. 3. Генеративное растение Callitriche cophocarpa: а — схема расположения
мужских и женских цветков; б — мужской цветок; в — женский цветок; г — схема
нарастания и отмирания побега, пунктиром обозначены отмершие части, стрелкой
— направление отмирания побега, I—III — обозначения порядка побегов.
Розеточные листья непогруженной водной формы — от эллип-
тических до почти округлых, имеют от 1 до 5 жилок, более или менее
выемчатые или слабо закругленные на верхушке. Подводные листья
растений длиннее и уже надводных (до 35 мм длиной), от линейных
116
до эллиптических по форме, заметно суженные к основанию. Они от-
личаются от надводных также развитым мезофиллом, вытянутыми
клетками эпидермиса без устьиц (Arber, 1963)— приспособлением к
жизни в воде и на воздуху Проводящие пучки листьев состоят из раз-
бросанных клеток с утолщенными стенками (Arber, 1963).
Подводные растения болотника болотного
— без розеток листьев, не цветут и не плодоносят.
Погруженные листья линейной формы, суженные
к основанию. У болотника короткоплодного такие
формы нами не обнаружены.
У обоих видов обнаружены женские и обое-
полые особи (рис. 5). Мужские особи мы наблюда-
ли только у болотника короткоплодного. Цветки
обоих видов — однополые, обычно с двумя пере-
пончатыми, серповидно изогнутыми прицветнич-
ками, расположены в пазухах листьев по одному.
В литературе существуют указания на возможность
формирования двух цветков в пазухе одного листа
(Arber, 1963), но пока не найдено подтверждения
этому. У всех женских особей пестики находятся с
одной стороны стебля (рис. 5). Редко встречались
пестики в пазухах обоих листьев узла, но и при этом
Б
Рис. 4. Строение
железок: а —
Callitriche palustris;
б — Callitriche
cophocarpa.
на одной стороне побега («предпочтительной») пестики были круп-
нее, чем на другой.
Пестичные цветки представлены синкарпным гинецеем, состоя-
щим из двух плодолистиков с двумя нитевидными стилодиями. Завязь
состоит из двух гнезд и двух лопастей, но в результате образования
ложной перегородки становится четырехгнездной. Каждое гнездо за-
вязи имеет один анатропный висячий тенуинуцеллятный семязачаток
с простым интегументом. Тычиночные цветки состоят из одной ты-
чинки. Пыльники имеют выстилающий слой — тапетум (Тахтаджян,
1966).
Расположение мужских и женских цветков на одном растении
имеет следующие особенности: еще A. Arber (1963) отмечала, что у
водной плавающей формы С. cophocarpa женские цветки на стеб-
ле обычно располагаются ниже мужских. Наши наблюдения, касаю-
щиеся обоих видов, соответствуют этим данным (рис. 3). Побеги II
порядка обычно несут только женские цветки. Материнский побег
несет мужские цветки лишь в области плавающей розетки и несколь-
117
ких примыкающих к ней снизу метамеров. Редки случаи, когда жен-
ские и мужские цветки встречаются в средней части материнского по-
бега на одном узле.
Зрелые плоды — дробные, распадаются на четыре эрема, реже
на два мерикарпия (Schulthorpe, 1967; Цвелев, 1975). Семена болот-
ников имеют хорошо развитый эндосперм с гаусториями, как у мно-
гих растений порядка Tubiflorae, и прямой зародыш (Тахтаджян, 1966).
Эндосперм содержит много жиров (Тахтаджян, 1987; Arber, 1963). Обо-
лочка семени нередко тонка (Schulthortpe, 1967).
Проведенные нами (при консультации Н.Р. Мейер) исследова-
ния пыльцы, подвергнутой ацетолизу, показали отсутствие у пыльце-
вых зерен борозд и пор. Пылинки обоих видов идентичны, имеют две
области с утонченными экзинами, которые несколько сходны с пора-
ми, но в отличие от настоящих пор не имеют сквозного отверстия.
Экзина — тонкая, до 1 мкм, с ячеистой скульптурой. Ячеи — округ-
лые, у пылинок обоих видов образованы 5—6 округлыми бугорками.
По форме пыльца — почти шаровидная, в очертании — круглая или
широко эллипсоидальная. Длина экваториального диаметра пылинки
23,5—25,7 мкм.
Онтогенез
Болотник короткоплодный и болотник болотный имеют один и
тот же возрастной ряд, представленный на рисунке 6. Диаспоры пред-
ставляют собой эремы, покрытые оболочкой. На прорастание семян
болотника короткоплодного оболочка, по-видимому, не влияет. Про-
растают семена как с неповрежденной оболочкой (контроль), так и
стратифицированные замораживанием. Стратификация не оказывает
влияния на прорастание семян этого вида, но, вероятно, влияет на про-
растание семян болотника болотного, у которого прорастают только
предварительно замороженные семена (по нашим опытам с проращи-
ванием). Проростки характеризуются наличием смешанного питания
— за счет запасных веществ семени и развивающихся семядольных
листьев. Последние приобретают размеры, форму и окраску, не отли-
чающиеся от таковых у наземных листьев взрослого растения (рис.
7).
Прорастание у болотников, вопреки мнению A. Arber (1963),
надземное, а не подземное. У проростков, появившихся из семян
в субстрате, рост побеговой и корневой систем происходит быст-
рее, чем у помещенных на фильтровальную бумагу. При прорастании
118
первым из семени появляется зароды-
шевый корешок. В следующие 2 дня
появляется гипокотиль, который к 3-
му дню достигает примерно 1,5 см
длины. К этому сроку зародышевый
корешок достигает длины 0,7 см и за-
глубляется в почву на 0,5—0,7 см.
Растущий гипокотиль выносит над
поверхностью почвы семядоли, кото-
рые удлиняются от 0,10—0,12 см в
день появления из семени до 0,40—
0,45 см к 6-му дню и приобретают
бледно-зеленую окраску в отличие от
светло-желтой вначале. На 7-ой день
после начала прорастания гипокотиль
достигает 4,0—6,5 см в длину. Семя-
дольные листья все еще несут семен-
ную кожуру, однако, уже приобрета-
ют зеленую окраску и фотосинтези-
руют. Их длина достигает 0,45—0,50
см. На 8-ой день гипокотиль уд-
линяется до 6,5—7,3 см. Семенная
кожура отделяется от семядолей позд-
нее.
У проростков появляются и
развиваются длинные бесцветные
корневые волоски на главном корне.
Они отходят от утолщения, которое
располагается чуть ниже условной
границы корня и гипокотиля (рис.
7). Расходящиеся во все стороны
корневые волоски образуют шаро-
образную зону всасывания моло-
Рис. 5. Женское растение наземной
формы Callitriche sp.
дого корня и достигают длины 0,001—0,1 см. Главный корень функ-
ционирует до перехода растения в ювенильное состояние. Он отмира-
ет обычно еще до появления второго узла (аксиллярного комплекса)
стебля. Редко встречаются случаи, когда главный корень функциони-
рует в течение всего ювенильного состояния, и его отмира-
ние происходит при переходе в виргинильное состояние. Пророст-
119
ки не имеют придаточных корней. С их появлением из узлов на стебле
или (что характерно для водных форм) из гипокотиля, растение пере-
ходит в ювенильное состояние. В состоянии проростков растения, как
мы предполагаем, находятся 2—3 недели.
Ювенильные растения болотников имеют несколько узлов
(обычно в пределах 2—5). Первый придаточный корень может раз-
виться из семядольного узла или из следующих выше по стеблю
узлов. К тому времени главный корень уже отмирает или в зоне услов-
ной границы с гипокотилем, или в области апекса. Случаи, когда он
сохраняется и достигает значительной длины, редки. Придаточные
корни быстро перерастают по размерам отмерший главный.
Очевидно, он служит лишь для закрепления проростка в почве. Далее
наземное растение начинает стелиться по субстрату, удерживаясь
придаточными корнями, а водное занимает наклонное положение
в воде. У водных форм, поэтому, главный корень до отмирания дости-
гает относительно большей длины, придаточные корни активно обра-
зуются в области гипокотиля, что заякоривает растение в субстрате.
Ювенильное состояние длится недолго, предположительно несколь-
ко недель. В пазухах листьев ювенильных растений начинают
закладываться вегетативные почки. С развитием из них боковых побе-
гов и с гибелью главного корня растение переходит в виргинильное
состояние.
Виргинильные растения обладают разветвленной побеговой сис-
темой. Она представлена моноподиально нарастающим материнским
побегом и побегами ветвления в пазухах листьев. Придаточные корни
обеспечивают определенную самостоятельность в питании и закреп-
лении в почве побегов ветвления, позже они играют важную роль в
вегетативном размножении болотников. Ветвление — базитонное:
большее число метамеров содержат именно самые нижние побеги ветв-
ления. Виргинильные растения наземной формы распластаны по суб-
страту и укореняются за счет придаточных корней, которые достига-
ют длины 0,1—7,5 см.
Растения водной формы с плавающими розетками «выносят»
верхушку материнского побега над уровнем воды. На ней формиру-
ются листья эллиптической формы. Побеги характеризуются гетеро-
филлией. Листья бывают следующих форм: нижние подводные—ром-
бические с 3-мя жилками; листья срединной формации — линейные,
длинные и более тонкие, с выемками на верхушках, с 1 жилкой; ли-
стья плавающей розетки — более мелкие, эллиптические, с 1—3
120
Рис. 6. Ряд онтогенетических состояний Callitriche sp.
жилками. В неглубоких водоемах плавающая розетка образуется и у
ювенильных растений. В отличие от виргинильных растений, розетки
которых уже имеют «видовое» количество листьев — от 10 до 18 (5—
9 пар) (Флора европ. части СССР, 1978), ювенильные особи обоих ви-
дов несут в розетке только 4—6 листьев. В этой фазе растения изучае-
мых видов трудно различить. Виргинильные растения водных форм
могут без ущерба перезимовывать в замерзающих водоемах (по на-
шим наблюдениям апреля 1995 г. в четырех водных популяциях Ка-
лужской области), тогда как проростки и ювенильные растения поги-
бают.
Репродуктивные органы формируются в пазухах листьев на ма-
теринском побеге и на побеге II порядка. Закладка репродуктивных
органов у растений всех жизненных форм происходит акропетально.
Женские цветки закладываются «ниже» по стеблю и начинают разви-
ваться раньше, чем мужские.
В генеративный период у растений наземной формы не проис-
ходит серьезных изменений вегетативной сферы. У водных форм рост
материнского побега в длину ограничен глубиной водоема. За лето у
растений формируется несколько метамеров, а в пазухах их плаваю-
121
щих листьев закладываются мужские цветки. Основания материнских
побегов водных форм осенью приобретают плагиотропное располо-
жение на поверхности дна с последующим заглублением в него. Этот
процесс происходит в октябре и сопровождается потерей плагиотроп-
ной частью побега листьев и приобретением бурой окраски. Верхние
части материнского побега, несущие плавающие розетки, не теряют
листьев. Верхушечная почка перезимовывает, и из нее развивается про-
должение побега в следующем вегетационном сезоне.
Период старения особей непродолжителен по времени и харак-
теризуется отмиранием внутрипочвенных побегов у водной формы,
увяданием и полным отмиранием у наземных форм. Процесс отмира-
ния у водных и наземной форм протекает от основания побегов к вер-
шине.
Сезонное развитие
Массовое прорастание семян однолетних наземных форм на-
чинается в среднем со II декады мая. Цветки у однолетников появля-
ются к концу мая, так что к началу июня большинство растений нахо-
дится уже в генеративной фазе, которая продолжается до конца октяб-
ря. Первые плоды были обнаружены нами в первых числах июня (на-
блюдения 1995 года). У однолетней формы диссеминация наиболее
интенсивно происходит, по нашим наблюдениям, в конце августа —
начале сентября. Отмирание растений наземной формы начинается в
конце августа и заканчивается к концу октября. Проростки, ювениль-
ные и виргинильные растения, появившиеся в конце лета, отмирают,
не дойдя до генеративной фазы.
В естественных условиях прорастание семян водных форм про-
текает в течение всего вегетационного сезона, с мая по октябрь и, ви-
димо, в очень большой степени зависит от количества осадков. Пере-
сыхание водоема, в котором обитают растения, отрицательно влияет
на прорастание. Цветение растений водной формы продолжается до
конца октября. К тому времени наземные формы уже отмирают. Вы-
сыхание водных местообитаний приводит к развитию у генеративных
растений болотника короткоплодного новых побегов ветвления, кото-
рые имеют «наземный облик» — более округлые листья, более длин-
ные придаточные корни. Болотник болотный таких побегов не образу-
ет.
В последних числах октября — начале ноября (наблюде-
ния 1995 г.) происходит погружение материнских побегов водных форм
122
на дно водоема, после чего их укрывает оседающий ил. Отмирание
сенильных растений водных форм протекает в течение всего вегета-
ционного сезона, но особенно интенсивно — осенью. Летом отмира-
ние ускоряется при обмелении водоема.
Способы размножения и распространения
Болотники относятся к явнополицентрической модели биомор-
фы (Ценопопуляции растений, 1976) и обладают большой вегетатив-
ной подвижностью. У генеративных растений водной формы легко про-
исходит травматическая партикуляция материнских побегов, связан-
ная с повреждениями животными, человеком и другими агентами. В
этих случаях побеги ветвления, имея придаточные корни в узлах, мо-
гут отрываться и развиваться самостоятельно, в основании такого «об-
рывка» образуются придаточные межузловые корни.
У близкого вида — С. stagnalis — наблюдали специализирован-
ное вегетативное размножение водных форм с помощью отделения от
растения плавающей розетки с придаточными корнями (Ridly, 1930).
Такой способ вегетативного размно-
жения теоретически возможен и для
наших видов, однако в природе пока
не наблюдался.
Вегетативное размножение по-
могает растению захватить большую
площадь за короткий срок. Нередко
из-за этого водяные звездочки счита-
ются водными сорняками (Ridly,
1930). Их расселение происходит об-
рывками вегетативного «тела» с при-
даточными корнями за счет движений
воды, переноса птицами или скотом,
на льдинах во время половодья (опи-
сано Ridly, 1930 для Callitriche
palustris, С. hamulata и С.
hermaphroditica), посредством транс-
порта (кусочки стеблей застревают на
колесах телег, тракторов).
Репродуктивное усилие назем-
ной и водной форм определялось
Рис. 7. Проростки Callitriche copho-
carpa: ск — семенная кожура; с —
семядоли; г — гипокотиль; к — глав-
ный корень с корневыми волосками.
в июле в период максимального раз-
123
вития репродуктивных органов. В таблице оно представлено как от-
ношение массы репродуктивных органов к общей массе растения. Ре-
зультаты рассчитаны как среднее из 20 образцов по 10 растений в воз-
душно-сухом состоянии (точность измерений — 0,5 мг).
Таблица
Репродуктивное усилие (РУ) разных жизненных форм
С. cophocarpa и С. palustris
Вид Форма Сообщество РУ
Callitriche cophocarpa водная Piceetum sphagnosum 1/11
наземная Piceetum sphagnosum 1/23
наземная Piceetum oxalidosum 1/19
Callitriche palustris водная Piceetum sphagnosum 1/169
наземная Piceetum sphagnosum 1/176
водная Piceetum oxalidosum 1/122
наземная Piceetum oxalidosum 1/91
У болотников наблюдается дихогамия в форме протогинии. Жен-
ские цветки у водных форм находятся полностью в воде. Они форми-
руются на растении и увеличиваются в размерах раньше, чем муж-
ские. Та же закономерность наблюдается и у наземных форм. Агентом
переноса пыльцы у водных форм болотников является вода, причем
пыльца, видимо, переносится как по поверхности воды, так и в ее тол-
ще.
У особей наземной формы к моменту диссеминации на расте-
ниях не было вскрывшихся пыльников, следовательно, пыльца этих
растений не принимала никакого участия в оплодотворении. Таким
образом, у болотников происходит перекрестное опыление.
В природе мы наблюдали наземные формы в пределах несколь-
ких микропопуляций, в которых при отсутствии собственной зрелой
пыльцы и приноса ее извне растения имели зрелые плоды. Речь идет
о тех случаях, когда микропопуляпии находились на расстоянии при-
мерно 1 км от других, полет пыльцы с других площадок под пологом
леса, вероятно, был затруднен. Мужские цветки растений находились
примерно в одной стадии развития — начальной (зачаточная тычинка
— бугорок под листьями, и пестик, скрытый в пазухе листа, менее
124
0,5 мм в диаметре). Таким образом, опыление внутри микропопуля-
ции было исключено. Был проведен опыт с кастрацией растений. В
ходе опыта оказалось, что почти во всех сформировавшихся и окон-
чивших рост завязях С. cophocarpa и С. palustris находились незрелые
семена. Размеры завязей без контакта с пыльцой увеличились в
среднем от 0,5 до 0,86 мм за четверо суток, максимальный размер за-
вязи — 0,95 мм. В наблюдаемом случае развитие завязи происходило
без оплодотворения пыльцой того же растения. Однако констатиро-
вать апомиксис все же нельзя, так как не использовались клас-
сические методы квалификации апомиксиса (Хохлов и др., 1978).
Поперечные срезы завязей, наблюдаемых в конце их развития, пока-
зали наличие плотных тел внутри четырех секций завязи — не-
зрелых семян, сформировавшихся на 4-й день опыта. Развитие заро-
дышевого мешка у С. cophocarpa, таким образом, происходит очень
быстро, скачком.
Распространение плодов и семян обычно происходит при по-
средстве водных течений или движений льда с примерзшими частями
вегетативного тела. Зрелые эремы могут перемещаться ветром по по-
верхности воды или почвы (описано Ridly, 1930 для С. stagnlis и С.
palustris), а также животными — на конских копытах, на лапах водо-
плавающих птиц вместе с обрывками стеблей (Ridly, 1930; Arber, 1963;
Schulthorpe, 1967).
Экология
С. cophocarpa произрастает в более широком спектре экологи-
ческих условий, чем С. palustris и обладает большим полиморфизмом.
Водные формы С. cophocarpa произрастают в пресных водоемах, вы-
носят слабое засоление. Форма с плавающими розетками предпочита-
ет слабое течение, но встречается как в водоемах со стоячей водой,
так и при сильном течении в руслах судоходных рек. Водные формы
близкого вида предпочитают слабое течение или стоячие водоемы, не
переносят высыхания водоема, хуже выносят засоление (Соколовская,
1930; Белавская, 1994; Arber, 1963).
Растения наземных форм обоих видов предпочитают неболь-
шие понижения рельефа, где достигают более крупных размеров. Од-
нако однолетники С. cophocarpa можно встретить и на повышенных
элементах рельефа (сухих кочках в поле, например), где растения при-
обретают карликовые размеры (листья длиной 1—2 мм, междоузлия
длиной несколько мм), но при этом развивают генеративные органы.
125
Наземные формы С. cophocarpa были отмечены даже на мелах,
где растения были представлены полноценными цветущими особями
(Ridley, 1967). По данным Spruse (1908; цит. по: Schulthorpe, 1967),
отдельные особи этого вида произрастают в Эквадоре на склонах го-
ры Чимборазо на абсолютной высоте 2400 м.
Скорость роста побегов и зарослей С. cophocarpa довольно ве-
лика. Эти показатели изучены в реках Дании с апреля по июнь (Sand-
Sensen, Madsen, 1992). Средняя скорость роста заросли оказалась 1,02
см/день в длину и 0,55 см в ширину. Было также обнаружено, что она
не зависит от первоначальных размеров заросли. Для С. palustris та-
ких исследований не проводилось.
В течение вегетационного периода происходит изменение пло-
щадей микропопуляций болотников, изменяется проективное покры-
тие видов и их биомасса. На эти параметры положительно влияет ко-
личество выпадающих осадков и их регулярность. Сокращение мик-
ропопуляций может быть связано с вытаптыванием побегов (для на-
земных форм) или с колебанием уровня воды, пересыханием водоема
(для водных форм). Последнее приводит к тому, что растения нередко
отмирают полностью или у них сохраняются только погруженные в
ил части стебля, не играющие роли в фотосинтезе.
На перезимовывание особей болотника болотного оказывают
влияние условия замораживания. Мы попытались воспроизвести их
при лабораторном замораживании в холодильнике для 30 растений вод-
ной формы в трех вариантах: в водной, воздушной и почвенной сре-
дах с последующим оттаиванием. Опыт состоял из 2-х этапов по 15
дней каждый: 10 дней замораживания и 5 дней оттаивания. В резуль-
тате оказалось, что наиболее благоприятно на растениях сказывается
замерзание в воде (из 30 живых особей осталось 27 после первого
замораживания и 8 после второго). При отсутствии воды жизнеспо-
собность растений снижается (при замерзании в воздушной среде и
почве после первого этапа осталось соответственно 22 и 3 живых осо-
би, а после повторного замораживания все они погибли). Болотник
короткоплодный, видимо, обладает сходными требованиями к усло-
виям перезимовки.
Фитоценология
Фитоценотическая роль болотников в сообществах водных
растений изучена мало. Видимо, они вполне могут быть в них доми-
нантами. Выделяется, например, сообщество Callitricho-Lemneetum
126
minoris Weber 1969 союза Lemnion minoris Tx. 1955 порядка Lemne-
talia Tx. 1955 класса Lemnetea (Korotkov et al., 1991).
Болотниковые нередко являются диагностическими видами в
сообществах водных растений (Schulthorpe, 1967). Callitriche hamulata
и С. plathycarpa произрастают в сообществах олиготрофных, эвтроф-
ных и мезотрофных зарастающих водоемов, относящихся к классам
водной растительности Potametea и Lemnetea. В альпийском и субаль-
пийском горных поясах в олиготрофных местообитаниях северо-за-
падной Европы произрастают сообщества ассоциации Callitricho-
Sparganietum, относящиеся к классу Littorelletea, где одним из диагно-
стических видов является Callitriche palustris (Ellenberg, 1986).
По нашим наблюдениям, наиболее часто водным и наземным
формам болотников в изученных лесных сообществах (ельник-зеле-
номошник, ельник-кисличник, ельник-черничник, ельник сфагновый,
сосняк сфагновый, разреженный березняк) сопутствовал Ranunculus
repens. Во всех водных сообществах (где произрастали только водные
формы болотников) им сопутствовала Lemna minor. В целом флори-
стический состав микрогруппировок с участием болотников опреде-
ляется условиями окружающих сообществ.
Консортивные связи и хозяйственное значение
Консортивные связи болотников изучены, по-видимому, не в
полной мере. Еще не полностью выяснен круг животных, использую-
щих болотники в пищу. Однако известно, что при слабом развитии
перифитона водоема двукрылое насекомое Paraponyx stratiotata L. по-
едает их листья. В пищевом рационе ондатры Ondatra zibetica L. все-
гда присутствует Callitriche sp. (Гаевская, 1966), листья и побеги по-
едают гуси и утки. Болотник болотный поедают брюхоногие моллю-
ски Limnaea stagnalis (L.) и Melanopsis dufouri Fex., чешуекрылое
Nymphula nymphaeata. Другой вид рода — Callitriche stagnalis—явля-
ется пищей для рачков Gammarus pulex L. и водяного ослика Asellus
aquaticus L. при отсутствии в стоячих водоемах основного материала
для питания — диатомей, нитчатых водорослей (Гаевская, 1966).
Выше уже отмечались переносчики плодов и вегетативных органов
болотников, которые способствуют их расселению — водоплаваю-
щие птицы и скот. В странах Западной Европы водяные звез-
дочки считаются водными сорняками, которые вызывают зарас-
тание русла судоходных рек за счет быстрого вегетативного размно-
жения (Ridly, 1930).
127
М.В. Марков, Н.Г Уланова, Н.В. Чубатова
РОД НЕДОТРОГА
Род Недотрога (Impatiens L.) — единственный представи-
тель семейства Balsaminaceae в России. Всего в этом семейст-
ве 4 рода. В роде Impatiens — около 900 видов, распространен-
ных преимущественно в горных лесах тропиков и субтропиков Аф-
рики и Азии. Немногие виды произрастают в умеренно теплых об-
ластях Евразии, Африки и Сев. Америки (Флора Восточной Европы,
1996). Центром происхождения рода считаются Гималаи и Индия
(Valentine, 1971). На территории бывшего СССР отмечено 9 ви-
дов, из них три вида (I. furcellata Hemsl., I. maackiiHook. ex Кот.,
I. textorii Miq.) произрастают только на Дальнем Востоке и один (I.
brachycentra Кат. & Kir.) — в Средней Азии (Черепанов, 1995). В
европейской части России встречаются 5 ттвидов — Impatiens
noli-tangere L., I. parviflora DC, I. glandulifera Royle, I. balsamina L., I.
uralensis A. Skvorts. В Московской области произрастают 3 первых
вида.
Виды Impatiens — это однолетние травы с немногими мясисты-
ми .сочными прозрачными стеблями и простыми, цельными, очеред-
ными, супротивными или мутовчатыми листьями. Цветки — в пазуш-
ных кистях или пучках, реже — одиночные, обоеполые, зигоморфные,
перевернутые на 180° (резупинатные). Чашелистиков — 3, редко — 5,
из которых более крупный нижний образует нектароносный, обычно
окрашенный, шпорец. Лепестков — 5, из которых верхний крупнее
других, а 4 боковых — попарно сросшиеся, так что венчик кажется
трехчленным. Тычинок — 5, нити тычинок короткие, в нижней части
свободные, наверху срастаются в трубку вокруг завязи и рыльца в ви-
де крышечки, которая опадает при созревании рыльца. Завязь — верх-
няя. Гинецей состоит из 5 плодолистиков с коротким столбиком, ры-
лец — от 1 до 5, семязачатки многочисленны. Плод — сочная локули-
цидная продолговатая коробочка, вскрывающаяся внезапно 5 створ-
ками, которые, спирально закручиваясь, разбрасывают семена. В зре-
лом семени недотроги имеется скудный эндосперм и крупный хорошо
развитый зародыш, дифференцированный на очень короткую осевую
часть с конусом нарастания и мясистые, занимающие почти весь объ-
ем семени, семядоли.
128
Ключ для определения видов, произрастающих
в Московской области
1. Цветки лилово-розовые. Impatiens glandulifera—Недотрога же-
лезконосная.
— Цветки желтые....................................... 2
2. Цветки крупные, до 30 мм дл., ярко-желтые, с крючковидно
загнутым шпорцем, поникающие. I. noli-tangere — Н. обыкно-
венная.
— Цветки мельче, до 15 мм дл., светло-желтые, с прямым шпорцем;
цветоносы прямостоячие или отклоненные. I. parviflora — Н.
мелкоцветковая.
Недотрога обыкновенная
Номенклатура и внутривидовая систематика
Impatiens noli-tangere L. 1753, Sp. Pl.: 938; Кауфман, 1866,
Моск, фл., изд. 1: 115; Цингер, 1885, Сборник свед.: 121; Шмальгау-
зен, 1895, Фл. Средн. иЮжн. Росс., 1: 201; Сырейщиков, 1907, Илл.
фл. Моск, губ., 2: 351; он же, 1927, Опред. раст. Моск, губ.: 183;
Назаров, 1940, в Маевский, Фл., изд. 7: 505; Победимова, 1949,
Фл. СССР, 14: 626; она же, 1964, в Маевский, Фл., изд. 9: 283;
В. Тихомиров, 1966, Опред. раст. Моск, обл.: 229; D.M. Moore, 1968,
Fl. Europ., 2: 240; Новиков, 1987, Опред. раст. Мещеры, 2: 10;
Черепанов, 1995, Сосуд, раст. России: 218; Мордак, 1996, Фл.
Вост. Европы, 9:391. — Недотрога обыкновенная, недотрога жел-
тая, не тронь меня, разрыв (семейство Balsaminaceae — Бальзамино-
вые).
Растения I. noli-tangere исключительно с клейстогамными цвет-
ками и мелкими листьями выделены Мейнсхаузеном в особую вариа-
цию var. minutiflora Meinsh. (Флора СССР, 1949). Эта форма описана
на северной границе апеала (Псков, Луга, р. Печора) на вырубках и
торфяных болотах. М.Г. Попов описал этот вариант как особый вид I.
cleistogama М. Pop. По мнению Е.Г. Победимовой (Флора СССР, 1949),
выделение особого вида, равно как и вариации, лишено оснований,
так как соотношение хазмогамных и клейстогамных цветков зависит
от экологических условий.
129
Географическое распространение
Недотрога обыкновенная—неморально-бореальный евразиат-
ский вид. Распространена по всей Европе (кроме Крайнего Севера и
некоторых южных районов), на Кавказе, в Западной и Восточной Си-
бири, на Дальнем Востоке, в Малой и Средней Азии, Монголии, Япо-
нии и Китае, изредка — в горах Средней Азии (Флора Восточной Ев-
ропы, 1996). Вид также указан для Аляски (Флора СССР, 1949). Ви-
карный вид в Северной Америке — I. capensis Meerb., с оранжевыми
цветками (Jouret, 1977). На тихоокеанском побережье Америки встре-
чается и другой близкий вид — I. occidentalis Rydb., причем, мнению
L. Abrams (1951), этот вид следует считать интродуцированной на ти-
хоокеанское побережье США I. noli-tangere L.
В России северная граница распространения вида проходит на
Кольском полуострове у Мурманска. Восточнее, в северной тайге, до-
вольно часто встречается по р. Ваге, йижнему течению Северной Дви-
ны, верхнему и среднему течению Мезени, верхнему и среднему тече-
нию реки Печоры, а также по Вычегде, Сысоле и Илычу (Флора севе-
ро-востока. .., 1976). Недотрога обыкновенная встречается по всей лес-
ной зоне. Южная граница ареала проходит по нижнему Дону и южно-
му Заволжью. Встречается также в лесах Крыма и Кавказа.
В Московской области недотрога обыкновенная встречается час-
то, во всех районах. Обитает, в основном, в сырых лесах и лесных
оврагах, по берегам рек и ручьев.
Морфологическое описание
Недотрога обыкновенная — травянистое однолетнее стержне-
корневое растение. Главный корень ветвится до корней III—IV поряд-
ка. В основании гипокотиля часто развиваются мясистые сочные при-
даточные корни до 5 см длиной. Стебель — цилиндрический, голый,
до 120 см высотой, 0,5—1 см в диаметре, утолщенный, несколько взду-
тый в узлах, просвечивающийся, светло-зеленый; основания его меж-
доузлий часто лиловато-розового цвета. Боковые побеги (I—III поряд-
ков) утолщены в основании и в узлах. Листья — супротивные или оче-
редные, короткочерешковые, с цельными продолговато-яйцевидными
(до 12—13 см длиной, 4—4,5 см шириной), крупнозубчатыми по краю
пластинками и без железок в основании. Жилкование перисто-петле-
видное (Рычин, 1948).
130
Рис. 1. Строение цветков Impatiens parviflora (A), I. noli-tangere (Б), I. glandulifera
(В); 1 — общий вид цветка, 2 — чашелистики (вид сзади), 3 — лепестки (вид
спереди).
Главный побег ветвится мезотонно до побегов III, редко до IV
порядка. В пазухах семядолей и первых шести-семи листьев образу-
ются побеги обогащения II порядка, длина которых в акропетальной
последовательности увеличивается от 5 до 35 см, затем уменьшается.
Начиная с 7—8 узла, в пазухах листьев формируются цветоносы, ко-
торые по мере роста поникают и загибаются под листья. Соцветие —
малоцветковое (3—5 цветков), короче листа. Побеги II порядка повто-
ряют структуру главного побега. Нижние 3—4 узла несут очень корот-
кие побеги III порядка, остальные — соцветия.
Цветки — крупные (до 35 мм длиной). Чашечка из 3 листочков
желтого цвета. Один из листочков, морфологически верхний, — во-
ронковидный, до 25 мм длиной, 12 мм в диаметре, имеет крючковид-
но загнутый шпорец (рис. 1, б). Два других — очень мелкие (до 5 мм
131
длиной), косо яйцевидные с заостренной верхушкой. Венчик состоит
из 5 желтых лепестков, 4 из которых срастаются попарно, так что вен-
чик кажется трехчленным. Непарный лепесток, супротивный чашели-
стику, — со шпорцем, широкояйцевидный, с заостренной отогнутой
наружу верхушкой, слегка килеватый. Боковые лепестки — неравно
двулопастные с красными точками в средней части.
Андроцей — из 5 тычинок с короткими нитями, свободными в
основании и сросшимися в трубку вокруг верхней части завязи, с си-
дячими продолговато-яйцевидными пыльниками. Пыльцевые зерна—
бледно-желтые, четырехбороздные, билатерально асимметричные, эл-
липсоидальные, длиной 33—36 мкм, диаметром 20 мкм (Куприянова,
Алешина, 1972). Гинецей— из 5 плодолистиков, с сидячим пяти-ло-
пастным рыльцем. Плацентация — центрально угловая, в завязи за-
кладывается до 7 семяпочек.
Часто на растениях наряду с яркоокрашенными полноценными
хазмогамными цветками появляются полураскрытые цветки с недо-
развитыми венчиками и невзрачные, не раскрывающиеся клейстогам-
ные цветки. Растения, растущие в тени, имеют только клейстогамные
цветки.
Плод — многосемянная коробочка, расширенная в основании
и суженная кверху, длиной до 2 см, вскрывается ламинально, будучи
еще зеленой и сочной. В плодах вызревает от 1 до 7 семян. Семена —
эллиптические, 3—4 мм длиной, 1,5 мм в диаметре, темно-коричне-
вые, матовые, с редкими извилистыми продольными гребнями. В се-
менах содержится 16,02% белка и 21,8% жира (Jones, Earle, 1966).
Число хромосом 2п = 20 (Хромосомные числа..., 1969). Только
в работе J. Skalinska с соавторами (1959) указан тетраплоид (2п = 40).
Онтогенез и сезонный ритм развития
Для недотроги характерен временный запас семян. Все семена,
осыпавшиеся летом и осенью, во время зимовки проходят стратифи-
кацию (Комарова, 1986), которая продолжается не менее 20—25 не-
дель при температуре не выше +5°С; после стратификации семена вес-
ной дают 100%-ное прорастание. В образцах почвы, взятых весной
после дружного появления всходов, семян недотроги не было обнару-
жено (Марков, 1991). После обсеменения и полного отмирания рас-
тений осенью удалось обнаружить очень небольшое количество
семян (24 семени/м2) только в самых верхних слоях почвы. В широко-
лиственных лесах юго-западной Норвегии (Austad, Skogen, 1990)
132
отмечался факт двух массовых генераций недотроги в мае—июне и в
августе. Возможно, что банк семян был реализован весной не сразу,
либо семена проросли без зимней стратификации.
Прорастание семян начинается в конце апреля (Серебряков,
1947) с удлинения гипокотиля. Никакого видимого зачатка главного
корня не образуется, из якореобразного расширения в нижней части
гипокотиля сразу же во все стороны вырастают 5—10 совершенно оди-
наковых придаточных корней (рис. 2, а), горизонтально идущих в са-
мых поверхностных слоях почвы (Марков, 1991).
Рис. 2. Стадии прорастания семян и проростки Impatiens noli-tangere (А),
I. parviflora (Б), I. glandulifera (В).
133
Рис. 3. Возрастные состояния Impatiens noli-tangere.
Удлиняющийся до 3—4 см гипокотиль выносит семядоли на по-
верхность. Последние имеют яйцевидные или широкояйцевидные пла-
стинки, 8 мм длиной и 6 мм шириной, с выемкой на верхушке, где
располагается гидатода. Их верхняя и нижняя поверхность — плоская,
зеленая, матовая с сизоватым оттенком. К моменту развертывания пер-
вых листьев пластинки семядолей достигают 10—12 мм длины и 10
мм ширины, их черешки — 10 мм длины, а на гипокотиле образуется
до 11 тонких неветвящихся придаточных корней до 30 мм длиной. На
формирование первой пары листьев требуется неделя. Одновременно
с развертыванием первой пары листьев возникают зачатки следующих.
Ювенильные растения кроме семядолей (атрибута проростков)
имеют два длинночерешковых листа с эллиптической или яйцевидной
зубчатой по краю пластинкой (рис. 3). Последующие листья — оче-
редные, короткочерешковые. Удлинение эпикотиля и начало фор-
мирования удлиненного главного побега условно можно рассматри-
134
вать как переход растения в имматурное состояние. Очень быстро в
пазухах нижних 6—7 листьев начинают формироваться побеги обога-
щения. У некоторых растений начинают отмирать семядоли. Листья
увеличиваются в размерах, но максимальных размеров они достигают
лишь у зрелых генеративных растений.
Переход в генеративное состояние происходит при достижении
растением 15—20 см высоты, когда главный побег состоит из 7—8
метамеров, где закладываются первые соцветия. Начало бутонизации
приходится на конец мая — начало июня. Максимальное развитие ли-
стьев отмечается в середине июля. Постепенно с развитием боковых
побегов вслед за семядолями отмирают и нижние листья на главном
побеге. Начало цветения отмечается в начале июля, массовое цвете-
ние наблюдается с середины июля до конца августа и даже до первых
заморозков в сентябре.
На рис. 4 приведены два вида фенологических спектров для по-
пуляции под пологом липового леса (Татария) в 1987 году (Марков,
1991). Одни из них (а) построены с учетом наличия или отсутствия
семядолей и перехода к репродукции, а другие (б) — с учетом стадий
роста, т.е. стадий двух, четырех и т.д. настоящих листьев. Обращает
на себя внимание более длительное сохранение семядолей и более ран-
нее наступление всех выделенных фенофаз и стадий у растений в су-
хом местообитании.
Рис. 4. Детализированные фенологические спектры для популяций Impatiens noli-
tangere из сухого (А) и влажного (Б) местообитаний: 1 с, 2с — наличие одной или двух
семядолей; в — вегетация; г — генеративное состояние; 2—18 — фазы двух—восем-
надцати листьев.
135
Первые плоды созревают в конце июля. Период диссеминации,
так же, как и цветение, растянут, и длится до конца сентября (Сереб-
ряков, 1947). Отмирает растение обычно с наступлением заморозков.
Однако под пологом леса, как, например, это было в сероольшанике
(Тверская область), уже к началу августа практически все растения
могут погибнуть в результате затенения Mercurialis perennis или
Aegopodium podagraria.
Способы размножения и распространения
У недотроги обыкновенной кроме хазмогамных цветков обыч-
ны и клейстогамные цветки. Мы наблюдаем у вида тот наиболее рас-
пространенный случай, когда самоопыление в форме факультативной
клейстогамии страхует, сводит к минимуму степень риска, а возмож-
ная в оптимальных условиях хазмогамия служит поддержанию гете-
розиготности. Вначале образуются клейстогамные цветки, позднее хаз-
могамные (Schemske, 1978), более ценные в энергетическом отноше-
нии (Грант, 1984). М. Masuda и Т. Yahara (1994), однако, обнаружили,
что в популяциях, где особи растут в наилучших условиях освещенно-
сти, хазмогамные цветки образуются раньше клейстогамных, что под-
тверждается нами в популяции, растущей на молодой вырубке (Твер-
ская область).
Рис. 5. Относительная скорость роста особей в популяциях Impatiens noli-tangere. W
— общий воздушно-сухой вес особи.
В хазмогамных цветках имеется ряд приспособлений, способ-
ствующих перекрестному опылению. Цветки строго протерандричны.
Тычинки в верхней части срастаются в трубку вокруг верхней части
136
завязи, образуя своеобразную крышечку, отпадающую к моменту со-
зревания рыльца, что обеспечивает эффективное перекрестное опы-
ление (Полянский, 1950). На дне шпорца верхнего чашелистика име-
ется неспециализированный нектарник, представляющий собой мел-
коклеточную нектароносную ткань. Цветки всегда висят на цветонож-
ках таким образом, что располагаются под листьями. Капли дождя,
скатываясь с листьев, не задевают цветков даже при очень сильных
ливнях.
Соотношение клейстогамных и хазмогамных цветков опреде-
ляется условиями освещенности (Masuda, Yahara, 1994). Высокая ос-
вещенность обеспечивает появление хазмогамных цветков. Они, как
правило, размещаются в пазухах верхних листьев на главном и боко-
вых побегах. В условиях затенения преобладают клейстогамные цвет-
ки. Практически во всех исследованных нами популяциях недотроги
под пологом леса в Московской и Тверской областях не было найдено
особей с хазмогамными цветками. Хазмогамные цветки есть только у
растений, растущих на полном свету — около ручьев, лесных речек,
на молодых вырубках, в окнах древесного полога. Факультативную
клейстогамию, по мнению Masuda и Yahara (1994), можно считать стра-
тегией жизни в сезонно меняющихся условиях среды. Тип цветка, спо-
собность к образованию полноценного открытого цветка определяет-
ся в конце формирования бутона и зависит от жизненности особи, ос-
вещения и влажности места произрастания.
Важным биологическим последствием сочетания двух спо-
собов опыления является различие веса семян клейстогамного
и хазмогамного происхождения. Клейстогамные цветки обычно
образуют мелкие семена. Так в популяции, изученной A. Hibatsuka
и О. Inoue (1988) в Японии, средний вес семян в хазмогамных цветках
составлял 68,0±18,6 мг, у клейстогамных — 58,1± 14,1 мг. Вес семян
в изученных нами популяциях под Казанью существенно меньше: от
2,4 до 6,9 мг (среднее — 4,6 мг). В Подмосковье (Звенигород) 100
семян, собранных у недотроги под пологом липы, весили 337,5 мг.
Вес семян представляет собой очень важный параметр стратегии ви-
да. От него зависят размеры семядолей и вместе с ними вероятность
выживания проростка, а впоследствии темпы накопления биомассы в
ходе роста (Falencka, 1983; Марков, 1991). Преимущество в высоте и
биомассе у особей, которые в фазе семян и всходов имели большие
размеры, сохраняется и даже нарастает в течение всей жизни расте-
ния (рис. 5).
137
Коробочка недотроги вскрывается путем образования неполных
продольно-кольцевых щелей в стенках плодолистиков от основания к
вершине. В нижней расширенной части плода, где обычно находится
2 семени, стенки оказываются наиболее тонкими. В верхней части ко-
робочки стенки утолщаются, и в них развивается особый слой клеток
с очень высоким осмотическим давлением (до 25 атм.) (Кожевников,
1950). В результате большого напряжения, которое создается в стен-
ках плода, при малейшем прикосновении створки разверзаются, за-
кручиваются улиткообразно и с силой выбрасывают семена (Левина,
1987). Дальность разбрасывания семян при автомеханохории прямо
коррелирует с высотой растения, силой толчка и массой семени и дос-
тигает нескольких метров. Семена из коробочек хазмогамного проис-
хождения разлетаются в среднем на 136,4 см, а клейстогамного — на
44,9 см (Hibatsuka, Inoue, 1988). Семена из клейстогамных цветков ин-
бридингового происхождения в результате оказываются на участках,
занятых материнскими растениями, гарантируя поддержание популя-
ции на уже завоеванном месте. Семена же из хазмогамных цветков,
образованные в результате аутбридинга, разлетаются далеко от мате-
ринского растения и имеют шанс прорасти в новых местообитаниях.
Эти семена часто попадают в водные потоки осенью и разносятся на
большие расстояния. Возможна также зоохория, когда животные, про-
ходя мимо зарослей недотроги, оказываются облеплены ее семенами
и разносят их на большие расстояния.
Плодовитость недотроги обыкновенной в природных популя-
циях — весьма низкая. Так, для популяции в ольховом заболоченном
лесу и популяции в дубово-грабовом лесу (Беловежская Пуща) приво-
дятся средние значения плодовитости в интервале от 1,4±0,9 до 4,7±2,6
семян/особь (Falencka, 1983).
По нашим данным из Татарии, в одной коробочке число
закладывающихся семязачатков (при анализе бутонов или едва начав-
ших развитие завязей) достигало 9 в отдельной завязи. Однако, как
потенции плода, так и реализация этих потенций, судя по нашим
данным, зависят от условий произрастания. В популяции из Казани
(Татария) в 194-х полностью сформированных плодах было от 1 до 6
семян, а в выборке плодов из дальневосточной популяции (в Уссу-
рийском заповеднике) плоды содержали от 1 до 11 семян. График рас-
пределения плодов по числу содержавшихся в них семян в обеих по-
пуляциях был положительно асимметричным с модальными значения-
ми, соответственно, 2 и 4. Средняя масса одного семени на плод с
138
увеличением числа семян в плоде достоверно снижается от 2,9 до 2,5
мг. Для Японии приводятся данные о достоверно различном среднем
количестве семян в коробочке хазмогамных (5,4±0,7 семян на плод) и
клейстогамных цветков (3,2±1,2) (Hibatsuka, Inoue, 1988).
Экология
Недотрога обыкновенная — наиболее теневыносливый и гиг-
рофильный вид в роде Impatiens (EliaS, 1987). Обитает на влажных, не
просыхающих почвах (7-я ступень шкалы Элленберга), в условиях от
влажно-лугового до болотного увлажнения (68—100 ступени шкалы
Раменского). Недотрогу обыкновенную обычно относят к гигромезо-
фильному типу трав. Вид не выносит отсутствия кислорода и длитель-
ного застоя воды.
Высокая влажность воздуха — фактор, также лимитирующий
распространение недотроги. Потеря воды на солнце при пересчете на
сырой вес составляет 4,73 г/г в сутки, что почти в два раза выше, чем
у Impatiens parviflora.
При достаточной влажности почвы и воздуха основным факто-
ром, определяющим темпы роста, накопления биомассы ив итоге мощ-
ность репродукции, является свет. По экологическим шкалам Эллен-
берга недотрога обыкновенная располагается между теневыми
и полутеневыми растениями (4-я ступень). Длиннопобеговое строе-
ние недотроги сопряжено со стратегией выносливости к дефициту
света (Марков, 1990). Листовая ассимиляционная поверхность
сосредоточена в основном в верхней части стебля, если растения про-
израстают под лесным пологом в условиях недостатка света. На
открытом же пространстве, например, на вырубках листовая поверх-
ность расположена более равномерно. Под пологом леса дифферен-
циация растений по высоте начинается с момента образования пер-
вых листьев.
С понижением освещенности под пологом леса наблюдается уве-
личение листовой поверхности, компенсирующее недостаток освеще-
ния, что приводит к снижению репродуктивного усилия. Это проявля-
ется в формировании только клейстогамных цветков, в подавлении се-
менного размножения. При критических уровнях затенения растение
может существовать только в вегетативном состоянии, близком к им-
матурному.
Недотрога обыкновенная относится к облигатным нитрофилам
(Работнов, 1940). При этом растения гипертрофически разрастаются
139
и накапливают в тканях нитраты при обилии их в почве. По экологи-
ческим шкалам Элленберга недотрога обыкновенная растет на почвах
с довольно высоким содержанием минерального азота (6-я ступень).
Корневая система расположена в поверхностном почвенном слое, не
глубже 10 см. При этом почва должна быть гумусирована и хорошо
аэрируема. Произрастает обычно на почвах, богатых мягким гумусом,
бурых, глееватых. На подзолистых почвах растет редко, только на
слабооподзоленных. На молодых вырубках растения приуроче-
ны к строго определенным местам: кучам валежа и порубочных
остатков, перемешанным с гумусом верхних горизонтов почвы.
В ветровальных западинах недотрога растет на комьях гумусово-
го горизонта и подстилки. Предпочитает почвы с нейтральной реак-
цией, но может расти на кислых и щелочных (7-я ступень шкалы Эллен-
берга).
Недотрога обыкновенная растет только там, где имеются хотя
бы незначительные нарушения травяного покрова. Режим этих нару-
шений может зависеть как от деятельности человека (вырубка леса,
колеи дорог, вытаптывание с нарушением почвы), так и от естествен-
ных факторов (временные потоки воды, западины ветровалов в лесу,
окна в пологе леса).
Фитоценология
Недотрога обыкновенная произрастает в приречных ивняках,
около стариц, ключей, ручьев, в зарослях лабазника, крупнотравных
ольшаниках, сырых травяных ельниках и смешанных лесах, в кана-
вах, нередко на сырых и затененных мусорных местах, близ жилья.
Она характерна для широколиственных лесов Восточноев-
ропейской, Маньчжурской, Сахалине - хоккайдской провинций
(Карписонова, 1985). В Центральной Европе недотрога обыкновен-
ная является характерным видом порядка Fagetalia silvaticae Pawl,
in Pawl, e.a 1928 в широколиственных лесах класса Querco-Fagetea, и
типичным для союза Alno-Padion Knapp 1942 (Matuszkiewicz, 1981).
В южной тайге и широколиственных лесах европейской части России
вид обилен в асе. Urtico dioicae-Alnetum incanae этого союза
(Коротков, 1991). Это высокотравные сухие сероолыпаники на плодо-
родных почвах, описанные для Валдая. Alnus incana образует древо-
стой вместе с A. glutinosa, Picea abies и Padus avium. В подлеске обыч-
ны Ribes spicatum и Rubus idaeus. В травяном ярусе обильны Anemone
nemorosa, Galeobdolon luteum, Stellaria nemorum, Aegopodium podag-
140
raria, Paris quadrifolia, Urtica dioica, Oxalis acetosella, Equisetum pre-
tense и другие. T.A. Работнов (1939) называл эти сероольшаники Alnus
incana-Urti-Impatiens.
Кроме этого она встречается в пойменных черноолыпаниках
союза Alno-Ulmion в ценозах Stellario-Alnetum и мезофитных широ-
колиственных лесах с ясенем Alno-Fraxinetum, с вязом Alno-Ulnetum,
с липой Ulmo-Tilietum (Elias, 1987). Она также характерна для сооб-
ществ союза Cardaminion (Maas 1959) Den Held et Westh. 1969 в клас-
се Montio-Cardaminetea Br.-Bl. et Tx. 1943 в Польше, Чехии, Литве
(Strazdaite-Baleviciene, 1988). Около речек и ручьев произрастает вме-
сте с Cardamine amara, Chrysosplenium altemifolium, Lysimachia nemorum
и Veronica montana.
В лесах южной тайги России недотрога обыкновенная типична
для ельников неморальных: ильмово-пролесниковых, реже кленово-
зеленчуковых и ельников травяно-болотных: черноольхово-таволго-
вых, травяно-папоротниковых (Карпов, Шапошников, 1983).
В.Н. Федорчук и Ю.И. Бурневский (1986) относят недотрогу
обыкновенную к эколого-ценотической группе Filipendula ulmaria вме-
сте с Geum rivale, Ranunculus repens, Viola epipsila, Galium palustre,
Cirsium oleraceum. В Ленинградской области эта группа видов являет-
ся индикаторной для приручейно-разнотравных ельников (таволгово-
кисличная серия на недостаточно дренированных местообитаниях с
проточным увлажнением) и болотно-разнотравных ельников с ольхой
серой и черной (таволговая серия на низинных торфах и слабодрени-
рованных местообитаниях с проточным увлажнением).
Недотрога обыкновенная — типичный эксплерент с чертами
патиентности. Вид быстро захватывает временно свободные от
других видов нарушенные территории за счет большой семенной
продуктивности, 100% всхожести семян, одновременного быстрого
интенсивного семенного возобновления, в результате чего рано
весной (раньше других видов) формируются популяции высокой
численности. Обычно это монодоминантные заросли, под пологом
которых могут обитать только виды лесного мелкотравья: Galeobdolon
luteum, Glechoma hederacea, Ajuga reptans, Asarum europaeum, Oxalis
acetosella, Stellaria nemorum, S. holostea, Ranunculus repens, Chrysosple-
nium altemifolium. Основными конкурентами, способными заглушить
недотрогу, можно считать Urtica dioica, Filipendula ulmaria, Aegopodium
podagraria, Chelidonium majus, Chamerion angustifolium (на вы-
рубках). Эти, в основном длиннокорневищные многолетники,
141
расползаясь по территории, резко сокращают численность побегов не-
дотроги, хотя она может прорасти и существовать в угнетенном со-
стоянии под пологом этих трав, проявляя патиентность. Сосущество-
вание с корневищными видами возможно, т.к. корневая система не-
дотроги расположена в основном в подстилке, а у корневищных рас-
тений — в гумусовом и ниже расположенных горизонтах. Прораста-
ние, формирование стебля и листовой поверхности у недотроги про-
ходит раньше, чем начинают формироваться надземные побеги у дру-
гих видов.
Консортивные связи
Недотрога обыкновенная опыляется в основном шмелями и, из-
редка, медоносными пчелами. Шмели садятся на нижние лепестки и
наполовину влезают внутрь воронковидного чашелистика, при этом
измазывая спинку в пыльце. Перелетая на цветки, у которых сросшие-
ся пыльники уже отпали, они дотрагиваются спинкой до созревшего
рыльца, обеспечивая перекрестное опыление. Неопыленные цветки
семян не дают.
На недотроге обыкновенной, в отличие от двух других видов,
обильно растет гриб Sphaerotheca balsaminea (Wullr.) Kari. (S. fusca (Fr.)
Blumer), вызывая мучнистую росу (Горленко, 1983). Гриб появляется
в середине июля, а в начале августа всегда в массе формируются клей-
стотеции. Мучнистая роса поражает недотрогу ежегодно и повсеме-
стно, что часто приводит к массовому отмиранию растений уже в ав-
густе.
Биологическая продуктивность
Продуктивность недотроги обыкновенной была изучена на тер-
ритории Звенигородской биостанции в ельнике липняковом на склоне
к р. Москве и в сероольшанике в притеррасной части одновременно
22 июня 1996 года. В ельнике липняковом выборка проводилась по
трем микрогруппировкам. Растения были выкопаны на площадках 1 х 1
м в 5-ти кратной повторности. Средние показатели для растений оп-
ределялись как средневзвешенные средние по классам жизненности.
Растения взвешивались сразу после выкапывания и после высушива-
ния (воздушно-сухой вес). Полиморфизм растений проявляется в зна-
чительных колебаниях всех показателей фитомассы в различных эко-
логических условиях (табл. 1). Однако аллокация фитомассы схожа.
Основной вклад в фитомассу вносят стебли, наименьший — репро-
дуктивные органы.
142
Таблица 1.
Средний вес одного растения Impatiens noli-tangere (мг)
в лесах Московской области (Звенигородская биостанция)
Фитоценоз Вес Листья Стебель Корни Соцветия Всего
Ельник липняковый, под пологом свежий 238 479 100 8 824
% 28,9 58,1 12,1 1,0 100
сухой 29 36 11 3 78
% 37,7 46,1 14,8 1,4 100
Ельник липняковый, окно свежий 1100 3550 510 80 5240
% 21,0 67,7 9,7 1,5 100
сухой 150 210 60 10 430
% 34,9 48,8 14,0 2,3 100
Ельник липняковый, ветроваль- ный бугор свежий 227 358 72 10 667
% 34,0 53,7 10,8 1,5 100
сухой 25 20 6 1 52
% 48,5 38,2 12,0 1,3 100
Сероолыпа- ник, под пологом свежий 340 1190 200 18 1748
% 19,4 68,0 11,4 1,1 100
сухой 44 50 23 6 123
% 35,8 40,6 18,7 4,9 100
Влажность растений составляет 90—93% (табл. 2). Общая про-
дуктивность недотроги очень сильно зависит от экологических усло-
вий, варьируя от 49 г/м2 в окне до 6 г/м2 на ветровальном бугре и в
ельнике липняковом, при одинаковой плотности популяции.
Динамика изменения фитомассы растений в течение сезона ве-
гетации изучена в липняке под Казанью (Марков, 1991). Накопление
биомассы шло в течение всего лета (табл. 3). В более сухих условиях
конечная биомасса оказалась выше, чем во влажных условиях.
Конечная биомасса особей достоверно положительно коррелировала
с размерами семядолей (г= от +0,69 до 0,84”’).
143
Таблица 2.
Характеристики популяций I. noli-tangere на пробных площадках
в лесах Московской области (Звенигородская биостанция)
Фитоценоз Особей на м2 Влаж- ность, % Фитомасса, г/м2
свежая сухая
Ельник липняковый, под пологом 281,4±22,5 90,5 231,54 21,92
Ельник липняковый, окно 114,2±15,4 91,8 597,36 49,02
Ельник липняковый, ветровал 119,1±Ю,3 92,2 79,42 6,19
Сероолыпаник 91,4±8,7 92,9 159,25 11,19
Таблица 3.
Динамика роста биомассы (сухой вес, г) средней особи в популяци-
ях г noli-tangere под пологом липового леса, Казань (Марков, 1991)
Местообитание 26.V.1989 2. VI 10.VI 26.VII
Вйажное 45,3±4,3 45,7±3,4 139,9±8,9 306,6±43,9
Сухое 45,4±3,4 57,4±6,4 89,3±9,5 398,4±57,5
У растений, принадлежащих к разным классам по общей био-
массе, выявляется различное размещение биомассы в пределах особи
(рис. 6) (при этом классы биомассы можно рассматривать как показа-
тель жизненного состояния). Так, у особей, имеющих малую биомас-
су, наибольшая часть биомассы (и энергии) вкладывается в создание
листовой поверхности, т.е. в вегетативные органы, а репродуктивное
усилие остается очень низким. С возрастанием общей биомассы (улуч-
шением жизненного состояния особи) увеличивается вклад в репро-
дуктивные структуры за счет уменьшения относительного веса
листьев или фотосинтетического усилия. Динамические аллокацион-
ные спектры для обеих популяций показывают, что у растений во влаж-
ном местообитании относительный вклад биомассы в корневую сис-
тему был в среднем на 5% выше, чем у растений сухого местообита-
ния. Фотосинтетическое усилие лишь в начале наблюдений, т. е. до
144
формирования затеняющего полога, было на 5—7% выше у растений
сухого местообитания, после чего разница сменилась на обратную,
достигнув к концу наблюдений 12%.
20.V 26.V 1.VI 9.V) 18.VI 2.VII 14.VII
Рис. 6. Динамические аллокационные спектры для популяций Impatiens noli-tangere
из сухого (А) и влажного (Б) местообитаний. Относительный вклад биомассы (%): 1
— в репродуктивные структуры; 2 — в листья; 3 — в стебли; 4 — в семядоли; 5 — в
корневую систему.
Численность особей в обеих популяциях изменялась только по
причине отмирания, так как появления новых всходов не наблюдалось.
Процессы естественного выпадения особей были замедлены, и уско-
рение их было связано с действием внешних нарушений. Плотность
на учетных площадках составляла от 60 до 84 особей/м2.
Хозяйственное значение
Свежие листья недотроги обыкновенной содержат горькие, вы-
зывающие рвоту вещества — impatiinid (Гроздов, 1963). Во всел орга-
нах растения содержатся гликозиды, алкалоиды и сапонины, которые
угнетают сердечную деятельность и центральную нервную систему и
понижают артериальное давление (Орлов и др., 1990). Скотом не по-
едается, ядовита для рогатого скота и овец. Цветы и листья могут ис-
пользоваться для получения желтой краски (Рычин, 1952).
145
Недотрога мелкоцветковая
Номенклатура и внутривидовая систематика
Impatiens parviflora DC. 1824, Prodr. 1:687; Шмальгаузен, 1895,
Фл. Средн, и Южн. Росс., 1: 201; Петунников, 1896, Крит, обзор, 1:
113; Сырейщиков, 1907, Илл. фл. Моск, губ., 2: 351; он же, 1927, Оп-
ред. раст. Моск, губ.: 183; Назаров, 1940, в Маевский, Фл., изд. 7: 505;
Победимова, 1949, Фл. СССР, 14:631; В. Тихомиров, 1966, Опред. раст.
Моск, обл.: 229; D.M. Moore, 1968, Fl. Europ., 2: 240; Новиков, 1987,
Опред. раст. Мещеры, 2: 10; Черепанов, 1995, Сосуд, раст. России:
218; Мордак, 1996, Фл. Вост. Европы, 9: 391. — Недотрога мелко-
цветковая, прыгун (семейство Balsaminaceae — Бальзаминовые).
Исследования Ю.К. Виноградовой (1992) показали, что евро-
пейская популяция недотроги мелкоцветковой практически не обла-
дает полиморфизмом, в отличие от исходной центрально-азиатской по-
пуляции. Отмечена лишь фенотипическая вариабельность интенсив-
ности окраски желтого околоцветника. Вероятно, это связано с одно-
кратным заносом растений в Европу в 1831 году.
В естественном ареале произрастает две цветовые формы не-
дотроги — с желтыми и с лиловыми цветками, рассматриваемые ра-
нее (Флора СССР, 1949) в качестве отдельных видов. Однако Вино-
градова установила, что Impatiens nevskii Pobed. (с лиловыми цветка-
ми) не отличается от I. parviflora ни по одному из изученных призна-
ков: ареалу, экологии, морфологии цветков, качеству семян, феноло-
гии растений в природе и при интродукции. Поэтому недотрогу Нев-
ского следует считать лиловоцветковой формой I. parviflora DC (Май-
тулина, 1988).
Географическое распространение
Недотрога мелкоцветковая — центрально-азиатский вид, ее ес-
тественный ареал охватывает, в основном, территории всех респуб-
лик Средней Азии бывшего СССР: юг Казахстана, юг Туркмении, юг
и восток Узбекистана, Киргизию, Таджикистан. В горных районах под-
нимается на высоту до 2500 м над ур. моря.
В Европе этот вид впервые появился в 1831 г. в ботаническом
саду Женевы, в 1837 г. культивировался в Дрездене. В течение сле-
дующих нескольких десятилетий отмечались единичные находки во
многих пунктах Европы, обычно возле ботанических садов: Женева,
146
Прага, Юрьев, Кенигсберг, Берлин, Дрезден (Coombe, 1956). В сере-
дине XX века ареал вида в Европе стал стремительно расширяться. В
1947 г. вид отмечен в южной Швеции (Riska, 1957—58). В это же вре-
мя началась натурализация вида в Дании, южной Финляндии, Поль-
ше, Чехословакии, Германии, Венгрии, Австрии, Голландии, Бельгии,
северной и восточной Франции. В Швейцарии вид стал обычен на вы-
соте 760 м над ур. моря, но поднимается и выше до 1860 м в Альпах
(Coombe, 1956). В Северной Америке вид отмечен как заносный в Бри-
танской Колумбии Канады около Ванкувера (Barkworth, 1973). Есть
данные о натурализации вида в Монреале (Zinov’eva-Stahevitch, Grant,
1984).
Процесс инвазии недотроги мелкоцветковой в России изучен
Ю.К. Виноградовой (1992). В конце XIX века она выращивалась в са-
дах и парках Львова, Санкт-Петербурга, Юрьева (Тарту), Риги, Пярну.
В Москве первый экземпляр недотроги собран А.Н. Петунниковым в
1884 г., а уже в 1889 г. Н.Н. Кауфман сообщал о том, что вид встреча-
ется во многих садах и парках. Число местонахождений недотроги в
Москве увеличивалось крайне медленно, и только через 40 лет Д.П.
Сырейщиков (1927) отметил быстрое распространение ее по сорным
местам близ жилья и по садам. Интенсивное расширение ареала не-
дотроги в России началось лишь с семидесятых годов (через 90 лет
после первого ее появления в европейской России).
Сейчас вид распространился в Белоруссии, Молдавии, на Ук-
раине. Самые северные точки его вторичного ареала находятся в Ка-
релии, в республике Коми, Вологде (Майтулина, 1984). Южная грани-
ца вторичного ареала проходит по Северной Осетии (Комжа, Попов,
1990). На Дальнем Востоке недотрога мелкоцветковая впервые была
найдена в 1966 г., однако здесь она интенсивно начала распростра-
няться только через 20 лет (Ворошилов, 1982). В пределах естествен-
ного ареала вид обитает в узких глубоких лощинах, ущельях, под кро-
нами деревьев (особенно часто под грецким орехом), по берегам ручьев
(Майтулина, 1988). Во вторичном ареале произрастает в посадках, по
берегам рек, в садах, влажных тенистых местах. Усиленно внедряется
в широколиственные леса Европы (Csoentos, 1986).
В Московской области недотрога мелкоцветковая стала массо-
вым видом, произрастая обычно в местах с антропогенными наруше-
ниями: во дворах, в садах и парках. В лесопарках и нарушенных лес-
ных массивах нередко образует сплошные заросли, вытесняя естест-
венную растительность.
147
Морфологическое описание
Недотрога мелкоцветковая — однолетнее травянистое растение
со смешанной корневой системой. Корни (главный и боковые) тон-
кие, располагаются в 10 сантиметровом слое почвы и ветвятся до кор-
ней 4 порядка. Очень быстро на гипокотиле развиваются более тол-
стые прозрачные розоватые придаточные корни. Гипокотиль достига-
ет в длину 13 см.
Стебель — цилиндрический, до 80—90 см высотой и диамет-
ром 0,5—1 см, плотный, зеленый с 8 темными полосками, сильно утол-
щенный, почти вздутый в узлах, простой или слабо ветвящийся до по-
бегов III порядка, полый внутри, голый. Длина междоузлий в акропе-
тальной последовательности сокращается от 10—15 см до несколь-
ких мм, так что большая часть листьев скучена на верхушке побега.
Листья—короткочерешковые (1,5—6 см длиной), с мелкими стебель-
чатыми волосками в основании. Листья первой пары — супротивные,
у последующих листьев — очередные. Листья — короткочерешковые
(6—1,5 см длиной) с мелкими стебельчатыми железками в основании.
Пластинки листьев — ланцетовидные, длиной от 5 до 18 см, шириной
от 3 до 8 см, голые, немного лоснящиеся, по краю с зубцами, притуп-
ленными и железистыми (Рычин, 1952). Жилкование — перисто-сет-
чатое с поперечными анастомозами.
Боковые ветви располагаются в пазухах первых двух листьев.
Они не превышают 20 см и включают до 8 метамеров, утолщены в
основании и направлены вертикально вверх. По архитектуре боковые
побеги сходны с главными. Цветоносы, формирующиеся в пазухах
верхних листьев, — прямостоячие или отклоненные, на 6—8 см воз-
вышающиеся над верхушкой побега с кистевидным соцветием из 4—
12 маленьких цветков, реже цветки — одиночные. Ветвление — мезо-
тонное.
Цветки — протерандричные. Длина цветка — 7—15 мм. Ча-
шечка — из 3 лепестковидных листочков (рис. 1, а). Два боковых ча-
шелистика — желтоватые, с зеленой полоской вдоль средней жилки,
треугольные, до 2 мм длиной. Верхний чашелистик с прямым шпор-
цем 5—10 мм длиной. Форма и длина шпорца — довольна изменчи-
вы. Встречаются короткие (3—5 мм) широко клиновидные шпорцы и
длинные (5—9 мм) узко клиновидные шпорцы, иногда булавовидные
на конце (Майтулина, 1988). Внутри расширенная часть чашелистика
окрашена в оранжевый цвет, на фоне которого выделяются красные
пятнышки.
148
Венчик—из 5 лепестков, 4 из них попарно срастаются, так что
венчик кажется трехчленным. Непарный лепесток, супротивный ча-
шелистику со шпорцем, — округлый, 5 мм длиной, желтого цвета с
зеленой полоской по килю. Боковые лепестки—асимметричные, име-
ют различную степень надрезанности: двулопастные, двулопастные с
неглубоким вырезом одной из лопастей, трехлопастные. Даже у одно-
го цветка правые и левые боковые лепестки могут различаться по сте-
пени надрезанности (Виноградова, 1992). Окраска лепестков — от свет-
ло-желтой в основании до ярко-желтой с красными пятнышками в сред-
ней части.
Тычинок — 5, нити тычинок — короткие, в нижней части сво-
бодные, наверху — сросшиеся в короткую трубку. Пыльники вскры-
ваются продолговатыми щелями еще в бутонах, и пыльца высыпается
в полость цветка. Пыльцевые зерна сходны по форме с пыльцевыми
зернами Impatiens noli-tangere, их размеры — 0,03 Зх 0,016 мм. Рыльце
созревает после растрескивания и опадания тычинок.
Гинецей — из 5 плодолистиков. Пестик — с коротким столби-
ком и цельным рыльцем. Завязь — одногнездная, с многочисленными
семязачатками, прикрепленными к центральной колонке. Цветки со-
храняются открытыми 1—2 дня.
Плод — сочная, торчащая или несколько отклоненная, продол-
говатая или цилиндрическая, локулицидная коробочка, длиной 15—
25 мм, несколько расширенная вверху. Плоды созревают в течение 18—
20 дней. Обычно в коробочке вызревают 1—4 семени, хотя число се-
мяпочек может достигать 7.
Семена — бурые, матовые, удлиненные или продолговато-эл-
липтические, сплюснутые, с ясным семенным швом (рис. 2, б). По-
верхность семян — с частыми продолговатыми короткими, полого из-
вилистыми ребрышками — морщинками, похожими на нити жемчуга
(Кац и др., 1965) или с бугорчатыми валиками в рядах (Артюшенко,
1990). Размер семени 4—4,6Х2,3—2,5Х 1,4 мм.
В европейских популяциях преобладают особи с количеством
хромосом 2п = 24,26 (Хромосомные числа..., 1969). Однако есть так-
же сообщения о 2п = 20 (Coombe, 1956). В Канаде (Монреаль) у не-
дотроги мелкоцветковой было зарегистрировано 2п = 26 (Ziniv’eva-
Stahovitch, Grant, 1984).
149
Онтогенез
Семена — со скудным эндоспермом и крупным зародышем.
Продолговатые семядоли жизнеспособного зародыша окрашены в си-
не-зеленый цвет. Прорастание семян — надземное, гипокотилярное.
Рост гипокотиля несколько опережает развитие главного корня. Очень
быстро вслед за формированием слабо выделяющегося главного кор-
ня на границе с гипокотилем закладываются 4—8 придаточных кор-
ней (рис. 2, б) (Марков, 1994). На ранних стадиях у проростков наибо-
лее интенсивно развитие протекает в подземной сфере. Удлиняющий-
ся до 20—30 мм гипокотиль выносит семядоли на поверхность поч-
вы, он способен сильно удлиняться в зависимости от глубины прорас-
тания. Вполне развитый проросток имеет две семядоли, гипокотиль,
главный и придаточные корни. Почечка не дифференцирована. Пла-
стинки семядолей — блестящие, мясистые, эллиптические, 8 мм дли-
ной и 4,6 мм шириной, короткочерешковые или почти сидячие, с вы-
емкой на верхушке, где располагается гидатода. Пластинки — темно-
зеленые с пурпуровым отливом, с вдавленной жилкой на верхней сто-
роне и слегка выступающей на нижней стороне. Гипокотиль — обыч-
но 20—30 мм длиной, диаметром 1—1,5 мм. Главный корень — 25 мм
длиной, слабо ветвится. Придаточные корни — утолщенные, неветвя-
щиеся, до 65 мм длиной. Высота проростков — в среднем 7 см, ам-
плитуда изменчивости — 4,5—10,5 см.
С развитием первой пары ассимилирующих листьев растение
переходит в ювенильное возрастное состояние (рис. 7). Листья — ко-
роткочерешковые продолговато-яйцевидные, пильчатые по краю. Од-
новременно несколько удлиняется гипокотиль, разрастаются семядо-
ли и почти одновременно закладываются два следующих листа. Их
развитие сопровождается удлинением эпикотиля. С этого момента на-
чинает формироваться главный побег, сохраняющий способность к от-
крытому росту в течение всего вегетационного сезона. В данной фазе
развития растение кроме семядолей имеет два супротивных, два весь-
ма сближенных очередных листа и зачатки 5-го и 6-го листьев. Парал-
лельно с развертыванием 4-го или 5-го листьев в их пазухах заклады-
ваются одиночные цветки или малоцветковые соцветия, а особи пере-
ходят в генеративное состояние. В пазухах опавших семядолей и
первой пары листьев развиваются побеги обогащения, повторяющие
в основных чертах структуру главного побега. Ветвление —
баритонное.
150
Рис. 7. Возрастные состояния Impatiens parviflora.
Жизненность цветущих растений зависит от экологических и
фитоценотических условий (табл. 4). Угнетенные экземпляры высо-
той до 20 см имеют 6—7 листьев и соцветие из 3—4 цветков. Растения
высокой жизненности могут достигать высоты 90 см, иметь побег, со-
стоящий из 13 метамеров с листьями до 9—18 см длиной и 5,5—6 см
шириной. На главном побеге развиваются боковые ветви одного—двух
порядков, и многочисленные соцветия, несущие 50—80 цветков.
Сезонный ритм развития
По нашим наблюдениям, в Подмосковье под пологом ельника
липнякового дружное прорастание семян отмечено в последних чис-
лах апреля — начале мая, что в значительной степени определяется
погодными условиями. В 1997 году массовые всходы появились 1—7
мая. Дружность прорастания и появления всходов нарушается в том
случае, если весной наблюдается засушливый период. Тогда первая
когорта всходов может «отделяться» от последующих единичных всхо-
дов некоторым промежутком времени (1—2 недели), что заметно про-
является в степени дифференцированности особей.
151
Таблица 4.
Средние морфологические показатели растений
Impatiens parviflora различной жизненности
Жиз- нен- ность Вы- сота, CM Число Размеры корневой системы, см N вы- выб- ки
вет- вей I nop. лис- тьев прида- точных корней соцве- тий на гл. по- беге
• *♦ im 8,8 — 4,0 2,5 — 5,0х2,6 16
gf 34,7 4,2 9,6 4,5 7,8 9,9x5,3 40
g2- 29,3 3,4 9,5 4,0 6,1 9,5x4,0 39
g2- 27,6 1,8 7,0 5,5 6,8 9,4Х3,7 20
g3" 21,8 0,2 6,6 4,7 5,5 8,9Х3,8 25
g3’ 21,1 0,3 7,6 3,5 5,0 6,6Х2,6 25
g4-‘ 17,5 5,4 2,5 5,0 8,8x3,5 26
*♦ g 5 14,5 4,9 2,0 з,о 5,2Х2,4 17
Примечание: im — имматурные растения, g — генеративные расте-
ния высшей жизненности, g5 — низшей жизненности,'—заросли кле-
на ясенелистного, Москва; " — ельник лотовый, Звенигород.
В течение недели после прорастания разворачиваются семядо-
ли и закладываются зачатки первых листьев. В ювенильное состояние
основная масса растений в 1997 г. перешла 10—20 мая, 3—4-й лист
сформировался к 20—31 мая (хотя в 1996 году в эти сроки большинст-
во растений уже находилось в стадии бутонизации). Первые цветки
раскрылись 13—17 июня, а массовое цветение наступило, как прави-
ло, во второй половине июля. В конце июня в лесах Подмосковья еще
можно найти всходы и ювенильные растения. Период диссеминации
наступает с начала июля. Цветение и плодоношение продолжается
вплоть до осенних заморозков в сентябре—октябре. В г. Москве все
фенофазы наступают на 7—10 дней раньше.
Фенология вида специально изучалась при осеннем экспери-
ментальном высеве семян из разных регионов России и Средней Азии
в 1984 году на участках в ГБС РАН, Москва (Виноградова, 1992). Еди-
152
ничные всходы начинали появляться во 2-й декаде апреля, массовые
всходы — в третьей декаде. Почти одновременно (в двадцатых числах
мая) растения вступили в фазу бутонизации, в первой декаде июня —
в фазу цветения. К середине июня все растения образовали плоды, а к
9 июля начали «разбрасывать» семена. Новые бутоны и цветки про-
должают образовываться у особей, находящихся уже в стадии плодо-
ношения и диссеминации. Часть особей после диссеминации начина-
ет отмирать уже в середине июля.
От прорастания семян до начала цветения особям недотроги
требуется в природных популяциях 8—9 недель, а в отапливаемом ве-
гетационном домике — 5—6 недель.
Способы размножения и распространения
Насекомые—опылители до начала июля появляются редко, а во
влажные и холодные дни не появляются вовсе, тем не менее, практи-
чески каждый цветок завязывает семена. Превалирует хазмогамия,
очень редко наблюдается клейстогамия (Coombe, 1956). Вероятно, этот
вид—амфимикт. Случаев перекрестного опыления и образования гиб-
ридов не обнаружено (Виноградова, 1992).
Семенная продуктивность сильно варьирует. В лесах и у мел-
ких особей в загущенных популяциях в коробочке обычно содержит-
ся 1 семя, а на одном растении созревает 1—5 семян. На богатых, ув-
лажненных почвах хорошо развитые экземпляры недотроги способ-
ны формировать 4—5 семян в коробочке, а одно растение образует до
10000 семян. Диссеминация может продолжаться до трех месяцев. В
клетках плодов недотроги тургорное давление достигает 26 атмосфер
(Ulbrich, 1928). Семена разлетаются на 1—2 м, могут распространять-
ся по рекам и дождевыми водами (Виноградова, 1992).
В лесах Татарии средний вес семян — 8,21 мг (Марков, 1990).
Средний вес 100 семян, собранных в пределах естественного ареала,
составляет 0,65 г (амплитуда изменчивости от 0,52 до 0,80 г) (Май-
тулина, 1988). Лабораторная всхожесть семян, заложенных на прора-
щивание через 1 месяц после сбора, в холодильнике при температуре
5°С составила 48%. Выбор такой температуры проращивания вызван
тем, что семена натурализовавшейся в Москве недотроги начинают
прорастать в середине апреля, когда среднесуточная температура воз-
духа в течение нескольких дней держится на отметке 5°С. В опы-
те семена начали прорастать уже на 10-й день, загнивших семян не
обнаружено. При весеннем посеве в конце апреля нестратифициро-
153
ванных семян к середине мая проросли лишь единичные экземпляры,
выросшие из семян, собранных в Таджикистане и Туркмении. Всхо-
жесть семян, высеянных осенью на грядки, составляет 100%.
Экология
Большинство местообитаний, где произрастает недотрога мел-
коцветковая, характеризуется той или иной степенью затенения. По
экологическим шкалам Элленберга (Ellenberg, 1991) вид располагает-
ся между теневыми и полутеневыми растениями (4-я ступень).
Зависимость роста особей недотроги мелкоцветковой от осве-
щенности, фотопериодизма, температурных условий, условий ми-
нерального питания неоднократно изучена по темпам накопления био-
массы (Evans, Hughes, 1961; Hughes, Evans, 1962; Hughes, 1966;
Rackham, 1966; Peace, Grubb, 1982). В целом, недотрога обнаружива-
ет чрезвычайно высокую пластичность по отношению к затенению.
Ухудшение роста наблюдается при освещенности ниже 10% от полно-
го света. При колебании освещенности между 20 и 80% от естествен-
ного полного летнего освещения ее рост меняется очень слабо. При
освещенности выше 80% рост слегка подавляется из-за продолжаю-
щегося (по нарастающему градиенту освещенности) снижения общей
поверхности листьев (Hughes, 1966)
. Средняя интенсивность транспирации составляет в тени— 1,56
г/дм2 в сутки, а на солнце — 4,38. При пересчете на сырой вес потеря
воды на солнце очень мала (2,58 г/г в сутки), по сравнению с другими
лесными малолетниками (Impatiens noli-tangere—4,73, Alliaria petiolata
9,35 г/г в сутки). Листья у I. parviflora отмирают, когда теряют более
44% содержащейся в них воды и/или 40% их сырого веса (Levitt, 1980).
Вид отличается меньшей гигрофильностью, по сравнению с двумя дру-
гими рассматриваемыми видами. Наблюдения над недотрогой мелко-
цветковой, проведенные в Главном ботаническом саду Москвы, по-
зволяют считать вид холодостойким: он не повреждается морозами и
заморозками (Карписонова, 1985).
Растет на самых разных, но достаточно увлажненных, почвах
при условии, что они хорошо аэрируются и не подтопляются. Пред-
почитает почвы, умеренно или очень богатые основаниями (но не обя-
зательно карбонатами), с pH корнеобитаемого слоя от 4,5 до 7,6.
По экологическим шкалам Элленберга недотрога мелкоцветковая
растет на свежих почвах (5-я ступень), с содержанием минерального
азота между средним и высоким (6-я ступень), индифферентна к ре-
154
акции почвы. Почвы местообитаний недотроги обычно богаты мяг-
ким гумусом. Однако, при достаточно высокой освещенности недот-
рога может длительно существовать и на очень бедной почве, занимая
только свободные от других растений участки (Hughes, 1966).
Недотрога мелкоцветковая обычно растет на нарушенных поч-
вах. Это преимущественно полуестественные лесные сообщества, пар-
ки или отмели рек с облесенными берегами. Например, она встреча-
ется в донских приречных ивняках (Тихомиров, 1987). На подзоли-
стых почвах произрастает исключительно по нарушениям (Coombe,
1956).
Фитоценология
Недотрога мелкоцветковая характерна для широколиственных
лесов Средней Азии (Туркестанской и Джунгаро-тяныпанской провин-
ции) (Карписонова, 1985). В этих лесах недотрога растет по речным
долинам, по выходам грунтовых вод, где развиваются наиболее бога-
тые леса из грецкого ореха с примесью ясеня согдийского и несколь-
ких видов тополей. В этих сомкнутых насаждениях подлесок развит
слабо, но разрастается густой многоярусный травостой, в состав кото-
рого входят влаголюбивые виды: Impatiens parviflora, Lamium album,
Ferula foliosa, Arum korolkowii и др.
В Центральной и Восточной Европе недотрога типична для ши-
роколиственных лесов порядка Fagetalia silvaticae Pawl, in Pawl, e.a
1928 в классе Querco-Fagetea (Ellenberg, 1991). В Чехии может быть
доминантом в травяном покрове ассоциации Querco-Populetum
Neuhauslova-Novotna 1965 (Slavic, 1995). В Западной Европе распро-
странена в лесном экотоне Querco petraeae — Carpinetum pannonicum
и Mercuriali-Tilietum, положительно реагируя на сокращение площа-
ди, занятой покровом из геофитов. В этих условиях (например, при
снижении покрытия аборигенных видов до 10% и ниже) недотрога
может стать доминантом и сохранять лидирующее положение, по край-
ней мере, в течение всего лета (Csontos, 1986).
Недотрога мелкоцветковая — диагностический вид союза
Chelidonio-Robinion в классе Robinietea Jurco em Hadan et Sofron 1980
(Moravec et al., 1995). Недотрога типична для нитрофильной расти-
тельности на нарушенных богатых почвах в кустарниковых зарослях,
в парках, на пустырях, произрастая вместе с Chelidonium majus, Роа
nemoralis, Solidago gigantea, Urtica dioica, Glechomahederacea, Geranium
robertianum, Scrophularia nodosa.
155
Недотрога мелкоцветковая может существенно потеснить не-
дотрогу обыкновенную в местах совместного произрастания: во влаж-
ных лесах, в лесопарках и пойменных фитоценозах около населенных
пунктов. Этот адвентивный натурализовавшийся вид оказался более
конкурентоспособен, чем недотрога обыкновенная, так как имеет бо-
лее широкую экологическую амплитуду (Weise, 1968; Csontos, 1986).
Это можно объяснить также морфологическими особенностями вида
и, в частности, архитектурой корневой системы в ее онтогенетической
динамике (Марков, 1994). Недотрога мелкоцветковая не так резко сни-
жает относительный вклад биомассы в корневую систему, начиная с
конца июня, чем недотрога обыкновенная.
Биологическая продуктивность
Исследования популяций недотроги мелкоцветковой в природе
(Татария) и в эксперименте, проведенные М.В. Марковым, показали,
что биомасса растения однозначно связана с его высотой. Показано,
что первоначальные размеры семядолей (семядольный индекс, опре-
деляемый как произведение длины наибольшей семядоли на ее шири-
ну) достоверно положительно коррелировали с массой семядолей, и
от обоих этих показателей сильно зависели все остальные абсолют-
ные морфобиологические показатели, включая массу органов репро-
дуктивной сферы. Величина семядольного индекса (как коррелят мас-
сы семени), таким образом, определяет конечную биомассу каждого
растения (табл. 5).
Таблица 5.
Взаимосвязь конечной биомассы особей Impatiens parviflora с величи-
ной семядольного индекса в разных вариантах плотности модельных
популяций
Плотность популяций (экз./сосуд) Классы семядольного индекса
1 5 10
1,0—1,5 245,3±75,Г 127,6±25,7”’
1,6—2,1 1078,0±212,2 412,3±140,7' 222,2±22,6‘"
2,2—2,7 563,8±131,8* 235,0±38,8"
Уровни значимости для средних:' р<0,05; ’* р<0,01; р<0,001
156
Ниже приведены количественные данные анализа двух популя-
ций недотроги: в Москве под зарослями клена ясенелистного на пус-
тыре между домами и в Подмосковье (Звенигородская биостанция
МГУ) под пологом ельника с примесью серой ольхи на склоне древ-
ней террасы р. Москвы (табл. 6).
Таблица 6.
Средний вес (г) одного растения I. parviflora и продуктивность
популяции (Р) в различных условиях произрастания
Фито- ценоз Вес Ли- стья Сте- бель Кор- ни Соц- ветия Все- го Р, г/м2
Заросли свежий 2,54 5,44 0,85 0,18 9,01 311,7
клена, % 28,2 60,4 9,4 2,0 100
Москва сухой 0,31 0,28 0,15 0,02 0,76 26,4
% 40,8 36,9 19,7 2,6 100
Ельник, свежий 0,63 0,95 0,21 0,04 1,83 189,7
Звени- % 34,4 51,9 11,5 2,2 100
город сухой 0,21 0,03 0,02 0,01 0,27 28,4
% 77,8 П,1 7,4 3,7 100
Примечания: выборка N = 100 для каждого участка; сухой вес = воз-
душно-сухой вес.
В различных экологических условиях продуктивность популя-
ций (воздушно-сухая масса на м2) оказалась весьма близкой, но алло-
кация фитомассы по органам растений резко отличается. В затенен-
ном ельнике 77,8% сухой биомассы сосредоточено в листьях, а в бо-
лее осветленных и сухих условиях города большая доля фитомассы
приходится на стебли и корни.
Консортивные связи
Исследования Р.А. Карписоновой (1985), проведенные в Глав-
ном ботаническом саду РАН Москвы показали, что недотрога мелко-
цветковая болезнями и насекомыми не повреждается. Известно, что
вид выступает в качестве одного из промежуточных хозяев паразити-
157
ческого ржавчинного гриба Puccinia komarovii, который образует эци-
дии исключительно на нижней части стебля, иногда на семядолях, но
никогда — на листьях. Уредоспоры же образуются на листьях. В слу-
чае произрастания недотроги на бедных почвах на ее корнях развива-
ет микоризу гриб типа Endogone (Peace, Grubb, 1982).
Хозяйственное значение
На родине в Средней Азии местные жители используют недот-
рогу в качестве красителя (Жизнь растений, 1981). Разводится как де-
коративное растение в садах и парках Европы, несмотря на невзрач-
ные цветки.
Недотрога железконосная
Номенклатура
Impatiens glandulifera Royle, 1834, Ill. Bot. Himal. Mount.: tab.
28, fig. 2; В. Тихомиров, 1966, Опред. раст. Моск, обл.: 229; D.M. Moore,
1968, Fl. Europ., 2: 240; Новиков, 1987, Опред. раст. Мещеры, 2: 10;
Черепанов, 1995, Сосуд, раст. России: 218; Мордок, 1996, Фл. Вост.
Европы, 9: 392. — I. roylei Walp. 1842, Repert. Bot. Syst., 1: 475;
Сырёйщиков, 1914, Илл. фл. Моск, губ.: 183; Назаров, 1940,
в Маевский, Фл., изд. 7: 505, in adnot.; Победимова, 1949, Фл. СССР,
14: 633; она же, 1964, в Маевский, Фл., изд. 9: 283, in adnot. —
Недотрога железконосная (семейство Balsaminaceae — Бальзамино-
вые).
Географическое распространение
Естественный ареал I. glandulifera находится в Гималаях и Ин-
дии — центре происхождения рода, где обитает еще 400 видов рода
Impatiens (Valentine, 1971). Вид встречается от Кашмира до Гархвала
в интервале высот 1800—3000 м, по берегам рек, вдоль дорог, на ме-
жах полей и в нарушенных влажных лесах. Отмечали его и на боль-
ших высотах вплоть до 4000 м (Polunin, Stainton, 1984).
Вторичный ареал вида очень широк и охватывает Северную
Америку и Евразию. В качестве декоративного растения недотрога
железконосная выращивалась в садах Европы с 1823 г. К 1923
году (через 100 лет после введения в культуру) она встречалась лишь в
158
пятидесяти пунктах Средней Европы, так как плохо закреплялась на
одном месте: семена недотроги не успевали полностью Bbi3peTb(Hegi,
1923). В начале 60-х годов началась бурная экспансия этого вида во
всех странах Средней Европы. Он широко распространен сейчас в 18
странах Европы между 30 и 64° северной широты: в Ирландии, Анг-
лии (кроме Шотландии), на юге и на восточном побережье Швеции,
юге Норвегии и Финляндии, в Дании, Германии. Продолжается инва-
зия вида в Чехии и Словакии (Pysek, Prach, 1995), а также в Швейца-
рии, где вид уже широко распространился в поясе до 800 м н.у.м. Как
обычное растение недотрога железконосная отмечена во Французских
Пиренеях, в Голландии, Германии, Австрии (особенно в Альпах), Поль-
ше, Венгрии, Югославии (Beerling, Perrins, 1993).
Недотрогу железконосную можно охарактеризовать как вид, хо-
рошо натурализовавшийся и способный к быстрой инвазии (Виногра-
дова, 1992). В России недотрога культивировалась с конца XIX века.
До 1953 г. были отмечены только 3 случая одичания недотроги (Побе-
димова, 1953). В конце 50-х годов семена недотроги как декоративно-
го растения пользовались в Москве большим спросом. Однако в евро-
пейской части России бурная натурализация недотроги началась толь-
ко в 60-х годах. В 1956 и 1967 гг. недотрога была найдена в Ле-
нинградской области (Гусев, 1973), в 1971 г. — в Московской области,
в 1975 г. — в Рязанской области (Тихомиров, 1975), в 1979 — во
многих местах Тверской области (Малышева, 1979), в 1980 г.
— в Липецкой области (Вьюкова, 1983), в 1987 — в поймах рек
Воронеж и Дон (Вьюкова, 1987), в 1983 г. — в Брянской области (Бо-
сек, 1986). Массовая натурализация наблюдается в Удмуртии и Ма-
рийской АССР. Единичные Находки отмечены в Сибири, на Кавказе
около г. Кисловодска в 1979 г, в окрестностях г. Орджоникидзе в 1984
г., в Алагирском ущелье в 1983 г. (Комжа, Попов, 1990). В окрестно-
стях г. Владивостока собрана В.Н. Ворошиловым (1982) как сорное в
1960 г., а в 1974 г. найдена около г. Южно-Сахалинска и в Корсаков-
ском районе (Воробьев, Небайкин, 1984, цит. по: Виноградова, 1992).
Современная северная граница распространения вида в России про-
ходит в Карелии, Мурманской, Архангельской областях и республике
Коми.
В Московской области в настоящее время встречается повсе-
местно в населенных пунктах, на засоренных местах, вдоль дорог, рек,
около ключей. На протяжении десятков километров отмечена на
берегах р. Москвы в Можайском, Рузском и Одинцовском районах
159
(Игнатов и др., 1990). Местами заходит и на ключевые болота, образу-
ет обширные заросли на заболоченных лугах у водоемов.
Морфологическое описание
Недотрога железконосная—однолетнее травянистое растение.
Корневая система — смешанная, состоит из системы главного корня и
более толстых розовых, прозрачных, развивающихся в основании ги-
покотиля придаточных корней. Корни проникают до глубины 10—15
см (Ennos et al., 1993). Гипокотиль достигает 8—15 см длины и 1,5—5
см в диаметре.
Стебель — прямостоячий, цилиндрический, простой или вет-
вистый, от 0,8 до 2,5 м высотой; зеленый, с красноватыми продольны-
ми полосками, слегка прозрачный, от 5 до 50 мм в диаметре, с сильно
утолщенными, почти вздутыми узлами (до 30—50 мм), полый внутри,
голый. Длина междоузлий в акропетальной последовательности умень-
шается с 50—55 см до 11—20 см. В пазухах нижних листьев форми-
руются побеги обогащения, часто вегетативные, до 10 см длиной, с
очередными листьями. Ветвление — мезотонное.
Листорасположение — мутовчатое, с 3 (редко 4) листьями на
узле, реже — супротивное. Листья — от яйцевидно-ланцетных до
узколанцетных, с заостренной верхушкой, клиновидным основанием,
с коротким (3—4,5 см) слегка крылатым черешком, основание
которого покрыто стебельчатыми железками. Пластинки листьев —
5—18 см длиной, 2,5—7,0 см шириной, по краю на каждой стороне
несут от 18 до 50 мелких острых зубцов. Устьица расположены на верх-
ней и на нижней поверхности листьев (Beerling, Perrins, 1993). Жил-
кование — перисто-петлевидное с поперечными анастомозами. Раз-
меры листьев возрастают к 5—7 узлу, затем постепенно уменьшают-
ся.
Из пазух верхних сближенных листьев выходят 5—15-цветко-
вые соцветия — зонтиковидные кисти на длинных цветоносах.
Цветки — крупные, длиной 2,5—4,0 см, лиловато-розовые, протеран-
дричные. Околоцветник — двойной. Чашечка состоит из 5 лепестко-
видных яркоокрашенных листочков (рис. 1, в). Морфологически верх-
ний чашелистик (расположенный после перекручивания в нижней
части цветка) — мешковидный, 12—20 мм длиной и 9—17 мм
шириной, резко суженный в коленчато согнутый, короткий нектаро-
носный шпорец 2—7 мм длиной. Два боковых чашелистика — отно-
сительно мелкие (10 мм длиной и 7 мм шириной), косо яйцевидные с
160
заостренной верхушкой, два нижних — очень мелкие (до 3 мм дли-
ной), яйцевидные (Troll, 1957). Венчик — крупный, состоит из 3 бо-
лее светлых розовато-лиловых лепестков. Морфологически нижний
лепесток — широкояйцевидный, отогнут к цветоножке, боковые —
неравно двулопастные, вдоль свернутые, так что меньшие лопасти, на-
легая друг на друга краями, образуют козырек над тычинками и зевом,
предохраняя их от дождевой воды. После цветения весь околоцветник
опадает. Клейстогамные цветки неизвестны.
Окраска цветков варьирует от лиловых до бледно-розовых и бе-
лых. На юге Польши в натурализовавшихся популяциях особи несут
пурпурные или темно-розовые цветки, тогда как в Голландии домини-
рует бледно-розовая окраска цветков. В Великобритании наблюдается
5 вариаций окраски цветков: пурпурные, розовые, лиловые, белые и
голубые. Однако природа генетической изменчивости окраски цвет-
ков до сих пор неясна (Valentine, 1971). В России растения с белыми и
бледно-розовыми цветками встречаются только в культуре. Натурали-
зуются растения лишь с темно-пурпурными, лиловыми и розовыми
цветками (Виноградова, 1992).
Андроцей — пятичленный. Тычинки с коническими пыльника-
ми и короткими нитями образуют колпачок, прикрывающий рыльце.
Тычинки вскрываются после раскрытия цветка. Пыльцевые зерна —
бледно-желтые, 4-бороздные, билатерально симметричные, эллипсои-
дальные. Гинецей — из 5 плодолистиков. Пестик — с коротким стол-
биком, цельным рыльцем и верхней одногнездной завязью. Плод —
удлиненная, продолговато-обратнояйцевидная коробочка, длиной до
2,5 см и диаметром 4 мм. Она эластично лопается и разбрасывает се-
мена. На колонке в коробочке закладывается 4—16 семяпочек.
Семена—обратно широкояйцевидные, сплюснутые, с попереч-
ной перетяжкой в основании, бежевые, с коричневыми пятнами и кра-
пинками. Они имеют светлые бугорчатые Валики, такие же, как у I.
parviflora, но выровненные коричневые участки между валиками у I.
glandulifera расположены более плотно. Длина семени—4—7 мм, ши-
рина 2—4 мм. Сухая масса одного семени — 2—35 мг, среднее из 100
семян — 21,8±1,2 мг (Beerling, Perrins, 1993).
В естественном ареале (Джамму и Кашмир) число хромосом
2п = 20 (Khoshoo, 1957). Во вторичном ареале недотрога железконос-
ная существует, по меньшей мере, в двух цитотипах — 2n = 18 и 20
(Хромосомные числа..., 1969). Первая форма встречается только во
вторичном ареале.
161
Онтогенез и сезонный ритм развития
Зародыш в семени — крупный, прямой, дифференцирован на
короткую осевую часть и крупные мясистые округлые семядоли с вы-
емкой на верхушке, в которых локализованы питательные вещества.
Семенам недотроги железконосной (как и другим видам рода) для про-
растания необходима холодная стратификация (Jouret, 1976). Недот-
рога отнесена к группе видов растений с необычной реакцией на су-
хое хранение семян: свежесобранным увлажненным семенам для про-
растания требуется как минимум 45-дневный период охлаждения до
+4°С или 30-дневный период охлаждения до +5°С (Grime et al., 1981).
Однако длительное сухое хранение семян при 20°С приводит к экспо-
ненциальному снижению времени, необходимому на стратификацию.
Например, семенам после 200 дней сухого хранения достаточно для
прорастания двухдневного охлаждения до 4°С. Набухшие семена при
20°С сохраняют жизнеспособность по крайней мере в течение 3 лет.
Постоянный банк семян в почве для недотроги железконосной не ха-
рактерен.
В природе семена прорастают в конце апреля. Прорастание —
гипокотилярное, первым появляется главный корень, на котором очень
быстро в области границы с гипокотилем закладываются и отрастают
многочисленные придаточные корни (рис. 2, в). Затем появляется ги-
покотиль, который в течение 7 дней удлиняется до 4 см и выносит
семядоли на поверхность.
Всходы несут светло-зеленые глянцевые мясистые семядоли, с
вдавленной средней жилкой на верхней стороне и слабо выступающи-
ми жилками снизу. Семядоли — широкояйцевидные, 7—12 мм дли-
ной, 10—15 мм шириной, с широкой выемкой на верхушке, где распо-
лагается крупная гидатода. В основании гипокотиля образуется до 10
слабо ветвящихся придаточных корней. Гипокотиль по мере разраста-
ния семядолей утолщается до 2,5 мм. Постепенно на конусе нараста-
ния закладываются листовые зачатки.
К концу первой декады мая на первом узле формируется 2—4-
членные мутовки листьев (рис. 8). Одновременно происходит удлине-
ние (до 20 см) эпикотиля. Растение быстро переходит в имматур-
ное состояние. Основание эпикотиля и междоузлий утолщается.
На последующих узлах располагаются, как правило, супротивные ли-
стья или их трехчленные мутовки. К концу мая у наиболее крупных
экземпляров побег насчитывает 5—6 метамеров, достигая 30—45 см
в длину. Семядоли увеличиваются до 20 мм в длину и 25 мм в шири-
162
Рис. 8. Возрастные состояния Impatiens glandulifera.
ну, листовые пластинки — эллиптические или ланцетные, до 70—140
мм в длину, 40—65 мм в ширину. Придаточные корни на гипокотиле
заметно утолщаются и ветвятся до корней III—IV порядков. В пазухах
листьев нередко формируются вегетативные побеги обогащения, а на-
чиная с 5—6-го узла — прямостоячие соцветия.
В фазу бутонизации растение вступает в конце июня — начале
июля. Первые цветки раскрываются в середине июля. Зрелые генера-
тивные особи достигают высоты 2,5 м, главный побег насчитывает
5—6 удлиненных метамеров и 3—4 укороченных верхних. В пределах
побега длина междоузлий уменьшается в акропетальной последова-
тельности с 50 см до 5 см. В пазухе верхних листьев располагается
основная масса соцветий. Соцветия — прямостоячие, возвышаются
над верхушкой побега на 7—10 см. Массовое цветение приходится на
август, плодоношение — на август—сентябрь. В Англии цветет с ию-
ля до октября (Grime et al., 1988). Жизненный цикл недотроги желез-
коносной, как правило, заканчивается с наступлением заморозков.
Фенотип вида меняется в зависимости от влажности местооби-
тания. Высокие, маловетвистые растения обитают в довольно влаж-
ных условиях, низкие растения с обильно ветвящимися стеблями, ко-
роткими междоузлиями и мелкими листьями характерны для нарушен-
ных лесных участков с непостоянным увлажнением.
163
Способы размножения и распространения
Цветки—протерандричны, поэтому опыление их облигатно пе-
рекрестное, хотя возможно и опыление между цветками с одного и
того же растения (Valentine, 1971; Виноградова, 1992). Нектар нахо-
дится в шпорце чашелистика. Секреция нектара начинается после от-
крытия цветка, прекращается ночью и возобновляется на следующий
день (Burquer, Corbet, 1991), максимальное содержание нектара при-
ходится на полдень. Вид—энтомофильный, опыляется шмелями. Цве-
ток — достаточно большой, чтобы шмель полностью залез в мешко-
видный чашелистик, касаясь при этом спинкой тычинок или рыльца
(Daumann, 1967; Valentine, 1978).
Плодовитость одной особи варьирует в зависимости от плотно-
сти популяции. Особи среднего размера (при плотности 20 особей на
м2) продуцировали в Великобритании 700—800 семян (образованных
140 плодами при 5,7 семян на плод) (Salisbuiy, 1961). Увеличение плот-
ности популяции до 36 особей на м2 влекло за собой снижение сред-
ней семенной продуктивности до 500 семян на особь (образованных
95 плодами при 5,3 семян на плод). D.J. Beerling (1990) приводит урав-
нение, связывающее плодовитость с плотностью популяции недотро-
ги:
SP = 1886-2PD(’°>367); г2= +0,963,
где SP — плодовитость растения, PD — число особей на м2.
В Германии у недотроги железконосной отмечена более высо-
кая плодовитость: при плотности 20 особей на м2 общая семенная про-
дуктивность одного растения превышала 1600 (учтены семена, содер-
жавшиеся в 250 плодах при 6,4 семян на плод), а в зарослях с Urtica
dioica семенная продуктивность особи повышалась до 2500 семян на
особь (учтены семена, содержавшиеся в 390 плодах при 6,4 семян на
плод) (Koenies, Glavac, 1979, цит. по: Beerling, Perrins, 1993). Масса
100 семян варьирует в популяциях от 1,05 г в Ленинграде до 1,59 г в
Вологде (Виноградова, 1992).
Распространение семян—гидро-, антропо-, и автохорное. Дис-
персия семян на короткие расстояния осуществляется автомехано-
хорно, причем семена могут разлетаться на расстояние до 5 м в
зависимости от высоты расположения плода над землей, силы и на-
правления ветра, преобладающего в период диссеминации. Максималь-
ное число семян падает на почву в радиусе 1—2 м от материнского
растения. D.J. Beerling и J.M. Perrins (1993) подсчитали, что скорость
164
распространения популяции в одном местообитании составляет 2,47 м в
год. Однако описаны отдельные случаи, когда семена оказывались в
результате диссеминации на расстоянии до 10 м от материнского рас-
тения. Исследователи пытаются объяснить эти факты участием в рас-
пространении семян лесной мыши Apodemus sylvaticus.
В переувлажненных местах набухшие семена недотроги желез-
коносной погружаются в воду и способны прорастать под водой
(Lhotska, Kopecky, 1966). Вместе с наносами сезонных паводков се-
мена могут переноситься вдоль рек на большие расстояния.
Синхронное прорастание семян весной приводит к формирова-
нию густых травостоев из одновозрастных особей. Гибель пророст-
ков зависит от их плотности (Beerling, Perrins, 1993). При начальных
плотностях 70,140,284 особи на м2, смертность составила, соответст-
венно, 28, 56 и 60%, и в итоге были сформированы локусы с плотно-
стями 50—70 взрослых особей на м2. В других условиях, при началь-
ной плотности 350 особей на м2, конечная плотность взрослых особей
составила 30—40 на м2. Смертность проростков может зависеть так-
же от относительного времени их появления.
Экология
По экологическим шкалам Элленберга (1991) недотрога желез-
коносная отнесена к полутеневыми растениями (4-я ступень). Несмотря
на относительную теневыносливость, рост ее ухудшается, когда мощ-
ность светового потока становится меньше 30% от полного дневного
света (у I. parviflora — при 10%) (Beerling, Perrins, 1993). Особи в за-
тененных местах временно могут демонстрировать более высокую,
чем на открытых местах, продуктивность как результат возрастания
листового индекса (LAR).
В качестве факторов, которые могут ограничивать инвазию не-
дотроги железконосной, могут выступать: требовательность к высо-
кой влажности почвы, ее чувствительность к низким температурам
(слабая морозоустойчивость). Недотрога чувствительна к засухе,
вызывающей завядание, так как для поддержания тургора в тканях этого
высокого сочного растения необходим постоянный приток воды: влаж-
ность листьев — 83—85%, стеблей — 92—94%, корней — 73—77%.
Однако, если засуха непродолжительна, то растения могут выживать,
вероятно, благодаря тому, что обычные местообитания этого
вида даже в очень сухие годы обеспечены влагой в достаточной степе-
ни.
165
Поздневесенние и раннеосенние заморозки могут вызывать ги-
бель, соответственно, проростков и взрослых особей недотроги. В Ги-
малаях коренная популяции толерантна, по-видимому, к морозам, т.к.
произрастает высоко в горах до 4000 м над уровнем моря. Высотная и
северная граница ее распространения в Великобритании соответству-
ет суммарной температуре вегетационного сезона в 2505—2198°С
(Beerling, Woodward, 1994). На основании теоретически высчитанной
величины суммарных температур, наблюдаемых в ныне освоенных не-
дотрогой районах, была составлена карта, которая прогнозирует даль-
нейшее распространение вида на север при условии глобального по-
тепления на 1,5° и 4,5°С.
Этот вид отмечен как на равнинных участках, так и на крутых
(>40°) склонах, при условии умеренного затенения. Способен произ-
растать на почвах самого разного гранулометрического состава и встре-
чается как на тонком, так и на грубом аллювии, морской гальке, хоро-
шо дренируемых минеральных почвах, отвалах каменного угля и тор-
фах (Beerling, Perrins, 1993). По экологическим шкалам Элленберга
(Ellenberg, 1991) недотрога железконосная растет на богатых мине-
ральным азотом почвах (7-я ступень), предпочитая почвы с нейтраль-
ной реакцией, но может расти на кислых и щелочных (7-я ступень).
Величины pH при этом варьируют от 4,5 до 7,3—7,7. Обитает на поч-
вах от влажных, не просыхающих, до сырых, часто плохо аэрируемых
(8-я ступень). Для успешного приживания проростков требуются уме-
ренная нарушенность растительного покрова (вывалы деревьев, упав-
шие ветки деревьев, зимнее подтопление берегов) и наличие голых
участков грунта.
В середине лета отдельные популяционные скопления (локу-
сы) недотроги железконосной имеют в центре мощные растения вы-
сотой свыше 2 м, в то время как на периферии таких скоплений преоб-
ладают очень мелкие растения (<10 см) с характерной антоциановой
окраской, свидетельствующей, вероятно, о недостатке влаги и/или эле-
ментов минерального питания.
Фитоценология
Недотрога железконосная образует обычно сплошные заросли
вдоль русел рек на речном гравии, аллювии и щебне. Видов, которые
произрастают вместе с недотрогой, даже при низком проективном по-
крытии, относительно немного. Может встречаться на низинных бо-
лотах. В Великобритании произрастает в сообществах Phragmites
166
australis — Urtica dioica, среди зарослей Phalaris arundinacea c Urtica
dioica, Convolvulus sepium, Petasites officinalis, Poa trivialis (Beerling,
Perrins, 1993). Impatiens glandulifera является диагностирующим ви-
дом многих ассоциаций союзов Phalaridion arundinaceae Кореску 1961,
Convolvulion sepii Тх. apud Oberd. 1949, Filipendulo-Petasition Br.-Bl.
1947, Salion albae Moor 1958 (Lhotska, Kopecky, 1966).
Вид может образовывать скопления на мезотрофных лугах с заи-
ленными грунтами. В Англии обычен для райграсовых лугов с Myrrhis
odorata (L.) Scop. (Beerling, Perrins, 1993). Может расти в высокотрав-
ных сообществах с Chamerion angustifolium, Epilobium hirsutum и
Petasites hybridus. Произрастает по опушкам, реже под пологом моло-
дых посадок деревьев и смешанных древостоев любых пород на ран-
них стадии сукцессии: Alnus—Fraxinus—Lysimachia vulgaris, Fraxinus
— Acer campestre — Mercurialis и Alnus glutinosa — Urtica dioica. В
Центральной Европе обычно встречается в сообществах порядка
Calystegialia в классе Artemisietea (Ellenberg, 1991).
Недотрога железконосная — самое высокое из однолетних рас-
тений. Она часто обгоняет по высоте Urtica dioica и, развивая ветвя-
щийся главный побег, достигает господства в наземных сообществах.
В Великобритании этот вид, проявляя «рудерально-доминантную»
стратегию (Grime, 1979), добивается перевеса в конкуренции на реч-
ных берегах благодаря синхронному прорастанию крупных семян из
временного почвенного банка и быстрому накоплению биомассы, дос-
таточной для подавления соседних видов. Проявлением лишь одной
регенеративной стратегии (синхронного прорастания семян) без спо-
собности к вегетативному возобновлению и без создания постоянного
почвенного банка можно объяснить приуроченность вида к местооби-
таниям с такими регулярными нарушениями, как весенние разливы,
отложение ила, размывание берегов. Несмотря на высокое содержа-
ние воды в тканях, растительные остатки недотроги железконосной
сохраняются неразложившимися до следующей весны (возможно, бла-
годаря высокому содержанию целлюлозы) и помогают подавлять кон-
курирующие проростки других видов.
Консортивные связи
Недотрога железконосная может иметь везикулярно-арбускуляр-
ную микоризу (Grime et al., 1988). Вид опыляется шмелями, в основ-
ном Bombus lucorus L. В Англии опылителями являются также Apis
mellifera L. и Vespa vulgaris L. (Beerling, Perrins, 1993). В Германии
167
подробно изучены не только насекомые опылители, но и фитофаги не-
дотроги (Schmitz, 1991, 1994).
На корнях недотроги могут обитать тли Aphis impatiphila
Pashtshenko и A. impatiradicis Pashtshenko, которые питаются соком
исключительно этого вида (Пащенко, 1993).
Биологическая продуктивность и хозяйственное значение
Максимальная биомасса (сухой вес) у недотроги железконос-
ной в Англии (южный Уэльс) накапливается в августе, при этом в за-
тененных местах она составляет 80 г/м2, а на свету—40 г/м2 (Beerling,
Perrins, 1993). Максимальная общая продуктивность в августе в цен-
тральной Англии составляет 11,5 кг/га (с корнями) и 10,5 кг/га (над-
земная биомасса).
Недотрога железконосная — повсеместно культивируется как
декоративное растение и пользуется большой популярностью у цвето-
водов.
168
Н.П. Савиных
ВЕРОНИКА ЛЕКАРСТВЕННАЯ
Номенклатура
Veronica officinalis L. 1753, Sp. Pl.: 11; Кауфман, 1866, Моск.
фл.,изд. 1: 349; Цингер, 1885, Сборник свед.: 327; Шмальгаузен, 1897,
Фл. Средн, и Южн. Росс., 2: 274; Сырейщиков, 1910, Илл. фл. Моск,
губ., 3: 146; он же, 1927, Опред. раст. Моск, губ.: 232; Иконников-Га-
лицкий, 1940, в Маевский, Фл., изд. 7: 647; Борисова, 1955, Фл. СССР,
22: 449; она же, 1964, в Маев.ский, Фл., изд. 9: 446; Ворошилов, 1966,
Опред. раст. Моск, обл.: 291; Walters a. Webb, 1972, FL Europ., 3: 247;
Еленевский, 1981, Фл. европ. части СССР, 5:250; Киселева, 1987, Оп-
ред. раст. Мещеры, 2: 72; Черепанов, 1995, Сосуд, раст. России: 936 —
Вероника лекарственная (семейство Scrophulariaceae — Норични-
ковые).
Географическое распространение
Ареал вида охватывает все страны Европы, а также ряд стран
Азии (Турцию, Сирию, Ливан, Иран), западные и восточные штаты
США, Канаду. Северная граница в России проходит по верхней Мезе-
ни, верхней Печоре, среднему течению Камы, Уральскому хребту в
пределах Свердловской области. На юге распространена до Лисок, Там-
бова, Чистополя, Миасса; встречается в горном Крыму, в большинст-
ве горных районов Кавказа; отсутствует в степной части Украины (Фло-
ра СССР, 1955; Флора Кавказа, 1967; Вульф, 1969; Еленевский, 1978;
Флора европейской части СССР, 1981). В Московской области встре-
чается повсеместно (Ворошилов и др., 1966).
Морфологическое описание
Вероника лекарственная—вечнозеленый вегетативно подвиж-
ный ползучий лесной хамефит с открытыми почками. Отдельные
особи, особенно генеративные, имеют разное строение. Одни пред-
ставлены системами из разного числа побегов IV—V порядков ветв-
ления: первичными или парциальными кустами, куртинами; другие
169
— отдельными или слабо разветвленными укореняющимися побега-
ми (Савиных, 1978, 1979а).
Главный корень отмирает у раннегенеративных особей, поэто-
му в зрелом генеративном возрастном состоянии корневая система
представлена системой стеблеродных придаточных корней. Корни —
толстые, 0,5—0,9 мм диаметром у основания. У раннегенеративных
особей они отходят от узлов на второй год жизни побега, у более взрос-
лых особей—в год развития годичного побега. От каждого узла отхо-
дят по 2—4 корня. Они имеют светло-коричневую окраску, по всей
длине ветвятся до II—III и изредка до IV порядка. Длина корней—до
30—40 см. Основная масса корней располагается в подстилке
и приповерхностном слое почвы, лишь отдельные корни углубляются
до 20 см (Рысин, Рысина, 1987). Обычная длина корней II порядка
составляет 5—8 см при диаметре 0,2—0,3 мм, корней III порядка —
1—2 см и 0,2—0,3 мм, корней IV порядка — до 0,5 мм и 0,1 мм соот-
ветственно. Длина корневых волосков — 0,1—0,8 мм, толщина—5—
13 мкм.
Побеги — многолетние олигокарпические. Верхушечная почка
побегов состоит из двух—трех пар зачатков листьев и конуса нараста-
ния (см. рис.). Листья — супротивные, без прилистников, короткоче-
решковые, с овальной, по краю зубчатой листовой пластинкой. Соцве-
тие — вставочное, интеркалярное (по терминологии А. Имса, 1964):
двойная фрондозная открытая кисть, расположенная между двумя ве-
гетативными участками годичного побега. Парциальные соцветия —
пазушные брактеозные открытые кисти. Число кистей 1—5, в зависи-
мости от фазы онтогенеза.
Цветки — некрупные, 6—7 мм в диаметре. Чашечка—желези-
сто опушенная, четырехраздельная, с ланцетными долями. Вен-
чик — лиловый, спайнолепестный, состоит из трех широкояй-
цевидных туповатых долей и одной продолговатой, которая
вдвое уже остальных. Тычинок — две, длинных, превышающих
венчик. Гинецей — синкарпный, из двух плодолистиков. Плод —
сплюснутая обратно треугольная многосемянная коробочка около 4—
5 мм, ее длина равна ширине; основание — суженное, на верхушке
заметная выемка. Семена—плосковыпуклые, шириной в 1 мм. Число
хромосом, по данным разных авторов, различно: 2п = 18 (Hitchcock et
al., 1959; Walters, Webb, 1972), 2n = 36 (Афанасьева, Мешкова, 1961),
2n = 36 (Hitchcock et al, 1959; Fisher, 1969; Мешкова, 1965, цит. по:
Еленевский, 1978).
170
Онтогенез
Большой жизненный цикл вероники лекарственной относится
к полночленному типу с длительным генеративным периодом без яр-
ко выраженного сенильного состояния (Савиных, 1978). Прорастанию
семян способствует свет (Kinzel, 1913). Проростки появляются глав-
ным образом весной, имеют две овальные толстоватые семядоли с за-
метной средней жилкой, на верхушке — усеченные. Семядоли выно-
сятся на поверхность почвы изогнутым гипокотилем. На первых эта-
пах развития проростка почечка скрыта черешками семядолей. Пер-
вая пара настоящих листьев имеет трехзубчатые пластинки, покрытые
редкими белыми волосками. Семядоли желтеют в августе—сентябре
и в таком виде сохраняются до весны следующего года. Обычно с по-
желтением семядолей в конце вегетационного периода ортотропный
побег полегает. К этому времени он состоит из 3—5 метамеров и вер-
хушечной почки. В пазухах каждого листа находятся почки, состоя-
щие из двух пар зачатков листьев и конуса нарастания. Это — юве-
нильные растения. Побеги из пазушных почек таких растений разви-
ваются, как правило, на второй год жизни. Поэтому трехлетние расте-
ния представлены в надземной части уже разветвленным побегом. Эта
фаза онтоморфогенеза соответствует имматурному и взрослому веге-
тативному возрастным состояниям. Иногда в таком виде растение мо-
жет впервые зацвести (см. рис.).
В результате моноподиального нарастания и ветвления всех
побегов 4—5-летние особи приобретают вид куртин. Они состоят из
нескольких разветвленных цветущих побегов (подобных изображен-
ному на рис.) и относятся к раннегенеративным. Главный критерий
для выделения этого возрастного состояния — морфофизиологиче-
ская целостность организма и наличие главного корня. У 5—6-лет-
них особей начинается дезинтеграция: отмирают главный корень
и первые междоузлия боковых побегов в месте сочленения их
с главным побегом. Это — начало образования клона и перехода рас-
тения в зрелое генеративное состояние. Главный критерий отнесения
к этому возрастному состоянию — строение и развитие годичных
побегов. Они состоят из 4—5 вегетативных метамеров, 1—3 метаме-
ров, в пазухах листьев которых располагаются парциальные соцветия,
и еще 7—8 вегетативных метамеров. Такие годичные побеги развива-
ются в течение 2—3 лет. В целом развитие побега вероники лекарст-
венной идет по следующей схеме. Сначала развивается вегетативный
годичный побег, затем 2—3 вегетативно-генеративных, позднее —
171
A
Veronica officinalis в раннегенеративном состоянии (А) и верхушка ее побега (Б) в
августе. Обозначения на рис. А: 1 — зеленые листья, 2 — отмершие листья, 3 —
соцветие, 4 — гипокотиль, 5 — система главного корня, 6 — придаточные корни, 7
— граница годичных побегов; на рис. Б: 1 — продольный разрез, 2, 3 — разверну-
тые зеленые листья, 4 — сомкнутые зеленые листья, 5, 6 — зачатки листьев в почке,
6 — с конусом нарастания.
снова вегетативный. Затем происходит «перевершинивание»: верху-
шечная почка или вся верхушка побега из нескольких последних ме-
тамеров отмирает, и дальнейшее нарастание системы побегов и веге-
тативное расселение растения осуществляется за счет нескольких
(обычно 1—2) боковых силлептических побегов.
В этом возрастном состоянии растения различаются по интен-
сивности ветвления побегов, размерам особи, числу и размерам
отмерших участков стебля. Поэтому все морфологическое разнооб-
разие зрелых генеративных особей можно свети к следующим основ-
ным типам. 1) Разв етвленный цветущий побег; исходный побег нарас-
тает моноподиально, боковые побеги — цветущие, не ветвятся.
2) Куртина вегетативного происхождения из 2—3 разветвленных цве-
тущих побегов. Исходный побег у одних особей моноподиально на-
растает и в составе куртины имеет свою систему, у других — отмира-
ет, а небольшой участок его служит в качестве связующего зве-
172
на у нескольких разветвленных цветущих побегов. 3) Одноосный цве-
тущий побег. 4) Система из одноосных цветущих побегов, связанных
между собой остатками исходного побега из нескольких метамеров.
Некоторые особи вероники в составе клона могут временно не
цвести, на что указывал в своих работах Т.А. Работнов (1945). По на-
шим наблюдениям, они имеют разнообразное строение. Обычно в над-
земной части это — разветвленные или одноосные побеги. Процесс
отмирания захватывает у них только базальную часть исходного побе-
га. Условный возраст таких особей не превышает 3—4 лет (Кренке,
1940). В дальнейшем эти особи развиваются двумя путями.
У одних растений моноподиальное нарастание сменяется акро-
симподиальным, боковые побеги зацветают, и вегетативное растение
превращается в систему цветущих боковых укореняющихся побегов.
Перерыв в цветении таких особей, по-видимому, связан с тем, что ис-
ходный побег уже завершает цикл своего развития, определяемый дли-
тельностью моноподиального нарастания, и поэтому не цветет, а бо-
ковые побеги еще не перешли в фазу формирования вегетативно-ге-
неративного годичного побега.
В другом случае циклы развития исходного и боковых побегов
совпадают, все они нарастают моноподиально, у всех на следующий
год образуются вегетативно-генеративные годичные побеги, и особи
приобретают вид разветвленных цветущих побегов. Временно нецве-
тущие особи вероники встречаются, как правило, одиночно или
группами из нескольких растений. Перерыв в цветении можно объяс-
нить цикличностью развития вторичных особей (Жукова, 1976) в
составе клона. Образовавшиеся после дезинтеграции дочерние особи
могут не цвести в течение 1—2-х лет, в дальнейшем они зацветают
вновь.
Одноосные особи в составе клона также развиваются не одина-
ково. Одни из них не ветвятся, нарастают моноподиально в течение
нескольких лет, затем цветут. Вегетативно-генеративные побеги у них
соответствуют таковым у зрелых генеративных особей. Такие расте-
ния также могут временно не цвести.
У других особей соцветия образуются поздно летом, семена,
как правило, не вызревают, второй вегетативный участок в строении
годичного побега состоит из 1—2 метамеров, верхушечная почка со-
держит только одну пару зачатков листьев и конус нарастания. Такие
особи относятся к позднегенеративным. Они могут быть не только од-
ноосными, но и состоять из нескольких выше описанных побегов.
173
Сенильные особи имеют вегетативное происхождение: они пред-
ставляют собой «отпад», образующийся при дезинтеграции путем от-
членения верхушек побегов или небольших боковых побегов, образо-
вавшихся из спящих почек зрелых и позднегенеративных растений.
Обычно особи этого возрастного состояния состоят из 1—2-х годич-
ных вегетативных побегов, они — одноосные, без живых пазушных
почек, со слабо развитой корневой системой; верхушечная почка у них,
как и у позднегенеративных особей, содержит лишь одну пару зачат-
ков листьев и конус нарастания. Для отдельных особей в этом возрас-
тном состоянии свойственно частое «перевершинивание». Сенильные
растения могут быть не только с плагиотропными, но и с ортотропны-
ми побегами. В последнем случае они представлены боковым по-
бегом из нескольких метамеров, отходящим от исходного побега, от-
мершего с обоих концов. Таким образом, особи вероники лекарствен-
ной в ходе онтогенеза претерпевают сложные изменения; большая
часть онтогенеза приходится на фазу клона; естественного старения
клона наблюдать не удается вследствие вегетативной подвижности
особей.
Сезонное развитие
Вегетация вероники лекарственной в условиях средней полосы
России начинается в конце апреля — начале мая. Заложение новых
метамеров продолжается в течение всего вегетационного периода. В
пазухе каждого листа содержатся почки, состоящие из двух пар зачат-
ков листьев и конуса нарастания. Цветет вероника с июня по сентябрь.
Массовое цветение — в июне—июле. Семена созревают в июле—ав-
густе и тут же высыпаются из коробочек.
Длина вегетативно-генеративного годичного побегав зрелом ге-
неративном возрастном состоянии достигает 25 см. Образование ме-
тамеров в конце вегетативного периода не сопровождается удлинени-
ем междоузлий. В результате формируется открытая верхушечная поч-
ка: края третьей пары листьев, считая от конуса нарастания, плотно
смыкаются, и почка оказывается между черешками двух пар ранее
сформированных листьев (см. рис., б). Она не имеет специализиро-
ванных почечных чешуй, но оказывается достаточно защищенной от
неблагоприятных воздействий среды в осенне-зимний период. Под снег
растение уходит с зелеными листьями. Отмирают они только летом
следующего года.
174
Способы размножения и расселения
Расселение вероники лекарственной осуществляется с помощью
семян, которые, как правило, прорастают вблизи материнского
растения. Энергия семенного размножения зависит от возрастного
состояния особи, условий произрастания, числа семян в коробоч-
ке, ее положения на оси. Для вероники лекарственной характер-
на энтомофилия: в трубке венчика есть нектар в виде крошеч-
ной капли (Верещагина, 1970). Цветки раскрываются в широком
диапазоне температур (от 17,2° до 27,3°) и относительной влаж-
ности (от 59 до 96%) в наиболее светлое время суток: с 7 до 14 ча-
сов.
Опыление осуществляется двукрылыми, пчелами, шмелями.
Продолжительность жизни цветка — 10—12 часов. Если перекрест-
ное опыление не произошло, то при начинающемся увядании проис-
ходит самоопыление. Венчики осыпаются, соскальзывая по пестику с
еще способным к опылению рыльцем. При этом они поворачиваются
на 180°, т.к. верхняя часть венчика с широким лепестком более тяже-
лая. В результате тычинки касаются пыльниками неподвижного рыль-
ца. Если пыльники не заденут рыльца, то на него попадет пыльца, вы-
сыпавшаяся на венчик и столбик и удержанная волосками у входа в
узкую трубку венчика.
Автогамия возможна и при распускании цветка: липкое рыльце
удерживает еще невскрывшийся пыльник, который впоследствии вы-
сыпает на него пыльцу, но такое встречается редко. Эффективность
опыления у вероники — 91,5%. Насекомые в типичных местах обита-
ния встречаются не часто. Поэтому можно считать, что опыление на-
секомыми у вероники лекарственной встречается редко, скорее это ти-
пичное автогамное растение.
Более активное расселение происходит за счет ежегодного
нарастания (до 25 см) плагиотропных побегов, которые достигают
в длину 80—90 см. Нередко отдельные особи приблизительно че-
рез 7 лет почти совсем «уходят» с мест, занятых ими первоначаль-
но (Барсукова, Пятковская, 1977). Вегетативное размножение про-
исходит в виде полной ранней неспециализированной морфологи-
ческой дезинтеграции (по терминологии Работнова, 1969; Ценопо-
пуляции растений, 1976). Это обусловлено спецификой развития
клона.
175
Экология и фитоценология
Вероника лекарственная — полутеневое растение (5-я ступень
шкалы Элленберга, 3-я ступень шкалы Ландольта). Хотя вероника —
растение полутеневое, она чаще растет в относительно осветленных
местах: вдоль дорог, в лесных «окнах» на месте срубленных или упав-
ших деревьев, в старых изреженных лесах. Может выносить и полное
освещение, поэтому часто встречается на открытых местах: на выруб-
ках, гарях, лесных опушках, лугах на месте сведенных лесов. Свет
влияет на степень ветвления и развитие особей, особенно на началь-
ных этапах онтогенеза. На открытых местах и при умеренном увлаж-
нении куртины — компактны, состоят из укореняющихся цветущих
побегов I—III порядков ветвления, площадь, занимаемая ими, не пре-
вышает 0,5 м2. Побеги направлены в разные стороны от главного кор-
ня.
В лесу, даже на более освещенных местах (вдоль дорог, при от-
сутствии хорошо выраженного подлеска), куртины образуются редко.
Здесь они отличаются меньшей компактностью, имеют преимущест-
венно однонаправленный рост. Длина куртины достигает 1 м, а пло-
щадь, занимаемая ею, — 1,5 м2. Соцветия образуются только на побе-
гах I и II порядка. В условиях сильного затенения, густого напочвен-
ного покрова побеги II порядка отмирают или совсем не образуются.
Тогда раннегенеративное растение представлено в надземной части
лишь цветущим главным побегом. Освещенность и степень увлажне-
ния влияют на длину годичного побега: на освещенных и относитель-
но сухих местах побеги могут быть от 3 до 11 см; на влажных затенен-
ных — до 25—27 см (Андриенко, Прядко, 1975).
Вероника лекарственная обитает на почвах сухих, избегая очень
сухих и очень влажных (2-я ступень шкалы Ландольта, 3-я ступень
шкалы Элленберга), кислых (2-я ступень шкалы Ландольта: срН=3,5—
5,6); лучше развивается на почвах, богатых гумусом (4-я ступень шка-
лы Ландольта). В Ленинградской области растет преимущественно на
дренированных супесях и песках, суглинках и двучленных наносах (Фе-
дорчук, Бурневский, 1986).
Вероника лекарственная обитает в хвойных, смешанных и ши-
роколиственных лесах. Диагностический вид кисличной группы
типов леса в Ленинградской области, в которую входят также
Melica nutans, Carex digitata, Pyrola rotundifolia, Viola riviniana и Aego-
podium podagraria. В.Н. Федорчук и Ю.И. Бурневский (1986) не об-
176
наружили случаев доминирования вероники лекарственной в травя-
ном покрове изученных лесов. В зависимости от условий обитания
вид выполняет разные роли в сообществе. При достаточном освеще-
нии, в сосняках, ее относят к высоко константным видам (Леса Вос-
точного Подмосковья, 1979). На 10 см2 насчитывается до 4—11 побе-
гов; растение обильно цветет и плодоносит (Рысин, 1969). Вероника
прекрасно растет в нарушенных местообитаниях: на вырубках, гарях,
а также в возникших на их месте насаждениях (Kujala, 1926). В этих
условиях вероника лекарственная — типичный эксплерент. При мень-
шем освещении, в сосняках с липой, она встречается с небольшим
обилием наряду с неморальным широкотравьем (Рысин, 1975). В ель-
никах (Речан и др., 1993) вид отмечается как редкое растение с не-
большой интенсивностью ветвления и возможными перерывами в цве-
тении. Поэтому в этих условиях вероника лекарственная имеет стра-
тегию жизни патиента. Обычно имеет небольшое обилие (проектив-
ное покрытие— 1—5%), растет в основном пятнами (Барсукова, Пят-
ковская, 1977).
В широколиственных лесах Южного Урала встречается в дуб-
няке наперстянково-коротконожково-тростниковидновейниковом (Гор-
чаковский, 1972). На Украине (Киевская и Житомирская области) — в
сосновых, сосново-дубовых, дубовых, березово-сосновых лесах, но
отсутствует в грабовых лесах, а также в лесах с хорошо развитым яру-
сом подлеска, густых молодых посадках (Андриенко, Прядко, 1975).
В Молдавии распространена на полянах и в лесах: дубовых, липово-
ясеневых, дубовых с липой, ясенем, грабом, буком и черешней (Коре-
нева, 1989).
Биологическая продуктивность
Количество семян и плодов варьирует у разных особей. На од-
ном растении может быть от 17 до 45 коробочек. Число семян в плоде
зависит от расположения плода на оси: в нижней части — 12—20, в
верхней — 8—14. Средний вес 1000 семян составляет 0,0932 г (Гусев,
1976). Средняя семенная продуктивность на один побег — 150—230
семян (Луговые травянистые растения, 1990). Биологическая продук-
тивность зависит от условий произрастания. В сосново-дубовых ле-
сах сухая надземная масса составляет 4,0±0,7 г/м2; в сосновых лесах
— 5,3±0,8 г/м2; в дубовых 2,5±0,4 г/м2; на открытых местах — 2,3±0,5
г/м2 (Андриенко, Прядко, 1975).
177
Хозяйственное значение
Вероника лекарственная не имеет кормового значения, скотом
не поедается. Видовое название растения свидетельствует о давнем
использовании его для лечения ряда заболеваний: легочных и простуд-
ных, истощения, язвы желудка, заболеваний мочевого пузыря. Исполь-
зуется и как желчегонное средство, наружно — для лечения ран; вхо-
дит в состав грудного чая (Лесные травянистые растения, 1988; Луго-
вые травянистые растения, 1990). Вегетативные побеги вероники со-
держат 12 веществ флавоновой природы, 8 фенолкарбоновых кислот
(в том числе — кофейную, хлорогеновую, неохлорогеновую, хинную,
феруловую), а также — холин, витамины, сапонины, эфирное масло
(Гусев, 1976). Флавоновые вещества локализуются преимущественно
в эпидермисе и палисадной ткани, иридоиды — в палисадной ткани
листа. Свободные полифенолы активны в отношении золотистого ста-
филококка. Высказано предположение о целесообразности изучения
этого растения для введения его в медицинскую практику. Для исполь-
зования вероники в качестве лекарственного сырья следует собирать
надземную часть в период цветения и плодоношения, оставляя не ме-
нее пятой части растения для сохранения и воспроизведения в буду-
щем.
178
В.IL Викторов
КОЛОКОЛЬЧИК ШИРОКОЛИСТНЫЙ
Номенклатура и внутривидовая систематика
Campanula latifolia L. 1753, Sp. Pl.: 165; Кауфман, 1866, Моск,
фл., изд. 1: 307; Цингер, 1885, Сборник свед.: 283; Шмальгаузен, Фл.
Средн, и Южн. Росс. 2: 178; Сырейщиков, 1910, Илл. фл. Моск. губ. 3:
224: он же, 1927, Опред. раст. Моск, губ.: 247; Б. Федченко, 1940, в
Маевский, Фл., изд. 7: 689; Ан. Федоров, 1957, Фл. СССР, 24: 191; он
же, 1964, в Маевский, Фл., изд. 9: 530; id. 1976, Fl. Europ. 4: 88; он же,
1978, Фл. европ. части СССР, 3: 222; Ворошилов, 1966, Опред. раст.
Моск, обл.: 311; Октябрева, 1987, Опред. раст. Мещеры, 2:90; Черепа-
нов, 1995, Сосуд, раст. России: 296 — Колокольчик
широколистный (семейство Campanulaceae — Колокольчиковые).
На обширной площади своего ареала колокольчик широколист-
ный характеризуется высоким уровнем внутривидовой изменчивости.
Еще в 1831 г. С.А. Мейер описал var. macrantha Меу. Несколько позд-
нее, в 1879 г., Э.Р. Траутфеттер описал еще 3 формы: var. leiocarpa
Trautv., var. intermedia Trautv., var. canescens Trautv. На основе крити-
ческого анализа представителей семейства Campanulaceae на Кавказе
М.Э. Оганесян (1996) в пределах данного вида выделяет два подвида:
Campanula latifolia subsp latifolia и C. latifolia subsp. megrelica (Manden.
& Kuthath.) Oganesian.
Географическое распространение
Евро-западноазиатский, равнинно-субальпийский вид, ареал его
охватывает практически всю Европу, кроме Крайнего Севера, а также
Западную Азию вплоть до Гималаев (Флора СССР, 1957; Флора
европ. части СССР, 1978; Flora Europaea, 1976). На территории
России колокольчик широколистный встречается во всех районах ев-
ропейской части России, за исключением Арктики (Флора европ. час-
ти СССР, 1978), на Кавказе (Гроссгейм, 1949; Косенко, 1970), в Запад-
ной Сибири (зап. часть Верхнетобольского района, Алтай), на Сред-
нем Урале и в районах южного Предуралья; через горы в Сибирь не
179
переходит (Говорухин, 1937). В Московской области произрастает во
всех ботанико-географических районах (Ворошилов и др., 1966).
Морфологическое описание
С. latifolia — многолетнее, травянистое, поликарпическое, ко-
роткокорневищное растение с кистекорневой системой и симподиально
возобновляющимися, ортотропными, монокарпическими, моноцикличе-
скими, полурозеточными побегами (Викторов, 19916, 1993).
В тенистых лесах длина системы корневищ не превышает 6 см
и ежегодно на ней образуется только один генеративный побег (очень
редко — два). Возраст отдельных систем корневищ, как показывает
подсчет пеньков от отмерших генеративных побегов, достигает 10 лет,
при условии, что ежегодно формируется только один побег (рис. 1, а).
На опушках и вырубках общая длина системы корневищ увеличивает-
ся до 10—15 см (рис 1, б). От корневищ, как правило, ежегодно отхо-
дит несколько (2—3, до 5) надземных побегов (Викторов, 1993). У
взрослых особей от корневища в стороны и вниз отходят 20—30 до-
вольно длинных (до 20—25 см) светло-серых придаточных корней,
несколько утолщенных в основании и ниже ветвящихся. Длина круп-
ных корней II порядка—до 20 см (диаметр 1—2 мм), длина корней III
порядка — до 5 см (0,3 мм), IV порядка — до 1 см (0,2 мм), V — до 5
мм (0,1 мм). Ветвление усиливается ближе к окончаниям крупных кор-
ней, которые на некотором удалении веретеновидно утолщаются до
7—8 мм в диаметре, выполняя запасающую функцию. Глубина корне-
вой системы не превышает 15—20 см, ширина простирания — 50—
60 см (Рысин, Рысина, 1987).
Надземные побеги — ортотропные, довольно крупные, 115—
150 см высотой. Биометрические их показатели значительно варьиру-
ют в локальных популяциях Московской области (Тихонова и др.,
1989). Стебли — зеленые, гладкие, довольно толстые (до 1,5 см в диа-
метре), на поперечном срезе округлые, с неясно выраженными ребра-
ми. Ветвление наблюдается только в области соцветия в хорошо осве-
щенных местообитаниях. Листорасположение — очередное.
Листья — простые, цельные, без прилистников, опушены мяг-
кими одноклеточными волосками; на поверхности пластинки волос-
ки сосредоточены, в основном, по жилкам. Листовые пластинки—яй-
цевидно-продолговатые с округлыми основаниями, двоякопильчатым
краем и острой верхушкой. Размеры листовых пластинок и длина че-
180
Рис. 1. Подземные органы
колокольчика широколистного.
решка постепенно уменьшаются к
верхушке побега: нижние листья —
длинночерешковые (до 18—20 см),
верхние — сидячие.
Соцветия — фрондулезные,
кистевидные, закрытые, монотеличе-
ские (Troll, 1964). Степень ветвления
зависит от условий местообитания. У
растений, произрастающих в лесных
фитоценозах, формируются простые
соцветия; на опушках и вырубках —
сложные. Большое значение при ха-
рактеристике соцветий колокольчиков
имеет анализ порядка раскрывания
цветков (Викторов, Соколова, 1994).
Так, в простых соцветиях С. latifolia
первым распускается самый нижний
цветок или цветок, расположенный в нижней трети соцветия. Если пер-
вым зацветает самый нижний цветок, дальнейшее распускание идет
акропетально. Если первым зацветает цветок в нижней трети соцве-
тия — дальнейшее распускание идет в двух направлениях. Верхушеч-
ный цветок всегда распускается несколько раньше 2—3(4)-х ниже рас-
положенных.
В сложных соцветиях первым распускается верхушечный цве-
ток самого нижнего паракладия, или расположенного в нижней трети
объединенного соцветия. Если первым зацветает верхушечный цве-
ток самого нижнего паракладия, то вторым распускается верхушеч-
ный цветок выше расположенного паракладия и далее распускание
идет акропетально. Если первым зацветает верхушечный цветок па-
ракладия, расположенного в нижней трети синфлоресценции, то в даль-
нейшем распускание идет в двух направлениях. Практически одно-
временно распускаются верхушечные цветки паракладиев, располо-
женных выше и ниже по оси объединенного соцветия. Параллельно с
распусканием верхушечных цветков паракладиев наблюдается распус-
кание базальных цветков паракладиев, расположенных ниже и у кото-
рых верхушечный цветок к этому времени уже заканчивает цветение.
Верхушечный цветок в сложных соцветиях, как и в простых, несколь-
ко опережает в цветении 2—3(4)-й ниже расположенный одиночный
цветок.
181
Цветок—правильный, обоеполый, с двойным околоцветником,
на цветоножке. Зубцы совершенно голой чашечки — длинно заост-
ренные, к основанию расширенные, по краю — пильчатые, короче вен-
чика. Венчик — крупный (5,0—6,0 см), колокольчатый или воронко-
видный, синего, почти фиолетового, голубого или даже почти белого
цвета с волосками внутри трубки и обычно с оттянутыми зауженными
зубцами.
Плод — коробочка, поникающая, овальная, с тремя порами у
основания. Семена колокольчика широколистного — светло коричне-
вые, яйцевидные, несколько уплощенные, 1,6—2,0 мм длиной и 0,6—
1,0 мм шириной. Масса 100 семян варьирует от 25 до 42 мг. В эндос-
перме заключен прямой бесцветный двусемядольный зародыш длин-
ной около 1 мм (Рысина, 1973).
На территории Подмосковья нами, совместно с сотрудниками
ВНИИ природы, изучена изменчивость некоторых морфологических
показателей генеративного побега (Тихонова и др., 1989). Статисти-
ческий анализ вариабельности 13-ти параметров в 7 локальных попу-
ляциях позволил сделать вывод о высоком уровне гетероморфности в
пределах географической популяции (Викторов, 1996). Выявленные
различия между популяциями наблюдаются, по видимому, в результа-
те существенных изменений в соотношении аллелей. Проведенный
сравнительный анализ биометрических измерений растений в трех фи-
тоценозах в пределах одной популяции показал, что условия освещен-
ности существенно влияют на морфологию генеративного побега (табл.
1). Наибольшие значения показателей отмечены у растений, обитаю-
щих на опушке леса с боковым затенением. При уменьшении осве-
щенности закономерно уменьшаются все показатели генеративного по-
бега.
Диплоидный набор хромосом 2п = 34 (Sugiura, 1938; Gadella,
1962); 2n = 34+3—5В (Gadella, 1964).
Онтогенез
Г.П. Рысина (1973) отмечает, что всходов, как от свежесобран-
ных семян, так и от хранившихся в течение 6 лет в лабораторных ус-
ловиях, получить не удалось. Однако согласно М.Г. Николаевой с со-
авторами (1985), семена колокольчика широколистного имеют неглу-
бокий физиологический тип покоя. Авторы указывают, что семена —
светочувствительные: при 15°С в темноте проросло 4%, на свету —
75% семян.
182
Таблица 1.
Морфологические показатели генеративных побегов С. latifolia
в разных экологических условиях
Показатели Опушка леса Осиновый лес Широколи- ственный лес
Освещенность, лк 85x105 4,5x105 0,8x105
Длина побега, см 148,3±1,6 137,0±2,2 94,0±2,0
Длина соцветия, см 40,5±1,2 27,4±0,9 12,6±0,7
Число узлов 53,0±1,5 43,4±1,1 32,1±0,7 -
Число цветков 26,3±1,4 16,0±0,2 8,6±0,4
Длина венчика, мм 55,6±0,9 50,4±0,9 42,2±0,3
Диаметр венчика, мм 51,0±1,1 51,4±1,1 43,5±1,2
Длина черешка, мм 70,8±3,2 52,7±2,7 59,5±2,5
Длина пластинки, мм • 157,0±3,2 167,7±4,6 120,7±7,4
Ширина пластинки, мм 78,3±1,8 77,3±2,4 68,8±1,3
Примечание: число узлов дано только на удлиненной части ге-
неративного побега, а параметры листьев соответствуют 12—13 узлу.
Результаты проращивания семян в разных режимах (табл. 2),
полученные нами совместно с В.Л. Тихоновой и И.Ю. Макеевой,
показывают, что семена этого вида очень плохо прорастают в лабора-
торных условиях. Наибольшая всхожесть семян отмечена при исполь-
зовании переменного режима: 16 часов в холодильнике (+4-5°)
и 8 часов в термостате (+30°). При изучении лабораторной всхожести
хорошие результаты могут быть получены при использовании экспресс-
метода (Разумова, Поздова, 1987), заключающегося в использовании
замачивания семян в гиббереловой кислоте (ГКЗ) в концентрации 500
мг/л в течение 48 часов и дальнейшего их проращивании при подсве-
чивании. Лабораторная всхожесть семян, собранных в разных
популяциях, при использовании экспресс-метода за 60 дней соста-
вила от 42±9 до 94±4%. Однако часть семян по-прежнему не про-
росла, оставаясь жизнеспособной (так называемые «свежие не про-
росшие семена» согласно изданию: «Международные правила...,
1984»).
183
Таблица 2.
Результаты проращивания семян С. latifolia при разных режимах, в %
Режим проращивания Всхожесть Жизненность
Срок проращивания (месяцев) 13 13
Комната (+18-20°), свет 1 9
Комната (+18-20°), темнота 0 29
Термостат (+30°) 0 16
Холодильник(+4-5°) 4 36
Холодильник (+4-5°) — 16 часов, термостат (+30°) — 8 часов 64 77
Холодильник (+4-5°) —8 часов, 0 40
комната (+18-20°), свет— 16 часов
Холодильник (+4-5°) — 8 часов, термостат (+30°) — 16 часов 0 37
Изучение ранних этапов онтогенеза в условиях, близких к при-
родным, проведено Т.П. Рысиной (1973). Она, указывая на длитель-
ность виргинильного периода, отмечает, что растения четвертого года
жизни имели розеточные побеги с 2 листьями ювенильного типа и сла-
бо развитой корневой системой. Морфогенез садовой формы С. latifolia
var. macrantha С. Меу. изучен Е.А. Судаковой (1976, 1983). Автор от-
мечает ускорение развития растений в условиях культуры’ Особи пе-
реходили к цветению на 2-й год жизни, а на 3—4-й год наблюдалась
их партикуляция и «вырождение насаждений».
Нами (Викторов, 1990) в условиях питомника ГИЗЛ «Горки Ле-
нинские» подробно изучен онтоморфогенез и выделены возрастные
состояния колокольчика широколистного. Исходным материалом по-
служили семена, собранные близ г. Икша (Икшинский овраг). Их раз-
меры составляли: 1,85+0,02 мм в длину и 0,81+0,02 мм в ширину, мас-
са 100 семян — 27,4±0,3 мг. Лабораторная всхожесть за 60 дней —
80±3%. В питомнике при осеннем посеве всходы появляются в начале
июня. Полевая всхожесть — около 15%. Прорастание семян над-
земное. Первым появляется главный корень, а затем в воздушной
среде развертывается укороченный побег розеточного типа. Семядо-
184
Рис. 2. С. latifolia. Возрастные состояния
прегенеративного периода: р — проросток;
/ — ювенильное; im — имматурное; v —
молодое вегетативное; v — взрослой
вегетативное.
ли—светло-зеленые, яйцевид-
ные (4—6 мм длиной и 3—4
мм шириной) с выемкой на
верхушке пластинки и череш-
ками до 2,5—3,0 мм длиной.
Розеточный побег проростков
имеет 2—3 длинночерешковых
(2—3 см) листа, опушенных
редкими одноклеточными во-
лосками. Листовая пластинка
их — широкояйцевидная
(1,3—2,0 см длиной и 1,5—2,2
см шириной) с острой верхуш-
кой, волнистым краем и серд-
цевидным основанием. Гипо-
котиль проростка несколько
утолщен (1—2 мм в диаметре),
заметно варьирует по длине
(0,7—1,5 см). От него, как пра-
вило, отходит 1—2 тонких при-
даточных корня. Корневая сис-
тема — стержневая по форме
и смешанная по происхожде-
нию (рис. 2).
Ювенильные растения (рис. 2) имеют розеточный побег с 1—2
листьями того же типа, что и проростки, но несколько больших разме-
ров: черешок;—2,5—4,5 см длиной, пластинка— 1,5—3,0 см длиной
и 1,7—2,5 см шириной. Подсемядольное колено утолщается до 25—
30 мм в диаметре, от него отходит несколько придаточных корней. Кор-
невая система — смешанная. Главный корень интенсивно ветвится,
достигает 5—6 см длины и продолжает занимать лидирующее поло-
жение.
Розеточный побег имматурных растений (рис. 2) имеет 3—5 ли-
стьев, из них 1—2 — ювенильного типа. Листовые пластинки
«переходного» типа, округлые или широкояйцевидные (2,0—3,5 см в
диаметре) с тупой верхушкой, сердцевидным основанием и городча-
тым краем, опушены одноклеточными волосками. Черешок достигает
3,0—4,5 см длины. Корневая система — смешанная. Придаточные
корни отходят как от гипокотиля, так и от основания укороченного
185
побега. Главный корень по-прежнему занимает лидирующее положе-
ние (10—12 см длиной).
Для С. latifolia в пределах виргинильного возрастного состоя-
ния мы выделили молодые и взрослые вегетативные особи. Молодые
вегетативные растения (рис. 2) имеют розеточный побег с 3—7 длин-
ночерешковыми (4—7 см) листьями. Пластинки их — широкояйце-
видные или яйцевидные (3—5 см длиной и 3,0—4,5 см шириной), с
тупой или острой верхушкой, сердцевидным основанием и городча-
тым краем. Корневая система — смешанная. Главный корень занима-
ет лидирующее положение и достигает длины 15 см. На второй год из
верхушечной почки развивается, как правило, удлиненный побег. Он
может быть генеративным или оставаться вегетативным (побег с не-
полным циклом развития). Растения, имеющие удлиненный вегетатив-
ный побег, мы относим к взрослому вегетативному состоянию (рис.
2). Их побеги достигают 6—27 см и несут 3—8 длинночерешковых
(1,5—6,0 см) листьев. Листовые пластинки — широкояйцевидные или
яйцевидные (3,5—8,5 см длиной и 2,0—4,5 см шириной) с неравно-
пильчатым краем, острой верхушкой и сердцевидным основанием. Кор-
невая система — смешанная. Главный корень, как правило, по разме-
рам еще выделяется среди интенсивно развивающихся придаточных
корней.
, В условиях питомника отдельные особи зацветают на 2-й год
жизни. Растения, перешедшие к цветению, характеризуются неболь-
шими размерами. Их побеги имеют длину от 23 до 66 см (42,5±1,2) с
заметно меньшим числом узлов — от 9 до 14 (9,9±0,3) и цветков от 1
до 4 (2,1±0,2). Цветки же были незначительно мельче, по сравнению
со средневозрастными генеративными растениями, произрастающие
в оптимальных для вида условиях: длина венчика составляла от 4,0 до
6,0 см (4,7±0,9). Выращенные нами в Измайловском питомнике особи
имели несколько более крупные побеги. Их длина колебалась от 48 до
73 см (58,2±1,1). Количество узлов и цветов также было больше (соот-
ветственно: 13,9±0,4 и 5,0±0,4). На третий год жизни биометрические
показатели вегетативной сферы также несколько уступали растениям, дос-
тигшим своего максимального развития. Массовый переход в генератив-
ный период наблюдается на 3—4-й год. В питомнике колокольчик широ-
колистный в генеративном состоянии имеет кистекорневую жизненную
форму, которая сохраняется, по крайней мере, до 5 лет (до окончания сро-
ка наших наблюдений). При этом уже на третий год у одной особи обра-
зуется, как правило, более одного генеративного побега.
186
В природных условиях данный вид переходит в генеративный
период не ранее, чем на 7—10-й год жизни. Отмирание более старых
участков корневища происходит, по-видимому, после 20-летнего воз-
раста. При этом в благоприятных условиях, как было отмечено выше,
наряду с симподиальным нарастанием наблюдается его ветвление.
В дальнейшем, постепенное отмирание старых участков приводит
к партикуляции и, соответственно, к вегетативному размножению
особи.
Сезонное развитие
В условиях Подмосковья колокольчик широколистный — ве-
сенне-летнезеленое растение. Вегетирует с середины апреля до осен-
них заморозков (КарпиСонова, 1985), цветет в июле месяце. Семена
созревают в августе. После плодоношения надземные побеги начина-
ет постепенно отмирать. В конце вегетационного сезона (октябрь) поч-
ки возобновления содержат по 30—35 метамеров (из 40—50), т. е.
сформированы не полностью. В пазухах нижних 2—4(6) чешуевид-
ных листьев уже заложены почки следующего порядка. Только 1—2
из них на следующий год сформируются как почки возобновления,
остальные останутся спящими. Генеративные побеги развиваются по
моноциклическому типу. Вторичное цветение для данного вида не ха-
рактерно.
В питомнике Ботанического сада БИН (Санкт-Петербург) в
1972 г. особи зацвели 5 мая, при среднесуточной температуре 12,6°С.
С завершением роста генеративного побега и его листьев в середине
июля рост растения завершается полностью, ветвления побега не про-
исходит. Почки возобновления формируются во второй половине лета
(Шулькина, 1975).
Способы размножения и распространения
Для колокольчика широколистного характерно как семенное, так
и вегетативное размножение. В пределах сформировавшихся популя-
ций особи размножаются, в основном, в результате ветвления и по-
следующей партикуляции корневищ. Проникновение вида в новые ме-
стообитания происходит исключительно семенным путем. По спосо-
бу распространения, как и все представители рода, он относится к груп-
пе растений-баллистов. С.С. Хохлов с соавторами (1978) указывают
на наличие в цветках колокольчика широколистного небольшого про-
цента дефектной пыльцы.
187
Колокольчик широколистный характеризуется высоким уровнем
реальной семенной продуктивности (табл. 3). Ее показатели сущест-
венно зависят от эколого-фитоценотических условий произрастания
растений и возраста особи (Викторов, 1989).
Таблица 3.
Семенная продуктивность Campanula latifolia
Место сбора семян и возраст особей Число семян в 1 коробочке Число ко робочек на 1 побеге Реальная се- менная про- дуктивность
Опушка леса 370±11 23±1 8510±433
Осиновый лес 115±11 12±1 1380±175
Широколиственный лес 80±5 6±1 430±85
Питомник (2 года) 157±13 2±1 338±78
Питомник (4 года) 447±18 18±1 7958±453
Экология
Колокольчик широколистный не очень требователен к влаге
(55—56-я ступени шкалы Раменского, 6-я ступень шкалы Элленберга)
и произрастает на относительно богатых почвах (до 12-ой ступени шка-
лы Раменского, 8-я ступень шкалы Элленберга). По мнению
Л.П. Рысина и Т.П. Рысиной (1987), вид предпочитают богатые и влаж-
ные почвы. Он встречается на довольно кислых почвах (8-я ступень
шкалы Элленберга), являясь слабым ацидофилом — индикатором поч-
вы с pH 4,5—6,0 (Викторов, Ремезова, 1988). Проведенный нами ана-
лиз эдафических условий 9 местообитаний в разных районах Москов-
ской области (табл. 4) несколько «расширяет» эдафические границы
распространения вида. Колокольчик широколистный произраста-
ет в несколько затененных местообитаниях (4-я ступень шкалы
Элленберга).
Фитоценология
Колокольчик широколистный произрастает в лиственных, сме-
шанных, темнохвойных лесах и на опушках. Изучая поведение видов
в синузии широкотравья, О.В. Смирнова (1987) отнесла его к толе-
188
Таблица 4.
Физикохимические характеристики почв в местообитаниях
Campanula latifolia в Московской области
Показатель min значение max значение
Механический состав (суглинок) легкий тяжелый
pH водн. 5,50 7,66
pH сол. 4,62 6,92
Обменная кислотность: AljT следы 0,74
Н+ следы 0,05
Гидролитическая кислотность 0,66 9,08
Поглощенные основания Са2+ 4,38 38,06
Mg2+ 0,80 4,98
РЛ 1,46 28,90
8,19 39,73
N общ. 0,175 0,374
Гумус 1,75 10,38
рантам (патиентам). Плотность зрелых популяций колокольчика ши-
роколистного в Подмосковье относительно высока и колеблется от 1,4
до 8,5 генеративных побегов на 1 м2. Плотность вегетативных побегов
— очень низкая (от 0,1 до 1,1 на 1 м2). Общее проективное покрытие в
изученных местообитаниях колеблется от 1 до 50%. Наиболее часты-
ми спутниками колокольчика широколистного являются следующие
неморальные виды: Aegopodiumpodagraria, Galeobdolon luteum, Asarum
europaeum, Pulmonaria obscura, Ranunculus cassubicus, а также Urtica
dioica.
Колокольчик широколистный резко отрицательно реагирует на
вытаптывание и связанное с ним механическое повреждение надзем-
ных побегов, почек возобновления и уплотнение почвы. При усиле-
нии рекреационной нагрузки выпадает из природных сообществ (Ры-
син, Рысина, 1987).
189
Консортивные связи и биологическая продуктивность
Колокольчик широколистный — растение энтомофильное. На-
секомых (шмелей, пчел и мух) привлекает нектар и пыльца. Коровы,
козы и овцы довольно охотно поедают его молодые побеги.
Биологическая продуктивность колокольчика широколистного
достаточно высока. Средний вес одного генеративного побега в
различных фитоценозах колеблется от 95 до 250 г. Продуктивность
одних популяций не превышает 450 г/м2, других — может достигать
1900 г/м2.
Хозяйственное значение и охрана вида
Побеги колокольчика широколистного в период цветения очень
декоративны и интенсивно обрываются на букеты. С середины
XVI века используется в декоративном цветоводстве для групповых
посадок (Крупина, 1954). Он принадлежит к группе медоносов
(Хребтов, 1941; Благовещенский, 1994) и перганосов (Кулиев, 1952);
используется на корм для крупного и мелкого рогатого скота
(Ларин и др., 1956).
Корни, молодые побеги и листья пригодны в пищу (Гроссгейм,
1952). В листьях содержится большое количество витамина С (Панко-
ва, 1949). В надземной части обнаружены сапонины и алкалоиды (Ане-
ли и др., 1967), а также небольшое количество каучука— до 0,5% в
стеблях и корнях и до 1% в листьях (Карягин, 1936).
Колокольчик широколистный охраняется почти по всему ареа-
лу: включен в «Красные книги» Белоруссии (1981) и Карелии (1985),
взят под охрану в Удмуртии (Растительный мир Удмуртии, 1980), в
Горьковской, Псковской (Растения Псковской области..., 1977), Ива-
новской, Владимирской (Шилов, 1982), Московской (Скворцов, Тихо-
миров, 1986) и других областях.
Методов «пассивной» охраны явно недостаточно для сохране-
ния генофонда этого очень декоративного вида. В связи с этим пред-
приняты попытки (Тихонова, Викторов и др., 19896, 1991; Викторов,
Тихонова, 1994) сохранения генетического материала в виде диаспор.
Семена в герметически закупоренных сосудах хранятся при разных
температурных режимах: низкие положительные, невысокие отрица-
тельные и сверхнизкие (в жидком азоте). Ежегодно проводится мо-
ниторинг за изменением лабораторной всхожести. Анализ получен-
ных данных показывает, что единственным способом долговремен-
но
ного сохранения семян является их замораживание в жидком азоте в
течение 1 месяца (табл. 5). Сравнение результатов эксперимента с кон-
тролем по критерию Стьюдента показало отсутствие достоверной раз-
личий (Р=95%) всхожести и жизнеспособности семян колокольчика в
опыте и контроле.
Таблица 5.
Влияние замораживания семян С. latifolia на их всхожесть
и жизнеспособность, %
Опыт Всхожесть Жизнеспособность
лабораторная полевая
Контроль 59±5 35±3 98±2
Жидкий азот 67±5 40±5 99±1
Выращивание растений в питомнике из замороженных семян
показало, что хранение семян в жидком азоте не привело к образова-
нию уродливых экземпляров и изменению жизненной формы. Боль-
шинство биометрических показателей (за исключением длины черешка
и листовой пластинки) практически не отличались от контрольных
(Викторов, Тихонова, 1995).
В лесопарках Москвы и Подмосковья проведены эксперимен-
ты по созданию искусственных популяций колокольчика широколист-
ного с использованием семян и растений разного возрастного состоя-
ния (Викторов, 1991а, 1995 и др.). Наилучшие результаты получены
при пересадке в природу растений молодого генеративного состояния
и минимального нарушения растительного покрова. Пересаженные
растения цвели, плодоносили, т.е. являлись источником семян для соз-
дания полночленных популяций. Биометрические показатели их по-
бегов практически не отличались от растений из природных популя-
ций.
191
М.В. Марков, Н.М. Ключникова
РОД ЧЕРЕДА
Bidens L., Sp. pl. (1753); Gen. Pl. ed. 5, No. 840. 1754 Череда. Род
из трибы Heliantheae семейства Asteraceae. Лектотип рода — Bidens
tripartita L. Название рода произошло от латинских слов bis — дважды
и dens — зуб по числу остей на семянках. В роде Bidens насчитывает-
ся около 230 видов, встречающихся в обоих полушариях, но большая
часть видов обитает на территории Америки. На территории бывшего
СССР встречается И видов р. Bidens (Черепанов, 1995).
Виды рода—длинно побеговые однолетние и многолетние тра-
вы, кустарники. Побеги — прямостоячие, полегающие или приподни-
мающиеся. Стебли в поперечном сечении — округлые или угловатые,
с опушением или без него. Листья — супротивные, простые, цельные,
раздельные или рассеченные. Соцветия — корзинки, располагающие-
ся терминально на главном и боковых побегах. Обертка соцветий —
двурядная; листочки наружного ряда — зеленые, травянистые, т. е.
подобны листьям, а внутренние — пленчатые. Ложе соцветия —
плоское или слегка коническое, несущее чешуи. В корзинках боль-
шинства видов имеются цветки двух типов: стерильные язычковые,
расположенные по краю соцветия и часто отсутствующие, и обоепо-
лые трубчатые цветки. Семянки несут на верхушке 1—8, но, большей
частью, 2—4 ости или зубца, которые усажены шипиками, направлен-
ными вниз.
В трибе Heliantheae, к которой относится род Bidens, 65% ви-
дов — гетеродиаспоричны. На наличие полиморфных семянок у ви-
дов череды указывал монограф рода Е. Sherff (1937) и многие другие
исследователи рода Bidens (Lhotska, 1968; Gillett, Lim, 1970; Dacshini,
Aggarwal, 1974; Forsyth, Brown, 1982; Klein et. al., 1992), работавшие в
различных точках земного шара.
У видов рода Bidens полиморфные плоды формируются из од-
нотипных обоеполых трубчатых цветков. В пределах корзинки и осо-
би строго очерченные морфотипы семянок выделить крайне сложно.
Семянки, занимающие центральное положение в корзинке, связаны с
краевыми семянками гаммой переходных форм при постепенном из-
менении размеров и формы семянок вдоль по парастихам (рис. 1, 4).
Поскольку именно этот вариант гетерокарпии наиболее часто ветре-
192
чается у сложноцветных, и притом в разных трибах, логично исполь-
зовать для его названия таксономически нейтральный термин «граду-
альная* гетерокарпия» (Ключникова, Марков, 1994).
1 см
empt 6(3)
Рис. 1. Изменение размеров и формы семянок у В. tripartita от края корзинки к цен-
тру (слева направо). Размерные классы семянок: empt — семянки с неразвившимся
зародышем, для трехсотых семянок (I) 6(3), 8(3), 7(3), для двухсотых семянок (II) —
6(2), 8(2), 7(2). Объяснение в тексте.
Виды череды часто обитают близ воды — по берегам рек, озер
болотит. д. (Sherff, 1937; ФлораСССР, 1959;FloraEuropaea, 1976). В
Московской области встречается 4 вида этого рода.
Ключ для определения видов, встречающихся
в Московской области
1. Листья цельные, ланцетные, по краю расставленно пильчато-зуб-
чатые, сидячие, почти сросшиеся основаниями. Семянки трех-че-
тырех-гранные с 3—4 равными между собой остями, которые в 2
раза короче семянок. В. cernua L. — Ч. поникшая.
— Листья тройчатые или перистые, с черешками. Семянки сплюс-
нутые, клиновидные, с 2 остями (иногда бывает 3—4 ости, но в
таких случаях 2 из них длиннее остальных)................ 2
2. Сегменты листьев (по крайней мере, верхушечные) с хорошо вы-
раженными черешочками. Семянка по всей поверхности опуше-
на прижатыми волосками. В. frondosa L. — Ч. олиственная.
— Сегменты листьев без черешочков. Семянка голая, но по краям и
остям несет щетинки, обращенные вниз..................... 3
3. Корзинки пло ские, их диаметр в 2 раза больше высоты. Наруж-
ные листочки обертки листовидные, в числе 10—14, значитель-
* От латинского «gradatio» — постепенность перехода от одного к другому.
193
но превышают внутреннюю обертку и окружают ее в виде лучей.
Семянка около 4 мм длиной, ее ости по длине равны семянке или
чуть короче ее. В. radiata Thuill. — Ч. лучистая.
— Корзинки не плоские, их диаметр приблизительно равен высоте.
Наружные листочки обертки менее крупные и в меньшем числе.
Семянки крупнее, 6—7 мм длиной, их ости в 2 раза короче семян-
ки. В. tripartita L. — Ч. трехраздельная.
Череда трехраздельная
Номенклатура и внутривидовая систематика
Bidens tripartita L. 1753, Sp. Pl.: 831; Кауфман, 1866,
Моск, фл., изд. 1: 248; Цингер, 1885, Сборник свед.: 231;
Шмальгаузен, 1897, Фл. Средн. иЮжн. Росс., 2: 57; Петунников, 1900,
Крит, обзор, 2: 42; Сырейщиков, 1910, Илл. фл. Моск, губ., 3: 257;
он же, 1927, Опред. раст. Моск, губ.: 257; Ильин и др., 1940,
в Маевский, Фл., изд. 7: 715; Васильченко, 1959, Фл. СССР, 25: 557;
Кирпичников, 1964, в Маевский, Фл., изд. 9: 556; Ворошилов, 1966,
Опред. раст. Моск, обл.: 324; Tutin, 1976, Fl. Europ., 4: 140; Октябрева,
В. Тихомиров, 1987, Опред. раст. Мещеры, 2: 108: Протопопова, 1994,
Фл. европ. части СССР, 7: 38; Черепанов, 1995, Сосуд, раст. России:
87. — Череда трехраздельная (семейство Asteraceae — Сложноцвет-
ные).
Вид полиморфный. Монограф рода Е. Sherff (1937) выделил у
Bidens tripartita следующие вариации: 1) var. repens (D. Don) Sherff,
2) var. hirta (Jord.) Sherff, 3) var. orientalis (Velen.) Sherff, 4) var.
cemuaefolia Sherff.
Географическое распространение
Вид В. tripartita распространен в Восточном полушарии.
В Евразии он обитает на территории северной Ирландии, Норвегии,
Швеции, Финляндии, Франции, Италии, Турции, Ирана, западной
Индии, европейской части России (кроме Крайнего Севера), Закав-
казья, Средней Азии (кроме Туркмении), на юге Дальнего Востока и
южной части полуострова Камчатка. Встречается на Филиппинских
островах, юго-востоке Австралии, а так же в Северной Африке. В Азии
известен в азиатской части Турции к югу до Палестины, в западной
194
Индии, Японии, на различных участках между западной Монголией и
юго-восточной частью Сибири. Отсутствует в Южном полушарии
(Sherff, 1937; Флора СССР, 1959; Атлас ареалов и ресурсов лекарст-
венных растений, 1976; Flora Europaea, 1976).
На территории бывшего СССР вид встречается везде в Евро-
пейской части, а также на Кавказе, в Западной и Восточной Сибири,
на Дальнем Востоке, в Средней Азии (Флора СССР, 1959).
В Московской области встречается повсеместно.
Морфологическое описание
Череда трехраздельная — длиннопобеговое незимующее одно-
летнее растение с закрытой моноподиальной системой побегов. Тип
корневой системы взрослого растения связан с особенностями эколо-
гических условий. При умеренном увлажнении почвы формируется
стержневая корневая система, причем главный корень сохраняется в
течение всей жизни и хорошо различим среди небольшого числа тон-
ких придаточных корней. В условиях избыточного увлажнения на ниж-
них узлах побега образуется большое число придаточных корней, а
главный корень отмирает. Благодаря контрактильной активности кор-
ней нижняя часть главного и боковых побегов, отходящих от первого
и второго узлов (а иногда и расположенных выше — третьего, четвер-
того), погружена в почву. При этом нижние боковые побеги повторя-
ют структуру главной оси и часто почти равны ей по высоте. Сбли-
женные базальные узлы образуют на главном побеге зону, сходную с
зоной кущения у злаков.
Главный побег — ортотропный, от 3 до 100 см высотой. При
ветвлении несет супротивные побеги II и III (иногда и IV) порядков.
Стебли — голые или редко волосистые. Листья — темно-зеленые или
зеленые, супротивные (иногда самые верхние — очередные) с зубча-
тым краем. Обычно листья—раздельные, с 3—5 ланцетными или про-
долговато-ланцетными долями, или рассеченные, с более крупной вер-
хушечной долей. Последняя может быть зубчатой, надрезанной или
раздельной с боковыми долями, при основании черешковидно сужен-
ными. У мелких угнетенных экземпляров листья могут быть цельны-
ми, продолговато-овальными или ланцетными.
Прямостоящими корзинками увенчаны главный и боковые по-
беги. Диаметр соцветий равен или почти равен высоте и составляет
6—15 мм (иногда больше). Наружные листочки обертки (их 5—8) —
зеленые, продолговатые или продолговато-линейные, к основанию су-
195
женные, по краю с короткими шипиками, равны корзинке или превы-
шают ее иногда в 2—3 раза. Внутренние листочки обертки — буро-
желтые, овальные, более короткие. Прицветники — широколинейные,
равные по длине цветкам, а при плодах — семянкам (не считая остей
последних). Язычковые цветки обычно отсутствуют, и цветки бывают
все трубчатыми, желто-коричневыми (Флора СССР, 1959).
Трубчатые цветки — около 0,5 см длиной. Чашечка видоизме-
нена в чешуи, венчик — сростнолепестный, актиноморфный, пяти-
членный. Гинецей—ценокарпный. Пестики в цветках—с одним дву-
раздельным наверху столбиком. Завязь — нижняя, одногнездная, у ос-
нования с одним семязачатком. Тычинки, обычно в числе 5, прикреп-
лены к трубке венчика. Нити тычинок — свободные, а пыльники бо-
ковыми сторонами слипаются между собой, образуя пыльниковую
трубку, через которую проходит столбик.
Семянки — клиновидной формы, сжатые с боков, на плоских
поверхностях посередине обычно с одним продольным ребрышком,
5—8 мм длиной, 2—3 мм шириной в верхней части. По краю плодов
расположены светлые обращенные вниз щетинки. На верхушке на-
ходятся две—три ости, которые вдвое короче семянок. Е. Salisbury
(1942), изучавший популяции В. tripartita на прибрежных участках в
Великобритании, выделил 2 типа плодов — центральные (длинные и
узкие) и краевые (короткие и широкие). Среди них в равной степени
встречались как двухостые, так и трехостые семянки.
Из-за постепенности изменения размеров и формы семянок
вдоль по парастихам (градуальная гетерокарпия) лишь число остей
позволяет разделить семянки это га вида на два морфотипа (рис. 1).
В изучаемых популяциях В. tripartita (Московская обл., Истринский
р-н) частота встречаемости трехостых семянок (около 30%) была су-
щественно ниже частоты встречаемости двухостных при довольно
сходном распределении по классам длины семянок в пределах каждо-
го из двух морфотипов. Анализ процентного распределения семянок
различной длины в популяциях показал преобладание двухостых цен-
тральных семянок (класс — 7). Их было около 30% от общего числа
плодов.
Средний вес одной семянки у В. tripartita — 3,07±0,1 мг. В
верхушечных корзинках В. tripartita находится от 16 до 83 семянок,
среднее — 41,6±3,0 (Марков, Ключникова, 1994). Диплоидный набор
хромосом 2п = 48 (Хромосомные числа..., 1969; Flora Europaea,
1976).
196
Онтогенез и ритм сезонное развитие
Прорастание свежесобранных семянок В. tripartita в лаборатор-
ных условиях затруднено. Оно не наступает при температуре +20° и
требует воздействия дополнительных факторов: высоких температур
(не ниже +35°), обработки 1—2 % раствором тиомочевины (Николае-
ва и др., 1985). Опытным путем выявлено положительное влияние вы-
соких переменных температур (22—40°) и света на предварительно
стратифицированные и скарифицированные семянки (Rollin, 1956;
Попцов, Буч, 1969; 1970).
В рекомендациях по возделыванию В. tripartita как лекарствен-
ного растения содержатся указания на то, что нестратифицированные
семена имеют всхожесть не более 10% (Бенько, 1986), семянки же,
подвергнутые предпосевной обработке (стратификации, скарифика-
ции), обладают всхожестью 80—90%, при этом в полевых условиях
прорастание начинается при температуре +3 — +4°, а оптимальная тем-
пература прорастания — +15 — +20° (Кодаш и др., 1987).
Наши опыты с проращиванием зародышей, извлеченных из све-
жесозревших семян, показали ингибирующее влияние перикарпия и
семенной кожуры на прорастание семянок В. tripartita (Ключникова,
Марков, 1994). Принимая во внимание малую механическую прочность
перикарпия, и тем более, семенной кожуры, мы вслед за Р. Rollin (1956)
предполагаем, что ингибирование имеет биохимическую природу. Судя
по нашим данным, для резкого повышения потенциальной всхожести
семянок достаточно естественной холодной стратификации, в тече-
ние одной зимовки. Массовое прорастание семянок В. tripartita в при-
роде в мае—июне (в условиях Московской области) свидетельствует
о благоприятной ситуации (Работнов, 1986), при которой внешние фак-
торы устраняют ингибирующее влияние веществ, содержащихся в пе-
рикарпии и семенной кожуре, и стимулируют интенсивное развитие
зародыша.
Материал для изучения особенностей прорастания в природе и
запаса семянок в почве, собранный в мае, июне 1993,1994 гг. на зато-
пленных и свободных от воды участках луга в Московской области
(Истринский р-н), показал, что численность 3—4-х дневных пророст-
ков на участках, свободных от воды, подвержена существенным коле-
баниям.
Существует потенциальная способность В. tripartita к форми-
рованию второй когорты проростков при условии гибели первой. Воз-
можно, что ведущим фактором здесь является освещение находящих-
197
ся на поверхности почвы семянок. Свет ослабевает под пологом пер-
вой когорты всходов, а опыты доказывают значительную светочувст-
вительность (а возможно и чувствительность к спектральному соста-
ву света) семянок этого вида.
Полученные данные позволяют предположить наличие у видов
череды такого типа банка семян, для которого характерно присутст-
вие временного компонента и постоянного компонента относительно
меньших размеров. По-видимому, наличие постоянного компонента
обусловлено в какой-то степени гетерокарпией и задержанным про-
растанием одного из типов семянок. Ранее М. Leek (1989) описал сход-
ный тип банка семян у американского вида Bidens laevis (L.) B.S.P.,
относящегося к той же секции Platycarpaea (Sherff, 1937), что и В. cemua
и В. tripartita.
Построение возрастных спектров для оценки темпов развития
отдельных особей в популяциях возможно не только для многолетних
растений, но и для популяций однолетних растений. В частности, И.И.
Гуреева (1992) описала возрастные состояния В. tripartita в популяци-
ях Кузнецкого Алатау и показала сезонную и погодичную динамику
их возрастных спектров.
Однако фигурирующие в спектрах возрастные (онтогенетиче-
ские) состояния, оказываются на деле отражающими не биологиче-
ский возраст особей, а возникающую достаточно рано в онтогенезе
размерную дифференциацию, которая отчасти является следствием ге-
терокарпии (рис. 2). Наш опыт с В. tripartita продемонстрировал зави-
симость размеров (общей биомассы) 18-дневных проростков от раз-
меров семянок, из которых они появились, причем наиболее сущест-
венно различались проростки из краевых и центральных плодов (Мар-
ков, Ключникова, 1995а).
Даже на самых начальных этапах онтогенеза череды трехраз-
дельной И.И. Гуреева (1992) была вынуждена подразделять возрас-
тные периоды на подпериоды, выделяя среди проростков, ювениль-
ных, имматурных и виргинильных растений по 3 подгруппы, а среди
генеративных — 4. При этом, судя по описаниям, основным критери-
ем для их выделения было число узлов на главной оси и интенсив-
ность ветвления. Нами было показано (Ключникова, Марков, 1994),
что прибавление каждого нового фитомера на оси главного побега у
особей В. tripartita ведет к заметному сдвигу в количественных значе-
ниях всех наиболее важных биоморфологических показателей, но не
влияет на темпы развития растений. Ускорение темпо вразвития
198
Рис. 2. Онтогенетические состояния В. tripartita.
напрямую не связано с каким-либо изменением числа фитомеров на
главной оси и числа боковых побегов. Наоборот, чаще ускоренными
темпами развиваются неотенические модификанты, у которых на глав-
ной оси развивается всего 2—4 фитомера (Васильченко, 1965; Гурее-
ва, 1992; Марков, Ключникова, 1994, 19956).
Фитомер — единое структурное образование (Жмылев и др.,
1993), но каждая из составляющих его структур вносит свой вклад в
различия фитомеров на главной оси. Так как побеговое тело малолет-
них растений содержит полный «пластический слепок онтогенеза», от-
ражающий в своей пространственной структуре временную ритмиче-
скую последовательность образования метамеров (Юрцев, 1976), то
по строению однолетнего растения в конце онтогенеза можно судить
о ритме его роста и формообразования. Как показатель, демонстри-
рующий ритм деятельности верхушечной меристемы, можно рассмат-
ривать изменение длины междоузлий на главной оси. По свидетельст-
ву И.Г. Серебрякова (1952), длина междоузлий на побеге отражает ин-
тенсивность и ритм еги роста.
В природных (объем выборки — 162 особи) и модельных (объ-
ем выборки — 131 особь) популяциях (сентябрь, 1993 г.) у плодонося-
щих растений во всех вариантах наибольшие по длине междоузлия
располагались в средней части главного побега и смещались выше
199
вдоль по главной оси с увеличением числа узлов на ней. Число узлов
на главной оси у череды трехраздельной коррелирует с мощностью
растений. По нашим данным, связь логарифма общей биомассы (InW)
с числом узлов (Nnod) на главном побеге линейна и демонстрирует
тесную положительную корреляцию (для природной популяции:
InW = -0,834+0,941-Nnod; г = +0,859; р < 0,001; для модельной попу-
ляции: InW = 0,546+0,809-Nnod; г = +0,810; р <0,001). Чем больше
биомасса особи, тем выше находится максимальное по длине
междоузлие.
Наблюдения за маркированными особями в природных попу-
ляциях показали, что после формирования верхушечного соцветия вы-
сота растений может увеличиваться на 1—5-см при высоте главного
побега В. tripartita от 11 до 78 см. Такое незначительное изменение
высоты главного побега после формирования верхушечного соцветия
(не более 7% от конечной высоты побега), очевидно, является следст-
вием растяжения клеток верхних междоузлий.
Роль онтогенетических изменений формы листьев в изменчи-
вости фитомеров в целом вдоль по главной оси побега мы изучали на
маркированных особях в природных популяциях, регистрируя разме-
ры обоих листьев каждого узла через 2 недели в течение вегетацион-
ного сезона. Листья наибольших размеров располагались в верхней
половине побега во все сроки наблюдений, но никогда не формирова-
лись на верхних узлах. По мере появления новых листьев ярус с мак-
симальным значением размеров сдвигался вверх по главному побегу.
Обращает на себя внимание тот факт, что в нижних ярусах (узлах) ли-
стья отмирали при переходе растений к цветению и плодоношению.
Исследование роли листьев различных ярусов в снабжении других час-
тей растения ассимилятами у ряда длиннопобеговых малолетних сис-
тематически близких растений из трибы Heliantheae сем. Compositae
(у различных сортов подсолнечника — Четверикова, 1965; Василев-
ская, Ермолаева, 1970; Казарян, 1990, у Cosmos bipinnatus Cav. — Ва-
силевская, Борисовская, 1968) показали, что при отмирании нижних
листьев в генеративной фазе передвижение продуктов их ассимиля-
ции происходит в основном в терминальную корзинку и верхние ли-
стья. Можно предположить наличие аналогичного перераспределения
ассимилятов и у изучаемых нами видов.
При закономерном тождестве основной генетически закреплен-
ной программы роста (архитектурной модели) в любой популяции
Bidens tripartita (длиннопобеговых однолетников с закрытой монопо-
200
диальной системой побегов) представлено большое число архитектур-
ных вариантов или типов, отличающихся интенсивностью ветвления,
числом соцветий, уровнем отхождения боковых побегов, порядком их
формирования во времени и т.п.
Для определения путей формирования архитектурных типов в
природных популяциях В. tripartita были использованы наблюдения за
маркированными особями на постоянных площадках с анализом ди-
намики и последовательности заложения и развития боковых побе-
гов, заканчивающихся соцветиями. Основными критериями для вы-
деления архитектурных типов служили: наличие или отсутствие побе-
гов II порядка и характер их размещения на главной оси.
У череды трехраздельной выявлено три последовательности раз-
вития боковых побегов (паракладиев) из пазушных почек главного по-
бега. Они появлялись, либо, начиная с самых нижних узлов, включая
семядольный, с последующим охватом все более высоко расположен-
ных узлов (акротонно), либо начиная с верхних узлов, а затем спуска-
ясь вниз (базитонно), либо начиная со средних (3-го, 4-го) узлов и рас-
пространяясь вверх и вниз (дивергентно). В популяциях В. tripartita
боковые побеги появлялись у растений с тремя фитомерами на побеге
I порядка. Во всех изученных популяциях преобладали случаи обра-
зования боковых побегов из пазух двух супротивных листьев: от 77,3
до 91,4%. Максимальное число побегов III порядка у обнаруженных
нами наиболее мощных особей — 63, а IV порядка — 20. Размеры
кроющих листьев оказывают влияние на длину образовавшихся из по-
чек побегов. Сопоставление кривых, отражающих изменения средних
размеров листовой пластинки и кривых изменения средней длины по-
бегов II порядка вдоль по главной оси, показывает значительное соот-
ветствие размеров листьев и длины паракладиев, отходящих от одних
и тех же узлов.
Схемы (рис. 3) демонстрируют основные варианты образова-
ния побегов II порядка. Выявлено четыре основных варианта распо-
ложения паракладиев. Варианты I—III формируются у особей,
главная ось которых включает не менее 10 фитомеров. При этом вари-
ант II возникает в онтогенезе двумя путями. Один из них связан с тем,
что из нижних узлов (с 1-го по 4-й) вообще не развиваются боковые
побеги, а в другом случае начинающие развиваться из этих узлов па-
ракладии отмирают при переходе растения в генеративное состояние.
Обращает на себя внимание наличие некоторой зоны «неблагоприят-
ной для ветвления» в области 2—3-го узла: если боковые побеги и
201
Рис. 3. Схема образования побегов II порядка у особей В. tripartita.
развиваются из этих узлов, то длина их, как правило, меньше, чем у
побегов из 1-го и вышерасположенных узлов (см. вариант I и III). Это
находит отражение в средних величинах длины паракладиев в наблю-
даемых популяциях, хотя амплитуда изменчивости этих величин у от-
дельных особей довольна значительна. При формировании варианта
II первоначально появившиеся из нижних узлов (с 1-го по 3-й или 4-
ый) побеги отмирают при переходе особей к цветению. На базе I вари-
анта происходит развитие наиболее мощных особей в популяциях. Та-
кие особи мы наблюдали в благоприятных (по метеорологическим ус-
ловиям) 1993 и 1994 гг. на зарастающем берегу нового русла р. Руден-
ки в популяционных локусах с низкой плотностью (16—20 особей на
м2). В предыдущие годы на данных участках берега особи череды от-
сутствовали. Благодаря контрактильным корням нижняя часть глав-
ного и боковых побегов, отходящих от 1-го и 2-го узлов (а иногда и
расположенных выше — 3-го, 4-го), погружена в почву. При этом ниж-
ние боковые побеги повторяют структуру главной оси и часто почти
равны ей по высоте.
Изучение особенностей ветвления позволило выделить 4 основ-
ных архитектурных типа. Они различаются по числу побегов II поряд-
ка и их пространственному размещению. Первый тип составляют
неветвящиеся особи. Для второго характерно наличие одного—двух
паракладиев, развивающихся из пазушных почек верхнего узла на
202
главной оси. В третий тип объединены особи с тремя—шестью боко-
выми побегами II порядка, как правило, сосредоточенными в верхней
половине главного побега. Последний — четвертый тип составляют
особи, имеющие более шести боковых побегов. Растения этого типа
развиваются в результате I—III вариантов ветвления. У особей чет-
вертого типа порядок расположения и длина паракладиев может варь-
ировать вдоль по главной оси.
Биомасса малолетних растений дает наиболее полную инфор-
мацию о жизненном состоянии особей, как на промежуточных этапах
роста, так и в конце онтогенеза (Марков, 1992). Поэтому общую био-
массу, как интегрированный показатель мощности растения, исполь-
зовали при определении границ классов жизненных состояний для ка-
ждого из 4-х архитектурных типов в популяциях череды (табл. 1).
В выборках из природных популяций В. tripartita преобладают
особи первых двух архитектурных типов. Значения биомассы в наи-
большей степени варьируют у особей первого типа (максимальное зна-
чение превосходит минимальное в 120 раз). Пластичность у неветвя-
щихся особей — результат изменения числа и размеров фитомеров на
главной оси. В целом значительное перекрывание верхних и нижних
границ значений биомассы соседних классов жизненного состояния
говорит о том, что особи могут достигать одинаковой мощности на
основе различных архитектурных решений.
Представление о пластичности будет неполным, если не учи-
тывать изменчивость соцветий, которые тоже выступают как метаме-
ризованные структуры, способные к миниатюризации и грандизации
за счет изменения числа слагающих их субъединиц. Несмотря на то,
что цветение и плодоношение у В. tripartita растянуты (первыми начи-
нают распускаться терминальные соцветия на побеге I порядка, затем,
последовательно, на побегах II и следующих порядков сверху вниз
вдоль по главной оси, а созревание семянок происходит в той же по-
следовательности), связь между массой соцветия и числом семянок в
его составе оказалась прямой и однотипной для соцветий разных по-
рядков. По мере увеличения массы корзинки число семянок в ней
росло, асимптотически приближаясь к своему максимальному зна-
чению, а сила связи между этими положительно коррелирующими по-
казателями составила для соцветий I, II и III порядков от 0,825 до 0,897
(р < 0,001). Следовательно, по массе генеративных структур (кор-
зинок) можно судить и о плодовитости особей. Сравнивая в
популяциях особи различных архитектурных типов по массе ге.
203
Таблица 1.
Связь спектров архитектурных типов и жизненного состояния
особей в популяциях В. tripartita L.
Архи тект. типы N по- бегов II ИОр. N узлов на глав, оси Общая биомасса W, мг % ОСО- бей в классе
min среднее max
Природная популяция, 1992 год
I 0 3—9 9 77,8 235 36,5
II 1—2 4—13 29 201,8 545 38,9
III 3—6 4—8 118 402,9 820 17,4
IV >7 7—10 840 1128,7 1843 7,3
Природная популяция из проростков второй когорты, 1993 год
I 0 3—8 17 165,9 720 51,9
II 1—2 7—8 285 541,7 895 23,5
III 3—6 7—9 510 1047,6 1545 15,4
IV >7 7—9 1305 1811 2645 9,2
Модельная популяция, 1993 год
I 0 5—6 25 38 51 1,5
II 1—2 6—9 45 225,8 430 9,9
III 3—6 6—9 160 980 3530 26,7
IV >7 6—12 1370 9732 35576' 61,9
неративных структур (W ), можно видеть, что на фоне роста среднего
значения границы максимальной и минимальной генеративной массы
существенно перекрываются. Пластичность проявляется и на уровне
соцветий. В популяциях были обнаружены как неотенические моди-
фиканты, у которых главная ось была составлена 2—3 фитомерами, а
в единственной корзинке были сформированы 1—2 семянки, так и осо-
би, имевшие до 91 корзинки, при среднем числе семянок в них до
82,2±3,5.
204
Череда поникшая *
Номенклатура
Bidens cernua L. 1753, Sp. Pl.: 832; Кауфман, 1866, Моск, фл.,
изд. 1:248; Цингер, 1885, Сборник свед.: 231; Шмальгаузен, 1897, Фл.
Средн, и Южн. Росс., 2: 58; Петунников, 1900, Крит, обзор, 2: 42; Сы-
рейщиков, 1910, Илл. фл. Моск, губ., 3: 258; он же, 1927, Опред. раст.
Моск, губ.: 256; Ильин и др., 1940, в Маевск. Фл., изд. 9: 715; Василь-
ченко, 1959, Фл. СССР, 25: 555; Кирпичников, 1964, в Маевский, Фл.,
изд. 9:556; Ворошилов, 1966, Опред. раст. Моск, обл.: 324; Tutin, 1976,
Fl. Europ., 4: 140; Октябрева, В. Тихомиров, 1987, Опред. раст. Меще-
ры, 2: 108; Протопопова, 1994, Фл. европ. части СССР, 7: 38; Черепа-
нов, 1995, Сосуд, раст. России: 87. — Череда поникшая (семейство
Asteraceae — Сложноцветные).
Географическое распространение
Вид распространен в Северном полушарии. В Северной Аме-
рике — от Британской Колумбии к югу до Калифорнии, Нью-Мекси-
ко, Теннеси и Северной Каролины. В Европе встречается в Албании,
Австрии, Бельгии, Люксембурге, Великобритании, Болгарии, Чехосло-
вакии, Дании, Финляндии, Франции, Германии, Греции, Ирландии,
Швейцарии, Нидерландах, Испании, Венгрии, Италии, Югославии,
Норвегии, Польше, Румынии. Вид отсутствует на Крайнем Севере и в
большей части Средиземноморского региона (Sherff, 1937; Флора
СССР, 1959; Атлас ареалов и ресурсов лекарственных растений, 1976;
Flora Europaea, 1976).
На территории бывшего СССР встречается в европейской час-
ти (все районы), на Кавказе (Предкавказье, Западное и Южное Закав-
казье), в Западной Сибири (до 60° с.ш.), Восточной Сибири, на Даль-
нем Востоке, в Средней Азии (Флора СССР, 1959). В Московской об-
ласти встречается повсеместно.
Морфологическое описание
Череда поникшая—длиннопобеговое незимующее однолетнее
растение с закрытой моноподиальной системой побегов. Тип корне-
вой системы взрослого растения, так же как у В. tripartita, связан с
* Разделы «Способы размножения и распространения; кология, фитоценология, био-
логическая продуктивность, хозяйственное использование, консортивные связи» да-
ны в конце статьи по двум видам В. tripartita и В. cernua одновременно.
205
1 CM
Рис. 4. Изменение размеров и формы семянок у В. cemua от края корзинки к центру
(слева направо). Размерные классы семянок: empt — семянки с неразвившимся за-
родышем , 4d, 7, 6, 5, 41,3, 2. Объяснение в тексте.
особенностями экологических условий. При умеренном увлажнении
почвы формируется стержневая корневая система, главный корень со-
храняется в течение всей жизни и хорошо различим среди небольшо-
го числа тонких придаточных корней. При произрастании в условиях
избыточного увлажнения почвы на нижних узлах побега образуется
большое число придаточных корней, а главный корень отмирает.
Побеги — удлиненные, ортотропные, от 3 до 100 см высотой.
Стебель—голый или рассеянно железисто-волосистый. Листья — си-
дячие, ланцетные или продолговато-ланцетные, на верхушке длинно
заостренные, желтовато-зеленые, от 3—4 до 10—12 см длиной, осно-
ваниями обычно немного срастающиеся. Край листовой пластинки—
раставленно пильчато-зубчатый. Корзинки—на длинных ножках, пло-
ские, поникающие, довольно крупные и широкие (10—20 мм). Наруж-
ные листочки обертки — зеленые, в числе 5—9, продолговато-линей-
ные, по краю — короткореснитчатые, значительно длиннее продолго-
вато-овальных буро-зеленоватых внутренних, по длине почти равны
цветкам. Цветок — такого же строения как у Bidens tripartita.
Семянки—обратно-пирамидальные, внутренние в поперечном
разрезе — ромбические, внешние — треугольные, на верхушке с че-
тырьмя остями, которые наполовину короче семянок. По ребрам се-
мянки усажены вниз обращенными волосками и иногда более или ме-
нее бугроватые (Васильченко, 1959).
Изучение особенностей варьирования плодов в популяциях Мо-
сковской области (Истринский р-н) показало наличие и у В. cemua
градуальной гетерокарпии. Выделено 7 классов плодов, условный клас-
совый интервал выбран в 1 мм (без учета длины остей). Семянки с
длиной 4 мм подразделены на 2 класса: 41 — клиновидные, узкие в
206
верхней части и 4d — плоские, широкие в верхней части (рис. 4). В
популяциях наиболее высок процент семянок переходных форм (класс
— 5). Их около 40% от общего числа измеренных плодов. Средний
вес семянки у В. cemua (W = 1,08±0,07мг). В верхушечных корзинках
находилось от 3 до 253 семянок, среднее — 133,67+12,45 (Марков,
Ключникова, 1994).
Диплоидный набор хромосом 2п = 24 (Хромосомные числа...,
1969; Flora Europaea, 1976).
Онтогенез и ритм сезонного развития
Биологические свойства семянок В. cemua и В. tripartita очень
сходны. Свежесобранные семянки В. cemua так же, как и семянки
В. tripartita, в лабораторных условиях прорастают очень плохо. При
проращивании на свету для повышения процента проросших семянок
также нужна дополнительная их обработка высокими переменными
температурами (22—40°С) при условии предварительной стратифи-
кации и скарификации (Rollin, 1956; Попцов, Буч, 1969, 1970).
В опытах с проращиванием изолированных зародышей показа-
но торможение прорастания из-за биохимических свойств перикар-
пия и семенной кожуры (Hogue, 1976). Естественная стратификация
холодом, происходящая в течение одной зимовки, сопровождается мас-
совым прорастанием семянок в природе, которое в условиях Москов-
ской области происходит в мае—июне. Так же как и у В. tripartita в
популяциях формируется III тип почвенного банка семян (Thompson,
Grime, 1979).
У В. cemua, так же как у В. tripartita, выделены только онтоге-
нетические (а не возрастные) состояния (рис. 5). Выборку (149 экз.)
из природной популяции (Московская обл., Истринский р-н) обраба-
тывали по той же схеме, что и для В. tripartita. Наибольшие по длине
междоузлия были расположены в средней части побега, и смещались
вверх по главной оси при увеличении числа узлов на ней. При этом
смещение максимума длины междоузлий главной оси коррелировало
с мощностью растений. Зафиксирована линейная связь и высокая по-
ложительная корреляция логарифма общей биомассы (InW) с числом
узлов (Nnod) на главном побеге: InW = 0,388+0,657.Nnod; г = +0,787; р
<0,001.
В природных популяциях после формирования верхушечного
соцветия высота растений могла увеличиваться вследствие растяже-
207
Рис. 5. Онтогенетические состояния В. cemua.
ния клеток верхних междоузлий на 1—5 см (т. е. не более чем на 7% от
конечной высоты побега) при высоте главного побега у В. cemua от 38
до 111 см. Последовательность развития боковых побегов у особей
двух видов череды имеет большое сходство, но у В. cemua ветвление
начинается у особей, образовавших не менее 6 фитомеров на главной
оси. Во всех изученных популяциях преобладали случаи образования
боковых побегов из пазух двух супротивных листьев: от 65,2 до 81,2%.
Максимальное число побегов III порядка у обнаруженных нами наи-
более мощных особей — 15, побеги IV порядка у них отсутствовали.
Схемы (рис. 6) демонстрируют основные варианты образова-
ния побегов II порядка. Выявлено пять основных вариантов располо-
жения паракладиев. Варианты I—IV формируются у особей, главная
ось которых включает не менее 10 фитомеров. При этом вариант II
возникает в онтогенезе двумя путями. Один из них связан с тем, что из
нижних узлов (с 1-го по 4-й) вообще не развиваются боковые побеги,
а в другом случае начинающие развиваться из этих узлов паракла-
дии отмирают при переходе растения в генеративное состоя-
ние. Обращает.на себя внимание наличие некоторой зоны «неблаго-
приятной для ветвления» в области 3—5-го узла: если боковые побеги
и развиваются из этих узлов, то дайна их, как правило, меньше, чем у
208
Рис. б. Схема образования побегов II порядка у особей В. cemua.
побегов из 1-го, 2-го и вышерасположенных узлов (рис. 6, варианты
III и IV). На базе IV варианта происходит развитие наиболее мощных
особей в популяции.
Четыре основных архитектурных типа особей В. cemua были
выделены по тем же критериям, что и у В. tripartita. В выборках из
природных популяций В. cemua преобладали особи первых двух ар-
хитектурных типов — неветвящиеся или с малым (1—2) числом боко-
вых побегов (табл. 2). Варьирование количественных показателей как
биомассы (максимум больше минимума в 105 раз) так и генеративных
структур (W ) у череды поникшей аналогично с описанными выше у
череды трех^аздельной. Пластичность проявляется и на уровне соцве-
тий. Неотенические модификанты, найденные в популяциях В. cemua,
имели всего 2—3 фитомера, и одну единственную корзинку с 1—1
семянками, тогда как самые мощные особи несли до 33 корзинок при
среднем числе семянок в них до 187,2±2,3.
Способы размножения и распространения
Для обоих видов характерна гейтоногамия — для центральных
и краевых цветков и ксеногамия — для краевых цветков (Поляков,
1967) при участии насекомых (Sherff, 1937). Наряду с опылением
насекомыми отмечена и возможность самоопыления (Frank и Klotz,
1988).
209
Таблица 2.
Связь спектров архитектурных типов и жизненного состояния
особей в популяциях В. cernua L.
Архи те кт. типы N по- бегов II пор. N узлов на глав, оси Общая биомасса W, мг % ОСО- бей в классе
min среднее шах
Природная популяция, 1992 год
I 0 4—9 7 155,8 694 76,7
II 1—2 6—9 188 502,8 640 10,4
III 3—6 5—9 357 642,5 1995 8,6
IV >7 5—8 634 1641,0 3200 4,3
Природная популяция, 1993 год
I 0 8—11 120 341,4 740 19,5
II 1—2 9—12 580 1171,8 2020 34,2
III 3—6 10—12 790 2498 4150 22,8
IV >7 10—13 2440 6238 27075 23,5
. Модельная популяция, 1994 год
I 0 5—10 15 229,7 940 73,4
II 1—2 7—10 190 764,4 1600 17,2
III 3—6 7—11 465 904 1710 5,8
IV >7 8—12 890 1711 2280 3,6
У В. tripartita и В. cernua полиморфные плоды формируются из
однотипных обоеполых трубчатых цветков, причем П.П. Поляков
(1967) отмечает, что у представителей рода Bidens по краю корзинки
иногда встречаются рудименты пестичных цветков.
Наблюдения в популяциях обоих видов в течение трех вегета-
ционных сезонов (1992—1994 гг.) показали, что значительная часть
семянок опадает вблизи материнских растений. По крайней мере, та-
кой вывод можно сделать, оценивая семенную продуктивность, запас
семянок в почве и особенности их прорастания в природе. Следова-
210
тельно, сохранение территорий, которую занимают отдельные попу-
ляции В. tripartita и В. cernua осуществляется преимущественно пу-
тем автобарохории. Относительно высокий удельный вес семянок не
позволяет ветру относить их на значительные расстояния от материн-
ских особей.
Эпизоохория и антропохория семянок череды — хрестоматий-
ный пример во многих учебниках и пособиях. Но зачастую популяции
этого вида расположены на топких илистых участках, что резко сни-
жает возможность распространения семянок животными и человеком.
Наличие острых зазубренных остей у семянок обоих видов играет роль
не только в эпизоохорной дисперсии на дальние расстояния, но и спо-
собствует разносу диаспор путем флотационной гидрохории. Ости, зуб-
цы и волоски, покрывающие семянки, дают возможность плодам дос-
таточно длительное время удерживаться на поверхности воды. По дан-
ным Е. Salisbury (1942), это время может составлять несколько дней
или недель. По наблюдениям авторов, плавающие семянки, соединен-
ные между собой посредством остей и многочисленных зубцов, обра-
зуют группы (от 2 до 30 и более экз.). Якорная функция остей обеспе-
чивает своеобразный вариант синаптокарпии, когда агрегации семя-
нок формируются как в результате их совокупного опадания из одной
корзинки, так и вторично, за счет перемещения с токами воды и слу-
чайного столкновения плодов. Очевидно, что такая агрегированность
семянок лежит в основе формирования густых зарослей, характерных
для череды. Кроме того, распространение путем флотационной гид-
рохории повышает вероятность попадания диаспор на достаточно ув-
лажненные участки, что совсем немаловажно, учитывая экологическую
приуроченность этих видов.
Экология
В. cernua и В. tripartita — гигрофиты, весьма мало отличаю-
щиеся друг от друга по экологическим свойствам. Если В. tripartita
единично встречается в широком интервале — от влажнолуговых до
прибрежно-водных сообществ (ступени 72—107 шкалы Раменского),
но обильнее представлена в болотно-луговых и болотных сообще-
ствах (ступени 84—97), то экологический ареал В. cernua несколько
сдвинут в сторону более высокой влажности (соответственно, ступе-
ни 88—103 и 95—100) и сопряжен с более резко переменным режи-
мом увлажнения (ступень 13) и слабой аллювиальностью. При этом
211
по шкале Элленберга (Ellenberg, 1991) оба вида получают одинаковое
значение (9-я ступень), характеризующее приуроченность к сырым,
часто плохо аэрируемым почвам.
У X. Элленберга в. cernua — это растение, произрастающее
исключительно на богатых азотом почвах (9-я ступень), тогда как при-
сутствие В. tripartita диагностирует почвы с повышенным содержани-
ем азота (8-я ступень).
Фитоценология
Оба вида рода Bidens играют существенную роль в синантроп-
ных ценозах на территории бывшего СССР. Об этом свидетельствует
упоминание В. cemua и В. tripartita в списке диагностических видов
класса Bidentetea tripartiti Тх., Lohm. et Prsg. in Tx. 1950 (синантропные
сообщества переувлажненных мест) (Миркин и др., 1989). В. cemua и
В. tripartita образуют популяции высокой плотности на заболоченных
участках лугов и в прибрежных зонах водоемов. Компактность зарос-
ли, необходимое условие осуществления C-R стратегии, присущей
обоим видам, основана на заполнении объема пространства разно-
родными по архитектуре и размерам особями. При преобладании мо-
нокогортного варианта популяционной динамики основная ставка
в приспособлении видов к фитоценотической обстановке делается
на внутрикогортную дифференциацию особей по архитектуре и
размерам.
Консортивные связи
Цветки череды посещают насекомые: В. cemua—мухи из рода
Lucila и сирфиды из рода Syrphus (отряд Diptera), а на цветках
В. tripartita — сирфы и настоящие шмели (род Bombus из отряда
Hymenoptera) (Sherff, 1937).
Биологическая продуктивность
В дикорастущих зарослях В. cemua урожайность надземной час-
ти на Украине составляла 15 ц/га (Кондратенко, Чернов, 1988). Уро-
жайность В. tripartita в дикорастущих зарослях — от 13 до 1725 кг/га в
различных регионах; при культуре в специализированных хозяйствах
— 10—12 ц/га (Пучин, 1977; Кодаш и др., 1987; Растительные ресур-
сы СССР, 1993).
212
Хозяйственное значение
В. cemua и В. tripartita — ценные лекарственные растения. В
народной медицине надземные части В. cemua используют при рахи-
те, артрите, подагре, болезнях почек, кожи, цинге, нарушениях обме-
на веществ. В тибетской медицине растение применяют при анемии,
атеросклерозе, сибирской язве, в монгольской — при травмах черепа.
В. cemua обладает антибактериальными, антифунгальными, антигель-
минтными свойствами. Она успешно испытана в клинике кольпитов и
дерматомикозов. Спиртовой экстракт в эксперименте стимулирует
функцию коры надпочечников, оказывает желчегонное и противовос-
палительное действие (Растительные ресурсы СССР, 1993).
В. tripartita в народной медицине используют аналогично В. сег-
nua, кроме того, есть укдзания на применение надземных частей этого
вида (в тибетской медицине) при злокачественных опухолях и ожогах.
Череда трехраздельная (надземные части) была включена в отечест-
венную фармакопею 1—3-го изданий и рекомендована к применению
при диатезах у детей (водный настой для ванны), а также как диурети-
ческое и потогонное при простудных заболеваниях. В медицинской
практике В. tripartita включают в состав сборов, как тонизирующее,
иммуностимулирующее, десенсибилизирующее, жаропонижающее
при острых гастритах, хронических гепатитах, панкреатитах, гломе-
рулонефритах, циррозе печени, сахарном диабете и ряде кожных за-
болеваний. Клинические испытания дали положительные результаты
при лечении бронхиальной астмы, панкреатита у детей, папилломато-
за гортани у детей (Растительные ресурсы СССР, 1993).
В связи с широким использованием этих видов в медицине в
настоящее время подробно изучен их химический состав. Надземные
части В. cemua содержат: эфирное масло, углеводы и их производные
(D-галактозу, L-арабинозу, D-глюкозу, L-раминозу, D-ксилозу, D-галак-
тоуроновую кислоту), флавоноиды (лютеолин, цинарозид, кверцитрин),
халконы и ауроны. В составе надземных частей В. tripartita также бы-
ли обнаружены выше перечисленные химические соединения. Кроме
того, в побегах череды трехраздельной содержатся органические ки-
слоты, стероиды, витамины (аскорбиновая кислота, токоферолы, ка-
ротин), дубильные вещества, кумарины, умбеллиферон, скополетин,
высшие алифатические углеводы, воска, липиды, а в корнях — поли-
ацетиленовые соединения (Бельтюкова, 1964; Исакова, 1980а, 19806;
Исакова и др., 1986; Растительные ресурсы СССР, 1993).
213
ЛИТЕРАТУРА
Александров В.Г. О гермафродитизме цветковых растений // Труды Ботан. ин-
та им, Комарова. Л., 1962. Серия 7, вып. 5.
Алехин А.А. Дремлик болотный в условиях культуры // Охрана, обогащение,
воспроизводство и использование рациональных ресурсов. Ставрополь, 1990. С. 302—
303.
Алехин А.А. Предварительные итоги интродукции орхидных флоры СССР в
Ботаническом саду Харьковского университета//Вести. Харьковск. ун-та, 1992. Я» 364.
С. 63-66.
Андр1енко Т.Л. Scheuchzeria palustris L. на Укрш'ш // Укр. ботан. ж., 1975. Т. 32,
№5. С. 617—623.
Андриенко Т.Л. Охрана гидрофильных видов орхидей на Украине // Охрана и
культивирование орхидей. М., 1987. С. 32—33.
Андриенко Т.Л. Распространение, экология и ценология редких болотных ви-
дов сем. Orchidaceae на Украине // Охрана и культивирование орхидей. Таллин, 1980.
С. 57—58.
Андриенко Т.Л., Прядко Е.И. Экология, ценология и запас фитомассы верони--
ки лекарственной на Украине // Растительные ресурсы. 1975. Т. 2, вып. 3. С. 406—409.
Андронова Н.Н., Кученева Г.Г., Кученева А.Е. Виды семейства Orchidaceae в
ценозах Куршской косы // Охрана и культивирование орхидей. Таллин, 1980. С. 42—46.
Анели Н.А., Ментенашашвили Т.С., Лагашвили И.И. Лекарственные растения
Цхинвальского и Джавского района ГССР // Тр. Ин-та фармакохимии, Тбилиси, 1967.
Вып. 10. С. 309—318.
Артюшенко З.Е, Атлас по описательной морфологии высших растений. Л. 1990.
204 с.
Атлас ареалов и ресурсов лекарственных растений СССР. М., Медгиз. 1976.
• Балявичене Ю. Орхидные, включенные в Красную книгу Латвийской ССР //
Охрана и культивирование орхидей. Таллин, 1980. С. 31—34.
Барсукова А.В., Пятковская В.П. Влияние корневой системы ели на динамику
численности и жизненное состояние некоторых растений травяно-кустарничкового яру-
са ельника-зеленомошника И Вестник Моск, ун-та, сер. биол. 1977. № 4. С. 56—65.
Белавская А.П. Водные растения России и сопредельных государств // Тр. Бо-
тан. ин-та им. В.Л. Комарова, СПб. 1994, Вып. 11. С. 1—63.
Белавская А.П., Леонова Т.П К морфологии и биологии Typha angustifolia L. //
Вопросы сравнительной морфологии семенных растений. Л., 1975. С. 129—137.
Бельтюкова К.И. О действии эфирного масла из череды поникшей и сальвииа
нафитопатогенные бактерии// Фитонциды в народном хозяйстве. Киев, 1964. С. 269—
271.
Бенько Т.Н. Морфологические и биологические особенности череды трехраз-
дельной в естественных местообитаниях и в условиях культуры//Лекарственные и ядо-
витые растения и их значение в педиатрии. Л., 1986. С. 13—17.
Благовещенский М.В. Медоносные растения Ульяновской области. Ульяновск,
1994. 138 с.
БлиноваИ.В. Эколого-биологические особенности некоторых представителей
семейства Orchidaceae Мурманской области: Автореф.дис.... канд.биол. наук.М., 1995.
24 с.
Богдановская-Гиенэф И.Д. О происхождении флоры бореальных болот Евра-
зии // М-лы по истории флоры и раст-ти СССР. Вып. 2, 1946. С. 425—468.
214
Бордзьтовский E.I Сем. Orchidaceae // Флора УРСР. Киев, Т. 3, 1950.
Босек П.З. О новых и редких растениях Брянской области // Ботаи. ж. 1983. Т.
41, №1. С. 98—101.
Босек П.З. О распространении видов семействаОгсЫбасеае в Брянской облас-
ти//Ботан. ж. 1985. Т. 70, № 8. С. 1097—1100.
Бон М.С. О классификации болотной растительности (на примере сфагновых
топей Северо-Запада РСФСР)//Ботан. ж., 1986. Т. 71, №9. С. 1182—1192.
Быков Б.А. Доминанты растительного покрова Советского Союза. Алма-Ата,
1962. Т. 2. С. 6—10.
Валетов В.В., Иванович В.С., Кудин М.В. Продукционно-биометрическая оцен-
ка растительности заповедных болот. Минск, 1989. 127 с.
Василевская В.К., Борисовская ГМ. Влияние листьев и верхушечного генера-
тивного побега на пазушные почки Cosmos bipinnatus Cav. // Ботан. ж., 1968. Т. 53, № 9.
С. 1189—1195.
Василевская В.К., Ермолаева Е.Я. О взаимосвязи передвижения ассимилятов
из листьев разного яруса и строения проводящей системы подсолнечника // Вестник
ЛГУ, сер. биол., 1970. Вып. 1, № 3. С. 33—42.
Васильченко И.Т. Неотенические изменения у растений. М.-Л.: Наука. 1965.84
С.
Васильченко Т.И., Кунина Л И. Ценопопуляций колокольчика персиколистно-
го в условиях рекреационной дигрессии. Воронеж., 1985. 5 с. Рукопись депонирована
в ВИНИТИ 25.03.1986, № 1963—В.
Вахрамеева М.Г, Варлыгина Т.И. Вопросы устойчивости и охраны орхидных
на территории Московской области // Вести. МГУ. Сер. 16. Биол. 1996. № 3. С. 30-35.
ВахрамееваМ.Г, Варлыгина Т.И., Куликов П.В. Пыльцеголовник длиннолист-
ный // Биол. флора Московской области. М., 1996. Вып. 12. С. 48—59.
Вахрамеева М.Г, Варлыгина Т.И., Татаренко И.В., Литвинская С.А., Загульский
М.Н., Блинова И.В, Виды евразиатских наземных орхидных в условиях антропогенно-
го воздействия и некоторые проблемы их охраны // Бюл. МОИП. Отд. биол. 1977. Т. 102,
вып. 4. С. 35-43.
Вахрамеева М.Г, Денисова Л.В., Никитина С.В., Самсонов С.К. Орхидеи на-
шей страны. М., 1991. 224 с.
ВахрамееваМ.Г, ТатаренкоИ.В., БыченкоТ.М. Экологические характеристи-
ки некоторых видов евразиатских орхидных//Бюл. МОИП. Отд. биол. 1994. Т. 99, вып.
4. С. 75-82.
Верещагина В.А. О цветении и опылении вероники лекарственной (Veronica
officinalis L.) // Уч. зап. Пермского гос. ун-та. 1970. № 206. С. 85—87.
Викторов В.П. Влияние экологических условий на семенную продуктивность
двух видов рода Campanula L. (Campanulaceae Juss.). //Изучение, охрана и рациональ-
ное использование природных ресурсов. Ч. Г, Уфа., 1989. С. 104.
Викторов В.П. Онтоморфогенез охраняемых видов рода колокольчик
(Campanula L.) в условиях питомника// Труды III молодежной конференции ботаников
г. Ленинграда., Л., 1990. С. 125—145. Деп. в ВИНИТИ 14.11.1990, № 5701—В90.
Викторов В.П. Опыт создания искусственных популяций некоторых видов ро-
да колокольчик // Проблемы рационального использования, воспроизводства и эколо-
гического мониторинга лесов. Свердловск, 1991а. С. 27—29.
Викторов В.П. Охраняемые виды р. Campanula L. (Campanulaceae Juss.) Под-
московья: морфологические и биологические особенности, перспективы реинтродук-
215
ции. Автор, дис ... канд. биол. наук, М., 19916. 16 с.
Викторов В.П. Жизненные формы редких колокольчиков (CampanulaL.) Под-
московья//Жизненные формы: онтогенез и структура, М., Прометей, 1993. С. 41—44.
Викторов В.П. Внутривидовая изменчивость некоторых видов рода Campanula
L. в Подмосковье // Научные труды МПГУ им В.И. Ленина., Сер. Естеств. науки. М.,
Прометей., 1996, С. 237—247.
Викторов В.П., Соколова Т.Г. Морфологические особенности соцветий неко-
торых видов рода колокольчик (Campanula L.) // Успехи экологической морфологии рас-
тений и ее влияние на смежные науки. М., Прометей, 1994. С. 126—128.
ВикгоровВ.П., ТихоноваВ.Л. Влияние продолжительностихранения семян рода
Campanula L. на их жизнеспособность // Научные труды МПГУ им. В.И. Ленина, сер.
Естеств. науки. Ч. 1., М.: Прометей, 1994. С. 164—171.
Викторов В.П., ТихоноваВ.Л. Влияние криоконсервации семян видов рода ко-
локольчик на морфо-биологические особенности растений //Актуальные вопросы эко-
логической морфологии растений. М., Прометей. 1995. С. 83—92.
Викторов С.В., Ремезова Г.П. Индикационная ботаника. М, 1988. 168 с.
Виноградова Ю.К. Процессы микроэволюции у адвентивных и интродуциро-
ванных растений. Дис.... докг. биол. наук., М. 1992.
Воронов А. Г. О некоторых приспособлениях растений к изменениям уровня озер
//Ботан.ж., 1948. Т.28,№5.
Ворошилов В.Н. Определитель растений Советского Дальнего Востока. М.: Нау-
ка, 1982. 672 с.
Ворошилов В.Н., Скворцов А.К., Тихомиров В.Н. Определитель растений Мо-
сковской области. М. 1966. 367 с.
Вульф Е.В. Флора Крыма. Ялта, 1969. 384 с.
Вьюкова Н. А. Новые материалы к характеристике адвентивной флоры Липец-
кой области // Бюл. МОИП. Отд. биол., 1983. Т. 88, № 1. С. 128—133.
, Вьюкова Н.А. Приуроченность адвентивных растений к фитоценозам Липец-
кой области // Растительный покров Центр. Черноземья и его охрана. Воронеж., 1987.
С. 29.
Гаевская Н.С. Роль высших водных растений в питании животных пресных во-
доемов. М.: Наука, 1966. 328 с.
Глухов М.М. Медоносные растения. М. 1955. 288 с.
Говорухин В.С. Флора Урала. Свердловск, 1937. 536 с.
Голубев В.Н. К биоморфологии природных растений Подмосковья с запасаю-
щими органами побегового происхождения//Учен. зап. МОПИ. Тр. каф. бот., 1956а. Т.
41, Я» 1. С. 41—104.
Голубев В.Н. Контогенезу корневищ кистекорневых растений// Ботан.ж., 19566.
Т. 41, №2. С. 248—253.
Голубев В.Н. Основы биоморфологии травянистых растений центральной ле-
состепи//Тр. Центрально-черноземного заповедника им. В.В. Алехина, Воронеж, 1962.
Вып. 7. С. 1—510.
Голубев В.Н. Эколого-биологические особенности травянистых растений и рас-
тительных сообществ лесостепи. М. 1965. 286 с.
Голубев В.Н. Биологическая флора Крыма. Ялта, 1984.215 с. Деп. ВИНИТИ №
5770884.
Горбик В.П. Семенная продуктивность Typha angustifolia днепровских водо-
хранилищ// Гидробиол. ж., 1988. Т. 24, № 2. С. 59—63.
Горленко М.В. Мучнисторосяные грибы Московской области (семейство
216
Erysiphaceae). M.: Изд-во МГУ, 1983. 72 с.
Горчаковский П. Л. Широколиственные леса и их место в растительном покро-
ве Южного Урала. М., 1972. 146 с.
ГорышинаТ.К., АнтоноваИ.С., Самойлов Ю.С. Практикум по экологии расте-
ний. СПб.: Изд-во С.-Петербург, ун-та. 1992. 140 С.
Грант В. Видообразование у растений. М.: Мир, 1984. 528 с:
Гроздов Б.В. Лесные травы, их индикаторное, кормовое и лекарственное зна-
чение. М.: Гослесбумиздат, 1963. 71 с.
Гроссгейм А.А. Определитель растений Кавказа. М., 1949. 747 с.
Гроссгейм А.А. Растительные богатства Кавказа. М., 1952. 632 с.
Губанов И.А. Конспект флоры Внешней Монголии (сосудистые растения). М.,
1996. 136 с.
Гуланян ТА. Морфолого-анатомическое исследование жизненных форм неко-
торых лютиковых в онтогенезе. Дис. ... канд. биол. наук. М., 1976. 192 с.
Гуреева И.И. Онтогенез, структура и жизненность ценопопуляций Bidens
tripartita L. в Кузнецком Алатау (Кемеровская область) И Растительные ресурсы, 1992.
Вып. 3. С. 28—43.
Гусев Н.В. Вероники Кунгурского-Красиоуфимской лесостепи Предуралья и
перспективы их использования. Автореф. дис.... канд. биол. наук. Пермь, 1976.
Гусев Ю.Д. Дополнения к адвентивной флоре северо-западных областей евро-
пейской России И Ботан. ж., 1973. Т. 58, № 6. С. 904—909.
Девятов А.Г. Об отклонениях в числе стилодиев в цветках в трибе Lychnideae
Fenzl in Endlicher//Бюл. МОИП. Отд. Биология. 1988. Т. 93, вып. 2. С. 112—117.
Девятов А.Г. Морфолого-анатомические признаки генеративной сферы в сис-
тематике трибы LychnideaeFenzl.(Caryophyllaceae-Silenoideae)//Автореф. дис.... канд.
биол. наук. М., 1992.
Декоративное садоводство. М. 1949. 438 с.
Дементьева С.М. К экологии и биологии видов рода дремлик И Докл. МОИП.
Зоол. и ботан. 1988 г. Ресурсы живой природы, их использование и охрана. М., 1990. С.
69.
Джапаридзе Л.И. Пол у растений. Ч. 1. Тбилиси, 1963.
Доктуровский ВС. Межледниковый торфяник уд. Немыкары Западной облас-
ти И Бюл. МОИП. Нов. сер., 1935. Т. 43, отдел геол. Т. 13, № 1. С. 106—ИЗ.
Домбровская А.В., Шляков Р.Н. Лишайники и мхи севера Европейской части
СССР. Л.: Наука, 1967. 182 с.
Дудинский Я.А., Бажутина В.М. Особенности роста листьев Typha latifolia и
Sparganium polyedrum изначальных этапах их развития// Ботан. ж., 1976. Т. 61. С. 263—
266.
ЕвтюховаМ.А. Дикорастущие весенние цветы для садов и парков. М. 1968.126
с.
Елина Г.А., Кузнецов О.Л., Максимов А.И. Структурно-функциональная орга-
низация и динамика болотных экосистем Карелии. Л.: Наука, 1984. 128 с.
Жизнь растений. М.-Л., 1980. Т. 5(1).
Жмылев П.Ю., Алексеев Ю.Е., Карпухина Е.А. Основные термины и понятия
в современной биоморфологии растений. М.: Изд-во МГУ 1993. 148 С.
Жукова Л.А. Изменения возрастного состава популяции луговика дернистого
на окских лугах. Автореф. дис.... канд. биол. наук. М., 1976.
Жукова Н.А. Кариология некоторых видов Ranunculaceae в Полярио-Альпий-
ском ботаническом саду И Ботан. ж., 1961. Т. 46, № 3. С. 421—428.
217
Загульский М.Н. Орхидные Львовской области и их охрана//Охрана и культи-
вирование орхидей. Киев. 1983. С. 33—34.
Загульский М.Н. Влияние хозяйственной деятельности человека на ценопопу-
ляции орхидных в 3 ападных областях Украинской ССР // Биогеоценолог. исслед, на Ук-
раине. Тез. докл. 3 Республ. совещ. Львов, 1984. С. 124—125.
Загульский М .Н. Некоторые особенности репродуктивной биологии орхидных
в Западных регионах Украины//Теорет. и прикл. карпология. Тез. докл. Всесоюзн. конф.
Кишинев. 1989: С. 179—180.
Злобин Ю. А. Принципы и методы изучения ценотических популяций растений.
Казань, 1989. 145 с.
Игнатов М.С., Макаров В.В., Чичев А.В. Конспект флоры адвентивных расте-
ний Московской области // Флористические исследования в Моск, области. М.: Наука,
1990. С. 5—105.
Имс А. Морфология цветковых растений. М., 1964. 497 с.
Интродукция лекарственных, ароматических и технических растений. М.-Л.,
1965.425 с.
Исакова Т.И. Поиск желчегонных средств в растениях рода череда // Фарма-
ция. 1980а. №5. С. 31—35.
Исакова Т.И. Химическое и фармакологическое исследование флавоноидов и
полисахаридов растений рода череда: Автореф. дис. ... канд. фармац. наук. Харьков.
19806. 20 С.
Исакова Т.И., Сербин А.Г., Беликов В.В., Чушенко В.Н. Флавоноиды и полиса-
хариды видов р. Bidens L. //Растит, ресурсы, 1986. Т. 22, вып. 4. С. 517—523.
Каден Н.Н. Типы плодов растений средней полосы европейской части СССР /
/ Ботан. ж., 1965. Т. 50, № 6. С. 775—787.
Казарян В.О. Физиологические аспекты эволюции от древесных к травам. Л.:
Наука. 1990. 348 С.
Карписоиова Р.А. Травянистые растения широколиственных лесов СССР. М.:
Наука, 1985. 205 с.
Карпов В.Г., Шапошников ЕС. Еловые леса территории//Факторы регуляции
экосистем еловых лесов. Л.: Наука, 1983. 318 с.
Карташова Н.Н. Строение и функции нектарников цветка двудольных расте-
ний. Томск, 1965.
Карягин И.И. Результаты Карабахо-Зангезурской экспедиции по пересмотру
флоры на каучуконосность // Тр. Азерб. фил. АН СССР. Баку, 1936. Т. 2. С. 69—80.
Катанская В.М. Материалы для изучения продуктивности зарослей водных рас-
тений дельты Амударьи// Тр. Лаб. озероразведения АН СССР. 1960. Т. 10.
Катенин А.Е. Эндотрофная микориза у видов лилейных и лютиковых в Аркти-
ке И Ботан. ж., 1962. Т. 47, № 9. С. 1273—1282.
Кауфман Н.Н. Московская флора, или описание высших растений и ботанико-
географический обзор Московской губернии. М. 1866. 708 с.
Кац Н.Я., Кац С.В., Кипиани М.Г. Атлас и определитель плодов и семян, встре-
чающихся в четвертичных отложениях СССР. М. 1965. 367 с.
Кимеридзе К.Р. К изучению формации Scheuchzeria palustris L. в горах Сване-
тии // Сообщ. АН Груз. ССР. 1964. Т. 35, № 1. С. 171—178.
КиртокаВ.А. Новые для флоры Молдавии виды рода Epipactis Zinn (Orchidaceae)
//Изв. АН Молд. ССР. Сер. биол., хим. 1979. № 4. С. 23—25.
Киртока В.А., Истратий А.И. Орхидные государственного заповедника лесо-
218
охотничьего хозяйства «Реденский лес» Молдавской ССР // Охрана и культивирование
орхидей. М., 1987. С. 43—44.
Ключникова Н.М., Марков М.В. Гетерокарпия и некоторые другие особенно-
сти популяционной биологии двух видов из рода Bidens L. (Compositae) // Бюл. МОИП.
Отд. биол, 1994. Т. 99, вып. 1. С. 80—91.
Князева О.И. Интродукция видов семейства Orchidaceae Уральской флоры и
перспективы использования их для озеленения // Новые декоративные растения в куль-
туре на Среднем Урале. Свердловск. 1986. С. 31—38.
Кодаш А.Г., Пищулин А.Т., Спиридонова В.П., Бабий Ф.Р., Заугольников В.Б.,
Черныш В.Я. Череда трехраздельная И Возделывание лекарственных культур. 1987. №
4. С. 105—112.
Кожевников А.В. Весна и осень в жизни растений. М., 1950. 238 с.
Комарова Т.А. Семенное возобновление растений на свежих гарях (леса южно-
го Сихотэ-Алиня). Владивосток: Изд-во ДВНЦ СССР, 1986. 221 с.
Комжа А.Л., Попов К.П. Новые данные об адвентивной флоре Северной Осе-
тии И Ботан. ж., 1990. Т. 75, № 1. С. 108-110.
Кондратенко Б.С., Чернов Б.А. Ресурсы череды поникшей в Правобережной ле-
состепи // 2-я республ. конферен. по мед. ботанике. Киев. 1988. С. 61—62.
Кондратьева-Мельвиль Е.А, Развитие структуры в онтогенезе однолетнего дву-
дольного растения // Тр. Ленингр. о-ва естествоиспытателей. 1979. Т. 74, вып. 3.
Константинова Н А., Потемкин А.Д., Шляков Р.Н. Список печеночников и ан-
тоцеротовых территории бывшего СССР //Arctoa: A Journal of Bryology, 1992. Т. 1(1—
2). С. 87—127.
Коренева Н.Н. Разнообразие видов рода Veronica L. в Молдавии // Ботан. иссл.
АН МССР. 1989. Вып. 5. С. 92—99.
Коротков К.О. Леса Валдая. М.: Наука, 1991. 160 с.
Косенко И.С. Определитель высших растений Северо-Западного Кавказа и
Предкавказья. М, 1970. 613 с.
Косых В.М. Современное состояние популяций орхидных наюжном макроскло-
не Крымских гор и их охрана//Охр ан а и культивирование орхидей. Киев. 1983. С. 34—
35.
Красная книга Белорусской ССР. Минск, 1981. 281 с.
Красная книга Карелии. Петрозаводск, 1985. 183 с.
Красная книга РСФСР. М, 1988.
Красная книга СССР. М., 1984.
Краснова А. Н. Экология и фитоценология видов рода Typha L. // Гидробиол.
ж., 1988. Т 24. № 1.С. 8—11.
Краснова А.Н. К систематике рогоза узколистного натерритории Северо-запа-
даевропейской части России и Таджикистана// Деп. в ВНЦ «Верас» и Изв. АН Белорус-
сии. Минск. 09.06.1993. 14 с.
Крискович И.В., Старшова Н.П. Анализ различных типов половой дифферен-
циации на примере сорных гвоздичных // Успехи экологической морфологии растений
и ее влияние на смежные науки, М., 1994. С. 138—139.
Крупина М.Г. Колокольчики. М., 1954, 78 с.
Кулиев А.М. Задачи изучения медоносных и перганосных растений. М.-Л., 1952.
208 с.
Куприянова Л. А., Алешина Л. А. Пыльца и споры растений флоры европейской
части СССР. Л.: Наука, 1972. Ч. 2. 170 с.
219
Кучеров Е.В., Мулдашев А. А., Галеева А.К. Охрана редких видов растений иа
Южном Урале, М., 1987. 205 с.
Лавренко Е.М. О генезисе сфагновых болот в пределах степной зоны в бассей-
нах рек Буга, Днепра и Доиа // Сов. бот., 1936. Т. 3. С. 24—42.
Ларин И.В., Агабабян Т.А.,Работнов Т.А.,ЛаринаВ.К.,КасименкоМ.А.,Люб-
ская А.Ф. Кормовые растения сенокосов и пастбищ СССР. М.-Л., 1956. Т. 3. 897 с.
Лебедева Н.В. Связь растительного покрова с движением воды в болотных мас-
сивах// Ботан. ж., 1957. Т. 42, №4. С. 635—639.
Левина Р.Е. Способы распространения плодов и семян. Л. 1957.
Левина Р.Е. Морфология и экология плодов. Л.: Наука, 1987, 160 с.
ЛейслеФ.Ф.,МакароваН.А. Влияние различной влажности почвы нарост, раз-
витие и другие процессы двудомного растения Melandrium album // Труды Ботаи. ин-та
им. Комарова. Л. 1950, вып. 7. С. 221—237.
Леса Восточного Подмосковья. М.: Наука, 1979. 183 с.
Лесные травянистые растения. Биология и охрана: справочник. М., 1988. 223
с.
Лоопман А., Пайдла А. О лаидшафтно-экологических условиях в грядово-мо-
чажинных комплексах выпуклых олиготрофных болот // Изв. АН Эст. ССР. Биология.
1981. Т. 30, № 1. С. 62—73.
Лопатин В. Д. Зависимость значения pH и хода оттаивания от растительности и
микрорельефа на верховом болоте//Вести. ЛГУ Сер. геологии и географии, 1956. Вып.
2, № 12. С. 86—98.
Лопатин В.Д. Экологические ряды растительности болот И Структура расти-
тельности и ресурсы болот Карелии. Петрозаводск, 1983. С. 5—38.
Луговые травянистые растения. Биология и охрана: справочник. М., 1990. 183
с.
Любарский Е.Л. Роль длиннокориевищных видов в сложении травостоя в свя-
зи с влажностью почвы // Ботан. ж., 1963. Т. 48, № 3. С. 403—405.
Маевский П.Ф. Флора Средней России. М. 1892; изд. 2-е, 1895; изд. 3-е, 1902;
изд. 4-е, 1912; изд. 5-е, 1917; 8-е изд., 1954.; 9-е изд., 1964.
Мазинг В.В. О размножении и распространении растений верховых болот при
помощи семян // Ежегодник общества естествоиспытателей при АН ЭССР. Таллин,1955.
Вып. 48. С. 141—162.
Майтулина Ю.К. К биологии и систематике среднеазиатских видов рода Не-
дотрога//Бюл. ГБС АН СССР, 1988а. № 150. С. 59—64.
Майтулина Ю.К. Новые и редкие адвентивные растения Вологодской области
//Бюл. ГБС АН СССР, 1984. № 132. С. 45—46.
Майтулина Ю.К. Скорость микроэволюции биологических признаков недот-
роги железконосной//Проблемы микроэволюции. М.: Наука, 19886. С. 25—26.
Малышева В.Г. К адвентивной флоре Калининской области // Экология расте-
ний южной тайги. Калинин, 1979. С. 48—54.
Малышева Г.М. Введение в культуру дикорастущих многолетников из семей-
ства лютиковых в Томске// Растения природной флоры Сибири для зеленого строитель-
ства. Новосибирск. 1972. С. 73—77.
МарковМ.В. Алгоритм популяционно-ботанического анализамалолетних рас-
тений: архитектурная модель — жизненная форма — эколого-ценотическая стратегия
//Биол. науки, 1989. № 11. С. 90—104.
МарковМ.В. Популяционная биология розеточных и полурозеточныхмалолет-
220
них растений. Казань: Изд-во Казан ун-та, 1990. 186 с.
Марков М.В. Популяционная биология недотроги обыкновенной Impatiens noli-
tangere L. // Экология, 1991. № 1. С. 17—26.
Марков М.В. Структура и популяционная биология малолетних растений Цен-
тра Русской равнины: Дис. ... доктора биол. наук. М., 1992. 328 С.
Марков М.В. Сравнительная характеристика морфологии корневых систем у
двух видов Недотроги Impatiens noli-tangere L. и1. parvifloraDC. (Balsaminaceae) на ран-
них этапах развития //Флора и растительность Тверской области, Тверь, 1994. С. 64—
68.
Марков М.В., Ключникова Н.М. Опыт изучения связи архитектуры с пластич-
ностью у длиннопобегового однолетника Bidens tripartita L. // Успехи эколог, морфоло-
гии растений и её влияние на смежные науки. М.: Изд-во МПГУ им. В.И. Ленина. 1994.
С. 104—105.
Марков М.В., КлючниковаН.М. Биологические следствия гетерокарпииудвух
видов рода Bidens L. //Научные труды МПГУ им. В. И. Ленина. Сер. Естественные нау-
ки. М.: Прометей, 1995а. С. 7—16.
Марков М.В., Ключникова Н.М. Связь архитектуры с пластичностью у длин-
нопобеговых однолетников Bidens cemuaL. и Bidens tripartitaL. //Актуальные вопросы
экологической морфологии растений. Межвузовский сб. научных трудов. М.: Проме-
тей, 19956. С. 35—40.
Миркин Б.М., Соломещ А.И., Ишбирдин А.Р., Алимбекова Л.М. Список и ди-
агностические критерии высших единиц эколого-флористической классификации рас-
тительности СССР (оперативно-информационный материал). М. 1989. 46 С.
Михайлова Л.Н. К экологии и биологии Typha angustifolia (Рогоза узколистно-
го) в дельте р. Волги И Тр. Астраханского гос. заповедника. М., 1940. Вып. 3. С. 116—
137.
Мишкин Б.А. Флора Хибинских гор. М.-Л., 1953. 112 с.
Назаров В.В. Семенная продуктивность некоторых орхидей, произрастающих
в Крыму// Охрана и культивирование орхидей. М., 1987. С. 38—39.
Наугольных В. Н. О половом диморфизме двудомных растений // Изв. ДАН СССР,
1945. Т. 49.
НаугольныхВ.Н.,БурковаТ.Н. О физиологических отличиях двудомных расте-
ний на ранних фазах их развития И Известия АН СССР, 1951, вып. 4.
Нейштадт М.И. Определитель растений средней полосы европейской части
СССР. М. 1963. 640 с.
Николаева М.Г., Разумова М.В., Гладкова В.И. Справочник по проращиванию
покоящихся семян. Л.: Наука, 1985. 348 с.
Новикова Н.Г. Использование рогоза на крахмал // Природа. 1940. Т. 29., № 5.
С. 58—60.
Оганесян М.Э. Семейство Campanulaceae Juss. На Кавказе. Автореф. дис. ...
канд. биол. наук. Ереван, 1995. 50 с.
Определитель растений Мещеры. М.: Изд-во МГУ, 1986, 1987. Ч. 1,2.
Определитель растений Средней Азии. Ташкент, 1987.
Определитель сосудистых растений Среднего Урала. М.: Наука. 1994. 525 с.
Определитель сосудистых растений Центра европейской России. М., 1995.560
с.
Орлов Б.Н., Гелашвили Д.Б., Ибрагимов А.К. Ядовитые животные и растения
СССР. М.: Высш, шк., 1990. 272 с.
Пащенко Н.Ф. Тли рода Aphis (Homoptera, Amphidinea, Aphididae), живущие
221
на растениях семейств Apiaceae,Balsaminaceae,Caryophylaceae,ChenopodiaceaHa Даль-
нем Востоке России. 2. // Зоол ж., 1993. Т. 72, вып. 7. С. 63—79.
Первухина Н.В. Проблемы морфологии и биологии цветка. Л.: Наука, 1970.
Петунников А.И. Критический обзор Московской флоры. СПб, 1896, ч. III. 220
с.
Петунников А.Н. Критический обзор московской флоры. Gamopetalae//Tp. СПб
о-ва естествоиспыт. 1900. Т. 30, вып. 2 (отд. ботан.). С. 1—142.
ПлискоМ.А. Семейство Scheuchzeriaceae//Сравнительная анатомия семян. Л.,
1985. Т. 1: Однодольные. С. 47—49.
ПобедимоваЕ.Г. О новых видах рода Typha// Ботан. матер. Гербария Ботан. ин-
та им. В. Л. Комарова АН СССР. Л., 1949. Т. 11. С. 3—17.
Поддубная-Арнольди В .А. Характеристика семейств покрытосеменных расте-
ний по цитоэмбриологическим признакам. М.: Наука, 1982. 351 с.
ПолевойВ.В., СаламатоваТ.С. Физиология роста и развития растений. Л : Изд-
во ЛГУ 1991. 240 с.
Поляков П.П. Систематика и происхождение сложноцветных. Алма-Ата: Нау-
ка. 1967. 336 с.
Полянский И М. Ботанические экскурсии. М.: Учпедгиз, 1950. 292 с.
Попцов А.В., Буч Т.Г. Температурный фактор в прорастании семян некоторых
видов череды // Бюл. Главн. ботан. сада АН СССР, 1969. Вып. 72. С. 67—69.
Попцов А.В., Буч Т.Г. Затрудненное прорастание семян некоторых видов чере-
ды и температурный фактор //Бюл. Главн. ботан. сада АН СССР, 1970. Вып. 77. С. 100—
103.
Похилько А. А. Семейство Orchidaceae—орхидные // Биол. флора Мурманской
области. Апатиты, 1993. С. 35—47.
Прядка Е.И. Охрана орхидных в природных национальных парках Украинско-
го Полесья // Охрана и культивирование орхидей. М., 1987. С. 28—29.
, Пучин В.М. Некоторые вопросы производства череды трехраздельной в совхо-
зах Союзлекраспрома//Науч-техн. реферат, сб. Сер. Лекарственное растениеводство.
М. 1977. №1. С. 7—9.
Работнов ТА. Типы серо-ольховых насаждений северо-западной части Москов-
ской области //Ботан ж., 1939. № 1.
Работнов ТА. Наблюдения над содержанием нитр атов в луговых и лесных рас-
тениях И Сов. бот., 1940, № 1.
Работнов ТА. Ranunculaceae Juss. —Лютиковые // Кормовые растения сеноко-
сов и пастбищ СССР. М. 1951. Т. 2. С. 327—383.
Работнов ТА. Некоторые вопросы изучения ценотических популяций // Бюл.
МОИП. Отд. биол., 1969. Т. 74, вып. 1. С. 141—149.
Работнов ТА. Фитоценология. М.: Изд-во МГУ, 1983. 292 с.
Работнов ТА. Жизнеспособные семена в почвах природных биогеоценозов //
Бюл. МОИП. Отд. биол, 1986. Т. 91. С. 3—18.
Разумова М.В., Поздова Л.М. О возможности ускоренного проращивания не-
которых представителей рода Campanula L. И Вопросы обогащения генофонда в семе-
новодстве интродуцентов. М., 1987. С. 100—101.
Раменский Л.Г., Цаценкин И.А., Чижиков О.Н., Антипин Н.А. Экологическая
оценка кормовых угодий по растительному покрову. М., 1956. 471 с.
Растения Псковской области, подлежащие охране. Псков, 1977. 20 с.
Растительные ресурсы России и сопредельных государств: Цветковые расте-
ния, их химический состав, использование. Сем. Butomaceae — Typhaceae (ред. Соко-
222
лов П.Д.) Л., 1997. С. 155—159.
Растительные ресурсы СССР. Цветковые растения, их химический состав, ис-
пользование. Сем. Magnoliaceae — Limoniaceae. Л. 1984. 460 с.
Растительные ресурсы СССР: Цветковые растения, их химический состав, ис-
пользование. СПб: Наука. 1993.
Растительный мир Удмуртии. Ижевск, 1980. 71 с.
Растительный покров Национального парка Литовской СССР. Вильнюс: Мокс-
лас, 1988. 164 с.
Рубцов Н.И., Привалова Л. А. Опыт сопоставления флор горного Крыма и За-
падного Закавказья // Тр. Гос. Никитского ботан. сада. 1961. Т. 35. С. 5—63.
Рысин Л.П. Сложные боры Подмосковья. М.: Наука, 1969. 111 с.
Рысин Л.П. Сосновые леса европейской части СССР. М.: Наука, 1975. 212 с.
Рысин Л.П., Рысина Т.П. Морфострукгура подземных органов лесных травя-
нистых растений. М. 1987. 207 с.
Рысина Г.П. Ранние этапы онтогенеза лесных травянистых растений Подмос-
ковья. М. 1973. 216 с.
Рычии Ю.В. Флора гигрофитов. М. 1948. 448 с.
Савиных Н.П. Большой жизненный цикл и сезонное развитие Veronicaofficinalis
Ь.//Бюл. МОИП. Отд. биол., 1978. Т. 83, вып. 4. С. 123—133.
Савиных Н.П. Побегообразование и взаимоотношения жизненных форм в сек-
ции Veronica рода Veronica L.//Бюл. МОИП. Отд. биол. 1979а. Т. 84, вып. 3. С. 92—105.
Савицкий В. Д. Морфология, классификация иэволюция пыльцы семейства лю-
тиковых. Киев. 1982. 124 с.
Саксонов С.В., Терентьева М.Е. Орхидные Жигулевского заповедника// Охра-
на и изучение редких видов растений в заповедниках. М., 1992. С. 81—91.
СацыпероваИ.Ф. Народно-лекарственные растения из семейства лютиковых и
их антибактериальные свойства. Дис.... канд. биол. наук. Л. 1948.
Серебряков И.Г. О ритме сезонного развития растений Подмосковных лесов /
/ Вест. Моск, ун-та, 1947. № 6. С. 75—108.
Серебряков И.Г. Морфология вегетативных органов высших растений. М. 1952.
391с.
Серебряков И.Г. Экологическая морфология растений. М.: Высшая школа, 1962.
Серебряков И.Г. Жизненные формы высших растений и их изучение// Полевая
геоботаника. Л., 1964. Т. 3. С. 146—205.
Серебряков И.Г, Галицкая Г. М. К биологии сезонного развития болотных рас-
тений Подмосковья в связи с условиями их жизни и происхождения // Уч. зап. Моск,
гор. пед. ин-та, 1951. Т. 19, вып. 1. С. 19—47.
Сидорский А.Г. Генетическиеосновы половой дифференциации покрытосемен-
ных растений// Успехи совр. биологии, 1983. Т. 96, вып. 4(5).
Скворцов А.К., Тихомиров В.Н. Редкие, исчезающие и нуждающиеся в охране
виды сосудистых растений Московской области//Бюл. МОИП. Отд. биол., 1986. Т. 91,
вып. 6. С. 11—118.
Смирнова Е С. Морфология побеговых систем орхидных. М., 1990. 208 с.
СмирноваО.В. Структура травяного покрова широколиственного леса. М.: Нау-
ка, 1987, 208 с.
Собко В.Г. Opxluei Украши. Кшв, 1989. 192 с.
Собко В.Г., Нефедова О.Н. Epipactis palustris (L.) Crantz в природел первичной
223
культуре // Охрана и культивирование орхидей. Киев, 1983. С. 45—47.
Сокол А.П. Экологические шкалы болотных растений (Болота северо-восточ-
ной частиПечеро-Илычскогозаповедника)//Ботан. ж., 1981. Т. 66,№2. С. 227—233.
Соколовская А. Значение микроморфологических признаков в систематике не-
которых видов водных растений//Журн. русского Ботан. общества, 1930. Т. 15, № 1—
2. С. 35—61.
Солоневич Н.Г. Материалы к эколого-биологической характеристике болотных
трав и кустарничков // Растительность Крайнего Севера СССР и ее освоение. М.-Л.,
1956. Вып. 2. С. 307—497.
Сравнительная анатомия семян: Однодольные. Л., 1985. Т. 1. 317 с.
Стоянов Н., Китанов Б. Диви полезни растения в България. София. 1960.480 с.
Судакова Е.А. Морфогенез вегетативных органов колокольчика широколист-
ного //Научные труды акад. комм, хоз-ва, 1976. Вып. 126. С. 79—86.
Судакова Е.А. Онтогенетический морфогенез некоторых представителей раз-
ных жизненных форм Campanula L. в культуре // Изв. Тимир. сельск. акад., 1983. Вып.
3. С. 48—51.
Сырейщиков Д.П. Иллюстрированная флора Московской губернии: В 4-х ч. Ч.
1. М., 1906; Ч. 2. М., 1907; Ч. 3. М., 1910; Ч. 4. М., 1914.
Сырейщиков Д.П. Определитель растений Московской губернии. М., 1927.294
с.
Талла Б. Орхидные, включенные в Красную книгу Латвийской ССР // Охрана
и культивирование орхидей. 1980. Таллин. С. 31—34.
Татаренко И.В. Орхидные России: жизненные формы, биология, вопросы ох-
раны. М„ 1996. 207 с.
Тахтаджян А.Л. Система и филогения цветковых растений. Л., 1966. 611 с.
Тахтаджян А.Л. Система магнолиофитов. Л.: Наука, 1987. 439 с.
Тимченко I.A. Особливост! в!ково! структуры ценопопулящй корневищных
видтв родини Orchidaceae//Te3. доп. 1Хз’1зду Укр. ботан. т-ва. Ки!в, 1992. С. 167—168.
Тимченко I. А. Стан ценопопулящй видав роду Epipactis Zinn. (Orchidaceae) флори
Украши И Укр. ботан. ж., 1993. Т. 50, № 3. С. 102—107.
Тимченко I.A. Стаи вивченосп популящй вид!в родини Orchidaceae Juss. //Укр.
ботан. ж., 1992. Т. 49, № 3. С. 64—71.
Тимченко I.A. Epipactis palustris (L.) Crantz (Orchidaceae) в Украгш // Тез. доп.
конф. «Охорона генофонду рослин в Укра'пп. Донецьк, 1994. Т. 49. С. 64—65.
Тимченко И.А. Структура ценопопулящй вид!в трибы Neottieae Lindt.
(Orchidaceae Juss.) флори Украши i теиденцп п змши в умовах синантротзацп. Авто-
реф. дис.... канд. бюл. наук. Кшв, 1996. 21 с.
Тихомиров Б.А. О лесной фазе в послеледниковой истории растительности се-
вера Сибири и ее реликтах в современной тундре // Материалы по истории флоры и рас-
тительности СССР. М.-Л., 1941. Вып. 1. С. 315—369.
Тихомиров В.Н. Новые материалы к флоре Воронежской области // Биол. нау-
ки, 1987. Т. 6. С. 74—78.
Тихонова В.Л., Викторов В.П., Макеева И.Ю. Научио-методические рекомен-
дации по размножению охраняемых видов рода Campanula L. (колокольчик). М., 1989.
68 с.
Тихонова В.Л., Викторов В.П., Макеева И.Ю., Яшина С.Г. Влияние низких и
сверхнизких температур хранения на лабораторную всхожесть семян дикорастущих тра-
вянистых растений. П. Семена видов родаСатрапи1аЬ. (колокольчик). Проблемы крио-
биологии, 1991. № 1. С. 43—49.
224
ТихоноваВ. Л., Викторов В.П., Разумова М.В., Поздова Л,В. Внутривидовая из-
менчивость семян Campanula latifoliaL. в природных популяциях Подмосковья и в куль-
туре//Раст. ресурсы., 1989. Т. 27, № 1. С. 61—67.
Ткачик В.П. Видовой состав, распространение и модификационная изменчи-
вость видов Typha (Typhaceae) в Прикарпатье И Ботан. ж. 1993. Т. 78. С. 84—93.
Толмачев А.И. Семейство Orchidaceae//Арктическая флора СССР. М.-Л„ 1963.
Вып. 4. С. 73—85.
Фегри К., Ван дер Пейл Л. Основы экологии опыления. М. 1982. 379 с.
Федорчук В.Н., Бурневский Ю.И. Рекомендации по определению и использо-
ванию типов леса при лесоустройстве (на примере Ленинградской области). Л.: Лен-
НИИЛХ, 1986. 71 с.
Федосеев Г.Е. Сравнительная анатомия семенной кожуры и ультраструкгуры
поверхности семян некоторых видов родов Melandrium и Silene // Источники информа-
ции в филогенетической систематике растений. М.: Наука, 1986. С. 83—85.
Флора европейской части СССР. Л., 1974—1989. Т. 1—8.
Флора Западной Сибири. Томск, 1966. 105 с.
Флора Кавказа. Л. Т. 7,1967.
Флора северо-востока европейской части СССР. Л., 1976. Т. 2.
Флора Сибири. Новосибирск, 1987—1988. Т. 1—2.
Флора СССР. М.; Л., 1934. Т. 1; 1935. Т. 3,4; 1936. Т. 6; 1937. Т. 7; 1949. Т. 14;
1955, Т. 22; 1957, Т. 24; 1959. Т. 25.
Хохлов С.С., ЗайцеваМ.И., Куприянов П.Г. Выявление апомикгичных форм во
флоре цветковых растений СССР. Изд-во Саратовского ун-та, 1978. 306 с.
Хребтов А.А. Полезные и вредные растения Урала. Свердловск, 1941. 256 с.
Хромосомные числа цветковых растений. Л. 1969. 927 с.
Цвелев Н.Н. Заметка о роде CallitricheL. в СССР//Нов. сист. высш, раст., 1975.
№ 12. С. 237—238.
Ценопопуляции растений: основные понятия и структура. М., 1976. 214 с.
Ценопопуляциирастений: развитие и взаимоотношения. М.: Наука, 1977.134
с.
Цингер В.Я. Сборник сведений о флоре Средней России. М., 1885. 520 с.
Цыганов Д.Н. Экоморфы флоры хвойно-широколиствеиных лесов. М. 1976.60
с.
Цыганов Д.Н. Фитоиндикация экологических режимов в подзоне хвойно-ши-
роколиственных лесов. М. 1983. 196 с.
Черепанов С.К. Сосудистые растения России и сопредельных государств (в пре-
делах бывшего СССР). СПб, 1995. 87 с.
Черных П.Я. Историко-этимологический словарь современного русского язы-
ка. М„ 1994. Т. 2. 118 с.
ЧетвериковаН.И. О направленности продвижения ассимилятов из листьев раз-
ных ярусов в онтогенезе подсолнечника И Изв. Сибирского отд. АН СССР, 1965. Т. 4, №
1.С. 68—74.
Числа хромосом цветковых растений флоры СССР. Moraceae Zygophyllcceae.
СПб, 1993. 430 с.
Шмальгаузен И. Флора Средней и Южной России, Крыма и Северного Кавка-
за. Киев, 1895. Т. 1.
ШулькинаТ.В. Особенности сезонного развития некоторых видов р. Campanula
L. //Ботан.’ж., 1975. Т. 60, № 10. С. 109—117.
Щербаков А.В. Конспект флоры водоемов Московской области // Флористиче-
225
ские исследования в Московской области. М., 1990. С. 106—120.
Юдин Ю.П. Реликтовая флора известняков северо-востока европейской части
СССР//Материалы по истории флоры и растительности СССР. М.-Л., 1963. Вып. 4. С.
493—571.
ЮдинаВ.Ф., МаксимоваТ. А. Сезонное развитие растений болот. Петрозаводск,
1993. 168 с.
Юрцев Б. А. Жизненные формы: один из узловых объектов ботаники // Пробле-
мы экологической морфологии растений. М. 1976. С. 9—54.
Ядовитые растения лугов и пастбищ. М.-Л., 1950. 526 с.
Aario L. Vegetation und Postglaziale Geschichte Nurmijarvi-Sees // Ann. Soc. Bot.,
Zool. Bot. Fennicae «Vanamo». Helsinki, 1933. T. 3., N 2. S. 115—124.
Abrams L. Illustrated flora of the Pacific States. Stanford: Stanf. Univ. Press, 1951.
V. 3. 267 p.
Alm T. Floraen i Finmark: 4. Marihandfamilien {Orchidaceae) //Polarflokken. 1992.
V. 16, N 1. S. 89—148.
Ansola G., Fernandez C., de Luis E. Removal of organic matter and nutrients from
urban wastewater by using an experimental emergent aquatic macrophyte system //Ecol. Eng.
1995. V 5. P. 13—19.
Arber A. Water plants. A study of aquatic angiosperms. New York, 1963. 436 p.
Archzewska A. Wstepne stadia rozwojowe storczykow europejskich w kulturze za-
chowawczej in vitro//Acta Univ. Wratislaviensis. 1993. Pr. Bot. V. 57, N1515. P. 215—221.
Arditti J., Michaud J.D., Oliva A.P. Seed germination of North American orchids. 1.
Native California and related species of Calypso, Epipactis, Goodyera, Piperia and
Platanthera // Bot. Gaz. 1981. V 142, N 4. P. 442—453.
Ascherson P., Graebner P. Synopsis der mitteleuropflischen Flora. 1913, Bd. 2. Aufl.
1.
AschersonP., Graebner P.FamilieTyphaceae//Synopsis der mitteleuropflischen Flora.
1898. Bd. 1.S.270—278.
Austad I., Skogen A. Restoration of a deciduous woodland in Western Norway formerly
used for fodder production: effects on tree canopy and field layer// Vegetatio, 1990. V. 88, N
l.P. 1—20.
Backitus I. Aboveground production and growth dynamics of vascular bog plants in
Central Sweden // Acta phytogeogr. Suecica., 1985. V. 74. P. 1—98.
BakerH.G. Melandriumalbum(Mill.)Garke//J.Ecol., 1947.V.35,N 1,2. P. 274—
282.
Barkworth M.E. Impatiensparviflora in British Columbia/ZMadrodo, 1973. V. 22,
N l.P. 24.
Bartolo G. Epipactis palustris (L.) Crantz, nuovo reperto per la flora sicala // Arch,
bot. ital. 1991. V. 67, N 3—4. P. 121—127.
Batousek P. Epipactis muelleri ,E. helleborine,E. palustris na secundamim stanovisti
v okrese Gottwaldow // Zpr. Cs. bot. spolec., 1985, V. 20, N 3, P. 234—236.
Beerling D.J., Perrins J.M. Impatiens glandulifera Royle (Impatiens roylei Walp.) /
/J. Ecol., 1993. V. 81, N 177. P. 367—382.
Beerling D.J., Woodward F.I. Climate change and the British scene // J. Ecol., 1994.
V. 82. P. 391—397.
Bellamy D. J. Some observations on the peat bogs of the wilderness of Pisz // Przegl.
Geogr., 1962. V. 34. P. 691—716.
Bocquet G. The structure of the placental column in the genus Melandrium //
Phytomorphology. 1960. V. 9, N 3. P. 217—222.
226
Brantjes N.B.M. Riddles around the pollination of Melandrium album (Caryo-
phyllaceae) during the oviposition by Nadena bickuris Hufu. (Noctuidae, Lepidoptera) //
Proc. Konikl. Nederl. Akad. Wot. Ser. C. 1976. V. 79, N 2. P. 127—141.
В ringer K.G. Den lagalpina Dryas—hedem differenting och standortsekologi inom
Tometraskom radet // Sven. bot. tidskr. 1961. P. 55, N 2. S. 349.—375.
Britton N.L. Scheuchzeriaceae //North American Flora, 1909. V. 17, Pt. I. P. 42.
Brunel B., Janse J.D., Laanbroek H.J., Woldendorp J.W. Effect of transient oxic
conditions on the composition ofthe nitrate-reducing community from the rhizosphere of Typha
angustifolia // Microb. Ecol., 1992. V. 24. P. 51—61.
Brunton D.F. The helleborine, Epipactis helleborine (Orchidaceae), in Northern
Ontario//Can. Field-Natur., 1986. V. 100,Nl.P, 127—130.
Buchenau F. Scheuchzeria L. // In: A. Engler. Das Pflanzenreich. Regni vegetabilis
conspectus, Heft 16, 1903. P. 14—15.
BurgeffH. SamenkeimungderOrchideenundEntwicklungihrerKeimpflanzen. Jena,
1936.312 s.
Burques A., Corbet S.A. Do flowers reabsorb nectar? // Functional Ecology, 1991. V.
5,N3. P. 369—379.
Catling P.M. Autogamy in Easrem Canadian Orchidaceae: a review of current
knowledge and some new observation //Natur. Can., 1983. V. 110, N 1. P. 37-53.
CelakovskyL. Uber die Inflorescenz von Typha // Flora. 1885. N 35. S. 617—632.
ClaassenP.W. Typha insects: their ecological relationships. New York, 1921.P. 463—
508.
Clarkson R.B. Scheuchzeria palustris L. var. americana // Castanea, 1961. V. 26. P.
102—103.
Cook Cr.D.K. Aquatic plants endemic to Europe and the Mediterranean // Bot. Jahrb.
Sys., 1983. V. 103, N 4. P. 539—582.
CoombeD.E. Biological floraoftheBritish Isles. ImpatiensparvijloraVtC.il J. Ecol.,
1956. V. 44, N3. P.701—713.
Coops H., Boeters R., Smit H. Direct and indirect effects of wave attack on helophytes
// Aquatic Botany. 1991. V 41. P. 333—352.
Coops H., Van der Velde G. Seed dispersal, germination and seedling growth of six
helophyte species in relation to water-level zonation//Freschwat. Biol., 1995. V. 34.P. 13—
20.
Csontos P. Phytosociological description of a hilly country stand of Impatiens par-
viflora DC. H Stud. bot. hung., 1986. V 19. P. 115—118.
Dacshini K.M.M., Aggarwal S.K. Intracapitular cypsele dimorphism and dormancy
in Bidens bipinnata // Biol, plant., 1974. V. 16, N 6. P. 469—471.
Daumann E. Zur Bestaubungs- und Verbeitungsukologie dreier Impatiens-Arten //
Preslia (Praha), 1967. V. 39. P. 43—58.
Delforde P., Tyteca D. Europaische wildorchideen. Zurich-Koln, 1984.
Dickerman J. A., Wetzel R.G. Clonal growth in Typha latifolia'. Population dynamics
and demography ofthe ramets H J. Ecol., 1985. V. 73. P. 535—552.
Dierssen K, Reichelt H. ZurGleiderungdes Rhynchosporion albae W. Koch 1926 in
Europa//Phytocoenologia, 1988, 16(1). S. 37—104.
Dierssen K., Mierward U. Atlas der Flora Schleswig-Holsteins und Hamburgs.
Wachholz-Verlag, 1982. 654 S.
Doroczewska A. Tetrapioid Trollius europaeus L. H ActaSoc. bot. pol„ 1964. V 33,
N. l.P. 9—15.
Dowden A.O. To pollinate an orchid // Audubon, 1975. V. 77, N 5. P. 40—47.
227
Doyon D., Cayonette R. Epipactis helleborine (L.) Crantz au Quebec // Natural.
Canad., 1966. V. 93. P. 171—176.
Dvorak J. An example of pelationships between macrophytes, macroinvertebrates
and their food resources in a shallow eutrophic lake // Hydrobiologia, 1996. V. 39. P. 27—36.
EberE. Karpellbauund Plazentationsverhnjtuisse in derReihederHelobiae. Miteinen
Anhang bber die Verwandtscaftlichen Beziehungen zwischen Ranales und Helobiae // Flora
Neue Folge, 1934. Bd. 27. S. 273—330.
EliaS P. Size inequality in coenopopulations of a wood annual Impatiens noli-tan-
gere L. // Biologia (Bratislava), 1987. V. 42, N 9. P. 881—891.
Ellenberg H. Zeigerwerte der Gefnsspflanzen Mitteleuropas // Scripta geobotanica.
1974. Bd. 9.
Ellenberg H. Vegetation Mitteleuropas mit den Alpen in ukologischer Sicht. Stut-
tgart: Ulmer, 1986. 990 S.
Ellenberg H., Weber H E., Dull R., Volkmar W., Willg W., Dirk P. Zeigerwerte von
Pflanzen in Mitteleuropa // Scripta Geobotanica. 1991. Bd. 18. 248 S.
Ennos A.R., CrookM.J., GrimshawC. A comparative study ofthe anchorage systems
of Himalayan balsam Impatiens glandulifera and mature sunflower Helianthus annuus // J.
of Exper. Bot„ 1993. V. 44. P. 133—146.
Evans G.C., Hughes A.P. Plant growth and aerial environment. I. Effect of artificial
shading on Impatiens parviflora DC. //NewPhytol, 1961. V. 60, N 2, P. 150—180.
Falencka M. The effect of the size of cotyledons on the development of individuals
inlmpatiens noli-tangere L. population//EkologiaPolska, 1983. V. 31,N 1. P. 123—143.
Fernaid M.L. The American variety of Scheuchzeria palustris II Bhoiotz., 1923. V.
25. P. 177—179.
Flora Europaea. London, New York, Melbourne. 1976. V. 4; 1980. V. 5.
Forsyth C., Brown N.A.C. Germination of dimorphic fruits of Bidens pilosa L. //
New. Phytologist, 1982. V. 90, N 1. P. 151—164.
' Frank D., Klotz S. Biologisch — okologische daten zur flora der DDR // Unter
Mitarbeit von Werner WesthusMartin-Luther-universitat Halle-Witenbergwissenschaftliche
beitrage. 1988. Bd. 60. 35 S.
Fuller F. Epipactis und Cephalanthera H Orchideen Mitteleuropas. Leipzig: Neue
Brehm-Bucherei. 1986. Bd. 5.
Gadella T. W. J. Cytotaxomic studies in the genus Campanula I I Wentia, 1964. Nil.
S. 1—104.
Gelbrecht J., Hamel G. Eine bisher unbeschreiben Epipactis — Sippe // Gleditschia,
1987. V. 15,N2. P. 203—204.
Geze J.B. Etudes botaniques etagronomiques surles Typha et quelques autres plantes
palustres. Villefranche-de-Rouergue. 1912. 174 p.
Gillet G.W., Lim E.K.S. An experimental study ofthe Genus Bidens (Asteraceae) in
the Hawaiian Islands //Berceley and Los Angeles: University of California press. 1970. V. 56.
63 p.
Gltlck H. Pteriodophyten und Phanerogamen unter gleichzeitiger Berbcksichtigung
der wichtigsten Wasser — und Sumpfgewachse des ganzen Kontinents von Europa. Die
Sbsswasserflora Mitteleuropas 15, 1936.
Goebel K. Blbtenbildung und sprossgestaltung (authohladen und inflorescenzen).
Jena: G. Fischer, 1931. S. 219—220.
Grace J.B., Harrison J. S. The biology of Canadian weeds. 73 //Can. J. Bot., 1986. V.
66. P. 361—379.
Grace J.B., Wetzel R.G. Effects of size and growth rate on vegetative reproduction
228
in Typha H Oecologia, 1981a. V. 50. P. 158—161.
Graebner P. Typhaceae П In: A. Engler. Das Pflanzenreich... 1900. V. 2. N 6—8. S.
9—16.
GraebnerP., MedlewskaE.,ZinzA. Typha alsNutzpflanze//AngewandteBot. 1919.
Bd. 1. S. 30—48, 98—99.
Grime J.P. Plant strategies and vegetation processes. Chichester: J. Wiley, 1979.
Grime J.P., Mason G., Curtis A.V. et al. A comparative study of germination
characteristics in a local flora// J. Ecol., 1981. V. 69. P. 1017—1059.
HagerupO. Notes on some boreal polyploids//Hereditas, 1974. V. 30. P. 152—160.
Hawkes A.D. Economic importance ofthe Orchidaceae H Am. Orch. Soc. Bull. 1943.
V. 12. P. 412-415.
Hawkes A.D. Economic importance ofthe Orchidaceae //Am. Orch. Soc. Bull. 1944.
V. 13. P. 56-58.
Hegelmaier C.F. Monographie derGattung Callitriche. Stuttgart, 1864.
HegiG. Illustrierte Flora von Mittel-Europa. Mbnchen: Lehmann. 1909. Bd. 2,H. 1;
1923. Bd. 5.
Heiser C.B., Whitaker T.W. Chromosome number, polyploidy, and growth habit in
California weeds П Amer. J. Bot., 1948. V. 35. P. 179—185.
Henry H.K. Scheuchzeria in Western Pennsylvania//Rhodora, 1963. V. 65, N 761,
P. 34.
Hibatsuka A., Inoue O. Dispersability of dimorphic seeds in Impatiens noli-tangere
and I. textori (Balsaminaceae) //Ecological Review, 1988. V 21. P. 157—161.
Hogue E.J. Seed dormancy of nodding beggarticks (Bidens cernua L.) // Weed. Sc.,
1976. V. 24, N4.P. 375—378.
Hooker W. J. Flora Boreali-Americana; or the botany of theNorthem parts of British
America. London, 1840. V. 2. P. 168.
Hoppe D.H. Botanisches Taschenbuch. Regensburg, 1792. S. I—VIII, 1—248.
Hughes A.P. The importance of light compared with other factors affecting plant
growth // Light as an ecological factor. Oxford: Blackwell, 1966. P. 121—146.
Hughes A.P., Evans G.C. Plant growth and the aerial environment. II. Effects of light
intensity on Impatiens parviflora DC. //New Phytol., 1962. V. 61, N 2. P. 154—174.
IrmishN. BeitragezurBiologie undMorphologiederOrchideen. Leipzig, 1853.828
S.
Jones Q., Earle F.R. Chemical analyses of seeds II. Oil and protein content of 759
species // Econ. Bot., 1966. V. 20, N 1. P. 127—155.
Jordan T.E., Whigham D.F., Correll D.L. Effects of nutrient and litter manipulations
on the narrow-leaved cattail, Typha angustifolia L. // Aquatic Botany, 1990. V. 36. P. 179—
191.
Jouret M.F. Ideologic de la dormance sitminale et de la germination chez diverse
ifspeces du genre Impatiens L. // Bui. Soc. Roy. bot. Belg. 1976. V. 109. P. 213—229.
Jouret M.F. Relation entre la dormance sfiminale et la chorologie de diverses espiices
du genre Impatiens L. // Bull. Soc. Roy. bot. Belg., 1977. V 110. P. 119—128.
Judd W.W. Wasps, (Vespidae) pollinating helleborine, Epipactis helleborine (L.)
Crantz at Owen Sound, Ontario //Proceed. Entomol. Soc. Ontario. 1972. V. 102. P. 115—118.
Kaminski P., Sarosiek J. Considerations on the age structure of Epipactis palustris
(Mill.) populations//Acta Univ. Wratislawiensis. 1990, N 1055. P. 181—188.
Keddy P.A., Constabel P. Germination often shoreline plants in relation to seed size,
soil particle size and water level: An experimental study//J. Ecol. 1986. V. 74.P. 131—141.
229
Keddy P. A., Ellis Т.Н. Seedling recruitment of 11 wetland plant species along a water
level gradient: Shared or distinct responses? // Can. J. Bot. 1985. V 63. P. 1876—1879.
KhoshooT.N. Cytology of some Impatiens species//Caryologia. 1957. V. 10. P. 55—
74.
Kinzel W. Frost und Licht als beeinflusende Кгдйе bei der Samenkeimung. Stuttgart,
1913.
Kirchner O., Loew E., Schruter C. Lebensgeschichte der BlbtenpflanZen
Mitteleuropas. 1906. Bd. I, Abt. 1. Lf. 6. S. 559—571.
Klein A.L.,Zaidan L.V.P., FelippeG.M. Floweringand heterophylly in Bidens gardneri
Baker//Rev. Bras. Bot., 1992. V. 15.N2.P. 139—144.
Kliphuis E. Cytological observation in relation to the taxonomy of the orchids of the
Netherlands // Acta Bot. Need. 1963. V. 12, N 2. P. 172—194.
Korotkov K.O., Morozova O.V., Belonovskaja E.A. The USSR vegetation syntaxa
prodromus. M., 1991. 346 p.
Krattinger K. Genetic mobility in Typha // Aquatic Botany. 1975. V 1. P. 57—70.
Kronfeld M. Monographic der Gattung Typha Toum. (Typhinae Agdh., Typhaceae
Schur Engl.) // Verh. Zool. Bot. Ges. Wien, 1889 Bd. 39. S. 89—190.
KUgler H. Blbtenukologie. Jena. 1970. 345 S.
Kujala V. Untersuchungen bber die Waldvegetation in Std-und Mittelfinnland. Hel-
sinki, 1926.
Ktlnkele S. Bemerkungen zu den autogamen Epipactis-'svp’pen in Wurttemberg //
Jahresber. Naturwiss. Ver. Wuppertal. 1969. Bd. 23, S. 104—105.
Ktlnkele S. Bemerkungen zu den autogamen Sippen der Epipactis helleborine (L.)
em Mill.) in Wurttemberg // Mitt. Bl. Arb. Kreis Heim. Orchid Baden-Wurt. 1970. Bd. 1, N
3. S. 16—18.
Ktltzing F.T. Monographia callitricharum germanicarum // Linnaea. 1832. V. 7. P.
174—102.
, LandoltE. OkologischeZeigerwertezurSchweizerFlora//Ventff. Geobot.Inst. Rbbel,
Zbrich, 1977. H. 64.
Langlet O.F.J. Beitrage zurZytologie der Ranunculazeen // SvenskBot. Tidskr. 1927.
Bd. 21, N 1. S. 1—17.
Lawler L. Ethnobotany of the Orchidaceae // Orchid Biology: Reviews and Per-
spectives. III. Ithaca, London. 1984. P. 27-149.
Lebel J.E. Callitriche, Esquisse monographique//Mem. Soc. imp. Sci. nat. Cherbourg,
1863. V. 9. P. 129—176.
Leek M. Wetland seed banks //Ecology of soil seed banks. 1989. V. 5. P. 283—308.
Lee D. W. Population variation and introgression in North American Typha // Taxon,
1975. V 24. P. 633—641.
Levitt J. Responses of plants to environmental stresses. V. 2. New York: Academic
Press. 1980.
Lhotska M. Karpologie und Karpobiologie der tschechoslowakischen Vertreter der
Gattung Bidens // Rozpravy CSAV. Ser. matt, natur., 1968. Bd. 78, N 10. S. 1—85.
Lhotska M., КорескэК. Zur Verbreitungsbiologie und Phytozunologia von Impatiens
glandulifera Royle an den Flussystemen der Svitava, Svratka und oberen Odra//Preslia, 1966.
V. 38. P. 376—385.
Linkola K., Tiirikka A. bber Wurzelsysteme und Wurzelausbreitung der
Wiesenpflanzen auf verschiedenen Wiessenstandorten // Ann. Bot. Soc. Zool.-Bot. Fenn.
Vanamo, 1936. V 6, N 6. 207 p.
230
Linnaeus C. Centuria I. plantarum. Uppsala, 1755a.
Linnaeus C. Flora suecica. Ed. 2. Holmiae, 1755b.
Linnaeus C. Species Plantarum. Holmiae, 1753. V. 2.
LoewE. Gattung Typha Toum. Rochkolben. //Lebensgeschichte der Blbtenpflanzen
Mitteleuropas. Stuttgart, 1908. Bd. 1., Abt. 1. S. 348—374.
Love A., LOve D. Cytotaxonomical studies on boreal plants, Ш. Some new chromosome
numbers of Scandinavian plants//Arkiv Bot. 1944. V. 31A. P. 1—22.
Love A., LOve D. Chromosome numbers of central and northwest European plant
species//Opera Bot. Suppl. Ser. Bot. Not. 1961. V. 5. 581 p.
Mabberley D.J. The plant-book. Cambridge, 1989.
Manton J. Note on the cytology of Scheuchzeria from Rannoch Moor // Watsonia,
1949. V. 1, pt. l.P. 36.
Markgraf F. Blbtenbau und Vemandschaftbei den einfachsten Helobiae //Ber. Deutch.
Bot. Ges. 1936. Bd. 54, N 1—2. P. 191—229.
Markgraf F. Scheuchzeriaceae // In: G. Hegi. Illustrierte Flora von Mitteleuropa. 1981,
Bd. I., P. 257—260.
Martinsson K. Callitriche in Sweden: case studies of reproductive biology and
intraspecific variation in a semi-aquatic plant genus 1) Acta Univ. Uppsal. Comp. Summ.
Uppsala diss. Fac. Sci. 1991a. N 327. P. 1—24.
Martinsson K. Geographical variation in fruit morphology in Swedish Callitriche I
/Nordic! Bot., 1991b. V. 11, N 5. P. 497—512.
Martinsson K. Natural hybridization within the genus Callitriche in Sweden // Nordic
J. Bot., 1991c. V. 11, N2. P.43—151.
Masuda M., Yahara T. Reproductive ecology of a cleistogamous annual Impatiens
noli-tangere L. occurring under different environmental conditions // Ecological Research,
1994. V 9, N l.P. 67—75.
Matuszkiewicz W. Przewodnik do oznaczania zbiorowisk roslinnych Polski //
Warszawa: PWN, 1981. 299 p.
McNaughton S.J. Ecotype function in the Typha community—type/ZEcol. Monogr.
1966. V. 36. P. 297—325.
McNaughton S.J. R- and k- selection in Typha // Amer. Natur. 1975. V. 109. P. 251—
261.
Metsavainio K. Untersuchungen bber das Wurzelsystem der Moorpflanzen // Ann.
Bot. Soc. Zool.-Bot. Fenn. Vanamo, 1931. V. 1, N 1. P. 58—62.
Meusel H., Jager E., Weinert E. Vergleichende Chorologie der zentraleropaischen
Flora. Jena: Gustav Fisher, 1965. Bd. 1—2.
Moore J.J. The distribution and ecology of Scheuchzeria palustris in a raised bog in
Offaly // Irish Naturalists’ J., 1955. V. 11, N 12. P. 321—329.
Moore P.D., Webb J.A., Collinson M.E. Pollen analysis. Ed. 2, Blackwell, Oxford,
1991,216 р.
Moravec J. et al. PostlinnO spoleuenstva Ceske republiky a jejich ohrozenn.
Seven»Keskou pmirodou, 1995. P. 206.
Morong T. Studies in the Typhaceae // Bull. Torrey Bot. Club. 1888. V 15. P. 1—8.
MbllerD. Epipactis helleborine f. albina i Danmark//Bot. Tidsskr. 1974. v. 46. P.
413—414.
Mbller-Doblies D. Uber die Verwandtschaft von Typha und Sparganum im
Infloreszenz und Blbtenbau И Bot. Jahrb. Syst., 1970. Bd. 89. S. 451—562.
Nakano T. Some epiphytic algae growing on the spikes of Typha // J. Jap. Bot., 1985.
V. 60. P. 5—10.
231
Nieschalk A., Nieschalk Ch. Autogame Epipact is-Aiten in Nordhessen // Jahrber.
Naturwiss. Ver. Wuppertal. 1970. Bd. 23. S. 98—103.
Nilsson L. A. Pollination ecology ofEpipactis palustris (Orchidaceae) I I Bot. Notis.,
1978. Bd. 131, N3.
Nilsson S. Orchids of Nothem Europe. Middlesex, 1979.
Peace W.J.H., Grubb P. J. Interaction of light and mineral nutrient supply in the growth
of Impatiens parviflora DC. // New Phytol. 1982. V. 90, N 1. P. 127—150.
Penko J.M., Pratt D.C. The growth and survival of early instars of Bellura obliqua
(Lepidoptera: Noctuidae) on Typha latifolia and Typha angustifolia И Great Lakes Entomol.
1986. V. 19. P. 35—42.
PoluninO., Stainton A. Flowers ofthe Himalayas. Oxford: Oxford Univ. Press, 1984.
Posluszny U. Re-evaluation of certain key relationships in the Alismatidae: floral
organogenesis of Scheuchzeria palustris И Amer. J. Bot., 1983. V. 70. P. 925—933
PotmuekO. Die Epipactis-Aiten derCSSR//Jahresber. Naturwiss. Ver. Wuppertal.,
1990. Bd. 23. S. 83—85.
Prochazka F., VelHsek V. Orchideje nasi prirody. Praha, 1983. 284 S.
Procopova A. Uber die Veranclerung des Geschlehtsverha Ithnisses derNachkom-
men Schaft im Zussammen Hang mit dem Alter der Bluten bei M. album H Biol, plantarum,
1964. Bd. 6,2. S. 99—103.
Pysek P., Prach K. Invasion dynamics of Impatiens glandulifera — a century of
spreading reconstructed//Biol. Conserv., 1995. V. 74. P. 41—48.
Rackham O. Radiation, transpiration and growth in a woodland annual //Light as an
ecological factor. Oxford: Blackwell, 1966. P. 167—185.
Rasmussen H. The vegetative architecture of Orchids // Lindleana. 1986. V 1, N 1.
P. 42—50.
Richards A. J., Porter A.F. On the identity of aNorthumberland Epipactis I/ Watsonia
(Gr. Brit ). V. 14, N 2. P. 121—128.
Riska S. Impatiens parviflora i Grankulla // Mem. Soc. fauna et flora fenn., 1957.
Bd. 5$. S. 34—49.
Rohrbach P. bber die europnischen Arten der Gattung Typha // Verh. Bot. Ver. Prov.
Brandenburg. 1869 (1870). Bd. XI. S. 67—104.
Rollin P. Action de la lumiere et de la temperature sur la germination des akenes du
Bidens tripartitus L. // Compt. Rend. Acad. Sci. 1956. V 243, N 3. P. 300—302.
RoscoeM.V Cytological studies in the genus Typha H Hot. Gaz., 1927. V. 84. P. 392—
406.
Ruijgrok H.W.L. Cyanogenese bei Scheuchzeria palustris II Phytochemistry, 1974.
V. 13, NIP. 161—162.
Ruprecht F. Flora Ingrica. Petropoly, 1860. P. I—XXVI, 1—670.
Sale P.J ,M., Wetzel R.G. Growth and metabolism of Typha species in relation to cutting
treatments//Aquatic Botany, 1983 V. 15. P. 321—334.
Salisbury E. Weeds. London: Collins, 1961.
Salisbury E.J. The reproductive capacity of plants: studies in quantitative biology.
London. 1942. 244 p.
Salmia A. Chlorophyll-free form of Epipactis helleborine (Orchidaceae) in SE
Finland // Ann. Bot. Fenn., 1986. V. 23. P. 49—59.
Sanders D. J. Epipactis helleborine — an European import // Amer. Orchid Soc. Bull.
1978. V. 47. P. 426—427.
Sand-Sensen K., Madsen TV Patch dynamics of the stream macrophyta, Callitriche
232
cophocarpa П Freshwater Biol., 1992. V. 27, N 2. P. 277—282.
Sarosiek J., Krukowski-Zdanowicz J., Woroniecka G. The ecological organization
of Epipactis palustris population (Mill.)Cr. inAugustowForest//ActaUniv. Wratislawiensis.
1990. N 1055. P. 173—180.
Satterwhite M.B. Spectral luminescence of plant polen // IGRASS 90:10th Ann. Int.
Geosci. and Remote Sens. 1990. V. 3. P. 1945—1948.
Scartch G. W. Scheuchzeria palustris L., its occurrence as a British plant, with a fresh
record // Notes Roy. bot. garden Edinburgh. 1911. P. 57—66.
Schemske W. Evolution of reproductive characteristics in Impatiens (Balsaminaceae)'.
the significance of cleistogamy and chasmogamy //Ecology, 1978. V. 59, N3. P. 596—613.
Schmeidt O. Eestimaa orchideed. Tallinn, 1996. 144 p.
Schmitz G. Spectrum of flower visiting insects on indigenous and neophytic species
of Impatiens // Entomologische Nachrichten und Berichte, 1994. V. 38, N 1. P. 17—23.
Schmitz G. The use of the neophytes Impatiens glandulifera Royle and Impatiens
parviflora DC. by phytophagous insects in the area of Bonn // Entomologische Nachrichten
und Berichte, 1991. V. 35, N 4. P. 260—264.
Schnizlein A. Dissertatio botanica de Typhacearum familia naturali. 1845. S. 1—
26.
Schnizlein A. Iconographia familiarum naturalium Regni vegetabilis. 1846. V. 1.
Schultz C.F. Prodromus florae stargardiensis. Berolini, 1806.
Sculthorpe C.D. The biology of aquatic vascular plants. London, 1967. 610 p.
Segadas V.F. A phytosociological and ecological study of cattail stands in Oakland
county, Michigan //J. Ecol. 1951. V. 39. S. 316—329.
Senghas K. Ubersicht zur Systematik und Taxonomie der Gattung Epipactis //
Jahresber. Naturwiss. Ver. Wuppertal. 1970. Bd. 23. S. 26—37.
Cerbmesco-Jitariu G. Considilrations sur le gyniicile et le fruit de Scheuchzeria
palustris L. // Rev. roumaine biol. Silr.bot. 1966. 11(6). P. 435—439.
SharmaK.P., Gopal B. Seed germination and occurrence of seedlings of Typha species
in nature // Aquatic Botany. 1978. V. 4. P. 353—358.
Sherff E.E. The Genus Bidens. Chicago, U.S.A: Publication 389 Field Museum of
Natural History. 1937. Part I—II. 709 P.
Slavik B. Rod Impatiens v Heskft republice // Preslia, 1996. V. 67. P. 193—211.
Sledge W.A. The distribution and ecology of Scheuchzeria palustris L. // Watsonia,
1949. V. 1,pt. l.P. 24—35.
Smith S.G. Experimental and natural hybrids in North American Typha (Typhaceae)
//Amer. Midland Naturalist, 1967. V. 78. P. 257—287.
Soy R. Die Epipactis-Таха der pannonischen und Karpatischen Flora und ihre
Soziologische Rolle // Jahresber. Naturwiss. Ver. Wuppertal. 1970. Bd. 23. S. 75—82.
StenarH. Embryologische Beobachtungen bber Scheuchzeria palustris L. // Bot. Not.,
1935. S. 78—86.
Sugiura T. A list of chromosome numbers in angiospermous plants. V. // Proc. Imp.
Acad. Tokyo, 1938. V. 10, N 1—2. P. 391—392.
Sukopp H„ Markstein B. Changes of the reed beds along the Berlin Havel, 1962—
1987 //Aquatic Botany. 1989. V. 35. P. 27—39.
Summerhayes VS. Wild orchids ofBritain. London. 1951. 290 p.
Sundermann H.Europnische und mediterraneOrchideen. Hildesheim. 2 Aufl. 1975.
SwamyB.G.L.,ParameswaranN. The helobiae endosperm//Biol. Rev. V. 38,1963.
P. 1—50
233
Szostek M. Nowe stanowiska czadkich gatunkow roslin we florze synantropijnaj
wybranych dzielnic Sosnowscal przyleglych terenow Dabrowy Gomiczen // Zesz. pizyr. OTPN
Opolu. 1986. N 24. P. 21—29.
Tallis J.H., Birks H.J. The past and present distribution of Scheuchzeria palustris in
Europe // J. Ecol., 1965. V. 53, N2. P. 287—298.
Tanaka R., Kamemoto H. Chromosomes in Orchids: counting and numbers // Orchid.
Biology: Rev., Persp., III. Ithaca-London: Cornell Univ. Press, 1984. P. 323—412.
Tanaka R., Kamemoto H. List of chromosomenumbers in species ofthe Orchidaceae
П Orchids: scientific studies. N.-Y, 1974. P. 411—484.
Thompson K., Grime J.P. Seasonal variation in the seed banks ofherbaceous species
in ten contrasting habitats // J. Ecol. 1979. V 67, N 3. P. 893—921.
Troll W. Die Infloreszenzen. Jena, 1964. Bd. 1. 615 s.
Troll W. Praktische Einfthrung in die Pflanzenmorphologie. II. Die Blbhende Pflanze.
Jena. 1957. 420 S.
Tyteca D., Dufrem M. Biostatical studies of Westernn European allogamous
populations ofthe Epipactis helleborine (L.) Crantz species group (Orchidaceae) //Syst. Bot.
1994. V. 19, N3. P424—442.
Ulbrich E. Biologie der Frbchte und Samen (Karpobilogie). Berlin: Springer, 1928.
Valentine D.H. Flower-color polymorphism in Impatiens glandulifera Royle //
Boissiera, 1971. V 19. P. 339—343.
Valentine D.H. The pollination of introduced species, with special reference to the
British Isles and thegenus Impatiens //The pollination of flowers by insects. London: Academic
Press, 1978. P. 117—123.
Velenovsky J. Vergleichende Morphologic derPflanzen. Prag., 1907. V. 2. S. 311,
314—315.
Warming E. Botaniske Exkursioner. 3. Skarridso. Vidensk. Meddel. fra den naturk.
Foren. Kjobenhavn. 1897. S. 164—197.
, Warming E. Smaa biologiske og morphologiske bidrag // Bot. Tidsskr. Ser. 3,1.1876.
S. 84—110.
Weise B. Untersuchungen uber die Konkurrenzbeziehungen von Impatiens parvi-
flora mdlmpatiensnoli-tangerel/Ser. Arbeitsgemeinsch. Sachsisch. Bot. 1966—1967 (1968).
Bd. 8. S. 101—122.
Weisner S.E.B. Long-term competitive displacement of Typha latifolia by Typha
angustifolia in an eutrophic lake // Oecologia, 1993. V. 94. P. 451—456.
Wiefelspiitz W. Zur Verbreitung der europaischen allogamen Epipactis-Arten //
lahrber. Naturwis. Ver Wuppertal. 1970. Bd. 23. S. 38-42.
Wildhaber O. Beitrag zur Karpologie von Epipactis-Alien // lahrber. Naturwis. Ver
Wuppertal. 1970. Bd. 23. S. 109-113.
Willis J.C. A dictionary of the flowering plants and fems. Ed. 8. Cambridge, 1988.
Young D.P. Studies in the British Epipactis // Watsonia, 1948. V 1. N 2. P. 102—
113.
Young D.P. Studies in the British Epipactis // Watsonia. 1962. V. 5. N 3. P. 127—
135.
Ziegenspeck H. Orchidaceae //Lebensgeschichte der Blbtenpflanzen Mitteleuropas.
Stuttgart, 1936. Bd. 1. Lfd. 4. 840 S.
Zinov’eva-Stahevitch A.E., Grant W.F. Chromosome numbers in Impatiens // Can.
J. Bot., 1984. V 62, N 12. P. 2630—2635.
234
СОДЕРЖАНИЕ
Предисловие................................................ 3
Рогоз узколистный. Е.В. Мавродиев...........................4
Шейхцерия болотная. Т.Ю. Минаева......................... 30
Род Дремлик. М.Г. Вахрамеева, Т.И. Варлыгина,
А.Е. Баталов, И.А. Тимченко, Т.И. Богомолова.............. 50
Дрема белая. М.А. Туленкова, С.К. Пятунина................ 88
Купальница европейская. Р.П. Барыкина, Н.В. Чубатова...... 97
Род Болотник. О.А. Горбачевская........................... НО
Род Недотрога. М.В. Марков, НГ. Уланова, Н.В. Чубатова.... 128
Вероника лекарственная. Н.П; Савиных..................... 169
Колокольчик широколистный. В.П. Викторов............... 179
Род Череда. М.В. Марков, Н.М. Ключникова................ 192
Литература............................................... 214
CONTENTS
Introduction....................................................... 3
Typha angustifolia. E. V. Mavrodiev................................ 4
Scheuchzeria palustris. N. Yu. Minaeva............................ 30
Genus Epipactis. M.G. Vakhrameeva, T.I. Varlygina,
A.E. Batalov, LA. Timchenko, T.I. Bogomolova...................... 50
Melandrium album. M.A. Gulenkova, S.K. Pyatunina.................. 88
Trollius europaeus. R.P. Barykina, N. V. Chubatova................ 97
Genus Callitriche. O.A. Gorbachevskaya........................... 110
Genus Impatiens. M. V. Markov, N.G. Ulanova, N. V. Chubatova..... 128
Verinica officinalis. N.P. Savinykh............................. 169
Campanula latifolia. N.P. Viktorov............................... 179
Genus Bidens. M. V. Markov, N.M. Klyuchnikova.................... 192
Literature cited................................................. 214
Научное издание
БИОЛОГИЧЕСКАЯ ФЛОРА
МОСКОВСКОЙ ОБЛАСТИ
вып. 13
Редактор к.б.и. Н.Г. Уланова
Компьютерная верстка А.А. Маслов
Изд. лицензия ЛР.№ 010295 Подписано в печать 14.12.96 г. Формат 60х84/1б.
Бумага тип. № 1. Гарнитура «Times New Roman». Офсет. Уел. кр. отг. 15,0.
Тираж 500 экз. Заказ Xs 381
Нелидовская типография
172500 Россия, Тверская обл., г.Нелидово, пер. Школьный, д. 4
Тел. (08266) 3-16-84
НЕЛИДОВСКАЯ ТИПОГРАФИЯ
Весь комплекс полиграфических услуг:
дизайн, верстка, изготовление оригинал-макетов,
цветная и чернобелая печать книг, журналов,
буклетов, визиток.
Опыт работы, индивидуальный подход к
каждому клиенту, разумные цены
Звоните: тел: (08266) 3-16-84.