


















































































































































Author: Грачев Ю.Н.
Tags: sauropsida ящероптицы биология птицы охрана природы орнитология издательство наука кеклик академия наук ксср издательство алма-ата
Year: 1983




АКАДЕМИЯ НАУК КАЗАХСКОЙ ССР
Институт зоологии
Ю. Н. ГРАЧЕВ
(Биология, использование и охрана)
Издательство «НАУКА» Казахской ССР
АЛМА-АТА • 1983
УДК 598.617.57(574)
Грачев Ю. Н. Кеклик: (Биология, использование и охрана). — Алма-Ата: Наука, 1983.— 148 с.
Кеклик — одна из популярных промысловых птиц Казахстана. Оседлость большинства популяций кеклика дает возможность успешно решать задачи его охраны и рациональной эксплуатации, что имеет несомненное преимущество перед перелетными видами дичи.
Изучение биологии разных географических форм кеклика представляет интерес для выяснения эволюции вида и приспособления его к различным экологическим условиям. Особое внимание уделено гнездовой биологии кеклика — наименее изученного вопроса.
Книга рассчитана на орнитологов, преподавателей вузов аспирантов, специалистов охотничьего хозяйства и любителей природы.
Библиогр. 109 назв. Ил. 23. Табл. 22.
Ответственный редактор доктор биологических наук Э. И. ГАВРИЛОВ
21008—075
Г 407(05)_8з 72.83.2005000000
©Издательство «Наука» Казахской ССР, 1983
Предисловие
В настоящее время в связи с преобразованием естественных ландшафтов происходят существенные изменения в животном мире. Одних охотничье-промысловых птиц, таких, ка*к дрофы, рябки, некоторые другие, пришлось внести в «Красную книгу», численность других настолько уменьшилась, что добычу их или ограничили, или сократили сроки охоты. В частности, к таким видам относятся водоплавающие, весенняя охота на которых на большей части территории нашей страны с конца 60-х годов запрещена.
На фоне общего уменьшения традиционных в охотничьем отношении групп птиц особое значение приобретают виды, которые еще в недалеком прошлом составляли лишь небольшой процент от всей отстреливаемой пернатой дичи. В Казахстане к ним в первую очередь следует отнести кеклика (Alector.is chukar Gray). Добыча его за последние годы заметно возросла и можно ожидать дальнейшего ее увеличения, поскольку численность кеклика после депрессии зимой 1968/69 г. достигла высокого уровня и стабильно удерживается на протяжении последних десяти лет.
На майском (1$)82 г.) Пленуме ЦК КПСС, принявшем Продовольственную программу СССР, большое внимание уделено производству мяса птицы. Важным подспорьем в этом отношении на территории Юго-Восточного Казахстана могут быть некоторые виды пернатой дичи, в частности кек-лик: при соблюдении сроков и норм отстрела в годы с высокой численностью этой птицы можно ежегодно добывать
3
50—75 тыс. кекликов, что позволит рационально использо-вгать дичные ресурсы.
У кеклика есть ряд преимуществ перед некоторыми промысловыми видами птиц семейства фазановых (например, серая и бородатая куропатки, фазан) и важнейшее из них — способность быстро восстанавливать численность после неблагоприятных условий зимовки; оседлость же кеклика дает возможность решать задачу охраны и рациональной эксплуатации, что имеет несомненное преимущество перед перелетными видами дичи.
Кеклик может служить объектом некоторых теоретических разработок. Так, изучение биологии разных географических форм представляет интерес для выяснения его эволюции и приспособления к различным экологическим условиям.
Несмотря на большое количество работ, затрагивающих различные аспекты жизни кеклика, биология его известна лишь в общих чертах. Из специальных статей по отдельным вопросам экологии следует упомянуть работы А. Свиридова (1876), Н. А. Зарудного (1900), Н. В. Минина (1939), А. К. Рустамова (1945), М. Д. Зверева (1948), М. А. Кузьминой (1948), А. А. Федорова (1949), М. К. Газаньяна (1954), А. И. Ханмамедова (1955), М. Е. Кутубидзе (1957), М. М. Остапенко (1958, 1965), Ю. А. Лебе^ва (1976), В. А. Вырыпаева и Р. Р. Будриса (1976). Некоторые вопросы биологии освещены в фаунистических работах И. А. Долгушина (1948, 1951), В. В. Шевченко (1948, 1949), А. И..Иванова (1969), И. А. Абдусалямова (1964, 1971), А. Ф. Ковшаря (1966). Наиболее полные сведения по экологии кеклика представлены в работе М. А. Кузьминой (1955), материал для которой собирали в 1948—1951 гг. в отрогах Джунгарского Алатау и Чу-Илийских горах. Однако некоторые вопросы (размножение и изменение численности) еще недостаточно изучены.
Систематике и распространению вида посвящены работы Н. И. Яблонского (1904), Н. А. Зарудного (1914), Е. Л. Ше-стоперова (1929), Н. В. Павлова (1931), В. А. Селевина (1935), Г. И. Дементьева (1946), С. К. Даля (1947), А. К. Рустамова (1948), Л. С. Степаняна (1975), а из зарубежных авторов —G. Watson (1962), Ch. Vaurie (1965). Общие сведения по кеклику приведены в сводках «Птицы Советского 4
Союза», «Птицы СССР» и в региональных сводках «Птицы Казахстана», «Птицы Киргизии», «Птицы Таджикистана» и «Птицы Туркменистана». Наконец, некоторые данные о способах отлова, добыче и краткие сообщения об образе жизни кеклика опубликованы в периодических изданиях, в основном в журналах, рассчитанных на широкий круг читателей.
О недостаточной изученности гнездовой биологии кеклика свидетельствует тот факт, что к настоящему времени на территории нашей страны найдено лишь немногим более 40 гнезд этой птицы (Бёме, 1975). Правда, эти сведения нисколько занижены, поскольку оказались неучтенными некоторые работы (Долгушин, 1948, 1951; Кузьмина, 1955; Остапенко, 1965 и др.). Литературные материалы содержат, как правило, лишь данные о сроках гнездования и, частично, о расположении гнезд, их строительном материале, величине полной кладки. Что же касается сведений о поведении птиц в брачный и предшествующий ему периоды, отходе яиц, судьбе гнезд, эффективности гнездования и некоторых других вопросов, то они остаются невыясненными.
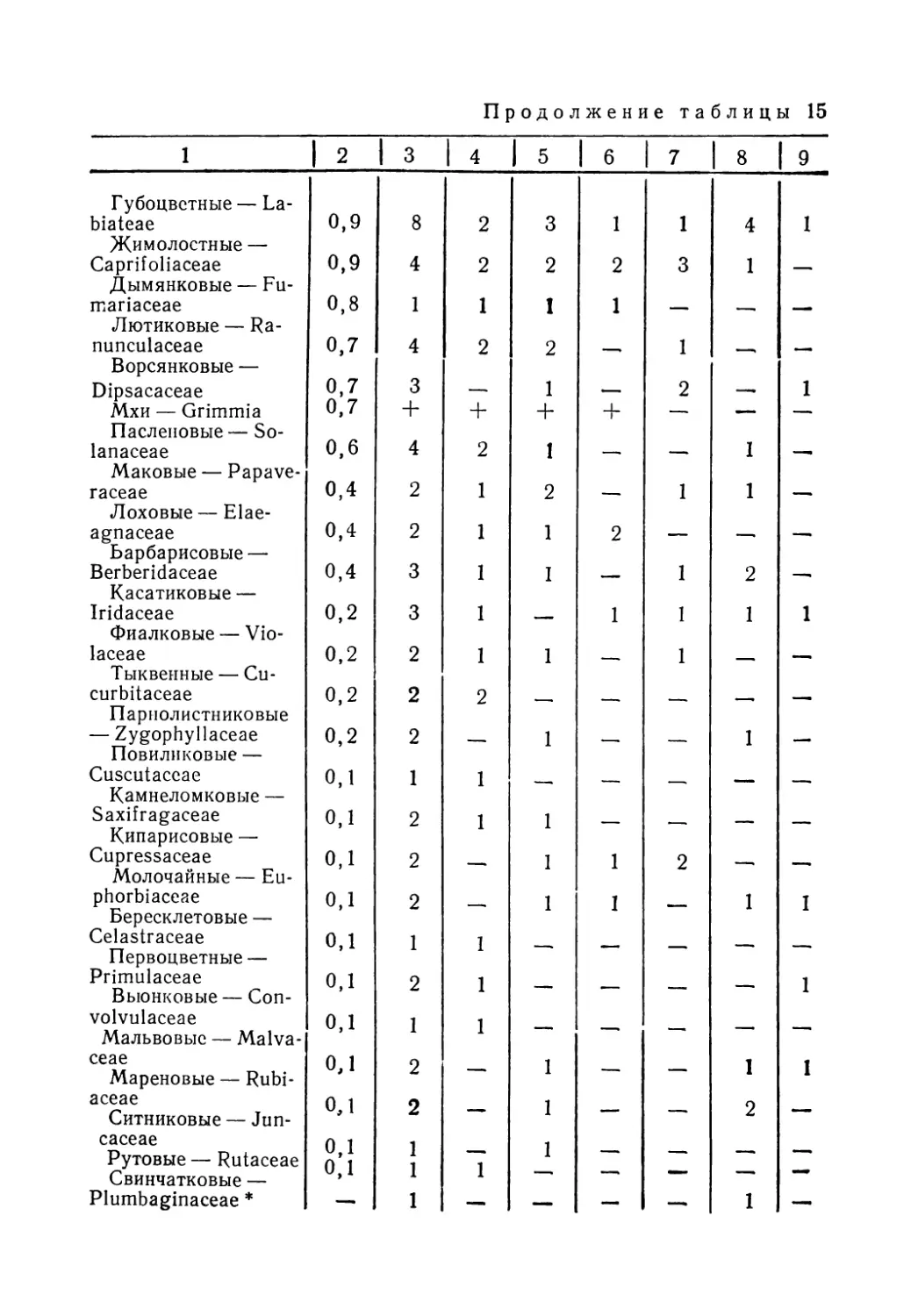
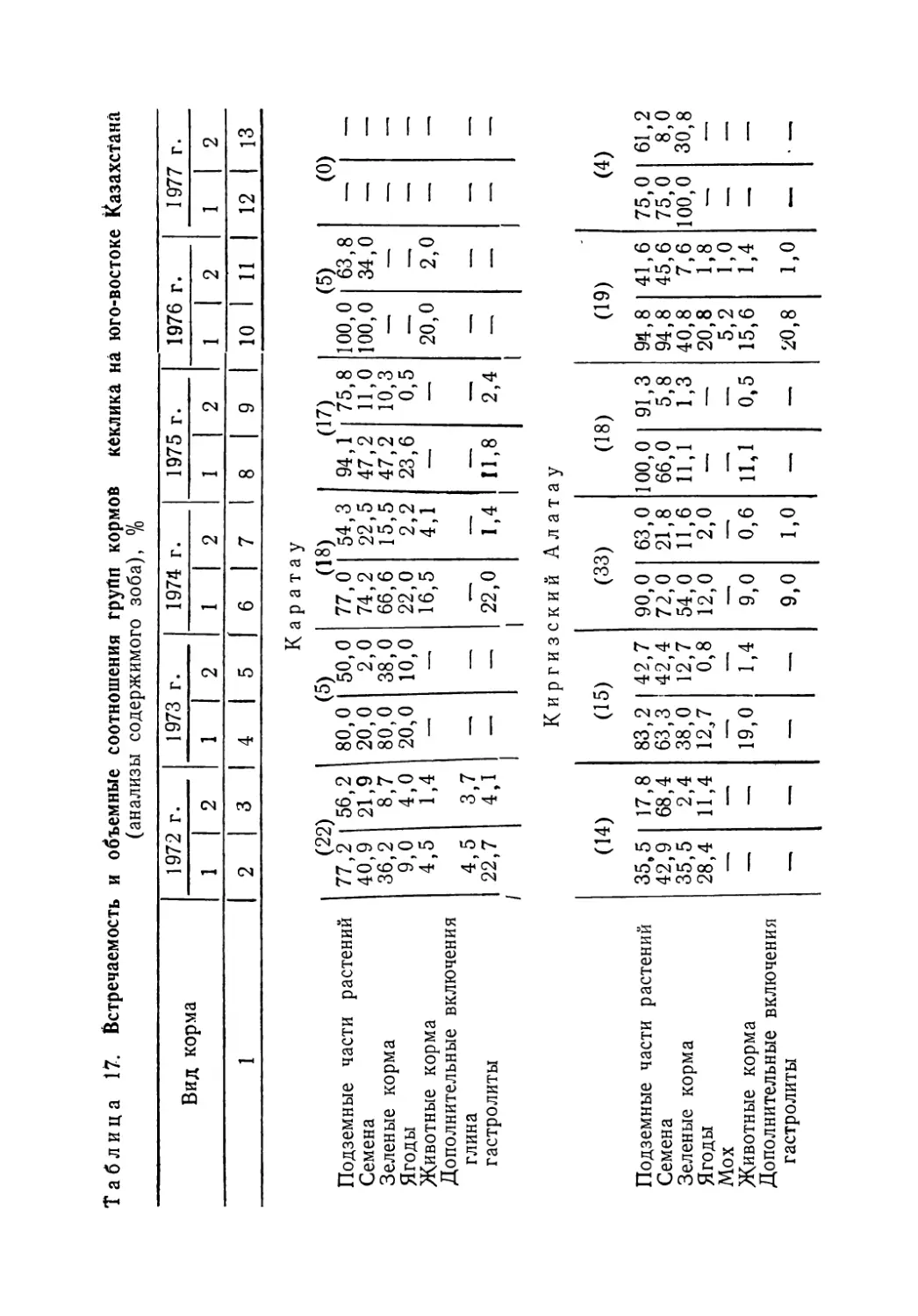
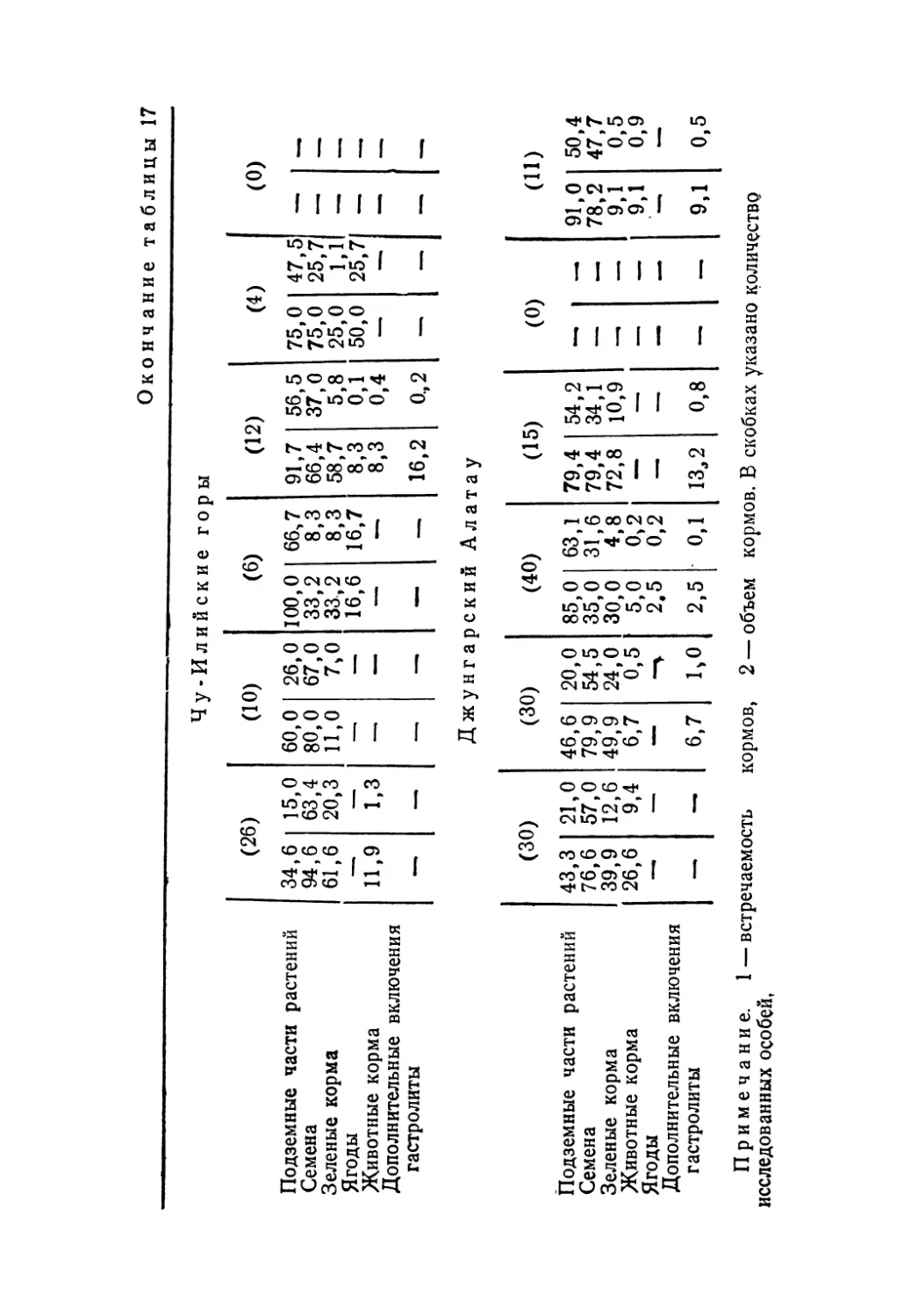
Лучше других вопросов биологии кеклика изучено его питание. В Казахстане наиболее полная информация собрана по Джунгарскому Алатау и хребтам Северного Тянь-Шаня, а Мангышлак, Устюрт, Тарбагатай и Алтай пока остаются «белыми пятнами». Нами предпринята попытка обобщить все имеющиеся данные о составе кормов кеклика в пределах СССР, учитывая отдельные статьи и материал региональных фаунистических сводок. В работе использованы публикации по следующим регионам: Джунгарский Алатау — горы Чулак и Чаган (Кузьмина, 1955); Северный Тянь-Шань — Заилийский Алатау, Чу-Илийские горы, Таучилик, Киргизский Алатау (Кузьмина, 1955; Пэк, Федянина, 1961); Западный Тянь-Шань — Таласский Алатау, Чаткальский хребет, хр. Нуратау (Остапенко, 1958, 1965; Ковшарь, 1966; Салих-баев и др., 1970); Памиро-Алай — Туркестанский, Гиссарский, Каратегинский и Зеравшанский хребты, Алай (Минин, 1939; Попов, 1959; Иванов, 1969; Абдусалямов, 1971); Копет-Даг и Кызылкумы (Рустамов, 1945; Федоров, 1949; Коган, 1950; Колесников, 1956); Кавказ (Даль, 1949; Ханмамедов, 1955)-
Обобщение литературных, а также* собранных нами материалов позволило составить наиболее полный список ъидо-вого состава кормов кеклика, проследить за сезонными из
5
менениями его рациона и определить встречаемость и количественные (объемные) соотношения основных компонентов пищи.
Учитывая степень изученности отдельных сторон экологии кеклика, мы обратили внимание на биологию размножения и выяснение причин колебания его численности. Эти разделы построены на оригинальном материале.
В работе постоянную помощь оказывал весь коллектив лаборатории орнитологии Института зоологии АН КазССР. Постоянное руководство работой, а также редактирование рукописи взял на себя заведующий лабораторией доктор биологических наук Э. И. Гаврилов. Ценные консультации по подготовке работы к печати получены от заведующего лабораторией проблем охраны диких животных доктора биологических наук А. Ф. Ковшаря и старшего научного сотрудника лаборатории млекопитающих кандидата биологических наук В. И. Капитонова. Большую помощь в сборе и обработке материалов оказала кандидат биологических наук М. А. Кузьмина — участник многих совместных экспедиций. В полевой сезон 1978 г. в работе принимал участие старший лаборант лаборатории орнитологии В. К. Захваткин. Насекомые, которые служат пищей кеклика, определены сотрудниками Института зоологии АН КазССР профессором П. И. Мариковским, кандидатами биологических ^наук В. Л. Казенасом, Р. Б. Асаповой, К. 3. Куленовой, Г. В. Николаевым, моллюски — кандидатом биологических наук К. К. Увалиевой, семена высших растений — научным сотрудником Главного ботанического сада АН КазССР М. С. Зориной. В оформлении иллюстраций принимали участие Л. М. Корелова, Е. В. Беседин, Р. Г. Пфеффер. Всем им автор выражает глубокую благодарность.
Сроки и места проведения работ. Материал и методика
Биологию кеклика изучали в течение 7 лет (1972— 1978 гг.) на маршрутах и в стационаре (табл. 1). Помимо сроков полевых работ, указанных в таблице 1, в сезоны охоты (ноябрь) совершались кратковременные выезды в ущелья Жаманты и Актерек (Заилийский Алатау), Чу-Илийские горы и горы Малай-Сары, где мы собирали данные о питании, поло-возрастном составе и упитанности птиц. Использовался также материал по кекликам, добытым охотниками. Таким образом, полевые работы охватили все сезоны года, а общая их продолжительность составила 609 дней.
Стационарные работы проводились в 1974—1978 гг. в горах Чулак — периферийных юго-западных отрогах Джунгарского Алатау (ущ. Кзылаус, 600—900 м над ур. м.), где изучались гнездовая биология, постэмбриональное развитие, поведение кекликов в разные сезоны года. Место было выбрано с учетом следующих соображений. Во-первых, оно отличалось высокой и стабильной численностью кеклика. Во-вторых, горы Чулак расположены на территории Капчагайского юхотвоспроизводственного хозяйства Главохоты Казахской ССР, где охота практически не ведется, а угодья не эксплуатируются. Это обстоятельство играет немаловажную роль при изучении вида, поскольку близко к условиям проведения работ в заповедниках. В-третьих, биотопы здесь представлены в основном осыпями, скалами и- открытыми склонами, что было использовано для наблюдений за отдельными парами, гнездами и стайками кекликов. Кроме того, в ущ. Кзылаус в 1949—1950 гг. М. А. Кузьмина собрала материал по пита-
7
Таблица 1. Сроки полевых работ
Год Стационарные ра-боты (горы Чула к) Маршрутные работы (учета)
Каратау Киргизский Алатау Чу-Илий-ские горы Горы Чулак А л тын-Эмель Малай-Сары
1972 — 26—27. VI И; 30.VIII; I.IX; 3.IX; 5-7.IX 12—13.IX; 15-21.IX 23—27. IX 12-13.Х 5-6.Х 8-10.Х
1973 — 29.VIH; 31.VIII- —2. IX; 5-6. IX 12-13.IX; 15— -16.IX; 18-20.IX 23-26.IX 4-6.Х 8-10.Х 12—16.Х
1974 8. IV—28. VI 28-29. VIII; ' 31.VIH-2.IX; 5-7.IX 12-14.IX 16-17.IX; 19-20. IX 23—25.IX 3-5.Х 9-I1.X 16.Х
1975 23.IV—15. VI; 8—23.XII 27.VIII; 29- -31.VIII; I.IX 14-21 .IX 23—24.IX 3-6.Х 10-12.Х 14.Х
1976 10.IV—13. VI; 15,11—I.III 8—11. IX 15-16.IX; 18— -19.IX 21-22.IX 28—30.IX 6-8.Х 10—11.Х 13.Х
1977 2—>21. II; 15. III— -15. VII —• — — — — —•
1978 29.IH-20.VI — — — — — —
Всего дней 449 29 37 17 15 13 11
нию кеклика, который оставался необработанным. После систематизации эти данные вошли в настоящую работу.
Маршрутные исследования велись в 1972—1976 гг. (конец августа — середина октября). В этот период собирали-материал по линьке, упитанности и питанию кеклика, проводили количественные учеты. Были обследованы следующие ущелья: на хребте Каратау — Актобе, расположенное на южном склоне, Алтынтау, Ран, Кумусты — на северном; в Киргизском Алатау — Сюгаты, Каракыстак и Мерке; в Чу-Илийских горах — Алмалы и Сарыбулак; на хребте Алтын-Эмель — Тюлькули; в горах Чулак — Кзылаус, Теректы, Унгур; в горах Малай-Сары — безымянные ущелья, расположенные в 10—20 км к северо-западу от железнодорожной станции Жоломан.
В работе применяли общепринятые методики полевых зоологических исследований (Новиков, 1953). Часть из них была, несколько изменена применительно к местным условййм и целям.
Гнезда искали, наблюдая за поведением отдельных пар в основных биотопах, преимущественно в утренние часы, во* время наибольшей активности кёкликов. Кроме того, в местах постоянных встреч пар тщательно осматривали основания кустов, ниши под камнями, куртинки травы. Для изучения поведения’ птиц при насиживании проводили учеты-наблюдения у гнезд в течение полного светового дня. Интервалы между бткладкой яиц, продолжительность инкубаций и сроки вылупления птенцов определяли при регулярной проверке гнезда, причем частота посещений зависела от целей и устанавливали ее для каждого случая индивидуальна. Гнёзда осматривали в основном во второй половине дня, блйжё к вечеру. Они получали порядковый номер и заносились в список проверок, где проставляли дату следующего посещения. Все гнезда с полными кладками измеряли и описывали, а после того как выводок покидал гнездо, его материал разбирали и взвешивали по фракциям. Яйца взвешивали и измеряли в заранее намеченных гнездах, когда там не было наседки, или использовали для этих целей брошенные кладки. Сведения о каждом из найденных гнезд заносили в гнездовую* карточку, форма которой была разработана проф. И. А. Долгушиным.
Для получения сведений о кочевках выводков и убыли1
&
птенцов в них с помощью сетей отлавливали наседок у гнезд. Пойманных кекликов окрашивали спиртовым раствором родамина (красный цвет — самцы) или пикриновой кислоты (желтый цвет — самки), метили цветными ножными метками и стандартными алюминиевыми кольцами. Система мечения позволяла избегать повторения комбинаций цветов ножных меток. Этот же способ мечения применялся при отлове кек-ликов зимой. В период насиживания кладок у части гнезд устанавливались «дворики» из мелкоячеистой сетки, натянутой на колья вокруг гнезда, что препятствовало уходу птенцов после вылупления (рис. 1). Молодняк, оказавшийся в
Рис. 1. «Дворик» из мелкоячеистой сетки
«двориках», взвешивали, измеряли и метили кольцами с подклеенным изнутри поролоном, что способствовало удержанию колец на цевках. Этим же способом метили птенцов в нелетных выводках.
В осенние месяцы (конец августа — середина октября) в горных районах юго-востока Казахстана проводили количе
10
ственные учеты кекликов. Основываясь на биологических особенностях вида, мы разработали методику учета на постоянных маршрутах, проходящих по дну ущелий и отщелков, где происходит концентрация птиц на кормежках и у водопоев. Каждый из двух учетчиков проходил один маршрут. Длина его обычно ограничивалась протяженностью щели или отщелка и определялась шагомером. Для более точного подсчета кекликов в группировках использовали восьмикратные бинокли. При прохождении маршрута в дневник записывали время встречи и количественный состав стай. После проведения учета заполняли специальные карточки, в которых отмечали дату и время учета, состояние погоды; здесь же описывали особенности маршрута (растительность, крутизна склонов, наличие осыпей и т. д.) и подводили результаты (длина учетной ленты и общее количество встреченных птиц). Для характеристики погодных условий в местах проведения учетов использовали данные метеостанций «Анархай», «Туркестан», «Мерке», «Луговая», «Кугалы», «Сары-Озек», «Ачисай».
В зимние периоды на стационаре в ущ. Кзылаус кекликов отлавливали тайниками, которые устанавливали на подкормочных площадках. Пойманных птиц измеряли, взвешивали и метили, после чего выпускали в местах отлова. Последующие наблюдения за ними и повторные отловы позволили определить участки, занимаемые отдельными стаями, выявить места их ночевок и кормежек. Мечение кекликов и повторный отлов дали возможность получить данные об изменении массы и упитанности отдельных особей.
Для изучения питания, сезонного изменения упитанности и линьки проводили отстрел кекликов. У добытых экземпляров при вскрытии определяли пол и возраст; гонады и фабри-циева сумка измерялись. Содержимое зоба и желудка извлекали, взвешивали и упаковывали для последующей обработки в лабораторных условиях. Состояние линьки крупного и мелкого пера оценивалось по шестибалльной шкале (Snow, 1970). Счет маховых перьев велся от наружного края в проксимальном направлении.
Летом 1978 г. на стационаре в ущелье Кзылаус в течение 6 недель изучали постэмбриональное развитие кеклика на ’10 птенцах, взятых из гнезда сразу после вылупления. Птен-щов поместили в вольеру, установленную среди естественного
11
травостоя. Первые 3 недели на ночь их помещали в коробку с подогревом. Со второго дня жизни выводок на 2—4 ч выйо-сили на прогулку на склоны гор или в долину ущелья/где* птенцы самостоятельно кормились; в качестве дополнительного питания они получали прямокрылых, реже — творог, ягоды эфедры, кукурузный комбикорм и мелко нарубленное мясо. Ежедневно утром (до кормления) каждого из птенцов взвешивали, измеряли и отмечали изменения, происходящие в, их перьевом покрове. Наблюдения за выводком в обстановке, приближенной к естественной, дали возможность проследить* за изменениями в поведении, усложнением локомоции и выяснить значение в их жизни звуковой сигнализации.
За период полевых работ собраны следующие материалы:: 1. Найдено и описано 80 гнезд с кладками.
2. Окольцовано 316 кекликов, в т. ч. 160 птенцов и 31 наседка.
3. Собрана коллекция из 136 тушек 2 подвидов кекликов различного пола и возраста, поступившая на хранение в Институт зоологии АН КазССР*.
4. Измерено и взвешено 1232 птицы различного возраста^ добытых преимущественно в сезон охоты.
5. Измерено 186 яиц, взвешено 53 свежих и 74 яйца с различной степенью насиженности.
6. Для выяснения состава кормов определено содержимое зобов и желудков 925 кекликов.
7. Для определения качеств кеклика как дичи исследовано 786 особей.
Систематическое положение, ареал
Каменные куропатки (Alectoris Каир., 1829)—гомогенный, хорошо выраженный род, объединяющий 7 рецентных видов. Современное распространение этого рода и известные палеонтологические находки в Южной Франции (Lambrecht, 1933) и окрестностях Одессы (Тугаринов, 1940) позволяют считать, что колыбелью этого рода является Средиземноморье. Однако G. Watson (1962а), изучая симпатрию и алло-патрию каменных куропаток, а также их поведение и морфологические отличия, приходит к выводу, что предковые формы современного, рода были очень широко распространены к востоку, вплоть до Гималаев. Автор предложил схему эволюции видов каменных куропаток с разделением их на 2 группы: древнюю реликтовую, куда включены A, graeca Meisnen A. magna Przew., A. melanocephala Ruppel, и более новую, динамичную, состоящую из A. chukar Gray, A. philbyi Bates, A. rufa L., A. barbara Bonn.
До 60-х годов текущего столетия самостоятельность 4 видов (A. rufa, A. graeca, A. barbara, A. melanocephala) была общепризнана, что находит отражение как в систематической сводке Y. L. Peters (1934), так и в последних работах по птицам Палеарктики (Vaurie, 1965; Бёме, 1975). Что же касается A. chukar, то до последнего времени он рассматривался либо как. самостоятельный вид, либо объединялся с европейской каменной куропаткой (A. graeca).
Впервые кеклика описал J. Falk в 1786 г., который по экземплярам из Бухары дал ему название Tetrao kakelik. Однако в середине первой половины XIX в. этот вид вновь был
13
описан по экземплярам из Индии и получил видовое название chukar (Gray, 1830). В 30-х годах текущего столетия в> систематике появилась тенденция укрупнения видов за счет объединения близких самостоятельных форм. Это коснулось и кеклика.. В списке птиц мира (Peters, 1934) кеклик теряет видовую самостоятельность и отнесен к европейской каменной' куропатке (A. graeca). Укрупнение видов в отечественной литературе не всегда находило единодушное одобрение, что-хорошо прослеживается на примере видовой самостоятельности кеклика. В сводке «Птицы СССР» кеклик считается самостоятельным видом — A. kakelik (Иванов, 1951), а в обширном труде «Птицы Советского Союза», где приводится биология птиц по отдельным подвидам, он отнесен к европейской каменной куропатке — A. graeca (Карташов, 1952). Авторитет этого крупного издания обусловил тот факт, что в дальнейшем в большинстве орнитологических статей и региональных сводок кеклика стали называть A. graeca, т. е. объединяли с европейской каменной куропаткой. Однако часть отечественных орнитологов продолжала считать кеклика самостоятельным видом (Иванов, 1940, 1969; Долгушин, 1948;. 1951; Попов, 1959; Кузьмина, 1962 и др.).
Полемику в вопросе разделения A. graeca на самостоятельные виды, куда помимо A. chukar вошли A. magna и A. philbyi, разрешил G. Watson (1962в), который указал на морфологические и экологические различия этих форм. В публикациях отечественных и зарубежных авторов, появившихся после исследований G. Watson (1962а, в, 1966), в том числе и в обширной систематической сводке Ch. Vaurie (1965), эти виды рассматриваются как самостоятельные. Необходимо отметить, что впервые A. magna как самостоятельный вид был описан Н. М. Пржевальским (1876). Позднее эту точку зрения подтвердил В. Л. Бианки (1907). Однако-Y. Peters (1934) считал A. magna подвидом A. graeca. Лишь позднее описанный Н. М. Пржевальским вид вновь признан самостоятельным (Watson, 1962в).
Одновременно с пересмотром видовой самостоятельности кеклика значительные изменения претерпевали и взгляды на подвидовое разделение, что выразилось в значительном сокращении форм, относящихся к A. chukar. В сводной работе С. А. Бутурлина и Г. П. Дементьева (1935)’ приводится 13 подвидов, в систематической сводке «Птицы СССР» (Иванов,
14
1951) — 16, в сводке «Птицы'Советского Союза» (Карташов, 1952)—9, и, наконец, в новейшей систематической работе «Птицы фауны СССР» (Степанян, 1975)—только 6.
В настоящее время в орнитологической литературе встречается 2 видовых названия кеклика — A. kakelik и A. chukar. Какое же из них нужно дать этой птице — решать Международной комиссии по зоологической номенклатуре; поскольку этот вопрос до сих пор не рассматривался, согласно существующим правилам, кеклика следует называть A. chukar.
Каменные куропатки населяют горные районы южной части Палеарктики и северо-восточной части Эфиопской области. Современные ареалы большинства видов ограничены. По Р. Л. Бёме (1975), ареал A. graeca охватывает Альпы от Франции и Италии до Австрии, отмечен также в Центральной Италии до Сицилии и от Югославии на юг до Греции, а на восток — до Западной Болгарии. A. rufa распространена от восточной Бретани к югу до Гибралтарского пролива; живет на Корсике и в^Северо-Западной Италии. A. barbara населяет Тунис, Алжир, Марокко и Канарские острова; A. magna — Северный Тибет от Цайдама на юг до истоков Хуанхэ, на восток до оз. Кукунор, к северу до Наныпаня. Еще 2 вида распространены в западных и южных районах Аравийского полуострова. Ch. Vaurie (1965) указывает, что A. melanocephala живет от Мекки к югу до Адена и Хадрамаута, а A. philbyi — от Таифа на севере до Асира и, вероятно, Северного Йемена на юге.
Область распространения A. chukar — единственного представителя рода каменных куропаток, встречающегося на территории нашей страны, — очень обширна. Она значительно' превышает ареалы других видов и охватывает горы юго-восточной части Балканского полуострова, Малой, Передней, Средней и Центральной Азии, Кавказа, Алтая. Северная граница проходит по Родопским горам, северному склону Большого Кавказа, Мангышлака, Кызылкумов, хребта Каратау, северных склонов Тянь-Шаня, Чу-Илийских гор, Джунгарского Алатау, Тарбагатая, Саура, Южного Алтая, Западного5 Танпу-Ола, Хангая; южная — охватывает восточные части Балканского полуострова, Малую Азию, Юго-Восточный Ирак, Южный Афганистан, Северо-Западную и Северную Индию, Северо-Западную Сычуань*и Шаньси (рис. 2). Следует отметить, что северная и северо-восточная границы
15
Рис. 2. Ареал рода Alectoris. Заштрихована территория распространения кеклика
ареала кеклика еще недостаточно ясны и требуют уточнения. Просмотр коллекций ЗИН АН СССР позволил предположить его нахождение на западном берегу Аральского моря к югу от Кератамака (сборы А. Р. Бурачека от 21.IV 1924 г., инв. № 10191) и в Саянах (сборы А. Я. Тугаринова от 24.IX 1908 г. у с. Тюрботы, 30 км от Кемчика, инв. № 23438; сборы С. И. Снигиревского с р. Абакан в марте 1936 г., инв. № 37383; экземпляры, полученные от Абаканской заготконторы в январе — апреле 1936 г.). До сих пор нуждается в уточнении вопрос об обитании кеклика в Мугоджарах (Зарудный, 1888), Улутау (Павлов, 1931) и окрестностях Семипалатинска, в Семейтау (Хахлов, Селевин, 1928).
Интересными представляются сведения о распространении кеклика в Крыму. Первая достоверная встреча здесь этой птицы описана А. Семеновым (1899); это послужило автору, поводом к пересмотру зоогеографического положения горного Крыма и возникновению теории происхождения фауны этой части полуострова. Однако Ю. В. Костин и В. В. Кормилици-на (1974) считают, что А. Семенов описал встречу потомков 3 пар кекликов, завезенных из Греции 1894 или 1895 г. В настоящее время в Крыму кеклик населяет безлесные и слабооблесенные южные склоны Главной гряды гор от Аюда-га на юго-западе до окрестностей Феодосии на востоке. Обитающие в этом районе особи — результат акклиматизационных работ (в 1947—1961 гг. из Казахстана и Средней Азии завезено и выпущено около 300 кекликов).
В Восточной Болгарии и Греции находится узкая зона контакта кеклика и европейской каменной куропатки, хотя экологически эти виды различаются по высотному распространению: A. graeca живет на высотах более 1000 м, a A. chukar— ниже указанной высоты. В области стыков ареалов не обнаружено гибридов или промежуточных форм между ними, но в вольерных условиях они получены, что говорит о их близком родстве (Watson, 1962). В восточной части ареала кеклик встречается с другим представителем рода — A. magna. Географическое перекрытие их ареалов в провинции Ганьсу (Китай) незначительно, но здесь, как и в предыду-' щем случае, отмечено высотное разобщение: A. magna обитает на высотах более 2600 м, a A. chukar — ниже 2300 м. Места обитания этих видов в указанном районе одинаковы и пред
2—24
17
ставляют собой каменистые склоны гор и ущелья с травой и мелким кустарником (Бёме, 1975).
В южной части ареала кеклик населяет сухие субтропики^ по климатическим условиям близкие к тем, в которых живут другие представители рода (Кузьмина, 1977). Однако на значительной территории он живет в районах с продолжительными зимами, которые периодически оказывают существенное воздействие на его численность, вызывая массовую гибель.
Благодаря акклиматизационным работам, проводящимся: в различных частях Земного шара, кеклик значительно расширил границы своего естественного ареала. Расселению этого вида способствовало исключительно легкое приручение к содержанию в неволе.
Первое упоминание об акклиматизации кеклика датируется 1588 г. (о. Св. Елены, центральная часть Атлантики), куда он был завезен из Индии или Ирана. Обитающие здесь в настоящее время птицы отличаются сероватой окраской оперения и чрезвычайно коротким крылом. Эти отличия скорее всего появились в результате эволюции на изолированном острове или, что менее вероятно, являются фенотипическими (Watson, 1966).
В Северной Америке кеклика начали акклиматизировать с 1828 г., сюда он был завезен из питомника в Индии. Успешное разведение этой птицы на фермах Калифорнии позволило1 в 1932—1955 гг. выпустить 52 184 особи в 54 графствах. Они успешно прижились в пустынных и полупустынных районах,, где годовая сумма осадков не превышает 250 мм. В лесистой, более влажной местности отмечен отрицательный результат (Harper е. а., 1958). Успешная акклиматизация осуществлена также в штатах Аризона, Невада и других, благодаря чему этот вид стал здесь объектом спортивной охоты.
Таким образом, кеклики, ввезенные в США, прочно уко-, ренились на большом горном пространстве от Калифорнии да Колорадо и от Нью-Мексико до Орегона (Wissel е. а., 1966).
В Новой Зеландии попытки акклиматизации предпринимались с 1920 г. Первого успеха добились в 1926 г., и с тех пор кеклики, завезенные из Индии, быстро расселились по Южному острову, где держатся на высоте 250—2000 м над. ур. м. С 1934 г. на этом острове на них разрешена охота..
18
Попытки расселить кеклика на Северном острове окончились неудачей (Marples, Gurr, 1955).
Если в североамериканских штатах и на островах Новой Зеландии кеклик получил «права гражданства», то в. Европе опыты по его расселению были редки, что, видимо, связано с распространением других видов каменных куропаток (A. graeca и A. rufa). Известен лишь случай расселения кеклика в Болгарии, где, по данным П. Драгоева (1971), за 1965—1969 гг. с этой целью выпущено 40 000 птиц, что должно было увеличить общее поголовье до 550 000 экземпляров, при ежегодной добыче 150 000. A. Menzdorf (1976), проводя краткий анализ попыток расселения каменных куропаток в ФРГ, указывает, что кеклик по сравнению с другими видами наиболее перспективен для акклиматизации в этой стране.
Из 6 подвидов кеклика, встречающихся на территории Советского Союза (Степанян, 1975), в Казахстане обитает три: A. ch. shestoperovi — мелкая светлая форма, населяющая Мангышлак и Устюрт, A. ch. falki — крупная темная форма, распространенная в Каратау, Таласском, Киргизском, Заилийском Алатау и Чу-Илийских горах, и, наконец, A. ch. dzungarica, населяющая Джунгарский Алатау, Тарбагатай, Саур и Алтай.
Наши материалы собраны в районах распространения двух последних форм.
Места обитаний и повадки
Кеклик — типичная птица пересеченной местности. Его обширный ареал охватывает районы, отличающиеся по геоморфологии, климатическим условиям и растительности, что подчеркивает исключительно высокую экологическую пластичность этого вида. В пределах Советского Союза кеклик населяет районы от равнин Туркмении, где поселяется на глинистых холмах с обрывами (Дементьев, 1952) до альпийских лугов Памира на высоте до 4000 м над ур. м. (Степанян, 1969). Наиболее обычны и даже многочисленны кеклики в диапазоне высот 500—2000 м. Гнезда из альпийского пояса пока неизвестны. Таким образом, наиболее излюбленными для этого вида являются невысокие горные группы, расположенные в пустынях и степях, и степной, лесо-лугостепной и субальпийский пояса крупных горных массивов.
При исключительном разнообразии местообитаний кеклик все-таки предпочитает ущелья с выходами скал и склоны, чередующиеся с каменистыми осыпями и открытыми травянистыми участками, иногда поросшими кустарником. Наличие водопоев (горные реки, ручьи, ключи) играет важную роль в жизни кекликов; в исключительных случаях птицы пьют горько-соленую воду. Иногда кеклики отмечались в совершенно несвойственных им условиях: на равнинах и среди песков, на значительном расстоянии от ближайших гор (Сержпин-ский, 1925; Молчанов, 1932; Шнитников, 1949; Ишадов, 1970).
Интересными представляются встречи кекликов близ озе-р'а Сорбулак (60 км к северо-западу от Алма-Аты). Здесь в полупустынной зоне с пологими холмами, где фоновая расти-
20
гельность в основном представлена несколькими видами полыней, кеклики встречены 1 ноября 1978 г. (3 особи), 27 марта 1979 г. (одиночка) и 12 сентября 1982 г. (из стайки 15 птиц добыта одна молодая). В 1979—1981 гг. в сентябре — октябре стайки птиц неоднократно отмечались близ Куртин-ского водохранилища. Несмотря на то что гнезд в этих районах пока не найдено, можно предположить спорадичное гнездование в некоторые годы.
В характере местообитаний кеклика существует множество различий, которые обусловлены природными факторами.
Кавказский кеклик (A. ch. kurdestanica) населяет разнообразные биотопы от сухих склонов гор и предгорий до 3500 м над ур. м., где живет по россыпям вблизи ледников (Сатунин, 1907). По безлесным каменистым участкам гор этот подвид спускается почти до равнины, где встречается на гнездовье по кустарникам вместе с турачом. Однако охотнее поселяется на крутых склонах предгорий или в ущельях горных рек с выходами скал и каменистыми осыпями, покрытых редкой ксерофитной растительностью. Реже занимает луговины северных склонов с арчовым редколесьем, избегая, как правило, сырых участков с богатой растительностью.
Места обитания копет-дагского кеклика (A. ch. shestope-rovi) занимают особое место среди других подвидов. Распространение его связано с небольшими ксерофитными горными группами, сухими хребтами и даже промоинами и обрывами. В Копет-Даге он встречается в небольших ущельях на высотах 500—600 м, иногда в местах, где имеются водопои, поднимаясь до 2000 м. Нижняя граница его распространения лежит в полынно-эфемеровой полупустыне, верхняя связана с участками нагорных ксерофитов. Кроме того, кеклик встречается и на участках, занятых арчовниками, зарослями трагакантовых астрагалов, .среди кустарников, растущих у осыпей и скал, зарослями дикого винограда, ежевики и различных плодовых (Федоров, 1949). Населяет он и обрывистые песчаные берега Узбоя в районе пресных озер, где встречается сре^ ди саксаула и песчаной акации ( Молчанов, 1932).
Кзылкумский кеклик (A. ch. koroviakovi) распространен от хребта Гязь-Гядык к востоку до Кугитанга и Байсунско-го хребтов (Степанян, 1975). Сведения о местах обитания его очень скудны. А. И. Иванов (1940) сообщает, что этот подвид держится в совершенно голых, безлесных, а местами и без
21
водных горах (Ходжа-Хозяин). Лишь на Бабатаге проникает в зону фисташки, а на Кумтаге поднимается в арчовники.
Наиболее высокогорным в Советском Союзе является кара-корумский кеклик — A. ch. pallescens, распространенный в южной половине Западного Памира. В долине Шах-Дары верхняя граница его распространения проходит на высоте около 4000 м. По данным Л. С. Степаняна (1969), здесь кеклик встречается по всему бассейну реки, но численность его невелика. Наибольшая плотность отмечена на высотах 2300—2600 м. В этих условиях птицы ведут оседлый образ жизни и держатся в период гнездования и^зимой на каменистых склонах и моренах среди россыпей.
Места обитания тянь-шаньского кеклика (A. ch. falki) по сравнению с другими подвидами наиболее разнообразны, что обусловлено особенностями климата в различных частях ареала и широким диапазоном вертикального распространения (300—3600 м над ур. м.). Населяет хребты Северного, Центрального и Западного Тянь-Шаня. Северная граница распространения проходит по хребту Каратау и Чу-Илийским горам; южная достигает хребтов Юго-Западного Таджикистана, Памиро-Алая, долины Вахша. На сыртах 4 в котловине Сонкуля и других высокогорных долинах Центрального Тянь-Шаня не найден. Отсутствие его в этих местах, по-видимому, связано с частыми летними снегопадами и похолоданиями (Янушевич и др., 1959).
В Зеравшанском, Туркестанском и Гиссаро-Каратегин-еком хребтах тянь-шаньский кеклик населяет скалистые склоны с зарослями кустарников, реже держится на безлесных скалах и осыпях, иногда — на травянистых склонах. Пределы высотного распространения в этих хребтах— 1200—3500 м. Особенностью биологии следует считать значительные сезонные' миграции, что приводит к четко выраженной смене разнообразных биотопов (Ковалев, Попов, 1980).
В'Таласском Алатау он встречается от культурного пояса до субальпийского (1000—3000 м). Населяет сухие каменистые склоны с разреженной травянистой растительностью и редким кустарником; обычен в арчовом редколесье. Иногда гнездится у подножья гор на лишенных каменистых обнажений участках с растительностью скорее лугового, чем степного характера (Ковшарь, 1966).
На северных склонах Киргизского хребта, водораздель-$2
ная линия которого достигает 4195 м над ур. м., кеклик распространен от подножья до вечных снегов, где был отмечен в небольшом числе (Спангенберг, Судиловская, 1959). В казахстанской части Киргизского Алатау, ледников нет, но на высотах свыше 3500 м снег удерживается все лето и снежные поля достигают значительных размеров. Полноводные реки (Мерке, Каракыстак и др.), начинаясь в пригребневой части, рассекают склоны глубокими ущельями и в предгорьях разбираются на орошение. В растительности Киргизского Алатау хорошо выражена вертикальная поясность. Предгорные шлейфы заняты степями, которые используются в земледелии. Начиная с 1000 м над ур. м. склоны ущелий покрыты зарослями кустарников: таволгой, жимолостью, шиповником; барбарисом и дикими плодовыми — яблоней, урюком, боярышником. С высоты 1500 м появляется высокоствольная арча, которая образует куртины редколесья, переходящие затем в стелющиеся заросли. В пределах 2500—3000 м отчетливо выражены нагорные типчаковые степи, а выше расположен пояс альпийских лугов. В ущельях по берегам рек, которые служат кеклику местами водопоя, отдыха и кормежки, встречаются тугайные заросли с преобладанием ивы (рис. 3—5).
В Киргизском Алатау численность кеклика довольно высока и стабильна, что объясняется наиболее благоприятными условиями существования по сравнению с Другими районами юго-востока Казахстана. Особенно высока плотность его в среднем поясе гор (900—1500 м над ур. м.), где он населяет склоны с кустарником и выходами скал, каменистыми осыпями. Зима здесь непродолжительна. Холодный период со среднесуточной температурой ниже 0° составляет лишь 3— 3,5 месяца. Средняя многолетняя температура января —5°. Продолжительность снежного периода 100—120, в особо теплые зимы—'всего 30—40 дней. Обширные горные склоны южной экспозиции обычно бывают бесснежны. Это обстоятельство помогает кекликам перенести наиболее трудное для них время года. Гнездятся в зарослях шиповника на опушках лиственного леса, среди скал и осыпей. В осеннее время стаи чаще встречаются в кустарниковых зарослях по склонам гор или берегам рек.
В Заилийском Алатау тянь-шаньский кеклик встречается ют предгорий до субальпийского пояса (3000 м), где отмечается лишь в очень небольшом количестве. В поясе листвен-
аз
Рис. 3. Тугайные заросли в ущелье Каракыстак (Киргизский Алатау, 1000 м над ур. м.) —места водопоев кеклика
г И
Рис. 4. Каменистые склоны в ущелье Сюгаты — типичное местообитание кеклика
Рис. 5. Древесно-кустарниковая растительность близ речки (ущелье Сюга-ты) — местообитание кеклика
ного леса (1100—1500 м) населяет участки склонов с выходами скал и осыпей, чередующихся территориями, поросшими таволгой, барбарисом, шиповником, урюком и яблоней. В поясе хвойного леса (1500—2700 м над ур. м.) встречается на открытых луговых участках с небольшими выходами скал. Наибольшая плотность птиц отмечена на высотах 900— 1600 м. В западной части хребта в некоторые годы кекликов
25
много в нижних участках гор, где они населяют остепненньщ склоны с кустами таволги и небольшими осыпями. /
В хребте Каратау кеклик распространен повсюду, за исключением самой северной отдельной группы Актау (Долгушин, 1948). По данным этого автора, кеклик встречается повсюду, где имеются скалы и каменистые выходы (рис. 6).
Рис. 6. Типичное местообитание кеклика в Каратау (ущелье Кумусты, -600— 800 м над ур. м.)
Иногда выходит кормиться на совершенно ровные места без камней. В Джувалинског! долине отмечен на посевах. По сведениям Д. Н. Кашкарова (1932), в 40-х годах в Каратау кекликов было несметное множество. Каратау — крайний северо-западный отрог Тянь-Шаня, простирающийся на 400 км. Наивысшие точки хребта находятся в центральной его части в горном узле Мынжилке, высота которого достигает 2176 м над ур. м. Этот узел разделяет хребет на две части, которые отличаются растительностью и климатическими условиями. Северо-западная характеризуется резко континентальным засушливым климатом: зима продолжительная, снежный покров лежит 100—130 дней, средняя температура января —10°, ле
26
то сухое и жаркое, с большой амплитудой суточных' температур (13—17°). В растительности основной фон создают различные полыни и засухоустойчивые злаки, в частности луковичный мятлик (Роа bulbosa). По речкам и у родников ►сосредоточены кустарники — тамариск, чингил, шиповник. Местами здесь можно встретить заросли тростника. Древесная растительность представлена ивами, растущими по беретам рек и родников. Во всем облике растительного покрова сказывается влияние *пустыни Муюнкум.
В юго-восточной части хребта преобладает мезофильная растительность, с обилием разнообразных плодовых, среди которых отмечается и ряд южных форм. Основной фон растительного покрова — мезофильные злаки: различные виды ячменя. (Hordeum crinitum, Н. mundrum, Н. bulbosum), костра (Bromus squamosum, В. tectorum), зонтичные. Склоны гор покрыты кустарниками: таволгой (Spiraea hypericifolia), кур-Чавкой (Atrafaxis pyrifoli), вишней (Cerasus tianschanica), а также фисташкой (Pistacia vera) и миндалем (Amygdalus petunnikovii). По дну ущелий, у берегов речек, развита уре-ма из многочисленных кустарников и деревьев: клена (Acer semenovii), боярышника (Crataegus turkestanica, С. ponti-са), разнолистной груши (Pirus regelii), яблони (Malus sie-versii) и различных жимолостей. Местами в уреме доминирует ясень (Fraxinus petamophila). Плоские вершины хребта (джоны) заняты полынно-типчаковыми ассоциациями. Лишь в пределах узла Мынжилке на высотах 1700—1800 м встречается лужайки с разнотравьем.
После джута зимой 1968/69 г. восстановление численности кеклика в хребте Каратау шло очень медленно и до 1977 г. юна оставалась самой низкой из всех обследованных хребтов Юго-Восточного Казахстана; первые стайки в Боролдае (у перевала Чокпак) появились лишь в 1972 г.
В Чу-Илийских горах кеклик является фоновым видом и встречается повсеместно, что объясняется однообразием биотопов, одинаково пригодных для его жизни. Однако чаще встречается на участках с выходами скал и осыпей.
Эти горы представляют собой сложную комбинацию отдельных невысоких хребтов, приподнятых на массивном плоскогорье, частью размытом эрозионными процессами. С запада они ограничены р. Чу, а с севера — пустыней Бетпак-Да-,ла, в которую горы заходят лишь отдельными отрогами-
27
Протяженность массива составляет около 250 км, макси-' мальная высота 1164 м над ур. м. На востоке к Чу-Илийским горам примыкает орографически связанное с ними плоскогорье Карой.
Ввиду сравнительно невысокого положения и значительной удаленности от морей Чу-Илийские горы получают всего1 250—300 мм осадков в год. Выпадают они очень неравномерно: весной до 70% годовой суммы, осенью — около 20, летом и зимой — немногим более 10%. Речки, имеющие истоки, в этих горах, маловодные и наполняются только во время таяния снега; едва достигая окраины возвышенностей, они, фильтруются в собственных наносах. Уже в начале лета многие водные источники пересыхают, и только в некоторых ущельях встречаются родники, густо заросшие травой и тростником. Однообразие почвенных условий и малые перепады высот обусловливают бедный видовой состав растительности. Исключение представляют речные долины, где в растительном покрове доминируют полыни, различные эфемероиды и кустарники. На щебнистых склонах много лука (Allium tra-chyscordum), местами занимающего значительные площади, а на пологих участках — ковыля, образующего сплошной ковер. Видовой состав кустарников беден. Среди них наиболее распространена таволга, реже — тамариск, чингил, эфедра, курчавка и кизильник. По берегам ручьев и речек растут чий и мята. Кроме того, в обводненных ущельях встречаются заросли тростника, местами на сотни метров покрывающие-дно долины. Древесная растительность представлена неболь-. шими группами ив, растущих у родников.
Климат Чу-Илийских гор резко континентальный, с большими перепадами температур даже в течение суток. Зима малоснежная, снеговой покров держится 90—100 дней. Продолжительность холодного периода со среднесуточной температурой ниже 0° 4 месяца; средняя температура зимних месяцев —4—7°. Лето жаркое и сухое; среднемесячная температура самого жаркого июля 25—27°.
Относительно небольшая высота Чу-Илийских гор и почти полная изолированность их от других хребтов накладывает отпечаток на жизнь кекликов, которые в течение года встречаются в одних и тех же районах, придерживаясь зимойг бесснежных участков склонов.
Джунгарский кеклик (A. ch. dzungarica) в пределах СССР
28
населяет Джунгарский Алатау, Тарбагатай, Саур, Западный
Южный Алтай, Западный Танну-Ола. В Джунгарском Алатау кеклик наиболее многочислен на высотах 500—1500 м, тде населяет пустынный, степной и лесолугово-степной пояса. В большом числе встречается по периферийным юго-западным отрогам Джунгарского Алатау, в горах Чулак, Матай, Ма-лай-Сары, Катутау и хребте Чаган.
В горах Чулак, протянувшихся вдоль северного побережья Капчагайского водохранилища почти на 100 км, наибольшая численность кеклика отмечена в нижней и средней частях гор (600—900 м над ур. м.). Нижняя граница их на высоте 600 м, обрываясь, переходит в пологий шлейф, представленный каменистой пустыней. Наиболее высокие точки гор достигают 1600—1700 м над ур. м. Орография гор Чулак представляет довольно пеструю картину: широкие долины у подножий перемежаются высокими крутыми скалами. Глубже в горы ущелья становятся теснее и местами образуют непроходимые цирки с вертикальными стенами. В верхней части иногда встречаются широкие долины, вытянутые вдоль хребта.
Гидрографическая сеть представлена ключами, водный баланс которых подвержен значительным колебаниям: в засушливые годы небольшие родники полностью пересыхают, а в полноводные воды становится значительно меньше.
Нижняя часть гор Чулак лежит в поясе пустынь. Сумма годовых осадков здесь составляет 300—330 мм; причем они выпадают в основном весной. Холодный период года со среднесуточной температурой ниже 0° составляет 4 месяца, но снежный покров сохраняется 90—100 дней. Зима достаточно суровая (средняя температура января —8—10°). Лето жаркое и сухое (средняя температура июля 23—24°). Растительный покров гор беден. Древесная растительность представлена лишь группами ив, растущих на обводненных участках ущелий, и небольшими рощицами или одиночными деревьями каркаса (Celtis caucasica) и боярышника (Crataegus sanguinea). В недалеком прошлом каркас в горах Чулак встречался чаще, но позднее был частично вырублен. Фоновыми видами щебнистых склонов являются боялыч (Salsola arbus-cula) и кохия; менее каменистые участки заняты полынями. Ранней весной склоны гор покрыты ковром эфемеров и эфемероидов, среди которых основное место занимают лукович-
29
Рис. 7. Места обитания кеклика в горах Чулак: а — ущелье Кзылаус; б — южная часть ущелья Унгур
ный мятлик, маки, тюльпаны и скалигерия. На обводненных участках долин местами встречаются тростник (Phragmites, communis), ежевика (Rubus caesius), мята. На северных склонах и дне ущелий растут кусты таволги, местами занимающие значительные площади, а также курчавка, барбарис и чин-гил. На высоте 800—900 м над ур. м. появляется хвощевидная эфедра (Ephedra intermedia), которая в некоторых ущельях становится фоновым видом.
Излюбленные места обитания кеклика — открытые участки склонов с небольшими выходами скальных пород, спускающихся к берегам ручьев (рис. 7, 8). Гораздо реже их мож-
Рис. 8. Кеклики на каменистом склоне ущелья
но встретить на крутых, лишенных растительности скалах, перемежающихся с обширными осыпями. В гнездовой период пары в основном держатся по мягким увалам с редкой кустарниковой растительностью. В июле — октябре кеклики чаще отмечаются на дне ущелий и примыкающих к ним нижних частях склонов. С выпадением глубокого снега, стайки в поисках пищи появляются на участках, где снежный покров
ЗГ
незначителен и имеются хорошо прогреваемые места. По данным М. А. Кузьминой (1955), зимой в нижних частях гор кекликов становится гораздо больше, чем осенью, что объясняется прикочевкой птиц из вышерасположенных районов Джунгарского Алатау. Мечение кекликов в горах Чулак показало, что часть особей в нижней части гор живет оседло.
В останцовых горах правобережья реки Биже и невысоком хребте Чаган наибольшая численность кеклика отмечена на высотах 1200—1700 м над ур. м. Здесь невысокие гряды с пологими склонами прорезаются глубокими оврагами и ущельями, в которых выступают обнажения коренных пород. Растительность довольно бедная. Склоны покрыты полынями, эфемерами и эфемероидами; в скалистых ущельях растут шиповник, таволга, кизильник. Зимой кеклики иногда спускаются в предгорную степь, где держатся по лессовым обрывам; на кормежке отмечены по кромкам убранных полей зерновых.
В сходных условиях живут кеклики и на хребте Алтын-Эмель — одном из юго-западных отрогов Джунгарского
Рис. 9. Типичные места обитания кеклика. Хребет Алтын-Эмель (ущелье Тюлькули, 1400 м над ур. м.)
32
Алатау, имеющим протяженность около 80 км. Северо-западные склоны этого хребта обращены в Кугалинскую долину, а на юге он ограничен обширной Конур-Уленской впадиной. Наибольшая высота хребта достигает 2920 м над ур: м. Ущелья глубокие, с отвесными склонами и уходящими под вершины осыпями. Периферийные части имеют более плавные очертания — преобладает плоскогорный и волнистый рельеф. От основных массивов Джунгарского Алатау хребет отделен хребтами Кояндытау и Токсанбай. Растительный покров, особенно в глубоких ущельях, где обычно развивается урема, благодаря обилию берез и черемухи, приобретает северный колорит( рис. 9). В верховьях ущелий степное разнотравье сменяется лугами и небольшими кочковатыми болотами. По мере подъема чаще встречаются заросли сте-лящейся арчи, в сочетании с лужайками они напоминают субальпийские луга. Зима продолжительная. Холодный период года со среднесуточной температурой ниже 0° длится более 4 месяцев, безморозный — НО—120 дней. Снег лежит 120—130 дней; в наиболее снежные годы высота его достигает 1 —1,6 м. Максимум осадков выпадает в весенние месяцы (250—370 мм). Лето теплое; среднесуточная температура июля составляет 20—23°.
Численность кеклика на хребте Алтын-Эмель подвержена большим колебаниям. После суровой многоснежной зимы 1968/69 г. она восстанавливалась медленно. На учетных маршрутах осенью 1972 г. здесь не встретили ни одной птицы. В последующие 4 года кеклик отмечался в небольшом количестве по открытым склонам, в заросших ущельях и арчов-никах.
В горах Малай-Сары кеклик встречается повсеместно. Эта горная группа представляет собой плоскогорье, приподнятое над окружающей степью. Высшие точки достигают 1300 м над ур. м. Склоны довольно крутые и изрезаны сетью неглубоких, в большинстве случаев безводных ущелий. На климат гор большое влияние оказывают окружающие степи и полупустыни, что обусловливает малое количество осадков и создает резкие колебания температур. Продолжительность холодного периода с среднесуточной температурой ниже 0° составляет 4 месяца, устойчивый снежный покров держится 80—100 дней; средняя температура января —9—11°; в аномальные годы она может снижаться на 12—14°. Лето жаркое
3—24
33
и сухое. Сумма осадков в июне—августе 15—35 мм; средняя температура июля 24—25°. Растительный покров беден и однообразен. Склоны гор в основном покрыты полынями, солодкой (Glycyrrhiza glabra), чием (Lasiagrostis splendens). На более пологих участках и долинах растут кусты таволги и курчавки.
Почти полная безводность гор Малай-Сары не сказывается на распределении здесь кеклика. Стайки птиц в летний период встречались в 15—20 км от ближайших водных источников. Осенью и зимой кеклики отмечены на плато, где они собирали оставшееся после уборки урожая зерно.
Исключительное разнообразие местообитаний свойственно кеклику не только как виду, но и отдельным его популяциям. Совершая вертикальные кочевки, стаи птиц в течение года сменяют ряд биотопов, избирая наиболее благоприятные условия на разных высотах. Высокая плотность птиц и меньшие колебания численности отмечаются в нижних поясах крупных горных хребтов в пределах до 2000 м. Здесь кеклики находят наиболее подходящие условия для существования.
По внешнему виду кеклик напоминает бородатую куропатку, вместе с которой живет в некоторых местах Казахстана и Средней Азии. Птица выглядит очень эффектно. Клюв, кольцо вокруг глаза и ноги красные. Радужина коричневая. На подбородке и у основания подклювья небольшие черные* пятна. От основания надклювья через глаз к уху н далее* под углом вниз проходит черная полоса, которая окаймляет светло-охристое горло и, опускаясь к зобу, образует V-об-разный «галстук». Кроющие уха рыжевато-коричневые. Верхняя сторона тела от оливково-коричневатого до серого цвета, на плечах и темени с винным оттенком. Маховые бурые (первостепенных 10, второстепенных 12); часть перьев с, охристыми наружными опахалами. Плечевые серо-голубые, оконтуренные широкой ярко-коричневой полосой; кроющие крыла бурые. Грудь серо-голубоватая*. Брюхо желтовато-охристое,
* Н. А. Зарудный (1900) отметил у кекликов, добытых в Северо-Восточной Персии, развитие на груди черных пятен, достигающих 1 см, свойственных преимущественно самцам. По нашим наблюдениям, эта особенность присуща некоторым кекликам из Юго-Восточного Казахстана. Однако черные пятна мы находили у взрослых и молодых птиц, самцов п самок,, особей разных подвидов, что не может служить критерием для определения половозрастных признаков и подвидовой принадлежности кекликов.
34
подхвостье охристое. Бока тела украшены перьями с широкими черными и каштаново-коричневыми полосами. Рулевые (12 шт.) коричнево-рыжие, за исключением центральной пары, имеющей такой же цвет, как и надхвостье.
Размеры. Самцы (п=225): крыло 152—168 (в среднем 161) мм, хвост 74—93 (83), цевка 43,4—50,2 (46,5), клюв 20,4—23,4 (21,7) мм. Самки (п=186): крыло 146—161 (в среднем 152) мм, хвост 69—90 (76), цевка 40,4—48,5 (43,5), клюв 19,6—22,8 (20,5) мм.
Масса. Самцы (п=324): 412—686 (в среднем 527) г; самки (п = 278) 381—528 (в среднем 457) г.
Половой диморфизм в окраске оперения у кекликов не выражен, и самцы отличаются от самок только по наличию «шпор» — рогового образования на тыльной стороне цевки.
Кеклики — общественные птицы и большую часть года проводят в стаях. Лишь в период размножения держатся парами. По окончании брачного периода особи, не участвующие в насиживании кладок и воспитании молодняка, объединяются в стаи. После вылупления птенцов выводки ведут самостоятельный образ жизни, или объединяются.
Ограниченная видимость в условиях пересеченной местности затрудняет общение между отдельными стаями, и поэтому основой общения у кекликов являются звуковые сигналы. Там, где птиц много, голоса их слышны в течение всего дня. Лишь в период насиживания и в первое время воспитания молодняка они становятся молчаливыми. При появлении опасности кеклики быстро убегают вверх по склону или улетают; в некоторых случаях затаиваются.
Кеклики очень живые, подвижные птицы, проводящие на земле всю жизнь. В сложных условиях горного рельефа они свободно передвигаются среди каменистых осыпей и участков отвесных скал. Вниз по склону бегают редко, но так же уверенно, как при ходьбе вверх. При появлении человека ведут себя по-разному. Если он находится на склоне ниже стайки — стараются убежать вверх или вдоль склона (скорость бегущей птицы может достигать 3 м/с). В случае, если опасность возникла сверху, — улетают. Взлетая со склона, птица переходит на стремительное скольжение. Взлет начинается с частых взмахов, которые чередуются со скользящим полетом на неподвижных крыльях. По данным А. В. Попова (1960), максимальное расстояние, которое могут пролететь кеклики,
35
составляет около 2 км. С вертикальной скалы к подножью слетают почти вертикально, вытянув ноги и часто взмахивая крыльями; при приземлении на склон иногда разворачиваются в воздухе и тормозят хвостом. По рыхлому глубокому снегу передвигаются с трудом и поэтому часто становятся легкой добычей разных хищников, особенно лисиц. В местах, где кекликов постоянно преследуют, они проявляют особую осторожность. Однако, если птиц не беспокоить, становятся доверчивыми и могут жить рядом с постройками человека.
Кормятся на земле, употребляя в пищу семена, листья, подземные части растений, беспозвоночных ^животных. Способы добычи корма довольно разнообразны. Зеленые части растений и плоды сначала захватывают клювом, а затем отрывают, делая резкое движение головой. Семена и мелких насекомых склевывают с поверхности земли, а расположенные в верхнем слое почвы подземные части растений предварительно раскапывают, разгребая попеременно лапами. Луковицы и клубни, находящиеся глубже, достают, выдалбливая клювом в земле вертикальные отверстия до 8— 10 см глубиной. Мелкие луковицы иксиолириона, клубни скалигерии и гераней извлекают целиком, а луковицы тюльпанов расклевывают прямо в лунках. Саранчовых, после того как они взлетают, преследуют, совершая стремительные броски, и настигают в момент приземления.
В период размножения кеклики держатся парами в районе гнезд, но встретить их рядом с ними можно лишь случайно, поскольку самка в сопровождении самца появляется здесь только для откладки очередного яйца. После появлении птенцов в гнездах выводки в основном встречаются на открытых участках склонов. Зарегистрировано 5 встреч меченых на гнездах наседок с выводками.
Большое значение в жизни кекликов имеют водопои, которыми они в основном пользуются в июле — сентябре. Частота их посещений во многом зависит от погодных условий и времени года. Весной, когда в пище кекликов преобладают зеленые части растений, птицы у воды встречаются редко. Летом, в связи с их переходом на питание семенами, водопои посещаются гораздо чаще. В зимний период кеклики обходятся без воды: потребность в ней они возмещают, склевывая снег.
36
Суточная жизнь кекликов в течение года однообразна. Зи-мой птицы весь день заняты добыванием корма. Летом и осенью, когда пищи много, добывание корма чередуется с посещением водопоев и продолжительными периодами отдыха. Ночуют в средних и верхних частях склонов, в распадках среди камней и кустов, но избегают оставаться на ночь на дне ущелий. Часто для ночевок используют одни и те же места. Это обычно небольшие пологие площадки под навесом скал или среди кустов, которые бывают покрыты слоем помета.
Биология кеклика
Брачный период. Календарные сроки разбивки зимних стай кекликов и образование пар во многом обусловлены географическим положением местности, ее высотой над уровнем моря и ходом весны. В горах Чулак с наступлением первых теплых дней среди стай кекликов наблюдается оживление, которое выражается повышенной двигательной и вокальной активностью. С восходом солнца громкие крики птиц раздаются на склонах и дне ущелий; часто особи из одной стаи перелетают в другую, иногда слышится писк самок, преследуемых самцами. Такое поведение кекликов мы отмечали 20 февраля 1977 г., первые встречи пар работники охотхозяйства зарегистрировали 8 марта 1974 г., 11 марта 1975 г., 5 марта 1976 г. и 10 марта 1977 г. По наблюдениям М. А. Кузьминой (1955), перезимовавшие стаи постепенно распадают^ ся: сначала на мелкие стайки, затем — на пары (первая встречена 7 марта 1949 г.). В феврале 1977 г. кеклики держались небольшими стайками и охотно посещали подкормочные площадки, где их отлавливали при помощи тайника, метили и тут же выпускали. Еще 3, 6, 9 и 13 апреля встречались стайки из 6—8 особей, но уже с середины марта здесь отмечались и пары.
Данные, полученные в результате мечения кекликов, позволили установить, что у некоторых особей проявляется привязанность к определенным районам, где их отмечали как зимой, так и в период размножения.
Помимо приведенных в таблице 2 сведений в апреле 1978 г. егерем Капчагайского охотхозяйства в ущелье Тал-дысай, в 8 км от места отлова, добыт меченый кеклик.
38
Таблица 2. Возврат меток кеклика в горах Чулак
Возраст особи при кольцевании Дата ата с уска,
Серия и номер кольца выпуска и [ОВТОрНОЙ встречи Срок возвр; начала вып дни Расстояние от места предыдущей встречи
1 2 3 4 5 1 6
Е Е Е Е Е Е Е Е Е Е Е ’£ 824101 824102 824103 824104 824105 824106 824109 824111 824113 824114 824115 824117 Взрослая > Молодая » > » Взрослая Молодая > 12. 12. 12. 12. 18. 14. 17. 17. 17. 18. 18. 18. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. II 1976 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. 16. 17. 20. 21. 22. 14. 16. 17. 20. 14. 16. 17. 20. 21. 22. 17. 20. 25. 10. 19. 15. 21. 15. 11. 11. 16. 20. 21. 21. 27. 19. 21. 24. 25. 26. 28. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. II 1976 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. XII 1975 г. 11 1976 г. V 1976 г. II 1976 г. IV 1976 г. IV 1976 г. V 1976 г. II 1977 г. II 1977 г. 11 1977 г. II 1977 г. II 1976 г. II 1976 г. II 1976 г. II 1976 г. 11 1976 г. И 1976 г. II 1976 г. 11 1976 г. II 1976 г. 4 5 8 9 62 2 4 5 8 2 4 5 8 9 10 5 8 7 82 67 120 126 150 422 396 401 405 66 65 71 63 65 68 69 70 72 * * 300 м * 300 м * * * 300 м * * * 300 м * 300 м * 300 м * 500 м (в группе из 3 птиц) * 1000 м (в паре) 1000 м > 300 м (в группе из 5 птиц) * * * * * * * * * * * *
Продолжение таблицы 2*
1 1 2 1 3 1 4 5 1 6
Е 824120 Взрослая 18. XII 1975 г. 26. II 1976 г. 70 ♦
Е 824 1 21 Молодая 18. XII 1975 г. 21. II 1976 г. 65
Е 824122 18. XII 1975 г. 19. II 1976 г. 63 ♦
27. 11 1976 г. 71 «
Е 824126 19. XII 1975 г. 21. II 1976 г. 64 ♦
Е 824127 19. XII 1975 г. 21. II 1976 г. 64 «
24. II 1976 г. 67 ♦
20. IV 1976 г. 123 100 м
Е 824132 18. II 1976 г. 23. IV 1976 г. 65 20fr м (в паре)’.
Е 824138 Взрослая 19. II 1976 г. 14. II 1977 г. 361 *
Е 824142 Молода» 19. II 1976 г. 27. II 1976 г. 8 300 м
Е 824143 19. II 1976 г. 27. II 1976 г. 8 300 м
Е 824144 19. II 1976 г. 26. II 1976 г. 69 «
Е 824145 19. II 1976 г. 3. I 1977 г. 319 1500 м (найде-
на лапка с
кольцом)
Е 824155 ВзрЬслая 23. II 1976 г. 11. II 1977 г. 354 —
16. II 1977 г. 359 ♦
Е 824156 Молодая 18. II 1976 г. 24. II 1976 г- 6 ♦
25. II 1976 г. 7 ♦
7. V 1976 г. 79 900 м (встрече-
на в паре)
Е 824175 Взрослая 8. II 1977 г» 16. II 1977 г. 8 •
31. III 1977 г. 51 800 м (в паре)
18. VI 1977 г. 130 500 м (с вы-
водкбм 3-4-
Дн. птенцЬв),
Е 824177 9. II 1977 г. 12. II 1977 г. 3 •
16. II 1977 г. 7 «
27. IV 1977 г. 138 400 м (с вы- ш
ВОДЙОМ; 12
птенцов 25-дн.
возраста)
Е 824188 11. II 1977 г. 18, II 1977 г. 7 •
Е 824195 12. II 1977 г. 20. 1Г 1977 г. 8 500 м
Е 824197 12. II 1977 г. 16. II 1977 г. 4. ♦
18. II 1977 г. 6 1000 м
Е 964103 12. II 1977 г. 20. II 1977 г. 8 700 м
Е 964105 12. II 1977 г. 20, II 1977 г. 8 700 м
Е 964108 12. II 1977 г. 20, II 1977 г. 8 500 м
Е 964115 16. II 1977 г. 5. V 1977 г. 78 : 1500 м (в паре)-
Е 964137** 3. VI 1977 г. 2. VII 1977 г. 30; 250 м (с вывод-
ком 2—-3-дн.
птейцов)
Е 964151** 14. V 1978 г. 18. VI 1978 г. 35 400 м (2 взрос-
лых и 15 птен-
1 цов)
Окончание таблицы 2.
1 2 3 1 4 5 6
Е 964155** Взрослая 19. V 1978 г. 5. VI 1978 г. 17 500 м (1 взрослый и 10 6-дн. птенцов)
Е 964157** » 22. V 1978 г. 12. VI 1978 г. 20 400 м (2 взрослых и 20—23 птенца)
Е 964158** 25. V 1978 г. 17. VI 1978 г. 23 500 м (2 взрослых и 21 птенец) 350 м (1 взрослый и 7 9-дн. птенцов) 700 м (2 взрослых и 15—17 птенцов)
Е 964159** 28. V 1978 г. 9. VI 12. VI 1978 г. 1978 г. 12 15
* Птицы, встреченные в месте поимки. ** Наседки, меченные на гнез^ дах.
А. И. Янушевич и др. (1959) и Г. П. Дементьев (1952} указывают, что кеклики — типичные моногамы. Однако> М. А. Кузьмина (1955) в Чу-Илийских горах в брачный период наряду с парами иногда отмечала группы из 3 птиц. По ее мнению, они состояли из самца и 2 самок. По данным R. Mackie и Н. Buechner (1963), у интродуцированных в штате Вашингтон (США) кекликов тоже отмечались случаи (10%), когда при самце было 2 самки. В апреле — мае 1974—1978 гг. в горах Чулак мы встретили 166 пар кекликов. и 8 (4,6%) групп (самец и 2 самки); в одной из них зарегистрирован случай спаривания самца с обеими самками. Эти наблюдения дают возможность заключить, что небольшая часть самцов, видимо доминантов, имеет 2 самок, т. е. налицо полигамия. В большинстве же случаев в период размножения кеклики держатся парами; самец постоянно находится рядом, с самкой и зорко следит, чтобы к ней не приблизился соперник. Допустимое расстояние между самцами, когда они спокойно переносят соседство, во многом зависит от сложности рельефа и густоты растительности. На открытых участках» оно составляет не менее 40—50 м. Вспугнутые человеком,, партнеры обычно улетают в одном направлении, иногда раз-
41.
метаются на противоположные склоны. В последнем случае, как только опасность минует, самец летит к месту приземления самки или преодолевает это расстояние пешком.
Образ жизни кекликов в марте — апреле весьма однообразен. Большую часть дня пары проводят на склонах или дне ущелий, где кормятся, приводят в порядок оперение и отдыхают. В этот период из 180 ч наблюдений за поведением самок в парах в разные часы дня ими было затрачено на кормежку 46 ч 40 мин (25,9%): утром они кормились 26 ч 10 мин (56,1%), днем — 7 ч 26 мин (15,9%), вечером 13 ч 4 мин (28,0%). Самцы из 202 ч наблюдений кормились лишь 23 ч 40 мин (11,7%): утром — 10 ч 32 мин (44,5%), днем — 6 ч 41 мин (28,2%) и вечером — 6 ч 27 мин (27,3%). Таким образом, продолжительность кормежек самок в этот период вдвое больше, чем у самцов.
Первое время образовавшиеся пары малозаметны и самцы в них не проявляют агрессивности по отношению друг к другу. Напротив, пары в некоторых случаях объединяются во временные стайки. Так, 3 апреля 1978 г. к паре кекликов присоединилась другая, а затем еще 4 особи. В течение 2 чони медленно передвигались по склону, иногда отдыхали под кустиками таволги и к концу наблюдений вновь разбились на пары и разошлись в разные стороны.
По устному сообщению В. А. Вырыпаева и Р. Р. Будриса, в условиях Чаткальского хребта сильные самцы начинают отделяться от стай в начале марта, а к концу этого месяца заканчивается распределение между ними гнездовых участков. К середине апреля у самцов устанавливаются стабильные иерархические взаимоотношения.
По нашим наблюдениям, в горах Чулак наибольшая вокальная активность у самцов отмечается в середине апреля. Первые голоса их слышны в ранние утренние сумерки. Поют обычно сидя на камне или на вершине скалы, что позволяет вовремя заметить приблизившегося к самке соперника. Песня состоит из хорошо известного квохтанья и заканчивается «скирканьем»: «скрю-ю-у», которое повторяется несколько раз подряд. Иногда в конце песни добавляются звуки «кро-о, кро-о», напоминающие вороньи. У поющей птицы перья на шее топорщатся, туловище вытянуто вверх и немного вперед. Вокальная активность к полудню ослабевает, и во второй половине дня, как, впрочем, в дождливые и ветреные дни,
42
песни слышатся редко. На кормежке особи в парах переговариваются между собой тихим, приятным для слуха щебетаньем, которое можно услышать с расстояния до 20 м. Во время наблюдений за отдельными парами без труда можно определить пол каждой птицы. Для этого достаточно кратковременного наблюдения: самцы подвижны и крикливы, самки малозаметны, и голоса их, отличающиеся высотой тона, слышатся гораздо реже.
В некоторых орнитологических работах приводятся указания, что разбивка стай кекликов сопровождается драками самцов. Так, А. К. Рустамов (1945) пишет: «Брачный период у кекликов в Туркмении начинается с первой половины марта. Птицы начинают разбиваться на пары, что сопровождается большими драками между самцами из-за самок. 11 марта 1942 г. в Копет-Даге, в горах Хунча (Роборовское) удалось наблюдать бой двух самцов. Увлеченные дракой, они подпустили меня на 10 шагов. Два самца Долбили друг другу головы, которые уже были красными от крови. Кроме клюва время от времени они пускали в ход также ноги и крылья» (с. 45—46). Для гор Каратау сведения о драках кекликов приводит И. А. Долгушин (1951). По мнению автора, причина возникновения драк заключается в гибели самок и попытках самца, оставшегося без подруги, отбить самку у другой пары. О драках между самцами из-за самок в Чат-кальском заповеднике, наблюдавшихся в марте, сообщает М. М. Остапенко (1965). А. В. Попов (1959) в Гиссаро-Кара-тегине за 12 лет лишь однажды, 16 апреля 1948 г. в ущелье р. Варзоб, наблюдал дерущихся самцов. В работах Н. А. За-рудного (1896), В. Н. Шнитникова (1949), А. Ф. Ковшаря (1966) сведений о драках кекликов в брачный период не приводится. В горах Чулак в период разбивки стаек драк у кекликов не наблюдалось, но во время откладки яиц они иногда бывают ожесточенными (Кузьмина, 1955).
За время полевых работ в горах Чулак нам 6 раз приходилось видеть схватки самцов. Одна из них в конце апреля 1978 г. по своему накалу превосходила все наблюдавшиеся. Дерущиеся самцы били друг друга крыльями, хватали клювами за затылок, подпрыгивали и, столкнувшись грудью, катились вниз по склону, поднимая пыль, иногда летели перья. Скатившись по склону на 20 м, кеклики, издавая потрескивающие звуки, поднялись к вершине, и схватка во
43
зобновилась с прежним ожесточением. В короткие перерывы между боем соперники издавали потрескивающие звуки, заканчивающиеся продолжительном «ко-коо», и в возбуждении «точили» клювы о камни. Схватка продолжалась в общей сложности 38 мин. Столь ожесточенные драки — явление довольно редкое, но преследование самцами друг друга нам приходилось видеть едва ли не ежедневно. Отмечено, что если неподалеку от пары кекликов появляется одиночный самец или другая пара, то на это самец моментально реагирует: бежит к сопернику, который спасается бегством или улетает. Иногда преследуемый кеклик затаивается и предпринимает попытку вновь приблизиться к самке, но во всех известных, нам случаях эти попытки решительно пресекались и кончались бегством пришельца.
Такое поведение самцов продолжается до второй декады мая, когда начинается массовое насиживание кладок. Наблюдения за кекликами в брачный период показывают, что ожесточенные драки, видимо, случаются только между самцами, занимающими одинаковое положение в иерархии. В. А. Вырыпаев и Р. Р. Будрис (1976), изучавшие поведение самцов в гнездовый период в Чаткальском заповеднике с помощью звуковых сигналов, воспроизводившихся через магнитофон, установили, что у кекликов четко выражена иерархия: доминанты своим голосом подавляют звуковую сигнализацию более слабых самцов и занимают наиболее удобные гнездовые участки.
Гнездовый период. Во второй половине апреля кеклики приступают к строительству гнезд. Г. П. Дементьев (1952) сообщает, что в устройстве гнезд принимают участие и самка и самец. М. А. Кузьмина же пишет (1955): «Принимает ли участие при устройстве гнезд самец, сказать трудно,, ясно лишь то, что у места постройки гнезда всегда рядом находятся самец и самка» (с. 129). Д. Н. Кашкаров (1932) сообщает о строительстве гнезд самцами, которые выстилают их мхом, листьями, травой и не только сторожат, но, когда самка отправляется пить или кормиться, заменяют ее на гнезде.
Мы располагаем следующими материалами о строительстве гнезд кекликами. 25 апреля 1974 г. из-под одного из редких кустов боялыча на склоне южной экспозиции вылетела помеченная нами самка. Самец в это время находился
44
в 5—7 м от нее. Осмотр места показал, что самка находилась в приготовленной для строительства гнезда ямке, в которой было несколько прошлогодних травинок и тонких веточек бо-ялыча. В другой раз, 29 апреля 1976 г., наблюдая за парой кекликов, удалось заметить, как самка зашла под куст. Спустя 5 мин она появилась и начала поспешно собирать сухие травинки, отбрасывая их к кусту. Затем, пройдя 10— 15 м, полетела вдоль склона, сопровождаемая самцом. Наконец, 5 мая 1978 г. при поиске гнезд нам удалось выгнать из-под куста самку; самец находился в 5—8 м от нее. При осмотре была обнаружена свежая ямка еще без строительного материала. На основании этих данных можно высказать предположение, что строит гнездо лишь самка.
Старые гнезда для устройства новых кеклики используют чрезвычайно редко. С таким явлением мы столкнулись лишь трижды. В ущелье Кзылаус 9 мая 1976 г. найдено гнездо с кладкой из 9 яиц, насиживаемых самкой; 26 мая оно оказалось разоренным. Разбирая строительный материал, мы обнаружили под его слоем мелкие белые частички скорлупы, которые находились в гнезде длительное время. В другом случае в июле 1977 г. в этом же ущелье неподалеку от берега ручья под валуном найдено гнездо со скорлупками, которое выводок покинул несколько дней назад; 29 мая 1981 г. под этим же валуном отмечен кеклик, насиживающий кладку. Наконец, 10 апреля 1978 г. найдено прошлогоднее .гнездо, края которого поросли молодой травой, а в лотке находилось яйцо. При проверке гнезда 15 апреля яйца в нем не оказалось. Неоднократные проверки найденных в прошлые годы гнезд показали, что ни одно из них не было вновь использовано для гнездования. Кроме того, в некоторых из них мы забирали весь строительный материал и оставшиеся после ухода выводка скорлупки. И в этих случаях ни одна из ямок не использовалась кекликами для строительства нового гнезда.
В большинстве случаев кеклики устраивают гнезда в скрытых местах, что значительно усложняет их поиск. Схема расположения гнезд представлена на рисунке 10. Из 80 жилых гнезд 19 (23,8%) обнаружено при наблюдениях за парами, 55 (68,7%) —при активном поиске и 6 (7,5%) —случайно. Кроме них удалось найти также свежие ямки без строительного материала и законченные гнезда (общее чис-
45
Рис. 10. Расположение гнезд кеклика в горах Чулак (ущелье Кзылаус, 600—900 м па ур. м.)
ло 31), в которых впоследствии яйца не появились. По сведениям М. А. Кузьминой (1955), в Чу-Илийских горах найдено* 5 гнезд без яиц; только в одном из них впоследствии появилась кладка.
Поскольку 7 недостроенных гнезд были найдены в после-гнездовой период, можно полагать, что кеклики строят больше гнезд, чем используют для откладки яиц.
Гнезда кекликов по своему устройству однотипны. Для их сооружения птицы предварительно выкапывают ямку у основания кустика или камня (169 случаев); реже гнезда располагаются открыто (6). В выкопанную ямку укладывается строительный материал, который обычно не отличается разнообразием и часто в состав его входят сухие веточки кустарников, прикрывающие гнездо. В 63 разобранных нами гнездах он состоял из сухих веточек боялыча (40 случаев),, ферулы (43), злаков (28), сухой прошлогодней травы (37), эфедры (31), курчавки (17), дикой вишни (3) и соцветий 'сложноцветных (1). Масса гнезд колеблется от 18,1 до 171,8г (в среднем 38,8 г). Самое массивное гнездо обнаружено на склоне восточной экспозиции, под кустом эфедры; наименьшее— в средней части склона южной экспозиции под чахлым кустиком боялыча. На склонах северной экспозиции масса гнезд (11) составила 21,3—76,2 г, на южных склонах (18) — 18.1—77,1 г.
Таким образом, масса строительного материала гнезд’ лишь в слабой степени зависит от экспозиции склона. Верхний край гнезд обычно несколько приподнят и образует невысокий валик из земли вокруг лотка. Г. П. Дементьев^ (1952)’ и А. И. Иванов (1969) высказывают предположение, что назначение валика — защита гнезда от сбегающей со склонов воды. Однако, как справедливо отмечает А. В. Попов (1959), в действительности валики по краям гнезда об-пазуютсь чисто механически при выбросе земли из углублений в почьи. ; zTe сооружаются специально. Это замечание подтверждается^ нашими наблюдениями: валики обнаружены и у гнезд, расположенных на дне ущелий, где стекающая со склонов вода угрожать им не может.
Кеклики устраивают гнезда обычно на склонах (124, или 85,5%); реже (15, или 10,3%)—в непосредственной близости от вершин и, как исключение, — на дне ущелий (6, или 4,2%). Наиболее часто (83, или 47,4%) они встречаются на склонах
47
южной, юго-западной и юго-восточной экспозиций; на север <яых склонах обнаружено 38 (21,7%) гнезд (табл. 3).
Таблица 3. Расположение гнезд кеклика на склонах разной экспозиции
Экспозиция склона Число гнезд %
Южный 63 36,0
Северный 27 15,4
-Западный 26 14,9
Восточный 18 10,3
Юго-западный 10 5,7
Юго-восточный 10 5,7
Дно ущелий 7 4,0
Северо-восточный 7 4,0
Северо-западный 4 2,3
Перевал 2 1,1
Вершина холма 1 0,6
Всего 175* | 100,0
* Только жилые гнезда, использовавшиеся для вывода птенцов.
Гнезда обычно располагаются на открытых участках склонов с редким кустарником, что создает наседке хороший обзор окружающей местности; подойти незамеченным к гнезду ие представляется возможным. Только одно из найденных гнезд находилось в густых кустах таволги, и насиживаю цая птица могла увидеть приближающегося человека с расстояния не более 3 м.
Большинство гнезд кеклики устраивают у основания кустов. Значительно реже они располагаются под навесом кам-нЯ, на уступе скалы, в щели между камнями или среди травы (рис. 11). Явное предпочтение при выборе места для гнезда отдается боялычу, под которым обнаружено 61 гнездо (35,3%), и курчавке — 35 (20,2%). Под кустамц эфедры найдено 7 (4%), терескена — 6 (3,5%), таволги — 5 (2,9%), вишни (Cerasus tianschanica)—2 (1,2%) гнезда. Под прикрытием колючих растений (карагана и др.) они встретились только 6 раз (3,5%). Несколько чаще гнезда устраиваются у основания ферулы (8, или 4,6%) и в куртинках травы (7, или 4%). В 8 случаях (4,6%) гнезда обнаружены под навесом камней (рис. 12). Остальные 28 гнезд (16,2%) распо-48
Рис. 11. Места размещения гнезд кеклика
4-24
лагались между двумя или тремя видами указанных выше растений.
Для кеклика отмечено и совершенно нетипичное гнездование. О. Сопыев и М. Караев (1979) сообщают о нахождении в Кугитанге гнезда в дупле арчи на высоте 1 м от земли. Это единственный приводимый в литературе случай. Нам с подобным явлением сталкиваться не приходилось.
Зачастую гнезда кекликов находятся на значительном расстоянии от водных источников, иногда в 2—3 км, поскольку в период их строительства, откладки и насиживания яиц.
59
Рис. 12. Гнезда кеклика: а — под навесом камня; б — в куртинке травы; в—в основании куста
птицы, довольно редко посещают водопои. По-видимому, поедаемые в этот период зеленые части растений (листья, стебли) полностью удовлетворяют потребность их организма во влаге. Только 2 гнезда располагались неподалеку от воды (10 и 30 м).
Начхало откладки яиц зависит от состояния погоды весной. Как показали наблюдения, появление первых яиц совпадает с установлением положительной ночной температуры, которая отмечается обычно в начале апреля (табл. 4).
Таблица 4. Сроки откладки первого яйца у кекликов в горах Чулак
Год
Месяц и декада
апрель | май
I I II I III I I I II
Всего гйезд
Наиболее ранняя откладка 1-го яйца
18. IV
16. IV
4. IV
3/IV
8. IV ,
В период откладки яиц самцы кекликов часто преследуют самок. Обычно после песни самец приближается к самке и начинает бегать вокруг нее. Голова низко опущена к земле и составляет прямую линию с вытянутой шеей; крыло приспущено и касается земли. Брачные игры часто заканчиваются спариванием;' оно продолжается 4—7 с. В некоторых случаях копуляция происходит без предварительного ухаживания самца. В горах Чулак брачные игры, заканчивающиеся спариванием, мы наблюдали с 14 по 29 апреля 8 раз, с 4 по 12 мая — 6 раз.
R. Mackie и Н. Buechner (1963) отмечают, что семенники у самцов кеклика достигают максимальной массы в мае (в 70 раз больше, чем зимой), а у самок, в сравнении с периодом покоя, масса яичников увеличивается в 155, яйцеводов — в 64 раза, независимо от возраста птицы (данные по размножению кекликов, акклиматизированных в США). По на-52
шим материалам, во второй половине апреля — начале мая семенники у самцов имели средний размер 18,4X9,6 мм (среднее по 6 измерениям). У самок в этот же период яичники по^ 5 измерениям имели средние размеры 39X21 мм, яйцевод был сильно вздут и приобрел молочный цвет.
В это время большую часть дня кеклики проводят в стороне от гнезда, чаще в 50—150 м; иногда в течение дня пары встречаются в одних и тех же местах. Приближаясь к гнезду, проявляют чрезвычайную осторожность и, заметив, опасность, затаиваются или улетают.
Расстояние между гнездами во многом зависит от весенней численности птиц. Нам ни разу не приходилось находить гнезда ближе 90 м одно от другого; среднее расстояние колебалось от 380 м в 1978 г. до 460 м в 1974 г. М. А. Кузьмина (1955) для Чу-Илийских гор приводит сведения о гнездах со свежими яйцами, которые располагались на расстоянии 100— 150 м; на противоположных склонах одной горы оно иногда сокращалось до 30 и даже 10 м.
Яйца кеклика бледно-охристые, по основному фону разбросаны коричневые или темно-бурые пятна. Индивидуальная изменчивость в окраске яиц наблюдается даже в одной кладке. В случае длительного пребывания яиц в гнезде (без насиживания) скорлупа выцветает. По наблюдениям за одной из брошенных кладок поверхность обращенных к свету яиц полностью обесцветилась за 50 дней, но к 20 дню яйца еще сохраняли первоначальную пигментацию.
Отношение длины яйца к его поперечнику составляет 1,3: Размеры (186): длина 37,0—43,6 мм (М=39,9+0,08), поперечник 27,6—31,7 мм (М=30,6+0,06). Масса свежего яйца (53): 18,7—21,6 г (М=20,1+0,6). В процессе насиживания она уменьшается на 10,95%^ и на 18—23 день равна 15,2— 20,3 г (М= 17,9+0,2). Таким образом, к концу насиживания масса яйца в среднем составляет 89,05% от массы свежего.
По данным R. Mackie и Н. Buechner (1963), рост отдельного фолликула у самки кеклика занимает 5 дней, а формирование яйца в яйцеводе—1 сутки. Однако откладка яиц происходит в среднем через 1,3 дня (первых семи через 1,6 дня), что, по мнению авторов, зависит от откладки части яиц вне гнезд, а также бросания или разорения части кладок.
По нашим материалам, средний интервал между отклад
53
кой каждого из яиц (наблюдения за 11 гнездами) составляет 32 ч. Только в одном гнезде они появлялись ежесуточно, в остальных интервал составлял 29—34 ч. Такой большой промежуток времени мы объясняем откладкой яиц в чужие гнезда и их потерей (нам неоднократно приходилось находить свежие яйца прямо на грунте).
При проверке гнезд было установлено, что в некоторых из них за сутки прибавляется более чем по одному яйцу. Так, в одном гнезде 28 апреля в 10 ч 15 мин было 15 яиц, в тот же день в 17 ч 50 мин—17, 1 мая в 16 ч — 21 яйцо, т. е. за трое суток было отложено 6 яиц. В другом гнезде 5 мая 1976 г. в 6 ч 30 мин кладка содержала 9, а к вечеру того же дня 17 4 40 мин) — 11 яиц. В третьем гнезде 1 мая 1978 г. в 10 ч 5 мин было 8 яиц, а 4 мая в 19 ч 40 мин количество их возросло до 15 (за 3,5 суток — 7 яиц). Еще в 3 гнездах, найденных в разные годы, удалось отметить появление яиц с интервалом откладки менее чем в одни сутки. Таким образом, в некоторых гнездах яйца откладываются разными самками, однако во всех известных нам случаях кладку в них насиживала только одна птица, что установлено мечением наседок и дальнейшими наблюдениями за ними.
В 1977 г. нами отмечен случай откладки яиц меченой самкой в два гнезда. Одно из них найдено 12 апреля с 3 яйцами; последнее 14-е яйцо кладки появилось в нем 27 апреля (откладка проводилась в среднем через 32,7 ч). Неоднократные проверки этого гнезда (до 23 июня) показали, что кладка оказалась брошенной. Спустя 3 суток (1 мая) у этой же птицы нашли второе гнездо с 3 яйцами, а 14 мая в нем было 9 яиц (интервал откладки последних 6 яиц составил в среднем 52 ч); самка приступила к насиживанию их, а 15 и 16 мая снесла еще 2 яйца. Таким образом, промежутка между откладкой яиц в разные гнезда не было, а первая кладка предназначалась самцу, который не стал ее насиживать.
Известно, что скорость откладки яиц во многом зависит от возраста и физиологического состояния самки. Как видно из приведенного выше примера, интервал между откладкой яиц одной и той же особью может колебаться от 24 до 52 ч.
В период откладки яиц кеклики ведут себя у гнезд очень осторожно. Прежде чем приблизиться к нему, птицы осматриваются и, только убедившись, что опасности нет, подходят. Обычно самка садится на гнездо, а самец находится в 5—
54
20 м. Отложив яйцо, она покидает гнездо. На кладке находится 6—21, в среднем (6 случаев) 12 мин, большую часть времени затрачивая на укладку строительного материала: птицы берут клювом находящиеся рядом с гнездом стебли сухой травы, злаков, мелкие веточки кустарников и укладывают в лоток :
Таблица 5. Величина кладки кеклика в горах Чулак
Год Количество гнезд с числом
9 1 10 I 1 11 | 12 I 1 13 I 1 14 1 15 1 16
1974 1 I 3 3
1975 —, —, 2 1 — 1 1 —.
1976 1 1 — 1 2 2 3 —
1977 1 2 —. 1 2 3 2
1978 1 1 2 3 6 3 3 —
Всего 3 3 6 5 10 11 • 13 2
% 4,3 4,3 8,6 7.1 14,3 15,7 18,6 2,8
Большинство кекликов заканчивают кладку в последних числах апреля — первой декаде мая. Величина полной кладки сильно колеблется — от 9 до 21 яйца, средняя величина ее по годам варьирует в пределах 12,6—14,9 яйца при средней многолетней 14,4 (табл. 5).
По данным Р. Л. Беме (1975), в различных частях ареала количество яиц в насиженных кладках составляет 8—21, в среднем 12,1. Согласно предположению К. А. Айрумяна (1965), рассматривающего лимитирующие величину кладки факторы у куриных, в частности кеклика, взрослые особи при выводке могут обеспечить необходимым теплом не более 15 птенцов, остальные обречены на гибель. Биологический смысл этого явления заключается в том, что под крыло родителей попадают наиболее сильные птенцы, вытесняя слабых, которые могут погибнуть и от многих других причин, т. е. происходит естественный отбор. Поэтому смертность в выводке будет тем выше, чем он крупнее.
Как видно из наших материалов, средняя величина кладки у кеклика близка к оптимальному количеству птенцов, которых может обогреть взрослая птица. При благоприятных погодных условиях отход птенцов, даже в близких к максимальному выводках, может быть незначительным и молодняк
55
достигает возраста, при котором перестает нуждаться в обогреве. Так, в июне 1974 г, встречено 2 выводка (16 и 18 птенцов месячного возраста), каждый из них водила одна взрослая птица, а в июне 1978 г. — три (19, 20 и 21 птенец) .
К насиживанию кеклики приступают чаще в день откладки последнего яйца (13 гнезд из 18), или на следующий! день.
(Джунгарский Алатау)
яиц в кладке Всего гнезд Средняя величина полной кладки
17 18 I 19 I 20 21
. 1 —. __ 9 14 J
—. —— —. —. — 5 12,6
1 1 1 13 14,5
1 2 —. 1 15 ' 14,9
3 3 2 1 — 28 14,5
4 7 2 2 2 70 14,4
5,7 10,2 2,8 2,8 2,8 100 —
Рис. 13. Кеклик на кладке
1(3 гнезда); в 2 гнездах после начала инкубации добавилось по 2 яйца.
56
Сроки начала насиживания установлены в 26 гнездах. В 1974 г. кеклики начали насиживать 1, 6, 10 и 12 мая; в>-1975 г. — 30 апреля и 1 мая; в 1976 г. — 30 апреля, 1, 1, 6, 7,8,. 10 и 10 мая; в 1977 г. — 2, 10, 14 и 15 мая и в 1978 г. — 30, 30 апреля, 2, 3, 3, 4, 6 и 24 мая. За исключением последнего’ случая к массовому насиживанию кеклики приступают в последние дни апреля и в первую половину мая.
Насиживая кладки, зачастую подпускают человека на 1 м, а иногда не сходят с гнезда даже когда до них дотрагиваются веточкой (рис. 13). В процессе инкубации кеклики покидают гнезда, улетают на кормежку за 150—200 м, иногда присоединяясь к стайкам взрослых птиц. Кормятся они даже в день вылупления птенцов (табл. 6).
И. А. Долгушин (1951) высказывает предположение, что кеклики насиживают кладки только утром и вечером, а также в пасмурную и дождливую погоду и совершенно оставляют гнездо на день. Согласно нашим материалам, они могут покидать кладку практически в любое время, с 7 ч 30 мин до 19 ч 10 мин, причем за день бывает до 4 отлучек общей продолжительностью 1 ч 40 мин — 3 ч 30 мин (см. табл. 6).
В жаркие часы дня у сидящей на гнезде наседки часто отмечается полипноэ — учащенное дыхание, связанное с регуляцией температуры тела: клюв приоткрыт, горло заметно вибрирует. Во время насиживания кеклики переворачивают яйца. Так, 8 мая 1978 г. с 6 до 18 ч наседка переворачивала их 5 раз и 27 раз меняла положение тела. По данным Д. Са-' гынбекова (1976), в Киргизском Алатау за 11 ч насиживания самка отлучалась от гнезда на 13—15% времени, на переворачивание яиц затрачивала 8—10%, производя его каждые’ 40—50 мин, что, по мнению автора, обеспечивает равномерное распределение температуры и влажности среди яиц и предотвращает гибель эмбрионов.
От длительного пребывания 'на гнезде во время насиживания у наседок в кишечнике скапливаются каловые массы. Во время отлучек на местах кормежки часто можно встретить «гнездовой» помет, размером и формой напоминающий яйцо кеклика. Первые находки его помогают установить начало насиживания^ кладок.
Срок насиживания яиц у кеклика в горах Чулак, по нашим данным, составляет 23—25 дней. В 3 гнездах птенцы вылупились спустя 23 дня после начала насиживания, в
57
Таблица 6. Распределение отлучек у кеклика в период насиживания кладок при наблюдениях с 4 до 19 ч
1 Пол'насиживаю-щей особи Экспозиция склона Да Га наблюдения День йасижйва-ния Число прогулок _в сутки Время отсутствий птицы на гнезде Общая продолжительность отлучек
Время %
Самец Дно от-щелка 8. V 1978 г. 7 1 15ч 28мин— 17 ч 38 мин t 2 ч Юмин 9,0
Самец 15. V 1978 г. 14 2 11 ч Юмин— 13 Ч 19 МИН; 18 ч 05мий— 19ч 10 мин 3 ч 9 мин 13,1
СЬмец 27. V 1978 г. 22 2 10 ч 51 мин— 11 Ч 36 МИН; 17 ч Обмий— 17 ч 58 мин 1 ч 39мин 6,9
Самка Восточный 20. V 1978 г. 12 4 7 ч 43мин— 8ч 12 МИН; 10 ч 35мин — 11 Ч 00 МИН; 12 ч 45мин— 13 ч 17 МИЙ; 15 ч 57мин— 16 ч 29 мин 2 ч 7 мин 9 8,9
Самка 12. V 1978 г. 4 2 9 ч 51мин — 11 ч 02’МИН; 16 ч 10 мин— 17 ч 07 мин 2 ч 8 мин 8*9
Самка Дно of-щелка 16. V 1974 г. 5 1 16 ч 30 мин- 18 ч 11 мий 1 ч 41 мин 7,0
Самка СеверЬ-западный 11. V 1976 г. 5 2 11 ч 03 мин— 12ч 25 МИН; 16ч 35мий— 18 ч 45 мин 3 ч 32мин 14,7
Самка Западный 5. V 1976 г. 3 2 8 ч 12 мин— 9 4 07 МИН; 15 ч 57 мин- 17 ч 12 мин 2 ч Юмин 9,0
Самец Южный 18. V 1977 г. 10 3 7 ч 35мин— 8 4 52 МИН; 12 ч 14мин— 13 ч 08 МИН; 17 ч 05 мин- 18 ч 07 мин 3 ч Юмин 13,3
трех — через 25, в пяти—через 24 дня. О продолжительности насиживания одной из кладок в течение 23 дней в горах Чудак сообщает М. А. Кузьмина (1955). Возможно, что на длительность инкубации яиц влияет частота и продолжительность отлучек насиживающей птицы и погодные условия.
С момента появления первого птенца до вылупления последнего проходит 6—8 ч, а начиная с проклева первого яйца и кончая уходом выводка—1—2 суток (наблюдения за 15 гнездами). Еще находясь в гнезде под наседкой, птенцы иногда покидают его на короткое время, удаляясь на расстояние до 0,5 м (рис. 14).
I
Рис. 14. Гнездо кеклика после ухода выводка
По данным вскрытия однодневных птенцов из 3 гнезд (36 экз.) в горах Чулак удалось установить соотношение полов, которое оказалось 1:0,89 (19 самцов, или 52,8%, и 17 самок, или 47,2%). Если принять во внимание, что гибель молодняка от различных причин воздействует на обе половые группы одинаково, можно полагать, что в природе среди кекликов в различные сезоны года в целом сохраняется такое же соотношение полов. Это предположение подтвердилось
59
результатами вскрытия кекликов, добытых в осенние периоды 1972—1978 гг. на юго-востоке Казахстана: из 925 экз. 486 (52,6%) оказались самками и 439 (47,4%) — самцами. Таким образом, по нашим материалам в различных популяциях кекликов наблюдается небольшое, но устойчивое преобладание самцов.
Естественный отход яиц в гнездах кеклика сравнительно невелик и составляет 2,3— 12% (табл. 7).
Таблица 7. Естественный отход яиц в гнездах кеклика
Год Чйсло яиц в гнез-дах Отход яиц Отход яиц Птейцы, покинувшие гнездо
Неоплодот-воренйые Погибшие эмбрионы | 1 Прбчие причины
Число % ЧйсЛо) % 1 Чисдо| % Число) % Число) %
1974 43 0 0 1 2,3 1 2,3 42 97,7
1975 25 0 0 2 8,0 1 4,0 3 12,0 22 88,0
1976 92 1 1,1 0 0 3 з,з 4 4,4 88 95,6
1977 164 6 3,7 2 1,2 4 2,4 12 7,3 152 92,7
1978 252 7 2,8 2 0,8 7 2,8 16 6,4 236 93,6
Всего 576 14 2,4 7 1,2 15 2,6 36 6,2 540 93,8
Следует заметить, что в 15 случаях наседки уводили выводки, оставляя в гнездах наклюнутые яйца с живыми, вполне сформированными птенцами, отставшими в развитии всего на несколько часов. Из таких яиц, помещенных на теплую грелку, нам удавалось выводить птенцов, которые впоследствии нормально росли и развивались.
По наблюдениям за 36 гнездами, в которых вывелись птенцы, только в 14 (38,8%) выход составил 100%. В остальных 22 (61,2%) после ухода выводка в гнездах оставались неоплодотворенные и с погибшими эмбрионами яйца. Что же касается судьбы кладок, то процент покинутых выводками гнезд колеблется от 37,5 в 1974 г. до 61,1 в 1978 г. (табл. 8).
Гибель гнезд происходит в большинстве случаев в первые 5—10 дней после начала насиживания. Из 18 разоренных гнезд только однажды яйцо исчезло на следующий день после появления его в гнезде. Во всех остальных случаях гибли полные кладки.
В орнитологических работах встречаются мнения, чта
60
кеклики могут выводить птенцов дважды в год (Зарудный, 1896; Рустамов, 1951; Дементьев, 1952). Однако М. А. Кузьмина (1955) совершенно исключает такую возможность, обращая внимание на длительный период насиживания, крайнюю истощенность наседок к моменту^ вылупления птенцов, продолжительную линьку с одновременной заботой о потомстве.
Таблица 8. Гибель кладок и вылупление птенцов в гнездах кеклика
. Гбд
Кол-во кладок БрРшены Разорены Птенцы вылупились
Числд % Число % Число' %
1974 8
1975 4
1976 13
1977 18
1978 33
Всего 76
1 12,5
1 25,0
1 7,6
3 66,7
9 27,3
15 19,7
4 50,0 3 37,5
1 25,0 2 50,0
6 46,2 6 46,2
4 22-, 2 11 61,1
4 12,1 20 60,6
19 25,0 42 55,3
На наш взгляд, эти выводы вполне убедительны. Даже простые расчеты показывают, что период размножения и воспитания молодняка у кекликов занимает не менее 3,5—4 месяцев. Следовательно,- ко второму репродуктивному циклу они могут приступить только в сентябре, а молодняк в таких выводках появится спустя 1,5 месяца, т. е. в конце октября — начале ноября. Появление выводков в такие поздние сроки у кеклика не зарегистрировано, что доказывает существование у этого вида в году только одной нормальной кладки. Однако в случае гибели кладок на ранней стадии насиживания кеклики могут гнездиться повторно. Так, М. А. Кузьмина (1955) 31 мая в горах Чулак добыла самку с наседным пятном, у которой в яйцеводе обнаружено яйцо. Этот факт, а также приводимые в литературе данные о поздних выводках подтверждают случаи повторного размножения.
Двойное гнездование — это постройка 2 гнезд одной парой птиц, кладки в которых насиживает каждый партнер самостоятельно. У птиц это явление встречается исключи
61
тельно редко и известно пока для небольшого числа видон. Оно описано для куликов — белохвостого песочника (Коханов, 1973) и кулика-воробья (Кищинский, Флинт, 1973).
Прежде чем перейти к изложению наших материалов, остановимся на литературных данных. В 1924 г. L. М. Dewar опубликовал работу, в которой отметил, что еще Аристотелю было известно о двойном гнездовании каменных куропаток, поскольку он в «Истории животных» в главе 6 книги IX писал: «Куропатки высиживают и тот и другой, причем самец и самка выращивают каждый своих» (цит. по М. А. Кузьминой, 1955, с. 127). По отношению к европейской каменной куропатке (A. graeca) имеются следующие сведения о двойном гнездовании. М. Portal (1924) сообщает, что в 1920 г. ему привезли с о. Крита пару куропаток. Птицы с подрезанными крыльями были помещены в загон (0,2 га) с кустами и травой. Во второй декаде июня самец водил 12 птенцов. Поскольку он был хромой, не вызывало сомнения, что птенцы были именно с этой птицей, а 6 июля отметили самку с выводком (7 или 8 молодых). Впоследствии на огороженном участке было обнаружено 2 гнезда. В одном находилось 3 яйца и 12 скорлупок, в другом — 9 скорлупок. Этот же автор сообщает о двойном гнездовании другого представителя рода каменных куропаток — красной куропатки (А. ги-fa). По кеклику имеются сведения С. Wissel и др. (1966). Авторы сообщают о двойном гнездовании кеклика в Индии, но не приводят конкретных материалов. М. Д. Зверев (1948), М. А. Кузьмина (1955) и Р. Л. Бёме (1975) добывали от вы-водков самцов с наседными пятнами. Эти факты можно расценивать и как участие самцов в насиживании яиц попеременно с самками, и как самостоятельное высиживание ими кладки, т. е. как двойное гнездование. Однако И. А. Долгушин (1948) пишет: «Сведения западноевропейских орнитологов о том, что горные куропатки устраивают 2 гнезда, делают две кладки, одну из которых насиживает самец, а дру-гую — самка и т. п., к кеклику неприменимы» (с. 145). К такому же выводу приходит К. А. Айрумян (1965). Основанием для такого заключения ему послужили 67 взрослых самцов без наседных пятен, добытых в репродуктивный период. Кроме того, автор отмечает, что «...для откладки 20 яиц в два гнезда, при наличии перерывов в откладке, потребуется около 20—26 дней. Следовательно, первые 40 яиц в течение 12—
62
26 дней будут лишены как подогрева, так и периодического переворачивания, без которого желток яйца с зародышевым диском всплывет и прилипнет к подскорлуповой оболочке, т. е. произойдет присыхание зародыша» (с. 84). Таким образом, по мнению этого автора, устройство двух гнезд кекли-ком теряет всякий биологический смысл.
По нашим наблюдениям, в горах Чулак некоторые пары кекликов устраивают по 2 гнезда (Грачев, 1979). Насколько широко распространено это явление у кекликов, можно заключить из того, что по данным отлова наседок на 31 гнезде, кладки в них насиживали 12 (38,7%) самцов и 19 (61,3%) самок. Кроме того, наблюдениями за 19 гнездами, в которых сопоставлялись даты окончания откладки яиц с началом насиживания, установлено, что 18 (94,7%) кладок насиживали самки и 1 (5,3%) самец. Таким образом, можно сделать вывод, что в насиживании кладок участвуют все самки и треть самцов. Установлена также принадлежность 8 гнезд четырем парам птиц. Так, 16 апреля 1976 г. найдено гнездо с 10-ю яйцами; 22 апреля в нем появилось последнее, 15-е, яйцо. С 25 апреля за этим гнездом установили наблюдение. На следующий день в 8 ч 45 мин самка посетила его (самец в это время находился в 2 м), а в 9 ч 25 мин эта пара, пройдя вдоль склона около 100 м, остановилась у небольшой ложбинки. В течение 5 мин птицы осматривались, затем самка пошла к одиночному кусту курчавки, расположенному в 7 м, и скрылась у его основания. В 9 ч 40 мин кеклики покинули этот район и скрылись за хребтом. При осмотре места обнаружено гнездо, содержащее 4 яйца. Таким образом, перерыва в откладке яиц не было, закончив первую кладку, самка уже на следующий день отложила яйцо во второе гнездо. В первом гнезде самец начал насиживать кладку 4 мая, а 28 мая покинул его с выводком. Во втором самка продолжала откладку яиц после того, когда самец приступил к насиживанию в первом гнезде. Кладка яиц в нем закончилась лишь 10 мая (13 яиц), и в этот же день она начала насиживание (спустя 5 дней его разорила лисица).
В другом случае 26 апреля 1977 г. нашли гнездо с 15 холодными яйцами на склоне северо-восточной экспозиции в , 40 м от дна отщелка. Наблюдая за ним 28 апреля в 9 ч 30 мин, мы заметили неподалеку пару кекликов. Через несколько минут самка в сопровождении самца спустилась на дно отщ£л-
63.
ка и начала подниматься по противоположному склону юж-лой экспозиции. Пройдя 70 м, юна скрылась в кусте курчавки, растущем на выступе скалы (самец в это время находился в 20 м). Через 9 мин самка появилась в поле зрения наблюдателя и, оглядевшись, полетела через отщелок к первому гнезду. Самец сразу же последовал за ней. Приземлившись, кеклики прошли 5 м, отделявшие их от гнезда, и сначала самка, а затем самец заходили в него на 10—15 с. В 10 ч 20 мин птицы, пройдя 3—4 м, улетели в скалы. Таким образом, в течение 50 мин пара кекликов посетила 2 гнезда. При проверке их в первом, как и в прошлые дни, оказалось 15 яиц, во втором— одно. На следующий день, 29 апреля, единственное яйцо во втором гнезде исчезло, а в первом самец приступил к насиживанию 6 или 7 мая и вечером 30 мая с выводком покинул гнездо.
В том же году зарегистрирован еще один случай двойного гнездования. 12 апреля найдено гнездо с 3 яйцами. Регулярные проверки показали, что 27 апреля кладка в нем была закончена и содержала 14 яиц, а наблюдения за этим гнездом дали возможность установить принадлежность еще одного этой же паре птиц (в течение 40 мин кеклики посетили 2 гнезда, расположенные на противоположных склонах ущелья, в 120 м одно от другого). В первом гнезде кладка оказалась брошенной, а во втором была закончена 16 мая и содержала 11 яиц (к насиживанию самка приступила 15 мая). Вечером 7 июня выводок с наседкой покинул гнездо.
Наконец, утром 25 апреля 1978 г. в результате наблюдений. за парой кекликов выяснилось, что им принадлежат 2 гнезда, которые они посетили в течение 45 мин. Гнезда располагались в 150 м одно от другого на склоне южной экспозиции. При их проверке оказалось, что в первом кладка содержала 19 яиц, а во втором — 3. При проверке 3 мая в первом отмечено насиживание; во втором кладка по-прежнему содержала только 3 яйца. Спустя двое суток (5 мая) первое гнездо оказалось разоренным, а во втором, как и в последующие дни, было 3 яйца (кладка брошена).
При наблюдениях за гнездами кекликов мы пришли к выводу, что ежегодно некоторые пары устраивают по 2 гнезда. Кладку в первом из них насиживает самец, во втором — самка. В некоторых случаях самцы приступают к насиживанию уже спустя 3—4 дня после появления последнего яйца в первом
64
гнезде, когда самка продолжает нестись во втором. Возможно, при этом она спаривается с другим самцом; если это так, то здесь мы сталкиваемся с явлением частичной полиандрии.
Регулярные наблюдения за гнездами показали, что яйца кекликов обладают повышенной жизнестойкостью. С одним из гнезд, кладка в котором 25 апреля "состояла из 3-х яиц и оказалась брошенной, мы провели следующий эксперимент. Днем 26 апреля яйца были пронумерованы и в определенном порядке расположены в гнезде. Спустя 14 дней, убедившись, что за это время гнездо не посещалось кекликами, их подложили в другое гнездо с 10 яйцами, где в этот день началось насиживание; 3 июня из подложенных яиц вылупились птенцы. Как видно из проведенного опыта, длительное нахождение яиц в состоянии покоя не повлияло на выведение птенцов.
О повышенной жизнестойкости яиц кеклика можно судить и по другому примеру. Днем 7 мая 1976 г. нашли гнездо с кладкой из 15 яиц, которые насиживала самка, а 26 мая оно было брошено, что установили путем неоднократных проверок гнезда. Через 7 дней, 2 июня, несмотря на длительное отсутствие наседки на гнезде (за это время дважды выпадали дожди, а днем яйца освещались солнцем) мы вскрыли 3 яйца из этой кладки. Одно из них было неоплодотворенным, а в двух оказались живые эмбрионы.
Проверки гнезд, кладки в которых насиживали самцы (яйца в них не насиживались 4—12 и более дней), показали, что из 150 яиц в 9 гнездах вывелся 141 птенец, а неоплодо-творенные яйца и эмбриональная смертность составили всего 6% (9 яиц).
В некоторых случаях кеклики после окончания откладки яиц в первое гнездо посещают его и переворачивают яйца. Так, 24 апреля 1975 г. обнаружено гнездо с 15 яйцами. Ежедневные проверки показали, что до 1 мая кладка не насиживалась, но 28, 29 и 30 апреля его посещали кеклики. Этот факт удалось установить путем сравнения расположения пронумерованных яиц. Таким образом, в случае двойного гнездования одни пары переворачивают в первом гнезде, другие нет, что, однако, существенно не повлияло на вывод птенцов.
Отлов насиживающих кладки самцов показал, что все они были не старше одного года, т. е. впервые принимали участие в размножении.
5-24
65
Эффективность гнездования у кеклика по годам? колеблется от 37,8 до 62% (табл. 9).
Сохранность гнезд кеклика во многом зависит от маскировки. От хищников чаще всего гибнут открыто расположенные гнезда. В некоторых случаях птицы бросают гнезда с неполными (10, или 29,4%) и полными (3, или 8,8%) кладками, что, однако, может быть связано с гибелью одного из членов пары.
Таблица 9. Эффективность гнездования кеклика (% птенцов, покинувших гнезда)
Год Число гнезд Количество Эффективность гнездования
яиц в гнездах птенцов, покинувших гнездо
1974 8 111 42 37,8
1975 4 48 22 45,8
1976 13 189 88 46,5
1977 18 245 152 62,0
1973 33 423 236 55,8
Всего 76 1016 540 53,1
К моменту появления первых выводков, а в некоторые годы на 10—15 дней раньше, не участвующие в насиживании взрослые кеклики начинают собираться в группы по 3—7 особей. Первые встречи их в горах Чулак отмечены 26 мая 1974 г., 11 мая 1975 г., 10 мая 1976 г., 25 мая 1977 г. и 17 мая 1978 г. Образовавшиеся стайки очень осторожны и предпочитают держаться на склонах с густой кустарниковой растительностью. Постепенно количество птиц в них увеличивается за счет особей, потерявших кладки и повторно не приступивших к размножению. Во второй половине июня стайки взрослых кекликов чаще состояли из 9—18 особей (максимальное число птиц в ' одной стае составило 35 особей). В 1974—1976 гг. в конце мая — начале июня из 4 стаек было добыто 5 птиц. Одна из них оказалась самкой, у которой начало зарастать наседное пятно, 4 — самцами (один из них тоже с зарастающим наседным пятном). М. А. Кузьмина (1955) в горах Чулак во второй половин мая — первой 66
половине июня добыла И линяющих самцов (один с йа-седным пятном) и И линяющих самок (шесть с наседными пятнами). Можно предположить, что самки без наседных пятен— особи из прошлогодних, поздних выводков, еще не приступившие к размножению.
Объединившиеся в стаи взрослые кеклики, видимо, разбиваются на пары только весной следующего года. По данным зимнего отлова на подкормочных площадках в феврале 1978 г. в ущелье Кзылаус, было отловлено 6 из стайки в 8 и 5 из 7 кекликов. Определение возраста пойманных птиц по разработанной нами методике (Грачев, 1978) показало, что все они были старше одного года.
Постэмбриональное развитие (рост и развитие, поведение, формирование оперения и линька). Постэмбриональное развитие птиц начинается после освобождения организма от яйцевых оболочек и заканчивается установлением окончательных пропорций тела, прекращением роста и наступлением половой зрелости. Изучение этого периода жизни позволяет проследить за сменой оперения, изменением массы, поведением, а иногда уточнить вопросы систематического положения. В этом отношении группа выводковых птиц наиболее трудна для изучения, поскольку спустя несколько часов после вылупления выводок покидает гнездо и получить необходимые сведения, касающиеся изменения массы, хода смены оперения и измерения различных частей тела у каждого из птенцов не представляется возможным, тем более, что получение таких данных требует ежесуточного контакта исследователя с изучаемым объектом.
Постэмбриональное развитие кеклика изучено слабо. Лишь в работах М. Д. Зверева (1948), М. А. Кузьминой (1955) и М. Н. Денисовой (1958) приводятся отрывочные сведения по изменению массы, поведению птенцов и развитию их* оперения.
Собранные нами материалы освещают постэмбриональное развитие кеклика с момента вылупления до 6-недельного возраста. Данные по росту и развитию, формированию оперения и, частично, поведению молодняка получены при наблюдении за 10 птенцами, которые находились на полувольном содержании. Для получения морфологического материала произведено вскрытие 28 птенцов (возраст до 38 дней). Сведения о линьке, изменении соотношений индексов органов и
поведении более старших возрастных групп получены при отстреле кекликов и наблюдениях за ними в естественных условиях.
У вылупившихся птенцов в первые дни жизни рассасывается желточный мешок, увеличивается индекс сердца, появляются пеньки маховых перьев, интенсивно развивается скелет и мускулатура плечевого пояса (табл. 10).
Таблица 10. Индексы массы внутренних органов кекликов различного возраста (отношение к массе тела, %)
Возраст/ дни Большая грудная мышца Подключичная* мышца Грудная мускулатура (в целом) Мышцы ног Органы пищеварения Желточйый мешок Сердце**
Перед вылуплением 0.80 0,24 1,04 8,88 7,28 13,60 6,4
1 0,76 0,23 0,99 7,78 11,15 12,46 I Г,5
7 2,46 0,91 3,37 9,96 15,50 9,6
14 9,80 2,73 12,59 11,63 13,49 — 6,9
21 8,04 2,3010,37 11,24 12,61 — 5,9
28 10,18 3,17 13,35 12,00 10,18 5,0
35 10,71 3,62 14,33 12,90 12,90 —. 4,5
42 10,00 3,68 13,68 13,68 11,54 — 6,6
4 мес. 17,60 5,50 23,10 16,55 9,30 — 4,6
Взрослые 16,90 5,10 22,10 18,25 9,36 — 4,2
* Мышцы локомоторных органов с двух сторон тела, ** Индекс сердца, °/оо.
Возрастание индекса сердца почти в 2 раза — результат становления процессов дыхания и кровообращения, связанных с началом активной деятельности. Прирост внутренних органов в первые дни осуществляется в основном за счет питательных резервов желточного мешка.
Масса только что вылупившихся птенцов кеклика варьирует в пределах 11,0—14,65 г, составляя в среднем (56 экз.) ,14,5 г. Среди птенцов одного выводка разница может достигать 2,9 г. В первые часы жизни птенцы имеют хорошо сформированные задние конечности (благодаря чему они способны активно передвигаться) и очень слабые плечевой пояс и
68
Рис. 15. Темп роста частей тела у птенцов кеклика. Обозначения: 1 — развернутое крыло; 2 — крыло от кистевого сгиба до вершины; 3— цевка; ^4 — средний палец; 5 — длина головы; 6 — клюв
грудину. Большой желточный мешок, составляющий после вылупления свыше 13% массы тела, и развитая подкожная жировая клетчатка особенно необходимы для жизнедеятельности организма в течение первых 2—3 дней после выхода птенцов из гнезда. Как показали вскрытия, эти образования сохраняются до 5-дневного возраста.
В первую неделю жизни особенно отчетливо выражена глубокая перестройка всего организма, обусловленная возрастающей активностью птенцов и усложнением их поведения. Это выражается в преимущественном развитии летательного аппарата (рост крыла и его оперейие, рост грудины, развитие грудных мышц), что способствует раннему подъему птенцов на крыло (рис. 15).
В первые 2 дня (а у отдельных особей и на третий) исчезает яйцевой «зуб», выполнивший свое прямое назначение — освобождение птенца от скорлупы. Переход к активной кормежке приводит к усложнению движений (бег, прыжки, лазанье по горизонтальным веткам низеньких кустиков), для чего используются лапы. Часто движения птенцов сопровождаются взмахами крылышек, что облегчает им перемещение и является постоянной тренировкой плечевого пояса. В результате отмечается интенсивный прирост зачаточных грудных мышц, масса которых к концу первой недели жизни возрастает почти в 7 раз (0,11 г при вылуплении, 0,76 г спустя 7 дней). За этот же период без малого в 2 раза увеличивается длина грудины (10,5—20,0 мм), что обеспечивает увеличение площади прикрепления грудных мышц. Почти в 2 раза возрастает длина развернутого крыла, в основном за счет прироста его дистальной части — кисти и первостепенных маховых перьев.
Таблица 11. Средняя масса и ее прирост у птенцов кеклика за неделю, % от предыдущей недели
Возраст, дни Масса, г Среднесу- | точный при-; рост, г 1 Прирост за неделю, %
При вылуплении 14,5
7 20,8 0,9 43,4
14 33,9 1,9 62,9
21 57,3 3,4 69,9
28 86,0 4,0 50,0
35 123,0 5,5 43,9
ТО
В перъые 2—3 дня жизни масса птенцов в среднем падает на 1,2 г, но к концу недели достигает 20,8 г, что в среднем составляет привес 0,9 г в сутки (табл. 11).
Наиболее характерная черта второй недели жизни птенцов — подъем на крыло. На 8-й день отмечены первые прыжки— перелеты (до 50 см); на 13-й — первое планирование над склоном на расстояние до 10—15 м. Полет вызывает повышенный расход энергии, что хорошо прослеживается на среднесуточных привесах. Спустя сутки после первого подъема на крыло (9-й день) среднесуточный привес составил всего 0,3 г; на 14-й день, после первого продолжительного планирования,— 1,2 г; среднесуточный привес за неделю — 1,9 г. В этот же период идет интенсивный прирост грудных мышц: на 4,3 г, или в 6,6 раза по сравнению с предыдущей неделей, а прирост мышц ног увеличивается на 4,07 г, или только в 2 раза.
Во вторую неделю прирост грудины, крыла и цевки замедляется. Это проявляется как в абсолютных величинах прироста, так и в снижении или лишь незначительном увеличении индексов линейных размеров (табл. 12). Следует от-
Таблица 12. Индексы линейных размеров отдельных органов кеклика различного возраста по отношению к длине туловища * на скелете, %
Возраст, дни
1
7
14
21
28
35
42
4 мес. Взрослые
38,2
63,8
61,4 151 ,'4
71,1 190,6
72,7 188,6
62,2
143,7
70,5185,5 72,0 1 68,0
85,7il80,0
83,7,180,0
102,2 192,3
237,8
262,0
253,0
282,0
260,7 223,0 237,0
57,3 54,7 61,3 54,9
56,5 50,2 п55,4
50,9
50,1
49,9
46,9 45,3
59,0
61,1
59,0
59,7
55,1
583,0
894,5
983,0
1212,8 1009,1
899,3
102,2
111,7
205,1
250,0
199,9 28; 6
217,51----
25,5
28,7
28,1
29,6
24,0
917,6,167,1 24,0
1006,9 225,6 23,7
1194,5 257,6 26,8
25,5
55,8
66,8
85,3
93,5
93,5
114,0
106,6
* Длина туловища — расстояние от переднего края последнего шейного позвонка до спинного края мозгового канала на задней поверхности таза.
71
метить, что за вторую неделю заметно увеличивается прирост туловища на скелете (на 10 мм, или на 36,1%). Вместе с тем прослеживается следующая закономерность: сначала идет интенсивный рост скелетных элементов, а позже — увеличение соответствующих мышц (интенсивный рост грудины в первую неделю жизни и грудных мышц — во вторую). Преимущественное развитие грудины резко изменяет соотношение ее мышц с мышцами задних конечностей. Если у птенцов в первый день жизни грудные мышцы составляли менее 1% массы тела и в 8 раз уступали в этом отношении мышцам ног, то в 2-недельном возрасте они достигают 12,5% массы тела и несколько превышают по этому показателю мышцы задних конечностей. К концу второй недели устанавливаются соотношения весовых и линейных размеров большинства органов, которые лишь с небольшими изменениями сохраняются до 6-недёльного возраста.
В течение третьей недели птенцы быстро растут и интенсивно оперяются. Общее увеличение их массы происходит в основном за счет роста скелета и внутренних органов: длина грудины увеличивается на 30,4, крыла — на 19,9, кишечника— на 85%, слепых отростков — на 20%. Рост мускулатуры локомоторных органов в этот период, наоборот, несколько приостанавливается: привес мышц ног составляет 19,3, грудных мускулов — только 11,8%, что приводит к некоторому снижению их индексов.
Для четвертой недели характерна интенсивная смена пухового наряда, дальнейшее развитие мускулатуры локомоторных органов. Масса мышц ног увеличивается на 45, грудных мышц — на 90%. Такой высокий темп возможен лишь благодаря увеличению линейных размеров грудины в предыдущую неделю. Начиная с четвертой недели происходит постепенное снижение темпа роста птенцов (прирост за неделю в %), хотя среднесуточный привес и продолжает возрастать.
В течение 5—6-й недели заметных изменений в соотношении размеров отдельных органов не происходит. Отмечается лишь снижение индекса крыла, что связано с окончанием роста юношеских первостепенных маховых перьев.
В возрасте 4 месяцев молодняк уже практически не отличается от взрослых кекликов по размерам и индексам линейных и весовых показателей (табл. 13, 14).
72
Таблица 13. Увеличение массы органов у кеклика за 4 месяца развития
Возраст Тело Грудная мускулатура Мускулатура ног Сердце Органы пищеварения
1 2 1 1 2 1 J 1 2 1 2 1 1 2
При вылуплении 13,0 0,14 1,11 0,15 1,45
2 недели 40,4 3,1 5,06 36,0 4,70 4,5 0,27 1,8 5,55 ЗЛ
6 недель 195,7 4,8 26,80 5,5 26,80 5,7 1,30 4,8 22,70 4,1
4 месяца 474,0 2,4 4 105,00 3,9 0 75,00 2,8 5 2,12 1,6 1 42,2011,8
Прирост за весь период (число 36, 750,' 67, 14, 29,1
раз)
Примечание, 1 — масса, г; 2 — прирост (число раз).
Таблица 14. Прирост линейных размеров у кеклика за 4 месяца развития
Элемейты измерений Возраст Прирост за весь период
й « и ч S к S S с ч я 2 недели 6 недель 4 месяца 1
Длина, мм
Крыло от кистевого сгиба 17,2 64,5 126,0 153,0
Грудина Длина туловища на 10,5 25,3 51,0 72,0 —
скелете 27,4 41,2 74,8 84,2
Цевка 15,5 23,3 44,0 50,3 —
Палец (третий) 15,0 20,7 37,0 39,5
Клюв 7,0 П,б 18,0 20,0
Кишечник 160,0 405,0 680,0 900,0 —
Слепые отростки 28,0 85,0 135,0 190,0 —
Прирост (число раз)
Крыло от кистевого сгиба 3,7 2,0 1,1 8,8
Грудина Длина туловища на —— 2.4 2,1 1,3 6,8
скелете —• 2,4 1,8 13 3*0
Цевка —. 1,5 1,6 1,1 3,2
Палец (третий) —. 1,4 1,8 1,1 2,1
Клюв — 1,6 1,5 1,1 2,8
Кишечник —. 2,5 1,7 1.3 5,5
Слепые отростки — 3,0 1,6 1,4 6,8
Первые звуки птенцы издают, еще не появившись на свет: в наклюнутом яйце можно уловить едва слышный писк. По данным В. Д. Ильичева и А. В. Тихонова (1979), становление звуковой сигнализации у эмбрионов обусловлено развитием их дыхательной системы. Звукоиздающая система у эмбрионов куриных, как отмечают авторы, достигает стадии функциональной готовности к моменту перехода на легочное дыхание (93—94% инкубационного периода). В зависимости от дыхательных движений эмбриона звуки выражаются одиночным писком и щелкающими сигналами дискомфорта и комфорта, что обеспечивает координацию движений и дружное вылупление. В конце насиживания резко возрастает звуковая активность самки, в частности ее призывные сигналы— «пищевой зов». Однако привязанность к кладке у насиживающих кекликов настолько велика, что если наклюнутые яйца в гнезде заменить на свежие, то птица будет продолжать насиживание.
У вылупившихся птенцов четко выражены безусловный рефлекс (добывание корма) и реакция на призывный и тревожный голос родителей. В поведении птенцов, даже в первые часы их жизни, отчетливо проявляются адаптивные особенности этого вида. Нам неоднократно приходилось наблюдать, как едва обсохшие птенцы, еще обогреваемые наседкой, удалялись от гнезда на 10—20 см и что-то склевывали на земле. Если потревожить такой выводок, наседка, притворяясь раненой, убегает с тревожными криками, а птенцы затаиваются в гнезде или поблизости от него между неровностями почвы, камнями, корнями кустов; окраска делает их совершенно незаметными. Как только опасность минует, наседка возвращается к птенцам, созывая их тихим квохтаньем. Затаивание— единственная защита нелетных птенцов. В более старшем возрасте такое поведение отмечается реже, хотя наблюдается и у взрослых птиц. Однако если у взрослых особей затаивание — непосредственная реакция защиты от врагов, полученная от зрительного восприятия, то у птенцов — реакция на тревожный звуковой сигнал родителей, заметивших опасность.
Поведение птенцов кеклика мы наблюдали при полуволь-ном содержании выводка, который воспитывался в тесном контакте с человеком до 43-дневного возраста. С момента, когда наклюнутые яйца, были взяты из гнезда, нарушился
74
звуковой контакт эмбрионов с наседкой. Поэтому можно считать, что издаваемые в дальнейшем птенцами звуки (сигналы тревоги, общение между собой) являются врожденными, поскольку перенять их было не у кого. Вместо созывающего выводок крика взрослой птицы в первые 2 дня жизни птенцы удвоили единственный служивший им общением с человеком собирающий позыв «цып-цып», привыкнув к которому они совершенно не реагировали на призывный сигнал взрослых кекликов. Подтверждает этот факт следующее наблюдение. В возрасте 2 недель наш выводок на прогулке встретился и соединился с выводком того же возраста, при котором находилась взрослая птица. Однако к призывному сигналу ее птенцы, воспитываемые человеком, относились равнодушно и спешили на подаваемый человеком позыв «цып-цып». Так происходили в природе и все последующие встречи прирученных птенцов с другими выводками.
Наши наблюдения показывают, что звуковые сигналы у штенцов кеклика включают пищевой позыв, сигнал сбора и .позыв отставшего от выводка птенца. Возможно, что при вос-
Тис. 16. Эмбриональный пух на различных частях тела птенца кеклика: J — пушинка брюшка; 2 — пушинка спины; 3 — пушинка темени; 4 — пушинка больших кроющих крыла
75
питании птенцов в тесном контакте с человеком звуковое общение между птенцами было несколько упрощено, поскольку при отсутствии родителей они не имели примера для подражания.
Только что вылупившийся птенец кеклика покрыт эмбриональным пухом (рис. 16). Верх головы (лоб, темя, затылок) коричневато-бурые, в неясных мелких пестринках; щеки и пятно за глазом желтовато-охристые; от глаза к затылку проходит узкая черная полоска, ограничивающая «шапочку». Спина пестрая: темный буровато-коричневый пух образует 3 продольные полоски более интенсивной окраски, чем верх головы; между ними находятся участки, покрытые длинным' серовато-серебристым пухом. На крыле удлиненные, слегка охристые пушинки. Подбородок, горло, бока шеи почти белые, со слабым желтоватым оттенком. Остальная часть нижней стороны тела покрыта длинным белым пухом с желтоватым оттенком. Клюв у вершины светлый, у основания коричневатый: яйцевой «зуб» белый. Ноги чуть розоватые.
Эмбриональные пушинки имеют вид кустика. По мере роста птенца эмбриональный пух выталкивается пеньком дефинитивного юношеского пера. Некоторое время пушинки остаются на вершине опахала, а затем отпадают. На участках тела, где оперение развивается позднее (брюшко), пух сохраняется дольше.
Первые перья юношеского наряда появляются на крыле. У птенца в возрасте одного дня уже хорошо видны пеньки 7 первостепенных и 6 второстепенных маховых. В первые дни жизни отклонения в темпах развития оперения составляют не более одного дня, а в возрасте 6 недель они могут достигать 5 дней. На 3—4-й день появляются пеньки больших кроющих перьев крыла. На 5-е сутки они, развернув кисточки, прикрывают на 7з—Vs маховые, скрывая их очины, и образуют вместе с ними сплошную поверхность крыла. Одновременно* появляются пеньки средних и малых кроющих крыла, плечевых и рулевых перьев. На 6-й день у части первостепенных маховых развертываются опахала; кроющие крыла превращаются в кисточки; первое второстепенное маховое представлено пеньком, остальные (2—10)—кисточками. Около глаз-к этому времени намечается темная полоска. Ноги становятся красновато-оранжевыми. В недельном возрасте опахала первостепенных маховых почти полностью развернуты, причем
76
наиболее длинные—10 и 9. В“ дальнейшем рост этих перьев приостанавливается, а дистальные продолжают интенсивно расти. В это же время пеньки рулевых развертывают кисточки. Остальная часть тела птенца покрыта эмбриональным пухс!м. Клюв становится темно-бурым, с буровато-желтой вершиной (рис. 17).
Рис. 17. Пятидневный птенец кеклика. Горы Чулак, июнь 1975 г.
В течение второй недели продолжается интенсивный рост оперения крыла и развивается дефинитивное перо, кроющее тело. На 8-й день появляются пеньки крылышка (alula) и отмечается значительный прирост первостепенных маховых; у части особей появляется пенек первого третьестепенного махового. Увеличивающаяся поверхность крыла вместе с интенсивным развитием грудных мышц и скелета плечевого пояса обеспечивают птенцам к этому времени возможность совершать первые небольшие подлеты. На 11-й день на боках тела появляется много пеньков, а на 12-й они отмечаются на передней части спины и верхней части шеи; в это же время
77
появляется пенек 3-го первостепенного махового. В возрасте* 13—14 дней отмечается быстрый рост оперения на большей части тела: развертываются кисточки на боках шеи, передней части спины, бедрах и появляются пеньки в области зоба. Пуховой наряд сохраняется лишь на голове, брюшке, нижней части спины и крестце. Охристая вершинная полоса больших кроющих крыла и светлые каплевидные пятна на других: кроющих создают пестрый рисунок крыльев, прикрывающих большую часть спины. Клюв в основании становится почти черным, в вершинной части — охристо-буроватым. Ноги заметно краснеют.
В третью неделю при максимальном привесе и совершенствовании полета у птенцов интенсивно растут перья крыла и хвоста, обеспечивающие увеличение несущей поверхности. Продолжается рост юношеского пера, расширяются покрытые пеньками участки в области зоба, на груди и спине. На 18-й день появляются пеньки по средней линии брюшной птерилии. К концу этой недели все маховые развертывают опахала, за исключением третьестепенных, представленных кисточками, и двух дистальных первостепенных, отрастающих позднее.
В течение 4-й недели идет дальнейшее развитие оперения туловища и начинается линька головы. Увеличиваются охваченные линькой участки, развертываются кисточки на пеньках средней части спины. В возрасте 22 дней появляются пеньки ушных кроющих, которые спустя 2 дня развертываются в кисточки; в 23 дня появляется пенек 3-го третьестепенного махового. В 26 дней отмечены пеньки на лбу, а к концу недели кисточки и пеньки образуют уже широкую темную полосу, проходящую от лба к затылку. В это же время появляются пеньки на горле. В крыле отмечаются следующие изменения: прекращают рост 10-е и 9-е первостепенные маховые (наиболее длинные в этот период 6-е и 7-е). Заканчивается развертывание опахал кроющих крыла. На 25-й день появляется зачаток пенька второго первостепенного махового.
Известно, что у куриных 2 дистальных первостепенных маховых при юношеской линьке не сменяются и имеют окраску взрослого наряда, лишь незначительно отличаясь от них несколько большей заостренностью и рисунком. Эти отличия дают возможность до первой полной линьки взрослого наряда отличать взрослых птиц от молодых. Более поздцие ис
78
следования показали, что -у молодых фазанов сменяются все 10 первостепенных маховых (Грачев, 1960), а у уларов не линяют 3 дистальных маховых (Кузьмина, 1960; Базиев, 1966). Необходимо отметить, что запоздалое развитие крайних дистальных первостепенных маховых и заостренность их вершин дали основание отнести эти перья к юношескому наряду. Эта точка зрения существует до сих пор. Однако, во-первых, эти перья отрастают одновременно с появлением отдельных перьев взрослого наряда и в это же время начинается ювенальная линька 10-го и 9-го первостепенных маховых перьев. Во-вторых, по окраске они не имеют ничего общего с ювенальным оперением и лишь с трудом отличимы от других первостепенных маховых взрослого наряда. В-третьих, расстояние между бородками первого порядка (10 измерений) вторых первостепенных маховых у взрослых и перелинявших во взрослое оперение молодых особей одинаково и составляет в среднем 0,2 мм, в то время как у доросшего 3-го первостепенного махового пера птенца оно в среднем оказалось равным 0,26 мм. Таким образом, изучение формирования и линьки юношеского оперения у кеклика показало, что эти перья правильнее отнести к взрослому наряду.
К концу 4-й недели в наряде птенцов частично сохраняется пух (на голове и брюшке), продолжается рост и развитие юношеского оперения на большей части тела и появляются первые перья взрослого наряда.
С 5-й недели начинается линька ювенального пера и появляется пенек 1-го первостепенного махового взрослого-наряда. Выпадение 10-го юношеского первостепенного махового пера отмечено в возрасте 29 дней, а к концу недели их остается только шесть (3—8-е). В это же время заканчивается формирование юношеского оперения третьестепенных маховых и идет быстрое развитие дефинитивного пера, кроющего тело. Пух сохраняется только на боках головы и задней части брюшка.
Для шестой недели характерен интенсивный рост ювенального пера (развертываются кисточки и опахала). Одновременно сокращаются покрытые пухом участки и заметно отрастают перья взрослого наряда. К 42 дню пух остается только на щеках и над глазами. Перья взрослого наряда появляются на задней части шеи, верхней части спины, зобе и голове (рис. 18).
79
Возраст Б днях П ер8осте пенные маходые Второстепенные маховые Третьест маходые
125456789 10 1 2545 67 89 10 1 2 3
1 2 3 4 5 6 7 8 5 10 11 12 73 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 • •••••• ••••••• • • н н + 1 1 i 4 i 4 i i 4 I + i 4 + + 4 4- 4 44+ 4+4+444 44444+4 + 44-1-444 •444+444 •444444+ i+4444+4 +44+4+4+ + +4+44+4 I+++++++ ++++++++ 4+44++++ 44444444 44444444 4444444+ 4+4444+4 +44+4444 A+++++444 A+++4++44 Л4444++44 A+44+4+44 AA+++++44O AA44++4++A AA+44444+A A 1+ + + -I—Ы—h 1 A ♦ "I—* +444+ i t 1 +4-4+44 OX 1 14+444+AX 4 1+4 + 44+ | X 4 444 + 444 4 X 4 4 44 + 4 + 4 4 X 4 4 +4 + 4 4 4 4 X 4 4 4 + 444+4 X 4 4+++44+lx 1+4+4+444 X 4 ++4 + 4+О 4 X •••••• • • • • • • {•••••• ill{}•••• H H H H t •HHiHH • ++++++ + i i I ++ ++•++ + + 1 1 +++++-H-++ t++++ + ++-++ 1+++++++++ ♦ + +++++-++ + 1+ + +-+++++ + ++++++++++ + + ++-+++•++ + + +++ + +-+++ + ++++++++++ ++ + +++-+ + + + ++- + + + + +++ + + + ++ + + 4-1-+ -t-++++++++++ ++++++++++ ++ + ++-+++ + + ++++++++++ -1—F+++ + + + ++ + + +++++-++ + ++++++++++ ++++++++++ + 4-+ + + + + + + + ++++++++++ +++-++-++++ + ++++++++++ ++++++++++ + -I- + ++ + + + + + -1—F + + + + +-+-1—f-++++++++++ +++++-(-+++ + + + +-+ +++ + + + ++++++++++ ++++++++++ ++ + + 4-++++ + +-F+ + + ++++ + • • I i • ♦ • 1 • i + + t I I f + ♦ + ♦ 1 + ♦ + t + ♦ + t • + i + + t f + + t + + + + + t + + i + + + + + + + + I + +i + + + + + + + ++• + + + + + + + + + + + + + + + + 4- +-
Юношеское оперение Оперение Взрослого наряда
• -a I-J" +-8 о-г k-a f-zf У.-3
Рис. 18. Рост и смена маховых перьев у птенцов кеклика (возраст 1— 43 дня). Юношеское оперение: а — пенек; б — кисточка; в — развернутое •опахало; г — выпавшее перо. Взрослое оперение: а — пенек; б — кисточка;
в — развернутое опахало
Приведенные данные показывают, что четко разграничить возрастные наряду у птенцов кеклика невозможно. Полный пуховой наряд сохраняется лишь 1—2 дня (до появления пеньков маховых), а с появлением 1-го и 2-го первостепенных маховых (25 и 29 дни) перьевой покров птенца состоит уже из трех возрастных нарядов. Необходимо отметить, что поздно линяющие участки тела (верх головы, задняя часть брюшка, крестец), минуя стадию юношеского, покрываются перьями взрослого наряда *.
Для уточнения дальнейшего хода линьки растущих молодых кекликов осмотрено 268 особей, добытых в августе — первой половине ноября в разные годы. Детальное изучение их показало, что смена юношеского оперения на взрослое и окончание роста последнего заканчиваются у большинства птиц в 4-месячном возрасте (середина ноября). Началом смены двух нарядов следует считать появление пенька 2-го первостепенного махового окончательного наряда и выпадение 10-го юношеского первостепенного махового. Почти одновременно начинают встречаться отдельные пеньки взрослого наряда на шее и верхней части спины. Линька первостепенных маховых перьев идет медленно — от 10-го первостепенного к дистальной части крыла и полностью заканчивается сменой 3-го юношеского пера вместе с третьестепенными маховыми в самом конце общей линьки.
Второстепенные маховые юношеского наряда начинают сменяться значительно позже, когда 10—7-е первостепенные достаточно отрастут. Линька их начинается с 4-го пера и идет в дистальном и проксимальном направлениях. Смена второстепенных происходит значительно быстрее, чем первостепенных; особенно быстро линяет проксимальная часть крыла, начиная с 5-го до третьестепенных. В результате ускоренной смены второстепенных маховых линька всего крыла заканчивается одновременно.
Смена рулевых идет от центральных перьев к крайним. Начало ее совпадает со сменой 8—7-го первостепенных маховых. Завершается линька хвоста вместе с окончанием общей линьки контурного оперения и крыльев.
Молодые особи отличаются от взрослых лишь более за
* В литературе приведены ошибочные указания (Остапенко, 1965), что у птенцов кеклика отмечается 3 линьки (смена пухового наряда и двух юношеских).
6-24
81
остренными 1-м и 2-м первостепенными маховыми перьями, а при вскрытии — слабо окостеневшей metasternuni грудины и наличием фабрициевой сумки, которая еще в сентябре имеет средние размеры 13,3X8,6 (98 экз.), в октябре 12,0X7,6 (113), в ноябре— 11,2X6,5 (51), в декабре — 9,1X4,1 (21). К началу размножения фабрициевая сумка полностью редуцируется: у 3 исследованных в июне птиц ее не было.
В линьке взрослых птиц остается еще много неясного. Судя по литературным данным, смена оперения у них не ограничивается летне-осенним периодом, а отмечается и в зимнее время (Дементьев, 1952; Карташов, 1955). К такому же выводу приходит М. Л. Кузьмина (1955), по данным которой в Чу-Илийских горах в феврале из 50 просмотренных экземпляров линька отмечена у пяти.
По мнению Г. П. Дементьева (1952), есть два объяснения этого явления: зимняя линька — нормальный факт и смена пера продолжается весь год за исключением периода активности гонад; линяющие зимой кеклики — родители вторых кладок или поздних выводков. Однако материалы М. А. Кузьминой (1955), собранные в Чу-Илийских горах, не подтверждают предположение, что активность гонад может задерживать линьку. Так, в апреле у 40 просмотренных птиц в различной степени функционировала половая система: у самцов вплоть до сперматогенеза, у самок — до сформированных яиц. У 28 (70%) особей в области шеи и верхней части спины обнаружены пеньки и кисточки. Таким образом, зависимости линьки от степени развития гонад отметить не удалось.
Что касается предположения Г. П. Дементьева о линяющих зимой кекликах, то можно отметить: эти особи не могут быть родителями вторых кладок, поскольку у кекликов в году только один нормальный репродуктивный период, а мнение об особях из поздних выводков, линяющих зимой, пока не находит подтверждения.
У взрослых особей сроки летне-осенней линьки зависят от участия в насиживании кладок. Самцы и самки, потерявшие кладки, не участвующие в насиживании сам щи и больные особи объединяются в стаи и у них сразу же наступает линька. Особи, насиживающие кладки и воспитывающие птенцов, начинают линять через 10—15 дней после вылупления птенцов, когда молодняк уже начинает подлетывать. У этих птиц на средней линии брюшка появляется широкая
82
полоса пеньков; затем — пеньки на боках шеи, спине, груди; почти одновременно наступает линька маховых и рулевых перьев. Смена первостепенных маховых идет от 10-го к 1-му, которое меняется последним. Указание о линьке первостепенных маховых после смены второстепенных (Остапенко,. 1965) ошибочно. Второстепенные маховые у взрослых особей начинают сменяться лишь после того, как 10—7-е первостепенные уже достаточно отрастут. Считают, что смена второстепенных маховых начинается с почти одновременным выпадением 4-го и 10-го перьев, от них линька распространяется в1 обе стороны, а последними сменяются 6—8-е маховые (Кузьмина, 1955).
Однако такая последовательность имеет значительные индивидуальные отклонения. Как показал осмотр 19 особей,, линька может начинаться не только с 4-го, а с любого из 4 дистальных второстепенных маховых. Начало линьки с 1-го или с 1—2-го пера отмечено у 2 особей (10,5%); со 2-го или; 2—3-го — у 6 (31,5%), с 3-го или 4-го — у 3 особей (15,8%). У 8 особей перья настолько отросли, что установить порядок смены оказалось невозможным: у 5 птиц (26,4%) 2-е, 3-е и 4-е оказались одинаковой длины, а у 3 (15,8%) наиболее длинными были 1-е, 2-е и 3-е второстепенные маховые. Таким образом, смена второстепенных маховых начинается с любого из-4 дистальных перьев. Линька проксимальных второстепенных, наступает несколько позже; чаще она начинается с 10-го пера, но иногда и с 9-го, и идет, как правило, в двух направлениях— дистальном и проксимальном. Отмечены случаи, когда линька третьестепенных маховых несколько задерживается.
Основное отличие линьки юношеского наряда — строгая последовательность смены второстепенных маховых, тогда как у взрослых особей порядок их смены подвержен значительным индивидуальным отклонениям (рис. 19, 20).
Процесс формирования юношеского оперения кеклика и линька на взрослый наряд существенно отличаются от развития перьевого покрова очень близкого вида — европейской каменной куропатки (A. graeca). У птенцов этого вида четко разграничивается окончание формирования юношеского оперения от начала появления взрослого наряда (Menzdorf, 1975). У кеклика, как указывалось выше, такого разграничения нет.
83
У европейской каменной куропатки развитие ювенального оперения, по сравнению с кекликом, идет более замедленными темпами и полностью заканчивается^ к 56—57 дню, одновременно с появлением пеньков дистальных маховых. Таким образом, в течение 10—12 дней птенцы европейской каменной куропатки имеют только юношеский наряд. У кеклика рост юношеского оперения до начала линьки идет более ускоренными темпами и смена маховых начинается уже в возрасте 29 дней, или на 27 дней раньше, причем четкого раз-
Рис. 19. Линька маховых перьев у молодых кекликов: а — перо юношеского наряда; б — перо взрослого наряда; в — направление смены пера
Рис. 20. Линька маховых перьев у взрослых кекликов: а — перо старого наряда; б — перо нового наряда; в — перья, смена которых подвержена индивидуальным отклонениям; г — направление смены пера
34
граничения возрастных нарядов у него нет. Однако если рост ювенального пера у европейской каменной куропатки идет медленнее, чем у кеклика, то юношеская линька протекает значительно быстрее. У кеклика выпадение 3 проксимальных первостепенных маховых (8—10-е) растянуто на 2 недели, а у европейской каменной куропатки занимает лишь 6 дней. Такие глубокие различия в темпах формирования перьевого покрова лишний раз подчеркивают видовую самостоятельность кеклика, еще недавно объединяемого в один вид с европейской каменной куропаткой.
Послегнездовая жизнь. Птенцы у кекликов появляются в конце мая — начале июня. По наблюдениям в горах Чулак уход птенцов из гнезд зарегистрирован в следующие даты: 1974 г. — 5 июня; 1975 г. — 21 и 28 мая; 1976 г.— 24, 28, 31 мая, 1, 4 и 13 июня; 1977 г. —30 мая, 1, 1, 1, 2, 4, 6, 7, 10 и 17 июня; 1978 г. —23, 26, 26, 26, 26, 27, 28, 29, 30, 31 мая, 1, 1, 3, 12, 12, 17 и 18 июня. Первые встречи выводков отмечены 1 июня 1974 г., 26 мая 1975 г., 27 мая 1977 г. и 25 мая 1978 г.
К моменту вылупления птенцов по сухим склонам гор и на дне ущелий появляется большое количество мелких беспозвоночных животных, служащих основным кормом для молодняка. Первые выводки чаще всего отмечались на пологих склонах с чахлой растительностью, среди которой преобладали полыни и злаки.
В первые дни жизни молодняк водопои не посещает. Только в недельном возрасте выводки появляются у берегов речек, ручьев, родников. В ущелье Чолбыр (горы Чулак) 22 июня 1974 г. неподалеку от пустующей кошары, в бетонированной колоде для водопоя овец обнаружено 7 трупов птенцов 6—10-дневного возраста: попав в колоду, они не в силах были выбраться. Чем старше становятся птенцы, тем чаще появляются у водопоев, что связано с изменением питания: в нем возрастает количество кормов с низким содержанием влаги (подземные части растений, семена). Частота появления выводков у водных источников во многом зависит от состояния погоды и часто не совпадает во времени в разные дни.
Определить число птенцов в нелетном выводке почти невозможно, поскольку при приближении человека молодняк в считанные секунды успевает затаиться. Взрослые особи при
'85
выводках ведут себя по-разному. При внезапном появлении опасности они, имитируя подранков и тревожно крича, отводят человека, давая возможность птенцам затаиться. Заметив опасность издали, убегают или отлетают от выводка на 40—60 м и выбирают позицию, откуда одинаково хорошо видно место затаивания выводка и подходящего к нему человека. Постоянно издаваемый тревожный крик их птенцы воспринимают как сигнал о близкой опасности и не двигаются до тех пор, пока его слышат. Только когда человек удалится на 150—200 м, родители перестают кричать и начинают молча приближаться к выводку, то и дело останавливаясь и осматриваясь. Птенцов созывают тихим, едва слышным с 20—30 м, голосом. В 2-недельном возрасте птенцы способны пролететь вдоль или вниз по склону 20—70 м и искусству затаиваться чаще предпочитают активный бег или полет.
Начиная с середины июня мы неоднократно встречали выводки с 2 взрослыми птицамй*. Мечение наседок на гнездах и последующие встречи их с выводками подтвердили эти наблюдения (см. табл. 2).
Касаясь вопроса о выводках, в которых отмечалось по 2 взрослых птицы, И. А. Долгушин (1951) пишет: «Самцы после окончания линьки присоединяются к выводкам вновь. Разыскивает ли при этом самец именно свою самку или присоединяется к первому попавшемуся выводку — судить не беремся» (с. 86). По устному сообщению В. А. Вырыпаева и Р. Р. Будриса, в условиях Чаткальского заповедника большая часть самцов, самки которых приступили к насиживанию, остаются в районе своих гнездовых участков и позднее присоединяются к выводкам. Другая, меньшая часть, объединяется в стайки по 4—7 особей и поднимается к верхней границе древесно-кустарникового пояса.
По нашим наблюдениям, выводки с 2 взрослыми птицами встречаются значительно реже, чем с одной. Так, в 1974 г. в июне встречено 105 выводков и только в 10 из них (9,5%) зарегистрировано по 2 взрослые птицы; в 1975 г. 5 (14,7%) из 34 выводков, в 1976 г. 5 (25%) из 20, в 1977 г. 25 (29,8%) из 84 и в 1978 г. 10 (15,4%) из 65. Таким образом, из 208 встреч 55 (26,4%) выводков были с 2 взрослыми птицами. Можно высказать предположение, что объединившиеся выводки— результат двойного гнездования, но материалов, подтверждающих этот факт, пока нет.
S6
Помимо встреч выводков с 2 взрослыми птицами в редких случаях при них отмечалось по 4—6 взрослых: за весь период работ отмечено только 16 таких встреч. Мы склонны расценивать такие факты как временные соединения выводков с группами взрослых птиц, не участвующих в насиживании кладок. Иногда в объединившихся выводках наблюдались разновозрастные птенцы.
После подъема молодняка на крыло проводился учет птенцов в выводках. По материалам 1977 г., в среднем покинуло гнездо 13,8 птенца (11 гнезд), а летных в выводке с одной взрослой оказалось 12,4 (59 выводков), т. е. в среднем число молодых, спустя 15—20 дней, сократилось на 1,4 (10,1%). По данным А. И. Иванова (1969), количество птенцов в выводках снижается очень быстро. Если пуховичков в выводках бывает 11 —12, то птенцов, достигших половины величины взрослых, в выводке сохраняется не больше 6—8.
В летний период с восходом солнца выводки часто встречаются на кормежке, а в жаркие часы дня отдыхают в тени кустов или под навесом скал. С наступлением вечерней прохлады активность их вновь возрастает: в поисках корма выводки перемещаются на открытые участки склонов, постепенно поднимаясь к вершинам, где в сумерки устраиваются на ночевку. Выпадение дождя значительно влияет на активность птиц. Ненастье кеклики пережидают в кустарнике, а с прекращением непогоды обычно кормятся неподалеку от этих мест.
Наблюдения за птенцами при полувольном содержании показали, что у них ^активная деятельность часто чередуется с отдыхом и сном; чем старше становятся птенцы, тем менее продолжителен у них отдых.
В конце августа — начале сентября на склонах и дне ущелий, где кеклики находят обильный корм, нередко встречаются стаи до 100 особей. Потревоженные, они легко распадаются, что свидетельствует о непрочности таких объединений. Обособившиеся в июне стайки взрослых особей, состоящие из самцов и самок, потерявших кладки, и самцов, не участвующих в воспитании молодняка, распадаются лишь к следующей весне, что подтверждается отловом кекликов в зимнее время. Число птиц в таких стайках невелико, чаще 8—12 особей.
Характерной чертой жизни кекликов в конце лета и нача
87
ле осени является регулярное посещение водопоев. С рассветом стайки спускаются со склонов к родникам и речкам (чаще перелетают). Водопои играют существенную роль в распределении кекликов. В обводненных ущельях Киргизского и Заилийского Алатау стайки распределяются более или менее равномерно, а в Чу-Илийских горах и горах Ма-лай-Сары в связи с дефицитом водных источников кеклики обычно держатся неподалеку от них. Для водопоев птицы выбирают открытые участки берега или полого спускающиеся к воде осыпи, избегают мест с густыми зарослями кустарников, скрывающих русло (рис. 21).
88
- В жаркие дни после водопоя кеклики часто устраиваются на отдых в ближайших кустах, где «купаются» в пыли,, оставляя в таких местах характерные «купальни», напоминающие лотки гнезд без строительного материала.
По наблюдениям в Киргизском Алатау 19 сентября 1975 г. стайка из 12 кекликов в течение дня 3 раза посещала водопой (6 ч 15 мин, 10 ч 40 мин и 14 ч 20 мин). В перерывах птицы поднимались на склон (200—250 м), где отдыхали и корми-
Рис. 21. Типичные места водопоя кекликов в Каратау: а — нижняя часть ущелья Ран, б — верхняя
89
лись. Во второй половине дня, ближе к вечеру, стайки птиц у водопоев встречались гораздо реже.
К концу ноября кеклики накапливают отложения подкожного и внутреннего жира, который необходим для перенесения трудных условий зимовки.
Зимняя жизнь. Зима — наиболее тяжелый период в жизни кекликов. Выпадение глубокого снега ограничивает передвижение птиц и значительно сокращает территорию кормовых угодий. По данным М. А. Кузьминой (1955), весовая нагрузка у кекликов колеблется в пределах 43—51 г на 1 см2 поверхности лап, что значительно затрудняет передвижение по рыхлому снегу. Чтобы добыть корм, птицы вынуждены перемещаться на склоны южной экспозиции, где снежный покров менее глубок и имеются участки, быстро освобождающиеся от снега. Частые снегопады с продолжительными морозами приводят к падежу кекликов от бескормицы.
В годы нашей работы в горах Чулак зимы особой суровостью и многоснежностью не отличались. Только зимой 1976/77 г. наблюдался частичный падеж кекликов. По сообщению чабанов, в эту зиму они несколько раз находили замерзших птиц. Необходимо отметить, что в ущельях Талды-сай, Унгур, Тайгак, Кзылаус и Чулак-Джиде еще с осени 1976 г. была организована подкормка кекликов, что, несомненно, сократило падеж птиц. В ущелье Кзылаус кеклики держались стайками по 8—15 особей на немногочисленных открытых от снега участках склонов. На дне ущелья, где глубина снежного покрова достигала более 30 см и образовался наст, мы их ни разу не встречали. Одна из стаек на протяжении всей зимы постоянно ночевала в поленнице дров на хоздворе. По сообщению егерей Капчагайского охотхозяйства В. И. Позднякова и Н. Ф. Рудометова, во время обильных снегопадов кеклики иногда появлялись на северном берегу Капчагайского водохранилища, в 7 км от ближайших гор. С окончанием ненастья птицы исчезали, несмотря на то, что их пытались подкармливать.
Наблюдения и результаты мечения кекликов в горах Чулак показали, что в зимний период у них прослеживается территориальный консерватизм. Так, в декабре 1975 г. и в феврале 1976 и 1977 гг. на подкормочных площадках было отловлено 128 кекликов, которых после мечения выпускали
90
в'этих же местах. Из 30 повторно отловленных кекликов 15 пойманы в тех же местах с интервалом в 2—10 дней, 13 отмечались или отлавливались спустя 62—422 дня, а 2 переместились на расстояние 300 и 1500 м.
У некоторых особей удалось установить привязанность к конкретным местам зимовок. Например, молодой самец 17 и 20 декабря 1975 г. отлавливался на подкормочной площадке в ущелье Биесимас. Первый раз он был выпущен там же, а во второй — в 2 км от площадки. Спустя 5 дней (25 декабря) его вновь поймали на прежнем месте.
Отмечены также случаи обмена особями в зимующих стаях. Так, 18 февраля 1975 г. отловлено 9 кекликов из стайки в 12 особей, которые после мечения выпущены там же. На следующий день 5 из них встречены на месте поимки в составе стайки из 14 птиц (9 особей не мечены), т. е. произошло соединение особей из разных стай.
Дневная жизнь кекликов зимой в основном сводится к добыванию корма. Только в ясные, относительно теплые дни их можно видеть неподвижно сидящими на вершинах скал. В декабре 1975 г. и в феврале 1976 и 1977 гг. некоторые стайки в течение дня не покидали участки размером 200X300 м.
Еще задолго до восхода солнца с мест ночевок раздаются голоса кекликов, «перекличка» через несколько минут обычно прекращается. С восходом солнца среди стаек наблюдается оживление и они начинают слетать к подножью склонов и на дно ущелий, где есть свободные от снега места. Здесь птицы проводят большую часть дня. К вечеру стайки поднимаются к вершинам и устраиваются на ночевку.
Интересные наблюдения за зимней жизнью кекликов провел Р. Г. Пфеффер (устн. сообщ.). Зимой 1974/75 г. в Алма-атинском заповеднике (ущ. Правый Талгар) стайка из 10— 15 птиц в течение 1 —1,5 месяца в дневные часы отмечалась у подножья склона среди высохших стеблей татарника. Площадка 15X15 м под слоем снега была изрыта ходами. Иногда при подходе удавалось выпугивать птиц, вылетающих из-под снега. По мнению Р. Г. Пфеффера, кеклики сделали в снегу ходы в поисках семян татарника.
Водопои кеклики в зимнее время посещают редко, потребность в воде возмещается склевыванием снега. Однако там, где по берегам ручьев и ключей есть зеленая трава, птицы охотно посещают такие места как кормовые.
91
Сильные снегопады птицы пережидают в кустарнике или под навесом скал; в случае длительного ненастья они вынуждены голодать по 2—3 дня. По данным М. А. Кузьминой (1955), перед снегопадом или в его начале кеклики поспешно кормятся пока снег не покроет оголенные участки земли.
В местах концентрации на кормежке отдельные стаи состоят из 100—150 особей. Инс^гда стайки посещают пустующие животноводческие фермы, где находят остатки зерна и комбикорм. Если кекликов не Преследовать, они становятся доверчивыми.
К концу февраля, когда обнаженных участков земли становится больше, крупные стаи не встречаются, кеклики чаще держатся по 10—15 особей, а в начале марта постепенно разбиваются на пары.
Питание
Кеклик — типичный эврифаг. В его пище зарегистрировано 311 видов высших растений 53 семейств, кроме того, мхи и водоросли, которые следует отнести к случайным кормам. Среди животной пищи основную массу составляют 132 вида насекомых, относящихся к 36 семействам (10 отрядов). Гораздо реже отмечаются моллюски (10 видов 4 семейств), и как исключение пауки (табл. 15).
По встречаемости, разнообразию видового состава и объему растительная пища в рационе кеклика значительно преобладает над животными кормами. В целом соотношение растительных и животных кормов выражается следующими цифрами: только растительная пища обнаружена у 1076 особей (77,3%), смешанная — у 309 (22,1%); исключительно животные корма встречены у 8 (0,6%). Среди растительных кормов можно выделить представителей 9 основных семейств: злаки, розоцветные, лилейные, амариллисовые, бурачниковые, сложноцветные, зонтичные, бобовые и гречишные, которые составляют около 2/з видов растений, отмеченных в пище кеклика в пределах СССР. Встречаемость их различна и достигает до 12,5% гречишных и до 58,3% злаков.
Злаки — основа питания кеклика. Различные части растений (семена, листья, стебли и прикорневые части) используются птицами во все сезоны года. Особенно часто в зобах птиц встречаются прикорневые части луковичного мятлика (Роа bulbosa), который встречен у 26% исследованных особей. Из других видов злаков отмечены семена нескольких видов костра (Bromus) и рисовидки (Piptatherum songoricum) —
93
Таблица 15. Состав кормов кеклика в различных регионах СССР
Корм Встречаемость, % Чйсло видов, отме. ченных в пище Джунгарский Алатау, A. ch. dzunga-rica (677) Северный Тянь-Шань, A. ch. falki (632) Западный Тянь-Шань, A. ch. falki (окбло 250) Памиро-Алай, А. ch. falki (около 120) Копет-Даг, A. ch. shestoperovi (более 300) Кавказ, A. ch. kur-destanica (64)
1 1 1 2 | 3 1 4 1 5 6 1 7 8 1 1 9
Растения (семе ! й с т в a)
Злаки — Gramineae Розоцветные — Ro- 58,3 35 19 16 Ю 14 9 5
saceae Лилейные — Lilia- 26,0 26 10 10 15 10 4 —
ceae Амариллисовые — 25,6 21 6 9 10 10 8 —
Amaryllidaceae Бурачниковые — Bo- 23,7 2 1 1 1 2 — —•
raginaceae Сложноцветные — 15,2 26 10 19 8 3 4 4
Compositae Зонтичные — Umbel- 13,1 36 15 16 6 7 8 3
liferae Бобовые — Papilio- 13,1 14 4 7 2 5 2 —
naceae Гречишные — Poly- 12,7 33 12 11 4 14 11 11
gonaceae Маревые — Cheno- 12,5 8 4 6 3 2 2 1
podiaceae Гераниевые — Gera- 8,2 11 9 7 2 — 2 —•
niaceae Крестоцветные — 7,9 3 2 2 2 2 1 —
Cruciferae Эфедровые — Ephed- 7,2 10 6 9 1 4 4 4
raceae Ильмовые — Ulma- 6,9 2 2 2 — 1 — —
ceae Норичниковые — 3,0 1 1 1 1 1 1 —
Scrophulariaceae Гвоздичные — Ca- 2,1 4 2 1 1 1 1 —•
ryophyllaceae Коноплевые — Can- 1,6 9 4 3 1 4- 2 2
nabaceae Амарантовые — 1,4 1 1 1 — —• — —
Amaranthaceae Осоковые — Cype- 1,3 1 1 1 — —- — —
raceae 1,0 4 3 2 1 2 2 —
Продолжение таблицы 15
1_______| 2 | 3 I 4 | 5 |б | 7 | 8 | 9
Г убоцвстные — La-biateae 0,9 8 2 3 1 1 4 1
Жимолостные — Caprifoliaceae 0,9 4 2 2 2 3 1
Дымянковые — Fu-mariaceae 0,8 1 1 1 1
Лютиковые — Ra-nunculaceae 0,7 4 2 2 1
Ворсянковые —
Dipsacaceae 0,7 3 — 1 — 2 —• 1
Мхи — Grimmia Пасленовые — So- 0,7 0,6 + + + — — —
lanaceae 4 2 1 —. —. I ——
Маковые — Papave- 0,4
raceae 2 1 2 — 1 1 —-
Лоховые — Elae-agnaceae 0,4 2 1 1 2 —.
Барбарисовые — Berberidaceae 0,4 3 1 I 1 2
Касатиковые — Iridaceae 3 1 1 1 1 1
0,2
Фиалковые — Vio-laceae 2 1 1 1
0,2
Тыквенные — Cu-curbitaceae 2 2
0,2
Парнолистниковые — Zygophyllaceae 2 1 1
0,2
Повиликовые — Cuscutaccae 1 1
0,1
Камнеломковые — Saxifragaceae 2 1 1
0,1
Кипарисовые — Cupressaceae 2 1 1 2
0,1
Молочайные — Eu-phorbiaceae 2 1 I 1 I
0,1
Бересклетовые — Celastraceae 1 1
0,1
Первоцветные —
Primulaceae Вьюнковые — Con- 0,1 2 1 — — — — 1
volvulaceae 0,1 1 1
Мальвовые — Malva- 1
ceae Мареновые — Rubi- од 2 — 1 — — 1 1
aceae Ситниковые — Jun- 0,1 2 — 1 — — 2 —
caceae 0,1 0,1
Рутовые — Rutaceae i 1 1
Свинчатковые — 1 1 —
Plumbaginaceae * —• 1 — — — — 1 —
Продолжение таблицы 15
1 2 3 1 1 4 5 1 6 1 7 1 8 1 9
Маслинные — О1е-асеае * 1 1
Волчниковые — Thymelaceae * —. 1 — —- — — - 1
Крушиновые — Rhamnaceae * 1 —— __ 1
Ладанниковые — Cistaceae * 1 — 1 __
Виноградные — Vi-taceae * 1 1
Ароидные — Ага-сеае * —-. 1 —. __
Хвощовые — Equise-taceae * —. + — — + — —
Тутовые — Мога-сеае * —. 1 — — 1 — — —
Подорожниковые — Plantaginaceae * — I — —> 1 — —. —
Водоросли (ближе не опр.)* + — —. + — —. —
Общее число видов растений — 311 134 146 79 92 79 38
Жив. Прямокрылые — Orthoptera Саранчовые — Acri-didae Э T H Ы I 9,2 г (от] 16 ряды, 8 семе 6 5ЙСТВ 3 a) 3
Кузнечики — Tetti-goniidae 0,6 3 1 2 2 — — —
Богомолы — Man-todea 0,5 1 1 1 1 — —- —,
Сверчки — Gryllidae 0,1 1 1 — —. —• —• —.
Жесткокрылые — Coleoptera Слоники — Curculi-onidae 2,5 23 20 3 6 1
Листоеды — Chry-somelidae 2,3 14 11 5 2 ___ + —-
Пластинчатоусые — Scarabaeidae 0,7 6 5 1 + —_
Усачи — Cerambyci-dae 0,4 4 4 1 + —. —.
Чернотелки — Te-nebrionidae 0,2 2 2 1 2
Щелкуны — Elateridae 0,2 2 2 — 2 — —
Коровки — Coccinel-lidae 0,2 1 1 —> — — — —-
Продолжение таблицы 15
1 2 3 1 4 I 5 1 6 1 7 I 8 1 9
Златки — Buprestidae 0,2 1 1 1
Жужелицы — Сага-bidae 0,1 1 1 + +
Приутайки — Byrrhi-
dae 0,1 + + —. —. — — —
Жуки, ближе не 1 0
опр. Полужесткокрылые — Hemiptera
Щитники — Pcnta-tomidae з,о 17 12 5 5 1
Наземники — Ly-gaeidae 0,4 3 2 1 1
Клопики — Micro-phisidae 0.3 1 1
Краевики — Corei-dae 0,1 4 . 2 2
Булавники — Rho-palidae 0,1 1 1 1
Слепняки — Miridae Черепашки — Scu- 0,1 2 1 1 — 2 —. —
telleridae Myodochli- 0,1 2 1 — 2
dae 0,1 1 1
Перепончатокрылые — Hymenoptera Муравьи — Formici-dae 10 6 5 2
2,7
Наездники — Ichne-
umonidac 0,1 1 1 1
Пчелы — Apidae Моллюски — Mol- 0,1 1 1 — — — —
lusca
Булиминиды — Bu-liminidae 0,3 3 3 1
Брадиобенидки — Bradybaenidae 3 3 1
0,2
Пупиллидки — Pu-
pilliadae 0,2 3 2 1
Слизни — Limacidae Моллюски, ближе 0,1 1 1 — *— — —
не опр. 1,9
Равнокрылые хоботные — Homoptera
Певчие цикады — Cicadidae 0,5 2 1 i 1
Листоблошки — 1 1
Psyllodea 0,1 1 I
Цикадки — Cicadel- —•
lidae 0,1 0,1 2 2
Тли — Aphidodea 1 1 —.
7—24
Окончание таблицы IS
1 I 2 3 I 4 | 5 I 6 | 7 | 8 | 9'
Чешуекрылые — Le-pidoptera
Нимфалиды — Nymphalidae
Совки — Noctuidae
Бабочки, ближе не опр.
Комары — Culicidae
Ripidae
Уховертки — Der-maptera Forficulidae
Двукрылые — Dip-tera
Мухи — Muscidae
Мелкие части насекомых, ближе не опр.
Красноклопы —
Pyrrhocoridae *
Эмпузы — Empusi-dae *
Пауки —Aranei
Общее число видов животных
0,1 0,1
0,4
0,1
0,1
3,1
Примечание. В скобках число особей. Знаком «+» отмечены жи-вотные и растения, определенные только до семейства; * семейства животных и растений, включенные в таблицу только по литературным источникам.
одного из самых ранних растительных кормов птенцов. Культурные злаки (пшеница, ячмень, рожь, кукуруза, овес, просо) отмечены у 43% особей и охотно поедаются кекликами в тех районах, где эти птицы живут в непосредственной близости от полей.
Розоцветные по встречаемости занимают в пище кеклика второе место и не обнаружены в его кормах только на Кавказе. В основном поедаются ягоды кустарников и деревьев, которые в некоторых случаях составляют значительную часть рациона. Необходимо отметить, что значение розоцветных несколько ниже, чем о них можно судить по содержимому желудков, поскольку костянки различных ягод, в отличие от другой пищи, практически не перевариваются и надолго за*
98
держиваются в желудках, дополнительно выполняя роль гастролитов.
Лилейные в рационе кеклика представлены более чем 20 видами. Весной в основном поедаются листья, летом и осенью — луковицы и семена, зимой — только семена. Чаще; других в пищу употребляются луковицы тюльпанов (Tulipa). и гусиных луков (Gagea). В Чу-Илийских горах, где в массе произрастает Allium trachyscordum, кеклики охотно поедают его семена (13,2% встречаемости) и луковицы (10,3%).
Для такого сравнительно немногочисленного семейства луковичных, как амариллисовые, отмечено поедание только Ixiolirion tataricum и Ungernia severzovii. Наличие первого из них зарегистрировано в пище кеклика повсеместно, за исключением Копет-Дага и Кавказа; второго как основного-корма — в Туркестанском хребте (Минин, 1939). По пищевому значению амариллисовые практически не отличаются от лилейных, так как кеклики употребляют различные части растений во все сезоны года. Если учесть почти одинаковую встречаемость лилейных и амариллисовых в кормах кеклика при более богатом (в 10 раз) видовом составе первых, можно констатировать, что амариллисовые являются массовым кормом. По частоте встречаемости Ixiolirion уступают лишь луковичному мятлику (23,7 и 26,0% соответственно). В Джунгарском Алатау, где эти растения встречаются в изобилии, кеклик явно предпочитает Ixiolirion, а где он реже — в большом количестве поедает луковичный мятлик.
Бурачниковые отмечены в кормах кеклика во всех рассматриваемых регионах и представлены в его питании более чем 20 видами. Наибольшее количество видов зарегистрировано в Джунгарском Алатау и Северном Тянь-Шане. В пи-* ще в основном встречаются семена и, гораздо реже, зеленые части побегов. Нередко в зобу одной особи отмечается несколько видов бурачниковых, а в некоторых случаях они составляют большую часть корма. Наиболее часто поедаются семена Rachela (6 видов), Lithospermum (3 вида) и Lappula (2 вида). В зимнем питании кеклика представители этого семейства редки.
Сложноцветные по разнообразию видового состава (36 видов) занимают в пище кеклика первое место, хотя встречаемость их (13,1%) значительно уступает таковой представителей ранее рассмотренных семейств. В основном кеклики
ЭД
поедают семена, реже — корзинки и зелень. Часто в кормах отмечаются 3 вида латука (Lactuca tatarica, L. viminae, L. serriola), обнаруженных в зобах кекликов в Копет-Даге, Туркестанском хребте, Северном Тянь-Шане и Джунгарском Алатау. В Западном, Северном Тянь-Шане и Джунгарском Алатау осенью и зимой в зобах птиц постоянно находили семена татарника (Onopordon acanthium), которые в этот период составляют существенную часть кормов. Цветы одуванчиков (3 вида) отмечены в Туркестанском хребте, Западном и Северном Тянь-Шане. Зеленые части полыней (4 вида) и тысячелистника (Achillea milleifolium) обнаружены в кормах в Северном Тянь-Шане и Джунгарском Алатау.
Зонтичные отмечены в пище кеклика во всех приводимых регионах, исключая Кавказ. В Северном Тянь-Шане и Джунгарском Алатау в кормах встречаются клубни скалигерии (Scaligeria setacea), в Туркестанском хребте — клубни Ви-nium capus и В. chaerophylloides (Минин, 1939). У остальных видов этого семейства кеклики поедают лишь семена, отдавая предпочтение феруле и шренкии.
Бобовые по разнообразию видового состава уступают лишь сложноцветным и злакам. Повсеместно в пищу используются семена горошков (Vicia sp., V. kokanica) и ползучего клевера. Наиболее разнообразен видовой состав бобовых в рационе кеклика в Туркестанском хребте, где наряду с указанными видами поедаются нут (Cicer songoricum и С. гпасгасап-thum) и остролодочник (Oxytropis macrocarpa), из 7 видов астрагалов, отмеченных в кормах, здесь используется шесть. Совершенно не обнаружены в пределах Туркестанского хребта виды чины (Lathyrus), часто встречающиеся в горных районах Юго-Восточного Казахстана. Состав кормов бобовых в Копет-Даге беднее: из астрагалов встречено только 4 вида, как и на Кавказе, здесь отмечено поедание 2 видов эспарцетов (Onobrychis Lipskyi, О. micrantha). Культурные растения представлены в небольшом числе — это фасоль и горох. При большом видовом разнообразии бобовых растений встречаемость их в кормах кеклика довольно низкая и каждому географическому району свойствен свой видовой состав.
Сравнительно небольшое количество видов гречишных в кормах кеклика объясняется трудностью определения их видов. Встречаемость этого семейства лишь немногим уступает бобовым, и они отмечены в зобах во всех рассматриваемых
100
регионах, что позволяет считать их важными кормовыми объектами. Наиболее разнообразен видовой состав гречишных в пище кеклика в Джунгарском Алатау и Северном Тянь-Шане, где в рационе отмечены семена курчавки (Atraphaxis lanceo-lata), птичьей гречихи (Polygonum aviculare) и луковички живородящей гречихи (Р. viviparum).
Второстепенные корма представлены в пище кеклика 12 семействами (57 видов), которые отмечены на ограниченных территориях, и встречаемость их составляет от 8,2% (маревые) до 0,8% (жимолостные). Среди этих кормов обнаружены семена маревых, норичниковых, гвоздичных, коноплевых, амарантовых, губоцветных, крестоцветных. Клубни (3 вида) поедаются только у гераниевых, но встречаемость их довольно высокая (7,9%). Возможно, значение последнего семейства в питании кеклика несколько выше, поскольку значительную часть мелких клубней определить не удалось. Ягоды в кормах кеклика зарегистрированы в летне-осенний период и представлены семействами эфедровых и жимолостных; зелень отмечается очень редко. Остальные растительные корма следует рассматривать как редкие, и они представлены 45 видами (22 семейства). Встречаемость их составляет около 1%. Чаще других в этой группе отмечаются семена, затем — ягоды (6 видов древесно-кустарниковых растений) и только у 3 видов в пищу используются клубни.
Необходимо подчеркнуть, что иногда второстепенные и даже редкие корма становятся массовыми. В первую очередь это относится к ягодам, которые в конце лета — начале осени могут преобладать над другой пищей. Так, в Копет-Даге осенью главным кормом становится виноград (Федоров, 1949). В хребте Нуратау в сезон созревания ягод тутовника кеклики в основном используют их (Салихбаев, Кашкаров, Шарипов, 1970). В Таласском Алатау в качестве основной пищи осенью употребляются ягоды облепихи (Ковшарь, 1966), а в Чу-Илийских горах в этот же сезон года мы отметили поедание в большом количестве ягод кизильника.
Все корма кеклика можно разделить на 5 групп:
1. Семена и плоды травянистых растений и кустарников (стручки, коробочки, орешки, семена).
2. Зеленые корма (листья, стебли, цветы).
3. Подземные части растений (луковицы, клубни, корни., корневища).
101
4. Ягоды.
5. Беспозвоночные животные (членистоногие, моллюски).
Из 4 групп растительных кормов, зарегистрированных в нище кеклика, по частоте встречаемости и разнообразию видового состава важнейшее место занимают семена (178 видов 35 семейств). Основу этой группы корма составляют семена злаков (25 видов), бурачниковых (21), крестоцветных (10), лилейных и маревых (по 9). Остальные семейства представлены меньшим числом видов.
Семена — высококалорийный корм, который включает растительные белки, жиры, углеводы. В течение всего года они служат основной пищей кекликов. Исключение составляют весенние месяцы: в это время сохранившихся после зимы семян мало и кеклики переходят на питание молодой зеленью. Особенно большое количество семян поедается во второй половине лета и осенью, когда линяющим птицам пос-ле^периода размножения требуются особенно питательные и разнообразные корма. Длительная сохранность семян в значительной степени обеспечивает выживаемость птиц в зимний период, и в этом их большое значение.
Обычно в зобах кекликов семена встречаются вместе с другими видами кормов. Реже пища состоит только из нескольких видов семян и, как исключение, — из одного вида. В последнем случае корм чаще бывает представлен зернами культурных злаков, собираемых на полях после уборки урожая.
Подземные части растений. В рационе кеклика отмечены луковицы, клубни, прикорневые луковицы, корневища и корни травянистых растений. Следует отметить, что среди представителей куриных нашей фауны кеклик наиболее широко использует эту группу корма, которая характеризуется высоким содержанием углеводов и крахмала, и это вместе с другой пищей обеспечивает накопление в организме запасов жира на зимний период. Кеклик поедает подземные части 25 видов растений (9 семейств) и особенно различных луков и тюльпанов. Однако по частоте встречаемости и обилию подземных частей первое место принадлежит прикорневым луковицам злаков (луковичных, мятлика и ячменя) и луковицам амариллисовых (иксиолирион и унгерния). Довольно часты в пище кеклика клубни гераниевых и зонтичных (скалиге-рия). Зимой добывание глубоко расположенных луковиц и
102
клубней затруднено из-за промерзания почвы и птицы используют лишь прикорневые луковицы мятлика живородящего.
Зеленые корма в рационе кеклика довольно разнообразны и представлены 61 видом (18 семейств) высших растений. Основную массу зеленых кормов составляют злаки (определенные до семейства), а также сложноцветные (15 видов, из них 5 видов полыней, одуванчики), бобовые (13 видов: 4 вида астрагалов, ползучий клевер, люцерна, горошки, 2 вида солодок и др.). Кроме того, встречаются представители маревых, зонтичных, бурачниковых.
Весной (март — апрель) основной корм кеклика — зелень и цветы эфемеров и эфемероидов. Среди этой группы особо следует выделить различные виды лилейных (Allium, Gagea, Tulipa), шафран, луковичный мятлик и иксиолирион. Поедание молодой зелени объясняется высоким содержанием различных витаминов, в которых особенно нуждаются птицы в этот период (Цюрупа, 1933). Поедая эти высокопитательные корма, кеклики за короткий период становятся упитанными: у них развивается подкожная жировая клетчатка и образуются отложения жира в полости тела; это способствует подготовке организма к размножению. За репродуктивный цикл самки расходуют на откладку яиц в среднем 300 г белков, жиров, извести, что составляет более половины собственной массы их тела. Этот расход веществ компенсируется главным образом за счет свежей сочной зелени, которую они используют в пищу.
Ягоды по сравнению с другими группами растительных кормов в рационе кеклика играют небольшую роль. Ограниченный видовой состав, относительно низкая встречаемость в пище и короткий период использования их дают основание отнести ягоды к второстепенным кормам. Всего в пище кеклика отмечено 25 видов ягод (И семейств). Во всех регионах встречены ягоды розоцветных (10 видов); чаще других поедаются плоды вишни, боярышника, шиповника, кизильника. Ягоды других семейств (лоховые, пасленовые, тутовые, жимолостные, барбарисовые, виноградные) отмечены на сравнительно ограниченных территориях. В период обилия ягоды иногда становятся массовым кормом, вытесняя на короткое время другие корма. В ягодах содержатся различные кислоты, витамины, сахара и жиры, которые придают им со
юз
вершенно отличную от других кормов питательную ценность. По сравнению со взрослыми птицами в рационе молодых в июле — августе ягоды встречаются гораздо чаще (рис. 22).
Животные корма по своей питательной ценности в рационе кеклика занимают особое место. Во-первых, это источник легко усвояемых высококалорийных белков животного происхождения. Во-вторых, поедание весной моллюсков способствует восполнению потерь кальция, необходимого для образования скорлупы яиц в период размножения.
Основные животные корма представлены отрядами прямокрылых (22), жесткокрылых (56) и полужесткокрылых
104
(33 вида). Прямокрылые, в основном саранчовые (16 видов), встречаются в пище наиболее часто. Представители другого отряда — жесткокрылые — самая разнообразная группа, причем в зобах и желудках кеклика часто обнаруживаются не определенные остатки жуков. Чаще других поедаются слоники (23 вида) и листоеды (14). Полужесткокрылые встречаются реже представителей предыдущих отрядов, а по видовому составу уступают лишь жесткокрылым. Однако в местах обильных скоплений клопы в массе поедаются кекликом. Так, в ноябре — декабре 1978 г. в Заилийском Алатау (ущелье Жаманты) этот вид корма был отмечен у 17 (28%) особей
105*
из 61, причем в ноябре клопы встречены у 32% (Ю особей из 32), в декабре — у 24,1% (7 из 29). Основная масса клопов >была представлена Dolicoris penicillatus, которые у отдельных птиц составляли 40—85% содержимого зоба. Кроме них здесь же отмечены Euridema ornata, Emlethis sp., Corizus hyoscyami, Corioneris histicornis, Sciocoris deltocephalus.
На юго-востоке Казахстана животные корма в пище кеклика (встречаемость 13,4%) отмечены с августа по январь (наиболее часто в августе и декабре) и обычно встречались в небольшом количестве, но у некоторых особей составляли 50—85%, а в одном случае — даже 100% содержимого зоба. Из насекомых наиболее часто отмечались саранчовые (30%), второе место занимали клопы (27,7%), которые отмечались в кормах с октября по декабрь, но в наибольшем количестве— в ноябре и декабре. По-видимому, в это время кеклики находили их в массовых скоплениях на местах зимовок.
Рис. 22. Растительные корма кеклика: а — ягоды эфедры; б — ягоды черноплодного кизильника; в—лук шероховатый
106
Особенно часто встречался Delicoris penicillatus, отмеченный Ф 16 зобах из 22, в которых обнаружены клопы, причем в 8 присутствовал только этот вид. В зобе одного кеклика найдено 15 D. penicillatus, что составило 85% пищевых объектов.
Жуки в пище кеклика отмечены с сентября по ноябрь. Их встречаемость составляет 24,4%, и наиболее часто они поедаются в октябре. Муравьи обнаружены в небольшом количестве с августа по октябрь (12,2% случаев); моллюски поедаются весной и в сентябре — ноябре. У 16 особей осенью обнаружены мелкие остатки хитина, крылышки и лапки насекомых, не поддающиеся определению.
Таблица 16. Изменения разнообразия кормов кеклика в осенне-зимний период на юго-востоке Казахстана •
Бид корма Август (п=26) Сентябрь (п=185)
1 2 1 1 1 2
Подземные части
^растений 4 15,3 30 16,2
Семена — —- 8 4,3
Зелень — —. —.
Ягоды —. --> 2 1.1
- Животные корма — — —
В с е г b 4 15,3 40 21,6
Октябрь (п=205) Ноябрь (п = 131) Декабрь (п=36) Январь (п=8)
1 1 2 1 1 2 1 1 2 1 1 2
19 9,3 15 11,3 4 11,1
9 4,3 15 11.3 —. 2 25,0
10 4,8 21 16,1 9 25,0 2 25,0
1 0,5 — — — — — —
1 0,5 — — — — —• —
40 19,4 51 38,7 13 38,7 4 50,0
1 — число особей с однородным кормом; 2 — % их
Примечание.
<от общего числа.
107
Летом пища взрослых особей по сравнению с другими сезонами года наиболее разнообразна, обычно в каждом зобе обнаруживается по 2—3 группы кормов. Животные корма в этот период составляют сравнительно небольшую часть. Лишь в одном случае у насиживающей самки в зобу обнаружено 17 моллюсков Ponsadenia semenovi, что составило 30% пищи от общей массы 19 г. По-видимому, поедание самкой моллюсков вызвано необходимостью восстановить в организме большой расход кальция.
Различное соотношение групп кормов и разнообразие их видового состава обеспечивают кеклику полноценный рацион питания.
На юго-востоке Казахстана преобладающие в августе — сентябре в пище кеклика подземные части растений в более поздние сроки уступают место семенам и зелени — основной пище зимнего периода. Процесс замены кормов обусловлен не только обилием последних в данное время. Так, сокращение подземных частей растений в рационе наступает задолго до выпадения снега. При обилии пищи у кекликов проявляется хорошо выраженная избирательная способность к определенным кормам и нередко к определенным видам растений. Одновременно с изменением соотношения групп кормов пища кеклика ближе к зиме становится более однообразной и зачастую у некоторых особей представлена только одной группой корма или даже одним видом растений (табл. 16).
В зависимости от погодных условий в разные годы кеклики могут изменять рацион кормов. Сопоставляя состав пищи по годам в одном и том же районе с суммой осадков, выпадающих в апреле — августе, удалось установить, что в двух разобщенных хребтах Каратау и Киргизском Алатау, несмотря на существенные отличия рельефа, высоты над уровнем моря и климата, в питании кеклика прослеживаются общие черты в изменении состава кормов. Такие же результаты получены при сравнении питания кеклика в Чу-Илий-ских горах и горах Малай-Сары.
По данным метеостанций (1972—1975 гг.) годовые изменения суммы осадков в Каратау и Киргизском Алатау шли синхронно. В наиболее засушливый 1975 год при сокращении урожая семян кеклики в обоих хребтах переходили на питание подземными частями растений, а в наиболее влажный' 1972 год они поедали в основном семена. Питание подзем-
108
Таблица 17. Встречаемость и объемные соотношения групп кормов кеклика на юго-востоке Казахстана (анализы содержимого зоба), %
Вид корма 1972 Г. 1973 Г. 1974 г. 1975 г. 1976 г. 1977 г.
1 1 2 1 1 2 1 1 2 1 1 2 1 1 2 1 1 2
1 2 1 3 1 4 | 5 1 f 6 1 7 1 1 8 1 9 | 10 1 11 1 12 | 13
Каратау
(22) (5) (18) (17) (5) (0)
Подземные части растений 77,2 56,2 80,0 50,0 77,0 54,3 94,1 75,8 100,0 63,8 —. —.
Семена 40,9 21,9 20,0 2,0 74,2 22,5 47,2 н,о 100,0 34,0 —. —
Зеленые корма 36,2 8,7 80,0 38,0 66,6 15,5 47,2 10,3 —. — —. —1
Ягоды 9,0 4,0 20,0 10,0 22,0 2,2 23,6 0,5 — —. —. —.
Животные корма Дополнительные включения 4,5 1,4 — — 16,5 4,1 — — 20,0 2,0 — —
глина 4,5 3,7 —* —. У « —. —- —-- —. —. —— —.
гастролиты 1 22,7 (14 4,1 ) 1 К и (П р Г И 3 ( >) 22,0 2 К И Й (3^ 1,4 А л а т 11,8 а у (Н 2,4 9 (К 0 (4 )
Подземные части растений 35,5 17,8 83,2 42,7 90,0 63,0 100,0 91,3 94,8 41,6 75,0 61,2
Семена 42,9 68,4 63,3 42,4 72,0 21,8 66,0 5,8 94,8 45,6 75,0 8,0
Зеленые корма 35,5 2,4 38,0 12,7 54,0 11,6 11,1 1,3 40,8 7,6 100,0 30,8
Ягоды 28,4 П,4 12.7 0,8 12,0 2,0 —. 20,8 1,8 —. —.
Мох —. — —. —1 —- —. —. —1 5,2 1,0 .
Животные корма Дополнительные включения —• — 19,0 1,4 9,0 0,6 11,1 0,5 15,6 1,4 — —
гастролиты —• —• — — 9,0 1,0 —- —- 20,8 1,0 — • —
Окончание таблицы 17
Чу-Илийские горы
(26) (10) (6) (12) (4) (0)
Подземные части растений 34,6 15,0 60,0 26,0 100,0 66,7 91,7 56,5 75,0 47,5 —*
Семена 94,6 63,4 80,0 67,0 33,2 8,3 66,4 37,0 75,0 25,7
Зеленые корма 61,6 20,3 н,о л° 33,2 8,3 58,7 5,8 25,0 1,1 —•—-•
Ягоды —• — 16,6 16,7 8,3 0,1 50,0 25,7
Животные корма Дополнительные включения 11,9 1,3 — — — 8,3 0,4 — —• —• —
гастролиты — — — —• 16,2 О.,2 —• — —• —
Джунгарский Алатау
(30) (30) (40) (15) (0) (11)
Подземные части растений 43,3 21,0 46,6 20,0 85,0 63,1 79,4 54,2 91,0 50,4
Семена 76,6 57,0 79,9 54,5 35,0 31,6 79,4 34,1 78,2 47,7
Зеленые корма 39,9 12,6 49,9 24,0 30,0 4,8 72,8 Ю,9 —» — 9,1 0,5
Животные корма 26,6 9,4 6,7 0,5 5,0 0,2 —. —— — 9,1 0,9
Ягоды —-W — — —v 2.5 0,2 —. —.
Дополнительные включения гастролиты — — 6,7 1>о 2,5 0,1 13„2 0,8 — — 9,1 0,5
Примечание. 1— встречаемость кормов, 2 — объем кормов. В скобках указано количество исследованных особей,
ними частями растений во влажные годы и сокращение семян в кормах в засушливые отмечено и в двух других районах— в Чу-Илийских горах и Малай-Сары (табл. 17). Однако наибольшее количество подземных частей и минимум семян зарегистрировано в нище кеклика не в самый засушливый 1975 год, а в наиболее влажный 1974 год, что было вызвано следующими причинами. Сумма летних осадков в 1975 г. составила 58,0 мм, но в марте их выпало 33,1 мм. Мартовское количество осадков оказалось достаточным для урожая семян эфемеров и очень важного осеннего корма — семян лука (Allium trachyscordiim). Сумма летних осадков 1974 г. оказалась несколько выше, распределялись они по месяцам бо-> лее равномерно, что в результате обусловило низкий урожай семян.
Следует отметить, что семена имеют существенное значение в питании кеклика осенью и, по данным М. А. Кузьминой (1955), составляют четвертую часть зимнего рациона. В случае их низкого урожая в осеннее время кеклики переходят на питание зеленью и подземными частями растений. Однако засушливое лето с плохим урожаем семян может стать причиной массовой гибели кекликов в суровые многоснежные зимы с низкими температурами. Таким образом, летние засухи могут оказать серьезное влияние на зимовки.
Обширный ареал кеклика и широкий диапазон вертикального распространения (от пустынь до альпийских лугов) существенно сказываются на разнообразии кормов. В пределах СССР у взрослых особей средняя встречаемость за год семян, подземных частей растений и зелени примерно одинакова и более, чем в 2 раза превышает этот показатель по ягодам и животным кормам. Вместе с тем в зависимости от сезона года наблюдаются глубокие отличия в поедании различной пищи. Встречаемость семян и подземных частей в течение года изменяется в 4—5 раз, достигая максимума в июле — августе и минимума — в апреле. Зеленые корма, наоборот, чаще отмечены в марте — апреле и гораздо реже — в июне — июле. Встречаемость животных кормов изменяется в 12 раз (максимум в июне — августе); ягоды, при отсутствии их в январе, в августе отмечены более чем у 40% особей (табл. 18).
Высокая пластичность в использовании кормов позволяет кеклику жить как в пустынных районах, так и высоко в го-
111
pax, достигая альпийского пояса. Несомненно, что благодаря этой особенности стала возможной его акклиматизация в различных частях земного шара.
Таблица 18. Встречаемость разных групп кормов кеклика (п=1378) по месяцам (материалы из различных районов юго-востока Казахстана), %
Месяц Число исследованных птиц Растительный корм Животный корм
Семена Подземные части Зелень Ягоды и кбстйнки
Январь 9 66,6 33,3 66,6 11,1
•Февраль 42 59,5 59,5 54,2 11,9 9,5
Март 69 34,8 36,2 75,4 7,2 4,3
Апрель 66 15,2 18,2 84,8 1,5 22,7
Май 50 66,0 66,0 54,0 8,0 50,0
Июнь 63 74,6 58,4 27,0 36,5 36,5
Июль 14 71,3 78,5 28,5 28,8 44,4
Август 35 74,4 72,3 40,4 40,4 21,3
Сентябрь 331 72,2 73,7 32,6 32,9 21,5
Октябрь 332 68,6 59,3 45,0 28,9 16,8
Нбябрь 195 53,8 49,7 55,9 14,4 11,8
Декабрь 172 52,3 33,7 55,8 29,7 10,5
Средняя встречаемость корма за год 59,1 53,3 51,5 21,8 21,7
Питание птенцов кеклика на территории нашей страны изучено очень слабо. По существу, материалы, освещающие этот вопрос, исчерпываются данными М. М. Остапенко (1958), которая считает, что молодняк кекликов в ран-
нем возрасте питается исключительно животной пищей.
Летом 1974—1978 гг. в горах Чулак мы изучали содержимое зобов и желудков 36 птенцов в возрасте от 2 дней до 1 месяца (у 16 особей возраст определен с точностью до 1 дня). Выяснилось, что корма птенцов в первый месяц жизни весьма разнообразны. Из насекомых поедаются саранчовые (Acrididae), щитники (Pentatomidae), певчие цикады (Ci-cadidae), цикадки (Jassidae), коровки (Coccinellidae), златки (Buprestidae), долгоносики (Curciilionidae), чернотелки (Tenebrionidae), пластинчатоусые (Scarabaeidae), мухи (Mus-cidae), муравьи (Formicidae), моллюски (Eulotidae, Enidae).
112
Среди растительных кормов отмечены семена гречихи (Polygonum sp.), рисовидки джунгарской (Piptatherum songori-cum), донника (Melilotus albus), шренкии (Schrenkia sp.), a также ягоды эфедры (Ephedra equisetina) и вишни (Cerasus tianschanica), луковицы иксиолириона и частички зеленых листьев.
В первую неделю жизни у одного птенца (возраст 2 дня) в зобу обнаружены личинки саранчовых (Acrididae), мелкие частицы двукрылых и муравей (Messor clyvorum). Зоб второго птенца (возраст 2 дня) оказался пустым, а в желудке находились мелкие остатки семян растений, определить которые по удалось, и гастролиты. У 5-дневного птенца в зобу были только семена (вид растения не определен), а в желудке— перетертые остатки их. Во вторую неделю жизни пища птенцов становится разнообразнее (просмотрены зобы и желудки 8-дневного и 11-дневного птенцов). У первого кроме личинок саранчовых и двукрылых, отмеченных ранее, в зобу обнаружены цикадка, коровка, златка, долгоносик и чернотелка; в желудке — остатки животных и растительных кормов. У второго птенца встречены мелкие луковицы иксиолириона, клопы (Euridema fieberi) и семена не определенных растений. В возрасте 14—21 дня (исследованы корма у 5 птенцов) начинают поедать ягоды эфедры и дикой вишни, а из животных — крупных жуков (жужелиц). В зобах обнаружены также семена гречихи. К концу первого месяца жизни состав кормов продолжает количественно увеличиваться. Впервые в пище птенцов появляется зелень (частички листьев) и различные моллюски. Разнообразится список семян: донник (Melilotus albus), липучка (Lappula patula). Как и раньше, отмечены клопы, прямокрылые, жуки, ягоды эфедры и луковицы иксиолириона; эти корма употребляются в большом количестве. Для переваривания столь разнообразной пищи молодняк поедает гастролиты, средняя масса которых в одном желудке составляет 0,5 г.
В 1,5—2-месячном возрасте соотношение животных и растительных кормов в рационе кеклика постепенно выравнивается, а в 3-месячном — мало чем отличается от пищи взрослых особей.
8—24
113
Динамика численности
Изучение динамики численности животных — одна из важнейших, интенсивно разрабатываемых проблем экологии. Для каждого вида существует определенное соотношение между рождаемостью и смертностью (Лэк, 1957), что определяется его приспособленностью к конкретной обстановке. Одной из основных причин колебания численности является естественная убыль в популяции, которая неодинакова в разные годы. При благоприятных условиях больше сохраняется молодняка и значительная часть его вовлекается в воспроизводство. С ухудшением условий обитания повышается смертность особей, что в конечном итоге сказывается на численности популяции в целом.
На колебание численности животных влияет совокупность различных факторов, из которых наиболее важны следующие:
1. Биологические особенности вида (плодовитость, время достижения половой зрелости, поло-возрастной состав популяции, естественная смертность и экологическая пластичность, обусловливающая степень жизнестойкости вида).
2. Погодные особенности, создающие конкретные жизненные условия.
3. Враги, болезни, паразиты.
4. Антропогенные факторы.
Оценка влияния этих факторов на динамику численности требует многолетних детальных изучений на небольших территориях и, по-существу, приемлема лишь для заповедников. Изучение динамики численности на больших территориях^
114
как правило, ограничивается проведением многолетних количественных учетов, изменения в которых сопоставляются с погодными условиями, численностью врагов и влиянием деятельности человека.
Биологические особенности вида. Половой зрелости кеклики достигают в возрасте 10—10,5 месяца. Основное поголовье молодняка, вылупившегося в начале — середине июня, в апреле следующего года вместе с особями старшего возраста приступает к размножению. Данные кольцевания показали, что среди меченых сеголеток, встреченных весной следующего года, случаев прохолостания не наблюдалось. Таким образом, можно полагать, что все перезимовавшие кеклики вовлекаются в воспроизводство.
Как показали вскрытия кекликов различного возраста, соотношение полов в природе у этого вида близко 1 : 1, а отношение взрослых к молодым зависит от сезона года. По нашим материалам, полученным в весенние периоды 1974— 1978 гг. в горах Чулак, на каждую наседку, успешно закончившую насиживание, в среднем приходилось от 11,0 до 14,7 птенца. Однако такое отношение взрослых к молодым довольно далеко от истины, поскольку, как показали наблюдения, эффективность гнездования у кеклика составляет 37,8—62,0%, т. е. в среднем половина наседок теряет кладки.
По нашим расчетам, на каждые 100 взрослых птиц (при соотношении полов 1:1) должно быть 66 наседок — 50 самок и 16 самцов. Учитывая, что птенцы вылупляются из половины кладок, а на каждую наседку приходится в среднем 12,4 птенца (в 1974 г.— 10,5, в 1975 г. —11,0, в 1976 г.— 14,7, в 1977 г.— 13,8, в 1978 г.— 11,8), можно считать, что 100 взрослых птиц дают 400 молодых. Следовательно, отношение взрослых к молодым после периода размножения должно быть примерно 1:4 или несколько выше, поскольку мы не располагаем данными об эффективности размножения при повторном гнездовании. Приводимые расчеты в какой-то мере подтверждаются и статистикой добываемых кекликов в осеннее время. Так, по данным осенних отстрелов в Заилийском Алатау (ущелье Актерек) 13—14 ноября 1976 г. соотношение взрослых и сеголеток среди 51 добытой особи было 1:3,25, а в начале ноября 1978 г. (ущелье Жаманты) из 145 экземпляров 42 птицы оказались взрослыми (соотношение 1 :3,45). Интересно, что при повторном отстреле, спустя месяц, в этом
115
же месте из 96 добытых особей 39 оказались взрослыми (соотношение 1 : 1,5).
Таким образом, можно констатировать, что в местах с высоким прессом охоты в течение сезона в основном отстреливается молодняк, но отношение молодых кекликов к взрослым в добыче охотников не превышает истинного соотношения взрослых и молодых птиц в природе, что может привести даже к некоторому «омоложению» популяции.
Ю. А. Лебедев (1976) для арчовых лесов северного склона Туркестанского хребта приводит данные о гибели кеклика в послегнездовой период, которая составила 4,2—39,3% от общей численности в период с конца гнездования до начала зимы.
Интересные сведения приводит A. Dayani (1978), изучавший популяцию кеклика в Иране близ Караджа. По его данным, зимой 1973 г. количество молодых на 100 взрослых составило 67, а зимой 1974 г.— ИЗ. Такое соотношение взрослых и молодых птиц объясняется дождливым летом и снежной зимой 1973/74 г.
Погодные условия. О влиянии погодных условий на динамику численности кеклика сведений очень мало. Лишь В. Н. Шнитников (1949) указывает, что после катастрофической гибели, вызванной суровой зимой, поголовье кеклика восстанавливается лишь спустя 2 года. В качестве примера автор приводит зиму 1918/19 г., когда держались необыкновенно сильные и продолжительные морозы, а снег совершенно не таял даже на солнечных склонах. В результате кеклики были лишены возможности добывать корм и начали гибнуть в массе. По данным этого исследователя, подобное повторилось в более суровую зиму 1929/30 г. в Джунгарском и За-илийском Алатау. Однако уже осенью 1932 г. поголовье кекликов достигло такого уровня, что, по образному выражению автора, «все горы кишат ими» (Шнитников, 1949).
На катастрофические последствия суровой зимы 1927/28 г. в Таласском Алатау указывает С. А. Северцов (1929). Эта зима привела к тому, что в августе 1928 г. выводки кекликов почти не попадались. Гибель их в этом же хребте описывает В. В. Шевченко (1949). Автор сообщает, что в заповеднике Аксу-Джабаглы в конце декабря 1944 г. после бурана, продолжавшегося несколько дней, и низкой температуры (до —31°) отмечался массовый падеж птиц; их находили
1161
мертвыми даже в с. Новониколаевке. К февралю кеклики настолько ослабели, что их можно было ловить руками. Пойманный в марте самец весил всего 260 г, что вдвое меньше средней массы птиц в осенне-зимний период. Летом 1945 г. в верховьях р. Джабаглы на маршруте в 10—12 км встречено только 3 выводка.
В зимы 1943/44 и 1944745 гг. массовый падеж кекликов отмечался по всему югу и востоку Казахстана и в Северной Киргизии. Много птиц погибло в Западном Тянь-Шане, в долинах рек Угам, Чаткал, Аксу, а также на Машате и Каратау. В Западном Тянь-Шане, включая хребет Каратау, гибель их продолжалась в зимы 1945/46, 1946/47, 1948/49 и 1949/50 гг. (Слудский, 1965).
Зимой 1950/51 г. в Казахстане разразился джут, наряду с дикими животными в массе погибал домашний скот, в основном овцы. О том, насколько сократилась численность кеклика в эту зиму, можно судить по наблюдениям орнитологов, работавших тогда в Чу-Илийских горах (Анархай). Так, за год до джута, в феврале 1950 г. за 8-часовую экскурсию здесь насчитывали до 450 птиц. В феврале 1951 г. за такой же промежуток времени — только около 100; весной 1951 г. М. Н. Корелов в этих местах за 4 дня отметил всего 2 пары кекликов, а А. А. Слудский и Б. К- Штегман за несколько длительных экскурсий не встретили ни одной птицы (Кузьмина, 1955).
Следует отметить, что иногда массовая гибель кеклика наблюдается лишь в отдельных хребтах, в других случаях она захватывает громадные территории. Например, резкое сокращение этих птиц зимой 1968/69 г. отмечалось не только в Казахстане, но и в республиках Средней Азии. По данным А. К. Ковалева и А. В. Попова (1980), в эту зиму в Таджикистане зарегистрирована бедственная зимняя миграция кеклика, в результате которой все зимующие стаи ушли е площади 7 тыс. км2. В пути кеклики гибли от истощения, хищников и браконьеров.
В Киргизском Алатау (ущелье Мерке), по сообщению охотинспектора В. Волкова, весной 1969 г. на чердаке пустующего дома обнаружены трупы кекликов. Видимо, в поисках пищи птицы попали туда через слуховое окно. Зимой 1968/69 г. в ущелье Мамай (Киргизский Алатау) сторож колхозного стада поймал 8 ослабевших кекликов. В течение
117
йсей зимы их держали в сарае вместе с курами. С наступлением первых теплых дней их стали выпускать на волю, но на ночь стайка самостоятельно возвращалась в сарай. В начале марта кеклики все реже и реже появлялись около дома, а в конце месяца ушли.
До сих пор еще не определена даже примерная смертность кекликов в годы массовой гибели. А. А. Слудский (1965) ориентировочно отмечает сокращение численности в 2—10 раз, но тут же указывает на возможность полного исчезновения в некоторых районах.
Большой интерес при изучении динамики численности кеклика представляет способность вида восстанавливать численность после катастрофической гибели. В литературных источниках приводятся указания на то, что через 2—3 года после массовой гибели поголовье кекликов вновь восстанавливается. Имеющиеся в нашем распоряжении материалы подтверждают эти сведения. Однако следует учитывать, что гибель птиц и следующее за ней восстановление имеют в разных частях ареала свои особенности: многое зависит от числа сохранившихся особей, а также от погодных условий последующих лет.-Особого внимания в этом отношении заслужи-вает Западный Тянь-Шань, где гибель кекликов регистрировалась на протяжении 6 лет (зимы 1943—1950 гг., за исключением зимы 1947/48 г.). При таком положении восстановление поголовья через 2—3 года невозможно. Вместе с тем указания на быстрое восстановление поголовья подтверждаются нашими наблюдениями. Так, учеты кеклика, проведенные осенью 1972 г. в .различных горных районах юго-востока Казахстана спустя 3 года после массовой гибели, показали, что в большинстве районов численность этих птиц достигла -высокого уровня.
Враги. Список врагов кеклика довольно обширен и включает представителей 3 классов позвоночных: млекопитающих, птиц и рептилий. Среди пернатых хищников опаснейшими врагами кеклика, безусловно, следует считать ястреба-тетеревятника и беркута.
Способ охоты у ястреба довольно прост и заключается в подкарауливании добычи на местах водопоев и коряжек. В сентябре и октябре, когда кеклики активно посещают водопои, тетеревятники обычно караулят их, скрываясь в ветвях деревьев, расположенных по берегам речек и родников.
Г18
Нападают неожиданно, и поэтому охота почти всегда успешна. В случае неудачной попытки ястреб преследует взлетевшую птицу и, как правило, быстро настигает ее. Предварительно ощипав жертву, хищник поедает ее неподалеку от места поимки. У добытой взрослой самки ястреба в Чу-Илийских горах И ноября 1975 г. в зобу обнаружено только мясо кеклика (177 г), в желудке — его лапка и трубчатые кости.
По В. Аракеляпцу (устн. сообщ.), в Чу-Илийских горах осенью 1965 г. он наблюдал неудачную попытку ястреба поймать одного из кекликов в стае, которая после броска хищника затаилась. Кеклики совершенно не реагировали на выстрелы, продолжая прятаться в основании кустов. Взлетели они лишь тогда, когда человек подошел к ним вплотную.
Об охоте ястреба-тетеревятника сообщает Э. М. Ауэзов, который в этих же горах в сентябре 1976 г. наблюдал, как хищник загнал в кусты 2 кекликов. Одного из них ястреб растерзал (оказались выеденными внутренности), а второй -сидел, затаившись, рядом; не обнаружил он себя и после того, как ястреба вспугнули, и был пойман руками.
Панический страх кекликов перед тетеревятником человек издавна использовал при охоте на этих птиц. В. Н. Шнитни-ков (1949) дает описание такой охоты: «...найдя табунок кекликов, охотники открывали тетеревятника, поднимали его высоко кверху на руке, и этого было достаточно, чтобы немедленно заставить всех кекликов залечь там, где каждый из них в то время находился. После этого один из охотников начинал их стрелять одного за другим и выбивал весь выводок» (с. 206). Кроме описанного способа известна и охота с чучелом ястреба.
Учитывая, что основу питания ястреба-тетеревятника на юго-востоке Казахстана в осенне-зимний период составляют кеклики, этот хищник в иные годы может существенно влиять на сокращение их численности.
Другой опасный враг кекликов — беркут. В некоторых торных районах юго-востока Казахстана, где этот хищник зимует и численность его бывает довольно высокой, по орлам, кружащимся над склонами, без ошибки можно найти богатые кекликами урочища. В таких местах кеклик является ^основным объектом питания беркута на протяжении всей зи
119
мы. По данным А. К. Рустамова (1945), беркут способен брать кеклика и на лету.
Среди других представителей пернатых врагов следует упомянуть ястреба-перепелятника и пустельгу, которые ловят нелетных птенцов, и ворона, специализирующегося на поиске гнезд и разоряющего кладки. Нам приходилось наблюдать, как вороны «прочесывали» склоны, летая в одиночку в 2— 5 м над ними. Пролетев в одном направлении, ворон возвращался обратно немного ближе к вершине или основанию; обнаружив гнездо, на котором нет наседки, брал в клюв одно из яиц и улетал. Такие кладки обречены на гибель. Если же ворон находит гнездо с насиживающей птицей, он стремится согнать наседку, но, по-видимому, всегда безуспешно. За годы нашей работы в горах Чулак отмечено 2 случая разорения гнезд вороном и 4 раза наблюдали летящих птиц с яйцами кеклика в клюве.
Из млекопитающих урон гнездам наносят лисица и каменная куница (рис. 23); кроме того, эти хищники уничто-
Рис. 23. Яйцо кеклика, выеденное каменной куницей
120
жают птенцов и взрослых птиц. Вред, наносимый ими, может рыть довольно значительным. Так, в 1974—1978 гг. из 18 погибших от хищников гнезд 4 разорили каменные куницьг и 6 — лисицы. Вполне вероятно, что указанные цифры занижены, поскольку у части гнезд невозможно было определить, каким именно хищником оно разорено.
Потенциальными врагами кеклика следует считать волка и барсука. Однако их численность в горах Чулак невелика.
Причиной гибели кладок и нелетных птенцов могут быть и змеи (щитомордник, степной удавчик, разноцветный и узорчатый полозы). В июне 1976 г. мы наблюдали, как небольшой щитомордник заполз в гнездо кеклика и безуспешно пытался заглотить яйца. Только небольшие размеры змеш не позволили ей разорить кладку (табл. 19).
Таблица 19. Причины гибели гнезд кеклика
Год Число гнезд Кем разорено гнездо Брошено Всегб погибло гнезд, %
Каменная куница Лисица Ворон Человек Неизве-стн. ' Хищник
1974 8 1 3 1 62,5
1975 4 —. —. 1 —. 1 50,0
1976 13 —. 4 —. 1 1 1 53,8
1977 18 2 1 — —. 1 3 38,9
1978 33 1 1 1 — 1 9 39,4
Всего 76 4 6 2 1 6 15 44,7
% 100,0 5,3 7,9 2,6 1,3 7,9 19,7
В целом степень влияния различных врагов на численность кеклика во многом зависит от их поголовья. Особенно* ощутимо это воздействие в годы джута, когда обессилившие птицы становятся легкой добычей хищников. Так, по сообщению Ю. С. Лобачева, в марте 1969 г. в ущелье Чемолган (Заилийский Алатау) после суровой многоснежной зимы он в течение месяца лишь несколько раз слышал голоса кекликов, а на склонах во множестве находил их перья и следы лисиц.
Таким образом, по сравнению с обычными зимами воздействие хищников, в частности лисиц, в годы джута заметно возрастает и наряду с плохими условиями зимовки является
121
одним из факторов, оказывающих влияние на низкую численность кеклика весной.
Паразиты. Болезни. В Казахстане у кеклика об^ наружено 5 видов трематод, 7 цестод, 22 нематод (Гвоздев, 1958). Зараженность гельминтами достигает 100%. Однако, несмотря на высокую зараженность отдельными видами, существенных патологических явлений, приводящих к значительной смертности птиЦ, не отмечено. Лишь нематода Tet-rameres sp., паразитирующая под серозной оболочкой железистого желудка, при интенсивной инвазии вызывает истощение кекликов, а трематода Lyperosomum corrigia, локализующаяся в поджелудочной железе, — тяжелое заболевание, иногда кончающееся смертью.
Среди эктопаразитов большую роль играют клещи Наеша-phisalis punctata, Jxodes persulcatus, Argas reflecus. Личинки и нимфы иксодовых клещей в массе воспитываются на кекли-ке в течение всего лета. В результате интенсивного заражения перья на голове и частично шее выпадают и под кожей образуются крупные твердые наросты, заметно изменяющие форму головы из-за механического раздражения. Особого внимания заслуживают аргасовые клещи как переносчики тяжелых заболеваний, в частности спирохетоза. При этом заболевании отмечается высокая температура тела, лихорадочное состояние, воспаление глаз. Нарушается подвижность (птицы сидят нахохлившись или лежат), появляется сонливость, понос и отсутствие аппетита. По данным лаборатории протозоологии Института зоологии АН КазССР, гибель от спирохетоза в экстремальных условиях среди кекликов достигает 70—80% (в основном погибает молодняк). В некоторых случаях наблюдается гибель кекликов от смешанных инвазий. Так, в конце марта 1979 г. из 40 кекликов, доставленных из гор Чулак, зараженность составила 52,5, а в июле—100%. Начиная с апреля отмечался падеж птиц, который с апреля по июнь составил 35, в июле — августе — 32,5, в сентябре — октябре— 15%. Всего пало 33 особи (82,5%).
В естественных условиях падеж кекликов от различных болезней — явление, видимо, довольно редкое. Только однажды нам удалось столкнуться со случаем летального исхода взрослой особи. В июне 1978 г. в горах Чулак встречен выводок 2-дневных птенцов со взрослой птицей, которая, как выяснилось после поимки ее руками, оказалась слепой. По
122
мещенная вместе с выводком в вольеру, она на следующее утро оказалась мертвой. Внешний осмотр и вскрытие трупа показали, что никаких внешних и внутренних повреждений у этой особи не было. О нахождении мертвого кеклика рядом с выводком живых 3—4-дневных птенцов в горах Чулак сообщает М. А. Кузьмина (1955). Как и в предыдущем случае, у мертвой птицы никаких патологических изменений не отмечено.
Таким образом, смертность кекликов от различных болезней в природе, несомненно, имеет место и в некоторой степени может влиять на их численность. Однако масштабы гибели от заболеваний не могут идти в сравнение с таковой от таких факторов, как суровые многоснежные зимы, вызывающие массовый падеж птиц.
Антропогенные факторы. Человек своей многогранной деятельностью оказывает на численность кеклика как прямое (охота), так и косвенное воздействие (выпас скота, сбор лекарственных трав, постройка поселков, дорог и т. д.).
Особенно интенсивная в последние годы охота оказывает в некоторых районах существенное влияние на снижение численности кеклика. Большой вред продолжает наносить браконьерство. Пользуясь тем, что многие угодья лишены элементарной охраны, браконьеры отстреливают кекликов еще задолго до начала открытия охотничьего сезона. Такие явления широко распространены.
В хребтах Каратау, Киргизском и Заилийском Алатау, где практикуется летний выпас скота, гнезда и молодняк кекликов часто попадают под копыта скота, в основном овец.
Заметно повлияло на численность кекликов и освоение территорий. Наглядным примером этого могут служить ближайшие окрестности Алма-Аты. Еще в 30—40-х годах кеклик был обычен в Малом Алматинском ущелье. Городские охотники добывали его неподалеку от границ города. Освоение горных районов под садоводство и расширяющееся жилищное строительство привели к тому, что в настоящее время кеклик сохранился здесь лишь в небольшом количестве.
Заметное уменьшение численности кеклика за последние 30 лет прослеживается и в Чу-Илийских горах. Так, в 1950 г. в этом районе населения практически не было, лишь в зимнее время угодья посещали чабаны, выпасающие отары овец.
123
По данным М. А. Кузьминой (1955); в феврале 1950 г. на маршруте в 8 км здесь учтено 450 кекликов— 56 особей на 1 км маршрута. В 1972—1‘976 гг., по данным осенних учетов,, максимальное поголовье этих птиц отмечено в 1974 г. — лишь 18,4 особи на 1 км маршрута. Именно к этому времени Чу-Илийские fopbi связались сетью асфальтированных дорог с населенными пунктами, что увеличило приток охотников в угодья. Кроме того, значительно возросло поголовье выпасаемого скота, увеличилось количество кошар и колодцев; горы часто посещают различные экспедиции. Словом, интенсивное освоение этого района привело к снижению поголовья кеклика.
Наряду с отрицательными антропогенными факторами, в той или иной степени воздействующими на численность кеклика, следует упомянуть и о положительном влиянии деятельности человека. Например, в горах Малай-Сары на пологих холмах ежегодно проводится посев ячменя. В зимнее время, когда кеклики особенно остро ощущают нехватку кормов, они выходят к оставшимся остожьям соломы подбирать зерно, что помогает птицам перенести наиболее тяжелый период в их жизни. Создание заповедников, заказников и воспроизводственных охотничьих хозяйств обеспечивает условия для воспроизводства кекликов, а акклиматизация этих птиц способствует расширению их ареала.
Результаты учетов. К настоящему времени разработано несколько методик учета кекликов. Впервые попытку проведения таких работ предпринял М. К. Лаптев (1936), который применил следующую формулу для определения плотности кекликов:
*=1007^7’
где о — количество особей на 1 км2, а — число зарегистрированных при обследовании птиц, V — скорость движения учетчика, t — продолжительность учета, г—^ширина обзора» (на каком расстоянии особь отмечалась), k — активность в момент подсчета (в какой степени особи данного вида обнаруживают себя). В результате работ в конце августа — начале сентября 1934 г. в предгорьях Копет-Дага на маршруте в 320 км
124
’было подсчитано 3650 кекликов, 11—21 мая того же года учтена 171 особь (128 км). На основании этих работ общее количество кекликов в Центральном Копет-Даге было определено в 121 000 птиц, или 8 100 выводков.
В последние 10 лет учеты кекликов проводились в Таджикистане (Ковалев, Попов, 1980), Узбекистане (Кашкаров, Павленко, 1975; Нам, 1975). Следует отметить, что наиболее эффективны эти работы в охотничьих хозяйствах, так как дают возможность определить численность птиц перед сезоном охоты и разработать план добычи с учетом дальнейшего воспроизводства.
Работы по учету кеклика на больших территориях нами впервые проводились в 1972—1976 гг.; цель их — проследить за колебаниями численности птиц в различных районах юго-востока Казахстана. Общую численность кеклика в обследованных хребтах мы не определяли, поскольку учетная площадь была очень мала.
Многолетнее изучение динамики численности кекликов начали в 1972 г., спустя 3 года после их катастрофической гибели в зиму 1968/69 г. К этому времени во многих горных районах юго-востока Казахстана поголовье этих птиц восстановилось, а местами, как, например, в Киргизском Алатау, стало высоким.
Учеты кеклика (конец августа — середина октября) проводились в горных районах, существенно отличающихся по физико-географическим условиям и отстоящих друг от друга на значительном расстоянии (рис. 24).
Так, наиболее удаленные точки учетов в хребтах Каратау и Алтын-Эмель находились на расстоянии по прямой около 1250 км, а расстояние между крайней южной точкой учета (Киргизский Алатау, ущелье Мерке и Чу-Илийскими горами) составило 450 км. В Каратау учеты проводились в ущельях Актобе (50 км к северо-задапу от г. Туркестана), Кумусты, Алтынтау и Ран (северо-восточные склоны). В 1972—1976 гг. здесь пройдено 49 маршрутов общей протяженностью 402,5 км (табл.20).
Обследование хребта Каратау показало, что поголовье кеклика после резкого сокращения в зиму 1968/69 г. в целом характеризовалось ежегодным приростом. Исключение составил лишь 1973 год, который оказался самым «неурожайным» за все годы учетных работ в этом районе. По данным
125
Рис. 24. Размещение пунктов учета кекликов на юго-востоке Казахстана в 1972—1976 гг.
Таблица 20. Численность кекликов на юго-востоке Казахстана
Год Маршруты Учтено пТиц Численность, % к предыдущему году
Число Длина, км Всего На 1 км маршрута
Каратау
1972 10 68,5 305 4,5
1973 10 79,5 221 2,8 62,2
1974 12 101,0 580 5,7 203,6
1975 9 88,0 595 6,8 119,3
1976 8 65,5 641 9,8 144,1
Киргизский Алатау
1972 8 55,5 844 15,2
1973 13 70,5 1862 26,4 173,7
1974 13 73,5 1541 21,0 79,5
1975 13 77,0 1416 18,4 87; 6
1976 13 77,0 1377 17,9 97,3
Чу-Илийские горы
1972 5 44,5 598 13,4
1973 7 48,0 768 16,0 119,4
1974 4 32,5 599 18,4 115,0
1975 3 27,0 354 13,1 71,2
1976 5 39,0 249 6,4 48,9^
Горы Чулак
1972 3 21,0 312 14,9
1973 6 30,6 1220 39,9 267,8
1974 7 41,5 1269 30,6 76,7
1975 7 44,0 568 12,9 42,2
1976 6 44,0 568 12,9 100,0
Хр е б е т А лтын-Эмель
1972 3 21,0 0 0
1973 5 22,8 227 10,0
1974 5 22>,5 225 10,0 100,0
1975 5 25,0 178 7,1 71,0
1976 4 25,0 393 15,7 221,1
Г оры Малай-Сары
1972 4 36,0 274 7,6
1973 5 42,0 779 18,6 244,7
1974 1 16,0 331 20,7 111,3
1975 2 30,0 211 7,0 33,3
1976 2 30,0 225 7,5 107,1
"метеостанции «Ачисай», зима 1972/73 г. в Каратау отличалась низкими температурами и многоснежьем. Начиная с декабря глубина снежного покрова постепенно увеличивалась и достигла максимума в третьей декаде января (112 см). Одновременно с накоплением снега отмечалось понижение температуры, минимум которой зарегистрирован в середине января (—22,3°). В феврале среднемесячная температура составила 0,9°, минимальная—14,8°. Снег начал уплотняться и достиг высоты 81 см. Еще более высокие температуры отмечены в марте (среднемесячная 2,2°, минимальна!. —7,9°). Высота снежного покрова в это время держалась на отметке 39 см, но уже к концу месяца была только 2 см. Лето 1973 г. отличалось малым количеством осадков и высокими температурами (со второй декады июня до конца августа выпало только 12,1 мм осадков), что сказалось на урожайности семян диких растений. Таким образом, неурожайность семян в летний период и неблагоприятные условия зимовки явились, no-видимому, основной причиной снижения численности кеклика в этот год.
Следует отметить, что в последующем восстановление поголовья в различных ущельях хребта Каратау происходило неравномерно. Так, в 1974 г. в ущелье Актобе численность птиц увеличилась на 340%, в ущелье Ран—на 104,3%, в ущелье Кумусты — на 122,7% и лишь в ущелье Алтынтау она сохранилась на уровне 1973 г. (96,8%). В 1975 г. в ущельях Кумусты и Ран отмечено снижение поголовья кеклика соответственно до 63,3 и 61,7%, а в ущельях Актобе и Алтынтау шло дальнейшее увеличение численности (соответственно на 131,8 и 70,0%). В следующем 1976 г. в ущелье Алтынтау этот показатель увеличился на 47,1%, а в ущелье Кумусты — на 35,5%; в ущелье Ран остался прежним. Возможно, изменение численности связано с перемещениями кекликов из одних ущелий в другие в поисках более кормных мест.
В большинстве обследованных ущелий хребта Каратау охота на кеклика в связи с удаленностью угодий от населенных пунктов практически не ведется и, таким образом, изменение численности в основном зависит от метеорологических условий.
Киргизский Алатау. Учеты проводились в нижних частях ущелий Сюгаты (50 км к юго-востоку от г. Джамбула), Ка-ракыстак (40 км к югу от ст. Луговая) и Мерке (20 км от .128
одноименного населенного пункта). Общая протяженность (И маршрутов составила 353,5 км (см. табл. 20).
К осени 1972 г., когда стали проводить учеты, численность кеклика в обследованных ущельях была самой высокой, несмотря на то, что в зиму1968/69 г. отмечалось значительное сокращение его поголовья, вызванное суровой зимой. По данным метеостанции «Мерке» в зиму 1968/69 г. морозы начались в середине декабря и держались до конца февреля. В янь эе минимальная температура доходила до —34,8°, в феврале — до —33,8°. Декабрь и большая часть января были малоснежными, и только в последней декаде января снежный покров достиг 33 см. Примерно на этом уровне он удерживался до середины марта. Такие условия зимы резко сократили численность кеклика. По словам местных охотников, к весне 1969 г. кеклик в ущельях Киргизского Алатау стал большой редкостью. Осенью 1972 г. он здесь стал обычным, а в 1973 г. — многочисленным.
В последующие годы, в связи с летней, засухой, поголовье кеклика несколько сократилось, но продолжало оставаться довольно высоким.
Такое состояние численности кеклика в Киргизском Алатау во многом объясняется малым воздействием антропогенных факторов. Сложный рельеф ущелий только местами пригоден для выпаса овец, а значительные площади, занимаемые осыпями и кустарником, создают благоприятные условия для жизни и воспроизводства кеклика.
Чу-Илийские горы. Учет численности проводили в ущельях Алмалы и Сарыбулак (горная группа Анархай) —местах, типичных для этого района. Здесь общая протяженность 24 маршрутов составила 191 км (см. табл. 20).
После многоснежной зимы 1968/69 г., когда произошла массовая гибель кеклика и он стал чрезвычайно малочисленным, следующие зимы были гораздо мягче. Если зимой 1968/69 г., по данным метеостанции «Анархай», снежный покров сохранялся с начала декабря до конца марта и особенно высоким был в конце января, когда высота его достигала 32 см, а минимальная температура —30,7°, то наиболее суровая из последующих трех зим (1970771 г.) характеризовалась неглубоким снежным покровом (2—12 см) и более высокими температурами (минимальная —18,6° во второй половине февраля). Благоприятные условия зимовки способство
9—24
129
вали увеличению численности кеклика, и к осени 1972 г. он вновь стал обычной птицей Чу-Илийских гор.
Результаты учетов 1972—1976 гг. показали, что максимальное поголовье кекликов было в 1974 г., а в последующие 2 года постепенно уменьшалось; и к 1976 г. оказалось самым низким за учетный период.
Уменьшение поголовья кекликов осенью 1975 г. и особенно 1976 г. явилось следствием сильной летней засухи: исчезли родники, которые служили водопоями, снизился урожай дикорастущих семян. Положение усугубилось и тем, что кеклики отстреливались браконьерами. Зачастую добыча их производилась с автомашин, что позволяли многочисленные дороги, пересекающие Анархай в различных направлениях. Возрастающее действие антропогенных факторов дает основание полагать, что Чу-Илийские горы, некогда славившиеся обилием кеклика, постепенно теряют свое значение как охотничьи угодья.
Горы Чулак. Учеты производили в ущельях Кзылаус, Те-ректы и Унгур. Протяженность 29 маршрутов составила 181,1 км (см. табл. 19).
Весенние учеты, проводившиеся на постоянных маршрутах в апреле 1974—1978 гг., показали, что численность кекликов в типичных местах обитания в период размножения изменялась следующим образом: в 1974 г. число птиц в среднем на 1 км маршрута составило 4,9, в 1975 г. — 4,5, в 1976 г. — 5,6, в 1977 г. — 2,2, и в 1978 г. — 5,3 особи.
В сентябре 1972 г. численность кеклика в горах Чулак была относительно высокой. К осени 1973 г. она возросла более чем в 2 раза и оказалась самой высокой за все годит учетов.
Увеличению поголовья способствовали благоприятные условия зимовки 1972/73 г. и летнего периода, что обусловило выживаемость птиц зимой и обеспеченность кормами молодняка летом. Однако в последующие годы поголовье кеклика стало снижаться и осенью 1975 и 1976 гг. оказалось даже ниже, чем к началу проведения работ. По сравнению с самььм «урожайным» 1973 г. численность кеклика за эти годы сократилась более чем в 3 раза. Причины снижения его поголовья заключаются в недостаточном обеспечении кормами, вызванными сильными летними засухами и сложными условиями зимовки 1975/76 г.
130
Следует отметить, что в отличие от других мест учета, являющихся угодьями общего пользования, на территории гор Чулак в конце 50-х годов было организовано приписное охотничье хозяйство «Динамо», которое в 1973 г. передано Казахскому зоокомбинату. В зимы 1973/74 и 1974/75 гг. штатными охотниками этой организации отловлено более 500 кекликов, которые доставлялись в научно-исследовательские институты для проведения различных экспериментов, а также в охотничьи хозяйства с целью пополнения численности местного поголовья. Начиная с 1975 г. эта территория закреплена за Капчагайским охотничье-воспроизводственным хозяйством и в настоящее время кеклик здесь практически не добывается.
Хребет Алтын-Эмель. Учеты производились в ущелье Тюль-кули (1400 м над ур. м.). Здесь пройдено 22 маршрута общей протяженностью 116,3 км (см. табл. 19).
К началу проведения работ мы знали, что осенью 1968 г. кеклик в этих местах был довольно обычен; более поздних сведений о его численности нет. Результаты осенних учетов 1972 г. показали, что за прошедшие 3 года поголовье кекликов резко сократилось и оказалось самым низким из всех обследованных точек юго-востока Казахстана. Так, 5 и 6 октября 1972 г. в ущелье Тюлькули и его окрестностях пройдено 3 маршрута (21 км). Однако ни на одном из них кекликов или следов их пребывания не встречено. Только 5 октября отмечена стайка из 15 особей, прилетевшая на водопой к лагерю. Таким образом, поголовье кеклика осенью 1972 г. после катастрофической гибели в зиму 1968/69 г. в обследуемом районе оставалось на крайне низком уровне.
Низкая численность кеклика, безусловно, вызвана тяжелыми условиями зим 1969—1972 гг., которые характеризовались высоким снежным покровом и устойчивыми низкими температурами. Так, в зиму 1969/70 г. снежный покров лежал с конца декабря до начала апреля. Максимальная его высота (57 см) отмечена в последней декаде января, в феврале она колебалась в пределах 34—43 см, а в марте — 32—38 см. Минимальная температура (—24,7°) отмечена в первой декаде января, а среднедекадная была около —14°. Приблизительно такие же температуры и высота снежного покрова наблюдались в зиму 1970/71 г. Суровые зимы, конечно, ограничивали восстановление численности кеклика. В последние
131
годы поголовье его благодаря благоприятным условиям зимовок несколько возросло и достигло максимума к осени 1976 г.
Следует отметить, что в связи с удаленностью охотничьих угодий хребта Алтын-Эмель от крупных населенных пунктов эксплуатируются они незначительно. В летний период в ущельях производится выпас овец и крупного рогатого скота, что, несомненно, сказывается на распределении кеклика. Однако изменение численности этого вида по годам в основном зависит от условий зимовок.
В горах Малай-Сары общая протяженность 14 маршрутов составила 154 км. Осенью 1972 г. численность кекликов в этом районе была невысокой (см. табл. 19). Спустя год она возросла в 2,5 раза и удерживалась на этом уровне до осени 1974 г., а в 1975 и 1976 гг. поголовье птиц здесь вновь снизилось до уровня 1972 г. Увеличению поголовья кеклика в 1973 и 1974 гг. способствовали благоприятные условия зимовки: зима 1973/74 г. была значительно мягче предыдущей. Постоянный снежный покров образовался лишь в последней декаде декабря, максимума достиг только в первой декаде февраля (14 см), но уже в середине марта полностью исчез. Среднемесячные температуры зимнего периода были несколько выше обычных для данного района и составили в декабре —3,6, январе —12,1, феврале —14°. Такие условия дали возможность кекликам перенести зимовку, и численность их осенью 1974 г. оказалась самой высокой за все годы проведения учетов. Однако в 1975 и 1976 гг. в горах Малай-Сары отмечались летние засухи. Так, в 1975 г. сумма выпавших за июнь осадков составила лишь 4,4, в июле — 7,5, в августе — 27,2 мм, а среднемесячные температуры — соответственно 21,0, 24,6 и 19,9°. Примерно такие же условия наблюдались здесь летом 1976 г. Таким образом, неблагоприятные летние сезоны 1975 и 1976 гг. усложнили положение с кормами. В результате численность кекликов осенью 1975 и 1976 гг. находилась на крайне низком уровне.
Горы Малай-Сары имеют малые относительные высоты и слабую изрезанность склонов, что создает хорошие условия для охоты. В годы с высокой численностью кекликов (1973 и 1974) здесь добывали не менее 1,5—2 тыс. птиц, а в 1975 и 1976 гг. отстрел практически не проводился: было мало дичи.
13'2
Ежегодные осенние учеты в хребтах Каратау, Киргизском и Джунгарском Алатау, в горах Малай-Сары, Алтын-Эмель и Анархай позволили проследить динамику численности кекликов в отдельных ущельях. Удалось установить, что после массовой гибели в зиму 1968/69 г. восстановление поголовья во всех обследованных районах шло крайне неравномерно (табл. 21). Это объясняется такими причинами, как верти-
Та блица 21. Численность кеклика на юго-востоке Казахстана
Район учетов 1972 г. 1973 г.
Длина маршрута, км Учтено птиц Длина маршрута, км Учтено птиц
всего на 1 км маршрута всего на 1 км марш-рута
Каратау 68,5 305 4,5 79,5 221 2,8
Киргизский Алатау 55,5 844 15,2 70,5 1862 26,4
Чу-Илийскйе горы 44,5 598 13,4 48,0 768 16,0
Горы ЧуЛай 21,0 312 14>9 30,6 1220 39,9
Горы Малай-Сары 36,0 274 7,6 42,0 779 18,6
Горы Алтын-Эмель 21,0 0 0 22,8 227 10,0
Всего 246,5 2333 9,5 293,4 5077 17,3
1974 г. 1975 г. 1976 г.
Длина маршрута, км Учтено птиц Длина маршрута, км Учтено птиц Длийа маршрута, км Учтено птиц
всего на 1 км маршрута всего на 1 км маршрута всего на 1 км маршрута
101 580 5,7 88 595 6,8 65,5 641 9,8
73,5 1541 21,0 77 1416 18,4 77 1377 17,9
32,5 599 18,4 27 354 13,1 39 249 6,4
41,5 1269 30,6 44 568 12,9 44 568 12,9
16,0 331 20,7 30 211 7,0 30 225 7,5
22,5 225 10,0 25 178 7,1 25 393 15,7
287 4545 15,8 . 291 3322 П,4 280,5 3453 12,3
кальиое распространение, условия обитаний, антропогенные воздействия и погодные влияния.
Самая низкая численность кеклика отмечена на хребте Каратау, где на протяжении 5 лет (1972—1976 гг.) она колеба
133
лась в пределах от 2,7 особи на 1 км маршрута в 1973 г.г до 9,8 — в 1976 г. Необходимо отметить, что ущелья и склоны этого хребта лишь в небольшой степени испытывают влияние антропогенных факторов. Охота на кеклика здесь практически не проводится, меньше выпасается скота, чем в других районах исследований.
Наиболее высокая численность кеклика отмечена в Киргизском Алатау, где несмотря на значительное влияние антропогенных факторов (массовый выпас скота в апреле — сентябре и существенный пресс охоты) поголовье птиц хотя и претерпевало колебания, но на протяжении 5 лет удерживалось на высоком уровне (максимальная численность 26,4 особи на 1 км маршрута в 1973 г., минимальная—15,3 в 1972 г.). После достижения пика численности в 1973 г. в последующие 3 года отмечалось постепенное снижение поголовья, что объясняется летними засухами, вызвавшими низкий урожай кормов.
В Чу-Илийских горах численность кекликов по годам характеризовалась подъемами в 1972—1974 гг., после чего осенью 1975 и 1976 гг. произошел некоторый спад. Максимальное количество птиц отмечено в 1974 г. (18,4 особи на 1 км маршрута), минимальное — в 1976 г. (6,4). Заметное уменьшение поголовья кеклика осенью 1975 г. и особенно 1976 г. явилось следствием значительной летней засухи, когда большинство родников и речек пересохло и повсеместно выгорела растительность. Положение усугубилось и тем, что в этой изолированной группе велась интенсивная охота на кеклика, а в зимнее и весеннее время выпасалось большое поголовье овец и лошадей.
В горах Чулак численность кеклика в 1972 г. составила 14,9 особи на 1 км маршрута, а осенью 1973 г. была максимальной (39,9). В 1974—1976 гг. шло постепенное снижение поголовья, которое в 1975—1976 гг. оказалось с^иым низким за все годы учетов в этом районе (12,9 особи на 1 км маршрута). Падение численности кеклика в последние 2 года объясняется летней засухой, которая привела к сплошному выгоранию растительности и пересыханию водных источников, что резко снизило численность насекомых, в частности прямокрылых, и существенно отразилось на выживаемости птенцов.
Горы Малай-Сары — богатые охотничьи угодья. Однако?
134
численность кеклика здесь подвержена значительным колебаниям, которые во многом зависят от погодных условий, высокого пресса охоты (в годы «урожая» кеклика) и использования территории как зимне-весеннего пастбища.
На хребте Алтын-Эмель численность кеклика, по данным осенних учетов в 1972—1976 гг., находилась на низком уровне. Только в 1976 г. здесь отмечен некоторый прирост поголовья (максимальный за 5 лет). Низкая численность кеклика в этом районе объясняется неблагоприятными условиями зимовки. Следует отметить, что охотничьи угодья хребта испытывают меньшее, чем в других районах, антропогенное влияние.
Динамика численности кеклика — наиболее трудный вопрос экологии, и успешно решен он может только при проведении учетных работ в течение длительного времени. Судя по литературным данным, депрессия численности этой птицы иногда происходит через 10—15 лет.
Проведенные осенью 1972—1976 гг. на юго-востоке Казахстана учеты показали, что поголовье кеклика подвержено значительным колебаниям из-за погодных условий, в результате чего происходят изменения как в структурах, так и в количественных соотношениях популяций.
Наши работы были начаты спустя 3 года после катастрофической гибели кеклика в зиму 1968/69 г. В это время в ряде районов он стал обычным, а в Киргизском Алатау даже многочисленным. Однако мы не располагаем данными о том, как шло восстановление его численности в 1969— 1971 гг., неизвестна степень воздействия врагов и болезней на различные популяции птиц. Эти и некоторые другие вопросы экологии можно решить лишь путем многолетних стационарных исследований в заповедниках, что поможет научно обосновать колебания численности кеклика .и прогнозировать их.
Охрана и рациональное использование кеклика
,, Охота на кеклика в горных районах Юго-Восточного Казахстана издавна была популярной. В 30—40-х годах в Казахстане заготавливали эту ценную дичь, которая поступала не только на внутренний, но и на внешний рынок (Руданов-ский, Насимович, 1933; Шнитников, 1949).
По М. М. Кондратенко и Ю. А. Смирнову (1973), среди представителей отряда куриных фауны Казахстана кеклик в. 1962—1963 гг. занимал по добыче второе место после серой куропатки, а в 1965 г. — первое. В охотничий сезон 1962 г. бъ1ло добыто 20650 особей (или 12,4% от общей добычи куриных)^ 1963 г.— 16090 (17,4%) ив 1965 г. —53490 (36,1%). Следует отметить, что это лишь приблизительные цифры, поскольку в анкетном опросе в силу ряда причин приняло участие только около 30% членов общества охотников. Кроме того, много дичи отстреляно сверх нормы. Не вызывает сомнения, что в последние годы, несмотря на отсутствие данных о количестве добычи, удельный вес кеклика в общем объеме добываемой дичи неуклонно возрастает.
В настоящее время плановых заготовок кеклика не проводят и многие безружейные способы добычи утратили свое значение, уступив место ружейной спортивной охоте.
Охотники обычно обнаруживают кекликов по далеко слышным голосам. Двигаясь в направлении крика, они зачастую подходят к стайке на близкое расстояние и производят выстрел. Даже разлетевшись, птицы в скором времени начинают перекликаться, что дает возможность стрелку, ориентируясь по голосам, вновь отыскать их.
£36
Как показал опрос охотников, регулярно проводимый .в 1974—1978 гг., в сезоны охоты (ноябрь) ими было добытр 458 кекликов, а 169 (27%) не попали в руки охотников ( подранки, потеря птиц). Таким образом, при ружейной охоте до трети добываемой дичи пропадает.
Безружейные способы охоты менее губительны для дичи. Они описаны в литературе (Бутурлин, 1932; Попов, 1956). однако многие из них в настоящее время не применяются? Исключение составляют тайники, с помощью которых кекликов отлавливают для расселения. Тайник очень эффективен и в местах большой концентрации птиц: один охотник может отловить 30 и более птиц в день.
Еще с осени, в сентябре — октябре, до выпадения снегаг ловец оборудует подкормочные площадки на дне ущелий, с которых удаляются камни, трава, корневища растений. В 7— 15 м от них сооружается скрадок из подручного материала (камни, ветки кустарников, солома и т. д.). На площадки выкладывается подкормка, в основном различные зерноотходы, устанавливаются тайники.
Перед рассветом охотник занимает скрадок и ждет появления кекликов. С восходом солнца стайки птиц следуют к местам подкормки и скоро появляются на площадках. Выбрав подходящий момент, ловец дергает шнур, и тайник накрывает кекликов. Птицы, не попавшие под тайник и находящиеся неподалеку от него, разлетаются на склоны и обычно в этот день здесь не появляются. На следующий день ловец занимает скрадок у другой подкормочной площадки, потом—* у третьей и т. д., давая возможность кекликам оправиться от испуга и вновь начать посещать места первых отловов.
Особенно активно кеклики посещают подкормочные площадки в глубокоснежные зимы, когда им трудно добывать корм. В этот период голод делает их менее осторожными, и особи одной стаи, не попавшие под тайник первый раз, могут уже через 2—3 ч снова появиться на подкормочной площадке. Этот способ отлова кекликов мы успешно применяли для мечения и он хорошо себя оправдал.
В последнее время начинает возрождаться охота на кеклика с помощью ястреба-тетеревятника. Этот способ находит все большее число поклонников и его следует всемерно развивать, поскольку он исключает возможность потери подран
137
ков и добытых птиц, что неизбежно случается при охоте с ружьем.
Наряду с изучением некоторых вопросов экологии кеклика нами собирался материал по упитанности этой птицы (табл. 22).
Таблица 22. Упитанность кекликов (по данным осенних отстрелов)
Месяц Добыто птиц Упитанность
низкая средняя выше средней высокая
абс. % абс. | % абс. % абс. %
Сентябрь ПО, 31 28,2 49 44,6 26 23,6 4 3,6
Октябрь 300 128 42,7 119 39,7 46 15,3 7 2,3
Ноябрь 294 30 10,2 40 13,6 132 14,9 92 31,3
Декабрь 32 11 13,4 13 15,9 40 18,8 18 21,9
Всего 786 200 25,4 221 28,1 244 31,1 121 15,4
Следует учесть, что высолю упитанность в сентябре и октябре имеют лишь взрослые самцы, раньше заканчивающие линьку. Однако число их невелико (11 экз., или 2,7%). Основную массу в этот период составляют молодые птицы, имеющие низкую и среднюю упитанность (327 экз., или 79,8%). В ноябре и декабре эти показатели существенно меняются: у 75% особей отмечена высокая и выше средней упитанность (см. табл. 22). С установлением постоянного снежного покрова кеклики начинают испытывать затруднения с кормами и упитанность у них резко падает. Особи, добытые в'конце декабря и январе, за небольшим исключением имели низкую и среднюю упитанность.
Исследования показали, что наиболее ценная в товарном отношении дичь отстреливается во второй половине ноября и первой половине декабря, когда средний весовой показатель взрослых самцов (24 экз.) составлял 613 г, при среднем выходе товарной продукции* 73,7%, взрослых самок (17) — 504 г. (71,5%), молодых самцов (56)—553 г. (71,1%) и молодых самок (44) —475 г. (71,6%). В этот же период средняя
* Масса ощипанной и выпотрошенной птицы без головы, лапок и внутренних органов.
138
масса пера одной птицы, включая маховые и рулевые, составляет 36,8 г (10 экз.), печени (11) —8,9 г и желудка (7) — 8,2 г.
Учитывая, что кеклик является основным видом пернатой дичи, добываемой в позднеосенний период на юго-востоке Казахстана, следует чрезвычайно бережно относиться к его поголовью. Биологические особенности кеклика (окончание линьки у 75% особей в середине ноября и наиболее высокая упитанность во второй половине ноября — первой половине декабря) дают основание считать, что оптимальными сроками охоты являются указанные месяцы. Такие же сроки охоты на кекликов в Казахстане предлагает Э. М. Ауэзов (1966). Действующие в настоящее время «Правила охоты в Казахской ССР» предусматривают охоту с 15 октября по 1 декабря и их необходимо пересмотреть, что позволит добывать более ценную в товарном отношении дичь.
Своеобразная гнездовая биология кекликов, не известная для других представителей отряда куриных нашей фауны, способствует поддержанию высокой численности их в благоприятные годы и, что особенно важно, увеличению поголовья после массовой гибели. В малоснежные зимы с небольшими морозами необходимо наладить охрану мест концентрации кеклика и его подкормку, что поможет птицам перенести неблагоприятный период. После суровых многоснежных зим следует полностью запрещать охоту не менее чем на 2 года. Такие мероприятия позволят сократить гибель кеклика и быстрее восстановить численность этой ценной промысловой птицы.
* * *
Таким образом, ареал кеклика охватывает огромную территорию и в несколько раз превышает ареалы других, вместе взятых, представителей рода Alectoris. Широкое вертикальное распространение указывает на исключительную экологическую пластичность вида, который живет как в пустынной местности, так и в альпийском поясе гор. Благодаря этой особенности удалось успешно акклиматизировать кеклика за пределами естественной области распространения (США, Новая Зеландия и др.).
В результате изучения биологии кеклика удалось уста
139
новить, что наряду с известной для этого вида моногамией у части самцов (4,6%) выявлена полигамия и установлено явление двойного гнездования: некоторые пары ежегодно устраивают по 2 гнезда. Кладку в первом из них насиживает самец, во втором— самка. В насиживании кладок вовлекаются все самки и треть самцов (не старше одного года). Случаев смены партнеров на гнездах не зарегистрировано. Вполне вероятно, что двойное гнездование распространяется на весь род каменных куропаток, поскольку оно известно еще •для двух представителей рода—A. rufa и A. graeca.
Полные кладки*кекликов содержат 9—21 яйцо (по литературным данным, 7—24), которые откладываются в среднем через 32 ч. Насиживание обычно начинается в день откладки последнего’ яйца, реже в предыдущий или на следующий день; как исключение некоторые особи приступают к насиживанию за 2 дня до окончания кладки.
Естественный отход яиц в гнездах кеклика (неоплодотво-ренные и с погибшими эмбрионами) составляет 2,3—12%, а средняя эффективность гнездования (процент птенцов, покинувших гнезда, от общего числа отложенных яиц) составляет 53,1%.
Период эмбрионального развития длится 23—25 дней. Данные вскрытия 36 однодневных и 925 особей, добытых осенью в разные годы, позволили выявить небольшое, но устойчивое преобладание самцов (1 :0,89).
В линьке молодняка четкого разграничения возрастных нарядов нет: в месячном возрасте перьевой покров птенца состоит из пухового, юношеского и взрослого нарядов. У взрослых особей сроки начала линьки зависят от участия в насиживании кладок. Потерявшие кладки особи и не участвующие в насиживаниии самцы объединяются в стайки и начинают линять на месяц раньше птиц, связанных с заботой о потомстве. Продолжительность линьки у взрослых особей составляет 120—135 дней.
В кормах кеклика из различных регионов СССР зарегистрировано 311 видов высших растений 53 семейств, а также мхи и водоросли; из животных кормов 132 вида насекомых (10 отрядов), 10 видов моллюсков и паукообразные. К основным кормам, учитывая встречаемость их в пище, относятся растения 9 семейств (злаки, розоцветные, лилейные, гречишные, бобовые, амариллисовые, бурачниковые, сложноцвет
140
ные, зонтичные) и представители 3 отрядов насекомых (прямокрылые, жесткокрылые и полужесткокрылые).
Важнейшими факторами, определяющими численность кеклика, следует считать погодные условия, в зависимости от которых происходят изменения в структурах и количественных соотношениях популяций. Особое воздействие на численность кеклика оказывают катастрофические условия зимовки. Например, зима 1968/69 г. вызвала массовый падеж кекликов на огромной территории — охватила Казахстан и республики Средней Азии. Восстановление численности птиц во многом зависит от оставшегося поголовья; в некоторых районах уже через 2—3 года она достигает высокого уровня, при: котором становится возможным открытие охоты.
Наиболее рациональными сроками проведения охоты на кеклика следует считать ноябрь и первую половину декабря. В этот период добывается самая ценная в товарном отношении дичь, которая имеет высокую упитанность: средняя масса одной особи составляет 540 г, а тушки без крыльев, внутренних органов, головы и лапок— 390 г, или 72% от общей массы птицы.
Закон СССР «Об охране и использовании животного мира», принятый в 1979 г., подчеркивает необходимость «рачительного, хозяйского и гуманного отношения к животному миру как к одному из важнейших компонентов природной среды нашей Родины». Следует чрезвычайно бережно относиться к поголовью кеклика. После суровых многоснежный зим, когда происходит падеж птиц, нужно полностью закрывать охоту на него не менее чем на 2 года.
Литература
Абдусалямов И. А. Птицы горного Зеравшана. Душанбе, 1964. 248 с.
Абдусалямов И. А. Фауна Таджикской ССР. Душанбе, 1971. Т. 19. 392 с.
(Айрумян К. А. Факторы, лимитирующие величину кладки у куриных и сохранность их птенцов. — Изв. АН АрмССР. Биол. науки, 1965, -№ 11, с. 82—87.
Ауэзов Э. М. Нужно ли запрещать охоту на кекликов? — Охота и охотн. х-во, 1966, № 8, с. 5.
Базиев Ж. X. Линька кавказского улара. — Зоол. журн., 1966, т. 45, № 1, с. 99—108.
Бёме Р. Л. Птицы гор Южной Палеарктики. М., 1975, с. 1—180.
Бианки В. Л. Материалы для авифауны Монголии и Восточного Тибета: (Труды экспедиции РГО, совершенной в 1899—1901 гг. под руков. П. К. Козлова). Спб., 1907. Т. 5.
Бутурлин С. А. Настольная книга охотника. М., 1932. 272 с.
Бутурлин С. А., Дементьев Г. П. Полный определитель птиц СССР: Гагаровые, веслоногие, цапли, пластинчатоклювые, куриные, пастушковые, триперстки. М.; Л., 1935. Т. 2. 279 с.
Вырыпаев В. А., Буд рис Р. Р. Поведение самцов кеклика в гнездовой период. — В кн.: Групповое поведение животных: Докл. II Все-союзн. конф, по поведению животных. М., 1976, с. 59—60.
Газаньян М. К. К биологии каменной куропатки в Азербайджане,— Труды Азерб. ун-та. Серия биол., 1954, № 6, с. 119—139.
Гвоздев Е. В. Паразитические черви куриных птиц Казахстана. Алма-Ата, 1958. 265 с.
Грачев В. А. О линьке семиреченского фазана (Phasianus colchi-cus mongolicus Brandt). — Труды Ин-та зоол. АН КазССР, 1960, т. 13, с. 176—179.
Грачев Ю. Н. О насиживании кладок самцами у кекликов.— В кн.: Биология птиц в Казахстане. Алма-Ата, 1978, с. 149—150.
Грачев Ю. Н. Половые и возрастные отличия у кекликов. — В кн.: Биология птиц в Казахстане. Алма-Ата, 1978, с. 151—152.
142
Грачев Ю. Н. Кеклик. — Охота и охотн. х-во, 1979, № 7, с. 20— 21.
Грачев Ю. Н. Насиживание и продолжительность инкубации яиц у джунгарского кеклика. — В кн.: Экология гнездования птиц и методы ее •изучения: Тезисы Всесоюзн. конф, молодых ученых. Самарканд, 1979, с. 54—55.
Даль С. К. Данные о. вариациях окраски каменной куропатки (Alectoris graeca Meisner) в Южном Закавказье. Ереван, 1947.
Даль С. К. Очерк позвоночных животных Айодзорского хребта. — Зоол. сборник, 1949, вып. 6, с. 21—37.
Дементьев Г. П. Новая форма каменной куропатки. — Изв. Туркм. фил. АН СССР. Ашхабад, № 3—4, 1946.
Дементьев Г. П. Птицы Туркменистана. Ашхабад, 1952, 438 с.
Денисова М. Н. Особенности роста и развития птиц в связи с условиями гнездования. — Уч. зап. Моск. обл. пед. ин-та, 1958, № 65, с. 165—179.
Долгушин И. А. О фауне птиц полуострова Мангышлак. — Изв. АН КазССР. Сер. биол., 1948, вып. 8, с. 131—160.
Долгушин И. А. К фауне птиц Каратау. — Изв. АН КазССР. Серия зоол., 1951, вып. 10, с. 72—117.
” Драгоев П. Възможности за по-широко разпространение на кеклика (Alectoris graeca L.). — Гороко стопанство, 1971, т. 27, № 5, с. 41— 45.
Зарудный Н. А. Орнитологическая фауна Оренбургского края.,— Зап. Акад, наук, 1888, т. 57, № 1, с. 17—55.
Зарудный Н. А. Орнитологическая фауна Закаспийского края ‘{Северной Персии, Закаспийской области и Хивинского оазиса и равнинной Бухары). — В кн.: Мат-лы к познанию фауны и флоры Рос. Империи. Отд. зоол. Спб., 1896. Вып. 2. 555 с.
Зарудный Н. А. Экскурсия по Северо-Восточной Персии и птицы этой страны. — В кн.: Импер. акад, наук и физико-математич. отд. Спб., 1900. Т. 10, № 1, с. 90—93.
Зарудный Н. А. К вопросу о формах каменной куропатки (Сас-cabis kakelik Falk.)—Орнитол. вести., 1914, № 1, с. 52—60.
Зверев М. Д. Заметки по биологии некоторых диких животных в Алма-Атинском зоопарке: К вопросу о размножении кекликов. — Труды Алма-Атинск. зоопарка, 1948, вып. 1, с. 94—106.
Иванов А. И. Птицы Таджикистана. М.; Л., 1940, с. 74—77.
Иванов А. И. Птицы СССР. М.; Л., 1951. Ч. I, с. 229—230.
Иванов А. И. Птицы Памиро-Алая. Л., 1969, с. 91—94.
Ильичев В. Д., Тихонов А. В. Биологические основы управления поведением птиц: I. Куриные (Galliformes). — Зоол. журн., 1979, т. 58, вып. 7, с. 1021—1032.
Ишадов Н. К распространению кеклика в Западных Каракумах. — В кн.: Животный мир Туркмении. Ашхабад, 1970, с. 182—183.
Карташов Н. Н. Птицы Советского Союза. М., 1952. Т. 4. с. 154— 173.
Кашкаров Д. Н. Животные Туркестана. Ташкент, 1932. 450 с.
Кашкаров Д. Ю., Павленко Т. А. Методические указания по учету численности охотничье-промысловых животных в условиях
143
Узбекистана. — Информ, сообщ. Ин-та зоол. и паразитол. АН УзССР^ 1975, №126, с. 7—9.
К и щи н ск ий А. А.,, Флинт В. Е. Случай сдвоенного гнездования у кулика-воробья. — В кн.: Фауна и экология куликов. М., 1973. Вып. 1, с. 56.
Ковалев А. К., Попов А. х В. Миграции кеклика в горах Центрального Таджикистана. — В кн.: Миграции птиц в Азии. Душанбе, 1980. с. 261—272.
Ков шарь А. Ф. Птицы Таласского Алатау. — Труды гос. запов. Аксу-Джабаглы, 1966, вып. 3. с. 1—425.
Коган! III. И. Материалы по питанию кеклика в Колет-Даге. — Изв. Турк. ФАН, 1950, № 4. с. 48—55.
Колесников И. И. Фауна наземных позвоночных Кюрен-Дага. — Труды Среднеаз. гос. универ., 1956, вып. 86. с. 168—169.
Кондратенко М. М., Смирнов 10. А; Опыт учета дичи, добываемой охотниками в Казахстане. Растительный и животный мир заповедников Казахстана. — Труды заповедников Казахстана, 1973, вып. Зг с. 190—200.
Костин Ю. В., Кормилицина В. В. Материалы по аккли? матизации кеклика и фазана в Крыму. — В кн.: Сб. научно-технической информации. Киров. Вып. 44—45, 1974, с. 49—51.
Коханов В. Д. Материалы по экологии белохвостого песочника-в Кандалакшском заливе Белого моря. — В кн.: Фауна и экология куликов. М, 1973. Вып. 1, с. 66—71.
Кузьмина М. А. Экология кеклика (Alectoris kakelik Falk) в Ка? захстане и его роль как хозяина иксодовых клещей. — Изв. АН КазССР. — Сер. зоол., 1948, вып, 5, с. 67—73.
Кузьмина М. А. Джунгарский кеклик, его экология и хозяйственное значение. — Труды Ин-та зоол. АН КазССР, 1955, т. 4, с. 97—165;
Кузьмина М. А. Материалы по размножению, питанию и линьке темнобрюхого улара. — Труды Ин-та зоол. АН КазССР, 1960, т. 13,. с. 133—141.
Кузьмина М. А. Отряд куриные. — В кн.: Птицы Казахстана. Алма-Ата, 1962. Т. 2, с. 389—487.
Кузьмина М. А. Тетеревиные и фазановые СССР. Алма-Ата, 1977. 264 с.
Кутубидзе М. Е. О размножении кавказского кеклика (Alecto* ris graeca caucasica Suschkin). — Сообщ. АН ГрузССР, 1957, № 5^ с. 18—20.
Лаптев М. К. Учет наземной фауны позвоночных методом маршрутного подсчета. — Труды Среднеазиатск. гос. ун-та. Серия VIII. зоология, 1930. Вып. 2. (Ташкент), с. 1—15.
Лебедев Ю. А. К сезонной динамике численности кеклика. — Изв». АН ТаджССР. Отд. биол. наук, 1976, № 2, с. 57—59.
Л э к Д. С. Численность животных и ее регуляция в природе. М»„ 19о7. 339 с. *
Минин Н. В. Некоторые закономерности в летнем питании кеклика (Alectoris graeca falki Hart.) на северном склоне Туркестанского' хребта. — Вопросы экологии и биоценологии, 1939, вып. 4. с. 112—127.
144
Молчанов Л. А. Охотничьи и промысловые птицы Средней Азии. Москва; Ташкент, 1932, 175 с.
Нам Н. Каменная куропатка в Акчинском хозяйстве. — В кн.: Охрана природы и воспроизводство дичи. М., 1975, с. 123—127.
Новиков Г. А. Полевые исследования по экологии наземных позвоночных. М., 1953. 490 с.
Остапенко М. М. К вопросу о питании каменной куропатки в юго-западных отрогах Чаткальского хребта. — Уз. биол. журн., 1958, № 1, с. 25—31.
Остапенко М. М. Материалы по экологии каменной куропатки в юго-западных отрогах Чаткальского хребта. — Труды Чаткальского горнолесного заповедника, 1965, вып. 2, с. 209—219.
Павлов Н. В. Комплексные степи и пустыни Карсакпая. М., 1931. 164 с.
Попов А. В. Орудия и приемы промыслового лова кеклика. — Изв. отд. общ. наук ТаджССР, 1956, вып. 8, с. 123—127.
Попов А. В. Птицы Гиссаро-Каратегина. — Труды Ин-та зоол. и паразитол. АН ТаджССР. Сталинабад, 1959. 176 с.
Попов А. В. Миграции кеклика в Таджикистане. — Труды пробл. и тематич. совещ. зоол. Ин-та АН СССР, 1969, вып. 9, с. 169—174.,
Пржевальский Н. М. Монголия и страна тангутов. — В кн.: Трехлетнее путешествие в восточной и нагорной Азии. Спб., 1876. 127 с.
Пэк Л. В., Федянина Т. Ф. Пища птиц Киргизии. — В кн.: Птицы Киргизии. Фрунзе, 1961. Т. 3, с. 59—119.
Рудановский В. Я., Насимович А. А. Боровая дичь: Заготовка, обработка, экспорт. М., 1933. 122 с.
Рустамов А. К. К биологии кеклика (Alectoris graeca Meisner) в Туркмении. — Изв. Туркм. филиала АН СССР, 1945, № 1, с. 45—47.
Рустамов А. К. Новая форма каменной куропатки с Устюрта. — Охрана природы, 1948, № 5.
С а г ы н б е ко в Д. Время переворачивания и перемещения насиживаемых яиц. — Труды Кирг. ун-та. Сер. биол. наук, 1976, № 15/2, с. 17—18. р Салихбаев X. С., Кашкаров Д. Ю., Шарипов А. Птицы, млекопитающие. — В кн.: Экология позвоночных хребта Нуратау. Ташкент, 1970, с. 42—100.
С а т у н и н К. А. Материалы к познанию птиц Кавказского края. — Зап. Кавк. отд. РГО, 1907, кн. 26, вып. 3. 114 с.
Свиридов А. Каменная куропатка. — Природа, 1876, кн. 2.
Селевин В. А. Новые данные по распространению птиц в Западном Алтае и его предгорьях.Бюл. Среднеаз. ун-та, 1935, вып. 21, № 13.
Семенов А. Несколько соображений о прошлом фауны и флоры Крыма по поводу нахождения там горной куропатки (Caccabis chukar G. R. Gray). — Зап. Импер. Акад, наук, 1899, т. 8, № 6.
Сержпинский А. Кеклик в сыпучих песках. — Среднеаз. охотник, 1925, № 1, с. 16.
Сопыев О., Караев М. Материалы по куриным птицам Куги-танга.— В кн.: Охрана природы Туркменистана. Ашхабад, 1979, вып. 5, с. 46—52, - ...
Слудский А. А. Роль стихийных бедствий в динамике численнос
10—24 145
ти птиц в Казахстане. — В кн.: Новости орнитологии. Алма-Ата, 1965^ с. 348—351.
Спангенберг Е. П., Судиловская А. М. Материалы по биологии и вертикальному размещению птиц в Киргизском Алатау. — Уч. зап. Моск. обл. пед. ин-та, 1959, т. 71, вып. 4, с. 142—150.
Степанян Л. С. Состав и распределение птиц фауны СССР: Неворобьиные Non-passeriformes. М., 1975, с. 136—138.
Степанян Л. С. Птицы Южного Бадахшана (в пределах бассей-на р. Шах-Дары): Фауна, экология и география животных. — Уч. зап. Моск. пед. ин-та им. В. И. Ленина, 1969, № 362, с. 176—333.
Тугаринов А. Я. Новые находки плиоценовой орнитофауны Одессы.—Докл. АН СССР, 1940, т. 26, № 3, с. 311—313.
Федоров А. А. К биологии горной куропатки Копет-Дага. — Природа, 1949, № 6, с. 64—65.
Флинт В. Е., К и щ и н с к и й А. А. Материалы по биологии? острохвостого песочника: Фауна и экология куликов. М., 1973, с. 100—105.
Хан мамедов А. И. К биологии кеклика (каменной куропатки} в Азербайджане. —Труды Ин-та зоол. АН АзербССР, 1955, т. 18, с. 117— 187.
X а х л о в В. А., С е л е в и н В. А. Список птиц окрестностей Семипалатинска.— Uragus, 1928, кн. 7, № 2, с. 19—34.
Цюрупа В. Д. Веточный корм. М.; Л., 1933. 105 с.
Шевченко В. В. Суровая зима 1944/45 г. и ее влияние на птиц предгорий Таласского Алатау. — Охрана природы, 1949, № 6, с. 36—41.
Шевченко В. В. Птицы Государственного заповедника Аксу-Джа-баглы. — Труды заповед. Аксу-Джабаглы, 1948, вып. 1, с. 36—71.
Шестоперов Е. Л. Материалы для орнитологической фауны Илийского края. — Бюл. Моск, об-ва испыт. природы, 1929, т. 38, вып. 1/2а, с. 154—204.
/ Яблонский Н. И. Охотничьи и промысловые птицы и звери Южного Алтая. — Природа и охота, 1904, № 2, с. 8—9.
Янушевич А. И., Тюрин П. С., Яковлева И. Д., К ы д ы-ралиев А., Семенова Н. И. Птицы Киргизии. Фрунзе, 1959, Т. 1» 222 с.
Falk J. Beitrage zur topographische Kenntnis des Russisches ReichsF 1786, v. 11, p. 55.
D a у a n i A. A population study of chukar (Alectoris chukar) near Karadj Iran. — Linz. biol. Beitr, 1978, v. 9, № 2, p. 279—286.
Gray J. E. Hardwicke. — Illustr. Indian Zool.. 1830, v. 1, p. 54.
Harper H., Harry B., Bailey W. The chukar partridge in California. Calif. — Fish and Game, 1958, v. 44, № 1, p. 5—50.
Lambrecht K. Handbuch der Palaeornithologie. Berlin, 1933, p. 1— 54.
Mackie R., Buechner H. The reproductive cycle of the chukar. — J. Wildlife Manag, 1963, v. 27. № 2, p. 246—260.
Marples B., Gurr L. The chukar in New Zeland. Emu, 1953, part 4, p. 283—291.
Menzdorf A. Zur jugendentwicklung des Steinhuhns Alectoris graeca Meisner 1804. — Sitzungsber, Osterr. Akad. Wiss. Math. — naturwiss KL”. 1975, Abt. 1, V. 183, № 8—10, p. 288—306.
146
Menzdorf A. Zur Moglichkeit der Einbiirgerung verschiedener Feld-liuhnarten in der Bundesrepublik Deutschland. — Ornithol. Mitt, 1976, v. 28*. № 11-12, p. 221—227.
Peters Y. L« Chek-List of birds of the World., v. 2. Cambridge,. 1934, p. 1—403.
Portal M. Breeding Habits of the Red-legged Partridge; British Birds XVII, 1924, p. 315—316.
S n о w D. W. A guide to moult in British birds. British trust for ornithology.— Field Gnide, 1970, N 11, p. 1—30.
V a u r i e Ch. The birds of the Palearctic fauna, Non-Passeriformes. London, 1965, p. 1—763.
Watson G. Sympatry in Palearctic Alectoris partridges. — Evolution*.. 1962a, v. 16, № 1, p. 11—19.
Watson G. Three sibling species of Alectoris partridge. — Ibis*. 1962 B, v. 104, N 3, p. 353—367.
Watson G. The chukar partridge (Aves) of St. Helena Island South Atlantic Ocean. — Proc. Biol. Soc. Wash, 1966, v. 79, p. 179—182.
W i s s e 1 G., S t e f a n i M., R a e t h e 1 H. Fasanen und andere Hiihner-vogel. Ein Handbuch fiir Ziebharber von Ziichter und Handler von Hiihner--vocln und Tauben. Radebenl und Berlin, 1966, p. 1—433.
Оглавление
Предисловие > Сроки и места проведения работ. Материал и методика 3 7
Систематическое положение, ареал . 13
Места обитаний и повадки . 20
Биология кеклика . 38
Питание 93
Динамика численности 114
Охрана и рациональное использование кеклика 136
Литература , • > 142
Юрий Николаевич Грачев
КЕКЛИК
(Биология, использование и охрана)
Утверждено к печати Ученым советом
Института зоологии Академии наук Казахской ССР
Рецензенты: доктор биологических наук А. Ф. Ковшарь, кандидаты биологических наук Б. М. Губин, В. А. Грачев
Зав. редакцией Д. М. Глазырина
Редактор А, Н. Ведерникова
Худ. редактор А. Б. Мальцев
Оформление художника Н. Ф. Чурсин^
Техн, редактор Е. М. Рахметова Корректор Н. Н. Горбатенко
ИБ № 1302
Сдано в набор 6.01.83. Подписано в печать 8.04.83. УГ10069.
Формат 60X84716- Бум. тип. № 1. Литературная гарнитура.
Высокая печать. Усл. п. л. 8,6. Уч.-изд. л. 8,7. Тираж 1800. Заказ 24.
Цена в переплете 1 р. 10 к., в обложке 1 руб.
Издательство «Наука» Казахской ССР.
Типография издательства «Наука» Казахской ССР« Адрес издательства и типографии: 480021, г. Алма-Ата, Шевченко, 28.
Цена 1 руб.